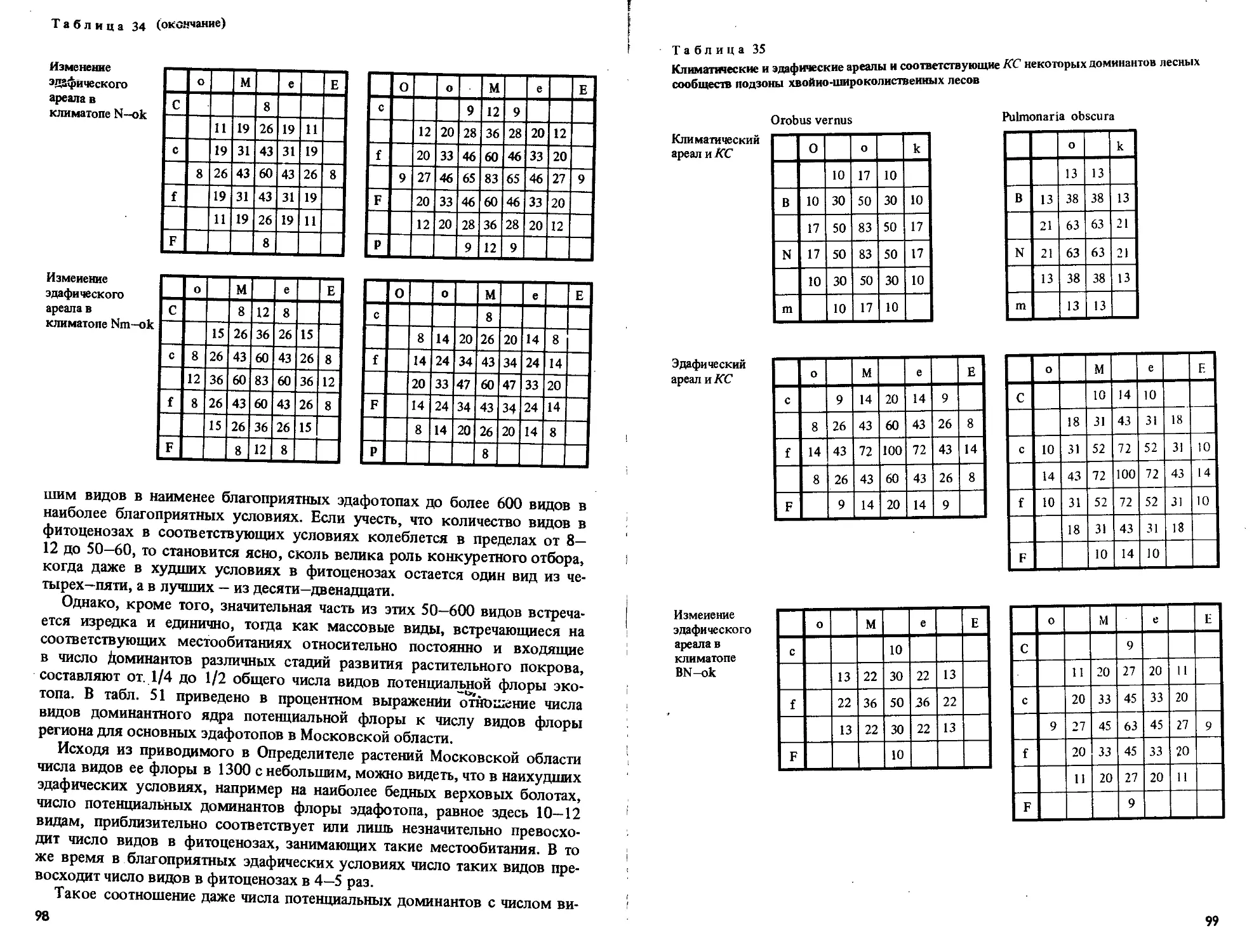
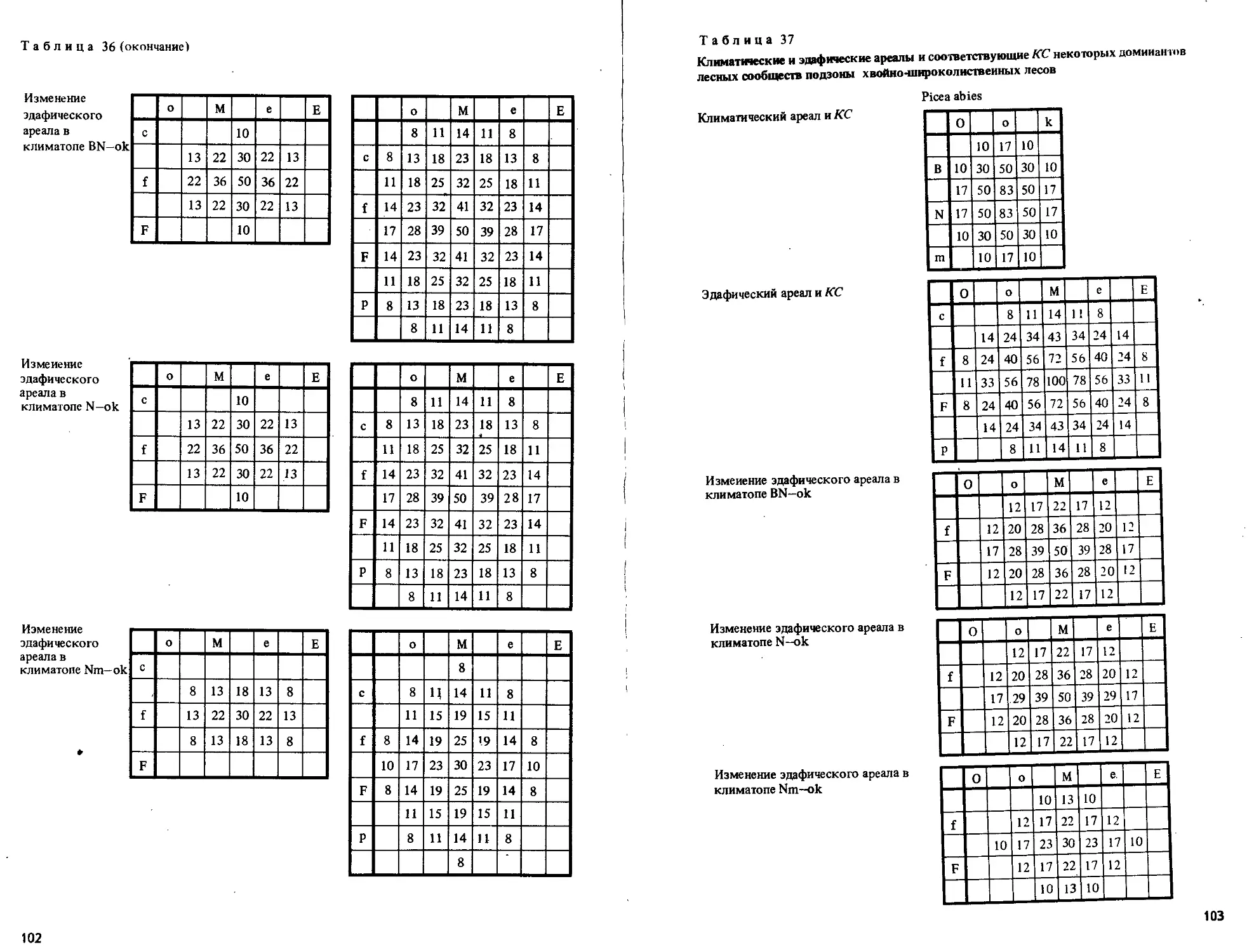
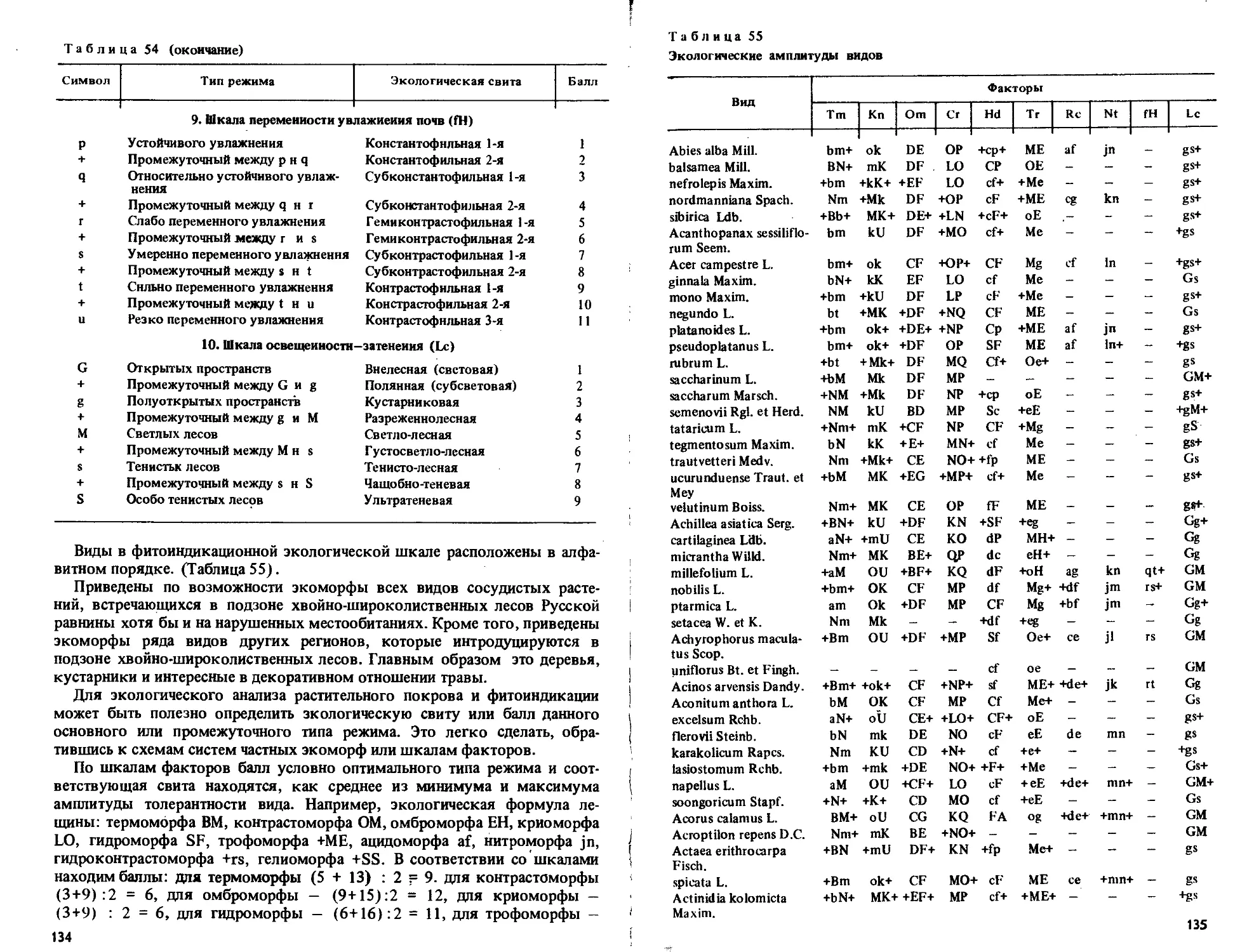
/















































































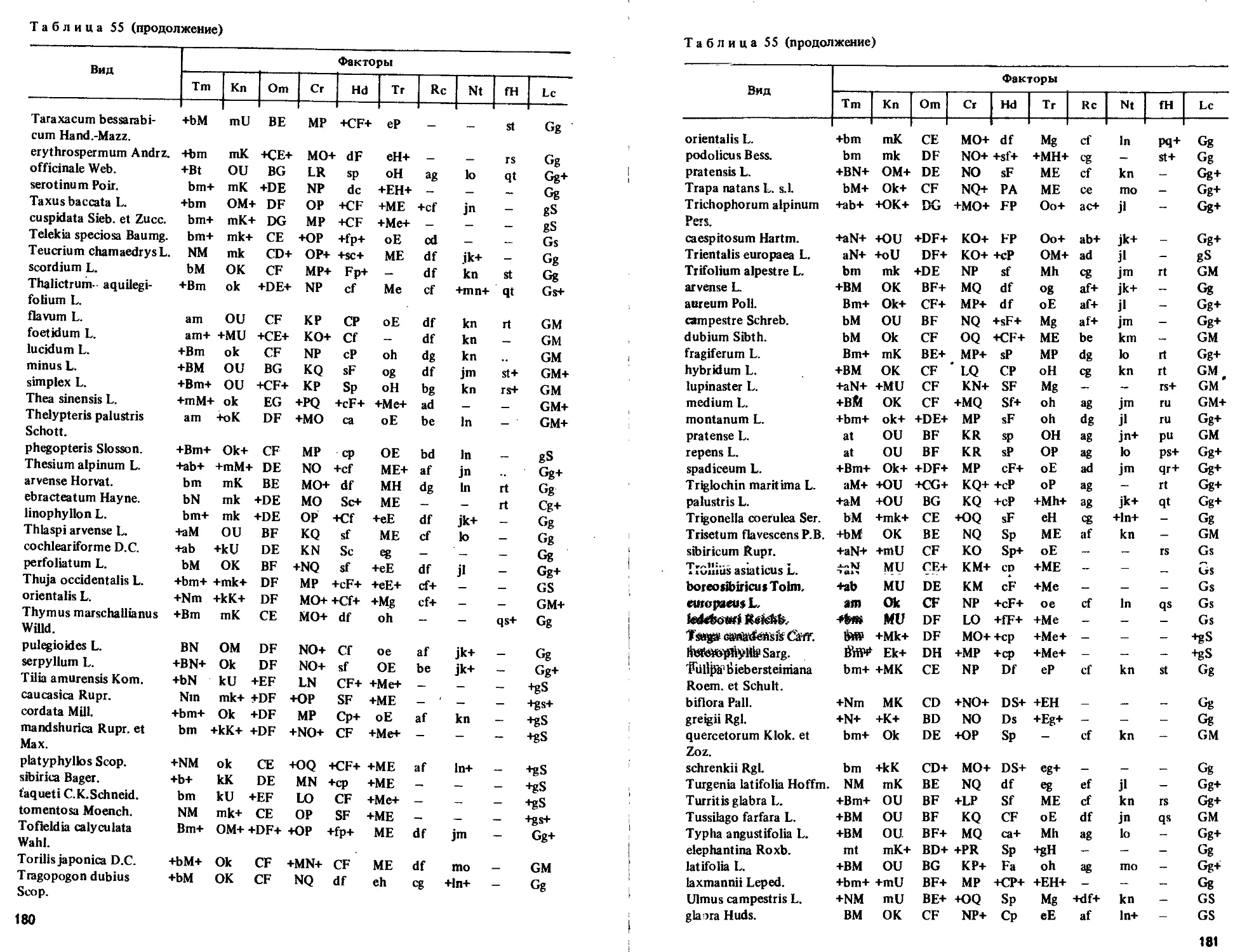
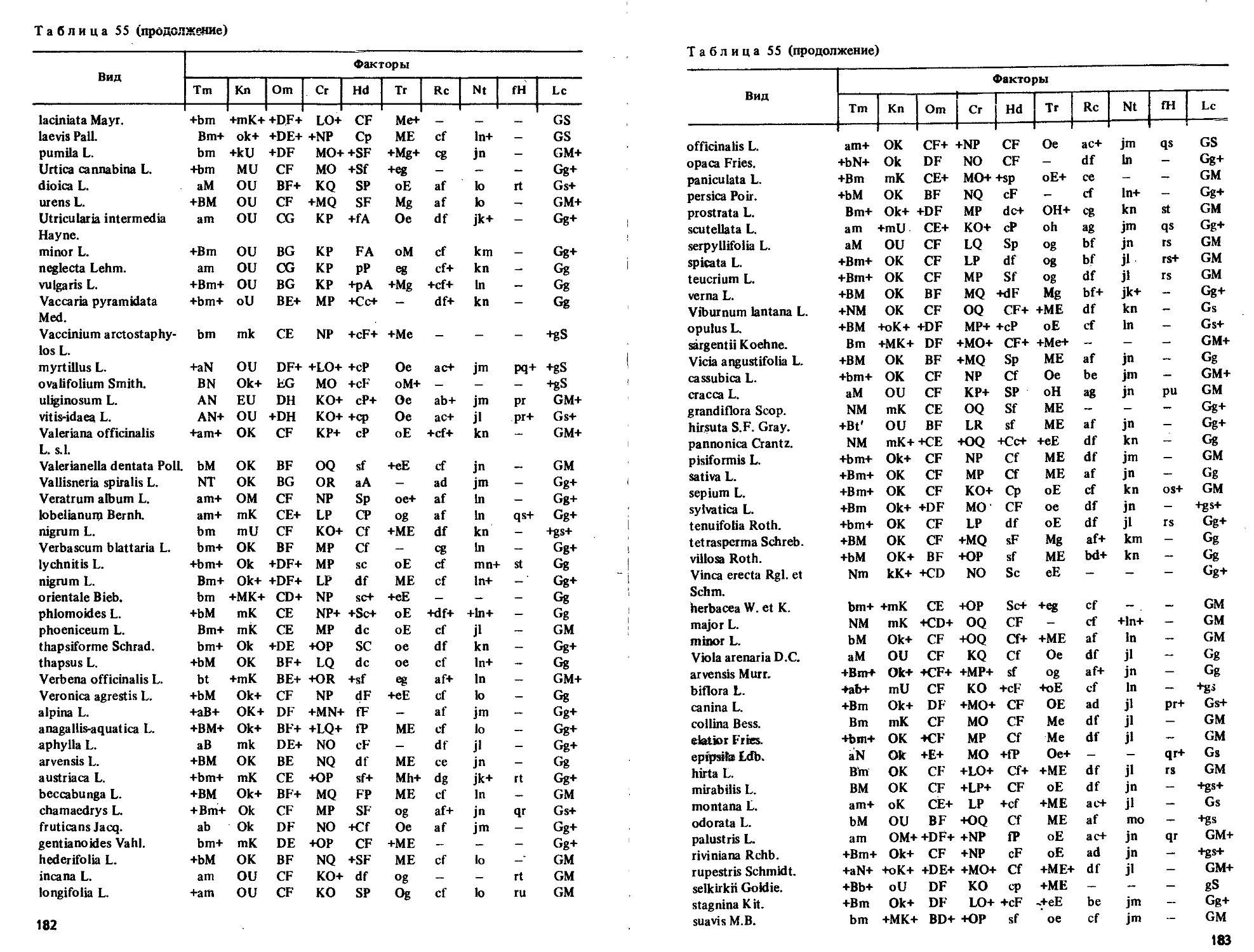












Текст
ФИТОИНДИКАЦИЯ
ЭКОЛОГИЧЕСКИХ
РЕЖИМОВ
В ПОДЗОНЕ ХВОЙНО-
ШИРОКОЛИСТВЕННЫХ
ЛЕСОВ
Аа
1 АВ ,
аВ t Ab
3 ab AN
, Bb aN Am
BN am AM
, Вт aM At
bm BM at
Nm bM Bt aT
m t NM bt BT
mM Nt bT
M mt NT
Mt mT
L T MT
Tt
bN
AT
АКАДЕМИЯ НАУК СССР
ЛАБОРАТОРИЯ ЛЕСОВЕДЕНИЯ
Д.Н.ЦЫГАНОВ
ФИТОИНДИКАЦИЯ
ЭКОЛОГИЧЕСКИХ
РЕЖИМОВ
В ПОДЗОНЕ ХВОЙНО-
ШИРОКОЛИСТВЕННЫХ
ЛЕСОВ
/ /
ИЗДАТЕЛЬСТВО ’’НАУКА
Москва 1983
УДК 581:522.4
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широ-
колиственных лесов. М.: Наука, 1983.
В книге рассматриваются принципы и методы фитоиндикации условий среды.
В основу индикационных признаков растительного покрова положены амплитуды
толерантности видов.
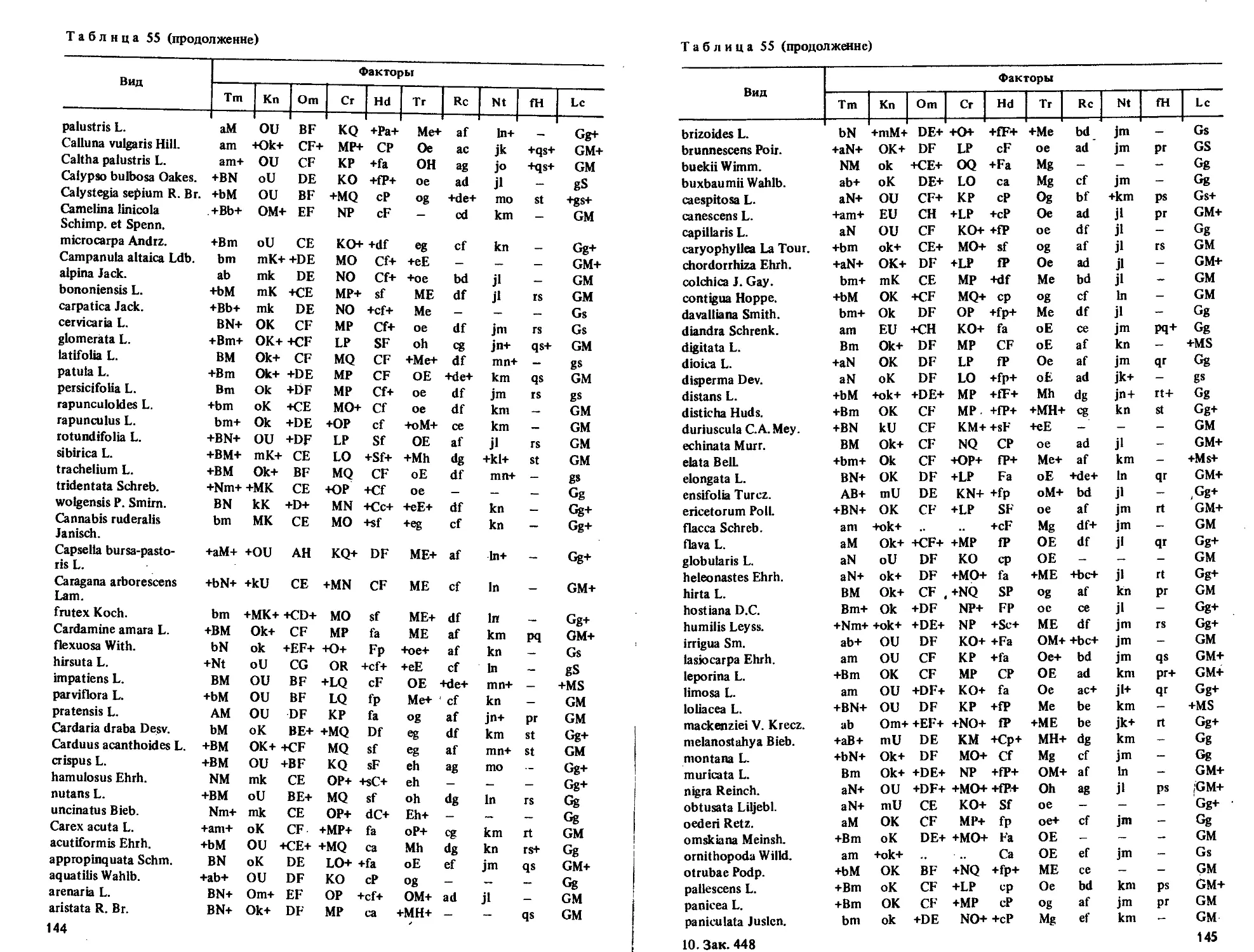
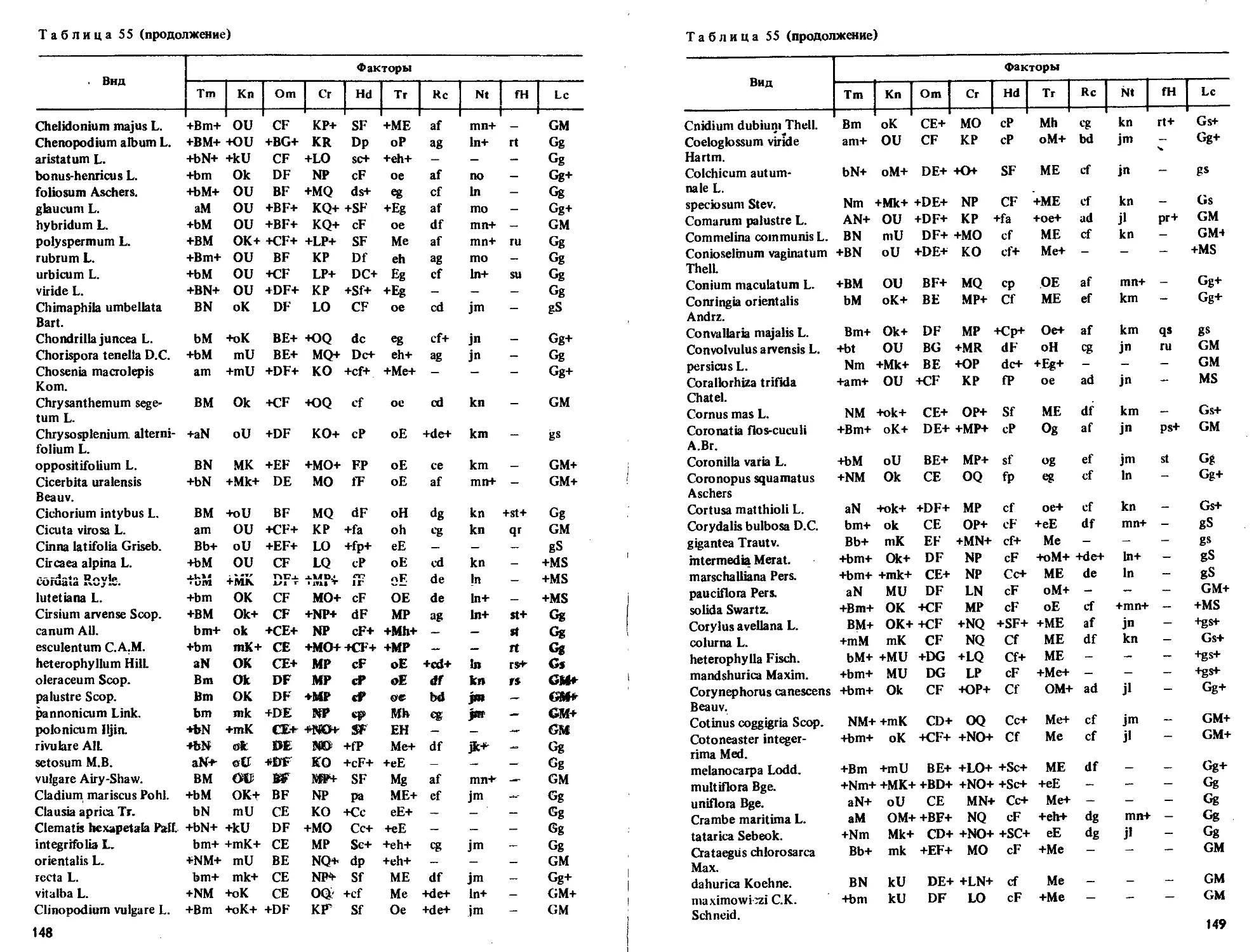
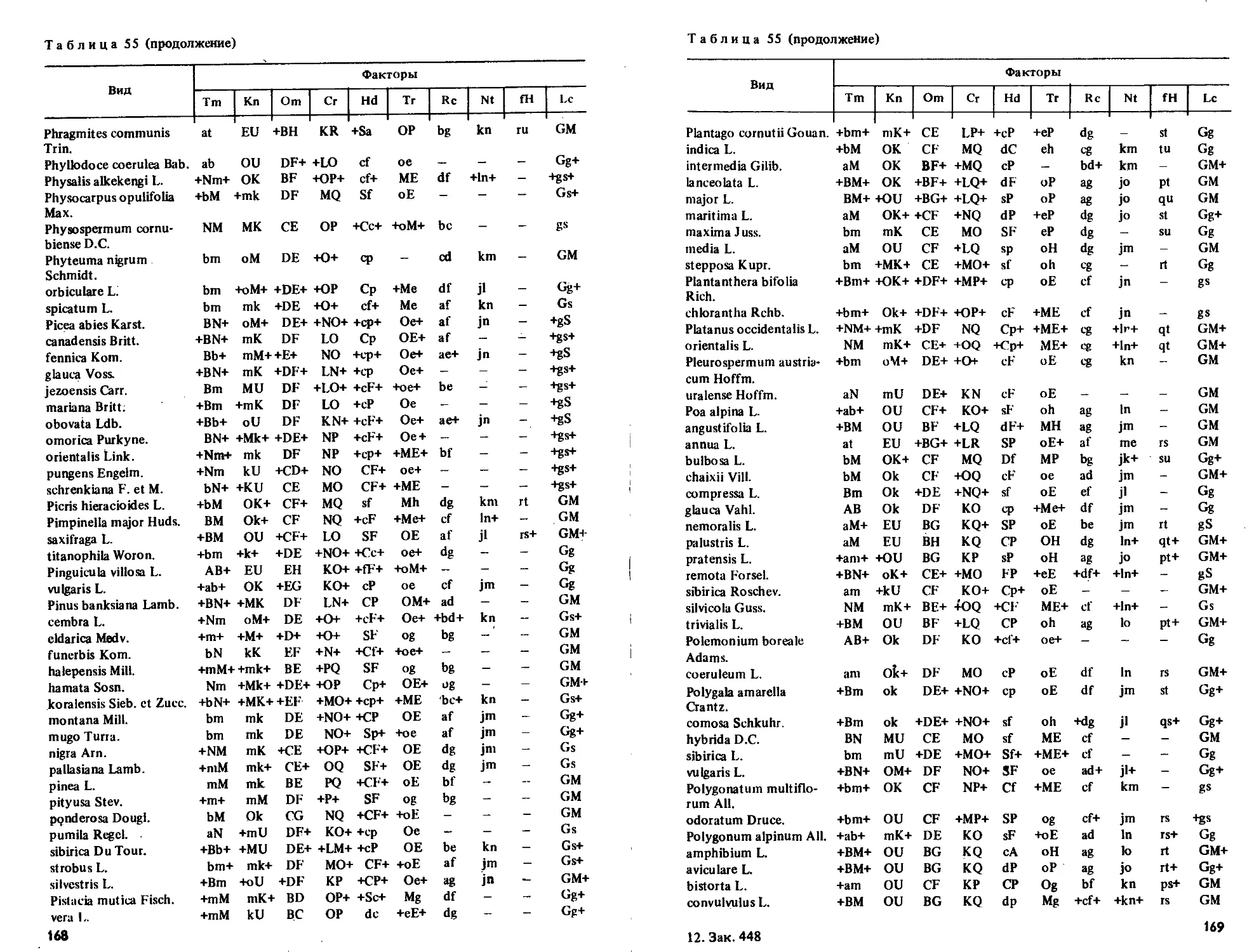
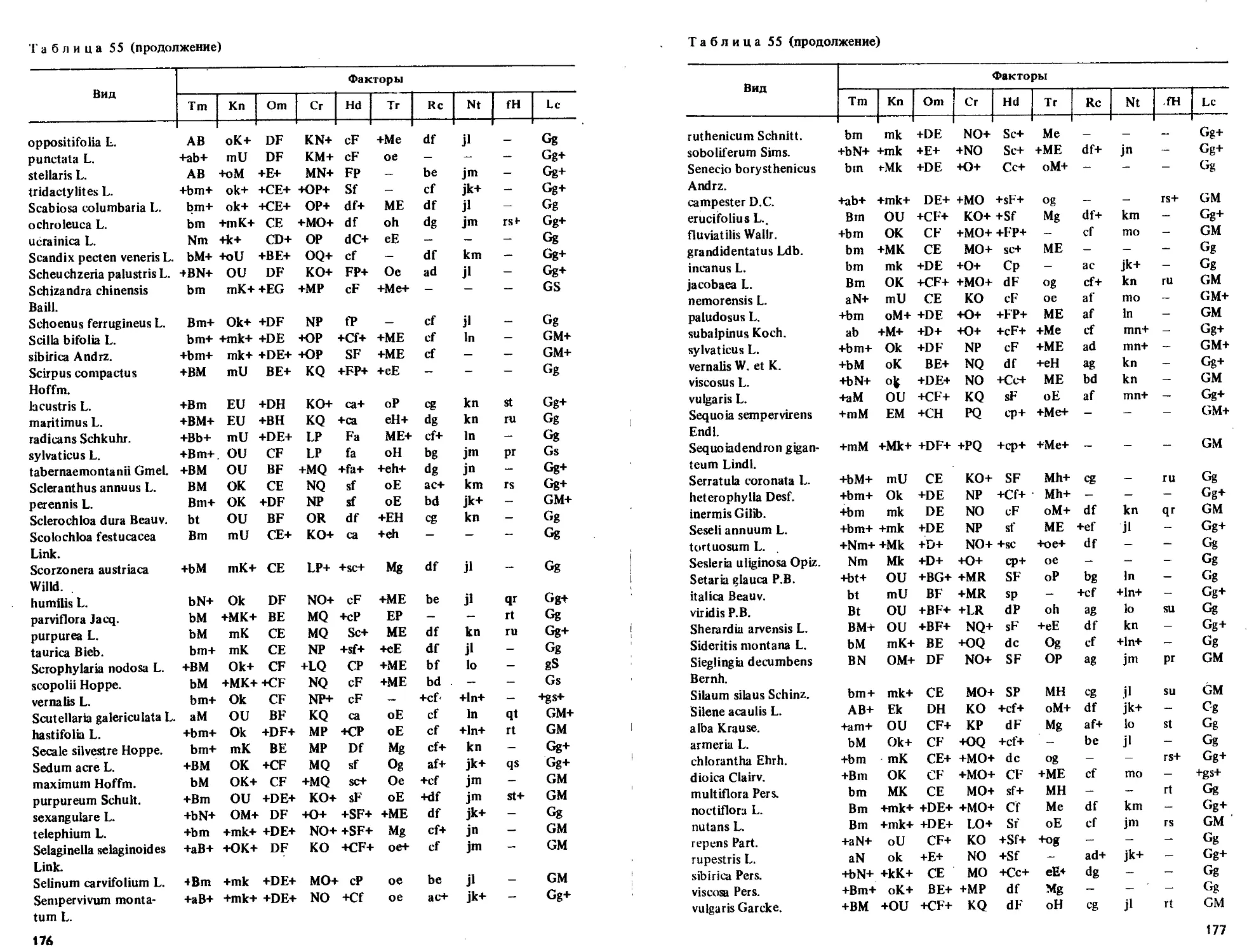
Приводятся таблицы амплитуд толерантности более 2000 видов по их отношению
к 10 экологическим режимам.
Для геоботаников, экологов, фитогеографов, лесоведов.
Табл. 55, ил. 7, библиогр. 69 назв.
Ответственный редактор
канд. биол. наук И.М. КУЛЬТИАСОВ
Ц
3903000000- 380
042(02) -83
340-83-III
©Издательство ’’Наука”, 1983 г.
ГЛАВА I
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ФИТОИНДИКАЦИИ
И РАЗВИТИЕ КОНЦЕПЦИИ Л.Г. РАМЕНСКОГО,
КАК ОДНО ИЗ ПЕРСПЕКТИВНЫХ ЕЕ НАПРАВЛЕНИЙ
Индикация — это, когда мы смотрим на то, что видим, и видим то, что
не видим. Такое шутливое, но принципиально верное определение сущ-
ности индикации любил повторять один из основоположников индика-
ционной геоботаники С.В. Викторов.
Б.В. Виноградов в книге ’’Растительные индикаторы и их использование
при изучении природных ресурсов” главу ’’Основы теории и практики
растительных индикаторов” начал следующим определением: ’’Раститель-
ными индикаторами могут служить как отдельные растения и фитоценозы,
так и особенности строения и состава растений, которые в силу тесной
их связи с различными элементами ландшафта указывают на характер,
распределение и динамику условий окружающей среды” [Виноградов,
1964, с. 5].
С своей стороны Б.А. Быков [1967] в Геоботаническом словаре дает
такие определения: ’’Индикаторы — растения, определяющие условия
внешней среды” и ’’Индикационная геоботаника — раздел геоботаники,
изучающий фитоценологические основы и практические способы исполь-
зования растительного покрова, как показателя условий внешней среды.
Во ’’Введении и фитоценологию” Быков [1970] говорит о фитоценоин-
дикации.
А.А. Корчагин [1971] предлагал особо выделять геоботаническую
индикацию, биогеохимическую индикацию, флористическую индикацию
и т.д., а для совокупности всех этих форм индикации применять объеди-
няющий их термин — фитоиндикация, или ботаническая индикация. Там
же он писал, что фитоиндикация не имеет своего особого фитоиндика-
ционного метода, своего предмета, а потому не может рассматриваться
как особая наука; что теоретической базой ее являются различные раз-
делы ботаники, но особое значение имеет экология растений; что успехи
фитоиндикации и уровень ее исследований зависят от успехов разработки
теоретических вопросов геоботаники, экологии, морфологии и других
разделов ботаники. Наконец, в очень конспективной, хотя и несколько
узкоспециализированной, современной сводке ’’Фитоценология” инди-
кация трактуется, как ’’определение по растительности дискретных клас-
сов среды” [Маркин, Розенберг, 1978].
Резюмируя изложенное выше, можно охарактеризовать фитоиндика-
цию как раздел экологии растений, включая и аутэкологию, и синэкологию,
имеющий задачей в прикладных целях конкретизировать связь определен-
ных ботанических объектов (видов, сообществ, экологических групп
видов, морфофизиологических состояний индивидов в популяциях и
т.п.) с определенными качественными и количественными параметрами
состояния среды, включая как косную, так и биотическую ее часть.
В сводках "Введение в индикационную геоботанику” [Викторов и др.,
1962] и ’’Растительные индикаторы” [Виноградов, 1964] истоки фито-
индикации, в смысле документально зафиксированных наблюдений о
приуроченности определенных растений и растительности к определен-
ным условиям среды, относятся еще к античному времени. Упоминаются
сочинения Теофраста, Катона, Плиния Старшего, Колумеллы, даже Вир-
гиния. Говорится о развитии представлений о растительности, как об
индикаторе вод и почв у народов древнего и средневекового Ближнего
и Среднего Востока. Логически осмысливая данные этнографии, архео-
логии и антропологии, не приходится сомневаться, что наблюдения такого
рода должны были накапливаться у человечества уже с времен палеолита.
Однако основоположниками научного подхода к проблеме фитоинди-
кации почв в России называют Ф.И. Рупрехта, а затем создателей почво-
ведения П.А. Костычева и В.В. Докучаева, а на западе - Хильгарда, Чем-
берлена, Контежана и др.
В дальнейшем вопросами, способствующими развитию фитоиндикации,
выявлением и уточнением закономерностей связи растений и растительнос-
ти с теми или иными элементами среды, занимались в той или иной мере
все ведущие и многие рядовые геоботаники, экологи и почвоведы. Осо-
бенно большую роль в развитии теории фитоиндикации сыграли работы
геоботаников и ботанико-географов, да иначе и не могло быть, поскольку
основой фитоиндикации является экология растений, а синэкология —
ведущий раздел и геоботаники, и ботанической географии.
В уже упоминавшейся сводке ’’Фитоценология” [Миркин, Розенберг,
1978] авторы рассматривают фитоиндикацию в качестве одного из трех
основных методических подходов к изучению связи растительности и
среды: ординации, координации и индикации. К задачам ординации от-
носится упорядочение видов или сообществ вдоль осей конкретных эко-
логических факторов (прямая ординация) или осей максимального
варьирования (непрямая ординация). К задачам координации относится
определение взаиморасположения в системе координат экологических
факторов единиц растительности, которые как бы уже разорвали непре-
рывное варьирование на дискретные звенья. К задачам индикации от-
носится определение по растительности дискретных классов факторов
среды.
Как мы видим, все эти методические подходы тесно связаны между
собой и являются, собственно, разными ракурсами одной сущности: связи
определенных сочетаний растений с определенными параметрами среды.
Поэтому, говоря об одном из этих методов, обязательно затронешь и при-
чинно-следственно связанные с ним оба других.
Действительно, если ординация означает раздельное определение ампли-
туд растительных общностей по градиентам отдельньииэкологических фак-
торов, то координация уже определяет их экологические ареалы в сис-
темах координат, построенных на осях этих факторов (т.е. является
следующей операцией, так как невозможна без предварительной орди-
нации), а индикация определяет, на какие ограниченные определенными
градациями факторов режимы указывает та или иная растительная общ-
ность (естественно, что она является уже третьей операцией, так как не-
возможна без первых двух).
4
Что касается видов индикации, то здесь существуют разные ее под-
разделения, основанные на различных принципах. Викторов [1971]
выделял гидроиндикацию (индикацию глубины залегания водоносных
слоев в аридных районах), геоиндикацию (индикацию разностей почв),
галоиндикацию (индикацию вида и интенсивности засоления) и инди-
кацию отдельных полезных ископаемых. Виноградов [1964] отдельно
рассматривал индикацию климатопа (климаиндикацию), индикацию
почв (педоиндикацию), индикацию природных вод (гидроиндикацию),
индикацию горных пород, полезных ископаемых и геологического строе-
ния (литоиндикацию), индикацию форм рельефа, геоморфологических
процессов и четвертичных отложений, индикацию стаций животных,
индикацию культурного ландшафта (антропогенных нарушений естест-
венной среды). Далее Викторов [1971] выделяет прогнозную индикацию
(прогноз будущего состояния тех или иных элементов природной среды),
стадийно-синхронную индикацию (индикацию современного их состояния)
и ретроиндикацию (индикацию истории развития до современного сос-
тояния тех или иных природных объектов и явлений), причем фитоин-
дикационные методы он считает пригодными лишь для стадийно-синхрон-
ной индикации, а для прогнозной индикации и ретроиндикации считает
приемлемыми методы ландшафтной индикации. Л.Н. Соболев [1978]
говорил о качественной (биоскопической) и количественной (биометри-
ческой) индикации, а также об одновидовой, многовидовой и ценотичес-
кой фитоиндикации.
Наконец, Миркин и Розенберг подразделяют фитоиндикацию на пять
основных направлений.
1) оценка среды по отдельным видам-индикаторам. (Этот вид фито-
индикации имеет очень узкую сферу применения - для оценки жесткой
связи почв или других явлений среды с растениями в условиях аридной
зоны или других, экстремальных по режимам тех или иных факторов,
регионов. Сюда же относится и индикация отдельных видов полезных
ископаемых. Вообще же этот метод авторы считают устаревшим.)
2) Оценка среды по растительным ассоциациям-индикаторам. (Этот
подход был свойствен школе Викторова, пока она почти полностью не
перешла на ландшафтные методы. Миркин осуждает и этот метод из-за
сложности выделения и особенно идентификации ассоциаций, а также
за трудность статистической обработки. Кроме того, и этот метод счи-
тается малопригодным при средних, не экстремальных режимах факто-
ров среды.)
3) Оценка среды по соотношению индикаторных групп видов. (Этот
подход авторы оценивают достаточно высоко, видя его преимущества
в том, что индикаторы выбираются из числа всех видов и сообщества
оцениваются по соотношению участия представителей разных индика-
торных групп. Здесь авторы повторяют одно из основных положений
школы Раменского, что в принципе видов, которые не могут быть ин-
дикаторами, нет и что каждый вид всего информативнее у границ своего
экологического ареала. К этому направлению относятся методы инди-
кации со шкальной или балловой оценкой среды и таким же выражением
экологических амплитуд видов: школы Раменского, Погребняка, Эппен-
берга и другие сходные с названными направления, а также математи-
5
ческий метод Джеглама — метод прямого учета среды и оценки моды
встречаемости и покрытия.)
4) Распознавание образов с использованием списков индикаторов.
Математический метод, требующий для реализации применения ЭВМ.
Возможны два варианта — с учителем и без учителя. Вариант с учителем
выполняется в два этапа: 1) нахождение видов-признаков, характери-
зующих принадлежность описания к классу среды, указанному учите-
лем, и 2) построение правила для такого разбиения градиента среды на
классы, чтобы качество расположения было наилучшим. В решении за-
дачи предлагаются три операции: 1) выбор индикаторных видов с пред-
варительным (методами градиентного анализа) и окончательным (мате-
матическими методами) отбором; 2) выбор алгоритма и обучение и 3) эк-
замен.
5) Распознавание образов с использованием групповых индексов. Тоже
математический метод, требующий применения ЭВМ. В этом подходе
указываются четыре этапа: 1) выделение индикаторных групп видов с
применением расчета средневзвешенной напряженности фактора и дис-
персии; 2) выведение синтетических формул-образцов классов обучения;
3) обоснование количества классов распознавания и 4) экзамен.
Кроме того, авторы особым параграфом рассматривают ’’некоторые
аспекты оптимизации распознавания среды по растительности”, куда вклю-
чают оптимизацию числа информативных видов, оптимизацию длины
обучающей последовательности и оптимизацию числа распознаваемых
градаций фактора.
В заключение же авторы приходят к довольно пессимистическому
выводу, что связь растительности и среды значительно менее прочная,
чем это казалось на этапе типического отбора материалов, и что по дан-
ным растительности среда оценивается в небольшом числе градаций и
поэтому геоботаническая индикация применима в ограниченном числе
случаев. В то же время авторы считают должным признать неоспоримым
преимуществом фитоиндикации тот факт, что растительность принимает
среду в своей естественной шкале, а такая интерпретация условий среды
косвенно объективизирует и решение задачи классификации раститель-
ности на экологической основе.
Особое внимание, уделяемое здесь сводке Миркина и Розенберга, объяс-
няется тем, что на настоящее время она является одним из наиболее ин-
формативных справочных руководств по сочетающему детальный кри-
тический раэбор материала с конспективностью его изложения. Неко-
торую настороженность вызывает лишь очень уж однозначная тенденция
авторов оценивать концепции и методы различных школ и направлений
только с позиций степени их удобства для математических обработок.
Между тем сложные математические методы обработки экологического
и фитоценологического материала, требующие применения ЭВМ, пока
еще доступны далеко не всем исследователям. В настоящее время в био-
ценологических и экологических исследованиях как в ботанике, так и
в зоологии оправдывают себя лишь элементарные статистические методы
обработки материалов, для которых достаточно возможностей настоль-
ных счетных машин. К сожалению, применение сложных математических
методов пока еще не раскрывает новых закономерностей формирования
6
растительных сообществ в условиях динамического взаимодействия с
элементами экологической среды, а лишь подтверждают в общем уже
известные положения геоботаники, экологии и других смежных дисцип-
лин. Иного не приходится и ожидать, так как математический анализ в
любой науке не может освободить нас от присущей этой науке на данном
этапе развития логики интерпретации накопляющихся факторов в свете
той или иной сформировавшейся в ней концепции. Это всего лишь один
из инструментов исследования, призванный облегчить выполнение тру-
доемких операций при обработке фактического материала, и применение
его должно быть оправдано экономией труда и времени, а не желанием
модернизировать методику исследований. Ни одна программа для ЭВМ,
ни одна математическая модель, ни одна схема не в состоянии подняться
выше уровня познаний своих составителей, и высшая квалификация в
математике не компенсирует недостатка эрудиции в биоценологии, эколо-
гии, флористике, биогеографии и тл.
Только что изложенные соображения побуждают считать, что развитие
теории фитоценологии, экологии и фитоиндикации возможно и без повсед-
невного использования ЭВМ, что карандаш и дневник еще долго будут
для фитоценолога привычнее и удобнее как орудия исследования. Много
еще в фитоценологии и экологии задач, проще и легче разрешимых, чем
это требуется для целесообразности использования ЭВМ. Одной из таких
задач, не требующей для решения особо сложных математических опера-
ций, является проблема фитоиндикации условий среды по соотношению
индикаторных групп видов, или зкоиндикации (такое название право-
мерно вдвойне, поскольку этот вид индикации предполагает в качестве
объекта индикации, или индиката, экотопы, или экологические режимы,
а в качестве агента индикации, или индикатора, — экологические объеди-
нения видов).
Наиболее разработанными методиками в решении задач такого рода
являются методики, разработанные и применяющиеся школами после-
дователей Раменского, Погребняка и Элленберга, сущностью которых
является сопоставление балловых или шкальных оценок градиента среды
с аналогичными оценками пределов толерантности видов. Близость к
этим методикам полярной ординации школы Уиттекера, равно как и
изначальная предназначенность самих этих методик к решению задач ор-
динации, лишний раз показывают, насколько тесно связаны между собой
ординация и индикация. В сущности, пытаться разделить три упомянутых
выше направления фитоценологии и экологии: ординацию, координа-
цию и фитоиндикацию, пытаться говорить об одной из них, почти не
затрагивая другие, — довольно неблагодарная задача, трудновыполни-
мая и практически ненужная. Целесообразнее говорить об экологическом
анализе, понимая под ним совокупность проблем и методов ординации,
координации и фитоиндикации. Тогда экологический анализ распреде-
ления растительного покрова (видов, их популяций, сообществ, флор)
вдоль отвлеченно построенных осей отдельных экологических факто-
ров или пофакторных экологических рядов будет представлять собой
прямую ординацию, как методический прием.
Непрямую ординацию, как методический прием, будет представлять
собой экологический анализ распределений растительного покрова вдоль
7
осей максимального варьирования или натуральных экологических рядов.
Координация, как методический прием, будет тогда представлена эколо-
гическим анализом распределения растительного покрова в системах
координат, построенных на осях отдельных экологических факторов
или на различных осях максимального варьирования. В зависимости от
числа осей можно различать плоскостную координацию (две оси), про-
странственную (три оси) и гиперпространственную (больше трех осей).
Так как единственно наглядной координацией является плоскостная,
о чем свидетельствует наличие хорошо разработанных двухфакторных
сеток (например, эдафические сетки Погребняка, Раменского, Зойоми
и др.), то в случае необходимости целесообразно строить не пространст-
венную систему координат (если факторов, которые нужно учесть, три)
и не абстрагировать материал в гиперпространстве (если факторов боль-
ше трех), а составлять иерархическую серию систем координат (например,
глобальную климатическую сетку, для каждой из ее ячей свою эдафи-
ческую сетку и т.д.). Кроме того, по аналогии с ординацией можно раз-
личать прямую координацию, когда системы координат построены на
осях отдельных факторов, и непрямую координацию, когда они построе-
ны на осях максимального варьирования, или натуральных экологических
рядах. Так как натуральные экологические ряды могут быть двоякого
рода: обобщенные, отражающие принципы распределения растительного
покрова по градиенту среды (например, схемы растительности в интер-
претации школы Уиттекера, отчасти эколого-фитоценотические ряды
Сукачева, точнее являющиеся переходными между прямой и обобщен-
ной непрямой координациями), и топографические, отмечаемые на топо-
графическом плане, то и непрямая координация может быть обобщенной
и топографической, последняя является в сущности экологическим кар-
тированием. Наконец, фитоиндикация, как прием, сводится тогда к по-
нятию экологической типологии земель, т.е. будет представлять собой
экологический анализ местностей по растительному покрову (разбиение
территории на мозаику экотопов).
В подходах экологического анализа предпочтение следует отдать мето-
дикам, использующим шкально-балловую оценку среды и соответственно
такую же оценку толерантности видов. Их преимущество перед методом
полярной ординации заключается именно в стандартном характере шкал
и баллов, тогда как при полярной ординации градации и даже располо-
жение самих осей устанавливается для каждого вновь исследуемого ре-
гиона заново. В шкально-балловых оценках среды и экологических ампли-
туд видов, как уже говорилось, наиболее развиты направления школ
Раменского, Погребняка и Элленберга, из которых, на наш взгляд, шка-
лы Раменского обладают тем неоспоримым преимуществом, что охва-
тывают весь возможный диапазон режимов таких важных свойств среды,
как богатство почв влагой и доступными растениям минеральными соля-
ми. Относительными преимуществами шкал Элленберга являются охват
большего числа факторов (включая климатические, отсутствие которых
составляет один из главных недостатков шкал Раменского), одинаковое
количество баллов во всех шкалах и связь с наиболее разработанной в
геоботанике флористической классификацией Браун-Бланке.
Сходными у направлений Элленберга и Раменского являются построе-
8
ния однофакторных экологических рядов: возможно большее число опи-
саний ранжируется по интенсивности проявления каждого фактора, на-
чиная с выделения описаний, соответствующих крайним проявлениям
его интенсивности в данном регионе (и уже это определяет соотноше-
ние масштабов шкал двух направлений, ибо Элленберг разрабатывал свою
методику для средней Европы, а Раменский — для почти всей Восточно-
Европейской равнины). Далее сторонники подхода Элленберга выделяли
лишь по пять градаций для каждого фактора: 1 - если так можно вы-
разиться, прожиточный минимум по данному фактору для самых вынос-
ливых видов; 2 - низкая обеспеченность по данному фактору; 3 - сред-
ние значения интенсивности данного фактора; 4 — относительно хорошая
обеспеченность по данному фактору; 5 — высшая интенсивность фактора
в диапазоне благоприятного для развития растительности режима, удов-
летворяющая потребности самых требовательных видов. Для оценки
толерантности вида использовались 6 баллов: 1 — виды, которые встре-
чаются исключительно при низких значениях данного фактора; 2 — виды
со сходным поведением, но с более широкой амплитудой; 3 — виды, осо-
бенно часто встречающиеся при средних значениях данного фактора; 4 -
виды довольно широких амплитуд, но тяготеющие к высоким значениям
данного фактора; 5 —, виды, встречающиеся почти исключительно при
высоких его значениях; 0 - виды, индифферентные к данному фактору
или имеющие очень широкую амплитуду по нему, перекрывающую все
градации.
Индикация режима фактора дается по преобладанию видов, характер-
ных для этой градации, а при сомнительном результате (нечетком) он
проверяется наибольшим произведением числа видов на сумму их зна-
чимостей (чаще всего индексов обилия-покрытия по шкале Браун-Бланке).
Что касается количества факторов, то разные авторы этого направления
в разных сводках приводят разные их сочетания. Всегда даются харак-
теристики видов по влажности, трофности и кислотностр почв, большей
частью даются числа по азоту почв, температуре и континентальности кли-
мата, часто - по отношению к затенению, к воздухопроницаемости почв
и др. Так, Элленберг [Ellenberg, 1950] дает характеристики видов сор-
няков по шести факторам, а в 1952 г. — характеристики луговых и паст-
бищных растений по пяти факторам (всего по семи факторам: свету,
теплу, влаге почв, кислотности почв, азоту, континентальности и воздуш-
ному режиму почв). Мраз и Самек [Mraz, Samek, 1966] дали характе-
ристики 370 видов лесных растений Чехословакии по восьми факторам:
влажности почв, трофности, кислотности, содержанию азота, порозности,
световому режиму, температурному режиму и континентальности кли-
мата. Зойоми и Пречни [Zolyomi, Precsnyi, 1964] дали сводку по TWR
(отношение к тепловому режиму, увлажнению и кислотности) для лесной
флоры Венгрии (1400 видов). Аналогичную сводку по числам TWR, а также
светолюбию и типам географического ареала 1000 видов лесной флоры
Румынии выпустили в 1977 г. Доница, Пурцеляну и др. [Donita et al.,
1977]. В ФРГ, кроме Элленберга, сводки по экологическим характерис-
тикам луговых трав выпустил Клапп [Klapp, 1971] и в ГДР Хундт [Hundt,
1966]. Сводка швейцарского исследователя Ландольта [Landolt, 1977]
содержит характеристики 3400 видов (с использованием также 5-балль-
9
ных шкал) по девяти факторам: влажности, трофности, кислотности,
гумусированности, засолению и порозности почв, световому режиму,
температурному режиму и континентальности. В 1974 г. Элленберг вы-
пустил новую сводку, где для 1600 видов среднеевропейской флоры дал
характеристики по 15 факторам, детализировав разбивку рядов факторов на
градации: 12 градаций в ряду фактора увлажнения почв и по 9 градаций в
рядах всех остальных. Следует отметить, что истоками европейских шкально-
балловых методик в экологическом анализе являются работы Погребняка и
Раменского (первая работа Погребняка, содержащая обоснование его мето-
дики, вышла в 1927 г., а первая работа Раменского, где он обосновал
метод рядовой координации, - в 1929 г., т.е. на 40 лет раньше аналогич-
ных работ Элленберга и других европейских исследователей).
В методике Раменского в обобщенных шкалах увлажнения, богатства
почв и других весь ряд фактора имеет диапазон, охватывающий все воз-
можные режимы и весь этот диапазон разбивается на возможно большее
еще различимое число ступеней. В этих ступенях выражаются и эколо-
гические амплитуды видов, отдельно по величинам обилия-покрытия.
Индикация экотопов (вернее, режима фактора) производится либо ме-
тодом засечек (наибольшее совпадение в описании определенных обилий-
покрытий видов с интервалами ступеней в шкалах, к которым эти оби-
лия-покрытия у данных видов приурочены), либо методом ограничений
(по совпадению или наибольшей близости начальных и конечных ступе-
ней шкал для имеющихся обилий-покрытий присутствующих видов).
Как показывает опыт, оба способа дают почти всегда одинаковый резуль-
тат, если же получается значительное расхождение, то это указывает на
дефекты в описании (неверное определение видов или их обилий-покры-
тий) или на мозаичность растительного покрова на пробной площади,
его экологическую неустойчивость (частые флюктуации) либо на переход-
ный его характер (положение на границе разных экотопов).
Школой Раменского, в основном уже после его смерти, выпущено семь
сводок, охватывающих всю территорию СССР, а также Карпаты и Балка-
ны, т.е. часть территории Болгарии, Румынии, Венгрии, Чехославакии и
Польши. В этих сводках приведены характеристики примерно 4000 видов
растений по 4—6 факторам (увлажнению почв, активному богатству почв,
переменности увлажнения, аллювиальности, пастбищной дигрессии и вы-
сотности) .
Принципиально близким к методике рядовой координации Раменского
можно считать лесотипологическое направление Погребняка, но в самой
технике индикации между ними наблюдается значительная разница. Как
отмечал Соболев [1978], разница наблюдается уже в выборе индикато-
ров: если в методике Раменского совместными индикаторами являются
все виды ценоза, да еще при учете их обилия-покрытия, то у Погребняка
такие виды находятся эмпирически исходя из опыта исследователя. Соот-
ветственно определение эдафотопа у школы Раменского идет по шкалам
с использованием всего списка видов, а у школы Погребняка — по списку
видов-индикаторов. С нашей же точки зрения, главным недостатком ме-
тодики школы Погребняка, как и методик Элленберга и других евро-
пейских экологов, является неполнота экологических рядов, в частности
ряда увлажнения и ряда солевого режима. К чему это приводит, можно
10
показать на очень простом примере. В статье Самойлова [1973] о резуль-
татах сравнения шкал Раменского со шкалами Элленберга, Хундта и Клап-
па отмечается, что их шкалы укладываются в интервал шкалы Раменского
от сухолугового (в крайнем случае лугово-степного) типа увлажнения
до болотного типа увлажнения. Как показывает построенная на основании
шкалы Раменского схема системы гидроморф [Цыганов, 1974, 1976],
при диапазоне охвата режимов фактора, равном шкале Элленберга, воз-
можно существование 65 гидроморф из общего числа 78, соответствую-
щего полной шкале. Однако, принимая шкалу Элленберга за полную,
можно выделить лишь 21 гидроморфу, так как число различаемых эко-
морф по данному фактору связано с числом различаемых его режимов
формулой
п
№(« + 1)— ,
где N — число экоморф, ан- число режимов фактора в шкале. (При
этом надо заметить, что мы приняли здесь шкалу Элленберга в масшта-
бе шкалы Раменского, посчитав вместо пяти номинальных типов режи-
ма шкал европейских экологов шесть. Как видно их схемы системы гид-
роморф, исходя из законченности шкалы европейских экологов, со сто-
роны ксерофильного крыла крайние экоморфы такой урезанной шкалы
будут соответствовать каждая пяти экоморфам полной шкалы, а со сто-
роны гидрофильного ряда — трем, что же касается гидроморфы, характе-
ризующей виды, индифферентные к данному фактору, т.е. способные су-
ществовать при всех различаемых его режимах, то такой гидроморфе
шкалы европейских экологов соответствуют 15 гидроморф полной шка-
лы. Если учесть, что видов с крайне узкой амплитудой толерантности по
каждому фактору, при достаточно подробном различении его режимов,
практически не обнаруживается, то все виды, встречающиеся в диапазоне
шкал европейских экологов, должны принадлежать к 59 гидроморфам
полной шкалы почвенного увлажнения, и тогда число индифферентных
видов, принадлежащих к 15 индифферентным гидроморфам, будет сос-
тавлять более 25%. Следует еще отметить, что виды, гидроморфы которых
не различаются шкалами европейских экологов и относятся к одной гид-
роморфе, со стороны ксерофильного крыла охватывают ио пяти эколо-
гических свит, со стороны гидрофильного — по три, а индифферентные —
семь экологических свит. Возникает вопрос: не слишком ли много ин-
формации мы теряем при таком подходе?
Учитывая, что ряд богатства почв все школы, кроме школы Рамен-
ского, рассматривают оторванно от ряда засоления, что только школой
Раменского осознано единство ряда солевого режима, так как острый
недостаток солей постепенно переходит в достаток и затем во все более
прогрессирующий избыток. Не приходится удивляться, что и на схеме
системы трофоморф обнаруживаются те же преимущества шкал Рамен-
ского. Например, трудно понять исходя из располовиненной шкалы, по-
чему одни виды широких амплитуд, охватывающие все типы режимов,
поместившихся на такой шкале, тяготеют к эвтрофным местообитаниям,
а другие — к мезотрофным. Так, например, амплитуду толерантности,
охватывающую и олиготрофные и эвтрофные, эдафотопы, имеют и ду-
11
шистый колосок, и иван-чай, и щучка, и кислица, и тысячелистник, и бело-
крыльник, и калужница, и донник, и кровохлебка, и клевер луговой,
и клевер ползучий, и овсяница красная, а между тем они принадлежат
к пяти трофическим свитам от мезотрофной до эвтрофной [Цыганов,
1976]. Исходя из половинной шкалы, трудно объяснить известный лу-
говодам факт,что овсяницу красную невозможно вытеснить из траво-
стоя никакими дозами удобрений. Сравнение шкал Раменского со шка-
лами Погребняка [Воробьев, 1967; Толчаин, 1980; Соболев, 1978] по-
казывает такую же фрагментарность последних по отношению к первой,
как и у шкал европейских экологов, ведущую к той же потере инфор-
мации и к затруднению построения на их основе такого удобного инстру-
мента экологического анализа, как схемы систем экоморф и экологи-
ческих свит растений [Цыганов, 1974,1976].
Все эти соображения, на наш взгляд, достаточно четко показывают
преимущество шкал Раменского для целей экологического анализа,
разработки его теории и техники практического применения.
Самым же ценным в этом смысле оказался сам принцип, построения
именно полных, а не фрагментарных экологических рядов. Именно на
этом принципе были основаны применяемые нами экологические шкалы
климатических и ценотических факторов, которые вместе с эдафическими
шкалами Раменского послужили основой для построения упомянутых
схем систем экоморф и экологических свит.
У автора этих строк всегда вызывало некоторое недоумение, почему
и сами создатели, и последующие модернизаторы шкал любой школы,
в том числе и школы Раменского, полностью удовлетворялись самим
фактором их создания, как будто шкалы суть только шкалы и самое
большее, что с ними можно еще сделать, - это построить на их основе
эдафические сетки. Правда, была в последние годы построена и клима-
тическая сетка [Лавриненко, 1978], впрочем тоже фрагментарная по
отношению к полной. Между тем на основании полных экологических
шкал по пяти ведущим прямодействующим факторам легко удалось
построить схемы систем экоморф, а путем их анализа была найдена фор-
мула, выражающая связь различимого числа экоморф по любому фак-
тору с числом выделенных в его непрерывном ряду градаций. Автор этих
строк склонен подозревать, что подобные построения не были произве-
дены ни Раменским, ни Погребняком, ни их непосредственными после-
дователями потому лишь, что им приходилось тратить слишком много
сил и времени на защиту созданных ими оригинальных направлений от
чересчур рьяных нападок приверженцев иных взглядов и почитателей
иных маститых авторитетов, может быть болезненно воспринимавших
возможность оспаривания универсальности их собственных концепций.
Так как всякая концепция обладает своей логикой развития, приняв
принцип амплитудных характеристик экоморф и придя через это к фор-
мализации нахождения их общего числа по каждому фактору, приходится
убедиться, что их набор для каждой градации соответствующей шкалы
тоже предопределен и находится по формуле
Nx = (п + 1)х -х2,
где Nx - число экоморф при режиме фактора, соответствующему гра; .
12
ции х; п — число выделяемых режимов фактора в соответствующей шкале,
а х — порядковый номер данного режима на этой шкале.
Из этой закономерности следует, что в каждом флопистическо.м регио-
не для ка»£0.г9 охарактеризованного сочетанием определенных режимов
основных прямодействующих факторов экотопа можно чисто теорети-
чески определить состав его потенциальной флоры. Более того, на осно-
вании таких соображений можно утверждать, что флора региона = S флор
его климатопов, флора климатопа = S флор его эдафотопов, флора эда-
фотопа = £ флор его ценотопов, а флора ценотопа = S видовых составов
фитоценозов. К выводу о положении фитоценозов, как совокупностей
популяций видов, на низшей ступени иерархии флор на основании совер-
шенно других построений приходили и другие авторы, в том числе в
последнее время Юрцев и Семкин [1980]. В свете их работы можно с
некоторыми оговорками рассматривать потенциальные флоры экото-
пов различного ранга, как различного ранга парциальные флоры.
Далее, из анализа координатных схем систем экотопов (климатопов,
эдафотопов) легко было сформулировать представление о различии их
принципиальных типов безотносительно к их факторному составу. И лег-
ко было увидеть, что каждый принципиальный тип экотопа имеет свой
набор принципиальных типов экологических ареалов, обусловливающих
различные способы их индикации.
Из анализа экологических ареалов видов пришлось прийти к представ-
лению о коэффициентах удовлетворительности условий среды (КС) для
вида в каждом из пригодных для него экотопов, а отсюда и о представле-
нии о совокупности видов с высокими значениями КС, как о домини-
рующем ядре потенциальной флоры экотопа, из числа которых и должны
происходить доминанты возможных в нем фитоценозов. С другой стороны,
давая экотопу характеристики, как качеству среды обитания для каждого
вида в форме КС, поневоле приходишь к мысли о возможности харак-
теристики его, как качеству среды, с точки зрения потенциальных воз-
можностей развития растительного покрова вообще, т.е. к определению
коэффициента общей комфортности экотопа (КК).
Все это, вместе взятое, дает возможность говорить об экологических
предпосылках конкурентных, взаимоотношений видов, об экологических
предпосылках успешности естественной или искусственной инвазии ви-
дов в то или иное местообитание, об экологических предпосылках удер-
жания видом той или иной занятой им территории, об экологических пред-
посылках нахождения способов помочь или помешать ему удержаться
на: ней, т.е. в целом об экологических предпосылках ассоциирования ви-
де» и направленности динамики определенных фитоценозов на опреде-
ленных типах местообитаний.
13
ГЛАВА II
ЛЕСОРАСТИТЕЛЬНЫЕ УСЛОВИЯ
И ЭКОЛОГИЧЕСКАЯ ТИПОЛОГИЯ ЗЕМЕЛЬ
Термин ’’лесорастительные условия” был принят в качестве одного из
двух необходимых фундаментальных понятий лесной типологии на сове-
щании по лесной типологии в 1950 г. На этом же совещании было принято
решение считать эдафическую сетку Алексеева—Погребняка удобной
основой для типологии лесорастительных условий.
Тем не менее, за исключением немногих случаев, последователи фито-
ценологического, а затем биогеоцеяологического направления в лесной
типологии эдафической сеткой почти не пользовались, предпочитая харак-
теризовать их традиционными методами соответствующих научных дис-
циплин (почвоведения, геоморфологии, гидрологии и др.).
Больше доверия оказывалось данным физико-химических исследова-
ний элементов среды, чем ее фитоиндикационной оценке. Между тем фито-
индикационная методика характеристики экологических условий среды
вообще и лесорастительных условий в частности имеет ’’неоспоримое
преимущество в том, что растительность принимает среду в своей есте-
ственной шкале” [Миркин, Розенберг, 1978, с. 92]. Кроме того, неоспори-
мо и положение, что данные физико-химических измерений отражают
статическое состояние среды, тогда как в растительности отражена ее
динамика [Вальтер, 1960].
Тем не менее определение физико-химических показателей, а лучше
их динамики имеет большое значение и для экологического анализа, но
лишь в соотношении с динамикой состояния растительного покрова, что
проще всего достигается соотношением динамики физико-химических
показателей с фитоиндикационно установленными градациями шкал соот-
ветствующих факторов, т.е. установлением амплитуд колебания физиче-
ских и химических показателей, соответствующих каждой градации эко-
логической шкалы.
В сущности, в градациях шкал можно характеризовать и через них
соотносить совершенно разнородные явления и элементы функциониру-
ющих экосистем, потому что экологические шкалы суть фитомеры каче-
ственного и количественного состояния экологической среды. Каждая
их градация соответствует определенному динамически равновесному
состоянию среды в отношении того или иного фактора. Поэтому в града-
циях экологических шкал могут быть представлены и амплитуды толе-
рантности видов, и амплитуды устойчивого и относительно устойчивого
равновесия сообществ, и амплитуды равновесного состояния почвенных
разностей, и амплитуды допустимых для сохранения данного типа режима
какого-либо фактора колебаний геологической и геоморфологической
среды, и амплитуды колебания отдельных составляющих режима соответ-
ствующего фактора и многое другое.
Например, можно выразить в единицах шкал, в пределах каких баллов
или ступеней шкал увлажнения и богатства-засоления возможно существо-
вание дерново-среднеподзолистых среднесуглинистых почв и в каких —
легкосуглинистых, какие балльные или ступенчатые параметры среды
14
допускает склон той или иной экспозиции и крутизны, сложенной теми
или иными породами. Разница будет лишь в том, что одни элементы (ви-
ды, сообщества, отчасти почвы) больше сами зависят от установившихся
помимо них режимов факторов, в то время как другие (геоморфологи-
ческие образования, геологические пласты) определяют установление
тех или иных режимов связанных с ними факторов. Но и в том и в другом
случае определенным параметрам состояния индикатора (легко наблюда-
емого элемента системы) должны соответствовать определенные пара-
метры состояния индиката (трудно наблюдаемого элемента системы).
Синонимом типа лесорастительных условий можно назвать тип условий
местопроизрастания [Сукачев, 1931; Сукачев, Зонн, 1961].
Принимая во внимание, что под типом лесорастительных условий сле-
дует понимать ’’объединение участков территорий, вызывающих однород-
ный лесорастительный эффект - однородный комплекс действующих
на растительность природных, климатических и почвенно-гидрологиче-
ских факторов” [Рысин, 1979, с. 40], справедливо будет подчеркнуть
принадлежность этого комплекса условий к свойствам косной, абиоти-
ческой среды. Но факторы косной среды и Раменский [1924], и Сукачев
[1931] подразделяли на прямые, физиологически действующие, физиче-
ские и химические свойства среды и косвенные, влияющие на раститель-
ность посредством изменения режимов прямодействующих факторов.
Сюда относятся: рельеф местности, литологический состав материнских
пород, характер надземной и подземной гидрологической сети, окружа-
ющие ландшафты и т.п.
Совокупности прямых или прямодействующих факторов - экотопы —
удобнее всего классифицируются путем построения экологических сеток,
тогда как классификации энтопиев, совокупностей косвенных, или кос-
вениодействующих факторов, пока еще не достигли хотя бы относительно
общепризнанной стадии обобщения, даже в смысле разработки самих
принципов их построения. Так, например, Л.Н. Соболев [1976] предлага-
ет такую классификацию энтопиев открытых и подгорных равнин:
1. Плакорные энтопии. Характеризуются зональными почвами и рас-
тительностью. Преобладает дренаж через породу.
2. Наклонные или присклоновые энтопии. Слабые уклоны (1—3°),
обнаруживаемые нивелиром; наблюдается некоторый перенос и сортиров-
ка почвенного субстрата водой. Экотопы зональные.
3. Пологосклоновые энтопии. Уклоны 3—7°. Повышенный поверх-
ностный дренаж, заметная транспортировка субстрата, некоторое иска-
жение нормального почвенного профиля.
4. Покатосклоновые энтопии. Уклоны 7—15°. Явный поверхност-
ный дренаж и смыв, замечается некоторое воздействие направления экс-
позиции. Сказывается влияние величины водосбора и положения в разных
частях склона.
5. Крутосклоновые знтопии. Уклоны 15—40°. Резкий дренаж, эрозия,
размыв почвы. Сильное воздействие направления экспозиции.
6. Обрывистые энтопии. Уклоны свыше 40°, Разорванный раститель-
ный покров, смытые почвы.
7. Конусовые энтопии. Встречаются на выходах временных или посто-
янных потоков из горных долин на равнину. Часто развиты гальконос-
15
ные почвы. Подвержены действию селевых потоков. В чисто равнинных
условиях этот тип энгопиев, очевидно в более смягченном и менее долго-
вечном варианте, развивается у выходов крупных оврагов с сильно повы-
шенных местоположений на нижележащие выровненные территории (пой-
мы или низкие террасы рек). При этом основной особенностью таких
местообитаний будет относительное накопление грубых делювиальных
отложений.
8. Пойменные энтопии. Большая группа разнообразных местоположе-
ний в речных долинах, расположенных в прирусловой, центральной и
притеррасной частях поймы, с несколькими террасными уровнями.
В качестве примера другого рода можно привести принципы определе-
ния лесорастительных условий, даваемые в книге ’’Леса восточного Под-
московья” [Рысин, 1979]. Там предлагается в пределах одного лесорасти-
тельного района, т.е. при одних и тех же климатических условиях, опреде-
лять тип лесорастительных условий следующими параметрами: 1) генети-
ческим типом рельефа; 2) формой мезорельефа; 3) механическим соста-
вом почвогрунта; 4) генетической разновидностью почв; 5) степенью
оглеения; 6) уровнем грунтовых вод; 7) карбонатностью. Как можно
видеть, все эти параметры характеризуют не экотопы, а именно энтопии.
Что касается этих двух подходов, то первый (подход Соболева) произ-
водит впечатление дедуктивно-формализованного, направленного в первую
очередь на систематизацию энтопиев с точки зрения выявления характера
и интенсивности процессов, влияющих на установление определенного
сочетания режимов прямодействующих факторов, в то время как во
втором подходе прослеживается стремление охарактеризовать каждый
элемент среды в классификационной традиции той специальной дисципли-
ны, которая его изучает (геоморфологии,гидрологии,почвоведения) .остав-
ляя увязывание этих характеристик с характером растительного покрова
различных местообитаний на долю эмпирических наблюдений и интуиции
исследователя.
Наиболее удачной, в смысле глубокой экологичности, классификацией
эктопических условий среды, комплекса косвенных факторов, в наиболь-
шей степени влияющих на режимы прямодействующих, является классифи-
кация типов лесных земель, опубликованная в книге ’’Экология лесных
земель” [Чертов, 1981].
Будучи почвоведом, автор, исходя из возможностей современного
почвоведения, не считает в настоящее время возможным определение
режимов прямодействующих факторов непосредственно и находит выход
в классифицировании типов земель по признакам доступных современ-
ным методам диагностики косвенных факторов, наиболее влияющих на
установление и поддержание режимов основных прямодействующих эда-
фических факторов — водного и солевого режимов почв. Хотя автор и за-
являет о своей солидарности с принципами экологической типологии
Раменского, в создании своей классификации он опирается на традицион-
ный подход лесного почвоведения, очевидно считая методы Раменского
слишком связанными с типологией сельскохозяйственных, луговых и
пастбищных угодий. Тем не менее типология лесных земель Чертова,
являясь на настоящее время, по крайней мере, одной из лучших типоло-
гий эктопических условий, может и должна послужить основой многих
16
исследований в области лесной типологии, общей типологии земель и
экологического анализа связи растительного покрова со своими условиями
существования.
Употребляя пятиступенчатую классификацию, автор выделяет класс
лесных земель по общности климатических условий, считая одним классом
все земли одной климатической провинции; группу лесных земель по
общности условий дренажа, отражающего влияние ведущего эдафического
фактора таежной зоны — режима увлажнения почв; подгруппу лесных
земель по общности дренажа и почвообразующих пород, определяющих
общий потенциал элементов солевого довольства растительности; тип
лесных земель по сходству дренажа, почвообразующих пород и рельефа
как фактора перераспределения тепла и влаги, а также определяющего
напряженность современных геологических процессов; вид лесных зе-
мель по типу гумуса лесных почв, отражающего направленность процес-
сов минерализации и гумификации опада, интенсивность биологического
круговорота и плодородие почв (ресурсы элементов питания, накопленные
в процессе биологической аккумуляции).
В этой классификации вызывает некоторое сомнение лишь тенденция
считать климат региона относительно однородным, тогда как это опреде-
ленно не соответствует действительности, чему подтверждением служит
явление экстразональности и в растительном покрове, и в почвенном
покрове, и как единственная причина этого экстразональность топокли-
матов и ценоклиматов определе!нных местоположений. В то же время
само выделение классов земель по признакам климата региона не вызыва-
ет сомнений, так как все топо- и ценоклиматы тоже зональны, если не для
своего, то для смежных регионов, а интразональные климатопы являются
лишь вариантами тех или иных зональных типов климата.
Особенно ценно, что типология всех остальных косвенных факторов,
играющих роль в диагностике типов земель, основана на относительно
легко доступных прямому наблюдению признаках, которые таким обра-
зом легко использовать в качестве параллельной контрольной индикаци-
онной системы, позволяющей сверять, а в некоторых случаях и дополнять
результаты фитоиндикации экотопов. В частности, типы дренажа выделя-
ются по признакам почвообразующих пород, положения в рельефе, а в
некоторых случаях и мощности органических горизонтов почв (например,
торфа) и, кроме того, сопоставлены с принятыми в лесоводстве града-
циями влажности. Систематизация почв по типам гумуса основана также
на легко наблюдаемых признаках верхних горизонтов: Ао, Ат, Ai и А2.
В построении классификации широко использованы разработки Благови-
дова [Благовидов, Бурков, 1959] и Таргульяна [Таргульян, 1971].
Сам Раменский предлагает целую систему признаков, которые следует
учитывать при расчленении исследуемой площади на энтопии, или экологи-
ческого диагноза энтопия.
1. Оценка условий натека поверхностных вод (в зависимости от укло-
на, положения на склоне, формы склона, развития микро- и нанорельефа,
проницаемости почвы).
2. Условия питания грунтовыми водами в зависимости от глубины
их залегания, проточности, вероятных колебаний уровня и капиллярных
свойств почвы и грунта.
2. Зак. 448
17
3. Подверженность весеннему затоплению и нерегулярным паводкам
(характер заливания — длительность, проточность, регулярность и т.д.).
4. Вероятное или видимое (по ряду признаков) развитие геологиче-
ских процессов — смыва, размыва, намыва, отложения аллювия, разве-
вания и т.д.).
5. Условия испарения в зависимости от залегания в рельефе (откры-
тость ветрам) и окружения (увлажняющего или иссушающего атмосферу
данного местоположения).
6. Термические и отчасти водные условия: пригревность или холод-
ность (и позднее весеннее развитие), подверженность заморозкам, тума-
нам, выпадению росы.
7. Литологические условия почвообразования — карбонатность, соле-
ность, глинистость, водоупорность или проницаемость, дренирующее влия-
ние подстилающей породы.
Однако, давая такое детальное перечисление условий, которые необ-
ходимо учитывать при диагностике энтопиев, и приводя примеры разно-
стороннего и содержательного анализа экологических условий местополо-
жений по эктопическим факторам, Раменский все-таки отмечал, что эко-
логические оценки, сделанные по условиям местоположений, являются
только сравнительными; большей частью довольно грубыми частью
гипотетичными [Раменский, 1938].
Решающее слово в экологической оценке среды он оставляет за ин-
дицируемым по растительности сочетанием экологических режимов пря-
модействующих факторов. Примерно в том же смысле высказывался
и Сукачев [1931], когда писал о понимании условий местопроизрастания
Морозовым: ”. .. по мнению Морозова, каждому типу леса строго свой-
ствен свой тип условий местопроизрастания.
Однако это положение во всем объеме будет справедливо только тог-
да, когда мы будем иметь в виду только комплекс прямодействующих
факторов, т.е. то, что мы условились называть средой, обязательно вклю-
чая в это понятие как экзогенные, так и эндогенные факторы. Морозов
при установлении типов не имел в виду комплекс только прямодейству-
ющих факторов, а, напротив, все условия местопроизрастания и прежде
всего искал связь типа с энтопическими факторами — рельефом, грунтом,
почвой в целом и т.п., т.е. с теми факторами, которые при обычном поле-
вом изучении леса легче всего распознаются, но сами не являются непосред-
ственно определяющими характер растительности, и поэтому между ними
и типами не всегда бывает строгая связь. И пользуясь ими как критерием
при установлении типов, можно ошибиться”. Вспомнив, что экзогенными
факторами называются прямодействующие факторы в том виде, как
диктует энтопий, а эндогенные — они же, но ® трансформированном влия-
нием фитоценоза виде, справедливо сделать заключение, что классифика-
ция лесорастительных условий должна быть прежде всего классификаци-
ей комплексов режимов прямых факторов, т.е. экотопов.
Классификации энтопиев играют второстепенную роль, являясь вслед-
ствие относительной легкости их наблюдения своего рода вспомогательной
системой индикационных признаков условий среды. Итак, если характери-
зовать экотопы по принципам, поддержанным и Сукачевым, и Раменским,
то эдафическая сетка Раменского является наиболее полной классифи-
18
кацией эдафотопов, если понимать их как, в первую очередь, сочетание
определенного режима увлажнения с определенным трофическим режи-
мом, полагая, что остальные режимы будут либо находиться в прямой
зависимости от сочетания этих режимов, либо в большей степени харак-
теризовать климатоп или ценотоп. Действительно, и эдафическая сетка
Погребняка, даже в наиболее дополненном виде [Лавриненко, 1978],
и сетки, которые можно построить на основании шкал Элленберга и других
европейских экологов, суть лишь фрагменты ее, охватывающие лишь
20-30% скоординированных в ней эдафотопов (рис. 1). Даже если отбро-
сить часть сетки Раменского, представляющую эдафотопы в режимах
засоления, который представители других школ не связывают с трофи-
ческим режимом и тогда охват ими сетки Раменского повысится лишь
до 40-60%. Но отрыв ряда засоления от ряда богатства почв принципиаль-
но не верен и объясняется скорее привычкой, чем достаточно вескими
аргументами. Если утверждают, что говорить просто о засолении нельзя,
так как очень важно, какие именно соли находятся в избытке, то и о тро-
фности высказывают аналогичное суждение, так как не менее важно,
каких именно соединений не хватает в тех или иных бедных почвах. И
обобщенная трофность, и обобщенное засоление представляют собой лишь
две ветви одной сущности — солевого режима почв.
В бедных почвах содержание солей понижено до минимума и за мини-
мум толерантности большинства видов, а в засоленных оно повышено
до максимума и сверх максимума большинства видов. В сущности, и ряд
увлажнения, будь то в шкале Раменского, Погребняка или Элленберга,
тоже имеют две ветви, потому что вода, как таковая, в избытке быть не
может и понижение качества среды при избыточном увлажнении вызвано
не избытком воды, а недостатком воздуха для дыхания корневых систем.
Поэтому ряд увлажнения в полном объеме правильнее рассматривать
в качестве ряда водно-воздушного режима почв. О том, что аэрация почв
находится большей частью в обратной зависимости к ее увлажнению, писал
и Сукачев [1938], но наиболее наглядно это изобразил в таблице Лаври-
ненко [1978] на своем варианте эдафической сетки Погребняка, где самые
благоприятные режимы располагаются в свежих и влажных эдафотопах
(при достаточно водном и хорошем воздушном довольстве и при хорошем
водном и достаточном воздушном довольстве). С возрастанием сухости
или влажности растения получают уже не имеющие значения повышение
обеспеченности удовлетворения одной из потребностей при необеспечен-
ности удовлетворения другой (излишняя обеспеченность влагой
при недостатке воздуха или прекрасная обеспеченность воздухом при
недостатке влаги). Но эдафическая сетка представляет собой лишь графи-
ческую схему системы эдафотопов, тогда как экотоп в целом — явление
более сложное, в характеристику которого входят и эдафические (водно-
воздушный и солевой режимы почв), и климатические (термический
режим, континентальность, аридность), и ценотические (режим затенения,
режим задернения) характеристики среды.
Об учете климата при характеристике условий среды, о недопустимости
рассмотрения эдафических режимов все связи с климатическими неодно-
кратно писали и Сукачев, и Раменский, и Погребняк, и все их последова-
тели. Школой Погребняка были предприняты попытки построения кли-
19
Рис. 1. Соотношение эдафических сеток, построенных по шкалам увлажнения и бо-
гатства почв различных авторов
1 — границы типов увлажнения и богатства почв по Раменскому; 2 — верхний край
эдафической сетки по Лавриненко; 3 — верхний край эдафической сетки по Эллен-
бергу; 4 — верхний край эдафической сетки по Воробьеву; 5 — нижний край эдафи-
ческих сеток школы Погребняка. (Все авторы, кроме авторов школы Раменского,
рассматривают ряд избыточного засоления отдельно от ряда богатства почв минераль-
ным питанием). Значение символов режимов увлажнения и солевого см. в тексте
магических сеток [Воробьев, 1967; Лавриненко, 1978], но, к сожалению,
обе эти сетки охватывают только те климатопы, которые представлены
в СССР, т.е. опять-таки не более 2/3 всего разнообразия климатопов. Что
касается ценотических факторов — прямо действующих факторов, завися-
щих от типа и состояния фитоценоза, занимающего данную территорию,
то для них пока относительна разработана лишь шкала светолюбия и самая
предварительная схема их системы пока сводится лишь к последователь-
ности элементарных режимов затенения.
В предлагаемой нами системе экотопов, базирующейся на1 экологиче-
ских рядах пяти основных прямодействующих факторов [Цыганов, 1976],
климатические факторы градуированы по природным зонам и поясам
континентальности, для эдафических факторов применяется градация
их но Раменскому, но не по отдельным ступеням, число которых для це-
лей систематизации слишком велико, а по типам режимов, фактор затене-
ния градуирован очень обобщенно в связи со слабой разработкой шкал
светолюбия. Таким образом, различая восемь градаций зонального режи-
ма тепла, четыре градации режима континентальности, 12 градаций водного
20
режима почв, 10 градаций солевого режима почв и три градации режима
затенения, можно получить климатическую сетку из 32 ячей, эдафическую
сетку из 120 ячей и отдельный ряд затенения. Так как климатические
факторы имеют глобальную по преимуществу природу, то высшей едини-
цей классификации экотопов целесообразно считать климатоп, тогда как
эдафотоп целесообразно считать единицей, подчиненной климатопу, по-
скольку эдафические факторы больше связаны с геоморфологическими,
гидрологическими и другими условиями регионального характера.
Низшей же единицей такой трехчленной иерархии является ценотоп,
так как факторы, ценотически обусловленные, зависят уже от состава,
структуры и состояния растительного покрова именно данного участка
местности.
В зависимости от целей исследования можно строить экологические
сетки и на сочетании шкал других пар факторов (например, зонального
режима тепла и режима увлажнения почв, увлажнения почв и затенения,
аридности и солевого режима почв и т.д.), но наиболее естественными
будут все же экологические сетки, построенные на осях факторов, мас-
штаб формирования режимов которых достаточно близок (глобально-
климатический, эдафотопологический, ценотический). \
Такие разнообразные сетки могут быть полезны при анализе совместно-
го влияния какой-либо пары факторов. Что касается классификации экото-
пов, то схематически она может быть представлена в виде глобальной
климатической сетки, в каждую ячею которой вписана эдафическая сетка
(точнее, фрагмент ее, охватывающий только те эдафотопы, существование
которых возможно в данном климатопе), а в каждую ячею эдафической
сетки вписан экологический ряд режима затенения.
Однако, кроме таких естественных пофакторных классификаций эко-
топов, могут быть полезны и общие принципиальные их типизации, когда
принципом разделения на типы будет положение их в средней части или
на концах экологического ряда, во внутренней или периферийной зоне
экологической сетки. Такая типизация экотопов и экологических режимов
помогает проанализировать и учесть особенности индикации экологических
режимов и экотопов, принципиально различных по своему положению
в экологических рядах и на экологических сетках.
Прежде всего, экотопы любого экологического ряда, любой экологи-
ческой сетки, будь то климатопы, эдафотопы или экотопы, построенные
на иных сочетаниях прямодействующих факторов, разделяются на нормаль-
ные экотопы, или ортотопы, характеризующиеся средними (или относи-
тельно средними, но не крайними) режимами данного фактора или данной
совокупности двух факторов, и экстремальные экотопы, или экстремото-
пы, характеризующиеся именно крайними (минимальными или макси-
мальными) режимами. Так, если мы обратимся к схеме системы климато-
пов (климатической сетке) или к схеме системы эдафотопов (эдафиче-
ской сетке), то увидим, что центральная часть любой из них (основной
массив клеток) представляет собой ортотопы, или нормальные экотопы,
а периферийный ряд экстремотопы, или экстремальные экотопы. Можно,
если позволяют размеры сетки (в смысле достаточного количества града-
ций каждого фактора), выделить еще второй от периферии ряд ячей, кото-
рые будут представлять собой субэкстремальные экотопы, или субэкс-
21
О О м е Е g h н р S
D Э г г г г г г г г э
d Г О 0 О О 0 О 0 0 г
В Г 0 О 0 0 О 0 О О г
S Г 0 0 0 О 0 О 0 0 г
с Г 0 0 0 0 0 0 0 0 г
с Г 0 0 0 0 О О О 0 г
f Г О 0 0 0 0 0 0 О г
F Г 0 0 О 0 0 О 0 0 г
Р г 0 0 0 О 0 0 О 0 г
Р г 0 0 0 0 О 0 О 0 г
а г 0 0 0 0 0 0 0 0 г
А э г г г г г г г г э
Рис. 2. Ортотопы и экстремотопы на примере эдафнческой (гидротрофнческой)
сетки
По верхнему краю снаружи сетки расположены символы основных типов солевого
режима почв, а по левому краю — символы основных типов режимов увлажнения
почв. Обозначения внутри сеток: о — ортотопы, или нормальные экотоны, у которых
оба режима неэкстремальные; г — гемиортотопы, или полу экстремальные экотоны,
у которых лишь один иэ факторов в экстремуме, э— экстремотопы (эуэкстремотопы)—
оба фактора в экстремуме. Значение символов режимов увлажнения и солевого см.
в тексте
тремотопы. Далее, если ортотопы представляют собой принципиально
мало разнящиеся явления, то экстремотопы в этом отношении гораздо
более разнообразны.
Как видно из рис. 2, разниться будут экотопы, расположенные по сторо-
нам экологической сетки, и экотопы, расположенные по ее углам, так как
у первых в экстремальном режиме находится лишь один из факторов,
тогда как у вторых — оба. Вероятно, целесообразно первые называть полу-
нормальными экотопами, или гемиортотопами, так как режим одного
из факторов у них не экстремальный, а вторые — собственно экстремо-
топами. При этом следует иметь в виду, что и они (собственно экстремо-
топы) группируются попарно, так что у одной пары режимы обоих фак-
торов гомометричны (оба минимальные или оба максимальные) а у дру-
гой пары — гетерометричны (минимум по одному фактору сочетается
с максимумом по другому, и наоборот).
По-видимому, целесообразно принципиальро выделить первые под
22
названием гомоэкстремотопов, а вторые — гетероэкстремотопов. Вообще
же на примере любой экологической сетки можно видеть, что для пары
факторов существуют четыре возможных принципиальных направления
изменения режимов: параллельно оси абсцисс — изменение режима х-фак-
тора при стабильном режиме .у-фактора; параллельно оси ординат — изме-
нение режима .у-фактора при стабильном режиме х -фактора; параллельно
диагонали, соединяющей гомометричные экстремальные режимы (гомо-
экстремотопы) — гомодинамическое изменение режимов обоих факторов
(одновременное уменьшение или одновременное увеличение их значимо-
стей) ; параллельно диагонали, соединяющей гетерометричные экстремаль-
ные режимы (гетероэкстремотопы) — гетеродинамическое изменение ре-
жимов обоих факторов (уменьшение значимости одного при одновремен-
ном увеличении значимости другого и наоборот). Кстати, осью гомоди-
намических изменений режимов пары факторов является ось ординат
эколого-фитоценотической схемы Сукачева (ось A—D), тогда как ось
абсцисс этой схемы (ось В-С) является осью гетеродинамических их
изменений.
Если же выделять на экологических сетках и субэкстремальные эко-
топы, то и среди них можно по аналогии с экстремотопами различать суб-
экстремальные гемиортотопы, гомосубэкстремотопы и гетеросубэкстре-
мотопы. Выделение экстремотопов и субэкстремотопов, как принципи-
альных типов экотопов любой природы (климатопов, эдафотопов, цено-
топов), имеет смысл по той причине, что к ним приурочен другой набор
экоморф с другими типами экологических ареалов видов, что обуслов-
ливает и другие приемы индикации условий среды.
Также безотносительно к физико-химической сущности факторов
каждый тип экотопа может быть охарактеризован, с одной стороны, с точ-
ки зрения его пригодности для отдельных видов, вернее видов одной эко-
морфы по отношению к режимам рассмариваемых факторов, а с другой
стороны, с точки зрения его общей комфортности, в смысле качества ус-
ловий для развития растительного покрова вообще, для создания за год
наибольшей фитомассы.
В первом случае (пригодность для отдельных видов) исходить надо,
очевидно, из экологических амплитуд видов по каждому из прямодей-
ствующих факторов, полагая, что в средней части экологической ампли-
туды вида или в центре его экологического ареала качество среды, с точки
зрения этого вида наивысшее (близко к 100%), а на концах амплитуды
или на периферии ареала — самое низкое (близко к 0). Во втором случае
(общая комфортность экотопа) исходить надо из всей фитометрической
шкалы каждого прямодействующего фактора, при этом надо только ре-
шить, какие из режимов отдельных факторов, какие из климатопов, какие
из эдафотопов могут претендовать на наименование наиболее общеблаго-
приятных для развития растительного покрова вообще.
Данные биогеографии и экологии, особенно же данные по продуктив-
ности растительного покрова различных зон и характерных местообитаний,
позволяют сделать вывод, что в отношении зонального режима тепла таким
является тепловой режим тропической зоны, в отношении фактора кон-
тинентальности — режим субокеанических климатов, в отношении атмо-
сферного увлажнения — режим гумидного типа климата, в отношении
23
I 0 0 к К
А 9 13 10 5
а 18 27 22 11
в 22 40 32 16
. N 36 54 43 22
m 45 67 54 27
м 53 80 64 32
t 63 94 75 38
т 67 100 80 40
II А В С D. Е F G Н
А 2 4 6 7 9 11 7 4
а 4 7 11 15 18 22 15 7
в 6 11 17 22 27 33 22 11
ь 7 15 22 29 37 44 29 15
N 10 18 28 37 46 56 37 18
m 11 22 34 45 56 67 45 22
м 13 26 39. 52 65 78 52 26
t 15 29 45' 60 74 89 60 29
т 17 33 50 67 83 100 67 33
Рис. 3. Комфортотопы и коэффициенты общей комфортности на климатических
сетках
I— климатическая сетка 1976 г: по оси ординат—зональные режимы тепла, по
оси абсцисс—пояса континентальности (и то и другое по Мёйэелю). Коэффициенты
комфортности климатопов даны в максимальном значении; II — климатическая
сетка 1981 г: по оси абсцисс — типы режимов аридности—гумидности климата, по
оси ординат — типы термического режима климата (по величинам годового радиа-
ционного баланса) . Коэффициенты комфортности климатопов даны в max значении.
Максимальные величины КК в каждом вертикальном или горизонтальном ряду соот-
ветствуют гемикомфортотопам, т. е. климатопам, комфортным лишь по одному из
факторов, а КК-100 соответствуют эукомфортотопам данной сетки, т. е. климатопам,
комфортным по обоим факторам (значение символов типа режима см. в тексте).
почвенного увлажнения — режим сыро-лугово-лесного типа увлажнения
(в трактовке школы Раменского), в отношении воздушного режима
почв - режим влажных песчаных почв (именно влажных потому, что
интенсивное дыхание корневых систем немыслимо без достаточного ув-
лажнения) , в отношении солевого режима почв — эвтрофный тип режима,
в отношении света — световой режим открытых пространств. Таким обра-
зом, наиболее комфортным климатопом можно считать экваториальный
субокеанический и гумидный, а наиболее комфортным эдафотопом -
влажно-лесо-луговой эвтрофный.
При анализе климатической и эдафической сеток (рис. 3 и 4) видно,
что полный комфортотоп (комфортный по обоим факторам) в любой
из них только один, а количество комфортных экотопов, в которых ре-
жим лишь одного соответствует среднеобщеоптимальным его значениям,
или гемикомфортотопов, равно сумме выделяемых градаций каждого
из факторов минус число этих факторов. Так, в климатической сетке
различаются две ветви термокомфортотопов (с комфортным термическим
режимом) — океаническая и континентальная, но лишь одна ветвь ом-
24
0 О М е Е g h Н Р S
D 3 7 10 13 15 13 11 8 5 3
d 4 14 20 27 31 28 22 17 11 6
8 10 20 30 41 46 41 33 25 17 8
S 14 27 41 55 62 56 45 33 22 И
с 17 34 51 68 77 69 55 42 28 14
с 20 40 61 81 92 83 66 50 33 17
f 22 44 66 88 100 90 72 54 36 18
F 20 40 59 79 90 81 65 49 32 16
Р 16 32 48 63 72 65 . 52 39 26 13
Р 12 24 36 48 54 49 39 29 19 10
а 8 16 24 32 36 32 26 19 13 6
А 4 8 12 16 18 16 13 10 6 3
Р и с.4.Комфортопы и коэффициенты комфортности на эдафической сетке
По оси ординат эдафической сетки расположены типы режима увлажнения почв
по оси абсцисс — типы солевого режима почв. Коэффициенты комфортности (КК)
для эдафотопов даны в максимальном значении. По вертикальному ряду Е (эвтроф-
ный тип солевого режима) и по горизонтальному ряду f (влажио-лесолуговой тип
режима увлажнения) располагаются гемикомфортотопы—экотопы, у которых лишь
один режим — общекомфортиый, а на пересечении этих рядов лежит эукомфрртотоп—
эдафотоп, у которого оба режима общекомфортны (значение символов типов режимов
см. в тексте)
брокомфортотопов (с комфортным режимом континентальности), по-
скольку тропический режиТл занимает крайнее положение в шкале зональ-
ных режимов тепла.
В связи с этим в климатической сетке все термокомфортотопы, вклю-
чая и эукомфортотоп (полный комфортотоп), являются в то же время
и экстремотопами. Аналогичная картина наблюдается в ряду фактора
затенения, где комфортный тип режима — режим открытых местообита-
ний — тоже является одновременно и экстремальным. В этом факте нет
никакой несообразности, потому что экстремальный в буквальном смысле
означает только крайний, но отнюдь не худший. Считать его синонимом
худшего нам стало привычным лишь потому, что в большинстве случаев
объекты, характеризуемые этими понятиями, совпадают по той причине,
что в большинстве шкал прямодействующих факторов комфортные режи-
мы находятся в середине ряда и оба края шкалы занимают экстремальные
дискомфортные типы режимов, а в оставшихся шкалах экстремальные
25
дискомфортные типы режимов занимают один край, т.е. для семи упомя-
нутых прямо действующих факторов 12 типов режима отдельного фактора
являются экстремальными дискомфортными, тогда как экстремальными
комфортными только два — тропический тип зонального режима и свето-
вой режим открытых пространств. Что касается эдафической сетки, то
на ней имеются две ветви гидрокомфортотопов — гликофильная и гало-
фильная и две ветви трофокомфортотопов — аэрально-ксерофильная
и анаэрально-гидрофильная.
Естественно, что комфортный тип режима должен характеризоваться
степенью комфортности к 100%, а дискомфортный — близкой к 0. Все
промежуточные между ними типы экотопов имеют свой коэффициент
комфортности (КК), выраженный в процентах от полной.
Коэффициенты комфортности экотопов на экологических сетках на-
ходятся способом нахождения процента от процента. Также находятся
КК подчиненной сетки. Операция может продолжаться, пока не будет
исчерпан весь набор учитываемых прямодействующих факторов. За то,
что зти подразделения и эти определения КК имеют не символический,
а реальный смысл свидетельствует то, что им пропорциональны величины
первичной продуктивности как зональной растительности, так и расти-
тельности типов эдафотопа, соответствующих тем, для которых взят КК.
Близость КК и величин первичной продуктивности в процентах к мак-
симальной, за которую взята ПП влажных тропических лесов (табл. 1).
Так как климатоп и эдафотоп не всегда можно было указать однознач-
но, то КК даются в виде допустимых, при соответствующем разбросе
возможных режимов разных факторов, пределов, иногда в виде средней
из нескольких вариантов сочетаний, допустимых для данной растительной
общности.
Как видно из табл. 1, РП в процентах к максимально возможной очень
близка к КК. Это указывает на прямо пропорциональное отношение годич-
ного прироста к КК экотопа.
Рассматривая схемы систем климатопов и эдафотопов, а особенно
серию систем эдафотопов для различных климатопов климатической
сетки, можно наглядно пронаблюдать зависимость качества среды вооб-
ще от качества экологических режимов отдельных факторов, особенно
в их дискомфортном значении. Качество среды в дискомфортных кли-
матопах настолько низкое, что хоть сколько-нибудь благоприятные для
развития растительного покрова эдафотопы там сосредоточены в самом
центре эдафической сетки, т.е. наличие растительного покрова в таких
климатопах уже само по себе, безотносительно к его видовому составу,
свидетельствует о самых благоприятных эдафических условиях.
Это звучит непривычно, по крайней мере в отношении режима богатства
почв, но иначе не может быть. Медленное разложение растительных остат-
ков в сочетании со слабопромывным, чаще аккумулятивным, режимом
делают эти почвы достаточно богатыми необходимыми зольными элемен-
тами, полностью удовлетворяющими потребности того низкопродуктивно-
го растительного покрова, существование которого допускается терми-
ческими условиями. В то же время обращает на себя внимание та особен-
ность преобразований эдафической сетки в разных климатопах, что в ком-
фортных и относительно комфортных климатопах якобы возможно су-
26
Таблица 1
Полный годичный прирост фитомассы (/W) и коэффициент комфортности эко-
топов (КК)
Растительная общность ц/га %Kmaxi- тит Климатоп Эдафотоп KK Maxi- mum
Арктическая тундра А c-f
о-в Большой Ляховский 10 3 К М 3
Кустарничковая тундра а c-f
Корякский автономный 25 8 к-К о—М 6-9
округ
Ельник-черничник а-В f-F
Мурманская обл. 65 20 о-к М 20-21
Ельник-зелёномошник, В c-f
Архангельская обл. 60 18 о-к М 17-20
Ельник-зеленомошник, B-N с—f
Архангельская обл. (юг) 79 24 о-к М 22-23
Липняк осоково-снытевый N с— f
Мордовская АССР 84 25 к М-е 23-27
Сложный ельник, B-N с—f
Великолукская обл. 90 27 о—к М-е 28-29
Березняк травный, N с—f
Московская обл. 111 33 о-к М-е 30-31
Дубравы, N—т S-c
Воронежская обл. 77 23 к е 20-25
Буковый лес, N С—f
Центральная Европа 130 39 О М-е 36-45
Луговая степь, т S-C
Курская обл. 104 31 к Е 29-36
Сухая степь т s—S
Аскаиия-Нова 87 26 к е-Е 19-29
Пустынная степь, т d-e
Монголия 24 7 К M-g 6,6
Северная полупустыня, т-т d—s
Волгоградская обл. 43 13 к-К e-g 11-14
Биюргуиовый полынник М D
Усгь-Урт 12 4 К e—h 3-4
Суккулентная пустыня, м D
КрасноВодская обл. 7 2 К e—h 2-4
Мятликовый полынник М D
Сирия (пустыня) 24 7 к e—h 6-8
Субтропическая пустыня М D
Сирия 7 2 К e—h 2-4
Сухая саванна, t S-C
Индия 73 22 К e-E 18-25
Тропическая саванна, t S—c
Гана 150 45 к e-E 41-52
Субтропический лес, М f-F
о-в Кюсю 245 74 О E 75-80
Влажнотропический лес, t-T F
Таиланд 320 96 о E 94—100
Экваториальный лес, Т F
Заир 330 100 О E 100
При м еч ан ие-РПвзяты из работы Ефимовой {1976]. Коэффициенты КК взяты
с учетом климатических и эдафических режимов.
Символику климатопов и эдафотопов см. в тексте.
Ществование почти всех ее эдафотопов, во всяком случае всех типов режима
каждого фактора: и водного режима, и солевого.
Между тем общеизвестно, что это не так. Это могло бы навести на мысль
о порочности самого метода составления экологических сеток, но следует
иметь в виду, что на сетках в виде изменений КК отражается лишь ха-
рактер изменения качества среды при взаимодействии спектра учитыва-
емых факторов при допущении, что сами режимы любого фактора авто-
номны по отношению к режимам других факторов. На самом деле это не
так, и каждый тип режима любого фактора имеет свою амплитуду суще-
ствования по отношению к режиму любого другого фактора. В случае
слабой зависимости одного фактора от другого эта амплитуда может быть
очень широкой, вплоть до охвата всех типов режима второго фактора, от-
раженных на его шкале. Такое положение наблюдается в отношении режи-
мов тепла к режимам других факторов, поскольку источник его ими не
контролируется, в отношении режимов почвенного увлажнения к трофи-
ческому режиму, к режиму затенения и т. д. В случае явной зависимости од-
ного фактора от другого амплитуда каждого типа его режима имеет более
или менее узкую амплитуду, охватывающую лишь какой-то отрезок шкалы
второго фактора.
Так, режимы почвенного увлажнения, особенно ксерофильной ветви,
имеют суженную амплитуду по отношению к фактору атмосферного увлаж-
нения, к фактору континентальности; режим солевого довольства имеет
суженную амплитуду по отношению к факторам и почвенного, и атмосфер-
ного увлажнения, и т.п.
Все эти амплитуды, естественно, также могут быть выражены в едини-
цах соответствующих шкал отдельных факторов, тех самых, на которых
отмечаются и амплитуды видов.
Имея в виду это обстоятельство, мы, конечно, не будем отметать в
сторону, как нонсенс, экологические сетки только за то, что на них может
быть показан КК эдафотопов пустынного и полупустынного типа увлаж-
нения почв для некоторых океанических или даже гумидных типов кли-
мата. Одно дело — принцип изменения КК при различных сочетаниях режи-
мов всего спектра прямодействующих факторов, а ограничение факти-
ческого набора этих сочетаний в силу взаимосвязи факторов — уже явле-
ние другого порядка.
Что касается определения КК экотопов, то вследствие условной дис-
кретности любого из типов экологических режимов на любой шкале, в
любой экологической сетке, следует различать KKmin, ККтях и A7Cmed
(минимальное, максимальное и среднее для экотопа значение КК). Все
они легко определяются по приведенным формулам
^>п=— ЮО; ^тах=— -ЮО; Kffied = ^--100,
«1 «1 2«i
где т — порядковый номер экотопа от края шкалы, а «1 — число экото-
пов, или, вернее, типов режима фактора от края шкалы до общеблагоприят-
ного (общекомфортного).
Переходя к вопросу о наборе экотопов в подзоне хвойно-широколист-
венных лесов (и климатопов, и эдафотопов), следует вспомнить правило
предварения и географической зональности, которое гласит: плакорный
28
вид или плакорный фитоценоз предваряется на юге или на севере в соот-
ветствующих условиях местообитания [Алехин, 1951].
Логически развивая эту мысль, можно сказать, что сами соответству-
ющие экологии такого вида или фитоценоза условия местообитания при-
урочиваются к особенностям среды данной местности, к ее энтопиям,
где создается сочетание режимов прямодействующих факторов, которое
более характерно для зоны плакорного распространения этого вида или
фитоценоза. Иными словами, климатопы и эдафотопы каждой зоны и под-
зоны, так же как виды и сообщества, могут быть экстр азональными.
Амплитуда колебаний здесь, конечно, невелика — вполне вероятно,
даже обязательно, присутствие экотопов, характерных для соседних зон
и регионов, но маловероятно нахождение экотопов, характерных для
более удаленных зон и регионов, не говоря уже о том, что наличие хотя
бы отдельных экотопов, характерных, скажем, для подзоны северной
тайги, невозможно в подзоне хвойно-широколиственных лесов. В связи
с этим климатопы подзоны хвойно-широколиственных лесов, вернее,
их топические и ценотические варианты следует подразделять лишь на
зонально-региональный и экстразональные, уклоняющиеся от зонального
в разной степени и в разных направлениях. Строго говоря, такое подраз-
деление климатопов справедливо для любой зоны, подзоны или региона.
Схематически оно может быть представлено в виде малой координат-
ной сетки, где варианты режимов каждого из двух образующих сетку
прямодействующих факторов подразделяются на три основных типа:
для фактора термического режима — это криофильный, термофильный
и зональный варианты термического режима; для фактора континенталь-
ности - это смягченный, региональный и контрастированный варианты ре-
жима континентальности.
В подзоне хвойно-широколиственных лесов, где зональным климато-
пом на Восточно-Европейской равнине является эунеморальный полукон-
тинентальный (он же — северонеморальный полуконтинентальный). В
малой климатической сетке, применяемой для выделения вариантов топо-
климатов и ценоклиматов, он находится в окружении следующих клима-
топов: по оси ординат — суббореоидного полуконтинентального и термо-
эунеморального полуконтинентального; по оси абсцисс — эунеморального
смягченно-полуконтинентального и эунеморального контрастированно-
полуконтинентального; по гомодинамической оси (оси AD) - субборео-
идного смягченно-полуконтинентального и термоэунеморального кон-
трастированно-полуконтинентального, а по гетеродинамической оси
(оси ВС) - термоэунеморального смягченно-полуконтинентального и суб-
бореоидного контрастированно-полуконтинентального. Название ’’суббо-
реоидный” относится к топо- и ценоклиматам, уклоняющимся в сторону
суббореального типа режима настолько, что, в сущности, стоят ближе
к нему, чем к своему зональному термоклимату.
Название ’’термоэунеморальный” относится к топо- и ценоклиматам,
аналогично уклоняющимся в сторону термонеморального (южнонемо-
рального) типа терморежима. Название ’’смягченно-полуконтинентальный”
относится к топо- и ценоклиматам, уклоняющимся от регионального
типа режима континентальности в сторону субокеанического типа. Назва-
ние ’’контрастированно-полуконтинентальный” относится соответственно
29
к топо- и ценоклиматам, уклоняющимся к субконтинентальному типу
режима континентальности.
Степень этого уклонения может быть различной и в каждом конкрет-
ном случае должна определяться фитоиндикационно, но в целях разграни-
чения при систематизации целесообразно считать экстразональными лишь
те климатопы (топоклиматы и ценоклиматы), уклонение которых от
зонально-регионального, хотя бы по режиму одного из факторов превыша-
ет 50%. Принимая условно, как это делается в излагаемой системе, индекс
эунеморального термического режима за 7, а полуконтинентального режи-
ма - за 4, мы относим к ним, собственно, все полученные индикационно
индексы порядка < 7,5 и >6,5 по режиму тепла и < 4,5 и >3,5 по режиму
континентальности. Тогда как индексы терморежима порядка >7,5 и
<6,5 относятся уже к экстразональным топо- и ценоклиматам, более
близким соответственно термонеморальному и суббореальному климато-
пам соседних зон. Также и в отношении режима континентальности ин-
дексы >4,5 и <3,5 характеризуют топо- и ценоклиматы, более близкие
соответственно субконтинентальному и субокеаническому типам режима
континентальности. В принципе подзона хвойно-широколиственных лесов
в пределах полуконтинентального типа климата индицируется относитель-
но равным участием (и те и другие выступают в качестве содоминантов)
бореальных хвойных и широколиственных пород в лесных фитоценозах
на плакорных местообитаниях.
Там, где преобладание на плакорах явно переходит к хвойным лесам,
а хвойно-широколиственные леса занимают местообитания с термофиль-
ным вариантом зонального режима тепла, начинается уже суббореальная
подзона, или подзона южной тайга. Где преобладание на плакорах перехо-
дит к широколиственных лесам, а хвойно-широколиственные отступают
на местообитания с относительно криофильным термическим режимом,
начинается подзона термонеморальных, или собственно широколиственных
лесов.
В подзоне хвойно-широколиственных лесов лесорастительные условия,
способствующие формированию хвойных лесов, наблюдаются в место-
положениях, обусловливающих относительно более криофильный тип
термического режима. Например, склоны достаточной протяженности
и крутизны северного сектора экспозиций, отрицательные формы мезо-
рельефа и макрорельефа, при условии, что они способствуют застою хо-
лодного воздуха, заболоченные участки с достаточно мощным развитием
мохового покрова, наиболее затененные темнохвойным насаждением
территории и т.п.
Лесорастительные условия, способствующие формированию широко-
лиственных лесов, возникают в местоположениях, обусловливающих
относительно более термофильный терморежим. Например, склоны доста-
точной протяженности и крутизны южного сектора экспозиций, отрица-
тельные формы мезорельефа и макрорельефа,закрытые для ветров север-
ных румбов и открытые к югу, некоторые хорошо прогреваемые, хорошо
дренированные положительные формы рельефа и тд. Те же закономер-
ности, только в других сочетаниях, наблюдаются во всех зонах и подзонах,
как и все явления природы, не ограниченные жесткими границами обо-
собленного материального тела. Например, организм, геологический
30
пласт, некоторые геоморфологические образования и т.п. Климатопы
существуют также в виде некоего гемиконтинуума — непрерывно меня-
ющегося по качественным и количественным характеристикам явления,
естественно разграничимого на относительно дискретные отдельности,
обусловленные пространственными или временными перепадами в интен-
сивности изменений. Пограничные зоны или периоды этих отдельностей
представляют собой те состояния явлений, где изменения наиболее интен-
сивны, а сами отдельности - те, где изменения менее интенсивны.
Практически это выражается в том, что климатопы в природе представ-
лены упорядоченными соответственно другим особенностям региона
совокупностями топоклиматов и ценоклиматов, между которыми располо-
жены более или менее узкие (относительно размеров топо- и ценоклима-
тов) переходные зоны. Также и в динамическом отношении, при неката-
строфических сменах топоклимата или ценоклимата одним или нес-
колькими другими, между ними проходит некоторый переходный
период.
Другой климатический фактор, используемый в предлагаемой климати-
ческой сетке, — фактор континентальности в принципе полностью следует
тем же закономерностям смен режимов, но вследствие менее подробной
градуировки его шкалы. Уловимые фитоиндикационно территориальные
проявления эффекта предварения его режимов растянуты на большие
пространства и в пределах Московской области практически не наблюда-
ются. В Московской области в основном представлены топо- и ценокли-
маты: суббореоидно-полуконтинентальный, эунеморально-полуконтинен-
тальный и термоэунеморально-полукоитинентальный — при амплитуде
колебаний порядка от 5,8 до 7,8 для зонального режима тепла и порядка
от 3,3 до 4,3 для режима континентальности, что в отношении последнего
фактора почти не выходит за пределы одного типа режима — собственно
полуконтинентального.
Чтобы представить, что собой представляют данные типы климата в
отношении их координатного положения, приводим характеристики зо-
нальных режимов тепла подзон южной тайги, хвойно-широколиственных
лесов, а также полуконтинентального режима континентальности, соот-
ветствующих условиям восточноевропейских вариантов южнотаежных,
хвойно-широколиственных и широколиственных лесов (табл. 2).
Как видно из табл. 2, отдельные данные физических измерений почти ниче-
го не объясняют в смысле различия подзон. Изотерма января имеет такой
большой разброс с запада на восток. На западе климат приближается к
субокеаническому, а на востоке — субконтинентальному. В центральной
части подзоны (Московская область и сопредельные районы) средняя Г
января —10° 4-11°. Наиболее показательны: сумма t за период >+10°,
количество безморозных дней и радиационный индекс сухости.
Переходя к рассмотрению эдафотопов подзоны хвойно-широколист-
венных лесов, прежде всего следует остановиться на некоторых общих за-
кономерностях формирования спектра эдафотопов зон, подзон и регионов.
В каждом регионе каждой зоны или подзоны всегда можно выделить
группировку зонально-региональных эдафотопов, встречающихся пре-
имущественно на плакорах, и группировки экстразональных, интразо-
нальных и азональных эдафотопов, приуроченных к особым энтопиям,
31
Таблица? ,,,
Некоторые климатические характеристики суббореальной (южиотаежной), эунемо-
ральной (хвойно-широколиствениых лесов) и термонеморальной (широколиствен-
ных лесов) подзон в условиях полуконтинентального типа климата
Климатическая характеристика Южная тайга Хвойно-широко- лиственные леса Широколиствен- ные леса
Средняя f июля +16°+ +19° +16° + +19° +18° + +20°
Средняя t° января -5° + -16° -5° + -16° -5° + -16°
Сумма/0 за период >+10° 1800-2000° 2000-2200° 2200-2500°
Количество безморозных дней 100-120 120-130 130-140
Количество осадков, мм 500-600 550-650 500-600
Коэффициент увлажнения 1 1 1
Высоцкого -Иванова Радиационный индекс сухости Будыко 2/3 2/3-1 1
обусловливающим отступление режимов прямодействующих факторов
от их зональных (региональных) типов.
При этом энтопии, условия среды которых формируют экстразональ-
ные эдафотопы, более или менее имитируют условия среды соседних зон,
подзон или регионов. Энтопии, формирующие интразональные эдафотопы,
в сущности характерны именно для данной зоны, но разбросаны в ней фраг-
ментарно, будучи специфичны по местоположению, характеру почвогрунтов
и т.д., к тому же преимущественно не на плакорах. Азональные эдафо-
топы формируются в энтопиях, одинаково обычных в целом ряде зон,
подзон или регионов, так как представляют собой совокупности косвен-
ных факторов незональной или преимущественно незональной природы.
Естественно, что в различных частях зоны, подзоны или региона на-
блюдается некоторое изменение спектров как зонально-региональных,
так и экстразональных эдафотопов. В меньшей степени такое изменение
спектра наблюдается у интразональных эдафотопов и почти не наблюдает-
ся у азональных. Ближе к соответствующей смежной зоне или смежному
региону изменение спектров эдафотопов на плакорах будет идти таким
образом, что все большее количество экстразональных эдафотопов соот-
ветствующего типа будут наблюдаться в положениях, все более приближа-
ющихся к плакорным, вплоть до полного выхода из плакор, и таким
образом включаться в зональный спектр.
В конечном итоге спектр эдафотопов плакорных положений станет
одинаково похож на зональные спектры эдафотопов обеих смежных
зон, что и будет соответствовать пограничной полосе между этими зона-
ми. Что касается интразональных и азональных эдафотопов, то они как
явления, относительно независимые от зональных факторов, и изменять
свой спектр от местности к местности должны относительно несвязанно
с изменением зональных факторов, т.е. безотносительно к зональным
границам. (Кстати, очевидно, интразональные эдафотопы или таксоны
растительности не самостоятельные явления, а представляют собой част-
ный случай азональных эдафотопов или таксонов растительности, выделя-
ющихся среди них лишь узкой амплитудой, укладывающейся в интервал
32
Тип водного режима почв Тип солевого режима почв
О О М | е Е g h
с эр эр эр эр эр эр : эо эо эр эр эр э р ЭО эо зо зр зр э р эо зо зо зо зр р 30 30 ЭО 30 зо р
с э 5
f ар а ап а
F ар ар ао ао ао зо зо ао ар ар ар ао ао ао ао ао ао ар ар ао ао ао ао ао ао ар а
Р р
Р р uv ас/ ас/ ас/ “ ао ао ао ао ао ао ар
а а о ао ао ао а ао ао ао ао ао ао 0
А
Рис. 5. Зональные, азональные и экстразональные эдафотопы в Московской обл.
Положение на эдафической сетке и частота встреч
эр — экстраэональный редкий; эо — экстраэональный обычный; зо — зональный
обычный; зр — зональный редкий; ар — азональный редкий; ао — азональный обычный
колебаний условий среды, ограниченный одной зоной или подзоной или
даже одним регионом). Кроме того, следует оговориться, что разделение
на зональные, экстразональные и азональные эдафотопов, или фитоцено-
зов, или других объектов не является строгим, а всегда допускает переход
части объектов из одной категории, в другую даже при незначительных
относительно перемещениях в географическом пространстве или во вре-
мени.
Часть же объектов, в том числе и эдафотопов, имеет промежуточный
в любых принятых критериях различия, характер; например, часть эда-
фотопов встречается в плакорных и во внеплакорных местоположениях.
Исходя из принятой нами системы эдафотопов в подзоне хвойно-ши-
роколиственных лесов теоретически могло бы существовать порядка
40—45 основных и порядка 110—115 дополнительных, или промежуточ-
ных, эдафотопов, в том числе наземных порядка 30—35 основных и поряд-
ка 90—95 промежуточных.
В действительности спектр эдафотопов подзоны хвойно-широколист-
венных лесов в пределах Восточно-Европейской равнины значительно
уже и составляет от 60 до 80% теоретического. Тем не менее поимённая
характеристика даже 60-70 эдафотопов заняла бы слишком много места
и была бы трудно читаема. Поэтому было сочтено целесообразным ограни-
читься характеристикой 14 типов режима увлажнения почв и И типов
солевого режима почв, а сами эдафотопы дать лишь в виде отметок на
координатной сетке с указанием на их зональность, экстразональность
3. Зак. 448 33
или азональность, на обычность, относительную редкость.или исключитель-
ность (рис. 5).
Характеристика режимов увлажнения почв и солевого режима почв
даются в том плане, как их давал Раменский, с учетом новых данных
и современных представлений, взятых по сводкам В.А. Ковды [1973],
М.А. Глазовской [1972], А.А. Роде и В.Н. Смирнова [1971], В.М. Фрид-
ланда [1972], А.И. Перельмана [1966], В.Р. Волобуева [1973].
ТИПЫ РЕЖИМА УВЛАЖНЕНИЯ ПОЧВ В ПОДЗОНЕ
ХВОЙНО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ НА ВОСТОЧНО-ЕВРОПЕЙСКОЙ
РАВНИНЕ
1. Среднестепной тип водного режима почв. Крайне
редок в подзоне в естественных условиях, где встречается иногда (пре-
имущественно на юге подзоны) на вершинах и в верхних частях крутых,
реже — сильно покатых склонов южного сектора экспозиций, в более
северных частях подзоны — при сочетании указанных выше местоположе-
ний с песчаными почвами. Чаще встречается как антропогенно-производ-
ное явление в непосредственной близости песчаных карьеров, по пусты-
рям среди городских застроек, по бровкам и в верхних частях откосов
насыпей при преимущественно южном секторе экспозиций.
Режим эрозионно-промывной, реже — промывной автоморфный, чаще
переходный между ними, поступление влаги исключительно с осадками.
На склонах при сильных ливнях наблюдается поверхностный сток, господ-
ствует инфильтрация через породу. Весной и в дождливые периоды зна-
чительна роль испарения, транспирация ослаблена, так как в растительном
покрове господствуют малопродуктивные сообщества типа луговых и
пустошных фитоценозов абсолютных суходолов. (Возможно заселение
этих местоположений преимущественно сосной, но тогда тип водного
режима изменяется в сторону менее ксерофильных условий).
В естественных условиях режим установившийся, компенсированный
подземным оттоком, как антропогенно-производный, нередко неустано-
вившийся, отрицательно декомпенсированный увеличением оттока грун-
товых вод.
2. С в е ж е с т е п н о й тип водного режима почв. Тоже
редок в подзоне хвойно-широколиственных лесов в естественных услови-
ях и также более обычен как антропогенно-производное явление. Встреча-
ется вместе с предыдущим типом как его обрамление, но чаще сам зани-
мает относительно аналогичные местоположения.
По вершинам и достаточно крутым склонам холмов южной экспози-
ции, также аналогично ориентированным бровкам достаточно крутых
склонов от вторых, реже — первых надпойменных террас, в верхних частях
таких склонов этот тип режима увлажнения приурочен не только к пес-
чаным почвам, - возможен и на супесях и даже суглинках. В случае пес-
чаных почв может на аналогичных формах рельефа занимать верхние части
склонов и не южного сектора экспозиций. Также и в антропогенном релье-
фе менее связан с южными экспозициями и легким механическим соста-
вом субстрата.
Режим также колеблется между эрозионно промывным и промывным
34
автотрофным (как и для предыдущего типа, характеристики местополо-
жений и типов режима даются лишь для условий подзоны хвойно-широко-
лиственных лесов — в зональных положениях они будут существенно
иными).
Поступление влаги исключительно с осадками, расход влаги через ин-
фильтрацию и поверхностный сток. Вследствие открытости и высокого
местоположения значительна роль испарения. Транспирация в пределах
возможностей растительности лугово-пустошного типа, характерного
для абсолютных суходолов. Как и в предыдущем случае, режим в есте-
ственных условиях установившийся, компенсированный подземным отто-
ком, но часто при значительном участии поверхностного стока. При ан-
тропогенных нарушениях на некоторый период режим становится неуста-
новившимся, отрицательно декомпенсированным усилением оттока.
3. Лугово-степной тип водного режима почв. До-
вольно обычный в подзоне тип на самых сухих местообитаниях, приуро-
ченных к вершинам и верхним частям склонов холмов, гряд, увалов,
бровкам и верхним частям склонов относительно крутых или покатых
берегов, террас, крупных оврагов и балок, чаще южного сектора экспо-
зиций.
При сочетании легких почв и южной экспозиции возможно установление
этого типа режима и в средних частях склона, где выше него будут нахо-
диться или один, или оба описанных выше типа.
Режим эрозионно-промывной при положении на склоне или промывной
автоморфный при положении на вершине холма, гребне увала или бровке
и прибровной полосе поверхности высшего уровня. Как и в первых двух
случаях (при обычных описательных обследованиях эти три типа режи-
мов в подзоне хвойно-широколиственных лесов даже не различаются),
грунтовые воды лежат глубоко, до 10 м и глубже. Верховодки не бывает.
Во второй половине лета и в начале осени ощущается острый недостаток
влаги, в засушливые годы — возможен с начала лета. (При описанных
типах режима увлажнения недостаток влаги уже в начале лета и даже в
конце весны — явление довольно обычное.)
Источник поступления влаги — осадки, а на склонах к ним добавляется
некоторый поверхностный сток, особенно если почвы более тяжелого
механического состава.
Расход влаги — грунтовое просачивание, в меньшей степени поверхно-
стный сток, транспирация и испарение. Водный режим возможен и на
лесопокрытых территориях (преимущественно в дюнных сосняках на
юге подзоны). Режим грунтовых вод установившийся, компенсированный
подземным оттоком.
4. Сублуговой тип водного режима почв. Еще более
обычный в подзоне тип режима увлажнения, приуроченный не только
к самым высоким элементам рельефа, но встречающийся и на песчаных
почвах местоположений плакорного типа при сильно развитой дрениро-
ванности территории. Возможен в особо сухих лишайниковых или остеп-
ненных сосняках, реже — в особо сухих ’’нагорных” дубравах по высоким
берегам.
При сведении или иного рода гибели леса переходит, в зависимости от
местоположения, в один из описанных выше типов режима увлажнения.
35
Режим уже преимущественно промывной автоморфный, эрозионно-про-
мывной сравнительно реже на склоновых местоположениях. Приход воды—
преимущественно осадки, поверхностный транзитный сток может наблю-
даться на склонах, если позволяет характер почвогрунтов.
Грунтовые воды глубоко—до 10 м и глубже. Верховодки не бывает.
Режим грунтовых вод установившийся, компенсированный подземным
оттоком.
5. Сухолуговой тип режима увлажнения почв. Обыч-
ный плакорный тип режима хорошо дренированных территорий при легком
механическом составе почв и подстилающих пород. Встречается как на
облесенных, так и на безлесных территориях. При сведении или гибели
леса происходит трансформация типа режима в сторону более ксерофиль-
ного. Полностью господствует промывной автоморфный тип режима,
но при склоновых положениях возможен промывной эрозионный делю-
виально-элювиальный.
Склоновые положения достаточной крутизны могут иметь и суглини-
стые почвы, а также относиться и к северному спектру экспозиций. Вод-
ный баланс складывается из поступления влаги преимущественно в виде
осадков (на плакоре это единственный ее источник), которые в склоновых
положениях дополняются транзитным поверхностным стоком и иногда
незначительным внутрипочвенным тоже транзитным стоком, и из расхода
влаги на инфильтрацию вниз, поверхностный сток, транспирацию и испа-
рение.
Грунтовые воды залегают относительно глубоко (5—7 м). На большин-
стве местоположений верховодки не бывает, на других она устанавлива-
ется на очень короткий срок по весне, реже — при продолжительных дож-
дливых периодах, но при прекращении осадков быстро иссякает, так как
по причине слабого развития водоупорных прослоек, их фрагментарности
и относительной проницаемости не накапливается в значительных коли-
чествах. Режим грунтовых вод установившийся, компенсированный под-
земным оттоком. Во второй половине лета и в начале осени наблюдается
дефицит влаги, а в сухие годы и гораздо раньше.
6; Свежелуговой тип водного режима почв. Зани-
мает обширные относительно хорошо дренированные пространства на пла-
корах, пологих склонах, террасах и поймах рек. Встречается как на поч-
вах легкого механического состава (при условии достаточного развития
водоупорных прослоек), так и на почвах тяжелого механического соста-
ва (при условии достаточного дренажа).
Режим переходного типа между промывным автоморфным и промыв-
ным гидроморфным, в случае склонового положения наблюдаются и
некоторые черты эрозионно-промывного режима (слабо выраженная
эрозия, некоторый перенос субстрата). Поступление влаги с осадками
при одновременной большей или меньшей доле поверхностного и внутри-
почвенного притока.
Расход влаги в виде вертикальной и боковой инфильтрации, в гораздо
меньшей степени — поверхностного стока, интенсивной транспирации
и более или менее ослабленного хорошо развитым растительным покровом
физического испарения. По весне и в начале лета возможна верховодка,
преимущественно в виде мозаики блюдец.
36
Грунтовые воды залегают на глубине 3—5 м, иногда несколько глубже.
Режим их установившийся, компенсированный внутрипочвенным отто-
ком и транспирацией.
7. Влажно-луговой тип водного режима почв. Зани-
мает обширные, относительно дренированные или относительно слабо
дренированные плакорные, террасные и пологосклоновые территории
с различного, но чаще тяжелого механического состава почвами. В этом
типе наблюдается двоякая тенденция изменения водного обеспечения
при утрате лесной растительности — наблюдается как некоторое иссушение,
так и некоторое переувлажнение, что частично связано с внутритипными
различиями свойств разных энтопиев, а частично с климатическими ано-
малиями лет, в которые это обезлесение произошло.
Режим этот промывной гидроморфный, или элювиально-гидроморф-
ный. Грунтовые воды —на глубене 1—3 м. Капиллярная кайма весь год
в пределах корнеобитаемого слоя, по крайней мере древесной и кустар-
никовой растительности и трав с относительно глубокой корневой систе-
мой. В весенний, раннелетний и все дождливые периоды почва полностью
насыщена влагой, наблюдается явление временно-избыточного увлажнения.
Внутрипочвенный сток — боковой, по дренирующим прослойкам.
Главные источники поступления воды — транзитная боковая инфиль-
трация и осадки, поверхностный натек почти не наблюдается. Расход воды—
боковой внутрипочвенный сток, транспирация и, в значительно меньшей
степени, испарение, вертикальное просачивание эпизодично и возможно
далеко не на всех местоположениях.
Режим грунтовых вод — установившийся, компенсированный подзем-
ным оттоком и транспирацией.
8. С ы р о в а т о - л у г о в о й тип режима почвенного
увлажнения. Приурочен к слабо дренированным плакорным, террас-
ным или пойменным, относительно выровненным местоположениям,
реже — к пологим склонам значительной протяженности, особенно на
участках, хотя бы с незначительным уменьшением угла уклона. Почвы
преимущественно тяжелого механического состава. Возможны процессы
оглеения и торфообразования. При сведении леса изменение режима про-
исходит в основном в сторону гидрофилизации. Режим промывной.
Грунтовые воды относительно близко — иногда глубина их залегания
даже менее 1 м и редко больше 2 м. Даже в обычные по выпадению осад-
ков годы капиллярная кайма почти не опускается ниже корнеобитаемого
слоя. Кратковременные периоды дефицита влаги возможны лишь в самые
засушливые годы, зато периоды временного переувлажнения, и подчас
довольно длительные, совершенно обычны. Во влажные годы вода настоль-
ко близка к поверхности, что создает впечатление временного заболачи-
вания. Как правило, относительно развит моховой покров.
Источники увлажнения — осадки и боковая инфильтрация. В. расходе
влаги велики доли боковой инфильтрации и транспирации, много меньше
доли испарения, инфильтрация вниз может наблюдаться только в засушли-
вые годы при небольших осадках, так как значительные осадки тотчас
восстанавливают подпор грунтовых вод.
Режим грунтовых вод — установившийся, компенсированный боковым
оттоком и транспирацией.
37
9. Сыро-луговой тип водного режима почв. Наблю-
дается в понижениях как на плакоре, так и на террасах и в поймах. Поч-
вы, как правило, тяжелого механического состава, но при условиях, обе-
спечивающих создание и сохранение ортзандовых горизонтов, могут быть
и супесчаными. Переходный от промывного гидроморфного к намывному
(амфибиальному) типу режима.
Грунтовые воды настолько близко, что капиллярная кайма находится
у самой поверхности. Хронически возникает состояние переувлажнения,
развиваются процессы оглеения и торфообразования. Относительное об-
сыхание, не достигающее дефицита влаги, возможно лишь в сухие годы.
Сами понижения, в которых формируется такой тип водного режима,
могут быть и бессточными западинами, и ложбинами временных водото-
ков. Поступление влаги в сырые годы преимущественно поверхностным
натеком и осадками, а в сухие — боковой инфильтрацией и осадками.
Расход воды в виде поверхностного стока (особенно в период выпаде-
ния осадков в форме временных водотоков), грунтового бокового стока
(главным образом в сухие годы), транспирации и испарения.
Режим грунтовых вод — установившийся, компенсированный глав-
ным образом транспирацией и испарением при эпизодическом участии
поверхностного стока и боковой инфильтрации.
10. М о к р о-л у г о в о й тип режима увлажнения почв.
В более резко выраженных западинах, блюдцах и ложбинах временных
водотоков, по низким берегам водоемов, в местах выклинивания водо-
носных горизонтов на относительно пологих склонах. Различного механи-
ческого состава почвы, хотя суглинистые и глинистые чаще, но обязательно
при близком залегании водоупоров.
Грунтовые воды застойного или проточного характера близко к по-
верхности, капиллярная кайма достигает горизонта Ао, так что он почти
постоянно напитан водой. Интенсивные процессы оглеения и торфообра-
зования. Часто мощно развитый моховой покров. Кратковременное осу-
шение возможно лишь в засушливые годы.
Тип режима намывной, амфибиальный. Приход влаги — поверхностный
натек, меньше осадки, еще меньше инфильтрация. Характер расхода вла-
ги зависит от застойности или проточности увлажнения: в первом случае
главная роль принадлежит транспирации и испарению, во втором — в от-
носительно равной мере поверхностному стоку, боковой инфильтрации,
транспирации и испарению.
Режим грунтовых вод в первом случае компенсирован лишь испарением
и транспирацией, а во втором — всеми четырьмя составляющими.
11. Болотно-луговой тип водного режима почв.
Различные депрессии рельефа с близким к поверхности залеганием водо-
упорных горизонтов, часто со слоем торфа большей или меньшей мощности,
аккумулирующим влагу.
Грунтовые воды у самой поверхности и частично (в виде разной кон-
фигурации мочажинок) кое-где выходят на поверхность. Часто развит
кочковатый микрорельеф. Поступление воды натеком и осадками, тогда
как расход преимущественно транспирацией и испарением. Ими обычно
и компенсирован режим грунтовых вод. Однако не редкость и частично
проточный режим вблизи русла постоянного или временного водотока.
38
12. Бол. о тный тип водного режима почв. Возможен в
виде собственного болотного типа с застойными грунтовыми водами. Ка-
пиллярная кайма выходит на поверхность. Большую часть года вода стоит
на поверхности в виде крупных мочажин или в понижениях между кочка-
ми. В засушливые годы возможно обсыхание поверхности вплоть до опус-
кания капиллярной каймы на некоторую глубину под поверхность.
Режим амфибиальный натечный, в некоторых случаях переходный
к субаквальному. Интенсивно проходит процесс торфонакопления. Ком-
пенсация режима грунтовых вод в собственно болотном типе осуществля-
ется транспирацией и испарением, а в грунтово-болотном — поверхностным
и частично внутрипочвенным оттоком, транспирацией и испарением.
13. Суб болотный тип водного режима почв. Является
промежуточным между болотно-луговым и болотным типами. Режим
натечный, амфибиальный. Также возможны варианты с застойным и про-
точным характером переувлажнения. Компенсация режима грунтовых
вод подчинена тем же закономерностям, что и у смежных типов. Отли-
чия от смежных типов почти чисто количественного характера: меньшая
обводненность и меньшая оторфованность по сравнению с болотным типом
и большая обводненность и большая оторфованность по сравнению с
болотно-луговым типом.
14. В о д н о-б о л о тный тип режима увлажнения почв.
Приурочен к почти постоянно залитым водой депрессиям рельефа. Пред-
ставляет собой сочетание обширных мочажин с крупными моховыми,
травяными кочками, деревьями на кочках и относительно глубокими
бочагами. Вода может быть как застойной, так и частично проточной.
Обсыхание поверхности случается очень редко, только в самые засушливые
годы.
Режим субаквального по преимуществу типа с некоторыми элемента-
ми натечного. Компенсация режима грунтовых и поверхностных вод осу-
ществляется полностью или преимущественно транспирацией и испарением.
ТИПЫ СОЛЕВОГО РЕЖИМА ПОЧВ В ПОДЗОНЕ ХВОЙНО-
ШИРОКОЛИСТВЕННЫХ ЛЕСОВ ВОСТОЧНО-ЕВРОПЕЙСКОЙ РАВНИНЫ
1. Олиготрофныйгликофильный тип солевого ре-
жима почв. Связан с промывным автоморфным, элювиальным типом
водного режима. Интенсивный вынос солей, кремнезема и железа, отчасти и
марганца, алюминия, органических соединений. Развитие кислотности. В
гидрофильной ветви экологического ряда режимов увлажнения развивает-
ся торфообразование, начиная примерно с сыровато-лугового типа увлаж-
нения. Материнские породы — суглинки, супеси и пески.
Почвы подзолистые, песчаные, выщелоченные, верховые торфяники.
Крайняя бедность солями: 34-74 мг/л, НСОз1, SO4" и С1' нет, жест-
кость воды болот 0,6—1,4, pH 3,0-4,3 для минеральных почв, а для тор-
фяников 2,5—5,0, зональность торфа 2—4%.
2. Суболиготрофный тип солевого режима. Также
связан с промывным автоморфным типом водного режима. Почвенные
процессы примерно те же, но несколько меньшей интенсивности. Как
и первый тйп, в подзоне хвойно-широколиственных лесов возможен только
39
на верховых болотах, и то в исключительных случаях. Также очень малое
количество солей: 75—82 мг/л, НСО3’ , S04” и С1’ нет, жесткость
воды болот 1,4—2,3, pH 3,0—5,0 для минеральных почв, для торфяников
pH 3,0—5,5, зольность торфа 3,6%.
3. Семиолиготрофный гликофильный тип солево-
го режима почв. Тоже связан с промывным автоморфным, элюви-
альным типом водного режима. В подзоне хвойно-широколиственных ле-
сов обычен на верховых болотах, на минеральных почвах всречается сравни-
тельно редко — главным образом на достаточно протяженных и достаточ-
ной крутизны склонах северного, реже — западного секторов экспозиций.
Почвы суглинистые и супесчаные, чаще подзолистые, реже подзолисто-
глеевые и торфяно-подзолистые, часто верховые и переходные торфяники.
Интенсивный вынос всех геохимических подвижных минеральных и ор-
ганических соединений, в том числе кремнезема, железа, марганца и алю-
миния. Бедность солями: 83—88 мг/л, зольность торфа 6—9%, НСО3’ ,
SO4” й Cl’ нет, жесткость воды болот 2,0— 4,5, pH 3,2-5,5 для ми-
неральных почв, для торфяников pH 3,5—5,7.
4. Субмезотрофный гликофильный тип солевого
режима почв. Встречается при промывном автоморфном, промывном
гидроморфном (реже) и эрозионно-промывном типах водного режима
почв. Из них промывной гидроморфный тип водного режима в сочетании
с данным типом солевого режима встречается почти исключительно на
болотах переходного типа.
Преобладают процессы выноса геохимических подвижных соединений,
однако устанавливающаяся временами связь с грунтовыми водами, пре-
имущественно верховодкой, обусловливает такое же временное разви-
тие гидрогенной аккумуляции вторичных минералов: глин и полуторных
окислов, сочетанию процесса оподзоливания с процессом оглеения.
На переходных болотах вынос большинства геохимически подвиж-
ных соединений (особенно кремнезема) сочетается с относительно ин-
тенсивной гидрогенной аккумуляцией окислов железа и марганца, гидро-
генным оглиением, и все это на фоне интенсивного торфообразовательного
процесса. Относительно большая обеспеченность солями': 88—120 мг/л,
зональность торфа 8—10%, НСО3’ , S04” и С1’ нет, жесткость воды
болот 4,0—6,0, pH 3,5-6,0 для минеральных почв, для торфа pH 4,0-6,0.
5. Мезотрофный гликофильный тип солевого ре-
жима почв. Встречается как при промывном автоморфном, так и при
промывном гидроморфном, реже—при эрозионно-промывном типах вод-
ного режима почв. В то же время промывной автоморфный тип режима
в чистом виде на местоположениях, где формируется данный тип солево-
го режима, практически не наблюдается, а существует в сочетании с про-
мывным гидроморфным, в периоды существования верховодки тип режи-
ма промывной гидроморфный, а в периоды, когда она иссякает, тип режи-
ма промывной автоморфный. Сочетание выщелачивания преимущественно
легко растворимых соединений с восстановительным процессом, накопле-
нием полуторных окислов.
Почвы в различной степени и в различных сочетаниях имеют признаки
подзолообразования, оглеения и торфообразования (первого со вторым,
первого со вторым и третьим, второго с третьим). Относительно удовлет-
40
верительная обеспеченность солями: 98—150 мг/л, зольность торфа 9—11%,
HCO3f , SO4"h С1' нет, жесткость воды болот 4,5—6,5, pH 4,0—6,5.
6. Пермезотрофный гликофильный тип солевого
режима почв. Связан с явным преобладанием гидроморфного про-
мывного режима, когда автоморфный режим может временно устанавли-
ваться только в наиболее сухие периоды года. Промывание большей частью
периодично и неинтенсивно — выщелачиваются лишь наиболее раствори-
мые соединения. Восстановительные процессы преобладают. Гидрогенная
аккумуляция окислов железа и марганца, глинных минералов. Становит-
ся возможной незначительная аккумуляция углекислого кальция.
Почвы с признаками дернового, подзолообразовательного (в ослаб-
ленном варианте), глеевого и торфообразовательного процессов. Преобла-
дание того или другого диктуется местоположением, водным режимом
и характером растительного покрова. Относительно хорошая обеспечен-
ность солями: 120—180 мг/л, зольность торфа 10—14%, НСОз' — до
1,4—5,0 мг/100 г почвы, SO4//h Cl1 нет, жесткость воды болот 5,0—
7,5, pH 4,5-7,5.
7. Семиэвтрофный гликофильный тип солевого
режима почв. В подзоне хвойно-широколиственных лесов связан
более с намывным амфибиальным типом водного режима и в меньшей
степени, главным образом на местонахождениях с улучшенным терми-
ческим режимом (например склоны южных экспозиций), с промывным
гидроморфным или переходным между ним и промывным автоморф-
ным типом водного режима почв. Может наблюдаться и сочетание эле-
ментов намывного и промывного режима, преимущественно гидроморф-
ного, но возможно и всех трех при периодических или эпизодических
чередованиях преобладания какого-либо из них. Промывание относитель-
но незначительно. Обычно накопление аллювия, делювия или пролювия.
Гидрогенная аккумуляция полуторных окислов и в меньшей степени —
углекислого кальция. В зависимости от соотношения оттока, транспи-
рации и испарения складываются разные сочетания механической, гео-
химической и биогенной аккумуляций.
Преобладающие местоположения: поймы рек, низины, нижние части
склонов. Хорошая обеспеченность солями: 160 мг/л, зольность торфа
12%, НСОз' 4,0—16,0 мг/100 г почвы, SO4” и С1₽ — следы, жесткость
воды болот 7,0—10,0, pH 5,5—7,5.
8. Субэвтрофный гликофильный тип солевого
режима почв. В подзоне хвойно-широколиственных лесов редок
и связан почти исключительно с намывным, амфибиальным типом вод-
ного режима, к тому же при условии исключительно проточного увлаж-
нения. Вследствие этого в почвах преобладают аэробные, окислительные
процессы.
В зависимости от соотношения оттока, транспирации и испарения скла-
дывается то или иное сочетание механической, биогенной и,геохимической
аккумуляции.
Почти исключительно в поймах рек и главным образом на юге подзоны.
Промывание и выщелачивание слабое — обеспечивает вынос только самых
легкорастворимых солей. Транспирация и испарение вызывают выпадение
менее растворимых соединений в осадок. Высокая обеспеченность солями:
41
200 мг/л, НСОз' 15,0—37,0 мг/100 г почвы, S04" — следы, С1Г — следы,
жесткость воды притеррасных болот 10,0, pH 6,0-7,8.
9. Эвтрофный гликофильный тип солевого режима
почв. В подзоне хвойно-широколиственных лесов крайне редок в естест-
венных условиях. Наиболее вероятно нахождение его только на юге под-
зоны в условиях намывного аллювиального типа водного режима. Более
вероятно формирование его как антропогенно-производного, на хорошо
ухоженных, обильно удобряемых обрабатываемых землях. Также возмож-
но формирование подобного ему временного солевого режима на некото-
рых осушенных территориях из-под эвтрофных болот с исходно богатым и
мощным гумусово-торфяным аккумулятивным горизонтом. В отдельных
случаях вероятно образование относительно близкого ему солевого режима
при сочетании намывного водного режима с близким залеганием мощных
слоев карбонатных пород.
Промывание и выщелачивание в более южных зонах обеспечивают
только незасоление, а в подзоне хвойно-широколиственных лесов практи-
чески многократно перевешиваются биогенной и гидрогенной аккуму-
ляцией. Преобладают глинистые и суглинистые почвы, изредка возможны
и супесчаные (при близком залегании карбонатных пород, при питании
карбонатными водами и т. п.). Наивысшая обеспеченность солями при
отсутствии признаков засоления: НСО3’ 30—50 мг/100 г почвы, SO/ и
С11 — следы, pH 6,5—8,0.
10. Пертрофный, или субгликофильный тип соле-
вого режима почв. В подзоне хвойно-широколиственных лесов
в естественных условиях вдали от морских побережий не встречается,
но, по-видимому, возможен в современных условиях на ограниченных
территориях как антропогенное явление. Как следствие регулярного или
сезонно-периодического поступления сульфатов, хлоридов, фосфатов и
других соединений в результате различного рода хозяйственной деятель-
ности промышленных, транспортных, коммунальных и сельскохозяйст-
венных организаций. В более южных зонах семиаридного и аридного кли-
мата распространен на местоположениях, обусловливающих переходное
состояние водного режима между промывными и испарительными, как
автоморфными, так и гидроморфными, или между намывным и испа-
рительным гидроморфным типами режима.
В лесных и более северных зонах этот тип солевого режима возможен,
естественно,.лишь на приморских лугах в полосе перехода от соленовод-
ного к пресноводному увлажнению. Обеспеченность солями выше обще-
благоприятной, но еще не затрудняющая (или практически не затрудняю-
щая) деятельность корневых систем и не нарушающая (или практически
не нарушающая) физиологических процессов. НСО/ ’40-60 мгЧОО г
почвы, SO4” и С1' 10 мг/100 г почвы, pH 7,5-8,0.
В заключение целесообразно остановиться на некоторых вопросах се-
зонной динамики водного режима почв как ведущего экологического
режима на ландшафтном уровне.
Явление сезонного промерзания почв в подзоне хвойно-широколиствен-
ных лесов, особенно в условиях полуконтинентального климата, доста-
точно хорошо выражено и вследствие этого экологически весьма значи-
мо. В частности, этим явлением обусловлено сходство начальных стадий
42
годового динамического цикла водных режимов почв почти всех место-
положений в пределах всего ареала распространения явления длительного
промерзания почв в сочетании с частичным или полным их оттаиванием
в летнее время.
Во всех этих регионах к началу периода вегетации устанавливается
надмерзлотный водный режим с близким залеганием надмерзлотных
грунтовых вод, до некоторой степени аналогичный испарительному гид-
роморфному типу водного режима, характерного для местоположений с
близкими грунтовыми водами в аридных районах. При этом в расходной
части баланса влаги господствуют испарение и боковая инфильтрация,
причем последняя относительно велика лишь на склоновых местополо-
жениях. Преобладают процессы гидрогенной аккумуляции.
Почвенные реакции носят преимущественно восстановительный ха-
рактер. В дальнейшем ход динамики режима принимает различное нап-
равление в зависимости от положения в рельефе и состава почвообразую-
щих пород, применительно к климатическим условиях каждой зоны, под-
зоны или региона. В подзоне хвойно-широколиственных лесов прежде
всего выходят из надмерзлотного водного режима относительно повы-
шенные место поле жения с почвами легкого механического состава, где
сразу же устанавливается промывной автоморфный тип режима увлаж-
нения со всем комплексом сопутствующих явлений: выносом всех геохи-
мически подвижных соединений, развитием кислотности, господством
аэробных процессов и др.
На всех других местоположениях, кроме местоположений, обусловли-
вающих формирование в качестве коренного намывного или эрозионно-
промывного типов водного режима почв, надмерзлотный режим сменяется
промывным гидроморфным, различающимся по степени собственно увлаж-
нения в зависимости от глубины залегания водоупорных горизонтов. В слу-
чае наличия верховодки режим существует до ее иссякания, а затем может
перейти в промывной автоморфный или, в случае относительно близкого
горизонта постоянных грунтовых вод, в другую модификацию того же
промывного гидроморфного режима.
В местоположениях, где коренным типом водного режима почв являют-
ся промывной гидроморфный, намывной амфибиальный или эрозионно-
промывной (смывной), смена надмерзлотного режима происходит непос-
редственно на коренной тип.
В осенний и предзимний периоды развитие нормально происходит в об-
ратном направлении. Таким образом, если не считать, что различные типы
промывного режима могут сменять друг друга несколько раз за безмо-
розный период в зависимости от режима выпадения осадков, общая тен-
денция динамики водного режима, по крайней мере большинства пла-
корных местоположений и равноценных им по гидрологическому эффекту,
заключается в последовательной смене надмерзлотного испарительного
гидроморфного типа режима промывным гидроморфным, а последнего -
промывным автоморфным, в конечном итоге — опять промывным гидро-
морфным, его мерзлотным сублимационным и, наконец, к началу нового
вегетационного периода—снова надмерзлотным.
На местоположениях с промывным гидроморфным, намывным или
смывным коренными типами режима цикл его сезонных смен сокраща-
43
ется, включая, помимо коренного, лишь надмерзлотный тип по весне
и мерзлотный в зимний период.
В районах распространения вечной мерзлоты господствует цикл лишь
из двух типов водного режима: надмерзлотного испарительного гидро-
морфного в летний период и мерзлотного сублимационного в зимний
период. Это не может не сказаться на разнообразии других режимов эда-
фотопов и в конечном итоге на разнообразии растительного покрова.
Пример динамического цикла водных режимов лишний раз подтвержда-
ет специфичность преломления характеристик среды в восприятии расти-
тельным покровом, трудно поддающимся расшифровке методами фи-
зико-химических определений. В самом деле, ни результаты инструмен-
тальных измерений, ни физико-химические объяснения сущности тех или
иных геохимических или гидрологических процессов не могут четко увя-
заться с состоянием и динамикой растительного покрова — растительность
воспринимает, среду в своей естественной шкале, интегрирующей все
многообразие динамики различных сочетаний режимов отдельных фак-
торов. Единственным, еще далеко не совершенным, но все же прибли-
жением к этой естественной шкале являются шкально-балловые методи-
ки оценки среды.
Что касается ценотопов, то основное подразделение их пока проводи-
лось по отношению к режиму факторов затенения, характеристике, силь-
но меняющейся в пределах очень мелких контуров, соразмерных не только
синузиям, но зачастую и отдельным экземплярам видов, составляющих
эдификаторный ярус сообщества. .
Ту же мелкоконтурность обнаруживают и другие факторы ценотической
природы, как, например, фактор занятости пространства, иначе фактор
задернения, или корневой сомкнутости. Таким образом, мелкоконтур-
ность конкретных пятен изменения режимов при относительной круп-
ноконтурности усредненных их характеристик связана с самой структу-
рой фитоценозов и является общей чертой всех экологических режимов,
зависящих от состава и состояния растительного покрова территории.
В сущности, поле влияния фитоценозов слагается из совокупности фито-
генных полей отдельных видов.
В принятой нами шкале различается пять основных типов режима за-
тенения, которые все встречаются в любом регионе, где возможно су-
ществование лесной растительности.
1. Световой режим открытых пространств. Возможен при отсутствии
в растительном покрове древесного, кустарникового и особо высоко-
травного ярусов.
2. Световой режим полян, опушек, относительно невысоких и мелко-
листных кустарниковых зарослей, некоторых высокотравных зарослей
(заросли тростника, бурьянного крупнотравья и т. п.). Режим опреде-
ляется эффектом бокового затенения (поляны), бокового, осветления
(редкие опушки), затенения маломощным верхним ярусом (кустарники,
высокотравья).
3. Световой режим светлых лесов. Наблюдается в насаждениях светло-
хвойных и мелколиственных пород низкой и средней сомкнутости и в
сильно разреженных древостоях широколиственных и темнохвойных
пород.
44
4. Световой режим тенистых лесов. Наблюдается в широколиственных
и темнохвойных лесах средней сомкнутости или в мелколиственных и
светлохвойных лесах высокой сомкнутости, или в них же при сильно
развитом ярусе высоких кустраников (лещины и др.).
5, Световой режим особо тенистых лесов. Наблюдается в насаждениях
темнохвойных, реже — широколиственных пород при высокой сомкнутос-
ти. Данные характеристики относятся к условиям затенения на уровне
нижних ярусов. Естественно, что на более высоких уровнях условия ос-
вещенности другие, благодаря чему при довольно сильном затенении на
уровне травостоя и даже кустарникового яруса в верхних ярусах могут
благополучно существовать светолюбивые породы, что при обычно зна-
чительной их долговечности длительное время обеспечивает возможность
заселения ими вышедших по той или иной причине из режима затенения
соседних территорий.
В целом же лесорастительные условия не представляют собой некое
особое явление в системе элементов среды и являются не экологическим
инебиогеоценологическом, а скорее хозяйственным термином. Объект,
трактуемый в лесоведении и лесоводстве как ,.лесорастительные условия”,
в экологии и биогеоценологии называется просто „условия местопроиз-
растания”, представляющие собой экотоп, несколько трансформирован-
ный в процессе взаимодействия с растительным покровом. Лесовод может
рассматривать его с точки зрения возможности развития на нем естествен-
ного или саженого древостоя и называть лесорастительными условиями,
луговод может рассматривать его с точки зрения условий для формиро-
вания травостоев той или иной продуктивности, полевод — с точки зрения
целесообразности использования под полевые культуры и т. д.
Таким образом, назовем ли мы данное явление лесорастительными,
лугорастительными или еще какими-то растительными условиями — это
все будет лишь данью той или иной отрасли хозяйства, а сущность его
останется одна — условия местопроизрастания, иначе экотоп, трансфор-
мированный взаимодействием с фитоценозом. Наука, призванная изучать
такие объекты, — экологическая типология земель, а основной метод
исследования у нее — фитоиндикация.
ГЛАВА III
ЛЕСОРАСТИТЕЛЬНЫЕ УСЛОВИЯ
НА ЛЕСОПОКРЫТЫХ И БЕЗЛЕСНЫХ ТЕРРИТОРИЯХ
Известно мнение, что принцип школы Браун-Бланке: главное внима-
ние при диагностике единства растительного покрова и среды уделять
травяному покрову — обусловлен отсутствием естественных древостоев
вследствие интенсивного способа ведения лесного хозяйства в Западной
Европе. В условиях выпаса скота в лесу при экстенсивном лесном хозяй-
стве главное внимание должно быть уделено особенностям формирования
именно древостоев, так как травяной покров в таких условиях изменен
сильнее [Вальтер, 1969]. Это справедливо лишь до тех пор, пока мы сто-
им на позициях предположения наличия жесткой связи определенных
параметров среды с определенными, фитоценотически или биогеоцено-
45
тически обусловленными сочетаниями видов растений. Пока мы пытаем-
ся утверждать, что условия среды индицируются лишь коренной расти-
тельностью.
Для целей фитоиндикации пригоден любой относительно разнообраз-
ный, в смысле количества видов, растительный покров, независимо от
степени его нарушенности, от того, является ли он коренным, длительно
производным, кратковременно производным или пионерным, даже ру-
деральным.
Сочетание режимов прямодействующих факторов можно определить
и в любом лесу, и на лугу, и на пустыре. Однако определенные фитоиндика-
ционно режимы, хотя бы факторов увлажнения почв и солевого богатства
почв, в высокополнотном насаждении, близком коренному типу, в рас-
строенном или производном лесу, на лугу и на пустыре, будут характе-
ризовать собой качественно различные состояния среды и идентифици-
ровать их по принадлежности к фитоиндикационно установленному од-
ному и тому же типу режимов этих факторов неправомерно.
Так, определив на некоторых участках, один из которых занят лесом,
другой лугом и третий пустырем, одно и то же сочетание режима увлаж-
нения почв, например свежелугового с солевым режимом пермезотроф-
ным гликофильным, будет все же неправомерно на этом основании иден-
тифицировать эти эдафотопы как одни и те же условия местопроизра-
стания.
Под лесом это будет некий эдафотоп, в некоторой степени измененный
воздействием лесной, в первую очередь древесной, растительности. Под
лугом и пустырем тоже некие эдафотопы, но измененные в какой-то степе-
ни разнонаправленной и разной интенсивности деятельностью человека.
Если обозначить данный лес как Л, данный луг — Т, пустырь — П, а све-
желуговой пермезотрофный эдафотоп — Э, то можно условно записать
следующие простейшие неравенства: 1) х+ Л = Э; 2) + Т = Э; 3) z +
+ П = Э. Не вызывает сомнения, что можно записать такие неравества:
1) Л =# Т; 2) Л¥=П; 3) Т¥=П. Но тогда и вторые слагаемые этой одной и
той же суммы не равны и справедливо записать другие неравенства:
]) х у; 2) х =£ z; 3) у =£ z. Иными словами исходные условия место-
произрастания, дающие один и тот же эффект (одно и то же сочетание
режимов прямодействующих факторов), при взаимодействии с явно
различными фитоценозами различны между собой.
Может представлять известный интерес вопрос о том, какое состояние
эдафотопа считать исходным: под лесом, лугом, пустырем или вообще
без растительного покрова. С точки зрения влияния энтопия в чистом виде
исходным состоянием следует считать экотоп такого типа на участке, вооб-
ще лишенном растительности, но с точки зрения естественности экотопа
это будет совершенно неприемлемо, так как в естественных условиях
все эдафотопы, состояние режимов факторов которых хоть в какой-то
мере пригодно для произрастания растений, обязательно покрыты расти-
тельным покровом.
С точки зрения естественности экотопа исходным, очевидно, следует
признать экотоп, находящийся под воздействием своего коренного расти-
тельного покрова. Однако в условиях прогрессирующей и уже очень далеко
зашедшей ашропогенной дигрессии фитоценозов в подавляющем боль-
46
шинстве ландшафтов определение коренного фитоценоза на каждом дан-
ном местоположении оказывается далеко не простым делом и любое
заключение по этому поводу чаще всего оспоривается разными исследо-
вателями с различных позиций.
Данные фитоиндикации тоже не могут в подавляющем большинстве
случаев дать однозначный ответ. Потенциальные флоры экотопов и их по-
тенциальное доминантное ядро указывают лишь, из каких видов может
формироваться фитоценоз и какие виды могут в нем доминировать, но чем
больше по числу видов потенциальная флора экотопа, тем больше разно-
образных их сочетаний будут обладать достаточной устойчивостью в усло-
виях данного экотопа, тем больше фитоценозов могут претендовать на
название коренных, или климаксовых, сообществ, тем труднее доказать
правильность субъективного выбора того или иного исследователя. От-
сительно сходны будут лишь мнения о коренных сообществах в диском-
фортных условиях среды, где потенциальные флоры и их доминантное
ядро предельно бедны видами.
Учитывая все сказанное выше практически целесообразно, по-видимому,
считать исходным состоянием любого экотопа вообще и эдафотопа в част-
ности сочетание его с травяным растительным покровом. Эта чисто услов-
ная посылка удобна тем, что травяной растительный покров распростра-
нен по всем зонам, по всем климатопам и практически по всем эдафо-
топам, так что в сравнении номинально одних и тех же эдафотопов из
разных зон и регионов влияние различной развитости травостоев будет
все же относительно одноразмерной поправкой, чего не скажешь о корен-
ных растительных сообществах.
Даже при сравнении лесорастительных условий возможно удобнее ис-
ходить из условия первичного состояния экотопа во взаимодействии с
травянистой растительностью, так как с этим состоянием его впервые стал-
кивается молодой лес. Кроме того, различные типы леса более контрастно
влияют на экотоп, чем различные травостои, да к тому же травостои быст-
рее приходят в равновесие с условиями среды, чем леса, потому что у них
короче сукцессионные смены: травостой становится все ближе к коренному
с каждым годом, тогда как древостой изменяется в этом направлении,
по крайней мере, в десятки раз медленнее.
Итак, условившись считать нормальным состоянием экотопа вообще
и эдафотопа в частности их сочетание с травяным растительным покро-
вом, на лесопокрытых территориях мы должны уже рассматривать ин-
дицируемые на них экотопы как трансформированные лесной раститель-
ностью. В этом случае основным становится уже вопрос о том, как сильно
и в каком направлении изменил тот или иной лес исходное состояние
экотопов, т. е. каким сочетаниям режимов прямодействующих факторов
под травяными сообществами соответствуют индицированные нами со-
четания этих режимов под лесом.
Вопросом изменения эдафических условий под влиянием леса занима-
лись многие исследователи, и в общих чертах направленность изменений
можно считать уже выясненной. Известное высказывание Высоцкого
[1932]: ’’Лес сушит равнины и увлажняет горы” - сохранило значение
и по сей час, будучи лишь детализировано и подправлено новыми иссле-
дованиями.
47
Вторая ее часть (’’увлажняет, горы”) остается без изменений, лишь с
тем разъяснением, что под ’’горами” здесь следует понимать любую силь-
но пересеченную местность. Первая же часть (’’сушит равнины”) подвер-
глась некоторому пересмотру и уточнению. Так, установлено, что эффект
взаимодействия с лесом для различных равнинных условий разный.
Высокопроизводительные древостои на суглинках расходуют влаги
на 40% больше безлесных пространств, а малопродуктивные на песках —
столько же или меньше, чем безлесные пространства [Рутковский, 1949]
Влияние леса на грунтовые воды зависит от глубины их залегания:
уровень близких, корнедоступных он понижает в результате сильной десук-
ции, тогда как уровень глубоких грунтовых вод повышает в результате
хороших фильтрационных свойств лесных почв [Воронков, 1970]. Лес
обусловливает существование верховодки в силу различной влагопро-
ницаемости генетических горизонтов лесных почв, горизонт В менее про-
ницаем, , чем горизонт А, отчего при обильных и продолжительных осад-
ках в последнем накапливается влага и устанавливается боковая инфиль-
трация [Роде, 1972]. В принципе же особенности динамики грунтовых
вод под лесом обусловлены большей, чем в безлесной местности, прони-
цаемостью лесных почв [Рахманов, 1971].
Отличие лесного ценоклимата от зонального типа климата или топо-
климата того или иного местоположения определяется составом и стро-
ением соответствующих лесных фитоценозов. Так, темнохвойные насаж-
дения высокой сомкнутости характеризуются постоянным режимом зате-
нения, стабильно ослабляющим прогревание почвы, постоянным режимом
задержания осадков на кронах, особенно в твердой фазе (снег), резко
уменьшающим толщину снегового покрова в насаждении, постоянным
режимом повышенной влажности воздуха под кронами, ослабляющим
эвапотранспирацию нижних ярусов растительности и уменьшающим су-
точные колебания температур под кронами древостоя.
Все это изменяет климатические условия в сторону более глубокого
промерзания почвы, более позднего ее оттаивания, более позднего схода
снежного покрова, меньшего прогревания почв и нижних слоев воздуха
в теплый период года, т. е. сокращения вегетационного периода, а в те-
чение его некоторого ослабления вегетации. Таким образом, ценокли-
мат имитирует условия более сурового климатопа, чем зональный кли-
матоп или топоклимат. В светлохвойных (сосновых) насаждениях вслед-
ствие гораздо большей свето- и влагопроницаемости крон описанные тен-
денции выражены гораздо слабее, и потому отклонение ценоклимата от
зонального, как правило, незначительно.
Ужесточение температурного режима почв и припочвенного воздуха
может наблюдаться лишь в условиях мощного развития мохового ков-
ра или в условиях особых местоположений, например достаточной кру-
тизны и протяженности склоны северных экспозиций, морозобойные кот-
ловины и т. п., но в последнем случае более суровым будет уже топокли-
мат, а не ценоклимат. В летнезеленых насаждениях вследствие перемен-
ности их радиационного режима и режима атмосферного увлажнения
тому же с гораздо меньшим перехватом осадков кронами даже в пе-
риод максимального развития листвы) тенденции отклонения ценокли-
мата от зонального климатопа или топоклимата уже качественно иные:
48
Режим затенения и соответственно уменьшение прогревания почвы, соз-
дание повышенной влажности приземного слоя воздуха и уменьшение
суточных колебаний температур наблюдаются только в теплый период,
точнее, только в тот его подпериод, когда полностью развита листва.
В период отсутствия листвы или ее неполного развития кроны почти
не ослабляют ни радиационного потока, ни выпадающих осадков. В то
же время слой опавшей листвы препятствует охлаждению почвы в хо-
лодный период года, этому же препятствует более мощный, чем на от-
крытых местах, снеговой покров. Иссушающее влияние ветра сильно
ослаблено.
Отсутствие листвы приводит к резкому увеличению нагреваемой по-
верхности (поверхность почвы, прикрытой опадом + поверхность ство-
лов и ветвей), что в сочетании с ее пониженной теплопроводностью при-
водит к дополнительному прогреванию нижних слоев воздуха. Все эти
явления уменьшают глубину и продолжительность промерзания почв,
смягчают суточные колебания температур и в летний (за счет затенения)
и в зимний (за счет некоторой аккумуляции тепла в стволах деревьев)
периоды, вносят дополнительный элемент неравномерности схода сне-
гового покрова (за счет дополнительного прихода тепла от нагретых ство-
лов) . Таким образом, ценоклимат листопадного леса или мало отличается
от зонального климатопа или топоклимата, или мягче его, но ни в коем
случае не холоднее и не контрастнее.
По наблюдениям в центральных районах подзоны хвойно-широколист-
венных лесов можно прийти к следующим выводам об отклонении цено-
климатов различных насаждений от зонально-регионального климатопа
или топоклимата определенного местоположения (табл. 3).
Что касается режима континентальности, то он, как уже говорилось,
в условиях центральных районов подзоны хвойно-широколиственных ле-
сов колеблется в нешироких пределах и всегда остается ближе к полу-
континентальному типу, чем к субокеаническому или субконтиненталь-
ному, хотя и сдвигается несколько под темнохвойными и широколист-
венными насаждениями в сторону субокеанического (в сторону субкон-
тинентального сдвигается на некоторых безлесных местоположениях).
Относительно влияния леса на режимы увлажнения почв прежде всего
следует отметить некоторые общие закономерности, заключающиеся в
том, что эрозионно-промывной тип режима лесная растительность исклю-
чает, переводя его в зависимости от глубины водоносного горизонта в
промывной автоморфный, или промывной гидроморфный, или переходный
между ними тип режима.
Промывной автоморфный режим лес несколько смягчает, большей ча-
стью переводя его в переменный автоморфно-гидроморфный промывной
тип режима. Промывной гидроморфный тип режима лес тоже несколько
смягчает, делая его ближе к переменному автморфно-гидроморфному.
Намывной амфибиальный тип режима лес достаточной протяженности
тоже трансформирует в промывной гидроморфный или застойно-боло-
тный в условиях гумидных климатов.
Кроме того, и осушительная, и регулирующая увлажнение роль леса
зависит от механического состава почв и почвообразующих пород, от глу-
бины залегания водоносных горизонтов и в сильнейшей степени от про-
4. Зак.448 49
ТаблицаЗ
Изменение терморежима древостоями разных типов
Травяные фитоценозы | Листопадные леса Светлохвойные леса Темнохвойные леса
Суббореоидаый Суббореоидаый и эунеморапьный Суббореоидаый М езо бореон дный
Эунеморапьный Эунеморапьный и термоэунемо- ральный Эунеморапьный Суббореоидаый
Термоэунемораль- ный Термоэунемораль- иый и термонемо- ральный Термоэунемораль- ный Эунеморапьный
ТаблицаД.
Изменение режима влажности в зависимости от механического состава почв
На почвах легкого механического состава (песках и супесях) На почвах тяжелого механического состава (суглинках и глинах)
Травянистые фитоце* нозы Лес Травянистые фитоце* нозы Лес
Средне степной Лугово-степной — —
Свежестепной Сублуговой — —
Лугово-степной Сухолуговой — —
Субпуговой Сухолуговой Сублуговой Сухолуговой
Сухолуговой Свежелуговой Сухолуговой Свежелуговой
Свежелуговой Влажио-луговой Свежелуговой Свежелуговой
Влажно-луговой Сыровато-луговой Влажно-луговой Свежелуговой
Сыровато-луговой Сыровато-луговой Сыровато-луговой Влажно-луговой
Сыро-луговой Сыровато-луговой Сыро-луговой Сыровато-луговой
Мокро-луговой Сыро-луговой Мокро-луговой Сыро-луговой
Болотно-луговой Мокро-луговой Болотно-луговой Мокро-луговой
Субболотный Болотно-луговой Субболотный Болотно-луговой
Болотный Субболотный Болотный Субболотный
Водно-болотный Болотный Водно-болотный Болотный
дуктивности насаждения. Но так как и продуктивность насаждений, и глу-
бина залеганйя грунтовых вод самым непосредственным образом связа-
ны с различными типами водного режима почв не только в смысле харак-
тера притока и оттока воды, но и в смысле величины увлажнения, то при
составлении таблицы изменения режимов увлажнения на облесенных
территориях по сравнению с необлесенными учитывается лишь связь этого
явления с механическим составом почв и почвообразующих пород. При-
веденная таблица ориентировочно показывает истинные аналоги режимов
увлажнения почв на облесенных и безлесных территориях в центральных
районах хвойно-широколиственной подзоны.
Как видно из табл. 4, общая тенденция фитоценотических изменений
режима увлажнения такова, что в любом случае в крайних типах ветви
недостаточного увлажнения участки под лесом будут более влажными,
50
чем безлесные, а в ветви так называемого избыточного увлажнения, нао-
борот, участки под лесом более сухие, чем безлесные. Разница между
участками с легкими и тяжелыми почвами сводится в основном к тому,
что переход от большего увлажнения облесенных территорий по сравне-
нию с безлесными территориями совершается при легких почвах пример-
но на уровне сыровато-лугового типа увлажнения, а при тяжелых — при-
мерно на уровне свежелугового типа увлажнения почв.
Тенденция изменения под влиянием леса режима солевого довольст-
вия состоит в основном в том, что, солевой режим довольно жестко со-
пряжен, с одной стороны, с водным режимом почв, . ас другой — с интен-
сивностью биологического круговорота веществ, с характером и интен-
сивностью биогенной аккумуляции. Вероятно, одним из лучших изобра-
жений связи солевого режима с увлажнением почв является так называе-
мый ’’сукачевский крест”, прекрасно и наглядно отражающий закон про-
точного увлажнения: ’’чем влажнее, тем богаче”, и закон застойного ув-
лажнения: ’’чем влажнее, тем беднёе”.
Сущность проточного увлажнения — промывные режимы, а сущность
застойного увлажнения — испарительные режимы. Крайние проявления
промывных режимов: с одной стороны, эдафотопы вересковых пусто-
шей на песчаных холмах в условиях гумидного климата при крайней
выраженности промывного автоморфного режима, а с другой стороны,
приморские сильно засоленные марши в условиях гумидного климата
при интенсивном промыве морскими грунтовыми водами в связи с при-
ливно-отливным колебанием их уровня. Крайние проявления испаритель-
ных режимов: с одной стороны, верховые болота в гумидных климатах,
когда поступающие осадки почти не стекают, а аккумулируются моховым
ковром и торфом и в бездождные периоды интенсивно испаряются, и,
с другой стороны, сухие солончаки в аридных зонах. В связи с этим можно
было бы предположить, что в случаях, когда лес изменяет водный режим
в сторону более влажного, он тем самым сдвигает и солевой режим в
сторону большего богатства солями, однако это не всегда так. Кислая
реакция разложения опада может не только уравновесить, но и перекрыть
уменьшение гидрогенного выщелачивания усилением геохимического
выщелачивания.
Зато когда лес уменьшает увлажнение, параллельное уменьшение соле-
вого довольства почти всегда сопутствует ему.
В связи со всем сказанным выше представляется более целесообразным
рассматривать изменение солевого режима под лесом в связи с его пород-
ным составом, определяющим в основном характер и интенсивность био-
логического круговорота. Приведенная таблица, так же как и все пре-
дыдущие, предназначена для центральных районов подзоны хвойно-широ-
колиственных лесов и отражает трансформацию солевого режима почв
насаждениями основных групп лесных формаций подзоны.
Тенденция изменения под лесом солевого режима почв аналогична
той, какую мы наблюдаем для водного режима. Бедные солями почвы
лес несколько обогащает, а богатые — несколько обедняет. Однако у раз-
ных групп формаций лесов разные рубежи перехода обогащающей роли
леса в обедняющую. У широколиственных лесов этот рубеж находится
в субэвтрофном типе солевого режима почв, у мелколиственных — в пер-
51
Таблица 5
Изменение солевого режима почв древостоями различного породного состава
Травяные фито- ценозы Широколист- венные леса Мелколиствен- ные леса Светло хвойные леса Т емно хвойные леса
Олиготрофный гликофильный - Суболиготроф- ный Олиготрофный -
Суболиготроф- ный — Семи олиготроф- ный Суболиготроф- ный Семиолиго троф- иый
Семиолиготроф- ный - Субмезотроф- ный Семиолиготроф- Субмезотрофный ный
Субмезотроф- ный Мезотрофный Мезотрофный Субмезотрофн.1. ный Мезотрофный
Мезотро-фный Пермезотроф- ный Пермезо троф- ный Мезотрофный 33
Пермезотроф- ный Субмезотроф- ный 33 33
Се ми эвтроф- ный Субэвтрофный Пермезотроф- ный Пермезотрофный
Субэвтроф- ный 3» Семиэвтроф- ный Семиэвтроф- ный Семи эвтроф- ный
Эвтрофный 33 Субэвтроф- ный Субэвтроф- ный Субэвтрофный
Пертрофный Эвтрофный Эвтрофный Эвтрофный Эвтрофный
мезотрофном, у светлохвойных и темнохвойных — в мезотрофном, но у
светлохвойных как бы нет обогащающей ветви, а есть как бы ветвь урав-
новешенного состояния, что, очевидно, объясняется слабой эдификатор-
ной способностью сосны (в лиственничниках вследствие иного химизма
опада и ежегодности смены хвои обогащающая роль насаждения на бедных
почвах выражена достаточно четко).
Предположительно можно сформулировать общий принцип обратного
влияния леса на среду так: лес смягчает условия среды, т. е. приближает
их к неким средним, но каждый вид (каждая порода) смещает их к своему
оптимуму и изменение направленности влияния насаждения на режим дан-
ного фактора, по-видимому, происходит на уровне оптимального для
вида режима.
Опираясь на приведенные выше закономерности изменения экологи-
ческих режимов под влиянием лесной растительности, можно достаточно
уверенно прогнозировать как изменение условий среды, так и изменение
потенциальных возможностей разных видов на данной территории при
сменах растительного покрова: травяного на лесной, лесного на травя-
ной и лесного одной группы формаций на лесной же другой группы фор-
маций. В случае насаждений переходного типа: хвойно-широколиствен-
ных, широколиственно-хвойных, суборевых, хвойно-мелколиственных
и т. Д. — приведенные закономерности тоже могут быть полезными, но
прогноз изменений среды и соотношений потенциальных возможностей
разных видов будет зависеть от верности: нашей оценки средоизменяю-
щей роли каждого из соэдификаторов в отдельности.
52
ГЛАВА IV
СИСТЕМА ЭКОЛОГИЧЕСКИХ ХАРАКТЕРИСТИК ВИДОВ
КАК ОСНОВА ФИТОИНДИКАЦИИ
Закон толерантности Шелфорда гласит: ’’Само присутствие и процвета-
ние организма в данном местообитании зависит от целого комплекса усло-
вий. Отсутствие или невозможность процветания определяется недостатком
(в качественном или количественном смысле) или, наоборот, избытком
любого из ряда факторов, уровень которых может оказаться близким к
пределам переносимого данным организмом”. Другая формулировка этого
закона: ’’Организмы характеризуются экологическим минимумом и эко-
логическим максимумом; диапазон же между этими двумя величинами со-
ставляет то, что принято называть пределами толерантности” [Одум, 1975,
с. 140].
Пределы толерантности, или пределы выносливости, естественно, различ-
ны у разных видов (в принципе и у разных индивидов), но также, естест-
венно, разнообразие этих пределов не может быть беспредельным. Вполне
возможны и закономерно существуют классификации видов по сходству
положений их пределов толерантности, или выносливости.
Объединения видов со сходными характеристиками толерантности в от-
ношении тех или иных факторов получили не совсем подходящее по смыс-
лу название экоморф, так как близость пределов выносливости отнюдь
не обязательно сопровождается морфологическим сходством. Конечно, бо-
лее подходящим было бы название экологический тип, распространив ана-
логичный термин внутривидового ранга ’’экотип” на объединения видов со
сходными амплитудами выносливости, оставив термин ’’экоморфа” для
объединений видов с морфологически и анатомически сходными приспо-
соблениями, обеспечивающими ту или иную экологическую приурочен-
ность. Однако существует множество терминов в самых различных науках,
не вполне соответствующих сущности вкладываемых в них понятий.
С точки зрения фитоиндикащш особенно важно знать именно пределы
толерантности. Систематизация типов толерантности видов к режимам пря-
модействующих факторов - основа основ всех фитоиндикационных по-
строений. Система экоморф по отношению видов к режимам основных пря-
модействующих факторов была предложена Бельгардом, где он пытался
сгруппировать виды по усредненным характеристикам их экологической
приуроченности: ксерофиты, мезофиты, олиготрофы, мезотрофы, эвтрофы,
гелиофиты, сциофиты и другие, используя терминологию, предложенную
еще Декандолем [Бельгард, 1950, 1971].
В этой и подобной ей системах, начало которых восходит к временам
Гумбольдта и Декандоля, еще нет даже попытки учесть амплитуды вынос-
ливости видов — характеристики видам даются однозначно.
Разработанные в 30—50-х годах шкалы режимов основных прямодейст-
вующих факторов [Погребняк, 1927, 1958; Раменский, 1929, 1938, 1956;
Ellenberg, 1950, 1952] позволили разработать систему экоморф, основан-
ную на учете полных экологических амплитуд видов [Цыганов, 1974, 1976].
Система экоморф разработана как совокупность пофакторных систем ча-
стных экоморф. В ходе разработки найдены формулы определения коли-
53
чества различимых экоморф в зависимости от числа различаемых градаций
фактора, понимаемых как основные, или элементарные, типы его режимов.
Экологический ряд терморежима, или зонального режима тепла дан в ви-
де шкалы из восьми основных типов режима фактора, определяемых как
термозоны широтного размещения, приведенные в работе Мейзеля ’’Срав-
нительная хронология центральноевропейской флоры” [Meusel, 1965].
Экологический ряд режимов континентальности взят по той же работе.
Экологические ряды увлажнения и солевого богатства почв даны по клас-
сам увлажнения (12 классов) и богатства-засоления (10 классов) в шка-
лах Раменского. Экологический ряд затенения дан в очень схематичном
виде. В нем выделены всего лишь три основных типа режима.
Схемы систем изображены в виде координатных фигур, по оси ординат
которых возрастают количественные оценки факторов, а по оси абцисс —
амплитуды экоморф. Общие экоморфы видов даны в виде экологических
формул, построенных в одной последовательности: термоморфа -омбро-
морфа (здесь так названа, вероятно не совсем удачно, экоморфа конти-
нентальности) — гидроморфа — трофоморфа — гелиоморфа.
С целью сделать такие формулы компактными введена символика, где
каждый символ в ряду какого-либо фактора обозначает одновременно и
один из основных типов его режима и монотопическую экоморфу, соответ-
ствующую этому типу режима данного фактора. Символика разработана
для схем систем частных экоморф.
Для схемы системы термоморф:
А — арктическая термозона и термоморфа — гекистотермы;
а — субарктическая термозона и термоморфа — субгекистотермы;
В — бореальная термозона и термоморфа — микротермы;
N — неморальная термозона и термоморфа — субмикротермы;
m — субсредиземноморская термозона и термоморфа — субмезотермы;
М — средиземноморская термозона и термоморфа — мезотермы;
t — субтропическая термозона и термоморфа — субмегатермы;
Т — тропическая термозона и термоморфа — мегатермы.
Для схемы системы омброморф:
О — океанический тип климата и омброморфа — океанисты;
о — субокеанический тип климата и омброморфа — субокеанисты;
к — субконтинентальный тип климата и омброморфа — субконтинента-
лы;
К — континентальный тип климата и омброморфа — континенталы.
Для схемы системы гидроморф:
D — пустынный тип увлажнения и гидроморфа — гиперксерофиты;
d — полупустынный тип увлажнения и гидроморфа — ксерофиты;
s — сухостепной тип увлажнения и гидроморфа — субксерофиты;
S — среднестепной тип увлажнения и гидроморфа — семиксерофиты;
С — лугово-степной тип увлажнения и гидроморфа — субмезофиты;
с - сухолуговой тип увлажнения и гидроморфа - мезофиты;
f — влажно-луговой тип увлажнения и гидроморфа — пермезофиты;
F - сыро-луговой тип увлажнения и гидроморфа — семигигрофиты;
р - болотно-луговой тип увлажнения и гидроморфа — субгигрофиты;
54
Р — болотный тип увлажнения и гидроморфа — гигрофиты;
а — прибрежно-водный тип увлажнения и гидроморфа — субгидрофиты;
А - водоемный тип увлажнения и гидроморфа - гидрофиты.
Для схемы системы трофоморф:
О — тип солевого режима особо бедных почв и трофоморфа — олиготро-
фы;
о — тип солевого режима бедных почв и трофоморфа — семиолиготро-
фы;
М — тип солевого режима небогатых почв и трофоморфа — мезотрофы;
е — тип солевого режима довольно богатых почв и трофоморфа — семи-
эв.трофы;
Е — тип солевого режима богатых почв и трофоморфа — эвтрофы;
g — тип солевого режима слабо засоленных почв и трофоморфа — суб
гликофиты;
h — тип солевого режима средне засоленных почв и трофоморфа — суб-
галофиты;
Н — тип солевого режима сильно засоленных почв и трофоморфа — га-,
лофиты;
Р — тип солевого режима резко засоленных почв и трофоморфа — пер-
гал офиты;
S — тип солевого режима злостных солончаков и трофоморфа — супер-
галофиты.
Для схемы системы гелиоморф:
G - тип светового режима открытых пространств и гелиоморфа — гелио-
фиты;
g - тип светового режима светлых лесов и гелиоморфа - семигелиофиты;
S — тип светового режима теневых лесов и гелиоморфа — сциофиты.
Учитывая приведенные выше формулы зависимости: числа зкоморф
в системе от числа различаемых режимов фактора; числа экоморф, сопря-
женных с каждым данным типом режима фактора, от числа различаемых
режимов и порядкового номера режима на шкале соответствующего эко-
логического ряда, — можно рассчитать, что:
различия 8 типов терморежима, получаем всего 36 термоморф, из чиста
которых при арктическом терморежиме могут существовать 8 экоморф,
субарктическом — 14, при бореальном — 18, неморальном — 20, субсреди-
земноморском — 20, средиземноморском - 18, субтропическом — 14, тро-
пическом — 8;
различая 4 типа режима континентальности, получаем всего 10 омбро-
морф, из числа которых при океаническом типе режима могут существо-
вать 4 омброморфы, при субокеаническом — 6, при субконтинентальном —
6, при континентальном — 4;
различая 12 типов режима увлажнения почв, получаем всего 78 гидро-
морф, из числа которых при пустынном типе гидрорежйма могут сущест-
вовать 12 гидроморф, при полупустынном - 22, при сухостепном — 30,
при среднестепном — 36, при лугово-степном — 40, при сухолуговом — 42,
при влажно-луговом - 42, при сыро-луговом - 40, при болотно-луговом -
36, при болотном — 30, при прибрежно-водном - 22, при водоемном - 12;
55
различая 10 типов солевого режима почв, получаем всего 55 трофоморф,
из числа которых на особо бедных почвах могут существовать 10 трофо-
морф, на бедных — 18, на небогатых — 24, на довольно богатых — 28, на бо-
гатых — 30, на слабо засоленных — 30, на средне засоленных — 28, на сильно
засоленных — 24, на резко засоленных — 18, на злостных солончаках — 10;
различая всего лишь 3 типа гелиорежима, получаем всего 6 гелио морф,
из которых на открытых пространствах могут существовать 3 гелиоморфы,
в светлых лесах — 4, в теневых лесах — 3.
Как видно из приведенных данных, наиболее бедны по составу экоморф
экстремальные типы режима, независимо от того, будут ли они диском-
фортными или комфортными, а наиболее богаты по составу экоморф —
средние в данном экологическом ряду типы режима.
Только что изложенная система экоморф оказалась весьма информатив-
ной и позволяющей делать далеко идущие выводы качественного и коли-
чественного порядка о связи видов растений со средой, о потенциальных
возможностях формирования растительного покрова территорий с опре-
деленным сочетанием режимов прямо действующих факторов.
Несмотря на это, система не лишена ряда существенных недостатков, из
которых особенно выделяются: недостаточное число градаций в рядах за-
тенения и континентальности, комплексный характер фактора континен-
тальности, отсутствие ряда значимых дополнительных характеристик (пе-
ременность режимов наиболее динамичных факторов, некоторые состав-
ляющие режимов, имеющие большое самостоятельное значение, как экстре-
мумы температур, баланс атмосферного увлажнения, pH, азот и т. д.), не
всегда достаточно точная корреляция подразделений климата по Мёйзелю
с принятыми у нас системами его классификаций и районирования. Все
только что изложенное предопределяет желание внести в изложенную си-
стему экоморф ряд корректив.
В отношении климатических амплитуд толерантности растений и ампли-
туд, хотя бы наиболее различимых формаций (иногда и ассоциаций) расти-
тельного покрова, представляет большой интерес концепция термо- и
гидрозон В.Р. Волобуева [1973], предназначенная, собственно, для райони-
рования почв, но вполне применимая и для целей диагностики климати-
ческих экоморф видов.
Выделение им термозон по величинам радиационного баланса и относи-
тельной увлажненности, находимой как отношение суммы осадков к сум-
марной испаряемости, так же может быть увязано с растительным покро-
вом, как и с почвами. Однако, кроме общего годового баланса тепла, в
распределении растений и растительности большую роль играет также вы-
раженность или не выраженность зимнего периода, особенно как периода
господства отрицательных температур.
В то же время всегда следует иметь в виду, что всякая перестройка систе-
мы может предприниматься только тогда, когда она оправдывается, с од-
ной стороны, гораздо большей информативностью, а с другой стороны, по
крайней мере не меньшим или большим удобством работы с новой систе-
мой, что достигается или особенностями самой системы или большей до-
ступностью материалов, на которых она базируется.
Система климатических экоморф, построенная на базе термо- и гидро-
56
зон, выделенных по изолиниям радиационного баланса и относитель-
ной увлажненности, несомненно, более понятна и менее дискусионна, чем
система климатических экоморф, построенная на базе зональности расти-
тельного покрова, по какому бы автору эта зональность ни давалась.
Но вместе с тем любая уже разработанная система обладает известной
инерцией и заменить ее сразу новой системой невозможно, поскольку
существуют уже многочисленные разработки смежных вопросов, опираю-
щиеся на старую систему. В силу этого, давая амплитуды толерантности
видов и схемы систем климатических экоморф и по прежней и по заново
разработанной системе климатических экоморф, приходится весь фито-
индикационный и иллюстрационный материал давать в основном по преж-
ней, уже опубликованной системе, несмотря на относительно меньшую
ее информативность, которая, однако, оказалась вполне достаточной для
принципиально верной оценки связи растений и растительности с клима-
тическими факторами.
Во вновь разработанной системе участвуют три категории климатичес-
ких экоморф: термоморфы, понимаемые как объединения видов с оди-
наковыми в принятом масштабе амплитудами толерантности по отноше-
нию к климатическому режиму тепла, выражаемому через величины радиа-
ционного баланса; криоморфы, понимаемые как объединения видов с
одинаковыми в принятых градациях амплитудами выносливости к су-
ровости зимнего периода, выражаемому через изотерму самого холод-
ного месяца; омброморфы, понимаемые здесь уже в истинном смысле
термина как объединения видов с одинаковыми в принятых градациях
амплитудами толерантности к климатическому режиму увлажненности, вы-
ражаемому через разности годовых сумм осадков и испаряемости.
Используя во вновь разработанной системе сам принцип выделения тер-
мозон и гидрозон в концепции Волобуева [1973], автор все же не счел
целесообразным отождествлять границы основных типов терморежима и
омброрежима с границами термозон и гидрозон Волобуева.
Во-первых, это сделано потому, что Волобуев избрал границами термо-
зон и гидрозон изолинии радиационного баланса и разности осадков и ис-
паряемости по принципу их наибольшей близости к зональному и регио-
нальному разграничению почвенного покрова. Между тем даже границы
зонального и провинциального разделения растительного покрова не всег-
да соответствуют таковым почвенного покрова, несмотря на неоспоримую
тесную связь этих объектов природы.
Во-вторых, неоспоримо, что всяческие зональные или региональные
границы и почв, и растительности континуальны и в пространственном, и
во временном, и в физическом смыслах, тогда как определять амплиту-
ды толерантности хоть в одном из смыслов (физическом) удобнее по
равномерной шкале.
В-третьих, шкалы, построенные на основе термозон и гидрозон Воло-
буева, были бы менее подробны и менее равноинформативны в различных
своих частях, чем это требуется для шкал, предназначенных к определе-
нию толерантности видов в целях экологического анализа вообще и фито-
индикации в частности.
57
Система термоморф в новой трактовке имеет следующую символику
и кодируемый ею набор понятий:
А — арктическая термозона (0—10 ккал/см2 . год), термоморфа — ге-
кистотермы;
а — субарктическая термозона (10—20 ккал/см2 . год), термоморфа —
субгекистотермы;
В — бореальная термозона (20-30 ккал/см2 • год), термоморфа — мик-
ротермы;
b — суббореальная термозона (30—40 ккал/см2 • год), термоморфа --
субмикротермы;
N — неморальная термозона (40—50 ккал/см2 • год), термоморфа —
субмезотермы;
m — субсредиземноморская термозона (50—60 ккал/см2 • год), тер-
моморфа — мезотермы;
М — средиземноморская термозона (60—70 ккал/см2 • год), термо-
морфа — пермезотермы;
t - тропическая термозона (70-80 ккал/см2 • год), термоморфа -
субмегатермы;
Т - экваториальная термозона (80 ккал/см2 . год), термоморфа — ме-
гатермы.
Для системы криоморф употребляется следующая символика и кодиру-
емый ею набор понятий
К — пояс очень суровой зимы (средняя t° самого холодного месяца
ниже —32°), криоморфа - гиперкриофиты;
L - пояс суровой зимы (средняя V самого холодного месяца от —24
до —32°), криоморфа — перкриофиты;
М — пояс довольно суровой зимы (средняя t° самого холодного меся-
ца от —16 до —24°), криоморфа — криофиты;
N — пояс умеренной зимы (средняя t° самого холодного месяца от —8
до -16°), криоморфа - субкриофиты;
О — пояс мягкой зимы (средняя t° самого холодного месяца от 0 до
—8°), криоморфа — гемикриофиты;
Р - пояс теплой зимы (средняя t° самого холодного месяца от 0 до
+8°), криоморфа — акриофиты;
Q — пояс очень теплой зимы (средняя t° самого холодного месяца от
+8 до +16), криоморфа — субтермофиты;
R — пояс невыраженной зимы (средняя t° самого холодного месяца
выше +16°), криоморфа — термофиты.
Система омброморф в новой трактовке получила следующую символи-
ку и кодируемый ею круг понятий:
А — экстрааридный пояс (Р-Е = -1600 мм в год и ниже), омбромор-
фа — гипераридофиты;
В — аридный пояс (Р—Е = —800 — 1600 мм в год), омброморфа - ари-
дофиты;
С - мезоаридный пояс (Р—Е = —400 - 800мм в год), омброморфа —
мезоаридофиты;
D — субаридный пояс (Р—Е = 0—400 мм в год), омброморфа — субари-
дофиты;
Е — субгумидный пояс (Р—Е = 0—400 мм в год), омброморфа — суб-
омброфилы;
F — гумидный пояс (Р—Е = 400—800 мм в год), омброморфа — мезо-
омброфилы;
С — пергу мидный пояс (Р—Е = 800—1600 мм в год), омброморфа —
омброфилы;
Н - гипергумидный пояс (Р-Е = 1600 мм и выше), омброморфа - ги-
перомброфилы.
(Р — осадки за год, Е — испарение за год.)
Все изолинии можно найти в Географическом атласе [1980].
Кроме переработки системы климатических экоморф, представляется
целесообразным дополнить систему эдафических экоморф введением до-
полняющих характеристик пределов толерантности видов по отношению к
pH почвенного раствора и к содержанию доступного азота в почве, а также
переменности режима увлажнения, трактуя эти дополнительные характе-
ристики как эдафоморфы вспомогательного значения: pH-морфы, нитро-
морфы и контрастоморфы.
Для системы pH-морф применяется следующая символика и кодиру-
емый ею набор понятий:
а - очень сильно кислые почвы (pH =3,5), pH-морфа - гиперацидофи-
лы;
b — сильно кислые почвы (pH = 3,5—4,5), pH-морфа — перацидофилы;
с — кислые почвы (pH = 4,5—5,5), pH-морфа — ацидофилы;
d — слабокислые почвы (pH = 5,5—6,5), pH-морфа — субацидофилы;
е — нейтральные почвы (pH = 6,5—7,2), pH-морфа — нейтрофилы;
f - слабощелочные (pH = 7,2-8,0), pH-морфа — субалкалифилы;
g - щелочные почвы (рН> 8,0), pH-морфа — алкалифилы.
Для системы нитроморф имеется следующая символика и кодируемый
ею набор понятий:
j — практически безазотные почвы, нитроморфа — анитрофилы;
к — почвы очень бедные азотом, нитроморфа — субанитрофилы;
1 — почвы бедные азотом, нитроморфы — геминитрофилы;
m — почвы, достаточно обеспеченные азотом, нитроморфа — субнитро-
филы;
п — почвы, богатые азотом, нитроморфа — нитрофилы.
Для системы контрастоморф (экоморф переменности увлажнения почв)
употребляется’ следующая символика и кодируемый ею набор понятий:
р — высоко обеспеченное постоянное увлажнение, контрастоморфа —
константофиты;
q - средне обеспеченное постоянное увлажнение, контрастоморфа -
субконстантофиты;
г - переменно-обеспеченное увлажнение, контрастоморфа — геми-
константофиты;
s — умеренно переменное увлажнение, контрастоморфа — субконтра-
стофиты;
t — сильно переменное увлажнение, контрастоморфа — контрастофи-
ты;
59
u — резко переменное увлажнение, контрастоморфа — гиперконтрасто-
фиты.
Наконец, переработаны в плане увеличения числа градаций и соот-
ветственно набора экоморф шкала затенения и система гелиоморф,
для которых в новой редакции дана следующая символика и коди-
руемый ею набор понятий:
G — тип светового режима открытых пространств (всяческих травяных,
моховых, лишайниковых, реже кустарничковых и переходных
между всеми ними фитоценозов, а также участков без раститель-
ного покрова), гелиоморфа — гелиофиты;
g — тип светового режима полуоткрытых пространств (кустарниковых
зарослей, опушек, полян, редколесий и т. п.), гелиоморфа — суб-
гелиофиты;
М — тип светового режима светлых лесов (светлохвойных, мелколист-
венных, сильно расстроенных темнохвойных и широколиствен-
ных) , гелиоморфа — семигелиофиты;
s — тип светового режима тенистых лесов (темнохвойных и широко-
лиственных средней сомкнутости), гелиоморфа — субсциофиты;
S - тип светового режима особо тенистых лесов (некоторых особо
высокосомкнутых темнохвойных и широколитсвенных лесов: пих-
тачей, реже ельников, часто бучин, каштанников и т. д.), гелиомор-
фа — сциофиты.
Как уже отмечалось ранее, экоморфы могут объединяться как по коли-
чественному признаку — амплитуде пределов толерантности, так и по ка-
чественному признаку — положению в соответствующем экологическом ря-
ду медианы этой амплитуды. Объединение экоморф с амплитудами, охва-
тывающими 1, 2, 3 и т. д. типов режима фактора, получили название клас-
сов амплитуд [Цыганов, 1976] и выражают гамму перехода от крайних сте-
нотопов к крайним эвритопам, от достоверных моноиндикаторв к видам
без индикационного значения относительно данного фактора. Гораздо более
информативно с точки зрения фитоиндикации объединение экоморф по
одинаковым медианам амплитуд выносливости — экологическая свита.
Принятая для обозначения частных экологических свит нумерация их по-
следовательными числами от одного до другого края соответствующего эко-
логического ряда [Цыганов, 1974, 1976] позволяет при индикации режима
данного фактора производить с ними те же действия, что и с баллами в ме-
тодике Элленберга.
Каждая частная экоморфа связана с режимом данного фактора, соответ-
ствующим ее экологической свите (той, в которую она входит по положе-
нию медианы своей амплитуды), тем теснее, чем уже ее амплитуда.
Это является теоретическим обоснованием того давно известного факта,
что стенотопные виды — хорошие индикаторы, а эвритопные — плохие.
Объединяя частные характеристики видов в их экологических форму-
лах так, как это говорилось выше, получаем основную экологическую
формулу вида (формулу его общей экоморфы) и дополнительную эколо-
гическую формулу вида (формулу его общей экологической свиты). Раз-
бив эти формулы на климатическую, эдафическую и ценотическую части
соответственно сущности режимов отдельных факторов, получим клима-
морфу и климатическую свиту, эдафоморфу и эдафическую свиту и цено-
60
морфу и ценотическую свиту. Причем в координатных системах климато-
пов и эдафотопов климатическая и эдафическая свиты будут соответство-
вать определенному климатопу и эдафотопу, а климаморфа и эдафоморфа
будут характеризоваться некими климатическим и эдафическим ареалами.
Типы и свойства этих ареалов будут различны у разных климаморф и эда-
морф в зависимости от положения, которое они занимают на климатичес-
кой или эдафической сетке.
В то же время экологические свиты как объединения видов, хоть и об-
ладающих тождественными экологическими оптимумами по данному фак-
тору или данному сочетанию факторов, но самых различных по амплитудам
толерантности, могут искажать результаты фитоиндикации условий место-
произрастания. В этом отношении более показательными могут быть эколо-
гические группы, или экогруппы, — объединения видов по преимуществен-
ной приуроченности к определенному, достаточно узкому диапазону режи-
мов отдельного фактора или сочетания факторов. Подход к разделению
системы экоморф на системы экогрупп может быть двоякого рода: или
глобально-теоретический, или регионально-прикладной. Первый более уме-
стен при различных видах геоботанического или экологического райониро-
вания обширных территорий, включающих несколько климатических зон и
регионов, а второй — при геоботаническом, типологическом или экологи-
ческом обследовании ограниченных территорий, полностью лежащих в пре-
делах одного климатического региона.
Принцип и способы выделения экогрупп на схемах систем экоморф и в
том и в другом случае будут практически одинаковы, но во втором случае
нехарактерные для региона экоморфы объединяются в более широкие
группы, так как индицируют экстремальные для региона условия среды с
гораздо меньшей подробностью, чем обычные зонально-региональные усло-
вия. Способы разделения схем систем экоморф на экогруппы в глобально-
теоретическом плане приведены в табл. 6—10, а пример регионально-при-
кладного разделения дан для центральных районов подзоны хвойно-широ-
колиственных лесов.
Так как термический режим местообитаний индицируется с небольшой
степенью подробности: зональный, холоднее зонального и теплее зональ-
ного, то 20 термоморф, принадлежащие восьми термоклиматическим сви-
там, присущие подзоне хвойно-широколиственных лесов, целесообразно
разделить лишь на семь, а не распределять по 15 экогруппам, как это име-
ло место при глобально-теоретическом подходе. Термоморфы AN, aN, Am и
BN, принадлежащие к арктобореальной, мезобореальной и суббореальной
свитам, объединяются в экогруппу бореальных растений; термоморфы ат
и AM, принадлежащие к суббореальной свите, — в экогруппу неморо-бо-
реальных растений; термоморфы Вт и аМ, принадлежащие к эунемораль-
ной или северонеморальной свите, — в экогруппу бореонеморальных расте-
ний; термоморфы Nm и ВМ, принадлежащие к термонеморальной, или
южнонеморальной свите, — в экогруппу неморальных растений; термомор-
фы NM и Bt, принадлежащие к криомеридиональной, или субсредиземног
морской, свите, — в экогруппу субтермофильных (для данной подзоны)
растений; термоморфы Nt, ВТ и NT, принадлежащие к мезомеридиональ-
ной и эумеридиональной свитам, - в экогруппу термофильных растений, а
термоморфы At, at, АТ и аТ, широкоамплитудные экойорфы эунемораль-
61
Таблицаб
Схема системы термоморф (по отношению видов к зональным режимам тепла) и
зкогруппы по отношению видов к терморежиму климата
Термоклиматическая зона Термоморфа Схема системы термоморф
Арктическая Гекистотермы А Аа
Субарктическая Су бгекистотермы а АВ аВ AN
Бореальная Микротермы В aN Am BN am AM
Неморальная Субмикротермы N Bm aM At Nm BM at AT
Субсредиземноморская Субмезотермы m NM Bt aT mM Nt BT
Средиземноморская Мезотермы M mt NT M t mT
Субтропическая Тропическая Субмегатермы Мегатермы t MT tT T
Примечание. Экогруппы: гекистотермы (А, Аа, а) , микротермы (В, BN, N) ,
мезотермы (т, тМ, М), мегатермы (t, tT, Т), ортотермы гипоарктические (АВ,
аВ, AN, aN), эвортотермы (Bm, Nm, BM, NM), ортотермы эпитропические (mt, Mt,
mT, МТ), политермы внетропические (Am, am, AM, аМ) , политермы внеарктические
(Bt, Nt, ВТ, NT) , эвритермы (At, at, AT, aT) .
Таблица?
Схема системы контрастоморф (омброморф) и зкогруппы по отношению видов к
коитииентальности климата
Пояс континентальности Контрастоморфа Схема системы контрасто- морф
Океанический Океанисты О Оо
Субокеанический Субокеанисты о Ок ок ОК
Субконтинентальный Континентальный. Субконтиненталы Континенталы к оК кК К
Примечание. Экоморфы: океанисты (О, Оо, о), континенталы (к, кК, К),
океанические политопы (Ок) , ортополитопы (ок) , континентальные политопы (оК),
эвритопы (ОК).
ной, термонеморальной и криомеридаональной свит — в экогруппу эври-
термных растений.
В подзоне хвойно-широколиственных лесов преобладание видов бореаль-
ной экогруппы определенно указывает на суббореоидаый тип ценоклимата
или топоклимата. Преобладание видов неморальной экогруппы тоже указы-
62
Таблица8
Схема системы гидроморф и экогруппы по отношению видов к водному режиму
почв
Тип увлажнения Гидроморфа Схема системы гидроморф
Пустынный Гипер ксерофиты D
Dd
Полупустынный Ксерофиты d Ds
ds DS
Сухостепной Субксерофиты s dS DC
sS dC De
Среднестепной Семиксерофиты S sC de Df
SC sc df DF
Лугово-степной Субмезофиты C Sc sf dF Dp
Cc Sf sF dp DP
Сухолесолуговой Мезофиты c Cf SF sp dP Da
cf CF Sp sP da DA
Влажно-лесолуговой Пермезофиты f cF Cp SP sa dA
fF cp CP Sa sA
Сыро-лесолуговой Семи гигрофиты F fp cP Ca SA
Fp fP ca CA
Болотно-ле солуговой Субгигрофиты p FP fa cA
pP Fa fA
Болотный Гигрофиты P pa FA
Pa pA
Прибрежно-водный Субгидрофиты a PA
aA
Водный Гидрофиты A
Примечание. Экогруппы: ксерофиты (D, Dd, d, Ds, ds, DS), субксерофиты
(s,dS,sS,S), мезоксерофиты (DC, dC, De, sC, de, Df, SC, sc,df,DF), ксеромезофиты
(Sc,sf,dF,Sf,sF,SF), мезофиты (C, Cc, c, Cf, cf, CF, f,cF, fF, F),гигромезофиты (Ср, ср,.
CP, fp, cP, Ca), мезогигрофиты (Fp, fp, ca, CA, FP, fa, cA, Fa, fA, FA), гигрофиты (p
pP, P, pa) , гидрофиты (Pa, pA, a, PA, aA, A) , мезополифиты (Dp, dp, sp, dP, Sp, sP, SP,
sa, Sa, SA), эвриполифиты (DP, Da, da, DA, dA, sA).
вает на тот же тип цено- или топоклимата, но менее определенно, а опреде-
ленно лишь в сочетании со значительным участием видов бореальной эко-
группы. Преобладание видов бореонеморальной и неморальной экогрупп
указывает на зональный тип терморежима. Преобладание неморальной
экогруппы в сочетании со значительным участием субтермофильных и тер-
мофильных видов указывает на термоэунеморальный тип топо-или ценокли-
мата. Эвритермные виды из анализа исключаются как неинформативные с
точки зрения индикации терморежима. Что касается омброморф как эко-
морф континентальности климата, то их в подзоне хвойно-широколиствен-
ных лесов целесообразно разделить на четыре экогруппы: субокеанических
видов (омброморфа — Ок), полуконтинентальных видов (омброморфа —
ок), субконтинентальных видов (омброморфы — оК и кК)и континенталь-
нопатиентных видов (омброморфа — ОК).
Аналогично, как можно видеть из прилагаемых схем, можно распреде-
лять виды по экогруппам и в глобально-теоретическом плане, и в регио-
нально-прикладном, также и во вновь разработанных схемах систем термо-
морф (см. табл. 10) ,климаконтрастоморф (табл. 11), омброморф (табл. 12)
и криоморф (табл. 13).
63
Таблица)
Схема системы трофоморф и экогруппы по отношению видов к солевом) режиму
почв
Тип солевого режима почв Трофоморфа Схема системы трофоморф
Особо бедных почв Бедных почв Олиготрофы Семиолиготрофы о Оо о ОМ
Небогатых почв Мезотрофы оМ Ое М ое ОЕ
Довольно богатых почв Семиэвтрофы Me оЕ Og е ME og Oh
Богатых почв Эвтрофы еЕ Mg oh ОН E eg Mh oH OP
Слабо засоленных почв Субгликофиты Eg eh MH oP OS g Eh eH MP oS
Средне засоленных почв Субгалофиты gh EH eP MS h gH EP eS
Сильно засоленных почв Галофиты hH gP ES H hP gS
Резко засоленных почв Пергалофиты HP hS P HS
PS Злостных солончаков Супергалофиты S Примечание. Экогруппы: олиготрофы (О, Оо, о), меэотрофы (М, Me, е),
мегатрофы (Е, Eg, g) , галофиты (h, hH, Н) , пергалофиты (Р, PS, S) , мезоолиготро-
фы (ОМ, оМ, Ое, ое) , мезомегатрофы (ME, еЕ, Mg, eg) , мезогалотрофы (Eh, gh, ЕН,
gH), мегагалотрофы (hP, HP, hS, HS) , мезополитрофы (OE,oE, Og, og) , мегаполитро-
фы (Mh, eh, MH, eH), галополитрофы (EP, gP, ES, gS), галогликотрофы (Oh, oh, OH,
oH) , гликогалотрофы(МР,еР, MS, eS) , звритрофы (OP, oP, OS, oS) .
Для эдафоморф разница между глобально-теоретическим и регионально-
прикладным подходами к выделению экогрупп сводится в основном к ог-
раничению числа экоморф при регионально-прикладном подходе исклю-
чительно теми, которые совместимы с климатическими условиями данно-
го региона. Что касается менее дробного выделения экогрупп для регио-
нально дискомфортных эдафотопов, то вследствие менее четкой связи
эдафических режимов с условиями климатопов это не всегда целесообразно.
Таким образом для хвойно-широколиственной подзоны выделяются
следующие экогруппы: мезоксерофиты (гидроморфы: DC, dC, De, sC, de,
Df), ксеромезоамфифиты (гидроморфы: SC, sc, df, DF), ксеромезофиты
(гидроморфы: Sc, sf,dF, Sf,sF, SF), эпимезофиты (гидроморфы: Cc,Cf), ор-
томезофиты (гидроморфы: cf и CF), гипомезофиты (гидроморфы: cF и
fF), ксеромезополифиты (гидроморфы: Dp, dp, sp, dP), ортомезополифи-
ты (гидроМорфы: Sp и sP), гигромезополифиты (гидроморфы: SP, sa, Sa,
SA), эвриполифиты (гидроморфы: DP, Da, da, DA, dA, sA), гигромезофи-
ты (гидроморфы: Ср, ср, СР, fp, cP, Са), мезогигроамфифиты (гидромор-
фы: Fp, fP, са, СА), мезогигрофиты (гидроморфы: FP, fa, сА, Fa, fA, FA),
гигрофиты (гидроморфы: рР и ра), гигрогидрофиты (гидроморфы: Ра и
рА), гидрофиты (гидроморфы: РА и аА) — по обобщенному водному ре-
жиму; олиготрофы (трофоморфа: Оо), мезоолиготрофы (трофоморфа
64
Таблица 10
Схема системы термоморф (по соотношению ареалов с изолиниями радиационного
баланса) и экогруппы по отношению видов к обобщенному терморежиму климата
Термоэона Термоморфа Схема системы термоморф
Арктическая Геки сто тер мы А Аа
Субарктическая Су бгеки стотермы а АВ аВ АЬ
Бореальная Микротермы В ad AN Bb aN Am
Суббореальная Субмикротермы b BN am AM bN Bm aM. At
Неморальная Субмезотермы N bm BM at AT Nm bM Bt aT
Субсредиземноморская Мезотермы m NM bt BT mM Nt bT
Средиземноморская Пермезотермы M mt NT Mt mT
Тропическая Субмегатермы t MT tT
Экваториальная Мегатермы T
Примечание. Экогруппы: арктические виды (А, Аа, а) , гипоарктические ви-
ды (АВ, аВ, Ab), бореальные виды (В, Bb, b, Вп) ; трансбореальные виды (ab, AN,
aN) , криопатиентные виды (Am, am, AM, аМ) , неморальные виды (N, bN) , трансне-
моральные виды (Вт, bm, ВМ) . эвритермные виды (At,at, АТ,аТ) .термонемораль-
ные виды (Nm, ЬМ, NM) , субтермофильные виды (Bt,bt, ВТ) .средиземноморские
виды (m, тМ, М) , термофильные виды (Nt, ЬТ, NT), субтропические виды (mt, Mt,
mT, МТ) , тропические виды (t, tT, Т).
ОМ), олигомезотрофы (трофоморфа: оМ), олигополитрофы (трофомор-
фа Ое), мезополитрофы (трофоморфы: ое, ОЕ), мегаполитрофы (трофо-
морфа оЕ), мегамезотрофы (трофоморфа Me), мезомегатрофы (трофо-
морфа ME), мегатрофы (трофоморфа еЕ), стеногалогликофиты (трофо-
морфы: Og, og, Mg, oh, Mh), гликогаломегатрофы (трофоморфы: eg, Eg,
eh, Eh), амфитрофы (трофоморфы: MH и oP), галогликопатиенты (трофо-
морфы: Oh, OH, oH, OP), стеногликогалофиты (трофоморфы: еН, ЕН, еР,
ЕР, ES), гликогалопатиенты (трофоморфы: MP, oS, MS, eS), евритрофы
(трофоморфа OS) — по обобщенному солевому режиму.
Как видно из схем систем гидроморф и трофоморф, в одну экогруппу,
как правило, входят виды не только разных экоморф, но и разных, хотя
и близких, экологических свит. Также и наоборот, виды и экоморфы од-
ной экологической свиты могут принадлежать разным экогруппам. Это вы-
текает из различия принципов выделения экогрупп и экологических свит.
В экологические свиты объединяются экоморфы с одним и тем же опти-
мальным режимом данного фактора, но с различными амплитудами толе-
рантности: от самой узкой до самой широкой при данном положении опти-
мума. В экогруппы объединяются экоморфы с близкими, но не всегда
совпадающими оптимальными режимами по данному фактору, зато с
близкими же амплитудами толерантности, когда совмещающиеся их части
гораздо больше несовмещающихся.
5. Зак. 448
65
Таблица 11
Схема системы климакоитрастоморф (переработанный вариант) * и экогруппы по
отношению видов к континентальности климата
Пояс континентальности Контрастоморфа Схема системы климаконтрас- томорф
Эксграокеанический Океанический Экстр аокеанисты Мезоо кеанисты Е ЕО 0 Ео
Морской Гемиокеанисты Оо Em о От ЕМ
Субматериковый Субконтиненталы от ОМ Ek т оМ Ok ЕК
Материковый Мезоконтиненталы mM ok OK EU М тк оК OU
Субконтинентальный Орто континенталы Мк mK oU к МК MU
Континентальный Эуконтиненталы kK MU К kU
Ультраконтинентальный Ультраконтиненталы KU и
* В основу переработанной шкалы положены пояса континентальности Н.Н. Ива-
нова [1959].
Примечание. Экогруппы: океанические стенобионты (Е, ЕО, О) , океаничес-
кие полибионты(Ео, Оо, Em, От), субокеанические стенобионты (о, от, т) , суб-
океанические полибионты (ЕМ, ОМ, Ek, Ok) , ортополибионты (оМ, mM, ок, тк) ,
зврибионты (EK, OK, ED, OLJ), материковые стенобионты (М,Мк, к),материковые
полибионты (оК, mK, oU, mU,), континентальные полибионты (МК, kK, MU, kU),
континентальные стенобионты (К, KU, U) .
Таблица 12
Схема системы омброморф и экогруппы по отношению видов к режиму влажности
климата
Тип режима аридности Омброморфа Схема системы омброморф
Экстрааридный Гипераридофиты А АВ
Эуаридный Эуарндофиты В АС ВС AD
Мезоаридный Me зоаридофиты С BD АЕ CD BE AF
Субарндный Субаридофиты D CE BF AG DE CF BG AH
Субгумидный Субомброфилы E DF CG BH EF DG CH
Гумидный Мезоо мброфилы F EG DH FG EH
Пергу мидный Эуомброфилы G FH GH
Гипергумидный Гиперомброфилы H
Примечание. Экогруппы: аридные стенобионты (А, АВ, В) , аридные поли-
бионты (АС, ВС, AD, BD), субаридные стенобионты (С, CD, D) , субаридные поли- бионты (АЕ, BE, AF, BF) , ортополибионты (СЕ, DE, DF, DF), эврибионты (AG, BG,
АН, ВН), субгумидные стенобионты (E.EF.F), субгумидныеполибионты (CG, DG,
CH, DH) , гумидные полибионты (EG, FG, ЕН, FH) , гумидные стенобионты (G,GH,H)
66
Таблица 13
Схема системы криоморф и экогруппы по отношению видов к суровости зим
Криозона Криоморфа Схема системы криоморф
Очень суровых зим Гиперкриофиты к KL
Суровых зим Перкриофиты L КМ LM KN
Довольно суровых зим Криофиты М LN КО MN LO КР
Умеренных зим Субкриофиты N МО LP KQ NO MP LQ KR
Мягких зим Гемикриофиты О NP MQ LR OP NQ MR
Теплых зим Акриофиты Р OQ NR PQ OR
Очень теплых зим Невыраженных зим Субтермофиты Термофиты Q PR QR R
П римечание. Экогруппы: стенобионты суровых зим (К, КО, L) , полибион-
ты суровых зим (КМ, LM, KN.LN), стенобионты умеренных зим (М, MN, N) , поли-
бионты умеренных зим (КО, КР, LO, LP) , ортополибионты (МО, NO, MP, NP), эври-
бионты (KQ, LQ, KR, LR) , стенобионты мягких зим (О, OP, Р), полибионты мяг-
ких зим (MQ, NQ, MR, NR) , полибионты безморозных зим (OQ, PQ, OR, PR),стено-
бионты безморозных зим (Q, QR, R) .
Таблица 14
Схема системы ацидоморф и экогруппы по отношению видов к кислотному режи-
му почв
Тип режима pH почв Аци до морфа Схема системы рН-морф
Крайне кислых Гиперацидофилы а
ab
Сильно кислых Перацидофилы b ас
be ad
Кислых Ацидофилы с bd ае
cd be af
Слабо кислых Субацидофилы d ce bf ag
de cf bg
Нейтральных Нейтрофилы e df eg
ef dg
Слабощелочных Субалкалифилы f eg
fg
Щелочных Алкалифилы g
Примечание. Экогруппы: перацидофильные станобионты (a, ab, Ь), пера-
цидофильные полибионты (ас, be, ad, bd), мезоацидофильные стенобионты (с, cd,
d), мезоацидофильные полибионты (be, се, cf) , субэврибионты (ае, bf, eg), эври-
бионты (af, ag, bg) , нейтрофильные стенобионты (de, е, ef), алкалифильные полиби-
онты (df, dg, eg) , алкалифильные стенобионты (f, fg, g) .
67
Таблица 15
Схема системы нитроморф и экогруппы по отношению видов к азотному режиму
почв
Схема азотного режима почв Нитроморфа Схема системы нитроморф
Безазотных Очень бедных азотом Бедных азотом Анитрофилы Субанитрофилы Геминитрофилы j jk k jl kl jm 1 km jn
Достаточно богатых Субнитрофилы Im kn jo m In ko
азотом Богатых азотом Избыточно богатых азотом Нитро фи л ы Меганит рофилы mn lo n mo no 0
Примечание. Экогруппы:анитрофильныестенобионты(j, jk, к),темииитрофиль-
ныеполибионты (JI, kl, jm, km), мезонитрофильные стенобионты (1,1m, т),эврибионты
(jn, kn, jo, ко) , нитрофильные полибионты (In, mn, lo, mo) , нитрофильные стено-
бионты (п, по, о) .
Таблица 16
Схема системы гидроконтрастоморф и экогруппы по отношению видов к режиму
переменности увлажнения почв
Тип режима перемен- ности увлажнения Гидрокоитрастоформа Схема системы гидрокоит- растоморф
Постоянный Относительно посто- янный Слабо переменный Умеренно переменный Сильно переменный Резко переменный Константофиты Субкоистантофиты Гемиконтрастофиты Субконтрастофиты Контрастофиты Гиперконтрастофиты р РЧ q рг qr ps г qs pt rs qt pu s rt qu ru t su st tu u
Примечание. Экогруппы: константофиты-стенобионты (р, pq, q), гемикон-
трастофиты-полибионты (pr, qr, ps, qs), субконтрастофиты-стенобионты (г, rs, s) ,
зврибионты (pt, qt, pu, qu) , контрастофиты-полибионты(г1, st, ru, su) .контрастофи-
ты-стенобионты (t,tu, u) .
На прилагаемых дополнительных схемах систем экоморф можно ви-
деть принцип выделения pH-групп (табл. 14), нитро групп (табл. 15), групп
переменности увлажнения почв (табл. 16).
Для ценоморф пока целесообразно выделять зкогруппы лишь для ге-
лиоморф (табл. 17). Для подзоны хвойно-широколиственных лесов, как,
впрочем, и для всех почти зон, кроме зоны полярных пустынь, зоны тундр
и частично зоны пустынь, целесообразно различать следующие зкогруппы
режима затенения: гелиофиты (гелиоморфа Gg), гипогелиофиты (гелиомор-
68
Таблица 17
Схема системы гелиоморф и экогруппы по отношению видов к режиму затенения
Тип режима затенения Гелиоморфа Схема системы гелиоморф
Открытых пространств Гелиофиты G Gg
Полуоткрытых пространств Субгелиофиты g GM
gM Gs
Светлых лесов Семигелиофиты М gs GS
Ms gS
Тенистых лесов Субсциофиты S MS
sS
Особо тенистых лесов Сциофиты S
Примечание. Экогруппы: гелиофиты (G, Gg, g) , гипогелиофиты (GM, Gs) ,
гемисильванты (gM, gs, М, Ms), гелиосциофиты (GS), сильванты (MS, gS), сциосиль-
ванты (s, sS, S).
фы GM и Gs), гемисильванты (гелиоморфы gM, gs, Ms), сильванты (ге-
лиоморфы: MS и gS), сциосильванты (гелиоморфа sS), гелиосциофиты
(гелиоморфа GS).
Как видно из изложенного выше, основанием для отнесения вида к той
или иной экоморфе, экологической свите или экогруппе является характер
его экологической амплитуды, или амплитуды толерантности по отноше-
нию к существующему фактору, т. е. положение этой амплитуды на шкале
элементарных типов режима данного фактора.
Пользуясь координатным методом, строя соответствующие экологичес-
кие сетки (климатические, эдафические, ценотические, а также основные,
построенные на осях ведущих факторов климатической, эдафической или
ценотической природы, и дополнительные, построенные на осях одного из
ведущих факторов и второстепенного фактора той же природы), можно
рассматривать совместно амплитуды толерантности вида по двум и более
факторам, которые в соответствующих координатных сетках будут очер-
чивать плоскостные, объемные или абстрактные многомерные фигуры
экологических ареалов видов. Более удобно вместо этого, очевидно, рас-
сматривать экологические ареалы видов на иерархически условно сопод-
чиненных сериях двумерных (т.е. двухфакторных) экологических сеток.
Однако при всех этих методических приемах рассмотрения экологичес-
ких сущностей видов следует ни на минуту не упускать из виду следующих
элементарных истин. Вычленяя какой-либо фактор, говоря об амплитуде
толерантности вида по отношению к режимам этого фактора, мы имеем
в виду некий непрерывный ряд величин, характеризующих интенсивность
данного фактора, в пределах которых возможно существование данного
вида. Вполне закономерно поэтому графическое расположение режимов
данного фактора, вообще пригодных для существования растений, в виде
линейной шкалы последовательных градаций, обозначающих различаемые
нами режимы, на которых амплитуды толерантности видов имеют вид не-
которых непрерывных отрезков шкалы.
При одновременном изображении амплитуд видов даже всего лишь по
69
двум факторам мы уже более уходим в область отвлеченных построений,
ибо независимость факторов друг от друга относительна, а координатные
схемы предполагают полную независимость одного фактора от другого.
Любой фактор, даже космического или техногенного происхождения, на-
ходясь в поле влияния других факторов, изменяет в той или иной степени
интенсивность и направленность своего воздействия, применительно к
особенностям сред и поверхностей в которых и на которых наблюдается
эффект его взаимодействия со всем комплексом режимов других факто-
ров вместе или какого-либо из них преимущественно.
Чем больше факторов мы пытаемся вместить в схему, отражающую
закономерности распределения видов и сообществ по градиентам среды,
тем более мы отвлекаемся от реальности. Путаясь в математических изощре-
ниях списания никогда не существовавших гиперпространств, мы ни на
микрон не приближаемся к познанию истинной связи организмов, сообществ
и факторов, ибо она осуществляется не в гиперпространстве, а в единствен-
но реально существующем трехмерном пространстве. Если же нас интере-
сует зависимость распределения на местности наземных видов и сообществ
от сочетания режимов факторов среды, то для достаточно наглядного мо-
делирования такой зависимости вполне возможно пользоваться и двухмер-
ным пространством.
В сущности карты ареалов видов и геоботанические карты и являются
такими моделями. Сложность заключается главным образом в анализе
карт, как моделей, в нахождении логически последовательного и практи-
чески удобного пути вычленения из многофакторной схемы, какой являет-
ся карта, простых двухфакторных координатных схем и однофакторных
рядов.
Что касается принципиальной классификации экологических амплитуд и
экологических ареалов (рассматриваются двухфакторные ареалы в коор-
динатных схемах), то первые удобно классифицировать по трем призна-
кам: отношение к экстремальным типам режима, положение относительно
оптимума и размерность, а вторые по сочетанию характеристик обеих
амплитуд толерантности или по положению на соответствующей координат-
ной схеме экотопов.
По отношению к экстремальному типу режима данного фактора ампли-
туды толерантности видов делятся на нормальные, не захватывающие экст-
ремальных режимов, и экстремальные, захватывающие хотя бы один из
экстремальных режимов. По положению относительно оптимума экологи-
ческие амплитуды делятся на адаптивные, целиком лежащие при больших
или меньших, чем в оптимальном режиме, значениях фактора, и анадаптив-
ные с амплитудой толерантности, включающей и оптимальный тип режима
данного фактора. По размерности амплитуд толерантности можно выде-
лять стенотопные виды (амплитуда < 1/4), политопные виды (амплитуда от
1/4 до 3/4) и эвритопные виды (амплитуда >3/4 полной шкалы данного
фактора). Таким образом, если обратиться к схемам систем частных эко-
морф (табл. 18—25), то можно видеть, что число принципиальных типов
амплитуд толерантности зависит от дробности шкалы элементарных типов
режимов данного фактора и положения на этой шкале оптимального типа
режима.
Что касается экологических ареалов, то их целесообразно рассматривать
70
Таблица 18
Схема системы термоморф и принципиальные типы амплитуд толераитиостн
Термозона Схема системы термоморф
Арктическая А,
Аа,
Субарктическая а3 АВ,
аВ3 АЬ2
Бореальная В3 ab, AN,
Bb3 aN4 Am,
Суббореальная b3 BN, am, AM,
bN3 Bm, aM, Ats
Неморальная N3 bm, BM, at, AT6
Nm3 bM, Bt, aT,
Субсредиземноморская m3 NM, bt, ' ,
mM3 Nt, bT,
Средиземноморская M3 mt, NT7
Mt3 mT,
Тропическая t3 MT,
tT,
Экваториальная T8
Примечание. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный; 2 -
экстремально-адаптивно-политопный; 3 — нормально-адаптивно-стенотопный; 4 —
нормально-адаптивно-политопный; 5 — экстремально-адаптивно-эвригопный; 6 — эк-
стремально-анадаптивно-эвритопный; 7 — экстремально-анадаптивно-политопный; 8 -
экстремально-анадаптивно-стенотопный.
Таблица 19
Схема системы климаконтрастоморф и принципиальные типы амплитуд толерант-
ности
Пояс континентальности Схема системы климаконтрастоморф
Экстр аокеанический Е,
ЕО,
Океанический 0, Ео3
Оо, Ет3
Морской о, От5 ЕМ3
от, OMS Ек3
Субматериковый т2 oMs Oks ЕК6
тМ, ok5 OKS EU6
Материковый М2 mk, oKs OU6
Mk2 тК, oU3
Субконтинентальный k2 MK, mil,
kK, MUa
Континентальный K, kU,
KU,
Ультраконтинентальный u,
Примечание. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный;
2 — нормально-адаптивио-стенотопный; 3 — экстремально-анадаптивно-политопный;
4 — нормально-анадаптивно-стенотопный; 5 — нормально-анадаптивно-политопный;
6 — экстремально-анадаптивно-эвритопный; 7 — нормально-адаптивно-политопный;
8 — экстремально-адаптивно-политопный.
71
Таблица 20
Схема системы омброморф и принципиальные типы амплитуд толерантности
Тип режима аридности Схема системы омброморф
Э кстр ааридный А,
ABt
Эуаридный В3 АС,
вс3 ad,
Меэоаридный с3 bd4 ае,
CD3 ВЕ4 AFs
Субаридный D3 СЕ„ BF6 AG7
DE3 CF6 BG8 AH,
Субгумидный E3 DF6 CG6 BH,
ef8 dg8 ch5
Гумидный f8 eg6 dh5
fg8 eh5
Пергумидный g8 fh5
GH,
Ги пе р г у ми дный H,
Примечание. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный;
2 — экстремально-адаптивно-политопный; 3 — нормально-адапти'вно-стенотопный;
4 — нормально-адаптивно-политопный; 5 — экстремально-анадаптивно-политопный;
6 — нормально-анадаптивно-политопный; 7 — экстремально-анадаптивно-эвритопный;
8 — нормально-анадаптивно-стенотопный.
Таблица 21
Схема системы криоморф и принципиальные типы амплитуд толерантности
Криозона Схема системы криоморф
Очень суровых зим К,
КЦ
Суровых зим L3 км.
LM3 KN,
Довольно суровых зим М3 LNS KO, .
NN3 LOS KP,
Умеренных зим N3 мо5 LPS KQ,
NO3 MP8 LQS KR8
Мягких зим о3 np5 MQS LR6
op3 NQS. MR,
Теплых зим Р3 oq5 NR,
PQ3 OR,
Очень теплых зим Q3 PR,
QR8
Невыраженных зим . R8
Примечание. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный;
2 — экстремально-адаптивно-политопный; 3 — нормально-адаптивно-стенотопный;
4 — экстремально-адаптивно-эвритопный: 5 - нормально-адаптивно-политопный;
6 — экстремально-анадаптивно-эвритопный; 7 — экстремально-анадаптивно-политоп-
ный; 8 — эк стр емально-анадаптивно-стейотопный.
72
Таблица 22
Схема системы гидроморф и принципиальные типы амплитуд толерантности
Тип режима увлажнения Схема системы гидроморф
Пустынный D,
Ddi
Полупустынный d3 DS;
ds3 DS2
Сухостепной s3 dS3 DC2
sS3 dC, Dc2
Среднестепной S3 sC3 de, Dfs
SC3 sc, df, DF,
Лугово-степной C3 Sc3 sf, dF, Dp,
Cc3 Sf, sF, dp, DP,
Сухолесолуговой c3 Cf, . SF. sp, dP, Da,
cf. CF. Cp, sP, da, DA,
Влажно-лесолуговой f, cF3 Cp, SP, sa, dA,
fF„ cp, CP, Sa, sA,
Сыро-лесолуговой F3 fp, cP, Ca, SA,
Fp3 fP, ca, CA,
Болотно-лесолуговой p3 FP3 fa, cAs
pP3 Fa, fA,
Болотный P3 pa3 FA2
Pa, pA2
Прибрежно-водный a3 PA,
aA,
Водный A,
Примечание. Типы амплитуд: I — экстремально-адаптивио-стенотопный;
2 — экстремально-адаптивно-политопный; 3 — нормально-адаптивно-стенотопный;
4 — нормально-адаптивно-политопный; 5 — экстремально-анадаптивно-политопный;
6 — нормально-анадаптивно-политопный; 7 — экстремально-анадаптивно-эвритопный;
8 — нормально-анадаптивно-стенотопный; 9 — нормально-анадаптивно-эвритопный.
на экологических сетках, построенных на осях ведущих факторов различ-
ной природы: климатической, эдафической или ценотической, а при необхо-
димости и на дополняющих экологических сетках, построенных с учетом и
относительно второстепенных факторов климатической или эдафической
природы-. Такие двухфакторные экологические ареалы типизируются тоже
относительно просто и по тем же признакам, что и экологические амплиту-
ды, т. е. как совокупность амплитуд по обоим факторам. Так, например,
эдафические ареалы, рассматриваемые на эдафической сетке, построенной
на осях-шкалах обобщенного водного и обобщенного солевого режимов
почв, подразделяются по отношению к экстремальным эдафотопам на: ксе-
ро-трофоэкстремальные, ксеро-галоэкстремальные, гидро-трофоэкстре-
мальные, гидро-галоэкстремальные, ксероэкстремальные, гидроэкстремаль-
ные, трофоэкстремальные, галоэкстремальные и амфинормальные; по от-
ношению к осям гидрокомфортотопов и галокомфортотопов на: ксеро-
трофоадаптивные, ксерогалоадаптивные, гидротрофоадаптивные, гидро-
галоадаптивные, ксероадаптивные, гидроадаптивные, трофоадаптивные,
галоадаптивные и амфианадаптивные; по размерности амплитуд Толерант-
ности относительно обоих сопряженных факторов на: стеногидротопные,
стеногалотопные, стеноэдафотопные (стеногидро-галотопные), полигидро-
73
Таблица 23
Схема системы трофоморф и принципиальные типы амплитуд толерантности
Типы солевого режима почв Схема системы трофоморф
Особо бедных О, Оо,
Бедных °э оМ3 ом, Ое,
Небогатых М3 ое4 ОЕ,
Ме3 °Е, Og,
Довольно богатых ез еЕ, МЕ, og. Oh, Mg, oh, OH,
Богатых е, Eg, eg, Mh, oH, OP, eh, MH, oP, OS,
Слабозасоленных Eh, eH, MP, oS,
gh9 EH, eP, MS,
Среднезасоленных h3 hH3 gH4 EP, eS, gP, ES,
Сильнозасоленных н3 HP3 hP4 gS hS,
Резко засоленных Злостных солончаков Рз PS, S. HS,
Примечание. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный;
2 — экстремальио-адаптивно-политопный; 3 — нормально-адаптивно-стенотопный;
4 — нормально-адаптивно-политопный; 5 — экстремально-анадаптивно-политопный;
6 — нормально-анадаптивно-политопиый; 7 8 — нормально-анадаптивно-стенотопный. - экстремально-анадаптивно-эвритопный;
Таблица 24
Схема системы ацидоморф и принципиальные типы амплитуд толерантности
Тип режима pH почв
Схема системы ацидоморф
Крайне кислых а> ab, 1
Сильно кислых Ь, bc3 ac, adj
Кислых сз «1з bd. be« ae5 af,
Слабо кислых 4» ce« bf,
de, cf, bg,
Нейтральных е. ef8 df. dS5 eg,
Слабощелочных f, egS
Щелочных почв g»
Примечанне. Типы амплитуд: 1 — экстремально-адаптивно-стенотопный;
2 — экстремально-адаптивно-политопный; 3 — иормально-адаптивно-стенотопный;
4 — нормально-адаптивно-политопный; 5 - экстремально-анадаптивно-политопный;
6 — нормалыю-анадативно-политопный; 7 — зкстремально-анадаптивно-эвритопный;
8 — нормально-анадаптивно-стеиотопный.
74
Таблица 25
Схема системы гелиоморф и принципиальные типы амплитуд толерантности
Тип режима затенения Схема системы гелиоморф
Открытых пространств G, Gg2
Полуоткрытых пространств Светлых лесов Тенистых лесов Особо тенистых лесов П римечание. Типы амплитуд: 1 g3 gm2 gM4 Gs2 M3 gs4 GSs Ms4 gS6 s3 MS6 sS6 s7 — экстремал ьно-анадаптивно-стенотопный
2 - экстремально-анадаптивно-политопный; 3 — нормально-адаптивно-стенотопный;
4 — нормально-адаптивно-политопный; 5 — экстремально-анадаптивно-эвритопный;
6 — экстремально-адаптивно-политопный; 7 — экстремально-адаптивно-стенотопный.
топные, полигалотопные, полиэдафотопные, эвригидротопные, эвригалотоп-
ные и эвриэдафотопные (рис. 6).
Эти 24 типа эдафических ареалов, конечно, не исчерпывают всего их раз-
нообразия. Однако их вполне достаточно для наглядной иллюстрации прин-
ципов их диагностики: экстремальные типы ареалов охватывают экстре-
мальные типы режимов факторов, нормальные — целиком лежат вне их;
адаптивные типы ареалов не охватывают комфортотопов, а анадаптивные —
включают их в себя; стенотопные типы ареалов занимают не более 1/4 шка-
лы фактора, политопные — не более 3/4, а эвритопные — более 3/4 шкалы.
Знаком + обозначены комфортотопы.
Климатические ареалы в связи с принципиально иным расположением
на климатической сетке осей комфортотопов имеют и принципиально иной
набор типов. По отношению к экстремальным климатопам климатические
ареалы подразделяются на: арктоокеанически экстремальные, арктоконти-
нентально экстремальные, тропоокеанически экстремальные, тропоконти-
нентально экстремальные, арктоэкстремальные, тропоэкстремальные, океа-
нически экстремальные, континентальноэкстремальные и амфинормаль-
ные; по отношению к осям комфортотопов на: океанически-термоадаптив-
ные, континентально-термоадаптивные, монотермоадаптивные и амфиана-
даптивные; по размерности амплитуд толерантности относительно режимов
обоих сопряженных факторов на: стенотермотопные, стеноконтрастотоп-
ные, стеноклиматопные, политермотопные, поликонтрастотопные, поли-
климатопные, эвритермотопные, эвриконтрастотопные, эвриклиматопные.
Если же обратиться к вновь разработанной климатической сетке, где кли-
матические терморежимы сопряжены с режимами аридности — гумидности
климата, то типизация ареалов будет принципиально та же, с той лишь раз-
ницей, что вместо океанически и континентальности будут фигурировать гу-
мидность и аридность, а вместо контрастотопов — омбротоп- >ис. 7).
Эти 15 видов климатических ареалов, конечно, не исчерпывают всего их
разнообразия. Если возвратиться к табл. 18 и 20, то легко подсчитать, что
шкала терморежимов, при учете положения на ней общекомфортног- типа
75
Трофотопы
1
II
Р и с. 6. Эдафнческая сетка и принципиальные типы эдафических ареалов
I — недостаток солен; П — избыток солей. Типы эдафических ареалов: 1 — ксеро-
экстремальный, трофоэкстремально-амфиадаптивно-амфнполитопный; 2 — ксеро-
экстремальный, трофонормально-амфиадаптивно-амфистенотопный; 3 — ксероэк-
стремальный трофонормально-ксероадаптивный, трофоанадаптивно-амфиполитопный;
4 ' — ксероэкстремальный, галоэкстремально-ксероадаптивный, галоанадаптивно-
амфиполитопный; 5 — ксероэкстремальный, галоэкстремально-амфиадаптивно-амфи-
политопный; 6 — ксеронормальиый, трофоэкстремально-амфиадаптивно-амфопо-
литопный; 7 — амфинормально-ксероадаптивный, трофоадаптивно-амфиполитопный;
8 — амфинормально-ксероадаптивный, трофоанадаптивно-амфиполитопный; 9 — амфи-
нормально-амфнанадаптивно-амфиполитопный; 10 — амфинормально-ксероадаптив-
ный, галоадаптивно-амфиполитопный; 11 — ксеронормальиый, галоэкстремально-
амфнадаптивно-амфиполитопный; 12 — гидронормальный, трофоэкстремально-гидро-
анадаптивный, трофоадаптивно-амфиполитопный; 13 — амфинормально-ксероанадап-
тивный, трофоадаптивно-амфиполитопный; 14 —амфинормально-гидроанадаптивный,
галоадаптивно-амфиполитопный; 15 — гидронормальный, галоэкстремально-гидроана-
даптивный, галоадаптивно-амфиполитопный; 16 — гидронормальный, трофоэкстре-
режима, допускает существование 8 принципиальных типов амплитуд толе-
рантности по этому фактору, а шкала омброрежимов — 10. Таким образом,
можно было бы ожидать существование 80 принципиальных типов клима-
тических ареалов по совокупности этих двух факторов. В действительно-
сти, учитывая, что сочетание некоторых типов амплитуд толерантности, по
крайней мере, маловероятно, практически значимое число принципиальных
типов климатических ареалов сократится до 60—65. Но и 60 типов поме-
стить на одной таблице — значит сделать ее нечитаемой. В то же время и 15
типов климатических ареалов более чем достаточно для иллюстрации прин-
ципов диагностики: экстремальный тип ареала охватывает и экстремаль-
ные типы режимов факторов, нормальный — целиком лежит вне их, адап-
тивный тип ареала целиком лежит в экотопах, худших для большинства ви-
дов, чем общекомфортный режим данного фактора, анадаптивный — охва-
тывает и комфортотопы, стенотопный тип ареала охватывает более 1/4
шкалы фактора, политопный — не более 3/4 шкалы, а эвритовый — более
3/4. Знаком + обозначены комфортотопы.
Типизация экологических амплитуд и экологических ареалов полезна
той наглядностью, с которой она отображает различные индикационные воз-
можности видов, принадлежащих различным экоморфам.
Как и экологическая (климатическая, эдафнческая, ценотическая, основ-
ная и дополняющая) сетка в целом, экологический ареал вида или эколо-
гический ареал ряда видов, объединяемых одной экоморфой, как разнока-
чественное в различных своих частях поле взаимодействия режимов двух
факторов, обладает восемью основными типами изменения характеристик
среды: улучшение ее состояния при смещении от периферии к центру и
ухудшение - при смещении от центра ареала к периферии в направлении
стабильного состояния режима фактора оси х и изменения режимов факто-
ра оси j', в направлении стабильного состояния режима фактора оси и сме-
ны режимов фактора оси х, в направлении однонаправленной смены режи-
мов обоих факторов и в направлении разнонаправленной смены режимов
обоих факторов.
Как и в случае оценки комфортности экотопов (климатопов, эдафото-
пов или ценотопов) для растительного покрова вообще, качество среды для
данного вида или для некоего набора видов, объединяемого принадлежно-
стью одной экоморфе, может быть выражено количественно в виде некото-
рого процентного коэффициента, изменяющегося от 100% в центре эколо-
гического ареала до 0 на его границе.
Этот коэффициент, в отличие от коэффициента комфортности (КК), це-
лесообразно именовать коэффициентом удовлетворительности условий сре-
ды (КС). Формулы определения КС (коэффициентов удовлетворительности
мально-гидроадаптивный; трофоадаптивно-амфистенотопный; 17 — амфинормально-
гидроадаптивный, трофоадаптивно-амфиполитопный; 18 — гидроэкстремальный,
трофонормально-гидроадаптивный, трофоадаптивно-амфиполитопный; 19 - амфи-
нормально-гидроадаптивный, галоадаптивно-амфиполитопный; 20 — гидронормаль-
ный, галоэкстремально-амфнадаптивно-гцдростенотопный, галополитопный;
21 — гидроэкстремальный, трофоэкстремально-амфиадаптивно-амфиполитопный;
22 — гидроэкстремальный, трофонормально-амфиадаптивно-амфнполитопный; 23 —
гидроэкстремальный, галонормально-гидроадаптивный, галоанадаптивно-гидростено-
топный, галополитопный; 24 — гидроэкстремальный, галоэкстремально-амфидаптив-
но-амфиполитопный
77
Омброклиматический пояс
Р и с. 7. Климатическая сетка и принципиальные типы климатических ареалов
I — аридный тип климата; II — гумидный тип климата. Типы климатических ареа-
лов: 1 — арктоэкстремальный, аридоэкстремально-амфиадаптивно-амфиполитопный;
2 — арктоэкстремальный, аридно-нормально-амфиадаптивно-амфиполитопный; 3 —
арктоэкстремальный, омбронормально-арктоадаптивный, омброанадаптивно-амфипо-
литопный; 4 — арктоэкстремальный, гумидоэкстремально-амфиадаптивно-амфистено-
толный; 5 и 6 — термонормальные, аридоэкстремально-амфиадаптивно-амфиполитоп-
ные; 7 — амфинормально-амфиадаптивно-амфистенотопный; 8 — тропоэкстремальный,
омбронормально-амфианадаптивно-термоэвритопный, омброполитопный; 9 — термо-
нормальный, гумидоэкстремально-амфиадаптивно-термополитопный, омбростенотоп-
ный; 10 — амфинормально-термоадаптивный, омброанадаптивно-термополитопный,
омбростенотипный; 11 — амфинормально-амфиадаптивно-амфиполитопный; 12 — тро-
поэкстремальный, гумидоэкстремально-амфианадаптивно-амфиполитопный; 13 — тро-
поэкстремальный, аридоэкстремально-термоадаптивный, омброадаптивно-амфиполи-
топный; 14 — тропоэкстремальный, омбронормально-термоанадаптивный, омброадап-
тивно-амфиполитопный;; 15 — тропоэкстремальный, гумидоэкстремально-термоаиа-
даптивный, омброадаптивно-амфистенотопный
условий среды) аналогичны формулам определения КК (коэффициенте!
комфортности экотопов):
KCmin = 2’ ЮО; КСтеЛ, ЮО; КСтак = — 100 ,
где х — порядковый номер данного типа режима фактора от края амплиту
ды толерантности вида по этому фактору; а — число элементарных режи
мов фактора, охватываемых амплитудой толерантности вида. КС для сово-
купности режима двух и более факторов вычисляется по принципу нахож-
дения процента от процента [/СС климатопа = КС термоклиматопа • КС
омброклиматопа : 100; КС эдафотопа = КС гидроэдафотопа • КС трофоэда-
фотопа : 100; КС эдафотопа в условиях данного климатопа (хоротопа) =
= КС климатопа • КС эдафотопа : 100; КС экотопа = КС эдафотопа в усло-
виях данного климатопа (хоротопа) • КС Ценотопа : 100]. В качестве при-
мера приводятся климатические и эдафические ареалы некоторых доминан-
тов лесных фитоценоэов подзоны хвойно-широколиственных лесов, на кото-
рых показано размещение КС этих видов в границах их экологических
ареалов (табл. 26—41).
В верхних табличках-схемах (см. табл. 26—41) приведены КС разных
климатопов в пределах климатических ареалов данных видов, в следую-
щих за ними - КС разных эдафотопов в пределах эдафических ареалов дан-
ных видов, на трех остальных — КС разных хоротопов в пределах трансфор-
мированных названным климатопом эдафических ареалов данных видов.
Название хоротоп представляется целесообразным применять для обозна-
чения класса экотопов, характеризуемых совокупностью только клима-
тических и эдафических режимов, аналогично тому, как мы называем кли-
матопом класс экотопов, характеризуемый только по совокупности клима-
тических режимов, и эдафотопом — только эдафических.
Из сказанного выше о КС и рассмотрения схем экологических ареалов
следует, что низкие значения КС по какому-либо фактору обусловливают
необходимость высоких значений их по другим факторам, что чем дальше
от оптимума режим одного из факторов, тем меньшее отклонение от опти-
мума возможно для режимов других факторов.
При несоблюдении этих условий само существование любого вида в та-
ком местообитании невозможно. Собственно говоря, об этой же закономер-
ности, только менее определенно, свидетельствует и одно из дополнений к
закону толерантности: ’’Если условия по одному экологическому фактору
не оптимальны для вида, то может сузиться и диапазон толерантности к
другим экологическим факторам” [Одум, 1975, с. 141]. Эмпирический
анализ схем двухфакторых экологических ареалов показывает, что преде-
лом устой>швого существования популяций видов является КС порядка
7-8% (0,07—0,08). Элементарные расчеты показывают, что КС= 90% (0,9)
могут одновременно иметь 24 фактора,КС = 80% (0,8) одновременно иметь
12 факторов, что КС = 70% (0,7), 7 факторов, КС = 60% (0,6) - 5 факто-
ров, КС = 50% (0,5) - 3 фактора,КС = 40% - только 2 фактора, КС = 30% -
только 2 фактора, КС = 20% — лишь 1 фактор и КС 10% — тоже 1 фактор.
Так как разные виды (принадлежащие разным экоморфам) обладают
различными амплитудами толерантности, т. е. могут успешно произрастать
при различном наборе режимов каждого фактора, в том числе при охвате
79
Таблица 26
Климатические и эдафические ареалы и соответствующие КС
некоторых номинантов лесных сообществ хвойно-широколиственных лесов
Климатический
ареал иКС
Acer platanoides
0 О к
в 10 17 10
10 30 50 30 10
N 17 50 83 50 17
17 50 83 50 17
m 10 30 50 30 17
10 17 10
Corylus avellana
о О к
В 8 14 8
9 26 43 26 9
N 14 43 72 43 14
20 60 100 60
m 14 43 72 43 14
9 26 43 26 9
м 8 14 8
Эдафический
ареал и КС
М е Е
10 10
С 19 31 31 19
11 32 52 52 32 11
f 15 43 72 72 43 15
15 43 72 72 43 15
F И 32 52 52 32 11
19 31 31 19
Р 10 10
М е Е
7 7
S 14 22 22 14
8 23 37 37 23 8
С 11 32 53 53 32 11
14 41 68 68 41 14
С 17 50 83 83 50 17
14 41 68 68 41 14
f И 32 53 53 32 11
8 23 37 37 23 8
F 14 22 22 14
7 7
соответствующими амплитудами различного количества элементарных ти-
пов режима, то возможна и различная оценка качества среды по данному
фактору с точки зрения потребностей видов разных экоморф. Иными сло-
вами, возможны различные бонитировочные шкалы, в которых один и
тот же элементарный тип режима каждого фактора оценивается по-разно-
му в зависимости от уровня удовлетворения потребностей интересующих
нас видов. В руководствах по лесной таксации также указывается, что од-
80
Таблица 26 (окончание)
Изменение эдафического М е Е
ареала в климатопе с 9 9
BN-ok 10 16 16 10
f 13 22 22 13
13 22 22 13
F 10 16 16 10
9 9
Изменение эдафического
м е Е
ареала в климатопе N—ок С 10 16 16 10
16 26 26 16
f 8 22 36 36 22 8
8 22 36 36 22 8
F 16 26 26 16
10 16 16 10
6. Зак. 448
Изменение
эдафического
ареала в
климатопе
Nm-ok
М е Е
С 10 16 16 10
16 26 26 16
f 8 22 36 36 22 8
8 22 36 36 22 8
F 16 26 26 16
10 16 16 10
м е Е
10 10
С 8 14 14 8
11 18 18 И
С 13 21 21 13
11 18 18 И
f 8 14 14 8
10 10
М е Е
9 9
С 10 16 16 10
14 23 23 14
С 18 29 29 18
22 36 36 22
f 18 29 29 18
14 23 23 14
F 10 16 16 10
9 9
М е Е
S 8 13 13 8
14 22 22 14
С 19 32 32 19
8 25 41 41 25 8
с 10 30 50 50 30 10
8 25 41 41 25 8
f 19 32 32 19
14 22 22 14
F 8 13 13 8
81
Таблица 27
Климатические и эдафическне ареалы и соответствующие КС некоторых доминан гов
лесных сообществ подзоны хвойно-широколиствеиных лесов
Климатический ареал и КС
О О к
В 8 14 8
9 26 43 26 9
N 14 43 72 43 14
20 60 100 60 20
ГЛ 14 43 72 43 14
9 26 43 26 6
м 8 14 8
Эдафический ареал и КС
О О М е Е
С 8 И 14 11 8
14 24 34 43 34 24 14
С 8 24 40 56 72 56 40 24 8
11 33 56 78 100 78 56 33 11
f 8 24 40 56 72 56 40 24 8
14 24 34 43 34 24 14
F 8 11 14 11 8
Изменение эдафического
ареала в климатопе BN—ок
0 О М е Е
9 11 9
С 10 15 19 15 10
9 15 20 26 20 15 9
f 10 15 19 15 10
9 11 9
82
Таблица 27 (окончание)
Изменение эдафического
ареала в климагопе N—ок
О О М е Е
10 15 18 15 10
С 10 17 24 31 24 17 10
14 24 34 43 34 24 14
f 10 17 24 31 24 17 10
10 15 18 15 10
Изменение эдафического
ареала в климатопе Nm-ok
О О М е Е
С 8
8 14 20 26 20 14 8
С 14 24 34 43 34 24 . 14
20 34 47 60 47 34 20
f 14 24 34 43 34 24 14
8 14 20 26 20 14 8
F 8
но и то же местообитание может оцениваться разным бонитетом в зави-
симости от породы, для которой производится бонитировка.
Возможно создание множества бонитировочных шкал: в зависимости
от величин амплитуд толерантности разных видов, от разработанности
шкал по тому или иному фактору, от набора учитываемых факторов, от
способа градации бонитировочной шкалы (словесными оценками, циф-
ровыми баллами, в процентном выражении от ведущего признака, по ко-
торому диагностируется бонитировочный балл по качественным или ко-
личественным оценкам данной популяции вида, или по индикационным
оценкам среды данного местоположения, или по другим критериям),
но целесообразно создавать только такие шкалы, которые могут быть
достаточно просто и удобно сопоставимы друг с другом, градации которых
достаточно просто и удобно могут быть переведены друг в друга либо не-
посредственно, либо при помощи некой третьей шкалы. В этом смысле не
подлежит сомнению, что бонитировочные шкалы разных видов должны
быть увязаны с КС (коэффициентами удовлетворительности условий сре-
ды) этих видов, а через них с экологическими шкалами, экологическими
сетками и КК (коэффициентами общей комфортности) экотопов.
83
Таблица 28
Климатические иэдафические ареалы и соответствующие КС
некоторых домииаитов лесных сообществ подзоны хвойно-широколиственных лесов
Convallaria majalis
Климатический ареал и КС
О О к
8 14 8
в 9 26 43 26 9
14 43 72 43 14
N 20 60 100 60 20
14 43 72 43 14
ГЛ 9 26 43 26 9
8 14 8
Эдафический ареал и ТСС
О О М е Е
9
С 9 15 21 27 21 15 9
15 25 35 45 35 25 15
С 21 36 50 64 50 36 21
9 27 46 64 82 64 46 27 9
f И 33 56 78 100 78 56 33 11
9 27 46 64 82 64 46 27 9
F 21 36 50 64 50 36 21
15 25 35 45 35 25 15
Р 9 15 21 27 21 15 9
9
Примеры бонитировочных шкал, оценивающих режимы отдельных
факторов с позиций удовлетворения потребностей разных видов, при-
ведены на табл. 42.
Как видно из табл. 42. для дуба по режимам климатических факто-
ров возможна лишь трехбалльная шкала бонитировки со значениями:
КС: 0— 33%, 34 — 66 и 67 — 100%; для сосны по режимам климатичес-
84
Таблица 28 (окончание)
Изменение эдафического ареала в климатопе BN—ок О О М е Е
с 9 12 9
11 15 19 15 и
с 9 15 21 28 21 15 9
12 20 28 35 28 20 12
f 14 24 34 43 34 24 14
12 20 28 35 28 20 12
F 9 15 21 28 21 15 9
11 15 19 15 11
Р 9 12 9
Изменение эдафического
ареала в климатопе N—ок
О О М е Е
С 9 13 16 13 9
9 15 21 27 21 15 9
С 13 22 30 38 30 22 13
16 28 38 49 38 28 16
f 20 34 47 60 47 34 20
16 28 38 49 38 28 16
F 13 22 30 38 30 22 13
9 15 21 27 21 15 9
Р 9 13 16 13 9
Изменение эдафического
ареала в климатопе Nm—ок
О О М е Е
С 9 12 9
11 15 19 15 11
С 9 15 21 28 21 15 9
12 20 28 35 28 20 12
f 14 24 34 43 34 24 14
12 20 28 35 28 20 12
F 9 15 21 28 21 15 9
11 15 19 15 11
Р 9 12 9
85
Таблица 29
Климатические и эдафическне ареалы и соответствующие /«'некоторых доминантой лесных
сообществ подзоны хвойно-иифоколиственных лесов
Asarum europeum
Климатический
ареал и КС
О К
8 14 8
в 9 26 43 26 9
14 43 72 43 14
N 20 60 100 60 20
14 43 72 43 14
ш 9 26 43 26 9
8 14 8
Ajuga rep tans
о О к К
9 12 9
В 16 27 38 27 16
9 27 45 62 45 27 9
N 12 38 63 88 63 38 12
12 38 63 88 63 38 12
П1 9 27 45 62 45 27 9
16 27 38 27 16
м 9 12 9
Эдафический
ареал и КС
О М е Е
С 9 14 20 14 9
8 26 43 60 43 26 8
f 14 43 72 100 72 43 14
8 26 43 60 43 26 8
F 9 14 20 14 9
О М е Е
С 9 14 20 14 9
8 26 43 60 43 26 8
f 14 43 72 100 72 43 14
8 26 43 60 43 26 8
F 9 14 20 14 9
Изменение
эдафического
ареала в
климатопе
BN—ок
О М е Е
С 10 14 10
19 31 43 31 19
f 10 31 52 72 52 31 10
19 31 43 31 19
F 10 14 10
О М е Е
С 9 12 9
16 27 37 27 16
f 9 27 45 62 45 27 9
- 16 .27 37 27 16
F 9 12 9
ких факторов и для дуба по солевому режиму почв — четырехбалльная
шкала со значениями: КС: 0 — 25%, 26 — 50, 51 — 75, 76 — 100%; для
дуба по водному режиму почв и для сосны по солевому режиму почв —
пятибалльная шкала со значениями КС: 0 — 20%, 21 — 40, 41 — 60, 61 —
80, 81 - 100%; для сосны по водному режиму почв — семибалльная шка-
ла со значениями КС: 0 - 14%, 15 - 28, 29 - 42,43 - 57, 58 - 71, 72 - 86,
87 — 100%. Таким образом, видно, что размерность бонитировочных шкал
режимов отдельных факторов жестко задается возможностями шкалы
86
Таблица 29 (окончание)
Изменение
эдафнческого
ареала в
климатопе
Nm- ok
О М е Е
С 8 12 18 12 8
23 38 53 38 23
f 12 38 63 88 63 38 12
23 38 53 38 23
F 8 12 18 12 8
О М е Е
С 10 14 10
19 31 43 31 19
f 10 "31 52 72 52 31 10
19 31 43 31 19
F ю’ 14 10
О М е Е
С 8 12 18 12 8
23 38 53 38 23
f 12 38 63 88 63 38 12
23 38 53 38 23
F 8 12 18 12 8
режимов данного фактора (ее дробностью) и амплитудой данного вида.
В то же время для сочетаний режимов нескольких факторов (клима-
топов, эдафотопов, хоротопов и т. д.) размерность бонитировочной шка-
лы можно выбрать произвольно. Кроме того, на табл. 42—47 показана
возможная бонитировка климатопов и эдафотопов на основе КС неко-
торых лесообразователей европейской части СССР. Такая бонитировка,
неплохо согласуясь с общепринятой лесоводственной в отношении кли-
матических факторов и в отношении ряда почвенного увлажнения, хуже
согласуется с ней в отношении ряда солевого богатства почв.
На табл. 43—47 баллы бонитировки и КС даны для условий зональных
и региональных климатопов и хоротопов. Естественно, что топоклиматы,
отличные от зональных, с теми же эдафотопами образуют другие, не зо-
нальные тоже, хоротопы, где КС и соответственно бонитировочные оцен-
ки будут иными.
Лесоводственные бонитировочные критерии, основанные на увеличе-
нии линейного и объемного прироста, не соответствуют характеру отри-
цательных изменений древесных индивидов при избыточном солевом
режиме почв. Очевидно, истинный высший бонитет среды для растения
должен представлять собой наивыгоднейшие условия для оптимального
сочетания определенных величин скорости роста с долговечностью, высо-
кой репродуктивной способностью и устойчивостью к заболеваниям. С
этой точки зрения условия среды, лесоводственно определяемые для
данной породы I и 1а, в действительности не являются оптимальными
для нее.
Лссоводственная оценка бонитета, как и любая другая хозяйственная
«7
Таблица 30
Климатические и эдафические ареалы и соответствующие КС некоторых домииаитов
лесных сообществ подзоны хвойио-широколиственных лесов
Calamagrostis arundinacea
Климатический
ареал и КС
о О к
В 10 17 17 10
10 30 50 50 30 10
N 17 50 83 83 50 17
10 30 50 50 30 10
m 10 17 17 10
Эдафический
ареал и КС
О О М е Е О э
S 8
10 15 19 23 19 15 10
С 10 17 24 31 38 31 24 17 10
15 24 35 44 54 44 35 24 15
С 19 31 44 57 69 57 44 31 19
8 23 38 54 70 85 70 54 38 23 8
f 9 27 45 64 82 100 82 64 45 27 9
8 23 38 54 70 85 70 54 38 23 8
F 19 31 44 57 69 57 44 31 19
15 24 35 44 54 44 35 24 15
Р 10 17 24 31 38 31 2 4 17 10
10 15 19 23 19 15 10
Р 8
оценка объектов живой природы, имеет в виду лишь скорейшее получе-
ние наибольшей выгоды человеком и далеко не во всех случаях совпа-
дает с наиболее выгодными условиями для самих организмов и популя-
ций. Кроме того, очевидно, далеко не однозначно для организма, нахо-
дится он при режиме какого-либо фактора, близком к минимуму или
близком к максимуму, хотя за пределами и того и другого существова-
88
Таблица 30 (окончание)
Изменение
эдафического
ареала в
климатопе N—ок
0 О М е Е g
8 12 16 19 16 12 8
С 8 14 20 26 31 26 20 14 8
12 20 29 36 45 36 29 20 12
с 16 26 34 47 57 47 34 26 16
19 31 45 58 70 58 45 31 19
f 22 37 53 68 83 68 53 37 22
19 31 45 58 70 58 45 31 19
F 16 26 34 47 57 47 34 26 16
12 20 29 36 45 36 29 20 12
Р 8 14 20 26 31 26 20 14 8
8 12 16 19 16 12 8
Изменение
эдафического
ареала в
климатопе BN—ок
и Nm—ок
0 О М е Е
8 10 12 10 8
С 9 12 16 19 16 12 9
8 12 18 22 27 22 18 12 8
С 10 16 22 29 35 29 22 16 10
12 19 27 35 43 35 27 19 12
f 14 23 32 41 50 41 32 23 14
12 19 27 35 43 35 27 19- 12
F 10 16 22 29 35 29 22 16 10
8 12 18 22 27 22 18 12 8
Р 9 12 16 19 16 12 9
8 10 12 10 8
89
Таблица 31
Климатические и эдафические ареалы и соответствующие
КС некоторых доминантой лесных сообществ подзоны хвойно-широколиственных лесов
Calamagrostis epigeios
Климатический
ареал и КС
о О к К
9 12 9 1
а 16 27 38 27 16
9 27 45 62 45 27 9
в 12 38 63 88 63 38 12
12 38 63 88 63 38 12
N 9 27 45 62 45 27 9
16 27 38 27 16
m 9 12 9
Эдафический
ареал и КС
0 О М е Е g h Н Р
10 12 14 16 18 16 14 12 10
S 12 15 19 22 26 29 26 22 19 15 12 8
12 17 22 27 32 36 41 36 32 27 22 17 12
S 10 15 22 28 34 41 47 53 47 41 34 28 22 15 10
12 19 27 34 42 50 57 65 57 50 42 34 27 19 12
с 14 22 32 41 50 59 68 77 68 59 50 41 32 22 14
16 26 36 47 57 68 77 88 77 68 57 47 36 26 16
с 18 29 41 53 65 77 88 100 88 77 65 53 41 29 18
16 26 36 47 57 68 77 88 77 68 57 47 36 26 16
f 14 22 32 41 50 59 68 77 68 59 50 41 32 22 14
12 19 27 34 42 50 57 65 57 50 42 34 27 19 12
F 10 15 22 28 34 41 47 53 47 41 34 28 22 15 10
12 17 22 27 32 36 41 36 32 27 22 17 12
Р 12 15 19 22 26 29 26 22 19 15 12
10 12 14 16 18 16 14 12 10
ние его равно невозможно. По-видимому, при недостатке каких-либо благ:
тепла, влаги, света, солей и т. д. — имеет место состояние голода, характе-
ризующееся в пределах переносимого организмом главным образом за-
медлением роста и развития, без существенных нарушений присущего
виду характера обмена веществ, тогда как при избытке каких-либо благ
90
Таблица 31 (продолжение)
Изменение
эдафического
ареала в
климатопе
BN-ok
О О М е Е g h Н Р
9 11 12 14 16 14 12 11 9
S 11 13 17 19 23 26 23 19 17 13 И
11 15 19 24 28 32 36 32 28 24 19 15 11
S 9 13 19 25 30 36 41 47 41 36 30 25 19 13 9
11 17 24 30 37 44 50 57 50 44 37 30 24 17 11
с 12 19 28 36 44 52 60 68 60 52 44 36 28 19 12
14 23 32 41 50 60 68 77 68 60 50 41 32 23 14
с 16 26 36 47 57 68 77 88 77 68 57 47 36 26 16
14 23 32 41 50 60 68 77 68 60 50 41 32 23 14
f 12 19 28 36 44 52 60 68 60 52 44 36 28 19 12
11 17 24 30 37 44 50 57 50 44 37 30 24 17 11
F 9 13 19 25 30 36 41 47 41 36 30 25 19 13 9
11 15 19 24 28 32 36 32 28 24 19 15 И
Р 11 13 17 19 23 26 23 19 17 13 11
9 11 12 14 16 14 12 11 9
Изменение
эдафического
ареала в
климатопе N—ок
О О М е Е g h Н Р
9 10 11 10 9
S 9 12 14 16 18 16 14 12 9
11 14 17 20 22 25 22 20 17 14 11
S 9 14 17 21 25 29 33 29 25 21 17 14 9
12 17 21 25 31 35 40 35 31 25 21 17 12
с 9 14 20 25 31 37 42 48 42 37 31 25 20 14 9
10 16 22 29 35 42 48 .54 48 42 35 29 22 16 10
с 11 18 25 33 40 48 54 62 54 48 40 33 25 18 11
10 16 22 29 35 42 48 54 48 42 35 29 22 16 10
f 9 14 20 25 31 37 42 48 42 37 31 25 20 14 9
12 17 21 25 31 35 40 35 31 25 21 17 12
F 9 14 17 21 25 29 33 29 25 21 17 14 9
11 14 17 20 22 25 22 20 17 14 11
Р 9 12 14 16 18 16 14 12 9
9 10 11 10 9
91
Таблица 31 (окончание)
Изменение
эдафического
ареала в
климатопе
Nm—ок
0 О М е Е g h Н Р
S 8 10 11 10 8
8 10 12 14 16 14 12 10 8
S 8 И 13 16 18 20 18 16 13 11 8
10 13 16 19 22 25 22 19 16 13 10
с 8 12 16 19 22 26 29 26 22 19 16 12 8
10 14 18 22 26 29 33 29 26 22 18 14 10
с 11 16 20 25 29 33 38 33 29 25 20 16 11
10 14 18 22 26 29 33 29 26 22 18 14 10
f 8 12 16 19 22 26 29 26 22 19 16 12 8
10 13 16 19 22 25 22 19 16 13 10
F 8 11 13 16 18 20 18 16 13 11 8
8 10 12 14 16 14 12 10 8
Р 8 10 И 10 8
имеет место уже состояние отравления, характеризующееся в первую
очередь именно нарушением обмена веществ, тогда как замедления роста
и развития может какое-то время и не наблюдаться, если степень отравле-
ния не слишком велика, и даже, наоборот, какое-то время может наблю-
даться гипертрофированное ускорение роста и развития, но всегда имеет
место ослабление сопротивляемости организма неблагоприятным воз-
действиям и, как неминуемое следствие этого, сокращение срока жизни.
Человек же, хозяйственно заинтересованный в ускорении именно роста и
развития, в скорейшем наращивании биомассы, оценивает состояние среды,
соответствующее такому гипертрофированному росту и развитию, как
оптимальное.
Все изложенное выше об экоморфах, экогруппах, экологических сви-
тах, о системах экоморф, об экологических амплитудах и ареалах, о КС
подводит нас к тому, что в каждом экотопе может существовать только
определенный набор видов, принадлежащих определенным экоморфам.
Поскольку набор экоморф, существование которых возможно при
каждом типе режима того или иного фактора, предопределен и легко мо-
жет быть найден по соответствующей схеме системы экоморф, то и каж-
дый климатоп, эдафотоп и ценотоп обладает своей потенциальной фло-
рой, в той или иной степени отличной от потенциальных флор других кли-
матов, эдафотопов и ценотопов. Практически это означает, что своей по-
тенциальной флорой обладает каждый географический регион каждое
местоположение и каждое местообитание. Для данного географического
региона такая флора будет включать в себя все виды, потребностям кото-
рых удовлетворяют климатические условия региона и проникновению ко-
92
Таблица 32
Климатические и эдафические ареалы и соответствующие КС некоторых домииантов лесных
сообществ подзоны хвойно-широко лиственных лесов
Carex digitata
Климатический
ареал и КС
0 О k К
В 9 14 29 14 9
8 26 43 60 43 26 8
N 14 43 72 100 72 43 14
8 26 43 60 43 26 8
m 9 14 29 14 9
Carex pilosa
О к
12 20 12
N 12 36 60 36 12
20 60 100 60 20
П) 12 36 60 36 12
12 20 12
Эдафический
ареал и КС
О М е Е
С 10 14 10
18 31 43 31 18
С 10 31 52 72 52 31 10
14 43 72 100 72 43 14
f 10 31 52 72 52 31 10
18 31 43 31 18
F 10 14 10
М е Е
С 12 20 12
12 36 60 36 12
f 20 60 100 60 20
12 36 60 36 12
F 12 20 12
Изменение
эдафического
ареала в
климатопе
BN-ok
О М е Е
С 8
11 19 26 19 И
с 19 31 43 31 19
8 26 43 60 43 26 8
f 19 31 43 31 1П
11 19 26 19 и
F 8
М е Е
С
12
f 12 20 12
12
F
93
Таблица 32 (окончание)
Изменение
эдафического
ареала в
климатопе N—ок
О М е Е
с 10 14 10
18 31 43 31 18
с 10 31 52 72 52 31 10
14 43 72 100 72 43 14
f 10 31 52 72 52 31 10
18 31 43 31 18
F 10 14 10
М е Е
С 12
22 36 22
f 12 36 60 36 12
22 36 22
F 12
Изменение
эдафического
ареала
в климатопе
Nm-ok
О М е Е
С 8
11 19 26 19 11
С 19 31 43 31 19
8 26 43 60 43 26 8
f 19 31 43 31 19
11 19 26 19 11
F 8
М е Е
С 12 20 12
12 36 60 36 12
f 20 60 100 60 20
12 36 60 36 12
F 12 20 12
торых в регион не мешают географические или иного характера преграды.
Для данного местоположения потенциального флора будет включать в се-
бя все виды флоры региона, потребностям которых удовлетворяют клима-
тические и эдафические условия данного местоположения.
Для данного местообитания потенциальная флора представлена видами
флоры региона, потребностям которых удовлетворяют не только клима-
тические и эдафические, но и ценотические условия данного местообита-
ния. Учитывая сказанное выше об условиях устойчивого существования
популяций, следует считать принадлежащими потенциальной флоре экото-
па любого ранга все те видь;, КС которых по совокупности всех, учиты-
ваемых при характеристике данного экотопа, режимов отдельных факто-
ров не менее 7—8%.
При этом всегда следует помнить о законе минимума, который при-
менительно к излагаемой системе должен быть выражен так: ’’Никакое бла-
гоприятнейшее сочетание режимов других факторов не может повысить
КС среды выше КС по фактору, находящемуся в наименее благоприят-
94
Таблица 33
Климатические и эдафические ареалы и соответствующие КС некоторых доминаитов лесных
сообществ подзоны хвойно-широколиственных лесов
Климатический
ареал и КС
Festuca gigantea
о О к
15 25 15
N 15 45 75 45 15
15 45 75 45 15
m 15 25 15
Milium effusum
о О к
В 12 20 12
12 36 60 36 12
N 20 60 100 60 20
12 36 60 36 12
m 12 20 12
О. М е Е
С 10 14 10
18 31 43 31 18
f 10 31 52 72 52 31 10
14 43 72 100 72 43 14
F 10 31 52 72 52 31 10
18 31 43 31 18
Р 10 14 10
Изменение
эдафического
ареала в
климатопе
BN-ok
ном режиме”. Элементарные расчеты показывают, что для того, чтобы КС
по совокупности режимов учитываемых факторов был не ниже 8%, тре-
буется, чтобы по всем остальным факторам он был равен 100%, если у
одного фактора равен 8%, или у двух — по 28%, или у трех — по 43%, или у
четырех - по 53%, или у пяти - 60% и т. д. При этом на линейной шкале
95
I а б п и u a 33 (окончание)
О М е Е
С 8
11 19 26 19 11
f 19 31 43 31 19
8 26 43 60 43 26 8
F 19 31 43 31 19
И 19 26 19 11
Р 8
Изменение
здафического
ареала в
климатопе
Nm—ок
м е Е
12 19 12
С 19 32 19
9 27 45 27 9
f 19 32 19
12 19 И
О М е Е
11 15 11
f 11 18 25 18 11
15 25 36 25 15
F 11 18 25 12 11
11 15 11
амплитуды толерантности вид теряет соответственно 7, 27, 42, 52, 59%
охватываемых ею типов режима фактора, а на двухмерной экологической
сетке — 16, 38, 55, 64% экотопов своего экологического ареала по данной
паре факторов. Зачастую именно этим объясняется явное ’’нежелание”
вида расти на многих местообитаниях, где по каждому фактору условия
вроде бы подходящие (широко известны случаи, когда вид находили в
виде процветающей популяции при условиях, гораздо худших по любому
из данных факторов в отдельности), а в совокупности дают слишком
низкий КС, не обеспечивающий достаточной конкурентоспособности и
даже достаточной выживаемости вида в условиях данного местообита-
ния. Это же явление в 1,5—4,5 раза ограничивает состав потенциальных
флор экотопов по сравнению с тем, который можно рассчитать, исходя
из схем систем экоморф и флоры региона.
В табл. 48—50 показано отношение численности видов во флорах ос-
новных наземных эдафотопов подзоны хвойно-широколиственных ле-
сов, найденное отвлеченно теоретически исходя только из анализа схем
систем гидроморф и трофоморф (см. табл. 48), найденное теоретически
с учетом еще и реального состава эдафоморф флоры региона (см. табл. 49),
найденное эмпирически исходя из видового состава флор эдафотопов и
флоры региона. Разница в характере распределения видов по эдафото-
96
Таблица 34
Климатический и эдафический ареалы и соответствующие КС некоторых доминаитов лесных
сообществ подзоны хвойно-широколиствениых лесов
Климатический
ареал и КС
Galeobdolon luteum
О О к
В 12 12
22 31 31 22
N 12 36 60 60 36 12
17 50 83 83 50 17
m 12 36 83 83 36 12
22 31 31 22
м 12 12
Oxalis acetosella
о О к
12 12
В 22 31 31 22
12 36 60 60 36 12
N 17 50 83 83 50 17
12 36 60 60 36 12
m 22 31 31 22
12 12
О О М е Е
С 8 11 14 11 8
14 24 34 43 34 24 14
f 8 24 40 56 72 56 40 24 8
11 33 56 78 100 78 56 33 И
F 8 24 40 56 72 56 40 24 8
14 24 34 43 34 24 14
Р 8 И 14 11 8
0 О М е Е
С 8
8 14 20 26 20 14 8
£ 14 24 34 43 34 24 14
20 33 47 60 47 33 20
F 14 24 34 43 34 24 14
8 14 20 26 20 14 8
Р 8
пам между табл. 48 и табл. 49 и 50 легко объясняется обычностью или
редкостью эдафотопов определенной ориентации в климатических усло-
виях региона. Характер распределения видов по эдафотопам на табл. 49 и
50 довольно близок и относительно близки даже числовые величины в
них, хотя одна из них исходит из теоретических посылок, а другая по-
лучена эмпирически.
Таким образом, исходя из флоры Московской области, можно видеть,
что количество видов в потенциальных флорах колеблется от 50 с неболь-
7.Зак.448 97
Таблица 34 (окончание)
Изменение
эдафического
ареала в
климатопе N-ok
О М е Е
С 8
11 19 26 19 11
с 19 31 43 31 19
8 26 43 60 43 26 8
f 19 31 43 31 19
11 19 26 19 11
F 8
О О М е Е
С 9 12 9
12 20 28 36 28 20 12
f 20 33 46 60 46 33 20
9 27 46 65 83 65 46 27 9
F 20 33 46 60 46 33 20
12 20 28 36 28 20 12
Р 9 12 9
Изменение
эдафического
ареала в
климатопе Nm—ок
О М е Е
С 8 12 8
15 26 36 26 15
С 8 26 43 60 43 26 8
12 36 60 83 60 36 12
f 8 26 43 60 43 26 8
15 26 36 26 15
F 8 12 8
О О М е Е
С 8
8 14 20 26 20 14 8
f 14 24 34 43 34 24 14
20 33 47 60 47 33 20
F 14 24 34 43 34 24 14
8 14 20 26 20 14 8
Р 8
шим видов в наименее благоприятных эдафотопах до более 600 видов в
наиболее благоприятных условиях. Если учесть, что количество видов в
фитоценозах в соответствующих условиях колеблется в пределах от 8—
12 до 50—60, то становится ясно, сколь велика роль конкуретного отбора,
когда даже в худших условиях в фитоценозах остается один вид из че-
тырех-пяти, а в лучших — из десяти—двенадцати.
Однако, кроме того, значительная часть из этих 50—600 видов встреча-
ется изредка и единично, тогда как массовые виды, встречающиеся на
соответствующих местообитаниях относительно постоянно и входящие
в число Доминантов различных стадий развития растительного покрова,
составляют от. 1/4 до 1/2 общего числа видов потенциальной флоры эко-
топа. В табл. 51 приведено в процентном выраженйи отношение числа
видов доминантного ядра потенциальной флоры к числу видов флоры
региона для основных эдафотопов в Московской области.
Исходя из приводимого в Определителе растений Московской области
числа видов ее флоры в 1300 с небольшим, можно видеть, что в наихудших
эдафических условиях, например на наиболее бедных верховых болотах,
число потенциальных номинантов флоры эдафотопа, равное здесь 10—12
видам, приблизительно соответствует или лишь незначительно превосхо-
дит число видов в фитоценозах, занимающих такие местообитания. В то
же время в благоприятных эдафических условиях число таких видов пре-
восходит число видов в фитоценозах в 4—5 раз.
Такое соотношение даже числа потенциальных номинантов с числом ви-
98
Таблица 35
Климатические и эдафические ареалы и соответствующие КС некоторых доминантов лесных
сообществ подзоны хвойно-широколиственных лесов
Orobus vernus
Климатический
ареал и КС
0 О к
10 17 10
В 10 30 50 30 10
17 50 83 50 17
N 17 50 83 50 17
10 30 50 30 10
ш 10 17 10
Puhnonaria obscura
О к
13 13
В 13 38 38 13
21 63 63 21
N 21 63 63 21
13 38 38 13
m 13 13
Эдафический
ареал и КС
О М е Е
С 9 14 20 14 9
8 26 43 60 43 26 8
f 14 43 72 100 72 43 14
8 26 43 60 43 26 8
F 9 14 20 14 9
О М е Е
С 10 14 10
18 31 43 31 18
С 10 31 52 72 52 31 10
14 43 72 100 72 43 14
f 10 31 52 72 52 31 10
18 31 43 31 18
F 10 14 10
Изменение
эдафического
ареала в
климатопе
BN-ok
О м е Е
С 10
13 22 30 22 13
f 22 36 50 36 22
13 22 30 22 13
F 10
0 М е Е
С 9
И 20 27 20 11
С 20 33 45 33 20
9 27 45 63 45 27 9
f 20 33 45 33 20
11 20 27 20 11
F 9
99
Таблица 35(окончание)
Изменение
эдафического
ареала в
климатопе
N-ok
О М е Е
С 12 17 12
22 36 50 36 22
f 12 36 60 83 60 36 12
22 36 60 36 22
F 12 17 12
О М е Е
С 9
11 20 27 20 И
С 20 33 45 33 20
9 27 45 63 45 27 9
f 20 33 45 33 20
11 20 2” 20 11
F 9
Изменение
эдафического
ареала в
климатопе
Nm-ok
О м е Е
С 10
13 22 30 22 13
f 22 36 50 36 22
13 22 30 22 13
F 10
О М е Е
12 16 12
С 12 20 27 20 12
16 27 38 27 16
f 12 20 27 20 12
12 16 12
дов в фитоценозах дает основание полагать (и это в более чем 90% случаев
подтверждается фактическим материалом), что фитоценозы, как прави-
ло, состоят из обычных, массовых видов, способных уже в незначительно
изменившихся условиях дать вспышку численности и войти в число до-
минантов. Даже в экстремальных экотопах, в условиях ослабленной кон-
куренции фитоценоз, как правило, составлен видами, входящими в число
потенциальных доминантов экотопа.
В различных фитоценозах, находящихся в экстремальных или нормаль-
ных условиях по тому или иному фактору, в комфортных или диском-
фортных, близких коренным или явно производных, относительно моно-
тонных или с выраженной синузиальностью, по-видимому, всегда, отмеча-
естя еще одна интересная закономерность — КС любого массового вида,
любого доминанта, во всяком случае, не ниже, а, как правило, значительно
выше КК данного экотопа, если и тот и другой взяты по совокупности
всех основных прямодействующих факторов. Табл. 52, составленная на
основе анализа 108 произвольно выбранных описаний, главным образом
лесных, но также еще луговых и болотных, фитоценозов, показывает,
что КС массовых, или доминирующих, видов может быть то выше, то ниже
КК экотопа (хотя чаще все же выше), когда дело касается режимов от-
дельных факторов или совокупности режимов нескольких из них, но
всегда выше КК экотопа по совокупности режимов всех пяти основных
100
Таблица 36
Климатические и эдафические ареалы и соответствующие КС некоторых доминантов лесных
сообществ в иодэоне хвойночшфоколиственных лесов
Paris quadrifolia
Климат
ческий
ареал и КС
Эдафический
ареал и КС
О М е Е
С 9 14 20 14 9
8 26 43 60 43 26 8
f 14 43 72 100 72 43 14
8 26 43 60 43 26 8
F 9 14 20 14 9
0 О М е Е
С 9
9 15 21 27 21 15 9
С 15 25 35 45 35 25 15
21 36 50 64 50„ 36 21
f 9 27 46 64 82 64 46 27 9
11 33 56 78 100 78 56 33 11
F 9 27 46 64 82 64 46 27 9
21 36 50 64 50 36 21
Р 15 25 35 45 35 25 15
9 15’ 21 27 21 15 9
Р 9
прямодействующих факторов: зонального режима тепла, режима конти-
нентальности климата, водного режима почв, солевого режима почв и
светового режима в основных синузиях фитоценоза. (Оговорка о све-
товом режиме в основных синузиях фитоценоза необходима по той при-
чине, что говорить об общем световом режиме фитоценозов, обладающих
развитым ярусным строением, неправомерно, так как в них участки,
отличающиеся друг от друга по крайней на ступень, в принятой нами гра-
дации чередуются в сложном переплетении как в вертикальном, так еще
более в горизонтальном направлении.)
Представления о потенциальных флорах и еще более о потенциальных
доминантах экотопов особенно ценны тем, что дают возможность с го-
раздо большей степенью достоверности производить оценку условий среды
и устойчивости наблюдаемого сообщества еще в поле по хорошо заметным
101
Таблица 36 (окончание)
Изменение
эдафического
ареала в
климатопе BN-ok
О м е Е
С 10
13 22 30 22 13
f 22 36 50 36 22
13 22 30 22 13
F 10
О М е Е
8 11 14 11 8
С 8 13 18 23 18 13 8
11 18 25 32 25 18 11
f 14 23 32 41 32 23 14
17 28 39 50 39 28 17
F 14 23 32 41 32 23 14
11 18 25 32 25 18 И
Р 8 13 18 23 18 13 8
8 11 14 11 8
Изменение
эдафического
ареала в
климатопе N—ок
О м е Е
С 10
13 22 30 22 13
f 22 36 50 36 22
13 22 30 22 13
F 10
О М е Е
8 11 14 И 8
С 8 13 18 23 18 13 8
11 18 25 32 25 18 11
f 14 23 32 41 32 23 14
17 28 39 50 39 28 17
F 14 23 32 41 32 23 14
11 18 25 32 25 18 11
Р 8 13 18 23 18 13 8
8 11 14 11 8
Изменение
эдафического
ареала в
климатопе Nm—ок
О М е Е
С
8 13 18 13 8
f 13 22 30 22 13
8 13 18 13 8
F
О М е Е
8
С 8 И 14 11 8
11 15 19 15 11
f 8 14 19 25 19 14 8
10 17 23 30 23 17 10
F 8 14 19 25 19 14 8
11 15 19 15 11
Р 8 11 14 11 8
8
102
Таблица 37
Климатические и эдафические ареалы и соответствующие КС некоторых доминанте
лесных сообществ подзоны хвойно-широколиственных лесов
Picea abies
Климатический ареал и КС 0 0 k
10 17 10
В 10 30 50 30 10
17 50 83 50 17
N 17 50 83 50 17
10 30 50 30 10
ш 10 17 10
Эдафический ареал и КС 0 0 M e E
с 8 11 14 11 8
14 24 34 43 34 24 14
f 8 24 40 56 72 56 40 24 8
11 33 56 78 100 78 56 33 1)
F 8 24 40 56 72 56 40 24 8
14 24 34 43 34 24 14
Р 8 11 14 11 8
Изменение эдафического ареала в климатопе BN—ок
0 0 M e E
12 17 22 17 12
f 12 20 28 36 28 20 12
17 28 39 50 39 28 17
F 12 20 28 36 28 20 12
12 17 22 17 12
Изменение эдафического ареала в климатопе N-ok
0 0 M e E
12 17 22 17 12
f 12 20 28 36 28 20 12
17 29 39 50 39 29 17
F 12 20 28 36 28 20 12
12 17 22 17 12
Изменение эдафического ареала в климатопе Nm-ok
0 0 M e. E
10 13 10
f 12 17 22 17 12
10 17 23 30 23 17 10
F 12 17 22 17 12
10 13 10
103
Таблица 38
Климатические и эдафические ареалы и соответствующие КС некоторых домииаитов
лесных сообществ подзоны хвойно-широколиственных лесов
Picea obovata
климатический ареал и КС О к К
а 8 14 14 8
8 25 42 42 25 8
в 14 42 69 69 42 14
14 42 69 69 42 14
N 8 25 42 42 25 8
8 14 14 8
Эдафический ареал и ТСС О О М е Е
8 11 14 11 8
С 14 24 34 43 34 24 14
8 24 40 56 72 56 40 24 8
f И 33 56 78 100 78 56 33 11
8 24 40 56 72 56 40 24 8
F 14 24 34 43 34 24 14
8 И 14 11 8
Изменение эдафического ареала в климатопе BN-ok 0 О М е Е
8 10 8
С 10 17 23 30 23 17 10
17 28 39 50 39 28 17
f 8 23 39 54 69 54 39 23 8
17 28 39 50 39 28 17
F 10 17 23 30 23 17 10
8 10 8
Изменение эдафического ареала в климатопе N-ok О О М е Е
С 10 14 18 14 10
10 17 24 30 24 17 10
f 14 24 33 42 33 24 14
10 17 24 30 24 17 10
F 10 14 18 14 10
Изменение эдафического ареала в климатопе Nm—ок О О М е
8 10 8
f 8 11 14 11 8
8 10 8
104
Таблица 39
Климатические и эдафические ареалы и соответствующие КС некоторых домюшгтов
лесных сообществ подзоны хвойно-широколиственных лесов
Климатический
ареал и КС
Pinus sylvestris
о О к К
а 10 14 10
18 31 43 31 18
. в 10 31 52 72 52 31 10
14 43 72 100 72 43 14
N 10 31 52 72 52 31 10
18 31 43 31 18
m 10 14 10
Эдафический
ареал и КС
О О М е Е
8
С 8 13 18 23 18 13 8
13 21 30 38 30 21 13
С 18 30 42 54 42 30 18
8 23 39 54 69 54 39 23 8
f 9 28 48 66 85 66 48 28 9
11 33 56 78 100 78 56 33 11
F 9 28 48 66 85 66 48 28 9
8 23 39 54 69 54 39 23 8
Р 18 30 42 54 42 30 18
13 21 30 38 30 21 13
Р 8 13 18 23 18 13 8
8
105
Таблица 39(окончание)
Климатоп BN-ok является климатическим оптимумом сосны и потому эдафический ареал
в нем остается тот же, что приведен выше, то есть максимальный потенциально-возможный,
так как КС этого климатопа - 100%
Изменение эдафического
ареала в климатопе N—ок
О О М е Е
С 9 13 17 13 9
9 15 22 27 22 15 9
с 13 22 30 ? 30 22 13
17 28 39 50 39 28 17
f 20 35 48 61 48 35 20
8 24 40 56 72 56 40 24 8
F 20 35 48 61 48 35 20
17 28 39 50 39 28 17
Р 13 22 30 39 30 22 13
9 15 22 27 22 15 9
Р 9 13 17 13 9
Изменение
эдафического ареала
в климатопе Nm-ok
О О М е Е
С 8 10 8
9 13 16 13 9
С 8 13 18 23 18 13 8
10 17 23 30 23 17 10
f 12 21 28 37 28 21 12
14 24 34 43 34 24 14
F 12 21 28 37 28 21 12
10 17 23 30 23 17 10
Р 8 13 18 23 18 13 8
9 13 16 13 9
Р 8 10 8
106
Таблица 40
Климатмеские и эдафические ареалы и соответствующие КС некоторых домннаитов лесных
сообществ подзоны хвойно-широколиственных лесов
Климати
ческий
ареал и КС
Quercus robur
О о к
10 17 17 10
N 10 30 50 50 30 10
17 50 83 83 50 17
т 10 30 50 50 30 10
10 17 17 10
Tilia cordata
о О к
В 10 17 17 10
10 30 50 50 30 10
N 17 50 83 83 50 17
10 30 50 50 30 10
т 10 17 17 10
Эдафический
ареал и КС
М е Е g
С 10 10
13 21 29 29 21 13
с 21 35 49 49 35 21
9 30 48 69 69 48 30 9
f 12 38 62 88 88 62 38 12
9 30 48 69 69 48 30 9
F 21 35 49 49 35 21
13 21 29 29 21 13
Р 10 10
О • М е Е
С 9
12 19 27 19 12
'С 19 32 45 32 19
9 27 46 64 46 27 9
f 11 35 59 82 59 35 11
14 43 72 100 72 43 14
F И 35 59 82 59 35 11
9 27 46 64 46 27 9
Р 19 32 45 32 19
12 19 27 19 12
Р 9
Изменение
эдафического
ареала в
климатопе
BN-ok
М е Е
С 8 8
8 12 12 8
f 11 15 15 11
8 12 12 8
F 8 8
О М е Е
10 14 10
С 10 16 23 16 10
14 23 32 23 14
f 18 30 41 30 18
22 36 50 36 22
F 18 30 41 30 18
14 23 32 23 14
Р 10 16 23’ 16 10
10 14 10
107
Таблица 40 (окончание)
Изменение эдафи* М е Е g 0 M e E
ческого ареала в 11 <1-5 15 И 10 16 22 16 10
климатопе N—ок с 11 18 25 25 18 11 С 16 27 37 27 16
15 24 35 35 24 15 22 38 53 38 22
f 19 31 44 44 31 19 f 9 29 49 68 49 29 9
15 24 35 35 24 15 12 36 60 83 60 36 12
F 11 18 25 25 18 11 F 9 29 49 68 49 29 9
11 15 15 11 22 38 53 38 ,22
P 16 27 37 27 16
10 16 22 16 10
Изменение эдафи- ческого ареала в климатопе Nm-ok
М е Е g 0 M e E
С 8 8
И 17 23 23 17 11 10 14 10
С 17 29 41 41 29 17 c 10 16 23 16 10
25 40 57 57 40 25 14 23 32 23 14
f 10 31 51 73 73 51 31 1С f 18 30 41 30 18
) .. 25 40 57 57 40 25 22 36 50 36 22
F 17 29 41 41 29 17 p 18 30 41 30 18
И 17 23 23 17 11 14 23 32 23 14
Р 8 8 p 10 16 23 16 10
10 14 10
визуально признакам: сочетанию и распределению массовых и хорошо за-
метных видов. С другой стороны, знание состава потенциальных флор и
потенциальных доминантов экотопов помогает избежать одной из рас-
пространеннейших ошибок, не опирающихся на систему экологического
анализа эмпирически создавшихся взглядов фитоценологов и биогеоце-
нологов — искусственного разделения подобного и искусственного соеди-
нения разнородного.
Резюмируя сказанное, следует еще раз подчеркнуть, что для проведения
качественного и количественного экологического анализа фитоценозов
в их динамической связи со средой, без чего уже немыслимо поднятие
геоботанических исследований и даже обследований на новый, отвечаю-
щий общему состоянию современной науки уровень, необходимо так или
108
Таблица 41
Климатические и эдафические ареалы и соответствующие КС некоторых цеминантов лесных
сообществ подзоны ХВОЙНО-Широколиствеииых лесов
Vaccinium myrtillus
Климати-
ческий
ареал и КС
о О к К
А 10
13 22 30 22 13
а 22 36 50 36 22
10 30 50 70 50 30 10
в 13 39 65 90 65 39 13
13 39 65 90 65 39 13
N 10 30 50 70 50 30 10
22 36 50 36 22
m 13 22 30 22 13
10
Vaccinium vitis— idaea
о О к К
А 8 11 8
14 24 33 24 14
а 8 24 40 56 40 24 8
11 34 56 78 56 34 11
в 14 43 72 100 72 43 14
11 34 56 78 56 34 11
N 8 24 40 56 40 24 8
14 24 33 24 14
m 8 11 8
Эдафи ческий
ареал и КС
О О М е
10
с 13 22 30 22 13
22 36 50 36 22
г 30 50 70 50 30 10
13 39 65 90 65 39 13
F 13 39 65 90 65 39 13
10 30 50 70 50 30 10
Р 22 36 ,5® |36 tt
13 22 30 '22' 13
Р 10
О О М е
10 10
с 13 21 29 29 21 13
21 35 49 49 35 21
f 9 30 48 69 69 48 30 9
12 38 62 88 88 62 38 12
F 9 30 48 69 69 48 30 9
21 35 49 49 35 21
Р 13 21 29 21 13
10 10
иначе иметь в виду систему экологических характеристик видов, основ-
ные понятия которой автор пытался изложить и обсудить в настоящей
главе. Такими понятиями, согласно изложенному, являются следующие.
1. Экологическая амплитуда (амплитуда толерантности) вида — отрезок
шкалы режимов фактора, в пределах которого возможно само существо-
вание вида.
2. Экологический ареал — ограниченное совокупностью экологических
амплитуд по всем основным или нескольким избранным факторам жиз-
109
Таблица 41 (окончание)
Изменение
эдафического
ареала в
климатопе
BN-ok
О О М е
9
С 12 20 27 20 12
20 32 45 32 20
f 9 27 45 63 45 27 9
12 35 59 81 59 35 12
F 12 35 59 81 59 35 12
9 27 45 63 45 27 9
Р 20 32 45 32 20
12 20 27 20 12
Р 9
О О М е
8 8
С 10 16 23 23 16 10
16 27 38 38 27 16
f 23 37 54 54 37 23
9 30 48 69 69 48 30 9
F 23 37 54 54 37 23
16 27 38 38 27 16
Р 10 16 23 23 16 10
8 8
Изменение эдафического ареала в климатопе N-ok О О М е
С 9 15 21 15 9
15 25 35 25 15
f 21 35 44 35 21
9 27 45 63 45 27 9
F 9 27 45 63 45 27 9
21 35 44 35 21
Р 15 25 35 25 15
9 15 21 15 9
О О М е
С 12 16 16 12
12 20 27 27 20 12
f 17 27 39 39 27 17
21 35 49 49 35 21
F 17 27 39 39 27 17
12 20 27 27 20 12
Р 12 16 16 12
Изменение
эдафического
ареала в
климатопе
Nm—ок
О О М е
с 11 15 11
11 18 25 18 11
f 15 25 35 25 15
20 33 45 33 20
F 20 33 45 33 20
15 25 35 25 15
Р 11 18 25 18 11
11 15 11
О О м е
С 10 10
11 16 16 12
f 10 16 23 23 16 10
13 20 29 29 20 13
F 10 16 23 23 16 10
12 16 16 12
Р 10 10
110
Таблица 42
Пример бонитировки основных климатических н эдафических режимов с позиций
сосны лесной и дуба черешчатого
Фактор и тип режима Оценка качества данного типа режима для сосны для дуба
Субарктический Аркто-бореальный Бореальный Суббореальный Эунеморальный Термонеморальный Криотермальный Субтермальный Зональный режим тепла Малоудовлетворительный - Удовлетворительный - Хороший - Лучший Малоблагоприятный Хороший Среднеблагоприятный Удовлетворительый Благоприятный Малоудовлетворительный Среднеблагоприятный - Малоблагоприятный
Эуокеанический Мезоокеанический Субокеаническнй Полуконтинентальный Субконтинентал ьиый Мезоконтииентальный Ультраконтинеитальный Режим континентальности Малоудовлетворительный Малоблагоприятный Удовлетворительный Среднеблагоприятный Хороший Благоприятный Лучший Среднеблагоприятный Хороший Малоблагоприятный удовлетворительный - Малоудовлетв орительный
Свеже степной Влажно-степной Сублесолуговой Сухолесолуговой Свежелесолуговой Влажно-лесолуговой Сыровато-лесолуговой Сыро-лесолуговой Мокро-лесолуговой Болотно-лесолуговой Субболотный Болотный Водно бол отный Водный режим почв Малоудовлетворительный - Удовлетворительный Малоудовлетворительный Весьма удовлетворительиыйУдовлетворительный Средний Средний Относительно хороший Хороший Хороший Лучший Лучший Хороший Хороший Средний Относительно хороший Удовлетворительный Средний Малоудовлетворительный Весьма удовлетворительный - Удовлетворительный — Малоудовлетворительный -
Олиготрофный Су болиготрофный Семиолиготрофный Субмезотрофный Мезотрофный Пермезотрофный Семиэвтрофиый Субэвтрофный Эвтрофный Пертрофный Галоэвтрофный Солевой режим почв Малоудовлетворительный - Удовлетворительный - Средний - Хороший Малоудовлетворительный Лучпшй Удовлетворительный Хороший Хороший Средний Лучший Удовлетворительный Лучший Малоудовлетворительный Хороший - Удовлетворительный - Малоудовлетворительный
111
Таблица 43
Средние бонитеты различных хоротопов в полу континентальном климате
различных широтных зон с позиций сосны лесной
Зона лесотундры
Зона северной тайги
О О M e E
С
с Va
Va Va Va
f Va Va Va
Va Va Va Va Va
F Va Va Va
Va Va Va
Р Va
Р
0 0 M e E
C Va Va Va
Va Va V Va Va
c Va Va V V V Va Va
Va V V IV V V Va
f Va V V IV V V Va
Va V IV III IV V Va
F Va V V IV V V Va
Va V V IV V V Va
P Va Va V V V Va Va
- Va Va V Va Va
P Va Va Va
Зона средней тайги
Зона южной тайги
0 О M e E
C Va Va V Va Va
Va V V V V V Va
c Va V IV IV IV V Va
V V IV III IV V V
f V IV Ш II Ш IV V
Va V IV III I Ш IV V Va
F V IV HI П Ш IV V
V V IV HI IV V V
P Va V IV IV IV V Va
Va V V V V V Va
P Va Va V Va Va
0 0 M e E
Va
C Va Va V V V Va Va
Va V IV IV IV V Va
c V IV IV III IV IV V
Va V IV III П DI IV V Va
f Va V III II I II III V Va
Va IV in I la I Ш IV Va
F Va V III II I II III V Va
Va V IV III II III IV V Va
P V IV IV III IV IV V
Va V IV IV IV V Va
P Va Va V V V Va Va
Va
ценное поле вида может быть представлено на двухфакторных или поли-
факторных схемах или сериях схем, а также на картографическом ма-
териале, где в силу сложности сочетаний и взаимодействия режимов раз-
личных факторов в географической среде ареалы приобретают широко
известные из хорологической литературы формы: сплошные различных
конфигураций, дизъюнктивные различного вида, а в сущности при доста-
112
Таблица 43 (окончание)
Зона хвойио-широколисгвенных лесов Зона широколиственных лесов и лесостепи
0 0 M e E
C Va Va V Va Va
Va V V V V V Va
c Va V IV IV IV V Va
V V IV III IV V Va
f V IV III П in IV V
Va V IV III I III IV V Va
F V IV П1 П П1 IV V
V V IV III IV V V
P Va V IV IV IV V Va
Va V V V V V Va
P Va Va V Va Va
0 0 M e E
С Va Va Va
Va Va V Va Va
с Va Va V V V Va Va
Va V V IV V V Va
f Va V V IV V V Va
Va V IV III IV V Va
F Va V V IV V V Va
Va V V IV V V Va
Р Va Va V V V Va Va
Va Va V Va Va
Р Va Va Va
Таблица 44
Средние бонитеты разлшных хоротопов в полукоитинеитальиом климате
разных широтных зон с позедий ели европейской
Зона южной тайги
Зона хвойно-широколиственных лесов
0 0 M e E
c
Va V V V Va
f Va V V IV V V Va
V V IV in IV V V
F Va V V IV V V Va
Va V V V Va
О 0 M e E
c
Va V V V Va
f Va V V IV V V Va
Va V IV III IV V Va
F Va V V IV V V Va
Va V V V Va
8. Зак. 448
113
Таблица 44 (окончание)
Зона широколиственных лесов и лесостепи
О О M e E
С
Va Va Va
f Va V V V Va
Va V V IV V V Va
F Va V V у Va
Va Va Va
Таблица 45
Средине бонитеты различных хоротопов в полукоитиненталыюм климате разных
широтных эон с познтий ели сибирской
Зона лесотундры Зона северной тайги
0 0 M e E
c
Va Va Va
f Va Va Va Va Va
Va Va Va
F
0 О M e E
c Va V V V Va
Va V V IV V V Va
f Va V IV IV IV V Va
Va V V IV V V Va
F Va V V V Va
Зона средней тайги Зона южной тайги
0 0 M e E
Va Va Va
c Va V V IV V V Va
V V IV III IV V V
f Va V IV III В HI IV V Va
V V IV IB IV V V
F Va V V IV V V Va
< Va Va Va
О О M e E
Va Va Va
c Va V V IV V V Va
V V IV III IV V V
f Va ; V IV III II III IV Va
,v У IV in IV V V
:F !V» I V V IV iV iv Va
Va Va Va
Зона хвойно-широколиственных лесов Зона широколиственных лесов и лесостеп
0 0 ; M * e E
c Va V V V Va
Va V V IV V V Va
f Va V IV IV IV V Va
Va V V IV V V Va
F Va V V V Va
•0 0 M e E
c
Va Va Va
f Va Va Va Va Va
Va Va Va
F
114
Таблица 46
Средние бонитеты различных хоротопов в полуконтииентальном климате
разных широтных зон с позиций дуба черешчатого
Зона южной тайги
О м е E
С
с Va Va
Va Va Va Va
f Va V V Va
Va Va Va Va
F Va Va
Зона широколиственных лесов и лесостепи Зона степей
Зона хвойно-широколиственных лесов
0 M e E
C Va V V Va
Va V V V V Va
c V V IV IV V V
V IV III III IV V
f V V IV IV V V
Va V V V V Va
F Va V V Va
0 M e E ?
C Va Va
Va V V V V Va
c V IV IV IV IV V
V IV III III IV V
f Va IV III I I III IV Va
V IV III III IV V
F V IV IV IV IV V
Va V V V V Va
P Va Va
0 M e E
C
Va V V Va
c Va V V V V Va
V V IV IV V V
f V IV III III IV V
V V IV IV V V
F Va V V V V Va
Va V V Va
P
точной детализации всегда мозаичные с различной плотностью мозаики
в разных частях.
3. Экоморфа — объединение видов с одинаковыми в данной системе
отсчета экологическими амплитудами по одному или нескольким факто-
рам (различного порядка частные экоморфы: термоморфы, омброморфы,
гидроморфы, трофоморфы, гелиоморфы, а также климаморфы, эдафо-
морфы, ценоморфы) или по всем основным факторам (общие экоморфы).
4. Экологические свиты — объединения экоморф с тождественно рас-
положенными медианами разновеликих экологических амплитуд (также
могут быть частными и общими).
5. Экологические группы (экогруппы) — объединения экоморф с не
всегда тождественным положением медиан экологических амплитуд, хотя
и близким, но зато с большим совпадением самих амплитуд, когда их
перекрывающиеся части значительно больше неперекрывающихся.
6. КС (коэффициент удовлетворительности условий среды экотопа
115
Таблица 47
Средние бонитеты различных хоротопов в полукоитинеитальиом климате
разных широтных зон с позиций липы сердцевидной
Зона средней тайги
О м e E
С Va
Va Va Va
f Va Va Va
Va Va Va
F Va Va Va
Va Va Va
Р Va
Зона южной тайги
0 M e E
Va Va Va
c Va V V V Va
Va V IV V Va
f V IV IV IV V
V IV III IV V
F V IV IV IV V
Va V IV V Va
P Va V V V Va
Va Va Va
Зона хвойно-широколиственных лесов
0 M e E
Va V V V Va
c V V IV V V
V IV in IV V
f Va IV III II III IV Va
Va IV II I II IV Va
F Va IV III II III IV Va
V IV HI IV V
P V V IV V V
Va V V V Va
Зона широколиственных лесов и лесостепи
0 M e E
Va Va Va
c Va V V V Va
Va V IV V Va
f V IV IV IV V
V IV III IV V
F V IV IV IV V
Va V IV V Va
P Va V V V Va
Va Va Va
для данного вида) — процентное выражение относительной пригодаости
экотопа для существования популяций вида в зависимости от положения
данного экотопа на шкалах режимов отдельных факторов.
/CCmed ~ (2х — 1)/аг где® — количество типов режима данного5фактора,
охватываемых амплитудой вида, ах — расстояние данного типа режима от
ближайшего края амплитуды (его порядковый номер от этого края).
7. Потенциальная флора экотопа — совокупность видов флоры региона,
КС которых в данном экотопе не меньше 7—8%.
8. Потенциальные доминанты экотопа (доминантное ядро потенциальной
флоры экотопа) — виды потенциальной флоры экотопа, способные играть
роль доминантов или субдоминантой либо иметь относительно многочис-
116
Таблица 48
Процентное соотношение числа видов в различных эдафотопах неморального полу-
коитииентальиого климатопа (по отношению к общему числу видов климатопа),
рассчитанное теоретически, по схемам систем гидроморф и трофоморф
Основной тип гидрорежима почв Основной тип солевого режима почв
О ° 1 М е 1 Е
С 25 56 65 68 69
С 25 56 65 68 69
f 25 56 65 68 69
I 25 56 65 68 69
р 24 54 64 66 67
р 22 49 58 60 60
Таблица 49
Процентное соотношение числа видов в различных эдафотопах иеморальио-полу-
коитинеитальиого климатопа (по отношению к общему числу видов региона), рас-
считанное теоретически, ио с учетом состава экоморф флоры региона
Основной тип гидрорежима почв Основной тип солевого режима почв
О 1 ° М е Е
с 3 21 34 41 38
с 5 25 43 49 44
f 5 28 48 53 48
F 5 27 43 45 41
Р 4 20 32 32 28
Р 3 16 24 25 22
Таблица 50
Процентное отношение числа видов в различных эдафотопах неморальио-полукои-
тинентального климатопа в Московской области (отношение числа видов потенци-
альных флор эдафотопов к числу видов во флоре области)
Основной тип гидрорежима почв Основной тип солевого режима почв
О О М е Е
с 31 33
с 24 42 46
f 26 46 48
F 4 23 37 38 32
Р 15 23 23 19
Р 16 16 14
117
Таблица 51
Процентное отношение числа видов доминантного ядра потенциальных флор раз-
личных эдафотопов в Московской области
Основной тип гидрорежима почв Основной тип солевого режима почв
О О м е Е
С 10 15
С 8 15 22
f 8 17 23
F 1 7 13 17 15
Р 4 7 9 8
Р 4 5 5
Таблица 52
Процент случаев: КС вида > КК экотопа (режима отдельного фактора, комплекса
режимов части факторов) для 108-произвольно взятых описаний лесных, болотных
и луговых фнтоценозов
Режим отдельных факторов или экотопы Случаев КС > КК, %
Термический режим климата 99,1
Режим континентальности климата • 61,3
Климатоп в целом 95,1
Обобщенный водный режим почв 48,5
Обобщенный солевой режим почв 70,7
Эдафотоп в целом 60,9
Хоротоп в целом 91,9
Экотоп в целом при усредненном режиме затенения
в фитоценозе 86,2
Экотоп в целом при режиме затенения в синузиях
с высоким покрытием вида 100,0
ленную популяцию как в коренных, так и в производных сообществах
данного экотопа, по-видимому, обязательно должны иметь КС в данном
экотопе не ниже его КК (коэффициента общей комфортности).
В заключение следует добавить относительно потенциальных флор зко-
топов, что они по своей сущности являются парциальными флорами по
Юрцеву [Юрцев, Семкин, 1980].
118
ГЛАВА V
ИНДИКАЦИЯ ЭКОТОПОВ, КАК ИНДИКАЦИЯ
КОМПЛЕКСИРУЮЩИХСЯ В НИХ ЭКОЛОГИЧЕСКИХ РЕЖИМОВ
ОТДЕЛЬНЫХ ПРЯМОДЕЙСТВУЮЩИХ ФАКТОРОВ
Часто в очень полезных и полных ценными мыслями и выводами обзор-
ных сводках упорно провозглашаются и совершенно неверные положения.
Примером такого принципиально неверного тезиса служит заявление, что
’’связь растительности и среды значительно менее прочная, чем это казалось
на этапе типического отбора материала, и потому возможности оценки
среды по растительности ограниченны”. Далее в том же духе заявляется,
что ”по данным растительности среда оценивается в небольшом числе
градаций, причем последние не имеют равного интервала, а в соответствии
с природой биологических закономерностей часто характеризуются лога-
рифмически возрастающим интервалом. Все это дает основания полагать,
что геоботаническая индикация имеет практическое значение в ограни-
ченном числе случаев”. Как дополнительный довод в пользу такого заклю-
чения авторы ссылаются на мнение Кюхлера, который ’’предостерегает про-
тив увлечения принципом адекватности границ растительности и почв и
приводит примеры индифферентности характера растительности характеру
почв в условиях тропиков” [Миркин, Розенберг, 1978, с. 91]. Относитель-
но несовпадения требований растительных общностей к почвам с класси-
фикационными воззрениями почвоведов, вероятно, не стоит много гово-
рить. Во всяком случае, в настоящее время вопрос об экологизации под-
хода к почвам не только поставлен, но уже есть и у нас и за рубежом ис-
следования и методики такого рода [Чертов, 1981]. Что же касается пред-
ставлений о нечеткости, неопределенности связи растительности с условия-
ми среды, то единственный их источник в нечеткости и расплывчатости на-
ших представлений о требованиях растений как к среде в целом, так и к
режимам отдельных факторов, с одной стороны и в незкологичности на-
ших оценок параметров и состояний самой абиотической среды с другой.
Дело даже не в уточнении и детализации наших знаний о требованиях
видов к режимам тех или иных факторов, хотя и в этом отношении наши
познания оставляют желать лучшего. Решение загадки характера взаимо-
связи растительных объектов (на уровне таксонов, популяций или син-
таксонов) с отдельными элементами среды или со средой в целом зави-
сит от совершенствования самой системы экологического анализа, от ее
непрерывного совершенствования, ибо всякая догматизированная схема
или программа очень скоро перестает удовлетворять и исследователей и
практиков.
Ключ к решению лежит в расшифровке системы отношений друг к другу
экологических характеристик, и самих растительных объектов, и условий
их существования, а не в разбиении видов на такие отдельности или тако-
го сужения радиуса применимости индикационной схемы, чтобы макси-
мально сузить и ужесточить привязку индикатора к индикату (в идеале
это значит, что каждый такой таксон или синтаксон должен сочетаться
только с каким-то одним узким интервалом характеристик среды). Однако
строгое совпадение объектов различной природы, выделяемых и подраз-
119
деляемых по не связанным друг с другом системам признаков, как пра-
вил, чаще объясняется случайным совпадением, чем какой-либо законе-,
мерностью. В этом смысле эмпирически устанавливаемые совпадения
немногого стоят, если нет системы, объясняющей их связь и позволяющей
прогнозировать встречу с такими совпадениями в аналогичной обстанов-
ке. Об этом хорошо сказал С.В. Мейен: ’’Установление совпадений, не под-
крепленных знанием более глубоких причин, видимо, не стоит называть
закономерностью, а лучше сопоставить с народными приметами” [Мейен,
1981, с. 101].
Наконец, необходимо еще и еще раз подчеркнуть, что индикация усло-
вий среды немыслима без строго экологичного подхода и к индицируе-
мым объектам, и к индикаторам, и к их причинно-следственным связям.
Конкретнее это значит, что индикаторами являются хорошо различимые
виды, подвиды, расы и т. п. не как таксономические категории, а как опре-
деленного типа зкобиоморфы. Они индуцируют не инструментально изме-
ряемые физические и химические величины, выражаемые в системе CGS
или какой-либо подобной ей, а обобщенные экологически значимые ре-
жимы прямодействующих факторов. Мы вынуждены сосредоточивать
свое внимание на индикации режимов отдельных факторов в силу не-
совершенства своих аналитических способностей, заставляющего нас по-
стоянно прибегать к мысленной изоляции исследуемого явления от всех
крмплексирующихся с ним иных явлений, тогда как растения-индикато-
ры дозволяют нам такой подход в силу того, что режимы различных фак-
торов влияют на различные процессы их жизнедеятельности.
Но так как существование организмов и популяций обеспечивает лишь
весь комплекс процессов их жизнедеятельности, то и при анализе связи
видов и популяций с режимом какого-либо из факторов необходимо
постоянно не упускать из виду, что эта связь осуществляется всегда на
фоне и во взаимодействии со всем комплексом факторов среды.
Все это давно осознано, обо всем этом много написано, но положение
остается таким, как будто, приступая к сбору и осмысливанию конкрет-
ного материала, исследователь тотчас попадает в тенета эмпиризма и опять
превращается в коллекционера фактов и их совпадений, в качестве анали-
тического инструмента располагающего лишь непознаваемой силой инту-
иции и более или менее догматизированными высказываниями автори-
тетов.
В результате выявляются закономерности без объяснения (или с откро-
венно гипотетичным объяснением) факта их наблюдения. Логическим
следствием этой необъясненности (или предположительности) становится
стремление из осторожности настолько сузить возможности экстраполя-
ции полученных выводов, что в любом, хоть сколько-нибудь удаленном,
другом пункте аналогичные исследования приходится проводить заново.
Причиной этого является то, что в экологии пока еще не разработана об-
щепризнанная система понятий, как этой имеет место в точных науках, а
существует лишь некий набор понятий, отдельные группы которых раз-
личные исследователи пытаются свести в некие частные системы, руко-
водствуясь при этом различными принципиальными положениями.
При этом в одних случаях абсолютизировались и даже догматизиро-
вались частные явления. Например, хорошая корреляция между распреде-
120
пением видов по местообитаниям узаконила представления об эдификато-
рофилах и эдификаторофобах в фитоценологии и о не существующих в
природе свитах ели, дуба и других лесообразователей, выделенных фор-
мально на основании сходства их экологических и географических ареа-
лов, тогда как истинная причина таких корреляций в соответствии эколо-
гических потребностей данных видов условиям среды, создающимся под
влияние^ эдификатора данной или близкой ей экобиоморфы. В других
случаях установление общих закономерностей затемнялось формальным
коллекционированием частных эмпирических наблюдений. Например,
общность жизненных потребностей вида по всему его ареалу затемня-
лась фиксированием отличий сопутствующей ему экологической обста-
новки в разных регионах, что в действительности объясняется измене-
ниями в комплексе экологических режимов и в наборе конкурентов, а
не в изменении потребностей вида.
Пожалуй, одним из наиболее досадных экологических ляпсусов явля-
ется упорно переписываемое из книги в книгу расползшееся по всем учеб-
никам и сводкам утверждение о частичной взаимозаменяемости факторов.
Так, утверждается, что частичное замещение одного фактора другим — ши-
роко распространенный факт. В подтверждение этого положения указы-
вается на общеизвестную связь между обеспеченностью азотом и устойчи-
востью к засухе [Миркин, Розенберг, 1978]. В другом случае утверждает-
ся, что существует частичная заменяемость основных экологических фак-
торов и вместе с тем их полная незаменимость. Основанием вывода о су-
ществовании частичной заменяемости факторов служит факт получения
сходного биологического эффекта ’’при частичной замене действия одного
фактора другим”: в данном примере замене увеличения освещенности по-
вышением концентрации СОг [Горышина, 1979].
Истинная же причина существования фактов, якобы подтверждающих
ограниченную замещаемость факторов, заключается в ~ом, что приближе-
ние к оптимальному режиму любого из факторов улучшает условия су-
ществования индивидов и популяций, повышает для них КС (коэффициент
удовлетворительности условий среды). Однако зто повышение КС улучше-
нием режима одного из факторов лимитируется состоянием режимов ос-
тальных факторов, и прежде всего фактора, находящегося в наименее
благоприятном режиме. КС вида по режиму наименее благоприятного фак-
тора является тем потолком, выше которого мы не можем поднять общий
КС, оптимизируя хоть вплоть до оптимума режимы всех остальных фак-
торов. Поэтому в каждом климате, на каждом местоположении сущест-
вует свой потолок урожайности определенной культуры, а более обоб-
щенно — свой потолок продуктивности каждого вида. Очень наглядно
это можно представить на координатных схемах, построенных на осях
интересующих нас факторов. Если мы вернемся к рассмотрению табл. 26—
41 (климатические и эдафические ареалы и соответствующие КС некото-
рых доминантов хвойно-широколиственных лесов), то сможем просле-
дить, как изменяет КС улучшение режима одного из факторов (например,
одного из эдафических факторов) при стабильном состоянии другого и
как даже оптимизация обоих эдафических режимов не может увеличить
КС экотопа выше КС данного климатопа. В то же время никогда не вред-
но лишний раз оговориться, что в природе изменение режима одного из
121
факторов в силу принципа их взаимодействия никогда не бывает изоли-
рованным, что наиболее тесно связанные между собой факторы (например,
увлажнение и солевой режим почв, тепловой и водный режимы атмосфе-
ры и т. д.) изменяются почти синхронно. В этом смысле размещение основ-
ных эдафических координат в схеме Сукачева более натуралистично, а в
сетке Погребняка — более умозрительно.
Неточно в экологическом смысле и утверждение, что ’’вид меняет отно-
шение к фактору Л в зависимости от напряженности фактора В” [Миркин,
Розенберг, 1978]. Хотя речь идет о ’’взаимодействии фитоценотических
оптимумов” (очень небрежное выражение), приводимый пример — рас-
ширение эдафического ареала типчака под влиянием выпаса — говорит о
влиянии снятия конкуренции для выявления подлинного экологического
ареала вида, но никак не об изменении его отношения к одному фактору
под влиянием другого. В принципе же потенциальная экологическая амп-
литуда вида по отношению к режимам данного фактора — та амплитуда,
которую можно наблюдать только при оптимальных режимах остальных
факторов. В естественных условиях она, как правило, сужена в той или
иной степени, поскольку КС ее режимов в сочетании с КС режимов других
факторов дает слишком низкий общий КС среды по мере удаления от
оптимума вида как режима данного фактора, так и режимов всех осталь-
ных (включая сюда и конкуренцию с другими видами, так как влияние
конкуренции практически осуществляется через ухудшение режимов
тех или иных факторов). Однако этот факт ни в коей мере не означает
изменения требований вида к режимам одного фактора под влиянием
состояния режимов других факторов. Он означает лишь ограничение по-
тенциальных возможностей удовлетворения потребностей вида при каж-
дом из пригодных для него типов режима данного фактора под влиянием
ухудшения жизненного состояния этого вида недостаточно благоприят-
ным сочетанием режимов других факторов. По-видимому, не требуется
доказывать, что это не одно и то же.
Выразительные примеры изменения эдафических амплитуд видов (ам-
плитуды по режиму влажности почв) в лесостепной и степной зоне Сибири
по сравнению с аналогичной зоной европейской части СССР приведены в
работе Цаценкина, Савченко и Дмитриевой [1978]. Весьма приближенно
к объяснению этого явления авторами можно резюмировать, что факты,
приведенные в названной работе, отражают изменение КС хоротопов (кли-
матоп X эдафотоп) для данных видов и их конкурентного окружения в
условиях более континентального климата Сибири.
Наконец, прежде чем перейти к изложению приемов фитоиндикации
условий среды, основанной на применении изложенной в предыдущей
главе системы экологических характеристик видов, нужно остановиться
еще на одном досадном примере живучести традиционных представлений.
Было высказано сомнение в точности шкалы богатства-засоления Рамен-
ского на том основании, что индицируемое по растительности почвенное
богатство на удобренном NPK типе луга в отдельные годы оказывалось
на 0,5 ступени ниже, а в другие на 0,5—1,0 ступень выше, чем на неудоб-
ренном. При этом обеспеченность азотом по шкале Элленберга колебалась
в пределах на 0,1—0,6 балла выше на удобренном варианте, чем на неудоб-
ренном [Работнов, 1979].
122
Здесь прежде всего обращает на себя внимание, что шкала Элленберга
и составлена специально для определения фитоиндикационным путем
содержания азота в почвах, тогда как шкала Раменского служит для опре-
деления общего солевого богатства, что не одно и то же. Представление о
том, что внесение минеральных удобрений имеет абсолютное положитель-
ное влияние на трофический режим, устарело, так как ведет к истощению
органического комплекса в почвах и через это к изменению физической
структуры почв, в свою очередь неблагоприятно отражающуюся на фильт-
рационных и водоудерживающих их свойствах, а кроме того, еще и за-
грязняет почвы труднорастворимыми токсичными для растений примесями
солей тяжелых металлов [Рамад, 1981]. Наконец, относительно азота
известны данные, что внесенный рано весной он очень быстро использу-
ется растениями, в связи с чем в поверхностном слое почвы возникают
условия достаточно выраженной переменности в обеспечении растений
азотом [Работнов, 1973]. Кроме того, изменение конкурентной напря-
женности как под влиянием нарушающих ход сезонной динамики соле-
вого режима почв внесений удобрений, так и в силу климатических осо-
бенностей разных лет не могло не повлиять на результаты фитоиндикации,
особенно если иметь в виду и тесную взаимосвязь солевого режима с ре-
жимом увлажнения.
Все эти соображения заставляют с осторожностью относиться к выводу
о ненадежности шкалы богатства-засоления только на основании несовпа-
дения ее показаний с традиционно ожидаемым эффектом внесения ми-
неральных удобрений не только на непосредственно следующий урожай,
но и на изменение трофического режима почв.
Взятые нами примеры из работ ведущих авторитетов современной
экологической мысли показывают, насколько живучи в экологии растений
и фитоценологии давние выводы традиционного эмпирического подхода.
Между тем в настоящее время уже невозможно надежно оринетироваться
в океане фактов без жестко логически связанной системы экологических
понятий и зависимостей. Любая схема, любая теория, включающая в себя
интуитивные построения, может быть признана лишь более или менее удоб-
ной на определенном уровне накопления фактов рабочей гипотезой. И в
этом нет ничего крамольного, так как удачные рабочие гипотезы всегда
были и будут залогом успеха любой исследовательской работы, тогда
как устоявшиеся теории являются основой практической деятельности.
’’Принимать же гипотезу за нечто окончательно установленное — нс вы-
ражение принципиальности и не научная смелость, а лишь выражение са-
моуверенности” [Мейен, 1981, с. 157].
Предлагаемая здесь рабочая гипотеза исходит от концепции Раменско-
го об экологической индивидуальности растений и континууме расти-
тельного покрова. В этой связи в фитоиндикационных построениях ис-
пользуются понятия экологических амплитуд видов, экологических аре-
алов видов, экоморф видов, экологических свит, экогрупп, экологичес-
ких режимов прямодействующих факторов, экотопов разного ранга (кли-
матопов, эдафотопов, хоротопов, ценотопов и общих экотопов, как их
совокупности), коэффициентов удовлетворительности условий среды —
КС (по режиму отдельного фактора или экотопа того или иного ранга),
потенциальных флор и потенциальных доминантов экотопов.
123
Основные понятия, из которых следуют все остальные, — экологичес-
кие режимы прямодействующих факторов и экологические амплитуды —
установлены на основании критического сопоставления шкал Раменского,
шкал. Элленберга и некоторых принципиально сходных с ними, с учетом,
по возможности, сопоставимых с ними данных по экологии видов авто-
ров других направлений, а также на основании расположения географи-
ческих ареалов относительно некоторых изолиний климатических карт.
Принимая концепцию генетически обусловленного постоянства требо-
ваний видов к режимам прямодействующих экологических факторов,
изла пемая рабочая гипотеза относит изменения экологических амплитуд
видов в различных регионах главным образом на счет изменения жизнен-
ного состояния видов при различных сочетаниях режимов прямодействую-
щих факторов. В то же время такая концепция отнюдь не отвергает до-
статочно широкой распространенности различий генотипического состава
популяций видов в разных регионах и даже на разных местоположениях.
Смысл кажущегося противоречия этих двух подходов заключается в том,
что такое различие генотипического состава не может утверждаться апри-
ори, и потому боле осторожным с фитоиндикационной точки зрения будет
ориентироваться на общность требований видов к условиям среды по все-
му кх ареалу, внося поправки лишь на изменение жизненного состояния
вида под влиянием различных КС экотопов в разных регионах.
Вся система фитоиндикации условий среды (в том числе и лесорасти-
тельных условий) основана на сопоставлении экологических свойств
(потребностей) растений с экологически значимыми качествами среды,
в первую очередь с режимами прямодействующих факторов.
Предлагаемая методика фитоиндикации лесорастительных условий
и условий среды вообще предназначена в основном для стадийно-синхрон-
ной индикации, т. е. индикации состояния экотопа на настоящее время,
но при использовании дополнительно понятий: потенциальная флора,
потенциальные доминанты, КС и КК экотопа, ее данные могут служить
и для целей прогнозной индикаций и ретроиндикации.
Исходными для большинства предлагаемых приемов фитоиндикации
условий среды были техника Раменского и техника Элленберга.
В принципиальном отношении различаются: индикация по присутствию
и индикация по отсутствию. Первая (индикация по присутствию) наиболее
всем знакома и применима в нормальных (не экстремальных) экотопах.
Вторая (индикация по отсутствию) означает заключение о принадлежности
режима фактора к экстремальному или близкому ему типу на основании
отсутствия на данной территории видов, принадлежащих экоморфам опре-
деленного типа. Индикация по отсутствию применима исключительно или
в экстремальных, или в близких к ним по режиму того или иного фактора
экотопах. Так, мы не можем методами индикации по присутствию опреде-
лить, относится ли данный экотоп по солевому режиму к олиготрофному,
суболиготрофному или семиолиготрофному типу, потому что видов, ко-
торые указывали бы нам, что данный тип режима не богаче олиготрофно-
го и суболиготрофного, нет, а видов, которые бы указывали, что данный
тип солевого режима не богаче семиолиготрофного, практически нет.
С другой стороны, есть достаточно видов, которые могут указывать,
что данное местообитание имеет солевой режим почв не беднее, чем мезо-
124
трофный, субмезотрофный и семиолиготрофный. Не логично ли заключить,
что отсутствие первых и вторых (например, принадлежащих экоморфам
Me, +*Ме, ME, + ME и др.) свидетельствует о солевом режиме семнолиго-
трофном гликофильном, а отсутствие третьих (например, принадлежащих
экоморфам оМ, ое, оЕ, + оМ, + ое и т. п.) — о суболиготрофном и олиго-
трофном солевом режиме? В то же время индикация по присутствию в та-
ких местообитаниях может показать лишь, что солевой режим здесь не
богаче субмезотрофного, а то и мезотрофного (нередки и случаи, когда
в таких местообитаниях присутствуют только виды с трофическими ампли-
тудами лишь типа Ое и ОЕ). Тот же порядок рассуждений можно приме-
нить и в отношении других прямодействующих факторов.
Что же касается индикации условий среды по сообществам, то здесь
главная трудность пока заключается в том, что выделение и отграничение
сообществ друг от друга вс^ еще не вышло из стадии относительного рав-
новесия дискуссионных мнений. Поэтому фитоценоиндикации в своих
разработках мы предпочитали не затрагивать.
Переходя к характеристике приемов индикации по присутствию, сле-
дует вспомнить, что виды-индикаторы подразделяются на пассивные (обыч-
ные индикаторы настоящего состояния среды) и активные (виды с сильны-
ми средоизменяющими способностями) индикаторы [Викторов, 1971],
а также на моноиндикаторы (индицируют данный тип режима фактора в
одиночку) и кониндикаторы (индицируют его лишь в сочетании друг с
другом, в силу того что лишь в нем могут быть найдены вместе) [Цыганов,
1975,1976].
Используя изложенную выше систему экологических характеристик
видов, можно предложить следующие несколько методов фитоиндика-
ции режимов прямодействующих факторов.
I. Метод определения режима фактора по видам-кониндикаторам. Прием
этот аналогичен до некоторой степени методу ограничения ступеней в тех-
нике Раменского. Сущность его заключается в том, что одни из видов
своим присутствием указывают на максимальную границу режима данного
фактора в данном местообитании, а другие — на минимальную. Так, соче-
тание в одном сообществе майника, рамишии и василистника водосборо
листного, с одной стороны, а коротконожки и овсяницы гигантской — с
другой, указывает, что ценоклимат данного сообщества представляет собой
основной вариант эунеморального (севернонеморального) климата, так
как первые три вида указывают, что он не может быть теплее, а вторые
два — что он не может быть холоднее по терморежиму (Мытищинский
ЛИХ, Яузский ЛП, кв. 54, 3, дубняк лещиновый злаково-зеленчуковый).
Другой пример: сочетание в одном сообществе колокольчика персиколист-
ного и лютика кашубского указывает на тип режима увлажнения — пере-
ходный между свежелесным и влажно-лесным, так как амплитуда по
этому фактору у колокольчика — Cf, а у лютика + fF1, (колокольчик ука-
зывает не влажнее влажно-лесного, а лютик — не суше свежелесного)
(Клязьминский ЛИХ, Лобненский ЛП, кв. 2, ельник снытево-кисличный).
Еще пример: ряд видов сообщества указывают на режим почвенного увлаж-
1 Знак ”+” при символе экоморфы значит, что амплитуда захватывает переходный
тип режима.
125
нения не влажнее сыро-лесного (сныть, живучка, осока волосистая, май-
ник и др.), а осока плевельная на тип режима — не суше влажно-лесного и
лютик кашубский - не суше свежелесного. Таким образом, этим методом
в данном примере индицируется не определенный тип режима увлажнения,
а его возможный диапазон. Такие случаи и в отношении режима этого
фактора, и в отношении режимов других факторов отнюдь не редки, а,
пожалуй, даже более часты, чем случаи возможности точной индикации
таким методом (Клязьминский ЛПХ, Лобненский ЛП, кв. 16, ельник
крупнопапоротниково-кисличный).
Приведенные примеры показывают, с одной стороны, малую трудо-
емкость при некотором навыке индикации режимов факторов этим ме-
тодом, а с другой стороны, ограниченность его точности во многих очень
обычных ситуациях. Дело в том, что виды-кониндикаторы встречаются
лишь при малоблагоприятных для обоих индикационных подгрупп усло-
виях по данному фактору, к чему они, естественно, не стремятся. По этой
же причине режим каждого из факторов индицируют, как правило, разные
виды (сочетания двух и более неблагоприятных режимов дают слишком
низкий КС экотопа для данного вида, слишком понижаются его жизнен-
ность и конкурентоспособность). Кроме того, этот фитоиндикационный
прием действительно доступен лишь специалистам с квалификацией выше
средней даже для геоботаников, не говоря уже об агрономах, лесоводах
и землеустроителях, потому что требует хорошего знания флоры, хотя бы
данного региона.
П. Метод определения режима фактора по преобладанию видов соответ-
ствующей экологической свиты. Этот метод напоминает метод засечек в
технике Раменского, только там определенные обилия видов индицируют
определенные интервалы ступеней шкалы фактора и затем по сопостав-
лению указанных разными видами интервалов выбирается определенная
ступень, а здесь тип режима определяется сопоставлением числа видов в
различных свитах. В сущности, этот прием гораздо проще метода засечек,
ибо его принцип: есть такой-то индикатор — индикат определенный ( в дан-
ном случае: преобладает по числу видов такая-то свита - тип режима
тот же).
Метод дает однозначный ответ в том случае, если одна из свит явно
преобладает по числу видов, а число видов в других свитах постепенно
убывает по мере удаления от нее. Такие случаи не редкость, однако столь
же обычны случаи, когда трудно сказать, которая из соседних, а порой и
не соседних свит преобладет. В первом случае (относительно равновес-
ны по числу видов соседние свиты) можно предполагать или промежуточ-
ный тип режима, или его сукцессионную смену, а во втором (относитель-
но равновесны по числу видов удаленные друг от друга свиты) вероят-
нее предположить сильно переменный режим с частными разногодичными
флюктуациями. Например, больше всего видов в суббореальной свите —
6 (ель, сосна, белокрыльник, вейник седоватый, вахта, черника), за ней
следует мезобореальная свита — 5 (береза пушистая, сфагнум головчатый,
осока черная, кизляк кистецветный, брусника), а в следующей по числу
видов свите их лишь 2 (пушица многоколосковая и плевроций) — это
эунеморальная свита.
Таким образом, принимая во внимание положение объекта в эунемо-
126
ральной зоне, мы можем заключить, что ценоклимат данного местооби-
тания — суббореоидный (экстраэональный, более близкий суббореаль-
ному, чем северонеморальному). Далее, больше всего видов в болотно-
лесолуговой свите — 5 (пушица многоколосковая, вейник седоватый,
сабельник, вахта, кизляк кистецветный), затем идет мокро-лесолуговая —
4 (белокрыльник, калужница, сфагнум головчатый, клюква), третья по
числу видов — 3 — сыро-лесолуговая (береза пушистая, осока черная,
черника). Видов-кониндикаторов здесь практически нет: их показания
указывают интервал от влажно- до болотно-лесолугового типа режима.
Принимая во внимание, что, кроме названных свит, представлены еще
влажно-лесолуговая и сыровато-лесолуговая свиты (3 и 2 вида), резонно
предположить, что режим местообитания все же суше болотно-лугового,
скорее всего мокро-лесолуговой, но не устойчивый (Мытищинский ЛИХ,
Алексеевский ЛП, кв. 20, сосняк вейниковосфагновый). Таким образом,
как видно из примеров, этот прием тоже не всегда дает четкий ответ и
может служить скорее дополнением к предыдущему, хотя и в этом случае
возможно порой лишь предположительное решение задачи. Кроме того, и
этот метод требует четкого знания флоры, как и метод кониндикаторов.
Ш. Метод определения режима фактора нахождением среднего балла.
Аналогичный метод применяется последователями Элленберга и отлича-
ется от предлагаемого лишь номенклатурой и способом оценки экологи-
ческих характеристик видов. В основном это сводится к тому, что Эллен-
берг дает видам по большей части однозначные балловые оценки, тогда
как предлагаемая система исходит из амплитуд толерантности и в ней
баллы соответствуют порядковым номерам частных экологических свит
в полных шкалах режимов этих факторов, которые одновременно явля-
ются и порядковыми номерами соответствующих свитам режимов (кста-
ти, баллы Элленберга тоже олицетворяют собой порядковые номера ре-
жимов в шкале фактора). Сущность метода заключается в суммировании
показаний всех видов и выведения из их суммы среднеарифметического.
Показанием вида служит произведение его балла на определенный коэф-
фициент, характеризующий его значимость в растительном покрове данно-
го участка. Западноевропейские экологи выводят этот коэффициент из
балловой оценки обилия-покрытия шкалы Браун-Бланке: 5 = 5, 4 = 4,
3 = 3, 2 = 2, 1 = 1, + и г = 0,1. Такая система оценки значимости вида в прин-
ципе может быть приспособлена и к другим шкалам оценки обилия-по-
крытия (но только не обилия отдельно!), даже к оценкам, основанным на
переадресованной обилию-покрытию шкале Друде, которую в последнее
время столь определенно осудили за субъективность, как будто шкала
Браун-Бланке гарантирует объективность (наш опыт показал, что есть
лица, совершенно неспособные оценить обилие-покрытие вида, если он
не встречается на каждом шагу, отчего весь список, за исключением мак-
симум четырех-пяти видов, характрезиуют значком + в шкале Браун-Блан-
ке или — sol. в шкале Друде). Определение типа режима фактора этим при-
емом может быть выражено формулой
__к i + ^2X3 + ... +А: хи . Sfcx
к I +к2 + - +кп
где х - балл типа режима фактора; xt, х2, ...,х„ — баллы свит, к которым
127
принадлежат виды: fc>, к2, .... кп— коэффициенты значимости видов;
Skx - сумма показаний видов; — сумма значимостей видов.
В случае достаточно дифференцированного по баллам обилия-покрытия
списка видов описания возможно упрощенное определение режимов факто-
ров этим методом, при котором используются лишь показания массовых
видов, т.. е. только тех видов, коэффициент значимости которых > 1.
Этот прием делает такой принцип индикации несколько более доступным,
потому что массовые виды знают и специалисты средней квалификации.
Насколько отличаются результаты фитоиндикации по среднему баллу при
использовании всего списка видов или только массовых видов, видно на
основании табл. 53.
Как можно видеть из табл. 53, расхождение между результатами инди-
кации по показаниям всего состава и показаниями одних массовых видов
не столь уж велики: обычно не превышают 0,5 балла, лишь в отдельных
случаях, главным образом там, где имеют место сильные антропогенные
нарушения растительного покрова и условий его существования, разница
может иногда значительно превышать 0,5 балла. Обращает на себя внима-
ние и более дифференцированная оценка режима, чем это допускают оба
предыдущих метода, хо.я предел точности, очевидно, касается лишь деся-
тых долей, а сотые доли создают только иллюзию точности и полезны как
детерминанты десятых долей. Все же даже возможность индицировать тип
режима до десятых долей балла означает возможность различать более
100 вариантов зональных режимов тепла, более 50 вариантов режимов
континентальности, более 200 вариантов режима увлажнения почв, более
150 вариантов солевого режима почв и более 20 вариантов режима зате-
нения, даже при самой примитивной шкале режимов этого фактора. В то
же время, принимая за основу индикации лишь массовые виды, нельзя
претендовать на большую точность, чем 0,5 балла. Таким образом, и этот
прием индикации дает лучшие результаты при условии наличия специалис-
тов достаточно высокого класса. В то же время его сокращенный вариант,
доступный специалистам среднего уровня, дает результаты, относительно
более дифференцированные и более независимые от случайностей перерас-
пределения видов по территории, чем первые два метода.
Однако, как и первые два метода, и этот прием фИтоиндикации по
присутствию непригоден в экстремальных и близких к ним типах режимов
факторов. Это связано с общими закономерностями распределения по
градиентам среды и эволюцией экологических амплитуд видов.
Анализ состава экоморф флоры подзоны хвойно-широколиственных ле-
сов, а также сопоставление его с составом экоморф флор некоторых дру-
гих регионов, позволяют утверждать, что наиболее распространены эко-
морфы с амплитудами толерантности по отношению к режимам отдельных
факторов, охватывающими 5—6 основных типов режима (это соответствует
11—13 типам режима, включая переходные), чуть реже встречаются эко-
морфы с охватом 3—4 основных типов режима (5—7, включая промежу-
точные) , экоморфы, амплитуды которых охватывают только 2 основных
типа режима (3, включая переходный между ними), редки и приурочены
почти исключительно к наиболее благоприятным (общекомфортным)
экотопам, тогда как стенотопы, узкоприспособленные к одному типу
режима, практически не встречаются, по крайней мере вне тропиков.
128
Таблица 53
Различия в балловой оценке режимов основных прямодействующих факторов при
учете показаний всех видов и только массовых видов
Балловая оценка типов режима
Тип леса и его местона- хождение тепловой режим климато- па континен- тальность водный режим почв солевой режим почв. режим затенения
Ольхо-березняк 5,87 3,80 15,51 5,78 2,67
дернистоосоково- таволговый. Мы- тищинский ЛПХ, Лосиноостровский ЛП, кв. 4,4. 5,85 3,42 15,11 5,77 2,46
Лило-дубняк ле- 7,52 3,46 12,91 6,54 3,71
щиновый снытево- волосистоосоковый Москворецкий ЛПХ, Подушкинский ЛП, кв. 31 7,60 3,44 12,84 6,60 3,36
Ельник рябинно- 6,53 3,41 13,22 5,77 3,21
бузинный зелен- чуковый, руде- рально-дигрес- сивный. Клязь- минский ЛПХ, Хлебниковский ЛП, кв. 96 7,19 3,33 12,57 5,77 3,24
Примечание. Числитель — по всем видам, знаменатель — по массовым видам.
Исключение могут составлять лишь экоморфы, выделенные на основе
очень слабо разработанной шкалы фактора, например гелиоморфы в трех-
балльной шкале. Поэтому в крайних условиях, если ни не являются одно-
временно комфортными (тропический тип терморежима, световой режим
открытых пространств), не всегда можно встретить даже экоморфу с ам-
плитудой в три основных типа режима, не говоря уже о более узкоампли-
тудных. Ясно, что в олиготрофных, например, или суболиготрофных усло-
виях можно встретить лишь немногие виды, оптимум которых по солево-
му режиму оценивается баллом 3, а у большинства он будет равен 4, а то
и 5, исключая тем самым возможность объективной индикации режима
по присутствующим видам. Очевидно, можно сформулировать основной
закон фитоиндикации следующим образом: индикация по присутствую-
щим видам тем лучше удается, чем ближе данный тип режима к общебла-
гоприятному (общекомфортному), чем дальше данный тип режима от
общеблагоприятного, тем больше возрастает значение индикации по от-
9. Зак.448
12»
сутствию видов определенных экоморф, так что в экстремальных и бли- t
жайших к ним одном-двух типах режима последний тип индикации стано-
вится единственно возможным.
Все изложенное выше позволяет прийти к выводу, что фитоиндикация
возможна везде, где есть растительный покров. Случаи одновидовых за-
рослей относительно редки, и, как правило, даже в них при более внима-
тельном поиске обнаруживается примесь хотя бы единичных экземпляров
других видов, если не сосудистых растений, то хотя бы мохообразных.
Ограничение другого рода связано с антропогенными причинами и заклю-
чается в том, что все посеянное и посаженное естественно должно быть
исключено из анализа, хотя естественная гибель, а также состояние таких
искусственных популяций и могут иметь индикационное значение.
Вместе с тем индикация с выходом в экологическое прогнозирование,
равно как и в раскрытие основных особенностей прошлого развития рас-
тительного покрова и экологической обстановки, посильна лишь специ-
алистам высшего класса, хорошо разбирающимся не только в принципах
и методах экологического анализа растительного покрова и непосредст-
венно влияющих на него факторов среды, но также ив динамике косвенных
явлений, влияющих на формирование режимов прямодействующих фак-
торов.
Связь растительности и среды значительно более прочная, чем это может
казаться при отсутствии единной системы экологических характеристик
растительных, объектов и экотопов.
ГЛАВА VI
ФИГОИНДИКАЦИОННАЯ ЭКОЛОГИЧЕСКАЯ ТАБЛИЦА
В фитоиндикационной экологической таблице приведены частные зко-
морфы 2129 видов сосудистых растений, выраженные через их амплитуды
толерантности по отношению к режимам 10 факторов: общему терморе-
жиму климата (Тт), континентальности климата (Кп), влажности климата
(От), морозности климата (Ct), увлажнению почвы (Hd), обобщенному
солевому режиму почв (Тг), кислотности почв (Rc), богатству почв азо-
том (Nt), переменности увлажнения почв (fH) и режиму затенения (Lc).
Подробнее об основных типах режимов этих факторов, экоморфах и эко-
логических свитах сказано во II и IV главах, Здесь же необходимо в виде
краткой справки привести шкалы этих факторов, экологические свиты
и определение усредненного балла амплитуды толерантности (табл. 54).
Последнее необходимо при фитоиндикации режимов методом нахожде-
ния среднего балла.
Буквенные обозначения символизируют крайние основные режимы
амплитуды толерантности по данному фактору. Знак + означает, что с
этой стороны амплитуда заканчивается режимом, промежуточным
между обозначенным буквой и предшествующим ему или следующим за
ним на шкале фактора.
130
Таблица 54
Шкалы факторов
Символ Тип режима Экологическая свита Балл
1. Термоклиматичес» сая шкала
А Арктический Полярная 1
4- Промежуточный между Айа Мезоарктическая 2
а Субарктический Субарктическая 3
+ Промежуточный между а и В Арктобореальная 4
в Бореальный Эубореальная 5
+ Промежуточный между В и Ь Мезобореапьная б
в Суббореальный Суббореальная 7
+ Промежуточный между в и N Бореонеморальная 8
N Неморальный Эунеморальная 9
+ Промежуточный между Num Термонеморальная 10
m Субсредиземноморский Субсредиземноморская 11
+ Промежуточный между т и М Мезосредиземноморская 12
м Средиземноморский Эу средиземноморская 13
+ Промежуточный между М и t Субтропическая 14
t Тропический Тропическая 15
+ Промежуточный между t н Т Субэкваториальная 16
т Экваториальный Экваториальная 17
2. Шкала континентальности климата (Кп)
Е Экстраокеан ический Экстраокеаническая 1
+ Промежуточный между Е и О Океаническая 1-я 2
О Океанический Океаническая 2-я 3
+ Промежуточный между Оно Субокеаннческая
о Морской Морская 5
+ Промежуточный между о и m Приморская 6
m Субматернковый Субматериковая 7
+ Промежуточный между ти М Материковая 1-я 8
М Материковый Материковая 2-я 9
+ Промежуточный между Мик Полуконтин витальная 10
к Субконтинентальный Субконтинентальная И
+ Промежуточный между к и К Мезоконтинентальная 12
К Континентальный Континентальная 1-я 13
+ Промежуточный между К и U Континентальная 2-я 14
и Ультраконтинентальный Ультраконтинентальная 15
3. Омброклиматическая шкала аридности-гумидности (Ош)
А Экстр ааридный Экстрааридная 1
+ Промежуточный между А и В Пераридная 2
В Эу аридный Эуаридная 3
+ Промежуточный между В и С Мёзоаридная 1-я 4
С Мезоаридный Мезоаридная 2-я 5
+ Промежуточный между С и D Субарндная 1-я 6
D Субаридный Субаридная 2-я 7
+ Промежуточный между D и Е Семиаридная 8
Е Субгумидный Субгумидная 1-я 9
+ Промежуточный между Е и F Субгумидная 2-я 10
F Гумидный Мезогумидная 11
131
Таблица 54 (продолжение)
Символ Тип режима Экологическая свита Балл
3. Омброклиматическая шкала арндности-гумидности (От)
+ G + Н Промежуточный между F н G Пергу мидный Промежуточный между G и Н Гипергумидный Эугумидная Пергумидная 1 -я Пергумидная 2-я Гипергумидная 12 13 14 15
К 4. Криоклиматическая шкала (Сг) Очень суровых зим Гиперкриотермиая 1-я 1
+ Промежуточный между К н L Гиперкриотермиая 2-я 2
L Суровых зим Перкриотермная 1-я 3
+ Промежуточный между L и М Перкриотермная 2-я 4
М Довольно суровых зим Крнотермиая 1-я 5
+ Промежуточный между М и N Криотермная 2-я 6
N Умеренных зим Субкриотермная 1-я 7
+ Промежуточный между N и О Субкриотермная 2-я 8
О Мягких зим Гемикриотермная 1-я 9
+ Промежуточный между О и Р Гемикриотермная 2-я 10
Р Теплых зим Акриотермиая 11
+ Промежуточный между Р н Q Субтермофильная 1-я 12
Q Очень теплых'зим Субтермофильная 2-я 13
+ Промежуточный между Q и R Термофильная 1-я 14
R Невыраженных зим Термофильная 2-я 15
5. Шкала увлажнения почв (Hd)
D Пустынный Сухопустынная 1
+ Промежуточный между Dh d Среднепустынная 2
d Полупустынный Полупустынная 3
+ Промежуточный между d и S Пустынно-степная 4
S Сухостепной Субстепная 5
+ Промежуточный между s н S Сухостепная 6
S Среднестепной Среднестепная 7
+ Промежуточный между S и С Свежестепная 8
С Лугово-степной Влажно-степная 9
+ Промежуточный между С и с Сублесолуговая 10
с Сухолесолуговой Сухолесолуговая . 11
+ Промежуточный между с н f Свежелесолуговая 12
f Влажно-лесолуговой Влажно-лесолуговая 13
+ Промежуточный между f и F Сыровато-лесолуговая 14
F Сыро-лесолуговой Сыро-лесолуговая 15
+ Промежуточный между F и р Мокро-лесолуговая 16
Р Болотно-лесолуговой Болотно-лесолуговая 17
+ Промежуточный между р и Р Субболотная 18
Р Болотный Болотная 19
+ Промежуточный между Р н а Водно-болотная 20
а Прибрежноводный: Прибрежноводная 21
+ Промежуточный между а и А Мелководная 22
А Водный Водная 23
132
Таблица 54 (продолжение)
Символ Тип режима Экологическая свита Балл
6. Шкала солевого ре жима почв (Тг)
О Особо бедных почв Г ликоолнготрофная 1
+ Промежуточный между Ойо Гликосуболиготрофная 2
0 Бедных почв Гликоссмиолиготрофная 3
+ Промежуточный между о и М Г ликосу бмезотрофная 4
м Небогатых почв Гликомезотрофная 5
+ Промежуточный между Мне Гликопермезотрофная 6
с Довольно богатых почв Гликосемиэвтрофная 7
+ Промежуточный между е и Е Глнкосубэвтрофная 8
Е Богатых почв Гликоэвтрофная 9
+ Промежуточный между Е и Д Псртрофная 10
g Слабозасоленных почв Галоэвтрофиая 11
+ Промежуточный между g и h Галосубэвтрофная 12
h Среднезасолеиных почв Галосемиэвтрофная 13
+ Промежуточный между h и Н Галопермезотрофная 14
н Снльнозасолеиных почв Г аломезотрофная 15
+ Промежуточный между Н и Р Г алосубмезотрофная 16
р Резко засоленных почв Галосемнолиготрофная 17
+ Промежуточный между Р и S Г алосуболнготрофная 18
S Злостных солончаков Г алоолнготрофная 19
7. Шкала кислотности почв (Rc)
а Очень кислых почв Гиперацидофильная 1-я 1
+ Промежуточный между а и b Гиперацидофильная 2-я 2
ь Сильно кислых почв Перацидофильная 1-я 3
+ Промежуточный между b н с Перацидофнльная 2-я 4
с Кислых почв Мезоацидофильная 1-я 5
+ Промежуточный между с н d Мсзоацидофильная 2-я 6
d Слабокислых почв Субацидофнльная 1-я 7
+ Промежуточный между due Субацидофильная 2-я 8
е Нейтральных почв Нейтрофильная 9
+ Промежуточный между е н f Субалкалифильная 1-я 10
f Слабощелочных почв Субалкалифильная 2-я 11
+ Промежуточных между f н g Мезоалкалифнльная 12
g Щелочных почв Алкалифильная 13
8. Шкала богатства почв азотом (Nt)
j Безазотных почв Анитрофильная 1
+ Промежуточный между j н к Субанитрофильная 1-я 2
к Очень бедных азотом почв Субанитрофильная 2-я 3
+ Промежуточный между к и 1 Геминнтрофильная 1-я 4
1 Бедных азотом почв Гемнннтрофильная 2-я 5
+ Промежуточный между 1 н m Субнитрофильная 1-я 6
m Достаточно обеспеченных азотом Субнитрофнльная 2-я 7
ПОЧВ
+ Промежуточная между шип Ннтрофильная 1-я 8
n Богатых азотом почв Нитрофильная 2-я 9
+ Промежуточная между п и о Ннтрофильная 3-я 10
о Избыточно богатых азотом почв Нитрофильная 4-я 11
133
Таблица 54 (окончание)
Символ Тип режима Экологическая свита Балл
р 9. Шкала переменности ув Устойчивого увлажнения лажиеиия почв (fH) Константофнльная 1-я 1
+ Промежуточный между р н q Константофнльная 2-я 2
q Относительно устойчивого увлаж- Субконстантофильная 1-я 3
+ нения Промежуточный между q н г Субконстантофильная 2-я 4
г Слабо переменного увлажнения Гемиконтрастофильная 1-я 5
+ Промежуточный между г и s Гемиконтрастофильная 2-я 6
S Умеренно переменного увлажнения Субконтрастофильная 1-я 7
+ Промежуточный между s н t Субконтрастофильная 2-я 8
t Сильно переменного увлажнения Контрастофнльная 1-я 9
+ Промежуточный между t н и Констрастофильная 2-я 10
U Резко переменного увлажнения Контрастофнльная 3-я 11
G 10. Шкала освещенности Открытых пространств —затенения (Lc) Внелесная (световая) 1
+ Промежуточный между G и g Полянная (субсветовая) 2
g Полуоткрытых пространств Кустарниковая 3
+ Промежуточный между g и М Разреженнолесная 4
м Светлых лесов Светло-лесная 5
+ Промежуточный между М н s Г устосветло-лесная 6
S Тенистьк лесов Тенисто-лесная 7
+ Промежуточный между s н S Чащобно-теневая 8
S Особо тенистых лесов Ультратеневая 9
Виды в фитоиндикационной экологической шкале расположены в алфа-
витном порядке. (Таблица 55).
Приведены по возможности экоморфы всех видов сосудистых расте-
ний, встречающихся в подзоне хвойно-широколиственных лесов Русской
равнины хотя бы и на нарушенных местообитаниях. Кроме того, приведены
экоморфы ряда видов других регионов, которые интродуцируются в
подзоне хвойно-широколиственных лесов. Главным образом это деревья,
кустарники и интересные в декоративном отношении травы.
Для экологического анализа растительного покрова и фитоиндикации
может быть полезно определить экологическую свиту или балл данного
основного или промежуточного типа режима. Это легко сделать, обра-
тившись к схемам систем частных экоморф или шкалам факторов.
По шкалам факторов балл условно оптимального типа режима и соот-
ветствующая свита находятся, как среднее из минимума и максимума
амплитуды толерантности вида. Например, экологическая формула ле-
щины: термоморфа ВМ, контрастоморфа ОМ, омброморфа ЕН, крио морфа
LO, гидроморфа SF, трофоморфа +МЕ, ацидоморфа af, нитроморфа jn,
гидроконтрастоморфа +rs, гелиоморфа +SS. В соответствии со шкалами
находим баллы: для термоморфы (5 + 13) : 2 f 9. для контрастоморфы
(3+9) :2 = 6, для омброморфы — (9+15):2 = 12, для криоморфы —
(3+9) : 2 = 6, для гидроморфы — (6+16) :2 = 11, для трофоморфы -
134
Таблица 55
Экологические амплитуды видов
Вид Факторы
Tm Kn Om Cr Hd Tr Rc | Nt fH Lc
Abies alba Mill. bm+ ok DE OP +cp+ ME af jn — gs+
balsamea Mill. BN+ mK DF LO CP OE — — — gs+
nefrolepis Maxim. +bm +kK+ +EF LO cf+ +Me — — — gs+
nordmanniana Spa ch. Nm +Mk DF +OP cF +ME eg kn — gsi-
sibirica Ldb. +Bb+ MK+ DE+ +LN +cF+ oE — — gs+
Acanthopanax sessiliflo- rum Seem. bm kU DF +MO cf+ Me - - — +gs
Acer campestre L. bm+ ok CF +OP+ CF Mg cf In — +gs+
ginnala Maxim. bN+ kK EF LO cf Me — — — Gs
mono Maxim. +bm +kU DF LP cF +Me — — — gs+
negundo L. bt +MK +DF +NQ CF ME — — — Gs
platanoides L. +bm ok+ +DE+ +NP Cp +ME af jn — gs+
pseudoplatanus L. bm+ ok+ +DF OP SF ME af ln+ — +gs
rubrum L. +bt + Mk+ DF MQ Cf+ Oe+ — — — gs
saccharinum L. +bM Mk DF MP — — — — — GM+
saccharum Marsch. +NM +Mk DF NP +cp oE — — gs+
semenovii Rgl. et Herd. NM kU BD MP Sc +eE — — +gM+
tataricum L. +Nm+ mK +CF NP CF +Mg — — — gs
tegmentosum Maxim. bN kK +E+ MN+ cf Me — — — gs+
trautvetteri Medv. Nm +Mk+ CE NO+ +fp ME — — — Gs
ucurunduense Traut. et Mey +bM MK +EG +MP+ cf+ Me — — — gs+
velutinum Boiss. Nm+ MK CE OP fF ME — — — gat-
Achillea asiatica Serg. +BN+ kU +DF KN +SF +<% — — — Gg+
cartilaginea Ldb. aN+ +mU CE KO dP MH+ - — — Gg
micrantha Willd. Nm+ MK BE+ QP de eH+ — — — Gg
millefolium L. +aM OU +BF+ KQ dF +oH ag kn qt+ GM
nobilis L. +bm+ OK CF MP df Mg+ +df jm rs+ GM
ptarmica L. am Ok +DF MP CF Mg +bf jm — Gg+
setacea W. et K. Nm Mk — — +df +eg — — — Gg
Achyrophorus macula- tus Scop. +Bm OU +DF +MP Sf Oe+ ce jl rs GM
uniflorus Bt. et Fingh. — — — — cf oe — — — GM
Acinos arvensis Dandy. +Bm+ +ok+ CF +NP+ sf ME+ +de+ jk rt Gg
Aconitumanthora L. bM OK CF MP Cf Me+ — — — Gs
excelsum Rchb. aN+ oU CE+ +LO+ CF+ oE — — — gs+
flerovii Steinb. bN mk DE NO cF eE de mn — gs
karakolicum Rapes. Nm KU CD +N+ cf +e+ — — — +gs
iasiostomum Rchb. +bm +mk +DE NO+ +F+ +Me — — — Gs+
napellus L. aM OU +CF+ LO cF + eE +de+ mn+ — GM+
soongoricum Stapf. +N+ +K+ CD MO cf +eE __ — — Gs
Acorus calamus L. BM+ oU CG KQ FA og +de+ +mn+ — GM
Acroptilon repens D.C. Nm+ mK BE +NO+ - — — — — GM
Actaea erithrocarpa Fisch. +BN +mU DF+ KN +fp Me+ — — — gs
spieata L. +Bm ok+ CF MO+ cF ME ce +mn+ — gs
A ct in id ia kolomicta Maxim. +bN+ MK+ +EF+ MP cf+ +ME+ — +gs
135
Таблица 55 (продолжение)
Вид Факторы
Tm 1 Кп 1 От 1 на Тг 1 Re Nt fH Lc
Adenophora iiliifolia bm oK CE MO+ Cf+ OE df km gS+
Bess.
tricuspidata A.D.C. —- — — — +cf+ +eg — — Gs
Adianthuni capillus- +NM OK +CF NQ +fp+ Me eg +kl+ — gs
veneris L.
pedatum L. bm MU DF+ +MO+ +fF +Me +Ms+
Adonis aestivalis L. NM OU BF MQ de eg df +kl+ GM+
sibiricus Patr. BN MU +DE LN Cf ME — — Gg+
vernalis L. bm OK CD+ +MP df Mg +de+ jm su Gg+
wolgensis Stev. bni mk +CE +NO+ +sC+ eE+ — — Gg+
Adoxa moschatellina L. +aM+ OK+ CF KQ cF ME +de+ mn+ +MS
Aegilops cylindrica Host. bM mU BE +NP+ DC+ eg+ +ce+ +lm+ — Gg
Aegopodiuni podagraria L. Bm+ oK+ CF+ MP CF OE +de+ mn+ qs Gs+
Aesculus hippo casta- +mM+ MK CE OP cp ME ce+ kn +MS
num L.
octandra Marsch. +mM+ +Mk+ DE +OP+ —
pavia L. mt +Mk DE OQ — — — —
Aethusa cynapium L. Bm+ Ok+ CF NP cF oE df +mn+ — GM+
Agrimonia eupatoria L. BM OK BF MP+ SF Mg df km st gs
pilosa Ldb. +bm+ +MU CF LP Cf ME — — gs
Agropyron intermedium bM +oU BE +NQ +sc+ +eH+ dg kn GM+
P.B.
folio ides Rochev. bN +MK+ +DE +MO +C+ eE — — Gg+
orientale Roem. et +NM +MU BD MQ Ds+ EH+ __ Gg
Schult.
pectiniforme Roem. et bm MK+ BD+ MO+ df oH — st Gg
Schult.
pungens Roem. et bN Om+ +EF CP cF Mg cf ln+ — Gg
Schult.
repens P.B. +aM+ OU +BG+ KQ +da+ oP af mn+ +ru GM
triticeum Gaertn. bM +MU BD+ MQ+ DS EP — , — — Gg
Agrostemma githago L. +BM oU BE KQ sF Mg af jn — Gg
linicola Terech. bN+ +Mk+ CE NO SF ME — — — Gg
Agrostis canina L. am Ok+ CF +NP Sa OE ad jl pr+ GM
ciavata Trin. +BN oU DF KO cF+ +eE — — — Gg
gigantea Roth. aM+ OU BF +LQ CP +oP dg In qt+ GM
stolonifera L. aM OU +CF KP+ +Ca +oP ag kn qt+ Gg
straminea G. Hartm. ab ok +E+ MO cF+ +gh +fg km qs Gg
syreistschikowii P. Sm. +bm+ OK+ CF +MP+ CP og+ — — GM
tenuis Sibth. aM OK+ CF +LP+ SF Og ad jm ps+ Gg+
Ajuga chia Schreb. NM mk CD OP de og — — — Gg
genevensis L. +bM+ OU CF +NQ df Mg df jl rs GM
pyramidalis L. +Bb+ +mk+ — — +cF+ oe ac jl — GM
reptans L. +BM OK CF +NP cF oE af In — gs
Alchemilla acutiloba +Bb+ ok +Ё+ MO cp+ Me Gg+
Opiz.
baltica G. Sam. aB+ ok+ +E+ +MN+ — — — — — —
breviloba Lindb. +b+ +M+ DE+ +N+ — — — — —
conglobata Lindb. Bb +M+ +E+ +N+ — — — — — —
136
Таблица 55 (продолжение)
Вид Факторы
Tm Kn 1 Om | Hd 1 ТГ | Rc | Nt 1 fH Lc
cymatophylia Juz. +Bb » oM+ +E+ 1 ON
filicaulis Bus. aN Om+ EF +NP cf oe ce+ Jm GM
glabra Neygent. +bN+ oM +E+ NP fP +Me ce+ kn rs gs
glabricaulis Lindb. +b+ +mk DE +NO
glaucescens Wallr. gibberubsa Lindb. +Bb +b+ +mk+ - mM++E+ NO CF Me ce jn - Gg+
glomeratus Bus. ab OM EF MO
hebescens Juz. +b+ mK DE MO
heptagona Juz. +b+ oM +E+ NO
hirsuticaulis Lindb. Bb+ ok +E+ MO Cc+ Oo-
leiophylla Juz. +b+ +M+ +E+ +N+
lindbergiana J uz. +b+ mk +E+ MO
lit wino wii Juz. +b+ +M+ +E+ +N+
micans Bus. +BN ok DE MO +cp+ Me GM
monticola Opiz. +BN ok EF MP cF +Me* Gg+
murbeckiana Bus. ab oK +DE+ MN
obtusa Bus. +b+ +mK+ +DE+ NO +fF +M+
plicata Bus. BN +mM +E+ NP
propinqua Lindb. +b+ mk +E+ NO
psiloneuta Juz. t-b+ +M+ +E+ +N+
sarmatica Juz. +BN ok DE +NO
schistophylla J uz. rt>+ +M+ +E+ +N+
semilunaris Alech. rb+ +mk +E+ NO
stellaris Juz. +Bb +Mk +E+ +N+
subcrenata Bus. +Bb mk DE MO
vulgaris L. s. str. +BN Om EF NP +cF +Ы- ce In es
vulgaris L. s.1. aN+ OK+ +DF +MP sP OE af In ps+ GM
Aldrovanda vesicu- losa L. bT OK BH +OR Fa+ oe ce jm
Alhagi pseudalhagi Desv. bm+ +kK+ BD MO+ Df +eH+ Gg Gg+
Alisma lanceolatum With. +BM+ OU BF NQ +pP +e+ ce kn -
loeselii Gorski BM oM BE+ LQ +pa
plantago-aquatica L. +BM+ OU BG KQ cA oP+ ag kn qt
wahlenbergii Holmb. Bb +mM++E+ NO __
Alliaria petiolata Cavara et Grande. +BM+ OK+ BF +NQ CF eE de +ПО - gs
Allium angulosum L. +Bm oK+ CE LO+ SP MH eg jm ru
flavescens Bess. +bm mk+ DE MO +dC+ eg Gg
inaequale Jank. Nm MK +CD+ NO+ +S+ Gg FgS
oleraceunt L. +bN+ ok+ CE NP +Cc+ ME ce kn 4
paniculatum L. bM oK+ BE NP+ <-dc+ oH eg kn
rotundum L. +bM +mK++CE (-NP+ SC eE dg kn Gg+ Gg+
schoenoprasum L. +aM oU +BE+ KQ K'P +og ce kn rt
sphaerocephalum L. bM +mK BE NQ Cf dg jm Gg+
strictum Schrad. BN mU CE KO SF +ME ce jm Ge+
ursinum L. bM Ok+ t CF -» OP+ Cf eE dg ln+ rt +Ms+
victorialis L. BM OU CF LQ Cf+ +ME +dg kn GM
137
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
Alnus barbata C.A.M. Nm+ mK CF +OP cp Me+ GM+
fruticosa Rupr. ab+ +MU +DE+ KN cp oE — — — GM+
glutinosa Gaertn. +Bm Ok+ +DF +NP fa °g cf jn qr GM+
incana Moench aN+ ok+ DE+ MO+ cP OE df jn qr Gs
viridis D.C. +Bb+ oM DE NO+ cp oe+ cd jn — Gg+
Alopecurus agrestis L. bM OU BF +OQ dF eh dg ln+ — Gg
aequalis Sobol. aM+ +OU BG KQ +fa Mh+ ag +mo — Gg
arundinaceus Poir. aM+ OU BF+ KQ sa MP — — rt Gg
geniculatus L. +BN Ok+ +DF NP +fp+ og+ de ln+ — Gg
pratensis L. aM OU BG KQ SP MH+ eg ln+ +ru GM
Althaea officinalis L. bm +MK+ +CD+ +NP +Cp oh+ cf In su Gg
Alyssum alyssoides L. +bM OU BF NP+ +sc +eE df jl — Gg
desertorum Stapf +bM+ +mU BE LQ Dc+ Mh+ — — — Gg
gmelinii Jord, et Fourr. bm mk DE NO+ +dc eh de jl — Gg
Amaranthus albus L. +NM oU BD+ MQ +sf +Eh ag ln+ — Gg
blitoides S. Wats. +mt +MK BD +OQ Cf Mg+ ag mo — Gg
hybridusL. +bt OU AF MR +cf+ Og af ln+ — Gg
lividus. L. bt mU AE MR +cf+ Mh ag mn+ — Gg+
retroflexus L. +bM+ mU BE MQ Sc eg af mo — Gg
Amelanchier canaden- bM mM DF NP+ — — —
sis Medic. rotundifolia Dum.- +NM+ oK+ BF +OQ Cf +Me+ eg km GM
Cours. spicata C. Koch BM mK CF NP+ Cf+ oE af jm GM
Ammophila arenaria +bN+ Om EF OP +cf+ Me+ de kn — Gg
Link. Amorpha fruticosa L. NM mK CE OQ Sf Mg+ GM
Amygdalus commu- +mM++kU +BD+ +OQ+ +Cf+ +Me+ cf +lm+ — GM
nis L. папа L. bm +MK+ CE MO+ de ME Gg+
Anagallis arvensis L. +BM OU BF NQ df eE af In — Gg
Anchusa ochroleuca +mM ok — — Cf +oM+ +ce+ io — Gg
M.B. officinalis L. +bm+ +mk+ +DE NP +Cc+ oe cf kn Gg
Andromeda polifolia L. AN EU DN KP SP OE ab+ jl — GM
Androsace elongata L. bm — — — sc oE ce — — Gg
filiformis Retz. ' +aN+ +mU CF KO SP oE — — — Gg+
septentrionalis L. aM OU CF KO sf oE cf kn — Gg+
Anemone altaica Fisch. ab mU DE +LN cf Me — — — GM
coerulea D.C. +b+ kK+ DE +M+ — — — — —
crinita Juz. BN kU DE LN cf Me — — — GM
dichotoma L. +Bb+ MU +DE+ KN+ cF+ ME — — GM
jenisseensis Kryl. aB+ mk+ +E+ KM cf Me — — — GM
narcissiflora L. +ab+ mU DE +MN CF oE +de+ km — GM
nemorosa L. Bm Ok DF NP Cp oE ce+ jn — GM
ranunculoides L. Bm +oK CE+ +NP+ cf ME df ln+ — GM
sylvestris L. +am+ mU CE KP sf Mg ce+ jm — GM
Anethum graveolens L. NM mk — — Cf Me ce+ kn — Gg
138
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt j fH Lc
Angelica palustris r-bm +mk+ CE +MO fp +oM-» rs Gg
Hoffm. syIvestris L. am oU DE KO+ cp oE af jn qs GM
Antennaria dioica hrM +OU +CF+ KQ Sf Oe ad jm +rs Gg
Gaertn. Anthemis arvensis L. BM Ok+ CF NQ sf ME ad In —. Gg
cotula L. bN mk — — df +Eg af kn — GM
ruthenica M.B. bm mk +DE NP+ +sf eh — — — Gg
tinctoria L. BM 0M+ CF NP+ SC oE ce km — Gg
Anthericum ramosum L. +bm+ ok+ +DE+ -ЮР+ Sf ME de km — Gg+
Anthoxathum odora- aM OU CF +MP+ SF OE cd jn rs GM
turn L. Anthriscus longirostris +NM mK BE +OP+ Cf eE GM
Bertol. silvestris Hoffm. aN+ Ok+ CE+ +NOh CF ME af mn+ rs Gs
Anthyllis polyphylla Bm +mM+ CE+ +NOh Sf oH — — rs GM
W. et. K. vulneraria L. s. str. Bb om +E+ NO+ +Sf OE cf jm — Gg
Antirrhinum oron- mM om — — +cF+ oe+ +bc+ ln+ — Gg
tium L. Apera spica-venti +Bm+ OU CF KP sf Og bd jn — Gg
Beauv. Apium graveolens L. NM OK BF OQ +fp +Me-t eg lo qs Gg
Aquilegia vulgaris L. +bm+ ok +DE+ NP+ +cf+ Me de km — Gg+
Arabidopsis thaliana +aM OK CF. MP+ cf — cd km — Gg+
Heynh. Arabis gerardii Bess. BM Ok+ CF +N+ — — — — Gg
hirsuta Scop. Bm oU +DE+ KP +Sf+ ME df jn Gg
penduia L. bN+ mU CE KO +CF eE — — — Gg
Aralia mandshurica +bm kU +EF +MN+ cf Me — — — GM+
Rupr. et Maxim. Archangelica litoralis +Bb +om +E+ NP cp Mh — — rt Gg
Agardh officinalis Hoffm. am +mk+ CE MO+ cP Mh ag mo rt GM
Arctagrostis latifolia Ab +oU DF KO cF+ oe — — Gg
Griseb. Arctium lappa L. +bM+ OU BG +MQ+ CF Mg de mo — GS
minus Bernh. +bm +mk DF NO+ cF eE df mo — Gg+
nemorosum Lej. +bm Ok CF NP fp — af mo — gs
tomentosum Mill +bm+ OU BF KPh CF eg ef mo — GM
Arctostaphylos uva- +Bb+ OU DF KO Cf+ Oe af jl — GM
ursi Spreng. Arctophila fuiva Anders. +aB +oU DE+ KO cP+ +Me+ — — — Gg
Arctous alpina Medenzu. AB oK DF KO cF OM+ af jl — GM
Arenaria koriniana bN kK CE MN dC +Eg — — — Gg
Fisch. longifolia M.B. +bm +MK+ CE MO dc+ Mg — — — Gg
saxatilis L. Bm oK CE KO dc+ Mg — — rs Gg
serpyllifolia L. +Bm+ oK BE MP Df Mh ag jn rt Gg
139
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn Om 1 Cr Hd Tr Rc Nt fH Lc
Aristida pennata Trin. +Nm+ kU BD+ +NO+ Dd +Eh+ — rs Gg
Aristolochia clemati- tis L. bm oK CE+ NP Sf ME df П1П+ — Gg
Armeniaca manshurica Skvortz. ,bN+ kK+ DF MN+ +Cf ME — — — GM
sibirica Lam. bm kU CE+ +MO Sf +ME — — — Gg+
vulgaris Lam. Nm+ kU BD+ NO+ Cf ME cf kn — GM
Armeria alpina Wilkl. Ab MK — — Cf eM+ ad jm — Gg
maritima Wilkl. AB+ OU DH KP +cf+ Og cd jm — Gg
Armoracia rusticana Gaertn. +bN+ ok +DE+ +NO+ cP ME af mo qt Gg
Arnica montana L. +bm +mM+ DE +O+ cF oe ad jl — Gg
Aronia melanocarpa Elliott bt +Mk+ DF +NQ +cf+ +Me+ — — - GM
Arrhenatherum elatius J. et C. Presl. +bm+ OK CF MQ +CF ME +de+ +ln+ — Gg+
Artemisia abrotanum L. +BM +mK+ CE +MP+ +sP MH+ eg kn — Gg
absinthium L. +BM+ OU BF +MQ dF Mg af mn+ rs GM
annua L. +Nm+ +MU BD+ +OP df eH eg mo — Gg
armeniaca Lam. bm +MK CE MO dc+ +eg — — rs+ Gg
austriaca Jacq. +BM oK CE MP+ Df+ oP eg — st+ Gg
campestris L. Bm+ OK +CF MP dF OH eg jl — Gg
cina Berg, et Poljak. Nm +K+ BC +NO+ ds+ eg — — — Gg
dracunculus L. +bm +kU +CD+ +LO+ +sf+ MH eg +ln+ +st+ Gg
glauca Pa IL bN+ kU +CD+ +MO+ +Sc+ eg — — rt Gg
latifolia Ldb. +bm +MU CE +MO +sf +eg — — rs Gg
lercheana Web. +N+ kK +CD+ MO — — — — —• Gg
maritima L. t-bm+ oM CE +OP +dF oP ag kn — Gg
monogyna W. et K. +N+ +kK CD+ NO Dp+ +EP — — — Gg
santolinifolia Turcz. bN kU CE KN+ +sf +<® — — — Gg
scoparia W. et K. +BM OU BF KP Df +eH — — — Gg
sericea Web. bN+ +MU CE KO Sf oE — «st Gg
sieversiana Wilkl. Bm +kU CD KO +Sf+ eg — — — Gg
vulgaris L. aM OU +CF KP SF og af mn+ rs+ Gg
Arum macuiatum L. bm+ OM +DF OP+ Sp Mg +de+ mn+ — gs
Aruncus vulgaris Raf. bm om+ DE +O+ cF+ ME af' mn+ +gs
Ar undo donax L. mT oU BG OR +SP Mg cf+ +ln+ — Gg+
Asarum europaeum L. +bm+ ok DE MO+ cF oE df in +MS
Asparagus caspius Hohen. Nm+ mK CD+ NP dC+ Eg — — — Gg
officinalis L. +bm OK CF MP dF MP eg +lm+ su GM
Asperugo procum- +BM bens L. oU BF +LQ cf ME df mo - GM
Asperula arvensis L. +NM +MK — - +Sf eE+ ef km — Gg+
cynanchica L. «bm+ MK CE NP FSC+ ME df jm — GM
galioides M.B. bm mK CE +NP sc+ ME+ — — — Gg
odorata L. Hbm+ Ok+ DF+ +NP CF oE af kn - +MS
tinctoria L. +bM +ok+ CE +NP Sf Mh+ eg jm rs gs
140
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt | fH | Lc
Asplenium ruta-mura- +bM OU +CF LQ Cf Me df jl GM
ria L. septentrionales Hoffm. +BM 0K+ +CF MQ Cf oe ac jl — Gg
trichomanes L. +bM OK CF +NP +cf+ Me af km — gs
Aster alpinus L. ab mk+ +DE MO sF +ME+ +de+ j' — Gg
amelloides Bess. Nm+ mk+ • — — +sc+ Mh — — rs Gg
amellus L. +bm+ Ok+ CF MP Cc ME ef jm — Gg*
novi-belgii L. bM +ОП1+ — — fF Mh ag mo — Gg
salignus Wilkl. NM oK — — fF Mh dg mo — GM
tripolium L. am oU BF LP CP oP ag ln+ st Gg+
Astragalus arenarius L. Bm OK CE+ NP Sf+ oe — — qt Gg*
austriacus L. bM +mK CE MP+ +sf +oH eg — — Gg
cicer L. bm+ Ok+ CF NP+ Sf Mh eg +ln+ — GM
danieus Retz. BN+ OU CF LP sf oh eg jl rs+ GM
falcatus Lam. bm mk CE NO+ — — — — — Gg
glycyphyllus L. BM Ok+ CF MP+ Cf ME +de+ km — GM
onobrychis L. +bM OK CE +MP+ Wf Mg df jm rt Gg
sulcatus L. +bm mK CE +MO sf eh+ — — st Gg
Astrantia major L. +bN mM DE +O+ cF Me df kn — Gs
Athyrium crenatum +Bb mU DF +MN+ +fp ME — — — gs
Rupr. distentifolium Tausch. aB +o+ +E+ NO +IT+ +oM+ cd mn gs
filix-femina Roth. aM OU DF+ LQ cP oE af In — gs
Atragene alpina L. +Bb+ +mk+ +E+ +O+ cF ME af jm — GM+
ochotensis PalL +BN mK +EF +LO +cf +Me+ — — — GM+
sibirica L. aN MU DE KN CF oE — — — GM+
Atriplex сапа C.A.Mev. bN MK CD+ MO Ds* EP — — —
hastata L. +BM +oU BE+ MQ CP piu ag mo su Gg
littoralis L. hm +oU BE+ MQ +dp+ EH+ ag mo — Gg
nit ens Schkuhr. bM +oU BE+ MQ +SF+ +EH+ eg +ln+ — Gg
patulaL. +BM OU +CF KQ Cp eE +de+ jn — Gg
bM mU BE +MQ DF EP ag In su Gg
At гора fteffarfow*» L. +Nm+ mK CD+ OP cf+ eE df mn + — GM
Avena fatua L. +BM OU +BF+ KQ de eg +df+ jn — Gg
Axyris amaranthoidcs +bN+ MU CE KN+ Cc* eE — — — Gg
L, BaBota nigra L. +bM OK+ +CF NP+ +SF °g af mn+ — Gg+
Barbarea srticta Andrz. +Bm+ +mK+ CE LP +sP+ oE +ce+ In qt Gg
vulgaris R.Br. +Bm Ok+ CF LP dF eh ag In qt Gg
Batrachium aquatile +NM Ok CF +OQ +a+ — cd In r- GM
Dum. divaricatum Schur. ab+ oK CF +LO+ Pa* — GM
foeniculaceum V.Krecz. BN+ OU CF KP +FP oh — — — GM
trichophyllum Bosch. Bm OU +DF KP +a+ — af jn — GM
Beckmannia eruciformis BM +mK BE+ MQ cP MP — — st+ Gg
Host. syzigachne Fern. am+ mU CF KO+ cP +Mg Gg
Bellis perennis L. NM OK BF +OQ CF ME af kn — GM
141
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn | Om Cr | Hd Tr Rc | Nt fH 1 Lc
Bellidiastrum michelii aB +mk cp Me df jm rs+ GM+
Gass.
Berberis amurensis +bN+ MU DF LO cf+ Me — — — GM
Rupr.
sibirica Pall. bN kU CE +MN+ Cf oe — — rs Gg+
vulgaris L. +bm+ +mk+ CE +OP+ Sf +ME df jm — GM
Bergenia crassifolia aN mU DE KM+ sf+ oe cf +lm+ — GS
Fritsch.
Bert его a incana D.C. Bm+ oK CE +MP df oH ag km +rs GM
Betonica officinalis L. +Bm+ +mk+ +DE+ MP sF oE af jm qs GM+
Betuia cajanderi Suk. +B+ KU DE KL+ +fp oM+ — — — GM
costata Trautv. +bN+ kK+ EF +MN+ cf+ +Me — — — Gs
dahurica Pall. +bN+ MU DF LO Cc+ Me+ — — — GM
ermani Cham. aN+ +mU DF+ +LO+ cf+ Me — — — GM
exilis Sukacz. +ab+ mU DF KN cP Oe — — — Gg+
fruticosa Pall. +B+ +KU DE KM cF+ oe — — — GM
gmelini Bge. +b+ kU DE +M+ Cf+ ME — — — GM
humilis Schrenk. Bb+ oK +DE +LO+ +fP+ +oe —. — pr GM
lutea Michx. bm+ mK +DF MO+ +cp+ +oE — — Gs
middendorffii Trautv. ab mU DF KN cF+ +oe —. — GM
et Mey.
nana L. +ab+ 0k+ +EF +LO+ +fa OM+ ac jl — Gg
papirifera Marsh. am+ +mK CF +LO+ CP OE af jm — GM+
pendula Roth. +Bm+ 0K+ +DF +LF CP OE af jn +qr+ GM+
platyphylia Sukacz. +Bb+ kU DF KM+ +cp+ +Me+ — — — GM+
pubescens Ehrh. aN+ OU DF KO+ +cP+ OE af jm — Gs
rotundifolia Spach. +bN kU +DE +LN +cf+ oe — — — Gg
schmidtii RgL bm MK EF MO +Cf oe —. — — Gg+
Bidens cernua L. BM OU CF +MP+ FP oh ag mo rt Gg
radiata ThuilL aM OK+ BF MQ FP — ag mn+ +rt Gg
tripartita L. +BM+ OU BG KQ ca+ oP ag mn+ +st+ Gg
Blechnum spicant Roth. +N+ Ok+ DF +OP fF OM ac jm — +MS
Blysmus compressus +BM OU BF +NQ +SP MH dg jm — GM+
Panz.
rufus Link. +BN+ OU CF +MN+ +1F+ oh+ ag km Gg+
Bolboschoenus compac- +BM oU BG KQ cP+ eh+ — — Gg
tus Drob.
maritimus Pall. +BT OU +BG+ KR +Sa+ +EH+ dg kn — Gg+
Borago officinalis L. +BM mU BE MQ +Cf +Me+ +ce+ +ln+ — Gg+
Botriochloa ischaemum +NM+ mU +BD+ +NQ de eE+ df jm — Gg+
К eng.
Botrichium lanceolatum +ab Ok EF+ +LO Cf oe ad jm — Gg+
Angstr.
lunaria Sw. +aM+ OU BF KQ cF Oe af jm . — GM
multifiduni Rupr. +Bm+ Ok+ +DF MP +cF+ Me bc+ im GM+
simplex E. Hitchc. ab OM DF MO+ fp +Me t-bc jm — Gg+
virginianum Sw. BN+ oK +E+ +LO fp+ +Me df kn — GM+
Brachypodium pinna- BM Ok BF LP+ sF oE tde+ km IS gs+
turn Beauv.
142
Таблица 55 (продолжение)
Факторы
ОПА, Tm Kn Om Cr Hd Tr Rc Nt fH Lc
sylvaticum Beauv. +bM+ OU BF+ +MQ SF ME +ce+ In — +gs+
Brassica campestris L. aM OU BF KQ df ME cf +ln+ — Gg
juncea Czern. bt oU AF LQ +cF +Mg +ce+ In — Gg
nigra Koch. +Nt oU BF NQ +cp +ME +ce+ In IS Gg
Briza media L. +bN+ Ok DF NP CF oE af jl -HJS Gg
Bromus arvensis L. +Bm+ OK +CF MP +sF ME cf +lm+ — Gs
benekenii Trim. +bm+ OK CF MP cf ME cf+ kn — gS
commutatus Schrad. +bM OU BF MQ +sF+ og +cf+ +lm+ — Gg+
erectus Huds. bm OM+ +CF OP Cf Me df jm — Gg+
hordeaceus L. +bN+ Om +EF +OP SP oE af jn — GM
inermis Leyss. am+ OU +CF LP dP MP dg kn ru GM
japonicus Thunb. +bM OU BF MQ +dc+ +eP+ eg +lm+ — Gg+
mollis L. +bM Ok+ +CF NP sF oE +cf +k> — Gg+
racemosus L. bm+ OK BF OP +Sp+ +eg cd kn — Gs
ramosus L. bm OM+ +DF OP CF ME cf In — Gs+
* riparius Rehm. bm +mk+ +CE +NO+ +sF Mh — — — GM
secalinus L. aM Ok+ +CF +NP+ SP +Me al' rs Gg+
squarrosus L. +bM OU BF MQ +dc+ eH+ eg kn — Gg
sterilis L. +NM OU BF +OQ +Sf+ +eE af kn — Gg
tectorum L. bM OU BF MQ De oh dg km st Gg
Bryonia alba L. +bm+ mK+ +CE+ NP sC Me +de+ in — +gM+
dioica Jacq. +bM mU BE NP+ cF — df In — +gM+
Bunias orientalis L. Bm +ok+ CF MP CF ME df kn rs Gg
Bupleurum aureum Fisch. BN+ mU CE LO Cf+ ME - - GM+
falcatum L. bm mk+ CE NP +Sc+ ME ef jm — Gg
multinerve D.C. +bN mU DE +MO Sf ME — — — Gg
rotundifolium L. bm+ OK CF MP +Cc+ Me ef km — Gg+
Butomus umbellatus L. BM OU BF +LQ cA oP ag mo st+ Gg+
Buxuz colchica Pojark. +niM mK CF +OP cF+ ME+ df jn — +MS
sempervirens L. NM mk CE+ OQ CF+ ME df jn — gs
Cacalia hastata L. +Bb+ +MU +DF KN +cF+ +ME — — rs gs+
Cachrys odontalgica ран. Nm MK CD+ NP Ds+ Eg - - - Gg
Cakile maritima Scop. +Bm Они BF+ +OQ fF — ag mo — Gg
Cabmagrostis arundina- cea Roth. Bm+ OK+ DF +MP+ SP Og +cf kn -HjS gs
canescens Roth. BN+ OK DE MP fa OE be kn +qs+ GM+
epigeios Roth. aM OU BF+ +LQ dP OP ag ln+ ru GM+
Гаррота Hartm. ab oU DF KM++cP +oe+ — — — Gg+
neglecta Gaertn. Mey. et Schreb. +aN+ (MI CF KO+ ca Og df jm qs Gg
obtusata Trin. Bb+ MU DF KM+ •cF Oe+ — — — gs+
purpurea Trin. +ab+ MU DF KM+ +cP oE — — — GM+
Calendub officinalis L. +mM mk+ BE OQ Cf ME cf +ln+ — GM
Calla palustris L. BN +oU +DF+ KO+ ca OH eg km qr GS
Callitriche cophocarpa Sendtn am+ oK CE LP +Pa+ Me+ bf jm — Gg+
hermaphrodita Juslen. am oU CE KP — - — — — Gg+
143
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы
Тш Кп 1 1 Сг на Тг Re Nt fH Lc
palustris L. aM OU BF KQ t-Pat- Met- af ln+ - Ggt-
Calluna vulgaris Hill. am +Ok+ CFt MP+ CP Oe ac jk +qs+ GM+
Caltha palustris L. am+ OU CF KP +fa OH ag j° +qs+ GM
Calypso bulbosa Oakes. +BN oU DE KO +fP+ oe ad jl gs
Calystegia sepium R. Br. +bM OU BF +MQ cP og +de+ mo st +gs+
Camelina linicola Schirnp. et Spenn. +Bb+ OM+ EF NP cF cd km - GM
microcarpa Andrz. t-Bm oU CE KO+ +df eg cf kn — Ggt-
Campanula altaica Ldb. bm mKt- +DE MO Cft- t-eE GM+
alpina Jack. ab mk DE NO Cd- +oe bd jl GM
bononiensis L. -t-bM mK +CE MP+ sf ME df jl rs GM
carpatica Jack. +Bb+ mk DE NO +cf+ Me — Gs
cervicaria L. BN+ OK CF MP Cf+ oe df jm rs Gs
glomerata L. +Bm+ OK+ +CF LP SF oh eg int- qs+ GM
latifolia L. BM Okt- CF MQ CF +Me+ df rant- gs
patuia L. t-Bm Okt- +DE MP CF OE +de+ km qs GM
persicifolia L. Bm Ok +DF MP Cft- oe df jm rs gs
rapunculokies L. +bm oK +CE MO+ Cf oe df km GM
rapunculus L. bm+ Ok +DE +OP cf +oM+ ce km GM
rotundifolia L. +BN+ OU +DF LP Sf OE af jl rs GM
sibirica L. +BM+ mKt- CE LO t-Sft- -t-Mh dg -t-klt- st GM
trachelium L. +BM Okt- BF MQ CF oE df mnt gs
tridentata Schreb. +Nm+ +MK CE +OP +Cf oe Gg
wolgensis P. Smim. BN kK +D+ MN +Cc+ +eE+ df kn Ggt-
Cannabis ruderalis Janisch. bm MK CE MO +sf +eg cf kn - Ggt-
Capselia bursa-pasto- ris L. +aM+ +OU AH KQ+ DF ME+ af Int- - Gg+
Caragana arborescens Lam. +bN+ t-kU CE +MN CF ME cf In - GM+
frutex Koch. bm +MK+ +CD+ MO sf ME+ df In __ Ggt-
Cardamine amara L. +BM Okt- CF MP fa ME af km pq GM+
flexuosa With. bN ok +EF+ +O+ Fp +oe+ af kn Gs
hirsuta L. t-Nt oU CG OR +cf+ +eE cf In gs
impatiens L. BM OU BF +LQ cF OE +de+ mnt- +MS
parviflora L. +bM OU BF LQ fp Met- cf kn GM
pratensis L. AM OU DF KP fa og af jn+ pr GM
Cardaria draba Desv. bM oK BE+ +MQ Df eg df km st Ggt-
Carduus acanthoides L. +BM OK+ +CF MQ sf eg af mn+ st GM
crispus L. +BM OU +BF KQ sF eh ag mo Ggt-
hamulosus Ehrh. NM mk CE 0P+ +sC+ eh — Gg+
nutans L. +BM oU BE+ MQ sf oh dg In rs Gg
uncinatus Bieb. Nmt- mk CE 0P+ dC+ Eht- — __ Gg
Carex acuta L. +am+ oK CF +MP+ fa oP+ eg km rt GM
acutiformis Ehrh. t-bM OU +CE+ +MQ ca Mh dg kn rs+ Gg
appropinquata Schm. BN oK DE L0+ +fa oE ef jm qs GM+
aquatilis Wahlb. +ab+ OU DF KO cP og — Gg
arenaria L. BN+ Omt- EF OP tcf+ 0M+ ad jl GM
aristata R. Br. BN+ Okt- DF MP ca +MH+ - qs GM
144
Факторы
Вид Tm 1 Kn I Om 1 Cr | Hd | Tr Rc Nt fH Lc
brizoides L. bN tmMt DE+ +O+ +fF+ +Me bd. jm - Gs
brunnescens Poir. +aN+ OK+ DF LP cF oe ad jm pr GS
buekii Wimm. NM ok +CE+ OQ +Fa Mg — — — Gg
buxbaumii Wahlb. ab+ oK DE+ LO ca Mg cf jm — Gg
caespitosa L. aN+ OU CF+ KP cP Og bf t-km PS Gs+
canescens L. +am+ EU CH +LP +cP Oe ad jl pr GM+
capillaris L. aN OU CF KO+ +fP oe df jl — Gg
caryophyllea La Tour. t-bm okt- CE+ MO+ sf og af jl rs GM
chordorrhiza Ehrh. +aN+ OKt- DF t-LP fP Oe ad jl — GM+
colchica J. Gay. bm+ mK CE MP +df Me bd jl — GM
contigua Hoppe. +bM OK +CF MQ+ cp og cf In — GM
davalliana Smith. bm+ Ok DF OP +fp+ Me df jl — Gg
diandra Schrenk. am EU +CH KO+ fa oE ce jm pqt- Gg
digitata L. Bm Okt- DF MP CF oE af kn —
dioica L. +aN OK DF LP fP Oe af jm qr Gg
disperma Dev. aN oK DF LO +fp+ oE ad jkt- — gs
distans L. t-bM +ok+ +DE+ MP +fF+ Mh dg jn+ rt+ Gg
disttcha Huds. t-Bm OK CF MP +fP+ +MH+ eg kn st Gg+
duriuscula C.A. Mey. +BN kU CF KM+ +sF +eE — — — GM
echinata Murr. BM Okt- CF NQ CP oe ad jl — gM+
elata Bell. t-bmt- Ok CF +OP+ fP+ Met- af km — +Ms+
elongata L. BN+ OK DF t-LP Fa oE +de+ In qr GM+
ensifolia Turcz. AB+ mU DE KN+ +fp oM+ bd jl ,Gg+
ericetorum Poll. +BN+ OK CF t-LP SF oe af jm rt gM+
flacca Schreb. am +ok+ +cF Mg df+ jm — GM
flava L. aM Ok+ +CF+ +MP fP OE df jl qr Ggt-
globuiaris L. aN oU DF KO cp OE — — — GM
heleonastes Ehrh. aN+ okt- DF +MO+ fa +ME +bc+ jl rt Ggt-
hirta L. BM Okt- CF , t-NQ SP og af kn pr gM
hostiana D.C. Bmt- Ok +DF NP+ FP oe ce jl — Ggt-
humilis Leyss. t-Nmt- +ok+ +DE+ NP +Sc+ ME df jm rs Ggt-
irrigua Sm. ab+ OU DF KO+ t-Fa OM+ +bc+ jm — gM
tasiocarpa Ehrh. am OU CF KP +fa Oet- bd jm qs GM+
leporina L. t-Bm OK CF MP CP OE ad km pr+ GM+
limosa L. am OU +DF+ KO+ fa Oe ac+ jl+ qr Ggt-
loliacea L. +BN+ OU DF KP +fP Me be km — +MS
mackenziei V. Krecz. ab Omt- +EF+ +NO+ fP +ME be jk+ rt Ggt-
melanostahya Bieb. +aB+ mU DE KM +Cp+ MH+ dg km — Gg
montana L. +bN+ Ok+ DF MO+ Cf Mg cf jm — Gg
muricata L. Bm Ok+ +DE+ NP +fP+ OM+ af In — GM+
nigra Reinch. aN+ OU +DF+ +MO+ +fP+ Oh ag jl PS [GM+
obtusata Liljebl. aNt- mU CE KO+ Sf oe — — — Ggt-
oederi Retz. aM OK CF MP+ fp oe+ cf jm — Gg
omskiana Meinsh. +Bm oK DE+ +MO+ Fa OE — — — GM
ornithopoda Willd. am +ok+ Ca OE ef jm — Gs
otrubae Podp. t-bM OK BF +NQ +fp+ ME ce — — GM
pallescens L. t-Bm oK CF t-LP cp Oe bd km PS GM+
panicea L. +Bm OK CF t-MP cP og af jm pr GM
paniculata Juslen. bm ok +DE NO+ t-cP Mg et km — GM
145
10. Зак. 448
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
pauciflora Light. aN oU DF KO +fP Oe ac jk+ — Gg
pediformis С.А.М. +BN+ oU +DE+ KO sf oE cf In rs Gs+
physodes Bieb. Nm +KU BC+ NP Ds eH — — — Gg
pilosa L. +bm ok+ +DE+ +NO+ +cF +ME cd+ kn — +MS
praecox Schreb. +Bm OU CF KP da oP ag jm ru GM
pseudocyperus L. BM+ OK+ +CF MQ fa oE ce kn — GM
rariflora Sm. +aB+ 0k+ +EF+ KO +fF+ oM+ — — — GM
remota L. +bM 0K+ BF NQ fP ME af jn — +Ms+
rhynchophysa C.A.M. aN +mU DF KO +fP oE — — — GS
riparia Cart. +bM OU BF +MP+ ca MP dg km — GM+
ro strata Stokes. am OU +CF+ KP +fa OE ad jm q« Gg+
stans Drej. AB+ OU DF KN cP oe — — — Gg
stenophylla Wahlb. bm mK DE MO Df +oH+ eg jm — Gg+
subspathacea Warmsk. AB+ ok +EF+ KN+ fP +gh+ — — Gg
supina Wahl ab oU DF KM+ +dc+ +Mh+ dg jl rt GM
sylvatica Huds. +bm Ok+ +DF +NP cF OE +de+ kn — +MS
tomentosa L. bm +oK CE+ +MO+ Cp ME ef jn st GM+
umbrosa Host. bN mM +E+ Ю+ cF OM+ cd jn — +MS
vaginata Tausch. +aN+ OK DF +LP cF OE ac+ jm rs GM+
vesicaria L. am OK CF LP +fa Og ce kn rt+ GM
vulpina L. Bm OK CF +MP ca oH ag kn rt Gg+
wiluica Meinsh. ab юи +DE+ KO cP oe+ — — qr GM
Carlina acaulis L. bm oM CE+ +OP Cf +Me af jl GM
biebersteinii Bernh. +bm Ok+ CF +MP Cf oe — — GM
vulgaris L. +bm OK BF NP Cf oE af jm — Gg+
Carpinus betulus L. bM Ok+ +DF OP+ CF oE af jn — +gs
caucasica Grossh. NM mk+ CF +OP +Cf+ +Me — — — +gS
oriental! s Mill. NM mU CE+ +OP+ sf ME df jn — Gs
Carum carvi L. +BM OU BF+ KQ Cp oh ag In qt Gg+
Cassiope ericoides D. Don. ab +kU DF KM cF oM+ — — — Gg
tetragona D. Don. AB +mU +E+ KN cF+ oe+ — — Gg
Castanea sativa Mill. NM +mK BF NQ SP ME bd+ J'n — +gS
Catabrosa aquatica Beauv. aM OU BF+ LQ CP oh eg +ln+ — GS
Cedrus deodara Loud. +mM mK CE+ OQ +cp+ oE af jn — Gs
Cenolophium fischeri Koch. +Bm oU CE+ KO Cp w — su Gg
Centaurea cyanus L. +Bt oK BF +MQ+ sf ME af j» — Gg
diffusa Lam. +NM mk+ +CD+ +OP+ +sC+ eH+ eg Gg
jacea L. +Bm+ Ok+ CF +NP+ Cp oh ag jn ps+ GM
maculosa Lam. bm ok +DE +O+ sc oE df In — Gg+
marschalliana Spreng. +Nhs mk +D+ +O+ de og — — — Gg+
micranthos Gmel. bm +Mk+ CD +N0* sc+ OE cf — — Gg
montana Baumg. ab ok DE NO +cF+ oe eg +lm+ — GM
phrygia L. +BN ok DE +MO Cf oe — — rs GM+
pseudophrygia C.A.Mey. +bm mk +DE MO SF Me cd km — GM+
ruthenica Lam. bm +MK CE MO+ dc+ eh+ — rt Gg
scabiosa L. +Bm OK +DF MP dF og df jm rs+ Gg+
146
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om 1 СГ Hd 1 ТГ 1 Rc 1 1 fH 1 Lc
sibirica L. +b+ +kK +DE MN SC eE rt Gg
trichocephala Bieb. bm +Mk+ CE NO+ df+ MH — — — Gg
Centarium erythraea Rafn. ' +bM oU BF NP Cp ME +cet jn — Gg
pulchellum Druce. +bm+ OK CF +NP+ sF +Mh+ eg kn — Gg+
Centunculus minimus L. bM Ok+ CF NP+ fP oe bd jm — Gg+
Cephalanthera damas- sonia Druce. +bm+ Om+ DF OP Cf +oM+ +de+ km - MS
longifoiia Fritsch. bM+ OU BF NQ Cf ME df jm — gs
rubra Rich. +bM OU BF +NP+ cf +Me df jm — +MS
Cephalaria uralensis Schrad. Nm +mK CE MO+ Cf oM+ +dg In - Gg
Cerastium alpinum L. AB+ Ok +EF +MO cf oe be jm Gg
arvense L. aN+ OU CF KP df oE be+ km — GM
dachuricum Fisch. Bm+ +MU +CE+ MP Cf+ ME — — — GM
glomeratum ThuilL +bM+ OK CF NQ sf+ +eE cd kn — GM
holosteoides Fries. at OU BF KR Sp OE cf kn ps GM
pauciflorum Stev. +bN+ +kU CF +LN+ CF Me+ — — — GM
semidecandrum bM OK +CF NP+ sf Mg af jn — Gg+
Cerasus austera Roem. +Nm+ mK +DE+ OP +Cf+ +ME+ — — GM+
avium Moench. NM mK+ CF NQ cF ME cf kn — Gs
collina Ley. et Court. bM oK CE +OP+ SF +ME+ — — — GM+
fruticosa Woron. bm mK CE +NP Sc+ +Mg df+ jn rs Gg+
vulgaris МШ. +bM oK CF +NP+ +Cf+ Mg cf kn — GM+
Ceratocarpus arenari- us L. bm MK+ +CD+ MO+ DC eH — — st Gg
Ceratocephaia falcata Pers. bM mU BE MQ Ds +Eh+ - - — Gg
Ceratophyilum demer- sum L. +BM OU BF KP+ PA ME df mn+ - GM+
submersum Schlecht. bm OK +CF MP PA eE df ln+ — gs
Cerinthe minor L. bM +mU +CE +NP+ Cf oE ce+ — — Gg
Ceterach officinarum D.C. NM+ +oK+ BF +OQ Sc oe df jl — GM+
Chaenorrhinum minus Lange. +bm ok +DE+ NP cf OM+ df km — Gg+
Chaerophylium aroma- ticum L. BM ok +CE+ NP+ CF oE be — — GS
bulbosum L. +bm oK CE+ +NO+ Sf ME df П1П+ Gs
hirsutum L. bm ok +DE +OP +fp Me af ln+ — GM+
prescottii D.C. am oK+ CE+ +MO+ SF eE — rt Gs
temulum L. Bm ok CE+ NP +cf+ ME af mn+ —. Gs
Chamaedaphne calyculata ab+ Moench. EU DH KO +fP Oe ad jl pq GM+
Chamaenerion angusti- folium Scop. aM+ EU + BG+ KQ Sp+ OE ad +ln+ qt GM+
latifolium Th. Fries. +aB+ oU DE KN cf +Me dg jl Gg
Chamaepericlymenum suecicum Graebn. ab+ Ok EH +MO+ +fP oe ad jl - GM+
147
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
Chelidonium majus L. +Bm+ OU CF KP+ SF +ME af mn+ GM
Chenopodium album L. +BM+ *OU +BG+ KR Dp oP ag ln+ rt Gg
aristatum L. +bN+ +kU CF +LO sc* +eh+ — — — Gg
bonus-henricus L. +bm Ok DF NP cF oe af no — Gg*
foliosum Aschers. +bM+ OU BF +MQ ds* eg cf In — Gg
glaucum L. aM OU +BF+ KQ* +SF +Eg af mo — Gg*
hybridum L. +bM OU +BF+ KQ* cF oe df mn+ — GM
polyspermum L. +BM OK+ +CF+ +LP+ SF Me af mn+ ru Gg
rubrum L. +Bm+ OU BF KP Df eh ag mo — Gg
urbicum L. +bM OU +CF LP+ DC+ Eg cf ln+ su Gg
viride L. +BN+ OU +DF+ KP +Sf+ +Eg — — Gg
Chimaphila umbellata BN oK DF LO CF oe cd jm — gs
Bart. Chondrilla juncea L. bM +oK BE+ +OQ de eg cf+ jn — Gg*
Chorispora tenelia D.C. +bM mU BE+ MQ* De* eh+ ag jn — Gg
Chosenia macrolepis am +mU +DF+ KO +cf+ +Me+ — — — Gg*
Korn. Chrysanthemum sege- BM Ok +CF +OQ cf oe cd kn GM
turn L. Chrysosplenium. alterni- +aN oU +DF KO+ cP oE +de+ km — gs
folium L. oppositifolium L. BN MK +EF +MO+ FP oE ce km GM*
Cicerbita uralensis +bN +Mk+ DE MO fF oE af mn+ — GM*
Beauv. Cichorium intybus L. BM +oU BF MQ dF oH dg kn +st+ Gg
Cicuta virosa L. am OU +CF+ KP +fa oh eg kn qr GM
Cinna latifolia Griseb. Bb+ oU +EF+ LO +fp+ eE — — — gs
Circaea alpina L. +bM OU CF LQ cP oE cd kn — +MS
coruata Reyle. +MK _lMD- • mi “ fir nF. de In — +MS
lutetiana L. +bm OK CF MO+ cF OE de ln+ — +MS
Cirsium arvense Scop. +BM Ok* CF +NP+ dF MP ag ln+ st* Gg
canum AU. bm* ok +CE+ NP cF+ +Mh+ — — st Gg
esculent um C.A.M. +bm mK+ CE +MO++CI + +MP — — rt Gg
heterophyllum HilL aN OK CE* MP cF oE +cd+ In rs* Gs
oleraceum Scop. Bm Ok DF MP cP oE df kn rs GM*
palustre Scop. Bm OK DF +MP cP oe bd jm — GM*
pannonicum Link. bm mk +DE NP «Р Mh eg — GM*
polonicum Iljin. +bN +mK CE* *NO* SF EH — — GM
rivulare AIL *bN ok № NO HP Me+ df jk+ — Gg
setosum M.B. aN* oCJ +DF KO *cF+ +eE — — —- Gg
vulgare Airy-Shaw. BM OU BF' mp+ SF Mg af mn+ — GM
Cladium mariscus Pohl. +bM OK* BF NP pa ME+ ef jm —- Gg
Ciausia aprica Tr. bN mU CE KO +Cc eE+ — — — Gg
Clematis hcxapetala PalL +bN+ +kU DF +MO Cc+ +eE — — — Gg
integrifolia L. bm* +mK+ CE MP Sc* +eh+ eg jm — Gg
orientalis L. +NM+ mU BE NQ+ dp +eh+ — — — GM
recta L. bm* mk* CE NP* Sf ME df jm — Gg*
vitalba L. +NM +oK CE OQ *cf Me +de+ ln+ — GM*
Clinopodium vulgare L. +Bm +oK+ +DF KF Sf Oe +de+ jm — GM
148
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn 1 Om Cr Hd Tr Rc Nt fH Lc
Cnidium dubiuni Thell. Bm oK CE+ MO cP Mh eg kn rt+ Gs+
Coeloglossum viride Hartm. am+ OU CF KP cP oM+ bd jm 4 Gg+
Colchicum autum- nale L. bN+ oM+ DE+ +O+ SF ME cf jn - gs
speciosum Stev. Nm +Mk+ +DE+ NP CF +ME ef kn — Gs
Comarum palustre L. AN+ OU +DF+ KP +fa +oe+ ad jl pr+ GM
Comntelina coinmunis L. BN mU DF+ +MO cf ME cf kn — GM4
Conioselfnum vaginatum Thell +BN oU +DE+ KO cf+ Me+ - - - +MS
Conium maculatum L. +BM OU BF+ MQ cp OE af mn+ — Gg+
Conringia orientalis Andrz. bM oK+ BE MP+ Cf ME ef km - Gg+
Convallaria ntajalis L. Bm+ Ok+ DF MP +Cp+ Oe+ af km qs gs
Convolvulus arvensis L. +bt OU BG +MR dF oH eg jn ru GM
persicus L. Nm +Mk+ BE +OP dc+ +Eg+ — — — GM
Corallorhiza trifida Chat el. +am+ OU +CF KP fP oe ad jn - MS
Comus mas L. NM +ok+ CE+ OP+ Sf ME df km — Gs+
Coronatia flos-cuculi A.Br. +Bm+ oK+ DE+ +MP+ cP Og af jn ps+ GM
Coronilla varia L. +bM oU BE+ MP+ sf og ef jm st Gg
Coronopus squamatus Aschers +NM Ok CE OQ fp eg cf In - Gg+
Cortusa matthioli L. aN +ok+ +DF+ MP cf oe+ ef kn Gs+
Corydalis bulbosa D.C. bm+ ok CE OP+ cF +eE df mn+ — gs
gigantea Trautv. Bb+ mK EF +MN+ cf+ Me — — — gs
intermedia Merat. +bm+ Ok+ DF NP cF +oM+ +de+ ln+ — gs
marschalliana Pers. +bm+ +mk+ CE+ NP Cc+ ME de In —. gs
pauciflora Pers. aN MU DF LN cF oM+ — — — GM+
solida Swartz. +Bm+ OK +CF MP cF oE cf +mn+ +MS
Corylus avellana L. BM+ OK+ +CF +NQ +SF+ +ME af jn — +gs+
colurna L. +mM mK CF NQ Cf ME df kn — Gs+
heterophy Ila Fisch. bM+ +MU +DG +LQ Cf+ ME — — — +gs+
mandshurica Maxim. +bm+ MU DG LP cF +Me+ — — — +gs*-
Corynephorus canescens Beauv. +bm+ Ok CF +OP+ Cf OM+ ad jl - Gg+
Cotinus coggigria Scop. NM+ +mK CD+ OQ Cc+ Me+ cf jm —. GM+
Cotoneaster integer- rima Med. +bm+ oK +CF+ +NO+ Cf Me cf jl — GM+
melanocarpa Lodd. +Bm +mU BE+ +LO+ +Sc+ ME df — Gg+
multiflora Bge. +Nm+ +MK+ +BD+ +NO+ +Sc+ +eE — — — Gg
uniflora Bge. aN+ oU CE MN+ Cc+ Me+ — — — Gg
Crambe maritima L. aM OM+ +BF+ NQ cF +eh+ dg mn+ — Gg
tatarica Sebeok. +Nm Mk+ CD+ +NO+ +SC+ eE dg jl — Gg
Crataegus chlorosarca Max. Bb+ mk +EF+ MO cF +Me — — — GM
dahurica Koehne. BN kU DE+ +LN+ cf Me — — — GM
maximowi tzi C.K. Schneid. +bm kU DF LO cF +Me — — — GM
149
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
monogyna Jacq. bm+ ok+ CE+ OP sF ME df jm — GM
oxyacantha L. bm OM DF OP CF ME cf kn — GM
pentagyna Waldst. et Kit. t-Nm+ +mk+ CE OP +€c+ oe de — Gg+
pinnatifida Bge. bm kU DF LN+ +cf t-eE — — Gg+
pontica C. Koch. NM MU BF NP sc+ ME — — — Gg+
sanguinea Pall. BN MU +DE+ KN+ Sf ME — — — GM
Crepis biennis L. Bm Ok DF NP +cf+ Me+ ce kn — Gs
capillaris Walk. +bm+ OM+ +DF ЮР sf ME cd jm — GM
paludosa Moench. +Bm Ok+ DF MP cP oe+ df jn — gs
pannonica C. Koch. bm+ +mK+ CD+ +NP tCc+ oM+ ce — — Gg
praemorsa Tausch. bN+ +mU +DE+ +LO+ tCf+ Me+ ef jm — Gs
sibirica L. am mU CE+ t-LO+ Cf+ ME+ — — rs Gg
tectoium. L. am OU CF t-LP dF oh ag In rt Gg
Crocus biflorus Mill NM MU +CE OP SC+ teE df kn — Gg+
moesiacusKer. tmM +MK+ CE OP Cf ME — — — Gg+
reticulatus Stev. Nm+ mk CE+ OP tCc+ ME — — — Gg
Cryptomeria japonica +mM++M+ Don. EG t-P+ FfF+ ME - — - GM+
Cucubalus baccifer L. +bM +mK CF MP t-cp-t- eE df ln+ — Gs
Cupressus semperivi- rens L. tmM+ mU BE+ f-PQ SF+ oE — — — Gs
Cuscuta epllinum +bm+ +mU Weiche. +CE+ +MO+ SP af jn - GS
epithymum Murr. Bm+ OK +CF MP Sp — af jl — GS
europaea L. +bM OU BF KQ fp — af ln+ — GS
Cyclamen coum Mill. +mM mk t-DE+ OP t-cF+ oE df kn — +MS
Cymbalaria muralis Gaertn. bM mk CE Q+ cF - df jm - GM
Cynanchum vincetoxi- cum Pers. +bN+ Ok+ DF +NP SF oE cf jm — Gs+
Cynodon dactylon Pers. +Nt+ OU AF+ OR dF MH+ ag kn st Gg+
Cynoglossum offici- nale L. t-Bm mK CE+ +MO+ Sf Me +de+ mn+ — Gg+
Cynosurus crist at us L. tBm-t Ok Cf NP+ CF oE af km PS Gg
Cyperus fuscus L. +bM OU BF LQ fa oe af In — Gg
Cypripedium calceolus L. tBm+ OU CF KP CF ME df jm — gs
guttatum Sw. BN mU +DF KO cf +Me — — — Gs+
macranthum Sw. BN MU +DF KO cf+ +Me — — — Gs+
Cystopteris filix-fragilis Borbas. +am+ OU BG KP cF ME cf kn - gs
montana Desv. t-ab+ mK DE+ LO cF oe df kn — GS
sudetica A. Br. +Bb+ mU DE+ KO t-fF +Me eg +km — Gs+
Cytisus austriacus L. bm mK DE Ю+ tCc+ t 4)M+ cf km — Gg+
nigricans L. bm mk +DE NO+ +Cf oE cf jm — GM+
ruthenicus Fisch. Him +mk+ +DE+ MO+ sf oE af jm — GM
Dactylis glomerata L. +BM OK+ +CF LP+ sF+ og af +ln+ rs GM+
polygama Horvat. +bm+ +mk +DE+ +OP cF oE be kn — Gs
Daphne altaica Pall. bN kU CE MN Cf ME — — — GM
150
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn | Om Cr 1 Hd Tr Rc | Nt fH Lc
cneorum L. bm+ Ok+ CF OP 1 cf OM+ df jl — Gg+
julia K-Pol. +N+ Mk DE NO cf +oe dg jl — Gg+
mezereum L. am Ok+ +DF MP+ cF oE cf kn — gs
sophia Kalen. +N+ Mk DE NO Cf oe+ dg km — GM
Datura stramonium L. +bM OK+ BF MQ CF ME af mn+ — Gg+
Daucus carota L. +bm+ OK BF +NP+ df Mh ag km su GM
Delphinium consoiidaL. +Bm+ ok CE+ MP+ sf ME df kn — Gg
cuneatum Stev. bN+ +Mk+ +DE +NO SC oe — — — GM
elatum L. ab+ mU +DE+ KO+ CP ME df +ln+ -r- GM+
puniceum Pall +N+ kK CD NO Cf ME dg — — Gg
Dentaria bulbifera L. bm+ +oM+ +DF +OP cF ME +de+ In — gS
glandulosa W. et K. +Nm+ mk CE +O+ CP ME — — gs
quinquefolia M.B. bm+ mK CE NP Cf oE — — — Gs
Deschampsia caespitosa Beauv. +aM OU BF KQ CP Og af+ jm pt GM+
flexuosa Trin. +am+ OK CF LP +cF+ Oe ac jl qr Gs+
Descurainia sophia Webb. aM OU BF KQ DF Mg af In — GM+
Dianthus andrzeiowskia- nusKulcz. bm mk+ +DE+ MO +sS+ eE+ — — — Gg
arenarius L. +Bm mk DE+ NO Sc OM+ ad jl — GM+
borbasii Vandas. bm mk+ CE+ MO+ dc+ Oe+ ad jm — GM+
carthusianorum L. bm+ mM+ CF+ OP -tCf ME cf jl — Gg,
chinensis L. Nm+ MK+ +DE+ NP +cF oe — — — Gg+
delthoides L. +BN+ ok+ +DE+ MO+ sf og ad jl — Gg+
fischeri Spreng. +bN mk DE+ +NO SC ME cf jm — GM+
polymorphus M.B. +Nm +MK CE +O+ dc+ OM+ - — Gg
pseudoarmeria Bieb. Nm++Mk CE OP Sc oe — — — Gg
superbus L. +aN+ oU +DE+ KO+ CF oE df jl st GM+
versicolor Fisch. bN +MK+ DE MO +sf og — — — Gg
Diapensia lapponica L. ab •OM EF +MO cf+ oM+ ac jk+ — Gg
obovata Nakai. +ab+ MU DF KO cF. oe+ ad jl — Gg
Dicentra peregrina Fedde. +ab+ mK +EF+ LN Cf Oe ad jk+ Gg+
Dictamnusalbus L. bM OU BF MQ Sf+ oE df jl — GM+
caucasicus Fisch. +Nm+ +MK+ CE+ NP +Sf+ oE cf jl — GM+
Digitalis grandiflora Mill. +bM Ok CF MP+ CF oE +cd+ +in - GM+
lanata Ehrh. bM mK CE+ +OP+ Sc +oE cf — — Gg+
purpurea L. bm OM DF +OP cF +ME ad In — GM+
Digitaria ischaemum Muehl. +bM OU BF +MQ +Cf+ oe+ ac+ jm - GM
sanguinalis Scop. +bM OU BE +NQ Cf oe cd km — GM
Diplotaxis muralis D.C. +Nm+ Ok +DF +OP+ +cf+ — df kn — Gg+
tenuifolia D.C. bM Ok+ CF OP+ Cf — af km — Gg+
Dipsacus fullonum L. +NM OU BF OQ cp — df kn — Gg
pilosus L. bM OK +CF MP+ +cp+ +eE +de+ In — . GM+
strigosus Willd. bM MU BE +OP+ +fF+ +eE — — — Gs
Doronicum austriacum +bm+ Ok+ CF OP Cp oe +de+ ln+ — gs
Jacq.
151
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn Om Cr Hd l ТГ Rc Nt 1 fH | Lc
Draba incana l.. ab 0M+ EF NP Sf oe df In Gg+
nemorosa L. +Bm oU BF KO df Mg ad kn — Gg
sibirica Thell. +aN oU CE+ KO Sf ME — — — Gg+
Dracocephalum nutans L. BN +mU CE+ LO Cc+ +eE - — — Gg
ruyschiana L. iBmt oU CE+ KO+ Sf ME cf jm qt GM
thymiflorum L. Bm+ oK+ CE+ +LO+ df oE — — — Gg
Drosera anglica Huds. +BN+ OU DG KP +Fa OM+ ad jl — Gg
rotundifblia L. aN+ OU DH KP +fa OM+ ac jk+ pq Gg
Dryas octopetala L. +ab +oU CF KO cf+ Oe df km — Gg
punctata Juz. +ab oU DF KN cF oe+ — — — Gg
Dryopteris austriaca Woyn. am OU +EF+ +MP cP oE af ln+ — gs
cristata Л. Gray. +Bm Ok+ DF+ +MP fP OE cd jn rs gs
carthusiana H.P. Fuchs. am Ok+ DF+ +MP cP OE bd jm gs
filix-mas Schott. aM Ok+ CF+ +MQ cF oE be In — gs
fragrans Schott. Ab+ mU DF KN+ cf+ oM+ — — +gs
Echinochloa crus-galli P.B. +bT EU AH MR dP eg af mn+ st GM
Echinops ritro L. +bM mK+ BE+ +MP df Mg df ln+ rs Gg
sphaerocephalus L. +bM mK +CE+ +MP SC eE df to — Gg
Echiuni rub rum Jacq. bm+ mK BE NP df eE — — — Gg
vulgare L. Bm+ mK+ CE +MP sf ME af km — Gg
Elaeagnus angusti- folia L. bM mK+ BE MP +dp +eH+ — - — GM
Elat ine alsinastrum L. <-Bm+ OK CF +MP+ FP — cd kn — Gg+
hydropiper L. +Bm+ Ok+ DF NP FP — ac+ jm — gs
Eleocharis acicu laris Roem. et Sch. taM+ EU CG KQ ca ME af jl rt Gg
mamillata Lindb. +BN mK DE+ +MO cP eE +cf jm rt Gg+
palustris R. Br. am OK CF +MP Ca oH ag jm, su GM
pauciflora Link. aM Ok+ BF NQ FP ME df jm — Gg+
uniflumis Schult. aM OU +CF+ +LQ cP+ +eP eg kn su GM
Eleuterococcus senti- cosus Max. bm MU DF+ +MO+ cF Me — — +gs+
Elodea canadensis Michx. t-bt EU AH MR aA +ME af ln+ - +gs
Elymus gjganteus Vahl. Nm mK +CD +NO+ +dc+ +eg+ cf kn — Gg
Empetrum hermaphrodi- tum Hagerup. Ab Om+ EF LO+ +cF OM ad — — Gg+
nigrum L. Ab+ OM EF LP +cP Oe af jl pq GM+
Ephedra distachya L. bm+ +MK BD+ MP De +eH dg jm — Gg
Epilobium collinum Gmel. Bm+ Ok CF NP+ cF +oM+ ac+ jl — Gg+
hirsutum L. H>M OU CF NP+ +cF+- +eE df +ПО qs GM
montanum L. am+ Ok+ CF MP cF oE ce In — GM+
obscurum Roth. bM Ok CE OP +fF+ +oM+ be km — GM
palustreL. tam+ OU CF KP cP +og af jm Gs
roseum Pers. +bm+ Ok CF NP FP +eE df mn+ — GM
152
Таблица 55 (продолжение)
Вид Факторы
Тш Kn I От Сг 1 Hd Тг Rc Nt fH Lc
parviflorum D.C. +bM OK CF NQ +fP oE df kn — GM
tetragonum L. +bM OK CF MQ +fP oE+ be kn qt GM
Epipactis atrorubens +Bm+ Ok+ CF NP Cf OM+ df jl — Gs
Schult.
helleborine Crantz. +BM OU +CF +MOh h CF ME cf kn — gs
palustris Crantz. +Bm+ OK CF MP fP Oe df jl — GM+
Epipogium aphyilum Bm+ OU CF +MP cF oM+ be jm — MS
Sw.
Equisetum arvense L. Am+ EU CH KP+ Cp oh ag jm qt Gg+
fluviatile L. am+ EU CH KP +fa Oh ag In qt GM
hiemale L. am OU CF +LP+ cF +ME cf In rt Gs+
palustre L. am+ OU CH KP cP og af jm qt GM+
ramosissimum Desf. bM +MU BE MQ dP +eg df jm It Gg
scirpoides Michx. +ab+ OU DG KO +FP Oe — — — gs
sylvaticum L. +am+ OU CH KP cp Oe+ ad km qs +MS
variegatum Schleich. AN EU DH KO+ cF oe df jl — Gg
Eragrostis poaeoides bM+ +mU BF MQ Df oE af kl — Gg+
P.B.
piiosa P.B. +bM+ +mU BF KQ df eh+ ag kn — Gg+
Eremopyrum orientale
Jaub. et Spach. bM+ +MU +BD+ +NQ+ DS+ eH+ — — — Gg
tritceum Nevski. bm+ mK+ BE MP+ DC+ EH+ — — — Gg
Eremurus inderiensis +Nm+ +KU BD +NP+ Ds +Eh Gg
Rgl-
robustus Rgl. +N+ kK+ +C+ +NO +sc+ eE — — — Gg
Erica carnea L. +Bm • t-Mk +DE NP Cf oe df jm — GM
tetralix L. BN+ Em +EG NP +Fp+ Oe ac jl Gg+
Erigeron acet L. +Bm+ OU CF+ KP SF og cf jm pq Gg
annuus Pers. +Bm ok +DE+ MO+ Cf ME cf +ln+ Gg+
canadensis L. +BM OU BF +MQ df oE ce+ kn — Gg
Er iophorum gracile aN+ OU DF X!O+ +fa Oe be jl Gg
Koch.
latifolium Hoppe. +Bm+ Ok+ CF+ MP fP oE df jl qs+ GM
polystachyon L. Am EU CH KP +fa OE df jm pr+ Gg
russeolum Fr. +aB+ OU DF+ KN+ fP+ +oe — — Gg
scheuchzeri Hoppe. AB OU +DF+ KO +FP+ Oe bd jl — Gg
vaginatum L. AN+ OU DG KO+ cP Oe ac+ jk+ pq GM
Eritrichium villosum +ab oU DF KN+ Cf +ME+ — — Gg
Bge.
Erodium cicutarium aM OU BF MQ De Mg af jn — Gg+
L’Herit.
Erophila verna Bess. +Bm+ OK+ BF +NP+ +df+ +eg af jl — Gg+
Erucastrum galiicum Nm+ oM+ +DE OP cf — df km — Gg
Schultz.
Eryngium campestre L. bM mK CE NP +df+ eg+ df kn — Gg
maritimum L. bM Ok+ +CF OP+ CF eg jn — Gg
planum L. bm+ mK CE MO dF Mh — — — Gg
Erysimum cheiranthoi- aN+ OU DF KO+ SF ME +de+ ln+ — Gg
des L.
rcpandum L. NM mK++CE NP+ +SC+ eE+ cf +ln+ - Gg
153
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы Lc
Tm 1 Kn От 1 СГ Hd Tr Rc 1 Nt fH
Erythronium caucasi- cum Wot. Nm ч-Мч- +EF OP +cF+ +Me+ - - - gs
dens-canis L. +Nm+ mk +DE+ +ОР +cF+ oe+ cf . kn gs
sibiricum Kryl. bN kK+ DE MN CF+ oe — gs
Euonymus europaea L. bm+ mk+ СЕ +OQ CF ME df kn — Gs+
latifolia Mill. Nm+ mK+ СЕ +OQ cp ME df km — 4gS
maackii Rupr. +bN+ MK+ +EF+ ч-МО CF +Me+ — — GM+
папа M.B. Nm +МК+ DE +O+ cF+ oe df GM
verrucosa Scop. Ьтч- оК+ СЕ +NO CF oE df jm Gs+
Eupatorium cannabi- +bM num L. 0U BF NP+ +SP +eE +de+ mn+ - GM+
Euphorbia cyparissias L. bm Ок+ CF +OP sf og af jm — Gg
esuia L. H>m+ Ок +DF +OP+ 4-sf Mh+ dg jn ru Gg+
helioscopia L. ч-ЬМ OU BF NQ dS eg +de+ ln+ Gg
lucida W. et K. bm тк DE +O+ cp oe dg Gg
palustris L. гЬтч- Окч- CF MP4- fa Mh dg jn +ru GM
peplus L. +bM Окч- CF NQ CF oE af mn+ — Gg+
segueriana Neck. bm тК BE +NO+ +dc+ чеНч- dg jk rs Gg
semivillosa Prokn. Kbm ч-Мк +DE +NO+ Sf Mg rt GM
uralensis Fisch. bm кК СЕ MO +sF MH — — su Gg+
virgata W. et К. ч -Bm+ тК +DE+ ч-МОч- dF MH eg Ч-1ПЧ- su Gg
Euphrasia hirtelia Jord. BM ОК+ CF LP+ Cf Oe be jl Gg+
montana Jord. +bN Ок DF NP fP oe be ji Gg+
officinalis L. s.1. aM OU BF KQ dp Og af jm ps GM
rostkoviana Hayne. Bm Ок +DE+ NP CF ч-Ме af jm GM
stricta Host. bm ОМ+ DF NP Cf +Me af jl Gg+
tatarica Fisch. +bM mU ВЕч- LQ +sF og — rs Gg
Fagopyrum sagittatum ч Gilib. -Ьтч- mU +СЕ+ +MO+ ч-cF ч-МЕ cf Ч-1ПЧ- Gg+
tataricum Gaertn. ч -Ьтч- mU +СЕ+ +MO++CF+ oE cf +1ПЧ- Gg+
Fagus orientalis Lipsky ч-NM ч-тк CF +OP +CF oE +be+ — — gS
silvatica L. ч-ЬМ +оМ+ +DE+ +ОРЧ- +CF+ oE af jn gS
Falcaria vulgaris Bernh. ч-ЬМ ч-тКч- ВЕч- MQ +df+ MH eg jm st GM
Festuca altissima AIL +bm+ ОК CF +MP+ cf ME ad In — gs
arenaria Osbec. ab+ +от+ t-EF-t NP CF oM+ cf jm Gg+
arundinacea Schreb. ЬМ mU BE MQ sp MP eg km rt Gg
gigantea VilL BM ОКч- +CF +MQ CF ME ce In — gs
ovina L. aN OU DF KO+ SF Og af jn ps GM+
pratensis Huds. +Bm+ ч-OU BF+ +LP+ CP oh ag In pt Gg+
rubra L. +aM +OU BF+ KQ SP OP ag jo pt GM
sulcata Nym. bm ч-МК СЕ MO Df oP eg jl+ rs+ GM
polesica Zapal. bN+ mk+ +DE+ MO dc4- ч-МЕч- — GM
trachyphylla Krajina. +bN4- Ок DF +OP SC ME — — — GM
valesiaca Gand. ч-ЬМ mU ВЕч- +MQ Df+ oP eg jl rt GM
Fucaria verna Huds. +Bm ОК CF MP CF Mg bg lo — GM+
Filago arvensis L. BM oU BF+ MQ Df oH ag jm qs Gg
minima Pers. +bm+ Ок CF NP+ Dc+ ч-Mg bd jl — Gg
l ilipendula kamtscha- +Bb+ tica Maxim. тк EF MO ср ч-Меч- - - GM+
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt I fH Lc
palmata Maxim, ulmaria Maxim. BN+ +mU DF CF4- KO КРч- cp CP+ Me+ og af km pu Gg+ GM+
am +OU
vulgaris Moench. +bm+ Окч- CF ч-МРч- sF og df jl qs+ Gg+
Fragaria moschata Duch. +bm+ +OM+ +CF+ NP4- SF oe cf +ln+ — GM+
vesca L. ч-Втч- OU 4CF4- KQ SF OE af In rt Gs+
viridis Duch. Bm ЮК+ CF KP sf og df jm rs+ GM
Franguia alnus Mill. +Bm+ ч-оКч- CF MP CP Oe ac+ jm pr gS
Fraxinus excelsior L. ч-ЬМ Ок CF NP +cp+ +ME+ cf+ ln+ — GM+
mandshurica Rupr. ч-bm +MU DF LO-t cF Me+ — — GM+
omus L. NM тК СЕ OQ Cf oe df jm — GM
pennsylvanica March. ЬМ тК CF +MP CP +ME — — - GM+
Fritillaria meleagris L. bm ОМ DE4- OP fp+ +oe+ cf kn — GM+
meleagroides Roth. bm ч-тК СЕ MO +dP +eH+ — — Gg
ruthenica Wickstr. bm тК ч<Еч- МОч- Cf Me — — — Gg
Fumaria officinalis L. ч-ВМ ч-тК СЕч- MP cf ' eE ce ln+ — Gg
Gagea erubescens Roem. ч-bm ок 4-DE4- 4-NO4- Cf Me+ — — — GM
et Sch.
lutea Ker-Gawl. Bm OU 4-DF4- MP cF +eE cf ln+ fa GM
minima Ker-GawL Втч- ч-ткч- ч-ОЕч- MP +cf+ +eE df +ln+ pr GM
Galanthus nivalis L. +Nm+ оК СЕч- ОРч- Sp oe cf ln+ — gs
plicatus M.B. ч-тч- +М+ чСч- ч-Рч- Cf Me+ — — — gs
Galatella punctata Nees. Bm тК СЕч- МОч- dp +eH+ — -- Ft GM
Galeobdolon luteum Втч- Окч- CF 4-NP4- CF oE cf kn — gS
Huds.
Galeopsis bifida Boenn. ат oU CF КОч- cf ME ad +ln+ — GM
iadanum L. ат ОКч- CF МОч- CF ME df jm Gg
pubescens Bess. ч-bN оМч- DE4- ч-Оч- CF Me af kn — GM
speciosa МШ. aN+ Ок DF +LO4- Sp ME cf mo — Gg+
tetrahit L. aN+ Ок DF NO+ cP +ME af ln+ — GM+
Galium aparine L. ч-ВМ OU CF4- KQ sF ME ce mo — GM
boreale L. ч-атч- OU CF4- КР SF oh dg jl qu GM
cruciata Scop. ЬМ тК+ СЕ 4OQ +CF+ oE cd In — GM
hercynicum Weigel. BN От EF OP cF — асч- jm — GM
mollugo L. +ВМ Окч- CF MQ SF oP ag jo qs Gs
palustre L. ч-Втч- ОК 4<F LP+ ca Oh ag km pt Gs+
rivale Griseb. Ьт Ок — — CP Mh — — — GM
rubioides L. Ьт тМ DE +Оч- Cp Mh eg +ln+ ru Gs+
ruprechtii Pobed. ч-Вт oU СЕч- К0+ Fa ME — — — Gg+
schultesii Vest. +Ьт ок DE4- +О+ cF ME cf km — gS
spurium L. +ВМ oU BF KQ +sP ME df kn qt GM
tricorne Stokes. ч-ЬМ OU BF +OQ sf eE df jm — GM+
triflorum Michx. +BN+ oU DF КО +fF+ +Me — — — gs
uliginosum L. ат ои CF4- K0+ cP Og af jn ps+ +gs+
vernum Scop. Ьтч- оК CE +NP SF+ +ME ce Im — GM
verum L. аМ ои CF KQ dp oH eg jm qu Gg+
Genista anglica L. Ьтч- ОМч- CF +P+ cF — bd jl — GM
germanica L. Ьтч- Ок CF NP Cf oe ac+ jm — GM
pilosa L. Вт Ок ч-DF NP Sp +ME +bc+ jk+ — Gg+
155
154
Таблица 55 (продолжение)
Факторы
Вид Tm | Kn Om Cr Hd l ТГ Rc 1 Nt 1 fH Lc
tinctoria L. +bm Ok +DF MP sF oE bd jl rt GM
Gent iaпа amarella L. aN OK CF LP Cf ME +cf jm — Gg
asclepiadea Scop. +bm+ Ok +CF+ OP+ cp OM+ cf jn — GM
campesttis L. am+ OM CF OP+ cF oe af jm — Gg+
clusii Petr, et Song. bN om+ DE +O+ CF oe ef jl — Gg
cruciata L. BM OK CF MP Sf Me df jm rt GM
pneumonanthe L. +bm+ OK+ CF +MP CP OE af jm rt GM
punctata L. bm+ ok+ CE+ +OP+ cF oe ad jm — Gg*
verna L. ab Ok DF +NO+ +cf+ oe +de+ jl — Gg+
Geranium bohemicum L. Bm ok CE+ +NO+ Cf oe* bf +ln+ — Gs+
coUinum Steph. +Nm+ oK+ BE +NO+ dP +eP — — rt+ Gg
divaricatum Ehrh. +bM OK+ BF MP+ Cf oE cf +ln+ — Gg
molle L. bM OK CF OP+ df Mh+ eg km — GM
palustre L. +Bm ok+ CE+ NO+ cP oE df mn+ qr GM+
pratense L. am OU CF KP CF oh dg ln+ qu Gg+
pseudosibiricum BN +kU CE KN +CF ME — — — Gg
J. Mayer.
pusillum L. +bM OK+ BF +NQ df eE af ln+ — GM
pyrenaicum L. +bm+ Ok+ CF NP+ CF Me af in — Gg+
robertianum L. +bM OK+ CF +NQ dP ME af io — gs*
rotundifolium L. +bM OK+ BF +NP+ +sf+ °g af km — gs
sanguineum L. bm+ Ok CF NP* Sf oE df jm — Gs+
sibiricum L. +bN+ mU CE+ LO +CF +Mg cf +ln+ — Gg+
sylvaticum L. am OK+ CF LP CF og cf ln+ rs gs
Geum aleppicum Jacq. +bm +mU CE+ KO+ SF +ME — — — Gs
rivale L. am OK CF LP CP OE af km ps* GM+
urbanum L. BM OU BF +MQ CP oE af lo pr Gs
Gladiolus imbricatus L. +bm+ +mk+ CF NP* Cp oH eg jm rs GM
palustris Gand. bN+ oM +E+ +O+ cp +ME df kn — Gg+
Glaux maritima L. +BM OU BF+ KQ Sp eP ag kn su GM+
Glechoma hederacea L. am+ OU +CF+ KP+ Sp oh ag b rt+ gs
hirsuta W. et K, +bm mk DE NP CF eE cf kn — gs+
Gieditschia triacan- bt mk CE+ NQ +CF +Mg cf+ kn — GM
thos L.
Glyceriai fluitans R.Br. +Bm Ok+ DF +NP fa og af kn qs Gg+
lithuanica Lindni. BN oU DE+ +LO +Fp+ ME cf kn — Gs
maxima Holmb. Bm Ok+ CF MP +fa Mh dg b — Gg+
nemoralis Uechtr. bm mk+ CE+ NO+ fP +ME cf — — Gs+
plicata Fr. Bt OK+ +BF+ MQ+ FP Mh dg mo — GM
Glycyrrhiza glabra L. +Nm+ mU BE NQ +dF eP bg kn tu GM+
Gnaphalium supinum L. ab OK CG MO +Cp+ oE ad jm — GM
sylvaticum L. Bm+ OU CF+ KP Sf oe ac+ In rs +gs
uliginosum L. +BN+ OU DH +LO+ cp +oe+ +cd+ kn — GM
Goodyera repens R. Br. +Bm+ OU CF+ KP cF Oe af jl — gs
Gratiola officinalis L. +bM OK+ CF MP+ Sa Mg af kn su GM
Grossuiaria reclinata +bM+ Ok+ +CF +OQ +cf+ ME af In GM+
Mill
Gymnadenia conopsea am+ OU CF +LP+ CP oe df jm — GM
R.Br.
156
Таблица 55 (продолжение)
Факторы
Вид Tm Kn | Om Cr Hd 1 ТГ Rc Nt fH Lc
odoratissima Rich. bm+ Ok+ 'cF OP CF oE ef jl — GM
Gymnocarpium dryopte- am OK CF+ MP cp oE bd kn — gs
ris Neun.
robertianum Chryst. +bm OU DF +MP cF Me df jn — gS
Gypsophila altissima L. bm mK DE MO 4SC4- oe-4 eg jk4- rt Gg
fastigiata L. +BN oM +E+ +NO KC4- OM+ df jkt- — GM
muralis L. +Bm+ OK CF MP Dp Mg ad jl st Gg-h
panicuiata L. H>m OK CF MO4-+dc4- ME4- bf im — Gg+
Halocnemum strobila- +Nt MU AD NQ4- Df +gs eg — st Gg
ceum M.B.
Haloxylon aphyllum NM kU AC +NQ Ds EH4- eg — Gg+
lijin.
persicum Bge. NM KU AC +NQ+ Dd-н Eh eg — — Gg
Harrimanella hypno ides 4-ab4- Ok +EF 4 -LO4- cp Oe ag — — Gg4-
Cov.
Hedera helix L. b.M OM +CF OQ Cp +ME af jn — GS
Hedysarum aipinum L. ab+ oU DF +MN+ +cp 4-ME — — — GM
grandiflorum Pall +bm +kK DE +NO SC4- 4-eE — — — Gg
Helianthemum canum bm+ Ok CF OP4- Sf4- +eE+ ef jl — Gg-h
Baumg.
nummularium Mill +Bm+ +mk+ CE+ NP Cf oe +de+ jk4- — Gg
ovaturn Dun. <-bm+ ok+ CE+ OP 4SC4- 4-ME ef jk+ — Gg4-
Melianthus annuus L. +NM +MK BE -i OQ CF ME 4-cf 4-1П4- — Gg4-
tuberosus L. +bM +MK CE 4 OQ CF +Me+ cf in — GM
Helichrysum arenarium bm+ oU +CE4- 4-MP de oE cd jk4- — Gg4-
Moench.
Helictitrichon deserto- bm mK CD-h MO HSC4- 4-ME4- — — — Gg
rum Pilger.
pratense Pilger. bN OM 4-E4- 4 OP Cf oe cf jm — Gg
pubescens Pilger. Bm+ OU CF LP4- dF oE 4df4- kn TS4- Gg-h
schellianum Kitagava +bN+ +kU CE KO Sf 4-ME — — rs+ Gg
Helleborus caucasicus Nm+ Mk+ CE4- 4 OP CF +ME — — — gs
A.Br.
niger L. bm+ mk CE4- +OP Kf+ +ME df km — gS4-
Hemerocallis flava L. tbmt +MU CD4- 4 LP+ Cf4- +ME cf kn — GM
Hepatica nobilis MiiL +bM ok +CG NP4- CF oE cf jm qs+ gS4-
transsilvanica Gilib. tNm +Mk 4-D4- 4 0+ +cf+ 4-ME cf — — gS4-
Heracleum dissectum aN MU 4-DF KN t-cf+ 4-ME — — — GM+
Ldb.
sibiricum L. am ok+ 4-DE4- MP CP oH — — rt+ Gs+
sosnowskyi Manden. +BN+ +Mk+ CE4- NO+ 4-cF-h 4-ME — — — Gs
spondylum L. BN oM+ DE+ NO4- cF Me af lo — GM+
Her minium monorchis t-bm-e OU +CF+ KP4- Sp +Me+ df jm GM+
R.Br.
Herniaria besseri Fisch. +NM mU BE NQ dS4- 4-eE 4-cd4- jl — Gg4-
glabra L. Bm+ OK BF MP4- sf og cf km rs GM
poiygama J. Gray. bm niK CE MO DC Me — — — Gg4-
Hesperis matronalis L. t-bM mK+ CE MQ 4-cf4- ME cf 4-1П4- — gs
Hieracium aipinum L. ab+ Ok 4-EF 4-NO4- 4-CF+ oMt- ac jk+ — Gg+
aurantiacum L. t-Bb+ ok+ DF +NO t-cF-H oe bd il — Gg-H
157
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
auricula L. Bm+ oM+ CE+ NP +Cp OE ad jm Gg
bauhinii Bess. +bm+ +mk+ CE+ +NP +Sc+ OE df jm — Gg
bifidum Kit. BN Om+ EF NO+ +cf+ +oM+ df jl — Gg+
су mo sum L. Bm+ mk+ CE MP SC oe df jm — Gg
echioides Lumn. +bm+ mK BE+ MP+ de Og cf jk+ rs Gg
lachenallii Gmel. +Bm+ Ok+ CF MP Cf oM+ bd jl — GM
laevigatum Wilkl. +Bm+ mk+ CE +NP+ cF OM+ ac+ jl — GM
murorum L. am Ok+ CF MP +cf+ oe cf kn — gs
pilosella L. +Bm+ +ok+ CE+ MP sF OE af jl ps GM
pratense Tausch. BN +ok+ +DE+ +MO+ cF Me — — pr+ Gg+
sabaudum L. BM OK BF NQ +cf+ +Me bd kn — GM
umbellatum L. aM OU +BF+ KQ SF OE bd jl rt+ GM
virosutn Pall. aN oK CE +NO+ sf +eg — — — GM
Hierochloe australis +bN+ oM DE+ +O+ +Cc+ +oM+ df In — GM
Roetn. et Schult, odorata Wahlb. aN+ OU CF KO+ sP og bd kn su GM
Hippocrepis comosa L. bm+ Ok CF +OP+ +Sf+ ME cf jl __ Gg+
Hippophae rhamnoi- bM oU BE+ +MP+ +Sf +cE+ df jl — Gg+
des L. Hippuris vulgaris L. +aM OU +CF+ KQ fa+ Mg df kn GM+
Holcus lanatus L. Bm+ Ok+ CF NP SP ME af km qs GM
mollis L. +bm+ OM+ CF +OP+ cf+ Me ad jm Gs
Holoschoenus vulgaris bM mK+ BE NQ SF+ eE+ df jm — Gg+
Link. Hordelymus europaeus bm+ Ok CF OP cF ME df kn gs+
Harz. Hordeum jubatum L. am+ EU +CH KO+ +Sf+ +oe+ cf kn Gg
vulgare L. BM OU +BF+ KQ SF oE cf +ln+ — Gg
Hottonia palustris L. +bm+ ok+ CE+ NP Pa ME cd km — GM
Humulus lupulus L. +bM +oK BE+ MP+ cP oE +ce+ +mo — gs
Huperzia selago Bernh. AN+ EU CH KO+ ca og ad kn — gs
Hyacinthella leucophaea Nm+ MK CD+ OP +sC+ +eE+ — — — Gg
Schur. Hydrocharis rnorsus- +Bm+ OK CF+ MP+ pA oE ce lo st Gg+
ranae L. Hyoscyamus niger L. BM OU BF LP+ dF eg cf mo — GM+
Hypericum elegans +bm+ ok+ CE MP +Sc+ Me+ dg In st Gs
Steph. hirsutum L. +bN+ OU +CF +MP sf ME df ln+ Gs
maculatum Crantz. BN oK DE +MO+ cF Oe+ ad jl ps GM+
montanum L. bm+ +mk+ CE +OP cf Me ce jm — GM
perforatum L. BM+ +mU BF LQ dF OE af jn rs GM
Hyfxrchoeris radicata L. +bM Ok CF +OP+ +CF +ME bd jm — Gg+
Hypopitys monotropa Bm+ OU CG +LP+ Cf+ oe bd jm — gs
Crantz. Hyssopus officinalis L. bM OK BE MQ +Cc+ +eE cf jl Gg+
Impatiens glandulifera Bt mK CF NQ fP +ME cf lo — Gs+
Royl. noli-tangere L. +BM+ EU CH KQ cP +ME cf In rs gs+
158
Таблица 55 (продолжение)
Вид Факторы
Tm 1 Kn 1 От 1 Ст 1 Hd | Tr 1 Rc | Nt 1 fH 1 Lc
parviflora D.C. +bM+ OK +CE MQ Sf +ME af In Gs+
Inula aspera Poir. +bM +MU BE MP+ sf eg — — — Gg
britannica L. am+ +OU BF+ KP+ dP MP dg kn ru GM
germanica L. +bm+ mK CE MO+ +sc+ oE be — Gg
helenium L. BM OK+ +CF+ MP+ cF+ oE bf +ln+ Gg
hirta L. Bm+ OK CF +MP +Sf MH+ dg jm tr GM+
ocuius christi L. bM mU BE NP+ sc oE ce — Gg
salicina L. +BM OU BF KP+ Sp Mh dg jm st GM
Iris aphylla L. bm+ mk+ CE NP +sf eh+ — — GM
germanica L. +bt oK +BE+ +OQ Cf+ +eE df kn — Gg+
halophila Pall. bm +kK+ CD+ MO SF eH+ — — — Gg
pseudacorus L. +bM Ok+ CF+ MP+ ca oh ag ln+ rt GM
pumila L. +NM mk+ CD+ NP +dC+ eg — — Gg
sibirica L. Bm+ mK СЕч- MP sp +eg df ji st Gg
Isatis tinctoria L. bM mK СЕ NP Cf — df jm — Gg+
Isopyrum thalictroi- des L. bM Ok+ CF OP+ +cf+ +Me+ cf kn - gs
IsoStes lacustris L. ab+ +Ok +EG+ MO+ aA — ad jk+ — GM+
Ixiob'rion tataricum Roem. et Schult. bni kU BD+ MP Dc+ +eh - - Gg
Jasione montana L. +bm Ok DF NP Sf oE ad jl — Gm+
Jug Ians magdshurica Maxim. +bN+ +MU DF +MN+ cF +Me+ - - GM+
nigra L. NM+ +MK DF NQ +CF +Me+ — — — GM+
regia L. +mM +MU BE NP CF +Me+ df+ +ln+ — GM+
Juncus alpinus Vill. +am +OU DG KO+ +fp+ Me df jm qt Gg+
articulatus L. aM OK+ BG KQ cP og af jl+ qt GM
atratus Krocker. +bm OK CF+ +MO+ +fP og+ af kn qt Gg
atrofuscus Rupr. ab+ oU CE+ KN cF +eE — — Gg+
balticus Willd. +Bb+ om +E+ +NO+ +Fp+ — ac+ jk+ — Gg+
bufonius L. +at OU BG KR SP +Mg ad jn — Gg
compressus Jacq. +Bm OU +DF LO+ cP og cf kn qs+ Gg
conglomeratus L. +Bm Ok +DF+ NP ca oe bd jn qs Gg+
effusus L. +Bm OK CF MP cP Og ad jm qr GM
filiformis L. am OU +DF+ KO+ cP OE bd jm pr+ GM
gerardii Loisel. am OK+ BF MP+ +sP +eP ag km ru Gg+
inflexus L. bM +oK+ BF NP +fp+ Mg df km — Gg+
squarrosus L. +bN+ +om+ +E+ +O+ +fp+ OM+ ac jk+ — Gg+
tenuis Willd. bm ok +DE+ +O+ +fF-' — cd km — GM+
Juniperus communis L. am OU DF KP Sp OE af jn — GM+
exelsa M.B. +mM mk+ +CD+ OP+ Sf oe — — — GM
oxycedrus L. +mM mK+ +CE+ OP +Sf +Me+ — — — GM
sabina L. bm +MU +CE MO+ +sf +ME ad jm — Gg+
semiglobosa Rgl. Nm kU +CD+ NP Sc+ +eE — — GM
Serawschanica Korn. +NM kU +BD+ MO+ dc+ +eE — — — GM
sibirica Borgsd. +aN +MU CE+ KN+ +cF +oE — — — Gg+
turkestanica Korn. Nm kU BC NO Sf +eE+ — — — GM
virginiana L. bM+ +Mk+ +DF+ +NP+ +CF+ +og — — — GM+
Jurinea arachnoidea Bge. +Nm Mk+ +D+ +O+ dC+ +eg dg — - Gg
159
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
cyanoides Rchb.. t-bm mkt- CDt- NOt- +dC oE eg — — GM
Kalopanax ricinifolium Miq. +Nm+ mKt- DG t-NPt- t-cFt- t-ME — — — -tgst-
Knautia arvensis Coult. Bmt- OK+ CF MP sF oh ag jm rs Ggt-
Kochia prostrata Schrad. bM mU BE t-MQ DC+ eH eg t-lnt- — Gg
Koeleria cristata Pers. +BM OU t-MF+ LQ DFt- MH eg jl rt Ggt-
delavignei Czern. +bN+ +Mk+ t-DE MO+ SF oP bg — rt GM
glauca D.C. t-bm OK+ +DF LO+ DF Mg df+ jl — Gg
grandis Bess. BN +mM++E+ NO Cf oe — — — GM
pyratnidata Beauv. bNt- om t-E+ OP +Cc+ +oM+ ce jl — GM
Lactuca serriola Torner. +bM юи BFt- MQ dF eg af km su Gg
sibirica Benth. aN •-mU CE+ KO +cF+ Me — — — GM
tatarica C.A. Mey. Bt юи t-BF+ MR +dF eht- — — — Gg
La'mium album L. aM OU BG •-LQ+ sF t-ME t-cf t-mo — GM
amplexicaule L. +BM OU BF+ MQ dF t-eE af Int- — Gs
maculatum L. Bm+ oK CEt- +NO+ cF t-ME cf mn+ — gs
purpureum L. aM OK+ +CF+ KP CF ME cf jn — Ggt-
Lappula myosotis Moench. aM hoU t-CEt- KP+ DF eh eg t-lnt- — Ggt-
patula Aschers. bM +MK+ BD+ MP+ +dc+ eg — — — Gg
Lapsana communis L. BM OU CF MP+ Sf +ME af Int- — gst-
Larix cajanderi Mayr. aBt- t-MU t-EF KMt- t-cP Oe+ af — — GM
dahurica Turcz. aN +MU DF KN CP OE — — — GM+
decidua Mill. +bN mk DE NOt- CFt- oE af jm — GM+
gmelini Rupr. aN +kU DE KN Cpt- OE af — — GM+
laricina K. Koch. am mK fDF+ LO +cP Oe+ af — — GM+
polonica Racib. +bN+ oM HE+ Ю+ Cp oE af — — GM+
rossica Trautv. bBbt- okt- t-DEt- t-MN Cp oE af — — GM+
sibirica Ldb. +BN +MK+ DE t-LMt- CF+ oE af — — GM+
Laser trilobum Borkh. bm+ OK CF NP +cf+ oE — — — gs
Laserpitium hispidum Bieb. Nm Mk +DE Ю+ +cf+ oE cf — — GM
latifolium L. bM OK CF NP sp Met- df kn — GM
prutenicum L. bm+ Ok CF NP cF Mh dg jm — GM
Lathraea squamaria L. +bM OU BF NQ cF ME cf In — gs
Lathyrus aphaca L. bmt- OK BF OP +Cf +eE af jn — Gs
maritimus Bigel. taNt- OM+ EFt- KO+ t-cft- Mgt- eg jn — Ggt-
paluster L. +BM OK CF LP+ cP Mh dg jm qs GM
pisiformis L. Bm mU CF LO+ Sf ME — — — GM
pratensis L. bBt OU +BF+ HR SP °g cf In qtt- Ggt-
sativus L. bM mKt- BDt- +MP+ CF df kn — Gg
sylvester L. BM Ok CF NQ Cf oE +cf kn rs GM+
tuberosus L. +bM OK BF NQ sf ME df km st Gg+
Laurocerasus officinalis Roem. +mM +mM+ +DF OP+ t-cp t-ME cf kn — gs
Laurus nobilis L. +mM Hmk+ CE+ tPQt- SF oE eg in — Gs
Lavatera thuringiaca +bM oK CEt- MP+ dF Mg df rt GM
Ledum decumbens Lodd. HaBt- oU +EF KN CP oe - — - GM
160
Таблица 55 (продолжение)
Вид Факторы
Tm 1 Kn Om Cr Hd Tr Rc | Nt fH Lc
palustre L. +aN oU DE+ KO cP Oe ac jl pq GM
Leersia oryzoides Sw. +bm+ OK BF +NP Fa oE+ af mo — Gg+
Lem na gibba L. bM OU BF +OP PA — cf mo — GM
minor L. aM+ OU BF KP +pa Og af jn st GM
trisulca L. at+ EU +BG+ KR FA Mh eg In — GM
Lens orientalis Schmalh. NM mU AD +OQ+ SC+ eE+ df kn — Gg
Leontodon asper Poir. bm+ OK CF NP sc+ +eE — — — Gg
autumnalis L. am Ok+ +DF+ MO+ SF oH ag kn PS GM
hispidus L. +Bm+ Ok+ +DF+ NP sF OE af jm pr+ GM
Leontopodium aipinum ab mk+ +DE NO SF og df jm — Gg+
Cass. leontopodioides Beauv. ab kU DE+ +MN +Sf +eE — Gg+
ochroleucum Beauv. ab kU CE +LN SF ME — — — Gg+
Leonurus cardiaca L. bm+ Ok+ +DE +OP dF ME df mo — GM
glaucescens Bunge. Bm +kK+ CE MO +cf eE — — - Gg+
quinquelobatus Gilib. +Bm +mk+ CE MO+ Cf oE — — — Gg
Lepidium campestre +bm+ Ok CF +O+ +Sc+ eH eg In — Gg
R.Br. latifolium L. bM OU BF MQ +dp eP+ eg In ru Gg+
rud efale L. +BM OU BF LQ DF+ MH eg +ln+ — Gg
Leucanthemum vulgare +Bm OK+ CF LO+ CF og af jm ps Gg+
Lam. Leucojum aestivum L. NM OK+ DE OP cp ME cf In __ GM+
vernum L bm+ mk +DE+ OP fp ME cf ln+ Gs
Leymus arenarius aN OM +EF+ +NO+ cp eg cf In — Gg
Hochst. Libanotis intermedia Bm mK+ DE MO+ sf ME cf jl+ rt+ GM
Rupr. Ligularia sibirica Cass. BN MU CE LN+ +cP oE GM
Ligusticum scoticum L. ab OM+ EF +NO+ cp +oM+ — -- — Gg
Ligustrum vulgare L. bM Ok+ CF +OP+ Sp Met- df jn — GM+
Lilium dahuricum Ker- Bm mU DF+ KO+ Cf +eE — — — GM
Gawl. martagon L. r-bm+ mK+ +DE+ MP+ CF oe+ cf kni rs Gs
Limonium latifolium Nm+ MK +D+ +O+ dS+ eg — — — Gg
Kuntze. sareptanum Gams. t-Nm +kK +CD+ +NO+ +dc+ MH+ dg Gg
Limosella aquatica L. +aM+ +OU +CF+ KQ+ dP+ +EH ag kn — GM
Lina ria genistifolia Mill. bm mK CE+ NO +sf+ OH+ eg — — Gg
minor Ldb. Bm mk DE NO+ sf oe df In — Gg+
vulgaris Mill. am Ok+ +DF MO+ dF Og cf jm ps Gg+
Linnaea borealis L. ab+ OU +EF+ KO+ cp oe ac+ jl qr gs
Linosyris vulgaris Cass. bM Ok+ CF +OP+ dF oh dg jm — GM
Linum catharticum L. BM OU BF NQ Cp oE af jk+ qr+ Gg
flavum L. +-bm+ mk+ +DE NO+ Sc 'Me cf — — Gg
hirsutum L. +Nm+ +Mk +D+ +OP +Cc+ OM+ cf — — Gg
nervosum Wald, et Kit. Nm +Mk +D+ OP +Sc+ — — — Gg
perenne L. am oK+ +DE+ KP df+ *ME+ — — — Gg
1/2 11. Зак. 448
161
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы Факторы
Tm Kn I Om Cr I Hd | Tr Rc Nt fH Lc Tm | Kn | Om I Cr | Hd | Tr Rc Nt fH Lc
usitatissimum L. Bm OK+ DF +MO+ +Cf+ oe+ +cf +1П+ — Gg+ vulgaris L. +BM OU +CF+ KP+ ca og af jn qu Gs
Liparis loeseUi Rich. +bm+ OK CF MP +FP+ — ef jl — Gg+ Lythrum salicariai L. +BM OU +CF+ +LQ Ca +Mg cf jn qt Gg+
Listera cordata R.Br. am+ OU CF KP fF +ME a cl- jl — gs virgatum L. +bm +mK BE+ +LO+ +CP og cf In su Gg
ovata R.Br. +am+ OK CF MP CP oe ef kn pr+ gs Maackia amurensis Rupr. +bm+ mK+ DG +MP+ Cp +Me+ — — rt GS
Lithospermum arven- +bM mU BE+ EQ df OE cf km Gg Magnolia grandiflora L. +mt ok+ CE OR +cp+ ME cf +ln GS
se L. Mahonia aquifolia Nutt. BM +OK +CG NQ CF oE cf kn — Gs+
officinale L. +BM mK+ CE+ KQ +sp+ OE df in rs GM Majanthemum bifoliunL. +Bm oU DF LP cF Oe ad jm pr+ gs
purputeo-coeruleum L. Nm+ mK +DE+ OP Sf ME df jm — GM+ Malabaila graveolens bm mK CE NO+ +sc oE ce — Gg
Litorella uniflora +bM+ OM BF +OQ+ Pa — af jl — GM Hoffm.
Aschers. Malaxis monophyllos Sw. +Bm+ oU +DE+ KP CP Oe ac+ jm — +gs+
Lloidia serotina Rchb. Ab +oU DF KO Cf oE ad- jl Gg+ Malus baccata Borkh. bM MU +DF+ +NP+ +Cf ME+ — — — GM+
Lobelia dortmanna L. +Bb+ +Om+ EF+ NO+ Pa — ad- jk+ — GM mandshurica Kom. bN+ kU DF +MN+ +cf+ Met- — — Gs
Loiseleuria procum- ab Ok +EF MP cF oM+ ad jk+ — Gg orientalis Uglitzkich. NM mU +CF +OP +Cf+ ME — — — Gs+
bens Desv. pallasiana Juz. +bN+ kU CE+ LN+ +Cf Me+ — — — Gs
Lolium multiflorum +bM OU +CF+ +MP+ +cf+ oE af jn — GM praecox Borkh. bm +MK CE NP +Cf+ +eg cf+ kn — Gs
Lam. silvestris Mill +bN Ok +EF NP +Cf+ Mg cf+ kn Gs+
perenne L. +BM OU BF+ MQ dF +ME af ln+ — Gg Malva excise Rchb. +bN+ ok+ DE+ NO+ cF — df mn+ — Gg+
temulentum L. +BM+ OU +BF+ +NQ+ +cf+ — df jn — GM neglecta Walk. +bM+ OU +DF+ NQ+ dF . eg af +no — Gg
Lonicera altaica L. ab +MK +DE+ MN +cF oe df lo — Gs+ pusilia Sm. et Sow. +BM OU +CF+ LQ dF eE+ af kn — Gg
caprifolium L. NM mK CE OP CF ME df kn — Gs+ sylvestris L. +bM OU BF LQ +cf+ ME af mn+ — Gg+
edulisTurcz. +bN+ mU +DF+ LO cF +Me — — Gs+ Marrubium praecox Nm+ mk+ CE+ NP +sC+ eg+ — — — Gg
paljasii Ldb. ab+ +mU DE+ +LN cF og ae jm GS Janka.
tatarica L. bM kU BD+ LO Sf+ Mg cf kn GS vulgare L. bM mU BE+ +OQ Cf — df mn+ — Gg
xylosteum L. Bm+ oU BF MP cp oe+ cf jn GS Marsilea quadrifolia L. Nt OU +BF+ +OR +pa+ — af ln+ — Gg+
Lotus comiculatus L.s.L +Bt oU +BF+ NR dF oP eg jl qu Gg+ Matricaria inodora L. +Bm+ OK +CF+ MP Sp MH eg In qs+ Gg
uliginosus Schkuhr. bN+ Om+ +EF+ +O+ +fP oe+ bd km Gg maritima L. +BN+ +Om EF NP Cf Mh eg +mo — Gg
Lunaria annua L. NM MK +CE OP+ cf oE cf +ln+ gs matricarioides Porter. aM +mU +BF+ +LQ +cF +ME cf +mo — Gg+
rediviva L. +bm+ Ok CF +OP+ +fp ME cf +mo +gs recutita L. +Bm+ OU +CF+ KP+ sF Mh eg kn — Gg
Lupinus luteus L. NM oU CD+ +PQ+ Cf +oe+ cf kn — GM Matteuccia struthiopte- +Bm+ OU CF+ LP+ +fF+ ME cf lo — gs
poiyphyllus Lindl. Bm OK CF +MP Cf +oe+ ad kn — GM+ tis Todaro.
Luzula albida D.C. +bN+ oM+ +E+ +O+ cf Oe ad km — gs Matthiola fragrans Bge. bN+ +MK +CD+ MO sf oe+ df jm — Gg
campestris D.C. +bm+ Ok+ CF OP Cp Oet- ad jm ps Gg+ Medicago denticulate NM mK BF OQ dS eg — — — Gg+
muitiflora Lej. +am OU +DF+ K0+ CF Oe cd jm GM Willd.
pallescens Swartz. am+ +OU BF+ KP CF +oe bd km — GM+ falcata L. +bM OU +CF+ +LQ df oP eg jm rt+ Gg+
pilosa Willd. am +oK+ +DE+ LO cp Oe cd. km qs gs+ lupulina L. +bM+ OU +BF+ +MQ+ dF oh df jn rt+ Gg+
Lychnis chalcedonies L. +Bm mK CE+ MO Cf ME — — GM+ minima Grufb. bt OU +BF+ NQ+ df +eH+ dg jk+ — Gg
Lycopodium alpinum L. ab oU DF KO +cF +oe act- jl — Gg+ romanica Prod. +bm+ +mK+ BE+ +MP dc+ +eg — — — Gg
annotinum L. +Bm OU DF+ KP CF+ OM+ id jm — gs+ sativa L. +bm+ +mK BE+ MP df +ef km — Gg+
clavatum L. am OU DF+ KP CF OM+ ac+ jl — GS Meiampyrum arvense L. +bm mk+ +DE+ +NO+ sF+ +ME df ' jm — GM+
complanatum L. am+ OU +DF KP CF OM+ ac jm Gs cristatum L. • +BM OK CF LP Cf og df jm rs Gg+
inundatum L. Bm Ok DF NP +FP+ — +bc+ jl — GM nemorosum L. +Bm mk+ CE+ MP CF Oe ce km qr +gs
Lycopsis arvensis L. +Bm+ +mk+ +DE+ NP Sf+ — ac In — Gg pratense L. am+ Ok+ +DF+ LP Cp Oe ad jm pr +gS
orientalis L. bM mU BE NQ dC+ eg — — — Gg sylvaticum L. am mk +DE+ NP CF OM+ ac+ jl — Gs+
Lycopus europaeus L. BM OU BF +LQ ca oE af +ln qs GM Melica altissima L. +bm+ mK +CE+ MP +Cc+ ME ce — — Gg+
exahatus L. +bm+ +mK++CE+ MP +FP+ oe cf +ln+ st Gg nutans L. am+ OU CF LP CF OE cf jm rs gs
Lysimachia nummularia +BM Ok+ CF NP SP oh ag jn su GM picta C. Koch. bm +mk+ CE+ +OP Cf +eE ce — GM
L. transsilvanica Schur. +bm+ +mK CE+ MP +sf +eE ce jk+ Gg+
punctata L. +NM Ok CF OP+ +cF+ oE cf +ln+ - GM uniflora Retz. +bM Ok+ CF OP cF +Me ce jn — gS*
162 163
Таблица 55 (продолжение)
Таблица 55 (продолжение)
•Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
Melilotus albus Desr. t-Bm+ HnU BE+ LO+ dF og cf jm SU Gg
altissimus Thuill. bm+ Ok CF +NP+ cP cf jn Gg
officinalis Lam. aM OU BF MQ dF OP ag jn rt Gg
Meniocus linifolius D.C. +bM mU BE MQ DS eE eg Gg.
Mentha aquatica L. bM mK++CE+ +OP+ ca Mg cf km — GM
arvensis L. at OK BF+ KR CP oH ag jn Gs
longifolia Huds. +bM OK CF MQ SP Mh dg mn+ — GM
Manyanthes trifoliata L. +aM oU +DF+ KP fa OE af jl pr GM
Mercurialis annua L. bM +mk 4-BE OQ +cf+ — cf mo GM+
perennis L. tbm Ok 4-DF+ NP CF ME cf ln+ gS
Mespilus germanica L. amt OU +CF+ LP cp oE af jn Gs
Milium effusum L. am+ OU +CF+ LP cp oE cd jn — gs
Mimulus guttatus D.C. +bm ok DE+ NP +FP+ — af ln+ GM
Moehringia lateriflora Fenzl. +am OU CG KO+ cF oe+ - - GM+
trinervia Glairv. +BM OK+ CF MQ cP +ME ce In gs
Molinia coerulea +BM+ Moench. OK ♦CF+ MQ ca Og af jl pr GM+
Moneses uniflora Gray. am юи CF+ KP cF oM+ bd jl gs
Montia rivularis C.C. +Bm+ Gmel. Ok CF+ NP fa — cd km -. Gg+
Morus alba L. NM KkK+ +DE NP Sf+ Mg dg — — GM
nigra L. +mM mK++CE OP+ Cf Mg dg — GM
Muscari botrioides Mill. bm +mk+ +DE+ +O+ sF oE af jn — GM
racemosum Mill. bM Hnk+ CE+ +OP+ Cf +oM+ df jl GM
Mycelis muralis Dumort. +bM OK CF+ +NQ CF oE af In gs+
Myosotis alpestris Schm. AN hmU DE KO Cf oe cf kn — Gg+
arvensis НШ. aM OU H2F+ KQ SF Mg af In — Gg
caespitosa Schultz. +BM+ EU rBG+ KQ+ CP Mg ad kn GM
coUina Hoffm. +bM OK+ BE NQ sc+ +eE __ Gg+
micrantha Pail BM -i -mU +BE+ LQ +sc ME — — — Gg
palustris Lam. tam+ OU CF+ LP ca oh ag kn ps GM+
sparsiflora Mikan. hbm+ oK CE+ MO+ cF oE cf gs
suaveolens W. et K. bN+ oU CE +MO+ Sf og GM
sylvatica Hoffm. aN+ mk+ DE t-LO Sf oE af lo rs gs+
Myosoton aquaticum +Bm Moench. oK CE+ <~LO+ fP <-eE af mo - GM
Myosurus minimus L. hBm+ Ok DF NP Df teP eg jm Gg
Myrica gale L. • BN+ Om +EF NP FP — ad jl — Gg+
tomentosa Asch, et Graebn. BN+ ok+ EF+ +MO+ fp +Me — - Gg+
Myricaria alopecuroides Schrenk. NM mU BE +NQ fp oe+ df - - Gg
Myriophyllum alterni- folium D.C. aN+ +OM +EF+ KNO+ aA - ad jm - GM
spicatum L. +aM +OU BG KQ tPA HeE dg kn GM+
verticillatum L. aM OU H CG KQ PA eh eg ln+ GM+
Najas minor AU. bt oK BF NR aA — df km GM+
marina L. +BM+ 164 OK CF +NQ+ aA eH eg In - GM+
Вид Факторы
Tm Kn Om Hd Tr Rc Nt fH Lc
Narcissus poeticus L. bM mk CE OP+ Cp +ME af kn — Gg+
Nardosmia frigida Hook. AB OM DF KO +fp oe+ — — — Gg+
laevigata D.C. ab oK +E+ +LN+ FP +Me+ — — — Gg+
Nardus stricta L. aN+ Ok+ +DF+ MP Cp Oe ac+ jn pr+ GM
Nasturtium officinalis R.Br. bt OU BF NR +PA Me+ cf ln+ — GM
Naumburgia thyrsiflora Rchb. aN+ EU BH KO+ +fa OE af jm qs GM+
Neottia nidus-avis Rich. +Bm+ Ok+ CF +NP+ +cf+ +ME cf kn — gs
Neottianthe cucullata Schl. +bN+ mU +DE+ +MO Cf oe — — — gs
Nepeta cataria L. +bM oK BE MQ Sc+ ME cf In — Gg
nuda L. +bm+ mK CE MP+ sf ME+ df kn rs Gg
Neslia paniculata Desv. Bm mU CE KO+ +cf+ — df km — GM+
Nicandra physaloides Gaertn. bm mK BE MP +cf+ — af ln+ — Gg+
Nonea pulla D.C. Bm mK CE MO+ df oh dg In st Gg+
Nuphar lutea Sibth. et Sm. +BM +OU CF+ KQ FA oh eg jn st GM
pumila D.C. +BN+ mU +DE+ KO fa+ ME ac+ jl — Gg+
Novosieversia glacialis G. Bolle. AB mU EF KM+ cf+ +Me+ — — — Gg
Nymphaea alba L. BM Ok+ CF NQ PA — cf ln+ — Gg+
Candida Presl. +Bm oK BE+ LO+ PA Og bd kn — GM
tetragona Georgi. +BN+ +MU +DF+ KN+ PA ME — — — Gg
Nympho ides peltata Ktze. BM OU BF LQ PA — cf ln+ — Gg+
Odontites serotina Dumort. BM OU +CF+ KQ dp oh ag jn — Gg
Oenanthe aquatica Poir. +Bm OK CF MP Fa °g cf kn qs+ GM
Oenothera biennis L. +bm+ OK CF+ MP Sf oE af km — GM
Olea europaea L. mt mk BD PR Sf Oe+ af jm — Gg+
Omphalodes Scorpio ides Schr. Bm +mk DE+ NO+ +cF+ oE cf kn — +gs+
Onobrychis arena ria Ser. bN+ mM+ DE+ +O+ df Mg df jm — Gg+
sibirica Turcz. +bN +kU CE+ KN +Sf +cg — — — Gg
viciaefolia Scop. bm +mk +DE OP sf og df jm rs Gg
Ononis arvensis L. +bm+ +mk+ +DE+ MP SF MH — — st GM
repens L. +bm+ OM+ +DF +OP CF — cf jl — Gg+
Onopordum acanthium L. BM OK+ BF MQ de +ME cf mo — Gg
Onosma simplicissi- mum L. BN+ kK CE MO dc+ +eg df+ jm rt Gg
Ophioglossum vulga- tum L. +BM Ok BF+ +NQ cF Me af jn — GM
Ophrys apifera Huds. +NM OK+ BF OQ +cf+ oe ef jl — Gg+
insectifera L. +bm+ Ok+ CF OP+ cF oe df jl — GM+
Orchis coriophora L. +NM OK+ BF +OQ Cf ME df jm ps Gg+
165
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Rc | Nt 1 fH Lc
fuchsii Druce. +BN+ OU +DF+ KO+ cF ME+ df kn pr GM+
latifolia L. +Bm+ OK CF LP+ cP og df jm st GM
maculata L. aN Ok DF MO+ cP OE af jm ps GM+
mascula L. +bM OU BF NQ +cf+ — df jm pr Gg+
militaris L. BM OK+ +CF PL+ Cf ME ef jl qs GM
morio L. bm Ok +DF +O+ +SF OE +cf jm pr Ggt-
palustris Jacq. +NM OK+ +CF+ +OP+ dF+ Mg df jm su Ggt-
tridentata Scop. NM mK CE OQ +Cf OM+ df jm qs Gg+
ustulata L. Bm+ Ok+ CF MP+ +cf+ OM+ af jm qs Ggt-
Origanum vulgare L. +BM OK+ CF +MQ sF og af jm qs GM
Ornithogalum gussonei Ten. Nm+ mK CE +OP+ +sf oE ce — — Gg
umbellatum L. +bm+ Ok CF OP+ cF, ME cf kn — Gg+
Orobanche purpurea Jacq. bM OK+ +CF NP+ +cf+ — af jl — GM
Orobus gmelinii Fisch. +bN +kK+ CE +LN+ +cf+ +ME — — — GM+
laevigatusW. et K. bm mk +DE+ OP cF +oM+ df km — Gg+
niger L. +bM Ok CF NP+ cf Me af jm — gs
tuberosus L. bm Ok CF OP Cf — df kn — gs+
venetus Mill. +Nm+ mk +DE OP +cf+ +oM+ ad kn — gs+
vernus L. +BM oK+ CE+ LQ CF oE cf jn — gs+
Orthilia secunda House. am+ EU CH KP CF+ Oe af jl pr gs
Osmunda regalis L. bT OM DG OR +Fp+ oe+ be kn ps gs
Oxalis acetosella L. t-BM OK CF LP +cF+ +oe+ af Int- qs gs
corniculata L. NM mU BE OQ +cf+ — af In __ GM+
stricta L. +bM+ 0k+ CG +OQ+ cF oe+ be In — GM
Oxy coccus microcarpus Turcz. Ab+ OU DF+ KO +FP OM ab+ jk+ — Ggt-
quadripetalus Gilib. ab+ EU DH KO+ fP OM+ af jk+ pr GM
Oxyria digyna Hill. Ab oU +DF KO cF +Me ad jn — Ggt-
Oxytropis campanulata Vass. +Bb+ +MK DE +M+ Cf +ME — - - Gg
glabra D.C. BN kU CE KN+ +CF+ +eg+ dg — — Gg
pilosa D.C. Bm+ mK+ +DE+ MO+ +sc+ +eg+ eg jm — Gg
sordida Pers. +aB+ OU DE+ KN+ Cf +Me+ — — — Gg
Padusasiatica Korn. +Bm+ +MU DF+ KP +Me+ — — — Gs
maackii Korn. bm kK+ +EF +MP CF +Me+ — — — GM+
mahaleb Borkh. +NM oU BE OQ +Cf +ME df jl+ — GM
racemosa Gilib. +Bm+ OK +DF LP cP og cf +ln — Gs+
serotina Agardh. r-Nt mk+ +DF +NQ+ CF+ oE cf kn — Gs
virginiana Mill. BM mK CF LP CF oe+ — — — Gs+
Paeonia anomala L. aN +mU +DE+ KN +Cf+ Met- — — — Gg+
tenuifoiia L. bm+ +mk+ CE+ +OP SC++eE — — — Ggt-
Paliurus spina Christi Mill. NM mK BE NP+ dc+ eg+ eg jm — Gg+
Panicum miliaceum L. +bM+ mU AE LR SF +ME cf +ln+ — Gg
Pa pa ver aipinum L. +ab+ Ok DF NO cF — ef — — Gg
lapponicum Nordh. +aB Ok +E+ KO cf t-Me — — — Gg
polare Perf. Aa+ ok +E+ KN+ cf+ oe — — — Gg
radicatum Rottb. AB Ok DF KO cf+ oe — — — Gg
166
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn | Om Cr Hd Tr Rc Nt fH Lc
rhoeas L. bM OU CF MQ sf ME cf jn — Gg
somniferum L. bM mU CE+ LQ SF — +cf +1П+ — GM
Paris quadrifolia L. +Bm+ +oK+ CF KP cF oE cf in — gs
Parnassia palustris L. am+ OU CF KP+ cP Og cf jl ps GM
Parthenocissus quinque- folia Planch. bt mK CF NQ+ CF oe+ — — — GM+
tricuspidata Planch. bm+ Mk+ +EG NP tCf+ oE+ — — — GM
Pastinaca sylvestris Mill. +BM Ok+ CF LP+ CF+ +eg df kn — Gg+
Pedicularis capitata Adams. +aB mU +DE+ KM+ cp oe+ ef jm — Gg
dasystachys Schrenk. bN +kK+ CE MN tCF+ +eH+ — — st Gg
hirsuta L. AB oM DF KN+ cf+ oe — — — Gg
kaufmannii Pinzg. +bm+ mk+ +DE+ MO+ sF+ Mg cf kn rs+ Gg
lanata Chum, et Schlecht. AB+ ok+ DF KN cF oe - — — Gg
oederi VahL +ab+ OU DF KN+ cp+ +oe eg jm — Gg+
palustris L. aN+ OU DF KO ca OE af jl qr+ Gg+
phy so calix Bge. bN MK+ CE MO i-sC +eE df jn — Gg
sceptrum-carolinum L. +aN+ mU +DE+ KO HP oM+ df jn qr Gg+
sudetica Wilid. Ab oU DF KO cp+ oe — — Gg
Peplis portuia L. BM Ok+ CF +NQ fp ME ad — — Gg+
Petasites albus Gaertn. +bN+ Ok +DF+ +O+ +fp ME af jn tgs+
hybrid us Gaertn. bm ok +DE+ Ю+ Fa eE cf mo — Gg+
spurius Rgl. Bm oK +CE+ MO+ cF Me+ — — — Gg
Peucedanum alsati- cum L. +bm +MK CE MO+ +sf oH eg kn — GM
cervaria Cuss. bm+ +mk CE+ OP Sc+ ME cf jm — GM
latifolium D.C. +N+ Mk +D+ +O+ cF +gH+ fg — — Gg
oreoselinum Moench. +bm+ +mk CE+ NP SF oe af jl — Gs
palustre Moench. +Bm ok+ +DE+ MO+ +ca OE af km pr GM+
ruthenicum Bieb. +Nm mk+ +DE ЮР sf +eE — — — Gg
Phacelia tanacetifolia Bern. bm ok DE Ю+ Sf — cf +ln+ — Gg+
Phalaris canariensis L. +bM ok+ +DE+ MQ Cp — cf +1П+ — Gg
Phalaroides arundinacea Rausch. aM OU BG KQ CP MH eg lo rt Gg
Phellodendron amurense Rupr. +bm+ MU DG +MP+ t-cF +Me+ — - — — GM+
Philadelphus corona- rius L. +Nm+ +oM+ +DF NP+ SP +ME cf kn — GM+
Phyllitis scolopendrium Newn. bM Ok +DF+ OQ cF +oe+ df In — gs
Phleum aipinum L. +aB+ OU +DF+ KO+ CF oE+ af ln+ — Gg+
nodosum L. BM OK CF MQ +Cf df +ln+ — Gg*
phleoides Karst. bM OU +BF LQ df oH dg jn rs+ Gg
pratense L. aM OU +CF+ LQ SP oh ag In ps+ Gg+
Phlomis pungens Wilid. +NM mU BE NQ dc+ eg — — — Gg
tuberosa L. +bM +mU BE KQ dF Mh eg — rt Gg+
Phlox sibirica L. +BN +kU DE KN Sf +ME — — — Gg
167
Таблица 55 (продолжение)
Вид Факторы
Tm I Кп Om I ° Hd I Tr Rc Nt ffl Lc
Phragmites communis at EU +BH KR +Sa OP bg kn ru GM
Trin.
Phyllodoce coerulea Bab. ab OU DF+ +LO cf oe — — — Gg+
Physalis alkekengi L. +Nm+ OK BF +OP+ cfT ME df +ln+ — +gs+
Physocarpus opulifolia +bM +mk DF MQ Sf oE — — — Gs+
Max.
Physospermum cornu- NM MK CE OP +Cc+ +oM+ be — — gs
biense D.C.
Phyteuma nigrum bm oM DE +O+ cp — cd km — GM
Schmidt.
orbiculare L. bm +oM+ +DE+ TOP Cp тМе df jl — Gg+
spicatum L. bm mk +DE +O+ cf+ Me af kn — Gs
Picea abies Karst. BN+ oM+ DET +NO+ терт Oe+ af jn — +gS
canadensis Britt. tBNt mK DF LO Cp OE+ af — — +gs+
fennica Kom. Bb+ mM+ +E+ NO терт Oer ает jn — +gs
glauca Voss. +BN+ mK +DF+ LN+ +CP Oe+ - — — -tgs+
jezoensis Carr. Bm MU DF +LO+ +cF+ +oe+ be — — +gs+
mariana Britt; TBm +mK DF LO +cP Oe — — — +gS
obovata Ldb. +ВЫ- oU DF KN+ +cF+ Oe+ ae+ jn —
omorica Purkyne. BN+ +Mk+ +DE+ NP +cF+ OeT — — — +gs+
orientalis Link. +Nm+ mk DF NP +cp+ +ME+ bf — — +gs+
pungens Engelm. +Nm kU +CD+ NO CF+ oe+ — — +gs+
schrenkiana F. et M. bN+ +KU CE MO CF+ +ME — — — +gs+
Pieris hieracioides L. +bM 0K+ CF+ MQ sf Mh dg km rt GM
Pimpinella major Huds. BM Ok+ CF NQ +cF +Me+ cf ln+ — GM
saxifraga L. +BM OU +CF+ LO SF OE af jl rs+ GM+
titanophila Woron. +bm +k+ +DE +NO+ +Cc+ oe+ dg — — Gg
Pinguicula villosa L. ABT EU EH KO+ +fF+ +oM+ — — — Gg
vulgaris L. +ab+ OK +EG KO+ cP oe cf jm — Gg
Pinus banksiana Lamb. +BN+ +MK DF LN+ CP OMt ad — — GM
cembra L. +Nm oMt DE +O+ +cF+ Oe+ +bd+ kn — Gs+
eldarica Medv. +m+ +M+ +D+ +O+ SF og bg — — GM
funerbis Kom. bN kK EF +N+ +Cf+ тоет — — — GM
halepensis Mill. ттМт +mk+ BE +PQ SF og bg — — GM
hamata Sosn. Nm +Mk+ +DE+ TOP Cp+ OE+ og — — GMT
koralensis Sieb. et Zucc. +bN+ +MK+ +EF +MO+ терт +ME bc+ kn GsT
montana Mill. bm mk DE +NO+ TCP OE af jm — GgT
mugo Turra. bm mk DE NO+ Sp+ Toe af jm — GgT
nigra Arn. +NM mK +CE +OP+ +CF+ OE dg jm — Gs
pallasiana Lamb. TniM mk+ CE+ OQ SF+ OE dg jm — Gs
pinea L. mM mk BE PQ +CF+ oE bf — — GM
pityusa Stev. ттт mM DF +P+ SF og bg — — GM
ppnderosa Dougl. bM Ok CG NQ +CF+ ToE — — — GM
pumila Regel. aN +mU DF+ KO+ +cp Oe — — — Gs
sibirica Du Tour. +Bb+ +MU DE+ +LM+ +cP OE be kn — GsT
strobus L. bm+ mk+ DF MO+ CF+ +oE af jm — GsT
silvestris L. TBm ToU +DF KP +CP+ Oe+ ag jn — GMT
Pistacia mutica Fisch. TmM mK+ BD OPT +Sc+ Mg df — — GgT
vera L. TmM kU BC OP de TeE+ dg - — Gg+
168
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
Plantago cor nut ii Gouan. +bm+ mK+ CE LP+ +cP +eP dg — st Gg
indica L. +bM OK CF MQ dC eh eg km tu Gg
intermedia Gilib. aM OK BF+ +MQ cP — bd+ km — GM+
lanceolata L. +BM+ OK +BF+ +LQ+ dF oP ag j° pt GM
major L. BM+ +OU +BG+ +LQ+ sP oP ag j° qu GM
maritima L. aM OK+ +CF +NQ dP +eP dg jo st Gg+
maxima Juss. bm mK CE MO SF eP dg — su Gg
media L. aM OU CF +LQ sp oH dg jm — GM
stepposa Kupr. bm +MK+ CE +MO+ sf oh eg — rt Gg
Planta nt hera bifolia +Bm+ +OK+ +DF+ +MP+ cp oE cf jn — gs
Rich, chlorantha Rchb. +bm+ Ok+ +DF+ +OP+ cF +ME cf jn — gs
Platanus occidental L. +NM+ +mK +DF NQ Cp+ +ME+ eg +lr+ q‘ GM+
orientalis L. NM mK+ CE+ +OQ +Cp+ ME+ eg +ln+ qt GM+
Pleurospermum austria- +bm oM+ DE+ +O+ cF oE eg kn - GM
cum Hoffm. uralense Hoffm. aN mU DE+ KN cF oE — — — GM
Poa alpina L. +ab+ OU CF+ KO+ sF oh ag In — GM
angustifolia L. +BM OU BF +LQ dF+ MH ag jm — GM
annua L. at EU +BG+ +LR SP oE+ af me rs GM
bulbosa L. bM OK+ CF MQ Df MP bg jk+ su Gg+
chaixii Vill. bM Ok CF +OQ cF oe ad jm — GM+
compressa L. Bm Ok +DE +NQ+ sf oE ef jl — Gg
glauca Vahl. AB Ok DF KO cp +Me+ df jm — Gg
nemoralis L. aM+ EU BG KQ+ SP oE be jm rt gs
palustris L. aM EU BH KQ CP OH dg ln+ qt+ GM+
prat en sis L. +am+ +OU BG KP sP oH ag jo pt+ GM+
remota Forsel. +BN+ oK+ CE+ +MO FP +eE +df+ +ln+ — gS
sibir ica Roschev. am +kU CF KO+ Cp+ oE — — — GM+
silvico la Guss. NM mK+ BE+ +OQ +CF ME+ cf +ln+ — Gs
tr ivialis L. +BM OU BF +LQ CP oh ag lo pt+ GM+
Polemonium boreale AB+ Ok DF KO +cf+ oe+ — — — Gg
Adams. coeruleum L. am ok+ DF MO cP oE df In rs GM+
Polygala amarella +Bm ok DE+ +NO+ cp oE df jm st Gg+
Gantz. cornosa Schkuhr. +Bm ok +DE+ +NO+ sf oh +dg jl qs+ Gg+
hybrida D.C. BN MU CE MO sf ME cf — — GM
sibirica L. bm mU +DE +MO+ Sf+ +ME+ cf — — Gg
vulgaris L. +BN+ OM+ DF NO+ SF oe ad+ jl+ — Gg+
Polygonatum multiflo- +bm+ OK CF NP+ Cf +ME cf km — gs
rum All. odoratum Druce. +bm+ OU CF +MP+ SP og cf+ jm rs +gs
Polygonum alpinum All. +ab+ mK+ DE KO sF +oE ad In rs+ Gg
amphibium L. +BM+ OU BG KQ cA oH ag lo rt GM+
aviculare L. +BM+ OU BG KQ dP oP ag jo rt+ Gg+
bistorta L. +am OU CF KP CP Og bf kn ps+ GM
convulvulus L. +BM OU BG KQ dp Mg +cf+ +kn+ rs GM
12. Зак. 448
169
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Факторы
Вид Tm Kn Om Cr Hd Tr | Rc Nt fH Lc GM+ Gg GM Вид Факторы
Tm | Kn Om 1 Cr 1 Hd Tr | Rc 1 Nt fH Lc
dumetorum L. hydropiper L. minus Huds. BM BM BM+ OU OU OU BG BG « BG LP -LQ LQ cP cP +fP +Me+ og oe+ cf bf bd +ln+ kn mn+ qs+
intermedia L. longipes Ldb. «-Ы- +bN mk +kK DE CE NO MN 1 Cf «Cc+ ME «-eh«- cf kn st Gg Gg-t-
mite Schrank. bM Ok CF NP+ +Fp+ ME af ln«- — GM multifida L. +BM +kU BE+ KP+ +sF +eg cf«- -t-ln-t- — GM
nodosum Pers. « -bM+ OU BG LQ ca MP ag mo — Gg nivea L. ab +MU DE KN +Cf+ f-ME-b cf«- «-ln«- — Gg«-
patulum Bieb. bM OK+ BF MQ Df+ +MP eg Gg norvegica L. am oU DF KO CF ME cd kn — Gg
persicaria L. +bM+ OU BG ' LQ FP eg af+ ln+ — Gg recta L. +bM mK++CE+ +NQ dc+ oh eg kn qs GM
sachalinenses Schmidt. hb+ +Mk EF hNO+ CF+ +Me+ — GM reptans L. +bM+ OU BF +MQ+ sP og cf kn ru Gs+
scabrum Moench. +BM+ OU BF «LQ sa oh eg +ln+ — Gg stipular is L. AB +MU DE+ KM +cF+ «-ME — — — Gg«-
viviparum L. Ab OU DF KN+ +cP oe+ ad jn — Gg supina L. +bM+ OU +BF+ KQ+ CP eE df -t-ln-t- su Gg
Polypodium vulgare L. am OK CF MP Sp oE ac+ jn — Gs+ Poterium sanguisorbaL. bm +mk+ +DE +O+ sc+ ME — — - • Gg
PolystichumbrauniiFee. +bN+ Ok+ DF LP +fF+ oE be ln+ — gs Prenanthes purpurea L. bM OK CF +OQ cf+ «•Me af kn — +gs
lonchitis Roth. ab oK CF MO cF oe df kn — +gs Primula acaulis L. bm mk CE OP+ CF Me«- cf kn — Gg«-
Populus alba L. bM+ oU BF MQ cp Mg dg In rt GM auricula L. aN mk DE+ +O+ SF — df jl pr Gg-t-
baisamitera L. +Bm oK «DF«- +LO cp +oe+ — GM+ elatior Hill. bm+ mk +DE+ OP+ «-cF Me cf ln«- — GM
deltoides Marsch. +ЬМ « mK CF MQ CF oE — — GM+ farinosa L. ab +oK+ DF +LO cP oE«- ef jl — Gg«-
diversifolia Schrenk. NM+ MU Bd +OQ+ +dF +eh+ dg — rt Gg+ halleri Gmel. ab mk DE +0+ SF oe+ df jm — Gg-t-
maximowiczii Henry. +bN+ MU DF LO+ CF+ oE — rt GM+ macrocalix Bunge. «•bm MK +DE+ MO-t- +Cf+ Me+ — — GM
nigra L. +bM+ oU +BF MQ CF Mg eg lo ru GM minima L. +ab mk DE NO +cF+ +oe ac«- jm — Gg«-
piramidalis Pers. NM mU BE «OQ SF +eg dg — qs Gg+ veris L. +bm+ «-mk DE+ NP CF+ «•ME df jm rs GM«-
suaveolens Fisch. t-Bb mU DF KM+ +cf+ +Me+ — rt GM+ Prinsepia sinensis Kom. bm kK DF MO CF «•ME cf — — Gs+
tremula L. f-Bm+ OU +DF KP«- CP +og ag jn+ — GM+ Prunella grandiflora +bM +mk+ CE+ NQ sf Me df jm — GM«-
tremuloides Michx. +Bm+ +oK+ Portulaca oleracea L. +NM mU CG +LP+ BF +NO Cp +sf+ +oe+ eE+ cf+ ln+ — GM+ GM Jacq. vulgaris L. aM +oU BF KQ SP Og bf jn — GM+
Potamoeeton crispus L. +BT EU +BH KR Pa+ +eE cf In — GM Prunus divaricata Ldb. NM MU BF NP+ SF Mg eg — — Gs
eramineus L. am OU CH LP +FA ME cf jm — Gg+ domestica L. +NM oU +CF NP+ SF ME cf kn — Gs
lucens L. +BM OK+ BH MQ PA +eE cf mo st GM spinosa L. Bm+ OK+ +CF NQ sf+ og ag jn — GM
natans L. +BM+ OU BH LQ+ PA oe cf km st GM Pseudotsuga taxifolia Britt. Pteridium aquilinum Kuhn +bm+ Ek+ CH +NP+ +cF+ +oe«- «cf jn «gs-t-
nodosus Poir. obtusifolius Mert. bM t-bm+ OU OU BF+ CF+ MQ KP+ aA aA eE cf bf In In — GM+ GM+ BT EU +CH LR Cp Oe ad jm rs +gs«-
pectinatus L. aM юи BG KQ aA eg eg kn — GM+ Puccinellia distans Pari. «•BM OU BF+ «-MQ +dF+ «-eP eg fg ln«- su
perfoliatus L. at EU BH KR PA Mg eg km st GM, palustris Grossh. bm+ mK BE+ MP +sp+ «gP
praelongusWult. aN EU +DG+ +LO+ PA df km — Gg+ Pulcaria prostrata Asch. «-bM OK+ BF+ MQ SP ad mn+ st Gg
pusillus L. at EU BH MR PA eE cf +mo — bM+ Pulmonaria angusti- bm+ +mk+ CE NP +Cf+ oe«- ce jm GM«-
PotentiUa alba L. bm+ anserina L. +aM mk+ «-DE OU BF+ NP KQ Sf dP ME oP cd ag kn lo pt GM GM folia L. mollissima Kern. «-bm+ mK CE LP +cf+ «•Me+ df kn GM
arenaria Borkh. +bm hmk DE+ +O+ de oe+ df jm — GM obscura Dumort. Bm +ok + DF MP CF+ oE df ln«- rs GS
argentea L. +Bm+ +ok+ «DE+ LO+ dp oP ag jk+ +rs+ Gg+ Pulsatilla alpina Schrank. aN +mk DE +O+ cF +oM+ df jm Gg«-
b if urea L. BN i-Ku DE KN +dF MH — — st Gg flavescens Juz. «ВЫ- MU DE KN sF oE Gg«-
canescens Bess. t Bm+ +mK+ CE+ LP +Cc+ +eE cf jm — Gg nigricans Stork. bN+ mM DE +O+ Cf Oe
crantziiG.Beck. «ab OK DF LO cF «•Me df jm — Gg+ patens Mill Bm mk+ DE+ MO sf «•ME rs GM
egedii Wormsk. AB Ok DF KO «-fF +eH dg — — Gg pratensis Mill. +bN+ +mM+ DE+ NO+ Sc oe cf jl rs Gg«-
erecta Raeusch. am Ok+ DF+ MP dP Og af+ jl qs Gs turczaninovii Kryl. BN kU +DE +LM+ +sf +eE«- Gg+
frag iform is Willd. ab OK «-DF+ LN +cf +Me+ — — — Gg+ vernalis Mill. «-bm+ Ok «DF+ NP «-cf«- ad jl GM
fruticosa Rydb. «am«- юи CG KP +CF+ oE cf kn — Gg+ vulgaris Mill. «-bm OM «DF+ +OP +Cc«- ce jl GM«-
goldbachii Rupr. BN Hnk+ DE+ MO cf Me — — rs GM Punica granatura L. +mM++MU «-BE OQ+ ds«- eg+ eg jm GM+
heptaphy Ua Jusl. bm mk DE +O«- df og df+ jm rs Gg Pycreus flavescens P.B. «bt+ OU CG NR cP cf kn Gg
humifusa Willd. Bm +MK+ CE +MO+ dc+ «•eg — — rs Gg Pyrethrum corymbosum «-bm«- Ok+ CF NP Sf ME df jm rs Gs
171
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы
Tm | Kn | Om Cr 1 Hd Tr Rc Nt fH Lc
parthenium Smith. bM OU BF +MQ +Cf+ oe+ cf +ln+ — Gs
Pyrola inearnata Fisch. aN mU DF KN cF+ +oE — — — +gs
media Sw. +Bm +Ok +DF+ +LP cF Oe+ be jl — +gs
minor L. +am +OK+ +DF+ +LP cF+ oM+ ad jl — +gs+
renifolia Max. Bm +mK+ DF +MO+ +cF oe — — — +gS
rotundifolia L. +am +OU CG KP +cF+ OE be jm qr +gs+
virescens Schweigg. +Bm OU DF+ +MO+ CF OM+ be jl — +gs+
Pyrus caucasica Fed. Nm +Mk+ CF +OP +CF ME — — — Gs+
communis L. +bM OK +CF MQ +CF Mg dg kn Gs+
elaeagrifolia Pall. +mM +mK CF NP +Cf+ ME de — — Gs
rossica Danil. bN +Mk+ DE +O+ SF +ME — — — Gs
ussuriensis Max. bm+ MU +CF+ LO CF ME — Gs+
Quercus alba L. Nt +Mk DE+ NO +Cf+ +ME+ — Gs
borealis Michx. +bm+ +mk+ CF +NP CF+ oE be km — Gs
castaneifolia C.A.M. СОН- +O+ +cF+ ME — - — Gs
cerris L. +NM +mk+ СЕ OP +CF +ME af jn — Gs
dentata Thunb. r'Nm +MK+ DF NO+ +Cf+ ME — — — Gs
iberica Stev. NM mK +DF OP +CF ME — — — Gs+
ilex L. NM+ mk+ BE +PQ Sf+ +MF. ag jn GS
macrantheta Fisch, et +Nm mK +CD+ +NO SF +Me+ — - GM+
Mey. Gs
macrocarpa Michx. bM+ mK CF MQ +CF+ oE — — —
mongolica Fisch. +bm+ MU DF +LO+ CF +ME — — — Gs
palustris Moench. Nm +MK CE +OP+ +cp+ ME — — — Gs
petraea Liebl. H-bmH- OM+ CF +OP +CF+ +ME+ af jn — Gs
pubescens Wilid. +Nm+ mk+ CE +OP+ +Sf+ ME+ eg jn — GM
robur L. +bm+ +ok+ +DF +NP +Cp +Mg ag jn ps Gs+
rubra L. +mt mk+ +D+ OQ CF+ oE bf kn — Gs
suber L. mM mk BD+ PQ CF Oe+ bg — — GM+
Rad io la linoides Roth. bt Ok+ CF +OR cP — ad jl — Gg+
Ranunculus alpestris L. +ab +ok+ DE NO +fp+ — df km — Gg
acris L. am oK CE MP SP oP ag jn pt GM
ar vensis L. NM mU BD+ OQ +cf+ — df jn — Gg
auricomus L. +Bm Ok+ +DF+ MO+ cP oh eg +ln+ ps GM+
borealis Trautv. +aN oU +DF+ KN+ +cF+ +ME de In — gs+
bulbosus L. oK+ CE+ NP+ Cf +eE cf jm — Gg+
cassubicus L. Bm ok+ DE+ +NO+ +fF+ +ME cf kn qs gs+
flammula L. +Bm+ Ok+ +DF+ +NP fP oh +bg jm qs Gg
glacialis L. Ab OM DF NO +fF+ — ad jl — Gg
gmeliniiD.C. Ab +mU DE+ KN cP oe+ — — — Gg+
illyricus L. bM +mK +DE NP de OE cf — — Gg
lapponicus L. +ab oU DE+ KN+ fF+ oM+ — — — GM+
lingua L. +bm OK CF +LO+ pa+ °g +cf kn qs GM
montanus Wilid. ab+ +ok DE+ NO cp — df In — GM+
nemorosus D.C. bm mk DE +O+ cF oe+ ce jn — GM+
polyanthemus L. +Bm+ OK +DF+ +MP SF oH ag jm pt+ GM+
pedatusW. et K. bm mK CD+ MO dF+ eh+ dg — — Gg+ 1
repens L. am+ OK CF+ KP cP oH ag jn pu GM
Вид Факторы
Tm Kn Om Cr j Hd Tr Rc Nt fH Lc
reptans L. +ab+ OU DF KO pa — af Gg+
scleratus L. aM+ +OU +BG+ KQ+ +fP+ Og df mo pr Gg
stevenii Andrz. +Nm mk+ CE -Юн- +cF+ +eE GM+
Raphanus raphanistrum L. aM +OU +BF+ LQ sf oE bd kn - Gg
Rapistrum rugosum All. bM OK +BF+ NQ den- +Eh eg +ln+ Gg
Reseda lutea L. +bM mK BE NQ se eE df +km+ Gg
Rhamnus cathartica L. +bm+ oK +DE+ +NO+ sF ME df jn rs GM+
dahurica Pall. +bm +kU DF +MO Cf Me — GM+
Rheum compactum L. +Bb+ +MU +DE+ KM+ cf+ oe — — Gg+
tataricum L. bN kK CD +N+ DS EH eg Gg
Rhinanthus angusti- +Bm+ OK+ +DF+ +LO+ SP oh eg jm ps+ Gg+
folius C.C. Gmel.
minor L. Bm Ok+ +DF+ MO+ cF oE af jl rs Gg
Rhodiola rosea L. +am+ +mU CE KO +Sf+ oe ac+ jl ps Gg
Rhododendron aureum aN oU +DF+ KO cp +oM+ Gg+
Georgi.
caucasicum Pall. Nm+ mk +DF +OP +cp+ +oe bd — GM
dahuricum L. +bN+ +kU DF LN CF +oe+ — — — Gs
luteum Sweet. +NM mk+ +CF +OP+ cp+ oe+ — — — GM+
laponicum Wahlb. aB+ om +EF +NO +fp+ +oe — — Gg+
parvifolium Adams. am oU +DF+ KO+ +fp+ +oe — — — GM
ponticum L. NM mk +DF +OP CF+ +oe+ be — — Gs+
Rhynchospora alba h-BNh- OU +DF+ +MO+ Fa OM ac+ jl Gg
VahL
fusca Ait. bN Ok DF NO fa ac jl Gg+
Ribes aipinum L. bm+ +om+ +DE+ +OP+ Sp +ME df jn Gs+
aureum Putsch. bm+ OU CG +NP+ +SF +Mg eg — GM
dicuscha Fisch. ab +kU DE KM cf+ +Me — Gs
fragrans Pall. BN +KU +DE+ KM+ cf oe — — — GM+
nigrum L. aN+ oK+ CE+ LO cP oE be kn ps +gS
procumbens Pall. +bN+ MU DF +MN+ fp +Me — __ +gs+
pubescens Hedl. +bN+ ok DE +NO+ cP oE — — +gs+
rubrum L. ab oU +DE+ KN+ cP oe ce In — +«s
triste Pall. am oU CG+ KO+ H-cfH- oe — — — +gs+
Robinia pseudacacia L. +NM+ mK CE+ NQ SF+ +Mg ag mn+ — GM+
Roegneria canina +BM OK+ +CF+ +LP+ cF oE cf lo +gs+
Nevski.
Rorippa amphibia Bess. +Bm+ OU CF KP+ fa Mh eg lo st GM
austriaca Bess. +bm+ +mk СЕН- NP cP +eP eg +ln+ Gg+
brachycarpa Hayek. bN+ MK СЕ MO dP Mh+ eg +1ПН- su Gg+
islandica Borbas. +aM+ OU BG KQ+ SP+ +Mg af lo — GM
prostrata Schinz. +bm+ Ok+ +DF MP fF+ Оен- cf +ln+ ru Gg+
sylvestris Bess. BM OK CF NP SP +Mg df In — GM+'
Rosa acicularis Lindl. am mU CF KN Sp OE — — __ Ga
caesia Smith. +bm Ok DF +O+ Sf — ce km — Gs
canina L. +bM+ OK+ BF+ +OQ sP oE af jn — GM+
cinnamomea L. aN+ oK DE+ +MO CF og +cf kn rs Gs+
172
173
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr | Hd Tr Rc Nt fH Lc
corymbifera Borkh. bM OU BE +OQ CF — cf jn — GM
dahurica Pall. Bm kU DF +LO +Cf+ +Me+ — — — Gs
gallica L. bm+ +mk+ CE+ OP Sf ME cf km — GM+
pend ulina L. bm mk CF OP cF ME cf kn — GM
rugosa Thunb. Bm oK DF+ MO+ CF +Mg eg — — GM+
tomentosa Smith. bM Ok CF +OP+ SF oM+ df km — Gs+
villosa L. +Nm+ oK CE+ OP+ Cf — cf jl — GM
Rubacer odoratum +NM+ mk DF NP CF+ +Me+ — — — Gs+
Rydb. Rubia tinctoria L. bM OK BF ' +OQ Sf cf kn — Gg+
Rubus arcticus L. +ab+ oU DE+ KO cP Oe+ — — qr GM+
caesius L. +bM OU BF +MQ SP Mg +cf+ mo st Gs+
chamaemorus L. +ab +oU DF KO+ cP Oe ac+ jk+ pr GM+
humulifolius C.A.M. +BN+ MU DF KN+ +fF+ oM+ ad — +gs
idaeus L. +Bm+ OU CF MP cP oE af lo — GS
nessensis W. Hall. BN 0k+ DF NP fP +ME af kn — Gs
sachalinensis Lew. am +MU DF KO +cF oe+ — — — Gs+
saxatilis L. am oU DF KO+ CP +oe+ af km qs +gS
Rumex acetosa L. +aM +OU CF+ +LQ SP oE af kn ps+ GM
acetosella L. aM OU CF+ KQ SF oE ac jl qs GM
aquaticus L. am+ oU CF KP ca og cf lo — Gg
confertus Willd. +bm+ mU CF +MP Cp oH — — ru Gg
conglomeratus Murr. bM OK BF +OQ fp eE af mo — Gg+
crispus L. +Bm+ OK BF NP SP oH ag kn — Gs
hydrolapathum L. bm+ Ok+ CF NP+ Fa og cf lo — GM
maritimus L. +BM+ EU BH KQ+ cP +eg+ dg mo — Gg+
obtusifolius L. +bM Ok+ CF NQ cF +ME af mo — +gs+
pseudonatronatus Borb. aN mK CE LO +SF+ +eE — — — GM
sanguineus L. bM Ok+ CF +OQ SP ME cf ln+ — +gs+
stenophyllus Ldb. bm MK BE MO dP MH+ — — rt+ Gg
thyrsiflorus Fingerh. am oU CE+ KO+ SF og cf kn rs Gg
Ruppia maritima L. ЬМ EK BH MQ+ aA gP fg — — GM+
Ruscus hypoglossum L. +mM Mk CD+ OP CF+ +ME — — +gS
ponticus Woron. +mM Mk CE+ OP Sf +ME+ +cf+ jm — Gs
Sagina nodosa Fenzl. +aN oU DE+ KO fP og dg kn pr+ GM
procumbens L. aM OU CF MQ Sp og eg In — Gg
saginoides Karst. +ab oU DF KO cp — ad km — GM
Sagittaria sagitti- +Bm+ Ok+ DF MO pA Mh eg In st Gg+
folia L. natans Pall. +BN oU DF KN+ pa +eE — — Gg+
Sallcornia europaea L. at OU BH MR +dp+ +gs dg lo st Gg
Salix acutifolia Willd. +Bm mK CE+ +NO+ CP OM+ df km pt Gg+
alba L. +bM+ OU BF +MQ +fP Mg dg kn pt GM
aurita L. Bm Ok DF +NO+ fP Oe ad jm ps GM+
caprea L. am +OU +DF+ MP cP oe cf jn qs Gs+
cinerea L. +Bm+ OK CF+ +MP ca Oe be km qs+. Gs
dasyclados Wimm. aN mU DE+ KO +cp+ Oe+ df jn — GM+
fragilis L. BM OU +DF MP fp ME be In rt GM
glauca L. +aB oU DE+ KO cp oe — — — Gg+
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt | fH Lc
hastata L. aB+ oU DF KO +fp+ oe cf km Gg+
herbacea L. AB OM+ DF+ +LN+ ep oM+ ad jm — Gg+
lanata L. +aB OU DF KO +fF+ +oe — — — GM
lapponum L. ab +ok+ DE+ KO+ fP Oe — — — GM
myrsinifolia Salisb. ab+ Ok+ DF+ MO+ fp +Me+ df In rs GM
myrtilloides L. aN oU DE+ KO cP Oe ac jm qs Gg
pentandra L. +Bm+ OU +CF KP fP oE ce km rt GM+
phyiicifolia L. +ab+ Ok +EF+ +LO+ cP +oe — GM+
polaris Wahlb. AB oU +EF KN cF+ oe — — — Gg+
purpurea L. s. str. bM +ok+ +CF +OQ cF ME df jn rt GM
repens L. Bm +Om+ +EF+ +OP Sp — af jn rs Gg+
reptans Rupr. Aa+ ok+ +E+ KN+ +cp+ oe — — — Gg+
rosmarinifolia L. Bnt +mK+ CE +MO+ +ca +oe+ cf jm pr GM
starkeana Willd. BN ok+ +DE+ +NO+ +cP OE af jm qs GM+
triandra L. +Bm+ OU CF LP fP oE df kn rt GM
viminalis L. +am+ OU BF KP cF Oe df jn tu GM
Salsola collina Pall. +bN+ +kU CD+ MO Df+ eg+ — — — Gg
kali L. . +bN+ Om EF +OP Sp eh+ eg mo — Gg
ruthenica Iljin. +bM +oK+ BE+ +MQ de EH+ eg — — Gg
soda L. +NM mK CD+ +NQ Dlr EP fg — SU Gg
Salvia aethiopis L. +NM OK+ BE +OQ +sC+ oE cf +ln+ — Gg
glutinosa L. bM Ok+ BF NQ cF ME cf lo — Gs+
nemorosa L. +Nm+ mk+ CE OP dF oH bg In — GM+
nutans L. +bm +Mk+ +DE NO+ sC+ eE — — — Gg
pratensis L. +bm+ Ok CF +OP+ sf ME df km qs GM
sclarea L. NM +MU BD+ OQ +sc +eE cf kn — Gg
stepposa Schost. +bm MK CD+ MO dc+ MH eg — rs+ Gg
verticillata L. BM OK CE+ +NQ df Mg eg +ln+ rt GM
Salvmia natans AU. bt OK BE MR Pa og cf lo st Gg+
Sambucus ebulus L. bM OK BF+ +OQ sF ME df lo — GM+
nigra L. bM OK BF +OQ Cp +ME af mo — GS
racemosa L. +bm+ Ok DF +OP +fF oE be lo — +gs
sibirica L. Bm kU DF +LN+ +cF+ oE — — — +gs
Samolus valerandii L. bM OU BF +OQ CP +Eg af+ In — GM
Sanguisorba officinalisL. am+ oU +CF+ KP SP OH ag jm qt+ GM
Sanicula europaea L. +bM Ok+ CF +NP+ Cf ME df In — gs
Saponaria officinalis L. +bM mK CE NQ cF — cf kn — GM
Sarothamnus scoparius lim+ Ok CE+ OP+ +cF — ad jm — Gg+
Wimm. Saturea hortensis L. Nm oK BE +OP CF +cf +ln+ Gg+
montana L. NM mK +CD OQ Sf — df kn — Gg+
Saussurea alpina D.C. +aB+ OU CF KO +CF+ +oe cf kn — Gg
amara D.C. Bm kU BE KO +SF+ eh+ — rt Gg
Saxifraga aizoides L. AB +oM+ +E+ +LN+ cp oe+ df jm — Gg+
caespitosa L. AB +oK+ DE+ KN+ cf+ Me — — — Gg
granulata L. +bm+ ok+ CE NP cF oe be jm — Gs
hircuius L. AN OU DF KO cP oe ad jm — Gg+
nivalis L. aB+ OU DF KO+ Cf +Me — • — — Gg
175
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
oppositifolia L. AB oK+ DF KN+ cF +Me df jl — Gg
punctata L. +ab+ mU DF KM+ cF oe — — — Gg+
stellaris L. AB +oM +E+ MN+ FP — be jm — Gg+
tridactylites L. +bm+ ok+ +CE+ +OP+ Sf — cf jk+ — Gg+
Scabiosa columbaria L. bm+ ok+ +CE+ OP+ df+ ME df jl — Gg
ochroleuca L. bm +mK+ CE +MO+ df oh dg jm rsr Gg+
ucrainica L. Nm +k+ CD+ OP dC+ eE — — — Gg
Scandix pecten veneris L. bM+ +oU +BE+ OQ+ cf — df km — Gg+
Scheuchzeria palustris L. +BN+ OU DF KO+ FP+ Oe ad jl — Gg+
Schizandra chinensis Ba ill. bm mK++EG +MP cF +Me+ — — — GS
Schoenus ferrugineus L. Bm+ Ok+ +DF NP fP — cf jl — Gg
Scilla bifolia L. bm+ +mk+ +DE +OP +Cf+ +ME cf In — GM+
sibirica Andrz. +bm+ mk+ +DE+ +OP SF +ME cf — — GM+
Scirpus compactus Hoffm. +BM mU BE+ KQ +FP+ +eE — - — Gg
lacustris L. +Bm EU +DH KO+ ca+ oP eg kn st Gg+
maritimus L. +BM+ EU +BH KQ +ca eH+ dg kn ru Gg
radicans Schkuhr. +Bb+ mU +DE+ LP Fa ME+ cf+ In — Gg
sylvaticus L. +Bm+ OU CF LP fa oH bg jm pr Gs
tabernaemontanii Gmel. +BM OU BF +MQ +fa+ +eh+ dg jn — Gg+
Scleranthus annuus L. BM OK CE NQ sf oE ac+ km rs Gg+
perennis L. Bm+ OK +DF NP sf oE bd jk+ — GM+
Sclerochloa dura Beauv. bt OU BF OR df +EH eg kn — Gg
Scolochloa festucacea Link. Bm mU CE+ KO+ ca +eh — — — Gg
Scorzonera austriaca Willd. +bM mK+ CE LP+ +sc+ Mg df jl — Gg
humilis L. bN+ Ok DF NO+ cF +ME be jl qr Gg+
parviflora Jacq. bM +MK+ BE MQ +cP EP — — rt Gg
purpurea L. bM mK CE MQ Sc+ ME df kn ru Gg+
taurica Bieb. bm+ mK CE NP +sf+ +eE df jl — Gg
Scrophylaria nodosa L. +BM Ok+ CF +LQ CP +ME bf lo — gS
scopolii Hoppe. bM +MK+ +CF NQ cF +ME bd — — Gs
vernalis L. bm+ Ok CF NP+ cF — +cf +ln+ — +gs+
Scutellaria galericulata L. aM OU BF KQ ca oE cf In qt GM+
hastifolia L. +bm+ Ok +DF+ MP +CP oE cf +ln+ rt GM
Secale silvestre Hoppe. bm+ mK BE MP Df Mg cf+ kn — Gg+
Sedum acre L. +BM OK +CF MQ sf Og af+ jk+ qs Gg+
maximum Hoffm. bM OK+ CF +MQ set Oe +cf jm — GM
purpureum Schult. +Bm OU +DE+ KO+ sF oE +df jm st+ GM
sexangulare L. +bN+ OM+ DF +O+ +SF+ +ME df jk+ — Gg
telephium L. +bm +mk+ +DE+ NO+ +SF+ Mg cf+ jn — GM
Selaginella selaginoides Link. +aB+ +OK+ DF KO +CF+ oe+ cf jm GM
Selinum carvifolium L. +Bm +mk +DE+ MO+ cP oe be jl — GM
Sentpervivum monta- tum L. +aB+ +mk+ +DE+ NO +Cf oe ac+ jk+ — Gg+
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
ruthenicum Schnitt. bm mk +DE NO+ Sc+ Me — — — Gg+
soboliferum Sims. +bN+ +mk +E+ +NO Sc+ +ME df+ jn — Gg+
Senecio borysthenicus Andrz. bm i-Mk +DE +O+ Cc+ oM+ — — - Gg
campester D.C. +ab+ t-mk+ DE+ +MO +sF+ og — — rs+ GM
erucifolius L._ Bm OU +CF+ KO+ +Sf Mg df+ km — Gg+
fluviatilis Walk. +bm OK CF +MO+ +FP+ — cf mo — GM
grandidentatus Ldb. bm +MK CE MO+ sc+ ME — — — Gg
incanus L. bm mk +DE +0+ Cp — ac jk+ — Gg
jacobaea L. Bm OK +CF+ +M0+ dF og cf+ kn ru GM
nemorensis L. aN+ mU CE KO cF oe af mo — GM+
paludosus L. +bm oM+ +DE +O+ +FP+ ME af In — GM
subalpinus Koch. ab t-M+ +D+ +O+ +cF+ +Me cf mn+ — Gg+
sylvaticus L. +bm+ Ok +DF NP cF +ME ad mn+ — GM+
vernalis W. et K. +bM oK BE+ NQ df +eH ag kn — Gg+
viscosus L. +bN+ +DE+ NO +Cc+ ME bd kn — GM
vulgaris L. +aM OU +CF+ KQ sF oE af mn+ — Gg+
Sequoia sempervirens Endl. +mM EM +CH PQ cp+ +Me+ — — — GM+
Sequoiadendron gigan- teum Lindl. +mM +Mk+ +DF+ +PQ +cp+ +Me+ — — — GM
Serratula coronata L. +bM+ mU CE K0+ SF Mh+ eg — ru Gg
heterophylla Desf. +bm+ Ok +DE NP +Cf+ Mh+ — — — Gg+
inermis Gilib. +bm mk DE NO cF oM+ df kn qr GM
Seseii annuum L. +bm+ +mk +DE NP sf ME +ef jl — Gg+
tortuosum L. +Nm+ +Mk +D+ N0+ +sc +oe+ df — — Gg
Sesleria uliginosa Opiz. Nm Mk +D+ +O+ cp+ oe — — — Gg
Set aria elauca P.B. +bt+ OU +BG+ +MR SF oP bg In — Gg
italica Beauv. bt mU BF +MR sp — +cf +ln+ — Gg+
viridis P.B. Bt OU +BF+ +LR dP oh ag lo su Gg
Sherardia arvensis L. BM+ OU +BF+ NQ+ sF +eE df kn — Gg+
Sideritis mo nt ana L. bM mK+ BE +OQ de Og cf +ln+ — Gg
Sieglingia decumbens Bernh. BN OM+ DF NO+ SF OP ag jm pr GM
Silaum silaus Schinz. bm+ mk+ CE MO+ SP MH eg jl su GM
Silene acaulis L. AB+ Ek DH KO +cf+ oM+ df jk+ — Gg
alba Krause. +am+ OU CF+ KP dF Mg af+ lo st Gg
armeria L. bM Ok+ CF +OQ +cf+ — be jl — Gg
chlorantha Ehrh. +bm mK CE+ +M0+ de Og — — rs+ Gg+
dioica Clairv. +Bm OK CF +MO+ CF +ME cf mo — +gs+
multiflora Pers. bm MK CE M0+ sf+ MH — rt Gg
noctiflora L. Bm +mk+ +DE+ +M0+ Cf Me df km — Gg+
nutans L. Bm t-mk+ +DE+ L0+ Sf oE cf jm rs GM
repens Part. +aN+ oU CF+ KO +Sf+ +og — — — Gg
rupestris L. aN ok +E+ NO +Sf — ad+ jk+ — Gg+
sibirica Pers. +bN+ r-kK+ CE MO +Cc+ eE+ dg — — Gg
viscose Pers. +Bm+ oK+ BE+ +MP df Mg — — Gg
vulgaris Garcke. +BM +OU +CF+ KQ dF oH eg jl rt GM
176
177
Таблица 55 (продолжение)
Вид Факторы
Tm Kn Om Cr Hd Tr | Rc Nt fH Lc
Stnapis arvensis L. +aM+ +OU +BF+ +MQh df +ME df In — Gg
Sisymbrium altissi- mum L. aM OK+ BF+ +NQ +df+ +Eg+ ag jn — Gg+
loeselii L. +BM OU BF KQ +df+ +MH eg kn —- Gg+
officinale Scop. +BM OU CF KQ Cf +eE+ af ln+ — Gg+
orientate L. bM OK+ +CE+ OQ +sC+ +eg cf+ +ln+ Gg
strictissimum L. +bm+ mk+ CE NQ +fF+ ME df ln+ — GM+
Sium latifolium L. +Bm+ OK CF +MP fa oH mo — Gg+
sisaroideum D.C. +bm+ +MK+ BE MP fP +Eh+ — — — Gg+
Smilacina trifolia Desf. aN mU DF KO +fp oe — — — Gs
Solanum dulcamara L. Bm+ OK CF+ MP ca oE af mo +gs
nigrum L. +bM+ OK BF+ +MQ+ Cf +ME cf mo GM+
Soldanella carpatica Viern. ab mk DE +O+ cF+ +oe df jn — Gg
montana Wilid. ab Ok DF NP . cp +oe ac+ jl — Gg
Solidago canadensis L. +bm+ +mK CE MP Sp — af lo Gg+
virga-aurea L. +BM Ok+ +CF+ +MQ SF Oet- af kn qs+ Gs+
Sonchus arvensis L. at+ EU BH KR sP +MP eg jo ru Gg
asper Hill. Bt EU BH KR sF eh eg lo — Gg
oleraceus L. Bt+ EU +BH KR Cp eg df+ mo — Gg+
palustris L. +bM OK+ BF+ +MQ fP eg cf+ lo — Gg+
Sorbaria sorbifolia A. Br. BN+ mU DF KO cp+ oE — — ps Gs
Sorbus amurensis Koehne. +bm+ +kU DF +LP CF+ +Me+ — — — GS
aria Crantz. bm +om+ DE OP CF — cf jm — GS
aucuparia aM +ok+ CF +NQ Cp Og af+ jn — GS
caucasica Zinserl. Nm mk +DE+ +O+ +cP oe — — — GS
domestica L. NM mkt- BE OQ +Cf+ df jm — • GS
sambucifolia Roem. ab +mk+ +EF+ LO Cf+ oe — — — Gs
sibirica Hedl. +Bb+ +MU +DE+ KN +cF+ OE+ — — — GS
tianschanica Rupr. bm kU CE MO +Cf+ +ME — — — Gs
torminalis Crantz. Nm+ +Mk+ CE+ OP Cp +ME cf km GS
Spaxganium angusti- folium Michx. +ab+ OU +EG+ KO Pa+ — ad jk+ — Gg+
erectum L. +bm OK+ +DF+ KO+ Fa oH ag' kn rt Gg+
minimum Wallr. aN+ OU DF+ +LO+ Pa oe be jm — Gg+
neglectum Beeby. bM OU +CF+ NQ Paf +eE cf +ln+ — Gg
simplex Huds. at OU CF+ KQ+ Pa oE cf +ln+ — Gg+
Spergula arvensis L. +BM OK+ +CF+ +MQ cF Oe ac+ In — Gg
Spergularia campestris . Asch. +BN oK+ CE+ LO Cp oe ad km — Gg
Spiraea betulifolia Pall. BN oU DF +LO+ cF+ oet- — — GM
chatnaedryfolia L. +bN+ mK+ CE +MO CF +ME cf +ln+ — GM
crenata L. +bm+ oK+ CE MP+ dc+ +eg — — — Gg+
hypericifolia L. bm+ +kU BE MO+ Df Mh — — — GM
media Schmidt. am mU DF KO Cf+ oE — — — GM
salicifolia L. am+ +MU +DF KO+ +Cp+ oE cf +ln+ — GM
Spirodela polyrrhiza +Bt+ EU CH KR PA +eE af +ln+ — Gg+
Schleid.
178
Таблица 55 (продолжение)
Факторы
ОИЦ Tm Kn Om Cr Hd Tr Rc Nt fH Lc
Stachys annua L. +bm+ OK CF MO+ Cf df km — Gg+
germa nica L. bM Ok CF OQ +Sf +eE df jn — Gg+
lanata Jacq. bM +kU BD+ +OP+ +sc+ +eE — — Gg
palustris L. +BM 0K+ BF +LQ cP oH eg lo rt+ Gg+
recta L. +bm+ +Mk+ CE NP Sc og df jl — Gg
sylvatica L. +Bm+ +mK+ CE +MP cF +ME cf ln+ — gS
Stellaria alsme Grimm.. +Bm Ok CF +MP cp +ME bd km — Gg+
bungeana Fenzl. aN MU DF KN+ cf+ +Me+ — — — +gs+
crassifolia Ehrh. AN+ oU CF KO +fa +og — — — Gg+
diffusa Wilid. am oU DF KO+ FP ME ad jm — +gs
graminea L. am OU BF+ +LP sF oH bg jn pt GM+
holostea L. +Bm+ Ok+ CF +MP +cp+ oH eg kn qs +gs+
media VilL +at+ EU +BH KR dF+ +Mg cf4- lo — GM+
nemorum L. am+ Ok- CF NP +cF+ oE be ln+ — gs
palustris Retz. +aM OU +CF+ KQ ca +Mg bf+ jl qs Gg+
Stipa capillata L. b.M mU BE KQ Df Mg df+ jm rt Gg
lessingiana Trin. et Rupr. bm+ MK BD+ MP De eh — — — Gg
pennata L. bm+ +mK+ CE +MP df og cf+ jm rs Gg+
pulcherrima C. Koch. bM mK CF MP+ de ME df jm — Gg
rubens Smirn. bm +MK CD+ MO+ dc+ +eg — — qs Gg
sareptana A. Beck. bN+ kK+ BD+ +MO+ DC eH — — — Gg
sibirica Lam. bN+ +kU CD+ MO +Sc+ +eE+ — — — Gg
stenophylla Czern. bm+ mk+ CD+ MO de +eE — — — Gg
Stratiotes aloides L. +Bm+ Ok CE MP+ PA oE cf mo st Gg
Suaeda maritima Dum. BM Om CF OQ +dp+ 4gP+ eg k> st Gg+
Subularia aquatica L. ab+ Ok+ +EF MO+ FP+ — ad jl qs Gg+
Succisa praemorsa Bm+ Ok+ +DF+ MP cp OE af jl PS GM+
A schers.
Swertia perennis L. +bm+ Ok +DF OP cP oe df jm — Gg+
Swida alba Opiz. BN+ MU +DF+ KO +cP+ +ME cf kn — Gs+
sanguinea Opiz. b.M OM+ CF +OQ Sp+ +Me+ df jn — Gs+
stolonifera Pojark. Bm +mK+ CF +MP+ +fP — cf kn qs +gs+
Symphoricarpus racemo- +bm+ +mk+ +DF MO+ +CF+ oE df kn — GM+
sus Michx.
Symphytum asperum BN+ mk +DE+ +NO+ +cF+ cf +ln+ — GM
Lepech.
officinale L. +Bm mK CE+ MO ca oH af mo rt GM
tauricum Wilid. Nm Mk +D+ +O+ cF +Me+ — — — GM+
Syringa amurensis Rupr. Bm oU +DF+ +LO+ +CF+ +Me — — — Gs
josikaea Jacq. bm mM+ +D+ +O+ sF oE — — — GM+
vulgaris L. bm+ +Mk CD+ OP SF+ +ME cf kn — Gs
Tamarix gracilis Wilid. bN+ MK CD :MO+ DS +gH - — - Gg+
hispida Wilid. +Nm +KU BC+ +NO+ Df EP — — — Gg+
ramosissima Ldb. +Nm+ MU +BD+ MP+ DP eH+ dg — — Gg+
Tanacetum achillei- +Nm MK CD MO+ Ds+ Eh+ — — rt Gg
folium Sch. Bip.
millefolium Tzvel bm +kK CD MO++dC Eg — - — Gg
vulgare L. am+ OU CF KP SF oH ag kn rt Gg+
179
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Вид Факторы
Tm Кп От 1 Cr | Hd I Тг Rc 1 Nt | fH 1 Lc
Taraxacum bessarabi- cum Hand.-Mazz. +bM mU BE MP +CF-* eP - - St Gg
erythrospermum Andrz. +bm mK +CE+ MO+ dF eH+ rs Gg
officinale Web. +Bt OU BG LR sp oH ag lo qt Gg+
serotinum Poir. bm+ mK +DE NP de +EH+ — Gg
Taxusbaccata L. +bm OM+ DF OP +CF +ME +cf jn gS
cuspidata Sieb. et Zucc. bm+ тКи ► DG MP +CF +Me+ gs
Telekia speciosa Baumg. bm+ mk+ CE +OP +fp+ oE cd Gs
Teucrium chamaedrysL. NM mk CD+ OP+ +sc+ ME df jk+ Gg
scordium L. bM OK CF MP+ Fp+ — df kn st Gg
Thalictrum- aquilegi- +Bm ok +DE+ NP cf Me cf +mn+ qt Gs+
folium L. flavum L. am OU CF KP CP oE df kn rt GM
foetidum L. am+ +MU +CE+ KO+ Cf — df kn GM
lucidum L. +Bm ok CF NP cP oh dg kn GM
minus L. +BM OU BG KQ sF og df jm st+ GM+
simplex L. +Bm+ OU +CF+ KP Sp oH bg kn rs+ GM
Thea sinensis L. +mM+ ok EG +PQ +cF+ +Me+ ad GM+
Thelypteris palustris Schott. am +oK DF +MO ca oE be in - GM+
phegopteris Slosson. +Bm+ Ok+ CF MP cp OE bd In gS
Thesium alpinum L. +ab+ +mM+ DE NO +cf ME+ af jn Gg+
arvense Horvat. bm mK BE MO+ df MH dg In rt Gg
ebracteatum Hayne. bN mk +DE MO Sc+ ME rt Cg+
linophyllon L. bm+ mk +DE OP +Cf +eE df jk+ Gg
Thlaspi arvense L. +aM OU BF KQ sf ME cf 10 Gg
cochleariforme D.C. +ab +kU DE KN Sc eg Gg
perfoliatum L. bM OK BF +NQ sf +eE df jl Gg+
Thuja occidentalis L. +bm+ +mk+ DF MP +cF+ +eE+ cf+ GS
orientalis L. +Nm +kK+ DF MO++Cf+ +Mg cf+ GM+
Thymus marschallianus Willd. +Bm mK CE MO+ df oh - - qs+ Gg
pulegioides L. BN OM DF NO+ Cf oe af jk+ Gg
serpyllum L. +BN+ Ok DF NO+ sf OE be jk+ Gg+
Tilia amurensis Kom. +bN kU +EF LN CF+ +Me+ — +gS
caucasica Rupr. Ntn mk+ +DF +OP SF +ME +gs+
cordata Mill. +bm+ Ok +DF MP Cp+ oE af kn +gs
mandshurica Rupr. et Max. bm +kK+ +DF +NO+ CF +Me+ - - - +gs
platyphyllos Scop. +NM ok CE +OQ +CF+ +ME af In+ +gS
sibirica Bager. +b+ kK DE MN +cp +ME — +gS
t'aqueti C.K.Schneid. bm kU +EF LO CF +Me+ — +gS
tomentosa Moench. NM mk+ CE OP SF +ME +gs+
Tofieldia calyculata Bm+ OM+ +DF+ +OP +fp+ ME df jm Gg+
Wahl.
Torilisjaponica D.C. +bM+ Ok CF +MN+ CF ME df mo GM
Tragopogon dubius +bM OK CF NQ df eh eg +ln+ — Gg
Scop.
Вид Факторы
Tm j Kn Om | Tr Rc Nt fH Lc
Cr | Hd |
orientalis L. +bm mK CE MO+ df Mg cf In pq+ Gg
podolicus Bess. bm mk DF NO+ +sf+ +MH+ eg — st+ Gg
pratensis L. +BN+ OM+ DE NO sF ME cf kn — Gg+
Trapa natans L. s.l. bM+ Ok+ CF NQ+ PA ME ce mo — Gg+
Trichophorum alpinum +ab+ +OK+ DG +MO+ FP Oo+ ac+ jl — Gg+
Pers. caespitosum Hartm. +aN+ +OU +DF+ KO+ FP Oo+ ab+ jk+ — Gg+
Trientalis europaea L. aN+ +oU DF+ KO+ +cP OM+ ad jl — gs
Trifolium alpestre L. bm mk +DE NP sf Mh eg jm rt GM
arvense L. +BM OK BF+ MQ df og af+ jk+ — Gg
aureum Poll. Bm+ Ok+ CF+ MP+ df oE af+ jl — Gg+
campestre Schreb. bM OU BF NQ +sF+ Mg af+ jm — Gg+
dubium Sibth. bM Ok CF OQ +CF+ ME be km — GM
fragiferum L. Bm+ mK BE+ MP+ sP MP dg lo rt Gg+
hybrid urn L. +BM OK CF LQ CP oH eg kn rt GM,
lupinaster L. +aN+ +MU CF KN+ SF Mg — — rs+ GM
medium L. +BM OK CF +MQ Sf+ oh ag jm ru GM+
montanum L. +bm+ ok+ +DE+ MP sF oh dg jl ru Gg+
pratense L. at OU BF KR sp OH ag jn+ pu GM
repens L. at OU BF KR sP OP ag lo ps+ Gg+
spadiceum L. +Bm+ Ok+ +DF+ MP cF+ oE ad jm qr+ Gg+
Triglochin maritima L. aM+ +OU +CG+ KQ+ +cP oP ag — rt Gg+
palustris L. +aM +OU BG KQ +cP +Mh+ ag jk+ q‘ Gg+
Trigonella coerulea Ser. bM +mk+ CE +OQ sF eH eg +ln+ — Gg
Trisetum flavescens P.B. +bM OK BE NQ Sp ME af kn — GM
sibiricum Rupr. +aN+ +mU CF KO Sp+ oE — rs Gs
Trolnus asiaticus L. XnM MU CE+ KM+ CD +ME - — - Gs
boreosibiricus Tolm. +ab MU DE KM cF +Me — — — Gs
europaeusL, am Ok CF NP +cF+ oe cf In qs Gs
fedebowi StefcSrt». MU DF LO +fF+ +Me — — — Gs
Tsags canadensis C&ff. to +Mk+ DF MO+ +cp — — — +gS
fefe«>pliyiteSarg. Ek+ DH +MP +cp — — +gS
"Piilljiti> 6 iebersteiniana bm+ +MK CE NP Df eP cf kn st Gg
Roem. et Schult, biflora Pall. +Nm MK CD +NO+ DS+ +EH — — Gg
greigii Rgl. +N+ +K+ BD NO Ds +Eg+ — — — Gg
quercetorum Klok. et bm+ Ok DE +OP Sp — cf kn — GM
Zoz. schrenkii Rgl. bm +kK CD+ MO+ DS+ eg+ — — — Gg
Turgenia latifolia Hoffm. NM mK BE NQ df eg ef jl — Gg+
Turritis glabra L. +Bm+ OU BF +LP Sf ME cf kn rs Gg+
Tussilago farfara L. +BM OU BF KQ CF oE df jn qs GM
Typha angustifolia L. +BM OU BF+ MQ ca+ Mh ag lo — Gg+
elephantina Roxb. mt mK+ BD+ +PR Sp +gH — — — Gg
latifolia L. +BM OU BG KP+ Fa oh ag mo — Gg+
laxmannii Leped. +bm+ +mU BF+ MP +CP+ +EH+ - — — Gg
Ulmus campestris L. +NM mU BE+ +OQ Sp Mg +df+ kn — GS
glaora Huds. BM OK CF NP+ Cp eE af ln+ — GS
180
181
Таблица 55 (продолжение)
Таблица 55 (продолжение)
Факторы
Вид Tm | Kn Om Cr | Hd Tr Rc Nt fH | Lc Вид Tm Kn Cr Ьакто| Hd >ы Tr Rc | Nt fH Lc
laciniata Мауг. +bm +mK+ +DF+ LO+ CF Me+ GS
laevis Pall. Bm+ ok+ +DE+ +NP Co ME cf ln+ GS
pumila L. bm +kU +DF MO+ +SF +Mg+ eg jn GM+ officinalis L. am+ OK CF+ t-NP CF Oe ac+ jm qs GS
Urtica cannabina L. +bm MU CF MO +Sf +eg — Gg+ opaca Fries. +bN+ Ok DF NO CF — df In — Gg+
dioica L. aM OU BF+ KQ SP oE af lo rt paniculata L. +Bm mK CE+ MO+ +sp oE+ ce — — GM
urensL. +BM OU CF +MQ SF Mg af lo GM+ persica Poir. +bM OK BF NQ cF — cf ln+ Gg+
Utricularia intermedia am OU CG KP +fA Oe df jk+ Gg+ prostrata L. Bm+ Ok+ +DF MP dc+ OH+ eg kn st Gm
Hayne. scutellata L. am tmU CE+ KO+ cP oh ag jm qs Gg+
minor L. +Bm OU BG KP FA oM cf km Gg+ serpyllifolia L. aM OU CF LQ Sp og bf jn rs GM
neglecta Lehm. am OU CG KP PP eg cf+ kn Gg । spicata L. +Bm+ OK CF LP df og bf jl rs+ GM
vulgaris L. +Bm+ OU BG KP +pA +Mg +cf+ In Gg teucrium L. +Bm+ OK CF MP Sf og df jl rs GM
Vaccaria pyramidata +bm+ oU BE+ MP +Cc+ df+ kn Gg verna L. +BM OK BF MQ +dF Mg bf+ jk+ — Gg+
Med. Viburnum lantana L. +NM OK CF OQ CF+ +ME df kn — Gs
Vaccinium arctostaphy- bm mk CE NP +cF+ +Me +gS opulus L. +BM +oK+ +DF MP+ +cP oE cf In — Gs+
los L. sargentii Koehne. Bm +MK+ DF +MO+ CF+ +Me+ — — — GM+
myrtillus L. +aN OU DF+ +LO+ +cP Oe ac+ jm pq+ +gs ! Vicia angustifolia L. +BM OK BF +MQ Sp ME af jn Gg
ovalifolium Smith. BN Ok+ EG MO +cF oM+ +gS 1 cassubica L. +bm+ OK CF NP Cf Oe be jm — Gm+
uliginosum L. AN EU DH KO+ cP+ Oe ab+ jm pr GM+ cracca L. aM OU CF KP+ SP oH ag jn pu GM
vitis-idaea L. AN+ OU +DH KO+ +cp Oe ac+ jl pr+ Gs+ grandiflora Scop. NM mK CE OQ Sf ME — — — Gg+
Valeriana officinalis +am+ OK CF KP+ cP oE +cf+ kn GM+ hirsuta S.F. Gray. +Bt' OU BF LR sf ME af jn — Gg+
L. s.l. pannonica Crantz. NM mK++CE +OQ +Cc+ +eE df kn — Gg
Valerianella dentata Poll. bM OK BF OQ sf +eE cf jn GM pisiformis L. +bm+ Ok+ CF NP Cf ME df jm — GM
Vallisneria spiralis L. NT OK BG OR aA ad jm Gg+ । sativa L. +Bm+ OK CF MP Cf ME af jn — Gg
Veratrum album L. am+ OM CF NP Sp oe+ af In Gg+ sepium L. +Bm+ OK CF KO+ Cp oE cf kn os+ GM
lobelianum Bernh. am+ mK CE+ LP CP og af In qs+ sylvatica L. +Bm Ok+ +DF MO CF oe df jn — +gs+
nigrum L. bm mU CF KO+ Cf +ME df kn +gs+ ' tenuifolia Roth. +bm+ OK CF LP df oE df jl rs Gg+
Verbascum blattaria L. bm+ OK BF MP Cf eg In Gg+ ; tetrasperma Schreb. +BM OK CF +MQ sF Mg af+ km — Gg
lychnitis L. +bm+ Ok +DF+ MP SC oE cf mn+ st Gg i villosa Roth. +bM OK+ BF +OP sf ME bd+ kn — Gg
nigrum L. Bm+ Ok+ +DF+ LP df ME cf ln+ — Gg+ i Vinca erecta Rgl. et Nm kK+ +CD NO Sc eE — — — Gg+
orientale Bieb. bm +MK+ CD+ NP sc+ +eE Gg Schm.
phlomoides L. +bM mK CE NP+ +Sc+ oE +df+ +ln+ Gg herbacea W. et K. bm+ +mK CE +OP Sc+ +eg cf — . — GM
phoeniceum L. Bm+ mK CE MP de oE cf jl GM major L. NM mK +CD+ OQ CF — cf +ln+ — GM
thapsiforme Schrad. bm+ Ok +DE +OP SC oe df kn Gg+ minor L. bM Ok+ CF +OQ Cf+ +ME af In GM
thapsus L. +bM OK BF+ LQ de oe cf ln+ Gg Viola arenaria D.C. aM OU CF KQ Cf Oe df jl — Gg
Verbena officinalis L. bt +mK BE+ +OR +sf eg af+ In GM+ arvensis Murr. +Bm+ Ok+ +CF+ +MP+ sf og af+ jn — Gg
Veronica agrestis L. +bM Ok+ CF NP dF +eE cf lo Gg biflora L. +ab+ mU CF KO +cF +oE cf In — +gs
alpina L. +aB+ OK+ DF +MN+ fF af jm Gg+ canina L. +Bm Ok+ DF +MO+ CF OE ad jl pr+ Gs+
anagallis-aquatica L. +BM+ Ok+ BF+ +LQ+ fP ME cf lo Gg+ collina Bess. Bm mK CF MO CF Me df jl — Gm
aphylla L. aB mk DE+ NO cF df jl elatior Fries. +bm+ OK +CF MP Cf Me df jl — GM
arvensis L. +BM OK BE NQ df ME ce jn Gg epipsila Ldb. aN Ok +E+ MO +fP Oe+ — — qr+ Gs
austriaca L. +bm+ mK CE +OP sf+ Mh+ dg jk+ rt Gg+ hirta L. Bm OK CF +LO+ Cf+ +ME df jl rs GM
beccabunga L. +BM Ok+ BF+ MQ FP ME cf In GM mirabilis L. BM OK CF +LP+ CF oE df jn — +gs+
chamaedrys L. +Bm+ Ok CF MP SF °g af+ jn qr Gs+ montana L. am+ oK СЁ+ LP +cf +ME ac+ jl — Gs
fruticans Jacq. ab Ok DF NO +Cf Oe af jm Gg+ odorata L. bM OU BF +OQ Cf ME af mo — +gs
gentianoides Vahl. bm+ mK DE +OP CF +ME Gg+ palustris L. am OM+ +DF+ +NP fP oE ac+ jn qr GM+
hederifolia L. +bM OK BF NQ +SF ME cf lo GM riviniana Rchb. +Bm+ Ok+ CF +NP cF oE ad jn — +gs+
incana L. am OU CF KO+ df Og rt GM rupestris Schmidt. +aN+ +oK+ +DE+ +MO+ Cf +ME+ df jl — GM+
longifolia L. +am OU CF KO SP Og cf lo ru GM selkirkii Goldie. +ВЫ- oU DF KO cp +ME — — gs
stagnina Kit. +Bm Ok+ DF LO+ +cF ~+eE be jm — Gg+
182 suavis M.B. bm +MK+ BD+ +OP sf oe cf jm — GM
183
Таблица 55 (окончание)
Вид Факторы
Tm Kn Om Cr Hd Tr Rc Nt fH Lc
tricolor L. +Bm+ Ok+ +DF+ +MP+ SP og af in qs GM
uliginosa Bess. BN oM+ DE +O+ Fp Me — — GM
uniflora L. aN +kU DE KM+ cf+ Me cf in — GM
Viscaria vulgaris Bernh. Bm ok+ +DE+ NO+ Sf og af jl IS GM+
Viscutn album L. bM OK CF +OQ — — — — — GM
Vitis amurensis Rupr. bm kU DF LN+ cp +Me+ — — — Gs
silvestris Gmel. +NM oU BF OQ Sp Og +dg +ln+ — Gs
vinifera L. +NM+ OU +BG +OQ+ SF Mg dg In — GM
Weigela middendorf- fiana C. Koch. +bm+ +MK +EG +NP CF +Me+ — — — Gs
praecox Bailey. +Nm++kK +EF NP Sf+ +Me+ — — — GM+
Woodsia alpina S.F. Gray. +ab EU DH KO+ Sf +ME+ ad jm — GM+
ilvensis R.Br. +aN+ +MU DF+ KO+ +Cc+ ME ad jl+ — GM+
Xanthium spinosum L. NT OU BF OR sF eE+ cf In — Gg+
strumarium L. +bM OK+ BF +NQ +sp +Eh+ ag In st Gg+
Xeranthemum annuutn L. NM +Mk+ CD +OQ +sC+ OE+ cf jm — Gg
Zannichelia palustris L. BM EK CH MQ aA eh eg In — GM+
Zizania aquatica L. bT ok DF NR +Fa +ME — — — Gg+
latifolia Stapf. bt mU DG +MR +Fa +ME — — — Gg+
Zostera marina L. aM+ Ek BH NQ aA gH eg In — GS
(4+9) :2 = 6,5 (или 6—7), для ацидоморфы — (1+11): 2 = 6, для нитромор-
фы — (1 +9) :2 = 5, для гидроконтрастоморфы — (4+8) : 2 = 6, для гелио-
морфы — (2+9) : 2 = 5,5 (или 5—6).
Может быть, не лишне сказать несколько слов о некоторых, не имею-
щих большого значения, особенностях шкал. Во-первых, на предлагаемых
10 шкалах прослеживаются два принципа составления символики режимов
факторов: смысловой и алфавитный. Смысловой принцип использован в
шкалах, разработанных в настоящем или несколько ином виде еще в
1972 г. [Цыганов, 1974, 1976]: термоклиматической шкале, шкале кон-
тинентальности, шкале увлажнения почв, шкале солевого режима почв и
шкале освещенности-затенения.
Алфавитный принцип употреблен во вновь разработанных шкалах:
омброклиматической, криоклиматической, кислотности почв, богатства
почв азотом и переменности увлажнения почв, причем в первых двух
(климатических) в качестве символов использованы прописные буквы
латинского алфавита, а в остальных трех (эдафических) - строчные.
Во-вторых, самый способ построения шкал и схрм систем частных эко-
морф определяет то, что баллы основных типов режима каждого фактора
представлены нечетными числами, а промежуточных - четными.
Наконец, по-видимому, не лишне еще раз оговориться, что предла-
гаемая фитоиндикационная экологическая таблица основана на вновь
разработанной системе экологических характеристик видов. Почти без
184
изменений в ней сохранились лишь шкалы режима увлажнения почв и соле-
вого их режима, тогда как термоклиматическая шкала, шкала конти-
нентальности и шкала затенения сильно переработаны, а остальные пять
шкал разработаны вновь. В то же время большинство рассуждений,
формулировок закономерностей и иллюстративных таблиц в других гла-
вах построены на системе экологических характеристик видов, изложен-
ной в работах 1974—1976 гг. Следствием этого является некоторое раз-
личие в названиях и символике частных экоморф и экологических режи-
мов в других главах с предлагаемой фитоиндикационной экологической
таблицей.
ГЛАВА VII
ВОЗМОЖНОСТИ ПРИМЕНЕНИЯ ФИТОИНДИКАЦИИ
ДЛЯ ДИАГНОСТИКИ ТИПОВ ЛЕСА
Возможность перехода от фитоиндикации экологических режимов
местообитаний к диагностике как наличного, так и наиболее потенциаль-
но устойчивых в данных условиях типов леса не должна бы вызывать
сомнений. Так как фитоиндикационно установленные типы режимов фак-
торов легко могут быть скоординированы с амплитудами толерантности
любых видов, то несложно и рассчитать КС данного местообитания для каж-
дого из интересующих нас видов с целью дать сравнительную оценку их
ценозостроительным возможностям как в существующем сообществе,
так и в перспективе на будущее.
С другой стороны, применение упорядоченной системы фитомер, чем
в сущности является фитоиндикационная экологическая таблица, в
неопределенно большой степени интуитивно выделенным единицам типо-
логии лесов или классификации растительных сообществ вообще вызывает
сомнения у сторонников фитоиндикации в определенности и реальности
объекта индикации такого рода, а у сторонников вдохновенной интуи-
ции — в полезности фитоиндикационного метода.
Таким образом, вопрос сводится к тому, какие типы леса (в смысле
того, на каких критериях основано их выделение) могут быть диагно-
зированы де факто и в потенции на основании фитоиндикационных
построений. Для ответа на этот вопрос следует уяснить, во-первых, что
входит в диагноз типа леса: какие явления, какие связи между ними и,
во-вторых, что из этого можно индицировать по растительному покрову.
При самом примитивном взгляде на проблему, может возникнуть
вопрос: как это можно и зачем это нужно индицировать тип раститель-
ного покрова по растительному покрову? В действительности же инди-
цируется не наличный видовой состав растительного покрова данного
участка, а его потенции, его динамика, что означает в сущности инди-
кацию не собственно синтаксономической принадлежности растительного
покрова данного участка, а условий среды на данном участке, и уже в
сопоставлении с ними диагнозируются динамические потенции налич-
ного растительного покрова. Задача, таким образом, сводится к сопостав-
13.Зак.448
185
лению индицированного экотопа с экологическими потребностями видов,
популяции которых либо заходят на данный участок, либо имеют возмож-
ность сообразно характеру и способам своего расселения попадать на него
с окружающих или более отдаленных территорий.
В то же время еще в тридцатые годы В.Н. Сукачев писал, что тип
леса мог бы быть вполне достаточно определен составом растений и
комплексом прямодействующих факторов. Необходимость при лесотиполо-
гических исследованиях изучать и почвенно-грунтовые условия, и
рельеф он относил целиком на счет невозможности или, по крайней
мере, очень большой трудности непосредственного определения комп-
лекса прямодействующих факторов [Сукачев, 1938].
Практически то же говорится и в его определении типа леса уже на
уровне теории биогеоценологии: ’’Тип леса — это объединение участков
леса (отдельных лесных биогеоценозов), однородных по составу древес-
ных пород, по общему характеру других ярусов растительности, по фауне,
по комплексу лесорастительных условий (климатических, почвенно-грун-
товых и гидрологических), по взаимоотношениям между растениями и
средой, по восстановительным процессам и по направлению смен на этих
участках леса ...” [Сукачев, Зонн, 1961, с. 47].
Но из этих необходимых для диагноза типа леса признаков комплекс
режимов прямодействующих факторов, или комплекс лесорастительных
условий как раз и является основным объектом фотоиндикации, в смысле
изложенных в этой книге взглядов, а из биотической среды экосистем
наиболее удобен для прямого наблюдения и диагностически наиболее
значим растительный покров, потенциальная динамика которого опять же
может быть выведена из сопоставления экологических особенностей видов
с тем же комплексом режимов прямодействующих факторов.
Из изложенного выше можно сделать вывод о возможности применения
фотоиццикации.в диагностике типов леса, если исходить из признания воз-
можности фитоиндикационной оценки режимов прямодействующих фак-
торов. Однако идол непосредственного (инструментального) измерения
режимов прямодействующих факторов еще властвует умами большинства
исследователей (геоботаников, лесоводов и особенно почвоведов), хотя
отдельные авторитеты и допускают несколько крамольные высказывания:
’’Прямое измерение отдельных факторов стоит дорого, а отдельные фак-
торы без учета их взаимодействия мало что могут сказать об экологи-
ческих условиях” [Миркин, Шеляг-Сосонко, 1980, с. 9] 1
В то же время не вызывает споров тот факт, что измерить режимы пря-
модействующих факторов в их динамике и взаимосвязи хотя бы прибли-
зительно относительно возможно только в стационарных условиях при
многолетних наблюдениях. Для маршрутных и даже детально-маршруп^гх
исследований значение индикационных приемов в целом не отвергается
никем, но многими отвергается шкально-балльная оценка режимов
факторов.
В защиту шкально-балльной оценки, о ее преимуществах в смысле
большей ее экологичности по сравнению с инструментальными измене-
ниями физико-химических характеристик абиотической среды говорилось
в начале книги. Здесь же стоит добавить, что фитоиндикация может быть
с успехом применена в диагностике синтаксонов лишь при условии экбло-
186
гичности их классификации или типологии. Это значит, что такая типоло-
гия или классификация должна основываться на достаточно четких пред-
ставлениях об экологических потребностях видов, о сущности экологи-
ческих режимов, о принципах изменения этих режимов под влиянием
популяций видов, в достаточной мере обладающих эдификаторными
(средотрансформирующими) возможностями. Это значит, что все под-
меченные эмпирически закономерности могут быть сформулированы,
типизированы (классифицированы) только в случае установления их
экологической причинности.
Все еще не объясненное или объясняемое интуитивно может связы-
ваться с классификационными или типологическими схемами лишь пред-
положительно и приблизительно, и только в рабочем порядке. Никаких
выводов и утверждений на основании интуитивных предположений при
отсутствии их экологического обоснования делаться не должно. Интуи-
тивно можно решить, как, где и когда проводить наблюдения или экспе-
рименты, но толковать их результаты и выводить закономерности интуи-
тивно нельзя.
Как иллюстрация и довод в пользу сказанного выше приводятся при-
меры фитоиндикации режимов пяти основных прямодействующих эко-
логических факторов конкретных участков по растительному покрову и
диагностики, на основании сопоставления полученных характеристик
режимов с экологическими потребностями видов, типологической при-
надлежности наличного растительного покрова, а также видового состава
и типологической принадлежности потенциального коренного фитоценоза.
Пример предлагается заведомо сложный для диагноза потенциального
коренного типа леса на основании фитоиндикационных данных — участок
80—90-летней культуры ели с небольшой примесью сосны (по-видимому,
старшего возраста) и дуба, сформировавшей к настоящему времени прак-
тически равноценный естественному тип леса: ельник условно лещиновый
(фактически бузинно-рябиновый) зеленчуковый, находящийся в послед-
ние десятилетия под воздействием постоянных и интенсивных рекреа-
ционных нагрузок в связи с расположением рядом с кольцевой автодо-
рогой напротив быстро застраиваемого микрорайона Москвы (Хлебни-
ковский лесопарк Клязьминского леспаркхоза, кГв. 96, юго-восточный
угол).
Ниже приводится краткая характеристика состояния растительного
покрова на лето 1980 г.
Сомкнутость крон 0,5—0,7; полнота древостоя 0,7 ± 0,06; высота пер-
вого яруса: ель и сосна — 28—32 м, дуб — 23—25 м; сомкнутость второго
яруса 0,1—0,2; высота второго яруса 9—14 м; сомкнутость подроста
< 10%; сомкнутость подлеска 15—25%; общее проективное покрытие
травостоя около 70%; общее проективное покрытие мхов
< 10%. В первом ярусе древостоя: береза пониклая, дуб, рябина, бредина;
в подросте: ель, липа, дуб, осина; в подлеске: рябина, бузина, жимолость,
крушина, бредина, калина, волчье лыко, малина. В травостое присутст-
вуют: сныть, живучка ползучая, копытень, кочедыжник женский, бук-
вица, осока пальчатая, ландыш, скерда болотная, ежа, щучка, щитовник
мужской, щитовник игольчатый, овсяница гигантская, таволга вязолист-
ная, земляника лесная, зеленчук, герань лесная, гравилат алеппский, гра-
187
вилат речной, зверобой пятнистый, ожика волосистая, марьянник дубрав-
ный, мицелис, перловник, мятлик дубравный, калган, Черноголовка, лютик
кашубский, лютик ползучий, костяника, норичник шишковатый, золотая
розга, крапива двудомная, вероника дубравная, вероника лекарственная,
фиалка собачья, фиалка сверху голая, фиалка удивительная. В моховом
покрове преобладают: атрих волнистый, брахитеций шероховатый, эврин-
хий Зетгерштедта, мний близкий.
Режимы основных прямодействующих факторов были индицированы
методом определения их среднего балла. Факторы и их шкалы даются
по разработке 1976 г. -{Цыганов, 1976].
Тепловой режим ценоклимата 6, 53, т.е. промежуточный между суб-
бореоидным и северонеморальным. Ужесточение зонального североне-
морального типа терморежима произошло преимущественно под воз-
действием сомкнутого темнохвойного насаждения, способствующего
в силу значительного задержания снега на кронах более глубокому про-
мерзанию почвы в начале зимы, вследствие большого затенения, задер-
живающего стаивание снегового покрова и оттаивание почвы весной и
препятствующего ее прогреванию в летние месяцы.
Режим континентальности ценоклимата 3, 4, т.е. несколько ближе к суб-
океаническому, чем к полуконтинентальному. Этот режим несколько
смягчен по сравнению с региональным и опять-таки влиянием сомкнутого
темнохвойного насаждения, несколько уменьшающего годичные и суточ-
ные колебания температур воздуха и почв (особенно суточные колебания
и в большей степени почв, чем воздуха), а также способствующего сохра-
нению большой влажности воздуха под пологом древостоя.
Обобщенный режим увлажнения почв 13,24, т.е. близок к влажно-
лесному, но несколько влажнее его. Это означает преобладание в течение
вегетационного сезона промывного гидроморфного типа водного режима
почв. Капиллярная кайма почти весь год находится в пределах корнеоби-
таемого слоя деревьев, кустарников и бопыпинстр.2 ipss. 5 геСёККИп,
ранИёлетний и все дождливые периоды почва полностью насыщена влагой,
наблюдаются явления, характерные для временно избыточного увлажнения.
Внутрипочвенный сток боковой, по дренирующим прослойкам. Главные
источники поступления влаги — осадки и боковая инфильтрация. Расход
воды — боковой внутритючвенный сток, транспирация и в значительно
меньшей степени испарение. Режим грунтовых вод — устаювивщийся,
компенсированный подземным оттоком и транспирацией. Влияние дре-
востоя, тем более темнохвойного, в плакорных условиях при относительно
тяжелых почвах несколько осушающее. Поэтому разреживание елового
порога, пусть даже с частичной заменой его лиственным, приведет к сдвигу
режима увлажнения в сторону более влажного.
Обобщенный солевой режим почв 5,77, т.е. близок к пермезотрофному
гликофильному, но несколько беднее. Это означает преобладание восста-
новительных процессов над выщелачиванием, когда выщелачиваются
периодически лишь наиболее растворимые соединения и относительно
постоянно происходит гидрогенная аккумуляция окислов железа и
марганца, а также глинных минералов. Содержание солей приблизительно
порядка 110-170 мг/л. Возможна незначительная аккумуляция углекисло-
го кальция (содержание НСОз порядка 10-40 мг на 100 г почвы). Величина
88
pH обычно порядка 5,5-6,5, причем наблюдается следующий ряд ее нарас-
тания: участки под кронами елей окна -» участки под кронами дуба.
Тип гумуса влажный модермуллевый, с некоторым преобладанием грубых
фракций, но с тенденцией перехода к состоянию с преобладанием мулля.
Обобщенный режим освещенности-затенения 3,2, т.е. несколько тенис-
тее светлолесного. Это означает, что еловый по преимуществу древостой
в значительной мере расстроен, т.е. среди участков с нормальной сомкну-
тостью древостоя встречается много окон и редин, где тип режима зате-
нения не тенистолесной, как это должно быть в темнохвойном насаждении,
а представляет собой различные степени перехода от рединно-полянного
к светлолесному.
Коэффициенты общей комфортности данного участка (КК) составляют:
по терморежиму ценоклимата - 45%, по режиму континентальности —
78% (т.е. по климатопу в целом — 35%), по обобщенному режиму увлаж-
нения почв—93%, по обобщенному солевому режиму почв — 65% (т.е.
по эдафотопу в целом - 60%, а по хоротопу в целом - 21%), по режиму
освещенности-затенения в среднем — 55% и по экотопу в целом и среднем —
12%. Тенденция изменения режимов под влиянием изменения состояния
эдификаторной популяции состоит в постепенном смещении режимов
прямодействующих факторов: терморежима ценоклимата — к северо-
неморальному (повышение КК), режима континентальности ценокли-
мата - к полуконтинентальному (понижение КК), режима увлажнения
почв - к сыровато-лесному (незначительное повышение КК), солевого
режима почв — в сторону эвтрофикации (повышение КК) и режима зате-
нения — в сторону осветления (повышение КК). Таким образом,ККцено-
климата вследствие противоречивости изменения слагающих его режимов,
по-видимому, или незначительно повысится, или останется практически
без изменения, КК эдафотопа - повысится, КК хоротопа в целом - тоже
повысится и КК экотопа в целом также станет больше.
В зафиксированных status quo условиях породы, слагающие древесный
ярус, имеют следующие коэффициенты удовлетворительности условий
среды (КС):
ель — 85% по терморежиму, 84% по режиму континентальности (71%
по климатопу в целом), 78% по режиму увлажнения почв, 85% по солевому
режиму почв (66% по эдафотопу в целом и 47% по хоротопу в целом),
92% по режиму затенения и 43% по совокупности всех пяти режимов (по
экотопу в целом);
дуб — 41% по терморежиму, 84% по режиму континентальности (34%
по климатопу в целом), 95% по режиму увлажнения почв, 59% по соле-
вому режиму почв (56% по эдафотопу в целом и 19% по хоротопу в целом),
65% по режиму затенения и 12% по совокупности всех пяти режимов
(по экотопу в целом);
сосна — 85% по терморежиму, 83% по режиму континентальности (71%
по климатопу в целом), 88% по режиму увлажнения почв, 85% по солевому
режиму почв (75% по эдафотопу в целом и 53% по хоротопу в целом),
но < 0 по режиму затенения, и, следовательно, непригодны условия ио
экотопу в целом, хотя в участках, где световой режим приближается
к рединно-полянномуКС, по режиму затенения может повышаться до-
33 -66%, повышая тем самым общий КС местообитания до 17—35%;
189
осина — 99% по терморежиму,. 36% по режиму континентальности
(36% по климатопу в целом), 86% по режиму увлажнения почв, 96% по
солевому режиму почв (83% по эдафотопу в целом и 30% по хоротопу в
целом); так же как и для сосны, непригоден режим затенения, но в
участках, где световой режим приближается к рединно-полянному,
общий КС местообитания может быть повышен до 10—20%;
береза пониклая — 99% по терморежиму, 83% по режиму континен-
тальности (83% по климатопу в целом), 86% по режиму увлажнения
почв, 85% по солевому режиму почв (73% по эдафотопу в целом и 61%
по хоротопу в целом); режим затенения тоже в среднем непригоден,
но в больших рединах и окнах может улучшаться, повышая тем самым
общий КС до 20—40%;
липа — 81% по терморежиму, 84% по режиму континентальности
(68% по климатопу в целом), 86% по режиму увлажнения почв, 93% по
солевому режиму почв (80% по эдафотопу в целом и 54% по хоротопу
в целом), 92% по режиму затенения и 50% по совокупности всех пяти
режимов (по экотопу в целом).
Кроме того, учитывая рассмотренные тенденции изменения режимов,
следует иметь в виду, что условия местообитания будут меняться различ-
ным образом (в смысле улучшения или ухудшения) для разных пород.
Так, сдвиг терморежима в сторону более теплого зонального типа
несколько понизит КС по нему для ели и сосны, но несколько повысит
для дуба и липы. Сдвиг режима континентальности (контрастности)
ценоклимата в сторону полуконтинентального ведет к понижению соот-
ветствующих КС у дуба и липы при одновременном повышении их у
сосны, осины и березы. Для ели же, учитывая гибридный характер ее по-
пуляций в данном регионе, смещение режима континентальности может
иметь знг ение лишь в смысле изменения соотношения выгодности приз-
наков, унаследованных от ели европейской и ели сибирской. Тенденция
к некоторому повышению увлажнения почв направлена к повышению
КС по этому фактору у всех названных пород, кроме дуба. В то же
время некоторая эвтрофикация местообитания, помимо основной при-
чины (уменьшения эдификаторного значения популяции ели), сопря-
женная с увеличением увлажнения, повышает соответствующие КС
для дуба, липы и осины, но понижает для сосны, ели и березы.
Тенденция к некоторому осветлению на ближайшее время повышает
КС всех пород. В дальнейшем же режим затенения может относительно
стабилизироваться где-то вблизи оптимума теневыносливых пород или
прогрессировать дальше в сторону осветления, повышая тем самым ка-
чество среды для светолюбивых пород.
По величине КС для хоротопа в целом (совокупность местных клима-
тических и эдафических режимов без учета ценотически обусловленного
режима затенения) породы выстраиваются в такой ряд: береза (61), липа
(54), сосна (53), ель (47), осина (30), дуб (19).
По величине КС для экотопа в целом (с учетом среднего балла режима
затенения) ряд получается иной: липа (50), ель (43), дуб (12).
Светолюбивые породы из ряда выпадают.
Таким образом, можно заключить, что в естественных условиях ель и
липа на этом местообитании относительно равноконкурентоспособны
190
что, захватив территорию первой, и та и другая несколько трансформи-
руют режимы прямодействующих факторов в благоприятную для себя
сторону. В то же время, поскольку ель изменяет экологические режимы
сильнее, соответственно сильнее сказывается на состоянии экотопа и расти-
тельного покрова снижение эдификаторной роли ее популяции в процессе
изреживания насаждения.
Что касается позиций дуба, то он явно менее конкурентоспособен на
данном местообитании (как, впрочем, и в данном регионе вообще), но
все же его общий КС в лесных экотопах, как правило, не ниже КК данных
экотопов, что свидетельствует о его способности давать устойчивую при-
месь в коренных сообществах местообитаний такого типа и образовывать
достаточно устойчивые насаждения при устранении более конкурентоспо-
собных ели и липы.
Обращает на себя внимание высокий КС хоротопа такого рода для
березы,что заставляет считать ее породой № 1 по потенции вселения на
любой осветлившийся участок данной и подобных ей территорий и в
силу этого всегда присутствовать в виде некоторой примеси в коренных
насаждениях и чаще других пород формировать вторичные насаждения.
В меньшей степени в силу биологических особенностей хвойного дерева
способна выступать в качестве примеси или вторичной породы в
хоротопах такого типа сосна, тоже имеющая в них очень высокий
КС. Большей частью явление сосны в таких ролях обусловлено пироген-
ными или антропогенными причинами.
Достаточно высокий КС хоротопа для осины наряду с явно выражен-
ной тенденцией его повышения при существующих направленностях
сдвигов всех режимов дает основания считать ее также потенциаль-
ной постоянной примесью любых образующихся на данной территории
фитоценозов.
Наконец, стоит отметить, что клен имеет в подобных существующему
на данном участке экотопах общий КС даже выше, чем у липы, что поз-
воляет считать и его одним из участников древостоя коренного типа
леса. Отсутствие же его на данном конкретном участке объясняется,
скорее всего, причинами случайного характера.
В подлеске при существующем сочетании режимов и их ближайших по-
тенциях изменения наиболее устойчивые позиции имеют малина, жимо-
лость, калина и бредина. У этих видов наибольшие общие КС должны еще
повыситься вследствие благоприятности для этих видов сдвига режимов
по всем факторам. Что касается позиций остальных видов, то они несколь-
ко лучше у волчьего лыка (достаточно высокий КС и благоприятность
перспектив по большинству факторов) и бузины (благоприятность пер-
спектив почти по всем режимам), а у рябины и крушины относительно
хуже (неблагоприятный в сумме эффект наметившейся динамики режимов
или низкий КО. В то же время не следует упускать из виду, что все наз-
ванные выше виды, кроме бредины, являются орнитохорами и их расселе-
ние в большой степени зависит от характера посещения данных участков
птицами. Несколько слов следует сказать о позиции лещины, которая
отсутствует на данном участке также из-за того, что сойки не занесли ее
семена сюда. Учитывая то, что КС лещины здесь достаточно высок (16%,
явно выше общего КК) и из всех наших кустарников она обладает,
191
по-видимому, наибольшими эдификаторными потенциями, следует счи-
тать ее присутствие довольно обычным и даже закономерным для эко-
топов данного типа, чем и оправдывается название данного ельника
условно лещиновым. В то же время стоит оговориться, что по режиму
увлажнения почв лещина находится здесь вблизи границы своего эколо-
гического ареала и на пределе своих доминантных возможностей.
Что касается травостоя, то в нем в лучших условиях находятся попу-
ляции лютика кашубского, копытня, мятлика дубравного, живучки,
кочедыжника, осоки пальчатой, зеленчука, фиалки удивительной, норич-
ника, Черноголовки, щучки и вероники дубравной (высокие и относитель-
но высокие КС и положительные для данных видов сдвиги всех или
большинства режимов). Относительно устойчивы позиции крапивы, лан-
дыша, щитовника мужского, лютика ползучего, гравилатов, герани лесной
и щитовника игольчатого. В той или иной степени ухудшаются, но
обладают достаточным запасом устойчивости позиции земляники, сныти,
марьянника, скерды, перловника, овсяницы гигантской, зверобоя и
калгана.
Все изложенное позволяет определить коренной тип леса на данном
участке как биогеоценоз, имеющий фитоценотическую основу с липой
и елью в роли соэдификаторов, с участием дуба и клена в первом ярусе,
единичной примесью березы и осины, лещиной, жимолостью, калиной,
брединой и волчьим лыком, как основными видами подлеска, с гос-
подством в травостое мезофильного пермезотрофно-семиэвтрофного
неморального широкотравья, при участии заметного количества более
влаголюбивых трав, и слабо развитым моховым покровом, образованным
в основном видами семейства брахитециевых.
Таким образом, мы видим, что индицируется, как, впрочем, и опре-
деляется сочетанием режимов прямодействующих факторов, не какой-
то определенный тип леса как коренной для данного местообитания, а
некий диапазон экологически относительно равнозначных типов.
По-видимому, логично заключить, что это обстоятельство может рас-
сматриваться в качестве лишнего довода в пользу концепции поликли-
макса, как она изложена Уиттекером [Уиттекер, 1980].
Вновь поднимаемая на щит концепция монокпимакса, равно как и
постулат дискретности растительных ассоциаций, пока что находит наи-
более веские аргументы в свою пользу лишь в пристрастиях интуиции
отстаивающих их авторов. В этом смысле весьма ценная своим динами-
ческим подходом монография ’’Закономерности динамики биоценозов”
[Разумовский, 1981] оставляет сожалеть о приверженности автора посту-
лированию своих интуитивных пристрастий. Чего стоит хотя бы категори-
ческое отрицание автором континуума, признание которого якобы возвра-
щает нас к чистой аутэкологии и перечеркивает геоботанику в целом.
Также сомнительно разделение видов на ценофильные и ценофобные,
попытка догматизировать дискретность сообществ, претензии дать окон-
чательный список климаксовых ассоциаций. В сущности, стремление рас-
сортировать все многообразие растительного покрова по ящикам карто-
теки противоречит основной идее монографии — упорядочению теории
динамики биоценозов.
С нашей точки зрения, гораздо ближе к истине положения, что разли-
192
чаемость фитоценозов ’’зависит от особенностей изменения градиентов
среды и специфики самой растительности, где непрерывность и дискрет-
ность сочленяются в диалектическое единство”, что ’’число фитоценозов
зависит от нашего определения масштаба неоднородностей внутри фито-
ценоза”, что ’’фитоценозы не являются исторически сложившимися груп-
пами растений, так как отдельные виды, за исключением некоторых узких
специалистов-паразитов, эволюционируют независимо, а сам процесс
эволюции носит сеткообразный характер” и ’’любой наблюдаемый нами
фитоценоз — некое сечение через пучок временно сближенных (разные
виды на разное время!) трендов развития отдельных видов” [Миркин,
1981, с. 825].
Что же касается нашего примера, то более определенно, чем собст-
венно коренной тип леса, индицируется его жизненная форма [Крылов,
1974]: темнохвойно-широколиственный лес с гемигигрофильно-мезо-
фильным травяным покровом.
Более конкретно набор видов на всех стадиях сукцессий, включая и
климаксовую, ограничивается флорой региона и региональной потен-
циальной флорой эдафотопа данного типа. Ограничение выбора проис-
ходит по принципу совместимости видов с характером изменений среды
развитыми популяциями потенциальных здификаторов.
В рассмотренном примере потенциальные эдификаторы — в первую
очередь ель и липа. Климаксовыми сообществами здесь будут ельники
с липой и липняки с елью, и липо-ельники, реже — ельники или липня-
ки — все с примесью дуба и незначительным участием других пород,
включая и светолюбивые, с теневыносливыми гигромезофитами и мезо-
фитами в подлеске и покрове. Тот или иной вариант климакса зависит
от истории формирования растительного покрова данной местности и от
состояния в ней популяций отдельных видов в периоды интенсификации
изменений экологических режимов.
Из сказанного выше о потенциальных флорах экотопов следует, что
близкими КС, а следовательно, и близкими потенциями по формированию
устойчивых популяций обладают многие виды. Это заставляет нас считать,
что каждому местообитанию соответствует определенный набор климак-
совых сообществ, относящихся к одной или близким жизненным формам.
Количество таких сообществ зависит от общей комфортности место-
обитаний.
Чем меньше КК, чем ближе местообитание к экстремуму по какому-
либо фактору, тем меньше возможностей варьирования у климаксовой
растительности, тем больше она соответствует моноклимаксу.
Чем выше КК, чем дальше от экстремумов режимы факторов, тем
больше разнообразие климаксовых сообществ, тем более они соответст-
вуют поликлимаксу.
Для практики лесного хозяйства зто означает, что далеко не во всех,
а лишь в немногих случаях экологическая обстановка заставляет нас
вести хозяйство на одну породу. В большинстве случаев нам представля-
ется выбор из ряда экологически относительно равноценных видов.
В заключение стоит пожалуй еще коснуться списка видов в табл. 55. Ко-
нечно же он не укладывается в рамки флоры подзоны хвойно-широколист-
венных лесов и автор вполне отдает себе в этом отчет. Однако, считая прин-
193
ципы анализа растительного покрова изложенные в данной работе универ-
сальными, автор должен был дать возможность оценить и апробировать их
и исследователям работающим в других природных зонах. Кроме того,
автор имел в виду, что наибольший интерес его разработки вызвали у
специалистов, работающих в области интродукции растений. Именно поэто-
му в состав списка включено довольно много экзотов, в первую очередь
деревьев и кустарников.
Автор глубоко убежден, что именно фитоиццикационная оценка условий
среды, с учетом закономерностей ее изменений при замене одного расти-
тельного покрова другим, может быть наиболее надежным критерием
оценки пригодности того или иного местообитания или местоположения
для размещения, культивирования или охраны тех или иных видов. Это
одинаково справедливо как для хозяйственных, так и для научных или
эстетических целей.
Оценка экологического состояния и экологических возможностей лю-
бых местоположений или местообитаний — первейшее условие рационали-
зации любого использования их под естественным или искусственным рас-
тительным покровом.
Наконец, не лишнее может быть заметить, что ортодоксальное следова-
ние позициям лишь одной какой-либо научной школы не отвечает требова-
ниям современной науки. В отличие от политических партий научные
школы отражают не антагонистические противоречия различных классов
общества, а различные стороны одной и той же истины. Поэтому, игнорируя
взгляды и достижения какой-либо школы, мы неминуемо суживаем свой
кругозор.
ЛИТЕРАТУРА
Алехин В.В. Растительность СССР в ос-
новных зонах. М.: Сов. наука, 1951.
Бельгард А.Л. Лесная растительность
юго-востока УССР. Киев: Изд-во
Киев, ун-та, 1950.
Бельгард А.Л. Степное лесоведение.
М.: Лесн. пром-сть, 1971.
Благовидов Н.Л., Бурков Г.Л. Методи-
ческие указания к производству поч-
венных исследований и характерис-
тике условий местообитания леса.
Л., 1959.
Быков Б.А. Геоботаническая термино-
логия. Алма-Ата: Наука, 1967.
Быков Б.А. Введение в фитоценологию.
Алма-Ата: Наука, 1970.
Вальтер Г. Растительность земного шара.
М.: Прогресс, 1969. Т. 1.
Викторов С.В. Индикация природных
процессов. - В кн.: Теоретические
вопросы фитоиндикации. Л.: Наука,
1971.
Викторов С.В., Востокова Е.А., Вышив-
кин Д.Д. Введение в индикационную
геоботанику. М.: Изд-во МГУ, 1962.
Виноградов Б.В. Растительные индикато-
ры и их использование при изучении
природных ресурсов. Л.: Высш, шк.,
1964.
Волобуев В.Р. Система почв мира. Баку:
Элм, 1973.
Воробьев Д.П. Типы лесов европейской
части СССР. Киев: Изд-во АН УССР,
1953.
Воробьев Д.П. Методика лесотипологи-
ческих исследований. Киев: Урожай,
1967.
Воронков Н.А. Элементы влагооборота
лесных водосборов. - В кн.: Докл.
сов. учен, на Междунар. симпоз.
по влиянию леса на внешнюю среду.
М., 1970, т. 1.
Высоцкий Г.Н. Лес сушит равнины и
увлажняет горы. - На лесокуль-
турном фронте, 1932, № 2.
Географический атлас. М.: ГУГК, 1980.
Глазовская М.А. Почвы мира. М.; Изд-во
МГУ, 1972.
Горы шина Т.К. Экология растений. М.:
Высш, шк., 1979.
Ефимова Н.А. Карты продуктивности
природных растительных покровов
континентов. - Ботан. журн., 1976,
т. 61, №12.
Иванов Н.П. Пояса континентальности
земного шара. - Изв. ВГО, 1959,
т. 91, вып. 5.
Ковда В.А. Основы учения о почвах.
М.: Наука, 1973.
Корчагин А-4. Использование раститель-
ных сообществ как индикаторов
среды. - В кн.: Теоретические воп-
росы фитоиндикации. Л.: Наука,
1971.
Крылов А.Г. Жизненные формы лесных
фитоценозов Приморья: Комаров-
ские чтения. Владивосток: ДВНЦ
АН СССР, 1974. Вып. 22.
Лавриненко Д.Д. Основы лесной эко-
логии. Киев: Укр. СХА, 1978.
Мейен С.В. Следы трав индейских.
М.: Мысль, 1981.
Миркин БМ. Методологические воп-
росы классификации растительнос-
ти. - Журн. общ. биологии, 1981,
т. 17, №6.
Миркин БМ. , Розенберг Г.С. Фитоцено-
логия: Принципы и методы. М.:
Наука, 1978.
Миркин БМ., Шеляг-Сосонко Ю.Р. Об
экологических классификациях рас-
тительности. - В кн.: Вопросы геобо-
таники и луговедения в Башкирии.
Уфа: БФАН СССР, 1980.
Одум Ю. Основы экологии. М.: Мир,
1975.
Перельман А.И. Геохимия ландшафта.
М.: Высш, шк., 1966.
Погребняк П.С Люо-рослинш умови
Правобережнього Полюся Украши. -
Тр. 3 лис. доев, справи на У крапп,
1927.
Погребняк П.С. Общее лесоводство. М.:
Колос, 1958.
Работное Т.А. Влияние минеральных
удобрений на луговые растения и лу-
говые фитоценозы. М.: Наука, 1973.
Работное Т.А. О применении экологи-
ческих шкал для индикации эдафи-
ческих условий произрастания рас-
тений. - Журн. общ. биологии, 1979,
т. 15, № 1.
195
Разумовский С.М. Закономерности ди-
намики биоценозов. М.: Наука, 1981.
Рамад Ф. Основы прикладной экологии.
Л.: Гидрометеоиздат, 1981.
Раменский Л.Г. Основные закономер-
ности растительного покрова. -
Вест, опытного дела, Воронеж, 1924.
Раменский Л.Г. К методике сравнитель-
ной обработки исистематизацииспис-
ков растений н других объектов,
определяемых несколькими несход-
но действующими факторами. -
В ки.: Труды совещ. ботаников-лу-
говедов, созванного Гос. луговым
ин-том 15-20 января 1928 г. в Ле-
нинграде. М.: Сельхозгиз, 1929.
Раменский Л.Г. Введение в почвенно-
геоботаническое исследование зе-
мель. М.: Сельхозгиз, 1938.
Раменский Л.Г., Цаценкин И.А., Чижи-
ков О.Н., Антипин Н.А. Экологичес-
кая оценка кормовых угодий по рас-
тительному покрову. М.: Сельхозгиз,
1956.
Рахманов В.В. Влияние лесов на вод-
ность рек в бассейне Верхней Волги.
Л.: Гидрометеоиздат, 1971.
Роде Ли4., Смирное В.Н. Почвоведение.
М.: Высш, шк, 1972. *
Рутковский В.И. Гидрологическая роль
леса. М.: Гослесбумиздат, 1949.
Рысин Л.П. Типы лесов Восточного
Подмосковья. - В кн.: Леса Восточ-
ного Подмосковья. М.: Наука, 1979.
Самойлов Ю.И. Некоторые результаты
сравнения экологических шкал Ра-
менского, Элленберга, Хундта и Клап-
па.- Ботан. журн., 1973, т. 58, № 5.
Соболев Л.Н. Методика эколого-типоло-
гического исследования земель.
Фрунзе: Илим, 1978.
Сукачев В.Н. Руководство к исследо-
ванию типов лесов. М.; Л.: Сель-
хозгиз, 1931.
Сукачев В.Н. Дендрология с основами
лесной геоботаники. Л.: Гослестех-
издат, 1938.
Сукачев В.Н, Зонн С.В. Методические
указания к изучению типов леса.
М.: Изд-во АН СССР, 1961.
Таргульян В.О. Почвообразование и
выветривание в холодных гумндных
областях. М.: Наука, 1971.
Толчаин Г.А. Опыт экологической инди-
кации растительности в Брянской
и Тульской областях для целей
мелиорации земель. - В кн.: Эколо-
гические и географические основы
мелиорации земель в бассейне
р. Десны. М.: МФГО, 1980.
Тюрин И.В., Пономарева В.В. Материа-
лы по изучению гумуса лесных
почв. - Тр. ЛТА, 1940, вып. 56.
Уиттекер Р. Сообщества и экосистемы.
М.: Прогресс, 1980.
Фридллнд В.М. Структура почвенного
покрова. М.: Мысль. 1972.
Цаценкин ИЛ., Савченко И.В., Дмитрие-
ва С.И. Методические указания к оцен-
ке кормовых угодий тундровой и
лесной зон Сибири и Дальнего Восто-
ка по растительному покрову. М.:
ВНИИ кормов им. Вильямса, 1978.
Цыганов Д.Н. Экоморфы и экологичес-
кие свиты. - Бюл. МОИП, 1974,
т. 79, вып. 2.
Цыганов Д.Н. Системы экоморф и инди-
кация основных экологических
режимов местообитаний. - Эколо-
гия, 1975, №6.
Ць.ганов ДН. Экоморфы флоры хвой-
но-широколиственных лесов. М.:
Наука, 1976.
Чертов О.Г. Экология лесных земель.
Л.: Наука, 1981.
Юрцев Б.А., Семкин Б.И. Изучение кон-
кретных и парцеллярных флор с по-
мощью математических методов. -
Ботан. журн., 1980, т. 65, № 12.
Donifa _N.„ Purcelean S., Ceianu J., Bel-
die A. Ecologie forestiera. Bucuresti:
Editura Ceres, 1977.
Ellenberg H. Uncrautgemeinschaften als
Zeiger fur Klima und Boden. Land-
wirtschaftliche Pflanzensociologie, 1.
Stuttgart, 1950.
Ellenberg H. Wiesen und Weiden und
ihre standoitliche Bevertung. Land-
wirtschaftliche Pflanzensociologie, 2.
Stuttgart, 1952.
Ellenberg H. Zeigerwerte der Gefasspflan-
zen Mitteleuropas. Gottingen, 1974.
Hundt R. Okologisch-geobotanische Un-
tersuchungen an Pflanzen der M t-
teleuropaischen Wiesenvegetatio i
Leipzig, 1966.
Klapp E. Wiesen und Weiden. 4. Au I
Berlin-Hamburg, 1971.
Meusel H. Vergleichende Chorologie der zen -
raleuropaischen Flora. Jena, 196.
Landolt E. Okologische Zeigerwert:
zur' Sweizer Flora. - Veroff. Geobot.
Inst. ETH. Zurich, 1977.
Mraz K., Samek V. Lesny rostliny. Praha,
1966.
Zolyomi B., Precsnyi J. Metrode zur
okologischen Charakterisirerung der
Vegetationseinheiten und zum Verg-
leich der Standorte. - Acta bot. Acad,
sci. hung., 1964, B. 10, Fasc. 3/4.
ОГЛАВЛЕНИЕ
Глава I. Современные представления о фитоиндикации и развитие
концепции Л.Г. Раменского, как одно из перспективных
ее направлений................................................... 3
Глава II. Лесорастительныс условия и экологическая типология зе-
мель ............................................................14
Глава III. Лесорастительные условия на лесопокрытых и безлесных
территориях ..................................................... 45
Глава IV. Система экологических характеристик видов как основа
фитоиндикации....................................................5 3
Глава V. Индикация экотопов, как индикация комплексирующихся
в них экологических режимов отдельных прямодействующих
факторов........................................................119
Г л а в а VI. Фитоивдикационная экологическая таблица...........130
Глава VII. Возможности применения фитоиндикации для диагностики
типов леса......................................................185
Литература..........................................1^5
Дмитрий Николаевич Цыганов
ФИТОИНДИКАЦИЯ
ЭКОЛОГИЧЕСКИХ РЕЖИМОВ
В ПОДЗОНЕ ХВОЙНО-
ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Утверждено к печати
Лабораторией лесоведения
Академии наук СССР
Редактор издательства Е.К. Исаев
Художник Н.Н. Якубовская
Художественный редактор И.Ю. Нестерова
Технические редакторы И.И. Джиоева
Г.И. Астахова
Корректор Л.А. Агеева
ИБ № 27206
Подписано к печати 29.06.83. Т—10467
Формат 60x90 1/16. Бумага офсетная № 1
Печать офсетная. Усл.печ.л. 12,5
Усл.кр.-отг. 12,8. Уч.-издл. 15,9
Тираж 850 зкз. Тип.зак. 448
Цена 2 р. 40 к.
Издательство ’’Наука”, 117864 ГСП-7,
Москва, В-485, Профсоюзная ул., д. 90
Ордена Трудового Красного Знамени
1-я типография издательства ’’Наука”
199034, Ленинград, В-34, 9-я линия, 12