/

















































































Автор: Павлов В.Н.
Теги: география растений фитогеография флора географическое распространение растений
ISBN: 5-8125-0332-X
Год: 2003




Текст
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
Выпуск пятнадцатый
Москва
2003
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ
ОБЛАСТИ
Под редакцией
профессора В.Н. Павлова
Выпуск пятнадцатый
Москва
2003
УДК 581.9 (471.311)
Биологическая флора Московской области. Вып. 15. Под ред.
В. Н. Павлова.— М.: Изд-во «Гриф и К», 2003.— 224 с.
ISBN 5-8125-0332-Х
В книге обобщены итоги оригинальных наблюдений, экспериментальных работ и
литературные данные по систематике, географическому распространению, морфоло-
гии, онтогенезу, экологии, консортивным связям, фитоценологии, хозяйственному
значению и охране видов растений, произрастающих в Московской области. В книгу
вошли статьи; вейник седеющий, шпажник черепитчатый, неоттеанте клобучковая,
пололепестиик зеленый, лютик кашубский, стальник полевой, тмин обыкновенный,
первоцвет весенний, вербейник обыкновенный, вьюнок полевой.
Для студентов, аспирантов, преподавателей высших учебных заведений и широ-
кого круга специалистов-биологов, работников сельского и лесного хозяйства, деяте-
лей охраны природы.
Редакторы выпуска М. Г. Вахрамеева, Н. Г. Уланова
Рецензенты: доктор биологических наук А. П. Меликян,
кандидат биологических наук С. В. Никитина
Biological Flora of the Moscow Region. Part 15. Moscow: Grif and K,
2003.—224 p.
Chief editor — Prof. V. N. Pavlov
Edited by M. G. Vakhrameeva, N. G. Ulanova
Издание осуществлено при поддержке грантов: Президента РФ для под-
держки ведущих научных школ№ 00-15-97734, HU1-2125.2003.4, Научной про-
граммы Минобразования РФ «Университеты России» НИР Не УР.07.03.010
и ГНТП Биологическое разнообразие
077(02) — 2003 — заказное
ISBN 5-8125-0332-Х
© Кафедра геоботаники, МГУ, 2003
ПРЕДИСЛОВИЕ
Начиная с 1974 г., опубликовано 14 выпусков сборника «Биологиче-
ская флора Московской области», в которых охарактеризовано 282 вида
растений из 54 семейств. В настоящем выпуске принимали участие со-
трудники государственных университетов: Московского, Московского пе-
дагогического, Тверского, Полярно-альпийского ботанического сада и
Башкирского заповедника. Растения описаны в традиционном для данного
издания плане: номенклатура и систематическое положение, географиче-
ское распространение, морфологическое описание, онтогенез, способы
размножения и распространения, сезонное развитие, экология, фитоцено-.
логия, консортивные связи, биологическая продуктивность, хозяйствен-
ное значение. Раздел «номенклатура» написан вед.н.с. каф. геоботаники
И.А. Губановым.
В разделе «экология», как и в предыдущих выпусках, использованы
экологические шкалы Л.Г. Раменского (Раменский и др., 1956), Г. Эллен-
берга (EUenberg, 1991) и Э. Ландольта (Landolt, 1977). В тексте приведены
только фамилии авторов экологических шкал без ссылки на источники.
Краткий перевод последних двух шкал опубликован в 6-м выпуске «Био-,
логической флоры Московской области» (1980).
Авторы обработок таксонов во «Флорах» и «Определителях» приведе-.
ны в номенклатурных цитатах, а списке литературы даны только названия
источников. Во всех статьях сборника для обозначения возрастных состоя-
ний растений применяются следующие условные обозначения: s — семя,
р — проростки, j — ювенильные, im — имматурные, v — виргинильные
(молодые вегетативные), g — генеративные, ss — субсенильные, s — се-
нильные.
Основную работу по комплектации и редактированию выпуска'осуще-'
ствляли сотрудники кафедры геоботаники Московского государственного
университета — член-корр.РАН В.Н. Павлов, с.н.с. М.Г. Вахрамеева, доц.
Ю.Е. Алексеев, с.н.с. Н.Г. Уланова.
3
A. H. Демидова, Н. Г. Уланова
ВЕЙНИК СЕДЕЮЩИЙ
Номенклатура, систематическое положение
и внутривидовая систематика
Calamagrostis canescens (Web.) Roth, 1789, Tent. Fl. Germ. 2,1: 93; Цвелев,
1964, в Маевский, Фл„ изд. 9 : 795; Скворцов, 1966, Опред. раст. Моск,
обл.: 76; Цвелев, 1974, Фл. европ. части СССР, 1 : 222; он же, 1976, Злаки
СССР: 313; Clarke, 1980, Fl. Europ. 5 : 237; Губанов и др., 1995, Опред. со-
суд. раст. Центра Европ. России: 120,— Arundo canescens Web. 1780, in
Wigg. Primit Fl. Holsat.: 10.— Calamagrostis lanceolata Roth, 1788, Tent. Fl.
Germ. 1,1: 34; Кауфман, 1866, Моск, фл.: 555; Сырейщйков, 1906, Илл. фл.
Моск, губ., 1: 120; он же, 1914, цит. соч., 4 : 13; Рожевиц, 1934, Фл. СССР,
2: 203.— Вейник седеющий [семейство Gramineae (Роасеаё)— Злаки
(Мятликовые)].
Вейник седеющий гибридизирует с другими видами вейника, причем
особенно часто встречаются межсекционные гибриды (Цвелев, 1976). Гиб-
ридогенные клоны, как правило, стерильны, но длительно существуют и
часто производят впечатление самостоятельных видов. Наиболее часто
встречаются гибриды (Флора СССР, 1934): 1) С. neglecta s.l. х С. canescens
(С. х vilnensis Bess.; С. gracilescens Blytt; С. obscura Downar; C. canescens'
subsp. vilnensis (Bess.) H. Schulz), 2) C. canescens x C. epigeios s.l. (C. x rigens
Lindgr.; C. x neumaniana Torges), 3) C. canescens x C. villosa (C. x kotulae
Zapal.). 4) C. canescens x C. obtusata (Цвелев, 1965)’ 5) C. arundinacea x
C. canescens (C. x hartmanniana Fries, 6) C. canescens x C. langsdorfii,
7) C. stricta x C. canescens (Crackles, 1998). В северной и восточной частях
ареала вида преобладают популяции, являющиеся результатом интрогрес-
сивной гибридизации С. canescens х С. purpurea. Эти виды связаны широ-
кой полосой переходных по признакам популяций и клонов гибридного
происхождения. Значительное перекрывание ареалов этих видов вторич-
ное (Цвелев, 1965), оно возникло во время наступивших в четвертичном
периоде периодических похолоданий климата, когда С. purpurea (Trin.)
Trin. вместе с другими элементами темнохвойной тайги спустился с гор на
4
равнины, оттесняя к югу и к морским побережьям, существовавший здесь
с неогена С. canescens (Web.) Roth (на западе). Возможно, происхождение
в результате интрогрессивной гибридизации имеют и все популяции С.
canescens с хорошо развитым и длинноволосистым продолжением оси ко-
лоска над основанием цветка — var. rudimentigera (Litv. ex Roshev.) TzveL
comb.nova (= C. lanceolata var. rudimentigera Litv. ex Roshev. 1928, во Фл.
Юго-Вост. 2: 66). Р.Ю. Рожевиц (Флора СССР, 1934) приводит другие при-
знаки этой разновидности: волоски на оси колоска немного длиннее цве-
точной чешуи; волосистого продолжения оси нет, реже имеется едва за-
метный, точечный, бледный зачаток с немногими волосками.
Вейник седеющий — полиморфный вид, в пределах которого кроме
var. rudimentigera (Litv. ex Roshev.) Tzvel. различают еще несколько разно-
видностей (Флора СССР, 1934): var. pallida Lange: бледная окраска листь-
ев; var. parviflora Harz: колоски 4-5 мм длиной, реже 3—4 мм; var.
grandiflora Harz: колоски 5-6 мм длиной; var. hypocrathera Torges: ость си-
дит немного ниже выемки (редко).
Географическое распространение
Вейник седеющий имеет западно-палеоарктический ареал (Гроссгейм,
1939). Распространен почти на всей территории Европы, включая европей1
скую часть бывшего СССР, на юге Западной Сибири и в северной части
Казахстана. На Украине вид изредка встречается по всей республике, кро-
ме Крыма (Прокудин, 1977). В горах вид распространен до нижнего горно-
го пояса (Цвелев, 1976).
В России границы ареала вейника седеющего проходят от Кольского
полуострова на севере до Предкавказья (окрестности Ставрополя), от Бал-
тийского моря до Иртыша на востоке. По данным А.С. Королевой с соавто-
рами (1973), вид встречается и в Новосибирской области. В лесной зоне
умеренных широт вид обычен до 65 с.ш., севернее встречается редко (Ра-
менская, 1982). В Средней полосе европейской части распространен
повсеместно почти во всех областях, но на юге и юго-востоке реже (Ма-
евский, 1964). К северо-востоку С. canescefis (Web.) Roth сменяется
С. langsdorfii Trin. ex' Litw., который в среднем течении Печоры и в верх-
нем течении Камы уже господствует (Соколова и др., 1956). В Централь-
ной части Среднего Поволжья вейник седеющий встречается спорадичен
ски во всех областях (Благовещенский, 1984). В степной зоне он произра-
стает только по долинам больших рек (Прокудин, 1977).
В Московской области вид встречается часто и повсеместно (Вороши-
лов и др., 1966).
5
Морфологическое описание
Вейник седеющий—длиннокорневищный травянистый поликарпиче-
ский многолетник с безрозеточными и вневлагалищными побегами (Се-
ребрякова, 1971).
Корневища гипогеогенные прочные, соломенного цвета, диаметром
1,5-2 мм, но обычно не более 1 мм (Бельков и др., 1974). От парциального
куста в среднем отходят до 4-х молодых без придаточных корней корне7
вищ (отбегов) и 2-х (реже 3) старых корневищ. Ветвистое корневище обра-
зует рыхлые дерновины (Рычин, 1948). В каждом из узлов корневища фор-
мируется по одному трубчатому низовому чешуевидному листу и по од-
ной пазушной почке (Бельков и др., 1974). Придаточные корни
сосредоточены в узлах корневищ, по 8-15 в каждом из них. Корни тонкие,
крепкие, на всем протяжении ветвятся. Длина корней П-го порядка —
1-2 см, диаметр — 0,1 мм, длина корней Ш-го порядка— 3-6 мм. Корни
густо покрыты корневыми волосками, отчего их поверхность приобретает
войлочный характер. В зонах кущения вейника, где сосредоточено боль-
шое количество узлов в небольшом пространстве, часто образуются очень
густые мочки корней (Бельков, 1974). По данным Л.П. Рысина (1987), в со-
сняке бруснично-молиниевом длина корней обычно не превышала 30 см
при диаметре у основания 0,3-0,4 мм.
Растение серовато-зеленого цвета. Стебель тонкий, прямой, цилиндри-
ческий, почти доверху ветвистый и олиственный, гладкий или шерохова-
тый в своей верхней части (Рычин, 1948). По данным разных авторов высо-
та растения варьирует от 50. до 150 см (Рычин, 1948; Быков, 1962; Проку-
дин, 1977; Weymar, 1972, Губанов и др., 1995).
Парциальные кусты состоят из 1-6 (чаще 1-3) побегов. Стебли с
3(4)-6(7) узлами, из которых верхний располагается значительно выше их
середины (Цвелев, 1965).
Срединные листья состоят из влагалища и листовой пластинки, ниж-
ние—только из чешуевидных влагалищ. Нижние влагалища гладкие,
верхние шероховатые (Рычин, 1948). Влагалищно-пластиночные сочлене-
ния обычно без бородки волосков. Влагалища замкнутые лишь у самого
основания (Цвелев, 1965). Пластинки листьев узколинейные или ланцет-
ные, длинно-заостренные, в почкосложении скрученные, к основанию за-
метно суженные, обычно 10-20 см длиной и около 5(1,5-3,5) мм шириной.
Листья светло-зеленые, с сероватым оттенком, голые, или опушенные; по
краям и снизу вниз шероховатые, жилки на просвет все, или почти все, бе-
лые (Рычин, 1948).
Высота ребер на листовых пластинках равна их ширине или превыша-
ет ее (на поперечном разрезе через середину пластинки), ребра у развито-
го листа выпуклые, килеватые или плоские (Штрекер, 1931). Пластинка
листа на разрезе четковидная, с тупоугольным или закругленным килем
6
(Прокудин, 1977). Клетки верхнего эпидермиса тонкостенные, нижне-
го — с утолщенными боковыми и наружными стенками. На верхней сто-
роне эпидермиса имеются заостренные шипики и длинные тонкие волос-
ки, на нижней стороне — тупые шипики. Лист эпистоматический. Пла-
стинка листа на верхней стороне бороздчатая, с моторными клетками в
каждой борозде. Жилок 23—44, из них средняя и несколько боко-
вых — крупные, остальные мелкие. Особенности анатомического строе-
ния листа вейника седеющего достаточно подробно обсуждаются в рабо-
те Р.Т. Протасевича (1963).
Язычок около 1 мм высоты у вегетативных побегов, 4-8 мм — у гене-
ративных. Он пленчатый, клиновидный или несколько закругленный, ко-
ротко-бархатистый со спинки (голый или с очень редкими волосками), ба-
хромчатый по краю, часто расщепленный.
Соцветие — крупная мнбгоколосковая метелка 10-17 см длиной и
2-4 см шириной, сжатая или слабо раскидистая, до отцветания фиолето-
во-серо-дымчатой окраски, иногда поникающая. Веточки метелки длин-
ные, тонкие, шероховатые от многочисленных шипиков. Самые длинные
веточки обычно в 1,5-2,5 раза короче всей метелки. Колоски одноцветко-
вые, ланцетные, темно-пурпуровые, реже зеленые, фиолетовые или буро-
вато-фиолетовые, на ножках, 3,5-5 мм длиной. Цветки обоеполые, с 3 ты-
чинками и 1 пестиком с двумя перистыми рыльцами Ось колоска под цвет-
ком с длинными волосками. Рудимент оси колоска над цветком
отсутствует или очень короткий (до 0,3 мм длиной), голый, редко с оди-
ночными колосками (Маевский, 1964). Колосковые чешуи ланцетные,
длиннозаостренные, по килям с короткими шипиками, иногда и по осталь-
ной поверхности с рассеянными шипиками, мало заметными даже при
сильном увеличении. Верхняя цветковая чешуя на */3 короче нижней. Ниж-
няя цветковая чешуя 2,0-2,7 мм длиной, с каллусом, на верхушке с зубца-
ми, с остью выходящей между верхушечными зубцами или чуть ниже в
верхней трети чешуи. Ость гладкая, слаборазвитая, часто почти незамет-
ная, обычно едва превышает верхушку чешуй (Цвелев, 1965). Каллус по
бокам с пучком густых волосков. Волоски каллуса немного превышают
половину длины нижних цветковых чешуй и к времени созревания зерно-
вок, в сухую погоду, они отгибаются в стороны от зерновки, образуя «па-
рашютик». Пленки при плодах усажены нежными волосками.
Зерновка продолговатая, слегка яйцевидная, светло-коричневая.
Число хромосом 2п=28 (Цвелев, 1976; Соколовская, 1972; Nygren,
1946, 1962, Hubbard, 1954, цит. по: Хромосомные числа..., 1969), очень
редко встречаются 2п=42, 56 (Hubbard, 1954, цит. по: Прокудин, 1977).
7
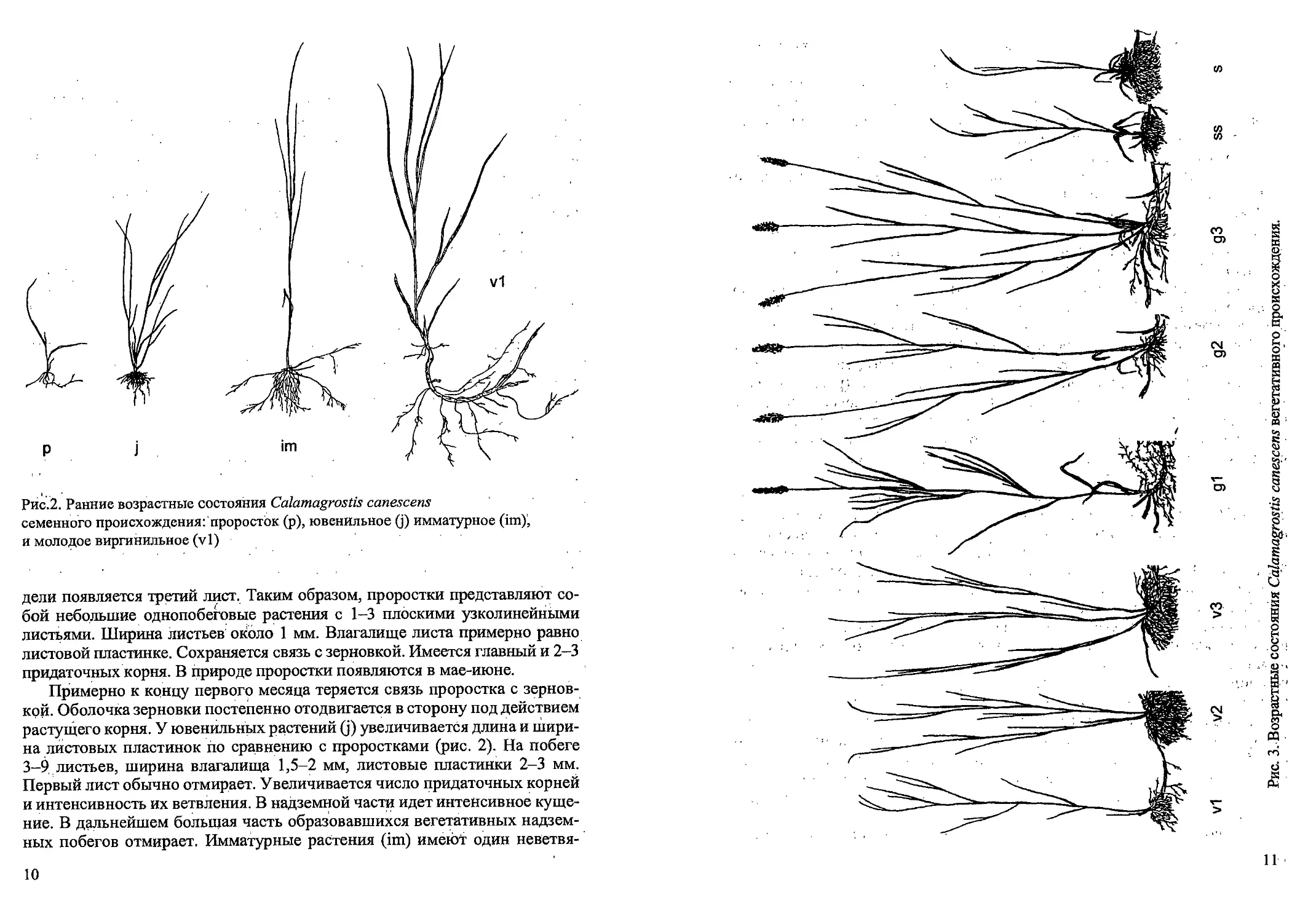
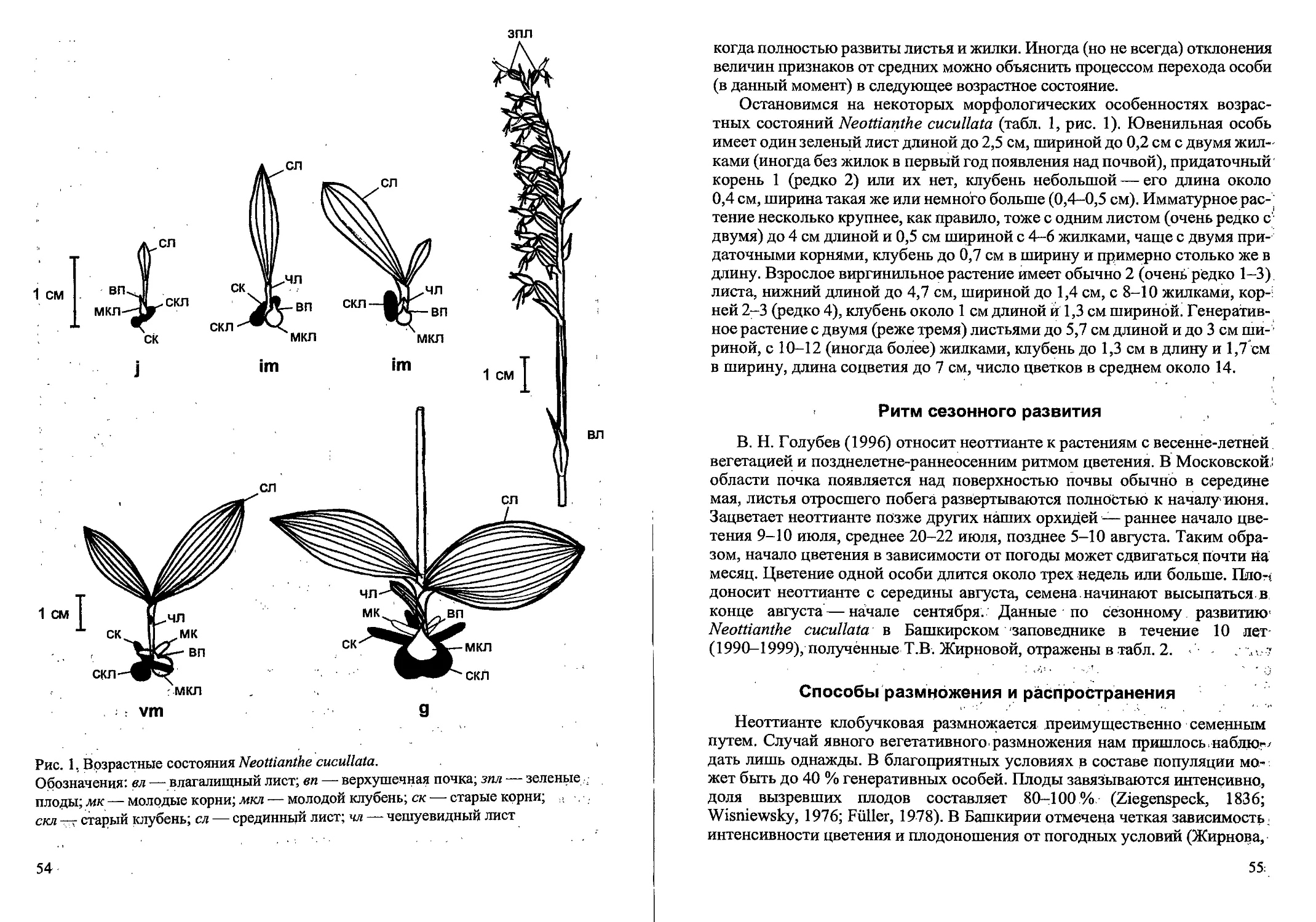
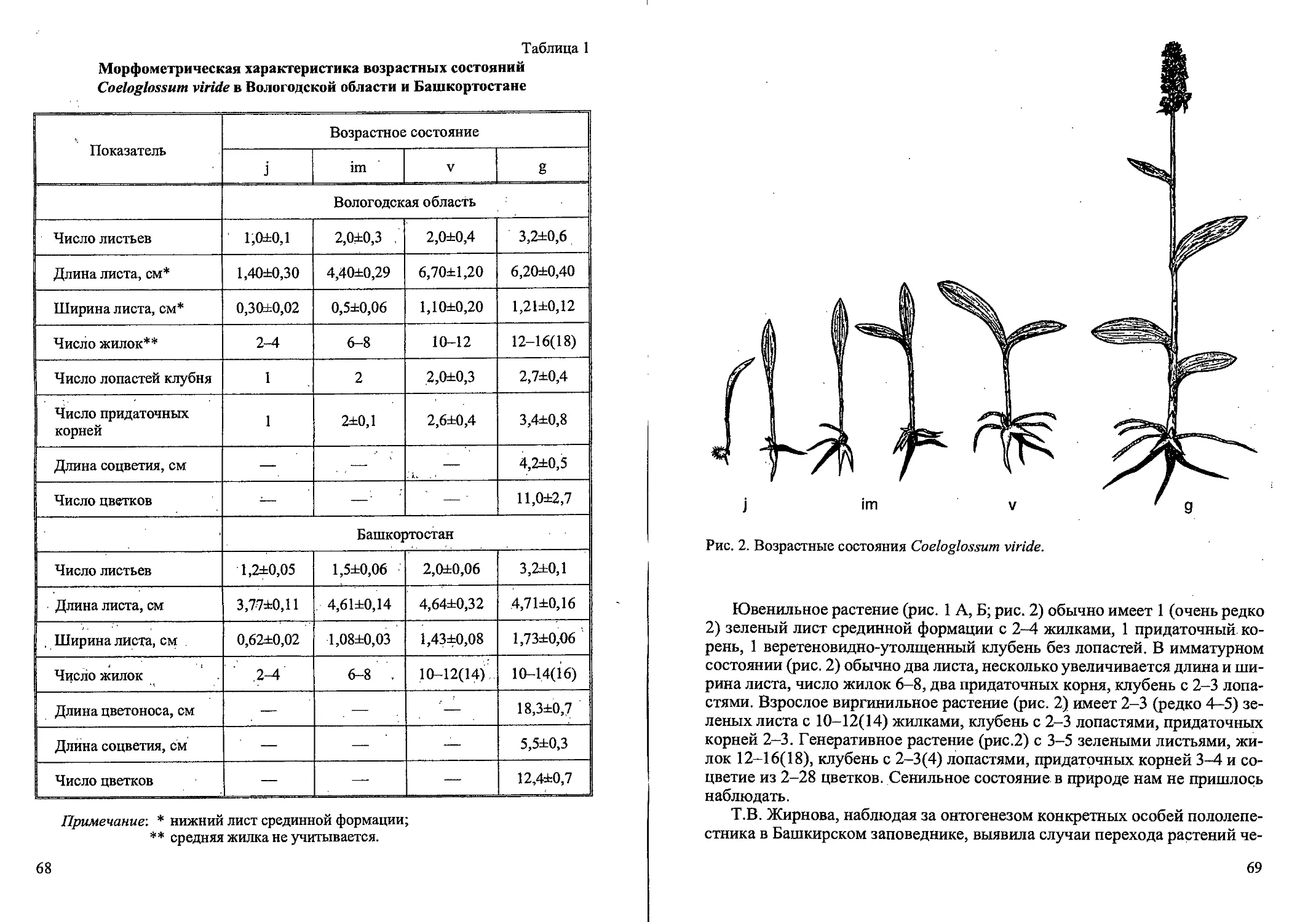
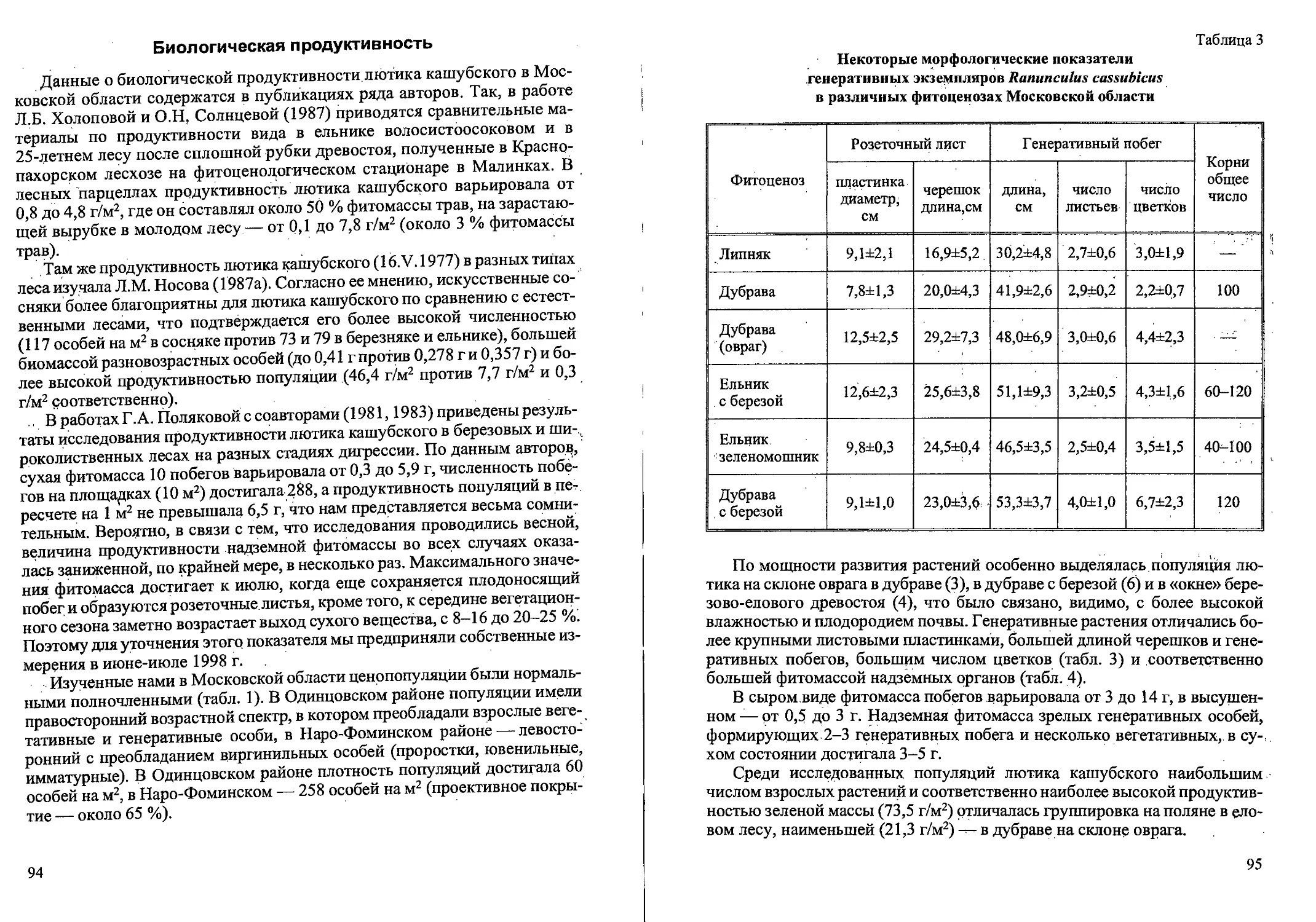
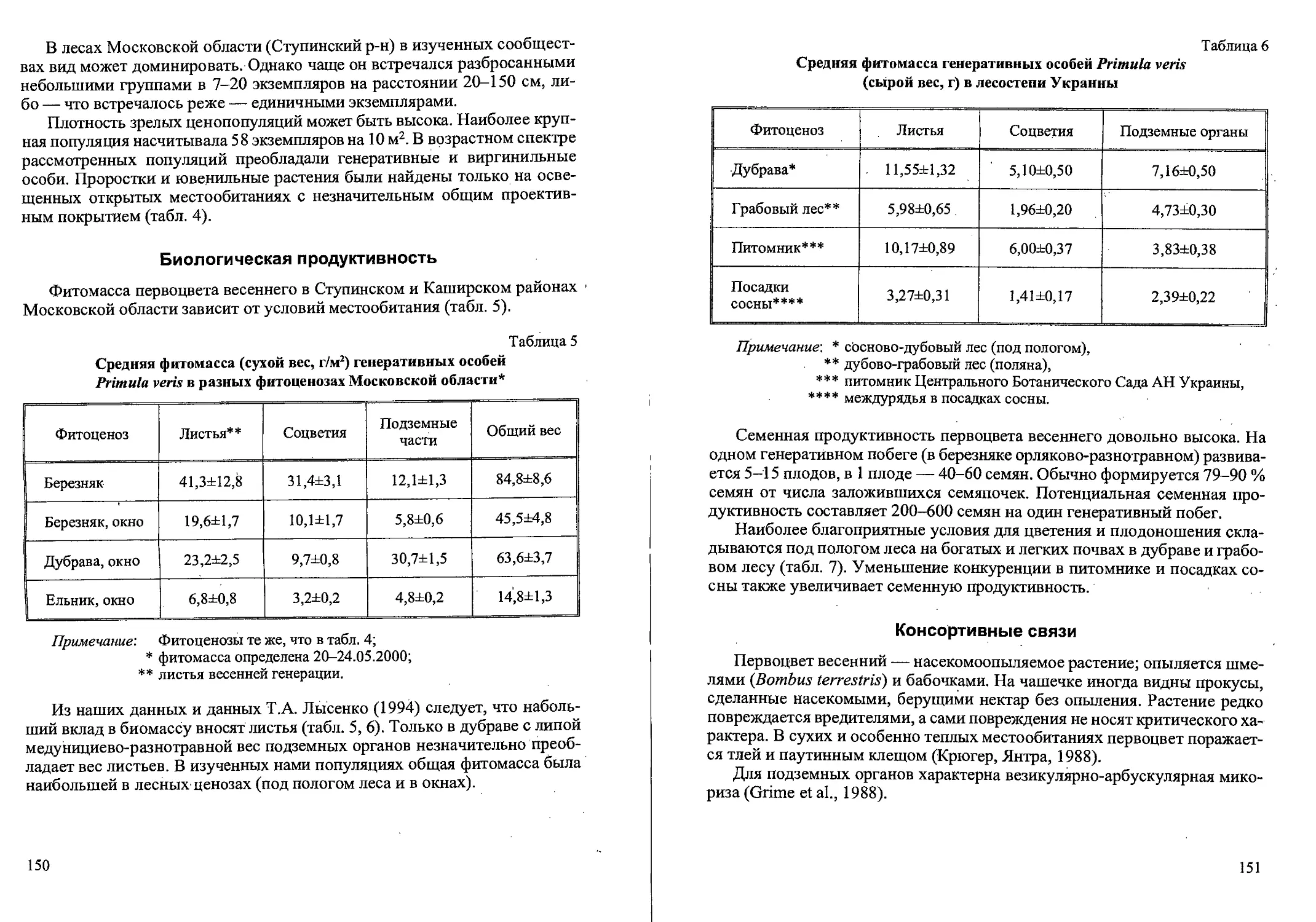
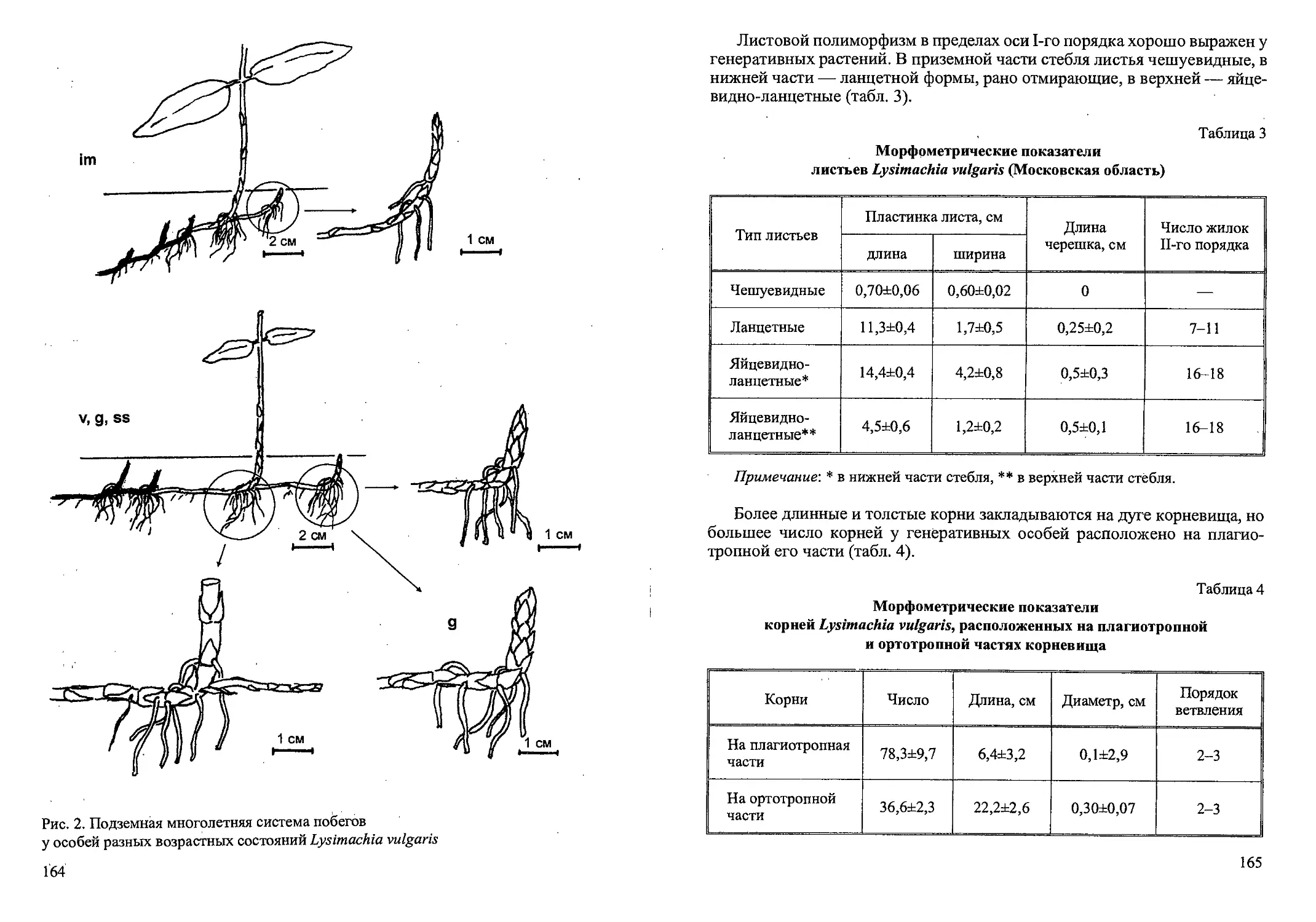
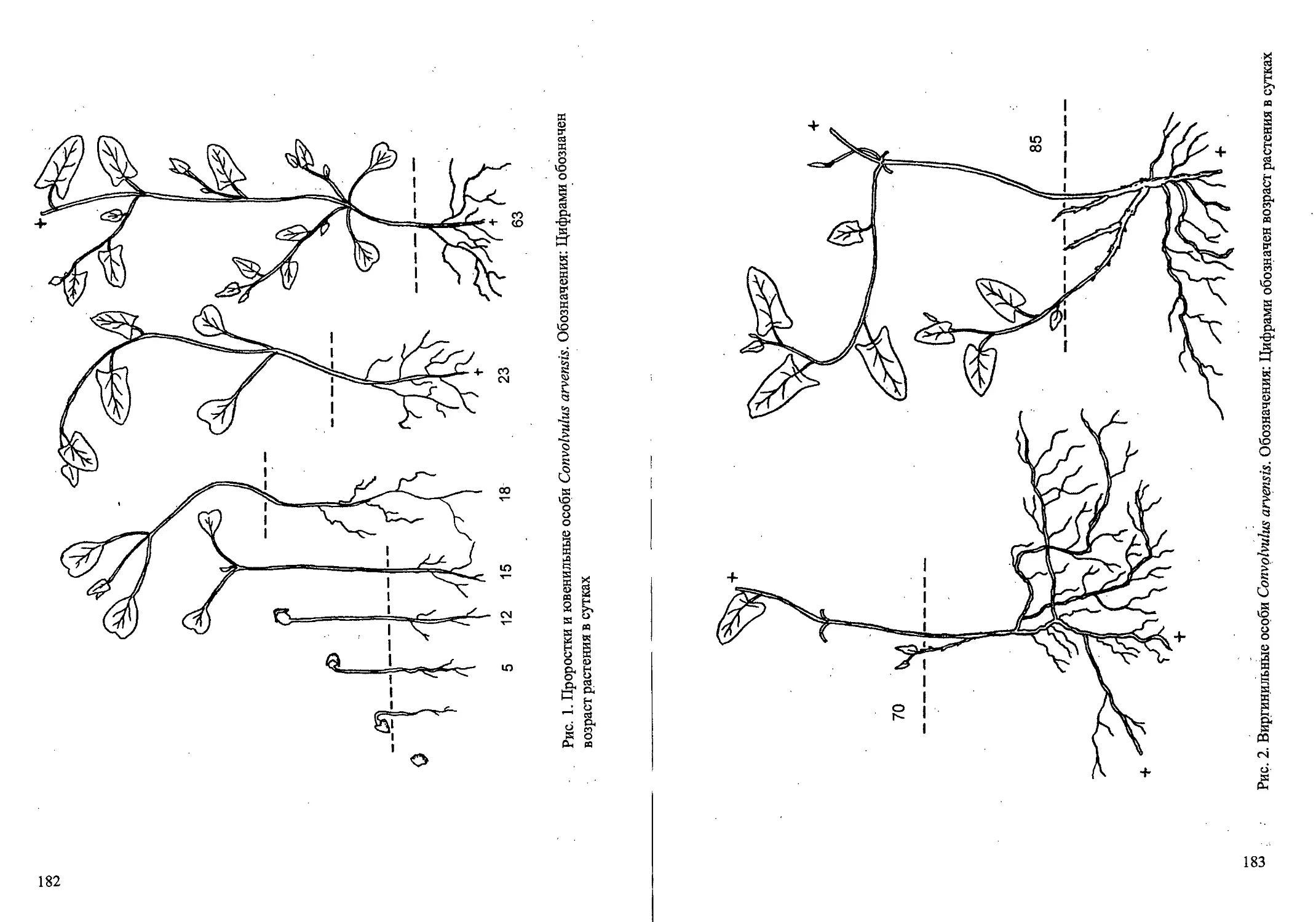
Рис. 1. Стадий прорастания семян и проростки Calamagrostis canescens
8
Онтогенез
Большой жизненный цикл вейника седеющего протекает с усилением
побегов последовательных порядков (Серебрякова, 1971): главная ось и
боковые побеги материнского семенного растения развиваются, но не про-
ходят полного цикла развития, они меньше и слабее побегов последующе-
го порядка. Только побеги в кустах П-Ш-го порядков могут пройти пол-
ный цикл развития.
В жизненном цикле вейника седеющего по особенностям возрастных
состояний можно выделить следующие фазы:
1) первичный прбег — этой фазе соответствует возрастное состояние
проростков, ювенильных и имматурных растений до начала кущения.
2) первичный куст — этой фазе соответствует виргинильный, генера-
тивный, субсенильный и сенильный периоды развития растения. Парци-
альный куст является основным структурным элементом взрослой особи
вейника. Развитие его начинается с появления вертикального побега из
терминальной почки плагиотропного корневища. Побеги последующих
порядков возникают на нем из пазушных почек нижних чешуевидных ли-
стьев выше или ниже поверхности почвы.
3) колония — образование системы парциальных кустов. Наиболее
продолжительная фаза в жизненном цикле вейника, характеризуется мак-
симальной жизненной активностью.
Семена вейника седеющего созревают в июле-августе. По нашим на-
блюдениям, свежесобранные семена при проращивании в лабораторных
условиях (на фильтровальной бумаге при 18-20°) в ноябре имели всхо-
жесть 63-72 %. После двухмесячной стратификации семян, собранных
осенью, их всхожесть не превысила 13 %. В природе проросшие семена,
проростки, а также взрослые растения семенного происхождения обнару-
жить очень сложно. Начальные стадии онтогенеза удалось проследить в
лабораторных условиях.
Наши наблюдения показали, что зерновки прорастают на 6-8-й день (4-й
после стратификации) при проращивании в чашках Петри на влажной
фильтровальной бумаге при температуре 18-20°. На 2-3-й день у пророст-
ков уже заметен первичный корешок и влагалищный лист (рис. 1). Верти-
кально стоящий первый лист появляется на 3-й день развития. Его длина со-
ставляет в среднем 3-5 мм. На 4-9-й день развития (редко на третий) начи-
нает формироваться корневая система. На 4-й день на поверхности
первичного корешка появляются небольшие бугорки. На 11—15-й день начи-
нается образование горизонтальных придаточных корней, в этот же период
появляется второй лист. У первого листа уже видны на просвет две жилки, и
длина его — 12-20 мм. Ежедневные измерения проростков показали нерав-
номерный рост корней и листьев в течение суток, и различную степень их
развития у одновозрастных проростков. В конце второй-начале третьей не-
9
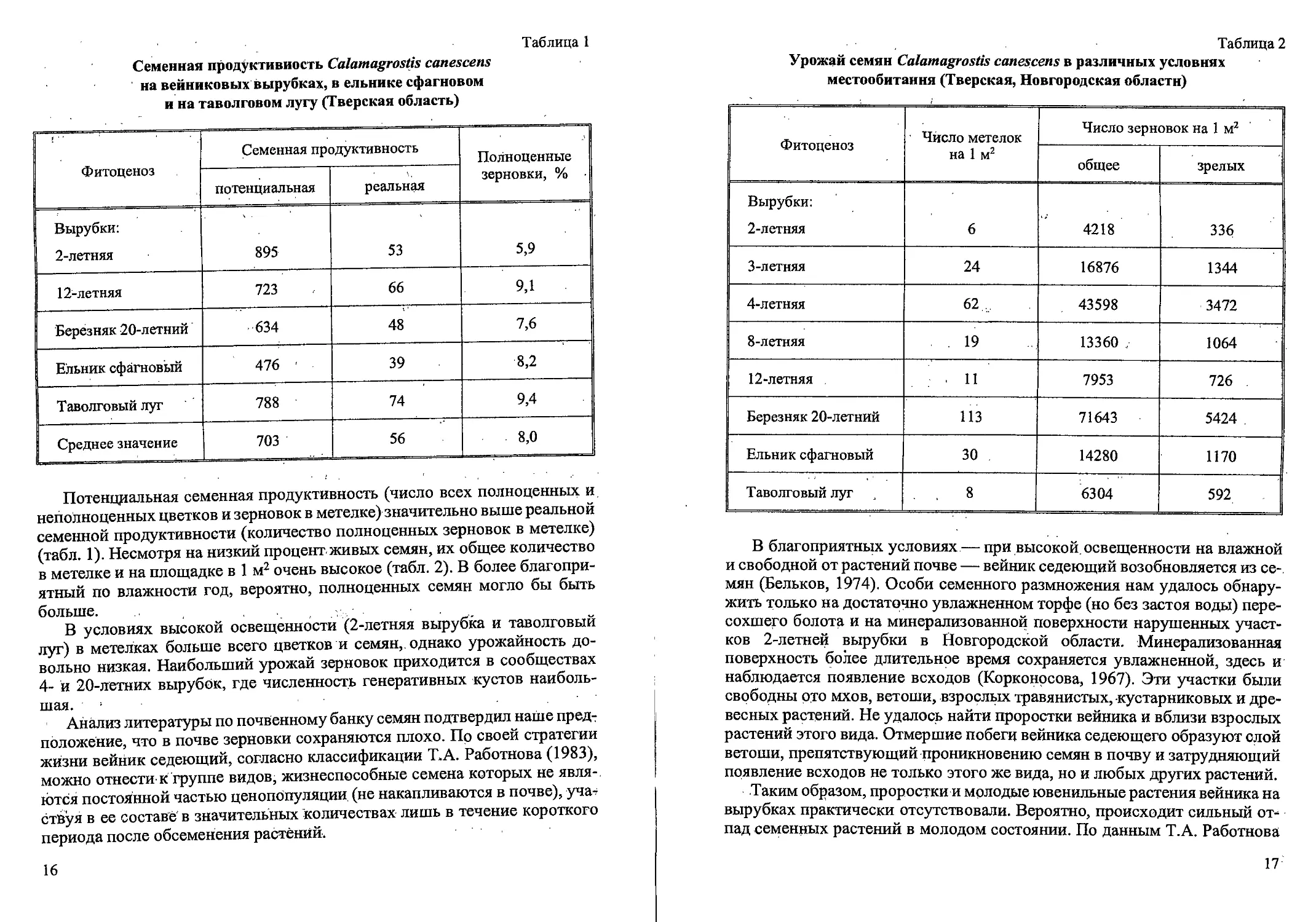
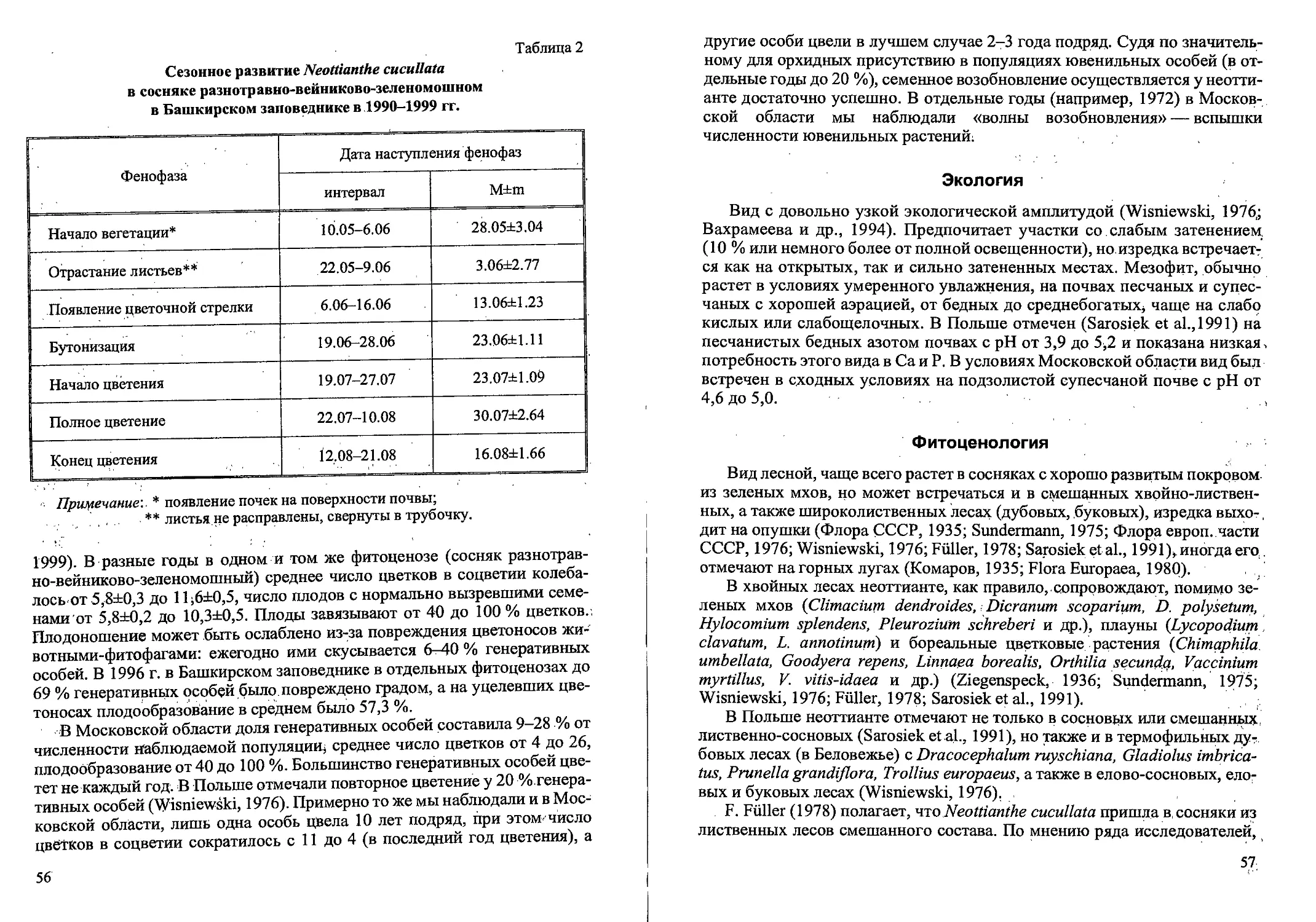
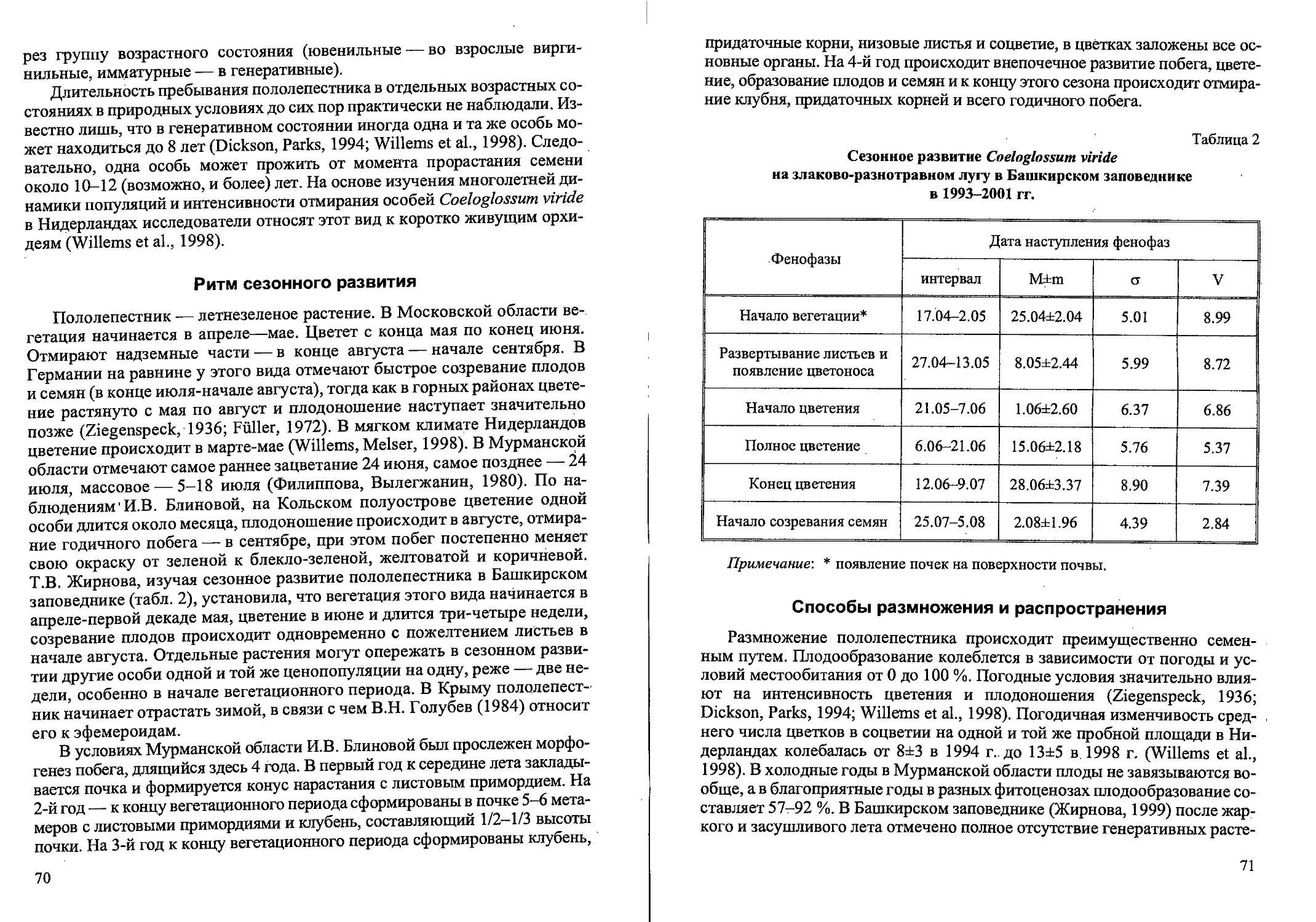
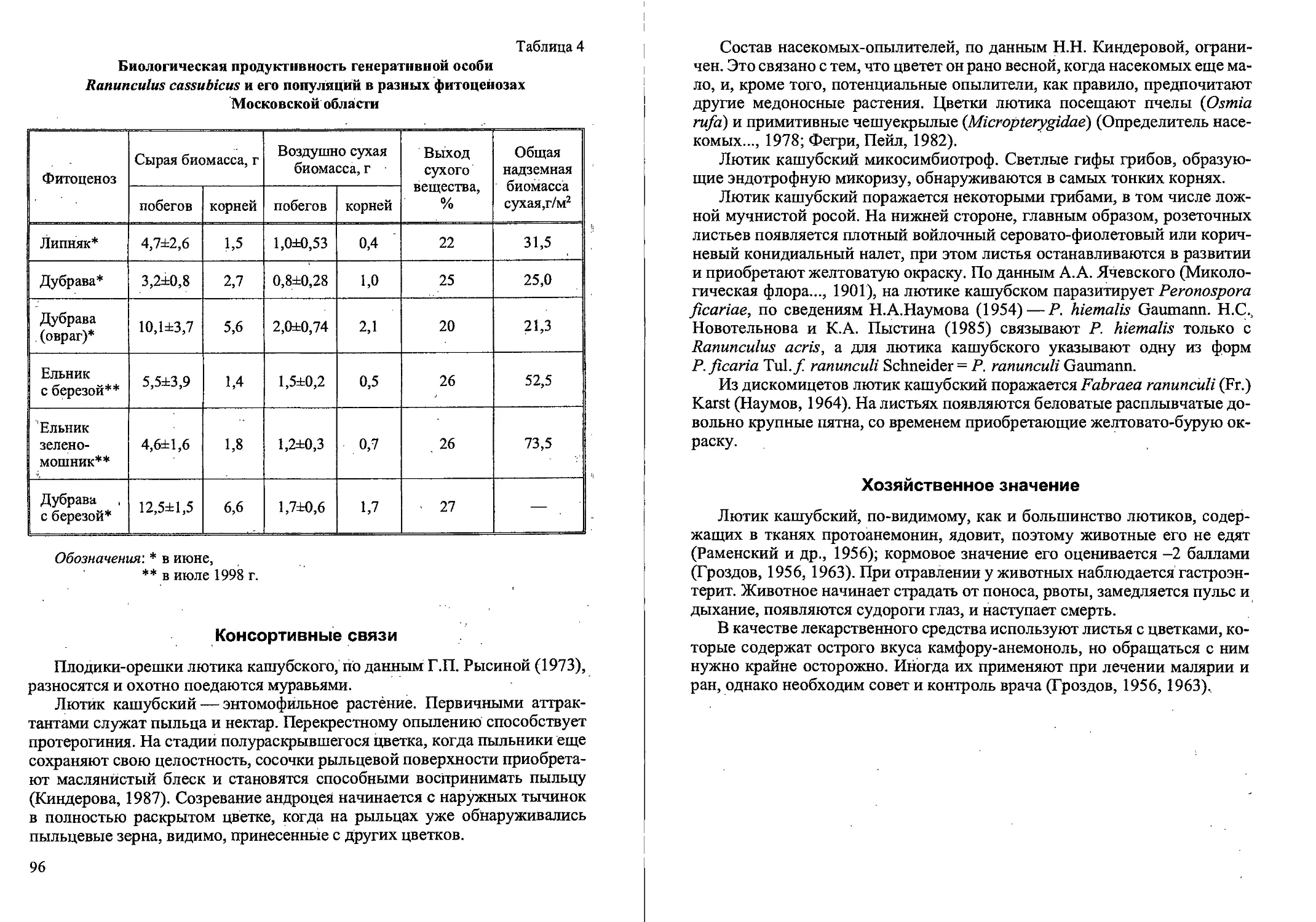
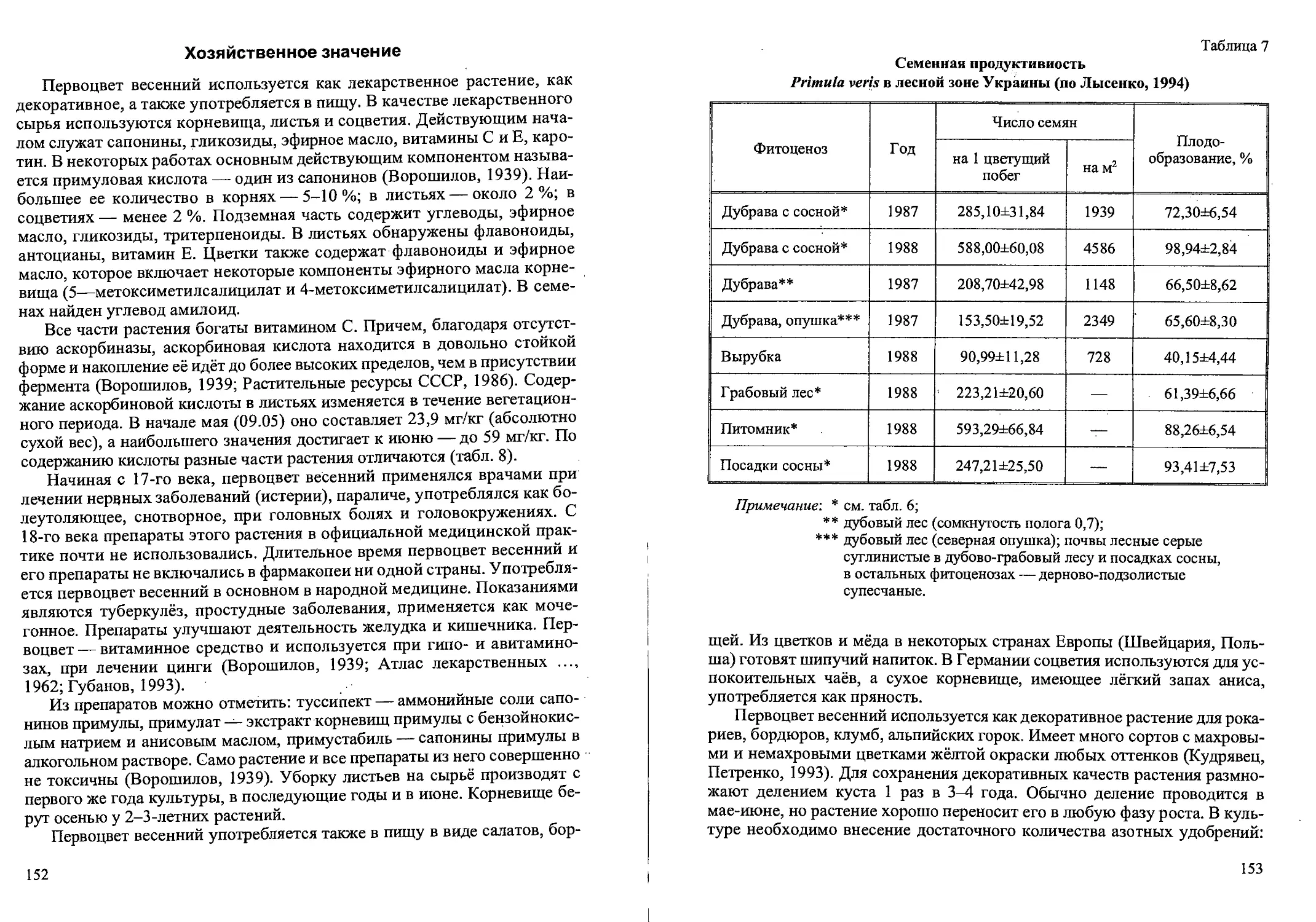
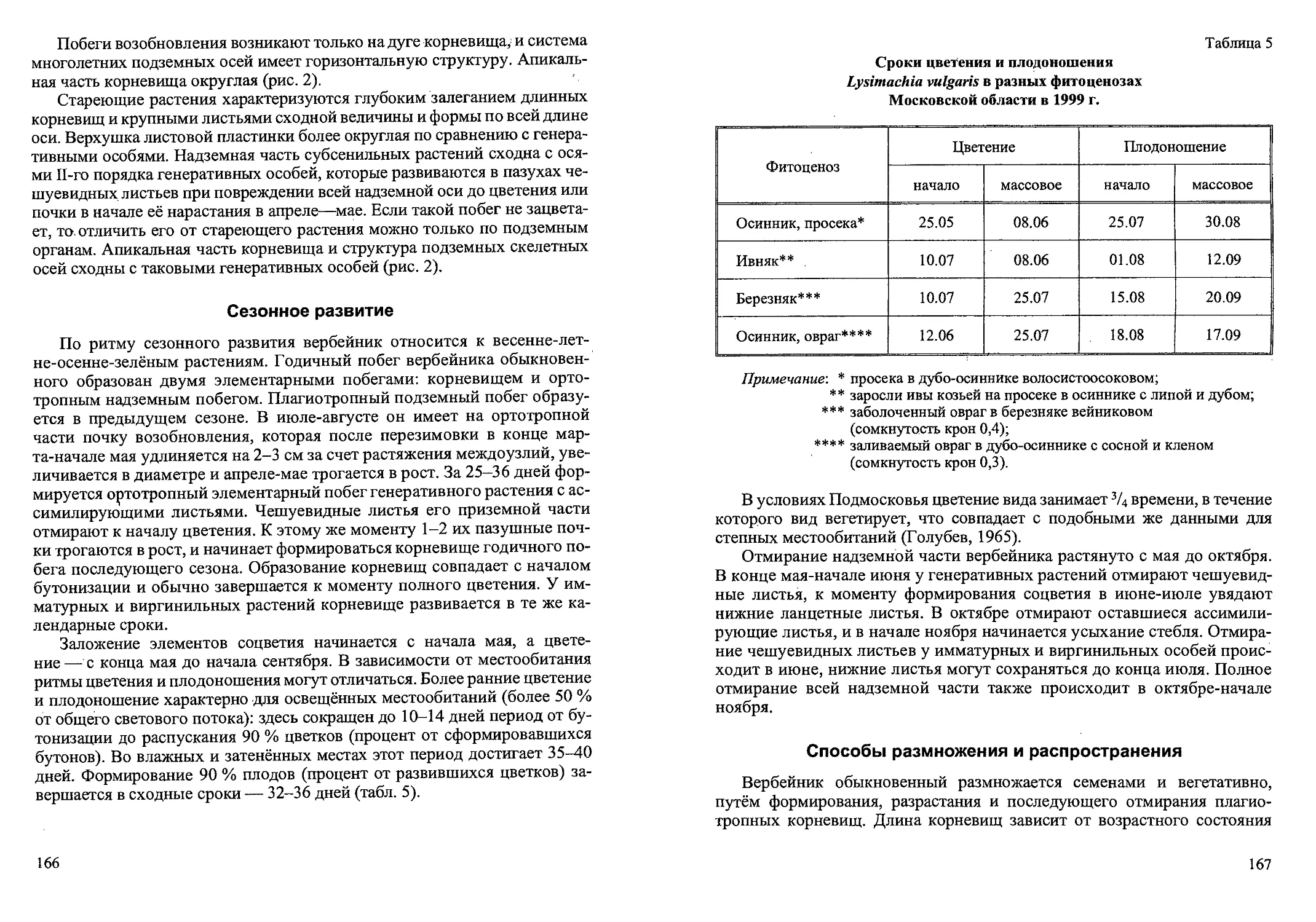
Рис.2. Ранние возрастные состояния Calamagrostis canescens
семенного происхождения: проросток (р), ювенильное (j) имматурное (im)’,
и молодое виргинильное (vl)
дели появляется третий лист. Таким образом, проростки представляют со-
бой небольшие однопобеговые растения с 1-3 плоскими узколинейными
листьями. Ширина листьев около 1 мм. Влагалище листа примерно равно
листовой пластинке. Сохраняется связь с зерновкой. Имеется главный и 2-3
придаточных корня. В природе проростки появляются в мае-июне.
Примерно к концу первого месяца теряется связь проростка с зернов-
кой. Оболочка зерновки постепенно отодвигается в сторону под действием
растущего корня. У ювенильных растений (j) увеличивается длина и шири-
на листовых пластинок по сравнению с проростками (рис. 2). На побеге
3-9 листьев, ширина влагалища 1,5-2 мм, листовые пластинки 2-3 мм.
Первый лист обычно отмирает. Увеличивается число придаточных корней
и интенсивность их ветвления. В надземной части идет интенсивное куще-
ние. В дальнейшем большая часть образовавшихся вегетативных надзем-
ных побегов отмирает. Имматурные растения (im) имеют один неветвя-
10
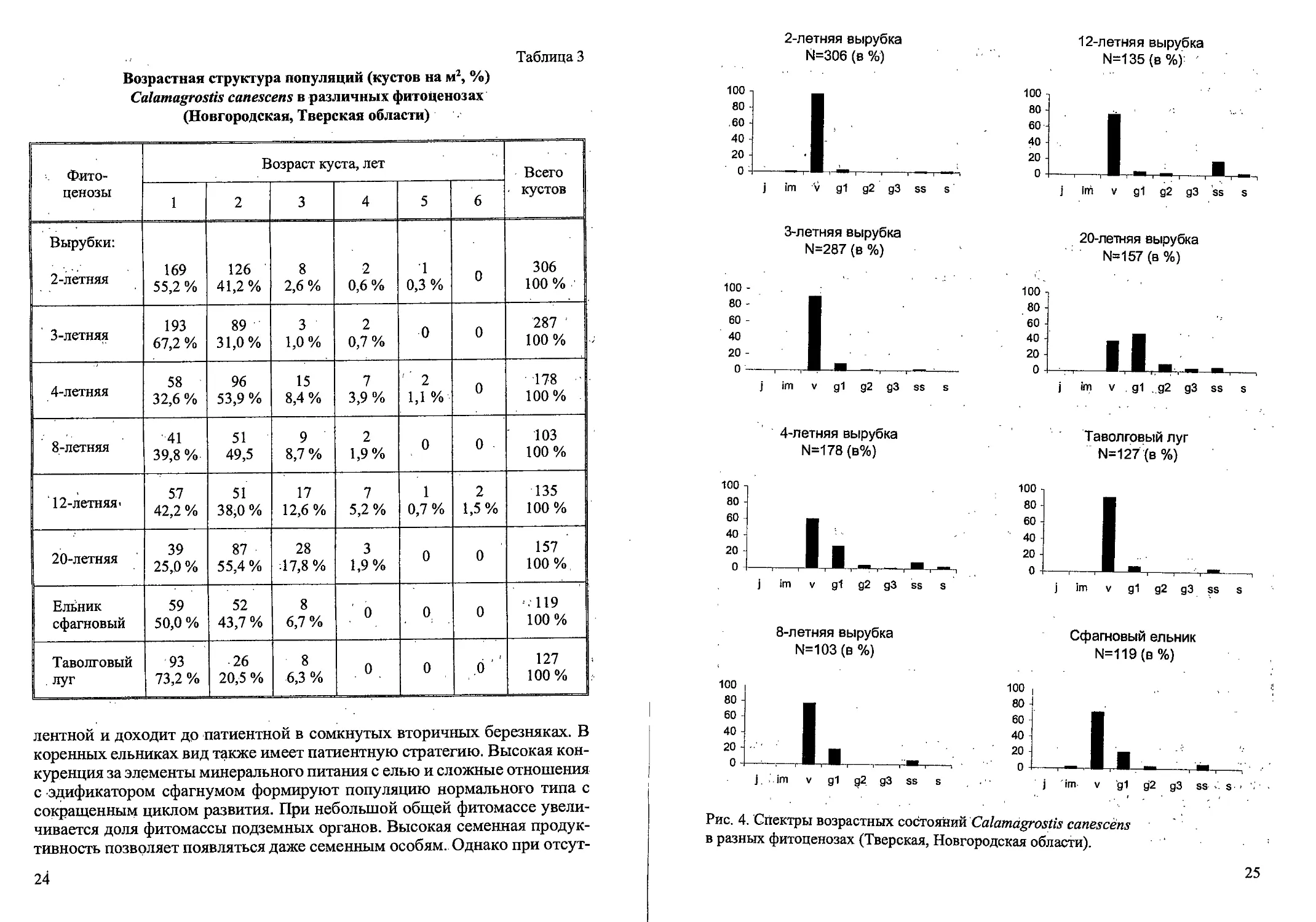
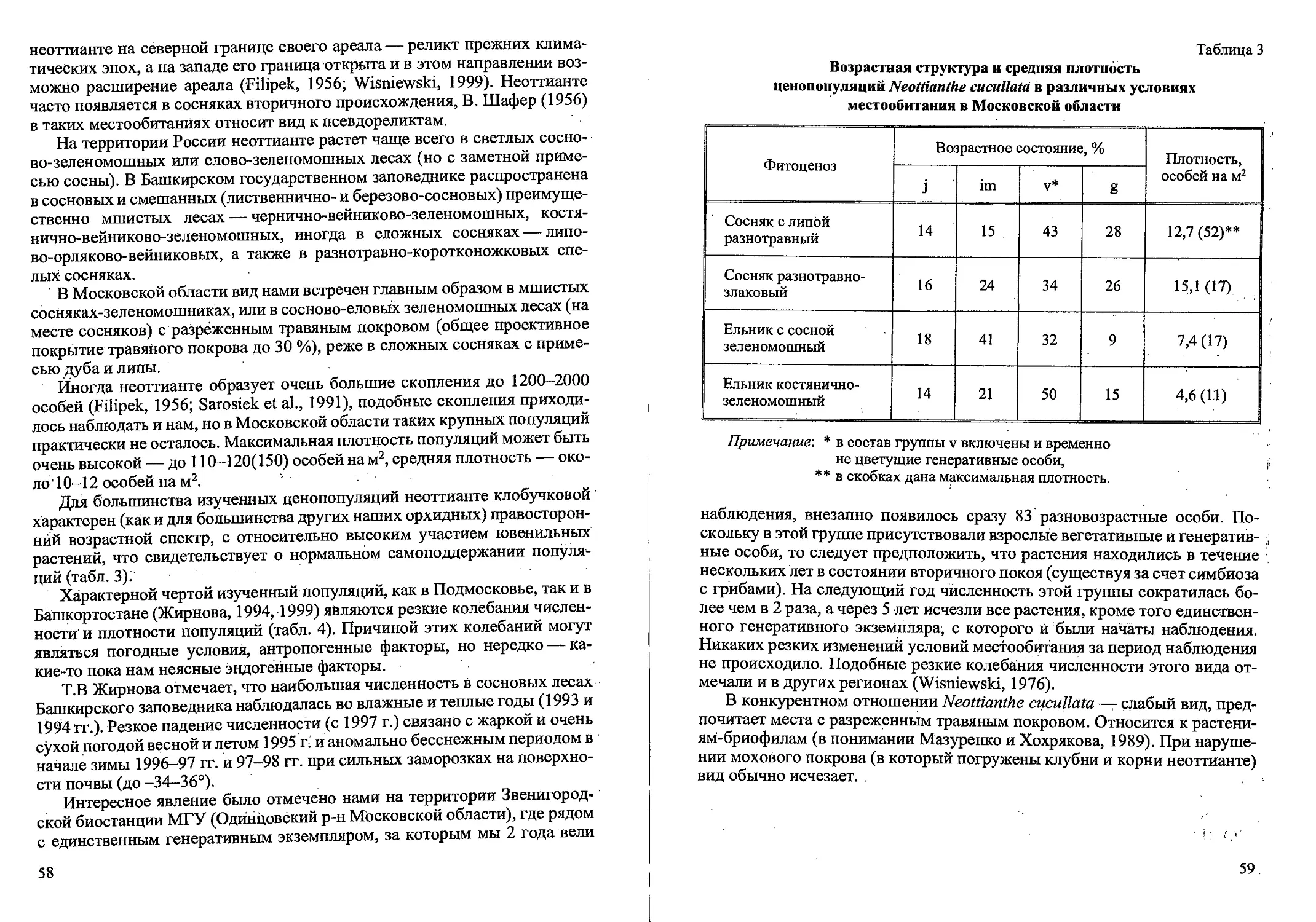
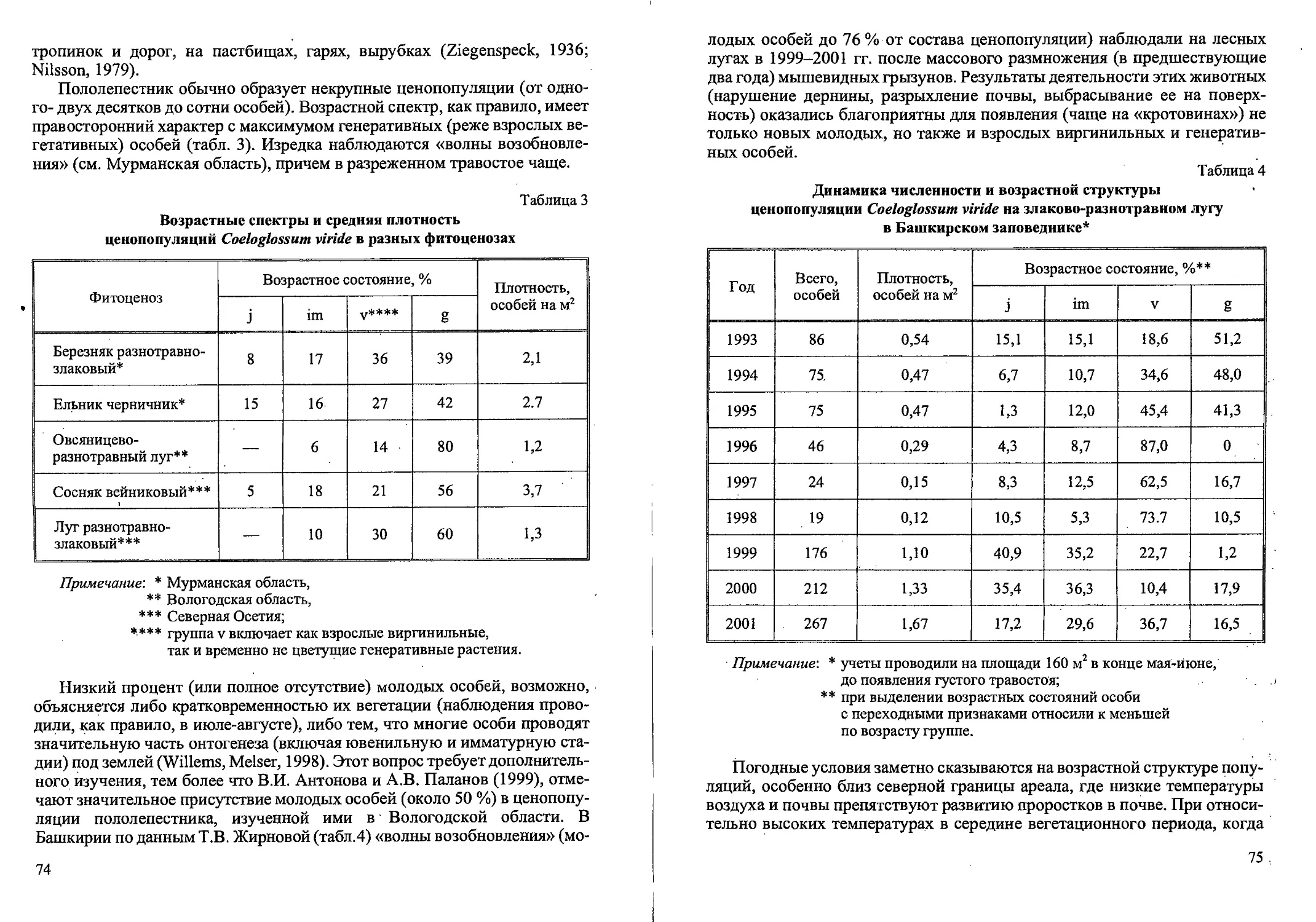
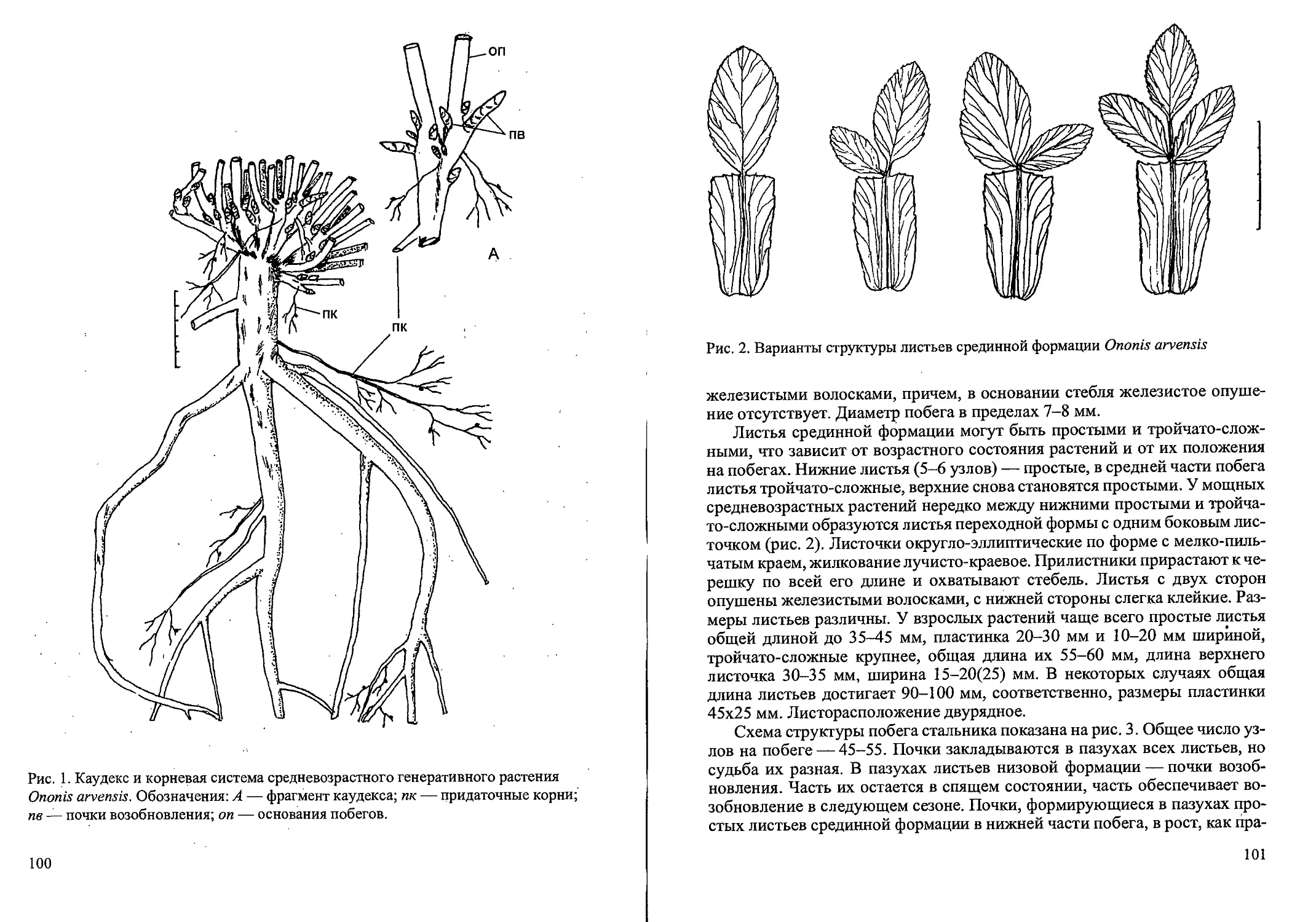
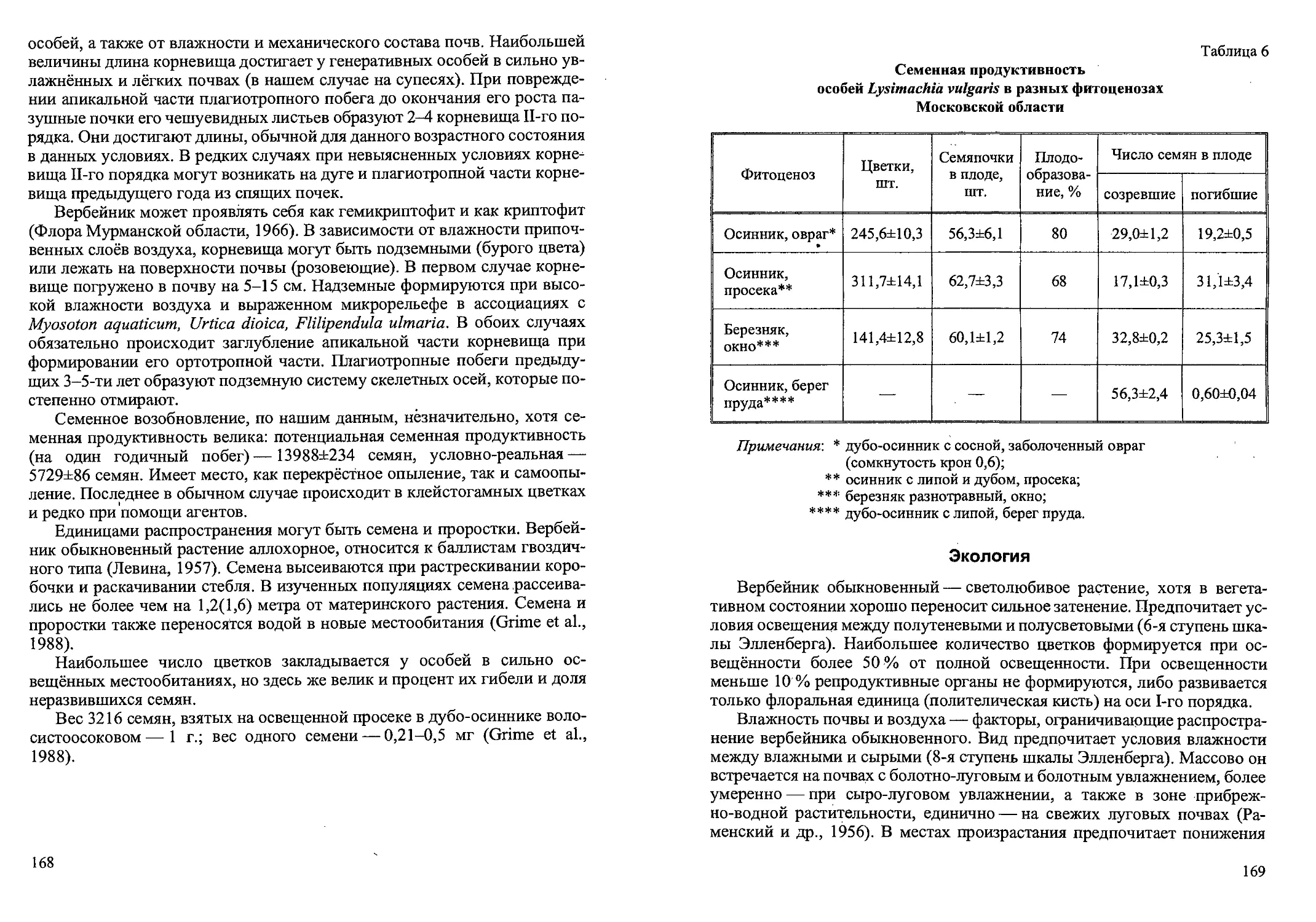
Рис. 3. Возрастные состояния Cdlamagrostis canescens вегетативного происхождения.
щийся вегетативный побег с 3-6 узлами, при этом появляется первое до-
чернее корневище. К июлю-августу первого года жизни растения
переходят в следующее, взрослое виргинильное состояние (Демидова и
др., 2000а, б). Встречаются особи и 2-летние, по габитусу напоминающие
имматурные растения 1-го года развития. Вероятно, некоторые растения
не переходят в виргинильное возрастное состояние в 1-й год жизни. Почка
возобновления у основания побега весной следующего года развивается в
полноценный вегетативный побег, и такие особи семенного происхожде-
ния погибают на 2-й год развития.
Виргинильные растения (V) состоят из 1-5 вегетативных побегов и не-
скольких засохших вегетативных побегов прошлых лет (рис. 2). Листья
вполне взрослые^ Побеги ветвятся, образуя вегетативные побеги II и Ш-го
порядков. В их зонах кущения идет интенсивное образование отбегов и до-
черних корневищ. Корневая система состоит из большого числа молодых
придаточных корней. Обычно в этой стадии растение пребывает 1-2 (реже
3) года. Экземпляры в молодом виргинильном состоянии (VI) имеют 1 (ре-
же 2) вегетативный побег с побегами II и Ш-го или только П-го порядка
(или без них). Если в кусте есть 2 вегетативных побега, то вегетативные
побеги II и Ш-го порядка малочисленны или отсутствуют (рис. 3). У осно-
вания побегов наиболее мощно развитых кустов VI к концу вегетационно-
гб сезона развиваются почки возобновления и уже сформировано 1-2 до-
черних корневища длиной 2-3 см. Такие особи дают начало дальнейшему
вегетативному размножению. Другая часть особей погибает в конце пер-
вого или второго года жизни.
Встречаются особи семенного происхожения высокой жизненно-
сти — (V2), они представляют собой 2-летний куст из 2 (реже 3) вегетатив-
ных побегов с побегами II и Ш-го (или только П-го) порядков (или без них)
и (V3) в 2-3-летнем кусте может быть от 3 до 6 вегетативных побегов с по-
бегами II и Ш-го порядка (или только П-го) или без них. Это мощно разви-
тые парциальные, кусты, однако высота этих экземпляров небольшая
(20-30 см), и они обычно погибают на 2-й год жизни.
Молодые генеративные растения семенного происхождения (gl)
представляют собой кусты с одним генеративным побегом, внешне сход-
ным с кустом вегетативного происхождения, однако вегетативные побе-
ги наблюдаются значительно реже. В виду того, что это возрастное со-
стояние наступает на 2-й год развития особи, обычно отчетливо заметен
сухой побег прошлого года (пенек). Корневая система достаточно разви-
та, преобладают молодые придаточные корни. Особи обычно погибают
на 2-й год развития. Крайне редко можно найти цветущую особь семен-
ного происхождения в 1-й год жизни. Это всегда один генеративный по-
бег с зачаточной метелкой, при этом вегетативные побеги П-го порядка
отсутствуют.
У молодых генеративных растений вегетативного происхождения в
12
кусте формируется один цветущий побег и 1-3 вегетативных побега (рис.
3). На вегетативных и генеративных побегах могут быть вегетативные
побеги II и Ш-го порядка. Сухие побеги прошлых лет единичны или от-
сутствуют. Общая мощность корневой системы возрастает, преобладают
молодые придаточные корни, подземные побеги и дочерние корневища.
Обычно растения вегетативного происхождения зацветают на 2-й год
жизни.
Средневозрастные генеративные растения (g2) морфологически близ-
ки молодым генеративным растениям, но в их кусте обычно 2-3 генератив-
ных побега, иногда с 1 (реже 2) вегетативным побегом (рис. 3). На вегета-
тивном побеге могут быть побеги II и Ш-го порядка, а также 1-2 сухих по-
бега прошлых лет. Корневая система состоит из многочисленных молодых
и старых придаточных корней, дочерних корневищ и подземных побегов.
В это состояние растения вегетативного происхождения могут переходить
на 2-3 год жизни.
Старые генеративные растения (g3) характеризуются наличием 3 гене-
ративных побегов и могут иметь 1-2 вегетативных побега (рис. 3). У осно-
вания куста обычны 2—4 сухих побега прошлых лет. Самые старые расте:-
ния могут формировать неполноценные метелки и переходить в субсе-
нильное состояние. Корневая система менее мощная, преобладают старые,
придаточные корни. В такое состояние; растения вегетативного происхож-
дения могут переходить в благоприятных условиях на 3-й год жизни
(рис. 3). ’
У субсенильных растений (ss) куст состоит из 1-2 живых неветвящихся
вегетативных и 3^1 сухих отмерших побегов прошлых лет (рис. 3). Генера-
тивных побегов нет. Интенсивность кущения слабая, при этом в зонах ку-.
щения образуется мало почек возобновления, часть из них отмирает; в этот
же год. Корневая система более поверхностная, при этом доля молодых,
придаточных корней мала. Обычно в это состояние растения переходят на
3-4-й (реже на 5) год жизни.
Сенильные растения (s) отличаются слабым развитием куста с одним
живым неветвящимся вегетативным побегом с листьями ювенильного,
типа (рис. 3). Отмирающий парциальный куст представляет собой систе-
му сближенных сухих отмерших 3-7 побегов прощлых лет и «служит» ,
для связи соседних более молодых парциальных кустов. Образование но-
вых ортотропных ассимилирующих побегов прекращается, отбеги в
большинстве случаев уже не образуются. В корневой системе преоблада-
ют старые постепенно разрушающиеся придаточные корни. Такие ста-
рые кусты могут жить 1-2 года, затем полностью отмирают, при этом
происходит старческая партикуляция колонии., Сенильные кусты встре-
чается редко, обычно у растений 4—5 (6-го) года жизни. На вырубке со-,
сняка сфагнового (Западнодвинский лесо-болотный стационар, Тверская
область) нами были обнаружены несколько экземпляров редко встречаю-
13
щиеся сенильные растения семенного происхождения. В наземной сфере
они сходны с сенильными особями вегетативного происхождения, одна-
ко растут обособленно. Их корневища (материнские, дочерние) не разви-
ты, однако корневая система из придаточных корней, большей частью
отмерших, очень мощная. Характерно наличие сухих отмерших побегов
прошлых лет (больше 3). Такие 4-летние особи встречаются крайне
редко.
Длительность жизни куста вейника седеющего вегетативного происхо-
ждения в среднем составляет 3^4 года. Редко кусты доживают до 6 лет.
Наши исследования различных аспектов популяционной структуры
вейника седеющего выявили поливариантность хода онтогенеза и реализа-
цию разных вариантов стратегии жизни в меняющихся условиях среды и
фитоценотического окружения (Уланова, Демидова, 2001).
Ритм сезонного развития
Весной листья начинают развиваться, по нашим наблюдениям в Мос-
ковской области, с апреля по май. В начале июня высота вегетативных по-
бегов составляла в среднем 50-70 см, отмечено начало кущения от второго
узла, некоторые кусты дали начало 1-2 отбегам длиной около 3 см. В ию-
ле—августе растения вейника достигают оптимальной для определенных
экологических условий высоты и максимальной биомассы. Новая генера-
ция листьев появляется со второй половины августа и в сентябре.
На вырубках в первый вегетационный сезон после рубки основная мас-
са растений вейника не цветет. Очень редко можно здесь встретить цвету-
щие побеги растений, которые произрастали, вероятно, в лесу на более ос-
вещенных участках. Развитие генеративных побегов и массовое цветение
наблюдается лишь со второго года после рубки.
Образование зачатков соцветий и цветков продолжается с мая до нача-
ла июля, цветение, по нашим данным,— с июня по август. Пик цветения
приходится на июнь-июль (Маевский, 1964). Основная масса семян созре-
вает в августе, а всходы появляются только в июне—июле следующего го-
да. Это анемофильное растение, «утренний злак». Раньше других зацвета-
ют популяции открытых местообитаний.
Ритм сезонного развития изучали В.П. Бельков с соавторами (1974) в
лесах Ленинградской, Псковской, Тульской областей. Летом после образо-
вания надземных побегов в пазухах низовых листьев в узлах корневищ или
базальной части надземных побегов трогаются в рост боковые почки во-
зобновления, образующие дочерние ветви корневища длиной от 4 до
40 мм, состоящие из 4-20 и более междоузлий. В зависимости от числа
междоузлий и их длины дочерние ветви корневища бывают очень корот-
кие (1,0-1,5 см) и длинные (30-40 см). К осени каждая ветвь корневища
меняет направление роста и растущим концом приближается к поверхно-
14
ста почвы. В переходной плагиотропно—ортотропной зоне корневища
при этом формируются укороченные междоузлия. Весной терминальная
почка каждой ветви выходит на поверхность почвы и образует терминаль-
ный генеративный или с неполным циклом развития (не доходящий до
цветения) вегетативный надземный побег с удлиненными междоузлиями.
Осенью генеративные и вегетативные надземные побеги отмирают почти
полностью (по нашим наблюдениям, в октябре-ноябре), за исключением
базальной части с укороченными междоузлиями. Эти сухие побеги полега-
ют, сохраняются под снегом до весны и спрессовываются в подстилку (ве-
тошь). Дальнейшее побегообразование осуществляется за счет аналогич-
ного вышеописанному развития боковых почек корневища или базальной
части надземных побегов, которые еще летом (в конце августа) трогаются
в рост. В базальной части отмирающего надземного побега в узлах укоро-
ченных междоузлий нередко трогается в рост несколько боковых почек..
Они образуют в следующем году соответствующее число дочерних над-
земных побегов с короткой плагиотропной частью и укороченными меж-
доузлиями в основании. У них в свою очередь в узлах зоны укороченных
междоузлий трогаются в рост боковые почки, образующие побеги третье-
го порядка и т.д. Таким образом, у вейника седеющего формируются пар-
циальные небольшие кусты. Взрослые особи представляют собой систему
парциальных кустов, соединенных в единое целое многочисленными и
часто длинными корневищами (Бельков и др., 1974).
Значительная часть боковых почек (до 70-90 %), ежегодно образую-
щихся у вейника в большом количестве, особенно из числа тех, которые за-
кладываются на плагиотропной части побегов (на корневищах), впадает в
состояние покоя на год или несколько лет (Бельков и др., 1974). Покоящие-
ся почки создают значительный резерв устойчивости вейника в покрове,
используемый при неблагоприятных или, наоборот, при улучшении усло-
вий существования.
Способы размножения и распространения
Вейник седеющий — облигатно ветроопыляемое и анемохорное расте-
ние.
Семенная продуктивность и урожайность вейника неодинаковы на мо-
лодых вырубках, на старой заросшей березой 12-летней вырубке, в освет-
ленном березняке на месте 20-летней вырубке, а также ельнике сфагновом
и на таволговом лугу (табл. 1). Семенная продуктивность вейника опреде-
лялась на каждой пробной площадке в середине августа до начала высыпа-
ния зерновок. Из всех полноценных метелок были выбраны по одной мо-
дельной метелке со средними морфологическими показателями (средняя
длина и ширина) и массы. Все цветки и зерновки подсчитаны и под микро-
скопом определены полноценные созревшие семена.
15
Таблица 1
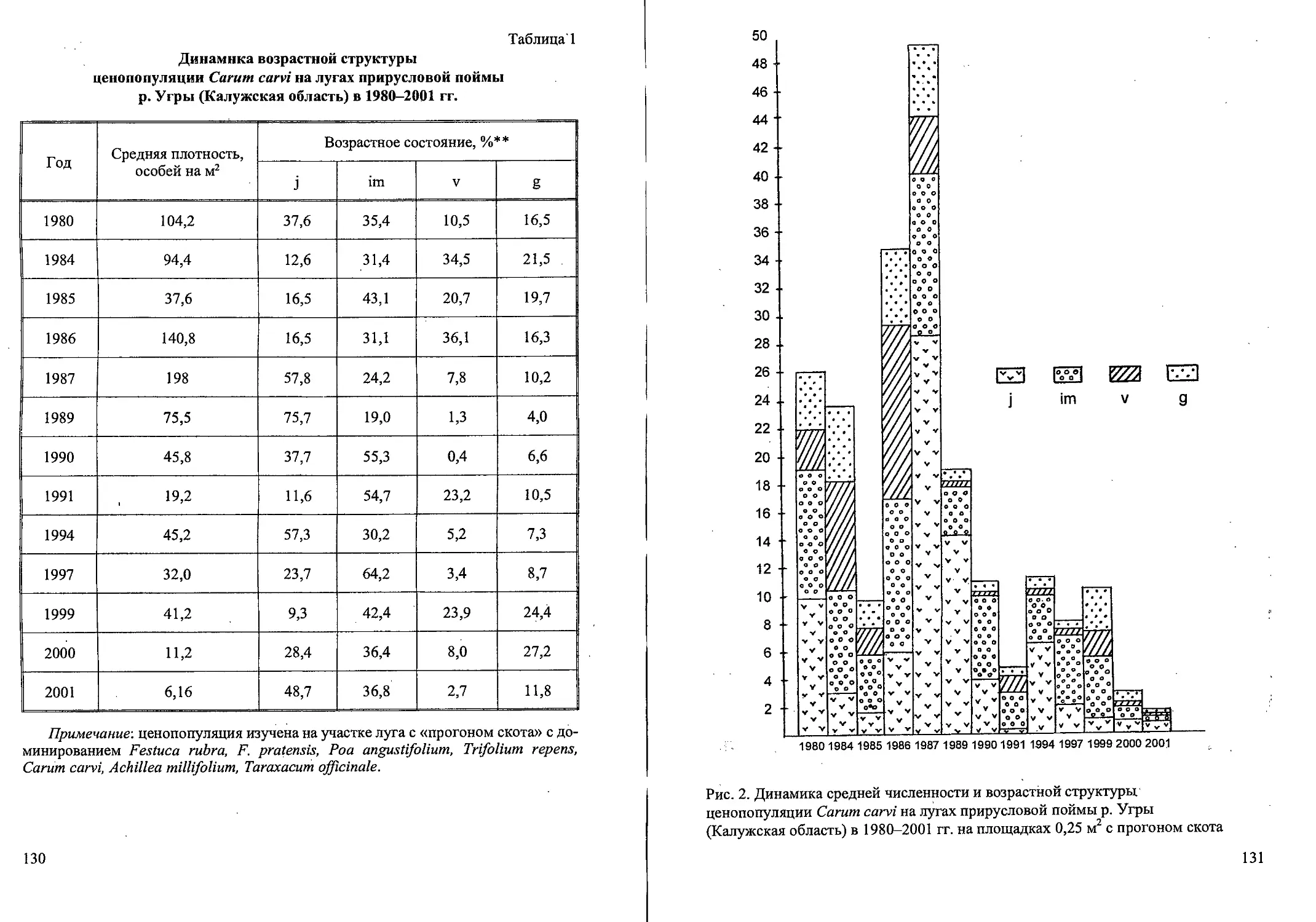
Семенная продуктивность Calamagrostis canescens
на вейниковых вырубках, в ельнике сфагновом
и на таволговом лугу (Тверская область)
Фитоценоз Семенная продуктивность Полноценные зерновки, %
потенциальная реальная
Вырубки: 2-летняя 895 53 5,9
12-летняя 723 66 9,1
Березняк 20-летний 634 48 7,6
Ельник сфагновый 476 ' 39 8,2
Таволговый луг 788 74 9,4
Среднее значение 703 56 8,0
Потенциальная семенная продуктивность (число всех полноценных и
неполноценных цветков и зерновок в метелке) значительно выше реальной
семенной продуктивности (количество полноценных зерновок в метелке)
(табл. 1). Несмотря на низкий процент живых семян, их общее количество
в метелке и на площадке в 1 м2 очень высокое (табл. 2). В более благопри-
ятный по влажности год, вероятно, полноценных семян могло бы быть
больше. .
В условиях высокой освещенности (2-летняя вырубка и таволговый
луг) в метелках больше всего цветков и семян, однако урожайность до-
вольно низкая. Наибольший урожай зерновок приходится в сообществах
4- й 20-летних вырубок, где численность генеративных кустов наиболь-
шая.
Анализ литературы по почвенному банку семян подтвердил наше пред-
положение, что в почве зерновки сохраняются плохо. Пр своей стратегии
жизни вейник седеющий, согласно классификации Т.А. Работнова (1983),
можно отнести к группе видов, жизнеспособные семена которых не явля-
ются постоянной частью ценопРпуляции (не накапливаются в почве), учат
стйуя в ее составё в значительных количествах лишь в течение короткого
периода после обсеменения растений.
16
Таблица 2
Урожай семян Calamagrostis canescens в различных условиях
местообитания (Тверская, Новгородская области)
Фитоценоз Число метелок на 1 м2 Число зерновок на 1 м2
общее Зрелых
Вырубки: 2-летняя 6 4218 336
3-летняя 24 16876 1344
4-летняя 62 .... 43598 3472
8-летняя .19 13360 .• 1064
12-летняя .11 7953 726
Березняк 20-летний ИЗ 71643 5424
Ельник сфагновый 30 14280 1170
Таволговый луг . . 8 6304 592
В благоприятных условиях — при высокой, освещенности на влажной
и свободной от растений почве — вейник седеющий возобновляется из се-
мян (Бельков, 1974). Особи семенного размножения нам удалось обнару-
жить только на достаточно увлажненном торфе (но без застоя воды) пере-
сохшего болота и на минерализованной поверхности нарушенных участ-
ков 2-летней вырубки в Новгородской области. Минерализованная
поверхность более длительное время сохраняется увлажненной, здесь и
наблюдается появление всходов (Корконрсова, 1967). Эти участки были
свободны ото мхов, ветоши, взрослых травянистых, кустарниковых и дре-
весных растений. Не удалось найти проростки вейника и вблизи взрослых
растений этого вида. Отмершие побеги вейника седеющего образуют слой
ветоши, препятствующий проникновению семян в почву и затрудняющий
появление всходов не только этого же вида, но и любых других растений.
Таким образом, проростки и молодые ювенильные растения вейника на
вырубках практически отсутствовали. Вероятно, происходит сильный от-
пад семенных растений в молодом состоянии. По данным Т.А. Работнова
17
(1964), среди всходов в естественных ценозах наблюдается большая
смертность, обычно лишь небольшая часть их достигает половозрелого со-
стояния (1-2 %).
Вид относится к вегетативно-подвижным растениям (Любарский,
1967). Его вегетативное размножение начинается с наступлением взросло-
го виргинильного состояния (с 1- или 2-го года) и продолжается вплоть до
субсенильного. Наиболее интенсивно оно в конце виргинильного и в гене-
ративном периоде.
Почка трогается в рост, развивая подземное корневище, растущее на
глубине около 5 см. Верхушка подземного побега загнута вверх, раздвигая
частицы почвы. В одной зоне кущения из почек у основания побега возни-
кает вневлагалищный побег, на котором формируется новая зона кущения.
После появления и отмирания генеративных и вегетативных побегов в
этой зоне кущения образуется новое корневище и возникает следующая
зона кущения; так продолжается до конца жизни растения. Осенью терми-
нальная почка выносится на поверхность, увеличивается в размерах, и в
таком состоянии зимует. Весной окончательно формируется ортотропный
побег, который дает начало парциальному кусту.
После цветения в основании вегетативных и генеративных побегов
трогаются в рост почки возобновления. Вейник принадлежит к группе рас-
тений с полностью сформировавшейся вегетативной сферой внутри почек
в предзимний период (Серебрякова, 1947). В конце августа — сентябре
длина укороченных молодых побегов, которые развиваются из почек во-
зобновления, составляет в среднем 3-5 см. Они имеют малиново-зеленова-
тую окраску; У некоторые из них в этот период уже заметен первый лист,
обычно с двумя отчетливыми жилками. Его длина составляет около 3^1
см. Вероятно, в таком состоянии они зимуют и развиваются уже весной
следующего года в полноценные вегетативные или генеративные побеги,
т.е. по классификации Т.П. Серебряковой (1971) побеги являются полуро-
зеточными.
В генеративном периоде возможны два варианта вегетативного раз-
множения: начало следующему парциальному кусту дают почки, располо-
женные в основании генеративного побега, или почки в пазухах пленчатых
листьев в основании вегетативных побегов данного парциального куста.
Это приводит к удлинению жизненного цикла и омоложению популяции.
О наличии омоложения свидетельствует усиление у дочерних особей про-
цессов роста и органообразования, проявляющееся в увеличении числа
растущих побегов, корней и почек возобновления.
Надземные побеги вейника седеющего способны к образованию при-
даточных корней при соприкосновении с почвой. Это явление особенно
широко распространено в тех условиях, где растения доходят до границы с
водным пространством (берега водоемов, на влажном песчаном или или-
стом субстрате), т.е. растение можно считать факультативноотводковым
18
(Шалыт,1960). Провести четкую грань между корневищами и надземными
побегами не всегда возможно.
На лугах вегетативное размножение преобладает над семенным. Под
пологом леса, по данным В.П. Белькова с соавторами (1974), вейник се-
деющий размножается почти исключительно вегетативно. На свежих вы-
рубках в первые годы он размножается преимущественно вегетативно, а
также за счет семян, заносимых ветром и опадающих с особей, росших под
пологом леса. В дальнейшем возобновление осуществляется главным об-
разом вегетативным путем. Вместе с тем в зарастании плужных борозд
значительную роль играет семенное размножение вейника, в то время как
на плужных пластах, минерализованных площадках и полосах он разрас-
тается в основном за счет вегетативного размножения и побегообразова-
ния от корневищ (Бельков, 1974 и наши наблюдения).
Экология
Вейник седеющий — светолюбивый вид, способный расти при относи-
тельной освещенности 10 %, но лишь при 40 % относительной освещенно-
сти может достигать 50 % покрытия (Вотсапнп, 1993). По нашим наблю-
дениям, наибольшего обилия вид достигает при полном освещении на вы-
рубках и лугах. Его относят его к группе промежуточной между
полусветовыми и полутеневыми растениями (6-я ступень шкалы Эллен-
берга). Любое затенение резко подавляет «генеративность» вейника, так
как не только ослабляет жизнедеятельность, но и направляет ее в сторону
вегетативного роста и размножения. При зарастании вырубок мелколист-
венными породами обилие вейника резко снижается именно из-за умень-
шения освещенности.
Вейник седеющий —гигромезофит, характерный злак болот и заболо-
ченных лесов и лугов. Он предпочитает сырые, часто плохо аэрируемые
почвы, являясь индикатором периодического избыточного увлажнения
(9-я ступень шкалы Элленберга). Может расти даже в литоральной поясе
при уровне воды на 0,2 м выше уровня почвы (Вотсаппп, 1993). По шкале
Л.Г. Раменского (1956), вейник относится к растениям влажно-луго-
вых прибрежно-водных местообитаний (70-105-я ступени шкалы), а по
шкале переменности увлажнения от высоко обеспеченного до сильно пе-
ременного (4-13-я ступени шкалы). Коэффициент эвапотранспирации вей-
ника седеющего в заболоченных ивняках в летние месяцы в среднем со-
ставляет 3,2 мм в день (Priban, Ondok, 1986). Он лучше развивается на рых-
лых, хорошо аэрируемых и достаточно влажных почвах, не подверженных
слишком резким колебаниям температуры и влажности в сдое, где распо-
ложены корневища и где возникают новые корни и побеги. Эксперимен-
тально показано, что улучшение аэрации почвы после фрезерования спо-
собствует разрастанию вейника (Вотсаппп, 1993). Может произрастать в
19
условиях слабой аллювиальности (0,3-0,5 см наилка) (Раменский, 1956). В
березняках с доминированием вейника капиллярная кайма грунтовых во-
ды может быть даже на глубине 15-18 см (Ипатов, 1960).
Вейник седеющий произрастает на глеевых и торфяных почвах разной
степени подзолистости (дерново-глеевых, торфяно-подзолисто-глеевых,
перегнойно-подзолисто-глеевых, болотно-подзолистых, пойменных луго-
во-болотных, пойменных болотных, иловато-перегнойно-глеевых, торфя-
нистых и торфяных, торфяно-глеевых, пойменных иловато-торфяных)
(Методические указания..., 1978). Строгая евтрофность местообитаний
вейника седеющего не очевидна (Powelson, 1992). Проведенный нами кла-
стерный анализ описаний фитоценозов вырубок в Тверской и Новгород-
ской областях показал обособленное положение сероватовейниковых и та-
волговых фитоценозов, это мезо^- и эутрофные сообщества, часто на влаж-
ных с оглеением почвах (Демидова и др., 2001). Г. Элленберг (Ellenberg,
1991) относит его к группе видов тяготеющим к почвам со средним содер-
жанием минерального азота (5-я ступень) и обитающих на промежуточных
между слабокислыми и нейтральными почвах (6-я ступень). Показатель
pH водной вытяжки в разных типах луговых сообществ с вейником состав-
ляет 5,5-7,5 (Методические указания..., 1978).
Фитоценология
Вейник седеющий широко распространен на пойменных и водораз-
дельных, сырых торфянистых лугах, в кустарниках и ольшаниках, в сырых
и заболоченных лесах и просеках, по берегам рек и ручьев, на низинных и
переходных болотах, в мелководьях мелких ручьев, ложбинах стока, кана-
вах, кюветах, нередко в воде (Маевский, 1964; Миняев, Конечная, 1976;
Губанов и др., 1995; Морозова, 1999). По нашим наблюдениям, в лесах
вейник седеющий чаще встречается на открытых участках: по обочинам
дорог, на просека^, в окнах, на лесных заболоченных лугах, по опушкам.
Вейник седеющий — диагностирующий вид асе. Carici acutae-Comare-
tum Schelyag-Sosonko, Solomaha, Sipailova 1986 (Шеляг-Сосонко и др.,
1986) союза Magnocaricion elatae W.Koch 1926 порядка Magnocaricetalia
Pignatti 1953 класса Phragmiti-MagnocariceteaKlikaet Novak 1941 (Миркин,
Наумова, 1998). Это сообщества притеррасной части пойм Днепра, Десны
и других, более мелких Полесских рек лесной зоны Украины. Фитоценоти-
ческий оптимум вида отмечен также на открытых солнечных местах, где
он, господствуя вместе с тростником (асе. Phragmitetum calamagrostietosum
canescentis Weber 1978), образует густые заросли, обильно цветет и плодо-
носит (Weber, 1979). Подобные сообщества были изучены на озере Бальк-
зее в Нижней Саксонии под названием Peucedano-Calamagrostietpm
canescentis Weber 1978, и широко распространены в Северной и Централь-.
ной Европе на территории Швеции и Германии. С.С. Ганешин (1925) отме-
20
тил заросли вейника седеющего в пойме реки. Волхова в понижениях мик-
рорельефа с Carex cespitosa (реже с Carex vulpina), занимающие иногда до-
вольно значительные участки.
Вид обычен в покрове гипново-сфагновых и травяных болот. М.М.
Данилова (1964) выделяет среди растительных формаций болот на тер-
расах долины реки Камы в юго-западных районах Пермской области
группу евтрофных злаковых болот с вейником (1 и 2-е террасы). На осо-
ковых болотах обычно растет вместе с Carex acuta, С. vesicaria, С.
canescens, С. vulpina, С. cespitosa. Ю.Ц. Федотов (1999) на болотах Не-
руссо-Деснянского Полесья Брянской области описывает доминирова-
ние вейника седеющего на травяных, черноольховых и гипновых боло-
тах, реже на мезотрофных сфагновых. Сильно возрастает доминирова-
ние вейником на первых стадиях осушения травяных болот в
мелиоративных канавах.
Вейник седеющий является также одним из диагностирующих видов
союза Alnion glutinosae Malcuit 1929 порядка Alnetalia glutinosae R. Tx.
1937 класса Alnetea glutinosae Br.-Bl. Et Th. Ex Westhoff et. al. 1946 (Мир-
кин, Наумова, 1998). Это низинные эутрофные черноольховые и пушисто-
березовые заболоченные леса. Класс Alnetea glutinosae включает богатые
видами черноольшаники на гидроморфных почвах, часто развивающиеся
в поймах речек при условии довольно высокой проточности. Иногда мо-
жет быть доминирующим видом (Воробьев, 1967).
Ю.П. Федотов (1999) отмечал обилие вейника седеющего в чернооль-
шаниках с хорошо развитым покровом Sphagnum squarrosum в междуреч-
ных пространствах и западинах на 2 и 3 террасах рек Неруссо-Деснянско-
го Полесья. М.П. Шилов (1971) в лесах притеррасных пойм р, Клязьма
(Владимирская область) выделяет сероватовейниковый цикл —- конеч-
ную стадию формирования черноольховых и кустарниковых трясин на
лесных оглеенных, болотных и лугово-болотных почвах. Доминирование
вейника характерно для низких длительно поемных притеррасных участ-
ков, а также для приозерных западин, центральной и отчасти прирусло-
вой поймы на торфянистых иловато-глеевых почвах с избыточным ув-
лажнением.
Также вейник седеющий является диагностирующим видом лесного
союза Sphagno-Piceenion K.-Lund 1981 (порядка Piceetalia excelsae Pawlow-
sky in Pawlowsky, Socolowski et Wallisch 1928 класса Vaccinio-Piceetea
Br.-Bl. in Br.-Bl., Siss. etVlieger 1939). Это переувлажненные и слегка забо-
лоченные европейские еловые таежные леса на бедных кислых почвах с
развитым моховым покровом. Согласно Sukopp (1959) (цит. по Hroudova,
1988), стадия с вейником седеющим замещает стадию с доминированием
сосны в ассоциации Ledo-Sphagnetum после вырубок. В европейской части
России вейник седеющий обычно встречается в сосновых заболоченных
лесах: круглоосочково-черничных, сфагново-черничных, травяно-сфагно-
21
вых (Рысин, 1975). В.Н. Федорчук с соавторами (2002) считают вейник се-
деющий диагностическим видом серии травяно-сфагновых типов леса в
Ленинградской области вместе с Frangula alnus, Salix aurita, S. cinerea,
Carex lasiocarpa, Phragmites ausralis, Menyanthes trifoliata, Eriophorum
vaginatum, Comarum palustris, Sphagnum angustifolium, Sph. magellanicum.
Обычно это сосняКи или березняки на слабо дренированных равнинах й
окраинах болот на торфяно- и торфяно-перегнойных почвах с мощностью
торфяного слоя более 25 см.
Вейник ланцетный типичный доминант березняков на длительно пере-
увлажненных почвах в условиях переходного и низинного рядов заболачи-
вания центральной части южной тайги Русской равнины (Соколова и др.,
1956; Зворыкина, 1983). Так, В.С. Ипатов (1960) в Ленинградской области
выделял березняки вейниково-ланцетные — Betuletum lanceolatae
calamagrostosum. Это бедные по видовому составу сообщества с пятнами
сфагнума, где вейник тяготеет к приствольным повышениям и подкроно-
вым участкам. Вейник обилен в березняках заболоченных (Betuletum-
paludosum), с высоким стоянием грунтовых вод и интенсивным торфооб-
разованием.
Согласно нашим данным, среди совместно обитающих с вейником ви-
дов в ельниках, березняках и вырубках чаще встречаются: Deschampsia
cespitosa, Equisetum sylvaticum, Lysimachia vulgaris, Trientalis europaea,
Juncus effusus, Oxalis acetosella, Majanthemum bifolium; Stellaria holostea,
Solidago virgaurea, Gymnocarpium dryopteris, Dryopteris cristata, Luzula
pilosa, Typha latifolia, Galium uliginosum, Filipendula ulmaria.
В таежной зоне вейник седеющий — важнейший эдификатор расти-
тельности вырубок на месте ельников травяных и сфагновых. Структура
популяций вейника была изучена нами на вырубках разного возраста в
Новгородской (Крестецкий район) и Тверской (Нелидовский район) облас-
тях (Демидова, Уланова, 2000; Демидова и др., 2000 а, б; Демидова, Улано-
ва, 2001; Уланова, Демидова, 2001). Вейник быстро заселяет вырубки в
течение первых 3 лет, интенсивно разрастаясь за счет вегетативного раз-
множения, а на 4-й год становится доминантом и эдификатором раститель-
ности на вырубке. К 8 годам начинается его угнетение подростом берез и
ив и в результате вейник теряет свои позиции. В молодых сомкнутых
12-летних березняках это уже типичная угнетенная популяция, неспособ-
ная цвести из-за нехватки света. Однако проведение осветления вырубок и :
создание полос без древостоя позволяет вейнику вновь стать типичным до-
минантом этих сообществ. В коренном ельнике сфагновом вейник может
занимать большие пространства, однако он находится в угнетенном со-
стоянии (Демидова и др., 2000).
На лугах и вырубках, где вейник седеющий прочно удерживает терри-
торию, он выступает как сильный эдификатор. Под его влиянием происхо-
дит значительное изменение условий внешней среды, как в надземной, так
22
и в подземной сферах. Жесткие листья и стебли вейника малопитательны и
плохо поедаются животными, отмирая осенью, образуют слой войлока
толщиной 7-10 см. Остатки листьев медленно разлагаются, формируя
плотную подстилку толщиной до 2-3 см, которая препятствует появлению
всходов. Однако подстилка и войлок не препятствуют нормальному разви-
тию самого вейника, у которого весенние надземные побеги имеют ост-
рые, жесткие верхушки, свободно проходящие через слой почвы и под-
стилку. В подземной части вейник образует очень густую систему длин-
ных, многократно ветвящихся подземных плагиотропных корневищ на
глубине от 2 до 20 см (обычно 5-8 см) в зависимости от почвенных усло-
вий и режима увлажнения (Бельков, 1974; Рысин, 1987).
Таким образом, вейник седеющий является сильным конкурентом за
воду и питательные вещества, и в его зарослях могут произрастать очень
немногие виды.
Вейник седеющий нельзя отнести к какой-то определенной группе по
стратегии жизни. Он обладает в той или иной степени выраженной пати-
ентностью, виолентностью и эксплерентностью (Уланова, Демидова,
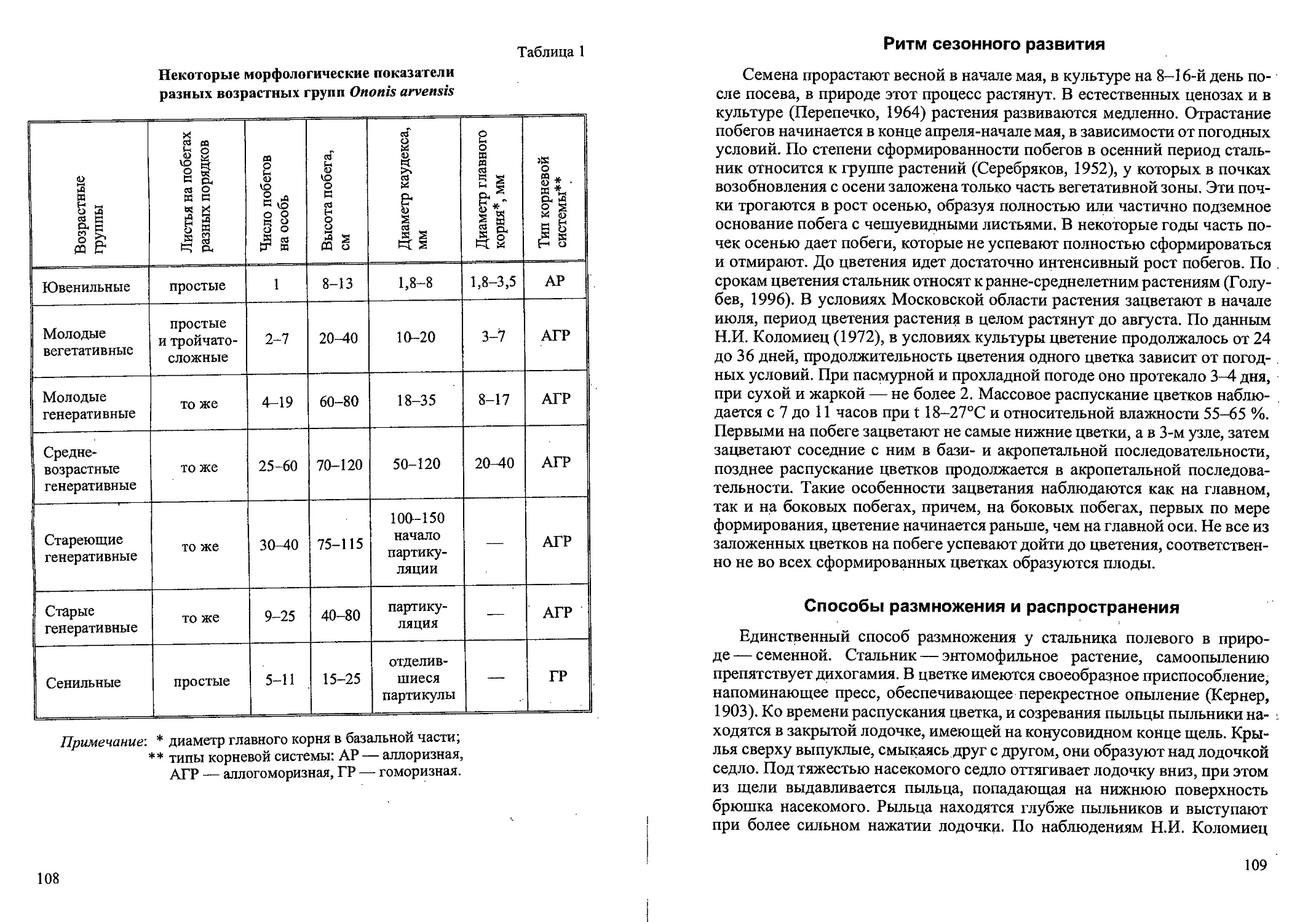
2001; Уланова и др., 2002). Стратегия поведения популяции вейника на 2-и
3-летних вырубках можно отнести к эксплерентному типу. После сильного
нарушения почвы и растительного покрова структура популяции имеет
инвазионный характер с преобладанием 1-2 летних виргинильных кустов
(табл. 3, рис. 4). Семян созревает мало, зато идет интенсивное вегетативное
размножение и популяция образует большую фитомассу. Стратегии пове-
дения популяции на 4-летней вырубке меняется в сторону виолентности.
Здесь вейник ведет себя как доминант и эдификатор лугового сообщества.
Структура популяции имеет нормальный полночленный тип с преоблада-
нием 2-летних виргинильных и генеративных кустов. Велика семенная
продуктивность и вклад генеративной сферы в общую фитомассу. Страте-
гию поведения популяции на 8- и 12-летних вырубках можно отнести к па-
тиентному типу. Особенно типична ситуация в 12-летнем березняке. Появ-
ление конкуренции с подростом деревьев за свет и элементы минерального
питания ведет к формированию двувершинности возрастного спектра, уд-
линению сроков жизни кустов в субсенильном состоянии и выпадению ге-
неративного состояния. Фитомасса популяции уменьшается. Семенная
продуктивность незначительна. Стратегию поведения популяции в освет-
ленном березняке на 20-летней вырубке можно отнести к конкурентному
типу. На открытых местах между рядами берез и елей популяция имеет
нормальную оптимальную полночленную структуру с преобладанием ге-
неративных кустов. Общая фитомасса большая со значительной долей
фракции генеративных побегов. Семенная продуктивность высокая при’
наибольшей количестве метелок на м2. Без конкуренции с древостоем Вид
может существовать в таком состоянии довольно долго. Следовательно, в
процессе демутации стратегия жизни меняется от эксплерентной к вио-
23
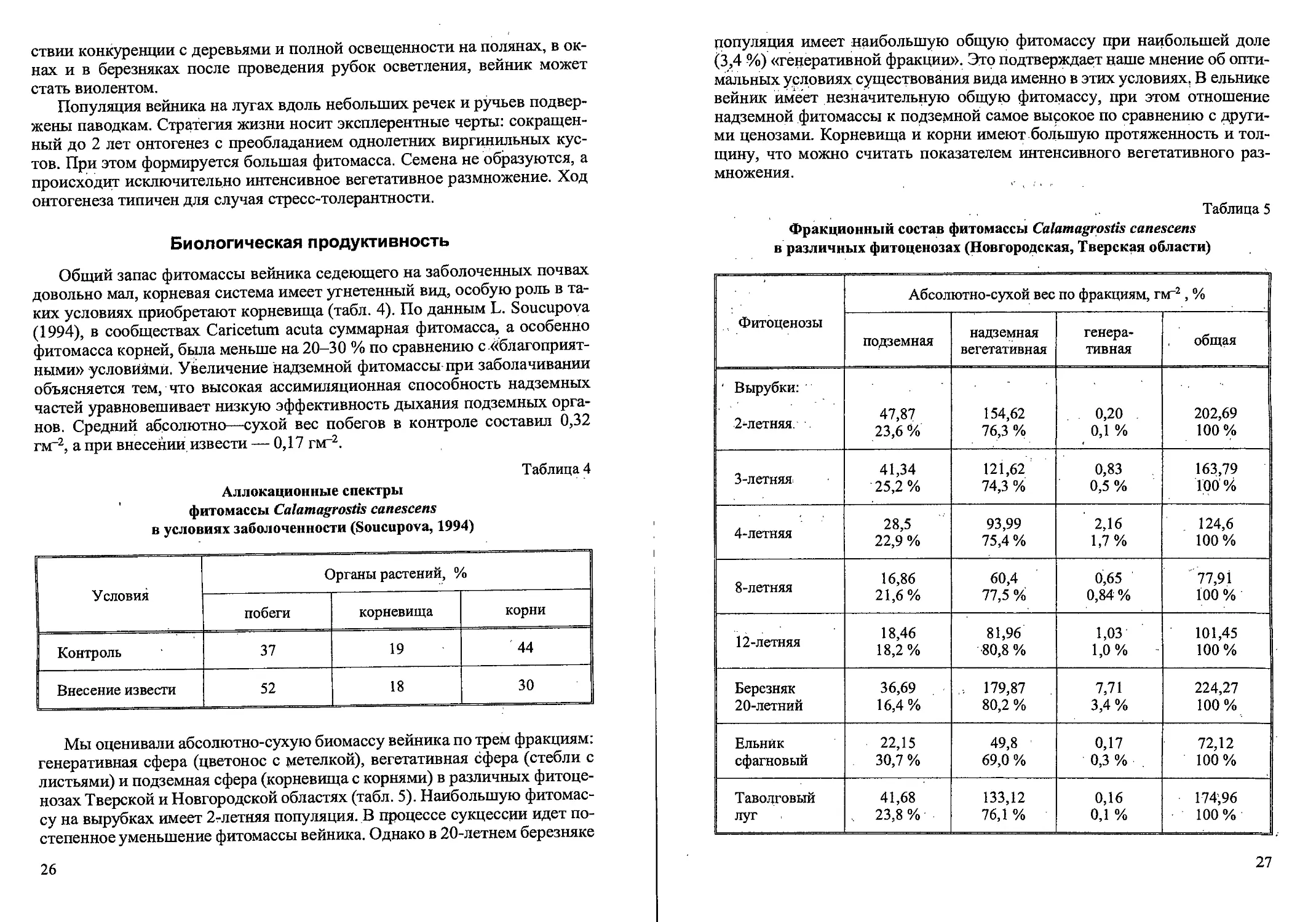
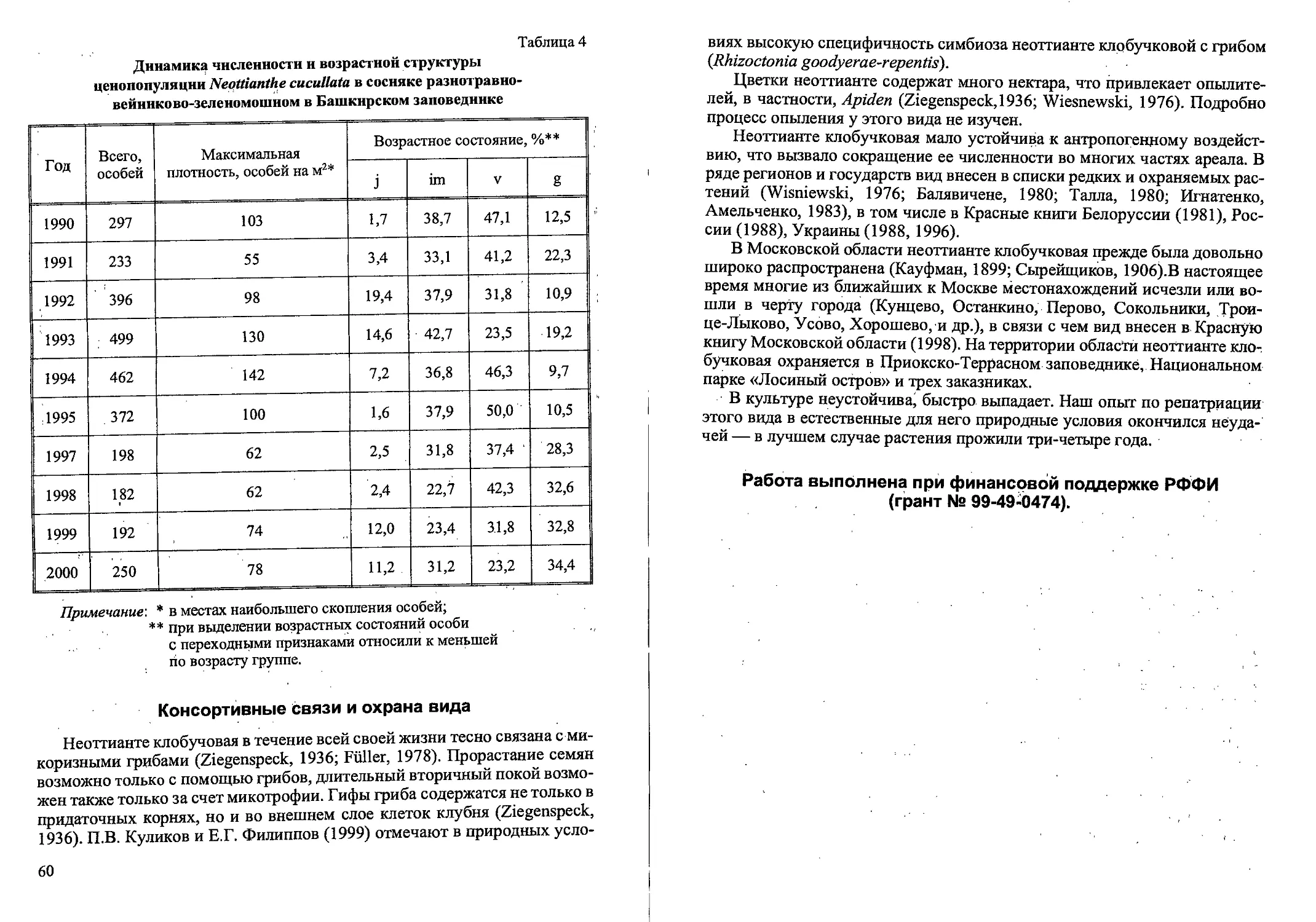
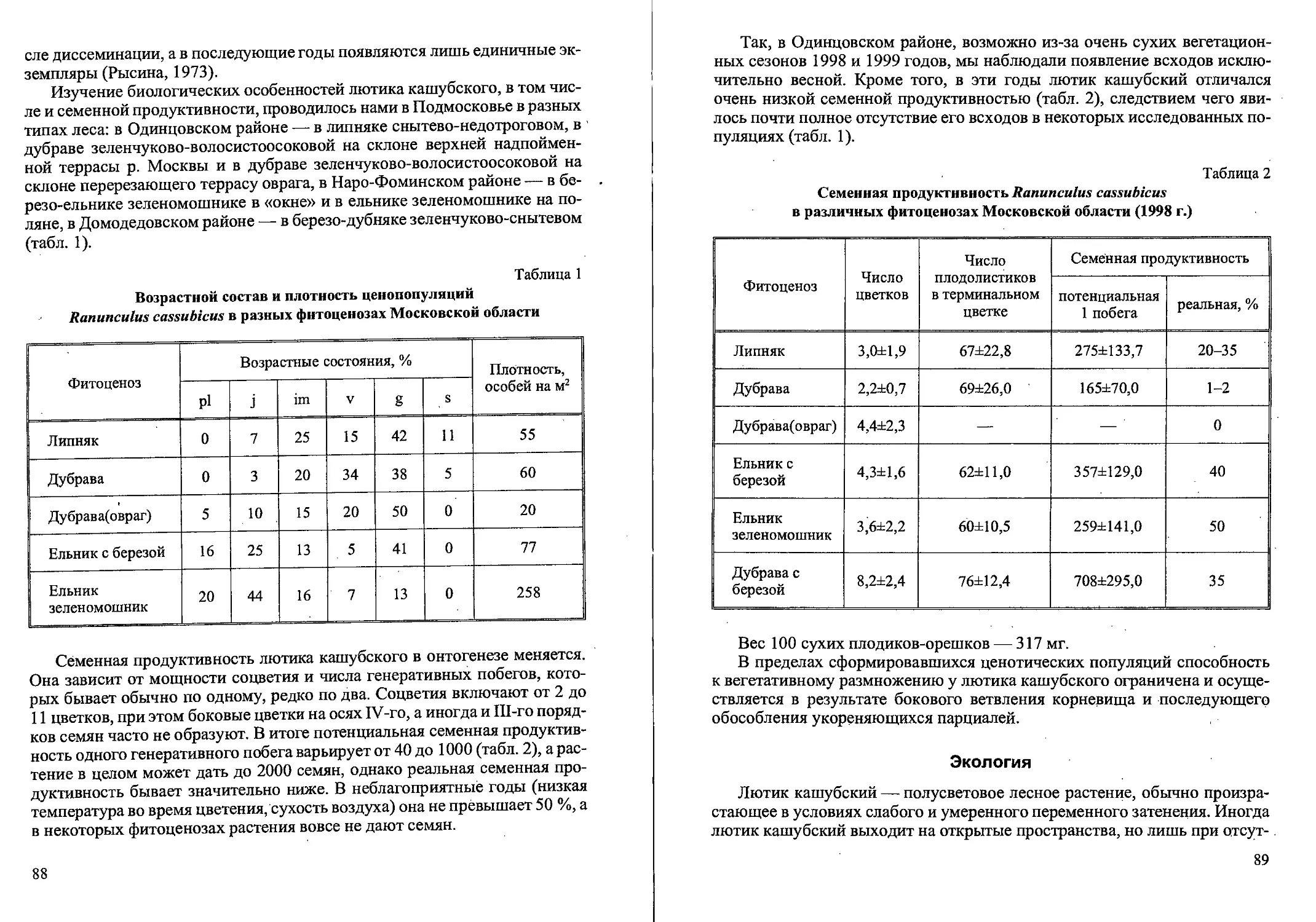
Возрастная структура популяций (кустов на м2, %)
Calamagrostis canescens в различных фитоценозах
(Новгородская, Тверская области)
Таблица 3
Фито- ценозы Возраст куста, лет Всего кустов
1 2 3 4 5 6
Вырубки: 2-летняя 169 55,2 % 126 41,2 % 8 2,6 % 2 0,6 % 1 0,3 % 0 306 100 %
3-летняя 193 67,2 % 89 31,0% 3 1,0% 2 0,7 % 0 0 287 ’ 100 %
4-летняя 58 32,6 % 96 53,9 % 15 8,4 % 7 3,9 % ' 2 1,1 % 0 178 100%
8-летняя 41 39,8 % 51 49,5 9 8,7 % 2 1,9% 0 0 103 100 %
12-летняя, 57 42,2 % 51 38,0 % 17 12,6 % 7 5,2 % 1 0,7 % 2 1,5 % 135 100 %
20-летняя 39 25,0 % 87 55,4 % 28 •17,8 % 3 1,9% 0 0 157 100%
Ельник сфагновый 59 50,0 % 52 43,7 % 8 6,7 % 0 0 0 .119 100 %
Таволговый . луг 93 73,2 % 26 20,5 % 8 6,3 % 0 0 .0 ' ' 127 100%
лентной и доходит до патентной в сомкнутых вторичных березняках. В
коренных ельниках вид также имеет патиентную стратегию. Высокая кон-
куренция за элементы минерального питания с елью и сложные отношения
с эдификатором сфагнумом формируют популяцию нормального типа с
сокращенным циклом развития. При небольшой общей фитомассе увели-
чивается доля фитомассы подземных органов. Высокая семенная продук-
тивность позволяет появляться даже семенным особям. Однако при отсут-
24
2-летняя вырубка
N=306 (в %)
12-летняя вырубка
N=135(B%y '
100 ,
80 -
60 -
40 -
20 -
0 -
j im v g1 g2 g3 ss s
3-летняя вырубка
N=287 (в %)
100 -
80 -
60 -
40
20 -
O'----,---(— .
j im v g1 g2 g3 ss s
4-летняя вырубка
N=178 (в%)
100 -|
80 -
j im v g1 g2 g3 ss s
8-летняя вырубка
N=103 (в %)
Im v g1 g2 g3 ss s
20-летняя вырубка
N=157 (в %)
Таволговый луг
N=127 (в %)
юо
80
60
40
20
О
im v g1 g2 g3 ss s
Сфагновый ельник
N=119(b%)
юо
80 -
60 -
40 -
20 -
О--
j i im v g1 g2 g3 ss s
Рис. 4. Спектры возрастных состояний Calamagrostis canescens
в разных фитоценозах (Тверская, Новгородская области).
25
ствии конкуренции с деревьями и полной освещенности на полянах, в ок-
нах и в березняках после проведения рубок осветления, вейник может
стать виолентом.
Популяция вейника на лугах вдоль небольших речек и ручьев подвер-
жены паводкам. Стратегия жизни носит эксплерентные черты: сокращен-
ный до 2 лет онтогенез с преобладанием однолетних виргинильных кус-
тов. При этом формируется большая фитомасса. Семена не образуются, а
происходит исключительно интенсивное вегетативное размножение. Ход
онтогенеза типичен для случая стресс-толерантности.
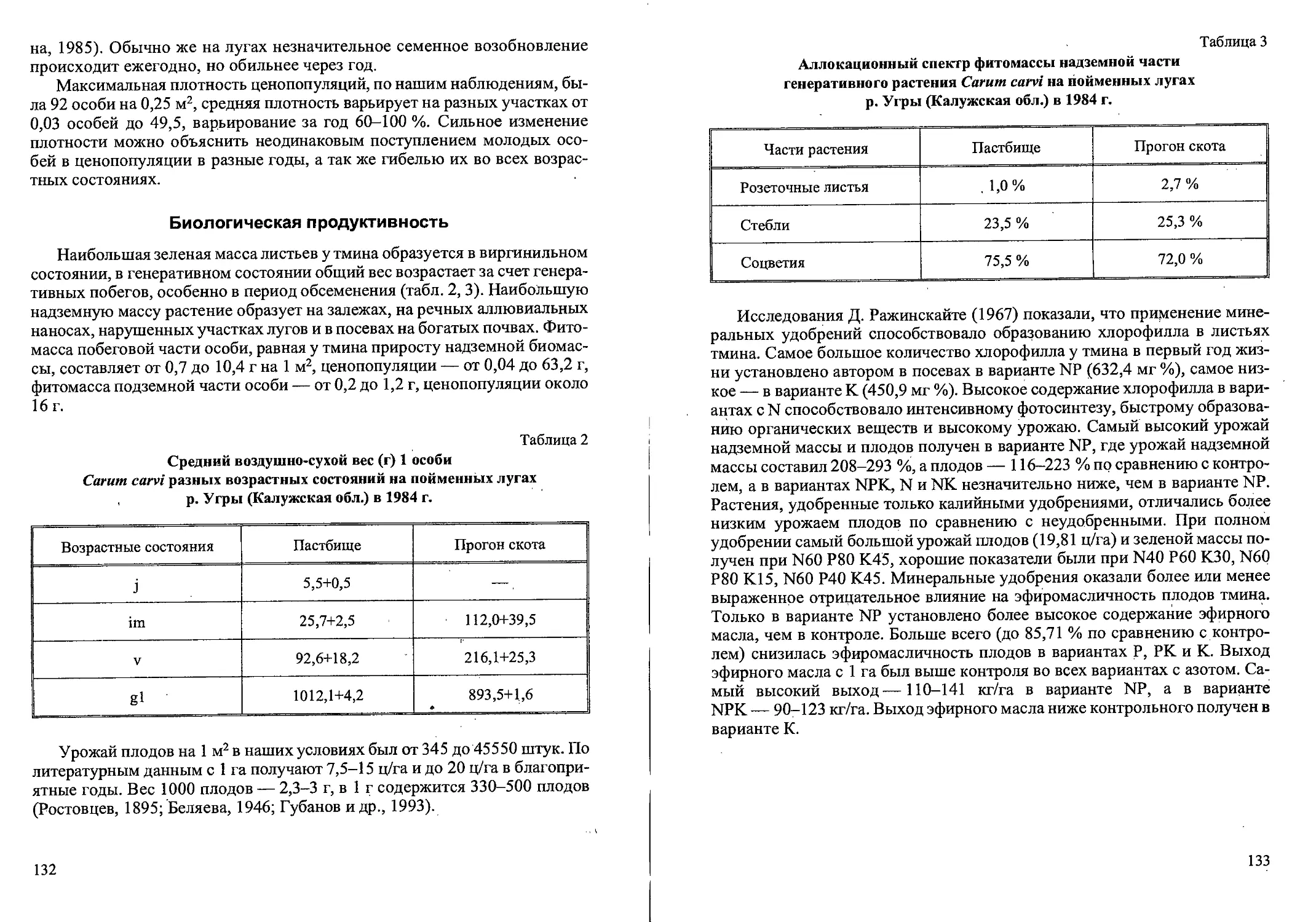
Биологическая продуктивность
Общий запас фитомассы вейника седеющего на заболоченных почвах
довольно мал, корневая система имеет угнетенный вид, особую роль в та-
ких условиях приобретают корневища (табл. 4). По данным L. Soucupova
(1994), в сообществах Caricetum acuta суммарная фитомасса, а особенно
фитомасса корней, была меньше на 20-30 % по сравнению с «благоприят-
ными» условиями. Увеличение надземной фитомассы при заболачивании
объясняется тем, что высокая ассимиляционная способность надземных
частей уравновешивает низкую эффективность дыхания подземных орга-
нов. Средний абсолютно—сухой вес побегов в контроле составил 0,32
гм-2, а при внесении извести — 0,17 гмг2.
Таблица 4
Аллокационные спектры
фитомассы Calamagrostis canescens
в условиях заболоченности (Soucupova, 1994)
Условия Органы растений, %
побеги корневища корни
Контроль 37 19 44
Внесение извести 52 18 30
Мы оценивали абсолютно-сухую биомассу вейника по трем фракциям:
генеративная сфера (цветонос с метелкой), вегетативная сфера (стебли с
листьями) и подземная сфера (корневища с корнями) в различных фитоце-
нозах Тверской и Новгородской областях (табл. 5). Наибольшую фитомас-
су на вырубках имеет 2-летняя популяция. В процессе сукцессии идет по-
степенное уменьшение фитомассы вейника. Однако в 20-летнем березняке
26
популяция имеет наибольшую общую фитомассу при наибольшей доле
(3,4 %) «генеративной фракции». Это подтверждает наше мнение об опти-
мальных условиях существования вида именно в этих условиях, В ельнике
вейник имеет незначительную общую фитомассу, при этом отношение
надземной фитомассы к подземной самое высокое по сравнению с други-
ми ценозами. Корневища и корни имеют большую протяженность и тол-
щину, что можно считать показателем интенсивного вегетативного раз-
множения.
Таблица 5
Фракционный состав фитомассы Calamagrostis canescens
в различных фитоценозах (Новгородская, Тверская области)
Фитоценозы Абсолютно-сухой вес по фракциям, гм-2 , %
подземная надземная вегетативная генера- тивная общая
' Вырубки:
2-летняя. 47,87 154,62 0,20 . 202,69
23,6% 76,3 % 0,1 % 100%
3-летняя 41,34 121,62 0,83 163,79
25,2 % 74,3 % 0,5 % 100%
4-летняя 28,5 93,99 2,16 124,6
22,9 % 75,4 % 1,7% 100%
8-летняя 16,86 60,4 0,65 77,91
21,6% 77,5 % 0,84 % 100 %
12-летняя 18,46 18,2% 81,96 80,8 % 1,03 1,0% 101,45 100%
Березняк 36,69 . 179,87 7,71 224,27
20-летний 16,4% 80,2 % 3,4 % 100%
Ельник 22,15 49,8 0,17 72,12
сфагновый 30,7 % 69,0 % 0,3 % 100%
Таволговый 41,68 133,12 0,16 174,96
луг 23,8 % 76,1 % 0,1 % 100%
27
Масса абсолютно сухого вещества вейника на вырубках березняков по
данным Л.С. Ермоловой (1981) варьировала очень сильно в течение 5 лет
наблюдений (от 3,05 до 56,29 гм-2). При этом наибольшая фитомасса фор-
мируется в августе. В сообществах с доминированием Thyphoides
arundinacea воздушно—сухая фитомасса вейника (проективное покрытие
2-5 %) может достигать 40 гмг2 (Балашев и др., 1988).
Консортивные связи
С.И. Ростовцев (1908) отмечает для вейника седеющего следующие
возбудители заболеваний: Ascochyta calamagrostidis Brun.— пятна на ли-
стьях; Epichloe typhina Pers.— беловатый или сероватый войлочный чехол
у основания междоузлий; Claviceps microcephala Wallr.— спорынья;
Fusoma triseptatum Sacc.— пятна на листьях; ржавчины: Puccinia
calamagrostidis Sydow. (Ured.), P. coronifera Klebhn., P. glumarum Schmidt.,
P. Rubigo veraDC., P. graminis Pers.; Septoria calamagrostidis Sacc.; голов-
ня: Tilletia calamagrostidis (Till.), T. striformis Westend, Ustilago hypodytes
Schlecht. (Ust.)
Нами были обнаружены поражения корней, побегов, семян, цветков
вейника седеющего. Черный налет на листьях образует гриб
Helmintosporium (кл. Deuteromycetos). При значительном поражении лист
сворачивается в трубочку. Вздутия оранжевого цвета на стеблях и корне-
вую гниль вызывает другой представитель Septoria кл. Deuteromycetos.
Массовоераспространение заболеваний было отмечено на низинном боло-
те. Процент пораженных растений в лесу был меньше, что, вероятно, свя-
зано с особенностями распространения грибов (для их развития требуется
температура 20-25).
Вейник седеющий — малоценное в кормовом отношении растение, но
зеленые листья поедаются гусями, а осенью и ранней зимой — зайцами,
(Растительное сырье СССР, 1957). На севере в большом количестве поеда-
ется оленями зимой из-под снега (Ларин, 1932). По нашим наблюдениям,
под слоем ветоши вейника на сухих участках складываются идеальные ус-
ловия для жизни мышевидных грызунов.
Хозяйственное значение
В лесном хозяйстве вырубки с доминированием вейник седеющий в та-
ежной зоне европейской части России считаются трудными для возобнов-
ления главных пород (Шутов и др., 1967). Связано это с сильной задернен-
ностью и неблагоприятными условиями, создаваемые вейниковым покро-
вом для развития самосева и посадок ели и сосны (Корконосова, 1967).
Естественный процесс возобновления леса на таких вырубках задержива-
ется на десятки лет.
28
Механические приемы борьбы с вейником седеющим (дискование,
рыхление почвы, окашйвание и т.п.) обычно малоэффективны и примене-
ние их- на лесокультурных площадях нецелесообразно (Бельков, 1974).
При толщине пласта от 10 до 20 см при плужной обработке молодые побе-
ги вейника в большом количестве начинают появляться и пробиваться на
поверхность почвы вскоре после вспашки. Наиболее надежным способом
борьбы с ним считается применение противозлаковых гербицидов/Семена
вейника на вырубках не погребены в почве, а прорастают на ее поверхно-
сти, что делает их всходы более уязвимыми к действию гербицидов (Шу-
тов и др., 1967). По данным В.П. Белькова с соавторами (1974) вейник се-
деющий чувствителен к триазинам (симазину и др.). При весенней обра-
ботке гербициды эффективно действует на вейник седеющий, не оказывая
отрицательного влияния на культуры ели, если ели находятся в момент оп-
рыскивания в состоянии покоя. Однако применять эти препараты в культу-
рах сосны и лиственницы нельзя.
Лекарственными являются подземные и надземные части вейника се-
деющего, так как содержат флавоноиды, углеводы, а также азотсодержа-
щие соединения. Растения обладают диуретическим, противокашлевым,
отхаркивающим, детоксикационным действием. Используется при болез-
нях почек и мочевыводящих путей и при асците.
Вейник седеющий — сенокосное растение, но малоценное в кормовом
отношении (табл. 6). К концу лета быстро грубеет, поэтому скашивать его
нужно до цветения (Кормовые растения ..., 1950).
Таблица 6
Химический состав Calamagrostis canescens
(Матвеева,1934; Попов и др., 1939)
Стадия развития Химический состав абсолютно сухого вещества, %
зола протеин белок жир клет- чатка прочие
Начало плодоношения 7,7 8,3 — 2,7 23,5 57,3
Плодоношение 5,5 7,1 6,6 2,5 35,1 49,8
Мнения о ценности вейника на пастбищах различны. Вследствие боль-
шой жесткости листьев и стеблей вейник относят к плохим или малоцен-
ным кормовым растениям (Кормовые растения ..., 1950; Раменская, 1960;
29
Быков, 1962; Нейштадт, 1963). Лишь в условиях Севера и высокогорий вид
приобретает существенное значение в качестве пастбищного растения
(Цвелев, 1976). Так, в пойме р. С.Двины вейник оценивается населением
как.хорошее кормовое растение (Кормовые растения..., 1950). В.В. Благо-
вещенский (1984) относит вейник седеющий к группе кормовых растений
для домашнего скота, зайцев и гусей (Дикорастущие полезные ..., 2001). В
Западной Сибири на пастбищах он плохо поедается крупным рогатым ско-
том (Кормовые растения ..., 1950). По наблюдениям И.В. Ларина (1932),
вейник .седеющий поедается весной, когда он дает сено среднего качества,
более ценное, чем у вейника наземного (Флора СССР, т. 2, 1934). Иногда
рассматривается как желательная примесь в сене (Кормовые растения ...,
1950). Устойчив к выпасу (Раменский, 1956).
Работа выполнена при финансовой поддержке
гранта INTAS № 01*0527
А. А. Нотов, Ю. В. Наумцев
ШПАЖНИК ЧЕРЕПИТЧАТЫЙ
Номенклатура, систематическое положение
и внутривидовая систематика
Gladiolus imbricatus L., 1753, Sp. Pl.: 37; Кауфман, 1866, Моск, фл.: 483;
Сырейщиков, 1906, Илл. фл. Моск, губ., 1: 247; Черняк, Фл. СССР, 4 : 583,
р.р.; Победимова, 1964, в Маевский, Фл., изд. 9 : 672; Скворцов, 1966, Оп-
ред. раст. Моск, обл.: 119; Цвелев, 1979, Фл. европ. части СССР, 4 : 310;
Hamilton, 1980, Fl. Europ. 5 : 101; Губанов и др., 1995, Опред. сосуд, раст.
Центра Европ. России : 193.— Шпажник черепитчатый [семейство
Iridaceae — Ирисовые (Касатиковые)].
Шпажник черепитчатый представляет род Gladiolus, объединяющий бо-
лее 250 видов, распространённых преимущественно в Африке (Рах, 1888;
Engler, 1936; Goldblatt, Manning, 1998). Центром видового разнообразия ро-
да является Капская область. В Европе и Азии встречается только 15 видов
рода Gladiolus. В последнем варианте системы ирисовых род Gladiolus от-
несён к трибе Ixieae подсемейства Ixioideae (Goldblatt, 1990,1991).
На Кавказе, в Западном Закавказье, Крыму встречается близкий вид
G. tenuis Bieb. (Черепанов, 1995), от которого G. imbricatus отличается бо-
лее крупными листочками околоцветника, большим числом листьев (3-5),
большей шириной листовых пластинок и крылатыми семенами (Цвелев,
1979). В настоящее время как G. tenuis идентифицируют некоторые опи-
санные ранее из Крыма и Кавказа виды — G. crispiflorus Herb., G. apterus
Klok., G. caucasicus Herb. (Цвелёв, 1979). Видовую самостоятельность
G. tenuis признают не все исследователи (Флора СССР, 1935; Тамберг,
1977). В последнем случае его соотносят с G. imbricatus.
Шпажник черепитчатый — достаточно полиморфный вид (Эристави,
1975, 1978, 1982). Выделяют разновидность var. libanoticus Boiss. (Там-
берг, 1977) с узкими листочками околоцветника, более жесткими и длин-,
ными прицветниками и более тонкими покровами клубнелуковицы. Расте-
ния с белыми цветками, имеющими пурпурные пятна, описаны как f.
albomaculata V. Krecz. (Флора СССР, 1935).
31
Географическое распространение
Евразиатский вид, распространённый в Средиземноморье, Средней и
Восточной Европе (Dietrich, 1832, FloraEuropaea, 1980; Atlas ..., 1986; Го-
лубев, 1996). Встречается в Прибалтике, на Украине, в Молдове и Белару-
си. Найден в северо-западном Казахстане (Мугоджарские горы) (Флора
СССР, 1935; Тамберг, 1977). В России известен от Кавказа до юга Ленин-
градской области (Маевский, 1964; Капинос, 1965; Тамберг, 1977; Цвелёв,
1979, 2000). Среди видов рода шпажник, G. imbricatus имеет самый боль-
шой ареал и встречается севернее всех остальных видов (Luther, 1961).
В Московской и смежной областях шпажник черепитчатый распро-
странен рассеянно, преимущественно на западе (Невский, 1952; Вороши-
лов и др., 1966; Опр. высш. раст. Северо-Запада..., 1981; Опр. сосуд, раст.
Мещеры, 1986; Опр. сосуд, раст. центра европ. России, 1995; Нотов, 1998,
2000; Цвелев, 2000).
Морфологическое описание
Травянистый клубнелуковичный монокормный (Голубев, Волокитин,
1986;,Борисова, 1992) поликарпик, геофит. И.Г. Серебряков (1962) относит
гладиолус к группе стебле-клубневых растений. Вегетативный малолет-
ник (однолетник по: Серебряков, 1954; Любарский, 1961) с симподиаль-
ным типом нарастания. Представляет симподиальную полурозеточную
модель побегообразования (Наумцев, 19996).
Корневая система представлена системой придаточных корней, кисте-
корневая (Голубев, 1996). Корни многочисленные, однотипные, желтова-
то-бурые, 1-1,2 мм в диаметре. Они располагаются горизонтально или на-
правлены дуговидно вниз. Ширина простирания 30-40 см, глубина про-
никновения 35 см (Голубев, 1962). Образуются придаточные корни на
нижнем метамере клубнелуковицы с сильнопаренхиматизированным меж-
доузлием (рис. 1). Функционируют они один вегетационный сезон (Сереб-
ряков, 1952; Голубев, 1962). Питание дочерней клубнелуковицы осуществ-
ляется за счёт придаточных корней прошлогодней материнской клубнелу-
ковицы. Придаточные корни шпажника черепитчатого обладают,
контрактильной способностью. Кора базальной части корня утолщена и
сильно обводнена. Верхушечная часть корня тонкая, покрыта корневыми
волосками, прочно укрепляется в почве. Благодаря сморщиванию и сплю-
щиванию базальной части происходит втягивание клубнелуковицы вглубь
почвы (Голубев, Г95бб). После этого корни функционируют как питаю-
щие. Обычно, клубнелуковицы залегают на глубине 6 см (Голубев, 1956а;
Рысйн, Рысина, 1987).
Основной структурной единицей системы побегов является монокар-
пический побег. Монокарпические побеги — однотипные, полурозеточ-
32
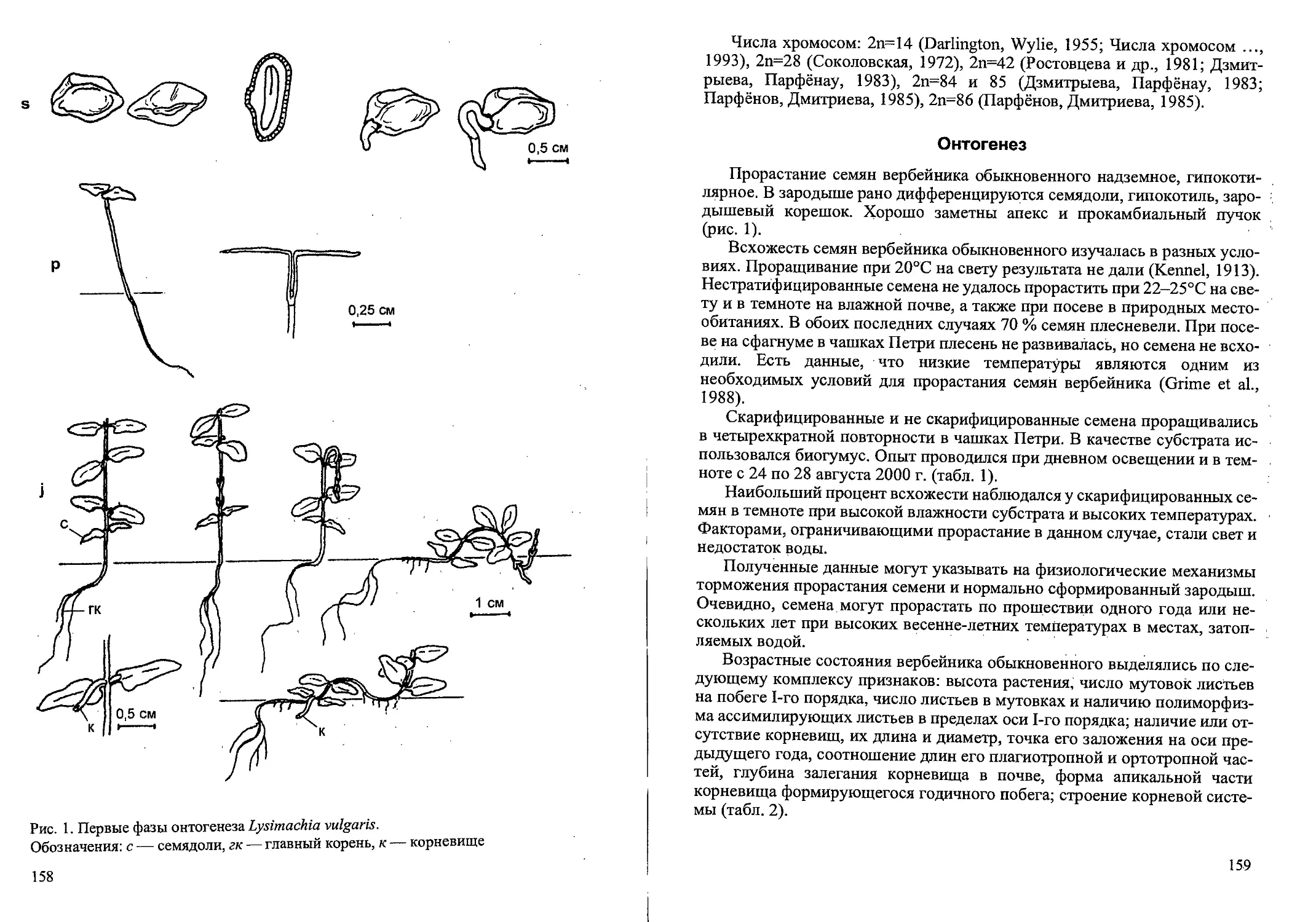
ные, моноциклические. Они состоят из удлинённой части, завершающейся
соцветием, и базального участка (рис. 1), который сохраняется в составе
многолетней части системы побегов не более двух сезонов.
Клубнелуковица образована, как правило, 8-ю метамерами (рис. 1).
Шесть нижних метамеров имеют укороченные, слабопаренхиматизиро-
ванные междоузлия и листья низовой формации. Следующие два метаме-
ра с укороченными сильнопаренхиматизированными междоузлиями и
листьями срединной формации. Чешуевидные листья четырех нижних
метамеров имеют форму колпачка. У листьев двух следующих метамеров
хорошо развита влагалищная часть. Почки, образующиеся в пазухах че-
шуевидных листьев, обычно остаются в спящем состоянии. Значительно
реже из них развиваются короткие столоны, завершающиеся клубнелуко-
вичками-детками. Столон состоит, как правило, из трех метамеров (рис.
1). Два из них с нежными быстроразрушающимися чешуевидными листь-
ями, а третий с плотным кожистым чешуевидным листом, который как
футляр окружает детку. Ось клубнелуковички-детки образована одним
сильнопаренхиматизированным междоузлием. На её конусе нарастания
можно обнаружить три листовых примордия. Розеточная часть базально-
го участка состоит из двух метамеров с листьями срединной формации.
Их междоузлия также сильно паренхиматизированы, больше утолщено
междоузлие, расположенное между верхним: влагалищным листом и
нижним листом срединной формации (рис. 1). В пазухах листьев средин-
ной формации образуются почки возобновления. Самая верхняя из них
дает начало монокарпическому побегу следующего порядка (из его ба-
зального участка развивается дочерняя клубнелуковица). Крайне редко
трогаются в рост две почки и развиваются два дочерних монокарпиче-
ских побега (Голубев, 1965а; Рысин, Рысина, 1987). Основания листьев
срединной формации материнского побега преобразуются в плотные по-
кровы. Они кожисто-перепончатые или перепончатые, окружают дочер-
нюю клубнелуковицу и выполняют защитную функцию. Сильной скле-
рификации жилок характерной для покровов некоторых представителей
семейства Iridaceae (Наумцев, 19996), и разрушения паренхимных участ-
ков, у шпажника черепитчатого не происходит.
На удлиненной части монокарпического побега (цветоносе) развива-
ются обычно 2 (реже 3) листа срединной формации и от 3 до 14 листьев
верховой формации (рис. 1). Листья срединной формации линейные или
узколанцетные, мечевидные (Рах, 1888; Кернер, 1899). Листовая пластин-
ка унифациальная.
Соцветие — однобокий, односторонний колос (Weimark, 1939; Фёдо-
ров, Артюшенко, 1979), из (3)5-10(14) цветков. Очень редко в пазухе верх-
него листа срединной формации развивается боковая кисть меньших раз-
меров с небольшим числом цветков. Такие кисти отмечены нами только на
монокарпических побегах, имеющих на удлиненной части 3 листа средин-
33
34
ной формации. Каждый цветок окружен кроющим листом ланцетной фор-
мы и профиллом.
Цветки зигоморфные, трёхчленные, пентамерные. Околоцветник про-
стой. Листочки его лепестковидные, овальные, с тупой верхушкой, розова-
тые или пурпурно-фиолетовые, в основании сросшиеся в короткую слегка
изогнутую трубку длиной до 1 см. Андроцей из трёх тычинок. Пыльники
линейные, интрорзные. Основания тычиночных нитей прикреплены вбли-
зи середины трубки околоцветника. Столбик с тремя рыльцами. Гинецей
синкарпный. Завязь нижняя, трехгцёздная. Плод трёхгнёздная локулицид-
ная коробочка, короткая, обратнояйцевидная, длиной 8-10 мм, ту-
потрёхгранная.
Число хромосом: 2п=60 (Цвелев, 1979; Retkeilykasvio, 1998). Евразиат-
ские виды гладиолусов — гекса- и октаплоиды (Громов, 1998). Различия в
числах хромосом препятствуют гибридизации G. imbricatus и африканских
видов (Тамберг, 1981).
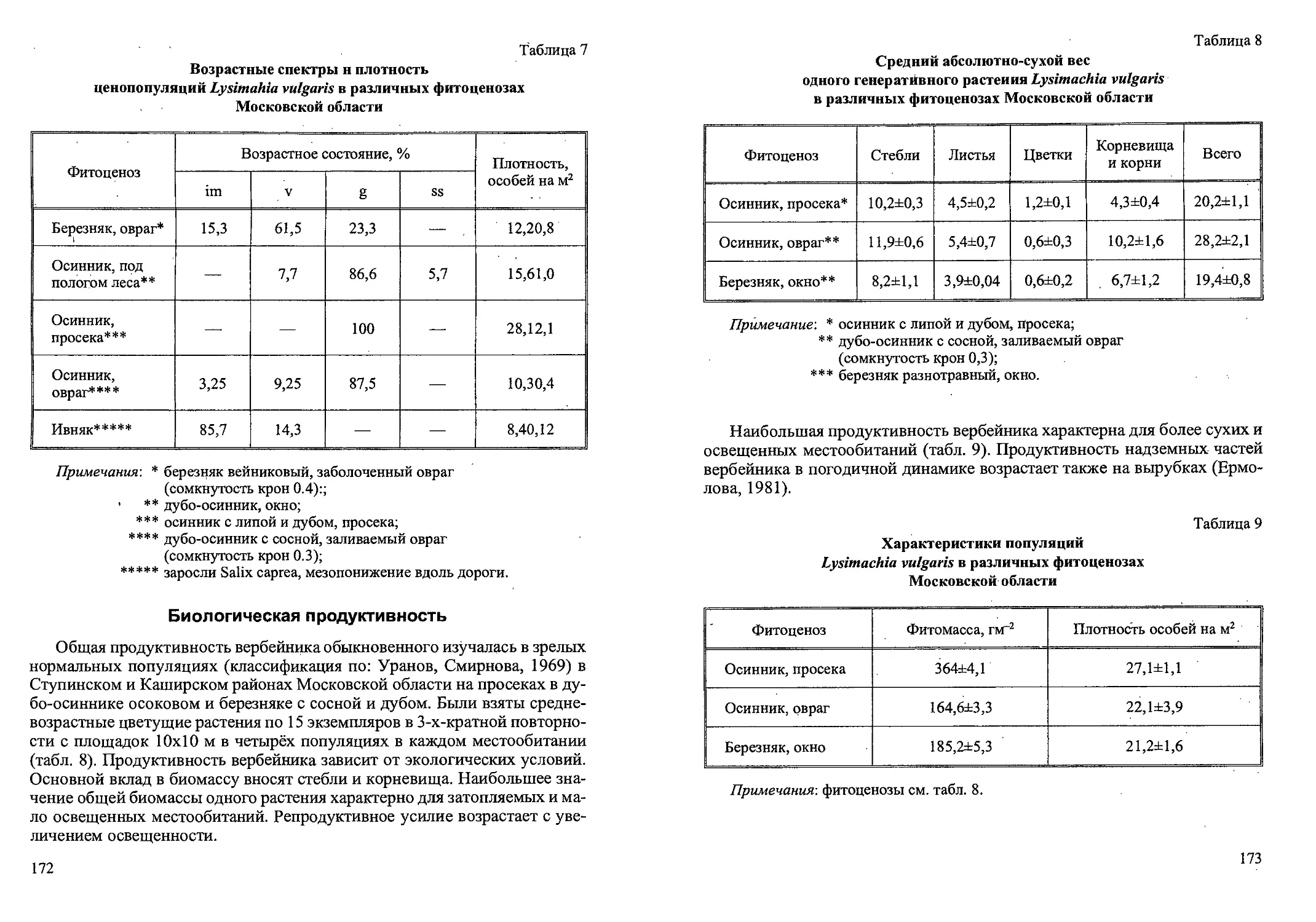
Рис. 1. Система побегов средневозрастного генеративного
растения Gladiolus imbricatus (20.08.1994 г.) (по: Наумцев, 1996а).
Обозначения: а — дочерняя клубнелуковица; б — материнская клубнелуковица;
в — столон; г — клубнелуковица детка; д — почка возобновления;
е — цветонос; ж — основание листьев срединной формации;
1,2,3 — порядковый номер метамера;
у ' — органы, функционирующие более одного сезона;
— органы, функционирующие не более одного сезона;
J — органы, разрушенные к моменту наблюдения;
— чешуевидные листья, имеющие форму колпачка;
— чешуевидные листья с развитой влагалищной частью;
— листья срединной формации;
— листья верховой формации;
ув — коробочки;
/7 — спящие почки;
° — придаточные корни;
— рубец от прошлогоднего цветоноса
35
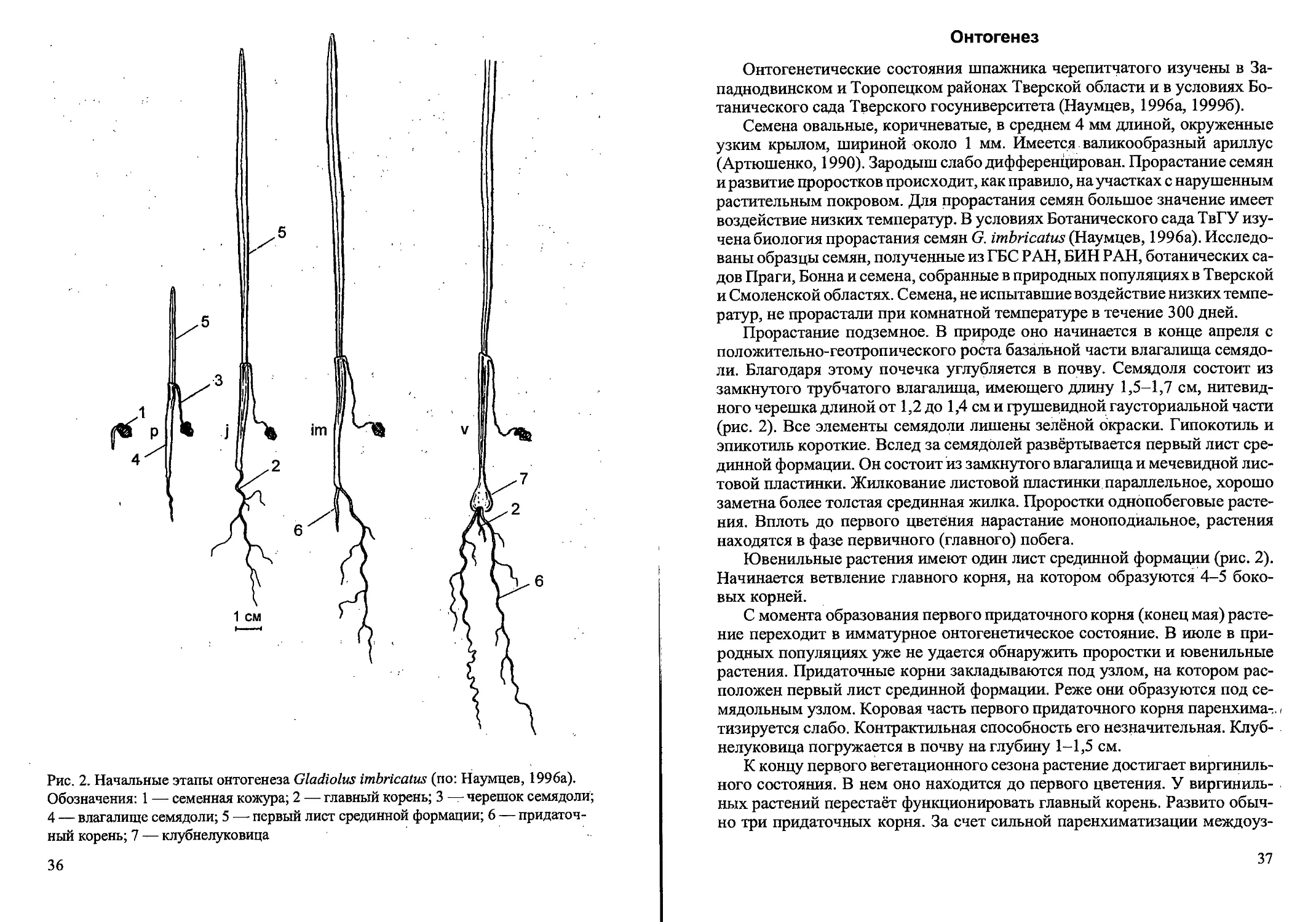
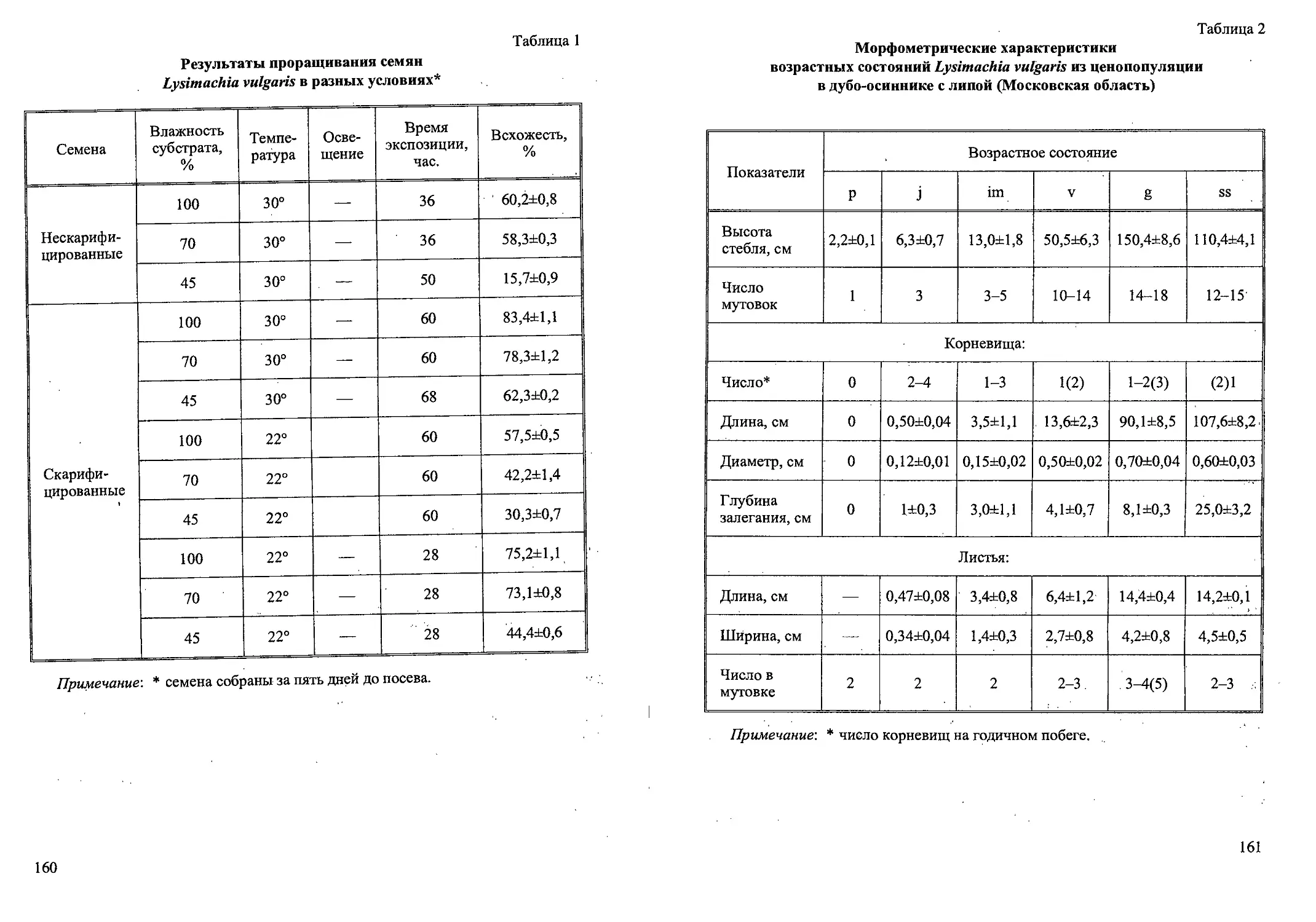
Рис. 2. Начальные этапы онтогенеза Gladiolus imbricatus (по: Наумцев, 1996а).
Обозначения: 1 — семенная кожура; 2 — главный корень; 3 — черешок семядоли;
4 — влагалище семядоли; 5 — первый лист срединной формации; 6 — придаточ-
ный корень; 7 — клубнелуковица
36
Онтогенез
Онтогенетические состояния шпажника черепитчатого изучены в За-
паднодвинском и Торопецком районах Тверской области и в условиях Бо-
танического сада Тверского госуниверситета (Наумцев, 1996а, 19996).
Семена овальные, коричневатые, в среднем 4 мм длиной, окруженные
узким крылом, шириной около 1 мм. Имеется валикообразный ариллус
(Артюшенко, 1990). Зародыш слабо дифференцирован. Прорастание семян
и развитие проростков происходит, как правило, на участках с нарушенным
растительным покровом. Для прорастания семян большое значение имеет
воздействие низких температур. В условиях Ботанического сада ТвГУ изу-
чена биология прорастания семян G. imbricatus (Наумцев, 1996а). Исследо-
ваны образцы семян, полученные из ГБС РАН, БИН РАН, ботанических са-
дов Праги, Бонна и семена, собранные в природных популяциях в Тверской
и Смоленской областях. Семена, не испытавшие воздействие низких темпе-
ратур, не прорастали при комнатной температуре в течение 300 дней.
Прорастание подземное. В природе оно начинается в конце апреля с
положительно-геотропического роста базальной части влагалища семядо-
ли. Благодаря этому почечка углубляется в почву. Семядоля состоит из
замкнутого трубчатого влагалища, имеющего длину 1,5-1,7 см, нитевид-
ного черешка длиной от 1,2 до 1,4 см и грушевидной гаусториальной части
(рис. 2). Все элементы семядоли лишены зелёной окраски. Гипокотиль и
эпикотиль короткие. Вслед за семядолей развёртывается первый лист сре-
динной формации. Он состоит из замкнутого влагалища и мечевидной лис-
товой пластинки. Жилкование листовой пластинки параллельное, хорошо
заметна более толстая срединная жилка. Проростки однопобеговые расте-
ния. Вплоть до первого цветения нарастание моноподиальное, растения
находятся в фазе первичного (главного) побега.
Ювенильные растения имеют один лист срединной формации (рис. 2).
Начинается ветвление главного корня, на котором образуются 4—5 боко-
вых корней.
С момента образования первого придаточного корня (конец мая) расте-
ние переходит в имматурное онтогенетическое состояние. В июле в при-
родных популяциях уже не удается обнаружить проростки и ювенильные
растения. Придаточные корни закладываются под узлом, на котором рас-
положен первый лист срединной формации. Реже они образуются под се-
мядольным узлом. Коровая часть первого придаточного корня паренхима-:. <
тизируется слабо. Контрактильная способность его незначительная. Клуб-
нелуковица погружается в почву на глубину 1-1,5 см.
К концу первого вегетационного сезона растение достигает виргиниль-
ного состояния. В нем оно находится до первого цветения. У виргиниль-
ных растений перестаёт функционировать главный корень. Развито обыч-
но три придаточных корня. За счет сильной паренхиматизации междоуз-
37
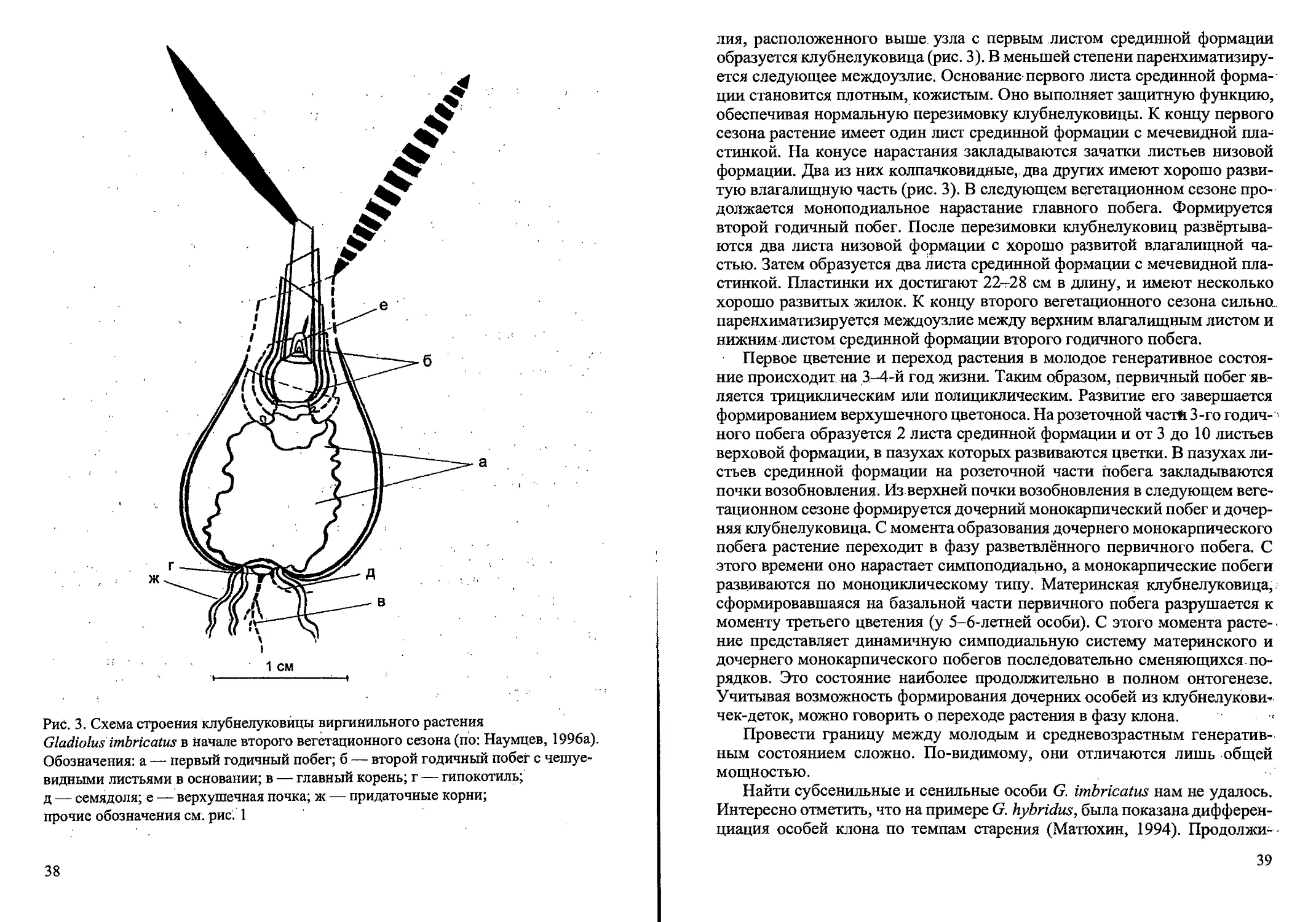
Рис. 3. Схема строения клубнелуковицы виргинильного растения
Gladiolus imbricatus в Начале второго вегетационного сезона (по: Наумцев, 1996а).
Обозначения: а — первый годичный побег; б — второй годичный побег с чешуе-
видными листьями в основании; в — главный корень; г — гипокотиль;
д — семядоля; е — верхушечная почка; ж — придаточные корни;
прочие обозначения см. рис. 1
38
лия, расположенного выше узла с первым листом срединной формации
образуется клубнелуковица (рис. 3). В меньшей степени паренхиматизиру-
ется следующее междоузлие. Основание первого листа срединной форма-
ции становится плотным, кожистым. Оно выполняет защитную функцию,
обеспечивая нормальную перезимовку клубнелуковицы. К концу первого
сезона растение имеет один лист срединной формации с мечевидной пла-
стинкой. На конусе нарастания закладываются зачатки листьев низовой
формации. Два из них колпачковидные, два других имеют хорошо разви-
тую влагалищную часть (рис. 3). В следующем вегетационном сезоне про-
должается моноподиальное нарастание главного побега. Формируется
второй годичный побег. После перезимовки клубнелуковиц развёртыва-
ются два листа низовой формации с хорошо развитой влагалищной ча-
стью. Затем образуется два листа срединной формации с мечевидной пла-
стинкой. Пластинки их достигают 22-28 см в длину, и имеют несколько
хорошо развитых жилок. К концу второго вегетационного сезона сильно.,
паренхиматизируется междоузлие между верхним влагалищным листом и
нижним листом срединной формации второго годичного побега.
Первое цветение и переход растения в молодое генеративное состоя-
ние происходит на 3-4-й год жизни. Таким образом, первичный побег яв-
ляется трициклическим или полициклическим. Развитие его завершается
формированием верхушечного цветоноса. На розеточной частй 3-го годич-'
ного побега образуется 2 листа срединной формации и от 3 до 10 листьев
верховой формации, в пазухах которых развиваются цветки. В пазухах ли-
стьев срединной формации на розеточной части побега закладываются
почки возобновления. Из верхней почки возобновления в следующем веге-
тационном сезоне формируется дочерний монокарпический побег и дочер-
няя клубнелуковица. С момента образования дочернего монокарпического
побега растение переходит в фазу разветвлённого первичного побега. С
этого времени оно нарастает симпоподиально, а монокарпические побеги
развиваются по моноциклическому типу. Материнская клубнелуковица,
сформировавшаяся на базальной части первичного побега разрушается к
моменту третьего цветения (у 5-6-летней особи). С этого момента расте-
ние представляет динамичную симподиальную систему материнского и
дочернего монокарпического побегов последовательно сменяющихся по-
рядков. Это состояние наиболее продолжительно в полном онтогенезе.
Учитывая возможность формирования дочерних особей из клубнелукови-
чек-деток, можно говорить о переходе растения в фазу клона.
Провести границу между молодым и средневозрастным генератив-
ным состоянием сложно. По-видимому, они отличаются лишь общей
мощностью.
Найти субсенильные и сенильные особи G. imbricatus нам не удалось.
Интересно отметить, что на примере G. hybridus, была показана дифферен-
циация особей клона по темпам старения (Матюхин, 1994). Продолжи-
39
тельность жизни клона зависит от характера соотношения особей с разной
динамикой старения. В случае преобладания и преимущественного выжи-'
вания быстростареющих особей продолжительность существования клона
может не превышать десяти лет (Андреева, 1977). Клоны, все особи кото-
рых находятся в состоянии медленного старения, функционируют не-
сколько десятков лет и, по- видимому, могут существовать неограниченно
долго.
В условиях интродукции отмечено ускорение темпов онтогенеза
шпажника черепитчатого (Наумцев, 19996). По нашим наблюдениям, в
культуре G. imbricatus переходит в генеративное состояние на второй год.
Ритм сезонного развития
Малый жизненный цикл монокарпического побега G. imbricatus скла-
дывается из трёх основных фаз: фаза внутрипочечного развития, фаза веге-
тации (включая цветение) и фаза резида (до полного отмирания многолет-
ней части побега) (Седова, 1972, 1976, 1982). В течение первой фазы про-
исходит активный органогенез. Вторая фаза в большей степени связана с
ростом зачатков побега и функционированием сформированных органов,
но при развёртывании монокарпического побега из почки продолжается
органогенез Генеративных органов. Фаза резида не связана ни с органоге-
незом, ни с ростом.
Данные о ритме развития G. imbricatus получены на основании наблю-
дений, проведённых в Ботаническом саду ТвГУ, и анализа материала, соб-
ранного в Западнодвинском районе Тверской области (Наумцев, 1996а,
19996).
Фаза внутрипочечного развития длится около одного года. Почки во-
зобновления закладываются в июне перед началом цветения материнского
монокарпического, побега. Внутрипочечное развитие дочернего монокар-
пического побега продолжается в июле после цветения. К концу августа на
конусе нарастания бывает сформировано, как правило, шесть листовых
примордиев. По терминологии В.А. Кукушкина (1983) почки морфологи-
чески развиты. С наступлением зимнего периода растение переходит в со-
стояние органического покоя. Таким образом, в почке возобновления зало-
жена только часть зачатков вегетативной сферы. Активный органогенез
возобновляется весной. В середине апреля в почке возобновления можно
обнаружить уже 8-9 примордиев. После формирования 10-го примордия
апекс переходит во флоральное состояние. Степень преформированности
побега у представителей рода Gladiolus разная, что определяет различия в
сроках их цветения (Наумцев, 19996). У G. palustris, также как у G. imbri-
catus, в почке возобновления заложена только часть зачатков вегетативной
сферы. У G. communis сформированы все зачатки вегетативной сферы. У
последнего вида наблюдается более раннее цветение
40
Фаза вегетации монокарпического побега захватывает только один ве-
гетационный сезон. Развертывание розеточных листьев начинается в кон-
це апреля-начале мая. Бутонизация наблюдается в середине-конце июня.
Цветение происходит с конца июня-начала июля до середины июля. Опре-
деленную роль в регуляции процесса цветения гладиолусов играет этилен
(Serek et al., 1994). Фенологические наблюдения, проведённые в Ботаниче-
ском саду ТвГУ в 1996-2000 гг., показали, что продолжительность цвете-
ния может изменяться от 10 до 24 дней (средняя — 19,8 дней). Наиболее
существенные отклонения от среднего значения обнаружены в 1999 и
2000 гг. В засушливое лето 1999 г. цветение началось раньше обычного
(15 июня). В этот сезон отмечена минимальная продолжительность цвете-
ния (от 10 до 15 дней у разных образцов) и наиболее, раннее созревание
плодов (19 июля-12 августа у разных образцов в зависимости от степени
освещенности местообитания). В более влажное, чем обычно, лето 2000 г.
отмечена максимальная продолжительность цветения (24 дня). С середи-
ны августа начинают созревать плоды. В конце вегетационного сезона на
оси материнской клубнелуковицы, представляющей базальную часть про-
шлогоднего монокарпического побега, можно видеть дочернюю клубне-
луковицу. Она образовалась на базальном участке моцокарпического по-
бега этого года в результате утолщения междоузлий. На дочерней клубне-
луковице сформирована почка возобновления, из которой в следующем
сезоне образуется новый монокарпический побег (рис. 1). Питательные ве-
щества материнской клубнелуковицы частично использует цветонос до-
чернего побега, поэтому к концу вегетационного сезона материнская клуб-
нелуковица сморщивается и уменьшается в размерах (Голубев, 1965). По-
сле отмирания надземной части дочернего монокарпического побега его
базальный участок становится материнской клубнелуковицей следующего
порядка, которая сохраняется в течение одного года.
Зачатки придаточных корней закладываются в конце вегетационного
сезона на дочерней клубнелуковице, но визуально они практически неза-
метны (Наумцев, 1996а, 19996). Придаточные корни формируются из них
в начале следующего вегетационного сезона. Таким образом, функциони-
рующие придаточные корни имеются только на материнской клубнелуко-
вице (рис. 1). За счёт них питается формирующаяся дочерняя клубнелуко-
вица. Позднее заложение и развитие придаточных корней у G. imbricatus-
приводит к гибели высаживаемых осенью клубнелуковиц, если при пере-.
садке удалена материнская клубнелуковица (Наумцев, >1996а, 19996).
В.Н Голубев (1962) относит G. imbricatus к группе .раннелетнецвету-
щих растений со средней продолжительностью вегетации, в Крыму и на
Кавказе он является поздневесенне-раннелетнецветущим видом (Эриста-
ви, 1984; Голубев, 1996). Установлено также более раннее наступление фе-
нофаз в условиях культуры в Болгарии (Протич, Евстатиева, 1994). Период
вегетации растения длится более 6 месяцев. Перерыв в вегетации связан с
41.
наступлением холодного зимнего сезона. По продолжительности, глубине
периода покоя и характеру его соотношения с перерывом вегетации
G. imbricatus можно отнести к растениям с устойчивым продолжительным
органическим покоем в течение неблагоприятного сезона, который сопро-
вождается длительным перерывом в вегетации (Наумцев, 19996). Физио-
логические аспекты периода покоя гладиолусов изучены Е.В. Танюшки-
ным (1991).
Способы размножения и распространения
Наличие крыла способствует анемохорному распространению семян.
В природных популяциях преобладает семенное размножение. Анализ
возрастных спектров позволил отнести большую часть изученных в Запад-
нодвинском и Жарковском районах Тверской области популяций
G. imbricatus к группе нормальных и инвазионных. Суммарная доля онто-
генетических групп прегенеративного периода в разных местообитаниях
достигала 79-83 %.
По мнению некоторых авторов (Цвелёв, 1979; Козловская, 1986) в при-
родных популяциях у G. imbricatus клубнелуковички-детки не образуются.
Однако возможность вегетативного размножения отмечена В.Н. Голубе-
вым (1956а), Т.О. Яницкой (Красная книга..., 1998). Ежегодное замещение
материнской клубнелуковицы Дочерней и втягивание клубнелуковицы
благодаря контрактильной способности придаточных корней способствуй
ет вытеснению деток ближе к поверхности почвы и распространению их
по территории (Голубев, 1956а).
В изученных нами популяциях в Западнодвинском районе Тверской
области клубнелуковички-детки отмечены у 11,1 % экземпляров (Наум-
цев, 1996а).
Детальный анализ клубнелуковиц на пробных площадках позволил об-
наружить в почве покоящиеся жизнеспособные клубнелуковицы, которые,
по-видимому, являются дополнительным резервом возобновления в попу-
ляциях, Аналогичные наблюдения сделаны в культуре на территории бота-
нического сада ТвГУ.
В условиях культуры происходит регулярное цветение и плодоноше-
ние G. imbricatus (Наумцев, 1999а, 19996). На слабозадернованных участ-
ках отмечен обильный самосев. В культуре увеличивается частота встреч
чаемости клубнелуковичек-деток (в некоторых случаях они образуются у
40 % растений). При этом чаще попадаются экземпляры, у которых сфор-
мировано более двух деток. Однако обнаружить у G. imbricatus разветв-
ленные столоны с несколькими клубнелуковичками-детками, которые
можно наблюдать у G. hybridus, нам не удалось. У последнего вида детки
мотуг образовываться даже в пазухах семядоли и первого ассимилирую-
щего листа проростка (Андреева, 1971).
42
Экология
G. imbricatus является сциогелиофитом (Голубев, 1996). В работе
D. Frank и S. Klotz (1990) отнесен к «полусветовым» растениям, пред-
ставляющим 7-ю ступень шкалы. Он может произрастать в условиях за-
тенения вместе с растениями 6-й ступени, но не встречается в местооби-
таниях, где наблюдается освещенность, соответствующая 5-й и 9-й сту-
пеням. По мнению D. Frank и S. Klotz, он не растёт в условиях полного
освещения. По нашим наблюдениям, G. imbricatus может расти в услови-
ях полного освещения. Под пологом древесного яруса прекращается нор-
мальное цветение.
В.Н. Голубев (1996) характеризует G. imbricatus как мезофит и глико-
фит. По экологическим шкалам Л.Г. Раменского (1956) шпажник черепит-
чатый отнесен к растениям, предпочитающим экотопы с увлажнением све-
жих и влажных лугов и лесов (53-70-я ступень). Может расти в условиях
сыролугового (74^-83-я) и лугово-степного (47-52-я) увлажнения (Рамен-
ский и др., 1956). Хотя G. imbricatus встречается надостаточно увлажнен-,
ных почвах, он не переносит избыточного застойного увлажнения (Губа-
нов и др., 1990). ;
Шпажник предпочитает довольно богатые почвы (9-12-я ступень), ре-
акция pH которых находится в интервале от слабокислой до нейтральной
(6,0-7,5). Отмечен также на бедных (5-6-я ступень) и богатых (14-16-я
ступень) почвах. Может расти в условиях слабого и среднего засоления
(Раменский и др., 1956; Красная книга ..., 1995). Встречается на луговых,
подзолистых лесостепных и суглинистых почвах.
В 1997-1999 гг. А.А. Нотовым и О.В. Романовым изучены ценопопуля-
ции G. imbricatus в Западнодвинском и Жарковском районах Тверской об-
ласти, расположенные в разных местообитаниях (Романов, 2000). Исполь-
зован метод экологической оценки экотопа по растительному покрову, ис-
пользуя шкалы Л.Г. Раменского (1956), D. Frank и S. Klotz (1990).
Оптимальным для G. imbricatus оказалось следующее сочетание характе-
ристик экотопа: увлажнение 70,54 балла, богатство почв 8,93 балла, осве-
щенность 6,96 балла. В местообитании с такими параметрами отмечен
наиболее высокий средний балл жизненности (3,27). Участие групп особей
разного жизненного состояния следующее: 9 % — особи 2-го уровня жиз-
ненности, 55 % — особи 3-го й 36 % — 4-го уровня жизненности. В усло-
виях повышенного затенения (6,55) и в достаточно сухих местообитаниях
(64,82) отмечены особи только 1-2(3)-го уровней жизненности. Средний с
балл жизненности от 1,13 до 2,45 (Романов, 2000).
Интересно сопоставление корреляций признаков, полученных при био-
метрическом анализе ценопопуляций в исследованных экотонах (Романов,
2000). В условиях оптимального для G. imbricatus сочетания характери-
стик экотопов чётко выражены корреляционные плеяды признаков вегета-
43
тивной и генеративной сфер. Отмечены многочисленные взаимосвязи ме-
жду признаками вегетативной и генеративной сфер. Обнаружена значи-
тельная взаимозависимость параметров 2-4-го листьев срединной
формации и связи их с признаками генеративной сферы. Значимые связи
между признаками 1-го листа срединной формации и признаками других
элементов монокарпического побега не установлены. При этом параметры
первого листа связаны между собой достаточно сильными связями. Опи- -
санный характер корреляции признаков свидетельствует о достаточно вы-
сокой целостности ростовых и морфогенетических процессов в пределах
удлиненной части монокарпического побега. Взаимозависимость призна-
ков 1-го листа и отсутствие связей с другими признаками, по-видимому,
обусловлено преформированностью зачатка этого листа и некоторой обо-
собленностью процесса его развертывания. В условиях, отличающихся от
оптимальных, отмечено уменьшение силы взаимосвязей между генератив-
ной и вегетативной сферой, меньшая степень выраженности корреляцион-
ных плеяд, меньшая взаимозависимость признаков листьев срединной
формации. Всё это свидетельствует об уменьшении целостности ростовых
и морфогенетических процессов, большей обособленности процессов фор-
мирования соцветия, розеточной, и удлиненной частей монокарпического
побега. В условиях недостаточного увлажнения отмечена большая взаимо-
зависимость признаков 1-го листа и сильные связи с генеративной сферой.
Последнее свидетельствует, по-видимому, об особой роли 1-го розеточно-
го листа в процессе формирования цветоноса в условиях дисбаланса вод-
ного режима.
Фитоценология
G. imbricatus произрастает на сырых и болотистых лугах, лесных поля-
нах и опушках, в разреженных березняках, среди кустарников (Голубев,
1956а, 1965, 1996; Рысин, Рысина, 1987; Красная книга ..., 1998). Указан
для союза Molinion (Frank, Klotz, 1990). В долинах рек может доминиро-
вать на злаково-разнотравных лугах (Губанов и др., 1990), В степной зоне,
на территории Центрального Черноземного заповедника, нередко встреча-.
ется у подножия северных склонов логов, реже растет по дну и на север-
ных склонах логов, на лесных полянах (Голубев, 1962). Отмечен в горах на
субальпийских лугах и у ледников на высоте до 1500 м. Изредка встречает-
ся как сорное по пустырям, залежам, на полях (Флора СССР, 1935; Luther,
1961).
В условиях Тверской области мы наблюдали шпажник черепитчатый
в Западнодвинском, Торопецком, Жарковском, Андреапольском районах
на пойменных и суходольных лугах разной степени увлажнения. Реже он
встречается на низинных болотах, развивающихся в поймах рек и по бе-
регам озер, на крутых береговых склонах и на опушках смешанного леса.
44
Наиболее часто G. imbripatus произрастает в разнотравно-злаковых,
злаково-разнотравных, разнотравно-осоково-злаковых ассоциациях. Не-
редко встречается в ассоциациях с Potentilla erecta (мелкозлаково-калга-
новая, калганово-злаковая). Шпажник черепитчатый отмечен также в
ассоциациях с Calamagrostis canescens (молиниево-вейниковая, таволго-
во-вейниковая, калганово-вейниковая, разнотравно-вейниковая и серова-
то-вейниковая), в белоусово-калгановой и таволгово-осоково-злаковой
ассоциациях.
Наиболее часто вместе с G. imbricatus произрастают Agrostis tenuis,
Festuca rubra, Anthoxanthum odoratum, Deschampsia cespitosa, Briza media,
Phleum pratens, Betonica officinalis, Potentilla erecta, Ranunculus acris, виды
^oac.Alchemilla, Hypericum maculatum, Centaurea jacea, Achillea millifolium,
Gentiana pneumonantha, Geranium palustre, Vicia cracca, Pimpinella saxi-
fraga, Luzula multiflora, Filipendula ulmaria, Molinia coerulea.
Изучение характера расположения особей в ценопопуляциях G. imbri-
catus, выполненное по стандартным методикам, показало случайное рас-
пределение особей, отсутствие ярковыраженных обособленных скопле-
ний. В некоторых экотопах, вне экологического оптимума G. imbricatus,
отмечены небольшие компактные группы особей, которые образуются в
результате вегетативного размножения. Сходная пространственная струк-
тура ценопопуляций отмечена у некоторых других клубнелуковичных рас-
тений (Черемушкина, Попова, 1976).
В травостое G. imbricatus может сохраняться до 100 и более лет (Губа-
нов и др., 1990). По данным D. Frank и S. Klotz (1990) шпажник черепитча-
тый является стресс-рудералом, урбанофобом. По нашим наблюдениям
G. imbricatus обладает невысокой конкурентной способностью. В этой свя-
зи полное прекращение сенокошения на лугах, приводящее к их зараста-
нию, может обусловить исчезновение вида из травостоя. Такая динамика
отмечена в окрестностях деревень Плавёнки, Чичаты Жарковского района
и Бибирево Западнодвинского района Тверской области. В 80-х гг. XX в. в
окрестностях указанных населённых пунктов численность популяций
шпажника черепитчатого достигала 200 и более особей. Полное прекраще-
ние сенокошения привело к значительному увеличению роли видов с вы-
сокой конкурентной способностью, зарастанию лугов ивами. В 1997 г. от- .
мечено исчезновение шпажника. Интенсивное сенокошение также приво-
дит к ослаблению популяции. В окрестностях деревни Чичаты
установлено выпадение шпажника на лугах в результате интенсивного се-
нокошения. Большое значение имеют также сроки сенокошения. Раннее
сенокошение препятствует нормальному семенному возобновлению вида.
Отмечено исчезновение шпажника в некоторых местонахождениях в бли-
жайшем Подмосковье (Красная книга .... 1998). Лимитирующие факто-
ры — уничтожение и нарушение местообитаний, распашка лугов, выпас
скота, сбор растений на букеты.
45
Биологическая продуктивность
На основе анализа морфометрических показателей в популяциях
шпажника черепитчатого в Тверской области предложена шкала жизнен-
ности для средневозрастных генеративных особей (табл. 1). Общие разме-
ры монокарпических побегов G. imbricatus варьируют от 27,5 до 133,7 см.
Таблица 1
Шкала жизненности средневозрастных
генеративных особей Gladiolus imbricatus
Баллы Высота, см Число цветков Число коробочек Длина коробочки, см
I 27,5-49,0 2-3 1-2 0,9-1,2
II 49,1-70,0 4-6 3-4 1,3-1,5
III 70,1-92,1 7-9 5-6 1,6-1,8
IV 92,2-13'3,7 10-11 7-8 1,9-2,1
Данные о биологической продуктивности получены нами в Андреа-
польском районе Тверской области. Материал собран 7 августа 1998 г. на
склоне берега озера Ореховское между деревнями Орехово и Лутки в раз-
нотравно-злаковой ассоциации. Исследовано 15 экземпляров средневозра-
стных генеративных растений, жизненное состояние которых соответство-
вало Ш-му баллу. Измерена фитомасса надземной части и клубнелуковиц
в сухом состоянии. Средняя фитомасса сухих клубнелуковиц вместе с при-
даточными корнями и покровами составила 0,68±0,4 г, варьируя в интерва-
ле от 0,35 до 1,6 г. Средняя фитомасса надземной части— 1,5±0,52 г варь-
ируя от 9 до 3,2 г. Учитывая, что особи в популяции были размещены со
средней плотностью 4 экземпляра на 1 м2, средняя фитОмасса надземных
частей в изученном местообитании составляет 5,7 г на 1 М2.
Исследования генеративной сферы растений данной выборки показа-
ло, что число коробочек на цветоносе варьирует от 4 до 13, в среднем обра-
зуется 7-8 коробочек. В коробочке может формироваться до 56 полноцен-
ных семян. Средняя семенная продуктивность — 20,3±15,0 семян. Уста-
новлена тенденция к недоразвитию коробочек и уменьшению средней
реальной семенной продуктивности в коробочках верхней части цветоноса
(табл. 2). Общая реальная продуктивность особи варьировала от 45 до 342,
в среднем она составила 149,2±59,9 семян.
46-
Семенная продуктивность Gladiolus imbricatus
(Тверская область)
Табдица2
Порядковый номер коробочки Средняя реальная семенная продуктивность Образования коробочек, %
полноценные недоразвитые и полноценные
1 32,5±14,5 93,3 100
2 29,7±8,1 100 100
3 27,9±12,3 93,3 100
4 25,9±12,2 93,3 100
5 15,3±12,0 60,0 93,3
6 9,1±11,4 33,4 86,7
- 7 '8,4±12,4 20,0 66,7
8 6,0±9,0 13,3 53.3
9 7,0±7,0 13,3 26,7
10 7,0±7,0 6,7 13,3
И 0 0 13,3
12 0 0 6.7
13 0 0 6,7
Консортивные связи
G. imbricatus опыляется пчёлами (Кернер, 1903; Нейштадт, 1954). В ос-
новании трубки околоцветника.на внутренней стороне имеется железистая
ткань, вырабатывающая нектар. Столбик и рыльца плотно прижаты к кры-
шеобразной части цветка, образованной верхними листочками околоцвет-
ника. Процессу опыления способствует перемещение тычинок и столбика.
47
В момент проникновения пчелы внутрь цветка благодаря движению тычи-
ночных нитей опускаются пыльники, и пыльца высыпается на спинку на-
секомого. Лучшему восприятию пыльцы способствуют многочисленные
сосочки, покрывающие край рыльца. Эффективное опыление осуществля-
ется благодаря перемещению столбика (Кернер, 1903). У шпажника чере-
питчатого отмечено изменение окраски от розового до пурпурно-лилового
цвета на разных стадиях развития цветка (Нейштадт, 1954). Однако этот
процесс, по-видимому, не играет той роли в опылении, которую он выпол-
няет у некоторых африканских гладиолусов (Vogel, 1954; Ingram, 1967;
Goldblatt et al., 1998).
В природе отмечены повреждения листьев, коробочек и соцветий лис-
тогрызущими насекомыми. По-видимому, некоторые из них образуются в
результате деятельности обыкновенной уховёртки (Forficula auricula-
ria L.). В Андреапольском районе Тверской области найдены клубнелуко-
вицы с ходами, сделанными личинками жуков-щелкунов, и погрызами мы-
шевидных грызунов.
В культуре, на территории Ботанического сада ТвГУ, мы наблюдали
некоторые повреждения и заболевания, отмеченные в Литературе для
G. hybridus (Вредители и болезни..., 1987). Однако они встречаются гораз-
до реже, чем у последнего вида. Отмечены листья и соцветия, поврежден-
ные тлями, слизнями, трипсами, гусеницами. Коркообразные бурые пятна
появляются на листьях, цветках и луковицах в результате деятельности
гладиолусовых трипсов (Taeniothrips simplex Moris). Отмечены клубнелу-
ковицы, повреждённые личинками жуков-щелкунов (Agriotes obscurus L.,
A. lineatus L., A. gurgistanus Fald., Athous niger L.). Личинки могут повреж-
дать также проростки и всходы. Отмечены фузариозы, вызываемые
Fusarium pxysporum Schl. f. gladioli (Mass.) Sn. et Hans, и склеротинозы
(возбудитель Sclerotinia gladioli (Mass.) Dray). Иногда в результате пора-
жения Botrytis gladiolorum Timm, развивается коричневая сердцевинная
гниль, при которой на листьях, соцветиях и стеблях образуются округлые
бурые пятна, покрытые серым пушистым налётом.
Хозяйственное значение и охрана вида
В средневековье высушенные и размолотые клубнелуковицы гладио-
луса добавляли к муке и использовали в хлебопечении. В XVI-XVIII вв. из
клубнелуковиц G. imbricatus готовили лекарственное средство от зубной
боли (Geelhaar, 1984; Grunert, 1989). Считали, что амулеты с сухими че-
шуями, которые покрывают снаружи клубнелуковицу, защищают от уши-
бов и ран (Кернер, 1903). Клубнелуковицы солдаты использовали как та-
лисман (Geelhaar, 1984; Grunert, 1989).
В.Н. Голубев (1996) характеризует G. imbricatus как декоративное, ле-
карственное и пищевое растение. Имеются сведения о съедобности клуб-
48
нелуковиц гладиолусов и о содержании в листьях витамина С (Гроссгейм,
1952). Отмечено значительное сходство состава белковых комплексов се-
мян евразиатских гладиолусов. Основой является солерастворимая фрак-
ция белков (39,3-43,0 %). Характерно преобладание глобулинов над аль-
буминами. Выявлено значительное, по сравнению с другими ирисовыми,
содержание спирторастворимых белков (12,9-18,1 %) (Соколова, Алек-
сандрова, 1990).
С XVI в. евразиатские гладиолусы выращивают в Европе в качестве де-
коративных растений. G. imbricatus впервые отмечен в культуре в 1604 г. К
80-м гг. XX в. он был интродуцирован в Кировске, Санкт-Петербурге, Тар-
ту, Риге, Брянске, Минске, Москве, Ставрополе и Ереване (Тамберг, 1977).
По данным, полученным в конце 90-х it. G. imbricatus выращивали в бота-
нических садах Ялты, Донецка, Киева, Санкт-Петербурга, Москвы, Мин-
ска, Баку, Риги и Сыктывкара (Каталог..., 1997). Культивируется в Болга-
рии (Протич, Евстатиева, 1994).
G. imbricatus обладает рядом особенностей, позволяющих рассматри-
вать его как перспективное декоративное растение (Наумцев, 1999а,
19966). Высокая морозостойкость исключает необходимость зимнего хра-
нения клубнелуковиц в помещении и укрытия растений на зиму. G. imbri-
catus можно культивировать в течение нескольких лет без пересадки. Он
устойчив к затенению, легкому и среднему задернению, достаточно силь-
ному увлажнению, нетребователен к почвам, в меньшей степени, чем
G. hybridus поражается болезнями и вредителями. В Ботаническом саду
ТвГУ отмечена достаточно высокая газоустойчивость G. imbricatus. в усло-
виях города. Шпажник черепитчатый можно использовать в ландшафтных
композициях с низкорослыми деревьями и кустарниками, на многолетник
цветниках, горках и рокариях, в посадках около водоемов, в парковых по-
садках — на опушках и полянах (Наумцев, 19966, 1999а).
G. imbricatus декоративен, поэтому его нередко собирают для букетов.
Однако в силу своих биоморфологических и фитоценотических особенно-
стей шпажник достаточно уязвим в природных ценозах. В связи с сокраще-
нием численности внесен в Красные книги Московской области (1998), Та-
тарстана (1995) и рекомендован к охране в других областях центра и севе-
ро-запада России (Редкие.и исчезающие виды ..., 1983; Сорокин, 1984;
Потов, 1998, 2000; Цвелев, 2000). Целесообразны специальные наблюде-
ния за состоянием популяций и контроль динамики их численности. •.
Работа выполнена при финансовой поддержке РФФИ
(грант № 98-04-48469)
М. Г. Вахрамеева, Т. В. Жирнова
НЕОТТИАНТЕ КЛОБУЧКОВАЯ
Номенклатура и систематическое положение
Neottianthe cucullata (L.) Schlechter, 1919, Feddes Repert. 16:292; Невский,
1935, Фл. СССР, 4 : 645; Скворцов, 1966, Опред. раст. Моск, обл.: 126;
Смольянинова, 19.76,'Фл. европ. части СССР, 2 : 33; Webb, 1980, Fl. Europ.
5 : 331; Губанов и др., 1995, Опред. сосуд, раст. Центра Европ. России :
201.— Orchis cucullata L., 1753, Sp. Pl.: 939.— Gymnadenia cucullata (L.)
L.C.M.Richard, 1818, Mm. Mus. Hist. Nat. (Paris), 4 : 57; Кауфман, 1866,
Моск, фл.: ; Сырешциков, 1906, Илл. фл. Моск, губ., 1 : 258; Голицын,
1964, в Маевский, Фл., изд.9 : 683.— Неотгианта клобучковая [семейст-
во Orchidaceae — Орхидные (Ятрышниковые)].
Вид филогенетически древний (Wisniewski, 1976). Мало изменчив, из-
вестны лишь формы с белыми цветками, а также с тремя хорошо развиты-
ми зелеными листьями. Сведений о гибридизации с другими видами нет.
Географическое распространение
Евразиатский вид, ареал которого охватывает Среднюю и Восточную
Ейропу и значительную часть Азии. В Европе неоттианте встречается в
Польше, Австрии, странах Балтии, на Украине, в Белоруссии, Европей-
ской части России, в Азии в Малой Азии, Северном Казахстане, Монго-
лии, Северном Китае, на полуострове Корея и Японии (Флора СССР, 1935;
Ziegenspeck, 1936; Wisniewski, 1969,1976; Флора европ. части СССР, 1976;
Fuller, 1978; Flora Europaea, 1980).
В России ареал неоттианте тянется полосой через значительную часть
лесной зоны. Северная граница проходит по 56-58° с. ш. (Красная книга
РСФСР, 1988) через Калининградскую, Ярославскую, Ивановскую облас-
ти, республики Мари-Эл и Удмуртскую, затем Кировскую, Пермскую,
Екатеринбургскую, Тюменскую, Омскую, Новосибирскую области, юг
Томской области, Красноярский край, Иркутскую область, юг Якутии и
Хабаровского края, далее граница проходит через Южный Сахалин и Юж-
50
ные Курилы. Южная граница проходит по Белоруссии, Воронежской, Са-
ратовской, Оренбургской областям, Башкортостану и Челябинской облас-
ти (Флора СССР, 1935; Флора Сибири, 1987; Красная книга РСФСР, 1988;
Сосудистые растения Дальнего Востока, 1996).
В Московской области изредка встречается' почти по всей террито-
рии— Сергиево-Посадский, Мытищинский, Красногорский, Одинцов-
ский, Люберецкий, Орехово-Зуевский, Воскресенский, Раменский, Серпу-
ховской, Луховицкий районы, на севере — особенно редок (Красная книга
Московской области, 1998).
Морфологическое описание
Многолетнее травянистое растение с шаровидным или почти шаровид-
ным клубнем, покрытым густым войлоком из буроватых волосков, цель-
ным или слабовыемчатым, 0,5-1,2 см в длину и 0,7-1,8 см в ширину. Одно-
временно присутствуют два клубня — старый, слегка сморщенный, буро-
ватый, функционирующий в текущем году, и молодой, более светлый и
плотный, с почкой возобновления побега будущего года. Как и другие
клубневые орхидные этот вид можно считать вегетативным однолетником
(в понимании Е.Л. Любарского, 1961,1994). По И.В. Татаренко (1996), не-
оттианте клобучковая:— вегетативный однолетник со сферическим стеб-
лекорневым тубероидом на коротком столоне.
Корни немногочисленные короткие и толстые, до 0,3 см в диаметре,
0,6-1,9 см длиной, расположены близко к поверхности почвы. Стебель тон-
кий, ребристый, голый, в диаметре 1-3 мм, 6,5-28 (35) см высотой, в основа-
нии с двумя сближенными хорошо развитыми срединными листьями. Ниж-
ний лист более крупный, эллиптический — 2,2-8,1 см длиной и 0,9-4,8 см ,
шириной, верхний более узкий, ланцетный, заостренный, длиной 2,0-7,5 см '
и 0,7-3,4 см шириной. Иногда бывает развит третий лист — до 3,2-4,7 см
длиной и 1,2-1,5 см шириной. Выше по стеблю расположены 1-3 (иногда их j
нет) мелкие узколанцетные длиннозаостренные влагалищные листья.
Соцветие — довольно рыхлая однобокая кисть до 8 см длиной, из
6-24 фиолетово-розовых цветков. Изредка встречается форма с бледно-ро- ?
зовыми или чисто белыми цветками. Прицветники линейно-ланцетные,
длинно заостренные равные или немного длиннее завязи. Цветок зиго-
морфный с простым околоцветником из 6 листочков, расположенных в два ;
круга. Листочки наружного круга ланцетные, длинно заостренные (6-8 мм ‘
длиной и 4-5 мм шириной) слипаются вместе с двумя листочками внутрен-
него круга и образуют неширокий шлем. Третий листочек внутреннего
круга —губа до 11 мм длиной, глубоко трехраздельная, направленная
вниз, густо покрыта мелкими сосочками; шпорец с нектаром почти равен
завязи, слегка согнутый, на конце шаровидно расширенный до 7 мм дли-
ной и 2 мм шириной. Пыльник продолговато-яйцевидный, к основанию су-
51
женный. Поллинии булавовидные с двумя полукруглыми железками;
рыльце вогнутое; завязь нижняя, почти сидячая скрученная. Плод —коро-
бочка 0,6—1,4 см длиной и до 0,34 см шириной (Флора СССР, 1935; Fuller,
1978; Флора Сибири, 1987; Сосуд, раст. Дальнего Востока, 1996).
Число хромосом: 2n=42 (Tanaka, 1965).
Онтогенез
Онтогенез вида, как и особенности его биологии, изучен очень мало
(Ziegenspeck, 1936; Fuller, 1978). Известно, что семена его прорастают
только в присутствии грибов. Первые годы растение развивается подзем-
но. Длительность подземного развития неизвестна. Наши наблюдения на
территории Московской области дают лишь самое приблизительное пред-
ставление о длительности большого жизненного цикла и отдельных ста-
дий онтогенеза (что отчасти связано с тем, что этот вид склонен к внезапно
наступающему и длительному вторичному покою). По предварительным
данным, общая продолжительность жизни особи (без учета постсеменного
подземного развития и вторичного покоя) от первого надземного появле-
ния побегов и до полного отмирания особи составляет от 12 до 20 лет, из
них ювенильное состояние длится 2-3 года, имматурное 2-3 (и более),
взрослое виргинильное 1 (редко 2-3) год, генеративное (включая временно
не цветущее состояние) 5-10(11) лет.
Таблица 1
Морфометрические характеристики возрастных (онтогенетических)
состояний Neottianthe cucullata в Московской области и Башкортостане
Показатель Возрастное состояние
j im V g
1 2 3 4 5 ;
Московская область
Число листьев 1 1—(2) (1)2(3) 2(3)
Длина листа, см * 2,20±0,18 3,50±0,41 4,50±0,20 5,30±0,40
Ширина листа, см 0,18±0,01 0,40±0,03 1,30±0,10 2,90±0,80
Число жилок** 2(0) 4-6 8-10 10-12(14)
Число корней (0)1-2 2(1) 2-3(4) 3(4-5)
52
1 2 3 4 5
Длина клубня, см 0,35±0,02 0,60±0,04 0,95±0,02 1,30*0,07
Ширина клубня, см 0,40±0,05 0,70*0,03 1,30±0,05 1,60*0,11
Длина соцветия, см — — — 6,3*0,43
Число цветков — — — 13,5*2,7
Башкирский заповедник
Число листьев 1(2)' 1-2. (1)2(3) (1)2(3) |
Длина листа, см 1,85±0,13 2,88±0,15 4,00*0,10 4,88*0,14
Ширина листа, см 0,25±0,02 0,51±0,02 1,46*0,04 2,76*0,08
Число жилок 0-2 4—6 8-10 12-14
Длина клубня, см 0,15-0,4 0,2-0,5 0,4-0,9 0,5-1,0
Ширина клубня, см 0,15-0,45 0,2-0,6 0,4-1,5 0,7-1,7
Число корней 0-1 (0)1-3 2-4(5) 2-4(5)
Длина соцветия — — — 5,83*0,40 1
Число цветков • — -• — —• 11,6*0,5
Примечание-. * нижний лист (срединной формации);
** число жилок, не считая средней.
Сравнение размеров одной и той же возрастной группы (табл. 1) в раз-
ных частях ареала не выявляет значительных различий. В Московской об-
ласти несколько крупнее листья, особенно у генеративных растений, что,
возможно, связано с более благоприятными здесь условиями увлажнения.
Следует особо подчеркнуть, что выделение возрастных групп у этого ви-
да нередко встречает значительные трудности в связи с очень высокой ва-
риабельностью признаков, особенно это касается числа и размеров листьев,
иногда бывает сложно подсчитать число жилок. Подобные затруднения с*1
определением возрастного состояния отмечают и у других видов орхидных, '
например, у Calypso bulbosa (Виноградова, 1998) и Malaxis monophyllos.
Легче определить возрастное состояние растений во второй половине лета,
53 '
зпл
Рис. 1, Возрастные состояния Neottianthe cucullata.
Обозначения: вл — влагалищный лист; вп — верхушечная почка; зпл — зеленые
плоды; мк — молодые корни; мкл — молодой клубень; ск — старые корни; .
скл старый клубень; сл — срединный лист; чл — чешуевидный лист
54
когда полностью развиты листья и жилки. Иногда (но не всегда) отклонения
величин признаков от средних можно объяснить процессом перехода особи
(в данный момент) в следующее возрастное состояние.
Остановимся на некоторых морфологических особенностях возрас-
тных состояний Neottianthe cucullata (табл. 1, рис. 1). Ювенильная особь
имеет один зеленый лист длиной до 2,5 см, шириной до 0,2 см с двумя жил-
ками (иногда без жилок в первый год появления над почвой), придаточный
корень 1 (редко 2) или их нет, клубень небольшой — его длина около
0,4 см, ширина такая же или немного больше (0,4-0,5 см). Имматурное рас- '
тение несколько крупнее, как правило, тоже с одним листом (очень редко с'
двумя) до 4 см длиной и 0,5 см шириной с 4—6 жилками, чаще с двумя при-
даточными корнями, клубень до 0,7 см в ширину и примерно столько же в
длину. Взрослое виргинильное растение имеет обычно 2 (очень редко 1-3)
листа, нижний длиной до 4,7 см, шириной до 1,4 см, с 8-10 жилками, кор-
ней 2-3 (редко 4), клубень около 1 см длиной и 1,3 см шириной. Генератив-
ное растение с двумя (реже тремя) листьями до 5,7 см длиной и до 3 см ши-
риной, с 10-12 (иногда более) жилками, клубень до 1,3 см в длину и 1,7 см
в ширину, длина соцветия до 7 см, число цветков в среднем около 14.
Ритм сезонного развития
В. Н. Голубев (1996) относит неоттианте к растениям с весенне-летней
вегетацией и позднелетне-раннеосенним ритмом цветения. В Московской;
области почка появляется над поверхностью почвы обычно в середине
мая, листья отросшего побега развертываются полностью к началу июня.
Зацветает неоттианте позже других наших орхидей •— раннее начало цве-
тения 9-10 июля, среднее 20-22 июля, позднее 5-10 августа. Таким обра-
зом, начало цветения в зависимости от погоды может сдвигаться почти на
месяц. Цветение одной особи длится около трех недель или больше. Плот
доносит неоттианте с середины августа, семена , начинают высыпаться в
конце августа — начале сентября. Данные по сезонному развитию'
Neottianthe cucullata в Башкирском заповеднике в течение 10 лет
(1990-1999), полученные Т.В. Жирновой, отражены в табл. 2. > . ?
Способы размножения и распространения
Неоттианте клобучковая размножается преимущественно семенным
путем. Случай явного вегетативного.размножения нам пришлось.наблюл
дать лишь однажды. В благоприятных условиях в составе популяции мо-
жет быть до 40 % генеративных особей. Плоды завязываются интенсивно,
доля вызревших плодов составляет 80—100 % (Ziegenspeck, 1836;
Wisniewsky, 1976; Fuller, 1978). В Башкирии отмечена четкая зависимость;
интенсивности цветения и плодоношения от погодных условий (Жирнова,
55:
Таблица 2
Сезонное развитие Neottianthe cucullata
в сосняке разнотравно-вейниково-зеленомошном
в Башкирском заповеднике в 1990-1999 гг.
Фенофаза Дата наступления фенофаз
интервал М±ш
Начало вегетации* 10.05-6.06 28.05±3.04
Отрастание листьев** 22.05-9.06 3.06±2.77
Появление цветочной стрелки 6.06-16.06 13.06±1.23
Бутонизация 19.06-28.06 23.06±1.11
Начало цветения 19.07-27.07 23.07±1.09
Полное цветение 22.07-10.08 30.07±2.64
| Конец цветения 12,08-21.08 16.08±1.66
Примечание'.. * появление почек на поверхности почвы;
** листья не расправлены, свернуты в трубочку.
1999). В разные годы в одном и том же фитоценозе (сосняк разнотрав-
но-вейниково-зеленомошный) среднее число цветков в соцветии колеба-
лось от 5,8±0,3 до 11;6±0,5, число плодов с нормально вызревшими семе-
нами от 5,8±0,2 до 10,3±0,5. Плоды завязывают от 40 до 100 % цветков..
Плодоношение может быть ослаблено из-за повреждения цветоносов жи-
вотными-фитофагами: ежегодно ими скусывается 6-40 % генеративных
особей. В 1996 г. в Башкирском заповеднике в отдельных фитоценозах до
69 % генеративных особей было повреждено градом, а на уцелевших цве-
тоносах плодообразование в среднем было 57,3 %.
В Московской области доля генеративных особей составила 9-28 % от
численности наблюдаемой популяции; среднее число цветков от 4 до 26,
плодообразование от 40 до 100 %. Большинство генеративных особей цве-
тет не каждый год. В Польше отмечали повторное цветение у 20 %.генера-
тивных особей (Wisniewski, 1976). Примерно то же мы наблюдали и в Мос-
ковской области, лишь одна особь цвела 10 лет подряд, при этом число
цветков в соцветии сократилось с 11 до 4 (в последний год цветения), а
56
другие особи цвели в лучшем случае 2-3 года подряд. Судя по значитель-
ному для орхидных присутствию в популяциях ювенильных особей (в от-
дельные годы до 20 %), семенное возобновление осуществляется у неотти-
анте достаточно успешно. В отдельные годы (например, 1972) в Москов-
ской области мы наблюдали «волны возобновления» — вспышки
численности ювенильных растений;
Экология
Вид с довольно узкой экологической амплитудой (Wisniewski, 1976,;
Вахрамеева и др., 1994). Предпочитает участки со слабым затенением
(10 % или немного более от полной освещенности), но изредка встречает?
ся как на открытых, так и сильно затененных местах. Мезофит, обычно
растет в условиях умеренного увлажнения, на почвах песчаных и супес-
чаных с хорошей аэрацией, от бедных до среднебогатых, чаще на слабо
кислых или слабощелочных. В Польше отмечен (Sarosiek et al., 1991) на
песчанистых бедных азотом почвах с pH от 3,9 до 5,2 и показана низкая,
потребность этого вида в Са и Р. В условиях Московской области вид был
встречен в сходных условиях на подзолистой супесчаной почве с pH от
4,6 до 5,0. >
Фитоценология
Вид лесной, чаще всего растет в сосняках с хорошо развитым покровом,
из зеленых мхов, но может встречаться и в смешанных хвойно-листвен-
ных, а также широколиственных лесах (дубовых, буковых), изредка выхо-,
дит на опушки (Флора СССР, 1935; Sundermann, 1975; Флора европ. части
СССР, 1976; Wisniewski, 1976; Fuller, 1978; Sarosiek et al., 1991), иногда его,,
отмечают нагорных лугах (Комаров, 1935; Flora Europaea, 1980).
В хвойных лесах неоттианте, как правило, сопровождают, помимо зе-
леных мхов (Climacium dendroides,: Dicranum scoparium, D. polysetum,
Hylocomium splendens, Pleurozium schreberi и др.), плауны (Lycopodium,
clavatum, L. annotinum) и бореальные цветковые растения (Chimaphila
umbellata, Goodyera repens, Linnaea borealis, Orthilia secunda, Vaccinium
myrtillus, V. vitis-idaea и др.) (Ziegenspeck, 1936; Sundermann, 1975;
Wisniewski, 1976; Fuller, 1978; Sarosiek et al., 1991).
В Польше неоттианте отмечают не только в сосновых или смешанных,
лиственно-сосновых (Sarosiek et al., 1991), но также и в термофильных ду-
бовых лесах (в Беловежье) с Dracocephalum ruyschiana, Gladiolus imbrica-
tus, Prunella grandiflora, Trollius europaeus, а также в елово-сосновых, ело-
вых и буковых лесах (Wisniewski, 1976),
F. Fuller (1978) полагает, что Neottianthe cucullata пришла в, сосняки из
лиственных лесов смешанного состава. По мнению ряда исследователей,
57
неоттианте на северной границе своего ареала — реликт прежних клима-
тических эпох, а на западе его граница открыта и в этом направлении воз-
можно расширение ареала (Filipek, 1956; Wisniewski, 1999). Неоттианте
часто появляется в сосняках вторичного происхождения, В. Шафер (1956)
в таких местообитаниях относит вид к псевдореликтам.
На территории России неоттианте растет чаще всего в светлых сосно-
во-зеленомошных или елово-зеленомошных лесах (но с заметной приме-
сью сосны). В Башкирском государственном заповеднике распространена
в сосновых и смешанных (лиственнично- и березово-сосновых) преимуще-
ственно мшистых лесах — чернично-вейниково-зеленомошных, костя-
нично-вейниково-зеленомошных, иногда в сложных сосняках — липо-
во-орляково-вейниковых, а также в разнотравно-коротконожковых спе-
лых сосняках.
В Московской области вид нами встречен главным образом в мшистых
сосняках-зеленомошниках, или в сосново-еловых зеленомошных лесах (на
месте сосняков) с разреженным травяным покровом (общее проективное
покрытие травяного покрова до 30 %), реже в сложных сосняках с приме-
сью дуба и липы.
Иногда неоттианте образует очень большие скопления до 1200-2000
особей (Filipek, 1956; Sarosiek et al., 1991), подобные скопления приходи-
лось наблюдать и нам, но в Московской области таких крупных популяций
практически не осталось. Максимальная плотность популяций может быть
очень высокой — до 110-120( 150) особей на м2, средняя плотность — око-
ло 10-12 особей на м2.
Для большинства изученных ценопопуляций неоттианте клобучковой
характерен (как и для большинства других наших орхидных) правосторон-
ний возрастной спектр, с относительно высоким участием ювенильных
растений, что свидетельствует о нормальном самоподдержании популя-
ций (табл. 3).
Характерной чертой изученный популяций, как в Подмосковье, так и в
Башкортостане (Жирнова, 1994, 1999) являются резкие колебания числен-
ности и плотности популяций (табл. 4). Причиной этих колебаний могут
являться погодные условия, антропогенные факторы, но нередко — ка-
кие-то пока нам неясные эндогенные факторы.
Т.В Жирнова отмечает, что наибольшая численность в сосновых лесах
Башкирского заповедника наблюдалась во влажные и теплые годы (1993 и
1994 гг.). Резкое падение численности (с 1997 г.) связано с жаркой и очень
сухой погодой весной и летом 1995 г. и аномально бесснежным периодом в
начале зимы 1996-97 гт. и 97-98 гг. при сильных заморозках на поверхно-
сти почвы (до -34—36°).
Интересное явление было отмечено нами на территории Звенигород-
ской биостанции МГУ (Одинцовский р-н Московской области), где рядом
с единственным генеративным экземпляром, за которым мы 2 года вели
58
Таблица 3
Возрастная структура и средняя плотность
ценопопуляций Neottianthe cucullata в различных условиях
местообитания в Московской области
Фитоценоз Возрастное состояние, % Плотность, особей на м2
j im у* g
Сосняк с липой разнотравный 14 15 . 43 28 12,7(52)**
Сосняк разнотравно- злаковый 16 24 34 26 15,1 (17)
Ельник с сосной зеленомошный 18 41 32 9 7,4(17)
Ельник костянично- зеленомошный 14 21 50 15 4,6 (1.1)
Примечание'. * в состав группы v включены и временно
не цветущие генеративные особи,
** в скобках дана максимальная плотность.
наблюдения, внезапно появилось сразу 83 разновозрастные особи. По-
скольку в этой группе присутствовали взрослые вегетативные и генератив-
ные особи, то следует предположить, что растения находились в течение
нескольких лет в состоянии вторичного покоя (существуя за счет симбиоза
с грибами). На следующий год численность этой группы сократилась бо-
лее чем в 2 раза, а через 5 лет исчезли все растения, кроме того единствен-
ного генеративного экземпляра, с которого й были начаты наблюдения.
Никаких резких изменений условий местообитания за период наблюдения
не происходило. Подобные резкие колебания численности этого вида от-
мечали и в других регионах (Wisniewski, 1976).
В конкурентном отношении Neottianthe cucullata — слабый вид, пред-
почитает места с разреженным травяным покровом. Относится к растени-
ям-бриофилам (в понимании Мазуренко и Хохрякова, 1989). При наруше-
нии мохового покрова (в который погружены клубни и корни неоттианте)
вид обычно исчезает.
59
Таблица 4
Динамика численности н возрастной структуры
ценопопуляцни Neottianthe cucullata в сосняке разнотравно-
вейннково-зеленомошном в Башкирском заповеднике
Год Всего, особей Максимальная плотность, особей на м2* Возрастное состояние, %**
j im V g
1990 297 103 1,7 38,7 47,1 12,5
1991 233 55 3,4 33,1 41,2 22,3
1992 396 98 19,4 37,9 31,8 10,9
1993 499 130 14,6 42,7 23,5 19,2
1994 462 142 7,2 36,8 46,3 9,7
1995 372 100 1,6 37,9 50,0 10,5
1997 198 62 2,5 31,8 37,4 ' 28,3
1998 182 62 2,4 22,7 42,3 32,6
1999 192 74 12,0 23,4 31,8 32,8
2000 250 78 11,2 31,2 23,2 34,4
Примечание-. * в местах наибольшего скопления особей;
** при выделении возрастных состояний особи
с переходными признаками относили к меньшей
по возрасту группе.
Консортивные связи и охрана вида
Неоттианте клобучовая в течение всей своей жизни тесно связана с ми-
коризными грибами (Ziegenspeck, 1936; Fuller, 1978). Прорастание семян
возможно только с помощью грибов, длительный вторичный покой возмо-
жен также только за счет микотрофии. Гифы гриба содержатся не только в
придаточных корнях, но и во внешнем слое клеток клубня (Ziegenspeck,
1936). П.В. Куликов и Е.Г. Филиппов (1999) отмечают в природных усло-
60
виях высокую специфичность симбиоза неоттианте клобучковой с грибом
(Rhizoctonia goody erae-repentis).
Цветки неоттианте содержат много нектара, что привлекает опылите-
лей, в частности, Apiden (Ziegenspeck,1936; Wiesnewski, 1976). Подробно
процесс опыления у этого вида не изучен.
Неоттианте клобучковая мало устойчива к антропогенному воздейст-
вию, что вызвало сокращение ее численности во многих частях ареала. В
ряде регионов и государств вид внесен в списки редких и охраняемых рас-
тений (Wisniewski, 1976; Балявичене, 1980; Талла, 1980; Игнатенко,
Амельченко, 1983), в том числе в Красные книги Белоруссии (1981), Рос-
сии (1988), Украины (1988, 1996).
В Московской области неоттианте клобучковая прежде была довольно
широко распространена (Кауфман, 1899; Сырейщиков, 1906).В настоящее
время многие из ближайших к Москве местонахождений исчезли или во-
шли в черту города (Кунцево, Останкино, Перово, Сокольники, Трои-
це-Лыково, Усово, Хорошево, и др.), в связи с чем вид внесен в Красную
книгу Московской области (1998). На территории области неоттианте кло-.
бучковая охраняется в Приокско-Террасном заповеднике, Национальном
парке «Лосиный остров» и трех заказниках.
В культуре неустойчива, быстро выпадает. Наш опыт по репатриации
этого вида в естественные для него природные условия окончился неуда-
чей — в лучшем случае растения прожили три-четыре года.
Работа выполнена при финансовой поддержке РФФИ
(грант № 99-49-0474).
М. Г Вахрамеева, И. В. Блинова,
Т. И. Богомолова, Т. В. Жирнова
ПОЛОЛЕПЕСТНИК ЗЕЛЕНЫЙ
Номенклатура, систематическое положение
и внутривидовая систематика
Coeloglossum viride (L.) C.Hartm. 1820, Handb. Scand. FI.: 329; Кауфман,
18(56, Моск, фл.: 472; Сырейщиков, 1906, Илл. фл. Моск, губ., 1: 260; Нев-
ский, 1935, Фл. СССР, 4 :647; Голицын, 1964, в Маевский, Фл., изд. 9 : 682;
Скворцов, 1966, Опред. раст. Моск, обл.: 124; Смольянинова, 1976, Фл. ев-,
роп. части СССР, 2 : 32; Moore, 1980, Fl. Europ. 5 : 333; Губанов и др., 1995,,
Опред. сосуд, раст. Центра Европ. России : 201.— Satyrium viride L., 1753,
Sp. PL: 944.— Пололепестник зеленый [семейство Orchidaceae — Орхид-
ные (Ятрышниковые)].
Согласно новейшим биохимическим исследованиям (Bateman et al.,
1998) род Coeloglossum предложено объединить с родом Dactylorhiza. В
последней сводке по орхидным (Genera Orchidacearum, 2001, v. 2, р. 1)
Coeloglossum viride упоминается как Dactylorhiza virides.
Вид полиморфный, ряд исследователей (Schultze, 1894; Hegi, 1936;
Peitz, 1962 и др.) выделяют три разновидности: var. bracteata (с длинными
прицветниками); var. islandicum (мелкие растения, не превышающие 10 см
в высоту, с 2 зелеными листьями); var. vaillantii (с золотисто-желтой губой
и красными или зеленоватыми зубчиками околоцветника). Другие иссле-
дователи считают их подвидами и выделяют, кроме выше указанных, еще
subsp. viride (сюда обычно включают var. vaillantii), а также subsp.
kaschmirum. Для Северной Америки указывают var. virescens (Nilsson,
1979). В. Г. Собко (1989), описывая орхидные Украины, выделяет две раз-
новидности — var. viride и var. vaillantii. В пределах указанных разновид-
ностей и подвидов выделяют около 10 форм, различающихся по величине
цветков, рассеченности губы, и длине прицветников (Fuller, 1972;
Prochazka, Velisek, 1983; Собко, 1989).
В Московской области помимо обычной формы указан var. bracteata
Led. с прицветниками, в 2-2,5 раза превышающими длину цветков (Маев-
ский, 1964; Аверьянов, 2000).
62
Вид нередко образует межродовые гибриды с видами рода Dactylorhiza
(D.fuchsii, D. incarnata, D. maculata, D. majalis.D. sambucina, D. purpurella,
D. umbrosa), а также видами родов Gymnadenia, Nigritella, Orchis,
Platanthera (Schulze, 1894; Hegi, 1936; Peitz, 1962; Sod, Borsos, 1966; Fuller,
1972; Schmid et al., 1974; Sundermann, 1975; Wennenberg, 1978; Lang, 1980;
Dressier, 1981; Prochazka, Vehsek, 1983).
Географическое распространение
Евразиатско-североамериканский, преимущественно голарктический
вид. Распространен практически по всей Европе — от Исландии, Британ-
ских островов и Скандинавии до Центральной Испании, Италии, Греции,;
Южной Болгарии и Крыма; на значительной части Азии — в Малой и Цен-
тральной Азии, Монголии, Тибете, Китае, Японии; в Северной Америке
распространен от Западной части Аляски до Ньюфаундленда (Флора
СССР, 1935;. Ziegenspeck, 1936; Summerhayes, 1951; Clapham et al., 1957; ;
Luer, 1975; Флора европ. части СССР, 1976; Flora Europ., 1980; Губанов,
1996). В северных районах это преимущественно равнинный вид, в юж-
ных— горный. t
В России пололепестник растет в основном в тундровой и лесной зо- d
нах. В европейской части России этот вид встречается от Кольского п-ова
по всей Средней полосе (редок в черноземных областях) и до Краснодар-
ского и Ставропольского краев, где растет в горах. В азиатской части Рос-
сии распространен на территории всей Сибири (от Ямала до Тувы и Дау-
рии) и Дальнего Востока (от Чукотки до южного Сахалина и Курильских
островов) (Флора СССР, 1935; Орлова, 1954; Аркт. Флора, 1963; Флора ев-
роп. части СССР, 1976; Flora Europaea, 1980; Алексеева, 1983; Флора Си-
бири, 1987;Опред. сосуд.раст...., 1995;Губанов, 1996; Аверьянов,2000).
В Московской области вид распространен широко, но обычно встреча-?,
ется рассеянно и в небольшом числе — в Клинском, Дмитровском, Серги-
ев-Посадском, Мытищинском, Можайском, Серпуховском районах; не от- .•
мечен в Заокском и восточных районах области (Красная книга Моск, обл.,
1998). Многие известные ранее местонахождения исчезли.
Морфологическое описание
Многолетнее травянистое растение с продолговатым пальчато-рассе-
ченным клубнем. По И.Г. Серебрякову (1962,1964), пололепестник следу- .
ет относить к классу наземных травянистых поликарпических растений с
ассимилирующими побегами несуккулентного типа, подклассу клубнеоб-
разующих травянистых многолетников, группе корнеклубневых много-
летников. Е.С, Смирнова (1990) относит такие растения к форме клубне-
вых растений с удлиненным побегом. Обычно у растения имеются одно-
63
временно два клубня: один, функционирующий в текущем году, несколько
сморщенный, с надземным ортотропным побегом, и другой — молодой,
плотный, с почкой возобновления (из которой разовьется побег в следую-
щем году). Морфологическую природу такого подземного утолщенного
образования трактуют по-разному. Обычно указывается на его корневое
происхождение. Однако, в ряде исследований (Sharman,1939; Ogura, 1953;
Kumazava, 1956, 1958; Тихонова, 1972,1983) было установлено, что такой
клубень является сложным образованием, включающим элементы как по-
бегового, так и корневого происхождения. Р. Дресслер (Dressier, 1981)
предложил для обозначения этого органа термин «стеблекорневой туберо-
ид», который используется в ряде современных работ (Аверьянов, 1991;
Татаренко, 1996; Блинова, 1996), но пока не стал общепринятым. Поэтому,
мы считаем возможным использовать для названия этого органа прежний
широко распространенный термин «клубень». И.В. Татаренко (1996) опре-.
деляет жизненную форму пололепестника как «вегетативный однолетник
(в понимании Е.Л. Любарского, 1961,1994) с пальчато-раздельным стебле-
корневым тубероидом».
Корневая система — кистекорневая (Голубев, 1984), образованная
придаточными корнями, отходящими от основания побега и закладываю-
щимися еще в период его внутрипочечного развития.
Стебель 6-35(40) см высотой, одетый при основании низовыми буры-
ми чешуевидными влагалищными листьями. Листья срединной форма-
ции —с хорошо развитой простой, цельнокрайней, ланцетной пластин-
кой, с четко выраженным влагалищем, обычно в числе 3-5, нижние с кры-
латым черешком, эллиптически-яйцевидные, эллиптически—-ланцетные
или реже ланцетные, 2,5-8(11) см длиной и 1-5 см шириной.
Соцветие 2-10(15) см длиной, колосовидное, негустое, из (3)5-25(31)
сидячих цветков. Прицветники линейно-ланцетные или ланцетные, тупо-
вато-заостренные, у типичной формы нижние длиннее цветков, до
2,5-3(4) см длиной; верхние короче цветков. Цветки зигоморфные, жел-
товато- или буровато-зеленые, со слабым медовым запахом. Околоцвет-
ник простой, венчиковидный, из 6 листочков, расположенных в двух
трехчленных кругах. Все листочки, кроме губы (среднего листочка внут-
реннего круга) обращены кверху и собраны в шлем, зеленоватые, иногда
слегка окрашенные в красновато- иди пурпурно-коричневый цвет. Лис-
точки наружного круга околоцветника яйцевидно-туповатые, 3-6(9) мм
длиной, 2,5-3,5(5) мм шириной; средний вогнутый, боковые — неравно-
бокие. Средний листочек наружного круга сближен с внутренними, боко-
вые наружные несколько расходящиеся. Два боковых листочка внутрен-
него круга линейные, около 1 мм шириной, туповатые, почти такой же
длины, как наружные. Губа со шпорцем, вниз обращенная, обратно кли-
новидная, 5-8(12) мм длиной, близ конца 3-3,5(5) мм шириной, плоская;.'
окрашена в желтоватый или желтовато-коричневый цвет, неглубо-
64
ко-трехлопастная; боковые лопасти параллельны и, обычно намного
длиннее, чем средняя. Шпорец очень короткий, около 2 мм длиной и 1 мм
шириной, мешковидный, тупой, зеленовато-беловатый. Андроцей реду-
цирован до одной фертильной тычинки, сросшейся нитью со столбиком в
короткую колонку (гиностемий), две другие тычинки преобразованы в
стаминодии и расположены по бокам колонки в виде бугорков. Пыльник
прикреплен спинкой, двугнездный, обратно яйцевидный, гнезда внизу
немного расходящиеся. Пыльцевые зерна склеены в булавовидные пол-
линии; каждый из двух поллиниев снабжен ножкой, с помощью которой
он прикрепляется к крупной клейкой округло-яйцевидный голой железке
(прилипальцу). Поллиний, ножка и прилипальце вместе составляют пол-
линарий. Гинецей образован тремя плодолистиками. Рылец 3, два из ко-
торых срастаются, образуя вогнутый диск, а третье — стерильно и преоб-
разовано в клювик, отделяющий пыльник от рыльца и служащий местом
прикрепления поллинариев. Клювик маленький, треугольный, слегка вы-
дающийся между основаниями гнезд пыльника. Завязь, нижняя, сидячая,,
скрученная, одногнездная, с тремя постенными семяносцами и много?,
численными семяпочками. Плод — коробочка, раскрывающаяся, при со-
зревании шестью продольными щелями. Семена очень мелкие, без эндос-
перма, веретеновидные, с прозрачной сетчатой оболочкой и шаровидным,
недифференцированным зародышем (Флора СССР, 1935; Burgeff, 193,6;
Ziegenspeck, 1936; Флора европ. части СССР, 1976; Флора Сибири, 1987;
Аверьянов, 2000).
Указывают различные наборы хромосом для этого вида: 2п=40
(Clapham et al., 1957; Флора Сибири, 1987; Reinhard et al.,. 1991;), 2n=40,42,
80 (Хромосомные числа.., 1969; Sundermann, 1975; Флора европ. части,
1976; Lid, 1994), 2n=40, 41, 42 (Аверьянов и др., 1982).
Онтогенез
Онтогенез вида изучен до сих пор недостаточно, и сведения, приводи-
мые в литературе, довольно противоречивы (Ziegenspeck, 1936; Блинова,
1996; Willems et al., 1998). Мелкие семена с недоразвитым зародышем
прорастают только при участии микоризообразующих грибов. Продол-
жительность периода прорастания семени в природных условиях неиз-,
вестна. В лабораторных условиях (Smereciu, Currah, 1989) семена пололе-
пестника в зависимости от состава среды и вида, внесенного в нее гриба,
прорастают в течение 2-27 недель. Установлено, что грибы Rhizoctonia
anaticola, Ceratobasidium obscurum, Ceratobasidium cereale стимулируют,
развитие протокормов. Однако, Rhizoctonia solani, обнаруженная в кор-
нях этого вида в природе, в лабораторных условиях подавляет прораста-
ние семян (Harvais, Hadley, 1967). Выяснено, что оптимальная температу-
ра для прорастания семян составляет 20-25°, хотя процесс прорастания
65
может происходить и при температуре 15-20°, Освещение подавляет
прорастание семян (Stoutrnire, 1974; Arditti et al., 1982). Отмечена воз-
можность прорастания семян и в асимбиотической среде (Linden, 1980).
При добавлении в питательную среду сахаров из протокормов могут раз-
виваться нормальные растения (Smerciu, Currah, 1989). При прорастании
семени (как'и у других видов орхидных) образуется так называемый
«прбтокорм» — округлое биполярное бесхлорофилльное тело, имеющее
в базальной части всасывающие волоски, содержащие гифы микоризооб-
разующего гриба. В апикальной части протокорма закладывается конус
нарастания будущего надземного побега, а при дальнейшем подземном
его развитии начинает формироваться побеговая часть и придаточный'
корень. Размеры протокорма могут значительно варьировать, даже в од-
ном и том же местообитании. До сих пор не существует общепринятого
термина для названия следующей за протокормом фазы развития орхид-
ных. Во многих немецких и англоязычных работах следующую за прото-
кормом фазу развития называют «микоризомом». Микоризом представ-
ляет собой стадию формирования первичного подземного побега, суще-
ствующего, главным образом, за счет микосимбиотрофии (Баталов,
1998). В отечественной литературе этот термин мало употребим. Вероят-
йо, следует согласиться с точкой зрения А.В. Баталова (1998) и понимать
под проростком орхидных совокупность протокорма (стадии послесе-
мейного развития спброфита по Т.Б. Батыгиной и В.Е. Васильевой, 1983)
и микоризома (стадии первичного подземного побега, существующего за
счет микосимбиотрофии).
И.В.Блинова (1998) предлагает различать «протокорм» (как образова-
ние одного ростового цикла в течение одного вегетационного периода)
и «проросток», который может быть и подземным, и надземным и состоит
из 2-3-сегментов, соответствующих разным вегетационным периодам
(рис. 1 А). Такой «подземный» проросток, очевидно, соответствует поня-
тию микоризом. Если протокорм расположен близко к поверхности почвы,
то он на следующий год может дать надземный побег, состоящий из не-
многих метамеров и, как правило, с единственным зеленым листом сре-
динной формации. В течение вегетационного сезона образует первый при-
даточный корень и первый клубень. На этой стадии, по мнению И.В. Бли-
новой (1998), растение переходит в ювенильное состояние.
Выделение «надземных» и «подземных» проростков орхидных не яв-
ляется общепринятым, что отчасти объясняется трудностью обнаружения
подземно существующих растений, а также опасностью нарушить или
уничтожить ценопопуляцию и без того редкого вида растения. В связи с
этим, вероятно, целесообразно объединить протокорм и «подземный про-
росток» (микоризом), оставляя за ними название протокорм, а «надзем-
ные» проростки отнести к ювенильным растениям. Особенно это оправда-
но при полевых популяционных исследованиях.
66
Рис. 1. Ранние стадии онтогенеза Coeloglossum viride. ’
А — подземные «проростки»; Б — ювенильные растения разных этапов развития
В условиях Средней Европы (Ziegenspeck, 1936; Summerhayes, 1951)
подземное развитие пололепестника длится 2-3 года, в конце второго года
образуется первый придаточный корень, на 4-й год формируются два че-
шуевидных листа и первый зеленый лист, а в конце этого сезона и малень-
кий клубень — т. е. растение переходит в ювенильное состояние. По мне-
нию Й. Виллемса с соавторами (Willems et al., 1998), период подземного
развития довольно длителен, поскольку они наблюдали отдельные расте-
ния пололепестника, зацветающие уже при их первом появлении над зем-
лей. Авторы полагают, что растение к этому времени благодаря симбио-
трофии накопили значительную биомассу, что позволило им «пропустить»
ювенильную, имматурную и взрослую виргинильную стадии. До сих пор.
такого ускоренного прохождения начальных стадий онтогенеза у других
наземных бореальных орхидных не отмечено. И.В. Блинова (1998), изучая
пололепестник зеленый в Мурманской области, высказывала совершенно
иную точку зрения. Она полагает (на основе косвенных наблюдений), что
развитие протокормов до ювенильной фазы может происходить в течение
месяца.
Остановимся на краткой характеристике отдельных возрастных (онто-
генетических) состояний пололепестника зеленого (табл. 1).
67
Таблица 1
Морфометрическая характеристика возрастных состояний
Coeloglossum viride в Вологодской области и Башкортостане
Показатель Возрастное состояние
j im V g
Вологодская область
Число листьев 1,0±0,1 2,0±0,3 . 2,0±0,4 3,2±0,6
Длина листа, см* 1,40±0,30 4,40±0,29 6,70±1,20 6,20±0,40
Ширина листа, см* 0,30±0,02 0,5±0,06 1,10±0,20 1,21±0,12
Число жилок** 2-4 6-8 10-12 12-16(18)
Число лопастей клубня 1 2 2,0±0,3 2,7±0,4
Число придаточных корней 1 2±0,1 2,6±0,4 3,4±0,8
Длина соцветия, см — — — 4,2±0,5
Число цветков — —* — • 11,0±2,7
Башкортостан
Число листьев 1,2±0,05 1,5±0,06 2,0±0,06 3,2±0,1
Длина листа, см 3,77±0,11 4,61±0,14 4,64±0,32 4,71±0,16
. Ширина листа, см 0,62±0,02 1,08±0,03 1,43±0,08 1,73±О,О6 '
Число жилок 2-4 6-8 , 10-12(14). 10—14(16)
Длина цветоноса, см — . — '— 18,3±0,7
Длина соцветия, см — — — 5,5±0,3
Число цветков — — — 12,4*0,7
Примечание-. * нижний лист срединной формации;
** средняя жилка не учитывается.
68
Рис. 2. Возрастные состояния Coeloglossum viride.
Ювенильное растение (рис. 1 А, Б; рис. 2) обычно имеет 1 (очень редко
2) зеленый лист срединной формации с 2-4 жилками, 1 придаточный ко-
рень, 1 веретеновидно-утолщенный клубень без лопастей. В имматурном
состоянии (рис. 2) обычно два листа, несколько увеличивается длина и ши-
рина листа, число жилок 6-8, два придаточных корня, клубень с 2-3 лопа-
стями. Взрослое виргинильное растение (рис. 2) имеет 2-3 (редко 4—5) зе-
леных листа с 10-12(14) жилками, клубень с 2-3 лопастями, придаточных
корней 2-3. Генеративное растение (рис.2) с 3-5 зелеными листьями, жи-
лок 12-16(18), клубень с 2-3(4) лопастями, придаточных корней 3-4 и со-
цветие из 2-28 цветков. Сенильное состояние в природе нам не пришлось
наблюдать.
Т.В. Жирнова, наблюдая за онтогенезом конкретных особей пололепе-
стника в Башкирском заповеднике, выявила случаи перехода растений че-
69
рез группу возрастного состояния (ювенильные — во взрослые вирги-
нильные, имматурные — в генеративные).
Длительность пребывания пололепестника в отдельных возрастных со-
стояниях в природных условиях до сих пор практически не наблюдали. Из-
вестно лишь, что в генеративном состоянии иногда одна и та же особь мо-
жет находиться до 8 лет (Dickson, Parks, 1994; Willems et al., 1998). Следо-
вательно, одна особь может прожить от момента прорастания семени
около 10-12 (возможно, и более) лет. На основе изучения многолетней ди-
намики популяций и интенсивности отмирания особей Coeloglossum viride
в Нидерландах исследователи относят этот вид к коротко живущим орхи-
деям (Willems et al., 1998).
Ритм сезонного развития
Пололепестник — летнезеленое растение. В Московской области ве-
гетация начинается в апреле—мае. Цветет с конца мая по конец июня.
Отмирают надземные части — в конце августа — начале сентября. В
Германии на равнине у этого вида отмечают быстрое созревание плодов
и семян (в конце июля-начале августа), тогда как в горных районах цвете-
ние растянуто с мая по август и плодоношение наступает значительно
позже (Ziegenspeck, 1936; Fuller, 1972). В мягком климате Нидерландов
цветение происходит в марте-мае (Willems, Melser, 1998). В Мурманской
области отмечают самое раннее зацветание 24 июня, самое позднее — 24
июля, массовое — 5-18 июля (Филиппова, Вылегжанин, 1980). По на-
блюдениям'И.В. Блиновой, на Кольском полуострове цветение одной
особи длится около месяца, плодоношение происходит в августе, отмира-
ние годичного побега — в сентябре, при этом побег постепенно меняет
свою окраску от зеленой к блекло-зеленой, желтоватой и коричневой.
Т.В. Жирнова, изучая сезонное развитие пололепестника в Башкирском
заповеднике (табл. 2), установила, что вегетация этого вида начинается в
апреле-первой декаде мая, цветение в июне и длится три-четыре недели,
созревание плодов происходит одновременно с пожелтением листьев в
начале августа. Отдельные растения могут опережать в сезонном разви-
тии другие особи одной и той же ценопопуляции на одну, реже — две не-
дели, особенно в начале вегетационного периода. В Крыму пололепест-
ник начинает отрастать зимой, в связи с чем В.Н. Голубев (1984) относит
его к эфемероидам.
В условиях Мурманской области И.В. Блиновой был прослежен морфо-
генез побега, длящийся здесь 4 года. В первый год к середине лета заклады-
вается почка и формируется конус нарастания с листовым примордием. На
2-й год — к концу вегетационного периода сформированы в почке 5-6 мета-
меров с листовыми примордиями и клубень, составляющий 1/2—1/3 высоты
почки. На 3-й год к концу вегетационного периода сформированы клубень,
70
придаточные корни, низовые листья и соцветие, в цветках заложены все ос-
новные органы. На 4-й год происходит внепочечное развитие побега, цвете-
ние, образование плодов и семян и к концу этого сезона происходит отмира-
ние клубня, придаточных корней и всего годичного побега.
Таблица 2
Сезонное развитие Coeloglossum viride
на злаково-разнотравном лугу в Башкирском заповеднике
в 1993-2001 гг.
Фенофазы Дата наступления фенофаз
интервал М±т СТ V
Начало вегетации* 17.04-2.05 25.04±2.04 5.01 8.99
Развертывание листьев и появление цветоноса 27.04-13.05 8.05±2.44 5.99 8.72
Начало цветения 21.05-7.06 1.06±2.60 6.37 6.86
Полное цветение 6.06-21.06 15.06±2.18 5.76 5.37
Конец цветения 12.06-9.07 28.06±3.37 8.90 7.39
Начало созревания семян 25.07-5.08 2.08±1.96 4.39 2.84
Примечание: * появление почек на поверхности почвы.
Способы размножения и распространения
Размножение пололепестника происходит преимущественно семен-
ным путем. Плодообразование колеблется в зависимости от погоды и ус-
ловий местообитания от 0 до 100 %. Погодные условия значительно влия-
ют на интенсивность цветения и плодоношения (Ziegenspeck, 1936;
Dickson, Parks, 1994; Willems et al., 1998). Погодичная изменчивость сред-
него числа цветков в соцветии на одной и той же пробной площади в Ни-
дерландах колебалась от 8±3 в 1994 г., до 13±5 в 1998 г. (Willems et al.,
1998). В холодные годы в Мурманской области плоды не завязываются во-
обще, а в благоприятные годы в разных фитоценозах плодообразование со-
ставляет 57-92 %. В Башкирском заповеднике (Жирнова, 1999) после жар-
кого и засушливого лета отмечено полное отсутствие генеративных расте-
71
ний, а в благоприятные годы они составляют значительную часть
популяции, при этом число цветков в соцветии колеблется от 12,22±0,97 до
16,25±0,83. В Вологодской области среднее число цветков на побег состав-
ляло 12,03±1,3, среднее число плодов 8,04±0,9, среднее плодообразование
66,7±9,2 %.
Число семян в плодах колеблется от 2300 в Нидерландах (Willems,
Melser, 1998) до 300 в Мурманской области. По наблюдениям И.В. Бли-
новой, в Мурманской области в зависимости от условий местообитания и
погоды число семян в плоде колеблется от 314 до 500. Наибольшее число
семян отмечено в плодах из нижней и средней части соцветия. Повторное
цветение одной и той же особи отмечается редко. Так за 9 лет наблюде-
ний в Шотландии (Dickson, Parks, 1994) из 184 генеративных растений
40 цвели 2 раза, а 8 растений — 3 раза. За 9 лет исследований в Башкир-
ском заповеднике Т.В. Жирновой установлено, что из 128 генеративных
растений 35 цвели 2 раза, 7 растений — 3 раза, а 4 и 5 раз цвело лишь по
одной особи.
При изучении пололепестника в Московской, Вологодской областях и
в Северной Осетии не было выявлено случаев вегетативного размножения,
это же отмечают и в Нидерландах (Willems, Melser, 1998). Г. Цигеншпек
(Ziegenspeck, 1936) полагает, что вегетативное размножение у этого вида
возможно. И.В. Блинова, изучая пололепестник в Мурманской области, от-
мечает случаи одновременного развития двух побегов у одной особи и
считает, что на севере ареала данного вида это не является редкостью.
Экология
Вид с довольно широкой экологической амплитудой (Summerhayes,
1950; Вахрамеева и др., 1994). Растет преимущественно в условиях полно-
го освещения, но может существовать и при незначительном затенении на
лесных полянах и опушках леса (4-я ступень шкалы Ландольта, 8-я сту-
пень шкалы Элленберга), сциогелиофит (Голубев, 1984). Может встре-
чаться в тенистых буковых и еловых лесах на более или менее осветлен-
ных участках после разреживания или частичной вырубки (Ziegenspeck,
1936). Оказываясь в условиях сильного затенения в густом лесу, переходит
к вторичному покою (Ziegenspeck, 1936).
Мезофит, предпочитает почвы промежуточные между сухими и свежи-
ми (3-я ступень шкалы Ландольта, 4-я ступень шкалы Элленберга), избега-
ет как очень сухие, так и заболоченные участки. В.Г. Собко (1989) в рав-
нинных условиях относит его к мезофитам, а в горных — к психромезо-
фитам.
Пололепестник растет на почвах, бедных минеральным азотом (2-я
ступень шкалы Элленберга), различного гранулометрического состава,
предпочитая, однако, мелкопесчаные (4-я ступень шкалы Ландольта). Ча-
72
ще встречается на почвах, богатых известью (Summerhayes, 1951;
Sundermann, 1975; Nilsson, 1979), на севере больше приурочен к таким поч-
вам, на юге — меньше (Ziegenspeck, 1936). Предпочитает слабокислые
почвы (3-я ступень шкалы Ландольта, 4-я ступень шкалы Элленберга) с pH
от 4 до 8 (Sundermann, 1975). В Северной Осетии вид встречен на почвах с
pH 6,1-6,8. Пололепестник часто находят на почвах, верхний горизонт ко-
торых имеет кислую реакцию, а ниже расположенный — реакцию близ-
кую к нейтральной (Ziegenspeck, 1936).
Пололепестник встречается как на равнине, так и (чаще) в горах, под-
нимаясь до 3000 и даже 4000 м над уровнем моря (Eichner, 1979; Nilsson,
1979).
Фитоценология
Обладая обширным ареалом, пололепестник не отличается привязан-
ностью к какому-либо типу фитоценозов. На севере ареала растет в тунд-
рах различных типов (вороничной, ерниковой и др.), нередко встречается
на начинающих зарастать песках (Флора Северо-востока, 1976). Южнее
растет в лесах хвойных (сосновых, еловых, лиственничных), мелколист-
венных (березовых, осиновых, ивовых), смешанных хвойно-лиственных,
широколиственных (буковых, липовых, дубовых). В горах Южного Урала
отмечен в остепненных лиственнично-сосново-можжевеловых редколесь-
ях, в горных луговых степях и на пограничных участках между лесом и
степью, а также на суходольных лугах (Жирнова, 1999). Нередко его мож-
но встретить на пойменных и материковых (суходольных и низинных), а
также горных лугах. На высокогорных лугах Кавказа, например, в Север-
ной Осетии поднимается до их верхней границы, где может встречаться в
несомкнутых фитоценозах в числе немногих видов (Betula litwinowii, Salix
caucasica, Dryas caucasica и др.). Здесь, практически по границе со Сказ-
ским ледником, все найденные особи пололепестника были хорошо разви-
ты, и большинство из них цвело.
В Московской области встречается в еловых, осиновых, березовых, се-
роольховых лесах, по сыроватым лугам, среди кустарников, на опушках и
лесных полянах (Ворошилов и др., 1966; Красная книга Московской облас-
ти, 1998).
Пололепестник зеленый — диагностический вид порядка Nardetalia
Prsg 1949 класса Nardo-Callunetea Prsg 1949 (Rothmaler, 1972; Matusz-
kiewicz, 1981). Б.М. Миркин и Л.Г; Наумова (1998) относят порядок
Nardetalia Oberd. ex Prsg 1949 к классу Calluno-Ulicetea Br.-Bl. et R.Tx. ex
Klika et Hadac 1944. Это луга на бедных кислых почвах равнинных и низко-
горных регионов Европы.
Пололепестник — слабый в конкурентном отношении вид, предпочи-
тает фитоценозы с негустым травяным покровом, часто встречается вдоль
73
тропинок и дорог, на пастбищах, гарях, вырубках (Ziegenspeck, 1936;
Nilsson, 1979).
Пололепестник обычно образует некрупные ценопопуляции (от одно-
го- двух десятков до сотни особей). Возрастной спектр, как правило, имеет
правосторонний характер с максимумом генеративных (реже взрослых ве-
гетативных) особей (табл. 3). Изредка наблюдаются «волны возобновле-
ния» (см. Мурманская область), причем в разреженном травостое чаще.
Таблица 3
Возрастные спектры и средняя плотность
ценопопуляцнй Coeloglossum viride в разных фитоценозах
Фитоценоз Возрастное состояние, % Плотность, особей на м2
j im у**** g
Березняк разнотравно- злаковый* 8 17 36 39 2,1
Ельник черничник* 15 16 27 42 2.7
Овсяницево- разнотравный луг** — 6 14 80 1,2
Сосняк вейниковый*** 5 18 21 56 3,7
Луг разнотравно- злаковый*** — 10 30 60 1,3
Примечание: * Мурманская область,
** Вологодская область,
*** Северная Осетия;
**** группа v включает как взрослые виргинильные,
так и временно не цветущие генеративные растения.
Низкий процент (или полное отсутствие) молодых особей, возможно,
объясняется либо кратковременностью их вегетации (наблюдения прово-
дили, как правило, в июле-августе), либо тем, что многие особи проводят
значительную часть онтогенеза (включая ювенильную и имматурную ста-
дии) под землей (Willems, Melser, 1998). Этот вопрос требует дополнитель-
ного изучения, тем более что В.И. Антонова и А.В. Паланов (1999), отме-
чают значительное присутствие молодых особей (около 50 %) в ценопопу-
ляции пололепестника, изученной ими в Вологодской области. В
Башкирии по данным Т.В. Жирновой (табл.4) «волны возобновления» (мо-
74
лодых особей до 76 % от состава ценопопуляции) наблюдали на лесных
лугах в 1999-2001 гг. после массового размножения (в предшествующие
два года) мышевидных грызунов. Результаты деятельности этих животных
(нарушение дернины, разрыхление почвы, выбрасывание ее на поверх-
ность) оказались благоприятны для появления (чаще на «кротовинах») не
только новых молодых, но также и взрослых виргинильных и генератив-
ных особей.
Таблица 4
Динамика численности и возрастной структуры
ценопопуляции Coeloglossum viride на злаково-разнотравном лугу
в Башкирском заповеднике*
Год Всего, особей Плотность, особей на м2 Возрастное состояние, %**
j im V g
1993 86 0,54 15,1 15,1 18,6 51,2
1994 75. 0,47 6,7 10,7 34,6 48,0
1995 75 0,47 1,3 12,0 45,4 41,3
1996 46 0,29 4,3 8,7 87,0 0
1997 24 0,15 8,3 12,5 62,5 16,7
1998 19 0,12 10,5 5,3 73.7 10,5
1999 176 1,10 40,9 35,2 22,7 1,2
2000 212 1,33 35,4 36,3 10,4 17,9
2001 267 1,67 17,2 29,6 36,7 16,5
Примечание-. * учеты проводили на площади 160 м2 в конце мая-йюне,
до появления густого травостоя;
** при выделении возрастных состояний особи
с переходными признаками относили к меньшей
по возрасту группе.
Погодные условия заметно сказываются на возрастной структуре попу-
ляций, особенно близ северной границы ареала, где низкие температуры
воздуха и почвы препятствуют развитию проростков в почве. При относи-
тельно высоких температурах в середине вегетационного периода, когда
75
закладываются генеративные почки, число генеративных особей в следую-
щем году увеличивается, а ранние осенние заморозки оказывают обратное
действие и приводят к снижению численности цветущих растений.
Изученные нами ценопопуляции пололепестника в разных частях его
ареала свидетельствуют о том, что вид предпочитает участки с разрежен-
ным и невысоким травяным покровом, где он, как правило, образует попу-
ляции с невысокими численностью и плотностью (1-6 особей на м2), с пра-
восторонним возрастным спектром (максимум генеративных и взрослых
вегетативных особей) и небольшим числом ювенильных особей (кроме лет
с «волнами возобновления»).
Консортивные связи
В жизни пололепестника зеленого существенную роль играет микори-
за. Прорастание семян в природных условиях возможно только при содей-
ствии микоризообразующих грибов, хотя в лабораторных условиях он
иногда может прорастать и асимбиотически (Warcup, 1973; Linden, 1980).
И.В. Татаренко (1996) относит этот вид к сильно Микотрофным видам и от-
мечает высокую интенсивность микоризной инфекции (40-90 %). Гифы
гриба содержатся практически во всех корнях. С возрастом, однако, зави-
симость от гриба ослабевает (Ziegenspeck, 1936). Некоторые грибы стиму-
лируют, другие — подавляют развитие протокорма (см. онтогенез).
X. Бургефф (Burgeff, 1936) изолировал из корней пололепестника гриб, на-
званный им Rhizoctonia anomala, впоследствии его видовая принадлеж-
ность была уточнена —Moniliopsis anomala (Currah, Smereciu, 1990). Поз-
же из корней пололепестника были выделены также Leptodontidum
orchidicola, Eulorhiza anaticula, Ceratorhiza goodyerae-repentis (Currah et
al., 1990). Для определения грибов-микосимбионтов орхидных составлен
ключ (Currah, Zelmer, 1992). И.В. Блинова отмечала в Мурманской области
начало инфицирования грибами молодых придаточных корней в конце ав-
густа-сентябре. После зимы микоризная инфекция распространяется по
всей длине корней, осенью микоризный гриб отмирает вместе с придаточ-
ными корнями.
Пололепестник зеленый — энтомофил, опыление чаще осуществляет-
ся насекомыми: мелкими двукрылыми, а также жуками и наездниками, ко-
торых привлекает медовый запах цветков (Ziegenspeck, 1936;
Summerhayes, 1951; Fuller, 1972). К числу возможных опылителей относят
виды родов Cantharis, Acalyptus, Ichneumon (Ziegenspeck, 1936). В Нидер-
ландах отмечали присутствие на цветках ос, мелких жуков, иногда наблю-
дали автогамию (Catling, 1981; Willems, Melser, 1998). О. Hagerup (1952)
отмечал особенности строения цветка пололепестника — вход в шпорец
закрыт тонкой мембраной, через которую может проникнуть насекомое с
сильно развитой мандибулой, но не бабочки и мелкие двукрылые. И.В. Та-
76
таренко (1996) относит пололепестник к слабоспециализированным энто-
мофилам.
В Башкирии (Жирнова, 1999) отмечено поедание клубней мышевидны-
ми грызунами, а также повреждения листьев листогрызущими насекомы-
ми. Молодые особи нередко погибают под земляными муравейниками.
Хозяйственное значение и охрана
Пололепестник зеленый, хотя и имеет обширный ареал, но числен-
ность его популяций обычно очень низка. Кроме того, в последние десяти-
летия многие местонахождения исчезли. Еще Г. Цигеншпек (Ziegenspeck,
1936) отмечал сокращение числа его местонахождений в Германии под
воздействием хозяйственного освоения территорий. Указывают на его ис-
чезновение и в других европейских странах — Германии (Kiinkele, Vogt,
1973; Hamel, 1981; Grieger, 1988), Шотландии (Dickson, Parks, 1994). На-
пример, в Нидерландах с 1950 исчезло 88 % местонахождений — из 55 ос-
талось 2 (Willems, Melser, 1998). В некоторых странах Европы (Нидерлан-
ды, Швейцария и др.) вид внесен в списки охраняемых растений (Reinhard
et al., 1991; Stewart, 1992).
Вид не внесен в Красную книгу России, поскольку в ряде регионов его
численность еще значительна. В Московской области пололепестник отне-
сен к охраняемым видам, численность которых сокращается — II катего-
рия (Красная книга Московской области обл., 1998). За последние десяти-
летия он исчез из многих известных ранее местонахождений, часть из ко-
торых ныне вошла в территорию Москвы (Воробьевы горы, Раменки,
Косино и др.). Главными лимитирующими факторами в Московской об-
ласти считаются мелиорация и торфоразработки.
Есть сведения о сохранении вида в культуре (Собко, 1989), при этой
его можно размножать ризорёституционным методом. О.И. Князева (1986)
сообщает об интродукции пололепестника зеленого в Ботаническом саду
Екатеринбурга, где вид успешно размножается делением замещающего
клубня. В Ботанических садах Москвы пололепестник пока не интродуци-
рован.
Р. П. Барыкина, Н. В. Чубатова
ЛЮТИК КАШУБСКИЙ
Номенклатура, систематическое положени
и внутривидовая систематика
Ranunculus cassubicus L., 1753, Sp. Pl.: 551; Сырейщиков, 1907, Илл. фл.
Моск, губ., 2: 160; он же, 1914, цит. соч., 4: 80; Овчинников, 1937, Фл.
СССР, 7 : 379; Turin, 1964, Fl. Europ. 1: 232; Маценко, 1964, в Маевский,
Фл., изд. 9: 85; Ворошилов, 1966, Опред. раст. Моск, обл.: 170; Turin а.
Akeroyd, 1993, Fl. Europ., ed. 2,1: 280; Губанов и др., 1995, Опред. сосуд,
раст. Центра Европ. России: 266; Цвелев, 2001, Фл. Вост. Европы, 10:
150.— R. auricomus L. В. cassubicus (L.) Kauftnan, 1866, Моск, фл,: 29.—
Лютик кашубский (семейство Ranunculaceae — Лютиковые).
Сложность изучения лютика кашубского связана с расплывчатостью
некоторых'его диагностических признаков. Во Флоре СССР (1937) вид ха-
рактеризуют как крайне варьирующий по степени надрезанности прикор-
невых и стеблевых листьев, в результате чего бывает трудно провести гра-
ницы между ним и близкими к нему видами; кроме того, отмечают сущест-
вование ряда переходных (неясного происхождения) форм к R. auricomus
L., R. allemanii Вг.-В1. и R. megacarpus W. Koch.
M.A. Розанова (1932) и W. Koch (1933) независимо друг от друга от-
крыли у лютика кашубского псевдогамию, что вскоре было подтверждено
цитологическими работами (Hafliger, 1953; Rutishauser, 1954). По их мне-
нию, вид представлен рядом стабильных апомиктических биотипов, не-
редко трактуемых в качестве самостоятельных таксонов (видов или подви-
дов). М.А. Розанова (1932) выделяет в пределах R.cassubicus 5 подвидов, а
L. Fagerstrom (1967) добавляет еще 4. Хромосомные наборы биотипов од-
нородны (Marklund, Rousi,1961), причем преобладают тетраплоиды. Л.Е.
Курлович (1982а) также различает несколько биотипов лютика кашубско-
го. Согласно автору, морфометрические признаки их варьируют в широ-
ких пределах. Однако, по нашему мнению, выделенные биотипы по сути
представляют собой морфотипы двух типов: лютика кашубского и лютика
золотистого.
78
С точки зрения Н.Н. Цвелева (1996), вторичные, главным образом, апо-
миктические виды, возникли в результате сложных гибридизационных
процессов между немногими первичными видами секции Ranunculus, и
собственно Ranunculus cassubicus s. 1. представляет собой сложный агрегат
видов с цельными прикорневыми листьями. Наличие у некоторых особей
прикорневых листьев с более или менее рассеченной пластинкой рассмат-
ривается им как результат аномального развития листа по типу стеблевых
листьев.
Таким образом, таксономический статус вида и внутривидовая систе-
матика все еще остается дискуссионными.
П.Ф. Маевский (1964) и В.Н. Ворошилов с соавторами (1966) лютиком
кашубским называют растения с цельными «прикорневыми» листьями. В
данной работе мы принимаем трактовку вида последних авторов. По наше-
му мнению, в целях диагностики вида необходимо четко различать розе-
точные листья дициклйческбго монокарпического побега и надрезанные
листья, иногда появляющиеся в основании генеративного побега.
Географическое распространение
Лютик кашубский широко распространен в лесной зоне Средней и
Восточной Европы, а также Скандинавии. Имеются сведения (Крылов,
1931; Флора СССР, 1937) о произрастании вида и в Западной Сибири (Ал-
тай). В новейшей сводке (Флора Сибири, 1993) для этой территории указы-
вается его восточный подвид Ranunculus cassubicus ssp. kemerovensis
Kvist., отличающийся тройчато-лопастными прикорневыми листьями
(Kvist, 1987). Это заметно отодвигает восточную границу ареала лютика
кашубского, карта которого в пределах европейской части СССР была
опубликована Л.М. Носовой (1987а). По данным автора, северная граница
ареала, пересекая 60 параллель, на западе немного отклоняется к северу, на
востоке — к югу. Восточная граница распространения лютика кашубско-
го, согласно Л.М. Носовой (1987а), проходит по пунктам: станция Баранча,
Верхотурье, село Я лани, Екатеринбург. С. А. Овеснов (1997) отмечал его и
для южных районов Пергской области.
Пределы распространения лютика кашубского на юг до сих пор не ус-
тановлены с достаточной точностью. С.И. Коржинский (1892), указывал
на нахождение его в северной части бывшей Самарской, Симбирской и
Тамбовской, а также в Харьковской, Полтавской, Киевской и Подольской
губерниях. Существуют старые, нуждающиеся в проверке, указания на на-
хождение его в северной части Ростовской области (Флора Нижнего Дона,
1984).
В Московской области лютик кашубский встречается повсеместно
(Ворошилов и др., 1966).
79
Рис. 1. Возрастные состояния Ranunculus cassubicus.
Условные обозначения: sm — внешний вид семени,р — проросток,/ — ювениль-
ное, im — имматурное, v — взрослое вегетативное, g — молодое генеративное
растения; с — семядоля, гп — гипокотиль, гк — главный корень, прл — прикор-
невой, чл — чешуевидный и ел — стеблевой листья генеративного побега,
рл — первый розеточный лист побега возобновления, орл — отмерший розеточ-
ный лист, кщ — корневище, пк — придаточный корень
80
Морфологическое описание
Лютик кашубский — короткокорневищный кистекорневой многолет-
ний травянистый гемикриптофит (Черняковская, 1986).
Генеративные побеги высотой 30-60 см, ортотропные, удлиненные,
толстые, бороздчатые, в верхней части ветвистые, при основании часто с
волокнами отмерших черешков прошлогодних розеточных листьев, 2-3
перепончатыми чешуевидными листьями, иногда с 1 длинночерешковым
прикорневым зеленым листом с расчлененной В той или иной степени ок-
ругло-почковидной пластинкой, 2-3 сидячими стеблевыми листьями и не-
многочисленными цветками, собранными в цимозное соцветие. Листья
срединной формации пальчаторассеченные, сегменты их, за исключением
самых верхних листьев, к основанию клиновидно сужены, кверху расши-
ренные, наверху и по краям равномерно более или менее остро пильча-
то-зубчатые, снизу более бледные, почти голые, сверху с короткими рассе-
янными волосками (особенно по жилкам). Листья, обычно описываемые
как прикорневые (Флора СССР, 1937), представляют собой предлистья ро-
зеточного бокового побега возобновления, который зацветает лишь в сле-
дующем году.
Пластинки прикорневых и стеблевых листьев заметно отличаются по
анатомической структуре. Согласно Е.Ф. Черняковской (1986), стеблевые
листья по ряду характеристик (толщина пластинки— 194±10 мкм, пали-
садной паренхимы — 57±3 мкм, коэффициент палисадности — 29%,
встречаемость устьиц — 64±7 на 1 мм2 в верхнем и 143±16 на 1 мм2 в ниж-
нем эпидермисе, густота жилок — 452±31 мм на 1 см2) можно отнести к
световым, прикорневые листья, имеющие более низкие значения тех же
показателей (соответственно 178±5 мкм, 50±5 мкм, 27 %, 42±5 на 1 мм2 и
94±11 на 1 мм2, 336±28 мм на 1 см2) — к теневым.
Побег возобновления развивается в пазухе верхнего чешуевидного
листа или катафилла удлиненного материнского цветоносного побега (Се-
ребряков, 1952). Таким образом, монокарпические побеги у лютика ка-
шубского формируются по двулетнему типу. У наиболее крупных экземп-
ляров в терминальной почке розеточного побега одновременно с формиро-
ванием метамеров генеративного побега в пазухе верхней его чешуи
закладывается монокарпический побег следующего порядка, который раз-
вивается, минуя фазу розетки, по типу моноциклического. В этом случае
новыми почками возобновления становятся пазушные почки отмирающе-
го листа розетки, а также катафилла бокового генеративного побега второ-
го порядка.
У ослабленных или стареющих особей монокарпические побеги ино-
гда развиваются как полициклические, тогда укороченный побег возоб-
новления в течение первых двух-трех сезонов развертывает по одному или
по два почковидных листа, лишь затем переходя к цветению.
81
Розеточные листья крупные, часто до 12-16 см шириной, до 9 см дли-
ной, в очертании округло-почковидные или округлые, при основании узко
сердцевидно-выемчатые со сближенными, иногда налегающими краями,
по краю равномерно пильчато-зубчатые; зубцы неравновеликие, широкие,
кверху суженные, на кончике островатые или притупленные; пластинка
прижато рассеянноволосистая, снизу с более длинными волосками; чере-
шок со слабо отстоящими редкими волосками или голый.
Корневая система мочковатая из этажированно расположенных длин-
ных (25-30 см), ветвистых придаточных корней, отходящих от короткого
(до 3-3,5 см), косого, симподиально нарастающего корневища, достигаю-
щего 0,6 см в диаметре.
Соцветие — сложный плейохазий, ветвящийся до побегов III (IV) по-
рядка и включающий от 2 до 11 цветков. Цветоножки без ясных бороздок.
Цветки 2-3 см в диаметре, актиноморфные, гемициклические с двой-
ным околоцветником. Цветоложе сильно выпуклое, густо волосистое. Ча-
щечка из 5 свободных желтовато-буровато-зеленых, более или менее ши-
рокоэллиптических (7-8 мм длиной), слабо опушенных прижатых квенчи-.
ку чашелистиков. Венчик из 5 желтых, с внутренней стороны блестящих,
обратно-яйцевидных, наверху расширенно-округлых, цо краю городчатых
лепестков, в основании с нектарной ямкой, прикрытой чешуйкой.
Тычинки многочисленные, спирально расположенные, с тонкими ни-
тями примерно такой же длины, что и неподвижные, овальнопродолгова-
тые пыльники (2-3,5 мм длиной); во время цветения тычинки превышают
пестичную «головку». Пыльники вскрываются экстрорзно, что связано с
положением нектарников. Пыльцевые зерна продолговато-сфероидальные
или сфероидальные (Р/Е 1,02), 3(4-6)-бороздные, в полярном положении
лопастные, в экваториальном — округлые или широкоэллиптические,
29-36 (в среднем 32,5) мкм в диаметре (Clarke et al., 1991). По уточненным
данным (устное сообщение Е.Э. Северовой, 1999) средний размер поляр-
ной оси (Р) пыльцевого зерна колеблется от 26,88 до 30,78 мкм (в среднем
29,18-30,45 мкм), экваториального диаметра (Е) — от 24,66 до 33,33 мкм
(в среднем 28,82-31,43 мкм). Борозды узкие, длинные, неглубокие. Края
борозд четкие. Поверхность экзины неровная, мелкошиповатая. Шипики
невысокие, многочисленные, разных размеров.
Пестиков много (до 150), они мелкие, в очертании округло-треуголь-
ные, с прямыми, бурыми, наверху слабо крючковатыми стилодиями и опу-
шенными завязями.
Плод — многоорешек. Плодики-орешки имеют овальную форму, не-
сколько сжаты с боков, 4—4,5 мм длиной, с длинным (до 1,5-1,7 мм) крюч-
коватым стилодием, густо опушены короткими одноклеточными отстоя-
щими волосками.
Число хромосом сильно варьирует. Отмечены образцы с 24, 32, 40, 44,
64 хромосомами (Хромосомные числа..., 1969; Числа хромосом .... 1993).
82
Онтогенез
Онтогенез лютика кашубского был подробно изучен Е.Ф. Черняков-
ской (1986). Созревшие орешки опадают с материнского растения в ию-
не-июле. В свежесобранных семенах можно видеть небольшой, состав-
ляющий пятую часть от длины эндосперма, зародыш, морфологически
расчлененный на корешок с корневым чехликом, гипокотиль и 2 семядоли,
что не подтверждает сведения Г.П. Рысиной (1973) о слабой дифференциа-
ции зародыша лютика кашубского в зрелых семенах. Недифференциро-
ванной остается лишь почечка. Осевая часть зародыша (гипокотиль и глав-
ный корень) короче семядолей примерно вдвое. Зародыш обращен кореш-
ком к основанию орешка. В обильном эндосперме содержатся жиры и
запасной белок.
По данным Е.Ф. Черняковской (1986), свежесобранные семена в лабо-
раторных условиях (при 18-22°С) начинают прорастать на 40-50-й день,
при этом всхожесть их не превышает 10—17 %. При высеве семян через
9-12 месяцев после сбора проростки появляются на 30—40-й день, а всхо-
жесть возрастает до 25 %. В опытах Г.П. Рысиной (1967, 1973), свежесоб-
ранные семена лютика кашубского, посеянные в лаборатории в июне, про-
растали лишь в сентябре. Автор связывает это с потребностью семян в не-
котором периоде дозревания.
Предварительная полутора-двухмесячная стратификация замоченных
семян в холодильнике (при 4°С), по данным Е.Ф. Черняковской (1986),
увеличивает всхожесть до 30 % и заметно ускоряет начало прорастания,
что согласуется с результатами, полученными ранее И.Ф. Сациперовой
(1972). Сухие семена после двухлетнего хранения при комнатной темпера-
туре полностью утрачивают всхожесть.
В природе семена при обилии влаги способны начать прорастание, как
весной, так и осенью (Рысина, 1967, 1973), а всходы появляются, главным
образом, в конце апреля-начале мая. Прорастание надземное, гипокоти-.
лярно-котиледонарное. Первым разрастается главный корень, вскрываю-
щий орешек, затем удлиняются (до 1см) гипокотиль и черешки семядолей,
извлекающие пластинки из семенной кожуры и выносящие их над поверх-
ностью почвы (рис. 1).
Главный корень (длиной до 4,5 см) на большем протяжении снабжен
корневыми волосками, слабо ветвится. Семядоли в числе двух (реже од-
ной, трех), длинночерешковые (до 1 см). Их пластинки, развертывающиеся
на 7-11-й день после начала прорастания и достигающие максимальных
размеров на 16-20-й день, округлые (5x5 мм), с небольшой выемкой на
верхушке; жилкование перисто-сетчатое.
В последней декаде мая с появлением первого ассимилирующего листа
растение вступает в ювенильную фазу развития. Первые листья (один или
два), сближенные в прикорневую розетку, почковидные до 0,6-2 см в дйа-
83
метре, по краю городчатые, длинночерешковые (до 4 см), с широким осно-
ванием. Гипокотиль часто полегает, в его базальной части и на семядоль-
ном узле развиваются 2-5 придаточных корней. Главный корень вскоре ос-
танавливается в росте и становится плохо различим среди придаточных
корней, которые развиваются в течение лета и на узлах розеточного побе-
га, проникая в почву на глубину 4-7 см.
Семядоли сохраняются до второй половины июня. В июле-августе ли-
стья розетки отмирают, а стеблевая часть розеточного побега, благодаря
контрактильности корней, втягивается в почву, превращаясь в эпигеоген-
ное корневище. Терминальная почка его зимует в поверхностном слое поч-
вы. Она защищена двумя почечными чешуями и расширенным основани-
ем зачатка ассимилирующего листа. Длительность пребывания в ювениль-
ном возрастном состоянии зависит от условий обитания и варьирует от 2
до 6 лет.
У имматурных растений побег остается розеточным, моноподиально
нарастающим. Ежегодно развертывается 2-3 чешуевидных и 2-3 длинно-
черешковых,(длина черешка до 5 см) листа. Листовые пластинки сохраня-
ют форму ювенильных, но увеличиваются в диаметре до 4—8 см. В течение
этой фазы развития происходит отмирание главного корня и базальной
части гипокотиля. Корневая система гоморизная, мочковатая из моно-
морфных стеблеродных корней, отходящих горизонтально от короткого
вертикального эпигеогенного корневища и ветвящихся близ верхушки до
корней II—Ш-го порядка. Продолжительность имматурного состоя-
ния —1—2 года.
Растения приобретают облик взрослых вегетативных в 4-8 лет. Сохра-
няется рбзеточность побега, 2-5 длинночерешковых листьев приобретают
форму и размеры дефинитивных. Они имеют округло-почковидные (7—12
см в диаметре), Опушенные с обеих сторон редкими одноклеточными жи-
выми волосками пластинки. При основании годичного побега развиваются
2-5 чешуевидных бело-лиловых листа. Подземная часть растения пред-
ставлена коротким вертикальным корневищем, включающим до 8 годич-
ных приростов с 50-65 придаточными корнями;
Растения, выросшие из семян, зацветают на 5-9-м году жизни. Генера-
тивный побег удлиненный, в основании его обычно развертываются ли-
стья низовой формации, но возможно появление 1-2 зеленых листьев пе-
реходного типа с удлиненным основанием и более или менее расчленен-
ной пластинкой (Гуленкова, Красникова, 1986). Листья срединной
формации сидячие, 3-8-пальчаторассеченные на линейно-ланцетные сег-
менты. Моноподиальное возобновление побегов после первого цветения
сменяется симподиальным. Монокарпические побеги последующих по-
рядков дициклические. Корневище «среднегенёративных» особей дости-
гает в длину 1,5-2 см и содержит то же число годичных приростов (до 7-8),
что и у взрослых вегетативных растений, число же придаточных корней
84
увеличивается до 92-115. У более мощных особей может одновременно
раскрыться 2(3) почки возобновления, что приводит к ветвлению корневи-
ща.
В исследованных нами популяциях сенильные особи встречались ред-
ко. Они имели розеточный побег, схожий по размерам листьев с побегом
имматурного растения, и участок полуразрушенного корневища.
На основании результатов изучения онтоморфогенеза лютик кашуб-
ский можно охарактеризовать как короткокорневищный, летнезеленый,
мезофильный гемикриптофит с дициклическими монокарпическими побе-
гами и гоморизной корневой системой из мономорфных стеблеродных
корней.
Сезонное развитие
По своему феноритмотипу (Митина, Антонова, 1974) лютик кашуб-
ский — летневегетирующее растение с ранним началом цветения и вегета-
ции (Г орышина, 1975).
Среди подмосковных лютиков лютик кашубский зацветает одним из
первых — в конце апреля-начале мая, вместе с эфемероидами, но в отли-
чие от них продолжает вегетировать до поздней осени. Эфемероидным
ритмом обладает лишь генеративный побег, т. е. монокарпический побег
на завершающей стадии своего морфогенеза.
В суточном ритме цветения максимальное количество распустившихся
цветков приходится на утренние часы. Однако из-за резких колебаний тем-
ператур и влажности воздуха в мае пик распускания цветков часто сдвига-
ется на послеполуденные часы (Киндерова, 1987). Продолжительность
жизни распустившегося цветка 6-7 дней. Цветки раскрываются однократ-
но. На ночь и во влажную погоду, а также во время дождя листочки около-
цветника полностью не смыкаются. Цветки поникают, тычинки и пестики,
прикрытые чашелистиками и лепестками, оказываются защищенными от
влаги.
Цветение продолжается до конца мая (начала июня). С двадцатых чи-
сел мая начинают завязываться плоды. Зрелые плоды появляются в сере-
дине июля, диссеминация продолжается до конца июля. После опадения
семян генеративный побег подсыхает в июле-сентябре (Серебряков, 1947,
1954; Голубев, 1965; Lukasiewicz, 1967), иногда в октябре (Шик, 1953). Од-
новременно с весенним развитием цветоносного побега начинает развора-
чиваться почка возобновления, дающая розеточный побег с 1, иногда 2-3,
зелеными длинночерешковыми листьями (нередко их ошибочно принима-
ют за прикорневые листья генеративного побега), которые функциониру-
ют до глубокой осени. Уже с апреля формируется его терминальная почка,
которую в литературе часто называют почкой возобновления (Серебряков,
1947); в апреле-мае на конусе нарастания ее появляются зачатки чешуе-
85
Рис. 2. Побег возобновления Ranunculus cassubicus в июне.
Условные обозначения: А — листья и почечные чешуи нормальной верхушечной
почки побега возобновления; Б — побег возобновления с вторичным приростом;
1 и 1’ — нормальные розеточные листья побега возобновления, 2,3, 4, 5 — нор-
мальные почечные чешуи, 2', 3' — розеточные листья и 4', 5' — катафиплы, воз-
никшие в результате реметаморфоза почечных чешуй; 6 и 6'— зачатки стеблевых
листьев
86
видных, в июне-июле — ассимилирующих листьев. Начиная с конца ию-
ля, закладывается соцветие, формирование цветков которого продолжает-
ся до середины апреля следующего года. В пазухе верхней чешуи почки
появляется зачаток следующей почки возобновления. К концу вегетацион-
ного сезона терминальная почка розеточного побега увеличивается в дли-
ну с 7-10 до 25-35 мм и содержит зачатки 6-7 листьев и соцветия (Сереб-
ряков, 1947, 1952).
Таким образом, в верхушечной почке бокового розеточного вегетатив-
ного побега к осени бывает полностью сформирован удлиненный побег
будущего года, а также зачаток следующей почки возобновления как буду-
щего побега Ш-го порядка. Зимует лютик кашубский без зеленых листьев.
В течение зимы длина терминальной почки увеличивается с 3-5 см до 6-8
см (Серебряков, 1947).
Иногда в конце июля, согласно И.Г. Серебрякову (1952), у лютика ка-
шубского после некоторого периода покоя возможно образование второго
элементарного побега (терм.: Грудзинская, 1960), новая розетка ассимили-
рующих листьев которого формируется в результате реметаморфоза по-
чечных чешуй (рис. 2); сам же генеративный побег остается в почке, не
трогаясь в рост.
В годы с обычным увлажнением почвы энергичное корнеобразование,
по наблюдениям О.В. Смирновой (1966), идет в первой половине июня, а
второй его период приходится на конец июля-начало августа; во влажные
годы корнеобразование, хотя и медленно, продолжается в течение всего
лета. Е.К. Горышина (1975) также отмечает два периода корнеобразования
у данного вида — в первой половине июня и в августе.
Согласно же И.Г. Серебрякову (1947), корни формируются лишь с мая
по июнь. По нашим наблюдениям в Наро-Фоминском районе Московской
области, мочки вновь образующихся молодых корней добавляются к уже
функционирующим в середине мая.
В лесопарковой зоне Москвы весеннее пробуждение лютика кашуб-
ского наступает на неделю-две раньше (Карписонова, 1967). Вегетация на-
чинается 15-20 апреля, с 20 апреля становятся заметны бутоны, а 10-15
мая раскрываются первые цветки. Весной 2001 г. цветение началось 20 ап-
реля. Заканчивается цветение в 20-х числах мая. Созревание плодов длится
до 10—15 июня. Диссеминация заканчивается 20-25 июня.
Способы размножения и распространения
Размножается лютик кашубский, главным образом, семенами. По спо-
собу распространения семян он относится к группе растений-баллистов.
Орешки опадают по одному вблизи материнского растения, после чего не-
редко растаскиваются муравьями (Рысина, 1973).
Основная масса семян прорастает в первую же осень и (или) весну по-
87
еле диссеминации, а в последующие годы появляются лишь единичные эк-
земпляры (Рысина, 1973).
Изучение биологических особенностей лютика кашубского, в том чис-
ле и семенной продуктивности, проводилось нами в Подмосковье в разных
типах леса: в Одинцовском районе — в липняке снытево-недотроговом, в
дубраве зеленчуково-волосистоосоковой на склоне верхней надпоймен-
ной террасы р. Москвы и в дубраве зеленчуково-волосистоосоковой на
склоне перерезающего террасу оврага, в Наро-Фоминском районе — в бе-
резо-ельнике зеленомошнике в «окне» и в ельнике зеленомошнике на по-
ляне, в Домодедовском районе — в березо-дубняке зеленчуково-снытевом
(табл. 1).
Таблица 1
Возрастной состав и плотность ценопопуляций
Ranunculus cassubicus в разных фнтоценозах Московской области
Фитоценоз Возрастные состояния, % Плотность, особей на м2
Р1 j im V g s
Липняк 0 7 25 15 42 11 55
Дубрава 0 3 20 34 38 5 60
Дубрава(овраг) 5 10 15 20 50 0 20
Ельник с березой 16 25 13 5 41 0 77
Ельник зеленомошник 20 44 16 7 13 0 258
Семенная продуктивность лютика кашубского в онтогенезе меняется.
Она зависит от мощности соцветия и числа генеративных побегов, кото-
рых бывает обычно по одному, редко по два. Соцветия включают от 2 до
11 цветков, при этом боковые цветки на осях IV-ro, а иногда и Ш-го поряд-
ков семян часто не образуют. В итоге потенциальная семенная продуктив-
ность одного генеративного побега варьирует от 40 до 1000 (табл. 2), а рас-
тение в целом может дать до 2000 семян, однако реальная семенная про-
дуктивность бывает значительно ниже. В неблагоприятные годы (низкая
температура во время цветения, сухость воздуха) она не превышает 50 %, а
в некоторых фитоценозах растения вовсе не дают семян.
88
Так, в Одинцовском районе, возможно из-за очень сухих вегетацион-
ных сезонов 1998 и 1999 годов, мы наблюдали появление всходов исклю-
чительно весной. Кроме того, в эти годы лютик кашубский отличался
очень низкой семенной продуктивностью (табл. 2), следствием чего яви-
лось почти полное отсутствие его всходов в некоторых исследованных по-
пуляциях (табл. 1).
Таблица 2
Семенная продуктивность Ranunculus cassubicus
в различных фитоценозах Московской области (1998 г.)
Фитоценоз Число цветков Число плодолистиков в терминальном цветке Семенная продуктивность
потенциальная 1 побега реальная, %
Липняк 3,0±1,9 67±22,8 275±133,7 20-35
Дубрава 2,2±0,7 69±26,0 165±70,0 1-2
Дубрава(овраг) 4,4±2,3 — — ' 0
Ельник с березой 4,3±1,6 62±11,0 357±129,0 40
Ельник зеленомошник 3,6±2,2 60±10,5 259±141,0 50
Дубрава с березой 8,2±2,4 76±12,4 708±295,0 35
Вес 100 сухих плодиков-орешков — 317 мг.
В пределах сформировавшихся ценотических популяций способность
к вегетативному размножению у лютика кашубского ограничена и осуще-
ствляется в результате бокового ветвления корневища и последующего
обособления укореняющихся парциалей.
Экология
Лютик кашубский — полусветовое лесное растение, обычно произра-
стающее в условиях слабого и умеренного переменного затенеция. Иногда
лютик кашубский выходит на открытые пространства, но лишь при отсут-
89
ствии конкуренции с другими видами (2-я ступень шкалы Ландольта
(Landolt, 1977)), при этом все листья его приобретают световую структуру.
При этом генеративные и вегетативные побеги функционируют в разных
по уровню освещенности условиях. Цветоносные побеги, появляющиеся
ранней весной еще до распускания листьев на деревьях и кустарниках, по-
лучают 50-80 % от освещенности открытых мест. Вегетативная часть мо-
нокарпического побега с развертыванием листвы на деревьях постепенно
оказывается в тени. Теневая фаза с освещенностью от 1-6 до 30 % от пол-
ной освещенности продолжается большую часть вегетационного сезона
вплоть до осеннего листопада. Фитомасса лютика кашубского в ельнике
максимальна при ФАР 2-4 % от полной освещенности (Солнцева, Холопо-
ва, 1987). В сосняке лютик кашубский хорошо себя чувствует при еще бо-
лее низкой освещенности (1 %), а также при освещенности более 10 %. В
лиственных лесах его продуктивность максимальна при ФАР 8-12 %. Та-
ким образом, диапазон толерантности лютика кашубского по отношению
к свету достаточно широк: от 0 до 18 % ФАР, в пределах которого он имеет
два фитоценотических оптимума: 1-4 % и 12-16 %.
Лютик кашубский — мезофит (Рысина, 1973), произрастает на почвах
от влажно- до сыролесолуговых (66-87-я ступени шкалы Раменского, 1956,
4w ступень шкалы Ландольта). Он предпочитает почвы средние по богат-
ству питательными веществами (особенно азотом) (7-12-я ступени шкалы
Раменского, 3-я ступень шкалы Ландольта), слабо кислые (pH 5,5-6,5) или
нейтральные. В Московской области лютик кашубский хорошо развивает-
ся на дерново-подзолистых, слабоподзолистых, бурых лесных почвах с до-
вольно легким механическим составом — супесчаных и суглинистых.
Диапазон толерантности лютика кашубского в отношении влажности поч-
вы от 10 до 32 %, а фитоценотический оптимум соответствует 20-25 %
(Солнцева, Холопова, 1987).
Лютик кашубский растет в лесах на хорошо аэрированных почвах с
мощной подстилкой, защищающей почки от вымерзания. Запас лесной
подстилки, по данным О.Н. Солнцевой и Л.Б. Холоповой (1987) достигает
1600 г/м2, а его фитоценотический оптимум находится в диапазоне от 320
до 640 г/м2.
В нормальных условиях связь продуктивности травянистых растений
с поступающей радиацией в два раза выше, чем связь с эдафическими
факторами, однако в условиях дефицита света или на открытых про-
странствах на первое место выходят такие факторы, как влажность суб-
страта и масса подстилки (Солнцева, Холопова, 1987). Солнечная радиа-
ция и влага — основные факторы, обусловливающие накопление фито-
массы. О.Н. Солнцевой (1987) приводятся результаты изучения
некоторых физиологических процессов у лютика кашубского, в том чис-
ле транспирации и эффективности использования им солнечной радиа-
ции и влаги при фотосинтезе. О.Н. Солнцева отмечает присущие данному
90
виду довольно большие колебания интенсивности транспирации от 100
до 500, а в некоторых типах леса до 600 мг/г/час, при этом сокращение
транспирации происходит только в периоды длительных засух, что с точ-
ки зрения автора, характерно для некоторых ксеромезофитных дубрав-
ных видов. Лютику кашубскому свойствен довольно высокий (380-760)
транспирационный коэффициент, показывающий низкую эффективность
использования воды в процессе роста.
Л.Е. Курлович (1982 б, в, 1984) была предпринята попытка связать
некоторые морфометрические показатели разнообразных форм лютика
кашубского с теми или иными характеристиками местообитания. Ис-
следовались 9 популяций в разных типах леса на территории Москов-
ской и Тульской областей. Была выявлена зависимость высоты стебля,
длины и ширины листовых сегментов первого стеблевого листа, а также
длины и ширины пластинки розеточного листа от освещенности, влаж-
ности и температуры воздуха и почвы в исследованных фитоценозах.
Опыты по культивированию образцов, взятых из природы, показали,
что изменение условий приводит к изменению основных морфометри-
ческих показателей. Таким образом, по мнению Л.Е. Курлович, морфо-
логическое разнообразие лютика кашубского объясняется, прежде все-
го, его широкой экологической амплитудой. Вместе с тем выявленные
морфологические различия, как считает автор, не являются чисто моди-
фикационными, а свидетельствуют о существовании в пределах рода
различных биотипов.
Лютик кашубский лучше других, сопутствующих ему растений, вы-
держивает вытаптывание и уплотнение почвы (Полякова и др., 1983). Он
способен сохраняться в фитоценозах при довольно значительной нагрузке
и высокой степени деградации. На малонарушенных участках широколи-
ственного леса лютик кашубский не обилен, но при увеличении антропо-
генной нагрузки летом и осенью его участие несколько увеличивается. Да-
же на IV и V стадиях дигрессии он остается довольно обилен, лишь макси-
мальные весенние воздействия его полностью угнетают и возобновление
происходит только на 3-й год. В березняках лютик кашубский достигает
максимальной численности на участках с П-й степенью деградации. Одна-
ко, при больших нагрузках, особенно весной, он полностью выпадает из
травостоя.
Фитоценология
Лютик кашубский — неморальный лесной вид, растет в широколист-
венных (дубовых, липовых) лесах, смешанных (хвойно-широколиствен-
ных) и в ельниках на влажных богатых почвах, реже в мелколиственных
лесах. Обычно разрастается в лесу в наиболее освещенных участках: в ок-
нах, на полянах и вырубках.
91
В Центральной Европе лютик кашубский — характерный вид широ-
колиственных лесов порядка Fagetalia silvaticae Pawl, in Pawle. a. 1928
класса Querco-Fagetea и типичный для союзов Carpinion betuli Oberd.
1953 и Alno-padion Knapp 1942 (Sokolowski, 1980), кроме того, он явля-
ется характерным видом ассоциации Tilio-Carpinetum союза Carpinion
(Matuszkiewicz, 1984). По данным К. О. Короткова с соавторами
(Korotkov et al., 1991), лютик кашубский — характерный вид ассоциа-
ции Trollio-Quercetum roburis Korot. этого же порядка, распространен-
ных в северной части Валдайской возвышенности. Согласно Т. Lippmaa
(1938), лютик кашубский — постоянный компонент травяного покрова
богатых по составу смешанных и лиственных лесов Средней Европы и
Скандинавии, которые автор объединил в союз Galeobdolon-Asperula-
Asarum.
На территории России лютик кашубский распространен в восточно-ев-
ропейских южнотаежных, смешанных и широколиственных лесах. В юж-
ной тайге он произрастает преимущественно в еловых лесах с включением
широколиственных пород и неморальных элементов в травяном покрове.
Чаще всего встречается в ельниках сложных: волосистоосоковых, сныте-
вых и зеленчуковых (Рысин, Савельева, 2002). Единичные экземпляры его
встречаются в пихтово-еловых неморально-травяно-кисличных лесах в
бассейнах рек Вятки, Ветлуги и Камы (Растительность европ. части СССР,
1980).
В подзоне смешанных лесов лютик кашубский распространен во влаж-
ных широколиственных, широколиственно-еловых, реже в еловых лесах,
произрастает по склонам речных долин и логам (Конспект флоры Псков-
ской области, 1970).
В северной части Московской области на Клинско-Дмитровской гряде
этот вид обычен в мелколиственных, а также смешанных широколиствен-
но-еловых лесах. Часто растет на вырубках и по берегам ручьев. Изредка
встречается на лугах (Киселева, 1965). На южных склонах Клинско-Дмит-
ровской гряды, на Верейско-Звенигородской равнине лютик кашубский
обилен (copl) в ельниках с липой и кленом лещиново-пролесниковых и
ельниках кленовых кислично-снытевых. Редко встречается в ельниках с
дубом лещиново-кислично-зеленчуковых и кочедыжниково-волосисто-
осоковых, в ельниках с сосной волосистоосоково-кисличных и кислич-
но-папоротниковых, в сосняках с дубом лещиново-пролесниковых (Кур-
наев, 1968; Ильинская и др., 1982; Речан и др., 1993).
К югу от Москвы лютик кашубский обитает в различных типах слож-
ных ельников и широколиственных лесов: липо-ельнике зеленчуково-во-
лосистоосоковом и зеленомошно-волосистоосоковом, дубо-ельнике воло-
систоосоковом, дубо-липняке волосистоосоковом и хвощево-снытевом,
осинниках дубо-лещиново-широкотравном, вейниково-таволговом, а так-
же в теперь уже редких вязово-липово-дубовых лесах, характерных для
92
ландшафтов с близким залеганием известняков (Бязров и др., 1971). В юж-
ной части Московской области (зона широколиственных лесов) лютик ка-
шубский — обычный компонент травяного яруса дубрав й липняков зе-
ленчуковых, медуницевых, а также смешанных широколиственных лесов
с участием ясеня, клена, осины, реже сосны с типичным неморальным тра-
востоем, где он достигает высокой степени обилия (сор 1-2) (Курнаев,
1980; Леса южного Подмосковья, 1985)^ Южнее, в дубравах лесостепи, лю-
тик кашубский встречается, как правило, по склонам лесных оврагов и
единичными экземплярами в лесу на плато (Левицкий, 1957; Курнаев,
1968).
Из искусственных лесонасаждений лютик кашубский йредпочитаёт ле-
щиново-снытевые дубравы на плоскохолмистой равнине северного Под-
московья, где встречаемость его достигает 98 % (Речан и др., 1993) и лещи-
ново-лютиковые сосняки, приуроченные, как правило, к пологим склонам
местных водоразделов южного Подмосковья (Носова, 19876).
В лесах Ленинградской области В.Н. Федорчук и с соавторами (2002)
относит лютик кашубский к индикаторному виду группы Pulmonaria'
obscura вместе с Lathyrus vernus, Siellaria holostea, St. nemorum, Actaea
spicata, Asarum europaeum, Galium odoratum, Viola mirabilis, Myosotis
sylvatica, Fraxinus excelsior (Федорчук и др., 2002). В северном Подмоско-
вье наиболее частыми спутниками лютика кашубского выступают
Galeobdolon luteum, Pulmonaria obscura, Ajuga reptans, Convatlaria majalis,
Maianthemum bifolium, Mercurialis perennis, Aegopodium podagraria, Carex ,
pilosa, Oxalis acetosella, Stellaria holostea, Gymnocarpium dryopteris,'
Pteridium aquilinum, Caltha palustris, Geum rivdle, G urbahum, в южном
Подмосковье — зоне смешанных и широколиственных лесов — кроме на- ',
званных неморальных и неморально-бореальных элементов Anemone
ranunculoides, A. nemorosa, Carex sylvatica, Viola mirabilis, Festuca altissi-
ma, Galium odoratum, Campanula trachelium, C. latifolium, из таежных видов
остаются Maianthemum bifolium, Gymnocarpium dryopteris, Phegopteris
connectilis (Карписонова, 1967; Курнаев, 1968; Леса южного Подмосковья,
1985).
Как типичный ассектатор лютик кашубский не оказывает большого
влияния на фитосреду в коренных естественных фитоценозах. Однако, в
нарушенных местообитаниях (посадки, сады, парки, лесные просеки и др.)
он, благодаря высокой устойчивости к вытаптыванию, сохраняется в тра-
востое в числе немногих растений (сныть), часто служа индикатором ан-‘
тропогенного воздействия на разных стадиях дигрессии. Здесь этот вид
выступает как субдоминант или доминант растительных сообществ, давая
в весеннее время красочный аспект. В таких местообитаниях ему часто со-
путствует недотрога мелкоцветковая.
93
Биологическая продуктивность
Данные о биологической продуктивности лютика кашубского в Мос-
ковской области содержатся в публикациях ряда авторов. Так, в работе
Л.Б. Холоповой и О.Н. Солнцевой (1987) приводятся сравнительные ма-
териалы по продуктивности вида в ельнике волосистоосоковом и в
25-летнем лесу после сплошной рубки древостоя, полученные в Красно-
пахорском лесхозе на фитоценодогическом стационаре в Малинках. В
лесных парцеллах продуктивность лютика кашубского варьировала от
0,8 до 4,8 г/м2, где он составлял около 50 % фитомассы трав, на зарастаю-
щей вырубке в молодом лесу — от ОД до 7,8 г/м2 (около 3 % фитомассы
трав).
Там же продуктивность лютика кашубского (16.V. 1977) в разных типах
леса изучала Л.М. Носова (1987а). Согласно ее мнению, искусственные со-
сняки более благоприятны для лютика кашубского по сравнению с естест-
венными лесами, что подтверждается его более высокой численностью
(117 особей на м2 в сосняке против 73 и 79 в березняке и ельнике), большей
биомассой разновозрастных особей (до 0,41 г против 0,278 г и 0,357 г) и бо-
лее высокой продуктивностью популяции (46,4 г/м2 против 7,7 г/м2 и 0,3
г/м2 соответственно).
., В работах Г.А. Поляковой с соавторами (1981, 1983) приведены резуль-
таты исследования продуктивности лютика кашубского в березовых и ши- ,
роколиственных лесах на разных стадиях дигрессии. По данным автород,
сухая фитомасса 10 побегов варьировала от 0,3 до 5,9 г, численность побе-
гов на площадках (10 м2) достигала 288, а продуктивность популяций в пет
ресчете на 1 м2 не превышала 6,5 г, что нам представляется весьма сомни-
тельным. Вероятно, в связи с тем, что исследования проводились весной,
величина продуктивности надземной фитомассы во всех случаях оказа-
лась заниженной, по крайней мере, в несколько раз. Максимального значе-
ния фитомасса достигает к июлю, когда еще сохраняется плодоносящий
побег и образуются розеточные, листья, кроме того, к середине вегетацион-
ного сезона заметно возрастает выход сухого вещества, с 8-16 до 20-25 %.
Поэтому для уточнения этого показателя мы предприняли собственные из-
мерения в июне-июле 1998 г.
Изученные нами в Московской области ценопопуляцйи были нормаль-
ными полночленными (табл. 1). В Одинцовском районе популяции имели
правосторонний возрастной спектр, в котором преобладали взрослые веге-
тативные и генеративные особи, в Наро-Фоминском районе — левосто-
ронний с преобладанием виргинильных особей (проростки, ювенильные,
имматурные). В Одинцовском районе плотность популяций достигала 60
особей на м2, в Наро-Фоминском — 258 особей на м2 (проективное покры-
тие — около 65 %).
94
Таблица 3
Некоторые морфологические показатели
генеративных экземпляров Ranunculus cassubicus
в различных фитоценозах Московской области
Фитоценоз Розеточный лист Генеративный побег Корни общее число
пластинка диаметр, см черешок длина,см длина, см число листьев ЧИСЛО цветков
Липняк 9,1±2,1 16,9±5,2 30,2±4,8 2,7±0,6 3,0±1,9 —
Дубрава 7,8±1,3 20,0±4,3 41,9±2,6 2,9±0,2 2,2±0,7 100
Дубрава (овраг) 12,5±2,5 29,2±7,3 48,0±6,9 3,0±0,6 4,4±2,3
Ельник с березой 12,6±2,3 25,6±3,8 51,1±9,3 3,2±0,5 4,3±1,6 60-120
Ельник зеленомошник 9,8±0,3 24,5±0,4 46,5±3,5 2,5±0,4 3,5±1,5 40-100
Дубрава с березой 9,1±1,0 23,0±3,б 53,3±3,7 4,0±1,0 6,7±2,3 120
По мощности развития растений особенно выделялась популяция лю-
тика на склоне оврага в дубраве (3), в дубраве с березой (6) и в «окне» бере-
зово-елового древостоя (4), что было связано, видимо, с более высокой
влажностью и плодородием почвы. Генеративные растения отличались бо-
лее крупными листовыми пластинками, большей длиной черешков и гене-
ративных побегов, большим числом цветков (табл. 3) и соответственно
большей фитомассой надземных органов (табл. 4).
В сыром виде фитомасса побегов варьировала от 3 до 14 г, в высушен-
ном — рт 0,5 до 3 г. Надземная фитомасса зрелых генеративных особей,
формирующих 2-3 генеративных побега и несколько вегетативных, в су-,
хом состоянии достигала 3-5 г.
Среди исследованных популяций лютика кашубского наибольшим
числом взрослых растений и соответственно наиболее высокой продуктив-
ностью зеленой массы (73,5 г/м2) отличалась группировка на поляне в ело-
вом лесу, наименьшей (21,3 г/м2) — в дубравена склоне оврага.
95
Таблица 4
Биологическая продуктивность генеративной особи
Ranunculus cassubicus и его популяций в разных фитоценозах
Московской области
Фитоценоз Сырая биомасса, г Воздушно сухая биомасса, г Выход сухого вещества, % Общая надземная биомасса сухая,г/м2
побегов корней побегов корней
Липняк* 4,7±2,6 1,5 1,0±0,53 0,4 22 31,5
Дубрава* 3,2±0,8 2,7 0,8±0,28 1,0 25 25,0
Дубрава (овраг)* 10,1±3,7 5,6 2,0±0,74 2,1 20 21,3
Ельник с березой** 5,5±3,9 1,4 1,5±0,2 0,5 26 52,5
Ельник зелено- мошник** 4,6±1,6 1,8 1,2±0,3 0,7 26 73,5
Дубрава с березой* 12,5±1,5 6,6 1,7±0,6 1,7 27 —
Обозначения'. * в июне,
** в июле 1998 г.
Консортивные связи
Плодики-орешки лютика кашубского, по данным Г.П. Рысиной (1973),
разносятся и охотно поедаются муравьями.
Лютик кашубский — энтомофильное растение. Первичными аттрак-
тантами служат пыльца и нектар. Перекрестному опылению способствует
протерогиния. На стадии полураскрывшегося цветка, когда пыльники еще
сохраняют свою целостность, сосочки рыльцевой поверхности приобрета-
ют маслянистый блеск и становятся способными воспринимать пыльцу
(Киндерова, 1987). Созревание андроцея начинается с наружных тычинок
в полностью раскрытом цветке, когда на рыльцах уже обнаруживались
пыльцевые зерна, видимо, принесенные с других цветков.
96
Состав насекомых-опылителей, по данным Н.Н. Киндеровой, ограни-
чен. Это связано с тем, что цветет он рано весной, когда насекомых еще ма-
ло, и, кроме того, потенциальные опылители, как правило, предпочитают
другие медоносные растения. Цветки лютика посещают пчелы (Osmia
rufa) и примитивные чешуекрылые (Micropterygidae) (Определитель насе-
комых..., 1978; Фегри, Пейл, 1982).
Лютик кашубский микосимбиотроф. Светлые гифы грибов, образую-
щие эндотрофную микоризу, обнаруживаются в самых тонких корнях.
Лютик кашубский поражается некоторыми грибами, в том числе лож-
ной мучнистой росой. На нижней стороне, главным образом, розеточных
листьев появляется плотный войлочный серовато-фиолетовый или корич-
невый конидиальный налет, при этом листья останавливаются в развитии
и приобретают желтоватую окраску. По данным А. А. Ячевского (Миколо-
гическая флора..., 1901), на лютике кашубском паразитирует Peronospora
ficariae, по сведениям Н.А.Наумова (1954) — Р. hiemalis Gaumann. Н.С.,
Новотельнова и К.А. Пыстина (1985) связывают Р. hiemalis только с
Ranunculus acris, а для лютика кашубского указывают одну из форм
P.ficaria Tul./ ranunculi Schneider = P. ranunculi Gaumann.
Из дискомицетов лютик кашубский поражается Fabraea ranunculi (Fr.)
Karst (Наумов, 1964). На листьях появляются беловатые расплывчатые до-
вольно крупные пятна, со временем приобретающие желтовато-бурую ок-
раску.
Хозяйственное значение
Лютик кашубский, по-видимому, как и большинство лютиков, содер-
жащих в тканях протоанемонин, ядовит, поэтому животные его не едят
(Раменский и др., 1956); кормовое значение его оценивается -2 баллами
(Гроздов, 1956, 1963). При отравлении у животных наблюдается гастроэн-
терит. Животное начинает страдать от поноса, рвоты, замедляется пульс и
дыхание, появляются судороги глаз, и наступает смерть.
В качестве лекарственного средства используют листья с цветками, ко-
торые содержат острого вкуса камфору-анемоноль, но обращаться с ним
нужно крайне осторожно. Иногда их применяют при лечении малярии и
ран, однако необходим совет и контроль врача (Гроздов, 1956,1963),
Л. В. Петухова. О. Б. Черноброва
СТАЛЬНИК ПОЛЕВОЙ
Номенклатура, систематическое положение
и внутривидовая систематика
Ononis arvensis L., 1753, Syst. Nat, ed. 10, 2 : 1159; Муравьева, 1941, Фл.
СССР, 11 : 96; Борисова, 1964, в Маевский, Фл., изд. 9 : 147; Тихомиров,
1966, Опред. раст. Моск, обл.: 210; Ivimey-Cook, 1968, Fl. Europ. 2 : 147,
р.р.; Васильченко, 1987, Фл. европ. части СССР, 6 : 176; Губанов и др.,
1995, Опред. сосуд, раст. Центра Европ. России : 327.— О. hircina Jacq.
1770, Hort. Bot. Vindob. 1 : 40, р.р.; Кауфман, 1866, Моск, фл.: 123; Сырей-
щиков, 1907, Илл. фл. Моск, губ., 2:288.— Стальник полевой [семейство
Fabaceae (Papilionaceae, Leguminosae) — Бобовые (Мотыльковые)].
Выделяют две разновидности: О. arvensis var. spinescens Ledeb., имею-
щий колючки и О. arvensis var. inermis Ledeb., без колючек (Спасская,
1980). Род Ononis по одним оценкам включает 70 видов (Sirjaev, 1932), по
другим— 112 (Cannon, 1981), распространенных в Средиземноморье, За-
падной Европе, Западной Азии и в Африке (Эфиопия).
Научное название происходит от греческого слова Onos — «осел»,
считается, что эти растения представляют любимую пищу ослов (Ней-
штадт, 1963).
Географическое распространение
Стальник полевой — евразиатский вид, дизъюнктивный ареал которо-
го захватывает южную часть Скандинавии, Западную и Среднюю Европу,
Балканский полуостров, Малазию, Иран, в Восточной Европе встречается
на территории Белоруссии, Эстонии, Латвии, Молдавии, Украины, вклю-
чая Крым, Кавказ. На территории Европейской части России встречается
во всех районах, кроме Карелии, Мурманской области, Двинско-Печерско-
го бассейна, на Урале граница ареала проходит вдоль р. Белой до 60 вос-
точной долготы и захватывает южную часть южного Урала. Часто встреча-
ется в Ставропольском и Краснодарском краях (Середин, 1984). Отдель-
98
ные фрагменты ареала имеются на юге Западной Сибири, в Алтайском
крае, Восточной Сибири. Известен в некоторых северо-восточных районах
Казахстана, Киргизии, Таджикистана (Флора СССР, 1941; Спасская, 1980;
Флора европ. части СССР, 1987).
В Московской области отмечается довольно часто по всей долине р.
Оки и в низовьях ее притоков, изредка по откосам железно-дорожных на-
сыпей, вне речных долин встречается как заносное (Ворошилов и др.,
1966; Опред. растений Мещеры, 1986).
Морфологическое описание
Стальник полевой — стержнекорневой с многоглавым каудексом тра-
вянистый поликарпик, симподиально возобновляющийся. По терминоло-
гии К.Раункиера типичный протогемикриптофит, или безрозеточный ге-
микриптофит. Главный корень хорошо развит, длиной до 100-120 см
(Спасская, 1980) и, видимо, еще длиннее. В условиях культуры в первый
год вегетации проникает на глубину 160-180 см, а на 2-3-й год — до 210
см (Коломиец, 1972). Диаметр корня в основании достигает 35-45 мм.
Боковых корней, составляющих скелетную основу, 2-3, у взрослых
растений они отходят на глубине 5-8 см, незначительно отличаясь по диа-
метру от главного. Интенсивное ветвление наблюдается при длине главно-
го и боковых-корней свыше 50 см. В условиях культуры основная масса
корней (60-80 %) сосредоточена в слое 0-30 см (Коломиец, 1972). Кроме
системы главного корня на каудексе возникают придаточные корни, они
недолговечны, быстро обновляются, их диаметр 1-1,5 мм, наиболее тол-
стые достигают диаметра 3,1-3,5 мм. По мнению Н.И. Коломиец (1972),
мелкие придаточные корни в конце вегетационного периода отмирают, но-
вые появляются в апреле-мае. При партикуляции растений некоторые при-
даточные корни сильно утолщаются, производя впечатление стержневых,
формируя вторично-стержневую систему (Михайловская, 1981).
Побеги удлиненные, монокарпические и моноциклические. По терми-
нологии Т.П. Серебряковой (1971), их можно отнести к безрозеточным с
укороченным основанием, которое составлено 6-7 метамерами с чешуе-
видными листьями низовой формации, общей длиной 15-30 мм. В пазухах
этих листьев формируются почки возобновления.
В конце сезона вегетации побеги отмирают до укороченных основа-
ний, из которых образуется многоглавый каудекс (рис. 1). Почки возобнов-
ления стимулируют деятельность камбия в этой зоне. У взрослых растений
диаметр каудекса в зависимости от числа побегов возобновления разраста-
ется до 10-20 см. На одном растении может быть до 60 и больше побегов.
Высота генеративных побегов от 60 до 120 см (40-80, Спасская, 1980).,
Стебли цилиндрические, зеленые, иногда с антоциановым окрашиванием,
в конце вегетации буроватые в основании. Они густо опушены простыми и
99
Рис. 1. Каудекс и корневая система средневозрастного генеративного растения
Ononis arvensis. Обозначения: А — фрагмент каудекса; пк — придаточные корни;
пв — почки возобновления; on — основания побегов.
100
Рис. 2. Варианты структуры листьев срединной формации Ononis arvensis
железистыми волосками, причем, в основании стебля железистое опуше-
ние отсутствует. Диаметр побега в пределах 7-8 мм.
Листья срединной формации могут быть простыми и тройчато-слож-
ными, что зависит от возрастного состояния растений и от их положения
на побегах. Нижние листья (5-6 узлов) — простые, в средней части побега
листья тройчато-сложные, верхние снова становятся простыми. У мощных
средневозрастных растений нередко между нижними простыми и тройча-
то-сложными образуются листья переходной формы с одним боковым лис-
точком (рис. 2). Листочки округло-эллиптические по форме с мелко-пиль-
чатым краем, жилкование лучисто-краевое. Прилистники прирастают к че-
решку по всей его длине и охватывают стебель. Листья с двух сторон
опушены железистыми волосками, с нижней стороны слегка клейкие. Раз-
меры листьев различны. У взрослых растений чаще всего простые листья
общей длиной до 35-45 мм, пластинка 20-30 мм и 10-20 мм шириной,
тройчато-сложные крупнее, общая длина их 55-60 мм, длина верхнего
листочка 30-35 мм, ширина 15-20(25) мм. В некоторых случаях общая
длина листьев достигает 90-100 мм, соответственно, размеры пластинки
45x25 мм. Листорасположение двурядное.
Схема структуры побега стальника показана на рис. 3. Общее число уз-
лов на побеге — 45-55. Почки закладываются в пазухах всех листьев, но
судьба их разная. В пазухах листьев низовой формации — почки возоб-
новления. Часть их остается в спящем состоянии, часть обеспечивает во-
зобновление в следующем сезоне. Почки, формирующиеся в пазухах про-
стых листьев срединной формации в нижней части побега, в рост, как пра-
101
Рис. 3. Структура монокарпического побега Ononis arvensis.
Обозначения: чл — чешуевидные листья зоны возобновления; вп — верхушечная
почка, пазушные вегетативные почки; пл — простые листья в зоне торможения;
трл — тройчато-сложные листья зоны обогащения; цв — цветки
102
вило, не трогаются, составляя зону торможения. Пазушные почки
тройчато-сложных листьев дают побеги обогащения. В пазухах следую-
щих за ними простых листьев закладываются цветки (15-25 узлов). Выше
них верхушечная почка имеет 10-15 зачатков простых листьев, в пазухах
которых вегетативные почки, т. е. закладывается второй вегетативный
участок, который, однако, не успевает развернуться, верхушка часто отми-
рает во время цветения, хотя на боковых побегах может пролиферировать.
Подобный тип цветорасположения для многолетних осей назван интерка-
лярным (Кузнецова, 1987), в нашем случае после цветения ось соцветия от-
мирает целиком. На боковых побегах 1-го порядка расположение зон соот-
ветствует главному. Первые листья простые, затем следуют тройча-
то-сложные, пазушные почки которых могут дать боковые побеги
следующего порядка, а выше их — простые листья с пазушными цветка-
ми, что соответствует соцветию. Побеги Ш-го порядка не имеют своей зо-
ны обогащения, имеют только простые листья, в пазухах которых заклады-
ваются цветки.
Цветки на коротких цветоножках, располагаются по два в пазухах ли-
стьев. Соцветие стальника называют кистью (Нейштадт, 1963), густой ко-
лосовидной кистью (Коломиец, 1969, 1972), густым колосовидным соцве-
тием (Спасская, 1980). По нашему мнению, соцветие у стальника фрондоз-
ная, псевдотерминальная (Кузнецова, 1987) кисть. Боковые побеги зоны
обогащения можно назвать паракладиями, а синфлоресценцию — фрон-
дозной метелкой.
Цветки мотылькового типа, розовые или белорозовые, венчик в два
раза длиннее чашечки. Чашечка длиной около 10 мм, густо опушена про-
стыми и железистыми волосками, колокольчатая, рассеченная на 5 линей-
но-ланцетных зубцов. Андроцей однобратственный из 10 тычинок. Гине-
цей апокарпный из одного плодолистика. Пестик с верхней одногнездной
завязью, длинным, тонким, загнутым кверху столбиком и головчатым
рыльцем. В завязи — 5-6 семязачатков.
Плод — боб, короткий (Артюшенко, Федоров, 1986), широкояйцевид-
ный, железистоопушенный, немного вздутый. Длина боба около 7 мм, ши-
рина — 5-6 мм, при этом боб короче остающейся при нем чашечки. Семе-
на округло-почковидные, буровато-зеленоватые, мелкобугорчатые, по 2^4
в бобе. Вес 1000 семян 3,2-3,8 г в природных популяциях и до 4,4-5,2 г. в
культуре (Коломиец, 1969,1972).
Число хромосом 2п=24, 30, 32 (Флора европ. части СССР, 1987).
Онтогенез
Семена стальника не имеют глубокого эндогенного покоя. В фазе мо-
лочной спелости имеют всхожесть от 86 до 90 % с энергией прорастания
(определяемой на 7-й день) — 70-80 %. Однако у созревших семян, взятых
103
Рис. 4. Проросток (1) и ювенильные растения
первого (2) и второго (3) года жизни Ononis arvensis.
104
с этих же растений, после хранения в течение месяца энергия прорастания
снижается до 22 % (Коломиец, 1969). Зрелые семена обладают твердока-
менностью, т. е. имеют экзогенный механический покой (Николаева и др.,
1999). Скарификация семян повышает их всхожесть до 79-82 % (Коломи-
ец, 1972). По данным Г.И. Короткевич и О.А. Ароновой (2000), в связи с
твердосемянностью всхожесть зрелых свежесобранных семян ниже, чем
после их хранения в течение 6-8 месяцев. Максимальная всхожесть зрелых
семян — 80 %, после двух лет хранения семена имеют всхожесть 5-10 %.
Прорастание семян надземное. На первых этапах растения развивают-
ся медленно. Проростки имеют главный корень до 4 см длины и 1 мм в диа-
метре, гипокотиль короткий, 4-5 мм длиной и 1-1,1 мм в диаметре. Семя-
доли мясистые, овальные по форме, почти без черешка, пластинка шири-
ной 4,5-5 и длиной 5-5,5 мм. Основания семядолей срастаются, образуя
короткую (0,8-1мм) трубочку. В пазухах семядолей закладываются почки.
Первый лист проростка простой, с округлой пластинкой (5,5x6 мм), редко
зубчатым краем (по 4 зубца с каждой стороны и такое же число жилок при
перисто-краевом жилковании). Второй лист имеет по 5 ветвящихся боко-
вых жилок, соответственно увеличивается число зубцов, жилкование ста-
новится лучисто-краевым. Постепенно у последующих листьев удлиняют-
ся листовые пластинки, увеличивается число и степень ветвления боковых
жилок, край становится мелкопильчатым.
Ювенильные растения первого года имеют единственный относитель-
но невысокий (до 8 см) удлиненный, неветвящийся побег, состоящий из
4-7 фитомеров, которые несут простые листья с прилистниками (рис. 4).
На главном и боковых корнях — сформированные бактериями клубеньки.
К концу первого сезона вегетации побег отмирает. Сохраняющаяся базаль-
ная часть включает в себя семядольный узел и 1-2 следующих за ним узла
с почками возобновления. Главный корень обладает контрактильной спо-
собностью, благодаря чему почки возобновления оказываются погружен-
ными в почву.
Со 2-го года жизни растение переходит к симпо диаль ному возобновле-
нию. Обычно развивается один побег, реже — два. К концу вегетационно-
го периода надземная часть побегов отмирает, остается укороченная ба-
зальная часть, которая состоит из 4-7 узлов с почками возобновления. На-
чинает формироваться многолетняя основа растения — каудекс. В
ювенильном состоянии особи могут пребывать на протяжении 1-3 лет.
Календарный возраст виргинильных растений можно определить пу-
тем подсчета пеньков (сохраняющихся базальных частей) отмерших побе-
гов предыдущих порядков. Как правило, в этом возрастном состоянии рас-
тения имеют от 2 до 7 побегов, причем, побеги могут ветвиться. В зоне
ветвления на побегах 1-го порядка листья тройчато-сложные, на боко-
вых — простые. Корневая система становится аллогоморизной, наряду с
главным образуется 1-2 тонких придаточных корня диаметром 0,5-1 мм.
105
Рис. 5. Возрастные состояния Ononis arvensis
106
Зацветают растения на 3-5-й год, в культуре — на 1-й год. У молодых
генеративных растений увеличивается общее число побегов (табл. 1), пар-
тикуляция отсутствует. С возрастом наблюдается постепенное усиление
побегов, увеличивается число составляющих их метамеров (рис. 5). Все
побеги возобновления потенциально генеративные, однако не все они до-
ходят до цветения, значительная часть их остается в вегетативном состоя-
нии. Наряду с тонкими корнями появляются утолщенные до 3-х мм прида-
точные корни, хотя по-прежнему значение их невелико.
С возрастом диаметр каудекса увеличивается, у средневозрастных ге-
неративных растений все биометрические показатели достигают макси-
мальных значений. Побеги ветвятся до Ш-го порядка, часто трогаются в
рост верхние почки зоны торможения, однако эти побеги никогда не дохо-
дят до цветения. Они имеют тройчато-сложные листья на главном и боко-
вых побегах (побеги П-го порядка), на побегах Ш-го порядка все листья
простые. При большом числе генеративных побегов часть из них не дохо-
дит до цветения, хотя цветки закладываются. Соотношение вегетативных
и генеративных побегов зависит от условий произрастания.
Стареющие генеративные растения характеризуются началом партику-
ляции, центральные части каудекса разрушаются, в центре образуется дуп-
ло (рис. 5). Процессы некроза затрагивают и главный корень. Возрастает
значение придаточных корней. Они формируются не только на каудексе,
но и на главном корне. Общее число побегов у растения уменьшается, од-
нако все побеги доходят до цветения. Побеги интенсивно ветвятся лишь в
верхней части растения, побеги Ш-го порядка не образуются, зона тормо-
жения длиннее зоны обогащения. На боковых побегах формируются толь-
ко простые листья.
Старые генеративные особи представлены отделившимися партикула-
ми. При этом партикулы долго остаются морфологически связанными
проводящими пучками в зоне каудекса, имея при этом собственный вто-
ричный стержневой корень. Создается впечатление нескольких растений,
растущих рядом, и только при внимательном рассмотрении можно устано-
вить их единое происхождение. У старых особей значительная часть побе-
гов остается в вегетативном состоянии.
Сенильные особи по надземным органам практически не отличаются
от молодых вегетативных. Это отделившиеся партикулы, еще сохранив-
шие жизнеспособность. Имеют слабые неветвящиеся побеги с простыми
листьями, до цветения побеги не доходят. Следует отметить, что сениль-
ный период у стальника выражен слабо, чаще всего способность к цвете-
нию сохраняется, по-видимому, до конца жизни. Общая продолжитель-
ность жизни растений по нашим предположениям до 40-50 лет (Петухова,
1996). В оптимальных условиях существования жизненный цикл проходит
значительно быстрее.
107
Некоторые морфологические показатели
разных возрастных групп Ononis arvensis
Таблица 1
Возрастные группы Листья на побегах разных порядков Число побегов на особь Высота побега, см Диаметр каудекса, мм Диаметр главного корня*, мм Тип корневой системы**
Ювенильные простые 1 8-13 1,8-8 1,8-3,5 АР
Молодые вегетативные простые и тройчато- сложные 2-7 20-40 10-20 3-7 АГР
Молодые генеративные то же 4—19 60-80 18-35 8-17 АГР
Средне- возрастные генеративные то же 25-60 70-120 50-120 20—40 АГР
Стареющие генеративные то же 30-40 75-115 100-150 начало партику- ляции — АГР
Старые генеративные то же 9-25 40-80 партику- ляция — АГР
Сенильные простые 5-1! 15-25 отделив- шиеся партикулы — ГР
Примечание: * диаметр главного корня в базальной части;
** типы корневой системы: АР — аллоризная,
АГР — аллогоморизная, ГР — гоморизная.
108
Ритм сезонного развития
Семена прорастают весной в начале мая, в культуре на 8-16-й день по-
сле посева, в природе этот процесс растянут. В естественных ценозах и в
культуре (Перепечко, 1964) растения развиваются медленно. Отрастание
побегов начинается в конце апреля-начале мая, в зависимости от погодных
условий. По степени сформированное™ побегов в осенний период сталь-
ник относится к группе растений (Серебряков, 1952), у которых в почках
возобновления с осени заложена только часть вегетативной зоны. Эти поч-
ки трогаются в рост осенью, образуя полностью или частично подземное
основание побега с чешуевидными листьями. В некоторые годы часть по-
чек осенью дает побеги, которые не успевают полностью сформироваться
и отмирают. До цветения идет достаточно интенсивный рост побегов. По ,
срокам цветения стальник относят к ранне-среднелетним растениям (Голу-
бев, 1996). В условиях Московской области растения зацветают в начале
июля, период цветения растения в целом растянут до августа. По данным
Н.И. Коломиец (1972), в условиях культуры цветение продолжалось от 24
до 36 дней, продолжительность цветения одного цветка зависит от погод-.
ных условий. При пасмурной и прохладной погоде оно протекало 3-4 дня,
при сухой и жаркой — не более 2. Массовое распускание цветков наблю-
дается с 7 до 11 часов при 118-27°С и относительной влажности 55-65 %.
Первыми на побеге зацветают не самые нижние цветки, а в 3-м узле, затем
зацветают соседние с ним в бази- и акропетальной последовательности,
позднее распускание цветков продолжается в акропетальной последова-
тельности. Такие особенности зацветания наблюдаются как на главном,
так и на боковых побегах, причем, на боковых побегах, первых по мере
формирования, цветение начинается раньше, чем на главной оси. Не все из
заложенных цветков на побеге успевают дойти до цветения, соответствен-
но не во всех сформированных цветках образуются плоды.
Способы размножения и распространения
Единственный способ размножения у стальника полевого в приро-
де — семенной. Стальник — энтомофильное растение, самоопылению
препятствует дихогамия. В цветке имеются своеобразное приспособление,
напоминающее пресс, обеспечивающее перекрестное опыление (Кернер,
1903). Ко времени распускания цветка, и созревания пыльцы пыльники на- ;
ходится в закрытой лодочке, имеющей на конусовидном конце щель. Кры-
лья сверху выпуклые, смыкаясь друг с другом, они образуют над лодочкой
седло. Под тяжестью насекомого седло оттягивает лодочку вниз, при этом
из щели выдавливается пыльца, попадающая на нижнюю поверхность
брюшка насекомого. Рыльца находятся глубже пыльников и выступают
при более сильном нажатии лодочки. По наблюдениям Н.И. Коломиец
109
(1972), пыльца оказалась наиболее активной к прорастанию в течение
1-2-го дня после растрескивания пыльников.
Плоды образуются из цветков, расположенных в нижней части соцве-
тия, сформированных первыми. В условиях культуры завязывание плодов
составляло 16,9-64,3 % от нормально развитых цветков. Среднее количест-
во плодов на растении 165, в каждом плоде 2-4 семени (Коломиец, 1972).
Учитывая число формирующихся цветков на растении, потенциальная се-
менная продуктивность одного побега около 1000 семян. У средневозраст-
ных растений в среднем 20 плодоносящих побегов, таким образом, потен-
циальная семенная продуктивность одного средневозрастного растения
20000 семян. В условиях культуры, по данным Н.И. Коломиец (1972), уро-
жай семян растений 1-го года составил 130-184 кг/га, на 2-3 год — 212-286
кг/га. Уборку проводили механизированным способом при побурении
60-80 % бобов. По нашему мнению, стальник по способу распространения
семян следует отнести к аллохорам, баллистам (Левина, 1957).
Вегетативное размножение стальника в условиях культуры возможно
частями каудекса с придаточными корнями (Перепечко, 1964). В природе
при партикуляции также может увеличиваться число особей, однако такое
увеличение связано с глубоким старением растения, продолжительность
жизни обособившихся партикул мала, поэтому партикуляция не может
считаться размножением.
Экология
Стальник — светолюбивое растение, гелиофит (Голубев, 1996). Зани-
мает промежуточное положение между световыми и полусветовыми рас-
тениями (8-я ступень шкалы Элленберга). При затенении побеги распро-
стертые, часто не доходят до цветения.
Стальник полевой — мезофит или ксеромезофит (Голубев, 1996). Про-
израстает на почвах от влажностепных до влажнолуговых (46-86-я ступе-
ни шкал Раменского), но обилен на свежих лугах (49-66-я ступени), реже
обитает при условиях умеренно-переменного до сильно переменного ув-
лажнения (10-14-я ступени). Предпочитает почвы между сухими и свежи-
ми (4-я ступень шкалы Элленберга). Имеет достаточно широкую амплиту-
ду по отношению к кислотности почв. Растет на нейтральных почвах, реже
на кислых и щелочных почвах (7-я ступень шкалы Элленберга). Часто про-
израстает на обнажениях известняков. Голубев В.Н. (1996) по отношению
к засолению почв относит стальник к гликофитам, может расти даже на за-
соленных (от слабо-солончаковых до сильно-солончаковых) почвах (сту-
пени 12-23-я шкал Раменского).
Обилен на богатых и довольно богатых почвах (ступени 12-13-я,
14-16-я шкал Раменского). Однако по шкалам Элленберга относится к рас-
тениям мало требовательным к богатству почвы минеральным азотом, рас-
110
тущим на бедных почвах (2-я ступень). По мнению Н.П. Перепечко (1964),
в культуре стальник достаточно требователен к плодородию почв, предпо-
читает хорошо проницаемые среднесуглинистые почвы, но хорошо растет
и на легких суглинках и супесчаных почвах, достаточно богатых перегно-
ем. Предпочитает слабо аллювиальные почвы с мощностью наилка 0,1-0,3
см, единично встречается на умеренно аллювиальных с мощностью наилка
0,5-2,0 см (по шкалам аллювиальности 3-7-я ступени шкал Раменского).
Фитоценология
Стальник растет на пойменных и суходольных лугах, по берегам водо-
емов, среди кустарников, на лесных опушках и полянах, на горных скло-
нах, на залежах и обочинах дорог (Спасская, 1980). В Московской области
произрастают на заливных лугах, по кустарникам, обнажениям известня-
ков (Ворошилов и др., 1966). По берегам Волги встречается одиночными
экземплярами или небольшими группами, иногда образует сплошные за-
росли на небольших площадях. По нашим наблюдениям, по берегам Волги
произрастает в цикорево-васильковой ассоциации вместе с такими вида-
ми, как Melilotus officinalis, Achillea millefolium, Lotus comiculatus,
Medicago lupulina, Pimpinella saxifraga, Trifolium pratense, Prunella
vulgaris, из злаков — Роа pratensis vi Festuca pratensis. По берегам рек час-
то встречается в нарушенных ценозах в разнотравных ассоциациях.
Стальник полевой считается характерным видом ксеротермных суб-
континентальных травяных сообществ союза Cirsio-Brachypodium pinnati
Hadac et Klika in Klika et Hadac 1944 порядка Festucetalia valesiacae Br.-Bl.
et Tx. 1943 класса Festuco-Brometea Br.-Bl. et Tx. 1943 (Ellenberg, 1991).
Б.М. Миркин и Л.Г. Наумова (1998) относят этот союз к порядку
Brometalia W. Koch 1926 em Br.-Bl. 1936 того же класса.
Спектр возрастных состояний ценопопуляции стальника был изучен в
Тверской области по правому берегу Волги на участке от устья р. Нижняя
Старица до устья р. Холохольня, где стальник образует небольшую за-
росль. Было обнаружено 595 разновозрастных особей (Петухова, 1996).
Преобладают генеративные особи (370), на долю которых приходится
62,2 %, виргинильные особи составили в совокупности 37,1 % (221 особь).
Сенильные особи составили 0,7 % от общего количества, хотя следует от-
метить, что по надземным частям не всегда можно четко отделить вирги-
нильные и сенильные особи. Таким образом, наши наблюдения показали,
что в популяции стальника преобладают генеративные особи. По класси-
фикации А.А. Уранова и О.В. Смирновой (1969) исследованную популя-
цию следует отнести к зрелым нормальным, поскольку в ней преобладают
генеративные особи, а виргинильных особей в ней больше, чем сенильных..
По мнению названных авторов, зрелые нормальные популяции составля-
ют устойчивую основу коренных фитоценозов.
1.11
Биологическая продуктивность
Биомасса надземных и подземных частей особи стальника увеличива-
ется в течение жизни, достигая наибольших показателей в. средневозраст-
ном генеративном состоянии, когда формируется максимальное число по?,
бегов у растения. По нашим данным, сухой вес одного побега в цветущем
состоянии составляет в среднем 12 г, сухой вес подземных частей при дли-
не корней 40см — 184 г. По данным Н.П. Перепечко (1964), средний вес
корней 100 г, выход сухой массы — 40 %. Продуктивность посевов зави-
сит от густоты стояния растений и плодородия почвы. Урожай сухой мас-
сы корней в конце первого года вегетации на недостаточно плодородных
почвах составил 9 ц/га, при повышении плодородия — от 17 до 30 ц/га
(Перепечко, 1964). По данным Н.И. Коломиец (1972), урожай воздуш-
но-сухих корней стальника в первый год вегетации составил от 12 до 20
ц/га, на 2-3 год жизни масса увеличивается на 27-45 %, но ухудшается ка-
чество в связи с одревеснением.
Урожай семян в первый год составил 180-184 кг/га, на 2-3
год — 216-286 кг/га. При механизированном способе рекомендуется про-
изводить уборку при побурении 60-80 % бобов (Коломиец, 1972).
Консортивные связи
В южных районах, по данным Н.И. Коломиец (1972), всходы и юве-
нильные растения поражаются песчаным медляком (Opatrum sabulosum),
клубеньковым долгоносиком рода Sitona, большим люцерновым слоником
(PhytonomeS variabilis). Взрослые поедаются совкой люцерновой (Heliothis
viriplaca), кроме того, им вредит бобовая тля (Aphis fabae), обычная й в Мо-
сковской области, и толстоножка (Bruchophagus roddi).
Из грибных болезней на стальнике встречается ржавчина Uromyces
ononidis (Guyot, 1951), а по нашим наблюдениям и мучнистая роса Erysiphe
communis.
Стальник опыляется перепончатокрылыми.
Хозяйственное значение
Стальник полевой — лекарственное растение. Вся потребность в его
сырье удовлетворяется за счет культурных посадок (Спасская, 1980). В
корнях стальника полевого содержатся флавоноиды (формононетин, даид-
зеин, онозид), птерокарпаны (трифолиризин), тритерпеноиды (оносерин),
дубильные вещества, эфирные масла, смолы, крахмал, органические ки-
слоты. В надземных частях алкалоиды, фенольные соединения, флавонои-
ды (трифолии и формононетин), в цветках из флавоноидов содержится ас-
трагалин (Растительные ресурсы СССР, 1987; Растительные ресурсы Рос-
112
сии, 1996). С лекарственными целями используются отвары и настойки
корней, обладающие послабляющим и кровеостанавливающим действием,
эффективны при геморрое (Турова, 1974; Спасская, 1980; Гаммерман и др.,
1983; Минаева, 1991). В гинекологии используются галеновые препараты
при метроррагиях, особенно в климактерический период. Экстракт обла-
дает антиоксидантными свойствами (Растительные ресурсы России, 1996).
В народной медицине настои и отвары используются для лечения по-
дагры, асцита, при головной боли, ревматизме, как диуретическое, пото-
гонное, обезболивающее, противовоспалительное средство, при мочека-
менной болезни, нарушениях функции желудочно-кишечного тракта. Из
отваров травы делают ванночки при экземе (Минаева, 1991). Следует от-
метить, что препараты стальника нетоксичны.
В Молдавии листья, отваренные в соленой воде, используются в пищу
(Растительные ресурсы СССР, 1987).
Надземные части используются для окрашивания тканей, с квасцами
дают желтую окраску, с железным купоросом — зеленую (Роллов, 1908а;
Рытов, 1918).
Во время цветения выделяет липкий, дурно пахнущий сок (Ларин,
1951). О кормовой ценности сведения противоречивы. А.Х. Роллов (19086)
указывал, что в молодом возрасте стальник представляет приятный корм
для всех видов животных, овцы поедают его с жадностью. М.И. Нейштадт
(1963) отмечает, что это любимая пища для ослов. Однако, по наблюдени-
ям И.В. Ларина в Казахской ССР и Т.А. Работнова в пойме реки Оки (Ла-
рин, 1951), стальник почти не поедается скотом.
Вытяжки растения и измельченный тонкий порошок сухой травы чрез-
вычайно ядовиты для рыб, что объясняется действием ононина (Ларин,
1951).
Стальник полевой считается хорошим медоносом.
Работа выполнена при финансовой поддержке РФФИ
(гранты № 98-04-48469, 99-04-48086).
И. М. Ермакова
ТМИН ОБЫКНОВЕННЫЙ
Номенклатура, систематическое положёни
и внутривидовая систематика
Carutn carvib., 1753,Sp. Р1.:263; Кауфман, 1866, Моск, фл.: 198;Сырейщи-
ков, 1907, Илл. фл. Моск: губ., 2 : 401; Шишкин, 1950, Фл. СССР, 16 : 386;
он же, 1964, в Маевский, Фл., изд.9 : 383; Тихомиров, 1966, Опред. раст.
Моск, обл.: 248; Tutin, 1968, Fl. Europ. 2 : 354; Губанов и др., 1995, Опред.
сосуд, раст. Центра Европ. России : 385.— Тмин обыкновенный [семей-
ство Umbelliferae (Apiaceae) — Зонтичные (Сельдереевые)].
Выделены «осыпающиеся» и «неосыпающиеся» формы вида. Формы с
осыпающимися плодами возникли раньше в естественных условиях как
приспособление к распространению семян. Вторые появились позже в ре-
зультате мутации. У «осыпающихся» форм при основании плода с ранних
стадий его развития появляетсяразделительный слой, идет «расхождение»
эпидермиса, которое приводит к созданию особой перетяжки в месте осы-,
пания плодов. У «неосыпающихся» форм, наряду с отсутствием явно вы-
раженного разъединительного слоя, эпидермис не образует перетяжки и не
прерывается в основании плода, а представляет одно целое с эпидермисом
плодоножки. Возможно отличие этих форм уже в фазе бутонизации, что
важно в хозяйстве (Савченко, 1938; Яковлев, 1938; Roon, Bleijenberg,
1964).
Разделяются скороспелые, средне- и позднеспелые формы по длине ве-
гетационного периода (Кузьмич, 1977) и географические формы: образцы
из Калининской обл., Чехословакии и Нидерландов по сравнению с образ-
цами из Узбекистана, Казахстана и Монголии на второй год жизни интен-
сивнее растут, достигают большей высоты, быстрее развиваются и явля-
ются более скороспелыми (Курлянчик, 1988).
Название рода Carum от греческого Karos или Karon — у Диоскорида
название зонтичных, плоды которых имели медицинское применение. По
Плинию название произошло от местности Кария в Малой Азии, где расте-
ние разводилось и где растет лучший тмин. По другим объяснениям назва-
114
ние происходит от слова Kara — голова, по соцветию или от арабского
Karwaia, Karvia — тмин. Видовое название carvi — измененное арабское
Karvia, Karwaia — тмин (Флора СССР, 1950, Каден и Терехова, 1975). На-
родные названия: гуньба, кмин, ганус, тимон, анис полевой или дикий,
чернушка (Роллов, 1908; Рычин, 1952; Подымов, Суслов, 1990).
Географическое распространение
Тмин обыкновенный в основном евроазиатский вид с дизъюнктивным
ареалом (Meusel, 1978; Атлас ареалов..., 1980; Губанов, 1996). Он распро-
странен в Средней и Атлантической Европе, Средиземноморье, в Север-
ной Африке, на Балканах, в Малой Азии, провинции Синь-Цзянь Китая, в
Монголии и Гималаях. Занесен в Северную Америку и Новую Зеландию.
Широко распространен в Латвии, Литве и Эстонии, а также в Белоруссии,
северной части Украины (южная граница проходит через гг. Каменец-По-
дольский, Тульчин, Винницу, Харьков), на самом севере Молдавии. В
Крыму встречается как заносное растение. На Кавказе распространен в го-
рах очень широко, поднимаясь до 800 м, но не встречается на Кура-Арак-
синской и Колхидской низменностях, редок на Армянском нагорье, В За-
падном Казахстане тмин отмечен близ г. Уральска, в верховьях Утвы, Руд-
ном Алтае; фрагменты ареала этого вида имеются в Джунгарском Алатау и
на северо-востоке Казахского мелкосопочника. Изолированное местона-
хождение известно близ г. Туркестана. Среднеазиатская часть ареала вида
охватывает горные системы Тянь-Шаня и Памиро-Алая. Изолированные
участки его ареала отмечены в нижней части долины р. Пяндж и на Цен-
тральном Копетдаге.
В России основная часть ареала простирается от западных границ до
западного Забайкалья в лесных и лесостепных районах. Его северная гра-
ница идет вдоль береговой линии Кольского полуострова, по правому бе-
регу Северной Двины, пересекает верховья Печоры (изолированное место-
нахождение имеется в долине Усы под Северным полярным кругом), Се-
верный Урал и достигает долины р. Оби около 63° с. ш. Далее к востоку
граница несколько отклоняется к югу и до долины р. Лены идет по
58-60° с. ш. По правому берегу р. Оби граница проходит до устья р. Кеть,
откуда отклоняется к северу и вблизи устья р. Кас пересекает р. Енисей,
идет вдоль широтного отрезка р. Ангары на верховья р. Лены, достигая
устья р.. Киренги. Изолированные местонахождения имеются вблизи
р. Нижней Тунгуски (60° с. ш.) и в окрестностях г. Ленска.
Восточная граница евро-сибирского фрагмента ареала проходит близ
северо-восточной оконечности оз. Байкала, пересекает среднюю часть до-
лин р. Баргузин, Уда, Хилок, Чикой и вдоль левого берега последнего ухо-
дит за пределы России. В Читинской области имеются изолированные ме-
стонахождения на р. Шилке около пос. Усть-Черная и в верховьях р. Уров.,
115,
Кроме того, этот вид собран южнее с. Оймякон — на территории севе-
ро-восточной Якутии.
Южная граница проходит чуть южнее широты Харькова, пересекает в
среднем течении р. Хопер (идет к гг. Саратову, Оренбургу) и р. Урал. Обо-
гнув Южный Урал в районе г. Магнитогорска, граница на 55° с. ш. пересе-
кает Тобол, а близ г. Омска, огибает с севера оз. Чаны, а с юга Рудный Ал-
тай. В Саянах тмин встречается редко, как заносное растение.
В Приморье произрастает в юго-западной части (Приханкайская низ-
менность) и на восточном побережье, около пос. Рудного. На Камчатке его
ареал состоит из трех самостоятельных частей: северо-западной (побере-
жье Охотского моря), центральной (долина р. Камчатки) и юго-восточной
на побережье Тихого океана (Роллов, 1908; Флора СССР, 1950; Атлас ареа-
лов..., 1980; Meusel, 1978).
Тмин обыкновенный обычен по всей Московской области (Ворошилов
и др., 1966).
Морфологическое описание
Тмин обыкновенный -— стержнекорневой монокарпический травяни-
стый дву-, реже одно- или малолетник. Главный корень хорошо развит,
уходит глубоко в почву (Рысина, 1973), веретенообразный или цилиндри-
ческий, мясистый, до 1 см в диаметре; светло-бурый, на срезе белый. Утол-
щенная часть корня имеет длину 10-20 см и обладает втягивающей способ-
ностью. Боковые корни тонкие, волосовидные, расположены малочислен-
ными пучками по всей длине главного корня. Крупные боковые корни
единичны, встречаются на глубине 4—14 см. Корневище вертикальное до 1
см длины и 1 см в диаметре.
Побеги розеточные вегетативные и полурозеточные генеративные.
Одиночные или малочисленные стебли прямостоячие, в верхней части вет-
вистые (иногда ветвление начинается почти у основания), 30-80 (115) см
высотой, округлые, голые, бороздчатые, зеленые, во время плодоношения
светло бурые, 1-2 мм в диаметре, длина междоузлий 3-6 см. В посевах ко-
личество боковых побегов достигает 50 и более (Глущенко, 1963).
Листья очередные, простые, нижние на длинных, до 10 см, черешках,
верхние сидячие или на коротких черешках, расширенных при основании
во влагалище, по краям бело- или розово-перепончатые. Листовые пла-
стинки в очертании продолговатые, дважды- или триждыперистые, с глу-
бокозубчатыми или надрезанными долями (сегментами) Ш-го порядка; ко-
нечные дольки линейные или линейно-ланцетные, у прикорневых листьев
короткие, у стеблевых листьев, чем выше, тем все более тонкие и длинные,
до нитевидных; сегменты 1-го порядка сидячие 3-4 мм длиной и 1-1,5 мм
шириной. При основании влагалищ стеблевых листьев, особенно верхних,
развиты как бы прилистнички (за счет смещения нижней пары сегментов
116
листа). Проводящие пучки в черешке без склеренхимной обкладки (Тихо-
миров и др., 1996).
Объединенное соцветие (синфлоресценция), формирующаяся из од-
ной верхушечной почки возобновления, полителическое метельчатое.
Таких синфлоресценций у особи может быть от одного до несколько, и
каждая состоит из одного-нескольких соцветий — сложных зонтиков.
Они 4-8 см в диаметре, с 5-16 неодинаковыми голыми лучами; обертка
отсутствует или она из 1-2 листочков. Простые зонтики без обертки в по-
перечнике около 1 см, с 15-28 цветками. Цветки актиноморфные обоепо-
лые или женские (гинодиэция), зубцы чашечки почти незаметны. Венчик
свободнолепестный из 5 лепестков. Лепестки белые или розовые, обрат-
нояйцевидные, глубоковыемчатые, с загнутой внутрь верхушкой, около
1,5 мм длины. Тычинок 5, они светло-желтые. Пыльцевые зерна блед-
но-желтые, трехбороздно-оровые, эллипсоидальные. В очертании с по-
люса округло-треугольные, с экватора эллиптические; полярная ось
26,4-27,6 мкм. Борозды узкие с неровными краями, мембрана борозд
гладкая. Оры экваториально вытянуты, эллиптические. Скульптура мел-
кобугорчатая, бугорки сглаженные, густо расположены, покров толстый,
почти равен по толщине стерженьковому слою, стерженьки тонкие (Ку-
приянова, Алешина, 1972). Завязь полунижняя, двугнездная; столбиков
2, с отвороченными головчатыми рыльцами; подстолбие (нектарники,
стилоподий) светлое, почти белое, сплюснутое, подушковидное с волни-
стым краем, лежит открыто.
Плод — серовато-коричневый продолговатый (эллиптический),
сплюснутый со спинки вислоплодник (двусемянка), легко распадается на 2
полуплодика, длиной 3-5 мм и 1-2 мм шириной с сильным ароматом и
своеобразным пряным вкусом. Карпофор на верхушке двураздельный. По-
луплодики голые цилиндрические, слабо изогнутые с 5 примерно равными
светло-серыми ребрами и широкими красновато-коричневыми ложбинка-
ми; три ребрышка проходят по спинке, а два других отодвинуты к краям;
секреторные канальца в ложбинках одиночные; на коммиссуральной сто-
роне их 2. Эндосперм с внутренней стороны почти плоский (Флора СССР,
1950; Маевский, 1964; Рысина, 1973; Атлас ареалов.., 1980).
Хромосомные числа: 2п=20 или 22 (Хромосомные числа..., 1969). Од-
нако отмечены и гаплоидные числа хромосом в Приморском крае (Гурзен-
ков, Горовой, 1971), на Алтае (n=l 1), в Новосибирской обл., в Хакасии и
Туве (п=10) (Ростовцева, 1979,1982). Е.Л. Кордюм (1967) обнаружила под
Киевом на разных фазах мейоза 10-11 хромосом. При этом в материнских
клетках микроспор одного и того же гнезда может быть 10 и 11 хромосом
(Schulz- Gaebel, 1930). Кроме того, в теплице у тмина было п=10, а в поле-
вых условиях n=l 1. Получены также тетраплоидные формы (Zderkiewicz,
1964, 1964а).
117
Онтогенез
Семена тмина могут прорастать сразу после созревания на 5-7-й день в
лабораторных условиях (Рысина, 1973), потом на 2-3 месяца процент про-
растания снижается, далее происходят незначительные его колебания.
Прорастает в течение года — 58-88 % семян (Formanowiczowa, Kozlowski,
1961). Всхожесть семян из разных географических пунктов варьирует от
70 до 96 % (Курлянчик и др., 1989). В полотняных и бумажных мешочках
способность прорастать сохраняется в течение 3-х лет. Всхожесть после 10
лет хранения составила 29 % за 21 день, но за 7 дней (время выявления
энергий прорастания) не проросло ни одно семя. Семена прорастают на
свету (Formanowiczowa, Koztowski, 1961). На всхожесть семян оказывает
влияние температура воздуха: при 1-3° всхожесть — 28 %, при 12-15 —
40 %, при 30-35° — 72 %.
Всхожесть зависит от экологических условий сбора и районов выращи-
вания (Fonnanowizowa, Kozlowski, 1961), от места образования плодов в со-
цветии, так как период формирования плодов растянут и определяет вес и
размер плодов (Ражинскайте, 1964, 1967). Растянутость цветения и созрева-
ния плодов в зонтиках разных ярусов приводит к различиям в них биохими-
ческих и физиологических процессах. Кроме того, меняются метеоусловия в
столь длительный срок. В результате семена из разных ярусов не только от-
личаются морфологически, но у них различаются всхожесть, энергия про-
растания и интенсивность роста проростков. Мелкие плоды (абсолютный
вес 1000 шт.— 1,7019 г) из зонтиков нижних ярусов имели лабораторную
всхожесть 74;3 % при энергии прорастания 61 %, начало прорастания про-
исходило в лабораторных условиях на шестой день, в полевых — на 22-й
день. Эти показатели у средних (абсолютный вес 1000 шт.— 2,3561 г) и
крупных (2,5283 г) плодов гораздо выше: лабораторная всхожесть 89-90 %,
энергия прорастания 67-77 %, начало прорастания в лаборатории происхо-
дило на 4— 5-й день, в полевых условиях на 17-18-й день. К сходным выво-
дам еще раньше пришел А.А. Хотин (1957): наивысшим качеством облада-
ют плоды, сформированные на центральном и зонтиках 1-го порядка верх-
них ярусов ветвления. С центрального зонтика абсолютный вес 1000 плодов
был 3,18 г, энергия прорастания 97 % и общая всхожесть 73 %; с зонтиков
П-го порядка соответственно 1,8 г, 67 % и 67,5 %. В фазе молочной спелости
семена не прорастают. Предпосевная обработка (стратификация, тепловой
обогрев и ферментирование) улучшает посевные качества семян.
В природных условиях часть семян сохраняет всхожесть в течение осен-
не-зимнего периода. По нашим данным, при хранении в лаборатории в апре-
ле-мае в чашках Петри за 18 дней проросло в среднем 35 % семян, в после-
дующие 8 дней еще 26 % от первоначального количества. На делянках при
летнем посеве прорастание произошло за 18 дней. Семена могут прорастать
уже при 9°, при 15-18° всходы появлялись на 18-20 день (Шилова, 1956).
118
Рис. 1. Возрастные состояния Canon carvi
Прорастание семян надземное: первым появляется корешок, затем про-
долговатые узкие семядоли. Первый настоящий лист формируется через
9-11 дней после появления всходов, а каждый следующий через 7-8 дней
(Ражинскайте, 1967).
Онтогенез тмина может быть разделен на 2 периода (виргинильный и
генеративный) и 5 возрастных состояний (Ермакова, 1983).
Проростки — растения 1,5-2,5 см высотой с розеточным побегом 1-го
порядка. Семядоли продолговатые до 1,5-2,5 см длиной, на верхушке ту-
пые, книзу постепенно суженные в черешки, при основании расширенные
в небольшое влагалище и слегка разросшиеся. Первые 1—2 листа широко-
яйцевидные или обратноширокояйцевидные пальчато-трехраздельные
или пальчато-трехрассеченные на длинных черешках. Главный корень
тонкий, менее 1 мм в диаметре у основания. Гипокотиль слабо выражен. В
посеве это состояние длится 30-40 дней (рис. 1).
У ювенильных растений семядоли отсутствуют. Главный побег — ро-
зеточный, с 2-4 простыми яйцевидными или узкояйцевидными непарно-
перистыми, дважды рассеченными листьями, с 2-3 парами сегментов 1-го
порядка. Главный корень 1,3-2,0 мм в диаметре. На лугах это состояние
длится от 3 до 12 месяцев.
У имматурных особей розеточный побег 1-го порядка с 3-5 яйцевидны-
ми или узкояйцевидными непарноперистыми, трижды рассеченными ли-
стьями с 4—5 парами сегментов 1-го порядка. Есть отмершие листья. Диа-
метр главного корня 2,5-5 мм. Продолжительность возрастного состояния
на лугах 1,5-3 месяца.
У виргцнильных растений продолжает функционировать главный
розеточный побег с 3-8 продолговатыми непарноперистыми трижды рас-
сеченными листьями о 6-7 и более пар сегментов. Главный корень утол-
щен до 3-9 мм. Образуется вертикальное эпигеогенное корневище до
0,2-0,5 см длины с хорошо выраженной зоной втягивания. Продолжитель-
ность возрастного состояния от 1 месяца до 2 лет. Задержка в развитии на-
ступает при неблагоприятных условиях и может продолжаться несколько
лет (Серебрякова, 1953; Рысина, 1973). Весь виргинильный период при за-
гущенном подсеве на лугах продолжался 7 и более лет (Работнов, 1966).
Генеративные растения формируют прямостоячий ветвящийся генера-
тивный побег. Прикорневые розеточные листья, если сохраняются, про-
долговатые дважды или трижды перисторассеченные. Главный корень
утолщен до 1 см. Вертикальное корневище 0,5-1 см длины. Продолжи-
тельность возрастного состояния — 1 месяц.
Общая продолжительность жизни по нашим наблюдениям 2 года,
гораздо реже до 4 лет. По данным Т.П. Серебряковой (1953) вид в культу-
ре живет 2 года, в травостое тмин может вегетировать несколько лет до
цветения.
120
Ритм сезонного развития
Проростки тмина появляются на лугах во второй половине лета (Сереб-
рякова, 1953), по нашим данным — как в августе, сентябре, так и в начале
мая. Проростки в массе гибнут. Через 30-40 дней выжившие переходят в
ювенильное состояние, через 3-12 месяцев ювенильные превращаются в
имматурные. При весеннем посеве проростки появляются через 8-20 дней
(Лебедева, 1992).
У растений, развившихся из летних проростков (наблюдения Т.П. Се-
ребряковой, 1956) на Дединовских пойменных лугах р. Оки в Московской
обл., с осени в центральной розетке бывает сформирован весь побег буду-
щего года: розетка или генеративный побег. В случае развития вегетатив-
ной розетки у нее с весны до июня развертывается от 5 до 10 листьев. Ко-
личество их зависит от возраста розетки и условий ее обитания; у хорошо
развитых растений в благоприятных условиях листьев значительно боль-
ше. Эти развернувшиеся листья составляют примерно половину количест-
ва перезимовавших в почке листовых зачатков. Остальные 6-12 листовых
зачатков остаются пока внутри почки. Листья из них развертываются в те-
чение августа-сентября, параллельно с начинающимся еще в конце июля
процессом заложения новых листовых зачатков для побега будущего года.
К октябрю розетка имеет уже столько развернувшихся листьев, сколько их
было заложено в почке с осени прошлого года (12-22). Из них только
3-5 нижних листьев засыхают в течение лета, остальные зимуют зелены-
ми. Некоторая часть их гибнет в течение зимы, так что весной из-под снега
выходят живыми от 4 до 10 листьев укороченного побега.
К октябрю же заканчивается формирование в почке листовых зачатков
будущего года. Если побег переходит в генеративную фазу, то зачатки со-
цветия формируются в почке тоже в октябре. В это время на конусе нарас-
тания имеется зачаток верхушечного сложного зонтика с наметившимися
бугорками простых зонтиков и несколько гладких округлых зачатков па-
зушных зонтиков. Зацветает тмин рано: бутонизация наступает в начале
июня, полное цветение начинается в середине июня. Собственно стебле-
вых листьев, расположенных на вытянутой части стебля, с удлиненными
междоузлиями, бывает 6-8; все они несут в пазухах боковые цветоносные
побеги (нижние о 2-3 листьях, верхние с 1 листом). Ниже расположенные
листья относятся к розетке. В начале июля цветение кончается, к концу
июля осыпаются зрелые плоды, и отплодоносившие растения нацело засы-
хают. Перезимовавшие розеточные листья засыхают в течение весны, до
цветения.
Органогенез тмина обыкновенного по методике Куперман изучен
А. А. Курлянчик с соавторами (1989) в посевах в Рязани. Выявлено 3 пе-
риода формирования всходов и 12 этапов морфогенеза. Во второй декаде
апреля в почке происходит формирование лучей сложных и простых зон-
121
тиков. В третьей декаде происходит «стеблевание»; формирование эле-
ментов цветков в зонтиках идет в акропетальной последовательности: вна-
чале закладываются чашелистики, потом одновременно зачатки лепестков
и тычиночные бугорки и позже всех гинецей. Во второй декаде мая в со-
цветии уже есть зеленые и окрашенные бутоны, в них формируются муж-
ские и женские гаметы, завершается рост всех частей цветка и подготовка
к цветению. В первой декаде июня все цветки цвели. На этом этапе проис-
ходит оплодотворение яйцеклеток в зародышевых мешках. В середине ию-
ня в некоторых семяпочках формируется семя, зеленые плоды интенсивно
растут. В третьей декаде июня в плодах происходит накопление питатель-
ных веществ — фенофаза восковой спелости, плоды светло-желтые. В это
же время заканчивается рост побегов растений в длину. У раннеспелых
растений к середине июля происходит превращение питательных веществ
в запасные. Плоды приобретают желто-бурую окраску, становятся воз-
душно-сухими.
У «поздних» форм экземпляров тмина плоды созревают к концу июля,
в августе (Сорные растения..., 1934; Курлянчик и др., 1989). Плоды легко
осыпаются в сухое время суток (барохория). Цветение тмина происходит
одновременно на сенокосах и пастбищах, при разной их нагрузке. Вторич-
ное цветение наблюдается в конце августа на пастбищах после перегона
скота на сенокосы. При этом у тмина образуются короткие генеративные
побеги, на уровне прикорневой розетки (Конюшков, 1930).
Тмин обыкновенный — гемикриптофит (Ражинскайте, 1967), зимует с
зелеными листьями, которые к концу сентября приобретают желтоватый
оттенок и опускаются к земле.
Переход растений в репродуктивное состояние возможен при форми-
ровании в первый год жизни особи определенного числа листьев на побе-
ге. Но этот показатель неодинаков в разных частях ареала. В условиях
Литвы (Ражинскайте, 1967) на второй год в посеве вегетация начинается
в середине апреля, когда сходит снег. Через 30-34 дня после начала от-
растания начинается рост стебля и формирование генеративных органов.
Бутоны покрыты влагалищами верхних стеблевых листьев. На 11-13-й
день после появления стебля бутоны центрального зонтика освобожда-
ются от влагалищ и через 4-5 дней раскрываются первые цветки. Один
зонтик цветет 5-7, а все растение — 27-33 дня. От завязывания плода до
полного созревания проходит 30-35 дней. Весь период цветения и созре-
вания плодов тмина очень растянут и занимает в Литве 40-47, под С.-Пе-
тербургом в Детском Селе — 50-55 дней (с начала июня до начала авгу-
ста) (Бабарыкина, 1992). Образование плодов тмина на зонтиках разных
порядков происходит с неодинаковой скоростью: на центральных зонти-
ках они формируются в течение 27-30 дней, на зонтиках 1-го порядка
22-25 и П-го порядка 16-19 дней (Хотин, 1948). В условиях Литвы на 2-м
году вегетации плодоносили растения достаточно развитые, образовав-
122
шие в первый год не менее 8 листьев, корень длиной 17 см и набравшие
общий вес 5 г (при посеве не позднее 1-ой половины мая), при более позд-
них посевах плодоносили только 3-50 % особей. Сходные критерии дос-
таточности развития в первый год для зацветания на второй приводит
А.А. Хотин (1959) для Краснодарского края: розетка из 8-10 листьев и
корень диаметром не менее 6—8 мм. В зависимости от климатических ус-
ловий минимальные размеры растения, обеспечивающие цветение на
следующий год, меняются: в Болгарии достаточно в первый год образо-
вать розетку из 5 листьев и иметь вес 0,31-0,38 г (Янкулов, 1960 цит. по:
Ражинскайте, 1964). О том, что на втором году жизни тмин не всегда пло-
доносит, свидетельствуют исследования и А. Дастикайте (1976). Плодо-
ношение, по автору, зависит от дифференциации конуса нарастания на
первом году жизни.
Способы размножения и распространения
Тмин обыкновенный размножается исключительно семенным путем.
Его особи женские или обоеполые (гинодиэция) (Knuth, 1899, Демьянова и
др., 1989). Это явление у зонтичных редкое, зарегистрированное лишь у
0,8 % видов и 5 % родов. Тмин опыляется многочисленными насекомыми
(мухами, жучками). Для тмина, как и для многих других зонтичных, харак-
терна протерандрия — смена мужской и женской фаз, обусловленная син-
хронностью или очередностью цветения соответствующих зонтиков (По-
номарев, 1960). А.Н. Бекетов (1889) допускал у тмина гейтоногамию в пре-
делах одного побега, исходя из того, что вторичные зонтики (П-го
порядка) у него возвышаются над главным. Как показали наблюдения
А. Н. Пономарева (1961), в действительности этого нет, т.к. зонтики П-го
порядка зацветают в мужской фазе лишь после того, как будут завершена
женская фаза главного зонтика у данного побега. Но если у особи от стерж-
невого корня отходят несколько генеративных побегов (т. е. имеется не-
сколько синфлоресценций), то они ведут себя независимо, как отдельные
особи, и тогда возможна гейтоногамия. (Пономарев, 1961; Ермакова,
2000). Однако, Е.И. Демьянова с соавторами (1989) считает, что дихогамия
у тмина охватывает всю особь в целом, и возможно только перекрестное
опыление.
Цветения тмина в условиях коллекционного участка Троицкого заказ-
ника Пермской области, выращенного из семян, собранных в 20 км в при-
родной популяции продолжалось около 30 дней (с 24 мая по 25 июня)
(Демьянова и др., 1989). Первыми зацвели зонтики 1-го порядка женской
формы, а через 1-2 дня начали цвести зонтики этого же порядка обоеполой
формы. Обоеполые особи тмина цветут с многократной сменой тычиноч-
ной и рыльцевой фаз у зонтиков всех порядков. Одна и та же особь много-
кратно и последовательно выступает то в тычиночной, то в рыльцевой фа-
123
зе. У обоеполой формы цветение отдельных зонтиков I—IV-ro порядка на-
чиналось с периферических зонтичков и цветков и постепенно
распространялось к центру зонтичков и зонтиков. Распускание обоеполых
цветков сопровождается постепенным выдвижением тычинок, раскрыва-
нием лепестков и выделением нектара на стилоподии. Экспонирование
столбиков и рылец начинается после увядания и опадения лепестков.
Рыльцевая фаза у обоеполых цветков заканчивалась с прекращением выде-
ления нектара, полным расхождением столбиков и опадением лепестков.
Зонтики 1-го порядка обоеполой формы цвели в течение 4—5 дней, на-
ходясь в тычиночной фазе, 2-3 дня — в рыльцевой; П-го порядка: 4—5
дней — в тычиночной и 3-4 дня — в рыльцевой; Ш-го порядка — 3-4 дня
в тычиночной и 3-4 — в рыльцевой. Цветки в зонтиках IV-ro порядка цве-
ли 4—5 дней, причем они почти полностью оказались мужскими (тычиноч-
ными). По-видимому, их назначение — поддерживать необходимый
пыльцевой режим для зонтиков Ш-го порядка, находящихся в это время в
рыльцевой стадии.
Женские особи тмина обыкновенного «процветали» быстрее обоепо-
лых, поскольку цветение здесь протекало только в одной стадии — рыль-
цевой. Цветки женских особей начинали цветение с выдвижения рылец; в
это время на стилоподии выделялся нектар. При развертывании лепестков,
которое здесь не бывает никогда таким полным, как у обоеполых цветков,
становились заметными короткие тычинки со стерильными пыльниками.
В целом цветки и соцветия женских особей выглядят менее крупными, но
более компактными. Цветение женских особей происходило в следующем
порядке: зонтики 1-го порядка цвели 5-6 дней, П-го — 6—7 дней,
Ш-го — 5-6 и IV-ro порядка — 5-6 дней. Таким образом, обоеполые рас-
тения, имевшие зонтики I—IV-ro порядков, цвели в среднем 31-32 дня, а
женские — 28-29 дней.
Несмотря на разную продолжительность цветения особей обоеполой и
женской формы, пики (максимумы) цветения зонтиков разных порядков у
них совпадают. Подобный характер цветения можно рассматривать как
одно из приспособлений к перекрестному опылению, поскольку в такой
ситуации женским особям обеспечен оптимальный пыльцевой режим на
протяжении всего периода их цветения. Более того, как отмечают авторы,
число зонтиков обоеполой формы, находящихся в тычиночной стадии,
всегда выше в сравнении с зонтиками женской формы, пребывающих в
рыльцевой стадии. И, наконец, самое важное — фазы цветения двух поло-
вых форм совпадали во времени.
По абсолютному числу выполненных плодов женская форма продуктив-
нее обоеполой, хотя эта разница невелика: процент семинификации (ПС)
35,8 у обоеполых и 40,1 у женских особей. ПС у зонтиков с увеличением по-
рядка уменьшается от 76 у обоеполых и 69 у женских 1-го порядка до 23—27
у зонтиков Ш-го порядка соответственно. Вес семян зонтиков 1-го порядка
124
больше у обоеполой формы. Семена, сформированные на зонтиках П-го и
Ш-го порядков, по весу примерно одинаковы. Проращивание семян тмина в
лабораторных условиях выявило их невысокую всхожесть (30-53 %) и от-
сутствие разницы по этому признаку у обеих форм. Лучшее качество семян
обнаружено в зонтиках Ш-го порядка у обоеполых и женских особей.
Существует биологическая неравноценность растений, выращенных из
семян, сформировавшихся в разных местах соцветий (Глущенко, 1954). Так
семена, формирующиеся в нижней части особи, наиболее быстро прораста-
ют и дают наиболее интенсивно растущие проростки и наибольшее число,
однолетних и незимостойких экземпляров. Семена, формирующиеся на пе-
риферийных побегах, замедленно прорастают, обладают ослабленной энер-
гией роста проростков и заметно выраженной тенденцией к многолетности.
Семена, формирующиеся около верхушки, в центральной части особи, наи-
более полно повторяют материнские свойства и дают наиболее жизненные,
зимостойкие и урожайные формы. Различия, проявившиеся в первом поко-
лении в зависимости от места формирования семян в исходных особях, зна-
чительно сглаживаются во втором поколении, если не производить такого
же отбора семян, как в урожае первого поколения. При осенних посевах уси-
ливаются различия, вызванные неодинаковыми условиями формирования
семян в разных местах особи. Этим способом можно преобразовывать двух-
летние формы в однолетние (озимые). Однолетний тмин при ежегодном
плодоношении приблизительно в 2 раза увеличивает продуктивность по
сравнению с двухлетним. Автору удалось получить однолетние формы, ко-
торые сохраняли однолетность в течение 7 лет опыта (7 поколений).
Протерандрия тмина (Бабарыкина, 1992), имеет 4 фазы цветения: плот-
ного бутона, тычиночную, пестичную и усыхания цветка. Нектарный диск
в тычиночную фазу подушкообразный, вздутый, края приподняты; в пес-
тичную фазу поверхность диска имеет густые сосочки и становится боль-
ше, края опускаются; в фазу усыхания цветка нектарный диск съеживает-
ся, поверхность его становится морщинистой.
Каждый экземпляр дает от 969 до 15000 семян (Сорные растения...,
1934; Доброхотов, 1961; Демьянова и др., 1989). По нашим данным, на
пойменных лугах р. Угры реальная семенная продуктивность (РСП) особи
и побега (в особи часто бывает 1 побег) колебалась от 100 до 5176, потен-
циальная (ПСП) — от 542 до 7400. Эти показатели отличались на разных
участках и в разные годы. Составляющие семенной продуктивности тми-
на: в одном цветке завязываются 1 двуплодник, в одном зонтичке от 2 до
28 таких плодов, зонтичков в 1 зонтике от 7 до 14, зонтиков на 1 побеге бы-
вает от 1 до 36, побегов в особи от 1 до 4. Например, в 1987 г. на прогоне
скота в среднем у особи было 7,9 общих зонтиков, в каждом по 22,7 част-
ных, у особи;— 179,3 частных зонтиков. В 1 частном зонтике образовалось
19,2 плода, в общем зонтике — 436,4, на особи — 3446,7. Процент плодо-
образования был в разные годы на разных участках от 32,6 до 84,6; про-
125
цент пустых и поврежденных плодов от 15,4 до 68,4. Урожай семян на 1 м2
был от 2543 до 45550 семян в разные годы и на разных участках.
По данным А.Н. Бабарыкиной (1992), в посеве у растения насчитывает-
ся до 27-30 сложных зонтиков. В начале цветения число цветков в зонти-
ках от 136 до 168, в период массового цветения — от 78 до 121, а в конце
цветения от 42 до 78. На одном растении за период цветения бывает от
1878 до 2594 цветков. В Пермской обл. среднее число зонтиков на особь
было от 5,47 до 9,59, в каждом от 6,71 до 11,34 зонтичков, среднее число
цветков в зонтичках от 15,14 до 17,77, среднее число выполненных плодов
в зонтичках от 3,33 до 11,56; ПСП особи от 2500 до 27007, РСП особи 969,7
до 1003,4 (Демьянова и др., 1989).
Семена осыпаются вокруг материнской особи, но могут, по-видимому,
разноситься полыми водами, а также птицами (жадно поедаются певчими
дроздами) (Кернер, 1903).
Экология
Вид не требователен к климатическим условиям (Серебрякова, 1953).
Начало вегетации под Москвой по нашим данным — 15-19 апреля, в те же
сроки — в Литве (Ражинскайте, 1967). Всходы появляются при температуре
почвы 4-6° градусов (Лебедева, 1992). К октябрю заканчиваются активные
формообразовательные процессы в почках, наружные листья в розетках
желтеют и отмирают, внутренние приобретают желто-зеленую окраску и
опускаются к земле. Морозоустойчив (Фроленко, 1993). Резкие колебания
температуры виду не вредны (Ростовцев, 1895). Рост и развитие его хорошо
протекают гфи среднесуточной температуре 15-18°. Повышение температу-
ры до 25-27° ускоряет развитие, а при более высоких температурах проис-
ходит отмирание. Снижение температуры и увеличение влажности воздуха
в период созревания плодов приводят к увеличению содержания эфирного
масла, так как уменьшается его испарение (Хотин, 1957).
Тмин обыкновенный произрастает на открытых и полуоткрытых уча-
стках (Цыганов, 1983). Н. Ellenberg (1991) относит вид к промежуточной
группе световых-полусветовыми растений (8-я ступень шкалы). На лугах
часто оказывается в тени более высоких растений, но обилен на нарушен-
ных лугах с малым покрытием и низким травостоем — там его генератив-
ные органы оказываются в 1-ом горизонте, а прикорневая розетка во 2-ом
или 3-ем. Световую стадию проходит на длинном дне.
Растение мезофит (Серебрякова, 1953а) или ксеромезофит (Тюрина,
1978), предпочитает свежие почвы (5-я ступень шкалы Элленберга). По
Л.Г. Раменскому с соавторами (1956) этот вид произрастает на почвах от
сухих луговых до болотно-луговых (61-90-я ступени), но обилен на све-
жих и сухих лугах (ступени 61-66-я), при средне- и сильно-переменном ув-
лажнении (ступени 6—13). По Д.Н. Цыганову (1983) произрастает на поч-
вах от лугово-степных до болотно-лесолуговых (9-17-я ступени). Сильно
126
изрезанные листья, небольшая общая масса, по-видимому, позволяют тми-
ну произрастать в широком диапазоне влажности почвы. Однако, при про-
растании семян и развитии проростков, а также в период с начала стеблева-
ния до формирования плодов на всех зонтиках, необходима влажная поч-
ва, но без излишней сырости, так как это стержнекорневое растение
требовательно к хорошей аэрации почвы (Ростовцев, 1895; Чехонина-Три-
фонова, 1954; Хотин, 1957; Фроленко, 1993).
Тмин имеет широкий экологический диапазон богатства почвы. Н.
Ellenberg (1991) относит его к группе растений, растущих на почвах проме-
жуточных между средними и богатыми минеральным азотом (6-я ступень
шкалы), Е. Landolt (1977) — на почвах от среднебедных до среднебогатых
(3-я ступень). Распространен в диапазоне от бедных до среднезасоленных
почв (5—21-я ступени по Раменскому), но очень обилен на довольно бога-
тых почвах (10-12-я ступени); по Д.Н. Цыганову (1983) встречается в диа-
пазоне от бедных до богатых азотом почв (5-8-я ступени). В эксперимен-
тах с внесением удобрений показано, что рост растений более всего стиму-
лирует N и NP; NPK и NK в меньшей степени; К — тормозит рост
(Ражинскайте, Славенас,1963; Ражинскайте, 1967). При N60 Р80 К45 полу-
чен в опытах самый большой урожай зеленой массы и плодов. При сокра-
щенном световом дне на подзолистой почве лучше растет при внесении
аммиачного азота, так как на нитратном азоте в этих условиях отстает в
росте и продукции, но на не известкованной подзолистой почве больший
урожай был получен при внесении нитратного азота. При внесении нитра-
та натрия и хлористого калия (а не сернокислого), содержание эфирного
масла в плодах выше (Хотин, 1957). Азот в вариантах N, NP, NK и NPK
стимулирует рост растений и образование хлорофилла, Р и К в вариантах'
Р, К и РК тормозят синтез хлорофилла. Влияние удобрений на эфиромас-
личность не велико: выше контроля она была на вариантах N, NK и NP (Ра-
жинскайте, Славенас, 1963).
Тмин индифферентен к кислотности почв (по шкалам Элленберга),
произрастает и на очень кислых и на щелочных почвах (1-13-я ступень по
Цыганову), однако оптимум находится на почвах в диапазоне от слабокис-
лых до субацидофильных (7-я). По данным А.А. Глущенко (1963), тмин от-
рицательно реагирует на кислую реакцию почвы. Предпочитает хорошо
аэрируемые, мелкопесчаные почвы (4-я ступень шкалы Ландольта). Для
Мариийской АССР указывается приуроченность к глинистым почвам и
мергелям (Подымов, Суслов, 1990). В Приокско-Террасном заповеднике
предпочитает более тяжелые, хорошо увлажненные почвы на заливных лу-
гах р. Оки (Смирнов, 1958). В культуре хорошо растет на суглинистых и
супесчаных почвах. Способен заселять речные песчаные наносы и голую
почву у полей и огородов, галечные обнажения долин рек. Выдерживает
присыпание песком до 10 см (наши данные). Растет по дорогам и нарушен-
ным местам (Смирнов, 1958).
127
Фитоценология
Тмин обыкновенный произрастает в лесной и лесостепной зонах, в го-
рах по долинам рек, изредка на лугах степной зоны. Встречается на пой-
менных, суходольных, субальпийских лугах, в разреженных лесах, на
опушках и лесных полянах, нередко по склонам и дну оврагов, на дегради-
рованных пастбищах, вдоль дорог, на залежах.
Тмин обыкновенный является диагностическим видом порядка
Arrhenatheretalia R. Тх. 1931 класса Molinio-Arrhenatheretea R. Тх. 1937 ет
R. Тх. 1970 (Matuszkiewicz, 1981), это мезофильные вторичные послелес-
ные луга умеренной зоны Евразии, формирующиеся на месте широколист-
венных лугов на хорошо дренированных минеральных почвах. Оптималь-
ными условиями для его произрастания являются суходольные и влажные
луга на богатых почвах с переменным увлажнением и слабым или умерен-
ными аллювиальными отложениями. На Дединовских пойменных лугах р.
Оки тмин в больших количествах произрастал на лугах высокого и средне-
го уровня в прирусловой и центральной пойме (Серебрякова, 1953; Атлас
ареалов..., 1980; Растительные ресурсы, 1988). На пойменных лугах реки
Угры в Калужской области его особенно много на пастбищах в прирусло-
вой пойме среднего и высокого уровня. Он входит как ассектатор в различ-
ные варианты разнотравно-злаковых и злаково-разнотравных лугов сено-
косов и смешанных угодий и как содоминант вместе с ним на прогоне ско-
та господствовали Festuca pratensis L., F. rubra L., Trifolium repens L.,
Taraxacum officinale Wigg.
Тмин обыкновенный в лесной и лесо-степной зонах входит в состав
травостоев мезофитных злаково-разнотравных и разнотравно-злаковых
лугов, где кроме злаков {Phleum pratense L., Festuca pratensis Huds, Poa
pratensis L. и Agrostis gigantea Roth.) большую роль играет мезофитное
разнотравье {Achillea millefollium L., Geraniumpratense L., Ranunculus acris
L., Rumex acetosa L., Trifolium medium L., Lathyruspratensis L. и др.). На бо-
лее увлажненных лугах состав разнотравья формируют Polygonum bistorta
L., Sanguisorba officinalis L., Veronica longifolia L., Geum rivale L., Seseli
libanotis (L) Hoch и др. (Атлас ареалов..., 1980). В Прибалтике выделена
обыкновеннотминная формация среди гликомезофитных формаций в
группе формаций мелкоразнотравных в классе формаций настоящие луга
(Матвеева, 1960, 1965).
В горных областях Средней Азии тмин обыкновенный встречается в
сухостепном, субальпийском и альпийском поясах, достигая высоты 3500
м над уровнем моря. В долинах рек входит в состав луговой растительно-
сти надпойменных террас. Низкие аллювиальные террасы в сухостепном
поясе Тянь-Шаня, где встречается тмин, периодически затопляются при
подъеме воды в реках в результате таяния снегов и ледников. Нередко
тмин встречается на поливных сенокосах.
128
На Кавказе он встречается от среднего горного до субальпийского поя-
са. Растет налесных опушках, травяных склонах, вдоль ручьев и на сорных
местах.
В Средней Азии, на Карпатах и на Кавказе массовое распространение
тмина в горных районах связано с сильно нарушенными выпасом пастби-
щами.
Средообразующая способность тмина невелика: его низко располо-
женные розеточные листья, ажурные листья и некрупные соцветия генера-
тивных побегов могут затенять только всходы и растения самого низкого
яруса.
Тмин не обладает эдификаторными свойствами, часто выступает в ро-
ли ассектатора, хотя в отдельные годы на сильно нарушенных местах обра-
зует микрогруппировки с высоким обилием или даже является содоминан-
том в сообществах. По типу поведения может быть оценен как эксплерент
или, по классификации Т.А. Работнова (1978), как ценофлуктуент. Высо-
кая семенная продуктивность, массовое прорастание семян и хорошая
приживаемость всходов в благоприятных условиях делает возможным бы-
строе размножение вида и захват пустых мест, которые могут образоваться
при отмирании других растений по разным причинам. По нашим данным,
при длительном сохранении благоприятных условий доминирование бы-
вает продолжительным: на прирусловой пойме р. Угры на прогоне скота
тмин сохранял высокое обилие в течение 20 лет. На культурном пастбище
в Эстонии, созданном на месте семенников тмина, этот вид сохранял высо-
кое обилие (13 % урожая) до 18 лет (Тоомре, 1966).
Возрастные спектры ценопопуляций тмина левосторонние, с абсо-
лютными максимумами на одной из групп прегенеративного периода или
редко на молодых генеративных (табл. 1, рис. 2). Погодичная и посезон-
ная динамика заключается в перемещении максимума с одной возрас-
тной группы на другую (коэффициент вариации возрастности неве-
лик— 35). Чаще всего максимум приходится на ювенильные или имма-
турные растения, что объясняется значительной гибелью особей в этих
возрастных состояниях: смертность j — 50-80 %, im— 50 %, т. е. в вир-
гинильное состояние переходит небольшая часть ценопопуляции (около
20 %), из которых отмирает до 50 %. Если принять все маркированные
особи за весь срок наблюдений за 100 %, то в молодом состоянии (j+im)
отмирает до 80 %, остальные 20 % во взрослом состоянии (v, gl). Период
полужизни тмина (время, за которое численность ценопопуляции умень-
шается в два раза (Harper, 1977) варьирует от 1,8 до 12 месяцев. Преобла-
дание молодых генеративных растений можно наблюдать редко, только в
течение одного месяца цветения и там, где инвазия происходила одномо-
ментно: на залежах, сильно нарушенных местах. После отцветания про-
исходит гибель всей или почти всей ценопопуляции, а на следующий год
или. через несколько лет совершается новая инвазия (Ермакова, Сугорки-
129
Таблица!
Динамика возрастной структуры
ценопопуляции Carum carvi на лугах прирусловой поймы
р. Угры (Калужская область) в 1980-2001 гг.
Год Средняя плотность, особей на м2 Возрастное состояние, %**
j im V g
1980 104,2 37,6 35,4 10,5 16,5
1984 94,4 12,6 31,4 34,5 21,5 .
1985 37,6 16,5 43,1 20,7 19,7
1986 140,8 16,5 31,1 36,1 16,3
1987 198 57,8 24,2 7,8 10,2
1989 75,5 75,7 19,0 1,3 4,0
1990 45,8 37,7 55,3 0,4 6,6
1991 19,2 11,6 54,7 23,2 10,5
1994 45,2 57,3 30,2 5,2 7,3
1997 32,0 23,7 64,2 3,4 8,7
1999 41,2 9,3 42,4 23,9 24,4
2000 11,2 28,4 36,4 8,0 27,2
2001 6,16 48,7 36,8 2,7 11,8
Примечание: ценопопуляция изучена на участке луга с «прогоном скота» с до-
минированием Festuca rubra, F. pratensis, Роа angustifolium, Trifolium repens,
Carum carvi, Achillea millifolium, Taraxacum officinale.
130
50
1980 1984 1985 1986 1987 1989 1990 1991 1994 1997 1999 2000 2001
Рис. 2. Динамика средней численности и возрастной структуры
ценопопуляции Carum carvi на лугах прирусловой поймы р. Угры
(Калужская область) в 1980-2001 гг. на площадках 0,25 м2 с прогоном скота
131
на, 1985). Обычно же на лугах незначительное семенное возобновление
происходит ежегодно, но обильнее через год.
Максимальная плотность ценопопуляций, по нашим наблюдениям, бы-
ла 92 особи на 0,25 м2, средняя плотность варьирует на разных участках от
0,03 особей до 49,5, варьирование за год 60-100 %. Сильное изменение
плотности можно объяснить неодинаковым поступлением молодых осо-
бей в ценопопуляции в разные годы, а так же гибелью их во всех возрас-
тных состояниях.
Биологическая продуктивность
Наибольшая зеленая масса листьев у тмина образуется в виргинильном
состоянии, в генеративном состоянии общий вес возрастает за счет генера-
тивных побегов, особенно в период обсеменения (табл. 2, 3). Наибольшую
надземную массу растение образует на залежах, на речных аллювиальных
наносах, нарушенных участках лугов и в посевах на богатых почвах. Фито-
масса побеговой части особи, равная у тмина приросту надземной биомас-
сы, составляет от 0,7 до 10,4 г на 1 м2, ценопопуляции — от 0,04 до 63,2 г,
фитомасса подземной части особи — от 0,2 до 1,2 г, ценопопуляции около
16 г.
Таблица 2
Средний воздушно-сухой вес (г) 1 особи
Carum carvi разных возрастных состояний на пойменных лугах
р. Угры (Калужская обл.) в 1984 г.
Возрастные состояния Пастбище Прогон скота
j 5,5+0,5 —
im 25,7+2,5 112,0+39,5
V 92,6+18,2 216,1+25,3
gl 1012,1+4,2 893,5+1,6
Урожай плодов на 1 м2 в наших условиях был от 345 до 45550 штук. По
литературным данным с 1 га получают 7,5-15 ц/га и до 20 ц/га в благопри-
ятные годы. Вес 1000 плодов — 2,3-3 г, в 1 г содержится 330-500 плодов
(Ростовцев, 1895; Беляева, 1946; Губанов идр., 1993).
132
Таблица 3
Аллокационный спектр фитомассы надземной части
генеративного растения Carum carvi на пойменных лугах
р. Угры (Калужская обл.) в 1984 г.
Части растения Пастбище Прогон скота
Розеточные листья . 1,0% 2,7 %
Стебли 23,5 % 25,3 %
Соцветия 75,5 % 72,0 %
Исследования Д. Ражинскайте (1967) показали, что применение мине-
ральных удобрений способствовало образованию хлорофилла в листьях
тмина. Самое большое количество хлорофилла у тмина в первый год жиз-
ни установлено автором в посевах в варианте NP (632,4 мг %), самое низ-
кое — в варианте К (450,9 мг %). Высокое содержание хлорофилла в вари-
антах с N способствовало интенсивному фотосинтезу, быстрому образова-
нию органических веществ и высокому урожаю. Самый высокий урожай
надземной массы и плодов получен в варианте NP, где урожай надземной
массы составил 208-293 %, а плодов — 116-223 % по сравнению с контро-
лем, а в вариантах NPK, N и NK незначительно ниже, чем в варианте NP.
Растения, удобренные только калийными удобрениями, отличались более
низким урожаем плодов по сравнению с неудобренными. При полном
удобрении самый большой урожай плодов (19,81 ц/га) и зеленой массы по-
лучен при N60 Р80 К45, хорошие показатели были при N40 Р60 КЗО, N60
Р80 К15, N60 Р40 К45. Минеральные удобрения оказали более или менее
выраженное отрицательное влияние на эфиромасличность плодов тмина.
Только в варианте NP установлено более высокое содержание эфирного
масла, чем в контроле. Больше всего (до 85,71 % по сравнению с контро-
лем) снизилась эфиромасличность плодов в вариантах Р, РК и К. Выход
эфирного масла с 1 га был выше контроля во всех вариантах с азотом. Са-
мый высокий выход—110-141 кг/га в варианте NP, а в варианте
NPK — 90-123 кг/га. Выход эфирного масла ниже контрольного получен в
варианте К.
133
Консортивные связи
У тмина в условиях Центрального Предуралья в корнях обнаружена эн-
дотрофная микориза, образованная эндотрофным грибом, относящимся к
классу фикомицетов сем. Endogonacea и роду Endogone (Крюгер, 1961).
Тмин опыляют многие насекомые (91 вид), так как открытые цветки с
доступным нектаром привлекают неспециализированных насекомых опы-
лителей (Демьянова и др., 1977). Среди них преобладали двукрылые
(38 видов): сем. Syrpidae (12 видов), Tachinidae (11 видов) и Caliphoridae
(4 вида). Отряд Hymenoptera был представлен 49 видами, относящимся к
13 семействам. Из жесткокрылых отмечено 4 вида из трех семейств.
Медоносное растение (Бабарыкина, 1992): в условиях северо-запада
Нечерноземной зоны России сахаропродуктивность цветков тмина неоди-
накова на протяжении периода цветения. Наиболее высок этот показатель
в фазе начала и массового цветения, к концу цветения нектаровыделение
сокращается. За период цветения сахаропродуктивность тмина колебалась
от 30,3 до 32,2 кг/га, в пересчете на медопродуктивность — от 37,9 до
40,2кг/га.
Опасным и главнейшим вредителем тмина является тминная моль
Depresaria daucela Den et Schiff. В условиях лесостепной Украины, где по-
тери от нее составляют 40-59 %, моль откладывает яйца с конца апреля до
начала мая на внутренней стороне черешков листьев. Появившиеся через
3-4 недели гусеницы выедают небольшие углубления в черешках и стеб-
лях тмина. Достигнув старших возрастов, они мигрируют вверх по стеблю,
прячутся во влагалищах листьев, потом переселяются в зонтики, обвола-
кивают его паутиной, отъедают цветки, лучи зонтика и плоды. Соцветия
темнеют и засыхают. Одна гусеница способна повредить несколько зонти-
ков. Начало окукливания совпадает с окончанием цветения тмина. Гусени-
цы прогрызают отверстия в стеблях и там окукливаются. Через 3—4 недели
в конце июня-начале июля вылетают бабочки и разлетаются в поисках
мест зимовки. Они зимуют под отставшей корой деревьев, в скирдах соло-
мы, в неровностях стен, под черепицей крыш и т.п. (Чумак и др., 1977).
Разработан метод долгосрочного прогноза численности тминной моли
и найдены биологические способы борьбы с ней с помощью 7 видов энто-
мофагов паразитов из сем. Ichneumonidae, личинки которых высасывают
содержимое пораженных яиц моли. Специализированные формы
Trichogramma pintoe, выпущенные в начале яйцекладки моли, снижают ко-
личество яиц последних на 46-89 %. Затраты на эти приемы борьбы окупа-
ются в 10-15 раз (Назаров, 1985а, б; 1987). Автор метода пришел к важно-
му выводу, что при численности гусениц тминной моли 24—39 экземпля-
ров на м2 при 5 % уровне допустимых потерь урожая, проведение каких
либо защитных мер экономически не оправдано. При очень сильной зара-
женности поля в краевой полосе до 100 м шириной, куда слетаются бабоч-
134
ки весной из зимних укрытий, следует применять такие инсектициды как
фазалон и базудин, которые менее токсичны для энтомофагов и пчел и
обеспечивают высокую гибель гусениц. Обработку проводят ночью за 1-2
дня до начала цветения тмина. Кроме тмина тминная моль повреждает
анис, фенхель, пастернак, сельдерей и другие зонтичные. Но полный цикл
развития моль проходит только на тмине (Чумак и др., 1977).
Другие вредители тмина — зонтичная моль {Depressaria depresselba
Hb.) и тминный клещ {Eriophyes peucedanii Carw). Клещ зимует в почках и
пазухах розеточных листьев, весной расселяется на посевы, накапливается
на старых полях тмина. Поврежденные им листья курчавятся, цветки зеле-
неют и приобретают вид плотных, махровых скоплений и не образуют
плодов. Клещ заносится ветром, водой, животными, орудиями обработки и
насекомыми (зонтичным клопом, тлями) и опылителями тмина. Меры про-
филактики: 1) новые посевы размещать, возможно дальше от его старых
плантаций; 2) возделывать на одном поле не более 2—х лет; 3) тщательно
запахивать стерню и корни тмина. Для уничтожения клеща опыляют гек-
сахлораном (Глущенко, 1963; Obarski, 1965).
Может поражаться грибными заболеваниями: мучнистой росой, аль-
тернариозом, антракнозом, церкоспорозом, вызываемыми грибами
Erysiphe polygoni, Mycocentrospore acerina, Cercospoza carvi (Zacha, 1965;
Patel, Desai, 1971; Mathur, Masih, 1971; Evenhuis и др., 1997). Корни тмина
сильно повреждаются мышевидными грызунами.
Тмин в молодом возрасте хорошо поедается на пастбище крупным ро-
гатым скотом, лошадьми, овцами. Отлично поедаются на пастбище листья
и соцветия в период цветения (Тоомре, 1966). В свежем виде включают в
корм кроликам, особенно растущему молодняку. Отлично поедается мара-
лом; хорошо — пятнистыми оленями в кормушках, в небольшом количе-
стве — рябчиком. Семена тмина являются хорошим кормом для голубей и
куропаток, диких гусей. Тмин переносит умеренный выпас (Ларин, Пала-
марчук, 1949; Агабабян, 1956; Паламарчук, .196О; Иванов, 1975). По дан-
ным Н.С. Конюшкова (1930), тмин встречается на пастбищах всех ступе-
ней выпаса. По сравнению с сенокосом при слабой интенсивности выпаса
весовое обилие, покрытие и встречаемость увеличиваются, достигают
максимума при средней и уменьшаются при сильной. Тмин отлично ми-
рится с вытаптыванием (Смирнов, 1958) и выдерживает многолетнее 5-7,
разовое стравливание на культурных пастбищах (Тоомре, 1966).
Хозяйственное значение
Надземные части тмина в фазах цветения и начала плодоношения со-
держат 9,67 % протеина, 5,85 % белка, 2,03 % жира, 35,52 % клетчатки,,
46,17 % безазотистых веществ, 6,61 % золы, 2,36 % окиси кальция и 0,78 %
окиси фосфора (Сабардина, 1952). Листья до бутонизации имеют в своем
135
составе 9,30 % гигроскопической воды, 18,12 % протеина, 13,37 % белка,
2,66 % жира, 16,62 % клетчатки, 52,59 % беззазотистых веществ, 10,01 %
золы, 0,1 % фосфора и 1,39 % кальция от абсолютно сухого веса (Матвее-
ва, Знаменская, 1960). По данным Р.И. Тоомре (1966), тмин содержит 19 %
клетчатки и 18,1 % протеина. В 1 кг сухого вещества находится 401 мг ка-
ротина и 0,74 мг кобальта. Тмин накапливает кобальта больше, чем даже
бобовые (Матвеева, Знаменская, 1959, 1960а), поэтому может служить
обогатителем кормов этим элементом, снижение которого в кормах до
0,05-0,06 мг/кг вызывает эпидемические заболевания скота, ведущие к
смерти (Берзинь, 1952 цит. по: Матвеева и др., 1964; Пейве, 1954 цит. по:
Матвеева, Знаменская, 1960а).
Из травы и цветков выделены флавоноиды — кверцетин, кемпферол,
изорамненин и полиины (ацетиленовые соединения) (Horhammer et al.,
1958; Bohlman et al., 1961; Crowden, 1969 цит. по: Атлас ареалов ..., 1980).
Плоды тмина содержат 2,6-7,67 % эфирного масла (основные его компо-
ненты — карвон и лимонен), 18,4—21,18 % жирного масла с высоким содер-
жанием петроселиновой кислоты и дубильные вещества (Зарайская, Бори-
сюк, 1956; Гроссгейм, 1967; Вульф, Малеева, 1969 цит. по: Атлас ареалов ...,
1980; Растительные ресурсы..., 1988). В последнем источнике приводится
наиболее полное описание химического состава всех органов тмина.
Динамика эфирного масла изучена Д. Ражинскайте (1967, 1968): на
втором году жизни максимум содержания эфирного масла в листьях при-
ходится на время массового цветения — 305,01 мм3/100 г сухого веса; в
стеблях в фазе бутонизации — 206; в бутонах в фазе бутониза-
ции — 287,35; в цветках в начале цветения — 835,226, а во время массово-
го цветения — 540,3; в плодах в молочной спелости — 11246,73. В созре-
вающих плодах содержание эфирного масла падает, но в фазе твердой спе-
лости, когда плоды буреют и начинают осыпаться, содержание его
несколько увеличивается. Но так как темпы накопления сухого вещества
превышают темпы образования эфирного масла, создается видимость бес-
прерывного уменьшения содержания эфирного масла, к тому же некоторая
часть его улетучивается. С созреванием плодов меняется и качество эфир-
ного масла: увеличивается содержание окисленных компонентов, главным
образом карвона. Поэтому убирать плоды лучше в фазе восковой спелости
до их осыпания (Ражинскайте, 1967) или через 3-4 дня от начала молочной
спелости (Ражинскайте, 1965) или за 3-5 дней до начала побурения семян
на центральном зонтике (Хотин, 1948) или в начале побурения плодов на
центральных зонтиках (Губанов и др., 1993).
В раскрытых цветках центрального зонтика и зонтиков I—II порядка
верхних ярусов эфирного масла обнаружено больше, чем в цветках II—IV
порядка ветвей нижнего яруса, цветущих в период созревания плодов в
верхних ярусах (Ражинскайте, 1968).
У тмина в листья обнаружена аскорбиновая кислота обратимо-окис-
136
ленной формы в количестве 5339,5 мг на 1кг сухого вещества, что не по-
зволило исследователям отнести этот вид к высоковитаминным (Муравь-
ев, Баньковский, 1947).
Плоды тмина действуют раздражающе на вкусовые рецепторы, реф-
лекторно повышают функцию пищеварительного аппарата, усиливают его
тонус и перистальтику, снижают процессы гниения и брожения в кишеч-
нике, облегчают переваривание и усвоение жирных и тяжелых блюд. В на-.
учной медицине настой плодов используется при гипофункции желудка и
кишечника, метеоризме. Тмин входит в фармакопеи многих стран: в тибет-:
ской медицине применяется при неврозах, нарушении обмена веществ, ин-
токсикациях, опухолях, болезнях глаз; в китайской и индийской медицине
используется для лечения болезней органов желудочно-кишечного тракта,
хроническом холецистите, как лактогенное. В фармакопею СССР (IX изд.,
N 213) введены вполне зрелые плоды (Соболевская, 1972).
В народной медицине применяется как седативное, отхаркивающее,
диуретическое, при метиоризме; плоды входят во многие сборы: аппетит-
ный, желудочный, ветрогонный, слабительный, седативный; используют-
ся при бронхиальной астме, асците, лихорадках, экземе и как антигель-
минтное (Роллов, 1908; Халматов и др., 1984). В монгольской ветеринарии
применяется при диарее у молодняка, тимпатите, лихорадке, кашле (Дико- t
растущие полезные..., 1985).
Эфирный экстракт в эксперименте проявлял антибактериальную ак-
тивность, эфирное масло оказывало противотуберкулезное действие.
Эфирное масло из плодов используется для ароматизации лекарственных
препаратов (Агабабян, 1956; Атлас ареалов..., 1980; Растительные ресур-
сы..., 1988; Губанов, 1993; Губанов идр.,1993; Фроленко, 1993).
Плоды тмина широко применяются в пищевой промышленности при
хлебопечении, изготовлении кондитерских изделий, сыров, колбас, при
квашении капусты. Используют их также в ликерном и парфюмерном про-
изводствах, мыловарении, в табачном махорочном, в домашнем хозяйстве
при засолке капусты, помидор, и огурцов.
Молодые витаминизированные листья употребляют как салаты и пря-
ность. Корни однолетних растений отваривают, тушат, обжаривают, до-
бавляют в гарниры и супы как пряность, сырыми в салаты; на зиму их заго-
тавливают с уксусом и медом или сахаром. Молотые семена добавляют в
салаты, сыры, паштеты, а в целом виде во все жирные блюда для облегче-
ния пищеварения (Роллов, 1908; Беляева, 1946; Агабабян, 1956; Синяков,
1990; Лебедева, 1992).
После извлечения эфирного масла из плодов жмых содержит до 19 %
протеина и до 2,3 % жирного масла. Жирное масло используется для тех-
нических целей, жмых — в качестве концентрированного корма для скота.
Дистилляционные остатки тмина прекрасно поедаются животными: коэф-
фициент переваримости протеина 60 %, жира 97 %, безазотистых веществ
137
76 %, клетчатки 85 %. Отрицательной особенностью этого корма является
резкий запах, который передается молоку (Болотин и Зафрен, 1939).
Тмин — хорошее кормовое растение, приравниваемое в ранние перио-
ды развития по питательности к луговому клеверу. В сене хорошо поедает-
ся. Примесь его в сене повышает удои коров (Дмитриев, 1904). При высоком
содержании протеина и белка имеет очень небольшое количество клетчат-
ки, мало увеличивающееся по мере перехода в более зрелое состояние. От-
рицательная особенность — осыпание листьев при сушке (Матвеева,
1960а). Тмин считается желательным в виде небольшой примеси (до 5 %) на
лугах и пастбищах, как растение, способствующее поеданию трав, пищева-
рению и обмену веществ скота. Семенами его кормят молочный скот для по-
вышения удойности (Агабабян, 1956; Шилов, 1978). Семена тмина рекомен-
дуется добавлять при посевах кормовых смесей, особенно клевера, так как
снимается опасность вздутия живота у коров при поедании влажного моло-
дого красного клевера (Беляев, 1905); а так же включать в состав травостоя
низинных лугопастбищных угодий (Trzaskos, 1992).
В листьях и плодах содержится ничтожное количество дубильных и
красящих веществ (Шлыков, 1932). Из семян тмина получен ингибитор
роста талент, используемый для борьбы с прорастанием клубней картофе-
ля при хранении (Diepenhorst, 1995).
Медоносное и перганосное растение (Глухов, 1950; Демьянова и др.,
1989; Бабарыкина, 1992). Тмин введен в культуру задолго до нашей эры в
Малой Азии, с IX века нашей эры возделывается в Европе. Промышлен-
ные посевы в ближнем зарубежье есть на Украине, в Белоруссии, Латвии и
Эстонии, в России: в Осетии, Воронежской, Ростовской, Ярославской, Ор-
ловской, Курской и Тульской областях. Но большая часть плодов в России
собирается в природных популяциях в Архангельской, Вологодской, Вят-
ской, Пермской и Воронежской областях. До 1914 года из России вывози-
ли до 2500 центнеров семян (Буравлев, 1932).
Разработаны технологии промышленного выращивания тмина (Бурав-
лев, 1932; Беляева, 1946; Агротехнические указания..., 1950; Глущенко,
1963; Ларин, 1978; Гайдамака, 1986)., на приусадебных участках и тепли-
цах (Кардашина, 1984; Лебедева, 1992; Фроленко, 1993). Выводятся сорта
тмина (Кузьмич, 1986). Посевы тмина возможны по всей подзолистой зоне
с гарантированным урожаем не ниже 6-7 ц/га, но на черноземах можно по-
лучать больше (Хотин, 1948, 1957).
Работа выполнена при финансовой поддержке
Федеральной целевой программы «Интеграция»
проект 5.1. № 439; РФФИ и администрации
Калужской области (грант № 01-04-96006).
И. Б. Седых
ПЕРВОЦВЕТ ВЕСЕННИЙ
Номенклатура и внутривидовая систематика
Primula veris L., 1753, Sp. PL: 142; Сырейщиков, 1910, Илл. фл. Моск, губ.,
3 : 28; Ан. Федоров, 1952, Фл. СССР, 18 : 145; он же, 1964, в Маевский, Фл.,
изд. 9 : 404; Скворцов, 1966, Опред. раст. Моск, обл.: 261; Valentine a. Kress,
1972, Fl. Europ. 3:16; Ан.А. Федоров, 1981, Фл. европ. части СССР, 5 : 68;
Губанов и др., 1995, Опред. сосуд, раст. Центра Европ. России : 400.—
Р. officinalis (L.) Hill, 1765, Veg. Syst. 8 : 25; Кауфман, 1866, Моск, фл.:
403.— Первоцвет весенний (семейство Primulaceae — Первоцветные).
Согласно одной из современных точек зрения (Федоров, 1981) вид
Primula veris L. включает два подвида: subsp. veris L. (чашечка после цвете-
ния разрастается мало) и subsp. macrocalix Bge. (чашечка сильно разрос-
шаяся, как бы вздутая). Последний подвид распространен в южных и вос-
точных районах Русской равнины, а также в Сибири, на Кавказе и в Кры-
му. За пределами бывшего СССР —в северном Иране и Афганистане.
Взаимоотношения данных подвидов нуждаются в дальнейшем изучении.
Ниже охарактеризован типовой подвид.
Географическое распространение
Элемент голарктической флоры, центр его происхождения — Цен-
тральный Китай (Невский, 1937). Северная граница ареала проходит около
30° с. ш., южная — вблизи 43° ю. ш. Ареал занимает 74 % территории Ев-
ропы, в основном вид распространен в странах западной и средней Европы
(Tutin, 1952; Flora Europaea, 1972; Heller, Heyn, 1986), а также в Дании,
Швеции, Норвегии и Финляндии (Федоров, 1981). Восточная часть ареала
первоцвета весеннего охватывает лесную и лесостепную зоны России, Ук-
раины, Молдавии, Белоруссии и Прибалтики (Флора Ленинградской об-
ласти, 1965; Раменская, Андреева, 1982; Гейдеман, 1986; Табака, 1988).
Ареал вида доходит до Урала, где типовой подвид subsp. veris замещается
subsp. macrocalix. Согласно Флоре СССР (1952) и Флоре Европейской час-
139
ти СССР (1981) первоцвет весенний распространен в Ладого-Ильменском,
Верхне-Днепровском, Верхне-Волжском, Волжско-Донском и Ниж-
не-Донском районах (Федоров, 1952; Попов, 1959; Атлас лекарственных
растений СССР, 1962).
Как заносной, вид встречается в Канаде: южный Онтарио, Квебек, Ван-
кувер (Scoggan, 1979).
В Московской области распространен во всех районах (Ворошилов и
др., 1966), однако становиться редким, и нуждается в охране (Воликов,
Тукчев, 1992).
Морфологическое описание
Первоцвет весенний — многолетнее, травянистое, короткокорневищное
розеточное растение. Корневище образовано короткими округлыми годич-
ными побегами (0,47x0,2 мм в диаметре) и почти полностью покрыто плот-
ными клиновидными остатками листьев, отчего выглядит шиповатым, а
также несколько извилистым благодаря симподиальному нарастанию.
Корневая система взрослого растения образована придаточными кор-?
нями. Корни (10)20-45(53) см длиной и 0,23±0,07 см в диаметре у основа-
ния; белые, прочные и ветвятся до IV-V-ro порядков. Обычно проникают
на глубину до 45-50 см. Корни формируются как узловые (наиболее круп-
ные) и надузловые; в последнем случае корень либо оказывается между
листьями, либо проходит через плотный черешок листа.
Листья первоцвета весеннего очередные, с крылатым трехгранным че-
решком и широкой яйцевидно-овальной или округлой пластинкой, отде-
ленной от черешка более или менее выраженным сужением. Пластинка
листа цельная, слабо городчато-зубчатая, морщинистая, весьма рельефная,
с сильно выступающими жилками на нижней стороне. Листья имеют гида-
тоды с одним отверстием (Эзау, 1969).
Соцветие — терминальный открытый фрондозный зонтик, расположен-
ный на безлистной цветочной стрелке. Цветочная стрелка безлистная, около
24,0±2,1 см высотой, светло-зеленая, опушенная короткими, неветвистыми
волосками. Соцветие состоит из 8-15 цветков, расположенных по спирали и
более или менее отклоненных в одну сторону. Цветки закладываются и раз-
виваются акротонно в пазухах прицветников. Прицветники светло-зеленые,
цельнокрайние, ланцетовидные, 0,5см длиной, в основании 0,1 см шириной.
Встречаются двухъярусные соцветия, что имеет место и у других видов при-
мул (например, у Р. japonica). Вероятнее всего такой тип возникает благода-
ря удлинению одного из средних междоузлий укороченной части цветоноса.
Такие соцветия обычно формируются под пологом леса.
Чашечка светло-зеленая, гранистая, рассеченная на треть, с острыми
зубцами, вздувается после цветения. Венчик пятичленный, спайнолепест-
ный, трубчатый; желтого цвета с оранжевым пятном у основания долей, с
140
вогнутым плоским отгибом, диаметр — 1,2±0,6 см. Андроцей из пяти ты-
чинок, противолежащих долям венчика. Завязь верхняя, столбик один с го-
ловчатым рыльцем. Гинецей лизикарпный, плацентация относится к верх-
нему колончатому типу.
Плод коробочка, которая вскрывается сутурально; вскрывание не-
полное, зубцами. Коробочка вдвое короче чашечки. Семена мелкие
(1,3±0,8 мм в диаметре), черно-коричневые, округлые, слегка сплюснутые,
по краю чуть бугорчатые, с эндоспермом; зародыш прямой.
Диплоидный набор хромосом 2п=22 (Хромосомные числа ..., 1969).
Онтогенез
Первоцвет весенний плодоносит с июня по август. Попавшие в почву се-
мена прорастают весной следующего года, прорастание надземное. Лабора-
торная всхожесть семян изучалась при разных режимах (табл. 1). Семена
были собраны 26.07 в вейниковом березняке за 3-5 дней до проведения экс-
перимента. Проращивались на крупнозернистом песке в чашках Петри.
Таблица 1
Всхожесть семян Primula veris при разных режимах
Темпе- ратура, С* Время экспозиции, дней Дата посева Дата появления всходов Число дней** Всхожесть, %
+22° 0 26.07 — — 0
30 30.08 14.10 41 1,2
50 05.10 22.11 48 0,8
70 15.11 25.12 40 4,7
-5° 30 30.08 05.10 36 46
50 05.10 26.10 21 44
70 15.11 08.12 23 58
Примечание-. * температура, при которой выдерживались семена
до посева;
** число дней от времени посева до появления
массовых всходов.
141
Рис. 1. Возрастные состояния Primula veris.
Обозначения: г — главный корень, с — семядоли
142
Нертратифицированные семена прорастают через 40-48 дней, причем
всхожесть невелика. В случае же стратификации при низкой температуре
значительно возрастает процент всхожести семян, и уменьшаются сроки
их прорастания.
Сведения о жизнеспособности семян в природе различны. По одним
данным (Кудрявец, Петренко, 1993), семена теряют всхожесть через год.
По другим (Рысина, 1973) — опавшие семена прорастают не одновремен-
но и в почве сохраняют всхожесть до 2-3 лет.
Проростки — небольшие растения с двумя семядолями, бесцветным
гипокотилем 1,2±0,3 см в длину, главным корнем и несколькими боковы-
ми ветвящимися корнями (рис. 1). Главный корень белый, слегка зеленею-
щий. Семядоли зеленые, округло-овальные, цельнокрайние, внезапно су-
жаются в короткий черешок.
Сформировавшиеся ювенильные листья имеют тонкий черешок и ок-
ругло-яйцевидную, с зубчатым краем пластинку (рис. 1). Ювенильное рас-
тение обычно несет 4—6 листьев и в лабораторных условиях сохраняет
именно такое их количество в течение 10-15 дней. Уже на стадии пророст-
ка и, особенно у ювенильных растений начинают формироваться прида-
точные корни, но главный корень сохраняется. Первые 3-5 придаточных
корней образуются в основании черешков первых ювенильных листьев.
Также у этих растений происходит удлинение и утолщение главного и
придаточных корней.
Имматурные растения имеют розетку из 3-6 листьев переходной фор-
мы. Листья с продолговатой, овально-округлой пластинкой, слегка мор-
щинистой, с зубчатым краем, плавно преходящей в крылатый черешок.
Имматурные особи сохраняют систему главного корня, но он начинает от-
мирать.
Виргинильные растения имеют листья сходные с листьями генератив-
ных особей, но они с более округлой листовой пластинкой. Система глав-
ного корня отмирает.
У молодых генеративных особей корневище без следов отмирания и
розетка листьев с несколько заостренной верхушкой. Через 4—5 лет расте-
ние переходит в средневозрастное генеративное состояние. Средневозра-
стные генеративные особи отличаются наибольшей толщиной корневища
и для них характерно начало его отмирания. У этих растений длина и тол-
щина корней и корневища по сравнению с растениями других возрастных
состояний наибольшие, листья с более округлой верхушкой. Корневище
может ветвиться за счет пазушных почек побега текущего года. Розетки
П-го порядка также образуют терминальные цветоносы. В силу этого,
средневозрастные генеративные особи могут образовывать 2—3(4) цвето-
носа на одной особи. У старых генеративных особей ветвление корневища
происходит очень редко, оно становится более тонким и уменьшается вы-
сота розетки. Эти растения почти всегда имеют один цветонос.
143
Таблица 2
Морфометрические характеристики
возрастных состояний Primula veris (Московская область)
Показатели Возрастное состояние
Р j im V g ss s
Корневище:
Длина, см — • 0,3*0,05 1,8*0,13 4,2*0,2 5,1*0,4 4,7*0,7 2,1*0,3
Диаметр, см — 0,3*0,02 0,2*0,06 0,3*0,07 0,4*0,07 0,4*0,03 0,2*0,06
Корень:
Главный, длина, см 3,7*0,2 2,3*0,1 0,7*0,1 — — - — —
Придаточный, число — 4,1*0,2 10,4*0,4 31,4*2,3 14,7*1,3 18,0*3,2 10,2*1,7
Придаточный, диаметр, см — 3,6*0,5 6,3*1,4 10,2*3,1 44,7*2,8 14,0*1,4 9,7* 1,1
Листья:
Число 2 4,0*0,8 5,1*0,8 5,7*0,6 9,8*1,1 8,2*0,3 3,2*0,5
Длина пластинки, см 0,6*0,07 0,7*0,08 4,6*0,4 12,4*1,2 15,7*2,8 11,4*1,3 5,1*0,3
Длина черешка, см 0,2*0,04 2,7*0,6 2,3*0,4 2,2*0,7 4,2*1,1 2,7*0,5 1,1*0,2
Длительность возрастного состояния, мес. 0,5-1 5-12 2-3 2-36 60-108 ? ?
144
Субсенильные растения представлены отдельными партикулами. При
переходе в это возрастное состояние корневище становиться еще тоньше,
годичные побеги имеют толщину как у молодых генеративных растений,
корней становиться меньше и они более тонкие. Листья имматурного типа.
Длительность субсенильного состояния не выяснена, однако большинство
растений имели корневища, сформированные 6-7 многолетними частями
годичных побегов, что приблизительно указывает на продолжительность
этого этапа.
Сенильные растения имеют розетки, образованные 2-4 листьями юве-
нильного типа. Отличаются от ювенильных растений подземной частью
(темный, слабый годичный побег со следами отмирания). Эти растения по-
гибают через 1-3 года.
Некоторые морфометрические параметры растений разных возрас-
тных состояний представлены в таблице 2.
Ритм сезонного развития
Годичный побег первоцвета весеннего начинает развиваться как ось
П-го порядка на побеге предыдущего года. В пределах годичного побега
можно выделить три типа листьев, которые формируются в разные кален-
дарные сроки. Они отличаются по форме и величине (табл. 3).
Таблица 3..
Морфометрические характеристики листьев
генеративных особей Primula veris (Московская область)*
Листья Черешок, см Пластинка, см Число листьев**
длина . ширина длина ширина
Весенние 4,2±1,1 0,6±0,4 15,7±2,8 5,2±0,3 3-6
Летние 4,2±1,4 0,6±1,1 26,2±1,8 6,1±0,5 4-5
Осенне- зимние 1,4±0,8 0,8±0,2 2,6±0,4 1,8±0,5 2-4
Примечание-. * березняк вейниково-разнотравный, 1997-2000 гт.,
** число листьев данного типа в системе годичного побега.
145
В конце апреля-начале мая формируется заложенное с осени терми-
нальное соцветие. Рост пазушной оси идет параллельно его развитию. По-
беговая система вида нарастает симподиально за счет пазушной почки
ближайшего к цветоносу листа. Новая розетка образована летним типом
листьев. Иногда в близкие сроки трогаются в рост пазушные почки ниже-
лежащих листьев, образуя 1-3 цветущие розетки, сформированные весен-
ним типом листьев.. В дальнейшем эти розетки либо гибнут, либо одна из
них (или единственная) также нарастает симподиально, что приводит к
ветвлению корневища.
Молодые листья побега П-го порядка копьевидны, с завернутыми на
нижнюю сторону краями листа, которые отличаются весьма равномерной
городчатостью. Несмотря на различную высоту (от 0,1 до 4 см в зависимо-
сти от степени развития), соотношение длин черешка и листовой пластин-
ки одинаково — приблизительно 1:4. Высота пазушной розетки на момент
цветения обычно составляет 4,5±0,8 см. После отцветания в июне (реже в
июле-августе) отмирают листья побега 1-го порядка и он замещается побе-
гом И-го порядка с летними листьями. В основании первых 4—5 листьев в
июле обычно формируются самые крупные корни — до 0,15 см в диамет-
ре. Благодаря контрактильной функции, они втягивают осенью растение в
почву. Вышерасположенные корни имеют меньший диаметр. Придаточ-
ные корни живут у растений 6-7 лет (Пошкурлат, 1962); далее они посте-
пенно отмирают и самые первые годичные побеги могут совсем не иметь
корней.
В конце сентября — октябре закладывается соцветие. В ноябре листья
летнего типа отмирают. Растение в конце ноября — начале декабря затор-
маживается в росте и уходит под снег. Почка здесь защищена осенне-зим-
ними листьями, сходными с чешуевидными (0,4±0,03 см в высоту). Лист в
данном случае неразвит и представлен в основном черешком. Когда вес-
ной растение трогается в рост, листья этого типа формируют округлую
листовую пластинку. Осенне-зимние сохраняются на растении до мая-ию-
ня. Зимой побег продолжает нарастать моноподиально. Было отмечено
максимальное удлинение апикальной почки на 1,3 см. Обычно ее длина в
середине декабря составляет в среднем 1,3 см, а в марте-2,5 см. По данным
Н.А. Прозоровского (1940), в период подснежного роста в степных услови-
ях Центрально-Черноземного заповедника длина листьев почки может
увеличиваться на 1-1,5 см.
Начиная с апреля-мая, начинается интенсивный рост побега и образу-
ется розетка, образованная листьями весеннего типа. По ритму развития
листового аппарата первоцвет весенний — зимне-зеленое растение (Шик,
1953).
146
Способы размножения и распространения
Первоцвет весенний может размножаться как семенным путем, так и
вегетативно, но преобладает последний способ. Он заключается в ветвле-
нии и последующем делении корневища. Обычно в этом случае развива-
ются почки двух ближайших к цветоносу листьев и формируются полно-
ценные цветущие розетки. Новые корневища располагаются под острым
углом друг к другу и первые 2-3 года нарастают вертикально. При пере-
гнивании соединяющих участков корневища или гибели старого новые
корневища обособляются, образуя клон. Очень редко встречались побеги,
которые возникли из спящей почки, заложившейся за 3-^4 года до этого.
Подобный способ был отмечен только для генеративных особей с сильным
повреждением верхушечной розетки.
Семенное возобновление первоцвета весеннего может происходить
только на открытых нарушенных местообитаниях (Grime et al., 1988; Лы-
сенко, 1994). По нашим данным, всхожесть семян в природных условиях в
лесных и луговых сообществах невысока. Высеянные семена прорастали
на второй год на открытых местообитаниях с легкими почвами и достаточ-
ным увлажнением. Проективное покрытие травостоя на таких участка бы-
ло низкое, а также почти отсутствовали мертвые остатки. Всхо-
жесть— 6-15 %. Установлено отрицательное влияние лесной подстилки
на прорастание семян первоцвета и последующее развитие проростков
(Лысенко, 1994). Семена высевались по ненарушенной подстилке, а также
в почву после полного удаления подстилки. В первом случае всхожесть со-
ставила 1,4 %, во втором — 11,8 %; выживаемость проростков — 7,1 % от
общего их числа и 70 % соответственно. Таким образом, семенное возоб-
новление первоцвета весеннего под пологом леса, а также в сформирован-
ных луговых сообществах затруднено. В то же время жизненность особей
гораздо выше именно в лесных сообществах, где обеспечено достаточное
затенение и влажность почвы и воздуха.
Для первоцвета весеннего характерна гетеростилия, при которой число
завязавшихся семян зависит от особенностей опыления. Если пыльца
длинных тычинок попадает на длинный пестик, то образуется почти в два
раза больше семян, чем при попадании пыльцы длинных тычинок на ко-
роткой пестик. В первом случае образуется 100 семян, во втором — 65
(Фегри, 1982).
Первоцвет весенний — баллист гвоздичного типа. Семена высыпаются
при раскачивании упругого прямостоячего цветоноса. Поскольку коробоч-
ка первоцвета вдвое ниже чашечки, последняя является барьером для про-
извольного высыпания семян, то есть играет регулирующую роль в диссе-
минации (Левина, 1957).
147
Экология
Первоцвет весенний растет в затененных и на открытых местообитани-
ях. Относится к полусветовым растениям (7-я ступень шкалы Элленберга).
В условиях хорошей освещенности, при достаточном увлажнении у перво-
цвета формируются наиболее крупные розетки (25-30 см высоты) с двумя,
реже тремя цветоносами. В местах со слабой освещенностью количество
особей на единицу площади меньше и жизненность их ниже. На первых
возрастных этапах (проростки, ювенильные и имматурные особи) потреб-
ность в свете невелика и растения лучше развиваются при затенении.
Первоцвет весенний предпочитает почвы между сухими и свежими
(4-я ступень шкалы Элленберга), избегает заболоченные почвы и места с
длительным затоплением (Grime et al., 1988). Находясь на открытых мес-
тах, растение располагается в небольших понижениях, западинах, вблизи
кустарников, на склонах оврагов.
Первоцвет весенний в Европе растет в основном на почвах бедных ми-
неральным азотом (3-я ступень шкалы Элленберга). Это могут быть каме-
нистые пустоши, скалы по берегу моря, песчаные дюны и вдоль насыпей
железных дорог (Grime et al., 1988). В России может расти и на богатых
почвах: типичных, выщелоченных и оподзоленных черноземах (Афанась-
ева, Голубев, 1962). Все изученные нами популяции были приурочены
только к богатым почвам. В Подмосковье вид растет на серых лесных
средне-оподзоленных и темно-серых слабооподзоленных глинистых поч-
вах. Высокая жизненность растений на дерново-подзолистых почвах так-
же была отмечена Т.А. Лысенко (1994).
Вид предпочитает хорошо аэрируемые нейтральных почвах (8-я сту-
пень шкалы Элленберга) с кислотностью от слабокислых (pH 5,0) до сла-
бощелочных (pH 8,4). Оптимальная кислотность — pH 6-7 (Grime et al.,
1988). В Подмосковных местообитаниях кислотность корнеобитаемого
слоя составляла рНВ0Д. =4,7-5.
Хорошо переносит морозы до -35 °C, для семян же низкие температу-
ры— необходимое условие прорастания. Растение хорошо развивается
только на прохладных почвах. Перегрев вызывает бледную окраску цвет-
ков, мелколистность, замедляет развитие новых розеток и сокращает сроки
цветения. Оптимальная температура приземных слоев воздуха —
10°-15°С (Ильинская, 1993).
Первоцвет весенний — стресс-толерантный вид (Grime et al., 1988), пе-
реносит умеренное сенокошение (Раменский и др., 1956).
Фитоценология
Первоцвет весенний — бореально-неморальный вид, произрастает в
лесной и лесостепной зонах. Преимущественно встречается в смешанных
148
Таблица 4
Возрастной состав популяций
Primula veris в разных фитоценозах Московской области
Фитоценоз Возрастное состояние, % Плотность, особей на м2
Р j im V g ss s
Березняк* — — 3 10 86 1 —' 12
Березняк, опушка** 0,5 1 2 1 94 1,5 — 8
Дубрава*** — 3 18 10 68 — 1 16
Ельник**** — — — — 100 — — 4
Сосняк***** — — ' — 8 85 1 6 14
Примечание'. * березняк орляково-разнотравный (сомкнутость полога 0,4);
** березняк вейниково-разнотравный (опушка вдоль дороги);
*** дубрава с липой медунициево-разнотравная (окно);
**** ельник-кисличник (окно);
***** сосняк с березой разнотравный (сомкнутость полога 0,3).
и лиственных лесах — в широколиственных, хвойно-широколиственных и
мелколиственных.
Некоторые авторы считают Primula veris в центральной Европе харак-
терным видом класса Querco—Fagetea Br.-Bl. et Vlieger in Vlieger 1937, не-
которые дифференцирующим видом союза Carpinion betuli Issler 1931 em
Mayer 1937 порядка Fagetalia silvaticae Pawl, in Pawl. e.a. 1928 того же клас-
са (Moravek, 1983; Neuhauslova-Novotna, 1964 цит. по Коротков, 1991). Это
мезофильные широколиственные и смешанные леса центральной и запад-
ной Европы.
Реже вид принимает участие в формировании лесостепных фитоцено-
зов в качестве содоминанта и иногда доминанта. Может создавать аспект,
например, в степи с Adonis vemalis (Растительный покров СССР, 1956). Ча-
ще встречается в виде разбросанных единичных растений или удаленных
небольших групп по 3-5 экземпляров. Произрастает в основном в разно-
травно-злаковых сообществах. На сухих местообитаниях растет вместе с
Campanula glomerata, Plantago lanceolata, Pimpinella saxifraga, Trifolium
montanum, на влажных — c Melampyrum nemorosum, Trollius europaeus,
Centaurea phrygia (Алексеев и др., 1988).
149
В лесах Московской области (Ступинский р-н) в изученных сообщест-
вах вид может доминировать. Однако чаще он встречался разбросанными
небольшими группами в 7-20 экземпляров на расстоянии 20-150 см, ли-
бо — что встречалось реже — единичными экземплярами.
Плотность зрелых ценопопуляций может быть высока. Наиболее круп-
ная популяция насчитывала 58 экземпляров на 10 м2. В возрастном спектре
рассмотренных популяций преобладали генеративные и виргинильные
особи. Проростки и ювенильные растения были найдены только на осве-
щенных открытых местообитаниях с незначительным общим проектив-
ным покрытием (табл. 4).
Биологическая продуктивность
Фитомасса первоцвета весеннего в Ступинском и Каширском районах
Московской области зависит от условий местообитания (табл. 5).
Таблица 5
Средняя фитомасса (сухой вес, г/м2) генеративных особей
Primula veris в разных фитоценозах Московской области*
Фитоценоз Листья** Соцветия Подземные части Общий вес
Березняк 41,3±12,8 31,4±3,1 12,1±1,3 84,8±8,6
Березняк, окно 19,б±1,7 10,1±1,7 5,8±0,6 45,5±4,8
Дубрава, окно 23,2±2,5 9,7±0,8 30,7±1,5 63,6±3,7
Ельник, окно 6,8±0,8 3,2±0,2 4,8±0,2 14,8±1,3
Примечание'. Фитоценозы те же, что в табл. 4;
* фитомасса определена 20-24.05.2000;
** листья весенней генерации.
Из наших данных и данных Т.А. Лысенко (1994) следует, что наболь-
ший вклад в биомассу вносят листья (табл. 5, 6). Только в дубраве с липой
медунициево-разнотравной вес подземных органов незначительно преоб-
ладает вес листьев. В изученных нами популяциях общая фитомасса была
наибольшей в лесных ценозах (под пологом леса и в окнах).
150
Таблица 6
Средняя фитомасса генеративных особей Primula veris
(сырой вес, г) в лесостепи Украины
Фитоценоз Листья Соцветия Подземные органы
Дубрава* . 11,55±1,32 5,10±0,50 7,16±0,50
Грабовый лес** 5,98±0,65 1,96±0,20 4,73±0,30
Питомник*** 10,17±0,89 6,00±0,37 3,83±0,38
Посадки сосны**** 3,27±0,31 1,41±0,17 2,39±0,22
Примечание'. * сосново-дубовый лес (под пологом),
** дубово-грабовый лес (поляна),
*** питомник Центрального Ботанического Сада АН Украины,
**** междурядья в посадках сосны.
Семенная продуктивность первоцвета весеннего довольно высока. На
одном генеративном побеге (в березняке орляково-разнотравном) развива-
ется 5—15 плодов, в 1 плоде — 40-60 семян. Обычно формируется 79-90 %
семян от числа заложившихся семяпочек. Потенциальная семенная про-
дуктивность составляет 200-600 семян на один генеративный побег.
Наиболее благоприятные условия для цветения и плодоношения скла-
дываются под пологом леса на богатых и легких почвах в дубраве и грабо-
вом лесу (табл. 7). Уменьшение конкуренции в питомнике и посадках со-
сны также увеличивает семенную продуктивность.
Консортивные связи
Первоцвет весенний — насекомоопыляемое растение; опыляется шме-
лями (Bombus terrestris) и бабочками. На чашечке иногда видны прокусы,
сделанные насекомыми, берущими нектар без опыления. Растение редко
повреждается вредителями, а сами повреждения не носят критического ха-
рактера. В сухих и особенно теплых местообитаниях первоцвет поражает-
ся тлей и паутинным клещом (Крюгер, Янтра, 1988).
Для подземных органов характерна везикулярно-арбускулярная мико-
риза (Grime et al., 1988).
151
Хозяйственное значение
Первоцвет весенний используется как лекарственное растение, как
декоративное, а также употребляется в пищу. В качестве лекарственного
сырья используются корневища, листья и соцветия. Действующим нача-
лом служат сапонины, гликозиды, эфирное масло, витамины С и Е, каро-
тин. В некоторых работах основным действующим компонентом называ-
ется примуловая кислота — один из сапонинов (Ворошилов, 1939). Наи-
большее ее количество в корнях — 5-10 %; в листьях — около 2 %; в
соцветиях — менее 2 %. Подземная часть содержит углеводы, эфирное
масло, гликозиды, тритерпеноиды. В листьях обнаружены флавоноиды,
антоцианы, витамин Е. Цветки также содержат флавоноиды и эфирное
масло, которое включает некоторые компоненты эфирного масла корне-
вища (5—метоксиметилсалицилат и 4-метоксиметилсалицилат). В семе-
нах найден углевод амилоид.
Все части растения богаты витамином С. Причем, благодаря отсутст-
вию аскорбиназы, аскорбиновая кислота находится в довольно стойкой
форме и накопление её идёт до более высоких пределов, чем в присутствии
фермента (Ворошилов, 1939; Растительные ресурсы СССР, 1986). Содер-
жание аскорбиновой кислоты в листьях изменяется в течение вегетацион-
ного периода. В начале мая (09.05) оно составляет 23,9 мг/кг (абсолютно
сухой вес), а наибольшего значения достигает к июню — до 59 мг/кг. По
содержанию кислоты разные части растения отличаются (табл. 8).
Начиная с 17-го века, первоцвет весенний применялся врачами при
лечении нервных заболеваний (истерии), параличе, употреблялся как бо-
леутоляющее, снотворное, при головных болях и головокружениях. С
18-го века препараты этого растения в официальной медицинской прак-
тике почти не использовались. Длительное время первоцвет весенний и
его препараты не включались в фармакопеи ни одной страны. Употребля-
ется первоцвет весенний в основном в народной медицине. Показаниями
являются туберкулёз, простудные заболевания, применяется как моче-
гонное. Препараты улучшают деятельность желудка и кишечника. Пер-
воцвет — витаминное средство и используется при гипо- и авитамино-
зах, при лечении цинги (Ворошилов, 1939; Атлас лекарственных ...,
1962; Губанов, 1993).
Из препаратов можно отметить: туссипект — аммонийные соли сапо-
нинов примулы, примулат — экстракт корневищ примулы с бензойнокис-
лым натрием и анисовым маслом, примустабиль — сапонины примулы в
алкогольном растворе. Само растение и все препараты из него совершенно
не токсичны (Ворошилов, 1939). Уборку листьев на сырьё производят с
первого же года культуры, в последующие годы и в июне. Корневище бе-
рут осенью у 2-3-летних растений.
Первоцвет весенний употребляется также в пищу в виде салатов, бор-
152
Таблица 7
Семенная продуктивность
Primula veris в лесной зоне Украины (по Лысенко, 1994)
Фитоценоз Год Число семян Плодо- образование, %
на 1 цветущий побег на м2
Дубрава с сосной* 1987 285,10±31,84 1939 72,30±6,54
Дубрава с сосной* 1988 588,00±60,08 4586 98,94±2,84
Дубрава** 1987 208,70±42,98 1148 66,50±8,62
Дубрава, опушка*** 1987 153,50±19,52 2349 65,60±8,30
Вырубка 1988 90,99±11,28 728 40,15±4,44
Грабовый лес* 1988 ‘ 223,21±20,60 — 61,39=1=6,66
Питомник* 1988 593,29±66,84 — 88,26±6,54
Посадки сосны* 1988 247,21 ±25,50 — 93,41±7,53
Примечание: * см. табл. 6;
* * дубовый лес (сомкнутость полога 0,7);
* ** дубовый лес (северная опушка); почвы лесные серые
суглинистые в дубово-грабовый лесу и посадках сосны,
в остальных фитоценозах — дерново-подзолистые
супесчаные.
щей. Из цветков и мёда в некоторых странах Европы (Швейцария, Поль-
ша) готовят шипучий напиток. В Германии соцветия используются для ус-
покоительных чаёв, а сухое корневище, имеющее лёгкий запах аниса,
употребляется как пряность.
Первоцвет весенний используется как декоративное растение для рока-
риев, бордюров, клумб, альпийских горок. Имеет много сортов с махровы-
ми и немахровыми цветками жёлтой окраски любых оттенков (Кудрявец,
Петренко, 1993). Для сохранения декоративных качеств растения размно-
жают делением куста 1 раз в 3-4 года. Обычно деление проводится в
мае-июне, но растение хорошо переносит его в любую фазу роста. В куль-
туре необходимо внесение достаточного количества азотных удобрений:
153
Таблица 8
Содержание аскорбиновой кислоты (мг/кг)
в разных частях Primula veris в период цветения 21-23 мая
(Рябова, 1939)
. Часть растения Сырой вес Абсолютно-сухой вес
Соцветие 3,550 22,326
Корневище 1,052 7,907
Листья 8,997 51,122
селитры — 20 г/м2 или мочевины -10-15 г/м2 (Ильинская, 1993; Кудрявец,
Петренко, 1993; Лисовская, 1994).
Как кормовое, первоцвет весенний имеет низкое кормовое достоинство
(Раменский и др., 1956). В начале цветения поедается овцами, к концу ле-
та — гусями. В Закавказье наряду с первоцветом крупночашечным служит
кормом маралам (Растительные ресурсы СССР, 1986; Алексеев и др.,
1988).
И. Б. Седых
ВЕРБЕЙНИК ОБЫКНОВЕННЫЙ
Номенклатура и систематическое положение
Lysimachia vulgaris L., 1753, Sp. PL: 146; Кауфман, 1866, Моск, фл.: 406;
Сырейщиков, 1910, Илл. фл. Моск, губ., 3 : 24; Штейнберг, 1952, Фл.
СССР, 18 : 266; Ан.Федоров, 1964, в Маевский, Фл., изд.9 : 407; Скворцов,
1966, Опред. раст. Моск, обл.: 263; Ferguson, 1972, Fl. Europ. 3 : 26; Ан.Фе-
доров, 1981, Фл. европ. части СССР, 5 : 82; Губанов и др., 1995, Опред. со-
суд. раст. Центра Европ. России : 402.— Вербейник обыкновенный (се-
мейство Primulaceae — Первоцветные).
Географическое распространение
Вербейник обыкновенный — евразийский вид. Северная граница ареала
проходит между 64° и 64°5', южная — между 32° и 40° с. ш. Вид распро-
странён в южной и средней Скандинавии, обычен в центральной и атланти-
ческой Европе, становится более редким в северной и исчезает в северной
Шотландии и на Фарерских островах. Южная граница в Европе проходит в
средней Италии, Хорватии и южной Греции. На островах Средиземного мо-
ря вид отсутствует, но отмечен в северной Африке на севере Алжира (Флора
СССР, 1952; Clapham et al., 1952; Флора Киргизской ССР, 1959; Boissier,
1963; Flora Europaea, 1972; Флора европ. части СССР, 1981; Pignatti, 1982;
Флора Таджикской ССР, 1984). В Турции вербейник распространен в Анта-
лии и на островах Эгейского моря. Встречается на севере Ирана, северо-за-
паде Ирака, в северной части Афганистана и занимает северо-западные рай-
оны Китая (Флора Таджикской ССР, 1984; Heller, Неуп, 1986).
Вербейник обыкновенный широко распространён на большей части
территории бывшего Советского Союза кроме Заполярья, Дальнего Восто-
ка и пустынь Средней Азии (Флора СССР, 1952; Флора Казахстана, 1964;
Флора европ. части СССР, 1981; Растительные ресурсы СССР, 1986). Он
занимает всю европейскую часть кроме арктической области. Его северная
граница проходит по линии Реболы — Кемь — г. Онега — дельта р. Се-
верной Двины — Зимний берег — верховья Кулоя — Мезень — Ух-
155
та — устье р. Сыни (Раменская, 1960; Флора северо-востока СССР, 1977).
Отсутствует в Ненецком национальном округе и лишь изредка встречается
в районах верхней Печоры, Ухты, Вычегды и на южном побережье Белого
моря (Флора Мурманской области, 1966; Флора европ. части СССР, 1981;
Раменская, Андреева, 1982). На юге отмечен в степном и горном Крыму, на
Кавказе. В Средней Азии занимает Арало-Каспийский, Прибалхашский,
Сыр-Дарьинский подгорный, Памиро-Алайский и Тянь-Шаньский флори-
стические районы (Гроссгейм, 1967; Определитель высших растений Кры-
ма, 1972; Растительные ресурсы СССР, 1986). Как пантаёжный вид вербей-
ник распространён по всей территории Западной и Центральной Сибири.
На востоке граница ареала проходит в Вилюйско-Верхнеленском и Анга-
ро-Саянскомрайонах (Говорухин, 1937; Попов, 1959; Флора Сибири, 1997;
Флора Центральной Сибири, 1979). Как заносное отмечен в США и Канаде
(Scoggan, 1979).
В Московской области обычен во всех районах (Ворошилов и др., 1966;
Губанов, и др., 1995).
Морфологическое описание
Вербейник обыкновенный — травянистый длиннокорневищный мно-
голетник. Скелетная основа годичного побега состоит из плагиотропного
гипогеогенного корневища, переходящего в надземную ортотропную
часть. Корневище залегает на глубине 5-12 см, бурое или розовато-бурое,
30-140 см длиной и 0,5-0,8 см в диаметре. Несёт чешуевидные листья
(0,2-0,5 см Длиной и 0,3-0,4 см шириной). Зона перехода от плагиотроп-
ной к ортотропной части (дуга корневища) обычно заглублена на 3-5 см
больше, чем плагиотропная часть.
Листорасположение очередное на 2/3 длины корневища, выше стано-
вится супротивным. Оно хорошо выражено в части, переходной к орто-
тропному побегу, Длины междоузлий при очередном расположении ли-
стьев непостоянны: чередуются длинные (6-12см) и короткие (1-3 см).
Корневая система вербейника образована придаточными корнями. На
корневище их распределение не имеет какой-либо закономерности. На ду-
ге и нижней части ортотропного побега корни формируются по два в узлах
чешуевидных листьев по бокам их, либо в пазухе листа над пазушной поч-
кой. В междоузлиях, в последнем случае, корень прорастает через листо-
вую пластинку.
Стебель округлый или округло-четырёхгранный. Высота
70-215(300)см, диаметр средней части 0,3-0,8 см. В нижней половине сла-
бо, в верхней более густо опушён, как и листья, простыми, неветвистыми и
редкими железистыми волосками.
Листорасположение мутовчатое, на коротких участках оси супротив-
ное и очередное, в мутовках (2)3-5 листьев. Число листьев колеблется в
156
пределах оси: в нижних мутовках на 1—2 листа меньше. В мутовках листья
расположены по спирали и расстояние между ними в мутовке — от 0,01 до
8 см. Число мутовок колеблется от (13)14-16(18). В пазухах листьев 8-11-й
мутовки расположены побеги обогащения оси Ш-го порядка закладывают-
ся в 5-7-й мутовке. Длина междоузлий варьирует от 7 до 17 см в зависимо-
сти от соотношения высоты надземной части и числа мутовок.
Срединные листья продолговато-ланцетные, цельнокрайние, черешко-
вые, жилкование брохидодромное. В области соцветия листья постепенно
уменьшаются и переходят в брактеи.
Листья содержат смолистые вещества, экскретируемые в межклетники
и образующие там гранулярный слой вдоль клеточных оболочек (Эзау,
1969). Места скопления смол образуют точечный рисунок листьев, чётко
выраженный у молодых растений и мало заметный у средневозрастных
особей (мало точек на единицу площади).
Соцветие — полителическая кисть. В нижней части фрондозное, в
верхней — брактеозное. Образовано системой паракладиев (побегами
обогащения); флоральная единица — открытая ярусная кисть. Распуска-
ние в пределах осей Ш-го порядка и у флоральных единиц происходит ак-
ропетально, у всей системы соцветия — базипетально.
Цветки обоеполые, актиноморфные, с двойным околоцветником.
Чашечка пятираздельная, доли её зелёные, ланцетовидные, за-
острённые, по краям с коричневым окаймлением в местах накопления смо-
листых веществ. Чашечка густо опушена преимущественно железистыми,
а также простыми волосками. Почкосложение свёрнутое.
Венчик колесовидный, пятичленный, спайнолепестный, диа-
метр— 0,7-1,8 см, с короткой трубкой (длина 0,2-0,4 см.), ярко-жёлтый.
Иногда в зеве с красноватым пятном. Доли венчика яйцевидные, продолго-
ватые, заострённые; длина свободной части долей 0,6-1,5 см, шири-
на — 0,5-0,8 см. Внутри венчика до половины длины густо расположены
железистые волоски (Флора СССР, 1952).
Андроцей из пяти тычинок. Тычиночные нити равные, вдвое короче
венчика, железистые, внизу расширенные и сросшиеся. Длина пыльников
до 1,5 мм. Мужской гаметофит относится к 2-х клеточному типу. Линей-
ные размеры спермиев 10:3 микрона (Руденко, 1956).
Гинецей образован верхней завязью. Плацентация колончатая, столбик
один, с головчатым рыльцем, покрыт по всей длине железистыми волоска-
ми. Величина генеративной клетки 38:4 микрона (Руденко, 1956).
Плод — одногнёздная многосемянная коробочка, вскрывается суту-
рально пятью зубцами (Левина, 1987).
Семена трехгранные, с обильным беловатым налётом, мелкоячеистые.
Длина 1-1,5 мм, ширина 0,8-1мм, толщина 0,6-0,8 мм. Грани ромбические
или эллиптические, с широкой оторочкой по краю. Рубчик семени на
брюшной стороне между гранями (рис. 1).
157
Рис. 1. Первые фазы онтогенеза Lysimachia vulgaris.
Обозначения: с — семядоли, гк — главный корень, к — корневище
158
Числа хромосом: 2n=14 (Darlington, Wylie, 1955; Числа хромосом ...,
1993), 2п=28 (Соколовская, 1972), 2п=42 (Ростовцева и др., 1981; Дзмит-
рыева, Парфёнау, 1983), 2п=84 и 85 (Дзмитрыева, Парфёнау, 1983;
Парфёнов, Дмитриева, 1985), 2п=86 (Парфёнов, Дмитриева, 1985).
Онтогенез
Прорастание семян вербейника обыкновенного надземное, гипокоти-
лярное. В зародыше рано дифференцируются семядоли, гипокотиль, заро-
дышевый корешок. Хорошо заметны апекс и прокамбиальный пучок
(рис. 1).
Всхожесть семян вербейника обыкновенного изучалась в разных усло-
виях. Проращивание при 20°С на свету результата не дали (Kennel, 1913).
Нестратифицированные семена не удалось прорастить при 22—25°С на све-
ту и в темноте на влажной почве, а также при посеве в природных место-
обитаниях. В обоих последних случаях 70 % семян плесневели. При посе-
ве на сфагнуме в чашках Петри плесень не развивалась, но семена не всхо-
дили. Есть данные, что низкие температуры являются одним из
необходимых условий для прорастания семян вербейника (Grime et al.,
1988).
Скарифицированные и не скарифицированные семена проращивались
в четырехкратной повторности в чашках Петри. В качестве субстрата ис-
пользовался биогумус. Опыт проводился при дневном освещении и в тем-
ноте с 24 по 28 августа 2000 г. (табл. 1).
Наибольший процент всхожести наблюдался у скарифицированных се-
мян в темноте при высокой влажности субстрата и высоких температурах.
Факторами, ограничивающими прорастание в данном случае, стали свет и
недостаток воды.
Полученные данные могут указывать на физиологические механизмы
торможения прорастания семени и нормально сформированный зародыш.
Очевидно, семена могут прорастать по прошествии одного года или не-
скольких лет при высоких весенне-летних температурах в местах, затоп-
ляемых водой.
Возрастные состояния вербейника обыкновенного выделялись по сле-
дующему комплексу признаков: высота растения, число мутовок листьев
на побеге I-ro порядка, число листьев в мутовках и наличию полиморфиз-
ма ассимилирующих листьев в пределах оси 1-го порядка; наличие или от-
сутствие корневищ, их длина и диаметр, точка его заложения на оси пре-
дыдущего года, соотношение длин его плагиотропной и ортотропной час-
тей, глубина залегания корневища в почве, форма апикальной части
корневища формирующегося годичного побега; строение корневой систе-
мы (табл. 2).
159
Таблица 1
Результаты проращивания семян
Lysimachia vulgaris в разных условиях*
Семена Влажность субстрата, % Темпе- ратура Осве- щение Время экспозиции, час. Всхожесть, %
Нескарифи- цированные 100 30° — 36 ' 60,2±0,8
70 30° — 36 58,3±0,3
45 30° — 50 15,7±0,9
Скарифи- цированные t 100 30° — 60 83,4±1,1
70 30° — 60 78,3±1,2
45 30° — 68 62,3±0,2
100 22° 60 57,5±0,5
70 22° 60 42,2±1,4
45 22° 60 30,3±0,7
100 22° — 28 75,2±1,1
70 22° — 28 73,1±0,8
45 22° — 28 44,4±0,6
Примечание: * семена собраны за пять дней до посева.
160
Таблица 2
Морфометрические характеристики
возрастных состояний Lysimachia vulgaris из ценопопуляции
в дубо-осиннике с липой (Московская область)
Показатели Возрастное состояние
Р j im V g SS
Высота стебля, см 2,2±0,1 6,3i0,7 13,0±1,8 50,5i6,3 150,4i8,6 110,4±4,l
Число мутовок 1 3 3-5 10-14 14-18 12-15
Корневища:
Число* 0 2-4 1-3 1(2) 1-2(3) (2)1
Длина, см 0 0,50±0,04 3,5±1,1 13,6±2,3 90,li8,5 107,6i8,2
Диаметр, см 0 0,12±0,01 0,15i0,02 0,50i0,02 0,70i0,04 0,60i0,03
Глубина залегания, см 0 1±0,3 3,0il,l 4,l±0,7 8,l±0,3 25,0i3,2
Листья:
Длина, см — 0,47±0,08 3,4i0,8 6,4±1,2 14,4±0,4 14,2±0,l
Ширина, см — 0,34±0,04 l,4±0,3 2,7i0,8 4,2i0,8 4,5i0,5
Число в мутовке 2 2 2 2-3 3-4(5) 2-3
Примечание: * число корневищ на годичном побеге.
161
Проростки и ювенильные растения выращивались в лабораторных ус-
ловиях при постоянной температуре 22°С и при освещении в течение
13 часов в сутки. Полностью развитые проростки образуются на 12-15-й
день после появления молочно-белого полупрозрачного главного корня.
Его длина—1,5-2 см. Семядоли цельнокрайние, яйцевидные, с за-
острённой верхушкой, черешковые. Они могут быть асимметричными, а
также разной величины. Длина черешка до 0,3 см, пластинки лис-
та — 0,3-0,4 см. Черешки семядолей расположены вертикально и парал-
лельно друг другу, а пластинка листа — под углом 90° к ним (рис. 1). Че-
решки защищают крупную (до 0,1 см в диаметре) верхушечную почку.
Длительность этапа — 18-24 дня.
Ювенильные особи начинают формироваться с моментароста верхушеч-
ной почки и образования 1-2 боковых корней. Побеги удлиненные, эпико-
тель укорочен (0,2-0,6 см) по сравнению с другими междоузлиями (1-2,5 см).
Листья черешковые, овально-яйцевидные, цельнокрайние, листорасположе-
ние супротивное. По линейным размерам все листья близки: 1-я пара — дли-
на 0,4-0,6 см, ширина — 0,3-0,4 см; 2-я и 3-я пары —длина 0,6-0,8, шири-
на — 0,3-0,5. Однако в пределах оси одной особи величина листьев, их число
и длина междоузлий могут быть различны. Первая (и вторая) пара листьев
округлой формы может иметь мелкие размеры (длина 0,2-0,3 см, ширина
0,1-0,2 см), при этом черешок составляет 0,5 длины листовой пластинки. Об-
щее число пар листьев возрастает до 4—5. Первые этапы онтогенеза характе-
ризуются вариабельностью путей развития (рис. 1). В одних случаях через
18-20 дней после начала формирования ювенильного растения в пазухах се-
мядолей начинают развиваться корневища розовато-зелёного цвета, с 2-3 че-
шуевидными листьями. При полегании стебля узел с семядолями оказывает-
ся лежащим на субстрате и корневище заглубляется.
В других случаях, если происходит задержка развития корневищ в па-
зухах семядолей, верхушечная почка проявляет положительный геотро-
пизм и формирует плеть с 4—5 парами мелких листьев (длина междоузлий
0,4-0,7 см), которая заглубляется в почву. Нормальный рост восстанавли-
вается после начала развития корневищ в пазухах семядолей или вышерас-
положенных мелких листьев. Общее число пар листьев возрастает здесь до
10-12. Развитие корневищ в пазухах семядолей тормозится и в случае
очень короткого (0,1 см) эпикотиля. В этом случае они развиваются в пазу-
хах ювенильных листьев.
Корневая система в течение 25-30 дней представлена главным корнем
(длина до 3,5-4 см) с 3-4 боковыми корнями П-го й Ш-го порядков. Если
длина эпикотиля не менее 0,3 см, то на нем обязательно закладывается
один придаточный корень. Он сохраняет вид бугорка до полегания расте-
ния на субстрат. В последнем случае в местах соприкосновения оси с поч-
вой развиваются придаточные корни. На одну особь приходится 5-10 кор-
ней: длина их — 0,2-0,4 см, толщина — менее 0,1 см.; по оси корни рас-
162
пределены без видимой закономерности. Через 30-35 дней начинается
отмирание главного корня и семядолей. Длительность этапа не выяснена.
Имматурные растения имеют гипогеогенное корневище с 2-4 парами че-
шуевидных листьев и чётко не выраженной плагиотропной частью. Подзем-
ный побег обычно сразу начинает расти вверх под углом 10-15°. На песча-
ных заливаемых почвах его плагиотропная часть вообще отсутствует, и кор-
невище растёт вертикально вверх, прилегая к оси материнского растения.
Надземная часть ортотропного побега несёт рано отмирающие чешуевидные
и ассимилирующие супротивные листья. Последние в пределах оси в при-
родных условиях имеют одинаковую форму (овально-яйцевидные, с за-
острённой верхушкой) и близки по линейным размерам. Придаточные корни
в основном сосредоточены в области перехода к ортотропной части побега.
Если растения в начале августа перенести в лабораторные условия (при
дневном освещении с постоянной температурой 22-24°С на обильно ув-
лажненный субстрат из сфагнума при влажности воздуха 90-95 %), то че-
рез 30-35 дней они начинают развивать 3-й элементарный побег (пер-
вый — корневище, второй — ортотропная часть растения). Он отделён от
предыдущего укороченным (0,5-1 см) междоузлием. В первой паре оба
листа могут иметь мелкие размеры (длина 0;5-1,3 см, ширина 0,3-0,6 см)
или один из них имеет нормальные размеры. Третий элементарный побег
несёт 3-5 пар листьев. В пазухах первых 2-3 пар могут формироваться
корневища — длина их не более 2 см, диаметр — 0,3-0,4 см. Выше закла-
дываются только побеги обогащения. У 15 % особей в начале октября воз-
никает 4-й элементарный побег. В природных условиях подобных явлений
не обнаружено. При погружении точки роста имматурных растений под
воду или в сильно обводнённый субстрат она растёт симподиально в тече-
ние 2-3 лет, образуя плеть 7-15 см длиной с мелкими супротивными ли-
стьями. Корневища имматурных растений образуются как в области пере-
хода к ортотропной части побега, так и на ортотропном побеге. В послед-
нем случае надземный побег погружён в подстилку на 3-6 см. В силу
подобной особенности скелетная система предыдущих лет имеет «верти-
кальную» структуру: корневища разных лет не лежат на одной линии, как
у растений последующих этапов, а расположены на разных линейных го-
ризонтах (рис. 2). Апикальная часть корневища имеет острую верхушку.
Длительность этапа не менее 3 лет.
У виргинильных растений появляется листовой полиморфизм в пределах
оси 1-го порядка годичного побега. Листья в нижних 2-3 мутовках более мел-
кие (длина 1,5—3 см, ширина 0,7-1,5 см), чем в вышерасположенных мутов-
ках. Их листовая пластинка имеет овально-округлую форму с суженой вер-
хушкой. Остальные листья по размерам и форме близки к таковым генера-
тивных растений. Корневища формируются только на дуге, и многолетняя
система скелетных осей расположена на одной линии («горизонтальная»).
Апикальная часть корневища имеет заострённую верхушку (рис. 2).
163
Листовой полиморфизм в пределах оси 1-го порядка хорошо выражен у
генеративных растений. В приземной части стебля листья чешуевидные, в
нижней части — ланцетной формы, рано отмирающие, в верхней — яйце-
видно-ланцетные (табл. 3).
Таблица 3
Морфометрические показатели
листьев Lysimachia vulgaris (Московская область)
Рис. 2. Подземная многолетняя система побегов
у особей разных возрастных состояний Lysimachia vulgaris
Тип листьев Пластинка листа, см Длина черешка, см Число жилок П-го порядка
длина ширина
Чешуевидные 0,70±0,06 0,60±0,02 0 —
Ланцетные 11,3±0,4 1,7±0,5 0,25±0,2 7-11
Яйцевидно- ланцетные* 14,4±0,4 4,2±0,8 0,5±0,3 16 18
Яйцевидно- ланцетные** 4,5±0,6 1,2±0,2 0,5±0,1 16-18
Примечание: * в нижней части стебля, ** в верхней части стебля.
Более длинные и толстые корни закладываются на дуге корневища, но
большее число корней у генеративных особей расположено на плагио-
тропной его части (табл. 4).
Таблица 4
Морфометрические показатели
корней Lysimachia vulgaris, расположенных на плагиотропной
и ортотропной частях корневища
Корни Число Длина, см Диаметр, см Порядок ветвления
На плагиотропная части 78,3±9,7 6,4±3,2 0,1 ±2,9 2-3
На ортотропной части 36,6±2,3 22,2±2,6 0,30±0,07 2-3
165
164
Побеги возобновления возникают только на дуге корневища, и система
многолетних подземных осей имеет горизонтальную структуру. Апикаль-
ная часть корневища округлая (рис. 2).
Стареющие растения характеризуются глубоким залеганием длинных
корневищ и крупными листьями сходной величины и формы по всей длине
оси. Верхушка листовой пластинки более округлая по сравнению с генера-
тивными особями. Надземная часть субсенильных растений сходна с ося-
ми П-го порядка генеративных особей, которые развиваются в пазухах че-
шуевидных листьев при повреждении всей надземной оси до цветения или
почки в начале её нарастания в апреле—мае. Если такой побег не зацвета-
ет, то. отличить его от стареющего растения можно только по подземным
органам. Апикальная часть корневища и структура подземных скелетных
осей сходны с таковыми генеративных особей (рис. 2).
Сезонное развитие
По ритму сезонного развития вербейник относится к весенне-лет-
не-осенне-зелёным растениям. Годичный побег вербейника обыкновен-
ного образован двумя элементарными побегами: корневищем и орто-
тропным надземным побегом. Плагиотропный подземный побег образу-
ется в предыдущем сезоне. В июле-августе он имеет на ортотропной
части почку возобновления, которая после перезимовки в конце мар-
та-начале мая удлиняется на 2-3 см за счет растяжения междоузлий, уве-
личивается в диаметре и апреле-мае трогается в рост. За 25-36 дней фор-
мируется ортотропный элементарный побег генеративного растения с ас-
симилирующими листьями. Чешуевидные листья его приземной части
отмирают к началу цветения. К этому же моменту 1-2 их пазушные поч-
ки трогаются в рост, и начинает формироваться корневище годичного по-
бега последующего сезона. Образование корневищ совпадает с началом
бутонизации и обычно завершается к моменту полного цветения. У им-
матурных и виргинильных растений корневище развивается в те же ка-
лендарные сроки.
Заложение элементов соцветия начинается с начала мая, а цвете-
ние — с конца мая до начала сентября. В зависимости от местообитания
ритмы цветения и плодоношения могут отличаться. Более ранние цветение
и плодоношение характерно для освещённых местообитаний (более 50 %
от общего светового потока): здесь сокращен до 10-14 дней период от бу-
тонизации до распускания 90 % цветков (процент от сформировавшихся
бутонов). Во влажных и затенённых местах этот период достигает 35-40
дней. Формирование 90 % плодов (процент от развившихся цветков) за-
вершается в сходные сроки — 32-36 дней (табл. 5).
166
Таблица 5
Сроки цветения и плодоношения
Lysimachia vulgaris в разных фитоценозах
Московской области в 1999 г.
Фитоценоз Цветение Плодоношение
начало массовое начало массовое
Осинник, просека* 25.05 08.06 25.07 30.08
Ивняк** 10.07 08.06 01.08 12.09
Березняк*** 10.07 25.07 15.08 20.09
Осинник, овраг**** 12.06 25.07 18.08 17.09
Примечание'. * просека в дубо-осиннике волосистоосоковом;
** заросли ивы козьей на просеке в осиннике с липой и дубом;
*** заболоченный овраг в березняке вейниковом
(сомкнутость крон 0,4);
**** заливаемый овраг в дубо-осиннике с сосной и кленом
(сомкнутость крон 0,3).
В условиях Подмосковья цветение вида занимает 3/4 времени, в течение
которого вид вегетирует, что совпадает с подобными же данными для
степных местообитаний (Голубев, 1965).
Отмирание надземной части вербейника растянуто с мая до октября.
В конце мая-начале июня у генеративных растений отмирают чешуевид-
ные листья, к моменту формирования соцветия в июне-июле увядают
нижние ланцетные листья. В октябре отмирают оставшиеся ассимили-
рующие листья, и в начале ноября начинается усыхание стебля. Отмира-
ние чешуевидных листьев у имматурных и виргинильных особей проис-
ходит в июне, нижние листья могут сохраняться до конца июля. Полное
отмирание всей надземной части также происходит в октябре-начале
ноября.
Способы размножения и распространения
Вербейник обыкновенный размножается семенами и вегетативно,
путём формирования, разрастания и последующего отмирания плагио-
тропных корневищ. Длина корневищ зависит от возрастного состояния
167
особей, а также от влажности и механического состава почв. Наибольшей
величины длина корневища достигает у генеративных особей в сильно ув-
лажнённых и лёгких почвах (в нашем случае на супесях). При поврежде-
нии апикальной части плагиотропного побега до окончания его роста па-
зушные почки его чешуевидных листьев образуют 2-4 корневища П-го по-
рядка. Они достигают длины, обычной для данного возрастного состояния
в данных условиях. В редких случаях при невыясненных условиях корне-
вища И-го порядка могут возникать на дуге и плагиотропной части корне-
вища предыдущего года из спящих почек.
Вербейник может проявлять себя как гемикриптофит и как криптофит
(Флора Мурманской области, 1966). В зависимости от влажности припоч-
венных слоёв воздуха, корневища могут быть подземными (бурого цвета)
или лежать на поверхности почвы (розовеющие). В первом случае корне-
вище погружено в почву на 5-15 см. Надземные формируются при высо-
кой влажности воздуха и выраженном микрорельефе в ассоциациях с
Myosoton aquaticum, Urtica dioica, Flilipendula ulmaria. В обоих случаях
обязательно происходит заглубление апикальной части корневища при
формировании его ортотропной части. Плагиотропные побеги предыду-
щих 3—5-ти лет образуют подземную систему скелетных осей, которые по-
степенно отмирают.
Семенное возобновление, по нашим данным, незначительно, хотя се-
менная продуктивность велика: потенциальная семенная продуктивность
(на один годичный побег)— 13988±234 семян, условно-реальная —
5729±86 семян. Имеет место, как перекрёстное опыление, так и самоопы-
ление. Последнее в обычном случае происходит в клейстогамных цветках
и редко при помощи агентов.
Единицами распространения могут быть семена и проростки. Вербей-
ник обыкновенный растение аллохорное, относится к баллистам гвоздич-
ного типа (Левина, 1957). Семена высеиваются при растрескивании коро-
бочки и раскачивании стебля. В изученных популяциях семена рассеива-
лись не более чем на 1,2(1,6) метра от материнского растения. Семена и
проростки также переносятся водой в новые местообитания (Grime et al.,
1988).
Наибольшее число цветков закладывается у особей в сильно ос-
вещённых местообитаниях, но здесь же велик и процент их гибели и доля
неразвившихся семян.
Вес 3216 семян, взятых на освещенной просеке в дубо-осиннике воло-
систоосоковом— 1 г.; вес одного семени — 0,21-0,5 мг (Grime et al.,
1988).
168
Таблица 6
Семенная продуктивность
особей Lysimachia vulgaris в разных фитоценозах
Московской области
Фитоценоз Цветки, шт. Семяпочки в плоде, шт. Плодо- образова- ние, % Число семян в плоде
созревшие погибшие
Осинник, овраг* • 245,6±10,3 56,3±6,1 80 29,0±1,2 19,2±0,5
Осинник, просека** 311,7±14,1 62,7±3,3 68 17,1±0,3 31,'1±3,4
Березняк, окно*** 141,4±12,8 60,1±1,2 74 32,8±0,2 25,3±1,5
Осинник, берег пруда**** — — — 56,3±2,4 0,60±0,04
Примечания’. * дубо-осинник с сосной, заболоченный овраг
(сомкнутость крон 0,6);
** осинник с липой и дубом, просека;
*** березняк разнотравный, окно;
**** дубо-осинник с липой, берег пруда.
Экология
Вербейник обыкновенный — светолюбивое растение, хотя в вегета-
тивном состоянии хорошо переносит сильное затенение. Предпочитает ус-
ловия освещения между полутеневыми и полусветовыми (6-я ступень шка-
лы Элленберга). Наибольшее количество цветков формируется при ос-
вещённости более 50% от полной освещенности. При освещенности
меньше 10 % репродуктивные органы не формируются, либо развивается
только флоральная единица (полителическая кисть) на оси 1-го порядка.
Влажность почвы и воздуха — факторы, ограничивающие распростра-
нение вербейника обыкновенного. Вид предпочитает условия влажности
между влажными и сырыми (8-я ступень шкалы Элленберга). Массово он
встречается на почвах с болотно-луговым и болотным увлажнением, более
умеренно — при сыро-луговом увлажнении, а также в зоне прибреж-
но-водной растительности, единично — на свежих луговых почвах (Ра-
менский и др., 1956). В местах произрастания предпочитает понижения
169
рельефа. Встречается в канавах, на заболоченных пустошах, влажных
поёмных лугах, по берегам водоёмов. Распространён и в более сухих ме-
стообитаниях в разреженных лесах, на опушках, просеках, в окнах. Для
Урала отмечена подводная бесплодная форма, развивающаяся на глубине
60-70 см, со слабым неветвящимся стеблем — L. vulgaris var. submersa
Gluck (Говорухин, 1937). Подобная форма отмечена для средней Европы
(Hegi, 1912).
Вербейник имеет широкий экологический диапазон по отношению к
трофности и кислотности почв (Ellenberg, 1991). Может расти и на бед-
ных и на богатых почвах (Лесные травянистые растения, 1988). Предпо-
читает почвы от слабокислых до слабощёлочных — pH 5,5-8,0 (Grime et
al., 1988). Наши исследования в Ступинском и Каширском районах Мос-
ковской области (20-29.07.99) показали, что pH водной вытяжки корне-
обитаемого слоя на лесных просеках был близок к 5, а в заболоченных ов-
рагов— 4-4,5. Гранулометрический состав почв в изученных случаях
был разный: от песчаных до суглинков. К аэрации почв вид нетребова-
телен.
Данные о высотном распределении вербейника обыкновенного недос-
таточны. В гористых районах Средней Азии вид приурочен к их равнин-
ным частям (Флора Казахстана, 1964; Флора Таджикской ССР, 1984). Од-
нако в средней и северной Европе вид поднимается на небольшие высоты:
до 1840 м в Баварских Альпах, до 1180 м в Южном Тироле, на 1150 м в кан-
тоне Валлис в Швейцарии (Hegi, 1912).
Выпас не оказывает губительного действия на заросли вербейника (Ра-
менский и др., 1956).
Фитоценология
Вербейник обыкновенный произрастает в лесной, лесостепной и степ-
ной зонах Евразии. Является диагностическим видом класса й порядка
Salicetea-Purpureae Moor 1958, в пойменных ивняках и ивово-тополевых
лесах умеренной и бореальной зоны. Отмечен как диагностический вид
союза Anemonido-Salicion albae Denisova in Iljina et al. 1988 того же класса
в поймах рек Восточной Сибири (Миркин, 1985; Korotkov et al., 1991).
В лесах класса Querco-Fagetea Br.-Bl. et Vlieger in Vlieger 1937 порядка
Fagetalia silvaticae Pawl, in Pawl. e.a. 1928 вербейник характерен для союза
Alno-Padion Knapp 1942. Это европейские мезофитные широколиственные
леса. Является дифференцирующим видом в ассоциации Urtico
dioicae-Alnetum incanae Korot. 1986 (высокотравных ольшаников). Как ди-
агностический вид вербейник обыкновенный указан в ассоциации
Filipendulo ulmariae-Betuletum Dumina 1989 (порядок Fagetalia silvaticae
Pawl, in Pawl. e.a. 1928), описанной в пойме p. Оби (Миркин и др., 1998;
Korotkov et al., 1991; Миркин и др.,2001)
170
Вербейник обыкновенный считается диагностическим видом порядка
Molinietalia Koch 1926 класса Molinia-Arrhenatheretea R.Tx. 1937 лугов
Башкортостана (Влажные луга 2002). Это вторичные луга умеренной
зоны на месте пойменных влажных и заболоченных лесов после вырубок,
существующие благодаря регулярному сенокошению или выпасу скота.
В наиболее переувлажненных условиях, граничащих на экологическом и
топографическом градиенте с осоковыми болотами, вербейник является
диагностическим видом ассоциации Lysimachio vulgaris-Filipenduletum
ulmariaeBal.-Tul. 1978 союза Caltion R.Tx. 1937 em. Bal.-Tul. 1978 того же
класса и порядка. Это сообщества влажных лугов понижений пойм либо
полян пойменных лесов. Доминирующее положение занимают Fili-
pendula ulmaria, Lysimachia vulgaris и Urtica dioica. Ассоциация широко
распространена в Центральной Европе. В Восточной Европе выявлена в
поймах рек Брянской, Курской и Орловской областей России и Башкор-
тостане.
На просеках и полянах в смешанных мелколиственных лесах южного
Подмосковья (Ступинский, Каширский и Серпуховской р-ны) вербейник
обыкновенный произрастает совместно с Calamagrostis arundinacea, Роа
trivialis, Carex pilosa, Hypericum perforatum, Geum rivale, Fragaria vesca,
Potentilla erecta, Juncus conglomeratus, Stellaria lanceolata и некоторыми
видами рода Alchemilla. На болотистых местообитаниях вид произрастает
вместе с Urtica dioica, Sonchus palustris, Scirpus maritimus, S. sylvaticus,
Myosotis scorpioides, Filipendula ulmaria, Equisetum palustre. В луговых це-
нозах вербейник растёт совместно с Filipendula ulmaria, Polygonum
bistorta, Urtica dioica, Geranium palustre (Барыкина, Чубатова, 1997).
Участие в травостое различно. Проективное покрытие может колебать-
ся от 1-2 % (ельники) до 90-95 % (просеки смешанных мелколиственных
лесов).
Все рассмотренные популяции неполночленные, большая часть их
представлена виргильными и генеративными особями. Очень редко имма-
турные растения составляют значительную часть популяции (85,7 %). Та-
кой спектр был отмечен только для популяций инвазионного типа в нару-
шенных и сильно увлажненных местах под пологом леса. Стареющие осо-
би также весьма редки. Наибольшая плотность популяции отмечена для
сухих, открытых местообитаний.
171
Таблица 7
Возрастные спектры н плотность
ценопопуляций Lysimahia vulgaris в различных фитоценозах
Московской области
Фитоценоз Возрастное состояние, % Плотность, особей на м2
im V g ss
Березняк, овраг* 15,3 61,5 23,3 — 12,20,8
Осинник, под пологом леса** — 7,7 86,6 5,7 15,61,0
Осинник, просека*** — — 100 — 28,12,1
Осинник, овраг**** 3,25 9,25 87,5 — 10,30,4
Ивняк***** 85,7 14,3 — — 8,40,12
Примечания: * березняк вейниковый, заболоченный овраг
(сомкнутость крон 0.4):;
** дубо-осинник, окно;
*** осинник с липой и дубом, просека;
**** дубо-осинник с сосной, заливаемый овраг
(сомкнутость крон 0.3);
***** заросли Salix саргеа, мезопонижение вдоль дороги.
Биологическая продуктивность
Общая продуктивность вербейника обыкновенного изучалась в зрелых
нормальных популяциях (классификация по: Уранов, Смирнова, 1969) в
Ступинском и Каширском районах Московской области на просеках в ду-
бо-осиннике осоковом и березняке с сосной и дубом. Были взяты средне-
возрастные цветущие растения по 15 экземпляров в 3-х-кратной повторно-
сти с площадок 10x10 м в четырёх популяциях в каждом местообитании
(табл. 8). Продуктивность вербейника зависит от экологических условий.
Основной вклад в биомассу вносят стебли и корневища. Наибольшее зна-
чение общей биомассы одного растения характерно для затопляемых и ма-
ло освещенных местообитаний. Репродуктивное усилие возрастает с уве-
личением освещенности.
172
Таблица 8
Средний абсолютно-сухой вес
одного генеративного растения Lysimachia vulgaris
в различных фитоценозах Московской области
Фитоценоз Стебли Листья Цветки Корневища и корни Всего
Осинник, просека* 10,2±0,3 4,5±0,2 1,2±0,1 4,3±0,4 20,2±1,1
Осинник, овраг** 11,9±0,6 5,4±0,7 0,6±0,3 10,2±1,6 28,2±2,1
Березняк, окно** 8,2±1,1 3,9±0,04 0,6±0,2 6,7±1,2 19,4±0,8
Примечание: * осинник с липой и дубом, Просека;
* * дубо-осинник с сосной, заливаемый овраг
(сомкнутость крон 0,3);
* ** березняк разнотравный, окно.
Наибольшая продуктивность вербейника характерна для более сухих и
освещенных местообитаний (табл. 9). Продуктивность надземных частей
вербейника в погодичной динамике возрастает также на вырубках (Ермо-
лова, 1981).
Таблица 9
Характеристики популяций
Lysimachia vulgaris в различных фитоценозах
Московской области
Фитоценоз Фитомасса, № Плотность особей на м2
Осинник, просека 364±4,1 27,1±1,1
Осинник, овраг 164,6±3,3 22,1±3,9
Березняк, окно 185,2±5,3 21,2±1,6
Примечания: фитоценозы см. табл. 8.
173
Консортивные связи
Вербейник обыкновенный опыляется пчёлами (Aris mellifera L.) и му-
хами-журчалками (Syrphus ribesii L.). Самоопыление с участием агента мо-
жет происходить благодаря личинкам трипса отличного (Haplothrips
distinguendus Uz.), которые питаются выделениями железистых волосков
на внутренней стороне венчика и репродуктивных органах. Для Англии
отмечены два вида пчел, опыляющих вербейник: Macropis еигораеа и
М. filpipes. Пчелы посещают цветки для сбора масел, выделяемых трихо-
мами венчика и репродуктивных органов. Пчелы садятся на цветок так,
что пыльники трутся о нижнюю часть брюшка и задние лапки. Масла в
трихомах долгое время могут оставаться в жидком состоянии, но при со-
прикосновении с пыльцой приобретают вид кристаллической водонерас-
творимой массы. Пчелы переносят именно эту смесь масел и пыльцы. Дос-
таточное количество смеси пчела собирает за 60 посещений, совершаемых
в течение 10-15 минут. Масла обеспечивают водонепроницаемость ячеек
сот, а в целом смесь используется для питания личинок (Kirk, 1996).
На открытых освещённых местах на вербейнике часто живет слюнявица
обыкновенная (Philaenus leucophthalmus L.). В европейской части России
листьями вербейника питается жук листоед Lythraria salicariae Pk. (Опреде-
литель насекомых ..., 1965). Листья повреждаются также двумя видами
моллюсков: Bridibaena truticum и Oxiloma elegans. Вирусных заболеваний
зелёных частей растения не отмечено ни для одного местообитания.
Корни имеют везикулярно-арбускулярную микоризу (Grime et al.,
1988).
Хозяйственное значение
В медицинской практике все части вербейника обыкновенного исполь-
зуются в виде отваров, водных настоев и спиртовых вытяжек, которые об-
ладают антикоагуляционными свойствами, а также проявляют антивирус-
ную и антифаговую активность. Вербейник употребляется при желтухе,
болезнях желудочно-кишечного тракта, гинекологических заболеваниях.
Отвар и настои применяются при коньюктивитах, как противосудорожное;
компрессы — при ушибах, грыже, артритах. Семена входят в состав сбо-
ров, употребляемых при циститах. Благодаря высокому содержанию вита-
мина С в зелёных частях растения (158-260 мг %), свежий сок из листьев
используют при лечении цинги, анемии и астении. Препараты из надзем-
ных частей растения обладают выраженным гомеостатическим свойством
и используются как кровоостанавливающее при внутренних и наружных
кровотечениях (Роллов, 1908; Энциклопедический словарь ..., 1951; Рас-
тительные ресурсы СССР, 1976; Манолова, Максимова, 1988; Раститель-
ные ресурсы ..., 1996).
174
Вербейник использовали для уничтожения некоторых насекомых и их
личинок. Так в ветеринарии порошком высушенного корневища присыпа-
ли раны скота от червей, а дым и пепел надземных частей токсичен для мух
(Роллов, 1908).
Вербейник обладает красящими свойствами, что используется при кра-
шении шерстяных тканей, а ранее употреблялся в косметике (краска для
волос). Растение даёт жёлтую и зелёную краску (вытяжки по железу) и, в
зависимости от протрав, чёрную и коричневую. Благодаря наличию ду-
бильных веществ, служит для дубления кож (Флора СССР, 1952; Лекарст-
венные растения ..., 1976).
Сведения о наличии нектара у вербейника обыкновенного различны.
Ф.Х. Роллов (1908) считал вербейник медоносом, который дает небольшое
количество нектара. Однако другие авторы (Kirk, 1996) указывают на от-
сутствие нектарников и не относят вербейник к медоносным растениям.
Большинство источников указывают, что вербейник обыкновенный аб-
солютно не поедается скотом из-за содержащихся в нём сапонинов. И-
только некоторые авторы (Кормовые растения ..., 1950, 1951, 1956) отме-
чают его как кормовое для лося и пятнистого оленя.
Вербейник разводится в садах как декоративное растение (Роллов,
1908; Scoggan, 1979).
М. А. Новикова, Ю. Е. Алексеев.
ВЬЮНОК ПОЛЕВОЙ
Номенклатура, систематическое положение
и внутривидовая систематика
Convolvulus arnensis L., 1753, Sp. Pl,: 153; Кауфман, 1866, Моск, фл.: 330;
Сырейщиков, 1910, Илл. фл. Моск, губ., 3 :44; Григорьев, 1953, Фл. СССР,
19 : 29; Победимова, 1964, в Маевский, Фл., изд.9 : 421; Тихомиров, 1966,
Опред. раст. Моск, обл.: 266; Stace, 1972, Fl. Europ. 3 : 81; Смольянинова,
1981, Фл. европ. части СССР, 5 : 95; Губанов и др., 1995, Опред. сосуд,
раст. Центра Европ. России : 411.— Вьюнок полевой (семейство Convol-
vulaceae — Вьюнковые).
Вьюнок полевой сильно изменчивый вид, возможно имеющий не-
сколько экотипов и географических рас, которые еще не стали предметом
специальных исследований. Тем не менее, уже давно отмечен полимор-
физм венчика этого вида (Розен, 1916), который наблюдается в пределах
региональных и локальных флор.
Изменчивость касается высоты (1,6-3,5 см)идиаметра (2,2-3,5см) вен-
чика, формы его края (ровный или с выемками), и границ лепе.стков (жил-
ки упираются в выемку или в углубление), особенностей пигментации. По
нашим данным, в области средней России встречаются следующие основ-
ные морфы (морфодемы) вьюнка полевого, которые характеризуются ком-
плексом признаков венчика:
• var. alba nom. provis.— венчик внутри белый, снаружи белый или ро-
зоватый с розовыми полосками;
• var. rosella nom. provis.— внутренняя и внешняя поверхность имеет
розовые полоски;
• var. purpurea nom. provis.— обе поверхности густо розового или пур-
пурного цвета.
Существенных закономерностей в вариации венчика и их связей с ме-
стообитанием не выявлено. По предварительным данным на пойменных
лугах (по р. Оке и Дону) доминирует var. alba, тогда как var. purpurea поч-
ти отсутствует. С другой стороны на осенних водораздельных лугах и в лу-
176
говых степях (верховья Дона в Липецкой обл.) var. purpurea встречается
намного чаще и составляет около 10 % от численности всех форм. Кроме
того, обнаружено, что в одной отдельной популяции обычно встречается
не больше двух морф вьюнка полевого. Изменчивость вида нуждается в
специальном исследовании.
Географическое распространение
Первичный ареал вида охватывал обширную территорию Евразии в
умеренном и субтропическом поясах Земли. Благодаря распространению в
агроценозах он сейчас занимает огромный регион между 60° с. ш. и
45° ю. ш. в Африке, Северной и Южной Америке, Юго-восточной Азии,
Австралии и островах Океании (Holm et al., 1977). В Северную Америку
вьюнок полевой попал из Европы в 1739 г., в штат Вирджиния. Его даль-
нейшая экспансия была связана первоначально с железными дорогами
(Weaver, Riley, 1982), но в современный период он распространен в Север-
ной Америке повсеместно кроме северных районов Канады и южных пус-
тынных районов США. В 80-е годы текущего столетия вьюнок был зареги-
стрирован как сорняк в посевах 332 сельскохозяйственных культур на тер-
ритории 44 стран (Holm et. al., 1977). Можно предположить, что в
настоящее время эти показатели должны быть увеличены. В России в свя-
зи с антропогенными изменениями растительного покрова также происхо-
дит постепенное расширение ареала Convolvulus arvensis и в современный
период. Он распространен в умеренной зоне по всему земному шару, по
всей Европейской части до Кавказа, за исключением Архангельской обл.,
Сев. Коми, Карелии и Мурманской обл. (Смольянинова, 1981). В Сибири
встречается в Тюменской (Тобольский флористический р-н), Омской,
Томской, Новосибирской, Курганской и Кемеровской областях, в Алтай-
ском крае, Хакассии, верховьях Енисея, Туве, Иркутской обл. (включая
Саяны, Приленско-Хатангский р-н, бассейн Нижней Тунгуски), Бурятии,
Читинской обл., в Якутии в большинстве районов включая тундровую зо-
ну (Фризен, 1997). Вьюнок также распространен в южных районах Дальне-
го Востока. Северные границы ареала этого вида находятся в стадии фор-
мирования, поскольку происходит интенсивный занос его семян в новые
районы. В 80-е годы этот вид впервые обнаружен в Мурманской области
на усадьбе Лапландского заповедника (Берлина, Костина, 1999). Прибли-
зительно в этот же период он встречен как заносный вид в приколымских и
приохотских районах Магаданской области (Хохряков, 1989), на Нижнем
Амуре около Софийска, Архангельского (Шага, 1989). В Московской об-
ласти вьюнок полевой встречается во всех районах (Ворошилов и др.,
1966).
177
Морфологическое описание
Вьюнок полевой — корнеотпрысковый многолетник, криптофит-ге-
микриптофит — травянистая лиана с однолетними вьющимися побегами;
его жизненный цикл охватывает весь вегетационный период.
У взрослых растений корневая система аллоризная, состоящая из длин-
ных горизонтальных и вертикальных геотропных корней. Корни прямые
или изогнутые, шнуроподобные, коричневые, ветвящиеся обычно до
ГУ—V-ro порядка. В аридных регионах корни вьюнка уходят в глубину до
3 м, но обычно основная масса горизонтальных корней, дающих корневые
отпрыски, сосредоточена на глубине 0,6-1 м (Утеулина, 1969; Брутян,
1970; Абдуллаев, 1982).
У молодых растений развит главный корень со многими боковыми.
Позже часть последних отмирает и сохраняется несколько многолетних
горизонтальных корней. Придаточные почки появляются на горизонталь-
ных боковых корнях и развиваются в корневые отпрыски, становящиеся
вертикальными корневищами, которые на всем протяжении формируют
боковые почки (Kennedy, Grafts, 1931, цит. по Weaver, Riley, 1982). Корне-
вища достигают поверхности почвы и в разных экологических условиях
варьируют по длине от 2 до 60 см. Закладка придаточных почек эндоген-
ная, происходит в межпучковом камбии первичного сердцевинного луча.
Боковой корень формируется из перицикла на стадии первичного строения
основного корня. Кроме того, в перицикле корня против первичного луча
ксилемы происходят межклеточные деления, и формируется межпучко-
вый камбий, который образует почку. Таким образом, почка формируется
на стадии вторичного строения корня.
Ветвление и формирование корневой системы зависит от структуры и
влажности почвы. В американских популяциях вьюнка полевого главный
корень достигал длины 3 м, а вся система имела площадь до 6 м в диаметре
и 9 м в глубину. При этом от 50 до 70 % массы корней залегало на глубине
60 см (Holm et al.,1978, цит. по: Weaver, Riley, 1982). Горизонтальные боко-
вые корни на некотором расстоянии от стержневого круто изгибаются и
уходят в вертикальном направлении в нижележащие слои. На изгибах кор-
ней закладывается большое количество почек, из которых впоследствии
развиваются новые побеги со своей корневой системой.
Корни имеют пентархную структуру, обладая 5-лучевой первичной
ксилемой. Бугорчатые образования в одних случаях содержат зачатки бо-
ковых корней с одним тяжем ксилемы, в других — придаточные почки с
двумя ксилемными тяжами. Закладка придаточных почек эндогенная, про-
исходит в межпучковом камбии первичного сердцевинного луча (Тогеу,
1958).
Стебли достигают 1-3 м в длину, вьющиеся или стелющиеся. Они час-
то обвивают побеги других растений, а также и другие собственные побе-
178
ги, образуя спутанные «колтуны» и «канаты». Последние образуются из
отпрысков как одного, так и нескольких растений.
Стебли цилиндрические или слегка ребристые, гладкие, голые, либо
покрытые волосками. Тонкостенный мелкоклеточный эпидермис стебля
сильно кутинизирован, внешняя поверхность кутикулы сморщена в про-,
дольном направлении. Лигнин образуется слабо — много тканей с целлю-
лозными оболочками. В коре почти нет склеренхимы, много паренхимы,
которая не мешает стеблю растягиваться. Особенности строения стебля
вьюнка полевого позволяют ему успешно конкурировать с другими вида-
ми на лугах и в агроценозах (Камилова, 1982; Weaver, Riley, 1982).
На свободных от растительности участках побеги растут горизонталь-
но и достигают 100 см в длину (Abdallah, 1967). Боковые побеги очеред-
ные, но в семядольном узле супротивные. Боковой побег имеет два пред-
листа, расположенных латерально. При встрече с растением-опорой, пол-
зущий побег взбирается на него, приобретая спиральную форму.
Листья очередные, простые, цельнокрайние, гладкие, опушенные, 2-5
см длиной, различные по форме и размеру, треугольные либо овально-уд-
линенные с ладьевидными, сердцевидными или стреловидными базальны-
ми лопастями. Поверхность листа неровная, состоит из тонкостенных сла-
бо кутинизированных эпидермальных клеток. Листья имеют сильную фе-
нотипическую изменчивость по форме и размеру в зависимости от ряда
экологических факторов: освещения, влаги, в том числе от условий ирри-
гации (Weaver, Riley, 1982).
Соцветие интеркалярное. В пазухах листьев цветки расположены по
одному, либо по 2-3, образуя цимозные соцветия.
Цветки правильные, пентамерные, спайнолепестные, обоеполые, сидя-
щие на длинных цветоножках, с двумя очередными, ланцетовидными при-
цветниками 1-10 мм длиной, расположенными на 1,2—2,5 см ниже цветка.
Чашелистиков пять, 3-5 мм длиной, внутренние длиннее наружных,
овальные или обратнояйцевидные, на верхушке округленные или тупые,
иногда выемчатые или с небольшим остроконечием, по краю с пленчатой
каймой до 0,4 мм ширины или с волосками до 0,2 мм длиной, зеленые, при
плодах коричневые.
Венчик актиноморфный, воронковидный или колокольчатый, 1,5-2 см в
длину и около 2 см в диаметре, чаще белый с розовыми продольными полос-
ками (Смирнова, Каден, 1971). Венчик в почкосложении скручен. Андроцей
из 5 тычинок неравной длины, две короче остальных, прикрепленных около
основания венчика. Тычиночные нити с железистыми короткими, светлыми
волосками. В основании тычиночных нитей имеется углубление в виде «ло-
жечки», покрытое прозрачными головчатыми выростами по всей поверхно-
сти, кроме углубленной части. На поверхности пыльника также имеются
бесцветные скульптурные образования, расположенные в 1-2 ряда. В осно-
вании пестика находятся цельный нектарный диск, ткани которого выделя-
179
ют нектар. Мы наблюдали два варианта его строения: оранжевый лопастной
или желтый. Завязь верхняя, двугнездная, овально-шаровидная. Столбик
около 5-8 мм длиной, наверху с двумя удлиненными рыльцами.
Плод — коробочка, около 6-7 мм длиной, яйцевидная, остроконечная,
желтоватая или светло-коричневая, двугнездная раскрывается створками
(гинецей синкарпный); у основания коробочки почти всегда остается око-
лоцветник с плодоножкой. В норме в коробочке 4 семени, но нередко неко-
торые из них недоразвиты. Наиболее часто встречаются коробочки с 1 и 3
семенами, но обнаруживались и 10-семенные плоды. В зависимости от
числа семян в плоде, варьирует их форма: если семян несколько, то их при-
легающие стороны бывают уплощены, а если семя одно, то оно приобрета-
ет почти сферическую форму. Вес семян 10-14 мг. Кожура семян плотная,
мало проницаемая (Смирнова, Каден, 1971; Weaver, Riley, 1982).
Семя обратнояйцевидное, овальное или грушевидное, до 4 мм дайной,
в сечение округло-треугольное, на спинке округленное, на брюшной сто-
роне с двумя сходящимися под углом гранями, бугорчатое, голое, корич-
невое или почти черное, крупно-шероховатое. Одно растение способно
сформировать до 550 семян (Доброхотов, 1960).
Число хромосом 2п=48 установлено в Европе и Азии, а 2п=50 у популя-
ции из США (Wolcott, 1937, цит. по: Weaver, Riley, 1982).
Онтогенез
Всхожесть и жизнеспособность семян варьируют в зависимости от ус-
ловий, однако свежесобранные семена имеют сходные средние показате-
ли: проросшие семена — 5-25 %, «непроницаемые» — 60-80 %, жизне-
способные— 87-99 % (Weaver, Riley, 1982). Прорастание «непроницае-
мых» семян может быть стимулировано обработкой концентрированной
серной кислотой в течение 1 часа, чистым спиртом в течение 20 часов, ли-
бо механическим надрезанием. По данным В.Н. Доброхотова (1968), после
скарификации — только 1-2 % семян давали всходы.
Бороздка рубчика семени (место входа пыльцевой трубки) у семян дей-
ствует как гигроскопичная створка, которая открывается и закрывается в
зависимости от относительной влажности воздуха. Она состоит из двух
слоев палисадных макросклереид. Размягчающая обработка (например,
спиртом) вызывает сокращение самого внутреннего слоя створки, и бо-
роздка открывается. Серная кислота растворяет ткани вокруг самой бо-
роздки. Механическое надсекание кожицы приводит к многочисленным
трещинам в семенной кожуре, облегчающим проникновение воды внутрь.
Надсеченные семена прорастают при широком диапазоне температур
(5-40°С), при оптимуме 20-3 5°С. Экспонирование на свету несколько по-
вышает всхожесть свежесобранных семян (на 5-10 %). Поочередное дей-
ствие температур 40°С и 5°С также стимулирует прорастание семян. У пе-
180
резимовавших в почве семян проницаемость покровов возрастает более
чем на 30 %. (Brown, Porter, 1942).
Э. Сулайманов (1981) изучал прорастание семян в условиях темпера-
турного оптимума. Низкую всхожесть (11-21 %) семян, автор объяснял
«свежестью» семян, не прошедших стадию покоя, а также целостностью
покровов семени и неповрежденностью кожуры. Низкая всхожесть, по-ви-
димому, приводит к консервации покоящихся семян в почве, и обеспечи-
вает возможность семенного возобновления в течение десятков лет.
По нашим данным, в комнатных условиях скарифицированные семена
прорастают быстро: в течение первых суток формируется главный корень с
развитыми корневыми волосками зоны всасывания (рис. 1). На пятый день
гипокотиль выносит семядоли в кожуре семени за пределы почвы. На 10-12
день, когда гипокотиль вытягивается до 5 см и более, сбрасывается кожура,
и семядоли начинают раздвигаться. Через 15 дней семядоли полностью
«раскрыты» и начинается развитие первого листа. Примерно на 18-й день
развития, когда главный побег достигает длины 3 см, его гипокотиль изгиба-
ется, и стебель начинает совершать вращательные движения. Менее чем че-
рез месяц (25-27 дней), когда уже на побеге развиты 4-5 листьев, растение
достигает опоры (при наличии таковой) и начинает обвиваться вокруг нее.
Длина стебля в этот период уже превышает 20 см и быстро увеличивается, и
одновременно у отдельных растений семядоли начинают желтеть и отми-
рать (полностью они засыхают на 60-й день). Развитие побегов П-го порядка
из семядольных пазух происходит примерно на 50-й день развития. Если на
побегах 1-го порядка выше семядолей формируются срединные листья
обычной формы, то у побегов П-го порядка в основании расположены мел-
кие редуцированные листья. На 60-й день возможно появление побегов П-го
порядка и на последующих узлах главного побега (рис. 2). В то же время на-
блюдается также утолщение корня, появление на нем бугорчатых образова-
ний, а затем почек и корневых отпрысков. Примерно на 70-й день происхо-
дит отмирание верхушки побега 1-го порядка (при благоприятных условиях
произрастания), а доминировать начинают побеги П-го порядка. Растения
90-дневнего возраста уже имеют корневые отпрыски, которые вырастают на
10 см над поверхностью почвы и имеют 5-6 листьев. В семядольных пазу-
хах наблюдается рост боковых побегов и, помимо этого, прорастание 3-4
почек. Примерно у 30 % наблюдаемых нами растений побег пригибался к
почве (не укореняясь), а от семядольной зоны отрастало 6-7 боковых побе-
гов. Подобное полегание наблюдается, уже начиная с 60-го дня. При этом в
почве располагается мощная зона кущения, из которой развиваются (на этой
стадии) 10 побегов и начинают прорастать (или потенциально готовы к про-
растанию) около 10 почек, а семядольная зона образует своеобразную «вто-
ричную зону кущения». На данном этапе развития молодого растения суще-
ствует как бы две зоны кущения. Одна, надземная, находится в пазухе семя-
долей, а другая, подземная, размещается на главном или боковых корнях.
181
Рис. 1. Проростки и ювенильные особи Convolvulus arvensis. Обозначения: Цифрами обозначен
возраст растения в сутках
Рис. 2. Виргинильные особи Convolvulus arvensis. Обозначения: Цифрами обозначен возраст растения в сутках
Отдельные побеги, берущие начало от корневых отпрысков чаще имеют
лишь одну, подземную зону кущения. По нашим наблюдениям, такая форма
присуща побегам, произрастающим в условиях возделываемых полей, лу-
гов, железнодорожных насыпей и других местообитаний, где нет большой
механической нагрузки на почву. В тех же местах, где вытаптывание интен-
сивно (вдоль дорог, тропинок), корнеотпрысковые побеги вьюнка также об-
разуют вторичные надземные зоны кущения. Таким образом, признаки фор-
мы жизни гемикриптофита в основном проявляется у проростков первого
года жизни, а также в условиях большой механической нагрузки на почву.
Цветочные почки закладываются уже на ранних стадиях, когда длина глав-
ного побега достигает 50 см, к этому времени появляются побеги Ш-го по-
рядка, корни интенсивно ветвятся, семядоли отмирают и растут побеги из
почек на гипокотиле.
Возрастные изменения особи вьюнка полевого и периодизацию онто-
генеза условно можно охарактеризовать следующим образом.
Стадия проростка — растение имеет только развернувшиеся семядоли.
Ювенильная стадия —растение формирует главный побег с несколь-
кими листьями.
Имматурная стадия — у растения на главном побеге формируются бо-
ковые побеги как в пазухах семядолей, так и выше расположенных листь-
ев; возможно отмирание семядолей.
Виргинильная стадия — отмирание верхушки главного побега, энер-
гичное развитие боковых побегов П-го порядка; заложение придаточных
почек и развитие корневых отпрысков.
Генеративная стадия — формирование цветков и системы интеркаляр-
ного соцветия; на протяжении генеративного периода, периодизация кото-
рого затруднена, происходит отмирание системы главного корня и образо-
вание клона.
Сенильная стадия — ослабление роста корней и снижение числа кор-
невых отпрысков; конкретные параметры данного периода и его продол-
жительность не установлены,
Взрослые особи вьюнка полевого имеют обширную систему горизон-
тальных корней, которые нарастают с одного конца и постепенно отмира-
ют с другого (старого). Придаточные почки закладываются уже на одно-
летних корнях. Часть из них развивается в корневые отпрыски, часть отми-
рает или остается в спящем состоянии. Конкретная роль внутренних и
внешних (экологических) факторов в различной «судьбе» почек остается
неизвестной. У взрослых растений на горизонтальных корнях придаточ-
ные почки расположены тесно.
Таким образом, особенностью корневой системы вьюнка полевого явля-
ется гетероризия, т. е. существование корней двух типов. Они различаются
размерами и морфолого-физиологической специализацией. Если главный
корень растет вертикально вниз, то боковые корни вьюнка, возникшие на
184
главном корне, обычно растут горизонтально. Однако, нередки случаи, ко-
гда через некоторое время кончики боковых корней изменяют направление
роста, и начинают расти вертикально вниз, т. е. геотропно, как и главный ко-
рень. Гетероризия отчетливо проявляется у взрослых особей. Последние
имеют длинные горизонтальные корни, которые несут придаточные почки и
корневые отпрыски. Такие корни обычно называют корнями размножения.
Они имеют более тонкие боковые корни, которые являются питающими.
Корневые отпрыски, со своей стороны, имеют почти исключительно тонкие
питающие корни (придаточные по своему происхождению). Но, кроме того,
изредка у них формируются и толстые, длинные придаточные корни с новы-
ми корневыми отпрысками. Эти корни также можно квалифицировать как
корни размножения. Как у многих корнеотпрысковых растений, участки
корня в месте отхождения отпрыска имеют неравный диаметр: участок кор-
ня, направленный в сторону растущего конца корня, всегда тоньше, чем
смежный участок корня, направленный в сторону старой корневой системы.
Основной срок жизни корневого отпрыска составляет два года. Из его
боковых почек на второй год симподиально формируется в среднем 3-4
боковых побега П-го порядка. Они могут быть простыми или ветвистыми,
т. е. образуют побеги Ш-го порядка, нередко из самых нижних почек на
уровне почвы или под ним.
У растущего под землей отпрыска междоузлия удлиненные. На уровне
почвы у отпрыска формируются укороченные междоузлия, образуется зо-
на кущения и происходит образование нескольких (обычно 5) радиально
отходящих побегов, звездообразно распластанных по поверхности почвы.
Отпрыски не всегда достигают поверхности почвы в первый год своего
развития, и их верхушечная почка часто отмирает. Апогеотропный рост на
2-й год инициируется одной из верхних боковых почек.
Изолированные корни вьюнка, выращенные в питательном растворе
при 25°С, демонстрируют заметное доминирование верхушки и вытягива-
ются в среднем на 25 мм в день (Тогеу, 1958). Сегменты корня имеют чет-
кую полярность, формируя боковые корни на дистальном конце и побе-
ги — на проксимальном. Свет необходим для комплексного развития сис-
темы побегов, и корневым почкам не удается вытянуться более чем на 10
мм, когда рост происходит в темноте. Изолированные сегменты корня, со-
держащиеся в питательном растворе, сохраняют способность формиро-
вать эндогенные почки в течение нескольких лет (Тогеу, 1958).
Опыты укоренения отрезков вертикальных корневищ показали неспо-
собность их укореняться (Котт, 1956). Медленно образующиеся на них мо-
лодые побеги дают только новые 1-2 пары листьев и в таком виде живут до
тех пор, пока не израсходуют всего запаса питательных веществ. Несколь-
ко лучше приживаются отрезки настоящих корней, хотя и у них прижива-
ние проходит медленно, и возможности полного закрепления в пахотной
земле нет.
185
Как выше отмечено, на уровне почвы у отпрыска формируются укоро-
ченные междоузлия, а из почек зоны кущения, возникающих также на
уровне почвы, образуются один или несколько побегов. Наземным ветвя-
щимся годичным побегам условно свойственна базитония. Боковые побе-
ги П-го порядка развиваются из почек 2-4 околоземных узлов побега 1-го
порядка. Примечательно, что часть боковых побегов имеет редуцирован-
ное строение, листья на них уменьшены (примерно в 5 раз) по сравнению с
листьями побегов 1-го порядка.
Побеги П-го порядка (если отпрыск считать побегом 1-го порядка) ха-
рактеризуются тем, что у них пробуждаются боковые почки, из которых в
свою очередь образуются побеги Ш-го порядка. Следовательно, годичные
побеги на двулетнем отпрыске могут быть как простыми, так и формиро-
вать систему годичных побегов, поскольку у проростков побеги П-го по-
рядка могут формироваться из боковых почек, закладывающихся в пазу-
хах семядолей. Эта особенность позволяет считать, что переход из стадии
проростка в стадию «ювенильная особь» может происходить уже в тече-
ние первого года жизни особи.
Свойственные вьюнку полевому круговые нутации (вращательные
движения побегов) являются одним из типов активного движения расте-
ний, возникающего за счет растяжения клеток, ведущего к удлинению осе-
вого органа. Но если ростовые настии и тропизмы являются необратимым
процессом, идущим под действием осмотических сил, то круговые нута-
ции обратимы за счет чередования растяжения клеток в разных участках
осевого органа. В настоящее время известен механизм «роста растяже-
ния», который описан в ряде работ (Полевой, Саламатов, 1985, и др.) и ко-
торый мы здесь не рассматриваем. Отметим только, что основную роль в
регуляции роста растения выполняет индолил-3-уксусная кислота, которая
взаимодействует с рецепторами, локализованными в мембране и цито-
плазме. Биохимические реакции, в конечном счете, приводят к разрыхле-
нию клеточных стенок и увеличивают их растяжимость.
Как отметил еще Ч. Дарвин (1941), закручивание вьющихся стеблей
представителей рода Convolvulus происходит против часовой стрелки. При
этом виды одного рода вьются с различной скоростью. Эта скорость,
по-видимому, не определяется толщиной побегов, так как у некоторых
древесных растений стебли вьются быстрее, чем у травянистых. Молодые
междоузлия вьются медленнее, чем тогда, когда они становятся старше.
По нашим наблюдениям, в комнатной культуре около окна, до тех пор,
пока побеги не достигли длины 15-20 см, вращения или изгиба верхушки Не
происходит. У побегов длиной более 17 см начинала отклоняться верхушка,
и наблюдалось вращение. У побегов достигших 20-25 см наблюдалось уже
два изгиба: верхушки и участка стебля на уровне 4—5 листа. Выявлено две
формы движения побега: вращение срединной части стебля и верхушки. У
обоих, стебля и верхушки, скорость движения по теневой стороне выше, чем
186
по световой, а амплитуда отклонения меньше. Верхушка вьюнка делает
один оборот (двигаясь против часовой стрелки) в среднем за 2 часа 50 минут.
Скорость вращения видоспецифична, например, скорость вращения
стебля у травянистой лианы Humulus lupulus 2 часа 8 минут и имеет на-
правление по часовой стрелке. Кроме круговых движений побега (круго-
вая нутация — по Саксу) у вьющихся растений происходит перекручива-
ние самого побега, не связанное напрямую с вращением стебля. Например,
у хмеля после 37 оборотов стебля, последний оказался перекручен только
3 раза. Осевое перекручивание уменьшало гибкость стебля, увеличивая
его прочность, когда стеблю приходилось совершать круговые движения
без подпорки. До соприкосновения с опорой круговые движения соверша-
ются быстрее, чем после закрепления побега на опоре (Дарвин, 1941).
Постепенное увеличение шага спирали у вьюнка полевого происходит
выше места первоначального закрепления на подпорке. Побег, сильно пе-
рерастающий подпорку, опускается вниз под влиянием силы тяжести.
Подпорки большого диаметра затрудняют обвивание, так побеги вьюнка
полевого были неспособны обвить опору диаметром 5-6 сантиметров.
По нашим наблюдениям, не обнаружено четкой зависимости в выборе
направления роста побега в зависимости от ориентации по сторонам света.
Наблюдалось лишь небольшое преимущество в выборе северо-западного
направления. Связь с затенением практически отсутствует.
В естественных условиях вьюнок обвивал любые близко расположен-
ные опоры — как живые, так и неживые, независимо от диаметра (наи-
больший был у дерева и каменной изгороди — около 5 см) и характера по-
верхности. Тонкие стебли злаков (до 0,2 см) также использовались в каче-
стве опоры, причем вьюнок иногда обвивал сразу несколько стеблей и
листьев растений разных видов. В этом качестве чаще выступают Phleum
pratense, Dactylis glomerata, виды родов Festuca, Taraxacum. Один побег
может переходить с одной опоры на другую, нередко оплетая до 4-5 опор
одной плетью и поднимаясь на высоту до 72 см от поверхности почвы. При
отсутствии опоры вьюнок обвивает собственные побеги, которые скручи-
ваются в канат, всегда против часовой стрелки. Иногда опоры оплетены
именно таким «канатом».
Цветки закладываются на побегах, начиная в среднем с 7-го узла его
надземной части. В пазухе одного листа обычно закладывается один цве-
ток или дихазиальное соцветие; в последнем случае нижние цветки имеют
несколько меньшие размеры.
Ритм сезонного развития
В природе прорастание семян начинается весной уже при 4-10°С, оп-
тимальный интервал температуры 20-28°С. Лучшая всхожесть при заглуб-
лении семян на 4—5 см. При размещении семян в почве на глубине свыше
187
20 см всходы практически не образуются. Семена вьюнка прорастают в те-
чение всего культивационного периода при условии, что достигается дос-
таточное увлажнение почв, однако пик прорастания приходится на позд-
нюю весну и раннее лето (Утеулина, 1969; Брутян, 1970).
Семена вьюнка, надсеченные и посаженные в Айове (США) с интерва-
лами в 2 недели с 25 апреля до 26 сентября всходили и давали проростки
вскоре после каждой посадки (Brown, Porter, 1942). Пики прорастания при-
ходились на весну (60-75 %) и позднее лето (43-57 %), а спад наблюдался
в середине лета (1-10 %). Растения, развившиеся весной и ранним летом,
укоренялись к ноябрю до глубины 1,3—1,7 м, в то время как всходы августа
и сентября только на 25-50 см.
Вьюнок полевой — летне-зеленое растение, его вегетация охватывает
весь вегетационный период. Он имеет продолжительный период роста побе-
гов и также продолжительный период цветения (в среднем с июня по сен-
тябрь). Вследствие наличия интеркалярного соцветия, элементы которого
находятся на многих междоузлиях, репродуктивные органы отдельного по-
бега в каждой период времени находятся в нескольких фазах развития. Рас-
тение переживает зимние условия в виде системы корней и корневищ, спо-
собных выдержать охлаждение до -6°С. Их гибель наступает при промерза-
нии почвы ниже -8°С. Зимостойкость корней повышается в течение осени.
Подкопанные ранней осенью, корни обладают пониженной способностью к
перезимовке (погибают при температуре около 2°С) (Weaver, Rily, 1982).
Поздней осенью из-за низких температур надземная часть побегов от-
мирает. Во второй и последующие года новые отпрыски возникают из эн-
догенных корневых почек, образованных на зимующих вертикальных и
некоторых боковых корнях. Латентные почки на более крупных корнях на-
чинают расширяться и развивают корневища ранней весной. Быстрый рост
корневищ и побегов начинается, как только дневная температура достига-
ет 14°, а ночная 2°С.
Интенсивный рост вьюнка в Азербайджане наблюдался уже со второй
половины марта, цветение с середины мая до начала июня (Абдуллаев,
1982).
Рассада, выросшая в теплице, получала способность возобновляться из
придаточных почек корня (когда надземная часть была удалена) только че-
рез 19 дней после прорастания.
В Канаде наблюдалось отрастание 25 побегов от корневого фрагмента
длиной 5 см в течение 4 месяцев после посадки, причем ближний дочерний
побег отходил от него на 45 см, а самый дальний на 1,3 м (Best,1963, цит.
по: Weaver, Riley, 1982). А после 15 месяцев вегетации новые побеги воз-
никали на расстоянии 2,85 м от материнского участка корня. В природной
популяции на культивируемых почвах Техаса численность побегов, дос-
тигших 15 см высоты от поверхности, составила в среднем 285 на 1 м2
(Wiese, Rea, 1962).
188
Отмечена сезонная динамика активности корневых почек. Максимум
формирования числа побегов из сегментов боковых корней приходится на
раннюю весну. Эндогенные почки корня, в отличие от пазушных почек,
могут оставаться в покое, либо развиваться в побеги и корни. Они находят-
ся под контролем ауксина из побегов и цитокинина из корней (Weaver,
Riley, 1982).
Процентное содержание крахмала в подземных органах достигает мак-
симума в августе или сентябре и с этого времени быстро идет на убыль,
превращаясь в сахар — его содержание максимально в октябре, мини-
мально в мае. Максимум содержания азота наблюдается в середине апре-
ля — до побегообразования, а затем быстро снижается, достигая миниму-
ма в мае (Barr, 1940 цит. по: Weaver, Riley, 1982).
Способы размножения и распространения
Вьюнок полевой имеет генеративный и вегетативный способы размно-
жения. Обычно вегетативное размножение корневыми отпрысками преоб-
ладает в локальном распространении, тогда как семена играют важную
роль при распространении вида на обширные территории.
Цветение длится так долго, как позволяют условия. В первый год жиз-
ни растения вьюнка редко образуют семена, даже если они были выраще-
ны в теплице и цвели в течение 6 недель после прорастания семян (Weaver,
Riley, 1982).
Цветки обычно функционируют только в течение одних суток и опыля-
ются насекомыми. Цветение начинается ранним утром и сопровождается
выработкой нектара в нектарнике у основания венчика (Waddington, 1976,
цит. по: Weaver, Riley, 1982). Появление нектара, вероятно, сопровождает-
ся наибольшей интенсивностью запаха цветка (Faegri, Pijl, 1971).
Вид, возможно, обладает гинодиэцией. В его популяциях женская фор-
ма составляла около 7 % (30 из 432 изученных особей) (Пономарев, Демья-
нова, 1975). Указанные авторы относят вьюнок полевой к видам, характе-
ризующимся ложной гинодиэцией. Для нее характерен более мелкий по
размеру функционально женский цветок.
Существуют различия в длине тычинок и рыльца со столбиком. Эти
органы могут располагаться приблизительно на одном уровне или ты-
чинки могут быть короче лопастей рыльца. У Ipomoea purpurea в цветках
разных особей также наблюдали различные уровни расположения тычи-
нок и рылец (Brown, Porter, 1942). При этом E.O.Brown и R.H.Porter
(1942) не связывали это с явлением гинодиэции, не отмечая какой-либо
связи между окраской венчика и относительными размерами тычинок и
столбика. Однако исследователи не изучали процент фертильной пыль-
цы у Ipomoea purpurea. Между тем А.Н. Пономарев и Е.И. Демьянова
(1975) установили, что у физиологически женских особей Convolvulus
189
arvensis фертильная пыльца составляет около 80 %. Этот показатель яв-
ляется очень высоким (норма) и поэтому можно поставить вопрос, на-
сколько сильно дифференцированы обоеполые и женские особи у вьюка
полевого. Кроме того, по нашим данным, в пределах одной популяции
вьюнка обоеполые и условно женские особи не имеют заметных разли-
чий в размерах венчика. Между тем А.Н. Пономарев и Е.И. Демьянова
показывают на рисунке существенные различия этого признака у обоепо-
лых и женских экземпляров. А именно, у женских экземпляров венчик по
высоте и диаметру приблизительно в два раза меньше, чем у обоеполых
растений. По нашим данным, обширные популяции вьюнка сравнитель-
но однородны по окраске венчика. Обычно в них можно обнаружить одну
или две морфы. Очень часто популяции представлены единственной мор-
фой, что можно объяснить особенностями вегетативного размножения
вьюнка. Вероятно, генетический состав отдельных популяций вьюнка
сравнительно однообразен. Но для полного ответа на этот вопрос необхо-
димы дополнительные исследования.
Созревающие семена приобретают способность к прорастанию уже
спустя 10-15 дней после опыления при 80% содержании влаги внутри
них. Непроницаемость семян наступает на 23-25 день после опыления, ко-
гда содержание внутренней влаги падает до 13 %. Семенная кожура полно-
стью дозревает спустя 30 дней (Sriplieng, Smith, 1960). Жизнеспособность
семян в почве сохраняется свыше 20 лет (Timmons, 1949), а при комнатной
температуре — свыше 50 лет (Brown, Porter, 1942). Семена теряют всхо-
жесть после 72 лет (Brown, 1960, по Брутяну, 1970).
Созреванцю семян благоприятствует сухая, солнечная погода, а также
произрастание на сухих известковых почвах. Но оно везде может замед-
ляться в дождливые периоды и, особенно на бедных, сильно истощенных
почвах. На регулярно возделываемых почвах растения не успевают проду-
цировать семена.
Нередко развитые семенные коробочки содержат неполное число се-
мян. Таких плодов может быть от 4 до 40 % (Brown, Potter, 1942, цит. по
Weaver, Riley, 1982). Одно растение способно дать в среднем 470 семян
(Абдуллаев, 1982). В отдельных районах вьюнок продуцирует от 50 тыс.
до 20 млн. семян на гектар (Brown, Porter, 1942). С помощью семян этот ру-
деральный и космополитный вид осуществляет широкую экспансию.
Семена распространяются животными и человеком, в частности, один
из главных путей попадания вьюнка в агроценозы — через засоренный по-
севной материал. В распространении вьюнка на пустырях, насыпях дорог
и в городских фитоценозах играет определенную роль и перемещение
грунта при строительных работах. Семена обычно осыпаются вблизи мате-
ринского растения, но могут разноситься путем эндозоохории и водой.
Важный распространитель — перелетные птицы, в их пищеварительном
тракте семена сохраняют жизнеспособность до 144 часов (Proctor, 1968).
190
Семена могут десятилетиями покоиться в почве, увеличивая способность к
повторной инвазии этого вида.
Вид размножается вегетативно посредством эндогенных корневых по-
чек, из которых развиваются корневые отпрыски. Поскольку последние
живут 2 года, то они становятся подземными корневищами, имеющими па-
зушные почки, а также собственные придаточные корни. Корни, отрастаю-
щие от корневищ, делают отпрыски относительно независимыми от роди-
тельского растения. Если отпрыск имеет хотя бы одну почку, он способен
в естественных условиях произвести новое растение (Брутян, 1970).
Вышесказанное позволяет считать наиболее характерными параметра-
ми вида — обильное цветение и плодоношение, низкую всхожесть семян,
слабую конкурентноспособность проростков и интенсивное вегетативное
размножение.
Экология
Совокупность биоморфологических признаков вьюнка свидетельству-
ет о его приспособлении к открытым травяным фитоценозам на суховатых
и свежих почвах. Это плюризональный вид. Он светолюбив, растет на лу-
гах, в степях, лесостепи и лесолуговых, реже полупустынных экотопах
(Раменский, 1956). Обычен и во вторичных местообитаниях, особенно на
разнообразных сельскохозяйственных землях и залежах. Н. Ellenberg
(1991) относит вьюнок полевой к полусветовым растениям (7-я ступень).
Е. Landolt (1977) отмечает, что он растет чаще на полном свету, но иногда
при некотором затенении.
Вьюнок хорошо растет на сухой, либо умеренно увлажненной почве
(6-я ступень шкалы Элленберга) и может переносить длительный период
засухи, благодаря своей разветвленной корневой системе (Weaver, Riley,
1982). Выдерживает резкую переменность увлажнения (Раменский, 1956).
Предпочитает в основном нейтральные, но растет также на кислых и ще-
лочных почвах (7-я ступень) (Ellenberg, 1991) с pH 5,5-8,0 (Landolt, 1977).
Индифферентен к содержанию в почве доступного азота (Ellenberg, 1991),
но чаще растет на богатых, плодородных почвах (Раменский, 1956), реже
на бедных, каменистых и песчаных (Weaver, Riley, 1982) и даже на слабо и
сильно засоленных почвах (Раменский, 1956).
В разных экологических условиях изменяются некоторые параметры
жизненной формы вида. К их числу относятся: расстояние между соседни-
ми корневыми отпрысками, расположенными на одном корне; число мета-
меров на подземной части корневого отпрыска, т. е. на вертикальном кор-
невище; число подземных побегов, образующихся в зоне кущения корне-
вого отпрыска, расположенной на границе двух сред—почвенной и
воздушной; образование прямостоячих и вьющихся или горизонтальных
(диагеотропных), лежащих на поверхности почвы побегов. Вариабель-
191
Рис. 3. Схема строения парциальных кустов (корневых отпрысков)
Convolvulus arvensis в разных экологических условиях: 1 — залежь без травостоя,
2 — старая залежь, 3 — луг, 4 — щебенка дорожного полотна.
192
ность всех этих признаков в принципе не изменяет существа жизненной
формы растения (его взрослых особей), но очень существенно влияет на
общую архитектуру систем корней и побегов и, безусловно, имеет адап-
тивный характер (рис. 3).
Фитоценология
Вьюнок полевой входит в состав как естественных, так и искусствен-
ных фитоценозов: возделываемых полей, пастбищ, садов, пустырей, вдоль
автомобильных трасс и железных дорог. В агроценозах существенно засо-
ряет виноградники, фруктовые сады, плантации сахарной свеклы, чая,
хлопчатника и овощей. Растение часто формирует небольшие заросли на
щебенке полотна железной дороги, где в грунте развивает своеобразные,
сильно разветвленные побеги и надземные стебли с мелкими листьями,
способно поселяться на стенах городских построек (Lisci, 1997).
С. arvensis — диагностический вид союза Panico-Setarion Siss. in Westh.
et al. 1946 порядка Polygono-Chenopodietalia (R.Tx. et Lohm. in R. Ttx. 1950)
J. Tx. in Lohm. et al. 1962 класса Chenopodietea Br.-Bl. 1951 em Lohm., J et R.
Tx. 1961 ex W. Matusz. 1962 (Миркин, Наумова, 1998). Это сообщества ого-
родов и залежей в южных засушливых областях России. В сводке К.О. Ко-
роткова с соавторами (Korotkov, 1991) приведена ассоциация Convolvulus
arvensis-Amaranthetum retroflexi Abrm. et Sach. in Mirk, et al. 1986 из того же
союза для предуральских степей и Воронежской области.
Вьюнок полевой — обычный участник ранних этапов сукцессии лугов,
пашен и пустырей. На заключительных этапах сукцессий и в коренных со-
обществах его роль очень мала, нередко он полностью выпадает из траво-
стоя. В связи с тем, что условия освещенности на всех этапах сукцессий ос-
таются практически неизменными, можно предположить, что лимитирую-
щим фактором в сохранении вьюнка в составе фитоценозов является,
прежде всего, корневая межвидовая конкуренция. Известно, что на позд-
них фазах сукцессий увеличивается роль многолетних видов, имеющих
развитую корневую систему, которая эффективно осваивает различные го-
ризонты почвы и увеличивается общее проективное покрытие растениями.
По-видимому, эти обстоятельства существенно затрудняют нормальное
развитие корневых отпрысков вьюнка. К тому же последние обычно не
имеют собственных придаточных корней, которые бы способствовали
усилению жизненности этих отпрысков. Наконец, очень часто корневые
отпрыски в первый год своего формирования растут только в почве и не
выходят в воздушную среду.
Успешная конкуренция вьюнка на ранних стадиях сукцессий связана с
разветвленной корневой системой, которая позволяет ему быстро захваты-
вать новые территории и подавлять рост других растений. Корни вьюнка
оплетают корни других растений, понижая кислотность почвы и затрудняя
193
доступ к влаге (Бржезицкий, Эфендиева, 1961; Брутян, 1970; Weaver, Riley,
1982).
Растение часто образует большие скопления вместе с другими одно-,
дву- и многолетними недолгоживущими сорняками. Обнаружена высокая
отрицательная аллелопатия С. arvensis по отношению к другим растениям,
в частности к сорняку Amaranthus retroflexus L. (Reynders, Duke, 1979).
Консортивные связи
Консортивные связи вьюнка изучались главным образом в агроцено-
зах. Большинство животных связано с вьюнком трофическими связями. В
степной и пустынной зонах Узбекистана и России Б.М. Горенштейн (1982)
и В.А. Слободянюк (1987) обнаружили фитофагов, поражающих вьюнок
полевой: долгоносики рода Alcides', зерновка Euspermophagus sericeus
Geofr., которая повреждает генеративные органы, в частности, ее личинки
прогрызают незрелый эндосперм; блошки рода Longitarsius, имаго кото-
рых перфорируют листья, а личинки грызут корни; златка Trachys minute
L., минирующая листья; Discestra trifolii Rott., Xylena exoleta L., Spodoptera
exigua Hb., Loxostega sticticalis L., а также Melangromyza aeneiventis из дву-
крылых. Гусеницы бражника Herse, пальцекрылки Alucitapentadactylea L.,
совок Acontia, Erastria в условиях Узбекистана иногда очень сильно объе-
дают листья вьюнка.
В средней полосе России выделены олигофаги, узкоспециализирован-
ные на вьюнке: Nepticula freyela Heyd., Emmelia trabealis Scop., Pterophorus
monodactylus L., Longitarsus longipennis, L. pellucidus, которые могут быть
потенциальными агентами для биологической борьбы с вьюнком. Но наи-
более эффективным в этом качестве считается вид-олигофаг щитоноска
вьюнковая Hypocassida subferruginea Schrank, повсеместно поражающая
вьюнок полевой. Она безопасна для основных культивируемых растений,
но сильно повреждает вьюнок, особенно в период бутонизации и цветения
(Радченко, Клепикова, 1977).
В Америке листьями Convolvulaceae кормятся щитоноски Аргуса
(Chelymorpha cassidea Fabr.), при этом они совершенно не повреждают
рожь и кукурузу, а также хризомелида Galerica rufa German, родом из Юж-
ной Европы (Selleck, 1979). Подробно изучены насекомые, обитающие в
популяциях Convolvulus arvensis в Канаде и США (Weaver, Riley, 1982).
Среди сопутствующих видов также описаны нематоды Meloidogyne sp.,
Pratylenchus penetrans и вирус Annulus orae Holmes (Weaver, Riley, 1982).
У вьюнка полевого перекрестное опыление осуществляют насекомые.
Mulligan и Findlay (цит. по: Weaver, Riley, 1982) сообщают об опылитель-
ной самонесовместимости вьюнка. Цветки посещают Halictidae, медонос-
ные пчелы и шмели, бабочки и моли, чаще всего Rhizoccus falcifer Kunk,
Paratrioza cockerelli Sulk., Macrosteles fascifrons Stal. (Weaver, Riley, 1982).
194
По нашим наблюдениям, в городских фитоценозах Москвы цветки вьюнка
посещают муравьи, трипсы, цикады, пчелы. Были также отловлены и опре-
делены насекомые-хищники: муравьи Lasius niger, Myrmica ruginodis, кло-
пы Reduvius sp., жуки Ctenicera sp. Вероятно, их консортивные связи кос-
венные — это охота на фитофагов вьюнка. Цветки, по-видимому, привле-
кают насекомых не только окраской венчика, но и ароматом, который
ощущается в течение нескольких часов после распускания. Энтомофауна
опылителей была относительно бедна из-за влияния городской среды. Бед-
ность энтомофауны сказывается и на практическом отсутствии фитофагов
вьюнка в городских условиях — на территории МГУ практически отсутст-
вуют поражения листьев насекомыми.
Вьюнок может вступать в консортивные связи со специфическими гри-
бами. В американских популяциях вьюнка Conners (цит. по: Weaver, Riley,
1982) описал следующие виды грибов: Leptosphaeria doliolum (Pers.) de
Not., Pistillaria micans Pers, ex Fr., Puccinia convolvuli Cast., Ramularia
sepium Deam. et. Bisby, Septoria concolvuli Desm., 5. flagellaris Ell. et. Ev.,
Trinacrium subtile Reiss. Кроме того, определены виды Albugo ipomoeae-
panduratae (Schw.) Swing, Cercospora tuberculella (Schw.), Coleosporium
ipomoeae (Schw.) Burr., Diplodia convolvuli Deam. et. House, Parodiella
paraguayensis Speg., Physalospora obtusa (Schw.) Cke., Rhabdospora sp.,
Rhizoctonia solani Kuehn, Septoria calystegiae West, 5. septulata Beach,
Stagonospora convolvuli Deame et. House. Экземпляр вьюнка полевого из
Подмосковья оказался поражен грибом Sphaerotheka convolvuli (определил
В.П. Прохоров кафедра микологии и альгологии МГУ). Информация о
симбиотических (микоризных) отношениях с грибами отсутствует
(Weaver, Riley, 1982).
В распространении семян вьюнка участвуют грызуны, которые исполь-
зуют их в пищу. В частности, около норок полевки (Microtus socialis) не-
редко встречаются побеги вьюнка, выросшие из принесенных семян. По-
следние также были обнаружены в зимних запасах серого хомячка
(Cricetulus migratorius'). (Левина, 1957).
Продуктивность и хозяйственное значение
Вьюнок полевой считается одним из главных сорняков сельскохозяй-
ственных культур. Определение вьюнка полевого как особо вредного сор-
няка в Америке закреплено законодательно (Federal Seeds Act, 1980 цит.
по: Weaver, Riley, 1982). Существует даже опубликованная библиография
по этому виду (Вьюнок полевой..., 1954, 1967).
Наибольшее влияние вьюнок оказывает на сельскохозяйственные
культуры как конкурент за влагу. Помимо этого вьюнок препятствует сбо-
ру урожая, вызывает полегание посевов, является резерватом вредителей
культурных растений (Горенштейн, 1982) и альтернативным хозяином фи-
195
топатогенов, в частности вирусов, вызывающих болезни «X» картофеля,
табачную мозаику и другие (Holmetai., 1977,цит. по: Weaver, Riley, 1982)
Из-за сходства формы и размера, семена вьюнка засоряют семенной
материал пшеницы и других зерновых (Иванов, 1953). Засорение семян
приносит большой ущерб на плантациях чая: лианы могут настолько густо
оплетать кусты, что полностью скрывают чайные листья. А при ядовито-
сти вьюнка, даже небольшая примесь его листьев в собранном чае делает
его непригодным. Своими корнями вьюнок оплетает корни чайных кустов,
делает почву более сухой и увеличивает ее pH, что еще более ухудшает ус-
ловия для произрастания чая (Бржезицкий, Эфендиева, 1961). Констатиро-
вано, что вьюнок вызывает отравление лошадей (Иванов, 1956).
У вьюнка есть и некоторое позитивное хозяйственное значение — его
иногда используют в фармакогнозии для изготовления слабительных и
нейроактивных средств. В Средиземноморском регионе его с удовольст-
вием поедают коровы (Saad, 1967, цит. по Weaver, Riley, 1982). Однако за-
мечено, что его ткани имеют низкое содержание аскорбиновой кислоты
(до 50 мг %; сборы Цыганковой (1971) на берегу р. Урал; анализ по Тиль-
мансу). Иногда вьюнок разводится в декоративных целях, подобно другим
вьюнковым.
Самые эффективные программы по борьбе с вьюнком сочетают в себе
культивацию и чередование культур с использованием гербицидов. Ин-
тенсивная обработка земли было одно время ведущим средством борьбы.
Культивация уменьшает длину корней и количество запасенных углево-
дов. Постоянная глубокая обработка почвы в течение 5 лет позволяет исто-
щить корневые ресурсы и эффективно ограничивать популяцию вьюнка на
поле (Wiese, Rea, 1962). Среди культур существенную конкуренцию вьюн-
ку оказывают люцерна и озимая пшеница — последняя за счет быстрого
роста ранней весной, когда вьюнок растет слабо и не конкурирует за поч-
венную влагу (Wiese, Rea, 1962). Озимая пшеница, кормовые травы и дру-
гие конкуренты помогают существенно снизить инвазию вида на поля че-
рез 3-5 лет (Wiese, Rea, 1962).
Для борьбы с вьюнком полевым из гербицидов чаще всего применя-
лись аминная соль 2,4-Д и кротилин. Обнаружены межпопуляционные ва-
риации по восприимчивости к гербициду 2,4-Д (Weaver, Riley, 1982). Ше-
роховатые кутинизированные покровы стебля и листьев делают растение
более устойчивым к обработке гербицидными средствами (Weaver, Riley,
1982).
Биологические средства борьбы, в частности, использование щитоно-
ски, осложнены мобильностью насекомых. Для ее снижения предлагаются
способы по предварительной деформации крыльев (Слободянюк, 1987).
ЛИТЕРАТУРА
Абдуллаев Г.Д. Биоэкологические особенности вьюнка полевого и меры борь-
бы с ним // Флорист, анализ различных групп растений в низменной части АзССР.
Баку, 1982. С. 45-46.
Аверьянов Л.В., Аверьянова Е.Л., Лавренко А.Н. Кариосистематическое иссле-
дование орхидных (Orchidaceae) на территории Коми АССР // Ботан. ж. 1982. Т. 67,
№11. С. 1491-1499.
Аверьянов Л.В. Основные пути морфологической эволюции в семействе
Orchidaceae // Ботан. ж. 1991а. Т. 76, № 7. С. 921-935.
Аверьянов Л.В. Происхождение и некоторые особенности эволюции, биологии
и экологии орхидных (Orchidaceae) И Ботан. ж. 19916. Т. 76, № 10. С. 1345-1359.
Аверьянов Л.В. Орхидные (Orchidaceae) Средней России // Turczaninowia.
2000. № 3 (1). с. 30-53.
Агабабян Ш.А. Тмин обыкновенный // Кормовые растения сенокосов и паст-
бищ. 1956. Т. 3. С. 121-124.
Агротехнические указания по возделыванию тмина. Сост. ВНИИ эфиромас.
культ. Мин. с-х СССР. М., 1950. 4 с.
Алексеев Ю.Е., Вахрамеева М.Г., Денисова Л.П., Никитина С.В. Лесные травя-
нистые растения. М., 1988.
Алексеева Л.М. Флора острова Кунашир. Владивосток, 1983. 128 с.
Андреева И.И. Морфогенез вегетативных органов шпажника гибридного
(Gladiolus х hybridus) в первые годы жизни // Бюлл. Главн.Бот. сада. 1971. Вып. 79.
С. 80-87.
Андреева И.И. Жизненный цикл шпажника гибридного Gladiolus х hybridus
hort. (Iridaceae) // Ботан. ж. 1977. Т. 62, № 8. С. 1183-1195.
Антонова В.И., Папанов А.В.Структура популяций некоторых орхидных на
территории национального парка «Русский Север» // Охорона i культивування
орхщей. Кшв, 1999. С. 27-28.
Арктическая флора. М.-Л.: Наука, 1963. Т. 4.
Артюшенко З.Т. Атлас по описательной морфологии высших растений: Семя.
Л.: Наука, 1990. 204 с.
Артюшенко З.Т., Федоров Ф.Ф. Атлас по описательной морфологии высших
растений. Плод. Л.: Наука, 1986. 392 с.
Атлас ареалов и ресурсов лекарственных растений СССР. М., 1980. 339 с.
Атлас лекарственных растений СССР. М.: Медгиз, 1962, 703 с.
197
Афанасьева Е.А., Голубев В.Н. Почвенно-ботанический очерк Стрелецкой сте-
пи. Центрально-Черноземный заповедник им. проф. В.В. Алехина, Курск, 1962,
68 с.
Бабарыкина А.Н. Тмин — медоносное растение // Пчеловодство. 1992, № 3-4.
С. 12-13.
Балашев Л.С., Сипайлова Л.М., Соломаха В.А., Шеляг-Сосонко Ю.Р. Типоло-
гия лугов Украины и их рациональное использование. Киев: Паукова думка, 1988.
240 с.
Балявичене Ю. Семейство ятрышниковые (Orchidaceae) в Литве // Охрана и
культивирование орхидей. Таллинн, 1980. С. 32-34.
Баталов А.Е. Биоморфология, экология популяций и вопросы охраны орхидей
Архангельской области. Автореф. дис. ... канд. биол. наук. М., 1998. 16 с.
Батыгина Т.Б., Васильева В.Е. Развитие зародыша и проростка у некоторых ор-
хидных // Охрана и культивирование орхидей. Киев, 1983. С. 73-75.
Бекетов А.Н. О протерандрии зонтичных // Тр. СПБ общ-ва естествоиспытате-
лей, отд. бот. 1889. Т. 20. С. 11-14.
Бельков В.П., Омельяненко А.Я., Мартынов А.Н. Регулирование травяного по-
крова в лесу. М.: Лесная промышленность, 1974. 112 с.
Беляев В.Г. Критический свод русских данных по кормовым растениям. М.:
Изд-во Моск, сельхоз. ин-та, 1905. Т. 11. 153 с.
Беляева В. А. Пряно-вкусовые растения их свойства и применение; М.: Госторг-
издат, 1946. 107 с.
Берлина Н.Г., Костина В.А. Антропогенные изменения видового состава сосу-
дистых растений Лапландского заповедника / Флора и растительность Мурман-
ской обл. Аппатиты, 1999. С. 55-61.
Благовещенский В.В., Пчелкин Ю.А., Раков Н.С., СтариковаВ.В., Шустов В.С.
Определитель растений Среднего Поволжья. Л.: Наука, 1984. 392 с.
Блинова И.В. Особенности морфологического строения и побегообразования
ряда орхидных на северном пределе их распространения // Бюлл. МОИП. Отд.
биол. 1996. Т. 101, вып. 5. С. 69-80.
Блинова И.В. Особенности онтогенеза некоторых корнеклубневых орхидных
(Orchidaceae) Крайнего Севера//Ботан. ж. 1998. Т. 83. С. 85-94.
Болотин Е.А., Зафрен С.Е. Кормовые отходы промышленности и их использо-
вание. 1939. Ч. 2.
Борисова И.В. Монокормные поликарпические травы // Ботан. ж. 1992. Т. 77,
№ 10. С. 39-46.
Бржезицкий М., Эфендиева Ш. Вьюнок полевой (Convolvolus arvensis L.), его
биологические и экологические особенности // Уч. Зап. Азерб. ун-та. 1961. № 3.
С. 15-18.
Брутян А.С. Вьюнок полевой (Convolvolus arvensis L.) и меры борьбы с ним в
условиях Араратской равнины Армянской ССР. Автореф. дис. ... канд. с.-х. наук.
Ереван, 1970. 32 с.
Буравлев Г.Н. Тмин и его возделывание. Воронеж: Коммуна, 1932. 20 с.
Быков Б.А. Доминанты растительного покрова Советского Союза. Алма-Ата:
Издательство АН Казахской ССР, 1962. Т. 2. 434 с.
Бязров Л.Г., Дылис Н.В., Жукова В.М. и др. Основные типы широколиствен-
198
но-еловых лесов и их производных Малинского лесничества Краснопахорского
лесхоза Московской области // Биогеоценологические исследования в широколи-
ственно-еловых лесах. М.: Наука, 1971. С. 7-150.
Вахрамеева М.Г., Татаренко И.В., Быченко Т.М. Экологические характеристи-
ки некоторых видов евразиатских орхидных // Бюл. МОИП. Отд. биол. 1994. Т. 99,
вып. 4. С. 75-82.
Виноградова Т.Н. Проблема выделения возрастных состояний у орхидных на
примере калипсо луковичной (Calypso bulbosa (L.) Oakes) // Бюл. МОИП. Отд.
биол. 1998. Т. 1-3, вып. 1. С. 47-55.
Влажные луга Республики Башкортостан: синтаксономия и вопросы охраны /
Григорьев И.Н., Соломещ А.И., Алимбекова Л.М., Онищенко Л.И. Уфа: Гилем,
2002. 157.
Воликов С.Н., Тукачев А.Г. Лекарственные растения северо-запада России. М.:
ТИМР, 1992. 159 с.
Воробьев Д.В. Методика лесотипологических исследований. Киев: Урожай,
1967. 388 с.
Ворошилов В.Н., Скворцов А.К., Тихомиров В.Н. Определитель растений Мос-
ковской области. М.: Наука, 1966. 365 с.
Вредители и болезни цветочно-декоративных растений / Ю.В. Синадский, И.Т.
Корнеева, И.Б. Доброчинская и др. М.: Наука, 1987. 592 с.
Вьюнок полевой. Библиографический список литературы за период 1890-1953
гг. в колич. 22 назв., и иностранных за период 1934—1954 гг. в колич. 64 названий.
Сост. справ.—библ. отд. ЦНСХБ. 1954. 94 с.
Вьюнок полевой. Библиографический список литературы за период 1954—1966
гг. в колич. 23 русских и 26 иностранных названий. Сост. справ.-библ. отд. ЦНСХБ.
1967.76 с.
Гайдамака И.П. Опыт выращивания тмина // Масличные культуры. 1986, № 3.
С. 36.
Гаммерман А.Ф., Кадаев Г.Н., Яценко-Хмелевский А.А. Лекарственные расте-
ния (Растения — целители). Справочное пособие. М.: Высш. Школа, 1983. 400 с.
Ганешин С.С. Растительность поймы р. Волхова // Материалы к исследованию
реки Волхова и его бассейна. Почвенные и ботанические исследования в поймах
оз. Ильменя и р. Волхова (предварительный отчет). Л.: Тип. Росс. Гидрол. Ин-та
ВСНХ, 1925. С. 27-58.
Ганюшкин Е.С. Особенности физиологии покоя клубнелуковиц Гладиолусов //
Бюлл. Главн. Бот. сада. 1991. Вып. 162. С. 78-83.
Гасанов А.М. К экологической анатомии вьюнка полевого (Convolvolus
arvensis L.) // Извест. АН АзССР. Сер. биол. и мед. наук. 1962. № 5. С. 9-16.
Гейдеман А.Ф. и др. Определитель высших растений Молдавской ССР. Киши-
нев, 1986, 263с.
Глухов М.М. Медоносные растения. М.: Сельхозгиз, 1950. 543 с. .
Глущенко Н.Н. Свойства потомства ворсянки и тмина в зависимости от места
формирования семян на кустах и сроков посева. Автореф. дис....канд. биол. наук.
М., 1954. 17 с.
Глущенко Н.Н. Тмин обыкновенный // Эфиромасличные культуры. М., 1963.
С. 359.
199
Говорухин В.С. Флора Урала. Свердловск: Свердловское обл. изд-во, 1937.
536 с.
Головач А.Г. Лианы, их биология и использование. Л.: Наука, 1973. 260 с.
Голубев В.Н. Биологическая флора Крыма. 2-е изд. Ялта: Изд-во Никит, гос.
бот. сада, 1996. 85 с.
Голубев В.Н. Биологическая флора Крыма. Ялта. 1984. Деп. ВИНИТИ № 5770
В 84.
Голубев В.Н. К биоморфологии природных растений Подмосковья с запасаю-
щими органами побегового происхождения // Уч. зап. Москов. обл. пед. ин-та. М.,
1956а. Т. 41,вып. 1. С. 41-104.
Голубев В.Н. О биологическом значении геофилии у травянистых растений //
Ботан. ж. 19566. Т. 41, № 2. С. 236-242.
Голубев В.Н. Основы биоморфологии травянистых растений центральной ле-
состепи // Тр. Центр.-Чернозем. гос. заповедника им. В.В. Алехина; Вып. 7, Воро-
неж: Изд-во Воронеж, ун-та, 1962. 601 с.
Голубев В.Н. Эколого-биологические особенности травянистых растений и
растительных сообществ лесостепи. М.: Наука, 1965. 286 с.
Голубев В.Н., Волокитин Ю.С. Методические рекомендации по изучению ан-
тэкологических особенностей цветковых растений. Морфологическое описание
репродуктивной структуры. Ялта: Изд-во Никит, гос. бот. сада, 1986. 43 с.
Горенштейн Б.М. Насекомые-узкоспециализированные фитофаги вьюнка по-
левого (в УзССР) // Научн. докл. высш, школы. Биол. науки. 1982. № 1. С. 68.
Горышина Е.К. Экология травянистых растений лесостепной дубравы. Л.:
Изд-во Ленингр. ун-та, 1975. 128 с.
Гроздов Б.В. Лесные травы. Брянск, 1956. 45 с.
Гроздов Б.В. Лесные травы, их индикаторное, кормовое и лекарственное значе-
ние. М.: Госдесбумиздат, 1963. 61 с.
Громов А.Н. Гладиолус — Gladiolus L. (биология, адаптивность, исходный ма-
териал, селекция): Автореф. дис. ... докт. с.-х. иаук. СПб., 1998. 71 с.
Гроссгейм А.А. Растительные богатства Кавказа. М.: Изд-во МОИП, 1952.
631с.
Гроссгейм А.А. Флора Кавказа. Баку: Изд-во Аз ФАН, 1939. Т. 1. 402 с.
Гроссгейм А.А. Флора Кавказа. Л.: Наука, 1967. Т. 7. 493 с.
Грудзинская И.А. Летнее побегообразование у древесных растений и его клас-
сификация // Ботан. ж. 1960. Т. 45, № 7. С. 968-978.
Губанов И.А. Лекарственные растения. М.: Изд-во МГУ, 1993. 273 с.
Губанов И.А. Конспект флоры Внешней Монголии (сосудистые растения). М.,
1996. 136 с.
Губанов И. Пищевые растения России. М.: ABF, 1996. 556 с.
Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. Луговые травяни-
стые растения. Биология и охрана: Справочник М.: Агропромиздат, 1990. 183 с.
Губанов И.А., Киселева К.В., Новиков В.С. Дикорастущие полезные растения.
М.: Изд-во МГУ, 1993. 300 с.
Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. Определитель со-
судистых растений Европейской России. М.: Аргус, 1995. 560 с.
200
Туленкова М.А., Красникова А.А. Летняя полевая практика по ботанике. М.:
Просвещение, 1986. 175 с.
Гурзенков Н.Н., Горовой П.Г. Числа хромосом Umbelliferae Дальнего Востока
//Ботан. ж. 1971. Т. 56, № 12. С. 1805-1814.
Данилова М.М. Болота юго-западных районов Пермской области // Ученые за-
писки Пермского гос. ун-та им. А.М.Горького. 1964. Т. 114. С. 79-91.
Дарвин Ч. Движения и повадки лазящих растений // Ч. Дарвин. Сочинения.
Т. 8. М.-Л.: АН СССР, 1941. 544 с.
Дастикайте А. Особенности развития тмина // Сб. науч. тр. Литовской с-х ака-
демии. Кн. 1. Каунас, 1976 (1977). С. 217.
Демидова А.Н., Уланова Н.Г. Динамика популяций вейника седеющего
(Calamagrostis canescens (Web.) Roth) на вырубках Новгородской области И
Тез. VII Мол. конф, ботаников в С.-Петербурге. С.-Петербург: Буслай, 2000.
С. 281-182.
Демидова А.Н. Уланова Н.Г. Динамика популяций Calamagrostis canescens
(Web.) Roth на вырубках Новгородской и Тверской областей // Труды междун.
конф, по фитоценологии и систематике высших растений, посвящ. 100-летию
А.А. Уранова, Москва. М., 2001. С. 51-52.
Демидова А.Н., Уланова Н.Г., Борисова О.В. Возрастная структура популяций
вейника седеющего (Calamagrostis canescens (Web.) Roth на вырубках Новгород-
ской области // Исследования на охраняемых природных территориях Северо-За-
пада России. Великий Новгород, 2000а. С. 230-236.
Демидова А.Н., Уланова Н.Г., Борисова О.В. Динамика популяций вейника се-
деющего (Calamagrostis canescens (Web.) Roth при демутации растительности вы-
рубок // Уч. зап. Академии с-х и природных ресурсов Нов ГУ. Великий Новгород,
20006. Вып. 2. С. 77-83.
Демидова А.Н., Уланова Н.Г., Борисова О.В. Растительность вырубок севе-
ро-запада Новгородской области // Тез. школы конференции «Актуальные пробле-
мы геоботаники. Современные направления иследований в России: методологии,
методы и способы обработки материалов». Петрозаводск, 22-26 октября 2001 г.
Петрозаводск, 2001. С. 86-87.
Демьянова Е.И., Лыков В.А., Логинова Е.А. К антэкологическому изучению
тмина обыкновенного (Carum carvi L.) в условиях интродукции // Экология цвете-
ния и опыления растений. Пермь, 1989. С. 58-69.
Дзмитрыева С.А., Парфёнау В.И. Храмасомныя лш некаторых вщау флоры
Беларус! // Весц! АН БССР, сер. б!ял. навук, 1983. № 2. С. 7-12.
Дикорастущие полезные растения России / Отв. ред.А.Л. Буданцев, Е.Е. Леси-
овская. СПб: Изд-во СПХФА, 2001. 663 с.
Дикорастущие полезные растения флоры Монгольской Народной Республики.
Л.: Наука, 1985. 235 с.
Дмитриев А.М. Луга Холмогорского района. СПб., 1904.
Доброхотов В.Н. Семена сорных расений. М.: Сельхозгиз, 1961. 414 с.
Ермакова И.М. Тмин обыкновенный (Carum carvi L.) // Диагнозы и ключи воз-
растных состояний луговых растений. Ч. 3. М.: Изд-во МГПИ им В.И. Ленина,
1983. С. 19-21.
201
Ермакова И.М. Синеголовник плосколистный // Биологическая флора Москов-
ской области, М.: Гриф и Ко, 2000. Вып. 14. С. 143-159.
Ермакова И.М., Сугоркина Н.С. Динамичность и устойчивость ценопопуляций
на естественных лугах Калужской области И Динамика ценопопуляций растений.
М.: Наука, 1985. С. 63-82.
Ермолова Е.В. Определение максимальной и полной продуктивности надзем-
ной части травянистого покрова в лесах Подмосковья с разной степенью вырубки //
Земледелие и растениеводство. М., 1969. С. 180-188.
Ермолова Л.С. Динамика травяного покрова на вырубках в связи с лесовозоб-
новительными процессами. М.: Наука, 1981. 140 с.
Жирнова Т.В. Обзор видов растений Красных книг СССР и РСФСР в Башкир-
ском заповеднике // Растения Красных книг в заповедниках России. Сб. научн.
тр. ЦНИЛ. М., 1994. С. 57-76.
Жирнова Т.В. Орхидные Башкирского заповедника (Южный Урал) // Изучение
природы в заповедниках Башкортостана. Миасс, 1999. С. 141-160.
Жуков И.Н. Биологические особенности вьюнка полевого и меры борьбы с ним
в условиях степи и лесостепи Омской области. Автореф. дис. ... канд. с.-х. наук.
Омск, 1961. 20 с.
Забелкин Н.А. Жизненные формы травянистого многолетника (Chamerion
angustifolium (L.) Holub.) // Успехи экол. морф, растений и ее влияние на смежные
науки. Межвузовский сб. науч, трудов. М.: Прометей, 1994. С. 97-98.
Зворыкина К.В. Эколого-ценотическая характеристика некоторых представи-
телей рода Calamagrostis Adans, (на примере южнотаежных березняков) // Эколо-
го-ценотические и географические особенности растительности. М., 1983.
С. 85-98.
Иванов В.В. Отравление [лошадей] вьюнком//Ботан. ж. 1956. Т. 41, № 5. С. 48.
Иванов В.П. Сорные растения и меры борьбы с ними. М.: Изд-во АН СССР,
1953. С. 47—48.
Иванов ДА. Повышение продуктивности сенокосов и пастбищ. Л.: Колос,
1975. 288 с.
Игнатенко Н.А., Амельченко В.П. Орхидные Томской области // Охрана и
культивирование орхидей. Киев, 1983. С. 18-19.
Изучение структуры и взаимоотношения ценопопуляций. М: Изд-во МГПИ
им. В.И. Ленина, 1986. 73 с.
Ильинская О.П. Примула — дивная незнакомка // Цветоводство. 1993. № 2.
С. 25-26.
Ильинская С.А., Матвеева А.А., Речан С.П. и др. Типы леса // Леса Западного
Подмосковья. М.: Наука, 1982. С. 20-150.
Ипатов В.С. Березняки восточных районов Ленинградской области // Уч. зап.
Ленингр. гос. ун-та им. А.А. Жданова. Биолого-почвенный факультет. Л.: Изд-во
Ленингр. ун-та, 1960. Вып. 48, № 290. С. 156-169.
Каден Н.Н.,Терентьева Н.Н. Этимологический словарь латинских названий
растений, встречающихся в окрестностях агробиостанции. М.: Изд-во МГУ, 1975.
203 с.
Камилова Ф.Г. Некоторые структурные особенности сорнячающих вьюнков //
Науч. тр. Ташк. СХИ. 1982. Вып. 101. С. 56.
202
Капинос Г.Е. Биологические закономерности развития луковичных и клубне-
луковичных растений на Апшероне. Баку: Изд-во АН АзССР, 1965. 240 с.
Кардашина Л.А. Конвеерное выращивание тмина в теплице // Тр. Уральского
НИИ с-х, 1984. Т. 41. С. 76-79.
Карписонова Р.А. Дубравы лесопарковой зоны Москвы. М.: Наука, 1967.104 с.
Каталог цветочно-декоративных растений ботанических садов СНГ и стран
Балтии // Совет бот. садов России; Центр, бот. сад АН Беларуси. Минск: Э.С. Галь-
перин, 1997. 475 с.
Кауфман Н.Н. Московская флора, или описание высших растений и ботани-
ко-географический обзор Московской губернии. М., 1866. 2-е изд. М., 1889.
Кернер А.М. Жизнь растений: В 2 т. СПб.: Просвещение, 1899-1903.
Киндерова Н.Н. Ритм цветения и стадии распускания цветка двух видов люти-
ка (Ranunculus acris L. и R .cassubicus L.) // Морфогенез и ритм развития высших
растений. М.: Изд-во МГПИ им. В.И. Ленина, 1987. 121 с.
Киселева К.В. Осинники Клинско-Дмитровской гряды и их динамика // Ботан.
ж. 1965. Т. 50, № 4. С. 155-163.
Князева О.И. Интродукция видов семейства Orchidaceae Уральской флоры и
перспективы использования их для озеленения // Новые декоративные растения в
культуре на Среднем Урале. Свердловск, 1986. С. 31-38.
Козловская Н.В. Загадки белорусской флоры. Минск: Выш. шк., 1986. 222 с.
Коломиец Н.И. Биологические основы введения в культуру Ononis arvensis L. //
Раст, ресурсы. 1969. Т. 5, вып. 1. С. 100-108.
Коломиец Н.И. Введение в культуру и возделывание стальника полевого в ле-
состепи Украины. Автореф. дис... канд с-х наук. М., 1972. 18 с.
Конспект флоры Псковской области. Л.: Изд-во ЛГУ, 1970. 280 с.
Конюшков Н.С. Влияние выпаса на растительный покров луга // Бюлл. Ин-та
лугов и пастбищ. М., 1930. № 5. С. 542-561.
Кордюм Е.Л. Матер1али до вивчення чисел хромосом зонтичных —
Umbelliferae // Допов1да АН УРССР. Сер. Б. 1967. № 1. С. 89-93.
Коржинский С.И. Флора Востока в Европейской России в ее систематических
и географических отношениях. Томск, 1892. 227 с.
Корконосова Л.И. К вопросу формирования вейниковых вырубок на Европей-
ском Севере // Вопросы таежного лесоводства на Европейском Севере. М.: Наука,
1967. С. 101-113.
Кормовые растения сенокосов и пастбищ. М.—Л.: Гос. изд-во с-х лит-ры, 1950,
Т. 1. 850 с.; 1951. Т. 2. 948 с.; 1956. Т. 3. 879 с.
Королева А.С., Красноборов И.М., Пеньковская Е.Ф. Определитель растений
Новосибирской области. Новосибирск: Наука, 1973. 367 с.
Короткевич Г.И., Аронова О.А. Особенности латентного периода некоторых
лекарственных растений сем. Fabaceae и Lamiaceae при их интродукции в Ленин-
градской области // Тезисы VII Мол. конф, ботаников в С-Петербурге. СПб.: Бус-
лай, 2000. С. 230.
Коротков К.О. Леса Валдая. М.: Наука, 1991. 160 с.
Красная книга Белорусской ССР. Минск, 1981.281 с.
Красная книга Московской области / Отв. ред. В.А. Зубакин, В.Н. Тихомиров.
М.: Аргус: Рус. ун-т, 1998. 560 с.
203
Красная книга республики Татарстан: Животные, растения, грибы. Казань:
Природа, 1995. 452 с.
Красная книга РСФСР. М.,1988. 591 с.
Красная книга УРСР. Киев, 1988.
Крылов П.Н. Флора Западной Сибири. Вып.У. Aisoaceae— Berberidaceae.
Томск: Изд-во Томск. Русск. бот.об-ва, 1931. 1227 с.
Крюгер Л.В. Эндотрофная микориза травянистой растительности некоторых
фитоценозов Центрального Предуралья// Ботан. ж. 1961. Т. 46, № 5. С. 617 627.
Крюгер У., Янтра И. 1000 прекрасных растений в вашем доме. М.: Бетельсманн
Медиа Москау АО, 1996. 379 с.
Кузнецова Т.В. Методы исследования соцветий. Проблема интеркалярного со-
цветия// Бюлл. МОИП. Отд. биол. 1987. Т. 92, выл. 1. С. 81-96.
Кузьмич Н.К. Результаты изучения коллекции тмина // Тр. ВНИИ эфиромас-
личных культур. Симферополь, 1977. Т. 10. С. 57-62.
Кузьмич Н.К. Новый сорт тмина Подольский-9 // Тр. ВНИИ эфиромасличных
культур. Симферополь, 1986. Т. 17. С. 29-33.
Кукушкин В.А. Морфогенез клубнепочек гладиолуса гибридного при разных
температурных режимах хранения // Бюлл. Главн. Бот. сада. 1983. Вып. 129.
С. 76-79.
Куликов П.В., Филиппов Е.Г. Симбиотическое прорастание некоторых нетро-
пических орхидных in vitro // Охорона i культивування орхщей. Юев. 1999.
С. 57-58.
Куприянова Л. А., Алешина Л. А. Пыльца и споры растений флоры Европейской
части СССР. Л: Наука, 1972. Т. 1. 184 с.
Курлович Л.Е. О морфологическом разнообразии лютика кашубского в Мос-
ковской и Тульской областях // Бюлл. Главн. Бот. сада. 1982а. Вып. 123. С. 41-44.
Курлович,Л.Е. Исследование корреляций между факторами среды и морфоло-
гическими признаками лютика//Бюлл. Главн.Бот. сада. 19826. Вып. 125. С. 40-44.
Курлович Л.Е. Об изменчивости морфологических признаков лютика в питом-
нике и природных условиях // Бюлл. Главн. Бот. сада. 1982в. Вып. 126. С. 41-45.
Курлович Л.Е. Сравнительное изучение сеянцев и родительских форм биоти-
пов лютика кашубского и лютика золотистого // Бюлл. Главн. Бот. сада. 1984. Вып.
132. С. 46-48.
Курлянчик И.А. Органогенез тмина обыкновенного // Новые аспекты исследо-
вания биологии флоры и фауны СССР. М., 1988. С. 97-99.
Курлянчик И.А., Скокова А.А., Бобков С.Н.. Особенности роста и развития
тмина обыкновенного. Рязань, 1989. 12 с. Деп. ВИНИТИ № 2936 В 89.
Курнаев С.Ф. Основные типы леса средней части Русской равнины. М.: Наука,
1968.354 с.
Курнаев С.Ф. Теневые широколиственные леса Русской равнины и Урала. М.:
Наука, 1980. 313 с.
Ларин И.В. Избранные труды. М.: Колос, 1978. 432 с.
Ларин И.В. Кормовые растения сенокосов и пастбищ СССР. Т. II. Двудольные.
М-Л., 1951. С. 548-549.
Ларин И.В., Паламарчук И. А. Введение в изучение кормовых растений марало-
204
водческих совхозов Алтайского края // Тр. Пушкинского с-х ин-та, 1949. Т. 19.
С. 63-75.
Лебедева В.И. Тмин — салатная, пряная культура // Картофель и овощи. 1992.
№ 5-6. С. 22.
Левина Р.Е. Способы распространения плодов и семян. Л.: Изд-во МГУ, 1957.
358 с.
Левина Р.Е. Морфология и экология плодов. Л.: Наука, 1987. 160 с.
Левитский Г. А. Цитология растений: избранные труды. М., 1976. 351 с.
Левицкий С.С. Список сосудистых растений Центрально-Черноземного госу-
дарственного заповедника // Тр. Центр.-Черноз. гос. заповедника им. В.В. Алехина.
1957. С. 110-173.
Леса южного Подмосковья. М.: Наука, 1985. 281 с.
Лисовская А.В. Цветоводство открытого грунта. Йошкар-Ола, 1994. 240 с.
Лысенко Т.А. Эколого-биологические особенности Primula veris в лесостепи
Украины // Растительные ресурсы. 1994. Т. 30, вып. 3. С. 15-24.
Любарский Е.Л. Об эволюции вегетативного возобновления и размножения
травянистых поликарпиков // Ботан. ж. 1961. Т. 46, № 7. С. 957-968.
Любарский Е.Л. Экология вегетативного размножения высших растений. Ка-
зань: Изд-во Казан, ун-та, 1967. 180 с.
Любарский Е.Л. Вегетативные однолетники // Успехи экологической морфоло-
гии растений. М., 1994. С. 10-11.
Маевский П.Ф. Флора средней полосы европейской части СССР. 9-е изд. 9, М.:
Колос, 1964. 880 с.
Мазуренко М.Т., Хохряков А.П. Бриофилы — своеобразная экологическая
группа растений И Бюл. МОИП. Отд. Биол. 1989. Т. 94, вып. 4. С. 64-73.
Манолова А., Максимова В. Лечебни растения инхибитори на вирусы. София,
1988.148 с.
Матвеева Е.П. Кормовые достоинства дикорастущих растений природных лу-
гов северо-западной части лесной зоны СССР // Организация кормопроизводства
по зонам СССР. М., 1960а. С. 245-246.
Матвеева Е.П. Фитоценотическая классификация и производственная типоло-
гия травянистой растительности сенокосов и пастбищ Советской Прибалтики //
Геоботаника. М.-Л., 19606. Вып. 12. С. 229-252.
Матвеева Е.П. Сравнительный анализ природной и культурной травянистой
растительности сенокосов и пастбищ Советской Прибалтики. Авт. дис... докт.
биол. наук. Л., 1965. 41 с.
Матвеева Е.П., Гагарин П.К., Знаменская Л.А. Биохимический состав некото-
рых растений группы лугового разнотравья// Ботан. ж. 1964. Т. 49, № 6. С. 875-877.
Матвеева Е.П., Знаменская Л.А. Содержание кобальта в дикорастущих луго-
вых растениях // Ботан. ж. 1959. № 7. С. 978-987.
Матвеева Е.П., Знаменская Л.А. Содержание кобальта в дикорастущих луго-
вых растениях // Тр. Бот. ин-та АН СССР, сер. 3. Геоботаника. 1960а. № 12.
С. 162-170.
Матвеева Е.П., Знаменская Л.А. Химический состав и кормовая характеристи-
ка растений, преобладающих на сенокосах и пастбищах Ленинградской обл. //
Тр. Бот. ин-та АН СССР, сер. 3. Геоботаника. 19606. № 12. С. 128-161.
205
Матюхин Д.Л. Возрастные изменения в клонах гладиолуса гибридного, полу-
ченых in vitro. Автореф. дис. ... канд. биол. наук. М., 1994. 18 с.
Методические указания по классификации сенокосов и пастбищ равнинной
территории Европейской части СССР. М.: Всес. академия с-х наук им. В.И. Лени-
на, ВНИИ кормов им. В.Р. Вильямса, 1978. 148 с.
Микологическая флора Европейской и Азиатской России. Т. 1. ПероноспорО-
вые // Мат-лы к познанию фауны и флоры Российской империи. Отд. ботан. 1901.
Вып. 4. 230 с.
Минаева В.Г. Лекарственные растения Сибири. Новосибирск: Наука. Сиб. отд.,
1991.431 с.
Миняев Н.А., Конечная Г.Ю. Флора Центрально-лесного государственного за-
поведника / Отв. ред. В.Г. Карпов. Л.: Наука, 1976. 104 с.
Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное
состояние основных концепций). Уфа: Гилем, 1998. 413 с.
Миркин Б.М., Соломещ А.И., Ишбирдин А.Р., Алимбекова Л.М. Список и ди-
агностические критерии высших единиц эколого-флористической классификации
растительности СССР (оперативно—информационный материал). М., 1989. 49 с.
Митина М.Б., Антонова Л. А. Сезонное развитие широколиственной дубравы //
Биологическая продуктивность и ее факторы в лесостепной дубраве. 1974.
С. 74-92.
Михайловская И.С. Корни и корневые системы. М., 1981. 136 с.
Морозова О.В. Леса заповедника «Брянский лес» и Неруссо-Деснянского По-
лесья (синтаксономическая характеристика). Брянск, 1999. 98 с.
Муравьев В.И., Баньковский А.И. Исследование растений, применяющихся в
народной медицине, на содержание аскорбиновой кислоты // Тр. ВИЛАР. М: Мед-
гиз, 1947. Вып. 9. С. 39-118.
Назаров В-Г. Долгосрочный прогноз численности тминной моли // Биологиче-
ская и химическая защита растений от вредителей, болезней и сорняков в УССР.
Киев, 1985а. С. 117-120.
Назаров В.Г. Роль энтомофагов в ограничении численности тминной моли и
эффективность примеииения биологических средств борьбы // Биологическая и хи-
мическая защита растений от вредителей, болезней и сорняков в УССР. Киев,
19856. С. 105-107.
Назаров В.Г. Тминная моль Depresaria daucela Den et. Schiff, и интегрирован-
ные приемы борьбы с ней в условиях лесостепи Украины. Авт. дис.... канд. с.-х. на-
ук. ЦСХА. Киев, 1987. 23 с.
Наумов Н.А. Флора грибов Ленинградской области. М.-Л.: Изд-во АН СССР,
1954. Вып. 1. Архимицеты и фикомицеты. 182 с.; 1964. Вып. 2. Дискомицеты. 257 с.'
Наумцев Ю.В. О биоморфологических особенностях и онтоморфогенезе
шпажника черепитчатого (G. imbricatus L.) в связи с его интродукцией // Флора и
растительность Тверской области. Тверь: Изд-во ТвГУ, 1996а. С. 43-54.
Наумцев Ю.В. О перспективах использования некоторых евроазиатских видов
рода Gladiolus в ландшафтном озеленении городов Средней России // Тр. 1 Всерос.
конф, по ботан. ресурсоведению. СПб.: Изд-во БИН РАН, 19966. С. 174.
Наумцев Ю.В. Интродукция Gladiolus imbricatus L. в ботаническом саду ТвГУ
206
в связи с вопросами его охраны // Материалы 1 науч.-практич. конф, студентов и
аспирантов вузов г. Твери. Тверь: Буквица, 1999а. С. 79-80.
Наумцев Ю.В. Итоги первичной интродукции представителей сем. Iridaceae
Juss. в Ботаническом саду Тверского госуниверситета. Дис.... канд. биол. наук. М.,
19996. 133 с.
Невский М.Л. Флора Калининской области. Т. 2. Калинин: Обл. кн. изд-во,:
1952. С. 309-1032.
Невский С.А. Материалы к флоре Кугитанга и его предгорий // Флора и систе-,
матика высших растений. М.-Л.: Изд-во АН СССР, 1937. Т. 4. С. 199-346.
Нейштадт М.И. Определитель растений средней полосы Европейской части
СССР. М.: Учпедгиз, 1954. 495 с.; 1963. 640 с.
Николаева М.Г., Лянгузова И.В., Поздова Л.М. Биология семян. СПб: Изд-во
НИИ химии, 1999. 230 с.
Новотельнова Н.С., Пыстина К.А. Порядок Peronosporales И Флора споровых
растений. Т. 11. Грибы (3). Л.; Наука, 1985. 363 с.. • .
Носова Л.М. География и экология Ranunculus cassubicus L. // Динамика цено-
популяций травянистых растений. Киев: Наукова думка, 1987а. С. 106-113.
Носова Л.М. О горизонтальной структуре сосновых культур на дерново-подзо-
листых почвах // Динамика естественных и искусственных лесных биогеоценозов
Подмосковья. М.: Наука, 19876. С. 5-25.
Нотов А.А. Материалы к флоре Тверской области. Ч. 1. Высшие растения.
Тверь: Изд-во ТвГУ, 1998. 99 с.
Нотов А.А. Материалы к флоре Тверской области. Ч. 1: Высшие растения. 3-е
изд., [Электрон; ресурс]. Тверь, 2000. 122 с. http://garden.tversu.ru/publ/flora/
spisok_2
Овеснов С.А. Конспект флоры Пермской области. Пермь: Изд-во Пермского
университета, 1997. 252 с.
Определитель высших растений Северо-Запада европейской части РСФСР
(Ленинградская, Псковская и Новгородская области). Л.: Изд-во ЛГУ, 1981. 376 с.
Определитель насекомых европейской части СССР. М.—Л.: Наука, 1965. Т. 2;
1978. Т. 4.
Определитель растений Мещеры / Под ред. В.Н. Тихомирова: В 2 ч. М.: Изд-во
МГУ, Ч. 1. 1986. Ч. 2. 1987.
Определитель сосудистых растений Центра европейской России. М., 1995. .
560 с.
Орлова Н.И. Orchidaceae II Флора Мурманской области. М.-Л., 1954.
С. 214-238.
Паламарчук И.А. Основные результаты десятилетних работ по изучению паст-
бищ мараловодческих совхозов Алтая // Вопросы сенокосно-пастбищного хозяйст-
ва. М„ 1960. С. 466-495.
Панкова И.А. Травянистые С—витаминосы // Тр. Бот. ин—та АН СССР, сер 5,
1949. Вып. 2. С. 472—478.
Парфёнов В.И., Дмитриева С.А. Кариологическая дифференциация у видов
флоры Белоруссии и её роль в формообразовании // Докл. АН БССР. 1985. Т. 29,
№ 6. С. 554-557.
207
Перепечко Н.П. Новое лекарственное растение — стальник пашенный Ononis
arvensis // Ботан. ж. 1964. Т. 49, № 4. С. 592-594.
Петров В.А. Критический обзор вьюнков флоры СССР // Бюлл. МОИП. Отд.
биол. и почв. 1935. Т. 44, вып. 3. С. 74.
Петухова Л.В. К биоморфологии стальника полевого // Флора и растительность
Тверской области. Тверь, 1996. С. 22-31.
Подымов А.И., Суслов Ю.Д. Лекарственные растения Марийской АССР. Йош-
кар-Ола: Марийское кн. изд-во. 1990. 192 с.
Полевой В.В., Саламатова Т.С. Эволюция способов движения растений // Эво-
люция функций в растительном мире / Тр. биол. науч.-исслед. инет. (Ленингр. гос.
унив.). Л.: ЛГУ, 1985. № 36. С. 188-200.
Полякова Г.А., Малышева Т.В., Флеров А.А. Антропогенное влияние на сосно-
вые леса Подмосковья. М.: Наука, 1981. 144 с.
Полякова Г.А., Малышева Т.В., Флеров А.А. Антропогенные изменения широ-
колиственных лесов Подмосковья. М.: Наука, 1983. 117 с.
Пономарев А. Н., Демьянова Е.И. К изучению гинодиэции у растений // Ботан.
ж. 1975. Т. 60, №1. С. 58.
Пономарев А.Н: О протерандрии у зонтичных//Доклады АН. 1960.Т. 135,№3.
С. 750-752.
Пономарев А.Н. О протерандрии зонтичных //Уч. Зап. Перм. ун-та. 1961. Т. 18,
вып. 3. С. 27-31.
Попов М.Г. Флора Средней Сибири. М.—Л.: Изд—во АН СССР, 1959. Т. 2. 918
с.
Пошкурлат А.П. Анализ ритма развития первоцвета весеннего // Ботан. ж.
1962. Т. 47, № 2. С. 262-267.
Щюзоровский Н.А. Наблюдения над осенним и зимним состоянием степных
растений // Тр. Центрально-Черноз. гос. заповедника. Воронеж, 1940. Вып. 1.
С. 273-301.
Прокудин Ю.Н., Вовк А.Г., Петрова О.А., Ермоленко Е.Д., Верниченко Ю.В.
Злаки Украины (анатомо-морфологический, кариосистематический и эколого-фи-
тоценотический обзор). Киев.: Наукова думка, 1977. 517 с.
Протасевич Р.Т. Особенности анатомического строения некоторых видов рода
Вейник // Ботаника. Исследования, вып. 5. Минск: изд-во АН БССР, 1963.
С. 102-103.
Протич Н., Евстатиева Л. Петльов гребен (Gladiolus imbricatus L.) в условията
на култура // Растен. науки.1994. Т. 31, № 1-2. С. 60-61.
Работнов Т.А. Методы изучения семенного размножения травянистых расте-
ний в сообществах (Всесоюзный институт кормов им. В.Р. Вильямса) // Полевая
геоботаника / Ред. Е.М.Лавренко, А.А. Корчагина. М.-Л.: Наука, 1964. Т. 2.
С. 20-40.
Работнов Т.А. Об эколого-биологическом и ценотическом своеобразии видов
на примере луговых растений // Тр. МОИП. Отд. Биол. 1966. Т. 27. С. 139-153.
Работнов Т.А. Фитоценология. М.: Изд-во МГУ, 1978. 382 с.; 2-е изд., 1983.
296 с.
Радченко Ю.Д., Клепикова П.И. Значение осотовой и вьюнковой щитоносок в
208
подавлении роста осота розового и вьюнка полевого // Тр. ВНИИ защиты растений.
1977. Т. 5. С. 164.
Ражинскайте Д.К. Предпосевная обработка семян и сроки посева тмина обык-
новенного (Carum carvi L.) // Тр. АН Литовской ССР, серия В, 1964. Т. 33, № 1.
С. 47-53.
Ражинскайте Д.К. Изменение эфирного масла плодов тмина в зависимости от
фазы зрелости и условий минерального питания // Тр. АН Литовской ССР, серия В.
1965. Т. 36, №1. С. 85-96.
Ражинскайте Д. Тмин обыкновенный и кориандр посевной, их эфиро-маслич-
ность и перспективы выращивания в Литве // Автореф. дис. ... канд. биол. наук.
Вильнюс, 1967. 20 с.
Ражинскайте Д. Изменение содержания эфирных масел в органах тмина и ко-
риандра посевного в онтогенезе растений // 4-й Межд. конгресс по эфирным мас-
лам. Т. 2. Тбилиси, 1968/1972. С. 147-150.
Ражинскайте Д.К., Славенас И.Ю. Влияние минеральных удобрений на рост и
развитие тмина обыкновенного и накопление эфирного масла // Тр. 1-й научн.
конф, по исследованию и обогащению раст. ресурсов Прибалтийских республик и
Белоруссии. Вильнюс, 1963; С. 165-176.
Раманаускене К. Почвенные водоросли в посеве тмина // Интенсификация с-х
производства. Каунас, 1978. С.91-92.
Раменская М. Л. Определитель высших растений Карелии. Изд—во Карельской
АССР, 1960.485 с.
Раменская М.Л., Андреева В.Н. Определитель высших растений Мурманской
области и Карелии. Л.: Наука, 1982. 435 с.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая
оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 471 с.
Растительное сырье СССР (под ред. М.М. Ильина). Т. 2. Л.: Изд-во АН СССР.
Бот. ин-т им. В.Л. Комарова. Отдел растительных ресурсов, 1957. 581 с.
Растительность европейской части СССР. Л.: Наука, 1980. 426 с.
Растительные ресурсы России и сопредельных государств. Ч. 1-2, доп. к 1-7 т.
СПб: Мир и семья-95., 1996. 571 с.
Растительные ресурсы СССР. Л., 1984. Т.1. 328 с.
Растительные ресурсы СССР: цветковые растения, их химический состав и ис-
пользование. Л.: Наука, 1986. Т. 2; 1987. Т. 3; 1988. Т. 4.
Редкие и исчезающие виды флоры СССР, нуждающиеся в охране / Ред.
А Л. Тахтаджяна. Л.: Наука, 1981. 568 с.
Речан С.П., Малышева Т.В., Абатуров А.В., Меланхолии П.Н. Леса северного
Подмосковья. М., 1993. 316 с.
Розанова М.А. Опыт аналитической монографии conspecies Ranunculus aurico-
mus Korsh. // Тр. Петергоф, естеств.-науч. ин-та, 1932. № 8, С. 19-138.
Розен В.В. Список растений, найденных в Тульской губернии до 1916 г. // Изв.
Тульск. об-ва любит, естествозн. 1916. Вып. 4. 282 с.
Роллов А.Х. Красильные растения Кавказа // Вест. Тифлис, бот. сада. 1908а.
Вып. 10. С. 41-58.
Роллов А.Х. Дикорастущие растения Кавказа, их распространение, свойства и
применение. Тифлис, 19086. 599 с.
209
Романов О.В. Структура ценопопуляции Gladiolus imbricatus L. в западных
районах Тверской области: Дипломная работа / Твер. гос. ун-т. Тверь, 2000. 99 с.
Ростовцев П.В. Мята, анис и тмин // Промышленные растения. 1895. Т. 4.29 с.
Ростовцев С.И. Список паразитных грибов, расположенных по растениям хо- ,
зяевам, 1908. 250 с.
Ростовцева Т.А., Красноборов И.М., Красникова С.А. Числа хромосом некото-
рых видов флоры Сибири // Новые данные о фитогеографии Сибири. Новосибирск:
Изд—во АН СССР, 1981. С. 215-220.
Ростовцева Т.С. Числа хромосом ряда видов семейства Apiaceae на юге Сибири
И Ботан. ж. 1976. Т. 61, № 1. С. 93-99.
Ростовцева Т.С. Числа хромосом некоторых видов семейства Apiaceae // Ботан.
ж. 1982. Т. 67, № 2. С. 206-210.
Руденко Ф.В. Строение мужских гамет покрытосемянных растений // Науч,
зап. Ужгородского гос. ун—та, 1956. Т. 23. С. 3-36.
Рысин Л.П. Сосновые леса европейской части СССР. М.: Наука, 1975. 210 с.
Рысин Л.П., Рысина Т.П. Морфоструктура подземных органов лесных травяни-
стых растений. М.: Наука, 1987. 207 с.
Рысина Т.П. Биология прорастания семян и развитие всходов некоторых лес-
ных травянистых растений // Бюлл. Главн. Бот. сада. 1967. Вып. 64. С. 48-55.
Рысина Т.П. Ранние этапы онтогенеза лесных травянистых растений Подмос-
ковья. М.: Наука, 1973. 216 с.
Рытов М.В. Русские лекарственные растения. Птр., 1918. Т. Г 256 с.
Рычин Ю.В. Флора гигрофитов. Определитель по вегетативным признакам со-
судистых растений водоемов и сырых и влажных местообитаний центральной час-
ти европейской территории СССР. М.: Советская наука, 1948. 447 с.
Рычин Ю.В. Сорные растения. М.: Учпедгиз, 1952. 278 с.
Рябова О.В. Primula officinalis как источник витамина С // Фарамация. 1939.
Вып. 11. С. 19-21.
Сабардина Г.С. Дикорастущие кормовые растения Латвийской ССР И Сб. тр.
ин-та Зоотехники и зоогигиены АН Латв. ССР. 1952. Т. 2. С. 17-42.
Савченко М.И. Материалы к анатомии осыпающихся и неосыпающихся форм
тмина (Carum carvi L.) // Ботан. ж. 1938. Т. 23, № 5-6. С. 397-402.
Сацыперова И.Ф. Биологические особенности семян лютиковых И Тр. Бот.
ин-та АН СССР. Сер. 5, Растительное сырье. 1972. Вып. 16. С. 136-147.
Седова Е.А. Особенности органогенеза и жизненного цикла луковичных и
клубнелуковичных геофитов // Экспериментальный морфогенез цветковых расте-
ний: опыт исследования различных жизненных форм. М.: Изд-во МГУ, 1972.
С. 194-207.
Седова Е.А. Закономерности органогенеза луковичных и клубнелуковичных
геофитов. М.: Изд-во МГУ, 1976. 30 с.
Седова Е.А. К вопросу о жизненных циклах многолетних травянистых расте-
ний типа луковичных и клубнелуковичных геофитов // Вестник МГУ. Сер. биол.
1982. №4. С. 28-34.
Серебряков И.Г. О ритме сезонного развития растений подмосковных лесов //
Вестник МГУ. 1947. № 6. С. 75-107.
210
Серебряков И.Г. Морфология вегетативных органов высших растений. М.:
Сов. наука, 1952. 391 с.
Серебряков И.Г. О методах изучения ритмики сезонного развития растений в
стационарных геоботанических исследованиях И Уч. зап. Моск. гор. пед. ин-та
им. В.П. Потемкина. М., 1954а. Т. 37, вып. 2. С. 3-20.
Серебряков И.Г. Биолого-морфологический и филогенетический анализ жиз-
ненных форм покрытосеменных // Уч. зап. Моск. гор. пед. ин-та им. В.П. Потемки-
на. М., 19546. Т. 37, вып. 2. С. 21-84.
Серебряков И.Г. Экологическая морфология растений; жизненные формы по-
крытосеменных и хвойных. М.: Высш, шк., 1962. 379 с.
Серебряков И.Г. Жизненные формы высших растений И Полевая геоботаника.
М.-Л., 1964. Т. 3. С. 146-205.
Серебрякова Т.И. Побегообразование и ритм сезонного развития растений за-
ливных лугов Средней Оки. Дис....канд. биол. наук. М., 1953.
Серебрякова Т.И. Побегообразование и ритм сезонного развития растений за-
ливных лугов Средней Оки // Уч. зап. Моск. гос. пед. ин-та. 1956. Т. 97, вып. 3.
С. 43-120.
Серебрякова Т.И. Морфогенез побегов и эволюция жизненных форм злаков.
М.: Наука, 1971. 359 с.
Середин Р.Н. Лекарственные растения // Растительные ресурсы: Природные
ресурсы и производительные силы Кавказа. Ростов-на-Дону, 1984. Ч. 2. С. 194-236.
Синяков А.Ф. Полезные свойства овощных растений // Картофель и овощи.
1990. № 1. С. 26-27.
Слободянюк В.Я. Насекомые-фитофаги вьюнка полевого и осота розового ле-
состепи Центрально-Черноземного района РСФСР и перспективы их использова-
ния в биологической борьбе. Автореф. дис... канд. с.-х. наук. Киев, 1987. 18 с.
Смирнов Б.М., Мамонов Н.Н. Борьба с молоканом и вьюнком полевым в усло-
виях орошения // Земледелие. 1954. № 9. 86 с.
Смирнов П.А. Флора Приокско-террасного государственного заповедника//
Труды Приокско-Террасного гос. заповедника. М., 1958. Вып. 2. 246 с.
Смирнова Е.С. Морфология побеговых систем орхидных. М., 1990. 208 с.
Смирнова О.В. Ритм роста корневых систем некоторых травянистых растений
дубрав //Бюлл. МОИП. Отд. биология. 1966. Т. 71, вып. 2. С. 54-63.
Смирнова С.А., Каден Н.Н. Семейство Convolvulaceae — Вьюнковые // Мор-
фология плодов и семян некоторых сорных растений СССР / Уч. записки Моск,
обл. пед. ин-та им. Н.К. Крупской. М., 1971. Т. 292, вып. 5. С. 12-29.
Смольянинова Л.А. Семейство Вьюнковые // Флора Европейской части СССР,
Л.; Наука, 1981. Т. 4. 380 с.
Собко В.Г. Орхще! Украши. Ки1в, 1989. 190 с.
Соболевская К. А. (отв. ред.) Полезные растения Западной Сибири и перспекти-
вы их интродукции. Новосибирск: Наука, 1972. 380 с.
Соколова Л.Е., Шиффере Е.В., Родин Л.Е., Лукичева А.Н. Луга и травяные бо-
лота // Растительный покров СССР: пояснительный текст к геоботанической карте
СССР. М.-Л.: Изд-во АН СССР, 1956. Т. 1. С. 520-563.
Соколова С.М., Александрова Е.Г. Биохимическая эволюция семейства ирисо-
вых//Бюлл. Главн. Бот. сада. 1990. Вып. 155. С. 40-46.
211
Соколовская А.П. Кариологическая характеристика представителей флоры Ле-
нинградской области//Вестник ЛГУ, сер. биол. наук. 1972. Т. 21, вып. 2. С. 56-63.
Солнцева О.Н. Транспирация травяного яруса и эффективность использования
им радиации и влаги в лесных биогеоценозах // Динамика естественных и искусст-
венных лесных биогеоценозов Подмосковья. М.: Наука, 1987. С. 97-105.
Солнцева О.Н., Холопова Л.Б. Роль экологических факторов в формировании
травяного покрова // Динамика естественных и искусственных биогеоценозов Под-
московья. М.: Наука, 1987. С. 88-97.
Сорные растения СССР. Л.: Изд-во АН СССР, 1934. Т. 3. 447 с.
Сорокин А.С. Растения Калининской области, нуждающиеся в особой охране //
Взаимоотношения компонентов биогеоценозов в южнотаежных ландшафтах. Ка-
линин: Изд-во КГУ, 1984. С. 41-48.
Сосудистые растения Советского Дальнего Востока. С.-Пб., 1996. Т. 8. 382 с.
Спасская Н.А. Стальник полевой (стальник пашенный) — Ononis arvensis L.
(Ononis hircine Jacq., Ononis intermedia C.A. Mey. ex Becker) // Атлас ареалов и ре-
сурсов лекарственных растений СССР. М., 1980. С. 306.
Сулайманов Э. Строение корневых систем вьюнков линейнолистного и ястре-
бинколистного // Некоторые исследования по вопросам биологии растений и жи-
вотных. М.: Наука, 1981. 58 с.
Сырейщиков Д.П. Иллюстрированная флора Московской губернии: в 4 ч. М.,
Ч. 1. 1906. Ч. 2. 1907. Ч. 3. 1910. Ч. 4. 1914.
Сырейщиков Д.П. Определитель растений Московской губернии. М., 1927.
294 с.
Табака Л., Гаврилова Г., Фатаре И. Флора сосудистых растений Латвийской
ССР. Рига: Зинатне, 1988. 195 с.
Талла Б. Орхидные, включенные в Красную книгу Латвийской ССР // Охрана и
культивирование орхидей. Таллинн, 1980. С. 32-34.
Тамберг Т. Г. Род Gladiolus L. // Декоративные травянистые растения для от-
крытого грунта СССР. Л.: Наука, 1977. Т. L С. 177-199.
Тамберг Т. Г. Южноафриканский вид Gladiolus carneus (Iridaceae) в условиях
Ленинграда//Ботан. ж. 1981. Т. 66, № 11. С. 1623-1625.:
Татаренко И.В. Орхидные России: жизненные формы, биология, вопросы охра-
ны. М., 1996. 206 с.
Тихомиров В.Н., Яницкая Т.О., Пронькина Г.А. Зонтичные Средней России.
Определитель по вегетативным органам. 2 изд., М.: Аргус, 1977., 88 с.
Тихонова М.Н. Строение и формирование побегов некоторых представителей
семейства орхидных. Автореф. дис. .. .канд. биол. наук. Л., 1972. 19 с.
Тихонова М.Н. К вопросу о природе подземных органов некоторых представи-
телей трибы Orchideae И Ботан. ж. 1983. Т. 68, № 5. С. 648-652.
Тоомре Р.И. Долголетние культурные пастбищам.: Колос, 1966. 400 с:
Турова А.Д. Лекарственные растения СССР и их применение. М.: Медицина,
1974.424 с.
Тюрина Е.В. Интродукция зонтичных в Сибири. Новосибирск: Наука, 1978.
240 с.
Уланова Н.Г., Демидова А.Н. Популяционная биология вейника седеющего
212
(Calamagrostis canescens (Web.) Roth) на вырубках ельников южной тайги // Бюлл.
МОИП. Отд. биол. 2001. Т. 106, вып. 5. С. 51-58.
Уланова Н.Г., Демидова А.Н., Клочкова И.Н., Логофет Д.О. Структура и дина-
мика популяций вейника седеющего Calamagrostis canescens-. модельный подход И
Журнал общ. биол. 2002. Т. 63, № 6. С. 509-521.
Уранов А.А., Смирнова О.В. Классификация и основные черты развития попу-
ляций многолетних растений // Бюлл. МОИП. Отд. биол. 1969. Т. 74, № 1.
С. 119-134.
Утеулина Р.Г. Вьюнок полевой и меры борьбы с ним в условиях сухой степи
Целиноградской области. Автореф. дис.... канд. с.-х. наук. Казань, 1969. 26 с.
Фегри К., Ван дер Пейл Л. Основы экологии опыления. М., 1982. 379 с.
Фёдоров А.А., Артюшенко З.Т. Атлас по описательной морфологии высших
растений. Соцветие. Л.: Наука, 1979. 296 с.
Федорчук В.М., Бурневский Ю.И. Рекомендации по определению и использо-
ванию типов леса при лесоустройстве (на примере Ленинградской области). Л.:
Лен. НИИЛХ, 1986. 71 с.
Федорчук В.Н., Егоров А.А., Гаубервиль К., Чернов И.М. Краткий определи-
тель типов леса Ленинградской области. С-Пб, 2002. 36 с.
Федотов Ю.П. Болота заповедника «Брянский лес» и Неруссо-Деснянского по-
лесья (флора и растительность). Брянск, 1999. 106 с.
Филиппова Л.Н., Вылегжанин Т.Н. К вопросу о цветении трех видов орхидных
// Биолого-флористические исследования в связи с охраной природы в Заполярье.
Апатиты, 1980. С. 60-66.
Флора Восточной Европы. Л.: Наука, 2001. Т. 10.
Флора европейской части СССР. Л.: Наука, 1974. Т. 1; 1976. Т. 2; 1979. Т. 4;
1981. Т. 5; 1987. Т. 6.
Флора Казахстана. Алма—Ата: Изд—во АН Каз ССР, 1964. Т. 7.
Флора Киргизской ССР. Фрунзе: Изд—во АН Кирг ССР, 1959. Т. 8.
Флора Ленинградской области. Л.: Наука, 1955. Т. 1.
Флора Ленинградской области. Л.: Изд-во Ленингр. ун-та, 1965.
Флора Мурманской области. М.: Наука, 1966. Т. 5.
Флора Нижнего Дона. 4.1. Ростов: Изд-во Ростовского университета, 1984. 280
с.
Флора северо—востока Европейской части СССР. Л.: Наука, 1976. Т. 2; 1977.
Т.4.
Флора Сибири. Новосибирск: Наука, 1987. Т. 1; 1993. Т. 6; 1997. Т. 11.
Флора СССР Л.: Изд-во АН СССР, 1934. Т. 2; 1935. Т. 4; 1937. Т. 7; 1941. Т. 11;
1950. Т. 16; 1952. Т. 18; 1953. Т. 19.
Флора Таджикской ССР. Л.: Наука, 1984. Т. 7.
Флора Центральной Сибири. Новосибирск: Наука, 1979. Т. 2. 1045 с.
Фроленко А.И. Такой обыкновенный тмин // Приусадебное хоз-во. 1993, № 3.
С. 37.
Халматов Х.Х., Харламов И.А., Алимбаева П.К., Карыев М.О., Халтов И.Х. Ос-
новные лекарственные растения Средней Азии. Ташкент, 1984. 199 с.
Холопова Л.Б., Солнцева О.Н. Растительный и почвенный покров через 25 лет
213
после сплошной рубки древостоя // Динамика естественных и искусственных лес-
ных биогеоценозов Подмосковья. М.: Наука, 1987. С. 52-62.
Хотин А.А. Плодообразование и накопление эфирного масла у тмина // Докл.
ВАСХНИЛ. 1948. № 12. С. 17-21.
Хотин А.А. Биологические основы культуры эфиромасличных растений // Ав-
тореф. дис....докт. с-х наук. 1957. 24 с.
Хотин А.А. О стадийном развитии некоторых эфиромасличных растений // Аг-
робиология, 1959. № 2. С. 231-235.
Хохряков А.П. Сорная и заносная флора Магаданской области // Проблемы
изучения адвентивной флоры СССР, Матер, совещ. 1-3 февр. 1989 г. М.: Наука,
1989. С. 92-94.
Хромосомные числа цветковых. Л.: Наука, 1969. 926 с.
Цвелев Н.Н. К систематике рода вейник (Calamagrostis Adans.) в СССР И Ново-
сти систематики высших растений. М.-Л.: Наука, 1965. Т. 2. С. 5-50.
Цвелев Н.Н. Злаки СССР. М.-Л.: Наука, 1976. 787 с.
Цвелев Н.Н. О лютиках (Ranunculus L., Ranunculaceae) секции Ranunculus в
Восточной Европе // Новости систематики высших растений. С.-Пб: Мир и се-
мья-95, 1996. Т. 30. С. 53-78.
Цвелев Н.Н. Определитель сосудистых растений Северо-Запада России (Ле-
нинградская, Псковская и Новгородская области). СПб.: Изд-во СПХФА, 2000.
781 с.
Цингер В.Я. Сборник сведений о флоре Средней России. М., 1985. 520 с.
Цыганкова Т.А. К изучению некоторых витаминоносных растений Северного
Прикаспия // Матер, по флоре и раст-ти Сев. Прикаспия. Л., 1971. Вып. 5, ч. 2.
С. 160-181. '
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-ши-
роколиственных лесов. М.: Наука, 1983. 196 с.
Цыганов Д.Н. Экоморфы флоры хвойно-широколиственных лесов. М.: Наука,
1976. 60 с.
Черемушкина В.А., Попова Н.А. Связь возрастного состава ценопопуляций с
ходом онтоморфогенеза (на примере корневищных луков и клубнелуковичных) //
Ботан. ж. 1976. Т. 61, № 2. С. 187-196.
Черепанов С.К. Сосудистые растения СССР. Л., 1981.
Черепанов С.К. Сосудистые растения России и сопредельных государств
(в пределах бывшего СССР). СПб.: Мир и семья-95, 1995. 990 с.
Черняковская Е.Ф. Морфолого-биологическая характеристика представителей
рода Ranunculus L. средней полосы европейской части СССР. Дис.... канд. биол. на-
ук, М., 1986. 153 с.
Чехонина-Трифонова М.В. Эколого-биологическая характеристика главней-
ших растений основных типов лугов южной части Карело-Финской ССР. Авт.
дис.... кан. биол. наук. Петрозаводск, 1954. 17 с.
Числа хромосом цветковых растений флоры СССР. Асегасеае — Menyantha-
сеае. Л.: Наука, 1990. 510 с.
Числа хромосом цветковых растений флоры СССР. Т. 2. Могасеае — Zygophy-
Пасеае. C-Пб.: Наука, 1993. 427 с.
214
Чумак В.А., Бондаренко Р.В., Панчук Г.И. Тминная моль и опыт борьбы с ней //
Тр. ВНИИ эфиромасличных культур, 1977. Т. 10. С. 121-123.
Шага А.Н. Адвентивная флора Нижнего Амура И Проблемы изучения адвен-
тивной флоры СССР. Матер, совещ. 1-3 февр. 1989 г. М.: Наука, 1989. С. 105-108.
Шалыт М.С. Методика изучения морфологии и экологии подземной части от-
дельных растений и растительных сообществ // Вегетативное размножение и во-
зобновление высших растений и методы его изучения. Крымский пед. ин-т, 1960.
С. 369-447.
Шафер В. Основы общей географии растений. М., 1956.
Шеляг-Сосонко Ю.Р., Соломаха В.А., Сипайлова Л.М. Сообщества пойм лес-
ной зоны Украины // Классификация растительности СССР с использованием фло-
ристических критериев). М.: Изд-во Моск, ун-та, 1986. с. 50-51.
Шик М.М. Сезонное развитие травяного покрова дубравы // Уч. зап. Моск. гос.
пед. ин-та им. В.И. Ленина. 1953. Т. 73, № 2. С. 159-250.
Шилов М.П. Растительные сообщества и их комплексы в пойме р. Клязьмы
(в пределах Владимирской области) // Уч. зап. Владимирского гос. пед. ин-та
им. П.И. Лебедева-Полянского. Владимир: Тип. им. 50-летия Октября, 1971.
С. 63-112.
Шилов М.П. Использование и охрана растительности кормовых угодий // Есте-
ственные кормовые ресурсы СССР и их использование. М., 1978. С. 161-167.
Шилова С.Н. Тмин // Сад и огород. 1956. № 5. С. 33-34.
Шлыков Т.Н. Дубильные растения СССР. Л.-М.: Сельхозгиз, 1932. 190 с.
Шмальгаузен И.Ф. Флора Средней и Южной России, Крыма и Северного Кав-
каза. Киев, 1897. Т. 2.
Штрекер В. Луговые злаки. Определитель луговых злаков в цветущем и нецве-
тущем состоянии, с указанием значения их для лугов и пастбищ. М.-Л.: Сельхоз-
гиз, 1931. 311 с.
Шутов И.В., Козлова Л. М., Бельков В.П., Самгин П.А., Мартынов А.Н. Приме-
нение гербицидов при лесовыращивании. М.: Лесн. пром-ть, 1967. 188 с.
Эзау К. Анатомия растений. М.: Мир, 1969. 564 с.
Экзерцева В.В., Вахрамеева М.Г., Денисова Л.В., Никитина С.В. Особенности
структуры ценопопуляций орхидных на северной границе ареала // Охрана и куль-
тивирование орхидей. М., 1987. С. 14-15.
Энциклопедический словарь лекарственных эфирно-масличных и ядовитых
растений. М.: Гос. изд-во с.-х. лит-ры, 1951. 486 с.
Эристави М.И. Заметки о виде Gladiolus imbricatus L. // Заметки по систематике
и географии растений. Тбил. Бот. ин-т, 1975. Вып. 31. С. 70-72.
Эристави М.И. Кавказские представители рода Gladiolus L.: Автореф. дис. ...
канд. биол. наук. Тбилиси, 1978. 26 с.
Эристави М.И. Изучение биологии развития проростков видов р. Gladiolus в
связи с таксономией И Заметки по систематике и географии растений. Тбил. Бот.
ин-т, 1982. Вып. 38. С. 37-40.
Эристави М.И. Ритм развития некоторых дикорастущих шпажников в услови-
ях Тбилиси // Интродукция растений и зеленое строительство. АН ГССР, Центр,
ботан. сад. Тбилиси, 1984. Вып. 15. С. 137-144.
215
Яковлев М.С. Ранняя диагностика осыпающихся и неосыпающихся форм тми-
на // Ботан. ж. 1938. Т. 23, № 5-6. С. 403^108.
Abdallah S., F. Et Z. The Convolvulus species ofthe Canary isles, the Mediterranean
region and the Near and Middle East. 1967. 288 p.
Arditti J., Clements M., Fast G., Hadley G., Nishinura G., Ernst R. Orchid seed
germination and seedling culture — a manual // Orchid biology. Review and perspec-
tives. Ithaca. N.-Y., 1982. B. 2. P. 243-270.
Atlas of Norht European vascular plants north of the tropic of cancer. Koeltz Sci.
Books, 1986. V. 1-3.
Bateman R.M., Pridgeon A.M., Chase M.W. Phylogenetic of subtribe Orchidinae
(Orchidoideae, Orchidacea) based on nuclear ITS sequences. 2. Intrageneric
relationships and reclassification to achieve monophyly of Orchis sensu stricto //
Lindleana. 1997. V. 12. P. 113-141.
Boissier E. Flora orientalis. V. 4. Coralliflora, Monochlamydae, 1879. Reipression
facsimile. A. Asher & C.O., 1963. 1266 p.
Bomkamm R., Meyer G., Marcan K. Uber die Wirkung von KronenschluB und
Dungung auf die Entwicklung von Waldbodenarten eines Kiefemforstes in Berlin —
Spandau // Phytocoenologia. 1993. № 23. S. 601-610.
Brown E.O., Porter R.H. The viability and germination of seeds of Convolvolus
arvensis L. and other perenial weeds // Iowa agric. exp. sta. res. bull. Iowa: Ames, 1942.
№ 294. P. 475-504.
Burgeff H. Samenkeimung der Orchideen und Entwicklung ihrer Keimpflanzen.
Jena. 1936.312 s.
Cannon P.F. Systematic studies in the genus Ononis L. 1981.
Catling P.M. Autogamy in Eastern Canadian Orchidaceae-. a review of current
knowledge and some new observations //Natur. Can. 1983. V. 110, № 1. P. 37-53.
Clapham A.R., Tutin T.G., Warburg E.F. Flora of the British Isles. Cambridge:
University press, 1952-1957.
Clarke G.C.S., Punt W., Hoen P.P. The northwest European pollen flora, 51.
Ranunculaceae // Review of Palaeobotany and Palynology. 1991. V. 69. P. 117-271.
Crackles F. E. Calamagrostis stricta (Timm) Koeler, C. canescens (Web.) Roth and
their hybrids in S.E. Yorks, v.c. 61, northern England J// Watsonia. 1998. V. 20, № 1.
P. 51-60.
Currah R.S., Smreciu E.A., Hambleton S. Mycorrhizae and mycorrhizal fungi pf
boreal species of Platanthera and Coeloglossum (Orchidaceae) II Can. J, Bot. 1990. №
68.P. 1171-1181.
Currah R.S., Zelmer C. A key and notes for the genera of fungi mycorrhizal with
orchids and a new species in the genus Epulorhiza H Rept. Tottori Mycol. Inst. 1992,
V. 30. P. 43-59.
Darlington C.D., Wylie A.P. Chromosome atlas of flowering plants. London: George
Allen & Unwin L.T.D., 1955. 524 p.
Dickson C. A., Parks W. Ten years of population counts of orchids at Dumbrock Loch
meadow, Stirlingshire, and problems of management // Glasgow Naturalist. 1994. V. 22,
№ 4. P. 349-360.
216
Diepenhorst P. Talent, a natural sprout inhibitor for potatoes // Veroff. der
Arbeitsgemeinshaft Kartoffelforschung e.v. Detmold, 1995. Bd. 17. S. 89-94.
Dietrich A. Uber die europaischen Arten der Gattung Gladiolus // Jahresberiche uber
die Konigliche Realschule. Berlin, 1832. S. 1-13.
Dressier R.L. The Orchids. Natural history and classification. Cambridge, 1981.
332 p.
Eichner Ch. Orchideen im Alpenraum// Sitzungsber. Ges. 1979. № 19. S. 49-69.
Ellenberg H. Zeigerwerte der Gefasspflanzen Mitteleuropas П Scripta Geobotanica.
1974. Bd. 9. 98 s.
Ellenberg H., Weber H.E., Dull R., Wirth V., Werner W., Paulisen D. Zeigerwerte
von Pflanzen in Mitteleuropa // Scripta Geobotanica, 1991. Bd. 18. 248 s.
Engler A. Syllabus der Pflanzenfamilien: Eine Ubersicht uber das gesamte
Pflazensystem / Bearbeit. L. Diels. 11. Aufl., iiberarb. Berlin: Gebruder Bomtraeger,
1936.419 s.
Evenhuis A., Verdam B., Zagoks J.C. Splash dispersal of conidia of Mycocentro-
spora acerina in the field // Plant Pathol. 1997. V. 46, № 4. P. 459-469.
Faegri K., van der Pijl L. The principles of pollination ecology. 2 ed. Oxford: Perga-
mon Press, 1971. 294 p.
Fagerstrom L. Neue Sippen des Ranunculus auricomus — Komplexes aus des
Sowjetunion / Acta soc. fauna et flora fenn. 1967. V. 79, № 2. P. 1-15.
Filipek M. Neottianthe cucullata (L.) Schlechter w Puszy Augustowskieg // Spraw.
Poznansk. Towarz. Przyjac. Nauk. 1956, № 3. P. 48-53.
Flora Europaea / Ed. T.G. Tutin et al. Cambrige: Cambrige Univ. Press, 1968. V. 2;
1972. V. 3; 1980. V. 5; 1980. V. 6; 1993. V.l.
Formanowiczowa H., Kozlowski J. Biologia Kietkowania i ocena laboratoryjna
nasiou roslin leczniczych jako materia tu siewnego vie. Nasiona uprawianych. W. Polsca
gatunkow z rodziny baldaszkowa tych. (Umbelliferae) — Kminen zwyczajny (Carum
carvi L) // Herba polon. rok. 1966. T. 12, № 4. S. 284—290.
Formanowiczowa H., Kozlowski J. Obserwacje Kielkowania nasion roslin leczni-
czych po dziesisciu latact przechowywania // Wiad. bot. 1961. T. 5, № 3. S. 254—256.
Frank D., Klotz S. Biologisch-okologische Daten zur Flora der DDR // Wissen-
schaftliche Beitrage. Martin-Luther-Univ. Halle-Wittenberg, 1990. Bd. 32 (41). 168 s.
Frazier I.C. Nature and rate of development of root system of Convolvolus arvensis //
Bot. Gazette. 1943. V. 194, № 3. P. 417—425.
Fuller F. Alpine und nordisch-alpine Orchideen. Die Orchideen Deutschlands. 1972,
№10. 80s.
Fuller F. Platanthera, Gymnadenia, Neottianthe. Neue Brehm-Bucherei. 1978. T. 8.
Geelhaar H. Gladiolen im Garten. Berlin, 1984. 154 s.
Godblatt P. Phylogeny and classification of lridaceae H Ann. Missouri Bot. Gard.,
1990. V. 77, №4. P. 607-627.
Godblatt P. An overview of the systematic, phylogeny and biology of the African
Iridaceae П Bolus Herbariorum. 1991. V. 13. P. 1-79.
Godblatt P., Manning J.C. Gladiolus in Southern Africa: systematics, biology and
evolution. Cape Town: Femwood, 1998. 320 p.
Godblatt P., Manning J.C., Bernhardt P. Adaptive radiation of bee-pollinated Gladio-
2\T
lus species (Iridaceae) in southern Africa// Ann. Missouri Bot. Garden. 1998. V. 85, №
3. P. 492-517.
Grieger H. Zur situation der heimischen Orchideen in Kreis Schwarzenberg // Mitt.
Arbeitskr. Heimische Orchideen. 1988, V. 17. S. 47-51.
Grime J.P., Hodson J.G., Hunt R. Comparative plant ecology (a functional approach
to common British species). London, 1988. 742 p.
Griinert C. Gartenbliimen von A bis Z. Leipzig; Radebeul: Neumann, 1989. 608 s.
Guyot A.L., Massenot M., Saceas A. Les Rouilles des Ononis // Encyclopedie
mycologique. Uredineana, 1951. T. 3. S. 112-119.
Hafliger E. Zytologisch-embryologische Untersuchungen pseudogamen Ranunculus
der Auricomus-Gruppe // Ber. Schweiz, bot. Ges. 1953. Bd. 10. S. 491-512.
Hagerup O. The morphology and biology of some primitive orchid flowers //
Phytomorphology. 1952, № 2. P. 134—138.
Hamel G. Die Gefahrdungsgrad der heimischen Orchideen in der DDR // Mitt.
Arbeitskr. «Heimische Orchideen». 1981, V. 10. S. 67-73.
Harvais G., Hadley G. The relation between host and endophyte in orchid mycorrhiza
// New Phytologist. 1967, V. 66, № 2. P. 205-215.
Hegi G. Illustrierte flora von Mittel-Europa. Munchen: J.F. Lehmanns Verlag, 1912.
Bd. 3,H. 3.; 1936. Bd. 2. H. 2.
Heller D., Heyn C.C. Conspectus Florae orientalis // An annotated catalogue of the
flora of the Middle East, fasc. 3. Jerusalem: Israael academy of scinces and humanities,
1986.160 р.
Holm L.G., Plunknett D.L., Pancho J.V., Herberger J.P. The world’s worst weeds.
Honolulu, Hawaii: East-West Center Book, Univ. Press of Hawaii, 1977. 609 p.
Hroudova Z. Littoral vegetation of the Rozmberk fishpond and its mineral nutrient
economy. Praha: Vydala Academia nacladatelstvi Ceskoslovenske akademie, 1988.
108 p.
Ingram C. The phenomenal behaviour of a South American Gladiolus И J. R. hort.
soc., 1967. V. 92. P. 396-398.
Kinzel W. Frost und Licht als beeinflussende Krafte oei der Samenkeimung.
Stuttgart, 1913. 170 s.
Kirk W.D.J. Yellow Loosestriff// Bee World (IBRA. The international link between
beeykeeping science and practice). 1996. V. 77, № 4. P. 197-199.
Knuth P. Handbuch der Bliitenbiologie. Leipzig: Verlag von Wilhelm Engelmann,
1899. Bd. 2.705 s.
Koch W. Schweizerische Arten aus der Verwandtschaft der Ranunculus auricomus
L. // Ber. Schweiz, bot. Ges. 1933. Bd. 42. S. 740-753.
Korotkov K.O., Morozova O.V., Belonovskaja E.A. The USSR vegetation syntaxa
prodromus. M., 1991. 346 p.
Kumazava M. Morphology and development of the sinker in Pecteillis radiata
(Orchidaceae) // Bot. mag. Tokyo. 1956, № 69. P. 455-461.
Kumazava M. The sinker of Platanthera and Perularia, it’s morphology and
development// Phytomorphology. 1958, V. 8. № 1-2. P. 137-145.
Kiinkele S., Vogt A. Zur Verbreitung und Gefahrdung der Orchideen in Baden-
Wurttemberg. 1973, Bh. 1. S. 8-72.
218
Kvist G. Ranunculus cassubicus subsp. kemerovensis subsp. nova from South Siberia
I I Ann. bot. fenn. 1987. V. 24, № 1. P. 79-80.
Landolt E. Okologische Zeigerwerte zur Schweizer Flora // Veroff. geob. Inst. ETH,
1977. H. 64. S. 1-208.
Lang D. Orchid of Britain. 1980. 213 p.
Lid J., Lid D.T. Norsk Flora. Oslo, 1994. P. 723-741.
Linden B. Aseptic germination of seeds of northern terrestrial orchids // New Phytol.,
1980, V. 72. P. 387-392.
Lippmaa T. Areal und alterbestimmung einer Union Galeobdolon-Asperula-
Asarum-U. sowie das problem der Charakterarten und der Konstanten // Acta Inst, et
Horti bot. Univ. Tartuensis. 1938. № 6. S. 1-152.
Lisci M. Flora vascolare dei muri in aree urbane della Toscana centro-meridionale //
Webbia. 1997. V. 52, № 1. P .,43-66.
Luer C.A. The native orchids of United States and Canada excluding Florida. N.-Y.,
1975. P. 35-349.
Lukasiewicz A. Rytmica rozwojonia bylin (ze szczegolnum uwzglednic nien feno-
logii organow nadziemnych) // Prace Komis. Biol. 1967. T. 31, № 6. S. 309-461.
Luther H. Om Gladiolus imbricatus synantropa forekotnst speciellt in Finland //
Memorande Soc. fauna et flora fennica. 1961. Vi 36. S. 21-27.
Marklund G., Rousi A. Jutlines of evolution in the pseudogamuos Ranunculus
auricomus group in Finland И Evolution. 1961. V. 15, № 4. P. 510-522.
Mathur R., Masih B. Evaluation of fungicides against powdery meldew disease of
cumin (cuminum cyminum) caused by Erysiphe polygoni // Indian Phytopathol. 1971.
V. 24, № 4. P. 796-798.
Matuszkiewicz W. Die Karte der Potentiellen Naturlichen Vegetation von Polen //
Braun-Blanquetia. Recueil de Travaux de Geobotanique, Camerino, 1984. V. 1.
Meusel H., Jager E., Weinert E. Vergleichende Chorologie der zentraleuropaishen
Flora. Jena, 1978. S. 418.
Nilsson S. Orchids of Nothem Europe. Middlesex. Penguin books Ltd, 1979. 146 p.
Obarski J. Szkodniki Kminku // Ochrona Rosl. 1965. V. 9, № 6.
Ogura Y. Anatomy and morphology of the subterranean organs in some Orchidaceae
// J. Fac. Sci. Univ. Tokyo. Sect. 3. 1953, V. 6. P. 135-157.
Patel R.M., Desai M.V. Altemaria blight of cuminum cyminum and its control //
Indian Phytopathol. 1971. V. 24, № 1. P. 16-22.
Pax F. Iridaceae II Die naturlichen Pflanzenfamilien, T. 2. Abt. 5. Leipzig, 1888.
S. 137-158.
Peitz E. Coeloglossum viride 11 Dhauner Echo, 1962. № 11. P. 17-18.
Pignatti S. Flora D’ltalia. Bolonia: Via Emilia Levante, 1982. P. 7. 732 p.
Powelson R. A., Lieffers V.J. Effect of light and nutrients on biomass allocation in
Calamagrostis canadensis И Ecography. 1992. № 15. P. 31-36.
Priban K., Ondok J.-P. Evapotranspiration of a willow carr in summer // Aquatic
botany. 1986. V. 25, № 2. P. 203-216.
ProchazkaF., Velisek V. Orchideje nasi prirody. Praha, 1983. 284 s.
Proctor V.W. Long-distance disersal of by retention in digestive tract of birds //
Science. 1968. V. 160. P. 321-322.
219
Reinhard H.R., Golz P., Peter R., Wildermuth H. Die Orchideen der Schweitz und
angrenzender Gebiete. 1991. 348 s.
Retkeilykasvio / L. Hamet-Ahti, J. Suominen, T. Ulvinen, P. Uotila. 3. painoksen
pohijalta. Helsinki: Luonnontiellienen Keskusmuseo, Kasvimuseo, 1998. 656 p.
Reynders M.M., Duke W.B. Utilization of axenic culture in allelopathic studies //
Proc. N.E. Weed Control Conf. 33. 1979. 122. p.
Roon van E., Bleijenberg H.J. Breeding caraway for nonshattering seed // Euphytica.
1964. V. 13, №3. P. 281-293.
Rothmaler W. Exkursionflora for die Gebiete der DDR und BRD. Berlin, 1972.
613 s.
Rutishauser A. Entwicklungserregung der Eizelle bei pseudogamen Arten der
Gattung Ranunculus // Bull, schweiz. Akad. med. Wiss. 1954; Bd. 10. S. 491-512.
Sarosiek J., Kolon K., Giza G., Wojcik G. Ecological characteristic of populations of
Neottianthe cucullata (L.) Schlechter in Augustow Forest // Acta Univ. Wratislav. 1990.
№ 1055. P. 95-102.
Schmid W., Reinhard H., Golz P. Coeloglossum viride (L.) Hartm. X Dactylorhiza
maculata (L.) Soo s.l.—erst fond in der Schweitz // Orchidee. 1974. V. 25, № 2.
S. 69-74.
Schultze M. Die Orchidaceen Deutschlands, Deutsch-Oesterreiches und der
Schweiz. 1894.
Schulz-Gaebel H. Entwicklungsgeschichtliche — zytologische Studien an der
Umbelliferen-Unterfamilie der Apioideen // Beitr. Biol. Pflanzen., 1930. Bd. 18, № 3.
S. 345-398.
Scoggan H.I. The flora of Canada. Ottawa: Nat. Museum of Nat. Sc., 1979. V. 4.
1225 р.
Serek M., Jones R.B., Reid M.S. Role of ethylene in opening and senescence of
Gladiolus spj flowers // J. Am. Soc. Hortic. Sc. 1994. V. 119, № 5. P. 1014-1019.
Sharman B.C. The development of sinker of Orchis mascula // Linn. Soc. J. Bot.
1939. V. 59, № 342. P. 145-158.
Sirjaev G. Generis Ononis L. revisio critica // Beih Bot. Centrabl. 1932. Bd. 49, № 2.
S. 381-665.
Smerciu E.A., Currah R.S. Symbiotic germination of seeds of terrestrial orchids of
North America and Europa // Lindleyana. 1989. № 1. P. 6-16.
Sokolowski A.W. Zbiorowiska Jesne polnocno-wschodniej Polski // Monographiae
botanicae. Warszawa, 1980. V. 60. 205 s.
Soo R., Borsos O. Geobotanische Monographic der pannonischen und karpatischen
Flora// Ann. Univ. Sci. Budapest. Sect. Biol. 1966. V. 8. P. 315-336.
Soucupova L. Allocation and modular structure in clonal graminoids in response to
waterlogging // Folia Geobot. Phytotax. Praha, 1994. № 29. P. 227-236.
Sriplieng A., Smith F.H. Anatomy of the seed of Convolvolus arvensis L. // Amer. J.
Bot. 1960. V. 47, № 5. P. 386-392.
Stewart J. The conservation of European orchids. Nature and environment. V. 57.
Strassbourg: Council of Europe Press., 1992. 64 p.
Stoutmire W. Terrestrial orchid seedlings // The Orchids. Scientific Studies. N.Y.,
1974. P. 101-129.
Summerhayes V.S. Orchids of Britain. London, 1951. 290 p.
220
Sundermann H. Europaishe und mediterrane Orchideen. Hidelsheim. 2 Aufl. 1975.
243 s.
Timmons F.L. Duration of viability of bindweed seed under field conditions and
experimental results in the control bindweed seedlings // Agron. J. 1949. V. 41, № 3.
P. 130-133.
Torrey J.G. Endogenous bud and root formation by isolated roots of Convolvulus
grown in vivo // Plant Physiol. 1958. V. 33. P. 258-263.
Trzaskos M. Some aspects of sowing Carum carvi into lowland swards 11 Proc, of the
14-th general meet of the Europ. grassland federation, Lahti, 1992. P. 549-551.
Tutin T.G. Primula 11 Flora of the British Isles. Cambridge: Universiti press, 1952.
1590 р.
Vogel S. Bliitenbiologische Typen als Elemente der Sippengliederung // Bot. Stud.
1954. Bd. 1. S. 1-338.
Warcup J.H. Symbiotic germination of some Australian terrestrial orchids // New
Phytol. 1973. №72. P. 387-392. <
Weaver S.E., Riley W.R. The biology of Canadian weeds. 53. Convolvolus arvensis
L. // Can. J. Plant Sci. 1982. № 62. P. 461^472.
Weber H.E. Zur soziologischen Stelluhg von Calamagrostis canescens (Weber)
Roth. //Phytocoenologia. 1979. № 6. S. 252-258.
Weimark H. Types of inflorescences in Aristea and some allied genera // Bot.
Notiser. 1939. № 40. P. 616-626.
Wennerberg A. Ett fynd av Coeloglossum viride x Dactylorhiza maculata И Sv. Bot.
Tidskr. 1978. V. 72, H. 2. P. 101-102. ,
Weymar H. Buch der Graser und Binsengewachse. Neumann: Verlag, 1972. 359 S.
Wiese A.F., Rea H.E. Factors affecting the toxity of phenoxy herbicides to fields
bindweed//Weeds. 1962. V. 10. P. 58-61.
Willems J.H., Melser C. Population dynamics and life-history of Coeloglossum
viride (L.) Hartm.: an endangered orchid species in the Netherlands // Bot. J. Linn. Soc.
1998. V. 126. P. 83-93.
Wisniewski N. Einige Bemerkungen zur Neottianthe cucullata (L.) Schlechter I I
Mitt. Arbeitskr. zur Beobachtungen und zum Schutz heimische Orchideen. 1969. H. 5.
S. 52-57.
Wisniewski N. Zum Verhalten von Neottianthe cucullata (L.) Schlecht. und der
Westgrenze ihrer Verbreitung in Europa // Feddes Rep. 1976. Bd. 87, H. 9-10.
S. 587-609.
Zacha V. Preco tohto roku na niektorych miestach odimiera rasca // Za vysoku Urodi.
1965. V. 13, №8. P. 302.
Zderkieiwicz T. Badania di- i tetraploidalnych Kminku zwyczainego Carum carvi L.
//Roczn. Nauk. roln., ser. a. 1964. V. 88, № 3. S. 683-710.
Zderkiewicz T. Kminek tetraploidalny {Carum carvi L.) (Doniesienie tymczasowe)
// Acta agrobotanica. Warszawa, 1964. V. 15. S. 169-171.
Ziegenspeck H. Orchidaceae // Lebensgeschichte der Blutenpflanzen Mitteleuropas.
Stuttgart, 1936. Bd. 1. Lf. 4. 840 s.
СОДЕРЖАНИЕ
Предисловие............................................... 3
Вейник седеющий. А.Н. Демидова, Н.Г. Уланова.............. 4
Шпажник черепитчатый. А.А. Потов, Ю.В. Наумцев............ 31
Неотгеанте клобучковая. М.Г. Вахрамеева, Т.В. Жирнова ...... 50
Пололепестник зеленый. М.Г. Вахрамеева, И.В. Блинова,
Т.И. Богомолова, Т.В. Жирнова...................... 62
Лютик кашубский. Р.П. Барыкина, Н.В. Чубатова . . . ...... 78
Стальник полевой. Л.В. Петухова, О.Б. Черноброва....... . . . 98
Тмин обыкновенный. И.М. Ермакова...............'..........114
Первоцвет весенний. И.Б. Седых.................... . . . 139
Вербейник обыкновенный. И.Б. Седых....................... 155
Вьюнок полевой. М.А. Новикова, Ю.Е. Алексеев ........... 176
Литература...............:............................... 197
CONTENTS
Introduction......................................................... 3
Calamagrostis canescens. A.N. Demidova, N.G. Ulanova................. 4
Gladiolus imbricatus. N.A. Notov, Yu.V. Naumtsev.................... 31
Neottianthe cucullata. M.G. Vakhrameeva, T.V. Zhirnova.............. 50
Coeloglossum viride. M.G. Vakhrameeva, I.V. Blinova,
T.I. Bogomolova, T.V. Zhirnova................................. 62
Ranunculus cassubicus. R.P. Barykina, N.V. Chubatova................ 78
Ononis arvensis. L.V. Petukhova, O.B. Chemobrova.................... 98
Carum carvi. I.M. Ermakova......................................... 114
Primula veris. LB. Sedykh.......................................... 139
Lysimachia vulgaris. I.B. Sedykh................................... 155
Convolvulus arvensis. M.A. Novikova, Yu.E. Alekseev................ 176
Literature cited................................................... 197
Научное издание
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
вып. 15
Редактор к. б. н. Н. Г. Уланова
Компьютерная верстка Н. Г. Уланова
Подписано в печать 10.07.03 г. Формат 60x90*/i6. Бумага тип. № 1. Гарнитура «Таймс».
Печать офсетная. Усл. печ. л. 14,0. Тираж 500 экз. Заказ № 122.
ИПП «Гриф и К»
г. Тула, ул. Октябрьская, 81-а.
Тел. (0872) 49-76-96