/



































































































































































Текст
экология
НАСЕ КОМЫХ
Н. С. АНДРИАНОВА
ЭКОЛОГИЯ НАСЕКОМЫХ
КУРС ЛЕКЦИЙ
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА
1970 ।
УДК 595.7—15
Печатается по постановлению
Редакционно-издательского совета
Московского университета
2-10—6
179—69
ВВЕДЕНИЕ
Экология насекомых — основа для решения многих теоре-
тических и практических проблем энтомологии. На экологиче-
ской основе развивается сельскохозяйственная, лесная, меди-
цинская энтомология. Без знания экологии невозможна ра-
циональная борьба с вредителями, использование многочис-
ленных полезных видов.
Массовые вспышки размножения насекомых, приносящие
колоссальные убытки народному хозяйству, значение насеко-
мых как переносчиков разнообразных болезней человека, жи-
вотных и растений и многие другие вопросы заставляют зани-
маться экологией насекомых в разных ее аспектах. Огромное
разнообразие видов насекомых, удивительная приспособлен-
ность морфологических структур, поведения и других черт
организации к жизни в различных средах делают возможным
использование отдельных групп и видов насекомых как инди-
каторов среды и незаменимыми объектами для решения мно-
гих биологических проблем.
Огромна роль насекомых в биогенном круговороте веществ
и энергии в жизни биогеоценоза и в целом биосферы. Отдель-
ных публикаций по экологии насекомых очень много. Есть
и обобщающие руководства советских и зарубежных авторов.
Но задачи экологии настолько разнообразны, теории много-
численны и часто противоречивы, а изучение многих вопросов
только начато, что автор «Экологии насекомых» считает по-
лезным дать краткое изложение современного состояния
основных ее разделов, надеясь, что это поможет быстрее
ориентироваться в сложном круге экологических проблем.
Автор не ставит задачи полного изложения разделов эко-
логии, так как многие из них перерастают в самостоятельные
дисциплины.
Например, лабораторно-экспериментальная, или физиоло-
гическая, экология имеет свой круг вопросов, изучая влияние
физико-химических условий среды обитания насекомых на их
жизненные функции (рост, развитие, размножение, поведение
и т. п.). (
3
Несколько иные задачи популяционной экологии, изучаю-
щей численность отдельных видов, вспышки размножения
вредителей, закономерности размещения видов по территории,
изменения популяций во времени.
Экология видов (аутэкология) перерастает в экологию со-
обществ разных видов — биоценологию, а последняя — в био-
геоценологию, поскольку биоценозы могут существовать лишь
в единстве с абиотической средой.
Биогеоценозы почти не затронуты исследованиями, хотя
именно они представляют ту элементарную структурную еди-
ницу биосферы земли, в которой осуществляется круговорот
форм, веществ и энергии. При изучении биогеоценозов жела-
тельно применение комплексного метода исследования, с уча-
стием специалистов разных профилей: почвоведов, ботаников,
зоологов, микробиологов. Роль насекомых в жизни биогеоце-
нозов велика, поэтому участие энтомологов в их изучении обя-
зательно. Все разделы экологии насекомых тесно связаны и
направлены в конечном итоге на выяснение их жизни и значе-
ния в биогеоценозах.
Курс лекций по экологии насекомых, положенный в осно-
ву книги, читался студентам-энтомологам Московского универ-
ситета, уже достаточно ориентированным в вопросах морфо-
логии, физиологии и систематики насекомых, знакомых с ме-
дицинской, сельскохозяйственной и лесной энтомологией,
а также с рядом общебиологических дисциплин. Однако не
все дисциплины, необходимые экологу, отражены учебным
планом. Автор считает полезным для студентов ознакомиться
с учебными пособиями по почвоведению, геоботанике, биогео-
графии, климатологии.
Автор выражает признательность коллективу кафедры
энтомологии МГУ и ее заведующему проф. Е. С. Смирнову за
поддержку работы, проф. Б. В. Добровольскому, ст. науч, со-
труднику О. А. Черновой и доктору биол. наук Г. А. Викторо-
ву за ряд ценных критических замечаний при работе над ру-
кописью.
Автор глубоко благодарен О. Т. Помаленькой за большую
помощь при подготовке рукописи к печати.
Глава I
ПРЕДМЕТ И ЗАДАЧИ ЭКОЛОГИИ
Со времен Э. Геккеля, впервые предложившего термин
«экология» (1869 г.), ее считают наукой, изучающей взаимо-
связи живых организмов со средой их обитания. В определе-
нии Геккеля экология — это наука об отношениях животного
к окружающей его органической и неорганической среде, в ча-
стности— к тем животным и растениям, с которыми оно вхо-
дит в прямой контакт.
Связи живых организмов с естественной средой их обита-
ния чрезвычайно сложны, разнообразны и специфичны для
разных групп организмов. Поэтому современная экология
представлена разными направлениями, разными разделами,
и определение содержания и задач экологии дается различ-
ное. Это говорит о том, что экология — развивающаяся и бы-
стро дифференцирующаяся наука, включающая экологию
растений, экологию животных, экологию микроорганизмов.
Изучение закономерных связей живых существ с условиями
их жизни осложняется разнообразием и самой среды жизни —
жизни в воде, на суше, в воздухе, в почве, внутри других орга-
низмов. Гидробиология, педобиология, аэробиология, парази-
тология— науки экологические по своему содержанию.
Экологические исследования у ботаников, зоологов, гидро-
биологов, почвоведов, биогеографов шли, в значительной мере,
своими независимыми путями. Однако ряд выводов оказался
общим. Главнейший из них — признание закономерного раз-
мещения организмов по территории в зависимости от физико-
химических условий жизни и объединения их в единые целост-
ные системы популяций и биоценозов.
Вместе с тем в трактовке ряда вопросов у экологов нет
единого мнения. Так, например, ботаники предметом экологии
считают лишь взаимоотношения растений с мертвой средой,
отношения же между самими растениями — предметом особой
5
науки — фитоценологии, а отношения животно-раститель-
ных сообществ — биоценологией (Шенников, 1950).
С фитоценологией отождествляют геоботанику, изучающую
строение, экологию, развитие, систематику и географию рас-
тительных сообществ (Шенников., 1964).
В зоологии сущность экологических исследований была
сформулирована задолго до появления Термина «экология»
в известных положениях о единстве организмов с условиями
их жизни К. Ф. Рулье (1850 г.) и И. М. Сеченова (1861 г.).
По Рулье, ни одно органическое существо не живет само по
себе, каждое вызывается к жизни и живет только постольку,
поскольку находится во взаимодействии с относительно внеш-
ним для него миром. По Сеченову, существование организма
без окружающей его среды невозможно. Много сделал для
развития экологии в нашей стране Д. Н. Кашкаров (1945). Он
считал необходимым изучать «проекцию поведения вида на
условия его местообитания», так как каждый организм «жи-
вет в определенной среде, из которой вырвать его безнаказан-
но невозможно и вне которой рассматривать его ненаучно».
В отличие от ботаников, по Кашкарову, экология есть наука
об отношениях организма к среде не только физической, но и
биотической. Мы не можем рассматривать организм или вид
изолированно, только в его физическом окружении, ибо его
окружает и биотическая среда. Мы должны рассматривать
всякий вид как член «биоценоза». При этом можно заняться
изучением отдельного вида (аутэкологией) или отношениями
к условиям местообитания всего биоценоза (синэкология).
Современная экология включает экологию особей, эколо-
гию популяций и экологию сообществ. При этом, поН.П. Нау-
мову (1963), «экология имеет дело лишь с той стороной взаи-
модействия организмов со средой, которая обусловливает
развитие, размножение и выживание особей, структуру и ди-
намику образуемых ими популяций видов, структуру и дина-
мику сообщества разных видов и исторически сложившиеся на
их основе: 1) специфические приспособления видов; 2) вну-
тривидовые отношения и специфическую структуру вида;
3) сообщества популяций разных видов, различные на разных
участках земной поверхности, их взаимные приспособления,
обеспечивающие биогенный круговорот веществ». Основная
проблема экологии — проблема численности видов.
Энтомологи, определяя содержание и задачи экологии, под-
черкивают обязательность изучения особенностей жизни как
отдельных видов, так и биоценозов в естественной обстановке,
изучение насекомых «как части природы». По Н. А. Холод-
ковскому, к области экологии относится изучение видовой
жизни насекомых, охватывающей те явления, в которых насе-
комое выступает не как отдельная особь, а как вид, вступая
6 .
в те или иные взаимоотношения с другими видами насекомых
или других организмов, а также с физической средой. При
рассмотрении индивидуальной жизни мы имеем в виду насе-
комое как нечто данное отдельно от окружающих условий.
В главе о видовой жизни нам придется обращать внимание на
те отношения, в которых насекомое находится к остальной
природе, и на те результаты, которые следуют для насекомого
из этих отношений.
Б. П. Уваров (1927) сущностью экологической точки зре-
ния считал изучение насекомых как производное и в то же
время как составную часть единого чрезвычайно сложного
организма природы. Предмет экологии, по Уварову, — «изу-
чение взаимоотношений животных с естественными условиями
обитаемых ими стаций».
На необходимость изучения насекомых в связи с естест-
венноисторическими условиями обитания (типами леса) обра-
щали внимание специалисты по лесной энтомологии (Старк,
1931). Как часть природы насекомых рассматривает Вебер
(Weber, 1933), считая основным вопросом экологии — вопрос
о сохранении вида, изучение противоречивых факторов, влия-
ющих на жизнь вида.
По В. Тишлеру (1960), экология — «наука об организмах
в природе как целом». Экологические исследования, по его
мнению, можно вести в трех направлениях: 1) изучать отдель-
ные виды на любом числе особей с их специфическими требо-
ваниями и возможностями, их зависимостью от условий осве-
щения, температуры, влажности, структуры почвы, питания
и т. д.; 2) изучать популяцию, изменение ее численности в про-
странстве за ряд поколений; 3) изучать систему связей живых
организмов с окружающей неживой природой (экосистема,
или биоценоз). Изучению особи вида, популяции и экосисте-
мы, по Тишлеру, соответствуют три узловых раздела эколо-
гии: аутэкология, или физиологическая экология, экология
популяций и синэкология, не всегда четко разграничиваемые.
В основе же — это только три разных аспекта изучения взаи-
мосвязей организмов в природе. С 1930 г. исследования по
защите растений начали перерастать в экологию сельскохо-
зяйственных территорий, или культурных ландшафтов.
Советские экологи считают, что основным содержанием
экологии является: 1) изучение видовых приспособлений как
основы для понимания многообразия взаимосвязей организ-
мов и среды; 2) изучение закономерностей образования и раз*
вития популяций как формы существования вида, их диффе-
ренцировки и динамики, их численности; 3) изучение законо-
мерностей формирования и развития биоценозов как выраже-
ния взаимоотношений организмов в конкретных условиях мес-
тообитания.
Таким образом, центральным объектом экологии признает-
7
ся жизнь вида в природной обстановке и жизнь биоценоза
в единстве с абиотической средой (биогеоценоз, экология
культурных ландшафтов).
МЕСТО ЭКОЛОГИИ СРЕДИ ДРУГИХ НАУК
Изучение связи организмов со средой составляет теоре-
тическую основу современной биологии. Поэтому данные эко-
логии стали необходимы для морфологов, физиологов, систе-
матиков и специалистов многих других биологических дисцип-
лин. Возникли «экологическая морфология», «экологическая
физиология» и т. д., что свидетельствует о тесной связи эко-
логии с рядом смежных наук. По Н. П. Наумову (1965), при
современном уровне знаний можно вести исследования ми-
кро-, мезо- и макросистем, изучать жизнь на разных уровнях
(молекулярном, клеточном, тканевом, органном, организмен-
ном, видовом и т. д.). Макросистемы (популяции, сообщества,
биосфера) изучает популяционная биология, включаяющая
систематику, биогеографию, экологию, биогеохимию и др. Со-
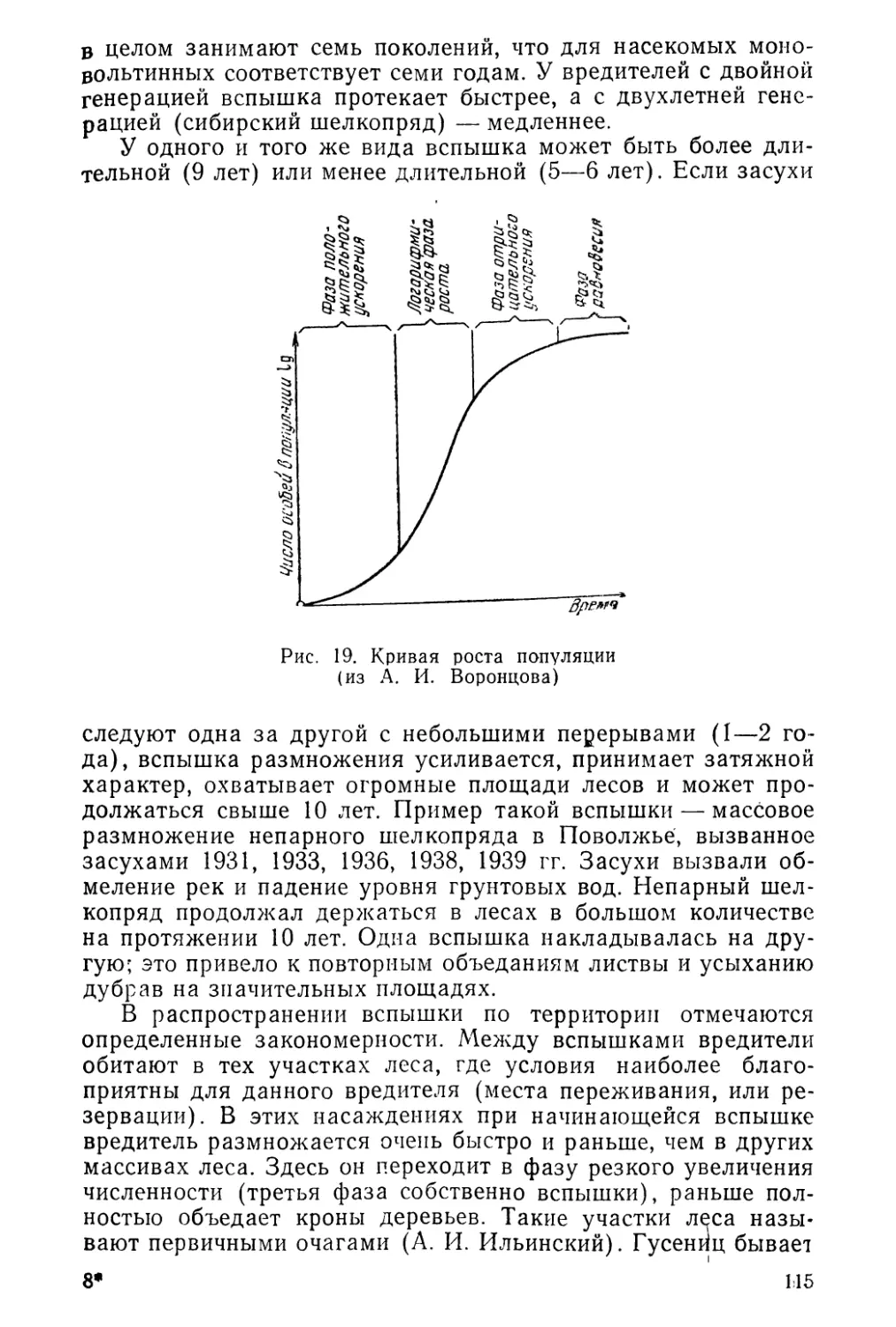
отношение этих наук иллюстрирует рис. 1.
Виды существуют в форме популяций, популяции в осо-
бях, и, следовательно, экология особей представляет собой
часть экологии популяции, а биоценоз невозможно понять без
изучения популяций отдельных видов. Все «три раздела эко-
логии» тесно связаны и представляют лишь последовательные
этапы исследования жизни биоценоза. Рассматривать физио-
логию или экологию и т. д. отдельной особи без учета состоя-
ния популяции нельзя, так как особи существуют в конкрет-
ной популяции, а не «сами по себе»; нет и абстрактных попу-
ляций, вырванных из конкретного биоценоза.
Для уточнения места экологии среди других наук нам ка-
жется полезно вспомнить классификацию, предложенную
В. Н. Беклемишевым (1949). Все биологические науки он де-
лит на два больших отдела. К одному из них Беклемишев,
относит науки, занимающиеся изучением отдельных предста-
вителей животных и растений — протозоология, гельминтоло-
гия, ихтиология, орнитология, энтомология и т. д. На наш
взгляд, возможны и еще более дробные подразделения: коле-
оптерология, лепидоптерология и т. п. Это предметные (таксо-
номические) науки. Среди них появились и новые, например,
вирусология, акарология. Вместе с тем каждый объект пред-
метных наук может изучаться в его различных жизненных
проявлениях. Этим занимается группа наук, которые Бекле-
мишев обозначил как частные науки (в настоящее время они
известны как науки фундаментальные). К ним он относил
морфологию, физиологию, экологию, систематику, биогеогра-
фию, биохропологию. К этому отделу, по-видимому, следует
отнести биофизику, биохимию, молекулярную биологию, гене-
8
тику. Как и предметные науки, частные науки могут быть бо-
лее дробными. Морфология, например, может подразделяться
на анатомию, гистологию, цитологию и др.
Схему деления биологических наук, сходную по принципу
с классификацией Беклемишева, приводит Е. П. Одум (Odum,
1957) (рис. 2).
Рис. 1. Положение экологии и ее Рис. 2. Схема деления наук (из Е. Odum
отношение к другим наукам (по с изменениями)
Н. П. Наумову)
Ьтл согра/рил
а а сеонтцлог ия
биохимия
duoipujuHQ
’ л ^аменталоные
чаукц
(частные)
Lu. '! с машина —
Морфологи» _
оология -
Экология стоит в одном ряду с морфологией, физиологией
и другими науками, имеющими дело со всеми «таксономиче-
скими подразделениями», отражающими специфику организма
(зоология, ботаника, бактериология и более узкие — миколо-
гия, энтомология и т. д.). Из основных разделов биологии
экология теснее всего связана с физиологией и генетикой. Но
экологи рассматривают индивидуум как мельчайшую единицу
исследования, а для физиологов это — наикрупнейшая едини-
ца, обычно же они заняты изучением механизма работы серд-
ца, эндокринной системы и других органов, тканей, клеток.
Генетика интересует механизм наследственности, а эколога —
поведение, распространение, выживание и т. д., и факторы,
воздействующие на экологическую изменчивость.
Энтомология опирается, как и другие предметные науки,
па достижения частных наук. Например, чтобы понять вред,
причиняемый комаром, и предотвратить его, нужно уметь
отличить комара от других насекомых, малярийного от иема-
лярийных. Этому учит нас систематика. Далее мы должны
знать образ жизни взрослого комара и его личинок, места,
где они встречаются, пищу их, условия, при которых они на-
падают на человека и т. д. Обо всем этом говорит экология
комаров. Но и систематика и экология опирается на знание
9
технический, лесохозяйственный и другие методы борьбы
основаны на знании экологии.
Роль насекомых в общем балансе природы еще далеко не
изучена. Лишь недавно стало известно, что, например, личин-
ки комара Bibio обогащают лесную почву азотом, а стаи са-
ранчи, достигающие массы до 50 000 т, ежедневно могут уни-
чтожать пищу 5 млн. людей. Подсчеты показывают, что 99%
насекомых полезны или нейтральны для человека и следовало
бы поставить вопрос об охране многих из них (Steinchaus,
1964). В ряде случаев и «хозяйственно вредные» виды могут
иметь большое положительное значение в общем балансе при-
роды. Термиты, например, уничтожая отмирающие деревья,
являются полезными компонентами биоценоза. Выяснение
основных взаимосвязей насекомых с окружающей природой и
человеком остается важнейшей задачей экологии. Большое
теоретическое и практическое значение имеет проблема массо-
вых размножений насекомых. Наконец, по строению насеко-
мых можно определить условия, в которых оно обитает. Тако-
го рода исследования имеют значение для разных отраслей
знаний и практики, в том числе и палеоэкологии. Находки
ископаемых остатков насекомых дают возможность судить
о реально существовавших условиях жизни и, следовательно,
об экологических и климатических картинах геологического
прошлого. Проблема организма как индикатора среды доста-
точно обширна.
Учитывая особенности этих организмов, задачами эколо-
гии насекомых следует признать изучение:
1. Микроусловий, в которых живут насекомые, и их значе-
ние для разных фаз онтогенеза и распределения видов по
местообитаниям.
2. Систем адаптивных признаков'как отражения длитель-
ного влияния среды. Выявление морфо-экологических типов
и жизненных форм и использование их как индикаторов
среды.
3. Особенностей популяций отдельных видов, их качествен-
ного состояния и численности и их изменений в зависимости
от времени и условий жизни.
4. Роли отдельных видов насекомых в биоценозах, связей
с популяциями других видов животных, микроорганизмов и
растений.
5. Участия насекомых в биогенном круговороте форм, ве-
щества и энергии в биогеоценозах, их значения в становлении
и состоянии вторичных биоценозов и сельскохозяйственных
ландшафтов.
Глава II
СРЕДА ОБИТАНИЯ НАСЕКОМЫХ
СВЯЗЬ СО СРЕДОЙ ЧЕРЕЗ ОБМЕН ВЕЩЕСТВ
Отношения насекомых со средой обитания подчинены не-
которым общим закономерностям. Окружающая среда служит
для организма основным условием его бытия: она дает тепло,
воду, воздух, свет, пищу и т. д. Воздействие среды на орга-
низм вызывает систему соответствующих ответных реакций.
Найти определенные связи в этих взаимоотношениях, выявить
влияние элементов среды и их совокупностей на отдельные
стороны жизни насекомого необходимо как начальный этап
изучения экологии вида.
Связи со средой осуществляются через обмен веществ,
сложные саморегулирующиеся физико-химические процессы—
гза счет энергии, получаемой из внешней среды. В настоящее
уремя считают, что организмы могут существовать в трех со-
стояниях: биоза, мезобиоза и анабиоза.
• Состояние биоза (жизни) характеризуется двух-
сторонним обменом веществ, ассимиляцией и диссимиляцией,
неразрывным единством структуры и функции. Возможно вре-
менное снижение интенсивности обмена (сон, спячка, оцепе-
нение, диапауза) как экологическое приспособление к перене-
сению неблагоприятных условий жизни.
Состояние промежуточное (мезобиоз) харак-
теризуется наличием только процессов диссимиляции, одно-
сторонним функционированием ферментативных систем жиз-
неспособных структур.
Состояние анабиоза, или абиоза, характери-
зуется отсутствием обмена и незначительностью процессов
разрушения. Жизнеспособные структуры сохраняются в не-
функционирующем состоянии: нет обмена, нет роста, разви-
тия, раздражимости. Но это и не мертвое тел^, поскольку со-
13
строения и развития комаров, которые составляют предмет
морфологии. Понять образ жизни комара невозможно без зна-
ния его жизненных отправлений — дыхания, пищеварения,
размножения и т. д., о которых говорит физиология. Частные
науки связаны друг с другом и вместе с тем сохраняют свои
особенности. Все это говорит о необходимости комплексного,
всестороннего изучения объекта, особенно необходимого для
эколога.
Экология опирается на данные физиологии при изучении
связей организмов с физико-химическими условиями среды,
обеспечивающими обмен веществ. Этими вопросами занимает-
ся физиологическая экология. При этом физиолога интересует
не жизнь вида, а процессы, происходящие в организме. При-
способительные особенности видов, их жизненные формы не-
возможно понять без проекции исторического влияния среды
на морфологию тех или иных видов. Объяснить закономерно-
сти размещения видов по территории невозможно без привле-
чения данных биогеографов, геоботаников, ландшафтоведов,
почвоведов, климатологов. Понять биогенный круговорот ве-
ществ, форм, энергии в биоценозах, закономерности их сложе-
ния и существования возможно лишь учитывая биогеохимию
ландшафтов, почвообразовательные процессы, особенности
фитоклимата и растительного покрова.
Итак, экология представляет собой науку синтетическую,
связанную с рядом биологических и небиологических наук,
сохраняющую в то же время свои специфические задачи и
объекты исследования. Экологу необходимо пользоваться син-
тезом различных научных дисциплин, особенно необходимым
при изучении биогеоценозов, при решении вопросов рацио-
нального использования и организации сельскохозяйственных
территорий.
ОСОБЕННОСТИ ЭКОЛОГИИ НАСЕКОМЫХ
Насекомые представляют собой один из наиболее разнооб-
разных классов животного царства. Современные насекомые
представлены более чем 30 отрядами, 1000 семействами, де-
сятками тысяч родов и более чем миллионом видов. Имеются
все основания считать, что такое разнообразие форм было
присуще насекомым давно. Обилие систематических единиц
в той или иной группе организмов — следствие ее долгой и
сложной истории (Родендорф, 1957). Многочисленность и раз-
нообразие форм насекомых указывают на присущую их орга-
низму удивительную гибкость, способность приспосабливаться
к изменяющимся условиям среды. Влияние среды отражено
в особенностях морфологии, физиологии, развития, поведения.
Господствующие на суше насекомые могут быть по праву на-
званы как «дети воздуха и света». У насекомых мы еще мно-
10
тому можем научиться. Это хорошо знают специалисты по
бионике, использующие насекомых в качестве моделей для
технических устройств.
Процветанию насекомых способствуют небольшие размеры
их тела (от 0,2 до 30 см) и возможность использования всех
сред жизни, их «вездесущность». Насекомых можно встретить
в воздухе до высоты 15 км, в холодных льдах Арктики и в го-
рячих источниках, в воде и в почве, в корнях растений, в пло-
дах, внутри стеблей и листьев, под корой и в древесине, в дру-
гих насекомых и животных и т. д. Все это вызывает необходи-
мость изучать микроусловия жизни насекомых в больших
масштабах, чем это требует экология других, более крупных,
животных и создает специфику экологических исследований
в энтомологии. Особенности метаморфоза насекомых, дивер-
генция развития, сопровождающаяся резкой сменой условий
жизни и гетеротопностью, вынуждают изучать экологию. от-
дельных фаз развития. Личинка, куколка, взрослое насекомое
имеют разную морфологию, разные функции и разные требо-
вания и связи с внешней средой. Это усложняет экологиче-
ские исследования.
В. Н. Беклемишев в классической монографии «Эколо-
гия малярийного комара» показал, что экология комара рас-
падается на экологию водных фаз, экологию взрослого насе-
комого и экологию вида в целом, поскольку на разных фазах
жизненного цикла комары заселяют разные биотопы. На при-
мере малярийного комара Anopheles он разработал учение
о жизненном цикле насекомых, согласно которому все ана-
томические, физиологические и экологические признаки вида
представляют единую систему приспособлений, направленных
на его самосохранение.
В процессе жизнедеятельности насекомые производят ра-
боту космического значения, перерабатывая огромную массу
органических и минеральных веществ, непрерывно синтезируя
новые органические и неорганические вещества.
Огромно значение насекомых в процессах почвообразова-
ния. Они резко усиливают деятельность бактерий. Общая мас-
са беспозвоночных животных в почве составляет 3—4 т/га.
Велика практическая значимость экологии насекомых. По
К. Фридериксу (1932), «прикладную биологию, имеющую хо-
зяйственное значение, вообще говоря, можно было бы назвать
«прикладной экологией», так как методы ее большей частью
тождественны с экологическими методами, а предмет ее со-
ставляет часть предмета экологии». Чтобы рационально вести
борьбу с вредителями сельскохозяйственных и лесных куль-
тур, охранять здоровье человека и животных или использовать
полезных насекомых, мы должны знать образ их жизни и
связь насекомых с окружающей средой. Биологический, агро-
11
храняются жизнеспособные структуры, способные функциони-
ровать при соответствующих условиях. Это «как бы смерть
для сохранения жизни». Для насекомых состояние анабиоза
хорошо известно. Активные связи со средой осуществляются
в состоянии биоза.
Специфической особенностью живых тел является то, что
реакции синтеза и распада образуют в них единую систему.
В основе этого единства лежит ферментативная система ката-
литических белков. В организме насекомых также имеется
ряд ферментов, активность которых меняется как от внутрен-
них условий, так и от воздействий окружающей среды. Коор-
динация обмена определяет характерный для данного орга-
низма тип обмена веществ, сложившийся в процессе истори-
ческого развития, поддерживаемый и направляемый взаимо-
действием с внешней средой.
Для различных организмов характерны свои особенности
связи с условиями внешней среды.
Насекомые отличаются от растений подвижностью, нали-
чием нервной системы и органов чувств, а от теплокровных
животных — слабой способностью теплорегуляции, что в ряде
случаев сближает их с растениями. Разные виды насекомых
живут в различных условиях, отличаясь активной избиратель-
ностью к среде. Для каждого вида характерен свой экологи-
ческий стандарт, постоянство требований к среде обитания.
Одни виды теплолюбивы и сухолюбивы, другие — влаголюби-
вы; одни любят солнце, другие — тень; одни активны днем,
другие — только ночью. Благодаря подвижности и развитым
органам чувств насекомые активно отыскивают благоприят-
ные условия и уходят от неблагоприятных. Постоянство тре-
бований к условиям среды у каждого вида складывалось
в процессе эволюции и оно может войти в видовую характе-
ристику, как, например, связь с кормовыми растениями у тлей,
листоблошек, некоторых щитовок.
Разные виды насекомых по-разному переносят амплитуду
колебаний условий среды. По выносливости (валентности)
К воздействию отдельных элементов, или факторов, среды и
их комплексов различают насекомых стено- и эврибионтных.,
стено- и эвритопных, стено- и эвритермных и т. п. Пределы
способности насекомых переносить те или иные колебания
условий жизни, их пластичность поясняет рис. 3.
Пластичность видов меняется в зависимости от места, вре-
мени, состояния организма. Отношение насекомых к условиям
среды резко меняется на разных этапах онтогенеза и в разные
сезоны года. Наряду с постоянством выбора экологических
условий, или экологическим стандартом, у насекомых развита
способность менять отношение к факторам среды; в ответ на
изменчивость условий жизни меняется направленность физио-
логических процессов. Реакция насекомого на воздействие
14
одного и того же фактора среды может оказаться противо-
положной в зависимости от состояния организма, его возраста
и стадии развития, связанной с сезонной цикличностью. На-
пример, для жука-притворяшки Ptinus fur L., по нашим на-
блюдениям, в период его активной жизни оптимальны темпе-
ратуры 20—24°, а осенью 1—5°. Соответственно период актив-
ной жизни отличается иным ходом физиологических процес-
сов и соотношением обменных реакций.
КЛАССИФИКАЦИЯ ФАКТОРОВ СРЕДЫ И УСЛОВИЙ ЖИЗНИ
Среда, в которой проходит жизнь тех или иных видов на-
секомых, представляет сложную систему совместно действую-
щих элементов этой среды, или экологических факторов. На-
секомое, каки другие организмы,— система не изолированная,
а открытая, подверженная влиянию окружающего мира.
В этом окружении многое для организма может быть безраз-
личным или почти безразличным или еще очень мало изучен-
ным, но многое необходимо. Необходимые для жизни элемен-
ты среды называют условиями существования, или условиями
жизни. Другие факторы не составляют необходимых условий
жизни, оказывая на процесс жизнедеятельности лишь косвен-
ное влияние или представляя нейтральный фон. Одни авторы
средой обитания считают всю биосферу (В. Н. Беклемишев),
Другие — лишь то, что окружает организм. По Г. В. Николь-
скому (1955), «средой организма являются только те природ-
ные тела и явления, с которыми организм находится в прямых
приспособительных связах».
По характеру и способам воздействия на организм факто-
ры среды подразделяют на: 1) климатические, или климато-
географические; 2) эдафические, т. е. почвенные — механиче-
ские, физические, химические свойства почв, включая и
15
рельеф; 3) биотические — влияние организмов друг на друга,
4) антропогенные, или «сверхорганические» (К. Фридерикс,
Д. Н. Кашкаров и др.). В данном случае подчеркивается за-
висимость групп факторов и отдельных факторов друг от Дру-
га, их комплексное действие и доминирующее значение клима-
тических факторов.
В последнее время (1958, 1961 гг.) вопросу классифика-
ции факторов среды много внимания уделяет А. С. Мончад-
ский, пользуясь преимущественно энтомологическим материа-
лом. Экологическим фактором он предлагает считать любой
из меняющихся элементов среды обитания, вызывающий
активную приспособительную реакцию. В своей классифика-
ции Мончадский факторы среды пытается оценить по ответным
приспособительным реакциям организмов, по их влиянию на
распространение, колебание численности, жизненные циклы,
т. е. по явлениям, еще недостаточно изученным.
Большинство экологов в своих классификациях учитывают
естественные взаимосвязанные группы факторов, влияние ко-
торых на организм различно, как, например, влияние климати-
ческих факторов и биотических. Влияние факторов среды на
отдельные индивидуумы определяется прежде всего их влия-
нием на обмен веществ, на поддержание жизни особи. А влия-
ние факторов среды на состояние вида, его популяций опреде-
ляется, кроме того, и взаимоотношениями с другими особями
своего вида и популяциями других видов.
Принципиальные различия в отношениях с абиотической и
биотической средой хорошо отражены в делении факторов
среды на факторы, представляющие прямые условия метабо-
лизма, непосредственно определяющие возможность жизни
особей, и факторы, действующие на процессы метаболизма
через внутривидовые и межвидовые отношения (Наумов,
1963). К первым Н. П. Наумов относит пищу (состав, каче-
ство, обилие, доступность, сезонность) и физико-химическую
обстановку (температура, свет, кислород, осмотическое давле-
ние и другие факторы), действующие в природе как микро-
климат или экоклимат. Для позвоночных животных на первое
место по экологическому значению он ставит пищу. Для на-
секомых как животных пойкилотермных, по Н. Я. Кузнецову,
на первое место следовало бы поставить условия абиотиче-
ские, особенно температуру. Но для таких оценок необходим
анализ значения отдельных факторов среды в разные перио-
ды онтогенеза насекомых.
ФИЗИКО-ХИМИЧЕСКИЕ УСЛОВИЯ жизни
Физико-химические условия жизни насекомых создают:
солнечная радиация, температура, влажность воздуха и суб-
страта, обеспеченность кислородом, осмотическое давление и
16
пугие факторы, действующие в природе. Не менее важную
категорию условий жизни составляет пища как источник энер-
гии и химических веществ, необходимых для обменных
реакций.
! Значение солнечной радиации. Источник энергии для жиз-
ни на земле — солнце. В конечном итоге вся энергия, связан-
ная в органических соединениях, есть энергия солнца.
Рис. 4. Схема прохождения сол-
нечных лучей через атмосферу (по
А. Ф. Клешнину)
Распределение солнечной энергии неравномерно и зависит
от многих причин, в результате которых попеременно сильнее
нагревается то северное, то южное полушарие, что приводит
к перемещению воздушных масс и в конечном итоге — форми-
рованию различных климатических зон. Солнечная радиа-
ция— одна из главных причин тепловых явлений в атмосфе-
ре, биосфере и литосфере, биофизических и биохимических
процессов, протекающих в организмах.
Известно несколько типов солнечного излучения: 1—элек-
тромагнитное (радиоволны); 2 — инфракрасное (тепловое);
3 — видимый, свет, составляющий 50% лучистой энергии; 4 —
ультрафиолетовое излучение (3—5% лучистой энергии); 5 —
жесткое излучение — х (рентгеновское) и у-лучей. Жесткое
излучение гибельно для животных. Наиболее изучена экологи-
ческая роль видимой части солнечной энергии — света.
Свет оказывает на организмы смешанное влияние; лучи
красной части спектра обеспечивают тепловое, синей и фиоле-
товой частей — химическое действие. Спектральный же состав
света зависит от состояния атмосферы и угла падения лучей.
Чем меньше угол падения лучей, тем больший путь проходят
они через атмосферу и тем сильнее меняется их спектральный
состав (рис. 4).
В полдень лучи проходят наиболее короткий путь, утром
и вечером — длиннее. На экваторе день равен 12 ч. В тропи-
ках в году разница в длине дня — 1—2 ч; в субтропиках — 3—
4 ч. На юге нашей страны утро и вечер очень короткие, быстро
наступает ночь, а на севере сумерки бывают длинные. От вы-
соты солнца зависит интенсивность и количество света.
Разные лучи спектра поглощаются и рассеиваются раз-
лично. Вечером и утром (при низком стоянии солнца), когда
лучи проходят более длительный путь (рис. 5), почти совсем
гасятся короткие волны и наблюдается максимум длинновол-
2 Заказ 193 '17
новых лучей. В полдень, при высоте стояния солнца 60—90%
отмечается максимум сине-зеленых и синих лучей. Прохожде-
ние луча в атмосфере имеет основное влияние на спектраль-
Рис. 5. Изменение спектрального состава солнечной радиации в зави-
симости от высоты солнца над горизонтом (по А. Ф. Клешнину):
1 — ультрафиолетовые лучи (меньше 400 ммк); 2 — фиолетовые лучи
(400—440 ммк); 3 — синие лучи (440—490 ммк); 4 — зеленые лучи
(490—565 ммк); 5 — желтые лучи (565—595 ммк); 6 — красные лучи
(595—760 ммк); 7 — инфракрасные лучи (760 ммк)
заметно снижается (в два раза) количество красных, желтых
и зеленых лучей. В зависимости от высоты солнца сильно ме-
няется процент ультрафиолетовых, фиолетовых и синих лучей:
при высоте солнца 90° их 12%, при 10° — 4,4%, при 5° —
1,7%.
Количество и качество радиации меняется и в течение се-
зона. Наиболее изменчив световой режим в высоких широтах.
Дни весеннего и осеннего равноденствия, дни самых длинных
и самых коротких ночей отличаются как разным режимом фо-
топериода, так и преобладанием в спектре тех или иных лучей.
18
Сточные и сезонные изменения светового режима имеют
мное значение в жизни организмов, способных улавлп-
°ГР° малейшие изменения в этом режиме. Световосприиимаю-
БагьОрГаны насекомых очень разнообразны: простые глазки,
лгсеточные глаза, теменной орган, светочувствительные уча-
Фа коЖИ, особые пигментные пятна. Для видов, обитающих
СТпочве, живущих в пещерах, характерна простая светочув-
Втвительность. У многих личинок имеются простые глазки,
пигментные пятна и другие световоспринимающие структуры.
У златки пожарищ обнаружены специальные рецепторы, улав-
ливающие инфракрасные лучи малой энергии. Огромно значе-
ние глаз, способных к цветовому восприятию света, восприя-
тию ультрафиолетового излучения.
Чувствительность к свету велика и разнообразна. Одни
насекомые светобоязливы, другие любят свет и активны днем.
Различны реакции на свет у дневных, сумеречных и ночных
видов. Насекомые могут реагировать и на лунный свет. Фо-
топериодическая реакция отмечена при освещенности 0,1 —
5 лк и даже 0,02 лк (у комарика Metriocnemus).
Многие вечерние и ночные насекомые появляются в строго
определенные часы. Например, восточный майский хрущ
(Melolontha hippocastani F.) в районе песчаных террас Сред-
него Дона начинает летать с 8 ч вечера (московское время)
при освещенности 378 лк, массовый лет отмечен в 20 ч 30 мин
при освещенности 200—150 лк. Июньский хрущ (Amphimallon
solstitialis L.) обычно летал после захода солнца — с 19 ч
35 мин до 20 ч 45 мин, а мраморный хрущ (Polyphylla fullo
L.) — с 20 ч 30 мин, когда лет июньского хруща заканчи-
вается. Только в течение 30 мин перед заходом солнца летает
волжский хрущ (Amphimallon volgensis Fisch.) в районе Ка-
мышина. Очень чувствительны к свету насекомые, живущие
при слабом освещении или при отсутствии света (в почве, дре-
весине, в пещерах и т. д.): почти все они избегают освещения.
Однако большинство насекомых, развивающихся без света,
имагинальную фазу проводят на свету. Если личинки хрущей
отрицательно фототропичны, то имаго вылетают из почвы
только при определенной силе света.
Свет влияет на активность и поведение насекомых: у одних
насекомых он стимулирует, у других — подавляет активность.
Реакция на свет связана со средой обитания. Активность на-
секомых, живущих при слабом свете, уменьшается, если свет
усилить, и, наоборот, активность светолюбивых насекомых
тормозится при ослаблении света. Так, например, личинки
Anopheles maculipennis Mg. заселяют незатененные, прогро
ваемые водоемы, и движение к свету для них совпадает с воз-
вращением на поверхность водоема, реакция на свет у них
положительна. Личинки Anopheles plumbeus ^teph. живут
2*
19
в сильно затененных дуплах и яркий свет для них необычен;
их реакция на свет отрицательна.
Помимо длительности и интенсивности освещения большое
значение имеет качество света — его спектральный состав.
Способность различать цвета большое значение имеет для
насекомых, посещающих цветы. Бабочки предпочитают цветы
яркоокрашенные, двукрылые — частично белые и голубые.
Красный цвет не видят бражники Sphingidae и жужжала
Bombyliidae, но видят шмели Bombus, белянки Pieridae, кома-
ры Culicidae, осы Vespidae. Ко всем лучам чувствительны зла-
ковые щелкуны Agriotes, пчелы Apis различают желто-зеле-
ный, сине-зеленый, сине-фиолетовый, ультрафиолетовый и не
различают желтый и зеленый, голубой и фиолетовый цвета,
а красный воспринимают как темноту. Муравьев Formica раз-
дражают желтые лучи.
Реакции насекомых на свет ярко проявляются в лёте има-
го на источник света, особенно ночных насекомых. Это многие
чешуекрылые Lepidoptera, ручейники Trichoptera, жужелицы
Carabidae, пластинчатоусые Scarabaeidae, чернотелки ТепеЬ-
riopidae, щелкуны Elateridae, стафилиниды Staphilinidae, ци-
кадки lassidae, комары Culicidae, мокрецы Heleidae, мошки
Simulidae, долгоносики Curculionidae, наездники Ichneumo-
nidae, златоглазки Chrysopidae, тараканы Blattodea и др.
Основную массу насекомых, летящих ночью на свет, состав-
ляют представители водной фауны и прибрежных биотипов
и насекомые, ведущие сумеречный и ночной образ жизни.
Дневные формы — слепни (Tabanidae), скакуны (Cicindeli-
dae), тлевые коровки (Coccinnelidae), пчелы (Apidae) и дру-
гие— летят на желто-зеленые и синие лучи, но по сравнению
с ними на ультрафиолетовые лучи насекомых прилетает
в 20 раз больше (Мазохин — Поршняков, 1967). Действует
источник света на расстоянии от 10 ж до 5—7 хж, что зависит
от рельефа и мощности лампы.
Летом время лёта смещается в зависимости от широты и
высоты места и условий температуры. В степной и лесостеп-
ной зонах Украины массовый лёт на ,свет отмечен при 20°,
в предгорьях Карпат — при 15°, в горных районах Карпат —
при 6—7°. Пороговые температуры смещаются в разных ши-
ротах: на Украине—10—11°, в Голландии — 8°, Финлян-
дии— 4°. Ранневесенние и позднеосенние виды летят на свет
при температурах, близких к нулю и даже при падающем
снеге. Лёт усиливается перед грозой; причины этого явления
не изучены; возможна связь с насыщенностью электричеством
и уменьшением атмосферного давления.
Реакция насекомых на свет меняется с их половым созре-
ванием, что часто сопровождается изменением окраски, па-
пример у стадной формы саранчи (Schisiocerca gregaria
Forsk.). Прослежена реакция на свет зеленой дубовой листо-
20
тки (Tortrix viridana L.). До откладки яиц бабочки кон цен-
ве£руются в кронах дубов, где освещенность равна 30—
ТЕ тыс. лк. Но при откладке яиц они выбирают уже более
атененные места с освещенностью не выше 10 тыс. лк. Так
з е меняется реакция на свет и садового листогрыза (Luperus
longicornis F), ясеневого пилильщика (Tomostcthus nigritus
р ) Бабочки дубовой листовертки совершают в кроне дубов
суточные вертикальные миграции. Рано утром они сидят не-
подвижно и начинают летать лишь при увеличении силы света
до 500 лк, а в вечерние часы, перед заходом солнца, вновь опу-
скаются с вершины кроны в среднюю и нижнюю части. При
силе света 90—100 лк в воздухе не остается ни одной бабоч-
ки. Наибольшая активность листовертки отмечена в 11 —15 ч
при освещенности 30—50 тыс. лк. Активность может меняться
в разные сезоны. Южные пустынные муравьи зимой бывают
активны в середине дня и всего 6 ч в сутки; весной и осенью
они деятельны до 12 ч, преимущественно вечером или в нача-
ле ночи; летом активность возрастает до 16 ч в сутки, но на-
секомые деятельны ночью. У насекомых лесной зоны отме-
чаются все типы суточной активности, в пустынях же преоб-
ладают ночные формы, в тундре — дневные или круглосуточ-
но активные.
Значение фотопериодизма. Для многих видов насекомых
(наземных, водных, растительноядных, хищных и паразитиче-
ских, открыто живущих и живущих скрытно при очень слабой
освещенности) доказано влияние на цикличность их развития
чередования дня и ночи (фотопериода). Уменьшение длины
дня к осени, за пределы критических для насекомых величин,
ведет к прекращению роста и морфогенеза и переходу в со-
стояние физиологического покоя — диапаузе.
Переход в состояние покоя можно вызвать эксперимен-
тально, действием короткого дня в весенний период. Так, гу-
сениц китайского дубового шелкопряда (Anthereaea pernyi,
Guer) весной выкармливали в темной комнате и на свет вы-
носили ежедневно лишь на 8 ч. При таком коротком дне гу-
сеницы завили коконы в последних числах июня. Через месяц
из контрольных куколок, находившихся при нормальном осве-
щении, начался вылет бабочек. Из куколок опытной партии
через месяц вылетело всего 1,6% бабочек. Через 70 дней
-8,5% куколок оставалось в состоянии глубокой диапаузы,
а из 70% куколок бабочки вылетали с разной продолжитель-
ностью задержки — от нескольких до 40 дней. Часть куколок
находилась в диапаузе 9 месяцев (при температуре 20—23°
11 нормальной влажности), вполне развитые бабочки выле-
ТдЛИ из этих куколок лишь в апреле следующего года
(Андрианова, 1948). В настоящее время диапауза, вызванная
фотопериодом, известна у 130 видов в разных отрядах насеко-
мых (Данилевский, 1961).
21
Р. С. Ушатинская (1957) различает разные формы физио-
логического покоя, разной продолжительности и устойчиво-
сти, что делает возможным существование пойкилотермных
насекомых в меняющейся среде с ритмически повторяющими-
ся благоприятными и неблагоприятными условиями темпера-
туры, влажности, длины дня. По ее данным, для колорадского
жука характерны четыре категории покоя — разной глубины,
продолжительности и биологической значимости: летняя диа-
пауза; диапауза, переходящая с наступлением зимних холодов
в зимнюю спячку; повторная диапауза; затяжная (многолет-
няя) диапауза, сопровождающаяся глубоким торможением
процессов основного метаболизма, с преобладанием анаэроб-
ных процессов. Итак, множественность типов физиологическо-
го покоя гарантирует выживание части особей при неблаго-
приятных условиях в разные сезоны года: летом — при засу-
хах, зимой — при низких температурах. Многолетняя диапауза
гарантирует, видимо, переживание многолетних периодиче-
ских колебаний климата и других неблагоприятных условий.
Диапауза проявляется в прекращении роста и морфогене-
за, физиологически — в торможении обмена веществ (чрез-
вычайно экономное расходование энергетических резервов и
защита от дегидратации). Эти явления обнаруживаются при
наличии определенных условий (температуры, влажности,
освещения, пищи), необходимых для нормального роста и раз-
вития. Диапауза наступает задолго до осеннего понижения
температур, когда длина светового дня сокращается до 14—
16 ч. Насекомые перестают питаться, размножаться, несмотря
на обеспеченность пищей и оптимальные условия температу-
ры и влажности. Диапауза продолжается часть лета, осень и
часть зимы. Окончание диапаузы не приурочено к концу хо-
лодного периода и повышению температуры; заканчивается
она еще зимой, и насекомое переходит в состояние ванужден-
ного покоя: в состояние оцепенения, характеризующееся по-
вышенной морозоустойчивостью. Выходу из диапаузы способ-
ствует увеличение длины дня и воздействие холодом.
У разных видов диапауза наступает на разных фазах раз-
вития. Различают диапаузу эмбриональную, личиночную, ку-
колочную и имагинальную. У одного и того же вида гусенич-
ная диапауза может наступать в разных возрастах. Действие
определенного фотопериода для возникновения диапаузы не-
обходимо лишь на какой-то одной светочувствительной фазе
развития, которая предшествует диапаузе. Диапауза кровосо-
сущих комаров определяется, например, световым режимом
жизни старших личинок и куколок, а диапауза куколок китай-
ского дубового шелкопряда—действием короткого дня на гу-
сениц 4—5-го возрастов. Действие коротким днем на молодых
гусениц (1—3-го возраста) китайского дубового шелкопряда
диапаузы не вызывает.
22
Разные виды и даже популяции одного вида насекомых
еходят в состояние физиологического покоя при разных по-
пе₽аХ и разных оптимумах светового режима. Моноцикличе-
Р°г вИДы приспособлены к узким пределам фотопериода.
Пиапауза у них не наступает при длине дня 18—20 ч\ при
склонениях от этого оптимума возникает диапауза. Для по-
лициклических видов характерны относительно широкий пре-
дел фотопериода для развития и относительно узкий — для
возникновения диапаузы (табл. 1).
Таблица 1
Влияние фотопериодических условий на процент диапаузирующих особей
(по Гейспиц, 1953, с изменениями)
Световых Вид полициклический Виды моноциклические
часов
в сутки капустная белянка желтогузка ивовая волнянка
(Pieris brassicae L.) (Euproctis similis F.) (Stilpnocia salicis I.)
24 0 88,3 97,3
22 0 — 93,4
20 0 2,7 53,4
18 0 0 96,7
17 — 0 —
16 0 100 100
15 0 100 —
14 76,8 100 100
12 100 100 100
Темнота 0 100 100
В опытах по фотопериоду исследователи выяснили влияние
на развитие насекомых суммы часов света или темноты и рит-
мичности их чередования. Установлено, что наступление диа-
паузы может определяться не абсолютной величиной дневных
п ночных часов, а постепенным сокращением дня, поэтому
сроки наступления диапаузы в разных климатических зонах
различны.
У насекомых установлены разные типы реакций на длину
дня. Длиннодневные виды преобладают в умеренных широтах.
Длинный день со светлым периодом 14—17 и более часов бла-
гоприятен для развития этих видов без диапаузы. Так, напри-
мер, развиваются капустная и репная белянки (Pieris brassi-
сае L., Р. rapae Е.,), луговой мотылек (Loxostege sticticalis
L.), малярийные комары, колорадский жук (Leptinotarsa de-
cemlineata Say.). Короткодневные виды у нас немногочислен-
ны, характерны для южных широт; без диапаузы развиваются
при коротком дне (менее 13—16 ч в сутки). Пример коротко-
дневных видов насекомых — тутовый шелкопряд (Bombyx
wort L.), цикадка (Stenocranus minutus F.). ।
23
Обращает внимание сходство фотопериодических реакции
у насекомых и растений и синхронность развития насекомых-
фитофагов с фазами развития кормовых растений. Активность
насекомых совпадает с началом сокодвижения, распусканием
почек, ростом растений, а состояние диапаузы — с состоянием
покоя. Растения длинного дня — это растения северных шп-
рот, растения короткого дня — это растения преимущественно
южные. Короткодневным растениям необходимо больше ко-
ротковолновых лучей. Длиннодневным растениям осенью и
весной нужен свет малой интенсивности, обилие длинноволно-
вых и отсутствие коротковолновых лучей, а летом — коротко-
волновая радиация, большая интенсивность света, более вы-
сокое солнцестояние. Можно, по-видимому, ожидать и для
насекомых большого значения в фотопериодических реакциях
не столько суммы и чередования дневных и ночных часов,
сколько изменений в качестве и интенсивности света.
Сходство диапаузы насекомых и физиологического покоя
у растений проявляется не только в сходстве влияния фотопе-
риодов, но и в направленности физиологических процессов.
Состояние покоя у растений, как и у насекомых, проявляются
в остановке роста и морфогенеза, но в скрытой форме в пе-
риод покоя протекают сложные физиологические процессы,
подготавливающие растение к активной жизни. Период по-
коя— период, когда растение не способно расти несмотря на
наличие соответствующих для роста условий (Генкель, 1964).
В период покоя у растений увеличивается содержание ве-
ществ, задерживающих рост, их распада не происходит. Уве-
личение длины дня весной способствует расщеплению ингиби-
торов роста и их тормозящее действие снимается. У насекомых
процессы, приводящие к торможению и прекращению роста,
сложнее и связаны с совместной деятельностью нервной и гор-
мональной систем. Тормозящие рост гормоны и активаторы
контролируются головным мозгом. Процессы торможения
роста сменяются реактивацией, которую можно рассматри-
вать, как и у растений, подготовкой к бурной весенней актив-
ности. Периоды активной жизни всегда чередуются с перио-
дами остановки или задержки формообразовательных процес-
сов— диапаузой, которую считают приспособлением к био-
логически неэффективным температурам (5—10°), недостатку
влаги (летняя диапауза) и отсутствию пищи. Так как все фак-
торы, снижающие обмен веществ у насекомых, способствуют
увеличению сопротивляемости их низким температурам,
а условия, стимулирующие обмен веществ, связаны с паде-
нием сопротивляемости холоду, прямым следствием диапаузы
является повышение холодостойкости (Ушатинская, 1957).
Наступление диапаузы, эффект сокращенного дня может
стимулироваться или тормозиться воздействием термических
условий. Пороговая длина дня, характеризующая фотоперио-
24
ескую реакцию, зависит от температуры, и уменьшается
Димерно на 1 —1,5 ч при повышении температуры на 5°. Это
наделяет сроки наступления диапаузы в природе соответст-
оПР температурным условиям сезона. При понижении тем-
веоатуры диапауза наступает быстрее, повышение температу-
П{ способствует активному развитию. Слишком высокие и
Тишком низкие температуры фотопериодическую реакцию на-
Сущают; нормально она протекает лишь при естественных ко-
лебаниях температуры, когда день теплее ночи. Весной низкие
температуры диапаузу не вызывают, осенью же даже при
20_22° наступает полная диапауза. Холодные вечера усили-
вают действие короткого дня, а теплые вечера задерживают
наступление диапаузы (Горышии, 1955).
Различен и температурный оптимум фотопериода у разных
видов насекомых. У северных форм оптимум фотопериода
сильно сдвинут в сторону высоких температур. У видов длин-
нодневных высокая температура подавляет действие фотопе-
риода, исчезает оно и при низких температурах. Сложной си-
стемой фотопериодических реакций регулируется сезонное
развитие у хвойных шелкопрядов Dendrolimus. Диапауза
у них вызывается действием короткого осеннего дня. Однако*
диапауза прекращается без холодовой закалки, поэтому воз-
никновение зимующих стадий сдвинуто к периоду устойчивого
осеннего похолодания. Ведущее значение в этом процессе
имеет замедление темпов формирования диапаузы при длин-
ном летнем дне и возможность ее появления в любом возрасте
гусениц. С этим связаны сроки ухода гусениц на зимовку, раз-
новозрастность зимующих гусениц, сдвиги циклов развития от
двухлетнего к однолетнему и другие особенности фенологии
хвойных шелкопрядов (Гейспиц, 1965).
С периодичностью освещения и его спектральным составом
связаны и другие особенности роста и развития насекомых.
Известна зависимость появления крылатых форм тлей от дей-
ствия света. Так, у Macrosiphutn крылатые особи исчезают
при повышении температуры и увеличении длины дня. В при-
роде крылатое поколение этих тлей появляется осенью, когда
длина дня равна примерно длине ночи. Появление мигрирую-
щих форм и полового поколения связано с действием фототер-
мопериодов. Высокие температуры и длинный день способст-
вуют появлению девственниц, снижению численности самцов,
а понижение температуры и короткий день — обоеполого по-
коления и яйцекладущих самок. Низкие температуры и корот-
кий день вызывают появление летне-осенних сезонных форм
насекомых, отличающихся размерами, длиной надкрылий,
изменением структуры полового аппарата. Осенние формы на-
секомых более темные, мелкие, короткокрылые. Перемещение
пигмента отмечено у стадной формы саранчи как приспособ-
ление к сильному свету, а у одиночной — к слабЬму.
25-
Действие света сказывается на скорости роста, качестве
коконов, плодовитости. Известно, например, что гусеницы ту-
тового шелкопряда при развитии в темноте бывают более мел-
кими, а коконы — на 20% легче, чем при воспитании на свету;
то же отмечено и для китайского дубового шелкопряда (Ан-
дрианова, 1948). Прирост в весе гусениц шелкопряда оказался
Рис. 6. Изменение веса гусениц в те-
чение суток (июнь 1939 г., Крым)
•связанным с наступлением сумерек; для них установлено за-
кономерное падение веса в первые часы наступления темноты
(рис. 6).
Изменение прироста в весе под влиянием разного освеще-
ния подтверждают наблюдения К. Ф. Гейспиц за сосновым
шелкопрядом (Dendrolimus pint L.). При круглосуточном
освещении средний вес гусеницы оказался 289 а, при чередо-
вании света (9 ч) и темноты (15 ч) — 112,9 г. При чередова-
нии обычного света и красного гусеницы весили 126,5 а, а при
желтом свете—134,7 а. По заключению А. С. Данилевского,
«в отличие от гидротермических и пищевых условий свет не
является необходимым для основных процессов жизнедеятель-
ности». Приведенные выше данные противоречат этому заклю-
чению.
О большом значении света в основных процессах жизне-
деятельности насекомых говорят также изменения плодовито-
сти насекомых. Например, воспитание гусениц китайского
дубового шелкопряда в темноте привело к снижению плодо-
витости бабочек на 33% (Андрианова, 1948). Мало плодовито
осеннее обоеполое поколение тлей. Изменение окраски насеко-
мых в связи с половым созреванием и фазовой изменчиво-
стью с одновременным изменением фототаксиса известно для
многих видов, известно и резкое колебание плодовитости у на-
26
паЮщих массовые размножения. Следовало бы обра-
«секомых, д на СВЯзь этих явлений с действием интепсивно-
тить внима_ солнечной радиации в разные годы и сезоны,
S разных погодных уе.ювнях
пр р нообразие фоторецепторов у насекомых, чувствитсль-
десятым и даже сотым долям люкса, активный лёт
ность т и другие особенности поведения насекомых, четкие
На "ии на спектральный состав света, огромное значение
Ре3 а в появлении диапаузы, влияние света на рост, скорость
Авития плодовитость, сезонный диморфизм и пигментацию
насекомых свидетельствуют о большом значении света в жиз-
ни насекомых. Ведущее значение света установлено в циклич-
ности развития. Влияние света, нарастание или уменьшение
длины дня усиливается или ослабляется одновременно дей-
ствующими термическими условиями.
'Значение температуры. Тепловой обмен считают основным
и ведущим энергетическим процессом в отношениях организ-
ма и среды. Температура определяет агрегатное состояние тел
и все важнейшие явления жизни. У насекомых температура
тела и происходящие в нем химические реакции зависят от
температуры окружающей среды, от поглощения и отражения
лучистой энергии солнца. Температура тела насекомых под
действием радиации может меняться в десятки раз. Так, по
опытам И. Д. Стрельникова (1934) с гусеницами крапивницы,
температура их тела в тени была близка к температуре воз-
духа (12° — температура воздуха, 16° — тела гусениц). Если
гусеницы оказываются на солнце, то через несколько минут
температура их тела резко поднимется, достигая 48°, а при
помещении в тень — снова быстро падает и становится близ-
кой к температуре воздуха. Для насекомых характерна пря-
мая, пассивная зависимость от температуры среды, так как
способность регулировать температуру тела у них очень огра-
ничена.
Температура тела насекомых связана с особенностями по-
глощения и отражения солнечных лучей покровом тела. По-
глощению солнечного тепла способствует темная, часто чер-
ная окраска тела. Она чаще встречается у горных и аркти-
ческих насекомых. Сильное нагревание темных экземпляров
отмечено у итальянского пруса, разница температуры тела
^емных экземпляров достигает 5—6° по сравнению со светлы-
ми. Блестящая металлическая поверхность часто встречается
У жуков, держащихся на ярком свете. Температура тела у этих
видов на солнце бывает ниже, чем у насекомых с другими
типами окраски. От перегревания спасает воздушная прослой-
- -Д элитРамп’ характерная для многих пустынных видов
Резкое повышение температуры тела насекомых связано
их активной мышечной работой. У хороших и сильных лету-
27
нов (бражники, стрекозы, шмели) при полете температура1
тела в течение нескольких минут повышается на 15—20°.
У летящей азиатской саранчи Locusta migratoria L. при тем-
пературе воздуха 17—20° температура тела достигает 30—37°,
а у сидящей — не отличается от окружающего воздуха. У вин-
ного бражника (Pergesa elpenor L.) при движении крыльев
температура тела поднимается с 19,6 до 30,3°. В высокогорье
Кавказа на высоте 2300 м при температуре воздуха 4—8°
температура летящих шмелей была 38—44°. Высокая темпера-
тура тела шмелей при полете сохранялась даже при снежных
метелях и температуре воздуха 2°. У плохих летунов (лу-
говой мотылек) температура тела при полете не поднимается
выше 2—5° (Стрельников, 1940).
Теплоотдача в небольших пределах может регулироваться
через испарение воды с поверхности тела и при дыхании. Жи-
вые жуки чернотелок (Adesmia) на солнце нагреваются мень-
ше, чем мертвые; разница в температуре достигает 2—9°. Гу-
сеницы хлопковой совки благодаря испарению с поверхности
тела и охлаждению от контакта с растениями не нагреваются
более чем на 5—8Q (при радиации в 1 —1,2 кал! см2 в мин).
В пределах нескольких градусов насекомые могут менять тем-
пературу тела, меняя его положение. Например, если саранча
сидит на стебле растения, то солнечные лучи скользят парал-
лельно телу и оно имеет температуру 38,3°, а саранча, сидя-
щая на земле и получающая лучи, падающие вертикально,
нагревается до 41,7°.
С этим явлением у насекомых связаны различного рода
миграции, подъемы в воздух, заползание на кусты и деревья.
Личинки азиатской саранчи ночью при температуре воздуха
10—15° неподвижно сидят на поверхности почвы или на расте-
ниях, утром при недостатке тепла собираются в небольшие
группы — солнечные кулижки, тела личинок тесно соприка-
саются, при этом поверхность лучеиспускания резко умень-
шается, что замедляет охлаждение тела личинки. При повы-
шении температуры тела примерно до 40° солнечные кулижки
распадаются, а при падении температуры ниже этого уровня
кулижки снова образуются.
Темноокрашенные стадные личинки саранчи имеют темпе-
ратуру тела выше одиночных. В жаркие часы дня личинки
передвигаются большими кулигами, тело личинок перегре-
вается, п у них наступает тепловое оцепенение. При пасмур-
ной погоде передвижений в виде больших кулиг и теплового
оцепенения не происходит, личинки остаются в кулижках.
Окрылившаяся саранча избегает перегрева, поднимаясь стая-
ми в воздух. Для охлаждения в воздух поднимаются роющие
осы (ВетЬех), когда песок нагревается выше 50°, и добраться
до более глубоких и прохладных слоев бывает трудно. В этом
отношении интересно поведение муравьев. Некоторые муравьи
28
специальные земляные своды—холмики, свободные от
СТстений. Такие холмики нагреваются сильнее, чем окружаю-
Ра гнездо почва, покрытая травой. Дерновой муравей Lasius
Шместах, прогреваемых солнцем, живет в земле и надземных
Е'ружений не строит, а в затененных местах строит земляные
^олмики, в зависимости от степени затенения и от высоты
читальная зона
JOfia актиВности
Пессимум
внешней
среды
Нижняя гжница
ытивности
Ц0° +15° +20и +23° +30°\+35° +30°
Оптимум s______________
+53°
Верхняя
v ----------^граница
Пи ос и мум аь т ибн о с пт
Рис. 7. Изменение активности гусениц шелкопряда-монашенки в зависимо-
сти от температуры среды
травяного покрова. В густом лесу, где солнечные лучи слабо
прогревают поверхность, муравьи гнездятся в гнилой древеси-
не, которая теплее почвы. Известна регулировка температуры
в пчелином улье. Если зимой температура падает ниже 13°,
то пчелы начинают усиленно двигаться, питаться, образуют
клубки и таким образом поддерживают в гнезде температуру
около 25°.
Зависимость жизнедеятельности насекомых от температу-
ры среды легко проследить по изменению их активности. Для
каждого вида насекомых имеются свои оптимальные границы
активности и пассимумы. Имеется зона смертельно-высоких
и смертельно-низких температур, зона теплового и холодового
оцепенения и зона нормальной активности. В состояние оце-
пенения насекомые впадают за пределами действия оптималь-
ных температур (рис. 7).
Отношение разных видов к температуре среды различно:
одни из них эвритермны, другие стенотермны, одни криофиль-
НЬ1, другие термофильны. Бабочки зимней пяденицы Орего-
Phtera brumata L. активны поздней осенью при 5—8°, другие
виды при таких температурах впадают в оцепенение. Гусени-
цы соснового шелкопряда выходят из мест зимовки при темпе-
ратуре почвы 4°, ледничники (Boreas), некоторые жужелицы,
личинки мягкотелой бывают активны даже зимой при —1—3°.
29
Оптимальные температуры, как и пессимальные, в значи-
тельной мере зависят от тех условий, в которых насекомое по-
стоянно живет. Средний верхний предел находится около 50°.
Водяные клопы — Notonecta glauca L. и Nepa cinerea L.—
встречены в теплых источниках при 45°, личинки Chirono-
mus— при 49—51°, личинки мух береговушек (Ephydridae)
остаются живы при 65°, личинки жуков-златок — при темпера-
туре 52°. Постоянные обитатели найдены в источниках с тем-
пературой не выше 45°. Тепловое окоченение комнатной мухи
наступает при 45—46,5°, жигалки (Stomoxis) —при 43,5°. Не-
которые бабочки могут жить при температуре 40—45°.
Действие низких температур менее губительно. Холодовое
оцепенение у разных насекомых также наступает при разных
температурах. Пчелы, например, не переносят охлаждения и
погибают уже при 0°, рыжие лесные муравьи (Formica ги-
fa L.)—при —1,5°, черный таракан (Blatta orientalis) при.
температуре —5° погибает через час. В то же время гусеницы
лугового мотылька выносит температуру — 30°. У совки-гаммы
куколки выдерживают —18°, гусеницы младшего возраста —
—12°, а гусеницы IV и V возраста — всего —4°. Яйца азиат-
ской саранчи выдерживают —30°, а личинки струйчатого
заболонника — даже —53°. Холодостойкость обычно выше
у тех видов, которые в природе живут при более низких тем-
пературах.
Для насекомых характерны сезонные колебания холодо-
стойкости. Например, клопа Perillus bioculatus F. зимой мож-
но переохладить до температуры —17° и даже до —26°,
а в марте и позднее уже при температуре —10° наступает за-
мерзание и смерть.
Холодостойкость различна у разных экологических групп
насекомых. Насекомые, живущие и зимующие в стволах де-
ревьев, зимой более холодоустойчивы, чем весной. Содержа-
ние насекомых при более высоких температурах делает их ме-
нее холодоустойчивыми. Холодостойкость водных насекомых
постоянно невысокая. У них не отмечаются периодические
колебания стойкости к низким температурам, но они могут
противостоять долгим периодам низкой температуры. Другая
группа насекомых способна выносить сильные понижения тем-
пературы. Так, личинки Synchroa punctata, живущие в дубах,
в июне, июле переносят температуру —2,8°, в декабре —12,0°,
в январе —17,(У, а в феврале —22,0°. При этом содержание
воды (в процентах к весу тела) у них падает с 54% в августе
до 31% в феврале. Холодостойкость выше у насекомых, зи-
мующих открыто или в ненадежных укрытиях. Очень чув-
ствительны к холоду пчелы, личинки майских жуков и другие
виды, обитающие в среде с малым колебанием температуры.
'В недеятельном состоянии насекомые умеренных широт про-
водят большую часть года. В оцепенении или в состоянии диа-
зо
паузы они зимуют в различных укрытиях. В местах зимовки
многих видов колебания температуры сглажены. В наших
шпротах под снегом у поверхности почвы температура близка
к 0°, в почве на глубине 10—25 см даже в холодные зимы тем-
пература бывает не ниже —2------4°. Ряд обитающих в почве
личинок насекомых мигрирует вслед за инверсией температур,
Рис. 8. Миграции личинок восточного майского хруща (по Г. Г. Гарунову)
не впадая в состояние диапаузы. Хорошо, например, изучены
миграции личинок восточного майского хруща (рис. 8).
Рассмотрим влияние на насекомых сверхнизких
температур. Экспериментальные работы по измерению1
температуры тела насекомых с помощью термоэлектрического
термометра позволили установить, что ниже 0° наступает пе-
реохлаждение, которое может привести к замерзанию соков
тела. Температура замерзания соков называется «критической
точкой». Она изменчива у разных видов насекомых и зависит
от физиологического состояния насекомого. Переохлаждение
до температуры критической точки не сопровождается обра-
зованием льда в организме. После прохождения критической
точки начинается образование льда. Лед разрывает нежные
ткани, что приводит к смерти организма. Свободная вода пре-
вращается в лед быстрее, чем связанная с коллоидами, поэто-
му насекомые с большим содержанием свободной воды (вод-
ные, личиночные фазы, весенние формы) менее холодоустой-
чивы. Медленное охлаждение способствует переходу в пере-
охлажденное состояние на более длительный срок. Постепен-
ное снижение температур осенью в ряде случаев спасает на-
секомых от промерзания и образования кристаллов льда.
Устойчивы к низким температурам диапаузирующие насеко-
мые.
У насекомых удалось наблюдать состояние анабиоза и
оживление даже после их замерзания. Л. К. Дозипа-Лозин-
31
<жий (1956) замораживал диапаузирующих гусениц кукуруз-
ного мотылька (Pyrausta nubilalis Hb.), яблочной плодожор-
ки (Carpocapsa pomonella L.), малинного шелкопряда (Ма-
crothylacia rubi L.) до —80°, выдерживал их при этой темпе-
ратуре в течение суток. Гусеницы становились твердыми, зве-
нели при падении, как стекло, но после оттаивания половина
гусениц прожила больше месяца. Предварительная подготов-
ка гусениц к замораживанию (постепенный перевод в усло-
вия все более низких температур) позволила ему сохранить
гусениц кукурузного мотылка жизнеспособными даже при
действии температур, близких к абсолютному нулю (—253J
в жидком водороде, —269° в жидком гелии) (Лозина-Лозин-
ский, 1964). Состояние анабиоза при замораживании жидким
гелием или жидким водородом возможно благодаря переходу
жидкостей тела в стекловидное состояние без образования
кристаллов льда и сохранению неповрежденной тончайшей’
структуры тканей и клеток. В опытах этого исследователя
часть гусениц кукурузного мотылька, охлажденная до —219°,
после выхода их из состояния анабиоза оставалась живыми
около двух недель.
Остановимся на в л и я н и н температуры на разви-
тие. Активная жизнь насекомых, рост и морфогенез, начи-
нается при достижении «порога развития», «биологического
нуля». Температурный порог развития различен у разных ви-
дов, на разных фазах развития, при разном физиологическом
состоянии. У термофилов он выше, у криофилов — низок и
варьирует в пределах от 4-15° до —7°. Обычным показателем
действия температур служит скорость развития, прирост веса,
процент смертности, плодовитость. В опытах с постоянными
температурами установлено, что скорость развития в пределах
оптимума пропорциональна повышению температуры. Ско-
рость развития и продолжительность развития — величины
обратно пропорциональные (рис. 9).
Скорость развития v определяют в процентах за единицу
юо
времени по формуле: v = —, где х — длительность развития
х
в часах или сутках при данной средней температуре (все вре-
мя развития принимают за 100). Такой метод вычисления по-
зволяет при известном ходе температуры дать оценку воз-
можной скорости развития па разных фазах роста насеко-
мого.
Установлено, что для завершения каждой фазы развития
необходимо определенное количество тепла, или сумма темпе-
ратур. Зависимость скорости развития от температуры в пре-
делах, близких к оптимальным, выражают произведением
эффективных температур на продолжительность развития.
Учитывают температуры дня, или «градусы-дни». Исключают
32
них температуры ниже порога развития и таким образом
излучают эффективные температуры. Произведение эффектив-
п0 температур на продолжительность развития считают ве-
Ничиной постоянной, характерной для отдельных видов. Зави-
симость выражают формулой С=(Г—i)D, в которой Т — тем-
JLnaTVDa развития, /“ — температура порога развития, D —
время развития.
.1______lr<J----J----1----!----J----Ю
Q 75 10 15 20 25 30 35
Температура, °C
Рис. 9. Развитие малярийного кома-
ра от яйца до окрыления (по В. Н.
Беклемишеву):
а — скорость развития; б — про-
должительность развития
Для примера проследим развитие плодовой мухи (Ceratitis
capitata Wied.). При температуре 26° ее развитие продолжает-
ся 20 дней, при температуре 19,5° — 41,7 дня. Отсюда порог ее
развития вычисляем по формуле: (26 — t°) -20= (19,5° — t°) •
•41,7=13,5°. Эту величину подставляем в формуле (Т — /°) •
D = C. Получаем сумму эффективных температур — С = 250°.
Длительность развития обратно пропорциональна температу-
ре. Чем выше температура, тем быстрее идет развитие, но об-
щая потребность в сумме тепла остается одинаковой.
Сумма тепла, необходимая для полного развития, различ-
на у разных видов, для разных фаз развития. Для теплолюби-
вых видов она выше, чем для холодолюбивых. Например, для
рыжего таракана (Blatella germanica L.) нужно 1895,4°, для
капустной моли (JPlutella maculipennis Curt.) —371°, для раз-
вития гусениц озимой совки (Agrotis segetum Schiff.) — 582,2°,
а Для куколок — 216,2° (Кожанчиков, 1961). Однако эти циф-
ры получены на основании опытов с постоянными температу-
рами, отсутствующими в природе, и весьма приблизительны.
Температурные константы во многих случаях оказываются
различными. Восточная азиатская саранча, например, при
оптимально высоких температурах растет быстрее, чем при
низких температурах, и общая сумма тепла нужна ей разная.
Для личинок, развивающихся при 26—32°, эффективная сум-
580^еМПеРаТУР Равна 460°, ПРИ 20—28° — 530°, при 18—24° —
3 Заказ 193 33
Суммой эффективных температур пользуются для расчетов
г
возможного числа генераций: — = U, где С — сумма тепла
Ci
для данного года или многолетняя средняя для данного места,
Ci — сумма тепла, необходимая для полного развития данного
вида, и И — индекс вероятного развития. Если индекс <1, то
успешное развитие вида в данном месте, в данном году мало-
вероятно или невозможно. Если индекс >1, то возможно раз-
витие одной генерации, >2 — двух генераций и т. п. Расчеты
показывают, что, например, яблонная плодожорка в районах
с суммой эффективных температур (выше 10°), равной 930°,
дает одну генерацию, а на Украине, где сумма температур
равна 1870°, развиваются три генерации (Васильев, 1950).
В ряде случаев число генераций остается постоянным при лю-
бых высоких температурах. В опытах повышение температуры
может привести к их тепловому оцепенению, но второго поко-
ления вывести не удается. Это насекомые строго моновольтин-
ные, имеющие в цикле развития глубокую диапаузу, появле-
ние которой определяется фотопериодом.
В естественных условиях насекомые постоянно подвергают-
ся действию переменных температур; это меняет и скорость
развития, и число генераций. По данным Шовена, азиатская
саранча при развитии в условиях постоянной температуры
(30° — для личинок и 35° — для взрослых) дает не более трех
поколений, тогда как при колебаниях температуры (26—30° —
для личинок и 20—37° — для взрослых) можно получить
неограниченное число поколений.
Установлено, что при переменных температурах многие на-
секомые развиваются быстрее. Так, развитие яиц яблонной
плодожорки при нормальных колебаниях дневной температу-
ры ускоряется на 7%, гусениц — на 8%, куколок — на 7%.
Личинки из яиц саранчевых выводились через 5 дней при по-
стоянной температуре 32°, а при чередовании температур в 12°
и 32° для развития личинок понадобилось 2 дня. В ряде слу-
чаев отмечено стимулирующее влияние холода. Яйца саранче-
вых, подвергшихся действию температуры 0°, развивались за
11 —16 дней, а при 27—37° для этого понадобилось 26—
46 дней. Действие низких температур ускоряет выход из со-
стояния диапаузы. Температурные границы, в пределах кото-
рых возможно размножение, нередко оказываются очень узки-
ми. У самок саранчи яйца не созревают, если дневная и ноч-
ная температура колеблется между 30—20°. Блохи не откла-
дывают яип при температуре ниже 25°, а малярийные кома-
ры—при температуре воздуха 7—9° и воды в лужах 11 —13°
(В. Н. Беклемишев) и т. д. Термические условия в период со-
зревания половой продукции и откладки яиц ускоряют или
тормозят размножение насекомых.
34
Ограничивающее влияние термических условий сказывает-
ся и на географическом распространении насекомых. Считают,
что распространение капустной белянки в Сибири ограничи-
вают зимние и весенние холода: открыто зимующие куколки
этого вредителя здесь вымерзают. На юг бабочка идет далеко,
по горам она проникает в Среднюю Азию и Индию, но в жар-
кие равнины не спускается. По Боденгеймеру, капустная бе-
лянка в зимние месяцы залетает в Палестину, дает до пяти
генераций, но в июле, когда температура поднимается выше
26°, все яйца и гусеницч погибают. Азиатская саранча не рас-
пространяется к северу от средней изотермы теплового перио-
да года в 13,6°, так как к северу от этой линии саранча не до-
стигает половой зрелости. Кровяная тля (Eriosoma lanigerum
Hausm.) не распространяется к северу от январской изотермы
в 3—4°, будучи вредителем яблони в Средней Азии, в Крыму
и на Кавказе. Не идет она и в горы, где температура ниже.
Распространение яблоневой и плодовой молей (Hyponomeuta
malinellus Zell, и Н. padellus L.) ограничивают температурные
минимумы ниже —38°; такие температуры приводят к массо-
вой гибели гусениц. В горных районах Средней Азии, Кавка-
за, Закавказья и Дальнего Востока зимние температуры не
падают ниже —30°, поэтому моли размножаются здесь еже-
годно. В южных районах распространение обоих видов моли
ограничивают высокие летние температуры (достигающие
45°) в сочетании с пониженной влажностью воздуха (40%).
Этим объясняется отсутствие молей в жарких районах плодо-
водства: в долинах Узбекистана, Туркмении, Таджикистана
и других полупустынных зонах поливного земледелия. Моль,
завозимая сюда с гор, вымирает (Петров, 1955).
Как и свет, температурные условия меняются в течение
суток и сезонов. В природе их действие осуществляется со-
вместно. Поглощению тепла способствует темная окраска на-
секомых, которая в свою очередь связана с влиянием корот-
кого дня. Появление сезонных форм можно вызвать как дей-
ствием температур, так и фотопериодом. При действии высо-
ких температур насекомые бывают светлее; более темная
окраска связана с низкими температурами. Фотопериодиче-
ская же реакция усиливается или тормозится действием тем-
ператур.
Значение воды в жизни насекомых. Без воды нет жизни.
Живых существ, не содержащих воды, мы не знаем. Все про-
цессы обмена веществ — питание, дыхание, выделение — тре-
буют участия воды. Интенсивность обмена связана с содер-
жанием воды в организме. У разных видов оно колеблется
в пределах от 45 до 92%. Воды меньше у насекомых с более
толстыми и крепкими покровами тела и больше у личинок
Q мягкими покровами (например, у амбарного долгоносика
Calandra granaria L. воды в теле 46—47%, а у гусенйц бабо-
чек 90—92%). У личинок насекомых воды содержится 74—
80%; смерть наступает уже при 35—68%. Минимальное со-
держание воды у пустынной саранчи (35%), у клопа Rhodnius
(60%). У молодых гусениц воды в теле 82%, у десятиднев-
ных— 79%; у молодых куколок — 86%, у десятидневных —
78%, у взрослых бабочек — 60%. Личинки японского хруща
(Popillia japonica Motsch.) воды содержат 78—81%,
куколки — 74%, имаго — 66%. Со старением содержание воды
в теле уменьшается. Зависит оно также от влажности возду-
ха, субстрата и влажности пищи.
Насекомые, защищая себя от высыхания, содержание во-
ды в организме регулируют разными способами. Многие пче-
лы, мухи, бабочки, гусеницы пьют воду. Например, гусеница
китайского дубового шелкопряда в старшем возрасте может
выпить 500 мг воды при нормальном питании, температуре
воздуха — 22—23° и влажности воздуха—70% (Андрианова,
1948).
Насекомые могут впитывать воду и непосредственно через
покровы тела. У водных насекомых кутикула легко проницае-
ма для воды. То же отмечается у личинок, обитающих в почве,
в сырой древесине, где воздух насыщен водяными парами.
Выделяют воду насекомые при дыхании, испарении через
покровы и с экскрементами. У ряда насекомых (палочников,
многих жуков, перепончатокрылых, гусениц, личинок муравьи-
ного льва) транспирации не происходит, слабо выражена она
у сверчков и очень значительна у всех сосущих насекомых.
У кузнечика Pholidoptera транспирация бывает настолько ве-
лика, что на поверхности тела образуются капельки росы;
самка саранчи при 35° за час выделяет через покровы около
10 мг воды.
Избыток воды может удаляться из организма и через
мальпигиевы сосуды. Таким образом интенсивно выделяют
воду личинки цикадок-пенниц; на безводном рационе они не
могут существовать больше суток. Червецы, наоборот, могут
удерживать воду и жить на растениях с малым ее содержа-
нием. Испарение воды идет и через дыхальца. У насекомых
пустынь они очень малы, продолжительное время закрыты,
снабжены различного рода отростками, щетинками и другими
приспособлениями, помогающими регулировать испарение.
При открытых дыхальцах испарение удваивается.
Экономное расходование воды очень нужно при жизни на
суше. От испарения предохраняет водонепроницаемый слой
эпикутикулы, богатый восковыми веществами. Он характерен
для насекомых, живущих в сухой среде. Расход воды умень-
шается и путем изменения продуктов выделения: у водных на-
секомых продуктом обмена служит аммиак, его выделение
требует большого расхода воды, у сухопутных насекомых вы-
деляется мочевина, сохраняющая запасы воды. Дефицит воды
36
компенсируется метаболической водой, получающейся при
расщеплении резервных веществ и особенно жира (полное
сгорание 100 г жира дает 105г воды). Благодаря наличию ме-
таболической воды многие ксерофилы (Derinestes, Tribolium,
Ephestia и др.), живущие в сухом субстрате и питающиеся су-
хой пищей, могут жить в очень сухих условиях при влажности
всего 5 % •
Другие виды при недостатке влаги мигрируют в поисках
более влажных условий и сочного корма. Быстро реагируют
на изменение влажности .личинки хрущей, проволочники и
другие обитающие в почве насекомые, направляясь в места,
где влажность близка к насыщению; для них влажность ниже
98—99% уже создает угрозу высыхания. Не чувствительны
к изменениям влажности воздуха насекомые, высасывающие
соки растений.
Приспособления к защите от высыхания М. С. Гиляров де-
лит на морфологические, физиологические и экологические.
К морфологическим приспособлениям относятся особенности
структуры покровов, наличие эпикутикулы, строение дыхалец.
Физиологические приспособления — связывание воды коллои-
дами, выделение метаболической воды, выделение мочевины
вместо аммиака. Экологические приспособления представляют
особенности поведения, обеспечивающие выбор мест, защи-
щенных от высыхания, и соответствующий период суток, ми-
грации в более влажные глубокие слои почвы, зарывание
в подстилку, активную жизнь ночью и т. д.
При недостатке воды развитие может задержаться или
даже приостановиться и насекомое переходит в состояние оце-
пенения. Дегидратация — один из наиболее важных моментов,
ограничивающих жизнь как при воздействии засухи, так и
низких температур. Состояние покоя при засухе, как и при
холодовом оцепенении, ведет к снижению всех жизненных
процессов. Летняя диапауза известна у многих насекомых:
гессенского комарика (Mayetiola destructor Say), совок (Try-
phaena ravida Schiff, Euxoa basigramma Stgr., Euxoa villiersi
Gn.) и других видов, особенно в засушливых районах юго-
востока. Куколки совок здесь лежат в колыбельках на глуби-
не 20—25 см в течение 1,5—2 месяцев — до периода осенних
Дождей. Насекомые засушливых мест более стойки к недо-
статку влаги за счет использования метаболической воды и
приспособительного поведения.
Известен случай анабиоза при высыхании. Личинки кома-
ров Dasychelea geleiana Mg. (Ceratopogonidae) живут в не-
больших, часто пересыхающих лужах среди скал в горах
Венгрии. При высыхании лужи личинки зарываются в детрит,
перестают двигаться, а потом, когда детрит пересыхает и ста-
новится пылью, личинки превращаются в сухие сморщенные
мумии. В высушенном состоянии личинки могут находиться
37
до 3 и даже до 5,5 месяцев. При непродолжительном пребыва-
нии в высушенном состоянии они оживают через несколько
минут после попадания в воду.
Большой влажности для развития требуют яйца многих
насекомых. Вес яиц саранчи за счет адсорбции воды увеличи-
вается от 6 до 14 мг; содержание ее поднимается с 52 до 80%.
Рис. 10. Влияние влажности на ги-
грофилов (а) и ксерофилов (б) (ио
Н. Andrewartha, L. Berch):
1 — долговечность; 2 — плодови-
тость; 3 — скорость развития; 4 —
смертность
Адсорбция воды яйцами пустынной саранчи происходит лишь
из влажного субстрата (песок) и невозможна из влажного
воздуха. На плодовитость насекомых положительно влияет
влажный корм, дожди, особенно весной. Весенние дожди мо-
гут быть причиной высокой численности второго поколения
кукурузного мотылька. Прямое и косвенное действие выпадаю-
щих дождей, снега, росы может приводить к гибели насеко-
мых или способствовать их размножению. Годы с небольшим
количеством осадков особенно гибельны в пустынях. Гибель
насекомых вызывают и ливни, иногда полностью прекращая
массовое размножение вредителей. В тропических странах
частые летние дожди отражаются на численности, составе и
приспособлениях насекомых. Например, многие тли здесь не-
полноциклые, имеют плоское тело, сильно покрыты восковыми
выделениями.
Переживанию насекомыми суровых зим помогает обильный
снеговой покров. Необычно морозная зима 1955/56 г., когда
в течение 22 дней температура не поднималась выше —20°,
привела к значительной гибели (67%) яблоневой запятовид-
38
лой щитовки и других вредителей садов. Сохранились лишь
насекомые, зимовавшие под снегом.
Общие закономерности влияния влажности на смертность,
скорость развития, плодовитость насекомых ксерофилов и ги-
грофилов сведены на рис. 10.
Действие термических условий зависит от влажности. Низ-
кая влажность при высоких температурах оказывает иное
Рис. 11. Влияние температуры и влажности на смерт-
ность гусениц монашенки
воздействие, чем та же влажность при низких температурах.
Поэтому в экологии принято оценивать совместное действие
температуры и влажности. Для этой цели широко пользуются
графическим методом анализа. Обычно на график наносят
показатели выживаемости и смертности насекомых, получен-
ные экспериментально при разных комбинациях температуры
и влажности. В качестве примера на рис. 11 показана смерт-
ность гусениц монашенки (Porthetria monacha L.) в разных
гидротермических условиях.
Из диаграммы видно, что оптимальные условия для этого
вредителя наступают при температуре 17—23° и влажности
Аналогичные данные о витальных границах комбинирован-
ного влияния температуры и влажности на развитие яйца и
гусеницы сосновой совки (Panolis flammea Schiff.) показаны
на рис. 12.
Исследования показывают, что колебания температуры и
влажности играют большую роль в появлении вспышек мас-
сового размножения насекомых, а также в понижении раз-
множения и гибели насекомых. Для изучения зависимости
размножения насекомых от погодных условий пользуются
графическим методом биоклимограмм. Для построения кли-
39
мограмм необходимо иметь средние многолетние данные по
температуре и осадкам и данные анализируемого года по де-
кадам. По ним строят график (рис. 13), позволяющий срав-
Рис. 12. Влияние температуры и влаж-
ности на развитие сосновой совки
Рис. 13. Биоклимограмма
(по А. И. Ильинскому)
1 — многолетние данные; 2 — дан-
ные за один год
нить для каждой декады отклонение показателей температуры
и влажности.
Климограмма сопоставляется с данными о состоянии фаз
развития вредителя, также обозначенные на графике. Они
подтверждают, что погодные условия 1936 г. были благоприят-
ны для развития совки; в первой половине вегетационного се-
зона стояла засушливая погода с высокой температурой, низ-
кой влажностью и безоблачным или малооблачным небом.
40
Биоклимограмму можно получить и другим спосооом
/пис. И). Центральную линию принимают за ось многолетних
редких температур и осадков, кверху от нее наносят отклоне-
ния от средней для данного года положительные, а книзу —
отрицательные температуры. Вдоль центральной оси простав-
ляют показатели фаз развития поднадзорного вредителя.
Рис. 14. Биоклимограмма (по А. И. Ильинскому):
1 — осадки, 2 — температура
Относительные показатели температуры дополняют абсолют-
ными данными.
В природе влажность воздуха и субстрата, осадки, темпе-
ратура и солнечная радиация действуют одновременно, оказы-
вая разнообразные влияния на все стороны жизни насеко-
мых
КЛИМАТ И МИКРОКЛИМАТ
Конкретное влияние климатических факторов на насеко-
мых осуществляется через состояние погоды и образование
микроклиматов. На протяжении климатической зоны (напри-
мер, тайги, степей) формируются многолетние режимы погоды
со своими особенностями атмосферной циркуляции. Над евро-
пейской частью СССР различают несколько типов погоды. За-
падный {тип циркуляции приносит тепло зимой и холод летом,
восточный тип зимой приносит холод, а летом тепло. Погода
1 Методы измерения влажности и соответствующие приборы описаны
в книге И. В. Кожанчикова (1961) и Н. А. Горышина (1966), а методы
анализа погоды, необходимые для прогнозов о состоянии вредителей, крат.
изложены у А. И. Ильинского (1965). Принятая в СССР для при-
кладного использования шкала условных обозначений фенологии насеко-
МЫх разработана Б. В. Добровольским (1961, 1969).
41
может установиться ясная, малооблачная, теплая и сухая или
облачная, дождливая и ветреная.
Естественно, что при разных типах атмосферной циркуля-
ции условия жизни насекомых в разные периоды сезона и
в разные годы складываются либо благоприятно для перези-
мовки, размножения, питания, либо наоборот, что ведет к из-
менению численности популяций.
С переменой погоды связаны миграции насекомых, коле-
бания их численности. Так, периодическое вторжение аркти-
ческого воздуха наблюдается в притропических пустынях, где
обитает пустынная саранча. Вторжение арктического воздуха
приводит к обильным дождям и мощному развитию расти-
тельности, что в свою очередь ведет к нарастанию численности
саранчи и ее массовым вылетам за пределы первоначальных
очагов (Щербиновский, 1952). С годами засух связывают раз-
множение ряда других вредителей (сибирский шелкопряд —
Dendrolimus sibiricus Tschw., непарный шелкопряд — Porthet-
ria dispar L., сосновая совка и др.). В засушливых степях
Волгограда необычно дождливый 1952 г. вызвал массовое
появление тлей, совки-карадрины (Laphygma exigua Hb.),
степного кистехвоста (Orgyia dubia Tausch.), капустной моли,
зеленой дубовой листовертки и ряда других видов. Ливневые
дожди, ветры, заморозки весной, оттепели зимой, сильные мо-
розы могут прекратить размножение насекомых.
Очень велико и еще мало изучено влияние климата и пого-
ды на пищу насекомых и их многочисленных врагов — пара-
зитических и хищных энтомофагов.
Климатические факторы определяют географический ареал
видов, зону их постоянного и неустойчивого вреда, особенно-
сти поведения и расселения. В районах, где сильные постоян-
ные ветры, насекомых мало и деятельны они в часы затишья.
С воздушными течениями насекомые переносятся на 1600—
1800 км. Вместе с циклонами совершают миграции стаи са-
ранчи, перемещаясь из зон высокого давления в области
с низким давлением. В теплом секторе циклона происходит
взлет бабочек лугового мотылька, а направление его полета
совпадает с направлением ветра. Бабочки перелетают на
400—600 км и оседают вместе с дождем там, где холодный и
теплый фронт воздуха надвигается друг па друга и ветер за-
тихает.
Огромное количество бабочек непарного шелкопряда было
занесено теплым юго-западным ветром вместе с грозой
в Москву и ее окрестности в июле 1957 г. Бабочки кружились
на площадях и улицах города, залетали в дома и даже откла-
дывали яйца на цветочные горшки и другие предметы \ Мас-
1 Более 2000 самок непарного шелкопряда отложили яйца под цоко-
лем здания биолого-почвенного факультета МГУ на Ленинских горах
(по нашим подсчетам, проведенным на четвертый день после залета ба-
бочек 20. VII 19*57 г.).
42
сОвое появление вредителя создало серьезную угрозу лесным
насаждениям.
В массовых миграциях большое значение имеет законо-
мерное перемещение циклонов и антициклонов. В лесах Ка-
нады установлена зависимость появления западного кольча-
того шелкопряда Malacosoma pluviale с движением центра
циклона в районах с северными широтами 52,5—55° и запад-
ными долготами 100—115°. Массовое появление шелкопряда
в этих районах сопровождается вторжением центрального
циклона с юго-запада (Wellington, 1964).
Для выяснения действия климатических факторов непо-
средственно на те или иные виды насекомых метеорологиче-
ских данных оказывается часто недостаточно. Действие кли-
мата преломляется через гидротермический и световой режи-
мы разнообразных местообитаний, которые характеризуются
своим экологическим климатом, или «экоклиматом». Термин
был предложен Б. П. Уваровым для характеристики особен-
ностей климата леса, болота, песчаных дюн, лугов, полей и
других местообитаний. Это понятие играет большую роль
в экологии. Нередко бывает так, что два вида живут рядом,
как будто в одинаковых условиях, а более внимательный ана-
лиз дает глубоко различные показатели.
Под кронами деревьев и на поляне, например, показатели
температуры, влажности, света различны. В лесу колебания
температуры выражены меньше, влажность выше, нет ветра,
зимой почва меньше промерзает. Густая крона деревьев за-
держивает много солнечных лучей, изменяя их спектральный
состав. В зависимости от состояния полога его проницаемость
для радиации различна. В период облиствления количество
радиации, проникающей сквозь полог, падает с 60% (начало
апреля) до 8% (конец мая). Летом в период максимального
развития полога сомкнутые насаждения высокой полноты
пропускают лишь 4—8% солнечной энергии. В условиях лесо-
степи в период максимального развития полога при одинако-
вом размещении и количестве растений на единйце площади
липа пропускает 4,7%, дуб 13,6%, ясень 18,4% радиации.
В подмосковном лесу первый ярус из осины и березы задер-
живает 20% радиации. В июне — августе под полог насажде-
ния проникает 10—13% радиации (Созыкин, Кожевникова,
1.965). Свои особенности освещения, температуры, влажности
складываются в лесополосах, в зависимости от их возраста,
состава пород, структуры. Лесная полоса снижает скорость
ветра в 5—10 раз, повышает влажность воздуха, задерживает
снег, способствует промачиванию почвы. В зависимости от
этих изменений меняется и фауна насекомых.
Большое влияние растений на климатические факторы под-
черкивается термином «фитоклимат». Но при изучении эколо-
гии насекомых во многих случаях и знание фитоклимата бы-
43
вает недостаточно. Необходимы сведения об условиях жизни
не только под пологом того или иного типа леса, лесополосы
и т. п., но и под корой дерева, затененного или освещенного,
в разных ярусах травяного покрова и других небольших уча-
стках, отличающихся своим микроклиматом.
Провести четкую грань, где кончается экоклимат, а где на-
чинается микроклимат, бывает трудно. Принято считать (Нау-
мов, 1963), что экоклпмат — это климат относительно крупных
участков, покрытых однотипной растительностью, а микрокли-
мат— частные особенности климата отдельных небольших
участков. Микроклимат характеризуется абсолютными пока-
зателями тепла, влаги, давления, отличаясь степенью общей
устойчивости и степенью контрастности сезонов, как и клима-
ты различных зон. В каждой зоне существует огромное разно-
образие микроклиматов. Суточные и сезонные изменения ми-
кроклимата различны на склонах разной экспозиции, в раз-
ных ярусах растительного покрова, на теневой или освещен-
ной стороне дерева и т. п.
Так, например, в жарких районах температура на теневой
стороне дерева была 2°, на солнечной — 33°. В воронке му-
равьиного льва (Eureoleon nostas) восточный склон днем на-
гревается больше, чем другие склоны на 3—5,5°, и меньше,
чем поверхность почвы рядом на 13,7° (24° в воронке, 37,7° —
рядом). Личинки муравьиного льва в соответствии с положе-
нием солнца ориентируют тело утром в направлении запад —
восток, в полдень — север — юг, после полудня — восток —
запад.
Значительные различия температуры на поверхности коры
и под корой при разной экспозиции зафиксированы на стволе
лиственницы (Петренко, 1965) (табл. 2).
Таблица 2
Температура бревен лиственницы
(в 16 ч при t° воздуха 14,5°)
Участок бревна Освещенная сторона Затененная сторона
ср. макс. мин. ср. макс. МИН.
Поверхность коры Под корой по- 21,9 25,2 19,8 14,4 15,0 13,8
верхность дре- весины 18,2 19,6 17,4 13,4 15,2 12,4
Нагревание деревьев лиственницы на опушке и в куртине
также весьма различно (табл. 3).
Установлена отчетливая связь микроклимата с силой сол-
нечной радиации, силой ветра и другими показателями клима-
44
Таблица 3
Температура деревьев лиственницы в зависимости от экспозиции
(в 12 ч при t° воздуха 26° на высоте 1,5—2 м)
Местоположение лиственницы Южная экспозиция Северная экспозиция
кора щели коры кора щели коры
На опушке dm — 32 см 38,1 35,0 26,8 25,1
В куртине dm—30 см 27,1 23,0 27,4 22,6
та в зависимости от сроков и густоты посева, направления
рядков и полос и от высоты растений, что позволяет активно
менять условия жизни насекомых. Направление сосновых ку-
лис с севера на юг ведет к усилению утреннего и вечернего
освещения с преобладанием длинноволновых лучей, что бла-
гоприятно для расселения насекомых северных широт; то же
происходит и на посевах сельскохозяйственных растений. Свои
микроклиматические условия создают разные виды и сорта
растений, гнездовые и рядовые посевы дуба, одиночно стоя-
щие деревья и кусты, особенно в полупустынных и пустынных
условиях. Все это резко сказывается на видовом составе и
обилии насекомых. Например, считается, что в верхнем слое
1 га цветущего люцерника в погожие дни обитает 1,5 млн.
полезных видов перепончатокрылых, а вместе с пчелами об-
щая сумма паразитических и хозяйственно полезных насеко-
мых на 1 га более 2 млн.
Особые условия многие насекомые создают сами, прогры-
зая мины в листьях, побегах и плодах, делая из листьев сверт-
ки, устраивая чехлики и коконы, а также образуя плотные
скопления, меняющие температуру и влажность мест обитания.
СЕЗОННАЯ РИТМИЧНОСТЬ ЖИЗНИ НАСЕКОМЫХ
Цикличность размножения насекомых приспособлена к се-
зонным изменениям погоды и наличию корма. Ритм жизнен-
ных процессов соответствует ритму сезонных изменений кли-
мата; если такое соответствие нарушено какими-либо причи-
нами, то это приводит к гибели насекомых. У непарного шел-
копряда и шелкопряда-монашенки зимуют кладки яиц. В не-
которые годы с теплой осенью вылупляются гусенички, но зи-
мовать они не могут и погибают. Массовая гибель потомства
наблюдалась и у некоторых короедов, когда холода и замо-
розки застигали их в фазе куколки или личинки. Цикличность
развития насекомых тесно связана с цикличностью развития
45
кормовых растений и вызывается одними и теми же климати-
ческими факторами. Поэтому наблюдается синхронность опре-
деленных фаз развития насекомых и растений. С повыше-
нием температуры и началом сокодвижения и набухания по-
чек связано весеннее пробуждение многих насекомых. В этот
период в крону деревьев поднимается казарка (Rhynchites
bacchus L.), яблонный цветоед (Anthonomus' pomorum L.),
начинают питаться гусеницы боярышницы (Aporia crategi L.),
непарного шелкопряда (Porthetria dispar L.), червецы (Coc-
cidae), тли (Aphididae), откладывает яйца грушевая медяница
(Psylla pyri L.), а личинки яблонной медяницы (Psylla mail
L.) собираются на верхушках лопающихся почек и т. д.
Зависимость сезонных явлений в мире животных и расте-
ний от повторяющихся изменений погоды изучает фенология.
Фенодаты меняются в зависимости от широты, долготы и вы-
соты над уровнем моря. Например, перезимовавшие гусеницы
яблонной моли выходят из-под щитков в период распускания
почек яблони. На высоте 800 м в Средней Азии это происхо-
дит в первой декаде мая, а на высоте 1600 м— во второй де-,
каде июня. Соответственно и зимующие гусеницы на высоте
800 м появляются в первой декаде августа, а на высоте
1600 м— в конце сентября (Петров, 1955).
Насекомые отличаются неустойчивым уровнем обмена ве-
ществ, меняющимся в зависимости от смены микроклиматиче-
ских режимов. В течение сезона закономерно меняются абсо-
лютные показатели действия света, тепла, влаги. Обычно по.
условиям жизни противопоставляют весенне-летний период,
благоприятный для активной жизни, развития и размножения
насекомых, и осенне-зимний — с неблагоприятными условиями
жизни для них. Правильнее оценивать значение кламатиче-
ских факторов по их периодически меняющейся направленно-
сти действия в разные сезоны года. В наших широтах можно
выделить четыре сезонных цикла с разным сочетанием дей-
ствующих факторов, что находит отражение в особенностях
жизни насекомых и в смене их требований к условиям жизни.
Характеризуются они следующими особенностями.
1. Период нарастания длины дня (с 22/ХП) при низких
температурах сопровождается зимним спектром радиации
с преобладанием длинноволновых лучей и температурой ниже
порога развития. Продолжительное действие низких темпера-
тур, увеличение фотопериода способствуют реактивации диа-
паузирующих насекомых и их переходу в холодовое оцепене-
ние. Зимние оттепели ведут к высокой смертности зимующих,
но уже реактивированных насекомых неустойчивых к положи-
тельным температурам. Насекомые к концу этого сезона бы-
вают подготовлены к быстрому переходу к активной жизни
и этот момент определяется лишь временем перехода темпе-
ратур через порог развития.
46
2. Период резкого нарастания длины дня (с 23/111}
с одновременным повышением температуры и влажности.
Зимний спектр радиации меняется на весенний — увеличивает-
ся ультрафиолетовое и коротковолновое излучение. 23/Шдень.
становится равным ночи, температура в ряде районов перехо-
дит пороговые величины, что вызывает массовое «пробужде-
ние» насекомых и переход их к активной жизни. Поздние ве-
сенние заморозки, весенние суховеи могут вызвать гибель
насекомых. От таких невзгод насекомых спасает растянутость
лёта, обеспечивающая выживание какой-то части популяции,
а также способность использовать всевозможные укрытия.
3. Период постепенного сокращения длины дня (с 22/VI)
при высоких нарастающих температурах. Спектральный со-
став лучей и их интенсивность меняются, влажность воздуха
снижается. Активная жизнь насекомых продолжается до по-
роговых величин фотопериода (15—1472 ^), наступление ко-
торых приводит к постепенному прекращению роста, морфо-
генеза, размножения. Усиливается торможение всех активных
процессов жизнедеятельности, насекомые переходят в состоя-
ние диапаузы и термоустойчивости.
4. Период заметного сокращения длины дня, сопровож-
дающийся снижением температур вплоть до пороговых вели-
чин. 23/IX день снова равен ночи, но направленность измене-
ний светового, температурного режимов и режима влажности,,
а также качества пищи иные, чем в период весеннего равно-
денствия. Состояние диапаузы, вызванное непродолжитель-
ным фотопериодом, усиливается действием низких температур
и переходит в глубокий осенний покой.
Таким образом в течение года у насекомых период актив-
ной жизни сменяется периодом перестройки и подготовки
к осенне-зимнему покою, а последний — заблаговременной
подготовкой к активной жизни. Утверждение, что у насекомых
имеются глубокие адаптации лишь к переживанию неблаго-
приятных условий бескормицы и холодов, неверно. Не мень-
шее значение имеют приспособления к максимальному исполь-
зованию благоприятных для развития и размножения весен-
них условий.
ПИЩА КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Насекомые питаются готовыми органическими вещества-
ми. Они могут быть фитофагами (более 50% насекомых), эн-
томофагами, гематофагами, сапрофагами, некрофагами, коп-
рофагами, ксенофагами. Есть насекомые, питающиеся воском,
клетчаткой, кератином волос, перьев, рогов, копыт. Личинка
музейного кожееда (Afithrenus museorum L.) развивается на
чистом шелке, а личинка Psilopa petrolei живет в нефтяных
Источниках. Во многих случаях насекомые питаются смешан-
47
ной пищей: и растительной, и животной. Для многих насеко-
мых характерна резкая смена режима питания у личинок и
имаго, сопровождающая смену среды обитания. Многие энто-
мофаги (двукрылые, перепончатокрылые) во взрослом состоя-
нии переходят на питание пыльцой и нектаром цветов, многие
кровососы в личиночный период жизни питаются планктоном
и т. д. Различают насекомых стенофагов и эврифагов. Бывает,
что личинка — стенофаг, а имаго того же вида — эврифаг (на-
пример, личинки желудевого долгоносика Curculio glandiutn
Marsch. питаются желудями, а жуки — на очень многих де-
ревьях и кустарниках). Насекомых делят также на моно-,
олиго-, поли- и пантофагов. Деление это условно. Ряд много-
ядных видов имеет излюбленные растения. Перелетная саран-
ча предпочитает злаки, особенно тростник, а итальянский
прус (Calliptamus italicus L.)—полыни. Непарный шелко-
пряд в одних районах предпочитает дуб, в других — березу.
Особенности питания в разных отрядах разнообразны.
Прямокрылые Orthdptera, термиты Isoptera, тараканы Blatto-
dea, уховертки Dermatoptera преимущественно многоядны;
специализация питания встречается редко. Например, из са-
ранчевых саксауловая горбатка (Dericoris annulata Fieb.) пи-
тается только саксаулом, а пустынница (Sphingonotus halo с-
nemi Uv.)—сарсазаном. Отряд богомолов Mantodea пред-
ставлен многоядными хищниками. Среди сетчатокрылых Neu-
roptera, тоже хищников, наблюдается специализация: Mantis-
pa питаются пауками, a Chysopa — тлями.
Среди жуков Coleoptera много многоядных хищников, мно-
гоядных фитофагов. Наряду с этим есть немало и стенофагов
(узкотелые златки Agrilus, долгоносики — ринхиты Rhynchites,
семееды Tychius, зерновки Bruchidae и т. п.). Широкой поли-
фагией отличаются виды, обитающие в почве в личиночный
период жизни (пластинчатоусые, щелкуны, чернотелки, пыль-
цееды). Многим из них свойственна сапрофагия, дающая воз-
можность длительное время обходиться без живой раститель-
ной пищи. Особенности передвижения в почве затрудняют
выбор специальной пищи, что и ведет к полифагии.
В отряде чешуекрылых преобладают фитофаги разной сте-
пени специализации — от полифагов до монофагов. Последних
много среди минеров, листовертов. Хищничество развито сла-
бо. Многоядны гусеницы подгрызающих совок.
Очень разнообразно питание полужесткокрылых (Hemipte-
га). В этом отряде развита и фитофагия, и зоофагия. Наряду
с обширными семействами слепняков (Mieridae), щитников
(Pentatomidae), кружевниц (Tingitidae) и других, большин-
ство видов которых питаются соками растений, имеется ряд
семейств хищников (Reduviidae, Nabidae, Anthocoridae, Gerri-
dae и др.).
Представители нескольких семейств питаются кровью теп-
48’
локровных животных и человека. Для водных клопов (Noto-
nectidae, Belastomatidae) постоянной пищей служат мальки
рыб, головастики, а для наземных хищных клопов — насеко-
мые, их яйца, гусеницы и т. д. Клопы-фитофаги питаются гри-
бами-трутовиками, соком деревьев, листвы, хвои, семян, пло-
дов, лишайников и мхов, папоротников, злаков, плавающих
водных растений.
В противоположность клопам в отряде равнокрылых (Но-
moptera) все виды фитофаги. Из них червецы обычны на тра-
вянистых растениях, а щитовки — на деревьях и кустарниках.
Большую группу образуют одноядные червецы. Встречаются
не только виды, но и роды червецов и щитовок и даже семей-
ства стенофагов. Например, все представители сем. Кеггпо-
coccidae живут на дубах, виды рода Leucaspis—на соснах.
Специализированное питание распространено и в других груп-
пах Homoptera. Только на хвойных живут тли рода Cinara.
С постоялыми видами растений связаны многие цикадки и
особенно листоблошки Psyllodea; это использовали как видо-
вой диагностический признак в своих работах А. К. Мордвил-
ко и другие систематики.
Определенных хозяев, придерживаются эктопаразиты
в отрядах вшей (Anoplura), пухоедов (Mallophaga), блох
(Aphaniptera). При отсутствии специфического хозяина блохи
могут переходить и на других животных; это имеет большое
эпидемиологическое значение.
Очень разнообразно питание двукрылых Diptera. Наряду
с фитофагами, например злаковыми мухами (Chloropidae),
распространены энтомофаги (тахины — Tachinida, серые мяс-
ные мухи — Sarcophagidae и др.), гематофаги (комары, слеп-
ни, мокрецы, мошки), паразиты животных — оводы (Oestri-
dae). В отряде перепончатокрылых (Hymenoptera) разнооб-
разны фитофаги (пилильщики Tenthredinidae, рогохвосты —
Siricidae, орехотворки — Cinipidae и др.), паразиты — энтомо-
фаги (наездники Ichneumonidae, Chalticidae, Braconidae
и др.), хищники, запасающие парализованную добычу для
потомства (роющие осы — Sphecidae и др.), нахлебники-ку-
кушки (осы блестянки Chryzididae и др.), антофилы, питаю-
щиеся пыльцой и нектаром (пчелиные).
Специализация питания у насекомых проявляется не толь-
ко в выборе предпочитаемых растений, но и определенных их
органов и тканей — ксилофаги, плодожорки, корнегрызы, ли-
стогрызы и т. д.
Особенности питания отразились на строении ротового ап-
парата, форме тела, способах движения, особенностях пове-
дения. В ряде случаев насекомые обладают специфическим
набором ферментов, позволяющим перерабатывать и усваи-
вать клетчатку, шерсть, воск, нефть. Насекомые, выедая лишь
определенные ткани, глубоко меняют биохимический состав
4 Заказ 193 '49
потребляемого растения. Ряд филлофагов не переваривают
клетчатки, но хорошо усваивают белки клеток, поглощая ка-
ротиноиды, хлорофилл. Минеры, питающиеся палисадной па-
ренхимой, поедают много углеводов. В губчатой ткани вы-
едаются и белки, и углеводы. В результате размножения тех
или иных минеров в листьях изменяется химический состав и
направление обмена веществ. Активные изменения в химизме
потребляемой пищи производят насекомые инъекцией фер-
ментов, вырабатываемых слюнными железами. Соответст-
вующие защитные реакции растений зависят от того, каким
частям, органам и тканям нанесено повреждение, при каком
состоянии растения и каким видом вредителя.
Типы повреждения разнообразны и постоянны. Насекомые
с грызущим ротовым аппаратом производят скелетирование,
сплошное объедание, изъязвление, объедание окошечное и фи-
гурное, разнообразное минирование, прогрызание ходов внут-
ри стеблей, стволов, ветвей, корней, выедание почек, генера-
тивных органов и т. д. Характер повреждения, рисунок хода
служит часто более легким диагностическим признаком вида,
чем его морфологические признаки. Насекомые с колюще-со-
сущим типом ротового аппарата производят интерцеллюляр-
ный укол без нарушения целостности клеток или интрацсл-
люлярный прокол с сильным нарушением растительных тка-
ней. Многие виды при уколе инъицируют ферменты. В резуль-
тате повреждений колюще-сосущими насекомыми наблюдает-
ся изменение окраски вокруг укола, образование вздутий на
листьях, их скрученность, засыхание листьев, искривление
веток, цветоножек, опадение бутонов и цветков, образование
галлов. Повреждения могут приводить к отставанию в росте,
снижению холодоустойчивости.
Выбор предпочитаемой пищи зависит как от свойств на-
секомого, так и особенностей растений. В ряде случаев одна
предпочитаемая пища сменяется другой, в зависимости от
сложившихся условий. Шестизубый короед (Ips sexdentatus
Воегп.) на Кавказе предпочитает заселять ель, в европейской
части СССР — обыкновенную сосну, в Сибири — сибирский
кедр, а в Приморском крае — кедр корейский. Короед-типо-
граф (Ips typographus L.) на Кавказе сильно повреждает
обыкновенную сосну, в Сибири — сибирскую ель, в Примор-
ском крае — саянскую ель. Обычный вредитель ели—большой
еловый лубоед (Dendroctonus micans Kug.) — в сильно забо-
лоченных местах предпочитает сосну. Известно, что насекомые
встречаются на свойственных им кормовых растениях в ме-
стах, типичных для их произрастания. Короеды (Ipidae) очень
чувствительны к влажности, сокодвижению и температуре в
местах размножения. Поэтому деревья они заселяют неравно-
мерно; разные виды селятся в различных частях ствола, кро-
ны, корней.
50’
Большое значение для заселения того или иного дерева
имеет его состояние. Короеды, усачи (Cerambicidae), златки
(Buprescidae) предпочитают ослабленные деревья. Но в ряде
случаев ослабление дерева достигается предварительным на-
падением на него массы короедов. Первые партии нападаю-
щих жуков гибнут, но последующие — благополучно разви-
ваются, так как дерево теряет защитные функции. Ослабление
деревьев достигается также дополнительным питанием жуков
в кронах и в ветвях. Так, короеды-садовники (Blastophagus pi-
niperda L., В. minor Hart.), втачиваясь в сердцевину тонких
веточек на вершинах здоровых сосен, вызывают обламывание
ветвей (иногда до 75%), что ведет к ослаблению всего дерева,
особенно при повторных нападениях жуков. Непригодность
древесной породы для питания насекомых обусловливается
содержанием эфирных масел, степенью кислотности соков,
ядовитостью, обилием выделяемой смолы и соков, их давле-
нием, механическими свойствами древесины.
Некоторые виды насекомых специализированы на питании
растениями, выделяющими млечный сок, содержащими эфир-
ные масла (гусеницы молочайного бражника — Celerio
euphorbia L., махаона — Papilio machaon L.). Ядовитый сок
обезвреживается выделениями особой железы. Жуки-листо-
еды (Metasoma saliceti Ws.), живущие на ивах, имеют кож-
ные железы, выделяющие салициловую кислоту.
Выбор растений происходит активно в период откладки
яиц. На избирательную способность влияют разные причины.
Снабженные массой хеморецепторов насекомые с поразитель-
ной тонкостью выбирают определенную пищу, необходимую
для той или иной фазы их развития. Например, колорадский
жук бывает многочислен на одном виде картофеля (Solanum
tuberosum) и избегает другие виды. Жуки-щелкуны в период
дополнительного питания сначала питаются на одних расте-
ниях, потом переходят на другие. Бронзовый щелкун (Selato-
somus aeneus L.) сначала питается пыльцой сосны, а потом
переходит на пшеницу.
Для яйцекладущих самок злаковых мух большое значение
имеют анатомо-морфологические особенности сортов пшениц:
форма и плотность прилегания к стеблю листового язычка,
особенности кущения, опушение, а также фенологические осо-
бенности растения.
Мало еще изучено влияние микроклиматических условий,
создаваемых разными видами и сортами растений (густота
стояния, форма кроны, особенности кущения, обилие плодов,
затененность и т. п.), на избирательную способность насеко-
мых. Установлена связь развития насекомых с органогенезом
Цветка костра безостного и костра прямого. Питание личинок
пшеничного комарика (Sitodiplosis mosellana Gehin.) неопло-
Дотворенной завязью вызывает многогенерациоцный тип раз-
4* 51
вития, питание оплодотворенной завязью приводит к личиноч-
ной диапаузе, длительностью от нескольких месяцев до двух
лет.
Переход к новой пище, смена питания у насекомых обычна
в онтогенезе и бывает очень резкой. Общеизвестны смены
кормовых растений половых и бесполых поколений у тлей, от-
мечаются перелеты с травянистых растений на хвойные у не-
которых листоблошек. Смены пищевого режима своеобразны
в онтогенезе чешуекрылых. Гусеницы многих бабочек пи-
таются растениями с богатым содержанием белков, а бабочки
используют нектар цветов, богатый углеводами. Жуки часто
при дополнительном питании используют пыльцу цветов,
обильную белками и витаминами. Личинки комаров соскабли-
вают и заглатывают при фильтрации разнообразные микро-
организмы, а самки комаров обязательно должны напиться
крови, иначе у них не будут развиваться яичники.
Насекомые иногда переходят на питание необычным для
данного вида растением, что может привести к появлению но-
вого вредителя. Таких примеров известно много. На питание
подсолнечником с сорняков перешла подсолнечниковая огнев-
ка (Homoeosoma nebulella Hb.)\ картофельный жук на кар-
тофель— с диких пасленовых; мальвовая моль (Gelechia mal-
vella Hb.) стала вредить хлопчатнику; местный хальцид (Pros-
paltella aurantii) стал развиваться на калифорнийской щитовке
(Diaspidiotus perniciosus Comst.); то же произошло с тлевы-
ми коровками (Chilocorus bipustulatus L., Ch. renipustulatus
Scr.); филоксера (Phylloxera vastatrix Pianch.) перешла с ди-
ких видов винограда; кровяная тля, став вредителем яблони,
уже не может вернуться на основное растение — американ-
ский вяз.
Практика и история сельскохозяйственной энтомологии
показывают, что в каждой данной местности специализиро-
ванными вредителями новых культурных растений становятся
те виды насекомых, которые были связаны с родственными
видами дикорастущих растений. «Общей закономерностью
является то, что вводимые культурные растения получают
массовых вредителей из состава местной фауны в тех случаях,
когда в составе местной флоры оказываются растения, систе-
матически (и биохимически) близкие к ввезенному растению
и, кроме того, сходные с ним морфологически, фенологически
и экологически» (Гиляров, 1955).
Для полевых культур экологически наиболее близки степ-
ные растения. Массовым вредителем пшеницы в целинном
крае стала зерновая совка (Hadena sordida Bkh.). В юго-во-
сточной Азии на рис перешли местные стеблевые огневки
(Schoenobius inertellus Walk., Chilo simplex Butler, и др.).
В Северной Америке прослежен переход клопа Psallus seria-
tus Reut. на хлопок с растений целинных земель. В Южной
52
Африке на хлопок с мальвовых перешел красный коробочный
червь (Diparopus castanea).
В степях юго-востока европейской части СССР на новые
лесные полосы переходят в основном вредители с местной
древесной и кустарниковой растительности. Не повреждаются
или мало повреждаются виды древесных пород, завезенные
из других районов, и наиболее заселяются виды, близкие к
местной флоре. В лесополосах под Волгоградом массовыми
вредителями вяза перистоветвистого стали виды, обычные в
балках на бересте,— берестовый листоед (Galerucella luteola
Mull.), ильмовый ногохвост (Exareta ulmi Schiff.), вязовая ци-
кадка (Empoasca haupti Rib.), заболонник кирша (Scolytus
kirschi Seal.). На ясень зеленый и жимолость татарскую пере-
брался сиреневый бражник (Sphinx ligustri L.), по-видимому,
из местных садов. С терна на иргу перешла в полосы и на
вишню в сады черная златка (Capnodis tenebrionis L.). Сла-
бо заселяются чуждые местной флоре ясенелистный клен, бе-
лая акация.
Свекловичная муха (Pegomyia hyosciaini Pz.) может раз-
виваться и на свекле, и на беладонне, но мухи, выведенные
на свекле, предпочитают свеклу, а на беладонне — бе-
ладонну. На яблоню перебралась с черники пестрокрылка
(Rhagoletis pomonella Walsch.), дав новую форму с новым
образом жизни; то же произошло с клопом Plericoris rugicol-
lis, обычно живущим не на яблоне, а на ивах.
Смена кормового растения, привыкание к новому пищево-
му режиму — один из путей эволюции, ведущий к образова-
нию новых наследственных особенностей у насекомых. Экспе-
риментальными работами (В. И. Кузнецов, Е. С. Смирнов,
И. А. Рубцов, Г. В. Самохвалова, Г. X. Шапошников) на гу-
сеницах, тлях, щитовках показана возможность образования
новых наследственных свойств. Вынужденное питание стано-
вится постоянным. Новая избирательность к корму в опытах
с античной волнянкой (Orgyia antique L.) проявилась уже
через десять дней, у кольчатого шелкопряда (Malacosoma
neustria L.) —в пятом возрасте, у пядениц (Lycia) —лишь в
следующем поколении. В опытах с тлями адаптация к новому
корму появилась лишь через ряд поколений, после восьми
поколений способность жить на старом растении для тлей бы-
ла потеряна; появились новые признаки, позволившие гово-
рить о появлении нового вида.
У многоядных видов смена питания совершается непре-
рывно, что развивает и укрепляет многоядность.
Влияние пищи на рост и развитие насекомых. Для нор-
мального развития насекомых необходимо поступление с пи-
Щей белков, углеводов, воды, витаминов, различных зольных
веществ. Их содержание в потребляемой пище различно на
протяжении онтогенеза и в разные периоды сезрна, что ока-
53
зывает разнообразное влияние на процессы роста и развития.
Влияние качества п-ищи изучалось на многих видах насеко-
мых. Известно, например, что гусеницы непарного шелкопря-
да гибнут при постоянном питании листьями липы и хорошо
развиваются на листьях дуба. Сибирский шелкопряд лучше
всего развивается на хвое лиственницы и хуже — на хвое
сосны. Плодовитость бабочек в партиях, выкармливающихся
лиственницей, была 826 яичек, а сосной — всего 12,6.
Питание, несвойственное данному виду насекомых, экспе-
риментально вынужденное, приводит к высокой смертности и
резкому сокращению размеров тела и плодовитости. Так,
оранжерейная тля (Neomyzus circumflexus Buck.) при корм-
лении неблагоприятным для нее редисом произвела незначи-
тельное количество потомков карликовых размеров. Другие
крестоцветные растения действовали так же угнетающе на
развитие тлей, как и редис, что авторы связывают с наличием
в этих растениях специфического вещества — синигрина. Для
белянок же, постоянно питающихся крестоцветными, запах си-
нигрина оказался привлекающим.
Наиболее подробно влияние химического состава пищи
изучено на шелкопрядах и пчелах. Шелководы считают, что
гусениц тутового шелкопряда в каждом возрасте нужно кор-
мить листом соответствующего качества. Возраст гусеницы
должен соответствовать возрасту листа. Весной узким местом
в корме может явиться недостаточное содержание белков,
а для более поздних выкормок — содержание воды. Наиболее
ценен как корм сочный и богатый белками весенний лист
шелковицы.
Сходные результаты получены при выкормке гусениц ки-
тайского дубового шелкопряда листьями разных видов дуба.
В период вегетации сопоставлялись показатели роста гусениц
с биохимическими изменениями состава листьев дуба. На ро-
сте и развитии гусениц резко сказывается снижение содер-
жания воды и белков в корме. Недостаток воды особенно
неблагоприятен для молодых гусениц; он ведет к задержке
развития и высокой смертности. Влажный корм в последнем
возрасте способствует повышению плодовитости бабочек, од-
нако снижает вес шелковой оболочки. Добавление воды к
корму повышало плодовитость до 40%. Гусеницы младших
возрастов отличаются большой интенсивностью роста, что ве-
дет к повышенной потребности в белках. У гусениц последнего
возраста недостаток белков тормозит рост бурно развиваю-
щейся в это время шелкоотделительной железы (Андрианова,
1948).
Велико значение коллоидно-связанной воды, поступающей
с пищей. Недостаток влажной пищи приводит у насекомых
к уменьшению числа образующихся половых клеток, сниже-
нию плодовитости, а в ряде случаев — и к бесплодию. У насе-
54
комых афагов потенциальная плодовитость определяется ко-
личеством резервных жиро-белковых отложений и соответст-
венно метаболической водой. Недостаток воды, поступающей
с пищей, особенно в жаркую сухую погоду, вынуждает насе-
комых питаться необычной пищей; многие сапрофаги перехо-
дят на питание живыми сочными растениями.
Для многих насекомых отмечены переходы на питание раз-
ной пищей в зависимости от их половозрелости. Так, земляные
блошки (Chalticini) весной после зимовки грызут многие ра-
стения, затем переходят к питанию лишь определенной груп-
пой растений, богатой легко усваивающимися белками, что
обеспечивает накопление запасных веществ; в период актив-
ного размножения блошки становятся монофагами и питают-
ся лишь на растениях, обеспечивающих созревание половых
продуктов.
Значение отдельных элементов пищи меняется на разных
этапах онтогенеза. В ряде случаев имаго как бы восполняет
недостающие элементы пищи в личиночный период развития.
Например, рост личинок восточного майского хруща идет тем
интенсивнее, чем больше в корнях углеводов, а жуки, наобо-
рот, требуют больше белковой пищи. Углеводов больше в кор-
нях березы и сосны, а азота — в листьях дуба. Указанными
особенностями питания объясняется наличие постоянных оча-
гов размножения майского хруща там, где произрастают сос-
на, береза, дуб.
Плодовитость ячменной мушки (Oscinella pusilia Mg.)
резко падает, если этот вредитель злаков не может менять
кормовое растение хотя бы в течение нескольких поколений.
При однотипном питании в течение пяти поколений (только
на ячмене или только на яровой пшенице) количество потом-
ков резко снижается, а в последнем поколении выживае-
мость сводится к нулю. Переменное питание (три поколения
на яровой пшенице, одно на пырее и одно на озимой пшени-
це) приводит к резкому увеличению числа выживших потом-
ков. Личинки ячменной мушки развиваются только в укоро-
ченных вегетативных побегах; их число возрастает весной и
осенью; к этому времени возрастает число самок мушки со
зрелыми яйцами (Нарчук, 1955).
Биохимический состав корма, как и другие факторы сре-
ды, разнообразен. Зависит он от колебаний погодных усло-
вий, сезона, географического положения, условий произраста-
ния растений, микроклимата. В освещенном листе больше
растворимых углеводов и сухого вещества, чем в затененном.
На химизм растений оказывает влияние водный режим, со-
держание азотистых и зольных веществ в почве. Длительные
паводки изменяют биохимический состав дуба в благоприят-
ную для размножения непарного шелкопряда сторону. Под-
сев люпина в сосновых культурах улучшал азотистое питание
55
сосны, что неблагоприятно сказывалось на личинках майского
хруща. Обеднение почв азотом и зольными веществами (старо-
пахотные участки) изменяет водный режим, снижает защит-
ные функции деревьев и способствует размножению ряда
лесных вредителей. Искусственная дефолиация деревьев и их
окультуривание показали, что имеется оптиум биохимическо-
го состава (определенное соотношение азота, углеводов, воды
и других веществ), благоприятный для развития насекомых
(Положенцев и Ханисламов, 1954, 1962; Гримальский, 1958,
1961). Активность ферментативной системы и содержание пи-
тательных веществ понижается в окольцованном дубе, что
благоприятно для развития непарного шелкопряда, а прежде-
временная дефолиация ведет к неблагоприятным изменениям.
Отклонения от оптимума в химизме растений приводят к ги-
бели гусениц непарного шелкопряда (Амирханова, 1962).
Смерть гусениц наступала при содержании белка в листьях
дуба 12% (от веса абсолютно сухого вещества); смертельный
исход наступал и при избытке азотистых веществ (23%).
Меньше нормы белковых веществ имеют листья сосны, ели,
ясеня, вяза, лиственницы. На всех этих породах гусеницы раз-
виваются плохо. Оптимальное содержание протеина (17—
18%) установлено в листьях дуба, осины, березы, боярышни-
ка. С этими породами связаны очаги размножения вредителя.
Г еографическая изменчивость питания. Биохимический со-
став растений в разных климатических зонах различен. Гео-
графическая зависимость химизма растений ведет к зональ-
ной зависимости питания насекомых. В разных зонах кормо-
вые растения закономерно меняются.
Значительная часть «дубовой» фауны Средней Европы у
нас фигурирует как «березовая» (Фридолин, 1936). Семейство
белянок связано с генетическим рядом пищевых растений —
от примитивного комплекса (каперцовые — крестоцветные —
резедовые) к более высоко стоящему комплексу (розоцвет-
ные— мотыльковые) с параллельными ветвями (крушинные
и вересковые) (Кузнецов, 1930). То же отмечено для голубя-
нок. Гусеницы пестрянки (Zygaena), голубянки (Lycaena),
желтушки (Colias) в лесной и пустынно-степной зонах живут
на мотыльковых, а в открытых тундро-альпийских ландшафтах
переходят на каменоломковые и вересковые (Фридолин,
1936). Махаон по всей Евразии живет на зонтичных, а в об-
ластях с реликтовыми третичными растениями (Алжир, Уссу-
рийский край) — на рутовых. По нашим наблюдениям,
в бугристо-колковом ландшафте песчаных террас Сред-
него Дона (Арчеда) гусеницы махаона питаются только
полынью Маршалла, а не зонтичными, что, видимо, связано
с особенностями истории ландшафта. Перламутровки (Л г gin-
nis) питаются на розоцветных и особенно на фиалках в от-
крытых ландшафтах северного и альпийского типов, а на тор-
56
фяниках— еще и на вересковых. Обыкновенная свекловичная
блошка (Chaetocnema concinna Marsch.) в Западной Европе
живет только на гречишке и щавеле, в европейской части Со-
ветского Союза — и на маревых, а в Западной Сибири пред-
почитает маревые; в сухих и жарких условиях она может
питаться дубом. Непарный шелкопряд в Западной Сибири
предпочитает березу, а в Нижнем Поволжье — дуб.
Распространение многих монофагов ограничено кормовыми
растениями. Распространением дуба и сосны определяются
границы ареалов большинства древоядных насекомых. С на-
личием крестоцветных растений связано распространение ка-
пустной моли, капустной белянки, жуков-баридов (Baris),
с цитрусовыми связаны цитрусовый (Pseudococcus gahani
Green) и желобчатый (Iceria purchasi iMask.) червецы.
Стенофагия обычна среди представителей тропических ле-
сов с постоянным запасом пищи и более устойчивым клима-
том. Обилие однородной пищи способствует узкой специали-
зации питания и сопровождается более полным и эффектив-
ным усвоением корма. Использование другой пищи ограничи-
вается или даже исключается (Наумов, 1963). Бедность же
кормовой базы, редкие сезонные и годовые изменения ее ка-
чества и запасов ведут к многоядностп.
Стенофагия и эврифагия — два различных эволюционных
пути приспособлений насекомых к использованию пищи в ме-
няющихся условиях среды, к богатству и различной устойчи-
вости кормовой базы.
ОСОБЕННОСТИ ДЕЙСТВИЯ ФИЗИКО-ХИМИЧЕСКИХ
ФАКТОРОВ В РАЗНЫХ СРЕДАХ
Особенности жизни в воде по сравнению с жизнью на су-
ше определяются прежде всего обеспеченностью кислородом.
Способность воды растворять воздух — важнейшее условие
жизни в ней. Насекомые с трахейными жабрами (поденки,
веснянки, ручейники, вислокрылки, сетчатокрылые, стрекозы,,
вертячки) и личинки, дышащие через покровы тела (молодые
личинки комаров, веснянок, поденок и других), используют
воздух, растворенный в воде.
Содержание кислорода в воде зависит от глубины водо-
ема, обилия водной растительности, скорости течения, разло-
жения органических веществ и других факторов. В воду кис-
лород поступает из атмосферного воздуха и за счет фотосин-
теза растений: там, где много водорослей и зарослей
различных растений, кислорода больше. Но если поверхность
водоема зарастает так, что нарушается контакт поверхности
воды с воздухом, то кислорода становится мало. Неблагопри-
ятные условия для дыхания создаются в мелких водоемах с
энергично протекающим разложением органических остатков
летом, а также зимой при толстом ледяном покрове.
57
Потребность в кислороде у насекомых различна и зависит
от интенсивности процессов обмена, подвижности, возраста,
физиологического состояния, температуры среды. Недостаток
кислорода водные организмы переносят тяжелее, чем недо-
статок пищи (С. Н. Скадовский). Виды, чувствительные к не-
достатку кислорода (личинки мошек, многих веснянок, поде-
нок), приспособлены к жизни на быстринах (.реофилы) и в
стоячих водоемах погибают.
Солевой режим обеспечивает поступление воды в тело
насекомых через покровы тела. Насасывание воды регули-
руется осмотическим давлением. Лишняя вода активно выво-
дится через мальпигиевы сосуды, и концентрация солей внут-
ри тела всегда поддерживается на постоянном уровне — вы-
ше, чем в воде. Солевой режим водоемов очень различен, кон-
центрация солей в пресных водоемах, наиболее заселенных
насекомыми, колеблется от 0,01 до 16 г на 1 л.
Растворы различных солей, кислот и оснований, распа-
даясь на ионы, создают определенную щелочную или кислую
реакцию среды. Щелочную реакцию имеют водоемы, богатые
углекислыми солями (рН = 8—9); болота же типа сфагновых
и моховых отличаются кислой реакцией (pH = 6,9 до 1). Реак-
ция воды колеблется в разное время суток, разные сезоны,
зависит и от глубины водоема. pH воды закономерно на-
растает и падает при ослаблении или усилении фотосинтеза,
влияет на pH внутренней среды организмов и изменяет про-
ницаемость покровов насекомых, их водно-солевой обмен. Тре-
бования насекомых к pH среды также различны. Лишь не-
многие виды переносят ее колебания в широких пределах.
Личинки комара Chironomus plumosus L. выдерживают
кислую реакцию до pH = 2—3, они же могут жить в щелочной
среде (pH =12). Для личинок малярийного комара оптималь-
на слабокислая реакция, а для личинок мошек — слабоще-
лочная.
Минеральные вещества встречаются не только в раство-
ренном виде, но и в форме взвешенных в воде частиц. Ими
могут быть частицы глины, кварца, органо-минеральные ча-
стицы разложившихся организмов.
Пищевой режим в воде отличается тем, что ряд насекомых
питается только органическими взвесями, фильтруя воду. Без-
выборная фильтрация может привести к заполнению кишеч-
ника минеральными частицами, что мешает нормальному
усвоению пищи. Избыток минеральных частиц неблагоприя-
тен для ряда личинок насекомых. Так, рисовые поля, зали-
ваемые мутной речной водой, личинки малярийного комара
не заселяют. Молодые личинки комаров могут отфильтровы-
вать частицы размером 22,8—34,2 мк, а взрослые личинки за-
глатывают частицы размером 68—165,3 мк. Поэтому в воде,
имеющей лишь мелкодисперсные органические вещества, ли-
58
чинки могут развиваться до третьего возраста, личинки стар-
ше третьего возраста в таких условиях погибают от голода.
Другие способы питания в воде аналогичны питанию на
суше. Среди водных насекомых есть детритофаги, фитофаги,
много хищников-энтомофагов. Как и на суше, разнообразные
пищевые связи составляют основу биоценозов. В водоемах
создаются различные режимы температуры, освещения, насы-
щения кислородом, растворами различных солей, пищей. Все
это ведет к определенному распределению насекомых в гидро-
сфере, к образованию водных биоценозов.
Температурный режим водоемов более постоянен, чем
воздуха. Вода охлаждается и нагревается медленно. При
замерзании она расширяется, лед всплывает и благодаря ма-
лой теплопроводности льда вода под ним не замерзает. Жизнь
в водоеме продолжается и зимой. Но часть насекомых зимует
в толще льда, где температура, даже при —30° в воздухе,
остается равной —Г (С. А. Зернов). В таких условиях под
Москвой отмечена зимовка 26 видов насекомых.
По температурному режиму различают три группы водо-
емов: 1) водоемы тропические, температура в них не бывает
ниже 4°; 2) водоемы полярных областей и высокогорные озе-
ра, температура которых не поднимается выше 4°; 3) водо-
емы умеренного пояса, хотя бы ненадолго покрывающиеся
льдом и хотя бы ненадолго прогревающиеся выше 49. Годо-
вые колебания температуры глубоких озер этого пояса велики
и сложны, а суточные колебания незначительны. Небольшие
водоемы прогреваются насквозь; ветер перемешивает в них
воду, и различий температур в разных слоях не наблюдается;
температура в таких водоемах обычно выше, чем в воздухе.
Летом мелкие водоемы могут высыхать, зимой — промерзать
до дна. Прогреванию водоемов мешают водные растения, пре-
граждая путь солнечным лучам, а также массовое развитие
планктона.
Световой режим в воде также иной, чем на суше. Интен-
сивность света в водоемах значительно меньше. Большая
часть лучей отражается поверхностью воды и в воду не про-
никает. Особенно это сказывается на косых лучах, падающих
в утренние и вечерние часы, поэтому ночь в водоемах насту-
пает раньше и длится дольше, чем на суше. Чем больше глу-
бина водоема, тем короче в нем день и длиннее ночь. Вода
поглощает больше красных лучей, меньше синих. Слабее про-
никает свет в загрязненные водоемы с мутной водой; макси-
мум в них сдвигается в сторону желто-зеленых лучей. Корот-
коволновые, желтые и синие лучи видимого спектра прони-
кают глубже, чем лучи красной части спектра. Очень сильно
поглощаются ультрафиолетовые лучи.
К действию света водные насекомые очень чувствительны.
Вместе с зоопланктоном они совершают ритмические верти-
59
кальные миграции. Миграции наблюдаются даже в мелких
водоемах, глубина которых не превышает 30—40 см. Основ-
ной причиной вертикальных миграций живых организмов в
океанах и пресных водах считают способность при опреде-
ленных условиях двигаться ориентированно на свет. Насеко-
мые, живущие в илу и на поверхности грунта, обычно отри-
цательно фототаксичны, но при кислородном голодании и они
становятся положительно фототаксичными (Скадовский,
1955). Как и на суше, реакции на свет различны у разных
видов, в разные периоды развития и в значительной степени
зависят от условий температуры и других факторов среды.
ОСОБЕННОСТИ ЖИЗНИ В ПОЧВЕ
Почва состоит из твердых частиц, воды и воздуха (имеет
трехфазную структуру). От размеров почвенных частиц зави-
сит объем воды и воздуха в почве, а также возможность пе-
редвижения в ней насекомых.
Наибольшее количество насекомых связано с легкими поч-
вами, в которых складывается наиболее благоприятный гидро-
термический и воздушный режим. Чем влажнее почва, тем
меньше в ней воздуха. Наполнение пор водой может резко
нарушить дыхание насекомых. Но оптимальная влажность
воздуха для почвенных насекомых близка к насыщению; к вы-
сыханию они чувствительны почти в такой же степени, как и
водные организмы. Открытая трахейная система и проницае-
мость покровов для воды — показатели приспособленности
почвенных насекомых к существованию только во влажной
среде, в атмосфере с высоким содержанием паров воды. На
дефицит влаги насекомые отвечают активными миграциями
в более глубокие и влажные слои почвы, иногда до 2 м (бо-
гарный щелкун Pleonomus tereticollis Men.) и глубже. Глубина
миграций зависит от свойств почвы и условий климата. На
глинистых, засоленных почвах в полупустынных степях юго-
востока, по нашим наблюдениям, более глубокие слои почвы
так уплотнены н сухи, что миграции личинок происходят в
поверхностных слоях горизонтально в сторону отдельных
групп растений и кустарничков, дающих тень и сохраняющих
в почве больше влаги. Высокую влажность воздуха имеют
песчаные почвы: в них уже на глубине 8—10 см в жаркие и
сухие дни она близка к насыщению.
В почве спасаются от высыхания многие насекомые. Она
благоприятна для окукливания, откладки яиц. Яйца насеко-
мых при контакте с влажной почвой способны сильно набу-
хать, адсорбируя воду. Адсорбция может идти против осмо-
тического градиента, даже из раствора глюкозы с осмотиче-
ским давлением 14,6 атм. Личинки, живущие во влажных
почвах, не имеют воскового слоя (эпикутикулы) и других
60
приспособлений, регулирующих потерю воды. Для них опасен
дефицит почвенной влаги и потеря воды при испарении. Де-
фицит влаги в верхних горизонтах почвы бывает в период
засух и в засушливых районах, когда почва теряет всю влагу,
кроме гигроскопической, прочно связанной с частичками поч-
вы и недоступной живым организмам. Избыток же воды,
заполняющей скважины, вытесняет воздух, и насекомые мо-
гут погибнуть от удушения.
Воздушный режим в почве определяется постоянным
притоком кислорода из атмосферы. Чем влажнее почва, тем
труднее доступ кислорода. С глубиной становится меньше
кислорода и больше углекислоты. В верхних слоях (до 15 см)
свободный воздух заполняет промежутки почвы. По составу
он близок к атмосферному, но содержит в 10 раз больше СО2.
Состав его непостоянен и меняется в течение суток, сезонов,
глубины, активности организмов, влажности. Повышение тем-
пературы ведет к расширению воздуха и его выходу из почвы,
а при охлаждении — наоборот. Почва, как говорят, «дышит».
Для мелких форм (Protura, Collembola) характерно кож-
ное дыхание за счет растворенного в воде воздуха. Через ку-
тикулу идет газообмен у личинок долгоножек, проволочников
и других насекомых, имеющих трахеи. Поэтому они долго
могут быть в воде и не погибать, особенно при низких темпе-
ратурах. При разливах Днепра личинки щелкунов (Agriotes,
Athous) и других насекомых переносили затопление в течение
40 дней, если температура воды была не выше 10°, в период
же летних паводков при температуре 17—20° личинки в массе
гибли (А. Ф. Кришталь). Очень чувствительны к недостатку
кислорода насекомые в период активной жизни. Наличие кис-
лорода в почве в свободном и растворенном состоянии при
насыщенности воздуха водяными парами создает условия ды-
хания, промежуточные между водной и воздушной средой.
Температурный режим в почве определяется ее
механическим составом, влажностью, положением в рельефе,
степенью развития травяного покрова и подстилки, цветом
почвы. Влажная почва и нагревается, и охлаждается медлен-
нее, чем сухая. Глинистая почва нагревается меньше, чем пес-
чаная. На поверхности почвы колебания температуры резче,
чем в глубине и в воздухе. Под Волгоградом, например, свет-
ло-каштановые почвы нагревались до 55—62° при температу-
ре воздуха 40°, ночью температура падала до 15—20°. Суточ-
ные колебания температур сказываются до глубины 1 м.
Днем, с глубиной, температура падает, ночью возрастает.
С суточными изменениями температур связывают соответ-
ствующие миграции насекомых на небольшую глубину
(10 см). Осенью и весной наступают температурные реверсии.
Миграции насекомых в эти периоды более значительны. На-
пример, личинки восточного майского хруща на песках Сред-
61
него Дона осенью постепенно опускаются вглубь (до 90 см),
вслед за температурой 7—9°. В верхних слоях ко времени
осенней миграции температура ниже (6—5°). Основная масса
личинок зимует на глубине 40—70 см при температуре 2°—3°,
а весной начинается быстрый подъем личинок при про-
гревании верхнего горизонта почвы до 8—10°. Действие темпе-
ратуры в почве зависит от влажности, аэрации, зональных
условий.
Световой режим в почве изучен недостаточно. Из-
вестно, что лучи видимой части спектра проникают слабо.
Многие насекомые живут в верхних слоях или периодически
поднимаются к поверхности, где не исключено прямое влия-
ние света. Личинки в почве избегают света, но в фазе имаго
они очень чувствительны к свету. Наличие у личинок органов
зрения, прекращение суточных миграций в пасмурные дни,
миграции в глубь почвы, когда нет угрозы от высыхания, да-
ют основание предполагать, что суточные и сезонные пере-
движения обитателей почвы подобны ритмическим миграциям
водных организмов, вызываемых светом.
Особенности питания выражаются в эврифагии.
Многим видам свойственна сапрофагия, заглатывание субст-
рата с растительными остатками и гумусом, что сближает
питание почвенных насекомых с насекомыми бентоса. Чем су-
ше почва, тем больше насекомых питается живыми сочными
корнями растений. Во влажной почве личинки долгое время
могут обходиться без пищи. Вред, приносимый личинкой, тем
выше, чем суше и беднее перегноем почва.
В ряде случаев отмечается избирательность к пище. Ко-
марик Neosciara medesta Staeg., широко распространенный в
подстилке дубрав, как правило, питается опавшими листьями
кустарников. Многие личинки щелкунов предпочитают куку-
рузу и корневищные злаки, майский хрущ — корни сосны, не-
которые проволочники — посевы лоха, абрикоса и др.
Особенности среды обитания паразитических энтомофагов
и фитофагов мало исследованы. Жизнь в полостях и тканях
своеобразна и ведет к разнообразным приспособлениям для
движения, дыхания, ’размножения. Дыхание эндопаразитов
осуществляется за счет кислорода, растворенного в тканевых
жидкостях, использования воздуха трахей и контакта с атмо-
сферным воздухом. Формы личинок напоминают организмы,
живущие в воде — циклоповидные, с выростами, похожими
на плавники, и т. п. Интересны морфо-физиологические адап-
тации у некоторых энциртид. Дыхальца их личинок в первых
стадиях прикрепляются к воздухоносной полоске хориона яй-
ца, что и обеспечивает энциртид атмосферным воздухом. Не-
которые личинки яйцеедов имеют длинную хвостовую нить,
позволяющую взбалтывать яйцо хозяина, не давая ему раз-
виваться.
62
Личинки, живущие внутри листьев, в древесине и других
органах и тканях, приспособлены, в зависимости от мягкости
поедаемых частей, либо к активному прокладыванию ходов,
либо к раздвиганию рыхлых частей, как при движении в.
почве.
Во многих случаях насекомые активно изменяют среду,
выделяя ростовые или токсические вещества, разнообразные
ферменты, что приводит к образованию галлов, тератозов, из-
менению роста клеток. Например, в галлах яблоковидной оре-
хотворки найдено ростовое вещество — индолуксусная кисло-
та, выделяемая личинкой. Изучением взаимоотношений фито-
фагов с растениями-хозяевами, выяснением природы ответных
защитных реакций в форме новообразований клеток, тканей,
органов растений занимается цецидология. Биологический
смысл образования галлов для растений — ограничение миг-
раций возбудителей галлов и их токсинов, ограничение пато-
логических процессов. Изучение микросреды жизни паразити-
ческих насекомых — энтомофагов и фитофагов — необходимо
развивать.
ОБЩИЕ ЗАКОНОМЕРНОСТИ ДЕЙСТВИЯ ФАКТОРОВ СРЕДЫ
Анализ значения отдельных физико-химических факторов
среды показывает, что: 1). Факторы среды действуют в при-
роде не изолированно, а взаимосвязаны и представляют систе-
му факторов, и организм связан не с отдельными элементами
среды, а с их системой. 2). Действие фактора или комплекса
взаимосвязанных факторов в процессе онтогенеза насекомых
и в разные сезоны года различно, поэтому необходимо их зна-
чение выяснить отдельно для личинок, куколок, имаго и в раз-
ные сезоны. 3). Влияние среды сложно и действует на орга-
низм через изменения в направленности обмена, изменения
скоростей отдельных биохимических реакций. Насекомые ак-
тивно осваивают среду, приспосабливаются к ней сообразно
наследственным особенностям. Каждый вид реагирует на
влияние факторов среды различно, строго по-своему. 4). Эко-
логическое значение одних и тех же факторов различно, если
Другие факторы выступают в иных сочетаниях. Например,
песчаная почва в холодном и влажном климате бывает суше
и теплее глинистой почвы, а в сухом и жарком климате пески
служат аккумулятором влаги, оправдывая наблюдения иссле-
дователей пустынь: «где пески, там жизнь». 5). Важно знать
особенности действия факторов в разные периоды онтогенеза,
в разные сезоны года и разные годы в зависимости от мест-
ных условий. Если, например, сравнивать средние температу-
ры января в Архангельске и Астрахани, то они окажутся
близкими, однако годовой режим их в этих местах будет раз-
личен и в целом условия жизни также глубоко ^различны;
63
6). Экологические факторы в основном незамещаемы, но эф-
фект их действия может быть сходен. Так, свет не может быть
заменен избытком тепла, но, действуя низкой температурой,
в некоторых случаях можно получить диапаузу, как и при
действии короткого дня. Однако основным фактором, вызы-
вающим диапаузу, остается короткий день, и замещение фак-
торов ограничено. Недостаток тепла не заменить обилием
холодного света. Все факторы, необходимые для процессов
обмена, необходимы в равной мере, хотя ведущее значение
каждого из них в разные периоды жизни насекомых может
быть различно. 7). Имеет значение продолжительность и по-
вторность действия факторов; кратковременные воздействия
не вызывают существенных изменений в организме. 8). Мно-
гие факторы физической среды еще ждут своих исследований.
Мы еще мало знаем о влиянии на жизнь насекомых давления,
земного магнетизма, космических лучей, радиоволн, звуковых
волн, спектрального состава света, жесткого излучения и дру-
гих факторов.
ЗАКОНОМЕРНОСТИ ТЕРРИТОРИАЛЬНОГО РАЗМЕЩЕНИЯ
НАСЕКОМЫХ
Распределение климатических и других факторов среды
закономерно, что находит выражение в зональности климата,
почв, растительности и животного мира. К различным физико-
географическим условиям приурочены определенные виды на-
секомых и других организмов, их комплексы. Выбор мест оби-
тания составляет характерное свойство вида, а сами место-
обитания рассматриваются как суммированный показатель
условий среды (Г. Я. Бей-Биенко). В выборе тех или иных
местообитаний проявляется связь насекомых с соответствую-
щим комплексом абиотических факторов. Насекомые подобно
многим другим организмам входят непременными компонен-
тами природных и культурных ландшафтов.
В каждой зоне, районе, местности встречаются сходные
сочетания рельефа, почв и других условий среды. Ландшаф-
товеды называют их микроландшафтом, или фацией, а эколо-
ги — экотопом, или биотопом. Ландшафтоведы пришли к вы-
воду, что однородные участки территории характеризуются не
только сходными физическими условиями среды, но и соста-
вом растений (фитоценозом) и животных (зооценозом).
А экологи, изучая сосуществование различных видов организ-
мов, считают, что их естественные группировки занимают
участки территории с однородными физико-географическими
условиями — экотопы или биотопы. Биотоп как микроланд-
шафт представляет элементарную территориальную единицу
изучения условий существования сообществ организмов. Ко-
гда одно и то же явление природы описывается представи-
64
целями разных наук, то естественно возникает много сходных
по содержанию терминов. Их обилие связано также с неяс-
ностью классификации деления территории на различные по
объему природные комплексы.
Среда обитания насекомых очень разнообразна, поэтому
термином «биотоп» обозначают часто разные по содержанию
понятия. Иногда «биотоп» не связывают с территорией, назы-
вая биотопом нору грызуна или гнездо птицы. Обычно же
под биотопом подразумевают участок среды обитания жи-
вотных, характеризующийся однородными экологическими
условиями или участок земной поверхности, отличающийся
от окружающих его участков по тем условиям существования,
который он предоставляет для населяющих его организмов.
Как примеры биотопов можно привести: однородные участки
луга, песчаные участки, осоковое болото. Каждый биотоп су-
ши характеризуется сходными особенностями рельефа, почво-
грунта, гидротермического режима, почвенного и раститель-
ного покрова и определенной группировкой животных. Био-
топ— это участок территории, сходный по характеру
жизненных условий. Свойственный же биотопу комплекс ра-
стений и животных называют биоценозом. Оба эти понятия
тесно связаны. Биотоп определяют и как место жизни био-
ценоза, тем самым подчеркивая, что в природе не существует
незаселенных биотопов, как и биоценозов, без участка терри-
тории. Для целей анализа, однако, бывает необходимо поль-
зоваться и понятием биотоп, и понятием биоценоз. Реально
же существующее в природе единство животно-растительных
сообществ (биоценозов) с абиотическими условиями их жиз-
ни (биотопом или экотопом) правильнее называть биогеоце-
нозом (термин, предложенный В. Н. Сукачевым). Биотоп —
участок однородной территории с комплексом свойственных
ему физических и химических факторов среды, соответствую-
щий элементарной географической единице — микроланд-
шафту.
Микроландшафт в своем типичном проявлении должен
представлять один элемент рельефа, сложенный одной поро-
дой или наносом и‘покрытый в данный момент определенным
растительным сообществом. Изучение местности следует начи-
нать с расчленения ее на элементарные ландшафты (Поли-
нов, 1953). Каждый элементарный ландшафт (микроланд-
шафт) слагается из более мелких структурных единиц — де-
талей ландшафта. Предельным структурным элементам
микроландшафта будут соответствовать части биоценоза —
микроценозы — с соответствующими элементами биотопа —
микростациями. Примерами последних можно назвать куст,
дерево, кочку на болоте, нору грызуна, шишку сосны и т. п.
При анализе особенностей распределения насекомых по
территории следует различать места обитания отдельных ви-
5 Заказ 193
65
дов и целых биоценозов, часть которых составляют сообщест-
ва насекомых — энтомоценозы.
При характеристике особенностей территориального раз-
мещения отдельных видов насекомых пользуются термином
стация обитания, или местообитание. По Г. Я- Бей-Биенко,
«стацией, или местообитанием вида, называют участок терри-
тории, характеризующийся определенным .комплексом усло-
вий, заселяемый тем или иным видом животного. Местообита-
ние характеризуется определенными почвенными, микрокли-
матическими условиями, а также определенной группировкой
растений, чаще всего ассоциацией». В этом случае понятия
биотоп и стация сходны. Но биотоп — место жизни биоценоза,
а стация — место жизни отдельного вида.
В других случаях стацией обитания называют места, в ко-
торых проходит лишь часть цикла развития вида. Например,
выделяют стацию питания личинок и имаго, стацию размно-
жения. Места обитания личинок и взрослых форм насекомых
часто бывают очень различны.
Н. П. Наумов стацию рассматривает как часть местооби-
тания вида. Она может изменяться в разные сезоны года,
в дневные и ночные часы. Местообитание же — это совокуп-
ность условий существования, обеспечивающая весь цикл раз-
вития вида.
Местообитание может распространяться на несколько зон,
а может ограничиваться одним организмом — хозяином.
Пустынная саранча в годы массового размножения успешно
развивается не только в зоне пустынь Индии, но и далеко за
ее пределами. А такие представители насекомых, как пухо-
еды или вши, весь цикл развития проходят на каком-либо
виде теплокровных животных, а иногда на какой-то части тела
хозяина.
Одни виды насекомых очень ограничены в выборе мест
обитания — с т е н о т о п н ы, другие — э в р и т о п н ы — с успе-
хом заселяют разные биотопы. Многие виды меняют биотопы
в процессе онтогенеза, а многие меняют стации обитания по
зонам и на границах ареалов своего распространения. Знание
экологических условий, определяющих ареалы распростране-
ния отдельных видов и особенности их размещения по биото-
пам, имеет большое значение для выявления зон и очагов
вредности, причин их возникновения.
Расселяясь по территории, каждый вид избирает свойст-
венные ему условия среды, проявляя известного рода консер-
ватизм. В центральных частях ареала вид находит наиболее
благоприятные условия развития, встречается здесь чаще.
На границах области распространения виды становятся сте-
нотопными, встречаются реже. При продвижении на север
своего ареала виды занимают более сухие, прогреваемые уча-
стки, а продвигаясь к югу — более влажные и затененные
66
биотопы. Песчаные участки на севере, хорошо прогреваемые
склоны южной экспозиции — те биотопы, по которым идет
продвижение более южных видов на север, а вдоль рек по
более затененным и влажным местам северные виды прони-
кают к югу в более сухие зоны. Следуя за своими стациями,
отдельные виды часто заходят далеко за пределы основного
центра распространения, за пределы типичных для них при-
родных зон.
Географическая зона включает три отдельных группы био-
топов: плакорные, интразональныс и экстразональные (Воро-
нов, 1964). Плакоры заняты типичными для данной зоны поч-
вами и растительными формациями и заселены зональными
группировками насекомых. По интразональным биотопам
(поймы рек с заливными лугами, песчаные террасы, болота,
солонцы) многие северные виды насекомых распространяются
далеко к югу, а теплолюбивые южные виды — далеко к севе-
ру. Экстразональные биотопы — вышедшие за пределы своей
зоны плакорные участки. Они занимают незональное положе-
ние к югу и к северу от зоны, где они типичны на плакоре.
Таковы байрачные леса по глубоким балкам на юге степей,
степные участки на южных склонах в зоне тайги, пятна раз-
нотравно-ковыльной степи по западинам в полупустыне. В не-
зональных биотопах складывается иной гидротермический
режим, иной микроклимат, чем в зональных условиях, и это
приводит к закономерной смене стаций обитания многих ви-
дов при их переходе в другие географические зоны.
Г. Я. Бей-Биенко сформулировал впервые выявленное им
изменение местообитаний у саранчевых как правило зональ-
ной смены стаций обитания. Примеров, подтверждающих это
правило, много. Так, итальянский прус на севере степной зоны
живет на сухих песчаных почвах, а в Средней Азии — на
орошаемых участках. Июньский хрущ в степной зоне вредит
полевым культурам на целине и залежах, а в Средней Азии
он — преимущественный вредитель садов. Причину зональной
смены стаций обитания Бей-Биенко видит в изменении темпе-
ратурного режима. Если тот или иной вид в разных зонах
заселял бы внешне однородные стации (например, песчаные
участки), то фактически он оказался бы в резко различных
условиях гидротермического режима. В лесной зоне темпера-
тура поверхности песчаной почвы поднимается до 35—40е,
а в пустынях она бывает выше 65°.
Анализируя распределение саранчевых в разных зонах,
Бей-Биенко все стации их обитания объединил по степени
увлажнения, прогревания и густоты травяного покрова в три
экологические группы: ксерофитные, мезофптные и гигрофит-
ные. Ксерофилы лесной зоны в более южных зонах (в степях,
полупустынях) переходят в мезофитные стации, мезофилы —
в гигрофитные стации. Гигрофилы во всех зона^с приурочены
5*
67
х местам с избыточным увлажнением и пониженному
рельефу.
Дальнейшие исследования показали, что правилу зональ-
ной смены стаций подвержен каждый вид насекомых. В об-
щей форме С. И. Медведев (1954) выразил это правило в сле-
дующих положениях: 1). В северных пределах ареала каждый
вид по сравнению с типичными условиями данной зоны при-
урочен к биотопам, которые характеризуются более высокой
температурой и пониженной влажностью. 2). В оптимальных
условиях для данного вида, в центральных частях ареала,
наблюдается приуроченность к типичным условиям данной
зоны (а для видов с широкой экологической валентностью —
нахождение также и в ряде других биотопов). 3). У южных
границ ареала каждый вид приурочен к биотопам, характе-
ризующимся пониженной температурой и повышенной влаж-
ностью сравнительно с типичными условиями данной зоны.
Отношение отдельных видов насекомых к температуре и
влажности характеризуется их приуроченностью к господст-
вующим (типичным) условиям определенной ландшафтно-гео-
графической зоны. Ксерофилами в этом случае нужно считать
обитателей плато в зонах с недостаточным увлажнением и вы-
сокими летними температурами, т. е. пустынь, полупустынь и
засушливых степей. Мезофилы — обитатели плато, но уже
разнотравно-ковыльных степей с суммой годовых осадков
450—500 мм, луговых степей и широколиственных лесов.
В более сухих подзонах мезофилы будут заселять уже не
плато, а понижения, а в более влажных подзонах — сухие
и лучше прогреваемые участки. Среди ксерофилов С. И. Мед-
ведев выделяет пустынных, полупустынных и степных.
Для примера рассмотрим размещение насекомых по био-
топам в подзоне засушливых степей. Подзона характеризует-
ся преобладанием светло-каштановых почв, располагающихся
в условиях равнинного рельефа и формирующихся под полын-
но-типчаковой растительностью. Заселены они ксерофилами —
степной кобылкой (Asiotmethis muricatus Pall.), копьеуской
пустынной (Myrmeleotettix pallidas Вг.), чернотелками (Теп-
tijria nomas Pall., Opatrum sabulosum L.), совкой (Euxoa ba-
sigramma Stgr.) и другими видами. На фоне типичных светло-
каштановых почв встречаются небольшие участки с почвами
черноземовидными. Они занимают понижения глубиной более
30 см, где задерживается больше воды и развита разнотрав-
ная растительность. В таких блюдцеобразпых западинах с
черноземовидными почвами и разнотравно-злаковой степной
растительностью распространены насекомые — мезофилы раз-
нотравно-ковыльных степей: чернотелки (Pedinus femoralis L.,
Blaps halophila F.—W., Asida lutosa Sol.), пыльцееды (Po-
donta daghestanica R., Omophlus proteus R.), щелкун широкий
(Selatosomus latus F.), июньский хрущ, крестовичка (Docio-
68
staurus brevicollis Ev.), степной конек (Euchortippus pulvina-
tus F.—W.) и другие. Растительность западин отличается гу-
стым и высоким травостоем, растения остаются зелеными на
протяжении всего сезона, температура поверхности почвы на
2—3° ниже, чем на светло-каштановых почвах, где преобла-
дает типчак, ромашник и ковыль Лессинга. В течение сезона
по мере выгорания растительности в степи многие виды пря-
мокрылых мигрируют в западины. Перемещение во все более
влажные места происходит постепенно. В западины мигриру-
ют чернополосая кобылка (Oedaleus decorus Ger.), крестовая
кобылка (Pararciptera microptera F.), травянка (Stenobothrus
eurasius F.), кузнечик эверсмана (Platycleis eversmanni Kitt.),
итальянский прус и другие виды. В основе сезонных миграций
лежит та же причина, что и в зональной смене стаций обита-
ния. Весной необходимые условия прямокрылые находят в
одних биотопах, ранним летом — в других, поздним летом —
в третьих. Миграции — приспособление к быстро меняющимся
условиям среды.
Проявлением правила смены стаций обитания считается
переход к жизни на корнях в аридных условиях короеда-са-
довника (Blastophagus piniperda L.), уход на зимовку из крон
в подстилку в районах с сильными морозами боярышницы.,
изменения сроков размножения, привязывающих виды к оп-
ределенным биотопам. Смена стаций обитания наряду с про-
явлением консервативности вида ставит вид в несколько от-
личные условия и ведет к изменению его особенностей. Пере-
летная саранча на севере представлена особым подвидом —
среднерусской саранчой (Locusta migratoria rossica Uv.);
крестовая кобылка (Par аг су pt er a microptera F.—W.) на юге
замещена другими подвидами, живущими среди более густой
растительности; сибирская кобылка (Gomphocerus sibiricus L.)
в степной зоне фитофил, а в зоне тайги — геофил.
Глава III
ЖИЗНЕННЫЕ ФОРМЫ НАСЕКОМЫХ
При изучении многообразного мира насекомых очень ярко
вскрывается соответствие двух систем: организма и среды.
Оно проявляется в особенностях морфологии, физиологии,
особенностях поведения, питания, выборе мест обитания, свя-
зях с другими организмами. Разнообразные адаптивные сис-
темы признаков проявляются как в крупных систематических
категориях, так и в пределах родов и видов, определяя их
жизненную форму. По Н. П. Наумову (1955), «жизненная
форма — это экологическая характеристика вида или любой
другой систематической группы».
Естественная классификация животных строится на фило-
генетически сложившихся признаках, отражающих длительное
влияние среды. Экологическая классификация организмов
-совпадает с их естественной системой. В отдельных система-
тических группах животных отношения к окружающей среде
своеобразны и зависят от особенностей строения и поведения
животного, от типа его приспособлений, всей его предшество-
вавшей эволюции. Классы, отряды, семейства отличаются как
морфологически, так и экологически.
Особую жизненную форму представляет насекомое как тип
сухопутно-воздушного животного. Многие отряды и семейства
насекомых хорошо очерчены экологически. Например, бого-
молы — типичные подстерегающие хищники; тараканы — му-
сорщики, боящиеся света, со сплющенным и мягким телом
и другими особенностями, позволяющими жить в укрытиях
и узких щелях; многочисленные саранчевые — жители откры-
тых мест. Во многих случаях русские названия семейств мо-
гут служить их экологической характеристикой: плавунцы,
водолюбы, листовертки, мертвоеды, короеды, древоточцы, оре-
хотворки, листоблошки, кожееды и т. п. Эти примеры под-
тверждают правильность положения о совпадении классифи-
кации жизненных форм с их естественной системой.
70
Вместе с тем известно не мало случаев, когда сходные
черты присущи представителям разных систематических кате-
горий. Существуют приспособления различных типов органи-
зации к однородной среде, что ведет к аналогичным измене-
ниям, к появлению сходного комплекса приспособительных
структур. Они служат хорошим способом оценки сходного
влияния внешней среды на совершенно различные организмы
и получили название «жизненные формы» и «биологические
типы». В некоторых странах Европы под экологией понимают
исследования именно таких жизненных форм.
Признаки, по которым выделяют жизненные формы, и спо-
собы их выделения различны и часто несопоставляемы. В од-
них случаях жизненная форма принимается как совокупность
морфологических, физиологических и экологических свойств
вида, выработавшихся в течение его филогенетической исто-
рии в соответствии с внешней средой; это находит выражение
в морфо-экологических типах. В других случаях жизненную
форму выделяют по какому-то частному бросающемуся в гла-
за признаку. Им может быть форма тела, приспособленная
К движению в средах различной плотности, и способ передви-
жения: ползающий, прыгающий, летающий, плавающий и т. д.
Выделяют жизненные формы хищников — свободных, засад-
ников, скользящих, плавающих и др. В качестве примера на-
зывают сходную форму хищника в семействах Mantidae, Ne-
pidae, Reduviidae с характерным типом передних ног. Такого
же типа строение имеет Mantispa из отряда сетчатокрылых.
Сходные типы строения имеют многие паразиты, например
кровососка (Hippobosca), бобровая блоха (Platypsyllus),
вошь (Haematopinus) и пухоед (Gyropus). Сходны «проволоч-
ники» — личинки семейств щелкунов, чернотелок, пыльцеедов,
приспособленных к жизни в почве.
У самых различных насекомых могут быть сходные реак-
ции на действие отдельных факторов среды или их совокуп-
ности. Выделяют насекомых стено- и эвритермных, стено- и
эвритопных и т. д. По местообитанию различают группы ор-
ганизмов дендробионтов, тамнобиоитов, хортобионтов, герпе-
тобионтов, геобионтов, псаммофилов, фитофилов и т. п.
Группы организмов несут на себе черты влияния особенно-
стей ландшафтов. Поэтому есть все основания говорить о жиз-
ненных формах лесных, степных, пустынных. Это дает воз-
можность оценить особенности целых фаун. Определенные
приспособительные черты характерны, например, для насеко-
мых пустынь. Широко распространенные насекомые заселяют
здесь наиболее влажные экстразональные местообитания, они
переходят к жизни в почве. В пустыне много скрытоживущих
фитофагов, видов эфемеров с весенней активностью, ночных
форм. Характерны приспособления, снижающие расход воды
у насекомых. i
71
Для насекомых известны классификации жизненных форм
муравьев (К. В. Арнольди), прямокрылых (Г. Я. Бей-Биенко),
поденок (О. А. Чернова), стрекоз (Л. Н. Притыкина), чешуе-
крылых (А. М. Герасимов, Г. А. Мазохин), личинок жужелиц
(И. X. Шарова), личинок стафилин (В. А. Потоцкая), личинок
древоядных жуков (Б. М. Мамаев).
ЖИЗНЕННЫЕ ФОРМЫ САРАНЧОВЫХ
Саранчовые как целая группа насекомых в отряде прямо-
крылых характеризуются фитофагией, многоядностью и тес-
ной зависимостью в распределении по территории от расти-
тельного покрова. Специфические свойства саранчевых ярко
проявляются в выборе ими местообитания. Густота раститель-
ного покрова часто оказывает решающее влияние на распре-
деление саранчовых потому, что степень покрытия почвы
растениями определяет степень прогревания поверхности поч-
вы и прилегающего слоя воздуха солнечными лучами, его про-
ветривание и относительную влажность. Следовательно, гу-
стота растительного покрова определяет микроклимат данного
участка территории и является выражением воздействия взаи-
мосвязанных факторов среды.
Г. Я. Бей-Биенко при выделении жизненных форм саран-
човых особое внимание уделяет их местообитанию, связи с
характером растительного покрова. Особенность местообита-
ния — «суммированный показатель условий существования».
Сопоставление обычных видов саранчовых дает представле-
ние не только об их морфолого-систематической дифференциа-
ции, но и о резких различиях в экологии и поведении, свя-
занных с внешним обликом насекомых.
Всех саранчовых Г. Я. Бей-Биенко делит на следующие
жизненные формы:
I класс. Обитатели растений, или фитофилы.
1) Обитатели толщи травяного покрова (хортобионты) —
Locusta, Chortippus, Omocestus, Aiolopus, Охуа и др.
2) Обитатели кустарников и деревьев (тамнобионты) —
Dericorys, Anacridium, Euprepocnemis и др.
II класс. Обитатели поверхности почвы, или геофилы.
1) Открытые геофилы, т. е. обитатели открытых участков
на поверхности почвы — Sphingonotus, Oedipoda, Bryodema,
Asiotmethis и др. Среди них могут быть специализированные
песколюбы (псаммобионты), приспособленные к жизни и пе-
редвижению на рыхлых песках, а также камнелюбы (петро-
бионты) — обитатели каменистого субстрата.
2) Скрытоживущие геофилы (герпетобионты) — обитатели
поверхности почвы, покрытой растительным покровом, опав-
шей листвой и другими растительными остатками. Существен-
ная особенность этих саранчовых — жизнь в затененных ме-
72
стах и требовательность к влаге. Питаются почвенными во-
дорослями и лишайниками — Tetrix.
Большинство видов, вредных для сельского хозяйства, от-
носится к хортобионтам. Они приспособлены к свободному
передвижению и жизни в толще травяного покрова и являют-
ся характерными обитателями открытых степей, лугов, саванн
и полей. Их тело стройное, гладкое, без шероховатостей, цвет
обычно зеленоватый или желтоватый, между коготками ног
хорошо развиты присоски, помогающие удерживаться на ра-
стении. Геофилы отличаются шероховатой скульптурой покро-
ва тела (морщинистой, бугорчатой), землистой окраской, при-
соски развиты слабо. Эти формы свойственны преимуществен-
но пустыням и полупустыням.
Приведенные выше жизненные формы саранчовых охва-
тывают только наиболее типичные и резко выраженные слу-
чаи. Между ними возможны разнообразные переходы.
Г. Я. Бей-Биенко отмечает консервативность жизненной
формы, устойчивость. Однако это наследственно закрепленное
свойство не закрывает путей для дальнейшего развития и при-
способления вида и даже выработки признаков новой жиз-
ненной формы. Необходимость морфо-экологической пере-
стройки вида возникает при его расселении и проникновении
в другие зоны, что ведет к изменению характера местооби-
таний.
Наиболее консервативными признаками жизненной формы
Бей-Биенко считает морфологические, а наиболее гибкими1
и способными к перестройке — экологические. Поэтому пере-
стройка вида начинается с изменений его физиологических
качеств, т. е. требований к окружающей среде. Этим объяс-
няются случаи, когда тот или иной вид или даже группа род-
ственных видов обладает морфологическими признаками од-
ной жизненной формы, а экологические — уже перестроились
на другую жизненную форму.
Классификация жизненных форм саранчовых позволяет
отметить, что морфологические особенности жизненной формы
отражают ее наиболее глубокие наследственные свойства и
должны быть использованы при выделении жизненных форм,,
особенно в крупных систематических категориях. Удобный
признак выделения жизненных форм—также местообитание
вида как показатель влияния системы факторов среды (пи-
щи, микроклимата, почвы).
МОРФО-ЭКОЛОГИЧЕСКИЕ ТИПЫ ВОДНЫХ НАСЕКОМЫХ
Типы водных насекомых наиболее полно разработаны на
личинках поденок в работе О. А. Черновой. В отличие от са-
ранчевых, поденки — типичные обитатели водоемов. Кроме
того, они гетеротопны и свою короткую имагинальную жизнь
73
проводят в воздушной среде, выполняя биологические функ-
ции размножения и расселения. Личинки поденок, обитающие
в разнообразных водоемах, живут долго, имеют различное
строение и ведут разный образ жизни. Изучение фауны поде-
нок позволяет сделать вывод, что «громадное большинство
родовых и семейственных категорий легко может характери-
зоваться по особенностям личинок и лишь в немногих случаях
личинки различных семейств обнаруживают между собой по-
верхностное сходство» (Чернова, 1952).
При классификации жизненных форм насекомых возника-
ют трудности, связанные со сменой среды обитания в онто-
генезе. Поэтому чрезвычайно важно либо выделить одну из
фраз развития как ведущую при оценке жизненной формы,
либо найти критерии, позволяющие оценить жизненную фор-
му с учетом особенностей онтогенеза. По Веберу, перенос
«центра тяжести жизни» на определенную стадию развития —
особая форма приспособления, при которой полностью исполь-
зуются возможности двух сред обитания. В отряде поденок
«центр тяжести жизни», длительный контакт с разнообразны-
ми условиями среды падает на личиночный период развития.
Исследования Черновой подтверждают ведущее значение ли-
чинок при выделении жизненных форм в этом отряде.
Классификация морфо-экологических типов поденок по-
строена на основе особенностей морфологии, поведения и ме-
стообитания личинок. В основном она отражает естественную
систему поденок и одновременно дает возможность оценить
случаи параллельного развития личинок в разных семействах.
Кроме того, морфо-экологическая классификация поденок,
разработанная Черновой, показывает, что по приспособлениям
личинок к обитанию в разных условиях, по строению их ор-
ганизма можно определять условия, в которых они живут,
судить о характере водоемов по обнаруженным в нем поден-
кам.
Все разнообразие личиночных форм поденок Чернова объ-
единяет в следующие типы и подтипы:
I тип. Крупножаберные личинки зарослей:
1-й подтип. Узкожаберные личинки равнинных ру-
чьев.
2-й подтип. Широкожаберные личинки стоячих вод.
3-й подтип. Ползающие личинки прибрежий рек.
II тип. Крышечножаберные личинки глинистых грунтов.
III тип. Длиннокоготковые и со «щетками» на голове ли-
яинки песчаных грунтов.
IV тип. Мелкожаберные, быстринные, активно плавающие
личинки.
V тип. Уплощенные или с цепкими ногами, быстринные ли-
чинки.
74
1-й подтип. Уплощенные личинки нижней стороны
камней.
2-й подтип. Личинки с цепкими ногами верхней сто-
роны камней.
3-й подтип. Малоподвижные, щитневидные личинки.
VI тип. Грунтовые, закапывающиеся личинки с бивнями.
1-й подтип. Закапывающиеся личинки с бегательны-
ми ногами.
2-й подтип. Роющие личинки с копательными нога-
ми, делающие ходы в грунте.
Большинство личинок — стенобионты; часто в массе слу-
жат пищей рыбам и могут быть использованы в качестве ука-
зателя кормовых стаций промысловых рыб. При этом важно
учитывать не только характер местообитаний и другие осо-
бенности экологии, но и особенности приспособительных мор-
фологических структур, а также систематическое положение
группы и отдельных видов.
Сходная по принципу классификация разработана для ли-
чинок стрекоз (Притыкина, 1964).
БИОЛОГИЧЕСКИЕ ТИПЫ ЧЕШУЕКРЫЛЫХ
При выделении жизненных форм, или биологических типов,
у чешуекрылых затруднения возникают из-за различия биоло-
гических функций личинок (преимущественно питание) и има-
го (преимущественно размножение и расселение).
А. М. Герасимов (1952) дает экологическую классифика-
цию только гусениц. Он делит их на две большие группы:
ведущих открытый образ жизни и ведущих скрытый образ
жизни. Переходное положение занимают гусеницы, живущие
в чехликах, что свойственно первичным гусеницам семейств
Adelidae, Incurvariidae, Psychidae и др. Свободно живущих
гусениц он подразделяет на травоядных и древесных. Послед-
ние характеризуются бородавчато-волосистым покровом, свой-
ственным обитателям деревьев (семейство Lasiocampidae, Ly-
mantriidae, Attacidae, Notodontidae). Скрыто живущие гусе-
ницы представлены разнообразными семействами, но преиму-
щественно Microfrenata. Герасимов выделяет группы гусениц
подземных (семейство Noctuidae, подгрызающие совки), лис-
товертов (Tortricidae и другие семейства), плодожорок, буриль-
щиков-ксилофагов (Cossidae, Aegeriidae), минеров, галлооб-
разователей и водных. Многочисленные минеры представлены
разными семействами, филогенетически часто не связанными.
Однако их больше среди Microfrenata. Очень редки они на
ранних фазах развития Hesperiidae, Lycaenidae. Минирующий
образ жизни распространен в отряде Diptera, реже — у Ну-
menoptera и Coleoptera. Мины, свойственные гусеницам, легко
отличимы от мин, характерных для других отрядов. Мины
75
служат для распознавания видов. Биологические типы гусе-
ниц, по классификации Герасимова, полностью не совпадают
с систематическими категориями, установленными по имаго.
Морфологическая характеристика биологических типов
изучена недостаточно. Г. А. Мазохин-Поршняков (1954) по-
пытался классифицировать биологические типы чешуекрылых
по особенностям всех фаз развития в онтогенезе. Термин
«жизненная форма» он предложил для отдельных фаз разви-
тия (яйца, гусеницы, куколки, имаго), а «биологический
тип» — для совокупной оценки особенностей онтогенеза по
важнейшим биологическим функциям (питания, размножения,
расселения). Для чешуекрылых, живущих в лесу, им выделе-
но девять биологических типов: гепиалоидный, нептикулоид-
ный, тортрикоидный, окнериоидный и т. д. Папилиоидный тип
он, например, характеризует следующим образом: представ-
лен всеми семействами Rhopalocera, многими представителя-
ми Sphingidae, Zygaenidae, Geometridae, Noctuidae, гусеницы
открыто живущие, куколки в почве, имаго живут долго, полет
быстрый, темп расселения высокий, проявляется забота о по-
томстве, овладение воздушно-сухой средой совершенно. При
выделении биологических типов анализа специализации пи-
тания гусениц и многих других экологических и морфологиче-
ских особенностей автор не проводит.
Биологические типы чешуекрылых, выделенные Мазохи-
ным-Поршняковым, и биологические типы гусениц, по Гераси-
мову, при сопоставлении во многом совпадают. Так,
гепиалоидный тип соответствует ксилофагам-бурильщикам,
нептикулоидный — минерам, тортрикоидный — листовертам и
плодожоркам, психеоидный — чехлоносам, папилиоидный —
свободно живущим гусеницам на травянистых растениях, три-
феноидный — подгрызающим совкам.
По-видимому, как и в отряде поденок, при выделении жиз-
ненных форм чешуекрылых ведущее значение имеет личиноч-
ный период развития. Обе системы биологических типов не
соответствуют таксономической системе чешуекрылых, по-
строенной на признаках имаго.
Приведенные примеры классификации жизненных форму
морфо-экологических типов и биологических типов у насеко-
мых показывают, что проблема организма как индикатора
внешних условий имеет огромное значение и решается по-раз-
ному. Наиболее глубокое и правильное ее решение может
быть достигнуто на базе экологической характеристики систе-
матических категорий разных рангов и анализа отклонений
от системы в случаях изоморфизмов как проявления частных
особенностей системы или более глубоких специализаций, вы-
званных эволюцией разных исходных форм в однотипных
условиях среды. При таком анализе, как нам думается, воз-
можны поправки к существующим классификациям.
Глава IV
БИОТИЧЕСКИЕ ФАКТОРЫ СРЕДЫ
Животные и растения неизбежно вступают в разнообраз-
ные связи друг с другом. Отдельные виды организмов распре-
делены в пределах своих ареалов неравномерно, образуя изо-
лированные скопления особей — популяции. Популяции одно-
го вида разбросаны по разным биоценозам, а биоценозы
состоят из популяций разных видов. Виды всегда существуют
в форме отдельных популяций, а биоценозы — в форме сосу-
ществующих популяций разных видов. Таким образом, попу-
ляция— структурная единица и вида, и биоценоза (М. С. Ги-
ляров). Отношения особей и популяций одного вида друг к
другу и к популяциям других видов неизбежны; они связы-
вают животно-растительный мир с остальной природой как
географическим ландшафтом.
Отношения популяций одного вида направлены и подчи-
нены задаче самосохранения этого вида в изменчивых усло-
виях среды, а межвидовые связи — на длительное существо-
вание биоценоза, в котором сосуществование популяций раз-
ных видов, сложившееся в процессе длительной эволюции,
обеспечивает биогенный круговорот форм, вещества и энергии.
Жизнь вида как самостоятельного сочлена биоценоза имеет
свои особенности, подчиненные интересам вида.
ВНУТРИВИДОВЫЕ ОТНОШЕНИЯ
В. Н. Беклемишев рассматривал вид как определенное
биологическое единство входящих в него особей и популяций,
характеризующееся сходством морфологических, физиологи-
ческих и экологических свойств, а основным жизненным на-
значением вида считал задачу самосохранения. Вся внутри-
видовая жизнь подчинена ее решению. Существование вида
обеспечивается связями между его популяциями, содгавляю-
77
щими основу внутривидовой структуры. Большинство эколо-
гов под популяцией понимает группу особей, которая населяет
определенную территорию, входит в состав существующего
там биоценоза и представляет форму существования вида в
конкретных ландшафтных условиях. Это — естественные про-
странственно расчлененные группировки особей.
У насекомых с обширными ареалами и большой изменчи-
востью видовое население очень разнообразно.
Так, популяции сибирского шелкопряда {Dendrolimus su-
perans sibiricus Rosh.), самого вредоносного подвида — боль-
шого хвойного шелкопряда (Dendrolimus superatis Butl.) -
явно предпочитают определенный вид пищи: они связаны ли-
бо с лиственницей, либо с кедром, либо с пихтой. Экологиче-
ские популяции разных племен сибирского шелкопряда отли-
чаются темпами роста, сроками размножения, размерами,
плодовитостью, выбором местообитаний, циклами развития,
годами массового лёта (А. С. Рожков, 1965).
У других видов насекомых экологические популяции выра-
жены также хорошо и отличаются друг от друга по феноло-
гии и стациям обитания. Популяции стенотопных и малопод-
вижных видов насекомых могут быть разобщены даже в пре-
делах небольшой территории. Например, популяция одуван-
чикового долгоносика (Ceuthorrhynchus optator Faust.) на
южном склоне развивается на 15—20 дней раньше, чем на се-
верном, в силу чего скрещивания между этими популяциями
не происходит (М. С. Гиляров). Бахчевая чернотелка (Tenty-
ria nomas Pall.) в степных популяциях на солонцах и на пес-
ках разобщена настолько, что их представители морфологи-
чески различны (К- В. Арнольди, 1941). У жука-короеда (Са1-
ligraphus) особи популяции, обитающие на липе и вязе,
крупнее, чем на кизиле. Чем более разобщены биоценозы, тем
больше отличий может возникнуть в популяции одного вида.
Возможность постоянного скрещивания, характерная для
внутривидовых отношений, обеспечивается постоянными миг-
рациями насекомых, перемешиванием различных популяций.
Популяции могут быть и независимыми от притока особей
извне, а могут быть зависимыми, когда собственный приплод
не покрывает смертность и без притока особей извне популя-
ции вымерли бы (Беклемишев, 1961).
Для многих видов насекомых характерна стенотопность,
узкая специализация. Поэтому способность к расселению для
них особенно существенна.
Активное расселение (полет) свойственно большинству
насекомых. Для многих бабочек, саранчи, стрекоз, жуков
установлены дальние миграции. Многие виды перелетают на
очень большие расстояния. Например, олеандровый бражник
(Delephila nerii L.) и мертвая голова (Acherontia atropos L.)
за 2—3 недели пролетели пространство более 5000 км, летя
78
только ночью и не по прямой. Бабочки чертополоховой угло-
крыльиицы (Pyrameis cardui L.) за три дня пролетели более
700 км (от Армении до Абхазии), пробыли там 3—4 дня и по-
летели дальше на север (Миляновский, 1956). Массовый лёт
этих бабочек и переползание гусениц наблюдались нами в
1964 г. на Цимлянском песчаном массиве. Бабочек было мно-
го; во многих местах на цветах их собиралось по 60—70 экз.
на 1 м2. В том же году массовый залет бабочек был зарегист-
рирован в Англии, предположительно — с юго-востока нашей
страны. Перелеты насекомых длятся от нескольких часов до
нескольких суток. Перелетная саранча иногда летит в течение
трех суток, покрывая расстояние до 2000 км (Andersen, 1964).
Полет имеет направленный характер даже у таких насекомых,
как тли.
Функции расселения и размножения для многих насеко-
мых падают на имаго; в ряде случаев имаго совсем утрачива-
ют функции питания.
Многие насекомые расселяются пассивно с помощью ветра,
воды и других организмов. При пассивном расселении вероят-
ность попадания особи в благоприятные условия подчиняется
закономерностям случайного распределения. Поэтому для та-
ких организмов выгодно иметь меньший вес и большее число
расселяющихся особей. Меньший вес облегчает возможность
перелетов. Если бы насекомые не обладали способностью к
расселению различными способами, то при изменении условий
существования за пределы пластичности данного вида это
привело бы к вымиранию вида. У пассивно расселяющихся
видов вырабатываются особые способы размножения, веду-
щие к увеличению расселяющихся особей. Таковы бескрылые
партогенетические самки у тлей, дающие крылатых рассели-
тельниц; полиэмбриония у мелких паразитических перепонча-
токрылых.
Разные жизненные функции личиночной (преимущественно
питание) и имагинальной фаз развития (размножение, рассе-
ление), гетеротопность многих видов приводят к большой
разнородности одновидовых популяций насекомых. Она уси-
ливается различиями половыми, возрастными, генетическими,
суточными, сезонными и многолетними. Меняется интенсив-
ность обмена, состав гемолимфы, возбудимость, требования
к условиям среды.
Разнокачественность особей популяции — специфически
видовое приспособление к изменчивым условиям среды — за-
кономерное и постоянное явление, подчиненное интересам ви-
да, страхующее вид от вымирания.
Свойства популяции не остаются постоянными и во време-
ни, что особенно ясно можно наблюдать у видов, дающих
вспышки массовых размножений и фазовую изменчивость.
79
ФАЗОВАЯ ИЗМЕНЧИВОСТЬ НАСЕКОМЫХ
Расселение животных почти всегда совершается в годы их
высокой численности, обычно путем массовых миграций. Аре-
ал вида резко увеличивается. Во многих случаях расширение
ареала и миграции насекомых приходятся на годы массовых
размножений и перехода от одиночного образа >жизни к стад-
ному.
Впервые фазовая изменчивость была установлена
Б. П. Уваровым на саранчевых. У перелетной, пустынной, ма-
роккской и итальянской саранчи в зависимости от плотности
популяции появляются одиночные или стадные формы. По-
добная же изменчивость наблюдается у многих чешуекрылых,
жуков и, по-видимому, она вообще широко распространена
в природе.
Подробно фазовая изменчивость изучалась на стадных са-
ранчевых. По Г. Я. Бей-Биенко и Л. Л. Мищенко (1951),
сущность ее сводится к следующему. Среди саранчевых
имеются виды стадные и нестадные. Обычное состояние стад-
ных видов — одиночное рассеянное существование, при кото-
ром плотность популяции невелика и особи не раздражают
друг друга. Они менее активны, окрашены в зеленые или
желтые тона, имеют меньший показатель отношений длины
надкрыльев к длине бедра, высокий гребень переднеспинки,
часто — меньшие размеры тела. При изменении условий су-
ществования в местообитаниях отдельных популяций (нерав-
номерное высыхание растительности или увеличение количест-
ва влаги, ведущее к образованию густого растительного
покрова) личинки саранчи начинают собираться на более бла-
гоприятных для них участках (богатых растительностью, на
возвышениях и буграх). Скученность для них становится при-
вычной, возникает условный рефлекс стадности: особи не рас-
сеиваются, передвигаются плотными кулигами или стаями
даже в том случае, если их пытаются разогнать. Для стадной
саранчи характерна повышенная возбудимость нервной систе-
мы, активность движений, усиление обмена веществ, положи-
тельный фототаксис, что также связано с изменением пигмен-
тации. Окраска становится черной, усиливается поглощение
солнечны?: лучей; это ведет к повышению температуры тела
и увеличивает возбудимость. Появляются и морфологические
отличия (высота гребня становится ниже, индекс пропорции
крыла и бедра — больше). Стадная саранча начинает совер-
шать сначала массовые переходы, а затем (после окрыле-
ния) — мш рации огромными стаями на далекие расстояния,
нанося страшные опустошения.
Между одиночной и стадной фазой имеются переходы, об-
разующие фазы рассеивания, или фазы скучивания, как по-
казано на схеме:
80
Фаза скучивания
Т 4
Фаза одиночная Фаза стадная
Т !
Фаза рассеивания
Стадная фаза имеет важное биологическое значение, так
как в этом состоянии саранча совершает дальние миграции.
Миграции же служат основным способом заселения новых
мест обитания и восстановления старых очагов.
По наблюдениям за размножением саранчи в естествен-
ных очагах, для превращения одиночной фазы в стадную при
всех необходимых условиях требуется два года. Этот процесс
может быть прерван изменениями погоды, биотических фак-
торов и особенно — применением активных мер борьбы. Раз-
реживание популяций саранчи активными мерами борьбы
приводит к образованию одиночной фазы и недопущению раз-,
лета. Многие опыты показали, что фазовая изменчивость оп-
ределяется прямым воздействием среды.
Стадная и одиночная фазы отличаются также особенно-
стями развития гонад, плодовитостью, весом отрождающихся
из яиц личинок и рядом физиологических особенностей (Ken-
nedy, 1961). Изучение хромометрическими методами показало,
что потемнение стадной фазы вызывается увеличением содер-
жания омохрома и окрашенных в коричневый цвет белков.
У одиночных фаз меланин отсутствует. У Locusta pardalina
Walk, установлено, что переход в стадную фазу и меланиза-
ция сопровождаются резким увеличением частоты кроссинго-
вера (Nolte, 1965). Влияние света на окраску и плодовитость
насекомых позволяют предполагать связь фазовой изменчи-
вости через изменение деятельности ферментов с изменениями
солнечной радиации. Работы в этом направлении необходимо
продолжать.
У других насекомых, для которых характерно массовое
размножение, есть много общего с тем, что установлено для
саранчи. Так, в работе А. Г. Шарова (1953) по фазовой из-
менчивости ильмового ногохвоста установлено наличие гусе-
ниц четырех типов. Первый тип свойствен периоду массового
размножения. Гусеницы подвижны, активны днем, прожорли-
вы, имеют интенсивный желто-зеленый цвет с рисунком из
темных полос. Черный пигмент сосредоточен в кутикуле, жел-
то-зеленый— в гиподерме. Гусеницы имеют хорошо развитое
жировое тело и кишечник. Питаются круглые сутки. Гусеницы
другого типа монотонно серые с мраморным рисунком или
зеленые. Черного пигмента (меланина) почти нет. Гусеницы
сидят неподвижно на листьях или обхватив веточки дерева;
этому способствуют длинные волоски, расположенные по бо-
кам брюшка. Днем гусеницы не питаются, сидят неподвижно,
6 Заказ 193
81
тесно прижавшись к веточкам, с которыми они сливаются по
окраске. При сотрясении ветки гусеницы не падают, подобно
первым, а продолжают оставаться неподвижными, крепко
прицепившись к ветке. Между этими крайними типами гусе-
ниц наблюдается тип с переходными признаками. По мере
затухания массового размножения ногохвоста соотношение
типов гусениц менялось: подвижных, активных листовых гу-
сениц становилось меньше, а малоподвижных — больше. Если
сопоставить данные из работы Шарова по годам, то соотно-
шение типов гусениц в одном и том же насаждении береста
таково:
Годы наблюдений Стадный тип Одиночный тип Переходный тип
1951 100% 0 0
1953 39% 16% 45%
Наблюдения, проводившиеся нами за массовым размно-
жением этого вредителя в лесных полосах под Волгоградом,
позволяют дополнить данные Шарова. Массовое размножение
ильмового ногохвоста отмечено в 1949—1951 гг. В эти годы
во всех насаждениях преобладали гусеницы стадного типа.
В 1953 г. численность ногохвоста резко снизилась; в значи-
тельном количестве появились гусеницы одиночного типа.
В 1954—1955 гг. с большим трудом можно было находить
только гусениц одиночного типа, крепко сидящих на ветках.
Эксперименты Шарова по воспитанию гусениц показали,
что массовая форма получалась при совместном содержании
гусениц, при уплотнении популяции. Совместное воспитание
сказывалось лишь в том случае, если гусеницы не старше кон-
ца четвертого возраста. При разреживании популяции возни-
кают переходные и одиночные формы. Как и у саранчевых,
у ногохвоста существует непрерывный ряд переходов от мас-
совой формы к одиночным. У ногохвоста одиночные формы
гусениц также окрашены под цвет окружающего фона, но в
отличие от саранчевых имеется не одна, а две одиночные
формы: светло-зеленая, окрашенная под цвет листвы, и серая,
окрашенная под цвет коры. У массовой формы ногохвоста в
кутикуле также откладывается в большом количестве мела-
нин, отсутствующий у одиночных форм. В районах, где мас-
совое размножение ногохвоста не наблюдалось, не обнаруже-
но и стадной формы гусениц.
Фазовая изменчивость, установленная у других чешуекры-
лых: совка карадрина, L. exempta Walk., Spodoptera abissitiia
Guer., совка гамма (Plusia gatntna L. и др.) —характеризует -
82
ся также тем, что стадные формы имеют темную, меланистиче-
скую окраску, а одиночные — светлую. Темные, гусеницы пре-
обладают в годы массовых размножений, в то время как свет-
лые встречаются в периоды депрессий этих вредителей. Пере-
ход одиночных форм в массовые во всех случаях совершается
быстрее, чем переход массовых форм в одиночные.
Веллингтоном (Wellington, 1957) установлены разные ти-
пы поведения в популяциях гусениц кольчатого шелкопряда
(Malacosoma pluviale) в период его массового размножения
в лесах Канады. По его наблюдениям, имеются четыре формы
гусениц. Одна из них характеризуется положительным фото-
таксисом, быстрым активным передвижением, большей про-
жорливостью, характером повреждений листвы, вытянутой
формой паутинных гнезд и активностью имаго. Таких гусениц
было меньше в старых очагах размножения и много в новых
зонах вреда. Вторая крайняя форма гусениц отличалась мед-
ленными движениями, иным типом повреждения листьев и
слабым их поеданием, отрицательным фототаксисом при вы-
сокой температуре, более медленным развитием и большим
числом линек, физиологической ослабленностью, треугольной
формой гнезда, его большей плотностью и неактивным пове-
дением имаго. Автор статьи допускает возможность того, что
преобладание первого и второго типов гусениц будет связано
с изменением плотности популяций в годы массового размно-
жения шелкопряда. В период наблюдений среди гусениц,
взятых непосредственно в лесу, активные формы составляли
38%.
Сравнивая эти данные с тем, что нам известно по фазовой
изменчивости насекомых, мы вправе предположить, что Вел-
лингтон имел дело со стадной и одиночной фазами кольчатого
шелкопряда. Исследователь считает, что благодаря полимор-
физму гусениц шелкопряда вид поддерживает свое существо-
вание в неблагоприятных условиях среды. Активность одних
гусениц и пассивность, малоподвижность других передается
имаго. Наиболее активные бабочки мигрируют, а оставшиеся
локальные популяции со временем становятся менее жизнен-
ными и могут вымирать, если не пополнятся иммигрантами.
Разнокачественность популяции и ее изменчивость во времени
в зависимости от условий среды, безусловно, полезна для су-
ществования вида.
Изменения, сходные с фазовой изменчивостью, регистри-
руются у многих видов, дающих резкие колебания числен-
ности. Наблюдения за качественным состоянием популяции
в период вспышки размножения и в годы депрессии числен-
ности у непарного, кольчатого, сибирского шелкопрядов
и других вредных видов показывают, что состояние это ме-
няется (А. И. Ильинский, В. О. Болдаруев, А. С. Рожков).
При нарастании численности появляются темноокрашенные
6*
83
гусеницы и бабочки, что свидетельствует о повышении энер-
гии обмена веществ. Крупные и более тяжелые экземпляры
с высокой плодовитостью вначале размножения заменяются
формами более мелкими, увеличивается подвижность, меняет-
ся соотношение полов в- популяциях с изменением плотности.
Появляется инстинкт разлета и миграции из населенных оча-
гов. Бабочки из кучнокладущих становятся раздельнокладу-
щими, рассеивающими свои яички по насаждениям. Как и у
стадных саранчевых, изменяется поведение, интенсивность об-
мена, состав гемолимфы, фототаксис, наряду с резкими изме-
нениями плотности популяций. Массовому размножению
обычно сопутствует массовое расселение, предотвращающее
перенаселение очагов и способствующее расширению ареала.
Однако лишь немногие виды насекомых дают вспышки раз-
множения. Обычно для каждого вида характерна определен-
ная оптимальная плотность популяции, резкое нарушение
которой приводит к понижению плодовитости, скорости раз-
вития и продолжительности жизни особей. Для вида небла-
гоприятны отклонения от оптимума и в сторону перенасе-
ленности, и в сторону недонаселенности.
Наблюдая за жизнью насекомых, легко заметить, что она
подчиняется правилу: «вместе жить легче». Насекомые обыч-
но образуют разного рода скопления — временные или по-
стоянные. Отродившиеся гусеницы капустной белянки, злато-
гузки, кольчатого шелкопряда, непарного шелкопряда, личин-
ки саранчевых и многих других видов держутся кучно, соби-
раются в «зеркала». Подобные скопления бывают часто
связаны с постройкой общих паутинных гнезд, как, например,
у яблоневой моли. Совместная жизнь защищает от дождей,
ветра, прямых солнечных лучей, многих врагов, облегчает пи-
тание, заселение еще здоровых деревьев. Очень дружно гры-
зут лист плотной компанией гусенички серебристо?! лунки
(Phalera bucephala L.), личинки сосновых пилильщиков
(Diprion pini L.), многих других насекомых, особенно в мо-
лодом возрасте. Известно, что короеды часто нападают на
здоровые деревья целым роем. При этом часть гибнет (зали-
тые смолой), но новые рои продолжают атаку до тех пор,
пока дерево не будет ослаблено и короеды его заселят, оста-
вив многочисленное потомство. Сосновые тли из рода Cinaria,
по наблюдениям В. К. Гавриловой, питаются группами, опу-
стив хоботки в одну точку, собравшись плотным кольцом на
стволе сосны. Это облегчает прокол плотной коры и приток
пищи. Летние личинки тли Periphyllus aceris L. при недостат-
ке сока в листьях остаются без пищи и в этом случае соби-
раются группами; их длинные волоски, торчащие вверх, обра-
зуют над ними войлок, удерживающий слой воздуха, что по-
нижает испарение и дает возможность переживать неблаго-
приятный период времени (Weber, 1933). Легче пробивать по-
84
верхностную пленку воды и получить доступ к воздуху ли-
чинкам комаров, когда их популяция не разрежена.
Экспериментальные данные показывают, что изолирован-
ное воспитание неблагоприятно сказывается на росте и раз-
витии. Дрозофила (Drosophila) и малый мучной хрущак
(Tribolium confusum Duv.) в культурах с малым числом осо-
бей размножаются медленно. Плохо поодиночке растут гу-
сеницы златогузки; в конце концов, они погибают. То же слу-
чается с сеноедами (Liposcelis divinatorius Mull.); для их
жизни необходима известная массовость, наличие других
особей.
Огромные скопления насекомые образуют в период раз-
множения. Общеизвестно роение пчел, термитов, муравьев,
во время которого большие массы насекомых перемещаются
на далекие расстояния, занимая новые территории. Большие
стаи образуют многие поденки, стрекозы, бабочки, веснянки.
Известны скопления насекомых (таких, как вредная черепаш-
ка Eurygaster integriceps Put., свекловичный долгоносик
Bothynoderes punctiventris Germ, и многих других) в местах
зимовок. Плотные скопления помогают активно изменять
среду.
У пчел, ос, муравьев, термитов колониальная жизнь со-
провождается глубоким полиморфизмом с образованием каст,
проявлением сложных инстинктов, высоко развитой системой
сигнализации при общении друг с другом. Сложная жизнь
улья или термитника общеизвестна. Во многих случаях поли-
морфизм поддерживается специальным пищевым режимом.
Колониальная жизнь общественных насекомых обеспечивает
защиту от врагов, добывание пищи, усиленное размножение
и возможность поддержания постоянного гидротермического
режима в гнезде. Уход за молодью, кормление личинок рабо-
чими свидетельствует о высоко развитой у общественных на-
секомых заботе о потомстве. Забота о потомстве у насекомых
имеет и другие разнообразные формы. Она проявляется в
способах размещения яиц: в укрытия, в ткани растений, в спе-
циальные камеры с запасом консервированной пищи. Часто
самки устраивают свертки из листьев, куда и откладывают
яички, катают навозные шары и т. д. В некоторых случаях
(Orgyia и другие) гусенички пожирают тело своей матери.
Самка откладывает яйца в кокон, где остается вместе с по-
томством, обеспечивая его готовой высококалорийной пищей
собственного тела. Внутривидовые отношения, возникнув в про-
стых формах, в процессе филогенеза все более усложняются
и достигают наибольшего совершенства у общественных на-
секомых. Однако у других видов насекомых повышение плот-
ности популяции за пределы нормы ведет к замедлению роста
и развития, повышению смертности и другим неблагоприят-
ным для вида последствиям.
85
Экспериментальные работы показывают, что в ряде случа-
ев численность популяции колеблется с четкой и постоянной
периодичностью во времени. За отклонением в одну сторону
неизбежно следует отклонение в другую сторону, выявляются
механизмы саморегуляции численности.
УРОВЕНЬ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Каждому виду насекомых свойствен свой уровень числен-
ности. Подавляющее число видов насекомых постоянно редки
и встречаются лишь в очень ограниченных местах обитания
и массовые размножения их не известны. Другие насекомые
обычны и изредка дают увеличение численности популяции.
Есть и такие, которые постоянно встречаются в большом ко-
личестве или дают очень резкие вспышки размножения, уве-
личиваясь от 2—3 экз. до многих тысяч на гектаре, возрастая
по сравнению с наименьшей численностью в 10 000 раз, а ино-
гда и больше. Одни виды дают вспышки размножения очень
редко, другие чаще, принося колоссальные убытки. Виды,
дающие массовые размножения, отличаются очень изменчивой
плодовитостью — от десятка до нескольких сотен и даже
тысяч яиц.
Современный уровень численности видов — качество, исто-
рически сложившееся. Его следует рассматривать как одно
из свойств вида, как приспособление к условиям жизни.
Плодовитость у разных видов также различна, а ее из-
менчивость есть также приспособление вида к изменениям
условий среды. Изучена она у насекомых недостаточно. Пло-
довитость зависит как от наследственных свойств, так и от
экологических условий, места и времени. Некоторые живоро-
дящие мухи рождают всего одну личинку, а самка термитов
может откладывать до 7000 яиц в день. Яйца, богатые желт-
ком, более крупные, но их меньше. Мелких яиц откладывается
больше, но и гибнет их также больше. Генерации весенние
обычно более плодовиты. Так, капустная тля (Brevicoryne
brassicae L.) весной откладывает 40 яичек, а осенью — всего
2—3 яйца, но более крупных. Плодовитость кровяной тли вес-
ной— 100—150 личинок, а осенью — 10—20. Высокая плодо-
витость характерна для многих паразитических насекомых со
сложным циклом развития. Высокая плодовитость свойствен-
на видам, потомство которых подвержено большой случайной
смертности. Сюда относятся паразиты (Bombyliidae, Cyrtidae,
Larvaevoridae, Meloidae, Mantispidae), некоторые виды черве-
цов, совок, мешочниц, поденок, веснянок. Высокая плодови-
тость не связана с обилием этих видов в природе. Высокая
плодовитость (>1000 яиц) отмечена для паразитических пе-
репончатокрылых — Braconidae (Apanteles), Ichneumonidae
(Euceros). Плодовитость выше у эндопаразитов, чем у экто-
86
паразитов. Перепончатокрылые, длительное время сохраняю-
щие контакт с потомством (ВетЬех, Xylocopa и др.), яиц от-
кладывают мало (6—8); половое созревание у них очень мед-
ленно. Яйца пчел древогрызов (Xylocopa aripennis, X. orich-
lycea), гнездящихся в стеблях бамбука, имеют самые крупные
размеры, длиной более сантиметра (Jamata Kunio, 1964).
Таким образом, уровень плодовитости связан с образом жиз-
ни насекомого.
Вместе с тем на плодовитость большое влияние оказывает
микроклимат, режим освещения, биохимический состав кор-
ма, особенно содержание воды и белков.
Влияние среды на потенциальную плодовитость и ее кон-
кретную реализацию у многих видов регулируется через из-
менение плотности популяции. Колебания плотности популя-
ций определяются изменениями плодовитости и выживанием
потомства, представляя суммированный итог взаимодействия
наследственных свойств вида с условиями среды, меняющейся
во времени. Каждому виду свойственны свои пределы откло-
нений от оптимальной плотности популяций в ряду поколе-
ний. У многих насекомых (например, у соснового подкорного
клопа, майского хруща, сибирского шелкопряда и других)
поколения многочисленные чередуются с поколениями мало-
численными. Величину подобных отклонений считают видо-
вым свойством (Никольский, 1961). По нашим наблюдениям,
в сосновых культурах на песках Среднего Дона (Арчединский
лесхоз Волгоградской обл.) в 1962 г. обилие четного (массо-
вого) поколения («колена») превосходило обилие нечетного
(малочисленного) у подкорного соснового клопа в 8—9 раз,
а у восточного майского хруща в тех же культурах обилие ли-
чинок лётного года превосходило обилие нелётного года в
7 раз.
Причины чередований разных по численности поколений
недостаточно изучены. По-видимому, немаловажное значение
у многих видов имеет дополнительное питание имаго, так как
в период лёта происходит созревание яиц, определяется их
количество и жизнеспособность нового поколения. У афагов
потенциальная плодовитость и ее реализация в значительной
мере зависят от условий питания личинок старших возрастов,
когда идет интенсивное накопление резервных питательных
веществ. Изменить плодовитость афагов возможно действием
фотопериода, изменением качества корма в период развития
гусениц последнего возраста, как это экспериментально по-
казано нами на гусеницах китайского дубового шелкопряда.
У насекомых, питающихся в фазе имаго, недостаток нужной
пищи (сахаров, белков, воды) приводит к снижению плодо-
витости и даже полному бесплодию, например у бабочек лу-
гового мотылька. На значение обильной и сочной пищи в год,
предшествующий массовому размножению ряда насекомых,
87
обращал внимание Н. М. Кулагин. По его мнению, массовое
размножение совпадает с годом сухой и теплой погоды, если
такая погода следует за годом влажным, богатым сочным
кормом.
Колебания численности имеют разный характер. Они мо-
гут быть нерегулярными (флуктуации), а могут закономерно
отклоняться от средней величины, напоминая качание маят-
ника (осцилляции — по Макфедьену, 1963). Эксперименталь-
ное изучение осцилляций показало большое значение плотно-
сти популяций как саморегулирующего фактора. Так, при ис-
кусственном разведении падалы-юй мухи (Lucilia cuprina
Wied.) увеличение числа личинок в садке от 10 до 40 ведет
к увеличению выходящих из них мух. Но дальнейшее увели-
чение плотности популяции личинок приводит к снижению
числа имаго, а при плотности популяции 200 личинок на са-
док выход имаго прекращается. Переуплотнение личинок при-
водит к уменьшению их размеров, вплоть до прекращения
метаморфоза: личинки перестают окукливаться. Это происхо-
дит в том случае, когда число имаго в садке становится боль-
ше 300. Если же имаго мало, яиц откладывается меньше, то
личинки ускоренно переходят к окукливанию, компенсируя
недостаток имаго. Подобного рода компенсаторную реакцию
допускают в природе при проведении истребительных меро-
приятий (Макфедьен, 1963).
Количественные изменения популяции ведут к качествен-
ным переменам в ее составе и структуре. Искусственное пере-
население замкнутых популяций у насекомых вызывает за-
держку роста и развития отдельных особей. На изменения
условий жизни популяция отвечает изменением численности
особей: изменяется плодовитость, жизнеспособность потомст-
ва, мельчают или увеличиваются в размерах и весе особи,
меняются сроки и темпы размножения, поведение и другие
особенности популяции, вскрытые при фазовой изменчивости.
Изменчивость форм в популяциях одного вида, полиморф-
ность,— выражение специфичности внутривидовых отношений.
Физиологическая и генетическая неоднородность популяции,
меняющаяся во времени, дает возможность виду различно
реагировать на внешние факторы, что приводит к сдвигам
численности популяции и ее качественного состояния.
СВЯЗИ НАСЕКОМЫХ В БИОЦЕНОЗЕ
В природе нет организмов, изолированных от воздействия
других видов. Все насекомые существуют за счет потребления
готовых органических веществ и уже в силу этого неизбежно
вступают в связи с другими видами растений и животных.
Эти связи чрезвычайно разнообразны. Существование насеко-
мого-фитофага без кормовых растений невозможно. Невоз-
88
можно и существование насекомого-паразита без хозяина, за
счет которого он живет. Растения и многие насекомые не мо-
гут существовать без почвы как среды обитания и источника
пищи. В свою очередь невозможно и образование самой поч-
вы без деятельности растений, микроорганизмов и многих
животных.
В биоценозах организмы представлены создателями орга-
нических веществ (продуцентами), потребителями этих ве-
ществ (консументами) и организмами-разрушителями тех и
других и продуктов их жизнедеятельности (редуцентами). Со-
вокупное действие всех трех групп организмов обеспечивает
биогенный круговорот веществ и возможность существования
биоценозов и в целом биосферы Земли. Насекомые составля-
ют весьма существенное звено в биоценозах и по обилию
форм и массы живого вещества, и по разнообразию связей.
Во многих случаях эти связи имеют огромное значение
для человека. Их расшифровка помогает вести борьбу с мно-
гочисленными вредителями, защищая посевы и здоровье че-
ловека, помогает использовать и не менее многочисленных
друзей из мира насекомых. Изучение связей насекомых в био-
ценозах лежит в основе биометода и других методов борьбы
с вредителями, в основе учения об очаговости трансмиссивных
заболеваний. Основные участники триады очагов — возбуди-
тель, переносчик и животные-доноры (носители возбудите-
ля) — являются сочленами биоценоза определенных биотопов
известного географического ландшафта страны (Павловский,,
1948).
В. Н. Беклемишев (1951) подчеркивает, что в биоценозах
важны связи, складывающиеся не между отдельными особя-
ми, а между популяциями. Это связи длительные; при извест-
ных условиях они могут изменить соотношение популяций
разных видов в биоценозах. Если плавунец съел головастика,
то это пример лишь возможной кратковременной связи. Если
же популяция плавунцов связана с популяцией лягушек дли-
тельное время, это приводит к изменению численности и воз-
растного состава в популяции лягушек.
Популяционные связи могут быть разными по интенсив-
ности или степени воздействия. Экстенсивные связи отличают-
ся более широким охватом при меньшей степени воздействия.
Связи, возникающие в биоценозах, могут быть обязательными
(облигатными), например связь монофага с хозяином, необя-
зательными (факультативными), затрагивающими лишь часть,
популяции, случайными, односторонними и двусторонними^
простыми и очень сложными. Примером сложной связи мож-
но назвать связь пчелы с ее паразитом. Так, пчела питается
на ромашке, переносит се пыльцу и может, кроме того, встре-
тить на цветке личинку жука — триунгулину (сем. нарывни-
ки— Meloidae), паразитирующую в гнездах пче^.
89
Основными межвидовыми связями Беклемишев считает
связи пищевые (трофические); по месту обитания (топиче-
ские) ; с использованием построек, убежищ, строительного ма-
териала других видов (фабрические) и связи по транспорти-
ровке на других видах (форические связи).
Трофические связи насекомых разнообразны, как разнооб-
разны и сами способы добывания пищи. Поедать объект пи-
тания насекомые могут целиком, заглатывая его вместе с
илом или почвой, или при специальной охоте за добычей. Ра-
стение они, как правило, обгрызают или высасывают частич-
но, или съедают лишь специально предназначенную часть
(выросты семян чистотела, плодовые тела шляпных грибов —
поедаются личинками двукрылых, муравьями). Питаться
насекомые могут не телом организмов-производителей, а лишь
различными их выделениями (нектаром, падью, слизью, по-
том, гноем). Поедаются мертвые остатки — трупы, древесина,
лесной опад, гумус. Разнообразны и косвенные трофические
связи. Например, облегчается доступ к пище кожеедам, жи-
вущим в гнездах орлов; кожееды питаются остатками пищи,
доставляемой в гнездо орлами. Шелкопряд, объедая хвою
сосны, облегчает ее заселение короедами, энтомофаги спаса-
ют растение от фитофагов и т. п.
Топические связи насекомых выражаются в использовании
тех условий среды, которые создаются присутствием других
видов организмов. Навозники связаны с домашними живот-
ными, лосями и другими позвоночными; блохи и прочие ви-
ды — с норами грызунов; многие жужелицы в засушливых
условиях — с растениями, дающими тень и влагу. Своеобраз-
ные связи создаются через использование свертков листьев,
мин, ходов усачей, короедов и других древогрызов. Например,
тля Hoplohaitophorus zachvatkini Aiz. встречается только в
свертках дубовых листьев, сделанных гусеницами дубовой ог-
невки (Salebria atrifasciella Rag.), а в ходах усачей поселяют-
ся пчелы-плотники (Xylocopa), пчелы-листорезы (Megachile),
муравьи. Косвенные топические связи возникают, например,
между оводами и мухами (Calliphora), откладывающими
яйца на кожу, поврежденную личинками оводов; между коро-
едами-типографами на ели и короедами — обитателями сос-
ны, поскольку сосна в тени ели расти не может и фауна сосны
оказывается в косвенной зависимости от врагов ели.
Фабрические связи возникают в связи с постройкой спе-
циальных ловчих сооружений (ловчие сети хищных личинок
ручейников — Hydropsiche, воронки муравьиных львов), со-
оружений, предназначенных для защиты потомства (коконы,
норки пчел и т. д.), хранением запасов пищи (гнезда пчел),
защитой симбионтов (грибные сады и колонии тлей, охраняе-
мые термитами и муравьями). Насекомые вступают в разно-
образные связи при добывании строительного материала —
90
пуха, шерсти, листьев и пр. Это пчелы-шерстобиты (Anthi-
dium), пчелы-листорезы (Megachile). Листорезы связаны с де-
ревьями и кустарниками, из листьев которых они вырезают
кружочки, и с усачами, в ходы которых эти кружочки укла-
дывают.
Форические связи распространены широко. Чаще насеко-
мые используются как переносчики другими организмами, осо-
бенно разнообразными клещами. Форезия обеспечивает рас-
селение и миграции многих видов. Форические связи сопро-
вождаются связями топическими, доставкой в места,
пригодные для размножения. Транспортируемые насекомыми
организмы часто способны к длительной диапаузе, анабиозу,
снабжены защитными оболочками. Пчелы разносят пыльцу,
часто являясь единственными опылителями культурных расте-
ний (например, люцерны); пыльцу разносят и многие другие
насекомые (двукрылые, осы, бабочки). Ряд ржавчинных гриб-
ков издают запах, привлекающий насекомых. Падальных мух
привлекает трупный запах некоторых грибов. Мясистые пло-
довые тела грибов можно рассматривать как приспособле-
ние к переносу спор насекомыми. Споры грибов разносятся
грибными комариками (Fungivoridae), мухами (Muscidae),
некоторыми жуками. Пухоеды для расселения используют
Кровососок, муха (Dermatobia) прикрепляет свои яйца к ко-
марам и во время сосания крови личинки перебираются на
животных. Самки специализированных яйцеедов (Telenomus,
Anastatus и др.) забираются под чешуйки тела, прикрепляют-
ся к волоскам на брюшке бабочек сибирского шелкопряда
и вместе с ними мигрируют в новые места.
НАСЕКОМЫЕ И ВЫСШИЕ РАСТЕНИЯ
Вследствие многочисленности фитофагов, разнообразия
повреждаемых растений, способов питания и ответных защит-
ных реакций растений на повреждения трофические связи на-
секомых с растениями также чрезвычайно разнообразны и
представляют основу взаимосвязей насекомых в биоценозах.
Эти связи не остаются постоянными. Устойчивость растений
К повреждениям насекомыми зависит от особенностей расте-
ния, характерных для разных фаз его развития, от состояния,
возраста и вида насекомого, а также от меняющихся условий
абиотической среды. Эти отношения не ограничиваются толь-
ко повреждениями насекомыми каких-либо тканей и частей
растений. Широко распространено использование нектара и
пыльцы многими насекомыми для питания.
У растений выделение нектара и обилие пыльцы служит
приманкой насекомых-опылителей, барьеры же из волосков,
выделение клейких веществ не допускают нежелательных на-
секомых. Яркая окраска и аромат цветов также привлекают
91
насекомых. Не все насекомые в равной степени приспособле-
ны к посещению цветов. Одни из них повреждают органы
цветка (многие жуки, их личинки, муравьи, трипсы); другие
мало способствуют опылению: их движения беспорядочны
(жуки, клопы, прямокрылые, многие двукрылые, осы). Насе-
комые с лижущим и лижуще-сосущим ротовым аппаратом,,
густым опушением для сбора пыльцы и ловкими движениями
приспособлены к посещению цветов значительно лучше (ко-
роткохоботные пчелы, мухи-бомбилиды, мухи-журчалки, мно-
гие чешуекрылые).
Наиболее специализированные насекомые-опылители име-
ют сильно опушенное тело, длинный хоботок, способность пи-
таться, не садясь на цветок (бражники).
По степени взаимной адаптации к насекомым и специали-
зации цветка (по Кузнецову, 1951) растения подразделяют-
ся на:
1) . Цветы перепончатокрылых — преимущественно крас-
ной, синей и фиолетовой окраски, со скрытым нектаром и
пыльцой, со сложным механизмом расправления и с площад-
ками для прилетающих насекомых (губоцветные, мотылько-
вые). Есть среди них и «осиные» цветы с более поверхностно
лежащим нектаром.
2) . Цветы чешуекрылых — с белой или светлой окраской
для дневных бабочек и с сильным запахом для ночных видов,,
с нектаром в глубоких трубочках, с открыто лежащей пыль-
цой.
3) . Цветы двукрылых — белые или синие, открытые, с
легко доступным нектаром; тусклоокрашенные, с вздутыми
ловушками, в которых мухи задерживаются, пока не произой-
дет опыление. Цветы с запахом падали привлекают падальных;
и мясных мух.
4) . Цветы инжира опыляются специальными хальцидами
(Blastophaga).
5) . Цветы юкки оплодотворяют бабочки юкковой моли
(Pronuba), которые после откладки яиц в цветок переносят
и оставляют пыльцу на рыльце пестика.
6) . Муравьиные растения имеют различные полости внутри
стеблей, клубней, шипов; в этих полостях живут и питаются
муравьи.
Пищевые связи с растениями исторически опредилили раз-
витие насекомых, обусловили особенности строения ротовых
частей и другие приспособления, связанные с добыванием пи-
щи. Одновременно в направлении приспособления к насеко-
мым шло эволюционное развитие и растений. Однако значе-
ние растений в жизни насекомых не ограничивается пищевы-
ми связями. Растения дают убежище от многочисленных вра-
гов, дождя и ветра, служат местом для откладки яиц, для
зимовки и т. д.
92
НАСЕКОМЫЕ И НИЗШИЕ РАСТЕНИЯ
Многие насекомые тесно связаны с грибами и бактериями.
Эти связи часто взаимно полезны: различают эктосимбиоз и
эндосимбиоз.
Пример эктосимбиоза — питание насекомых грибка-
ми: грибки способствуют перевариванию целлюлозы и древе-
сины. Ряд насекомых готовят субстрат для грибка, разводят
грибные сады. Муравьи листорезы (Atta) в ходах своих жи-
лищ культивируют гриб Rhozites, термиты — гриб Xylaria,
короед (Anisandrus) — гриб Monilia. Известны и многие дру-
гие подобные связи насекомых с грибами.
Эндосимбиоз — случаи нахождения микроорганизмов,
преимущественно бактерий, в кишечнике и тканях тела насе-
комых. У многоядных насекомых эндосимбиоз характерен для
тараканов и муравьев. Значительно чаще он встречается у на-
секомых-минеров, у потребителей древесины и семян. У терми-
тов в расширениях прямой кишки живут жгутиконосцы,
у бронзовки (Potosia), жука-носорога (Oryctes), долгоножки
(Tipula) бактерии населяют мешковидные расширения задней
кишки. Сахоромицеты встречены у личинок жуков-притворя-
шек (Ptinidae), некоторых усачей и других насекомых. Живут
бактерии в кишечнике у клопов, у равнокрылых-хоботных,
в гемолимфе червецов, в специальных мицетомах тлей, цикад.
Имеются симбионты и у кровососущих насекомых (мухи
це-це, кровососки, постельные клопы). Предполагают, что
симбионты участвуют в обмене веществ хозяина, выделяя
ферменты и витамины.
В некоторых случаях бактерии вызывают свечение. Так,
гусеница Mamestra oleracea L. светится от Bacterium haemo-
phosphoreum. От бактерий, по-видимому, светятся некоторые
поденки и другие насекомые. При откладке яиц , насекомые
обмазывают их содержимым кишечника с симбионтами, моло-
дые личинки съедают скорлупу яйца, обеспечивая дальней-
шее существование симбионтов.
МИКРООРГАНИЗМЫ - ВОЗБУДИТЕЛИ БОЛЕЗНЕЙ НАСЕКОМЫХ
Насекомые подвержены многим болезням, вызываемым
бактериями (бактериозы), вирусами (вирозы), грибами (ми-
козы), простейшими (протозоонозы), нематодами (гельмин-
тозы). Заболевания принимают часто эпидемический характер
и приводят к вымиранию популяций массовых видов насеко-
мых на больших площадях. Много наблюдений проведено по
заболеваниям вредителей леса: сибирским и непарным шелко-
прядом, монашенкой, сосновой совкой, сосновыми пилильщи-
ками, хрущами и другими видами. Особенности заболевания
связаны с экологией вредителя, природой возбудителя болез-
93
ни и условиями среды. Для одних видов характерны вирозы,
для других — микозы или бактериозы. Наибольшее значение
имеют смешанные заболевания, вызываемые одновременно
разными вредителями (например, полиэдренным вирусом и
микроспоридией, или вирусом и энтомофторовым грибом).
При смешанных заболеваниях эпизоотии протекают быстро и
сказываются на потомстве. Болезни насекомых изучены недо-
статочно !.
Бактериальные болезни (бактериозы) давно известны пче-
ловодам и шелководам. Заболевание пчелы «европейский гни-
лец» распространено повсеместно и вызывается спороносной
бактерией Bacillus alvei Ches. Повсюду личинки пчел болеют
американским гнильцом; возбудитель — спороносная палочко-
видная бактерия Bacillus larvae White. Общеизвестно заболе-
вание тутового червя — флашерия; возбудитель — микроб Ba-
cillus bombysepticus. Бактериальные возбудители обнаружены
при заболеваниях майского и июньского хрущей, японского
жука. Болезни личинок жуков известны под названием «мо-
лочные болезни»; японский жук — главный хозяин возбудите-
ля этой болезни (Bacillus popilliae Dutky). В. popilliae исполь-
зуют для борьбы с японским жуком. Известны и многие
другие бактериозы — у гусениц совок, кукурузного мотылька;
саранчевых и др. Распространены болезнетворные споровые
бактерии, действующие на насекомых токсическими вещества-
ми. Это бактерии типа Bacillus cereus Frankl., В. thuringensis
Berk Они выделены во время эпизоотий соснового и сибирско-
го шелкопрядов, яблонной плодожорки, черепашки, пилиль-
щиков и многих других насекомых и используются для борь-
бы с вредителями.
Эффективность бактериологического метода борьбы с на-
секомыми определяется многими факторами: вирулентностью
возбудителя, его агрессивностью, устойчивостью возбудителя
к метеорологическим условиям, плотностью вредителя-хозяина
в местах локализации, миграцией бациллоносителей, ослабле-
нием сопротивляемости насекомого.
Возбудитель заболеваний сибирского шелкопряда — спо-
роносная палочковидная шелкопрядная бацилла Bacillus den-
drolimi Tai. — вызывает острую форму эпизоотии у гусениц
этого насекомого в старших его возрастах. При известных
условиях применение препарата дендробациллина, изготов-
ленного из спор бактерии и наполнителя, приводит к полной
гибели гусениц. Эффективность препарата зависит от степени
сопротивляемости гусениц; применять его желательно ко вре-
мени выхода гусениц из лесной подстилки, после их пере-
зимовки.
Важнейшим условием интенсивного развития патогенных
1 Наиболее полная сводка по патологии насекомых принадлежит
Штейнхаузу (1952, 1963).
94
микроорганизмов в теле насекомых является физиологическое
ослабление насекомых. Оно может быть вызвано неблагопри-
ятными условиями питания, влажности, температуры, ослаб-
лением газообмена, а также влиянием человека — воздействи-
ем на них слабыми дозами ДДТ и гексахлорана. Применение
бактериологического метода борьбы в сочетании с инсектици-
дами повышает гибель насекомых (Н. А. Теленга, 1955). Для
получения промышленных бактериальных препаратов в основ-
ном используют Bacillus thuringensis Berl.
Грибные заболевания (микозы) часто вызывают гибель на-
секомых. Грибы включают различные типы организмов, кото-
рые часто обозначаются как плесени, дрожжи, актиномицеты
и т. д. Представителями грибов, вызывающих болезни насеко-
мых, являются главным образом грибы-водоросли (Phycomy-
cetes) и несовершенные грибы (Ascomycetes).
Наиболее важные из них — грибы из семейства энтомофто-
ровых (Entomophthoracea), особенно Empusa, Entomophtora
и Massospora. На насекомых паразитируют более 60 видов
энтомофторовых грибов. Эпизоотии, вызываемые этими гри-
бами, отличаются массовым характером. Заражаются ими
двукрылые, полужесткокрылые, чешуекрылые и жуки. Извест-
но заражение комнатной мухи грибом Empusa inuscae Cohn.,
многих саранчевых — Empusa grylli Fres., сосновой совки —
Empusa aulicae Reich., капустной белянки — грибком Ento-
mophtora sphaerospherma Fressen и т. д. Энтомофторозы ча-
сто поражают златогузку, хрущей, подкорного соснового кло-
па, кольчатого шелкопряда и др.
Другая группа грибных болезней — мускардинозы. Вызы-
ваются они несовершенными грибами порядка гифомицетов,
например Beauveria bassiana Bals. Мускардина известна у
тутового шелкопряда, кукурузного мотылька, японского жу-
ка, яблонной плодожорки, пилильщиков, златогузки и многих
других насекомых. Мускардиноз наблюдается при физиологи-
ческом ослаблении насекомых. Распространению болезни спо-
собствуют условия высокой влажности и умеренной темпера-
туры. Положительные результаты при борьбе с вредителем
получены от применения биопрепарата и химических средств,
обеспечивающих физиологическое ослабление вредителя.
Опудривание семян сахарной свеклы биопрепаратом зеленой
мускардины и внесение гексахлорана в почву в смеси с супер-
фосфатом повышает заболевание мускардиной. Применение
против яблонной плодожорки водной суспензии биопрепарата
белой мускардины и дуста ДДТ позволило снизить числен-
ность зимующих гусениц на 80% (Н. А. Теленга).
В ряде случаев применение патогенных микроорганизмов
вызывает развитие других возбудителей болезни. Так, при
применении против свекловичного долгоносика белой мус-
кардины личинки погибают главным образом от зеленой мус-
95
кардины; применение белой мускардины вызывает гибель
зимующих гусениц плодожорки от гриба спикария.
Среди возбудителей микозов известны высокоспециализи-
рованные облигатные паразиты (Coelomomyces) комаров,
а также представители сумчатых грибов из рода Cordyceps,
паразитирующие преимущественно на личинках многих насе-
комых.
Вирусные заболевания (вирозы) широко распространены
и у насекомых, и у растений. С вирусами насекомых давно
сталкиваются в практике шелководства и пчеловодства. Ви-
русные заболевания наносят здесь огромный ущерб, вызывая
массовую гибель гусениц шелкопрядов и личинок пчел.
Среди насекомых известны вирозы нескольких типов: по-
лиэдренные, характеризующиеся образованием в теле насе-
комых включений в виде многогранных телец — полиэдров;
гранулезы, с образованием включений в виде овальных те-
лец — гранул, или капсул, и вирусные болезни, не связанные
с включениями.
Полиэдренные болезни бывают ядерного типа и цитоплаз-
менные. Вирусы ядерной полиэдрии относятся к роду Borreli-
navirus, а цитоплазменной (сферической) — к роду Stnithia-
virus.
Наиболее распространена ядерная полиэдрия: она извест-
на более чем у 100 видов чешуекрылых и немногих видов
перепончатокрылых, жуков, двукрылых. Полиэдроз известен
у многих вредителей леса, таких, как непарный и кольчатый
шелкопряд, монашенка, боярышница, златогузка, сосновая
совка, сосновая пяденица, пилильщики и др. Характерна по-
лиэдренная болезнь для личиночных фаз, встречается у ку-
колок и редко у бабочек.
Цитоплазменная полиэдрия известна у нескольких десят-
ков видов насекомых. Вирус размножается преимущественно
в цитоплазме клеток эпителия средней кишки. Это заболева-
ние известно у непарного шелкопряда, зимней пяденицы, аме-
риканской белой бабочки и др.
Через яички вирусы передаются следующим поколениям.
Возбудитель ослабляет эти поколения и надолго задерживает
размножение жизнеспособного потомства. Вирозы носят резко
выраженный эпизоотический характер и приводят к вымира-
нию популяций.
Для разных видов насекомых установлен ряд специфиче-
ских вирусов. Указывают на связь вирусных болезней с энто-
мофагами. Там, где появляется полиэдренная болезнь, на
больных гусениц перестает паразитировать тахина, распро-
странение вирусной болезни вынуждает многих тахин пере-
селяться в менее зараженные области (Штейнхауз, 1952).
По другим данным, паразитические саркофаги больше напа-
дают на больных гусениц. Вирусные заболевания зависят от
96
условий температуры, влажности и состояния гусениц. Мно-
гие исследователи указывают, что для развития вирусных эпи-
демий необходима высокая плотность популяции хозяина.
В ряде случаев вирусные заболевания приводят к полной
гибели вредителя. Часто вирус остается в латентном состоя-
нии. Изучение ядерного полиэдроза, цитоплазменных вирусов,
латентности вирусной инфекции и факторов, активизирующих
заболевание, показало возможность практического их исполь-
зования для борьбы с вредителями. В ряде стран вирусные
биопрепараты применяются против пилильщиков.
Велико значение насекомых в биоценозах как переносчи-
ков вирусов растений и животных. У растений вирусы вызы-
вают заболевания мозаикой и желтухой. Мозаики передаются
сосущими насекомыми во время их питания на растении, при
прививках через сок больных растений и другими путями.
Желтухи, например, такие, как скручивание листьев хлопчат-
ника, столбур пасленовых и другие, переносят только насеко-
мые, особенно тли, цикады, листоблошки, некоторые клопы,
трипсы. Каждый вид тли может переносить несколько виру-
сов, а некоторые виды вирусов переносятся многими видами
тлей. Например, персиковая тля, питаясь на различных ра-
стениях, переносит 21 вирус, а желтуху лука переносят 53 раз-
личных вида тлей. Цикады, как правило, строго специфичные
переносчики: каждый вид переносит какой-то один вирус.
Установлен сложный цикл развития вирусов; часть этого
цикла проходит в теле насекомого. Например, в кишечнике
цикады обнаружен вирус закукливания овса в кристалличе-
ском виде. Отсюда вирус попадает в слюнную железу, а во
время питания насекомого проникает в ткани растения. Все
вирусы в теле цикад проходят инкубационный период, во вре-
мя которого вирус размножается.
Распространение вирусов зависит от условий погоды, на-
личия источника заражения, переносчика. И патология насе-
комых, и патология растений могут быть поняты лишь при
учете влияний как абиотических факторов среды, так и связей
между организмами в биоценозах.
Нематодные болезни (гельминтозы) вызываются энтомо-
гельминтами. В качестве хозяев нематод известно около
400 видов насекомых из 100 семейств 21 отряда. Подавляю-
щее число видов-хозяев с большим числом гельминтопарази-
тов известно в отрядах двукрылых, жуков, бабочек, хоботных,
прямокрылых, тараканов, перепончатокрылых, блох. Значение
гельминтов в жизни насекомых не меньше, чем вредных гель-
минтов в жизни человека и домашних животных. Они вызы-
вают бесплодие, болезни и гибель насекомых.
Общее количество видов энтомогельминтов превышает
1000. Наибольшее значение имеют нематоды, паразитирующие
в полости тела и в тканях насекомых. В одном насекомом мо-
7 Заказ 193 97
жет быть от 1 до 1500 нематод во всех фазах развития. Они
встречаются и снаружи: в складках груди и брюшка, на но-
гах, под крыльями и на других частях тела.
Питаются энтомогельминты кровью и жировым телом хо-
зяина, часто нарушая развитие половых клеток. Известны
инвазии, приводящие к массовой гибели насекомых.
В настоящее время разработаны методы культивирования
нематод на искусственных средах. Перспективные результаты
получены при применении для борьбы с вредителями немато-
ды рода Neoaplectana. Neoaplectana высоко патогенна, пере-
носит бактериальные инфекции, размножается на искусствен-
ных средах, способна образовывать латентных личинок и мо-
жет расселяться с насекомыми. Паразитирует в жуках, ба-
бочках, двукрылых, перепончатокрылых (Положенцев, Артю-
ховский, 1963). Известно более 10 видов гельминтов этого
рода.
Многие насекомые имеют огромное значение как перенос-
чики гельминтов человека и животных. Для группы энтомо-
биогельминтов насекомые служат лишь промежуточными
хозяевами. Так, комары, мошки, мокрецы, слепни передают
человеку через уколы несколько видов филярий. Многие жуки,
особенно копрофаги, передают многочисленных возбудителей
разнообразных гельминтозов домашних и диких животных
(Положенцев, Негробов, 1967). Мероприятия по оздоровлению
пастбищ не могут быть решены без учета роли насекомых —
переносчиков заболеваний. Установлена зональность распро-
странения гельминтов, зависимость путей и источников зара-
жения от условий среды обитания зародышевых форм гель-
минтов, питания и возраста их хозяина — насекомого. Изуче-
ние роли энтомогельминтов в подавлении численности
популяции вредных насекомых и роли насекомых-переносчи-
ков гельминтозов животных — необходимо продолжать.
Протозоонозы — болезни насекомых, вызываемые простей-
шими — в динамике численности насекомых имеют большое
значение. Простейшие передаются от зараженных насекомых
в следующее поколение и снижают жизнеспособность популя-
ции. Патогенными для насекомых могут быть амебы, грегари-
ны, гемоспоридии и микроспоридии. В качестве паразитов
насекомых описано свыше 100 видов микроспоридий. Боль-
шинство из них принадлежит к роду Nosema.
Протозойные заболевания насекомых, в отличие от бакте-
риальных и вирусных, развиваются очень медленно, прини-
мая хронический характер. Общеизвестно длительное заболе-
вание шелковичных червей — пебрина и заболевание пчел,
вызываемое Nosema apis Zander. Микроспоридиозы известны
у лугового мотылька, златогузки, сибирского и непарного
шелкопрядов, комаров и других насекомых. У непарного шел-
копряда и златогузки паразитирует Nosema polyvora и Pits to-
te
phora schubergi Zvoef. Одновременно в одном насекомом мо-
жет быть до семи различных видов простейших.
Часто простейшие из классов Sporozoa, Mastigophora,
Sarcodina, Ciliophora развиваются в теле насекомых как про-
межуточных хозяев и безвредны для самих насекомых. Из-
вестен сложный цикл развития в теле комаров возбудителя
малярии — плазмодиума. Разнообразные кровососы переда-
ют человеку при сосании крови трипанозом, лейшманий и дру-
гих возбудителей болезней, но сами от них не страдают.
Роль простейших в изменении численности насекомых изу-
чена недостаточно.
НАСЕКОМЫЕ КАК ПЕРЕНОСЧИКИ БОЛЕЗНЕЙ
Огромно значение насекомых как переносчиков разнооб-
разных болезней.
Насекомые передают возбудителей болезней человеку и
животным разными способами. Ряд болезней — трансмиссив-
ные — передаются только через переносчика. Многим этим бо-
лезням (чуме, туляремии, энцефалитам и другим) свойствен-
на природная очаговость. Возбудитель болезни, его перенос-
чик, животное-носитель возбудителя (хозяин-донор) — все
являются сочленами определенных биоценозов. Природные
очаги связаны с норами животных (например, грызунов),
с пещерами, гнездами птиц и другими элементами биотопов.
Инфекционные болезни, поражающие человека и животных
(бруцеллез, сап, туляремия, чума, бешенство и многие дру-
гие), называют зоонозами. Зоонозы существуют только там,
где живут переносчики и их хозяева, где возможен непре-
рывный контакт возбудителя через паразита с хозяином. Та-
кие места Е. Н. Павловский и называет «природными очагами
заболеваний». В основе природного очага лежат тесные пище-
вые связи. При равномерном расселении носителей инфекции
и их высокой численности могут существовать очаги с по-
стоянными или периодически возникающими эпизоотиями.
Периодичность вспышек заболеваний обычно совпадает
с массовым появлением и расселением переносчиков или ос-
новных хозяев. При неравномерности распределения живот-
ных природные очаги приурочены к плотно заселенным ме-
стам. Например, южноазиатские очаги чумы приурочены к
местам, где на крысах обильны блохи Xenopsylla cheopis Both.
(Н. П. Наумов).
При циркуляции возбудителя в природе он может быть
связан с несколькими видами хозяев и переносчиков, что уве-
личивает возможность его размножения. Множественность
хозяев и переносчиков обеспечивает устойчивость очагов. На-
пример, туляремию переносят клещи, вши, блохи, слепни, ко-
7* 99
мары, мухи-жигалки. В каждом очаге существуют основные
виды, обеспечивающие его существование, но участие других
делает очаг более устойчивым.
Контакт переносчика, возбудителя и хозяина обеспечивает-
ся и передвижениями животных по определенным «кочевым
путям». Расположенные на таких путях норы грызунов почти
всегда заняты основными хозяевами или пришельцами.
Изучение природных очагов заболеваний, путей циркуля-
ции возбудителей болезней вместе с их переносчиками-насе-
комыми, требует комплексности исследований. Понять приро-
ду заболеваний можно, лишь раскрыв связи в биоценозах
и взаимные влияния природных очагов на человека и обрат-
но— в связи с его сознательной и бессознательной деятель-
ностью. Биоценотические исследования в сочетании с изуче-
нием аутэкологии возбудителей, переносчиков и животных,
страдающих от них, дают надежную основу для построения
рациональных систем борьбы с трансмиссивными паразитар-
ными болезнями человека (Е. Н. Павловский). Экологиче-
ские — эпидемиологические — исследования должны основы-
ваться на паразитологии в такой же мере, как и на микро-
биологии. Учение о насекомых-паразитах одинаково важно,
как и учение о патогенных микробах. Роль специфического
переносчика в эпидемиологии трансмиссивных болезней не
менее важна, чем значение самого возбудителя. Порознь они
безразличны для человека.
Энтомофаги насекомые — паразиты и хищники — играют
существенную роль в биоценозах.
Насекомые-энтомофаги многочисленны, включают хищни-
ков, экто- и эндопаразитов. Хищники ведут свободный образ
жизни, уничтожая яйца, личинок, куколок и взрослых насе-
комых. К их числу относятся представители жуков, верблюд-
ки (отряд Rhaphidioptera), двукрылые, перепончатокрылые,
ряд полужесткокрылых и пр. Хорошо известны красотелы
(род Calosoma) и тлевые коровки и другие хищные жуки.
Личинка зеленого красотела съедает до 40, а взрослый жук —
до 270 гусениц. До 80% яйцекладок непарного шелкопряда
поедают кожееды (сем. Dermestidae). Деятельно уничтожают
вредителей муравьи, особенно Formica polyctena Foerst. В те-
чение дня муравьи одного большого гнезда собирают до
20 000 насекомых. Для борьбы с картофельным жуком стали
использовать хищных клопов. Паразитические энтомофаги
также истребляют насекомых на всех фазах развития. Ис-
числяются они десятками тысяч видов, составляя около 70%
общего числа известных видов паразитических животных.
По В. А. Догелю, паразитических насекомых насчитывалось
не менее 60 000, к сегодняшнему дню эта цифра значительно
возросла. Известны паразиты из 87 семейств, но особенно бо-
гаты ими перепончатокрылые (сем. Ichneumonidae, Braconi-
100
dae, Chalcidoidea, Aphidiidae, Scoliidae и др.), двукрылые
(Larvaevoridae, Sarcophagidae, Phasiinae, Bombilyidae и др.).
Яйца паразитические энтомофаги откладывают либо
внутрь тела хозяина, прокалывая покровы яйцекладом, либо
на покровы и внутрь тела вбуравливается уже личинка, либо
на растение и в тело хозяина личинки и яйца паразита попа-
дают с пищей. Одни виды заражают только яйца, другие —
молодых личинок, третьи и личинок, и куколок, и даже имаго.
Развитие паразита может пройти в одной из этих фаз хозяи-
на либо, начавшись в яйце или личинке, завершится в кукол-
ке и имаго. Паразиты могут нападать только на одного
хозяина, а могут переключаться на питание другими видами
хозяев, что способствует сохранению численности паразита.
Нападать паразиты могут на разных хозяев в одно время и
в одном месте, а могут менять хозяев по мере исчезновения
одних и появления других.
Отношения паразитов с хозяевами усложняются наличием
сверхпаразитов разного порядка, живущих за счет паразитов
первичных, вторичных и т. д., до пятого и даже шестого по-
рядка. Нередки случаи сопаразитизма, когда в одной особи
хозяина живут разные виды паразитов. Исход такого сопара-
зитизма бывает различный: один паразит выживает, другой
гибнет; остаются жить оба паразита, не страдая друг от дру-
га, или одновременно ослабляются, задерживаются в росте и,
наконец, оба паразита погибают из-за недостатка пищи и
преждевременной гибели хозяина.
Формы и случаи паразитизма разнообразны, и не всегда
легко разграничить хищников и паразитов. Паразитизмом
принято считать все случаи питания одного животного за счет
другого. Обычно считают, что хищные насекомые убивают
добычу сразу, а паразитические — питаются ею долгое время.
Для хищного насекомого требуется несколько особей жерт-
вы, а паразитическое насекомое успевает закончить развитие
в одной особи. Так, личинка семиточечной коровки за сутки
съедает до 270 тлей, а яйцеед трихограмма развивается в од-
ном яйце какой-нибудь бабочки. В одном яйце может одно-
временно развиваться и по нескольку особей паразита. Так,
например, яйцеед Trichogramma откладывает в каждое яйцо
хозяина разное число яиц, что зависит от размеров яиц хо-
зяина. В яйце сосновой совки развивается до 8 особей трихо-
граммы, а в яйце сибирского шелкопряда—до 39. Но многие
паразиты откладывают лишь по одному яйцу. В некоторых
случаях паразитирование идет за счет яиц, личинок и запасов
пищи, заготовленных другими насекомыми, что особенно рас-
пространено у перепончатокрылых. Осы блестянки (Chryzidi-
dae), пчелы-кукушки (Nomada, Psythyrus и др.), осы семейст-
ва Mutillidae откладывают яйца на чужие запасы пищи; их
развивающиеся личинки поедают .яйца., и. личинок, хозяина
. 101
вместе с их пищей. Широко распространено у насекомых «во-
ровское сожительство». Например, муравей-вор (Solenopsis
fugax Latr.) живет в гнездах более крупных муравьев и тас-
кает у них пищу.
Специализация питания энтомофагов разнообразна и у ли-
чинок, и у имаго. Одни виды развиваются на одном хозяине
и только на одной из фаз его развития, другие паразитируют
на широком круге хозяев. Например, Telenomus gracilis Mayr.
и Ooencyrtus pinicola Mats, специализированы на питании
яйцами сибирского шелкопряда; Aphelinus mali Hald. зара-
жает только кровяную тлю; Rodolia cardinalis Muis, истреб-
ляет червеца ицерию, a Trichogramma evanescens West, спо-
собна уничтожать яйца 84 видов бабочек, хищник криптоле-
мус одинаково охотно поедает и мучнистых червецов, и
пульвинарий. За счет бобовой огневки живет 41 вид парази-
тов, соснового шелкопряда — 42 вида и т. д. Очень часто в
первом поколении паразит живет на одном вредителе, а во
втором поколении — на другом. Иногда один и тот же вид
энтомофага является и первичным, и вторичным паразитом.
Например, многоядный Dibrachys cavus Wek. развивается и
в куколках соснового шелкопряда, и в куколках его парази-
тов (Meteorus versicolor Wesm., Rhogas esenbeski Htg.).
Сложные взаимоотношения разнообразных энтомофагов со
своими хозяевами и между собой дополняются связями с кор-
мовыми растениями при дополнительном питании имаго.
Большинство имаго различных энтомофагов любят солнечную
теплую погоду, деятельны днем. Питаются на цветах пыльцой
и нектаром, на листьях деревьев медвяной росой. Для многих
наездников дополнительным питанием служит гемолифма хо-
зяина. Они прокалывают тело хозяина и слизывают выступаю-
щую жидкость. На цветах часто питаются тахины, как прави-
ло,— нектаром зонтичных. На цветах тамарикса собираются
тахины, наездники, сколии. Питание взрослых паразитических
форм изучено недостаточно.
Сложность взаимоотношений между насекомым-фитофа-
гом и его паразитами можно показать на примере бобовой
огневки. В Волгоградской обл. бобовая огневка постоянно в
массе повреждает семена желтой акации в лесных полосах.
По Г. А. Викторову (1956), на ней живет 41 вид энтомофагов.
Среди них выделяется пять экологических группировок:
1). Яйцееды, развитие которых проходит в яйцах (Tricho-
gramma sp., Liophanurus sp.). Во втором поколении они унич-
тожают до 18% яиц. 2). Яйце-гусеничные паразиты; зара-
жают яйца, но продолжают развиваться в гусеницах до их
коконирования: Один из паразитов указанной группы (Phane-
rotoma rjabovi Voin.— Кг.) специализирован на питании бо-
бовой огневкой и развивается синхронно с ее развитием; они
вместе, впадают и в диапаузу. 3). Гусеничные паразиты —
102
заражают гусениц бобовой огневки в период питания в пло-
дах и заканчивают развитие до ухода гусениц в почву для
коконирования. Наиболее многочисленны среди членов этой
группы эктопаразитические формы из семейства Ichneumoni-
dae (Epiurus ventricosa Fshek; E. offinis Haberm.), Braconidae
(Bracon piger Wesm. u Habrobracon brevicornis Wesm.) и
Chalcididae (Habrocytus). Несколькими видами представлены
эндопаразиты (Apanteles apellator Haberm., A. longicaudis
Wesm. и др.). Наибольшее значение среди гусеничных парази-
тов имеют Epiurus ventricosa, Bracon piger и Elasmus sp.
4). Гусенично-пронимфальные паразиты; самки откладывают
яйца в кормящихся гусениц, а личиночное развитие заканчи-
вается после их коконирования. Сюда относится большое ко-
личество эндопаразитов из семейств Ichneumonidae, Braconi-
dae, Larvivoridae. Наиболее эффективен из них Litnneria fus-
clcarpus Thoms. 5). Пронимфальные паразиты; нападают на
уже закоконировавшихся гусениц. Обычен из них Cryptus vi-
duatorius и Mesostenus abbinotatus.
Многочисленные паразиты приурочены к разным место-
обитаниям: 1) ксерофильным (на южных склонах и плато
полупустыни); 2) мезофильным (в верховьях долин и на всех
склонах, кроме южных); 3) гигрофильным (на влажных лу-
гах, в долинах балок). В каждой из перечисленных стаций
преобладают определенные виды энтомофагов. По сравнению
с бобовой огневкой ее паразиты менее пластичны: далеко не
все переходят за ней с диких бобовых растений в лесные по-
лосы. Так, из двух гусеничных паразитов один (Epiurus vent-
ricosa Tshek.) господствует в условиях повышенного увлаж-
нения, а другой (Bracon piger Wesm.) — в ксерофильных ста-
циях. Для обоих видов характерна специализация в отноше-
нии кормовых растений бобовой огневки: на плодах мышиного
горошка они заражают около 30% гусениц бобовой огневки,
а на ракитнике в этих же условиях совсем не встречаются.
Крайне малочисленны они и в посадках желтой и белой ака-
ции, что связано с плотностью стенок бобов. Ксерофильные
виды паразитов не идут в глубину насаждений, хотя бобовой
огневки бывает там много.
Сложность взаимоотношений, существующих между бобо-
вой огневкой, ее кормовыми растениями и паразитами-энтомо-
фагами, показана на рис. 15.” •
Энтомофаги из мира позвоночных животных также играют
большую роль в истреблении насекомых. Необыкновенной
прожорливостью, способностью быстро находить скопления
насекомых отличаются насекомоядные птицы. Они склевыва-
ют насекомых как здоровых, так и зараженных паразитами
и болезнями. Коконы наездников и насекомых, поражённых
микозами, птицы не трогают, способствуя- сохранению пато-
генных грибов. Ряд птиц,, /грачи, . воро.бьц); истреб-
103
ляют насекомых лишь при выкармливании птенцов. В одних
случаях птицы резко снижают численность вредных насеко-
мых, в других — их влияние на биоценозы сказывается мало.
Многих полезных птиц, в том числе и домашних (куры, утки),
используют для борьбы с вредителями, особенно в садах и
лесах. Одна курица за день съедает до 1500 клопов-черепа-
Рис. 15. Биоценотические связи бобовой огневки и ее паразитов
(по Г. А. Викторову).
Сплошными кругами обозначены два поколения бобовой огневки
(I, II) и ее кормовые растения; пунктирными кружками — ее основ-
ные паразиты. Толщина стрелок соответствует проценту повреждения
плодов и зараженности паразитом
шек, до 1400 долгоносиков. Мало изучено влияние на числен-
ность насекомых ящериц, лягушек, летучих мышей, землероек,
кротов, ежей и других животных. Зоологи Туркменской ака-
демии наук ставят вопрос об охране уникальной колонии ле-
тучих мышей Захарденской пещеры. Здесь их насчитывают
30 тыс., и каждую ночь, вылетая на плантации у подножий
.Копет-Дага, летучие мыши уничтожают до 120 кг насекомых-
вредителей.
СПОСОБЫ ОБЩЕНИЯ И ЗАЩИТА ОТ ВРАГОВ
У насекомых способы общения и защита от врагов разно-
образны. Обилие контактов друг с другом, с объектами пита-
ния, с представителями биоценозов привели к выработке раз-
104
нообразных способов общения, распознавания, защиты. Хищ-
ники могут преследовать добычу, подстерегать, выслеживать,
фитофаги должны различать, узнавать нужные кормовые ра-
стения, тонко реагируя на их состояние. От нападения врагов
насекомые защищаются быстрыми оборонительными движе-
ниями, спасаются в трещинах, полостях, чехликах, свертках,
паутинных гнездах и других укрытиях. Защите помогают
быстрый бег, полет, прыжки, иногда неподвижность (насеко-
мые падают «притворяются» мертвыми). Широко используют-
ся такие органы, как жало, вооруженные шипами ноги. Боль-
шое значение имеют разнообразные ядовитые железы, волос-
ки, выбрызгивание ядовитых и отпугивающих веществ. Яйца
паразитов подвергаются в теле хозяина фагоцитозу и инкап-
сулированию.
В процессе эволюции у насекомых возникла и совершенст-
вовалась выработка биологически активных веществ, специ-
фически действующих на других особей своего вида и на осо-
бей другого вида. Эти вещества называются телергонами
(Киршенблат, 1962). Вещества, действующие на особей дру-
гих видов (гетеротелергоны-феромоны), могут быть защитны-
ми или отпугивающими, предохраняющими от нападения,
парализующими или убивающими добычу, возбуждающими
(наркотическими) у сожителей и специальными для воздейст-
вия паразитов и комменсалов на своих хозяев. Вырабатывают
телергоны особые одноклеточные и многоклеточные железы.
Телергоны очень разнообразны по химическому строению и
физиологическому действию. По биологической активности
они не уступают гормонам и витаминам. Необходимо их даль-
нейшее, более глубокое изучение.
Глава V
МАССОВЫЕ РАЗМНОЖЕНИЯ НАСЕКОМЫХ
Насекомых, дающих вспышки размножения, относительно
немного, но значение их в жизни природы и человека огромно.
Проблема динамики численности животных по праву считает-
ся центральной проблемой современной экологии, и для ее
решения необходимо знать и значение абиотической среды,
и специфику внутривидовых отношений, и связи в биоцено-
зах. Эта проблема объединяет экологов всех направлений.
Фактических данных о размножениях вредных насекомых
накоплено немало, но нерешенных вопросов остается еще
больше. Поскольку насекомые, дающие массовое размноже-
ние, очень разнообразны и по таксономическому положению,
и по особенностям экологии и распространения, то и законо-
мерности колебания численности популяций, причины вспышек
также различны. Занимались изучением массовых размноже-
ний экологи разных направлений, на разных объектах, в раз-
ных странах и в разных условиях. Это одна из причин, поро-
дивших обилие теорий и много споров.
Вспышки размножения у разных видов имеют разную про-
должительность, широко распространены или ограничены тер-
риторией, периодически повторяются или периодичность не
проявляется. Причину массовых размножений одни авторы
усматривают в случайных сочетаниях разнообразных факто-
ров среды, благоприятных или неблагоприятных для данного
вида; другие признают существование и большое значение ре-
гуляторных механизмов, компенсирующих случайные откло-
нения плотности популяции за пределы нормы. Регулирующие
факторы часто связывают с плотностью популяции и в ее из-
менениях видят механизм регуляции, плотности популяции.
Факторы регулирующие в отличие от факторов, вызывающих
случайные отклонения (модифицирующие факторы), дейст-
вуют по принципу отрицательной обратной связи. В роли мо-
106
дифицирующих факторов выступают преимущественно клима-
тические факторы, в роли регулирующих — внутривидовые и
межвидовые связи. Способность действовать по принципу от-
рицательной обратной связи некоторые экологи (например,
Викторов, 1967) признают только за биотическими факторами
среды.
Многочисленные теории динамики численности насекомых
в основном соответствуют разнообразию факторов, прямо или
косвенно влияющих на численность насекомых.
«Климатические» теории преимущественное значение при-
знают за влиянием погодных условий, их благоприятном или
гибельном действии. Значение прямого влияния радиации,
температуры, влаги, осадков, режима погоды в разные сезоны
года и в разные годы для развития, выживания, плодовитости
насекомых хорошо известно. Большинство сторонников «кли-
матических» теорий признают действие не отдельных факто-
ров, а комплекса погодных условий, типа атмосферной цирку-
ляции. Ряд авторов связывают периодичность вспышек раз-
множения с периодическими колебаниями погоды, последние
с активностью солнца.
Большое влияние погодные условия оказывают на числен-
ность вредителей через пищу. Оптимум биохимического со-
става корма фитофагов зависит от светового режима, режима
влажности, уровня грунтовых вод и других условий абиоти-
ческой среды. Физиологическое состояние дерева, его защит-
ные реакции (смоловыделение, осмотическое давление, фитон-
цидность) зависят от погодных условий. Ослабление деревьев
стимулирует, а иногда и задерживает размножение вредите-
лей. Качество пищи меняется и под воздействием самих фито-
фагов, разной степени объедания листвы (дефолиации), инъ-
екции слюны сосущими насекомыми. Например, у непарного
шелкопряда после двухлетнего объедания крон начинаются
активные миграции гусениц, вызванные биохимическими изме-
нениями листвы. Листва на деревьях становится непригодной
для нормального развития насекомых и вызывает вымирание
популяции. Кроме того, объедание листвы ведет к изменению
гидротермического и светого режимов и в целом микрокли-
мата, что в свою очередь отражается на популяции фитофага.
Некоторые данные показывают, что изменение микроклима-
тических условий, вызванное деятельностью фитофагов, при-
водит к резкому сокращению их численности и даже вымира-
нию. Так, численность листоедов Chrysomela gemellata Pk.,
Ch. hyperica Forst, увеличивается до тех пор, пока они не
съедят зверобой, на котором питаются. Это, в свою очередь,
ведет к изменению и микроклимата: лишает жуков необходи-
мых укрытий (тени), и популяция жуков вымирает. Заросли
зверобоя восстанавливаются раньше, чем восстановится плот-
ность популяции листоедов (Э. Макфедьен, 1963).
107
Внутривидовые особенности видов, дающих массовые раз-
множения, значение плотности популяций в возникновении
стадного поведения и массовых миграций породили теорию
саморегуляции численности и миграционную теорию массовых
размножений. Замечено, что уменьшение плотности популя-
ции наступает до истощения пищевых ресурсов, предотвращая
перенаселенность. Глубокие изменения в качественном состоя-
нии популяции (изменение пигментации, энергии метаболиз-
ма, измельчание особей, снижение плодовитости, появление
миграционного инстинкта и др.) нами рассмотрены выше. Ка-
чественные сдвиги в состоянии популяции в процессе нара-
стания и затухания вспышки размножения имеют большое
значение для понимания массовых размножений, и работы
в этом направлении должны быть усилены.
Биоценотические теории отражают влияние на численность
насекомых межвидовых связей в биоценозах.
Многими авторами отмечается существенная, а иногда
единственная роль в подавлении массовых размножений мно-
гочисленных энтомофагов и разнообразных возбудителей бо-
лезней. Энтомофаги способны быстро реагировать на измене-
ние численности популяций насекомого-хозяина. Фиксируется
также увеличение численности вирусов, грибов, бактерий с
ростом плотности популяций вредителя. Возникла «паразитар-
ная» теория массовых размножений. Сторонники этой теории
по-разному оценивают значение тех или иных врагов насеко-
мого-вредителя. Одни отводят ведущую роль энтомофагам,
другие — бактериальным возбудителям, третьи — вирусам.
Эпизоотии также связывают с плотностью популяций, считая,
что скученность часто сопровождается потерей устойчивости
К заболеваниям. Значение энтомофагов оценивается по-разно-
му. Одни авторы решающую роль в подавлении вспышки
размножения отводят специализированным энтомофагам, дру-
гие — полифагам. Многоядные энтомофаги отличаются узкой
зоной активности и важную роль играют в периоды низкой
численности насекомых. Специализированные энтомофаги
способны реагировать на численность популяции хозяина в ее
более широких пределах (Викторов, 1965). Паразиты, хищ-
ники, возбудители разнообразных болезней связаны с хозяи-
ном как с источником пищи, поэтому обилие этой пищи и ее
качество имеют для них такое же значение, как и состояние
растения для фитофага. Изменение реакции гемолимфы, жи-
рового тела, активности, восприимчивости к заболеваниям
и т. п. отражаются на состоянии энтомофагов, усиливая или
ослабляя их деятельность. Разнообразные количественные от-
ношения между «хищником и жертвой» были предметом экс-
периментального изучения, математического моделирования.
В схематической форме суть этих взаимоотношений можно
иллюстрировать, рис. 16.
108
Увеличение плотности популяции хозяина (жертвы) ведет
к увеличению численности хищника (паразита), а последнее—
к уменьшению численности жертвы и т. д., получается замкну-
тый круг; колебание численности около средней величины.
Рис. 16. Зависимость между плотностью популяций хищника и жертвы
1 — жертва; 2 — хищник
В природе связи неизмеримо сложнее. Вмешательство
лишь еще одного сочлена биоценоза (а их в действительности
значительно больше) может полностью изменить исход отно-
шений «хищник—жертва». При анализе взаимоотношений
между энтомофагами и их хозяевами (пищей) следует учиты-
вать ряд обстоятельств: энтомофаг размножается при высоких
плотностях популяции хозяина быстрее, чем при низких, по-
этому его плодовитость зависит от популяции хозяина. При
всех условиях должно быть достаточно пищи для дополни-
тельного питания имаго энтомофага (цветущих растений,
медвяной росы и др.), чтобы обеспечить созревание яичников.
При низких плотностях популяции хозяина паразит находит
других хозяев, смертность энтомофага должна быть низкой.
Все более осознается значение комплексного действия фак-
торов среды. Сложность такого рода действия Швердфегер
(F. Schwerdtfeger) пытался изложить в своей теории градо-
цена. Согласно этой теории учитываются влияния как абио-
тических, так и биотических факторов, но допускается поло-
жение о стремлении популяции к равновесию, что некоторые
авторы считают ошибочным (Викторов, 1955; Яхонтов, 1964).
Сущность теории градоцена иллюстрируется схемой (рис. 17).
Приведенная схема, конечно, не исчерпывает всю слож-
ность взаимосвязей в биоценозе, но ряд существенных черт,
влияющих на численность отдельных популяций, отражает.
Безусловно, что и значение факторов не может быть равно-
ценно: оно меняется в зависимости от особенностей вида и
от соотношений действующих факторов, места, состояния вре-
Ц)9
Дителя и времени. Ведущее значение отдельных факторов так-
же меняется. Схема градоцена не отражает значения качест-
венной изменчивости популяции.
Как следует из проведенного нами раньше анализа значе-
ния физико-химических условий жизни (микроклимата, пищи
и др.), анализа внутривидовой жизни и связей межвидовых,
Рис. 17. Схема градоцена
(по F. Schwerdtfeger):
Р — популяция насекомого вреди-
теля; W — погода; N — пища;
R — паразиты; S — хищники;
К — болезни; А — другой вид
причины массовых размножений насекомых и действие регу-
лирующих систем лежат во взаимодействии условий физико-
химических с особенностями популяций отдельных видов и
межвидовыми (биоценотическими) связями. Если рассматри-
вать значение каждой из этих трех групп взаимодействующих
факторов отдельно, то это приведет к обоснованию климати-
ческих теорий, теории саморегуляции и миграций, теориям
биоценотическим в разных их вариациях. Каждая из теорий
базируется на определенной группе факторов, но при игнори-
ровании других данных будет страдать односторонностью.
Практически влияние разнообразных факторов среды на вре-
дителя необходимо рассматривать не изолированно, а в их
совместном действии, что не исключает в каждом конкретном
случае выделение ведущего фактора.
Рассмотрим конкретные примеры.
ОСОБЕННОСТИ МАССОВЫХ РАЗМНОЖЕНИЙ САРАНЧИ
Наибольшее количество сведений имеется об изменении
численности перелетной саранчи (Locusta migratoria L.) и
пустынной саранчи (Schistocerca gregaria F.) (Захаров, Щер-
биновский).
110
По данным Бей-Биенко, область массовых размножений
в СССР этого вида представлена двумя резко отличными зо-
нами: северной, приуроченной к южной окраине нечернозем-
ной полосы европейской части СССР, и южной, приуроченной
к пустынно-степным территориям.
В северной зоне перелетная саранча живет на песчаных
землях; больших перелетов не делает; вспышки происходят
редко, после двух-трех жарких и сухих лет. В этой зоне са-
ранча представлена особым подвидом (Locusta migratoria ros-
sica Uv. et Zol.). Наибольшее количество саранчи в нечерно-
земной полосе наблюдали в 1892—1893 гг., 1899 г. и в 1922—
1923 гг. Исследованиями С. А. Предтеченского установлено,
что годы перед массовым появлением характеризуются повы-
шенной температурой (на 13,6° сравнительно с многолетней
средней) и пониженным количеством осадков (320 мм), а го-
ды после максимума численности отличались пониженной тем-
пературой и повышением количества осадков. В теплые и су-
хие годы саранча отрождается раньше, быстрее развивается,
имаго появляются также раньше, живут дольше и откладыва-
ют больше яиц. В сырые и холодные годы отрождение и окры-
ление происходит поздно. При холодной осени половые про-
дукты у большинства особей не развиваются, и саранча по-
гибает, почти не отложив яиц. В зоне пустынно-степной
(южной) перелетная саранча заселяет сырые болотистые уча-
стки с зарослями тростника, расположенные вдоль рек и по
берегам озер. В этих районах саранча представляет основной
подвид, или типичную азиатскую перелетную саранчу Locusta
migratoria migratoria L. Массовое размножение ее наблюдает-
ся чаще, и вылетает она далеко за пределы основных очагов.
Многолетние наблюдения Л. 3. Захарова и анализ литера-
турных данных о вспышках размножения саранчи на протя-
жении 130 лет на юго-востоке РСФСР и Северном Кавказе
показывают, что вспышки размножения саранчи закономерно
повторяются и в некоторые годы охватывают огромные терри-
тории. Л. 3. Захаров выделяет большие и малые периоды
размножения. Малые, или короткие, периоды — от момента
возникновения до наступления упадка длятся 7—9 лет. Два
или три малых периода вместе образуют большие, или длин-
ные, периоды, продолжающие 19—21 год. Во время больших
периодов наблюдается усиленное размножение саранчи на
огромной территории (Северный Кавказ, юго-восток, Средне-
Русская равнина и даже Средняя Азия).
Одновременность вспышек размножения на огромной тер-
ритории Захаров объясняет действием засух, также периоди-
чески захватывающих одновременно большие территории.
Влияние засухи на азиатскую саранчу выражается в следую-
щем: 1) частичное высыхание территории по бассейнам рек,
где гнездится саранча, дает возможность шире расселиться
Ш
и образовать новые очаги; 2) соответственно увеличивается
кормовая площадь; 3) условия питания личинок и имаго
улучшаются, и плодовитость увеличивается; 4) реки слабо
разливаются, и яйца саранчи менее продолжительное время
находятся под водой; 5) воздействие хищников и паразитов
уменьшается, так как насекомые переселяются на новые уча-
стки. Прямое влияние засухи сказывается в том, что, повы-
шенная температура сокращает эмбриональное и личиночное
развитие, имаго живут дольше, ускоряется половое развитие,
больше откладывается яиц, быстрее идет превращение в стад-
ную форму. По расчетам Захарова, 100 экз. одиночной саран-
чи, живущей на площади в 1 га, через три года превращаются
в кулигу на площади в 30 га с плотностью 100 экз. на 1 м2.
Максимума численность достигает на 5—б-й год, а на
7—9-й год (иногда и раньше) обычно резко снижается. При-
чинами резкого падения численности являются главным об-
разом, сильные разливы рек, затопляющие места выплода ли-
чинок, эмиграции личинок и имаго, истребительные меро-
приятия.
Накоплению саранчи в плавнях по берегам нижнего тече-
ния Волги способствуют засухи (средняя температура за веге-
тационный период 21,8° и относительная влажность 25—50%),
сильные сухие ветры и богатая растительность по берегам
водоемов. В таких условиях одиночная фаза саранчи через
2—3 генерации превращается в стадную фазу со всеми ее
особенностями. Наиболее излюбленные места для стай саран-
чи — заросли тростника, лугово-степная растительность, посе-
вы кукурузы и хлопчатника и сорняки. Растительность опре-
деляет распределение откладываемых кубышек и способствует
усиленному размножению. При наличии обильного сочного
корма саранча откладывает при теплой сухой осени до трех
кубышек с полным количеством яичек (80—110). При недо-
статке корма или его сухости откладка яиц уменьшается до
40—20, кубышки бывают непрочны и легко распадаются.
В ряде случаев паразиты саранчи заметного влияния на ее
численность не оказывают. От заражения паразитами саран-
чу предохраняют ежегодные перемещения залежей кубышек.
В других южных гнездилищах саранчи, также связанных
с плавнями рек, основной фактор, регулирующий численность
саранчи,— водный режим рек.
У других видов саранчевых закономерности размножения
могут быть другими. Для мароккской саранчи (Dociostaurus
maroccanus Thnb.) решающим фактором становятся осадки,
особенно весенние. Ареал этого вида связан с областями, где
весной мало дождей, количество выпадающих осадков около
100 мм; их резкое повышение или понижение гибельно отра-
жается на саранче. При повышении осадков в массе разви-
ваются паразитные грибы (Fusarium sp.), уничтожающие 50__
112
100% яиц. Резкое уменьшение осадков приводит к высыханию
яиц и гибели личинок от недостатка корма.
Н. С. Щербиновский (1960) и др. связывают периодичность
больших и малых циклов размножения азиатской и пустын-
ной саранчи с периодичностью колебаний солнечной радиации.
РАЗМНОЖЕНИЕ ВРЕДИТЕЛЕЙ ЛЕСА
В лесах СССР массовые размножения свойственны 100 ви-
дам вредителей хвои и листьев. Более 20 видов из них отно-
сятся к группе межзонального значения, они дают вспышки
массового размножения наиболее часто и на огромных терри-
ториях (пандемические вспышки), остальным видам (преиму-
щественно листогрызущим) свойственны локальные вспышки,
ограниченные отдельными областями и районами. Виды меж-
зонального значения (сибирский шелкопряд, непарный шелко-
пряд, монашенка, сосновая совка и другие) изучены довольно
подробно рядом авторов (А. И. Ильинский, А. И. Воронцов,
М. Г. Ханисламов, Д. Ф. Руднев, В. О. Болдаруев, А. С. Рож-
ков и др.). Значительно меньше данных имеется о размноже-
нии стволовых вредителей, вредителей семян, корней.
Общая картина хода вспышки размножения (градацион-
ная кривая) непарного и сибирского шелкопрядов рисуется
в следующем виде. По мнению ряда авторов (А. И. Ильин-
ский, В. О. Болдаруев и др.), возникновению вспышки способ-
ствует солнечная, теплая и засушливая погода, особенно в пе-
риод питания личинок среднего и старшего возрастов, вызы-
вающая изменения в биохимическом составе корма и
создающая оптимальные условия для жизни и развития личи-
нок. Они быстро заканчивают развитие и дают имаго с высо-
кой плодовитостью.
Поколение, развившееся при солнечной теплой и сухой по-
годе, относят к первой, или начальной, фазе вспышки. Чис-
ленность популяции вредителя в начальной фазе увеличивает-
ся немного — в 2—4 раза. Численность двух следующих поко-
лений (у непарного шелкопряда, занимающих два календар-
ных года, а у сибирского — с двухлетней генерацией — 4 го-
да) увеличивается, но она остается еще небольшой, и повреж-
дения в глаза не бросаются. Это вторая фаза, или фаза роста
(продромальная). Гусеницы в этой фазе имеют большие за-
пасы жировых и белковых веществ, что ведет к увеличению
плодовитости питающихся гусеницами энтомофагов и их быст-
рому размножению. При переходе в третью фазу, или фазу
собственно вспышки (эруптивную), численность вредителя
резко увеличивается, и вспышка принимает качественно иной
характер: кроны деревьев бывают объедены сильно или даже
полностью, в перенаселенных местах гусеницы активно пол-
зают, частью гибнут, среди них распространяются^ болезни,
8 Заказ 193
113
плодовитость имаго снижается, возрастает численность хищ-
ников, паразитов и сверхпаразитов. Третья фаза чаще всего
распространяется на два поколения. Нарастающая числен-
ность энтомофагов и болезни в третью фазу также дают скач-
кообразный подъем, и вспышка переходит в свою четвертую,
последнюю фазу (фазу кризиса), качественно отличную от
Рис. 18. Схема градационной кривой
(по А. И. Ильинскому)
всех других. Плотность популяции резко идет на убыль. Вы-
сокая плодовитость сменяется низкой, резко возрастает зара-
женность паразитами, болезнями, хищниками. Численность
вредителя падает до минимума, вспышка заканчивается. В ти-
пичных случаях последняя фаза включает два поколения.
Последующие годы численность вредителя держится на низ-
ком уровне (рис. 18).
Изменение плотности отдельных популяций многих видов
на протяжении вспышки имеет сходную картину. Вначале
плотность популяции увеличивается медленно, затем нара-
стание ее ускоряется, переходит в фазу логарифмического
(экспоненциального) роста и, наконец, замедляется (фаза
отрицательного ускорения) и заканчивается фазой относитель-
ного равновесия (рис. 19).
Между вспышками численность вредителя держится на
низком, но непрерывно колеблющемся уровне. Плодовитость
близка к средней, свойственной виду. Все периоды вспышки
114
в целом занимают семь поколений, что для насекомых моно-
вольтинных соответствует семи годам. У вредителей с двойной
генерацией вспышка протекает быстрее, а с двухлетней гене-
рацией (сибирский шелкопряд) — медленнее.
У одного и того же вида вспышка может быть более дли-
тельной (9 лет) или менее длительной (5—6 лет). Если засухи
Рис. 19. Кривая роста популяции
(из А. И. Воронцова)
следуют одна за другой с небольшими перерывами (I—2 го-
да), вспышка размножения усиливается, принимает затяжной
характер, охватывает огромные площади лесов и может про-
должаться свыше 10 лет. Пример такой вспышки — массовое
размножение непарного шелкопряда в Поволжье, вызванное
засухами 1931, 1933, 1936, 1938, 1939 гг. Засухи вызвали об-
меление рек и падение уровня грунтовых вод. Непарный шел-
копряд продолжал держаться в лесах в большом количестве
на протяжении 10 лет. Одна вспышка накладывалась на дру-
гую; это привело к повторным объеданиям листвы и усыханию
дубрав на значительных площадях.
В распространении вспышки по территории отмечаются
определенные закономерности. Между вспышками вредители
обитают в тех участках леса, где условия наиболее благо-
приятны для данного вредителя (места переживания, или ре-
зервации). В этих насаждениях при начинающейся вспышке
вредитель размножается очень быстро и раньше, чем в других
массивах леса. Здесь он переходит в фазу резкого увеличения
численности (третья фаза собственно вспышки), раньше пол-
ностью объедает кроны деревьев. Такие участки леса назы-
вают первичными очагами (А. И. Ильинский). Гусениц бывает
8* 115
так много, что они, не закончив своего развития, предприни-
мают массовые странствования в поисках корма, образуя
новые миграционные очаги. Миграционные очаги могут фор-
мироваться также путем активных и пассивных массовых пе-
релетов имаго. Перелеты предпринимают самки с пониженной
плодовитостью или уже успевшие частично отложить яйца.
Медленнее, чем в первичных очагах, развивается вспышка
в насаждениях, где условия хуже оптимальных. В таких на-
саждениях — вторичных очагах — меньше исходное число
вредителя, так как больше хищников, паразитов и болезней,
ниже плодовитость бабочек. Оголение крон гусеницами про-
исходит с запозданием на одно поколение по сравнению с пер-
вичными очагами. Вторичные очаги бывают менее заселен-
ными, объедание листвы сильное, но не полное.
Все остальные насаждения, наименее пригодные для раз-
множения вредителя, относят к третичным очагам. Повреж-
даются они слабо, размножение вредителя идет медленно и
не достигает хозяйственного значения.
В процессе развития вспышки размножения поблизости от
первичных и вторичных очагов возникают миграционные оча-
ги, путем разлета бабочек, переползания гусениц или пере-
носа их ветром. Миграционные очаги возникают в любых
насаждениях, где есть кормовые растения вредителя. В неко-
торых случаях возникают самостоятельные миграционные оча-
ги в силу перелетов имаго на далекие расстояния от первич-
ных очагов.
Развитие вспышки и формирование очагов зависит от
(свойств насаждений, и, если они изучены, можно предвидеть,
в каких массивах леса и его отдельных биотопах разовьются
очаги вспышки. Во всех трех категориях очагов продолжи-
тельность вспышки охватывает чаще всего девять поколений.
По массиву в целом возможно ее увеличение до 10—11 лет
или уменьшение до 7—8 лет. На территории СССР наиболее
частые вспышки листогрызущих лесных вредителей происхо-
дят в степной и лесостепной зонах. Эти зоны А. И. Ильин-
ский относит к зоне первичных вспышек. Именно в них часто
бывают засухи.
Территориальное развитие вспышки и формирование ее
очагов после засухи 1936 г. показаны на рис. 20.
Судить о численности вредителя в насаждениях и о ее
росте можно как по количественным, так и по качественным
показателям.
К количественным показателям относятся абсолютная за-
селенность насаждений, коэффициент размножения и коэффи-
циент нарастания вспышки. Абсолютная заселенность — число
особей вредителя, приходящееся на 1 м2 поверхности почвы
или на одно дерево в насаждении. Коэффициент размноже-
ния— отношение абсолютной заселенности вредителем насаж-
116
дений в данном году к заселенности в предшествующем году.
Если коэффициент размножения выше единицы, численность
вредителя растет, ниже единицы — снижается. Коэффициент
нарастания вспышки вычисляется как отношение абсолютной
заселенности вредителем насаждения за данный год к абсо-
лютной заселенности в год перед вспышкой.
Рис. 20. Очаги вспышки
(по А. И. Ильинскому):
1 — первичный; 2 — вторичный; 3 —
третичный
Для примера А. И. Ильинским приводятся количественные
показатели вспышки сосновой совки (табл. 4).
Из таблицы видно, что численность совки на пробной пло-
щади за три года массового размножения выросла в 1300 раз
(2,47 : 0,0019= 1300). Для первичных очагов создалась угроза
полного объедания крон.
Качественными показателями вспышки могут быть плодо-
витость вредителя, окраска, отношение между полами, дея-
тельность паразитов, хищников и распространение болезней.
Плодовитость в процессе вспышки меняется: в начале
вспышки она очень велика, в конце — незначительна, а мно-
гие самки могут быть вообще бесплодными, особенно в пер-
вичных очагах. Например, плодовитость непарного шелкопря-
да в начале вспышки в среднем 500—750 яиц, в конце вспыш-
ки— 100—500 яиц; златогузки — соответственно ^00—500 и
117
Таблица 4
Показатели вспышки сосновой совки (по А. И. Ильинскому)
Год Фазы вспышки Абс. заселен- ность на 1 м2 (куколок) Коэффици- ент размно- жения Коэффици- ент нара- стания вспы- шки
1935 последний год перед вспышкой 0,0019 — —
1936 год начала вспышки 0,0050 2,6 2,6
1937 1-й год фазы нарастания 0,1057 21,1 55,6
1938 2-й год фазы нарастания 2,47 23,4 1300,0
1939 1-й год фазы собственно вспышки 37,11 15,0 19531,6
1940 2-й год фазы собственно вспышки (5-й год вспышки) 0,3 0,0008 15,2
40—60 яиц; кольчатого шелкопряда — 300—350 и 50—100 яиц.
При развивающейся вспышке размножения наблюдается
изменение окраски. В период первых трех фаз появляются
темноокрашенные личинки, что свидетельствует о повышении
энергии обмена веществ. Иногда становятся темными и ба-
бочки.
Вспышки размножения некоторых вредителей наблюдают-
ся одновременно на огромных территориях. Непарный шелко-
пряд, например, в период 1951 —1956 гг. в массе размножался
в центральных и южных областях европейской части СССР,
в Западной Сибири, в США и в Западной Европе. Повторяе-
мость вспышек размножения в разных районах различна:
так, например, в Крыму — через 9 лет, в Башкирии —
11 лет, в Москве очаги малоустойчивы и быстро затухают,
в небольших количествах непарный шелкопряд встречается
ежегодно. Что касается других лесных бабочек, то некоторые
из них (златогузка, кольчатый шелкопряд и др.) часто сопут-
ствуют непарному шелкопряду или появляются в большом ко-
личестве за один-два года до массовой вспышки шелкопряда,
как это отмечалось в центральных областях европейской ча-
сти СССР для зеленой дубовой листовертки. Сосновый шелко-
пряд в лесах Западной Европы дает вспышки размножения
очень редкие (за 80 лет — 2 раза), шелкопряд-монашенка за
150 лет 6 раз. У сосновой пяденицы за последние 40 лет часто-
та градаций выросла, но угрожающего лесам подъема чис-
ленности не наблюдалось. Массовое размножение сибирского
шелкопряда наступает после засушливых лет, особенно в июне
и июле. Одновременно в большом количестве размножаются
и другие бабочки (например, ивовая волнянка, непарный шел-
копряд, боярышница). Первичные очаги сибирского шелко-
118
пряда существуют много лет (примерно 8 лет) и лишь незна-
чительно изменяют свой радиус. И только при достижении
максимальной плотности появляются более мелкие бабочки,
разлетающиеся за пределы первичных очагов. Вредитель за-
полняет все окрестные насаждения в радиусе до 30 км и бо-
лее, образуя множество вторичных очагов. Массовое размно-
жение сибирского шелкопряда продолжается в среднем
16 лет. При достижении значительной плотности гусениц
(в третьей фазе вспышки) появляется инстинкт разлета (ми-
грационный инстинкт). Во время разлета у бабочек меняется
способ откладки яиц: из кучнокладущих они становятся раз-
дельнокладущими (бабочки рассеивают свои яйца по насаж-
дениям) .
Причинами угасания вспышки служат внешние факторы;
решающее значение часто играют паразиты и хищники. Их
массовое размножение начинается вслед за размножением
хозяина. На сибирского шелкопряда (по В. О. Болдаруеву)
в годы затухания вспышки один за другим производят мас-
совые нападения следующие специализированные паразиты:
тахина Masicera zimini, Koi. — выводит из строя 25—60% ку-
колок шелкопряда; яйцеед Т eleno tnus gracilis Mayz. — 70—
90% яиц и наездник Rhogas dendrolimi Mats. — 65—90% гу-
сениц. В последующие годы паразиты полностью уничтожают
популяцию шелкопряда, и территория практически очищается
от вредителя.
Анализ размножения сибирского шелкопряда за 100 лет
показывает, что в большинстве случаев очаги возникают и
развиваются одновременно в различных частях ареала, в том
числе и наиболее отдаленных. Раз в 10 лет они возникают
сразу во многих районах. Чередуются «большие» и «малые»
вспышки размножения. За последние 50 лет общая депрессия
для всего ареала не наблюдалась.
По А. С. Рожкову, повышенная плотность вредителя, гро-
зящая вспышкой, может длиться много лет и не всегда за-
вершается вспышкой. Переход в очаговый образ жизни сопро-
вождается активной концентрацией гусениц, нарастанием чис-
ленности с последующими миграциями.
Очаг — это качественно иное состояние популяции. Он
претерпевает две фазы развития: эруптивную и кризиса. Про-
дромальное же состояние (1-я и 2-я фазы вспышки, по
А. И. Ильинскому) — предвестник вспышки, но не начальная
фаза развития очага. Продолжительность развития очагов
7—9 лет. Эруптивная фаза распространяется на два-три, а
фаза кризиса — на одно поколение. Миграционные очаги
обычно кратковременны, так как вместе с бабочками в новые
места доставляются Telenomus и другие паразиты (форезия),
обеспечивающие быстрое заражение яиц в миграционных
очагах. i
119
В очагах активные перемещения совершают гусеницы, пе-
реползая даже безлесные участки (иногда на расстоянии до
1,5 км). Волна миграций идет от центра очага к периферии.
Меченые гусеницы, по А. С. Рожкову, вынесенные из плотных
очагов на 200—500 м, через 10 дней приползли обратно в зо-
ну очагов (80% гусениц), а гусеницы, взятые из мест с невы-
сокой плотностью и выпущенные в плотных очагах, уползали
из очагов.
Колебания плотности популяций вредителя на большой
территории обусловлены климатическими факторами; они же
вызывают концентрацию особей в определенных стациях. Ес-
ли концентрация происходит в период общего подъема чис-
ленности вредителя, она принимает форму массовой, с обра-
зованием первичных очагов. Этому могут способствовать по-
жары, уничтожающие подстилку вместе с яйцеедами;
вымерзания (по данным Н. Г. Коломийца, у Telenomus’a и у
шелкопряда отношение к температуре различное) и другие
причины, снижающие активность энтомофагов.
Условия погоды, вызывающие увеличение плотности попу-
ляции, для разных видов различны. Так, выходу из депрессии
монашенки в сосновых лесах Башкирии (Ханисламов и Ла-
тышев, 1962) способствует погода с высокими гидротермиче-
скими коэффициентами мая и июня в сочетании с большой
суммой отрицательных температур ноября. Прохладная дожд-
ливая погода вызывает длительное цветение сосны, что обес-
печивает гусениц необходимым питанием ее пыльцой. Муж-
ские «соцветия» сосны дают оптимальную пищу гусеницам
в младших возрастах. При обилии сухих солнечных дней в
начале вегетации мужские «соцветия» освобождаются от по-
кровных чешуек раньше отрождения гусениц, а при преобла-
дании пасмурной погоды — опережает вредитель. Образова-
ние очагов связано с продолжительностью питания молодых
гусениц пыльцой сосны; сосна цветет дольше (на 2—3 недели)
при обильных дождях.
На монашенке зарегистрировано 26 видов энтомофагов,
но их эффективность незначительна. Основная причина огра-
ничения численности монашенки полиэдренная болезнь, разви-
тие которой во многом зависит от трофических условий.
Деятельность энтомофагов в ряде случаев снижается на
полях монокультур. Так, численность вредной черепашки огра-
ничивают яйцееды и другие энтомофаги. В условиях крупного
зернового хозяйства (по Викторову, 1967) энтомофаги утра-
чивают регулирующее значение и становятся лишь одним из
модифицирующих факторов. По К. В. Арнольди, колебания
численности вредной черепашки в зоне хронических массовых
размножений определяются в основном действием погодных
условий, которые, в свою очередь, влияют на численность ее
естественных врагов.
120
Ко всему изложенному следует добавить, что вспышки
размножения разнообразных видов насекомых протекают по-
разному в зависимости от особенностей биоценозов, их поло-
жения (биотопа), структуры (возраста и состояния растений
и др.), а также деятельности человека, вносящей существен-
ные изменения в условия абиотической и биотической среды
(обработка почв, акклиматизация новых энтомофагов, приме-
нение химических средств защиты и др.).
Глава VI
СТРОЕНИЕ БИОЦЕНОЗОВ
В определении понятия «биоценоз» до сего времени нет
полного единства мнений. Из многочисленных определений, по
Э. Макфедьену (1965), следует три главных представления
о биоценозе: 1) совместное присутствие определенных видов
растений и животных; 2) повторяемость видов в пространстве
и времени; 3) динамическая стабильность. По мнению ряда
экологов, эти качества биоценоза определяются саморегуля-
цией биоценотических процессов, осуществляющихся только
в единстве с абиотической средой. Такое единство присуще
экосистеме (Т. А. Балог, Э. Макфедьен, Е. Одум и др.), или
биогеоценозу (В. Н. Сукачев), в которых происходит круго-
ворот форм, вещества и энергии. Биоценоз мыслится только
в конкретной физико-географической обстановке, определяю-
щей облик биоценоза и его пространственную ограниченность.
Биоценоз не мыслим без биотопа, как и биотоп без биоценоза,
и в этом смысле «биоценоз» — синоним биогеоценоза (К. и
Л. Арнольди, 1963). Биоценоз — понятие и биологическое,
и географическое. Жизнь биоценоза определяется как особен-
ностями связей организмов, населяющих биоценоз, так и осо-
бенностями физико-географической среды, меняющейся под
воздействием организмов (Воронов, 1963).
Биогеоценоз, по В. Н. Сукачеву (1964), включает взаимо-
действие экотопа (-биотопа) и биоценоза, а последний право-
мочно рассматривать состоящим из фитоценоза, зооценоза и
микробоценоза (рис. 21).
Внимание большинства экологов сосредоточено преимуще-
ственно на изучении отдельных компонентов биоценоза, от-
дельных его сторонах. Самостоятельно исследуются почвы,
фитоценозы, зооценозы, энтомоценозы, паразитоценозы, а еще
чаще — виды и их группировки. В целом биоценозы почти не
исследованы.
722
Сложные биоценотические связи, обеспечивающие биоген-
ный круговорот веществ и длительное существование биоцено-
зов, изучают в трех разных направлениях. В большей сте-
пени выявлены статические, или физиономические, особенности
биоценозов. При изучении биоценозов в их статике выясняют-
Рис. 21. Схема биогеоценоза (по В. Н. Сукачеву)
ся структурные особенности, пространственное размещение и
типология биоценозов. Второе направление — трофическое —
развивалось преимущественно на основе изучения пищевых
связей в биоценозах и колебаний численности популяций, их
биомассы. В последнее время дается энергетическая трактов-
ка трофических взаимоотношений компонентов экосистем. Ос-
новные закономерности превращения энергии в процессах
биотического круговорота универсальны, но для изучения
возможны только на основе анализа специфических биоло-
гических особенностей отдельных видов. Энергетическое п
трофическое направления изучают биоценозы в 'их динамике.
123
Самая существенная черта биоценоза — наличие типично-
го состава организмов, сохраняющее относительное постоян-
ство во времени и пространстве. Большинству природных био-
ценозов присуща стабильность связей между популяциями,
составляющими основу сообщества, и вместе с тем подвиж-
ность, изменчивость этих связей. Для каждого биоценоза ха-
рактерен определенный набор разнообразных организмов,
в том числе и насекомых, и определенные соотношения между
ними. Популяции видов, сосуществующих в биоценозе, при-
способлены как к условиям физической среды данного биоце-
ноза, так и друг к другу. Изменение численности одного из
членов биоценоза может привести к изменениям во всем
составе биоценоза. Для биоценоза характерно наличие по-
стоянных видов, наиболее прочно связанных прежде всего пи-
танием. Одни из них остаются малочисленными, но показа-
тельными для биоценоза, другие оказываются многочисленны-
ми, составляющими ядро биоценоза.
Связи в биоценозе не остаются постоянными. Один доми-
нирующий вид может быть заменен другим. Числ'енные соот-
ношения популяций непостоянны: они меняются в течение су-
ток (днем преобладают одни виды, ночью—другие) и в те-
чение сезонов. Одни виды, заканчивая развитие, переходят а
недеятельное состояние или перекочевывают в новые биоцено-
зы; другие, наоборот, приходят из соседних биоценозов или
переходят в активное состояние.
Состав биоценозов непрерывно изменяется, одновременно
сохраняя относительное постоянство, состояние подвижного
равновесия. Отдельные популяции биоценозов то численно
разрастаются и занимают большую территорию, как это бы-
вает при вспышках размножения, то уменьшаются, оставаясь
на небольших участках территории. Биоценозы, как говорят,
пульсируют, отражая изменчивость условий среды. Выявить
типичные черты биоценоза поможет оценка смены сезонных
форм насекомых.
Облик биоценозов в течение сезона заметно меняется.
По С. И. Медведеву (1964), в плакорной степи Аскании Но-
ва смена основной массы доминирующих видов изменяет эн-
томологическую физиономию биоценоза и определяет наступ-
ление новой фазы. Энтомологические сезонные аспекты по
срокам наступления очень близки к ботаническим и имеют
характерный состав активных форм насекомых. Им выделены
виды, встречающиеся в течение всего сезона, поэтому удобные
для сравнительной оценки разных биоценозов. К таким фор-
мам отнесены семейства жужелиц и чернотелок. При изучении
видового состава биоценозов сборы важно проводить в одно'
и то же время, при определенной погоде и в возможно корот-
кий срок.
Для биоценоза характерен набор доминирующих растений-
124
эдификаторов, определяющих особенности биоценоза и су-
ществующих за их счет потребляющих видов — предоминан-
тов. По своей роли в биоценозе выделяются виды доминан-
ты, господствующие по численности, и содоминанты. Ими
могут быть и растения, и насекомые. Ведущее положение,
естественно, занимают доминанты и прежде всего растения-
эдификаторы, создающие структурную основу биоценоза, его
облик, его тип. Видов-эдификаторов, как правило, немного.
С ними тесно связаны существующие за их счет фитофаги.
Обычно их тоже немного. Изучение видового состава насеко-
мых в биоценозе необходимо начинать с выделения наиболее
многочисленных (доминирующих) видов, определить их оби-
лие как одну из важнейших величин.
В пределах биоценоза наблюдаются не только различные
соотношения численности популяций разных видов, но и раз-
личного рода более тесные группировки, приуроченные или к
растениям-эдификаторам или к каким-либо элементам био-
ценоза: микроценозам, более мелким его подразделениям или
частям (мероценозам, по зарубежным экологам, от «меро» —
часть). Биоценоз синузиален, пятнист. Синузия — топографи-
чески и экологически более или менее обособленная структур-
ная часть биоценоза, пространственно оформленная группи-
ровка организмов, связанных между собой более тесно, чем
с другими организмами, или объединенных каким-нибудь ха-
рактерным субстратом, например лесной подстилкой. Микро-
ценозы показательны для пространственной структуры биоце-
ноза. Ими могут быть живое дерево со всем комплексом насе-
ляющих его организмов и группировкой насекомых, гниющий
пень, шишка сосны, нора грызуна, гнездо муравья и т. п.
Среди экологических группировок биоценоза, его синузий,
выделяют очень важную структурную единицу — консорцию.
Это группировки, адаптированные к основному виду биоцено-
за, без которого вся группировка существовать не может. Кон-
сорцию определяют как сочетание разнородных организмов,
тесно связанных друг с другом в их жизнедеятельности (Ра-
менский, 1952). Приспособление в консорциях либо взаимное,
выраженное в различных связях, либо одностороннее. Без вы-
деления и изучения консортивных группировок наше знание
биоценозов не будет полным.
Консорцию можно определить также как «сочетание попу-
ляции вида высшего растения в данной растительной ассоциа-
ции со связанными с этим высшим растением популяции низ-
ших растений и популяций животных. Связи эти могут быть
как длительные, так и сезонные. То обстоятельство, что во
главу угла ставится высшее растение, должно быть понятно,
так как на поверхности суши фотосинтезирующие высшие ра-
стения являются как бы энергетической установкой биоцено-
за» (Лавренко, 1959).
125
Примером консортивных связей в биоценозах можно на-
звать сосну и связанных с ней монофагов и олигофагов в
сосновых лесах; дерновинные злаки с населяющими их насе-
комыми и организмами в сухих степях; группировку специа-
лизированных опылителей и опыляемые растения; «хозяина»
и его паразитов; центральные организмы в консорциях одно-
временно представляют доминанты биоценоза.
Разнокачественность популяций массовых видов, гетеро-
цикличность (чередование «мощных» и «слабых» поколений)
меняют значение популяций в структуре и динамике биоцено-
за. Различия в пищевой специализации, способах, сроках и
местах размножения, смена стаций обитания, скопления и
миграции особей приводят к разнообразию структуры и видо-
вого населения биоценозов.
Популяции отдельных видов в биоценозе занимают опреде-
ленное место в связях с другими видами, свою нишу. Если
местообитание вида рассматривать как его местожительство
(адрес), то ниша — значение вида, его место в биоценозе, его
«профессия» (Одум). Одни виды определяют облик биоцено-
за. В биоценозе они занимают ведущее положение как основ-
ные продуцирующие виды, поэтому группировки насекомых,
связанные с доминирующими растениями-эдификаторами, не-
обходимо выделять в первую очередь. Чтобы описать эколо-
гическую нишу какого-либо вида, нужно знать, чем он питает-
ся, кому служит пищей, его поведение, движение, воздействие
на другие организмы и неживые элементы среды. Решение
этой сложной задачи облегчает наличие в биоценозах кон-
сорций.
Биоценозам свойственна определенная ярусность строе-
ния— стратиграфия. С ярусами растений связаны такие спе-
цифические группировки насекомых и других организмов, как
стратоценозы. Ими могут быть филлобиоиты, дендробионты,
заселяющие верхний ярус, тамнобионты — жители кустарни-
кового яруса, хортобионты — жители травяного яруса. Свои
особенности имеют герпетобионты — жители мохового покро-
ва, подстилки, поверхности почвы и геобионты, населяющие
почвенный ярус, повреждающие корневую систему — ризосфе-
ру. Ярусность выражена различно в лесных и степных биоце-
нозах. Она во многом определяет внешний облик биоценоза
и другие его особенности.
Методы полевого изучения биоценозов разнообразны. На-
чинают с выявления видового состава. Число видов в
биоценозе различно. Установить их важно, но еще важнее
выявить соотношение численности популяций видов, значение
отдельных видов в биоценозе.
В. Н. Беклемишев (1961) рекомендует следующий порядок
изучения биоценоза. Следует выделить границы однородного
пятна растительности (ассоциации), описать его положение
126
в рельефе, экспозицию, почву, микрорельеф, растительность^
окружающую местность. Изучить микроклимат с его суточ-
ными и сезонными изменениями. С этой целью измерения
необходимо проводить в разные часы суток, лучше в 13—15 ч,
и повторять их в течение года. Наблюдения необходимо про-
водить при определенном типе погоды и в возможно короткий
срок. Не стремиться к полному выявлению видов. Фиксиро-
вать внимание на выявлении многочисленных видов и видов
редких, но постоянно встречающихся, показательных для изу-
чаемого биоценоза, видов-индикаторов.
Определив основной состав видов в биоценозе, следует
установить численность — общее число особей, со-
ставляющих данную популяцию или какую-либо единицу на-
селения.
Очень важный показатель учета — обилие, или плот-
ность. Обилие — среднее число особей данного вида или
данной группы видов, приходящееся на единицу учета. Еди-
ницей учета может быть площадь в данном биоценозе (обычно
1 л/2), объем субстрата, определенный период времени при
вылове сачком, ловушка-сутки, особь хозяина, нора, дерево,
растение или погонный метр при учете населения крон де-
ревьев или рядков растений и т. п. При вычислении показа-
телей обилия принимаются во внимание все пробы, включая
пустые. Отношение численностей двух видовых популяций,
входящих в состав одного и того же биоценоза, равно отно-
шению их обилий. Выбор единицы учета определяется особен-
ностями изучаемого вида.
Встречаемость вида — число проб, в которых обна-
ружены особи этого вида, выраженное в процентах от общего
числа исследованных проб. Число особей, обнаруженных в
той или иной пробе, при вычислении встречаемости не прини-
мается во внимание.
Доминирование вида выражают в процентах к
суммарному обилию всех сравниваемых видов данного био-
ценоза или его подразделения. Сумма показателей домини-
рования всех видов равна 100%. Индекс доминирования —
величина вспомогательная и не может заменить основные по-
казатели (индексы встречаемости и обилия).
Индекс верности, или степени приурочен-
ности вида, популяции к данным условиям.
Он позволяет судить о распределении популяции по различ-
ным единицам среды. Вычисляется для каждого из сравнивае-
мых биотопов по обилию, по численности. Например, обилие
эктопаразита на одной особи полевки было 0,05, а в одном
гнезде той же полевки — 2,15. Верность хозяину равна 4,5%,
верность гнезду — 95,5%. Обилие другого паразита на хозяи-
не— 0,44, а в его гнезде 0,78; верность хозяину — 36,1%, вер-
ность гнезду — 63,9%- Степень приуроченности к хозяину од-
127
иого вида паразита по сравнению с другим видом оказывается
в 8 раз выше.
Индекс общности видового состава между
двумя биоценозами, или микроценозами, мера сходства фау-
ны. Например, в одной норе грызуна найдено 12 видов насе-
комых, в норе другого грызуна — 8 видов, из них общих 4 ви-
да. Значит в двух норах было учтено 20 + 8—4= 16 видов. Че-
4-100 0 ~
тыре вида общих для обеих нор составляет —— =2э7о-^то
16
и есть показатель общности по встречаемости.
Выявляя видовой состав и другие особенности биоценозов,
следует обратить внимание на большое индикаторное значе-
ние почвенной фауны насекомых. Изучать почвенную фауну
необходимо на одних ключевых участках с геоботаниками и
иочвоведами. При этом можно ограничиваться только важ-
нейшими для характеристики биоценозов группами насеко-
мых. Слабая миграционная способность почвенных насекомых
делает их хорошими индикаторами биотических условий, как
и растительный покров. Насекомые и другие беспозвоночные
животные могут быть даже более надежными показателями
почвенных условий, чем растения, особенно когда раститель-
ный покров нарушен человеком.
Количественный учет, необходимый для выявления чис-
ленности доминантов и соотношений различных групп насеко-
мых, не может быть стандартным. Он определяется размера-
ми объектов, их особенностями, численностью, почвенными
условиями и задачами учета. Размеры и число проб зависят
от объектов. Обычно, чем мельче насекомое, тем больше его
численность и тем меньше нужно проб и меньше их размеры
(Гиляров, 1964).
Для учета крупных насекомых, легко видимых невооружен-
ным глазом (личинки хрущей, щелкунов, жужелиц, ктырей
и т. п.), универсален и доступен метод ручной разборки выка-
пываемых почвенных проб по 0,25 м2 во влажный период и по
1 м2 и даже 2 м2 — в сухое время и в сухих местообитаниях.
Пробы берут на глубину встречаемости насекомых. В одно-
родном сообществе — не менее 4 проб по 0,25 м2, а на стацио-
нарных участках — не менее 12 проб для каждого варианта
с распределением их по‘сетке. При выраженной комплексно-
сти среды пробы лучше брать в разных условиях. Например,
в типчаково-ковыльной степи часть проб — под типчаком,
часть — под ковылем.
В центре внимания сначала должны быть фитофаги, свя-
занные с эдификаторами,— наиболее массовые вредители,
влияющие на состояние растительности. Оценка того, как на-
секомые используют растение, имеет немаловажное значение.
Связь с растением может быть специфична для определенных
128
видов и носить исторически сложившийся консервативный ха-
рактер, а может быть неспецифична, без явного выбора, с ис-
пользованием опада, надземных и подземных частей. Связи
первого типа характеризуют специфику биоценоза, а связи
второго типа важны для понимания динамики и круговорота
органических веществ в биоценозе, его производительности.
Отсюда вытекают два основных направления работы эко-
лога в полевой обстановке (по Л. В. Лриольди): 1) оценка
консортивных и неконсортпвных связей; 2) оценка раститель-
ного покрова и его отдельных компонентов в жизни энтомо-
логической части ценоза.
Общие сборы (кошение) необходимо проводить по одно-
родным куртинам с одним, либо двумя-тремя видами расте-
ний, но обязательно далеких систематически (например, зла-
ки, сложноцветные, зонтичные), чтобы выявить связь насеко-
мого с одним из видов растений. Одновременно ведут сборы
напочвенно-растительных видов, дополняющих список видов
биоценоза. Общие сборы дают ориентировку в распределении
видов по растениям и выполняются за несколько дней. После
этого переходят к тщательному анализу отдельных видов
растений.
Для разных групп насекомых требуется применение раз-
личных приемов учета. Одни растения оказываются заселен-
ными очень слабо, насчитывается всего 2—3 вида насекомых,
на других регулярно встречается более 10 видов, большинство
обычных. Но и один вид растения может быть заселен по-
разному в разных вариантах сообщества. Для растений с бед-
ной фауной насекомых нужно просматривать большее число
экземпляров. Иногда, чтобы обнаружить редкие случаи, необ-
ходимо просмотреть не менее 100 растений. Растения с обиль-
ной энтомофауной дают хорошую картину распределения на-
секомых по его органам (в среднем на 30 экз.). А1еньше заселе-
ны бывают злаки (нужно просматривать больше 30 растений).
Насекомых дерновин учитывают по 30 кустам, а находящиеся
внутри стеблей златки (Cylindromorphus) повреждают только
около 1 % побегов ковыля, и, чтобы их обнаружить, нужно
просмотреть несколько сот побегов.
Наиболее сложен учет насекомых в лесах, особенно с мно-
гоярусной структурой. Своих приемов учета требуют листо-
грызущие и хвоегрызущие виды, короеды, усачи, златки, раз-
рушители пней, население мохового покрова, подстилки,
почвы.
ТИПОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ БИОЦЕНОЗОВ
Основным фактором, определяющим облик биоценоза, его
структуру, тип, как в естественных, так и во вторичных био-
ценозах, выступают растения. Растительный пркров имеет
9 Заказ 193
1,29
большое индикаторное значение при оценке почв, рельефа,,
водного режима и других особенностей абиотической среды,
а его физиономическая выраженность дает возможность клас-
сифицировать биоценозы на ботанической основе. Геоботани-
ческая карта — основа и для выделения границ биоцензов.
Комплексные исследования биологических группировок, изу-
чение животных компонентов ценозов в единстве с раститель-
ными ассоциациями, а тех и других совместно в их взаимо-
связях с почвенными условиями и с абиотическими факторами
среды показали возможность выделения типов биоценозов
(В. П. Сочава, 1961). Типология и картирование биоценозов—
путь познания и систематизации разнообразных соотношений
между почвенным покровом, животным и растительным ми-
ром.
Опыт сопряженного изучения растений и насекомых на
равнинных плакорах подтвердил, что наиболее рациональной
основой для выделения биоценозов является геоботаническая
карта (Л. В. Арнольди, А. А. Юнатов). К настоящему времени
выяснено, что для каждой растительной ассоциации — исход-
ной единице ботанической классификации — типичен свой эн-
томологический комплекс, соответствующий определенному
биоценозу.
Классифицировать биоценозы наиболее целесообразно,
взяв за исходную единицу растительную ассоциацию, не соз-
давая особых таксономических единиц (А. Г. Воронов, К. Ар-
нольди, В. Б. Сочава). Ассоциация характеризуется одинако-
вым видовым составом растений и животных, одинаковым
набором ярусов, консорций, синузий и местообитанием. Тип
леса, широко используемый в лесной энтомологии, соответ-
ствует ассоциации и биогеоценозу. Лесная типология положе-
на в основу лесной биогеоценологии В. Н. Сукачевым. Ряд
последовательных таксономических единиц, принятых геобо-
таниками и ландшафтоведами, следующий: ассоциация,
группа ассоциаций, формация, группа формаций, класс фор-
маций. Примерами таксономических единиц различного ран-
га можно назвать: ассоциация — еловый лес-зеленомощник с
черникой или еловый лес-зеленомошник с брусникой. Группа
ассоциаций — еловые леса-зеленомошники: черничники, брус-
ничники, кисличники. Формация — еловые леса-зеленомошни-
ки: лишайниковые, мертвопокровные. Группа формаций—тем-
ноховойная тайга — леса еловые, елово-пихтовые, пихтовые.
Самую крупную таксономическую единицу — класс форма-
ций — представляют леса темнохвойные и светлохвойные (сос-
новые, лиственничные).
В природе биоценозы разных формаций обычно чередуют-
ся, образуя комплексность покрова, что является одним из
основных свойств биоценозов. Биоценоз испытывает влияние
соседствующих с ним биоценозов других формаций и всей зо-
130
ны. По положению в каждой зоне встречаются типично зо-
нальные биоценозы, приуроченные к плакорам, интразональ-
ные (луга, болота, пески) и экстразональные. Внимание ис-
следователей больше привлекалось к типично зональным
биоценозам. Однако азональные биоценозы заслуживают бо-
лее пристального внимания, так как отличаются повышенной
биологической продуктивностью (Бей-Биенко, 1966).
В ряде случаев азональные ландшафты представляют конт-
растное сочетание биоценозов различных зон. Примеры таких
биоценозов можно видеть на песчаных террасах рек. Так, ле-
вобережная надпойменная терраса р. Арчеды (приток р. Мед-
ведицы в районе среднего течения Дона), по данным экспе-
диции МГУ, представляет ландшафт полого бугристых пес-
чаных степей с березовыми колками по понижениям.
Метод профилей при картировании песков, применяемый
проф. А. Г. Гаелем, дает наглядную картину комплексности
биоценозов, позволяет выделить их границы, вскрывает исто-
рию формирования, первичность и вторичность сложившихся
здесь биоценозов.
На пологих буграх развиты связнопесчаные почвы средней
мощности. Растительность степная (ковыль, иоанна, типчак
беккера, келерия сизая, полынь маршалла, лапчатка, тысяче-
листник гербера и Др.). Это исходный тип древних почв и
растительности. Почвенная фауна насекомых здесь, по нашим
данным, представлена псаммофилами юго-восточного проис-
хождения, распространенными в полупустыне: Anatolica ab-
breviata Gelb., Microdera convexa Tausch., Anomala errans F.,
Anisoplia deserticola Fisch., Chioneosoma pulvereum Knoch.,
Ceratophyes polyceros Pall.
Из прямокрылых обычны Callipiamus barbarus Costa,
Bryodema tubercilatum F., из муравьев — Cataglyphis aenes-
cens Nyl., Lasius alienus Fieb. Обращает внимание необычная
приуроченность гусениц махаона к полыни маршалла.
В понижениях между буграми почвы заилены, связнопес-
чаные, дерново-луговые, лугово-болотные и торфянисто-болот-
ные. Понижения заняты лесными колками — березово-осино-
выми, иногда с участием дуба. В колках обычны Melolontha
hippocastani F., Serica brunnea L., Hoplia parvula Kryn, Pros-
ternon tesselatum L., Lacon murinus L., Athous haemorrhoidalis
F., Melanotus rufipes Hb., Selatosomus aeneus L., Elater sp.
и другие виды, характерные для лесной и лесостепной зон.
Чернотелки в колках не обнаружены. Общих видов с песчаной
степью, расположенной рядом на буграх,— нет. Между песча-
но-степным биоценозом и лесным колком проходит полоса лу-
гово-степной растительности на лугово-степных почвах. Здесь
обычны виды лесостепной и степной зон: Polyphylla fullo L.,
Amphimallon solstitialis L., Anomala aenea Dag., Selatosomus
latus F., Decticus verrucivorus L., Omocestus ventralis Zett.
9*
Г31
Рис. 22. Профиль через участок бугристо-колкового ландшафта надпойменной террасы р. Арчеды (по А. Г.
Г аелю):
/ — современный эоловый нанос песка; 2 — эоловый нанос раннепастбищной фазы дефляции; 3 — глубоко-
гумусированная связнопесчаная почва; 4 — среднегумусированная почва на песках древней фазы дефляции; 5 -
псевдофибры; 6 капиллярная зона; 7 —зона насыщения грунтовых вод
и др. (рис. 22). Коэффициент общности фауны насекомых пес-
чано-степного биоценоза и лугово-степного равен 10%, что
свидетельствует о самостоятельности лугово-степного биоце-
ноза наряду с общностью некоторых форм.
Классификация биоценозов на типологической основе име-
ет большое значение при изыскании путей хозяйственного ос-
воения территории, в том числе облесения песков.
Глава VII
ДИНАМИКА БИОЦЕНОЗОВ
Каждый биоценоз всегда находится в изменении, динами-
ке. Подвижность структуры биоценозов — их характерная
черта. Она проявляется в смене доминирующих видов, изме-
нении численности популяций отдельных видов, в суточных,
сезонных и многолетних колебаниях состава биоценозов и
всех других его свойств. Биоценоз представляет арену очень
сложных, разнообразных и противоречивых процессов. Боль-
шей частью изменения в биоценозах протекают медленно, по-
этому биоценозы кажутся неизменными. Другие процессы
периодически повторяются и также выражают относительную
стабильность биоценозов. Но ряд процессов в биоценозах но-
сит необратимый характер и приводит к смене одних биоце-
нозов другими — к сукцессии. Во всех случаях существование
самого биоценоза, его обратимые и необратимые изменения
возможны лишь при непрерывном круговороте форм, вещест-
ва и энергии. Все перемещения, все сложные процессы в био-
ценозе связаны с переносом химических веществ и энергии,
их накоплением, превращением и циркуляцией в природе.
Жизнь возможна лишь как непрерывный биогенный кругово-
рот, осуществляющийся в конкретных биоценозах.
БИОЛОГИЧЕСКИЙ КРУГОВОРОТ ВЕЩЕСТВ
«Под биологическим круговоротом понимается поступле-
ние элементов из почвы и атмосферы в живые организмы,
превращение в них поступающих элементов в новые сложные
соединения и возвращение в почву и атмосферу в процессе
жизнедеятельности с ежегодным опадом части органического
вещества или полностью организмами, входящими в состав
биогеоценоза» (Ремезов и др., 1963). Биологический кругово-
рот, его устойчивость в биоценозах обеспечивается наличием
организмов — производителей органического вещества (фото-
134
^синтезирующих и хемосинтезирующих растений), потребите-
лей (насекомые и другие животные) и разрушителей (микро-
организмы). Связи биоценоза с абиотической средой его жиз-
ни включают как понятие обитания, так и понятие питания
как химического процесса между биосферой, атмосферой и
литосферой (В. Ю. Фридолин).
Через живые организмы постоянно осуществляется круго-
ворот углерода, азота, воды, фосфора и других элементов, а
также и энергии. Подсчитано, что над каждым гектаром по-
верхности земли находится углерода (в форме СО2) 2,5 т,
а в растениях сахарного тростника на 1 га посевов — 8 т уг-
лерода. Если бы углерод не возвращался через дыхание, гние-
ние, брожение и другие процессы жизни, то весь его запас был
бы исчерпан за 35 лет. Подобные же процессы происходят
с нитратами почвы и воды, водой, фосфором, кальцием, маг-
нием и другими химическими веществами, потребляемыми
растениями, а затем и животными. Передача и превращение
веществ и энергии являются важной стороной биологического
круговорота. Осуществляется она через цепи питания разных
трофических уровней. Популяции животных, входящих в био-
ценоз, это не только определенное число особей, но и количе-
ство протоплазмы, участвующей в обмене химическим вещест-
вом и накапливающее в себе вещества и энергию за счет
других видов. Биомасса и1 метаболизм, интенсивность биохи-
мических процессов — это тоже определенные признаки попу-
ляции. Скорость создания органического вещества определяет
продуктивность живых систем. Продуктивность отдельных по-
пуляций учитывают по числу особей и по биомассе.
Главную часть биомассы составляют массовые виды, доми-
нанты. Значение их в круговороте веществ велико. Среди фи-
тофагов из мира беспозвоночных доминируют насекомые.
Поедая растение, они трансформируют его вещество, которое
поступает в почву в форме выделений и трупов. Эта транс-
формация может пройти через ряд трофических уровней (фи-
тофаг, хищник, паразит, паразит второго порядка и т. д.).
Многие насекомые аккумулируют в себе ряд химических ве-
ществ, элементов, в том числе радиоактивных. Радиоактивные
вещества в малых дозах стимулируют азотфиксирующую дея-
тельность микроорганизмов. Микроорганизмы продуцируют
очень много веществ, нам еще плохо известных (стимуляторы
роста, антибиотики, гормоны, витамины и пр.). Установлено,
что на 1 см2 поверхности почвы адсорбированных частиц мик-
роорганизмов встречается до 140 млн., а общая бактериаль-
ная масса на 1 га поверхностного слоя достигает 5—7 т. Мно-
гие из микроорганизмов поступают в кишечник почвообитаю-
щих насекомых с частичками почвы. Часть из них насекомые
разносят. Мелких немикроскопических членистоногих (клещи,
.ногохвостки и другие) на 1 м2 пахотного слоя цочвы насчи-
135
тывается десятки и сотни тысяч, а в лесных почвах — нередко
миллион. Общая масса почвенных беспозвоночных равна 3—
4 т/га (Гиляров, 1949). В процессе жизнедеятельности насе-
ление почвы производит биохимическую работу космического*
значения, а в миграциях химических элементов решающее
значение имеют биоценотические связи особенно многочислен-
ных и подвижных насекомых.
Большое значение в круговороте веществ играют личинки
двукрылых. В дубравных лесах лесостепи и степи опад листь-
ев некоторых пород превращают в гумифицированную массу
исключительно личинки комариков Bibio, Neosciara, Scatopse
и др. (Зражевский, 1957). Наиболее распространенный Neos-
ciara modesta Staeg. развивается на свежих опавших листьях
многих древесных пород, встречаясь очагами. Личинки про-
являют избирательность к опаду листьев разных пород, но
предпочитают листья кустарников. В экскрементах личинок
обнаружен аммиак, нитраты; в некоторых случаях азота
обнаружено в несколько раз больше, чем было в листьях до
опыта. Личинок садовой мошки Bibio hortulans L. в степных
насаждениях дуба на темнокаштановых почвах встречается
до 12 000 экз. на 1 м2 и перерабатывать они могут до 2 т/аа
подстилки. В насаждениях белой акации при том же количе-
стве личинок на 1 га они перерабатывают до 5,6 т/га подстил-
ки. Обнаружено, что в экскрементах личинок содержалось до
20,5% гумусовых веществ (Травлеев, 1961). Роль насекомых
в механической переработке органических остатков, активи-
зации бактерий и грибов велика. Они выступают существен-
ными ускорителями почвообразовательного процесса. Пленоч-
ные почвы населяют Collembola, Diptera, Dermaptera, Coleop-
tera, участвуя в формировании первичных почв (Стебаев^
1958).
Очень мало данных о роли в круговороте веществ листо-
грызущих насекомых. Подсчеты таких изменений за семилет-
ний цикл размножения непарного шелкопряда сделаны
П. М. Рафесом (1964). Запас весенней листвы на дубах со-
ставлял 9,3 т/га. В первый год размножения деятельность не-
парного шелкопряда оставалась незаметной. На второй год
гусеницы съели 0,07 т/га листвы, в почву с экскрементами
поступило 0,03 т/га. На четвертом году размножения было
съедено 5,5 т/га, на пятый год — 6,1 т/га. Увеличилось и ко-
личество поступающих органических веществ в почву (рис. 23).
Основное значение массового размножения для метаболи-
ческих процессов состоит в резком ускорении минерализации
поступающего в почву органического вещества и изменении
его состава. Значение его для почвообразовательных процес-
сов, минерального питания растений и, следовательно, всего
биогеоценоза требует глубоких исследований. Количество ве-
ществ, перерабатываемых почвенной фауной насекомых, за-
,136
висит не только от их обилия и биомассы, но и от интенсив-
ности процессов жизнедеятельности и избирательности пищи.
Имеет место качественная и количественная дифференциация
мезофауны почвы в разных биоценозах. Разложение опада и
подстилки зависит от свойств доминирующих видов. Механи-
ческое размельчение органического опада производят, как.
Рис. 23. Поступление органического вещества на 1 га почвы за период
вспышки непарного шелкопряда (по Рафесу, 1964)
правило, личинки представителей подкласса крылатых насе-
комых (Pterygota), а к гумусообразователям относят Aptery-
gota (Козловфая, 1959).
Из изложенного выше есть все основания считать, что
роль насекомых и других беспозвоночных в ускорении процес-
са гумификации отмирающих растений весьма велика.
СМЕНА БИОЦЕНОЗОВ
Процессы, происходящие в биоценозах, имеют разное зна-
чение для жизни биоценоза. Одни изменения непродолжитель-
ны и легко восстанавливаются, другие приводят к замене од-
ного биоценоза другим (к сукцессии). Различают сукцессии
вековые, длительные, быстрые и катастрофические; вызывают-
ся они разными причинами (как эндогенными, так и экзоген-
137
ними). Чаще всего сукцессии необратимы, но иногда можег
восстановиться биоценоз исходного типа. Сукцессии вызы-
ваются изменениями климата, условиями геоморфологически-
ми, внедрением в биоценозы новых видов, усилением значения
какого-то одного вида, массовым размножением вредителя
(фитофага) пожарами, вырубкой лесов и другими причинами.
Благодаря замедленности происходящих в биоценозе измене-
ний может создаться впечатление, что биоценоз неизменен,
но в действительности в нем непрерывно идут процессы, при-
водящие к смене его другим биоценозом.
Многолетние наблюдения за жизнью биоценоза позволяют
заметить глубокие сдвиги в численном соотношении популя-
ций разных видов растений и животных и многие другие от-
клонения. Эти изменения могут затрагивать или отдельные
микроценозы, или даже весь биоценоз. Так, луговой мотылек,
объедая только двудольные растения и не трогая злаков, спо-
собствует выравниванию типчаковых степей; садовый хрущик,
выедая мягкие злаки, приводит к появлению мохового покро-
ва; вследствие постоянного размножения яблонной моли в
орехово-плодовых лесах Южной Киргизии яблоню вытесняет
клен, а сибирский шелкопряд, вызывая усыхание кедра, спо-
собствует замене кедровых лесов другими хвойными порода-
ми. Ель хорошо возобновляется там, где почва разрыхлена
муравьями и имеется много выбросов дождевых червей.
В лесах очень характерны процессы сукцессии микроце-
нозов пней и постепенное превращение их в кочки. Наблюде-
ния за этапами разложения древесины показывают, что сук-
цессии беспозвоночных — разрушителей древесины — могут
иметь разную направленность и связаны с деятельностью гри-
бов (рис. 24).
Восстановление леса после вырубки или пожара происхо-
дит путем смены разных биоценозов. Так, вместо елового ле-
са на месте вырубки или гари появляются прежде всего сор-
ные растения. Они разрастаются, но позднее их вытесняют
разрастающиеся кустарники и поросль березы и осины. Де-
ревья быстро растут, и кроны их смыкаются. Прежняя травя-
нистая растительность сменяется более теневыносливыми
растениями лесного типа. Таким образом, вместо елового леса
вырастает березово-осиновый со своим миром насекомых. Че-
рез 10—20 лет лиственные леса становятся светлее, и под их
пологом начинает расти ель. Появление ее изменяет почвенно-
грунтовые условия в лесу; ель создает много тени, благодаря
чему светолюбивые береза и осина возобновляться уже не мо-
гут, и через 60—80 лет после вырубки ели березово-осиновые
леса снова заменяются еловыми. Происходит изменение и жи-
вотного населения биоценозов.
Смена биоценозов может принять и. другое направление.
Пастьба скота приводит к уничтожению кустарников и под-
'L38
роста деревьев; на месте вырубки может образоваться не лес,
а суходольный луг.
Биоценозы елового леса, березового леса, суходольного
луга и вырубок, занятых бурьяном, являясь отдельными
звеньями в цепи последовательных превращений, сохраняют
каждый свою самостоятельность и относительное постоянство.
Рис. 24. Сукцессия беспозвоночных — разрушителей древесины
(по Мамаеву)
Хорошо известно, что каждому типу леса (биоценозу) соот-
ветствует набор определенных видов насекомых, среди кото-
рых можно выделить ведущих. Например, сосновые лубоеды
и четырехзубчатый короед типичны для сосновых боров-зеле-
номошников, короеды типограф и гравер — для ельников-зе-
леномошников, березовый заболонник — для березовых лесов
и т. д. Энтомофауна как составная часть биоценоза зависит
от типа леса и изменяется вместе с последним. Массовое раз-
множение первичных вредителей, объедающих листву, ведет
к сильному ослаблению древостоя. Обнажение крон изменяет
режим освещения: под полог леса проникает много света.
Это ведет к преобладанию светолюбивой травянистой расти-
тельности, изменению гидротермического режима , к проник-
новению степных видов. Изменение состояния леса резко ска-
зывается на составе энтомофауны. В лесу, ослабленном ли-
стогрызущими вредителями или пожарами, в массе размно-
жаются стволовые вредители: короеды, златки, усачи. Появ-
ление новой энтомофауны, в свою очередь, по-разному ска-
зывается на насаждении.
139
Рассмотрим еще пример. Так, по В. Н. Беклемишеву, вновь
образованный водоем не имеет на дне органических отложе-
ний, в нем мало питательных веществ, поэтому население его
бедно. В таком водоеме особенно мало животных организмов,
в том числе насекомых. Первыми поселяются жгутиковые во-
доросли и инфузории. При наличии водорослей в водоеме мо-
гуть жить личинки комаров. Уже на второй год водоем засе-
ляется водными растениями, с их поселением создаются
благоприятные условия для ряда насекомых. В последующие
годы водоем обогащается продуктами жизнедеятельности ор-
ганизмов— органическими веществами, солями. Появляются
новые типы водных растений, заросли становятся густыми,
нередко они становятся многоярусными. Образуется надвод-
ный ярус растений, нейстический ярус (плавающие растения)
и погруженный. Намечается зональность распределения ра-
стений от берега к центру водоема. Дно таких зарастающих
водоемов покрыто слоем черного органического ила. В воде
идет интенсивный фотосинтез высшими растениями и взве-
шенными водорослями, что сказывается на газовом режиме.
Биоценозы таких водоемов становятся все более разнообраз-
ными. Из года в год водоем больше засоряется мертвыми
растительными остатками и постепенно мелеет, свободной по-
верхности воды становится меньше, и, наконец, она исчезает.
Водоем превращается в болото низинного типа с соответст-
вующим типом растительности и животного населения.
Динамичность биоценоза определяется: 1) меняющимися
взаимодействиями организмов, составляющих биоценоз;
2) вселением новых организмов; 3) заносом минеральных и
органических веществ (с пылью и водой); 4) выносом мине-
ральных и органических веществ. Процесс внутренних взаи-
модействий никогда не прекращается, но постепенно замед-
ляется. Общая тенденция к замедлению сукцессии привела
зарубежных экологов к представлению об устойчивой, заклю-
чительной фазе,— климаксу. Устойчивость растительности в
соответствии с климатом соответствует нашему представле-
нию о зональности. Климакс определяют как конечную ста-
дию в развитии растительности, которая занимает данную
территорию до тех пор, пока остается неизменным климат.
Для климакса характерно состояние динамического равнове-
сия между климатом, геоморфологией, почвой и раститель-
ностью. Но поскольку влияние растительности, животного
мира и микроорганизмов на другие компоненты биоценоза
не может прекратиться, то процесс изменения биоценоза ни-
когда не останавливается. Он все время приводит к смене
одних биоценозов другими, что исключает возможность со-
стояния климакса.
Смена биоценозов может замедлиться на любом этапе.
Таким, образом, любой из «временных» биоценозов может ока-
140
заться на положении «завершающей формации». Но и «за-
вершающая формация» не останавливается в своем поступа-
тельном развитии и может стать «начальной стадией» разви-
тия новых биоценозов. Постоянство в природе в любой его
форме (кратковременное или длительное) относительно.
Изменение биоценозов можно изучать при постоянных
многолетних учетах на стационарных участках. Однократные
непродолжительные исследования при самых детальных опи-
саниях фиксируют лишь отдельный этап, выхваченный из
общей цепи развития.
Насколько изменчивы состав и численность насекомых в
формирующемся биоценозе, показывают наши наблюдения
за массовыми видами в лесополосах, выращенных в сухих
степях юго-востока (под Волгоградом) (табл. 5).
Таблица 5
Изменения в видовом составе насекомых в лесополосе
Виды 1950 г. 1951г. 1 1952 г. J1953 г. 1954 г. 1956г.
Opatrum sabulosum L. 1,3 M2 0,2 л2
Pedinus femoralis L. 2,4 M2 — — 0,1 м2 — .—
Agriotes sputator L. 0,3 M2 — — — — —
Selatosomus latus L. 0,1 M2 — — — — —
Hister bipustulatus Schr. — — — 11 м2 —
Carabidae (Amara Har- 4,1 M2 — — — 22 л/2 —
palus)
Laphygma exigua Hb. — до 80 м2 ед. — —
VII—IX V-VI
Orgyia dubia Tausch. + + н—I—H + 4 +
Gracilaria alchimiella Sc. — + 44 + 4 ед. ед.
Galerucella luteola Mull. — ++ Н—h + 4 + '4~ + +
Sphenoptera cuprina — + 4
Motsch. Agrilus constant ini Obenb 4
Empoasca haupti Rib. —. 4 4 4-4 4 4 44 4 +
Aphis craccivora Koch. — 4~-|- 4 444 ед. ед.
Terioaphis tenera Aizenb. — ++
Hayhurstia tatarica Aizenb. — 444 4 4 4 + + 4 +++ 4-4 +
Tettigonia caudata Ch. — — 4 4 ед. ед.
Metrioptera eversmanni —- — + 4 ед. ед.
Kitt Cynipidae | —- — ед. ед. ед. -|~ +
Примечание: ед. — единичен,
4------------------обычен,
-|—|--в значительном количестве,
+++— в массе.
Обращает внимание массовое появление ранее отсутство-
вавших видов и их быстрое исчезновение: Hister — в 1954 г.;
Laphygma — ь 1952 г.; Orgyia — в 1952—1953 гг.; Aphis —
141
в 1951, 1952 гг.; Ter ioaphis — в 1953 г. 1952 год был годом мас-
совой вспышки размножения совки-карадрины (Laphygma
exigua Hb.) на огромной территории (в районе наблюдений
карадрина не отмечалась с 1936 г.). В последующий за
вспышкой год (1953 г.) гусеницы карадрины встречались еди-
нично, а затем пропали. 1952 год отличался обилием дождей
и теплой погодой. В полосах появились кузнечики, в массе
размножился степной кистехвост (Orgyia dubia Tausch.), ми-
нирующая моль-пестрянка (Gracilaria alchimiella Sc.).
В следующие годы численность вредителей резко снизилась,
а гусениц кистехвоста отыскать вообще не удалось. Для
1954 г. характерно было большое количество жуков-карапу-
зиков (Hister) и некоторых жужелиц — типичных обитателей
лесной подстилки в лесных балках (Cymindis, Amar а), ранее
отсутствовавших в полосе. Люцерновая тля (Aphis medicagi-
nis Koch.), в массе встречавшаяся в 1952 и 1953 гг. на акации,
после влажного 1952 г. стала единичной, но появился другой
вид тли, однако на непродолжительный срок. Изменение чис-
ленности каждого из фитофагов сказывалось на энтомофагах
и на состоянии самой полосы.
Резкие колебания видового состава и численности насеко-
мых, заселяющих лесные полосы, в необычных для леса за-
сушливых условиях полупустыни, по-видимому, черта, харак-
терная для новых биоценозов. Эта особенность соответствует
неустойчивому водному режиму, неравномерному выпадению
и распределению осадков. Ряд видов стали постоянными оби-
тателями лесополосы, такие, как ильмовый листоед, ильмовая
цикадка, моль-пестрянка, орехотворки, некоторые листовертки
и другие дендрофильные виды, преимущественно моно- и оли-
гофаги.
Итак, сукцессии биоценозов разнообразны; происходят они
в относительно непродолжительные сроки, нередко носят ка-
тастрофический характер, а многие сукцессии проходят за
длительный период и связаны со сменой целых ландшафтов
(например, смена еловых плакоров на березово-сосновые леса
на Русской равнине, проходившая в голоцене— 10 000 лет
назад). Много изменений связано с деятельностью человека.
Глава VIII
ЭКОЛОГИЯ КУЛЬТУРНЫХ ЛАНДШАФТОВ
Человечество уже тысячелетиями воздействует на природу
Распашка земель, вырубка лесов, выпас скота, сенокошение
и другие формы хозяйственной деятельности человека привели
к замене первичных естественных биоценозов вторичными
биоценозами, или агробиоценозами, на большей части терри-
тории земли. Во многих странах мира целина сохранилась
только в заповедниках. Человек, используя природу в своих
практических целях, сознательно, а чаще невольно способст-
вует коренному преобразованию и разрушению первичных при-
родных биогеоценозов и целых ландшафтов. Исчезает пер-
вичная фауна и флора, создаются новые ценотические связи.
Поля, сады, огороды, сенокосы, лесные посадки — вторичные,
или культурные, биоценозы (агробиоценозы).
Агробиоценозы отличаются от первичных биоценозов. По
структуре они приближаются к экстремным биоценозам, ха-
рактеризующимся небольшим числом видов и их высокой чис-
ленностью. Комплекс насекомых в агробиоценозах опреде-
ляется характером растительного покрова — возделываемыми
культурами. Однако четких граней между агро- и биоценоза-
ми нет, поэтому нет оснований отрицать наличие в агробио-
ценозах признаков, свойственных биоценозам (Гиляров, 1963).
Вторичные биоценозы — аналоги сукцессионных рядов естест-
венных биоценозов. К характерному отбору видов и упроще-
нию агробиоценозов приводит ежегодная обработка почвы.
Остаются виды, способные к факультативной сапрофагии,
всеядные, эвритермные и эвритопные виды с высоким потен*
циалом размножения или легко переносящие неблагоприят-
ные условия. Поверхностно обитающие насекомые, специали-
зированные вредители полевых культур, способны быстро
передвигаться, развиваться и расселяться.
Простота структуры вторичных биоценозов и возможность
широкой их регуляции позволяют экспериментально решать
143
биоценотические проблемы при работах по прикладной энто-
мологии и при других исследованиях. Удаление или введение
отдельных звеньев агробиоценоза можно достичь обработкой
почвы, снятием урожаев, интенсивностью выпаса, химической
борьбой с вредителями, выпуском энтомофагов. Экологиче-
ские исследования выливаются в конечном счете в экологию
сельскохозяйственных ландшафтов, которая рассматривает
комплексные взаимосвязи отдельных частей ландшафта.
В этом аспекте должны идти работы по защите растений и
охране природы, ибо только знание экологических взаимосвя-
зей в природе позволяет понять историю образовавшихся
ландшафтов, формирующихся на протяжении тысячелетий, по-
нять и те изменения, которые связаны с деятельностью чело-
века (Медведев, 1959).
Изучение закономерностей формирования вторичных био-
ценозов имеет первостепенное теоретическое и практическое
значение. Сравнение особенностей комплексов обитателей
культурных земель в разных местностях позволяет рассмат-
ривать их как закономерно складывающиеся группировки,
развивающиеся по тем же законам, что и естественные био-
ценозы. Но их устойчивость зависит от степени и повторяемо-
сти изменений, вносимых человеком.
В агробиоценозах лишь единственный вид — возделывае-
мое растение (в смешанных культурах два или несколько ви-
дов) — является его константой и доминантой; остальные не-
многие виды представлены сорными растениями и в идеале
их не должно быть. Устойчивость растительного покрова в
агробиоценозе достигается лишь постоянной заботой человека,
а регулярное изъятие биологической продукции в виде урожая
восполняется внесением удобрений и соответствующей агротех-
никой. Смена агробиоценозов также определяется человеком.
Но основное свойство биоценоза—устойчивость состава
растительного покрова, хотя и сведенного к одному домини-
рующему виду, сохраняется и в агробиоценозе. Относительная
устойчивость видового состава характерна и для фауны и в
целом определяет самую существенную черту любого биоце-
ноза— наличие в нем типичного состава организмов, сохра-
няющего постоянство во времени и пространстве.
Анализ фауны целинной степи и пшеничного поля (Бей-
Биенко, 1961) показывает, что целинная степь по видовому
составу вдвое богаче, нежели пшеничное поле (330 видов на
целине и 142 вида на пшеничном поле). Однако среднее оби-
лие на пшеничном поле почти вдвое превышает плотность
населения на целине (351 особь на 1 м2 на пшеничнохМ поле
и 199 особей на целине). Число показательных видов (доми-
нанты и константы) на целине вдвое больше, но на пшенич-
ном поле они резко преобладают над остальными видами
(94,2%). Доминанты пшеничного поля — пшеничный трипс
144
(Haplotlirips tritici Kurd.), ячменная тля (Brachycolus noxius
Mordw.), поедающая ее божья коровка (Adonia variegata
Gr.), дерновый муравей (Lasius alienus Forst.) и серая зерно-
вая совка (Аратеа Sordida Bkh.). Целина имеет более раз-
нообразный видовой состав, но ни один вид не выделяется
резко по численности, тогда как на посевах пшеницы основу
населения составляют лишь немногие господствующие виды.
При распашке целины часть видов исчезает, часть видов рез-
ко уменьшается по обилию, а часть видов на посевах пшени-
цы дают резкую вспышку — численность их возрастает в де-
сятки и сотни раз (хлебная цикадка — Deltocephalus collinus
Dahlb., пшеничный трипс — Haplothrips tritici Kurd., полоса-
тая хлебная блоха — Phyllotreta vittula Redt., серая зерновая
совка — Hadena sordida Bkh. и муравей — Cataglyphis cursor
aenescens Nyl.). Освоение целинных земель под посевы при-
водит к полной перестройке фауны: вымиранию доминантных
целинных видов и к необычайному возрастанию численности
отдельных, прежде второстепенных, видов. Эти виды в агро-
биоценозах становятся своеобразными сверхдоминантами,
трофически связанными с культурой растения.
Изучение агробиоценозов позволяет установить, какие осо-
бенности экологии вредных насекомых позволяют им стать
постоянными обитателями культурных полей. По М. С. Ги-
лярову, ведущее значение приобретают приспособления к рез-
ким и быстрым сменам растительного покрова: многоядность
и моно- и олигофагия при способности к расселению и корот-
ком жизненном цикле. Если личиночное развитие протекает
долго и за это время успевают снять урожай и заменить куль-
туру, то при таких условиях могут выжить только многоядные
насекомые, способные переключиться на питание другим
растением (проволочники, чернотелки, хрущи и др.). Моно-
фаги и олигофаги должны иметь короткий период развития
и высокую способность к расселению. Если цикл развития на-
секомого захватывает период больший, чем развитие расте-
ния, или если у насекомого цикл развития короткий, но имаго
не способны к расселению, то такие насекомые не могут жить
на полях.
Многие биологи считают, что частые катастрофические
нарушения численности отдельных видов (массовые размно-
жения вредителей)—следствие нарушений каких-либо свя-
зей, исторически сложившихся в биоценозах. В естественных
биоценозах не отмечают массовых размножений отдельных
видов; богатые видами биоценозы имеют более постоянное
«уравновешенное около средней нормы» соотношение числен-
ности видов.
Известны многочисленные факты массовых «нашествий»
различного вида животных и растений чужеземного проис-
хождения, завезенные человеком в несвойственные им биоце-
10 Заказ 193
145
нозы. В ряде случаев такие виды на своей родине массовых;
размножений не дают.
Интересная сводка дана английским экологом Ч. Элтоном
в книге «Экология нашествий животных и растений». Элтон
приводит пример пандемии малярии в Бразилии 1938—
1939 гг., вызванной завозом из Африки Anopheles gambiae
Gil., пример стремительного размножения колорадского кар-
тофельного жука в Европе, размножений многих лесных вре-
дителей в лесах США и Канады, завезенных из лесов Европы
(непарный шелкопряд, златогузка и др.). Известны печаль-
ные результаты завоза в Европу филлоксеры и многих других
карантинных объектов.
Появление новых видов в биоценозе вызывает сдвиги в1
местной природе; аборигенные ценозы изменяются, соотноше-
ние численности их популяций катастрофически нарушается,
что и приводит к перестройкам в биоценозах и появлению
вредителей. Так, например, Tetropium gabrieli Weise не вредит
в районах естественного ареала, а там, где лиственница интро-
дуцирована и с ней завезен жук, он становится вредителем.
Предполагают, что более частым вспышкам размножения
отдельных видов и постоянному вреду на культурных расте-
ниях способствует обеднение видового состава энтомофагов.
Популяции вредителей в отдельных биоценозах выходят из-
под биологического контроля паразитов и хищников.
К упрощению биоценозов и увеличению численности от-
дельных видов приводит обработка почвы, внесение ядохими-
катов, неумеренный выпас скота, ветровая и водная эрозии
почв. Химическая обработка, проводимая в целях борьбы с
вредителями и сорными растениями, сопровождается гибелью
многих других видов биоценоза, в том числе и полезных. Но
изучено это недостаточно, и данные противоречивы. Так, одни
авторы отмечают губительное действие гексахлорана, ДДТ и
ряда других химикатов на полезную фауну и флору, накопле-
ние ядов в организмах, перенос их при питании другим орга-
низмам, что приводит к «цепной реакции смерти». Многие ядо-
химикаты не имеют избирательного действия и, уничтожая
вместе с вредителем его энтомофагов, дают новые возможно-
сти размножения вредителя, что, в свою очередь, требует все
большего применения ядов. Это вызывает появление устойчи-
вых к ядам форм. Идут поиски новых избирательного дейст-
вия инсектицидов, новых приемов борьбы. Например, в Ан-
глии на яблонях паразитировало более 100 видов насекомых;
за последние 50 лет их число снизилось до 5, но оставшиеся
пять видов стали чувствительны лишь к очень сильным ядам,
и стоимость обработки садов повысилась (Ч. Элтон, 1960).
Обработка садов ГХЦГ и ДДТ привела к сильному размно-
жению паутинного клещика (Metatetranichus), так как яды
истребили его паразитов и хищников (Тишлер, 1960). По дру-
146
гим данным, мнение о нарушении оаланса в природе при при-
менении современных сильнодействующих инсектицидов пре-
увеличено. Анализ последствий применения ядохимикатов в
лесах Дании (Bejer—Petersen Broder, 1964) показывает, что
вред от них незаметен (ДДТ теряет свою активность через
1—2 недели); случаи отравления птиц и млекопитающих не
известны. Имеются сведения, что 0,15%-ный водный раствор
хлорофоса при норме расхода 0,75 кг]га при борьбе с капуст-
ной молью вызывает даже рост численности энтомофагов.
ДДТ для энтомофагов губителен, но их численность вырав-
нивается через месяц после обработки.
Однако в последнее время все чаще звучат советы обра-
тить основное внимание на рациональную организацию ланд-
шафта, охрану природы, охрану полезной фауны, пытаться из-
бегать химических средств защиты, особенно в лесных биоце-
нозах. Но для этого необходимо знать особенности естествен-
ных биоценозов и историю формирования вторичных биоцено-
зов. По В. TI. Сукачеву (1964), «атмосфера, материнская гор-
ная порода, вода атмосферы, почвы и подпочвы являются
как бы первичным материалом данного биогеоценоза, а расте-
ния, животные и микроорганизмы по преимуществу служат
трансформаторами и аппаратами обмена веществ и энергии».
Но особое значение имеют почва и лесная подстилка. Они
суммируют в себе результаты сложного биогеоценотического
процесса и представляют наглядное и наиболее полное выра-
жение итогов этих процессов. К каким последствиям для
судьбы биогеоценозов, к чему ведет разрушение почв, легко
проследить на опыте комплексной песчано-эрозионной экспе-
диции МГУ, работавшей ряд лет под руководством проф.
А. Г. Гаеля на песчаных массивах Дона и его притоков. Лег-
кие песчаные и супесчаные почвы легко поддаются ветровой
эрозии, поэтому нарушение растительного покрова распашкой
и выпасом скота быстро приводит к их разрушению, образо-
ванию котловин, эоловых наносов, превращению в бугристые
пески. На речных песчаных террасах в зоне засушливых сте-
пей имело место неоднократное перевевание почв, их разру-
шение и формирование новых почв на эоловом наносе.
В районе Среднего Дона (Доно-Арчединский песчаный
массив) экспедицией вскрыто три яруса дерново-степных почв
разного возраста. Глинисто-песчаные, дерново-степные глубо-
ко- и среднегумусированные почвы встречаются преимущест-
венно на левобережной второй надпойменной террасе. Они
развились на мелкозернистых древних песках афитогенных
фаз дефляции в период послеледникового климатического оп-
тимума. На небольших участках эти почвы сохранились и для
них характерна типичная целинная песчаная степь с преобла-
данием ковыля иоаниа, овсяницы беккера, келерии сизой с
примесью разнотравья — полыни маршалла, тысячелетника
ю*
147
гербера, лапчатки серебристой, цмина, чебреца и ракитника
(рис. 22). Доминирующие виды насекомых_____Anatolica abbre-
viata Gebl., Microdera convexa Tausch., Anomala errans F.,
Chioneosoma pulvereum Knoch., Anisoplia deserticola Fisch.,
Tentyria nomas Pall., Calliptamus barbarus Costa, Cataglyphis
aenescens Nyl., Lasius alienus Fieb., Platyope leucogramma
Pall.; типичны Bryodetna tuberculatum F. и Papilio machaon L.
В бронзовый и железный века эти почвы местами были
разбиты выпасом скота, но за 3—0,5 тыс. лет разбитые пески
заросли, и на них сформировались новые маломощные почвы,
рыхлопесчаные. Для них в настоящее время характерна так-
же песчано-степная растительность, но более разреженная и с
большой примесью чебреца, василька гербера, ракитника. На-
секомых в почве меньше, преобладают: Chioneosoma pulvere-
um Koch., Anomala errans F., Cataglyphis, Myrmeleon. Разру-
шение древних песчаных почв (рис. 25) вместе с их биоцено-
зами и последующее образование маломощных рыхлопесча-
ных почв привело к обеднению первичных биоценозов и преоб-
ладанию некоторых псаммофилов.
Наконец, за последние 150—100 лет вблизи рек и озер пес-
чаные пастбища были еще раз разбиты выпасом скота. В та-
ких местах рельеф принял дюнно-барханные формы. Но поч-
венные разрезы вскрывают здесь три яруса разновозрастных
дерново-степных почв: 1) древних мощных и среднемощных,
более пли менее разрушенных; 2) маломощных — на песках
раннепастушеской фазы дпфляцин; 3) сыпучих песков, лишен-
ных почв или с примитивными песчаными почвенными образо-
ваниями (гумусом прокрашены 5—10 см). Растительный по-
кров на полуразвеваемых песках современной фазы дефляции
пионерно-псаммофитный (пырей пушистоцветковый, осока
колхидская, колосняк, полынь песчаная, ракитник, казацкий
можжевельник). Почвенная фауна жуков крайне бедна, еди-
нично встречены личинки Anisoplia deserticola Fisch., Anatoli-
ca angustata Stev., Platyope leucogramma Pall. Обильны ти-
пичные псаммофилы — личинки муравьиного льва, осы-бем-
бексы. Исходные типы биоценозов не восстановились даже за
1000 лет.
Создание сосновых насаждений на песках привело к ново-
му изменению первичных и вторичных песчано-степных биоце-
нозов. В культурах сосны на склонах и березовых колках, где
сохранились остатки реликтовых форм, абсолютно доминирует
восточный майский хрущ, расселившийся из березовых колков
вместе с сосной. На более высоких, сухих местах распростра-
нены другие виды хрущей (Lasiopsis, Amphimallon).
Приведенные примеры показывают, сколь велика роль че-
ловека в разрушении исходных типов биоценозов и как труд-
но и даже невозможно их восстановление. Но человек может
и должен воздействовать па природу, избегая нежелательных
148
Рис. 25. Профиль через участок бугристых песков с погребенными почвами (из А. Г. Гасля):
/ — бывшая поверхность глубоко гумусированной связнопесчаной почвы; 2 — песок современной фазы
дефляции — с поверхности слабо окрашенный гумусом на глубину 3—5 см; 3 — песок раннепастушеской фа-
йл дефляции с гумусовым горизонтом до 20 см; 4— остатки древней глубоко гумусированной связнопесча-
ной почвы; 5— прослои аллювиального суглинка; 6—капиллярная кайма над уровнем грунтовых вод; 7 —
зона насыщения нижнего зеркала грунтовых вод; 8 — скважины и шурфы; 9 — можжевельник; 10 — осина;
11 — ракитник; 12 — келерия, типчак, ковыль; 13— колосник, вейник, осока колхидская; 14 — чебрсц,
цмин; 15 — молочай, песчаная полынь, полевая полынь, василек
последствий, создавая новые биоценозы. Мощным фактором
преобразования природы, создания культурных ландшафтов
выступает лесная растительность. Она предохраняет почвы от
водной и ветровой эрозии. Бережное к ней отношение, созда-
ние лесных полос — в руках человека. Умеренный выпас ско-
та, пастбищные севообороты, подсев кормовых трав, улучшаю-
щих луга, подбор устойчивых к повреждениям пород и сортов
растений, повышение их устойчивости путем разнообразной
обработки почвы, внесения удобрений, борьбы за влагу в поч-
ве, обогащение культурных биоценозов полезными видами
насекомых и других животных — все это пути далеко нами не
использованы. Они требуют к себе больше внимания, знания
жизни и значения отдельных видов и группировок насекомых
в конкретных биогеоценозах.
Во многих случаях причиной деградации лесных культур
и снижения урожайности сельскохозяйственных культур яв-
ляется не повреждение вредителем, а снижение природной ус-
тойчивости насаждений ввиду несоответствия их структуры
почвенно-климатическим условиям. Биоценозы наших полей и
лесных насаждений, как и естественные биоценозы, форми-
руются в зависимости от абиотических условий среды, свойств
биотопа, его почвенно-грунтовых и других особенностей, т. е.
как вторичные биогеоценозы, и об этом забывать нельзя. Мо-
нокультуры в разных местах могут оказаться заселенными по-
разному, и значение одного вида вредителя тоже будет раз-
личным и зависимым от состояния растения.
Агробиоценозы, или вторичные биоценозы, закономерности
их формирования, особенности структуры, их фауна изучены
еще очень слабо, а вне биоценозов существование любого ор-
ганизма в природе невозможно. Поэтому изучение закономер-
ностей формирования вторичных биоценозов составляет перво-
степенную теоретическую и практическую задачу современной
экологии. Решать эту задачу можно лишь комплексным мето-
дом исследования, базируясь на хорошем знании всех сторон
жизни отдельных видов в биоценозе, их биотической и абиоти-
ческой среды, меняющейся во времени и пространстве.
ОСНОВНАЯ ЛИТЕРАТУРА
АрнольдиК. В., А р н о ль ди Л. В. О биоценозе как одном из
'•основных понятий экологии, его структуре и объеме. «Зоол. журн.», 1963,
т. 42, вып. 2.
А р н о л ь д и Л. В. Учет насекомых в степных и пустынных сообщест-
вах. В кн.: «Полевая геоботаника», т. 3. М.—Л., «Наука», 1964.
Беклемишев В. Н. Экология малярийного комара. М., Медгиз,
1944.
Беклемишев В. Н. Учебник медицинской энтомологии. М., Мед-
гиз, 1949.
Беклемишев В. Н. О классификации биоценотических (симфизно-
логических) связей. «Бюлл. Моск, об-ва испыт. природы», отдел биол.,
1951, т. 56, № 5.
Бей-Биенко Г. Я. Общая энтомология. М., «Высшая школа», 1966.
Вернадский В. И. Биосфера. М., «Мысль», 1967.
Викторов Г. А. Проблема численности насекомых (на примере
вредной черепашки). М., «Наука», 1967.
Воронцов А. И. Биологические основы защиты леса. М., «Высшая
школа», 1963.
Воронцов А. И. Лесная энтомология. М., «Высшая школа», 1967.
ВороновА. Г. Биогеография. Изд-во МГУ, 1963.
Вопросы экологии. По материалам 4-й экологической конференции,
т. 4, Киев, 1962.
Герасимов А. М. Насекомые чешуекрылые, т. I, вып. 2. Гусеницы,
ч. I. Фауна СССР. М—Л., Изд-во АН СССР, 19512.
Гиляров М. С. Особенности почвы как среды обитания и ее зна-
чение в эволюции насекомых. М., Изд-во АН СССР, 1949.
Гиляров М. С. Вид, популяция и биоценоз. «Зоол. журн.», 1954,
т. 28, вып. 4.
Гиляров М. С. Современные представления о вторичных биоцено-
зах. 5-е совещание ВЭО. Ташкент, 1963.
Гиляров М. С. Зоологический метод диагностики почв. М., «Нау-
ка», 1965.
Г о р ы ш и н Н. И. Техническое оснащение экологических исследова-
ний в энтомологии. Изд-во ЛГУ, 1966.
Данилевский А. С. Фотопериодизм и сезонное развитие насеко-
мых. Изд-во ЛГУ, 1961.
Добровольский Б. В. Фенология насекомых вредителей сель-
ского хозяйства. М., «Высшая школа», 1961.
Ильинский А. И. и Тропин И. В. Надзор, учет и прогноз
массовых размножений насекомых в лесах СССР. М., «Лесная промыш-
ленность», 1965.
Кашкаров Д. Н. Основы экологии животных. Л., Учпедгиз, ,1945.
151
Кожанчиков И. В. Методы исследования экологии насекомых. Л.,
«Высшая школа», 1961.
М а з о х и н-П о р ш н я к о в Г. А. Основные приспособительные типы
чешуекрылых (Lepidoptera). «Зоол. жури.», 1954, т. 33, вып. 4.
Макфедьен Э. Экология животных. Цели и методы. М., «Мир»,
1965.
Наумов Н. П. Экология животных. 2-е изд М., «Высшая школа»,
1963.
Павловский Е. Н. Руководство по паразитологии человека, ч. 2,
М., Медгиз, 1948.
Павловский Е. Н. О синтезе учения о природной очаговости бо-
лезней и теории паразитоценозов. «Журн. микробиол. эпидемиол. и имму-
нол.», 1957, т. XI.
Правдин Ф. Н. Растительноядные насекомые как паразиты. «Ус-
пехи современной биологии», 1950, т. 29, вып. 1.
Роден дорф Б. Б. Многообразие органического мира и его истори-
ческое развитие. «Журн. общ. биологии», 1965, т. 26, № 4.
Рожков А. С. Массовое размножение сибирского шелкопряда и-
меры борьбы с ним. М., «Наука», 1965.
Руднев Д. Ф. Влияние физиологического состояния растений на'
массовое размножение вредителей леса. «Зоол. журн.», 1962, т. 41, вып. 3.
Старк В. Н. Короеды. Фауна СССР. Насекомые жесткокрылые, т. 31.
М., Изд-во АН СССР, 1952.
Сукачев В. Н. и др. Основы лесной бпогеоценологии. М., «Наука»,
1964.
Тишлер В. Общие проблемы экологии (преимущественно по агро-
экологическим исследованиям). «Журн. общ. биол.», 1960, т. 21, № 5.
Ушатинская Р. С. Основы холодостойкости насекомых. М., Изд-во-
АН СССР, 1957.
Фасулати К. К. Полевое изучение наземных беспозвоночных. М.„
«Высшая школа», 1961.
Чернова О. А. Поденки (Ephemeroptera) бассейна Амура и приле-
гающих вод и их роль в питании амурских рыб. «Тр. Амурск, ихтиол, эк-
спедиции 1945—1949 гг.», 1952, т. 3.
Шенников А. П. Введение в геоботанику. Изд-во ЛГУ, 1964.
Штейнхауз Э. Микробиология насекомых. М., ИЛ, 1950.
Штейнхауз Э. Патология насекомых. М., ИЛ, 1952.
Элтон Ч. Экология нашествий животных и растений. М., ИЛ, I960:
Яхонтов В. В. Экология насекомых. М., «Высшая школа», 1964.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Амирханова С. Н. Химизм растений и выживаемость непарного'
шелкопряда. «Научн. конференц, по вопросам массовых размножений вре-
дителей леса» (рефераты докладов). Уфа, 1962.
Андрианова Н. С. Влияние различного качества корма на рост
гусениц китайского дубового шелкопряда. В сб.: «Культура дубового шел-
копряда в СССР». М., ОГИЗ, 1948.
Андрианова Н. С. Влияние света на рост и развитие дубового
шелкопряда. В сб.: «Культура дубового шелкопряда в СССР». М., Сель-
хозгиз, 1948.
Ар ноль ди К. В. Жизненные формы муравьев. ДАН СССР, 1937,
т. 16, № 6.
А р н о л ь д и К. В. К вопросу об экологической дивергенции видовых
популяций у жуков Tentyria nomas Pall. (Coleoptera, Tenebrionidae). «Тр.
Зоол. ин-та АН СССР», 1941, т. VI, вып. 4.
А р н о л ь д и К. В., Г и л я р о в М. С., О б р а з ц о в Б. В. Животный
мир в условиях степного лесоразведения. «Научн. вопр. полезащитного-
лесоразведения», т. I. М., Изд-во АН СССР, 1951.
152
АрнольдиК. В. О некоторых закономерностях сложения энтомо-
комплексов биоценозов при степном лесоразведении. «Зоол. журн.», 1951.
т. 30.
А р н о л ь д и К. В. К выяснению зональных закономерностей образо-
вания новых группировок насекомых и заселение лесопосадок ксерофиль-
ными видами при степном лесоразведении. «Зоол. журн.», 1952, т. 31,
вып. 3.
Беклемишев В. Н. Возбудители болезней как члены биоценозов.
«Зоол. жури.», 1956, т. 35, вып. 12.
Беклемишев В. Н. Термины и понятия, необходимые при коли-
чественном изучении популяций эктопаразитов и индиколов. «Зоол. журн.»,
1961, т. 40, № 2.
Бей-Биенко Г. Я. Смена местообитаний наземными организмами
как экологический принцип. «Журн. общ. биол.», 1966, т. 27, № 1.
Бей-Биенко Г. Я. и Мищенко Л. Л. Саранчевые фауны СССР
и сопредельных стран, т. I. М.—Л., Изд-во АН СССР, 1951.
Болдаруев В. О. Плодовитость и пищевая специализация сибир-
ского шелкопряда. «Зоол. журн.», 1955, т. 34, вып. 4.
Вейзер Я. Патология насекомых, ее настоящее и будущее. «Энто-
мол. обозр.», 1959, т. 38, № 2.
Викторов Г. А. Роль паразитических насекомых в размножении
бобовой огневки (Etiella zinckenella Тг.). «Зоол. журн.», 1956, т. 35, № 1.
Викторов Г. А. Колебания численности насекомых как регулируе-
мый процесс. «Журн. общ. биол.», 1965, т. 36, № 1.
Г е й с п и ц К. Ф. Реакция моновольтинных чешуекрылых на продол-
жительность дня. «Энтомол. обозр.», 1953, т. 33.
Гейспиц К. Ф. Фотопериодические и температурные реакции, опре-
деляющие сезонное развитие хвойных шелкопрядов. «Энтомол. обозр.»,.
1965, т. 44, № 3.
Гиляров М. С. Параллелизм в формировании энтомоцеиозов злако-
вых полей в Восточной Европе и Северной Америке. ДАН СССР, 1943,.
т. 38, № 1.
Гиляров М. С. Основные особенности вредных насекомых, приспо-
сабливающихся к полевым севооборотам. ДАН СССР, 1945, т. 47, № 3.
Гиляров М. С. Закономерности формирования комплексов насеко-
мых при освоении целинных земель. «Журн. общ. биол.», 1955, т. 16, № 6.
Гиляров М. С. Основные направления приспособлений насекомых,
к жизни в пустыне. 5-е совещание ВЭО. Ташкент, 1963.
Гиляров М. С. Исследования почвенной фауны и некоторые на-
блюдения над насекомыми при геоботанических исследованиях. В кн.:
«Полевая геоботаника», ч. 3. М.—Л., Изд-во АН СССР, 1964.
Гримальск-ий В. И. Причины устойчивости сосновых насаждений
к хвоегрызущим вредителям. «Зоол. журн.», 1961, т. 40, вып. И.
Гр ямальский В. И. Устойчивость сосновых насаждений против,
хвоегрызущих вредителей. М., «Лесная промышленность», 1964.
Г о р ы ш и н Н. И. Соотношение светового и температурного факто-
ров в фотопериодической реакции насекомых. «Энтомол. обозрен.», 1955,
т. 34.
Длусский Г. М. Муравьи рода формика. М., «Наука», 1967.
Зражевский А. И. Дождевые черви как фактор плодородия почв.
Киев, 1957.
Киршенблат Я. Д. Телергоны как средства воздействия животных
на биотические факторы окружающей среды. «Вопр. экол.», т. 4. Изд-во
Киевского ун-та, 1962.
Кузнецов Н. Я. Связь географического распространения белянок
с распространением их кормовых растений и с химизмом последних. «Еже-
годн. зоол. музея АН СССР», 1930.
Кузнецов Н. Я. Класс насекомых (Insecta, или Hexapoda). Руко-
водство по зоологии, т. 3, ч. 2. М., «Советская наука», 19511
153.
Лавренко Е. М. Полевая геоботаника т I М — Л., Изд-во АН
СССР, 1959. ’
Латышев Н. К. Динамика численности шелкопряда-монашенки в
Башкирии и ее прогнозирование. В кн.: «Исследование очагов вредителей
леса в Башкирии», т. 2. Уфа, 1962.
Л о з и н а-Л о з и н с к и й Л. К. Организм и условия жизни. М., Учпед-
гиз, 1956.
Лозина-Лозинский Л. К. Устойчивость насекомых к глубокому
охлаждению. В кн.: «Клетка и температура». М.—Л., «Наука», 1964.
Л э к Д. Численность животных и ее регуляция в природе. М., ИЛ,
1957.
М а з о х и н-П о р ш н я к о в Г. А. Зрение у насекомых. М., «Наука»,
1965.
Мамаев Б. М. Зоологическая оценка стадий естественного разру-
шения древесины. «Изв. АН СССР», сер. биол., 1962, № 4.
Медведев С. И. Особенности расространения некоторых экологи-
ческих форм насекомых в различных ландшафтно-географических зонах
Украины. «Зоол. журн.», 1954, т. 33, вып. 6.
Медведев С. И. Основные черты изменения энтомофауны Украины
в связи с формированием культурного ландшафта. «Зоол. журн.», 1959,
т. 33, вып. 1.
Медведев С. И. О сезонных аспектах аэтомофауны типчаково-
ковыльной степи юга Украины. «Вопросы генетики и зоологии». Харьков,
1964.
Ми л яновский Е. С. Миграции насекомых. «Зоол. журн.», 1956,
т. 35, вып. 8.
М о н ч а д с к и й А. С. Экологические факторы и принципы их клас-
сификации. «Журн. общ. биол.», 1962, т. 23, № 5.
Наумов Н. П. Популяционная экология и ее основные проблемы.
«Вопр. экол.», т. 4. Изд-во Киевск. ун-та, 1962.
Наумов Н. П. Методологические проблемы и задачи биологии.
Изд-во МГУ, 1965.
Наумов Н. П., Никольский Г. В. О некоторых общих законо-
мерностях динамики популяций животных. «Вопр. экол.», т. 4. Изд-во
Киевск. ун-та, 1962.
Никольский Г. В. О содержании, теоретических основах и ос-
новных задачах экологии животных. «Зоол. журн.», 1955, т. 31, вып. 1.
Н а р ч у к Э. П. Ритмика размножения и кормовые растения Oscinella
pusilia Mg. «Зоол. журн.», 1955, т. 34, вып. 5.
Павловский Е. Н. Задачи зоологической науки в свете решений
XIX съезда КПСС. «Зоол. журн.», 1952, т. 31, вып. 6.
Петренко Е. С. Насекомые — вредители лесов Якутии. М., «Нау-
ка», 1965.
Петров А. И. Яблоневая и плодовые моли в Средней Азии. Алма-
Ата, 1955.
П о л ы н о в Б. Б. Учение о ландшафтах. «Вопр. географии», вып. 33,
1953.
Поливанова Е. Н. Причины, определяющие численность хлебных
клопов (Pentatomidae) в южных зерновых районах европейской части
СССР. ДАН СССР, 1957, т. 112, вып. 3.
Положенцев П. А. и Хан исламов М. Г. О возможности
применения «живичного индикатора» в сосновых молодняках для опреде-
ления их жизнеспособности. «Тр. Башкирского с.-х. ин-та», т. 6'. Уфа, 1954.
Положенцев П. А., Хан исламов М. Г. История и перспекти-
вы зависимости градаций насекомых от физиологического состояния де-
ревьев. «Научи, конференц, по вопросам массовых размножений вредите-
лей леса» (рефераты докладов). Уфа, 1962.
Положенцев П. А., АртюховскийА. К. Об использовании не-
матод Rhabditis и Neoaplectana в борьбе с вредными насекомыми. «Мат-лы
154
конференции об-ва гельминтологов 9—12 декабря 1963», ч. 2. М., Изд-во
АН СССР, 1963.
Положенцев П. A., Her робов В. П. О насекомых-хозяевах па-
разитических червей человека и животных. В кн.: «Вредные и полезные
насекомые». Воронеж, 1967.
Правдин Ф. Н. Растительноядные насекомые как паразиты. «Ус-
пехи современной биологии», 1950, т. 29, вып. 1.
Приты кин а Л. Н. Материалы к морфо-экологической классифика-
ции личинок стрекоз. «Энтомол. обозр.», 1965, т. 44, № 3.
Раменский Л. Г. О некоторых принципиальных положениях со-
временной геоботаники. «Бот. журн.», 1952, т. 37, № 2.
Р а ф е с П. М. Массовые размножения вредных насекомых как осо-
бые случаи круговорота веществ и энергии в лесном биоценозе. Сб. «За-
щита леса от вредных насекомых». М., «Наука», 1964.
Ремезов Н. П. и др. Методические указания к изучению биологи-
ческого круговорота зольных веществ и азота наземных растительных сооб-
ществ в основных природных зонах умеренного пояса. «Бот. журн.», 1963,
т. 18, № 6.
Родендорф Б. Б. Палеоэнтомологические исследования в СССР.
«Тр. палеонтол. ин-та», 1957, т. 66.
Рубцов И. А. Биологический метод борьбы с вредными насекомыми.
М.—Л., Сельхозгиз, 1948.
Рустамов А. К. К вопросу о понятии «жизненная форма» в эколо-
гии животных. «Зоол. журн.», 1955, т. 34, вып. 4.
Рывкин Б. В. Энтомофаги и защита леса. Минск, 1963.
Сочава В. Б. Вопросы классификации растительности, типологии
физико-географических стадий и биогеоценозов. «Тр. ин-та биол. Уральск,
фил. АН СССР», вып. 27. Свердловск, 1961.
Смирнов Е. С. и ЧувахинаЗ. Ф. Наследование приобретаемых
свойств и проблема возникновения вредителей. «Вест. МГУ», 1953, № 5.
Созы кин Н. Ф., Кожевникова С. А. Поглощение коротковол-
новой радиации лесным пологом. «Мат-лы по лесной гидрологии и био-
геофизике». М., «Лесная промышленность», 1965.
Старк В. Н. Вредные лесные насекомые. М., Сельхозгиз, 1931.
Старк В. Н. Устойчивость лесных насаждений к повреждениям их
вредителями. В сб.: «Защита лесов от вредителей и болезней». М., 1961.
Стебаев И. В. Население прямокрылых насекомых ландшафта глав-
ного водораздела северных Ергеней. «Зоол. журн.», 1957, т. 34, вып. 8.
Стебаев И. В. Животное население первичных наскальных почв и
его роль в почвообразовании. «Зоол. журн.», 1958, № 10.
Стебаев И. В., Наплекова Н. И, Гукасян А. Б. Саранче-
вые (Acrididae) и чернотелки (Tenebrionidae) как стимуляторы микробио-
логических процессов в почвах сухих степей Тувинской АР. «Почвоведе-
ние», 1964, № 9.
Стрельников И. Д. Свет как фактор в экологии животных (Дей-
ствие солнечной радиации на температуру тела некоторых пойкилотерм-
ных животных). «Изв. науч.-исслед. ин-та им. Лесгафта», 1934, тт. 27—28.
Стрельников И. Д. Значение теплопродукции при движении и
под действием солнечной радиации в экологии высокогорных насекомых.
«Зоол. журн.», 1940, т. 19, вып. 3.
Суитмен X. Биологический метод борьбы с вредными насекомыми
и сорными растениями. М., «Колос», 1964.
Талалаев Е. В. Микробиологический метод борьбы с вредными на-
секомыми. М., «Наука», 1963.
Теленга Н. А. Биологический метод борьбы с вредными насекомы-
ми сельскохозяйственных и лесных культур. Киев, 1955.
Трав леев А. П. Роль личинок садовой мошки в разложении лесной
подстилки в степи. «Научн. докл. высш, школы», биол. науки, 1961, № 1.
Третья экологическая конференция по проблеме массовых размноже-
ний животных и их прогнозы, чч. 1, 2, 3. М., 1954. 1
155
Уваров Б. П. Саранча и кобылки. Библиотека хлопкового дела.
М.» 1927.
Ф р и д е р и к с К. Экологические основы прикладной зоологии и энто-
мологии. М., 193'2.
Фридолин В. Ю. Биоценотика на химическом основании. Сб. «Ака-
демику В. И. Вернадскому». М., Изд-во АН СССР, 1936.
ХанисламовМ. Г. О ведущих условиях начала вспышки хвое- и
листогрызущпх вредителей. «Вопр. лесозащиты», 1963, 2.
Ханисламов М. Г., Векшина Р. С. Зависимость затухания оча-
гов непарного шелкопряда от физиологического состояния деревьев.
«Научная конференция по вопросам массовых размножений вредителей-
леса». Уфа, 1962.
X а н и с л а м о в М. Г., Л а т ы ш е в Н. 1\. О значении питания мона-
шенки мужскими соцветиями сосны. В кн.: «Научная конференция по во-
просам массовых размножений вредителей леса». Уфа, 1962.
Шаров А. Г. Фазовый характер изменчивости гусениц ильмовогс-
ногохвоста. ДАН СССР, 1953, т. 88, № 5.
Шарова И. X. Морфоэкологические типы личинок жужелиц (Сага-
bidae). «Зоол. журн.», 1960, т. 39, вып. 5.
Шапошников Г. X. Возникновение и утрата репродуктивной изо-
ляции и критерий вида. «Энтомол. обозр.», 1966, т. 49, вып. 1.
Шапошников Г. X. К вопросу о переходе тлей с одних растений
на другие. «Труды Зоол. ин-та АН СССР», 1955, т. 21.
Шенников А. П. Экология растений. М., «Сов. Наука», 1950.
Ш о в е н Р. Физиология насекомых. М., ИЛ, 1953.
Шовсн Р. Жизнь и нравы насекомых. М., Сельхозгиз, 1960.
Щерби но в с кий Н. С. Пустынная саранча шистоцерка. М., Изд-во
с.-х. литерат., 1952.
Щербиновский Н. С. Солнечно-обусловленная цикличность мас-
совых размножений вредных насекомых и других организмов. «Астроном,
сбор.», Львовский ун-т, вып. 3—4, 1960.
Щ е п е т и л ь н и к о в а В. А., Чумакова Б. М. Современные проб-
лемы акклиматизации энтомофагов в СССР. «Тр. Всес. ин-та защиты рас-
тений», 1964, вып. 21.
Andrewartha Н. G. and Birch L. С. The distribution and abun-
dance of animals. The University of Chicago Press., 1954.
Andrewartha H. G. Introduction to the Study of animal populati-
ons. London, 1961.
Balogh T. A. Zooconologie alapjai (Grundziige der Zooconologie),
Budapest, 1953.
Balogh T. A. Diskussionsbeitrag Reaiitat, Abgrenzung und Ordnung-
sprinzipien der Biozonosen. «Z. ang. Entomol.», Bd. 47, № 1, 1960.
Bajer-Petersen Broder. Om pesticiden i Danmarks skove. «Na-
tur. verden», 1964.
Dudich E., Balogh T., Loksa T. Produktionbiologische Unter-
suchungen fiber die Arthropoden der Waldboden. «Acta Biol.», Bd. Ill, f. 3.
Budapest, 1952.
Elton Ch. «Animal ecology». London, 1960.
Kaldewey Harold. Wachstumsregulatoren aus JPflanzengallen und
Larven der Gallenbewohner. «Вег. Dtsch. bot Ges.», 1965, 78, Nr. 2.
Kennedy T. S. Continuos polymorphism in locusts. «Sympos. Roy.
Entomol. Soc.». London, 1961, No. 1.
Nicholson A. I. Dynamics of insect populations. «Annual Rev. Ento-
mol.», vol. 3, Palo Alto, Calif., 1958.
Nolte D. T. The pigmentation of locusts. «S. Afr. Sci.», 61, No. 4,
1965.
Odum E. P. Fundamentals of Ecology. London, 1957.
Richards O. W. The theoretical and practical study of natural in-
sect populations. «Annual Rev. Entomol.», vol. 6, Palo—Alto, Calif., 196L
156
Steinchaus E. When an insect dies. «Bull. Entomol. Soc. America»,
1964, No. 3.
Schwenke W. Grundzuge der Populationsdynamic und Bekampfung
•der gemeinen Kiefernbuschhorn-Blattwespe. Diprion pini L. «Z. angew. Ento-
niol.», 1964, B. 54, Nr. 1—2.
Schwerdtfeger F. Okologie der Tiere. В. 1. Autokologie. Hamburg
und Berlin, 1963.
Scliwerdfeger F. Uber das Minimum-Gezetz in der angewandten
Entomologie. «Z. angew. Entomol.», 1964, 54, Nr. 1—2.
T i s c h 1 e г W. Synokologie der Landtiere. Stuttgart, 1955.
Uvarov В. P. Insects and Climate. «Trans. Entom. Soc.», London,
1931, B. 79.
Weber H. Lehrbuch der Entomologie. Jena, 1933.
Wellington W. G. The Synoptic approach to studies of insects and
•climate. «Annual Rev. Entomol.», Palo Alto, Calif.», 1957. vol. 2.
ОГЛАВЛЕНИЕ
введение......................................... . . * * 3»
"лава I. Предмет и задачи экологии.............................. 5
Место экологии среди других наук............................ 8
Особенности экологии насекомых...............................10
'лава II. Среда обитания насекомых я . . « 13-
Связь со средой через обмен веществ..........................13
Классификация факторов среды и условий жизни . 13
Физико-химические условия жизни..............................16
Климат и микроклимат ....................................... 41
Сезонная ритмичность жизни насекомых в 45
Пища как экологический фактор................................47
Особенности действия физико-химических факторов в разных
средах....................................................57
Особенности жизни в почве....................................60
Общие закономерности действия факторов среды.................63
Закономерности территориального размещения насекомых . . 64
'лава III. Жизненные формы насекомых...........................70«
Жизненные формы саранчовых...................................72
Морфо-экологические типы водных насекомых....................73
Биологические типы чешуекрылых..............................75'
'лава IV. Биотические факторы среды ........ 77
Внутривидовые отношения......................................77
Фазовая изменчивость насекомых...............................80
Уровень численности популяций................................86
Связи насекомых в биоценозе..................................88
Насекомые и высшие растения..................................91
Насекомые и низшие растения..................................93
Микроорганизмы — возбудители болезней насекомых .... 93
Насекомые как переносчики болезней...........................99
Способы общения и защита от врагов..........................104
лава V. Массовые размножения насекомых.........................106
Особенности массовых размножений саранчи.....................НО
Размножение вредителей леса.................................113
лава VI. Строение биоценозов...................................122
Типологическая классификация биоценозов.....................129
'лава VII. Динамика биоценозов.................................134
Биологический круговорот веществ............................134
Смена биоценозов............................................137
лава VIII. Экология культурных ландшафтов..................... 143
Основная литература ....................................... 151
Дополнительная литература............................. . 152’
Андрианова Нина Семеновна
ЭКОЛОГИЯ НАСЕКОМЫХ
Курс лекций
Тематический план 1969 г. № 179
Редактор О. Т. Помаленъкая
Технический редактор Н. А. Рябикина
Корректоры: В. П. Кададинская,
С. Ф. Будаева
Сдано в набор 5/VI 1969 г.
Подписано к печати 14/1 1970 г.
Л-34994 Формат 60X90716
Бумага офсет. № 2 Физ. печ. л. 10,0
Уч.-изд. л. 10,26 Изд. № 711
Заказ 193 Тираж 6000 экз.
Цена 28 коп.
Изд-во Московского университета
Москва, Ленинские горы
Административный корпус
Тип. Изд-ва МГУ (филиал).
Москва, проспект .Маркса, 20