













































































































































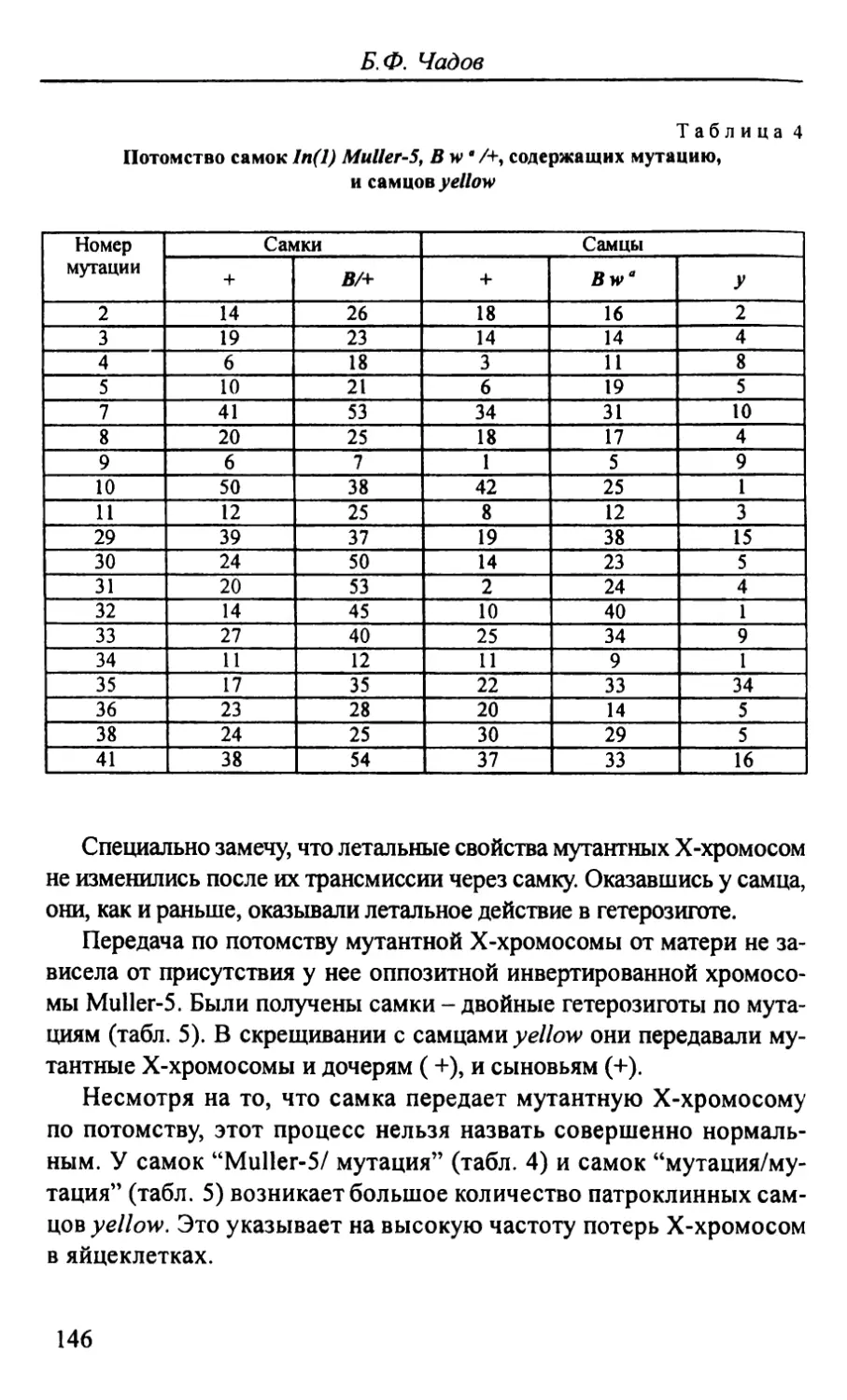








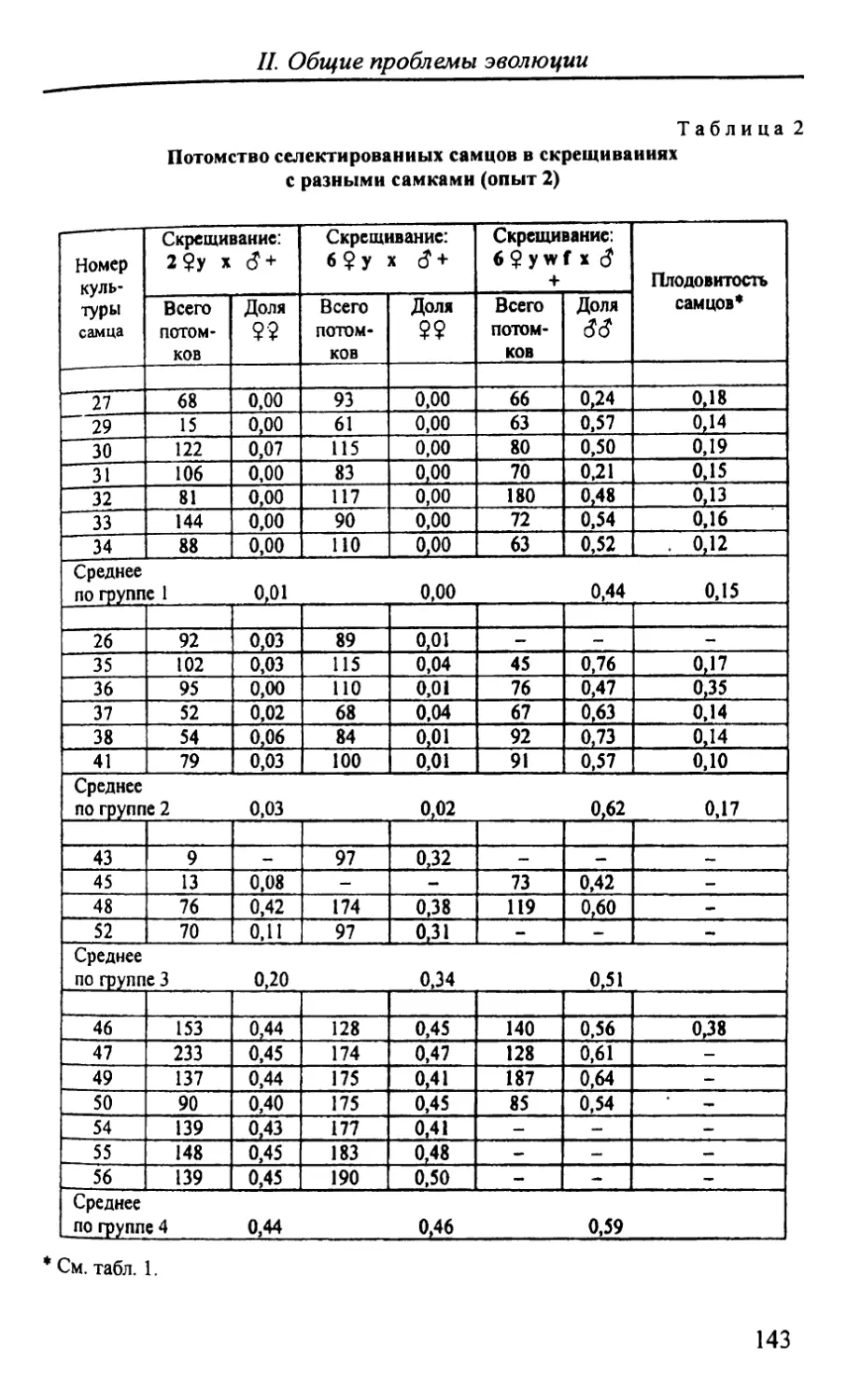


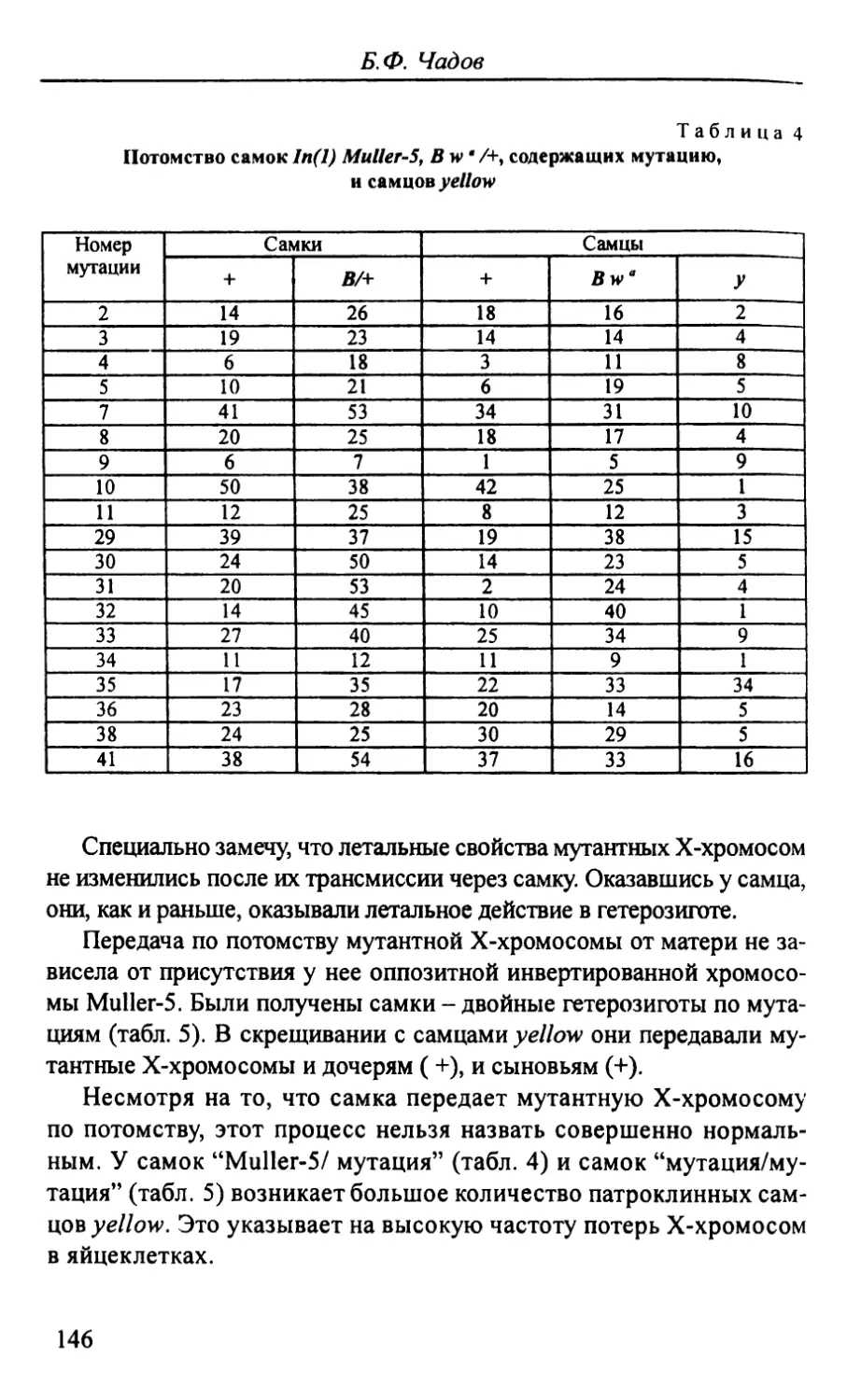















































































































































































































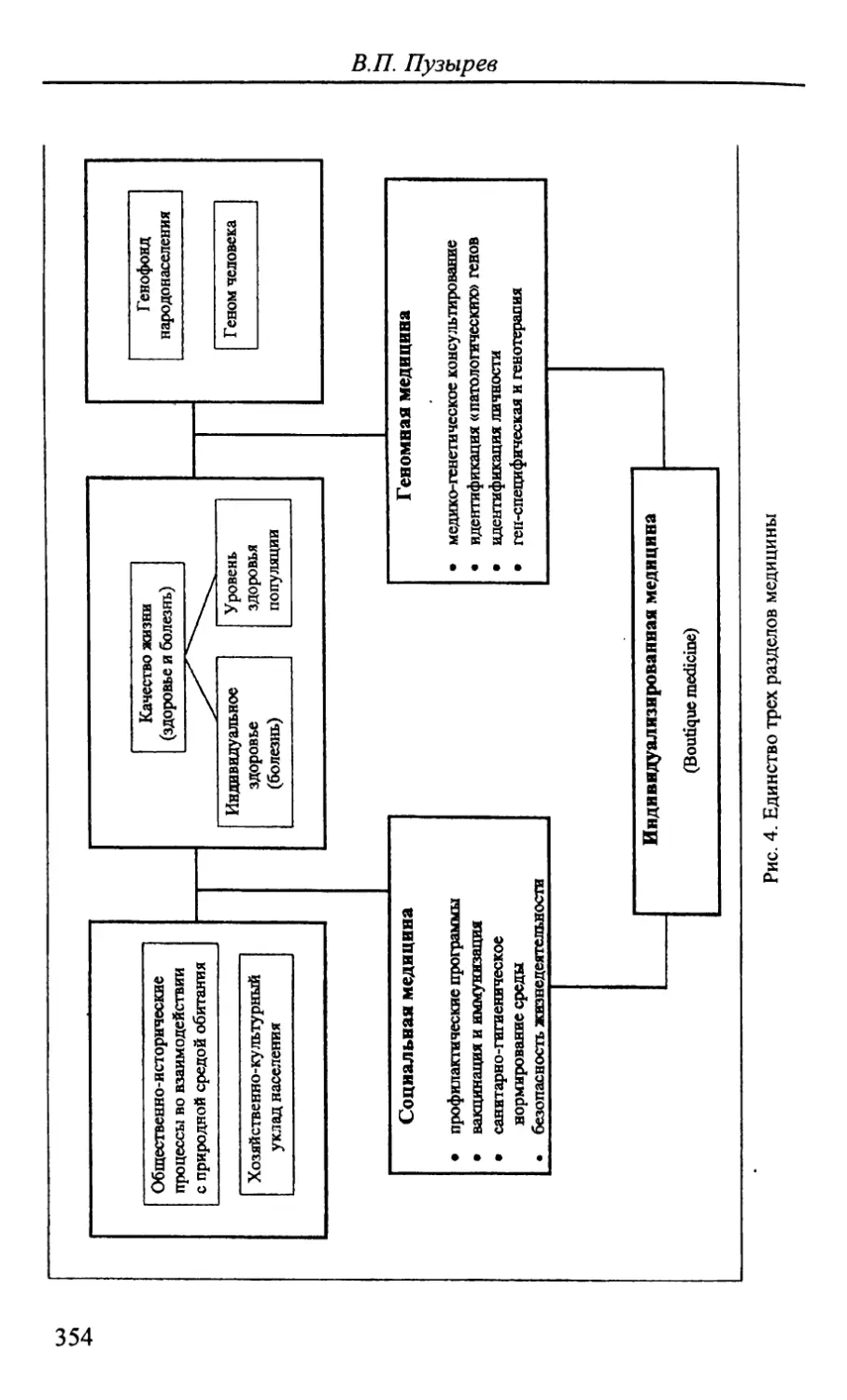












































Автор: Стегний В.Н.
Теги: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез развитие органического мира (филогенез) биология эволюционный процесс эволюция
ISBN: 5-94621-001-7
Год: 2001




Корочкин
Леонид
Иванович,
чл.-корр. РАН,
Институт биологии
развития РА!!,
г. Москва
Малолетко
Алексей
Михайлович,
д.г.н., профессор,
Томский
государствен н ы й
университет
Назаренко
Сергей
Андреевич,
д.б.н., профессор,
НИИ медицинской
генетики ТНЦ СО РАМН,
г. Томск
Островерхова
Галина
Петровна,
д.б.н., профессор,
Томский
государственный
университет
2
Положий
Антонина
Васильевна,
д.бл., профессор,
Томский
государственный
университет
Северцов
Алексей
Сергеевич,
д.бл., профессор,
Московский
государственный
университет
Сергеев
Михаил
Георгиевич,
д.бл., профессор,
Новосибирский
государстве ины й
упнвсрси тег
Стегний
Владимир
Николаевич,
д.б.п., профессор,
Томский
государственный
унивсрситег
Томский государственный университет
Научно-исследовательский институт биологии и биофизики
Научно-исследовательский институт медицинской генетики
ТНЦ СО РАН
ЭВОЛЮЦИОННАЯ
БИОЛОГИЯ
Том 1
Материалы конференции
«Проблема вида и видообразование»
г. Томск, 3-6 октября 2000 г.
Томск - 2001
УДК 575.8
ББК 28.02
Э 15
Э 15 Эволюционная биология: Материалы конференции
«Проблема вида и видообразование» / Под ред.
В.Н. Стегния. Томск: Томский государственный
университет, - Т. 1. - 2001. - 396 с.
ISBN 5-94621-001-7
В настоящем издании представлены работы ряда ведущих специалистов по эволю-
ционной биологии. Рассматриваются механизмы сальтационного видообразования, про-
блемы исторических и современных представлений о виде, проблемы соотношения мик-
роэволюционных и макроэволюционных преобразований у растений, животных и чело-
века. Представлены данные по молекулярной биологии, цитогенетике, иммунологии,
эволюционной морфологии, экологии, палеонтологии, матемагической биологии.
Кроме того, приводятся избранные главы из работы выдающегося эволюциониста
С.И. Коржинского «Гетерогенезис и эволюция. К теории происхождения видов» (1899)
Книга представляет интерес для биологов и очень полезна для молодых ученых,
студентов, аспирантов, так как в ней представлены самые современные данные по про-
блемам эволюции.
УДК 575.8
ББК 28.02
Редакционная коллегия:
В.Н. Стегний, А.В. Положий, И.И. Гуреева,
А.К. Сибатаев, Т.В. Ананьина, О.П. Брагинец
ISBN 5-94621-001-7
© Томский государственный университет, 2001
Посвящается памяти выдающегося
биолога-эволюциониста,
профессора Томского университета
Сергея Ивановича Коржинского
ПРЕДИСЛОВИЕ
В настоящем издании публикуются материалы докладов-лекций
ряда ведущих ученых-эволюционистов, представленных на конферен-
ции «Проблемы вида и видообразования» (3-5 октября 2000 г., Томс-
кий госуниверситет). По спектру эволюционной проблематики дан-
ная книга, как и сама конференция, очень разнообразны: история эво-
люционной биологии, эволюционная морфология и экология,
систематика, палеонтология, цитогенетика, молекулярная биология и
другие направления. Обширен и охват исследованных жизненных
форм: микроорганизмы, растения, различные группы животных, че-
ловек. Следует отметить, что в основном в статьях освещаются про-
блемы эволюционного развития жизни на Земле в целом, а также от-
дельных таксономических групп и намечаются направления научного
поиска. В середине XX века сложилось мнение, что эволюционное
учение Дарвина - единственная парадигма и ее можно только подтвер-
ждать новыми данными. Была создана так называемая «синтетичес-
кая теория эволюции» - апофеоз неодарвинизма. В то же время крити-
ческое осмысление дарвинизма, начавшееся с момента появления уче-
ния, было очень продуктивным для развития других эволюционных
взглядов, и здесь следует отметить особую роль отечественных уче-
ных (С.И. Коржинский, Л.С. Берг, Д.Н. Соболев, Ю.А. Филипченко,
А.А. Любищев).
Это издание, как и сама конференция, посвящены памяти профес-
сора Томского университета Сергея Ивановича Коржинского, который
в 1899 г. опубликовал свою теорию скачкообразного видообразования
на основе гетерогенных вариаций. Только спустя 2 года, в 1901 г., выш-
ла знаменитая «мутационная теория» Г. де Фриза, в которой излага-
лись аналогичные взгляды на видообразование. Ю.А. Филипченко в
своей книге «Эволюционная идея в биологии» отметил: «Справедливо
3
Предисловие
считать его (Коржинского) наравне с Г. де фризом, создателем мута-
ционной теории, которая играет теперь столь важную роль во всех
наших представлениях об изменчивости и эволюции организмов»
(с. 163). В настоящем издании воспроизводятся избранные главы из
работы С.И. Коржинского «Гетерогенезис и эволюция. К теории про-
исхождения видов», которая и сегодня, спустя 100 лет, представляет
не только исторический, но и научный интерес. При этом приводятся
только первая и последняя главы работы, так как в промежуточных
главах даны подробные примеры подтверждения концепции гетероге-
неза, а в последней - в сжатой форме изложена ее суть.
Предполагается в ближайшее время подготовить и выпустить 2-й том
издания на основе П конференции «Проблемы вида и видообразова-
ния», которая состоится в Томске на базе Томского госуниверситета в
ноябре 2001 г.
В.Н. Стегний
I. ИСТОРИЯ эволюционной
БИОЛОГИИ
ЭВОЛЮЦИОННЫЙ И РАЗВИТИЙНЫЙ подход
В ТРУДАХ РУССКИХ БИОЛОГОВ
В.В. Бабков
Институт истории естествознания и техники им. С И. Вавилова РАН
103012, Москва, Старопанский пер., 1/5
E-mail: babkoff@ihst.ru
Борьба и гармония
Вопросы видообразования и изменчивости видов перешли с перифе-
рии в центр дискуссий после совместного сообщения Ч. Дарвина и
А. Р. Уоллеса о характере и механизмах видообразования на заседании
Линнеевского общества летом 1858 г. и первого издания ‘'Происхожде-
ния видов” осенью 1859 г. В России теоретические вопросы зоологии
обсуждались к тому времени в рамках направления К. Бэра и школы
К. Рулье. Изучение флоры России к середине века приняло особую ок-
раску: на первый план выступили вопросы о растительных формациях, о
взаимной связи между растительностью, климатом и почвой.
В теоретической конструкции Дарвина конкуренция является исход-
ным моментом, а эволюция выводится из борьбы за жизнь. Напротив,
для русской традиции характерно представление о целостности приро-
ды. Говоря языком кибернетики, природа трактовалась как сложная са-
моорганизующаяся система, поэтому подчеркивалась идея её гармонии,
баланса. Развивая это умонастроение, Н.А. Северцов заложил основы
экологии животных. Здесь же основа традиции рассматривать взаимо-
помощь как фактор эволюции. Во взаимодействии учения Дарвина и
русской биологии важной темой стала дилемма гармонии и борьбы.
В своей речи при учреждении Русского энтомологического обще-
ства в мае 1860 г. его первый президент К. Бэр утверждал, что наука,
5
В. В. Бабков
основанная на идеалистической онтологии, дает более адекватную
картину мира, чем материалистическая наука. Бэр понимал природу
как сложную целостную систему, а ее развитие - как непрерывное
творение: все существа “низмыслены на Землю’’. Этот взгляд не ис-
ключает трансформизма: "вечно продолжающаяся смена жизни есть
не что иное, как вечное развитие, эволюция”. Нет застывших форм:
действительно и вечно одно постоянное изменение - в виде развития.
(О том же в 1876 г. С.А. Усов писал: “Организм есть особь status quo,
то есть особь в разрезе настоящего времени”; в середине XX в. эту
мысль В.Н. Беклемишев выразил формулой “организм есть морфоп-
роцесс”). По Бэру, природа развивается в своем масштабе времени,
ином, чем у ее составных частей. В ходе развития возникают момен-
ты конкуренции; они подчиняются законам системы, каковые суть
законы равновесия природы.
В очерке “Гармония в природе” А.Н. Бекетов [1], основатель гео-
ботанической школы, рассматривал закон корреляции на двух уров-
нях: органов в рамках организма и организмов в окружающей среде.
Изменение строения организмов является следствием мировой гар-
монии: “Причина строения, наружного вида и всей сущности каждого
существа заключается в окружающих его условиях, в зависимости его
от этих условий, короче сказать, в гармонии”. Впоследствии его пози-
цию называли предвосхищением идеи борьбы Дарвина. Однако у Бе-
кетова борьба возникает как побочный результат перехода от одного
равновесия к другому, и тем сохраняется гармония в природе.
В рецензии [2] к немецкому переводу “Происхождения видов” Петр
Лавров предложил отличать способность выжить от способности дать
потомство (так отвечая на замечание переводчика Г. Бронна об отсут-
ствии промежуточных форм в геологической летописи); эта тема ге-
нетики популяций случайно была названа в XIX в.
Будучи сторонником идеи взаимопомощи И.И. Мечников к числу
недостатков книги относит “неверное обобщение мальтусова закона и
сообщение особого значения принципам естественного избрания и
вымирания” и утверждает, что понятие преимущества и понятие со-
вершенства организации “должны быть строго разграничены” [3].
В своем предисловии к французскому переводу «Происхождения
видов» (1862) К. Руайе предложила новый аспект обсуждения дарви-
низма- применительно к развитию общества. Н.Н. Страхов резко кри-
6
/. История эволюционной биологии
тиковая попытку Руайе вывести “основания против учения о полити-
ческом равенстве людей” (по Руайе, такое равенство “невозможно,
вредно и противоестественно”). Расовые, физические, психические
различия не являются открытием, аргументировал Страхов: “Люди
равны между собой именно как люди, а не как животные. Это одина-
ково всем принадлежащий признак человеческого достоинства...” [4].
Попытка отвергнуть “сентиментальную философию” Страхова в пользу
“веры в науку” Руайе [5] не привлекла внимание русской публики: ей
были чужды идеи социал-дарвинизма.
Московский ботаник С.А. Рачинский, выпустивший первый рус-
ский перевод “Происхождения видов”, предварил его появление очер-
ком “Цветы и насекомые”, адресованным широкой публике: дарвинов-
ские приспособленность (успех в конкуренции и вытеснение сопер-
ника) и неприспособленность (вытеснение соперником) у Рачинского
означают соответственно взаимоприлаживание и выпадение из баланса
природы [6]. Перевод Рачинского вышел в начале 1864 г. и служил
32 года. Рецензенты сравнивали новое учение с гипотезой Коперника
(М. Антонович), с законом тяготения Ньютона (Дм. Писарев), чуть ли
не с Творцом (К.А. Тимирязев).
Система Дарвина как программа исследований
Зоогеограф и систематик Н.А. Северцов в июле 1867 г. утверждал:
“Лучшая из существующих гипотез есть, конечно, Дарвинова, но и она
во многих частностях неточна и подлежит проверке, и при всей гени-
альности ее логического развития она еще далеко не имеет достаточ-
ного фактического основания”. Достоинство книги Дарвина он усмат-
ривает именно в систематическом подходе к проблеме: “Это превос-
ходная программа исследований”. “Истинное научное значение” труда
Дарвина необходимо отметить, так как “у нас очень многие считают
его гипотезу окончательным решением вопроса о происхождении ви-
дов, чего нет” [7]. Однажды приняв “гипотезу Дарвина” в качестве
программы исследования, Северцов решил наполнить ее фактическим
содержанием. В монографии 1873 г. он говорил об изменении бара-
нов (увеличение размеров горных форм и т.п.) как реальном примере
естественного отбора [8] (хотя его можно трактовать в духе школы
7
В. В. Бабков
Рулье). В статье “Аркары” он описал случаи, когда "‘видовой или даже
родовой признак, постоянный у одних диких животных, у других ди-
ких же форм характеризовал только непостоянные изменения или даже
личные различия” [9]. Тем временем его ландшафтно-экологический
подход к фаунистическому районированию имел коренное отличие от
уоллесовского генетического подхода, который ассоциировался с дарви-
низмом. Северцовское определение фауны “сложность животных, живу-
щих в естественной области и имеющих строго определенные взаимо-
отношения” напоминает не о конкуренции, но о гармонии.
Задача Александра Ковалевского - построить солидный фактичес-
кий фундамент для эволюционного принципа - подразумевала демон-
страцию сходства эмбриологического развития у всех групп ныне су-
ществующих животных. Задача была весьма дерзкой: в ту пору рабо-
ты мирового авторитета К. Бэра доказывали, что эмбриологическое
развитие в животном царстве идет по четырем самостоятельным пла-
нам, соответствующим четырем типам другого мирового авторитета,
Ж. Кювье. Его результаты приветствовали и Дарвин, и Бэр, сказавший,
что работа по истории развития ланцетника “открыла нам такое стран-
ное и неожиданное сходство между первыми процессами развития
организмов, не близких между собой” [10]. Но и не будучи трансфор-
мистом можно искать такие связи в развитии: “Сравнительная исто-
рия развития имеет дело с фактами, из которых она делает непосред-
ственные выводы, не входя в рассуждение о путях происхождения раз-
личных видов” [11]. А.О. Ковалевский не был увлечен филогенети-
ческим направлением. Не занимаясь вопросом, какой способ образо-
вания энтодермы первичный, более древний (в этом сущность спора
между сторонниками идеи гастреи и идеи паренхимеллы), он считал
разные способы гаструляции гомологичными, легко переходящими
друг в друга, что соответствует выводам экспериментальной эмбрио-
логии XX в., выраженным афоризмом Д.П. Филатова: “орган консер-
вативнее, чем способ его становления”.
Из 40 лет жизни Владимира Ковалевского 4 года (1871-1874) были
заняты написанием немногих работ по палеонтологии, которые сдела-
ли бессмертным его имя. Исходным моментом было намерение объе-
динить учение Кювье с трансформизмом (в форме учения Дарвина).
Для нас особенно интересны выявленные им общие принципы про-
цесса эволюции.
8
I. История эволюционной биологии
В.О. Ковалевский отмечал иррадиации линий копытных от некое-
го первичного типа; здесь речь идет об адаптивной стратегии рода,
состоящей в заполнении всех ниш. Тема направленности в эволюции
возникла в связи с вопросом, что такое прогресс (Ковалевский разли-
чает общий прогресс и прогресс-специализацию). Принцип направ-
ленности эволюционных траекторий проявляется в параллелизмах в
ходе редукции конечностей и в развитии зубной системы у различных
групп копытных, и в том, что эволюция может идти только по несколь-
ким определенным направлениям, но отнюдь не в каждом мыслимом
направлении. Из этого последнего принципа, в соединении с принци-
пом координации, следует положение о необратимости эволюции.
В общей картине эволюции копытных, построенной В.О. Ковалев-
ским, важным моментом эволюционного изменения является резкое
ускорение на небольшом участке эволюционной траектории (это объяс-
няет отсутствие переходных форм, но не согласуется с принципом гра-
дуализма Дарвина). Ковалевский обсуждал среди прочего проблему
преадаптации и более общую проблему смены функций (впоследствии
тема А.Н. Северцова, а также А. Дорна). Некоторые из упомянутых
принципов Ковалевского предвосхитили punctuational model эволюции
- альтернативу градуалистической модели. Ковалевский связывал при-
знаки роскоши (рога простые и рога оленьи) с дошедшим до конца
прогрессом-специализацией: эти признаки появляются лишь тогда,
когда вполне разрешены все жизненные вопросы, связанные с редук-
цией скелета. Он не привлекал к объяснению признаков роскоши пред-
положений об их возможном адаптивном значении и не ссылался на
теорию полового отбора Дарвина. В XX в. подобные примеры стали
материалом для формирования концепций косвенного отбора
В.С. Кирпичникова и стабилизирующего отбора И.И. Шмальгаузена.
Ведущая проблема эволюционной теории XX века - коррелятив-
ная филогенетическая изменчивость органов - была поставлена Вла-
димиром Ковалевским: ‘‘При переходе одной формы в другую все ча-
сти изменяются строго коррелятивно”. Ковалевский различает два типа
редукции конечностей у копытных и вообще два типа развития: адап-
тивная эволюция - коррелятивна; неадаптивная эволюция (приводя-
щую группу в эволюционный тупик) - не коррелятивна. Эта тема не
касается адаптивного видообразования, темы Дарвина. Напротив, она
перекликается с корреляциями Кювье: от принципа статических кор-
9
В. В. Бабков
реляций Кювье Ковалевский переходит к принципу динамических кор-
реляций, рассматривая филогенетические линии изменения форм и
отдельных органов на чрезвычайно богатом материале. В.О. Ковалев-
ский никогда не обсуждал дарвинизм и не упоминал естественный
отбор (однажды упомянул борьбу), но занимался автономной задачей.
Она не включает видообразования, но поставленные им проблемы были
в центре эволюционных дискуссий XX в.
А.Н. Северцов подчеркивал самостоятельное значение “теорети-
ческого исследования эволюционной морфологии” и поэтому стремил-
ся не связывать своих взглядов с признанием того или иного теорети-
ческого объяснения причин эволюционного процесса, “то есть Ламар-
кизмом или Дарвинизмом, или другой из распространенных в
настоящее время теорий эволюции” [12]. В своем основном труде
“Морфологические закономерности эволюции” [ 13] А.Н. Северцов дал
анализ разнообразных модусов, или принципов, филогенетических
изменений. Для эволюционной морфологии А.Н. Северцова характер-
но, что морфология понимается как самостоятельная наука о форме, не-
зависимая от физиологии; эволюция подразумевает ограниченную и оп-
ределенно направленную изменчивость формообразования; филогенез
неравномерен и включает периоды радикальных реорганизаций.
Зачаток конкурирующей программы
Карл Бэр, стремившийся к полной и целостной концепции, изло-
жил ее принципы в речи “Самый общий закон природы во всяком ее
развитии” [14], произнесенной в начале 1834 г в Кёнигсберге. В неко-
торых моментах его концепция противоположна дарвинской. Д ля дар-
винова эволюционизма, основанного на понятии материи и причины,
а не мирового разума или Бога, главной трудностью всегда было поня-
тие ценности, производное от понятия цели. Под “развитием” обычно
понимается предсказуемое необратимое изменение; под “эволюцией”
- накопление исторической информации, то есть индивидуация. Под-
ход Дарвина в “Происхождении видов” подразумевает необходимость
объяснить, каким образом получается так много различных форм орга-
низмов на Земле. Каждый вид уникален; стало быть, следует объяс-
нить множественность исторических уникальностей [15]. Напротив,
10
I. История эволюционной биологии
подход Бэра основан на самом общем законе природы, который про-
является в развитии различных организмов и видов, в развитии живой
природы Земли и жизни во Вселенной. С позиций Бэра дарвинизм не
может ответить, как возникла жизнь на Земле и как появились первые
виды. Понимание приспособления, развития и наследственности тре-
бует связи с телеологией, а не причинностью. Принцип отбора не объяс-
няет, каким образом одна организация может превратиться в другую,
поэтому принцип отбора нельзя распространить на развитие в приро-
де в качестве универсального закона. Бэр убежден, что именно пред-
сказание является делом естественных наук, поэтому их теоретичес-
кая основа должна быть не эволюционной, но развитийной. Действи-
тельно, в фаунистике и систематике Н.А. Северцова, эмбриологии
А.О. Ковалевского и И.И. Мечникова, палеонтологии В.О. Ковалевс-
кого, морфологии А.Н. Северцова содержатся не только эволюцион-
ные, но и некоторые важные развитийные моменты.
Три взгляда на дарвинизм
Дарвинизм воспринимается разными группами общества по-раз-
ному. Согласно одному взгляду, дарвинизм является открытой систе-
мой: это “систематически формулированная теория перерождения”
(В. Ковалевский) и “превосходная программа новых исследований”
(Н. Северцов). Этот широкий взгляд характеризовал наиболее свобод-
но мыслящих и одаренных, ориентированных на собственное творче-
ство русских биологов. Такой подход позволил им построить на осно-
ве богатого самостоятельно добытого фактического материала широ-
кие обобщения, сравнимые по оригинальности с теорией естественного
отбора. Их обобщения содержали моменты, противоречащие тем или
иным основам учения Дарвина, но все они внесли выдающийся вклад
в мировой эволюционизм.
Другой смысл: дарвинизм есть мировоззрение. Социальная функ-
ция дарвинизма состояла в замещении иудейско-христианско-мусуль-
манского мифа творения, и колоссальная популярность дарвинизма
обязана этой роли. В России дарвинизм ассоциировался с нигилизмом,
материализмом, атеизмом и оказывал им поддержку, а в 1930-е гг. он был
взят на вооружение Агитпропом. Вследствие характера радикальной
11
В. В. Бабков
интеллигенции (как и большевиков) дарвинизм, в истолковании того
или иного ее вождя, превращался в самодовлеющую партийную докт-
рину. В рамках этого взгляда оставалось место либо для пропаганды и
защиты дарвинизма, либо же для идеологической критики и атак на
дарвинизм.
Еще одно понимание - дарвинизм есть законченное объяснение
причин эволюции, замкнутая система - соответствует логическому и
экспериментальному анализу его ключевых моментов (борьба за жизнь,
естественный и половой отбор).
Учение Дарвина имело сильный идеологический элемент. Это об-
стоятельство сделало первое ознакомление русской публики с дарви-
низмом настороженным и напряженным, а последующее принятие
дарвинизма (а позже отход от него) - чрезмерно экзальтированным и
выходящим за рамки биологии.
В начале 1900-х гг. И.В. Сталин высказал мнение о перспективах
эволюционных концепций и предсказал большую будущность неола-
маркизму. Очерк был напечатан на грузинском языке в провинциаль-
ной подпольной газете; в т. 1 “Сочинений” (1946) Сталин поместил
перевод, назвав эти соображения мыслями незрелого марксиста. Поз-
же взгляды Сталина на причины эволюции расширились и включили
внутривидовую борьбу (по аналогии с внутриклассовой). Когда
Т. Д. Лысенко стал отрицать внутривидовую борьбу и уклонился от вер-
сии Сталина, тот распорядился критиковать это отклонение. ЦК пору-
чил деятелю президиума АН СССР Н.Д. Иванову написать для “Бота-
нического журнала” критическую статью [16]. Она попала на рецен-
зию к Н.В. Турбину, грамотному генетику и эволюционисту, перешед-
шему на службу к обскурантам; он надолго задержал статью Иванова
и успел написать свою на ту же тему [17], так что обе они открыли
дискуссию по вопросам эволюции.
Полемика
Идеологическая подоплека новой книги Дарвина “Происхождение
человека и подбор по отношению к полу” (три перевода в 1871 г.),
расколола общество на тех, кто восторженно приветствовал новое воз-
зрение на мир и человека, и тех, кто принять его не мог.
12
1. История эволюционной биологии
В 1873 г. появился очерк А.П. Лебедева “Учение Дарвина о проис-
хождении мира органического и человека” [18], который был ориен-
тирован на университетскую публику, но нарушил неписаный закон
научного сообщества: называть в учении Дарвина те или иные силь-
ные моменты и умалчивать о слабых, к критике которых научное со-
общество тогда не было готово. Неявным ответом на него стала речь
А.С. Фаминцына на акте Санкт-Петербургского университета 8 фев-
раля 1874 г. “Дарвин и его значение в биологии”, представляющая ти-
пичное отношение члена русского научного сообщества периода боль-
ших ожиданий к учению Дарвина. Автор говорит о Дарвине с боль-
шим пиететом, но четко обозначает слабые или неверные моменты
учения. “Высокое ученое значение Дарвина обусловливается ... при-
менением строгого метода исследования”; “... главный вопрос, вопрос
о происхождении человеческого рода, невозможно в настоящее время
считать разъясненным”; “... он сам в последнее время выяснил истин-
ное значение естественной подборки, признав ее, в то же время, недо-
статочной для доказательства генетической связи между низшими и
высшими формами животных и растений” [19].
К. Бэр подверг критике данную Дарвином картину процесса эво-
люции и защищал общие принципы: что виды не могут выйти за пре-
делы типа в своих изменениях, что в рамках каждого типа есть виды
творения и другие, возникшие в результате эволюции, что все транс-
мутации обусловлены взаимодействием форм жизни со средой и что
география - единственный фактор эволюции. Бэр подчеркивал эти-
ческие последствия пренебрежения телеологией, замененной слепой
силой естественного отбора. Его аргументы направлены против пред-
полагаемого устремления Дарвина стать Ньютоном биологии посред-
ством распространения механического принципа на область жизни.
Проблемы жизни, которые Дарвин трактует в ньютонианском духе,
отличаются от области механики: приспособление, развитие и наслед-
ственность - проблемы иного порядка, так как их понимание требует
связи с телеологией, а не причинностью. Вывод Бэра состоит в том,
что теорию Дарвина, нарушающую принципы научной методологии,
необходимо отвергнуть [20].
Первый том самой знаменитой антидарвиновской сводки “Дарви-
низм” Н.Я. Данилевского, соратника К. Бэра, вышел в свет в ноябре
13
В. В. Бабков
1885 г. [21]. Во введении Данилевский не оставляет сомнений в при-
чинах, побудивших его предпринять разбор теории естественного от-
бора Дарвина. Это вопрос о разуме в природе. Дарвин уходит от рас-
смотрения разумного мироздания. Главная ошибка дарвинизма, по Да-
нилевскому, кроется в абсолютной случайности, положенной в его
основу. Поскольку свойствами изменчивости, удовлетворяющими дар-
винизму, должны быть “постепенность, неопределенность, безгранич-
ность, мозаичность”, то из дарвинизма нельзя вывести удивительную
целесообразность природы. Данилевский указывает, что “системати-
ческие группы животного царства не имеют ничего общего с прино-
ровленностью, творческим или критическим влиянием среды”, и на
этом основании отделяет таксономию и систематику от принципа от-
бора. Он приводит примеры, из которых заключает, что “в строении
организмов сторона морфологическая есть главное и сущест- венное”,
а что “адаптивная сторона есть уже нечто второстепенное”. Из реально-
сти искусственного отбора и борьбы за существование не выводится
какой-либо отбор в природе, и отсюда - заключение: “...так называе-
мый естественный подбор - не реальный природный деятель или фак-
тор, а не более, как фантазм, мозговой призрак...”.
Н.Н. Страхов спровоцировал К.А. Тимирязева на полемику вокруг
дарвинизма и книги Данилевского. Низкий стиль дискуссии, имевшей
предметом учение Дарвина, не мог оставить равнодушным академи-
ческую публику. Н.А. Холодковский напечатал в 1888-1889 гг. боль-
шой очерк “Теория Дарвина, ее критики и ее дальнейшее развитие”
([22] переиздание). Выбор тем (идеи А. Кёлликера, К. Нэгели, А. Вей-
смана) указывал, что для плодотворного развития эволюционной тео-
рии необходимо учитывать все новейшие достижения биологии. От-
вечая на полемику, А.С. Фаминцын отверг мнение, что теория Дарви-
на “господствует в ученом мире” [23]. Впоследствии Ю.А. Филипченко
отметил, что некоторые возражения Данилевского Дарвину, не нашед-
шие отклика в атмосфере 1880-х годов, могли бы быть важны для дис-
куссий 1920-х [24]. Однако к концу 1920-х, в обстановке Культурной
революции, вопрос решился просто: работы Данилевского и Страхо-
ва, среди прочих, были запрещены.
14
I. История эволюционной биологии
К.А. Тимирязев и М.А. Мензбир
Тимирязев не был специалистом в эволюционной теории и даже не
интересовался ею (так, в библиотеке музея-квартиры Тимирязева нет
книг по эволюции). Он трактовал дарвинизм не как теоретическое по-
строение, подлежащее проверке, но как важную часть обязательного
миросозерцания прогрессивной интеллигенции. Тимирязев полагал,
что учение Дарвина полностью завершено и нуждается лишь в незна-
чительной отделке. Занимаясь дарвинизмом в качестве hobby, Тими-
рязев в 1890-е ив 1910-е годы оставался в этой области на уровне
1860-х годов. Тимирязев принимал систему Дарвина не во всех ее ос-
новных моментах. Он желал заменить метафору “естественный отбор”
понятием elimination и ссылался в этом отношении на Конта, но не
мог себе этого позволить в период всеобщего восхищения дарвиниз-
мом. (В начале XX в. такую замену произвел английский позитивист
биометрик Карл Пирсон.) Хотя Тимирязев считал, что перенаселение
необходимо для успеха эволюции, он хотел бы отказаться от метафо-
ры “борьба за существование”. После 1900 г. Тимирязев должен был
учитывать менделизм и мутационизм. Он однажды назвал менделизм
достижением: с его помощью “кошмар Дженкина, испортивший
столько крови Дарвину, рассеивается без следа”. Но Тимирязев трак-
товал наследственность как физиологическую проблему, свойство це-
лого организма. Поэтому менделизм стал лишь подробностью среди
типов наследования, которые он принимал.
М.А. Мензбир отличался от Тимирязева спокойным темперамен-
том и уравновешенным тоном аргументов, которые никогда не выхо-
дили за рамки академических приличий. Он проводил взгляд, что тео-
рия Дарвина закончена, не подлежит проверке и изменениям и допус-
кает лишь некоторую коррекцию. Мензбир представлял научную школу
Н.А. Северцова, однако он имел совершенно иной темперамент: всю
жизнь он провел в университетском кабинете, разбирая коллекции
Северцова и публикуя их, тогда как Северцов провел жизнь в путеше-
ствиях. Создав научное направление, он распространял учение Дар-
вина - в своем избирательном и персональном понимании - среди не-
скольких поколений учеников. Мензбир чутко реагировал на любые
уклонения от своего понимания дарвинизма и отсекал их спокойно,
осторожно, систематично.
15
В. В. Бабков
Комбинация тактики Мензбира с тактикой Тимирязева оказалась
чрезвычайно эффективной, и не только в пропаганде упрощенного
дарвинизма. Она послужила моделью при реформе университетов в
начале 1920-х и большевизации Академии наук в конце 1920-х годов;
она же использовалась Т.Д. Лысенко и И.И. Презентом при монопо-
лизации биологии. При этом новый творческий дарвинизм, хотя и был
пародией на дарвинизм Тимирязева и Мензбира и понимание наслед-
ственности как свойства целого организма Тимирязева, но уже не имел
никакого отношения к науке.
Сообщества
Русская традиция в теоретических вопросах биологии, с предпоч-
тением гармонии, была естественным союзником “самого общего за-
кона природы” Бэра, а не “борьбы за жизнь” Дарвина. Поэтому мно-
гие естествоиспытатели, желающие развивать учение Дарвина, выде-
ляли одни моменты и принижали другие; придавали произвольный
смысл понятию отбора и вводили новые направления исследований, в
которых борьба за существование получала более широкое, и вообще
иное, звучание. По этой же причине русские биологи естественным об-
разом заложили основы экологии, фитосоциологии, биогеоценологии.
Сообщая в 1863 г. о скором появлении русского перевода “Проис-
хождения видов”, С.А. Рачинский умолчал о Мальтусе. А Страхов в
1862 г. отверг мальтузианскую модель эволюции. Тимирязев в рецен-
зии 1864 г. прямо поддержал идею о необходимости перенаселения
для дарвиновской эволюции. Статья А.Н. Бекетова, опубликованная в
1873 г., обычно приводится в доказательство его взглядов о необходи-
мости перенаселения для борьбы за существование. Однако Бекетов
писал: “Борьба за существование в широком дарвиновском смысле про-
исходит от совместного действия двух причин, но из них самая жизнь
является инициатором явления, а ограниченность пространства и
средств к жизни представляются второстепенною причиною, поддер-
живающей и обостряющей борьбу”. Как видно, он подчеркивает здесь
роль внутренних причин; он отрицает мальтузианскую эволюцию и
соглашается принять “борьбу” в качестве “риторической фигуры”.
Борьба ведет к гибели, напротив, “равновесие или гармония природы
16
I. История эволюционной биологии
выражается именно в многочисленных приспособлениях организмов
к окружающей среде” [25].
И.И. Мечников в очерке “Борьба за существование в обширном
смысле” [26] оспаривает заключение Дарвина, что борьба за существо-
вание и вытекающий из нее естественный отбор ведут к усовершен-
ствованию организации. Связь между борьбой за существование и об-
разованием видов не всеобща: “Напротив, можно утверждать, что во
многих случаях они бывают разобщены, то есть что есть виды, при-
знаки которых фиксировались помимо борьбы за существование, и, с
другой стороны, что последняя не необходимо ведет к образованию
форменных признаков, составляющих видовые отличия”.
А.С. Фаминцын отмечал, что все доводы в пользу прогрессив-
ного характера эволюции носят косвенный характер и нет “ни од-
ного непосредственного наблюдения над превращением низшей
формы в высшую” [27]. В ранних опытах С.А. Фаминцын и И. Ба-
ранецкий пришли к выводу, что лишайники суть комплексы гри-
бов с водорослями [28]. Термин “симбиоз” был введен в науку че-
рез 12 лет после этих опытов, и Фаминцын его принял; он дал свое
решение вопроса о возникновении более сложной организации из
простой в представлении о роли симбиоза в эволюции [29 и др.].
Идеи о симбиозе как основном (но не исключительном) способе
образования видов разрабатывал К.С. Мережковский [30], а позже
Б.М. Козо-Полянский (впоследствии заново этот круг идей успеш-
но разработала Линн Маргулис).
Серьезным исключением в отношении русских радикалов к дар-
винизму оказался самый влиятельный мыслитель того времени
Н.Г. Чернышевский. В 1888 г. он отверг дарвинизм по моральным и
научным основаниям [31]. Его аргументация сводилась к следующе-
му. Борьба есть зло; зло в применении к понятию организации есть
дезорганизация, поэтому сказать, что в результате борьбы может по-
лучиться благо, значит думать, что вредное полезно.
К.Ф. Кесслер [32] и П.А. Кропоткин [33] оспорили всеобщую при-
ложимость учения о борьбе за существование и выставили противо-
положный принцип - взаимной помощи. Ихтиолог и географ
Л.С. Берг посвятил этому вопросу небольшую книжку “Борьба за су-
ществование и взаимная помощь” [34]. Вопреки мнению Кропоткина
Берг не признает взаимную помощь в качестве фактора эволюции:
17
В. В. Бабков
эволюция организмов идет по строго определенным законам, вне
зависимости от взаимопомощи и борьбы за существование”. Но борь-
ба и взаимная помощь суть факты, с которыми должен считаться есте-
ствоиспытатель: “В отношении прогресса моральные соображения
Кесслера и Кропоткина сохраняют силу”.
В ходе знаменитых Полтавских экспедиций 1888-1891 гг. по изу-
чению русских черноземов В.В. Докучаев демонстрировал справед-
ливость “закона содружества, мировой сопомощи и любви”, должен-
ствовавшего дополнить “закон великого Дарвина - закон борьбы за
существование”. В своей знаменитой книге “Наши степи прежде и те-
перь” (СПб, 1892) и в ряде речей и статей он повторял вывод, сделан-
ный в процессе изучения почв как природных геобиологических об-
разований: “В мире, кроме жестокого, сурового, ветхозаветного зако-
на постоянной борьбы, мы ясно усматриваем теперь закон содружества,
любви...”Он подчеркивал различие областей, где действует тот или
другой закон: “Закон Дарвина вполне применим к небольшому полю
наблюдений: чем ближе друг к другу сталкивающиеся существа, тем
борьба между ними сильнее. Если же окинем взором обширные Зо-
нальные пространства, то увидим, что на протяжении тысяч верст чер-
нозем, сурки, ковыли и пр. превосходно уживаются вместе и дополня-
ют друг друга”. Поэтому Докучаев призывал “обратить внимание на
вековечную зависимость, генетическую и всегда закономерную связь,
какая имеется между силами, телами и явлениями, между мертвой и
живой природой, между растительными, животными и минеральны-
ми царствами - с одной стороны, человеком, его бытом и даже духов-
ным миром - с другой” [35]. Учение о биосфере, созданное В.И. Вер-
надским, вполне созвучно мыслям его учителя.
С.И. Коржинский искал решения знаменитого “степного вопроса”
[36] - почему нет леса в степях - за пределами влияний климата и
особенностей почвы. Он подчеркивал, что “фито-географические ус-
ловия еще не выполняют собой всей жизненной обстановки растений:
еще остается целый мир социальных отношений к другим организ-
мам”. Именно эти воздействия ответственны за наблюдаемую извили-
стость границы леса и степи, которую нельзя объяснить лишь клима-
том и почвой: “... смена как отдельных видов, так и целых формаций и
глубокие изменения характера растительности могут происходить со-
вершенно самостоятельно, помимо всяких изменений климата”.
18
I. История эволюционной биологии
Работы Коржинского привели к формированию особой научной дис-
циплины - фитоценологии, которую он основывал на принципе взаи-
мопомощи, не придавая значения идее борьбы за жизнь.
С.А. Северцов основал эволюционную экологию, дополняя иссле-
дованиями популяционной динамики в полевых условиях теорию про-
грессивной эволюции Алексея Северцова и учение Дарвина (напр.,
[37]). “Мы можем надеяться, что дальнейшие исследования в области
эволюционной экологии, синтез экологии с эволюционной морфоло-
гией, а в дальнейшем и с генетикой позволит создать настоящую науч-
ную теорию эволюции, вскроет законы, пользуясь которыми мы смо-
жем направить как процесс формообразования, так и численность
животных на пользу человека” [38]. В середине 1930-х его работы по
установлению корреляций между плодовитостью и продолжительно-
стью жизни животных были раскритикованы сталинским идеологом
И.И. Презентом, после чего работы по применению математических
методов в биологии оказались невозможными.
В 1940-е годы геоботаник, фитоценолог, лесовед и болотовед
В.Н. Сукачев ввел в науку понятие о биогеоценозе и основал комплек-
сную науку биогеоценологию, связавшую экологические биологичес-
кие дисциплины с географией, почвоведением, биогеохимией [39,40].
К определению понятия “биогеоценоз” логически можно прийти дву-
мя путями: по восходящему пути от особи и вида, основных объектов
исследований биологов, и по нисходящему - от представлений о био-
сфере, развитых В.И. Вернадским. Если рассматривать биосферу как
пронизанную “живым веществом” оболочку Земли, изменяемую мощ-
нейшим геологическим фактором - жизнью, то биогеоценоз предста-
ет как элементарная эволюционирующая единица биосферы. На этих
основаниях Н.В. Тимофеев-Ресовский заложил в 1960-е годы прин-
ципы радиационной биогеоценологии [41,42]. Благодаря Тимофееву-
Ресовскому идеи Вернадского и Сукачева воспринял и включил в свои
работы С.С. Шварц [43].
Номогенетическая линия
В 1920-х появилось сразу несколько сочинений по теории паралле-
лизма, чисто номогенетических концепций, относящихся к бэрианс-
кой, или развитийной, традиции. По закону гомологических рядов
19
В. В. Бабков
Н.И. Вавилова (различные редакции [44]) многообразие признаков, воз-
можных в пределах вида, рода, семейства, строго упорядочено и огра-
ничено. Множество признаков в различных видах и родах данного
семейства точно повторяют друг друга, поэтому гомологические ряды
Вавилова позволяют предсказывать существование форм, наделенных
совершенно определенными признаками (предсказания неизменно
сбывались). Если теория естественного отбора предполагала накопле-
ние мелких непрерывных уклонений, то закон гомологических рядов
- выбор из ограниченного числа предсуществующих дискретных форм.
Филогенетическое развитие основано в первом случае на последова-
тельной дивергенции, во втором - на заполнении клеток таблицы, по-
добной таблице Менделеева.
Тогда же появились сочинения Л.С. Берга [45, 46], основанные на
данных сравнительной анатомии животных, и вышла работа Д.Н. Со-
болева, где он развивал идею цикличности филогенеза и дополнял ее
принципом постоянства видов (Линней), принципом градаций (Ламарк)
и законом прерывистости (катастроф) [47]. Принцип параллелизма
применялся к строению тканей организмов, принадлежащих к нерод-
ственным группам, в работе А.А. Заварзина, где тонкая архитектони-
ка расположения и соединения нейронов друг с другом в брюшной
цепочке стрекозы и спинном мозге высших позвоночных оказалась до
мелочей тождественной [48].
Одна из наиболее интересных работ Берга по теории эволюции - не-
большой доклад на Съезде по генетике [49]. “Влияние географического
ландшафта на организмы двоякое: преобразующее и отбирающее, - пи-
сал Берг. - Непосредственно воздействуя на всю массу организмов, лан-
дшафт преобразует их. С другой стороны, он отбирает формы, приспо-
собленные для данного ландшафта: при этом неприспособленные или
гибнут, или переселяются в другой, более соответствующий ландшафт”.
Берг выводит, что “одни и те же признаки появляются в одном и том же
ландшафте пачками - у целой группы растительных видов”. Механизм
воздействия ландшафта пока скрыт от нас, но ясно, что “секрет формо-
образующего влияния изоляции, на котором настаивают многие авторы,
коренится именно в воздействии определенного географического ланд-
шафта...’’Другая сторона действия географического ландшафта - рас-
пределяющая: “Естественный отбор и ландшафт распределяют формы
по лику Земли, но, понятно, не создают эти формы”.
20
I. История эволюционной биологии
Хотя открыто выражать убеждение в направленности эволюции на-
чиная с 1930-х годов было невозможно, но и позже появлялись некото-
рые выдающиеся, номогенетические по характеру, работы. К ним отно-
сятся главный труд А.Н. Северцова “Морфологические закономерности
эволюции” [13], а также принцип олигомеризации В.А. Догеля [50], до-
полнивший список модусов филогенетических преобразований; промор-
фология, учение о симметрии В.Н. Беклемишева [51], ставшая основой
архитектонического объяснения организма. Номогенетический характер
носит сформулированная М.С. Гиляровым мысль о роли параллелизмов
и конвергенций в эволюции различных филогенетических ветвей члени-
стоногих [52]. Номогенетические моменты играют выдающуюся роль в
теории стабилизирующего отбора И.И. Шмальгаузена и представлениях
о дестабилизирующем отборе Д.К. Беляева.
Некоторые взгляды на видообразование
Занимаясь лесной энтомологией, Н.А. Холодковский внес ясность в
вопрос о параллельных рядах распространенного вредителя леса, жел-
того хермеса. Считалось, что это две группы особей, происходящих парте-
ногенетически от одной самки-родоначальницы: одна мигрирует на ли-
ственницу, другая остается на ели. Холодковский установил, что две внеш-
не неразличимые группы суть два разных вида [53]. На этой основе он
построил представление о видах-двойниках [54], впоследствии играв-
шее существенную роль в генетике популяций.
Н.В. Цингер описал две расы сорняка, засоряющего посевы льна
[55]; одна из рас имеет семена того же (большого) размера, что и се-
мена льна, так что они не отделяются при предпосевной обработке;
эта ситуация в целом сходствует с феноменом ярового и озимого об-
раза жизни у растений, с расами рыб [56], с моно- и бивольтинизмом
у шелкопряда.
С.И. Коржинский отвергал эволюционное значение внешних вли-
яний, мелких индивидуальных отклонений, борьбы за жизнь. “Все
виды, раз сформировавшись, остаются неизменными, но временами
отщепляют от себя новые формы путем гетерогенезиса”. Происхож-
дение высших форм из низших зависит от внутренней тенденции про-
гресса, связанной с тенденцией изменчивости [57].
21
В. В. Бабков
Ю.А. Филипченко, один из основателей русской генетики и пос-
ледователь Бэра, строго отделял происхождение высших таксонов
(макроэволюцию) от происхождения видов (микроэволюции) и ис-
кал решение проблемы макроэволюции за пределами генетики и дар-
винизма: в плазмоне, а не генотипе и отборе.
Н.К. Кольцов, один из основателей экспериментальной биологии
в России, занимался поисками физико-химического объяснения фор-
мы живых образований: клеток (принцип клеточного скелета), моле-
кул (матричный принцип воспроизведения “наследственных моле-
кул”), возникновения новой формы в индивидуальном развитии (ко-
торое он трактовал в терминах силового поля), организмов. Кольцов
критиковал взгляд о безграничном прогрессе в ходе формообразова-
ния (и схемы филогении); он подчеркивал роль регрессов и неотений
в процессе эволюции (и строил схему “мангровой заросли”). Опира-
ясь на опыты Е.И. Балкашиной с гомеозисными мутациями, он выд-
винул гипотезу об эволюции путем дупликации генов: “Запас не про-
являющихся в развитии генов, которые могут мутировать в гены, про-
являющиеся в развитии уже неотенической формы, влечет за собой
высокую изменчивость последней и позволяет ей иногда обнаружить
в дальнейшем пышный расцвет прогрессивной эволюции” [58].
И.А. Рапопорт дал экспериментальное подтверждение концепции [59],
которая позже стала известной в независимой постановке Сусуму Оно.
Природа и поддержание изменчивости в популяциях
Эту проблему поставил С.С. Четвериков. Он проводил различие
между индивидуальными и географическими (полигонными) вариа-
циями: значение для понимания процессов видообразования имеет
географическая изменчивость, “а из мутаций непосредственно, сразу
никакого вида не получается”. Он различал половую изоляцию (суще-
ственную для видообразования) и пространственную и ставил вопрос
о симпатрическом видообразовании в связи с экологической изоляци-
ей. Программа построения новой дисциплины, эволюционной гене-
тики, была изложена в классической работе 1926 г. “О некоторых мо-
ментах эволюционного процесса с точки зрения современной генети-
ки” [60]. Будучи натуралистом и дарвинистом и освоив эксперимен-
22
I. История эволюционной биологии
тальную генетику, Четвериков желал устранить их несовместимость и
разрешить основное затруднение дарвинизма того времени: найти во
внешне однородных диких популяциях достаточное для эволюции
путем отбора количество “геновариаций”. В первом анализе генофон-
дов диких популяций дрозофилы 1923-1928 гг. он нашел скрытый ре-
зерв наследственной изменчивости, “из которого отбор может свобод-
но черпать всегда, как только возникнет необходимость”. Четвериков
пришел к выводу, что “приспособленность не является ключом к по-
ниманию всех проблем эволюции”: он различал адаптивную эволю-
цию на основе отбора и неадаптивную эволюцию - “процесс видооб-
разования, процесс разнообразия органического мира”, который про-
исходит в результате изоляции. Он выдвинул проблему целостности
генотипа в форме концепции генотипической среды [61].
По материалам первого исследования генофондов диких популя-
ций Д.Д. Ромашов и Е.И. Балкашина заключили, что “изменчивость
отдельных признаков не представляет случайного, беспорядочного
процесса, но является “определенно направленной” и ограниченной”;
они выделили три категории в фонде популяционной изменчивости и
сделали предположение об эволюционной роли каждой из них [62];
идея Ромашова о роли случайных процессов в эволюции [63], осно-
ванных на ошибках выборки, обязанных менделевской стохастике, и
усиливаемых “волнами жизни”, повела к представлениям об “эффек-
те горлышка бутылки” и “эффекте родоначальника”. Е.А. и Н.В. Ти-
мофеевы-Ресовские впервые нашли летальные мутации в процвета-
ющих популяциях [64]. Н.В. Тимофеев-Ресовский сформулировал си-
стему основных элементарных понятий микроэволюции, которая
оформила представления школы Четверикова о новой биологической
дисциплине [65, 66]; среди прочих крупных достижений, он сформу-
лировал принцип усилителя в биологии, трактующий дискретность
как условие жизненных явлений и включающий в качестве частного
случая принцип отбора Дарвина [67].
Серия работ Д.Д. Ромашова 1929-1948 гг. по феногенетике массо-
вой популяционной изменчивости и цикл работ Н.К. Беляева 1930-х, а
также В.П. Эфроимсона 1940-х гг. по изучению коррелированной из-
менчивости конституциональных признаков (объединивших подходы
генетики, индивидуального развития, систематики, эволюции, эколо-
23
В. В. Бабков
гии), привели к постановке вопроса об эволюционно значимой измен-
чивости и дали основу для пересмотра генетики популяций (большая
часть этих работ не опубликована; их материалы см. в [68]).
Цитология и проблема вида
Среди достижений Б Л. Астаурова выделяются экспериментальные
доказательства возможности клонирования животных и возникнове-
ния в природе полиплоидных рас: гипотеза непрямого (опосредован-
ного партеногенезом) происхождения естественной полиплоидии у
бисексуальных животных [69]. (На растениях Г.Д. Карпеченко еще
раньше преодолел бесплодие гибридов между капустой и редькой пу-
тем искусственной полиплоидии.) Н.Н. Соколов исследовал влияние
плазмы при гибридизации (нарушение митоза и проч.) и, в частности,
показал, что случаи, которые Р. Гольдшмидт трактовал как интерсек-
сы, наделе оказались мозаиками [70]. Наиболее перспективным здесь
является направление В.Н. Стегния [71], сочетающее выяснение трех-
мерной организации хромосом, зоогеографии, системных мутаций и
включающее ряд развитийных моментов.
Развитийные концепции эволюции
Идея генотипической среды Четверикова дала возможность объе-
динить рассмотрение процессов наследственной передачи и формо-
образовательных процессов в рамках одной теоретико-биологической
концепции. Е.И. Балкашина нашла у дрозофилы мутации aristopedia и
tetraptera (выводящие мух из класса и отряда) и всесторонне изучила
первую из них, дающую лапку вместо кисточки усика [72]. Рихард
Гольдшмидт на основе ее результатов выдвинул идею “перспективно-
го уродца” и макромутаций. В анализе мутации tetraptera, где левое и
правое крылья “второй пары” развивались независимо друг от друга,
Б.Л. Астауров поставил вопрос о возникновении устойчивости разви-
тия в процессе эволюции.
Заново вопрос об эволюции онтогенеза, включавший вопрос об
эволюционном возникновении устойчивости и представление об эво-
24
I. История эволюционной биологии
люции как побочном продукте процесса, ведущего к поддержанию и
возрастанию автономности биологической системы, был поставлен
И.И. Шмальгаузеном. В концепции стабилизирующего отбора он объе-
динил принцип трансформизма и принцип типа, идеи гармонии при-
роды и борьбы, тихогенеза и номогенеза [73]. Важным моментом кос-
венного отбора В.С. Кирпичникова было представление об автоном-
ной изменчивости частей организма и возрастании автономности
онтогенеза в восходящем ряду форм [74].
35-летние опыты Д.К. Беляева по одомашниванию серебристо-чер-
ных лисиц, поставленные поначалу ради чисто утилитарной цели (по-
лучить приплод несколько раньше стандартного срока), привели его к
мысли о дестабилизирующем отборе. Он вел отбор по поведению,
иерархически высокому признаку, поэтому отбор затронул системы
центральной регуляции комплекса адаптивных признаков. Картина
модельной эволюции (дестабилизация и канализация формообразова-
ния) совпадает с тем, что говорил Берг о двояком воздействии геогра-
фического ландшафта (“одним из самых мощных преобразователей
ландшафта является человек”) [75]. Эти соображения могут пролить
свет и на редчайший случай образования новой видовой формы в экс-
перименте ГХ. Шапошникова [76].
Вместо заключения
Материалы очерка показывают, что нет царской дороги не только в
геометрию (как говорил Аристотель ученику Александру Македонс-
кому), но и в теорию вида. Занимаясь выяснением видообразования
на одном объекте и с одним подходом, следует помнить о существова-
нии многих других. Имея в виду перспективу построения работаю-
щей общей картины процесса эволюции, будет полезно стремление к
некоему развитийному фундаменту, центр кристаллизации которого
содержится в Бэрианской традиции в широком смысле.
Литература
1. Бекетов АН. Гармония в природе // Русский вестник. 1860. Ноябрь. С 197-241.
Декабрь. С. 534-558.
2. Обзор иностранной литературы И Отечественные записки. 1861. Февраль.
25
В. В. Бабков
3. Несколько слов о современной теории происхождения видов И Мечников И.И. Из-
бранные биологические произведения. М.: Изд-во АН СССР, 1950.
4. Страхов Н.Н. Дурные признаки И Время. 1862. Ноябрь.
5. Бибиков П.А. Критические очерки. СПб., 1865.
6. Рачинский С.А. Цветы и насекомые // Русский вестник. СПб. 1863. № 1.
7. Северцов Н. Орнитология и орнитологическая география Европейской и Азиатской
России. СПб., 1867. Вып. 1.
8. Северцов Н. Вертикальное и горизонтальное распределение туркестанских живоз-
ных. М., 1873.
9. Северцов Н. Аркары // Природа. 1873. Кн. 1.
10. Давыдов К.Н. А О. Ковалевский как человек и ученый // Труды Института истории
естествознания и техники. М.: Изд-во АН СССР, 1960. Т. 31.
11. Мечников И.И. Современное состояние науки о развитии животных // Журн. мин.
нар. просвещения. 1869.
12. Северцов А Н. Этюды по теории эволюции. Киев, 1912.
13. Северцов А Н. Морфологические закономерности эволюции. М.; Л.: Изд-во АН
СССР, 1939.
14. Бэр К.Э.ф. Избранные работы / Сост. и пер. Ю.А. Филипченко. Пг., 1924.
15. Salthe S. Comparing Darwinism with possible renewed developmental discourse. MS.
1992.
16. Иванов Н.Д. О новом учении Т.Д. Лысенко о виде И Ботанический журнал. 1952.
№6. С. 819-842.
17. Турбин Н.В. Дарвинизм и новое учение о виде И Ботанический журнал. 1952. № 6.
С. 798-818.
18. Лебедев А П. Учение Дарвина о происхождении мира органического и человека:
Философско-критические этюды И Русский вестник. 1873. Июль. С. 118-163. Ав-
густ. С. 429-509.
19. Фаминцын А С. Дарвин и его значение в биологии //Отечественные записки. 1874.
№З.С. 129-150.
20. Baer К. Uber Darwins Lehre И Reden... SPb, 1876. II T. S. 235-480.
21. Данилевский Н.Я. Дарвинизм. Критическое исследование. СПб.. Комаров. 1885.
Ч. 1.519 с. Ч. 2. 148 с.
22. Холодковский Н.А. Биологические очерки. М.; Пг.: Госиздат, 1923.
23. Фаминцын С.А. Н.Я. Данилевский и дарвинизм. (Опровергнут ли дарвинизм Дани-
левским?)//Вестник Европы. 1889. Февраль. С. 616-643.
24. Филипченко Ю.А. Эволюционная идея в биологии. Пг.: Сабашников, 1923.
25. Бекетов А Н. О борьбе за существование в органическом мире И Вестник Европы.
1873. № 10. С. 558-593.
26. Мечников И И. Борьба за существование в широком смысле И Вестник Европы.
1878. №7, 8.
27. Фаминцын А С. Современное естествознание и психология. СПб., 1898.
28. Famintzin A., Baranetzky I. Zur Entwicklungsgeschichte der Gonidien und Zoosporenbildung
der Flechten // Mem. Acad. sci. SPb., 1867. Ser. VII. Til. № 9. S. 1-6.
29. Фаминцын А.Н. О роли симбиоза в эволюции организмов // Зап. Ими. акад наук.
1886. Сер 7.Т. 20. №2. С. 1-14.
30. Мережковский К С. Теория двух плазм как основа симбиогенезиса, нового учения о
происхождении организмов. Казань, 1909.
26
I. История эволюционной биологии
31. Чернышевский Н.Г Происхождение теории благотворности борьбы за жизнь // Рус-
ская мысль. 1888. Сентябрь.
32. Кесслер К.Ф. О законе взаимной помощи // Труды СПб. Общества естествоиспыта-
телей. 1880. Т. 11 С. 124-136.
33. Кропоткин 11.А. Взаимная помощь как фактор эволюции СПб., 1907.
34. Берг Л.С. Борьба за существование и взаимная помощь. Петербург: Время, 1922.
35. Докучаев В.В. Избранные сочинения. М.: Сельхозгиз, 1954. С. 338, 330.
36. Коржинский С И Северная граница черноземно-степной области восточной поло-
сы Европейской России в ботанико-географическом и почвенном отношении. Ка-
зань Ч. 1. 1888. 4.2. 1891
37. Северцов С.А Динамика населения и приспособительная эволюция животных.
М.; Л.: Изд-во АН СССР, 1941.
38. Северцов С.А. Дарвинизм и экология // Зоологический журнал. 1937. Т. 16. Вып. 4.
С. 591-613.
39. Сукачев В.Н. Биогеоценология и фитоценология И ДАН СССР. 1945. Т. 47.
40. Сукачев В Н. О соотношении понятий географический ландшафт и биогеоценоз
//Вопросы географии. 1949. Т. 16.
41. Тимофеев-Ресовский Н.В. Некоторые проблемы радиационной биогеоценологии
// Проблемы кибернетики. М.: Физматгиз, 1964. Вып. 12. С. 201-232.
42. Тимофеев-Ресовский Н.В. О некоторых принципах классификации биохорологи-
ческих единиц // Труды Института биологии УФ АН СССР. Свердловск, 1961.
Вып. 27. С. 23-29.
43. Шварц С.С Экологические закономерности эволюции. М.: Наука, 1980.
44. Вавилов Н И. Закон гомологических рядов в наследственной изменчивости.
Л.: Наука, 1987. 261 с.
45. Берг Л.С. Теории эволюции. Петербург. Academia, 1922. 120 с.
46. Берг Л .С. Номогенез, или Эволюция на основе закономерностей//Труды Географи-
ческого института Т. 1. Петроград, 1922. 306 с.
47. Соболев Д.Н. Начала исторической биогенетики. Харьков: Госиздат Украины, 1924.
203 с.
48. Заварзин А.А. Параллелизм структур как основной принцип морфологии И Изв.
Биол. ин-та при Пермском ун-те. 1923. Т. 2. № 4
49. Берг Л.С. Воздействие географического ландшафта на культурные растения и жи-
вотных И Труды Съезда по генетике. Л., 1930. Т 2. С. 107-112.
50. Догель В.А. Олигомеризация гомологичных органов как один из главных путей
эволюции животных. Л., 1954.
51. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. 3-е изд. М.,
1964 Т 1-2.
52 Гиляров М.С. Закономерности приспособлений членистоногих к жизни на суше.
М., 1970.
53 Холодковский Н А. К биологии и систематике рода Chermes И Труды Русск. энто-
мол. об-ва. 1890. Т. 24.
54 Холодковский Н А. О биологических видах // Изв. Имп. акад. наук. 1910. Сер. VI.
Т 4. № 10.
55. Цингер Н.В. О подвидах большого погремка Alectorolophus major. Вологда, 1928.
56 Берг Л.С Яровые и озимые расы у проходных рыб // Изв. Отд. мат. и ест. наук АН
СССР. Сер. биол. 1934. № 5. С. 711-732.
27
В. В. Бабков
57. Коржинский С.И. Гетерогенезис и эволюция. I. // Записки Имп. акад. наук.
1899. Сер. VIII. Т. 9. №2.
58. Кольцов Н.К. Проблема прогрессивной эволюции // Биологический журнал. 1933.
Т. 2. Вып. 4-5. С. 475-500.
59. Рапопорт И. А. Многократные линейные повторения участков хромосомы и их эво-
люционное значение // Журн. общей биологии. 1940. Т. 1. № 2. С. 235-270.
60. Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения
современной генетики //Журн. эксп. биол. Сер. А. 1926. Т. 2. № 1. С. 3-54.
61. Четаериков С.С. Проблема общей биологии и генетики. Новосибирск: Наука, 1993.273 с.
62. Ромашов Д.Д., Балкашина Е.И. Материалы по генетике Drosophila funebris
// Журн. эксп. биол. 1929. Т. 5. № 2. С. 102-146.
63. Ромашов Д.Д. Об условиях “равновесия” в популяциях // Журн. эксп. биол. 1931.
Т. 7. Вып. 4. С. 442-454.
64. Timofeeff-Ressovsky Е.А., N.W. Genetische Analyse einer freilebenden Drosophila
melanogaster-Population//Roux’Arch. Entwicklungsmech. Organ. 1927. Bd. 109.
H. 1. S. 70-109.
65. Timofeeff-Ressovsky N.W. Mutations and geographic variations // The New Systematics
/ J.S. Huxley, ed. L.: Oxford Univ. Press. 1940. P. 73-136.
66. Тимофеев-Ресовский H.B. Микроэволюция. Элементарные явления, материал и
факторы микроэволюционного процесса// Ботанический журнал. 1958. Т. 43. № 3.
С. 317-336.
67. Тимофеев-Ресовский Н.В., Ромпе Р.Р. О сгатистичности и принципе усилителя в
биологии И Проблемы кибернетики. М.: Физматгиз, 1959. Вып. 2. С. 213-228.
68. Бабков В.В. Московская школа эволюционной генетики. М.: Наука, 1985. 216 с.
69. Астауров Б.Л. Экспериментальная полиплоидия и гипотеза непрямого (опосредо-
ванного партеногенезом) происхождения естественной полиплоидии у бисексуаль-
ных животных // Генетика. 1969. Т. 5. № 7. С. 129-140.
70. Соколов Н.Н. Взаимодействие ядра и цитоплазмы при отдаленной гибридизации
животных. М.: Изд-во АН СССР, 1959.
71. Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск:
Изд-во НГУ, 1993. 110 с.
72. Балкашина Е.И. Случай наследственного гомеозиса (геновариация “Aristopediа”)
у Drosophila melanogaster // Журн. эксперим. биол. 1928. Сер. А. Т. 4. Вып. 2.
С. 93-106.
73. Шмальгаузен И.И. Организм как целое в индивидуальном и историческом разви-
тии. М ; Л.: Изд-во АН СССР, 1938. 144 с.
74. Кирпичников В.С. Роль наследственной изменчивости в процессе естественного
отбора (гипотеза о косвенном отборе) // Успехи соврем, биол. 1944. Т. 18. Вып. 5.
С. 775-800.
75. Бабков В.В. Системный стиль в изучении естественного отбора // Системные ис-
следования. Ежегодник 1980. М.: Наука, 1981. С. 404-420.
76 Шапошников Г.Х. Возникновение и утрата репродуктивной изоляции и критерий
вида// Энтомологическое обозрение. 1966. Т. 45. № 1. С. 3-33.
ИСТОРИЯ ФОРМИРОВАНИЯ понятия
О ВИДЕ ВЫСШИХ РАСТЕНИЙ
А.В. Положим
Томский государственный университет
634050, г.Томск, пр. Ленина, 36
E-mail: herb@bio.tsu.ru
Термин «вид» был введен в науку в XVII столетии. Английский
естествоиспытатель Джон Рей назвал видом совокупность тождествен-
ных друг другу организмов, способных давать подобное себе потом-
ство. Но вид как таксон в систематике получил широкое применение
только в XVIII столетии, после того, как К. Линней доказал, что вид
представляет собой универсальную форму существования всех орга-
низмов. Основываясь на идее сотворения и постоянства видов, он по-
нимал вид как определенный тип структуры (species - внешность).
Единственным критерием вида считался комплекс морфологических
признаков, по которым различаются виды.
На определенном этапе типологическая концепция вида Линнея иг-
рала положительную роль в систематике высших растений, позволила
привести в систему накопленный обширный фактический материал.
Важнейшее значение имели введение в систематику единого латинского
языка и разработанная Линнеем бинарная номенклатура. Виды, прини-
маемые Линнеем, позднее получили название «линнеонов».
Появление эволюционного учения обосновало новые представле-
ния о виде и видообразовании. Эволюционное учение доказывало ис-
торичность, изменчивость видов - виды зарождаются, достигают зре-
лости, дают начало новым видам, старые со временем вымирают.
Систематики растений в теоретических работах разделяли взгля-
ды эволюционного учения, в том числе и на проблемы вида, но прак-
тически продолжали описывать виды только по морфологическим при-
знакам. Накапливались факты о внутривидовом разнообразии, о су-
ществовании промежуточных форм между видами. Это привело к
созданию так называемой политипической концепции вида. Вид стал
29
А. В. Положий
рассматриваться как система, состоящая из соподчиненных таксонов:
подвидов, разновидностей, форм. Основным критерием вида считал-
ся четко выраженный гиатус между видами, а промежуточные формы
представляют собой внутривидовые единицы.
Политипическая концепция была принята во флористических тру-
дах систематиков немецкой школы: А. Энглера, П. Ашерсона, П. Греб-
нера, а также многих французских, итальянских и русских системати-
ков. По мере использования политипической концепции активно ра-
ботающие ботаники стали обращать внимание на господствующий
субъективизм в оценке признаков, отличающих виды от внутривидо-
вых таксонов. Приходили к выводу, что одних морфологических при-
знаков в качестве критерия вида недостаточно.
Новый этап познания вида у высших растений начался во второй
половине XIX столетия. Австрийские ботаники А. Кернер, Р. Веттш-
тейн выдвигали мысль о том, что в понятие вида следует включать его
географическое распространение. Эту точку зрения поддерживали и
некоторые другие зарубежные ученые.
Из российских ботаников идея эколого-географического подхода к
пониманию вида была обоснована С.И. Коржинским. В предисловии
к труду «Флора востока Европейской России» (1892) он пишет, что
реально в природе существуют расы (proles), они обладают морфоло-
гическими отличиями и занимают обособленный ареал. Эти расы пред-
ставляют собой стадии развития вида. Видами С.И. Коржинский счи-
тает вполне сформированные расы с вымершими промежуточными
формами и не смешивающиеся между собой на участках их общего
обитания, хотя и способные спорадически образовывать гибридные
формы. Подвидами он называет не вполне сформированные расы, скре-
щивающиеся между собой при совместном распространении и обра-
зующие разные промежуточные формы. Таким образом, С.И. Коржин-
ский впервые обращает внимание на нескрещиваемость видов, хотя и
не считает это свойство облигатным.
Дальнейшее развитие идея Коржинского получила в трудах
В.Л. Комарова. Во время своих многочисленных экспедиций от евро-
пейской части России до Дальнего Востока и при обработке собран-
ных материалов он убеждался в том, что представления о критериях
вида у ботаников того времени крайне субъективны. В.Л. Комаровым
опубликован ряд статей по проблеме вида, позднее его представления
30
/. История эволюционной биологии
о виде у растений обобщены в монографии «Учение о виде у расте-
ний» (1940). Он доказывал, что каждый вид занимает определенное
место в природе. У высших растений это выражается в том, что каж-
дый вид имеет собственный эколого-географический ареал.
В.Л. Комаровым разработана монотипическая концепция вида.
Сущность этой концепции состоит в том, что вид понимается как пле-
мя, имеющее общее происхождение, стойкие морфологические, фи-
зиологические, биохимические особенности, генетическую целост-
ность и собственный ареал. К важнейшим критериям вида В.Л. Кома-
ров относил генетическую целостность его, что неразрывно связано с
обособленным ареалом. Особый непрерывный ареал вида обеспечи-
вает возможность перекрестного опыления, при котором генотипы
внутривидовых форм объединяются в единый генный фонд вида.
Монотипическая концепция вида получила признание у многих сис-
тематиков, главным образом в России. Зарубежные ботаники большей
частью придерживались политипической концепции. Монотипическая
концепция была принята за основу при составлении фундаментального
труда - «Флоры СССР». Однако автором монотипической концепции была
допущена крайность в смысле переоценки монотипности каждого вида.
В отличие от С.И. Коржинского В.Л. Комаров не признавал выделение
подвидов, переводя их в ранг видов. Еще дальше в этом направлении
пошли некоторые другие авторы обработки материалов для составления
«Флоры СССР» и некоторых региональных «Флор». Без достаточных
оснований широкоареальные виды делились на комплексы мелких мо-
нотипных видов. Это привело к искусственному видодробительству и
дискредитации монотипической концепции в целом.
Возникновение политипической и монотипической концепций от-
ражает определенные пройденные ступени изучения вида ботаника-
ми. На современном уровне развития биологии стало неоспоримым,
что видовые критерии должны прежде всего основываться на позна-
нии вида в эволюционно-генетическом аспекте.
Изучение вида с генетических позиций было начато в двадцатых
годах XX столетия работами С.С. Четверикова (1926), показавшими,
что каждый вид обладает большими возможностями изменчивости,
это обеспечивается богатством его генофонда.
В сороковых годах под влиянием популяционно-генетических пред-
ставлений Ф. Добржанского, автора книги «Генетика и происхожде-
31
А. В. Положий
ние видов» (1937), сформировалась концепция «биологического вида».
Основными идеологами концепции являются американские биологи
Э. Майр и В. Грант. Сущность этой концепции детально изложена при-
менительно к высшим растениям в монографии В. Гранта «Видообра-
зование у растений» (1984). Биологический вид рассматривается как
система скрещивающихся популяций, репродуктивно изолированная
от других систем популяций. Биологические виды не всегда различа-
ются по морфологическим признакам, могут быть виды-двойники.
Единственным критерием биологического вида считается его стопро-
центная репродуктивная изоляция от других видов. Гибридизация про-
исходит только между полувидами, у которых нескрещиваемость с
другими видами не достигла 100%.
Согласно концепции биологического вида в природе существуют
только биологические виды и микровиды. Понятие «биологический
вид», как отмечает сам автор, относится только к бипарентальным (дву-
родительским видам). Микровиды представляют собой апомикты и
популяции, возобновляющиеся вегетативным способом. Таксономи-
ческие виды, по мнению сторонников этой концепции, выделялись
искусственно, они в природе не существуют за исключением некото-
рых случаев, когда таксономический вид соответствует критериям
биологического вида.
С критикой этой концепции выступали как зарубежные, так и отече-
ственные биологи (из ботаников - А. Кронквист, А.Л. Тахтаджян и др.).
А.Л. Тахтаджян в предисловии к русскому изданию монографии
В. Гранта обращает внимание на то, что концепция «биологического
вида» не универсальна, поскольку относится только к бипаренталь-
ным видам, и то не ко всем. Он приводит примеры аллопатических,
пространственно удаленных видов, которые при введении в культуру
успешно скрещиваются (к ним, в частности, относятся некоторые виды
Platanus, Catalpa, сформировавшиеся и произрастающие на разных
континентах).
Совершенно другие критерии применяются к «микровидам», ско-
рее морфолого-географические, но довольно неопределенные.
Вопрос о таксономическом ранге апомиктов дискуссионен. Вероят-
но, что апомикты, так же как и полиплоиды и гибриды, могут служить
материалом для видообразования, но большей частью представляют со-
бой внутривидовые формы или (гибриды) видовые комплексы.
32
I. История эволюционной биологии
Понятие «полувид» очень неопределенно. Автор считает полуви-
ды результатом постепенного видообразования, т.е. молодыми вида-
ми, в то же время он рассматривает некоторые полувиды как результат
естественной гибридизации. Гибридизация очень широко распрост-
ранена в некоторых группах (родах) цветковых растений. На какой
возрастной стадии формирования вида она происходит, установить
можно только путем специальных экспериментальных исследований.
Виды-двойники, не отличающиеся по морфологическим признакам,
но не скрещивающиеся между собой, вероятно, существуют, но со-
ставляют исключение.
Концепция биологического вида полностью игнорирует историю
познания вида в природе, многолетние исследования систематиками
структуры флор. Выдающиеся знатоки природного растительного по-
крова, основывающиеся на позициях эволюционного учения,
С.И. Коржинский, В.Л. Комаров и многие другие ботаники доказали,
что в природе реально существуют морфолого-эколого-географичес-
кие расы, которые получили название видов. Такое понимание приня-
то в систематике высших растений во всем мире. В соответствии с
этим вид принят как основной таксон, номенклатура таксонов разра-
батывается и утверждается на международных конгрессах.
Ботаники, безусловно, понимают, что сущность вида составляет его
генетическая структура, но генный фонд вида представляет собой про-
грамму, которая определяет весь комплекс признаков вида, в том чис-
ле и его морфологические признаки. Связь между генетической струк-
турой вида и его фенотипом неоднократно отмечали известные био-
логи Н.В. Тимофеев-Ресовский (1961), А.В. Яблоков (1980).
Фенотипы, или морфологические признаки, вида как и все другие
характеризуют вид в целом. Генофонд вида обеспечивает широкие
возможности внутривидовой изменчивости, но ограничивает преде-
лы этой изменчивости. Существует не только межпопуляционная, но
и внутрипопуляционная изменчивость, каждая особь имеет какие-то
отличия, поэтому признаки вида нельзя установить по одной или не-
скольким особям. Должен быть исследован обширный многолетний
материал, собранный из разных популяций вида с учетом возрастных
состояний. Кроме того, используются богатые коллекции гербариев.
Таксономические виды (расы) по мере развития науки и техничес-
ких приемов подвергались все более разностороннему изучению.
33
А. В. Положий
Современная систематика использует большое количество мето-
дов: кроме внешней морфологии и эколого-географической характе-
ристики используются анатомические признаки, в необходимых слу-
чаях - особенности микроструктуры (морфология пыльников, пыль-
цы, хромосом и др.), особенности фитохимии и физиологии, гене-
тические связи, а также, по возможности, данные генетики.
Современному пониманию вида у растений и его критериев в наи-
большей степени отвечает определение вида, которое дано А.Л. Тахтад-
жяном (1984). Он определяет вид как обособившуюся в процессе эволю-
ции систему популяций, объединенную общим происхождением, мор-
фологическими, экологическими, физиологическими, биохимическими
признаками, собственным ареалом и изолирующим, в разной степени,
репродуктивным барьером от других видов. Репродуктивная изоляция
вида служит одним из важнейших, но не единственным критерием вида.
Нельзя не учитывать, что виды в разных группах организмов, даже
среди высших растений, существенно различаются по объему, струк-
туре, степени выраженности морфологических отличий. Различия ви-
дов по объему и структуре зависят от эволюционного уровня группы,
к которой они относятся, от территории, на которой происходило их
формирование, от способа видообразования.
У голосеменных растений лесной зоны преобладают виды боль-
шого объема с широким ареалом, большей частью имеющие подви-
ды. У цветковых растений, находящихся на вершине эволюционного
ствола высших растений, темпы эволюции несравненно выше, чем у
голосеменных, спектр местообитаний очень разнообразен в разных
растительных зонах и поясах. В связи с этим у них преобладают виды
меньшего объема, с более локальными ареалами, иногда и монотип-
ные, особенно в горах.
Как полагает А.Л. Тахтаджян, важное значение имеет так называе-
мое «взвешивание», т.е. оценка видового значения признаков с учетом
происхождения, филогенетических связей вида.
Таксономический вид - реальная категория. Вид как таксон необ-
ходим для познания богатства и структуры флор, мониторинга за их
состоянием, познания филогенеза, в частности высших растений, для
экологических и ботанико-гео графических исследований.
Долгое время ботаники-систематики были убеждены в том, что у
высших растений существует только постепенное, градуальное видо-
34
L История эволюционной биологии
образование. Это согласуется с представлением о господствующем
аллопатическом видообразовании у растений. Если возникает барьер,
изолирующий одну или несколько популяций от ареала исходного вида,
они путем мелких мутаций образуют новый подвид или вид.
Однако с позиций градуального видообразования трудно было
объяснить существование резко обособленных видов, узких эндеми-
ков, не обнаруживающих связи с какими-то исходными видами, иног-
да они выделялись систематиками даже в особые монотипные роды.
В начале XX столетия голландским биологом Де-Фризом и
С.И. Коржинским обосновывалась возможность возникновения мак-
ромутаций и, следовательно, вероятность сальтационного видообра-
зования. Но эти идеи не были восприняты систематиками растений.
Значительно позднее А.Л. Тахтаджян (1983) пришел к выводу о
существовании макрогенетических скачков в эволюции высших рас-
тений. По его мнению, процесс микроэволюции, основанный на на-
коплении мелких мутаций, представляет собой основную форму эво-
люции высших растений, но на этом фоне происходят макрогенети-
ческие скачки и сальтационное видообразование. Многие макро-
мутанты вымирают. В.Н. Стегнием (1993) разработана теория систем-
ных мутаций, которая убедительно доказывает существование и при-
роду сальтационного образования видов.
Появление макромутантов, как отмечают А.Л. Тахтаджян и
В.Н. Стегний, происходит в экстремальных условиях - на границах
ареалов, в горах, видимо, также на территориях, подвергавшихся гео-
логическим и климатическим преобразованиям. Именно к таким рай-
онам приурочены распространением резко обособленные виды. Их
сальтационное происхождение не вызывает сомнения.
В.Н. Стегний приходит к выводу о том, что системные мутации
являются непременным условием формирования любого вида эука-
риот и доказывает принципиальную невозможность градуального
видообразования.
Происходит ли видообразование исключительно сальтационным пу-
тем или в некоторых группах организмов, в частности у высших расте-
ний, сочетается с градуальным, остается вопросом дискуссионным.
У высших растений наиболее вероятны оба способа видообразования.
Исследования генетиков внесли и вносят крупный вклад в позна-
ние вида и видообразования. Приходится только сожалеть о том, что
до сих пор отсутствует тесная связь в исследованиях генетиков и си-
35
А. В. Положий
стематиков по этой сложной проблеме. На отрыв экспериментальных
исследований генетиков от исследований систематиков, что приво-
дит к односторонним подходам в познании вида, за последние годы
обращали внимание некоторые ученые (В.М. Медников, 1989;
Р.В. Камелин, 2000).
Хочется надеяться, что более тесный контакт в работе генетиков и
систематиков позволит сблизить понятия биологического и таксоно-
мического вида и разработать обоснованные и доступные для систе-
матиков критерии вида.
Литература
Грант В. Видообразование у растений. М.: Мир, 1984. 520 с.
Камелин Р.В. Систематика сосудистых растений в России (Вехи истории) И Бот. журн.
2000. Т. 85. № 6. С. 2-18.
Комаров ВЛ. Учение о виде у растений. М.; Л.: Изд-во АН ССС Р. 1940. 212 с.
Комаров ВЛ. Флора полуострова Камчатка.Ч. I. М.: Л.: Изд-во АН СССР, 1951.
С. 47-49.
Коржинский С.И. Флора востока Европейской России. Т. I. Томск, 1892. С. 21-27.
Медников Б.М. Современное состояние проблемы вида и видообразования. Вавиловс-
кое наследие в современной биологии. М., 1989. С. 133-147
Стсгний В.Н. Архитектоника генома, системные мутации и эволюция. Томск: Изд-во
Том. ун-та. 1993. 107 с.
Тахтаджян АЛ. Предисловия редактора русского перевода монографии В. Гранта "Ви-
дообразование у растений”, М.: Мир, 1984. С. 5-9.
Тахтаджян А.Л Макроэволюционные процессы в истории растительного мира // Бот.
жури. 1983. Г. 68. № 12.593 с.
Тимофеев-Ресовский Н.В. О некоторых принципах классификации биохорологичсс-
кнх единиц//Труды ин-та биологии Урал фил. АН СССР 1961. Вып. 27. С. 27-29.
Четвериков С.С. О некоторых моментах эволюционною процесса с точки зрения со-
временной генетики//Бюл МОИН. Биология. 1965 Вып 4 С. 33
Яблоков А.В. Фаготика. Эволюция, популяция, признак. М. Наука, 1980. 135 с.
Dobzhansky Th. Geneties and the Origin of Species. New York. Columbia University Press,
1971.364 p.
II. ОБЩИЕ ПРОБЛЕМЫ ЭВОЛЮЦИИ
ИНВАЗИЯ МОБИЛЬНЫХ ЭЛЕМЕНТОВ -
ПРИЧИНА ВЗРЫВНОГО САЛЬТАЦИОННОГО
ВИДООБРАЗОВАНИЯ
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К.И. Пятков,
В. В. Вел и код ворс кая, Н.Г. Шостак, Г.Т. Лезин
Институт молекулярной биологии РАН
117984. г Москва, ул. Вавилова. 32
E-mail. evgenev((/ genome.eimb.relarn ru
Как известно, перемещение мобильных элементов (МЭ) является
одним из главных факторов, вызывающих как точечные, так и хромо-
сомные мутации [1]. Значение таких мутаций для функционирования
отдельных генов и всего генома в целом хорошо известно [2, 3]. Зна-
чительно менее изучена роль МЭ в эволюции эукариотических гено-
мов, крайним случаем которой является появление репродуктивно изо-
лированных форм - видообразование. В последние десятилетия на-
капливается все больше разнообразных фактов, свидетельствующих о
том, что быстрая, взрывная эволюция служит не исключением, а ско-
рее правилом, по крайней мере, на первых этапах, когда происходит
расщепление древней предковой формы на целый веер производных.
Ясно, что в дальнейшем естественный отбор отбраковывает неудач-
ные и поддерживает успешные варианты. Если случаи взрывного, саль-
тационного формообразования хорошо документированы, то причи-
ны таких взрывов далеко не ясны и широко дискутируются. Активиза-
ция различных семейств МЭ и последующая хромосомная революция
неоднократно назывались в ряду таких причин [4, 5]. Описание раз-
личных типов гибридного дисгенеза у дрозофилы [6, 7], сопровождаю-
щихся множественными перестройками, служит сильным аргументом в
пользу такого предположения. С другой стороны, в настоящее время опи-
сано всего лишь несколько случаев ассоциации природных хромосом-
ных перестроек с МЭ [8]. Еще хуже обстоит дело с доказательством того,
37
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К. И. Пятков и др.
что хромосомные перестройки, приведшие к видообразованию или со-
провождающие это явление, были ассоциированы с активностью транс-
позонов. Дело гут в том, что МЭ, по которым могли произойти разрывы
при хромосомных перестройках, приведших к видообразованию, по-ви-
димому, утрачиваются за время последующей эволюции вида.
Для проверки постулируемой последовательности событий: инва-
зия МЭ - хромосомная революция репродуктивная изоляция - видо-
образование необходима удобная экспериментальная модель. «Virilis»
- группа близкородственных видов дрозофилы - отвечает всем требо-
ваниям для того, чтобы служить такой модельной системой.
Во-первых, филогенетические взаимоотношения между 12 видами
группы хорошо изучены [9]; во-вторых, видообразование в этой группе
сопровождалось возникновением хромосомных перестроек (инверсий
и робертсоновских транслокаций); в-третьих, у видов этой группы опи-
сано 6 семейств мобильных элементов, которые активируются при дис-
генных скрещиваниях между линиями D. virilis [10, 11 ] и, наконец,
в-четвергых, D. virilis находится в настоящее время в процессе инвазии
одного из этих МЭ Пенелопы [12]. Интересно, что в отличие от всех
остальных видов группы спонтанные хромосомные перестройки у
D. virilis никогда не были обнаружены ни в лабораторных линиях, ни в
природных популяциях этого вида, несмотря на то, что хромосомы
D. virilis интенсивно изучались [9]. Ранее нами было показано, что Пе-
нелопа является старым компонентом генома во всех остальных ви-
дах группы «virilis» и копии этого семейства локализуются у всех ви-
дов группы (кроме D. virilis) в основном в гетерохроматиновом хро-
моцентре (рис. I, а).
Рис. 1.1 нбрндшация содержащею Пенелоп) клона, а с хромосомами I), texana
(метка локализована в гетерохроматиновом хромоцентре) и б-с инверсией по-
лученной в результате дпегенных скрещиваний линий I) virilis (стрелками oiме-
чены ТР, совпадающие с 11снелоной)
38
II. Общие проблемы эволюции
В настоящей работе мы разработали экспериментальный подход
для получения хромосомных перестроек у предкового вида всей груп-
пы «virilis» - D. virilis. Была изучена связь этих новых перестроек,
полученных у D. virilis в потомстве от дисгенных скрещиваний, с раз-
личными МЭ, а также установлено совпадение точек разрыва этих
перестроек с местами разрыва у перестроек, описанных ранее у дру-
гих видов группы «virilis».
Материалы и методы
Линии Drosophila virilis. Мух содержали на стандартной среде при
25°С. Мы использовали линии из нашей собственной коллекции, под-
держиваемой в Институте биологии развития (Москва), собранные в
природе в различное время, а также полученные из других лаборато-
рий, включая Bowling Green Drosophila Species Center. Номера, обо-
значающие линии, соответствуют номерам линий либо в Bowling Green
Center, либо в коллекции линий Института биологии развития. Линия
9 дикого типа Drosophila virilis была использована для создания ге-
номной библиотеки. Эта линия была заложена в 1972 г. из популяции,
обитающей в окрестностях Батуми (Грузия). Она представляет собой
типичную М-подобную линию и используется в качестве материнской
в дисгенных скрещиваниях. Мы также использовали старую линию
Drosophila virilis (160) из Японии, несущую маркеры b, gp, cd, ре и gl.
Эта типичная P-подобная линия содержит более 40 копий Пенелопы и
неоднократно использовалась в дисгенных скрещиваниях в качестве
отцовской [13]. Из нее также была получена геномная библиотека.
Кроме того, для получения геномных библиотек мы использовали ли-
нию дикого типа (201) D. lummei.
Методы работы с ДНК. Геномные библиотеки из ДНК дрозофил
видов группы «virilis», упомянутых выше, были приготовлены частич-
ным гидролизом Sau3A с последующим лигированием в сайт узнава-
ния рестриктазы BamH 1 вектора X Dash (Stratagene). Рекомбинантные
ДНК фага X были упакованы in vitro при помощи упаковочных экст-
рактов Gigapack Gold extracts (Stratagene). Фаговые реплики снимали
на нитроцеллюлозные мембраны (Millipore) и гибридизовали с 32Р-ме-
неными зондами. Все геномные библиотеки, за исключением библио-
теки, полученной из линии (160) D. virilis, гибридизовали в течение
39
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К. И. Пятков и др.
ночи в мягких условиях (6XSSC, 55°С). Геномную библиотеку, полу-
ченную из линии 160 D. virilis, гибридизовали в течение ночи при 67°С
в 4XSSC. Вставки рекомбинантных Х-клонов субклонировали в век-
тор pUC19 и проводили их рестрикционное картирование. Секвени-
рование клонов, содержащих копии Пенелопы, проводили с использо-
ванием набора реактивов Sequenase II kit (Stratagene).
Зонды, использованные для гибридизации in situ и скрининга
геномных библиотек. В большинстве экспериментов по гибридиза-
ции в качестве зонда мы использовали клон рб, содержащий полно-
размерные копии Пенелопы и около 1 т.п.о. фланкирующих геномных
последовательностей [14]. В некоторых экспериментах мы выделяли
из агарозы внутренний фрагмент Пенелопы, ограниченный сайтами
узнавания рестриктазы Xhol размером 2,6 т.п.о., который представля-
ет полноразмерную копию данного элемента, и использовали его в
качестве зонда для гибридизации in situ.
Цитологический анализ и гибридизация in situ на политенных
хромосомах. Для анализа были использованы как самцы, так и самки
упомянутых выше видов группы «virilis». Слюнные железы извлекали
из личинок третьего возраста в 45%-ной уксусной кислоте и готовили
давленые препараты по методу Лима [ 15]. Для экспериментов по гибри-
дизации in situ личинок выращивали при температуре 18°С и за два дня
до сбора в корм добавляли раствор живых дрожжей. В каждой повторно-
сти использовали не менее 5 личинок. Меченые зонды для гибридиза-
ции in situ получали методом ник-трансляции с использованием биоти-
на- 14-dATP в основном, как описано Лимом. Для определения сайтов
гибридизации использовали фотографическую карту хромосом D. virilis.
Результаты
Распределение Пенелопы и Улисса в хромосомах D. virilis.
Как видно из табл. I, различные географические линии D. virilis
отличаются по числу копий Пенелопы. Некоторые линии вообще не
имеют этого элемента. Мы проанализировали локализацию Пенелопы
и Улисса, мобилизующихся в результате дисгснных скрещиваний в хро-
мосомах 12 изученных линий D. virilis. Всего было отмечено 181 сайт
гибридизации Пенелопы и 144 сайтаУлисса, которые распределились
по 1359 подсекциям хромосом D. virilis. Анализ распределения пока-
40
1L Общие проблемы эволюции
зывает, что наблюдаемое распределение не случайно. Статистический
анализ показал, что только пятая часть всех подсекций, по-видимому,
Таблица 1
Число копий Пенелопы в различных линиях D. virilis
Линии несущие более 20 копий Пенелопы Линии несущие менее 20 копий Пенелопы Линии свободные от Пенелопы
160, 138, 149, 112, 146, 140, Чили, Аргентина, Мексика, Нью-Мексико, Китай, Сендай (Япония), Траки (США) Умань (Украина), 110,142 Магарач (Крым), Ялта (Крым), Краснодар, 101 (Япония), Баку, Берлин, Пасадена, Сейшельские острова, Мцхета, Ереван, Батуми
доступна для инсерции этих МЭ. Замечательно, что наблюдается по-
ложительная корреляция между локализацией Пенелопы и Улисса
(табл. 2). Иными словами, эти элементы статистически достоверно
предпочитают одни и те же районы хромосом для транспозиции.
Анализ распределения сайтов Пенелопы и Улисса по хромосомам
D. virilis позволяет заключить, что отдельные подсекции представля-
ют собой «горячие точки» для внедрения этих элементов. Нам не уда-
лось найти никаких корреляций между этими «горячими точками» и
цитогенетически значимыми районами в хромосомах этого вида, та-
кими как гены, кодирующие 5S, тРНК слабые точки и т.д. С другой
стороны, когда мы сопоставили места инсерции Пенелопы и Улисса в
хромосомах D. virilis с точками разрыва (ТР) хромосомных перестро-
ек, описанных ранее у других видов группы, то обнаружили статисти-
чески высоко достоверную положительную корреляцию (см. табл. 2).
Так, 80 сайтов инсерции Пенелопы и 57 сайтов инсерции Улисса
совпали с точками разрыва инверсий, описанных у других видов груп-
пы «virilis». Возникает парадоксальная ситуация: с одной стороны, мы
наблюдаем высокодостоверное совпадение сайтов инсерции МЭ, ак-
тивируемых при дисгенезе с группами хромосомных перестроек у
других видов группы, а с другой стороны, никто никогда не наблюдал
спонтанных хромосомных аберраций в самом виде D. virilis. Для объяс-
нения кажущегося противоречия мы предположили следующую пос-
ледовательность событий. Древняя предковая форма, давшая начало
всей группе «virilis», подверглась инвазии мобильного элемента Пене-
41
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К.И. Пятков и др.
лопы, что вызвало активизацию других МЭ, множественные хромо-
сомные перестройки и послужило в конечном итоге пусковым меха-
низмом видообразования. Подтверждением такой последовательнос-
ти событий служит обнаружение МЭ на границах перестроек у видов
группы «virilis» [16]. Вид D. virilis в настоящее время находится в про-
цессе инвазии Пенелопы [12], и поскольку транспозиции МЭ неслу-
чайно распределяются по длине хромосомы, мы наблюдаем совпаде-
ние современных транспозиций МЭ с границами инверсий, возник-
ших в результате предыдущей инвазии (или инвазий) Пенелопы.
Остается, однако, неясным почему мы не наблюдаем аберрации у
D. virilis в настоящее время, если этот вид в самом деле находится в
процессе инвазии Пенелопы.
Таблица 2
Неслучайное распределение Пенелопы (Пен.), Улисса и точек
инверсионных разрывов (ТР) в геноме D. virilis
Сравниваемые величины Снаб Сожид Достоверность р Совпадение р
Пен. V? ТР 80 18,78 <<0,0001 0,50
Улис vs ТР 57 14,94 *<0,0001 0,33
Пен. vs Улис 82 19,18 «0,0001 0,50
1 Пен&Улис vs ТР 3) 56 8,51 «0,0001 0,65
Примечание. Ciut6 - реально наблюдаемое число совпадений; Са/кш) - ожидае-
мое число совпадений исходя из того, что общее число дисков в гюлитенных хромосо-
мах D. virilis равно 1359.
Пен. vs Улисс отражает число совпадающих сайтов инсерции этих элементов (82).
Общее число инверсионных точек разрыва у видов группы «virilis» равно 141, а число
сайтов инсерции Пенелопы и Улисса у D. virilis равно 181 и 144 соответственно.
Достоверность - р и коэффициент совпадения - У рассчитывали согласно ранее
описанным процедурам.
Хромосомные перестройки в потомстве
дисгенных скрещиваний
Для поиска хромосомных перестроек у D. virilis мы скрещивали
самок линии 9, не имеющих Пенелопу, с самцами линии 160, несущи-
ми более 30 копий этого элемента, т.е. проводили типичное дисгенное
скрещивание [13]. Потомство Fj от таких скрещиваний было скреще-
но inter se, и в дальнейших поколениях F2 и F3 мы отбирали особей с
выраженными морфологическим аномалиями. Этих особей скрещи-
42
1L Общие проблемы эволюции
вали с пятью самцами или самками линии 9 и анализировали слюн-
ные железы 10-15 личинок от таких скрещиваний.
В результате такого анализа нам удалось выделить 16 хромосом-
ных перестроек, включая 12 инверсий, две транслокации и две деле-
ции (табл. 3). Наш метод не позволяет точно определить частоту воз-
никновения перестроек в потомстве дисгенных форм, поскольку оче-
видно, что возникающие аберрации не всегда приводят к феноти-
пическим изменениям, обнаруженные ненормальные фенотипы часто
отличались стерильностью. Проведенный анализ (см. табл. 3) показал,
что значительная часть разрывов этих дисгенных перестроек совпадает
с местами локализации Пенелопы и/или Улисса у родительских линий.
Отклонения от случайности высокодостоверны. В большинстве случаев
МЭ сохранялся в обеих точках разрыва (табл. 4, рис. 1, 6). Такой харак-
тер совпадения напоминает аналогичную картину, наблюдаемую при
возникновении перестроек в случае Р-m и Hobo типов дисгенеза, ког-
да перестройки происходят благодаря эктопическому контакту между
предсуществующими (у родительской линии) сайтами МЭ.
По-видимому, перестройки у D. virilis происходят по такому же меха-
низму. Важным отличием является то, что перестройки, полученные в
результате дисгенных скрещиваний у D. virilis, несут на своих границах
не один (как в случаях гибридного дисгенеза у D. melanogaster), а по край-
ней мере два (Пенелопа и Улисс) элемента, активизируемых при дисге-
незе.
Т а б л и ц а 3
Перестройки, полученные в потомстве от дисгенных скрещиваний линий
D. virilis
Тип перестройки Общее число Присутствие Пенелопы или Улисса в ТР Совпадение с перестройками, описанными ранее у видов группы «virilis»
Транслокация 2 1 1
Делсция 2 1 —
Инверсия 15 10 11
В данной работе мы провели также сравнение границ разрывов
«свежих» дисгенных инверсий, полученных у D. virilis, с границами
инверсий, описанных ранее у других видов группы. Данные, приве-
денные в табл. 4, показывают, что и в данном случае наблюдается вы-
43
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К.И. Пятков и др.
соко достоверное совпадение. Так, 12 точек разрыва “свежих” инвер-
сий совпали с границами “старых” перестроек при ожидаемой вели-
чине 3,29 (при случайном распределении).
Таблица 4
Характеристика хромосомных перестроек, полученных в потомстве дисгенных
скрещиваний D. virilis
Тип перестройки Располо- жение дистальной Т Располо- жение прокси- мальной TP Пенелопа в линии 160 Улисс в линии 160 или 9 Инверсионные TP, ранее описанные у видов группы «virilis» +
Инверсия XDk Xie
Xia XTf XTf(N,T,A)
XRc XXa
2Bj* 2Gd 2Bj
2Ij* 2Za* ?Ij, 2Za 2Ij (Mo, La, Ezo, Lu); 2Za (Mo, La, Li, Ezo, Ka)
2Si/Ta 2Zc 2Si/Ta (Ezo, Lu); 2Zc (Ka)
3FeS# 3Z# 3Fe, 3Z 3Fe (Li); 3Z (N, A, Mo, La)
3Hk* 3Wf* 3Hk, 3Wf 3Hk (Mo, La)
3Nhi 3Ri 3Nhi (A, N)
4Mhi* 4Sa* 4Mhi, 4Sa
4Od 4Sa* 4Sa 4Od (Mo)
51ab# 5Pn# 5 lab, 5Pn 5Pn (Ezo, Ka)
Транслока- ция XHki 5Ma
2Eh* 4Ga* 2Eh, 4Ga 4Ga (Li)
Деления 5Ab 5Ci 5Ci
2Eh 2Fa
Всего 7/16 7/16 11 4 12
* Совпадение с локализацией Пенелопы в родительской линии 160.
# Совпадение с локализацией Улисса в родительских линиях 160 или 9.
Сокращенные обозначения видов N: D. novamexicana; Т: D. texana; A: D. americana,
Mo: D. montana; La: D. lacicola; Ezo: D. ezoana; Lu: D. lummei; Li: D. littoralis; Ka:
D. kanekoi. Li: D. littoralis.
Примечание. Совпадение между расположением ТР дисгенных инверсий (32) и
ТР инверсий, описанных ранее у других видов группы “virilis” (141), не случайно:
12 случаев совпадений против ожидаемых 3,2, исходя из наличия 1359 дисков.
44
II. Общие проблемы эволюции
Полученные результаты являются прямым доказательством наших
представлений о ключевой роли инвазии мобильных элементов в ин-
дукции хромосомного полиморфизма (хромосомная революция), ко-
торая при определенных внешних условиях может привести к репро-
дуктивной изоляции.
Неслучайное распределение инсерций МЭ, а также границ пере-
строек по длине хромосомы можно объяснить следующим образом:
во-первых, возможно, что интеграза (эндонуклеаза) Пенелопы, обес-
печивающая транспозиции, может иметь определенную специфичность
узнавания на уровне нуклеотидов, во-вторых, возможно, что специ-
фичность разрывов определяется структурой хроматина в тот доста-
точно узкий период онтогенеза, когда МЭ находится в активном со-
стоянии. Иными словами, предпочтительной мишенью инсерций МЭ
являются гены, функционирующие (диффузный хроматин) в момент
активации МЭ. Следуеттакже помнить, что мы можем наблюдать транс-
позиции МЭ и перестройки только в тех случаях, когда они не наруша-
ют жизненно важные локусы. Таким образом, ясно, что лишь часть
генома “открыта” для инсерций МЭ и перестроек.
Для того чтобы выяснить, насколько специфичны на нуклеотид-
ном уровне инсерции Пенелопы, мы клонировали один и тот же хро-
мосомный локус, содержащий этот элемент, из D. virilis и близкого вида
D. lummei.
Структура сайта инсерции Пенелопы у D. virilis и D. lummei
Мы отобрали из геномной библиотеки D. lummei клон, гибридизу-
ющий с клоном р 6 (9) D. virilis, который содержит интрон “eyeless”
гена, несколько тандемно расположенных Pdv элементов [17J, а так-
же копии Пенелопы. При скринировании библиотеки D. lummei мы
пользовались уникальной частью клона рб. Гибридизация обоих кло-
нов in situ на политенных хромосомах линий D. virilis и D. lummei,
свободных от Пенелопы, показал, что данный сегмент ДНК располо-
жен в обоих видах в 49F подсекции хромосомы 4. Рис. 2 иллюстриру-
ет структуру гомологичных участков ДНК, выделенных из геномных
библиотек D. virilis и D. lummei.
Видно, что оба сегмента содержат копии Пенелопы, а также одни
и те же фланкирующие последовательности. Ясно также, что в обоих
45
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К.И. Пятков и др.
случаях копии Пенелопы находятся в разной ориентации по отноше-
нию к фланкирующим последовательностям, а в случае D. lummei при-
сутствуют лишь фрагменты копии Пенелопы. Можно заключить, что
в изучаемых видах инсерции Пенелопы произошли в один и тот же
район хромосомы, на уровне нуклеотидов сайты инсерции не совпа-
дают. Сравнение элемента Пенелопы, выделенного из генома
D. lummei, с копией D. virilis (рб) показало, что копия Пенелопы у
D. lummei содержит много делений, инсерций и замен на уровне ос-
нований и, очевидно, не функциональна. Идентичность на уровне нук-
леотидов между клонами Пенелопы обоих видов составляет лишь 82%.
1 kb
Рис. 2. Схематическое изображение Пенелопы, выделенной из геномных библиотек (/1)
D virilis, (Б) D. luinmei. Серым цветом обозначены последовательности Пенелопы в
обоих клонах, черным цветом - последовагсль нести гена eyeless, стрелки направле-
ние транскрипции, латинские буквы-сайты рестрикции (X-Xhol. N-Ndel, E-EcoR I,
В-ВаМН1)
Полученные результаты показывают, что наблюдаемая неслучай-
ность инсерций Пенелопы и других элементов, мобилизующихся при
дисгенных скрещиваниях у D. virilis, осуществляется не на уровне
нуклеотидов. Клонирование сайтов разрывов новых дисгенных хро-
мосомных перестроек и старых, давно существующих инверсий, опи-
санных у других видов группы «virilis», покажет, насколько точно они
совпадают, и поможет ответить на вопрос: могут ли идентичные хро-
мосомные перестройки повторно возникать в процессе эволюции у
одного и того же или близкородственных видов. Описание и характе-
ристика хромосомных перестроек у D. virilis позволяет по-новому
представить себе ход эволюции в группе «virilis», который, возмож-
46
//. Общие проблемы эволюции
но, если не универсален, то весьма распространен. Полученные ре-
зультаты можно объяснить следующим образом. Инвазия Пенелопы
произошла по крайней мере однажды на ранних этапах истории этой
группы. Эта инвазия, возможно произошедшая горизонтально, при-
вела к активации ряда не родственных семейств транспозонов, что
повлекло к возникновению множества хромосомных перестроек -
«транспозиционному взрыву», послужившему материалом для видо-
образования. Следы этой первой инвазии мы обнаруживаем в виде
дивергированных копий Пенелопы и Улисса на границах видоспеци-
фических инверсий у видов этой группы [17]. Подобная древняя ко-
пия Пенелопы, выделенная из генома D. lummei, описана в этой рабо-
те. В настоящее время вид D. virilis находится в процессе вторичной
инвазии Пенелопы, которая способна вызывать мобилизацию других
семейств МЭ в дисгенных скрещиваниях, что приводит к хромосом-
ным перестройкам, часто совпадающим с аберрациями, описанными
у других видов группы. Чрезвычайно важно для постулируемой цепи
событий было бы обнаружение хромосомных перестроек в природ-
ных популяциях D.virilis, претерпевших недавнюю инвазию Пене-
лопы.
Литература
1 Arkhipova 1 R., Lyuboimrskaya N.V, Ilyin Y V. (1995) Drosophila retrotransposons. R.G
Landes Company, Austin, Texas.
2 . Lim J K. & Simmons M J (1994) Bioessays 16. 269-275
3 Mathiopoulos K.D . della Torre A , Predazzi V, Petrarca V & Coluzzi M (1998) Proc Natl
Acad Sci USA. 95 12444 -12449
4 Craig N L (1997) Annu. Rev. Biochem. 66. 437 -474.
5 . Engels W R. & Preston C R (1984) Genetics 107. 657-678
6 . Busseau 1 A., Pelisson A & Bucheton A (1989) Mol. Gen Genet. 218. 222-228.
7 Kidwell M G , Kidwell.! E & Sved J A (1977) Genetics. 36. 813-833
8 Lyttle Г W , Haymer D.S. (1992) Genelica 86 113-126
9 . Patterson .1 T & Stone W.S. (1952) Evolution in the Genus Drosophila (The Macmillan
Co , New York)
10 Vieira J. Vieira CP, Hartl DL and Lozovskaya E.R. (1998) Genet. Res 71. 109-117
11 Petrov. D A , Schutzman J L., Hartl D L & Lozovskaya E R (1995) Proc Natl. Acad
Sci 92 8050- 8054
12 Evgen'ev M , Zelentsova IL, Mnjoian L, Poluectova 11 & Kidwell M.G (2000)
Chromosoma 110 375-380
13 . Lozovskaya E R , Schemker VS. & Evgen'ev MB. (1990) Genetics 126 619-623
47
М.Б. Евгеньев, Е.С. Зеленцова, Е.С. Полуэктова, К.И. Пятков и др.
14 . Evgen'ev MB , Zelcntsova 11, Shostak N., Kozitsina M , Barsky V, Lankenau D -I I &
Corccs VG (1997) Proc. Nall. Acad. Sci. USA. 94 196-201.
15 Lim J K. (1993) Dros Inf Serv 72 73-77.
16 Zelcntsova H , Poluectova II., Mnjoian L , Lyozm G , Vcleikodvorskaja V., Zhivotovsky
L, Kidwell M G & Evgen'cv M B (1999) Chromosoma 108
17 Zelcntsova E.S , Vashakidze R.P., Kraev A S. & Evgen’cv M B (1986) Chromosoma 93
469-475
ГЕНЫ, ОНТОГЕНЕЗ И ПРОБЛЕМЫ
ЭВОЛЮЦИОННОГО РАЗВИТИЯ
Л.И. Корочкин
Институт биоло! ии гена PA! I, Институт биологии развития им И.К. Кольцова PAI I
117334, Москва, ул Вавилова, 34/5
E-mail: likor^nix.ibg.relarn.rii
Проблема вида и видообразования всегда замыкается на общеэво-
люционную концепцию, венцом которой принято считать синтетичес-
кую теорию эволюции (СТЭ). В этой теории (а вернее - гипотезе)
была предпринята попытка совместить с дарвиновскими градуализ-
мом и естественным отбором классическую генетику, первоначально
достаточно резко с ним расходившуюся (Грант, 1991). К числу созда-
телей СТЭ принадлежат выдающиеся российские ученые Н.С. Чет-
вериков и Ф.Г. Добжанский. Следует отметить крупный вклад в раз-
витие «ортодоксальных» сторон СТЭ наших замечательных биологов
А.Н. Северцова, И.И. Шмальгаузена, Д.К. Беляева, А.В. Яблокова,
Л.П. Татаринова, Э.В. Воробьевой, В.А. Ратнера, А.С. Северцова и др.,
результаты исследований которых имеют огромную ценность и дол-
жны учитываться при построении любой эволюционной схемы.
В то же время как в зарубежной, так и в отечественной науке скла-
дывались взгляды, противополагаемые СТЭ (или, во всяком случае,
существенно ее модифицирующие, не-
редко на уровне философско-биологи-
ческом). В зарубежной эволюционис ги-
ке наиболее распространены гипотеза
системных мутаций Р. Гольдшмидта, те-
ория прерывистого равновесия Стэнли-
Элдриджа и гипотеза нейтралистской
эволюции Кимуры, Джукса и Кинга.
Рис. 1. Лев Семенович Верг. Великий российс-
кий ученый - географ и биолог, создатель ори-
гинальной эволюционной концепции, предвос-
хитивший многие современные взгляды на те-
чение филогенеза
49
П.И. Корочкин
В отечественной биологии можно выделить три этапа становления
недарвиновских взглядов на процессы эволюционного развития (под-
робнее см.: Назаров, 1991).
Первый - концепция номогенеза Л.С. Берга (рис. 1), сформулиро-
ванная в 20-е годы. Суть ее заключается в постулировании иных движу-
щих факторов эволюционного процесса, нежели формулировал Дарвин
и СТЭ: вместо монофилии - попифшшя, вместо градуальности - скачко-
образность, вместо случайности -- закономерность (см.: Берг, 1977).
Второй этап (60-70-е годы) - работы Ю.П. Алтухова и Н.Н. Во-
ронцова. Первый [на Западе ему вторит Carson (1975)] подразделил
геном на полиморфный и мономорфный и выдвинул гипотезу, соглас-
но которой полиморфизм и обеспечивающая его полиморфная часть
генома способствуют сохранению постоянства вида, но не его «транс-
формации» в другой вид, расширяют приспособительные возможнос-
ти вида и соответственно ареал его распространения. Видообразова-
ние же происходит за счет скачкообразного изменения мономорфной
части генома (Алтухов, 1983).
Рис. 2 Николай Николаевич Воронцов Выдающий-
ся зооло!-эволюционис'1. реформировавший сишс-
тичсскую теорию эволюции, внес крупный вклад в
развитие отечественной экологии
Н.Н. Воронцов (рис. 2) сформулировал
концепцию мозаичной эволюции и глубо-
ко разработал учение о хромосомном ви-
дообразовании, роли макромутаций и сей-
смических факторов в филогенезе (Ворон-
цов, 1999).
Третий этап (80- 90-е годы) знамену-
ется выдающимся, с моей точки зрения, о г-
крытием томского генетика В.Н. Стегния
(1991), продемонстрировавшего видоспецифич кость точек прикреп-
ления политенных хромосом насекомых к ядерной мембране и уста-
новившего отсутствие полиморфизма по данному признаку. Следова-
тельно, видообразование по постулированному ранее СТЭ принципу
постепенного изменения генных частот в данном случае исключается
и должно происходить путем скачка, макромутации.
50
II. Общие проблемы эволюции
Будучи сторонником гипотезы о «макромутационной эволюции»,
я всегда придавал существенное значение взаимоотношению, или, если
хотите, единству исторического и индивидуального развития (Ко-
рочкин, 1999; Korochkin, 1993).
Об этом единстве заговорили сразу после создания эволюционно-
го учения. Это и понятно, поскольку эволюционные преобразова-
ния не могли начинаться иначе как через изменения программы
индивидуального развития.
Первоначально это единство выражали в так называемом БИОГЕ-
НЕТИЧЕСКОМ ЗАКОНЕ. Основываясь на работах Меккеля и Дарвина,
немецкий биолог Фриц Мюллер еще в 1864 г. высказал идею о тесной
связи предков с эмбриональным развитием потомков. Эта идея была пре-
образована в биогенетический закон известным дарвинистом Эрнстом
Геккелем. В 1866 г. он сформулировал этот закон следующим образом:
«Онтогенез является коротким и быстрым повторением филогенеза, по-
вторением, обусловленным физиологическими функциями наследствен-
ности (воспроизведения) и приспособленности (питания)».
Следует сказать, что наиболее выдающиеся эмбриологи того време-
ни критически восприняли идеи Мюллера-Геккеля. Свое негативное
отношение к ним выразили А. Келликер, В. Гис, Карл Бэр, Оскар Герт-
виг, А. Седжвик. Последний заметил, в частности, что глоточные дуги
и щели эмбриона млекопитающих гомологичны жаберным дугам и
щелям рыб, однако нет свидетельств в пользу того, что они выполняли
в прошлом жаберную функцию. Без таких свидетельств нельзя утвер-
ждать, что жаберные щели и дуги доказывают происхождение млеко-
питающих от рыбообразных предков.
Новое в онтогенезе возникает не за счет прибавления новых ста-
дий к онтогенезу предков, как думал Геккель, но главным образом за
счет такого изменения хода эмбриональных стадий, которое преобра-
зует процесс онтогенеза в целом. Клайненберг (Kleinenberg, 1886)
высказал мысль, что такие, казалось бы, лишенные функции эмбрио-
нальные структуры, как хорда или трубчатая закладка сердца у позво-
ночных, считавшиеся простыми примерами рекапитуляции, возмож-
но, имеют жизненно важное значение для развития зародыша, прини-
мая участие в формировании более поздних структур. С. Уитман
(С. Whitman), один из основателей американской эмбриологии, про-
рочески писал в 1895 г., что наши глаза похожи на глаза наших пред-
ков не вследствие генеалогических связей, а в связи с тем, что
51
Л. И. Корочкин
молекулярные события, лежащие в основе морфогенеза глаз, про-
исходили в сходных условиях.
Наконец, давно известно такое явление, как ПРЕАДАПТАЦИЯ. Еще
Карл Бэр (рис. 3) писал, что если бы биогенетический закон был прави-
лен, то в развитии некоторых животных не наблюдалось бы в проходя-
щем состоянии образований, которые остаются навсегда лишь у выше-
Рис 3 Карл Бэр - отец современной эмбрио-
логии. Выступил категорически против дар-
виновской эволюционной гипотезы
стоящих форм: у всех мле-
копитающих в первое
время развития челюсти
так же коротки, как у чело-
века, мозг птиц в течение
первой трети эмбриогене-
за гораздо более близок к
мозгу млекопитающих,
чем во взрослом состоя-
нии. А российский палеон-
толог А.П. Павлов еще в
1901 г. показал, что моло-
дые особи некоторых ам-
монитов обладают призна-
ками, которые исчезают в
зрелом возрасте, но впос-
ледствии снова обнаружи-
ваются у более высокосто-
ящих в системе форм, у ви-
дов, появляющихся в более
новых отложениях. Следовательно, предковые, как говорят, анцестраль-
ные формы, в молодом состоянии как бы предвещают то, что со време-
нем будет свойственно взрослым особям выше в эволюционном отно-
шении стоящих форм (см.: Назаров, 1991).
В 20-30-е годы критика биогенетического закона была продолже-
на учеником Седжвика Ф. Гарстангом, утверждавшим, что онтогенез
не повторяет филогенез, а творит его. В его поддержку высказались
Л. Берталанфи, Т. Морган и др. Морган, в частности, отметил, что в
ходе эволюции сами эмбриональные стадии могут изменяться и те-
рять сходство с соответствующими эмбриональными стадиями более
ранних форм, а потому, если теория рекапитуляции является «зако-
52
II. Общие проблемы эволюции
ном», то он имеет так много исключений, что становится бесполез-
ным и часто ошибочным. Понимая всю серьезность такого рода воз-
ражений и стремясь тем не менее «спасти» биогенетический закон,
выдающийся российский биолог А.Н. Северцов (1967) выдвинул тео-
рию филэмбриогенеза, под которым он понимал эмбриональные из-
менения, связанные с филогенетическим развитием взрослого орга-
низма. Он выделил три типа филэмбриогенеза: 1) АНАБОЛИЯ - над-
ставка конечной стадии (например, развитие челюстей у саргана);
2) ДЕВИАЦИЯ - изменение пути развития (развитие чешуи у акуловых
рыб и рептилий); 3) АРХАЛЛАКСИС - изменение первичных зачатков.
Однако работы А.Н. Северцова не положили конец критике пред-
ставлений Геккеля-Мюллера. Негативное отношение к ним продемон-
стрировали палеонтолог Шарль Депере, зоолог А.А. Любищев, эмб-
риологи Д. Дьюор (Dewar), С.Г. Крыжановский, физиолог И. А. Аршав-
ский и др.
Дьюор, например, заметил, что пищеварительный канал эмбриона
некоторое время замкнут в себе и отделен от рта и анального отвер-
стия, а это не может представлять какую-то предковую стадию: разве
можно представить такую фазу эволюции, когда примитивный пище-
варительный канал имеет смысл в качестве слепой трубки? Закладка
однопалой конечности лошади с самого начального момента обнару-
живает четкую специфичность - латеральные пальцы в ходе эволю-
ции не рекапитулируют в онтогенезе, утраченные в эволюции пальцы
редуцированы в самой ранней эмбриональной закладке, что противо-
речит биогенетическому закону (Dewar, 1931).
Сходные противоречия обнаруживаются и в ходе сравнительно-
эмбриологических исследований. Как уже отмечалось, становление
плана строения тела различных организмов в онтогенезе обусловлено
изменениями паттерна экспрессии генов сегментации и гомеозисных
генов. Стадия, на которой эмбрионы одного филума обнаруживают
наивысший уровень морфологического сходства друг с другом, обо-
значается как ФИЛОТИПИЧЕСКАЯ стадия. Стадия, на которой уста-
навливаются различия в плане строения тела животных разных филу-
мов, связанные с функционированием гомеозисных генов, обознача-
ется как ЗООТИПИЧЕСКАЯ стадия.
Например, хордовые проходят стадию развития, на которой они
имеют сходное устройство нервной трубки, нотохорды и сомитов. Эта
53
Л. И. Корочкин
стадия является той филотипической точкой, на которой устанавлива-
ется региональная идентичность паттерна экспрессии гомеозисных
генов. При этом, несмотря на консерватизм филотипической и зооти-
пической стадий, специалисты по генетике развития определяют, что
начальные стадии эмбриогенеза внутри каждого филума характеризу-
ются значительным разнообразием. Например, в то время как эмбрио-
ны человека, цыпленка и рыбки полосатая данио (zebrafish) выглядят
сходно на филотипической стадии, их более ранние стадии развития
морфологически совершенно РАЗЛИЧНЫ, что находится в противо-
речии с биогенетическим законом (см.: Корочкин, 1999).
Спрашивается, отражают ли эти морфологические и морфогене-
тические различия какую-либо соответствующую этим различиям
молекулярно-генетическую специфичность? Имеющийся фактичес-
кий материал позволяет предположить, что молекулярно-генети-
ческая «машина» является во всех случаях сходной, а морфоло-
гические различия обусловлены сдвигами во ВРЕМЕННОЙ ПОС-
ЛЕДОВАТЕЛЬНОСТИ одних и тех же молекулярных событий.
Именно разнообразие временного развертывания сходных моле-
кулярно-генетических процессов детерминирует различия в мор-
фогенезе разных таксонов.
Эти особенности можно проследить на примере эволюции насеко-
мых. Например, у дрозофилы «полноценный» паттерн сегментов тела
устанавливается уже к концу стадии бластодермы. Эмбрионов таких
насекомых (мухи, пчелы) называют «зародышами с длинной заклад-
кой» (long-germ).
У таких животных, как кузнечик, синцитий и клеточная бластодер-
ма формируются, как и у дрозофилы, но только малая фракция бласто-
дермы, обозначенная как зародышевая закладка (germ aniage), вносит
вклад в развитие эмбриона. Остальная бластодерма дает начало экст-
раэмбриональным мембранам. Эксперименты показали, что план стро-
ения животного в этом случае не представлен полностью в зародыше-
вой закладке. Из нее возникает только головная область зародыша, все
остальные части эмбриона генерируются из зоны роста, которая про-
дуцирует соответствующий материал путем пролиферации клеток,
следующей за процессом гаструляции. Такие эмбрионы называют «за-
родышами с короткой закладкой» (short-germ). Есть эмбрионы с про-
межуточным типом развития, когда, например, голова и грудь разви-
54
II. Общие проблемы эволюции
ваются из зародышевой закладки, а брюшная область генерируется
позднее. Все явления такого рода нелегко согласовать с биогенетичес-
ким законом, а потому скептическое к нему отношение становится
преобладающим и понятным.
Собственно, в настоящее время этот закон всерьез принимается
лишь в отечественной литературе, в западных источниках по эмбрио-
логии и генетике развития он обычно не упоминается или отрицается.
Яркий тому пример - книга Р. Рэффа и Т. Кауфмана «Эмбрионы, гены,
эволюция» (1986).
Вот что пишут авторы по этому поводу: «В конечном счете роко-
вые слабости биогенетического закона заключались в его зависимос-
ти от ламарковской теории наследственности и в его непременном
условии, что новая эволюционная ступень может быть достигнута толь-
ко как добавление к взрослой стадии непосредственного предка. Вто-
ричное открытие и развитие менделевской генетики на рубеже двух
столетий покажет, что в сущности биогенетический закон - это всего
лишь иллюзия» (с. 30). И еще: «Последний удар биогенетическому
закону был нанесен тогда, когда стало ясно, что морфология и морфо-
логические адаптации имеют важное значение не только для взросло-
го организма, но и для всех стадий его онтогенеза... Если морфология
развивающегося организма имеет такое же важное, а может быть, и
еще более важное значение, чем его морфология во взрослом состоя-
нии, то это трудно согласовать с геккелевской моделью эволюции.
В совокупности менделевская генетика, обособленность клеток заро-
дышевой линии и важность морфологических признаков на всем про-
тяжении развития положили конец теории рекапитуляции...» (с. 31).
Тем не менее у нас нет оснований сомневаться в том, что индиви-
дуальное и историческое развитие организмов тесно друг с другом
связаны и, следовательно, несмотря ни на что, составляют некое ЕДИН-
СТВО. В чем же это единство заключается? На мой взгляд, при его
оценке следует исходить из того обстоятельства, что как индиви-
дуальное, так и эволюционное развитие основываются на одном
и том же материале, а именно ДНК, и потому им должны быть
присущи ОБЩИЕ закономерности.
Трудно представить, что в онто- и филогенезе наследственная ин-
формация, заключенная в ДНК, развертывается разным способом. Тем
не менее допущение этих различий является в настоящее время обще-
55
Л. И. Корочкин
принятым. Действительно, предполагается, что филогенез осуществ-
ляется на базе нецелесообразных, ненаправленных процессов и осно-
вывается на постепенном накоплении в популяции случайных, т.е. не-
направленных мелких мутационных изменений. Однако, исходя из
единства индивидуального и исторического развития, а также из
единства принципа, лежащего в основе того и другого, гораздо
разумнее и логичнее распространить экспериментально выявлен-
ные и доказанные особенности индивидуального развития на вто-
ричные и обусловленные ими эволюционные события, которые
сами по себе не поддаются, как правило, точной эксперименталь-
ной проверке, а потому формулируются как спекулятивные, «под-
тянутые» под ту или иную экспериментально неверифицирован-
ную концепцию.
При экстраполяции точных данных генетики развития на филоге-
нетические процессы следует опираться на следующие факты:
1. Индивидуальное развитие подчинено реализации развивающимся
организмом определенной «цели» - преобразованию во взрослый, де-
финитивный организм, следовательно, оно ЦЕЛЕСООБРАЗНО. От-
сюда следует предположение о целесообразности и эволюционного
процесса, коль скоро его осуществление зависит от того же самого
материала - ДНК.
2. Процесс онтогенеза НЕ СЛУЧАЕН, он протекает направленно
от стадии к стадии, так что прохождение одной стадии с необходимо-
стью влечет за собой переход на следующую, точно запрограммиро-
ванную стадию развития. Всякого рода случайности исключают точ-
ную реализацию плана нормального развития. Отчего же эволюция
должна основываться на случайных мутациях и идти неведомо куда
по «ненаправленному» пути? Собственно, посмотрев внимательно на
различные эволюционные ряды, на появление в этих рядах сходных
структурных образований (крылья у птиц, у летучих мышей, насеко-
мых, древних рептилий, подобие крыльев у некоторых рыб), поневоле
начинаешь подозревать наличие как бы предопределенного генети-
чески, ЗАКОНОМЕРНОГО, «запрограммированного» в самой
структуре ДНК (как и в случае индивидуального развития) фи-
логенеза, как бы направленного по некоему «преформированно-
му» каналу, о чем, собственно, высказывался уже Лев Семенович Берг
в своей теории НОМОГЕНЕЗА.
56
Ih Общие проблемы эволюции
3. В ходе онтогенеза фазы относительно «спокойного» развития
сменяются так называемыми критическими периодами, характеризу-
ющимися морфогенетической активностью ядер и активацией фор-
мообразовательных процессов. Очевидно, и в эволюции длительные
фазы стазиса, покоя, должны сменяться взрывами видообразова-
ния. Иными словами, эволюция носит не градуалистский, а пунк-
туалистский, СКАЧКООБРАЗНЫЙ характер.
Следует отметить, что среди эмбриологов давно наблюдалась тен-
денция рассматривать феномен эволюции не как результат накопле-
ния мелких мутаций под влиянием естественного отбора, постепенно
и медленно ведущих к формированию нового вида через бесконечные
ряды промежуточных форм, а как следствие внезапных и коренных
преобразований процессов онтогенеза, сразу вызывающих появление
нового вида. Еще Е. Рабо (Rabaud) в 1908 г. допускал, что видообразо-
вание может быть сопряжено с мутациями большой амплитуды, про-
являющимися на ранних этапах морфогенеза и нарушающими слож-
ную систему онтогенетических корреляций. Е. Гийено(Сиуеппо1, 1921)
отметил, что Ж. Бюффон был близок к истине, когда, описывая неле-
пое строение и форму клюва, характерные для некоторых видов птиц,
причислял их к тератологическим уклонениям, едва совместимым с
жизнью. Он подметил, что одни и те же уродства у некоторых групп
беспозвоночных (например, иглокожих) представляются то как слу-
чайные индивидуальные особенности, то как постоянные признаки ви-
дов, родов и семейств. Он предположил, что некоторые катастрофи-
ческие уродства являются очевидными следствиями макромутаций,
изменяющих ход онтогенеза. Это - исчезновение способности к поле-
ту у многих птиц открытых пространств (страусы, казуары, эпиорнис),
которое отнюдь не является следствием приспособления к бегу, но
принадлежит к числу уродств, обрекающих его носителей на един-
ственно возможный образ жизни в ограниченном биотопе. Усатые киты
- настоящий парадокс природы и живая коллекция уродств. Гийено
считает, что любое животное можно описать в терминах тератологи-
ческого языка. Так, передние лапы крота - пример ахондроплазии (на-
рушение окостенения длинных костей конечностей), среди китов на-
блюдается двусторонняя эктромелия (врожденное отсутствие конеч-
ностей), у змей - амелия. Организация человека уродлива по сравнению
с его предками. Таковы, например, анатомические особенности, свя-
57
Л. И. Корочкин
занные с вертикальным положением тела, отсутствие хвоста, сплош-
ного волосяного покрова и т.д.
В 1957 г. бельгийский эмбриолог Альберт Дальк (Dalcq, рис. 4)
предположил, что со времени кембрия посредством радикальных транс-
формаций самых ранних стадий эмбриогенеза должны были устано-
виться два-три десятка основных планов строения (архетипов). Рез-
кие преобразования строения, случись они у взрослого, обернулись
бы для него катастрофой и обрекли бы его на гибель. Дальк отмечал,
что они могут переноситься зародышем в силу его чрезвычайной пла-
стичности и присущей ему высокой регуляционной способности. Он
полагал, что основой эволюционного процесса является событие, кото-
рое он обозначил как «онтомутация» и которое заключается в резких,
глубоких, радикальных и в то же время жизнеспособных трансформаци-
ях в цитоплазме яйцеклетки как морфогенетической системы.
Однако особенно ясно положения о филогенетической роли рез-
ких уклонений эмбрионального развития были сформулированы Ри-
хардом Гольдшмидтом (рис. 5).
Рис 4. А. Дальк. Выдающийся
бельгийский эмбриолог. Одним
из первых выдвинул тезис о гом.
чю наибольшее эволюционное
значение имеют мутации, заггра-
гивающие самые ранние этапы
эмбриогенеза и, в частности,
процесс созревания яйцеклетки
Рис 5 Рихард Гольдшмид!
Один из основоположников
генетики и эксперименталь-
ной эмбриологии Автор )во-
люциопной концепции, ус-
пешно в настоящее время
конкурирующей с классичес-
ким дарвинизмом
58
//. Общие проблемы эволюции
Он разработал стройную концепцию макроэволюции, основыва-
ясь на принципах генетики развития. Его взгляды выражены в следу-
ющих постулатах:
1. Макроэволюция не может быть понята на основе гипотезы о
накоплении микромутаций. Она сопровождается реорганизацией ге-
нома, реорганизацией хромосом.
2. Изменения хромосомного «паттерна» могут вызывать значитель-
ный фенотипический эффект независимо от точковых мутаций.
3. Фенотипический эффект основывается на преобразовании сис-
тем межтканевых взаимодействий в процессе индивидуального раз-
вития и может иметь эволюционное значение, обусловливая появле-
ние так называемых «многообещающих уродов», значительно откло-
няющихся в своем строении от нормы. Они могут быть преадап-
тированы к существованию в определенных условиях среды и, попав
в эти условия, дать начало новым таксономическим единицам.
4. Системная реорганизация онтогенеза реализуется либо че-
рез эффекты генов-модификаторов, либо благодаря макромута-
циям, существенно изменяющим функционирование структур
типа эндокринных желез, продуцирующих различные гормоны и
оказывающих влияние на развитие организма в целом.
Гольдшмидт приводит в качестве примера такие фенотипические
эффекты, основывающиеся на действии гормонов, как акромегалия и
гигантизм, карликовость, ахондроплазия. С. Стоккард (С. Stockard)
связывает с функцией эндокринных желез многие расовые признаки у
собак. Д.К. Беляев продемонстрировал существенные сдвиги в функ-
ции эндокринных желез при доместикации лисиц.
Достаточно давно, в начале 30-х годов, были проведены интерес-
ные эксперименты на рыбке Peryophthalmus megaris, продемонстри-
ровавшие возможность значительной морфогенетической перестрой-
ки в условиях трехлетнего непрерывного введения тироксина. Оказа-
лось, что в этом случае удлиняются грудные плавники, которые при-
обретают внешнее сходство с конечностями амфибий, а рассеянные в
норме эндокринные элементы, продуцирующие тироксин, группиру-
ются в более компактные образования также наподобие свойственных
амфибиям. Изложенные факты дали основания Гольдшмидту сделать
вывод о значительном фенотипическом эффекте тех изменений гено-
ма, которые отражаются на состоянии механизмов гормонального конт-
59
Л. И. Корочкин
роля. Они могут быть ответственны за инициацию серии изменений
преадаптивного характера. Выдающийся российский зоолог Н.Н. Во-
ронцов, разделявший взгляды Р. Гольдшмидта, представил два бесспор-
ных фактора макромутационного возникновения безволосых видов
млекопитающих за счет единственной макромутации типа hairless,
противоречащих концепции облигатного градуализма (рис. 6). Ворон-
цов полагает, что одним из ведущих факторов, индуцирующих макро-
мутации, является сейсмическая активность (Воронцов, 1999).
Другим примером макромутаций могут служить гомеозисные му-
тации, выводящие признак на уровень нового таксономического ран-
га. В целом постулируется, таким образом, существование такого рода
адаптаций, возник-
новение которых не
требует ни ламар-
ко вс ко го, ни селек-
тогенетического
пути.
Рис 6 Макромутации по
НН Воронцову. А - без-
волосые муганты олень-
их хомячков Видны со-
хранившиеся вибриссы и
складки ороговевшего
эпителия. В норме особи
этого вида покрыты
обычным меховым по-
кровом, Б - молодой,
нормально пигментиро-
ванный хомячок - гомо-
зигот по мутации hair-
less. /У-молодой безволо-
сый хомячок-альбинос
(гомозигот по двум ре-
цессивным - hairless,
albino несцспленным
признакам). Г - безволо-
со сть как систематичес-
кий признак у цейлон-
ской бабирусы
Один из самых выдающихся палеонтологов современности Отго
Шиндевольф (рис. 7) также исходил из факта предварения филогении
60
//. Общие проблемы эволюции
Рис. 7 Отго Шиндсвольф -
один из крупнейших палеон-
тологов современности Сто-
ронник теории скачков в эво-
люции
онтогенезом и выдвинул теорию типостро-
физма. В ней он игнорировал процессы,
протекающие в популяциях, отверг эволю-
ционную роль случайности, признав носи-
телем эволюции отдельную особь. Он счи-
тал, что отсутствие промежуточных форм
в палеонтологических останках объясняет-
ся особенностями эволюционного процес-
са, который заключается в быстрых транс-
формациях форм, связанных с резкими из-
менениями уровня космической и
солнечной радиации. Шиндевольфу при-
надлежит крылатая фраза: «Первая птица
вылетела из яйца рептилии».
Сходные взгляды под названием «те-
ория прерывистого равновесия» испове-
дуют американские палеонтологи Нильс
Элдридж (Eldredge), Стив Стэнли (Stan-
ley) и Стефан Гоулд (Gould). Они прида-
ют также важное значение в эволюции педоморфозу, т.е. таким изме-
нениям онтогенеза, которые характеризуются полной утратой взрос-
лой стадии и соответствующим укорочением индивидуального
развития, когда последней становится стадия, бывшая прежде личи-
ночной. Животные приобретают способность размножаться именно
на этой стадии. Таким путем возникли, по-видимому, некоторые груп-
пы хвостатых земноводных (протеи, сирены), аппендикулярии, неко-
торые группы насекомых (гриллоблаттиды), паукообразных (ряд по-
чвенных клещей) (подробнее см.: Назаров, 1991).
Каковы, однако, те конкретные события и процессы, которые мо-
гут обусловливать преобразование типов онтогенеза? На мой взгляд,
это особый вид мутаций, точкой приложения которых являются вре-
менные параметры созревания взаимодействующих систем в разви-
тии. В сущности, весь процесс онтогенеза представляет собой цепь
эмбриональных индукций, взаимодействий индуктор - компетентная
ткань. Полноценное осуществление эмбриональной индукции зави-
сит от того, насколько точно соответствует в развитии время созрева-
ния индуктора таковому компетентной ткани. В нормальных условиях
61
Л. И. Корочкин
реагирующая компетентная система приобретает способность отвечать
формообразовательным процессом на стимулирующий импульс индук-
тора в тот момент времени, когда индуктор становится способным про-
дуцировать этот импульс. Рассогласования во времени созревания ин-
дуктора и компетентной ткани ведут к нарушению соответствующих мор-
фогенетических процессов. Мутации, вызывающие такие рассогласо-
вания, по-видимому, распространены достаточно широко. Так, у мышей
известны мутанты с нарушением индуктивных взаимодействий между'
выростом вольфова протока и метанефрогенной мезенхимой, которое
обусловлено временным рассогласованием в созревании этих тканей, в
результате чего нарушается развитие почки. При совмещении в орган-
ной культуре индуктора и компетентной системы от мутантных и нор-
мальных зародышей почечная ткань развивается нормально.
Становление пигментации у амфибий определяется взаимодействи-
ем эпидермиса, выступающего в качестве индуктора, и ткани нервно-
го гребня, который является источником меланобластов, мигрирую-
щих субэпидермально под влиянием индуктора. Одна из мутаций (d)
вызывает в гомозиготе (dd) резкое ослабление окраски аксолотля, так
что лишь спина животного слегка окрашена (так называемая белая
раса аксолотлей). Мною вместе с моей сотрудницей В.И. Богомоло-
вой было выяснено, что отсутствие окраски детерминируется не утра-
той эпидермисом индуцирующих свойств или меланобластами спо-
собности мигрировать и синтезировать пигмент, а рассогласованием
во времени созревания этих двух взаимодействующих закладок, со-
ставляющих, таким образом, единую корреляционную систему. В се-
рии экспериментов по трансплантации кусочков презумптивного эпи-
дермиса между зародышами аксолотлей белой расы разного возраста
мы обнаружили, что при некоторых сочетаниях возраста донора и ре-
ципиента в зоне трансплантата развивается пигментация (рис. 8).
И.И. Шмальгаузеном (рис. 9) и Д.К. Беляевым (рис. 10) была про-
демонстрирована доместикация - типичный случай, который характе-
ризуется распадом, дезинтеграцией такого рода корреляционных сис-
тем. Корреляционная система оказывается нарушенной во многих слу-
чаях, не нарушается лишь функционирование жизненно важных
органов. Например, в окраске домашних животных обычно отмечает-
ся неправильное распределение пятен различного цвета (у коров, со-
бак, кошек, морских свинок), чего не бывает у диких животных, у ко-
62
//. Общие проблемы эволюции
Рис 8 Результаты экспериментов
В.И. Богомоловой и Л И. Корочкина
но анализу генетической регуляции
пигментогснеза у аксолотлей. А -
контрольные эмбрионы аксолотлей
белой линии на стадиях развития
39-40. На их боковой поверхности
отсутствуют пигментные колетки; В
- результаты трансплантации пре-
зумптивного эпидермиса от эмбри-
онов белой линии на стадиях разви-
тия 34-35 к эмбрионам той же ли-
нии на сiалиях развития 25-26.
Эмбрионы зафиксированы на стади-
ях 40-41. В месте трансплантата
развилась пигментация (показана
стрелками)
Рис. 10 Дмитрий Константинович Бе-
ляев Крупнейший эволюционный ।сне-
тик, один из основоположников отече-
ственной генетики поведения. Один из
тех, кто боролся с лысснковщиной и
возрождал генетику в нашей стране,
создал мощный генетический центр в
новосибирском Академгородке
Рис 9. Иван Иванович Шмаль-
гаузен (.'формулировал iсе-
рию стабилизирующего отбо-
ра, автор классического груда
«Организм как целое в шщи-
видуалыюм и историческом
ратвит ин»
63
Л. И. Корочкин
торых имеется либо однотонная окраска, либо закономерное распре-
деление полос или пятен. Следует отметить, что генетический конт-
роль однотонной серой окраски достаточно сложен. Тем не менее
сложный корреляционный механизм легко разрушается. Мутации, про-
являющиеся в процессе доместикации животных (по-видимому, в сво-
ем большинстве накопленные ранее, но не распространявшиеся в по-
пуляциях в силу малой жизнеспособности мутантов в естественной
обстановке), в условиях их размножения под охраной человека дей-
ствуют на уровне корреляционных соотношений. При этом существен-
ные корреляции часто теряются, а взамен устанавливаются совершен-
но новые. Развитие хохла и перьев на ногах у кур, а также курдюка у
овец обусловлено действительно новыми корреляциями. Установле-
ние новых признаков связано с установлением новых корреляций.
Шмальгаузен рассматривает редукцию органов как распад корреля-
ционных систем, а атавизм - как локальную реинтеграцию, в основе
которых лежат сдвиги во времени наступления формообразователь-
ных реакций.
Детальный биохимический анализ развития личиночных и имаги-
нальных ткайей у разных видов Drosophila группы «virilis» позволил
выявить видовую специфичность времени биохимического становле-
ния исследованных тканей. Межлинейные различия в поведении ла-
бораторных животных в ряде случаев являются результатом различ-
ных скоростей накопления нейрохимических субстратов в ходе разви-
тия. Каковы возможные феногенетические основы выраженной
вариабельности формообразования, обусловленной сдвигами времен-
ных параметров созревания взаимодействующих тканей?
Предположим, что в развивающейся системе имеется два гена: At
и А2 (которые могут быть аллельными и неаллельными, что, в общем,
не принципиально для данного случая), осуществляющие контроль
соответствующих морфогенетических реакций (а1 и а2) через синтез
специфических веществ а1 и а2 (рис. 11). Очевидно, при многоуровне-
вом контроле осуществления признака сам факт транскрипции данно-
го локуса еще не означает, что контролируемый им признак будет вы-
ражен в фенотипе. Существуют многочисленные генетические конт-
ролирующие элементы, которые способны подавить проявление
признака на том или ином этапе его осуществления.
Допустим, что морфогенетическая реакция, контролируемая геном
А2, не имеет выхода в фенотип вследствие блока на том или ином уровне
64
1L Общие проблемы эволюции
регуляции, например, торможения синтеза вещества а2 или рассогла-
сования времени его синтеза со временем созревания реагирующей
системы. В таком случае осуществится лишь один морфогенетичес-
кий процесс а1,. Допустим теперь, что в одном из генов-модификато-
ров (М) произошла мутация, результатом которой является совмеще-
ние времени синтеза вещества а2 и созревания реагирующей системы,
а следовательно, и выход в фенотип признака, контролируемого геном
А В таком случае осуществляется также событие а2г Если между ре-
акциями а1 и а2 имеет место взаимодействие, то возможны различные
дополнительные, «промежуточные» формообразовательные процессы.
Поскольку на относительную выраженность каждой из этих реакций
в фенотипе будут влиять многочисленные гены-модификаторы, коли-
чество различных возникающих при этом фенотипических вариантов
практически безгранично. Следует также учитывать возможность того,
что ген М контролирует уровень того или иного гормона в развиваю-
щемся организме. Это, в свою очередь, существенно отразится на об-
щем гормональном балансе. Последний же играет важную роль в ре-
гуляции особенностей, в том числе и временных, фенотипического
выражения целого комплекса различных признаков и осуществления
разнообразных морфогенетических реакций.
Рис. 11. Гипотетическая схема
макромутации (М), влияющей на
морфогенетические процессы: а,
- продукт кодируется геном А, и
детерминирует реализацию мор-
фогенетической реакции а1 г -
продукт кодируется геном А2 и
принимает участие только под
влиянием гена-модификатора
(М). В этом случае он детерми-
нирует реализацию морфогенети-
ческой реакции а22 взаимодей-
ствия продуктов, обеспечивает
вариации в морфогенетических
событиях, контролируемых каж-
дым из них (Корочкин, 1999)
Изменения временных характеристик, например, преждевремен-
ная или задержанная пролиферация определенных групп клеток, спо-
65
Л. И. Корочкин
собны преобразить фенотипический «облик» организма. Хорошие
примеры такого «преображения» можно найти в работах д’Арси Том-
псона. Как показано на рис. 12, заимствованном из его работы, путем
простого преобразования координат достигаются значительные раз-
личия между органическими формами. Так, заменив координаты
рис. 12, А, изображающего морскую рыбу Scaurus, на изогнутую орто-
гональную систему, мы получим изображение не очень отдаленного
рода Pomacanthus (рис. 12, £)), которое по отношению к Scaurus впол-
не можно назвать «счастливым монстром» Гольдшмидта. По-видимо-
му, именно такие преобразования осуществляются в ходе морфо-
генетического процесса, измененного макромутацией.
Рис. 12. Трансформации д’Арси Томпсона. Рисунок демонстрирует, что измене-
ния (вариации) в направлении роста и клеточной пролиферации (линии коорди-
натной сетки) во время развития могут вызвать существенные сдвиги в фенотипе
животных (в данном случае рыбок). Макромутации, вызывающие такого рода из-
менения, способны продуцировать вариации видо-специфических, а то и родо-
специфических признаков, посредством, например, сдвигов в скорости митоти-
ческого цикла: А - Diodon; В - Scaurus; С - Pomacanthus; D Orthagoriscus
66
II. Общие проблемы эволюции
И в этом смысле сходство или различие органических форм и струк-
тур отнюдь не является доказательством их родственного происхож-
дения. Отношение в этом случае подобно известным в логике отноше-
ниям равносильности. Равносильные формулы - это те, которые неза-
висимо от значений истинности входных переменных характеризуются
идентичностью выходных значений истинности, например:
(р-* -ч) ~(pAq)
р q ~q (р-* ~q) р q pAq ~(pAq)
и и л Л и и и Л
л и л и л и л и
и л и и и л л и
л л и и л л л и
Разве не подобно отношениям равносильности развитие камерно-
го глаза у головоногих и позвоночных? Рис. 13 отчетливо демоне-
Рис. 13. Схема эмбрионального развития и строения глаза головоногих моллюсков
(л-d) и позвоночных (е-к). 1 - сетчатка, 2 - пигментная оболочка, 3 - роговица, 4 -
радужина, 5 - хрусталик, 6 - ресничное (эпителиальное) тело, 7 - сосудистая оболоч-
ка> Я - склера, 9 - зрительный нерв, 10 - покровная эктодерма, 11 - головной мозг
67
Л. И. Корочкин
трирует, как на основе совершенно различных морфогенетических
процессов формируются подобные органы. Именно таким путем мо-
жет быть осуществлено конвергентное развитие признаков у филоге-
нетически неродственных организмов. В основе событий, последова-
тельно строящих данную структуру, лежит, очевидно, генетически зап-
рограммированный план развития. Последовательное развертывание
этих событий регулируется сложным и точно настроенным генетичес-
ким механизмом, начало которому может положить одноразовая мак-
ромутация Гольдшмидта.
Что же, однако, заставляет гены изменять время экспрессии? Есть
основания предполагать, что существенная роль в этом принадлежит
гетерохроматиновым участкам хромосом, которые могут составлять
от 20 до 80% генома.
У дрозофилы гены, необходимые для обеспечения жизнеспособ-
ности и плодовитости самцов, были локализованы в Y-хромосоме и
центромерном гетерохроматине хромосомы 2. Гетерохроматиновые
области оказывают эффект на компенсацию дозы рДНК, экспрессию
кластера генов, тесно сцепленных в хромосоме 2, мейотическое пове-
дение хромосом, рекомбинацию, супрессию эффекта положения. Бо-
лее того, специфические области Y-хромосомы подавляют экспрессию
Х-сцепленных генов в семенниках.
Фенотипический эффект изменения количества гетерохроматина
часто проявляется в раннем эмбриогенезе, обусловливая, например,
снижение количества клеток в органе или сохранение фетальных ха-
рактеристик после рождения. Именно гетерохроматину и, в первую
очередь, входящей в его состав сателлитной ДНК приписывают роль
регулятора скорости клеточного деления и роста и, следовательно,
временных параметров индивидуального развития.
Гетерохроматин и сателлитная ДНК, возможно, оказывают эффект
на время экспрессии генов двояким способом: они могут ассоцииро-
ваться с определенным классом белков, которые влияют на структуру
хроматина, они могут также влиять на трехмерную организацию ин-
терфазного ядра. В приведенном выше примере с нарушением пиг-
ментации у аксолотлей детерминирующие этот процесс изменения во
времени созревания взаимодействующих тканей обусловлены, веро-
ятно, делецией кусочка гетерохроматина в области ядрышкового орга-
низатора. Так, у Drosophila littoralis получены лабораторные сублинии,
68
Н. Общие проблемы эволюции
отличающиеся наличием или отсутствием гетерохроматинового бло-
ка в районе G4 хромосомы 2, прилежащего к кластеру генов, кодиру-
ющих изоферменты эстеразы. Оказалось, что присутствие гетерохро-
матинового блока сдвигает время экспрессии изоферментов эстеразы
в различных органах дрозофилы в ходе онтогенеза.
Особенно интересны случаи, когда гетерохроматиновый блок ло-
кализуется вблизи района G5 хромосомы 2 D. littoralis, содержащего
гены, кодирующие три изофермента бета-эстеразы, в том числе эсте-
разы, расщепляющей ювенильный гормон (ЮГ-эстеразы). В этом слу-
чае особи, гомозиготные по гетерохроматиновому блоку, погибают на
стадии куколки. Характерно также и то, что не только задерживается
время появления изоферментов ЮГ-эстеразы, но и тормозится свой-
ственный нормальному развитию рост их активности. Предполагает-
ся, что сохранение низкого уровня активности ЮГ-эстеразы обуслов-
ливает дисбаланс в соотношении экдизон/ювенильный гормон, так что
сложившийся гормональный статус развивающейся дрозофилы не
позволяет завершить метаморфоз. Моя ученица И.Ю. Раушенбах (1990)
выдвинула гипотезу, согласно которой этот изофермент, являясь орга-
но- и тканеспецифичным, составляет целостную систему вместе с ней-
роэндокринными органами. Эта система регулирует адаптивную ре-
акцию дрозофилы. В результате селекции в данной фенотипической
системе отбираются комплексы генов-модификаторов, контролирующих
экспрессию ЮГ-эстеразы в критические моменты развития особей, спо-
собствуя сохранению или элиминации сложившихся генотипов в опре-
деленных условиях среды. В соответствии с этими представлениями ко-
лебания активности ЮГ-эстеразы являются частью реакции онтогене-
тической системы, принимающей участие в регуляции процесса
индивидуального развития и, возможно, играющей исключительную роль
в этой регуляции. Внезапные и глубокие наследственные перестройки
функционирования именно таких систем могут произвести на свет «мно-
гообещающих уродов» с эволюционным будущим.
Таким образом, можно себе представить, что перераспределение
гетерохроматина вызывает функциональную реорганизацию генома в
Целом, порою затрагивающую лишь отдельные признаки, а порою
достаточно глубоко преобразующую фенотипическое становление
систем таких признаков.
Особый интерес в связи с этим представляет организация кариоти-
па у разных видов Drosophila группы «virilis», обнаруживающих от-
69
ЛИ. Корочкин
четливые различия в количестве гетерохроматина на геном и отчасти
в его распределении по хромосомам. Эта группа дрозофилид включа-
ет по крайней мере 12 видов, объединяемых в филады по степени мор-
фологического, биохимического сходства, а также скрещиваемости
между собой. Различные филады очень четко отличаются по количе-
ству сателлитной ДНК, собранной преимущественно в гетерохрома-
тиновых районах хромосом.
Так, у D.virilis количество сателлитной ДНК составляет почти 50%
генома. Филада1ехапа(ВЛехапа, D.americana, D.novamexicana, D. lummei)
характеризуется значительным уменьшением количества гетерохрома-
тина по сравнению с D.virilis. В филадах littoralis и montana наблюдается
дальнейшее снижение количества гетерохроматина, так что значитель-
ная часть блока альфа-гетерохроматина полностью выпадает.
Дж.Голл (Gall) с сотрудниками обнаружили, что имеется три глав-
ных типа сателлитной ДНК у D.virilis - 25% генома, последователь-
ность нуклеотидов 5'АСАААСТ, 8% генома - 5'АТАААСТ, и 8% -
5'АСАААТТ. Была также продемонстрирована тканевая специфичность
в распределении и дифференциальной репликации разных фракций
сателлитной ДНК (рис. 14).
Рис. 14. Фракции сателлитной ДНК
70
II. Общие проблемы эволюции
Распределение небольших количеств сателлитной ДНК, содержа-
щихся в эухроматиновых районах, также различается у разных видов
Drosophila группы «virilis».
Стегний показал, что различия в количестве сателлитной ДНК со-
провождаются видоспецифическими различиями в трехмерной орга-
низации хроматина ядра, в спецификации точек прикрепления хромо-
сом к ядерному матриксу.
Но что за факторы вызывают в ходе эволюции перераспределение
гетерохроматина? Л.И. Корочкин (1983) и М.Б. Евгеньев (Evgenev,
1982) предположили, что такого рода события происходят благода-
ря подвижным генетическим элементам, как бы «растаскиваю-
щим» кусочки гетерохроматиновой ДНК по разным ячейкам ге-
нома, результатом чего и являются гольдшмидтовские макромутации.
Подвижные генетические элементы могут по крайней мере двояким
способом влиять на реализацию наследственной информации в разви-
тии. Во-первых, внедряясь в область структурного гена, они изменяют
скорость транскрипции, в результате чего концентрация кодируемого
этим геном белка изменяется в несколько раз. Так, в лаборатории аме-
риканского генетика Кэти Лаури (Lauri) было показано, что инсерция
ПГЭ в зону гена, который кодирует алкогольдегидрогеназу, вызывает
снижение активности фермента примерно в 4 раза. Если в подобной
ситуации окажется ген, кодирующий тот или иной фактор, формирую-
щий полярный градиент, это существенно скажется на развитии эмб-
риона. Во-вторых, ПГЭ способны изменять временной паттерн эксп-
рессии генов, что точно так же оказывает влияние на взаимодействие
тканей в развитии и соответственно на морфогенетические процессы.
Иными словами, происходящие в определенных точках генома эли-
минации, вставки и перераспределения блоков сателлитной ДНК, обус-
ловленные их «захватом» перемещающимися подвижными генетичес-
кими элементами (ПГЭ), могут быть механизмом реализации направ-
ленности эволюционного процесса, поскольку сайты вставок ПГЭ
расположены закономерно, а не разбросаны беспорядочно по геному.
Такого рода перемещения, по-видимому, способствуют «взрывам» раз-
личных инверсий и транслокаций, как правило, сопровождающих ви-
дообразование. В работах М.Б. Евгеньева четко продемонстрирована
корреляция в расположении сателлитной ДНК и ПГЭ у различных
видов Drosophila группы «virilis», что является косвенным подтвер-
ждением этой гипотезы.
71
Л. И. Корочкин
Как показал английский генетик Г. Довер (Dover, 1982), переме-
щения ПГЭ носят «массовый» характер и связаны с резким увеличе-
нием их количества на геном, что могло бы являться молекулярно-
генетическим механизмом сальтационного видообразования.
Большое значение в происхождении видообразовательных «взрывов»
придает ПГЭ и современный палеонтолог Дж. Валентайн (Valentine,
Campbell, 1975).
И все же основанные на данных генетики развития эволюционные
представления являются лишь гипотетическими, и решающее слово
пока что остается за палеонтологами.
Литература
Алтухов Ю.П. Генетические процессы в популяциях. М.: Наука, 1983.
Берг Л.С. // Труды по теории эволюции. Л.: Наука, 1977.
Воронцов Н.Н. Развитие эволюционных идей в биологии. М.: Прогресс, 1999.
Грант В. Эволюция организмов. М.: Мир, 1991.
Корочкин Л И. Эволюционное значение подвижных генетических элементов // Цито-
логия и генетика. 1983. Т. 17. С 67-75.
Корочкин Л И. Введение в генетику развития. М.: Наука, 1999.
Назаров В.И. Учение о макроэволюции. М.: Наука, 1991.
Раушенбах И.Ю. Нейроэндокринная регуляция развития насекомых в условиях
стресса. Новосибирск: Наука, 1990.
Рэфф Р., Кауфман Т. Эмбрионы, гены, эволюция. М.: Мир, 1986.
Северцов А.Н. Морфологические направления эволюционного процесса. М.: Наука, 1967.
Стегний В.Н. Архитектоника генома. Системные мутации и эволюция. Новоси-
бирск: Изд-во НГУ, 1993.
Carson Н. The genetics of speciation at the diploid level // Amer. Nat. 1975. Vol. 109.
P. 83-95.
Dewar D. Difficulties of the evolution theory. London I E. Arnold and Co, 1931.
Korochkin L. Hopeful monsters and jumping genes // Evol. biol 1993. Vol. 7. P. 153-172.
Thompson d’Arcy. On growth and form. Cambridge: Univ Press, 1917.
ПРИЧИНЫ ЭВОЛЮЦИОННОГО СТАЗИСА
А.С. Северцов
Московский государственный университет
119899, г. Москва, Воробьевы горы
E-mail: asevertsov@soil.msu.ru
Эволюционные исследования традиционно направлены на изуче-
ние прогрессивной эволюции, механизмов и закономерностей новых,
отсутствовавших у предков, адаптаций. И.И. Шмальгаузен (1946) пер-
вым поставил вопрос о причинах сохранения сформировавшихся адап-
таций, объяснив это явление действием стабилизирующего отбора.
Однако стабилизирующий отбор действует только при условии ста-
бильного соотношения признака и тех условий среды, к которым этот
признак обеспечивает адаптацию. Поэтому стабилизирующим отбо-
ром можно объяснить сохранение только отдельных признаков орга-
низма. Естественный отбор идет по функциональным признакам фе-
нотипа (Северцов, 1990), поэтому он и сохраняет те признаки, кото-
рые остаются адаптивными, хотя другие признаки изменяются под
действием движущего отбора.
Между тем экологическая ниша вида никогда не предоставляет ему
стабильных условий существования: колебания погоды и климата, изме-
нения численности пищевых ресурсов, конкурентов и хищников, т.е. из-
менения экологической обстановки. Перестройка экосистем нарушает
соотношение организации особей и среды их обитания. Эволюционные
изменения видов - компонентов биоценозов - заставляют коадаптиро-
вать-ся между собой и другими компонентами этого сообщества. Поэто-
му объяснить действием стабилизирующего отбора эволюционный ста-
зис - длительное существование в неизменном состоянии фенотипичес-
кого облика видов - не представляется возможным.
Со времени классической работы Симпсона (Simpson, 1953) и бла-
годаря многим исследованиям других авторов, посвященным изуче-
нию на палеонтологическом материале темпов эволюции, стало изве-
стно, что продолжительность существования вида у животных колеб-
лется от сотен тысяч до миллионов лет. У персистирующих форм это
время составляет десятки, и даже сотни миллионов лет. Объяснить
73
А. С. Северцов
столь длительную неизменность стабильностью условий существова-
ния, т.е. действием стабилизирующего отбора, невозможно. Таким
образом, возникает вопрос о причинах эволюционного стазиса, т.е. об
эволюционных механизмах поддержания эволюционной неизменнос-
ти популяций и видов.
По данным Гингериха (Gingerich, 1976), у плейстоценовых грызу-
нов скорость эволюции составила 222 новых вида за 1 млн лет. В то
же время средняя продолжительность существования уже возникшего
рода составила 5,9 млн лет. Продолжительность существования вида
у млекопитающих около 0,5 млн лет. Очевидно, что за время суще-
ствования видов, составляющих эти роды, их экологическое окруже-
ние существенно изменялось, а следовательно, должны были суще-
ствовать какие-то специальные механизмы, тормозившие их прогрес-
сивную эволюцию.
В связи с этим необходимо отметить, что концепция прерывистого
равновесия (Gold, Elderege, 1976), постулировавшая закономерное чере-
дование в эволюции видов периодов эволюционного стазиса и быстрого
видообразования, не объясняла причин существования стазиса. Равно-
весное соотношение организации вида и среды его обитания только по-
стулировалось. Дискуссия о прерывистом равновесии привела лишь к
отказу от представления о регулярном закономерном характере наруше-
ний равновесия, но не объяснила его существования.
Существуют, по-видимому, два основных механизма, обеспечива-
ющих эволюционный стазис на популяционно-видовом и внутрипо-
пуляционном уровнях: внутривидовое разнообразие и контрбаланс
векторов отбора.
Критерии эволюционной стабильности
В рамках синтетической теории эволюции сформировалось пред-
ставление о том, что наименьшее, элементарное, эволюционное изме-
нение представляет собой стойкое изменение частот (концентраций)
аллелей в генофонде эволюирующей популяции. В отечественной ли-
тературе наиболее четко это было сформулировано Н.В. Тимофеевым-
Ресовским (Тимофеев-Ресовский и др., 1977; Яблоков, 1987).
Однако такой подход, иногда работающий в популяционной гене-
тике и кариосистематике, совершенно не применим там, где генети-
ческие исследования не проводились или невозможны, например в
74
II. Общие проблемы эволюции
палеонтологии. Там, где о видовой принадлежности или о структуре
вида приходится судить по признакам фенотипа, действует, как извес-
тно, подход, основанный на степени и (или) дискретности различий
по фенотипическим полигенным признакам (Глиценко и др., 1983;
Мина, 1986). Только таким подходом могут пользоваться палеонтоло-
ги: вид остается неизменным до тех пор, пока сохраняются свойствен-
ные ему фенотипические признаки. Этот критерий использован и в
данном сообщении.
Внутривидовое разнообразие как причина эволюционной
стабильности
Сбалансированный генетический полиморфизм - широко распро-
страненное явление. Наличие двух или большего числа дискретных
или плавно переходящих друг в друга генетически детерминирован-
ных морф обеспечивает популяциям возможность существования в ко-
леблющихся условиях среды и (или) в условиях пространственной дис-
персии микроусловий. Этот экологический аспект полиморфизма хо-
рошо известен. Однако полиморфизм ответствен не только за
экологическую устойчивость, но и за эволюционную неизменяемость
видов. Один из наиболее бросающихся в глаза примеров адаптивного
полиморфизма демонстрируют синантропные сизые голуби Columba
livia. Адаптивное значение этого полиморфизма было изучено
Н.Ю. Обуховой и А.Г. Креславским на кафедре теории эволюции Био-
логического факультета МГУ. Две крайние морфы - типичная сизая и
меланистическая - связаны двумя рядами переходных морф - через
черно-крапчатую окраску, при которой меланин концентрируется на
концах кроющих перьев, и через равномерное потемнение окраски,
при котором меланин равномерно распределяется по всему опахалу
пера. Посемейный генетический анализ показал, что существуют два
гена меланизации, соответствующие каждому типу - Black и Dark, на
проявление которых влияет значительное количество генов-модифи-
каторов (около 6 на каждый ген с сильным действием). В результате,
при перекомбинации, возникает весь спектр окрасок, наблюдаемых в
любой стае сизых голубей.
Окраска сцеплена с поведением. Сизая морфа более активна. Голу-
би, принадлежащие к этой морфе, охраняют свой гнездовой участок,
75
А. С. Северцов
более активны в поисках корма. Меланисты толерантны к плотности
гнездования. Их гнезда могут располагаться практически вплотную
друг к другу. Они менее активны и в поисках корма. Все промежуточ-
ные формы и по окраске, и по поведению образуют непрерывный ряд
переходов между крайними формами.
Динамика полиморфизма в московских популяциях голубей за обо-
зримый период времени связана с динамикой численности популяции.
Во время Великой Отечественной войны почти все голуби погибли,
по-видимому, отчасти от бескормицы, а отчасти были съедены насе-
лением. Сохранились всего 2-3 малочисленные колонии, представлен-
ные светлой морфой. Численность голубей резко возросла после Все-
мирного фестиваля молодежи и студентов в 1957 г. благодаря подкормке
и одичанию массы домашних голубей. Как показал еще Ч. Дарвин, ок-
раска домашних голубей при их одичании через 2-3 поколения ревер-
сирует к дикому типу. В результате увеличения численности и плотно-
сти населения росла и концентрация меланистов, достигнув к концу
60-х годов примерно 80%. Толерантные к плотности меланисты и близ-
кие к меланистам промежуточные формы вытеснили своей массой
светлоокрашенных голубей на неудобные чердаки, где эта морфа мог-
ла поддерживать свои гнездовые территории. Не ясно, происходило
ли восстановление меланистов за счет сохранившихся аллелей или за
счет повторных мутаций. Можно, однако, утверждать, что иммигра-
ции из других популяций не было.
Точных данных за последнее десятилетие у нас нет. Но приблизи-
тельные визуальные прикидки показывают, что в период начала пере-
стройки, когда численность голубей резко сократилась, концентрация
сизой морфы резко возросла, а затем снова упала, когда уличная тор-
говля и сопровождающая ее масса отбросов восстановили кормовую
базу голубей.
Таким образом, на изменение экологической обстановки, в данном
случае - кормовой базы и лимита мест гнездования, популяция отве-
чает изменением численности морф, адаптивных к каждой из возмож-
ных ситуаций - снижение или увеличение плотности населения. Вид в
фенотипически узнаваемом состоянии сохраняется. Он более устой-
чив, чем если бы не было полиморфизма.
Аналогичный и, может быть, более убедительный результат был
получен сотрудником нашей кафедры В.Г. Гриньковым (2000) при изу-
76
II. Общие проблемы эволюции
чении полиморфизма окраски оперения самцов мухоловки-пеструш-
ки (Ficedula hypoleuca Pall). Полиморфизм представляет собой непре-
рывный ряд окраски оперения от контрастной черно-белой до неотли-
чимой от окраски самок. Условно в этом ряду выделяют семь морф,
хотя на самом деле это картина непрерывного изменения полигенного
количественного признака. Вместе с тем наследуемость окраски h2g=l.
Подход к полиморфизму как к количественному признаку позволил
разделить ряд на три группы морф: ярко окрашенные, промежуточ-
ные и светлые.
Условием стабильного полиморфизма является равноценность стра-
тегий приспособленности каждой из морф. Приспособленность оцени-
вали по репродуктивному успеху за жизнь. Выяснилось, что репродук-
тивный успех в Подмосковье (на Звенигородской биологической стан-
ции МГУ) равноценен у каждой из трех групп морф. В южной Карелии
две крайние группы морф оказались более приспособленными, чем про-
межуточные. Однако стратегии достижения этого результата различны.
Ярко окрашенные самцы прилетают первыми и первыми присту-
пают к размножению. Самку они привлекают, главным образом, де-
монстрируя себя на открытых присадах. Более раннее размножение
при коротком северном лете обеспечивает надежность прохождения
постгнездового периода, линьки и подготовки к осеннему перелету.
Однако при поздних и холодных веснах яркие самцы проигрывают.
Кроме того, яркие самцы и их гнезда больше страдают от хищников.
Хищники в Карелии разоряют около 16% гнезд, а в Подмосковье - до
50%. Кроме того, яркие самцы обладают одним, не вполне понятным,
свойством. Проиграв в размножении в первый свой сезон, они и затем
проигрывают в размножении - во все последующие сезоны размно-
жения своей жизни.
Светлоокрашенные самцы приступают к размножению позже, по-
этому их выводки реже страдают от весенних холодов. Неуспех пер-
вого года размножения для них не существен. Они могут наращивать
репродуктивный успех в течение жизни, видимо, по мере накопления
индивидуального опыта. К тому же они меньше страдают от хищни-
ков, т.к. демонстрируют самке не столько самих себя, сколько досто-
инства своего гнездового участка.
При наблюдениях в южной Карелии выявился проигрыш проме-
жуточных форм. В Подмосковье, при более продолжительном теплом
77
А. С. Северцов
периоде года и неустойчивой погоде, промежуточные формы облада-
ют приспособленностью, равной и ярким и светлым морфам. Проме-
жуточным хватает времени, чтобы подстроиться к условиям данного
сезона. Возможно, при более длительном периоде наблюдений в Ка-
релии приспособленность промежуточных форм оказалась бы и там
не ниже, чем у крайних. Наблюдения в Карелии продолжались два года,
и оба сезона были жаркими.
Для темы данного сообщения важно, что частота морф мухоловки-
пеструшки подвержена клинальной изменчивости. Частота ярко окра-
шенных самцов максимальна на северо-западе ареала: в Скандинавии,
Финляндии и Карелии. К югу она постепенно понижается, как и к во-
стоку, вплоть до Западной Сибири так, что в Центральной Европе пре-
обладают промежуточные формы. Таким образом, полиморфизм опос-
редованно обусловлен климатическими факторами. Он обеспечивает
устойчивость вида на ограниченной территории от Западной Европы
до Енисея и от Средиземного моря до Норвегии. При этом неустойчи-
вость локальных изменений экологической ситуации демпфируется
разными репродуктивными стратегиями групп морф.
К тем же результатам стабилизации существования вида во време-
ни приводит и более продвинутая стадия дифференциации - экологи-
ческие расы. У жука ивового листоеда Lochmea caprea L., детально
изученного А.В. Михеевым (1985), в Европейской России существует
две симпатричные экологические расы. Кормовыми растениями од-
ной из них служат широколиственные ивы, главным образом козья и
ушастая, а также осина. Другая раса питается в основном листьями
пушистой и, реже, бородавчатой березы.
Личинки и имаго ивовой расы не могут питаться листьями бе-
резы. В эксперименте при принудительном питании листьями бе-
резы гибель составляла 100%. В то же время березовая раса спо-
собна питаться листьями ивы. Осиновыми листьями питаются
жуки, мигрирующие с ив, по-видимому, при повышенной плотно-
сти. Аналогично жуки, питающиеся на бородавчатой березе, вы-
селяются с пушистой. В эксперименте жуки березовой расы выби-
рают пушистую в 93,5 случаев из 100. В природе, по результатам
массового мечения, обмен между жуками на иве и осине составля-
ет около 40%. Примерно такой же уровень обмена между жуками,
питающимися двумя видами берез.
78
IL Общие проблемы эволюции
Обмен между ивовой и березовой расами составляет от 5,0 до 0,3%
при том, что ивы и березы, заселенные соответствующими расами,
буквально соприкасаются ветвями, а миграция с зимовки до кормово-
го растения достигает 2-4 км. Другими словами, кормовая специали-
зация связана с поведением. Связана она и со слабыми экологически-
ми различиями. Жуки ивовой расы несколько более гигрофильны и
несколько более термофильны. Как видно из сказанного, дивергенция
рас ивового листоеда достигла довольно высокого уровня, по мнению
Д.В. Михеева (1985), - уровня подвидов.
Возникает вопрос: почему она не продолжается, т.е. почему в Ев-
ропейской России эти расы нигде не дивергировали сильнее. В Вос-
точной Сибири, по предварительным данным, дивергенция их слабее
и экология их иная, чем в европейской части.
Каждая из двух рас ивового листоеда связана с кормовыми расте-
ниями, представляющими собой закономерный и достаточно длитель-
ный этап демутационных сукцессий. Таковыми являются оба вида бе-
рез и осина. Ивы, согласно Разумовскому (1981), являются ценофоб-
ными видами. Они не входят в типичные сукцессионные ряды, но
растут в любых местах, подходящих им по условиям освещенности и
влажности. Обычно это различные нарушенные территории. Длитель-
ность этапов сукцессии, мозаичность демутаций и наличие ценофоб-
ных кормовых растений обеспечивает каждой из рас устойчивое су-
ществование на обширных территориях. Для поддержания популяций
достаточно локальных изменений плотности населения, происходящих
по мере возникновения и выпадения насаждений кормовых растений.
Стабильность более специализированной ивовой расы, как и единство
вида, поддерживаются асимметричным притоком генов из березовой
расы. Несмотря на то, что обмен генами составляет в среднем менее
3% на достаточно длительном промежутке времени, он, по-видимому,
эффективен. В эксперименте, при свободном выборе брачного парт-
нера, частота межрасовых скрещиваний колеблется от 8,6 до 20,9%,
причем потомство таких гибридов плодовито.
Таким образом, дифференциация и рас, и внутри каждой расы со-
здает условия, при которых даже значительные колебания экологичес-
ких условий на обширных территориях позволяют существовать каж-
дой из них. Мало того, даже если одна из двух рас вымрет при более
сильном изменении условий существования, это не приведет к исчезно-
вению вида Lochmea caprea L. как целого.
79
А. С. Северцов
Приведенные данные, которые можно было бы дополнить (см.,
например: Person, 1967; Сергаевский, 1987 а,б; Новоженнов, 1987 и
др.), свидетельствуют о двояком значении внутривидового разнообра-
зия. С одной стороны, любая степень внутривидовой дифференциа-
ции, от полиморфизма до хорошо обособленных экологических рас,
представляет собой различные этапы микроэволюции. С другой - они
же препятствуют дальнейшей дифференциации вида. Повышая его эко-
логическую устойчивость, внутривидовое разнообразие препятствует
прогрессивной эволюции.
Контрбаланс векторов естественного отбора как причина
эволюционного стазиса
На любую популяцию любого вида в природе действует множе-
ство векторов естественного отбора. Анализ их действия на популя-
циях двух видов бурых лягушек: травяной Rana temporaria L. и остро-
мордой Rana arvalis Nilss., основанный на материалах, собранных
Г.С. Суровой, С.М. Ляпковым, М.Б. Корниловой и мной, привел к кон-
цепции контрбаланса векторов движущего отбора, препятствующего
прогрессивной эволюции.
Эти данные опубликованы с различной степенью полноты (Север-
цов, 1986, 1990, 1995 и 1999), поэтому здесь не будут приведены циф-
ровые данные, обосновывающие эффективность каждого исследован-
ного вектора. Однако обосновывать эффективность отбора по иссле-
дованным признакам: темпам роста и развития, признакам с широкой
нормой реакции и нулевой наследуемостью, необходимо.
Многочисленные оценки аддитивной компоненты генетической
изменчивости темпов роста и темпов развития разных видов лягушек,
проведенные методом сибсов, показывают практически нулевую на-
следуемость этих признаков (Сурова, 1988;Berven, 1982; Mitchel, 1990;
Newmen, 1988; Travis, 1980, 1981 и др.). Основное значение для из-
менчивости темпов роста и развития головастиков имеет эффект груп-
пы: существует оптимальная плотность населения: 1 особь на 0,6-0,7 л
воды (Шварц и др., 1976). При этой плотности темпы онтогенеза мак-
симальны, а дисперсия по скорости роста и развития минимальна.
С повышением плотности населения средние темпы развития и роста
снижаются, а дисперсия по этим признакам увеличивается (рис. 1,2).
80
IL Общие проблемы эволюции
Этот эффект обусловлен, главным образом, экзометаболитами - веще-
ствами белковой природы, выделяемыми головастиками в воду и ин-
гибирующими их рост и развитие. Кроме экзометаболитов показано
аналогичное влияние зрительных и тактильных контактов между осо-
бями и влияние некоторых водорослей, которыми они питаются. Бо-
лее крупные головастики меньше страдают от эффекта группы, мел-
кие - сильнее. Поэтому по мере онтогенеза дисперсия по обоим при-
знакам увеличивается.
Известно, что чем ниже наследуемость признака, тем менее эф-
фективен идущий по нему отбор. Однако сравнение популяций травя-
ной лягушки из Подмосковья (Звенигородская биологическая станция
МГУ) и Соловецких островов Белого моря показывает, что он эффек-
тивен (рис. 1,2). Головастики соловецкой популяции почти не чувстви-
тельны к эффекту группы, и личиночный период онтогенеза проходит
у них существенно быстрее и с более низкой смертностью. Это обус-
ловлено адаптацией к более суровому климату.
Рис. 1. Темпы роста головастиков из звенигородской и соловецкой популяций
травяной лягушки при выращивании их в группах с оптимальной плотностью
1 особь/0,6 л воды (Л) и в группах с высокой плотностью 1 особь/0,13 л воды (Б)
81
А. С. Северцов
и I
О 10 20 30 40
сутхн развития
Рис. 2. Дифференциация по стадиям развития головастиков из зве-
нигородской и соловецкой популяций травяной лягушки при выра-
щивании их в группах с оптимальной плотностью 1 особь/0,6 л
воды (Л) и в группах с высокой плотностью 1 особь/0,13 л воды (Ь)
Можно предположить два механизма генетической фиксации по-
добных эволюционных изменений. Они могут происходить либо на
основе генокопирования модификационных изменений по схеме
Шмальгаузена-Уодинктона, либо на основе неаддитивной компонен-
ты генотипической изменчивости. Методы анализа этой изменчивос-
ти отсутствуют.
На наличие неаддитивной генетической изменчивости роста голо-
вастиков травяной лягушки может указывать следующий эксперимент.
Были взяты 379 головастиков, вылупившихся из одной кладки икры.
На стадии почки задней конечности они были разделены на четыре
группы. В первую группу были выделены наиболее мелкие особи, на-
ходившиеся на стадии ранней (округлой) почки задней конечности.
Во вторую - средние по размерам головастики на той же стадии ок-
руглой почки задней конечности. В третью группу были отнесены го-
82
IL Общие проблемы эволюции
ловастики тех же размеров, но более старшие, находившиеся на ста-
дии конической почки задней конечности. В последнюю, четвертую,
группу вошли более крупные особи с конической почкой задней ко-
нечности. В дальнейшем каждая из четырех групп развивалась в от-
дельной емкости при одинаковой плотности (5 особей на 1 л воды),
выше оптимальной в 3 раза. За 10 дней в результате эффекта группы
распределение головастиков по длине тела в каждой группе приобре-
ло характер, типичный для подобных экспериментов. Однако средние
и модальные значения у разных групп не совпадали, что указывает на
то, что для особей каждой группы существуют пределы модификаци-
онной изменчивости скорости роста. Наличие этих пределов может
свидетельствовать о существовании генетической составляющей их
изменчивости. Так или иначе, отбор в процессе эволюции бурых лягу-
шек эффективен, что позволяет рассматривать эволюционные послед-
ствия его действия, реконструируя на этой основе направления каждо-
го из векторов.
Среди бурых лягушек Европейской России R. temporaria наиболее
плодовита. На Звенигородской биостанции количество икринок в клад-
ках этого вида варьирует от 700 до 6700 шт. при модальном значении
в 1800 икринок. У симпатричной и в значительной степени синтопич-
ной остромордой лягушки лимиты числа икринок в кладке составля-
ют 280-2100 шт при модальном их количестве - 1100. Различия по
средним значениям еще более заметны - 3700 и 1190 соответственно.
Более высокая плодовитость травяной лягушки объясняется тем,
что она мечет икру очень рано, когда водоемы наполнены талыми во-
дами. Когда вода спадает, икра или головастики обсыхают и гибнут
Остромордая лягушка мечет икру на 7-10 дней позже, и ее потомство
гибнет от обсыхания существенно реже (Северцов, 1990). На связь
обсыхания икры с отбором на плодовитость обратил внимание еще
С.С. Шварц (1976). Относительно крупные размеры травяной лягуш-
ки, а она примерно в 2 раза крупнее остромордой, объясняются более
высокой плодовитостью. Большое количество икры требует более круп-
ных размеров родителей.
Но икра обсыхает ежегодно, причем гибнет ее при этом много: во
временных водоемах - до 80% от отложенной, а в постоянных - до
15% (Сурова, Северцов, 1985). Следовательно, возникает вопрос: по-
чему с конца плиоцена размеры травяной лягушки не увеличивались
83
А. С. Северцов
доя того, чтобы самки могли откладывать больше икры? Другими сло-
вами: почему отбор на плодовитость не эффективен?
Дело в том, что отбору на плодовитость препятствует увеличение
эмбриональной смертности, происходящее по мере увеличения коли-
чества икры в водоеме. Кладки травяных лягушек, помимо того, что
они сами большие, слипаются в плотные подушки, что способствует
аккумуляции тепла и тем самым обеспечивает нормальное эмбриональ-
ное развитие в холодной воде при раннем икрометании. Даже ночью
температура в скоплении икры на 1-2 градуса выше, чем температура
окружающей воды. Но эта адаптация имеет оборотную сторону: в скоп-
лении икры развивается гипоксия, и от нее гибнет в среднем около
30% эмбрионов. Иногда, например на фоне сероводородного отравле-
ния, эта смертность достигает 70% (Сурова, Северцов, 1985). Значит,
чем выше плодовитость, тем выше эмбриональная смертность.
Нельзя сказать, что у травяной лягушки не шел отбор, обусловлен-
ный смертностью от гипоксии. Чувствительность к недостатку кисло-
рода у нее ниже, чем у остромордой лягушки. Но если эмбриональная
смертность низка, из икры вылупляется много головастиков и начина-
ет сказываться эффект группы. Особенно сильно он проявляется, ког-
да головастики образуют плотные скопления на оболочках икры. За
первые дни личиночной жизни, пока головастики не расплылись по
водоему, гибнет около 30% от числа вылупившихся особей (Сурова,
1985). Затем личиночная смертность начинает уменьшаться, но диф-
ференциация генерации по темпам онтогенеза, обусловленная эффек-
том группы, продолжается.
Высокая смертность головастиков в результате повышения плот-
ности их населения в то время, когда они свободно плавают по водо-
ему, - явление довольно редкое, обычно связанное с усыханием водо-
ема, а не с переуплотнением популяции, вызванным обилием икры,
отложенной в пруд. Другими словами: смертность на эмбриональных
и постэмбриональных стадиях развития до расплывания головастиков
достаточно интенсивна, и эффект группы на более поздних стадиях
развития проявляется, прежде всего, дифференциацией генерации по
темпам онтогенеза и большими или меньшими средними для генера-
ции сроками развития в данном водоеме. Во всяком случае, корреля-
ция между численностью икры в водоеме и численностью выходящих
сеголеток низкая (порядка 0,3) (Ляпков, Северцов, 1994). Сроки выхо-
84
IL Общие проблемы эволюции
да сеголеток в большой степени зависят и от температурного режима
водоема: чем холоднее пруд, тем дольше предметаморфозное разви-
тие. Этот фактор часто перекрывает зависящее от плотности торможе-
ние темпов онтогенеза. Но так или иначе, эмбриональная и постэмб-
риональная смертность есть всегда, и она препятствует отбору на пло-
довитость (Ляпков, Северцов, 1994).
Тогда возникает вопрос: почему эта смертность не снижается в
результате отбора на снижение чувствительности к эффекту группы?
Тем более, что отбор в этом направлении возможен. Головастики со-
ловецкой популяции к нему гораздо менее чувствительны, чем под-
московные. Мало того, чем дольше головастики живут в воде, тем боль-
ше они экспонируют себя хищникам, в первую очередь, личинкам пла-
вунцов и крупных стрекоз. Пик численности последних приходится
на конец личиночного развития головастиков (Северцов, Сурова, 1979).
Поэтому существует вектор отбора на ускорение личиночного разви-
тия, то есть отбор против эффекта группы. Второй фактор, работаю-
щий против чувствительности к повышению плотности, - приспособ-
ленность сеголеток. С.М. Ляпков (1992) показал, что приспособлен-
ность сеголеток тем выше, чем они крупнее и чем раньше они метамор-
физировали.
Тем не менее эффект группы сохраняется. В естественных водо-
емах он выражается, кроме смертности, прежде всего в том, что рас-
тягивает сроки выхода сеголеток на сушу. Если икрометание у травя-
ной лягушки продолжается около недели, то метаморфоз генерации
занимает 1,5-2 месяца, а иногда, при высокой плотности головасти-
ков или холодном водоеме, часть генерации не успевает метаморфи-
зировать до ледостава, как это произошло в 1994 г. на двух прудах Зве-
нигородской биостанции.
Сохранение эффекта группы, т.е. чувствительности головастиков к
повышению плотности, обусловлено тем, что у сеголеток, вышедших
на сушу, существует высокая и к тому же плотностно-зависимая ги-
бель, определяемая хищниками. Действительно, вблизи водоема плот-
ность сеголеток на пике выхода на сушу составляет десятки особей на
1 кв.м. В результате они оказываются привлекательной добычей для
очень широкого спектра видов хищников. Их едят пауки-серебрянки,
тритоны, зеленые лягушки, поползни, сойки, вороны. Но основную
массу сеголеток выедают, по данным С.М. Ляпкова, землеройки.
85
А.С. Северцов
Так или иначе, за первые недели жизни на суше гибнет до 70% вышед-
ших сеголеток (Ляпков, 1992).
Если бы не было эффекта группы, плотность сеголеток вблизи во-
доема была бы еще выше, и эта высокая плотность сохранялась бы на
большей территории - на большем расстоянии от уреза воды. Други-
ми словами: эффект группы, снижая динамическую плотность сеголе-
ток вблизи водоема, делает их менее привлекательными для хищни-
ков. Когда пик выхода проходит, большинство хищников покидает ок-
рестности пруда, т.к. высокая плотность доступных жертв, как
привлекающий фактор, теряет свое значение.
Таким образом, отбор сеголеток хищником сохраняет эффект груп-
пы, а он, в свою очередь, препятствует отбору на плодовитость. Выяс-
няется, что в целом популяция травяной лягушки все время находится
под воздействием разнонаправленных и противодействующих векто-
ров отбора. Причем усиление отбора в одном каком-нибудь направле-
нии приводит к усилению отбора в противоположном направлении.
В результате, несмотря на то, что каждый вектор отбора действует не-
прерывно и достаточно интенсивно, эволюции в смысле направленно-
го изменения популяции не происходит, а суммарное действие всех
векторов обусловливает низкую выживаемость генерации. Эту ситуа-
цию нельзя считать частным случаем, характерным для единственной
изученной нами популяции Rana temporaria. В литературе удается об-
наружить достаточное число аналогичных примеров контрбаланса
векторов движущего отбора, которые просто до сих пор интерпрети-
ровали либо как стабилизирующий, либо как циклический отбор.
Напомню знаменитую работу Лэка (Lack, 1954) о величине кладки
у птиц, в которой написано, что отбору на плодовитость, т.е. на увели-
чение числа яиц в кладке, препятствует отбор на уменьшение числа
яиц в кладке, обусловленный трудностями выкармливания большого
количества птенцов.
Напомню известную работу О’Дональда (O’Donald, 1972) о корот-
кохвостом поморнике, в которой показано, что баланс темно- и свет-
лоокрашенных форм поддерживается, с одной стороны, высокой по-
ловой активностью темноокрашенных самцов, а с другой стороны -
повышенной рецептивностью светлоокрашенных самок.
Напомню известную работу Н.В. Тимофеева-Ресовского и
Ю.М. Свирежева (1966) о полиморфизме божьей коровки, где показа-
86
II. Общие проблемы эволюции
но, что баланс морф поддерживается тем, что темная форма более хо-
лодоустойчива, а красная - более плодовита.
Число примеров можно увеличить, но и приведенных достаточно,
чтобы сказать, что ситуация с травяной лягушкой не уникальна. Веро-
ятно, большую часть случаев циклического, плотностно-зависимого и
частотно-зависимого отбора следует трактовать как контрбаланс век-
торов движущего отбора.
Тогда возникает два вопроса. Во-первых: если контрбаланс отбора
стабилизирует популяцию, то какова ситуация, в которой происходит
прогрессивная эволюция? Во-вторых, насколько эффективно контр-
баланс стабилизирует популяцию?
Ответ на первый вопрос может звучать следующим образом: для
того, чтобы шла прогрессивная эволюция, т.е. чтобы популяция изме-
нилась, нужно либо нарушение баланса уже существующих векторов,
либо возникновение нового, не сбалансированного вектора отбора.
Первую из двух ситуаций иллюстрирует соловецкая популяция тра-
вяной лягушки. Контрбаланс в этой популяции изменен более жестким
климатом: с одной стороны, сокращение теплого периода стимулирует
отбор против эффекта группы, с другой - тот же фактор замедляет рост
метаморфизировавших лягушек. Они мельчают и откладывают меньше
икры. Величина кладки у соловецких лягушек в среднем 500 (от 106 до
990 шт.) икринок, приблизительно в три раза меньше, чем у подмосков-
ных. Это, в свою очередь, приводит к прекращению отбора на десинхро-
низацию метаморфоза, т.к. плотность выходящих сеголеток и без того
низка. В результате и утрачивается эффект группы.
Иллюстрацией второго случая - возникновения нового несбалан-
сированного вектора отбора - могут служить все примеры возникно-
вения резистентности насекомых к пестицидам. Понятно, что пести-
циды - новый фактор. Их массовое применение началось около 40 лет
назад, и сейчас более 1000 видов насекомых устойчивы к одному, а
часто и к нескольким хлорорганическим и фосфороорганическим со-
единениям. Как возникновение такого несбалансированного вектора
сказывается на балансе всех остальных, судить трудно.
Второй вопрос, касательно эффективности стабилизации популя-
ции, пока не имеет ответа. В этом отношении очень заманчиво иссле-
дование популяции вблизи границ ареалов видов. Известно, что гра-
ницы ареалов более или менее стабильны, и если меняются в сторону
87
А. С. Северцов
расселения, то обычно это связано с изменением экологической ситу-
ации в благоприятную для вида сторону. С другой стороны, большин-
ство факторов среды меняется по градиенту, т.е. в принципе к их изме-
нению возможна адаптация. Однако этой адаптации, как правило, не
происходит. Можно предположить, что именно контрбаланс векторов
отбора делает границы ареалов относительно стабильными, т.е. пре-
пятствует дальнейшей адаптации краевых популяций. Однако этот
вопрос требует отдельного изучения.
В заключение следует отметить, что нельзя себе представить попу-
ляцию под действием отбора таким образом, что на какой-то признак
действует вектор движущего отбора, а на другой - стабилизирующе-
го. Популяции любого вида существуют в сложной среде и сами они
сложно структурированы. Поэтому каждый фактор внешней среды,
вызывающий гибель или устранение от размножения части особей в
популяции, является причиной возникновения того или иного направ-
ления отбора. Этих направлений много, они противодействуют друг
другу, и состояние популяции, с эволюционной точки зрения, - это
состояние равновесия между противодействующими друг другу на-
правлениями отбора. А это, в свою очередь, значит, что состояние по-
пуляции по любому признаку, подвергающемуся отбору, отнюдь не
оптимально. Контрбаланс препятствует оптимизации адаптаций. Он
лишь уравновешивает их на том уровне, на котором выравниваются
интенсивности отбора в противоположных направлениях.
Следовательно, нецелесообразно считать, что особи, составляющие
популяцию, оптимально приспособлены к среде своего обитания, а под-
ход, основанный на расчетах эволюционно стабильных стратегий, веро-
ятно, интересен с точки зрения математики, но едва ли приложим к ре-
альным процессам эволюции популяции под действием отбора.
Наконец, из вышеизложенного следует, что структура взаимодей-
ствий особей в популяции влияет на характер естественного отбора
сильнее, чем наследуемость отбираемых признаков. Быть может, это -
частный результат, свойственный Rana temporaria, но он интересен для
понимания эволюции.
МИКРОЭВОЛЮЦИЯ И ФИЛОЦЕНОГЕНЕЗ
А.С. Северцов
Московский государственный университет
119899, г. Москва, Воробьевы горы
E-mail: asevertsov@soil.msu.ru
Вопрос об эволюции многовидовых сообществ дискутируется уже
более 80 лет. Он поставлен еще Ф. Клементсом [35] и В. Сукачевым
[24]. Однако мнения о том, что считать эволюцией сообществ, каковы
ее механизмы, и даже существует ли она как таковая, до сих пор явля-
ются дискуссионными.
С одной стороны, эволюция экосистем - палеонтологический факт.
Мезозойские фитоценозы голосеменных и кайнозойские сообщества
покрытосеменных существенно отличались друг от друга. Эвтрофи-
кация пресноводных водоемов также началась только после смены
мезофиты кайнофитом [7]. Однако какую долю в этих отличиях со-
ставляла дарвиновская эволюция видов, составлявших и составляю-
щих экосистемы, а какую - эволюция экосистемы, как многовидовой
системы, не ясно. Сукачев [24, 25] считал, что эволюция сообществ
происходит по мере эволюции видов, составляющих экосистемы, и
вместе с ней, что и выражено в предложенном им термине «филоце-
ногенез».
С другой стороны, хорошо известно, что биоценозы формируются
из уже существующих, «готовых» видов. А.М. Гиляров [4] обратил вни-
мание на то, что три основных компонента любого сообщества - про-
дуценты, редуценты и деструкторы - существуют с самых ранних эта-
пов эволюции биосферы. Формирование сообществ - результат ком-
поновки популяций уже существующих видов, а сама эта компоновка
происходит по неизменным правилам, таким, например, как полный
круговорот вещества и энергии, пирамида биомасс трофических уров-
ней и т.п. Неизменность правил формирования многовидовых сооб-
ществ позволяет отрицать само существование эволюции экосистем,
как это сделал, например, Р. Мэй [16].
Многие сторонники существования эволюции экосистем призна-
ют, что теория этого процесса до сих пор отсутствует [21,28-31 и др.].
89
А. С. Северцов
В.В. Жерихин [10] предложил считать элементарным событием эво-
люции экосистем изменение набора реализованных экологических
ниш, что равносильно изменению видового состава сообщества.
Сложности анализа эволюции экосистем на конкретном материале
породили неоднократные попытки механически экстраполировать фак-
торы микроэволюции, действующие на популяционном уровне на эво-
люцию многовидовых сообществ. Предлагалось, например, считать ма-
териалом филогенеза сумму генофондов всех популяции, составляющих
биоценоз. Ряд авторов рассматривали в качестве движущей силы биоце-
нотической эволюции отбор популяций и целых экосистем, т.е. вымира-
ние одних популяций и выживание других, или процессы необратимой
сукцессии типа заболачивания озера. Исчерпывающая критика подоб-
ных представлений дана Ю.И. Черновым [29,30]. К ней можно приба-
вить лишь одно: вымирание или переживание популяций, групповая эли-
минация, по И. Шмальгаузену [33], не ведет к прогрессивной эволюции
даже на популяционно-видовом уровне [33, 38].
Наиболее распространенным представлением о механизме эволю-
ции сообществ является концепция филогенеза [25], согласно которой
эволюционная перестройка сообществ представляет собой результат
микроэволюции разных видов, образующих сообщество.
С другой стороны, многие авторы, например Жерихин [8-10], рас-
сматривают в качестве эволюции экосистем их перестройку в резуль-
тате вхождения в них или выпадения из них тех или иных компонен-
тов. Поэтому Жерихин [9] подчеркнул, что эволюцией сообществ сле-
дует считать такие процессы, которые приводят к фиксированным,
стабильно воспроизводящимся изменениям структуры экосистем. Если
в результате перестройки состава сообщества возникает новая, ранее
не существовавшая структура экологических связей внутри него и эта
структура в дальнейшем сохраняется, - это эволюция.
Эта концепция отличается от концепции филогенеза. Филогенез не-
посредственно основан намикроэволюционных изменениях популяций,
составляющих сообщество. Перекомпоновка состава сообществ, созда-
ющая их новую структуру, связана с микроэволюцией лишь опосредо-
ванно. По Жерихину, эволюция экосистем представляет собой процесс
новообразования, а не адаптации. Однако обе концепции нельзя считать
взаимно исключающими друг друга, что, в свою очередь, предполагает
сосуществование двух различных механизмов эволюции экосистем.
90
II. Общие проблемы эволюции
Источники, из которых могут рекрутироваться новые компоненты
биоценозов, были рассмотрены Г. Длусским [5] и Ю. Черновым [29].
Это либо виды, сформировавшиеся в близких по структуре (конвер-
гентных [6]) экосистемах, но в географически удаленных регионах;
либо виды, сформировавшиеся в условиях интрастациальных [5], или
интразональных [28, 29], мест обитания, способные внедриться в но-
вые для них зональные сообщества. Длусский показал, что внедрение
нового вида (видов) в коадаптивный комплекс пустынных муравьев-
зоонекрофагов (капрофагов) более четко разграничивает ниши ком-
понентов этого комплекса и канализирует их дальнейшую эволюцию.
Прямых наблюдений или экспериментальных данных по эволю-
ции экосистем ожидать не приходится - процесс слишком медленный.
Данные палеонтологии также мало помогают в решении этой пробле-
мы. Накоплено много сведений о смене во времени локальных экоси-
стем (см. например, материалы конференции «Эволюция экосистем»
[34]). Однако доказать, что наблюдаемые изменения - именно эволю-
ционные, на палеонтологическом материале невозможно, т.к. даже в
случае доказательства непрерывности наблюдаемых локальных изме-
нений нет гарантии, что это эволюция, а не изменение границ экосис-
тем, например, под влиянием изменения климата. Поэтому палеонто-
логи стараются оперировать большим промежутком времени, порядка
периодов и геологических веков, и доступным материалом, либо палео-
зоологическим, либо палеоботаническим, что, естественно, не позволя-
ет рассматривать ни биоценоз в целом, ни факторы его эволюции.
Исходя из сказанного, приходится признать, что попытка анализа
эволюции экосистем может опираться, главным образом, на две кате-
гории данных: данные о структуре и динамике рецентных сообществ
и данные по механизму микроэволюции. В дополнение к ним должны
привлекаться любые как палеонтологические, так и неонтологичес-
кие сведения, относящиеся к обсуждаемой проблеме.
Традиционно изучение механизмов и закономерностей изменения
сообществ основано на рассмотрении этого процесса с точки зрения
самого сообщества как целостной системы [4, 7-10, 16 и др.]. Несмот-
ря на значительные успехи, достигнутые благодаря этому подходу, те-
ория эволюции экосистем отсутствует. Поэтому представляется целе-
сообразным взглянуть на проблему с другой стороны - с точки зрения
эволюции популяций, составляющих экосистему. Правомочность та-
91
А.С. Северцов
кого подхода обусловлена тем, что экосистемы слабо интегрированы
[29, 30], а следовательно, целесообразно выяснить, насколько микро-
эволюционные процессы, в первую очередь видообразование, влияют
на эволюцию многовидовых сообществ.
Любая биологическая эволюционная теория, в том числе и теория
эволюции экосистем, должна отвечать минимум на три вопроса. Ка-
ков механизм эволюции? Куда направлена эволюция рассматриваемой
экосистемы? Каковы темпы этой эволюции? Желательно также выде-
лить элементарную единицу эволюции на изучаемом уровне органи-
зации биологических систем. Попытке ответа на эти три вопроса и
посвящена настоящая статья.
Единица эволюции экосистемного уровня
В.В. Жерихин [10] предложил рассматривать в качестве единицы
эволюции многовидовых сообществ сукцессионную систему. Сама сук-
цессия сопоставима с онтогенезом многоклеточного организма, а ее
необратимая перестройка - с филогенезом. С общетеоретических по-
зиций этот подход безупречен, но трудно приложим к реальным ситу-
ациям. В качестве необратимой перестройки сукцессии можно пред-
ставить и заболачивание озера (десятки тысяч лет и десятки километ-
ров поверхности земли), и смену мезофита кайнофитом в середине
мелового периода - меловой биоценотический кризис (миллионы лет
и вся суша земного шара).
По своим размерам экосистемы образуют непрерывный ряд, от
самых малых, например, деструктивной сукцессии фекалий какого-
нибудь млекопитающего, до природно-климатических зон и биосфе-
ры в целом. Ни очень маленькие, ни очень большие экосистемы не
могут являться единицей эволюции: маленькие - эфемерны, большие
включают в себя автономные подсистемы.
Единицей микроэволюции считается локальная популяция. Исхо-
дя из этого, можно считать, что единицей эволюции экосистем долж-
но быть сообщество, в котором численность наиболее малочисленно-
го компонента более или менее соответствует размерам популяции
этого вида, т.е. обеспечивает его воспроизводство на данной террито-
рии. Плотность населения консументов 1-3-го порядков, особенно
крупных животных, меньше, чем мелких. Поэтому размеры экосис-
92
IL Общие проблемы эволюции
тем, способных к длительному существованию, а тем самым и к эво-
люции, должны быть сопоставимы с размерами территории популя-
ций крупных млекопитающих [1].
С другой стороны, в силу своей эврибионтности популяции круп-
ных травоядных и хищных млекопитающих не имеют строгой стаци-
альной приуроченности. Даже у микромаммалий одна и та же популя-
ция может являться компонентом одновременно нескольких сообществ.
То же самое относится и к растениям. Общеизвестно, что границы
ценопопуляций и границы локальных популяций редко совпадают. Тем
не менее, если выбирать в континууме размеров экосистем нечто со-
поставимое с размерами популяций, наиболее близким к искомому
представляется биоценоз в том объеме, в котором это понятие предло-
жил Сукачев [25]. Это достаточно крупный территориальный комп-
лекс, внутри которого происходит полный круговорот вещества и энер-
гии, и размеры его сопоставимы с размерами популяций консументов
1-2-го порядков. Жерихин [9] специально подчеркнул, что рассматри-
вать следует именно биоценоз, а не биогеоценоз, т.к. абиотические
условия в эволюции не участвуют, они лишь создают пределы допус-
тимых изменений биоты. Выделяя биоценоз как единицу эволюции
сообществ, надо еще раз подчеркнуть всю условность подобного вы-
деления. На местности границы биоценозов обычно определяются как
границы фитоценозов. С.М. Разумовский [19], во многом следуя в этом
отношении взглядам Сукачева, настаивал на дискретности границ ра-
стительных сообществ. Противоположную точку зрения, как извест-
но, развивали Л.Г. Раменский и его школа. Если же учесть, что биоце-
ноз -это не только фитоценоз, но и все остальные его компоненты, т.е.
и консументы, и деструкторы, то границы биоценозов на местности
станут совершенно условными. Можно выделить некое сообщество,
но нельзя провести его границы.
Механизм эволюции экосистем
И.И. Шмальгаузен [32] предложил рассматривать микроэволюцию,
как авторегуляторный цикл, в котором регулятором является биогеоце-
ноз, а регулируемым объектом - популяция, составная часть этого био-
геоценоза. По каналу прямой связи передается информация о составе
популяции. Этот канал работает на генетическом уровне. Гаметы про-
93
А. С. Северцов
шедших отбор особей, объединяясь в зиготы, дают начало следующему
поколению популяции, которое, по своим характеристикам оказывается
соответствующим условиям существования в данном биогеоценозе.
В процессе онтогенеза происходит перекодирование информации
с генетического на фенотипический уровень. Генетическая информа-
ция, закодированная линейными последовательностями нуклеотидов,
преобразуется в фенотипическую информацию, развертываемую во
всех четырех измерениях. Эпигенетическими аспектами онтогенеза
модель пренебрегает. Каналом обратной связи является борьба за су-
ществование, по Шмальгаузену - активность организмов по отноше-
нию к окружающей среде. В процессе борьбы за существование био-
ценоз получает информацию о соответствии популяции условиям дан-
ного сообщества. Отбор, таким образом, является процессом регу-
ляции, а размножение выживших особей - второй перекодировкой
информации. Полный цикл регуляции проходит за одно поколение.
Изложенная модель микроэволюции легко преобразуется в модель
филоце но генеза [21].
Действительно, любая популяция - компонент данного биоценоза,
одновременно является и регулируемым объектом, и частью регулято-
ра для любой другой популяции того же биоценоза. Сколько популя-
ций - членов сообщества, столько и прямых и обратных связей между
каждой данной популяцией (регулируемым блоком) и всем остальным
сообществом (регулятором). Следуя этой модели, можно сказать, что
микроэволюционный процесс, начавшись в той или иной популяции,
автоматически приведет к изменению условий регулирования для ос-
тальных популяций сообщества, тем самым - к их микроэволюцион-
ным изменениям, т.е. к филоценогенезу. Поскольку границы ценопо-
пуляций и локальных популяций не совпадают, микроэволюционные
изменения должны затронуть не только рассматриваемую экосистему,
но и те экосистемы, частями которых являются эволюирующие локаль-
ные популяции. В результате эволюционировать будет не только один
биоценоз, но и экосистемы более высокого уровня.
Изменения абиотических условий, стимулируя микроэволюцион-
ные изменения внутри биоценозов, одновременно и сдерживают эти
изменения. Изменения климата, такие, например, как ледниковые пе-
риоды и межледниковые потепления, не только стимулировали видо-
образование, но и приводили к смещению климатических зон по по-
94
II Общие проблемы эволюции
верхности Земли и, тем самым, к смещению входящих в эти зоны био-
ценозов, что сохраняло и сами сообщества, и их компоненты. Ледни-
ковые изменения ареалов палеарктических видов служат хорошим
примером подобных перестроек, тогда как расселение видов часто
связано с микроэволюционными процессами.
Причины, инициирующие прогрессивную эволюцию популяций,
могут быть разными: изменение абиотических условий; внедрение
новых видов, не характерных для данного этапа сукцессии; смена
доминантов из числа видов, уже сосуществующих в данном био-
ценозе. Условием начала филоценогенеза должна быть лишь дос-
таточная интенсивность этих воздействий. Они должны вызывать
дифференциальную смертность, т.е. стимулировать изменения на-
правления и интенсивности естественного отбора. При этом попу-
ляции не должны вымирать. Вымирание отдельных популяций, как
это следует не только из представления Шмальгаузена [33] о веду-
щем значении для эволюции внутривидовой борьбы за существо-
вание и из критики Е. Уилсоном [38] и его последователями кон-
цепции группового отбора, но и из многочисленных примеров унич-
тожения человеком тех или иных популяций или целых видов,
обычно не приводит к заметным перестройкам экосистем. Унич-
тожение европейского тура, тарпана и кулана в европейских сте-
пях не уничтожило степь как экосистему. Уничтожение лося, боб-
ра, соболя в таежных экосистемах не привело к заметным пере-
стройкам таежных экосистем.
Ограничения на действие модели
Изложенная модель очень далека от реальности. Она подразумева-
ет равноценность всех микроэволюционных связей внутри биоценоза
и высокую интегрированность биоценоза в целом. И то и другое не
соответствует действительности. Биоценоз - сложно структурирован-
ная система, в ней выделяются коадаптивные комплексы [5], экологи-
ческие связи внутри которых более жестки, чем между этими комп-
лексами. Более того, и сами популяции (и виды) обладают механизма-
ми, стабилизирующими их положение в окружающей среде. Поэтому
следует рассмотреть биоценотические и популяционно-видовые огра-
ничения на действие изложенной модели.
95
А.С. Северцов
Биоценотические ограничения
Микроэволюционное изменение какой-нибудь популяции затронет
прежде всего популяции внутри коадаптивного комплекса, заставляя
их коадаптироваться дальше, но не затронет, либо лишь слабо затро-
нет, другие такие же комплексы. Коадаптация в комплексе пустынных
муравьев-зоофагов лишь в слабой степени скажется на эволюции пус-
тынных муравьев-капрофагов [5].
Другим важным ограничением является интегрированность био-
ценозов, выражающаяся в слабости и неоднозначности экологичес-
ких связей в цепях питания и между популяциями разных видов, нахо-
дящихся на одном трофическом уровне. Эта слабость становится за-
метной в случаях реинтродукции или успешной интродукции видов,
что эквивалентно внедрению в биоценоз новых видов или выпадению
видов при их вымирании (или уничтожении). Реинтродукция бобра,
соболя, восстановление численности лося и целый ряд аналогичных
событий, так же как и предшествующее уничтожение этих видов на
огромных пространствах лесной зоны Евразии, не повлекли за собой
заметных перестроек других компонентов биоценозов. Расселение
кабана на север и восток Европейской России, интродукция ондатры в
Палеарктику, интродукция енотовидной собаки и ротана в европейс-
кую часть Евразии, зайца русака в Западную Сибирь - не вызвали се-
рьезных биоценотических последствий, хотя определенные сдвиги
численности конкурентов этих видов происходили. Однако ни времен-
ное сокращение численности лисицы при конкуренции с енотовидной
собакой, ни сокращение численности окуня в тех водоемах, в которых
он конкурирует с ротаном, не затронули эти виды настолько, чтобы
можно было ожидать микроэволюционных последствий.
Все вышеизложенное относится к достаточно эврибионтным ви-
дам. Перестройка ценотического окружения для стенобионтных ви-
дов, например для олиго- или монофагов, будет существенно более
серьезной. Считается [19], что жесткость экологических связей или,
что в данном случае то же самое, плотность упаковки экологических
ниш возрастает от начальных стадий сукцессии к ее климаксу. Другим
фактором, определяющим плотность упаковки экологических ниш,
является степень стабильности абиотических условий. В обоих этих
случаях, например в тропических влажных лесах или в дубравах зоны
96
II. Общие проблемы эволюции
широколиственных лесов, изменения состава биоценоза более суще-
ственны, чем на ранних этапах зарастания вырубки или гари. В то же
время структурные и (или) эволюционные изменения в популяциях
сообществ на ранних стадиях сукцессии едва ли будут иметь заметное
влияние на более поздних стадиях.
Оборотной стороной вышеизложенного, также определяемой низ-
кой плотностью упаковки ниш, т.е. эврибионтностью видов, является
то что регулятор (биоценоз как целое) «не различает» экологически
близкие виды. Примером могут служить травяная (Rana temporaria) и
остромордая (Rana arvalis) лягушки. На огромном пространстве от
восточной Франции на западе до Уральского хребта на востоке эти
виды симпатричны и в значительной степени синтопичны. Часто они
мечут икру в одни и те же водоемы, хотя эмбрионы остромордой ля-
гушки менее чувствительны к низким значениям pH, чем эмбрионы
травяной. В холодные весны их икрометание происходит одновремен-
но, хотя обычно травяная лягушка приступает к размножению на не-
делю раньше остромордой. Головастики всеядны, и пищевая конку-
ренция и между видами и внутри видов отсутствует. Взаимного или
одностороннего подавления головастиков этих видов не наблюдается.
Несмотря на то, что головастики остромордой лягушки более чув-
ствительны к действию экзометаболитов - белковых веществ, выделяе-
мых головастиками в воду и ингибирующих рост и развитие более мел-
ких особей, - ни один из видов не подавляет предметаморфозное разви-
тие другого, что обусловлено лишь частичным совпадением сроков
икрометания и предпочитаемых водоемов для размножения. Сеголетки
обоих видов также не конкурируют. По мере миграции от берегов водо-
емов их плотность быстро снижается, а пищевые ресурсы даже в непос-
редственной близости от берега недоиспользуются. Несколько большая
привлекательность для хищников более многочисленных сеголеток тра-
вяной лягушки нивелируется меньшей смертностью от хищников менее
многочисленных сеголеток остромордой лягушки.
Биотопические предпочтения взрослых лягушек в летний, актив-
ный период их жизни в значительной степени перекрываются. Лягуш-
ки встречаются в одних и тех же стациях. При этом за пищу они не
конкурируют. Пищевые ресурсы далеко недоиспользуются, а хищни-
ки не различают эти два вида. Таким образом, в зоне симпатрии они
не конкурируют ни на одном этапе онтогенеза. Пожалуй, наиболее
97
А.С. Северцов
веским аргументом в пользу того, что травяная и остромордая лягуш-
ки не только не конкурируют сейчас, но никогда не конкурировали в
прошлом, хотя они симпатричны со второй половины плиоцена, явля-
ется то, что все их экологические различия укладываются в три комп-
лекса, связанные не с биотическими, а с абиотическими условиями их
обитания. Остромордая лягушка, по сравнению с травяной, несколько
более сухоустойчива, несколько более термофильна, а ее икра выдер-
живает несколько более сильную кислотность воды. Очевидно, если
между этими видами возникнет конкуренция и они начнут дивергиро-
вать сильнее, чем они различаются в настоящее время, их ценотичес-
кое окружение не среагирует на эти изменения до тех пор, пока не
изменится их положение в цепях питания.
Другим примером подобной ситуации могут служить домовый и
полевой воробьи, не различающиеся ни по питанию, ни по отноше-
нию к хищникам и синтопичные во многих местообитаниях [11, 12].
Аналогичные виды-двойники Microtus arvalis и Microtus rossomeridio-
nalis в Европейской России экологически различаются лишь тем, что
второй вид несколько более плодовит и несколько более синантропен
[27]. Возможно, представление об экологической эквивалентности ви-
дов с точки зрения их ценотического окружения можно экстраполи-
ровать на гильдии близких видов.
Другими словами, микроэволюция тех или других видов - членов
биоценоза - еще не означает перестройки структуры сообщества, так-
же как не означают ее перестройки частные замены одних видов дру-
гими. Структура сообществ более консервативна, чем составляющие
эти сообщества виды [18]. Таким образом, микроэволюционные воз-
мущения структуры биоценоза затухают в нем, так же как затухают и
частные экологические возмущения.
Микроэволюционные ограничения
Вторым фактором, препятствующим филогенезу, является эволю-
ционная стабильность популяций и видов. В настоящее время выделе-
но два механизма, поддерживающие эту стабильность: внутривидо-
вое разнообразие [20] и контрбаланс векторов отбора [21, 22].
Внутривидовое разнообразие, т.е. разнообразие морф сбалансиро-
ванного полиморфизма, разнообразие экологических рас или (и) гео-
98
II. Общие проблемы эволюции
графических подвидов означает, что при ухудшении условий существо-
вания часть этого разнообразия вымирает или сокращает численность,
а часть либо выживает, либо даже получает преимущество.
Иллюстрацией роли полиморфизма в поддержании стабильности вида
может служить история московских сизых голубей (Columba livia) [17].
Светлая морфа сизого голубя более активна в поисках корма и охране
гнездовой территории. Меланистические формы более пассивны и по-
этому более толерантны к повышению плотности населения. Лимитиру-
ющим фактором является недостаток гнездовий. Пищевые отбросы -
основная пища голубей в Москве - не лимитированы. Во время Великой
Отечественной войны голуби в Москве были почти полностью истреб-
лены. Сохранились лишь две-три небольшие колонии, состоящие лишь
из светлых форм. Затем, особенно после Международного фестиваля мо-
лодежи в 1957 г., численность голубей восстановилась и преимущество
получили меланисты, тогда как первоначально преобладавшая светлая
морфа была вытеснена на неудобные чердаки, где население голубей
редко и можно сохранить охраняемый гнездовой участок.
Иллюстрацией роли экологических рас в эволюционной стабили-
зации видов может служить работа А.В. Михеева [15] по ивовому ли-
стоеду (Lochmea саргеа). В Подмосковье этот вид имеет две расы. Одна
из них питается на ушастой и козьей ивах и на осине, а другая - на
пушистой и бородавчатой березах. Расы почти полностью изолирова-
ны, т.к. личинки березовой расы могут питаться листьями ивы, выжи-
вают и превращаются в имаго, а личинки ивовой расы в 100% случаев
гибнут при питании листьями березы. Очевидно, что несмотря на же-
сткую изоляцию, исчезновение на данной территории либо ивовой,
либо березовой расы, например в ходе сукцессии, не приведет к выми-
ранию вида - вторая раса сохранится. При восстановлении леса в ходе
сукцессии после вырубки или пожара восстанавливается и исчезнув-
шая раса. Другими словами, внутривидовое разнообразие стабилизи-
рует вид даже при очень резких биоценотических перестройках.
Второй механизм эволюционной стабилизации популяций сводит-
ся к тому, что разнонаправленные векторы естественного отбора, дей-
ствующие на популяцию, взаимно препятствуют друг другу. Подроб-
нее смотрите в книге А.С. Северцова «Направленность эволюции».
Изложенный способ, а также ряд других аналогичных примеров
[21, 22] объясняют, почему даже существенные экологические пере-
99
А. С. Северцов
стройки экосистем не способны стимулировать микроэволюционные
изменения тех популяций, на которых сказываются эти перестройки.
Длительный, порядка миллионов лет, эволюционный стазис видов -
палеонтологический факт [36, 37].
Микроэволюционные изменения популяций происходят при наруше-
нии контрбаланса векторов отбора, т.е. в тех случаях, когда изменение
среды настолько существенно, что тот или иной вектор (векторы) отбора
оказывается не сбалансированным противодействующим ему вектором
(векторами) отбора, или в тех случаях, когда возникает новый, ранее от-
сутствовавший вектор отбора [21-23]. Однако когда начинается микро-
эволюция каких-либо популяций - членов сообщества, даже в тех случа-
ях, когда она заканчивается возникновением нового вида, в силу биоце-
нотических ограничений, это еще не означает филоценогенеза.
Темпы эволюции экосистем
Из вышеизложенного следует, что два процесса - процесс микро-
эволюции и процесс перестройки экосистем за счет «готовых» компо-
нентов - происходят в значительной степени автономно друг от друга.
При этом видообразование создает те компоненты, которые затем вклю-
чаются в экологическую структуру сообществ. Темпы видообразова-
ния, как известно, варьируют в очень широких пределах, но включе-
ние новых видов в биогеоценозы еще не означает существенных из-
менений этих ценозов.
Следовательно, филоценогенез, в понимании этого термина по
Сукачеву, не связан с видообразованием как таковым. Он должен за-
висеть в первую очередь от коадаптации видов внутри экосистемы.
Именно в этом случае должно выдерживаться соответствие модели
эволюции сообщества как авторегуляторного процесса. Однако коа-
даптация, обусловленная когерентной эволюцией, - процесс медлен-
ный. Видимо, еще медленнее меняется облик сообществ за счет пере-
компоновки входящих в них видов. Дело в том, что еще В.Н. Кашка-
ров [13] подчеркнул, что облик сообщества формируется не столько
входящими в него видами, сколько жизненными формами. С Кашка-
ровым солидарен и Чернов [30]. Новые жизненные формы возникают
в процессе филогенеза, с филогенетическими скоростями, т.е. за де-
сятки и сотни миллионов лет. На становление облика млекопитающих,
100
IL Общие проблемы эволюции
от примитивных тероморф до примитивных млекопитающих, ушло
около 70 млн лет от начала перми до середины триаса [26]. На станов-
ление амфибий - почти весь девон, около 50-60 млн лет [3]. Филоге-
нез лошадей от Palaeoterium до Hipparion длился с начала эоцена до
плиоцена, т.е. около 45 млн лет. Сказанное иллюстрирует то, что фи-
лоценогенез происходит обычно очень медленно.
Представление о медленности филоценогенеза как когерентной
коадаптивной эволюции и представление о медленности формирова-
ния в ходе филогенеза новых жизненных форм, появление которых
меняет облик экосистем, означает, что два рассматриваемых в литера-
туре механизма эволюции надвидовых сообществ на макроэволюци-
онных временах сливаются воедино. Действительно, филоценогенез
создает новые формы, а новые формы, меняя структуру сообществ,
стимулируют филоценогенез.
Темпы филоценогенеза могут существенно возрастать только в слу-
чае экологических кризисов, когда разрушаются старые сообщества, зна-
чительная часть составляющих их видов вымирает, а новые сообщества,
группировки видов - по терминологии Жерихина [9] возникают из ос-
татков старой, докризисной, ценофильной флоры и фауны интерстаци-
альных и (или) интерзональных местообитаний, т.е. ценофобных, по тер-
минологии Разумовского [19]. В группировках происходит быстрая коа-
даптивная эволюция видов, приводящая к возникновению новых
сукцессионных рядов. По-видимому, именно в случае экологических
кризисов, в период некогерентной эволюции выравниваются темпы мик-
роэволюции и темпы филогенеза, т.е. модель эволюции сообщества как
авторегуляторного процесса может воплощаться в реальных условиях.
Направленность эволюции экосистем
Наиболее распространенным является утверждение, что эволюция
экосистем идет по пути коадаптации их компонентов [5, 9]. Однако
коадаптацию популяций нельзя считать направлением эволюции эко-
систем в целом. Все компоненты экосистемы тундры (или любой дру-
гой экосистемы) более или менее коадаптированы, т.е. адаптированы
к обитанию в тундре. Однако экосистема тундры и есть тундра. Сама к
себе она не может быть приспособлена. Другими словами, экосисте-
мам нельзя приписывать свойства адаптации или коадаптации.
101
А.С. Северцов
Кроме того, эволюция популяций, составляющих биоценоз, не ис-
черпывается их коадаптацией. Коадаптация выступает на передний
план эволюционных изменений популяций лишь в случае жестких эко-
логических связей между ними, таких как жесткая конкуренция или
олиго- или монофагия и т.п. В других случаях на передний план выс-
тупает адаптация к абиотическим или биотическим факторам. После-
днее совершенно не обязательно сопровождается взаимным приспо-
соблением компонентов экологических взаимодействий. Семена ели
приспособлены к распространению ветром, но не защищены от по-
едания их птицами и млекопитающими.
Весьма существенным фактором, препятствующим коадаптации по-
пуляций, являются внутрипопуляционные (внутривидовые) отношения.
Внутривидовая борьба за существование - основная причина естествен-
ного отбора [33]. Из вышеприведенных данных по контрбалансу векто-
ров отбора видно, что все ценотические связи популяции преломляются
через внутрипопуляционные взаимодействия особей - их плотность и
эффект группы. Внутрипопуляционные взаимоотношения, как извест-
но, чрезвычайно многообразны, от химических взаимодействий одно-
клеточных водорослей до сложного поведения высших животных. С био-
ценотической точки зрения эволюция, вызванная внутрипопуляционны-
ми связями, автономизирует популяции от их ценотического окружения,
т.е. она противоположна коадаптации. Вероятно, благодаря ей, за исклю-
чительно редким случаем мутуализма, нельзя говорить о том, что по мере
филоценогенеза увеличивается интегрированность сообществ. Напро-
тив, морфофизиологический прогресс, эмансипируя организмы от окру-
жающей среды, должен был бы понижать целостность экосистем. Кос-
венным подтверждением последнего является слабая ценотическая при-
уроченность и высших позвоночных и высших беспозвоночных.
Другим препятствием повышению интегрированности экосистем
является несовпадение размеров ценопопуляций и локальных попу-
ляций. Поскольку локальные популяции часто являются компонен-
том нескольких биоценозов, их эволюционные изменения должны
удовлетворять требованиям коадаптированности разных сообществ.
В результате в каждом данном сообществе коадаптированность не
может быть высокой.
Таким образом, уровень коадаптации компонентов сообщества, и
тем самым степень его интегрированности, представляет собой комп-
102
II. Общие проблемы эволюции
омисс между отбором на коадаптацию, отбором на адаптацию попу-
ляций к разным экосистемам и отбором, обусловленным абиотичес-
кими факторами и внутрипопуляционными взаимоотношениями. В
силу этих причин за редким исключением симбиогенеза в широком
смысле, т.е. высокой коадаптированности между теми или иными не-
многими членами биоценоза, общий уровень интеграции экосистем
не повышается. Следствием низкой интегрированности экосистем и
является то, что они могут формироваться из уже существующих «го-
товых» компонентов.
Таким образом, эволюция сообществ не идет ни по пути адапта-
ции, ни по пути коадаптации. Единственное, что можно приписать
эволюции сообществ как многовидовых системнее поступательность.
В этом состоит кардинальное отличие эволюции экосистем от эволю-
ции составляющих эти экосистемы популяций и видов.
Низкая интегрированость экологических систем [29,30] - несомнен-
но, основной фактор, определяющий специфику их эволюции. Именно
низкая интегрированость допускает компоновку биоценозов из готовых
компонентов, особенно ярко выраженную в пионерных сообществах и
формировании группировок видов в ходе экологических кризисов.
Коадаптация популяций внутри сообщества тем выше, чем дольше
происходит их совместная эволюция и чем стабильнее абиотические
условия среды, допускающие стенобионтность видов, слабое перекры-
вание их ниш и тем самым - более плотную «упаковку» реализован-
ных экологических ниш популяций, составляющих экосистему. Чем
сильнее интегрировано сообщество, тем сложнее чуждым ему видам
внедриться в него, но тем глубже и экологическая и эволюционная пе-
рестройка, вызываемая такими внедрениями.
Исходя из этого, наиболее чувствительными к экологическим и
микроэволюционным изменениям должны быть поздние стадии сук-
цессии, включая климакс. Существуют и географические отличия,
определяемые стабильностью абиотических условий, например отли-
чия тропических лесов от лесов бореальных.
Если экологическая перестройка происходит, т.е. в сообщество вне-
дряется новый вид (виды) или из него выпадают те или иные компо-
ненты, или их замещают (вытесняют) более приспособленные потом-
ки, причем новая компоновка экосистемы остается устойчивой, мож-
но говорить о биоценотической эволюции.
103
А.С. Северцов
Микроэволюционный процесс как таковой, не влияя существенно
на структуру сообществ (за исключением некогерентной эволюции в
периоды экологических кризисов), формирует материал для компонов-
ки экосистем. Однако поскольку для их состава важны не столько но-
вые виды, сколько новые жизненные формы, филоценогенез идет мед-
леннее микроэволюции, со скоростью макроэволюции.
Таким образом, по своему механизму эволюция экосистем дуа-
листична - с одной стороны, она основана на эволюции популя-
ций компонентов сообщества, с другой - на компоновке «готовых»
жизненных форм.
Основным же отличием эволюции экосистем от эволюции популя-
ций и видов является неадаптивность эволюции надвидовых сообществ.
Литература
1. Антипина Е.Е. Облик и история фауны наземных млекопитающих и птиц равнин-
ного Крыма в позднем голоцене: Автореф. дис... канд. биол. наук. М., МГУ, 1993.
16 с.
2. Быков Б.А. Геоботаника. Алма-Ата: Наука, 1978. 259 с.
3. Воробьева Э.И. Проблемы происхождения наземных позвоночных. М.: Наука, 1992.
343 с.
4. Гиляров А.М. Эволюция на уровне экосистем И Журн. общ. биол. 1973. Т. 34, № 1.
С. 13-20.
5. Длусский Г.М. Муравьи пустынь. М.: Наука, 1981. 229 с.
6. Джиллер П. Структура сообществ и экологическая ниша. М.: Мир, 1993. 184 с.
7. Жерихин В.В. Развитие и смена меловых и кайнозойских фаунистических комплек-
сов. М.. Наука, 1978. 197 с.
8. Жерихин В.В. Природа и история травяных биоценозов // Степи Евразии - проблема
сохранения и восстановления. СПб.: БИН РАН, 1993. С. 29-49.
9. Жерихин В.В. Эволюционная биоценология: Проблема выбора моделей // Экосис-
темные перестройки в эволюции биосферы. М.: Наука, 1994. Вып. 1.С. 13-15.
10. Жерихин В.В. Основные закономерности филоценоге! этических процессов: Авто-
реф. дис... докт. биол наук. М., 1997.
11. Иваницкий В.В., Матюхин А.В. К изучению коллективных ночевок и структу-
ры популяции домовых воробьев (Passer domesticus) И Биолог, науки. 1990.
№ 12. С. 50-56.
12. Иваницкий В.В., Матюхин А.В. К популяционной биологии полевого воробья (Passer
montanus) в городских лесопарках // Зоол. журн. 1990. Т. 69. № 4. С. 78—88.
13. Кашкаров В.Н. Среда и сообщество. М.: Медгиз, 1933. 244 с.
14. Левченко В.Ф., Старобогатов Я.И. Сукцессионные изменения и эволюция экосис-
тем (некоторые изменения эволюционной экологии)//Журн. общ. экол. 1990. Т. 51.
№5. С. 619-631.
104
II. Общие проблемы эволюции
15 Михеев А.В. Симпагрическая дифференциация в природных популяциях жука ли-
стоеда Lochmea саргеа (Crisomelidae) в Подмосковье: Автореф. дис... канд. биол.
наук. М.: МГУ, 1986. 16 с.
16 Мэй Р.М. Эволюция экологических систем И Эволюция. М.: Мир, 1981.
С. 173-195.
17 Обухова Н. Ю. Изменчивость окраски оперения в популяциях сизых голубей и ме-
ханизмы ее поддержания: Автореф. дис... канд. биол. наук. М.: МГУ, 1987. 16 с.
18 Плотников В.В. Эволюция структуры растительных сообществ. М.: Наука, 1979.
275 с.
19. Разумовский С.М. Закономерности динамики биоценозов. М.: Наука, 1981. 382 с.
20. Северцов А.С. Внутривидовое разнообразие, как причина эволюционной стабиль-
ности // Журн. общ. экол. 1990. Т. 51, № 5. С. 79-89.
21. Северцов А.С. Направленность эволюции. М.: Изд-во МГУ, 1990. 272 с.
22. Северцов А.С. Популяция как объект естественного отбора // Экология популяций:
структура и динамика: Матер. Всерос. совещания. М., 1995. Т. 1. С. 42-63.
23. Северцов А.С., Сурова Г.С. Наследуемость темпов онтогенеза и скорость эволюции
чувствительности к эффекту группы у травяной лягушки И Зоолог, журн. 1995.
Т. 74. № 2. С. 74-84.
24. Сукачев В.Н. Растительные сообщества (введение в фитосоциологию). М.; Л.: Изд-
во АН СССР, 1928.232 с.
25. Сукачев В.Н Идея развития в фитоценологии. М.: Современная ботаника, 1942.
Т. 1-3. С. 5-17.
26. Татаринов Л.П. Морфологическая эволюция териодонтов и общие вопросы фило-
генетики. М.: Наука, 1976. 256 с.
27. Тихонов И.А., Тихонова Т.Н., Полякова Л.В. Виды-двойники Microtus arvalis и
Microtus rossomeridionalis (Rodentia, Cricetidae) на северо-востоке Московской об-
ласти II Зоол. журн. 1998. Т. 77. № 1.С. 95-100.
28. Чернов Ю.И. Проблема эволюции на биоценотическом уровне организации жизни
II Развитие эволюционной теории в СССР. Л.: Наука, 1983. С. 264-512.
29. Чернов Ю.И. Эволюционный процесс и историческое развитие сообществ И Фау-
ногенез и филоценогенез. М.: Наука, 1984. С. 5-23.
30. Чернов Ю.И. Эволюционная экология - сущность и перспективы II Успехи совре-
менной биологии. 1996. Т. 116. № 3. С. 277-292.
31. Шварц С.С. Эволюция и биосфера И Проблемы биоценологии. М.: Наука, 1973.
С. 213-228.
32. Шмальгаузен И.И. Кибернетические вопросы биологии. Новосибирск: Изд-во АН
СССР, 1968. 223 с.
33. Шмальгаузен И.И. Проблемы дарвинизма. Л.: Наука, 1969. 492 с.
34. Эволюция экосистем: Тез. конф. М.: Изд-во ПИН РАН, 1995. 155 с.
35. Clements ЕЕ. Plant successions. An analysis of the development of vegetation I I Carnegie
Inst. Wash. Publ. 1916. №242. 512 p.
36. Gingerich P.D. Paleontology and Philogeny: patterns of evolution at the species level in
early Tertiary mammals //Amer. J. of Sci. 1976. T. 276. P. 1-28.
37. Simpson G.G. The Major Features of Evolution. N.Y.: Columb. Univ. Press. 1953.434 p.
38. Wilson E.O. Sociobiology. The new sintesis. Cambridge: Beeknap press of Harvard Univ.
Press. 1975.366 р.
ВИДООБРАЗОВАНИЕ И ЭВОЛЮЦИОННАЯ
БИОГЕОГРАФИЯ
М.Г. Сергеев
Институт систематики и экологии животных СО РАН
630091, г. Новосибирск, ул. Фрунзе, И
E-mail: mgs@fen.nsu.ru
Законы, управляющие жизнью,
представляют замечательный
параллелизм во времени
и пространстве...
Ч. Дарвин
“Происхождение видов путем
естественного отбора”
Введение
Проблема происхождения видов - одна из ключевых в понимании
эволюционных процессов. Для ее решения необходимо найти ответы
по крайней мере на несколько вопросов: как, почему, когда и где?
К сожалению, далеко не на все эти вопросы есть приемлемые ответы.
Но на вопрос “где?” может помочь найти ответ биогеография.
Хорошо известно, что биогеография - наука, соединяющая биоло-
гию и географию. Основная ее цель - установление закономерностей
распределения биоты на поверхности Земли [1,2]. Именно поэтому
биогеография - в первую очередь наука о пространстве. Однако, есте-
ственно, она является и исторической, точнее эволюционной наукой.
В биогеографии обычны попытки реконструкции прошлого распреде-
ления живых организмов и их сообществ [3,4 и др.]. Именно поэтому
главная ее цель может быть сформулирована другими словами - опре-
деление местоположения биологических явлений и процессов (в том
числе эволюционных) на поверхности Земли.
Конечно, биогеография использует эволюционные представления
для решения своих задач. Но естественна и обратная связь.
106
II. Общие проблемы эволюции
Биогеография и формирование эволюционных представлений
Пространственные (в первую очередь географические) аспекты
эволюции живых систем были одной из основ современных эволюци-
онных воззрений. Ч. Дарвин в своей классической и блестящей книге
“Происхождение видов путем естественного отбора” [5] отмечал: “Ник-
то не станет удивляться тому, что в вопросе о происхождении видов и
разновидностей многое остается еще необъясненным, если только от-
дать себе отчет в нашем глубоком неведении в вопросе о взаимных
отношениях множества существ, нас окружающих. Кто может объяс-
нить, почему один вид широко распространен и многочислен, а дру-
гой, близкий ему вид имеет узкую область распространения и редок.
И тем не менее эти отношения крайне важны, так как они определяют
современное благосостояние и, как я полагаю, будущий успех и моди-
фикацию каждого обитателя Земли.” (с. 23-24).
Дарвин особо подчеркнул следующие важные для развиваемых им
представлений пространственные закономерности:
1) ярко выраженную изменчивость широко и наиболее равномер-
но расселенных видов;
2) временную (историческую) детерминированность многих осо-
бенностей распространения;
3) роль различных барьеров в разделении близких таксонов;
4) родство таксонов внутри одного географического региона;
5) формирование каждого вида первоначально в одном географи-
ческом районе и последующие его миграции;
6) своеобразие биоты океанических островов (выпадение ряда так-
сонов, повышенная степень эндемизма других групп, филогенетичес-
кие связи таксонов с обитателями ближайшего континента).
Соратник Дарвина А. Уоллес [6] отмечал, что одна из основных
задач биогеографии - поиск ответа на вопросы: почему вид распрост-
ранен здесь, а не там, почему в каких-то районах высоко разнообразие
таксонов? Но, как подчеркивает Л.П. Татаринов [7], вскоре после вы-
хода в свет самого знаменитого труда Дарвина биогеография в эволю-
ционных исследованиях была оттеснена на задний план.
Вместе с тем пространственные аспекты обсуждались и в работах
многих других эволюционистов, в том числе и противников теории
естественного отбора. Так, Л.С. Берг [8] отмечал определяющую роль
107
М.Г. Сергеев
географических ландшафтов и природных зон в изменении живых
существ и писал: “... географический ландшафт воздействует на орга-
низмы принудительно, заставляя все особи варьировать в определен-
ном направлении ...” (с. 238). Можно вспомнить работы Н.И. Вавило-
ва, Ф.Г Добржанского, А.С. Серебровского, Н.В. Тимофеева-Ресовс-
кого, в которых ставились различные проблемы эволюционной
биогеографии, в том числе и связанные с геногеографией.
К сожалению, обычно пространственные аспекты эволюционных
процессов обсуждаются на самом простом уровне [9]. Во-первых, не-
редко все ограничивается изоляцией и соответственно выяснением
роли разных географических барьеров. Во-вторых, часто рассматри-
ваются миграции в самом общем виде, в первую очередь как способ
обмена генетической информацией. В-третьих, обычно используются
крайне упрощенные (как правило, контурные) мелкомасштабные кар-
ты. Вместе с тем следует подчеркнуть, что географическое простран-
ство отнюдь не пустое. Реально это географическая оболочка (геосфе-
ра) Земли, представления о которой наиболее хорошо развиты в оте-
чественной географии (см. [10] и др.).
Несомненно, что пространственная специфика эволюционных про-
цессов требует привлечения обширных материалов по изменчивости
различных таксонов и структуре их ареалов. С сожалением приходится
констатировать, что географическим (особенно ландшафтно-географи-
ческим) аспектам эволюционного процесса, как правило, уделяется да-
леко не достаточное внимание. Так, Н.Н. Воронцов [11] отмечал, что связь
генетики с географией до сих пор не самоочевидна для многих предста-
вителей той и другой наук, хотя были и есть исследования на их стыке.
Однако интересно, что тот же Воронцов в своей недавней и значитель-
ной по объему книге “Развитие эволюционных идей в биологии” [12]
пространственные аспекты фактически не затрагивает и роль биогеог-
рафии в развитии эволюционных представлений не обсуждает.
Эволюционистами, да и вообще биологами, в первую очередь не-
доучитываются сложности организации географического пространства,
в том числе не только мало популярные за пределами физической гео-
графии закономерности и структуры (например, стоковые серии, мо-
заичность ландшафтов), но широко известные, такие как зональность
и секторность. Как правило, не принимается во внимание разная мас-
штабность пространства - планетарная, региональная, топологичес-
108
IL Общие проблемы эволюции
кая [Ю]. Между тем понятно, что для слона, грызуна и ногохвостки
пространства, в которых они существуют, a priori разные.
Биогеография: типология и регионализм
в решении эволюционных проблем
В последние два десятилетия вновь началось обсуждение, иногда
- в среде биогеографов - очень бурное, пространственных аспектов
эволюции и организации биоты. Это (по крайней мере частично) свя-
зано с новым развитием регионализма как одного из основных подхо-
дов, основанных на едином пространстве-времени Тейяра де Шарде-
на [13], и определенным кризисом в биогеографических исследовани-
ях [4, 14 и др.].
Современные и классические биогеографические исследования в
своей значительной части могут быть охарактеризованы как типоло-
гические. Сущность типологического подхода связана с выявлением
на поверхности Земли участков, занятых сходно организованными,
близкими или даже идентичными группами или явлениями. Обычно
такие участки не являются целостными, т.е. их единство основано не
на пространстве, а на структурном сходстве. Таковы, например, ареа-
лы, природные зоны.
Именно типологический подход обычно используется в эволюци-
онной биогеографии, так как она, как правило, направлена на реконст-
рукцию истории отдельных таксонов. Обычно описывается распреде-
ление каких-то групп - таксонов, популяций, сообществ. Затем пред-
принимается попытка воссоздать их историю на основе имеющихся
филогенетических реконструкций, палеобиологических данных, па-
леогеографических карт и т.п. [15-19]. Начиная с классических работ
Ф.Л. Склэтера [20], Н.А. Северцова [21], Уоллеса [22] и многих дру-
гих авторов, подобные исследования развивались очень интенсивно.
Но такой подход может помочь нарисовать только часть сложной кар-
тины биогеографии и пространственной эволюции. Следует помнить,
что биогеография сопряжена не только с биологией и геологией
(см. [17,23]), но и географией [1,2,24,25]. Конечно, она может объяс-
нить какие-то типы распределения таксонов или сообществ. Но все-
таки ее основная цель - выявление собственно биогеографических за-
кономерностей, в том числе эволюционных.
109
М.Г Сергеев
В установлении закономерностей распределения биоты региональ-
ный подход во многом противоположен типологическому. Его сущность
заключается в выявлении индивидуальных интегрированных частей (уча-
стков или регионов) поверхности Земли [10, 26 и др.]. Как правило, та-
кие участки организованы определенным образом и имеют относитель-
но четкие границы. Создание системы таких регионов - это процесс рай-
онирования [26]. В результате может быть создана их иерархическая
система. Каждый из регионов, естественно, имеет свою историю. Это
позволяет нам обсуждать исторический процесс формирования любого
региона как единого целого со свойственными ему границами.
Кроме того, следует подчеркнуть, что региональный подход не си-
нонимичен хорологическому. Последний обычно противопоставляет-
ся историческому [27]. Кроме того, он обычно ассоциируется с хоро-
логической концепцией А. Геттнера [28], связанной с идеями И. Нью-
тона о пространстве, которое исследователь заполняет какими-то
объектами или процессами [29]. Однако и хорологический, и регио-
нальный подходы в определенной степени основаны на идее геогра-
фического детерминизма [28, 30].
В биологии, в том числе в эволюционной теории, значимость реги-
онального подхода обычно недооценивается, и он не очень популярен
[25, 31, 32]. Однако этот подход основан на определенной географи-
ческой (пространственной) организованности поверхности Земли и
полностью соответствует пространственному духу биогеографии.
Фактически последовательное использование регионального подхода
во многом позволяет снять многолетние споры между сторонниками
исторических и экологических интерпретаций [25, 33-35]. В частно-
сти, это позволяет понять справедливость двух внешне противореча-
щих друг другу идей - Н. Элдриджа о том, что “процесс создает кар-
тину” [23], и Берга [8] о влиянии картины (фактически географичес-
кой оболочки) на процесс.
К сожалению, даже в богатой регионалистическими традициями
отечественной биогеографии биота часто отрывается от реального гео-
графического пространства и, по сути дела, рассматривается как эво-
люционирующая в каком-то пустом либо очень простом пространстве.
В последние годы часто весь пространственный анализ ограничивает-
ся элементарной привязкой к перемещающимся (в соответствии с мо-
билистскими представлениями) континентальным массивам [36 и др.].
ПО
II. Общие проблемы эволюции
Многие зарубежные биогеографы разделились на два лагеря - сторон-
ников викариантного и приверженцев дисперсалистского объяснений.
Но очевидно, что даже в истории одного таксона есть элементы и ви-
карирования, и расселения. К сожалению, биогеографы, особенно при
исторических реконструкциях и рассмотрении эволюционных проблем,
тоже нередко забывают об иерархической и мозаичной структуриро-
ванности географического пространства.
Каждый биогеографический регион всегда является результатом
исторического и пространственного взаимодействия между биотой и
географической средой. Каждый регион может быть охарактеризован
как географическое и как биологическое явление [37]. Первое позво-
ляет нам описать закономерности, общие для различных групп живых
существ и их группировок [38-40 и др.]. Второе дает возможность
подчеркнуть экологические и исторические различия между биотами
[39,41]. Выделение регионов и их групп, отражающее дифференциа-
цию биосферы, создает основу как для пространственного прогноза
[42], так и для установления картины их вероятного развития, опреде-
ляемого пространственной организацией эволюции.
Современная биогеография и развитие эволюционных
представлений
Не только классические, но и современные биогеографические
исследования могут способствовать развитию и пересмотру эволюци-
онных представлений. Во многом это определяется тем, что совре-
менная биогеография развивается в значительной степени благодаря
осознанию иерархической структурированности и комплексности гео-
графического пространства.
I. Сейчас очевидна сложная пространственная структурирован-
ность популяционной системы любого вида. Примеры работ, в кото-
рых это ярко продемонстрировано, - сводка по таежному клещу [43],
многочисленные работы по насекомым, например саранчовым [44],
мелким наземным позвоночным (см. [45]). Однако в тесно связанной
с синтетической теорией эволюции эволюционной генетике структу-
рированность географического пространства часто недооценивается.
Так, Н.П. Дубинин [46] подчеркивал значимость так называемого мик-
ротопографического распределения с генетической точки зрения, но
111
М.Г Сергеев
сразу же отмечал, что в таком распределении нет очевидных геогра-
фических закономерностей, а для изменения популяций наиболее су-
щественны генетико-автоматические процессы, т.е. дрейф генов. Вме-
сте с тем С.С. Шварц [47] писал, что “сложная топографическая струк-
тура популяции лишь в особых случаях приводит к дифференциации
популяций и к последующему формообразованию, но нам представ-
ляется, что по крайней мере во многих случаях она является мощным
фактором эволюции популяции как единого целого. Сложная топогра-
фическая структура популяции выступает не только как стабилизиру-
ющий и объединяющий, но и как творческий фактор” (с. 202).
Проведенные в последние годы исследования пространственной
структуры популяций саранчовых [44, 48-50] позволяют утверждать
следующее:
(А) В популяционной системе вида (фактически в его метапопуля-
ции) обычно можно выделить вполне определенные пространственные
группы сходных демов в пределах природных зон и их фрагментов или
же физико-географических регионов. Возможно, это отраженный в про-
странственном размещении результат их эволюционного родства.
(Б) В картине распределения популяций хорошо прослеживаются
четко выраженные барьеры. Причем это не только и не столько геогра-
фические структуры, явно могущие выступать в качестве препятствий
для обмена генетической информацией (например, для многих саранчо-
вых - крупные реки, хребты и т.п.), а структуры ландшафтной размерно-
сти, такие как границы между террасами. Для широко распространенно-
го и массового представителя саранчовых - короткокрылого конька
(Chorthippus parallelus)- нами показано, что практически на всем протя-
жении ареала четко разделены локальные популяции речных долин (пойм
и нижних террас) и водораздельных равнин (вместе с верхними терраса-
ми) [44]. Причем это четко прослеживается даже в оптимальных для вида
районах, где его популяции распределены непрерывно от нижних пойм
до водораздельных равнин. При этом равнинные популяции близки друг
к другу в пределах каждого физико-географического выдела, тогда как
долинные популяции на протяжении речной долины значительно разли-
чаются и обычно располагаются пятнами. Сходная картина прослежена
нами и для ряда других видов саранчовых.
Аналогичным барьером для многих видов являются горные хреб-
ты [51] и речные долины. Причем это проявляется не только для неле-
112
II. Общие проблемы эволюции
таюших форм, но и для видов, которые хорошо летают и могут пере-
сечь такую долину по воздуху (например, Bryodema tuberculatum).
На исследованном нами профиле в Центральном Алтае популяцион-
ные группировки В. tuberculatum на северном и южном степных скло-
нах, разделенных примерно 100-метровым луговым пространством
пойм и нижних террас, достоверно различаются по частоте встречае-
мости белого пятна на боковых килях переднеспинки [49].
Для ряда саранчовых установлено существование устойчивой и
хорошо прослеживающейся границы между поселениями горных и
равнинных форм каждого вида. Она соответствует границе между гор-
ными склонами передовых хребтов и подгорными равнинами и чаще
всего выражена в резкой смене характера ландшафтного размещения
этих поселений [52]. Последнее четко отражается и в тяготении гор-
ных и равнинных форм к разным ландшафтам. Особенно это типично
для Calliptamus italicus italicus (полупустыни) и С. italicus reductus (гор-
ные полусаванны) [53].
Во многом сходная ситуация описана для испанских популяций
С. barbarus [54], где горные и долинные поселения, обычно относи-
мые к этому виду, довольно четко различаются как по рисуночно-ок-
расочным, так и морфометрическим признакам. Замечательно, что эти
две группы практически не смешиваются.
Резко различаются локальные популяции короткокрылого конька в
горных долинах Кавказа [44]. По крайней мере одна из них была вы-
делена в самостоятельный подвид - Chorthippus parallelus aemulus (Ха-
мышки). Тогда, если быть последовательным, необходимо присвое-
ние подвидового статуса и популяциям из других горных долин.
(В) Очень часто переоценивается или, наоборот, недооценивается
способность особей вида к миграциям. Так, обычно считается, что
хорошо летающие виды саранчовых способны преодолевать суще-
ственные барьеры - до нескольких сотен метров шириной. Вместе с
тем наши эксперименты с В. tuberculatum (см. выше) показывают, что
это не совсем так [49]. В настоящее время нередко миграции являются
результатом человеческой активности и могут, как известно, приво-
дить к самым разнообразным последствиям. Так, например, бескры-
лая кобылка Podisma pedestris с ее очень низкой миграционной актив-
ностью может расселяться на значительные расстояния (много превы-
шающие радиус ее репродуктивной активности) в шерсти овец [55].
113
М.Г. Сергеев
Наши результаты в целом соответствуют данным уральских эколо-
гов, показавших значительные экологические различия между сосед-
ними популяциями грызунов и низкие возможности миграционных
обменов [47 и др.]. Шварц подчеркивал, что интенсивность обмена
особями между соседними популяциями подчиняется достаточно стро-
гим закономерностям.
Все это говорит о необходимости оценки эволюционной и эколо-
гической значимости разных частей популяционной системы каждого
вида, в том числе отдельных демов. Прослеживаемые различия между
локальными популяциями на ландшафтном и внутриландшафтном
уровнях показывают возможность их эволюционной дифференциации,
причем эта возможность реальна не только для краевых поселений
вида, но и для демов внутри оптимальной области ареала.
Кроме того, анализ распределения локальных популяций, частей
популяционных систем видов, установление их границ позволяют и
по-иному взглянуть на взаимоотношения биогеографических регио-
нов и границ между ними, в частности, снять (или обойти) известную
проблему викариантной биогеографии - обязательного присутствия
эндемичных видов в отдельно взятом биогеографическом выделе.
II. Классическая географическая модель видообразования в общем
виде проста и красива. Идея о роли географических преград в диффе-
ренциации видов и других таксонов имеет долгую историю. Вероят-
но, М. Вагнер [56] был первым исследователем, ясно сформулиро-
вавшим эту идею и подчеркнувшим роль пространственной изоля-
ции для дивергенции видов. Кроме того, он обратил внимание на то,
что близкородственные формы, в частности виды, обычно обитают
на смежных территориях.
Типичное аллопатрическое видообразование обсуждалось неодно-
кратно и здесь вряд ли стоит подробно останавливаться на этой про-
блеме. В модели, предложенной более полувека тому назад Э. Май-
ром [57], много рационального, и в самом общем виде она хорошо
описывает этот процесс.
Географическое видообразование может быть представлено как мно-
гостадийный процесс, следующий за географической изоляцией [58]:
1) Появление генетических различий между разделенными частя-
ми популяционной системы, затрудняющих обмен генетической ин-
формацией. Начало дивергенции по экологическим нишам.
114
IL Общие проблемы эволюции
2) При вступлении изолированных популяций в контакт (если реп-
родуктивная изоляция достаточна) естественный отбор может усили-
вать различия. Продолжение расхождения экологических ниш.
3) Продолжение независимой эволюции и дальнейшее расхожде-
ние видов за счет вхождения в состав разных сообществ.
Однако классическое представление об аллопатрическом видооб-
разовании основано на примерной эквивалентности исходных для каж-
дого нового вида частей первоначальной популяционной системы.
Вместе с тем понятно, что могут быть случаи формообразования за
счет небольшой части популяционной системы. Или же за счет не-
скольких таких частей может образовываться ряд видов, т.е. реально
множественное видообразование [27].
Одним из первых на возможность такого формообразования обра-
тил внимание Г.Я. Бей-Биенко [59] при обсуждении зональной смены
местообитаний. Он отметил своеобразие условий существования и
изолированность популяций на северных и южных окраинах ареала.
Майр [60] описал процесс отпочковывания нового дочернего вида
от небольшого периферического изолята обширного полиморфного
предкового вида. Позже Грант [61] предложил для обозначения такого
пути термин “квантовое видообразование”. Сам Майр использовал на-
звание “видообразование путем генетической революции”, а позже -
парапатрическое видообразование. В основе такого процесса лежат
изоляция, дрейф генов и близкородственное скрещивание, последнее
может привести к формированию и распространению новых феноти-
пов. Скорее всего, к этому же варианту видообразования относится и
так называемое стасипатрическое видообразование [62], связанное с
обособлением из полиморфного вида форм, гомозиготных по струк-
туре хромосом.
Ш. Современные биогеографические данные могут быть исполь-
зованы для реконструкции возможной эволюции той или иной группы
живых существ, в том числе для определения возможного района и
эколого-географических условий формирования. Естественно, это наи-
более актуально для ситуации (к сожалению, крайне обычной), когда
полностью или почти полностью отсутствуют палеобиологические
данные.
Для решения этой проблемы предлагались различные подходы [63]:
а) установление геометрического центра ареала (что в общем виде
115
М.Г Сергеев
связано с толерантностью); б) оценка распределения численности,
отражающей положение оптимума ареала; в) распределение размеров
особей внутри ареала; г) разнообразие разных признаков в пределах
ареала и д) распределение примитивных и прогрессивных признаков.
Естественно, что не все эти подходы реалистичны, а некоторые (осо-
бенно определение геометрического центра ареала) практически бес-
смысленны. Однако приспособления и характер зонально-ландшафт-
ного распределения многих видов внутри их ареалов, очевидно, отра-
жает условия их формирования [31,64 и др.]. Это позволяет на основе
принципа актуализма использовать современные данные по устрой-
ству популяционной системы вида для частичной реконструкции его
прошлого. Вместе с тем это применимо не для всех видов, поскольку
некоторые из них, особенно патиенты, уходят от конкуренции в спе-
цифические стации, что и определяет характер их распределения. В
любом случае такой подход позволяет реконструировать не реальное
положение района обособления вида, а связь вида с некоторыми ис-
ходными условиями.
Дифференциация близких видов в разных частях их общего ареала
идет различными путями. Это в значительной степени соответствует
выявленному нами ранее слабому перекрыванию пространственных
экологических ниш таких форм [48,52], а также подтверждает вероят-
ную роль в этом процессе мозаичности местообитаний [51].
Нередко можно наблюдать ситуацию, когда даже современное рас-
пределение ареалов родственных видов и родов проще всего может
быть интерпретировано с точки зрения викариантной биогеографии,
т.е. как дивергенция вслед за дифференциацией природных зон и ре-
гионов [65]. Вместе с тем совершенно очевидно, что такие виды и
роды, так же как и предковые формы, явно мигрировали, в том числе и
в связи с известными смещениями природных зон. Примечательно,
что дифференциация близких родов и видов нередко как бы повторя-
ется в пространстве. Это означает возможность экстраполяции такой
картины в пределах одного таксона.
Хотя сейчас очевидно, что центры или очаги видового разнообра-
зия крайне редко соответствуют тем районам, где произошло обособ-
ление рода или другого таксона, тем не менее выявление подобных
центров и их анализ много дают для понимания особенностей эволю-
ции таксона. Здесь возможно использование дифференциального си-
116
IL Общие проблемы эволюции
стематико-географического метода [66], ориентированного на опре-
деление центров формообразования и включающего:
1) Дифференциацию данного вида на так называемые линнеевские
виды и генетические группы при помощи разных методов - морфоло-
го-систематического, гибридологического, цитологического, парази-
тарного и т.д.
2) Определение ареалов этих форм, по возможности в отдаленное
время, когда сообщение было более затрудненным, чем сейчас.
3) Детальное определение состава разновидностей и рас (точнее,
наследственно варьирующих признаков) каждого вида и общей систе-
мы наследственной изменчивости.
4) Установление распределения наследственного разнообразия
форм данного вида по областям и странам и определение географи-
ческих центров скопления разнообразия. Области максимального раз-
нообразия, включающие обыкновенно также ряд эндемичных форм и
признаков, скорее всего, и будут центрами формообразования.
Анализ центров разнообразия (особенно с резко выраженной кладо-
генетической активностью) [17, 63,67 и др.], распределения эндемизма
[68] и распространения таксонов [69] - частные случаи этого метода.
Реальное распределение узкоэндемичных видов (т.е. видов с очень
небольшими ареалами), обитающих как на суше, так и в море, часто
демонстрирует их приуроченность к четко дифференцированному
пространству, например окраинным хребтам крупных горных систем
[70], шельфам у границ биогеографических выделов в океанах [71].
Заключение
Современная картина распределения живых существ и их сооб-
ществ на поверхности Земли - результат вполне определенного раз-
мещения видов и других таксонов - фактически терминальных ста-
дий филогенетических и фауно(флоро)генетических стволов и ветвей
(см. [72, 73]). Эта картина может быть охарактеризована как результат
решения диалектического противоречия между викариатом (т.е. видо-
образованием в пространстве), когда границы выступают как изоли-
рующие препятствия, и распространением (дисперсализм), когда гра-
ницы могут быть представлены как полупроницаемые мембраны.
Каждый биогеографический регион обладает своей биотой, кото-
рая представляет собой не просто мешанину из видов, а скорее, сбор-
117
М.Г. Сергеев
ную солянку, в которой каждый (или почти каждый) элемент важен и в
которой все элементы дополняют друг друга. Это не просто истори-
ческая комбинация эндемичных и широко распространенных видов,
но определенным образом организованное сообщество популяций,
закономерно распределенных в системе местообитаний [32, 52]. Эти
популяции формируют соответствующие сообщества в каждой стации,
а также на каждой стадии сукцессии.
Биогеографические данные дают возможность охарактеризовать
каждый таксон как целостную совокупность, эволюционирующую в
пространстве и времени [74, 75], а саму эволюцию - как единый про-
цесс, связанный с дифференциацией биосферы, и демонстрируют об-
щие закономерности на разных уровнях организации.
Региональная дифференциация биосферы в целом может быть сво-
его рода эволюционным организатором. По-видимому, можно развить
идею о своеобразии отбора в пределах каждого участка земной повер-
хности и о связанной с этим ограниченности случайности в разнооб-
разии и эволюции [63, 76, 77].
Литература
1. Бобринский НА. География животных. М.: Учпедгиз, 1951.384 с.
2. Jones R.L. Biogeography II Progr. Phys. Geogr. 1987. Vol. 11. P. 133-145.
3. Буко А. Эволюция и темпы вымирания. М.: Мир, 1979. 318 с.
4. Симпсон Дж. Великолепная изоляция. М.: Мир, 1983. 256 с.
5. Дарвин Ч. Происхождение видов путем естественного отбора. Л.: Наука, 1991. 539 с.
6. Wallace A.R. Island Life. London: Macmillan, 1880.
7. Татаринов Л.П. Предисловие редактора перевода // Симпсон Дж. Великолепная изо-
ляция. М.: Мир, 1983. С. 5-7.
8. Берг Л.С. Труды по теории эволюции. Л.: Наука, 1977. 387 с.
9. Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эво-
люции. М.: Наука, 1977. 297 с.
10. Сочава В.Б. Введение в учение о геосистемах. Новосибирск. Наука, 1978. 319 с.
11. Воронцов Н.Н. Генетика и география // Чтения памяти Н.В. Тимофеева-Ресовского.
Ереван, 1983. С. 200-236.
12. Воронцов Н.Н. Развитие эволюционных идей в биологии. М.: УНЦ ДО МГУ,
Прогресс-Традиция, АБФ, 1999. 639 с.
13. Тейяр де Шарден П. Феномен человека. М.: Наука, 1987. 240 с.
14. Thornton I.W.B. Vicariance and dispersal: confrontation or compatibility? 11 Geojoumal.
1983. Vol. 7. P. 557-564.
15. Uvarov B.P. A new method in biogeography // Nature. 1937. Vol. 139. P. 492-494.
16. Brundin L. Methods and principles of phylogenetic biogeography//Chironomidae: Ecol.,
Syst., Cytol. and Physiol. Proc. 7th Int. Symp. Oxford et al., 1980. P. 91-92.
118
II. Общие проблемы эволюции
17 Nelson G., Platnick N.I. A vicariance approach to historical biogeography // BioScience.
1980. Vol. 30. P. 339-343.
18 Еськов К.Ю. Дрейф континентов и проблемы исторической биогеографии // Фауно-
генез и филоценогенез. М.: Наука, 1984. С. 24-92.
19. Cracraft J. Deep-history biogeography: retrieving the historical pattern of evolving
continental biotas // Syst. Zool. 1988. Vol. 37. P. 221-236.
20. Sclater P.L. On the general geographical distribution of the numbers of the class Aves
//J. Proc. Linn. Soc. Zool. 1858. Vol. 2. P. 130-145.
21. Северцов Н А. Вертикальное и горизонтальное распределение туркестанских жи-
вотных. М.: Изд-во АН СССР, 1953. 270 с.
22. Wallace A.R. The geographical distribution of animals. London: Macmillan, 1876.
23. Ferris VR. A science in search of a paradigm? - review of the symposium «Vicariance
biogeography: a critique» // Syst. Zool. 1980. Vol. 29. P. 67-76.
24. Meadows M.E. Biogeography: a happy ending to the fairy tale? // S. Afr. Geogr. J. 1985.
Vol. 67. P. 40-61.
25. Smith C.H. Historical biogeography: geography as evolution, evolution as geography
//N. Z. J. Zool. 1989. Vol. 16. P. 773-785.
26. Алаев Э.Б. Социально-экономическая география. M.: Мысль, 1983. 350 с.
27. Балл И.Р. Планарии, множественность и биогеографические объяснения И Биосфе-
ра: Эволюция, пространство, время. М.: Прогресс, 1988. С. 351-367.
28. Hettner A. Die Geographie. Ihre Geschichte, ihr Wesen und ihre Methoden. Breslau:
F. Hirt, 1927. 463 S.
29. Мукитанов H.K. От Страбона до наших дней. М.: Мысль, 1985. 239 с.
30. Войлошников М.В. Судьба идеи геодетерминизма И Природа. 1991. № 11. С. 76-82.
31. Сергеев М П Закономерности распространения прямокрылых насекомых Север-
ной Азии. Новосибирск: Наука, 1986. 237 с.
32. Сергеев М.Г. Закономерности распространения прямокрылых насекомых азиатс-
кой части СССР: Автореф. дис. ... докт. биол. наук. СПб.: ЗИН РАН, 1991. 37 с.
33. Endler J. A. Problems in distinguishing historical from ecological factors in biogeography
//Amer. Zool. 1982. Vol. 22. P. 441-452.
34. Brooks D.R. Scaling effects in historical biogeography: a new view of space, time, and
form П Syst. Zool. 1988. Vol. 37. P. 237-244.
35. Gray R Oppositions in panbiogeography: can the conflicts between selection, constraint,
ecology, and history be resolved? 11N. Z. J. Zool. 1989. Vol. 16. P. 787-806.
36. Craw R.C. Biogeography and biogeographical principles //N. Z. Ent. 1984. Vol. 8. P. 49-52.
37. Матвеев С.Д. Географическое деление Балканского полуострова в целях био-
географического районирования // Изв. Всесоюз. геогр. об-ва. 1969. Т. 101.
№2. С. 125-137.
38. Емельянов А.Ф. Предложения по классификации и номенклатуре ареалов // Энто-
мол. обозрение. 1974. Т. 53. № 3. С. 497-552.
39. Крыжановский О.Л. К вопросу о предмете зоогеографии и методах зоогеографи-
ческих исследований /7 Журн. общ. биол. 1976. Т. 37. № 5. С. 762-768.
40. Юрцев Б.А. Флористический и фитоценологический подходы к растительно-
му покрову: соотношение, проблемы синтеза И Журн. общ. биол. 1988. Т. 49.
№ 4. С. 437-450.
41. Чернов Ю.И. Природная зональность и животный мир суши. М.. Мысль, 1975.222 с.
119
М.Г Сергеев
42. Киселев АН. Прогнозное биогеографическое картографирование (региональный
аспект). М.: Наука, 1985. 104 с.
43. Коренберг Э.И. Биохорологическая структура вида (на примере таежного клеща)
М.: Наука, 1979. 171 с.
44. Казакова И.Г., Сергеев М.Г. Пространственная организация популяционной систе-
мы вида у короткокрылого конька Chorthippus parallelus Zett. (Insecta: Orthoptera) /
I Жури. общ. биол. 1992. T. 53. № 3. С. 373-383.
45. Яблоков А.В. Популяционная биология. М.: Высшая школа, 1987. 303 с.
46. Дубинин Н.П. Эволюция популяций и радиация. М.: Атомиздат, 1966. 743 с.
47. Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 278 с.
48. Казакова И.Г., Сергеев М.Г. Закономерности распределения популяций прямокры-
лых (Orthoptera) в естественных и антропогенных ландшафтах горных котловин
юга Сибири И Энтомол. обозрение. 1992. Т. 71. № 4. С. 731-741.
49. Казакова И.Г, Сергеев М.Г О проблеме определения границ популяционных груп-
пировок саранчовых // Сиб. экол. журн. 1997. Т. 4. № 3. С. 315-321.
50. Сергеев М.Г, Валькова И.А. Зонально-ландшафтное распределение популяцион-
ных группировок итальянского пруса - Calliptamus italicus L. (Insecta, Orthoptera,
Acrididae) 11 Сиб. экол. журн. 1996. T. 3. № 3-4. С. 219-225.
51. Hewitt G.M. Limited introgression through orthopteran hybrid zones // Bol. San. Veg
Plagas (Fuera de Serie). 1990. N 20. P. 329-339.
52. Сергеев М.Г. Биологическое разнообразие прямокрылых насекомых Северо-Вос-
точной Палеарктики: распределение популяционных группировок //Сиб. экол. журн
1994. Т. l.№6. С. 547-554.
53. Sergeev M.G., Vanjkova I.A. The range structure of Calliptamus italicus and estimation
of its significance for natural and anthropogenic landscapes // Proc. VI Meeting on the
Project “Species and its Productivity in the Distribution Area”. St. Petersburg, 1993.
P. 270-271.
54. Clemente M.E., Garsia M.D., Presa J.J. Morphometric and pigmentary variation in
Calliptamus barbarus Costa, 1836 in relationship with the environment, and its taxonomic
value 11 Evolutionary biology of Orthopteroid insects. Chichester: Ellis Homwood Ltd.,
1987. P. 184-189.
55. Nichols R.A., Hewitt G.M. Population structure and the shape of chromosomal cline
between two races of Podisma pedestris (Orthoptera: Acrididae) // Biol. J. Linn. Soc.
1986. Vol. 29. N 4. P. 301-316.
56. Wagner M. Die Darwinische Theorie und das Migrationsgesetz der Organismen. Leipzig:
Dunckerand Humblot, 1868.
57. Mayr E. Systematics and the origin of species. New York: Columbia Unniversity Press,
1942. 334 p.
58. Левонтин P. Генетические основы эволюции. M.: Мир, 1978. 351 с.
59. Бей-Биенко ГЯ. К вопросу о зонально-экологическом распределении саранчовых
(Orthoptera, Acrididae) в Западно-Сибирской и Зайсанской низменностях // Тр. по
защите растений. Сер. энтомол. 1930. Т. 1. № 1. С. 51-90.
60. Mayr Е. Change of genetic environment and evolution // Evolution as a process. London:
alien and Unwin, 1954. P. 157-180.
61. Грант В. Эволюция организмов. М.: Мир, 1980. 407 с.
62. White M.J.D. Models of speciation П Science. 1968. Vol. 159. P. 1065-1070.
120
II. Общие проблемы эволюции
63 Красилов В.А. Эволюция и биостратиграфия. М.: Наука, 1977. 256 с.
64 Hengeveld R. Dynamic biogeography. Cambridge et al.: Cambridge University Press,
1990. xiv+249 p.
65 Сибирская кобылка (Aeropus sibiricus L.). Новосибирск. Наука, 1995. 175 с.
66 Вавилов Н.И. Происхождение и география культурных растений. Л.: Наука,
1987. 439 с.
67 Holloway J.D. The biogeography of Indian butterflies // Monogr. biol. 1974. Vol. 23.
P. 473-499.
68 Матюшкин E.H. Региональная дифференциация лесной фауны Палеарктики в про-
шлом и настоящем И Теоретические и прикладные аспекты биогеографии. М.:
Наука, 1982. С. 59-80.
69 Комаров В.Л. Введение к флорам Китая и Монголии // Тр. С.-Петерб. бот. сада.
' 1908. Т 29. №1. С. 1-176.
70. Sergeev M.G. Conservation of orthopteran biological diversity relative to landscape change
in temperate Eurasia// J. Insect Conservation. 1998. Vol. 2. N 3/4. P. 247-252.
71. Валентайн Дж.У, Яблонский Д. Видообразование в морском мелководье: общие
тенденции и проверка биогеографическими данными // Биосфера: Эволюция, про-
странство, время. М.: Прогресс, 1988. С. 175-196.
72. Cracraft J. Geographic differentiation, cladistics, and vicariance biogeography:
reconstructing the tempo and mode of evolution 11 Amer. Zool. 1982. Vol. 22.
P. 411-424.
73. Мейен C.B. Флорогенсз и эволюция растений // Природа. 1986. № 11. С. 47-57.
74. Черных В.В. Проблема целостности высших таксонов. М.: Наука, 1986. 143 с.
75. Bennett K.D. Evolution and ecology: The pace of life. Cambridge et al.: Cambridge
University Press, 1997. 241 p.
76. Васильев В.Н. Физико-географическая среда и видообразование. М.; Л.: Наука, 1966.
75 с.
77. Smith С.Н. A system of world mammal faunal region. II. The distance decay effect upon
inter-regional affinities // J. Biogeog. 1983. Vol. 10. P. 467^482.
ВИД В СИСТЕМАТИКЕ СЕГОДНЯШНЕГО ДНЯ
А.К. Скворцов1
Главный ботанический сад РАН
127276, г. Москва, ул. Ботаническая, 4
E-mail: seed@aha.ru
В настоящем сборнике представлен целый ряд интересных иссле-
дований, посвященных конкретным видам растений и животных.
Я же хочу высказать несколько замечаний по общим вопросам, кото-
рые, думается, еще в известной мере дискуссионны. Некоторые конк-
ретные виды растений упомяну только в качестве иллюстраций.
Слово «вид» может употребляться во множестве разных значений.
Но в биологии это слово имеет вполне определенный смысл и обозна-
чает основную систематическую единицу, основной «строительный
кирпич» систематики. Дня биолога вид - не логическая категория, не
условная классификационная единица, опирающаяся на какие-то из-
бранные признаки; вид - это эмпирически обнаруживаемое истори-
чески сформировавшееся природное образование. Это множество
живых существ, связанных между собой генетически, устойчиво вос-
производящее себя в ряду поколений и нашедшее свое специфическое
место в тех или иных экосистемах.
Феномен вида наиболее полно проявляется у организмов, в попу-
ляциях которых существует панмиксис - свободное перекрестное оп-
лодотворение. Благодаря панмиксису создается общий генофонд, или
коллективный генотип популяций и всего вида в целом, который обес-
печивает определенное единообразие свойств и характеристик вида
на всем его ареале, вместе с тем заключает в себе и огромные возмож-
ности адаптивной изменчивости.
К. Линней первый гениально угадал, что из всех таксономических
категорий только уровень вида обладает некими общими критериями,
позволяющими аргументированно опознать его, и положил вид в ос-
’Автор выражает признательность за поддержку его работы грантом РФФИ
№ 99-04-48202.
122
II. Общие проблемы эволюции
нову систематики. Говоря современным языком, вид - это наиболь-
шее множество организмов, объединяемых единым коллективным ге-
нотипом (генофондом), или - что то же самое в представлении натура-
листа - наибольшее множество, внутри которого вся изменчивость
носит характер постепенных градаций. Вместе с тем вид - это наи-
меньшее множество, генофонд которого четко обособлен от генофон-
дов других подобных множеств, что находит выражение и в существо-
вании гиатусов - перерывов в рядах изменчивости по ряду парамет-
ров. Совпадение этих двух критериев и дает нам (идеализированный,
конечно) уровень вида. Что же до установления рангов надвидовых
категорий, то уже со времен А. Жюсье почти всеми признается их от-
носительность; переходное положение можно признать за ближайшей
к виду категорией: это «ряд» у ботаников, «надвид» у зоологов.
Ограничения панмиксиса неизбежно нарушают и целостность кар-
тины вида. Существование географических или экологических пре-
пятствий для свободной миграции особей и диаспор со временем при-
водит к более-менее отчетливому разделению вида на подвиды, а за-
тем и на ряд молодых видов. Стало быть, в действительности мы далеко
не всегда встречаемся с реализацией идеальной модели вида, и таксо-
номические решения нам приходится принимать лишь с известной
мерой приблизительности. Нижеследующая схема поясняет соотно-
шение уровней идеальных моделей вида и близких к нему группиро-
вок с реальными ситуациями.
ряд (series) | |
вид | |
подвид | |
локальная популяция | ~ |
разнообразие
реальных
ситуаций
Совсем особая ситуация возникает при агамном размножении, ко-
торое преобладает у прокариот и нередко возникает в разных группах
эвкариот. Агамное размножение создает серии биотипов, которые об-
ладают частью характеристик вида (как устойчивое воспроизведение
себя в потомстве и определенная экология) и потому многими автора-
ми трактуются в ранге видов. Однако они лишены такого существен-
ного атрибута вида, как коллективный генофонд, обеспечивающий виду
123
А.К. Скворцов
широту адаптивных возможностей. Поэтому агамные биотипы можно
рассматривать и в качестве клонально размножившихся индивидов в
рамках агамного комплекса, принимаемого в этом случае за один вид.
А поскольку агамность животных и растений в большинстве случаев
имеет факультативный характер и возникают всевозможные комбини-
рованные ситуации, то и здесь систематику приходится принимать
приблизительные и даже весьма условные решения.
В приведенной схеме ниже подвида показана локальная популя-
ция. А где же разновидность? Красивый афоризм ранних эволюцио-
нистов «разновидность - зачинающийся вид» стал, увы, анахрониз-
мом. За зачинающийся вид может быть принята только изолированная
популяция. Разновидность же, еще числящуюся в Международном ко-
дексе ботанической номенклатуры в качестве таксономической кате-
гории, давно пора оттуда исключить. Разновидность - это не более
чем индивидуальная особенность, чем-то приглянувшаяся наблюдав-
шему ее автору. Еще Линней сказал, что разновидностей можно раз-
личить столько, сколько разных растений может вырасти из семян од-
ного вида («Философия ботаники»). Оставим рассмотрение разновид-
ностей морфологам. И еще более нуждается в передаче морфологам
«форма».
Среди подходов к отграничению и изучению видов основными и
пока что наиболее надежными для итоговых интегральных оценок
остаются морфолого-географический и эколого-генетико-популяцион-
ный. Они прямее всех других нацелены на определение места вида в
биосфере и познание его жизни в конкретных природных условиях.
Позволю себе несколько замечаний о других подходах.
Биометрия. В нашем деле она в основном может только подтвер-
дить то, что внимательный наблюдатель видит и простым глазом. Что
в расчеты вложишь, то из них и получишь. Возьмешь ничего не знача-
щие параметры или небрежно составленную выборку - толкового ре-
зультата ни из каких вычислений не получится. Например, на протя-
жении без малого ста лет не раз делались попытки с помощью биомет-
рии установить, являются ли пушистая и бородавчатая березы разными
видами или одним. Биометрия четких границ установить не смогла:
значит, либо вид один, либо (что, впрочем, фактически то же самое)
имеется некий гибридный комплекс. Но если найти более информа-
тивные признаки и исследовать более широкие материалы, то оказы-
124
II. Общие проблемы эволюции
вается, что названные виды не только четко различимы, но и принад-
лежат к разным рядам: у каждого из них свои родичи в Азии и в Север-
ной Америке. И вдобавок генетики утверждают, что пушистая и боро-
давчатая березы с трудом поддаются скрещиванию друг с другом.
С другой стороны, нередок и другой биометрический казус. Меж-
ду популяциями обнаруживаются достоверные биометрические раз-
личия, и исследователь уже готов придать им некое таксономическое
значение, но тут оказывается, что диапазон различий между отдель-
ными индивидами внутри популяции намного шире различий между
средними показателями популяций. Значит, никакой таксономии при-
тянуть нельзя.
Кариосистематика. Здесь особенно важно помнить тезис Линнея
(высоко оцененный и Дарвином): «не признаки определяют род, а род
определяет свои признаки»; это же можно отнести и к виду: не плоид-
ность определяет вид, а вид сам регулирует свою плоидность (а то
даже и общее число хромосом). Например, исследованиями Т.А. Ре-
тиной и С.В. Клицова в бытность их моими аспирантами было пока-
зано, что у Sanguisorba officinalis на фоне преобладающего в целом
по ареалу диплоидного уровня существуют отдельные очаги тетра-
плоидности, а у Lonicera coerulea - наоборот, на тетраплоидном фоне
есть очажки диплоидности. При отсутствии морфологических разли-
чий между ди- и тетраплоидами им нельзя придать статус разных ви-
дов; наиболее вероятно предполагать, что в обоих случаях основной
фоновый уровень - исходное состояние, а изолированные очаги вто-
ричны, производны. В то же время, как известно, многие другие виды
и даже роды столь же суверенно блюдут постоянство своей плоидно-
сти. У той же бородавчатой березы везде и всегда 2п=2х; а у пушис-
той 2п=4х.
Наконец, самый новый подход - молекулярный. В 1950-1970-х го-
дах один из идейных лидеров кариосистематики, ныне покойный Ас-
кел Лёве (A. Love) считал, что, изучая структуру хромосомного аппа-
рата, он непосредственно изучает генотип растений. Примерно в то
же время у нас А.Н. Белозерский, внесший большой вклад в биохи-
мию нуклеиновых кислот и оказавший существенную поддержку сис-
тематике, также считал, что ДНК и РНК - это и есть генотип, и для их
изучения в систематическом плане предложил название «геносисте-
матика»; еще более определенно эта мысль выразилась в употребляе-
125
А.К. Скворцов
мом и поныне школой А.Н. Белозерского обозначении «систематика
генотипов», противопоставляемом «систематике фенотипов», каковой
будто бы является классическая систематика. (И еще в те же годы в
английской литературе появилось выражение «фенетическая система-
тика» - по созвучию как будто синонимичное «систематике феноти-
пов». Однако по содержанию оно определяет собой антитезу не «сис-
тематике генотипов», а филогенетической систематике; далее его не
рассматриваю.)
Исследования хромосомного аппарата и еще более - ДНК и РНК
очень много дали для развития систематики. И даже не только потому,
что открыли целые пласты новых сведений о живых существах, а еще
и в силу психологического, лозунгового эффекта: «Новое направле-
ние, новый этап науки! - Дорогу ему, дорогу!» Этот эффект положи-
тельно сказался на систематике в целом. Но вместе с тем новые обо-
значения внесли в систематику изрядную путаницу. Фенотип и гено-
тип в организме еще более неразделимы, чем в монете орел и решка.
Нет ни одного качества, ни одного признака, которые сформировались
бы в организме без всякого участия генотипа. Поэтому говорить о «си-
стематике генотипов» - это все равно (или еще хуже), чем сказать «я
Вам заплачу решками».
И более того: нет других способов узнать генотип иначе как по его
реализации в фенотипе. За доказательством этой простой истины не
надо далеко ходить: ведь именно по признакам фенотипа (окраска се-
мян гороха) Грегор Мендель судил о генотипах сортов, и именно опи-
раясь на эти признаки установил фундаментальные законы генетики.
Генотип - не структура, а информация, это программа всего, на что
организм, обладающий этим генотипом, способен в течение всей сво-
ей жизни. А фенотип - это вполне осязаемое наличное состояние орга-
низма в любой момент или период его жизни.
Все это уже в самом начале XX века вполне отчетливо понимал и
изложил классик генетики и автор самих терминов «генотип», «ген»,
«фенотип» - В. Иогансен. Но, как ни странно, это понимание до сих
пор не стало среди биологов всеобщим. И еще более странно, что струк-
туры (хромосомы или нити ДНК) считают генотипом, тогда как для
этих кодирующих наследственность структур уже достаточно давно
существует обозначение «геном». Теперь, когда исследования геномов
стали быстро расширяться и для них появилось вполне удачное назва-
126
II Общие проблемы эволюции
ние «геномика», должно восстановиться и корректное употребление
терминов.
Следует ожидать, что развитие геномики поможет систематике под-
няться на совершенно новый уровень и также стать на путь быстрого
прогресса. Но заменить собой систематику геномика в принципе не
может. Во-первых, потому что объекты систематики - целые организ-
мы и образования надорганизменного уровня (популяции, колонии,
симбиотические формирования), геномика же изучает, хотя и важней-
шую, но все же только часть организма. Во-вторых, геномика изучает
молекулярные структуры как таковые, тогда как дело систематики -
осмысление структур в плане их значимости для основной задачи си-
стематики - выяснения степени генетического родства между разны-
ми группами живых существ. Сейчас еще трудно представить себе все
возможные варианты того, как будет выглядеть это осмысление. Пока
ясно только, что подсчеты нуклеотидных замен ни универсальным, ни
прямым и решающим критерием в систематике служить не могут; в
стремлении к естественности системы она последовательно идет по
пути деформализации критериев, а внесение грубого числового фор-
мализма было бы шагом к системе искусственной.
СИСТЕМНАЯ РЕОРГАНИЗАЦИЯ ГЕНОМА
ПРИ ВИДООБРАЗОВАНИИ
В.Н. Стегний
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
E-mail: steg@info.tsu.ru
Эволюционная идея в биологии базируется на осмыслении фунда-
ментальных философских принципов, а именно - значения и соотно-
шения «внутреннего и внешнего» в развитии или, по биологической
терминологии, - автогенеза и эктогенеза. Основатель эволюционно-
го учения гениальный Ж.Б. Ламарк впервые ввел эти два принципа в
биологическое мировоззрение и воплотил их в двух законах: 1) града-
ции (автогенез) и 2) приспособительной эволюции (эктогенез). Ч. Дар-
вин и его последователи, включая и современных «неодарвинистов»,
проповедуют лишь второй принцип (эктогенез), оценивая естествен-
ный отбор как главную причину эволюционного развития. Другой край-
ностью является абсолютизация «внутреннего» в эволюции, или авто-
генеза. Наиболее яркий приверженец этого подхода - Л.С. Берг.
С позиций современной биологии становится очевидным, что эво-
люционный процесс на всех этапах осуществляется при непрерывном
взаимодействии «внутреннего и внешнего» [1]. Именно это взаимо-
действие отвечает принципам системного подхода, когда система (внут-
реннее) функционирует или развивается только под контролем среды
(внешнее). В данной статье предполагается оценить проблему «внут-
реннего», то есть организации видовой системы, и прежде всего ее
генетической компоненты, на современном уровне развития биоло-
гии. Эволюционная судьба вида как генетической системы может быть
разной в зависимости от его структурно-функциональной организа-
ции. Системный подход предполагает наличие в видовой системе ин-
вариантной (мономорфной) структуры - видового генома и вариант-
ной (полиморфной) компоненты - видового генофонда. При этом
вводим постулат, что реорганизация видового генома может привести
к развитию системы или видообразованию, тогда как трансформация
видового генофонда является основой поддержания функциональной
128
IL Общие проблемы эволюции
стабильности вида (видового гомеостаза). По определению [2], осно-
вой функционирования системы являются гомеостатические (равно-
весные) отношения. Основой развития являются противоречивые (не-
равновесные) отношения. Если в процессе функционирования каче-
ство системы не меняется, то такой тип ее поведения требует сохра-
нения определенной устойчивой структуры.
Адаптивный генетический полиморфизм - основа устойчивого
функционирования вида
Видовой генофонд - полный набор вариантных состояний генов
(аллелофонд) и хромосомных структур (инверсионный полиморфизм
и др.), характерный для популяций в пределах видового ареала. При
этом не всегда генетический полиморфизм относится к адаптивному
типу, а лишь тогда, когда он обеспечивает стационарность видовой
системы в условиях экоклиматических градиентов среды обитания.
В настоящее время находит подтверждение точка зрения, оцениваю-
щая адаптивный генетический полиморфизм в терминах стационар-
ной видовой системы и популяционно-генетического гомеостаза, ис-
ключающих возможность градуальной трансформации видов [3]. Аль-
тернативная позиция большинства современных неодарвинистов [4, 5],
оценивающих пространственно-временные частотные флуктуации
полиморфных вариантов генов и хромосомных инверсий как проявле-
ние постепенной видовой дивергенции или филетической эволюции,
представляется нам ошибочной. При этом сторонники синтетической
теории эволюции обычно используют следующие аргументы:
1) в пространстве видового ареала происходят постепенные заме-
щения альтернативных аллельных и инверсионных вариантов (клины),
что, по их мнению, в конечном счете может привести к полной диз-
рупции; подобные процессы могут характеризовать и темпоральные
частотные сдвиги;
2) реальные частоты аллельных и инверсионных генотипов не со-
ответствуют распределению Харди-Вайнберга (наблюдается значитель-
ный дефицит гетерозигот);
3) обнаруживаются ассоциативные взаимодействия между разны-
ми инверсиями и аллелями (linkage disequilibrium). Необходимо отме-
тить, что подобная «эволюционная» интерпретация обычно является
следствием крайне редукционного подхода, когда отдельные аллели
129
В.Н. Стегний
или инверсии, обнаруживающие подобные тенденции, сознательно вы-
рываются из контекста оценки всего кариофонда или аллелофонда.
Напротив, комплексный и тщательный анализ генофонда и карио-
фонда вида показывает наличие у видов исключительной стационар-
ности адаптивного полиморфизма, отражающей высокую степень по-
пуляционно-генетического или, точнее, видового гомеостаза. При этом
выявляются популяционные системы, обладающие мощным интег-
рирующим эффектом на фоне дифференциальной частотной динами-
ки по отдельным аллелям или инверсиям. Было убедительно доказа-
но, что популяционно-генетический гомеостаз реализуется у видов с
адаптивным генетическим полиморфизмом через пространственно-
временную частотную динамику инверсий и аллелей и динамичность
их ассоциативных связей. При этом в генокомплексах (сочетаниях раз-
нохромосомных инверсий и аллелей) всегда существуют генотипи-
ческие варианты (отдельные гены или хромосомы), играющие роль
интеграторов генофонда, обеспечивающих видовое единство в тех
случаях, когда некоторые инверсионные варианты или аллели имеют
тенденции к дифференциации генофонда за счет ассоциативных свя-
зей или дефицита гетерозигот [6].
Несмотря на некоторые модификации взглядов неодарвинистов,
касающиеся предварительных условий и скорости видообразования,
все они базируются на постулате: при видообразовании реорганизует-
ся накопленный ранее генофонд вида. Градуализм зиждется на час-
тотном преобразовании полиморфной популяции с последующей фик-
сацией наиболее адаптированных аллельных или хромосомных вари-
антов. Этот методологически ошибочный, противоречащий систем-
ному подходу дарвиновский принцип «разновидность - суть возника-
ющий вид», подхваченный современными неодарвинистами в их обыч-
ной трактовке «от полиморфизма к видообразованию», не выдержи-
вает критики. Из теории систем следует, что абсолютное преоблада-
ние гомеостатических структур (адаптивного генетического полимор-
физма) приводит только к сохранению системы, но не к ее преобразо-
ванию (видообразованию).
После фундаментальных работ Ю.П. Алтухова и Ю.Г. Рычкова
[3, 7], в которых наряду с конструктивной критикой градуального ви-
дообразования были сформулированы принципы преобразования ви-
дов, основанные на реорганизации инвариантной (мономорфной) части
видового генома, в последние десятилетия XX века были получены
130
II. Общие проблемы эволюции
оригинальные данные, подтверждающие и развивающие эту концеп-
цию [6, 8, 9, 10, 11].
Идеология данного подхода базируется на возрождении типологи-
ческой концепции вида и макромутационистском толковании пробле-
мы видообразования. Убедительным аргументом этого являются ге-
номные мутации и, прежде всего, полиплоидия. Около 70% видов по-
крытосеменных растений возникло путем полиплоидии - резкого
скачкообразного преобразования видового генома [12]. Этот способ
видообразования, кроме растений, отмечен и в разных группах живот-
ных и является единственным в природе доказательным (и воспроиз-
водимым человеком) примером сальтационного видообразования.
Хорошо увязываются с сальтационным принципом видообразования
и скачкообразные модификации гетерохроматиновых компонентов
генома. К настоящему времени получено много данных относительно
реорганизации гетерохроматиновых блоков хромосом в близкород-
ственных видовых комплексах. Основной закономерностью при этом
является локализация гетерохроматина в прицентромерных блоках и
в микросомах у «стволовых» видов и «диспергирование» гетерохро-
матина по хромосомным плечам у видов, занимающих терминальные
положения в филогенетических ветвях. С сальтационным видообра-
зованием хорошо согласуются и данные по мобильным генетическим
элементам, обнаруживающим видовую специфику [10, 13].
Системные мутации как основной механизм видовой
реорганизации генома
Все известные с начала XX века мутации были подразделены на
три типа: генные, хромосомные и геномные. К настоящему времени
стало очевидным, что существует еще один тип - системные мутации.
В 1979 г. был выявлен феномен кардинальной реорганизации архи-
тектуры интерфазных ядер в онто- и филогенезе малярийных комаров
[8], затем подтвержденный на дрозофиле [14, 15]. Именно анализ архи-
тектуры ядер генеративных клеток позволил нам выявить наиболее су-
щественные закономерности: 1) принципиальное различие архитектуры
хромосом в клетках генеративной ткани и в соматических клетках; 2) суще-
ственные различия в трехмерной организации хромосом в самих генератив-
ных клетках между разными (в том числе и очень близкими) видами.
131
В.Н. Стегний
Видоспецифичность проявляется в следующих показателях: 1) на-
личие - отсутствие связей хромосом с ядерной оболочкой и локализа-
ция мест контактов на хромосомах; 2) морфология хромосомных уча-
стков прикрепления; 3) разобщенность локусов прикрепления гомео-
логичных хромосом у близких видов на ядерной оболочке; 4) наличие
- отсутствие локального или диффузного хромоцентра.
Взаимоотношения хромосом с ядерной мембраной облигатно инва-
риантны в пределах каждого вида малярийных комаров и четко различа-
ются между восемью изученными видами (рис. 1). У межвидовых гиб-
ридов проявляются видоспецифичные особенности каждого из видов,
причем значительная пространственная разобщенность зон прикрепле-
ния каждого из гомеологов в гибридных клетках свидетельствует о ви-
доспецифичности координат точек прикрепления на мембране.
На основании данных по видовым комплексам “D.melanogaster” и
“D.virilis” можно оценить основные закономерности реорганизации
хромосомной архитектуры в филогенетических направлениях от ство-
ловых (исходных) видов к терминальным (производным) видам [14,
15]. Общая схема такова: у стволовых видов (D.orena, D.virilis,
D.kanekoi, D.flavomontana) наблюдается концентрация гетерохроматина
в области хромоцентра, затем у производных видов хромоцентр визу-
ально исчезает, и хромосомы демонстрируют либо свободное (без при-
крепления к ядерной оболочке) состояние (D.mauritiana, D.teissieri,
D.yakuba), либо частичное (отдельные хромосомы) прикрепление
(D.novamexicana) (рис. 2).
Обнаруженный принцип хромосомной реорганизации рассматрива-
ется нами как явление мутационного ранга, не имеющего аналогов сре-
ди известных типов хромосомных мутаций. Таким образом, перестрой-
ку архитектуры интерфазного ядра можно оценить как новый тип мута-
ций, названный нами системными мутациями.
Системные мутации возникают в результате пространственной ре-
организации интерфазных хромосом в ядре за счет изменения хромо-
сомно-мембранных взаимоотношений. Системные мутации в своем
происхождении связаны с перестройкой хромоцентрального аппара-
та: изменяется структура хромоцентра (от локального до диффузного)
вплоть до визуального исчезновения последнего; появляется (или ис-
чезает) прикрепление центромерных, теломерных и других хромосом-
ных локусов к ядерной оболочке. Особенностью системных мутаций
132
LU
LU
Рис. 1. Схема эволюционного изменения архитектуры политенных хромосом трофоцитов яичников вось-
ми видов малярийных комаров палеарктического комплекса Anopheles maculipennis. Стрелками показа-
ны направления видообразования (сплошными - наиболее вероятные, пунктирными - менее вероятные)
II. Общие проблемы эволюции
В.Н. Стегний
Рис.2. Схема эволюционного изменения архитектуры первично по-
лигонных хромосом трофоцитов яичников восьми видов комплекса
Drosophila melanogaster. Стрелками показаны направления видо-
образования, основанные на комплексных данных [14]
является также то, что они: 1) четко выявляются только в клетках ге-
неративной ткани; 2) являются видоспецифичными и не обнаружива-
ют внутривидового полиморфизма; 3) проявляют «гетерозиготность»
только у межвидовых гибридов, геномы которых отражают различия
в архитектуре хромосом «отцовского» и «материнского» видов.
Таким образом, архитектура хромосомного аппарата генеративной
ткани представляет собой элементарную типологическую систему,
отражающую видоспецифичность признаков.
Происхождение системных мутаций. Как выше было отмечено,
перестройка архитектуры хромосомного аппарата, или системные
мутации, были обнаружены только в питающих клетках (трофоцитах)
яичников самок комаров и дрозофилы. Изучение соматических тка-
134
II. Общие проблемы эволюции
ней с политенными хромосомами (слюнные железы, мальпигиевы со-
суды) не выявило каких-либо очевидных различий в пространствен-
ной организации хромосом между видами. Для всех изученных видов
малярийных комаров в соматических тканях существует универсаль-
ная организация хромосом: они объединены центромерными района-
ми в единый хромоцентральный узел по принципу эктопической конъ-
югации. Для дрозофил также характерна однотипная система органи-
зации политенных хромосом слюнных желез, хотя и хромоцент-
ральное объединение центромерных участков представляет более диф-
фузное хроматиновое «облако», чем у комаров. Интересно, что у меж-
видовых гибридов F1 в клетках слюнных желез малярийных комаров
хромоцентр остается общим для гомеологов «отцовского» и «материнс-
кого» видов при отсутствии конъюгации между гомеологичными хромо-
сомами (частичном - у аутосом и полном - у половых хромосом).
Таким образом, учитывая происхождение питающих клеток, систем-
ные мутации относятся непосредственно к генеративной сфере. Действи-
тельно, исследования, проводимые уже более 80 лет (Э. Вильсон, X. Ра-
вен, R. King), убедительно и однозначно свидетельствуют, что трофоци-
ты у организмов с нутриментарным (алиментарным) типом оогенеза
являются сестринскими с собственно ооцитом. В генеративной ткани
самок первичные оогонии делятся несколько раз (у малярийных кома-
ров - 3 раза, у дрозофилы - 4 раза). В результате возникает 8 клеток у
комаров и 16 - у дрозофилы. Центральная по суперпозиции клетка ста-
новится ооцитом, остальные (7 - у комаров и 15 - у дрозофилы) - пита-
ющими (трофоцитами). R. King [16] показал, что ооцит связан со всеми
остальными сестринскими клетками (трофоцитами) цитоплазматичес-
кими мостиками. Впоследствии ооцит претерпевает мейотические пре-
образования, а питающие клетки остаются диплоидными. В начале пе-
риода созревания фолликула хромосомы трофоцитов интенсивно поли-
тенизируются и с высочайшей активностью синтезируют РНК-продукты,
практически полностью обеспечивая рост и развитие ооплазмы, и, что
самое главное, формируют ооплазматическую сегрегацию,
Системные мутации наиболее четко проявляются в период макси-
мального развития политенных хромосом у малярийных комаров.
У дрозофилы в норме в трофоцитах существуют так называемые первично
политенные хромосомы, размеры которых значительно меньше таковых у
комаров, и между тем системные мутации проявляются достаточно четко.
135
В.Н. Стегний
Методически пока невозможно оценить архитектуру хромосом в оого-
ниях и ооците. Однако есть все основания считать, что топологические
параметры взаиморасположения отдельных хромосом и их связи с ядер-
ной оболочкой в оогониях и ооците так же, как и у трофоцитов, носят
видоспецифичный характер. Наши данные показывают, что для ооцита
малярийных комаров характерно отсутствие общего локального хромо-
центра, что совпадает с данными по трофоцитам, следовательно, схема
хромосомной организации у них идентична. В пользу этого свидетель-
ствуют также «гибридные конфигурации» (F1) ядер ооцитов на стадии
пахитены (от скрещивания Anopheles maculipennis и A.subalpinus), пока-
зывающие отсутствие конъюгации гомеологов хромосом, как и в «гиб-
ридных» трофоцитах [9].
В заключение, касаясь принципов пространственной организации
хромосом половых клеток (яйцеклеток и спермиев), необходимо отме-
тить, что методические ограничения визуализации их хромосом не по-
зволяют пока детально оценить характер топологии. Было бы опромет-
чиво предполагать, что архитектура хромосом и трофоцитов идентична,
поскольку и структурно и функционально эти системы различаются, хотя
и имеют общее происхождение. Однако есть достаточно оснований счи-
тать, что и в ооците, и в оогониях, а также в спермиях и сперматогоникх
элементы видоспецифичной архитектуры хромосом идентичны трофо-
цитам. В этом отношении важные данные демонстрируют межвидовые
гибриды малярийных комаров. В трофоцитах самок F1 обнаруживаются
все специфичные элементы организации хромосом, характерные для
каждого из родительских типов. Впечатляет то, что посредством спер-
миев от родителя-самца передается полная информация относительно
видоспецифичной архитектуры хромосом в трофоцитах самок своего
вида. Это достаточно убедительный аргумент в пользу видоспецифич-
ности хромосомной архитектуры ядер в генеративной ткани в целом с
подобными же видоспецифичными особенностями в отдельных клеточ-
ных системах ткани. Клетки зародышевого пути от оогониев в гермарии
до зрелого ооцита (у самок) и от сперматогониев до сперматозоидов (у
самцов) определенно имеют видовую специфику в структурной органи-
зации ядерного материала, как это предполагал известный цитолог кон-
ца XIX века О. Гертвиг.
В следующей статье предполагается осветить взаимодействие ви-
довой системы с внешней средой и прежде всего вскрыть механизмы
трансформации видового генома в видообразовательном смысле.
136
IL Общие проблемы эволюции
Литература
1 Голубовский М.Д. Организация генотипа и формы наследственной изменчивости
эукариот// Успехи совр. биологии. 1985. Т. 100. Вып. 6. С. 323-339.
2 Садовский В.Н. Основания общей теории систем. М.: Наука, 1974. С. 274.
3 Алтухов Ю.П., Рычков Ю.Г. Популяционные системы и их структурные компонен-
ты. Генетическая стабильность и изменчивость // Журн.общ.биологии. 1970. Т. 31.
№ 5. С. 507-526.
4. Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
5. Dobzhansky Th. Genetics of evolutionary process. N.Y. L.: Columbia Univ. Press, 1971.
520 p.
6. Стегний В.Н. Популяционная генетика и эволюция малярийных комаров. Томск: Изд-
во Том. ун-та, 1991. 137 с.
7. Алтухов Ю.П., Рычков Ю.Г. Генетический мономорфизм видов и его возможное био-
логическое значение И Журн. общ. биологии. 1972. Т. 33. № 3. С. 281-300.
8. Стегний В.Н. Реорганизация структуры интерфазных ядер в онто- и филогенезе ма-
лярийных комаров // ДАН СССР 1979. Т. 249. № 5. С. 1231.
9. Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск:
Изд-во Новосиб. ун-та, 1993. 110 с.
10. Евгеньев М.Б., Ениколопов Е.Н., Пеунова Н.И. Транспозиция мобильных диспер-
гированных генетических элементов у дрозофилы // ДАН СССР. 1982. Т. 64. № 6.
С. 145.
11. Тахтаджян А.Л. Макроэволюционные процессы в истории растительного мира
И Ботан. журн. 1983. Т. 68. № 12. С. 1593-1603.
12. Грант В. Видообразование у растений. М.: Мир, 1980. 407 с.
13. Корочкин Л.И. Эволюционное значение генетических подвижных элементов // Ци-
тология и генетика. 1983. Т. 17. № 4. С. 67-78.
14. Стегний В.Н., Вассерлауф И.Э. Видовая архитектоника хромосом генеративной ткани
и проблемы филогенетических отношений в подгруппе melanogaster рода Drosophila
( Sophophora) И Генетика. 1994. Т. 30. № 4. С. 478-483.
15. Стегний В.Н., Вассерлауф И.Э., Ананьина Т.В. Взаиморасположение первичных
политенных хромосом яичников у 12 видов группы «virilis» рода Drosophila
(Sophophora) И Генетика. 1996. Т. 32. № 6. С. 750-754.
16. KingR. The meiotic behavior of the Drosophila oocyte //Intern. Rev. Cytol. 1970. Vol. 28.
P. 125.
МУТАЦИИ, СПОСОБНЫЕ ИНИЦИИРОВАТЬ
ВИДООБРАЗОВАНИЕ
Б.Ф. Чадов
Институт цитологии и генетики СО РАН
630090, г. Новосибирск, пр. Лаврентьева, 10
E-mail: chadov@bionet.nsc.ru
Введение
Мутации, с которыми имеет дело экспериментальная генетика, об-
ладают одним общим свойством: они не меняют видовой принадлеж-
ности индивидуума. Мутантные особи скрещиваются с особями свое-
го вида, оставляют плодовитое потомство, т.е. следуют правилам, “пред-
начертанным” особям в составе вида. Иные мутации и не могли бы
использоваться в генетической работе. Особую группу составляют
доминантные летали. О них нельзя сказать, сохраняют ли они видо-
вую принадлежность, поскольку, погибая на ранних стадиях развития,
в ранг мутантов они уже не попадают. Может ли мутация в отдельном
гене изменить видовую принадлежность особи или видовая принад-
лежность определяется группой генов и поэтому поиск отдельных,
“видообразующих”, мутаций обречен на неуспех - вопросы пока ис-
ключительно теоретического рассмотрения.
Здание синтетической теории эволюции построено на мутаци-
ях, с которыми реально имела дело генетика. Как сказано выше,
это - мутации, не меняющие видовой принадлежности. Согласно
синтетической теории вид образуется по мере накопления мута-
ций, создающих некий новый генный комплекс. Параллельно с этим
идет нарастание изоляции от особей исходного вида [1]. По сути
предполагается, что из элементов, не обладающих определенным
свойством (изоляция), строится система, обладающая этим свой-
ством. Виды предстают как различные конструкции, собранные из
одного строительного материала.
Разные конструктивные решения должны быть, в свою очередь,
генетически детерминированы. Дело только в том, что гены эти дру-
138
II. Общие проблемы эволюции
гие - это регуляторные гены. Можно предполагать, что видообразова-
ние идет путем мутирования регуляторных генов, причем той их час-
ти, которая еще не попала в поле зрения экспериментальной генетики.
В статье я расскажу об обнаружении необычных мутаций у
Drosophila melanogaster, полученных мной с сотрудниками в процес-
се работы, начатой в ноябре 1999 года. Первое сообщение о результа-
тах опубликовано в августе 2000 года в [2]. С момента подачи статьи
появились новые данные. Полученные мутации, на мой взгляд, обла-
дают характеристиками, необходимыми для мутаций, принимающих
участие в видообразовательном процессе.
Мой интерес к видообразованию возник под влиянием моего учи-
теля Н.В. Тимофеева-Ресовского. Его работы по микроэволюции из-
вестны. В 2000 году отмечалось 100 лет со дня рождения Тимофеева-
Ресовского, и эту статью я посвящаю его памяти.
Умозрительные посылки о существовании факультативных
летальных мутаций и изыскание способа их регистрации
В проблеме видообразования генетик и биолог находятся во влас-
ти разных парадигм. Генетическое исследование построено на изуче-
нии генетических вариантов, поэтому внимание генетика целиком со-
средоточено на внутривидовом разнообразии. Предположение о су-
ществовании части генома, общей для всех особей вида, непродуктивно
для генетика. Оно лишает его возможности пользоваться традицион-
ным генетическим методом и неосознанно вытесняется из рассмотре-
ния. Внимание биолога, особенно систематика, напротив, сосредота-
чивается на сходстве особей одного вида. Биологическая парадигма
разрешает биологу в качестве полноправного представителя вида ис-
пользовать любую особь этого вида.
Два взгляда на общность, называемую видом, логично объединить.
Принять, что видовой геном состоит из двух частей: части общей и
неизменной для особей вида и изменяемой части, которой разрешено
находиться в разных состояниях (аллелях). Такое объединение, при-
чем сделанное не умозрительно, а на основе изучения вариабильнос-
ти изоферментных спектров, было сделано Ю.П. Алтуховым с соавто-
рами еще в 70-х годах. Со всей определенностью было заявлено, что
процесс видообразования связан с изменением инвариантной, а от-
нюдь не вариабельной части видового генома [3]. Стремительное разви-
тие генетики морфогенеза: появление данных о мастер-генах, гомео-
139
Б.Ф. Чадов
называя тот же самый уровень дочерей в потомстве. Вопрос о причинах
разной экспрессии летальности у разных мутантов составляет отдель-
ную тему и пока рассматриваться не будет. Далее, говоря о получен-
ных мутантах, я буду иметь в виду прежде всего группу самых “стро-
гих” мутантов, которые не дают ни одной дочери в выборке не менее
чем из 200 потомков. Это мутанты групп 1 и 2.
Таблица!
Потомство селектированных самцов в скрещиваниях
с разными самками (опыт 1)
Номер культуры самца Скрещивание: Скрещивание. 6?у X (? + Скрещивание: * 6 $ у w f х (? + Плодо- витость самцов*
2$у х
Всего потом- ков Доля 9$ Всего потом- ков Доля 99 Всего потом- ков Доля <?<?
1 119 0,00 191 0,00 51 0,35 0,02
2 65 0,00 435 0,00 42 0,60 0,15
3 112 0,00 180 0,00 33 0,39 0,12
4 114 0,00 293 0,00 85 0,38 0,07
5 50 0,00 303 0,02 8 0,25 0,14
6 47 0,00 283 0,02 133 0,50 0,14
Среднее по группе 1 0,00 0,00 0,41 0,11
7 47 0,02 100 0,00 37 0,57 -
8 47 0,15 216 0,07 19 0,47 0,16
9 182 0,07 529 0,00 199 0,55 0,40
10 162 0,03 297 0,04 30 0,40 0,09
И 13 0,08 409 0,06 152 0,51 0,09
12 46 0,13 103 0,13 69 0,30 0,03
Среднее по группе 2 0,08 0,05 0,47 0,15
14 171 0,28 257 0,18 83 0,46 0,47
15 87 0,34 121 0,12 82 0,50 0,18
Среднее по группе 3 0,31 0,14 0,48 0,32
16 187 0,34 199 0,36 77 0,52 0,37
17 221 0,40 208 0,42 100 0,44 0,40
Среднее по группе 4 0,37 0,38 0,47 0,38
18 212 0,49 426 0,44 107 0,42 0,82
19 237 0,55 296 0,48 122 0,61 0,69
20 187 0,38 322 0,52 125 0,58 0,76
21 221 0,47 371 0,52 51 0,51 0,68
22 222 0,46 318 0,54 87 0,49 0,31
Среднее по группе 5 0,47 0,50 0,53 0,72
* Плодовитость самцов измерена отношением числа взрослого потомства, развившегося из
яиц, отложенных самками yellow в скрещивании с этими самцами, к числу отложенных яиц.
142
II. Общие проблемы эволюции
Таблица 2
Потомство селектированных самцов в скрещиваниях
с разными самками (опыт 2)
Номер куль- туры самца Скрещивание: 2 $у х 5 + Скрещивание: б$у х с? + Скрещивание: 6 ? уwf X 3 + Плодовитость самцов*
Всего потом- ков Доля ?? Всего потом- ков Доля $>? Всего потом- ков Доля <?<?
27 68 0,00 93 0,00 66 0,24 0,18
29 15 0,00 61 0,00 63 0,57 0,14
30 122 0,07 115 0,00 80 0,50 0,19
31 106 0,00 83 0,00 70 0,21 0,15
32 81 0,00 117 0,00 180 0,48 0J3 _
33 144 0,00 90 0,00 72 0,54 0,16
34 88 0,00 110 0,00 63 0,52 . 0,12
Среднее по группе 1 0,01 0,00 0,44 0,15
26 92 0,03 89 0,01 — - -
35 102 0,03 115 0,04 45 0,76 0,17
36 95 0,00 НО 0,01 76 0,47 0,35
37 52 0,02 68 0,04 67 0,63 0,14
38 54 0,06 84 0,01 92 0,73 0,14
41 79 0,03 100 0,01 91 0,57 0,10
Среднее по группе 2 0,03 0,02 0,62 0,17
43 9 - 97 0,32 - — —
45 13 0,08 — — 73 0,42
48 76 0,42 174 0,38 119 0,60 -
52 70 о.п 97 0,31 - - —
Среднее по группе 3 0,20 0,34 0,51
46 153 0,44 128 0,45 140 0,56 0,38
47 233 0,45 174 0,47 128 0,61 —
49 137 0,44 175 0,41 187 0,64 -
50 90 0,40 175 0,45 85 0,54 —
54 139 0,43 177 0,41 — — -
55 148 0,45 183 0,48 — — —
56 139 0,45 190 0,50 — — -
Среднее по группе 4 0,44 0,46 0,59
* См. табл. 1.
143
Б.Ф. Чадов
Отобранные мутанты и их потомки мужского пола не имели никаких
дефектов в экстерьере. Более того, они отличались высокой жизнеспо-
собностью и гиперсексуальностью, видимой без специального исследо-
вания. Указанные черты сочетались со сниженной плодовитостью сам-
цов (табл. 1,2). Уровень плодовитости, определенный в скрещивании с
yellow самками, отрицательно коррелировал с уровнем дочерей в потом-
стве, хотя не был им напрямую обусловлен.
2. Изоляцию мутантов можно преодолеть. Полученные мутан-
ты сохранялись в качестве культур только потому, что, используя са-
мок со сцепленными Х-хромосомами, можно передавать Х-хромосо-
му от отца к сыну. Обычный путь наследования Х-хромосомы от отца
к дочери был закрыт из-за гибели дочерей. Окажись мутантный самец
в природных условиях, мутация не получила бы распространения.
Оказалось, однако, что летальность мутации в гетерозиготе не абсо-
лютна. Она ослабляется и даже ликвидируется, если мутантного самца
скрещивать не с самками yellow, на которых проводился отбор леталь-
ных мутаций, а с другими самками. Летальность стала исчезать при скре-
щивании с самками D.melanogaster линий Berlin wild, Barnaul, Canton-S
и с самками, имеющими инвертированную хромосому Muller-5. В табл. 3
приведены результаты скрещиваний с самками yellow, Muller-5, Berlin
wild и Barnaul. В большинстве случаев нехватка дочерей в потомстве
мутантных самцов сохранялась. Для некоторых (“сильных”) мутантов
летальность Х-хромосомы в гетерозиготе не удавалось преодолеть и в
скрещивании с другими самками (номера 1,10, 31, к примеру). Вопрос
о роли перестроек я рассмотрю подробнее дальше, здесь же укажу на
практический аспект найденного факта. Получение гетерозиготы с ин-
вертированной Х-хромосомой позволяло, во-первых, изучить экспрес-
сию полученных мутаций у самки, во-вторых, построить методику полу-
чения подобных мутаций в аутосомах дрозофилы.
3. Мутации в Х-хромосоме передаются от матери ее дочерям и сы-
новьям. Скрещивание самок, получивших мутантную Х-хромосому и Х-хро-
мосому Muller-5, с самцами линии yellow показывает; что из генома самки
мутантная Х-хромосома без проблем передается потомству (табл. 4). У сам-
ки возникают и сыновья с этой хромосомой (+ и В ж“), и дочери (+), содержа-
щие мутантную Х-хромосому в гетерозиготе. Летальное действие мутаций в
гетерозиготе исчезло. В гомозиготе оно осталось: получить самок, гомози-
готных по мутациям, не удавалось. Таким образом, у самок полученные му-
тации вели себя как рецессивные летали.
144
II. Общие проблемы эволюции
Таблица 3
Доля дочерей в потомстве мутантного самца, скрещенного
с самками стандартных лабораторных линий
Номер куль- туры самца Типы самок
yellow Muller-5 Berlin wild Barnaul
Всего потом- ков Доля 9? Всего потом- ков Доля 99 Всего потом- ков Доля 99 Всего потом- ков Доля 99
1 191 0,00 — — — 0,00 91 0,02
2 435 0,00 23 0,22 7 0,17 143 0,03
3 180 0,00 36 0,25 18 0,17 146 0,04
4 293 0,00 156 0,31 — — 135 0,01
5 303 0,02 41 0,34 — — 143 0,19
6 283 0,02 161 0,50 212 0,31 138 0,03
7 100 0,00 — — — — - —
8 216 0,07 — — — — - —
9 529 0,04 160 0,55 128 0,29 303 0,28
10 297 0,00 63 0,06 — — — -
11 409 0,06 182 0,53 223 0,38 178 0,22
Средняя доля самок 0,01 0,35 0,26 0,10
27 93 0,00 62 0,29 84 0,08 112 0,10
29 61 0,00 53 0,53 64 0,20 113 0,26
30 115 0,00 76 0,36 102 0,33 135 0,П
31 83 0,00 77 0,18 87 0,08 129 0,07
32 117 0,00 62 0,53 140 0,34 80 0,30
33 90 0,00 59 0,39 149 0,28 48 0,27
34 110 0,00 68 0,31 99 0,24 117 0,27
Средняя доля самок 0,00 0,37 0,22 0,21
26 89 0,01 69 0,29 100 0,22 111 0,23
35 115 0,04 73 0,33 145 0,27 123 0,15
36 ПО 0,01 99 0,28 92 0,36 124 0,29
37 68 0,04 67 0,55 89 0,39 119 0,30
38 84 0,01 54 0,61 36 0,50 39 0,38
41 100 0,01 129 0,40 92 0,37 130 0,38
43 97 0,32 30 0,60 — - 57 0,49
45 - - - — 26 0,58 — —
48 174 0,38 131 0,67 172 0,53 232 0,57
52 97 0,31 27 0,59 — — 62 0,24
Средняя доля самок 0,13 0,48 0,40 0,34
46 128 0,45 108 0,60 145 0,46 191 0,50
47 174 0,47 130 0,54 194 0,45 237 0,57
49 175 0,41 111 0,55 209 0,59 196 0,51
50 175 0,45 ПО 0,67 213 0,38 225 0,49
53 111 0,42 43 0,53 60 0,55 79 0,56
54 177 0,41 92 0,52 157 0,50 159 0,52
55 183 0,48 111 0,45 34 0,57 184 0,56
56 190 0,50 83 0,53 134 0,44 189 0,52
25 83 0,55 85 0,48 104 0,30 131 0,47
Средняя доля самок 0,46 0,54 0,47 0,52
145
Б.Ф. Чадов
Таблица 4
Потомство самок ln(l) Muller-5, BW /+, содержащих мутацию,
и самцов yellow
Номер мутации Самки Самцы
4- В/+ + Bwa У
2 14 26 18 16 2
3 19 23 14 14 4
4 6 18 3 И 8
5 10 21 6 19 5
7 41 53 34 31 10
8 20 25 18 17 4
9 6 7 1 5 9
10 50 38 42 25 1
11 12 25 8 12 3
29 39 37 19 38 15
30 24 50 14 23 5
31 20 53 2 24 4
32 14 45 10 40 1
33 27 40 25 34 9
34 11 12 11 9 1
35 17 35 22 33 34
36 23 28 20 14 5
38 24 25 30 29 5
41 38 54 37 33 16
Специально замечу, что летальные свойства мутантных Х-хромосом
не изменились после их трансмиссии через самку. Оказавшись у самца,
они, как и раньше, оказывали летальное действие в гетерозиготе.
Передача по потомству мутантной Х-хромосомы от матери не за-
висела от присутствия у нее оппозитной инвертированной хромосо-
мы Muller-5. Были получены самки - двойные гетерозиготы по мута-
циям (табл. 5). В скрещивании с самцами yellow они передавали му-
тантные Х-хромосомы и дочерям (+), и сыновьям (+).
Несмотря на то, что самка передает мутантную Х-хромосому
по потомству, этот процесс нельзя назвать совершенно нормаль-
ным. У самок “Muller-5/ мутация” (табл. 4) и самок “мутация/му-
тация” (табл. 5) возникает большое количество патроклинных сам-
цов yellow. Это указывает на высокую частоту потерь Х-хромосом
в яйцеклетках.
146
II. Общие проблемы эволюции
Таблица 5
Потомство самок, содержащих мутацию в каждой из Х-хромосома
(скрещивание с самцами yellow)
Номер культуры самки Дочери + Сыновья 4- Сыновья у
6/41 97 78 3
6/8 58 62 3
6/38 29 22 1
6/29 96 84 —
6/7 50 38 —
6/3 57 50 6
6/10 33 27 8
6/11 40 29 5
6/4 5 1 3
6/9 67 78 1
6/35 28 35 24
6/5 13 12 3
3/11 11 14 —
3/8 38 25 19
9/10 29 32 25
9/5 1 6 1
9/11 24 24 20
Примечание, а - номер самки составлен из номеров мутаций.
4. Факультативные доминантные летальные мутации в ауто-
соме 2. Открытие способа ликвидации летальности с помощью хро-
мосомной перестройки дало возможность получить мутации в ауто-
сомах дрозофилы. Самцов дрозофилы облучили и скрестили с самка-
ми, имеющими перестроенную аутосому 2. Получили сыновей,
содержащих облученную и перестроенную аутосомы 2. Гарантией того,
что мутация в гетерозиготе не погибнет, была перестройка в оппозит-
ной аутосоме - инверсия In(2LR) Су Bl L4. Самцов индивидуально про-
веряли на наличие мутации в скрещивании с самками yellow. Показа-
телем наличия мутации было отсутствие в потомстве самца дочерей и
сыновей Су+. В результате одного опыта было получено 10 мутаций.
Из них было 4 “строгих” летали, 4 “просто” летали и 2 “нестрогих
летали”. Приведем результаты повторного тестирования мутаций
(табл. 6). Летальность проверялась опять на самках yellow. Самца с
деталью в одной хромосоме 2 и перестройкой Су - в другой скрещи-
147
Б.Ф. Чадов
вали с самками yellow. “Строгие” летали давали только Су потомство,
“просто летали” - 1-2 особи “не Су” на 100-150 Су, а “нестрогие”
летали давали потомства Су(+), только достоверно ниже, чем потом-
ства Су (табл. 6).
Таблица 6
Потомство самцов, содержащих доминантную деталь в хромосоме 2
(скрещивание: 9 у х 3 2* / Су Bl L4)
Номер куль- туры самца Особи + Особи CyBIL4 Всего потомства Доля особей +
?+ <?У ? + <?У
7а - - 116 110 226 0,00
37а — 1 124 130 254 0,00
44а — 1 125 109 234 0,00
53а — — 101 130 231 0,00
5а 11 6 146 140 303 0,06
8а 6 — 135 98 239 0,03
9а 9 9 96 84 198 0,09
62а 1 1 154 144 300 0,01
1а 56 55 83 94 288 0,39
61а 63 75 90 88 316 0,44
Конт- роль
42а 111 139 135 120 505 0.51
26а 128 107 106 106 447 0,53
Примечание ♦ - хромосома, содержащая мутацию.
Гомозигот по деталям среди “строгих” деталей не было. Среди “про-
сто деталей” было несколько случаев образования гомозигот. Полу-
ченные летали можно было считать рецессивными деталями, если бы
не их основное свойство: не давать с линией yellow потомства с мутан-
тной хромосомой 2 в гетерозиготе.
Доминантная летальность, как и в случае мутаций в Х-хромосоме,
проявлялась только при одном направлении скрещивания: когда жел-
тых самок скрещивали с мутантными самцами. Мутантные самки с
желтыми самцами давали обычное потомство, не обнаруживая при-
знаков наличия летали в хромосоме 2.
5. Материнский эффект хромосомных перестроек. Снятие доми-
нантной летальности с помощью хромосомной перестройки поднимает
старую проблему о связи видообразования и хромосомных перестроек.
148
IL Общие проблемы эволюции
Хорошо доказанная, но абсолютно непонятная связь между видообразо-
ванием и закреплением в гомозиготе перестроек как будто бы получает
объяснение. Ведь из полученных данных следовало, что дефект в регу-
ляторном гене можно оставить в популяции, если его совместить с хро-
мосомной перестройкой, иными словами, создать комплекс: мутация в
регуляторном гене + перестройка. Специально исследовали действие
хромосомных перестроек в Х-хромосоме и в аутосомах дрозофилы.
Изучили сначала, как будет сказываться присутствие перестроек в
аутосомах самок дрозофилы на проявлении доминантной летальнос-
ти Х-мутаций. В контроле мутантного самца скрещивали с самками
yellow, в опыте - с самками yellow Curly, содержащими в аутосоме 2
инверсию Су, с самками у; Dichaete, содержащими инверсию в ауто-
соме 3; и с самками у; Су; D, содержащими инверсии в обоих аутосо-
мах. Исследовали также влияние инверсии Рт в аутосоме 2. В табл. 7
для краткости представлены данные только по инверсиям Су и Рт.
У контрольных самок yellow, не несущих перестроек в аутосомах,
как и ранее, дочерей не возникало. Присутствие и Curly, и Plum выз-
вало появление дочерей. Перестройки в аутосомах явно снимали ле-
тальное действие Х-хромосомных мутаций. Но как? Отнюдь не сво-
им присутствием в зиготе. Если бы это было так, то исключительные
серые самки содержали бы только одну из аутосом - инвертированную.
Таблица 7
Результаты скрещиваний мутантных самцов с самками
у;+/+ , у; + 7 Су и у; +7 Рт
Но- мер куль- туры самца Самки у; +/+ Самки у; +7 Су Самки у; + 7 Рт
Си- + <?У 9 + <?У 9 + <?У
Су* Су Су* Су Рт* Рт Рт* Рт
1 0 230 0 0 178 163 0 0 107 57
2 0 230 14 13 127 134 4 3 70 72
4 0 270 9 4 185 159 1 7 86 81
5 2 197 23 21 80 95 6 4 47 48
27 4 167 1 0 102 ИЗ 2 1 53 65
29 0 163 32 27 71 56 26 24 55 20
30 0 184 15 13 81 76 9 12 60 47
31 0 242 32 20 127 102 5 4 28 29
32 0 197 22 10 90 77 9 17 36 32
~ 33 0 209 20 18 95 101 11 8 87 47
34 0 140 11 14 88 101 25 20 68 54
149
Б.Ф. Чадов
В опыте же они получали примерно поровну и инвертированную, и
нормальную аутосому. Совершенно ясно, что изменение летальности
происходило за счет действия перестроек еще до редукционного деле-
ния ооцита. В тот период, когда яйцеклетка созревала, наполняясь про-
дуктами трофоцитов, так же как и яйцеклетка, гетерозиготная по пе-
рестройкам.
Сказанное проиллюстрировано на рис. 2.
Яйцеклетка
Оплодотворение
Знгота
Рис. 2. Материнский эффект хромосомной перестройки. Созревающие ооциты +/+ и
1п(2) Су/+ окружены четырьмя питающими клетками, продукты которых поступают в
ооцит. В результате мейоза образуется один тип яйцеклеток у ооцитов +/+ и два типа -
у ооцита 1п(2) Су/+. После оплодотворения яйцеклеток спермием, содержащим мутан-
тную Х-хромосому, возникают зиготы XX; +/+ и XX; Су/+. В питающих клетках самок
1п(2) Су/+ образуется фактор (обозначен точками), благодаря которому выживают оба
типа зигот, несмотря на наличие доминантной летальной мутации в пагроклинной
Х-хромосоме. В питающих клетках самок +/+ такого фактора не образуется, и зиготы с
леталью в Х-хромосоме погибают. Благодаря действию 1п(2) Су до мейоза жизнеспо-
собными оказываются как зиготы с инверсией 1п(2) Су, так и зиготы без нее
Самка с инверсией Су образует два типа яйцеклеток: X; + и X; Су.
Выжившие дочери XX образовались из яйцеклеток обоих типов. До-
чери XX; +/+, возникшие у самок с инверсией Су, по генотипу ничем
не отличались от дочерей XX; +/+, возникших у самок без инверсии
Су. Однако они выживали, потому что содержали цитоплазму, образо-
вавшуюся в генотипе Су /+ (обозначена точками). Точно такие же до-
чери XX; +/+, получившие от самца мутантную Х-хромосому, гибли,
поскольку получили цитоплазму, образовавшуюся в генотипе +/+.
150
II Общие проблемы эволюции
Перестройки в Х-хромосоме действовали точно так же. Появление
серых дочерей в потомстве мутантных самцов происходило не только в
скрещиваниях самцов с самками, гомозиготными по инверсии Muller-5,
но и с самками Muller-5/+, гетерозиготными по этой же инверсии. Из
табл. 8 видно, что у мутантных самцов в скрещиваниях с гетерозиго-
тами по Muller-5 возникают не только сыновья, но и дочери. Причем
одни из них содержат инверсию Muller-5 (фенотип В/+), другие (+/+)
- не содержат.
Таблица 8
Потомство от скрещивания мутантных самцов (+) с самками
In (1) Muller-5, w-B/y
Номер культуры самца Фенотип потомства
Дочери Сыновья
+ В/+ У
1 1 7 127 46
2 11 21 47 89
4 5 12 77 167
5 22 31 36 71
6 9 26 58 62
30 31 41 86 109
31 16 33 65 79
32 42 51 60 81
33 23 23 61 88
34 6 11 44 72
36 21 25 58 108
Если мутантный самец сам содержал инверсию Су или Dichaete,
то инверсии нисколько не снижали летальное действие мутаций в его
Х-хромосоме. Мутантные самцы с инверсией Curly не имели дочерей
в потомстве точно так же, как их братья без инверсии Curly (табл. 9).
6. Модификация летальности: влияние собственно хромосомной
перестройки или генов в перестроенной хромосоме? Получив измене-
ние летального действия мутаций у гетерозигот по перестроенным хро-
мосомам, задались вопросом: это - эффект перестройки или специфи-
ческого набора аллелей в составе перестроенной хромосомы? Чтобы
ответить на него, получили перестройки в самой тестерной линии yellow.
Испытали две инверсии в Х-хромосоме и две транслокации, включаю-
151
Б.Ф. Чадов
боксах и гомеодоменах [4], со своей стороны, подводят к идее суще-
ствования инвариантной части в видовом геноме, предполагая, что это
должны быть гены, управляющие морфогенезом.
Признание существования инвариантной части видового генома с
генетической точки зрения означает следующее. У особей, составля-
ющих вид, гены инвариантной части должны находиться в гомозиго-
те. Никаких аллельных вариантов: ни отрицательно действующих на
жизнеспособность, ни нейтральных, ни положительных, не должно
существовать. Аллельные варианты должны элиминироваться весьма
необычно: без проверки их действия, только потому, что первичная
последовательность мутанта отличается от предковой. Мутация дол-
жна выглядеть как доминантная деталь, а причина летальности для
нас просто парадоксальна, это - гетерозиготность.
При наличии состояния гетерозиготности как причины летальности
можно предположить, что найдутся мутации, летальные в гетеро-, но не
в гомозиготе. Открывается возможность получить мутации, проявляю-
щиеся в двух, казалось бы, несовместимых формах: как жизнеспособная
особь (гомозигота, не деталь) и как доминантная деталь (гетерозигота).
Далее такие мутации я буду называть факультативными доминантны-
ми деталями. Следовало сначала получить мутантных особей, гомози-
готных по мутации (поэтому жизнеспособных), а потом испытать мута-
ции на летальность в гетерозиготе. В случае успеха можно было бы по-
лучить коллекцию доминантных летальных мутаций для последующего
изучения, как это делается в генетике для всех прочих мутаций. Дрозо-
фила с ее гетерогаметным определением пола позволяла осуществить
указанную процедуру в отношении мутаций в Х-хромосоме.
Методика получения доминантных летальных мутаций
в Х-хромосоме Drosophila melanogaster
Самцов дрозофилы облучили стандартным образом гамма-лучами и
скрестили с самками, содержащими сцепленные Х-хромосомы (рис. 1).
В результате получили выборку сыновей примерно в 3000 особей.
Каждая из них содержала облученный набор хромосом (X, 2, 3 и 4) от
отца и необлученный (У, 2, 3 и 4) от матери. Предполагали, что некото-
рые из самцов содержат в облученной Х-хромосоме искомую мутацию.
На рисунке - это Х-хромосома со звездочкой. Чтобы найти самца с мута-
цией, каждого из отобранных самцов скрестили с двумя самками линии
140
II. Общие проблемы эволюции
yellow. Самцы, не содержащие
jviyrauHH, имели в потомстве
серых дочерей (+/у) и желтых
сыновей (у), а самцы с мута-
цией - только желтых сыно-
вей. Последних ввели в куль-
туру, скрещивая с самками,
имеющими сцепленные X-
хромосомы. В настоящее вре-
мя коллекция мутаций, полу-
ченная в двух повторностях
опыта, насчитывает более
30 культур.
$ 3000 R
sxxv ® аш
««да». «хх ®
' «XX ® tfBtY
«XX ® tf<Y
Fl Норма:
дочери хЦ
*»*ри
Рис. 1. Схема обнаружения факультативных до-
минантных летальных мутаций в Х-хромосоме
Основные свойства мутаций в Х-хромосоме
1. Изоляция мутантов (мутантная Х-хромосома не передает-
ся от отца к дочери). Дважды проведенная селекция мутаций в
Х-хромосоме привела к выделению двух похожих групп мутантов. Дан-
ные по ним для сравнения представлены отдельно (табл. 1 и 2). По
основному критерию - отсутствию дочерей в потомстве - мутантные
самцы отличались друг от друга. Одни (номера 1-6 табл. 1 и номера
27-34 табл. 2) характеризовались полным отсутствием дочерей в по-
томстве, состоящем из нескольких сотен потомков, вторые имели
1-2% от положенных 50%, у третьих доля самок была еще выше и так
до самцов самой последней группы, у которых доля дочерей в потом-
стве лишь статистически достоверно была ниже 50%. Селекционная
процедура не предусматривала отбора мутантов со слабо выражен-
ным свойством, однако они оказались отобранными. Отбор самцов,
перспективных на содержание мутации, проводился рано, когда по-
томство в пробирке еще только начинало вылетать из куколки. После
подсчета всего вылетевшего потомства оказывалось, что в некоторых
пробирках то или иное количество дочерей все же образовывалось.
Вне зависимости от того, каким был уровень дочерей в потомстве, он
стабильно сохранялся в потомстве самца. В таблицах приведены данные
Двух испытаний мутантных самцов с самками yellow', первый - при от-
боре самца, второй - после введения мутации в культуру. В процессе
работы с мутантами некоторые были испытаны 4 и 5 раз, неизменно по-
141
Б.Ф. Чадов
казывая тот же самый уровень дочерей в потомстве. Вопрос о причинах
разной экспрессии летальности у разных мутантов составляет отдель-
ную тему и пока рассматриваться не будет. Далее, говоря о получен-
ных мутантах, я буду иметь в виду прежде всего группу самых “стро-
гих” мутантов, которые не дают ни одной дочери в выборке не менее
чем из 200 потомков. Это мутанты групп 1 и 2.
Таблица!
Потомство селектированных самцов в скрещиваниях
с разными самками (опыт 1)
Номер культуры самца Скрещивание: 2 $у х (J + Скрещивание: 6 $У х (? + Скрещивание: * 6 $ у W f X (? + Плодо- витость самцов*
Всего потом- ков Доля 99 Всего потом- ков Доля 99 Всего потом- ков Доля <?<?
1 119 0,00 191 0,00 51 0,35 0,02
2 65 0,00 435 0,00 42 0,60 0,15
3 112 0,00 180 0,00 33 0,39 0,12
4 114 0,00 293 0,00 85 0,38 0,07
5 50 0,00 303 0,02 8 0,25 0,14
6 47 0,00 283 0,02 133 0,50 0,14
Среднее по группе 1 0,00 0,00 0,41 0,11
7 47 0,02 100 0,00 37 0,57 —
8 47 0,15 216 0,07 19 0,47 0,16
9 182 0,07 529 0,00 199 0,55 0,40
10 162 0,03 297 0,04 30 0,40 0,09
И 13 0,08 409 0,06 152 0,51 0,09
12 46 0,13 103 0,13 69 0,30 0,03
Среднее по группе 2 0,08 0,05 0,47 0,15
14 171 0,28 257 0,18 83 0,46 0,47
15 87 0,34 121 0,12 82 0,50 0,18
Среднее по группе 3 0,31 0,14 0,48 0,32
16 187 0,34 199 0,36 77 0,52 0,37
17 221 0,40 208 0,42 100 0,44 0,40
Среднее по группе 4 0,37 0,38 0,47 0,38
18 212 0,49 426 0,44 107 0,42 0,82
19 237 0,55 296 0,48 122 0,61 0,69
20 187 0,38 322 0,52 125 0,58 0,76
21 221 0,47 371 0,52 51 0,51 0,68
22 222 0,46 318 0,54 87 0,49 0,31
Среднее по группе 5 0,47 0,50 0,53 0,72
* Плодовитость самцов измерена отношением числа взрослого потомства, развившегося из
яиц, отложенных самками yellow в скрещивании с этими самцами, к числу отложенных яиц.
142
II. Общие проблемы эволюции
Таблица 2
Потомство селектированных самцов в скрещиваниях
с разными самками (опыт 2)
Номер куль- туры самца Скрещивание: 2 $у х <? + Скрещивание: 6?у х 5+ Скрещивание: 6 $ у wf х (У Плодовитость самцов*
Всего потом- ков Доля 99 Всего потом- ков Доля 9? Всего потом- ков Доля 63
27 68 0,00 93 0,00 66 0,24 0,18
29 15 0,00 61 0,00 63 0,57 0,14
30 122 0,07 115 0,00 80 0,50 0,19
31 106 0,00 83 0,00 70 0,21 0,15
32 1 81 0,00 117 0,00 180 0,48 0,13
33 144 0,00 90 0,00 72 0,54 0,16
34 88 0,00 НО 0,00 63 0,52 . 0,12
Среднее по группе 1 0,01 0,00 0,44 0,15
26 92 0,03 89 0,01 — - -
35 102 0,03 115 0,04 45 0,76 0,17
36 95 0,00 110 0,01 76 0,47 0,35
37 52 0,02 68 0,04 67 0,63 0,14
38 54 0,06 84 0,01 92 0,73 0,14
41 79 0,03 100 0,01 91 0,57 0,10
Среднее по группе 2 0,03 0,02 0,62 0,17
43 9 - 97 0,32 - — —
45 13 0,08 — — 73 0,42 -
48 76 0,42 174 0,38 119 0,60 -
52 70 0,11 97 0,31 - - —
Среднее по группе 3 0,20 0,34 0,51
46 153 0,44 128 0,45 140 0,56 0Д8
47 233 0,45 174 0,47 128 0,61 —
49 137 0,44 175 0,41 187 0,64 -
50 90 0,40 175 0,45 85 0,54 —
54 139 0,43 177 0,41 — — -
55 148 0,45 183 0,48 — — —
56 139 0,45 190 0,50 — — -
Среднее по группе 4 0,44 0,46 0,59
* См. табл. 1.
143
Б.Ф. Чадов
Отобранные мутанты и их потомки мужского пола не имели никаких
дефектов в экстерьере. Более того, они отличались высокой жизнеспо-
собностью и гиперсексуальностью, видимой без специального исследо-
вания. Указанные черты сочетались со сниженной плодовитостью сам-
цов (табл. 1, 2). Уровень плодовитости, определенный в скрещивании с
yellow самками, отрицательно коррелировал с уровнем дочерей в потом-
стве, хотя не был им напрямую обусловлен.
2, Изоляцию мутантов можно преодолеть. Полученные мутан-
ты сохранялись в качестве культур только потому, что, используя са-
мок со сцепленными Х-хромосомами, можно передавать Х-хромосо-
му от отца к сыну. Обычный путь наследования Х-хромосомы от отца
к дочери был закрыт из-за гибели дочерей. Окажись мутантный самец
в природных условиях, мутация не получила бы распространения.
Оказалось, однако, что летальность мутации в гетерозиготе не абсо-
лютна. Она ослабляется и даже ликвидируется, если мутантного самца
скрещивать не с самками yellow, на которых проводился отбор леталь-
ных мутаций, а с другими самками. Летальность стала исчезать при скре-
щивании с самками D.melanogaster линий Berlin wild, Barnaul, Canton-S
и с самками, имеющими инвертированную хромосому Muller-5. В табл. 3
приведены результаты скрещиваний с самками yellow, Muller-5, Berlin
wild и Barnaul. В большинстве случаев нехватка дочерей в потомстве
мутантных самцов сохранялась. Для некоторых (“сильных”) мутантов
летальность Х-хромосомы в гетерозиготе не удавалось преодолеть и в
скрещивании с другими самками (номера 1, 10, 31, к примеру). Вопрос
о роли перестроек я рассмотрю подробнее дальше, здесь же укажу на
практический аспект найденного факта. Получение гетерозиготы с ин-
вертированной Х-хромосомой позволяло, во-первых, изучить экспрес-
сию полученных мутаций у самки, во-вторых, построить методику полу-
чения подобных мутаций в аутосомах дрозофилы.
3. Мутации в Х-хромосоме передаются от матери ее дочерям и сы-
новьям. Скрещивание самок, получивших мутантную Х-хромосому и Х-хро-
мосому Muller-5, с самцами линии yellow показывает; что из генома самки
мутантная Х-хромосома без проблем передается потомству (табл. 4). У сам-
ки возникают и сыновья с этой хромосомой (+ и В >₽“), и дочери (+), содержа-
щие мутантную Х-хромосому в гетерозиготе. Летальное действие мутаций в
гетерозиготе исчезло. В гомозиготе оно осталось: получить самок, гомози-
готных по мутациям, не удавалось. Таким образом, у самок полученные му-
тации вели себя как рецессивные летали.
144
II. Общие проблемы эволюции
Таблица 3
Доля дочерей в потомстве мутантного самца, скрещенного
с самками стандартных лабораторных линий
"Номер куль- туры самца Типы самок
yellow Muller-5 Berlin wild Barnaul
Всего потом- ков Доля 22 Всего потом- ков Доля 22 Всего потом- ков Доля 22 Всего потом- ков Доля 22
' 1 191 0,00 — — - 0,00 91 0,02
j 435 0,00 23 0,22 7 0,17 143 0,03
3 180 0,00 36 0,25 18 0,17 146 0,04
4 293 0,00 156 0,31 — — 135 0,01
5 303 0,02 41 0,34 - — 143 0,19
6 283 0,02 161 0,50 212 0,31 138 0,03
7 100 0,00 — — — — - —
8 216 0,07 — — — — — —
9 529 0,04 160 0,55 128 0,29 303 0,28
10 297 0,00 63 0,06 — — — -
11 409 0,06 182 0,53 223 0,38 178 0,22
Средняя доля самок 0,01 0,35 0,26 0,10
27 93 0,00 62 0,29 84 0,08 112 0,10
29 61 0,00 53 0,53 64 0,20 113 0,26
30 115 0,00 76 0,36 102 0,33 135 0.17
31 1 83 0,00 77 0,18 87 0,08 129 0,07
32 117 0,00 62 0,53 140 0,34 80 0,30
33 90 0,00 59 0,39 149 0,28 48 0,27
34 110 0,00 68 0,31 99 0,24 117 0,27
Средняя доля самок 0,00 0,37 0,22 0,21
26 89 0,01 69 0,29 100 0,22 111 0,23
35 115 0,04 73 0,33 145 0,27 123 0,15
36 НО 0,01 99 0,28 92 0,36 124 0,29
37 68 0,04 67 0,55 89 0,39 119 0,30
38 84 0,01 54 0,61 36 0,50 39 0,38
41 100 0,01 129 0,40 92 0,37 130 0,38
43 97 0,32 30 0,60 — - 57 0,49
45 - — - — 26 0,58 - —
48 174 0,38 131 0,67 172 0,53 232 0,57
52 97 0,31 27 0,59 — — 62 0,24
Средняя доля самок 0,13 0,48 0,40 0,34
46 128 0,45 108 0,60 145 0,46 191 0,50
47 174 0,47 130 0,54 194 0,45 237 0,57
49 175 0,41 111 0,55 209 0,59 196 0.51
50 175 0,45 ПО 0,67 213 0,38 225 0,49
53 111 0,42 43 0,53 60 0,55 79 0,56
54 177 0,41 92 0,52 157 0,50 159 0,52
55 183 0,48 111 0,45 34 0,57 184 0,56
56 190 0,50 83 0,53 134 0,44 189 0,52
25 83 0,55 85 0,48 104 0,30 131 0,47
Средняя доля самок 0,46 0,54 0,47 0,52
145
Б.Ф. Чадов
Таблица 4
Потомство самокIn(l) Muller-5, Bw‘/+, содержащих мутацию,
и самцов yellow
Номер мутации Самки Самцы
+ В/+ + Bwa У
2 14 26 18 16 2
3 19 23 14 14 4
4 6 18 3 11 8
5 10 21 6 19 5
7 41 53 34 31 10
8 20 25 18 17 4
9 6 7 1 5 9
10 50 38 42 25 1
11 12 25 8 12 3
29 39 37 19 38 15
30 24 50 14 23 5
31 20 53 2 24 4
32 14 45 10 40 1
33 27 40 25 34 9
34 11 12 11 9 1
35 17 35 22 33 34
36 23 28 20 14 5
38 24 25 30 29 5
41 38 54 37 33 16
Специально замечу, что летальные свойства мутантных Х-хромосом
не изменились после их трансмиссии через самку. Оказавшись у самца,
они, как и раньше, оказывали летальное действие в гетерозиготе.
Передача по потомству мутантной Х-хромосомы от матери не за-
висела от присутствия у нее оппозитной инвертированной хромосо-
мы Muller-5. Были получены самки - двойные гетерозиготы по мута-
циям (табл. 5). В скрещивании с самцами yellow они передавали му-
тантные Х-хромосомы и дочерям (+), и сыновьям (+).
Несмотря на то, что самка передает мутантную Х-хромосому
по потомству, этот процесс нельзя назвать совершенно нормаль-
ным. У самок “Muller-5/ мутация” (табл. 4) и самок “мутация/му-
тация” (табл. 5) возникает большое количество патроклинных сам-
цов yellow. Это указывает на высокую частоту потерь Х-хромосом
в яйцеклетках.
146
II. Общие проблемы эволюции
Таблица 5
Потомство самок, содержащих мутацию в каждой из Х-хромосом*
(скрещивание с самцами yellow)
Номер культуры самки Дочери + Сыновья + Сыновья у
6/41 97 78 3
6/8 58 62 3
” 6/38 29 22 1
6/29 96 84 —
6/7 50 38 —
6/3 57 50 6
6/10 33 27 8
6/11 40 29 5
6/4 5 1 3
6/9 67 78 1
6/35 28 35 24
6/5 13 12 3
3/11 11 14 —
3/8 38 25 19
9/10 29 32 25
9/5 1 6 1
9/11 24 24 20
Примечание, а - номер самки составлен из номеров мутаций.
4. Факультативные доминантные летальные мутации в ауто-
соме 2. Открытие способа ликвидации летальности с помощью хро-
мосомной перестройки дало возможность получить мутации в ауто-
сомах дрозофилы. Самцов дрозофилы облучили и скрестили с самка-
ми, имеющими перестроенную аутосому 2. Получили сыновей,
содержащих облученную и перестроенную аутосомы 2. Гарантией того,
что мутация в гетерозиготе не погибнет, была перестройка в оппозит-
ной аутосоме - инверсия In(2LR) Су Bl L4. Самцов индивидуально про-
веряли на наличие мутации в скрещивании с самками yellow. Показа-
телем наличия мутации было отсутствие в потомстве самца дочерей и
сыновей Су+. В результате одного опыта было получено 10 мутаций.
Из них было 4 “строгих” летали, 4 “просто” летали и 2 “нестрогих
летали”. Приведем результаты повторного тестирования мутаций
(табл. 6). Летальность проверялась опять на самках yellow. Самца с
Деталью в одной хромосоме 2 и перестройкой Су - в другой скрещи-
147
Б.Ф. Чадов
вали с самками yellow. “Строгие” летали давали только Су потомство,
“просто летали” - 1-2 особи “не Су” на 100-150 Су, а “нестрогие”
летали давали потомства Су(+), только достоверно ниже, чем потом-
ства Су (табл. 6).
Таблица 6
Потомство самцов, содержащих доминантную деталь в хромосоме 2
(скрещивание: $ у х с? 2* / Су Bl L4)
Номер куль- туры самца Особи + Особи Су Bl L* Всего потомства Доля особей +
?+ <?у 2 + ву
7а — — 116 но 226 0,00
37а — 1 124 130 254 0,00
44а — 1 125 109 234 0,00
53а - — 101 130 231 0,00
5а 11 6 146 140 303 0,06
8а 6 — 135 98 239 0,03
9а 9 1 9 96 84 198 0,09
62а 1 1 154 144 300 0,01
1а 56 55 83 94 288 0,39
61а 63 75 90 88 316 0,44
Конт- роль
42а 111 139 135 120 505 0,51
26а 128 107 106 106 447 0,53
Примечание. * - хромосома, содержащая мутацию.
Гомозигот по деталям среди “строгих” деталей не было. Среди “про-
сто деталей” было несколько случаев образования гомозигот. Полу-
ченные летали можно было считать рецессивными деталями, если бы
не их основное свойство: не давать с линией yellow потомства с мутан-
тной хромосомой 2 в гетерозиготе.
Доминантная летальность, как и в случае мутаций в Х-хромосоме,
проявлялась только при одном направлении скрещивания: когда жел-
тых самок скрещивали с мутантными самцами. Мутантные самки с
желтыми самцами давали обычное потомство, не обнаруживая при-
знаков наличия летали в хромосоме 2.
5. Материнский эффект хромосомных перестроек. Снятие доми-
нантной летальности с помощью хромосомной перестройки поднимает
старую проблему о связи видообразования и хромосомных перестроек.
148
II. Общие проблемы эволюции
Хорошо доказанная, но абсолютно непонятная связь между видообразо-
ванием и закреплением в гомозиготе перестроек как будто бы получает
объяснение. Ведь из полученных данных следовало, что дефект в регу-
ляторном гене можно оставить в популяции, если его совместить с хро-
мосомной перестройкой, иными словами, создать комплекс: мутация в
регуляторном гене + перестройка. Специально исследовали действие
хромосомных перестроек в Х-хромосоме и в аутосомах дрозофилы.
Изучили сначала, как будет сказываться присутствие перестроек в
аутосомах самок дрозофилы на проявлении доминантной летальнос-
ти Х-мутаций. В контроле мутантного самца скрещивали с самками
yellow, в опыте - с самками yellow Curly, содержащими в аутосоме 2
инверсию Су, с самками у; Dichaete, содержащими инверсию в ауто-
соме 3; и с самками у; Су; D, содержащими инверсии в обоих аутосо-
мах. Исследовали также влияние инверсии Рт в аутосоме 2. В табл. 7
для краткости представлены данные только по инверсиям Су и Рт.
У контрольных самок yellow, не несущих перестроек в аутосомах,
как и ранее, дочерей не возникало. Присутствие и Curly, и Plum выз-
вало появление дочерей. Перестройки в аутосомах явно снимали ле-
тальное действие Х-хромосомных мутаций. Но как? Отнюдь не сво-
им присутствием в зиготе. Если бы это было так, то исключительные
серые самки содержали бы только одну из аутосом - инвертированную.
Таблица 7
Результаты скрещиваний мутантных самцов с самками
у;+/+, у; + / Су и у; +/ Рт
Но- мер куль- туры самца Самки у; +/+ Самки у; +/ Су Самки у; + / Рт
СМ- + 5у 5 + <?У 9 + с?У
Су* Су Су* Су Рт* Рт Рт* Рт
1 0 230 0 0 178 163 0 0 107 57
2 0 230 14 13 127 134 4 3 70 72
4 0 270 9 4 185 159 1 7 86 81
5 2 197 23 21 80 95 6 4 47 48
27 4 167 1 0 102 ИЗ 2 1 53 65
29 0 163 32 27 71 56 26 24 55 20
30 0 184 15 13 81 76 9 12 60 47
31 0 242 32 20 127 102 5 4 28 29
32 0 197 22 10 90 77 9 17 36 32
33 0 209 20 18 95 101 11 8 87 47
Г~34 0 140 11 14 88 101 25 20 68 54
149
Б.Ф. Чадов
В опыте же они получали примерно поровну и инвертированную, и
нормальную аутосому. Совершенно ясно, что изменение летальности
происходило за счет действия перестроек еще до редукционного деле-
ния ооцита. В тот период, когда яйцеклетка созревала, наполняясь про-
дуктами трофоцитов, так же как и яйцеклетка, гетерозиготная по пе-
рестройкам.
Сказанное проиллюстрировано на рис. 2.
Яйцеклетка
Рис. 2. Материнский эффект хромосомной перестройки. Созревающие ооциты +/+ и
1п(2) Су/-»- окружены четырьмя питающими клетками, продукты которых поступают в
ооцит. В результате мейоза образуется один тип яйцеклеток у ооцитов +/+ и два типа -
у ооцита 1п(2) Су/+. После оплодотворения яйцеклеток спермием, содержащим мутан-
тную Х-хромосому, возникают зиготы XX; +/+ и XX; Су/+. В питающих клетках самок
1п(2) Су/+ образуется фактор (обозначен точками), благодаря которому выживают оба
типа зигот, несмотря на наличие доминантной летальной мутации в пагроклинной
Х-хромосоме. В питающих клетках самок +/+ такого фактора не образуется, и зиготы с
леталью в Х-хромосоме погибают. Благодаря действию 1п(2) Су до мейоза жизнеспо-
собными оказываются как зиготы с инверсией 1п(2) Су, так и зиготы без нее
Самка с инверсией Су образует два типа яйцеклеток: X; + и X; Су.
Выжившие дочери XX образовались из яйцеклеток обоих типов. До-
чери XX; 4-/4-, возникшие у самок с инверсией Су, по генотипу ничем
не отличались от дочерей XX; 4-/4-, возникших у самок без инверсии
Су. Однако они выживали, потому что содержали цитоплазму, образо-
вавшуюся в генотипе Су /+ (обозначена точками). Точно такие же до-
чери XX; +/+, получившие от самца мутантную Х-хромосому, гибли,
поскольку получили цитоплазму, образовавшуюся в генотипе 4-/4-.
150
II. Общие проблемы эволюции
Перестройки в Х-хромосоме действовали точно так же. Появление
серых дочерей в потомстве мутантных самцов происходило не только в
скрещиваниях самцов с самками, гомозиготными по инверсии Muller-5,
но и с самками Muller-5/+, гетерозиготными по этой же инверсии. Из
табл. 8 видно, что у мутантных самцов в скрещиваниях с гетерозиго-
тами по Muller-5 возникают не только сыновья, но и дочери. Причем
одни из них содержат инверсию Muller-5 (фенотип В/+), другие (+/+)
- не содержат.
Таблица 8
Потомство от скрещивания мутантных самцов (+7 с самками
In (1) Muller-5, w" В / у
Номер культуры самца Фенотип потомства
Дочери Сыновья
+ В/+ н?В У
1 1 7 127 46
2 11 21 47 89
4 5 12 77 167
5 22 31 36 71
6 9 26 58 62
30 31 41 86 109
31 16 33 65 79
32 42 51 60 81
33 23 23 61 88
34 6 И 44 72
36 21 25 58 108
Если мутантный самец сам содержал инверсию Су или Dichaete,
то инверсии нисколько не снижали летальное действие мутаций в его
Х-хромосоме. Мутантные самцы с инверсией Curly не имели дочерей
в потомстве точно так же, как их братья без инверсии Curly (табл. 9).
6. Модификация летальности: влияние собственно хромосомной
перестройки или генов в перестроенной хромосоме? Получив измене-
ние летального действия мутаций у гетерозигот по перестроенным хро-
мосомам, задались вопросом: это - эффект перестройки или специфи-
ческого набора аллелей в составе перестроенной хромосомы? Чтобы
ответить на него, получили перестройки в самой тестерной линии^в/Zow.
Испытали две инверсии в Х-хромосоме и две транслокации, включаю-
151
Б.Ф. Чадов
щие Х-хромосому и одну из аутосом. В табл. 10 представлены 3 пере-
стройки. Все 4 перестройки, находясь в гетерозиготе у самки, вызывали
заметное снижение летального эффекта большинства полученных мута-
ций. Резонно считать, что хромосомные перестройки сами по себе изме-
няют летальность. Следует отметить также, что причиной отсутствия
дочерей в потомстве мутантных самцов не является наличие собственно
мутации yellow у самок, с которыми мы их скрещивали. Гетерозиготные
самки с перестройками в Х-хромосоме также были самками yellow, но,
несмотря на это, имели дочерей в потомстве.
Таблица 9
Влияние комплексной инверсии Су в аутосоме самца
на пенетрантность летали в Х-хромосоме
Номер культуры самца Фенотипы потомства
Скрещивание: 2 у х <?+ Скрещивание: 2 У х <?+;Су
2 + <? У 2 + с?У
2 59 14
4 — 125 1 137
5 — 95 — 144
26 - 56 - 43
30 — 77 — 49
31 — 114 — 137
32 — 55 — 82
33 — 66 - 54
34 — .41 - 14
36 - 89 - 140
Гипотеза о механизме действия мутаций
Выделенные мутации по характеру их выражения представляют
собой условные летали. Мутации в Х-хромосоме у самца XY не де-
тальны, у самки - это типичные рецессивные летали, жизнеспособ-
ные в гетерозиготе и нежизнеспособные в гомозиготе. В том случае,
если мутантная хромосома включается в зиготу с геномом самца, му-
тация проявляет себя как доминантная деталь. Мутации в аутосоме 2
проявляют себя в зависимости от направления скрещивания как ре-
цессивные или как доминантые летали. Проявление доминантной ле-
тальности только в определенных условиях позволило назвать мута-
ции факультативными доминантными деталями.
152
Таблица 10
Потомство мутантного самца и самок yellow, содержащих перестройки в Х-хромосоме
№ ку- ль- туры с? Самки 1п(1)5/у Самки 1п(1)23/у Самки Т(1;2)19/у
5? <?<? Всего потом- ков Доля ?$ ? $ <?<? Всего потом- ков Доля ?? ?? <?<? Всего потом- ков Доля ??
I 10 159 169 0,06 1 86 87 0,01 4. 144 148 0,03
2 28 208 236 0,12 8 68 76 0,11 22 151 173 0,13
3 295 174 469 0,63 82 108 190 0,43 192 139 331 0,58
4 16 193 209 0,08 7 155 162 0,04 18 195 213 0,08
5 29 78 107 0,27 1 111 112 0,01 21 75 96 0,22
6 31 105 136 0,23 24 82 106 оДз 28 84 112 0,25
7 56 98 154 0,36 29 106 135 0,21 57 88 145 1 0,39
26 29 92 121 0,24 64 146 210 0,30 19 50 69 0,28
27 6 117 123 0,05 2 115 117 0,02 5 74 79 0,06
29 100 103 203 0,49 69 53 122 0,57 70 58 128 1 0,55
30 54 88 142 0,38 17 83 100 0,17 20 86 106 0,19
31 22 96 118 0,19 27 96 123 0,22 52 143 195 0,27
32 24 159 183 0,13 19 82 101 0,19 41 76 117 0,35
33 49 95 144 0,34 30 93 123 0,24 22 78 100 0,22
34 29 86 115 0,25 5 66 71 0,07 6 35 41 0,15
36 27 81 108 0,25 25 102 127 0,20 148 175 323 0,46
II. Общие проблемы эволюции
Б.Ф. Чадов
Основной формой проявления найденных мутаций следует все же
принять рецессивную летальность. Это состояние мутации демонст-
рируют в диплоидной зиготе при наличии гомологичной хромосомы
с активным нормальным аллелем. Состояния “нелетали” и доминант-
ной летали можно описать как варианты состояния рецессивной ле-
тальности.
Причина снятия летального действия мутаций в Х-хромосоме ока-
залась лростой: это присутствие Y-хромосомы. Скрещивание мутант-
ных самцов с самками С(1) у w f / О, содержащими сцепленные
Х-хромосомы без свободной Y-хромосомы, привело к тому, что му-
тантные самцы почти перестали появляться в потомстве (табл. 11). Без
наличия Y-хромосомы самцы в большинстве культур погибали.
Таблица 11
Влияние Y-хромосомы на летальное действие мутаций в Х-хромосоме
(скрещивание: ? С(1)у wf/O х <? +)
Номер культуры самца $>*'/ <?+ Всего потомков Доля самцов в потомстве
1 129 2 131 0,02
2 112 6 118 0,05
3 88 2 90 0,05
4 137 0 137 0,00
5 85 12 97 0,12
6 36 1 37 0,03
7 125 17 142 0,12
8 117 6 123 0,16
9 117 6 123 0,05
10 92 15 107 0,14
11 83 12 95 0,13
12 107 111 218 0,51
13 49 5 54 0,09
14 77 43 120 0,36
15 56 14 70 0,20
16 8Т 70 151 0,46
17 73 56 129 0,43
18 135 113 248 0,46
19 135 123 258 0,48
20 98 91 189 0,48
21 109 128 237 0,54
22 101 82 183 0,45
154
II. Общие проблемы эволюции
Превращение рецессивной летали в доминантную - основной фе-
номен данной работы, феномен новый и порождающий весьма нео-
бычные предположения. Доминантная летальность возникает при со-
четании мутации в отцовском геноме с “особым типом” яйцеклетки.
Изначально это были яйцеклетки линии yellow. Модификация доми-
нантной летальности перестройками в Х-хромосоме и аутосомах ясно
показала, что причиной такой формы летальности со стороны матери
является не гаплоидный хромосомный набор яйцеклетки, а весь дип-
лоидный геном матери. С большой долей уверенности можно считать,
что доминантная форма летальности обусловливается взаимодействием
цитоплазмы, привнесенной в зиготу яйцеклеткой, и гаплоидного от-
цовского генома. Я полагаю, что найденные мутации маркируют один
из самых первых этапов жизнедеятельности зиготы: начало работы
женского и мужского геномов, окруженных цитоплазматическими про-
дуктами материнского происхождения.
На рис. 3 показано объединение в зиготе (прямоугольник) гапло-
идных геномов яйцеклетки и спермия.
Одна из хромосом спермия содержит мутацию. В зиготе находятся
и многочисленные регуляторные продукты, синтезированные в комп-
лексе “яйцеклетка - трофоциты” в период созревания яйцеклетки.
Нормальным событием является посадка некоего транскрипционного
фактора на хромосому. На отцовскую хромосому он не может сесть
из-за изменения нуклеотидной последовательности в результате мута-
ции. Возникает вопрос, почему дефект отцовской хромосомы не по-
крывается нормальной работой соответствующего гена материнской
хромосомы? Ведь именно так работает диплоидный геном. Ведь имен-
но в этом состоит причина рецессивности.
Я полагаю, что на первых этапах развертывания активности мужс-
кого и женского геномов имеет место запрограммированная задержка
включения женского генома. В период времени t-t2мужской геном
находится в состоянии функциональной гаплоидии, хотя структурная
диплоидность зиготы имеет место с момента оплодотворения. В мо-
мент t( мужской геном находится в активном состоянии (отмечено тем-
ным цветом), а женский продолжает находиться в неактивном состоя-
нии (светлый цвет), как это было в яйцеклетке. В этот момент времени
регуляторный белок не может сесть ни на отцовскую хромосому, ни
на материнскую. В первом случае из-за мутантного искажения после-
155
Б.Ф. Чадов
довательности, во втором — из-за неактивности хромосомы. Результат
- мутация в отцовской хромосоме действует как доминантный летал ь-
ный фактор.
Рис. 3. Функциональная гаплоидия отцовского генома. Хромосомные наборы яйцеклет-
ки и спермия находятся в неактивном состоянии. Спермий содержит в одной из хромо-
сом летальную мутацию с факультативным доминантным проявлением. Яйцеклетка со-
держит в цитоплазме молекулы регуляторного белка, в норме садящегося на место мута-
ции. В зиготе (прямоугольник) в момент t, в активном состоянии находится только
отцовский набор хромосом (темный цвет). Несмотря на наличие обоих наборов, функци-
онально отцовский находится в гаплоидном состоянии. Молекулы регуляторного белка
не могут взаимодействовать с материнским набором из-за его неактивности, а с отцовс-
ким - из-за мутантного повреждения на месте посадки. В результате имеет место доми-
нантное проявление летальной мутации (факультативная доминантная летальность). Во
время Ц активируется материнский набор хромосом. Появляется возможность посадки
белкового фактора на материнскую хромосому. Доминантная форма действия летали прев-
ращается в рецессивную
Во время t2 материнский набор хромосом входит в состояние ак-
тивности и посадка регуляторного белка на соответствующее место
материнской хромосомы становится возможной. С этого момента та
же самая мутация в отцовской хромосоме перестает быть доминант-
ной деталью и становится рецессивной деталью.
С биологической точки зрения временная функциональная гапло-
идия мужской половины генома на ранней стадии развития имеет боль-
шое значение. Это состояние фактически означает проверку мужского
генома на соответствие стандарту. Стандартом является геном матери.
156
II. Общие проблемы эволюции
В группу проверяемых входят гены, активирующиеся самыми первы-
ми и поэтому ключевые с точки зрения развития. В это время прохо-
дит сверка как раз той инвариантной части генома, о которой шла речь
в начале статьи. Именно на данном этапе должны элиминироваться
мутации по генам этой группы.
О свойствах мутаций, делающих их перспективными
для видообразования
Перечислим свойства мутаций, имеющих отношение к процессу
видообразования.
1. Полученные мутации, судя по частоте образования, локализа-
ции в разных хромосомах и отличиям в проявлении, составляют це-
лый класс мутаций. Главной чертой мутаций является факультативная
доминантная летальность. Существование подобных мутаций согла-
суется с концепцией разделения генома на вариабильную и неизменя-
емую части. Факультативные доминантные летали, согласно этой кон-
цепции, являются мутациями инвариантной части видового генома.
2. Летали с факультативным доминированием обеспечивают изо-
ляцию особи самым эффективным способом: гибелью потомка на ста-
дии зиготы.
3. Существуют способы снятия изоляции. Одним из них является
использование самок, содержащих хромосомные перестройки. Спо-
собность перестроек снимать летальность мутаций объясняет давно
известную связь между видообразованием и наличием новой пере-
стройки у особей образовавшегося вида.
4. Снятие летальности обусловлено действием перестройки в пи-
тательных клетках, что согласуется с ранее обнаруженной связью меж-
ду актом видообразования и порядком расположения хромосом в яд-
рах питающих клеток [5].
5. Гены, по которым найдены мутации, относятся к генам раннего
развития. Последние определяют основные черты устройства особи -
черты, отличающие организмы разных таксонов от вида и выше.
6. Гены, по которым найдены мутации, судя по картине их проявле-
ния, являются регуляторными генами. Мутации по этим генам ставят
организм на грань жизни и смерти и инициируют процесс залечива-
ния образовавшегося дефекта.
157
Б.Ф. Чадов
7. Мутации одномоментно разрушают старую систему регуляции, не
создавая новой. Однако последняя может постепенно возникнуть в про-
цессе ликвидации образовавшегося дефекта регуляторной системы.
8. Метастабильное состояние регуляторной системы, вызванное
мутированием регуляторного гена, может явиться фактором, иниции-
рующим видообразование.
Модель видообразования
Существование сложных генетических программ индивидуально-
го развития с неизбежностью приводит к выводу о существовании
программного генетического обеспечения филогенеза. С биологичес-
кой точки зрения задача наращивания видового разнообразия, экспан-
сии живого не менее актуальна, чем создание самой сложной програм-
мы индивидуального развития. Растет количество данных о тесной
связи и переплетениях этих двух программ [4].
С эволюционно-генетической точки зрения геном организма мож-
но разделить на две части: вариабильную и консервативную. Мутаци-
онный процесс в первой приводит к образованию аллельных вариан-
тов, существующих в гетеро- и гомозиготном состоянии. Аллельные
изменения таковы, что они не меняют видовой принадлежности осо-
би. Мутационный процесс в этой части генома ведет к образованию
только внутривидового разнообразия. К процессу образования новых
видов он прямого отношения не имеет [3,5].
Консервативная часть генома включает в себя исторически сложив-
шуюся программу онтогенетического развития особей данного вида.
Сложность и взаимосвязанность этой программы такова, что одиноч-
ное мутационное событие всегда оказывается летальным. Это либо
рецессивная, либо доминантная деталь. Жизнеспособных вариантов,
аллелей, в обычном понимании этого слова, при мутировании этой
части генома не возникает.
Признаком повреждения в консервативной части генома является
образование факультативной доминантной летальной мутации. Судь-
ба такой мутации двоякая. Если она возникла в структурно-нормаль-
ном геноме или если мутант скрещивается с особями, имеющими струк-
турнонормальный геном, она исчезает из популяции, поскольку гете-
розигота по мутации гибнет. Таким путем сохраняется постоянство
158
IL Общие проблемы эволюции
вИда. Если мутация возникла в структурно перестроенном геноме или
получает возможность скреститься с партнером, имеющим структур-
но перестроенный геном, она получает шанс на выживание и даже
закрепление в популяции. Комплекс “доминантная деталь + перестрой-
ка” являет собой генетическую конструкцию, на основе которой мо-
жет пойти формирование нового вида.
Какое новое качество имеет этот комплекс по сравнению с обыч-
ными мутациями, чтобы его считать перспективным для видообразо-
вания? Глубокий дефект регуляционной системы, поставивший организм
на грань гибели. Столбовая дорога регуляции оказалась разрушенной в
результате мутации. Я полагаю, что серьезное нарушение, сохранившее-
ся, но поставившее организм на грань гибели, побудит систему к измене-
нию с перспективой найти новое устойчивое состояние.
Традиционно считается, что импульсом к видообразованию явля-
ется изменение условий среды. Несмотря на широкое хождение, этот
тезис не верен. Нет никаких доказательств того, что только стратегия
к изменению организмов вслед за изменениями среды является выиг-
рышной. Существование на данный момент времени иерархии видов
от простейших до сложнейших говорит о том, что простейшие виды
ничего не потеряли, оставшись равнодушными к имевшим место из-
менениям среды. Они так же существуют, и их биомасса не меньше, а
больше биомассы высших. Во всяком случае, изменения среды, имев-
шие место на Земле, не явились императивом к изменению организ-
мов. Если бы это было так, биосфера была бы представлена неболь-
шим числом видов и задача систематики явно упростилась бы.
Возникает вопрос, если не среда, то что толкает систему к измене-
нию? Я полагаю, что таким толчком является событие внутри самой
живой системы. Это - образование доминантной летальной мутации в
регуляторном гене, ставящей организм на грань жизни и смерти и в то
же время не элиминирующейся сразу из популяции.
Опыты с влиянием хромосомных перестроек на летальное действие
найденных мутаций показывают, что перестройки имеют первостепен-
ное значение для создания в популяции очагов видообразования. Совре-
менная литература располагает хорошо аргументированным доказатель-
ством причастности перестроек к видообразованию и в то же время не
может объяснить, как это происходит. По нашим данным, их роль состо-
ит в изменении работы материнского генома по формированию програм-
159
Б.Ф. Чадов
мы развития нового организма. Иными словами, хромосомные перестрой-
ки существенным (но пока неизвестно каким) образом меняют деятель-
ность генома питающих клеток, функция которых состоит в наполнении
яйцеклетки РНК и белками. В то время как присутствие перестройки
мало или совсем не сказывается на работе диплоидного генома в зиготе,
оно резко меняет работу диплоидного генома питающей клетки. Питаю-
щие клетки наполняют яйцеклетку регуляторными продуктами для обес-
печения раннего развития зиготы.
Полученные результаты с неожиданной стороны входят в контакт
с результатами и теорией видообразования В.Н. Стегния [5]. В его ра-
ботах была обнаружена связь между видообразованием у комаров и
изменением расположения хромосом в питающих клетках женской
генеративной ткани. Было предположено, что процесс видообразова-
ния связан с перестройками, изменяющими расположение хромосом
в питающих клетках. Такие перестройки Стегний назвал “системны-
ми мутациями”, поскольку они меняют старую систему регуляции на
новую.
Генетические данные, представленные выше, приводят к выводу о
важной роли хромосомных перестроек в работе генома питающих
клеток. Таким образом, данные Стегния и наши данные связывают
видообразование с событиями, происходящими в женской генератив-
ной ткани. Однако по гипотезе Стегния, изменения регуляции в пита-
ющих клетках достаточно для генерации вида. С моей точки зрения,
этого недостаточно. Должна быть мутация в регуляторном гене, при-
надлежащем к особой группе регуляторных генов. Она может быть и
“точковой”. Хромосомные перестройки, изменяя работу питающих
клеток, уберегают мутацию в регуляторном гене от немедленной эли-
минации. Сами по себе хромосомные перестройки - это в большин-
стве случаев “разрешенные” мутации, и инициировать видообразова-
ние они не могут.
Момент образования комплекса “мутация в регуляторном гене +
перестройка” - это только начало процесса видообразования. При бла-
гоприятном исходе генетическая конструкция путем повторных актов
мутирования может восстановить свою работу, “регенерировать”. По-
казателем окончания регенерации явится потеря летального эффекта
комплекса и его огомозигочивание. Вероятность того, что произойдет
восстановление прежней структуры и прежней системы регуляции,
160
II. Общие проблемы эволюции
очень мала хотя бы потому, что в процесс переустройства регуляции
оказались вовлеченными хромосомные перестройки, ранее не бывшие
обязательным элементом генома. Возникнет нечто иное. С большой
долей вероятности это будет вариант регуляторной системы, отлич-
ный от старого, иначе говоря, претендент на новый вид.
В.А. Ратнером и Л.А. Васильевой с сотрудниками разрабатывает-
ся концепция, согласно которой серьезная перестройка регуляторной
системы генома происходит в результате перемещения мобильных эле-
ментов [6]. Не исключено, что этот процесс способствует функцио-
нальному залечиванию дефекта, вызванного мутацией в регуляторном
гене.
Знаменательным моментом в процессе залечивания дефекта долж-
но быть образование гомозиготной самки. В скрещивании с гетерози-
готным самцом появятся гомозиготные потомки, которые смогут да-
вать потомство, скрещиваясь только внутри себя. Поскольку заготов-
ки в виде гетерозиготных вариантов будут еще находиться в популяции,
возможны повторные случаи появления гомозигот.
Принципиальными моментами предлагаемой модели видообразо-
вания являются скрытая сборка нового вида и представление его как
нового организма уже в готовом виде. Синтетическая теория предпо-
лагает создание вида по частям, каждая из которых в составе организ-
ма предварительно обкатывается отбором. В предлагаемой модели
стимулом к видообразованию являются определенные изменения в
самой живой системе, но не воздействия внешней среды, не ее изме-
нения, не приспособление организма к условиям среды, как это пред-
писывается синтетической теорией видообразования.
Новый вид представляется среде в виде группы особей, содержа-
щей указанную конструкцию в гомозиготном состоянии. После появ-
ления первых представителей нового вида начинается популяционный
и биосферный этап в его становлении. Образование нового вида мож-
но сравнить с вынашиванием плода в организме матери, где матерью
является вся популяция, а плодом - организмы, содержащие генети-
ческую конструкцию “мутация регуляторного гена + перестройка”.
На популяционном (биосферном) этапе взаимоотношения между
кандидатом на новый вид и средой явятся определяющими: либо кан-
дидат окажется удачливым реципиентом солнечной энергии в той или
иной ее форме и, размножившись, станет видом, либо погибнет. Успех
161
Б.Ф. Чадов
претендента на новый вид или иной таксон зависит от того, насколько
он способствует выполнению энергетической задачи, стоящей перед
биосферой в целом. Во всяком случае, процесс видообразования пред-
ставляет собой многоэтапный и эволюционно закрепленный способ
создания новых форм. В нем есть генетическая и биосферная части.
Представленная модель прописывает только несколько этапов в гене-
тической части видообразовательного процесса. Мое глубокое убеж-
дение состоит в том, что шаг за шагом мы сможем восстановить длин-
ную цепь генетических и биосферных событий, ведущих к образова-
нию нового вида. Задача генетиков - понять, какие классы генов
мутируют и в какой последовательности, чтобы одно стабильное со-
стояние регуляторной системы через фазу нестабильности могло пе-
рейти в другое стабильное состояние.
Автор благодарит Е.В. Чадову и Н.Б. Федорову за сотрудничество
в выполнении экспериментальной части работы и С.А. Копыла за по-
мощь при подготовке рисунков.
Работа поддержана грантом РФФИ.
Литература
1. Шмальгаузен И.И. Факторы эволюции (теория стабилизирующего отбора). М.: На-
ука, 1968. 451 с.
2. Чадов Б.Ф., Чадова Е.В., Копыл С.А., Федорова Н.Б. Новый класс мутаций у Drosophila
melanogaster И Доклады РАН. 2000. Т. 373. № 5. С. 714-717.
3. Алтухов Ю.П. Генетические процессы в популяциях. М.: Наука, 1989. 328 с.
4. Корочкин Л.И. Введение в генетику развития. М.: Наука, 1999. 254 с.
5. Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск:
Изд-во НГУ, 1993. 110 с.
6. Ratner V.A., Vasilyeva L. A., Bubenshchikova Е. V. Genetic diversity of interacting patterns
of mobile genetic elements (MGES) and polygenic systems: induction with stress factors,
response to selection and possible evolutionary consequences // Proc. Intern. Conf
Biodiversity and Dynamics of Ecosystems in North Eurasia, August 21-26. 2000
Novosibirsk, Russia. VI. Part 1. P. 97-98.
Ш. ЭВОЛЮЦИЯ РАСТЕНИИ
ПРОИСХОЖДЕНИЕ В-ХРОМОСОМ У РАСТЕНИЙ.
ТЕОРЕТИЧЕСКИЙ И ЭКСПЕРИМЕНТАЛЬНЫЙ
ПОДХОДЫ
В.А. Бердников, Ф.Л. Горель, О.Э. Костерин, В.С. Богданова
Институт цитологии и генетики СО РАН
630090, г. Новосибирск, пр. Лаврентьева, 10
E-mail: berdnik@bionet.nsc.ru
В-хромосомы отмечены у около 15% исследованных видов живот-
ных и растений [1]. Эти необязательные для жизни организма сверх-
численные хромосомы обычно присутствуют только у части особей
одной популяции и отличаются от хромосом основного набора (А-хро-
мосом) такими свойствами, как: 1) меньший размер; 2) гетерохрома-
тичность; 3) отсутствие спаривания с А-хромосомами в мейозе; 4) от-
сутствие генов с качественным эффектом; 5) слабое влияние на фено-
тип и приспособленность организма; 6) повышенная склонность к
попаданию в клетки зародышевого пути, что у растений чаще всего обус-
ловлено нерасхождением хроматид в одном из митотических делений
микроспоры; 7) нерасхождение в митозе соматических клеток, что ведёт
к отличиям в числе В-хромосом в разных клеточных линиях одного орга-
низма; 8) влияние на уровень рекомбинации А-хромосом.
В каждом конкретном случае сверхчисленная хромосома может
быть лишена некоторых из этих свойств, однако три из них всегда
сохраняются [1]: 1) необязательность присутствия в геноме; 2) отно-
сительная безвредность для организма; 3) отсутствие рекомбинации
с А-хромосомами.
Наиболее вероятно происхождение сверхчисленных хромосом от
каких-то элементов основного набора [2]. Особенно правдоподобным
представляется происхождение В-хромосом у многих животных от
гетероморфных половых Y- или W-хромосом [3], имеющих многие из
163
В.А. Бердников, Ф.Л. Горелъ, О.Э. Костерин, В.С. Богданова
отмеченных выше свойств сверхчисленных хромосом. Как и В-хро-
мосомы, они обычно малы и гетерохроматичны и, главное, обладают
очень небольшим числом генов, определяющих пол или фертильность
гетерогаметной особи. Хорошо известно, что присутствие одной
Y-хромосомы у самок Drosophila melanogaster практически не отра-
жается ни на фенотипе, ни на фертильности и скорости развития мух,
хотя добавление большего числа Y-хромосом снижает приспособлен-
ность [4].
Д. Грин с соавторами [5,6] убедительно показали, что у новозелан-
дской реликтовой лягушки Leiopelma hochstetteri многообразные сверх-
численные хромосомы произошли от унивалентной W-хромосомы за
время, истекшее после последнего оледенения. Столь же убедитель-
ным выглядит и почти мгновенное возникновение В-хромосом от
Х-хромосомы у насекомых из отряда Orthoptera [7].
Заметим, что, принимая происхождение сверхчисленных хромосом
от половых, мы переносим возникновение главных особенностей пер-
вых, таких, как отсутствие генов с качественным эффектом, гетерох-
роматичность и отсутствие рекомбинации с гомологичными сегмен-
тами нормального кариотипа, на этап дифференциации половых хро-
мосом. Предполагается, что изначально гомоморфная Y-хромосома
отличалась от Х-хромосомы лишь аллелями по малому числу генов,
определяющих пол. Далее должен был действовать какой-то механизм,
ответственный за дегенерацию только Y-хромосомы. Важно отметить,
что при наличии свободной рекомбинации с гомологичной Х-хромо-
сомой любая (даже повышающая приспособленность) нуль-мутация в
Y-хромосоме с высокой вероятностью будет замещаться нормальным
аллелем, находящимся в Х-хромосоме. В то же время если рекомбина-
ция между половыми хромосомами отсутствует, то накопление мута-
ций только в одной из них можно объяснить с помощью совершенно
нейтрального механизма, предложенного Мёллером [8]. Суть механиз-
ма “храповика Мёллера“ состоит в том, что в силу одного лишь дрей-
фа в каждом поколении популяции конечных размеров существует
определённая вероятность потери элитного класса Y-хромосом с наи-
меньшим числом вредных мутаций. Продолжительное действие этого
механизма способно привести в конце концов к генетической деграда-
ции Y-хромосомы [9]. Следует заметить, что реальный процесс эво-
164
IIJ. Эволюция растений
люции гетероморфной пары половых хромосом много сложнее, по-
скольку потеря функционирующих генов одной из них нарушает ген-
юяй баланс, что должно восполняться развитием механизма дозовой
компенсации [10]. Тем не менее, основным предусловием для возник-
новения генетически обеднённой хромосомы является подавление ре-
комбинации с её гомологом.
Для объяснения эволюции В-хромосом тоже прибегают к механизму
храповика Мёллера [3]. Анализируя кариотипы лягушки L. hochstetteri,
Грин с сотрудниками [5, 6] обнаружили в популяциях о. Северный
мноюобразные и многочисленные В-хромосомы, проявляющие явное
морфологическое сходство с W-хромосомой, определяющей женский
пол. Всего за 10 тысяч лет у этих лягушек произошла смена системы
детерминации пола с ZW к W0 с возникновением гипервариабельных
W- и В-хромосом. Повышенное разнообразие W-хромосом Грин с со-
авторами объясняют её рекомбинационной изолированностью, позво-
ляющей накапливать изменения под действием храповика Мёллера.
Однако следует заметить, что эффективность этого механизма довольно
низка, поскольку на один щелчок храповика уходит в среднем не ме-
нее тысячи поколений [11]. Для объяснения высокой скорости эволю-
ции кариотипа L. hochstetteri авторы прибегают к предположению о
присутствии в будущей В-хромосоме повторённых последовательно-
стей ДНК, способных быстро распространяться по хромосоме и тем
самым снижать гомологию с исходной W-хромосомой [5, 6]. Наибо-
лее важным представляется самый первый шаг возникновения В-хро-
мосомы, хотя её сходство с унивалентной W столь высоко, что для
превращения последней в В-хромосому было бы достаточно одной
мутации, инактивирующей ген, детерминирующий женский пол; пос-
ле такой инактивации просто возник бы самец с В-хромосомой [5].
Учитывая очень маленький размер унивалентной W-хромосомы и ее
внутри- и межпопуляционную гипервариабельность, весьма вероят-
но, что даже самки в потомстве такого самца будут обладать В-хромо-
сомой, исключенной из рекомбинации с самого момента своего воз-
никновения.
Возникает вопрос, какой механизм может обеспечить появление
сверхчисленных хромосом у растений, как правило, лишённых поло-
вых хромосом? Данная работа посвящена попытке ответить на этот
вопрос.
165
В.А. Бердников, Ф.Л. Горель, О.Э. Костерин, В.С. Богданова
Теоретический подход к проблеме
Если В-хромосомы происходят от А-хромосом того же вида, то на
первой фазе их эволюции должно наблюдаться переходное гиперплоид-
ное состояние генома. Оставив без рассмотрения менее вероятные, на
наш взгляд, варианты происхождения через фрагментацию А-хромосом
или межвидовой перенос [1,2], остановимся на двух: 1) первичная три-
сомия, когда одна хромосома нормального кариотипа присутствует в трёх
копиях; 2) третичная трисомия, когда к нормальному кариотипу добав-
лена обменная экстрахромосома, состоящая из сегментов, происходя-
щих от двух разных хромосом нормального кариотипа.
В литературе уже более полувека продолжается спор о значении
В-хромосом для организма: повышают ли они приспособленность или,
напротив, являются терпимыми внутриядерными паразитами [I].
Уклоняясь от дискуссии по этому вопросу, лишь отметим, что присут-
ствие в геноме одной В-хромосомы обычно нельзя обнаружить ни по
внешнему виду растений, ни по скорости их развития, ни по фертиль-
ности. В то же время трисомики, как правило, уступают нормальным
диплоидам по всем показателям приспособленности, при этом неред-
ко трисомия по каждой хромосоме набора приводит к совершенно
конкретному изменению фенотипа растения [12]. По-видимому, меж-
ду дозами некоторых генов, расположенных в разных частях генома,
должно выполняться строгое соотношение, и изменение этого балан-
са ведёт к серьёзным последствиям для процессов развития, как пра-
вило, уменьшая приспособленность организма. Ясно, что присутствие
в трисомной хромосоме генов, чувствительных к дозовому дисбалан-
су, представляет собой серьёзное препятствие для возникновения
В-хромосом, поскольку эволюция, направляемая естественным отбо-
ром, не может идти по пути, сопряженному с падением приспособлен-
ности. Отсюда следует, что фиксация трисомиков должна осуществ-
ляться с помощью селективно-нейтральных механизмов.
Первичная трисомия по любой хромосоме возникает вследствие
нерасхождения гомологов достаточно регулярно с частотой порядка
0,001 [13], однако трудно представить, чтобы экстрахромосомы пер-
вичных трисомиков могли лежать в основе возникновения В-хромо-
сом. С точки зрения рассматриваемой проблемы нужно, чтобы состо-
яние трисомии передавалось потомкам, и при этом одна и та же из
166
III. Эволюция растений
трёх хромосом была постоянно исключена из рекомбинационного про-
цесса. В принципе, это было бы возможно, если бы одна из хромосом
несла инверсию, а две другие - леталь. Однако вероятность совпаде-
ния во времени столь редких событий (трисомия, инверсия и леталь)
слишком мала, и трудно предложить сценарий последовательного свер-
шения этих событий.
Много предпочтительнее в качестве источника происхождения
В-хромосом выглядит третичная трисомия. В этом случае экстрахромо-
сома является продуктом соединения фрагментов двух негомологичных
хромосом. Третичные трисомики возникают с довольно высокой часто-
той (до 10-20%) в потомстве гетерозигот по транслокациям. Такие гете-
розиготы весьма обычны в природных популяциях растений [12,13,14].
Биологический смысл этого явления не до конца понятен, возможно, оно
является следствием склонности многих видов растений к обмену хро-
мосомными плечами, хотя не исключено, что для изолированных попу-
ляций гетерозиготность по транслокациям в какой-то степени снижает
инбредную депрессию. На стадии зиготены-пахитены мейоза спарива-
ние гомологичных районов четырёх вовлечённых в транслокацию хро-
мосом ведёт к образованию крестообразной фигуры с центром креста
вблизи точек транслокационных разрывов (ВР) (рис. 1).
Рис. 1. Схема возникно-
вения третичного три-
сомика из гетерозиготы
по транслокации: N1,
N2 - хромосомы с ис-
ходным порядком ге-
нов; Tl, Т2 - обменные
хромосомы. Штрихо-
вой линией обозначен
Н-район основного на-
бора, гомологичный эк-
страхромосоме. а - ис-
ходная гетерозигота; Ь,
с - гаплоидные гамето-
фиты; d- конкурентос-
пособный гиперплоид-
ный гаметофит; e-g -
Диплоидные зиготы; h -
трисомная зигота
167
В.Л. Бердников, Ф.Л. Горель, О.Э. Костерин, В.С. Богданова
Такой крест образован двумя хромосомами с исходным порядком
генов (Nl, N2) и двумя обменными хромосомами (Tl, Т2). В анафазе
1 нормальное (2:2) расхождение хромосом креста нередко заменяет-
ся на расхождение 1:3, что влечёт образование четырёх вариантов ги-
перплоидных гамет (Nl, N2, Tl), (Nl, N2, Т2), (Tl, Т2, Nl), (Tl, Т2,
N2), слияние которых с эуплоидными гаметами приведёт к возникно-
вению трисомных зигот. Если обменные хромосомы отличаются по
длине, то, как правило, чем короче обменная хромосома, тем выше
шансы её прохождения через стадию гаметофита и тем выше фер-
тильность и жизнеспособность соответствующего третичного трисо-
мика. В связи с этим мы ограничимся рассмотрением третичных три-
сомиков, имеющих в качестве экстрахромосомы малую обменную
хромосому (Т1). Кроме того, дополнительная хромосома порядком
своих генов отличается от хромосом основного набора, и в этом слу-
чае создаются определённые препятствия для вовлечения экстрахро-
мосомы в рекомбинацию. Таким образом, наибольший интерес пред-
ставляют трисомики, кариотип которых содержит неперестроенные
хромосомы основного набора плюс малая обменная хромосома (N1/
Nl, N2/N2, Т1). Такие трисомики будем называть далее перспектив-
ными трисомиками.
Для удобства изложения хромосомные сегменты основного набо-
ра, гомологичные малой обменной хромосоме, будем называть Н-рай-
оном генома. Замечательной особенностью транслокационных гете-
розигот является снижение рекомбинации вблизи ВР, что, вероятно,
объясняется локальным нарушением гомологичного спаривания. Если
в район слабого спаривания попадает пара рецессивных спорофитных
деталей (одна в N-, другая в Т-хромосомы), то гетерозиготность по
транслокации закрепляется, так как обе структурные гомозиготы по-
гибают. Однако кроме гетерозигот выживают и перспективные трисо-
мики с конституцией Nl/Nl, N2/N2, Т1; более того, эти трисомики
могут размножаться в чистоте.
Следует заметить, что в природных популяциях нескольких видов
Clarkia с высокой частотой встречаются растения со сверхчисленны-
ми хромосомами, причём встречаемость экстрахромосом коррелиру-
ет со встречаемостью транслокационных гетерозигот [14]. Третичные
трисомики С. unguiculata, спонтанно возникшие от самоопыления
168
III. Эволюция растений
транслокационных гетерозигот, были изучены X. Льюисом [15]. Экст-
рахромосомы трисомиков вели себя в мейозе так же, как сверхчислен-
ные хромосомы природных популяций [17].
Если в популяции с высоким уровнем гетерозиготности по транс-
локации часто происходят резкие сокращения численности, то с до-
вольно высокой вероятностью может происходить фиксация перспек-
тивных трисомиков, несущих в Н-районе спорофитную деталь. Такие
трисомики должны выживать и размножаться, поскольку деталь пере-
крывается нормальным аллелем экстрахромосомы. По своей приспо-
собленности трисомики должны несколько уступать породившей их
транслокационной гетерозиготе, однако при хорошей изолированнос-
ти популяции выжившему трисомику предоставляется некоторое вре-
мя для самостоятельной эволюции.
Теперь проследим за судьбой рецессивной мутации по гену, пред-
ставленному в клетках спорофита трисомика тремя копиями - две в хро-
мосомах Н-района и одна в экстрахромосоме. Сначала рассмотрим класс
гаметофитных мутаций (деталей и полулеталей), существенно снижаю-
щих жизнеспособность или конкурентоспособность гамет. Трисомик
производит два типа гаметофитов: 1) обычные гаплоидные (без экстрах-
ромосомы) и 2) гиперплоидные, в которых каждый ген Н-района пред-
ставлен своей копией в экстрахромосоме. Если новая гаметофитная му-
тация инактивировала один из генов Н-района, то, попав в обычный гап-
лоидный гаметофит, она не достигнет зиготы и будет потеряна
популяцией. Если же такая мутация попадёт в гиперпловдный гамето-
фит, содержащий неповреждённую копию того же гена в экстрахромо-
соме, то, пройдя через гамету, она попадёт в геном трисомного потомка.
В следующем поколении мутантный ген может снова попасть в гаплоид-
ный гаметофит и исчезнуть из популяции. Поскольку вероятность поте-
ри мутации на стадии гаметофита соизмерима с 50%, то гаметофитная
мутация по Н-району должна довольно скоро покинуть популяцию.
Иное дело, если гаметофитная мутация затронула ген, расположен-
ный в экстрахромосоме. В данном случае копия этого гена в Н-районе
остаётся неповреждённой, поэтому новая мутация не должна снижать
приспособленность гиперплоидного гаметофита, более того, скорее все-
го эта мутация её даже повысит, так как приблизит набор неповрежден-
ных генов гаметофита к оптимальному гаплоидному состоянию. Однако
169
В.А, Бердников, Ф.Л. Горель, О.Э. Костерин, В. С. Богданова
с помощью рекомбинации мутация, возникшая в обменной хромосоме,
может попасть в Н-район основного набора и вскоре после этого поки-
нуть популяцию. Если же кроссинговер между экстрахромосомой и
Н-районом подавлен, то ничто не препятствует накоплению в экстрахро-
мосоме гаметофитных мутаций с помощью храповика Мёллера.
Если новая рецессивная мутация строго спорофитная, то есть она
нарушает развитие растения только на фазе спорофита, совершенно не
затрагивая гаметофит, то легко пройдя через гамету, она попадёт в три-
сомную зиготу. Удаление такой мутации из перекрёстно-опыляющейся
популяции трисомиков весьма затруднительно, так как она перекрыта
нормальным аллелем экстрахромосомы. Однако известно, что на стадии
гаметофита экспрессируется значительная доля генов спорофита (по не-
которым оценкам не менее 60% [16]), поэтому большая часть спорофит-
ных мутаций должна отрицательно сказаться на жизнеспособности или
конкурентоспособности мужского гаметофита и довольно быстро уда-
ляться из популяции. Исключение составят лишь мутации, попавшие в
рекомбинационно изолированную экстрахромосому, поскольку их при-
сутствие в этом случае не приведёт к снижению конкурентоспособности
гиперплоидных гаметофитов. Таким образом, если скорость мутирова-
ния генов экстрахромосомы будет выше скорости удаления мутантных
генов за счёт кроссоверной рекомбинации, то в популяции трисомиков
должен идти процесс генетической дегенерации экстрахромосомы. Важно
отметить, что каждая мутация, связанная с потерей или инактивацией
гена в экстрахромосоме, должна в большей или меньшей степени повы-
шать жизнеспособность трисомного спорофита и/или конкурентоспо-
собность мужского гиперплоидного гаметофита, приближая их геномы
к эуплоидному состоянию, так что к храповику Мёллера присоединяет-
ся более мощный фактор - естественный отбор. Наиболее благоприят-
ными здесь должны оказаться мутации, связанные с инактивацией ге-
нов, чувствительных к генному дисбалансу.
Можно возразить, что область сниженной рекомбинации ограни-
чивается лишь небольшим районом, непосредственно примыкающим
к ВР, однако заметим, что в этом районе со временем следует ожидать
появления структурных мутаций, ослабляющих спаривание в сосед-
них, более удалённых от ВР районах хромосомы. В конце концов экст-
рахромосома на всём своём протяжении потеряет способность к спа-
риванию с Н-районом.
170
III. Эволюция растений
Мы провели компьютерную симуляцию процесса деградации эк-
страхромосомы. Величина перекрёстной популяции трисомиков рав-
^ядась 50 индивидуумам, экстрахромосома содержала 200 локусов. Как
й ожидалось, в условиях, когда скорость добавления мутаций в экст-
пахромосоме превышала скорость их удаления за счет рекомбинации,
наблюдалось накопление мутаций только в экстрахромосоме (рис. 2),
что подтверждает правильность приведенных выше рассуждений.
Рис. 2. Компьютерная модель
накопления мутаций в экстрах-
ромосоме трисомика при по-
давлении рекомбинации. G -
число поколений; п - среднее
число мутаций на хромосому.
Условия машинного экспери-
мента: число особей - 50; чис-
ло генов в экстрахромосоме -
200; частота мутирования -
10-4 на ген на поколение. Ре-
комбинационная длина экст-
рахромосомы - 0,01 сМ
Интересно, что такая особенность В-хромосом, как её преимуще-
ственное попадание в гаметы, может быть объяснена в рамках пред-
лагаемой модели. Дело в том, что значительную часть потомства пер-
спективных трисомиков составляют диплоиды, погибающие в ходе
развития, так как гомозиготны по спорофитной летали, зафиксиро-
ванной ещё предками, гетерозиготными по транслокации. Дефицит
плодовитости трисомиков может быть уменьшен при снижении доли
диплоидных зигот, возникающих от слияния гаплоидных гамет. В этой
ситуации естественный отбор будет способствовать появлению в эк-
страхромосоме мутаций, повышающих вероятность её попадания в
гамету. Когда популяция, развившаяся из трисомиков, войдёт в кон-
такт с популяцией того же вида, представленной диплоидами с нор-
мальным порядком генов, частично деградированная унивалентная
экстрахромосома, несущая мутации такого рода, может удержаться в
новом геноме, превратившись в В-хромосому.
171
В.А. Бердников, Ф.Л. Горелъ, О.Э. Костерин, В.С. Богданова
Экспериментальный подход к проблеме
Как показывает приведённый выше анализ, самым принципиаль-
ным пунктом в происхождении В-хромосомы является рекомбинаци-
онная изоляция экстрахромосомы перспективного трисомика от ос-
тального генома. Чтобы подвергнуть эту проблему эксперименталь-
ной проверке, мы получили у гороха на основе транслокации
Хаммарлунда жизнеспособные трисомики и провели генетическую
маркировку Н-района (т.е. района основного набора, гомологичного
экстрахромосоме) рецессивными мутациями. Мы выбрали посевной
горох в качестве объекта из двух соображений: во-первых, он относи
тельно неплохо изучен генетически, и, во-вторых, В-хромосомы не
отмечены ни у В sativum, ни у близкородственных видов, поэтому едва
ли в геноме гороха сохранились какие-то элементы, которые служили
ранее для поддержания сверхчисленных хромосом и могли бы спо-
собствовать успеху в синтезе и поддержании экстрахромосомы.
Открытая в 1927 г. транслокация Хаммарлунда является первой
генетически изученной транслокацией. Показано, что у гетерозигот
по этой транслокации кроссоверная рекомбинация между маркерами,
фланкирующими точку разрыва, уменьшена более чем на порядок [20].
Уже в работах 1939-1940 гг. [18, 19] было показано, что в потомстве от
самоопыления гетерозиготы по транслокации Хаммарлунда с довольно
высокой вероятностью (до 18%) возникают трисомики, кариотип кото-
рых представлен семью парами хромосом стандартного набора и малой
обменной хромосомой, состоящей из коротких плеч хромосом 2 и 5 (здесь
и далее номера хромосом совпадают с номерами групп сцепления). Хотя
по жизнеспособности эти трисомики уступают своим диплоидным со-
братьям, их нетрудно выращивать, и они вполне фертильны. Таким об-
разом, у нас были все основания считать эти трисомики перспективны-
ми. В 1992 г. мы получили трисомное растение TRIS-54, у которого обе
хромосомы 5 несут в своём Н-районе рецессивную мутацию cri, при этом
аллель дикого типа Cri находится в экстрахромосоме недалеко от точки
транслокационного разрыва ВР (рис. 3).
Мутация cri резко нарушает строение листьев, поэтому в потом-
стве трисомиков диплоидные растения с генотипом cri/cri можно было
уже на стадии проростков легко отличить от трисомиков с генотипом
cri/cri/Cri. Доля трисомиков в таких потомствах колебалась от 10 до
172
III. Эволюция растений
Рис. 3. Схема наследования эксграхромосомы в линии третичных трисомиков го-
роха TRIS. Цифрами обозначены: 2 - хромосома 2; 5 - хромосома 5; 52 - малая
обменная хромосома, состоящая из коротких плеч хромосом 2 и 5. Штриховой
линией обозначен Н-район основного набора, гомологичный экстрахромосоме.
а - исходный третичный трисомик; b - эуплоидный гаметофит; с - гиперплоид-
ный гаметофит; d-эуплоидный гамегофит, несущий рекомбинантную хромосому.
Кроссинговер прошел между точкой разрыва и маркером cri; г - нормальный дип-
лоид cri/cri\f- трисомик cri/cri/Cri, g - кроссоверный диплоид cri/Cri
30%. Дополнительным свидетельством трисомии были признаки, от-
меченные Е. Саттон [18]: сильно увеличенные прицветники, увеличен-
ное число цветков в соцветии, волнистые края прилистников и лис-
точков. В сомнительных случаях проводился анализ бивалентов в ма-
теринских клетках пыльцы (МКП). Трисомное потомство исходного
TRIS-54 было многократно размножено на протяжении нескольких
поколений, при этом среди 1052 диплоидов мы зафиксировали 4 крос-
совера с фенотипом Cri (остальные диплоиды имели фенотип cri), что
173
В.А. Бердников, Ф.Л. Горелъ, О.Э. Костерин, В. С. Богданова
свидетельствует о рекомбинационной дистанции между cri и ВР, рав-
ной 0,19 сМ.
Для введения дополнительных маркеров в Н-район мы скрестили
трисомик TRIS с диплоидным растением с генотипом cri U" / cri
Доминантный аллель Ust локуса U также находится в пятой хромосо-
ме в 20 сМ от cri [21], он не снижает приспособленности и вызывает
появление на семенной кожуре характерных антоциановых пятен, тог-
да как у семян растений с генотипом и/и таких пятен нет. У исходного
трисомика TRIS-54 все три копии этого локуса заняты рецессивным
аллелем и. Из F1 от скрещивания был отобран трисомик, ставший
основателем изогенной линии трисомиков, получившей название
TRUST.
Одно растение из F13 было размножено в течение нескольких по-
колений до нескольких десятков тысяч, все они - и дисомики и трисо-
мики - имели признак Ust, что, скорее всего, свидетельствовало о том.
что все три копии гена U представлены аллелем U5t. Доля трисомиков
в этой линии составляет около 24,0%, тетрасомиков в потомстве три-
сомиков обнаружено не было. В реципрокных скрещиваниях линии
TRUST с тестерной линией, маркированной рецессивными аллелями
cri, мы установили, что дополнительная хромосома передается с час
тотой около 25,5% через женский гаметофит (объем выборки N = 383)
и около 12,3% через мужской гаметофит (N= 983).
Заметим, что при таких частотах передачи экстрахромосомы через
оба зародышевых пути мы должны иметь при самоопылении 65,3%
диплоидных потомков с фенотипом cri вместо наблюдаемых в опыте
76,0%. Это расхождение (и отсутствие в потомстве тетрасомиков) ско-
рее всего объясняется разницей в уровне конкуренции между пыльце-
выми трубками с эу- и гиперплоидным геномами. По-видимому, при
искусственном опылении количество прорастающих сквозь пестик муж
ских гаметофитов соизмеримо с числом зародышевых мешков, что ис-
ключает конкуренцию пыльцевых трубок.
Если в скорости прорастания гиперплоидные гаметофиты (с экстрах-
ромосомой) уступают нормальным гаплоидным, то при самоопылении
в условиях острой конкуренции за зародышевые мешки доля диплоид-
ных зигот будет выше расчетной при отсутствии конкуренции. Более того,
близость долей диплоидных зигот и гаплоидных женских гаметофитов,
вероятно, указывает на то, что при самоопылении зиготы возникают прак-
174
III. Эволюция растений
тически только от слияния гиперплоидных яйцеклеток с гаплоидными
спермиями, лишенными экстрахромосомы.
Самая удивительная особенность линии TRUST заключалась в пол-
ном отсутствии диплоидов с фенотипом Cri, что свидетельствовало
об отсутствии кроссинговера между локусом cri и точкой разрыва
транслокации ВР. К настоящему времени изучено более 10 тысяч по-
томков, но нс обнаружено ни одного кроссовера по этому району. Фе-
нотип трисомиков линии TRUST по сравнению с исходным трисоми-
ком TRIS-54 несколько приблизился к фенотипу нормального диплои-
да - края листьев стали менее волнистыми, фертильность пыльцы и
озерненность бобов приблизились к норме. Тем не менее, неизменно
приблизительно 76% их потомства составляли диплоидные растения
cri, а в МКП, кроме семи бивалентов, неизменно обнаруживалась ма-
ленькая унивалентная экстрахромосома (рис. 4).
Мы попытались ввести в Н-район генома линии TRUST дополни-
тельные рецессивные мутации, которые проявлялись бы только у дип-
лоидных растений с фенотипом cri и не проявлялись бы у трисомиков.
Две тысячи семян, собранных с трисомиков, были обработаны 0,1%
ЭМС, на стадии проростков, расте-
ния с фенотипом cri были удалены,
и оставшиеся 500 трисомиков были
выращены в теплице. Завязывае-
мость большинства растений оказа-
лась очень низкой. Семена с 210
наиболее “урожайных44 растений
Ml были высеяны посемейно в теп-
лице, и от каждой семьи было ото-
брано по одному трисомному рас-
тению М2 немутантного фенотипа,
их потомки были выращены в поле.
Отбирая растения нормального
фенотипа, мы пытались отбросить,
хотя бы частично, мутации, не пе-
рекрываемые экстрахромосомой.
Из 207 семей поколения М3 в 41
были выявлены различные мутации,
но только три из них обнаружили
Рис. 4. Метафаза I в материнских
клетках пыльцы линии TRUST. Эк-
страхромосома отмечена стрелкой
175
В.А. Бердников, Ф.Л. Горелъ, О.Э. Костерин, ВС. Богданова
сильное сцепление с cri. Ojx&&,fas-3, имела фасциированный стебель,
а две другие - kil (killer) и rmv (remover) - были деталями, поскольку в
семьях, где они были обнаружены, вообще отсутствовали растения с
фенотипом cri. Мутация kil блокирует прорастание семян, a rmv оста-
навливает развитие на самых первых стадиях развития зародыша. Три-
сомные носители мутаций rmv и kil стали основателями линий TRUST-
R и TRUST-K соответственно.
Последующий генетический анализ показал, что все три мутации
находятся в хромосоме 5, причём в случае мутации кН удалось пока-
зать, что в диплоидном геноме расстояние между kil и cri составляет
около 6 сМ. Для оценки рекомбинации между рецессивной спорофит-
ной деталью и геном cri достаточно подсчитать частоту появления в
потомстве растений с фенотипом cri. Тем не менее, при просмотре
более тысячи потомков растений линии TRUST-R с генотипом cri rmv
/ cri rmv / Cri Rmv не было найдено ни одного растения cri, то есть
расстояние между cri и rmv меньше 0,05 сМ. Просмотр проростков,
появившихся из более тысячи семян линии TRUST-K с генотипом cri
kil/cri kil/Cri Kil, выявил всего одно растение cri Kil. Таким образом,
рекомбинационная дистанция между kil и cri в экстрахромосоме в со-
ставе трисомика составляет 0,05 сМ, что на два порядка меньше того
же расстояния в Н-районе диплоида. Линия TRUST-R замечательна
тем, что все ее семена прорастают и дают исключительно трисомиков,
тогда как у линии TRUST-K прорастают только семена с триплоидны-
ми зародышами, составляющие около 25% производимых семян.
Кроме этих линий мы получили ещё три с существенным измене-
нием доли трисомиков. В двух линиях, TRUST-6 и TRUST-138, доля
трисомиков составляет около 50%, а в линии TRUST-111 эта доля до-
стигает 90%. В настоящее время идёт изучение фокуса действия мута-
ций, лежащих в основе повышения доли трисомиков. Не исключено,
что некоторые из них могут быть связаны с повышением вероятности
попадания экстрахромосомы в гаметы.
Наконец, мы попытались проверить, действительно ли трисомик
линии TRUST несёт три копии аллеля Ust. Мы скрестили растения
этой линии с диплоидной линией CRIF с генотипом cri и/cri и. Расте-
ния F1 имели фенотип Cri Us>, а у трисомиков в F2 от самоопыления
наблюдалось выщепление растений и/и с частотой, близкой к 1/4, что
много выше, чем можно было ожидать от рекомбинации между гена-
176
III. Эволюция растений
мй удалёнными на стандартной карте на 20-30 сМ и не менее чем на
порядок ближе на рекомбинационной карте экстрахромосомы. Такой
пезультат соответствует предположению о том, что локус U в экст-
оахромосме занят рецессивным аллелем и. Отсюда следует, что экст-
рахромосома в линии TRUST, по-видимому, несмотря на 13 поколе-
ний изогенизации, продолжает сохранять в экстрахромосоме исход-
ный ген и.
К сожалению, трисомики, в потомстве которых все растения cri
имели фенотип fas-З, отличаются пониженной мощностью и семен-
ной продуктивностью, поэтому пока мы смогли оценить только 86 их
потомков cri, но все они имели фенотип fas.
Резюмируя информацию по линии TRUST и ее производным, можно
заключить, что по меньшей мере одно (правое) плечо экстрахромосо-
мы трисомика выведено из рекомбинационного процесса фактически
сразу же после возникновения трисомика. В настоящее время мы по-
лучаем трисомики, в которых Н-район, гомологичный левому плечу
экстрахромосомы, маркирован мутацией crd. Такой трисомик позво-
лит оценить степень рекомбинационной изоляции экстрахромосомы
на всём её протяжении.
Сравнение трисомиков TRUST с исходным TRIS-54 показывает, что
замена части генома, скорее всего Н-района хромосомы-5, привела к
рекомбинационной изоляции экстрахромосомы. Н-район хромосомы
5, несущий Ust, линии TRUST, происходит от многомаркерной линии
WL1238, широко используемой в генетическом анализе гороха. Син-
тез этой линии был осуществлён Лампрехтом на основе сортов евро-
пейской селекции, порядок генов в этой линии обычный.
Итак, на основе реципрокной транслокации Хаммарлунда мы по-
лучили обменную хромосому, которая обладает такими основными
свойствами В-хромосом, как небольшой размер, необязательное при-
сутствие, относительная безвредность и отсутствие рекомбинации с
хромосомами основного набора. Всё это хорошо согласуется с нашим
предположением, что именно третичная трисомия является первым
шагом в создании В-хромосом. В дальнейшем мы рассчитываем полу-
чить мутации, повышающие конкурентоспособность мужских гипер-
плоидных гаметофитов, и мутации, способствующие попаданию эк-
стра-хромосомы в гаметы.
177
В. А. Бердников, Ф.Л. Горель, О.Э. Костерин, В.С. Богданова
Литература
1. Beukeboom L.W. Bewildering Bs: an impression of the 1st В-Chromosome Conference
//Heredity. 1994. Vol. 73. P. 328-336.
2. Jones R.N. and Rees H.B. Chromosomes. London: Academic Press, 1982
3. Green D.M. Muller’s Ratchet and evolution of supernumerary chromosomes 11 Genome
1990. Vol. 33. P. 818-824.
4. Stern C. Dosage Compensation - Development of a Concept and New Facts // Canad J
Genet. Cytol. 1960. Vol. 2. P. 105.
5. Green D.M.,Zeyl C.W., SharbelT.F. The evolution of hypervariable sex and supernumeian
(B) chromosomes in the relict New Zealand frog, Leiopelma hochstetteri //J. Evol. Biol
1993. Vol. 6. P. 417-441.
6. Sharbel T.S., Green D M., I louben A. B-chromosome origin in the endemic New Zealand
frog Leiopelma hochstetteri through sex chromosome devolution // Genome. 1998
Vol. 41. P. 14-22.
7. Colombo P.C. and Remis M.I. On the origin of B-chromosomes: neo XY systems and
X-like supernumeraries in Orthoptera//Caryologia. 1997. Vol. 50. P. 151-162.
8. Muller H.J. The relation of recombination to mutational advance // Mutation Research
1964. Vol. LP. 2-9.
9. Felsenstein J. The evolutionary' advantage of recombination // Genetics. 1974. Vol. 78
P. 737-756.
10. Charlesworth B. Model for evolution of Y chromosomes and dosage compensation
// Proc. Natl. Acad. Sci. USA. 1978. Vol. 75. P. 5618-5622.
11. Gordo I. and Charlesworth B. The degeneration of asexual haploid populations and the
speed of Muller’s ratchet//Genetics. 2000. Vol. 154. P. 1379-1387.
12. Blakeslee A.F. New Jimson weeds from old chromosomes // J. Heredity. 1934. Vol. 25
P. 80-115.
13. Lewis H. Speciation in flowering plants П Science. 1966. Vol. 152. P. 167-172.
14. Lewis H. The origin of supernumerary chromosomes in natural populations of Clarkia
elegans // Evolution. 1951. Vol. 5. P. 142-157.
15. Wedberg H L., Lewis H. and Venkatesh C.S. Translocation heterozygotes and
supernumerary chromosomes in wild populations of Clarkia williamsonii // Evolution
1968. Vol. 22. P. 93-107.
16. Mascarenhas J.P. The male gametophyte of flowering plants // Plant Cell. 1989. Vol I
P. 657-664.
17. Mooring J.S. A cytogenetic study of Clarkia uniguiculata. II. Supernumerary chromosomes
//Amer. J. Bot. 1960. Vol. 47. P. 847-854.
18. Sutton E. Trisomics in Pisum sativum derived from an interchange heterozygotc
П J. Genet. 1939. Vol. 38. P. 459-476.
19. Pellew C. Genetical studies on the first reciprocal translocation found in Pisum sativum
//J. Genet. 1940. Vol. 39. P. 363-390.
20. Kosterin O.E., PukhnachevaN.V, Gorel F.L., Berdnikov VA. Location of the breakpoints of four
reciprocal translocations involving linkage group V and their influence on recombination distances
between neighbouing markers П Pisum Genet. 1999. Vol. 31. P. 13-20.
21. Weeden N.F., Ellis T.H.N., Timmerman- Vaughan G.M., Swecicki WK.., Rozov S.M., Berdnikox
VA A consensus linkage map for Pisum sativum // Pisum Genet. 1998. Vol. 30. P. 1-4.
ВИД С ТОЧКИ ЗРЕНИЯ ПРАКТИЧЕСКОГО
БОТАНИКА
А.Н. Куприянов
Алтайский государственный университет
656099, г. Барнаул, ул. Димитрова, 66
E-mail: altb@biogeo.dcn-asu.ru
Nomina si nescis
periit et cognito rerum
Linneus
Интерес к ботаническим знаниям заложен в биологической сущ-
ности человека, и в любые времена названия растений, их реаль-
ная или потенциальная полезность всегда его интересовала. Лю-
бое название всегда подразумевает вид растения. И каждый вид
должен отличаться от другого. Практическая ботаника занимается
тем, что тиражирует ботанические знания, составляя списки ви-
дов растений определенной территории, определители растений,
которые, в свою очередь, всегда находят своего читателя.
Несколько более отдаленно от практических задач работают
систематики, которые критически пересматривают объемы видов,
создавая новые комбинации названий часто одних и тех же расте-
ний, устанавливают родство между видами и определяют системы
растений, в которых виды группируются по их схожести и родству.
Этот процесс находится постоянно в развитии, даже когда мысль
ботаника возвращается к старым забытым комбинациям и вроде
бы занимается «повторением пройденного».
Для ботаника, занимающегося практическими вопросами, чрез-
вычайно важно знать точные критерии вида и его границы. Очень
важно, чтобы видов было столько, сколько их существует в приро-
де как объективных реальностей. Знание видовой принадлежнос-
ти того или иного экземпляра растения конкретной местности -
суть основа знаний любого ботаника*. И для него не безразлично,
* Вряд ли это представление найдет поддержку у многих ботаников, занимающихся
теоретическими вопросами определения вида, для которых отдельный экземпляр не
несет видовой значимости.
179
А.Н. Куприянов
что вкладывается в понятие “ботанический вид”. Но не является ли
поиск точности в определении вида поиском черной кошки в темной
комнате, которой там нет?
В 1855 г. на закате блистательной жизни в ботанике Альфонс де
Кандоль писал: «Самым тяжелым испытанием для натуралиста явля-
ется задача ясно выразить свои мнения о природе вида. Он знает, что
каждое слово его будет взвешено, что каждая мысль будет признана
еретичной и что все ошибочные представления об этой базе естествен-
ных наук бросят вполне заслуженную тень на его собственные описа-
тельные работы» [1].
Проблема вида всегда была краеугольной в ботанике, поскольку он
является мерой исчисления биологического разнообразия и несет черты
объективной реальности и субъективного подхода. Безусловно, про-
блема эта широко освещена, и достаточно отослать читателя к моно-
графическим обзорам [2, 3]. В шестидесятых годах была проведена
большая конференция по проблемам вида в ботанике, материалы ко-
торой вышли в 1958 г. под редакцией М.П. Баранова [4].
В настоящее время определений вида сделано немало. Каждый
исследователь пытается сформулировать собственное и часто делает
это как те слепые, которые пытались представить слона, ощупывая
каждый свою часть. Целостное всегда остается недоступным. Пони-
мая это, еще Ч. Дарвин в главе о виде писал, что определений вида
слишком много, чтоб все их воспроизводить. Почти такими же слова-
ми пишет В.П. Комаров [2] в своем «Учении о виде у растений».
И тем не менее попытаемся воспроизвести современную трактов-
ку вида, взятую из Словаря ботанических терминов [5] и отражаю-
щую биологическую концепцию вида: «Вид - совокупность особей,
образующих географические или экологические викарирующие попу-
ляции, обладающих общими морфо-физиологическими признаками,
способных в природных условиях скрещиваться друг с другом, в со-
вокупности занимающих общий сплошной или частично разорванный
ареал». Давно доказано, что скрещиваются и виды и даже роды, четко
разграниченные, как это происходит в семействе Rosaceae, и получа-
ются жизнеспособные виды. Согласно морфологической концепции
вид - это группы популяций, сильно похожих друг на друга и в мень-
шей степени - на другие. За этими попытками дать всеобъемлющий
портрет «слона» кроется односторонность специалистов, которые
180
III. Эволюция растений
«ощупывают» то популяционную сущность, то генетическую, то мор-
фологическую, и чем дальше будет развиваться наука, тем больше бу-
дет этих сущностей, за которыми уже сейчас не проглядывает целост-
ность вида.
Наиболее целостное представление о виде было у К. Линнея, счи-
тавшего вид только морфологической сущностью вне времени и про-
странства. Он первым среди ботаников использовал знание Логики
Аристотеля, заимствовав из нее понятия “вид” и “род”. А.К. Скворцов
считает, что к этим понятиям Линней пришел индуктивным методом в
результате попыток синтезировать собственные наблюдения. Так или
иначе, но вид у Линнея монотипичен. Это альфа и омега номенклату-
ры. «Видов столько, сколько создал их господь бог». Заменим слово
«бог» на слово «природа», и под этим подпишется любой ботаник. И,
очевидно, К. Линнея можно назвать основоположником типологичес-
кого подхода к определению объема вида. В дальнейшем типологи-
ческий подход оформился в морфологическую концепцию вида. На-
звание вида тесно связано с типом, который представляет гербарный
лист, по которому ботаник сделал это заключение. Но уже во времена
Линнея в стройном мире монотипичных видов появляется диссонанс
в форме внутривидовой изменчивости. Линней [6] тоже видел и фор-
мы, и разновидности, а в параграфе 158 «Философии ботаники» он
писал: «Разновидностей столько, сколько различных растений может
вырасти из семени одного вида». И тут же он замечает, что «разновид-
ности должны быть исключены из ботаники», и, наконец, в параграфе
310: «... ботаник не заботится о мельчайших разновидностях». Но сле-
дует помнить, что это он говорит о фенотипической изменчивости:
«Разновидность есть растение, измененное случайной причиной, кли-
матом, почвой, жарой, ветром».
Но невозможно отмести то, что видит глаз: в сумме популяций при-
мерно одной морфологической структуры всегда видна изменчивость
наследственного характера разной амплитуды, которая должна укла-
дываться в кривую нормального распределения. Дробление популя-
ции на клоны, серии, линии опять же даст изменчивость признаков,
которые также будут иметь нормальное распределение, и с заданной
вероятностью можно эти группы отделить от похожих. Причем вариа-
бельность может наблюдаться как на уровне морфологических, так и
анатомических, химических, генетических структур. Это создает пред-
181
А.Н. Куприянов
посылки для все большего и большего дробления видов. Поэтому уже
с середины прошлого века обособились два взгляда на вид: монотипи-
ческий и политипический. Монотипистами, или splitter «дробителя-
ми», были Августен Декандоль и Вильденев. Декандоль уже в 1813 г
выделял разновидности, как бы мы сейчас назвали фенотипические,
вызванные влиянием разных внешних условий. Но он же выделял раз-
новидности постоянные, наследственные «значения которых преуве-
личены культиваторами». Он их называл расами.
В противовес им lumper's - «укрупнители» - считали, что вид со-
стоит из различных подвидовых структур. К этим ботаникам относят-
ся К. Кох, М. Вильком, а из российских ботаников К. Ледебур. Они
стали отстаивать многоступенчатые подразделения видов. Например,
Помпани, монограф рода Artemisia, приводил вид, подвид, разновид-
ность и далее а, ₽, у ступени внутренней структуры вида. Надо ска-
зать, что укрупнительство не упрощает работу практическому ботани-
ку. Для практических нужд ботаники было бы удобнее пользоваться
видами примерно одинакового объема, а это возможно только при
монотипическом подходе.
Альфонс Декандоль, используя свой колоссальный опыт практи-
ческого ботаника, в 1867 г. вводит понятия вида, подвида, разновид-
ности (подразновидности), вариации (подвариации) в Закон ботани-
ческой номенклатуры. Им записано: «Многочисленные подразделения
вида должны служить во многих загадочных и спорных случаях, чтоб
избежать установления новых названий для новых видов». В посла-
нии Ботаническому конгрессу в 1901 г. С. Бейли предлагал руковод-
ствоваться следующим: «Вид таксономически неделим. Ниже вида нет
ничего кроме разновидностей. Термин разновидность не должен вхо-
дить в иерархический ряд номенклатурного закона как выражение не-
уверенности в суждениях относительно данной формы».
Вид - очень сложная система, однако практический ботаник изу-
чает его определенными методами. И главный из них - морфологи-
ческий. Этот метод сейчас, как и во времена Линнея, остается опреде-
ляющим. Поэтому морфологичность вида остается главным его свой-
ством. Именно это свойство создает виду необходимую стабильность
существования. Общепринято, что качественные морфологические
структуры имеют преимущество перед количественными, которые, как
правило, свидетельствуют о модификационной изменчивости. Хотя
182
III. Эволюция растений
есть и исключения. В систематике полыни есть примеры, когда коли-
чественные признаки становятся ключевыми. Например, у Artemisia
kasakorum N. Pavl. листья не превышают 3 см, а у распространенного
вида A. nitrosa Web. ex Stechm. листья 3-7 см. Основным морфологи-
ческим признаком, различающим Hieracium umbellatum L. и H.filifolium
Juxip., является ширина листовой пластинки. Морфологизм вида ни-
кем не оспаривается. Л.И. Малышев, выступая на VII съезде ВБО, от-
мечал важность сравнительно-морфологического метода для развития
ботаники в следующем столетии [7].
Ч. Дарвин в 1859 г. в работе “Прохождение видов” делает упор на
развитие вида во времени - историзм вида. Линнеевский таксономи-
ческий вид он признавал только как «полезный артефакт». Руковод-
ствуясь идеей развития, Дарвин полагал, что границы между видом,
подвидом, разновидностью условны. Разновидность есть заканчива-
ющийся вид, а вид - это определившаяся разновидность. Идея разви-
тия была продуктивна для практической ботаники. Руководствуясь ею,
ботаники стали пытаться обозначать естественные группы видов, ис-
пользуя различные построения надвидового ранга. Бейли предложил
категорию strips для объединения близкородственных видов. К.И. Мак-
симович предложил series - видовые ряды, что идентично strips. Труд-
но определить срок развития вида. Но бесспорно, что виды рождают-
ся, развиваются, умирают. Например, возраст Artemisia elenae, опи-
санной нами из южных ленточных боров, так же как и A. radicans,
сравнительно небольшой. Происхождение этих видов связано с появ-
лением крупнозернистых песков, возраст образования которых не бо-
лее 16-18 тыс. лет. Эволюция этих видов связана с формированием
ленточных боров. В отличие от палеоэндемов, жестко привязанных к
определенным экологическим условиям и обладающих консерватив-
ными мало изменяемыми морфологическими признаками, у неоэнде-
мов наблюдается большой полиморфизм и нестабильность в соблю-
дении условий произрастания. Историзм вида оказал огромное влия-
ние на построение естественных систем, в основе которых находится
понятие о монофилетической группе, где понятие “вид” исчезает, а
вместо него употребляется понятие «терминальный таксон», представ-
ляющий собой наименьшую монофилетическую группу, которая да-
лее уже не может быть разделена [8]. Подобное дробление может про-
ходить до конкретного индивидуума, что для практической ботаники
183
А.Н. Куприянов
не совсем удобно. С другой стороны, объединение терминальных так-
сонов в агрегации, что и делается во многих родах, и использование
этих псевдоукрупненных видов - единственно верный путь для прак-
тической ботаники.
В конце прошлого века стали появляться работы, раскрывающие
географизм вида (А. Кернер, Р. Веттштейн). Ареал, занимаемый ви-
дом, стал одним из важнейших свойств вида. В законченной форме
учение о виде, как географической расе проявилось у С.И. Коржинс-
кого. Р.В. Камелин по праву называет его пятым великим российским
систематиком после К.И. Максимовича, Э. Регеля, А.А. Бунге, Н.С. Тур-
чанинова [9]. Очевидно, эти идеи возникли у него в конце прошлого
века во время путешествия по Каркаралинскому уезду, поскольку кон
трастность условий низкогорий Центрального Казахстана порождает
широкий спектр внутривидовой изменчивости самых обыденных ви-
дов. Он писал: «Все формы, которые при обладании известными мор-
фологическими отличиями представляют особый ареал распростра-
нения, я считаю за отдельные расы (prolies). Эти расы - истинные си-
стематические и географические единицы. Они подлежат иссле-
дованию и как нечто действительно существующее. Между тем виды
и подвиды представляют нечто условное». В.Л. Комаров, не обладая
максимализмом Коржинского, но глубоко проникнув в идею вида как
географической структуры, сформировал понятие о виде как геогра-
фической расе, которое наиболее ярко выражено в афоризме: “Вид -
морфологическая система, помноженная на географическую опреде-
ленность”. Восприняв эти идеи от Коржинского, В.Л. Комаров рассмат-
ривает вид не просто викарирующей структурой, которая постепенно
меняет свой облик по мере широтного или долготного распространения,
но и как исторический феномен эволюционного развития. И здесь гео-
графические свойства вида смыкаются с историческими. Принципы
В.Л. Комарова были использованы для создания монументальной “Фло-
ры СССР”. Эти идеи, безусловно, плодотворны и сегодня, они создали
предпосылки для описания сотен мелких географических видов.
Для практических целей ботаника-флориста низводить вид до расы
и конкретной популяции неразумно. По сути это и есть монотипичес-
кая идея вида, доведенная до абсурда. Хотя с теоретической точки
зрения эта мысль безупречна. Реализацией этой идеи занимался
М.В. Клоков [10], которым было описано около 400 мелких видов -
184
III. Эволюция растений
географических рас. А.И. Кузмичев [11] считает, что, только прини-
мая концепцию вида как географической расы, ботаник-географ име-
ет ключ для анализа происхождения флоры и растительности. Отри-
цательной чертой применения географического подхода к пониманию
объема вида стало появление географических двойников. Например,
есть эндемик Алтае-Саянской горной страны Gymnospermium altaicum
(Pallas) Spach и есть его причерноморский аналог G. odessianum (DC.)
Takht. Есть вид с широчайшим ареалом Artemisia austriaca Jacq. и его
украинский аналог Л. repens Pall, ex Willd.
Существование вида во времени и пространстве полностью под-
держивалось Н.Н. Вавиловым. В 1931 г. в своем классическом труде
«Линнеевский вид как система» [12] он писал: «Виды, с которыми имеет
дело современный ботаник, возникают в пространстве и во времени,
без учета этих факторов даже углубленное на основе гибридологичес-
кого анализа знание вида не полно и не точно» (с. 173). Кроме того,
Вавилов считал, что вид - сложная система экотипов. В противовес
эволюционистическим отрицаниям вида он утверждает, что виды -
закономерные, действительно существующие комплексы подвижных
экосистем, которые могут охватывать категории разного объема и в
своем историческом развитии связаны со средой и ареалом.
Таким образом, экологичность вида является его третьим, важ-
нейшим свойством. Следует сказать, что многие ботаники, включая
В.Л. Комарова, не приняли множественную конструкцию вида. В «Уче-
нии о виде растений», критикуя Н.Н. Вавилова, Комаров не упомина-
ет его имени. Но возможно, это сделано не по концептуальным сооб-
ражениям, а по политическим. Экологическое викарирование широко
встречается в природе и имеет много ярких примеров. Например, ака-
демик В.Н. Сукачев еще в 1934 г. указывает, что сосна, растущая в лен-
точных борах, отличается от сосны других местообитаний не только
биоэкологическими свойствами, но и морфологическими особеннос-
тями. Так, ее шишки в 1,5-2 раза крупнее, с сильно развитыми щитка-
ми; хвоя удерживается на дереве до 8 лет, вместо обычных 5-6 лет.
Это позволило В.Н. Сукачеву выделить сосну ленточных боров в осо-
бый подвид - кулундинскую сосну (Pinus sylvestris L., ssp kulundensis
Suk.). Однако другие систематики не признают права сосны ленточ-
ных боров на статус подвида, считают ее экоморфой сосны обыкно-
венной.
185
А.Н. Куприянов
Примером экологического викарирования являются полыни род-
ства Artemisia frigida Willd., поселяющиеся на разных высотах.
A. frigida (низкогорный или среднегорный вид) замещается в высоко-
горьях A. argyrophylla Ledeb., который действительно в крайнем вы-
ражении сильно отличается от A. frigida как габитусом, так и темными
краями обверток. В переходной зоне отнесение конкретных экземпля-
ров к тому или иному виду практически невозможно и всегда несет
элементы волюнтаризма. Очень большое количество фактов влияния
среды на видообразование приводит В.Н. Васильев [13].
К сожалению, в систематике растений много видов описываются
по одному экземпляру. С одной стороны, это неизбежно, поскольку
всегда существует неполнота ботанических исследований, а с другой,
появляется возможность артефакта, выявления определенной морфо-
логической случайности, не подкрепленной ни множественностью, ни
ареалом, ни экологией. Например, на Ивановском хребте (Алтай)
Д.В. Чусовляновым был найден вполне жизненный экземпляр овся-
ницы группы Festuka kryloviana s.l., совершенно лишенный остей на
нижней цветковой чешуе. При существующей тщательности обработ-
ки сибирских видов рода Festuca напрашивается единственное реше-
ние - описание нового вида, но он кроме морфологической особенно-
сти не обладает никаким другим признаком вида. Поэтому существу-
ющий морфологический казус может быть отмечен как форма или
терата. Морфологические границы отдельных признаков вида теоре-
тически можно установить через доверительный интервал и заданный
уровень вероятности. Такой подход уже реализовался в российской
биологии Е.С. Смирновым, для которого вид «...есть элементарная ка-
тегория естественной иерархической системы; она характеризуется
наличием континиума особей, составляющих популяцию данного вида»
(цит. по [8]). Практически это делается посредством сравнения экзем-
пляра с диагнозом вида или с типовым экземпляром (куда должны от-
носиться и тип, и изотипы, и паратипы). Морфологическая концепция
наиболее важна для практической систематики, и по сути только она
удерживает ботанику от хаоса. Поэтому остается согласиться с мне-
нием Э. Майра [14] о том, что для типолога тип реален, а изменчи-
вость иллюзорна.
Трудность заключается в том, что тип для практического ботаника
недоступен, а диагноз зачастую не отражает действительную морфо-
186
III. Эволюция растений
логию признаков. И, конечно же, чрезвычайно важна работа с типом,
который должен пониматься не как один экземпляр, а как интегрирую-
щий образец типовых экземпляров с амплитудой изменчивости ключе-
вых признаков. И мы никак не можем согласиться с мнением Л.Н. Васи-
льевой [15], что «зависимость систематиков от типовых образцов об-
рекает их на деградацию мышления, на произвол и субъективизм
таксономических решений, на невозможность оценить действия друг
друга и амбициозное самомнение». Но вопрос о том, что нужно де-
лать, когда отдельный признак выходит за рамки диагноза, а весь га-
битус и остальные признаки вида соответствуют ему, остается слож-
ным. Всегда существует дилемма - описывать новый вид или обнаро-
довать изменение диагноза. Логичнее второе, но чаще всего ботаник
идет по первому пути или незаконно «вбивает» уклоняющийся обра-
зец в название как внутривидовой таксон. Внутривидовая изменчи-
вость отражает реальное изменение признаков в пределах диагноза и
объективно должна находиться в разных частях кривой нормального
распределения признаков. Она должна строиться на иерархии свойств
и их признаков.
Собственно, эта же идея заложена у Васильевой [ 15] в методе апо-
стериорного взвешивания признаков: признаки свойств она называет
«состоянием». В лаконичной форме у нее это выглядит так: «призна-
ки всегда отличают таксоны, а состояния всегда характеризуют» (с. 34).
В качестве свойств и признаков могут быть использованы самые раз-
личные уровни организации растений и самые различные методики.
В 60-х годах большое значение для определения видовой специфич-
ности придавалось химическим соединениям - маркерам (глобулины
- для злаков и лилейных; секвитерпеновые лактоны - для астровых;
кумарины - для сельдерейных; флавоноиды - для розоцветных и т.д.).
Химическим соединениям придавался ранг таксономического при-
знака, и делалась попытка с их помощью решать систематические воп-
росы [16]. В последнее время большое значение придается структуре
хромосом и содержания ДНК [17]. Оказалось, что количество ДНК
на ядро у разных видов растений может различаться в сотни раз. Ко-
личество ДНК - признак видоспецифический и стабильный, а посе-
му он также может рассматриваться как один из морфогенетических
признаков вида. Очень заманчиво выстроить систему видов по весу
ДНК.
187
А.Н. Куприянов
С развитием науки появятся новые методы изучения растений и
обнаружатся новые признаки вида, которые могут использоваться в
систематике растений. Тем не менее следует заметить, что для целей
практической ботаники эти признаки являются вторичными, к кото-
рым следует прибегать в особо сложных случаях, поскольку вид дол-
жен быть различим прежде всего габитуально, а для доказательства
этого отличия допустим весь комплекс методов, которыми располага-
ет наука. Вопрос о гибридогенных, апомиктических, амфимиктичес-
ких видах, скорее всего, не должен затрагивать практическую ботани-
ку, имеющую в основе анатомо-морфологические, эколого-географи-
ческие методы работы. Само применение эпитета к виду не правомерно,
поскольку понятие «вид» - с одной стороны, элемент логики, а с дру-
гой, элемент выбранной системы координат расчленения единого био-
логического разнообразия.
В настоящее время нет-нет, да и вспыхивает жаркая полемика между
сторонниками биологической и морфологической концепций вида. На-
конец появляются попытки примирения обеих сторон, что, собственно
говоря, и происходит при попытке дать обобщенное понятие ВИД. Это
попытка совместить не совместимое, поскольку вид как биологическая
категория - это сложнейшая система атомных, молекулярных, биохими-
ческих, анатомических и других уровней. И мы полностью согласны с
И.Я. Павликовым [18], что концепции биологического вида просто не
может быть, поскольку нет единой концепции биоты, в которой он суще-
ствует. Вид как таксономическая категория - это мельчайшая частичка
биологического разнообразия. Объем вида- понятие субъективное. При
его определении необходимо учитывать основные его свойства: морфо-
логическое отличие, географическую и экологическую обособленность.
Впрочем, и эти свойства суть предмет общей договоренности между
ботаниками или личной убежденности. Количество видов зависит от
уровня квалификации исследователя данной таксономической группы.
Образно говоря, видов столько, сколько различает их исследователь.
Иначе как объяснить, что на одной территории со времени выхода «Фло-
ры СССР» количество видов увеличилось с 17 до 21 тысячи [19]. Таким
образом, вид для практической ботаники - это единица биологи-
ческого разнообразия, обладающая множественностью особей или
популяций, морфологической индивидуальностью, ареалом и эко-
логической приуроченностью.
188
111. Эволюция растений
Литература
1 De Candolle А.Р. Geographic botanique raisonnee ou exposition des faits principaux et de
lois concernant la distribution geographique des plantes de 1’epoque actuelle. Paris;
Geneve, 1855. T. 1-2. 1365 p.
2 Комаров В.П. Учение о виде у растений. М.; Л., 1944. 245 с.
3 Завадский К.М. Вид и видообразование. М., 1971. 404 с.
4 Проблема вида в ботанике: Сб. научн. статей / Под ред. М.П. Баранова. М.; Л., 1958.
5 Словарь ботанических терминов. Киев, 1984. 308 с.
6. Линней К. Философия ботаники. М., 1989. 451 с.
7. Малышев Л.И. Систематика растений и перспективы ее развития // Актуальные воп-
росы развития ботаники в СССР. Алма-Ата, 1988.
8 Шагалкин А.И. К вопросу о таксономическом виде И Журнал общей биологии. 1983.
Т.44.№2. С. 172-186.
9. Камелин Р.В. Систематика сосудистых растений в России (вехи истории) И Ботани-
ческий журнал. 2000. Т. 85. № 6. С. 2-18.
)0. Клоков М.В. Географическая раса как историческое явление // Природная обста-
новка и фауны прошлого. Киев, 1974. Вып. 8. С. 105-111.
11. Кузмичев А.И. Гигрофильная флора юго-запада Русской равнины и ее генезис. СПб.,
1992. 208 с.
12. Вавилов Н.Н. Линнеевский вид как система// Избр. труды. М.; Л., 1965. Т. 5.
13. Васильев В.Н. Физико-географическая среда и видообразование. М.; Л., 1966. 73 с.
14. Майр Э. Зоологический вид и эволюция. М., 1968. 597 с.
15. Васильева Л.Н. Типологическая школа систематики // Методические проблемы
биологии и экологии. Владивосток, 1989. С. 26-43.
16. Адекенов С.М., Куприянов А Н., Кагарлицкий А.Д. Структурные особенности сес-
квитерпеновых лактонов и систематика рода Artemisia L. И Известия АН КазССР.
1986. № 10. С. 55-66.
17. Гриф Н.Г. Проблемы и перспективы кариологии и кариосистематики растений
//Ботанический журнал. 1997. Т. 82. № 2. С. 1-13.
18. Павлинов И.Я. Есть ли биологический вид, или в чем «вред» систематики И Жур-
нал общей биологии. 1992. Т. 53. № 5. С. 757-767.
19. Черепанов С.К. Свод дополнений и изменений к «Флоре СССР». Л., 1973. 668 с.
СИНДРОМЫ В СРАВНИТЕЛЬНОЙ
ФЛОРИСТИКЕ
Л.И. Малышев
Центральный Сибирский ботанический сад СО РАН
630090, г Новосибирск, ул. Золотодолинская, 101
E-mail: “Malyshev” <root@botgard.nsk.su>
Введение
Методы сравнительной или количественной флористики позволя-
ют выявить закономерности пространственного разнообразия таксо-
нов растений. Они основаны на сопоставлении исходных флористи-
ческих данных в таксономических списках или таблицах. Рубеж XX и
XXI столетий знаменует приблизительно 150 лет в развитии методов
сравнительной флористики. Основоположником этого направления бо-
таники был Альфонс Декандоль (годы жизни 1806-1893) - сын Огюста
Декандоля и отец Казимира Декандоля. Они образуют знаменитую ди-
настию швейцарских ботаников французского происхождения [1].
Флористика является старшей сестрой фитоценологии, которая
оформилась в недрах ботанической географии в последней четверти
XIX века. Европейские ботаники нередко отождествляют флористику
с геоботаникой, но русские ученые часто понимают под геоботаникой
только фитоценологию. Флористика развилась из ботанической гео-
графии еще в первой четверти XIX века, когда впервые были исполь-
зованы некоторые количественные характеристики флоры. De Candolle
[2] изложил основы количественного анализа флоры в сводке «Обо-
снованная география растений или изложение главных фактов и за-
конов о географическом распределении растений в настоящую эпо-
ху». Девизом сравнительной флористики может быть его высказыва-
ние: «Методы исследования - это то, что характеризует сущность
науки в каждую эпоху и что больше всего определяет ее прогресс»
[Les methodes sont се qui caracterise 1’etat de la science a chague epoque
et qui determine le plus ses progres].
190
III. Эволюция растений
Основой сравнительной флористики является общая статистика.
Декандоль, как основоположник сравнительной флористики, увлекался
статистикой еще в юности. Когда отец предоставил ему выбор - изу-
чать право или медицину для последующей защиты диссертации и
получения степени доктора, Альфонс выбрал право, так как в этом
случае он мог заниматься статистикой, которую очень любил [ 1, с. 14].
Некоторые суждения Декандоля [2] в области сравнительной флорис-
тики важны вплоть до нашего времени, особенно в его сводке глава
аО разнообразии форм растений в разных странах и мире в целом”
[De la variete des formes vegetales dans divers payes et dans le monde
entier] (p. 1270-1298). Декандоль указывал на абсурдность сравнения
численности видов в регионах, если они хотя бы приблизительно не
одинаковы по площади. Этому правилу пытался следовать Е.В. Вульф
[3] при составлении карты деления земного шара на растительные об-
ласти по количественному распределению видов.
Стимулом для успехов сравнительной флористики и фитоценоло-
гии было развитие методов биологической статистики в 20-е и осо-
бенно 30-е годы XX века. Значительным был вклад польских ученых:
С. Кульчинского, Д. Шимкевича, В. Шафера и др. [4-9]. В более по-
зднее время сравнительная флористика как отрасль ботанической гео-
графии была усовершенствована в методологическом отношении рус-
скими ботаниками. Благоприятным для этого было огромное простран-
ство бывшего СССР с большим разнообразием растительного покрова.
Большой вклад в сравнительную флористику внесли А.И. Толмачев,
Б.А. Юрцев и В.М. Шмидт [10-16]. По инициативе Толмачева (годы
жизни 1903-1979) и его преемника Юрцева стало традицией проведе-
ние рабочих совещаний по сравнительной флористике. Первое из них
состоялось в Ленинграде в 1971 г. под руководством Толмачева, вто-
рое - в г. Неринга (Литва) в 1983 г., третье - в Кунгуре в 1988 г., чет-
вертое - в Березинском биосферном заповеднике (Белоруссия) в 1998 г.,
наконец, пятое - в Ижевске в 1998 г. Результаты трех из этих совеща-
ний опубликованы в сборниках под редакцией Юрцева: “Теоретичес-
кие и методические вопросы сравнительной флористики”, “Актуаль-
ные проблемы сравнительного изучения флор”, “Изучение биологичес-
кого разнообразия методами сравнительной флористики” [17-19].
В последние десятилетия в России, как и в зарубежных странах, мощ-
ным стимулом для использования и развития методологии сравнитель-
191
Л.И. Малышев
ной флористики было внедрение в научный обиход компьютеров и со-
здание компьютерных программ для анализа базовых данных. Одновре-
менно становилось все более очевидным большое значение углублен-
ных флористических исследований для познания биологического разно-
образия в целом и для решения вопросов по охране окружающей среды.
За истекшие полтора столетия в сравнительной флористике, так же
как фитоценологии и экологии растений, совершенствовались методы
исследования. В то же время введение более информативных или более
корректных методов иногда сопровождалось непониманием их со сто-
роны части ботаников. Одновременно развивалась склонность загромож-
дать результаты исследования отдельных флор цифровыми показателя-
ми без критического осмысления их значения. Эти недостатки можно
назвать синдромами сравнительной флористики. В связи с этим уместно
напомнить, что за эпиграф статьи Толмачева [11, с. 3] “О количествен-
ной характеристике флор и флористических областей” было взято пре-
достережение Декандоля [2]: “/Zy a des chiffres bon a calculer et d’autres
parfaitement inutiles ou meme trompeurs” [Одни цифры вполне заслужива-
ют подсчета, а другие совершенно бесполезны и даже ошибочны].
Задача нашего исследования не сводится к критическому обзору
основных методов сравнительной флористики или к изложению исто-
рии их развития, для чего понадобилось бы написать книгу. Это уже
выполнил отчасти Шмидт [15-16] в сводке “Статистические мето-
ды в сравнительной флористике” (1980), а в учебном пособии “Ма
тематические методы в ботанике” (1984) отдельная глава посвяще-
на математическим методам в сравнительной флористике. Поэтом)
наше внимание сосредоточено на некоторых тенденциозных неточно-
стях и ошибках в методологии, которые стали очевидными лишь к
настоящему времени. Такими синдромами в сравнительной флорис
тике являются: 1) неравенство территории сравниваемых флор; 2) не-
равновеликость флористических списков при сопоставлениях; 3) ана-
лиз недостаточно информативных таксономических спектров.
1. Синдром неравенства территории сравниваемых флор
Опубликованные “Флоры” обычно составлены для разных по пло
щади территорий, иногда с неопределенными границами. Поэтом)
192
III. Эволюция растений
Декандоль [2: Ch. 24, § 1; р. 1270] рекомендовал идти не по пути на-
м омождения цифровых данных для сравнения, а по пути отбора:
этом вопросе речь идет не о соединении многих цифр, но о выборе
небольшого числа тех, которые являются ценными для того, чтобы
прийти к заключению” [Il ne s’agit pas dans cette question de reunir
beacoup de chiffres, mais de choisir le petit nombre de ceux qui offrent les
conditions voulues pour arriver a une conclusion], что “было бы нелепо
сравнивать относительно полного числа видов регионы, которые не
будут приемлемо равными по поверхности” [... il serait absurde de
comparer, au point de vue du nombre total des especes, de regions qui ne
serait pas sensiblement egales en surface].
Этому совету пытался следовать Вульф [3] в работе “Опыт деле-
ния земного шара на растительные области на основе количествен-
ного распределения видов ”. Для него наиболее трудной задачей оказа-
лось числовое сопоставление флор, различающихся по размерам тер-
ритории. Флоры в действительности были неравными по площади,
поэтому Вульф принимал приблизительное равенство территории в
слишком широком диапазоне. В связи с этим Толмачев [11] отметил
недостаточную обоснованность выводов Вульфа о распространении
растений и недостоверность карты областей земного шара по видово-
му богатству отдельных флор. Но исследование Вульфа все же было
первой после Декандоля попыткой количественного сопоставления
флор всего земного шара.
В противоположность учету флористического состава больших и
неизбежно разнородных по физико-географическим условиям терри-
торий, Толмачев [10,11] предложил метод конкретных или элементар-
ных флор (КФ) для сравнения районов при условиях, что эти районы
1) приблизительно одинаковые (равноценные) по площади; 2) невели-
ки по площади; 3) изучены с одинаковой, весьма значительной полно-
той. В частности, район КФ “должен быть достаточно мал для того,
чтобы обеспечить действительную универсальность для него данной
флоры, вместе с тем достаточно велик для того, чтобы охватить все
возможные в его условиях стации, так как в противном случае отли-
чия систематического состава разных сообществ затемнят чисто фло-
ристические особенности страны, и получившийся в результате работ
список флоры будет носить отчасти фациальный, а не региональный
характер” [11, с. 15]. Уместно напомнить, что первое и третье условия
193
Л. И. Малышев
были названы еще Декандолем и относятся не только к КФ, а также к
“сводным” или “коллективным” флорам в понимании Толмачева, со
свойственной им континуальностью (постепенностью) в сменах так-
сономического состава по мере нарастания территории.
Впервые метод КФ был применен Толмачевым [10] на Таймыре
при изучении флоры двух районов у низовий рек Ямутарида и Яму
Неры в 1928 г. Было сделано существенное заключение, что невоз
можно дать какие-либо общие обязательные рекомендации о размере
площади КФ, но допускается, что для выявления полного видового
состава достаточна площадь радиусом около 10 км в Арктике, а на
севере лесной зоны она может быть от 100 до 500 кв. км [13].
Метод КФ был использован для познания закономерностей про-
странственного разнообразия флоры на плато Путорана, где коллек-
тив ботаников в 1968-1972 гг. обследовал 16 мест [20]. Для выявле-
ния зональности флоры на Среднесибирском плоскогорье Н.С. Водо
пьянова [21] исследовалав 1976-1979 гг. 10 КФ по меридиану 108°в.д
- от Прибайкалья на юге до Хатангского залива у Ледовитого океана,
на профиле длиной 2120 км.
Метод КФ стал популярным в исследованиях бывшего СССР, но с
противоречивым пониманием сущности [14]. За рубежом он не нашел
признания, вероятно, в связи с относительно высоким там уровнем
общей изученности флоры. Но канадский ботаник Р. Бешель [22] обо-
сновал понятие о геоне для сопоставления флор по количественным
показателям. Геон - это представительный участок флоры размером
около 100 кв. км на севере, но может быть около 1 000 кв. км в тропи-
ческих странах. Понятия о КФ и геоне в основном совпадают, более
того, обе концепции основаны на примере северных флор, именно
Канадского арктического архипелага и Гренландии в случае геона.
Иногда исследователи необоснованно отождествляют метод КФ с
минимальной единицей флористического районирования, а не с эта-
лоном территории вокруг него.
Метод КФ - более продуктивный при изучении флоры трудно дос
тупных и мало изученных северных территорий. Но и там условие
детального обследования позволяет изучить в среднем лишь две КФ
за сезон. Поэтому даже стойкие приверженцы этого метода все же
иногда изучали “локальные” флоры вместо более определенных КФ
например, на Чукотке и Ямале.
194
III. Эволюция растений
Как условие использования метода КФ иногда оговаривается фи-
зико-географическая однородность территории. Но это условие лишено
смысла, когда требуется изучить особенности флоры в условиях эко-
тона, т.е. переходной полосы между биомами или предполагаемыми
вЬ1делами флористического районирования. На видовом составе не-
достаточных по площади участков, принятых в качестве КФ, сказыва-
ется дискретность (мозаичность) растительного покрова, а при значи-
тельном увеличении территории за пределы минимума проявляется
континуальность флоры в виде постепенных переходов. Как справед-
ливо отмечал Шмидт [15, с. 9], “флора одновременно и непрерывна и
дискретна”. Соглашаясь с этим, можно заключить, что деление флор
на однородные (гомогенные или конкретные) и разнородные (гетеро-
генные или континуальные) - условное и имеет значение в основном
при разработке схем флористического районирования.
Самое спорное в методе КФ условие - это требование прибли-
зительно одинакового размера (равноценности) площади обследо-
вания флоры при одновременном отказе учитывать размер обсле-
дованной территории, поскольку размер минимальной территории
будет увеличиваться при движении с севера на юг в связи с зональ-
ным ростом флористического богатства. Как выход из этого весь-
ма затруднительного положения предлагалось совсем не учитывать
размер площади при подборе для обследования однородной в бо-
танико-географическом отношении территории. Шмидт [16, с. 200]
писал: “Одна из главных особенностей конкретной флоры состоит
в том, что это понятие не требует привязки к определенной площа-
ди, хотя эта площадь и может быть установлена. Границы конкрет-
ной флоры обусловлены не квадратными километрами, а биоти-
ческими и историческими факторами, действовавшими в данном
районе.” В соответствии с этой парадоксальной концепцией,
Шмидт [16, с. 237-252] усреднил показатели о числе видов для
разных конкретных и “близких к ним” флорах европейской части
бывшего СССР, затем вычислил регрессию числа видов, родов и
семейств для интервалов географической широты. Эта попытка
интересна оригинальностью, но расхождение между фактически-
Ми и прогнозируемыми данными оказалось значительным
U6, с. 250-251, табл. 93]. Это можно объяснить прежде всего не-
равенством территорий и экологическим разнообразием.
195
Л. И. Малышев
В настоящее время несомненно, что флористическое богатство за-
висит от размера территории и экологических условий [23]. Для полу-
чения сравнимых данных необходимо вычислить прогнозируемое (ожи-
даемое) число видов на площади стандартного размера. Эта стандарт-
ная площадь не должна быть меньше минимального ареала флоры как
целостной системы, иначе будут получены абстрактные показатели.
В Арктике ареал-минимум флоры может быть около 100 кв. км или
даже меньше. Так, на п-ве Ямал репрезентативной оказалась флора на
площади около 30 кв. км из-за пространственного однообразия этого
сектора Арктики [24]. По мере продвижения к югу ареал-минимум фло-
ры увеличивается до 500-1000 кв. км. В полосе подтайги он равен 600
650 кв. км для Латвии [25] и 700-750 кв. км для Псковской и Новго-
родской областей [15, с. 26].
Для определения уровней флористического богатства путем экст-
раполяции пригодны формулы Аррениуса [26, 27] (1) и Глизэна [28]
(2); кроме того, формула сомнительной корректности предложена Еван-
сом [29] (3):
log5“ = [z * log А] - log 5; (I)
S“ = [/>*logJ]-S; (2)
S “ = [S * log (a + 1)] / (log A + 1) или S“ = SI log A. (3)
В них Su - ожидаемое число видов на единице площади стандарт-
ного размера (a), S - реальное число видов на обследованной площа-
ди, А - размер обследованной площади в стандартных единицах, кон-
станты z и b - индексы пространственного флористического разнооб-
разия.
При этом более корректные результаты могут быть получены при
экстраполяции по альтернативным формулам Аррениуса и Глизэна.
В отличие от них формула Евэнса не учитывает пространственное
флористическое разнообразие (z или Ь), и экстраполяция приводит в
этом случае к менее корректным прогнозным данным, которые зани-
жены для малых площадей и завышены для больших.
Ориентировочные значения z, так же как и Ь, вычислялись эмпири-
чески по регрессии “виды/площадь” для различных флор в пределах
отдельных регионов [30-31 ]. Величина z обычно варьирует от 0,11 до
0,19 в материковых флорах, возрастая от арктической полосы к тропи-
196
IIL Эволюция растений
ческой, а для являющихся изолятами островных флор обычно изменя-
ется в пределах от 0,22 до 0,44 [32]. Весьма примечательно, что пока-
затель z использован также в каноническом уравнении Ф. Престона
[33] для оценки разнородности или гетерогенности двух флор или фаун
относительно друг друга:
[а/(а + ед1/2+[а/(а + с)],/г=1,
где а - число общих видов, b - число специфических видов для одной
флоры и с - для другой. При значениях z больше 0,27 сравниваемые
флоры или фауны являются изолятами, а при меньших значениях -
лишь частями общей биоты. Этот индекс был предложен для сравне-
ния островных биот, но может быть использован также как коэффици-
ент совместимости (1-z) материковых флор или фаун [34, 35].
В качестве стандартной единицы площади для получения сопоста-
вимых данных о флористическом богатстве могут быть условно при-
няты 100, 1 000, 10 000 или 100 000 кв. км. При этом площадь в 100
или 1 000 кв. км приближается к размеру КФ, тогда как 10 000 или
100 000 кв. км соответствует сборным континуальным флорам. Не
могут быть рекомендованы в качестве стандарта 1 кв. км или 1 кв. ми-
ля, так как столь малые территории многократно меньше размера ре-
альной флоры как целостной системы и поэтому представляют абст-
ракцию, лишенную биологического смысла.
А. Кайё [36] был пионером оценки флористического богатства путем
пересчета реальных данных на площадь стандартного размера (richesse
areale), именно, на 10 000 кв. км. Т. Моно [37] за стандарт принял
100 000 кв. км при изучении бедной и однообразной пустынной флоры
Мавритании на севере Африки. При составлении карты флористическо-
го богатства районов Африки Лебрэ [38] использовал пересчет на
10 000 кв. км. Затем Бартлот и др. [39] составили цветную карту деления
земного шара на зоны по градиентам флористического богатства также
на 10 000 кв. км. Все названные исследователи-Моно, Лебрэ, Бартлот -
применяли для пересчета формулу Еванса, при этом без ограничения
пределов экстраполяции. Это снижало достоверность результатов.
Л. Малышев [32] использовал уравнение Аррениуса при составле-
нии карты богатства видами сосудистых растений флор земного шара в
пересчете на 100 000 кв. км. При этом допускалась экстраполяция не
197
Л. И. Малышев
более 10 раз от исходной площади. Составлена также серия карт ддя
флоры бывшего СССР с уровнями видового богатства сосудистых рас-
тений для 100,1 000, 10 000 и 100 000 кв. км. В каждом случае экстрапо-
ляция также не превышала 10-кратный размер изученной территории
[40-42].
Можно заключить, что при оценке флористического богатства син-
дром неравенства размера территории сравниваемых флор может быть
преодолен пересчетом на стандартную площадь, размер которой дол-
жен быть не ниже репрезентативного. При пересчете необходимо учи-
тывать градиент пространственного разнообразия флоры, определяе-
мый по уравнению регрессии. Наиболее корректной для пересчета
является формула Аррениуса.
2. Синдром неравновеликости флористических списков
при сопоставлениях
Количественное сравнение списков растений для определения сход
ства или различия практиковалось прежде всего в фитоценологии, а
позднее было принято также при исследовании флоры или фауны.
Швейцарский ботаник П. Жаккар [43] первым предложил формулу для
оценки флористического сходства участков фитоценоза по индексу
сопряженности (коэффициенту общности IJAC):
1МС= а/ (а+Ь+С)’
где а - число общих видов, b - число специфических видов для одного
участка или флоры и с - для другого. В этом случае число общих (со-
вместных) видов делится на все число видов (не сумму) в двух объе-
диненных участках или флорах.
К настоящему времени предложено до 40 различных индексов [15,
44-45 и др.]. Одни из них представляют сходство, другие - различие,
могут варьировать от 0 до +1 или же от -1 до +1. Но это не означает,
что диапазон от -1 до +1 более точный, чем от 0 до +1.
Не все индексы одинаково информативны. Большинство из них
учитывает три величины: а, b и с. Таковы индексы Жаккара, Чеканов-
ского, Стугрена и Радулеску и большинство других. Некоторые фор-
мулы учитывают только две величины: а и b либо а и с. Соответствен-
но, они менее информативные, так как оценивают только односторон-
нюю связь: меру включения меньшей флоры в большую (индекс
198
111. Эволюция растений
Щимкевича) или, наоборот, большей флоры в меньшую (индекс Бра-
ун-Бланке). Самые информативные индексы учитывают четыре вели-
чины: а, Ь, с и где d - число не общих видов, т.е. отсутствующих в
сравниваемой паре, но найденных в других флорах или фаунах в пре-
делах генеральной совокупности. Такими являются индекс Сокэла и
Снята по формуле 5 (7Ш) и аналогичный ему индекс Фи (1рн1) для че-
тырехсторонней корреляции [46-48]. Но они имеют значение при изу-
чении всех флор в генеральной совокупности. Так, индекс ISS5 исполь-
зован для флористического районирования Азиатской России на ос-
нове анализа 49 базовых выделов [49].
Синдром неравновеликости сравниваемых списков выражается в
продолжающемся использовании мало информативных либо некор-
ректных индексов сопряженности. Большинство формул (Жаккара,
Чекановского, Стугрена и Радулеску и др.) учитывает среднее ариф-
метическое сходство. Но это допустимо лишь при условии, когда фло-
ристические списки содержат почти одинаковое число видов. В про-
тивном случае необходимо вычислять среднее гармоническое сходство,
например, по формуле 2 Кульчинского [4] (JKUL)'-
4и= °>5 [а/(а+Ь) + а/(а+с)].
Еще более информативным при анализе неравновеликих списков
является индекс среднего геометрического сходства по формуле Отиаи
или Баркмана [50, 51] (1ОС1}У- _________
1ЖН= a/V(a+b)(a+c).
Альтернативными ему являются предложенный Сокэлом и Снитом
индекс по формуле 5 или же индекс Фи. Пригоден также показатель со-
вместимости флоры или фауны сравниваемых территорий, вычисляе-
мый по каноническому уравнению Престона как величина 1-z (см. выше).
Для целей районирования возможно использование также индекса
Шимкевича [5] (Jszy):
1SZY= а/ (а+с).
Эта формула оценивает не среднее сходство, а только односторон-
нюю связь: меру включения меньшей флоры (а+с) в большую (я+6),
т.е. при условии (Ь? с). Наоборот, меру включения большей флоры в
меньшую оценивает формула Браун-Бланке [52]
IB№ = a/(a+b)-
Этот индекс все же отражает нереальную для флористического рай-
онирования ситуацию, тогда как индекс Шимкевича завышает “под-
199
Л.И. Малышев
чиненность” бедной флоры [53]. Объединение обоих индексов (Шим-
кевича и Браун-Бланке) для оценки двусторонней связи равнозначно
среднему гармоническому сходству по формуле Кульчинского (/* ).
Вышесказанное призвано показать, что синдром неравновеликос-
ти сравниваемых флористических списков видов растений может быть
преодолен в наибольшей мере при использовании для флористическо-
го районирования индексов Отиаи (7^), Сокэла и Снита (7VW) или Фи
а также гомогенности объединенных флор (1-z). Они являются
альтернативными.
Из-за разнообразия арсенала индексов (коэффициентов общности или
различия) для сравнения флор по таксономическому составу желательно
использовать формулы с учетом приоритета авторов, которые их пред-
ложили. Нельзя объединять фамилии знаком соединения (дефис или тире),
если не имеется в виду автор со сдвоенной фамилией. Так, будет пра-
вильно говорить: “индекс Стугрена и Радулеску”, а не Стугрена-Раду-
леску, которые были лишь соавторами; “индекс Чекановского”, а не “ин-
декс Сёренсена-Чекановского”, так как приоритет принадлежит Чека-
новскому, к тому же этот индекс до Сёренсена использовал также Дайс;
“индекс Шимкевича”, а не “индекс Симпсона”, который применил его
значительно позже. Но допустимо использовать пояснения: “индекс
Шимкевича или Симпсона” для индекса Шимкевича; “индекс Чекановс-
кого или Дайса или Сёренсена” для индекса Чекановского.
3. Синдром недостаточной информативности
таксономических спектров
Структура флоры может быть выражена весовыми характеристи-
ками семейств, родов или видов по содержанию в них таксонов низ-
кого ранга. Тогда список таксонов, расположенных в порядке убыва-
ния богатства, составит таксономический спектр флоры. Он имеет
значение при сравнении отдельных флор для уяснения их особеннос-
тей и эволюции (генезиса). Наиболее контрастны “ведущие” по бо-
гатству таксоны, которые образуют головную (главную) часть спект-
ра. У других таксонов последовательно уменьшается богатство и од-
новременно возрастает их число с одинаково малыми весовыми
показателями. Графически это соответствует параболе для степенной
функции.
200
III. Эволюция растений
Стремление ограничить исследование головной частью спектра
облегчает подбор данных, но уменьшает репрезентативность спектра.
Толмачев (1970; 1974, с. 120-133) показал на многих примерах, что
репрезентативность 10 богатых семейств по содержанию видов боль-
ше в бедных флорах Арктики и пустынных флорах Средней Азии, где
достигает величины 65-76 и 63-71%. Она понижается до 48-61% в
бореальных и особенно в южных не аридных флорах иногда до 30-
41%. Примечательно также, что видовая репрезентативность 10 глав-
ных семейств в КФ также выше, чем во флоре в целом, частью кото-
рой является КФ, например, в соотношении 56,7 и 54,9% или 74,9 и
72,0% (Толмачев, 1974, с. 195). Показатели видовой репрезентативно-
сти совокупности главных семейств все же не имеют самостоятельно-
го ботанико-географического значения, так как зависят от нескольких
факторов: числа семейств в выборке и видового богатства флоры, а
оно - от размера территории и экологических условий. Такая репре-
зентативность мало информативна по сравнению с числом видов рас-
тений во флоре в целом с последующим пересчетом на стандартную
площадь для определения уровня видового богатства (см. выше).
Примечательно, что репрезентативность видо-семейственных и
родо-семейственных спектров, составленных для “главных” семейств,
гораздо выше свойственной видо-родовым спектрам из того же числа
членов. Поэтому видо-родовые спектры должны быть много длиннее
для достижения высокой репрезентативности таксономического бо-
гатства. Но это затрудняет сбор исходных данных и их анализ. Так, в
10 флорах южного Средиземноморья таксономическая репрезентатив-
ность 10-членных списков достигает для видо-семейственной и родо-
семейственной структуры 55-65 и 52-67%, а для видо-родовой лишь
11-16%; аналогично этому репрезентативность 10-членных спектров
для отдельных Курильских о-вов составляет 49-58 и 50-56% в видо-
семейственных и родо-семейственных спектрах и только 16-27% в
видо-родовых (Шмидт, 1980, с. 73-117).
Сравнение структуры таксономических спектров (не их репре-
зентативности) полезно для выявления особенностей отдельных флор
и определения их места в системе районирования. При этом структура
родового богатства семейств (родо-семейственные спектры) более
показательна при сравнении флористических областей, тогда как видо-
Родовые спектры больше отражают провинциальные особенности
201
Л.И. Малышев
флор. Зато видо-семейственные спектры соединяют таксономические
ранги вида и семейства в обход промежуточного ранга рода, что зату-
шевывает их флорогенетическое значение (Malyshev et al., 1999).
В этом отношении примечательно замечание Толмачева (1974, с. 122),
что “более показательными кажутся соотношения между численнос-
тью видов и родов растений, свойственные различным флорам”. Уме-
стно напомнить, что Szymkiewicz (1934, 1936, 1947; Шафер, 1956;
Szafer, 1964) первым применил родо-семейственные спектры для ста-
тистического анализа. Для разных флор земного шара он построил
нисходящие ряды от 10 до 18 главных семейств по числу не космопо-
литических родов, а для анализа использовал меру включения мень-
шей флоры в большую (индекс Шимкевича).
Использование неполных таксономических спектров, построенных
только из более богатых семейств или родов, неизбежно снижает их
информативность. А.И. Пяк и А.А. Зверев (1997) выявили, что денд-
рограммы кластерного анализа по коэффициентам ранговой корреля-
ции не совпадали для спектров из 12, 15 и 20 главных по видовому
богатству семейств в локальных флорах Томской области.
Изучение пространственного разнообразия родовой и семействен-
ной структуры в базовых флористических выделах (БФВ) Сибири по-
казало, что коэффициенты ранговой корреляции (Кендэла, Спирмена
или гамма) менее информативны в спектрах, чем весовые характери-
стики, например, выраженное в промилле (процентах или частях еди-
ницы) число видов в роде или же число родов в семействе относи-
тельно общего числа видов или родов в том или ином БФВ (Малышев
и др., 1998а, б; Malyshev et al.9 1999). Найдено также, что дендрог-
раммы видо-родовых спектров не вполне совпадали для всего соста-
ва родов и только для 42 главных, при репрезентативности 42,6% от
всей флоры. Дальнейшее сокращение спектров до 24 главных родов
(репрезентативность 32,5%) и особенно до 20, 15 и 10 главных родов
привело к существенному изменению конфигурации дендрограмм.
Аналогично этому в случае родо-семейственных спектров дендрог-
раммы оказались лишь приблизительно сходными для всего состава
семейств и для 30 главных семейств, при репрезентативности родо-
вого состава 78,6%.
С учетом сказанного выше нельзя согласиться с суждением
А.П. Хохрякова (1995,2000), что качественное сравнение видо-семей-
202
III. Эволюция растений
сТвенных спектров, построенных на учете лишь первых 6 семейств,
из которых первые три образуют наиболее важную триаду, позволяет
удовлетворительно классифицировать флоры без применения мате-
матических методов.
Рецидивы синдрома некорректности сравниваемых таксономичес-
ких спектров могут быть устранены использованием: 1) полных так-
сономических списков вместе, составленных только для главных так-
сонов; 2) весовых параметров таксонов (например, в промилле от об-
щего числа таксонов подчиненного ранга) вместо ранжированных;
3) родо-семейственной и видо-родовой структуры флоры для сопос-
тавлений регионального или провинциального уровня. Не может быть
рекомендовано использование видо-семейственных спектров взамен
родо-семейственных и видо-родовых.
Исследование выполнено при поддержке Государственной научно-
технической подпрограммы “Биологическое разнообразие” и РФФИ.
Литература
1. Микулинский С.Р, Маркова Л. А., Старостин Б. А. Альфонс Декандоль (1806-1893).
М.: Наука, 1973. 295 с.
2. De Candolle A. Geographic botanique raisonnee ou exposition des faits principeaux et de
lois concemant la distribution geograppique des plantes de 1’epoque actuelle. T. 1-2.
Paris: Masson / Geneve: Kessmann, 1955 (1956). 1365 p.
3. Вульф E. В. Опыт деления земного шара на растительные области на основе количе-
ственного распределения видов // Тр. прикл. бот., ген., селекции. 1937. Сер. 1(2).
С. 315-368, карта (препринт в 1934 г.).
4. Kulczynski S. Die Pflanzenassozionen der Pieninen // Bull. Acad. Polon. Sci. et Lettrres,
Classe Sci. Math. Natur. 1927 (1928). Ser. В. P. 57-203.
5. Szymkiewicz D. Une contribution statistique a la geographic floristique //Acta Soc. Bot.
Polon. 1934. T. XI. N. 3. P. 249-265.
6. Szymkiewicz D. Seconde contribution statistique a la geographic floristique // Acta Soc.
Bot. Polon. 1936. T. XIII. N. 4. P. 271-292.
7. Szymkiewicz D. Cinquieme contribution statistique a la geographic floristique // Bull.
Inst. Acad. Polon. Sci., Ser. Bot. Krakow, 1947. P. 1-29.
8. Шафер В. Основы общей географии растений. М.: ИЛ, 1956. 380 с.
9. Szafer W. Ogolna geografia roslin. Warszawa: PWN, 1964. 433 p.
10. Толмачев А.И. К методике сравнительно-флористических исследований. Понятие о
флоре в сравнительной флористике // Журн. РБО. 1931. T. 16. № 1. С. 111-124.
11 • Толмачев А.И. О количественной характеристике флор и флористических областей
//Тр. Сев. базы АН СССР. 1941. Вып. 8. 40 с.
12. Толмачев А.И. О некоторых количественных соотношениях во флорах земного шара
// Вест. ЛГУ. 1970. № 15. С. 62-74.
203
Л. И. Малышев
13. Толмачев А.И. Введение в географию растений. Л., 1974. 244 с.
14. Юрцев Б. А. Некоторые тенденции развития метода конкретных флор // Бог журн
1975. Т. 60. № 1.С. 69-83.
15. Шмидт В.М. Статистические методы в сравнительной флористике. Л.: Изд-во Ле-
нингр. ун-та, 1980. 176 с.
16. Шмидт В.М. Математические методы в ботанике. Л.: Изд-во Ленингр. ун-та, 1984
288 с.
17. Теоретические и методические проблемы сравнительной флористики (Ред. Юрцев
Б.А.). Л : Наука, 1987. 292 с.
18. Актуальные проблемы сравнительного изучения флор (Ред. Юрцев Б.А ). СПб., 1994
354 с.
19. Изучение биологического разнообразия методами сравнительной флористики (Ред.
Юрцев Б.А ). СПб., 1998. 356 с.
20. Флора Путорана (Ред. Малышев Л.И.). Новосибирск: Наука, 1976. 246 с.
21. Водопьянова Н.С. Зональность флоры Среднесибирского плоскогорья. Новосибирск
Наука, 1984. 157 с.
22. Бешель Р. Флористические соотношения на островах Неоарктики // Бот. журн. 1969
Т. 54. №6. С. 872-891.
23. Малышев Л И. Зависимость флористического богатства от внешних условий и ис-
торических факторов И Бот. журн. 1969. Т. 54. № 8. С. 1137-1147.
24. Ребристая О.В. Опыт применения метода конкретных флор в Западносибирской
Арктике (п-ов Ямал) // Теоретические и методические проблемы сравнительной
флористики. Л.: Наука, 1987. С. 67-90.
25. Табака Л.В. Некоторые итоги сравнительного изучения флоры различных природ-
но-территориальных подразделений Латвии // Теоретические и методические про-
блемы сравнительной флористики. Л.: Наука, 1987. С. 104-107.
26. Arrhenius О. Distribution of the species over the area // Medd. Vetenskaps. Nobelinst
1920. Vol. 4(1). N. 7. P. 1-6.
27. Williams C.B. Patterns in the balance of nature and related problems in quantitative
ecology. London & New York: Academic Press, 1964. 324 p.
28. Gleason H.A. On the relation between species and area// Ecology. 1922. Vol. 3. No. 1
P. 158-162.
29. Evans F.C., Clark P.J., Brand R.H. Estimation of the number of species presen on a given
area// Ecology. 1955. Vol. 36. No. 2. P. 342-343.
30. Dony J.G. The expectation of plant records from prescribed area // Watsonia. 1963
Vol. 5. No. 6. P. 377-385.
31. Dony J.G. Species-area relationships. Areport to the Natural Environment Research Council
(Britain), 1971. 73 p.
32. Малышев Л И. Количественный анализ флоры: пространственное разнообразие,
уровень видового богатства и репрезентативность участков обследования // Бот
журн. 1975. Т 60. № И. С. 1537-1550.
33. Preston F.W. The canonical distribution of commonness and rarity // Ecology. 1962
Vol. 43. P. 185-215,410-432.
34. Старобогатов Я.И. Фауна моллюсков и географическое районирование континен-
тальных водоемов земного шара. Л.: Наука, 1970. 372 с.
35. Малышев Л И. Количественная характеристика флоры Путорана// Флора Путора-
на. Новосибирск: Наука, 1976. С. 163-186.
204
IIL Эволюция растений
, CailleuxA. Biogeographic mondiale. Paris: Presses Universitares, 1953. 128 p.
' Monod T. Sur un cas exceptionnel de richesse areale: le sables de la Mauritanie orientale
//Rec. Trav. Univ. Montpellier, Ser. Bot. 1955. N. 7. P. 63-67.
qg Lebrun J. Sur la richesse de la flore de divers territoires africains //Acad. Roy. Sci. Outre-
Mer, Bui. seances, Nouv. ser. 1960. T. 6. N. 4. P. 669-690.
39 Barthlott W, Lauer W., Placke A. Global distribution of species diversity in vascular
plants: towards a world map of phytodiversity 11 Erdkunde, Kleve (Germany). 1996.
Vol. 50. P. 317-327, map.
40 Малышев Л.И. Биологическое разнообразие в пространственной перспективе
Ц Биологическое разнообразие, подходы к изучению и сохранению. СПб., 1992.
С. 41-52.
41. Малышев Л И. Флористическое богатство СССР // Актуальные проблемы сравни-
тельного изучения флор. СПб., 1994. С. 34-87.
42. Малышев Л И. Прогноз пространственного разнообразия и изученность флоры
Сибири//Биоразнообразие. Степень таксономической изученности. М.: Наука, 1994.
С. 42-52.
43. Jaccard Р. Distribution de la flore alpine dans la basin de Dranses et dans quelques regions
voisines // Bui. Soc. Vaudoise, Sci. natur. 1901. T. 37. N. 140. P. 241-272.
44. Тамарин П.В., Шмидт B.M. Сравнительный анализ некоторых коэффициентов сход-
ства//Тр. Ленингр. общ. естествоисп. Л.: Изд-во Ленингр. ун-та, 1975. Т. 72. Вып. 5
(Успехи биометрии). С. 45-53.
45. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических
исследованиях. М.: Наука, 1982. 287 с.
46. Sokal R.R., Sneath Р.Н.А. Principles of numerical taxonomy. San Francisco, 1963.
359 p.
47. Rohlf F.J. NTSYS-pc: numerical taxonomy and multivariate analysis system. New York,
1990. VIII+11. 5 p.
48. Norusis M.J. SPSS for Windows: base system user’s guide. Chicago, 1993. 828 p.
49. Малышев Л И., Байков К С., Доронькин В.М. Флористическое деление Азиатской
России на основе количественных признаков // Krylovia (Томск). 2000. Т. 2. № 1.
С. 3-16.
50. Ochiai A. Zoogeographical studies on the soleoid fishes found in Japan and its neighbouring
regions, 2 // Bull. Jap. Soc. Sci. Fish. 1957. Vol. 22. P. 526-530.
51. Barkman J. J. Phytosociology and ecology of cryptogamic epiphytes, including a taxonomy
survey and description of their vegetation units in Europe. Assen: Van Gorcum, 1958.
628 p.
52. Braun-Blanquet J. Plant sociology: the study of plant communities. New York: Mc-Graw
Hill, 1932. 439 p.
53. Малышев Л И. Основы флористического районирования // Бот. журн. 1999. Т. 84.
№1.С. 3-14.
54. Malyshev L.I., Baikov K-S , Doronkin V.M. Spatial diversity of the Siberian flora// Flora
(Jena). 1999. Vol. 194. N. 4. P. 357-368.
55. Пяк А.И., Зверев A.A. Опыт сравнительного анализа локальных флор с помощью
прикладного статистического пакета Biostat // Бот. журн. 1997. Т. 82. № 5. С. 64-75.
56. Малышев Л И., Байков К С., Доронькин В.М. Пространственное разнообразие ро-
довой структуры во флоре Сибири // Изучение биологического разнообразия мето-
дами сравнительной флористики. СПб., 1998. С. 34-44.
205
Л.И. Малышев
57. Малышев Л.И., Байков К С., Доронькин В.М. Таксономические спектры флоры
Сибири на уровне семейств // Бот. журн. 1998. Т. 83. № 10. С. 3-17.
58. Хохряков А.П. Основные типы флористических спектров Средней России // фло.
ристические исследования Центральной России (ред. Тихомиров В.Н ). М., 1995
С. 12-16.
59. Хохряков А.П. Таксономические спектры и их роль в сравнительной флористике
//Бот. журн. 2000. Т. 85. № 5. С. 1-11.
ПРОБЛЕМА ВИДА И СОВРЕМЕННАЯ
СИСТЕМАТИКА РАСТЕНИЙ
М.В. Олонова
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
E-mail: herb@bio.tsu.ru
1. Современные представления о видообразовании
Проблема вида, являющаяся одной из основных в современной
биологии, решается биологами уже не одно столетие, и, как известно,
до разрешения ее еще очень далеко. Систематическая обработка лю-
бого крупного и полиморфного рода растений, включающая установ-
ление таксономического ранга как отдельных особей, так и популя-
ций, уточнение границ видов, поиски родственных связей между ними,
а также попытки воссоздания эволюции изучаемой группы заставля-
ют коснуться таких аспектов проблемы вида, как современные пред-
ставления о видообразовании и концепции вида. Эти теоретические
вопросы обыкновенно относят к компетенции эволюционистов и ге-
нетиков, однако они являются теоретической основой и для решения
задач систематики. Как отмечает JI.H. Васильева (1989), если эволю-
ционистов в данном случае более занимает вопрос, как образуются
виды, то систематиков - что и из чего образуется и какое место в сис-
теме это новообразование занимает - его ранг и родственные связи.
Задачи систематики осложняются тем, что генетики пока не могут
прийти к единому мнению относительно путей видообразования. Су-
щественно осложняют проблему вида у растений такие явления, как
полиплоидия, гибридизация, апомиксис и вегетативная пролиферация,
иногда называемая вивипарией.
В настоящее время существуют две точки зрения на процесс видо-
образования - синтетическая теория и теория прерывистого равнове-
сия (прерывистой эволюции).
Синтетическая теория эволюции и видообразования возникла в
первой половине нашего века в результате слияния двух направлений
207
М.В. Олонова
- дарвинизма и менделизма. Ее возникновение было предопределено
работами С.С. Четверикова (1926), посвященными исследованию воз-
никновения, проявления и поддержания наследственной изменчивос-
ти в популяциях и анализу генетических механизмов видообразова-
ния. Многочисленные работы генетиков, посвященные генетике по-
пуляций и влиянию естественного отбора, подготовили теоретическую
базу для создания синтетической теории эволюции. Далее, в работах
Добжанского (Dobzhansky, 1937), видообразование рассматривалось
уже с генетико-популяционных позиций. Трудами Н.В. Тимофеева-
Ресовского и др. исследователей (Timofeeff-Ressovsky, 1940 а, б; Huxley,
1942; Майр, 1947; Симпсон, 1948; Stebbins, 1950) популяционно-гене-
тические принципы исследования были распространены на система-
тику растений и животных. В. Грант (1991) характеризует синтетичес-
кую теорию как популяционно-генетический подход к микроэволю-
ции. Сущность этой гипотезы состоит в основном в признании гео-
графического способа видообразования как процесса медленного пре-
вращения под действием изоляции и естественного отбора одной или
нескольких локальных популяций сначала в географические расы, за-
тем, с углублением дивергенции, в биологические виды и таксоны бо-
лее высокого порядка (градуалистический подход). Другими словами,
в результате изоляции популяции некоторого вида обмен генами меж-
ду ней и другими популяциями этого вида прекращается. Таким обра-
зом возникает группа растений, генетически представляющая собой
более или менее случайную выборку из первоначального генофонда.
С течением времени изолированная популяция адаптируется к новым
условиям, их генные частоты начинают различаться, и в конечном итоге
популяции становятся генетически разными настолько, что даже если
по той или иной причине изолирующие барьеры вдруг исчезнут, скре-
щивание между материнской и дочерней популяциями станет уже не-
возможным, другими словами - из дочерней популяции образуется
новый вид.
Как отмечалось В. Грантом (1991), синтетическая теория эволю-
ции достаточно гибкая и способна воспринимать новые идеи, даже
если они в чем-то не соответствуют ее изначальным принципам. Так,
многие последователи синтетической теории признают квантовую те-
орию видообразования, которая, в сущности, противоречит идее по-
степенного, градуального видообразования.
208
III. Эволюция растений
Квантовая теория видообразования была выдвинута в 1944 г.
Г Симпсоном (1948), и затем поддержана многими другими исследо-
вателями (Lewis, 1962; Carson, 1971, 1975; Грант, 1984 и др.) В отли-
чие от последовательного видообразования при рассмотренном выше
географическом способе, при квантовом происходит непосредствен-
ный переход от локальной расы к новому виду. От общего ареала не-
которого вида отпочковывается дочерняя, небольшая и пространствен-
но изолированная популяция. Благодаря сравнительно небольшим раз-
мерам в ней происходит инбридинг, который приводит к образованию
и закреплению новых гомозиготных сочетаний генов. Эти сочетания,
которые в большой предковой популяции были бы разрушены в ре-
зультате панмиксиса, служат источником фенетических изменений.
Большинство новых фенотипов, не обладая адаптивной ценностью,
вымирает, но те немногие, которые такой ценностью обладают, закре-
пятся в дочерней колонии в результате совместного действия инбри-
динга, дрейфа и отбора (Грант, 1991). Р. Hanelt и D. Mettin (1975) отож-
дествляют квантовую эволюцию и сальтацию. В.Н. Стегний (1993) в
обзоре работ сторонников синтетической теории эволюции отмечает,
что, несмотря на некоторые расхождения во взглядах, касающихся
предварительных условий и скорости видообразования, все они неиз-
менно принимают положение, утверждающее, что при видообразова-
нии реорганизуется уже имеющийся генофонд вида или популяции.
Градуализм опирается на частотное преобразование полиморфной
популяции с последующим закреплением наиболее благоприятных
аллельных или хромосомных сочетаний.
В противоположность сторонникам синтетической теории эволю-
ции последователи теории прерывистого равновесия (Eldredge, Gould,
1972; Stanley, 1975) считают, что видообразование - скачкообразный
(сальтационный) процесс, осуществляющийся за счет макромутаций.
В ходе эволюции кратковременные периоды быстрого изменения сме-
няются длительными периодами равновесия. Целесообразность же
возникает в результате отбора видов, а не особей (Кейлоу, 1986). Сто-
ронники этой теории длительные периоды застоя объясняют неспо-
собностью большинства мутационных изменений войти в состав фе-
нотипа, однако допускают, что время от времени одно из таких изме-
нений может оказаться совместимым с существующим адаптивным
комплексом. Возможен и иной путь: возникновение резкого сдвига в
209
М.В. Олонова
сторону от генофонда родительской популяции в результате образова-
ния неслучайной выборки на ранних стадиях изоляции (Кейлоу, 1986).
Как отмечает В.Н. Стегний, с генетических позиций наиболее полно идея
сальтационного видообразования была разработана Р. Гольдшмидтом в
его теории макрогенеза. Эта теория предполагает, что видообразование
носит быстрый характер и связано с реорганизацией регуляторной сис
темы генома. При этом мутации отдельных генных локусов (микромута-
ции) здесь не играют значительной роли. Напротив, крупные хромосом-
ные реорганизации (макромутации и системные мутации) имеют шансы
предопределять события видообразования (Стегний, 1993).
В. Грант, выступая с критикой сторонников этой теории, доказыва-
ет, что в настоящее время никто из последователей синтетической те-
ории эволюции не пропагандирует филетический градуализм в чис-
том виде и что идея быстрого и резкого эволюционного изменения
ими отнюдь не отвергается. При этом он обращает внимание на идею
квантовой эволюции, возникшую в рамках синтетической теории, и
утверждает, что гипотеза прерывистого равновесия не содержит ниче-
го принципиально нового. Сторонники же этой теории утверждаю!,
что ее принципиальное отличие состоит в том, что в данном случае
происходит не перераспределение имеющихся генов, а возникнове-
ние совершенно новых, и отбор идет на ином - видовом - уровне.
«Видообразование - не постепенный процесс перестройки генетичес-
кой структуры популяций, а скачкообразное событие, связанное с воз-
никновением системных мутаций» (Стегний, 1993). В отличие ол
Р. Гольдшмидта В.Н. Стегний считает, что системные мутации возни-
кают не единично, а кластером, что лучше позволяет им закрепиться.
При этом он подчеркивает высокую роль отбора на всех стадиях ста-
новления нового вида. Принципиальные отличия между градуалисти-
ческим и сальтационным видообразованием состоят в том, что в пер-
вом случае идет перераспределение имеющегося генетического мате-
риала, а во втором - создание принципиально новых генетических
структур, и отбор в этом случае идет на видовом уровне.
А.С. Кашин (1996, 1999), поддерживая идею сальтационного ви-
дообразования, отмечает, что при возникновении видов путем внезап-
ного изменения генотипа, сопряженного с репродуктивной изоляци-
ей, избежать нивелирования, вызываемого половым процессом, по-
могает апомиксис.
210
III. Эволюция растений
А.Л. Тахтаджян (1983) своей концепцией макрогенеза объединяет
достижения обеих теорий. Не отвергая возможности скачкообразных
(макрогенетических) изменений в процессе видообразования и эво-
люции, он, в отличие от Гольдшмидта, считавшего мутационный про-
цесс «двигателем эволюции», главную роль отводит отбору. Развивае-
мая им концепция макрогенеза не противопоставляется синтетичес-
кой теории эволюции, но дополняет ее. Не укладываясь в рамки
квантовой эволюции, она, тем не менее, не противоречит ей. А.Л. Тах-
таджян (1983, с. 1601) подчеркивает, что «постепенный ... градуалис-
тический процесс микроэволюции, основанный на накоплении мел-
ких мутаций, представляет основную и безусловно наиболее распрос-
траненную форму эволюции, на фоне которой время от времени
происходят редкие макрогенетические скачки. Это редкие события,
так как ... большинство макромутантов постигает неудача и лишь нич-
тожное меньшинство способно пройти через узкое «бутылочное гор-
лышко» естественного отбора и дать начало новой жизнеспособной
популяции». При этом макрогенез более присущ растениям, так как
растения гораздо менее интегрированы и поэтому резко выраженные
мутации для них менее детальны, поскольку легче корректируются в
онтогенезе.
П. Кейлоу (1986), отмечая в своем обзоре слабые стороны каждой
из теорий, считает, что главная проблема, стоящая перед сторонника-
ми синтетической теории эволюции, не скорость, с которой могут про-
исходить эволюционные изменения, а тот факт, что признаки могут
оставаться неизменными на протяжении длительных периодов геоло-
гического времени. Сторонникам же теории прерывистого равнове-
сия следует прежде всего решить проблему случайности в скачкооб-
разных изменениях. В заключение он пишет, что вряд ли удастся, опи-
раясь на фактические данные, найти ясное решение всех этих вопросов.
Очень возможно, что эволюция одних групп проходила в соответствии
с теорией прерывистого равновесия, а других - в соответствии с нео-
дарвинистской моделью (синтетическая теория эволюции), и малове-
роятно, чтобы одна какая-то модель оказалась применимой ко всем
случаям. В настоящее время многие исследователи, как ботаники, так
и зоологи, считают возможными оба пути видообразования, однако
конечную точку в этом вопросе должны поставить генетики. Если им
Удастся доказать, что виды образуются только сальтационным путем,
211
MB. Олонова
а совокупности особей, образующиеся при расхождении аллеломорф
в результате адаптивной дивергенции, не отвечают критериям вида, то
ранг видов, образовавшихся таким путем, будет понижен до внутри-
видового и будет отражать внутривидовое разнообразие.
2. Роль полиплоидии, гибридизации
и апомиксиса в видообразовании
При всем различии взглядов на видообразование сторонники обе-
их теорий признают видообразование путем полиплоидии и гибриди-
зации, однако определение ранга образующихся таким образом таксо-
нов также вызывает много недоразумений.
Н.Н. Цвелев (1967) полагает, что полиплоидия является достаточ-
но обычным способом образования эколого-географических рас в пре-
делах политипических видов в тех группах высших растений, в кото-
рых она распространена особенно широко. Особое значение имеет по
липлоидия, наступившая в результате гибридизации (аллополиплои-
дия), нередко приводящая к образованию серий популяций с разными
хромосомными числами. С развитием генетики как важный таксоно-
мический признак стало рассматриваться число хромосом, а именно
основное или базовое хромосомное число и уровень плоидности. Эти
данные помогают решать проблемы макро- и микроэволюции, уста-
навливать или, наоборот, исключать родственные отношения между
видами (Цвелев, 1972; Hunziker, Stebbins, 1986), однако ни числа хро-
мосом ни даже особенности их строения не могут быть приняты в
качестве универсального видового критерия, так как признак этот, как
и большинство морфологических признаков, нередко варьирует в са-
мых широких пределах. Хорошо известны случаи, когда одинаковые
числа хромосом встречаются у совершенно различных организмов и,
с другой стороны, нередко даже сложные хромосомные исследования
не выявляют видовых отличий (Кабанова, Карташова, 1972 и др.).
Известно, что одни систематические группы характеризуются на
редкость стабильным кариотипом, у других же он варьирует в самых
широких пределах. Зоологические исследования, основанные на боль-
шом статистическом материале, с достоверностью показали, что су-
ществует статистически достоверная корреляция между плодовитос-
212
III. Эволюция растений
Тью и изменчивостью кариотипа, причем у организмов, характеризу-
ющихся высоким коэффициентом плодовитости (насекомых, рыб, ля-
гушек), различия в структуре кариотипа не являются видовым крите-
рием (Орлов, 1974, цитируется по: Медников, 1989). Этот вывод мож-
но отнести к растениям, коэффициент плодовитости которых
чрезвычайно высок. Таким образом, хотя видообразование путем уве-
личения хромосомных чисел и является неоспоримым, различия по-
пуляций по числу хромосом сами по себе не являются основанием для
придания им видового статуса. По этой причине многие ботаники не
считают правомерным рассматривать кариотипические формы как
виды, предпочитая объединять их в полиплоидные комплексы, в осо-
бенности, когда между формами нет ни экологических, ни географи-
ческих различий (Грант, 1984).
Как известно, большую роль в эволюции растений в целом сыгра-
ло явление гибридизации.
Н.Н. Цвелев (1967), большое внимание уделявший проблемам гиб-
ридизации, отмечает, что наряду с образованием простых полиплоид-
ных рядов у высших растений, по-видимому, достаточно часты слу-
чаи, когда в результате гибридизации между членами нескольких по-
липлоидных рядов, сопровождающихся или не сопровождающихся
аллополиплоидией, члены этих рядов объединяются в один сложный
видовой агрегат, и установление таксономического ранга входящих в
этот агрегат единиц - задача настолько сложная, что ее не всегда уда-
ется разрешить даже при помощи самых детальных исследований.
О. и Д. Солбриг (1982) рассматривают гибридизацию наряду с поли-
плоидией, как частный случай мгновенного видообразования. По мне-
нию Н.Н. Цвелева (1973, 1975, 1996), гибридогенные таксоны не только
имеют значительно более широкие возможности для рекомбинации ге-
нов, чем первичные диплоиды, но и оказываются более или менее дес-
пециализированными, что облегчает их дальнейшую эволюцию.
Н.Н. Цвелев объясняет деспециализацию при гибридизации тем, что у
гибридов доминируют более древние признаки каждого из растений, а
более древние признаки обыкновенно являются и более примитивными.
Вместе с тем, за счет перегруппировки имеющихся генов у гибридов
могут возникнуть совершенно новые признаки (Цвелев, 19766). Как из-
вестно, глубокая специализация является эволюционным тупиком, а гиб-
ридизация способствует деспециализации (Lotsy, 1916; Stebbins, 1959).
213
М.В. Олонова
Н.Н. Цвелев (1995) считает, что темпы эволюции таксонов опреде-
ляются не столько временем их существования, сколько способнос-
тью к дальнейшей эволюции в данном направлении, и если такая эво-
люция возможна, то видообразование может пойти очень быстро (кван-
товое видообразование). Многие гибриды в результате стабилизации
тем или иным путем (амфиплоидия или интрогрессия) восстанавлива-
ют фертильность и избегают возможности расщепления в последую-
щих поколениях. Однако, как отмечает Stebbins (1959), залогом успе-
ха в большей мере является адаптивность новых форм по сравнению с
исходными и в меньшей - частота возникновения стабильных гибрид-
ных форм. Н.Н. Цвелев (19766) предполагает, что один, но перспек-
тивный гибридогенный таксон может выжить при прогрессивном ухуд-
шении окружающих условий и дать начало группам более высоких
таксономических рангов.
Наибольшее значение имела интрогрессивная гибридизация, в ре-
зультате которой происходило массовое проникновение генов одних
видов в генофонд других. Это явление также приводило к стиранию
морфологических различий между видами, образованию множества
форм неясного таксономического ранга, нередко стабилизировавших-
ся с помощью апомиксиса.
Апомиксис отмечается у многих высших растений и существенно
усложняет как определение видовых границ, так и проблему вида в
целом.
Согласно В.Л. Комарову (1934, 1940), апомикты либо составляют
особую группу видов, либо видовой критерий к ним не приложим вооб-
ще. По мнению В. Гранта (Grant, 1957), категория вида применима толь-
ко к бипарентальным формам, унипарентальные формы имеют только
клоны. В.Н. Тихомиров (1967) считает неправильным применение по-
нятия “вид” к облигатным апомиктам. По мнению А.Л. Тахтаждяна
(1984), категория вида приложима и к унипарентальным организмам,
однако вид у них, в отличие от бипарентальных, представляет собой си
стему клонов. Н.П. Дубинин (1966) также принимает универсальность
категории вида, однако у апомиктов вид приобретает другой характер,
поэтому критерии вида, разработанные для бипарентальных организмов
для них неприменимы. Однако, несмотря на то, что виды апомиктов пред-
ставляют собой группы параллельных линий, эти линии объединены эво-
люционным единством, связаны единым происхождением и единством
214
III. Эволюция растений
эколого-географических условий. Благодаря общности действия внут-
ривидового естественного отбора, эти линии еще сохраняют достаточ-
лую близость генных систем. Н.Н. Цвелев (1995) считает, что «микрови-
дь!» видовых комплексов или агрегатов заслуживают видового ранга не
меньше, чем многочисленные виды дрозофил или виды-двойники у зоо-
логов. Однако, по-видимому, здесь необходимы более осторожный под-
ход и детальное изучение, иначе количество описываемых видов превы-
сит все разумные пределы и существенно осложнит работу флористов
(Еленевский, 1998; Красников, 2000).
Анализируя мнения большинства исследователей об особенностях
вида у апомиктов, З.М. Рубцова (1989) считает, что у асексуальных
организмов ранг вида несколько отличается от такового у сексуаль-
ных. Однако вопрос о реальности вида у апомиктов приобретает ост-
роту лишь в том случае, когда апомиксис носит облигатный характер.
В этом случае, как отмечает Т. Добжанский (Dobzhansky, 1941), цело-
стная система вида нарушается и заменяется суммой репродуктивно
изолированных клонов, что в будущем приведет к исчезновению вида,
несмотря на то, что эти клоны могут сохранять и морфологическое
сходство, и сходную генетическую основу, и одинаковую экологичес-
кую приуроченность. Однако даже небольшой обмен генами позволя-
ет сохранить целостность популяции и вида.
Эволюционное значение апомиксиса заключается в том, что пере-
ход к нему позволяет сохраниться гибридам, возникшим в результате
скрещивания сексуальных видов. Эти гибриды могут стать родона-
чальниками новых линий. Как отмечает Стеббинс (Stebbins, 1941, цит.
по: Рубцова, 1989), этот процесс приводит к образованию большого
числа микровидов, сочетающих признаки родительских видов. Эти
микровиды являются генетически константными и отличаются от близ-
ких родственников мельчайшими деталями. Благодаря сохранению
апомиксисом сексуально стерильных гибридных производных стира-
ются генетические и морфологические различия между видами и об-
разуются структуры, названные Бэбкоком и Стеббинсом (Babcock,
Stebbins, 1938) агамными комплексами. Агамные комплексы характе-
ризуются очень высоким полиморфизмом, возникающим вследствие
Действия трех причин: гибридизации, полиплоидии и апомиксиса.
Авторы указывают, что эти комплексы обладают более сложным ти-
пом изменчивости, чем полиплоидные комплексы, и меньшим пере-
215
М.В. Олонова
рывом между группами, принимаемыми за виды. Как отмечает
З.М. Рубцова (1989), проанализировавшая литературу, касающуюся
апомиксиса, проблема вида может быть решена только частично. Ссы-
лаясь на работу В. Гранта (Grant, 1971, цит. по: Рубцова, 1989), она
считает, что апомиктические производные, имеющие на определен-
ной территории сходную морфологию, представленную отчетливой
комбинацией признаков, могут признаваться как таксономические
виды. Но в некоторых частях агамного комплекса видовая проблема
неразрешима. Если между сексуальными диплоидными и обширными
апомиктическими видами имеется огромное число производных, колеб-
лющихся от мелких биотипов до локализованных микровидов, то опре-
делить их таксономический ранг практически невозможно. Однако боль-
шинство систематиков склоняется к мысли, что в таких случаях целесо-
образно сохранить крупные виды в качестве видовых комплексов или
агрегатов (Heywood, 1963; Цвелев, 1967, 1976 а; Завадский, 1968).
Одним из частных случаев агамоспермии, также вносящим допол-
нительные сложности в систематику растений, является пролифера-
ция, когда над колосковыми чешуями возникает облиственный вегета-
тивный побег, или «бульбочка».
Большинство авторов указывают, что вивипарные популяции зла-
ков приурочены главным образом к высоким широтам и, реже
(Rajbhandari, 1988), к альпийскому поясу, при этом указывается
(Nannfeld, 1937), что вивипарные и невивипарные расы, морфологи-
чески отличающиеся только наличием бульбочек, нередко существу-
ют параллельно. Н.Н. Цвелев (1964, 1972а) отмечает, что они обыкно-
венно ведут себя как эколого-географические расы с вполне определен-
ным ареалом, который не всегда совпадает с ареалом основного вида, и
предполагает, что подобные вивипарные расы могли возникнуть в наи-
более холодное время четвертичного периода как адаптивное приспо-
собление, он считает целесообразным считать эти формы подвидами.
Вместе с тем исследования Munzig (1940) показали, что вивипарные
популяции тесно связаны с продуцирующими семена. Им были исследо-
ваны шведские популяции Р. alpina L., с 2п = 26 и 2п = 33, найденные
только в одном месте. Часть из них образовывала нормальные семена, а
часть - пролиферированные колоски, хромосомные же числа при этом
не изменялись, из чего был сделан вывод, что вивипарные клоны возни-
кают путем мутации из соответствующих семяпродуцирующих биотипов.
216
III. Эволюция растений
Многие исследователи склоняются к тому, что переход к пролиферации
осуществляется под воздействием особых экологических условий. Так,
Clayton (1965) высказывает мысль о том, что сочетание длинных и ко-
ротких дней должно было привести к гормональному дисбалансу у не-
которых генетически предрасположенных рас. Younger (1960, цит. по:
Vickery, 1970) провел экспериментальное исследование 30 австралийс-
ких популяций с разной степенью пролиферации в разное время года.
В результате было установлено, что некоторые растения имеют полнос-
тью пролиферированные метелки зимой, а метелки с почти всеми нор-
мальными колосками наблюдались поздней весной. Эксперименты по-
казали, что нормальные цветки развиваются при нормальной температу-
ре и долготе дня, а «бульбы» на их месте - под воздействием низких
температур и короткого дня. Позднее эту точку зрения поддержали мно-
гие исследователи. Nielsen (1941) добавил к списку факторов, способ-
ствующих пролиферации, повышенную влажность.
К. Flovic (1938), базируясь на цитологических исследованиях попу-
ляций пролиферированных Deschampsia, Festuca и Роа, заключил, что
пролиферация в Арктике - довольно обычное явление, обусловленное
сочетаниями следующих трех факторов: гибридизацией, полиплоидией
и экстремальными условиями. J.A. Nannfeld (1937, 1940), принимая во
внимание то, что пролиферированные растения являются, как правило,
полиплоидными и гибридогенными, высказал предположение, что это
явление продуцируется только в гибридогенных таксонах как способ
преодоления стерильности. Winkler (1908) рассматривал вегетативную
пролиферацию как частный случай апомиксиса.
Приведенные данные показывают, что, по-видимому, генетически
обусловлена сама способность тех или иных популяций образовывать
под воздействием соответствующих условий пролиферированные ко-
лоски. Причем эта способность может проявляться у одной части по-
пуляций вида и не проявляться у другой. При этом, по наблюдениям
Н.Н. Цвелева (1964), группа популяций, способная в определенных
условиях образовывать пролиферированные колоски, обладает сво-
им собственным ареалом, отличным от основного ареала вида. По-
этому, поскольку этот признак имеет определенную географическую
приуроченность и генетическую основу и связан, по мнению И.Е. Са-
рапульцева (1996), с генетическими рекомбинациями, считаем воз-
можным, следуя Н.Н. Цвелеву, придать популяциям, образующим
пролиферированные колоски, подвидовой ранг.
217
М.В. Олонова
Из вышеизложенного следует, что таксоны, образующиеся в резуль-
тате расхождения аллеломорф в популяциях, а также полиплоидизации
гибридизации и апомиксиса, имеют видовой уровень. Значительно чаще
результатом бывает повышение внутривидового полиморфизма.
3. Морфологическая и биологическая концепции вида
При решении вопроса о ранге образовавшейся совокупности прак-
тическая систематика вплотную приближается к другому аспекту про-
блемы - выбору концепции вида. Именно концепция вида, как указы-
вает А.С. Северцов (1988), является теоретической основой для выяв-
ления грани между подвидом и видом, именно от выбора концепции
зависит, считать ли 2 сравниваемые совокупности особей частями од-
ного и того же вида или принадлежащими к разным видам.
На протяжении нескольких веков со времен К. Линнея основным
критерием видовой самостоятельности признавалась морфологичес-
кая обособленность. В первой половине столетия работы С.И. Кор-
жинского (1892) и В.Л. Комарова (1934, 1940) послужили основой
создания так называемой монотипической концепции вида, которая в
России была принята большинством ботаников.
В сороковых годах нашего века сформировалась и получила широ-
кое распространение биологическая концепция вида, предложенная
Т. Добжанским (Dobzhansky, 1937), согласно которой вид является реп-
родуктивно изолированной совокупностью скрещивающихся между
собой популяций.
В.В. Гриценко, А.Б. Креславский, А.В. Михеев (1983), проанали-
зировав состояние проблемы вида на современном этапе, отметили
существование двух основных концепций - биологической и морфо-
логической, все остальные концепции в принципе можно свести к этим
двум. Согласно первой единство и целостность вида обеспечиваются
за счет обмена генами, а его дискретность обусловлена прекращением
этого обмена. Морфологическая концепция для разграничения видов
принимает весь комплекс фенотипических признаков. В морфологи-
ческой концепции основное внимание уделяется изменчивости коли-
чественных признаков и критерием для разграничения видов служит
наличие разрыва между значениями признаков.
218
III. Эволюция растений
Сторонники биологической концепции считали и считают репро-
дуктивную изоляцию основным критерием вида, отмечая при этом,
что истинные виды не способны давать при скрещивании плодовитое
потомство. Даже если два вида скрещиваются только в одной части
общего ареала, то и тогда их следует относить к одному виду. Наибо-
лее последовательным приверженцем этой концепции был Э. Майр
0974). Самой привлекательной чертой этой концепции является чет-
кий и объективный критерий, позволяющий ограничить рамки вида -
репродуктивная изоляция. Однако, как отмечает Е.Н. Панов (1993а),
если вид - строго генетически обособленная система, то вопрос о ре-
альности межвидовой гибридизации не может быть поставлен вооб-
ще, в то время как известно множество случаев, как среди животных,
так и среди растений, особенно среди злаков, когда плодовитое потом-
ство дают гибриды не только хорошо различающихся между собой
видов одной секции, но и представителей разных секций и даже ро-
дов. Тем не менее, по логике биологической концепции, как только
виды вступают в эффективную гибридизацию (дают жизнеспособное
и плодовитое потомство), их уже нельзя считать видами. Кроме того,
отмечает Е.Н. Панов со ссылкой на Г.А. Заварзина, у низших расте-
ний вполне обычным является обмен генетическим материалом меж-
ду таксонами, традиционно относящимися не только к разным видам,
но и к разным родам. Вместе с тем в литературе отмечаются случаи
репродуктивной изоляции между внутривидовыми формами (Грицен-
ко и др., 1983), а также особями разных популяций одного вида, не
имеющих морфологических отличий, которые сторонники биологи-
ческой концепции считают самостоятельными видами-двойниками
(Стегний, 1993). Если все эти случаи при желании можно рассматри-
вать как примеры неудачной систематики, то ограничения в примене-
нии биологической концепции являются серьезным ее недостатком.
Биологическая концепция неприменима не только к унипаренталь-
ным организмам, но, в отдельных случаях, по замечанию А.С. Север-
цова, и к бипарентальным. Это, во-первых, случаи разорванных ареа-
лов, когда изоляция заведомо абсолютная. Согласно биологической кон-
цепции каждая из частей разорванного ареала должна рассматриваться
как самостоятельный вид. Во-вторых, бипарентальные виды, у кото-
рых в качестве системы скрещиваний в отдельных популяциях преоб-
ладает инбридинг (некоторые беспозвоночные, самоопыляющиеся
219
М.В. Олонова
растения, апомикты). Вид в этом случае оказывается подразделенным
на систему чистых линий. В-третьих, при наличии гибридных зон меж-
ду бипарентальными видами. Кроме того, и поток генов, призванный
нивелировать возникающие в отдельных популяциях изменения, в на-
стоящее время признается лишь одним из механизмов, интегрирую-
щих вид, но механизмом не единственным и не обязательным (Север-
цов, 1988). По мнению же Г. Симпсона (Simpson, 1961), главный воп-
рос состоит не в том, происходит ли между двумя видами гибридизация
или нет, а если несмотря на гибридизацию эти виды не сливаются в
единое целое, то и в эволюционной перспективе они останутся отдель-
ными видами.
Снижает ценность этой концепции и признание сетчатой эволю-
ции не только у растений (Стеббинс, 1957; Цвелев, 1976а), айв неко-
торых группах рыб, амфибий и рептилий (Панов, 19936). Кроме того,
следует признать, что в отношении возможной гибридизации между
видами ботанические объекты в настоящее время изучены значитель-
но слабее, чем зоологические - практически очень трудно установить,
скрещиваются или не скрещиваются в природе те или иные популя-
ции, и уж совсем невозможно определить это на гербарном материа-
ле, большой объем которого приходится использовать при системати-
ческих исследованиях.
Широкое распространение у растений апомиксиса и гибридизаци-
онных процессов между отдаленно родственными группами, разор-
ванность ареалов многих видов, сетчатая эволюция, отмечающаяся во
многих группах, затрудняют использование биологической концепции,
но главное, что понижает ее ценность в глазах ботаника-систематика,
- это невозможность ее практического использования.
Морфологическая концепция возникла на базе типологической. Эта
концепция основана на дискретности видов, направлена на выявление
хиатусов по морфологическим признакам, а с развитием методов био-
химического анализа и молекулярных исследований принимает во вни-
мание и эти данные. Таким образом, главная роль здесь принадлежит
признакам. Классическая типология - это методы установления гомо-
логическим путем сравнения различных организмов с целью постро-
ения типа и его дальнейшего использования для построения класси-
фикации (Канаев, 1989). Как отмечает Л.Н. Васильева в этой же рабо-
те, в середине нашего столетия, в пору господства СТЭ, понятие типа
220
111. Эволюция растений
было сведено до представления о типовых музейных образцах (Майр,
1971; Грант, 1991), и типология из науки сравнительного анализа го-
мологических признаков в сознании многих исследователей превра-
тилась в науку стандартов и шаблонов, которой чуждо всякое разви-
тие. Однако в настоящее время типология вновь занимает утраченные
позиции. Решающая роль в разграничении видов вновь отводится при-
знакам, но поскольку фенетическая концепция, базирующаяся только
на количестве исследованных признаков, без оценки их значимости,
явно не оправдала себя, основное внимание последователи морфоло-
гической концепции уделяют исследованию признаков, изучению их
роли и оценке их значимости.
Согласно морфологической концепции, если группировки по сво-
им признакам дискретны относительно друг друга, они, по всей веро-
ятности, принадлежат к разным видам, а если не дискретны - то к
одному. Основная, однако, сложность состоит в том, что, как отмечает
А.С. Северцов (1988), далеко не всякая дискретность свидетельствует
о видовом статусе. Им приводится 5 случаев, когда виды были выде-
лены неправомерно несмотря на дискретность признаков. Среди них
- дискретный внутривидовой полиморфизм (выделение видов на ос-
нове кариологических различий), дискретный модификационный по-
лиморфизм, когда видовой ранг был присвоен разным цветовым мо-
дификациям бабочек, дискретный половой полиморфизм и дискрет-
ность последовательных стадий онтогенеза. Единственно значимой,
по мнению А.С. Северцова для выделения видов дискретностью явля-
ется дискретность по количественным непрерывно изменяющимся
признакам, определяемым полигенно. На последнее обстоятельство
также указывает и А.П. Кожевников (1987, 1996), отмечая многочис-
ленные случаи неправомерного придания видового статуса географи-
чески дифференцированным аллеломорфам. В.С. Фридман (1991) в
качестве недостатков также отмечает необходимость некоторых допу-
щений - об абсолютности обнаруженного хиатуса между формами, о
существенности выбранных систематиком признаков. Таким образом,
для успешного применения типологической концепции в систематике
требуется детальное изучение изменчивости признаков.
В.В. Гриценко, А.Б. Креславский, А.В. Михеев (1983) справедли-
во утверждают, что на данном этапе нет серьезных оснований считать
морфологическую концепцию устаревшей, подчеркивая, что «морфо-
221
М.В. Олонова
логическая концепция вида в настоящее время более точно отражает
объективные закономерности живой природы, нежели биологическая»
(Гриценко и др., 1983). Очень убедительные доказательства в пользу
морфологической концепции приводит в серии своих работ Л.Н. Ва-
сильева (1987, 1989а, б). В сущности, большинство возражений про-
тив морфологической концепции сводится к возражениям против опи-
сания видов по отдельным образцам без учета изменчивости призна-
ков в популяциях, но эта практика уже давно не находит одобрения у
ботаников.
При всех достоинствах типологической концепции вышеперечис-
ленные недостатки затрудняют ее применение и вызывают немало
нареканий. Л.В. Васильева (19896) указывает, что в том виде, в каком
она сейчас существует, типология не может стать лидером в система-
тике и привести к коренной перестройке. Дальнейшее развитие этой
концепции она связывает с филогенетической систематикой (кладиз-
мом, нашедшим широкое применение в зоологии, в настоящее время
приобретающим популярность и у ботаников).
Л.В. Васильева и Л.Я. Боркин (1986) причину близости кладизма и
классической типологии видят в том, что филогенетическая система-
тика зародилась в лоне морфологической типологии. При этом, как
указывает Л.В. Васильева, в современном кладизме естественно воз-
никла ветвь «типологических кладистов», идеи которых возвращают
систематике прочную основу. На подготовительном этапе классифи-
кации филогенетика использует почти те же самые приемы - распре-
деление организмов по состояниям одних и тех же признаков очень
напоминает гомологизацию различных частей в типологии. При этом
большинство различий, проводимых филогенетическими системати-
ками между плезиоморфными и апоморфными состояниями призна-
ков, фактически являются типологическими различиями. Это дает
право утверждать, что в основе филогенетической систематики лежит
морфологическая типология (Васильева, 19896). Развитие типологии,
по ее мнению, остановилось на первом этапе систематики - поиске
сходных черт с целью объединения близких организмов в группы.
Второй этап, представляющий для систематики наибольший инте-
рес, - распределение групп по рангам - методологически не разрабо-
тан, что приводит к полнейшему субъективизму в этой области, и во-
обще как нечто научное признается зависимость этого этапа от того,
222
III. Эволюция растений
является ли систематик «дробителем» или «укрупнителем». Крайние
сТОронники монотипической концепции стремятся разделить виды на
серию мелких, морфологически однородных видов. Но абсолютно
мономорфных видов, как полагал Н.И. Вавилов (1931), не существу-
ет. И важнейшее достижение эволюции - амфимиксис - как раз и со-
стоит в том, чтобы разные индивидуальные генотипы связать в еди-
ный генотип. Эта генетическая и морфологическая неоднородность и
служит основой экологической пластичности и устойчивости таксона
(Северцов, 1990). Однако субъективизм в этом вопросе уже породил и
продолжает порождать множество недоразумений и неудобств.
Для выхода из этой тупиковой ситуации, в которой оказалась со-
временная систематика, предлагается метод соотносительного взве-
шивания отличающих признаков на разных иерархических уровнях
(Васильева, 1987). Применение этого метода требует детального ис-
следования многих признаков самого разного характера, включая реп-
родуктивную изоляцию (поскольку эволюция идет не только по мор-
фологическим признакам), но представляется очень перспективным
для решения одной из основных задач систематики - определения так-
сономического ранга отдельных совокупностей.
В настоящее время многие сторонники биологической концепции
не считают репродуктивную изоляцию абсолютным видовым крите-
рием. Так, А.П. Расницын (1975) отмечает, что методика выявления
биологических видов комплексная и критерий скрещиваемости, взя-
тый изолированно, дает в этом отношении очень мало. В.С. Фридман
(1991), не отрицая возможности гибридизации, подчеркивает, что меж-
ду популяциями разных видов должен отсутствовать поток генов, в то
время как сама гибридизация в природе может происходить. Вопрос о
применимости биологической концепции к гибридизирующим фор-
мам, по его мнению, требует изучения интенсивности потока генов.
Однако в рамках биологической концепции гибридизация рассматри-
вается как редкое нарушение, не имеющее эволюционных перспек-
тив, в то время как морфологическая признает гибридизацию как один
из важнейших факторов видообразования.
А.К. Скворцов (1968) замечает, что степень генетической совмес-
тимости или несовместимости, определяющая способность к гибри-
дизации, - всего лишь один из возможных физиологических призна-
ков и, следовательно, ее таксономическая ценность в разных случаях
223
М.В. Олонова
может быть разной. Этого же мнения придерживается Л.Н. Васильева
(1987). Тем не менее, несмотря на некоторую схожесть взглядов по воп-
росу об оценке генетической совместимости, сближение этих концеп-
ций вряд ли возможно. Так, В.С. Фридман (1993), детально изучив сте-
пень согласованности дивергенции различных признаков и степень
приспособленности гибридов у птиц, установил, что все изученные
признаки можно разделить на 2 согласованно эволюционирующие
группы. Первая из них содержит селективно-нейтральные признаки
(различия в составе ДНК и белков, различия в наборе коммуникатив-
ных сигналов), а вторая - адаптивные признаки (морфологические и
экологические). Эти группы признаков, по мнению В.С. Фридмана, ли-
вергируют независимо друг от друга, при этом биологическая концеп-
ция выделяет виды по первой группе признаков, а типологическая
по второй. Поэтому если придерживаться первой концепции, то будут
выделены группировки, дискретные репродуктивно, но морфофунк-
ционально (в общем случае) не единые. Если же следовать второй - то
к одному виду будут отнесены особи, сходные по набору адаптаций и
эквифинальным фенотипам. При этом такие морфологически диск-
ретные виды могут интрогрессировать или скрывать в себе виды-двой-
ники. Если в основу выделения таксонов кладут единство происхож-
дения, то виды надо выделять в рамках биологической концепции, а
если утверждать, что каждый из таксонов обладает инвариантной для
него адаптивной зоной, то они должны выделяться на основании сход-
ства скоррелированных друг с другом адаптивных признаков.
Поскольку дивергенции признаков 1 и 2 групп не связаны между
собой, выделенные таким образом таксоны не будут монофилетичес-
кими, а дивергенция признаков, по которым будут выделяться таксо-
ны, с родством будет связана лишь случайно. Попытка объединения
этих подходов, которую предпринял в своей работе А.П. Расницын
(1983), по мнению В.С. Фридмана, приводит к ситуации, когда неиз-
вестно, в какой степени система отражает ветвление филума, а в какой
- адаптогенез. Справедливости ради следует отметить, что эта точка
зрения разделяется не всеми, в частности, Е.Н. Панов (19936) считает
выводы В.С. Фридмана интересными, но все же спорными. Тем не
менее по имеющимся литературным данным системы, составленные
с использованием молекулярных данных, нередко отличаются от сис-
тем, построенных традиционным способом на основе морфологичес-
224
Ill. Эволюция растений
ких признаков. А.С. Антонов (2000) объясняет это двумя причинами.
go-первых, молекулярные биологи обычно строят свои деревья на
основании изучения только одного из многих тысяч генов, образую-
щих геном, а такой объем явно недостаточен; во-вторых, иногда суще-
ствуют принципиальные отличия в механизмах молекулярной и орга-
низменной эволюции, особенно у таксонов низшего ранга, поэтому
феносистематика и геносистематика есть 2 различные и в разной сте-
пени разработанные части систематики, но попытки объединить их не
могут иметь успеха.
Современная систематика для решения своих задач использует дан-
ные различных фундаментальных наук по отдельным таксонам. Как
за рубежом, так и в нашей стране ботаниками проводятся успешные
работы по использованию биохимических признаков для целей систе-
матики. Эти методы позволяют с удивительной точностью установить
родственные связи между видами, особенно у гибридов и гибридоген-
ных видов. Однако, к сожалению, и они не универсальны, так как, во-
первых, случается, что одинаковые белки маркируют разные виды и
наоборот (Медников, 1989). Молекулярные методы позволяют рабо-
тать с точностью до отдельных популяций, поэтому разграничение
видов делается тоже достаточно субъективно. Главное же то, что для
успешного применения этого метода необходимо очертить границы
вида при помощи морфологического метода.
Заключение
В настоящее время большинство биологов признает, что абсолют-
ного критерия вида не существует, основные критерии вида, выдвига-
емые сторонниками биологической концепции, не носят универсаль-
ного характера и не могут использоваться в отрыве от методов класси-
ческой морфологии.
По-видимому, следует согласиться с мнением многих авторов, что
широкое привлечение данных биохимического и генетического ана-
лиза позволит избежать многих ошибок при определении границ ви-
дов, однако традиционный морфологический анализ и в настоящее
время сохраняет в систематике ведущую роль гибридологического или
биохимического методов, мы, тем не менее, всегда за основу берем
Морфологическое описание данной группы.
225
М.В. Олонова
С учетом всего изложенного для ботаников-систематиков более
приемлемой представляется морфологическая концепция, в которой
главная роль отводится морфологическим признакам с учетом данных
других специальностей, поскольку, хотя эволюция и идет по всем при-
знакам, отражением генотипа является именно фенотип.
Приведенный обзор литературы по проблеме вида у растений дает
основание считать, что несмотря на достижения эволюционной науки
и генетики определение объема вида и его границ остается в значи-
тельной мере субъективным. Поэтому ботаникам-систематикам для
повышения объективности своих заключений о видовых границах сле-
дует максимальное внимание уделить выбору и оценке признаков, на
основании которых строится систематика, причем изучать их измен-
чивость следует на всем протяжении видового ареала.
«Вид можно определить как обособившуюся в процессе эволюции
систему клонов или популяций, объединенную общими признаками
(морфологическими, экологическими, биохимическими, генетически-
ми, цитологическими и т.д.), общим происхождением, общим геогра
фическим ареалом и достаточно четко отделенную от близких видов
как совокупностью своих признаков, так и разными изолирующими
барьерами» (Тахтаджян, 1984, с. 8).
Автор выражает искреннюю благодарность профессору Томского
государственного университета А.В. Положий и старшему преподавате-
лю кафедры генетики Ю.М. Новикову за ценные советы и замечания.
Литература
Антонов А.С. О возможных причинах расхождения взглядов фено- и геносистемагиков на
филогению и систему высших растений И Боган, журн. 2000. T. 85. № 1. С. 3-11.
Боркин Л.Я. Через кладизм - к новой систематике (вместо предисловия) И Принципы и
методы зоологической систематики: Тр. Зоол. ин-та АН СССР. 1989. T. 206. С. 3-8
Вавилов Н.И. Линнеевский вид как система. М.; Л.: Сельколхозгиз, 1931. 32 с.
Васильева Л.Н. Пиреномицеты и локулоаскомицеты севера Дальнего Востока. Л.: На-
ука, 1987. 257 с.
Васильева Л.Н. Типологическая школа систематики И Методологические проблемы
биологии и экологии. Владивосток, 1989 а. С. 26-43.
Васильева Л.Н. Проблемы нетипологической систематики И Методологические про-
блемы биологии и экологии. Владивосток, 1989 б. С. 57-70.
Глотов Н.В. Необходимость эколого-генетического синтеза в теории микроэволюции
И Дарвинизм: история и современность. Л., 1988. С. 45-56.
226
III. Эволюция растений
г ант В. Видообразование у растений. М.: Мир, 1984. 528 с.
ЛитН Эволюционный процесс: Критический обзор эволюционной теории. М.: Мир,
j раш
1991. 488 с.
Гриценко В.В., Креславский А.Б., Михеев А.В. и др. Концепции вида и симпатричес-
кое видообразование. М.: Изд-во МГУ, 1983. 193 с.
Дубинин Н.П. Эволюция популяций и радиация. М.: Наука, 1966. 743 с.
Еленевский А.Г Рецензия на книгу «Флора Липецкой области» И Бюл. МОИП. 1998.
№ 5. С. 68.
Завадский К.М. Вид и видообразование. Л.: Наука, 1968. 404 с.
Кабанова В.М., Карташова Н.Н. Кариотипы кровососущих комаров рода Aedes (Dipfera,
Culucidae) // Генетика. 1972. Т. 8. № 3. С. 47.
Кашин А.С. Сальтационное видообразование и апомиксис у покрытосеменных
//IX Московское совещ. по филогении раст. М., 1996. С. 66-69.
Кашин А.С. Структура агамокомплексов и проблема сальтационного видообразования
у покрытосеменных // Ботан. журн. 1999. Т. 84. № 1. С. 15-29.
Кейлоу П. Принципы эволюции. М.: Мир, 1986. 128 с.
Кожевников Ю.П. Популяционно-генетическая изменчивость видов и ее отражение в
систематике растений // Ботан. журн. 1987. Т. 72. № 7. С. 874-887.
Кожевников Ю.П. Растительный покров Северной Азии в исторической перспективе.
СПб.: НПО «Мир и семья», 1996. 400 с.
Комаров В.Л. Предисловие редактора// Флора СССР. Л.: Наука, 1934. Т. 1. С. 1-12.
Комаров В.Л. Учение о виде у растений (страницы истории биологии). М.; Л.. Изд-во
АН СССР, 1940.212 с.
Коржинский С.И. Предварительные замечания // Флора востока Европейской части
России. Томск, 1892. С. 9-27.
Красников А.А. К вопросу разумного подхода к микровидам И Проблемы изучения
растительного покрова Сибири: Тез. докл. II Рос. науч, конф., поев. 150-летию со
дня рожд. П.Н. Крылова. Томск, 2000. С. 69-70.
Майр Э. Систематика и происхождение видов. М.: Изд-во Иностр, лит., 1947. 502 с.
Майр Э. Популяции, виды, эволюция. М.: Мир, 1974. 460 с.
Медников Б.М. Современное состояние проблемы вида и видообразованиия // Вави-
ловское наследие в современной биологии. М., 1989. С. 133-147.
Панов Е.Н. Предисловие // Гибридизация и проблема вида у позвоночных: Сборник
трудов. М., 1993. С. 3-5.
Панов Е.Н. Граница вида и гибридизация у птиц И Гибридизация и проблема вида у
позвоночных: Сборник трудов. М., 1993. С. 53-96.
Расницын А.П. К вопросу о виде и видообразовании // Проблемы эволюции. Новоси-
бирск, 1975. Т. 4. С. 221-230.
Расницын А.П. Филогения и систематика // Теор. проблемы современной биологии.
Пущино, 1983. С. 41-49.
Рубцова З.М. Эволюционное значение апомиксиса: историко-критическое исследова-
ние. Л.: Наука, 1989. 154 с.
Сарапульцев И.Е. Репродуктивная биология двух арктических видов мятлика
// Экология. 1996. № 5. С. 345-349.
227
М.В. Олонова
Северцов А.С. Современные концепции вида // Бюл. МОИП. Отд. биол. 1988
Т. 93. №6. С. 3-15.
Северцов А.С. Внутривидовое разнообразие как причина эволюционной стабильнос-
ти //Журн. общ. биол. 1990. Т. 51. № 5. С. 579-589.
Симпсон Дж. Темпы и формы эволюции. М.: Изд-во иностр, лит., 1948. 358 с.
Скворцов А.К. Ивы СССР. М.: Наука, 1968. 336 с.
Скворцов А.К. Сущность таксона и проблемы внутривидовой систематики растений
(некоторые соображения и предложения) // Бюл. МОИП. Отд. биол. 1971. Т. 7 6
Вып. 5. С. 72-81.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. М.: Мир, 1982. 488 с
Стеббинс Д.Л. О гибридном происхождении покрытосеменных // Боган, журн. 195^
Т. 42. №10. С. 1503-1506.
Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск
Изд-во Новосибирск, ун-та, 1993. 111 с.
Тахтаджян А.Л. Макроэволюционные процессы в истории растительного мира // Бо-
тан. журн. 1983. Т. 68. № 12. С. 1593-1603.
Тахтаджян А.Л. Предисловие редактора перевода // Грант В. Видообразование у расчс
ниЙ. М.: Мир, 1984. 528 с.
Тихомиров В.Н. Некоторые особенности систематики облигатных апомиктов на при-
мере рода Alchemilla L. И Совещ. по объему вида и внутривидовой систематике Л
1967. С. 3-55.
Фридман В.С. «Кризис» биологической концепции вида в орнитологии и таксономи-
ческий статус гибридизирующих и гибридогенных форм // Журн. общ. биол. 1991
Т. 52. № 4. С. 476-487.
Фридман В.С. Согласованность дивергенции гибридизирующих форм и состоятся!»
ность биологической концепции вида у птиц// Журн. общ. биол. 1993. Т. 54. № 1
С. 81-95.
Цвелев Н.Н. Роа L. - мятлик // Арктическая флора СССР. М.; Л., 1964. Вып. 2. С. 112-162.
Цвелев Н.Н. О таксономическом ранге членов полиплоидных рядов у высших расте-
ний И Совещ. по объему вида и внутривидовой систематике. Л., 1967. С. 59-60.
Цвелев Н.Н. О значении гибридизационных процессов в эволюции злаков (Роасеае)
И История флоры и растительности Евразии. Л., 1972. С. 5-16.
Цвелев Н.Н. О значении степени специализации таксонов для их дальнейшей эволю-
ции // Бюл. МОИП. Отд. биол. 1973. Т. 78. Вып. 2. С. 71-82.
Цвелев Н.Н. О возможности деспециализации путем гибридогенеза на примере эволю-
ции трибы Triticae семейства злаков (Роасеае) И Журн. общ. биол. 1975. Т. 36. № 1
С. 90-99.
Цвелев Н.Н. Злаки СССР. Л.: Наука, 1976а. 788 с.
Цвелев Н.Н. О происхождении арктических злаков // Ботан. журн. 19766. Т. 61, № Н)
С. 1354-1363.
Цвелев Н.Н. Вид как один из таксонов // Бюл. МОИП. Отд. биол. 1995. Т. 100. Вып. 5
С. 62-68.
Цвелев Н.Н. Новые тенденции в теории филогении высших растений ИIX Московское
совещ. по филогении раст. М., 1996. С. 151-153.
228
III. Эволюция растений
Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения совре-
менной генетики//Журн. эксперим. биол. 1926. Сер. А. Т. 2. Вып. 3. С. 54-95.
Ц1варн С.С. Экспериментальные методы исследования начальных стадий микроэво-
люционного процесса//Внутривидовая изменчивость наземных позвоночных жи-
вотных и микроэволюция: Тр. Всес. совещ. Свердловск, 1965. С. 21-32.
Babcock Е.В., Stebbins G.L. The american species of Crepis: their relationships and
distribution as affected by polyploidy and apomixis // Publ. Camegia Inst. Washington.
1938. № 504. P. 11-99.
Carson H.L. Speciation and the founder principle // Stadler Genet. Symp. 1971. Vol. 3.
P. 51-70.
Carson H.L. The genetics of speciation at the diploid level //Amer. Nat. 1975. Vol. 109.
№ 965. P. 1-83.
Dobzhansky Th. Genetics and the Origin of species. New York: Columbia Univ. Press, 1937b.
364 p.
Dobzhansky Th. Genetics and the origin of species. 2 ed. New York. 1941. 446 p.
Elredge N., Gould S.J. Punctuated equilibria: an alternative to phyletic gradualism
//Models in Paleobiology. San-Francisco, 1972.
Flovic K. Cytological studies of arctic grasses // Hereditas. 1938. Vol. 24. № 2. P. 265-376.
Grant V. The plant species in theory and practice // The species problem. Publ. Amer. Assoc.
Adv. Sci. 1957. № 50. P. 39-80.
Hanelt P, Mettin D. Spetiation process within the genus Vicia L. // Abstr. pap., presented at
the XII Int. Bot. Congr. Leningrad, 1975. Vol. 1. P. 14.
Hennig W. Philogenetic systematics. Urbana: Univ. Illinois press, 1966. 263 p.
Hunziker J.H., Stebbins G.L. Chromosomal evolution in the Gramineae // Grass Syst. and
Evolution: Int. Symp. Washington D.C. London, 1986. P. 179-187.
Huxley J.S. Evolution, the Modem Synthesis. London: Allen and Unwin, 1942. 654 p.
Lewis H. Catastrophic selection as a factor in speciation // Evolution. 1962. Vol. 16.
P. 257-271.
Lotsy J. Evolution by means hybridization. The Hague, 1916.
Muntzing A. Further studies on apomixis and sexuality in Poa// Hereditas. 1940. T. 26. № 2.
P. 131-154.
Nannfeldt J.A. On Poa jemtlandica (Alqn.)Riecht., its distribution and possible origin. A
criticism of the theory of hybridization as the cause of vivipary II Bot.nat. 1937. P. 22.
Nannfeldt J.A. On the polymorphy of Poa arctica R.Br., with special reference to its
Scandinavian forms // Symb. Bot. Uppsala. 1940. Vol. 4. P. 1-85.
Nielsen E.L. Grass studies Y Observations on proliferation // Bot Gaz. 1941. Vol. 103. P. 177-181.
Rajbhandari K.R. Three new species of Himalayan Poa, with a note on proliferation
//Acta Phytotaxon, et Geobot. 1988. Vol. 39. № 1-3. P. 55-66.
Simpson G.G. Principles of Animal Taxonomy. New York: Columbia Univ. Press, 1961.351 p.
Stanley S.M. A theory of evolution above the species level // Proc. Nat. Acad. Sci. U.S.A.
1975. Vol. 72. P. 646-650.
Stebbins G.L. Variation and evolution in plants. New York, 1950. 643 p.
Stebbins G.L. The role of hybridization in evolution // Proc. Amer. Phil. Soc. 1959. Vol. 103.
R 231-251.
229
М.В. Олонова
Timofeeff-Ressovsky N.W. Zur Analise des Polymorphismus bei Adalia punctata L
// Biol. Zentr.-Bl. 1940 a. B. 60. H. 3/4. S. 130-137.
Timofeeff-Ressovsky N.W. Mutation and Geographical variation // The New Systematics.
J.S.Huxley ed. Oxford: Clarendon Press, 1940 b. P. 13-136.
Vickery J. W. A taxonomic study of genus Poa L. in Australia// Contr. New South Wales Natl
Herb. 1970. Vol. 4. P. 145-243.
Winkler H. Uber Parthenogenesis und Apogamie im Pflanzenreiche // Progressus rei Botanicac
1908. B. 2. S. 293-454.
IV. эволюция животных
СТРАТЕГИИ ОТБОРА
М.И. Гордеев
Институт общей генетики РАН
E-mail: gordeev@vigg.ru
В теории эволюции важное место отводится проблеме адаптации
организмов. Адаптация (от лат. adapto - прилаживаю) - это совокуп-
ность морфофизиологических, поведенческих, популяционных и дру-
гих особенностей данного биологического вида, обеспечивающая воз-
можность специфического образа жизни в определенных условиях
внешней среды (Биологический энциклопедический словарь, 1995).
Под адаптацией понимают, с одной стороны, сохранение устойчивос-
ти биологических систем в данных условиях среды. С другой сторо-
ны, адаптация - это такое состояние системы, которое обеспечивает
ее развитие при изменении внешних условий. На популяционном уров-
не адаптация обеспечивается за счет создания и сохранения субпопу-
ляционной структуры и реализуется в процессе отбора.
Очевидно, существуют особые закономерности отбора в пери-
од, когда происходит формирование вида и освоение новой ниши.
Совершенно другие закономерности отбора характеризуют эволюци-
онный стазис - период сохранения популяционной структуры. И осо-
бые закономерности отбора присущи процессу деградации популяций
и вымиранию видов.
В генетике популяций элементарным эволюционным явлением счи-
тают устойчивое изменение частоты гена (аллеля). В 60-70-е годы
любое изменение частоты гена рассматривали как этап видообразова-
ния. В настоящее время мы отдаем себе отчет в том, что изменение
частоты само по себе не свидетельствует о закономерном преобразо-
вании популяционно-генетической структуры вида.
Строго говоря, отбор идет по фенотипам, и отбираются не отдель-
ные гены, а весь генотип в конкретном его проявлении. Фактически
231
М.И. Гордеев
отбору подвергаются особи, имеющие определенные комбинации скор-
релированных признаков. Группы таких особей и образуют субпопу-
ляционную структуру. Под адаптивной стратегией мы понимаем се
лекцию особей, характеризующихся системой скоррелированных при-
знаков и формирующих внутривидовые группировки с определенным
набором таких признаков. Само сочетание скоррелированных призна-
ков организма было названо оптимальной композицией (Rosen, 1967)
или адаптивным комплексом (Bartholomew, 1972; Пианка, 1981). Адап-
тивные комплексы создаются и поддерживаются характерной для дан-
ного местообитания системой векторов отбора (Северцов, 1990).
Характеристики адаптивных комплексов определяются тем, как
возникшие адаптации связаны с репродуктивной сферой, как они вли-
яют на трофические связи и энергетический обмен. Отправной точкой
для выделения адаптивных стратегий служит кривая роста популяции
в ограниченном объеме среды, которая описывается логистическим
уравнением Ферхульста-Пирла:
dN/dt = rN(K-N)/K,
где N - число особей; г - врожденная (специфическая) скорость есте-
ственного увеличения популяции; К - предельная плотность насыще-
ния (максимальная численность, которую может достигнуть популя-
ция в данном объеме среды). Согласно этому уравнению график роста
численности популяции представляет собой S-образную кривую, вы-
ходящую на плато при достижении предельной плотности насыщения
К (Пианка, 1981). Логистическая кривая дает основу для понимания
адаптивных характеристик популяционно-видовых систем.
Подходы, связанные с выделением типов адаптивных страте-
гий, развивались параллельно в фитоценологии и популяционной эко-
логии животных и в значительной степени пересекаются. В фитоце-
нологии адаптивные стратегии определяются как способы выживания
популяций растений в сообществах и экосистемах (Миркин и др., 1989)
Классификации адаптивных стратегий основываются либо на средо
образующей способности растений (Сукачев, 1928), либо на их конку-
рентоспособности в местообитаниях различного типа (Раменский.
1938; Grime, 1974, 1979).
232
IV. Эволюция животных
В популяционной экологии животных в основе классификации адап-
тивных стратегий лежит соотношение затрат на размножение и на под-
держание жизнедеятельности организмов. В разреженной среде наи-
лучшей репродуктивной стратегией считается максимальный вклад
вещества и энергии в размножение с целью получить наибольшее число
потомков в наиболее сжатые сроки. В насыщенной среде, где велико
действие зависящих от плотности факторов, оптимальной стратегией
будет расходование энергии в первую очередь на преодоление конку-
ренции, на повышение собственной жизнеспособности и продуциро-
вание более жизнеспособного потомства. Этот подход получил назва-
ние концепции r/K-отбора, в соответствии с двумя параметрами логи-
стического уравнения (MacArthur, Wilson, 1967).
Характерными признаками К-отбора считаются: высокое постоян-
ство или предсказуемость среды обитания; зависящая от плотности
смертность; относительно постоянный, близкий к предельной емкос-
ти биотопов размер популяции; острая внутри- и межвидовая конку-
ренция; высокая продолжительность жизни. Такой отбор должен бла-
гоприятствовать более медленному развитию особей, высокой их кон-
курентоспособности, позднему началу репродукции с многочислен-
ными повторяющимися в течение жизни актами размножения, мень-
шему числу более крупных потомков. К признакам r-отбора как аль-
тернативной стратегии соответственно относятся: неопределенный и
непредсказуемый характер изменений среды; ненаправленная, часто
катастрофическая, не зависящая от плотности смертность; неравно-
весный, обычно ниже предельной емкости биотопов размер популя-
ции; относительно слабая внутри- и межвидовая конкуренция; корот-
кая продолжительность жизни. Отбор благоприятствует: быстрому
развитию особей; высокой скорости роста популяции; более раннему
И единственному в течение жизни акту размножения; большому числу
мелких потомков. Подчеркивается, что r-стратегия свойственна ран-
ним стадиям сукцессии и направлена на более высокую продуктив-
ность популяций в биоценозах. К.-стратегия характерна для поздних,
климаксных стадий сукцессии и способствует повышению эффектив-
ности использования ресурсов. Реализованные ниши r-стратегов оп-
ределяются прежде всего физической средой, а у К-стратегов - пре-
имущественно действием биотических факторов (Пианка, 1981).
233
М.И. Гордеев
Отмечается относительность классификации видов на г- или К-стра-
тегов. Нет видов, подверженных исключительно г- или К-отбору: каж-
дый должен достигнуть определенного компромисса между двумя край-
ними вариантами адаптивных стратегий. О каждом организме можно
говорить как о r-стратеге или К-стратеге при сравнении с другими
организмами. Относительность классификации связана также с дина-
микой ниш. Давление отбора и ниша могут меняться в пространстве и
во времени. Например, в умеренной зоне ранняя весна - это время,
когда организмы с коротким жизненным циклом подвержены, в ос-
новном, r-отбору, тогда как позднее они оказываются в условиях К-
отбора. В пределах одного вида одни особи могут в большей степени
зависеть от r-отбора, чем другие, если они находятся в особых микро-
местообитаниях или частях видового ареала (Пианка, 1981).
Важнейшей задачей эволюционной экологии является определе-
ние свойств адаптивных комплексов, их генетической и метаболичес-
кой основы. Фундаментальное различие между г- и К-стратегами со-
стоит в относительном распределении ресурсов между анаболически-
ми и катаболическими процессами. Выделяют две фунциональные
стратегии адаптации в условиях стресса - резистентную и толерант-
ную (Кулинский, Ольховский, 1992). Резистентная стратегия характе-
ризуется активным противодействием внешней среде, максимизацией
функций основных физиологических систем и, в результате, сохране-
нием гомеостаза или даже явлением гиперкомпенсации. Биохимичес-
кой основой этой стратегии является сдвиг к преобладанию катабо-
лизма эндогенных и экзогенных питательных веществ, в первую оче-
редь углеводов. Стратегия резистентности - это гиперкатаболическая
калоригенная стратегия. Она неэкономична, но позволяет организму
преодолевать неблагоприятные факторы внешней среды за счет моби-
лизации энергетических ресурсов. Согласно нашим представлениям,
эта стратегия служит физиологической составляющей адаптивных
комплексов, поддерживаемых г-отбором.
Стратегия толерантности характеризуется большим “подчинени-
ем” условиям внешней среды, минимизацией функций, что в услови-
ях стресса неизбежно приводит к определенным нарушениям гомеос-
таза (Кулинский, Ольховский, 1992). Однако организм устойчив к та-
ким нарушениям, поскольку они не чрезмерны. Данная стратегия
направлена не на поддержание гомеостаза, а на сохранение жизни в
234
IV. Эволюция животных
неблагоприятных условиях. Например, разные формы гипобиоза уве-
личивают толерантность к травмам, гипоксии, интоксикациям. Мета-
болической основой стратегии толерантности является снижение ме-
таболизма эндогенных полимеров, энергозатрат и потребления О2.
Считается, что стратегия толерантности типична для более древних
животных, в том числе для низших позвоночных, которым свойствен-
ны: более низкий основной обмен, меньшее количество митохондрий
в тканях, слабая термоизоляция и, в результате, экзотермия. В выра-
женной форме это эволюционно закрепленное приспособление про-
является в виде периодического оцепенения (при перегреве; переох-
лаждении; пересыхании и т. д.). Стратегия толерантности наиболее
эффективна при ограничении потребления О2, при чрезмерном дей-
ствии повреждающих факторов или при недостатке пищи, которые
делают невозможной или невыгодной гиперкатаболическую калори-
генную стратегию. Это эволюционно закрепленная реакция опреде-
ленных групп организмов, которых мы относим к К-стратегам. Фи-
зиологические особенности организмов, сформировавшихся в усло-
виях г- и К-отбора, проявляются в первую очередь при пороговых
воздействиях факторов внешней среды, поскольку именно в этих ус-
ловиях удается зафиксировать несовпадение нормы реакции у срав-
ниваемых особей.
Способы идентификации г- и К-стратегов являются самостоятель-
ной проблемой, как и выявление генетических детерминант, опреде-
ляющих состав адаптивных комплексов. В этой связи представляет
интерес генетический анализ внутривидовых группировок, отличаю-
щихся по адаптивным характеристикам. Подобный анализ был прове-
ден у тихоокеанского лосося - нерки, имеющего сложную и своеоб-
разную внутрипопуляционную структуру (Алтухов, 1995). У этого вида
самцы крупнее самок, однако в нативных популяциях имеется неболь-
шая (доли процента) фракция мелких половозрелых самцов. Мелкие
самцы - молодые рыбы в возрасте до 3 лет, которые быстро растут и
рано созревают (r-стратеги). Крупные самцы имеют возраст 5-7 лет,
Долго растут и поздно созревают (К-стратеги). Самки 4-5 лет предпо-
читают спариваться с крупными самцами, однако в маловодные годы
и на мелководье преимущество в спаривании имеют мелкие самцы.
Установлено, что мелкие самцы максимально гетерозиготны по генам,
Кодирующим синтез изоферментов. Гетерозиготность крупных сам-
235
М.И. Гордеев
цов минимальна, а самки имеют средний (оптимальный) уровень ге
терозиготности. В результате интенсивного промысла наблюдается
резкое нарушение возрастной и генетической структуры популяций
нерки: резко возрастает доля мелких самцов.
В подразделенных популяциях существует свое исторически сло-
жившееся оптимальное соотношение К- и r-стратегов. Это соотноше-
ние поддерживается стабилизирующим отбором. Генетическое разно-
образие в такой популяции включает две компоненты: внутри- и меж-
популяционную. Если в результате антропогенного воздействия умень-
шается внутрипопуляционная компонента и, соответственно, возрас-
тает межпопуляционная, то это равнозначно действию инбридинга.
Популяция как бы распадается на дифференцированные субпопуля
ции. Такой процесс инадаптивен, необратим и ведет к вымиранию.
Если, напротив, возрастает внутрипопуляционная компонента и уве-
личивается средняя гетерозиготность особей, то это сопровождается
снижением продолжительности жизни, увеличением смертности и
некоторыми другими неблагоприятными эффектами. Однако этот про-
цесс обратим, и при восстановлении прежней среды субпопуляцион
ная структура восстанавливается.
Субпопуляционная структура может возникать и на основе хромо-
сомной изменчивости. Важную роль в процессе эволюции двукрылых
насекомых играют инверсии (инверсия - поворот участка хромосомы
на 180°). Инверсии препятствуют кроссинговеру внутри инверсион-
ной петли у гетерозигот, и в процессе отбора инвертированных после-
довательностей формируются коадаптированные генные комплексы.
Наиболее молодые в филогенетическом отношении виды малярийных
комаров сформировали полиморфизм высшего ранга - полиморфизм
по адаптивным стратегиям. Инверсионные варианты половой хромо-
сомы и аутосом, детерминирующие фенотипические признаки г- и К-
стратегов, объединяются в кариотипах полиморфных видов благодаря
этологической и экологической подразделенности внутривидовых
группировок. В группе A.maculipennis к полиморфным видам отно-
сится A.messeae, имеющий в своем кариофонде пять распространен-
ных парацентрических инверсий: XLp ХЦ, 2Rp 3Rp 3Lj (Стегний.
1991). Свойствами К-стратегов обладают комары A. messeae с “южны-
ми” хромосомными последовательностями XL0(XLl)-2R0-3R0-3L(f
свойствами г-стратегов - комары с “северными” сочетаниями инвср-
236
IV. Эволюция животных
сий XL/XL^Rj-SRj-SLj. В направлении с юго-запада на север и се-
веро-восток ареала происходит постепенное замещение “южных” хро-
мосомных сочетаний на “северные”, хотя на обширной территории
Восточной Европы, Урала и Сибири К- и r-стратеги в популяциях
д messeae встречаются совместно (Гордеев, Стегний, 1987, 1989; Гор-
деев, 1997а).
Внутривидовые группировки A.messeae, относящиеся к К-стра-
тегам, характеризуются более низкой плодовитостью, повышенной
конкурентоспособностью на личиночных стадиях развития, высокой
устойчивостью к голоданию, паразитам и бактериальным патогенам.
Они отличаются пониженной скоростью фильтрации и, соответствен-
но, способны более эффективно перерабатывать и усваивать поступа-
ющие в кишечник пищевые ресурсы, но менее приспособлены к кан-
нибализму и некрофагии. Благодаря более низкой подвижности ли-
чинки этих комаров меньше подвержены гибели от хищников и
способны развиваться в условиях высокой плотности преимагиналь-
ных стадий. Альтернативными свойствами обладают г-стратеги (Гор-
деев, Стегний, 1987, 1989; Гордеев, Сибатаев, 1995а,б, 1996; Гордеев,
Перевозкин, 1995; Гордеев, 19976; Гордеев и др., 1997).
Экспериментальный анализ взаимодействия г- и К-стратегов на
преимагинальных стадиях развития позволил установить принципи-
альное тождество индивидуального и различных форм группового
отбора (Гордеев, Бурлак, 1992; Гордеев, Перевозкин, 1997). По наше-
му мнению, групповые отношения способны модифицировать резуль-
тирующий вектор отбора и относятся к селективным факторам, опре-
деляющим эколого-генетическую структуру популяции, однако по спо-
собам действия и результатам они ничем не отличаются от других
селективных сил. Групповые эффекты создают преимущества для К-
стратегов, но к подобным результатам приводит, например, хищниче-
ство и паразитизм в полиморфных системах, включающих различные
виды или внутривидовые группировки. Индивидуальная и групповая
обработка личинок бактериальными патогенами вызывала однонап-
равленные сдвиги частот внутривидовых группировок у A. messeae,
хотя при инфицировании в группе эффективность отбора возрастала.
По-видимому, групповые селективные эффекты, в том числе кин-от-
бор (отбор родичей), играют важную роль при формировании адап-
тивных комплексов у К-стратегов.
237
М.И. Гордеев
Генетическая гетерогенность природных популяций и создава-
емая на ее основе дифференциация особей по составу адаптивных ком-
плексов обусловливают возможность селекции малярийных комаров
на резистентность к бактериальным инсектицидам (Гордеев, Бурлак,
1991, 1992, 1994; Гордеев, 1997а). Установлено, что генетическим ме-
ханизмом первичного селективного ответа на обработку личинок
A.messeae бактерией Bacillus thuringiensis subsp. israelensis является
увеличение доли особей с “южными” хромосомными сочетаниями
(К-стратегов). Более высокая устойчивость К-стратегов обнаружена и
при обработке патогеном особей разных видов. Очевидно, в условиях
К-отбора возникли фенотипы, обладающие неспецифической устой-
чивостью к действию бактериальных токсинов. По-видимому, такая
устойчивость обеспечивается за счет снижения скорости поступления
фрагментов d-эндотоксина энтомопатогенной бактерии в организм
насекомого. Возможности данного механизма повышения резистент-
ности ограничены, и для многократного увеличения устойчивости к
патогену необходима длительная селекция по данному признаку в ряду
поколений.
Сочетание популяционно-экологических и цитогенетических
подходов позволило выявить сходство наборов признаков, входящих в
состав соответствующих адаптивных комплексов, на внутривидовом
уровне и при межвидовых сравнениях. Внутри- и межвидовая измен-
чивость по структуре адаптивных комплексов рассматривается нами
как основа эколого-генетической системы гомеостаза в пределах од-
ной жизненной формы - в сообществах комаров. Эксперименты с бак-
териальными инсектицидами показали, что представители комаров
рода Culex способны модифицировать селективное действие патоге-
нов и способствуют сохранению гетерогенности в популяциях маля-
рийных комаров Anopheles (Гордеев, Бурлак, 1994). Взаимосвязь про-
цессов отбора у совместно обитающих видов является одним из усло-
вий развития сообществ и характеризует состояние экосистем. В этом
смысле устойчивость состава адаптивных комплексов у полиморфных
видов может служить индикатором стабильности и интегрированнос-
ти сообщества в целом.
В настоящее время в целом ряде географически удаленных место-
обитаний на юге Западной Сибири наблюдается перестройка кариоти-
пической структуры популяций A.messeae (Gordeev, 1996). Согласно на-
238
IV. Эволюция животных
щим данным, К-стратеги в период после 1985 г. повсеместно вытесняют
r-стратегов. Мы связываем этот процесс с изменением среды обитания
малярийных комаров в результате глобального потепления климата.
Наиболее показательна динамика хромосомного состава популя-
ции A.messeae в пос. Тегульдет на востоке Томской обл. В 1973 г. доля
“северных” гомозигот 2RH в этом местообитании составляла 100%
(Стегний и др., 1978). В июле 1974 г. в поселке было выпущено 2000
оплодотворенных самок A.messeae из популяции пос. Коларово (Стег-
ний, Кабанова, 1985). Выпущенная группа имаго на 52% состояла из
особей с “южными” гомозиготами 2R00 (К-стратегов). Через год в ли-
чиночных биотопах было обнаружено 28% гомозигот и всего лишь
3% гетерозигот 2R01 (п = 62). Однако уже в 1981 г. популяция верну-
лась к своему исходному состоянию. Доли “южных” гомозигот 2R00 и
гетерозигот 2R0l составили соответственно 0,4 и 4,6% (п = 280). Дан-
ный полевой эксперимент продемонстрировал высокую устойчивость
кариотипической структуры популяции и ведущую роль отбора в со-
хранении баланса внутривидовых группировок.
Через 18 лет после описанного эксперимента, 1992-1995 гг.,
нами был вновь зафиксирован рост частоты “южных” хромосомных
перестроек в популяции пос. Тегульдет (Гордеев, 1997а; Перевозкин,
Гордеев, 1997). Обследование мест выплода, выполненное в 1995 г.,
показало, что основная часть особей с “южными” гомозиготами 2R00
развивается непосредственно в жилой зоне поселка. В прилегающих
к поселку биотопах доля личинок с этими хромосомными сочетания-
ми была значительно ниже (61,0 ± 6,2 и 16,8 ± 6,2% соответственно;
п = 59 и 274). Вероятно, этот факт связан с экологическими предпочте-
ниями внутривидовых группировок. Комары с “южными” хромосомны-
ми вариантами преимущественно зимуют в подвалах жилых домов и
хозяйственных постройках. После вылета с зимовочных убежищ самки
откладывают яйца в близлежащие водоемы рядом с помещениями для
скота. В этих же водоемах происходит развитие комаров в течение лета.
Очевидно, однонаправленная перестройка кариотипической
структуры A.messeae в пос. Тегульдет и в других географически уда-
ленных местообитаниях Западной Сибири обусловлена климатичес-
кими изменениями, общими для всего региона. Главным фактором,
определяющим распространение малярийных комаров на север, явля-
ется температурный режим (Беклемишев, 1944). Зависимость хромо-
239
М.И. Гордеев
сомного состава A.messeae от температурного режима исследовали в
1974-1982 гг. в популяции пос. Коларово. Установлена положитель-
ная корреляция между среднегодовыми частотами “южных” хромо-
сомных последовательностей XL0, 2R0, 3R0, 3L0 и годовыми суммами
среднесуточных температур в период размножения (апрель-август)
(Новиков, 1983). В этом сообщении впервые было сделано предполо-
жение о том, что в результате прогрессирующего потепления климата
кариотипическая структура популяции пос. Коларово может стать по-
хожей на таковую на юге и на западе ареала.
По нашему мнению, основной причиной наблюдаемой нами
перестройки генетической структуры популяций является изменение
условий зимовки. Равновесное соотношение инверсионных часто! в
сибирских популяциях малярийных комаров устанавливалось в усло-
виях обратимых колебаний среднезимних температур вокруг опреде-
ленного уровня, присущего каждому конкретному местообитанию
Температурный оптимум для зимовки малярийных комаров лежит
около 0°С, в пределах от +2 до -2°С. Соответствующие условия име-
ются в подвалах жилых домов, а также в хлевах, пещерах и некоторых
других естественных убежищах. Но основная масса диапаузирующих
имаго зимует в природе, где температура опускается ниже оптималь-
ных значений. Сравнение численности уходящих на зимовку и пере-
зимовавших самок дает основание считать, что за зиму погибает не
менее 75-90% диапаузирующих комаров (Беклемишев, 1944). Различ-
ная жизнеспособность диапаузирующих имаго может быть связана с
экологическими предпочтениями при выборе зимовочных убежищ, а
также с особенностями энергетического обмена у комаров с разными
кариотипами. При понижении температуры в начале диапаузы в гемо-
лимфе насекомых сначала резко увеличивается концентрация глице-
рина, а при дальнейшем снижении температуры возрастает концент-
рация сорбита. В результате точка замерзания гемолимфы может сни-
жаться до -10°С (Storey et al., 1981). Вероятно, указанная температура
является критической для большинства зимующих насекомых. При
более низких ее значениях происходит замерзание гемолимфы, разру-
шение клеточных структур и гибель всего организма.
На территории бывшей Томской губернии (Томская, Новосибирс-
кая, Кемеровская обл. и Алтайский край) за 80 лет наблюдений с 1881
по 1960 гг. установлено, что средние январские температуры достига-
240
IV Эволюция животных
__lg5 -20°С, опускаясь в отдельные дни до -40, -45°С и ниже. За
указанный период в г. Томске и его окрестностях средняя температура
за ноябрь-февраль составляла-15,95°С (Справочник по климату СССР,
1965). По данным Томской метеостанции, с ноября 1982 по февраль
1995 г., когда наблюдалось изменение кариотипической структуры
A.messeae, среднемесячная температура повысилась до -12,23°С, т.е.
на 3,72°С. Особенно заметным был рост среднезимних температур пос-
ле относительно холодной зимы 1984-1985 гг. Все последующие зимы
были теплее нормы, и среднемесячная температура составила -11,80°С,
т.е. вплотную приблизилась к точке замерзания гемолимфы насеко-
мых. В связи с общим потеплением снизился и перепад зимне-весен-
них температур, что также может влиять на жизнеспособность кома-
ров с «южными» инверсиями на завершающем этапе зимовки.
Очевидно, имеющаяся хромосомная изменчивость у A.messeae слу-
жит механизмом стабилизации генетической системы вида. При обрати-
мых и кратковременных (в пределах нескольких поколений) климати-
ческих изменениях частоты инверсий колеблются вокруг равновесных
значений. Равновесие поддерживается за счет дифференциальной при-
способленности гомозигот на разных этапах сезонного цикла. Только при
длительной направленной трансформации среды обитания соотношение
генотипов может значительно отклониться от исходного уровня, вплоть
до полного замещения альтернативных хромосомных вариантов. Подоб-
ный процесс происходил в сибирских популяциях малярийных комаров
на протяжении последних 60 поколений. В данном случае климатичес-
кие изменения привели к увеличению концентрации «южной» инверсии
2R0 и сцепленных с нею последовательностей XL0, 3R0,3L0. Следует от-
метить, что комары с этими хромосомными последовательностями по-
тенциально являются более опасными переносчиками малярии, поскольку
зимуют и размножаются вблизи человеческого жилья.
В целом в условиях потепления климата на юге Западной Сибири
наблюдается разрушение исторически сложившейся системной орга-
низации популяций малярийных комаров. Происходит вытеснение
«северных» внутривидовых группировок и резкое сокращение, на за-
вершающем этапе, уровня внутрипопуляционной хромосомной измен-
чивости.
Результаты генетического мониторинга популяций малярийных
комаров позволяют зафиксировать изменение результирующего век-
241
М.И. Гордеев
тора отбора и отражают начальные этапы деградации таежных биоце-
нозов. Приведенный пример показывает, что выделение адаптивных
комплексов может служить основой для определения направленности
микроэволюционных процессов в популяциях полиморфных видов.
Все изложенное выше позволяет сформулировать программу по-
пуляционно-генетических исследований на современном этапе. Тако-
го рода исследования должны включать экологический анализ субпо-
пуляционной структуры, оценку внутри- и межпопуляционной компо-
нент изменчивости, реконструкцию исторически сложившегося
соотношения этих компонент в подразделенной популяции, выявле-
ние основных векторов отбора. Нам представляется важным изучение
и самих механизмов действия селективных сил.
Работа поддержана грантом РФФИ-рСибирь обл. № 98-04-03074.
Литература
Алтухов Ю.П. Внутривидовое генетическое разнообразие: мониторинг и принципы
сохранения //Генетика. 1995. Т. 31. № 10. С. 1333-1357.
Беклемишев В.Н. Экология малярийного комара. М.: Медгиз, 1944. 299 с.
Биологический энциклопедический словарь / Гл. ред. М.С. Гиляров. М.: Большая рос-
сийская энциклопедия, 1995. 864 с.
Гордеев М.И. Адаптивные стратегии в популяциях малярийных комаров: Автореф. дис...
докт. биол. наук. Томск, 1997а. 42 с.
Гордеев М.И. Устойчивость к голоданию как элемент адаптивной стратегии у личинок
малярийных комаров // Генетика. 19976. Т. 33. № 6. С. 844-851.
Гордеев М.И., Бурлак В. А. Инверсионный полиморфизм малярийного комара Anopheles
messeae. Сообщение X. Устойчивость личинок с разными генотипами к токсинам
кристаллообразующей бактерии Bacillus thuringiensis subsp. israelensis (серовар Hl4)
И Генетика. 1991. Т. 27. № 2. С. 238-246.
Гордеев М.И., Бурлак В. А. Инверсионный полиморфизм малярийного комара Anopheles
messeae. Сообщение XI. Групповой эффект при инфицировании личинок бактери-
ей Bacillus thuringiensis subsp. israelensis И Генетика. 1992. Т. 28. № 7. С. 82-88.
Гордеев М.И., Бурлак В.А. Влияние личинок комаров рода Culex на чувствительность
комаров рода Anopheles с различными кариотипами к энтомопатогенной бактерии
Bacillus thuringiensis subsp. israelensis И Генетика. 1994. Т. 30. № 3. С. 367-372.
Гордеев М.И., Перевозкин В.П. Стратегии отбора и устойчивость к асфиксии у личи-
нок малярийного комара Anopheles messeae с разными кариотипами И Генетика.
1995. Т. 31. №2. С. 180-184.
Гордеев М.И., Перевозкин В.П. Отбор родичей и скорость развития малярийных кома-
ров с разными кариотипами И Генетика. 1997. Т. 33. № 10. С. 1367-1373.
Гордеев М.И., Сибатаев А.К. Возрастной отбор и способность к каннибализму у личи-
нок малярийных комаров с разными кариотипами // Генетика. 1995а. Т. 31. № 7.
С. 939-946.
242
IV. Эволюция животных
ГорДеев Сибатаев А.К. Влияние хищного растения пузырчатки Utricularia vulgaris
на процессы отбора у личинок малярийных комаров // Экология. 19956. № 3.
С. 241-245.
Гордеев М.И., Сибатаев А.К. Цитогенетическая и фенотипическая изменчивость в цен-
тральных и периферийных популяциях малярийного KOMapaAnopheles messeae Fall.
(Diptera, Culicidae) // Генетика. 1996. T. 32. № 9. С. 1199-1205.
Гордеев М.И., Стегний В.Н. Инверсионный полиморфизм малярийного KOMapaAnopheles
messeae. Сообщение VII. Плодовитость и популяционно- генетическая структура
вида//Генетика. 1987. Т. 23. № 12. С. 2169-2174.
Гордеев М.И., Стегний В.Н. Инверсионный полиморфизм малярийного комара
Anopheles messeae. Сообщение VIII. Распределение личинок в биотопах в усло-
виях совместного обитания с Anopheles beklemishevi // Генетика. 1989. Т. 25.
№2. С. 283-291.
Гордеев М.И, Перевозкин В.П, Лукьянцев С.В. Генетические и экологические эффек-
ты охоты пауков-серебрянок Argyroneta aquatica на личинок комаров Anopheles и
Culex И Генетика. 1997. Т. 33. № 5. С. 704-709.
Кулинский В.И, Ольховский И.А. Две адаптационные стратегии в неблагоприятных
условиях. Роль гормонов и рецепторов // Успехи соврем, биол. 1992. Т. 112. № 5-6.
С. 697-714.
Миркин Б.М, Розенберг Г.С., Наумова Л.Г Словарь понятий и терминов современной
фитоценологии. М.: Наука, 1989. 223 с.
Новиков Ю.М. Микроэволюция популяции малярийного комара Anopheles messeae в
окрестностях Томска за девятилетний период // Молодые ученые и специалисты -
народному хозяйству: Биология. Томск: Изд-во Томск, ун-та, 1983. С. 31-32.
Перевозкин В.П., Гордеев М.И. Механизм вытеснения одних кариотипов другими у ма-
лярийного KOMapaAnopheles messeae. Анализ in flagrante delicto (на месте преступле-
ния) // Современные концепции эволюционной генетики: Магер, межд. конф., по-
священной 80-летию со дня рождения акад. Д.К. Беляева (Новосибирск,
9-12 сентября 1997 г.). Новосибирск: Ин-т цитологии и генетики, 1997. С. 254-256.
Пианка Э. Эволюционная экология. М.: Мир, 1981.400 с.
Раменский Л.Г. Введение в комплексное почвенно-геоботаническое исследование зе-
мель. М.: Сельхозгиз, 1938. 620 с.
Северцов А.С. Направленность эволюции. М.: Изд-во МГУ, 1990. 272 с.
Справочник по климату СССР. Томская, Новосибирская, Кемеровская обл. и Алтайс-
кий край. Температура воздуха и почвы. Л.: Гидрометеоиздаг, 1965. Вып. 20. Ч. 2.
396 с.
Стегний В.Н. Популяционная генетика и эволюция малярийных комаров. Томск: Изд-
во Томск. ун-та, 1991. 136 с.
Стегний В.Н, Кабанова В.М. Инверсионный полиморфизм малярийного комара
Anopheles messeae. IV. Локальный отбор и пространственная дифференциация ин-
версионных генокомплексов по плодовитости // Генетика. 1985. Т. 21. № 12.
С. 1970-1973.
Стегний В.Н., Новиков Ю.М., Плешкова Г.Н., Кабанова В.М. Инверсионный полимор-
физм малярийного комара Anopheles messeae. II. Межпопуляционная вариабель-
ность инверсионных частот// Генетика. 1978. Т. 14. № 6. С. 1016-1023.
Сукачев В.Н. Растительные сообщества. Введение в фитоценологию. 4-е изд. М.: Кни-
га, 1928. 232 с.
243
М.И. Гордеев
Bartholomew G. A. Body temperature and energy metabolism//Animal physiology: principles
and adaptations. Chapter 8. N. Y: Macmillian Press, 1972. P. 298-368.
Gordeev M.I. Karyotypic structure transformation of Siberian malaria mosquitoes (Diptera
Culicidae) as a result of global warming // Proceedings of XX International congress of
entomology (Firenze, Italy, August 25-31, 1996). Firenze. 1996. P. 244.
Grime J.P. Vegetation classification by reference to strategies // Nature. 1974. Vol. 250
P. 26-31.
Grime J.P. Plant strategies and vegetation processes. Chichester, N. G.: John Willey and
Sons, 1979. 222 p.
MacArthur R.H., Wilson E.O. The theory of island biogeography. Princeton, N. J.: Princeton
Univ. Press, 1967. 203 p.
Rosen R. Optimality principles in biology. N. Y: Plenum Press, 1967. 198 p.
Storey K.B., Baust J.G., Storey J.M. Intermediary metabolism during low temperature
acclimation in the overwintering gall fly larva, Eurosta solidaginis // J. Comp. Physiol
1981. Vol. 144. P. 183-190.
ДЕТЕРМИНИРОВАННЫЙ ХАОС В ДИНАМИКЕ
ПОПУЛЯЦИЙ КАК ЭВОЛЮЦИОННЫЙ ФАКТОР
Е.В. Евдокимов, А.В. Шаповалов
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
E-mail: evd@res.tsu.ru
Введение
В начале 70-х годов нашего столетия почти одновременно А.П. Ша-
пиро и Р. Мэй [1,2, 3, 4] обнаружили, что некоторые простые популя-
ционные модели с дискретным временем могут демонстрировать нео-
жиданно сложное динамическое поведение. Наиболее известными из
них являются модели популяций с неперекрывающимися поколения-
ми, динамика которых описывается отображениями типа
xn+=f(xn,R,K). (1)
Здесь хпи xn+l- численности популяции, соответственно, в n-й и
(п+1)-й сезоны, R - удельная скорость воспроизводства популяции
(мальтузианский параметр), К - параметр емкости лимитирующего
ресурса,/- нелинейная функция, имеющая один максимум в положи-
тельном квадранте и отражающая плотностно-зависимую динамику
размножения.
Виды с неперекрывающимися поколениями в биологии чаще изве-
стны как однолетние (растения, насекомые и т.п.), хотя более строго
их можно определить как виды, у которых родительские особи по тем
или иным причинам не доживают до репродуктивного возраста своих
потомков. Поэтому непрерывную ось времени в динамике таких по-
пуляций дискретизуют на интервалы, соответствующие жизни одного
поколения, а каждое поколение характеризуют численностью хп. Та-
кая дискретизация чаще всего имеет навязанный сезонный характер,
однако в некоторых случаях, например в популяциях тепличных насе-
245
Е.В. Евдокимов, А. В. Шаповалов
комых-вредителей, дискретизация динамики может явиться результа-
том самосинхронизации процессов размножения и короткого периода
жизни половозрелых особей [5].
В современной литературе в качестве отображения (1) наиболее
часто употребимы отображение Риккера [2, 6]:
х„+1 =f(R>K,xn )=х„ ехр{Я(1-х„/К)} (2)
и отображение Хассела [7]:
х,„=/(Л,К,х,) = х, К/(1-х,/КУ , 15|
а также их варианты. Такого типа отображениями описывается дина
мика популяций многих насекомых, рыб, однолетних растений и т.д.
[2, 6, 7]. Для удобства анализа часто вместо популяционных перемен
ныххп используют их нормированные значения х*п = хп/Х, предпола-
гая, что параметр К со временем не изменяется.
Главной особенностью таких систем является зависимость харак
тера их поведения от величины мальтузианского параметра/? [2, 7, 8].
При R меньше определенной величины поведение популяции харак-
теризуется ее стремлением к устойчивому стационарному состоянию
(рис. 1, слева). Этот вид популяционной динамики (типа логистичес-
кой кривой) наиболее знаком биологам и экологам.
Менее известным является циклический тип поведения популяции
(рис. 1, в центре), к которому она переходит, когда мальтузианский
параметр превысит определенную величину (в зависимости от типа
отображения, для отображения Риккера это R = 2), при этом с ростом
R период колебаний последовательно удваивается с сохранением в
спектре гармоники с предыдущим периодом. Важным здесь является
то, что периодические колебания численности популяции возникаю i
сами по себе без какого-либо влияния извне (хищничества, сезонных
колебаний и т.п.) как следствие нелинейности ее динамических свойств
Наконец, дальнейшее увеличение мальтузианского параметра при-
водит к существенному качественному изменению динамики: числен-
ность популяции начинает колебаться непредсказуемым образом
(рис. 1, справа). Такой режим был назван детерминированным хаосом
246
IV. Эволюция животных
Иля хаотического режима характерна очень большая амплитуда изме-
нений численности популяций, при этом в нижних точках численность
популяции может быть как угодно близка к нулю. Важно отметить, что
переходы из одного динамического режима в другой могут происхо-
дить при очень малых изменениях R, что отражает бифуркационный
характер динамики системы [8].
4000---------------
Устойчивое
|зооо
I
I
£2000
О
X
X
ф
О 1000
стационарное
состояние
Поколения
0
Рис. 1. Типы динамического поведения и переходы между
ними в популяциях с неперекрывающимися поколениями
Для наглядного представления динамики таких систем использу-
ют так называемую бифуркационную диаграмму (рис. 2), на которой
по оси абсцисс откладывают значения мальтузианского параметра, а
по оси ординат - соответствующие данному значению R состояния
системы хп для п больше некоторого номера N, имеющего смысл дис-
кретного времени, в течение которого завершаются переходные про-
цессы и динамика системы выходит на стационарный режим.
На рис. 2 видно, что для популяций, описываемых моделью Рикке-
ра, при R < 2 наблюдается первый тип динамического поведения (ус-
тойчивое стационарное состояние - одна линия на диаграмме), в ин-
тервале 2 < R < 2,69 с ростом мальтузианского параметра система ис-
пытывает каскад бифуркаций удвоения периода колебаний и в ней
реализуются различные предельные циклы, начиная с периода 2 (две,
четыре и более линий на диаграмме).
247
Е.В. Евдокимов, А.В. Шаповалов
Рис.2. Бифуркационная диаграмма динамики популяций, описывае-
мых отображением Риккера. По оси абсцисс - мальтузианский пара-
метр, по оси ординат - численность популяции
При дальнейшем увеличении R возникают хаотические режимы
(значения хп хаотически заполняют некоторый интервал), перемежае-
мые так называемыми /?-окнами, в которых наблюдаются предельные
циклы. Каждое /?-окно характеризуется своим базовым предельным
циклом, имеющим наименьший период, а число /?-окон составляет
счетное множество [8].
Теория перехода к хаосу в динамических системах, подобных рас-
смотренным выше, была разработана М. Фейгенбаумом [9]. Ее основ-
ная идея - потеря аттракторами (точкой, предельными циклами) сво-
ей устойчивости при изменении управляющего параметра R и соот-
ветствующий каскад удвоения периода реализуемого предельного
цикла при сильном нелинейном сжатии интервалов устойчивости на
каждом этапе. Это приводит к экспоненциальному росту числа гармо-
ник предельного цикла на малом интервале изменения параметра. Хаос,
по Фейгенбауму, наступает при их “бесконечном” числе. Такой меха-
низм генерации хаоса в динамических системах носит название “суб-
гармонический каскад” [8].
Таким образом, два фактора нелинейности, дискретность времени
и немонотонность функции размножения, приводят к резкому услож-
нению динамики популяций с неперекрывающимися поколениями,
248
IV. Эволюция животных
которое определяется значением управляющего мальтузианского па-
раметра.
Работы последних 30 лет показали, что детерминированный хаос -
не экзотический, редко встречающийся феномен, а широко распрост-
раненное свойство природных систем различных уровней организа-
ции, физических, химических, биологических и социальных. В этом
ряду - явление турбулентности в жидкостях и газах, нелинейные явле-
ния в процессах горения, электрического пробоя, генерация шума оп-
ределенного типа в нелинейных электрических цепях, макроскопичес-
кая стохастизация протекания цепных и автокаталитических химичес-
ких реакций, возникновение хаоса в нейронных сетях, демографи-
ческих и экономических моделях и многое другое [10, 11,12]. Из пос-
ледних работ, экспериментально подтверждающих теоретическую
модель Шапиро-Мэя-Хассела на достаточно высоком методическом
уровне, можно отметить исследования П. Филипса на нематодах [13]
и Р. Константино с соавторами на жуках [14].
С биологической точки зрения изменения среднего по популяции
мальтузианского параметра могут иметь как физиологическую детер-
минацию и вызываться внешними факторами (вариацией температу-
ры, влажности и т.д.), так и популяционно-генетическую, путем сме-
щения средней приспособленности в результате отбора в ходе микро-
эволюции. Ниже мы рассмотрим ряд полученных нами результатов
исследования влияния этих двух комплексов причин на проявление
детерминированного хаоса в динамике популяций.
1. Теорема Фишера, рост приспособленности
и возможные динамические последствия
В синтетической теории эволюции основополагающую роль игра-
ет теорема естественного отбора Р. Фишера, определяющая неизбеж-
ность роста приспособленности в популяциях в ходе отбора [15, 16, 17].
Эта теорема обычно формулируется следующим образом: “средняя
приспособленность в неравновесной популяции вследствие отбора
всегда возрастает, а скорость возрастания пропорциональна диспер-
сии приспособленностей в популяции” [15, с. 18]. Биологическое зна-
чение теоремы Фишера становится ясным, если учесть, что приспо-
249
ЕВ. Евдокимов, А.В. Шаповалов
собленность определяет темп увеличения численности популяции и
количественно равна среднепопуляционному значению удельной ско-
рости размножения [17].
Для одномерных моделей популяций с неперекрывающимися по-
колениями, описываемых отображением Риккера, приспособленность
зависит от двух параметров - мальтузианского R и ресурсного К. На-
пример, для отображения Риккера функция приспособленности имеет
вид [19, 20]:
- exp {Л(1-^)}
К.
Если в популяции возникает разнообразие по этим параметрам по
тем или иным причинам (мутации, рекомбинации и т.д.), то в ней с
необходимостью (по теореме Фишера) начинается процесс отбора на
увеличение средней приспособленности. Это, в свою очередь, вызы-
вает изменение среднепопуляционного значения мальтузианского па-
раметра. Поскольку динамический режим популяций с неперекры-
вающимися поколениями полностью определяется значением мальту-
зианского параметра, то в результате его изменения в процессе
микроэволюции популяции должны последовательно демонстрировать
все типы динамического поведения: “устойчивое стационарное состо-
яние -> предельный цикл -> хаос” и т.д. Смена динамических режи-
мов, таким образом, - обязательное следствие длительного направ-
ленного отбора в рассматриваемых популяциях. В силу бифуркаци-
онного характера переходов от режима к режиму их можно наблюдать
при изменении мальтузианского параметра на 1-2%, что достижимо в
селекционных экспериментах, а также, например, в естественных по-
пуляциях насекомых при их адаптации к инсектицидам.
Рассмотренная особенность микроэволюции в популяциях с непе-
рекрывающимися поколениями, “смена динамических режимов в ходе
отбора”, в неявном виде обсуждалась еще в ранних работах по детер-
минированному хаосу [2, 21], однако в явном виде, как новая законо-
мерность, присущая селекционной динамике и вытекающая из нели-
нейных свойств популяций, она была, по-видимому, сформулирована
Е. Фрисманом [22, 23]. Поскольку сложное динамическое поведение
с бифуркационными переходами между стационарным состоянием,
предельными циклами и детерминированным хаосом характерно не
250
IV. Эволюция животных
только дая популяций с неперекрывающимися поколениями, но и яв-
ляется общим свойством обширного класса нелинейных многовидо-
вЫХ систем [24], то всестороннее обоснование указанной закономер-
ности является, на наш взгляд, важным направлением в эволюцион-
ных исследованиях.
В данной работе мы рассмотрим полученные нами результаты по
численному моделированию процессов отбора в популяциях с непе-
рекрывающимися поколениями.
2. Смена динамических режимов в ходе отбора
в популяциях с неперекрывающимися поколениями
В соответствии с подходом Р. Мэя [2] рассмотрим двухпараметри-
ческую модель популяции с неперекрывающимися поколениями и бес-
полым размножением, динамика численности особей с различными
генотипами в которой описывается следующим отображением:
х;я=х;ехР[я'(1-5>'/Г)]. <4)
7
Здесь х!п и - численности особей /-го генотипа в и-й и (и+1)-й
моменты времени, R м К' - соответствующие данному генотипу значе-
ния мальтузианского и ресурсного параметров, j меняется от 1 до Z, где L
- число различных генотипов в популяции. Данная модель описывает
плотностно-зависимый отбор, определяемый лишь двумя фенотипичес-
кими признаками, скоростью размножения и глубиной использования
ресурса. Влияние иных конкурентных взаимодействий между особями
разных генотипов типа взаимоподавления или альтруизма оставлено за
рамками данной работы, поскольку их введение существенно усложняет
анализ, однако, как показали расчеты, не меняет качественную картину
хода микроэволюции в рассматриваемом контексте.
Численные эксперименты проводили путем итераций отображения
(4) в течение 500-5000 поколений при различных уровнях исходного
‘генетического” разнообразия в популяции. Значения R. выбирались
так, чтобы минимальное из них было в области стационарного состо-
яния, а максимальное - в области хаоса. Обобщенные качественные
Результаты исследования представлены в табл. 1.
251
Е.В. Евдокимов, А. В. Шаповалов
Таблица 1
Динамические режимы, реализуемые в ходе отбора в популяциях
с неперекрывающимися поколениями
Характеристика “генетического” разнообразия в исходной популяции Rj< Ri<Rl К,= Ki =KL Xio< XiO<XLO Rj- Ri-Rl Kj< К, <KL Xio< x,o<Xlo Ri< Ri<Rt Kt< Ki <KL X1O< XiO<XLO Я/> r,>rl Ki< к, <к, Хю< Xi0<Xui
Результат отбора Сосущество- вание всех генотипов Конкурентное исключение всех генотипов с K<KL Конкурентное исключение всех генотипов с K,<KL Конкурент" ное исклю- чение всех генотипов с K<KL
Наблюдаемые динамические режимы Один тип режима, определяемый среднепопу- ляционным значением R Один тип режима, определяемый значением R Последова- тельный переход от стационарного состояния к предельным циклам и, далее, к хаосу Последова- тельный переход о г хаоса к предельным циклам и. далее, к стационар- ному состоянию
Из анализа таблицы следуют три особенности динамики отбора в
исследуемых популяциях.
Во-первых, отбор всегда идет в сторону увеличения ресурсного
параметра К, если в популяциях есть соответствующее генетическое
разнообразие. При его отсутствии отбор, как таковой, в популяции не
наблюдается несмотря на наличие дисперсии по мальтузианскому па-
раметру R. Следовательно, для популяций с неперекрывающимися
поколениями с плотностно-зависимым контролем размножения харак
терен так называемый /С-отбор (по классификации Р. МакАртура [25]),
направленный на более полное использование ресурса. Аналогичный
тип отбора в популяциях микроорганизмов наблюдается в хемостак
[26]. Подобные результаты для несколько иной модели отбора были
получены М. Асмуссеном [19].
Во-вторых, в отсутствии дисперсии по К и при наличии разнооб-
разия по Я в популяции наблюдается сосуществование всех геноти-
пов. Их равновесное распределение устанавливается в течение пер-
252
IV. Эволюция животных
вых трех-четырех поколений и далее не меняется. Эффект сосуще-
ствования в популяции различных генотипов при определенных ус-
ловиях - известное и довольно необычное свойство популяций с не-
перекрывающимися поколениями, являющееся следствием существен-
но нелинейных свойств таких систем [19, 27, 28]. Оно отличает их от
популяций с невыраженной нелинейностью динамики, например от
проточных популяций бактерий, для которых характерен принцип
конкурентного исключения Гаузе [29].
В-третьих, при одновременном наличии в популяции разнообра-
зия по мальтузианскому и ресурсному параметрам течение отбора со-
провождается последовательной сменой динамических режимов от
стационарного состояния через предельные циклы к хаосу и наоборот
в зависимости от направления отбора по К. В популяции наблюдается
последовательная замена одного доминирующего генотипа другим, а
тип наблюдаемого при этом динамического режима определяется сред-
непопуляционным значением мальтузианского параметра.
На рис. 3 и 4 в качестве примера, иллюстрирующего последнее
свойство динамики ПНП, приведены данные о ходе отбора в попу-
ляции с пятью генотипами, отличающимися значением как R, так
и К. В ней в начале “опыта” доминировали генотипы с наимень-
шим значением К (1000 и 1010) и имеющие значение мальтузиан-
ского параметра R в стационарной области (1,5 и 1,9). Генотипы с
большим К (1020, 1025, 1030) и значением R из области циклов и
хаоса были представлены величинами, соответствующими типич-
ным частотам мутаций.
Как видно из сопоставления рис. 3 и рис. 4, динамика изменения
численности отдельных генотипов независимо от соответствующих
им значений R и К подчиняется режиму, свойственному для популя-
ции в целом. На первом этапе, до 245-го поколения, это гладкая дина-
мика, далее, до 530-го поколения, циклические колебания численнос-
ти всех генотипов, и затем наблюдается, опять же для всех, переход к
хаотическим колебаниям.
В среднем же эти кривые имеют характер, типичный для селекци-
онных экспериментов с конкурентным исключением [30]. Подобные
результаты для однолокусной двухаллельной модели популяции с по-
ловым размножением были получены Е. Фрисманом [23].
253
Е.В. Евдокимов, А. В. Шаповалов
Время, п
Рис. 3. Ход отбора в популяции, описываемой отображением (4). 1, 2, 3, 4, 5
- кривые изменения численности особей с генотипами, соответствующими
стационарной области (2, 2), области предельных циклов (3, 4) и области ха-
оса (5). Начальные условия: х;о=4О, х2=4, xJf=4*10~2, х^=4*10Л х^/М’ЧО-6
Время, п
Рис. 4. Смена динамических режимов в ходе отбора в популяции, описыва-
емой отображением (4). Кривая получена суммированием данных, соответ-
ствующих кривым 1-5 на рис. 3
254
IV. Эволюция животных
Подчинение динамики изменения численности каждого отдельного
генотипа общепопуляционной динамике свидетельствует о том, что
исследуемая популяция в ходе микроэволюции ведет себя как целос-
тная нелинейная система. Это наглядно демонстрирует рис. 5, на ко-
тором представлена зависимость между среднепопуляционным зна-
чением мальтузианского параметра <R> и соответствующей числен-
ностью популяции в ходе отбора в рассматриваемом численном
эксперименте. Как видно, эта корреляционная диаграмма носит яв-
ный бифуркационный характер, типичный для популяций, описывае-
мых отображением Риккера (см. рис. 2), роль управляющего параметра
в ней носит среднепопуляционное значение R.
3000
Среднепопуляционное значение мальтузианского параметра, п-1
Рис. 5. Взаимосвязь между среднепопуляционным значением мальтузианского па-
раметра <R> и численностью популяции в ходе отбора в численном эксперименте,
представленном на рис. 3 и 4. Для построения диаграммы в каждом поколении рас-
считывали <R> и откладывали против него соответствующее значение суммарной
численности популяции
Таким образом, предсказываемая из общих соображений смена
Динамических режимов “стационарное состояние - предельные цик-
лы - хаос” в ходе микроэволюции популяций с неперекрывающимися
поколениями подтверждается численными экспериментами с много-
мерным двухпараметрическим отображением Риккера. Необходимым
и достаточным условием для этого является наличие в популяции дис-
персии как по мальтузианскому, так и по ресурсному параметрам.
255
Е.В. Евдокимов, А. В. Шаповалов
3. Детерминированный хаос как фактор
биологической эволюции
Рассмотренное выше сложное динамическое поведение популя-
ций с неперекрывающимися поколениями с нелинейными свойства-
ми, проявляющееся как в реакции на действие внешних факторов,
так и в ходе отбора, может оказывать существенное влияние на ход
эволюционных процессов. В современной биологической литерату-
ре активно дискутируется этот вопрос и высказываются достаточно
аргументированные мнения о коррекции существующих эволюцион-
ных концепций и введении поправок на нелинейность и хаос в извес-
тные теоретические модели микро- и макроэволюции (см., например,
обзор [31]). Некоторые авторы даже высказываются о смене парадиг-
мы в теории эволюции [32]. Ниже мы представим ряд очевидных, на
наш взгляд, аспектов анализа эволюционных процессов, в которых
необходимо учитывать нелинейные свойства популяций и экосистем,
при этом особое внимание будем уделять проявлению детерминиро-
ванного хаоса в эволюционной динамике.
3.1. Особенности динамики генетической структуры популяций
с неперекрывающимися поколениями
Как было показано выше, динамика популяций с неперекрываю-
щимися поколениями характеризуется переходами “стационарное со-
стояние -> предельные циклы —> хаос”, управляемыми значением маль-
тузианского параметра. Предельно малая величина изменения маль-
тузианского параметра, необходимого для перехода из одного
динамического режима поведения популяции в другой, приводит к
следующим важным биологическим следствиям.
Различные популяции одного и того же вида, обитающие в разных
точках ареала, например в оптимуме и на окраине, могут существенно
отличаться по типу динамического поведения в силу различия в удель-
ных скоростях размножения, возникающих вследствие различия кли-
матических и иных экологических условий. Для перехода от предель-
ного цикла к хаотическому режиму (или наоборот) достаточно одно-
процентного (или даже меньше) изменения величины мальтузианского
параметра, что легко реализуется, например, при изменении средне-
годовой температуры на 1 градус или влажности на 1-2% и т.п.
256
IV. Эволюция животных
Медленные климатические тренды и колебания или другие длин-
нопериодные воздействия (антропогенные, например), способные из-
менять удельную скорость размножения (гибели) одной и той же по-
пуляции, могут также качественно модулировать ее динамику.
Существенные колебания численности популяции, перешедшей в
хаотический режим по вышеуказанным причинам, обязаны изменять
ее генетическую структуру, поскольку в минимуме численность попу-
ляции может составлять единицы процентов от максимума и менее.
Резкое понижение численности, как известно, имеет своим следстви-
ем случайную потерю тех или иных генных аллелей, “дрейф генов”
[16]. Особенно это касается редких аллелей, возникающих путем ген-
ных и хромосомных мутаций. Поскольку значительные минимумы
численности популяции в хаотическом режиме регулярно повторяют-
ся, то можно говорить о постоянной “генетической фильтрации” в та-
ких популяциях, результатом которой является гомозиготизация. Важ-
ным следствием этого является проблематичность протекания в таких
популяциях микроэволюционных процессов путем естественного от-
бора редко возникающих “положительных” мутаций. Можно, по-ви-
димому, утверждать, что популяции, находящиеся в режиме детер-
минированного хаоса, представляют собой “эволюционные запо-
ведники”.
По вышеуказанным причинам у видов, имеющих мальтузианские
параметры, близкие к /?-окнам, популяции в середине ареала и по его
краям обязаны отличаться по генетической структуре. Это должно
способствовать их дивергенции и, в конечном счете, видообразова-
нию, однако не путем естественного отбора, а вследствие случайного
“дрейфа генов” и последующей “генной фильтрации”.
Еще более сложная картина возникает на уровне метапопуляций,
состоящих из субпопуляций, способных к обмену особями путем миг-
раций. Если интенсивность миграций достаточно велика и линейно
зависит от градиента плотности особей по ареалу, то “перемешиваю-
щий” эффект миграций оказывает стабилизирующее действие на всю
метапопуляцию, переводя ее в целом в стационарное состояние не-
смотря на то, что значение мальтузианского параметра лежит в хаоти-
ческой области [33]. Однако при учете нелинейных свойств миграци-
онных процессов супрессия хаоса в метапопуляции наблюдается лишь
в определенной области характеризующих ее параметров [34, 35, 36].
257
Е.В. Евдокимов, А.В. Шаповалов
Другой важный экологический эффект миграций в метапопуляци-
ях, субпопуляции которых способны демонстрировать стационарное
состояние, предельные циклы и хаос, заключается в возможности ге-
нерации пространственно-временных структур типа спиральных волн,
концентрических колец с ревербераторами и др. [37]. Особенно слож-
ное поведение подобных структур свойственно метапопуляционным
системам типа “паразит-хозяин” [35, 36, 38].
3.2. Влияние внешних периодических факторов
В работе [39] нами было показано, что при периодическом возму-
щении мальтузианского и ресурсного параметров действием внешних
факторов в динамике популяций с неперекрывающимися поколения-
ми могут происходить существенные изменения. При соблюдении ряда
условий в них наблюдается резонансная супрессия хаоса и перевод
системы в режим предельного цикла, период которого совпадает с
периодом внешнего воздействия. Напомним, что амплитуда возмуще-
ния, необходимая для реализации этого феномена, может быть очень
мала. Рассмотрим возможные экологические и эволюционные след-
ствия данного свойства динамики ПНП.
Биосфера Земли и составляющие ее экосистемы существуют в пери-
одически изменяющейся среде. Это положение является общим местом
всего современного естествознания. В рассматриваемом аспекте важны
те циклические изменения биосферы, период которых превышает как
минимум в два раза цикл размножения в популяциях с неперекрываю
щимися поколениями и которые способны влиять на величину мальту-
зианского параметра. Особенно интересны циклы, порождаемые 11- и
22-летними изменениями солнечной активности, поскольку они имеют
глобальный общебиосферный характер. К настоящему времени с доста-
точной достоверностью установлено, что порождаемые этими циклами
периодические изменения климатических, геофизических и иных усло-
вий на Земле способны вызывать физиологические эффекты порядка 0,1
5% и тем самым ритмически менять величину мальтузианского пара-
метра [40]. Из этого следует, что все популяции различных организмов с
неперекрывающимися поколениями, мальтузианский параметр которых
находится на таком и меньшем расстоянии от 7?-окна с периодом 11 лет
или 22 года, должны перейти от хаотического режима изменения чис-
258
IV. Эволюция животных
ценности к 11- или 22-летнему циклу изменений, причем этот переход
должен был совершиться давно, немногим позднее момента формиро-
вания популяций данных видов. Численные эксперименты показывают,
что указанное множество значений мальтузианского параметра не явля-
ется очень уж малым. Оно представлено как минимум пятью интервала-
ми и находится в экологически вероятной, по крайней мере для насеко-
мых, зоне значений R (десятки яиц на одну самку в сезон). Следователь-
но синхронизация циклов изменения численности с солнечной
активностью должна наблюдаться довольно часто, особенно среди насе-
комых. Подобную же синхронизацию можно ожидать и для вспышек
различных эпидемий, динамика которых также описывается отображе-
ниями типа Риккера [32]. Гелиобиологические эффекты такого рода были
описаны еще А.Л. Чижевским [41].
С эволюционной точки зрения описанное явление интересно в двух
аспектах. Во-первых, периодические колебания численности какого-
либо вида неизбежно будут модулировать изменения численности ви-
дов, связанных с ним по экосистемным цепям, трофическим, ресурс-
ным и т.п. Затронутые таким влиянием виды будут передавать перио-
дическое возмущение своим соседям по трофической цепи, а также
видам, связанным с ними другими отношениями: конкуренцией, му-
туализмом и т.п. Таким образом, в экосистемах, содержащих виды,
подверженные резонансной дестохастизации, возникают своеобразные
генераторы колебаний с 11- и 22-летним периодом, влияние которых
более или менее распространяется на всю экосистему.
Отсюда следует второй важный аспект резонансной супрессии хао-
са: подобная синхронизация колебаний численности различных видов,
составляющих экосистему, вероятно, задает временной масштаб, харак-
терный для всей экосистемы в целом. Феномен дестохастизации может
выполнять роль механизма экологических часов и определять ритм эво-
люции и возможность коэволюции. Необходимо также отметить и опи-
санные выше генетические последствия, генный дрейф и фильтрацию.
3.3. Детерминированный хаос и различные
эволюционные стратегии
Выше было показано, что предсказываемый теоремой Фишера рост
приспособленности для популяций с неперекрывающимися поколе-
259
Е.В. Евдокимов, А.В. Шаповалов
ниями приводит к их попаданию в область хаотического режима. Та-
кая эволюционная неизбежность хаоса и ее последствия активно об-
суждаются в литературе в последние два десятилетия. Кратко рассмот-
рим основные направления дискуссии.
Главная особенность динамики популяций, находящихся в хаоти-
ческом режиме, это высокоамплитудные колебания численности, при
этом в низших точках количество половозрелых особей может падать
до критических для выживания популяции значений [2, 7]. Очевидно,
что в ходе эволюции различные группы организмов по-разному “ре-
шали” эту проблему и выработали свои эволюционные стратегии “борь-
бы с хаосом”.
Первая, достаточно очевидная стратегия, позволяющая уменьшит ь
риск коллапса популяции в минимуме численности, заключается в с>
щественном повышении плодовитости. Вероятно, это является одной
из причин, почему у видов с неперекрывающимися поколениями, на-
секомых, рыб и т.д., число яиц (икринок и др.) на одну самку достига-
ет величин порядка 102—104 [7].
Вторую, прямо противоположную стратегию, направленную на
уменьшение мальтузианского параметра и выведение таким образом
популяции из хаотической области R, аргументировали М. Добели и
Дж. Коэла [42].
На третью из них обратил внимание Дж. Ракстон [20]. Он показал,
что введение взаимодействия и “перемешивания” гамет в ходе поло-
вого размножения существенно сужает область хаотического поведе-
ния популяций по сравнению с популяциями с бесполым, вегетатив-
ным или ограниченно половым (типа самоопыления) размножением.
Эта работа дает еще один угол зрения на известное в биологии явле-
ние, когда популяции переключаются с вегетативного размножения на
половое в неблагоприятных условиях.
Четвертая стратегия “борьбы с хаосом” заключается во введении
фактора перемешивания путем миграций [43]. Она была, вероятно,
реализована у таких видов, как саранча, лемминги, некоторые виды
птиц, для которых свойственны иррегулярные, хаотические вспышки
численности.
Представленные тезисы не исчерпывают всю тематику дискуссии
о теореме Фишера и нелинейной популяционной динамике, однако
отражают ее характерные черты.
260
IV. Эволюция животных
В заключении отметим, что представленный выше обзор проявле-
ния нелинейных эффектов и детерминированного хаоса в экологии и
эволюционной теории не претендует на исчерпывающую полноту,
поскольку число публикаций по данной тематике постоянно растет.
Нашей целью было показать невозможность игнорирования детерми-
нированного хаоса в разработке эволюционных концепций и проде-
монстрировать привлекательность нелинейной динамики для анализа
эволюционных стратегий, а также планирования новых исследований.
Наконец, нельзя хотя бы вскользь не обратить внимание на методоло-
гические трудности в экологических исследованиях, которые проясняет
нелинейная теория динамики популяций. Речь идет об оценке экологи-
ческих последствий различных антропогенных факторов. Как правило,
в таких работах в качестве одного из главных показателей измеряют чис-
ленность популяций, подвергшихся воздействию. Повышение или по-
нижение численности берут за критерий оценки фактора (положитель-
ного или отрицательного). С точки зрения теории детерминированного
хаоса такой методический подход неправомерен, по крайней мере для
популяций с неперекрывающимися поколениями. Уменьшение или уве-
личение численности популяции может быть просто началом перехода в
другой динамический режим, колебательный или хаотический, экологи-
ческую оценку которому можно давать только после многофакторного
анализа последствий (изменения устойчивости, возможность появления
генетического дрейфа, синхронизация трофических цепей и т.д.).
Литература
1. Шапиро А.П. К вопросу о циклах и возвратных последовательностях И Управление и
информация. Владивосток: ДВНЦ АН СССР, 1972. Вып. 3. С. 96-118.
2. May R.M. Biological populations with non-overlapping generations: stable points, stable
cycles and chaos // Science. 1974. Vol. 186. P. 645-647.
3. May R.M. Simple mathematical models with very complicated dynamics //Nature. 1976.
Vol. 26l.P. 459-467.
4. Шапиро А.П., Луппов С.П. Рекуррентные уравнения в теории популяционной био-
логии. М.: Наука, 1983. С. 132.
5 Ризниченко Г.Ю., Рубин А.Б. Математические модели биологических продукцион-
ных процессов. М.: МГУ, 1993. С. 301.
6. Ricker W.E. Stock and recruitment // Theor. J. Fisheries Res. Bard. Can. 1954. Vol. 11.
N 5. P. 559-623.
7. Hassel M.P., Lawton J.H., May R. Patterns of dynamical behaviour in single-species
populations //J. Anim. Ecol. 1976. Vol. 45. P. 471-486.
261
Е.В. Евдокимов, А.В. Шаповалов
8. Schuster H.G. Deterministic Chaos: An Introduction, 2nd edn. VCH Weinheim. 1988.
9. Фейгенбаум M. // Успехи физ.наук, 1983. T. 141. С. 343-357.
10. Заславский Г.М., Сагдеев Р.З. Введение в нелинейную физику. От маятника до гур.
булентности и хаоса. М.: Наука, 1988. 368 с.
11. Берже П., Помо И., Видаль К. Порядок в хаосе. М.: Мир, 1991. 367 с.
12. Мун Ф. Хаотические колебания М.: Мир, 1990. 214 с.
13. Phillips PC. Distinguishing chaos from noise in Nematode population dynamics // Early
1995 International Worm Meeting, abstract 524.
14. Costantino R.F., Deshamais R.A., Cushing J.M., Dennis B. Chaotic Dynamics in an
Insect Population // Science. 1997. Vol. 275. P. 389-391.
15. Fisher R.A. The genetical theory of natural selection. Oxford: Clarendon Press, 1930
16. Ли Ч. Введение в популяционную генетику. М.: Мир, 1978. С. 411.
17. Пых Ю.А. Равновесие и устойчивость в моделях популяционной динамики М
Наука, 1983.
18. Свирижев Ю.М., Пасеков В.П. Основы математической генетики. М.: Наука, 1982
19. Asmussen М.А. Density dependent selection incorporating intraspecific competition 1
Ahaploid model // Journal of Theoretical Biology. 1983. Vol. 101. P. 113-127.
20. Ruxton G.D. Population models with sexual reproduction show a reduced propensit) to
exhibit chaos // Journal of Theoretical Biology. 1995. Vol. 175. P. 595-601.
21. Witten M. Fitness and survival in logistic models // Journal of Theoretical Biology 1978
V. 74. P. 23-32.
22. Фрисман ЕЯ. Изменение характера динамики численности популяции: механизмы
перехода к хаосу // Вестник ДВО РАН. Экология. 1995. № 4. С. 92-103.
23. Фрисман Е.Я. Эволюция характера динамики численности популяции: переход к
хаосу И Исследования по математической биологии / Под ред. Э.Э. Шноль. Пущи-
но: Изд-во Путинского НЦ РАН, 1996. С. 75-87.
24. Cavalieri L.F., Kocak Н. Intermittent transition between order and chaos in an insect pest
population // Journal of Theoretical Biology. 1995. Vol. 175. P. 231-234.
25. MacArthur R.H. Some generalized theorems of natural selection //
Proc.Natl.Acad.Sci.USA. 1962. Vol. 48. P. 1893-1897.
26. Печуркин H.C. Популяционная микробиология. Новосибирск: Наука, 1978. С. 278
27. Felsenstein J. г- and К-selection in a completely chaotic population model //American
Naturalist. 1979. Vol. 113. P. 499-510.
28. Tao Yi, Yang Qi-sen, Jiang Zhi-gang, Wang Zu-wang. Evolutionarily Stable Strategy
Stable State, Periodic Cycle and Chaos in a Simple Discrete Time Two-Phenotype Model
//Journal of Theoretical Biology. 1997. Vol. 188. P. 21-27.
29. Печуркин H.C., Брильков A.B., Марченкова ТВ. Популяционные аспекты биотех-
нологии. Новосибирск: Наука, 1990. С. 174.
30. Цоглин Л.Н., Владимирова М.Г., Семененко В.Е. Математическое и эксперимен
тальное моделирование процесса автоселекции микроводорослей в условиях про-
точного моделирования И Физиология растений. 1970. Т. 17. Вып. 6. С. 1129-1139
31. Earn D., Rohani Р. Complex dynamics in ecology II Trends in Ecology & Evolution
1999. Vol. 14. P. 43-44.
32. Philippe P. Chaos, population biology, and epidemiology: some research implications //
Hum. Biol. 1993. Vol. 65:4. P. 525-546.
33. Scheming I., Janosi I.M. When two and two make four: a structured population without
chaos//Journal of Theoretical Biology. 1996. Vol. 178. P. 89-97.
262
IV. Эволюция животных
_д paradis, Е. Metapopulations and Chaos: On the Stabilizing Influence of Dispersal //
journal of Theoretical Biology. 1997. Vol. 186. P. 261-266.
35 Rohani P., May R.M., Hassell M.P. Metapopulations and equilibrium stability: the effects
of spatial structure//Journal of Theoretical Biology. 1996. Vol. 181:2. P. 97-109.
36 Rohani P., Ruxton G.D. Dispersal-induced instabilities in host-parasitoid metapopulations
//Theor. Popul. Biol. 1999. Vol. 55:1. P. 23-36.
37 Hassel M.P., Comins H.N., May R. Spatial structure and chaos in insect population
’ dynamics //Nature. 1991. Vol. 353. P. 255-258.
38. Weisser W.W., Jansen VA., Hassell M.P. The effects of a pool of dispersers on host-
parasitoid systems //Journal of Theoretical Biology. 1997. Vol. 189:4. P. 413-425.
39. Евдокимов E.B., Евдокимов K.E., Крейдун Ю.А., Шаповалов А.В. Квазирезонансные
свойства периодически возмущенных однопараметрических логистически-подобных
отображений // Журнал физической химии. 1997. Т. 71. № 11. С. 2003-2008.
40. Материалы IV Международного симпозиума “ Корреляции биологических и физи-
ко-химических процессов с космическими и гелио-геофизическими факторами”.
Пущино, 1996.
41. Чижевский А Л. Земное эхо солнечных бурь. М.: Мысль, 1976. С. 368.
42. Doebeli М. and Koella J. Evolution of simple population dynamics // Proc. R. Soc. London
Ser. B. 1995. Vol. 260. P. 119-125.
43. Holt R D. and McPeek M.A. Chaotic population dynamics favours the evolution of
dispersal //Am. Nat. 1996. Vol. 148. P. 709-718.
ВИДОВЫЕ АДАПТАЦИИ НА ПРИМЕРЕ
ВЗАИМООТНОШЕНИЙ
РОДИТЕЛИ-ПОТОМКИ У МЛЕКОПИТАЮЩИХ
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
Институт систематики и экологии животных СО РАН
630090, г Новосибирск, ул. Фрунзе, 11
E-mail: ei@eco.nsc.ru
После возникновения редупликации крупнейшим “изобретением”
эволюции стало половое размножение. Половое размножение обеспе-
чивает постоянное совершенствование интегрированного генофонда
видов, т.к. способствует умножению в цепи поколений мутационною
резерва (Четвериков, 1926) и поддерживает за счет комбинативной из-
менчивости наследственное разнообразие, повышая тем самым устой-
чивость надорганизменных систем в изменчивой среде.
Половое размножение обеспечило становление и дальнейшую эво-
люцию онтогенетического уровня организации живой материи. Орга-
низм с его уникальным сочетанием генетических задатков и конечно-
стью существования становится в собственном смысле «особью», т.е.
чем-то особым, онтогенетически обособившимся из пространствен-
но-временного континуума жизни. Одновременно половое размноже-
ние, приведя к потере отдельным организмом значения самовоспро-
изводящейся единицы, увеличило взаимозависимость особей и озна-
меновало появление популяционного уровня, начало эволюции
популяций. Теперь размножение обеспечивается встречей двух индиви-
дуумов разного пола; в результате возникает нечто третье, генетичес-
ки отличающееся от обоих «прототипов». Единицей воспроизводства
становится триединая «ячейка» - семья. Присутствие тут младшего
члена символизирует преемственность репродуктивного (и эволюци-
онного) процесса, поскольку приспособленность потомка естествен-
ным образом приумножает или, наоборот, уменьшает адаптивную цен-
ность каждого из родителей.
264
IV. Эволюция животных
Организация жизни на Земле носит многоуровневый характер.
Выделение «уровней», несмотря на определенную условность, отра-
жает эту реальную сложность (Яблоков, 1987; Лидикер, 1999). На каж-
дом уровне возникают иерархически структурированные функциональ-
ные системы, состоящие из упорядоченно взаимодействующих ком-
понентов и обладающие качественно новыми («эмерджентными»)
свойствами, не сводимыми к сумме свойств частей. Понять сущность
биологических систем можно только с учетом их основополагающего
свойства, названного Э.С. Бауэром (1935) «принципом устойчивого
неравновесия». Называемое теперь динамическим равновесием, или
гомеостазом, свойство это заключается в стремлении живых систем
возвращаться после отклонения в исходное состояние с сохранением
основных параметров. И во всех случаях сохранение целостности био-
логической организации обеспечивается сложной эволюционно сло-
жившейся системой обратных связей, жертвующей «интересами» со-
подчиненных элементов ради сохранения всей системы.
Эволюция от агамных видов к видам с настоящим половым раз-
множением сопровождалась, с одной стороны, процессами дифферен-
циации и функциональной специализации полов, с другой, - возник-
новением и усилением механизмов интеграции на популяционно-ви-
довом уровне репродуктивных систем, обеспечивающих сохранение
целостности вида в чреде поколений (во времени). Отражением этой
интеграционной тенденции служит возрастание в эволюционном ряду
позвоночных зависимости потомков от родителей: появляется внут-
реннее оплодотворение; удлиняется период внутриутробного существо-
вания; у млекопитающих прогрессирует проницаемость плаценты; все
большее значение и продолжительность приобретает забота об отно-
сительно незрелом потомстве в постнатальный период.
О целостности видовых систем свидетельствует появление приспо-
соблений, полезных не отдельной особи, а группе представителей дан-
ного вида, т.е. «видовых адаптаций» (Завадский, 1967), не сводимых
ни к индивидуальным адаптациям, ни к их сумме, и относящихся, та-
ким образом, к эмерджентным свойствам вида. Можно различать не-
сколько групп видовых адаптаций, как это сделал С. А. Северцов (1951)
в учении о конгруэнциях. Конгруэнции - это соответствия в поведе-
нии, строении и функциях органов, возникающие между особями од-
ного вида (родителями и потомками; брачными партнерами; членами
265
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
колонии, стада и т.д.)- Польза таких соответствий для жизнедеятель-
ности и воспроизводства проявляется лишь при взаимодействии обо-
юдно заинтересованных индивидуумов, и в этом смысле конгруэнции
могут рассматриваться как видовые приспособления. В приложении к
млекопитающим первой из этих групп конгруэнций выделим соответ-
ствия в строении и функциях эмбриона и материнского организма.
Действительно, беременная самка у млекопитающих занимает по-
ложение на границе онтогенетического и популяционного уровней,
поскольку является одновременно и особью, и маленькой «популяци
ей» со специфическими внутриутробными взаимодействиями между
отдельными ее членами. В случае многоплодного вида самка потен
циально может стать основательницей «нормальной» популяции (Яб
локов, 1987). В некотором смысле беременную самку можно рассма.
ривать даже как «межвидовое» сообщество. Дело в том, что эмбрионы
воспринимаются материнской иммунной системой как чужеродна*
инвазия, да и сами они ведут фактически «паразитический» образ
жизни. Однако лишь эффективное взаимодействие в пределах систе-
мы мать-эмбрион служит залогом процветания вида, а необходимым
условием тому являются «взаимовыгодные» (мутуалистические) вза-
имоотношения. Если для эмбрионов выгода эта сиюминутна и конк
ретна, то для самки она имеет отсроченный, в эволюционном смысле
характер: ведь «качество» потомства определяет эволюционный успех
родителей в данном цикле воспроизводства (их адаптивную ценность)
Мать и потомки в период беременности являют нерасторжимо еди
ную систему, а адаптивные свойства представителей следующего по-
коления складываются в зависимости от условий их пренатального
развития. Малейшее несоответствие во взаимных реакциях может с г
рицательно сказаться на развитии потомка, поэтому его единственной
эволюционно-оправданной стратегией в условиях полной физиологи
ческой зависимости от матери, когда реализовать свои генетически
предопределенные потенции он способен лишь во взаимодействии с
ней, является поддержание с ней своеобразного «диалога» (Евсиков и
др., 1998) через посредство подаваемых сигналов о своем существо-
вании, состоянии, потребностях.
Многочисленные данные свидетельствуют, что важным фактором
определяющим пренатальное существование организма, является на-
личие генетических различий между ним самим и организмом мате-
266
IV. Эволюция животных
пи уровень которых для успешного функционирования репродуктив-
ной системы, впрочем, обычно не должен превышать внутривидово-
го Маркерами для опознавания иммунной системой матери генети-
ческих отличий ее плода от нее самой служат антигены гистосовмес-
тимости, по которым эволюционно достигнута поистине рекордная
изменчивость. Экспрессия этих белков на клеточных стенках зароды-
ша как раз и является тем первым сигналом, который он подает, всту-
пая в «заинтересованное общение» с матерью.
Однако еще за временными рамками существования потомка само
наличие генетических различий между его родителями определяет
«преадаптивный» характер его онтогенеза. У млекопитающих, для
большинства из которых главным коммуникативным каналом являет-
ся ольфакторный, антигенная индивидуальность особи закодирована
в определенных компонентах запаха, а специально поставленные опы-
ты свидетельствуют о действии механизмов запахового предпочтения
именно «чужеродных», отличающихся по антигенам, половых парт-
неров (см.: Yamazaki et al., 1999). К тому же существуют данные о раз-
личной фертилизации яйцеклеток спермиями в зависимости от нали-
чия у них (а значит, у потенциальных эмбрионов) антигенных отличий
от самки (Wedekind et al., 1996).
После оплодотворения, в ответ на возникающую угрозу иммунного
отторжения (Вербицкий, 1973; Говалло, 1987), эволюция выработала
механизмы, благоприятствующие сочетанию «несовместимого»; ведь это
служит залогом поддержания наследственного разнообразия в цепи смеж-
ных поколений. В частности, повышение уровня «гормона беременнос-
ти» - прогестерона - увеличивает иммунную защиту «чужеродных» за-
родышей, а морфогистологическая перестройка плаценты ведет к их луч-
шему снабжению белковыми соединениями и, соответственно, - лучшему
пренатальному развитию (Евсиков и др., 1998).
Еще в начале 70-х годов в ИМБиГ НАН Украины была выполнена
работа по межлинейным пересадкам эмбрионов, в результате которой
было установлено, что чужеродные «трансплантированные» мыши
обладают большими, чем в контроле, темпами не только пренатально-
го, но и постнатального развития (см.: Евсиков и др., 1998). Посколь-
ку сразу после рождения детеныши млекопитающих по-прежнему за-
висят от матери, обладая незрелой иммунной системой и нуждаясь в
молочном кормлении и воспитании, было интересно выяснить конк-
267
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
ретные механизмы ускоренного развития гетерогенных потомков в
период лактации. В недавних экспериментах показано, что у самок
мышей BALB/cLac (далее - С) после того, как они родили гибридов
от скрещивания с самцами C57BL/6J (В6), улучшается питательный
состав молока и увеличивается материнская забота о потомках. Уве-
личение материнского вклада приводит к тому, что детеныши (причем
даже приемные чистолинейные) развиваются более быстрыми темпа-
ми (Потапов, Евсиков, 2000). Таким образом, в лактационный период
продолжается действие запущенных еще во время беременности фи-
зиологических механизмов, способствующих эффективному развитию
генетически чужеродных потомков.
Между тем была продолжена и работа по межлинейным пересад-
кам эмбрионов, в результате которой выяснилось, что контрольные
самки В6 (стандартное разведение) предпочитают запах самцов В6,
«трансплантированных» на стадии ранних эмбрионов в матку самок
С, по отношению к запаху контрольных самцов В6 (Potapov et al., 1995)
Более того, в дальнейшем оказалось, что для повышения привлека-
тельности самцов В6 достаточно только вскармливания самкой С
Одним из возможных объяснений причин такого предпочтения может
быть следующее. Известно, что существует перенос иммунологичес-
ких факторов (как иммуноглобулинов, так и активных лейкоцитов) в
организм потомка через плаценту и с молоком (Вербицкий, 1973; Го-
валло, 1987; Goldman et al., 1994). Переход материнских лейкоцитов с
молоком, который получает все больше экспериментальных подтвер
ждений (Hanson, 2000), является причиной приобретения толерантно-
сти к материнским антигенам (матери вследствие этого - хорошие
доноры органов для своих детей). Таким образом, наши эксперимен-
тальные мышата становятся в некоторой степени «химерами» (этим
термином обозначают животных, состоящих из клеток с различной
генетической информацией, происходящих первоначально от двух или
более оплодотворенных яйцеклеток; см.: McLaren, 1976; Зубин, Веп-
ринцев, 1988). Если же антигены чужеродных лейкоцитов принимаю!
участие в формировании запаха (а такая возможность показана экспе-
риментально; см.: Yamazaki et al., 1999), то такие животные различи
мы соплеменниками, поскольку способности мышей улавливать тон-
чайшие различия запахов, связанных с антигенной индивидуальнос-
тью, широко известны. Конечно, предпочтения, отдаваемые «транс-
268
IV. Эволюция животных
^актированным» и «приемным» самцам, не являются доказательством
их химеризма (для этого требуется специальный анализ), однако в
любом случае свидетельствуют о большом значении взаимодействия
матери и потомков для формирования адаптивных черт последних.
Потенциальные эффекты перехода иммунологических факторов от
матери к детенышу через потребляемое им молоко не ограничивают-
ся только положительными: приобретенным иммунитетом и иммуно-
модуляцией (Goldman et al., 1994; Xanthou, 1998), поскольку мать им-
мунизирована антигенами зародыша во время беременности (класси-
ческий пример такой иммунизации - несовместимость по резус-
фактору у человека). В принципе, в период молочного вскармливания
возможно даже угнетение жизнеспособности потомков вследствие
перехода в их организм с молоком антизародышевых антител и разви-
тия РТПХ (реакции трансплантата против хозяина, т.е. вторгшихся
материнских иммунных клеток против тканей новорожденного; см.:
Beer, Billingham, 1975; Goldman et al., 1994). Известно, что с каждой
беременностью от одного и того же партнера (и даже с каждым вступ-
лением с ним в половую связь) в крови женщин нарастает титр анти-
тел против антигенов данного мужчины и увеличивается риск как для
течения беременности (бесплодие, привычные выкидыши), так и для
развития приплода (эритробластоз и лейкопения новорожденных; см.:
Вербицкий, 1973; Снелл и др., 1979; Говалло, 1987).
Чтобы убедиться в негативных последствиях повторной беремен-
ности от одного и того же «чужеродного» партнера, из числа единож-
ды рожавших самок В6 были сформированы две экспериментальные
группы, в каждой из которых были проведены гибридизующие скре-
щивания с самцами линии С. При этом в I группу вошли самки В6, в
предыдущий раз родившие чистолинейных мышат, во II группу - сам-
ки, в первый раз уже родившие гибридов (B6><C)F1. После того, как
Для полученных в опыте гибридных мышат закончился период вскар-
мливания (21-й день), определяли индекс их селезенки (отношение ее
массы к массе тела), который является показателем активации иммун-
ной системы и используется в качестве показателя РТПХ (Никитин,
1982). Выяснилось, что индекс селезенки гибридных потомков
(B6xC)Fl к моменту окончания вскармливания собственными мате-
рями превышает показатели для чистолинейных мышат (Потапов, Ев-
сиков, 2000), а в случае повторной гибридизации самок - еще более
269
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
возрастает (8,68± 1,05 мг/г (и=63) и 9,84±4,32 мг/г (и=67) для I и II групп
соответственно, р<0,05, /-критерий; здесь и далее приведены средние
± стандартное отклонение). Сам по себе индекс селезенки еще не гово-
рит о неблагополучии мышат, но нужно учесть, что он коррелирует со
смертностью (%) мышат в выводках (для гибридов (B6xC)Fl по несколь-
ким опытам: г=+0,64, и=162,р<0,001; отдельно для гибридов II группы
(повторная гибридизация): г=+0,81, и=67,р<0,001).
И все-таки надо сказать, что мы не наблюдали резкого снижения
жизнеспособности потомков даже при получении нескольких после-
довательных приплодов от таких «неудачных» репродуктивных пар
(самок В6 и самцов С). Очевидно, система мать-потомки имеет в сво-
ем арсенале достаточно мощные компенсаторные механизмы, способ-
ствующие успешной реализации репродуктивной функции. Тем нс
менее млекопитающие большинства видов предпочитают не «прове-
рять» надежность этих защитных гомеостатических механизмов; дру-
гими словами, им не свойственны моногамные отношения. При этом
самцы склонны к полигинии (многоженству) из-за стремления макси-
мизировать свой «вклад» в генофонд следующего поколения; самки
же, как мы предполагаем, стремятся к полиандрии, т.е. к брачному парт-
нерству в каждом следующем репродуктивном цикле с новым сам-
цом, генетически отличным не только от них самих, но и от предыду-
щих партнеров. При этом сохраняются все выгоды от подбора анти-
генно-отличных пар и предотвращаются негативные эффекты
повторной иммунизации.
На двух видах - беспородных домовых мышах (Mus musculus) и
джунгарских хомячках (Рhodopus sungorus) - мы проверили, насколь-
ко выражено у них стремление к перемене партнера. Репродуктивные
пары содержали до появления у самок явных признаков беременнос-
ти, после чего самцы были отсажены. Когда самки выкормиливывод-
ки и находились в эструсе, им были предложены запахи «своего» сам-
ца (отца детенышей) и «чужого», случайно выбранного из этой же эк-
спериментальной группы животных. На мышах опыты были проведены
дополнительно и в послеродовом эструсе, в котором самки грызунов
бывают сразу после рождения выводков. Аналогично, самцам предъяв-
ляли запахи «супруги» и незнакомой самки. Оказалось, что не только
самцы, но и самки того и другого вида грызунов предпочитают нового
партнера (рис. 1).
270
IV Эволюция животных
Рис. 1. Время исследования запаха предыдущего партнера («свой») и незна-
комого партнера («чужой») беспородными мышами и джунгарскими хомяч-
ками того и другого пола при нахождении самок в эструсе: послеродовом («п/
р») или по окончании выкармливания выводков («п/в»). Все отличия между
временем исследования своего и чужого партнера статистически достоверны
(отр<0,05 до р<0,0001, критерий Уилкоксона для парных сравнений)
И не только в экспериментах. Интересно, что, даже находясь под
контролем самца-доминанта (становящегося, как правило, отцом боль-
шинства потомков в демах - репродуктивных группах), самки мышей
иногда спариваются за пределами его участка (Potts et al., 1992). Появ-
ление нового партнера (и даже контакт с его запахом) может привести
к прерыванию беременности и восстановлению рецептивности - го-
товности к спариванию («эффект Брюс», названный так в честь от-
крывшей его в 1959 г. Хильды Брюс; см.: Соколов и др., 1986).
Для млекопитающих, с их длительным периодом беременности и
лактации, стоимость новой жизни высока, что повышает разборчивость
самок при формировании брачной пары. В случае «полюбовного» со-
четания партнеров наблюдается увеличение материнского вклада в
потомков в лактационный период (Потапов, Евсиков, 2000). Лучшее
развитие потомков к моменту отъема в значительной степени гаранти-
рует им больший репродуктивный успех: более быстрое половое со-
271
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
зревание и раннее вступление в размножение. Однако в неблагоприят-
ных условиях (при стрессе или при сочетании с «нежеланным» парт-
нером) у разных видов млекопитающих относительный материнский
вклад может компенсаторно повышаться за счет снижения плодовито-
сти из-за частичного отхода эмбрионов того или другого пола (Krackow,
1990; Pratt, Lisk, 1991). В основе дифференциации материнским орга-
низмом пола зародыша может лежать распознавание Y-антигена, име-
ющегося лишь у самцов (Schellinck et al., 1993). Наши эксперимен-
тальные данные по водяным полевкам (Arvicola terrestris) свидетель-
ствуют о способности данного вида к адаптивной регуляции пола
потомства (Evsikov et al., 1995; Назарова, Евсиков, 2000; Потапов, Ев-
сиков, 2000).
Мы провели изучение влияния подбора партнера на соотношение
полов в потомстве джунгарских хомячков. На основе результатов оль-
факторных тестов были сформированы две группы, в одной из кою
рых партнерами самок выступали наиболее предпочитаемые, а в дру-
гой - наименее предпочитаемые самцы. Выяснилось, что, как и у во-
дяных полевок, воспроизводство осуществляется более эффективно,
если самкам предоставлена возможность самим выбрать самца. В ре-
зультате же покрытий непривлекательными самцами рождается в сред-
нем меньше детенышей за счет сокращения числа самок (рис. 2). При
этом некоторое уменьшение числа детенышей в помете из-за перерас
пределения ресурса материнской заботы и кормления ведет к их луч-
шему постнатальному развитию, и в первую очередь - самцов. При
отсадке масса тела выкормленных самцов составила для, казалось бы,
«неблагоприятного» сочетания партнеров 17,0±1,9 г (и=21) по срав-
нению с 15,2± 1,1г (п= 19) для «благоприятного» (р=0,001, Г-критерий).
Получается, что лишенные выбора самки будто бы создают своим «пре-
емницам» в следующем поколении лучшие условия для подбора реп-
родуктивного партнера как в количественном, так и качественном пла-
не. С другой стороны, поскольку репродуктивный вклад крупных и
конкурентоспособных самцов может значительно превышать таковой
самок, половой состав выводков отражается на вкладе матерей в гено-
фонд следующих поколений.
Оказалось, что выявленная в опытах адаптивная регуляция соот-
ношения полов в потомстве реализуются и в природе (Evsikov et al,
2000). В популяции водяных полевок при усилении действия стресси-
272
IV. Эволюция животных
Шсамцы О самки
Число детенышей в выводке
Рис. 2. Число рожденных детенышей того и другого пола в выводках, принесен-
ных самками водяной полевки (по: Evsikov et al., 1995) и джунгарского хомячка
(оригинальные данные) после спаривания с предпочитаемыми («П») и непредпо-
читаемыми («Н») самцами. Отличия по числу самок и суммарному числу дете-
нышей в выводках статистически достоверны (/-критерий, медианный тест)
рующих факторов возрастает уровень эмбриональных потерь. С ним,
в свою очередь, положительно связана на межгодовом уровне доля
самцов среди эмбрионов (г=+0,56, и=14,р<0,05). Таким образом, мож-
но считать, что у млекопитающих, обладающих наиболее совершен-
ными механизмами сохранения жизни своего потомства, эмбриональ-
ная смертность в значительной мере носит адаптивный характер. Из-
менение уровня эмбриональной смертности лежит в основе
популяционного гомеостаза и обеспечивает популяциям млекопитаю-
щих адаптацию к меняющимся условиям внешней среды.
Очень интересной проблемой являются взаимоотношения с потом-
ками их отцов. Несмотря на то, что самцы у млекопитающих не слиш-
ком обременены заботой о потомстве, тут тоже существует изменчи-
вость. Скажем, у тех же домовой мыши и джунгарского хомячка есть
виды-двойники, сравнительно недавно получившие независимый си-
273
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
стоматический статус: курганчиковая мышь (Mus spicilegus) и хомя-
чок Кэмпбелла (Р hodopus campbelli), где связи отцов с потомками зна-
чительно прочнее. Есть мнение, что самец хомячка Кэмпбелла может
в какой-то мере участвовать в воспитании потомства (Reburn, Wynne-
Edwards, 1999), а курганчиковая мышь и вовсе склонна к моногамии
(Patris, Baudoin, 1998). Ну, а в Швейцарии живут себе парочками и
плодятся в своих подземных лабиринтах представители роющей фор^
мы водяных полевок (Arvicola terrestris scherman\ см.: Airoldi, 1976).
Но и у обычных домовых мышей, оказывается, тоже существусл
изменчивость по степени выраженности отцовской заботы о потом
стве, что расширяет возможности адаптивной стратегии вида в реали-
зации репродуктивного потенциала. Из мышей линии С путем предо
ставления самкам в каждом поколении самостоятельного выбора по-
лового партнера на основе исследования запаховых стимулов была
«отселектирована» сублиния ОСП («ольфакторный стимул предпочи-
таемый»). После 5 циклов «самоотбора» мыши ОСП отличаются от
стандартно разводимых контрольных как возросшей долей самцов,
демонстрирующих отцовскую заботу, - затаскивающих в гнездо уда-
ленных из него детенышей (56% заботливых отцов по сравнению с
27% в контроле, р<0,05, критерий с2), так и большей сохранностью
детенышей в выводках за период лактации (75% полностью сохранив-
шихся выводков по сравнению с 60% в контроле, р<0,05, критерий с2).
Во многих случаях благоприятствования со стороны родителей
развитию потомков (за счет увеличения уровня прогестерона, роди-
тельской заботы, питательного состава молока и т.д.), итогом чего яв-
ляется повышение адаптивной ценности не только последних, но и
самих родителей, важную роль может играть гормон пролактин (ПРЛ),
обладающий широким спектром биологических функций (Теппермен,
Теппермен, 1989; Goffin et al., 1998). У грызунов ПРЛ выполняет лю-
теотропное действие, регулируя продукцию прогестерона яичниками
и способствуя тем самым нормальному течению беременности (Green-
wald, Wang, 1994). Кроме того, ПРЛ влияет на родительское поведение
как самок (Bridges, 1994), так и самцов (Rebum, Wynne-Edwards, 1999),
а также на выработку молока у матери в ответ на акт сосания и сигна-
лы голода сосунков (в том самом «диалоге» с ней). При этом хороший
уровень сигнала, подаваемого потомком, гарантирует ему больший
материнский вклад в его развитие. ПРЛ играет также ключевую роль в
274
IV. Эволюция животных
синтезе материнского феромона, участвующего в миелинизации ЦНС
й формировании иммунорезистентности потомков (Новиков, 1988).
Интересно, что в данном случае своеобразной формой обмена сигна-
лами лактирующей самки и детеныша является взаимная копрофагия,
в результате которой регулируется метаболизм ПРЛ.
Половое созревание также является тем критическим для особей
периодом, где важны взаимоотношения между родительским и дочер-
ним поколениями. В первую очередь, речь идет об эффектах ускоре-
ния или замедления полового созревания и вступления в репродуктив-
ный процесс молодых самок и самцов в зависимости от половой при-
надлежности и физиологических характеристик взрослых животных,
с которыми (или с запаховыми стимулами которых) им приходится
сталкиваться. Описания классических работ У. Уиттена, Дж. Ванден-
берга, Л. Дрикамера и других исследователей приведены в моногра-
фии С.Н. Новикова (1988). Они находят свое дальнейшее развитие в
работах популяционных экологов, поскольку регуляция вступления
молодых особей в размножение является важным фактором демогра-
фических процессов.
Таким образом, приведенные примеры иллюстрируют приспособ-
ления вида, направленные на более полную реализацию его репродук-
тивного потенциала. Важнейшей видовой адаптацией, значение кото-
рой начинает проявляться на этапе образования брачной пары, захва-
тывая весь период взаимозависимости в системе родители-потомки и
определяя формирование триединой репродуктивной единицы - се-
мьи в последующих циклах воспроизводства, является полиморфизм
по тканевым антигенам. Взаимоотношения родителей и потомков ба-
зируются на постоянном обмене сигналами, загадочном диалоге по-
колений (Евсиков и др., 1998). Взаимодействующая система родите-
ли-потомки едина перед лицом естественного отбора. В этом един-
стве и взаимозависимости заложена сила семьи, ее способность
противостоять бурям и невзгодам существования.
В заключение обратимся только на одном примере к проблеме видо-
образования. Ведь, казалось бы, интеграция в системе родители-потом-
ки, приводящая к появлению «видовых адаптаций», есть «конечная стан-
ция» эволюции вида. Но зададимся вопросом: не может ли усиление вза-
имосвязей в этой системе само по себе выполнять функцию движущего
°тбора, задавать вектор эволюционно «прогрессивного» развития?
275
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
Ранее мы уже высказывали мысль о том, что формирование чело-
века современного типа сопровождалось (и, отчасти, определялось)
своеобразным «искусственным» отбором - «приручением» самками
предка человека своих самцов. Подобный внутривидовой отбор, в пол-
ном соответствии с тем, что наблюдается при доместикации других
видов, в конце концов мог привести к достижению чрезвычайной из-
менчивости фенотипического облика, инфантилизации поведения,
исчезновению сезонности размножения и т.д. (см.: Потапов, Евсиков,
2000). На эти любопытные аналогии обращал внимание еще академик
Д.К. Беляев. Одним из итогов действия такого отбора могло стать и
дальнейшее увеличение взаимозависимости в рамках воспроизводи-
тельной единицы - семьи, которая все больше приобретала черты мо-
ногамной, нетипичной для большинства других видов млекопитаю-
щих. Но, кроме этого, в эволюции человека интеграционный процесс
в системе мать-потомки зашел, пожалуй, слишком далеко и привел к
довольно неожиданным последствиям. Чтобы стало ясно, о чем идет
речь, напомним, что на лабораторных животных показано, что голая
кожа новорожденных входит в комплекс сигналов, провоцирующих
материнскую заботу в ранний лактационный период (Scribner, Wynne-
Edwards, 1994).
Так не прошли ли в свое время предки человека через отбор на
столь милый для материнского восприятия признак? Не послужила ли
затянувшаяся «детскость» (своеобразная «неотения») причиной бес-
прецедентного продления периода родительской опеки, создавшего в
том числе предпосылки культурной преемственности на основе на-
учения в семье (передачи навыков, включая новые, например, по изго-
товлению ставших необходимыми искусственных покровов)? Не без-
рассудная ли материнская любовь (отражающая «всего лишь» эволю-
ционную тенденцию к интеграции репродуктивных систем) стала той
силой, которая несет ответственность за закрепление в нашем облике
фенотипических черт тех из наших предков, кого прочие, «волосатые»,
соплеменники презрительно кликали до поры до времени «лягушата-
ми» (на их языке - «mowgli»)?
Исследования поддержаны РФФИ (гранты №№ 99-04-50013,
00-04-49210).
276
IV. Эволюция животных
Литература
Бауэр Э.С. Теоретическая биология. М.; Л., 1935. С. 43.
Вербицкий М.Ш. Особенности иммунологической регуляции эмбриогенеза у млекопитаю-
щих и человека// Основы иммуноэмбриологии. М.: Медицина, 1973. С. 184-280.
Говалло В.И. Иммунология репродукции. М.: Медицина, 1987.
Евсиков В.И., Герлинская Л.А., Мошкин М.П., Осетрова Т.Д., Потапов М.А. Генетико-
физиологические взаимоотношения мать-плод и их влияние на адаптивные при-
знаки потомков И Онтогенез. 1998. Т. 29. № 6. С. 405-417.
Завадский К.М. Вид как форма существования жизни. Структура вида. Видообразова-
ние//Современные проблемы эволюционной теории. Л.: Наука, 1967. С. 145-295.
Зубин М.Н., Вепринцев Б.Н. Химеры мышей. Методы, проблемы, перспективы. Пущи-
но, 1988.
Лидикер В. Популяционная регуляция у млекопитающих: Эволюция взгляда И Сиб.
экол. журн. 1999. Т. 6. № 1. С. 5-13.
Назарова Г.Г., Евсиков В.И. Влияние условий выкармливания на выживаемость потом-
ков, их репродуктивные характеристики и соотношение полов у водяной полевки
(Arvicola terrestris) И Зоол. журн. 2000. Т. 79. № 3. С. 58-63.
Никитин В.М. Справочник методов иммунологии. Кишинев: Штиинца, 1982.
Новиков С.Н. Феромоны и размножение у млекопитающих. Л.: Наука, 1988.
Потапов М.А., Евсиков В.И. Генетико-физиологические взаимоотношения мать-плод
и их влияние на адаптивные признаки потомков: Взгляд с третьей стороны И Со-
временные концепции эволюционной генетики. Новосибирск: ИЦиГ СО РАН, 2000.
С. 277-293.
Северцов С.А. О конгруэнциях и понятии целостности вида И Проблемы экологии
животных. М.: Изд-во АН СССР, 1951. Т. 1. С. 30-57.
Снелл Дж., Доссе Ж., Нэтенсон С. Совместимость тканей. М.: Мир, 1979.
Соколов В.Е., Лебедев В.С, Зинкевич Э.П. Нарушение беременности у грызунов под
воздействием обонятельных сигналов (Брюс-эффект) // Химическая коммуника-
ция животных: Теория и практика. М.: Наука, 1986. С. 329-338.
Теппермен Дж., Теппермен X. Физиология обмена веществ и эндокринной системы:
Вводный курс. М.: Мир, 1989.
Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения со-
временной генетики И Проблемы общей биологии и генетики (воспоминания, ста-
тьи, лекции). Новосибирск: Наука, 1983 (1926).
Яблоков А.В. Популяционная биология. М.: Высшая школа, 1987.
Airoldi, J.-P. Le terrier de la forme fouisseuse du campagnol terrestre, Arvicola terrestris
scherman Shaw (Mammalia, Rodentia) // Z. Saugetierk. 1976. Vol. 41. P. 23-42.
Beer A.E., Billingham R.E. Immunologic benefits and hazards of milk in maternal-perinatal
relationship //Ann. Intern. Med. 1975. Vol. 83. N 6. P. 865-871.
Bridges R.S. The role of lactogenic hormones in maternal behavior in female rats // Acta
Pediatr. Suppl. 1994. Vol. 397. P. 33-39.
Evsikov VI., Nazarova G.G., Potapov M.A. Female odour choice, male social rank, and sex
ratio in the water vole //Advances in the Biosciences. Oxford: Pergamon. 1995. Vol. 93.
P. 303-307.
277
В.И. Евсиков, М.А. Потапов, Е.А. Новиков, О.Ф. Потапова
Evsikov V.I., Nazarova G.G., Potapov М. A., Rogov V.G., Gerlinskaya L.A. Ecological factors
determine differential reproduction in mammals // Biodiversity and Dynamics of
Ecosystems in North Eurasia. Novosibirsk: IC&G. 2000. Vol. 1. P. 21-23.
Goffin V, Bouchard B., Ormady C.J., Weimann E., Ferrag F., Touraine P., Bole-Feysot C
Maaskant R.A., Clement-Lacroix P, Edery M., BinartN., Kelly P.A. Prolactin: a hormone
at the crossroads of neuroimmunoendocrinology 11 Ann. NY Acad. Sci. 1998. Vol 840
N l.P. 498-509.
Goldman A.S., Chheda S., Keeney S.E., Schmalstieg F.C., Schanler R.J. Immunologic
protection of the premature newborn by human milk H Semin. Perinatol. 1994. Vol. 18
N 6. P. 495-501.
Greenwald G.S., Wang M.W. Abortifacient effects in the pregnant hamsters of an antibody to
progesterone are reversed by exogenous prolactin // Biol. Reprod. 1994. Vol. 51, N 3
P. 504-508.
Hanson L.A. The mother-offspring dyad and the immune system // Acta Paediatr 2000
Vol. 89, N 3. P. 252-258.
Krackow S. Sex-specific embryonic mortality during concurrent pregnancy and lactation m
house mice // J. Exp. Zool. 1990. Vol. 256, N 1. P. 106-112.
McLaren A. Mammalian Chimaeras. L.: Cambridge Univ. Press. 1976. (Существует рус-
ский перевод: Мак-Ларен Э. 1979. Химеры млекопитающих. М.: Мир).
Patris В., Baudoin С. Female sexual preferences differ in Mus spicilegus and Mus musculus
domesticus: The role of familiarization and sexual experience 11 Anim. Behav. 1998.
Vol. 56, N 6. P. 1465-1470.
Potapov M.A., Yakushko O.F., Belogurova M.N. Long-term effects of interstrain embryo
transfer: Female olfactory preference in adult mice offspring // Advances in the
Biosciences. Oxford: Pergamon. 1995. Vol. 93. P. 313-316.
Potts W.K., Manning C.J., Wakeland E.K. МНС-based mating preferences in Mus operate
through both settlement patterns and female controlled extra-territorial matings 11 Chemical
Signals in Vertebrates VI. New York: Plenum Press. 1992. P. 183-188.
Pratt N.C., Lisk R.D. Role of progesterone in mediating stress-related litter deficits in the
golden hamster (Mesocricetus auratus)//J. Reprod. Fertil. 1991. V. 92, N 1. P. 139-146
Rebum C.J., Wynne-Edwards K.E. Hormonal changes in males of a naturally biparental and
uniparental mammals 11 Horm. Behav. 1999. Vol. 35, N 2. P. 163-176.
Schellinck H.M., Monahan E., Brown R.E., Maxson S.C. A comparison of the contribution
of the major histocompatibility complex (MHC) and Y chromosomes to the
discriminability of individual odors of mice by Long-Evans rats 11 Behav. Genetics. 1993
Vol. 23, N3. P. 257-263.
Scribner S.J., Wynne-Edwards K.E. Thermal constraints on maternal behavior during
reproduction in dwarf hamsters (Phodopus) // Physiol. Behav. 1994. Vol. 55, N 5.
P. 897-903.
Wedekind C., Chapuisat M., Macas E. Non-random fertilization in mice correlates with the
MHC and something else И Heredity. 1996. Vol. 77. P. 400-409.
Xanthou M. Immune protection of human milk 11 Biol. Neonate. 1998. Vol. 74, N 2
P. 121-133.
Yamazaki K., Singer A., Curran M., Beauchamp G.K. Origin, functions, and chemistry of H-
2 regulated odorants //Advances in Chemical Signals in Vertebrates. N.Y: Kluwer
Academic / Plenum Publishers. 1999. P. 173-180.
НЕКОТОРЫЕ АСПЕКТЫ ЭВОЛЮЦИИ
БИОЦЕНОТИЧЕСКИХ СВЯЗЕЙ
ДВУКРЫЛЫХ НАСЕКОМЫХ
С ВЫСШИМИ БАЗИДИАЛЬНЫМИ ГРИБАМИ
Г.П. Островерхова
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
E-mail: insect@bio.ru
Общеизвестно, что возникающие и развивающиеся в ходе эволю-
ции всевозможные биологические взаимодействия между видами яв-
ляются важными факторами, определяющими естественный отбор
среди их особей, резко усиливающими формообразовательные про-
цессы и в целом видообразование, умножая состав биоценозов и всей
биосферы. В результате эволюция создает определенные структуры
на уровне экологических систем - определенных сообществ, которые
обеспечивают саму возможность существования видов, их эволюци-
онные преобразования, в том числе и экологическую дифференциров-
ку. То есть в различных группах организмов формируются определен-
ные адаптивные типы, жизненные формы, способные выполнять ту
или иную биоценотическую функцию.
Как отмечал И.И. Шмальгаузен [1], в основе всех этих преобразо-
ваний лежит эволюция наиболее интегрированных систем организмов
как индивидуальных представителей видов. Поэтому несомненно, что
изучение биоценотических связей между различными группами орга-
низмов имеет непосредственное отношение к проблемам видообразо-
вания, структурно-функциональной организации биоценотической
формы жизни, а также к проблеме закономерностей исторического
преобразования сообществ, решение которой находится пока на на-
чальном этапе. Различные биоценотические совокупности взаимосвя-
занных организмов - это экологические объекты, и их изучение важно
Для решения проблем экологии и познания биологического многооб-
разия на биоценотическом уровне организации живого [2].
279
ГП. Островерхова
При современном разнообразии подходов к решению вопросов
системы органического мира и особом внимании при этом к ультра-
тонким структурным особенностям, биохимии, функциям клетки и ее
органоидов [3] экологический подход остается не менее актуальным.
Филогения представляет собой также и экологический процесс [4],
поскольку филогенетические отношения могут быть основаны на рас-
хождении в типе питания, и потому пищевую специализацию можно
рассматривать не только с экологических позиций [5, 6]. Таким обра-
зом, для выяснения филогенеза любого таксона необходимо рассмот-
рение его взаимоотношений со средой обитания и особенностей, ко-
торые определили его прогрессивное развитие. Анализ биоценотичес-
ких связей нередко оказывается полезным в решении сложных
вопросов филогении и системы некоторых сопряженно эволюциони-
ровавших групп организмов.
Особый интерес представляет коадаптивная эволюция различных
групп организмов с насекомыми - одним из наиболее многочислен-
ных классов животного мира, представители которого практически
вездесущи. Обширные данные о мутуалистическом взаимодействии
цветковых растений с опыляющими их насекомыми и об эволюцион-
ной роли этого взаимодействия; о взаимоотношениях между муравья-
ми и растениями, представляющими их жилье взамен защиты от рас-
тительноядных насекомых; плодов - с животными, распространяю-
щими их семена и т.д., являются прямым доказательством процесса
ускорения эволюции и формирования множества новых таксонов, воз-
никновения новых адаптаций.
Однако если коадаптивные связи покрытосеменных растений с
насекомыми достаточно полно изучены, прослежены их взаимные адап-
тации и влияния в ходе эволюции [7], то с гетеротрофными организ
мами, в частности с грибами, биоценотические связи насекомых изу-
чены далеко не полно. Однако, при современной неопределенности и
дискуссионное™ вопросов происхождения и филогении грибов [8,9],
сведения о их взаимосвязях с насекомыми, возможно, будут полезны-
ми. К тому же и сама микроэкосистема “гриб-насекомое” в плане со-
временного внимания к экосистемам как интегрирующим воздействия
среды весьма своеобразна.
Первые работы о связях насекомых с макромицетами относятся к
первой половине XX века [10-14]. Во второй его половине стали скла-
280
IV. Эволюция животных
даваться представления о грибе с его обитателями как некой системе
взаимосвязанных организмов. Так, указывая на роль грибных обита-
телей в распространении спор, В.Н. Беклемишев [15] в качестве при-
мера биоценологических (симфизиологических) связей приводил вза-
имоотношения плодовых тел шляпных грибов и их потребителей, на-
зывая их прямыми форическими связями. Он писал: “Плодовые тела
шляпных грибов, заселяющиеся многочисленными личинками двукры-
лых, жуками и прочими, поедающими ткань гриба и разносящими в
своем кишечнике его споры, имеют для жизни вида такое же значе-
ние, как мясистые плоды цветковых: плодовые тела грибов также пред-
ставляют живые части организма, специально предназначенные для
поедания их животными”. В.Н. Беклемишев отмечал при этом роль
форических связей насекомых с грибами в возникновении мясистых
плодовых тел шляпных грибов.
Другими авторами гриб со всеми его обитателями рассматривался
как “биоценотический комплекс” [ 16], как биотоп [17], отмечалась зави-
симость количественного и качественного состава двукрылых, поража-
ющих грибы, от морфологических особенностей их плодовых тел [18].
В отечественной литературе вопросы специфичности, сукцессии
мицетобионтов в грибах были рассмотрены в работах Г.П. Островер-
ховой [19, 20], а гриб с его обитателями представлялся как единое це-
лое, как экосистема (микроэкосистема), названная позже [21, 22] как
мицетоконсорция. В 60-80-е годы изучение грибных обитателей ин-
тенсифицировалось как в России, так и за рубежом.
В настоящее время из числа связанных с макромицетами насеко-
мых известно около 500 видов двукрылых, свыше 1100 видов жуков;
также в грибах отмечены чешуекрылые, перепончатокрылые, ногох-
востки, клопы и другие беспозвоночные (слизни, ракообразные, пау-
кообразные - клещи, пауки, нематоды и др.). Трофическая специали-
зация обитателей разнообразна - сапрофаги, зоофаги, но основная и
многочисленная группа - это мицетофаги, представленные группой
мицетофилоидных двукрылых. Биоценотическая связь “гриб-мицето-
фаг” а следовательно, мицетофагия - основа грибного сообщества
(мицето консорции).
Вообще мицетофагия - питание грибами - широко распростране-
на среди насекомых и известна в различных их отрядах. Однако в от-
личие от фитофагии - питания на высших растениях - о мицетофагии
281
ГП. Островерхова
исключительно мало сведений, особенно о специализированной ми-
цетофагии (эумицетофагии) - питании высшими базидиальными гри-
бами. Неспециализированная же мицетофагия - питание низшими
грибами - привлекает внимание многих авторов [23-30] как переход-
ное и промежуточное звено в эволюции других типов питания в раз-
ных группах насекомых, или как первичный (питание детритом и низ-
шими организмами - лишайниками, водорослями, грибами). Это го-
ворит о важной роли неспециализированной мицетофагии в эволюции
типов питания и филогении тех или иных таксонов.
Нередко в отдельных группах насекомых неспециализированная
мицетофагия сочетается с детритофагией или дает переходные типы
(фито-, зоомицетофагию), а в конечном итоге и соответствующие спе-
циализированные типы - фитофагию и зоофагию. Или, например, путь
к специализированной мицетофагии на трутовых грибах лежал через
древнюю мицетофагию, так же, как, вероятно, и путь к керато-, лихе-
но- и мусциофагии [28].
Появление мицетофагии - это своеобразный этап в эволюции био-
са, новый тип обмена веществ, когда источником пищи для насекомых
становятся первично гетеротрофные организмы - грибы. По данным
Э.К. Гринфельда [24], многие древние насекомые питались спорами и
пыльцой, и первая связь насекомых с генеративными органами расте-
ний возникала на базе питания пыльцой. Часто пыльца служила про-
межуточным кормом при переходе от примитивных режимов питания
- сапрофагии, детритофагии и хищничества - к фитофагии. Возмож-
но, что и для настоящей мицетофагии исходной была спорофагия.
Грибы - древняя группа, существовавшая еще до расхождения ра-
стений и животных [31]. С появлением насекомых, датируемым по-
разному: в девоне [32]; в перми, триасе, карбоне [33]; в карбоне - ран-
нем, среднем [34], возможно появление и мицетофагии. По данным
Панфилова [35], мицетофагия свойственна группам насекомых (пер-
вичнобескрылые, трипсы, сеноеды, сверчки), известным с девона, пер-
ми или триаса. Наибольшее развитие она могла получить в мезозое -
юра, мел [25] или во второй половине мезозоя - середина юры, когда
предполагается [30] появление трутовиков и грибов с крупными соч-
ными карпофорами.
Возникновение и становление мицетофагии достаточно хорошо
прослеживается на примере мицетофилоидных двукрылых (Diptera,
282
IV. Эволюция животных
jvfycetophiloidea), включающих девять семейств (Sciaridae,
Bolitophilidae, Ditomyiidae, Keroplatidae, Macroceridae, Diadocidiidae,
jvfanotidae, Sciophilidae, Mycetophilidae). Их личинки обитают в разла-
гающейся древесине, древесных и напочвенных грибах, во мху, под-
стилке; куколки - в исходных субстратах, или в почве; взрослые - в
темных, прохладных, с повышенной влажностью местах. Таксономи-
ческая гетерогенность надсемейства мицетофилоидов обусловлена
разнообразием мест обитания личинок и соответственно типов тро-
фики. Известные типы питания мицетофилоидных личинок (сапрок-
силофагия, мицетофагия неспециализированная, мицетофагия специ-
ализированная, зоомицетофагия, зоофагия, миксомицетофагия, брио-
фагия) свойственны личинкам, связанным с определенными субст-
ратами.
Происхождение сапроксилофагии мицетофилоидов связано с раз-
лагающейся древесиной - исходным субстратом для всех двукрылых
насекомых - бибиономорф [27]. Причем сапроксилофагия трудно от-
личима от ксиломицетофагии, поскольку в разлагающейся древесине
всегда имеются споры и мицелий низших грибов. Поэтому вполне за-
кономерно полагать, что и в этом случае потребление низших грибов
(мицелиальных пленок, нитей), то есть неспециализированная мице-
тофагия (ксиломицетофагия), явилась переходной от древней детри-
тофагии к сапроксилофагии. Вероятно, в разлагающейся древесине
трофическая эволюция мицетофилоидов первоначально шла по пути
становления двух типов питания - сапроксилофагии и мицетофагии
неспециализированной в виде ксиломицетофагии. Сапроксилофагия
в надсемействе не получила достаточного развития и представляет
узкую специализацию небольших и в значительной мере обособлен-
ных таксонов (Symmerus, Boletina). Судьба же неспециализированной
мицетофагии (ксиломицетофагии) различна. В немногих группах ми-
цетофилоидов (Diadocidiidae), обитающих на древесно-веточном опа-
де и питающихся разлагающими его низшими грибами, она осталась
таковой. А вообще же неспециализированная мицетофагия - это фак-
тор, на основе которого возникли другие типы питания мицетофилои-
дов как в пределах разлагающейся древесины, так и вне ее. Вероятно,
миксомицетофагия неспециализированная явилась исходной для ми-
цетофагии, бриофагии, зоомицетофагии, зоофагии, настоящей мице-
тофагии - эумицетофагии.
283
ГП. Островерхова
Миксомицетофагия в надсемействе свойственна немногим пред-
ставителям достаточно молодого семейства мицетофилид (Mycetophila,
Platurocypta, Phronia) [18, 36]. Бриофагия в основном ограничена се-
мейством сциофилид - Boletina [37], Viridivora, и мицетофилид -
Phronia [38]. Зоофагия отмечается у немногих представителей - ке-
роплатид, макроцерид, некоторых сциофилид, поедающих мелких бес-
позвоночных на поверхности или под отставшей корой разлагающей-
ся древесины, гименофорах трутовиков, на стенах пещер и других суб-
стратах. Известны, например, некоторые (Xenoplatyura) из
кувшинчиков непентеса [39], в которых они потребляют добычу, пред-
назначенную для непентеса, вступая, таким образом, в конкурентные
с ним отношения. Отмечено [40] паразитирование тасманийского рода
Planarivora, близкого к кероплатидам, в теле наземных планарий. Воз-
можно, обнаруженная у сциофилид зоомицетофагия [36] является пе-
реходным типом питания от неспециализированной мицетофагии к
настоящей зоофагии. Таким образом, в разлагающейся древесине на
разных стадиях ее разложения из древней детритофагии могли воз-
никнуть: сапроксилофагия, неспециализированная мицетофагия в виде
ксиломицетофагии, давшей в пределах этого же субстрата зоомицето-
фагию, зоофагию, миксомицетофагию, бриофагию и эумицетофагию.
Этот спектр трофических групп личинок мицетофилоидов и в настоя-
щее время представлен в полном объеме в разлагающейся древесине.
Эумицетофагия могла иметь своим началом спорофагию, способству-
ющую переходу и в ткань карпофоров грибов, как ксилобионтных, так и
филогенетически и экологически близких к ним напочвенных макроми-
цетов. Освоение карпофоров - разнообразной, мягкой, доступной пищи
- обеспечило успех в эволюционном развитии именно эумицетофагам,
которые на данном этапе - процветающая группа в надсемействе, в то
время как, есть предположение [35], в древних наземных биоценозах
грибы использовались менее интенсивно, чем в настоящее время. Ста-
новление эумицетофагии - генеральная линия экологической эволю-
ции надсемейства мицетофилоидов и основа коэволюционной связи
двух групп организмов - грибов и насекомых. Поскольку высшие ба-
зидиальные грибы представляют тупиковую ветвь [31 ], то и эумицето-
фагия - это вероятный эволюционный венец вообще в становлении
мицетофагии как типа питания. На современном этапе эумицетофагия
двунаправлена - на древесных и на напочвенных макромицетах, явля-
ясь основой существования соответствующих мицетоконсорций.
284
IV. Эволюция животных
Основная часть древесных грибов - это сапротрофы, представлен-
ные преимущественно низшими базидиомицетами. Известные из них
мицетофилоиды в таксономическом отношении являют группы, сто-
ящие в основе их филогенетического ствола. Трофическая их специа-
лизация - ксиломицетофагия (Ditomyia, Asioditomyia, Mycomyia,
Apolephtisa, Synapha, Rondaniella, Leia, Tetragoneura, Docosia, некото-
рые Exechia, Trichonta, Phronia) и зоофагия (Keroplatus, Cerotelion,
Mycomyia). Обращает внимание сходство родового, нередко видово-
го, а также состава трофических групп мицетофилоидов из древесных
сапротрофных грибов с таковыми из разлагающейся древесины. На-
пример, из 11 родов мицетофилоидов связанных с пориевыми гриба-
ми, 8 (Mycomyia, Neoempheria, Sciophila, Leptomorphus, Apolephthisa,
Tetragoneura, Trichonta, Dynatosoma) - - общих с разлагающейся древе-
синой. Или, 3 рода мицетофилоидов, связанных с кортициевыми, все
известны из разлагающейся древесины. Из 18 родов, известных из
полипоровых базидиомицетов, 6 родов - общих с разлагающейся дре-
весиной, а многие - общие с обитателями напочвенных грибов. С по-
липоровыми грибами связаны мицетофилоиды из группы разной эво-
люционной продвинутое™ и разной экологии (ксиломицетофаги и
зоофаги). Трофические группы, как в древесных грибах, так и в разла-
гающейся древесине - ксиломицетофаги, зоомицетофаги и зоофаги.
Совершенно иная картина наблюдается в напочвенных грибах по-
рядка Agaricales - группе эволюционно прогрессивной. В них обита-
ют преимущественно сциофилиды и мицетофилиды, трофическая спе-
циализация которых - эумицетофагия. Наибольшее количество родов
и видов мицетофилоидов отмечено для Boletaceae - 17 родов,
Russulaceae - 19, для Amanitaceae -11, Agaricaceae - 8, Hygrophoraceae
- 9, Paxillaceae - 6, Coprinaceae - 4, Gomphidiaceae - 4. Агариковые
грибы освоены в настоящее время молодым прогрессивным семей-
ством Mycetophillidae.
Таким образом, таксономический и трофический состав комплек-
сов мицетофилоидов, связанных с кортициевыми и с пориевыми дре-
весными грибами, близки таковому разлагающейся древесины. Ком-
плексы мицетофилоидов полипоровых грибов занимают как бы про-
межуточное положение между таковыми разлагающейся древесины
и напочвенных грибов. Комплексы мицетофилоидов напочвенных ага-
риковых грибов, хотя имеют сходство с таковыми древесных поли-
285
Г.П. Островерхова
поровых грибов, но достаточно своеобразны. Анализ мицетофилоид-
ных комплексов основных субстратов, с которыми связанны их ли-
чинки, а именно: разлагающаяся древесина, древесные грибы, напоч-
венные грибы, свидетельствуют о том, что становление специализи-
рованной мицетофагии и консортивных связей с высшими грибами
способствовали обеднению их трофической структуры. Так, с дре-
весными макромицетами связаны, в отличие от разлагающейся дре-
весины [41], немногие трофические группы, а именно мицетофаги и
зоофаги, а с напочвенными грибами - только мицетофаги. Вероятно,
питание напочвенными грибами - это решение основного противо-
речия в эволюции мицетофилоидов - недостатка личинок питания.
Состав мицетофилоидных комплексов, связанных с основными
экологическими группами грибов, позволяет предположить ход ста-
новления мицетоконсорций в ходе эволюции. Консортивные связи
мицетофилоидов на первых порах, видимо, устанавливались с древес-
ными сапротрофными афиллофоровыми грибами (Corticiaceae,
Thelephoraceae, Coniophoraceae, Poriaceae), о чем свидетельствует зна-
чительное таксономическое сходство их мицетофилоидных комплек-
сов с таковыми разлагающейся древесины. Паразитические ксилофиль-
ные макромицеты заселялись мицетофилоидами, вероятно, позже.
Из напочвенных грибов мицетофилоидные комплексы микоризо-
образующих грибов, особенно дающих смешанную микоризу и мико-
ризу с лиственными породами, более разнообразны таксономически и
в определенной степени специфичны. С подстилочными и гумусовы-
ми сапротрофами связаны широко адаптированные эволюционно мо-
лодые виды, что является возможным показателем более позднего воз-
никновения и далеко не завершенного формирования детерминиро-
ванных ими мицетоконсорций.
Становление эумицетофагии способствовало выработке различных
коадаптаций базидиомицетов и мицетофагов. Известно [8], что важным
направлением в эволюции грибов является выработка мощного фермен-
тативного аппарата, адаптация к которому, вероятно, была основной стра-
тегией перешедших к питанию грибами организмов и, в общем, в ста-
новлении эумицетофагии. Анализ комплексов мицетофагов некоторых
древесных макромицетов с разным ферментативным составом показал
определенные различия в их таксономическом составе.
286
IV. Эволюция животных
Так, для целлюлозоразрушающих грибов (Daedalea quercina,
Coriolus vaporarius, Polyporus squamosus) характерны представители
семейства Sciophilidae (роды Mycomyia, Leptomorphus, Sciophila,
Apolepthisa, Tetragoneura) и Mycetophilidae (роды Exechia, Brachypeza,
Trichonta, Dynatosoma, Mycetophila). Своеобразие этому комплексу
придают роды Leptomorphus, Apolepthisa, Tetragoneura - наиболее древ-
ние среди мицетофилоидов.
Для древесных грибов, разрушающих преимущественно лигнин
(Fomes fomentarius, Ganoderma applanatum, Pleurotes ostreatus, Phellinus
igniarius, Polystictus versicolor, Polyporus adastus), отмечены предста-
вители семейств: Sciaridae (роды Sciaria, Bradysia), Ditomyiidae (роды
Ditomyia и Asioditomyia), Sciophilidae (роды Mycomyia, Neoempheria,
Sciophila, Rondaniella), Mycetophilidae (роды Dynatosoma, Mycetophila).
Своеобразие этому комплексу придают виды рода Ditomyia.
Несмотря на наличие сходных семейств и родов мицетофилоидов
в целлюлозо- и лигнин-разрушающих грибах, виды их специфичны
для каждой ферментативной группы. При этом мицетофилоидные ком-
плексы целлюлозоразрушающих грибов наиболее сходны с таковыми
разлагающейся древесины.
Сравнение мицетофилоидных комплексов, связанных с напочвенны-
ми грибами, отличающихся по биохимическому составу [42], т.е. разру-
шающих как лигнин, так и клетчатку (в основном трихоломовые) и раз-
лагающих преимущественно лигнин (в основном руссулевые), показы-
вает в грибах первой группы из 52 известных видов: Sciophilidae
(35 видов), Mycetophilidae (6), а в грибах второй группы, наоборот, из
68 видов: Mycetophilidae (51), Sciophilidae (13). Возможно, это говорит о
более ранней связи мицетофилоидов с напочвенными грибами, разру-
шающими как лигнин, так и клетчатку (с трихоломовыми), в сравнении
с грибами, разлагающими лигнин (руссулевыми).
В группах грибов, выделенных по характеру действия на организм,
в мицетофилоидных комплексах только для грибов, действующих на
пищеварительную систему, отмечено 25 видов (Bolitophilidae - 3,
Sciophilidae - 1, Mycetophilidae - 21); только для группы грибов, дей-
ствующих на нервную систему, - 6 (Sciophilidae - 3, Mycetophilidae -
3); для группы грибов гемолитического действия - 1 вид Sciophilidae.
Среди мицетофилоидов особого внимания заслуживают сциофи-
лиды (Sciophilidae триба Sciophilini), в частности род Sciophila, для
287
Г.П. Островерхова
которых характерны все типы трофики, известные для надсемейства.
Другие же грибы этого семейства обнаруживают определенную при-
уроченность к тем или иным субстратам, менее дифференцированы в
трофическом отношении и менее продвинуты к эумицетофагии. Так,
наиболее примитивные гнористины (триба Gnoristini род Boletina) из-
вестны из разлагающейся древесины [36, 37]; из древесины и древес-
ных грибов (Poria vaporaria) - Apolephthisa [43]. Возможно, что они
могли стать исходными в направлении выработки сапроксилофагии и
неспециализированной мицетофагии. Микомиины (триба Mycomyiini,
роды Mycomyia, Neoempheria, Leptomorphus, Phtinia) широко представ-
лены в разлагающейся древесине, а также в древесных грибах (Myco-
myia, Leptomorphus, Allacotocera, Syntemna, Phthinia). Трофическая
специализация микомиин - специализированная мицетофагия на дре-
весных и напочвенных грибах с более твердой кожистой консистенци-
ей, а также отчетливо выраженные зоомицето - и зоофагия. Сходны с
микомиинами и Leiini. Таким образом, на примере Sciophilidae про-
слеживается становление всех типов питания, свойственных надсемей-
ству, и роль в этом различных субстратов. Это семейство представляет
как бы своеобразную эколого-филетическую линию, ближе связанную
с исходными субстратами - разлагающейся древесиной и древесными
грибами. Другое центральное семейство - Mycetophilidae (известные
как грибные комарики), более молодое, развитие личинок которого в
основном связано с напочвенными грибами и с преимущественным
представительством эумицетофагов.
Итак, рассмотренные биоценотические связи мицетофилоидов
группы, основным направлением экологической эволюции которых
является выработка эумицетофагии, с высшими базидиальными гри-
бами в аспектах: таксономическом, экологическом и биохимическом,
позволяют констатировать следующее. Исходным является мицетофи-
лоидный комплекс разлагающейся древесины, отличающийся значи-
тельной таксономической (родовой) и трофической дифференциров-
кой. Первыми детерминантами мицетоконсорций (первыми высшими
базидиальными грибами, с которыми установились биоценотические
связи мицетофилоидных двукрылых) могли быть афиллофоровые (по-
риевые или пориеподобные) целлюлозоразрушающие сапротрофные
макромицеты на хвойных. Об этом свидетельствуют некоторые осо-
бенности биоценотических связей мицетофилоидов с афилофоровы-
288
IV. Эволюция животных
ми трутовыми (Poriaceae, Polyporaceae) грибами. Во-первых, в этих
грибах преобладают мицетофилоиды более примитивных групп
(Ditomyiidae, Keroplatidae), а также переходных (Sciophilidae). Из про-
грессивного семейства Mycetophilidae наиболее характерны Trichonta,
phronia, Dynatosoma и некоторые Mycetophila. В целом таксономичес-
кий состав мицетофилоидов древесных грибов близок к таковому раз-
лагающейся древесины. Во-вторых, особое сходство с мицетофило-
идным комплексом разлагающейся древесины обнаруживает таковой
целлюлозоразрушающих сапротрофных грибов с хвойных пород. Так,
из Poriaceae общие с разлагающейся древесиной роды - Leptomorphus,
Apolephthisa, Tetragoneura; из сапротрофных Polyporaceae с листвен-
ных - Symmerus, Synapha. Трофическая специализация мицетофилои-
дов, связанных с древесными грибами, - мицетофагия (мицело-, спо-
рофагия и зоофагия).
Из напочвенных макромицетов вероятна более ранняя связь ми-
цетофилоидов с микоризообразующими грибами, из которых извест-
но 90 видов мицетофилоидов. Причем для микоризообразующих афил-
лофоровых грибов (Cantharellaceae, Hydnaceae) отмечены сциофили-
ды (Sciophila, Leia, Neoclastobasis, Docosia); для микоризообразующих
агариковых (Boletaceae, Tricholomataceae, Amanitaceae, Russulaceae)
наряду co сциофилидами отмечены мицетофилиды (Exechia, Rhymosia,
Tamania, Allodiopsis, Cordyla, Mycetophila), которые значительно пре-
обладают.
Из подстилочных сапротрофных афиллофоровых макромицетов
(некоторые Clavariaceae) и агариковых (некоторые Tricholomataceae)
отмечены Bolitophilidae (Bolitophila, Messala), Sciophilidae (Sciophila,
Rondaniella, Leia, Docosia) и Mycetophilidae (Exechia, Rymosia, Tamania,
Allodia, Cordyla, Trichonta, Mycetophila). Из гумусовых сапротрофных
агариковых грибов (Agaricaceae, Strophariaceae, Paxil laceae) отмечены
болитофилиды (Bolitophila), сциофилиды (Sciophila, Leia, Docosa),
мицетофилиды (Exechia, Rymosia, Alloidia, Mycetophila).
В общем, мицетофилоидные комплексы грибов (подстилочных и
гумусовых сапротрофов) во многом сходны количественно (по 36 ви-
дов из тех и других) и качественно (виды одних и тех же семейств -
Bolitophilidae, Sciophilidae, Mycetophilidae). Но значительная родовая
Дифференцировка (14 родов) мицетофилоидов, связанных с подсти-
лочными сапротрофами, в сравнении с таковой (8 родов) мицетофи-
289
ГП. Островерхова
лоидов из гумусовых сапротрофов, а также предполагаемая более ран-
няя связь мицетофилоидов с напочвенными грибами, способными раз-
лагать как лигнин, так и клетчатку, позволяют пока говорить о вероят-
ной более ранней связи мицетофилоидов с подстилочными сапротро-
фами. В целом в экологической линии напочвенных грибов состав
мицетофилоидов позволяет пока считать мицетоконсорций, основан-
ные микоризообразующими грибами, более ранними в сравнении с
мицетоконсорциями, детерминированными подстилочными и тем бо-
лее гумусовыми сапротрофами. Грибы порядков Tremellales и низшие
формы Thelephorales, положение которых в известных нам классифика-
циях [44] предполагается как первичное, - это афиллофоровые грибы и
преимущественно сапротрофы. Таксономический состав связанных с
ними мицетофагов соответствует их таксономическому статусу как пер-
вичных. Что касается первичности паразитизма или сапрофитизма у гри-
бов [9], то конкретные данные о комплексах мицетофилоидов сапрог
рофных и паразитных древесных макромицетов свидетельствуют боль-
ше о первичности сапрофитизма и вторичности паразитизма.
Общая же эволюционная направленность трофической специали-
зации макромицетов на основании данных о связанных с ними мице-
тофилоидных комплексах представляется таким образом: наиболее
ранним является сапрофитизм древесный - исходный для паразитиз-
ма и микоризообразования и более позднего сапрофитизма напочвен-
ных грибов (подстилочных и гумусовых).
Литература
1. Шмальгаузен ИИ. Кибернетические вопросы биологии. Новосибирск, 1968. 224 с
2. Емельянов И.Г. Разнообразие и устойчивость биосистем // Успехи соврем, биологии
1994. Т 114. № 3. С. 304-318.
3. Кусакин О.Г, Дроздов А.Л. Филемаорганического мира. 1. СПб.: Наука, 1994. 282 с
4. Wahlert G. von. Phylogenie als okologischer Prozess // Naturwiss. Rundschau. 1973
Bd. 26. № 6. P. 247-254.
5. Гиляров M.C. Экологические и энтомологические признаки в систематике и фило! е-
нетике насекомых И Журн. общ. биол. 1974. Т. 35. №1. С. 13-33.
6. Hengeveld R. Food specialisation in ground beetles: an ecological or a phylogenetic process9
//Netherl. J. Zool. 1980. Vol. 30. № 4. P. 585-594.
7. Тахтаджян А.Л. Вопросы эволюционной морфологии растений. Л., 1954. 214 с.
8. Гарибова Л.В. Обзор современной системы грибов, происхождение и место грибов в
органическом мире // Микология и фитопатология. 1975. 9. 2. С. 164-170.
9. Мирабдуллаев И.М. О происхождении и положении грибов в системе органического
мира//Успехи совр. биол. 1994. Т. 114. Выи. 1. С. 30-41.
290
IV. Эволюция животных
Ю Weiss Н.В., West Е. Notes on fungus - insect // Canad. Entom. 1922. 54. P. 198-199.
И Dieuzeide R. Les Repports ontre les Champignons et les Insects // Rev. Zool. Agric. E.
‘ Appl- 1923. 22. 10. P. 282-289.
j2 Okada J. Beitrag zur Kenntnis der Fungivoriden. Fauna Japans. II; Diadocidiina. 1936.
XI. 1-2 P 21-24.
13 Частухин В.Я. Значение плодовых тел у шляпочных грибов // Боган, журн. 1932. 2.
’ С. 102-114.
14, фридолин В.Ю. Животно-растительное сообщество горной страны Хибин // Тр.
Кольск. базы им. С.М. Кирова. 1936. 3. 215 с.
15. Беклемишев В.Н. О классификации биоценологических (симфизиологических) свя-
зей // Бюл. МОИП. Отд. биол. 1951. 56. Вып. 5. С. 3-30.
16. Benick L. Piizkafer und Kaferpilze //Acta. Zool. Fennica. 1952. 70. 250 p.
17. Eisfelder J. Beitrage zur Kenntnis der Fauna in hoheren Pilzen // Zeitschr. fur Pilzkunde.
1954. 22. P. 108-117.
18. Buxton P.A. British Diptera, associated with fungi. 3. Flies of al families reared from
abount 150 species of fungi // Entomol. Mon. Mag. 1954. 96. P. 73-95.
19. Островерхова Г.П. Съедобные грибы Томской области и вредящие им насекомые
И Заметки по фауне и флоре. Томск, 1966. Т. 19. С. 53-57.
20. Островерхова Г.П. О фауне насекомых белого гриба в Западной Сибири // Фауна и
экология членистоногих. Новосибирск: Наука, 1972. С. 76-78.
21. Островерхова Г.П. Комплексы мицетофилоидных двукрылых в лесных экосисте-
мах // Тр. IX сьезда ВЭО. 1984. С. 84.
22. Островерхова Г.П. Консортивные связи насекомых с грибами // Вопр. экологии бес-
позвоночных. Томск, 1988. С. 52-61.
23. Кузнецов Н.Я. Чешуекрылые янтаря. М.; Л., 1941. 136 с.
24. Гринфельд Э К. Происхождение и развитие антофилии у насекомых. Л., 1978. 208 с.
25. Родевдорф Б.Б. Историческое развитие двукрылых насекомых. М.: Наука, 1964. 315 с.
26. Мамаев Б.М. Эволюция галлообразующих насекомых - галлиц. Л.: Наука, 1968.
. 237 с.
27. Кривошеина Н.П. Морфо-экологические аспекты ранней эволюции двукрылых на-
секомых // Насекомые - разрушители древесины и их энтомофаги. М.: Наука, 1979.
С. 5-23.
28. Загуляев А.К. Пищевые связи и эволюция типов питания настоящих молей
(Lepidoptera, Tineidae) // Зоол. журн. 1972. Т. 51. Вып. 10. С. 1507-1516.
29. Нарчук Э.П. К эволюции фитофагии у двукрылых насекомых // Систематика и эво-
люция двукрылых насекомых. Л., 1977. С. 70-76.
30. Ковалев В.Г. Этапы и факторы исторического развития фауны Diptera// Фауногенез
и филоценогенез. М.: Наука, 1984. С. 138-153.
31. Горленко М.В. Курс низших растений. М.; Высшая школа, 1981. 520 с.
32. Мартынов А.В. Очерки геологической истории и филогении отрядов насекомых
(Pterygota) // Тр. ПИН АН СССР. 1938. 7. 4. С. 7-148.
33. Родендорф Б.Б. Эволюция крыла и филогенез длинноусых двукрылых И Тр. ПИН
АН СССР. 1946. Т. 13. Вып. 2. 107 с.
34. Расницын А.П. Происхождение и эволюция перепончатокрылых // Тр. ПИН АН
СССР. 1980. Т. 168.
35. Панфилов Д.В. О роли насекомых в древних и современных континентальных био-
ценозах// Зоол. журн. 1967. Т. 46. Вып. 5. С. 645-656.
291
ГП. Островерхова
36. Зайцев А.И. Направления морфологической специализации пищеварительного зрак-
та личинок высших мицетофилоидных Iderhsks (Diptera, Mycetophiloidea) // Биол
науки. 1984. № 1. С. 38-44.
37. Landrock К. Fungivoridae// Lindner Die Flicgen der palearktischen Region. 1927. 195
38. Lastovka P. Predbezna ekologicka kiasifikace celedi Mycetophilidae (Dipt.) // Shornik
Jihocskoho museaCeskych Budejovicich prirodni vedi. 1972. 12.2. P. 91-95
39. Matile L. Xenoplatyura beaveri n. sp. (Dipt., Mycetophiloidea) Keroplatidae nouveaux
de Malaisie in feode aux urines de Nepentes // Ann. Soc. ent. 1979. Fr. (N.S.). 15(1)
P. 31-35.
40. Hickman V.V. On Planarivora insignis gen. et sp. n. (Dipt., Mycetophiloidea) whose
lawal stages are parasitic in land planarians // Papers and Proc. Roy. Soc. Tasmania
1965. 99. P. 1-8.
41. Островерхова ГП. Мицетофилоидные двукрылые (Dipt., Mycetophiloidea) - типич-
ные представители лесных экосистем И Насекомые в экосистемах лесной зоны Си-
бири. Томск, 1984. С. 15-35.
42. Шиврина А Н. Биологически активные вещества высших грибов. М.; Л.: Наука,
1965.199 с.
43. Edwards F. W. British fungus-gnats (Diptera, Mycetophilidae)//Trans. Ent. Soc. London,
1925. P. 505-670.
44. Зеров Д.К. Очерк филогении бессосудистых растений. Киев. Наукова думка, 1972
315 с.
ВИДООБРАЗОВАНИЕ И ЭВОЛЮЦИЯ
ОРГАНИЗМОВ
(НА ПРИМЕРЕ ФОРАМИНИФЕР)
В.М. Подобина, Г.М.Татьянин
Томский государственный университет
634050, г Томск, пр. Ленина, 36
E-mail: decanat@dggf.tsu.tomsk.su
Палеонтологические данные являются одной из основ для восста-
новления закономерностей эволюции органического мира. Из них ав-
торами использованы мельчайшие организмы - фораминиферы. Это
наиболее многочисленный и разнообразный класс морских простей-
ших животных, претерпевший большой и сложный путь эволюцион-
ного развития, начиная с кембрийского периода до настоящего време-
ни. Каждому отрезку геологического прошлого отвечают характерные
ассоциации видов фораминифер, отличающиеся систематическим со-
ставом в разных экологических нишах и биохориях.
Вполне естественно рассматривать историческое развитие различ-
ных групп фораминифер как результат изменения видов, превращения
их в новые виды, отличные по строению от своих предшественников.
Фораминиферы, и особенно комплексы их видов, являются цен-
ными показателями возраста горных пород, условий осадконакопле-
ния. Они также используются для изучения фаций геологического про-
шлого, построения палеогеографических карт, а также установления
биогеографических связей между отдельными бассейнами.
В результате всестороннего исследования этих организмов сложи-
лось определенное представление об объеме основного их таксоно-
мического подразделения - вида - и о видообразовании. Вид пред-
ставляет соподчиненный роду таксон, обладающий рядом морфоло-
гических признаков, имеет определенную экологическую нишу, ареал
обитания и стратиграфический диапазон. Для его выделения, как и
Других таксонов, необходимо учитывать 5 основных критериев:
1. Морфофункциональный.
2. Онтофилогенетический.
293
В.М. Подобина, Г.М.Татьянин
3. Геохронологический.
4. Палеогеографический.
5. Палеобиогеографический.
Впервые эти критерии были разработаны А.В. Фурсенко (1978);
В.М. Подобиной (1998) добавлен последний - палеобиогеографичес-
кий критерий.
Первый из указанных пяти критериев - морфофункциональный
- является основным при исследовании всех групп фораминифер. По
степени морфофункциональной организации В.М. Подобиной (1998)
установлены 4 группы, которым соответствуют определенные высшие
таксоны (надсемейства и отряды) (табл. 1).
Таблица I
ГРУППЫ ТАКСОНЫ
1. Примитивная Astrorhizida, Ammodiscida
11. Усложненная Haplophragmiidea Textuiariida Ataxophragmiida
111. Сложная Miliolida Lagenida Rotaliida
IV. Сверхсложная Globigcrinida, Buliminida
Усложнение морфологического строения раковин высших таксонов
фораминифер, показанное по 4 группам, согласуется с общим повыше-
нием их организации и разнообразия на протяжении фанерозоя (Основы
палеонтологии: Общая часть. Простейшие, 1959; Фурсенко, 1978).
Основными морфологическими признаками первого критерия яв-
ляются:
1) строение раковин;
2) состав и строение стенки;
3) характер пористости стенки.
Все указанные признаки взаимосвязаны, однако для каждой груп-
пы один из них является ведущим.
Онтофилогенетический критерий, наряду с морфофункциональ-
ным, используется при исследовании как видов, так и более крупных
таксономических категорий (родов, семейств, надсемейств, отрядов).
Использование модусов филогенеза (архалаксис, девиация, анаболия)
294
IV. Эволюция животных
уточнило ранг и объем многих таксонов. Сравнительный анализ над-
семейства Haplophragmiidea с однорядным отделом позволил рассмот-
реть степень родства его двух соподчиненных семейств. Сохранение в
онтогенезе видов начального завитка у некоторых представителей се-
мейства Haplophragmiidae позволяет сделать предположение о проис-
хождении этого семейства от полностью спиральных гаплофрагмои-
дид. В данном случае формы с однорядным отделом отличаются от
полностью спиральных. Они возникли в результате появления новых
признаков на средних стадиях онтогенеза по способу девиаций, вслед-
ствие чего возник другой тип строения раковин (Северцев, 1949; Мас-
лакова, 1978; Подобина, 1978).
Кроме того, В.М. Подобиной (1978) выявлены ряды развития род-
ственных родов гаплофрагмоидид и прослежена определенная направ-
ленность в изменении их признаков. Так, в семействе гаплофрагмои-
дид, как и среди других групп фораминифер, наблюдается тенденция
создать более прочную инволютную раковину (Trochamminoides -
Labrospira - Haplophragmoides). В дальнейшем происходит утолще-
ние стенки раковины и более плотное свертывание некоторых гаплоф-
рагмоидид, что привело к происхождению новых родов
(Cribrostomoides и др.).
Третий - геохронологический критерий также является одним из
определяющих в систематике фораминифер, в том числе и в видооб-
разовании.
Установив некоторую зависимость между усложнением строения
раковин и степенью распространения их в разрезе для указанных че-
тырех групп фораминифер, мы на конкретном фактическом материа-
ле отмечаем значительные возрастные диапазоны для примитивно ус-
троенных фораминифер и их уменьшение соответственно для услож-
ненных и более сложных форм. Поэтому при описании наиболее
примитивно устроенных форм отмечается их относительно широкое
стратиграфическое распространение и ограничивается приуроченность
многих таксонов из усложненных (П группа), сложных (III группа) и
самых сложных (IV группа - Globigerinida и др.) форм к ярусным и
зональным подразделениям. Самые сложные по своей морфологии
ископаемые - планктонные фораминиферы - обладали высокой орга-
низацией в приспособлении к внешним условиям, что отразилось на
строении раковин и прежде всего на характере пористости. Планктон-
295
В.М. Подобина, ГМ. Татьянин
ные фораминиферы эволюционировали довольно быстро, и измене-
ние их родового и видового состава во времени послужило надежной
основой для эволюционных и биостратиграфических исследований.
Четвертый - палеогеографический критерий также имеет боль-
шое значение при определении объема вида, разработке систематики
фораминифер. По мнению авторов, сюда должны быть включены и
экологические особенности обитания видов фораминифер. Фациаль-
ное сходство вмещающих пород является зачастую определяющим при
сопоставлении различных комплексов видов фораминифер даже уда-
ленных регионов.
Несколько общих с западносибирскими палеоценовых видов про-
слежено в пределах Камчатки (Серова, 1966, 1987). Условия обитания
фораминифер здесь (в глубоководном бассейне) приближались, в не-
которой мере, к западносибирским, что обусловило наличие сходных
видов в палеоцене сравниваемых регионов.
Пятый - палеобиогеографический критерий имеет большое значе-
ние для выявления сходных комплексов видов фораминифер из разных
бассейнов в пределах общих биохорий. Определение простирания од-
ной биохории дает возможность проводить в ней более детальные сопо-
ставления с выявлением одновозрастных общих и викарирующих ви-
дов. Подобное прослеживание сходных комплексов фораминифер по-
могает более надежно отождествить составляющие их виды и роды,
уточнив систематическое положение данных таксонов и ареалы их рас-
пространения в пределах установленных биохорий.
Исследуя фораминиферы фанерозоя, а также учитывая развитие
других организмов по опубликованным работам, авторы предполага-
ют, что на эволюцию биоты оказали влияние следующие основные
факторы:
1. Космический (обращение Земли вместе с Солнечной системой
вокруг центра Галактики, изменение величины солнечной радиации,
падение астероидов, метеоритов, изменение эксцентриситета земной
орбиты, оси вращения Земли и др.).
2. Тектонический (орогенез, рифтогенез, образование глубоковод-
ных желобов, опусканий, поднятий и др.).
3. Геохронологический (геологическое время).
4. Палеогеографический (экосистемные перестройки: абиотичес-
кие и биотические изменения, взаимосвязь организмов).
296
IV Эволюция животных
5. Температурный (климатическая и вертикальная зональность:
уменьшение температуры к полюсам и с глубиной, повышение в от-
дельных местах температуры, связанной с эндогенными процессами).
В эволюции организмов возник 6-й - миграционный фактор, дей-
ствие которого стало особенно ощущаться при появлении активно или
пассивно перемещающихся нектонных, планктонных и некоторых бен-
тосных организмов в результате возникновения более разнообразных
климатических и других обстановок, что привело к ускоренной эво-
люции отдельных групп этих организмов (Подобина, Татьянин, 1997).
Скорость эволюции представителей биоты поэтому не оставалась
постоянной. На основании исследования некоторых отрядов форами-
нифер по скорости эволюции выделены три основные группы, кото-
рые могут быть прослежены и среди других органических форм
(табл. 2):
1 - ускоренной эволюции (планктон, нектон и частично подвиж-
ный бентос); 2 - умеренной эволюции (подвижный бентос); 3 - замед-
ленной эволюции (медленно передвигающийся и сидячий бентос).
В пределах каждой группы, в свою очередь, по скорости эволюции
могут быть выделены соподчиненные подгруппы, отличающиеся не-
которыми особенностями.
Среди исследованных фораминифер к первой группе могут быть
отнесены представители планктонных роталипор, глобигерин, глобот-
рункан, бентосных орбитоидид, нуммулитид и др. Для второй группы
характерны бентосные аномалиниды, булиминиды и др. Третья груп-
па включает преимущественно прикрепленные формы, среди которых
наиболее характерны астроризиды, ризамминиды, хиперамминиды и
другие морфологически сравнительно примитивно устроенные фора-
миниферы (рис. 1).
Нами выяснены причины, почему первая группа ускоренно эволю-
ционировала. Она охватывает планктон, нектон и частично подвиж-
ный бентос. Эти формы при своей миграции попадали в активные
зоны Земли - рифтовые зоны, разломы, где имелся повышенный ра-
диационный и тепловой фоны и другие катализаторы, что приводило
к различным мутациям. Далее, по Ч. Дарвину, шло приспособление
организмов к условиям внешней среды.
Одно из катастрофических вымираний организмов на границе мела
и палеогена коснулось, как известно, наиболее специализированных
297
298
Группы и подгруппы организмов с различными скоростями эволюции
Таблица 2
299
I. Группа
ускоренной
эволюции
II. Группа
умеренной
эволюции
Ш. Группа
замедленной
эволюции
Сем. Globigerinidae,
Сем. Globotruncanidae,
Сем. Heteropelicidae,
и др.
Сем. Discorbidae,
Сем. Anomalininidae,
Сем. Buliminidae,
и др.
Сем. Astrorhizidae,
Сем. Rhizamminidae,
Сем. Hyperamminidae
и др.
Рис. 1. Группы фораминифер по скорости эволюции
В.М. Подобии а, Г.М.Татъянин
форм, находящихся в большей степени на третьей стадии развития (1 -
появление, 2 - расцвет, 3 - угасание). Это преимущественно глобот-
рунканы, аммониты, белемниты, динозавры и др., которые к этому
времени были очень специализированы и исчерпали свой жизненный
потенциал. По скорости эволюции, как указывалось, они относятся к
первой группе. Большинство организмов второй и третьей групп, бо-
лее медленно эволюционирующих, в это время не столь резко специа-
лизированных и оказавшихся на стадиях появления или расцвета, про-
шли этот рубеж без заметных изменений. Для примера можно привес-
ти изученные агглютинированные с кварцево-кремнистой стенкой
гаплофрагмиидеи, которые продолжали без заметных изменений свое
развитие, сохранив в палеогене основные родовые таксоны, хотя ви-
довой состав почти полностью обновился.
В целом умеренно развивающиеся гаплофрагмиидеи представля-
ют сравнительно продвинутые в эволюции организмы, становление
которых началось с позднего палеозоя и ранней юры, расцвет - в по-
здней юре, мелу и палеогене, а угасание - с неогена до настоящего
времени (Подобина, 1978, 1989, 1998) (рис. 2).
Появление новых качественных видовых признаков, повышающих
организацию гаплофрагмоидид (ароморфоз), происходит сравнитель-
но кратковременно и обычно приурочено к началу этапов, сопряжен-
ных с рубежами геохронологических подразделений. Затем следует
более длительное время морфофункционального приспособления гап-
лофрагмоидид (идиоадаптация), растягивающееся на весь этап их раз-
вития, подготавливающий снова качественный скачок к появлению
новых таксонов.
В заключение хочется подчеркнуть, что именно изучение форами-
нифер дало существенный вклад в решение проблемы видообразова-
ния и в целом эволюции организмов. Оно явилось теоретической ос-
новой для разработки дробных стратиграфических схем и построения
палеогеографических карт, имеющих большое значение для прогноза
и поисков месторождений полезных ископаемых.
300
IV Эволюция животных
Рис. 2. Предполагаемая филогенетическая схема семейства Haplophragmoididae
(Mayns, 1952)
301
В.М. Подобина, ГМ. Татьянин
Литература
МаслаковаН.И. Глоботрунканиды юга европейской части СССР. М.: Наука, 1978. 166 с .
28 табл.
Основы палеонтологии: Общая часть. Простейшие. М.. Изд-во АН СССР, 1959. 367 с ,
13 табл.
Подобина В.М. Систематика и филогения гаплофрагмиидей // Томск: Изд-во ТГУ, 1978
91 с., 17 табл.
Подобина В.М. Фораминиферы и зональная стратиграфия верхнего мела Западной
Сибири. Томск: Изд-во ТГУ, 1989. 175 с., 35 табл., 25 рис.
Подобина В.М. Фораминиферы и биостратиграфия палеогена Западной Сибири. Томск
Изд-во НТЛ, 1998. 24 с.
Подобина В.М., Татьянин ГМ. Факторы эволюции организмов (на примере фанеро-
зойских фораминифер Западной Сибири) // Эволюция жизни на Земле: Материалы
1-го Международного симпозиума «Эволюция жизни на Земле», 24-28 ноября
1997 г., Томск. Томск: Изд-во НТЛ, 1997. С. 21-23.
Северцев А.Н. Морфологические закономерности эволюции (Собрание сочинений. Том
V.) М ; Л.: Изд-во АН СССР, 1949. С. 13-509.
Серова М.Я. Фораминиферы палеоценовых отложений Восточной Камчатки. М. На-
ука, 1966. 96 с., 12 табл.
Серова М.Я. Фораминиферы и биостратиграфия Северной Пацифики на рубеже мела
и палеогена. М.: Наука, 1987. 145 с.
Фурсенко А.В. Введение в изучение фораминифер. Новосибирск: Наука, 1978 243 с
(Труды ИГиГ СО АН СССР. Вып. 391).
ПРОБЛЕМА ВИДА И ВИДООБРАЗОВАНИЯ
В ОТРЯДЕ СТРЕКОЗ
А.Ю. Харитонов
Институт систематики и экологии животных СО РАН
630090, г. Новосибирск, пр. Фрунзе, 11
E-mail: ei@eco.nsc.ru
Стрекозы обладают высокой морфологической специализацией,
благодаря которой их обычно противопоставляют всем остальным
крылатым насекомым и выделяют в особый отдел или инфракласс
класса Insecta. Учитывая палеозойский возраст ископаемых стрекозо-
образных, современный отряд стрекоз считается одним из наиболее
древних по происхождению. Вместе с тем многим их таксонам свой-
ственны такие черты биологического прогресса, как широкое распро-
странение, видовое разнообразие (около 5500 рецентных видов), оби-
лие в водных и околоводных биоценозах умеренных и южных широт.
Своеобразие стрекоз, относительно крупные размеры и доступность
для наблюдения и сбора в природе привлекают к этому отряду насеко-
мых внимание многих специалистов. Об этом свидетельствует, в част-
ности, создание в 1971 г. Международного общества одонатологов,
которое объединяет в настоящее время более 800 исследователей из
70 стран мира, издает журналы и информационно-справочную лите-
ратуру, проводит регулярные симпозиумы, имеет ряд национальных
отделений и Международный институт одонатологических исследо-
ваний (Флорида, США). Только за последние 30 лет было опубликова-
но около 13200 работ, отреферированных в библиографическом раз-
деле журнала «Odonatologica» и посвященных прежде всего фаунис-
тике и систематике стрекоз.
При изучении видового богатства, построении системы и реконст-
рукциях эволюции отряда наряду с традиционной морфологией при-
меняются разнообразные методы исследований. Проводились работы
по изучению структуры и размеров геномов стрекоз разных таксоно-
мических групп, измерению доли гомологичных последовательнос-
303
А.Ю. Харитонов
тей нуклеотидов в их повторяющихся и уникальных фракциях, обна-
руживаемой при разных критериях проведения реакции гибридизации
и температурной устойчивости гибридных молекул (Алёшин, 1985),
Широкое применение находят цитологические методы исследований,
и стрекозы MOiyr считаться одним из наиболее цитологически изучен-
ных отрядов насекомых. Начиная с 1885 г., когда впервые были получе-
ны сведения о хромосомах европейских видов из родов Calopteryx и
Libellula (Camoy, 1885), многие исследователи из разных стран работали
над изучением особенностей кариотипа стрекоз (Oguma, 1930; Oguma.
Asana, 1932; Cumming, 1964;Kiauta, 1967, 1969,1975 и др.). К настояще-
му времени изучены хромосомные наборы 530 видов стрекоз, что со-
ставляет около 10% от числа описанных видов.
Благодаря благоприятным тафономическим особенностям отряда,
в частности, относительно хорошей сохраняемости в ископаемом со-
стоянии больших сетчатых крыльев стрекоз, эти насекомые предос-
тавляют для изучения богатый палеонтологический материал. В на-
стоящее время известно около 300 ископаемых видов стрекоз, относя-
щихся к 50 родам и 30 семействам. Хорошо изучено географическое
распространение отряда, для многих таксонов проанализирована из-
менчивость признаков в разных частях ареалов (Харитонов, 1990).
Все эти обстоятельства ставят стрекоз в ряд удобных модельных
объектов для изучения проблемы вида и видообразования. Однако даже
в рамках одного этого сравнительно компактного и систематически
хорошо очерченного отряда выявляется множество противоречивых
особенностей в трактовке видов и видообразовательных процессов.
В отряде стрекоз много таксономически и биологически «хороших
видов», не доставляющих особых хлопот систематикам при их иден-
тификации и определении места в системе. Многие виды характери-
зуются столь существенными морфологическими и иными отличия-
ми, что выделяются в особые монотипические роды. Даже в наиболее
молодом и эволюционно продвинутом семействе Libellulidae из 133
его родов 33% родов являются монотипическими (Белышев, Харито-
нов, 1985).
Обилие монотипических таксонов у стрекоз считается обычно след-
ствием древности отряда, благодаря которой до наших дней сохрани-
лись лишь единичные представители некогда богатых видами родов и
семейств, в результате чего рецентная одонатофауна носит реликто-
304
IV. Эволюция животных
вый, «осколочный» характер (Белышев и др., 1989). Однако если про-
анализировать географическое распространение представителей мо-
нотипических таксонов, то становится очевидным, что большинство
из них имеют островные ареалы, располагаясь в том числе и на геоло-
гически очень молодых островах, например Японских, что свидетель-
ствует о быстром формообразовании у этих таксонов в условиях ост-
ровной изоляции вопреки представлениям о низких темпах эволюции
у этих древних и консервативных насекомых.
Наряду с «хорошими видами» среди стрекоз немало видов, статус
которых остается неясным. Особенно много таких таксонов в семей-
стве Calopterigidae. Например, только среди переднеазиатских пред-
ставителей этого семейства насчитывается примерно полтора десятка
таксонов, систематическое положение которых проблематично и рас-
ценивается разными авторами в качестве или видового, или подвидо-
вого, или же они считаются внутривидовыми формами, не имеющими
таксономического статуса. Не внесли ясность в ситуацию с этими так-
сонами и попытки изучения их с применением цитогенетических и
молекулярно-биологических методов.
Положение с видами этого семейства усугубляется тем, что мно-
гие из них не обнаруживают существенных морфологических разли-
чий, имея в том числе идентичные по строению копулятивные аппара-
ты, но для них выявлены прекопуляционные механизмы репродуктив-
ной изоляции, заключающиеся в разных поведенческих репертуарах
(Харитонов, 1976; Неутег, 1973).
С подобными проблемами приходится сталкиваться и в других се-
мействах стрекоз. В частности, интересная ситуация выявлена в группе
таксонов, связанных с видом Sympetrum vulgatum L. из семейства
Libellulidae (Попова, 1999). В северной части своего ареала S. vulgatum
относительно однороден и представлен номинативным подвидом.
В южной же части ареала он распадается на ряд подвидов и обнаружива-
ет близкородственные связи с несколькими другими представителями
рода. Сравнительно-морфологический и зоогеографический анализ этих
таксонов показал, что все они, по-видимому, представляют монофилети-
ческую группу «vulgatum». Но степень родства и обособленности таксо-
нов внутри группы различна. Если видовая самостоятельность
& striolatum и S. haritonovi не вызывает сомнений, то 5. decoloratum,
S. imitans и 5. sinaiticum не утратили окончательно связь друг с другом и
305
А.Ю. Харитонов
с S. vulgatum, иногда образуя переходные формы в зоне контакта ареа-
лов. Их можно было бы принять за подвиды политипического вида 5.
vulgatum, но картина осложняется тем, что внутри них идет дифферен-
цировка, приведшая к образованию, так сказать, «подвидов второго по-
рядка», вполне таксономически значимых. Более того, по аналогии с уже
известными в эволюционной литературе «кольцевыми ареалами» попу-
ляции 5. vulgatum (север Средиземноморья) и 5. sinaiticum (юг Среди-
земноморья), достигнув с разных сторон Пиренейского полуострова,
симпатрично обитают здесь как подвиды разных видов -5. v. ibericum и
5. s. tarraconense. Возможно, в такой ситуации вид 5. vulgatum правиль-
нее было бы рассматривать как надвид, включающий в себя комплекс
формирующихся и идущих по пути обособления новых видов, тем бо-
лее, что по мнению некоторых авторов (Майр, 1968), надвиды - явление
совсем не исключительное и составляет постоянный, причем иногда до-
вольно высокий процент всякой фауны.
Вид S.striolatum морфологически и зоогеографически близок к
S.vulgatum, но его ареал сужен в широтном направлении, и в азиатс-
кой части он встречается только южнее 50 параллели. Вид политипи-
ческий, состоит из 4 подвидов, хотя некоторые авторы дополнительно
относят к нему на правах подвидов островные виды S.nigrifemur и
S.nigrescens, а также сближают с ним гималайский вид S.commixtum.
Вероятно, ситуация с этими таксонами близка к той, которая была опи-
сана для S.vulgatum, и S.striolatum также представляет собой сложный
таксон надвидового ранга, видообразовательные процессы в котором
находятся на разной стадии завершения. Интересно при этом, что гео-
графическая изменчивость S.striolatum и S.vulgatum обнаруживает
параллелизм. Так, в Средней Азии обесцвеченным формам S.vulgatum
flavum соответствует S.striolatum pallidum с аналогичной бледной ок-
раской и, наоборот, на крайнем востоке ареала совместно обитают
S.v.imitans (- S.imitans) и S.s.imitoides с ярко окрашенными крыльями.
Помимо аллопатрического видообразования у стрекоз выявлена
возможность дифференциации симпатрично обитающих популяций.
В частности, такие явления описаны нами у вида Aeshna viridis Ever, в
Западной Сибири. Ряд видов обнаруживает сезонный диморфизм, при
котором разные генерации имаго могут сильно различаться по своим
морфологическим признакам и разведены по занимаемым экологичес-
ким нишам (Борисов, 1987).
306
IV. Эволюция животных
Таким образом, отряд стрекоз несмотря на древность своего про-
исхождения и ожидаемую в связи с этим относительную стабилиза-
цию формообразовательных процессов демонстрирует многогранность
проблемы вида и путей видообразования. В пределах одного отряда
насекомых мы сталкиваемся здесь с разными темпами эволюции так-
сонов, возможностью замедления и резкого ускорения формообразо-
вания; аллопатрическим и симпатрическим видообразованием; разны-
ми стратегиями репродуктивной изоляции, способной идти по пути
как морфологической, так и этологической и экологической диффе-
ренциации; существованием наряду с оформившимися видами и под-
видами переходных стадий сложения таксонов, подтверждающих кон-
структивность применения понятия «надвид» в систематике насеко-
мых и в представлениях об их эволюции.
Литература
Алешин В.В. Филогенетические аспекты структуры ДНК некоторых беспозвоночных.
Автореф. дис. ... канд. биол. наук. М., 1985. 22 с.
Белышев Б.Ф., Харитонов А.Ю. Монотипические таксоны стрекоз (Insecta, Odonata)
Бореального фаунистического царства// Фауна Сибири. Новосибирск: Наука. Сиб.
отд-ние, 1985. С. 44-47.
Белышев Б.Ф., Харитонов А.Ю., Борисов С.Н. и др. Фауна и экология стрекоз. Новоси-
бирск: Наука. Сиб. отд-ние, 1989. 205 с.
Борисов С.Н. Фауна и экология стрекоз Таджикистана: Дис. ... канд. биол. наук. Ду-
шанбе, 1987. 275 с.
Майр Э. Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
Попова О Н. Стрекозы рода Sympetrum.. Автореф. дис. ... канд. биол. наук. Новоси-
бирск, 1999. 22 с.
Харитонов А.Ю. Основные направления эволюции репродуктивного поведения у рав-
нокрылых стрекоз и его таксономическое значение // Групповое поведение живот-
ных (Докл. участн. 2-й Всесоюзн. конф, по поведению животных). М.: Наука, 1976.
С. 409-412.
Харитонов А.Ю. Бореальная одонатофауна и экологические факторы географического
распространения стрекоз: Дис.... д-ра биол. наук. Новосибирск, 1990. 531 с.
Camoy B.J. La cytodierese chez les arthropodes. IV. Pseudo-Nevropteres // Cellule. 1885.
Vol. 1. P. 279-282.
Cumming R.B. Cytogenetic studies in the order Odonata // Thesis Univ. Texas. 1964.
P. 1-64.
Heymer A. Verhaltensstudien an Prachtlibellen //Z. Tierpsychol. (Beiheft). Berlin, 1973.
N 11. 100 S.
Kiauta B. Evolution of the chromosome complement in Odonata // Genen Phaenen. 1967.
Vol. 11. №4 P. 56-61.
307
А.Ю. Харитонов
Kiauta В. Sex chromosomes and sex determining mechanism in Odonata, with a review of
the cytological conditions in the family Gomphidae and references to the karyotypic
evolution of the order// Genetica. 1969. Vol. 40. № 2. P. 127-157.
Kiauta B. Cytotaxonomy of dragonflies. П. Lectures delivered at the Tribhuvan University
Nepal Res. Cent. 1975. 80 p.
Oguma K. A comparative study of the spermatocyte chromosome in allied species of the
dragonflies//}. Fac. Sci. Hokkaido Univ. 1930. Vol. 6. № 1. P. 1-32.
Oguma K., Asana J. Additional data to our knowledge on the dragonfly chromosome with a
note on the occurrence of X-Y chromosome in the ant-lion //J. Fac. Sci. Hokkaido Univ
1932. Vol. l.№4. P. 133-142.
V. ЭВОЛЮЦИЯ ЧЕЛОВЕКА
РАННИЕ МИГРАЦИИ И РАСОВАЯ ЭВОЛЮЦИЯ
HOMO SAPIENS
А.М. Малолетко
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
E-mail: decanat@dggf.tsu.tomsk.su
В решении вопроса о прародине человека разумного мы исходим
из парадигмы расширенного моноцентризма. Такой прародиной явля-
лись, по нашему мнению, бассейн Нила, включая область Великих
Озер, и ближнее Околосредиземнеморье. Процесс сапиентации чело-
века захватывает отрезок времени примерно 70-40 тыс. лет назад.
Возможно, наиболее рано сапиентация наших предков проявилась в
области Великих Озёр, колыбели человечества. Прямых доказательств
тому нет. Но именно здесь началось и интенсивно протекало биологи-
ческое развитие человека вследствие усиленной его мутации. Возмож-
но, эти мутации были спровоцированы флюидами, поступавшими из
земных недр по глубинным разломам Восточноафриканского грабена.
Процесс сапиентации волнообразно развивался в северном направ-
лении, где его следы особенно четко зафиксированы на Ближнем Во-
стоке. Сформировалась Околосредиземноморская ойкумена челове-
ка разумного.
Ранние миграции. В условиях присваивающего типа хозяйства
перенаселение изначальной ойкумены наступило рано, что вызвало
центробежное расселение человека разумного. Человек разумный миг-
рировал по трём маршрутам (рис. 1).
Аустрический (южный) путь. Для мигрантов он был относитель-
но легким, так как пролегал в широтном направлении, не выходя за
пределы родной субтропической зоны. Мигранты шли по побережью
Индийского океана до Индокитая. Часть мигрантов, осваивая просто-
ры Южной Азии, оставалась по пути следования [1]. Достигнув Юго-
Восточной Азии, миграционный поток разделился на две ветви. Одна
309
о
Рис. 1. Миграционные потоки и ранние популяции Homo sapiens: / - прародина человека разумного, пути миграции. 2 -
бореальный (северный). 3 - аустрический (южный). 4 - африканский, популяции аустрическая (5 - австралоиды, 6 -
восточные мошолоиды. ~ - американэиды). 8 - бореальная. 9 - африканская. /О - <'людоразде.1>». // -очаш монголизации
А.М. Малолетко
И Эволюция человека
из них через Зондские острова достигла Австралии и, позднее, остро-
вов Тихого океана. Другая по тихоокеанскому побережью Азии про-
шла до Камчатки и Чукотки и по Алеутским островам проникла в
Америку.
Предполагаемый путь человека в Америку через Берингов пролив,
по нашему мнению, был недоступным и гибельным для такого замыс-
ла. Заселение Америки началось в эпоху последнего оледенения. Аляс-
ка (не полуостров, а нынешний штат), находившаяся на широте По-
лярного круга, была занята ледником. Мигранты с большим успехом
могли использовать Алеутские острова (широта Новосибирска). Сама
по себе идея перехода первых людей в Америку по Алеутским остро-
вам не нова. В 1964 г. ее проверял Н.Н. Диков [2], который провел ар-
хеологическое обследование о. Беринга в группе Командорских ост-
ровов. Но следов древних культур он там не обнаружил. И неудиви-
тельно - в эпоху последнего вюрмского (висконсинского) оледенения
(60-10 тыс. лет назад) уровень Мирового океана был ниже современ-
ного, по разным подсчетам, от 80 до 160 м, археологические памятни-
ки того времени лежат на дне океана.
Бореальный (северный) путь. Человек современного типа начал
осваивать внеледниковые области Европы около 40 тыс. лет назад. Это
было холодное время. Заболоченные приледниковые области представ-
ляли собой тундру и редколесье. Однако обилие крупных травоядных
животных мамонтового комплекса привлекало сюда первобытных охот-
ников. На внеледниковые равнины Восточной Европы человек при-
шел позже: поселение Костёнки имеет дату 24 тыс. лет, могильник
Сунгирь 24430 ± 400 лет.
Из Восточной Европы палеоевропейцы перевалили через Урал и
распространились по предгорьям и низкогорьям Южной Сибири до
Байкала, где они основали 24 тыс. лет назад стоянки Мальта и Буреть.
Тип наземных жилищ Бурети и Мальты, исключительное богатство
костяного инвентаря, костяные женские статуэтки, каменный инвен-
тарь - все это, безусловно, западные, европейские аналоги [3].
А.П. Окладников и З.А. Абрамова [4] считали, что сибирские, в том
числе и байкальские каменные изделия этого времени несопоставимы
с дальневосточными. Последние генетически тяготеют к Юго-Восточ-
ной Азии (Япония, Индокитай). Предположение этих исследователей
подтвердил американский антрополог К. Тернер [5], который устано-
311
А.М. Малолетко
вил физическое родство жителей Мальты с палеоевропейцами (кро-
маньонцами) вообще и людьми Костёнок и Сунгиря, в частности. При
этом он категорически отвергал какое-либо родство мальтинцев с па-
леоиндейцами Америки и жителями Центральной Азии.
Очевидно, горы Южной Сибири и тихоокеанского побережья Азии
в позднем палеолите служили «людоразделом», который долгое время
разграничивал бореальную и аустрическую популяции. Он был пре-
одолен значительно позже.
Африканский путь. Первый «исход» человека разумного из ойку-
мены, очевидно, связан с миграцией населения района Великих Озер
на юг Африки. В Южной Африке (Флорисбад) самый древний череп
человека разумного имеет возраст 37 тыс. лет, но, в отличие от палес-
тинских, он уже был лишен неандертальских черт. Череп из Фуш-Хук,
близ Кейптауна, обнаруживает сходство с бушменским: он также име-
ет широкий и плоский нос на широком сильно уплощенном лице [6].
Очевидно, койсанские народы являются наиболее ранними из сапиен-
тных групп населения планеты.
Собственно негрская группа сформировалась в результате сравни-
тельно недавней (мезолит или неолит [7]) миграции околосредизем-
номорского населения. Предполагается их родство с ранними европе-
оидами [8, с. 177]. Однако биохимические сравнительные материалы
по европейцам и неграм как будто не подтверждают это родство, но
эти же исследования подтвердили генетическую самостоятельность
негров и бушменов.
Расовая эволюция. В преддверии верхнего палеолита заверши-
лась биологическая эволюция человека как вида. В дальнейшем чело-
век эволюционировал только в расовом отношении.
Антропологи, похоже, без особых разногласий пришли к выводу,
что наш общий предок имел признаки всех трех основных рас, то есть
он был слабо дифференцирован в расовом отношении, но в наиболее
типичном виде был близок к представителям австралоидной (южной,
экваториальной) расы. В этом нет ничего удивительного: формиро-
вался он в зоне субтропиков и на него издавна воздействовали те же
факторы природной среды, что и ныне воздействуют на негров Афри-
ки. В.П. Алексеев [9, с. 168] так описал нашего предка: «Он был вол-
нистоволос, довольно темнокож, имел выступающий нос. Все призна-
ки негроидной расы были выражены достаточно отчетливо. И сразу
312
И Эволюция человека
вслед за этим нас поразит сходство нарисованного силой воображе-
ния облика с реальным австралийцем». Этот прототип человека ра-
зумного получил названия протоавстралоид и евро-африканец.
Расовые различия были заложены в генах человека еще до его са-
пиентации, но реализовались значительно позже, когда человек разум-
ный мигрировал в иные природные зоны - в тундру, пустыни и на плос-
когорья умеренных широт. Расовые изменения обусловлены двумя
причинами: адаптационными (приспособление к иным природно-кли-
матическим условиям) и генетико-автоматическими. Первая при-
чина в большой ее сложности рассмотрена В.П. Алексеевым [8]. Бо-
лее ранние ламаркистские представления об адаптации (пример: мон-
гольский разрез глаз возник в результате частого прищуривания в
условиях пыльных бурь или ослепительного света при снежном по-
крове) ныне всерьез не воспринимаются. Речь может вестись в основ-
ном о появлении и отборе генетических мутаций, полезных в конкрет-
ных природных условиях. Вторая заключается в том, что гены орга-
низмов, в том числе и человека, по-разному проявляют себя в различных
условиях. Доминантные гены «работают» в центре видового ареала.
Здесь, как выразился В.П. Алексеев [9, с. 193], бурлит жизнь, созда-
ются все новые и новые формы, которые начинают преобладать. Гены,
которые были скрыты, подавлены, редки, в условиях изолята (горы,
пустыни), слабой заселенности или по периферии ареала (это тоже
своеобразный изолят, например, Кольский полуостров) начинают фор-
мировать и закреплять в потомстве новые признаки. Усиливаются те
или иные расовые признаки, расовая принадлежность человека стано-
вится более определенной.
Происхождение европеоидов. В составе европеоидов выделяется две
ветви - северная (скандинавы, исландцы, ирландцы и др.) и южная (юж-
ные французы и итальянцы, иранцы, турки, азербайджанцы, армяне и
др.). Различия между ними касаются в основном пигментации кожи, глаз,
волос. Между этими двумя ветвями располагаются народы, занимаю-
щие промежуточное положение. Сейчас, пожалуй, не оспаривается точ-
ка зрения, высказанная Н.Н. Чебоксаровым еще в ЗО-е годы, о том, что
южные европеоиды, промежуточные варианты и северные европеоиды
представляют собой результат последовательного процесса депигмента-
ции изначально темнопигментированного населения. Южные европео-
иды стоят к исходному типу ближе, нежели северные.
313
А.М. Малолетко
Человеку разумному пришлось приспосабливаться к суровым при-
родным условиям приледниковых областей Европы, чтобы избежать
переохлаждения. Сильное выступание носовой полости удлиняло путь
воздуха до дыхательных путей и способствовало его (воздуха) согре-
ванию. Ортогнатность лицевого скелета европеоидов предохраняла
глотку от охлаждения. Светлая кожа создает явные преимущества био-
химического порядка: она помогает аккумулировать витамин D при
воздействии света на кожу. У таких особей не развивается рахит в ус-
ловиях слабого солнечного облучения. Это сохраняет жизнь и продле-
вает репродуктивную способность, что приводит к увеличению числа
депигментированных особей.
В результате генетико-автоматических и адаптационных процес-
сов в приледниковой зоне Западной Европы сформировалась новая
антропологическая раса - европеоидная, стратотипный субъект кото-
рой находился в пещере Кро-Маньон в долине р. Везер (Франция),
отчего тип получил название кроманьонский (другие названия - па-
леоевропейский, древнеевропейский). Я.Я. Рогинский и М.Г. Левин [ 1 ]
следующим образом охарактеризовали кроманьонца. Специфически-
ми особенностями кроманьонца были сочетание длинного черепа с
очень широким лицом, заметное развитие надбровных дуг, очень низ-
кие угловатые глазницы, сильно выступающий в плоскости лица уз-
кий нос, прямой общий профиль лица. Он имел прямой свод черепа,
подбородок резко выступал, высота лица малая, как и ширина меж-
глазничного расстояния и ширина носа. Однако кроманьонца из пе-
щеры Кро-Маньон по современным меркам нельзя считать «чисток-
ровным» европеоидом. У него был мал носовой указатель, что при
невысоком переносье и альвеолярном прогнатизме (косо поставлен-
ные передние зубы), большой скуловой ширине и низкой черепной
коробке не характерно для европеоидов. В нем явно просматриваются
южные (австралоидные) черты.
Происхождение монголоидов. Типичный монголоид имеет смуг-
лый цвет кожи, прямые, нередко жесткие волосы, слабый или очень
слабый рост бороды. Характерны средняя ширина носа, низкое или
среднее по высоте переносье, слабо выступающий нос, средняя тол-
щина губ, уплощенное лицо, сильное выступание скул, крупные раз-
меры лица, наличие эпикантуса, косо поставленные глаза. Но лишь
немногие народы имеют полный «набор» этих признаков. К ним мож-
314
V. Эволюция человека
но отнести лишь якутов, монголов, бурят и некоторые группы тувин-
цев. Американские индейцы, включаемые в большую монголоидную
расу, имеют выступающий, даже по европейским нормам, нос, у бе-
зусловных «азиатов» - японцев и корейцев - отмечается увеличение
ширины носа и толщины губ, нередки волнистые волосы, что сближа-
ет их с представителями экваториальной расы. Бушмены и готтентоты
Южной Африки, включаемые в экваториальную (австралоидную) расу,
имеют скулы, более выдающиеся вперед, нежели у сибирских монго-
лоидов. Даже при беглом обзоре географического распространения
ныне живущих и исчезнувших монголоидных популяций обращает на
себя внимание разобщенность их ареалов, отсутствие связей между
отдельными группами. Не может быть и речи о едином центре возник-
новения монголоидов. Ныне можно говорить о нескольких разрознен-
ных и разновозрастных очагах (центрах) возникновения групп с тем
или иным «набором» монголоидных признаков.
Восточная Европа. В позднем палеолите часть палеоевропейцев
Западной Европы переместилась в восточном направлении, в сторону
Урала. Здесь они встретились с ранее пришедшими с юга (Ближний
Восток) евро-африканцами, которые имели черты европеоидные и
австралоидные. Смешанное население продвинулось в Северное Пре-
дуралье, которое в эпоху последнего оледенения не было занято лед-
никами. Именно здесь, на восточной периферии ойкумены, заявили о
себе гены, которые «отвечали» за монголоидные признаки. Несомнен-
но, сыграл свою роль так называемый дрейф генов, а также воздей-
ствие суровых для того времени природно-климатических условий
Северного Приуралья. Исследование черепов с Оленьеостровского
могильника (Онежское озеро, мезолит) показало, что слабая монголо-
идность, по существу, является обычной особенностью древнего на-
селения Восточной Европы [ 11 ] и сложилась она самостоятельно под
влиянием суровых условий жизни [12]. Такого рода монголоидность
сохранили некоторые финноязычные группы (коми, удмурты, некото-
рые группы карел и мордвы).
Таким образом, можно поставить вопрос о восточноевропейском
очаге монголообразования в завершающую фазу верхнего палеолита
на базе слабо дифференцированных антропологических типов. О ран-
нем появлении у населения Восточной Европы признаков монголоид-
ности свидетельствуют материалы памятника Сунгирь: восемь скеле-
315
А.М. Малолетко
тов дают представление о европеоидном восточнокроманьонском типе,
не дифференцированном в расовом отношении, но обладающем про-
томонголоидными и негроидными признаками [13]. Так возникла мон-
голоидность уральского типа. На самом Урале этот антропологичес-
кий тип имели носители шигирской культуры V-IV тыс. до н.э. [14].
По мнению М.М. Герасимова, среди неолитического населения лес-
ной полосы Восточной Европы и всей Центральной и Южной Сибири
нет ни одного черепа, сколько-нибудь приближенного по своим ос-
новным размерам к данной категории. Он был характерен только для
Урала. В пещере по ручью Тыткескень (левый приток Катуни) было
найдено неолитическое захоронение мужчины, антропологически близ-
кого к хантам и манси [15]. Очевидно, уральский антропологический
тип в неолите занимал более обширные территории, чем ныне.
Сибирь. Процесс монголизации усилился при переходе мигрантов
через Урал в Сибирь до Забайкалья (более 25 тыс. лет назад). Мигран-
ты вскоре достигли Забайкалья. Их следы известны у Красноярска
(Афонтова гора) и Иркутска (Мальта, Буреть). Здесь европейского типа
инвентарь сопровождался фигурками женщин с явно монголоидными
лицами. Особенно выразительны были глаза: узкие, раскосые, они
невольно вызывают в памяти черты, свойственные представителям
монгольской расы [3].
И.М. Золотарева [16] реконструировала облик ранних монголоидов
Восточной Сибири, представителей палеосибирской расы, которые, по
ее мнению, выглядели следующим образом. Они имели крупную голову,
широкое уплощенное лицо с умеренным выступанием скул, которое «ук-
рашал» широкий нос с прямой спинкой и невысоким переносьем. У них
была сильно развита складка верхнего века и эпикантус. Губы имели сред-
нюю толщину. Окраска волос и радужины глаз темная, волосы умеренно
тугие. Добавим, что лицо было низкое. При желании можно усмотреть и
кроманьонские реликты (широкое лицо) и южные (широкий нос). В этом
плане палеосибирские монголоиды были близки к жителям Костёнок и
Сунгиря. Исходя из нашей версии, можно сделать предположение, что
низколицые (северные или таежные) монголоиды Сибири - это потомки
выходцев из Европы и что исходной для них базой было население па-
леоевропейского (кроманьонского) типа.
Норландия. В Северной Норвегии, Северной Финляндии, на Коль-
ском полуострове сформировался своеобразный антропологический
316
V. Эволюция человека
тип, который принято называть лапаноидным. Носителями этого типа
являются саамы (лопари) и, какой-то мере, коми-пермяки, мордва-мок-
ша. По данным И.И. Гохмана [17, с. 41], антропологически саамы ха-
рактеризуются следующими особенностями. Коробка черепа корот-
кая и широкая, высота средняя. Лоб средней ширины и наклона, уме-
ренно выпуклый. Лицо низкое, довольно широкое, ортогнатное, однако
на некоторых черепах заметен альвеолярный прогнатизм при доволь-
но широком носе (признак австралоидной расы). Нос слабо выступа-
ющий, спинка вогнутая, кончик носа и основание приподняты. Воло-
сы прямые, но мягкие, пониженная растительность на лице и теле.
Пигментация умеренно темная.
Таким образом, у лапаноидов имеются признаки всех трех боль-
ших рас: европеоидной (ортогнатное лицо), южной (прогнатизм, ши-
рокий нос; признаки не всегда наблюдаемые) и монголоидной (плос-
кое лицо, выступающие скулы, иногда эпикантус, наклонное положе-
ние осей глаз).
По вопросу о происхождении физического типа саамов нет едино-
го мнения. По мнению одних исследователей (В.В. Бунак, В.П. Яки-
мов, Г.Л. Хить, В.Я. Шумкин, П. Хайду, К.Б. Виклунд), они являются
недифференцированной в расовом отношении группой древнейшего
населения Европы. По мнению других (Ю.Д. Беневоленский,
В.И. Хартанович, Г.Ф. Дебец, Н.Н. Чебоксаров, В.П. Алексеев), - это
метисная группа, сформировавшаяся на базе палеоевропейцев и при-
шлых из Центральной Азии через Сибирь монголоидов. Мы придер-
живаемся первой точки зрения и считаем, что саамы являются древ-
нейшим европейским населением, которое длительное время форми-
ровалось в Фенно-Скандии. Исходным «материалом» послужили
потомки палеоевропейцев (кроманьонцев), которые примерно 8 тыс.
лет назад пришли в Норландию и в условиях изоляции и под влиянием
адаптационных процессов сформировали свой антропологический тип.
Низколицесть саамов, альвеолярный прогнатизм (у некоторых инди-
видуумов) - это наследие палеоевропейцев.
Западная Европа. В палеолите население Околосредиземноморья
не имело явных монголоидных признаков, если не считать некоторую
уплощенность лицевого скелета на уровне назомалярных точек. Про-
исхождение такой уплощенности неизвестно Возможно, это следствие
большой ширины лица палеоевропейцев [10, с. 306]. Но на Офнетс-
317
А.М. Малолетко
кой стоянке (мезолит Баварии) наряду с черепами долихокранных
(длинноголовых) кроманьонцев найдены короткие (круглые) черепа с
уплощенным сводом, слабо развитым надбровьем, плосковатым ли-
цом, сильно выступающими вперед и в стороны скуловыми дугами,
широким и коротким носом с низким переносьем. Еще более отчетли-
во признаки монголоидности выражены на черепах из Бранденбурга
(Притцербергское озеро). Оба неолитические черепа обладают упло-
щенным лицом, сильно развитыми предносовыми ямками, слабо выс-
тупающим носом с низким переносьем. Но в отличие от офнетских -
здесь черепа длинные (долихокрания), массивные. Уплощенное лицо,
выступающие скулы, низкое переносье и другие монголоидные при-
знаки отмечены на черепах из неолитических памятников Домитца,
Дохлярмарки, Саферштеттена на Инне в Баварии [19]. Причиной мон-
голизации явились адаптационные (в условиях изоляции) и генетико-
автоматические процессы. Однако «бунт» рецессивных генов был по-
давлен, и начавшаяся было монголизация европеоидного населения
завершилась поражением.
Африка. Здесь древнейшее население - бушмены и готтентоты -
причудливо сочетает в своем физическом типе основные признаки двух
рас - негроидной и монголоидной. Они более светлокожи, чем негры,
но так же широконосы, толстогубы, прогнатны, курчавоволосы. К числу
признаков монголоидной расы относятся складка века, эпикантус, силь-
но развитые скулы, плоские лица и нос, еще более плоские, нежели у
сибирских монголоидов. Добавим, что бушменским женщинам свой-
ственна стеатопигия - отложение жира на ягодицах. Накопление жира,
но на лице, свойственно сибирским монголоидам.
В Южной Африке, близ Кейптауна, в пещере Скидергат найден
череп, сходный с бушменским. У него очень широкий и плоский нос,
очень малая высота и сильная уплощенность мезогнатного лица [6].
Этот протобушменоид не имеет сходства с палеоантропами, напри-
мер, неандертальцами из Брокен-Хилл той же Южной Африки. Оче-
видно, предки бушменов мигрировали из района Великих Озер.
Провинция Кам в Тибете. Часть ранних мигрантов, избравших путь
из Околосредиземноморья на восток по побережью Индийского океа-
на, уклонилась на север, вглубь материка. В суровых условиях плоско-
горья Тибет физический тип переселенцев изменился, усилились ев-
ропеоидные (выступающий нос, высокое переносье) и монголоидные
318
И Эволюция человека
(скуластость, эпикантус, жесткие волосы и пр.) признаки. Так возник
своеобразный тип населения провинции Кам (верховья рр. Наг-чу,
Меконг и Янцзы). В.П. Алексеев [7, с. 162] предполагал, что монголо-
идные признаки у тибетцев в прошлом были выражены менее резко. И
лишь в ходе своей последующей эволюции приобрели черты, которые
позволяют включить их в состав азиатской, а не американской ветви
монголоидов.
Центральная Азия. В послеледниковье сформировались современ-
ные природные зоны Северного полушария, в области высокого давле-
ния возникли крупные пустыни планеты, в том числе и Гоби. Специфи-
ческие особенности внутриконтинентальных пустынь (пыльные бури,
обилие яркого света летом, обжигающие морозы зимой) заставило на-
сельников выработать приспособительные морфофизические особенно-
сти (эпикантус, узкий разрез глаз, плосколицесть, лицевую стеатопигию
и пр.). Процесс адаптации заключался в появлении полезных мутаций,
которые закреплялись в потомстве. Индивидуумы, лишенные этих по-
лезных приспособительных признаков, постепенно исчезали из популя-
ции. Формировался физический тип. Гены, отвечающие за монголоид-
ность, в условиях изоляции стали доминирующими.
Голоценовый возраст «типичных» монголоидов подтверждается
памятником Чжалайнор (Внутренняя Монголия). Создатели этого па-
мятника имели очень плоское лицо, абсолютно широкое и высокое.
Носовые кости выступали слабо, клыковые ямки выражены не резко,
предносовые же развиты сильно.
Исходная популяция, которая в Центральной Азии претерпела столь
глубокую трансформацию, неизвестна. Черепа из Верхней пещеры близ
Пекина (возраст 18 тыс. лет [20]), несомненно, принадлежавшие че-
ловеку разумному, обладают так же ярко выраженными монголоидны-
ми особенностями. Мужской череп (№ 101) отличается значительной
массивностью коробки, крупными абсолютными размерами. Череп
резко долихокранный с огромным продольным диаметром и умерен-
ной шириной. Лоб сильно наклонный, надбровье мощное, глазницы
прямоугольной формы, абсолютно и относительно низкие. Лицо ис-
ключительно высокое и в то же время широкое. Но череп отчетливо
прогнатный - это явный признак экваториальной расы.
Монголоиды Восточной Азии. Восточное (тихоокеанское) побере-
жье Азии заселено группами, которые включаются в большую монго-
319
А,М. Малолетко
лоидную расу, но различаются по ряду признаков. Эти различия и лег-
ли в основу выделения малых монголоидных рас.
Представители южной малой расы (южные китайцы и малые наро-
ды Южного Китая) довольно темно пигментированы, но имеют слабо
уплощенное лицо, нередко волнистые волосы, относительно широкий
нос, заметно прогнатное лицо. Следовательно, наряду с явными при-
знаками монголоидной расы, южные китайцы имеют черты и южной
(толстые губы, прогнатность, волнистые волосы).
Дальневосточная раса включает северных китайцев, корейцев.
И те, и другие обладают явными монголоидными чертами (скуластость,
жесткие волосы, эпикантус и пр.) У корейцев более четко, чем у север-
ных китайцев, проявляются черты южной расы - губы у них толще,
ширина носа больше, третичный волосяной покров развит лучше.
По этим параметрам корейцы сближаются с южными китайцами.
Современное население Японии обладает смешанным антрополо-
гическим типом. По некоторым признакам японцы сближаются с ай-
нами, которые имеют веддо-полинезийских облик (поздний признак),
и южными монголоидами (относительно широкий нос, толстые губы,
малый рост).
Арктическая раса включает эскимосов, чукчей, коряков. Населе-
ние Северо-Востока имеет элементы, связанные с древним населени-
ем Восточной и Юго-Восточной Азии. У чукчей сохранились явные
признаки южной расы - большая ширина носа, толстые губы, чаще
встречается вогнутая спинка носа.
В эту же восточноазиатскую группу монголоидов можно включить
и нивхов. Наряду с явно монголоидными признаками (сильное разви-
тие эпикантуса, плоское лицо, низкое переносье) у них несомненны и
южные: сильное развитие бороды, заметный прогнатизм.
Все расовые группы тихоокеанского побережья Азии имеют сме-
шанные монголо-австралоидные черты, что их и объединяет. Их гене-
тическое единство подтверждается и ареальной непрерывностью. Всю
приморскую полосу Восточной Азии можно рассматривать как область
формирования особой группы монголоидов. Исходным «материалом»
послужило изначально австралоидное население южного (аустричес-
кого) миграционного потока. Нет оснований считать этих монголои-
дов метисами: до сих пор на побережье не найдены останки древних
«чистых» монголоидов. Известны только «чистые» австралоиды, при-
320
И Эволюция человека
шедшие по берегу Индийского океана, у которых где-то в Восточной
Азии возникли монголоидные признаки. Произошло это примерно 20
тыс. лет назад, но еще в неолите Юго-Восточный Китай был заселен
людьми полинезийско-меланезийского типа.
Американоиды. С современными азиатскими монголоидами аме-
риканоидов роднит желтовато-бурый цвет лица, прямые и очень жест-
кие волосы, слабый третичный волосяной покров, заметно выступаю-
щие скулы, высокие орбиты, очень часто встречающиеся лопатооб-
разные резцы верхней челюсти, эпикантус у детей. С европеоидами
их сближают высокое переносье, сильно выступающий нос и отсут-
ствие эпикантуса у взрослых. Большими размерами коренных зубов,
большой шириной рта, значительной шириной носа (до 42 мм), нали-
чием небольшого прогнатизма, удлиненностью предплечья индейцы
напоминают некоторые варианты австралоидной расы [1, с. 466]. Сле-
довательно, американоиды сохранили в своем обличье в разном соот-
ношении признаки всех трех основных человеческих рас. Это харак-
теризует древнего человека, еще слабо дифференцированного в расо-
вом отношении.
Все попытки найти «родственников» индейцев в Азии среди со-
временного населения Сибири, в частности кетов, не увенчались ус-
пехом и, как будто, оставлены за бесперспективностью. Признавая
безусловно правильной точку зрения о приходе населения Америки из
Азии, тем не менее, мы не можем найти в Сибири предков индейцев
не только среди современного населения, но и древнего.
Американоиды пришли в Новый Свет в «готовом» виде. Об этом
свидетельствует большое сходство в строении черепов современных
и древних индейцев.
Всех монголоидов можно разделить на две группы: низколицых,
сформировавшихся на базе бореальной (северной) популяции (лапа-
ноиды, уральцы и сибирские или таёжные монголоиды) и высоколи-
цых, сформировавшихся на базе аустрической (южной) популяции
(монголоиды Центральной и дальневосточной Азии, американоиды).
Обе группы имеют изначально самостоятельное происхождение.
Расовая эволюция народов Африки. Негрская группа является ти-
повой для австралоидной (негроидной, экваториальной) расы. У не-
гров мелкокурчавые волосы, очень слабый рост третичного волосяно-
го покрова (как у монголоидов), темная пигментация волос, глаз и кожи,
321
AM. Малолетко
толстые вывернутые губы, широкая глазная щель. По сравнению с ев-
ропейцами негры более длиннорукие и длинноногие. Такой тип сфор-
мировался поздно, в мезолите, возможно в неолите. Исходным расо-
вым типом был протоавстралоидный (евро-африканский).
Типичные негроидные признаки появились после ухода части на-
селения из Южного Околосредиземноморья в тропики и являются ре-
зультатом реакции организма на изменившиеся природные условия. В
основном они направлены на борьбу с перегревом тела. В итоге те
австралоидные признаки, которыми обладал предок, у негроидов Аф-
рики еще более усилились. Темный цвет (как следствие высокого со-
держания меланина) предохраняет кожу от ожога. Курчавые волосы
создают вокруг головы особую воздухоносную прослойку. Узкая, вы-
сокая и длинная голова, типичная для негров, нагревается медленнее,
чем широкая, низкая и короткая [21]. Широкий нос с крупными нозд-
рями, толстые губы с обширной поверхностью слизистой оболочки
усиливают теплоотдачу, как и большое количество потовых желез на
единицу поверхности тела, что характерно для всех негроидов.
Бушменская (койсанская, южноафриканская) малая раса является
древней, сохранившей с очень давних времен свои особенности, в том
числе и монголоидные признаки. Расовая эволюция бушменов была
направлена на усиление монголоидности, что вызвано было миграци-
ей их предков в пустыню Калахари. «Давление» пустыни на адаптаци-
онные процессы предков бушменов началось с момента прихода их в
пустыню (более 40 тыс. лет назад).
Носители эфиопской малой расы имеют глубокие местные корни.
Этот своеобразный расовый тип является автохтонным, и основой для
его оформления был восточносредиземноморский высоколицый ва-
риант евро-африканцев.
Расовая эволюция человека завершилась в неолите. С эпохи брон-
зы изменения физического типа тех или иных популяций происходило
лишь в результате процессов метисации, когда имело место глубокое
проникновение значительных групп населения в популяции с иным
антропологическим типом. Так, в эпоху раннего металла европеоиды
из Северного Прикаспия и Причерноморья прошли до Енисея, где жили
представители уральского антропологического типа. Волна за волной
шли в Сибирь европеоиды из Передней Азии. На рубеже эр централь-
но-азиатские высоколицые монголоиды вторглись в Сибирь, а позже
322
И Эволюция человека
они прошли до Восточной Европы. В средневековье массовое про-
никновение европеоидов в Америку привело к формированию мети-
сов, а завоз негров-рабов из Африки - мулатов.
Изменение внешней конституции человека в это время выражалось
также в приобретении древними матуризованными типами грациль-
ных черт и брахикефальных признаков (признаков круглоголовости).
В заключение следует сказать, что европеоидность и монголоид-
ность являются аномалиями в расовом развитии человека разумного.
Если бы этот человек длительное время не выходил за пределы род-
ной субтропической природной зоны и в пределах её завершил бы свою
расовую эволюцию, достигнув уровня специализированного расового
типа, население планеты было бы в расовом отношении единообраз-
ным, похожим на аборигенов Австралии. Человечество было бы из-
бавлено от тех коллизий в обществе, которые были порождены и по-
рождаются расовыми предрассудками.
Литература
1. Рогинский Я.Я., Левин М.Г Антропология. М.: Высш, школа, 1978.
2. Диков Н.Н. Археологические памятники Камчатки, Чукотки и Верхней Колымы. М.:
Наука, 1977.
3. Окладников А.П. Сибирь в древнекаменном веке. Эпоха палеолита И Древняя Си-
бирь (Макет I тома «Истории Сибири»). Улан-Удэ, 1964.
4. Окладников А.П., Абрамова З.А. Первоначальное освоение палеолитическим чело-
веком Сибири и Дальнего Востока И Первобытн. человек, его матер, культура и
природн. среда в плейст. и голоцене. М., 1974.
5. Тернер Кристи Г. Ребенок верхнепалеолитической стоянки Мальта И Изв. СОАН
СССР. Сер. истор., филос. и филол. 1990. Вып. 2.
6. Рогинский Я.Я. Антропологический состав населения Африки И Расы и народы. М.:
Наука, 1974. Вып. 4.
7. Алексеев В.П. География человеческих рас. М.: Мысль, 1974.
8. Алексеев В.П. Географические очаги формирования человеческих рас. М.: Мысль,
1985.
9. Алексеев В.П. В поисках предков. Антропология и история. М.: Сов. Россия, 1972.
10. Алексеев В.П. Историческая антропология и этногенез. М.: Наука, 1989.
11. Якимов В.П. О древней «монголоидности» в Европе И Кр. сообщ. Ин-та этногр.
1957. Вып. 28.
12. ОшибкинаС.В. Древнейшие этнокультурные образования Севера Восточной Евро-
пы // Indo-germanica. М., 1990. Ч. 2.
13. Бадер О Н. Человек, его культура и природная среда северного края европейской
ойкумены // Первобытн. человек, его матер, культ, и природн. среда в плейстоцене
и голоцене. М., 1974.
323
А.М. Малолетко
14. Герасимов М.М. Восстановление лица по черепу (современный и ископаемый че-
ловек). М.: Изд-во АН СССР, 1955.
15. Кирюшин Ю.Ф., Кунгуров А.А., Степанова Н.Ф. Погребение из Нижнетыткескенс-
кой пещеры 1 (Алтай) // Палеоэкол. и рассел. древн. человека в Сев. Азии и Амери-
ке. Красноярск, 1992.
16. Юкагиры. Историко-географический очерк. Новосибирск: Наука, 1975.
17. Гохман И.И. Происхождение центральноазиатской расы в свете новых палеоантро-
пологических материалов // Сб. Музея антропол. и этногр. 1980. Т. 36.
18. Шумкин В.Я. Этногенез саамов (археологический аспект) // Происх. саамов. М.:
Наука, 1991.
19. Чебоксаров Н.Н. Этническая антропология Германии И Кр. сообщ. Ин-та этногр.
1946. Вып. 1.
20. Ларичев В.Е. Верхнепалеолитические памятники Китая, датированные радиокар-
боном // Изв. СОАН СССР. Сер. общ. наук. 1978. № 1. Вып. 1.
21. Алексеева Т.И. Географическая среда и биология человека. М.: Мысль, 1977.
ФЕНОМЕН МИКРОСАТЕЛЛИТНОЙ
НЕСТАБИЛЬНОСТИ
КАК ВОЗМОЖНЫЙ ФАКТОР
ПРЕНАТАЛЬНОГО ОТБОРА У ЧЕЛОВЕКА
Т.В. Никитина, С.А. Назаренко
НИИ медицинской генетики ТНЦ СО РАМН
634050, г. Томск, Наб. р. Ушайки, 10
E-mail: snaz@img.tsu.ru
Особенностью человека как биологического вида является высо-
кая, по сравнению с другими млекопитающими, частота спонтанной
гибели зародышей. Так, до 50-70% зигот погибают уже в доимпланта-
ционном периоде, а из клинически распознаваемых беременностей 15-
20% заканчиваются спонтанными абортами [1]. Причинами такой
высокой смертности зародышей человека могут быть нейроэндокрин-
ные нарушения и инфекционные заболевания у матери, иммунологи-
ческие конфликты между матерью и плодом, неблагоприятное воздей-
ствие окружающей среды и другие причины. Однако в нарушениях
раннего эмбрионального развития наиболее существенная роль при-
надлежит генетическим факторам [2, 3].
Каждый раз развитие нового организма, возникающего в результа-
те слияния сперматозоида с яйцеклеткой, подвергается строгой про-
верке силами естественного отбора на способность к нормальному
автономному существованию. В результате такой проверки неудачные
новые “генетические конструкции” гибнут, причем чем серьезнее на-
рушение онтогенеза, тем раньше останавливается развитие. Грубые
цитогенетические аномалии обнаруживаются в среднем у 50% абор-
тусов [4, 5]. Причина гибели другой половины зародышей, с нормаль-
ным кариотипом, как правило, не выявляется.
Так как пренатальная смертность человека является не только на-
учной проблемой, но и тесно связана с практической медициной, по-
иски причин ранней внутриутробной гибели зародышей активно ве-
дутся во всем мире. В 1995 г. появилась работа греческих авторов,
325
ТВ. Никитина, С.А. Назаренко
обнаруживших мутации минисателлита, расположенного в 3’-конце
протоонкогена H-ras у 3 из 30 исследованных спонтанных абортусов
[6]. В 1998 г. эта же группа исследователей на основании изучения
ограниченной выборки из 35 спонтанных абортусов сообщила о связи
между эмбриональной гибелью у человека и повышенной скоростью
мутирования анонимных микросателлитных локусов, не связанных с
протоонкогенами [7]. Микросателлиты - это тандемно расположен-
ные повторы ДНК с длиной последовательности от 2 до 6 нуклеоти-
дов, принадлежащие к семейству повторяющихся некодирующих ДНК-
последовательностей, к которому относятся также сателлитные пос-
ледовательности и минисателлиты. Они получили также название
коротких тандемных повторов (short tandem repeats, STR) или простых
повторяющихся последовательностей (simple sequence repeats, SSR).
Феномен высокой мутабильности STR-последовательностей назы-
вается микросателлитной нестабильностью (МН). Впервые это явле-
ние было описано в начале 90-х годов. В ДНК, полученной из опухо-
левых клеток пациентов с наследственным неполипозным колоректаль-
ным раком, с частотой свыше 50% обнаруживались аллельные
варианты, не свойственные нормальной ДНК индивида [8, 9]. Различ-
ные формы наследственного неполипозного рака толстого кишечника
вызываются мутациями в генах MSH2, MLH1, PMS1 и PMS2. Все эти
гены кодируют белки, входящие в состав ферментативной системы
репарации ошибочного спаривания оснований ДНК (мис-матч репа-
рации). Именно нарушение функционирования системы мис-матч ре-
парации ДНК приводит к фатальному повышению частоты мутаций в
клетках-родоначальниках опухоли [10, 11]. После обнаружения МН в
ДНК клеток колоректального рака последовала целая серия исследо-
ваний, посвященных поиску МН и при других опухолях. Оказалось,
что МН с высокой частотой (до 50%) обнаруживается также у пациен-
тов с мелкоклеточным раком легкого, опухолями головы и шеи (29%)
и раком молочной железы (28%), что указывает на существенную роль
нестабильности генома в этиологии злокачественных новообразова-
ний [12-14]. Микросателлиты в данном случае являются индикатором
нарушения генетической стабильности.
Активное изучение микросателлитов генома человека началось в
последнее десятилетие благодаря их уникальным чертам: многочис-
ленности, широкому распространению в эухроматиновых областях
326
И Эволюция человека
генома, кодоминантному типу наследования и чрезвычайно высокому
полиморфизму. Сочетание этих достоинств с возможностью примене-
ния ПЦР-технологий благодаря тому, что повторы фланкированы уни-
кальными последовательностями ДНК, сделали микросателлиты от-
личным инструментом для генетического анализа. Они используются
как маркеры при локализации генов, в медико-генетическом консуль-
тировании семей с наследственными заболеваниями, в судебно-меди-
цинской экспертизе для идентификации личности и установления от-
цовства. Микросателлиты нашли также широкое применение как ге-
нетические маркеры для изучения проблемы генетической гетеро-
генности и особенностей этногенеза популяций человека, для созда-
ния филогенетического древа отдельных популяций с целью реконст-
рукции истории миграции и эволюции видов.
Оказалось, однако, что простые повторы ДНК генома далеко не
так просты. Во-первых, несмотря на активные исследования, до сих
пор неясны их эволюционное происхождение и функции. Классичес-
кой стала гипотеза японского эволюцониста и генетика Оно [15] об
эволюции генов путем дупликации участков ДНК с последующей их
дивергенцией. Дупликации также широко распространены в некоди-
рующей части генома, где тандемные повторы более или менее длин-
ных ДНК-последовательностей формируют высокоупорядоченную
организацию, для которых Оно предложил термин “генетический хлам”
(“junk DNA”). Во-вторых, STR не подвергаются действию естествен-
ного отбора в соответствии с теорией нейтральности молекулярной
эволюции, то есть подавляющая часть мутаций микросателлитов се-
лективно нейтральна [16]. Обладая очень высокой, по сравнению с
экспрессирующимися областями генома, скоростью мутирования, они
в некоторых случаях проявляют удивительную консервативность. На-
пример, аллельные спектры ряда STR не только сходны между раз-
личными человеческими популяциями, но и имеют аналоги у прима-
тов [17]. Для объяснения этого факта выдвигается предположение о
существовании некоего ограничивающего давления (constraints) на
микросателлитные последовательности [18]. Скорее всего, это действи-
тельно не давление отбора, в его классическом понимании, а внутрен-
ние механизмы регулировки размеров повторяющихся последователь-
ностей ДНК. Однако неконтролируемый рост длины последователь-
ности, характерный для тринуклеотидных STR, вызывает дисфункцию
327
ТВ. Никитина, С.А. Назаренко
сцепленных генов и подвергается действию отбора, так как связан у
человека с патологическими фенотипами (болезни экспансии тринук-
леотидных повторов). Другие вопросы, стоящие перед исследователя-
ми эволюции микросателлитов, заключаются в изучении видо-, локу-
со- и аллель-специфических скоростей мутирования, выяснении при-
чин преобладания мутаций отцовского происхождения и т.п.
Исследование мутаций микросателлитных последовательностей
ДНК у погибших эмбрионов, с одной стороны, позволит оценить воз-
можный вклад микросателлитной нестабильности в достаточно высо-
кую частоту пренатальной смертности человека, а с другой стороны,
накопление информации о мутациях в STR необходимо для понима-
ния особенностей генетических процессов в самих микросателлитах
и их эволюционной значимости.
Материал и методы
В 55 семьях с погибшими на ранних стадиях развития эмбрионами
с нормальным кариотипом исследовали характер наследования поли-
морфных участков ДНК, содержащих тетрануклеотидные STR. ДНК
абортусов выделяли из свежезамороженных экстраэмбриональных
тканей, а ДНК их родителей - из лимфоцитов периферической крови с
обработкой перхлоратом натрия и экстракцией хлороформом по стан-
дартной методике [19, 20]. Спонтанные абортусы 6-12 недель разви-
тия были представлены двумя формами патологии беременности -
анэмбриониями и неразвивающимися беременностями, которые объек-
тивно верифицируются с помощью ультразвукового обследования бе-
ременных женщин, что исключает вмешательство родителей в судьбу
эмбриона. В качестве микросателлитных повторяющихся последова-
тельностей ДНК были выбраны анонимные высокополиморфные (ге-
терозиготность не менее 0,80) тетрануклеотидные ДНК-маркеры:
D2S1242, D2S1788, D9S768, D9S252, D9S1119, D11S1304, D11S1983,
DI 1S488, D16S685, D16S2624, D16S751, D16S752, D19S601, D19S559,
D19S394, D20S161, D20S168, D20S431, D21S11, D21S1435 и
D21S1413. Таким образом, исследовали по 2-4 полиморфных локуса,
локализованных на 7 разных аутосомах, а общее число исследован-
ных локусов составило 21. Такой подход позволил не только оценить
328
V. Эволюция человека
сегрегацию ДНК-маркеров в семьях по каждому локусу с выявлением
мутаций de novo, но и с высокой точностью исключить случаи ложно-
го отцовства. Информация о маркерах была получена из базы данных
GDB (www.gdb.org.). Олигонуклеотидные праймеры были синтезиро-
ваны фосфитамидным методом на базе НИИ медицинской генетики
ТНЦ СО РАМН.
ПЦР проводили на программируемом термоциклере “Циклотемп-5”
(“СТМ”, Россия). Реакционная смесь, объемом 30 мкл, содержала 100 нг
очищенной ДНК, 30 пмоль каждого праймера, смесь четырех основных
дезокситрифосфатов в концентрации 0,1 мМ каждого, 1,5 ед. активнос-
ти ДНК-Тац-полимеразы (ТОО “Гель”, Новосибирск). Концентрация
хлористого магния подбиралась индивидуально. Продукты амплифи-
кации разделяли при помощи электрофореза в 7 и 8% нативных поли-
акриламидных гелях, с последующим окрашиванием в бромистом эти-
дии. Результаты анализа регистрировали с применением системы ком-
пьютерной видеосъемки на приборе “UV-VIS Imager-П” (США).
Нумерацию аллелей (числовую или буквенную) проводили в порядке
возрастания числа тандемных повторов ДНК. Числами обозначали
точно идентифицированные аллельные варианты, а буквами - аллели,
размер которых не был определен. При определении направленности
мутации руководствовались общепринятым положением, по которо-
му более вероятным является наименьшее из возможных изменение
числа тандемных повторов ДНК в новом аллельном варианте [21].
Значение вероятности отцовства в семьях вычисляли методом, ре-
комендованным экспертно-криминалистическим центром МВД Рос-
сии [22]. Статистическую обработку результатов проводили с исполь-
зованием программы “Statistica” (версия 5.0).
Результаты иссследования
Молекулярно-генетический анализ характера наследования спонтан-
ными абортусами аллелей микросателлитных повторов ДНК выявил 10
эмбрионов из 55 обследованных (18,2%), имевших аллели, отсутствую-
щие у обоих родителей. Анализ сегрегации хромосом по другим ДНК-
маркерам позволил с вероятностью не менее 99,97% исключить ложное
отцовство как причину несовпадения аллелей у родителей и потомков.
329
ТВ. Никитина, С.А. Назаренко
Случаи, когда можно было подозревать однородительскую дисомию (по
хромосомам 19 и 20) как причину неправильной сегрегации аллелей у
эмбриона, также были исключены при исследовании других локусов этих
же хромосом. Сочетание высокого значения вероятности отцовства и
присутствия у ребенка “нового” аллеля по какому-либо локусу указыва-
ет на мутацию, произошедшую либо в ходе гаметогенеза у родителей,
либо в ходе развития эмбриона (при появлении третьего “нового” алле-
ля). В табл. 1 представлены данные о 12 зарегистрированных мутацион-
ных событиях у десяти эмбрионов, включающих 10 мутаций гаметичес-
кого и 2 - соматического происхождения.
Из них у двух эмбрионов мутации были выявлены одновременно
по двум локусам.
Анализ направленности мутаций гаметического происхождения у
спонтанных абортусов показал увеличение числа тандемных повторов в
микросателлитах в двух случаях, уменьшение - в семи случаях и в од-
ном случае ее направление нельзя было точно определить. Кроме того,
обнаружено еще два события появления третьего аллеля - фрагмента
ДНК, отсутствующего у обоих родителей (рис. 1), что является следствием
соматической мутации, произошедшей в ходе эмбриогенеза. Во всех слу-
чаях, когда оказалось возможным определить родительское происхож-
дение мутации, она возникла в гаметогенезе отца эмбриона (табл. 1).
Всего в настоящей работе было изучено 1232 события передачи
аллеля от родителя потомку. Обнаружено 12 мутаций по 7 локусам
шести разных хромосом. Для локусов с мутациями микросателлитов
средняя частота мутирования составила 2,4-10’2 на локус на гамету на
поколение (табл. 2). По остальным 14 исследованным локусам мута-
ций не обнаружено. В среднем скорость мутирования всего комплекса
исследованных тетрануклеотидных повторов ДНК у спонтанных абор-
тусов составила 9,8-10"3 на локус на гамету на поколение.
Частота мутационных событий в нашем исследовании заметно ва-
рьировала между локусами. Из 21 исследованного STR-маркера они
были найдены в семи (35%). В табл. 2 показано соотношение скорос-
тей мутирования локусов, проявивших нестабильность. Наиболее ча-
сто мутирующим оказался локус D21S11, по которому обнаружены
три мутационных события. По две мутации найдены в локусах D9S768,
D19S601 и D20S168, и по одной - в локусах D21S1413, DI 1S1983 и
D2S1242. Не обнаружено достоверной корреляции между гетерози-
готностью локуса и его нестабильностью.
330
V. Эволюция человека
Таблица 1
Характеристика мутаций de novo в микросателлитных локусах,
выявленных у спонтанных а борту со в
Локус Генотип Направлен- ность мутации* Происхождение мутации
отца эмбриона матери
D2S1242 8,9 4,7 4,6 -1 Гаметич. (отц.)
D9S768 b, d а, b Ь, с -1 Гаметич. (?)
D9S768 а a, b, d с, d +1 Соматическое
D11S1983 4, 11 5, 12 5 + 1 Гаметич. (отц.)
D19S601 5 3** 3,6 -2 Гаметич. (?)
D19S601 5 2,4 2 -1 Гаметич. (отц.)
D20S168 b а, b В -1 Гаметич. (?)
D20S168 a, d Ь, с** Ь, с -1 Гаметич. (отц.)
D21S11 Ь, с а, b b,d -1 Гаметич. (?)
D21S11 с, d b, d a, d ±1 Гаметич. (?)
D21S11 а, b b, с а, b +2 Гаметич. (?)
D21S1413 5,7 5, 6,7 5 ±1 Соматическое
Примечание. Жирным шрифтом выделены новые аллели.
♦ Цифра означает, на сколько повторяющихся элементов изменился размер аллеля; знак
(+) или (-) - увеличение и уменьшение длины амплифицированного фрагмента ДНК
соответственно; (±) - равновероятное событие увеличения или уменьшения аллеля.
** Однородительская дисомия исключена по другому локусу этой же хромосомы.
А
В
Рис. 1 Примеры мутаций тетрануклеотидных повторов ДНК, выявленных у спонтан-
но погибших эмбрионов: А - мутация в локусе D2S1242 произошла в гаметогенезе
отца (слева - леддер); Б - соматическая мутация в локусе D21S1413 (справа - леддер)
331
ТВ. Никитина, С.А. Назаренко
Таблица 2
Характеристика локусов с мутациями микросателлитов
Генетический маркер Гетерози- готность Число передач родитель- потомок Число мутаций Скорость мутирования х10"2
D21S11 0,90 100 3 3,0
D20S168 0,80 68 2 2,9
D9S768 0,91 68 2 2,9
D19S601 0,81 80 2 2,5
D21S1413 0,88 54 1 1,8
D11S1983 0,90 62 1 1,6
D2S1242 0,94 106 1 0,9
Всего (среднее) 538 12 (2,4)
Обсуждение
Анализ наследования полиморфных аллелей микросателлитных
повторов ДНК у 55 спонтанных абортусов показал, что у десяти эмб-
рионов (18,2%) имеются аллели, отсутствующие у обоих родителей.
Характер сегрегации хромосом по другим ДНК-маркерам позволил с
высокой вероятностью исключить ложное отцовство и однородитель-
скую дисомию как возможные причины несовпадения аллелей у ро-
дителей и потомков. Этот результат указывает на мутацию числа тан-
демных повторов в микросателлитных последовательностях ДНК,
произошедшую в ходе гаметогенеза у родителей. Из десяти абортусов
с мутациями в микросателлитных повторах ДНК у двух зародышей
выявлены мутации одновременно по двум локусам. Мы обнаружили
также два случая появления третьего аллеля, отсутствующего у обоих
родителей, что является следствием соматической мутации, произо-
шедшей в ходе эмбриогенеза. Средняя скорость мутирования всего
комплекса исследованных тетрануклеотидных повторов ДНК у спон-
танных абортусов составила 9,8-10 3 на локус на гамету на поколение.
Известно, что скорость мутирования микросателлитов зависит от
нуклеотидного состава их последовательностей. Математические мо-
дели мутационного процесса предполагают, что с максимальной ско-
ростью мутируют динуклеотидные повторы, за ними следуют не свя-
заные с болезнями тринуклеотиды, а тетрануклеотидные STR мутиру-
ют в 1,5-2 раза медленнее динуклеотидных [23]. Однако эксперимен-
332
V. Эволюция человека
тальные данные часто не согласуются с этими оценками. Например, в
работе Вебера и Вонга [24], при исследовании скорости мутирования
28 STR-маркеров, локализованных на хромосоме 19 человека, сред-
няя скорость мутирования тетрануклеотидных локусов в четыре раза
превышала таковую для динуклеотидных. И хотя 8 из 24 обнаружен-
ных мутаций приходилось на два тетрануклеотидных локуса, исклю-
чение которых из анализа изменяло соотношение скоростей мутиро-
вания в пользу динуклеотидных повторов, в среднем скорость мути-
рования тетрануклеотидных STR хромосомы 19 в этом исследовании
составила 2,1 10~3 на локус на гамету на поколение [24]. Очень похо-
жие оценки скорости мутирования тетрануклеотидных локусов при-
водят и другие авторы: 2,1 10 3 на локус на гамету на поколение для
аутосомных маркеров [21] и 0,2% для маркеров Y хромосомы [25].
Эти цифры получены для нормальной скорости мутирования (при пе-
редаче от родителей здоровым детям). Обнаруженная в ходе настоя-
щего исследования частота мутаций de novo тетрануклеотидных STR
у погибших эмбрионов (9,8-10~3) практически в 5 раз превышает спон-
танный уровень их мутирования.
Частота мутаций может заметно варьировать даже между локуса-
ми одного типа. Среди тетрануклеотидов человека встречаются как
гипермутабильные локусы, например D21S1245 и DXS981, мутирую-
щие со скоростью (0,56-1,4)-10-2 и 1,5 • 10-2 соответственно [26,27], так
и чрезвычайно стабильные, скорость мутирования которых на 2-3 по-
рядка меньше [28]. Причины неодинаковой скорости мутирования от-
дельных тетрануклеотидных STR были изучены в работах Бринкман-
на с соавт. [21], где было показано, что частота мутаций в наибольшей
степени зависит от состава локуса: нестабильность экспоненциально
повышается с увеличением длины монотонно повторяющегося трак-
та. Вставки другого нуклеотидного состава оказывают стабилизирую-
щее действие на ДНК. По-видимому, это общая особенность всех по-
вторяющихся последовательностей ДНК, так как подобный феномен
обнаружен и при изучении болезней экспансии числа тринуклеотид-
ных повторов, а также для динуклеотидных STR [29, 30].
Нельзя исключить, что повышенная скорость мутирования в на-
шем исследовании получена в результате использования для анализа
высокомутабильных локусов. Однако тот факт, что в семьях с аборту-
сами мы изучали наследование по 21 различному маркеру, делает та-
333
ТВ. Никитина, С.А. Назаренко
кое объяснение маловероятным. Более того, самый мутабильный в
нашем исследовании локус D21S11, который обнаружил три мутаци-
онных события на 100 мейозов (табл. 2), в норме мутирует со скорос-
тью 1,810~3 на гамету на поколение [31]. К сожалению, нормальная
скорость мутирования остальных использованных в данной работе ло-
кусов неизвестна, и это является предметом нашего дальнейшего ис-
следования. Прямые сравнения скоростей мутирования STR-локусов
у погибших эмбрионов и в семьях со здоровым потомством позволят,
по-видимому, исключить влияние выбора микросателлитных марке-
ров, хотя оно и было случайным.
Интересно отметить, что появление третьего аллеля (соматический
мозаицизм) относительно часто встречается в ДНК лимфобластоидных
клеточных линий, трансформированных in vitro [24], однако практичес-
ки не регистрируется при исследовании нормальных индивидов. В лите-
ратуре мы встретили лишь два упоминания о подобных событиях [27, 32].
Соматический мозаицизм выявляется в ДНК онкологических больных
при сравнении образцов из здоровых и опухолевых тканей [33]. Подоб-
ную стратегию использовали Киарис с соавт. [34] при изучении микро-
сателлитной нестабильности у спонтанных абортусов: они сравнивали
образцы амплификации ДНК из тканевых срезов, взятых с диаметраль-
но противоположных сторон эмбриона. Результаты оказались весьма
неожиданными: из 30 спонтанных абортусов, обследованных по 12 мик-
росателлитным локусам, у 12 (40%) было выявлено несовпадение алле-
лей в образцах ДНК, выделенных из различных частей эмбриона [34].
Этот факт кажется еще более удивительным, если учесть, что простран-
ственная удаленность тканей не гарантирует их развития из далеких в
онтогенезе клеточных клонов. К сожалению, в качестве контрольной
группы авторы использовали только 5 медицинских абортусов, что не
позволяет считать их результаты бесспорными. Но в любом случае такой
высокий уровень соматических мутаций у погибших эмбрионов заслу-
живает дальнейшего изучения.
В нашей работе обнаружено два случая соматического мозаицизма
из 55 абортусов (3,6%). В исследовании была использована ДНК, вы-
деленная из тканей трофобласта, развивающегося из двух различных
зародышевых листков (плодные оболочки - из внезародышевой мезо-
дермы, а хорион - из трофоэктодермы). Для получения более полного
представления о масштабах соматической микросателлитной неста-
бильности в клеточных клонах погибших эмбрионов было бы инте-
334
V. Эволюция человека
ресно сравнить образцы ДНК экстраэмбриональных тканей с тканями
собственно зародыша. Но так как обследованная нами выборка заро-
дышей состояла из случаев анэмбрионий и неразвивающихся бере-
менностей ранних сроков, то операционный материал в подавляющем
большинстве случаев вообще не содержал тканей эмбриона.
Кроме соматических мутаций у погибших эмбрионов нами выявле-
но 10 мутации, произошедших в гаметогенезе родителей абортусов.
В двух случаях это было увеличение числа тандемных повторов, в семи
- уменьшение, и в одном случае направленность мутации определить
было невозможно. Соотношение событий потери и добавления повторя-
ющихся элементов в микросателлитных локусах обычно в пользу после-
дних [24, 32, 35]. Исследование гипервариабельного тетрануклеотидно-
го локуса у ласточек обнаружило положительный мутационный скос:
аллели втрое чаще мутировали в сторону увеличения размеров [32]. Та-
кая тенденция ведет к неограниченному росту повторяющихся последо-
вательностей в геноме, а следовательно, должен существовать и проти-
воположный механизм. Анализ ряда тетрануклеотидных STR человека
обнаружил, что в микросателлитах с непрерывной последовательностью
более длинные аллели мутируют чаще в направлении уменьшения числа
повторяющихся единиц, а мутации в коротких аллелях направлены пре-
имущественно в сторону увеличения [21, 36]. Интересно, что аналогич-
ная тенденция была обнаружена и при исследовании динуклеотидного
локуса у дрожжей: спектр мутаций различался между аллелями, причем
более длинные чаще подвергались протяженным делениям, то есть му-
тационное поведение микросателлитов может предотвращать неограни-
ченный рост ДНК последовательности [37].
Другой механизм противодействия экспансии STR - гипотезу “ча-
стичных мутаций” - предложили Палсболл с соавт. [38]. Анализируя
мутационные события в три- и тетрануклеотидных последовательнос-
тях ДНК китов, они обнаружили не только изменения целого числа
повторов, но и появление частичных повторов, которые прерывают
правильную последовательность, делая микросателлит гораздо менее
мутабильным. Накопление новых данных о мутационном поведении
микросателлитов различной структуры позволит более точно оценить
их эволюцию и вклад в формирование геномов организмов.
Анализ родительского происхождения мутаций у погибших эмб-
рионов в нашем исследовании показал, что они возникали преиму-
щественно в ходе гаметогенеза отца. По-видимому, преобладание “от-
335
ТВ. Никитина, С.А. Назаренко
цовских” мутаций - общая тенденция, характерная для микросател-
литных повторяющихся последовательностей ДНК (табл. 3).
Причиной преобладания мутаций отцовского происхождения, по-
видимому, является тот факт, что при созревании сперматозоид пре-
терпевает на порядок больше циклов репликации, чем яйцеклетка.
У женщин независимо от возраста до начала мейоза оогонии подвер-
гаются 22 митотическим делениям, тогда как у мужчин число претер-
певаемых сперматогониями клеточных делений зависит от возраста.
Например, у мужчины к 29-летнему возрасту спермин проходят в сред-
нем около 350 митозов [39], т.е. в 16 раз больше, чем ооцит. Если раз-
ница в числе мутационных событий в гаметогенезе действительно яв-
ляется следствием разности числа циклов репликации ДНК, то долж-
на наблюдаться зависимость частоты отцовских мутаций от возраста
отца. И действительно, обнаружен эффект отцовского возраста на ско-
рость мутирования микросателлитов: средний возраст отцов с мута-
циями оказался выше среднего возраста отцов в выборке [36]. Инте-
ресно, что подобное соотношение мутаций в гаметогенезе отца и ма-
тери наблюдалось и у ласточек [32], хотя у птиц самки являются
гетерогаметным, а самцы - гомогаметным полом по гоносомам W и Z
соответственно.
Таблица 3
Соотношение отцовских и материнских
мутаций в микросателлитах генома человека
Тип поспело- вательности Днк Мутации Ссылка
Всею Отцовские Материнские
Ди-, три-, тетра 24 15 4 Weber, Wong, 1993
Тетра- 4 2 1 Mahtani, Willard, 1993
Ди- 15 6 3 Amos et al., 1996
Тетра-, пента- 23 17 3 Brinlmann et.al., 1998a
Тетра- 11 10 - Sajantila et.al., 1999
Тетра- 10 4 - Настоящая работа
Всего 87 54 11
336
V. Эволюция человека
Мутации STR проявляются как сдвиг электрофоретической под-
вижности ПЦР-продукта за счет потери или добавления повторяющих-
ся единиц. Наиболее вероятными механизмами мутирования микро-
сателлитов являются:
1. Неравный кроссинговер. Обмен повторяющимися единицами
между гомологичными хромосомами приводит к образованию на них
разного числа повторов: один гомолог будет содержать делецию, а
другой - вставку.
2. Проскальзывание репликационной вилки (replication slippage).
ДНК-полимераза временно диссоциирует в репликационной вилке с
образованием неспаренного участка ДНК. Повторяющаяся единица
реассоциирует на некотором расстоянии, но уже не с аналогичным
повтором. В месте неправильного спаривания образуется “выпячива-
ние” из повторяющегося фрагмента ДНК, а ДНК-полимераза продол-
жает достраивать ДНК от места проскальзывания.
Большинство данных о мутациях микросателлитов говорите пользу
последнего механизма. Во-первых, в клеточных линиях с нарушения-
ми функций системы мис-матч репарации наблюдается резкий рост
микро сателлитной нестабильности [10, 11]. Во-вторых, проскальзы-
вание репликации - внутриспиральное событие и не должно зависеть
от частоты хиазм. Так и оказалось - мутации, элиминирующие реком-
бинацию, не влияли на скорость мутирования микросателлитов [41].
Кроме того, STR Y-хромосомы имеют сходную с аутосомными локу-
сами скорость мутирования, что было бы невозможно для процесса,
связанного с рекомбинацией [25]. В-третьих, существуют прямые до-
казательства, когда использование дополнительных маркеров, флан-
кирующих исследуемый локус, продемонстрировало у мутантных ал-
лелей и аллелей-предшественников идентичные гаплотипы по флан-
кирующим маркерам [27].
Причиной проскальзывания ДНК-полимеразы могут быть и кон-
формационные изменения структуры ДНК. Имеются сообщения о том,
что при нарушении метилирования ДНК в коротких повторяющихся
последовательностях образуются особые 4-нитчатые тугоплавкие
структуры и, как следствие, происходит их пропуск при репликации,
что приводит к изменению числа повторов. Нарушение процесса ме-
тилирования ДНК обнаружено и в клетках колоректального рака. От-
мечается существенная корреляция между генетической нестабильно-
337
ТВ. Никитина, С.А. Назаренко
стью при раке и метилированием ДНК [42, 43]. Таким образом, нару-
шение метилирования и сбои в работе системы репарации ошибочно-
го спаривания оснований ДНК могут вызвать сдвиг репликации, кото-
рый приводит к делеции или инсерции (indels) одного или нескольких
повторяющихся элементов. Возникновение МН может служить инди-
катором серьезных функциональных изменений в работе генетичес-
кого аппарата, и не удивительно, что клеточные линии, проявляющие
МН, демонстрируют и увеличение частоты мутирования экспрессиру-
емых генов [44], что приводит к фатальным последствиям для орга-
низма. Эти данные дают основание предполагать, что нестабильность
генома, выявляемая на уровне повторяющихся последовательностей
ДНК, может затрагивать не только генетически нейтральные локусы,
но и активные участки генома, играющие критическую роль в жизне-
способности эмбриона, что ведет к гибели клеток и прекращению раз-
вития зародыша.
В нашей работе 18,2% зародышей проявляли признаки микросател-
литной нестабильности. В исследовании Спандидоса с соавторами, про-
веденном на 35 спонтанных абортусах, мутации наблюдались у 8 из 35
обследованных эмбрионов, что составило 23% [7]. Доля семей с невы-
нашиванием беременности в анамнезе среди тех, в которых спонтанный
абортус проявлял признаки МН, в настоящем исследовании составила
50%, тогда как среди семей без признаков нестабильности генома у абор-
туса она составила только 27%. Хотя это отличие оказалось статистичес-
ки недостоверным (Р>0,05), возможно, вследствие недостаточного чис-
ла наблюдений, коэффициент корреляции оказался равным 0,53, что го-
ворит о средней силе связи между отягощенным акушерским анамнезом
и микросателлитной нестабильностью у абортусов. Эти данные согласу-
ются с результатами исследования [7], где была выявлена статистически
значимая ассоциация между МН у абортуса и отсутствием деторожде-
ния в анамнезе родительской пары. Вполне возможно, что в некоторых
семьях существуют наследственные факторы, обусловливающие невы-
нашивание беременности. Нам представляется, что одним из таких фак-
торов может быть гетерозиготное носительство родителями рецессив-
ных дефектных генов системы репарации или метилирования ДНК, ко
торые у некоторых эмбрионов переходят в гомозиготное состояние,
приводя к нарушению процесса репарации ДНК, нестабильности гено-
ма соматических клеток и гибели этих зародышей.
338
V Эволюция человека
Литература
1. Carr D.H. Cytogenetics of human reproductive wastage // Issues and Rewiews in Teratolody.
1983. Vol. l.P. 33-72.
2. Кулешов Н.П. Частота возникновения и судьба хромосомных аномалий у человека:
Автореф. дис. ... докт. мед. наук. М., 1979. 45 с.
3. Назаренко С.А. Изменчивость хромосом и развитие человека. Томск: Изд-во Томско-
го ун-та, 1993. 200 с.
4. Eiben В., Bartels I., Bahr-Porsch S. et. al. Cytogenetic analysis of750 spontaneous abortions
with the direct-preparation method of chorionic villi and its implications for studying
genetic causes of pregnancy wastage //Am. J. Hum. Genet. 1990. Vol. 47. P. 656-663.
5. Boue J., Boue A., Lazar P. Retrospective and prospective epidemiological studies of 1500
karyotyped spontaneous human abortions//Teratology. 1975. Vol. 12. P. 11-26.
6. Kiaris H., Ergazaki M., Spandidos D A. Instability at the H-ras minisatellite is associated
with the spontaneous abortion of the embryo I I Biochem. Biophys. Res. Commun. 1995.
Vol. 214. N3.P. 788-792.
7. Spandidos D.A., Koumantakis E., Sifakis S., Sourvinos G. Microsatellite mutations in
spontaneously aborted embryos // Fertility and Sterility. 1998. Vol. 5. P. 892-895.
8. Aaltonen L.A., Peltomaki P., Leach F.S. et al. Clues to the pathogenesis of familial colorectal
cancer//Science. 1993. Vol. 260. P. 812-816.
9. Thibodeau S.N., Bren G., Schaid D. Microsatellite instability in cancer of the proximal
colon//Lancet. 1993. Vol. 260. P. 816-819.
10 Bzapat B.V, Madlensky L., Temple L.K.F. et al. Family history characteristics, tumor
microsatellite instability and germline MSH2 and MLH1 mutations in hereditary colorectal
cancer//Human Genetics. 1999. Vol. 104. P. 167-176.
11. Liu B., Nicolaides N.C., Marcowitz S. et al. Mismatch repair gene defects in sporadic
colorectal cancers with microsatellite instability//Nature Genetics. 1995. Vol. 9. P. 48-55.
12. Chen X., Stroun M., Magnenat J.-L., Nicod L.P., Kurt A.M., Lyantey J., Lederrey C.,
Ancer P Microsatellite alterations in plasma DNAof small lung cancer patients //Nature
Medicine. 1996. Vol. 2. № 9. P. 1033-1035.
13. Nawroz H., Koch W., Ancer P, Stroun M., Sidransky D. Microsatellite alterations in
serum DNA of head and neck cancer patients // Nature Medicine. 1996. Vol. 2. № 9.
P. 1035-1037.
14. Rush E.B., Calvano J.E., van Zee K.J., Zelenetz A.D., Borgen P.I. Microsatellite instability
in breast cancer// Ann. Surg. Oncol. 1997. Vol. 4. № 4. P. 310-315.
15. Оно С. Генетические механизмы прогрессивной эволюции. М.: Мир, 1973.
16. Кимура М. Молекулярная эволюция: теория нейтральности. М.: Мир, 1985.
17. Garza J.C., Slatkin М., Freimer N.B. Microsatellite allele frequencies in humans and
chimpanzees with implication for constraintson allele size I I Mol. Biol. Evol. 1995.
Vol. 12. P. 594-603.
18. Nielsen R., Palsboll P.J. Single-locus tests 1 of microsatellite evolution: multi-step mutations
and constraints on allele size // Mol. Phylogenet. Evol. 1999. Vol. 11. N. 3. P. 477-484.
19. Johns M B.Jr., Paulus-Thomas J.E. Purification of human genomic DNA from whole
blood using sodium perchlorate in place of phenol //Anal. Biochem. 1989. Vol. 180.
N. 2. P. 276-278.
339
Т.В. Никитина, С.А. Назаренко
20. Current protocols in human genetics. New-York: John Wiley & Sons, Ink. 1998. Vol 2
Appendix 3B.1.
21. Brinkmann B., Klintschar M., Neuhuber F., Huhne J., Berkhard R. Mutation rate in
human microsatellites: influence of the structure and lenght of the tandem repeat 11 Am
J. Hum. Genet. 1998a. Vol. 62. N 6. P. 1408-1415.
22. ПерепечинаИ.О., Гришечкин С.А. Вероятностные расчеты в ДНК-дактилоскопии.
Методические рекомендации. М.: ЭКЦ МВД России. 1996. 16 с.
23. Chakraborty R., Kimel М., Stivers D.N., Davison L.J., Deka R. Relative mutation rates at
di-, tri-, and tetranucleotide microsatellite loci 11 Proc. Natl. Acad. Sci. USA. 1997
Vol. 94. P. 1041-1046.
24. Weber J.L., Wong C. Mutation of human tandem repeats // Hum. Mol. Genet. 1993
Vol. 2. №8. P. 1123-1128.
25. Heyer E., Puymirat J., Dieltiers P., Bakker E., De KnifTP. Estimating Y chromosome
specific microsatellite mutation frequencies using deep rooting pedigrees // Hum. Mol
Genet. 1997. Vol. 6. P. 799-803.
26. Talbot C.C. Jr., Avramoulos D., Gerken S., Chakravarti A. et al. The tetranucleotide repeat
polymorphism D2IS 1245 demonstrates hypermutability in germline and somatic cells
//Hum. Mol. Genet. 1995. Vol. 4. No. 7. P. 193-1199.
27. Mahtani M.M., Willard H.F. A polymorphic X-linked tetranucleotide repeat locus
displaying a high rate of new mutation: implications for mechanisms of mutation at
short tandem repeat loci // Hum. Mol. Genet. 1993. Vol. 2. N. 4. P. 431-437.
28. Chakraborty R., Stivers D.N., Zhong Y. Estimation of mutation rates from parentage
exclusion data: applications to STR and VNTR loci //Mutation Research. 1996. Vol. 354.
P. 41-48.
29. Heale S.M., Petes T.D. The stabilization of repetitive tracts of DNA by variant repeats
requires a functional DNA mismatch repair system // Cell. 1995. Vol. 83. P. 539-545.
30. Sturzeneker R., Haddad L.A., Bevilacqua R.A.U., Simpson A.J.G., Pena S.D.J Polarity
of mutations in tumor-associated microsatellite instability // Human Genetics. 1998
Vol. 102. P. 231-235.
31. Brinkmann B., Jungle A., Meyer E., Wiegand P. Population genetic diversity in relation
to microsatellite heterogeneity//Human Mutation. 1998. Vol. 11. P. 135-144.
32. Primmer C.R., Ellegren H., Saino N., Moller A.P. Directional evolution in germline
microsatellite mutations //Nat. Genet. 1996. Vol. 13. P. 391-393.
33. Boland R. Setting microsatellites free 11 Nature Medicine. 1996. Vol. 2. N. 9
P. 972-974.
34. Kiaris H., Koumantakis E., Ergazaki M., Spandidos D.A. Microsatellite instability in
aborted embryos // Molec. Hum. Reprod. 1996. Vol. 2. N. 1. P. 72-74.
35. Amos W., Sawcer S.J., Feakes R.W, Rubinsztein D.C. Microsatellites show mutational
bias and heterozygote instability//Nat. Genet. 1996. Vol. 13. P. 390-391.
36. Rolf B., Brinkmann B. Reply to Henke and Henke //Am. J. Hum. Genet. 1999. Vol. 64
P. 1473-1474.
37. Wierdl M., Dominska M., Petes T.D. Microsatellite instability in yeast: dependence on
the lenght of the microsatellite // Genetics. 1997. Vol. 146. P. 769-779.
38. Palsboll P.J., Berube M., Jorgensen H. Multiple levels of single-strand slippage at cetacean
tri- and tetranucleotide repeat microsatelite loci // Genetics. 1999. Vol. 151. P. 285-296
39. Фогель Ф., Мотульски А. Генетика человека. M.: Мир, 1989.
340
V. Эволюция человека
40. Sajantila A., Lukka М., Syvanen А-С. Experimentally observed germline mutations at
human micro- and minisatellite loci 11 Eur. J. Hum. Genet. 1999. Vol. 7. P. 263-266.
41. Levinson G., Gutman G.A. Slipped-strand mispairing:a maior mechanism for DNA
sequence evolution 11 Moi. Biol. Evol. 1987. Vol. 4. P. 203-221.
42. Lengauer C., Kinzler K.W., Vogelstein B. DNA methylation and genetic instability in
colorectal cancer cells I I Proc. Natl. Acad. Sci. USA. 1997. Vol. 94. P. 2545-2550.
43. Jones P.A., Gonzalgo M.L. Altered DNA methylation and genome instability. A new
pathway to cancer? // Proc. Natl. Acad. Sci. USA. 1997. Vol. 94. P. 2103-2105.
44. Loeb LA. Cancer cells exhibit a mutator phenotype //Adv. Cancer Res. 1998. Vol. 72.
P. 25-56.
ГЕНЕТИКА НАСЕЛЕНИЯ, ГЕНОГЕОГРАФИЯ
И ГЕНОМНАЯ МЕДИЦИНА
(к проблеме генофондов народонаселения)
В.П. Пузырев
НИИ медицинской генетики ТНЦ СО РАМН
634050, г Томск, Наб. р. Ушайки, 10
E-mail: vadimst@img.tsu.ru
Мы стоим у края необъятного моря. Тысячи раз-
личных драгоценных или вредных веществ - генов
растворены в этом море ... и море это волнуется. Не-
слышными взрывами ежеминутно взрываются в нем
мутации, даря нам новые ценности или отравляя это
море новыми ядами. Медленно расползаются диффу-
зионными процессами эти гены, захватывая все но-
вые и новые зоны. Сложными потоками переливают-
ся, смешиваясь и крутясь, разноцветные струи, рож-
дая новые комбинации генов, часто еще неведомые
человеку ... Имя этому морю - генофонд ... Познать,
понять и овладеть его взволнованной многосложной
жизнью - наша благородная задача!
А.С. Серебровский (1928)
Общественный интерес к успехам генетической науки огромен.
Достаточно вспомнить недавние обсуждения в средствах массовой
информации и дебаты в парламентах некоторых стран успехов в кло-
нировании животных (знаменитая овечка Долли) и перспектив клони-
рования человека. Первые клинические протоколы генотерапии не-
скольких десятков форм наследственных болезней с известными мо-
лекулярными дефектами, «открытие» генов широко распространенных
заболеваний (инфаркт миокарда, сахарный диабет, бронхиальная аст-
ма, шизофрения) и связанные с этим надежды на их первичную про-
филактику, полное секвенирование отдельных хромосом человека
(22-й и 21-й) и в ближайшее время завершение секвенирования всего
генома человека - все любопытно современному человеку.
342
V. Эволюция человека
Сегодня в центре общественного внимания - вопросы, относящи-
еся к проблеме сохранения и развития генофондов человеческих по-
пуляций. Однако термин «генофонд» по-разному воспринимается «обы-
денным» сознанием и фундаментальной наукой.
1. Семантический феномен понятия «генофонд»
Для обыденного сознания характерны категоричность суждений и им-
ператив рекомендаций по управлению состоянием генофондов популя-
ций человека, возвращающих нас во времена расцвета евгенического дви-
жения в нелучших рекомендательных вариантах. Кризисные явления в
обществе (депопуляция, деградация населения, деакцелерация) решитель-
но связываются с процессами, происходящими в генофонде, укрепляя
мысли о его вырождении и разрушении. Эти «де...» в обозначении попу-
ляционных характеристик населения в данном контексте представляют-
ся конгломератом значений понятия «генофонд», которые варьируют в
зависимости от политической конъюнктуры, различных предлагаемых
социальных программ или уже в ходе конкретных действий.
Для научного освещения сути понятия «генофонд» характерны ос-
торожность и осмотрительность, особенно, если это касается вопро-
сов прогноза направлений эволюции генофондов; констатация факта
недостаточности знаний о закономерностях генетических процессов
в народонаселении. На заре предложенного А.С. Серебровским тер-
мина «генофонд» (20-30-е годы и позднее) отечественные генетики
уточняли смысловое его содержание, отраженное в синонимах: гене-
тическая структура популяции (Тимофеев-Ресовский, 1927), запас (ре-
зерв) наследственной изменчивости (Четвериков, 1929), генетическое
строение населения вида (Балкашина, Ромашов, 1935), фонд наслед-
ственной изменчивости (Ромашов, 1935), резерв мобилизационной
изменчивости вида (Шмальгаузен, 1946), генный пул (Добржанский,
1955), аллелофонд (Тимофеев-Ресовский, 1973) [3].
Семантический феномен и заключается в том, что сложилась ситуа-
ция «двойного кодирования» термина «генофонд»: с одной стороны, он
эксплуатируется в тех случаях, когда необходимо усилить кризисность,
катастрофичность возникших новых явлений в природе (экология) или в
общественной жизни (перестройка сложившегося стиля жизни), предоп-
343
В.П. Пузырев
ределяя неизбежность негативных последствий для наследственности
живущих и будущих поколений; с другой - термин применяется в строго
научно определенном значении, а генофонд является предметом иссле-
дования специалистов. А это означает, что устанавливаемые взаимосвя-
зи между изменениями в природной и социальной среде обитания чело-
века с происходящими изменениями в генофонде народонаселения тре-
буют тщательного доказательства, оценки их значительности и
определения характера динамики генофонда во времени. Ведь без дока-
зательства декларируемых взаимосвязей невозможен прогноз.
2. Определение терминов «генофонд»
и «геногеография», состояние проблемы
Эти термины и понятия были введены А.С. Серебровским [18]. Он
писал: «Совокупность всех генов данного вида животных ... я назвал
генофондом, чтобы подчеркнуть мысль о том, что в лице генофонда
мы имеем такое же национальное богатство, как в лице запасов не-
фти, запасов золота, угля, скрытых в наших недрах». И далее: «Совер-
шенно так же мы должны говорить и о генофонде применительно к
человеку, и анализ происходящих в человеческом генофонде течений
и диффузий был бы, конечно, очень важным исследованием».
Геногеография по А.С. Серебровскому - это ветвь генетики, свя-
зывающая ряд генетических проблем с географией, изучающая рас-
пространение генов в данном пространстве. Геногеография домаш-
них животных (А.С. Серебровский), культурных растений (Н.И. Ва-
вилов) стали теоретической основой становления геногеографии
народонаселения, формировавшейся по мере «открытия» генов че-
ловека.
Дальнейшему развитию концепции генофонда и геногеографии
способствовали исследования Ю.Г. Рычкова и его учеников. Справед-
ливо замечено, что «понимание генофонда как фундаментальной про-
блемы популяционного уровня жизни сохранилось лишь в отечествен-
ной науке ...» [17], а идея А.С. Серебровского о том, что «современ-
ная география генов является результатом длительного исторического
процесса», по праву считается достоянием отечественной школы по-
пуляционной генетики человека [1].
344
V. Эволюция человека
Ю.Г. Рычков [16] основу связи генетики народонаселения с демог-
рафией и общественно-историческим процессом видит в процессе
воспроизводства, имеющего у человека двойственную (биологичес-
кую и общественную) природу, а также иерархичность данного про-
цесса в плане генетики (табл. 1).
Таблица 1
Иерархия процесса воспроизводства
Уровни организации Характер воспроизводства генетического материала
Молекулярный Воспроизводство генов (репликация)
Организменный Воспроизводство генотипов (индивидуальных сочетаний генов). Наследование от родителей к детям (степень родства членов семьи)
Популяционный Воспроизводство генофондов (совокупностей генов в группах населения). Наследование от родительских поколений к поколениям потомков (степень общности происхождения популяций)
Народонаселение (ндщтопуляпионная структура) Воспроизводство генетического разнообразия (исторически формируемая структура народонаселения)
Общие свойства генофондов народонаселения, выделяемые шко-
лой Ю.Г. Рычкова, могут быть суммированы в следующих положени-
ях [4, 17]:
- генофонд представляет не хаотическую массу генов, а стратифици-
рованную систему изменчивости генных частот (архитектонику), содер-
жащую информацию о многотысячелетних этапах развития генофонда;
- принцип эргодичности в понимании геногеографии как истории
генофонда: География как История в пространстве, а История как Гео-
графия во времени;
- изменчивость генных частот, связанная с селективной значимос-
тью генов, отражает адаптивные свойства генофонда и безграничную
генетически обусловленную множественность состояния здоровья;
- генофонд есть ключевой из природных ресурсов общества (раз-
витие производительных сил общества, проблемы социально-эконо-
мических преобразований, генетическое здоровье населения).
Так сложилось определение генофонда, конкретизированное при-
менительно к генофонду народонаселения. «Генофонд есть истори-
чески упорядоченная совокупность генов (и в целом любых реплици-
рующихся участков ДНК), свойственная народонаселению, удержи-
345
В.П. Пузырев
ваемая его самосознанием на протяжении поколений в границах, на-
ходящаяся в динамическом равновесии с состоянием вмещающей на-
селение и изменяемой им среды» [17].
Многие ключевые составляющие этого определения, стилистичес-
кая изящность и емкость, «вмещающая» многосложность феномена
Человека, перекликаются с определением термина «этнос», которое
дает выдающийся отечественный этнограф, географ, историк, фило-
соф, автор глобальной концепции формирования и развития этносов
в их неразрывной связи с природой Л.Н. Гумилев, в исследованиях
которого проблема человеческого генофонда всегда занимала замет-
ное место (известны его дискуссии по этому вопросу с Н.В. Тимофе-
евым-Ресовским). По Л.Н. Гумилеву (1991), « ... общечеловеческая
культура, одинаковая для всех народов, невозможна, поскольку все
этносы имеют разный вмещающий ландшафт и различное прошлое,
формирующее настоящее как во времени, так и в пространстве. Куль-
тура каждого этноса своеобразна, и именно эта мозаичность челове-
чества как вида придает ему пластичность, благодаря которой вид
Homo Sapiens выжил на планете Земля. Итак, этническая пестрота
это оптимальная форма существования человечества, хотя политичес-
кое объединение различных этносов обладает определенной устой-
чивостью во времени» [8].
Вклад в понимание проблемы генофонда населения этих трех вы-
дающихся отечественных ученых - А.С. Серебровского, Ю.Г. Рычко-
ва, Л.Н. Гумилева (рис. 1), обладающих редким даром единства энцик-
лопедических знаний, оригинальности концептуального мышления и
таланта увлекательного изложения своих идей, огромен и является
достоянием российской научной мысли.
Во всех случаях оценок состояния генофондов популяций человека в
прошлом и настоящем для прогноза их развития необходим анализ ос-
новных факторов эволюции - факторов динамики генофондов (мутации
- отбор генов, миграции, структура браков, стохастические процессы -
дрейф генов). Многие из них внимательно анализировались А.С. Сереб-
ровским [1]. Значительная роль в разработке генетико-математических
моделей, которые позволяют изучать и анализировать популяционно-ге-
нетические процессы, принадлежит «великому триумвирату популяци-
онной генетики» - Р Фишеру, Дж. Холдейну и С. Райту (рис. 2).
346
V. Эволюция человека
А.С. Серебровский
(1892 - 1948)
Ю.Г. Рычков
(1932- 1998)
Л.Н. Гумилев
(1911-1992)
Рис. 1. Российские основоположники концепций генофонда и этногенеза
Р Фишер
(1890 - 1962)
Дж. Холдейн
(1892- 1964)
С Райт
(1889- 1981)
Рис 2 Великий триумвират тюпуляционнной генетики
И Эволюция человека
3. Эволюционные последствия общественно-исторических
событий и круг нашего незнания в проблеме генофонда
Известный отечественный биолог-эволюционист Н.Н. Воронцов в
одной из своих публицистических статей [7] обсуждает чрезвычайной
важности вопрос - каковы эволюционные последствия войны, рево-
люции, застоя? Звучит в статье и ответ-намек: «Говорят о том, что в
результате репрессий был подорван генофонд нашего народа». Автор,
обозначающий периодом русской революции в широком смысле сло-
ва 1905-1955 гг., оценивает «урон», который понес «генофонд нашей
страны» от революции в 8 млн, Первой мировой войны - около 5 млн
и Второй мировой войны -от 18 до 27 млн человек.
Дополним, что недавние исследования по репрессивным принуди-
тельным миграциям [14] свидетельствуют о суммарном объеме депор-
тированного советского населения в период 20-50-х годов в 15 млн
человек: внутренние принудительные миграции затронули 6 млн, а
международные для гражданских лиц - около 9 млн человек (из СССР
- остарбайтеры 1941-1944 гг.; в СССР - репатрианты 1944-1952 гг. и
вестарбайтеры 1945 г.). Конечно же, департация населения привела к
вынужденному прерыванию мирного эволюционного процесса, и это
обостряет актуальность решения вопроса о судьбе генофонда.
Н.Н. Воронцов рассматривает еще один аспект - эволюционные
последствия политики 60-70-х годов нивелировки в образовании и
воспитании (он называет это «концепцией идентизма»), не учитыва-
ющей основной эволюционной особенности - генетической неодно-
родности популяций. Эта составляющая нашего общественного раз-
вития, наряду с вышеизложенными последствиями «революции в Рос-
сии 1905-1955 гг.», по мнению автора, сформировала, с одной
стороны, «людей с наиболее активным темпераментом, с наиболее
развитым чувством чести, с наибольшей ответственностью, с чувством
альтруизма», а с другой - «происходил искусственный отбор молчу-
нов, середняков, посредственностей». Несмотря на, казалось бы, ут-
вердительный ответ о присутствии связи негативных общественно-
исторических событий с процессами, происходящими в генофонде,
автор справедливо в итоге отмечает: «Мы все еще крайне далеки от
полного понимания того, к каким эволюционным последствиям при-
вели войны, революции и застойный период».
349
В.П. Пузырев
Вероятно, пришла пора в проблеме генофонда очертить круг на-
шего незнания, хотя сделать это нелегко - «мыслить известное» (Ге-
гель) всегда трудно. Проблема генофонда вытекает не только из от-
дельных частных научных дисциплин, но и из современных глобаль-
ных тенденций, порой негативных, развития человечества, приобретая
мировоззренческую окраску.
Один из вопросов - значение естественного отбора в формирова-
нии свойств народонаселения и запечатленности их в генофонде, т.е.
существовании «следа биологических факторов». Известна на этот счет
точка зрения В.П. Эфроимсона: «Надо вообще оставить мысль, что
естественный отбор за последние два тысячелетия существенно осла-
бел. Он только изменил свое направление и на него наслоился отбор
социальный, требующий самого пристального внимания» [20].
А.А. Любищев [11], внимательно проанализировавший взгляды
В.П. Эфроимсона на «войну и естественный отбор», «эволюционные
ограничения жестокости», «эволюционно-генетическое происхожде-
ние эстетических эмоций», отметив его широту и свободомыслие в
постановке вопросов, отличное от «густопсовых» ортодоксальных дар-
винистов, все-таки заметил, что «мы, несмотря на очень дружеские
отношения, принадлежим к разным лагерям ...». Он пишет: «Эта иде-
ологическая наследственность, совершенно не связанная с хромосо-
мами, играет у человека колоссальную роль, в частности в выработке
так называемого национального характера и длительных традиций
отдельных государств». И далее: «Идеологическая наследственность
играет ведущую роль в этике на всех уровнях: от индивидуального до
государственного, роль среды только как стимулятор, генетическая же
хромосомная наследственность играет совершенно ничтожную роль,
... что в первом приближении может полностью игнорироваться». Но
сейчас, похоже, селекционистские воззрения являются доминирующи-
ми в обсуждении проблем «генофонд и здоровье будущих поколений»,
«общественно-политический кризис - состояние генофонда - феноде-
виантное развитие общества в будущем» и прочее.
Другая область нашего незнания касается характера (динамики,
темпа, направления) изменений в состоянии генофонда как совокуп-
ности генов, следующих за изменениями в природной и социальной
среде. Анализ этих изменений сегодня основывается чаще на линей-
ных закономерностях эволюционного процесса, однако накапливают-
350
V. Эволюция человека
ся данные, что они могут носить нелинейный характер, а «синергети-
ческие» концепции, рожденные изучением физико-химических про-
цессов, проникают в методологию анализа популяционных, популя-
ционно-генетических и социальных явлений [10, 13].
Наконец, реальная угроза благополучию человеческого рода в бли-
жайшем будущем вследствие сегодняшних глобальных экологических и
демографических проблем вновь поднимает вопросы «евгенического
значения» (термин Н.И. Кольцова) общественно-исторических событий,
лечебно-профилактических мероприятий, отбора «лучших» в надежде
«подвинуть» существующий генофонд к оптимальному уровню. По это-
му поводу есть позиция, высказанная И. А. Захаровым [9]:«... проблема
биологической эволюции человека, протекающей в течение многих по-
колений и десятков и сотен тысяч лет, не может считаться сколько-ни-
будь актуальной, а возможность управления этой эволюцией не является
насущной научной задачей». Эта точка зрения не является бесспорной.
4. Евгеника - эволюционный абсурд?
С таким названием недавно опубликована небольшая работа в
Nature Genetics [23], в которой авторы дают утвердительный ответ на
обозначенный вопрос. Почему? Во-первых, отмечают они, без оши-
бок в репликации геномов и «продукции» генетических вариантов у
индивидов популяции не способны реагировать на новые средовые
«вызовы» (включая инфекции), ибо отсутствует основа для адаптации.
Во-вторых, по их мнению, эволюция «непредусмотрительна» (по
foresight) и в отношении Homo sapiens, для которого жизненное про-
странство, социальные и средовые условия коренным образом меня-
ются даже на протяжении нескольких поколений. Поэтому «искорене-
ние» признаков, рассматриваемых сегодня как нежелательные, может
быть напрасным, если не опасным (злополучным), в попытке «улуч-
шить» человеческую расу. Таких примеров преимущества гетерози-
готных носителей мутантных генов тяжелых рецессивных заболева-
ний перед появлением новых факторов среды достаточно много. На-
конец, было бы не эволюционно человеку самому конструировать в
пространстве обитания «горлышко бутылки», способствуя формиро-
ванию генетического единообразия и преждевременному вымиранию.
351
В.П. Пузырев
Эти соображения об эволюционной абсурдности широкого распрос-
транения евгенических взглядов не новы, если вспомнить обсуждение
проблемы «польза-вред» в блестящей работе А.С. Серебровского «Не-
которые проблемы органической эволюции» [19]. Однако коренные ос-
новы евгеники, заложенные в прошлом веке Ф. Гальтоном и независимо
от него в России (в одно и то же время - 1865 г.) В.М. Флоринским
(рис. 3), развиваемые и подтвержденные многочисленной плеядой из-
вестных биологов и медиков, среди которых лауреат Нобелевской пре-
мии Г. Меллер [12], так выпестовали формально-логическую структу-
ру евгенического значения предлагаемых социальных и медицинских
мероприятий и обросли громадой «фактов и наблюдений», что крити-
ческое отношение к евгенике, высказываемое перед широкой обще-
ственной аудиторией, редко находит поддержку. И этот психологичес-
кий момент удачно подмечен поэтом:
«К чему бесплодно спорить с веком?
Обычай - деспот меж людей».
(А.С. Пушкин)
Сложность ситуации состоит в том, что идейные основы евгеники
отдельными моментами вошли в некоторые научные направления: в
Британии евгеника дала основу математической генетике популяций,
в России - генетике человека, медицинской генетике и эксперимен-
тальной генетике популяций, во Франции - основу «пуэркультуре».
заботе о младенчестве и материнстве [2].
5. Эволюционно-генетический взгляд на геномную медицину
Недавно (1998 г.) появился термин “геномная медицина” - меди-
цина, которая предполагает рутинное использование генотипического
анализа, обычно в форме ДНК-тестирования, с целью улучшения ка-
чества медицинской помощи [21]. На новом этапе структурной гено-
мики каждый индивид и члены семьи могут быть прогенотипированы
одновременно по большому числу генов (аллелей, полиморфизмов).
Сегодня действительно формируется реальная основа для индивидуа-
лизированной медицины как осуществленного единения социальной
медицины и геномной (рис. 4) [21]. Более того, в практике решения
повседневных вопросов генетического здоровья становится доступ-
ной профилактика наследственной патологии (табл. 2) [5, 6].
352
И Эволюция человека
Ф. Гальтон
(1822-1911)
В.М. Флоринский
(1834-1899)
Рис. 3
Рис. 4. Единство трех разделов медицины
В.П. Пузырев
V. Эволюция человека
Таблица 2
Основные подходы к профилактике наследственной патологии
1. Управление экспрессией генов ("евфеника", по Н.К. Кольцову):
• периконцепционная профилактика
• перинатальная профилактика__________________________________
2. Элиминация эмбрионов и плодов с наследственной патологией:
• пренатальная диагностика
• прерывание беременности ____________________________________
3. Генная инженерия на уровне зародышевых клеток______________
4. Планирование семьи:
• отказ от деторождения при риске > 20%
• отказ от родственных браков
• отказ от браков гетерозиготных носителей
♦ окончание деторождения до 30-35 лет_________________________
5. Охрана окружающей среды:
• исключение мутагенных факторов из среды обитания
• исключение факторов, вызывающих
экогенетические патологические реакции
Ближайшая перспектива ДНК-тестирования по 10-100 тыс. генов и
их полиморфизмов/мугаций (технология биочипов) неизбежно соблаз-
няет оценить те или иные варианты гена или генные ансамбли как благо-
приятные (полезные, выгодные) или неблагоприятные (нежелательные,
негативные, рисковые) в отношении здоровья (болезни) индивидов и
членов их семей. И это действительно приходится делать при медико-
генетическом консультировании пациентов, хотя последние самостоятель-
но принимают решение о своем поведении (репродуктивном, реабили-
тационном и т.д.). Но с эволюционной точки зрения важно иметь в виду,
что наши оценки полезности (вредности) аллелей (генов) относятся толь-
ко к настоящему времени. Эволюционные аргументы положения о том,
что сохранение человеческого разнообразия «иммунизирует» нас от не-
предсказуемости будущего разнообразия нашей среды (природной и со-
циальной) обитания, должны убедить общество в необходимости забот-
ливого отношения к генетическому несовершенству индивида.
Заключение
Справедливо ли утверждать, что тревога о вырождении генофонда
(российского народа) в условиях кризиса, переживаемого обществом,
355
В.П. Пузырев
- патриотическое пустословие, а факты разрушения генофонда - пе-
строе собрание баек? Конечно, нет. Но, с другой стороны, следует
признать недостаточность наших знаний о динамике генофондов, о
взаимосвязи изменений в генофонде, в биологических характеристи-
ках населения с общественно-историческими событиями и меняю-
щейся природной средой обитания. Геногеография как один из наи-
более эффективных подходов в исследовании этих вопросов совсем
недавно из дескриптивных приобрела аналитические возможности
(современные статистические модели анализа, вовлечение в анализ
лингвистических, климато-географических, демографических, этно-
исторических, медицинских характеристик народонаселения, картог-
рафический анализ), дополнена огромным исследовательским ресур-
сом благодаря успехам молекулярной генетики, давшим возможность
работать со всей совокупностью генов, описываемых в структуре с
точностью до одного нуклеотида. Популяционная генетика стала ин-
теллектуальной основой понимания генофенотипических взаимосвязей,
закономерностей формирования конечного фенотипа индивида, опреде-
ления «сферы компетенции» отдельных генов и генных ансамблей.
Эволюционно-генетический ракурс рассмотрения проблем клини-
ческой медицины, вооруженной теперь генными технологиями (ге-
номная медицина), позволяет предполагать, что в интересах судьбы
генофонда народонаселения активность медико-генетического кон-
сультирования в обществе и возможные манипуляции с «болезнен-
ными» генами должны быть дифференцированными. Для подавляю-
щего большинства менделирующих форм патологии, особенно сни-
жающих качество жизни пациентов, индивидуально-семейный
уровень генетического консультирования вполне обоснован. Харак-
тер рекомендаций по репродуктивному поведению фенотипически здо-
ровых носителей мутантных генов должен быть синхронизирован с
высокой точностью методов пренатальной диагностики. Глобально-
евгенические рекомендации, распространяемые на популяции, осо-
бенно в отношении политенных форм патологии (широко распрост-
раненных заболеваний), в силу невысокой до сих пор надежности
обнаруживаемых ассоциаций генов и их комбинаций с клиническими
проявлениями болезней, мало обоснованы.
356
V. Эволюция человека
Литература
1. Алтухов Ю.П. И Генетика, 1992. Т. 28. № 1. С. 8-19.
2. Бабков В.В. И ВИЕТ. 1997. № 1. С. 76-94
3. Бабков В.В. Московская школа эволюционной генетики. М.: Наука, 1985. 216 с.
4. Балановская Е.В., Рычков Ю.Г. Геногеография (Гены человека на карте СССР).
М.: Знание, 1990. 64 с.
5. Бочков Н.П. Клиническая генетика. М.: Медицина, 1997. 288 с.
6. Бочков Н.П. // Вестн. РАМН. 1999. № 10. С. 3-5.
7. Воронцов Н. //Знамя. 1995. № 7. С. 176-181.
8. Гумилев Л.Н. И Наше наследие. 1991. № 3. С. 19-26.
9. Захаров И.А. И ВИЕТ. 1997. № 1. С. 65-67.
10. Кучер А.Н., Пузырев В.П., Санчат А.О., Эрдыниева Л.С. // Генетика. 1999. Т. 35.
№ 5. С. 688-694.
11. Любищев А.А. И Эфроимсон В.П. Генетика этики и эстетики. СПб.: Талисман, 1995.
С. 255-278.
12. Меллер Г. (Публикация письма к И.В. Сталину) И ВЕИТ. 1997. № 1. С. 68-76.
13. Моисеев Н.Н. Алгоритмы развития. М.: Наука, 1987. 304 с.
14. Полян П.М. И Природа. 2000. № 6. С. 3-12.
15. Пузырев В.П. И Вестн. РАМН. 2000. № 7. С. 28-33
16. Рычков Ю.Г. И Система знаний о народонаселении / Ред. Д.И. Валентей.
М.: Высш, шк., 1991. С. 210-222.
17. Рычков Ю.Г., Балановская Е.В. И Генетика. 1992. Т. 28. № 1. С. 52-75.
18. Серебровский А.С. И Научное слово. 1928. № 9. С. 22.
19. Серебровский А.С. Некоторые проблемы органической эволюции. М.: Наука, 1973.
168 с.
20. Эфроимсон В.П. Генетика этики и эстетики. СПб.: Талисман. 1995. 288 с.
21. Beaudet A.L. //Am. J. Hum. Genet. 1999. Vol. 64. P. 1-13.
22. Bloom B.R. //Nature. 1999. Vol. 402. Supp. 2. P. 63-64
23. Brosius J., Kreitman M. //Nat. Genet. 2000. Vol. 25. P. 253.
В ПОИСКАХ «АДАМА»: ЭВОЛЮЦИЯ
ГАПЛОТИПОВ Y-ХРОМОСОМЫ
В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА
В.А. Степанов, В.П. Пузырев
НИИ медицинской генетики ТНЦ СО РАМН
634050, г. Томск, Наб. р. Ушайки, 10
E-mail vadimst@img.tsu.ru
Вследствие того, что в каждом поколении часть линий передается
следующему поколению, а часть - нет, через определенное число поко-
лений все ныне существующие варианты Y-хромосомы можно свести к
одной исходной. Носителя или носителей этого исходного варианта и
можно условно назвать «Адамом» - именно от него мы все и ведем свое
происхождение. Аналогичная история и с мтДНК - точно так же все со-
временные линии можно свести к одной первоначальной, существовав-
шей некоторое число поколений назад. Носительницу или носительниц
этой линии также условно можно назвать «Евой».
Попробуем доказать это положение. Для этого нам придется сде-
лать несколько допущений. Предположим, что некоторое время назад,
скажем, 200 тысяч лет назад, все существующие тогда люди составля-
ли одну популяцию. Допустим, что эффективная численность такой
популяции была относительно небольшой - 10 тысяч человек - кста-
ти, эта оценка численности, по-видимому, недалека от истины. Допус-
тим также, что все члены этой популяции генетически отличались друг
от друга благодаря ранее накопленному разнообразию, в том числе по
Y-хромосоме. То есть в некий исходный момент существовало, услов-
но говоря, пять тысяч вариантов Y-хромосомы.
И еще допустим, что в течение длительного времени эффективная
численность популяции не изменялась (во всяком случае, не возраста-
ла), что также соответствует действительному положению вещей. Это
значит, что у каждой пары родителей в среднем только двое потомков
достигало репродуктивного возраста. При равном соотношении по-
лов в четверти семей должны были выживать два мальчика, в четверти
358
V. Эволюция человека
- две девочки и в половине - мальчик и девочка. В тех семьях, где
рождались или выживали только девочки, линия, или клон, Y-хромо-
сомы прерывалась в первом же поколении. Те семьи, где рождались
мальчик и девочка, передавали свой клон в следующее поколение в
той же доле, что и в предыдущем. Ну а семьи, где рождались или вы-
живали два мальчика, соответственно увеличивали долю своего клона
в следующем поколении. Тем самым вероятность утраты конкретной
линии Y-хромосомы в каждой конкретной семье в среднем равна 0,25.
То есть уже в первом поколении должно утрачиваться четверть всего
числа линий Y-хромосомы. Теперь представим себе, что за эти 200 ты-
сяч лет сменилось примерно 10000 поколений. Вероятность сохране-
ния для каждого исходного клона при этом практически равна нулю.
Конечно, реально средний темп исчезновения клонов был гораздо ниже,
чем четверть за поколение, из-за того, что некоторые клоны «расши-
рялись» и «сужались». Однако в какие-то моменты, когда предковая
популяция проходила через «горлышко бутылки» - резкое уменьше-
ние численности - темп, в силу эффектов генетического дрейфа, был
существенно выше.
Разумеется, не всегда резкое уменьшение численности было связа-
но с гибелью большей части членов популяции - чаще всего это была
просто изоляция части популяции, в результате которой в дальнейшем
антропогенезе участвовала только эта первоначально небольшая «от-
коловшаяся» популяция. Тем не менее, простые статистические моде-
ли показывают, что даже за гораздо меньшее время, чем 200 тысяч
лет, с вероятностью, практически равной единице, исчезнут все кло-
ны, кроме одного. Здесь вполне уместна аналогия с лотереей: вероят-
ность выиграть ничтожна, тем не менее, она реализуется и кто-то один
все-таки получает главный приз. Именно этого счастливчика и можно
назвать «Адамом». Это вовсе не значит, что в те времена он жил один
на Земле. Возможно, носителей уцелевшей линии Y-хромосомы было
гораздо больше, чем один. И уж совершенно точно одновременно с
нашим «Адамом» жило множество других мужчин, чьи Y-хромосом-
ные гены до нас не дошли.
Совершенно аналогичные рассуждения можно применить и к
мтДНК и «митохондриальной Еве». И еще. От «Адама» и «Евы» мы
получили лишь небольшую часть нашего генома - Y-хромосому и
мтДНК соответственно, причем уже далеко не в исходном виде, а из-
359
В.А. Степанов, В.П. Пузырев
мененную многочисленными мутациями. Большую же часть генов мы
унаследовали не от них, а от других и очень многочисленных предков.
Приведенные рассуждения, конечно, упрощают реальную ситуа-
цию. Мы не знаем ни эффективной численности «исходной» популя-
ции, ни характера ее динамики, ни степени изоляции палеопопуляций,
не учитываем селективных различий между различными линиями
Y-хромосомы, а они, несомненно, есть.
Конечно, термины «Адам» и «Ева» в отношении наших предков не
научны. В научной же литературе их принято называть MRCA (most recent
common ancestor) - наименее древний общий предок. «Мисс» MRCA,
или «митохондриальная Ева» - т.е. наименее древняя наша общая линия
мтДНК - имеет возраст в районе 200 тысяч лет и ведет свое происхожде-
ние из Африки. Возраст этой линии был установлен на основании темпа
накопления точечных мутаций в мтДНК, причем эти «молекулярные
часы» были проградуированы путем сравнения последовательности
мтДНК человека и шимпанзе (место жительства было установлено пу-
тем анализа генеалогического древа линий мтДНК в современных попу-
ляциях). В вопросе о возрасте и происхождении «Y-хромосомного Ада-
ма» полной ясности пока нет. Тем не менее, наиболее вероятный возраст
MRCA по Y-хромосоме оценивается примерно в 190-270 тыс. лет, а про-
исходит эта линия Y-хромосомы также, вероятнее всего, из Африки.
В «Библии» упоминается трое детей Адама и Евы - Каин, Авель и
Сет. Современные же генетические подходы выявили 18 линий мтДНК,
ведущих начало от MRCA по женской линии и 10 линий Y-хромосомы,
восходящих к мужскому MRCA. Линии мтДНК, определяемые по рест-
рикционным полиморфизмам и последовательности гипервариабельно-
го района молекулы, обозначаются латинскими буквами, а линии Y-xpo-
мосомы, определяемые по примерно полутора сотням мутаций (в основ-
ном, SNP), нумеруются арабскими цифрами. Три наиболее древние линии
мтДНК найдены в Африке. Производные от них 6 линий обнаружены в
Азии, 9 линий - в Европе и 5 линий (4 азиатских и 1 европейская) - в
Новом Свете. Три древнейших линии Y-хромосомы также найдены в
Африке. Семь линий, производных от них, присутствуют в Азии. Все
европейские линии являются вариантами африканских и азиатских, а все
североамериканские - производными от азиатских линий.
Теперь давайте попробуем более подробно разобраться с тем, как
прослеживаются линии Y-хромосомы и какую информацию относи-
360
V. Эволюция человека
тельно эволюции генофонда в популяциях человека можно извлечь из
такого анализа.
Для начала рассмотрим структуру Y-хромосомы. Большая часть
Y-хромосомы нерекомбинантна, т.е. не имеет гомологичных участков
в Х-хромосоме. Однако небольшие дистальные участки обоих плеч,
называемые псевдоаутсомными регионами (PAR), имеют гомологич-
ные районы в Х-хромосоме. В среднем размер Y-хромосомы человека
- примерно 60 Mb, из них 30 Mb занято эухроматиновыми доменами,
а оставшаяся часть - это гетерохроматин, длина которого может силь-
но варьировать у разных индивидов.
На Y-хромосоме сейчас картировано относительно небольшое чис-
ло генов, около 25. Большая часть из них имеет отношение к процес-
сам сперматогенеза, однако есть гены, которые экспрессируются и в
других тканях, в том числе «вездесущие» факторы транскрипции и
трансляции. Детальное физическое картирование Y-хромосомы про-
ведено с помощью панели делеционных клеточных линий. Было со-
здано 24 линии клеток, в каждой из которых делегирован определен-
ный участок Y-xp.
Такие специфические свойства Y-хромосомы, как передача только
отцовской линии и отсутствие рекомбинации, позволяют прослежи-
вать линии гаплотипов, которые представляют собой «запись» мута-
ционных событий в предыдущих поколениях. В этом отношении
Y-хромосома сходна с мтДНК. Однако есть и отличия. О размерах я
уже говорил - Y-хромосома примерно в 4000 раз больше, чем молеку-
ла мтДНК, которая, по сути, представляет собой один локус. Еще од-
ной специфической чертой Y-хромосомы, важной для популяционно-
генетического анализа, является низкая эффективная численность ее
пула - в 4 раза меньше, чем для аутосом, - т.е. на каждую Y-хромосо-
му, передающуюся в следующее поколение, приходится 4 аутосомы и
3 Х-хромосомы. В связи с этим Y-хромосома в большей степени под-
вержена эффектам генетического дрейфа и характеризуется большей
степенью межпопуляционной вариабельности и географической кла-
стеризации по сравнению с аутосомными и митохондриальными мар-
керами.
Наконец, в отличие от мтДНК, где преобладают точечные мута-
ции, Y-хромосома содержит маркеры различной природы - точечные
мутации (SNP), инсерции, делеции, микро- и минисателлиты, что по-
361
В.А. Степанов, В.П. Пузырев
зволяет решать эволюционные задачи разного масштаба. Современ-
ные подходы к анализу линий Y-хромосомы основаны на использова-
нии компаундных гаплотипов, построенных на основе комбинации ди-
аллельных и микросателлитных локусов.
С помощью бинарных полиморфизмов можно построить эволю-
ционное древо, отражающее последовательность накопления мутаций.
Узлы на этом древе представляют собой гаплотип («бинарную» гап-
логруппу), который характеризуется определенным аллельным состо-
янием: в данном случае 12 бинарных локусов. Соседние гаплотипы
связаны мутациями в одном бинарном локусе. Рекуррентные, или по-
вторные, мутации в одном и том же локусе приводят к образованию
замкнутых контуров на древе. Более детальные взаимоотношения меж-
ду гаплотипами внутри «бинарной» гаплогруппы выявляются с помо-
щью микросателлитов, для которых характерен гораздо более высо-
кий темп мутирования, чем для диаллельных локусов.
Распределение гаплотипов Y-хромосомы, как мы уже отмеча-
ли, характеризуется высокой популяционной специфичностью. У
африканских негроидов преобладают определенные гаплотипы, у
европеоидов - другие, у азиатов - третьи, а у американских индей-
цев - четвертые.
Популяционная специфичность характерна и для хромосомных пе-
рестроек в гетерохроматиновом районе длинного плеча У-хромосо-
мы. В 1985 г. С.А. Назаренко и В.П. Пузырев опубликовали в журнале
Human Genetics статью об интересной находке, обнаруженной в попу-
ляции северных хантов. У хантов с очень высокой частотой (более 20%)
встречалась субтотальная делеция гетерохроматинового сегмента У-
хромосомы. Авторы справедливо объяснили сей факт механизмом ге-
нетического дрейфа в изолированной хантыйской популяции. Это была
первая работа по Y-хромосоме, выполненная в нашем институте.
Мы в нашей лаборатории занимаемся конструированием гаплотипов,
используя 11 диаллельных локусов (YAP, Tat, DYF155S, DYS199 и др.) и
семь микросателлитов (DYS389 - DYS394). Задача, которую мы решаем
с помощью гаплотипов Y-хромосомы, а также с помощью и других сис-
тем генетических маркеров, о которых здесь речь идти не будет, - гене-
тическая реконструкция процесса заселения территории севера Евразии
современным человеком и выявление генетических взаимоотношений
между этническими группами, проживающими на этой территории.
362
V. Эволюция человека
Появление человека современного типа в Сибири датируется сред-
ним палеолитом. В последующие периоды Северная Евразия была
ареной многочисленных исторических миграций, включая перемеще-
ние европеоидного населения с запада Евразии на восток и более по-
здние миграционные волны монголоидов с востока на запад. Началь-
ные этапы этногенеза современных этносов, проживающих на терри-
тории Северной Евразии, этногрофы и антропологи датируют недавним
в историческом масштабе временем - 2-3 тыс. лет назад. Преемствен-
ность же генофонда должна просматриваться на гораздо более протя-
женных временных отрезках. Несмотря на обилие антропологичес-
ких, лингвистических и этнографических сведений об этносах Север-
ной Евразии, вопросы их генетических взаимоотношений (т.е. родства
и происхождения), вклада различных предковых групп в их генофонд,
датировки различных этногенетических событий, времени происхож-
дения их генофондов остаются неясными.
Первый срез, который мы сделали в нашей работе, - анализ рас-
пределения гаплотипов, построенных с помощью четырех диаллель-
ных полиморфизмов. Одним из наиболее интересных диаллельных
маркеров является транзиция Т-С в локусе Tat. Нас он заинтересовал
в связи с взаимоотношениями между алтайскими и финно-угорскими
народами и в связи с этногенезом русских. Мутантный и более редкий
аллель С встречается только на территории Северной Евразии, при-
чем распространен довольно широко у представителей алтайской и
уральской языковых семей. Встречается он и у восточных славян - с
частотой порядка 20% у русских и около 10% - у украинцев. Интерес-
но, что эволюционный предшественник линии YAP-/DYF-/TatC - гап-
лотип YAP-/DYF-/TatT - найден нами только у тувинцев и алтайцев и,
с небольшой частотой - у киргизов и узбеков. В литературе высказано
несколько гипотез происхождения ТатС. Одна из них связывает его с
древними континентальными монголоидами и территорией современ-
ной Монголии. Согласно другой гипотезе, ТатС возник у предков фин-
но-угорских народностей и распространился по территории Евразии
вследствие этнических контактов и миграций палеопопуляций. Отно-
сительно высокая частота аллеля С у русских может быть объяснена
при этом наличием финно-угорской и/или тюркской примеси. Русские
же получили хромосомы с аллелем TatC в результате ассимиляции
финно-угорских племен 1000-1200 лет назад. В те времена в Волго-
363
В.А. Степанов, В.П. Пузырев
Окском междуречье, куда мигрировали восточные славяне и где сло-
жилось ядро русского этноса, обитали многочисленые финно-угорс-
кие племена - ведь, мещера, меря и др., которые были ассимилирова-
ны русскими. Ну а собственно частота аллеля С у русских может кос-
венно свидетельствовать о доле финно-угорской крови в этом этносе
- по крайней мере, в «мужской» части его генофонда.
С учетом анализа дисперсии микросателлитных гаплотипов и оцен-
ки по ней возраста происхождения ТатС, на чем мы остановимся ниже,
наиболее вероятным сценарием распространения этой линии нам пред-
ставляется следующий. Транзиция ТатС произошла в популяции, пред-
ковой как по отношению к алтайцам, так и к финно-уграм, а затем в
результате эффектов дрейфа и миграций получила наблюдаемое ныне
относительно широкое распространение на севере Евразии.
Еще один интересный маркер - полиморфная инсерция мобильного
генетического элемента Alu (YAP). Мутантный аллель YAP+ с высокой
частотой представлен у африканских негроидов. На территории Евразии
он почти не встречается. До недавнего времени было известно лишь два
исключения - японцы и тибетцы, где частота YAP+ находится в пределах
3(М0%. Недавно мы обнаружили третью популяцию с высокой (около
20%) частотой YAP+ - бурят. Локус YAP в последнее время привлекал
особое внимание в связи с проблемой заселения Японского архипелага.
Общепризнано, что заселение Японии происходило как минимум двумя
миграционными волнами из континентальной Азии. Если со второй, бо-
лее поздней волной, которая датируется периодом 2300 лет назад, все
относительно ясно - эта волна, называемая культурой Яёй, шла с Корей-
ского полуострова, являясь одной из заключительных стадий миграций
древнего человека через Юго-Восточную Азию. Первая более древняя
волна (культура Йомон) имеет датировку 10000 лет назад. Существует
две гипотезы происхождения культуры йомон. Одна из них связывает
эту культуру с населением Юго-Восточной Азии, другая считает ее ис-
точником территорию Северо-Восточной Евразии. Считается, что пря-
мыми потомками йомон являются айны и жители островов Рюкю на юге
Японского архипелага. И у тех и у других была выявлена высокая часто-
та аллеля YAP, в то время как на территории Юго-Восточной Азии этот
аллель отсутствует. Позднее находка YAP+ в Тибете, как и наши данные
о частоте инсерции Alu-элемента у бурят, явно свидетельствуют в пользу
североазиатского происхождения йомон. Правда, какие-то более или ме-
364
V. Эволюция человека
нее обоснованные интерпретации, связывающие население Тибета, Бу-
рятии и Японии, пока невозможны в силу того, что нет данных о микро-
сателлитных гаплотипах у японцев и тибетцев.
Теперь давайте более подробно рассмотрим подходы к анализу
микросателлитных гаплотипов Y-хромосомы. В нашей лаборатории
для анализа микросателлитов (или STR) мы используем технологию
капиллярного электрофореза флюоресцентно меченых продуктов ПЦР,
что позволяет одновременно (в одном образце) анализировать все 7 ло-
кусов,составляющих микросателлитный гаплотип. Полученные таким
образом гаплотипы затем по принципу парсимонии (наименьшего чис-
ла мутационных шагов) объединяются в кластер, или древо.
Кластерный анализ выявил, что в большинстве популяций присут-
ствует более чем одна линия гаплотипов. В большинстве случаев «би-
нарные» гаплогруппы четко обособлены и на уровне микросателлит-
ных гаплотипов. Лишь у тувинцев наблюдается объединение в рамках
одного микросателлитного древа гаплотипов, принадлежащих разным,
но родственным бинарным гаплогруппам. По-видимому, это древо,
объединяющее линии DYF+/TatT, DYF-/TatT и DYF-/TatC, является
предковым для двух последних линий. У бурят три линии строго обо-
соблены, в том числе и линия YAP+/DYF-/TatC. Такая же картина на-
блюдается и у восточных славян, эвенков и алтайцев. У таджиков и
узбеков наблюдаются две разветвленных линии Y-xp. У киргизов же
практически все гаплотипы объединяются в одну очень протяженную,
а следовательно, древнюю линию.
«Разветвленность» древа гаплотипов, а точнее дисперсия микро-
сателлитов внутри этого древа, позволяет вычислять оценки возраста
линий Y-хромосомы в популяциях. При этом под «возрастом» пони-
мается время, за которое было генерировано наблюдаемое ныне раз-
нообразие. Для оценки возраста мы использовали такие показатели,
как число мутаций от предкового гаплотипа на локус на индивида в
пределах конкретного древа (лямбда), а также кумулятивные оценки
темпа мутирования Y-сцепленных микросателлитов. Наиболее древ-
ние линии присутствуют в популяциях Средней Азии - их возраст вос-
ходит к эпохе позднего палеолита и варьирует от 6500 до 12000 лет.
Мужской генофонд восточных славян чуть моложе. В исследованных
нами коренных этносах Сибири наиболее древние линии представле-
ны у тюркоязычных народов - алтайцев и тувинцев. Монголоязычные
365
В.А. Степанов, В.П. Пузырев
буряты и эвенки, принадлежащие к тунгусо-маньчжурской группе, ха-
рактеризуются относительной «молодостью» компонентов своего муж-
ского генофонда. Разумеется, эти расчеты в некоторой степени услов-
ны, тем не менее, они позволяют составить представление об относи-
тельном времени генерации генетического разнообразия в исследо-
ванных этнических группах и совершенно однозначно свидетельству-
ют, что преемственность генофонда прослеживается с времен, гораз-
до более ранних, чем выделяемые этнографами периоды этногенеза
того или иного этноса.
Всего мы наблюдали около 200 различных микросателлитных гапло-
типов. Интересно, что большую часть из них - 90% - можно объединить
в единое древо, что свидетельствует об общности (общем происхожде-
нии) пула Y-хромосом у современного населения Северной Евразии.
«Возраст» предкового гаплотипа для этого древа составляет 16000 лет с
95% доверительным интервалом от 9 до 30 тыс. лет, что находится в хо-
рошем соответствии с палеоантропологическими данными.
Интерес представляет и степень генетической дифференциации на-
селения Северной Евразии по гаплотипам Y-xp. Оценку генетической
дифференциации популяций проводили методом молекулярной диспер-
сии (AMOVA) с помощью пакета ARLEQUIN. Генетическая дифферен-
циация популяций Северной Евразии оценивалась нами на трех уровнях
иерархической структуры: подразделенность популяций внутри одного
этноса, дифференциация этнических групп внутри алтайской и индоев-
ропейской языковых семей и генетические различия между семьями.
В трех из четырех этнических групп, представленных более чем одной
популяциями (тувинцы, эвенки, киргизы), наблюдался относительно низ-
кий и недостоверный уровень генетической подразделенности по гапло-
типам Y-хромосомы. Лишь для бурятских популяций выявлена значи-
тельная дифференциация: около 35% генетического разнообразия обус-
ловлено межпопуляционными различиями (Р=0,000).
На уровне языковых семей наблюдается значительно большая сте-
пень структурированности генофонда. Так, по данным AMOVA, в эт-
носах алтайской языковой семьи 5,7% генетического разнообразия
определяются различиями между тремя группами алтайской семьи,
19,9% связано с различиями между популяциями внутри языковой
группы и лишь 74,3% молекулярной дисперсии можно отнести на счет
внутрипопуляционного разнообразия. Для включенных в работу по-
366
V. Эволюция человека
пуляций индоевропейской семьи около 4,4% генетической дисперсии
определяется межгрупповыми различиями. Доля молекулярной дис-
персии, определяемая различиями между алтайской и индоевропейс-
кой семьями, составила для исследованных популяций 2,0%.
Теперь давайте вернемся к началу нашего анализа - к «бинарным»
гаплогруппам, т.е. гаплотипам, построенным по диаллельным поли-
морфизмам.
Молекулярная дисперсия микросателлитных гаплотипов внутри этих
гаплогрупп также позволяет получить оценки их возраста - т.е. времени
возникновения мутаций, приведших к образованию нового «бинарного»
гаплотипа. Оказалось, что все выявленные гаплотипы DYF- и TatC
объединяются в единое древо. Анализ дисперсии микросателлитов в этих
линиях показал, что возраст делеции в локусе DYF оценивается пример-
но в 6900 лет, а возраст транзиции Т-С в локусе Тат - в 4200 лет с до-
вольно широкими пределами 95% доверительного интервала.
Что же касается возраста и происхождения ТатС, то наша оценка
совпадает с таковой, полученной Анти Саянтила с сотрудниками в Фин-
ляндии, которые при анализе гаплотипов у финно-угров оценили воз-
раст ТатС в те же 4 с небольшим тысячи лет. Следует заметить, что их
оценка базируется на дисперсии 6 микросателлитных локусов, а наша
- 7. Более того, предковый гаплотип ТатС, выявленный нами, имеет
практически ту же композицию микросателлитов, что и предковый
гаплотип ТатС у финно-угров. Тем самым финно-угорское происхож-
дение ТатС, равно как и «монгольское», вызывает сомнения. Вероят-
но, этот предковый гаплотип присутствовал в предковой по отноше-
нию к финно-уграм и тюркам популяции. Кроме того, возраст линии
ТатС у вост, славян также совпадает с возрастом предкового гаплоти-
па. Это вызывает сомнение в том, что русские получили ТатС при ас-
симиляции финно-угорских племен, которая произошла 1000-1200 лет
назад. Вероятно, эта линия Y-хромосомы присутствовала у предков
восточных славян гораздо раньше.
В результате нашего анализа вырисовывается следующая датиро-
ванная последовательность событий, приведшая к возникновению
наблюдаемых ныне в Северной Евразии линий Y-хромосомы. Транзи-
ция в локусе SRY3225 примерно 100 тыс. лет назад привела к образо-
ванию этого кластера гаплотипов. Затем инсерция Alu-элемента в ло-
кусе YAP, произошедшая ориентировочно 50 тыс. лет назад, раздели-
367
В.А. Степанов, В.П. Пузырев
ла его на два дочерних кластера. Дальнейшие события происходили
уже на территории Северной Евразии. Делеция в локусе DYF
(6-7 тыс. лет назад) привела к образованию следующего подкластера
гаплотипов. Наконец, следующим эволюционным событием была тран-
зиция в локусе Тат.
В качестве примера анализа гаплотипов Y-хромосомы в конкрет-
ной этнической группе и интерпретации генетических данных в связи
с этногенезом можно привести наши данные по линиям Y-хромосомы
у тувинцев. Предковым для кластера гаплотипов, найденных в Туве,
был выбран гаплотип 8. Такой выбор диктуется несколькими причи-
нами. Во-первых, этот гаплотип наиболее распространен и встречает-
ся во всех трех исследованных популяциях, что делает его наиболее
вероятным кандидатом на роль исходного гаплотипа. Во-вторых, гап-
лотип 8 имеет наименьшее среднее евклидово расстояние со всеми
остальными гаплотипами. В-третьих, галотип 8 состоит из наиболее
частых у тувинцев аллелей. В-четвертых, наиболее «компактная» то-
пология древа с минимальной длиной ветвей получается в случае, если
отталкиваться именно от гаплотипа 8.
Большая часть обнаруженных у тувинцев гаплотипов распадается
на 2 линии (древа А и В). Кластер А более представителен и содержит
22 гаплотипа, включая 4 наиболее частых. Всего кластер А включает в
себя 64% хромосом. Кластер В объединяет 16 гаплотипов и 24% хро-
мосом. В качестве предкового гаплотипа для линии В по изложенным
выше причинам можно рассматривать гаплотип 10 или гаплотип 16.
Кластеры А и В при наложенных на построение древа ограничениях
не объединяются. Минимальная дистанция между предполагаемыми
предковыми гаплотипами кластеров А и В составляет 5 мутаций в 3
микросателлитных локусах. Это свидетельствует, что линии А и В,
вероятно, являются независимыми и отражают вклад различных пред-
ковых групп в мужской генофонд современного тувинского этноса.
Кроме кластеров А и В имеются и 2 небольшие линии, дистанци-
рованные от обоих основных кластеров. Возможно, обособленность
этих линий указывает на присутствие в пуле Y-хромосом тувинцев ком-
понентов, отличных по происхождению от двух основных линий. Не
исключено и то, что дистанцированность этих линий объясняется тем,
что промежуточные гаплотипы не попали в проанализированную вы-
борку. Кроме того, три гаплотипа не попали ни в один из кластеров.
368
V. Эволюция человека
Эти гаплотипы имеют наибольшее среднее евклидово расстояние с
другими и, по-видимому, являются иммигрантными.
Для того чтобы проследить возможные этно-исторические корни
выявленных в тувинской популяции линий Y-хромосомы, мы исполь-
зовали информацию об их сходстве или различии с линиями в других
этнических группах и, оценив время, за которое было накоплено на-
блюдаемое разнообразие, рассчитав среднее число мутаций на локус
на индивида в пределах эволюционного кластера, мы получили следу-
ющие оценки возраста: для линии А - 2800 лет с 95% доверительным
интервалом от 1200 до 7300, для линии В - 4400 лет с 95% довери-
тельным интервалом от 1900 до 11500 лет.
Эти достаточно грубые оценки свидетельствуют, тем не менее,
о том, что истоки мужского генофонда современных тувинцев следует
искать в эпоху позднего палеолита или неолита. Появление человека
на территории современной Тувы датируют поздним палеолитом. Со-
гласно палеоантропологическим данным, в эпоху бронзы Тува была
населена племенами европеоидного типа. Заселение территории Тувы
европеоидным населением можно связать с обширной миграцией ев-
ропеоидов с запада на восток степной зоны Евразии в эпоху неолита.
Появление монголоидных черт в облике древних обитателей Тувы свя-
зывают с миграцией на территорию Тувы гуннов в так называемое
гунно-сарматское время (2200-1500 лет назад). Именно этот период
этнографы считают начальным этапом этногенеза тувинцев: начиная
с гунно-сарматского периода прослеживается этногенетическая и эт-
нокультурная преемственность между древним и современным насе-
лением Тувы. Преемственность же генофонда, даже по самым мини-
мальным оценкам возраста линий Y-хромосомы у тувинцев, просле-
живается с гораздо более ранних догуннских времен.
Предковый гаплотип линии А достаточно далеко дистанцирован
от наиболее распространенного у современных европеоидов гаплоти-
па - на 6 мутационных шагов в 4 локусах. Предполагаемый же пред-
ковый гаплртип линии В (16 или 10) значительно ближе к европейско-
му (Md = 3; 2 локуса). Линия В, если пользоваться оценками возраста
гаплотипов, древнее линии А, и верхняя граница 95% доверительного
интервала ее датировки предшествует гунно-сарматскому периоду.
Гунно-сарматское время входит в доверительный интервал датировки
лини А. Значит, можно предположить, что линия В отражает вклад в
369
В.А. Степанов, В.П. Пузырев
современный мужской генофонд тувинцев древнего европеоидного
населения территории Тувы. Более обширную и менее древнюю ли-
нию А можно связать с заселением территории современной Тувы
древними монголоидными тюркоязычными группами в гунно-сармат-
ское время. Небольшая линия С, чей предполагаемый предковый гап-
лотип (5) лишь на 1 мутацию отстоит от наиболее распространенного
гаплотипа у монголов, возможно, отражает вклад более поздних миг-
раций монгольских родоплеменных групп.
О том, что линия В - более древняя и имеет европеоидное проис-
хождение, свидетельствуют и точечные мутации. В частности, все гап-
лотипы линии В входят в одну гаплогруппу по бинарным полимор-
физмам, в которой представлены, как правило, более древние аллели.
Например, по локусу Tat все гаплотипы линии В несут аллель С. Вспом-
ните теперь возраст этой транзиции - 4200 лет - он как раз совпадает
с оценкой возраста линии В. Гаплотипы же предположительно «тюрк-
ской» линии А распадаются на несколько бинарных гаплогрупп, несу-
щих менее древние аллели, как по локусу Tat, так и по другим поли-
морфизмам.
Таким образом, в этой работе мы попытались проиллюстрировать
некоторые подходы, разрабатываемые современной популяционной и
эволюционной генетикой человека в отношении познания эволюции
генофонда человека с помощью Y-хромосомы. Эти подходы представ-
ляются весьма полезными для поиска ответов на вопросы о происхож-
дении популяций, этногенеза, генетической систематики этносов. Эт-
ническая история человечества является предметом различных облас-
тей науки, включая этнографию, антропологию, лингвистику,
археологию. Разумеется, генетические исследования сами по себе не
могут дать однозначных ответов, но они дают огромный массив фак-
тического материала, анализ которого в рамках мультидисциплинар-
ных подходов будет способствовать значительному прогрессу как в
понимании собственно этнической истории человечества, так и меха-
низмов, лежащих в основе этногенетических процессов.
Приложение
С. Коржинский
ГЕТЕРОГЕНЕЗИС И ЭВОЛЮЦИЯ
(избранные главы)
КОРЖИНСКИЙ СЕРГЕЙ ИВАНОВИЧ
(из книги: Профессора Томского университета: Биографический
словарь. Вып. 1. 1888-1917 / Отв. ред. С.Ф. Фоминых. Томск,
1996. С. 117-123)
КОРЖИНСКИЙ Сергей
Иванович (26 августа 1861 г.,
Астрахань - 18 ноября 1900 г.,
С.-Петербург) - экстраординар-
ный профессор по кафедре бо-
таники.
Его отец, Иван Иванович, из
потомственных дворян, служил
врачом в Астрахани и умер, ког-
да Серёжа был еще ребенком.
Воспитывала С.И. Коржинского
старшая сестра. Обучаясь в Ас-
траханской гимназии, которую
он окончил с золотой медалью
(1881 г.), увлекся ботаникой и
совершил ряд ботанических эк-
скурсий в район дельты Вол-
ги. Поступил на естественное отделение физико-математического, фа-
культета Казанского университета. Будучи студентом, С.И. Коржинс-
кий участвовал в ряде экскурсий и обработал собранные им еще в
гимназические годы материалы по флоре окрестностей Астрахани.
После окончания университета со степенью кандидата естественных наук
(1885 г.) он был оставлен при университете для подготовки к профессор-
скому званию по кафедре ботаники. 26 апреля 1887 г. С.И. Коржинский
защитил диссертацию «Материалы для географии, морфологии и био-
логии Aldrovandia vesiculosa L.» и был удостоен степени магистра бо-
таники. Осенью того же года он получил звание приват-доцента и стал
читать курс ботанической географии растений. Начиная с 1887 г.
С.И. Коржинский изучал южную часть Пермской губернии. Итогом
экспедиций стал труд «Флора востока Европейской России в ее систе-
372
матических и географических отношениях», который он начал изда-
вать в Томске (1892 г.). В Томск Сергей Иванович переехал после за-
щиты 15 мая 1888 г. в Казанском университете диссертации «Введе-
ние и ботанико-географический очерк Казанской губернии» на сте-
пень доктора ботаники. 1 июля 1888 г. он был назначен экстра-
ординарным профессором по кафедре ботаники Томского университе-
та и заведовал ею до 27 июля 1892 г. 1 сентября 1888 г. С.И. Коржин-
ский прочитал студентам первую в истории Томского университета
лекцию «Что такое жизнь?». На протяжении ряда лет он преподавал
курс ботаники.
В томский период научной деятельности Сергей Иванович совер-
шил ряд экспедиций. В 1889 г. он занимался изучением окрестностей
Томска, в 1890 г. - Барабинской и Кулундинской степей, посетил
оз. Балхаш. На следующий год по заданию Восточно-Сибирского От-
деления Императорского Русского географического Общества Коржин-
ский совершил экспедицию на Амур, где изучал местную флору и при-
годность края для колонизации. Поездка была организована на деньги
Общества и личные средства самого Сергея Ивановича. Во время этой
экспедиции он простудил почки. Обострение хронического воспале-
ния почек и явилось впоследствии причиной общего заражения крови
и смерти ученого. С.И. Коржинский стоял у истоков сибирской бота-
нической школы. Итог томского периода его деятельности - 13 науч-
ных работ.
В 1892 г. С.И. Коржинский переезжает в Петербург, где занимает
должность главного ботаника Императорского Ботанического сада
(1892-1899 гг.). В этом же году он совершает несколько ботанических
экскурсий в районы Средней России, в 1893 г. - отправляется в боль-
шое путешествие с целью рекогносцировочного ознакомления с рас-
тительностью юго-западной части России, в 1897 г. - путешествует в
Средней Азии, в 1898 г. - посещает Крым, где начинает опыты по вы-
ведению новых сортов винограда, в 1899 г. его направляют в команди-
ровку по Европе и в Алжир для ознакомления с ботаническими садами.
В 1898 г. Сергей Иванович завершил свою работу над капиталь-
ным трудом «Флора Востока европейской России...». В нем раньше и
независимо от Р. Веттштейна(1898 г.) было сформулировано понятие
расы как «истинной систематической единицы» и введен в системати-
ку растений географо-морфологический метод. Коржинский предло-
жил различать под суммой морфологических признаков (морфомой)
373
отражение внутренней биологической сущности, или «бионта» (тер-
мины Коржинского), опередив введение понятия о фенотипе и геноти-
пе (Иогансен, 1903). Эта работа получила высокую оценку ряда рус-
ских ученых. В опубликованной в русских и немецких научных жур-
налах 1-й части своей работы «Гетерогенезис и эволюция: К теории
происхождения видов» (1899) Коржинский создал мутационную тео-
рию эволюции («теория гетерогенезиса»), противопоставив ее дарви-
низму, что принесло ему мировую известность. В ней он первый раз-
работал понятия о чистых («чистый вид») и смешанных («гибридная
форма») линиях, опередив на 4 года В. Иогансена, а также понятия о
наследственных («гетерогенные вариации» мутации) и ненаследствен-
ных («модификации») изменениях, предвосхитив Де Фриза (1904).
За выдающиеся заслуги в науке Коржинский был избран адъюнк-
том Академии Наук по ботанике (1893 г.), экстраординарным (1897 г.)
и ординарным академиком Императорской Академии Наук (1898 г.).
Награды С.И. Коржинского: орден Св. Станислава 2 ст. (1896 г.), ор-
ден Св. Станислава 3 ст. (1891 г.), из иностранных орденов - Бухар-
ский Золотой звезды 1 ст.; медаль “В память царствования Императо-
ра Александра III”. В 1898 г. Сергею Ивановичу был присвоен чин
действительного статского советника.
С.И. Коржинский был женат на Ольге Михайловне Брылкиной
(1860-1930), по первому мужу Рудановская, из обедневшей дворянс-
кой семьи. С нею он познакомился в Нижнем Тагиле в 1890 г., где был
проездом из Томска. Она окончила Высшие (Бестужевские) женские
курсы и занималась музыкой у композитора М.А. Балакирева. Их сын,
Дмитрий Сергеевич (1899-1985), был одним из основоположников
физико-химической петрологии и минералогии, академиком АН СССР
(1953), Героем Социалистического Труда (1969), лауреатом Сталинс-
кой премии (1946), Государственной премии (1975), Ленинской пре-
мии (1958), Премии им. Карпинского (1949); награжден золотой меда-
лью им. В.И. Вернадского. Дочь, Ольга Сергеевна, занималась скульп-
турой, автор бюста отца.
374
запвсен юпттонжой шдшш m
:м±ал:огкЕа
ОВ КАСАПЙМШ 1МЙЙВШЖ DES SCIENCR8 DE CT,.P&TBRSBOUIW,
virrntaiB.
(К» *«Ш<ЬИАТ1аИГГВСК1ШГ отдмюимь «ЛЬЛИДО WUfWCOMAWfauWlt
Тоасъ1Х«Ж4Ж Volume IX. М Я.
ГЕТЕРОГЕНЕЗИСЕ И ЭВОЛЮЩЯ.
КЪ ТВОИМ ПРОИСХОЖДЕН1Я ВЙДОВЪ.
1
О, Коряшнскаго,
кйкй Амт
«МЙММ«4 яа Ч
С.-ПЕГЕРВУРГЬ. W. ST.-Pto5RSBOUB6.
Н]идмл« у «medQMMw ЯммкглтмгскоЯ « СошаЫ«вм)гй й ГЛлмймй iartauxt М
АыцгашНаугы 1 йймм;
L ГДжмц JL Эти* » Йж К JL Гмпн &вмммЦ Ж low* А СИ. еС й BWUr»HL-Hur^
«гь<?.-1(«г*рй]грг£, . Ыжпг,
L К^Шмйт «ъ (L-ffmpa, w*rt, Ндршмь ’ J. ДмйиЖаО ЫлРймтЬвд, Ммосм, Тш«*й е<
«кзд .
rbCHhwp5jr>rtj . Ьшъ
юуийиш «Ъ С- Птрб.,Mdrturt, HapttiMh ’ J. ДмЙшКМ! St...
надави*
гм
т-й«1«
375
Съ тйхъ поръ, какъ въ 1859 году появилось знаменитое сочинеше Дарвина: «О про-
исхожденш видовъ», мнопе ученые стали внимательно наследовать расиространен1е разно-
видностей и варьящй для того, чтобы проследить по нимъ процессъ Фориировав1я новыхъ
вкдовъ въ природе. Эти изейдоваюя обогатили науку большимъ количеством» Фактовъ,
осветили иксия загадочный и мало изученный явлешя, но относительно своей главной цели
далеко не дали ожидаемыхъ результатов^ Некоторые авторы не скрывали своего разоча-
рования (W. О. Focke), друпе же хотя и находили въ наблюдаемыхъ ими явлешяхъ соот-
ветствие съ теорией, но добытые ими Факты не были особенно убедительны. Замечательно,
что не смотря на большое число даровитыхъ горячить приверженцевъ учешя Дарвина,
Фактическая сторона собряиеняо дарвинизма (или трансмутацш), т. е. теорш происхождешя
вкдовъ путемъ подбора и накоилен1я ивдивхдуальныхъ признаковъ, осталась до енхъ поръ
почти въ томъ же состояши, какъ она была разработана самимъ авторомъ. А огромная
дарвинистическая литература, возникшая за последняя десятилетщ, состоять главвымъ
образомъ изъ теоретически» разеужденй, среди которыхъ положительно тонуть отдель-
ные разрозненные Факты, приводимые въ качестве прпгёровъ.
Съ самого начала моихъ научвыхъ занят!! я точно также съ особеннымъ интересомъ
изелкдовалъ всякая отклонея1я, отыскивалъ переходный Формы я вообще стремился изучить
следы постепенного развиты видовъ. Эта задача была всегда главной, затаенной целью
всехъ моихъ экскурс!! и путешеств!! въ Poccin и Сибири. Однако, чЬмъ более я изейдо-
валъ, тЬмъ глубже испытывалъ разочароваше. Факты решительно не находились въ соот-
ветствш съ теорией. ВсЬ явлешя, который мне удалось изучить, говорили за то, что виды
изменчивы;,1ю какъ происходить ихъ изменеше и происхождете новыхъ Формъ, оставалось
для меня попрежнему загадкой. Въ конце концовъ приходилось сознаться, что въ этой
Сфере теор!я Дарвина не осветила намъ явлешй, который остались столь же темными и не-
ясными, какъ были прежде. Невольно являлось сомнете, справедливо ли то объяснеше,
которое даетъ Дарвинъ процессу эволюцш; эта теор!я трансмутацш, столь остроумная и
заманчивая, согласна ли она на самомъ дШ съ действительностью?
Убедившись, что одними наблюден!ями надъ дикорастущими Формами нельзя разре-
шить возшжающихъ сомнки!!, я обратился къ изучение пронсхожден!я новыхъ Формъ въ
садоводстве. Какъ известно, вопросъ объ изменчивости животныхъ и растений въ культуре,
Вывш «щ.-Жп. Очи 1
376
2 С. К0РЖИНСК1Й. ГВТЕРОГЕНВЗЖОЪ И 8В0ЛЮЦ1Я.
есть коренной вопросъ дарвинизма. Ему посвятнлъ особенно много времени Дарвняъ и на
немъ, главнымъ образомъ, онъ построил свое учете. ТЪмъ не меийе мн± пришлось очень
скоро убедиться, что выводы къ которымъ пришел Дарвинъ относительно пронсхояценш
культурныхъ Формъ, основаны на нев^рномъ поннманш фвктовъ. По крайней irtpfe относи-
тельно садовыхъ растешй я могу смЪло утверждать, что никогда ни одянъ культиваторъ
для получен^ новыхъ расъ не оперировал съ индивидуальными признаками, и что никогда
не наблюдалось «накоплеюяэ этихъ послФднихъ. A set новыя разновидности (кромй помЪ-
сей), происхождение которыхъ намъ известно, возникли на самомъ д'Ьгё лутемъ внезапныхъ
отклонений отъ чистыхъ видовъ или гибридныхъ Формъ. Спрашивается теперь, не играют»
ли эти внезапныя отклонеи!я подобной же роли в въ природ^, и нельзя ли такинъ лутемъ
объяснить то несоотв£тств1е характера и распространена варьяцШ съ Teopieft Дарвина.
Существоваше внезапныхъ отклонен^ было хорошо известно Дарвину, который
однако придивалъ имъ мало значеюя, считая это явлеше, которое въ далыгЬйшемъ я буду
называть тапеуоияетосолл, какъ бы ненормальнымъ, всключительнымъ. По этой же прн-
чия$ большинство дарвянистовъ совершенно игнорировало его. Факты, которые изложены
мною въ настоящемъ сочинеши, покажутъ, какъ я надеюсь, достаточно ясно, что гетероге-
незисъ есть явленie, хотя к редкое, по вполн! нормальное, свойственное какъ животньшъ,
такъ и растительнымъ организмамъ и играющее огромную роль въ кхъ эволощи.
Первая часть моей работы посвящена характеристик^ самаго явления гетерогенеза,
во второй же, которую я представлю нисколько позднее, будетъ излагаться роль его въ
происхождети видовъ.
14-го явваря, 1899 голе.
С.-Петербургь-
377
I.
Пояяпе о гетерогенезие! введено въ науку, какъ laetcrHO, знаменитымъ гистологомъ
Кёлликеромъ въ 1864 году х). Именно, разбирая теорию Дарвина, названный ученый пы-
тался противопоставить ей свою собственную, которую опъ назвать «Theorie der heteroge-
nen Zeugong» и охарактеризовать въ стЦующихъ выражешяхъ:
«Основная мысль этой гипотезы заключается въ томъ, что, пода «мятежа общем за-
кона развитЬяуСуицжгы. производить изъ своихъ зародышей друпя, отклоняющаяся огь нихъ.
«Это можете произойти:
1) оттого, что оплодотворенный яйца при своемъ развили при особыхъ обстоятель-
ствахъ переходить въ высппя Формы;
2) оттого, что организмы безъ оплодотворетя изъ зародышей или яицъ даюгь начало
другимъ организмамъ».
Въ подтверждеше этихъ положена авторъ приводить слйдукнщя данныя:
1) См±ну покол-Ьпй, при которой, какъ известно, организмъ производить половымъ
или безполымъ путемъ Формы, совершенно на него не похожее. Такъ наприм^ръ, полипы и
медузы связаны у мЬкоторыхъ видовъ тЬсиыми узами и представляйте лишь стадш развила;
у другихъ же группъ эти Формы живуте и развиваются совершенно самостоятельно. Такюгь
образомъ здйсь полипы производить медузъ, который (по крайней Mipfc въ другихъ груп-
пахъ) могутъ жить самостоятельно. Факте гетерогеннаго развиля зд^сь на лицо.
2) Авторъ указываете, что оплодотворенный яйца розличныхъ животныхъ иногда
весьма сходны между собою, и потому онъ полагаете, что небольшого лзм£нен1я было бы
достаточно, чтобы произвести другую высшую Форму, Хотя «это не доказано ни однимъ
ирямымъ Фактомъ, но въ возможности такого процесса, конечно, нельзя сомневаться».
1) KQlHker, Ueber die Darvin'cche Schtyfungiiheorie. — ZelUchrift ftr vtaenicb. Zoologie, т. XIV
<1864), p. 174—180
1*
378
ГБТВРОГИНЕЗИСЪ I эводюци.
73
VIII.
Мы разсмотр’Ьли въ предыдущихъ главохъ различный категорш отклонен^, могу-
щихъ возникать гетерогеннымъ путемъ. Изъ собранныхъ данпыхъ прежде всего вытека-
етъ, что гетерогенезлсъ встречается совсЪмъ не такъ редко, какъ можно было думать, и
что его проявления бывають довольно разнообразны. Р&зберемъ же теперь ближе, на осно-
ваши всЬхъ извЬстныхъ Фактовъ, услов1я возникновентя и отличительный черты этого про-
цесса.
Сущность гетерогенезиса.'Хплпчная картина гетерогенезиса состоять въ томъ, что изъ
семянъ, полу чей и ыхъ огъ нормальныхъ экземнляровъ какого либо вида, выростаетъ средн
многихъ (сотенъ и тысячъ) сеянцевъ какой нибудь одинъ индивидуумъ, р£эко отлячающтйся
отъ всЬхъ остальныхъ тЬыъ или другвмъ прязнакомъ, пли ц1лымъ рядомъ этихъ послед-
пихъ. Такой экземпляръ нредставляетъ гетерогенную варьлцЬо, а его отличительные при-
знаки можно назвать гетерогенными. Вырастал, онъ производить потомство, наследующее
целпкомъ илл отчасти его свойства, и такимъ образомъ даетъ начало гетерогенной расе.
Отличгя гетерогенных* признаком. Для того, чтобы не смешивать гетерогеиныхъ
варьящйсъ индивидуальными и прочими изменениями, наблюдающимися въ организмахъ, не-
обходимо иметь въ виду следующее: гетерогенные признаки иредставляютъвсегда более или
менее резня уклопешя, тогда какъ индивидуальны л характеризуются всегда мелкими к не-
заметными отлич!ями. Конечно, это определено не можетъ назваться строгимъ, ибо попяпе
о мелкпхъ и крупныхъ признакахъ весьма относительно. Но вотъ уже более существенная
черта. Индивидуальная физ юном i я каждой особи состоять изъ комбин&щм мелкихъ призна-
ковъ, которые set повторяются у очень многихъ экземпляровъ даппаго вида, хотя и въ дру-
гихъ комбинащяхъ, Поэтому при нормальпыхъ условхнхъ, т. е. при отсутствы гибрцдиза-
щи и гетерогенеза, хотя каждый индивидуумъ данной расы и отличается несколько отъ
всякаго другого, но ни одинъ изъ пихъ не выделяется изъ массы всехъ остальныхъ. А
при гетерогенезе мы видимъ именно это последнее. Какой нибудь одинъ экземпляръ отли-
чается отъ всехъ остальныхъ появлешемъ такихъ свойствъ или признаковъ, которыхъ ветъ
вовсе у типпчныхъ представителей данной расы. Вкратце это различ1е можно Формулиро-
вать такимъ образомъ: все индивидуальный варьяцш заключаются въ пределахъ типа, гете-
рогенный же выходятъ изъ этихъ пред!ловъ и составляют^» нарушение типа.
Какъ гетерогенный, такъ и индивидуальный отклонения нужно отличать оть модифи-
каций, т. е. такихъ изменен^ организма, который находятся въ непосредственной зависи-
мости отъ вн1шнихъ услов1й: оть характера почвы, орошены, освещены, густоты или ред-
кости nocisa и т. п. Сюда же примыкаютъ изменены, эависяиця оть качества или возраста
Займа «м.-Мат. Отд. )0
379
74
0. КОРЖИИОК1Й.
сЪмянъ. ВсЪ подобный отклонены легко отличаются отъ варьящй другихъ категор1й.£во
первыхъ, оип простираются главнымъ образомъ на размеры органовъ, а не на ихъ специ-
Фическ1е признаки; во вторыхъ, они обнимаютъ обыкновенно все экземпляры, развиваю-
шдеся при данныхъ услов!лхъ, а не отдельные индивидуумы; в, въ третьихъ, эти изменен 1я
выражаются въ большей или меньшей степени, въ зависимости отъ интенсивности соответ-
ствующаго Фактора. Такъ иапримЪръ, если какая либо «нэрна въ тЬпя развиваетъ длинные
стебли и меньшее число, но более крупныхъ листьевъ, то при меньшей степени затЬвенш
rfc же самые признаки будутъ выражены въ болЪе слабой степени. И если изъ старыхъ
еймянъ (паприм^ръ б-лйтпихъ огуречныхъ) получаются слабый растешя, но ранее и обиль-
нее цвЪтущ1я, то изъ сЬмянъ но столь старыхъ (нанримйръ, 4-летнихъ) мы получимъ ра-
стения не столь слабыя в позже цвету Щ! я, т. е. приближающхяся къ экзенплярамъ, выро-
щепнымъ изъ свЪжихъ сЬмянъ. ВслЪдствйе такихъ градаций, причину пзменешй, который
мы называемъ модификащями, обыкновенно весьма легко констатировать уже простыми
наблюден!ями, а кроме того можно доказать и опытнымъ путемъ. Наконецъ, вей подобный
изменены не передаются по наследству J
Есть еще одинъ классъ изменешй, которыя необходимо строго отличать, чтобы избе-
жать эатемн!шя истиннаго смысла явлешй—это помеси. Семена, которыя ейетъ культвва-
торъ и которыя онъ собралъ отъ тяпячныхъ растений, нередко имЪютъ гибридное происхо-
ждеше, всл1дств1е заноса цвйтени отъ другого близкаго вида насекомыми или ийтромь.
Тогда въ поейвахъ среди типичныхъ растений онъ завгйчаетъ экземпляры, более или менее
сильно уклонявшиеся отъ типа, которые однако не гетерогеннаго происхождения.
Слйдуюпця данный позволяють констатировать гибридное происхожден!е отклонений.
Во первыхъ, если действительно произошло гибридное опыление цвйтовъ, то изъ еймяпъ
получается всегда несколько гибридовъ или даже довольно много. Редко и лишь въ виде
исключешя можетъ получиться только одинъ экземпляръ данной помеси. Затйнъ, если дей-
ствительно уклоняющееся экземпляры обязаны своимъ пронсхождешемъ скрещивашю, то
они должны представлять среднее признаки между своими родичами, г. е. между материн-
скшъ видомъ и тЬмь, которое дало пыльцу. Необходимо, следовательно, решить, къ ка-
кому виду приближаются полученныл отклонешя, и находится ли данный видъ где либо по
близости. При наличности этихъ услов1й есть полное основан!е предполагать гибридное
происхождение полученныхъ варьящй.
Но можетъ случиться, что изъ семянъ получается довольно пестрое поколеше, въ
которомъ находится много разнообраэныхъ отклонений отъ типичной Формы. TaKie Факты
наблюдались и наблюдаются нередко, но они происходить лишь тогда, когда семена взяты
отъ неустановившихся еще Формъ гибриднаго или гетерогеннаго происхождения. При этомъ
колебан!я призяаковъ всегда происходить въ нзвестныхъ тйсныхъ пределахъ, определяе-
мыхъ признаками родичей въ случае гнбридизацш, или признаками типа и разновидности,
въ случае гетерогеннаго ея происхождения. Расы же установивппяся, какого бы оне ни
были происхождения, никогда ле даютъ подобнаго снешаннаго потомства.
380
ГКТКРОПНЕВЮЪ и вводоцш.
75
Итакъ, чтобы наблюдать въ чистомъ виде явлешя гетерогенеза, необходимо искать
его среди потомства оть чистыхъ, т. е, не гибридяыхъ, и яормальныхъ, т. е. установив-
шихся, видовъ при услов^яхъ, устраняющих* возможность гибридацш. (Мшена, получен-
ный отъ такихъ растепш, даюгь обыкновенно совершенно однородное потомство, вполне
сходное съ своими родителями. Часто мнопе годы, даже десятки лЪгь ежегодныхъ посе-
вов* проходить безъ появлешя какяхъ бы то ни было отклонен^. Но иногда совершенно
неожиданно среди однородна™ потомства возникаете» какая либо гетерогенная варьяшя.
Въ гибридных* поколЪшяхъ мы вообще наблюдаемъ очень пестрое потомство, колеблю-
щееся въ своихъ признаках* между признаками родителей. Но квтл либо новыя черты,
отсутствующая у этихъ последних*, проявляются и здесь такъ же неожиданно, какъ у чи-
стыхъ видовъ, именно у какого нибудь одного экземпляра, отличающегося таким* образомъ
отъ всех* остальиыхъ, имеющих* то же происхождеше.
Редкость явлешя. Хотя случаевъ гетерогевеза въ общемъ известно довольно много,
но для каждого отдельного вида онъ все-таки составляете» чрезвычайно редкое явлете.
М nori я растешя, разводимый въ болыиомъ количестве и размножаемыл постоянно семе-
нами, въ течете целых* десятков* лЪтъ не даюгь никакяхъ варьяшй. Такъ Erytbrina Crista
galli была введена въ культуру въ 1771 г., по первая разновидность отъ нея была полу-
чена лишь въ 1844 г. Begonia semperflorens, введенная въ культуру въ 1829 году, дала
первую разновидность, отличающуюся розовыми цветами, лишь въ конце семидесятых*
годовъ, т. е. почти черезъ 50 л-Ьтъ. Еще черезъ десять лет* эта последняя дола варьящю
съ красными цветами, п кромЬ того была получена низкорослая Форма; скрещивате же
этихъ Формъ дало еще нисколько разновидностей, отличающихся различной комбинашей
тЬхъ же самыхъ прпзпаковъ. Crambe maritima въ течете бол!е пятидесяти л±тъ культуры
не дола никаких* варьящй. Правда, друг!я растетя начинают* варьировать очень быстро,
иногда уже въ первые год гл культуры, по таюе случаи довольно редки. Та же масса раз-
новидностей, которая получается отъ некоторых* видовъ въ немного л!тъ их* культуры,
обязана своимъ нроисхождешемъ нс гетерогенезу, но гибридизации съ другими видами.
Н£тъ ничего легче, какъ получить отъ двухъ близкихъ видовъ целый рядъ культурных*
варьящй, въ которьтхъ признаки родоначальников* будут* комбинироваться въ разпыхъ
группировках*. По новые признаки, не свойственные родоначальнымъ видамъ, происхо-
дить и въ такихъ гибридных* циклах* почти столь же редко, какъ и отъ чистыхъ видовъ.
Разсматривая различного рода отклонетя въ отдельности, мы найдемъ, что одни изъ
нихъ происходить р!же, друпя чаще. Частичный хлорозъ у сеянцев* некоторых* древе-
сныхъ породъ происходить, невидимому, очень нередко. Часто наблюдается также изме-
ueuie той или другой окраски венчика въ белый цв1тъ или по крайней wipe изменение от-
тЬнковъ окраски цветов*. Махровость цветов* считается одним* изъ обыкновеннейших*
изменений растешй въ культуре, но вот* катя данныя можно привести для того, чтобы
охарактеризовать точнее, насколько часто происходить это обыкновеннейшее явлете. Такъ
петунш разводятся въ садахъ съ двадцатых* годовъ истекающего столетия, во махровые
ю*
381
76
С. К0РХИНСК1Й.
цвйты были получены отъ нихъ лишь въ 1863 году. Cyclamen persicum находится въ
культурй съ 1731 года; махровые цвйты были замйчеяы у него въ пятидесятыхъ годахъ
въ Гентй и въ 1875 г. въ Варшавй. Ipomoea pnrpurea существуете. въ садахъ съ XVII
столйт!я; махровые цвйты у ней были замечены въ Парижй въ 1843 году, а затЬмъ въ
1895 г. Конечно никто не решится утверждать, что помимо зарегистрированныхъ случа-
евъ у этихъ растенш никогда не возникало варьяцш съ махровыми цветами, не замйчен-
ныхъ садоводами. Но нужно во всякомъ случай имйть въ виду, въ какомъ огронномъ
количествй разводятся сймепамп эти растения въ круиныхъ садовыхъ заведешяхъ, и съ
какой тщательностью отыскивають культиваторы всяк!я новинки, чтобы оцйнить ту степень
рйдкости возннкповешя даже такихъ обыкновенныхъ разновидностей. Изъ остальныхъ же
варьяцш большая часть или, лучше сказать, почти вей были лишь по одному разу занесены
въ лйтописп науки. Такъ обыкновенная бйлан акащя (Robinia Pseudacacia) съ половины
XVII столйпя (1640) разводится въ огромпомъ количествй ейменамп; но до 1858 г. никто
никогда не замйчалъ этого вида съ розовыми цвйтами, а между тймъ это такой признакъ,
который не ускользнулъ бы отъ вниман!я не только садовода, но и всякаго обыкновенная
смертнаго. Точно такъ*же, несмотря на обширные поейвы земляники, которые производятся
садоводами, никто, кромй Дюшена, ни раньше, пи поелй не паходилъ земляники съ простыми
листьямиг). Разъ какая либо новая Форма получена, она сохраняется, размножается и ста-
новится нерйдко очень обыкновенной. Ноне нужно забывать,что ея воз ник новен ie все-таки
было, такъ сказать, необычайнынъ собьгнемъ и, быть можете., составило единственный
въ своемъ родй, болйе пе повторившейся Факте».
Если о нйкоторыхъ варьящяхъ говорить, что онй получались неоднократно, то не
нужно думать, что дййствительно возникала вторично Форма, вполнй тождественная съ преж-
ней. На самомъ дйлй въ такихъ случаяхъ можете» быть рйчь лишь о вторичною» появлении
извйстнаго признака у одной изъ Формъ даннаго вида, но не о вторичномъ появленш
опредйленной Формы. Даже въ томъ случай, если однородныя отклоненйя происходить
отъ какого нибудь довольно постоянная вида, то все-таки внимательное изслйдован1е
откроете, или въ степени интенсивности главная признака или въ побочныхъ свой-
ствахъ много мелкихъ чертъ, позволяющихъ отличать одну отъ другой данный разновид-
ности, какъ напр. пурпуровый варьяцш Pninus cerasifera, изъ которыхъ одна возникла въ
Персии, а другая въ Гермаши. Въ большинствй же случаевъ вторичная появлен!я одно-
родной варьяцш, эта поелйдняя возникаете» совсймъ оть другой разновидности, но относя-
щейся къ тому же самому виду. Этотъ Факте» заслуживаете» пйкотораго внимашя съ чисто
научной точки зрйн!я; съ точки же зрйтя садоводства онъ не представляете» интереса, ибо,
разъ какой нибудь признакъ появился у одной разновидности, то, путемъ скрещивашя этой
1) Отъ одно! изъ крупноплодных* эемхявнкъ,
именно Frxi»ier docteur Nicaiee была получена однажды
въ шествдссятыхъ годахъ подобная же однолисточко*
ван Форма въ садояомъ оаведен1и Вильморена (Rev.
bort. 1867, р. 222). Неизвестно только, возникла дм она
самостоятельно или путемъ скрещиван1л съ Fragaria
monopbylla Дюшена, что ми1. кажется божЬе вероят-
ным*.
382
ГЕТХРОГМЕЗИСЬ Ж ВВ0ЛЮЦ1Я.
77
последней съ другими, его получаюсь во всехъ разновидностяхъ. Достаточно было полу-
чить одну лиловую петушю съ махровыми цветами, чтобы въ короткое время выростить
мах ров ыя петунш съ цветами всЪхъ существовавшихъ въ то время окрасокъ. Равнымъ
образомъ достаточно было выррстить одинъ экземпляръ петунш съ зубчатыми лепестками,
чтобы получить весьма быстро этотъ признакъ въ комбинацш со всеми окрасками и со
всЬми другими признаками различныхъ сортовъ даинаго цикла.
Надо заметить кроме того, что повторное возникповеше однородпыхъ варьящй, хотя
несомненно им^етъ мЬсто въ природе, по совершается далеко не такъ часто, какъ можно
было бы думать, судя по сообщешямъ садовыхъ журваловъ. Во всехъ случаяхъ, когда
дело идеть о новомъ возникяовенш варьяцш, однородном съ существовавшей уже раньше,
необходимо строгое раэследовате вопроса: 1) откуда были взяты семена, изъ которыхъ
выросла новая варьящя; и 2) не росла ли прежняя Форма где нибудь по близости въ томъ
же саду или въ окрестныхъ, и нс могла ли ся пыльца быть занесенной па материнское
растете новой варьяцш. И, если эти вопросы обойдены молчшпемъ или недостаточно разъ-
яснены, то мы вправе отнестись съ сомнешсмъ къ сообщаемымъ Фактамъ и не объяснять
ихъ гетерогепезисомъ, но скорее возможностью гнбрвднаго опылен in. Нередко стоить
только появиться какой нибудь оригинальной варьяцЫ, какъ раздаются занвлетя отъ раз-
ныхъ садоводовъ, что и они получили ту же Форму и даже на годь или два раньше. Къ та-
кимъ сообщетямъ надо вообще относиться съ недоверхемъ, если только они не сопровож-
даются описашемъ, какимъ образомъ и отъ какой Формы произошла данная варьящя, или
не подкреплены свидетельствомъ какого нибудь авторитетпаго лица.
Въ заключенхе надо коснуться еще одного вопроса, всегда ли гетерогенный варьяцш
появляются первоначально въ одномъ экземпляре или несколько ихъ можегъ возникнуть
сразу. На это надо ответить следующее: во всехъ случаяхъ, о которыхъ мы пмТ.емъ точ-
ный и авторитетный паблюдешя, всегда говорится о единствеппомъ индивидууме, отличаю-
щемся отъ всехъ прочихъ. Эго есть столь общее правило, что о числе пбдивидуумонъ обы-
кновенной не упоминается, но во всехъ случаяхъ, когда рЬчь идеть обт. одной новой разно-
видности, всегда подразумевается само собою, что опа получена въ одномъ экземпляре,
который навсегда и сохраняегъ назваюс «орпгпнальнаго». Но иногда встречаются упоми-
нания объ одповременпомъ появленхн 2—3 и даже болыпомъ числе индивидуумовъ, Bcf.
подобный сообщены однако очень сомнительны. Они всегда эаставляютъ подозревать, что
нибудь одно изъ двухъ: 1) не имПемъ ли мы передъ собою какой либо л ом to, или, если
этого нельзя предположить, 2) не произошло ли гетерогенное полвлеше дайной варьяцш по-
колешемъ раньше, что не было своевременно зам Пчено. Во всякомъ случае вполне точиыхъ
и внушающпхъ довЬр!е наблюденшнадъпоявлежемъкакой либо гетерогенной варьяцш более,
чемъ въ одномъ экземпляре одновременно, мне решительно неизвестно.
Вюъшмя услсипя гетерогенезиса. Спрашивается теперь, какхс же Факторы обусловли-
ваютъ гетерогенное появлеше новыхъ Формъ? На это пока возможепъ лпшь одинъ ответь,
что причина гетерогенезиса не заключается но нн[;шнпхъ условшхъ развитая. Да и нельзя
383
78
С. К0РЖИНСК1Й.
предположить непосредственное вл)яше на это явлеше каквхъ либо ввйшнихъ Факторовъ.
В!дь эти послйдше обнаруживали бы свое д!йств1е на вс! или по крайней мйрй на большую
часть индивидуумовъ. А между тйыъ изъ огромного числа ихъ, изъ сотенъ и тысячъ, ра-
стущихъ вмйстЬ, при совершенно одинаковыхъ услов!яхъ, лишь одинъ какой яибудь вдругъ
обнаруживаете гетерогенную варьящю. Очевидно, что не во внйшнихъ условшхъ нужно
искать причипъ этого явлешя, что зд!сь д!ло въ какпхъ то внутреннихъ процессахъ, въ
каквхъ то измйнентяхъ яйцевой клйтки, о сущности которыхъ впрочеиъ мы не можемъ еще
составить себ! никакого понятая.
Но если вн!шшя условш не могутъ считаться за непосредственную причину гетероге-
неза, то они во всякомъ случай могутъ играть роль предрасполагающего элемента. Въ этомъ
отношепш сходятся мнйшя почти вейхъ садоводовъ и культиваторовъ. Именно, они полага-
ютъ, что значительно способствуйте» полу чешю гетерогенныхъ варьящй елйдуюшдя условш:
1) Измененie условгё существовашя.
2) Условия культуры.
3) Повторенные поейвы въ возможно большихъ разм!рахъ.
На значеше пзмйнеппыхъ условШ существовашя указываютъмнопе садоводы. Поводъ
къ этому MHiiiiio дали наблюдешя надъ многими растениями, которыя, будучи введены
въ культуру, очень быстро начали варьировать. Однако этоть выврдъ слишкомъ односто-
ронне, ибо при этомъ предполагается, что въ дикомъ состоянии растешя не варьнруютъ
или варьнруютъ весьма мало. Бол!е тщательный наблюдешя ноказываютъ однако, что и
дикорастущ1я растешя очень часто даютъ отклонен!я съ тою лишь разницей, что ихъ от-
клонены обыкновенно вымираютъ, въ культур! же отмечаются и сохраняются. Кром! того
известны вйкоторыя растенш, которыя, будучи введены въ культуру, въ течете цйлыхъ
десятковъ л!тъ почти не варьировали, а потомъ вдругъ дали значительныя отклонения, какъ
напр. Primula sinensis, Cyclamen persicum и др. Зд!сь следовательно измйнеше условш
существовашя, сопряженное съ переходомъ изъ дикаго состояшя въ культурное, не ока-
зало вл1ян!я на изменчивость растешя; варьлщи же стали появляться, когда растете уже
много лйтъ находилось въ однихъ и т!хъ ясе услов!яхъ культуры.,
Относительно вл1яшл другихъ Факторовъ той же категорш, папр. перемйны почвы,
температуры, влажности л пр., въ садовой литератур! мн! не приходилось встречать ука-
заний. Притомъ же въ культур!; услов!я жизни вс!хъ растеши бол!е или нен!е однообразны,
а спещальныхъ опытовъ съ этой ц!лью, сколько мп! известно, не было производимо. Въ
природ! услов1я существовашя представляютъ огромное рознообраз!е, л Bjiaaic этихъ раз-
личай на измйнчивость признается очень многими учеными, хотя и зд!сь осязательныхъ
Фактовъ въ пользу этого воззрйшя до сяхъ норъ не было приведено. Во всякомъ случай
для культурныхъ растем# п!тъ никакихъ основащй признавать вл1яше иэмйнешя условий
жизни на появлеше гетерогенныхъ варьящй.
Условхямъ культуры, именно въ смысл! плодородной почвы, обильного орошены и
частой пересадки, лрвдаютъ огромное значеше въ д!л! молучешя новыхъ разновидностей
384
ггпрогвйиисъ 1 эволющя.
79
вс! садоводы, высказавшееся по атому поводу. Культиваторы полагаютъ,»что обильное пи-
танёе въ течете н!сколькихъ покол!шй производить какъ бы избыток* жизненно! энергш,
которая я выливается въ образование варьящи. Этотъ взглядъ имЬетъ, но моему мийшю,
солидное основаше, ио я долженъ зд!сь указать на некоторый стороны вопроса, который,
быть можете», заставляютъ садоводовъ преувеличивать значенёе этого Фактора. Во первыхъ,
обильное питан!о бываете» при тщательной культур!, которая предполагаете» заботливость и
вниманге со стороны культиватора, отм!чающаго при этомъ вс! появляющаяся уклонеп1я.
Плохое питашс, плохая культура есть обыкновенно результате небрежности садовода, при
которой появляюпцяся варьящи могутъ оставаться незамеченными. Во вторыхъ, обильное
питание уже само но себ! производить рядъ изм!нешй въ растеши и притонъ въ желатель-
номъ для садовода направление. Растепёе делается крупнее и красивее, листья бол!е сочными
и блестящими, цв!ты крупнее. Takin видоизменения часто даже обозначаются нозваншми но-
выхъ сортовъ, что впрочемъ д!лаютъ лишь культиваторы недостаточно опытные или недо-
бросовестные, такъ какъ подобный изменен!я суть просто модиФНкаши, не передающейся по
наследству и не имеющая отношены къ образована новыхъ расъ. Въ трстьихъ, такое весьма
распространенное и ценимое у садоводовъ отклонепёе, какъ махровость, необходимо требуете»
по самой своей природ! обильнаго нитан1я,хотя возникаете» всегда на почв! гетерогенеза. Въ
четвертыхъ, многёя вновь возникающей гетерогенный варьящи отличаются хилостью и сла-
бостью роста, почему при бол!е слабом* питанш могутъ погибнуть, не достигши цв!тенёя
или не давъ с!мянъ, между т!мъ какъ при бол!е тщательномъ уход! благополучно дости-
гаютъ плодоношения. Воте» эти данныя достаточно показывайте», почему вс! садоводы глу-
боко убеждены въ важномъ значешн условш хорошей культуры для получешя новыхъ
разновидностей. Но насколько влёяете эта хорошая культура па самый процсссъ появления
гетерогенныхъ варьяшй, все-таки остается мало доказанными,.
Наконецъ, что касается до посл!дняго Фактора, т. е. иовторныхъ пос!вовъ, то его
зваченёе столь ясно, что объ пемъ нечего и распространяться. Собственно говоря, его даже
нельзя и ставить въ число условёй, благопр1атствующихъ полученёю гетерогенныхъ варья-
щй, ибо само собою разумеется, что ч!мъ больще и чаще сеять какое либо растете, т!мъ
больше в!роятёя получить отклонешя, подобно тому, какъ въ лоттсре! каждый имеете т!мъ
бол!е шансовъ выиграть, ч!мъ больше купить билетовъ. Но тотъ Факте», что именно на
частыхъ и обширныхъ пос!вахъ такъ настаивайте вс! корифеи садоводства, какъ Виль-
моренъ, Декенъ, Пуато, Верло, Каррьеръ, что обширностью культуръ объясняйте такъ
часто усп!шное получение новыхъ Формъ, лучше всего указываете на независимость явле-
ния гетерогенеза отъ человека и на несостоятельность вс!хъ остальныхъ Факторовъ, не мо-
гущихъ значительно повысить вероятности получения гетерогенныхъ Формъ.
Направления изменчивости. Мы перечислили выше вс! главный направления измен-
чивости, по которымъ происходить гетерогенпыя отклонешя у находящихся въ культуре
растенёй. Къ вимъ следуете прибавить еще г! отклонения, который были мною упомянуты
въ качеств! побочных* признаков* гетерогенныхъ варьяцёй, но не разсматрявалжсь спе-
385
80
С. КОРЖИНСК1Й.
щально, какъ напр. большая или меньшая выносливость къ холоду или засухе, более ран-
нее или позднее зацвЪташе, обильное или скудное цв!теше, появление бохЬе крупныхъ цв1-
товъ или бол!е мелкихъ, пахучихъ или непахучихъ, окрашиваше стебля и т. п. Если шгёть
затЬмъ въ виду те культурный разновидности, относительно происхождения которыхъ н^тъ
точныхъ данныхъ, но которыл, какъ можно думать, также возникли лутемъ гетерогенеза
(напр. махровыя Аквилепн безъ шпорцевъ), то легко придти къ заключенно, что растения
могутъ давать гетерогенный варьяцш почти во вс!хъ возможпыхъ направлежяхъ.
Разсматривая эти отклояешя съ точки apinifl вероятного пропсхождешя данного вида,
можно принимать однЬ варьяцш за регрессивный, т. е. за проявлежя атавизма, друг!я,
наоборотъ, за прогрессивный, третьи, паконсцъ, съ точки эрЬшя эволюцш могутъ считаться
безразличными. Къ последней категорш можно отнести, напримЪръ, варьяцш роста (на-
ниэмъ и гигаптизмъ) я пзм^нен1я Формы кроны (образование ннрамидальпыхъ и плакучххъ
Формъ), потому что у иасъ и!тъ никакихъ основанхй считать более низкий или более высокй
ростъ за бол1е нормальный, иля какую либо Форму кроны за более примитивную, а другую
за болЪе усовершенствованную. Можно сказать вообще, что какъ плакучая, такъ и пира-
мидальная (въ условкомъ смысле, какъ у пирамидальнаго тополя) Форма кроны является не-
нормальной, такъ какъ въ природе мы видимъ главнымъ образомъ округлыя, продолгова-
тый или неправильной Формы кроны деревьевъ, за исключешемъ лишь н-Ькоторыхъ хвой-
ныхъ, которымъ свойственна конусовидная крона. Но въ другихъ случаяхъ мы можемъ
отличать признаки прогрессирующее отъ регрессивныхъ. Такъ напр. образование колючекъ
у растешй есть все-таки у с лож пен ie ихъ организации, следовательно прогрессъ. Обратное
же явлеше, т. е. исчезание колючекъ, можно считать за регрессъ, или, лучше сказать, за
атавизмъ, т. е. возвращеше къ типу нредковъ. У гетерогенныхъ Формъ мы часто наблю-
даемъ исчезание колючекъ, но появления колючекъ гетерогеннымъ путемъ у Формъ, не им4в-
шихъ ихъ, не наблюдалось ни разу. Впрочемъ огь Формъ колючихъ получались варьяц!и
еще съ большимъ рвзвтчемъ колючекъ, что нужно считать за некоторый прогрессъ.
Относительно листьевъ можно заметить следующее: изъ перистыхъ или тройчатыхъ
листьевъ нередко путемъ гетерогенеза получались простые, что, разумеется, можно раз-
сматривать за проявлено атавизма, ибо простые листья, какъ более примитивные, надо
считать за более древнюю Форму. Обратного перехода, т. е. изъ простыхъ листьевъ въ
перистые пли тройчатые, не наблюдалось ни разу. Но изъ тройчатыхъ листьевъ могутъ
получиться пятерные, что уже представляетъ известное усложнеше. Такой Фактъ наблю-
дался у одной разновидности земляники, описанной подъ именемъ Fragaria quinquefoliaх).
Но съ другой стороны въ сфере более мелкихъ отклонен^ мы видвмъ варьяцш, какъ
въ сторону прогресса, такъ и регресса. Въ силу гетерогенеза очень нерЬдко изъ листьевъ
цельнокрайныхъ получаются зубчатые, изъ зубчатыхъ падрезные или лопастные, а въ пе-
1) Rer. bort. 1660, р. 346. Эта «Орма была получена | Плоды «а близки къ «opat British Queen,
известны мъ ажглйскммъ культиваторомъ г. МуаЦ’оиъ. |
386
гвтитогвпвжгь и зволгоцш.
81
ристыхъ листьяхъ листочки делаются надрезными или лопастными. Обратные переходы въ
смысле уменьшены числа i глубины надреэовъ также происходить нередко.
Изъ раапчныхъ окрасокъ венчика можно считать аа самую простую а древнюю бе-
лый шг4ть. Растешя съ вЪнчиковгь, окрашеннымъ иначе, очень легко даютъ б4лоцв4тныя
варьяцш. Отъ Формъ же съ белыми цветами также происходить разновидности съ в4н-
чикомъ иной окраски, но реже. Изм4ненш остальныхъ окрасокъ (кроме белой) одна въ дру-
гую можно считать съ нашей точки зрйни безразличными.
Махровость цв4товъ обыкновенно считаютъ регрессивнымъ метаморфозомъ, но это
справедливо лишь съ чисто морфологической точки зр4шя. Съ точки зр4в!я зволюцш мах-
ровость можно считать безразлнчнымъ отклонешемъ, а если она состоять въ увелнченк
круговъ венчика, не въ ущербъ тычннкамъ, то скорее за прогрессивный признаю». Это
последнее явлено встречается нередко. Обратного перехода, т. е. нзъ нормально полу-
махровыхъ цнЬтовъ въ простые, неизвестно, безъ сомнЬнш, главнымъ образомъ потому,
что р&стенш, у которыхъ цветы нормально полумахровые, какъ НимФейныя и Магнолхевыя,
очень немного. Лжемахровость сложноцвЪтвыхъ, т. е. превращено трубчатыхъ цвйточковъ
въ язычковые нлн въ крупные трубчатые, съ точки эр4нш теории зволюцш безразлична,
такъ какъ обе Формы встречаются у данной группы сложноцв4тныхъ въ одномъ соцвйтш.
Превращеше ц4льнокрайныхъ лепестковъ въ зубчатые иля развгпе выростовъ на
лепесткахъ есть прнзнакъ явно прогрессивный, такъ какъ онъ составляет» осложнение
организацш. Равныиъ образомъ превращеше чашечки въ нйнчикъ, т. е. calycanthemia (у
Campanula, Mimulus Primula и пр.), есть признаю» прогрессивный. Обратный переходъ
венчика въ чашечку иногда наблюдается, но весьма редко *)•
Образоваше пелор!й, т. е. превращеше неправильного венчика въ правильный есть,
безъ сомнЬшя, атавизмъ, такъ какъ правильная Форма венчика является более простой н,
следовательно, бодйе древней. Обратныхъ переходовъ, т. е. правильней) венчика въ непра-
вильный, во всЬхъ цвйтахъ какого нибудь индивидуума никогда не наблюдалось ).
Превращеше раздельнолепестного венчика въ сростнолепестной нужно считать за
прогрессивный прнзнакъ; обратный переходъ, наблюдающ(йся чаще, за регрессивный.
Увеличеше числа завязей у миндальныхъ (вишня, миндаль) можно, повкдимому, раз-
сматривать за прогрессъ, т. е. усложнение организацш, хотя вполне определенно въ дан-
ною» случае трудно высказаться.
1) MmL Тег. р. 820.
2) Что тако! переходъ однако вдоляй возкожегъ,
уиазываетъ яа&иодеа1е Гмльдебрапда, ведано описав-
шее имъ въ Botan. Ceatralbl. (1899, ▼. LXXV1I, р. 177—
179). Именно ва одномъ акземплярй Fachiia coecinea
овъ жашелъ одмм цвйтокъ съ рйзко выражав но!
эхпжоро1е1. Этотъ цвйтохъ ве былъ опущегъ хъ визу,
во быть малравлевъ косо ввержъ. Изъ четырехъ ча«
вхеиствховъ два верхв!е были гораздо хрувжЪе в
Завмвж *аа.-Мм. Отд,
образовал в&что п род* шлема, два жвжв<е Maienxie
был обращевы ввмзъ. Изълеласткмъ одияъ aepuil
былъ очень врупжый и быль ваоравлавъ вмрхъ, ввж-
nil былъ очевь малъ, дм боковые средве! величавы.
Тычивокъ было нормальное число (8), но очень кеедв-
какоио! длины: верхм>е ддивнЬе, ннжше короче. Чрез*
вычайво интересно было бы проелйднть, получатся лн
сЫсеяа отъ этого цеЬтм, ж не окажутся л аакЪчен-
выя особенности намйдстмияынм.
11
387
82
С. ХОРЖИНОК1Й.
Резюмируя все сказанное, можно придти къ такому заключенно: въсилу гетерогенеза
происходить варьящи, какъ прогрессивны», представляюпця дальнейшее совершенствовав
aie, или по крайней мер! усложнение типа, такъ и регрессивный, обнаруживающая возврата къ
бол^е простымъ и по всей вероятности более древнимъ Формамъ; но во второмъ направле-
на, т. е. въ смысле атавизма, происходить более крупныя отклонешя (какъ напр. полуве-
ке простыхъ листьевъ изъ перистыхъ или правильныхъ в£нчиковъ изъ неправильныхъ),
который не могутъ возникать сразу въ прогрессивномъ направлены.
Свойства <етерогенкьт варъхщй. Фнзюлогичестя свойства Формъ, возннкшихъ пу-
темъ гетерогенеза, бываютъ весьма разнообразны: oni проявляются въ особенностяхъ
роста, эпохе и продолжительности цвететя я плодовошешя, отношение къ климатическимъ
Факторамъ и т. п. Каждое изъ этихъ Физюлогическнхъ свойствъ есть въ сущности такой
же призпакъ, какъ и всяюй морфологичесюй. Онъ точно также можеть возникать и насле-
доваться или изменяться, независимо отъ другихъ свойствъ растем я. Но есть одна черта,
которую, какъ кажется, нельзя подвести къ той же категорш, но, быть можетъ, справедли-
вее разсматрнвать, какъ общее свойство гетерогенныхъ варьящй, связанное съ самымъ
способомъ пхъ происхождешя.
Это свойство есть пониженная плодовитость, доходящая иногда до полной расшатан-
ности половой системы. Она выражается въ томъ, что некоторый гетерогенный Формы
вовсе не достигаютъ цвететя, какъ напр. Broussonetia papyrifera var. dissecta, ши цве-
тута чрезвычайно редко и скудно, какъ напр. Robinia Pseudacacia var. umbraculifera, Ti-
lia platyphyloe var. asplenifolia. Друпя же, хотя цвЬтутъ часто и обильно, но совсемъ не
даютъ семянъ, какъ папр. безшпорцевая neiopi» льнянки (Linaria vulgaris) или даютъ се-
мена, но слабыя я въ неболыпомъ количестве (Biota orientalis var. flagelliformis, Robinia
Pseudacacia v. monophylla, Fraxinus excelsior v. monophylla, Robinia Pseudocode var. De-
caisneana и др.).
Подобныхъ примеровъ можно бы привести еще довольно много, особенно относительно
древесныхъ породъ’). Точно такъ же и среди травявистыхъ растетй известны Формы съ
явно пониженной плодовитостью или даже безплодныя. Но съ другой стороны очень мнопя
гетерогенный варьящи и даже, быть можеть, большая ихъ часть обильно цвететь и при-
носить семена, не представляя явныхъ приэнаковъ уменьшенной плодовитости. Даже тах!я
варьящи, какъ махровыя разновидности, у которыхъ главный морфологически признакъ
неизбежно влечетъ за собою атроФ1ю половой системы, оказываются вполне плодовитыми,
если у нихъ метаморфозу подверглись не все органы размножешя. Въ виду этого является
вопросъ, действительно ли пониженная плодовитость многихъ гетерогенныхъ варьяц!й свя-
зана съ самымъ пропессомъ пхъ происхождешя? Не свойственна ли она скорее лишь нй-
которымъ Формамъ, к не составляете ли она ихъ отличительной черты, не имеющей общего
1) Ср. по атому поводу датам Каррмра отаосптмшо рашгомдвосте* Robinia (Rar bort 1879, р.
109—111).
388
птхроншввмсъ В ЭВОЛЮЦИ.
83
эиачешя, подобно другимъ физюлогичсскнмъ и морфологнческкмъ пркзнакамъ гетероген-
выхъ разновидностей?
Въ настоящее время дать на этотъ вопросъ вполн! точный ответе, обоснованный на
Фактахъ, нйть еще возможности. Въ пользу того, что уменьшенная плодовитость есть явле-
aie общее, связанное съ гетерогенезомъ, говорить главиымъ образомъ аналогия съ Фактами,
наблюдаемыми при гибридизац1и. Что у помйсей вообще понижена плодовитость, и что это
ихъ общее свойство, связанное съ самымъ происхождешемъ этихъ Формъ, есть Факте обще-
признанный. Но и зд£сь умепыпеше плодовитости выражается въ очень различной Степени
у разныхъ гибридовъ. Есть некоторые гибриды, которые даже нс могугъ достигнуть
цвйтешя или цвйтутъ весьма скудно и рЬдко. Друпе, и именно большая часть ихъ, цвйтутъ
обильно, но не прнносятъ сЪмянъ или прнносятъ ихъ въ маломъ количеств! и плохо разви-
тыхъ. Третьи, наковецъ, обильно цвгЬтутъ и прнносятъ сймена, такъ что понижение плодо-
витости у нвхъ почти или вовсе незаметно. Ч^мъ больше отличаются между собою родона-
чальныя Формы, гЬмъ бол!е сильное потрясение претерпйваетъ продукте ихъ скрещивашя,
т. е. пом!сь, и тймъ сильнее отличается она отъ материнского растешя. Въ такомъ случа!
помесь является вполн! безплодной или очень мало плодовитой. Чймъ ближе между собою
родоначальныя Формы, гЬмъ меньшее потрясете испытываете поыЪсь и соответственно съ
этимъ плодовитость ея страдаете мало или почти не страдаете. То же самое въ общихъ
чертахъ мы наблюдаемъ и при гетерогекезй: у варьящй, сильно отклоняющихся отъ типа,
плодовитость страдаете сильно, при бол!е же слабыхъ отклонешяхъ она понижается срав-
нительно слабо. Наконецъ нужно заметить, что въ слЪдующихъ покол!а1яхъ помЪсей пло-
довитость повышается по nipt того, какъ устанавливаются болйс или менйе постоянный
гибридныя расы. Есть основание думать, что и у гетерогенныхъ варьящй происходить то же
самое, и что первоначальный мало плодовитыя варьящй въ сл!дую1дихъ покол!н1яхъ, по
м!рЬ того, какъ устанавливается ихъ конституция, делаются бол!е плодовитыми. Намекъ
на это даете вамъ та разница въ плодовитости гетерогенныхъ Формъ отъ древесныхъ по-
родъ и травянистыхъ растенШ, которая очень сильно бросается въ глаза. Гетерогенный
разновидности древесныхъ породъ очень часто отличаются пониженной плодовитостью, при
чемъ нужно помнить, что обыкновенно мы им!емъ здЬсь д!ло съ частями оригинального
экземпляра, сохраняемого въ культу pi и размножаемого безполымъ путемъ. Отъ травя-
нистыхъ же растенкй гетерогенный разновидности обладаюте обыкновенно нормальной
плодовитостью, но зд!сь подъ разновидностью мы лонвмаемъ не оригинальную варьящю, а
сумму послйдующихъ поколШй, который, быть можете, значительно уже улучшили свою
плодовитость. Ибо известно много травянистыхъ гетерогенныхъ варьящй, который возни-
кали въ развое время, во затймъ безсл!дно пропадали, не оставввъ потомства.
Пониженная плодовитость помесей выражается нагляднее всего въ развили пыльцы.
У Формъ безплодныхъ очень часто пыльники сморщиваются и не заключимте въ себ! ни
одного зерна цвйтена. У Формъ съ пониженной плодовитостью въ пылышкахъ развивается
цвйтеяь, но на ряду съ нормальными пылинками находится много недоразвитыхъ. Эти no-
il*
389
84
0. К0РЖКИСХ1Й.
следшя встречаются въ цветет я плодовитыхъ гибридовъ въ размере, напримеръ, 10—
20%, указывая своимъ присутствуешь на известную расшатанность половой системы. У ге-
терогенныхъ варьяцзй мы находимъ то же самое. У Формъ вполне безплодныхъ все пыль-
ники бываютъ атрофированы, какъ напр. у беэшпорцевой пелорш льнянки (Linaria vulga-
ris), или лишь въ немногяхъ пыльнвкахъ развивается небольшое количество пыльцы, какъ
я убедился относительно Tilia asplenifolia. Но какъ обстоять дело относительно другихъ
гетерогенныхъ Формъ съ пониженной или нормальной плодовитостью, къ сожаленно, нйть
никакихъ наблюдений.
Надо заметить впрочемъ, что плохое состояние пыльцы еще не объясняете намъ без-
плошя пекоторыхъ помесей или гетерогенныхъ Формъ. Пыльцы образуется всегда довольна
значительное количество, и, если даже половина ея недоразвилась, то остального количе-
ства было бы вполне достаточно, чтобы обезпечить опылеше пестика. По всей вероятности
параллельно происходятъ глубокая изменения и въ женскомъ аппарате, именно въ яйцевой
клетке. Но въ чемъ состоять эти изменен!я, неизвестно. По этому поводу не существуете
никакихъ наследован!# ни относительно гетерогенныхъ варьящй, ни помесей.
На основании всего сказанного можно придти къ заключена, что угнетенность поло-
вой системы варьящй есть явлеше общее, связанное съ гетерогенезомъ, хотя и выражаю-
щееся въ различной степени подобно тому, какъ у помесей. Если точный наблюдения и
иэследован1я подтвердить эту аналопю, то можно будете придти къ определенному взгляду
и на причину этого понижения плодовитости, которое до сихъ поръ, сколько мне известно,
оставалось необъясненнымъ. Можно будете сказать, что всякое нарушеше наследствен-
ности, всякая, такъ сказать, расшатанность конститущи вида, будете ли она являться
слЬдств!емъ скрещивания или гетерогенеза, всегда влечете за собой угветея!е половой си-
стемы. Но когда въ последующихъ поколеншхъ, изъ гетерогенной ли варьящи иля помеси,
выработывается постоянная раса, въ которой, следовательно, все элементы приходить вновь
въ равновесие, тогда само собою разумеется, что и половая система постепенно приходить
въ нормальное состоян!е.
Изъ другихъ Физшлогическихъ свойствъ слйдовало бы обратить внимание на одну осо-
бенность, замечаемую главнымъ образомъ въ древесныхъ породахъ. Именно очень многш
разновидности этихъ последввхъ отличаются сравнительно съ типомъ слабостью своего
роста в меньшей выносливостью относительно мороза. Нечего к говорить про пестролист-
ныя Формы, которыя все гораздо слабее типичныхъ, медленнее растуть и отличаются боль-
шой нежностью. Эти свойства въ нихъ есть прямое следств1в недостаточного развит!я хло-
рофилла. Но и друпя разновидности, не имйющ!я, невидимому, никакого органического де-
фекта, очень часто оказываются слабее установившихся расъ. Такъ напримеръ, пурпурово-
листныя Формы отъ обыкновенной березы1 2), орешника1), ильма (Ulmns campeatrie1) я
1) Per. Сед. реет. I, стр. 142. I 3) Ibidem, стр. 100.
2) Per. Деядр. стр. 63. |
390
ППРОГЖНЫВСЪ I эволюци.
86
остролистнаго клева (Acer platanoides) ве выдерживаютъ климата Петербурга требують
ва зиму покрышки.
Изъ Формъ съ разоренными листьями Sambucus nigra var. laciniata еще ntatrte,
тЬгь типъ, в въ Петербург^ безъ покрышки совершенно вымерзаетъ ’). Также Acer pla-
tanoides var. dissoctnm, Tilia platy phylloa v. laciniata °), Ulmus campestris var. adianthifblia
a cucullata, Sambucus racemosa var. plumosan serratifolia”) не выдерживаютъ зимъ Петер-
бурга безъ покрышки, тогда какъ типы вполне устойчивы. Но по словамъ Э. Л. Регеля *),
A Inns glutiuosa var. quercifolia laciniata и incisa столь же выносливы, какъ атипичная ольха.
Безплодная разновидность обыкновенной калины ил такъ называемая Boule de neige
(Viburnum Opulus v. roseum, или v. sterile) гораздо uisate типа и требуегь на зиму при-
крыли (завязыванья соломой *). Meaie заметно это явлев(е на плакучпхъ или пирамидаль-
ныхъ разновидностях!.. ТЬмъ ве менЬе Ulmus montana v. pendula и fastigiate0) и Fraxinus
exelsior v. pendula7) требують прикрыли на зиму, ве говоря уже о пирамадальномъ сереб-
ра стомъ топол-Ь (Populus alba Bolleana) и итальянскоиъ тополЬ (Populus nigra pyramidalia),
которые растуть только въ южной половин^ Poccin.
МнЬше объ органической слабости разновидностей сравнительно съ типами очень рас-
пространено среди садоводовъ и культиватором», в мнопе авторы счигаютъ его общимъ
свойствомъ всЬхъ отклонен^’). Анализируя Факты, которые Mat удалось собрать по этому
поводу, я ве могу согласиться съ такнмъ заключещемъ. Но все-таки нужно признать, что
меньшая выносливость и слабость конституция составляюгь довольно часто отличительную
черту гетерогенныхъ варьящй, и, быть можетъ, являются нередко слЬдств!емъ самаго про-
цесса ихъ происхождения. Такъ какъ притомъ это свойство замечается именно у древес-
ныхъ породъ и многол!тниковъ, сохраняющихся въкулыурЬ въ орипшальвомъ экземпляра,
и не наблюдается у однолйтннхъ растений, то можно предположить, что у эткхъ посл£днихъ
оно сглаживается въ дальнЗДпихъ покол£шяхъ при размвожети семенами.
Наслпдстеенность гетерогенных» отхлонетй. Rd отклонения, возникала путемъ ге-
терогенезиса, не только вполнЬ постоянны при вегетативномъ размножен», но передають
свои признаки также н по наследству. Это общее ихъ свойство, чрезвычайно характерное
для даннаго явлешя, хотя проявляющееся въ различной степени у раэныхъ варьящй. Такъ
известны Формы, какънапр. Fragaria monophy Па, Chelidonium laciniatnm и Begonia eemperflo-
rens, var. rosea, которыя сразу по возникновенш оказались вполне постоянными при размноже-
ны семенами. Друпя даютъ лишь известный проценгь, то болышй, то менышй, Формъ, тож-
дественныхъ съ материнскямъ растешемъ. Наконецъ, были рЬдк!е случаи, когда возникла
варьяцш совершенно не воспроизводили свонхъ прнэнаковъ въ потомствй.
Причина такого разнообразия, повидимому, двоякаго рода. Во первыхъ, ed гетероген-
1) Ibidem, стр. 168.
2) По сообщению Р. Э. Рогом.
3) Per. я Косс. Кат. 1899, стр. 21 я 28.
4) Рог. Девдр. стр. 47.
б) Там-ъ же, стр. 1б&
6) Тамъ же, стр. 100.
7) По сообщая!» Р. Э. Регем.
8) Рог. Сод. я. I, стри 141.
391
86
C. KOFJKHHCKlI.
ныя Формы, по крайне! м!р! въ течете первыхъ покагЬтй после возникноветя, склонны
прв размножети семенами отчасти возвращаться къ типу. Даже самый устойчивыя расы,
какъ напркпгЬръ, Fragaria monophylla, все-таки даюта постоянно небольшой процента ci-
янцевъ, тождествеиныхъ съ типично! Формой. То же наблюдается и относительно другихъ
Формъ въ больше! или меньше! степени. Во вторыхъ, гетерогенный варьящй могутъ опы-
ляться цветенью типично! Формы, в тогда огь нихъ получается смешанное потомство, въ
которомъ одни экземпляры сходны съ матерннскимъ растешемъ, другие обнаруживать
возврата къ типу и третьи, иакопецъ, представляютъ среднее признаки между первыми и
вторыми. Если припомнить, что гетерогенный варьящй происходить всегда въ едннствен-
помъ экземпляр!, то легко понять, что при распространенности въ природ! перекрестнаго
опылешя гораздо больше в!роят1я получить отъ изучаемо! Формы смешанное потомство,
ч!мъ чистое. И то непостоянство результатовъ отъ посева с!мянъ, на которое мы им!ли
случай много разъ указывать выше, по большей части очень легко объясняется т!мъ,
росли ля по близости огь варьящй типичные индивидуумы или н!тъ, т. е. насколько велика
была вероятность опылешя разновидности цветенью типа.
Въ виду сказанваго удивнтельнымъ является скорее тотъ Факта, что некоторый геге-
рогенныя варьящй съ самаго начала оказались вполне постоянными, не давая сжйшаинаго
потомства и, невидимому, не вступая въ перекрестное оплодотвореше съ типической Формой.
Это заставляета предполагать, что некоторый варьяцш, возникшая путемъ гетерогенеза,
столь отличаются по свое! внутренне! коиституцш огь типа, что даже являются неспособ-
ными или мало способными къ скрещивание съ зтимъ посл!днимъ. Однако это предположен
Hie, чтобы быть доказаннымъ, должно быть проверено точными опытами и наблюдениями,
который, къ сожал!шю, еще ннкЬмъ не были сделаны.
Намъ неизвестно, почему, наприм!ръ, отъ Ulexeuropaeus v. inermis не удалось вывести
постоянную расу, но такче знатоки д!ла, какъ Верло и Ноденъ, объясняли неусп!хъ попы-
токъ лишь недостаточно! настойчивостью и неполпымъ изилировашемъ варьящй. Мы зна-
емъ кром! того, что ноеЬвы такихъ Формъ далеко не всегда бываютъ удачны. Однако,
если гетерогенная разновидность воспроизводится въ потомстве хотя бы въ ничтожною
процент!, то путемъ отбора наиболее* чистыхъ инддвцдуумовъ и ихъ строгая изолирования
отъ типа, всегда удается, если не сразу, то въ нисколько покол!шй, Фиксировать разновид-
ность, т, е. получить изъ нея постоянную при размножеши семенами расу.
Причина гетерогенеза. Наследственность изменчивость, отъ чего бы нк замедли
ихъ реальвыя причины, можно представлять себе какъ двЬ силы, скрытыя въ организме,
две тенденщи, находящаяся въ антагонизм!. При нормальныхъ условхяхъ, т. о. въ устано-
вившихся, не расшатанныхъ расахъ безусловно господствуета наследственность, опреде-
ляющая тождество сл!дующихъ одно за другвмъ покол!ш!. Что же касается до тевден-
щн изменчивости, то она не проявляется непрерывно. Въ течете многихъ покол!н1! она
должна, такъ сказать, накоплять энергио для того, чтобы наковецъ преодолеть силу наслед-
ственности и дать начало гетерогенно! pact.
392
ГИКРОППЗВСЪ И ЭВОЛЮЦ1Я.
87
Спрашивается теперь, как!е Факторы содействують этому накоплен^ энерпн измен-
чивости. Изъ вс!хъ виЬшнихъ условий, разобранныхъ нами выше, мы призвали, что лишь
одинъ Факторъ благопрхятствуетъ проявлешю изменчивости, это—хорошее питаше и вообще
процветите организма. Подобно тому какъ въ Mipe человека творчество, т. е. произведете
чего нибудь новаго, выходящаго изъ установденныхъ рамокъ, всегда требуетъ избытка
энергш; подобно тому, какъ для рождешя renin необходимо, какъ думаютъ, несколько поко-
xiaiH здоровыхъ и уранновешепныхъ людей, такъ, невидимому, и для проявлена гетеро-
генеза необходимо, чтобы несколько предыдущих! покодЫй пользовалось благопр1ятными
услов1лми для своего развипя. Въ этомъ сходятся единодушно мнения всЪхъ культиваторов!.
Что же касается до самаго механизма явлешя, т. е. до непосредственной причины
возникновешя гетерогенныхъ отклояешй,то,какъ можно думать, она заключается въ какихъ
нибудь иэменешяхъ, совершающихся въ половыхъ продуктах! материнскаго растен’ш, т. е.
въ пыльце или сЬмяпочке. Но предполагать, чтобы эти процессы совершались до оплодо-
творешя, довольно трудно, ибо тогда возникппя варьящи заключали бы въ себе лишь на
половину иэпгененную природу (т. е. если измененная семяпочка опыляется нормальной пы-
линкой или наоборотъ), чему противоречить цельный и устойчивый характеръ многихъ ге-
терогенныхъ варьяцш. Следовательно вероятнее, что изменения наступают! въ семяпочке
во время или после оплодотворстя. Но что же за причина этпхъ измененхй, и отчего, дей-
ствуя на одну семяпочку, она не простирает! своего влын1я на другую, находящуюся въ
той же завязи, остается совершенно непонятным!, какъ и многое другое въ этомъ таин-
ственяомъ явлешя.
393
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ...........................................3
I. ИСТОРИЯ ЭВОЛЮЦИОННОЙ БИОЛОГИИ 5
Бабков В.В.
ЭВОЛЮЦИОННЫЙ И РАЗВИТИЙНЫЙ подход
В ТРУДАХ РУССКИХ БИОЛОГОВ.............................5
Положий А.В.
ИСТОРИЯ ФОРМИРОВАНИЯ ПОНЯТИЯ
О ВИДЕ ВЫСШИХ РАСТЕНИЙ................................29
11. ОБЩИЕ ПРОБЛЕМЫ ЭВОЛЮЦИИ 37
Евгеньев М.Б., Зеленцова Е С., Полуэктова Е С., Пятков К.И.,
Великодворская В.В., Шостак Н.Г., Лезин ГТ.
ИНВАЗИЯ МОБИЛЬНЫХ ЭЛЕМЕНТОВ - ПРИЧИНА
ВЗРЫВНОГО САЛЬТАЦИОННОГО ВИДООБРАЗОВАНИЯ.............37
Корочкин Л И.
ГЕНЫ, ОНТОГЕНЕЗ И ПРОБЛЕМЫ
ЭВОЛЮЦИОННОГО РАЗВИТИЯ................................49
Северцов А.С.
ПРИЧИНЫ ЭВОЛЮЦИОННОГО СТАЗИСА.........................73
Северцов АС.
МИКРОЭВОЛЮЦИЯ И ФИЛОЦЕНОГЕНЕЗ........................89
Сергеев М.Г
ВИДООБРАЗОВАНИЕ И ЭВОЛЮЦИОННАЯ БИОГЕОГРАФИЯ.........106
Скворцов А.К.
ВИД В СИСТЕМАТИКЕ СЕГОДНЯШНЕГО ДНЯ................... 122
Стегний В.Н.
СИСТЕМНАЯ РЕОРГАНИЗАЦИЯ ГЕНОМА ПРИ ВИДООБРАЗОВАНИИ....128
Чадов Б.Ф.
МУТАЦИИ, СПОСОБНЫЕ ИНИЦИИРОВАТЬ ВИДООБРАЗОВАНИЕ.....138
III. ЭВОЛЮЦИЯ РАСТЕНИЙ................................
Бердников В.А., Горель Ф.Л., Костерин О.Э., Богданова В С.
ПРОИСХОЖДЕНИЕ В-ХРОМОСОМ У РАСТЕНИЙ.
ТЕОРЕТИЧЕСКИЙ И ЭКСПЕРИМЕНТАЛЬНЫЙ ПОДХОДЫ...........163
394
Куприянов А.Н.
ВИД С ТОЧКИ ЗРЕНИЯ ПРАКТИЧЕСКОГО БОТАНИКА.............. 179
Малышев Л.И.
СИНДРОМЫ В СРАВНИТЕЛЬНОЙ ФЛОРИСТИКЕ.................... 190
Олонова М.В.
ПРОБЛЕМА ВИДА И СОВРЕМЕННАЯ СИСТЕМАТИКА РАСТЕНИЙ........207
IV. ЭВОЛЮЦИЯ ЖИВОТНЫХ 231
Гордеев М.И.
СТРАТЕГИИ ОТБОРА........................................231
Евдокимов Е.В., Шаповалов А.В.
ДЕТЕРМИНИРОВАННЫЙ ХАОС В ДИНАМИКЕ ПОПУЛЯЦИЙ
КАК ЭВОЛЮЦИОННЫЙ ФАКТОР.................................245
Евсиков В.И., Потапов М.А., Новиков Е.А., Потапова О.Ф.
ВИДОВЫЕ АДАПТАЦИИ НА ПРИМЕРЕ ВЗАИМООТНОШЕНИЙ
РОДИТЕЛИ-ПОТОМКИ У МЛЕКОПИТАЮЩИХ........................264
Островерхова Г.П.
НЕКОТОРЫЕ АСПЕКТЫ ЭВОЛЮЦИИ БИОЦЕНОТИЧЕСКИХ СВЯЗЕЙ
ДВУКРЫЛЫХ НАСЕКОМЫХ С ВЫСШИМИ БАЗИДИАЛЬНЫМИ ГРИБАМИ .... 279
Подобина В.М., Татьянин ГМ.
ВИДООБРАЗОВАНИЕ И ЭВОЛЮЦИЯ ОРГАНИЗМОВ
(ИА ПРИМЕРЕ ФОРАМИНИФЕР)................................293
Харитонов А.Ю.
ПРОБЛЕМА ВИДА И ВИДООБРАЗОВАНИЯ В ОТРЯДЕ СТРЕКОЗ........303
V. ЭВОЛЮЦИЯ ЧЕЛОВЕКА 309
Малолетко А.М.
РАННИЕ МИГРАЦИИ И РАСОВАЯ ЭВОЛЮЦИЯ HOMO SAPIENS.........309
Никитина Т.В., Назаренко С. А.
ФЕНОМЕН МИКРОСАТЕЛЛИТНОЙ НЕСТАБИЛЬНОСТИ КАК ВОЗМОЖНЫЙ
ФАКТОР ПРЕНАТАЛЬНОГО ОТБОРА У ЧЕЛОВЕКА..................325
Пузырев В.П.
ГЕНЕТИКА НАСЕЛЕНИЯ, ГЕНОГЕОГРАФИЯ И ГЕНОМНАЯ МЕДИЦИНА
(к проблеме генофондов народонаселения).................342
Степанов В.А., Пузырев В.П.
В ПОИСКАХ «АДАМА»: ЭВОЛЮЦИЯ ГАПЛОТИПОВ Y-ХРОМОСОМЫ
В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА...................................358
ПРИЛОЖЕНИЕ
С Коржинский ГЕТЕРОГЕНЕЗИС И ЭВОЛЮЦИЯ (избранные главы).371
395
ЭВОЛЮЦИОННАЯ
БИОЛОГИЯ
Материалы конференции
«Проблема вида и видообразование»
Том 1
Под редакцией
В.Н. Стегния
Редактор К.Г. Шилько
Оригинал-макет В.К. Савицкого
Дизайн обложки А.Ю. Шиняковой
Лицензия ИД № 00208 от 20.12.99 г.
Подписано к печати 28.06.2001 г. Формат 84x108/32
Бумага офсетная. Гарнитура Times. Печ. л. 12,38.
Усл. печ. л. 20,8. Тираж 250 экз. Заказ № 049
Томский государственный университет
634050, г. Томск, пр. Ленина, 36
участок оперативной ризографии
и офсетной печати
Редакционно-издательского отдела ТГУ
Лицензия ПД № 00455 от 15.11.99 г.
Пузырев
Валерий
Павлович,
чл.-корр. РАМН,
НИИ медицинской
генетики ТНЦ СО РАМН,
г. Томск
Харитонов
Анатолий
Юрьевич,
Д.6.Н.,
Институт систематики
и экологии животных
СО РАН,
г. Новосибирск
Чадов
Борис
Федорович,
Д.6.П.,
Институт цитологии
и генетики СО РАН,
г. 11овосибирск
Подобина
Вера
Михайловна,
д.г.-м.н., профессор,
Томский
государственный
университет
4
Бабков
Василий
Васильевич,
Д.6.П.,
Институт истории
естествознания РАН,
г. Москва
Бердников
Владимир
Александрович,
д.б.н, профессор,
Институт цитологии
и генетики СО РАН,
г. Новосибирск
Гордеев
Михаил
Иванович,
д.б.н.,
Институт общей
генетики РАН,
г. Москва
Евгеньев
Михаил
Борисович,
д.б.н., профессор,
Институт молекулярной
биологии РАН,
г. Москва