/






















































































































































































































Похожие




Текст
ФА НА
СС
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ, № ИЗ
ФАУНА СССР
НАСЕКОМЫЕ ДВУКРЫЛЫЕ
Том VII, вып. 2
Н. Г. ОЛСУФЬЕВ
СЛЕПНИ
Семейство TABANIDAE
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
Ленинград 1977
УДК 595.772 : о7в.89
Слепни (сем. Tabanidae). Олсуфьев Н. Г. В серии: Фауна" СССР.
Насекомые двукрылые. Т. VII, вып. 2. 1977. Изд-во «Наука», Ленингр.
отд., Л. 1—435.
В книге на современном уровне обобщены данные по изучению слепней
в СССР, их строению, филогении, классификации, изменчивости,
географическому распространению, биологии, вредоносному значению, в том
числе как переносчиков ряда инфекционных болезней, методам
изучения и способам борьбы. Кратко освещена история изучения слепней
в нашей стране. В специальной части представлены определительные
таблицы и описания 195 видов взрослых насекомых фауны СССР и
сопредельных стран. Илл. — 358, табл. — 7, библ. — 510 назв.
Главный редактор
директор Зоологического института АН СССР
О. А. Скарлато
Редакционная коллегия:
И. М. Громов, О. Л. Крыжановский, И. М. Лихарев, А. А. Стрелков
(редактор тома), А. А. Штакелъберг
21008-550 „ ,, „„ 7„ © Издательство «Наука», 1977
Ф055(02)-77Б3-17-23-76
ПРЕДИСЛОВИЕ
Слепни представляют собой кровососущих насекомых. Они имеют крупное
экономическое значение как вредители животноводства и в особенности как переносчики
ряда инфекционных болезней, в том числе" опасных для человека. Интерес к изучению
слепней и разработке мер борьбы с ними демонстрируется возрастающим числом
публикаций как у нас в стране, так и за рубежом. Важным стимулом для этих
исследований могут служить монографические руководства, обобщающие основные итоги
изучения слепней. В 1937 г. в серии «Фауна СССР» Академии наук СССР была
опубликована наша монография по слепням СССР. В соответствии с общим планом издания
монография имела преимущественно фаунистическую направленность. Эта книга стала
библиографической редкостью и необходимость издания новой монографии ощущается
давно. В послевоенные годы были опубликованы региональные монографии по
слепням — В. В. Шевченко (1961) по Казахстану, Н. А. Виоловича (1968) по Сибири,
А. С. Лутты (1970) по Карелии, Р. В. Гребенюк и П. А. Чирова (1971) по Киргизии,
Г. В. Бошко (1973) по Украине, Р. Г. Соболевой (1974) по югу Приморского края,
М. Кадыровой (1975) по Узбекистану и др. Эти публикации оказались весьма
полезными для развития краевых исследований.
В предлагаемой вниманию читателя книге на современном уровне обобщены
доступные данные по изучению слепней СССР. В «Введении» представлены впервые
разделы по истории изучения слепней в СССР, филогении и классификации, а также по
борьбе с слепнями. Географическое распространение слепней рассмотрено широко,
впервые дается типология ареалов, показаны на ландшафтной основе количественные
аспекты фауны слепней СССР, дано зоогеографическое деление Палеарктики, а также
высказаны соображения о палеогенезе фауны слепней этой области. Разделы по
биологии и вредоносности слепней написаны с учетом зарубежной литературы. При
составлении специальной части мы учли обширные коллекции Зоологического института
Академии наук СССР, затем обработали коллекции Зоологического музея Московского
государственного университета, использовали собственные многочисленные сборы
и поступления от других лиц из разных мест СССР. Учтены по возможности все
доступные нам публикации. Данные о видовом составе слепней изложены на основе
современной таксономии и номенклатуры, подвергшихся некоторым изменениям в связи
с прогрессом надвидовои систематики и уточнением видовых названий в результате
изучения типовых экземпляров в коллекциях музеев Европы. В сравнении с
монографией 1937 г. в текст книги включены вновь 38 видов слепней и 32 подвида, всего 70
таксонов, что по видам составляет пополнение на 25% , а по подвидам — более чем в 5 раз.
Широкое применение внутривидовой таксономии, осуществленное нами за последние
годы, открывает перспективы в оценке направлений и темпов эволюции отдельных
таксонов применительно к определенным ландшафтам. Это отвечает современному
уровню биосистематики, характеризующемуся широким внедрением политипической
концепции вида (Майр, 1947; Майр и др., 1956). Далее нами уточнены сведения о
географическом распространении видов фауны СССР, причем по многим видам впервые
даны картограммы ареалов. Пересмотрены и уточнены описания видов. Строение тер-
миналий учтено в случаях, когда имеется необходимость в уточнении на этой основе
отдельных таксонов. Для видов, описанных нами до 1963 г., указаны голотипы и место
их хранения — Зоологический институт Академии наук СССР в Ленинграде.
Публикация этих данных при новоописаниях обязательна по современному Международному
кодексу Зоологической номенклатуры. Ранее мы этого правила не придерживались,
хотя типовые экземпляры были в коллекции обозначены соответствующими
этикетками. Рисунки деталей строения слепней, картосхемы и фотографии, как правило,
выполнены автором, тотальные рисунки сделаны (ныне покойным) И. В. Григорьевым;
для заимствованных фотографий или рисунков указаны их авторы. При использовании
для определения рисунков деталей строения слепней, в том числе терминалий, следует
иметь в виду, что рисунки сделаны с типичных экземпляров и не учитывают
изменчивости, весьма значительной у слепней.
В нашей работе над коллекциями в Зоологическом институте АН СССР и над
рукописью мы получали неизменную помощь и советы от проф. А. А. Штакельберга.
1*
4
ПРЕДИСЛОВИЕ
Ценное содействие нам оказано проф. К. Я. Груниным. Благодаря любезности А. Н. Же-
лоховцева и Л. В. Зиминой мы имели регулярный доступ к коллекциям Зоологического
музея Московского государственного университета. Существенную помощь в коллек-
цирнном материале мы получили от проф. К. В. Скуфьина (Воронеж), проф. В. В.
Шевченко (Алма-Ата), д-ра Н. А. Виоловича (Новосибирск), д-ра А. С. Лутты
(Петрозаводск) и др. Интересные коллекции для сравнительного изучения, а также
необходимую литературу нам прислали Моуха и Хвала (Moucha и Chvala, Чехословакия),
Леклерк (Leclercq, Бельгия), Филип (Philip, США), Пикьюмен (Pechuman, США),
МакАлпин (McAlpin) и Тескей (Teskey) (Канада). Ценные рекомендации мы получили
от редактора издания проф. А. А. Стрелкова. При оформлении рукописи большая
техническая помощь нам оказана В. Г. Дуденковой и И. С. Мещеряковой. В процессе
подготовки данного тома большая часть иллюстраций была перерисована
художником Б. А. Гирстуном.
Всем перечисленным лицам мы рады выразить свою глубокую признательность.
Мы будем также благодарны всем лицам, которые пришлют свои замечания и
предложения по поводу публикуемой монографии.
Н. Г. Олсуфьев
член-корреспондент АМН СССР
доктор биологических наук,
профессор.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ*
Сем. ТАВА NIDAE
1. Подсем. PANGONIINAE
1. Триба Р a n g о n i i n.i
1. Род Pangonius Latr.
Стр.
1. P. pyritosus Lw 127
2. Род Stonemyia Bren.
1. S. tigris Big 129
*2. S. caucasica Krob 129
II. Подсем. CHRYSOPSINAE
2. Триба Chrysopsini
3. Род Silvius Mg.
1. Подрод Silvius s. str.
1. S. (s. str.) vituli P 131
2. S. (s. str.) latifrons Ols 133
2. Подрод Nemorius Rond.
3. S. (N.) vitripennis Mg 135
4. S. (N.) caucasicus Ols 137
S. (TV) с molitor Bog. et Sam . . , 138
5. S. (N.) irritahs Ric '■. '138
3. Подрод Нeterosilvius Ols.
6. S. (H.) zaitzevi Ols ...... 138
4. Род Chrysops Mg.
1. Подрод Chrysops s. str.
1. Ch. (s. str.) japonicus Wied 145
2. Ch. (s. str.) sepulcralis F 145
3. Ch. (s. str.) nigripes Ztt 147
4. Ch. (s. str.) divaricatus Lw , . . . 148
5. Ch. (s. str.) anthrax Ols 150
6. Ch. (s. str.) makerovi PL 151
7. Ch. (s. str.) caecutiens L. ' 152
Ch. (s. str.) c. ludens Lw . . . 154
* Здесь и в тексте звездочкой помечены виды (6), не найденные в СССР, но
распространенные в смежных странах и с большой долей вероятности могущие быть
обнаруженными у нас.
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
8. Ch. (s. str.) pictus Mg 154
9. Ch. (s. str.) turanlcus 01s 156
10. Ch. (s. str.) validus Lw 157
11. Ch. (s. str.) rehctus Mg 158
12. Ch. (s. str.) angaricus Ols 161
13. Cfe. (s. str.) parallelogrammus Zell >~ 162
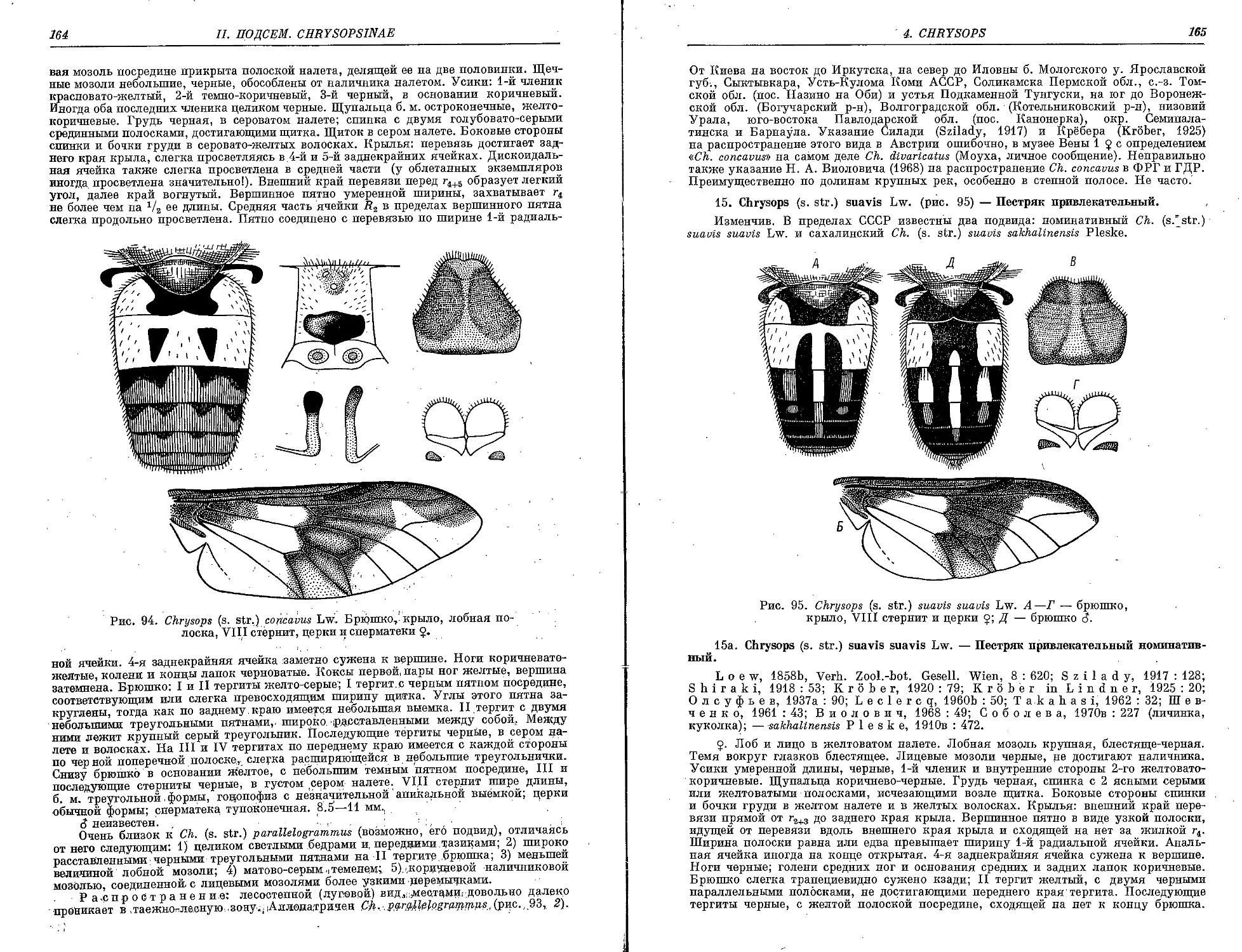
14. СЛ. (s. str.) concavus Lw 163
15. Ch. (s. Str.) suavis Lw 165
Cft. (s. str.) s. sakhalinensis PI 166
16. Ch. (s. str.) ricardoae PI 166
Ch. (s. str.) r. volgensis Ols 168
Cfe. (s. str.) r. jakutensis Ols 168
17. СЛ. (s. str.) dissectus Lw 169
18. Ch. (s. str.) та/ipes Mg. .'.'.' 170
2. Подрод Heterochrysops Krob.
19. Ch. (H.) sejunctus Szil. 173
20. Ch. (H.) italicus Mg 174
21. Ch. (H.) flavipes Mg 175
Ch. (H.) f. punctifer Lw. ......' .:' 177
22. Ch. (H.) vaderwulpi Krob. 178
23. Ch. (H.) mlokosiewiczi Big. . 179
24. Ch. (H.) oxianus Pi , . 180
3. Подрод Petersenichrysops M. et Chv.
25. Ch. (P.) buxtoni Aust. . ! 182
Ch. (P.) b. armeniensis Ols
4. Подрод Turanochrysops Stack.
26. Ch. (T.) stackelbergi Ols. . ; 183
27. Ch. (T.) hyalipennis Stack 183
III. Подсем. TABANINAE
3. Триба Diachlorini
5. Род Dasyrhamphis End.
1. D. umbrinus Mg 185
6. Род Philipomyla Ols.
*1. Ph. graeca F 187
2. Ph. aprica Mg 188
3. Ph. rohdendorfi Ols ' 189
7. Род Nanorhynchus Ols.
1. N. crassinervis Villen '. 190
4. Триба Tabanini
8. Род Tabanus L.
1. T. bifarius Lw 202
T. b. tarjukini Haus . 203
T. b. kurensis Djaf 203
2. T. lunatus F 203
3. T. bactrianus Ols 204
4. T: talyshi Ols 205
5. T. quatuornotatus Mg 206
T. q. araxis Ols 207
6. T. caucasius Krob 208
7. T. glaucopis Mg . 208
8.>Г. uccipiter Szil. '. 210
9t7\ subsdbuletorum Ols. 211
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 7
10. Т. sabuletorum Lw. 212
Т. s. gerkei Br 214
*11. T. stackelbergiellus Ols. . . 214
12. T. mistshenkoi Ols 215
13. T. freyi Szil . . . . 216
14. T. ansarii Abbas.-L. 216
T. a. badhysi Ols. et Shevtsh 216
15. T. ziraini Ols 217
16. T. filipjevi Ols 219
17. T. brunneocallosus Ols. . 220
18. T. leleani Aust 221
T. I. turkestanicus Ols 223
19. T. unifasciatus Lw ....;.. 223
20. T. smirnovi Ols 224
21. T. cordiger Mg 225
22. T. golovi Ols ' 226
T. g. mediaasiaticus Ols 228
23. T. fumidus Aust 228
24. T. kinoshitai Kono et Так 229
25. Т. rupium Br 230
26. T. maculicornis Ztt 230
27. T. geminus Szil '..... 232
28. T. raiki Br 233
T. m. colchidicus Ols ....... 235
T. m. australis Haus 235
29. T. indrae Haus. 235
T. i. vappa Bog. et Sam 236
T. i. montivagus Ols ........'. 237
30. T. infestus Bog. et Sam ....:.' 237
31. T. armeniacus- Krob . . . ; 238
32. T. regularis Jaeim .....";.. 238
33. T. bromius L > 239
T. b. flavofemoratus Strobl ■':'•': . . . 241
34. T. hissaricus Bar 242
35. T. semenovi Ols 242
36. T. laetetinctus Beck. . 243
T. 1. sordes Bog. et Sam 245
37. T. hauseri Ols 245
38. T. tergestinus Egg. . 246
39. T. armenicus Szil 248
40. T. argenteomaculatus Krob. 248
41. T. montiasiaticus Ols 249
42. T. schiva M. et Chv ." 251
43. T. shelkovnikovi Param 251
44. T. prometheus Szil 252
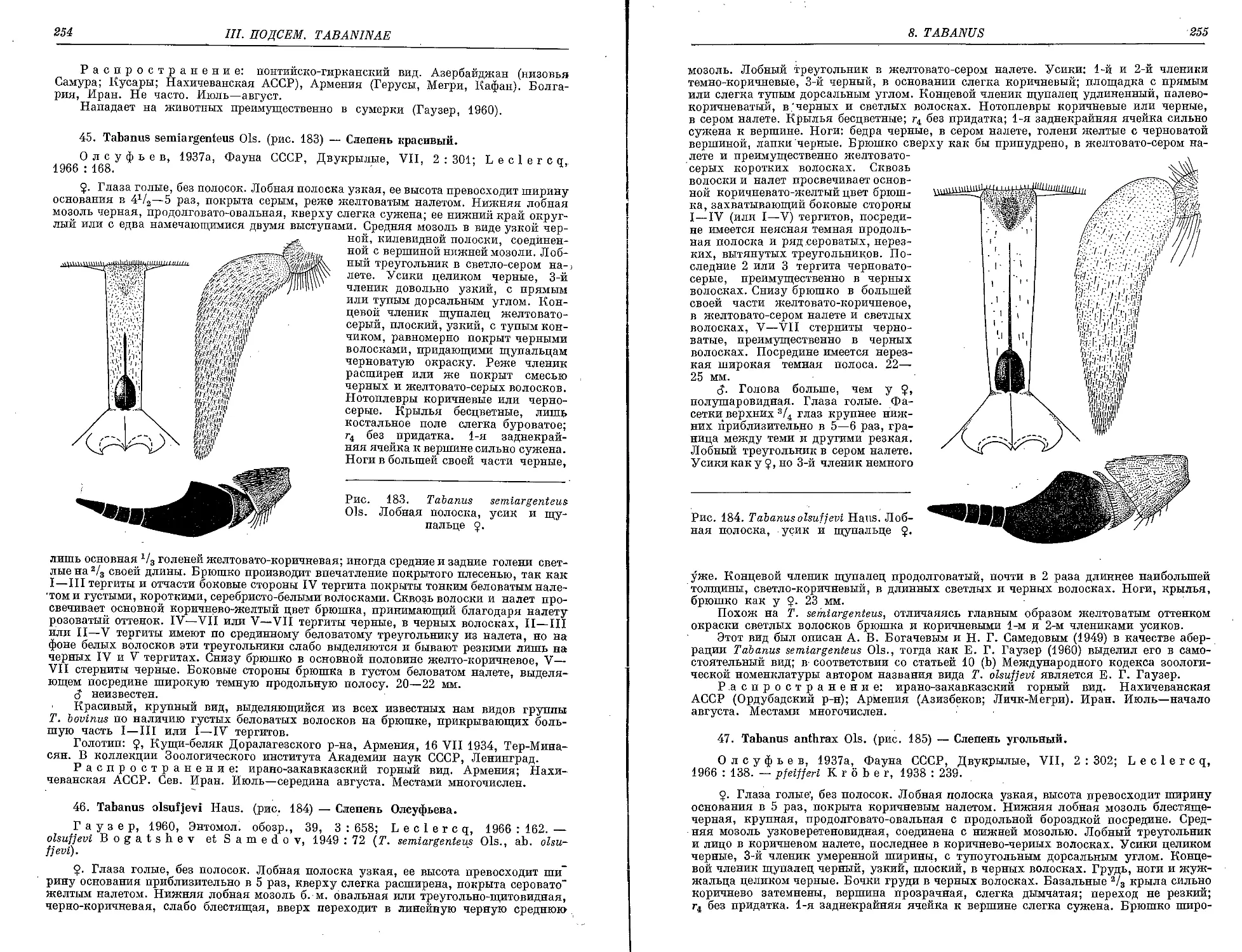
45. T. semiargenteus Ols 254
46. T. olsufjevi Haus 254
47. T. anthrax Ols - 255
48. T. subparadoxus Ols 25e
49. T. swiridowi Portsch 257
50. T. perelegans Ols 258
51. T. sudeticus Zell 259
T. s. verralli Oldr 261
52. T. portschinskii Ols . 261
53. T. spodopterus Mg 262
54. T. tinctus Wlk 263
55. T. bovinus L 265
56. T. pleskei Krob 267
57. T. sapporoenus Shir. 268
58. T. chrysurus Lw. 269
59. T. buddha Portsch. ■. 271
60. T. spectabilis Lw 272
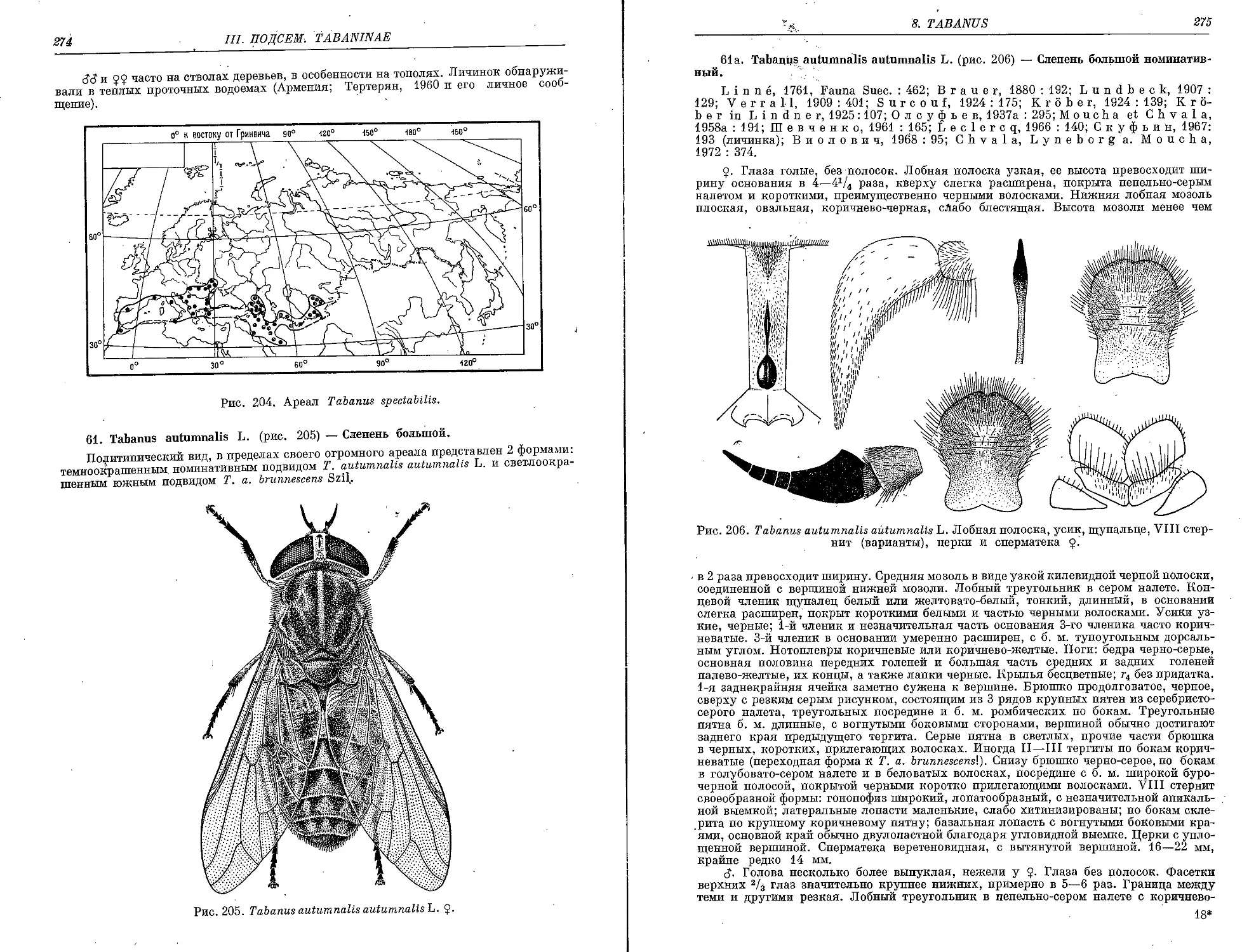
61. T. autumnalis L 274
T. a. brunnescens Szil 276
62. T. atropathenicus Ols 277
63. T. signatipennis Portsch 278
64. T. trigeminus Coq 279
65. T. amaenus Wlk 280
T. a. griseinus Phil 281
8
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
9. Род Atylotus О. S.
I. Подрод Atylotus s. str.
1. A. (s. str.) latistriatUs Br 284
2. A. (s. str.) fulvus Mg 285
4. (s. str.) /. aureus Haus 287
3. A. (s. str.) chodukini Ols 287
4. 4. (s. str.) loewianus Villen 288
5. A. (s. str.) horvathi Szil 289
6. A. (s. str.) miser Szil 290
7. A. (s. str.) pallitarsis Ols 291
8. 4. (s. str.) rustlcus L 293
9. A. (s. str.) proditor Bog. et Sam 294
10. A. (s. str.) quadrifarius Lw 295
11. A. (s. str.) flavoguttatus Szil 297
12. 4. (s. str.) pulchellus Lw 299
4. (s. str.) p. karybenthinus Szil 301
2. Подрод Surcoufiella Bequ.
13. A. (S.) plebejus Flln 301
A. (S.) p. sibiricus Ols 302
14. A. (S.) sublunaticornis Ztt 303
10. Род Therioplectes Zell.
1. Th. tricolor Zell. 305
Th. t. luminosus Ols ч 305
Th. t. pallidicauda Ols 306
2. Th. tunicatus Szil 306
3. Th. carabaghensis Portsch ' 307
Th. с orientalis Ols 308
4. Th. albicauda Ols 308
II. Род Hybomitra End.
1. Подрод Mauchaemyia Ols.
1. ff. (M.) caucasi Szil ....:.. 310
2. Подрод Hybomitra S; str.
2. ff. (s. str.) kaurii Chv. et Lyn 318
3. ff. (s. Str.) sexfasciata Hine 320
4. ff. (s. str.) stenopselapha Ols . 322
5. ff. (s. str.) olsoi Takah 323
6. ff. (s. str.) altaica Ols 324
7. ff. (s. str.) lapponica Wahlbg 325
8. ff. (s. str.) astuta O. S 327
9. ff. ,(s. str.) brevis Lw 329
10. ff. (s. str.) ussuriensis Ols 330
11. ff. (s. str.) arpadi Szil 332
12. ff. (s. str.) nigella Szil 333
13. ff. (s. str.) pavlovskii Ols • • 334
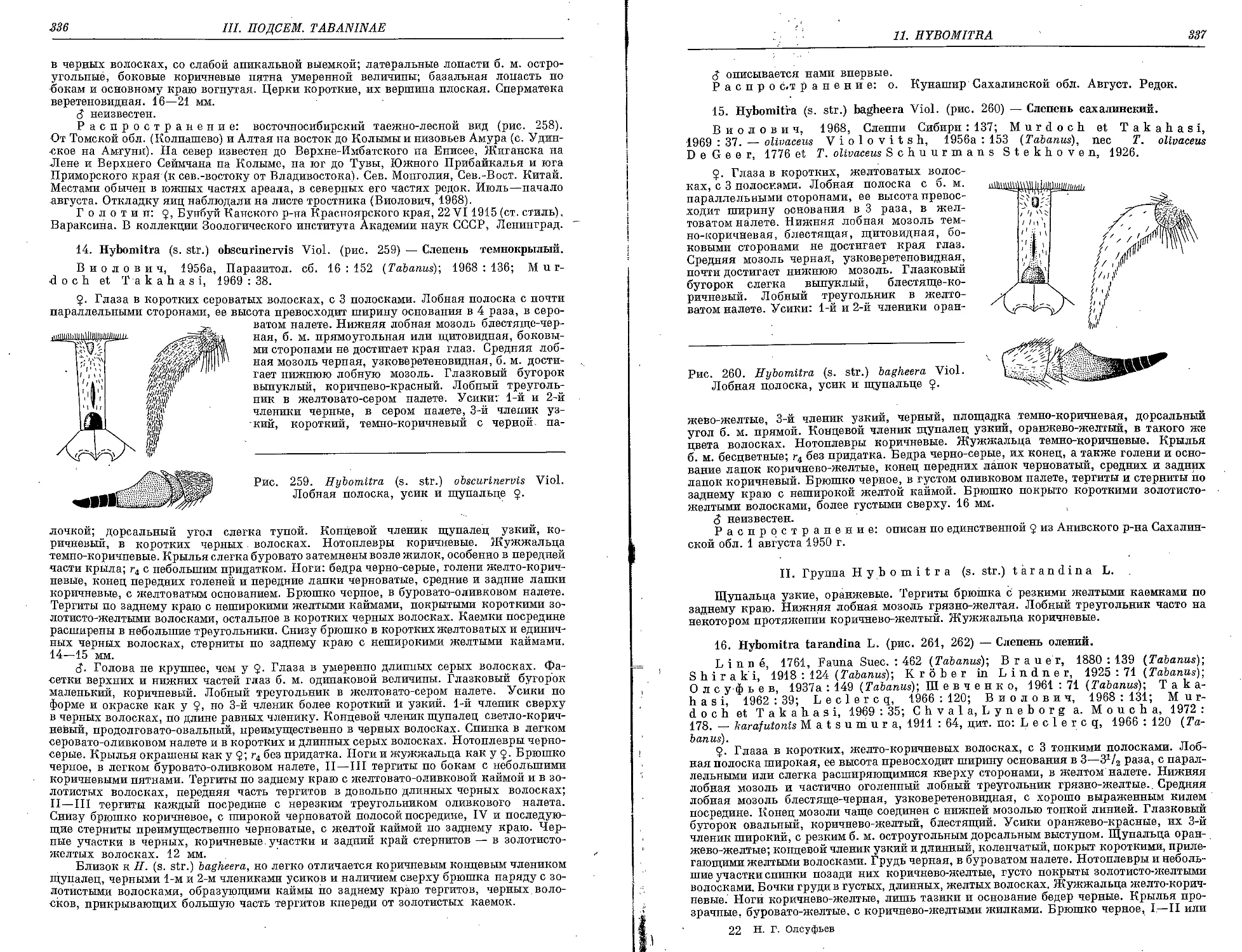
14. ff. (s. str.) obscurinervis Viol. 336
15. ff. (s. str.) bagheera Viol 337
16. ff. (s. str.) tarandina L 337
17. ff. (s. str.) tarandinoides Ols 339
18. #. (s. str.) aequetincta Beck 341
19. ff. (s. str.) stigmoptera Ols 343
20. Я. (s. str.) tatarica Portsch 344
21. ff. (s. str.) shevtshenkoi Ols 345
22. ff. (s. str.) zonata Szil 346
23. ff. (s. str.) astur Erichs 34?
24. Я. (s. str.) auripila Mg '. • . 349
*25. ff. (s. str.) aterrima Mg 350
*26. ff. (s. str.) micarcs Mg 350
27. ff. (s. str.) popovi Ols. . , 351
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 9
28. H. (s. str.) /wide Flln 352
29. H. (s. str.) nitidifrons Szil 354
ff. (s. str.) n. confiformis Chv. et M 355
30. H. (s. str.) sareptana Szil 356
31. ff. (s. str.) tschuensis Ols 358
32. ff. (s. str.) adachii Takag 359
33. ff. (s. str.) distinguenda Verr 360
ff (s. str.) d. contigua Ols 363
34. ff. (s. str.) ciureai Seg 363
35. ff. (s. str.) ukrainica Ols 365
36. ff. (s. str.) muehlfeldi Br 366
37. ff. (s. str.) bimaculata Macq. . . . ' 368
38. ff. (s. str.) caucasica Enderl 370
39. ff. (s. str.) hunnorum Szil 371
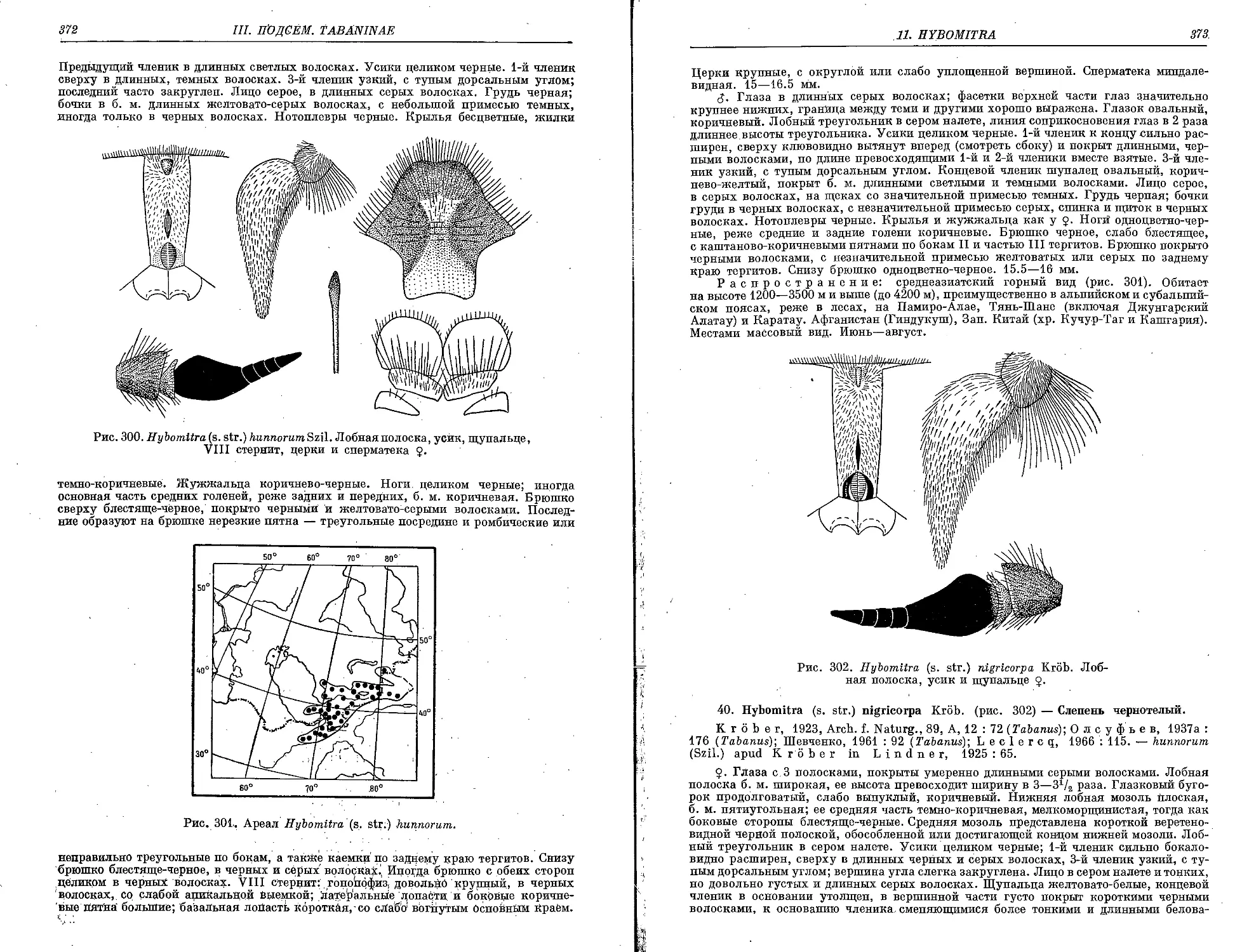
40. ff. (s. str.) nigricorpa Krob 373
41. ff. (s. str.) sogdiana Ols 374
42. ff. (s. str.) semipollinosa Ols 375
43. ff. (s. str.) shnitnikovi Ols 377
44. ff. (s. str.) turkestana Szil 378
45. ff. (s. str.) nigricornis Ztt 379
46. ff. (s. str.) lundbecki Lyn 381
ff. (s. str.) I. sibiriensis Ols 383
47. ff. (s. str.) montana Mg. 384
ff. (s. str.) m. morgani Sure 386
H. (s. str.) m. reinigiana End 387
ff. (s. str.) m. karatschafensis Skuf. 387
48. ff. (s. str.) expollicata Pand 387
H. (s. str.) e. orientalis Ols 389
49. ff. (s. str.) erberi Br 389
50. H. (s. str.) peculiaris Szil 392
51. ff. (s. str.) turanica Ols. 393
3. Подрод Sipala End.
52. H. (S.) acuminata Lw 394
5. Триба Haematopotini
12. Род Heptatoma Mg.
1. ff. pellucens F 396
ff. p. orientalis Ols. . 399
13. Род Haematopota Mg.
1. ff. grandis Macq. .,. . . 403
2. ff. pavlovskii Haus ^. . . 404
3. ff. caspica Abbas:-L 405
ff. с transcaucasica Ols 406
4. ff. гЫгса Mg 406
5. ff. bactriana Ols 408
6. ff. longeantennata Ols 409
* 7. ff. caenofrons Krob 410
8. ff. pollens Lw 411
9. ff. turkestanica Krob. . , 412
ff. f. orientalis Ols 414
10. ff. koryoensis Shir 414
11. ff. pallidula Krob 415
12. ff. desertorum Szil 416
13. ff. pluvialis L 417
ff. p. tristis Big. . .^ 419
ff. p. io//s Ols 420
14. ff. subcylindrica Pand ' 420
15. ff. scutellata Ols., M. et Chv 422
ff. s. pontica Ols 423
16. ff. crassicornis Wahlbg 423
17. ff. tamerlani Szil 425
18. ff. stackelbergi Ols 427
I. ВВЕДЕНИЕ
1. КРАТКАЯ ИСТОРИЯ ИЗУЧЕНИЯ СЛЕПНЕЙ В СССР
Исторический очерк целесообразно поделить на два периода: дореволюционный и
советский, поскольку они отличаются по направленности и объему исследований.
Мы ограничиваемся по возможности кратким изложением вопроса, опуская детали.
Первые указания на слепней нашей страны мы находим в известном труде
академика Палласа «Reise durch verschiedene Provinzen des Russischen Reiches» (1771—1776),
упомянувшего найденные им 4 вида: Tabanus tarandinus, Т. bovinus, Т. occidentalis
и Т. (Chrysops) caecutiens. Отдельные виды слепней были затем отмечены в списках
насекомых, опубликованных Цедерхельмом (Cederhielm, 1798) для Ингрии (примерно
территория б. Петербургской губ.), Двигубским (1802) для б. Московской губ., Гум-
мелем (Hummel, 1821—1829) для б. Петербургской губ., Эверсманном (Eversmann,
1834) для территории между Волгой и Уралом, Гиммерталем (Gimmerthal, 1847) для
различных мест России и т. д.
В 1842 г. австрийский энтомолог Целлер (Zeller) описал из «Южной России»
(очевидно, с Кавказа) эффектного Therioplectes tricolor. Это первое описание нового вида из
состава нашей фауны и одновременно первое «иностранное вмешательство». Ново-
описания видов слепней с территории нашей страны мы затем встречаем в публикациях
Эрихсона, Лёва, Биго, Фрея, Силади и Крёбера (Erichson, 1851; Loew, 1858a,
1858b, 1871; Bigot, 1880, 1892; Frey, 1915; Szilady, 1914-1926; Krober, 1920—1928).
Публикации двух последних авторов, относящиеся к 1917—1928 гг., основываются
на коллекционных материалах, поступивших в зарубежные музеи еще из
дореволюционной России, поэтому мы их рассматриваем в исследованиях дореволюционного
периода. Всего иностранными авторами описаны 37 видов * слепней фауны России.
В отечественной литературе первое новоописание мы встречаем в публикации
В. Мочульского (1859), но его постигла неудача: описанный им Chrysops bipunctatus
оказался синонимом Ch. dissectus Lw. Далее отметим новоописания в работах И. А. Пор-
чинского (1881—1892) и Ф. Плеске (1910а, 19106). BGero в дореволюционное время
отечественными авторами было описано 8 новых видов фауны России. Это более чем
скромная цифра в сравнении с многочисленными новоописаниями зарубежных авторов.
Для дореволюционного периода заслуживают еще упоминания следующие
фаунистические публикации по двукрылым, содержащие данные и о слепнях (но без ново-
описаний): Р. Остен-Сакена (1858) для окрестностей Петербурга, Б. Федченко (1892)
для б. Московской губ., И. А. Порчинского (1901) для б. Воронежской губ., В. Ярошев-
ского (1876) для окрестностей Харькова и др. Ряд фаунистических указаний для
разных мест России содержится в известной монографии Брауэра (Brauer, 1880) по пале-
арктическим Tabanus, в публикации Беккера (Becker, 1900) по фауне севера Сибири и
Фрея (Frey, 1911) —по фауне Финляндии, Карелии и Кольского п-ова.
Дореволюционная биологическая литература по слепням ограничивается
публикацией И. А. Порчинским в 1899 г. (и затем многократно переиздававшейся) небольшой,
но интересно составленной монографии. В ней наряду с данными о биологии слепней
и их вредоносности содержатся некоторые оригинальные рекомендации автора по
истреблению этих насекомых («лужи смерти», клеевые щиты). О слепнях как
переносчиках сибирской язвы имеются интересные эпизоотологические наблюдения в работе
В. Нагорского (1902). Строение ротового аппарата самки слепня было описано в
небольшой работе Ф. Штейнгеля (1897).
Для послереволюционного периода (и частью нескольких предшествующих ему
лет) отмечу прежде всего серию систематико-фаунистических работ А. А. Штакельберга.
Этим выдающимся советским диптерологом опубликованы в разное время (1916—1926)
* Из этой цифры, так же как из нижесообщаемых данных о количестве видов, вновь
описанных отечественными авторами, мы исключаем виды, отошедшие затем в
синонимы, сведенные в подвиды, ошибочно указанные для нашей территории и т. д.
ИСТОРИЯ ИЗУЧЕНИЯ СЛЕПНЕЙ
11
списки двукрылых, включавшие слепней, по б. б. Петроградской, Новгородской,
Черниговской губ., Армении и Средней Азии, в том числе описаны (1926b) 1 новый подрод
и вид Chrysops. Этот же автор положил начало отечественным определителям по
двукрылым, и для европейской части СССР в разное время их было издано три. А. А. Шта-
кельберг сыграл выдающуюся роль в создании коллекции двукрылых, в том числе
слепней, в Зоологическом институте Академии наук СССР. Эта коллекция,
включающая голотипы многих видов, является эталоном для фаунистических работ,
проводимых у нас в стране.
В послереволюционное время слепни привлекают внимание многих исследователей,
что в значительной мере связано с важной экономической ролью слепней в медицине
и ветеринарии.
Для довоенного периода из фаунистических работ упомянем публикации Ю. М.
Колосова и Л. В. Попова (1932) по Уральской обл., С. Я. Парамонова (1925—1933) по
Молдавии и Армении (с описанием нового вида), А. Дегтярева (1928) по б. Ярославской
губ., К. П. Самко (1929) по б. Тобольскому округу, Д. Н. Засухина с соавт. (1930)
по юго-востоку РСФСР, Е. Н. Павловского, Д. И. Благовещенского и Н. И. Алфеева
(1935) по Кустанайскому р-ну Казахстана, Е. Ф. Киселевой (1937) по Томской обл.,
Д. И. Благовещенского (1937) по Казалинскому р-ну Казахстана, В. Ю. Фридолина
(1936) по Кольскому п-ову, А. В. Гуцевича (1937, 1940) по Дальнему Востоку и
Забайкалью, Н. Я. Харитоновой (1939) по Пермской обл., К. В. Скуфьина (1938) по
Карачаево-Черкесской АО и Е. Г. Гаузера (1939, 1941) по Азербайджану (с описанием
нового вида). А. В. Гуцевич по довоенным исследованиям в Чувашской АССР
опубликовал еще статью в 1943 г. Аналогично Е. Г. Гаузер по исследованиям 1938 и 1939 гг.
в Азербайджане опубликовал еще две фаунистические работы — в 1953 и I960 гг.,
включающие некоторые новоописания, из них новыми можно признать лишь два вида.
Упомянем, наконец, публикации Эндерлейна (Enderlein, 1925, 1933), содержащие
некоторые описания новых видов из СССР, но в большинстве эти виды оказались уже
известными ранее (Олсуфьев, 1973).
Наши фаунистические исследования довоенного периода (1934—1941) по слепням
относятся к Ленинградской обл., Уралу, Западной Сибири, Смоленской обл.,
Казахстану, дельте Волги, Таджикистану и Кавказу; некоторые работы включают
новоописания, всего 10 видов. В серии «Фауна СССР» в 1937 г. опубликована наша
монография по слепням, включающая 153 вида, найденных к тому времени на территории
СССР, в том числе 24 вида описано впервые и установлен один новый род.* Основу
монографии составили результаты обработки обширных коллекций по слепням
Зоологического института Академии наук СССР (Ленинград), а также наших
многочисленных сборов в разных местах СССР и поступлений от отдельных исследователей.
В довоенный период получили некоторое развитие и другие направления в
изучении слепней. Отметим публикации по биологии слепней и гонотрофическому циклу
(Олсуфьев, 19356, 1940а), по строению половых органов самцов (Смирнов, 1923) и по
строению личинок слепней (Олсуфьев, 19366; Гаузер, 1939). Изучались влияние
нападений слёгшей на удойность коров (Дегтярев, 1928), патогенное действие слюны слепней
на кожу человека (Павловский, Штейн, Олсуфьев, 1935), роль слепней как
переносчиков трипанозомоза верблюдов «су-ауру» (Драчинский, 1927; Арбузов, 1941),
сибирской язвы (Олсуфьев и Лелеп,1935), туляремии (Олсуфьев и Голов, 1936; Сомов с
соавт., 1939; Боженко, 1941.) и инфекционной анемии лошадей (Родионов, 1936).
В послевоенные годы изучение слепней в СССР привлекло к себе внимание еще
большего числа исследователей и продолжало успешно развиваться.
Из фундаментальных работ этого периода прежде всего отметим интересные
многопрофильные исследования К. В. Скуфьина (1949—1968) по слепням Воронежской обл.
Им были изучены фауна, распространение, сезонная и суточная активность взрослых
насекомых, экология отдельных видов, места их выплода, преимагинальные фазы
развития, некоторые способы борьбы и т. д. Для отлова слепней автором разработана и
с успехом испытана чучелообразная ловушка, использованная затем в исследованиях
других авторов.
Заслуживает внимания серия статей и монография В. В. Шевченко (1953—1961) по
слепням Казахстана. Автором дана обстоятельная сводка по фауне, проделана большая
работа по изучению терминалий самок с целью видовой таксономии, выяснены
особенности географического распространения видов, обсуждены некоторые вопросы
генезиса фауны, отмечены особенности экологии слепней, места выплода, преимагинальные
фазы развития, паразиты и враги, вредоносное значение и т. д.
Далее отметим публикации Н. А. Виоловича (1956а) по слепням Сахалинской обл.
(с описанием двух новых видов) и его же (1965—1968) по Тувинской АССР и Западной
Сибири. Итоги этих работ, вместе с другими материалами и указаниями литературы,
обобщены автором (1968) в монографии по слепням Сибири. В ней представлены дан-
* В приведенные цифры не включены виды, сведенные позднее в подвиды,
синонимы или неправильно указанные для территории СССР.
12 ВВЕДЕНИЕ
ные по фауне, географическому распространению, сезонному ходу численности,
строению отдельных видов, включая терминалии самок, некоторым сторонам экологии,
вредоносности, а также высказаны отдельные соображения о палеогенезе бореальной
фауны Сибири.
Серия работ по слепням Карельской АССР и некоторых районов Средней Азии
выполнена А. С. Луттой (1950—1970), причем по слепням Карелии автором
опубликована отдельная монография. Изучались видовой состав слепней, географическое
распространение, сезонная и суточная активность, места выплода и обитания, биология
взрослых слепней, включая гонотрофический цикл, преимагинальные фазы развития,
паразиты слепней, меры борьбы и т. д.
Разнообразие направлений в изучении слепней должно быть также отмечено
в многочисленных исследованиях Р. Г. Петровой (Соболевой), начатые в Астраханской
и Московской областях (1955—1956) и продолженные в Приморском крае (1962—1974).
Эти исследования включают данные по фауне, распространению, строению отдельных
видов, сезонной и суточной активности, биологии взрослых слепней, включая
гонотрофический цикл, преимагинальные фазы развития, вредоносное действие на
животных и способы истребления.
Отметим, наконец, серию работ Г. В. Бошко (1950—1968) по слепням Украины,
завершившуюся опубликованием монографии (1973).
Наши публикации послевоенного периода (1949—1972) содержат описания
новых 1 рода, 2 подродов и 12 видов слепней фауны СССР, затем результаты изучения
терминалии самок некоторых видов Hybomitra, данные по фауне слепней Московской,
Волгоградской, Новосибирской областей, Алтайского края, Центрального и
Восточного Казахстана, горного Крыма и Западного Кавказа, некоторые биологические
наблюдения, в том числе по местам обитания личинок, вопросы палеогенеза отдельных
видов, а также результаты испытания некоторых отпугивающих веществ.
Значительное число преимущественно фаунистических исследований выполнено
в послевоенный период в разных местах Советского Союза. Не претендуя на полноту
обзора, отметим следующие, по нашему мнению, наиболее существенные публикации:
О. Н. Сазоновой (1949) и В. В. Попова (1962) по Тюменской обл., М..И. Волковой (1950)
по Татарской и Чувашской АССР, А. В. Богачева и Н. Г. Самедова (1948, 1949) (с
описанием трех новых видов) и Ш. М. Джафарова (1959, 1960) по Азербайджану,
Г. В. Бошко (1950—1968) по Украине и Молдавии, Ю. Г. Митрофановой (1951, 1955)
по Пермской обл., А. А. Штакельберга (1951, 1954) по Таджикистану и Ленинградской
обл., К. С. Сахибзадаева (1955—1958) по Северному Прикаспию, А. Г. Маевского (1956,
1960) по Белоруссии, X. Ремма (1959) по Эстонии, И. Г. Бей-Биенко (1957—1970)
по Белоруссии и Среднему Енисею, В. И. Алифанова (1955) и К. С. Растегаевой (1956—
1962) пд Омской обл., Е. В. Кучерова (1959—1965) по Башкирской АССР, X. А. Ай-
басова и М. С. Шакирзяновой (1958, 1960) по Джунгарскому Алатау и юго-западному
Алтаю, И. М. Олигера (1960, 1967) по Чувашской АССР, Т. Г. Мельниковой (1965) и
Ш. Баратова (1962—;1966) по Таджикистану (с описанием нового вида), Ф. Г.
Булановой (1963, 1967) по Удмуртской АССР, У. С. Куандыковой (1966, 1968) по бассейну
р. Или, В. А. Синелыцикова (1962, 1970) по Павлодарской обл. и Молдавской ССР,
П. А. Чирова (1968а,) Р. В. Гребешок и П. А. Чирова (1971) по Киргизской ССР,
А. Ф. Кателиной и Ю. А. Мясникова (1965) по Тульской обл., 3. М. Маршенкулова
(1967) по Кабардино-Балкарской АССР, В. А. Полякова (1965, 1.967, 1974) по
Таймыру и Магаданской обл., Д. Чарыкулиева (1967) по Туркмении, М. Кадыровой
и Э. И. Ган (1967) по Узбекистану, Т. Т. Васюковой(1967, 1968, 1972) поСреднему При-
ангарыо, Иркутской обл. и Якутской АССР, Ю. И.Запекиной-Дулькейт (1969) по
заповеднику «Столбы» Красноярского края, Ш. Исмаилова (1970) по Дагестану, Л. Н. Гур-
генидзе (1970, 1971, 1972а, 19726) по Грузии, П. П. Иваншцука (1973) по
Ивановской обл. и др.
Как общий итог фаунистических работ представляет интерес отметить, что за
советский период отечественными исследователями было описано 56 новых видов слепней*
фауны СССР и установлены 2 новых рода. Число известных видов доведено до 189.
По биологии слепней и преимагинальвым фазам развития, кроме уже упомянутых
выше, отметим исследования Н. А. Т.амариной (1951, 1956), Т. С. Чуписа (1954),
О. Ф. Колесовой (1966, 1968), П. П. Иванищука (1973), по гонотрсфическому циклу —
Р. В. Колычевой и Н.К. Паенко (1964), Н. К. Паенко (1966, 1968), Р. П. Павловой
(1965, 1972), И. Г. Бей-Биенко (1967), по сравнительной морфологии терминалии
самок — А. А. Штакельберга и А. Е. Тертеряна (1953) и самцов — А. Е. Тертеряна
(1972), по поведению слепней около животных и методам учета — К. А. Бреева (1948,
1950), А. А. Потапова (1961, 1964), А. Е. Тертеряна (1954) и др.
Вредоносное действие слепней на животных, а также вопросы защиты от нападений
этих насекомых изучали К. П. Андреев (1956 — 1966), К. С. Растегаева (1958),
И. Н. Гладенко и В. А. Фортушный (1952), С. Д. Павлов (1965), Н. Л. Симбирцев (1965 ,
* В эту цифру не включен 1 новый вид, описываемый в данной монографии.
СТРОЕНИЕ ВЗРОСЛОГО НАСЕКОМОГО
13
А. И. Черепанов (1966), Г. И. Янович (1958), Г. Н. В0лик(1966), В. А. Поляков (1967,
1974) и др.
Наконец, слепней как переносчиков инфекционных болезней изучали: П. С. Ива-
нова-Гобзем (1945) при трипанозомозе верблюдов, П. Д. Шатько (1952) при
эмфизематозном карбункуле, Г. В. Бошко (1963а) при туляремии, В. Н. Ягодинский (1962)
при клещевом энцефалите, О. Ф. Гробов (1961) при анаплазмозе крупного рогатого
скота, С. А. Аманжулов с соавт. (1965) при Ку-риккетсиозе, А. К. Анищенко и
П. В. Орехов (1967) при чуме свиней, А. Н. Алексеев с соавт. (1969) при листериозе
и т. д.
По слепням защищены 6 докторских и около 30 кандидатских диссертаций; только
в советский период опубликованы 10 монографий и свыше 500 статей.
2. ОБЩАЯ ХАРАКТЕРИСТИКА СЕМЕЙСТВА
Представители семейства Tabanidae на русском языке называются слепнями.
Это название закрепилось в научной литературе в целом за всем семейством и, кроме
того, специально применяется в отношении видов рода Tabanus и близких к нему
родов. Название «слепни» в корне противоречит их зрительным способностям, которые
у них прекрасно развиты. И. А. Порчинский (1915). объяснял его происхождение тем,
что во время сосания крови на животном насекомые теряют осторожность, становясь
как бы «слепыми», и могут быть в это время легко пойманы.
Слепней часто ошибочно смешивают с оводами, от которых первые легко отличаются
уже при поверхностном осмотре своими большими, обычно ярко окрашенными
глазами и наличием колющего хоботка. .
Семейство Tabanidae относят к подотряду короткоусых прямошовных двукрылых
{Diptera Brachycera Orthorrapha). Б. Б. Родендорф (1964) в недавно разработанной им
классификации включил большую часть этого подотряда в установленный им инфраот-
ряд Asilomorpha. Последний объединяет пять надсемейств, в том числе Tabanidea,
куда относится семейство Tabanidae.
Слепни представляют собой по большей части коренастых, средних или крупных
размеров мух, с поделенным на несколько сегментов (до 8) 3-м члеником усиков,
характерным жилкованием крыла (5 заднекрайних ячеек), развитыми закрыловыми
чешуйками (squamae), отсутствием на теле крепких щетинок и наличием между
коготками лапок хорошо развитого эмподия, т. е. срединной присоски, равновеликой
боковым присоскам-пульвиллам. По Б. Б. Родендорфу, надсемейство Tabanidea
объединяет 5 семейств: Tabanidae, Rhagionidae ( = Leptidae), Acanthomeridae (=Panto-
phthalmidae), Coenomyiidae (=Xylophagidae, Erinnidae) и Nemestrinidae. К этому
перечню надо добавить семейство Pelecorhynchidae. Последнее наиболее близко к
Tabanidae и сравнительно недавно выделено из его состава (Mackerras a. Fuller, 1942).
От всех перечисленных семейств Tabanidae отличаются хорошо развитыми
закрыловыми чешуйками. Rhagionidae дополнительно отличаются от слепней строением
3-го членика усиков, концевая часть которого щетинковидная и не поделена на колечки.
Coenomyiidae дополнительно отличаются от слепней наличием шипов на щитке или
жилкованием крыла: жилка гь упирается в вершину крыла, тогда как у Tabanidae
она впадает в косту позади вершины.
Наибольшие отличия можно отметить у Nemestrinidae: у них 3-й членик усиков
имеет щетинковидный конец, а жилки в вершинной половине расположены б. м.
параллельно заднему краю крыла и вливаются в косту впереди от его вершины.
Биологически Tabanidae выделяются среди других семейств надсемейства
Tabanidea тем, что самки большинства видов питаются кровью животпых. Слепни обладают
для этой цели сложно устроенными в виде хоботка ротовыми органами
колюще-режущего типа, посредством которых насекомые прокалывают кожу животных.
Указанная биологическая особенность Tabanidae, однако, не проводит резкой
грани между этим семейством и другими Tabanidea. Например, ряд представителей
сем. Coenomyiidae и Rhagionidae являются во взрослой фазе хищниками — нападают
на мелких насекомых; кроме того, известны некоторые Rhagionidae, которые питаются
кровью млекопитающих. С другой стороны, все самцы Tabanidae растительноядны,
известны также немногие виды слепней, самки которых не сосут кровь позвоночных.
3. СТРОЕНИЕ СЛЕПНЕЙ
ВЗРОСЛОЕ НАСЕКОМОЕ
Внешнее строение
Слепни имеют коренастое тело, относительно короткие ноги, более или менее"
прижатую к груди голову, широкое брюшко и хорошо развитые крылья (рис. 1). Облик
большинства видов свидетельствует об их первоклассных летательных способностях.
U / ВВЕДЕНИЕ
У самок это в известной мере связано с необходимостью разыскивания и преследования
быстро двигающихся животных, у самцов — преследования самок. Для этого же,
очевидно, им нужны хорошо развитые глаза. '
Половой диморфизм у слепней выражен весьма резко. Он присущ
многим двукрылым, но у слепней он усиливается различием в биологии самцов и
кровососущих самок.
Тело слепня обычно несколько сжато дорсовентрально. Хитиновые покровы
сравнительно тонкие, за исключением грудного отдела и передней части головы, где хитин
несколько утолщен. Опушение тела обычно умеренное. Оно сильнее развито
у бореальных и высокогорных форм и слабее у южных, особенно пустынных видов.
В окраске тела слепней преобладают блеклые тона — бурые, серые, желтые.
Бореальные и особенно горные формы обычно имеют темную окраску, наоборот,
южные и особенно пустыппыс виды характеризуются светлой окраской. Но темная, иногда
Рис. 1. Строение § слепня (схема), г — голова; у — усики; гр — грудной отдел
(спинка); нт — нотоплевра; щ — щиток; к — крыло; ж — жужжальце;. их,
и2) мз — передняя, средняя и задняя нога; бр — брюшко (1—7 его сегменты).
угольно-черная, окраска имеется у южных форм слепней, например, почти у всех
видов Dasyrhamphis, некоторых Pangonius и Haematopota фауны Средиземноморской
подобласти, не связанных обитанием с горами.
Следует учитывать, что у старых музейных экземпляров может произойти
выцветание хитина — черный цвет сменяется на коричневый и даже буро-желтый.
Выцветание наблюдается у мозолей на лбу и на лице, а также ног, особенно бедер, крыльев
и т. д.
В общей массе слепни довольно крупные мухи, в среднем от 10 до 20 мм дл. В
нашей фауне наиболее крупным является Tabanus chrysurus, достигающий 30 мм дл.,
и самой мелкой — дождевка Haematopota koryoensis, наименьшие размеры тела
которой 6 мм. Оба вида распространены на Дальнем Востоке.
Голова (рис. 2) большая, поперечная, спереди выпуклая, равная в ширину груди
или шире ее. У самцов чаще голова бывает крупнее, нежели у самок, и в таком случае
она имеет форму полушария. Сложные фасетчатые глаза занимают по
бокам большую часть головы, при жизни красиво и ярко окрашены в различные цвета
с металлическим оттенком: зеленый, коричневый, пурпурный, синий и т. д. Часто на
глазах располагаются более темные пятна или поперечные полоски, число и
расположение которых имеют известное таксономическое значение. У сухих экземпляров
раскраска глаз и полоски исчезают, глаза становятся тускло-черными. Но окраска может
быть восстановлена путем размачивания во влажной камере. У видов рода Atylotus
глаза бледной окраски — желтоватые, сероватые или зеленоватые, у сухих
экземпляров она обычно темнеет лишь немного.
Фасетки глаз у самок б. м. одинаковой величины, тогда как у самцов часто
фасетки верхних частей глаз значительно крупнее нижних, с резкой границей между
теми и другими. На юге преобладают виды с резким делением глаз на крупные и
мелкие фасетки, наоборот, в бореальном климате или в высокогорных областях .чаще
встречаются виды с слабо различающимися по величине фасетками. У некоторых ви-
СТРОЕНИЕ ВЗРОСЛОГО НАСЕКОМОГО Ц
дов слепней глаза бывают совершенно голые, тогда как у других покрыты короткими,
густыми волосками. Для того чтобы лучше выявить этот признак, необходимо глаза
слепня рассматривать в лупу сбоку против света. Волоски сидят между фасетками
перпендикулярно к поверхности глаза. Опушение на глазах сильнее развито у самцов,
нежели у самок. Помимо сложных глаз, у многих слепней (Chrysops, Silvius, Pangonius)
на темени имеются простые глазки (ocelli) в количестве трех; эти глазки
обычно расположены треугольно на плоском возвышении или бугорке. У других
слепней (Hybomitra) глазки редуцированы и заменены небольшим глазков ым
бугорком. Наконец, у ряда видов слепней (Haematopota, Tabanus и др.) темя лишено
каких-либо образований. У самок сложные фасетчатые глаза разделены лобной
л г/1 см
Рис. 2. Голова спереди § Hybomitra. г —
сложный фасетчатый глаз; гл — глазковый бугорок;
л — лобная полоска; см — средняя лобная
мозоль; нм — нижняя лобная мозоль; лтп —
лобный треугольник; у — усиковая ямка (усики
удалены); к — наличник; щ — щека; х —
хоботок; щп — щупальце (концевой членик).
полоской, или лбом, тогда как у самцов глаза соприкасаются; по этому признаку
' самцы легко отличаются от самок. Лобная полоска самки покрыта матовым налетом и
обычно несет отграниченные участки б. м. выпуклого и блестящего хитина —
лобные мозоли. В нижней части лба расположена нижняя лобная
мозоль, над ней часто бывает развита средняя лобная мозоль. У
различных видов слепней форма, окраска мозолей весьма разнообразны, что имеет важное
значение для видовой таксономии. Задняя сторона головы, включая узкую полоску .
позади глаз, носит название затылка. Верхний край затылка, называемый также
затылочной полоской, иногда бывает снабжен рядом длинных волосков, выступающих
над поверхностью глаз.
Усики (рис. 3) прикрепляются спереди к центральной части головы и имеют
вид двух торчащих вперед рожков. Они сближены основаниями, расположены обычно
на плоскости, в месте прикрепления каждого усика имеется небольшая, уплощенная .
усиковая ямка. Верхний край усиковых ямок обычно узкий, но у некоторых
Tabanus, реже Hybomitra, сильно расширен, составляя более трети высоты лобного
треугольника. Длина усиков различна, чаще они короче передне-задней ширины головы, ,
но известны виды, например Heptatoma, с более длинными усиками. Усики явственно
поделены на 3 членика. 3-й (т. е. концевой) членик усиков в свою очередь поделен на .
несколько сегментов, или колец, причем базальный сегмент обычно бывает
наиболее развит и носит наименование площадки, тогда как меньшие концевые
сегменты в совокупности образуют палочку (рис. 3, п). Строение усиков имеет ;
большое значение для надродовых и родовых группировок (рис. 53),. Так у Pangonii-
16
ВВЕДЕНИЕ
пае, например, 3-й членик поделен на 8 сегментов, постепенно уменьшающихся к концу.
Подобное строение несомненно нужно считать наиболее древним. У Chrysopsinae' и
Tabanint 3-й членик усиков состоит из 5 сегментов: 1-й членик в виде хорошо развитой
площадки, тогда как 4 концевых сегмента образуют палочку. Площадка образовалась
в результате слияния первых четырех сегментов. У Tabanus площадка в основании
обычно сильно дорсовентрально расширена и несет на верхней стороне
дорсальный угол; иногда угол настолько резкий, что он имеет вид выступа или острого
зубца. У Haematopota укороченная площадка срослась с вытянувшимся I сегментом
палочки, так что палочка стала трехчлениковой; границу этого срастания можно
обнаружить у многих видов благодаря наличию в чтом месте волосков, но особенно
хорошо она заметна у И. desertorum. Наиболее уклоняющиеся усики у Heptatoma: они шну-
ровидные, их 3-й членик поделен на 4 сегмента настолько резко, что усики легко могут
быть приняты за шестичлениковые.
Усики у слепней имеют довольно разнообразную окраску, используемую для
видовых различий. У самок усики всегда несколько более развиты, нежели у самцов.
Но у самцов волоеки на 1-м и 2-м члениках, особенно сверху, обычно бывают несколько
длиннее.
Рис. 3. Усик Tabanus. 1—3-й членики усика;
п — палочка; д — дорсальный угол (или
выступ).
Над местом прикрепления усиков расположен лобный треугольник.
У самок последний имеет форму трапеции и вершиной граничит с основанием лобной
полоски. Лежащая ниже места причленения усиков часть головы носит название
лица. Последнее состоит из слегка выпуклой средней части —.наличника и
резко отграниченных от него глубокими бороздками щек. Обычно у слепней лицо
покрыто матовым налетом и волосками, но у некоторых видов (Chrysops, Silrius, Pango-
reius)-Ha лице имеются хорошо'развитые участки выпуклого и блестящего хитина —
лицевые мозоли (рис. 70). У немногих видов налет может совершенно
отсутствовать, и в таком случае лицо целиком блестящее, как бы покрыто лаком.
Снизу к голове причленяются хорошо заметные ротовые органы, имеющие
вид массивного хоботка (см. стр. 21). Обычно длина хоботка не превышает высоту
головы, но у слепней Pangoniinae последний часто сильно развит и бывает равен
половине длины тела (рис. 56) и более. В основании к хоботку прилегают
нижнечелюстные щупальца, по одному с каждой стороны. Последние состоят
из двух члеников, причем у самцов концевой членик короткий, конусовидный,
яйцевидный или шарообразный, .тогда как у самюгон вытянут и заострен на конце.
Грудь широкая и массивная, с большим треугольным щитком позади. Как
и у всех двукрылых, она состоит из неясно разграниченных между собой трех колец:
передне-, средне- и заднегруди, из коих среднегрудь развита наиболее сильно. На
спинной стороне в передних углах бороздкой отграничены плечи. Кроме того, кпереди от
места прикрепления крыльев расположены хорошо заметные нотоплевры, по
одной с каждой стороны, представляющие собой треугольные образования на боках
спинки. От внутренних углов нотоплевр к центру спинки направляется с каждой
стороны по бороздке, исчезающей возле середины.
Боковые стороны груди (плевры) состоят из отдельных склеритов, из них наиболее
крупный носит название мезоплевры. По бокам передне- и заднегруди
расположены хорошо заметные, крупные стигмы.
Грудной отдел обычно покрыт налетом и б. м. густыми и тонкими волосками;
в таксономии некоторую роль играет их окраска, тогда как число и расположение не
имеют существенного значения.
К боковым сторонам среднегруди причленены крылья в числе 1 пары. У слепней
крылья широкие, с обильным, но б. м. однообразным в пределах семейства жилкованием
(рис. 4). Последнее хорошо удается рассмотреть в лупу при небольших увеличениях.
Различают следующие жилки: обходящую наружный край крыла костальную
жилку (е), затем субкостальную (sc), 4радиальных (гх, г2+3, г4, гь), 3
медиальных (ту— т3), 2кубитальных (сиу—си2) и 1 анальную (are). Между
СТРОЕНИЕ ВЗРОСЛОГО НАСЕКОМОГО 17
5-й радиальной (4-й по счету) и 1-й медиальной имеется короткая поперечная жилка,
носящая наименование радиомедиальной (гт). Кроме того, третья по счету
радиальная жилка (г4) в месте изгиба часто имеет небольшую, направленную в сторону
основания крыла придаточную жилку, или сокращенно — придаток г4.
Последний встречается независимо в самых различных группах слепней. В одних
случаях он постоянен и используется для диагностики, в других изменчив, и бывают
Рис. 4. Крыло Atylotus. Жилки: с — костальная, sc— субкостальная,
г — радиальные, т — медиальные, гт — радиомедиальная, си — ку-
битальные, an — анальная, ар — придаток г4. Ячейки: С — костальная,
S — субкостальная, R — радиальные, Р — заднекрайние, D — дискои-
дальная, В — базальные (Вг передняя и В2 задняя базальная ячейка),
А — анальная.
случаи, когда у одной и той же особи на одном крыле придаток г, имеется, на другом
отсутствует.
Перечисленные жилки ограничивают на крыле следующие ячейки:
костальную (С), субкостальную (Sc), 3 р а диальных (R±—R3), 2 б а з а л ь-
н ы х (передняя и задняя базальные ячейки) (Вх—В2), дискоидальную (D),
5 заднекрайних (Рг—Р5) и анальную (А).
В основании костальной жилки расположен небольшой отдельный склерит —
6 азикоста (рис. 5), опушение которой имеет важное значение для надродовой
Рис. 5. Основание крыла Philipomyia rohdendorfi (слева) и Tabanus su-
deticus (справа), т—тегула; б — базикоста; к—основание костальной
жилки.
таксономии (Philip, 1941). У Pangoniinae и Chrysopsinae базикоста голая (имеются
лишь микротрихии, видимые при сильном увеличении), у Tabanini и Haematopotini
она покрыта короткими волосками (щетинками), такими же, как основание (рукоятка)
костальной жилки. Крепление крыла к груди осуществляется при помощи небольшого
склерита — т е г у л ы.
В основании крыла снизу имеется хорошо заметное крылышко (alula) и пара
закрыловых пластинок, или чешуек (squamae). Последние хорошо
развиты.
У различных видов слепней крылья могут быть бесцветные или с темными
перевязями, пятнами или сетчатым рисунком (мраморные). Отличается способ складывания
крыльев, когда муха сидит спокойно. У Chrysopsinae крылья расставлены довольно
широко в стороны, но поверхность их находится в горизонтальной плоскости. У Та-
ЬапЩ крылья менее расставлены и поверхность располагается несколько наклонно.
У Haematopotini крылья складываются кровлеобразно вдоль, тела.
2 Н. Г. Олсуфьев
18
ВВЕДЕНИЕ
, Позади основания крыльев на груди расположены жужжальца, рудименты
задней пары крыльев. Жужжальца состоят из узкого, короткого стебелька и
булавовидной головки и окрашены в белый, желтый или черно-коричневый цвет.
Снизу к груди причленяются ноги (рис. 6). Последние умеренной длины,
покрыты волосками, без грубых щетинок. Каждая нога состоит из следующих частей,
считая от основания: тазик (утолщенный членик, соединяющий ногу с грудью),
затем в е р т л у г (маленький, плохо заметный членик), бедро, голень и пяти-
члениковая лапка. На конце последнего членика лапки имеется пара крепких к о-
г о т к о в и под ними три хорошо развитые б. м. равновеликие присоски: боковые —
пульвиллы и между ними —эмподий. У SS некоторых видов Hybomltra
(например, в группе Н. kaurii) на передних лапках внутренний коготок и
соответствующая ему пульвилла значительно укорочены (рис. 238). Иногда передние лапки на
дорсальной стороне несут длинные щетинки (Я. mi-
cans и др.) (рис. 266).
Обычно голени средней пары ног на самом конце
бывают снабжены парой сидящих рядом крупных
щетинок или шипов — так называемых шпор. Эти
же шпоры у ряда видов слепней имеются и на задних
голенях. Этот признак требует внимательного
рассмотрения и весьма существен, так как
характеризует подсемейства Pangoniinae и Chrysopsinae.
Брюшко широкое, сжато сверху вниз, его ба-
зальная или основная часть состоит из 7 пар
видимых полуколец: верхних тергитов и
нижних ■— стернитов. Впередилежащие
полукольца налегают своим задним краем на последующие.
Они соединены между собой интерсегментальной
перепонкой, которая способна растягиваться при
насыщении самки слепня кровью. На границе
полуколец, на интерсегментальной перепонке расположены
брюшные стигмы, по одной с каждой стороны
кольца (при наружном осмотре стигмы не видны).
У самцов брюшко на конце имеет коническую
форму, у самок — округлую. Терминальная или
концевая часть брюшка (сокращенно —
терминалии) обычно втянута в VII сегмент, она образована
4 концевыми сегментами (VIII—XI),
функционально модифицированными в дистальные части
Рис. 6. Задняя нога Chrysops. б.— бедро; г —
голень; л — лапка; к — коготок; п — пульвилла;
э — эмподий; ш — шпоры.
половых органов и анальные придатки. Описание их представлено ниже. Окраска
брюшка, играющая большую роль при определении ряда видов, состоит обычно
из темного (черного) фона, с более светлыми краспо-жслтыми боковыми
пятнами, в той или иной мере вытесняющими темную окраску. Рисунок дополняется
налетом и коротким волосяным покровом, образующим в комбинации каймы, ряды
различной формы пятен и т. д. Брюшко у самцов обычно окрашено темнее, чем у самок.
Окраска брюшка в одних группах довольно постоянна, например у Chrysops, в
других группах, например у Tabanus, она изменчива, причем у видов с широким
распространением южные популяции окрашены значительно светлее, чем северные (или
горные).
Терминалии брюшка составляют его концевую часть, у самок они обычно не видны,
у самца они часто выступают наружу. Строение терминалии слепней используют для
видовой и надвидовой таксономии (Collin, 1940; Штакельберг и Тертерян, 1953; Macker-
ras, 1954; Шевченко, 1960, 1961; Олсуфьев, 19626; Тертерян, 1972). В этом случае
обозначение «терминалии», принятое за рубежом, правильнее, чем «гениталии», поскольку
для диагностики учитывают также анальные придатки.
У самки дорсальная сторона терминалии (рис. 7) образована поперечным,
хорошо развитым VIII тергитом, двумя небольшими парными склеритами — IX и Хтер-
гитами, поделенными по продольной оси надвое (у Pangoniinae IX тергит цельный),
заканчивающимися парными церками. Парные склериты IX тергита чаще широко
расставлены, тогда как склериты X тергита обычно соприкасаются по медиальной линии.
XI тергит слит с церками и морфологически не выражен. Церки прикрывают сверху
анальное о .т в е р с т и е, они б. м. плоские и обычно имеют округлые очерта-
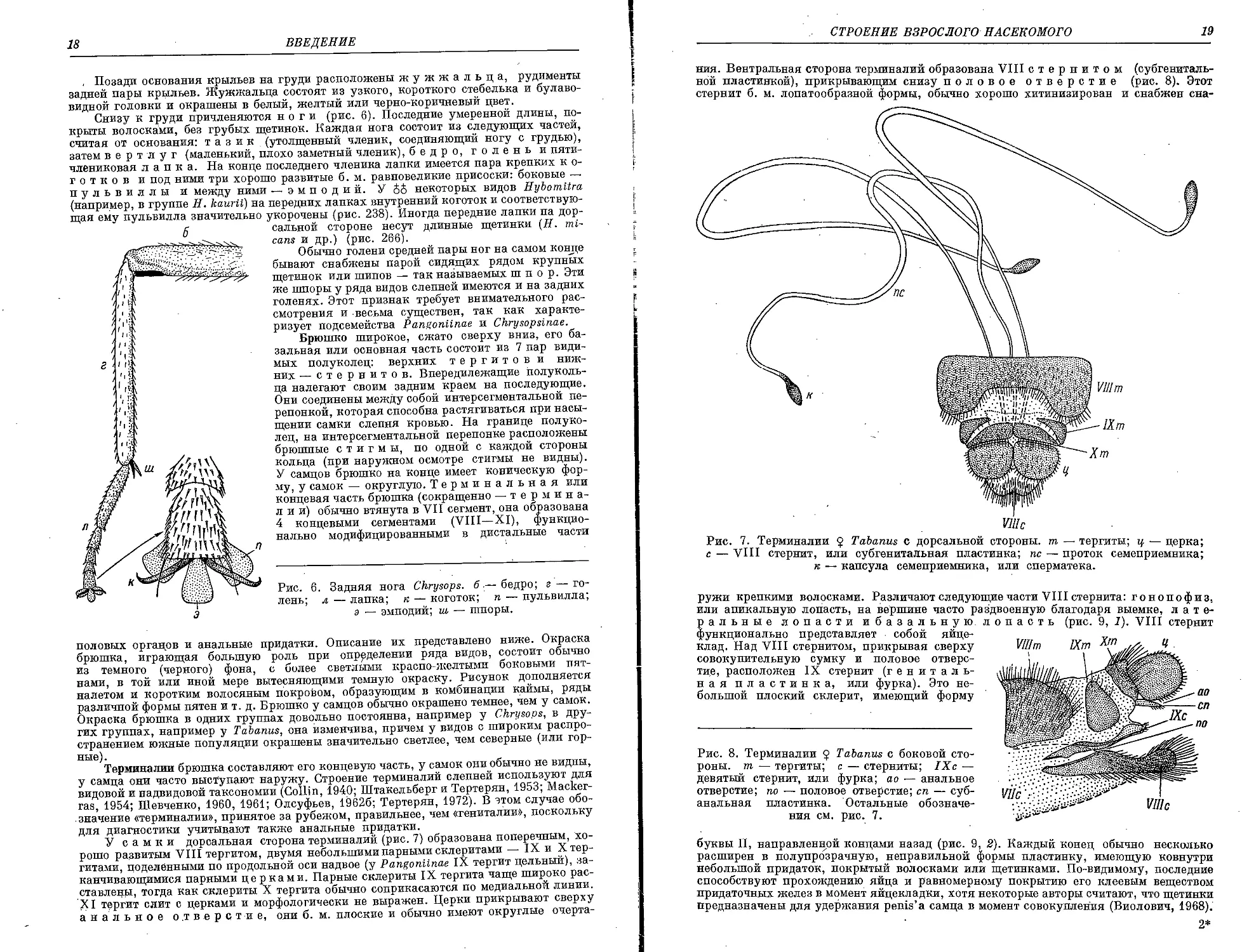
СТРОЕНИЕ ВЗРОСЛОГО НАСЕКОМОГО 19
ния. Вентральная сторона терминалии образована VIII стернитом (субгениталь-
ной пластинкой), прикрывающим снизу половое отверстие (рис. 8). Этот
стернит б. м. лопатообразной формы, обычно хорошо хитинизирован и снабжен сна-
VJlIc
Рис. 7. Терминалии § Tabanus с дорсальной стороны, m — тергиты; ц — церка;
с — VIII стернит, или субгенитальная пластинка; пс — проток семеприемника;
к — капсула семеприемника, или сперматека.
ружи крепкими волосками. Различают следующие части VIII стернита: гонопофиз,
или апикальную лопасть, на вершине часто раздвоенную благодаря выемке,
латеральные лопасти ибазальную. лопасть (рис. 9, 1). VIII стернит
функционально представляет собой
яйцеклад. Над VIII стернитом, прикрывая сверху
совокупительную сумку и половое
отверстие, расположен IX стернит (г е н и т а л ь-
ная пластинка, или фурка). Это
небольшой плоский склерит, имеющий форму
Рис. 8. Терминалии § Tabanus с боковой
стороны, m — тергиты; с — стерниты; 1Хс —
девятый стернит, или фурка; ао — анальное
отверстие; по — половое отверстие; сп —
субанальная пластинка. Остальные
обозначения см. рис. 7.
буквы П, направленной концами назад (рис. 9, 2). Каждый конец обычно несколько
расширен в полупрозрачную, неправильной формы пластинку, имеющую ковнутри
небольшой придаток, покрытый волосками или щетинками. По-видимому, последние
способствуют прохождению яйца и равномерному покрытию его клеевым веществом
придаточных желез в момент яйцекладки, хотя некоторые авторы считают, что щетинки
предназначены для удержания penis'a самца в момент совокупления (Виолович, 1968).
2*
SO ВВЕДЕНИЕ
Последнее объяснение сомнительно, если учесть, что у самца имеется значительно
более эффективное приспособление — гоностили для фиксации гоноподита в момент
спаривания.
VIIIc
1 2 3
Рис. 9. Детали строения терминалий $ Tabanus. 1 — VIII стернит (субгени-
тальная пластинка) с внешней стороны: г — гонопофиз, бл — базальная,
лл — латеральная лопасть; 2 — генитальные придатки: 1Хс — IX стернит
(фурка), прилегающий изнутри к VIII стерниту, м — муфта, пс — проток
семеприемника; 3 — анальные придатки (с вентральной стороны).
Остальные обозначения см. рис. 7—8.
В базальной части IX стернита с его дорсальной стороны прикреплены 3 с е м е-
приемника, ав апикальной части — 2 клеевых железы. Все 3
семеприемника одинаковы по величине и каждый из них представляет собой длинную,
Рис. 10. Терминалий 6 Tabanus с дорсальной
стороны, тп — тергиты; с — IX стернит, или
гоноподит; ц — церка; са — субанальная
пластинка; IX-{-Хт — эпандрий.
тонкую, согнутую пополам полупрозрачную трубку — семепроток,
оканчивающуюся на свободном конце продолговатой хитинизированной капсулой — сперма-
т е к о й. В базальной части семепроток снабжен хитинизированной муфтой
{рис. 9, 2). Обычно семеприемники располагаются в пределах VI и VII колец брюшка,
но, у некоторых видов (например, Philip omyia} они очень длинные и, их согнутая часть
Рис. 11. Терминалий в Tabanus Рис. 12. Терминалий в Tabanus, вид сбоку,
с вентральной стороны. Обозначения Обозначения см. рис. 10.
см. рис. 10.
Рис. 13. IX стернит (гоноподит) в Tabanus Рис. 14. Ротовые органы (in situ)
с дорсальной стороны (анальный сегмент уда- § Hybomitra. вг — верхняя губа;
лен), гк — гонококсит; гс — гоностиль; ф — нг — нижняя губа; щ — щупальце,
фаллус; пщ — придаточные щетинки; э — концевой членик; н — наличник,
чехол фаллуса, или эдеагус; ск — семеизвер-
гательный канал.
22 ВВЕДЕНИЕ
(петля) достигает II—III кольца. Над концевой частью IX стернита расположена
субанальная пластинка (X+XI стерниты), образованная двумя небольшими,
треугольной формы, слабо хитинизированными склеритами, соединенными в паре по
медиальной линии (рис. 9, 3).
Для видовой таксономии используют главным образом строение VIII стернита
и церок, для надвидовой систематики, помимо VIII стернита, важное значение Имеет
строение IX тергита, семеприемников и т. д.
У самца терминалии образованы теми же склеритами, что и у самки, но они
имеют несколько иную форму (рис. 10, 11, 12). VIII тергит представлен более узкой,
поперечной, изогнутой пластинкой. IX и X тергиты, так же как у самки, обычно
поделены по продольной оси пополам, но склериты IX тергита присоединены к таковым X
тергита, иногда граница между ними не выражена. Оба склерита вместе носят
название эпандрия. У Pangoniinae IX и X тергиты в виде цельной хорошо развитой
пластинки. Церки примерно как у самки.
На вентральной стороне терминалии расположен хорошо развитый трапециевидный
VIII стернит, за ним следует гоноподит (рис. 13). Последний представляет собой
видоизмененный IX стернит и сильно хитинизирован. От базальной хорошо развитой
части стернита по бокам отходят два каудально направленных пальцевидных выроста,
носящих наименование гонококситов или вальв. Последние целиком
срослись с стернитом. Каждый гонококсит на конце вооружен коротким, подвижным г о н о-
стилем или 2-м члеником вальв. Эти членики в паре представляют собой
щипцы, и их назначение защемлять конец брюшка самки в момент спаривания. В
средней части гоноподита дорсально расположен ф а л л у с, или penis, он хитинизирован
и имеет форму короткого кинжала. По бокам фаллуса, несколько дорсально, причле-
нена пара длинных, перекрещивающихся концами, крепких придаточных щетинок.
Фаллус и его придатки прикрыты особым хитиновым чехлом (эдеагусом),
образованным парой соединенных между собой боковых пластинок. Этот чехол имеет
форму конуса. Над гоноподитом расположена продолговатая субанальная
пластинка (X+XI стерниты), образованная парой соединенных по медиальной
линии удлиненных склеритов.
Строение терминалии самцов используют главным образом в надродовой
таксономии.
Строение ротовых органов слепня
Ротовые органы (рис. 14) слепня колюще-лижущего типа, причленены к нижней
стороне головы и имеют вид хоботка. Они слагаются из массивной, мягкой, темно-
окрашенной нижней губы, в жолоб которой вложен крепкий, хитиновый ко-
нч вч вг п иг еч . нч
Рис. 15. Ротовые органы в расчлененном виде § Tabanus. вг — верхняя губа+надгло-
точник; п — подглоточник; вч — верхняя челюсть; нч — нижняя челюсть; щ —
щупальце, концевой членик; нг — нижняя губа.
люще-режущий аппарат. Нижняя губа на свободном конце разделяется на две
широких лопасти (labellae), которые с внутренней стороны изборождены желобками,
лежащими параллельно друг другу. Колюще-режущий аппарат состоит (рис. 15) из
СТРОЕНИЕ ВЗРОСЛОГО НАСЕКОМОГО 23
шести тонких, стилетообразных, коричневоокрашенных пластинок, образующих
сосальце: желобчатой верхней губы — сросшейся с надглоточником и
прикрепленной основанием к наличнику, затем пары саблевидных или ножевидных верхних
Рис. 16. Внутренние органы § Tabanus. х — хоботок; еж — слюнные
железы; п — пищевод; а — зоб; пж — преджелудок; ж — желудок; м —
мальпигиевы сосуды; к — кишка; р .— ректум.
челюстей, пары тонких, в поперечном сечении трехгранных нижних
челюстей, снабженных двучленистыми нижнечелюстными
щупальцами, и узкого пятигранного языка или подглоточника. Последний на
всем протяжении пронизан слюнным протоком. Вершинная треть внутрен-
84
ВВЕДЕНИЕ
него края верхних челюстей, шиловидные концы нижних челюстей и кончик верхней
губы тонко зазубрены назад (рассматривать под микроскопом).
Рис. 17. Внутренние половые органы § Tabanus. я — яичник;
яв — яйцевод; с — семеприемник; кж — клеевая железа.
Колюще-режущий аппарат служит для прокалывания кожи животных и сосания
крови, тогда как нижней губой слепень подлизывает различные сладкие соки или
утоляет жажду.
У самцов верхние челюсти редуцированы,
что находится в связи с особенностями их
питания только растительными соками.
Внутреннее строение слепня
В общий чертах внутренняя анатомия
слепней имеет много общего с прочими
двукрылыми. Пищеварительный тракт (рис. 16)
начинается ротовымотверстием,
лежащим между основаниями надглоточника и
подглоточника. Ротовое отверстие переходит
в короткую глотку, окруженную сложной
системой мышц, насасывающих и
перегоняющих у самки кровь через следующий за ней
короткий пищеводвжелудок. В месте
соединения желудка с пищеводом в
последний открывается зоб, имеющий вид тонкой
трубки, на свободном конце расширенной в
двойной резервуар. Зоб служит резервуаром
для воды и сладкой растительной пищи,
откуда жидкость поступает затем в желудок.
Последний довольно длинный, состоит из двух
отделов: преджелудка и собственно ж е-
Рис. 18. Внутренние половые органы <J
Tabanus. с — семенник; ж — придаточная
железа; ск — семеизвергательный канал; ф —
фаллус; 1Хс — гоноподит.
л у д к а. Преджелудок имеет форму длинной трубки с рыхлыми стенками,
расширенной на переднем конце в два плоских крыла. Желудок представляет собой объемистый
мешок, растягивающийся от поступающей в него крови в несколько раз. В желудке
имеется очень тонкая перитрофическая мембрана, которая у самки отделяет
поступающую кровь от стенки желудка. Желудок на заднем конце переходит в з а д н ю ю
кишку; на их границе впадают 4мальпигиевых сосуда, соединенных
концами попарно. Задняя кишка переходит затем в короткую и расширенную
прямую кишку, снабженную шестью ректальными железами.
Анальное отверстие открывается на заднем конце брюшка и расположено
непосредственно над половым, они разделяются субанальной пластинкой.
ФАЗЫ РАЗВИТИЯ
25
В груди вентрально расположены парные слюнные железы, имеющие вид
прозрачных б. м. прямых трубок; через короткий общий слюнной проток
железы открываются в гипофаринк. (подглоточник). У самцов слюнные железы сильно
редуцированы и имеют вид очень тонких трубок.
В полости тела дорсально расположен сердечный сосуд,
представляющий собой тонкую, желтоватоокрашенную трубку. Задним концом сосуд прикреплен
на уровне границы VI и VII тергитов, тогда как его передний конец открывается в
голове; Центральная нервная система представлена крупным г о-
ловоглоточным ганглием, расположенным в. голове, и идущей от него
нервной цепочкой из шести сильно сближенных узлов. Цепочка лежит
вентрально в грудной полости и частью в первых двух брюшных кольцах; от нервной
цепочки во все стороны расходятся многочисленные нервные нити. В грудной полости
расположены мощные летательные мышцы, занимающие большую часть
ее объема.
Трахейная система представлена продольными трахейными
стволами, от которых отходят более мелкие боковые трахеи, пронизывающие всю
полость тела. Особенно густо оплетены трахеями желудок и половые органы самки.
В передней части брюшка лежит пара крупных воздушных мешков.
Внутренние органы, в особенности находящиеся в брюшной полости (кишечник,
мальпигиевы сосуды, половые органы), оплетены тонким слоем жирового тела,
строение которого слегка напоминает кружева.
Внутренние половые органы самки (рис. 17) обычного для
двукрылых строения. Бросаются в глаза довольно крупные, овальной формы
яичники, состоящие из множества яйцевых трубок с включенными в них яйцевыми
клетками. От яичников отходят короткие яйцепроводы, сливающиеся затем
в общий широкий яйцевод. Последний оканчивается половым
отверстием. В дистальную часть яйцевода, образованную IX стернитом, открываются
три семеприемника и несколько ниже — пара клеевых желез.
Последние сильно развиты, имеют вид малопрозрачных изогнутых трубок. Их окраска
варьирует от белых до желтых и коричневых цветов в зависимости от видовой
принадлежности.
Внутренние половые органы самца устроены сравнительно
просто (рис. 18). От продолговатых семенников отходят довольно тонкие
относящие протоки, которые далее впадают в небольшой резервуар. Этот резервуар
двойной, он выстлан довольно крупными клетками железистого характера и соответствует
придаточным железам прочих двукрылых. От резервуара отходит короткий парный
семеизвергательный канал, который далее сливается в один общий,
более узкий проток, открывающийся наружу на конце фаллуса.
ФАЗЫ РАЗВИТИЯ
Яйцо. Яйца откладываются слепнями в компактные кучки — в виде
пирамидки или округлой бляшки на листья и стебли различных растений (рис. 19). Обычно
кладки состоят из трех-четырех слоев склеенных между собой яиц, расположенных
один над другим (рис. 20), но у некоторых видов кладки бывают однослойными
(некоторые Chrysops, Hybomitra, Haematopota). Количество яиц в среднем 500—600, но
может доходить до 1000. Яйцо слепня длинное, узкое, цилиндрическое или слегка
суженное к переднему концу. На переднем конце сверху, на заднем снизу яйцо слегка
косо срезано и тупо закруглено. Размеры яйца зависят от вида, но для средней величины
слепней из рода Tabanus длина.составляет 1.8—2 мм, толщина 0.3—0.4 мм. Оболочка
яйца гладкая, бесструктурная, блестящая. Окраска колеблется в зависимости от
вида от буро-серых до угольно-черных цветов; только что отложенные яйца молочно-
белого цвета, но вскоре затем темнеют, принимая нормальную окраску. У некоторых
Tabanus яйца остаются белыми.
Личинка. Тело личинки (рис. 21) узко веретеновидное, с заостренными обоими
концами, в поперечном сечении круглое или слегка дорсовентрально сжатое, состоит
из 12 явственно разделенных сегментов и головы. Размеры личинок колеблются в
зависимости от возраста и вида слепня. У средней величины Tabanus только что
вылупившаяся личинка имеет длину 2.5—2.7 мм, при толщине 0.4—0.5 мм, тогда как
взрослая личинка достигает длины 25—30 мм, при толщине 4—5 мм. У крупных Tabanus
взрослые личинки достигают длины до 48 мм.
Окраска тела, личинки белая, бледно-зеленая, бурая или розово-коричневая,
часто имеются более темные поперечные кольцевые полосы или пятна, образующие
рисунок. У живой личинки сквозь кутикулу ясно просвечивают темный головной
скелет, оплетающее внутренние органы жировое тело, а с дорсальной стороны —
центральные трахейные стволы.
Тело покрыто упругой кутикулой, поделенной на голые и волосистые
участки; последние покрыты мелкими, густо расположенными волосками и опоясывают
26
ВВЕДЕНИЕ
сегменты тела. Голые участки обычно имеют продольно исчерченную (гофрированную)
поверхность, придающую кутикуле большую упругость. На границе брюшных
сегментов, по их переднему краю, расположены валикообразные утолщения и довольно
крупные, мягкие бородавчатые выступы, служащие для передвижения личинок.
Эти бугорки и утолщения покрыты короткими шипиками. На предпоследнем кольце
снизу у его основания расположено анальное отверстие, которое имеет
вид продолговатой щели с двумя полушаровидными утолщениями по бокам. Последнее
кольцо представляет собой узкую дыхательную трубку и несёт на конце
пару соединенных вместе стигм, расположенных в общей продольной щели. На
переднем конце тела, на границе I и II
колец, по бокам имеется еще пара
передних стигм, по одной с каждой стороны,
но эти стигмы очень сильно редуциро- ь '
Рис. 19. $$ Tabanus autumna- Рис. 20. Кладки яиц Hybomitra
lis, откладывающие яйца на на растениях,
растения.
Голова (рис. 22) личинки вытянута в трубку и сильно хитинизирована. На ее
переднем конце располагается ротовой аппарат, который состоит из
следующих частей: крючковатой, сильно сжатой с боков верхней губы, пары тонких,
изогнутых и сильно заостренных верхних челюстей, пары более широких
и менее хитинизированных нижних челюстейи раздвоенной на конце
нижней | г у б ы. По бокам головы возле основания нижних челюстей расположены
короткие усики, а ниже их более длинные двучлениковые
нижнечелюстные щупальца. Между основанием усиков и верхней губой имеется с каждой
стороны головы площадка, покрытая густо сидящими шипиками или щетинками.
В основании нижней губы имеется пара одночлениковых нижнегубных щ у п а-
л е ц. По бокам головной трубки в ее передней трети расположены простые
г л а з а в виде черных точек. Голова личинки очень подвижна и может поворачиваться
по продольной оси на 90° в ту или иную сторону, а также целиком втягиваться в
последующие кольца тела.
В спокойном состоянии ротовые органы (челюсти) у личинки втянуты в головной
конец, а обе площадки с щетинками ввернуты в особые складки кожи (рис. 22, А).
При нападении личинка резким движением выбрасывает челюсти вперед и вниз;
последние описывают полукруг и вонзаются в тело намеченной жертвы (рис. 22, В). При этом
челюсти издают характерный треск. Правая и левая пара челюстей могут выбрасываться
каждая порознь или совместно. Одновременно с челюстями выворачиваются наружу
ФАЗЫ РАЗВИТИЯ 87
площадки с щетинками, которые в вывернутом состоянии, вместе с челюстями,
образуют своеобразный якорь, посредством которого личинка сцепляется с телом своей
жертвы. У крупных личинок (например, Tabanus bovinus) сила удара челюстей такова,
что может причинить достаточно' болезненное ощущение коже ладони.
Личинки различных видов отличаются по величине, окраске тела, строению
головы, форме дыхательной трубки, размерам и числу бугорков на теле и т. д. Следует,
однако, иметь в виду, что систематика личинок слабо разработана, и поэтому наиболее
надежным для определения пока является выведение из них взрослой фазы.
Внутреннее строение личинок в основном следующее (рис.23): на переднем конце
головы, между верхней и нижней губой расположено ротовое отверстие,
которое ведет через хитинизированную г л о т к у в довольно длинный и тонкий п и-
щ е в о д. За пищеводом следует муфтообразный преджелудок,
представляющий собой сложно устроенный п и л о-
рический клапан. Далее следует
длинный желудок, поделенный пополам
перетяжкой в виде узкой трубки. Желудок
на заднем конце переходит в т о н к у ю
к и ш к у, на их границе впадают 4 м а л ь-
пигиевых сосуда. Последние
имеют вид длинных, извитых, тонких
трубок, окрашенных в желтовато-коричневый
цвет. Тонкая кишка переходит в более
широкую и довольно длинную прямую
кишку, заканчивающуюся анальным
отверстием.
В передней части тела расположены
очень длинные слюнные железы.
Они лежат по бокам желудка и имеют вид
полупрозрачных, извитых трубок,
согнутых пополам. Слюнные железы на
переднем конце переходят в длинные, тонкие
слюнные протоки, которые затем
сливаются в общий проток, открывающийся
в ротовую полость на нижней губе.
Органы пищеварения личинок слепня
на всем протяжении от заднего конца
пищевода до анальной части тела заключены
в особый длинный мешок, стенки которого
состоят из тонкого слоя жирового тела
кружевного строения. Над мешком с
внутренними органами дорсально расположены
центральные трахейные
с т в о л ы в количестве двух, которые у
многих личинок (Tabanus, Hybomitra и др.)
сильно расширены и играют роль гидро- р 21 Вполне развитые ЛИЧИНки Га-
статического приспособления, позволяю- Ьапш bovinus.
щего личинкам плавать на поверхности
воды. От центральных трахейных стволов
отходят боковые ветви, которые направляются в сторону кожно-мускульной оболочки и
к внутренним органам. На заднем конце оба трахейных ствола сложены вместе и
открываются наружу в общей щели. Между центральными трахейными стволами
расположен сердечный сосуд, представляющий собой длинную полупрозрачную
трубку. Последняя задним концом прикреплена к спинной стороне на границе X и
XI сегментов, а передним концом к головоглоточному ганглию.
Центральная нервная система представлена общим нервным узлом, состоящим
из гоповоглоточного ганглия и плотно слитых с ним последующих ганглиев нервной
цепочки. Центральный нервный узел лежит сразу же позади головной капсулы,
окружая кольцом пищевод. От центрального нервного узла отходят тонкие нервные
нити.
Из особенностей строения личинок Tabanidae упомянем орган Грабера
и хордотональные органы. Первый представляет собой, по-видимому,
статический орган. Он микроскопических размеров, расположен между центральными
трахейными стволами в пределах XI сегмента и имеет вид маленькой, полупрозрачной
груши с темными округлыми включениями в середине (рис. 24).
Хордотональные органы, похожие на таковые у Corethra, расположены латерально
на каждом сегменте вблизи кожной поверхности и присоединены к трахее; эти органы
компенсируют отсутствие специального тимпанального аппарата.
Куколка. Куколка слепня (рис. 25) напоминает до некоторой степени куколку
бабочки (pupa obtecta). Она цилиндрическая, длинная, слегка выгнута со спинной
28 ВВЕДЕНИЕ
стороны. Голова у большинства видов нашей фауны без шипов и резких выступов;
сверху на темени расположены 1—2 пары небольших бугорков, снабженных
волосками. Исключение составляют куколки Pangonius, у которых голова с резкими
выступами,— шипами. На лбу в верхней части имеются два двойных поперечных
округлых выступа; по сторонам лежат короткие конусовидные усики. Грудной
отделе хорошо намеченными крыловыми зачатками и ногами. На спинной стороне возле
заднего края головы помещается пара крупных грудных стигм; последние
Рис. 22. Голова личинки Tabanus. A — ротовые
органы во втянутом положении; Б — то же, ротовые
органы вывернуты наружу при нападении, вг —
верхняя губа; вч — верхняя челюсть; нч — нижняя
челюсть; нг — нижняя губа; у — усики; щ —
нижнечелюстные щупальца; нщ — нижнегубные щупальца;
п — площадка с щетинками; г — глаз.
имеют вид ушков. Брюшко состоит из 8 сегментов. Каждое кольцо брюшка, кроме
первого и последнего, возле заднего края снабжено густым рядом щетинок или
волосков, образующих вокруг кольца непрерывный круг. По бокам, на каждом
сегменте, кроме последнего, расположены хорошо заметные брюшные стигмы.
Последний сегмент короткий, на конце снабжен 6 крупными отростками, образующими
характерную розетку. Форма и расположение этих отростков различны у разных видов,
и поэтому строение розетки имеет большое-систематическое значение. Строение
последнего сегмента куколки несколько различается у самцов и самок. Куколка вначале
желтовато-белая или зеленоватая, но затем в последующие дни принимает более
темную окраску. Последняя варьирует в зависимости от вида, причем преобладают
коричневые, бурые или грязно-зеленые тона. К концу куколочного периода сквозь
хитиновую оболочку куколки начинают просвечивать пигментирующиеся части тела слепня
И прежде всего — глаза.
Рис. 23. Внутренние органы личинки Tabanus. г —
голова; еж — слюнные железы; и — нервный узел;
п — пищевод; пж — преджелудок; ж — желудок; м —
мальпигиевы сосуды; к — кишка; р — ректум; о —
анальное отверстие.
30
ВВЕДЕНИЕ
4. ФИЛОГЕНИЯ И КЛАССИФИКАЦИЯ
Немногочисленные ископаемые Tabanidae известны лишь из третичных отттожр
представителей близкородственных
слепням семейств позволили автору
высказать предположение о
происхождении слепней.
Первые представители надсемей-
ства Tabanidea* появляются в фауне
средней юры Каратау. Здесь
обнаружены виды ископаемых семейств
Eostmtiomyiidae, Rhagionempididae и
дожившего до современной эпохи
семейства Rhagionidae. По мнению
Б. Б. Родендорфа, Eostratiomyildae
дали начало семейству Tabanidae
и близкому к нему семейству
Acanthomeridae. К надсемейству
Tabanidea, кроме двух упомянутых
семейств, Б. Б. Родендорф
относит еще семейства Rhagionidae, Ne-
mestrinidae и Coenomyiidae. Надсемей-
ство Tabanidea, вместе с надсемей-
ствами Stratiomyiidea, Asilidea и
другими автор объединяет в инфраот-
ряд Asilomorpha.
Рис. 24. Орган Грабера. (По Sur-
couf, 1924).
Рис. 25. Куколка Tabanus.
Маккерас (Mackerras, 1954) основывает свои суждения о филогении Tabanidae
главным образом на сравнительной морфологии современных TabaZdeT* данных их
географического распространения. По аналогии с другими группами животные ч?^
геологическая история лучше известна, автор считТ/т, что тГсХмические группы
TabanofdTeJ}a° (MacbrraS' 1954> и «РУ1ие авт°Р* обозначают это надсемейство как
ФИЛОГЕНИЯ И КЛАССИФИКАЦИЯ
31
слепней с распространением на Гондване (т. е. в Австралии, южных Азии, Африке
и Америке) должны были развиться не позднее, чем в середине мезозоя. К этому
времени, по мнению автора, уже наметилось деление на подсемейства и выделились более
примитивные трибы. Формирование слепней как самостоятельного семейства автор
относит к началу мезозоя. Ближайшими родичами сем. Tabanidae автор считает
австралийско-американское семейство Pelecorhynchidae и затем сем. Rhagionidae.
Фэрчайльд (Fairchild, 1969), ссылаясь на ископаемые находки в юрских
отложениях других двукрылых, в частности Orthorrhapha и Cyclorrhapha, а также на
относительную примитивность строения слепней, заключает, что Tabanidae существуют как
семейство с триаса. Подсемейства, а возможно, и трибы сформировались перед началом
третичного периода, вероятно, в мелу.
Мы полагаем, что в мезозое слепни (самки) эволюционировали как кровососы
главным образом рептилий. В современной фауне известны виды, питающиеся кровью
крокодилов, варанов и крупных черепах. С появлением на жизненной арене
млекопитающих и вымиранием большинства крупных рептилий слепни сменили хозяев. Эта
смена, начавшаяся в мелу, сопровождалась интенсивным формообразованием и
появлением более специализированных групп. Интенсивное развитие в мелу (и позднее)
цветковых растений, подходящих для литания слепней (самцов и самок) углеводной
пищей, способствовало этому процессу. Своего расцвета и многообразия форм слепни,
видимо, достигли в третичном периоде. События четвертичного периода, в частности,
образование на материках северного полушария мощного ледяного покрова, вызвали
значительные территориальные перемещения фауны и, вероятно, гибель части более
теплолюбивых форм. Выжившие группы, особенно древние, сохранились
преимущественно в теплых частях земного шара — в Австралии, Южной Африке и Америке.
Здесь сосредоточено 2/3 видов современной фауны слепней. В северном полушарии
господствующей стала более молодая фауна, приспособленная к бореальным условиям
существования.
Надродовая классификация слепней имеет свою историю, которая вкратце
заключается в следующем. В качестве самостоятельного семейство Tabanidae установил
Лич (Leach) в 1817 г. Лёв (Loew, 1860) на основании наличия или отсутствия шпор на
задних голенях поделил семейство Tabanidae на два подсемейства: Pangoniinae и Та-
baninae. К началу текущего столетия, по данным Кертеса (Kertesz, 1908), глобальное
число описанных видов слепней достигало 1529, относящихся/к 33 родам. В
монографии Сюркуфа (Surcouf, 1921) уже указаны 2123 вида, относящиеся к 61 роду.
Естественно, это требовало дальнейшей разработки системы. Лутц (Lutz, 1909, 1913, цит. по:
Fairchild, 1961 : 234), сохранив деление семейства на две секции, дал им названия
Opisthacanthae и Opisthanoplae и подразделил каждую секцию на 3 подсемейства. Всего,
таким образом, было выделено 6 подсемейств. Эндерлейн (Enderlein, 1922, 1925),
использовав секции Лутца (Lutz), увеличил число подсемейств до 10 и ввел деление
на трибы, установив их в количестве 19. Но предложенная этим автором классификация
во многих отношениях оказалась неприемлемой из-за искусственности. Прогрессивным
было использование в таксономии триб. Бекворт (Bequaert, 1930) значительно упростил
надродовую систематику слепней. Он сохранил деление Лёва (Loew) на два
подсемейства и установил 5 триб: три в подсемействе Pangoniinae и две — в подсемействе Та-
baninae. Филип (Philip, 1947) повторил в общих чертах схему Бекворта (Bequaert):
он использовал деление слепней на два подсемейства и в подсемействе Pangoniinae
сохранил три трибы. Но подсемейство Tabaninae автор поделил на 5 триб, используя
опушение базикосты. Фундаментальную ревизию надродовой таксономии Tabanidae
осуществил Маккерас (Mackerras, 1954) на основании изучения терминалий брюшка
самцов и самок. Автор предложил деление семейства на 4 подсемейства и 9 триб.
Позднее Маккерас (Mackerras, см.: Fairchild, 1969) упразднил одно подсемейство (Scepsi-
dinae), понизив его до ранга трибы, а входившие в его состав, кроме номинативного,
еще два монотипичных рода, были выделены Диасом (Dias, 1958, 1962, см.: Moucha,
1969) в самостоятельные трибы.
По современной классификации семейство Tabanidae подразделяется на 3
подсемейства, 12 триб и 117 родов и насчитывает в своем составе 2992 вида (табл. 1).
Подсемейство Pangoniinae представлено 5 трибами и включает формы с наиболее
архаичными признаками: коротким, обычно поделенным на 6—8 сегментов 3-м
члеником усиков, хорошо развитыми глазками на темени (у трибы Philolichini могут
отсутствовать), концевыми шпорами на задних голенях, голой базикостои, IX тергитом
брюшка в виде цельной пластинки и т. д. Подсемейство охватывает распространением
главным образом страны южного полушария с теплым климатом, в частности,
Австралию, Южную Африку и Южную Америку; в Северной Америке пангонии обитают
главным образом в Сонорской подобласти, а в Евразии — в Южной Европе, Японии и
Восточном Китае.
Подсемейство Chrysopsinae благодаря несколько редуцированному строению
3-го членика усиков, состоящего не более чем из 5 сегментов, а также поделенному на
две пластинки IX тергиту брюшка, рассматривается как более специализированная
группа слепней. Но Chrysopsinae сохраняют глазки на темени, шпоры на задних голенях
32 ВВЕДЕНИЕ
Таблица 1
Количество родов и видов слепней
по подсемействам и трибам
Подсемейство
Pangoniinae
Chrysopsinae
Tabaninae
Всего . . .
Триба
Количество родов
17
6
2
1
2
8
10
6
1
44
16
4
117
Количество видов
185
234
90
1
2
119
52
289
1
648
978
393
2992
Примечание. Количество триб и родов указано по Моуха (Moucha, 1969), количество
видов по Фэрчайльд (Fairchild, 1969), с уточнением числа родов и видов Палеарктики
по Олсуфьеву (1969) и другим данным.
и голую базикосту, как у Pangoniinae. Наиболее древней в подсемействе Chrysopsinae
считается триба Bouvieromyiini, которая имеет преимущественно тропическое и
субтропическое распространение (главным образом в Австралии и в Африке) и лишь
немногие виды заселяют наиболее теплые части Голарктики. Своеобразная триба
Rhinomyzini обитает в тропических частях Африки и юга Азии. Наконец, триба
Chrysopsini, включая ряд относительно молодых форм, охватывает распространением наряду
с Южной Америкой и Африкой почти всю Голарктику, но практически отсутствует
в Австралии (1 вид) и малочисленна на юге Азии.
Подсемейство Tabanidae представляет собой относительно молодую группу
семейства. На это указывают значительное видоизменение 3-го членика усиков с дальнейшей
редукцией числа сегментов, наблюдаемое в некоторых группах; редукция глазков
и концевых шпор задних голеней и т. д.; IX тергит брюшка поделен на 2 пластинки,
а базикоста у большинства Tabaninae (кроме Diachlorini) покрыта короткими волосками
(щетинками). Подсемейство Tabaninae эволюционно является наиболее прогрессивным
— оно включает 2/3 видового состава семейства.
Более древняя в подсемействе Tabaninae триба Diachlorini в значительной мере
повторяет распространение Pangoniinae, т. е. является южной группой; в Палеарктике
она встречается только на юге Европы, на севере Африки, на Кавказе и частично
заходит в Среднюю Азию (род Nanorhynchus). Триба Tabanini имеет всесветное
распространение, но наиболее многочисленна в Палеарктике. Наконец, триба Haematopotini,
несомненно, филогенетически самая молодая ветвь подсемейства, распространена
почти исключительно в Африке и Евразии, немногие виды обитают в Северной
Америке.
5. ИЗМЕНЧИВОСТЬ СЛЕПНЕЙ
Вопросы внутривидовой изменчивости слепней впервые в нашей литературе были
рассмотрены В. В. Шевченко (1961) применительно к видам фауны Казахстана. Как
справедливо отметил автор, ни одна географическая форма, ни одна популяция не
бывает полностью гомогенной, и индивидуальные колебания признаков отдельных особей
являются правилом. Изменчивости подвержены размеры тела и его отдельных частей,
окраска, морфологические структуры, включая терминалии, и т. д. Изменчивость
может быть индивидуальной, встречаясь у особей одной и той же популяции, или
географической. Она в той или иной мере адекватна экологическим условиям
существования. Предполагается, что географическая изменчивость, в отличие от
индивидуальной, имеет наследственно закрепленный характер.
ИЗМЕНЧИВОСТЬ СЛЕПНЕЙ 33
ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ
Наибольшим колебаниям подвержены размеры тела. Видимо, это зависит от
условий питания и местообитания личинок, климатических воздействий и т. д. По данным
В. В. Шевченко (1961), исследовавшего 47 видов фауны Казахстана, относящихся
к 5 родам, длина тела у 99 разных видов варьирует в пределах от 13 до 50% по
отношению к средней величине этого цризнака. В меньшей степени колеблется длина
крыла — от 7 до 38%. По нашим данным, значительные колебания в размерах тела
встречаются у отдельных представителей всех изученных нами родов (табл. 2). Наибо-
Таблица 2
Индивидуальная изменчивость длины тела §$
в разных родах (крайние варианты)
Вид
Длина тела, в мм
6-10
7.5—12
7—11
8.5—15
11—16
17—24
20—26
14—22
8—14
13—18.5
10—16
12-18.5
9.5—13.5
6.5—12
8-13
Отклонение
в мм
4
4.5
4
6.5
5
7
6
8
6
5.5
6
6.5
• 4 .
5.5
5
в % к
наименьшему размеру
66
60
57
76
45
41
30
57
77
42
60
54
42
84
62
Примечание. Длина тела определяется путем измерения от фронтальной части головы
(без усиков) до кончика, брюшка.
лее значительные отклонения, вычисленные в процентах к наименьшей длине тела,
можно отметить у 99 Tabanus sabuletorum — 76%, Atylotus pulchellus— 77% и
особенно у Haematopota pluvialis — 84%. Они велики также у Silvius vitripennis — 66%,
Chrysops relictus—-60%, Tabanus bromius—45%, T. autumnalis—57%, Hybomitra
montana — 54% и др. Это все виды с очень широким распространением. У менее
изменчивых (или слабее изученных) видов, обычно имеющих меньшие ареалы,
отклонения в размерах тела составляют чаще 15—20%. Вероятно, в этих же пределах
колеблются размеры <J(J, но недостаток коллекционного материала затрудняет
соответствующий анализ. У одного и того же вида обычно длина тела у <J<J такая же, как у 99-
В отдельных случаях изменения размеров тела носят популяционный характер.
Например, 99 Chrysops relictus в дельте Волги немного мельче, чем в Тюменской обл.
(Березов) (Сазонова 1949), 9? Hybomitra peculiaris с о. Барса-Кельмес (Аральское
море) мельче, чем в среднем течении Или (Шевченко, 1961) и т. д.
Значительная индивидуальная изменчивость обнаруживается в окраске тела
слепней, особенно у 99- Некоторые повторяющиеся отклонения у отдельных особей в
популяции, но не имеющие географической локализации, обозначают варьететами.
У отдельных видов Hybomitra, например, Н. bimaculata, H. sareptana, H. tschu-
ensis, H. lurida, H. montana и других обнаруживается-меланизм в окраске брюшка —
коричневые пятна по его бокам вытесняет черная окраска. Обычно меланистические
особи составляют ничтожную часть популяции, но у некоторых видов местами они могут
преобладать, например, Н. montana montana var. flaviceps — на севере Европы, а
Н. bimaculata var. bisignata — на Дальнем Востоке. У горных Hybomitra с целиком
черным брюшком, например Н. hunnorum, H. caucasica, H. aterrima, может
варьировать окраска волосков тергитов брюшка, у одних особей они только"черные, у других —
с примесью беловато-серых. У Chrysops подвержен изменчивости темный рисунок II
тергита брюшка, именно черные пятна в его средней части, они могут быть крупнее
или мельче, а в некоторых случаях, например у Ch. dissectus и Ch. ricardoae, нацело
исчезать. Иногда изменчив также рисунок на последующих тергитах брюшка.
Известны меланистические особи Ch. sepulcralis var. maurus с черными волосками по бокам
груди. Индивидуальная вариабельность в окраске крыльев у Chrysops может
выражаться в изменении ее интенсивности, например, у Ch. concavus иногда обнаруживается
3 Н. Г. Олсуфьев
gi ВВЕДЕНИЕ
значительное просветление дискоидальной ячейки, что приводит к неправильному
определению этого вида как Heterochrysops. В южных популяциях Tabanus sabuletorum,
Hybomitra peculiaris и других видов у отдельных особей отмечается значительное по-
светлениё брюшка, бедер и т. д. Окраска глаз у слепней, в том числе количество и
расположение темных полосок или пятен, довольно константно. Однако у отдельных $$>
некоторых Hybomitra, например Н. expollicata expollicata, H. popovi, H. vaucasi,
их число варьирует в пределах 1—3, а иногда полоски не выражены вовсе.
Аналогичное колебание числа глазных полосок от 1 до 3 отмечено для Н. turkestana и Н. shnit-
nikovi (Шевченко, 1961).
Некоторые структурные признаки также подвержены индивидуальной
изменчивости. Практически у $$ всех Tabanini варьируют в известных пределах ширина
лобной полоски, очертания нижней и средней мозоли, в меньшей мере толщина концевого
членика щупалец, форма и окраска 3-го членика усиков и т. д. Своеобразна
изменчивость налета на лобном треугольнике у $ Hybomitra nitidifrons — от полного его
отсутствия (в таком случае склерит целиком блестящий) до сплошного покрытия. Весьма
вариабельна форма лобных мозолей у Silvius vitripennis, что привело к описанию
нескольких ложных видов. Число сегментов у 3-го членика усиков достаточно постоянно,
но у видов подрода Surcoufiella (род Atylotus) палочка усиков с варьирующим числом
члеников от 2 до 4. Придаток у жилки г4 крыла довольно константен у некоторых
групп слепней, например, у видов Atylotus s. str., Haematopota, некоторых Tabanus
и др. Эндерлейн (Enderlein) придавал этому признаку большое значение для
дифференциации видов и надвидовой таксономии. Но этот придаток может обнаруживаться,
чаще в виде рудимента, у отдельных особей видов, которым он не свойствен,
например, у Hybomitra astur, H. lurida, H. turkestana, H. nigricornis, H. montana и др.
Иногда придаток г4 бывает лишь на одном крыле, отсутствуя на другом. Известны и
другие отклонения в жилковании крыла. У Chrysops relictus около 1/3 особей имеют
незамкнутую анальную ячейку, тогда как обычно она замкнута. У Tabanus signatipennis
и Т. trigeminus 1-я заднекрайняя ячейка сильно сужена к вершине, но иногда она
замкнута. Строение глаз <$<$, в частности величина фасеток, довольно постоянный признак,
но у Т. bromius обнаруживаются колебания в размерах крупных фасеток, у одних
особей они больше, у других меньше. У <J<J T. sabuletorum в области крупных фасеток
иногда бывают заметны короткие волоски. В известных пределах подвержены также
изменчивости терминалии. У § VIII стернит может изменяться по ширине и длине, что
влияет на очертания гонопофиза, латеральных лопастей и т. д. Непостоянен темный
рисунок стернита — боковые коричневые пятна могут ограничиваться латеральными
лопастями или распространяться почти до центра склерита. Еще более изменчив
IX стернит (фурка), в том числе его слабо хитинизированные каудальные'части, кон- ,
фигурация которых в значительной мере зависит от техники препаровки. Значительная
изменчивость этого склерита ограничивает его использование для целей систематики.
Церки относительно постоянны по очертанию, но пропорции длины—ширины могут
индивидуально меняться. В небольшой степени изменчивы сперматеки, главным
образом их апикальная часть, изредка она бывает двойной (уродство!). Длина семеприемни-
ков б. м. постоянна.
ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Ряд видов слепней с широким распространением распадается на отдельные,
отличающиеся между собой формы, имеющие определенную географическую локализацию.
В местах стыка ареалов этих форм встречаются особи с промежуточными признаками
(интёрградация), что дает основание рассматривать эти формы как географические расы
или подвиды. Иногда ареалы подвидов бывают изолированными. Из 189 видов фауны
СССР можно отметить 37 политипических видов, для которых известны 2, а для
некоторых 3 и более подвида. Дальнейшее изучение, несомненно, пополнит число
политипических видов нашей фауны. В основном географическая изменчивость распространяется
на окраску тела, в первую очередь у $§ (<J<J хуже изучены!), в меньшей степени на
другие признаки. Вполне естественно, что ареалы политипических видов обычно
охватывают разные ландшафты, иногда значительно отличающиеся между собой. Очевидно,
этому способствует экологическая пластичность данных видов, что и находит отражение
в образовании географических рас. Для политипических видов нашей фауны можно
наметить следующие основные пространственные ориентации, в пределах и под
влиянием которых формируется их географическая изменчивость: широтная (север—юг),
• долготная (запад—восток) и вертикальная (горы юга). Говоря о пространственной
ориентации, мы не предрешаем, какая географическая (ландшафтная) локализация
i является исходной для родительской формы. В одних случаях исходной могла быть
южная форма, отделившая затем от себя бореальную форму, в других случаях события
могли развиваться наоборот.
Ярким примером формообразования в широтном направлении (север—юг) могут
служить Tabanus autumnalis и Т. bromius. Особи ($?) обоих видов, относимые к
номинативной форме, в северных и средних частях своего ареала имеют черно-серую окраску
ИЗМЕНЧИВОСТЬ СЛЕПНЕЙ 35
брюшка, темные усики, неяркие коричневато-зеленоватые глаза. На юге, например,
в Закавказье и Средней Азии, они замещаются светлыми подвидами Tabanus
autumnalis brunnescens и Т. bromius flavofemoratus с преимущественно коричневато-желтым
брюшком, светлыми усиками и ярко-зелеными глазами. Строение терминалии §
сохраняется, но VIII стернит окрашен значительно светлее, особенно у Т. autumnalis
brunnescens. Размеры тела у номинативной формы несколько больше, чем у южного подвида.
Любопытно, что в горах Кавказа и Средней Азии распространен номинативный подвид
Т. bromius bromius, а не южная форма, следовательно, его ареал характеризуется по-
литопностью. Примерно такие же цветовые различия имеются между таежно-лесным
Hybomitra montana montana и его степным подвидом Н. montana morgani, причем у
последнего посветление распространяется и на жужжальца — они беловатые в отличие
от коричневых у номинативной формы. Любопытно, что у § подвида Н. montana
morgani волоски на глазах значительно короче, чем у номинативной формы, имеются
небольшие различия и в строении церок. Но самым светлым подвидом является
описанный нами с юга Монголии Н. montana zaitzevi, у него же глаза практически голые.
Распространенный на Памире и Тянь-Шане горный подвид Н. montana reinigiana
обнаруживает значительное потемнение окраски брюшка и усиков, удлинение
волосков на глазах и уменьшение размеров тела, но белая окраска жужжалец и строение
терминалии сохраняются, как у Н. montana morgani. Примерами подвидообразования
в широтном направлении с посветлением окраски тела $$ могут быть еще Taianus
sudeticus verralli, Т. bifarius kurensis, T. quatuornotatus araxis, Atylotus pulchellus pul-
chellus, A. fulvus aureus (горы Кавказа), Chrysops caecutiens ludens и др.
Географическая изменчивость в долготном направлении (запад—восток) не имеет
таких четких закономерностей, как в широтном направлении. Остановимся на некото--
рых примерах. Так, номинативный подвид Hybomitra nitidifrons nitidifrons, распро*
страненный в Восточной Сибири и на Дальнем Востоке, отличается от европейско-за-г
падносибирского подвида Н. nitidifrons' confiformis беловато-серой окраской светлых
волосков брюшка; у последнего эти волоски золотисто-желтые. Восточносибирский под-^
вид Н. distinguenda contigua (§) отличается от номинативного европейско-западноси-
бирского подвида примесью черных волосков по бокам II тергита брюшка и несколько
измененной формой VIII стернита. Восточносибирский подвид Н. lundbecki sibiriensis
отличается от номинативного европейско-западносибирского подвида более светлыми
усиками. Н. expollicata orientalis, распространенная в Восточной Сибири, имеет в
сравнении с западным номинативным подвидом более темные бедра и 3 резких полоски на
глазах. Среднесибирский подвид Heptatoma pellucens orientalis отличается от
европейско-западносибирского номинативного подвида более светлыми (беловатыми)
волосиками тела. Наконец, у дальневосточного подвида Haematopota pluvialis tristis немного
изменен рисунок крыла в сравнении с таковым номинативного европейско-сибирского
подвида.
Географическая изменчивость по вертикали (вверх) наблюдается в горах юга,
особенно на Кавказе и в Средней Азии, выражаясь в потемнении окраски тела
(преимущественно брюшка), в некоторых случаях в уменьшении егоразмеровит. д. Примерамитемных
подвидов могут служить горные Tabanus miki colchidicus, Т. indrae montivagus, T. lae-r
tetinctus sordes, T. golovi golovi и др. Они имеют политопные ареалы, приуроченные
к отдельным горным системам. У популяций Hybomitra tatarica и Н. shertshenkoi,
обитающих на больших высотах Западного Тянь-Шаня, отмечается уменьшение размера
тела и изменение окраски глаз (Шевченко, 1961). Мы расцениваем эти'формы как варь-*
ететы, в ожидании накопления дальнейших материалов.
Заканчивая изложение, следует отметить, что проявления внутривидовой
изменчивости в зависимости от географической локализации имеют определенный
параллелизм с соответствующими проявлениями видового формообразования. Приведенные
выше примеры ясно показывают, что бореальные и горные подвиды обычно отличаются
более темной окраской тела в сравнении с южными и равнинными (или низкогорными)
подвидами, особенно обитателями аридного пояса. Иногда к цветовой изменчивости
присоединяются изменения в строении терминалии §§ и других признаков. Основная
масса видов слепней, обитающих на севере или в горах, имеет преимущественно темную
окраску, а на юге, особенно в пустынной зоне, светлую. Этот параллелизм в основном
касается окраски тела, но он может служить указанием на единые закономерности
образования подвидов и видов.
6. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И ВОПРОСЫ
ПАЛЕОГЕНЕЗА ФАУНЫ СЛЕПНЕЙ
ОБЩИЕ ДАННЫЕ О ФАУНЕ СЛЕПНЕЙ ПАЛЕАРКТИКИ И СССР
Наш опыт изучения слепней СССР показывает, что география этой группы насеко*-
мых в общем подчиняется закономерностям распределения наземных свободноживу-
щих животных, хотя взрослые слепни ($>§) представляют собой паразитов теплокров-
3*
36 ВВЕДЕНИЕ
ных, а личинки в значительной мере являются гидробионтами. Любопытно, что
отдельные виды слепней, птиц и млекопитающих имеют сходные очертания ареалов, не
обнаруживая специальных связей в биоценозах. Сходство ареалов указывает на то, что
эволюция этих животных находилась в тесной зависимости от общей эволюции
ландшафтов, которые они населяли.
Изучая зоогеографию слепней, можно выяснить закономерности, которые
окажутся общими для некоторых других групп животного мира.
Сем. Tabanidae является космополитным в своем распространении. Его
представители встречаются на всех континентах, кроме Антарктиды, а также на некоторых
океанических островах; слепни неизвестны в Гренландии и Исландии. Семейство по
своему происхождению является тропическим. Именно во влажных тропиках Старого
и Нового Света, изобилующих крупными млекопитающими, слепни достигли наиболее
Пышного расцвета в отношении численности и разнообразия форм. Так, на первом месте
по числу известных видов, в том числе более древних форм, стоит Южная Америка —
986, далее следует Эфиопская область — 587 видов. Австралия менее богата видами —
всего 234, но зато здесь многочисленны, древние группы слепней. Фауна слепней
Ориентальной области насчитывает 343 вида, причем преобладают молодые группы слепней
(Fairchild, 1969). Излагая данные о фауне Палеарктической области, мы полагаем, что
будет полезным произвести ее сравнение с фауной. Неарктической области, с которой
она наиболее близка. Это тем более необходимо, поскольку обе области часто
объединяют в единую Голарктическую область.
Фауна слепней Палеарктической и Неарктической областей насчитывает 860
видов, относящихся к 39 родам. Они объединены в 8 триб и 3 подсемейства: Pangoniinae,
Chrysopsinae и Tabaninae. В Палеарктической области * известен 501 вид, относящийся
к 3 подсемействам, 7 трибам и 19 родам, соответственно в Иеарктической области —
368 видов, относящихся к 3 подсемействам, 7 трибами 27 родам (табл. 3). Следовательно,
фауна Неарктики богаче фауны Палеарктики по количеству родов, но беднее по
количеству видов.
Из табл. 3 видно, что фауна Tabanidae Палеарктики и Неарктики полностью
совпадает в ранге подсемейств и почти полностью в ранге триб (расхождение по 1 трибе).
В ранге родов расхождение значительное — общими являются всего 7: Stonemyia,
Silvius, Chrysops, Tabanus, Atylotus, Hybomitra и Haematopota, тогда как
несовпадающих родов в Палеарктике 12 и в Неарктике 20. Несовпадающие роды в основном
относятся к филогенетически более древним группам, таким как подсемейство
Pangoniinae, или триба Вouvieromyiini в подсемействе Chrysopsinae, или триба Diachlorini
в подсемействе Tabaninae. Распространены они в обеих областях преимущественно
в условиях субтропического климата — в Палеарктике в Средиземноморской и в Вос-
точноазиатской подобластях, а в Неарктике — в Сонорской подобласти. Наоборот,
общие роды филогенетически относятся к более молодым группам, таким как триба
Chrysopsini в подсемействе Chrysopsinae и трибы Tabanini и Haematopotini в
подсемействе Tabaninae. Распространены они наряду с субтропиками в условиях умеренного
и холодного климатов, в Палеарктике — в Воревразийской и Центральноазиатской
подобластях, а в Неарктике — в Канадской подобласти. В ранге видов расхождение
фаун в рассматриваемых областях наибольшее: несовпадающих видов 851 и общих —
всего 9. Однако эти несовпадающие виды, особенно в Канадской подобласти Неарктики,
часто имеют большое сходство с таковыми Воревразийской подобласти Палеарктики.
И тут и там основную массу видов составляют Hybomitra и Chrysops s. str., а в северных
Частях Канадской подобласти обитают общие виды: Hybomitra astuta (syn. H. polaris),
И. uequeiincla (syn. H. flavipes), П. sexfasciala, II. lapponica, II. lurida, H. nitidifrons
(подвид Н. nitidifrons nuda), H. montana (подвид Н. montana frontalis), H. arpadi и
Chrysops nigripes.
Обращает на себя внимание большее разнообразие видов Tabanus, Atylotus,
Hybomitra и Haematopota в Палеарктике и видов Chrysops — в Неарктике. Последняя
особенно бедна видами Haematopota — их всего 5 против 58 в Палеарктике.
В целом фауна слепней Голарктики более молодая, чем фауна континентов южного
полушария (Mackerras, 1959).
Сравнительное изучение фаун слепней Палеарктики и Неарктики дает основание
заключить, что между Евразией и Северной Америкой существовали в прошлом два
пути обмена фаунами: более древний — через Атлантику, и более поздний — через
Берингию. Примером древних атлантических связей могут служить представители
подсемейства Pangoniinae (общий род Stonemyia), затем некоторые Silvius и Chrysops,
а также представители трибы Diachlorini и некоторые группы Tabanus. Нами,
например, установлено, что распространенные в Средней Азии Nanorhynchus crassinervis
Vill. и Chrysops (Turanochrysops) hyalipennis Stack, имеют ближайших родичей в CoHOpj
ской подобласти Неарктики (Олсуфьев, 1966, 1971). Примером более поздних связей
* На юго-востоке мы не включаем в Палеарктическую область территорию Китая
К югу от Янцзы, о. Тайвань, а также южные низкогорья Гималаев.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 37
Таблица 3
Количество родов и видов Tabanidae Голарктической надобласти
Подсемейство
Триба
Род
Количество видов
Палеарк-
тическая
область
Неарктическая
область
Pangoniinae
Chrysopsinae
Tabaninae
Р angoniini
Philolichini
Scionini
В ouvieromyiini
Chrysopsini
Diachlorini
Tabanini
Haematopotini
Pangonius . .
Stonemyia . .
Apatolestes . .
Zophina . . .
Brennania . .
Pilimas . . . .
Esenbeckia . .
Asaphomyia .
Ectinocerella
Goniops . . . .
Thaumastomyia
Gresittia . . .
Nagatomyia . .
Mericomyia . .
Silvius . . . .
Surcoufia . . .
Chrysops . . .
Neochrysops . .
Dasyrhamphis .
Philipomyia . .
Nanorhynchus .
Stenotabanus .
Anacimas . . .
Microtabanus .
Diachlorus . .
Chlorotabanus .
Bolbodimyia
Leucotabanus .
Holcopsis . . .
Tabanus . . .
Isshikia . . .
Atylotus . . .
Therioplectes
Whitneyomyia .
Hamatabanus .
Agkistrocerus .
Hybomitra
Haematopota
Heptatoma . .
30
5
5
13
1
2
3
12
1
1
1
1
—
20
1
46
—
12
4
1
—
—
—
—
—
—
—
—
182
2
30
7
—
—
—
96
58
1
—
—
^-
2
13
—
96
1
.—
—
—.
14
3
1
1
2
1
2
2
111
—
9
—
1
3
2
61
5
—
39
501
368
Примечание. Данные о фауне слепней Неарктической области сообщены Пикьюменом
(L. Pechuman, личное сообщение).
г
через Берингию могут служить представители Hybomitra, некоторые группы Tabanus,
Atylotus, Chrysops и Haematopota. Эти связи обнаруживаются благодаря наличию
в составе фаун обеих областей либо общих, либо близких видов; Но существовали и
древние фаунистические связи через Берингию, на что, например, указывает наличие
представителей трибы В ouvieromyiini в Восточноазиатской подобласти Палеарктики
и Сонорской подобласти Неарктики. Как известно по данным изучения позвоночных,
включая палеонтологические находки, атлантические связи фаун Евразии и Северной
Америки в третичном периоде датируются его началом или серединой (миоцен), тогда
как связи через Берингию определяются как многократные на протяжении третичного
периода, и даже в четвертичное время. Вероятно, также следует датировать и
палеогеографические связи фауны слепней Старого и Нового Света.
38 ВВЕДЕНИЕ
Фауна слепней Палеарктики обнаруживает некоторые связи с таковой Эфиопской
области. Не останавливаясь на деталях, укажем лишь, что в составе распространенных
в саваннах Восточной Африки Tabanus и Atylotus имеются виды, обнаруживающие
ясные родственные связи с пустынными видами Палеарктики, в том числе 4 вида
являются общими: Atylotus agrestis, Tabanus sufis, Т. gratus и Т. taeniola. Эти виды
распространены в Египте, а Т. sufis. — еще в Ираке, Иране и Афганистане. Общим видом
для Палеарктической и Эфиопской областей является также Haematopota coronata.
Вероятно, имеются связи между фауной слепней Восточноазиатской подобласти
Палеарктики и Ориентальной области. Такая связь хорошо известна по другим группам
Животных. Но сравнение фаун слепней затруднительно из-за отсутствия региональных
сводок по Китаю и Ориентальной области.
В пределах СССР слепни распространены весьма широко, встречаясь почти на
всей территории страны с севера на юг и с запада на восток. Пределы распространения
слепней на Крайнем Севере в точности не выяснены. В европейской части СССР, по-
видимому, в основном границей является береговая линия материка. В Азии слепни
(Hybomitra astuta) найдены в придельтовой части Енисея (Дудинка), Лены (Тикси), Яны
(Казачье), Колымы (Нижние Кресты), а на п-ове Таймыр северная граница обитания
слепней, по данным Н. П. Мезенева (1972), проходит между 72 и 73° с. ш., не достигая
береговой линии. В горах слепни распространены почти до границ вечных снегов,
поднимаясь иногда до высоты 4200 м над ур. м. (Hybomitra hunnorum в горах Средней
Азии). В пустынных местностях вдали от воды слепни отсутствуют, но встречаются
по долинам рек, в оазисах, возле озер и колодцев.
Всего на территории СССР обнаружено 189 видов, 39 подвидов * и 17 варьететов
слепней. Они относятся к 3 подсемействам: Pangoniinae, Chrysopsinae и Tabaninae.
Первое подсемейство представлено 2 родами — Pangonius (1) и Stonemyia (1), второе
также 2 родами — Chrysops (27) и Silvius (6), наконец, третье — 9 родами: Dasyrham-
phis (1), Philipomyia (2), Nanorhynchus (1), Tabanus (64), Atylotus (14), Therioplectes (4),
Hybomitra, (50), Heptatoma (1) и Haematopota (17), всего 13 родов; в скобках указано
количество видов каждого рода. Наибольшее число видов фауны СССР (89%) относится
к 5 родам: Chrysops, Tabanus, Atylotus, Hybomitra и Haematopota.
Выявление видового состава слепней фауны СССР мы не считаем завершенным.
Возможно нахождение видов известных в сопредельных странах, не исключено также
обнаружение новых видов. Это особенно касается Кавказа, юга Восточной Сибири,
Дальнего Востока и гор Средней Азии. Вероятно, в конечном итоге число видов
слепней фауны СССР будет доведено до 200. Нуждается в дальнейшей разработке и
внутривидовая таксономия.
ТИПОЛОГИЯ (КЛАССИФИКАЦИЯ) АРЕАЛОВ СЛЕПНЕЙ
В зоогеографии сложились 2 основных направления, из них каждое имеет свои
задачи и методы. Первое направление, называемое ареалогией (учение об ареалах),
имеет задачу выяснения области распространения тех или иных форм, что важно для
их таксономии (ареал — одно из свойств вида), экологии, палеогенеза, истории
расселения и т. д. Группы видов, имеющие сходные ареалы, обозначают фаунистическими
комплексами (Тупикова, 1969), объединяя их затем в типы фауны. Второе направление,
внимание к которому особенно привлек А. П. Кузякин (1962), изучает количественные
аспекты животного населения применительно к тем или иным условиям местообитания,
например, ландшафтным зонам. Синтез данных обоих направлений позволяет
наиболее полно обосновать зоогеографическое (фаунистическое) районирование территории.
Накопленные за ряд лет данные о распространении слепней на территории СССР
и сопредельных стран дают возможность определить для большинства видов их ареалы
и сопоставить их между собой. Это обобщение нами осуществляется впервые. Оно не
лишено недостатков изтза некоторой неполноты фаунистических материалов, особенно
по Восточной Сибири, отдельным участкам Средней Азии, Кавказа и Украины. Еще
более существенны пробелы в изучении распространения слепней в Передней и в
особенности в Центральной Азии. Картографирование ареалов слепней Палеарктики
имеется в монографии Леклерка (Leclercq, 19606, 1966), но для территории СССР оно
весьма несовершенно. По Казахстану картографические данные о распространении
слепней представлены в публикации В. В. Шевченко (1961).
Поскольку слепни в основном представляют собой свободноживущих насекомых,
их ареалы находятся в прямой зависимости от климата, типов растительности (лес,
кустарники), с которыми связаны обитанием и питанием взрослые насекомые,
водоемов, служащих местами развития личинок и т. д. Распределение по территории разных
видов млекопитающих, являющихся источниками питания кровью самок слепней,
вряд ли оказывает влияние на ареалы видов нашей фауны. Слепни мало разборчивы
* Tbanus ansarii badhysi, T. amaenus griseinus и Chrysops buxtoni armeniensis
учтены как виды вместо номинативных форм, не встречающихся на территории СССР.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 39
в объектах питания, лишь бы они были достаточных размеров и удобны для нападений.
Практически источники питания слепней кровью имеются повсеместно. Но численность
крупных позвоночных в той или иной местности может оказать существенное влияние
на обилие слепней.
Эколого-географическая характеристика того или иного вида основывается не
только на степени совпадения его ареала С границами той или иной ландшафтной
зоны, но и на степени его обилия в ее пределах. Это особенно важно для видов, ареалы
которых выходят за пределы какой-либо одной зоны. Несомненно, существенным
дополнением для такой характеристики могли бы служить сведения о местах выплода
(обитания) личинок тех или иных видов — водоемах различных типов: реках, ручьях,
озерах, болотах, лиманах и т. д. Но ввиду крайне скудных данных, которые
имеются, это задача будущего. Сейчас мы можем только говорить о приуроченности
ареала (и обилия) того или иного вида к той или иной ландшафтной зоне и соответственно
■обозначать виды как «лесные», «горно-лесные», «степные», «пустынные» и т. д.
Величина ареала весьма разнообразна. Наибольшие размеры ареала имеют виды,
связанные обитанием с равнинными ландшафтными зонами, например, с таежной или
лесной, имеющими в Палеарктике огромную протяженность. Некоторые виды с
большим ареалом, охватывающим разные ландшафты, в его пределах представлены
отдельными подвидами. Таковы политипические Hybomitra montana, Tabanus autum-
nalis, Т. bromius и др. Наименьшие ареалы имеют виды горных систем юга, например,
Т. smirnovi ограничен распространением лесами горного Крыма, или Hybomitra so-
gdiana и И. sheutshenkoi, известные из немногих частей гор Средней Азии. Обычно это
виды недавнего происхождения. Принято считать виды с небольшими ареалами,
особенно горной локализации, эндемиками, тогда как виды с большими ареалами,
таковыми не считаются, хотя по сути дела последние являются такими же эндемиками
соответствующих ландшафтных зон.
Большинство видов, охватывающих своим распространением значительные
пространства, бывают многочисленны лишь в определенном ландшафте, которому они
в основном свойственны, и вне его встречаются лишь в небольших количествах.
Следовательно, необходимо различать область распространения вида (ареал) и в его
пределах — зону обилия вида, которая является оптимальной в отношении условий его
существования. Так, Hybomitra distinguenda распространена от Англии до Японии,
но обычна лишь на Дальнем Востоке, тогда как в западных частях своего ареала
встречается не часто. Н. montana весьма многочисленна в Восточной Сибири, тогда как
в европейской части своего ареала относительно редка. Tabanus bromius и Hybomitra
•ciureai изобилуют в лесостепной зоне, многочисленны также в лесной зоне, но обычно
немногочисленны в таежной зоне, куда они частично проникают.
В представленном ниже анализе учтены ареалы 173 видов и 9 подвидов слепней,
распространенных в СССР. Ареалы 93 видов нами нанесены на карты, помещенные
в специальной части книги. На картосхемах места сбора того или иного вида
обозначены значками (точками, кружками и т. д.), причем, учитывая мелкий масштаб карт,
близко расположенные местонахождения объединены одним значком. Ареалы
обведены сплошной линией, но при недостатке данных границы показаны прерывистой
линией. Из рассмотрения мы исключаем виды, которые известны по немногим находкам
и характеристика их ареалов, а также экологической приуроченности, затруднительна.
Сюда относятся Hybomitra bagheera, H. obscurineruis, Tabanus talyshi, T. perelegans,
Chrysops anthrax, Haematopota koryoensis и некоторые другие.
Рассмотрение ареалов показывает, что каждому виду свойственно свое
неповторимое распространение. Это, очевидно, зависит от того, что каждый вид имеет свою
историю формирования и расселения и свои экологические требования. Вместе с тем можно
наметить группы ареалов, по очертаниям имеющие то или иное сходство,
причем такие ареалы могут быть как у видов, принадлежащих к одному и тому же роду,
так и у видов, таксономически отдаленных друг от друга. Наиболее близкородственные
виды в ряде случаев имеют аллопатрические ареалы, приуроченные к сходным
ландшафтным зонам. Все это указывает на сложность формирования фауны, проходившего
в рамках эволюции тех или иных ландшафтов в, разные геологические эпохи. Деление
ландшафта с видоизменениями его отдельных частей обычно приводило к делению
заселяющего его вида на самостоятельные формы.
Группу видов слепней, имеющих сходные ареалы, мы обозначаем фаунистиче-
ским комплек сом. В. В. Шевченко (1961) назвал такие группы «фаунистическими
элементами», или фаунулами (по П. П. Сушкину), тогда как термин «фаунистиче-
ский комплекс» он применил для обозначения того, что мы называем «количественный
аспект фауны». Под последним мы понимаем набор видов с выделением
господствующих и соподчиненных форм, населяющих ту или иную территорию или
ландшафтную зону (см. ниже стр. 54). Н. А. Виолович (1968) применил термин «фаунистический
комплекс» в том же значении, что и мы. В пределах фаунистического комплекса мы
выделяем его варианты, имеющие в очертании ареала те или иные отклонения. Группы
видов, ареалы которых более или менее типичны для комплекса, мы обозначаем номи-
40
ВВЕДЕНИЕ
нативным вариантом, тогда как виды с уклоняющимися ареалами выделяем в варианты,
соответствующие их географической локализации.
Сравнения между отдельными фаунистическими комплексами по их
таксономическим и ландшафтным связям позволяют произвести их .объединение в
субрегиональные типы фаун; региональным типом для них является палеарк-
Таблица.4
Типология (классификация) ареалов слепней фауны СССР
Субрегиональный тип фауны
I. Боревразий-
ский
II.
Средиземноморский
III. Афроевра-
зийский
аридный
ч
<
IV. Восточно-
азиатский
•
Фаунистический
комплекс
(группа ареалов)
1. Тундровый
2. Таежный
3. Европейско-си-
бирский лесной
4.
Восточносибирский лесной
5. Лесостепной
6. Среднеазиатский
горный
7. Южноевропей-
скип
8.
ско-среднеазиатский
9. Понтийско-гир-
канский
10. Бактрийско-иран-
ский
11. Степной
12. Пустынный
13. Японо-китайский
Вариант
А. Номинативный
Б. Тундрово-таежный
В. Горно-алтайский тундровый
А. Номинативный
Б. Таежно-лесной
В. Восточносибирский
таежный
А. Номинативный
Б. Европейский лесной
В. Европейский горно-лесной
А. Номинативный
Б. Дальневосточный лесной
А. Номинативный
А. Номинативный
Б. Западнотяньшанско-пами-
роалайский
В. Западнопамироалайский
А. Номинативный
Б. Прибрежный (литоральный)
А. Номинативный
А. Номинативный
Б. Балкано-кавказский
В. Кавказско-иранский
Г. Кавказский
Д. Крымский горный
А. Номинативный
А. Номинативный
Б. Южноукраинский
В. Восточноевропейско-севе-
роказахстанский
Г. Сибирско-монгольский
А. Номинативный
Б. Переднеазиатско-кавказ-
ско-туранский
В. Турано-монгольский
Г. Казахстано-монгопьский
Д. Туранский
А. Номинативный
Б. Китайский
В. Японский
Количество
видов ■
2
1
1
7
6
6
14
3
4
9
8
9
7
4
4
15
1
6
-
2
5
8
13
2
7
3
1
2
6
8 ■
4
1
1
6
6
1
2
185
Всего
видов
КО J
1
1
J
1
1
'
J
'
.
1
дплекса
► 4
19
'
1 21
! «
9
• 15
• 16
6
. 30
7
12
. 20
1 9
185
Примечание. Подвиды (13), замещающие в фаунистическом комплексе номинативную
форму, учтены как виды.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 41
тическая фауна в целом. Всего для фауны слепней СССР мы выделяем 4
субрегиональных типа: боревразийский, средиземноморский, афроевра-
зийский аридный и восточноазиатский. Они включают 13 групп
ареалов' или фаунистических комплексов. Последние подразделены на 36 вариантов,
включая номинативные (табл. 4). В Палеарктическом регионе имеется еще 5-й_тип —
центральноазиатский (или тибетский), но он не представлен в нашей фауне
и мы его рассматриваем лишь при зоогеографическом районировании (см. ниже, стр. 75).
Для ясности изложения по каждому фаунистическому комплексу мы представляем
в соответствующем месте текста картограмму, схематически изображающую его
распространение. При наложении друг на друга ареалов видов, относимых к какому-
либо одному "фаунистическому комплексу, выяснилось, что ареалы отдельных видов
иногда выходят далеко за пределы территории, которую охватывают распространением
все или большинство видов рассматриваемого комплекса и которая должна быть
признана для него наиболее типичной. Эту последнюю мы называем основным
ареалом комплекса. Обычно именно в его пределах популяции видов,
слагающих комнлекс, бывают наиболее многочисленными. Распространение (дисперсия)
за пределами основного ареала свойственно лишь немногим наиболее эвритопным
видам комплекса, причем они чаще бывают представлены малочисленными популяциями.
На составленных нами картограммах основной ареал комплекса показан
(схематически) вертикальной штриховкой, тогда как точками обозначены местонахождения
(дисперсия) отдельных видов за границами основного ареала. В немногих случаях
горизонтальной штриховкой показано распространение варианта рассматриваемого
комплекса. При составлении картограмм мы придерживались данных о
распространении видов, входящих в тот или иной комплекс. Ирландия оставлена на
картограммах без обозначений, поскольку сведения о фауне слепней этой страны
фрагментарны. В целях экономии места названия номинативных подвидов нами обычна
опущены после видового названия; не упоминаем мы и авторов видов.
Как и следовало ожидать, очерченные нами основные ареалы фаунистических
комплексов боревразийского и афроевразийского аридного типов фауны слепней более
или менее совпадают с принятым в отечественной биогеографии делением на равнинные
ландшафтные зоны. Но для выделяемых нами комплексов средиземноморского типа
фауны мы не нашли в доступной нам литературе соответствующих ландшафтных
разработок. Наши данные по комплексу слепней высокогорий Тянь-Шаня и Памиро-
Алая вполне четко указывают на самостоятельность этого комплекса, его
принадлежность боревразийскому типу фауны и чужеродность равнинной (аридной) фауне
Средней Азии, что совпадает с мнением отдельных исследователей других групп животных.
При сопоставлении основных ареалов комплексов выяснилось, что они в той или иной
мере налегают друг на друга и строгих границ между ними не существует. Особенна
это касается средиземноморских комплексов, отдельные части ареалов которых в
пределах СССР в значительной степени совпадают. Очевидно, это связано с гористостью
Средиземноморья, его южным положением и пестротой ландшафтов, а также
сложностью его истории. На Кавказе фауна усложняется присутствием в горах европей-
ско-сибирских лесных и таежно-лесных видрв.
Боревразийский тип фауны
Охватывает своим распространением северную и среднюю части Европы, всю
Сибирь, часть Северного Казахстана, верхние и частью средние пояса гор Средней
Азии (кроме Копет-Дага) и Северо-Восточный Китай; некоторые виды обитают также
в Крыму и на Кавказе. Фауна боревразийского типа может быть подразделена на.
следующие 6 фаунистических комплексов: тундровый, таежный, европейско-сибирский
лесной, восточносибирский лесной, лесостепной и среднеазиатский горный.
Тундровый фаунистический комплекс. Ареалы видов этого
комплекса в основном приурочены к крайнему северу Евразии в пределах зоны тундры,
но частично захватывают лесотундру и северную тайгу (рис. 26). Благодаря
особенностям орографии Восточной Сибири, именно наличию здесь' большого количества гор
с развитыми горными тундрами и лесотундрами, виды рассматриваемого комплекса
здесь мозаично распространены далеко к югу, достигая Алтая, южного Прибайкалья
и Приамурья.
К рассматриваемому комплексу относятся Hybomitra astuta и Н. aequetincta.
Ареал первого вида на западе достигает средней Швеции (видимо, изолированное
местонахождение) и Кольского п-ова, ареал второго вида заканчивается немного западнее
Печоры (Индига), на востоке оба вида распространены, включая Камчатку, а также
в Северной Америке.
В рассматриваемом комплексе представлены еще следующие варианты. Тундрово-
таежный вариант, насчитывающий всего 1 вид — Hybomitra sexfasciata, охватывает
распространением тундру и северную тайгу Голарктики, в Евразии от севера
Скандинавии на восток до Камчатки, в Сибири ареал отклоняется к югу и достигает Алтая,.
42 ВВЕДЕНИЕ
Приамурья и Сахалина. Второй вариант мы обозначаем как горно-алтайский, он также
представлен 1 видом И. altaica. Его ареал в основном включает высокогорные тундры
Алтая и частью Тувы (хр. Танну-Ола).
0° К ВОСТОКу ОТ ГриНВЙЧа 90° 120° 150° 180° «0°
0° 30° 60° 90° 120°
Рис. 26. Распространение тундрового фаунистического комплекса видов
боревразийского типа фауны слепней. Схема. 1 — основной ареал; 2 —
местонахождение отдельных видов комплекса вне основного ареала.
По своим таксономическим связям все виды тундрового комплекса
близкородственны представителям таежной и лесной фауны и, несомненно, являются ее
производной. Так, Н. aequetincta очень близка североамериканской Н. zonalis, H. astuta —
голарктической таежной Н. lapponica и палеарктической таежной — Н. nigricornis,
Рис. 27. Распространение таежного фаунистического комплекса видов
боревразийского типа фауны слепней. Схема. Обозначения см. рис. 26.
Горизонтальная штриховка — распространение восточносибирского варианта
таежного комплекса.
Л. sexfasciata — евразийской лесной Н. kaurii, наконец, Н. altaica имеет тесные
генетические связи с Н. sexfasciata. Поэтому нет оснований тундровый комплекс выделять
из боревразийской фауны в качестве самостоятельного арктического типа.
Таежный фаунистический комплекс. В типичном случае
(номинативный вариант) ареалы охватывают огромные пространства таежной зоны от
■Скандинавии на западе до Камчатки и Сахалина на востоке (рис. 27). На севере ареалы
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 43
■обычно включают лесотундру и даже тундру, на юге частично лесную зону. Многие
виды на западе своими ареалами захватывают Центральную Европу, но чаще в виде
локальных местонахождений и иногда только в горных районах. На востоке некоторые
,виды заходят в Северную Японию. К рассматриваемому комплексу относятся Chrysops
nigripes, Hybomitra lapponica, H. arpadi, H. nigricornis, H. tarandina и Atylotus sublu-
naticomis. Первые З вида распространены также в Сев. Америке. К группе таежных
видов, по-видимому, также относится Chrysops divaricatus, но недостаток данных о его
распространении в Восточной Сибири не позволяет высказываться более определенно.
Таежный комплекс видов дополняется еще 2 вариантами: таежно-лесным и
таежным восточносибирским. Виды таежно-лесного варианта имеют ареалы, уходящие
значительно далее на юг в лесную зону, на западе они достигают Франции и иногда
Англии. На севере и востоке граница их ареалов, примерно, та же, что и строго
таежных видов. Большая часть точек на картограмме показывает распространение видов
0° К ВОСТОку ОТ ГриНВИча 90° 120° 150° 180° 160°
Рис. 28. Распространение европейско-сибирского лесного фаунистического
комплекса видов боревразийского типа фауны слепней. Схема. Обозначения
см. рис. 26. Горизонтальная штриховка — распространение европейского
варианта комплекса.
этого варианта к югу и юго-западу от основного ареала комплекса. Большинство видов
таежно-лесного варианта локально распространено в горах Кавказа и восточных
пастей Казахстана. К рассматриваемому варианту относятся Hybomitra lurida, H. niti-
difrons, H. bimaculata, H. lundbecki, H. muehlfeldi и Н. montana. Первый вид, а также
■особые подвиды Н. nitidifrons nuda и Н. montana frontalis распространены в Сев. Америке.
'Следует отметить, что у некоторых видов этого варианта, например, Н. lundbecki
ш Н. nitidifrons намечается подвидовое формообразование с гранипей между западным
и восточным подвидами примерно по Енисейско-Байкальскому водоразделу.
Восточносибирский таежный вариант охватывает распространением значительную
территорию Восточной Сибири, на западе от Енисея (или Алтая), на востоке до
Магаданской обл. и Приморского края (иногда до Японии), на севере местами достигает
полярного круга. Практически ареал восточносибирского варианта почти полностью
налегает на восточную часть номинативного ареала комплекса (на картограмме эта
часть заштрихована в клетку). К этому варианту я отношу Chrysops makerovi, Ch. va-
lidus, Hybomitra olsoi и Н. pavlovskii, а также подвиды Н. nitidifrons nitidifrons и Н.
lundbecki sibiriensis. Первые 4 вида в значительной мере являются горно-таежными.
Всего таежный фаунистический комплекс насчитывает 17 видов (+2 подвида),
в том числе 4 вида Chrysops, 12 видов (+2 подвида) Hybomitra и 1 вид Atylotus. 6 видов,
в том числе 1 вид Chrysops и 5 видов Hybomitra, распространены также в Северной
Америке, т. е. являются голарктическими. Эти виды, а также остальные 11 видов так или
•иначе имеют родственные связи с одной стороны с представителями лесного
фаунистического комплекса, а с другой — с Неарктической (Канадской) фауной, что указывает
на общность происхождения с последней.
Европейск о-сибирский лесной фаунистический
■комплекс. В типичном случае (номинативный вариант) ареалы относящихся
■к этому комплексу видов охватывают лесную зону от Англии и Пиренеев на восток
до Алтая и Енисея (рис. 28). Наиболее эвритопные виды распространены и восточнее —
44 ВВЕДЕНИЕ
до Прибайкалья, 4 вида — еще и в Якутии, из них 2 вида в несколько измененном вида
охватывают распространением дальневосточные леса. Из эвритопных видов некоторые
имеют наибольшее в сравнении с другими распространение в северных частях Евразии,
проникая (главным образом по долинам рек) далеко в зону тайги и даже лесотундры.
Почти все виды рассматриваемого комплекса распространены на Кавказе и некоторые
виды — в восточных гористых частях Казахстана до Джунгарского Алатау
включительно. Многие виды спорадически распространены на крайнем юге Европы, в Малой
Азии и некоторые — в Сев.-Зап. Африке.
К европейско-сибирскому комплексу относятся следующие виды: Chrysops саеси-
tiens, Ch. pictus, Ch. sepulcralis, Tabanus maculicornis, T. miki, T. glaucopis, T. bovinus,
Hybomitra kaurii, H. distinguenda distinguenda, Atylotus fulvus, A. plebejus plebejusr
Heptatoma pelluceps, Haematopota pluvialis и H. crassicornis.
Рис. 29. Распространение восточносибирского лесного фаунистического
комплекса видов боревразийского типа фауны слепней. Схема. Обозначения
см. рис. 26. Горизонтальная штриховка — распространение
дальневосточного варианта комплекса.
Любопытно, что виды, распространенные до Алтая и Енисея, в ряде случаев алло-
патричны таксономически близкородственным видам восточносибирского лесного ком-:
плекса. Таковы пары: Tabanus bovinus — Т. pleskei, Т. maculicornis — Т. geminus,
Haematopota crassicornis — H. tamerlani. Наличие таких викарирующих пар указывает
на сравнительно недавнее их видообразование от общих предковых лесных форм,
по-видимому, в плейстоцене. Tabanus miki представлен в горах Кавказа, Крыма и
Карпат темной формой Т. miki colchidicus, а в горах Восточного Закавказья и Ирана —
подвидом Т. miki ausralis.
Помимо номинативной группы видов, в пределах рассматриваемого комплекса
имеются еще следующие 2 варианта: европейский лесной и европейский горно-лесной.
Европейский лесной вариант характеризуется ареалами, которые на западе б. м.
повторяют таковые лесных видов широкого распространения (с некоторым расширением
границ за счет Пиренейского п-ова и Сев.-Зап. Африки), но на востоке ограничены
Уральскими горами. На картограмме распространение видов этого варианта, почти
полностью налегающее на западную часть основного ареала комплекса, показано
штриховкой в клетку. Виды этого варианта обычно представлены на Кавказе после перерыва
распространения в южнорусских степях. Сюда мы относим Tabanus cordiger, T. sudeticus
и Haematopota italica (на Кавказе неизвестна).
Европейский горно-лесной вариант характеризуется ареалами, охватывающими
горы (и частично предгорья) Европы и изолированно Кавказа. К этому варианту
относятся Hybomitra caucasica, Tabanus rupium и Haematopota scutellata (на Кавказе
замещена подвидом pontica). Первый вид является преимущественно альпийским,
остальные — лесными видами. К этому же варианту мы относим Hybomitra auripila, но ее
ареал на востоке достигает лишь Карпат, Эстонии и Скандинавии, тогда как на Кавказе
этот вид отсутствует. Отметим, наконец, европейские лесные Н- micans и Н. aterrima,
охватывающие распространением Западную Европу, на восток до Польши; второй
вид является преимущественно горным.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 45
Всего европейско-сибирский лесной фаунистический комплекс в пределах СССР
насчитывает 21 вид, в том числе Chrysops — 3, Tabanus — 7', Hybomitra — 4, Atylotus— 2,
Heptatoma— 1 и Haematopota — 4. По происхождению этот комплекс разнороден.
Виды Hybomitra и 1 вид Atylotus (Surcoufiella) обнаруживают ясные связи с
представителями таежного комплекса Боревразии и Северной Америки. Виды Chrysops, Tabanus,
Haematopota и Atylotus s. str. близки видам средиземноморского и отчасти восточноази-
атского типов фауны. Но наибольшие связи имеются с видами восточносибирского
лесного фаунистического комплекса, о чем уже говорилось выше.
Восточносибирский лесной фаунистический
комплекс. Является аналогом европейско-сибирского лесного комплекса. Он
характеризуется ареалами, в типичном случае простирающимися по югу Средней и Восточной
Сибири от Алтая и Енисея (иногда западнее) на восток до низовий Амура, Приморского
края и Сев.-Вост. Китая (рис. 29). Виды этого комплекса в большинстве обнаружены
в Центральной Якутии, но здесь они обычно малочисленны и некоторые из них, видимо,
■обитают оторванно от основного ареала. Некоторые виды рассматриваемого комплекса
распространены на Сахалине и в Японии, в редких случаях найдены на
Корейском п-ове и в Вост. Китае.
К номинативному варианту восточносибирского лесного комплекса мы относим
Chrysops suavis, Hybomitra brevis, H. tarandinoides, H. astur, Tabanus geminus, T. pleskei
и Haematopota tamerlani. Эти виды чаще обитают в предгорно-горных условиях, видимо,
выплаживаясь в ручьях. К рассматриваемому комплексу принадлежат еще Hybomitra
distinguenda contigua и Atylotus plebejus sibiricus. Интересно отметить, что последний
подвид найден и на Камчатке.
В составе рассматриваемого комплекса выделяется дальневосточный лесной
(неморальной) вариант, имеющий ареал, уменьшенный в сравнении с таковым номинатив-
лого варианта. Виды дальневосточного варианта охватывают распространением юг
Приморского краяи Сев.-Вост. Китай, но 1 вид к северу достигает Хабаровска,3 вида —
нижнего и среднего течения Амура, а ареалы 3 видов простираются также в Японию.
К этому варианту принадлежат Chrysops dissectus, Hybomitra ussuriensis, H. nigella,
Л. stenopselapha, H. stigmoptera, Atylotus horvathi и Haematopota stackelbergi. Сюда же
мы относим Н. pluvialis tristis, своеобразный ареал которой охватывает низовья
Амура, о-ва Сахалин, Кунашир и некоторые др., а также Сев. Японию. Chrysops
■dissectus, возможно, является луголесным видом.
Всего восточносибирский лесной фаунистический комплекс в пределах СССР
насчитывает 14 видов и 3 подвида, в том числе Chrysops — 2, Hybomitra —• 8, Atylotus—2,
Tabanus — 2-й Haematopota — 3. Истинное количество видов, входящих в этот
комплекс, несколько больше, так как нами не упомянуты некоторые виды, найденные на
Дальнем Востоке, но ареалы которых остаются неизученными (Chrysops anthrax,
Haematopota koryoensis и др.). По количеству форм восточносибирский комплекс почти
равен европейско-сибирскому комплексу, насчитывающему 21 вид.
Почти все виды рассматриваемого комплекса имеют теснейшие таксономические
-связи с европейско-сибирскими лесными и даже таежными видами Боревразии. 2 вида —
Hybomitra astur и Н. stigmoptera родственны центральноазиатским видам Hybomitra.
Преобладание связей с боревразийской фауной дает нам основание восточносибирский
лесной комплекс вместе с его дальневосточным вариантом отнести к боревразийскому
типу фауны.
Лесостепной фаунистический комплекс. Основной ареал
этого комплекса в значительной мере повторяет таковой европейско-сибирского
лесного комплекса, но его северная граница смещена к югу. В типичных случаях ареалы
видов лесостепного комплекса простираются от Пиренеев на западе до Алтая и Енисея
на востоке (рис. 30). На севере их граница проходит близ 55-й параллели (на западе
около 50° с. ш.), на юге ареалы частично охватывают Средиземноморье или граничат
■с зоной пустыни. У некоторых видов ареалы простираются далеко на север и восток
(до Якутии и Прибайкалья), причем распространение обычно бывает приурочено к
долинам крупных рек, таких как Северная Двина, Обь, Енисей, Лена и т. д. Очевидно,
эти долины обладают более теплым микроклиматом в сравнении с таковым междуречий,
и на севере эти долины являются своеобразными резервациями для южных форм.
Некоторые лесостепные виды аналогичным образом по долинам рек
распространены далеко к югу, вклиниваясь в степную и даже пустынную зоны. Вместе с
лесостепными видами, но не так далеко на юг, бывают распространены также лесные и частично
таежные виды. На далекое проникновение к северу и югу по долинам крупных рек
животных лесостепи обращали внимание многие авторы, А. П. Кузякин (1962) даже
предложил эти долины присоединить к лесостепной (лесолуговой) зоне.
Возможно, что рассматриваемый комплекс правильнее называть европейско-си-
бирским лесостепным в отличие от восточносибирского лесостепного комплекса,
который, видимо, существует как аналог первого. Но недостаток данных о
восточносибирской фауне пока заставляет нас воздержаться от такого предложения.
К лесостепному комплексу мы относим 9 видов: Chrysops par allele grammus, Ch. con-
■cavus (эти виды, возможно, представляют собой западный и восточный подвид одного
ВВЕДЕНИЕ
вида!), Ch. relictus, Ch. rufipes, Hybomitra ciureai, Tabanus bromius, T. autumnalis,
Atylotus rusticus и Haematopota subcylindrica. Большинство этих видов ранее мы
относили к степным формам, однако на основании очертания ареалов их следует считать
лесостепными. В зоне лесостепи эти виды обычно наиболее многочисленны, причем
Рис. 30. Распространение лесостепного фаунистического комплекса видов,
боревразийского типа фауны слепней. Схема. Обозначения см. рис. 26..
некоторые виды явно тяготеют обитанием к озерам междуречий или старицам в речных
долинах.
Наиболее широкое распространение к югу имеют более эвритопные Hybomitra
ciureai, Tabanus bromius и Т. autumnalis — здесь они охватывают своими ареалами
Малую Азию, Кавказ и часть гор Средней
Азии. Наиболее далеко на север, иногда до
Полярного круга, распространены Chrysops
relictus и Hybomitra ciureai, эти же виды
обнаружены в Якутии. Однако почти всюду
на севере (кроме долины Оби и Иртыша) эти
виды малочисленны, что указывает на
несвойственную им экологическую
обстановку. Очень возможно, что
распространению на севере видов рассматриваемого
комплекса могла способствовать
хозяйственная деятельность человека, в частности,
вырубание лесов, расчистка речных долин
с формированием открытых берегов и
луговых участков и т. д. Но распространение
некоторых лесостепных видов, например,
Рис. 31. Распространение среднеазиатского
горного комплекса видов боревразийского
типа фауны слепней. Схема. Обозначения
см. рис. 26.
Chrysops relictus, далеко на севере, в том числе в^Якутии, по-видимому, представляет
собой естественную инвазию в ксеротермический период. К этому вопросу мы еще
вернемся ниже.
По своим таксономическим связям рассматриваемый фаунистичеекий комплекс
гетерогенен, очевидно, отражая гетерогенность ландшафта лесостепи, с которым он
связан. Часть видов, например, Hybomitra ciureai, Haematopota subcylindrica,
Chrysops relictus и другие имеют ясные родственные связи с боревразийскими лесными
видами, тогда как Tabanus bromius и Т. autumnalis, несомненно, родственны
представителям средиземноморского и восточноазиатского типов фауны. Эти оба вида (в виде
южных подвидов) широко охватывают ареалами Средиземноморскую подобласть, а по-
50° 60° 70° 80°
60° 70° 80°
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 47
следний вид— проникает в Афроевразийскую аридную подобласть. Atylotus rusticus
родствен степным A. miser, A. pallitarsis и пустынному А. quadrifarius.
Среднеазиатский горный фаунистичеекий
комплекс. Включает виды с относительно небольшими ареалами, приуроченными в
горных системах Памиро-Алая, Тянь-Шаня, Каратау и Тарбагатая преимущественно
к лесному,, альпийскому и отчасти высокогорному степному поясам (рис. 31). Наиболее
широкое распространение имеют Hybomitra shnitnikovi, H. semipollinosa и Н. turkestana,,
ареалы которых включают все перечисленные выше горные системы. Первые 2 вида;
охватывают ареалами также афганскую часть Памиро-Алая, тогда как Н. turkestana —
восточную (китайскую) часть Тянь-Шаня. Вероятно, здесь обитают и другие виды пере--
численной группы. К номинативному варианту мы относим также Hybomitra hunnorum,.
Н. tatarica, H. montana reinigiana (высокогорные степи) и Tabanus argenteomaculatus,
распространенные на Тянь-Шане и Памиро-Алае. Западнотяныпанско-памироалайский
вариант, как явствует из названия, имеет более узкий ареал и представлен 4 видами:
Hybomitra nigricorpa, Atylotus chodukini, Hybomitra sogdiana и Н. shevtshenkoi. Наконец,
западнопамироалайский вариант представлен 4 видами: Hybomitra zonata, Tabanus-
schiva, Т. hissaricus и Haematopota bactriana. Все виды, кроме Tabanus hissaricus,
охватывают своими ареалами также афганскую часть Памиро-Алая.
Всего, следовательно, среднеазиатский горный фаунистичеекий комплекс
насчитывает 14 видов и 1 подвид, в том числе Hybomitra — 10, Tabanus — 3, Atylotus — 1
и Haematopota — 1. Подавляющее большинство видов (9) обнаруживает тесные
родственные связи с видами лесного и даже таежного комплексов боревразийского типа
фауны. Популяции этих видов доминируют в соответствующих горных поясах
(см. ниже). Для Звидов — Hybomitra tatarica, H. shevtshenkoi и Н. zonata — несомненны,
таксономические связи с представителями фауны Центрально-Азиатской подобласти.
Наконец, 2 вида — Tabanus argenteomaculatus и Т. schiva ■— родственны видам,
распространенным западнее, в том числе в Иране, на Кавказе и в Малой Азии, и, таким
образом, могут рассматриваться как производные восточно-средиземноморской фауны.
Родственные связи своеобразного Т. hissaricus для нас неясны.
Преобладание видов боревразийского происхождения в составе среднеазиатского
горного комплекса дает нам основание отнести его к боревразийскому типу фауны.
Виды слепней, обитающие преимущественно в поясе орехоплодных лесов (и их
аналогов) жни по степным предгорьям и низкогорьям Западного Тянь-Шаня и Памиро-Алая,
мы относим к средиземноморскому типу фауны.(см. ниже).
Рассмотренные нами выше комплексы, относимые нами к боревразийскому типу
фауны и распространенные на территории СССР, включают 79 видов и 6 подвидов.
Средиземноморский тип фауны
Фауна этого типа охватывает распространением юг Европы, север Африки,
Переднюю Азию и Афганистан (без низменных пустынных частей), в СССР — горы Крыма,
часть Кавказа и юг Средней Азии (Копет-Даг, Паропамиз, предгорья, низкогорья и
средний пояс гор Гиссаро-Дарваза, Алая и Западного Тянь-Шаня). Отдельные
средиземноморские элементы проникают на Украину, Среднее и Нижнее Поволжье и на юге.
Средней Азии по предгорьям простираются на восток до Тарбагатая, Западного Алая
и даже Центральной Азии. Большинство видов обитает в горах или предгорьях, при-,
урочиваясь к лесному и степному поясам, иногда альпийскому; некоторые виды
населяют по преимуществу морские побережья.
Средиземноморский тип фауны слепней имеет значительные таксономические
отличия в надвидовых рангах от других фаунистических типов Палеарктики, что вполне,
оправдывает его выделение. Но он обнаруживает ясные генетические связи с восточно-
азиатским типом. К этому вопросу мы вернемся ниже (стр. 75 и 79).
В пределах СССР фауна слепней средиземноморского типа может быть подразде-.
лена на следующие фаунистические комплексы: южноевропейский, средиземноморско-
среднеазиатский, понтийско-гирканский и бактрийско-иранский.
Южноевропейский (северосредиземноморский)
фаунистичеекий комплекс. Виды этого комплекса охватывают
распространением Южную и частью Среднюю Европу, на западе от Иберийского п-ова и на востоке,
частично до Украины, в некоторых случаях до Урала, на юге включая Малую Азию,
и Кавказ (иногда также горную Туркмению и Иран), а некоторые виды проникают
в Северо-Западную Африку (рис. 32). В большинстве виды рассматриваемого комплекса
являются лесными, лесостепными или степными обитателями, но ландшафтно-эко-
логическая характеристика некоторых видов неясна. Особо выделяется прибрежный
(литоральный) вариант. К южноевропейскому комплексу принадлежат Chrysops cae-
cutiens ludens, Ch. jlavipes, Silvius vituli, Tabanus tergestinus, T. bifarius, T. quatuorno-.
tatus, T. unifasciatus, T. tinctus, Philipomyia aprica (два последних вида без Украины),,
Atylotus loewianus и Haematopota grandis. Сюда же могут быть отнесены Tabanus spodop-
terus (на Кавказе его замещает Т. portschinskii) и A tylotus latistriatus (на восток до Крыма}..
48 ВВЕДЕНИЕ
2 вида рассматриваемого комплекса — лесной Silvius vituli и лесостепной Chrysops
flavipes — к востоку распространены до Урала, т. е: значительно восточнее основного
ареала комплекса. Другие 2 вида — Tabanus quatuornotatus и Т. unifasciatus — в Иране
проникают по горной системе Загрос далеко к югу, а в Туркмении распространены
в Копет-Даге. К южноевропейскому комплексу мы относим еще Pangonius pyritosus,
0° К ВОСТОКУ ОТ ГриНВИЧа 90° Ш° «0° 180° 160°
Рис. 32. Распространение южноевропейского фаунистического комплекса
видов средиземноморского типа фауны слепней. Схема. Обозначения
см. рис. 26.
распространенного на Балканах и в Передней Азии, а у нас — на Украине, а также
Dasyrhamphis umbrinus, обитающего в южной и юго-восточной Европе, Передней Азии,
Закавказье и Туркмении (Копет-Даг). Оба вида могут быть обозначены как восточно-
средиземноморские формы.
Рис. 33. Распространение средиземноморско-среднеазиатского
фаунистического комплекса видов средиземноморского типа фауны слепней. Схема.
Обозначения см. рис. 26.
К литоральному варианту принадлежит Chrysops italicus, распространенный по
берегам Средиземного и Черного морей.
Средиземноморско-среднеазиатский фаунистиче-
с'к ий комплекс. В типичном случае распространен по остепненным предгорьям
и^низкогорьям юга Европы, затем в Малой (или Передней) Азии, Иране, Афганистане,
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 49
на Кавказе и на юге Средней Азии, на восток обычно до Каратау, редко восточнее
(рис. 33). Ареалы некоторых видов охватывают также Сев.-Зап. Африку. К этому
комплексу относятся Chrysops flavipes punctifer, Silvius vitripennis (на Кавказе его замещает
S. caucasicus), Tabanus spectabilis, T. bromius flavofemoratus, Т. autumnalis brun-
nescens и Т. leleani (на восток распространен по предгорьям до Западного Алтая и
Центральной Азии — Восточный Нань-Шань и хр. Алашань). Наиболее широко
распространен эвритопный Т. autumnalis brunnescens, который по долинам крупных рек
проникает в пустынную зону.
Основной ареал этого комплекса на западе и в средних частях (включая Кавказ)
в значительной мере накладывается на ареалы видов южноевропейского комплекса.
Понтийско-гирканский фаунистический комплекс.
Ареалы охватывают главным образом горные районы Кавказа и Северного Ирана,
некоторые виды распространены также на Балканах, в Крыму и в Малой Азии (рис. 34).
Рис. 34. Распространение понтийско-гирканского фаунистического
комплекса видов средиземноморского типа фауны слепней. Схема. Обозначения
см. рис. 26.
Представители этого комплекса обитают преимущественно в горнолесном поясе, но
некоторые виды заселяют остепненные предгорья или нагорья, и очень немногие —
альпийский пояс или равнины. Основной ареал комплекса почти целиком налегает
на восточную часть ареала южноевропейского комплекса. На Кавказе, особенно в его
южных частях, преобладают понтийско-гирканские виды.
Наиболее широкое распространение имеет Tabanus indrae, охватывающий своим
ареалом Балканы, Крым (горную часть), Кавказ, Иран (включая южные части хребта
Загрос) и часть гор Средней Азии (Копет-Даг, Гиссаро-Дарваз). Сходный ареал имеет
Т. laetetinctus, но на западе этот вид неизвестен на Балканах и в Крыму, а на востоке
он достигает по низкогорьям до Западного Тянь-Шаня. Балкано-кавказский вариант
этого комплекса имеет в своем составе 5 видов: Silvius latifrons, Tabanus prometheus
(заходит в Иран), Therioplectes tricolor (в Крыму его замещает подвид pallidicauda),
Th. tunicatus и Haematopota longeantennata. Кавказско-иранский вариант включает
значительную группу видов, распространенных в Закавказье и в Северном Иране:
Tabanus shelkovnikovi, Т. semiargenteus, Т. olsufjevi, T. anthrax (известен также в
Турции), Т. atrbpathenicus (на восток до Копет-Дага), Therioplectes carabaghensis (тоже
самое), Th. albicauda (Талыш и южное побережье Каспийского моря) и Haematopota
caspica (с подвидом Я. caspica transcaucasica). Еще более значительную группу
составляет кавказский вариант (эндемики Кавказа). Сюда относятся обитающие на Большом
и Малом Кавказе Silvius zaitzevi, S. caucasicus (преимущественно степные предгорья),
Philipomyia rohdendorfi (альпийский вид), Tabanus armeniacus, Т. portschinskii, T. sub-
paradoxus и Hybomitra caucasi, затем найденные только на Малом Кавказе Chrysops
sejunctus, Tabanus swiridovi, A tylotus proditor и Hybomitra popovi или только на Большом
Кавказе — Tabanus caucasius и Hybomitra montana karatschajensis.
pf. Наконец, упомянем крымский горный вариант, насчитывающий 1 вид и 1 подвид:
Tabanus smirnovi и Therioplectes tricolor pallidicauda. Эти виды имеют весьма
ограниченные по размерам ареалы, они охватывают лесные части южных и северных склонов
Крымских гор.
4 Н. Г. Олсуфьев
so
ВВЕДЕНИЕ
Почти все. виды рассмотренного нами понтийско-гирканского фаунистического
комплекса имеют явственные родственные связи с основными представителями
средиземноморского типа фауны и рассматриваются нами как производные формы
(дериваты) последнего. Лишь Hybomitra popovi, H. montana karatschajensis и Tabanus arme-
niacus представляют собой родственные формы видов боревразийского типа фауны.
Бактрийско-иранский фаунистический комплекс.
Виды этого комплекса охватывают распространением предгорья и горы Средней Азии
(чаще без Копет-Дага), на восток включая Западный Тянь-Шань (немногие виды
распространены и восточнее), к югу и юго-западу горы Афганистана (Гиндукуш) и Ирана
(Загрос и горы Белуджистана) (рис. 35). В горах виды обычно приурочены обитанием
к низкогорьям или среднегорному поясу, в частности, к орехоплодным лесам или их
аналогам. К этому комплексу относятся: Tabanus bactrianus, Т. semenovi, T. mistshenkoi,
Рис. 35. Распространение бактрийско-иранского
фаунистического комплекса видов
средиземноморского типа фауны слепней. Схема. Обозначения
см. рис. 26.
Т. montiasiaticus (на восток до Саура, но в Иране неизвестен), Т. ansarii, Т. accipiter
и Т. golovi (на восток до Джунгарского Алатау, но неизвестен в Иране). Последние
2 вида обитают преимущественно по степным предгорьям и низкогорьям, причем
Т. golovi (подвид Т. golovi mediaasiaticus) по долинам рек проникает в пустынную зону,
в частности, малочисленные популяции этого вида обнаружены в низовьях Амударьи
и Сырдарьи.
Значительный зоогеографический интерес представляет распространение большей
части видов этого комплекса (кроме Tabanus montiasiaticus и Т. golovi) в горах южного
и юго-западного Ирана, причем некоторые виды, например, Tabanus mistshenkoi,
Т. ansarii и Т. accipiter были собраны обследовавшей эти горы Аббасиан-Линтцен (АЬ-
bassian-Lintzen) значительными сериями. К сожалению, остается неясным, являются
ли эти местонахождения изолированными от среднеазиатского ареала, или же между
ними существует связь через горы Северного Ирана, оставшиеся мало обследованными.
Аббассиан-Линтцен (Abbassian-Lintzen) отмечает произрастание в горах Загроса в
местах сбора слепней грецкого ореха, сосны, тополя, платана, дикого миндаля, инжира,
винограда, граната и других растений. В горах Средней Азии, особенно в Гиссаро-
Дарвазе, отмечается сходный флористический состав в местах типичного
распространения бактрийско-иранского фаунистического комплекса слепней.
Виды рассматриваемого комплекса имеют ясные генетические связи с основными
представителями средиземноморского типа фауны. Tabanus mistshenkoi имеет большое
сходство с пустынным Т. sabuletorum, но оба они относятся к группе Т. cordiger,
объединяющей по преимуществу средиземноморские виды. Рассмотренные нами комплексы
средиземноморского типа фауны, обитающие на территории СССР, включают 53 вида
и 6 подвидов.
i
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
51
Афроевразийский аридный тип фауны
Ареалы представителей этого типа фауны охватывают равнинные пустынные
и степные пространства севера Африки, Передней Азии, Ирана, Афганистана, СССР,
Китая и Монголии. В пределах аридных территорий слепни приурочены обитанием
к берегам рек, озер, морским побережьям, рисовым полям, ирригационным каналам,
а также к колодцам, но отсутствуют на безводных пустынностепных участках плакора.
Так как последние доминируют, ареалы степных и особенно пустынных видов
представляются разорванными на отдельные части, иногда сильно удаленные друг от друга
в зависимости от очертаний гидросети («мозаичные» или «ленточные» ареалы).
Отдельные виды на западе местами локально распространены в Южной, реже
в Средней Европе, приурочиваясь чаще к солоноватым водоемам; видимо, эти виды
сохранились здесь от предшествовавшего ксеротермического периода, когда они имели
0° К ВОСТОКУ ОТ ГрИНВИЧа 90° 120° 150° 180° 150°
0° 30° 60° 90° 120°
Рис. 36. Распространение степного фаунистического комплекса видов афро-
евразийского аридного типа фауны слепней. Схема. Обозначения: см. рис. 26.
на западе более широкое распространение. В пределах СССР аридный тип фауны может
быть подразделен на 2 фаунистических комплекса: степной и пустынный.
Степной фаунистический комплекс. Ареалы наиболее
широко распространенных видов этого комплекса охватывают степные пространства от
Украины на восток до Сев .-Вост. Китая, на севере частично заходят в лесостепь и на
юге в зону пустыни. Отдельными пятнами эти виды встречаются в Зап. Европе (рис. 36).
Такое распространение имеют Hybomitra expollicata и Н. montana morgani, причем
первый вид от Тувы к востоку представлен подвидом Н. expollicata orientalis. H. montana
morgani изолированно распространена еще в остепненных местах Малого Кавказа и
Северного Ирана, куда она проникла, вероятно, в ксеротермический период.
Остальные виды этого комплекса имеют менее широкие ареалы. Южноукраинский вариант
представлен 1 видом — Hybomitra ukrainica, распространенным в северо-западном
Причерноморье.
Представители восточноевропейско-североказахстанского варианта охватывают
распространением территорию от Дона (или несколько западнее) до Иртыша (или Оби),
на севере частично проникая в зону лесостепи, на юге в зону пустыни. Сюда относятся
Hybomitra sareptana и Haematopota pallidula.
У видов степного сибирско-монгольского варианта ареалы в основном охватывают
степи Монголии, Сев-.Вост. и Вост. Китая, но у некоторых видов простираются в Зап.
Сибирь и 1 вида — на запад до Волги. На севере ареалы почти всех видов этого варианта
по степям заходят в Забайкалье, а у некоторых — в Предбайкалье (район Иркутска),
Туву и у 1 вида в Якутию (изолированное местонахождение). К этому варианту
относятся Chrysops ricardoae, Ch. angaricus, Atylotus pallitarsis, A. miser, Haematopota de-
sertorum и, видимо, Hybomitra tschuensis (горностепной Алтай). Последние 2 вида
являются викариатами Haematopota pallidula и Hybomitra sareptana, распространенных
западнее. Следует отметить, что популяции Chrysops ricardoae, распространенные оторг
ванно от основного ареала в низовьях Волги и Урала и затем в Центральной Якутии,
образуют локальные подвиды. Atylotus miser,, ката и его западный викариат A. rustir
cus, возможно, является в большей степени лесостепным, чем степным видом. Он
4*
52
ВВЕДЕНИЕ
весьма многочислен в южных частях Приморского края, обитая с некоторыми другими
видами степного комплекса.
Всего рассматриваемый комплекс насчитывает в пределах СССР 11 видов.
Последние гетерогенны по происхождению. Hybomitra и Chrysops обнаруживают связи с бор-
евразийскими лесостепными и частью таежно-лесными формами, Atylotus и Haematopota
в той или иной мере родственны пустынным видам.
Пу стынный фаунистический комплекс. Отличается
разнообразием видов и множеством вариантов ареалов, которые в будущем, возможно,
удастся выделить в самостоятельные комплексы.
Изученность ареалов ряда видов пустынного комплекса явно недостаточна, в
особенности за рубежом. По этой причине на картограмме (рис. 37) основной ареал рас-
матриваемого комплекса обозначен вертикальным штрихом лишь для территории СССР
(пустыни Средней Азии и Вост. Закавказья). За пределами СССР границы основного
Рис. 37. Распространение пустынного фаунистического комплекса видов
афроевразийского аридного типа фауны слепней. Схема. Обозначения
см. рис. 26. Ввиду недостатка данных границы основного ареала комплекса
за пределами СССР показаны прерывистой линией провизорно, исходя из
примерных границ пустынной зоны.
ареала пустынного комплекса помечены провизорно прерывистой линией, а
фактические местонахождения учтенных нами видов обозначены точками.
Многие виды рассматриваемого комплекса обитают в Сев. Африке и Передней
Азии, не достигая ареалами территории СССР. Из числа распространенных в СССР
видов наиболее широкий ареал имеет Atylotus pulchellus — от Сев. Африки через
Переднюю и Среднюю Азию до Вост. Китая. Широкое распространение, охватывающее почти
все пустынные территории Палсарктики (в том числе мозаично на юге Европы), имеют
A. quadrifarius, A., flavoguttatus (на восток до Прибалхашья), Hybomitra acuminata,
Л. erberi, H. peculiaris, Tabanus sabuletorum и Haematdpota turkestanica (последние 2 вида
на запад известны до северо-западного Причерноморья). Некоторые из перечисленных
видов, и Особенно Haematopota turkestanica, к северу от основного ареала довольно
далеко проникают в степную зону, но обычно здесь их популяции немногочисленны.
Всего номинативный вариант насчитывает 8 видов. Переднеазиатско-кавказско-туран-
'ский вариант представлен Tabanus zimini, Т. filipjevi (на восток до Монголии), Т. fu-
midus и Haematopota pollens. Турано-монгольский вариант представлен 1 видом —
'Chrysops mlokosiewiczi, казахстано-монгольский вариант также 1 видом — Tabanus
•brunneocallosus. Довольно многочислен туранский вариант (туранские эндемики),
включающий 6 видов: Chrysops oxianus (на запад до пустынных частей Закавказья),
Ch. stackelbergi, Ch. hyalipennis, Nanorhynchus crassinervis, Hybomitra turanica и Tabanus
■submbuletorum.
Рассматриваемый пустынный комплекс насчитывает в пределах СССР 20 видов.
Происхождение его сложно и, несомненно, формирование видов осуществлялось из
фазных источников. Виды рода Tabanus (подгруппа Т. sabuletorum группы Т. cordiger),
■Chrysops (подрод Heterochrysops ) и Atylotus обнаруживают родственные связи с
средиземноморскими и частью восточноафриканским'и видами, представители рода
Hybomitra — это видоизмененные родичи боревразийских Hybomitra, особенно подгруппы
Д. montana. Chrysops mlokosiewiczi весьма близок восточноазиатскому Ch. vanderwulpi.
ГЕОГРАФИЧЕСКОЕ. РАСПРОСТРАНЕНИЕ 53
Своеобразные по строению Nanorhynchus crassinervis и Chrysops hyalipennis
обнаруживают связи (несомненно, древние) с некоторыми североамериканскими видами.
Рассмотренные нами комплексы афроевразийского аридного типа фауны,
распространенные в СССР, всего включают 30 видов и 2 подвида.
\
Восточноазиатский тип фауны
Охватывает распространением юго-восток Палеарктики, главным образом в
пределах Китая, Корейского п-ова и Японии, где представлен значительным числом видов.
Вероятно, фауна рассматриваемого типа может быть подразделена на ряд комплексов,
но недостаток данных, особенно по Китаю и Корейскому п-ову, не позволяет нам этого
сделать. Все виды, которые своими ареалами захватывают территорию СССР,мыотш>-
сим к одному фаунистическому - комплексу— японо-китайскому, хотя экологически
они неодинаковы: одни виды, видимо, лесные, другие — степные.
Рассматриваемые нами виды, а их всего 9, распространены главным образом в
пределах Приморского, отчасти Хабаровского края и других территорий. Но они здесь
представлены малочисленными популяциями, тогда как доминируют и по количеству
видов, и по численности особей представители боревразийской фауны (см. ниже). Ос-
0° К ВОСТОКУ DT ГрИНВИЧа 90° 120° 150° 180° 150°
0° 30° 60° 90° 120°
Рис. 38. Распространение видов восточноезиатского типа фауны слепней.
Схема. Обозначения см. рис. 26.
новной ареал рассматриваемого комплекса расположен- за пределами СССР, причем
ареал типа охватывает восточную и юго-восточную часть Китая, Корейский п-ов и
Японию (без о. Хоккайдо) (рис. 38). Некоторые виды частично заходят в Ориентальную
(Индо-Малайскую) область. Наиболее широко распространены Chrysops japonicus,
Ch. vanderwulpi, Tabanus amaenus, T. trigeminus, T. signatipennis и Т. chrysurus. Они
известны как в Китае, так и в Японии.
Поразительно распространение Chrysops vanderwulpi. Этот характерный
представитель Врсточноазиатской фауны, ареал которого на юге простирается до Ю. Китая
(о-ва Тайвань и Хайнань), изолированными популяциями обитает в Центральной
Якутии, Прибайкалье и по долинам некоторых крупных рек Средней и Западной Сибири
(рис. 107, 5). Эти популяции, несомненно, представляют собой сохранившиеся «остатки»
от фауны предшествовавшего более теплого периода.
К китайскому варианту рассматриваемого комплекса мы относим Tabanus buddha,
к японскому варианту — Т. sapporoenus (о-ва Сахалин и Кунашир) и Т. kinoshitai
(о. Кунашир), широко распространенные в Японии и неизвестные в Китае.
Некоторые из перечисленных видов, например, Т. chrysurus, T. buddha, T.
amaenus и др. Н. А. Виолович (1968) определил как представителей индо-малайского типа
фауны (т. е. Ориентальной области). С этим нельзя согласиться, так как ареалы этих
видов в основном охватывают Восточноазиатскую подобласть Палеарктики.
Большинство видов рассмотренного нами комплекса имеет» несомненно,
родственные связи с более южной восточноазиатской фауной, а также отчасти с представителями
средиземноморской фауны.
54
ВВЕДЕНИЕ
КОЛИЧЕСТВЕННЫЕ АСПЕКТЫ ФАУНЫ СЛЕПНЕЙ
ЛАНДШАФТНЫХ ЗОН И ГОРНЫХ СИСТЕМ СССР '
Наряду с изучением типологии ареалов для зоогеографии важно также выяснение
численных соотношений популяций отдельных видов животных, заселяющих ту или
иную ландшафтную зону, иными словами структуры населения.
Практический смысл изучения количественных аспектов фауны заключается
в выявлении видов, имеющих наибольшее экономическое значение, а теоретический —
в выяснении биоценотических отношений и уточнении зоогеографического
районирования. В СССР изучение количественных аспектов фауны слепней наряду с выяснением
видового состава привлекло к себе внимание многих энтомологов, так как в отличие от
типологии ареалов оно вполне доступно в плане краевого исследования. За ряд лет
опубликовано большое число работ этого направления (свыше сотни) по разным
областям и краям СССР. Излагаемый ниже раздел по аспектам фауны слепней лишь сжато
суммирует накопившиеся данные (без таблиц), но в начале рассмотрения тех или иных
ландшафтных зон или горных систем нами сделаны ссылки на использованную
литературу.
Количественные аспекты видов слепней в отличие от простого их перечня
достаточно хорошо характеризуют отдельные ландшафты и могут вполне применяться в
качестве их показателей (индикаторов). В то же время слепни (взрослая фаза) мало
пригодны для характеристики отдельных стаций (биотопов), так как вследствие своей
подвижности они легко перемещаются из одной стации в другую, и поэтому видовой
состав их носит сравнительно смешанный характер. В горных местностях следует
считаться с возможностью сноса личинок вниз горными потоками и появления отдельных
взрослых особей горных видов в низкогорьях и даже предгорьях.
Ниже мы рассмотрим основные черты фауны слепней отдельных ландшафтных зон
и горных систем СССР, уделяя внимание «аспектным» или «фоновым» видам.
Количественное определение слепней основывается на учетных сборах самок в
момент их нападения на домашнее животное или человека. Учеты вокруг себя или около
животного могут дать несколько различные результаты особенно за счет крупных видов
типа Tabanus bovinus, которые нападают на человека с меньшей охотой. Учеты могут
также производиться специальными ловушками, например, ловушкой Скуфьина, или
отловом на ловчую лужу, покрытую керосином, но получаемые этим путем результаты
не всегда совпадают с данными учетов, на животном. Регулярно проводимые в течение
сезона сборы позволяют рассчитать в процентах соотношение (обилие) собранных
видов. Очень важно произвести сборы в течение всего сезона активности слепней,
например, в начале, середине и конце лета, так как в разные сроки соотношение популяций
разных видов может быть резко различным. Исследования показали, что
количественное соотношение популяций видов слепней в разные годы наблюдений обычно
изменяется мало, поэтому даже односезонные наблюдения дают ценные результаты для
Таблица 5
Количество видов и подвидов слепней
в равнинных зонах и горных системах СССР
Пространственные
группировки
Природная территория
Подвид
Всего
I. Равнинные зоны
II. Горные системы
Зона тундры
» тайги
» европейскосибирских лесов
» лесостепи
» - дальневосточных лесов . .
» степи
» пустыни
Карпаты
Горы Крыма
» Кавказа
Уральские горы
Горы Средней Азии
Алтайские горы
Горы Восточной Сибири ....
Сихотэ-Алинь
20
51
46
50
43
33
24
27
11
70
36
45
43
22
34
1
6
1
1
6
4
7
2
4
23
1
11
4
(неполные
данные)
2
21
57
47
51
49
37
31
29
15
93
37
56
47
36
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 55
характеристики аспектности фауны. Во всяком случае фоновые виды почти всегда будут
выявлены.
По степени обилия в сборах того или иного вида слепней К. В. Скуфьин (1949а)
предложил делить виды на 4 группы: доминирующие, или массовые
виды — 8% и более общей численности; субдоминанты, или
многочисленные виды — от 2 до 8%; малочисленные виды — от 0.5 до 2% я
редкие, или локальные виды — менее 0.5% от общей численности. В
дальнейшем К. В. Скуфьиным (1952а) было предложено выделить еще группу весьма редких
видов — менее 0.1% от общей численности, но нам представляется целесообразным
ограничиться подразделением степени обилия слепней на первые 4 группы. Степень
обилия, или аспектность того или иного вида в той или иной ландшафтной зоне,
В. В. Шевченко (19536) предложил определять не только по степени господства, т. е.
процентного отношения числа особей данного вида к числу особей всех видов, но и по
коэффициенту постоянства, т. е. процентному отношению числа встреч данного вида
к общему числу обследований (учетов) в данной зоне.
Аспектность фауны мы рассмотрим раздельно для равнинных ландшафтных зон
и для горных систем, имеющих вертикальную ландшафтную зональность (поясность).
Последние имеют фауну, отличающуюся от таковой окружающих их равнин, что
проявляется особенно резко в горных системах южного расположения. Некровососущие
виды мы исключаем из рассмотрения, поскольку методы их сбора и количественные
определения иные, чем кровососущих форм. Всего нами подвергнута анализу фауна
7 равнинных ландшафтных зон и 8 горных систем, по каждой природной территории
учтены все обнаруженные на ней виды и подвиды слепней, независимо от
принадлежности их к тому или иному фаунистическому комплексу (табл. 5). В пределах
равнинных зон наиболее разнообразна фауна в зоне тайги — 51 вид и 6 подвидов, из горных
систем — фауна Кавказа — 70 видов и 23 подвида.
Аспекты фаун равнинных ландшафтных зон
Тундровая зона. Охватывает полосой, местами шириной до 800 км,
север Евразии от Кольского п-ова до Чукотского п-ова и севера Камчатки. Фауна
слепней рассматриваемой зоны (и подзоны лесотундры) освещена в работах Ф. Синтениса
(1904), Н. Г. Олсуфьева (1934), В. Ю. Фридолина (1936), К. А. Бреева (1950), В. А.
Полякова (1965, 1974), Н. А. Виоловича (1968) и др.
В пределах всей зоны доминантами или субдоминантами
являются Hybomitra sexfasciata, H. arpadi и Н. montana (местами преобладает темная
форма Н. montana var. flaviceps); в европейской части в тундре многочислен
также (но не доминирует) Н. lundbecki, а в Сибири, начиная с Северного Урала и до
Чукотского п-ова абсолютно доминирует Я. aequetincta. Второй тундровый вид—
Н. astuta — обычно всюду редок, но В. А. Поляков (личное сообщение) обнаружил
в 1965 г. несколько более многочисленную популяцию этого вида (собрана серия 23 ?$>)
при отсутствии других видов в Таймырской тундре близ поселка Усть-Авам на р. Ду-
дыпте (71°20' с. ш.)-. Этот же исследователь собрал значительную серию (79??) этого
вида в июле 1971 г. в лесотундре верховьев р. Альденван, 150 км ссз пос. Омолон
Магаданской обл.
В зоне тундры малочисленны или редки другие таежные виды — Chrysops nigri-
pes, Hybomitra lapponica, H. nitidifrons eonfiformis, H. lurida и H. nigricornis.
Заслуживают упоминания единичные находки в зоне тундры европейской части СССР,
главным образом по долинам рек, лесных Chrysops sepulcralis, Ch. caecutiens, Haematopota
pluvialis, H. crassicornis и лесостепного Chrysops relictus (оз. Парусное на Канином п-ове).
По нашему мнению, это «остатки» популяций южных видов, сохранившиеся от
предшествовавшего ксеротермического периода.
В подзоне лесотундры, являющейся переходной между тундрой и расположенной
к.югу тайгой,-д оминантами и субдоминантами остаются виды, аспёкт-
ные в зоне тундры, но многочисленными становятся Chrysops nigripes
(местами обилие до 11—17% *), Hybomitra lurida (9—15%) и Н. nigricornis (2—23%),
последний вид многочислен лишь в Сибири. В Западной Сибири многочислен также
Н. nitidifrons eonfiformis. Малочисленными являются остальные виды, встречающиеся
в тундре, причем добавляются Н. bimaculata, H. tarandina, H. muehlfeldi, а в Восточной
Сибири — еще Н. olsoi (до 25%) и Chrysops makerovi (до 2%).
Та ежная зона. Охватывает широкую полосу- от Карелии и
Ленинградской обл. на восток до Камчатки и Сахалина (кроме его южной части). Фауну
слепней этой зоны изучали А. А. Штакельберг (1916, 1954), Н. Г. Олсуфьев (1934,
1936а), Ю. Г. Митрофанова (19,51, 1955), А. С. Лутта (1959, 1960, 1970), О. Н. Сазонова
* Здесь, как и в дальнейшем изложении, указываемый в скобках процент означает
отношение особей (?$) данного вида к числу особей всех видов слепней при массовых
сборах.
56 ВВЕДЕНИЕ
(1949), В. В. Попов (1962), Н. А. Виолович (1966, 1968), И. М. Олигер (1967), И. Г. Бей-
Биенко (1970), Ю. И. Запекина-Дулькейт (1969), В. Г. Фоминых с соавт. (1972),
Р. П. Павлова (1972), В. Б. Купрессова с соавт. (1972) и др.
В пределах рассматриваемой зоны в ее отдельных частях уже отмечается некоторая
неоднородность в аспектах фауны слепней.
В европейской части СССР в северных и средних частях тайги доминантами
являются таежные Hybomitra lundbecki, Н. bimaculata, H. tarandina, И. arpadi и
лесная Haematopota pluvialis, субдоминантами — таежные Hybomitra lapponica,
Н. nitidifrons confiformis, Н. muehlfeldi, H. montana и лесной Chrysops caecutiens (по
ручьям и речкам), относительно малочисленны таежные Hybomitra lurida, Н._ nigricornis,
Chrysops nigripes и Ch. divaricatus. В южных частях тайги наряду с перечисленными
видами многочисленны еще лесные Tabanus maculicornis, Atylotus fulvus
(местами), лесостепные Tabanus bromius, Hybomitra ciureai и в небольшом количестве
встречаются лесные Chrysops pictus, Ch. sepulcralis, Hybomitra kaurii, H. distinguenda,
Tabanus bovinus, Heptatoma pellucens, Haematopota crassicornis и лесостепные Atylotus rus-
ticus и Chrysops relictus, некоторые из этих видов проникают в среднюю и даже северную
тайгу. На крайнем юге таежной зоны единично встречаются лесные Tabanus glaucopis,
Т. cordiger, Т. miki, T. sudeticus и Haematopota italica.
Обращает на себя внимание, что по мере продвижения к востоку влияние лесной
фауны в южных частях тайги возрастает — например, в бассейне Ветлуги и Вятки
к северу от верхней Волги, по данным И, М. Олигера (1967), наряду с массовыми
эвритопными таежно-лесными видами Hybomitra bimaculata, H. lundbecki, H.
muehlfeldi, Н. nitidifrons и другими многочисленны Chrysops pictus (обилие до 21—37%),
Tabanus maculicornis (до 24—30%), Т. bromius (до 20—28%), Т. bovinus, Hybomitra
ciureai, Haematopota italica и др. В противоположность этому редки таежные
Hybomitra tarandina, H. arpadi, H. lapponica, H. lurida, H. nigricornis. Здесь же встречаются,
по обычно малочисленны такие южные элементы, как Silvius vituli (до Кирова!),
Chrysops flavipes (Казань), Tabanus autumnalis и Haematopota subcylindrica.
Такое расхождение между типом ландшафта — тайгой, и аспектом фауны
слепней — преимущественно лесным, надо объяснить относительно недавним наступлением
тайги на леса и степи в предшествовавший современному период формирования
ландшафтов европейской части СССР. Изменения в составе фауны слепней, не связанных
питанием с определенными растениями, несколько отстают от изменений лесного
покрова. Южные элементы в таежной зоне — это, видимо, реликты предшествовавшего
более теплого периода.
В таежной зоне Западной Сибири сохраняется сходный аспект фауны в сравнении
с таковой европейской части СССР. Доминантами и
субдоминантами являются Hybomitra lundbecki, H. bimaculata, H. arpadi, H. tarandina, H.
lapponica, Haematopota pluvialis (особенно в южной подзоне), местами Hybomitra montana,
Н. muehlfeldi и др. Но в отличие от тайги европейской части СССР значительно
возрастает по всей зоне численность Н. nitidifrons (3—16%) и Н. lurida (4—9%), в южной
подзоне местами становится много лесного Tabanus miki (3—6%), а по долинам рек —
лесостепных Hybomitra ciureai и Chrysops relictus, но уменьшается количество Tabanus
maculicornis, T. bromius и Chrysops caecutiens, которые становятся малочисленными или
редкими. Спорадичны лесные -Ch. pictus, Ch. sepulcralis и Haematopota crassicornis.
В южную подзону тайги проникают в небольшом количестве лесные Tabanus glaucopis,
Hybomitra distinguenda, Atylotus fulvus, Heptatoma pellucens orientalis (Томск и
восточнее), лесостепные ■ Tabanus autumnalis, Haematopota subcylindrica, Atylotus rusticus.
Местами (Тюменская обл.) довольно обычен Tabanus bovinus (до 6%).
Более заметно аспект фауны претерпевает изменения между Енисеем и Байкалом.
•В средней и южной тайге продолжают оставаться доминантами лишь
Hybomitra lundbecki, H. arpadi, H. bimaculata и на юге зоны — Haematopota pluvialis,
местами субдоминантами удерживаются Hybomitra montana, H. lurida, H.
muehlfeldi, Atylotus fulvus (средняя тайга) и Hybomitra ciureai, но переходят в группу
малочисленных или редких Н. tarandina, H. nitidifrons и исчезают лесные Tabanus
maculicornis и Т. bromius. Местами многочисленны Hybomitra nigricornis (до 12%),
Я. lapponica (до 25%), Chrysops divdricatus (северная и средняя'тайга — до 6%), Ch.
caecutiens (южная тайга — до 3%). Остальные виды сохраняются по преимуществу как
малочисленные или редкие. В небольшом количестве встречаются восточные таежные
виды — Chrysops makerovi, Hybomitra olsoi и Н. pavlovskii, а преимущественно в южных:
частях зоны — восточносибирские лесные виды — Н. astur (местами до 20%), Н.
brevis'(p,o 7.5%), Tabanus pleskei (до 2%) и малочисленные Т. geminus, Haematopota
tamerlani и Chrysops suavis. Однако на участке между Енисеем и Байкалом и далее до
верхнего Приамурья на долю этих видов суммарно обычно приходится 10—25% обилия,
тогда как основную массу составляют особи таежных видов (местами на юге резко
преобладает лесная Haematopota pluvialis). В северных частях зоны сильно влияние фауны
лесотундры, что выражается вдоминировании(до 27%) Hybomitra sexfasciata.
К доминантам относятся еще Н. lundbecki, H. olsoi, H. nigricornis, H. arpadi,
субдоминантам — Я. montana. Местами многочислен Chrysops nigripes. Осталь-
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 57
ные таежные и таежно-лесные виды малочисленны или редки. В небольшом количестве
отмечаются тундровые Hybomitra aequetincta и Н. astuta, а также лесные и лесостепные
Chrysops caecutiens, Atylotus fulvus, Hybomitra ciureai и Haematopota tamerlani. В
пределах Якутии и Магаданской обл. аспектность фауны слепней таежной зоны изучена
недостаточно. По разрозненным данным можно заключить, что фоновыми видами
здесь продолжают оставаться Hybomitra lundbecki (подвид Н. lundbecki sibiriensis)
и Н. arpadi, но к ним добавляются как доминанты или субдоминанты
Н. montana (обилие до 52%), Н. nigricornis, H. lurida, H. olsoi, Chrysops makerovi и
CH. nigripes, а на севере—Hybomitra sexfasciata и местами Chrysops divaricatus (Верхний
Сеймчан). Малочисленны Ch. validus, Hybomitra muehlfeldi, H. tarandina, H.
nitidifrons, H. bimaculata, H. lapponica и Н. pavlovskii.
Обращает на себя внимание наличие в Центральной Якутии многих лесных и
даже степных видов, популяции которых, видимо, обитают здесь в ряде случаев
изолированно от основного ареала. Так, здесь найдены западные лесные виды Chrysops
caecutiens, Haematopota pluvialis, Hybomitra distinguenda, лесостепные Chrysops relictus
и Hybomitra ciureai, восточносибирские лесные Hybomitra astur, H. brevis, H. tarahdi-
noides, Tabanus geminus, Haematopota tamerlani, степной Chrysops ricardoae (подвид
Ch. ricardoae jakutensis) и даже представитель восточноазиатского типа фауны Ch. van-
derwulpi. Но популяции большинства "перечисленных видов малочисленны и
присутствие их здесь мы объясняем распространением их в предшествующий ксеротермиче-
ский период.
Еще менее изучена аспектность фауны слепней таежной зоны в пределах
Амурской обл. и севера Хабаровского края. В сборах С. П. Расницына, А. П. Бурыкиной
и других в окр. Г. Зеи Амурской обл. абсолютно доминировала Hybomitra
lundbecki sibiriensis (46%) и в меньшей степени — Н. montana (14%) и Н. astur
(13%), многочисленны Я. lapponica (4%) и Я. montana, малочисленны (1—3%)
Я. arpadi, Я. brevis, H. olsoi, H. pavlovskii, Я. distinguenda contigua, Я. sexfasciata,
Я. nigricornis, Chrysops suavis и редки Hybomitra lurida, Chrysops makerovi, Ch. validus
и др. Налицо преобладание таежных форм с небольшой примесью представителей
восточносибирской лесной фауны.
Н. А. Виолович (1968) для побережья Охотского моря указывает в качестве д о-
минантов Hybomitra borealis (вероятно, Н. sexfasciata), H. arpadi, Я. astur и
Я. lurida, а несколько западнее— Chrysops nigripes и Ch. makerovi. На Камчатке,
также по разрозненным данным, дом и>н антами и субдоминантами
определяются Hybomitra montana, Я. lundbecki (вероятно, подвид Н. lundbecki
sibiriensis), Я. lapponica, H. arpadi, Chrysops nigripes, Hybomitra bimaculata, H. nigricornis,
немногочисленны Я. lurida, H. muehlfeldi, H. aequetincta, H. sexfasciata, H. astuta
и Chrysops makerovi. He найдены Hybomitra tarandina, H. nitidifrons, H. pavlovskii,
H. olsoi и Chrysops divaricatus, возможно, из-за недостаточного количества
исследований. Обращает на себя внимание отсутствие на Камчатке лесных видов слепней (иа
числа кровососущих форм). Найден лишь некровосос Atylotus plebejus sibiricus. На
Сахалине в пределах его средней части (север острова остался необследованным),
относящейся к таежной зоне, доминантами и субдоминантами
являются тундрово-таежный Hybomitra sexfasciata, таежные Н. lapponica, H. arpadi,
Я. tarandina, H. montana, H. nigricornis, Я. bimaculata, Chrysops makerovi и
дальневосточные или восточносибирские лесные Haematopota pluvialis tristis, Chrysops suavis-
и Hybomitra astur. Малочисленны или редки таежные Chrysops nigripes, Hybomitra
olsoi, H. lurida и лесной вид — Haematopota tamerlani. Фауна слепней юга Сахалина
и Курильских о-вов нами освещена ниже.
Всего на огромной территории, занимаемой таежной зоной, обнаружено 51 вид.
и 6 подвидов слепней. Наибольшее количество — 23 вида и 3 подвида — приходится на
лесные виды, широко проникающие с юга, тогда как таежные виды насчитывают 16 видов
и 2 подвида. Но по обилию особей господствующее положение занимают таежные виды.
С севера в зону тайги местами проникают 2 тундровых и 1 лесотундровый вид, с юга —
некоторые лесостепные (6 видов) и немногие более южные формы (3 вида и 1 подвид).
Но они чаще малочисленны по числу особей.
Европейско-сибирская лесная зона. В пределах СССР она
охватывает пространство от Прибалтийских республик и западных границ с Польшей
и Румынией на восток до среднего Приобья, далее, отдельными пятнами, до Канска за
Енисеем. Аналогом этой зоны является дальневосточная лесная зона.
В северных частях лесная зона на широком пространстве переходит в таежную зону,,
образуя подзону смешанных лесов, на юге — в сочетании со степями формирует зону
лесостепи (или островных лесов).
Данные о видовом составе и аспектах фауны слепней лесной зоны имеются в
публикациях А. А. Штакельберга (1922), А. В. Гуцевйча (1943), Н. Г. Олсуфьева (1949а),
Н. Г. Олсуфьева и Л. П. Мосолова (1964), М. И. Волковой (1950), Ю. Г. Митрофано-
вой (1951, 1955), Е. 3. Таскаевой (1951), Г. В. Бошко (1950), В. Ж. Алифанова (1955),
А. Г. Маевского (1956, 1960), И. Г. Бей-Биенко (1957), X. Ремма (1959), И. М. Олигера
(1960), О. Ф. Колесовой (1966), К. С. Растегаевой.(1960, 1962), В. В. Попова (1962),
58 ВВЕДЕНИЕ
Ф. Г. Булановой (1963, 1967), А. Ф. Кателиной и Ю. А. Мясникова (1965), Н. А. Вио-
ловича (1966, 1968) и др.
В европейской части СССР д оми пирующими видами лесной зоны
являются Tabanus maculicornis, Т. bromius, Hybomitra bimaculata, H. lundbecki и Нае-
matopota pluvialis, субдоминантами Hybomitra muehlfeldi, H. ciureai, H. ni-
tidifrons confiformis, H. montana, Chrysops caecutiens, Ch. pictus и Tabanus bovinus.
Малочисленны или редки таежные Chrysops divaricatus, Hybomitra lurida, лесные Chrysops
sepulcralis, Hybomitra distinguenda, H. kaurii (в Эстонии обилие 2.7%), Tabanus miki,
Т. cordiger, Т. glaucopis, Т. sudeticus, Atylotus fulvus (в окр. Иванова обилие 2.5%),
Haptatoma pellucens (в Тульской обл. обилие 3.3%), Haematopota crassicornis и Н. ita-
lica, лесостепные Chrysops relictus, Ch. concavus, Tabanus autumnalis, Atylotus rusticus
и Haematopota subcylindrica. Спорадически распространен средиземноморский лесной
вид Silvius vituli. В подзоне смешанных лесов в небольшом количестве встречаются
таежные Chrysops nigripes, Hybomitra lapponica, H. arpadi, H. tarandina, H. nigricornis.
Представляет интерес отметить обнаружение в Эстонии европейского лесного
(и частью горного) Hybomitra auripila, собранного единичными экземплярами.
Заслуживает внимапия обитание в Нижнем Прикамье немногочисленных популяций степных
видов Haematopota pallidula и Hybomitra expollicata. Здесь это несомненно реликты
ксеротермического периода.
В лесной зоне Западной Сибири доминантами являются Hybomitra
bimaculata (до 70%) и Н. ciureai (14%), субдоминантами Я. nitidifrons confiformis,
Н. lundbecki, Haematopota pluvialis, местами H. subcylindrica, Tabanus bromius, Т. miki,
Т. glaucopis, малочисленны или редки лесные Hybomitra distinguenda, H. kaurii,
Tabanus maculicornis, Т. bovinus, Atylotus fulvus, Chrysops caecuVens, лесостепные Tabanus
autumnalis, Atylotus rusticus, Chysops relictus и таежные Chrysops nigripes, Ch.
divaricatus, Hybomitra lapponica, H. arpadi (местами обычен), H. tarandina, H. lurida, H.
nigricornis, H. montana и Н. muehlfeldi.
Из лесостепной зоны единично проникают степные Hybomitra expollicata, Atylotus
pallitarsis и пустынная Haematopota turkestanica.
К востоку от Оби появляются в небольшом количестве восточносибирские лесные
виды — Hybomitra brevis, Haematopota tamerlani, Chrysops suavis, Tabanus geminus,
T. pleskei. В^неболыпом сборе слепней О. М. Мартыновой в окрестностях Новокузнецка
Кемеровской оби. на границе лесостепи и лесных предгорий оказались
представленными таежные Hybomitra lundbecki, H. bimaculata, H. muehlfeldi и Chrysops nigripes,
евронейско-сибирские лесные Chrysops caecutiens, Hybomitra distinguenda, Tabanus
bovinus и Haematopota pluvialis, лесостепной Chrysops relictus и восточносибирские
лесные Hybomitra tarandinoides и Н. brevis. Налицо преобладание таежных и лесных форм.
Всего на территории европейско-сибирской лесной зоны* обнаружено 46 видов
и 1 подвид слепней. Наибольшее количество — 23 вида — приходится на лесные виды
(из них 6 — дальневосточные формы), они же господствуют по численности особей.
С севера в зону проникают таежные 11 видов и 1 подвид, с юга — 7 лесостепных, 3
степных, 1 пустынный вид, а также 1 средиземноморский лесной вид.
Лесостепная зона. Расположена к югу от лесной зоны, охватывая
значительные пространства юга европейской части СССР и Западной Сибири.
Данные о видовом составе и аспектах фауны слепней имеются в работах Н. Г.
Олсуфьева (1936, 19496), Н. Г. Олсуфьева с соавт. (1958), К. В. Скуфьина (1952а),
Г. В. Бошко (1957), В. В. Попова (1962), Н. А. Виоловича (1966, 1968), В. Г. Федорова
(1971) и др.
В северных и средних частях лесостепной зоны в пределах европейской части
СССР аспекты фауны слепней заметно иные, чем в лесной зоне. Доминантами
являются лесостепная Hybomitra ciureai (до 30%), лесные Tabanus bovinus, Chrysops
pictus, Haematopota pluvialis и таежно-лесная Hybomitra bimaculata, на долю которых
приходится до 75% обилия. К субдоминантам относятся лесостепные Tabanus
bromius, T. autumnalis (местами), Chrysops relictus, Haematopota subcylindrica, Atylotus
rusticus (местами), лесной Tabanus glaucopis и таежно-лесная Hybomitra nitidifrons
confiformis. В сравнении с лесной зоной уменьшается до уровня малочисленных или
редких Tabanus maculicornis, Chrysops caecutiens, Hybomitra lundbecki и Н. montana.
Редко встречаются таежные Chrysops divaricatus, Hybomitra lurida, H. nigricornis и
лесные H. distinguenda, Tabanus miki, T. sudeticus, Atylotus fulvus, Heptatoma pellucens
и Haematopota crassicornis. В южных частях зоны появляются как редкие виды
лесостепные Chrysops concavus (на западе Ch. parallelogrammus), степные Hybomitra expollicata,
Haematopota pallidula и даже пустынные Atylotus quadrifarius, Hybomitra acuminata,
Haematopota turkestanica, H. pallens, Tabanus sabuletorum и Atylotus flavoguttatus. Для
лесостепи можно еще отметить средиземноморских Chrysops flavipes, Tabanus tergestinus,
Т. bifarius, Silvius vituli, а на Украине (преимущественно на западе) и в Молдавии —
Pangonius pyritosus, Tabanus spectabilis, Т. spodopterus, Т. quatuornotatus, Т. unifasciatus
и Haematopota grandis, но эти виды в общем редки за исключением Chrysops flavipes,
местами обычного по долинам рек. В долинах крупных рек (Волги, Дона, Днепра,
Днестра и др.) доминантами обычно являются Tabanus bromius, T. bovinus,
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 59
Т. autumnalis, Hybomitra ciureai, Chrysops relictus, Haematopota subcylindrica, Atylotus
rusticus и др. В большинстве это виды лесостепного комплекса.
В лесостепи Западной Сибири абсолютное доминироваиие переходит
к Hybomitra ciureai (до 79%), но еще местами встречается в массе Н. bimaculata (до 39%),
^субдоминантами являются Н. lundbecki, H. nitidifrons confiformis, H. montana,
'Atylotus rusticus, Haematopota pluvialis, местами Chrysops caecutiens (близ рек), Ch.
relictus, Haematopota subcylindrica, Tabanus bromius и Т. autumnalis. К малочисленным
или редким относятся таежные Chrysops nigripes, Ch. divaricatus, Hybomitra arpadi,
H. lurida, H. nigricornis, лесные Hybomitra distinguenda, Tabanus bovinus^ T. glaucopis,
Heptatoma pellucens и Atylotus fulvus, лесостепной Chrysops concavus и степные
Hybomitra sareptana, H. expollicata, Atylotus pallitarsis, Haematopota desertotum и пустынные
H. turkestanica, A. quadrifarius (в южных частях лесостепи). В долинах Оби и Иртыша
на большом протяжении к северу многочисленны те же виды, что и в долинах рек
европейской части СССР.
К востоку от Оби- в лесостепных предгорьях и низкогорьях Салаира (Тогучин-
-ский р-н Новосибирской обл.) доминантами и субдоминантами
в большинстве являются те же виды, что и в лесостепи Западно-Сибирской низменности
на Иртышско-Обском междуречье. Но обращает на себя внимание присутствие в
заметных количествах (1—2%) таежных Chrysops divaricatus, Hybomitra arpadi и Н.
muehlfeldi, лесного Н. brevis, и резкое сокращение до уровня редких — лесостепных видов
Haematopota subcylindrica, Atylotus rusticus, Tabanus bromius и Т. autumnalis.
Итого в пределах лесостепной зоны обнаружено 50 видов и 1 подвид слепней.
Лесостепных видов насчитывается 8, лесных — 13 (из них 1 вид проникает с востока,
остальные с севера) и степных — 5. Обычно господствуют по числу особей лесостепные
виды. Местами в северных частях зоны отмечаются таежные формы — всего 8 видов
и 1 подвид, на юге — 6 пустынных видов, а преимущественно на западе —10
средиземноморских видов, но в большинстве они представлены малочисленными популяциями.
Дальневосточная лесная зона. В пределах СССР охватывает
отдельные территории Приморского края, Сахалинской обл., юга Хабаровского края
и Приамурье к западу примерно до Благовещенска. Наличие на континентальном
Дальнем Востоке хребта Сихотэ-Алиня с его преимущественно горно-таежным ландшафтом,
я на равнинах — значительных лугостепных пространств (Суйфуно-Приханкайская
равнина, Прихасанские степи) сильно усложняют аспект фауны слепней
рассматриваемой зоны.
Фауну слепней дальневосточной лесной зоны изучали А. В. Гуцевич (1937, 1940),
А. С. Мончадский (1939), Д. И. Благовещенский (1946), А. В. Маслов * (1960), Р. Г.
Соболева (1962, 1965в, 1974) и др.-
В рассматриваемой зоне (в континентальной ее части) доминантами являются
восточносибирские и дальневосточные лесные виды Hybomitra brevis, H. stigmoptera (юг),
Atylotus horvathi, A. miser (степной или лесостепной вид), Tabanus pleskei и
Haematopota tamerlani, субдоминантами Hybomitra stenopselapha, H. distinguenda
contigua, Tabanus geminus, Chrysops suavis, затем таежные Hybomitra lapponica
(местами доминант), H. nitidifrons, H. lurida, H. tarandina, H. lundbecki sibiriensis и
степная Haematopota desertorum. Малочисленными или редкими являются
восточносибирские или дальневосточные лесные Hybomitra ussuriensis, H. nigella, H. tarandinoides,
Н. astur, Chrysops dissectus, Haematopota stackelbergi, таежные Chrysops nigripes,
Ch. validus, Ch. makerovi, Hybomitra bimaculata, H. montana, H. nigricornis, степные
H. expollicata orientalis, H. montana morgani и Atylotus pallitarsis. Упомянем еще
редких Chrysops anthrax, Haematopota koryoensis (оба вида на юге) и Hybomitra adachii,
ландшафтная приуроченность которых неясна. Особо следует отметить
представителей восточноазиатского типа фауны — Chrysops vanderwulpi, Ch. japonicus, Tabanus
chrysurus, T. buddha, T. amaenus (подвид griseinus), T. trigeminus и Т. signatipennis, но
все они малочисленны в рассматриваемой зоне.
Аспекты фауны дальневосточной лесной зоны сильно отличаются от таковых
европейско-сибирской зоны. Большинство видов этих зон не совпадает, общим является
* К сожалению, публикация А. В. Маслова по фауне слепней Дальнего Востока
(71 вид) в значительной мере дефектна, что лишает возможности ею пользоваться. Так,
автором ошибочно указаны для Дальнего Востока Tabanus sabuletorum, Hybomitra
tatarica, Atylotus'fulvus, A. rusticus. Сомнительны указания о распространении в
Арктической подобласти Hybomitra tarandinoides, H. brevis, Tabanus pleskei, Haematopota
desertorum, Chrysops vanderwulpi, а в Якутии — Tabanus buddha. Непонятно, почему
некоторые виды приведены автором дважды под разными названиями,
представляющими лишь синонимы, например, Chrysops vanderwulpi и Ch. striatus, Hybomitra arpadi
и H. stackelbergi, H. astur и Н. guttipennis и т. д. Автор ошибочно указывает Tabanus
ketonensis и Т. komurae для Приморского округа, поскольку оба вида японскими
авторами описаны с Сахалина. Как выяснено, первый вид идентичен Hybomitra nigricornis,
а второй вид — Н. lapponica; эти оба вида широко распространены на Дальнем Востоке
и упомянуты в работе А. В. Маслова. Имеются и другие ошибки.
60 ВВЕДЕНИЕ
лишь один лесной вид — Hybomitra distinguenda (и некровосос Atylotus plebejus),.
а также заходящие с севера из таежной зоны таежные Hybomitra lapponica, H. niti-
difrons, H. bimaculata, H. montana и др., а с юга из степной зоны — Н. expollicata,
Н. montana morgani, Atylotus pallitarsis (частично) и другие:
В южной части Сахалина, характеризующейся смешанным таежно-лесным
ландшафтом, резко доминирует лесная Haematopota pluvialis tristis (82% сборов!),
субдоминантами являются таежная Hybomitra montana и
восточносибирский лесной Chrysops suavis (подвид Ch. suavis sakhalinensis) (6%). Малочисленны или
редки лесная Hybomitra distinguenda contigua и таежные Chrysops makerovi, Ch. nigri-
pes, Hybomitra bimaculata, H. tarandina, H. lapponica, H. olsoi и H. lurida. Но здесь-
обитает японский вид Tabanus sapporoenus (местами в заметных количествах) и найден*
Hybomitra bagheera, известная пока по единственному экземпляру.
Обращает на себя внимание, что на Сахалине не найдены широко
распространенные в таежной зоне континента Hybomitra nitidifrons и Н. lundbecki, а также
характерные восточносибирские лесные виды — Н. brevis и Н. tarandinoides и дальневосточные-
лесные виды Н. ussuriensis, H. nigella и др. Вероятно, некоторые из перечисленных
видов в дальнейшем все же будут найдены на Сахалине. Сравнивая фауну слепней
Сахалина с таковой таежно-лесных ландшафтов континентального Дальнего Востока,,
следует отметить значительное обеднение первой. Так, на Дальнем Востоке
насчитывается 37 таежных и лесных видов, тогда как на Сахалине — 18 видов, из них
общих — 16 видов.
На о. Кунашир доминантами являются Haematopota pluvialis tristis и
Hybomitra montana, субдоминантами Н. tarandina и Chrysops suavis,
малочисленны и редки Ch. nigripes, Ch. makerovi, Hybomitra lapponica, H. obscurinervis (найдена
только на о. Кунашир), японские виды Tabanus sapporoenus, T. kinoshitai и восточно-
азиатский вид Chrysops vanderwulpi. В целом фауна о. Кунашир представляется
значительно обедненной в сравнении с таковой о. Сахалин, она насчитывает всего 11
видов, из них общих с фауной Сахалина — 8 видов. Наиболее аспектны виды лесного
комплекса фауны.
На Курильских островах, лишенных леса, отмечается наибольшее обеднение-
фауны слепней, но численность их местами значительная. Так, на островах Юрий
и Зеленый (Малая Курильская гряда) многочисленны Haematopota pluvialis
tristis и местами Hybomitra montana, последний вид обнаружен в небольших
количествах на островах Итуруп и Уруп Большой Курильской гряды.
Всего в дальневосточной лесной зоне насчитывается 43 вида и 6 подвидов слепней,
итого 49 таксонов. Из них лесных 14 видов и 3 подвида, таежных 11 видов и 1 подвид,
степных 3 вида и 2 подвида, дампированных 5 видов и представителей восточноазиат-
ского типа фауны 10 видов. Последние обычно малочисленны по количеству особей.
Степная зона. Охватывает непрерывным поясом значительные
пространства от юга Украины до верхнего Приобья, далее после перерыва участки степей
имеются в Хакассии, Туве, Забайкалье и южном Приморье. Степи имеются также
в Закавказье, но аспекты их фауны мы рассмотрим вместе с таковыми других его-
частей.
Фауну слепней рассматриваемой зоны изучали Н. Г. Олсуфьев (1936, 1937а, 1962),
Н. Г. Олсуфьев с соавт. (1958), К. С. Сахибзадаев (1955, 1957), М. С. Шакирзянова.
(1958), Г. А. Коренчевская (1960), В. В. Шевченко (1961), В. А. Синельщиков (1962),
Г. В. Бошко (1964, 1966), Н. А. Виолович и Н. П. Гомоюнова (1964) и др.
В пределах европейской части СССР, Западной Сибири и Северного Казахстана
по побережьям рек и озер (кроме соленых) степной зоны доминантами или
субдоминантами являются лесостепные Hybomitra ciureai, Tabanus bromius,
Т. autumnalis, Chrysops relictus, Haematopota subcylindrica и Atylotus rusticous.
От Волги и восточнее к ним присоединяются как субдоминанты (а
местами и доминанты) степные Hybomitra sareptana, H. expollicata и Н. montana morgani,
последние 2 вида распространены также на юге Украины, но малочисленны, а Н. sa^-
reptana к западу известна только до Дона (и Сев. Донца). Отметим еще для
рассматриваемой зоны лесостепного Chrysops concavus, средиземноморского Ch. flavipes
(на восток примерно до Урала) и степных Haematopota pallidula, Atylotus pallitarsis
(к западу до Урала) и Chrysops ricardoae (распространен оторванно от основного ареала
в Нижней Волге, по р. Урал и некоторым водоемам их междуречья, образуя подвид
Ch. ricardoae volgensis), но эти виды обычно малочисленны. В дельтовой части Дуная
многочисленна Hybomitra ukrainica. В степную зону, особенно в северную-
ее часть, проникают эврибионтные лесные виды — Tabanus bovinus, Haematopota-
pluvialis и некоторые другие и местами они могут быть многочисленными. На юге зоны
отмечаются пустынные виды —Hybomitra acuminata, H. erberi, H. peculiaris, Tabanus-
sabuletorum, Atylotus flavoguttatus, A. quadrifarius, Haematopota pallens (на восток
до р. Урал) и Н. turkestanica. В большинстве они малочисленны, но некоторые из них
местами встречаются в заметных количествах, например, Hybomitra acuminata, Atylo-
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 61
tus quadrifarius, Haematopota pallens или Н. turkestanica. По побережью Черного моря
■{и по лиманам) обычен и местами доминирует галофильный Chrysops italicus.
В Восточной Сибири (Тува, Забайкалье) и на юге Приморского края
доминантами или субдоминантами являются Hybomitra montana morgani, H.
expollicata orientalis и Haematopota desertorum. В Туве местами многочисленны
-еще пустынные Haematopota turkestanica, Tabanus sabuletorum и Atylotus quadrifarius,
& также-степной Atylotus pallitarsis. Кроме того, в небольшом количестве обнаружен
степной Chrysops ricardoae и пустыннай Hybomitra acuminata. Видимо, вследствие
близости гор и лесов в остепнённых котловинах Тувы многочисленны таежный
Chrysops nigripes и лесная Haematopota tamerlani, в небольшом количестве встречаются
также другие таежные и лесные виды. В Забайкалье многочисленны, кроме
вышеупомянутых доминантов, еще Haematopota turkestanica и Chrysops ricardoae.
На юге Приморского края в остепнённых местах многочисленны Atylotus
miser и A. pallitarsis.
Наряду с типичными степными видами в Забайкалье и южном Приморье местами
многочисленны другие виды, свойственные лесному и даже таежному
ландшафту, например, в Забайкалье — Hybomitra astur и Н. montana montana. Фауна
слепней степной части Хакассии изучена фрагментарно. В долине р. Абакан близ г.
Абакана в сборах ловушкой Скуфьина доминирующими оказались Chrysops sua-
■vis, Ch. ricardoae, многочисленными Ch. vanderwulpi, Hybomitra expollicata
й единичными — Chrysops felictus, Hybomitra ciureai, Haematopota pluvialis и Н.
turkestanica orientalis.
Всего в пределах степной зоны обнаружено 33 вида и 4 подвида слепней. Из них
8 видов и 2 подвида степных, 7 видов лесостепных, 5 видов лесных, 2 вида таежных,
£ видов и 1 подвид пустынных, 2 вида средиземноморских и 1 вид восточноазиатский.
Пустынная зона. В пределах СССР занимает крайнее южное положение,
она охватывает огромные равнинные пространства от Каспийского моря на западе
до Прибалхашья на востоке и на юге до гор Средней Азии; участки этой зоны имеются
в Восточном Закавказье в пределах Кура-Араксинской низменности.
Видовой состав и аспекты фауны пустынной зоны освещены в публикациях
Н. Г. Олсуфьева (19376, 1939, 19406), Н. Г. Олсуфьева и А. Н. Формозова (1953),
Д. И. Благовещенского (1937), А. С. Лутты (1950), К. С. Сахибзадаева (1957),
М. С. Шакирзяновой (1958), Ш. М. Джафарова (1960), В. В. Шевченко (1961),
"Т. Г. Мельниковой (1965), Д. Чарыкулиева (1967), М. Кадыровой и Э. И. Ган(1967),
М. Кадыровой (19726) и др.
Возле водоемов рассматриваемой зоны доминантами или
субдоминантами являются Hybomitra peculiaris, H. erberi, H. acuminata, Atylotus
pulchellus karybenthinus (кроме северного Прикаспия), A. pulchellus pulchellus (юг
^зоны), A. flavoguttatus (преимущественно в северных частях зоны и в Закавказье),
Tabanus sabuletorum и Haematopota turkestanica; последняя в сев.-зап. Прикаспии,
Закавказье и на юге Средней Азии замещена Н. pallens. Местами
многочисленны Tabanus zimini, Т. filipjevi, T. brunneocallosus (север зоны), Atylotus
quadrifarius, Chrysops mlokosiewiczi (юг и восток зоны) и средиземноморские Tabanus
autumnalis brunnescens и Т. bromius flavo.femoratus.
Из перечисленных видов слепней одни виды в той или иной мере галофильны,
например, Tabanus zimini, Т. filipjevi, Т. brunneocallosus, T. sabuletorum, Atylotus
quadrifarius, Haematopota turkestanica, H. erberi, H. acuminata, Atylotus pulchellus,
A. flavoguttatus, другие же виды, видимо, избегают солоноватых водоемов —
Hybomitra peculiaris, Tabanus autumnalis brunnescens и Haematopota pallens.
Соответственно это в значительной мере определяет их аспектность возле водоемов разных
типов.
Малочисленны и распространены лишь в южных частях зоны туранские виды —
Chrysops oxianus (известен также из Закавказья), Ch. stackelbergi, Ch. hyalipennis,
Tabanus subsabuletorum и Hybomitra- turanica, а на юге Туркмении, общий с Ираном
.Silvius irritans.
В рассматриваемую зону с юга (благодаря присутствию гор) частично
проникают, обычно в виде немногочисленных популяций, представители средиземноморского
типа фауны — Tabanus leleani (главным образом подвид Т. leleani turkestanicus),
Т. golovi (подвид Т. golovi mediaasiaticus), T. spectabilis, Silvius vitripennis, Chrysops
flavipes punctifer и др., а с севера — степные Hybomitra montana morgani, H.
expollicata и др. Для долины р. Араке (в пределах Нахичеванской АССР) отметим по
сборам Т. Худавердыева многочисленных Tabanus autumnalis brunnescens.
и Т. bromius flavofemoratus и более редких Tabanus zimini, T. leleani, A tylotus
quadrifarius, А. flavoguttatus, Haematopota pallens и H. pavlovskii.
Всего в пустынной зоне обнаружено 24 вида и 7 подвидов. В их числе пустынных
18 видов и 1 подвид, степных 1 вид и 1 подвид, средиземноморских 5 видов и 5
подвидов, последние главным образом на стыке со степными ландшафтами предгорий
среднеазиатских и кавказских гор.
62
ВВЕДЕНИЕ
Аспекты фаун горных систем
К а р п а т ы. О составе фауны и численном соотношении видов слепней на
Карпатах (в пределах СССР) имеются данные Г. В. Бошко (1957, 1963в), обследовавшего-
эту горную систему в пределах главным образом Закарпатской области. Им найдено
23 вида и 2 подвида. Эндемики отсутствуют. В лесном поясе доминантами
являются Haematopota subcylindrica, H. italica, Н. pluvialis, Tabanus glaucopis, Т. miki
(в высокогорьях подвид Т. miki colchidicus), Т. bromius и Chrysops pictus, в умеренном
количестве найдены Tabanus maculicornis, Т. cordiger, Т. bovinus, Т. autumnalis, Ну-
bomitra lundbecki, Silvius vituli, Chrysops caecutiens и A tylotus rusticus, редкими
оказались Tabanus sudeticus, Hybomitra ciureai, H. distinguenda, H. bimaculata, H. nitidi-
frons confiformis (найден в Ивано-Франковской обл.), Atylotus fulvus, Heptatoma pel-
lucens и Chrysops relictus. В альпийском поясе Карпат и других местах обнаружена
в небольшом количестве Hybomitra auripila, характерная для Средней и Северной
Европы.
В лесном поясе Карпат, следовательно, обитают почти все виды слепней,
свойственные равнинной лесной зоне Евразии, но в несколько иных соотношениях.
Обращает на себя внимание доминирование Tabanus glaucopis, T. miki, Haematopota italica
и редкость Hybomitra bimaculata или Н. ciureai. Под определением «Haematopota hi-
spanica» у Г. В. Бошко, по-видимому, были 2 вида: Н. subcylindrica и сходный с ним
Н. scutellata; последний вид мы обнаружили в сборах А. Н. Желоховцева на
Карпатах в Ивано-Франковской обл. (Олсуфьев, 1970). В этой же области (Тлумачский
р-н) недавно найден Tabanus indrae montivagus (Г. В. Бошко).
В остепненных предгорьях и низкогорьях Закарпатья обнаружен
средиземноморский вид Silvius vitripennis (местами в заметных количествах) ж Atylotus loewianus.
Представляет интерес сравнить фауну Советских Карпат с таковой Румынских
Карпат. По данным К. В. Скуфьина (19606), обследовавшего долину Быстрицы (в районе
Броштень и Баруар), наиболее многочисленными видамивиюле оказались
Silvius vitripennis и Haematopota pluvialis, обычными Chrysops caecutiens, Tabanus
bromius, T. maculicornis, T. sudeticus, Philipomyia aprica и Hybomitra auripila, редкими
или единичными Haematopota italica, Hybomitra lundbecki, H. ciureai, H. bimaculata,
Tabanus tergestinus, T. cordiger, T. glaucopis, T. miki, T. spodopterus ж Philipomyia.
graeca (многочислен в окр. Ясс). По этим сборам, несомненно, неполным, можно
отметить значительное сходство фауны слепней обоих участков Карпат. Однако
обращает на себя внимание, что в Румынских Карпатах обнаружены еще средиземноморские-
Philipomyia graeca, Ph. aprica ж Tabanus tergestinus, не найденные Г. В. Бошко в
Советских Карпатах.
Горы Крыма. Аспекты фауны слепней описаны главным образом в работе
Н. Г. Олсуфьева и Т. Г. Мельниковой (1962), а также Г. В. Бошко (19636). Всего в
горном Крыму найдено И видов и 4 подвида. Доминантами и
субдоминантами являются Tabanus smirnovi (местами обилие до 60%), Т. bromius, T. bifarius,
Т. autumnalis, Chrysops caecutiens ludens ж Haematopota pluvialis, малочисленны или
редки — Therioplectes tricolor■ pallidicauda, Tabanus indrae montivagus, T. miki
colchidicus, T. bovinus, Atylotus rusticus, Hybomitra ciureai (Алушта), Chrysops pictus-
ж Ch. flavipes. На южном побережье, частично проникая в горы, обитает в небольшом
количестве Atylotus latistriatus.
Из перечисленного состава 4 вида относятся к средиземноморскому типу фауны —
Tabanus bifarius, T. indrae montivagus, Chrysops caecutiens ludens ж Atylotus latistriatus,
так же как 2 эндемичных вида — Tabanus smirnovi ж Therioplectes tricolor pallidicauda,.
которые представляют собой лишь слегка измененные формы средиземноморских
видов. Остальные 9 видов являются боревразийскими лесными или лесостепными
обитателями. Следовательно, фауна слепней Крымских гор представляется смешанной,
с некоторым преобладанием видов боревразийского типа по их списочному составу
и видов средиземноморского типа — по численности популяций.
Популяции большинства видов обитают в горах Крыма оторванно от основного-
ареала, разделяясь значительными степными пространствами Крымского полуострова
и юга Украины. Обращает на себя внимание полное отсутствие в Крымских горах
таежных или таежно-лесных видов, хотя на Кавказе некоторые из них имеются.
По-видимому, эти виды были стерты (вытеснены) из Крымских гор в ксеротермический
период.
Горы Кавказа. Южное расположение, большие высоты, значительная
протяженность Кавказских гор, пестрота климата в сочетании со сложностью
формирования в прошлом флоры и фауны этого региона обусловливают разнообразие
ландшафтов и соответственно богатство видов и разнообразие аспектов' фауны слепней.
В горах и предгорьях Кавказа известны 70 видов и 23 подвида слепней, что составляет
более 1/3 фауны СССР. В эти цифры мы не включаем виды афроевразийского аридного
типа фауны, частично проникающие в предгорья.
Высок эндемизм фауны слепней Кавказа: на его территории (с частичным заходомх
в прилегающие районы Ирана и Турции) известны эндемичные 22 вида и 12 подвидов.•-
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
63
Видовой состав и аспекты фауны слепней Кавказа подвергнуты рассмотрению
в работах А. А. Штакельберга (19266), С. Я. Парамонова (1926, 1933), Н. Г. Олсуфьева
(1937а, 1941, 1964), К. В. Скуфьина (1938), К. В. Скуфьина и С. Р. Калиты (1959),
С. Г. Калугина (1945), Е. Г. Гаузера (1939, 1941, 1953, 1960), А. В. Богачева
и Н. Г. Самедова (1948, 1949), А. Е. Тертеряна (1954, 1969), Ш. М. Джафарова (1959,
1960), А. Ф. Сергеева (1962), 3. М. Маршенкулова (1967), Ш. И. Исмаилова (1970),
Л. Н. Гургенидзе (1970, 1971) и др. Однако изученность слепней Кавказа все еще
недостаточная, особенно в Грузии и Армении. Отсутствуют какие-либо сведения о фауне
слепней Колхидской низменности.
Фауна слепней Большого Кавказа (с предгорьями) насчитывает 46 видов и 12
подвидов.
В степных и лесостепных предгорьях северо-западной части Большого Кавказа
(Майкоп, Нальчик) доминантами и субдоминантами являются
Tabanus bromius, Chrysops pictus, Hybomitra ciureai, Tabanus autumnalis, T. bovinus,
T. tergestinus, местами Chrysops caecutiens ludens, Hybomitra muehlfeldi, Haematopota
pluvialis, H. subcylindrica ж Silvius caucasicus (Майкоп — до 7%). В большинстве это
виды боревразийского лесного или лесостепного фаунистических комплексов, но
немногие, например, Tabanus tergestinus ж Chrysops caecutiens ludens — лесные
средиземноморские виды, a Silvius caucasicus — эндемик Кавказа и преимущественный обитатель
лесостепных предгорий — является ближайшим родичем средиземноморского S.
vitripennis.
Заслуживает внимания присутствие на рассматриваемой территории 2 таежно-
лесных видов: Hybomitra muehlfeldi (местами многочислен) и Н. bimaculata (редок).
Из малочисленных видов отметим боревразийских лесных Hybomitra distinguenda,
Tabanus cordiger, T. miki, Atylotus fulvus (представлен кавказским подвидом A. fulvus
aureus), Haematopota crassicornis, лесостепного A. rusticus, средиземноморских лесных
и лесостепных Silvius vituli, Chrysops flavipes, Tabanus unifasciatus, T. bifarius, T. qua-
tuornotatus ж горно-лесных Philipomyia aprica ж Therioplectes tricolor.
В степных и лесостепных предгорьях северо-восточной части Большого Кавказа
(Дагестан; Куба-Хачмасская низменность Азербайджана), а также его южной части
(Закатали, Азербайджан) аспект фауны слепней значительно меняется в сторону
резкого преобладания средиземноморской фауны. Становятся доминантами эвритоп-
ные средиземноморские степные или лесостепные виды Tabanus bifarius, Т. quatuornota-
tus, Т. bromius flavofemoratus, T. autumnalis brunnescens, удерживается как
доминант средиземноморский Т. tergestinus ж как субдоминант— Chrysops caecutiens
ludens. В небольшом количестве распространены Silvius caucasicus, S. vituli,
Chrysops flavipes, Tabanus unifasciatus, Philipomyia aprica (местами многочисленна),
появляются отсутствующие в западных частях Большого Кавказа средиземноморские
Tabanus spectabilis, Т. regularis, T. leleani, Atylotus loewianus, Dasyrhamphisumbrinus,
Tabanus tinctus, Therioplectes tunicatus, затем генетически связанная с той же
средиземноморской группой Hybomitra (Mouchaemyia) caucasi. Но исчезают боревразийские
лесные Tabanus bovinus, Chrysops pictus, становятся малочисленными Haematopota
pluvialis, Hybomitra ciureai и др. Обращает на себя внимание обилие в лесистых низовьях
Самура (Куба-Хачмасская низменность) балкано-кавказских Tabanus prometheus ж
Therioplectes tricolor, а в Закавказье (Закаталы) эндемика Кавказа Haematopota caspica
transcaucasica, генетически связанной с средиземноморскими видами дождевок.
Фауна слепней горно-лесного пояса Большого Кавказа менее дифференцирована,
чем фауна предгорий. Повсеместно доминантами и субдоминантами
являются Tabanus bromius, Philipomyia aprica, Tabanus tergestinus, Т. cordiger, Chrysops
caecutiens ludens, Tabanus rupium, T. miki (с подвидом Т. miki colchidicus), Haematopota
crassicornis, местами H. subcylindrica; H.pluvialis, Tabanus bovinus (запад), Т. sudeticus
(с подвидом Т. sudeticus verralli), T. autumnalis, Т. armeniacus (преимущественно
высокогорья), Т. unifasciatus (центр и восток) и Т. caucasius, малочисленны Silvius vituli,
S. zaitzevi (южные склоны), Chrysops caecutiens caecutiens, Tabanus subparadoxus, T.
glaucopis, T. indrae (в более высоких частях гор замещен подвидом Т. indrae montivagus),
Hybomitra distinguenda, Therioplectes tricolor (в северо-западных частях местами
замещен подвидом Th. tricolor luminosus), Atylotus fulvus aureus, Haematopota scutellata ~\
pontica ж H. longeantennata (восток и южные склоны); местами в лесной пояс прони- -
кают немногочисленными популяциями Tabanus bifarius ж Т. quatuornotatus (оба вида
многочисленны в южном Дагестане). В Закаталах найден Т. portschinskii (многочислен
на Малом Кавказе), а из горных частей запада Грузии (Шови) нами описан подвид.
.Haematopota pluvialis ioffi.
На северных склонах Большого Кавказа встречается европейско-сибирский
лесостепной вид Atylotus rusticus, причем в центральных и восточных частях хребта
местами он весьма многочислен в субальпийском поясе. На северных склонах
центральной части Большого Кавказа на высокогорных остепненных участках
распространена (но немногочисленна) Hybomitra montana karatschajensis. Большой
интерес представляет обнаружение в юго-восточной части Большого Кавказа Therio-
64
ВВЕДЕНИЕ
plectes carabaghensis, основной ареал которого охватывает восточные части Малого
Кавказа, Талыш, Северный Иран и Западный Копет-Даг.
В целом фауна горно-лесного пояса Большого Кавказа, насчитывая в своем
составе 29 видов и 9 подвидов, представляется смешанной из элементов боревразийского
лесного и лесостепного комплексов — 14 видов и 2 подвида, а из представителей
средиземноморского типа фауны — 10 видов и 2 подвида, с присутствием 10 кавказских
эндемиков: Silvius zaitzevi, Therioplectes tricolor luminosus, Tabanus caucasius, T. sub-
paradoxus, T. portschinskii, T. armeniacus, Atylotus fulvus aureus, Haematopota
scutulata pontica, H. pluvialis iojfi и Hybomitra montana karatschajensis. Первые 5 форм
генетически связаны с средиземноморской фауной, остальные — с европейской лесной
■фауной, а Н. montana karatschajensis представляет собой несколько измененную форму
таежно-лесной Н. montana.
Заметим, что эндемизм фауны горно-лесного пояса Большого Кавказа (большая
часть видов распространена также на Малом Кавказе) значительно выше, чем
таковой лесостепных и степных предгорий. В лесном поясе эндемиков 10, тогда как в
предгорьях и низкогорьях известен только 1 — Silvius caucasicus.
Давая общую характеристику фауны слепней лесного пояса Большого Кавказа,
все же следует отметить, что на северных его склонах на восток до Дагестана аспект-
ность (по числу видов и обилию особей) в большей мере принадлежит видам
боревразийского типа фауны (лесного комплекса), тогда как по южным склонам (и в
восточной части северных склонов) видам средиземноморского типа фауны.
Фауна высокогорий Большого Кавказа в пределах альпийского пояса по числу
видов намного беднее таковой нижележащих поясов, но численность слепней здесь
местами очень высокая. Доминантами являются Hybomitra caucasica и Phi-
lipomyia rohdendorfi. Первый вид является европейским горно-лесным и альпийским
видом, второй вид — эндемик Кавказа. В альпийском поясе, но в большей мере в его
субальпийской части, местами в большом количестве встречаются
Tabanus rupium, T. miki colchidicus, Philipomyia aprica и Haematopota crassicornis и в
меньшем количестве Tabanus armeniacus, Silvius latifrons и некоторые др. В целом для
рассматриваемого пояса следует отметить почти равное соотношение боревразийских
горно-лесных видов (включая генетически связанных с ними), которых 4: Hybomitra
caucasica, Tabanus rupium, T. miki colchidicus и Т. armeniacus, и средиземноморских
видов (или их производных) — их 3: Philipomyia rohdendorfi, Ph. aprica и Silvius
latifrons. Эндемиков Кавказа всего 2: уже упоминавшийся Philopomyia rohdendorfi
и Tabanus armeniacus. Обращает на себя внимание преимущественно черная окраска
тела большинства высокогорных кавказских видов, в отличие от преимущественно
светлой окраски большинства видов, обитающих в предгорьях.
Фауна слепней Малого Кавказа несколько богаче таковой Большого Кавказа —
она включает 64 вида и 18 подвидов. Общими для обеих горных систем являются
43 вида и 7 подвидов.
В покрытых лесами предгорьях и низкогорьях западной части хребта
(Аджарская АССР), судя по сборам А. В. Соловьевой, доминируют Tabanus bromius
и Т. tergestinus, в небольшом количестве встречаются Т. rupium, T. armeniacus, T. 1е-
leani, Т. sudeticus, Т. maculicornis, Т. miki colchidicus, T. indrae, Haematopota
crassicornis, H. pluvialis, H. subcylindrica и Н. scutellata pontica; в соседнем Махарадзевском
районе Грузии обнаружены Chrysops pictus и Tabanus autumnalis. Почти все
перечисленные виды (кроме Т. maculicornis) обитают в лесном поясе Большого Кавказа,
имеется явное преобладание боревразийских лесных форм (или их производных) —
12 видов из 15, и только 3 вида — Tabanus tergestinus, T. leleani и Т. indrae относятся
к средиземноморским формам.
В степных предгорьях и лесостепных низкогорьях в пределах Азербайджана
(включая Нахичеванскую АССР) и Армении абсолютное доминирование
принадлежит Tabanus bromius flavofemoratus, местами массовыми видами
являются Т. quatuornotatus (в долине Аракса замещен подвидом Т. quatuornotatus araxis),
Т. bifarius (на юго-востоке местами замещен подвидом Т. bifarius tdrjukini, а в сев.-
аап. Азербайджане — подвидом Т. bifarius kurensis), в Нахичеванской АССР — еще
Т. laetetinctus, T. hauseri и Т. regularis. Вместе с ними распространены, но чаще
малочисленны, Т. spectabilis, T. tergestinus, T. atropathenieus, T. prometheus (главным
■образом в Нахичеванской АССР), Dasyrhamphis umbrinus, Chrysops flavipes (и подвид
Ch. flavipes punctifer), Ch. caecutiens ludens, Silvius caucasicus (в Армении и
Нахичеванской АССР замещен подвидом S. caucasicus molitor), S. vituli (Акстафа), Tabanus
indrae (с подвидом Т. indrae vappa), T. tinctus, Т. autumnalis brunnescens (обилен в
долине Аракса), Т. cordiger, T. lunatus, Hybom tra ciureai (Акстафа), Haematopota
subcylindrica, H. caspica transcaucasica, Chrysops buxtoni armeniensis (Западная Армения) и др.
Ш. М. Джафаров (1959) отметил Silvius zaitzevi для Мургабской степи, но это
указание нуждается в подтверждении.
В целом фауна степных предгорий и лесостепных низкогорий восточной части
Малого Кавказа в основных чертах повторяет таковую южных предгорий и низкогорий
Большого Кавказа. Преобладают виды средиземноморского типа фауны.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
65
Фауна горно-лесного пояса Малого Кавказа в его западных частях изучена
главным образом в окрестностях Бакуриани (сборы А. Г. Шарова и других). Здесь
доминантными видами являются Tabanus bromius, Philipomyia aprica, Hybomitra
popovi, Haematopota crassicornis и Н. subcylindrica, субдоминантами
Hybomitra distinguenda, Tabanus glaucopis, T. quatuornotatus, T. unifasciatus, T. cordiger,
T. portschinskii, T. miki (и подвид Т. miki colchidicus) и малочисленны или редки
Tabanus bifarius, Т. rupium, Т. tergestinus, Т. subparadoxus, T. armeniacus, A.
autumnalis, Atylotus fulvus aureus, Chrysops caecutiens (смешанно с подвидом Ch. caecutiens
ludens), Haematopota pluvialis, Silvius zaitzevi и Therioplectes tricolor. Особо следует
отметить обнаружение в районе Бакуриани немногочисленных популяций таежно-
лесных видов Hybomitra lurida, H. nitidifrons confiformis, H. bimaculata, H. muehlfeldi
и лесного вида Tabanus maculicornis. Первые 2 вида кроме Бакуриани нигде больше
на Кавказе не найдены (возможно, из-за недостаточного обследования), но Т.
maculicornis обнаружен еще в Аджарской АССР (см. выше) и на Талыше, a Hybomitra
bimaculata встречается в небольшом количестве также на Талыше и по северным низко-
горьям и предгорьям Большого Кавказа. Видимо, эти виды проникли на Кавказ
в плейстоцене вместе с лесными видами, в том числе европейскими горно-лесными
и альпийскими формами (Н. caucasica, Tabanus rupium и др.). Они пережили здесь
ксеротермический период и сохранились до наших дней.
В эту же группу плейстоценовых «вселенцев» надо отнести обитающего на северных
склонах Большого Кавказа (в пределах Карачаево-Черкесской АО и Кабардино-
Балкарской АССР) Hybomitra montana karatschajensis, а на Малом Кавказе — Н.
montana morgani; эти подвиды представляют собой несколько измененные формы
голарктического таежно-лесного вида Н. montana.
Судя по отдельным сборам из других частей лесного пояса Малого Кавказа в
пределах Грузии и Армении, видовой состав слепней примерно тот же, что
и в районе Бакуриани за исключением таежно-лесных видов и лесного Tabanus
maculicornis. Следует лишь отметить, что местами довольно многочисленны
Therioplectes tricolor, Hybomitra caucasi и в небольшом количестве встречаются Tabanus
bovinus, T. sudeticus (запад), Т. indrae, T. caucasius Silvius vituli и Chrysops sejunctus,
а по остепненным участкам — Hybomitra montana morgani и H. expollicata.
В восточных частях Малого Кавказа (Ханлар, Нагорный Карабах) аспекты
фауны слепней горно-лесного пояса в значительной мере повторяют таковые западных
частей хребта. Доминантами и субдоминантами являются Tabanus
bromius (вместе с подвидом Т. bromius flavofemoratus), Т. tergestinus, T. quatuornotatus,
Т. bifarius, Т. unifasciatus, T. portschinskii (главным образом субальпийский пояс),
Philipomyia aprica, Haematopota subcylindrica и Н. crassicornis (главным образом
субальпийский пояс), малочисленны Chrysops caecutiens ludens, Hybomitra caucasi, Tabanus
cordiger, T. glaucopis, T. rupium, T. miki colchidicus, T. indrae, T. spectabilis, T.
autumnalis, T. bovinus, T. atropathenieus, Therioplectes carabaghensis ш Atylotus fulvus aureus. Из
перечисленных 19 видов и 4 подвидов несовпадающими с фауной горно-лесного пояса
западных частей Малого Кавказа являются лишь 2 вида — Therioplectes carabaghensis,
замещающий западного Th. tricolorrm Tabanus atropathenieus. Доминантами и
субдоминантамив восточных частях Малого Кавказа являются за немногими
исключениями те же виды, что и в западных частях, однако в лесном поясе востока
не найдены наряду с упомянутым Therioplectes tricolor еще и другие западные виды,
в частности Hybomitra popovi, H. distinguenda, Chrysops caucutiens caecutiens, Silvius
zaitzevi, и группа таежно-лесных видов, обитающих возле Бакуриани.
В целом фауна горно-лесного пояса Малого Кавказа включает 37 видов и 6
подвидов. По своему составу и аспектности она в значительной мере повторяет таковую
горно-лесного пояса Большого Кавказа, особенно его восточной и южной частей.
В западной части Малого Кавказа добавляется эндемичный Hybomitra popovi (бореаль-
ного происхождения) и таежно-лесные реликты, а в восточной части — кавказско-
иранский Tabanus atropathenieus. Эти виды на Большом Кавказе неизвестны.
Но на Малом Кавказе не обнаружен Atylotus rusticus, распространенный на Большом
Кавказе,
Фауна горно-лесного пояса Малого Кавказа, равно как Большого Кавказа (см.
выше), может быть охарактеризована как смешанная боревразийская — 17 видов
и 3 подвида, и средиземноморская — 9 видов и 2 подвида, с присутствием 8 кавказских
и 2 кавказско-иранских эндемичных видов (Therioplectes carabaghensis и Tabanus
atropathenieus) и 1 подвида. 9 эндемичных видов генетически связаны с
средиземноморской фауной, а 1 вид и 1 подвид — с боревразийской лесной фауной. В лесном поясе
западных частей Малого Кавказа преобладают виды (и особи) боревразийского типа
фауны, в центральных и восточных частях — средиземноморского типа фауны.
Несколько измененный состав имеет фауна слепней ксерофитных нагорий и
островных лесов юго-восточной части Малого Кавказа в пределах Нахичеванской АССР
и юго-восточной Армении. Из многочисленной группы кавказских горно-лесных
видов здесь найдены только 13 видов и 4 подвида, из их числа доминирует
Tabanus bromius (с подвидом Т. bromius flavofemoratus) и Philipomyia aprica, многочисленны
5 Н. Г. Олсуфьев
66 ВВЕДЕНИЕ
Tabanus bifarius, Т. unifasciatus, Т. indrae (местами доминант наряду с подвидом
Т. indrae vappa) и Haematopota subcylindrica, малочисленны или редки Chrysops сае-
cutiens ludens, Tabanus glaucopis, Т. quatuornotatus, Т. rupium, Т. armeniacus, Atylotus
fulvus aureus и Haematopota crassicomis, а также Therioplectes carabaghensis и Tabanus
atropathenicus. Но наряду с этими видами на рассматриваемой территории обитает
значительная группа кавказско-иранских (или кавказских) эндемиков: Atylotus proditor
(Кавказ), Tabanus miki austraiis, T. infestus (Кавказ), Т. anthrax (найден еще в Турции),
Т. swiridowi (Кавказ), Т. perelegans, T. shelkovnikovi, T. olsufjevi, T. semiargenteus
(Кавказ). Первые 2 вида местами многочисленны, остальные малочисленны или редки.
Своеобразие фауны дополняется наличием местами многочисленных пон-
тийско-гирканских Tabanus prometheusm T. laetetinctus (местами с подвидом Т. laete-
tinctus sordes). Все эти виды за исключением Т. miki austraiis генетически связаны
с представителями средиземноморского типа фауны.
В итоге можно заключить, что в составе фауны рассматриваемого участка
Малого Кавказа, насчитывающей 23 вида и 6 подвидов, доминирование принадлежит
средиземноморским формам (или их производным) — всего 17 видов и 4 подвида, тогда
• как на долю боревразийских форм — 6 видов и 2 подвида. Почти половина форм —
11 видов и 3 подвида представляют собой эндемиков Кавказа (или ближайших стран).
Фауна альпийского пояса Малого Кавказа почти повторяет таковую Большого
Кавказа. Повсеместно распространена и доминирует Hybomitra caucasica,
многочисленна Haematopota crassicomis, местами встречаются' Philipomyta
aprica, Tabanus miki colchidicus, T. armeniacus, Chrysops sejunctus (эндемик Малого
Кавказа), в западных частях многочисленна Philipomyia rohdendorfi и, видимо, редок
Silvius latifrons. Все эти виды так или иначе проникают в субальпику и верхние части
лесного пояса, а некоторые, например, Т. miki colchidicus, T. armeniacus и другие,
больше свойственны последнему.
Фауна слепней Талыша насчитывает 29 видов и 4 подвида. Мы не учитываем
описанного с Талыша по единственному экземпляру 5 и нам неизвестного Tabanus агте-
nicus Szilady, отсутствующего в сборах советских исследователей и исследователей
фауны слепней Ирана. В низменных лесах Талыша доминирует Hybomitra
ciureai, H. distinguenda и Tabanus bromius (в смеси с подвидом Т. bromius flavofemoratus),
в меньшем числе встречаются Chrysops caecutiens (в смеси с подвидом Ch. caecutiens
ludens), Hybomitra bimaculata, Tabanus autumnalis brunnescens, T. tergestinus,
Therioplectes albicauda, Philipomyia aprica и Haematopota subcylindrica, и малочисленны
Chrysops flavipes, Tabanus miki colchidicus, T. spectabilis, T. portschinskii, T. talyshi,
T. maculicornis (в предгорьях), Atylotus loewianus и Haematopota caspica. В целом
преобладают виды европейской лесной (и лесостепной) фауны, в меньшем числе
представлены средиземноморские виды, имеется 1 общекавказский эндемик — Tabanus
portschinskii и 3 талышских (или гирканских) эндемика — Tabanus talyshi, Therioplectes
albicauda и Haematopota caspica; последние 2 вида, помимо Талыша, известны также
в Иране, где они обитают по южному побережью Каспийского моря. Генетически эти
эндемики связаны с средиземноморскими видами. Обращает на себя внимание
присутствие в лесах Талыша таежно-леснЬй Hybomitra bimaculata.
Фауна горно-лесного пояса Талыша несколько отличается от таковой низменных
лесов. Здесь доминируют средиземноморские Philipomyia aprica, Tabanus
unifasciatus, T. quatuornotatus и Т. spectabilis, в меньшем числе встречаются Chrysops
caecutiens, Tabanus tergestinus, T. bormius, Т. cordiger, Т. glaucopis и единичные
Chrysops flavipes, Silvius caucasicus, Tabanus leleani, T. regularis, T. rupium, T. armeniacus
и Haematopota subcylindrica. В ксерофитных нагорьях Талыша (Зуванд) фаунистиче-
ский состав слепней в основных чертах тот же, что и в горно-лесном поясе, но
обнаружены еще в небольшом количестве Therioplectes carabaghensis, T. indrae и Haematopota
pluvialis (указание Ш. М. Джафарова, нуждающееся в подтверждении), и становится
более многочисленным Tabanus leleani. Однако в Зуванде не найдены оба вида Chrsops,
Philipomyia aprica, Tabanus rupium и Т. quatuornotatus, но, по-видимому, это
случайные пропуски в сборах. Для лесных районов Талыша Ш. М. Джафаров (1959) еще
указывает как многочисленный вид «Tabanus aff. vappa Bog. et Sam.», не определенный
болер точно.
В целом фауна слепней Талыша в значительной мере повторяет таковую восточных
частей Малого Кавказа и является по преимуществу смешанной из средиземноморских
и боревразийских лесных (или лесостепных) видов. Но имеются 3 эндемика, и не
найдены некоторые виды, обитающие на Малом Кавказе: Tabanus bifarius T.
atropathenicus, Hybomitra caucasica, H. caucasi, Atylotus fulvus aureus, Haematopota crassicomis
и Chrysops sejunctus; вероятно, некоторые из перечисленных видов в дальнейшем
все же будут обнаружены на Талыше.
Уральские горы. Вытянутые в меридиональном направлении, имея
протяженность около 2000 км, Уральские горы пересекают следующие ландшафтные
зоны: на севере тундровую, в средних частях — таежную, а на юге — лесную и
лесостепь. Данные о видовом составе и численных соотношениях разных видов слепней
содержатся в публикациях Ю. М. Колосова и Л. В. Попова (1932), Н. Г. Олсуфьева
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 67
(1935в), Ю. Г. Митрофановой (1951, 1955), Е. В. Кучерова (1959, 1964), Б. В. Куче-
рова и Р. X. Еникеева (1965) и др.
Фауна слепней Урала в основном повторяет таковую соответствующих зон,
которые пересекает эта горная система, с некоторыми отклонениями, особенно заметными
в ее южных частях. Всего на Урале обнаружено 36 видов и 1 подвид слепней.
На Северном Урале в зоне каменистой тундры (и лесотундры) доминантами
являются Hybomitra aequetincta, H. sexfasciata и Н. lurida, в меньшем количестве
встречаются Н. nitidifrons confiformis, H. montana и единично найдены Н. astuta, H. lap-
ponica, H. lundbecki, Chrysops nigripes, Ch. caecutiens и Haematopota pluvialis.
На Среднем Урале (Красновишерский и Чусовский р-ны Пермской обл. и другие
места) доминантами и субдоминантами являются Hybomitra
lundbecki, Н. bimaculata, H. arpadi, H. tarandina, H. nitidifrons confiformis, Tabanus
maculicornis, Т. bromius, Haematopota pluvialis, местами Т. cordiger и Hybomitra «solstitialis)>
(? muehlfeldi), малочисленны или редки Н. montana, H. kaurii, H. nigricornis, H. lurida,
Н. distinguenda, H. lapponica, Tabanus bovinus, Т. glaucopis, Atylotus fulvus, Chrysops
caecutiens, Ch. pictus, Ch. divaricatus, Ch. nigripes, Heptatoma pellucens и др. На
таежный комплекс приходится 12 видов и^70—75% особей, тогда как на лесной и
лесостепной комплексы — 15 видов, но 25—30% особей (Митрофанова, 1951 и др.). В целом
аспект фауны горно-таежных частей Среднего Урала довольно хорошо совпадает
с таковым равнинных частей таежной зоны (ее средней полосы) в пределах
европейской части СССР. Лишь обращает на себя внимание повышенная численность лесного
Tabanus cordiger в горной тайге Чусовского р-на, тогда как в равнинных частях
таежной зоны (средней тайги) этот вид редок. На Южном Урале Т. cordiger еще более
многочислен (обилие 5%).
На Южном Урале доминант а м и и субдоминантами являются
Haematopota pluvialis, Н. italica, Tabanus bromius, Т. maculicornis, Т. cordiger, Т. su-
deticus, Chrysops caecutiens и Hybomitra lundbecki, т. е. в большинстве виды лесного
фаунистического комплекса. Малочисленны или единичны лесные Chrysops pictus,
Tabanus bovinus, T. glaucopis, Т. miki, Hybomitra distinguenda, H. kaurii, Atylotus
fulvus, Heptatoma pellucens, Haematopota crassicomis, таежные Hybomitra bimaculata,
H. nigricornis, H. montana, H. muehlfeldi, H. nitidifrons confiformis, H. tarandina (Бело-
рецк), лесостепные Н. ciureai^ Atylotus rusticus, Chrysops relictus, Haematopota
subcylindrica, Tabanus autumnalis и средиземноморский лесной вид Silvius vituli. Всего
на Южном Урале обнаружено 29 видов, в том числе на долю представителей лесного
комплекса приходится 15 видов и преобладание особей этих видов, тогда как на долю
таежных и лесостепных — 13 видов и меньшинство особей. Обращает на себя внимание
многочисленность на Южном Урале Haematopota italica (обилие 8.5%), Tabanus
cordiger (5%) и Т. sudeticus (4.2%), а также относительная частота средиземноморского
лесного вида Silvius vituli (1.5%); эти виды в равнинных местах лесной зоны
европейской части СССР обычно малочисленны или редки. Но на Южном Урале малочисленны
или редки Hybomitra ciureai, H. bimaculata, H. muehlfeldi, Tabanus bovinus и Chrysops
pictus, изобилующие в равнинных частях лесной полосы европейской части СССР.
По некоторым аспектам фауна слепней Южного Урала напоминает таковую
лесных Карпат. И тут, и там многочисленны Haematopota italica, в заметном количестве
встречаются Silvius vituli, Tabanus cordiger и малочисленны или редки Hybomitra
ciureai и Н. bimaculata. Но имеются и существенные различия — на горном Урале
многочислен Tabanus sudeticus, но относительно редки Т. glaucopis, T. miki, Chrysops pictus,
тогда как на Карпатах они обычны. Tabanus glaucopis многочислен (обилие 7.4%)
в западных предгорьях Урала в пределах Кунгурского и Кишертского районов
Пермской области, здесь также изобилует Haematopota italica (13.8%) и многочислен
Tabanus cordiger (2.7%) (Митрофанова, 1955).
Горы Средней Азии. Окаймляя с юга огромные аридные пространства
и имея большую протяженность с запада на восток, а также значительные высоты,
горы Средней Азии выделяются разнообразием ландшафтов. Последние резко
отличаются от прилегающих к ним пустынных равнин. Предгорья обычно бывают остепнен-
ными, горы покрыты лесами, в высокогорьях выражен альпийский пояс, а в особых
случаях — горные степи и даже полупустыни (Восточный Памир).
Видовой состав и аспекты фауны слепней гор Средней Азии изучались Н. Г.
Олсуфьевым (19376, 19396), Н. Г. Олсуфьевыми В. П. Боженко (1950), В. В. Шевченко
(1953а, 19536, 1955, 1961), X. А. АйбасовымиМ. С. Шакирзяновой (1960), Ш. Барато-
вым (1962а, 19626, 1966), П. А. Чировым (1968а, 19686), Т. Г. Мельниковой (1965),
У. С. Куандыковой (1966), Д. Чарыкулиевым (1967), М. Кадыровой и Э. И. Ган (1967)
и др.
В предгорьях и горах Средней Азии известно 45 видов и 11 подвидов слепней.
Сюда мы не включаем пустынные виды афроевразийского аридного типа фауны,
проникающие в предгорья и частично в горы. Эндемизм фауны гор (и предгорий) Средней
Азии выражен в. достаточной мере, на их территории (с частичным заходом в
прилегающие горы Западного Китая и Афганистана) обитают эндемичные 17 видов и 5 под-
5*
№
ВВЕДЕНИЕ
Твидов. В целом фауна слепней предгорий и гор Средней Азии намного беднее таковой
Кавказских гор, насчитывающей 70 видов и 2 s подвида. Общих видов 1 и подвидов 5.
Горы Средней Азии представлены несколькими самостоятельными горными
системами, различающимися по времени образования, высоте поднятия, широтному
и долготному расположению и т. д. Соответственно фауна слепней в них неоднородна.
Мы выделяем для самостоятельного рассмотрения следующие* горные системы: Тарба-
гатай (с хребтом Саур). Тянь-Шань (с хребтом Каратау), Памиро-Алай (с хребтами
Зеравшанском и Гиссарскнм! и Копет-Даг.
Фауна слепней Тарбагатая (главным образом Саура) и?.учена фра мзнта;) то.
Здесь обнаружено 13 видов и 1 подвид. Преобладаю, в гил ■ ^еечо.-о ита^ж ,о •:> .'[иуия-
стических комплексов, их найдено 8: Chrysops caeculi па, Hybm Ira lu:iu„ecki, H.
bimaculata, И. distinguenda, H. ciureai, Tajanus maiulic >, nis, -it'll tas ju'u i : и 11 i. muo-
pota pluvialis. По остенненным нпзкогорьпм об .1 i ia е.епная Hybomitra montana m r-
gani и здесь же встречается восточное етлче'.гн > :орскпй эвритопну.й вид Tab inus
leleani. Второй вид средиземноморского происхождения энлемнк гор Средней Азии —
Tabanus montiasiaticus— найден в небольшом количестве в горнолесном поясе Саура.
Здесь обитают еще 3 характерных вида среднеазиатского горного фаунпг.тпч. с .ого
комплекса: Hybomitra turkestana, II. shnltnikovi и Н. semipollinosa. Последние 2 вида,
а также Tabanus montiasiaticus имеют на Сауре северо-восточную границу ареала,
тогда как Hybomitra turkestana и Tabanus leleani частично распространены на юго-
западных склонах Алтая. Вероятно, на Тарбагатао обитают также лесостешкле. Пае-
matopota subcylindrica, Tabanus bromius и таежно-лесные Ilybnmitra nigricor/iis
и Н. muehlfeldi, найденные южнее на Джунгарском Алатау. Последний вид обитает
также еще южнее — по низкогорьям Заилийского Алатау Тянь-Шаня, a Tabanus
bromius широко распространен в горах Средней Азии.
Слепни Тянь-Шаня изучены значительно полнее. Здесь найдено 34 вида и 6
подвидов. В Джунгарском Алатау, составляющем северо-восточную оконечность Т>'нь-
Шаня, еще заметно присутствие широко распространенных боревразпйских видов,
обитающих в среднегорном лесном поясе и по низкогорьям. Здесь обнаружены та-
ежно-лесные Hybomitra bimaculata, H. muehlfeldi, II. nigricornis, лесные Chrysops cae-
cutiens, Hybomitra distinguenda, Tabanus maculicornis, Atylotus fuluus, Haematnpota ■■
pluvialis, лесостепные Hybomitra ciureai, Tabanus bromius (доминант), Т. autumnalis
(доминант) и Haematopota subcylindrica. Преимущественно к среднегориому лесному
поясу относятся также находки среднеазиатских эндемиков (средиземноморского
происхождения) Tabanus golovi (обитает также по предгорьям), Т. montiasiaticus и Т. аг-
genteomaculatus (единственная поимка в долине р. ТеректыП. По остепненным
предгорьям и низкогорьям обитают степные Hybomitra montana morgani, H. expollicata
и Chrysops ricardoae. В горно-лесном и альпийском поясах доминируют
характерные среднеазиатские горные виды: Hybomitra hunnorum, H. tatarica, H. turkestana
(массовый вид), И. semipollinosa и Н. shnitnikovi. Первые 2 вида более характерны для
альпийского пояса, остальные — для лесного пояса.
В Заилийском Алатау и Киргизском хребте, ограничивающими с севера
основной массив Тянь-Шаня, фауна слепней в значительной мере повторяет таковую
Джунгарского Алатау. Отличие заключается лишь в том, что здесь уже не
встречаются некоторыеборевразийскиеравнинныетаежно лесные i -.с. j. виды. Chrysopsсаеси-
tiens, Hybomitra bimaculata, H. nigricornis, H. distinguenda, Tabanus maculi ornis,
Haematopota pluvialis, не найдены также лесостепная Н. subcylindrica и степная Hybomitra
expollicata. Это обеднение не компенсируется появлением каких-либо видов
среднеазиатской фауны. В предгорьях и по низкогорьям доминируют Tabanus
bromius, Т. autumnalis, в меньшем количестве встречаются Hybomitra ciureai, II.
muehlfeldi, Tabanus leleani, Т. golovi, изредка Chrysops ricardoae и Hybomitra montana
morgani (восточнаячасть Заилийского Алатау). В горно-лесном поясе доминантами
и субдоминантами являются Hybomitra turkestana, И. shnitnikovi, H.
semipollinosa, более редки Atylotus fulvus и Tabanus montiasiaticus, в альпийском поясе
абсолютным доминантам является Hybomitra hunnorum, в меньшем количестве
встречается Н. tatarica.
К югу от этих хребтов в Центральном Тянь-Шане фауна слепней зпачительно
беднеет. В высокогорных его частях доминируют Hybomitra- hunnorum, H.
turkestana, местами многочисленна Н. tatarica, а но ксерофптным нагорьям
обычна Н. montana reinigiana и изредка встречается Tabanus leleani. В Иссык-Кульспой
котловине, кроме последнего вида, найдены еще обитающие з,п.есь изолированно
степные Chrysops ricardoae и Hybomitra montana morgani n пустынная Haematopota iurke-
stanica.
Фауна слепней Западного Тянь-С'аня (западная часть Таласского Алатау и сто
отроги, Ферганский хребет) и Каратау существенно отличаются ог тагююп групгх
частей Тянь-Шапя. По низкогорьям и предгорьям, запятим особыми «.эфемеровыми»
субтропическими степями, преобладают виды средиземноморского типа фауны. Д о-
минантами и субдоминантами здесь являются Sih-ius vilripenuis,
Chrysops flavipes punctifer, Tabanus autumnalis brunnescens, T. bromius flavofemoratus,
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
69
Т. leleani, Т. golovi (и подвид Т. golovi mediaasiaticus), в меньшем количестве
встречаются Т. laetetinctus, Т. spectabilis и изредка Т. accipiter (Леиинабад). В орехоплодных
лесах (и их аналогах) обитают почти все перечисленные виды, но, кроме того, здесь
многочисленны Т. bactrianus, T. semenovi (по П. А. Чирову, более обычен
в субальпийском поясе) и в меньшем числе встречается Т. montiasiaticus. Упомянутые
3 вида частично распространены также в остепненных низкогорьях. В предгорьях
и низкогорьях Западного Тянь-Шаня спорадически распространен эндемичный
Chrysops turanicus, ближайший родич европейского лесного вида Ch. pictus. В лесных
высокогорьях (местами покрыты арчевниками) обитают широко распространенные
виды среднеазиатского горного комплекса: Hybomitra hunnorum (преимущественно
в альпийском поясе), Н. turkestana, H. semipollinosa, H. tatarica и Н. shnitnikovi. Кроме
них в Западном Тянь-Шане распространены еще следующие представители того же
комплекса, но характеризующиеся более узкими ареалами: Н. nigricorpa, Atylotus
chodukini (викариат европейского лесного A. fulvus), а на крайнем западе Таласского
Алатау (и частью его отрогах) — Hybomitra sogdiana (альпийский пояс) и Ж. shev-
tshenkoi (преимущественно субальпийский пояс). Следует отметить отсутствие в
горном поясе и предгорьях Западного Тянь-Шаня боревразийских лесных и лесостепных
видов, распространенных восточнее по северным склонам Тянь-Шаня. Исключение
составляют лишь эвритопные Tabanus autumnalis и Т. bromius, распространенные в
номинативной форме в горном поясе, тогда как в низкогорьях и предгорьях они замещены
светлоокрашенными ере таземноморскими подвидами Т. autumnalis brunnescens и Т.
bromius flavofemoratus (или'промежуточными формами). Исчезают также степные виды
Hybomitra montana morgani и Chrysops ricardoae (крайняя находка последнего к
западу— на северном склоне Угамского хребта).
В высокой степени вероятно, что в Западном Тянь-Шане обитает горный Tabanus
argenteomaculatus. Он найден в Джунгарском Алатау (см. выше), на Алайском, Гис-
сарском и Зеравшанском хребтах, но везде малочислен.
В целом фауна слепней Тянь-Шаня носит смешанный характер. По предгорьям
и низкогорьям преобладают виды, боревразийского типа фауны, сменяемые в
Западном Тянь-Шане средиземноморскими видами. Любопытно, что сюда (т. е. до Западного
Тяш^Шаня) в почти неизмененном виде достигают такие типичные средиземноморцы,
как Silvius vitripennis, Chrysops flavipes punctifer, Tabanus spectabilis, T. autumnalis
brunnescens и Т. bromius flavofemoratus, ареалы которых на западе включают Испанию,
а у большинства, кроме Silvius vitripennis, также Северо-Западную Африку.
Остальные, виды, относимые нами к средиземноморскому типу фауны и распространенные
на Тянь-Шане (и по другим горным системам Средней Азии), представляют собой в той
или иной степени «переработанные» элементы, трансформировавшиеся в ряде случаев
в эндемиков гор Средней Азии. Но эти виды обнаруживают ясные генетические связи
с средиземноморской (и в чаети случаев с восточноазиатской) фауной.
Иной характер имеет фауна горно-лесного пояса и альпийских высокогорий.
В ней преобладают формы боревразийского типа фауны, в большинстве
превратившиеся в эндемиков гор Средней Азии. Сюда относятся Atylotus и почти все виды
Hybomitra. Для своеобразных мохнатых Н. tatarica и Н. shevtshenkoi прослеживаются связи
с центральноазиатским (тибетским) типом фауны.
Фауна слепней Памиро-Алая насчитывает 24 вида и 8 подвидов. Следовательно,
она несколько беднее таковой Тянь-Шаня. В западных частях Памиро-Алая
(обследованы преимущественно Гиссарский и Зеравшанский хребты) фауна в значительной ^
мере повторяет таковую западных частей Тянь-Шаня. По остепненным низкогорьям
(степи того же типа, что и в Западном Тянь-Шане) и в среднем поясе гор, включая
орехоплодные леса, доминируют или многочисленны Tabanus bromius
flavofemoratus (в более высоких частях гор замещен номинативным подвидом Т.
bromius), Т. leleani (в предгорьях замещен подвидом Т. leleani turkestanicus), Т. golovi
(в предгорьях и местами в низкогорьях замещен подвидом Т. golovi mediaasiaticus),
Т. laetetinctus (в горах близ верхнего предела распространения замещен более редким
подвидом Т. laetetinctus sordes), Т. autumnalis brunnescens, Т. semenovi, T.
montiasiaticus (оба вида также в лесном поясе) и Silvius vitripennis. По предгорьям в небольшом
количестве встречаются Tabanus spectabilis, T. accipiter и Chrysops flavipes punctifer,
спорадически в лесном поясе гор распространен Tabanus bactrianus. Возле Самарканда
найден Chrysops turanicus. В горном лесном поясе добавляются отсутствующие на Тянь-
Шане Tabanus, indrae vappa, T. mistshenkoi (оба вида также в предгорьях),
Haematopota bactriana, Tabanus schiva, T. hissaricus и Hybomitra zonata (хребты Петра Первого,
и Дарвазский). Первые 4 вида относятся к средиземноморскому типу фауны (его
дериваты), Н. zonata очень близка к Н. tatarica и, следовательно, генетически связана;
с центральноазиатским (тибетским) типом фауны. Первые 3 вида местами
многочисленны, остальные редки. Tabanus hissaricus известен только с Гиссарского хребта,
его родственные связи неясны. В горно-лесном и частью альпийском поясе обитают-
общие с Тянь-Шанем среднеазиатские горные виды — многочисленная Hybomitra:
shnitnikovi и более редкие Н. semipollinosa, H. turkestana, H. tatarica, H. hunnorum,.
70 ВВЕДЕНИЕ
Atylotus chodukini и Tabanus argenteomaculatus. Для северных склонов Алайского
и Туркестанского хребтов отметим Hybomitra nigricorpa. Фауна Памира (за
исключением его западных склонов, где найдены некоторые из вышеперечисленных видов)
значительно обеднена. В альпийском поясе распространены главным образом Н. hun-
norurn и Н. tatarica, тогда как в опустыненной восточной части обитает Н. montana
reinigiana.
В итоге можно отметить для фауны Памиро-Алая такой же смешанный характер,
как и для фауны Тянь-Шаня. По низкогорьям и среднегорному поясу (орехоплодные
леса и их аналоги) доминируют виды средиземноморского тина фауны, в высокогорьях
обитают виды среднеазиатского горного комплекса, являющиеся производными бор-
евразийского (7 видов), центральноазиатского (2 вида) и средиземноморского (2 вида)
типов фауны.
Фауна слепней Копет-Дага изучена фрагментарно.* По данным Д. Чарыкулиева
(1967), в горных долинах и предгорьях распространены Silvius vitripennis, Tabanus
acclpiter, T. spectabilis, Т. laetetinctus (с подвидом Т. laetetinctus sordes) и Dasyrhamphis
umbrinus. Позднее Д. Чарыкулиевым были найдены здесь же Tabanus atropathenicus,
Т. quatuornotatus и Т. indrae. В горной зоне и в горной части субтропической зоны
массовыми видами являются Dasyrhamphis umbrinus и Tabanus accipiter, за
20-минутный учет на животном максимально отлавливались соответственно 206—148 слепней
этих видов (Мамедниязов, 1971). В Каракале (и восточнее) обнаружены Therioplectes
carabaghensis orientalis, Tabanus unif'asciatus и Т. ansarii badhysi. По остепненным
предгорьям Копет-Дага доминирует Т. leleani (реже встречается подвид Т. leleani
turkestanicus), в меньшем количестве обитают Т. gqloui mediaasiaticus, T. bromius fla~
vofemoratus, T. autumnalis brunnescens и редок Chrysops flavipes punctifer. Виды пустын
ной фауны, частично заходящие в предгорья, мы опускаем из рассмотрения.
Всего на Копет-Даге найдено 10 видов и 8 подвидов слепней. Все они без
исключения принадлежат к средиземноморскому типу фауны. Обнаруживаются связи
С|Кавказом благодаря наличию в фауне Копет-Дага общих Therioplectes carabaghensis,
Tabanus atropathenicus и Т. laetetinctus, а также средиземноморских видов широкого
распространения: Dasyrhamphis umbrinus, Tabanus leleani, T. unifasciatus, Т.
quatuornotatus, Т. indrae, Т. spectabilis, Т. bromius flavofemoratus и Т. autumnalis brunnescens.
Виды, обнаруженные в Копет-Даге, обитают также в Иране. Исключение составляет
лишь Т. golovi, но он, несомненно, будет найден в Иране.
Горы Алтая и Тувы. Горы Алтая представляют собой массивную
систему хребтов, окаймляющих юг Средней Сибири и восток Казахстана, и имеют
значительные высоты. В Туве обследован главным образом хребет Танну-Ола (запад
и восток), с востока примыкающий к Алтаю. Горы покрыты преимущественно
хвойными лесами, а высокогорья заняты альпийскими лугами и тундрами. На Алтае
местами развиты высокогорные степи.
Сведения о фауне слепней Алтая имеются в публикациях Н. Г. Олсуфьева (1936а,
19376, 1962а), X. А. Айбасова и М. С. Шакирзяновой (1958), В. В. Шевченко (1961)
и Н. А. Виоловича (1968), а о фауне Тувы — в работах Н. А. Виоловича (1965, 1968).
В лесостепных предгорьях и лесных низкогорьях северных и западных склонов
Алтая преобладают европейско-сибирские лесные и лесостепные виды (12) с
значительной примесью таежно-лесных элементов (6). По численности популяций
преобладают первые. Доминантами являются Chrysops caecutiens, Hybomitra muehl-
feldi, H. bimaculata и Haematopota pluvialis, субдоминантами Hybomitra
distinguenda, II. ciureai и Tabanus mild, малочисленны или редки Chrysops nigripes,
Ch. relictus, Hybomitra nigricornis, H. lundbecki, H. montana, Atylotus rusticus, Tabanus
maculicomis, T. autumnalis, T. bovinus, T. bromius (обычен на юго-западных склонах)
и Haematopota crassicornis (крайне редка, найдена возле Кытманова и Барнаула).
Интересно обнаружение в низкогорьях юго-западных склонов Алтая спорадически
распространенных степного вида Hybomitra expollicata, пустынных Н. peculiaris и
и Haematopota turkestanica, а также средиземноморского Tabanus leleani (на север
до Змеиногорска). Эти виды, видимо, являются здесь реликтами ксеротермического
периода.
Фауна слепней горно-лесного пояса Алтая более сложна. - Упомянем прежде
всего значительную группу таежных и европейско-сибирских лесных видов, из них
доминантами и субдоминантами являются Chrysops caecutiens,
Ch. nigripes, Hybomitra lurida, H. bimaculata, H. nigricornis, H. lundbecki, H. "montana,
Tabanus maculicomis (по северным склонам до 20% обилия) и Haematopota pluvialis (no
северным склонам до 50% обилия), малочисленны или редки Chrysops divaricatus, Ch.
relictus (в нижней части пояса), Hybomitra lapponica, H. arpadi, H. tarandina, H. nitidif-
rons, П. confiformis, H. distinguenda, H. muehlfeldi, H. ciureai, Atylotus fulvus, Tabanus miki,
* Публикация Г. А. Бабаянца (1962) содержит ряд неправильных определений,
поэтому нами не используется.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 71
Т. autumnalis, T. bovinus и Heptatoma pellucens orientalis. В верхних частях
горнотаежного пояса местами многочислен лесотундровый вид Hybomitra sexfasciata
и единично встречается тундровый Н. aequetincta. В горной тайге Алтая
распространены также восточносибирские лесные виды, ареалы которых имеют на Алтае
западную границу. К этой группе относятся местами многочисленные Н. brevis,
Н. tarandinoides, И. astur, а также малочисленные Chrysops suavis, Tabanus geminus
и Haematopota tamerlani (Шебалино). Первые 2 вида обитают главным образом по более
влажным северным и западным склонам Алтая, Н. astur многочисленна в более сухих
юго-восточных частях Алтая. В горной тайге Алтая спорадически распространены
восточносибирские таежные виды -Chrysops makerovi, Hybomitra olsoi и II. pavlovskii.
На юго-западных склонах Алтая найдена в небольшом количестве Н. turkestana —
представитель среднеазиатского горного комплекса.
В высокогорном тундровом поясе в большом количестве обитает
эндемик Алтая (и Танну-Ола) горно-тундровый вид Н. altaica. Местами за
20-минутный учет сачком вокруг себя отлавливалось до 220 самок этого вида (Виолович, 1968).
В горной тундре многочисленны также лесотундровый Н. sexfasciata и
таежный Н. nigricornis, в небольшом количестве встречаются типичные тундровые Н.
aequetincta и Н. astuta (последний вид найден Н. А. Виоловичем на перевале Циган-Шибети
на стыке Алтая и хребта Танну-Ола), а также таежные Chrysops nigripes, Hybomitra
olsoi и Н. montana. Последний вид более характерен для альпийских лугов, где его
обилие достигает 20%. В тундровый пояс и на альпийские луга вклиниваются и
другие виды, обитающие в горной тайге, особо следует упомянуть Н. lundbecki sibiriensis,
местами весьма многочисленного на альпийских лугах юго-восточного
Алтая. Этот подвид имеет западную границу распространения на Алтае.
В горных степях юго-восточного Алтая многочисленны Н. montana mor-
gani, H. tschuensis (главным образом в Чуйской степи), местами Н. lundbecki
sibiriensis и Н. nigricornis, спорадически распространены и некоторые виды горно-таежного
пояса.
Всего в предгорном и горных ландшафтах Алтая найдено 43 вида и 4 подвида
слепней. Эндемизм в фауне Алтая незначительный — всего 1 вид (общий с Танну-
Ола) — Н. altaica. Фауна Алтая носит смешанный характер. По предгорьям и
низкогорьям преобладают европейско-сибирские лесные и лесостепные виды, в горном
поясе — таежные виды, наконец, в высокогорных тундрах — тундровые и некоторые
таежные виды; в горных степях обнаружены степные формы, в том числе
своеобразный вид Н. tschuensis, пока известный только с Алтая, но, вероятно, распространенный
и в Монголии. Особый интерес представляет наличие на Алтае восточносибирских
таежных и лесных видов, всего в количестве 9.
На Алтае находят предел распространения на восток некоторые
европейско-сибирские лесные и лесостепные виды, например, Tabanus maculicomis, T. bromius и
Haematopota crassicornis. Алтай, следовательно, является местом стыка ареалов некоторых
восточносибирских и европейско-западносибирских форм. Фауна слепней Кузнецкого
Алатау, примыкающего к северо-восточной части Алтая, практически не изучена.
Фауна слепней хребта Танну-Ола имеет много общего с таковой Алтая. Но
имеются и свои особенности. По предгорьям и таежным низкогорьям (до 1600 м) д о м и-
н и р у ю т или многочисленны Hybomitra lundbecki (подвид Н. lundbecki
sibiriensis), H. montana (в меньшем количестве Н. montana morgani), H. lurida, H. astur
и Chrysops suavis, малочисленны Ch. nigripes, Ch. caecutiens, Hybomitra arpadi, H.
nigricornis, H. ciureai, H. muehlfeldi, H. olsoi, H. sexfasciata, H. pavlovskii, Tabanus pleskei,
T. geminus и Haematopota tamerlani и редки Hybomitra bimaculata, H. distinguenda,
H. nitidifrons (вероятно, H. n. confiformis), H. aequetincta, H. lapponica, Atylotus fulvus,
A. rusticus (указание нуждается в подтверждении) и Haematopota pluvialis. Степные и
пустынные виды, местами встречающиеся по предгорьям и в Тувинской котловине, нами
обсуждены при описании степного фаунистического комплекса (см. стр. 61).
В верхних частях горной тайги и в горной тундре доминантами и
субдоминантами являются Hybomitra sexfasciata (до 55% обилия), Н. altaica,
Н. astur (преимущественно в горной тайге), Н. lurida, H. nigricornis и Н. lundbecki,
в меньшем количестве обнаружены Н. aequetincta, II. pavlovskii и единично Н. arpadi,
Н. olsoi, H. astuta (только в тундре) и Chrysops nigripes. Для альпийских лугов
Н. А. Виолович отмечает обилие степного вида Haematopota desertorum,
неизвестного на Алтае, второе и третье места занимают Hybomitra montana и Н. lundbecki
(вероятно, подвид Н. lundbecki sibiriensis).
Всего на хребте Танну-Ола обнаружено 26 видов и 3 подвида слепней, что намного
меньше, чем на Алтае. Обеднение фауны отмечается за счет европейско-сибирских
лесных и лесостепных видов, таких как Tabanus maculicomis, Т. bromius, f. miki,
Т. autumnalis, T. bovinus, Chrysops relictus, Heptatoma pellucens, Haematopota
crassicornis, не найдены таежные Hybomitra tarandina, Chrysops makerovi, Ch. divaricatus,
восточносибирские лесные Hybomitra tarandinoides и H. brevis, а также алтайский
горностепной вид Н. tschuensis. Но в фауне Тувы добавляется восточносибирский лесной.
72 ВВЕДЕНИЕ
Tabanus pleskei, аллопатричный европейско-сибирскому лесному Т. bovinus, а также
степной вид Haematopota desertorum.
Горы Восточной Сибири и Дальнего Востока. Сюда
относятся хребты: Саян, Становой, Верхоянский и другие, а также Сихотэ-Алинь.
К сожалению, специальные исследования аспектов фауны слепней этих горных систем
не проводились, за. исключением Сихотэ-Алиня. Упомянем лишь небольшой сбор
слепней Г. В. Сердюковой в Западном Саяне на р. Малый Кебеж в 120 км к
юго-востоку от Минусинска в зоне кедрово-пихтовой тайги. Доминировали Hybo-
mitra bimaculata, Н. lundbecki, H. lapponica и Tabanus pleskei, в небольшом количестве
были Hybomitra tarandina, Н. sexfasciata, Н. arpadi,. H. tarandinoides, Н. pavlovskii,
«Н. lolstitalis» (вероятно Н. muehlfeldi), H. distinguenda, Atylotus fulvus, Chrysops cae-
cutiens, Haematopota pluvialis и Н. tamerlani. Таким образом, аспект фауны был в
основном таежным. Имеющиеся фрагментарные данные но другим горным системам
дают основание заключить, что в их лесном (таежном) поясе господствуют виды
таежного фаунистического комплекса. Массовыми и многочисленными
видами являются Hybomitra montana, H. nigricornis, H. lundbecki sibiriensis, местами
Н. lurida, H. olsoi, H. arpadi, Chrysops makerovi и др. В горных тундрах и прилегающих
верхних частях лесного пояса обитают Н. sexfasciata, H. aequetincta и Н. astuta вместе
с некоторыми вышеперечисленными таежными видами. Наличие гор в Восточной
Сибири с развитыми тундрами отодвигает далеко на юг распространение тундровых видов
слепней.
По сборам В. А. Кирпичниковой (1972) и другим материалам, по предгорьям
и низкогорьям Сихотэ-Алиня (в пределах Приморского края) в зоне
широколиственных и кедрово-широколиственных лесов аспект фауны слепней примерно тот же, что
и в дальневосточной лесной зоне в целом (см. выше). Но кроме массовых и
многочисленных для зоны Hybomitra brevis, H. lapponica, H. lundbecki sibiriensis, H. montana
montana, H. distinguenda contigua, Tabanus pleskei, T. geminus, Chrysops, suavis,
Haematopota tamerlani и других видов, местами доминируют Hybomitra ussuriensis,
многочисленны Chrysops japonicus, а по восточным склонам хребта —
доминируют Hybomitra stigmoptera и Н. astur. Последние, два вида многочисленны также
в пихтово-еловом поясе тех же склонов, а также в лиственничных лесах северной части
хребта. Вместе с ними, но в меньших количествах, встречаются все типичные виды
дальневосточной зоны. Здесь же найдена в небольшом количестве тундрово-таежная
Н. sexfasciata. В высокогорном гольцовом (тундровом) поясе обнаружен единично
тундровый вид Н. aequetincta и в большом количестве — лесной Н. astur. Местами
в среднем и южном Сихотэ-Алине по западным склонам спорадически
распространены эффектные по величине и окраске Tabanus buddha и Т. chrysurus, а также
некоторые другие виды восточноазиатского типа фауны.
Итого на Сихотэ-Алине обнаружено 34 вида и 2 подвида слепней, всего 36 форм,
в том числе восточносибирских и дальневосточных лесных — 14, таежных — 13,
тундровых и тундрово-таежных — 2, степных и лесостепных — 2 и восточноазиат-
ских — 5.
ЗООГБОГРАФИЧБСКОЕ ДЕЛЕНИЕ ПАЛЕАРКТИКИ
Вопросы зоогеографического (фаунистического) районирования Палеарктиче-
ской области издавна привлекали к себе внимание отечественных исследователей.
Пионерами являются Н. А. Северцов (1877), М. А. Мензибр (1У14), П. П. Сушкин
(1925) и др., разработавшие деление Палеарктики на подобласти и провинции
главным образом в результате изучения географического распространения наземных
позвоночных. Были также предложены схемы, основанные на анализе фауны насекомых,
например жуков (Семенов-Тян-Шанский, 1936; Крыжановский, 1965). Эти схемы
близки схемам предыдущих авторов. Из работ, посвященных зоогеографическому
районированию пресноводных животных, упомянем фундаментальное исследование
Л. С. Берга (1949) по рыбами Я. И. Старобргатова (1970) по моллюскам. Однако
предложенные этими авторами деления Палеарктики сильно отличаются от зоогеографии
животных суши.
Ниже нами предлагается зоогеографическое деление Палеарктики и несколько
подробнее — территории СССР, основанное на анализе ареалов, а также
количественных аспектов фауны слепней. Такая попытка, основанная на изучении двукрылых
насекомых, делается впервые и, вероятно, она не лишена отдельных недочетов. Однако
мы надеемся, что по мере накопления новых данных детали этого деления будут
уточнены. Наше деление гораздо ближе к схемам районирования наземных животных,
нежели водных.
Палеарктическую область мы рассматриваем как часть Голарктической надоб-
ласти (отдела по А. П. Семенову-Тян-Шанскому, 1936; доминиона по О. Л. Крыжанов-
скому, 1965), куда входит еще Неарктическая область. О сходстве фаун слепней этих
областей говорилось выше.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 75
В соответствии с осуществленной нами типизацией фаун Палеарктическую
область мы подразделяем на пять подобластей (рис. 39). Они имеют одноименные
названия с таковыми субрегиональных типов фаун: Боревразийская, Средиземноморская,
Афроевразийская аридная. Восточноазиатская (Палеархеарктическая по А. П. Се-
менову-Тян-Шанскому, но без Сев.-Вост. Китая и Тибета) и Централыюазиатская
(Тибетская) подобласти. Мы применили здесь, как и на других картограммах,
линейное изображение границ, хотя отдаем себе отчет в том, что они имеют относительное
значение. Предлагаемое деление имеет ряд отличий от общепринятого. Так, мы не
выделяем Арктическую подобласть, присоединяя территорию, занятую фауной тундры,
к Боревразийской подобласти, и обозначая эту территорию как тундровую или
циркумполярную провинцию. Распространение в зоне туидры значительного количества
таежных и даже лесных видов и малого числа собственно тундровых форм (к тому же
генетически весьма близких таежным) вполне обосновывает отнесение фауны этой
зоны в ранге провинции к Боревразийской подобласти. Равнинные пустыни и сухие
степи Евразии и Северной Африки мы рассматриваем как самостоятельную Афроев-
разийскую аридную подобласть, не объединяя ее с нагорьями Центральной Азии.
Последние мы выделяем в другую подобласть. Горы (преимущественно высокогорья)
Средней Азии (кроме Копет-Дага) мы в ранге провинции объединяем с Боревразийской
подобластью. В Средиземноморскую подобласть мы включаем в Средней Азии не только
Копет-Даг и Паропамиз, но и предгорно-низкогорные части Памиро-Алая и
Западного Тянь-Шаня. Границу между Боревразийской и Восточноазиатской подобластями
мы проводим южнее, за пределами СССР.
Фауна Боревразийской подобласти представлена в основном видами родов НуЪо-
mitra и Chrysops s. str., относительно малочисленны виды Tabanus, Atylotus и Наета-
topota; имеется эндемичный монотипичный род Heptatoma. На западе из
Средиземноморской подобласти проникают 1 вид Pangonius и 1 вид Silvius.
Фауна Средиземноморской подобласти наиболее разнообразна. Преобладают
виды Tabanus, относительно многочисленны виды пангонии (3 рода июколо 40 видов),
Silvius, Haematopota, род Chrysops представлен преимущественно видами подрода
Heterochrysops, имеются эндемичные роды Dasyrhamphis, Philipomyia, Therioplectes,
Surcoufia. Немногочисленны виды Atylotus и Hybomitra, последний род представлен
главным образом видами эндемичного подрода Mouchaemyia.
Фауна Афроевразийской аридной подобласти в родовом отношении относительно
бедна. Здесь представлены главным образом Tabanus (в основном подгруппа Tabanus
sabuletorum из группы Т. cordiger), Hybomitra (подрод Sipala и группа Н. erberi),
Atylotus, Chrysops (эндемичный подрод Turanochrysops и некоторые Heterochrysops)
и немногие виды Haematopota. Имеется эндемичный монотипичный род Nanorhynchus.
Фауна Центральноазиатской подобласти изучена фрагментарно. Она представлена
многочисленными"и весьма своеобразными видами Hybomitra (в том числе эндемичным
подродом Tibetomyia), немногими Chrysops и Haematopota.
Наконец, фауна Восточноазиатской подобласти обнаруживает значительное
разнообразие, напоминающее таковое Средиземноморской подобласти. Родственные связи
с этой подобластью подчеркиваются наличием здесь нескольких видов пангонии (род
Stonemyia) и Silvius, отсутствующих в других подобластях Палеарктики.
Многочисленны виды Chrysops (в том числе имеются Heterochrysops), Tabanus и Haematopota,
но малочисленны Hybomitra и Atylotus. Имеются эндемичные роды Thaumastomyia,
Gresittia, Nagatomyia и Isshikia.
Территорию СССР по нашему делению охватывают три подобласти:
Боревразийская, Афроевразийская аридная и Средиземноморская. В пределах этих подобластей
применительно к территории СССР мы можем выделить провинции, а в некоторых
случаях округа и районы, используя разработанную нами типологию ареалов (рис. 40).
Границы между провинциями мы проводим, учитывая основные ареалы фауни-
стических комплексов, а также количественные аспекты фауны. Однако при этом
возникают известные трудности, поскольку основные ареалы комплексов частично
налегают друг на друга. Да и не всегда имеется достаточно фактического материала для
Рис. 40. Зоогеографическое деление территории СССР на основании распространения
слепней. Схема. Сплошная линия — границы подобластей; прерывистая линия—
границы провинций; точечная линия — границы округов. 1—6 —• Боревразийская
подобласть, провинции: 1 — тундровая; 2 — таежная, европейско-западносибирский
округ: 2А — восточносибирский округ, 2В — южносибирский округ; 3 — европейско-
сибирская лесная, европейский округ: ЗА — западносибирский округ, ЗВ —
кавказский округ; 4 — дальневосточная лесная; 5 — лесостепная; 6 — среднеазиатская
горная (деление на округа см. рис. 41). 7—8 — Афроевразийская аридная подобласть,
провинции: 7 — степная, европейский округ, 7А — казахстанский округ, 7В —
монгольский округ; 8 — пустынная. 9—10 — Средиземноморская подобласть, провинции:
9 — понтийско-гирканская, 9А —горнокрымский округ, 9Б —понтийский округ,
9В — копетдагский округ; 10 — бактрийско-иранская.
76 ВВЕДЕНИЕ
соответствующих уточнений, поэтому намеченные нами границы в значительной мере
схематичны. В проведении их мы в основном придерживались наших данных, а не
ботанических границ, как это часто делается.
Таблица 6
Зоогеографические (фаунистические) подразделения
Палеарктической области (преимущественно в пределах СССР)
на основании распространения слепней
Подобласть
Боревразийская
Афроевразийская
аридная
Средиземноморская
Центральноазиатская
Восточноазиатская
Провинция
Тундровая
Таежная
Европейско-сибирская лесная
Лесостепная
Дальневосточная лесная
Среднеазиатская горная
Степная
Пустынная
Понтийско-гирканская
Б актрийско-иранская
В СССР не представлена
То же
Округ
_
Европейско-западносибирский
Восточносибирский
Южносибирский '
Европейский
Западносибирский
Кавказский
—
—
Лесо альпийский
Горно-степной
Европейский
Казахстанский
Монгольский
—
Горнокрымский ■
Понтийский
Копетдагский
—
В общих чертах разработанное нами районирование (табл. 6) совпадает с
принятым в СССР делением на ландшафтные зоны. Исключение составляют лишь горы
Кавказа и особенно Средней Азии, где мы, основываясь на нашем материале,
осуществляем это районирование несколько по-иному.
Боревразийскую подобласть мы делим на 6 провинций: тундровую, таежную, ев-
ропейско-сибирскую лесную, лесостепную, дальневосточную лесную и
среднеазиатскую горную.
Тундровая провинция охватывает север Евразии и не имеет подразделений.
В пределах таежной провинции мы выделяем ее восточносибирский округ с
границей по Енисею и южносибирский округ, охватывающий участок от Алтая до
Нижнего Приамурья. Основанием для этого является наличие в тайге к востоку от Енисея
восточносибирских таежных видов, а на юге — восточносибирских лесных видов.
Но и те, и другие по численности популяций уступают видам основного таежного ком- -
плекса. Тундровые высокогорья Алтая мы выделяем в особый район. Это выделение
обосновывается наличием эндемичного горнб-алтайского тундрового вида НуЪотНга
altaica, популяции которого здесь многочисленны и доминируют. Участок таежной
провинции к западу от Енисея и Алтая мы обозначаем как западный (европейско-
западносибирский) округ.
Европейско-сибирскую лесную провинцию мы делим на округа: европейский
(западный) и западносибирский (восточный) с границей но Уральскому хребту.
Основанием для такого подразделения является отсутствие (или спорадичность) к востоку
от Урала некоторых лесных видов, широко распространенных на западе. К восточному
округу мы присоединяем Тарбагатай (точнее — обследованный Саур), на котором
господство принадлежит лесным и таежно-лесным видам.
В составе европейско-сибирской лесной провинции мы выделяем кавказский
округ. В него включаем западные части Большого и Малого Кавказа, северные склоны
Большого Кавказа на восток до Дагестана и провизорно — Колхидскую низменность.
Альпийские высокогорья Большого и Малого Кавказа ввиду особенностей
их фауны мы выделяем в особый район.
Лесостепная провинция расположена южнее европейско-сибирской лесной
провинции, с юга отграничивается степной провинцией Афроевразийской аридной под^
области. Делений на округа не предусматривается. Следует отметить, что некоторые
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 77
представители лесостепного комплекса фауны ленточно распространены (обычно вместе
с лесными видами) по предгорьям и частично низкогорьям северных склонов Большого
Кавказа, а также северных и частью западных склонов Тянь-Шаня (на запад до
Джамбула). Эти участки в дальнейшем могут быть подвергнуты более дробному
районированию.
Дальневосточная лесная провинция охватывает Приморский край (без
высокогорий Сихотэ-Алиня), юг Хабаровского края и^юго-восточную (приамурскую) часть
Амурской области. Сюда же мы относим южную часть о. Сахалин и о. Кунашир,
а за пределами СССР — северную и восточную часть Северо-Восточного Китая.
Вероятно, сюда же относится о. Хоккайдо (Япония), но для более точного решения этого
вопроса имеющихся в литературе данных недостаточно. Принадлежность рассматри-
Рис. 41. Зоогеографическое деление территории Средней Азии на основании
распространения слепней. Схема. 1 — Средиземноморская подобласть, ее
понтийско-гирканская провинция; 1А — бактрийско-иранская провинция той же подобласти. 2 —
Афроевразийская аридная подобласть, ее пустынная провинция. 3 — Боревразийская
подобласть, среднеазиатская горная провинция, ее лесоальпийский округ; ЗА — ее
горно-степной округ.
ваемой провинции к Боревразийской подобласти хорошо обосновывается
преобладанием видов (и их аспектностыо) боревразийского типа фауны. Встречающиеся в
рассматриваемой провинции виды восточноазиатского типа фауны представлены
ограниченным числом видов и популяции их малочисленны.
К среднеазиатской горной провинций мы относим высокогорья и частью средне-
горья Памиро-Алая и Тянь-Шаня. Включение этой провинции в Боревразийскую
подобласть обосновывается доминированием здесь видов, отчетливо связанных
происхождением с боревразийской фауной. Рассматриваемую провинцию мы делим на 2 округа:
северный — северо-западный (лесо-альпийский) и юго-восточный (горно-степной).
Последний охватывает восточный Памир, а на Тянь-Шане — остепненные высокогорья
(сырты) к югу от оз. Иссык-Куль. Возможно, что этот округ правильнее отнести к степной
провинции Афроевразийской аридной подобласти. Он характеризуется
распространением в нем почти в единственном числе (но местами многочисленной популяцией)
Hybomitra montana reinigiana. Ближайший родич этой формы Н. montana morgani
является одним из индикаторных видов степной провинции вышеназванной аридной
подобласти. В особый район следует выделить Иссык-Кульскую котловину, в которой
обитают наряду с некоторыми горными видами отдельные виды степной и пустынной
фаун, например, Chrysops ricardoae и Haematopota turkestanica.
Афроевразийская аридная подобласть в пределах СССР делится на 2 провинции:
степную и пустынную. Степную провинцию мы делим на 3 округа: европейский (на
восток до Волги), казахстанский и монгольский, последний с западной границей по
Алтаю и Тарбагатаю. Основанием для этого служат ареалы соответствующих вариантов
степного фаунистического комплекса.
78
ВВЕДЕНИЕ
Пустынная провинция, охватывающая в. пределах СССР равнинные части Средней
Азии и Казахстана, а также Восточного Закавказья, вероятно, может быть поделена
на 2 округа: северный и южный. Но недостаток материала затрудняет нас в
проведении границы между этими округами.
' Средиземноморская подобласть на территории СССР представлена своей восточной
окраиной. Анализ ареалов средиземноморского типа фауны позволил нам
сгруппировать их в 4 фаунистических комплекса. Однако ввиду значительных совмещений
их ареалов на территории СССР, особенно на Кавказе, мы по господствующему числу
видов выделяем в составе Средиземноморской подобласти лишь провинции: понтийско-
гирканскую (или восточносредиземноморскую) и бактрийско-иранскую. Первая
охватывает горы Крыма, значительную часть Кавказа и Колет-Даг. За пределам СССР
эта провинция включает Балканы, Переднюю Азию и северную часть Ирана. В
пределах этой провинции для СССР мы выделяем 3 округа: горнокрымский, поптийский
(кавказский) и копет-дагский. Бактрийско-ирапская провинция в пределах СССР
охватывает низкогорья и частью среднегорья западных частей Памиро-Алая,
западного Тянь-Шаня и Каратау, а в Иране — горы Загрос и др. Границы этой провинции
в СССР, имеющей сложные очертания, показаны на рис. 41.
При выделении понтийско-гирканской и бактрийско-иранской провинций мы
не выделяем в самостоятельные участки предгорья и низкогорья, по которым
преимущественно распространены виды средиземноморско-азиатского фаупистического
комплекса. Мы объединяем эти участки с соответствующими частями провинций.
Распространение этого комплекса приурочено к поясу своеобразных «эфемеровых»
субтропических степей, растительность которых ботаники сближают с таковой
Средиземноморья. По-видимому, в дальнейшем при более дробном районировании
соответствующее подразделение может быть сделано.
НЕКОТОРЫЕ ВОПРОСЫ ПАЛЕОГЕНЕЗА ФАУНЫ ПАЛЕАРКТИКИ
Ископаемые формы слепней немногочисленны, известны лишь следующие 14
находок (табл. 7). Обнаруженные виды, относящиеся к 5 родам, в большинстве вымерли,
но Tabanus sudeticus обитает на территории Европы и поныне.
Таблица 7
Ископаемые Tabanidae (Moucha, личное сообщение)
Эпоха
Олигоцен
Миоцен
Плиоцен (верхний)
Название
Silvius laticornis
Silvius sp.
Chrysops vectensis
Tabanus fulvescens
Tabanus—
Tabanus —
Tabanus —
Aemoaipus bornensis
Silvius merychippi
Tabanus hipparionis
T. parahippi
Tabanidae n. g. n. sp.
Hexatoma? oeningensis
Tabanus sudeticus
Автор
Loew
Helm
Cockerrell
Statz -
Keferstein
Serres
Goldfuss
Aymard
Cockerrell
»
»
Scudder
Heer
Durrenfeldt
Место
обнаружения
Европа
»
»
»
»
»
»
»
Сев. Америка
» »
» »
Европа
»
»
Из перечня видно, что уже в палеогене (олигоцен) в Европе были распространены
хорошо дифференцированные современные роды 2 подсемейств: Chrysopsinae —
Silvius, Chrysops и Tabaninae — Tabanus. В миоцене на территории Северной Америки
найденыше же роды, а в Европе предположительно еще род Hexatoma (=Heptatoma).-
Большой интерес представляет обнаружение на территории ФРГ в верхнем плиоцене
ныне существующего Tabanus sudeticus.
Естественно, что этих находок недостаточно для решения сложного вопроса
происхождения и формирования фауны слепней Палеарктики. Поэтому приходится
прибегнуть к сопоставлению зоогеографических и филогенетических данных по этой
группе насекомых с хорошо изученной палеонтологами эволюцией ландшафтов
Евразии в кайнозойскую эру. Разумеется, что эти сопоставления могут быть сделаны лишь
в самом обобщенном виде.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 79
Современная фауна слепней Средиземноморской и Восточноазиатской подобластей
является в основном субтропической и в наиболее южных частях — тропической.
Она включает ряд филогенетически более древних форм, таких как Pangoniinae,
а также представителей трибы Bouvieromyiini и рода Silvius подсемейства Chrysopsinae
и трибы Diachlorini подсемейства Tabaninae. Эта фауна уходит корнями в палеоген
и, вероятнее всего, в эоцен — олигоценовую эпоху. Наличие ископаемых Silvius и
Chrysops из олигоцена вполне это подтверждает. В Евразии в эоцене и олигоцене была
широко распространена разнообразная, но мало дифференцированная в зональном
отношении тропическая и субтропическая флора и фауна (Страхов, 1938). Она имела
много общего с таковой Северной Америки. В составе фауны уже было довольно много
средних размеров и крупных млекопитающих (носороги, тапиры и т. д.), на которых
слепни могли осуществлять кровососание.
Разумеется, обитавшие в эту эпоху виды слепней были лишь предшественниками
современных. С другой стороны, они являлись потомками более древних форм мезозоя,
о которых мы уже частично.говорили при рассмотрении филогении слепней (см. выше,
стр. 31). Важно подчеркнуть, что слепни палеогена уже относились к родовым
группировкам Tabanus, Chrysops, Silvius и другим, представленным и поныне как главным
в Средиземноморской и Восточноазиатской подобластях. Возможно, были и другие
родовые группировки, ныне вымершие.
В миоцене в Евразии сохранялся сходный с олигоценом тип флоры и фауны, по
уже с значительной примесью теплоумеренных форм. В северных частях Евразии
господство переходит к широколиственным листопадным древесным породам, развиваются
также хвойные деревья. Млекопитающие получают дальнейшее развитие, причем
появляется много разнообразных крупных форм — хоботные, оленьи и т. д. Видимо,
в этом теплоумеренном ландшафте получают развитие виды слепней Tabanus, Chrysops
и других родов, предшественники видов современных лесных фаунистических
комплексов. Первоначально единый на территории Евразии палеогеновый тип фауны из
наиболее теплолюбивых форм расчленяется на западную и восточную части, которые
начинают постепенно оттесняться к западу и востоку материка. С появлением саванн
формируются лесостепные, а затем и степные формы. По-видимому, в миоцене на фоне
начавшихся обширных горообразовательных процессов, появляются первоначально
в горах Hybomitra, получившие затем в плиоцене широкое развитие. На относительно
позднее появление Hybomitra указывает распространение видов этого рода, ныне
охватывающее в основном умеренный и холодный пояс Голарктики; многие виды
являются горными.
В верхнемиоценовое время осуществляются широкие связи Евразии с Северной
Америкой и обширные миграции млекопитающих с одного континента на другой.
Вероятно, в это время происходит очередной взаимный обмен фаунами слепней. В
начале плиоцена вместе с мощной инвазией из Африки крупных копытных (фауна Пи-
керми) в Евразию проникают Haematopotini. Относительно позднее формообразование
этой трибы, по всей вероятности в Африке, хорошо объясняет ее полное отсутствие
в Австралии и Южной Америке. В Северной Америке виды дождевок единичны и
проникли они сюда, очевидно, из Восточной Азии через Берингию.
В плиоцене по мере развития в северных и средних широтах Евразии бореального
климата происходит дальнейшая дифференциация ландшафтных зон. Н. М. Страхов
(1938) пишет: «Если бы мы попытались представить себе ландшафт тех огромных
континентальных пространств, которые раскинулись в конце неогена от Атлантического
океана до Тихого, то мы увидели бы здесь примерно те же флористические пояса, что
и теперь. Близ Средиземного моря растут еще лавры, мирты, пальмы, тропические
дубы и прочая вечнозеленая субтропическая флора. Севернее, в Западпой и Восточной
Европе и на юге Сибири (Уссурийский край), — лиственный листопадный и смешанный
лес. Остальная Сибирь покрыта хвойной тайгой. Наконец, в области современных
степных пространств юго-западной Сибири, Монголии, Китая располагаются степи,
возникшие, вероятно, еще с миоценового времени» (стр. 380). К сказанному добавим,
что на равнинах Передней, Средней и Центральной Азии в плиоцене развиваются
пустыни.
Имеются все основания полагать, что к концу плиоцена на территории Евразии
формирование 5 типов фаун слепней было в основных чертах закончено. Вокруг
средиземноморского бассейна и частично по горам Передней и Средней Азии локализовались
потомки палеогеновой фауны, в наибольшей степени сохранившие ее основные черты.
Такие же потомки, но в несколько видоизмененном и обедненном составе
локализовались на востоке Азии, ныне территории Китая, Кореи и Японии.* Сформировавшаяся
в неогене теплоумеренная и бореальная фауна заняла территории в средних частях
* На средиземноморский и восточноазиатский (палеархеарктический) центры
разнообразия более древних форм жуков указывает А. П. Семенов-Тян-Шанский
(1936) и уховерток — Г. Я. Бей-Биенко (1936). Оба автора отмечают генетическую
близость фаун этих центров и датируют их происхождение палеогеном.
80
ВВЕДЕНИЕ
и на севере Евразии. Поднявшиеся в Центральной Азии огромные горные системы
(Тибет и другие) с их крайними условиями существования вызвали образование особых
групп Hybomitra, в том числе подрода Tibetomyia.
Развитие равнинных степей и пустынь также повлекло за собой сложную
переделку фауны слепней и образование ее особого типа. Как справедливо указал В. В.
Шевченко (1961), формирование «пустынных» видов в значительной мере произошло за счет
переработки некоторых форм средиземноморской фауны. Но в этом приняли участие
и некоторые боревразийские элементы, затем выходцы из саванн Африки и т. д. Этот
вопрос нами уже обсуждался при рассмотрении типологии ареалов.
В плиоцене по мере поднятия Тянь-Шаня и Памиро-Алая и развития окружающих
равнинных пустынь средиземноморская фауна стала по преимуществу
концентрироваться по низкогорьям и среднегорьям, но немногие преобразовались в
высокогорные виды. С северо-востока (через Тарбагатай—Джунгарский Алатау) в
формирующиеся высокогорья проникли боревразийские формы, преобразовавшиеся затем в
эндемиков и ставшие господствующими. С востока по Куньлуню сюда проникли
немногие цептральноазиатские формы.
К концу плиоцена в Евразии, по-видимому, закончилось формообразование
многих, если не большинства ныне живущих видов. Косвенно на это могут указывать
особенности современного распространения некоторых видов, имеющих разорванные
ареалы. Так, весьма показательно обнаружение А. Г. Шаровым (1956) близ Баку-
риани на Малом Кавказе небольших популяций таежно-лесных Hybomitra lurida
и Н. nitidifrons вместе с некоторыми другими двукрылыми насекомыми (Dolichopus)
бореального типа. По справедливому заключению автора, инвазия этих видов на
Кавказ, очевидно, произошла в ледниковый период, когда теажно-лесной ландшафт
продвинулся далеко к югу. По нашему мнению, сформировались эти виды ранее, очевидно,
в плиоцене. Распространение указанных двух видов слепней на Кавказе прервано
от основных ареалов значительными пространствами степей, по крайней мере на
1000 км, что исключает возможность их случайного заноса в современный или близкий
к нему период.
Обитающие в высокогорьях Кавказа Hybomitra caucasica и Tabanus rupium
распространены также в высокогорьях Альп и Пиренеев. Эти виды, очевидно, также
сформировались до плейстоцена, а распространению их в столь удаленных друг от друга
горных системах способствовали ледники, которые на какое-то время вытеснили их
на равнины. Другие таежно-лесные виды, например, Hybomitra bimaculata и Н. muehl-
feldi, обитающие на Большом и Малом Кавказе в отрыве от основного ареала,
охватывающего равнинные таежную и лесную зоны Евразии, по-видимому, также
представляют собой инвазию ледникового периода. Очень интересен разрыв в распространении
Silvius latifrons — последний обитает в высокогорьях Большого и Малого Кавказа
и в горах Греции. Образование этого вида также произошло ранее плейстоцена.
Упомянем, наконец, находку в отложениях верхнего плиоцена (ФРГ) Tabanus sudeticus,
этого весьма характерного вида современной лесной фауны Европы. Здесь мы имеем
точное доказательство доплейстоценового видообразования.
В плейстоцене под влиянием мощных равнинных и горных оледенений в Евразии
произошли значительные смещения ландшафтных зон, а вместе с ними и фаунистиче-
ских комплексов слепней. О некоторых из них мы уже говорили. Эти смещения в
отдельных случаях сопровождались формообразованием на уровне видов. Сюда мы
относим, например, возникновение парных видов с аллопатрическим распространением:
Tabanus bovinus—T. pleskei, Т. rhaculicornis—T. geminus, Atylotus rusticus—A.^miser,
Haematopota crassicornis—H. tamerlani. Это виды в основном равнинной лесной зоны
(Atylotus — лесостепные или степные виды). Разрыв их ареалов на западную и
восточную части произошел, видимо, в Средней Сибири, где близость ледников в
плейстоцене.стерла почти нацело лесной ландшафт (Синицын, 1962). Под влиянием несколько
различных условий существования западных и восточных популяций этих видов они
превратились в самостоятельные, но очень близкие формы. Любопытно, что ^ареалы
этих видов в постплейстоценовое время «сошлись» на линии Енисей—Алтай, т. е.
в районе наибольшего ледникового разрыва.
Заслуживает внимания викариат аллопатричных степных видов Haematopota
pallidula—H. desertorum, возникновение которого, по-видимому, также связано^ с
разрывом ареала вида-предшественника, произошедшее в Средней Сибири в
плейстоценовое время.
Тундровые и лесотундровые виды, по-видимому, сформировались еще в
доледниковый период в горных условиях Сибири или Северной Америки, но в плейстоцене
получили широкое распространение вместе с равнинными тундрами и «мамонтовой»
фауной. Все виды тундрового комплекса являются циркумполярными. После
отступления ледников их основной ареал отодвинулся на север, но отдельные популяции
сохранились в горах Средней и Восточной Сибири далеко на юге. Алтайский
горнотундровый эндемик Hybomitra altaica, видимо, сформировался в плейстоценовый
период в результате какой-то местной изоляции в горах Алтая (возможно, в монгольской
его части). Этот вид, видимо, отделился от Н. sexfasciata.
БИОЛОГИЯ СЛЕПНЕЙ 81
Излагая наши соображения о палеогенезе бореальной фауны слепней Евразии,
мы должны отметить, что в некоторых частях они совпадают с высказываниями
Н. А. Виоловича (1968).
В постплейстоценовое (послеледниковое) время и, в частности, в ксеротермический
период, предшествовавший современному, в Евразии продолжались передвижения
ландшафтов и следы этих перемещений прослеживаются в современном
распространении слепней. На них мы неоднократно обращали внимание при изложении данных
о распространении отдельных фаунистических комплексов. Наиболее
демонстративными примерами являются: мозаичное распространение в Западной Европе пустынных
видов — Atylotus quadrifarius, Hybomitra erberi и др., наличие некоторых лесостепных
видов, например, Chrysops relictus, далеко на севере Европы, наконец, пятно
лесостепных и степных видов и даже одного вида восточноазиатского типа фауны ^
Ch. vanderwulpi — в Центральной Якутии. В послеледниковое (голоценовое) время
произошло ограниченное видообразование слепней, причем сформировавшиеся виды
лишь очень незначительно отличаются от исходных форм. В качестве примеров мы
можем привести обитающего в горах Крыма Tabanus smirnovi, произошедшего от
средиземноморского лесного Т. unifasciatus, а также заселяющего предгорья Западного
Тянь-Шаня и Зеравшанского хребта Chrysops turanicus, произошедшего от боревразий-
ского лесного Ch. pictus, проникшего сюда, видимо, в плейстоцене. Вероятно,
послеледниковым временем следует датировать происхождение некоторых очень близких
видов (или подвидов), распространенных в Европе (преимущественно в горах) и
изолированно на Кавказе. Таковы пары Tabanus paradoxus—Т. subparadoxus, T. spodo-
pterus—Т. portschinskii, Haematopota scutellata scutellata—H. scutellata pontica и др.
Очевидно, виды — предшественники этих форм проделали в плейстоцене под
влиянием ледников миграции на равнины и после изменения климата в голоцене их ареалы
оказались разорванными на европейскую и кавказскую части, при атом произошло
формообразование.
7. БИОЛОГИЯ СЛЕПНЕЙ
Характерными биологическими чертами слепней являются: 1) тесная связь
насекомых с влажными биотопами — развитие большинства личинок слепней происходит
в воде или прибрежных частях водоемов, а взрослые особи постоянно нуждаются
во влаге для утоления жажды; 2) паразитический образ жизни насекомых — самки
слепней нападают и сосут кровь главным образом крупных теплокровных животных.
Взрослые слегши появляются в природе с наступлением теплого периода, на юге
(Средняя Азия, Закавказье) — в середине апреля—начале мая, в средних широтах
(Московская обл., юг Пермской обл. и т. д.) — в последней декаде мая, на севере
(Карельской АССР) — в середине июня, на крайнем севере (Дудинский район
Красноярского края) — в конце 1-й декады июля и т. д. Самцы слепней появляются на
несколько дней раньше самок. Вылет слепней совпадает в средних широтах с цветением
черемухи. С установлением жаркой летней погоды лёт самок слепней становится
максимальным. В средних широтах и на юге слепни наиболее многочисленны в июне и июле
и исчезают в августе—сентябре, на севере слепни наиболее многочисленны в июле
и исчезают в августе и т. д. В средних широтах период лёта большинства слепней
не превышает 4—5 недель, причем в пределах этого срока самки одного и того же вида
встречаются в массе 10—15 дней, а затем довольно быстро исчезают. Лишь немногие
виды имеют более растянутые сроки лёта — до 2 месяцев и более, например,
Haematopota pluvialis, Chrysops caecutiens и др. Начюге период лёта у отдельных видов
слепней, например, Atylotus pulchellus, может продолжаться свыше 4 месяцев.
Длительность жизни самок слепней в лабораторных условиях обычно не превышает 1 месяца.
Нипгульц (Nieschulz, 1927) отмечает, что самки некоторых тропических видов слепней
у него жили в неволе до 70 дней. Длительность жизни самцов 4—6 дней, редко дольше
(Лутта, 1970).
Разные виды слепней характеризуются определенными сроками лёта, одни
появляются раньше, другие позже, сменяя друг друга в известной последовательности
(рис. 48). В соответствии с особенностями фенологии виды слепней можно поделить
минимум на 4 группы: 1) весенние, 2) раннелетние, 3) летние и 4) позднелетние виды.
В зависимости от ландшафта эти группы включают в себя различные виды слепней.
Так, в зоне европейских широколиственных лесов весенними видами являются
Hybomitra lurida и Н. nitidifrons, раннелетяими —■ Chrysops caecutiens, Hybomitra bimaculata,
H. lundbecki и т. д., летними — Haematopota pluvialis, -Tabanus bovinus, T. miki и т. д.,
и позднелетними — Atylotus rusticus и Tabanus glaucopis.
Виды слепней фауны СССР можно разделить на 2 экологические группы: 1) группу,
обитание которой связано с лесной или кустарниковой растительностью; 2) группу,
обитание которой связано с травянистой растительностью. Первые распространены
главным образом в лесной полосе и горах, вторые — в степной и пустынной зонах.
Самцы первой группы попадаются сравнительно редко, так как обычно держатся
на недоступной для наблюдателя высоте, тогда как самцы второй группы легко обна-
R Н. Г. Олсуфьев
82 ВВЕДЕНИЕ
руживаются на цветах и в траве. Связь с растительностью у слепней двоякая: с одной
стороны, слепни находят здесь стацию, удовлетворяющую их своим микроклиматом:
здесь имеется необходимая влажность, затем защита от прямых солнечных лучей,
от ветра, дождя, врагов и т. д., с другой стороны — растения (и их обитатели)
доставляют слепням необходимую пищу в виде нектара цветов, сока деревьев, выделений
тлей, листоблошек и т. д.
Большинство видов слепней представляет собой весьма теплолюбивых и
светолюбивых насекомых: максимум жизнедеятельности самцов и самок приходится на
теплые и солнечные часы дня. Это очень подвижные насекомые, которые значительную
часть своего времени проводят в воздухе в поисках пищи, необходимой влаги и т. д.
В средних широтах полеты слепней и нападения на животных начинаются при
температуре воздуха не ниже 15—16° и становятся интенсивными при 19—20° и выше.
Особенно настойчиво слепни нападают в жаркую, душную погоду, предшествующую
дождю. Ветреная погода, так же как холод или дождь, препятствуют полетам и
нападениям слепней. Дождевки могут нападать в пасмурную погоду и даже при слабом
дожде: температурный порог их активности лежит около 13—14°. Нив (Neave, 1912)
отмечает, что в тропической Африке местные виды дождевок всегда нападают в плохую,
влажную погоду и реже в солнечную жаркую погоду; в сырых тенистых лесах дождевки
могут быть назойливыми весь день. По наблюдениям в высокогорьях (Кавказ, Тянь-
Шань) и на крайнем севере (Дудинка) температурный порог активности слепней НуЪо-
mitra и Philipomyia — 11—12°, а при 15—16° и выше уже отмечается интенсивный лёт
насекомых (Скуфьин, 1938; Шевченко, 1961; Поляков, 1965). Среднегорные и
низкогорные слепни Tabanus в условиях'Тянь-Шаня имеют значительно более высокий
порог активности — 20—21° (Шевченко, 1961).
В средней полосе СССР лёт слепней в жаркий летний день начинается в 6—7 утра,
к 9 час. уже становится массовым и прекращается лишь к закату солнца. В то время
как самки заняты поисками добычи, самцы слепней держатся отдельно по лесным
опушкам, на полянках и т. д., поодиночке паря в воздухе.
В условиях жаркого климата юга слепни наиболее активны утром и во второй
половине дня, тогда как в самое жаркое время дня лёт слепней уменьшается; угнетающее
действие тепла на насекомых начинает сказываться при температуре воздуха выше
31—32° (Лутта, 1950; Скуфьин, 1952а, 1966а).
В отношении суточного режима среди слепней известны любопытные исключения.
Так; в тропических и субтропических частях Америки и Африки известны сумеречные
и частью ночные виды Tabanus и Chrysops, которые проявляют свою активность после
заката солнца или рано утром. К сумеречным (утренне-вечерним) видам относятся
Tabanus prometheus и Т. subparadoxus, обитающие в горах Кавказа (Гаузер, 1960;
Исмаилов, 1970) и Т. paradoxus, распространенный в горах Центральной Европы.
Имеется указание на то, что самцы и самки типично дневных видов слепней (Tabanus
bromius, T, bovinus, Chrysops relictus и др.) могут в теплые ночи лететь в светоловушку
(Бошко, 1954).
У слегшей высокая термофильность сочетается с большой чувствительностью к
недостатку влаги. Если продержать слепней несколько часов без увлажнения при
температуре 25—30°, то они вскоре все погибнут. В жаркие дни, когда слепни проявляют
максимальную жизнедеятельность, они постоянно нуждаются во влаге. Как это
установил еще И. А. Порчинский (1899), слепни для этого посещают мелкие водоемы,
с налета ударяются о поверхность воды и улетают затем дальше с захваченной
каплей воды. Из нашей фауны этим свойством обладают Hybomitra, Tabanus, Chrysops,
Silviusii Heptatoma. К водоемам летят в одинаковой мере самцы и самки. Особи с кровью
в желудке или с развитыми яичниками посещают водоемы наряду с несосавшими
кровь и неполовозрелыми самками.
Спаривание слепней обычно происходит в воздухе: самцы устремляются за
пролетающими мимо самками, быстро их настигают и копулируют на лету. Иногда
копулирующая пара располагается где-либо на растении, причем самец держится за него
ногами, а сцепленная с ним самка висит в воздухе вниз головой.
Самки слепней нашей фауны становятся способными к кровососанию только
после оплодотворения (Cameron, 1934; Лутта, 19646, 1970). Однако некоторые
тропические виды Tabanus в лабораторных условиях могут осуществлять кровососание
без оплодотворения.
Самцы слепней, у которых ротовые органы не приспособлены для прокалывания
кожи, никогда на животных не нападают: они питаются соком цветов, истечениями
деревьев, сладкими выделениями насекомых и т. д. Питание самок слепней имеет
двойственный характер: они, так же как и самцы, подлизывают различные растительные
соки, необходимые для жизнедеятельности, но наряду с этим нуждаются в
дополнительном питании кровью позвоночных животных, за счет которых главным образом
происходит развитие яйцеклеток в яичниках. В нашей фауне известны- лишь немногие
виды, например; Atylotus sublunaticornis, A. plebejus, Chrysops rufipes, Nanorhynchus
crassinervis, самки которых не сосут кровь позвоночных.
БИОЛОГИЯ СЛЕПНЕЙ 83
Имеются указания, что у заведомо кровососущих видов, например, у обитающего
в Японии Tabanus iyoensis, первая порция яиц в яичниках самки может развиваться
без питания кровью за счет потребления растительных соков, и уже затем после первой
яйцекладки самка становится кровососом (Saito, 1967; Watanaba a. Kamimura, 1971).
В эксперименте Кемерон (Cameron, 1926) наблюдал откладку яиц некоторыми
североамериканскими кровососущими Chrysops и Tabanus без предварительного питания
кровью; мухи были выведены из куколок. Автогенное развитие яиц на первом гоно-
трофическом цикле, без питания кровью, установлено в эксперименте и для отдельных
видов слепней нашей фауны, которое у Chrysops relictus и Tabanus bovinus завершалось
яйцекладкой; без оплодотворения личинки не вылуплялись (Павлова, 1972; Бошко
с соавт., 1972). На возможность автогенного развития яиц у Hybomitra borealis
(вероятно, Н. sexfasciata) и Haematopota pluvialis предположительно указывает И. Г. Бей-
Биенко (1970) на основании полевых наблюдений.
По-видимому, у ряда кровососущих видов слепней развитие яиц на первом гоно-
трофическом цикле при потреблении некровяной пищи (растительные соки и т. д.)
довольно закономерно и является своеобразной «страховкой» в обеспечении
потомства при затруднении с отысканием теплокровных животных. Вопрос этот заслуживает
дальнейшего тщательного изучения.
Слепни являются паразитами, не приуроченными строго к какому-либо
определенному хозяину, и могут производить нападения на многих млекопитающих
преимущественно крупных размеров и предпочтительно темных мастей. В некоторых
случаях слепни могут осуществлять питание на крупных колониальных птицах,
например, на пеликанах, а в тропиках известны слепни, нападающие на крупных
пресмыкающихся — крокодилов, варанов и т. д.
У некоторых пангоний наблюдается далеко зашедшая специализация хоботка
для добывания нектара из цветков, в таком случае хоботок иногда намного превосходит
длину тела. Мухи сосут нектар часто налету, т. е. так же, как некоторые бабочки.
В тропической Африке появление и лёт некоторых видов пангоний совпадает с
цветением определенных видов растений, на которых они питаются (Schwetz, 1919). Вместе
с тем эти пангоний являются кровососами. Интересно, что у длиннохоботных
пангоний колющие части хоботка значительно короче вытянутой нижней губы, в которую
они вложены, иногда больше, чем в 2 раза, и это заставляло многих авторов считать,
что эти формы не являются кровососами, а питаются только нектаром цветков. Но
наблюдения над Pangonius longirostris в Индии и некоторыми другими длиннохобот-
ными пангониями в Африке показали, что эти мухи также нападают и сосут кровь
животных, как и другие Tabanidae. При этом пангоний отгибают свою нижнюю губу
в сторону или кзади, между ногами, как хобот слона, обнажая колющие части.
Обладая хорошим полетом, самки слепней весьма настойчиво преследуют
животных. Особенно энергичны в этом отношении слепни рода Tabanus (в особенности
крупные виды —. Т. bovinus, Т. sudeticus, T. autumnalis и др.), которые при этом могут
преодолеть несколько километров. По некоторым наблюдениям, отдельные особи
■Hybomitra прилетают к объектам питания с расстояния до 5 км (Купрессова с соавт.,
19726). В Средней Азии отмечено удаление Hybomitra, Tabanus и Atylotus от мест
выплода на 25—30 км в полупустыню при преследовании насекомыми верховых
лошадей (Лутта, 1964г). Пестряки и дождевки обладают менее стремительным полетом
и обычно далеко не отлетают от мест выплода.
Слепни при отыскании добычи руководствуются в первую очередь зрением: их
привлекает любой предмет, выделяющийся на окружающем фоне. Особенно легко
слепни устремляются за движущимися объектами, в том числе за автомашинами.
В акте нападения слепней на животных определенную роль играет также обоняние.
Акт нападения слепня на млекопитающее (объект кровососания) представляет собой
сложный процесс, состоящий из нескольких фаз: 1) разыскивание объекта на
местности путем «прочесывающего» полета; 2) подлет к объекту; 3) полет вокруг объекта
и его преследование; 4) посадка на объект; 5) выбор места присасывания; 6) акт
кровососания (Скуфьин, 1952а). У дождевок и частью пестряков преобладает
«подстерегающий» тип нападения. Они часто садятся на объект без предварительных подлетов
и круговых облетов (Скуфьин, 1952а). У различных видов слепней при нападении
существует известная топографическая приуроченность к определенным частям тела
животных, которая выработалась еще на диких животных в связи с особенностями
распределения на их теле шерстного покрова, способностью к самообороне,
размерами слепней и т. д. Так, одни виды нападают на половые органы, вымя и грудь,
другие — на конечности и морду и т. д., тогда как наиболее крупные виды слепней
присасываются на спине, где кожные покровы животного достигают наибольшей
толщины. Дождевки менее разборчивы в выборе места для кровососания, чем Tabanus,
Hybomitra и Chrysops.
В природе слепням редко удается сразу же насосаться крови, так как атакуемые
ими животные, если они не ограничены в движениях, например упряжью, отбиваются
достаточно энергично. Обычно самкам приходится сделать ряд уколов, каждый раз
6*
84 ВВЕДЕНИЕ
меняя место или перелетая с одного животного на другое, прежде чем которой-нибудь
из них удается присосаться достаточно крепко и насытиться полностью. Прокол кожи
слепень осуществляет сложенными вместе хитинизированными частями хоботка
(сосальца), при этом нижнечелюстные щупальца, в покое прикрывающие хоботок сверху
и сбоку, раздвигаются в стороны (рис. 42), а нижняя (мягкая) губа сдавливается снизу
вверх по мере погружения хоботка в кожные покровы. Самка слепня, если ее не
тревожить, насасывается крови в течение от 1 1/2 до 5 мин. и более в зависимости от
величины кровеносного сосуда. Крупный слепень рода Tabanus или Hybomitra (например,
Т. autumnalis, H. tarandina) поглощает 180—200 мг крови, средних размеров слепень
(Я. muehlfeldi, Н. lundbecki) — 100 мг, Chrysops — около 50 мг (Олсуфьев, 19356,
19496; Петрова, 1955, и др.), а Т. bovinus — 300 мг (Паенко, 1968). Вес выпиваемой
слепнем крови обычно превышает вес насекомого, у Hybomitra и Tabanus максимально
в 1.5 раза, у Chrysops и Haematopota — в 3 раза (Паенко, 1968).
Прерывистое питание, равно как и способность нападать на свежие трупы
животных, имеет огромное значение в распространении слепнями различных
инфекционных заболеваний среди животных и человека (см. ниже).
Однократного полного насыщения кровью достаточно, чтобы обеспечить развитие
яиц в яичниках самки слепня (гонотрофическая гармония). В дельте Волги при
температуре 26—30° у Tabanus autumnalis,
Chrysops relictus и Haematopota subcylindrica
овогенез закапчивается в среднем через 3—4
суток после кровососания, а в отдельных
случаях даже через 2 суток (Олсуфьев, 1940а).
В условиях Карелии при температуре 24—28°
созревание яичников у слепней завершается
в течение 3.5—8 дней, снижение температуры
ниже 20° вызывает резкое замедление
развития яичников, а при 14—16° происходит
полная остановка процесса овогенеза и быстрая
гибель насекомых (Лутта, 19646). У Chrysops
relictus отмечалось созревание яйцевых
фолликулов при кормлении самок неполной
порцией крови, но число развившихся
фолликулов было сниженным (Скуфьин и Паенко,
1971). Однако в опытах с Tabanus leleani
выяснено, что самки этого вида при
неполном насыщении кровью не могут откладывать
зрелые яйца (Кадырова, 1972а).
Процесс развития и созревания яйцевых
фолликулов состоит из 7 морфологически
хорошо отличимых фаз (Лутта, 19646, 1970),
соответствуя таковым, по Кристоферсу и
Меру, у других кровососущих двукрылых.
После откладки яиц у самок в течение
1—2 суток сохраняется растяжение
яйцевых трубочек, после чего они сокращаются
до исходного состояния (Олсуфьев, 1940;
Скуфьин, 1959). В дистальной части трубочек
обнаруживается «желтое тело» — остаток питательных клеток яйца. Число
находимых в яйцевой трубке «желтых тел» соответствует числу яйцекладок и определяет
«физиологический возраст» самки слепня (Лутта, 19646, 1970; Павлова, 1965; Паенко,
1966; Бей-Биенко, 1967). Иногда в яичниках обнаруживаются застрявшие единичные
вполне сформированные яйца, свидетельствующие о том, что яйцекладка уже
осуществлялась (Олсуфьев, 1940).
Яйца откладываются компактной кучкой (кладкой) на стебли или нижнюю
поверхность листьев растений, расположенных над водой или близ нее (рис. 19). Для
яйцекладки используются также торчащие из воды палки, сухие ветки, деревья,
сваи и т. д. Обычно кладки имеют вид пирамидки, они состоят из 3—4 слоев
склеенных между собой яиц, расположенных один над другим (рис. 20), но у некоторых
видов кладки бывают однослойными (некоторые Chrysops, Haematopota, а также
Hybomitra lapponica). Окраска только что отложенных яиц молочно-белая, но затем они
темнеют и становятся в зависимости от видовой принадлежности буро-серыми или
блестяще угольно-черными. У некоторых видов, например Tabanus buddha, яйца
остаются белыми (Соболева, 1968а). Яйца откладываются в теплую, солнечную погоду
чаще в первой половине дня, для кладок обычно выбираются растения, хорошо
освещенные солнцем. В тропиках известны слепни, откладывающие яйца на камни,
выступающие из речных потоков. Самки Tabanus, Hybomitra, Atylotus откладывают
в среднем от 300 до 600 яиц, но некоторые виды до 1000 {Tabanus autumnalis, T. pleskei
и др.). Самки Chrysops caecutiens оставляют в кладке 300—400 яиц (Олсуфьев, 19356);
Рис. 42. $ Hybomitra в момент
кровососания: видны кончики
раздвинутых в стороны щупалец. Фото
И. М. Олигера.
БИОЛОГИЯ СЛЕПНЕЙ 85
Н. А. Виолович (Виолович и Евстигнеева, 1964) в Западной Сибири обнаруживал
в кладках этого вида~400—800 яиц. Местами яйцекладки чаще всего являются
выступающие из воды растения в прибрежных частях речек, озер, прудов, канав, болот
и т. д., расположенных по соседству с проезжей дорогой, пастбищем для
сельскохозяйственных животных или тропами диких копытных, т. е. недалеко от того места, где
слепни нападают на животных. Процесс яйцекладки длится 1—2 часа, иногда дольше.
Закончив ее, самка может снова насосаться крови и после этого вновь отложить яйца.
В естественных условиях по второму разу гонотрофический цикл осуществляет
значительная часть самок, а в условиях эксперимента выяснено, что число гонотрофи-
ческих циклов у одной самки Tabanus autumnalis может достигнуть 5 (Олсуфьев,
1940). В Карелии самки слепней в естественных условиях откладывают яйца не более
2 раз, крайне редко — 3 раза (определено по количеству «желтых тел») (Лутта, 19646,
1970), но в Приморском крае (Соболева, 19656, 1968а) и в Воронежской обл. в жаркое
лето (Паенко, 1968) в небольшом числе случаев у слепней отмечены 3 и- даже 4 гоно-
трофических цикла. До 4 кладок отмечено у Tabanus leleani и Т. autumnalis brunnescens
в Узбекистане (Кадырова, 1972а). С каждым проделанным самкой гонотрофический
циклом количество откладываемых яиц уменьшается (Олсуфьев, 1940; Бей-Биенко,
Рис. 43. Берег Дона близ Калача Волгоградской обл. В июле на листьях
водных растений обнаружено большое количество кладок яиц Chrysops
relictus и Ch. flavipes.
1967; Паенко, 1968; Скуфьин и Паенко, 1971). Отложив яйца, самка, если она
неспособна к новому, кровососанию, погибает.
Наиболее энергичная яйцекладка в средних широтах наблюдается в конце июня
и в июле, к концу массового лёта слепней. Число обнаруживаемых кладок различно
в зависимости от обилия слепней и распределения биотопов, удобных для яйцекладки
этих насекомых. При обилии мелких водоемов и заболоченностей, например в таежном
ландшафте, кладки слепней распределены в природе довольно дисперсно и лишь в
отдельных случаях наблюдается заметная их концентрация. Наоборот, при ограниченном
числе подходящих для яйцекладки биотопов происходит довольно значительная
концентрация кладок яиц слепней на растениях (рис. 43). В лесостепной части
Новосибирской обл. мы обнаруживали при однократном обследовании растительности
по берегу озера или старицы на каждые 100 шагов (приблизительно 70 погонных метров)
в среднем по 14—22 кладки яиц слепней, главным образм Tabanus и Hybomitra
(Олсуфьев, 19496). К. В. Скуфьин (19526) в Воронежской обл., обследуя один и тот же
пруд каждую пятидневку, собрал за лето с прибрежной растительности по 10 кладок
Chrysops relictus на каждый погонный метр береговой линии. При редком
расположении растений вдоль берега концентрация на них кладок яиц Ch. relictus бывает иногда
столь значительной, что последние сплошь покрывают снизу листовую пластинку,
на каждые 10 см листа осоки может приходиться 20—30 кладок (Тамарина, 1-951).
Эмбриональное развитие у слепней заканчивается в течение 4—8 дней, но может
затянуться до 3 недель или сократиться до 2 дней (на юге) в зависимости от окружаю-
86
ВВЕДЕНИЕ
щей температуры воздуха. Личинки вылупляются из всех яиц кладки почти
одновременно и, собравшись в комочек, падают в воду (или на влажную почву), линяют
и расползаются. Далее личинки живут поодиночке в берегах или прибрежных частях
водоемов, в поверхностных слоях моховых болот, во влажной почве и т. д. (рис. 44, 45).
Личинки слепней имеют характерное строение и легко узнаются по сигаровидному
упругому телу, заостренному на обоих концах и явственно поделенному на 12
сегментов. Личинки хищничают, уничтожая червей, мелких членистоногих или
моллюсков, или питаются органическими остатками. Некоторые виды Chrysops (например,
Ch. caecutiens) не хищны и питаются только различными органическими
остатками.
К осени личинки подрастают и зимуют в тех местах, где происходило их развитие
или вблизи них. Зимой глубина залегания личинок слепней старших возрастов
(например, Tabanus autumnalis) может достигать от 25 до 128 см, т. е. ниже промерзающего
слоя грунта, в то время как личинки более младших возрастов зимуют в
промерзающем слое (Бошко, 1950). По наблюдениям в Карелии личинки слепней (Hybomitra
и др.) осенью прекращают питание и мигрируют из водной среды в почву
(гидрофобия), где зимуют на глубине 5—10 см. Личинки обладают холодоустойчивостью, они
могут переносить длительное охлаждение до —11°; менее устойчивы личинки млад-
Рис. 44. Зарастающее озеро —место выплода слепней: весной в берегах
обнаружены личинки Hybomitra, Chrysops и Tabanus. Окрестности Вен-
герово Новосибирской обл.
шего возраста (Лутта, 19726). Весной с наступлением теплого периода
перезимовавшие личинки заканчивают метаморфоз. В наших широтах продолжительность жизни
личинок, если их развитие почему-либо не растягивается на второй год, в среднем
9 1/2—10 месяцев.
Количество возрастов у личинок слепней довольно велико и у одного и того же
вида часто непостоянно. Личинки Tabanus имеют 7—11 возрастов (Isaac, 1924, 1925),
Т. Meant — 8 возрастов (Кадырова, 1972а), Atylotus pulchellus karybenthinus — 8—
9 возрастов (Лутта, 1966а), Hybomitra lundbecki и Н. bimaculata — 9—11 возрастов
(Лутта, 1970), Haematopota pluvialis — 7—10 возрастов (Cameron, 1934), Chrysops
caecutiens— 6—7 возрастов (Лутта, 1970), наконец, Ch. relictus — 5 возрастов (Тама-
рина, 1951; Скуфьин, 19526). Длина вполне развитых личинок слепней колеблется
в зависимости от рода и вида от 11 до 48 мм (Скуфьин, 19526).
Стациальное распределение личинок слепней изучено недостаточно. По отдельным
данным можно заключить, что среда обитания личинок разных родов слепней может
сильно различаться. Личинки Chrysops — типичные водные формы (гидробионты),
они живут ниже уреза воды и только перед окуклением перемещаются в надводную
часть берега водоема, тогда как личинки Hybomitra, большинства Tabanus, некоторых
Haematopota — полуводные формы (амфибионты), они обычно обитают выше уреза
воды, хотя вначале они попадают в воду (Олсуфьев, 19356, 19496; Скуфьин, 1952а;
БИОЛОГИЯ СЛЕПНЕЙ 87
Лутта, 1970). Глубина залегания личинок весной и осенью 5—10 см, редко глубже.
Некоторые виды, например, Tabanus quatuornotatus, T. glaucopis оставляют кладки
яиц на растениях вдали - от воды в сравнительно сухих местах, и вылупившиеся
личинки падают и развиваются непосредственно в почве (почвенные формы или геоби-
онты).
Касаясь биотопов, заселяемых личинками отдельных видов, можно отметить
следующее. Личинки Chrysops caecutiens преимущественно обитают в лесных ручьях
или речках с относительно быстрым течением, личинки Ch. relictus предпочитают
непроточные водоемы с открытой водной поверхностью (пруды, пойменные озера и т. д.)
или реки с медленным течением, личинки Hybomitra bimaculata и Н. lundbecki чаще
заселяют лесные болота, заболоченные ямы, канавы и т. д. В условиях Карелии
личинки слепней заселяют низинные и переходные болота, берега рек, ручьев, озер
и т. д., их много на заболоченных участках в лесу и на лугах, в канавах и ямах на
пастбище, но личинки практически отсутствуют в верховых (сфагновых) болотах (Лутта,
1964а, 1970). В Центральной Якутии наиболее высокая плотность личинок и куколок
слепней отмечена по заболоченным берегам пойменных и аласных (плакорных) озер,
а также в лесных затопляемых котловинах. Выявлены сезонные миграции личинок
Рис. 45. Заболоченная канава на пастбище — место выплода слепней:-
на растениях, выступающих над водой, летом обнаружены
многочисленные кладки яиц Hybomitra. Окрестности Сиверской Ленинградской обл.
в зависимости от увлажнения и прогревания мест их обитания (Васюкова, 1972). В
Приморском крае личинки слепней обитают главным образом по берегам непроточных
водоемов, а также на заболоченных низинах, в почвах с различной влажностью и
составом, на глубине 5—30 см, от 10 см до 5 м от уреза воды (Соболева, 19656). В
Узбекистане личинок слепней находили в арыках, на рисовых полях, в поймах речек, под
камнями в горных ручьях и т. д. (Кадырова и Ган, 1967; Кадырова, 1972а).
Заслуживает внимания обнаружение вблизи г. Иванова большого количества личинок слепней,
в том числе Heptatoma pellucens, в прудах и канавах, загрязненных навозной жижей
из свинарников и коровников, а также находка личинок Tabanus bromius под коровьим
пометом на пастбище (Колесова, 1966, 1968) и Т. bovinus в стенке помойной ямы
(Лобанов, 1959). Частое обнаружение личинок и куколок слепней в «лепешках» коровьего
и конского навоза на лугах близ водоемов отмечает для Якутии Т. Т. Васюкова (1972).
Очевидно, в богатой разлагающимися веществами среде личинки слепней находят
обильную пищу, в том числе в виде мелких беспозвоночных. Отметим, наконец, галофильные
виды, например, Chrysops italicus, личинки которого обитают по морским побережьям,
или Tabanus sabuletorum, T. zimini и др., связанных обитанием с солоноватыми
водоемами.
Большой интерес представляют находки весной личинок слепней (до 5 экз. на 1 м2)
Tabanus bifarius и Т. tergestinus в почве безводной Ширакской степи (Гургенидзе,
19726).
88 ВВЕДЕНИЕ
Весной незадолго до окукливания концентрация личинок Chrysops caecutiens
может достигать 50—70 экз. на 1 м2 берега водоема (Олсуфьев, 19356), Ch. relictus —
100—160 экз. (Тамарина, 1951), Ch. pictus — 20—45 экз. (Скуфьин, 1952а), Tabanus
autumnalis — 33 экз. (Олсуфьев, 19496), Heptatoma pellucens—Ш экз. (Колесова, 1968)
и т. д. Однако гораздо чаще наблюдаются более низкие плотности личинок — в
среднем 5—7 экз. на 1 м2 (Олсуфьев, 19496; Скуфьин, 1952а, 19666; Запекина-Дулькейт,
1969; Иванищук, 1973) или меньше.
Часто личинки уползают далеко от места первоначального выплода, особенно
перед окуклением, которое обычно происходит в рыхлой почве, моховом покрове
и т. д., иногда вдали от воды. Фаза куколки длится от 5—7 дней до 2—3 недель и более
в зависимости от окружающей температуры. Перед вылетом куколка путем энергичных
движений брюшка поднимается к поверхности и высовывает наружу верхнюю
половину тела, после чего куколочная оболочка лопается и взрослый слепень вылезает
наружу.
Рис. 46. Яйцеед ■ Telenomus tabard Мауг. поражает кладку яиц
слепня.
В средних широтах цикл развития у слепней нормально заканчивается в один
год. Продолжительность отдельных фаз следующая:
Взрослый слепень — 10—30 дней
Яйцо — 4—20 »
Личинка — 9—10 месяцев (и больше)
Куколка — 5—28 дней
В отдельных случаях развитие личинки может затянуться до 2 (Олсуфьев, 19356,
Бошко, 1950; Скуфьин, 1952а, и др.), 3 и даже 4 лет в условиях Карелии, Московской,
Ивановской областей и Приморского края (Лутта, 1964а, 1970; Скуфьин, 19666;
Соболева, 19656; Иванищук, 1973). Задержка в развитии личинок слепней (с
повторными зимовками) наблюдается главным образом, в северных и средних широтах и
является приспособлением этих насекомых к климатическим колебаниям. Без этого
приспособления дождливые, холодные годы, неблагоприятные для лёта слепней
могли бы отрицательно отразиться на численности популяции в целом.
На юге СССР слепни, по-видимому, в большинстве заканчивают свое развитие
не дольше, чем за один год. Этот срок, например, указывает М. К. Кадырова (1972а)
для Tabanus leleani и Т. autumnalis в условиях Узбекистана. В тропиках и
субтропиках некоторые виды слепней могут иметь в год до 3 поколений (Isaac, 1924, 1925).
Наиболее обычными паразитами слепней являются яйцееды Telenomus tabani
(рис. 46), Т. oophagus и др. (в кладках яиц Tabanus и Eybomitra) и Trichogramma eva-
nescens (в кладках яиц Chrysops), которые заражают яйца в тот момент, когда они только
что отложены (Олсуфьев, 19356, 1938; Лутта, 1950; Скуфьин, 1952а, и др.). По
наблюдениям в Омской обл. (Растегаева, 1962), Карелии и Каракалпакии (Лутта, 1964в,
19666), яйцееды уничтожают в среднем 58—60% отложенных слепнями яиц. Сильнее
ВРЕДОНОСНОЕ ЗНАЧЕНИЕ СЛЕПНЕЙ 89
поражаются более доступные для яйцеедов однослойные кладки Chrysops, иногда
до 90%, и менее интенсивно — многослойные кладки Tabanus, Hybomitra, Atylotus
и Haematopota. Несмотря на столь большой урон, наносимый яйцеедами, численность
большинства видов слепней из года в год остается высокой за исключением некоторых
видов, размножение которых, возможно, лимитируется яйцеедами.
Кладки яиц слепней поедают личинки божьих коровок сем. Coccinellidae и жуки
сем. Staphylinidae (Бей-Биенко, 1963; Соболева, 1968а; Виолович, 1968). В Армении
личинки и куколки слепней довольно часто (до 15—20%) поражаются личинками
паразитической мухи Villa ventruosa из сем. Bombyliidae (Зайцев и Тертерян, 1966). Для
личинок и куколок слепней указаны паразиты — перепончатокрылое Trichoglena
complanatus (Чупис, 1947; Скуфьин, 1952а) и Diglachis sp. (Павлова, 1972), в личинках
слепней обнаруживали также другого перепончатокрылого — Trichopria sp. (Бей-
Биенко, 1963) и затем паразитических нематод из сем. Mermithidae (Скуфьин, 1952а).
Иногда этих нематод находили также в куколках и взрослых слепнях (Паенко, 1968).
В дельте Волги во взрослых слепнях Tabanus autumnalis нами в 1937 г. обнаружен
волосатик из сем. Gordiidae (определение Ж. К. Штрома); зараженность популяции
составила 5.6%, в одном слепне было от 1 до 6 паразитов. На взрослых слепнях
паразитируют личинки клещиков-краснотелок Trombidiidae. Из врагов слепней следует
отметить бембексов и других ос, которые охотятся за взрослыми слепнями и кормят
ими своих личинок. Слепни часто становятся добычей насекомоядных птиц, крупных
пауков, некоторых стрекоз, мух-ктырей и муравьев. Личинки и частично куколки
слепней могут поражаться паразитическими грибками, особенно Coelomomyces milkoi
и Metarrhizum anisopliae; на Украине в естественных условиях отмечено до 28%
пораженное™ личинок слепней, а в неволе до 75%. Грибки вызывают гибель личинок
(Коваль и Андреева, 1971).
Женкинс (Jenkins, 1960) в США указывает для слепней 37 видов паразитов, в том
числе перепончатокрылых — 20, двукрылых — 7, клещей — 3, нематод — 4 и
простейших — 3 вида.
8. ВРЕДОНОСНОЕ ЗНАЧЕНИЕ СЛЕПНЕЙ
Слепни составляют часть так называемого «гнуса», т. е. комплекса кровососущих
двукрылых, куда также входят комары, мошки, мухи-жигалки и т. п. Соотношение
этих групп по их численности не всегда и везде одинаково, но все же в большинстве
местностей, пораженных гнусом, слепням принадлежит одно из главных мест, причем
их вредоносное значение достаточно велико, особенно если принять во внимание
способность слепней распространять некоторые опасные болезни домашних животных
и человека.
Оценка значения слепней может быть сделана в результате рассмотрения
следующих ее аспектов: а) территории вредоносности, б) сезона вредоносности и в) степени
вредоносности. Если первые 2 аспекта освещают по преимуществу количественную
сторону проблемы, то третий характеризует ее качественную сторону.
ПОРАЖЕННОСТЬ РАЗЛИЧНЫХ ЛАНДШАФТНЫХ ЗОН СССР
Как уже указывалось, слепни весьма широко распространены на территории
СССР. Их можно встретить и в полярной тундре, и в оазисах жарких пустынь Средней
Азии, в равнинных лесах и на альпийских высокогорных лугах. Но не везде слепни
появляются в одинаковых количествах. Их численность зависит от наличия
благоприятных условий для их существования и размножения, за каковые в основном
нужно признать сочетание следующих элементов: 1) обилия различных водоемов
и болотных образований, 2) развитой лесной растительности и 3) наличия крупных
теплокровных животных (домашних или диких), являющихся источниками питания
самок слепней. В отношении второго условия для видов, распространенных на юге
по долинам рек, достаточно наличие крупной травянистой или кустарниковой
растительности, наоборот — для северных видов обычно необходимы высокоствольные леса.
В соответствии со сказанным, наиболее пораженным слепнями ландшафтом, именно
«царством слепней», является таежный пояс, который тянется через всю северную
половину СССР от Финляндии до берегов Тихого океана (рис. 47). Обилие в таежном
ландшафте болот и различных водоемов в сочетании с обширными пространствами,
занятыми лесами, обусловливают исключительное обилие слепней. В наиболее
жаркую часть летнего сезона слепни появляются здесь в таких количествах, что делают
совершенно невозможным пребывание скота на пастбищах или ведение на лошадях
полевых сельскохозяйственных работ в течение дневного периода. Насекомые
буквально тысячами облепляют животных и искусывают их до крови.
По мере продвижения на юг, тайга постепенно сменяется лесной зоной, а эта
последняя — лесостепью. Указанные ландшафты являются более сухими, а кроме
ВРЕДОНОСНОЕ ЗНАЧЕНИЕ СЛЕПНЕЙ 91
того, значительно обезлесенными. В соответствии с этим видовой состав здесь .иной,
и количество слепней в особенности в лесостепи значительно меньше, что допускает
в норме дневную пастьбу скота и сельскохозяйственные работы на лошадях.
На крайнем севере тайга через переходную зону, лесотундры примыкает к тундре.
Последняя, по-видимому, является слабо пораженной слепнями вследствие
отсутствия лесной растительности, но в переходной зоне лесотундры слепни местами
встречаются в изобилии, и здесь от них сильно страдает оленеводство.
Степной и пустынный ландшафты, широкой полосой охватывающие СССР с юга,
в основном свободны от слепней. Но здесь зато долины рек и побережья крупных
водоемов представляют прекрасные условия для массового выплаживания слепней, и
последние наблюдаются здесь в огромных количествах. В особенности в плавневом
ландшафте рек пустынной зоны с его образующей основной фон тростниковой
растительностью численность слепней столь велика, что местами мало чем уступает тайге, резко
отличаясь, однако, своим видовым-составом. Необходимо упомянуть, что в
Казахстане именно проблема гнуса в известной мере обусловила в свое время периодические
кочевки скотоводческого населения на лето от рек и озер в степи или горы, а на зиму,
когда кровососущие двукрылые исчезают, — обратно к рекам.
Некоторое представление о суммарной численности слепней могут дать
следующие цифры. В сезон массового лёта слепней за 1 учетный час на лошади (или корове)
удавалось поймать сачком следующее количество самок: в Карелии 500—800 и до 1000
(Лутта, 1959, 1970), в Московской обл. 400—500 (Олсуфьев, 1949а), в Воронежской
обл. 300—600 (Скуфьин, 1952а), в северной части Волго-Ахтубинской поймы 250—
500 (Олсуфьев с соавтор., 1958), в лесостепной части Новосибирской обл. 200—300
(Олсуфьев, 19496), в Кара-Калпакии 600—650 (Лутта, 1950) и т. д. В лесотундре
на п-ове Таймыр в дни и часы пика численность слепней на олене достигала 800 особей
за 30-минутный учет (В. А. Поляков, личное сообщение). Однако эти цифры не могут
считаться пределом. По наблюдениям. Р. Г. Петровой (1955), в одном из районов
Московской обл. в период массового лёта слепней на 1 корову в среднем нападало за 5 мин.
280 слепней и максимально 492 слепня.* По ее же данным, в сильно пораженных
слепнями районах Приморского края в период наибольшей активности этих насекомых
на 1 корову нападало за 15 мин. до 600—700 слепней (Соболева, 1965в).. Н. А. Виолович
(1965) в Новосибирской обл. в один из жарких дней начала июля за 20-минутный
вылов сачком около себя собрал 3216 слепней, что является своеобразным рекордом
численности слепней.
Торные ландшафты Кавказа и в особенности Средней Азии обычно умеренно
поражены слепнями; в массовых количествах последние наблюдаются лишь в зоне
субальпийских лугов и прилегающих частях лесной зоны.
Отметим влияние хозяйственной деятельности человека на места выплода
слепней и соответственно на численность этих насекомых. Так, широко проводимые
местами осушение болот, мелиорация и так далее приводят к снижению численности
слепней. На Украине отмечено, что при строительстве гидроэлектростанций после
затопления поймы рек слепни исчезают или становятся малочисленными в нижней
более глубокой части водохранилища, но в верхних мелководных, подвергающихся
заболачиванию частях слепни обычно остаются и их численность может быть
значительной. Установлено также формирование мест выплода слепней на некоторых
участках крупных оросительных каналов, где до их проведения слепней не было (Г. В. Бошко,
личное сообщение).
СЕЗОННАЯ ЧИСЛЕННОСТЬ СЛЕПНЕЙ
Мы уже отмечали, что взрослые слепни появляются в средних широтах
(Московская и другие области) в конце мая и держатся до конца августа. По мере
продвижения на север лёт слепней смещается на более поздние сроки, например, на Крайнем
Севере (Таймыр, Дудинский район) слепни появляются в конце первой и начале
второй декад июля и исчезают к началу второй декады августа. На юге лёт слепней может
продолжаться с мая (или с середины апреля) по сентябрь. Сводные данные по СССР
см.: Лутта, 1972а.
Соответственно общая продолжительность сезона лёта слепней на Таймыре
составляет около 30 дней (Поляков, 1967), в южной Карелии —.74 дня (Лутта, 1959), в
Московской, Ленинградской и Пермской (южная часть)4 областях — 90 дней (Олсуфьев,
1934, 1949а; Митрофанова, 1951), в Новосибирской обл. — 100 дней (Олсуфьев,
1949; Виолович, 1966), в Воронежской обл. — в среднем около 120 дней (Скуфьин,
1952а) и в Кара-Калпакии — 129—146 дней (Лутта, 1950).
Массовые нападения слепней на животных в средних широтах обычно
отмечаются в июне—июле, на юге также и в августе. В горах Кавказа и Средней Азии слепни
наиболее многочисленны в июле и в начале августа.
• * Сбор слепней одновременно производили 2 человека.
93
ВВЕДЕНИЕ
В пределах сезона лёта слепней выпадают прохладные и дождливые дни, когда
слепни неактивны. В средних широтах в июне—августе таких дней бывает 25—30%.
Если вычесть эти дни, а также периоды, когда слепни в природе вообще малочисленны
(в начале или конце их сезона), то массовый или многочисленный лёт слепней
насчитывает 25—30 дней (Олсуфьев, 1934). На юге соответственно более теплому и
сухому климату массовый лёт слепней охватывает большее число дней, например, в Кара-
К'алпакии около 60 дней (Лутта, 1950).
Как уже отмечалось, для каждой ландшафтной зоны характерен свой видовой
комплекс слепней, состоящий обычно из нескольких десятков видов. Из них лишь
небольшое число видов летает во множестве, являясь таким образом хозяйственно
вредными видами, прочие же виды немногочисленны или встречаются единичными
экземплярами. В продолжение сезона одни виды сменяются другими, эти в свою
очередь третьими и т. д. (рис. 48).
В разные годы численность взрослых слепней большинства видов довольно
постоянна (Олсуфьев, 1937а; Бошко, 1950; Скуфьин, 1952а, 1966а; Митрофанова, 1955)
и лишь у некоторых видов отмечались изменения их обилия по годам, например,
у Atylotus rusticus на юге Московской обл. (Олсуфьев, 1949а), у дождевок Haematopota
pluvialis и Н. italica в Пермской обл. (Митрофанова, 1955) и т. д.
Однако в средних и северных широтах в зависимости от различий в погоде сильно
меняется по годам число дней, когда слепни активны, и это создает впечатление
повышенной численности слепней в жаркие и сухие годы и малого их количества в
дождливые и прохладные годы. В северных широтах может наблюдаться преждевременное
уменьшение или прекращение массового лёта слепней вследствие резкого похолодания
и выпадения дождей, обусловливающих гибель значительной части куколок и
окрыленных особей (Лутта, 1959).
ПАТОГЕННОЕ ЗНАЧЕНИЕ СЛЕПНЕЙ
С целью кровососания слепни нападают на человека, а-также на различных
животных, преимущественно крупных копытных. Из домашних животных слепни
особенно интенсивно нападают на лощадей, верблюдов, крупный рогатый скот, буйволов
и северных оленей, из диких — на лосей, оленей, косуль, диких кабанов и т. д.
Уколы слепней болезненны. Помимо чисто механического повреждения кожи
в момент укола, слепень впрыскивает в ранку слюну, которая обладает ядовитыми
свойствами и препятствует свертыванию крови. В слюне слепней обнаружен фактор
диффузии — активная гиалуронидаза, деполимеризирующая гиалуроновую кислоту
соединительной ткани (Пегасова и Носкова, 1961). На месте укола слепня появляется
чувство жжения или резкая боль, развивается небольшая папула и покраснение,
а в некоторых случаях образуется весьма болезненная опухоль, удерживающаяся
несколько часов, а иногда несколько дней (Олсуфьев, 19356; Лутта, 1970). По данным
Е. Н. Павловского, А. К. Штейна и Н. Г. Олсуфьева (1935), внутрикожная инъекция
суспензии из слюнных желез Tabanus вызывает у человека сходную с естественной
(т. е. от укола слепня) реакцию кожи, заключающуюся в появлении на месте инъекции
чувства жжения, затем красноты, отечности кожи и т. д., причем в отдельных случаях
в реактивный процесс вовлекаются ближайшие лимфатические узлы и ощущается
легкое недомогание, а также повышается температура тела (до 37.6°).
В опытах Р. Г. Соболевой (Петровой) (1955, 1965а) внутрикожная инъекция
бычкам суспензии из слюнных желез Tabanus и Hybomitra вызывала появление
болезненной опухоли, повышение температуры тела, учащение пульса и дыхания и т. д.
Внутривенное введение бычку суспензии слюнных желез слепней обусловило его гибель
через 3 часа при явлениях геморрагического диатеза.
В местностях, изобилующих слепнями, число одновременно нападающих на
лошадь или другое крупное животное слепней может исчисляться десятками и даже
сотнями особей, а за час их может быть несколько сотен и даже тысяч. Количество
крови, поглощаемой слепнями, колеблется от 40 до 300 мг в зависимости от величины
насекомого. После укола слепня часто из ранки на коже животного выступает крупная
капля крови, а иногда кровь сочится некоторое время спустя. При массовом нападении
слепней отдельные участки кожи животных могут представлять собой сплошную
кровоточащую поверхность. Больше всего страдают от слепней лошади, особенно
находящиеся в упряжи, когда они лишены возможности отбиваться от атакующих насекомых.
Вследствие этого в слепневые дни использование лошадей становится почти
невозможным, что часто влечет за собой потерю рабочего времени. Слепни охотнее нападают
на животных темных мастей и сильно вспотевших.4 Большой урон слепни наносят
также крупному рогатому скоту. При массовом появлении слепней животные
прекращают еду, мечутся по пастбищу, забиваются в кусты, убегают в хлева, заходят в воду
и т. д. Имеются указания, что в Ярославской и Ленинградской областях удои молоки"
у коров снижаются на 12—15% при массовом нападении слепней (Дегтярев, 1928;
Олсуфьев, 1937а). По исследованиям в Московской, Тюменской, Омской,
Новосибирской областях, в Кизлярском р-не Дагестанской АССР и других местах,
g4 ВВЕДЕНИЕ
в период массовых нападений слепней удои у коров снижаются на 10—20—30% и более,
а привес у нагульного молодняка крупного рогатого скота — на 20—45% (Петрова,
1955; Растегаева, 1958; Андреев и Мяло, 1961; Растегаев, 1965; Виолович, 1966;
Велик, 1966).
По заключению К. П. Андреева (1966), в СССР в целом по поголовью крупного
рогатого скота, находящемуся в местностях, где слепни лётом массами нападают на жи
вотных, ежегодная потеря составляет полмиллиона тонн молока и 30 тыс. т живого
веса, или около 15 тыс. т мяса. Стоимость этой продукции по заготовительным ценам
выражается ежегодно в 640 млн рублей, теряемых народным хозяйством из-за
нападения слепней только на крупный рогатый скот.
На юге от слепней сильно страдают верблюды, а на севере — олени. На овец,
свиней и коз слепни нападают в меньшем числе, но остриженные овцы и козы
становятся для них более доступными.
Человек подвергается нападению главным образом со стороны мелких слепней,
в частности пестряков и дождевок. Последние бывают иногда весьма навязчивыми
и мучительными, в особенности в душную, жаркую погоду перед дождем. Очень охотно
слепни нападают на людей во время купания: смоченное водой тело особенно
привлекает слепней.
Слепни могут нападать на свежие трупы животных, что имеет большое значение
в распространении этими насекомыми отдельных инфекций. Мелкие слепни, например
некоторые виды пестряков (Chrysops relictus), могут нападать на зайцев и водяных крыс,
особенно на больных малоподвижных зверьков.
Не менее важной в эпидемиологическом отношении особенностью слепней
является утоление ими жажды в мелких водоемах, что в жаркие летние дни носит массовый
характер. Обычно слепни ударяются с налета о поверхность воды и уносят на хоботке
каплю, но иногда они просто садятся на воду и некоторое время держатся на
поверхности, всасывая хоботком воду. Дождевки могут утолять жажду, подлизывая влагу
с поверхности сырой почвы. Посещение слепнями загрязненной какими-либо
патогенными микробами пересыхающей лужицы может вызвать распространение заболеваний
среди животных и людей, на которых инфицированные слегши будут затем нападать.
Слепни имеют большое значение как переносчики возбудителей ряда
инфекционных болезней, в том числе туляремии, сибирской язвы, инфекционной анемии лошадей,
некоторых форм трипаносомозов и т. д. В отдельных местах Африки слепни (некоторые
виды пестряков) являются переносчиками глистного заболевания лоаоза.
Туляремия. Болезнь широко распространена в странах умеренного пояса
северного полушария и имеет природную очаговость, связанную в основном с грызунами,
зайцами и иксодовыми клещами. Возбудителем туляремии является мелкая бактерия
Francisella tularensis. Вполне доказана роль слепней в передаче туляремии в природных
очагах этой инфекции при так называемых трансмиссивных вспышках среди людей.
Источниками заражения слепней в первую очередь служат зайцы и водяные крысы.
Слепни инфицируются, пытаясь кормиться на находящихся в агонии больных
туляремией грызунах или трупах или при утолении жажды в мелких водоемах,
содержащих бактерии туляремии. Естественная зараженность микробом туляремии
установлена для ряда видов слепней, в том числе: Chrysops relictus, Ch. italicus, Tabanus autum-
nalis, T. bromius, Atylotus flavoguttatus, Haematopota pluvialis. В эксперименте перенос
туляремии от больного или павшего грызуна здоровому животному осуществлен
посредством слепней Tabanus (Т. bromius, Т. autumnalis), Hybomitra (Н. ciureai,
Н. turkestana, Н. erberi, H. peculiaris), Atylotus (A. flavoguttatus, A. pulchellus, A. qua-
drifarius) и дождевки Haematopota turkestanica, а также пестряков Chrysops relictus,
Ch. caecutiens и Ch. flavipes. Слепни сохраняли способность передавать инфекцию
уколом хоботка до 2—3 суток после инфицирующего кормления. Передача
осуществляется механическим путем. Из крупных животных посредством уколов инфицированных
слепней удалось заразить туляремией овцу. В эксперименте установлена также
возможность передачи слепням возбудителя туляремии от инфицированной воды
(которую слепни пили) восприимчивому животному.
В США экспериментально доказаны как переносчики туляремии Chrysops discalis,
Ch. noctifer, Hybomitra septentrionalis и Н. rupestris. В естественных условиях наиболее
частым переносчиком при заражении людей трансмиссивным путем американские
авторы считают Chrysops discalis. Недавно спонтанно зараженными возбудителем
туляремии были обнаружены Ch. fulvaster и Ch. aestuans.
Антракс (сибирская язва). Болезнь имеет всесветное распространение,
возбудителем ее является споровая палочка Bacillus anthracis. В таежно-лесной полосе нашей
страны, изобилующей слепнями, последние являлись в дореволюционный период
главными агентами распространения сибирской язвы среди лошадей и крупного
рогатого скота (Нагорский, 1902). Большая эпизоотия сибирской язвы среди лошадей
и крупного рогатого скота, обусловленная передачей главным образом слепнями,
наблюдалась в 1929 г. в Западной Сибири. Известны случаи заражения людей
сибирской язвой через уколы слепней.
..ВРЕДОНОСНОЕ ЗНАЧЕНИЕ СЛЕПНЕЙ
95
За рубежом слепни имеют важное значение как переносчики антракса среди
домашних животных в ряде стран, в том числе в некоторых южных штатах Америки
(Луизиана, Техас), в Индонезии, Индии, на Филиппинах, в Новой Каледонии, Южной
Африке и т. д. (обзор литературы см. Г. В. Колонии, 1969).
Описаны находки естественно инфицированных бациллами антракса слепней.
Культуры этого возбудителя были также выделены от Tabanus atratus, T. bromius
и Atylotus rusticus, пойманных в природе кормящимися на сибиреязвенном трупе. В
эксперименте способность передачи инфекции от больных животных (или трупов) здоровым
животным была доказана для ряда видов слепней, в том числе Tabanus striatus, Т. ги-
bidus, Т. рторе nigrovittatus, T. autumnalis, T. bovinus, T. bromius, Hybomitra ciureai
и др. В большинстве случаев передача осуществлялась методом прерванного
кормления, т. е. немедленного докармливания слепней на здоровом животном после их
инфицирования, но в некоторых опытах в отдельных случаях слепни передавали антракс
спустя 3 суток после заражения. Передача инфекции осуществляется слепнями
механическим путем. По данным Н. Г. Олсуфьева и П. П. Лелеп (1935), сибиреязвенные
бациллы могут сохраняться в хоботке слепней 5—7 дней. Этим же авторам удалось
уколами инфицированных слепней заразить сибирской язвой лошадь, а П. П.
Лелеп — овцу и бычка.
В эксперименте выяснена возможность передачи слепнями (Tabanus, Haematopota)
возбудителя антракса от инфицированной почвы (которую слепни подлизывали
хоботком) восприимчивым животным.
Трипаносомоз сурра, су-ауру — весьма опасное заболевание верблюдов, лошадей
и части других домашних животных, широко распространенное в странах Старого
Света. Возбудителем является Trypanosoma evansi. Первоначально сурру отличали
от североафриканского трипаносомоза (эльдебаб, мбори, сума), но Гоар и Кутелен
(Ноаге et Coutelen, 1933) считают все эти заболевания идентичными, вызываемыми
одной и той же Trypanosoma evansi, и сюда же относят су-ауру— трипаносомоз
верблюдов и лошадей, распространенный в наших среднеазиатских республиках и на юго-
востоке европейской части РСФСР.
По данным зарубежных исследователей, слепни являются главными переносчиками
возбудителя сурры (су-ауру), тогда как роль других кровососущих] двукрылых —
жигалки, комаров и др. — второстепенна. Передача осуществляется слепнями от
больных животных к здоровым механически в момент укола хоботком и никакого цикла
развития трипаносомы не проделывают в организме слепней. После кормления на
больном животном слепни сохраняют способность передавать трипаносом в
сравнительно небольшой срок, обычно в пределах нескольких часов, но как исключение
передача наблюдалась в опытах через сутки. Крупные виды слепней (Tabanus)
передают инфекцию лучше, чем мелкие (Chrysops, Haematopota).
В опытах П. Н. Арбузова (1941) передачу возбудителя су-ауру морским свинкам
удалось осуществить при помощи слепней Tabanus autumnalis, T. bovinus и Hybomitra
expollicata (syn. H. nigrivitta). '
Трипаносомоз крупного рогатого скота. Болезнь свойственна крупному рогатому
скоту, зубрам и буйволам. Возбудитель — Trypanosoma theileri. Заболевание
обнаружено во многих странах, в СССР оно встречается как на севере, так и на юге, но в виде
спорадических случаев. Течение болезни легкое. В зарубежной литературе имеются
указания, что переносчиками возбудителя являются слепни, в том числе Tabanus
и Haematopota. По Неллеру (Noller, 1925), трипаносомы проходят цикл развития
в организме слепней.
Инфекционная анемия лошадей. Эта болезнь, вызываемая фильтрующимся
вирусом и наносящая во многих странах большой урон коневодству, передается от больных
лошадей здоровым в условиях пастбища слепнями, главным образом видами рода
Tabanus. В опытах японской комиссии, а также И. М. Родионова (1936) при
содержании здоровых лошадей на пастбище изолированно в загородках и на некотором
расстоянии от больных животных, но так, чтобы слепни могли свободно нападать на тех и
других, через определенный срок среди здоровых лошадей начинались заболевания
инфекционной анемией. По сообщениям Скота (Scott, 1922), передачу вируса от больной
лошади к здоровой удалось осуществить через уколы слепня Hybomitra septentrionalis;
установить цикл развития вируса в слепнях не удалось, передача, видимо,
осуществляется механически.
Лоаоз (калабарский отек). Болезнь эндемична в зоне тропических лесов
экваториальной Африки (Гана, Либерия, Гвинея, Камерун, Ангола, Замбия и др.). Ее
возбудителем является филярия Loa loa. Специфическими переносчиками этой глисты
служат два местных слепня-златоглазика — Chrysops silacea и Ch. dimidiata, что было
установлено А. и С. Коннел (A. Connal и S. Connal, 1922) и др. Молодые личинки филя-
рии держатся в дневные часы в периферических сосудах больных людей и, будучи
поглощенными златоглазиком вместе с кровью, проникают из желудка в мышцы груди
мухи и здесь заканчивают свое развитие. Примерно к 10-му дню личинки мигрируют в
хоботок мухи, и с этого момента последняя становится способной производить
заражение при нападении на здоровых людей. В момент акта сосания крови личинки филярии
ВВЕДЕНИЕ
выходят из нижней губы златоглазика и проникают в ранку на месте укола. Попытка
получить заражение при посредстве других слепней — Tabanus, Haematopota, затем
жигалок и мух це-це — была безуспешной. Слепни-златоглазики, следовательно,
являются специфическими переносчиками этого филяриоза. Естественная зараженность
мух филяриями в природе достигает 5%.
Лоаоз обнаружен у диких обезьян в лесах экваториальной Африки и, таким
образом, может рассматриваться как природноочаговая болезнь. По мнению Дак и Крью
(Duke, 1955 и Crewe, 1954, цит. по: Oldroyd, 1957), переносчиками филярий среди
обезьян являются златоглазики Chrysops langi, Ch. centurionis и другие виды (цит. по:
Oldroyd, 1957).
Парафиляриоз. Заболевание лошадей и мулов, именуемое «сечением».
Возбудителем является мелкая глиста Parafilaria multipapillosa. В СССР болезнь широко
распространена, особенно на юго-востоке, и, как правило, достигает максимума
распространения в июне—июле, в период лёта слепней. Поражаются главным образом лошади
не моложе 3 лет. Предполагают, что переносчиками парафилярий являются
кровососущие двукрылые, в том числе слепни. У лошадей и мулов летом отмечается сходное
с «сечением» заболевание невыясненной этиологии, которое проявляется местными
подкожными опухолями — «шишками». Заболевание связывают с уколами слепней
(Благовещенский, 1937; Шевченко, 1961). Опухоли бывают различной величины,
от горошины до грецкого ореха и даже куриного яйца. Они локализуются
преимущественно в наиболее уязвимых слепнями местах — на шее, лопатках, на брюхе, реже
на спине и холке. На ощупь они болезненны. Опухоли исчезают сами по себе с
прекращением нападений слепней.
Другие болезни. На основании эпизоотологических наблюдений и
экспериментальных исследований слепней считают способными механически передавать
возбудителей пастереллеза буйволов и эмфизематозного карбункула.
Имеются указания о возможном участии слепней в передаче эризипелоида, некро-
бациллеза северных оленей, анаплазмоза крупного рогатого скота и пироплазмоза
северных оленей. Известны случаи выделения от слепней возбудителя Ку-риккетсиоза
и вируса из группы клещевого энцефалита. В эксперименте получена передача
слепнями вируса чумы свиней и листерий; для последнего возбудителя установлена
способность размножения в слепнях. В Северной Америке доказана роль слепней в передаче
филярий Elaeophora schneideri, вызывающих слепоту у лося и являющихся обычными
паразитами оленей и овец.
9. СБОР, ХРАНЕНИЕ И ОПРЕДЕЛЕНИЕ СЛЕПНЕЙ
Сбор взрослых слепней. С целью выявления фауны слепней
организуют регулярные сборы этих насекомых в различных частях области (края,
республики), __ отличающихся по своим ландшафтно-географическим особенностям. Сборы
слепней необходимо начать с момента их появления, т. е. с конца мая (на юге с конца
апреля—начала мая), и продолжать с 7-дневными промежутками (можно чаще) в
течение всего лета до момента исчезновения слепней, т. е. до конца августа—начала
сентября. Для полного охвата фауны той или иной местности сборы периодически про-,
водят в различных стациях — болото, берег реки, лесная поляна и т. д. Для сборов
нужно выбрать теплый, солнечный, маловетреный день и производить лов насекомых
по возможности в первой половине дня; самки слегшей прилетают для нападения
к человеку или животному и в этот момент могут быть легко пойманы обычным
энтомологическим сачком. Наиболее удобно слепней собирать сачком или просто руками
на лошадях, волах или верблюдах, находящихся в упряжи или привязанных к дереву
(рис. 49). В местностях, где слепней вообще мало, только около животных и можно
произвести более или менее полные сборы. В некоторых местах (например, в горах)
обильные сборы слепней иногда удается произвести на экспедиционной машине, кузов
которой обтянут брезентом. Для количественного учета слепней сбор на животном или
около себя производят энтомологическим сачком в течение 20 мин. Диаметр обруча
сачка 30 см, длина мешка сачка 75—80 см, конец мешка округлый. Для полного
определения количества слепней, нападающих на животных, могут применяться также
учетные сборы в течение 30 мин. или часа. А. Е. Тертерян (1954) рекомендует
отлавливать слепней с максимальным приближением сачка к поверхности тела животного,
а не «кошением» вокруг него. Этот автор (а также некоторые другие) проводил учеты
в течение 5 мин., но нам представляется, что сборы за такой короткий промежуток
времени не всегда точны. К моменту начала отлова происходит концентрация слепней
вокруг животного и она тем больше, чем позднее сбор начат, но затем по мере
вылавливания насекомых количество вновь подлетающих слепней снижается. Учет в течение
20 или 30 мин. охватывает ровнее нападающую часть популяции. С целью выявления
метеорологических условий, способствующих нападению слепней, регистрируют
во время проведения учетного сбора температуру и влажность воздуха, скорость ветра,
интенсивность инсоляции и т. д. Колокол Мончадского мало пригоден для учета слеп-
СБОР, ХРАНЕНИЕ И ОПРЕДЕЛЕНИЕ 97
ней, так как последние обычно кружатся вокруг добычи на расстоянии, которое не
захватывается колоколом в момент его опускания.
С целью характеристики степени пораженности территории слепнями К. В. Ску-
фьин (1952а) предложил следующую шкалу интенсивности лёта насекомых (количество
нападающих самок) (табл. 8).
Таблица 8
Шкала интенсивности лёта самок слегшей
(по Скуфьину, 1952а) (количество экз.)
Баллы интенсивности лёта
Массовый лёт (слегшей масса) . . .
Умеренный лёт (слепней много) . .
Слабый лёт (слепней мало)
Единичный лёт (слегшей очень мало)
Сбор сачком
за 4-часовую
экскурсию
Свыше 100
30—100
10-30
Менее 10
Сбор сачком на
себе за 20 мин.
на месте
Свыше 25
10—25
3—10*
1—3*
Сбор сачком на
лошади за час
Свыше 200
60-200
20—60
Менее 20
* Применение 20-минутных учетов на себе при слабом и единичном лёте слепней может быть
рекомендовано только в исключительных случаях, когда полностью отсутствует возможность
организовать сборы на животных.
Может быть выделена еще одна градация (балл) интенсивности лёта слепней —
«сверхмассовый лёт», когда за 1 час на лошади улавливается 600 и более слепней.
Для сборов и учета самок слегшей пригодна также чучелообразная ловушка
Скуфьина (см. ниже). Разработаны различные более портативные варианты этой-
Рис. 49. Сбор слепней энтомологическим сачком около лошади.
ловушки (Шевченко, 1961; Владимирова и Потапов, 1963), а также приборы для
автоматического их включения и фракционированного учета численности слепней
(Потапов, 1964; Потапов и Владимиров, 1962). Количество слепней, пойманных за
определенный отрезок времени, рассчитывают на 1 час. Следует иметь в виду, что данные
учетов 'ловушками могут расходиться с данными учетов на животных, например,
ловушкой Скуфьина лучше отлавливаются дождевки Haematopota pluvialis и хуже
Chrysops caecutiens и некоторые Hybomitra и Tabanus (Запекина-Дулькейт, 1969; Бей-
Биенко, 1959). В США предложена портативная" «шатровая» ловушка с привлечением
слепней (самок) углекислым газом, выделяемым подвешиваемым внутрь ловушки
сухим льдом; уловистость такой ловушки в несколько раз выше, чем ловушки без
7 Н. Г. Олсуфьев
98 ВВЕДЕНИЕ
сухого льда, и может достигать 1000 слепней за час в пик их численности (Catts, 1970;
Pechuman, личное сообщение). В жаркие дни слепни часто залетают в помещения
(конюшни, скотные дворы,веранды,станционные помещения и т.д.) и здесь скопляются
на окнах, где они могут быть легко собраны руками.
Пойманных слепней немедленно умерщвляют, для чего наиболее удобны морилки,
заряженные цианистым калием с гипсом на дне. Вместо этих веществ можно
использовать мелкие кусочки мягкой резины, пропитанные хлороформом. Их кладут на дно
морилки и прижимают кружком, вырезанным из картона. В. В. Шевченко (1961)
рекомендует морилку заряжать парадихлорбензолом. Ни в коем случае не следует
помещать сборы в одну морилку с различными жесткими насекомыми (жуками,
саранчовыми и др.) или в спирт, так как от этого очень сильно понижается качество
собранного материала.
Рис. 50. Извлечение слепней при помощи небольшой металлической
сетки на конце удилища из водоема, поверхность которого покрыта
соляровым маслом («лужа смерти»).
Указанными выше способами улавливаются лишь кровососущие самки, тогда*как
самцы слепней, питающиеся исключительно растительной пищей, требуют иных
способов поимки. Легче всего их ловить, пользуясь небольшими (1—5 м2) ловчими
водоемами («лужами смерти»), поверхность которых покрывают керосином или соляровым
маслом. Для этой цели могут быть использованы различные наполненные водой
рытвины, углубления в старых канавах или ямы со свободной (очищенной) от растений
поверхностью воды (рис. 50). Если поблизости пасется скот, то ловчий водоем
необходимо огородить. Погибших слепней извлекают из водоема в тот же день и тщательно
отмывают от керосина или солярового масла в нескольких порциях серного эфира
(бензина, хлороформа и т. п.). Полученные таким путем экземпляры почти не изменяют
своего вида. Нефтью или мазутом лучше не заливать водоемы, используемые для
сборов слепней в коллекцию, так как эти вещества трудно затем удалить с поверхности
тела насекомых.
С целью учета количество слепней, пойманных за день в ловчий водоем,
рассчитывают на 1 м2 поверхности последнего. Следует иметь в виду, что описываемым
методом не могут учитываться некоторые виды, например дождевки (Haematopota),
избегающие «лужи смерти».
Самцы слепней (и самки многих видов, в том числе некровососущих форм) могут
быть пойманы на цветах, стволах деревьев, изгородях, затем парящими в воздухе и т. д.
Но нахождение их требует специальных навыков и часто случайно.
Собранных насекомых затем накалывают на булавки. Для этого употребляют
специальные энтомологические булавки средних номеров (1 или 2). Слепня помещают
спинкой вверх между концами собранных щепоткой пальцев (большой и
указательный) левой руки и правой рукой вкалывают булавку в среднюю часть спинки (на
уровне основания крыльев) с таким расчетом, чтобы над насекомым осталось около
I
| СБОР, ХРАНЕНИЕ И ОПРЕДЕЛЕНИЕ 99
V3 ее длины, т. е. объект располагают на высоте 2/3 булавки. Наколотых слепней
немедленно снабжают этикеткой с указанием географического названия местности
или населенного пункта, точной даты поимки слепня и фамилии собирателя.
Географическое название указывают, ориентируясь на доступные карты. Наколотый
материал хранят в специальных энтомологических коробках с торфяным дном. Их
устройство, а также общие правила хранения коллекционного материала можно почерпнуть
в общих руководствах по этому вопросу. При невозможности немедленной наколки,
что обычно и бывает в условиях летних работ, собранный материал сохраняют на вате
(метод ватных сборов). Слепней раскладывают рядами спинками вверх на тонкие слои
ваты и сверху прикрывают белой бумагой, на которой отмечают необходимые для
этикетировки данные; слои ваты, переложенные листами бумаги, помещают по мере
заполнения материала друг на друга в соответствующую размерам слоев деревянную
или картонную коробку. Для защиты собранных насекомых от вредителей коллекций
внутреннюю поверхность коробки и покровные листы бумаги прочерчивают гекса-
хлорановым карандашом. В дальнейшем, если необходимо произвести наколку, ватные
слои кладут во влажную камеру (стеклянный сосуд с мокрым песком на дне и сверху
плотно прикрытый крышкой) примерно на 12—18 час, а затем с насекомыми
поступают, как со свежеумерщвленными.
Наружное строение слепней изучают на сухих, предварительно наколотых на
булавки объектах, рассматривая их в ручную 12—16-кратную лупу или бинокулярную
пупу (бинокуляр). Для исследования терминалий брюшка самцов или самок можно
использовать сухих насекомых. Концевую треть или половину брюшка отрезают
ножницами и кипятят в течение 4—5 мин. в 10%-м растворе едкого калия. Далее конец
брюшка промывают в кипяченой воде, вычленяют терминалий и делают из них
постоянные препараты в канадском бальзаме. В. В. Шевченко (1961) рекомендует терминалий
слепней кипятить в течение 10—,15 мин. в 15%-м едком калии и после промывки в воде
окрашивать в ацетокармине. Столь длительное кипячение в более крепком растворе
щелочи нам представляется нежелательным ввиду возможного обесцвечивания
отдельных частей терминалий.
Для определения физиологического возраста популяции самок прибегают к
подсчету желтых тел в яйцевых трубках. Извлечение яичников из тела слепня производят
методом задней тракции внутренних органов, т. е. вытягиванием их препаровальной
иглой из конца брюшка. Для обнаружения желтого тела очень тонкой
препаровальной иглой захватывают концевую нить и осторожно растягивают всю яйцевую трубку.
В момент растягивания желтые тела становятся хорошо различимыми и
поддающимися счету (Лутта, 19646).
Сбор кладок яиц. В местностях, изобилующих слепнями, сбор кладок
яиц не представляет особых затруднений: нужно только знать, когда и где их следует
искать. Наиболее успешны поиски в самое жаркое время лета, т. е. в конце июня
и в первой половине июля (на юге в конце июля), когда происходит массовая
яйцекладка. Кладки следует искать на растениях, произрастающих в воде или по берегам
ручьев, рек, озер, прудов, канав, заболоченных ям, по окраинам болот и т. д.
Некоторые слепни оставляют кладки вдали от воды, например на лугах. Легче всего найти
кладки в местах выпаса скота или же возле проезжих дорог. Обычно кладки бывают
расположены на нижней поверхности листьев (или на стеблях), хорошо освещаемых
солнцем растений на высоте от нескольких сантиметров от поверхности воды (или
почвы) до 2—3 м (на листьях кустарников и деревьев). Кладки сравнительно легко
заметны при осмотре растений вследствие своего обычно темного цвета и характерного
строения. Количество собранных кладок относят к длине обследованной береговой
линии водоема и степень их обилия рассчитывают на 1 погонный метр (Олсуфьев,
19496; Скуфьин, 1952а, и др.). Найденные кладки срезают вместе со стебельком или
листом, на котором они расположены, и помещают в банку с 70° спиртом. Кладки
некоторых видов при неосторожном соприкосновении легко отскакивают от
поверхности листа, к которому приклеены, и рассыпаются; поэтому надо избегать класть
в одну банку много материала.
Если из яиц надо вывести личинок, лист растения вместе с кладкой помещают
в открытую чашку Коха или какую-либо стеклянную банку диаметром 5—8 см, на дно
которой наливают воду. Кладка располагается с.таким расчетом, чтобы она была над
водой на высоте 1—1.5 см. Вылупившиеся личинки попадают в воду, линяют, после
чего их нужно рассадить и содержать в небольших сосудах. Вывести личинок из яиц
очень просто, но дальнейшее их воспитание представляет большие трудности. После
вылупления личинок кладку не выкидывать, а поместить на 3—4 недели в небольшой
сосуд, затянутый наглухо плотной материей для выведения паразитов — яйцеедов.
Воду на дно сосуда не наливают. Кладку яиц лучше всего исследовать под лупой
в часовом стекле в 70° спирте. Для рассмотрения отдельных яиц иглами осторожно
отделяют несколько штук и приготовляют из них постоянный препарат в канадском
бальзаме.
Сбор личинок. Сбор личинок легче всего производить весной, с
наступлением теплого времени (конец апреля—начало мая), когда перезимовавшие личинки
7*
100 ВВЕДЕНИЕ
заканчивают свое развитие и достигают наибольшей величины. Но если нужно
заготовить к зимовке живых личинок, то их можно с успехом собрать и в осеннее время —
в сентябре или в начале октября. Личинок добывают в местах их обитания, т. е. в
прибрежных частях различных водоемов (ручьев, рек, озер, прудов, канав и т. д.) путем
взятия и тщательного просмотра проб. Почвенные пробы берут лопатой, мох
извлекают металлическими граблями; взятую массу тут же тщательно измельчают руками
и просматривают. Пробу берут из поверхностных частей субстрата, на глубину до 10—
15 см. Размер проб 1U или V2 м2. Пробы со дна или из береговых частей водоема берут
лопатой (или небольшой драгой), помещают в сито с дном из металлической сетки
и промывают в воде (рис. 51). При весеннем обследовании, рассчитанном на более
крупных личинок, применяют металлическую сетку с ячеей 2 мм, при осеннем
обследовании, когда попадают мелкие личинки первых возрастов, применяют сетку с ячеей
--'C-Vs^ei *
■вК"'Ъ . ..." "
Рис. 51. Промывание в воде пробы грунта из
прибрежной части реки, для чего использован
небольшой деревянный ящик, дно которого
заменено металлической сеткой. На этом участке
весной добыты 230 личинок Chrysops caecutiens,
10 личинок Hybomitra sp. и 6 личинок Haemato-
pota sp. Окрестности Сиверской
Ленинградской обл.
1 мм. Нахождение личинок затруднено их одиночным образом жизни и дисперсией
в природе; однако тщательные и настойчивые поиски всегда вознаграждаются успехом.
Легче всего личинки обнаруживаются при исследовании растительной массы (мох,
ряска), взятой с поверхности воды из прибрежных частей небольших водоемов (пруд,
заболоченная яма возле проезжей дороги и т. д.).
Степень обилия личинок на обследуемой территории определяют путем
вычисления их среднего количества на 1 м2 (Олсуфьев, 19496; Скуфьин, 1952а).
Так как личинки многих видов в неволе проявляют каннибализм (кроме некоторых
Chrysops), то собранных во время экскурсий личинок лучше всего размещать
поодиночке в пробирки или небольшие баночки. Для дальнейшего сохранения личинок
консервируют какой-либо фиксирующей жидкостью, например, жидкостью Фрей-
линга или Дюбоска, дающими обычно хорошие результаты. При этом у крупных
личинок необходимо при фиксации проколоть в нескольких местах кожу. Очень
крупных личинок можно консервировать, облив их крутым кипятком, но этот способ все же
довольно груб. Фиксированный материал перекладывают для постоянного хранения
в 70° спирт.
Если нужно сохранить личинок живыми, их помещают поодиночке в небольшие
стеклянные сосуды (РЫИр, 1928; Олсуфьев, 19356; Скуфьин, 1952а). Для этой цели
удобны пилюльные баночки или колбы Эрленмейера емкостью 25—50 мл. Крупных
личинок можно содержать в более просторных банках емкостью 100—150 мл. На дно
сосуда кладут субстрат, в котором личинка была найдена (мох, ил, песок и др.), и
приливают немного воды; отверстие сосуда затягивают бязыо и последнюю закрепляют
посредством резинового кольца. Для мелких личинок используют небольшие пробирки
СБОР, ХРАНЕНИЕ И ОПРЕДЕЛЕНИЕ 101
с песком на дне. Хищных личинок по мере надобности подкармливают нарезанными
на куски дождевыми червями, моллюсками, личинками комаров и т. д. Замену корма
следует производить раз в 5 дней (можно чаще), а в случае загрязнения субстрата
необходима замена последнего и промывание сосуда. Собранные осенью личинки
могут жить месяцами, причем их кормление не обязательно; для этого надо личинок
держать при температуре, не превышающей комнатную, лучше даже немного ниже
(8—12°), и время от времени приливать на дно сосуда немного воды. В зимнее время
из достаточно крупных личинок можно получить при желании куколок и затем
взрослых слепней. С этой целью сосуды ставят в термостат с температурой около 28° и
личинок подкармливают.
Для знакомства с наружным строением личинок необходим спиртовый материал,
но для подробного изучения желательно иметь также живых личинок. Крупных
личинок (фиксированных) рассматривают в спирту или дистиллированной воде на часовом
стекле под лупой или бинокуляром. Мелких личинок (2—5 мм) обрабатывают едким
кали, окрашивают и заключают в канадский бальзам. У крупных личинок изучение
головы и ее придатков удобнее производить после того, как головной конец выпре-
парован из тела личинки и обработан щелочью. Исследование производят под
бинокуляром в часовом стекле в дистиллированной воде. Последующее заключение в
канадский бальзам позволяет рассмотреть ряд деталей строения головы под
микроскопом. К. В. Скуфьин (1952а) с успехом изучал морфологию личинок на живых
экземплярах с последующим доведением их до окрыленной фазы для целей диагностики.
Для иммобилизации мелких личинок автор заливал их теплым раствором 15—
20%-й желатины, заключая между двумя предметными стеклами. Более крупных
личинок автор помещал в склеенную из стекла плоскую ванночку, заливал более
густым раствором желатины и сверху прикрывал большой стеклянной
пластинкой.
Сбор куколок. Искать куколок слепней следует выше уреза воды в тех же
местах и теми же способами, что и личинок. Обычно куколки располагаются в несколько
более сухих поверхностных частях субстрата головным концом вверх. Несмотря
на плотную оболочку они очень нежны и легко гибнут от неосторожных надавливаний
на поверхность тела. Наилучшим временем для сбора является середина—конец мая,
когда их легче всего обнаружить.
Собранных куколок фиксируют крутым кипятком и перекладывают в 70° спирт.
Если нужно вывести взрослое насекомое, куколку помещают головным концом вверх
в какой-либо небольшой стеклянный сосуд — колбу Эрленмейера, пилюльную
баночку, чашку Коха и т. д. В качестве субстрата можно положить мох или песок,
который слегка увлажняют по мере надобности. В зависимости от окружающей температуры
воздуха фаза куколки длится от 5—7 дней до 3 недель. Для изучения строения
куколки пригодны спиртовый материал и сухие куколочные оболочки, из которых слепни
уже вышли. При лабораторном выведении слепней может быть совершенно точно
установлена видовая принадлежность куколки и ее пол. Пустую куколбчную
оболочку осторожно отмывают в воде кисточкой от приставших посторонних частиц
субстрата, подсушивают на фильтровальной бумаге и накалывают, лучше на одну и ту же
булавку вместе с вышедшим из нее слепнем, снабжая этикеткой. Куколку
рассматривают в часовом стекле в спирту под бинокуляром или лупой, тогда как куколочную
оболочку, наколотую на булавку, рассматривают в ручную лупу или также под
бинокуляром. Следует лишь иметь в виду, что головной и грудной отделы у пустых
оболочек несколько деформированы разрывами в момент выхода слепня.
Определение слепней. Определение до вида ведут по взрослым
насекомым. Фазы развития — яйца, личинки и куколки изучены недостаточно и
определить их до вида чаще не представляется возможным. В ряде случаев может быть
определена родовая и видовая принадлежность личинок (Скуфьин, 1967; Иванищук,
1970; Chvala a. Jezek, 1969; Соболева, 19706, 1971). Для определения слепней обычно
используют ручную лупу с -12—16-кратным увеличением или бинокулярную лупу.
Слепни должны быть хорошей сохранности, наколоты на энтомологические булавки
и иметь этикетки (см. выше). Всегда желательно иметь для определения серию одного
и того же вида, так как возможны некоторые уклонения от типичной формы. При
определении многих видов рода Tabanus необходимо предварительное 4—6-часовое
размачивание во влажной камере для выявления числа полосок на глазах (если этот
признак не был отмечен ранее при наколке или у живых особей в момент поимки).
Слепни довольно однообразны по своему строению, и поэтому их видовая
дифференциация представляет большие трудности, особенно для начинающих. При определении
самок некоторых Hybomitra (группа Н. bimaculata) и других родов иногда приходится
прибегать к исследованию терминалий. Помимо текста определительных таблиц,
совершенно обязательно сличение с описанием и рисунками. Но и это не всегда может
гарантировать от возможных ошибок, и поэтому в ряде случаев приходится прибегать
к сличению с точно определенными экземплярами или обращаться за консультацией
к специалисту.
102
ВВЕДЕНИЕ
10. БОРЬБА СО СЛЕПНЯМИ
Борьба со слепнями вследствие особенностей их образа жизни представляет
известные трудности. Все рекомендуемые способы могут быть разделены на
истребительные, защитные и предупредительные.
В целях истребления слепней рекомендуют на пастбище устраивать для них
«лужи смерти» (Порчинский, 1899). Для этого выбирают мелкие водоемы площадью
не более 10—20 м2, охотно посещаемые слепнями для утоления жажды, и заливают
поверхность нефтью (мазутом, автолом и т. д.). Подлетающие насекомые соприкасаются
с маслянистой пленкой и погибают почти моментально. В занефтеванные водоемы
улавливаются самцы и самки слепней, последние летят в водоемы и после насыщения
кровью на животных. Однократный расход нефти составляет 100 г на 1 м2 поверхности
водоема. Возобновлять нефтяную пленку необходимо через каждые 7—8 дней на
протяжении сезона массового лёта слепней. При отсутствии на пастбище подходящих
Рис. 52. Схема чучелообразной ловушки для слепней. (По Ску-
фьину, 19516).
для нефтевания мелких водоемов их можно создавать искусственно в местах
поверхностного залегания грунтовых вод. Искусственные водоемы могут иметь размеры
1—3 м2 при глубине 0.5 м. «Лужи смерти» необходимо огородить жердняком, чтобы
скот не мог в них напиться воды. В период массового лёта попадаемость на 1 м2 занеф-
теванного водоема составляет в среднем 50—100 слепней за день при максимуме до 500
(Олсуфьев, 19496; Скуфьин, 1952а). «Лужа смерти» в средних широтах за лето может
истребить 3000—5000 слепней и более на 1 м2 занефтеванной поверхности. На юге
Московской обл. В. И. Кузнецовым и нами летом 1939 г. в разгар лёта слепней (3-я
декада июня) был проведен опыт истребления слепней путем обработки нефтью 16
мелких водоемов общей площадью 62 м2. Водоемы были расположены цепочкой по дну
оврага на протяжении 1 км. За декаду в лужи попало около 30 000 слепней, при этом
через несколько дней от начала опыта в зоне действия «луж смерти» было отмечено
снижение в 3 раза количества слепней, нападающих на животных. Не все виды слепней
улавливаются водоемами, обработанными нефтью. Так, в них почти не попадают дож-
девки Haematopota и слепни Atylotus (например A. rusticus).
Дождевок, не попадающих в «лужи смерти», рекомендуют улавливать на черные
щиты, покрытые мушиным клеем (Порчинский, 1899), но применение на практике
данного способа борьбы затруднено, так как для этого требуются значительные
трудовые затраты: дождевки хорошо улавливаются только в присутствии человека,
несущего щит (Бошко, 1950; Маршенкулов, 1967).
Для истребления видов слепней, включая дождевок, может быть использована
чучелообразная ловушка (Скуфьин, 19516), которая представляет собой
приспособление (рис. 52), отдаленно напоминающее лошадь или корову. Из жердей изготовляют
каркас длиной 2 м и высотой 1.6 м, покрывают его грубошерстным материалом (два
■БОРЬБА СО СЛЕПНЯМИ
103
серых одеяла) и сверху приспосабливают мухоловку (35X35X50 см) с воронкой,
направленной внутрь каркаса. Ловушку ставят на пастбище, изобилующем слепнями.
Привлекаемые видом ловушки слепни подлетают к ней, и в том случае, когда пытаются
сесть на нижнюю затененную сторону чучела, насекомые устремляются вверх и
попадают в мухоловку. За лето ловушка может истребить 20 000—25 000 слепней (самок),
но в местах, особенно изобилующих слепнями, например в лесостепной Барабе,
дневной улов слепней ловушкой может достигать 13 000 экз. (Виолович, 1963). Чучело
может быть обработано контактным инсектицидом, и в таком случае будут погибать
от отравления и те насекомые, которые соприкоснулись лишь с внешней поверхностью
ловушки, но не залетели в нее. Обработку материи следует повторять через 1—2
недели (Скуфьин, 1952а).
К. В. Скуфьин (1952а) рекомендует истреблять слепней на пастбище комплексно,
применяя одновременно «лужи смерти» и чучелообразные ловушки.
С целью борьбы с взрослыми слепнями широкому испытанию были подвергнуты
ДДТ и гексахлоран, и установлена их токсичность для слепней (Бошко, 1950; Лутта,
1950; Скуфьин, 1952а; Андреев и Янович, 1954, и др.). Имеются сообщения о
возможности применения шашек типа НБК для истребления в природе взрослых слепней.
Но по данным А. И. Черепанова (1966), в Западной Сибири инсектицидный аэрозоль,
образуемый на смеси ДДТ и гексахлорана в дизельном топливе специальным мощным
генератором, не действовал на слепней, хотя комары от него погибали. Для защиты
крупного рогатого скота от нападения слепней и других кровососущих двукрылых
был применен полихлорпинен (Андреев и Закамырдин, 1960; Андреев и Мяло, 1961;
Павлов, 1965; Андреев, 1966, и др.). Этот препарат (в виде эмульсии) обладает
отпугивающим и инсекцитидным действием и в местах систематических обработок им
крупного рогатого скота отмечалось снижение численности слепней в 2—3 раза по
сравнению с таковой на контрольном участке. При этом уменьшался процент самок,
проделавших яйцекладку (Павлова, 1965).
Однако применение ДДТ и других полихлорорганических инсектицидов для
обработки скота или пастбищ теперь отменено, так как установлена способность этих
препаратов накапливаться в организме позвоночных животных и людей и вызывать
патологические изменения тканей. Вредны они и для пчел и других полезных
насекомых. С целью защиты животных от нападений слепней и других кровососов с хорошими
результатами испытаны хлорофос, гексамид (бензамин), карбоксид и другие вещества
(Андреев и Закамырдин, 1961; Тальдрик, 1965; Сперанская с соавт., 1965; Андреев,
1966, и др.). Для обработки крупного рогатого скота- против слепней в настоящее
время рекомендуют инсектициды растительного происхождения и соединения
фосфорной и карбаминовой кислот: пибутрин, диметилхлорвинилфосфат (ДДВФ), дибром,
байгон и др. Для массовой обработки животных устраивают специальные загоны
и применяют машинные опрыскиватели, при помощи которых за 10—15 мин. можно
обработать 120—150 голов (Павлов, 1965). Систематические (ежедневные по мере
необходимости) опрыскивания отдельных гуртов 0.5% эмульсиями ДДВФ или ди-
брома, а также 0.4% эмульсией байгона из расчета 500 мл на животное, позволили
в условиях Западной Сибири предотвратить снижение среднесуточных удоев коров
в период массового лёта слепней на 0.3—0.8 л. При этом общая численность слепней
на пастбищах опытных гуртов снижалась по сравнению с контролем в 2—4 раза
и более, а в первые 3-~4 часа после каждой обработки на животных в стаде нападали
лишь единичные насекомые (Павлов, 1973).
В оленеводческих хозяйствах с целью борьбы со слепнями с успехом применены
инсектициды ДДВФ, дибром, хлорофос и циодрин. Они использованы как
монопрепараты или в сочетании с репеллентами, например, гексамидом (инсектицидно-репел-
лентные эмульсии). Обработка стад такими эмульсиями при помощи опрыскивателей
«Север» или «Олень» быстро прекращала нападение слепней. В жаркий день при
обилии слепней стада приходилось обрабатывать в день дважды: утром и вечером
(Поляков, 1974).
Представляет интерес опыт борьбы со слепнями, проведенный Брюсом и Декером
(Bruce a. Decker, 1951) в штате Висконсин (США) на 25 стадах крупного рогатого
скота путем обработки животных раз в 3—4 дня синтетическим пиретрином в 0.1%
эмульсии в смеси с 1% piperonyl butoxide. Проведение этих защитных мероприятий
против слепней привело к повышению продуктивности молочного скота на 20—30%
(цит. по: Скуфьияу, 1952а).
Для защиты упряжных животных (лошадей, волов и т. д.) наряду с
вышеперечисленными препаратами могут быть применены масло оленьего рога, а также креолин
и лизол в форме эмульсий или мазей (на вазелине) и т. д. (Олсуфьев, 19496). Эти
вещества наносят на отдельные части тела животных, наиболее уязвимые для нападений
слепней. Действие этих препаратов продолжается в течение нескольких часов.
К. П. Андреев (1966) рекомендует использовать для защиты лошадей ленточные
покрывала, пропитанные в 10%-м растворе креолина или 10%-й эмульсии полихлор-
пинена.
104
ВВЕДЕНИЕ
Для защиты человека от нападения слепней может быть использован диэтил-
толуамид (Виолович, 1963; Черепанов, 1966). Против слепней Hybomitra
отпугивающее действие препарата, нанесенного на кожу, сохраняется до 8 час, но он слабо
действует против златоглазиков. Диэтилтолуамид можно применять для импрегнации
защитной сетки Павловского или капюшона. Диметилфталат не защищает от
нападений слепней.
Методы истребления личинок слепней в природе изучались недостаточно. В
лабораторных экспериментах отмечена токсичность ДДТ и ГХЦГ для личинок слепней,
особенно ранних возрастов (Лутта, 1950; Бошко, 1950; Скуфьин, 1952а). Но в полевых
условиях опыты обработки прибрежных участков, заселенных личинками слепней,
обильным смачиванием эмульсией ДДТ, дали малоудовлетворительные результаты,
так как часть личинок, ушедших в глубину грунта, не погибла (Гладенко и Фортуш-
ный, 1951; Лутта, 1950). Яйца слепней устойчивы к ДДТ и ГХЦГ, но личинки I
возраста погибают в момент вылупления из яиц, обработанных этими препаратами.
Заслуживают внимания появившиеся в США сообщения (Hansens, 1956; Jamnback
and Wall, 1957, цит. по: Расницын, 1963) об успешном истреблении личинок Tabanus
на атлантическом побережье в результате применения диэлдрина; препарат вносили
но 12.2 кг на 1 га вручную или с вертолета (цит. по: Расницын, 1963).
В США также делались локальные попытки использовать яйцеедов для
уничтожения слепней Tabanus и были получены положительные результаты (Parman, 1928).
Но дальнейшего внедрения этого метода в практику не последовало, видимо, из-за
его сложности.
С целью предохранения крупного рогатого скота, лошадей и других животных
от нападения слепней применяют ночные выпасы, тогда как днем животных держат
в стойлах. Эта мера довольно широко практикуется в сильно пораженных слепнями
местностях. В северной половине территории СССР ночные выпасы облегчены тем,
что в период массового нападения слепней — июнь и частью июль — стоят светлые
ночи. Днем животных необходимо обеспечить готовым зеленым кормом и
концентратами. Ночные выпасы можно практиковать лишь в местностях с ограниченной
численностью комаров. А. В. Федюшиным предложено устраивать для скота специальные
тенеубежища, где животных можно содержать в дневные часы, укрывая от нападения
слепней. Тенеубежище представляет собой затемненный загон, закрытый сверху
и с боков ветками ели, тростником, камышом и т. д. По наблюдениям в Омской обл.
К. С. Растегаевой (1958) применение противослепневого тенеубежища повысило в
опытной группе коров удой молока на 15.9% в сравнении с контрольной группой, не
пользовавшейся тенеубежищем. Предложены более совершенные тенеубежища для скота
в виде переносных теневых навесов, обрабатываемых отпугивающими слепней
веществами (Симбирцев, 1965).
На юге практикуют отгон животных на лето в более сухие местности, свободные
от слепней.
Радикальным способом борьбы со слепнями следует считать уничтожение мест
их выплода путем мелиорации речных пойм, осушки болот, засыпки мелких копанок
с водой и ненужных канав, поддержания в порядке прудов, осушительной сети и т. д.
Оздоровление местности от слепней путем мелиорации является мерой,
требующей капитальных затрат.
ЛИТЕРАТУРА *
Айбасов X. А. и Шакирзянова М. С. 1958. Материалы к фауне слепней
Юго-Западного Алтая. Тр. Инст. зоол. АН КазССР, 8 : 209—214.
Айбасов X. А. и Шакирзянова М. С. 1960. Материалы по слепням Джун-
гарского Алатау. Тр. Инст. зоол. АН КазССР, 11 : 173—179.
Алексеев А. Н>, Чиров П. А., Кадышева А. М. и • С в и р и-
д е н к о В. ф. 1969. Сохранение, размножение и выведение во внешнюю среду
возбудителей листериоза (Listeria monocytogenes) и безжелтушного лептоспироза
(Leptospira grippotyphosa) слепнями (Tabanidae) в экспериментальных условиях.
Мед. паразитол. и паразит, болезни, 38, 5 : 583—588.
Алифанов В. И. 1955. Слепни Омской области. Зоол. журн., 34, 5 : 1094—1098.
Аман ж улов С. А., Амосенкова Н. И. и Постричева О. В. 1965.
О нахождении риккетсий Бернета у слепней Tabanus staegeri. Мед. паразитол.
и паразит, болезни, 34, 5 : 613—614.
Андреев К. П. 1956. О мерах борьбы с гнусом. Ветеринария, 4 : 63—69.
Андреев К. П. 1966. Ветеринарная энтомология и дезинсекция. М.
Андреев К. П. и Закамырдин И. А. 1960. Эмульсия полихлорпинена
для защиты животных от гнуса. Ветеринария, 8 : 79—83.
* В список литературы включена лишь часть публикаций; более-подробные списки
литературы по слепням см. Н.А.Виолович. 1968. Слепни Сибири и А. С. Л у т т а.
1970. Слепни Карелии.
ЛИТЕРАТУРА 105
Андреев К. П. и Закамырдин И. А. 1961. Гексаметиленбензамид (гекса-
мид Б) — новый репеллент против слепней. Ветеринария, 6 : 68—69.
Андреев К. П. и М я л о И. И. 1961. Защита животных от гнуса. Благовещенск.
Андреев К. П. и Янович Г. И. 1954. Новые репелленты для защиты
сельскохозяйственных животных от гнуса. В кн.: Проблемы ветеринарной
дерматологии, арахнологии и энтомологии. М. : 195—198.
Анищенко А. К. и Орехов П. В. 1967. Слепни — хранители и переносчики
вируса чумы свиней. Тр. Белорусок, н.-иссл. ветер, инст., 6, Минск : 114—119.
Арбузов П. Н. 1941. Роль слепней в передаче трипанозомоза су-ауру.
Ветеринария, 5 : 32—33.
Арбузов П. Н. 1946. Роль слепней в передаче трипанозомоза су-ауру. Изв.
АН КазССР, серия паразитол., 33, 4 : 77—79.
Б а б а я н ц Г. А. 1962. Материалы к фауне слепней Туркменистана. Тр. Агахабадск.
н.-иссл. инст. эпидемиологии и гигиены, 5 : 243—248.
Баратов Ш. 1961. Новые для фауны Таджикистана виды слепней (Diptera,
Tabanidae). Тр. Инст. зоол. и паразитол. АН Тадж. ССР, XX : 181—188.
Баратов Ш. 1962а. Новые для Таджикистана виды слепней (Tabanidae).
Сообщение II. Докл. АН Тадж. ССР, 5, 3 : 40—42.
Баратов Ш. 19626. Новый вид слепня (Diptera, Tabanidae) из Таджикистана.
Докл. АН Тадж. ССР, 5, 6 : 44—45.
Баратов Ш. 1962в. Описания самцов слепней — Tabanus (Tylostypia) staegeri
reinigianus (End) и Т. (s. stir.) semenovi Ols. (Diptera, Tabanidae). Изв. отд. биолог,
наук АН Тадж. ССР, 2 (9) : 96—97.
Баратов Ш. 1966. О слепнях (Tabanidae) Гиссарской долины и Гиссарского
хребта (Таджикистан). Изв. отд. биолог, наук АН Тадж. ССР, 1 (22) : 71—82.
Баратов Ш. Б. и Мишкин К. В. 1972. О самцах малоизученных видов
слепней (Diptera, Tabanidae) из Таджикистана. Изв. АН Тадж. ССР, отд. биолог,
наук, 3 (48) : 61—64.
Бей-Биенко Г. Я. 1936. Насекомые жесткокрылые. Фауна СССР, нов. серия,
5. Изд. АН СССР, М.—Л.
Бей-Биенко И. Г. 1957. Материалы по фауне слепней Витебской области.
Энтомол. обозр., 36, 3 : 655—658.
Бей-Биенко И. Г. 1959. Двухлетние наблюдения над лётом слепней в
Витебской области. Тр. Инст. мед. паразитол. и тропич. мед. им. Е. И. Марцинов-
ского, М. : 332—341.
Бей-Бценко И. Г. 1963. К' фауне и биологии слепней (Diptera,, Tabanidae) тайги
Среднего Енисея. Энтомол. обозр., 42, 4 : 833—844.
Бей-Биенко И. Г. 1967. Особенности гонотрофического цикла слепней как
компонентов гнуса. Научная конференция профессорско-преподавательского
состава и аспирантов, посвященная 50-летию Советской власти. 21—24 марта.
Тезисы докл. секции защиты растений. Пушкин : 13—14.
Бей-Биенко И. Г. 1970. Слепни тайги бассейна Среднего Енисея. Дисс. канд.
Л.—Пушкин.
Белановский И. Д. 1950. К фауне двукрылых юго-западного Памира. Науков1
зап. Кшвськ. державн. ушв., IX, VI : 133—143.
Берг Л. С. 1949. Рыбы пресных вод СССР и сопредельных стран. Часть III. Изд.
АН СССР, М.—Л. : 929-1381.
Благовещенский Д. И. 1937. Материалы по фауне наружных паразитов
животных Казалинского и некоторых других районов Южного Казахстана.
Тр. Казахск. фил. АН СССР, 2 : 11—84.
Благовещенский Д. И. 1946. Клещи сем. Ixodidae и кровососущие двукрылые
низовья Амура. Паразитол. сб. 9 : 83—113.
Богачев А. В. и Самедов Н. Г. 1948. Материалы к познанию
распространения слепней (Tabanidae) в северо-восточной части Азербайджана. Изв. АН
Азерб. ССР, 6 : 74—80.
Богачев А. В. и Самедов Н. Г. 1949. Материалы к изучению паразитофауны
НахАССР (Tabanidae НахАССР). Изв. АН Азерб. ССР, 5 : 66—75.
Боженко В. П. 1941. Слепни Chrysops как передатчики туляремии. Журн. микро-
биол., эпидемиол. и иммунобиол., 12 : 21—25.
Бошко Г. В. 1950. Слепни Украинского Полесья, их биология, вредоносное
значение и меры борьбы. Дисс. канд. Киев.
Бошко Г. В. 1954. О возможности ночного лова слепней в светоловушки. В кн.:
III экологическая конференция, 4. Тезисы, Киев : 51—55.
Бошко Г. В. 1957. Матер!алы по фауни гедз1в захдаых областей Украши. Тр.
1нст. зоолог. АН СССР, 14 : 49—56.
Бошко Г. В. 1958а. Материалы по распространению слепней (Tabanidae) в УССР.
В кн.: Проблемы зоогеографии- суши. Львов : 25—31.
Бошко Г. В. 19586. Новий для фауни СРСР вид гедзя (Dipt., Tabanidae). Доп.
АН УРСР, 3 : 351—352.
106
ВВЕДЕНИЕ
Б о ш к о Г. В. 1962. Опис рашше невщомых самщв Tabanus tawicus N. Ols. 1952
(Diptera, Tabanidae). Доп. АН УРСР, 9 : 1251—1252.
Б о ш к о Г. В. 1963а. До можливого ешдемюлопчного значения золотоочок вшон-
чатых (Chrysops) Heterochrysops (italicus Mg.) на o-Bi Б1рючому. Доп. АН УРСР,
7 : 968—970.
Б о ш к о Г. В. 19636. Материалы к фауне и распространению слепней (Diptera,
Tabanidae) на территории Крымского полуострова. В кн.: Проблемы
паразитологии, 2. Киев : 182—194.
Б о ш к о Г. В. 1963в. Фауна и экология слепней (Diptera, Tabanidae) на
высокогорных пастбищах Украинских Карпат. В кн.: Флора и фауна Карпат, 2. М. :
192—207.
Б о шк о Г. В. 1964. Фауна и экология слепней (Tabanidae) на территории
Правобережной степи Украинской ССР. В кн.: Проблемы паразитологии, 3. Киев :
233—255.
Бошко Г. В. 1966. Слепни (Diptera, Tabanidae) Азово-Сивашского заповедно-
охотничьего хозяйства в пределах природного очага туляремии на п-ове
Бирючьем. В кн.: Паразиты, промежуточные хозяева и переносчики. Киев :
147—159.
Бошко Г. В. 1968. Экология пестряка окончатого (Diptera, Tabanidae). Вестн.
зоологии, 3 : 37—43.
Бошко Г. В. 1973. Гедз! Diptera, Tabanidae. Фауна Украши, 13, 4. Киев.
Бошко Г. В., Жданова Т. Г. и Андреева Р. В. 1972. Автогенне дозр1-
вання яэчншпв у гедз!в Chrysops relictus Meg. (Diptera, Tabanidae). В кн.:
Паразити, паразитози та шляхи 'ix лшвщащ, 1. Киев : 244—245.
Бошко Г. В. i Черногоренко Е. Б. 1963. До фауни гедз!в Tabanidae
Пруто-Дтстровского межир!ччя. Допов. АН УРСР, 10 : 1398—1400.
Б р е е в К. А. 1948. Активность нападения кровососущих двукрылых и оводов
на северного оленя и факторы ее регулирующие. Автореф. канд. дисс. Л.
Б р е е в К. А. 1950. О поведении кровососущих двукрылых и оводов при нападении
их на северного оленя и ответных реакциях оленей. Сообщ. I. Паразитол. сб.
12 : 167—198.
Буланова Ф. Г. 1963. О слепнях Удмуртской АССР. Зоол. журн., 42, 4 : 627—
628.
Буланова Ф. Г. 1967. Эколого-фаунистический обзор слепней (Diptera,
Tabanidae) Удмуртской АССР. Автореф. канд. дисс. Пермь.
Васюкова Т. Т. 1967. Фауна слепней Среднего Приангарья. В кн.: Итоги
исследования'! по проблеме борьбы с гнусом. Новосибирск : 197—202.
Васюкова Т. Т. 1968. О фауне слепней Центральной части Иркутской области.
Паразитол., 2, 4 : 365—368.
Васюкова Т. Т. 1972. Материалы по экологии личинок и куколок слепней
(Diptera, Tabanidae) Центральной Якутии. В кн.: Фауна и экология насекомых
Якутии. Якутск : 158—164.
Виолович Н. А. 1956а. Новые виды слепней (Tabanidae) дальневосточной фауны.
Паразитол. сб. 16 : 152—154.
Виолович Н. А. 19566. Слепни Сахалинской области. Тр. Дальневост. фил.
АН СССР, серия зоол., 3 (6) : 255—264.
Виолович Н. А. 1963. Репеллентное свойство диэтиламида метатолуиловой
кислоты по отношению к слепням. Мед. паразитол. и паразит, болезни, 32,
5 : 539—542.
Виолович Н. А. 1965. К фауне кровососущих двукрылых насекомых (Diptera)
Тувы. Энтомол. обозр., 44, 4 : 832—838.
Виолович Н. А. 1966. Слепни (Tabanidae). В кн.: Биологические основы борьбы
с гнусом в бассейне р. Оби. Новосибирск : 171—237.
Виолович Н. А. 1968. Слепни Сибири. Новосибирск.
Ви'олович Н. А. и Гомоюнова Н. П. 1964. К .фауне и экологии слепней
(Diptera, Tabanidae) степной зоны Западной Сибири. Изв. Сиб. отд. АН СССР,
4, 3 : 108—113.
Виолович Н. А., Гомоюнова Н. П. и' Евстигнеева Н. С. 1965.
Материалы по фауне и экологии слепней Барабинской лесостепи. В кн.:
Животный мир Барабы. Новосибирск : 220—228.
Виолович Н. А. и Евстигнеева Н. С. 1964. К экологии лесного пестряка
в лесостепной зоне Западной Сибири. Изв. Сибирск. отд. АН СССР, сер. биол.-
мед. наук, Новосибирск, 1 : 106—110.
Владимирова В. В. и Потапов А. А. 1963. Новые образцы ловушек
для слепней и мошек. Мед. паразитол. и паразит, болезни, 32, 1 : 83—87.
В о л и к Г. Н. 1966. Изучение гнуса и борьба с ним на кизлярских пастбищах
Дагестана. Автореф. канд. дисс. М.
ЛИТЕРАТУРА
107
Волкова М. И. 1950. К изучению слепней среднего Поволжья Татарской и
Чувашской республик. Уч. зап. Казанск. унив. им. В. И. Ульянова-Ленина,
110, 4 : 155—199.
В о njK о в а М. И. 1957. Эффективность действия фосфорорганических препаратов
на кровососущих двукрылых — комаров и слепней. В кн.: Химия и применение
фосфорорганических соединений. Казань : 420—430.
Г ату]з ер Е. Г. 1939. Очерк фауны слепней Ханларского района. Тр. Зоол. инст.
Азерб. фил. АН СССР, 10 : 135—159.
Г а у з е р Е. Г. 1941. К фауне слепней Нагорного Карабаха. Изв. Азерб. фил. АН
СССР, 2 : 70—74. '
Г a yja е р Е. Г. 1953. К фауне и фенологии слепней северо-западного Азербайджана.
Энтомол. обозр., 33 : 238—254.
Г а у~з ер Е. Г. 1960. О фауне и фенологии слепней Нахичеванской АССР. Энтомол.
обозр.,,39, 3 : 643—660.
Гладенко И. Н. и Фортушный В. А. 1951. Слепни (Tabanidae)
Нижнеднепровских плавней и меры борьбы с ними. Научн. тр. Укр. инст. экспер.
ветеринарии, 18 : 186—191.
Гладенко И. Н. и Фортушный В. А. 1952. Широкий опыт борьбы с
кровососущими двукрылыми насекомыми с помощью ДДТ и гексахлорана. Научн.
тр. Укр. инст. экспер. ветеринарии, 19 : 182—185.
Гребенюк Р. В. и Чиров П. А. 1971. Слепни Киргизии. Фрунзе.
Гробов О. Ф. 1961. Насекомые как переносчики анаплазмоза крупного рогатого
скота. Тр. Всесоюзн. инст. эксперим. ветеринарии, 24 : 160—169.
Гургенидзе Л. Н. 1970. О видовом составе и сезонной активности слепней
полупустынных и степных массивов Шираки-Эльдари (Грузинская ССР). Сообщ.
АН ГрузССР, 59, 1 : 197—200.
Гургенидзе Л. Н. 1971. 6 фауне и сезонной активности слепней Тушетии
(Грузинская ССР). Сообщ. АН ГрузССР, 61, 1 : 197—200.
Гургенидзе Л. Н. 1972а. Новые данные о распространении и поведении Silvius
(Heterosilvius) zaitzevi Olsufjev и описание самца этого вида. Сообщ. АН ГрузССР,
66, 1 : 201-203.
Гургенидзе Л. Н. 19726. О местах выплода слепней в Ширакской степи. В кн.:
Проблемы почвенной зоологии. Материалы IV Всесоюзного совещания, Баку.
М. : 44.
Гуцевич А. В. 1937. Материалы по изучению гнуса (кровососущие двукрылые
насекомые) на Дальнем Востоке. Тр. Военно-мед. акад. им. С. М. Кирова,
8 : 151—169.
Гуцевич А. В. 1939. Материалы по изучению кровососущих двукрылых
насекомых (гнуса) в Забайкалье. Тр. Военно-мед. акад. им. С. М. Кирова, 19 : 35—47.
Гуцевич А. В.' 1940. Материалы по изучению кровососущих двукрылых (гнуса)
североуссурийской тайги. Зоол. журн., 19, 3 : 428—444.
Гуцевич А. В. 1943. Кровососущие двукрылые насекомые Присурского лесного
массива (Чувашская АССР). Зоол. журн., 22, 1 : 19—26.
(Двигубский И.) Dwigubsky'I. 1802. Primitiae Faunae Mosquensis. Mos-
quae : 173—189.
Дегтярев А. 1928. Материалы к вопросу хозяйственного значения
кровососущих насекомых. Слепни (сем. Tabanidae) в условиях Ярославской губернии.
Производительные силы Ярославской губернии. Тр. II губ. конф. по изуч.
произв. сил Яросл. губ., Ярославль : 323—328.
Д ж а ф а р о в Ш. М. 1959. Слепни {Tabanidae, Diptera) Талыша и юго-восточной
части Муганской зоны. Тр. Инст. зоол. АН Азерб. ССР, 20 : 190—210.
Джафаров Ш. М. 1960. Слепни (Diptera, Tabanidae) Азербайджана. Зоол. журн.,
39, 5 : 714-722.
Драчинский СП. 1927. Методы и научное обоснование борьбы с трипанозомо-
зом верблюдов. Тр. I Всерос. вет. научн. орг. Съезда, Л., 2 : 384—386.
Евсеева Т. И. и Тимофеева А. Е. 1971. Об участии слепней в циркуляции
возбудителя эразипелоида на Сахалине. В кн.: Природа Сахалина и здоровье
человека. Изв. Сахалинск, отдела географ, общ. СССР, I. Южно-Сахалинск,
2 : 69—70.
Зайцев В. Ф. и Тертерян А. Н. 1966. Villa ventruosa Lw. (Diptera, ВотЪу-
liidae) паразит личинок и куколок слепней (Diptera, Tabanidae) в Армянской ССР.
Биол. журн. Армении, 19, 12 : 84—89.
Закамырдин И. А. 1961. Опыт защиты скота от кровососущих двукрылых
насекомых. Ветеринария, 6 : 70—73.
Запекина-Дулькейт Ю. И. 1969. Слепни и другие кровососущие
двукрылые заповедника «Столбы». Вопросы энтомологии, Тр. Гос. заповеди. «Столбы»,
7, Красноярск : 4—105.
108 ВВЕДЕНИЕ
Засухин Д. Н., Федоров Н. С, Боженко В. П. и Т и ф л о в В. Е.
1930. Материалы к познанию фауны Tabanidae (Diptera, Brachycera) Юго-Востока
РСФСР. Вести, микробиол., эпидемиол. и паразитол., 9 : 401—405.
Иванищук П. П. 1970. К морфологии и классификации личинок слепней
(Diptera, Tabanidae) Ивановской области. В кн.: Насекомые— переносчики
заразных заболеваний. Тр. Ивановск. гос. мед. инст., 46 : 137—153.
Иванищук П. П. 1973. Некоторые вопросы морфологии, экологии и фенологии
. преимагинальных фаз развития слепней (Diptera, Tabanidae). Дисс. канд.
Иваново.
Иванова-Гобзем П. С. 1945. Трипанозомоз верблюдов в Вахшской долине
Таджикской ССР. В кн.: Комплексные исследования по вредителям
животноводства в Таджикистане и по борьбе с ними. Тр. Таджикск. фил. АН СССР,
14 : 160—171.
Исмаилов Ш. И. 1970. Кровососущие двукрылые Дагестана и их распределение
по высотным поясам. Дисс. канд. М.
Истомина Л. П. 1963. О фауне слепней Семипалатинской области. В кн.:
Паразиты сельскохозяйственных животных Казахстана. Алма-Ата : 121—124.
Кадырова М. К. 1972а. К биологии Tabanus leleani Aust. (Diptera, Tabanidae)
в условиях Узбекистана. В кн.: Природная очаговость болезней и вопросы
паразитологии животных. Тр. VII Всесоюзн. конф. по прир. очаговости
болезней и общим вопросам паразитол. животных, 6, 2; 214—218.
Кадырова М. К. 19726. Слепни (сем. Tabanidae). В кн.: Паразитические
членистоногие Ферганской долины. Ташкент : 4—40.
Кадырова М. 1975. Слепни "Узбекистана. Ташкент.
Кадырова М. и Ган Э. И. 1967. Слепни (Diptera, Tabanidae). В кн.:
Переносчики возбудителей болезней человека и животных. Ташкент : 66—114.
Калугин С. Г. 1945. К изучению слепней центральной части северных склонов
Кавказского хребта. Работы Северо-Кавказской гидробиологической станции,
4, 2—3, Дзауджикау : 21—25.
Кателина А. Ф. и Мясников 10. А. 1965. Фауна слепней природных
очагов туляремии Тульской области. В кн.: Туляремия и сопутствующие
инфекции. Омск : 86—89.
Кирпичникова В. А. 1972. Фауна и экология слепней (Diptera, Tabanidae)
северной части Приморского края. Дисс. канд. Владивосток.
Киселева Е. Ф. 1937. К фауне слепней лесной зоны восточной части Западной
Сибири. Тр. Биол. н.-иссл. инст. Томск, унив., Томск, 4 : 93—97.
Коваль Е. 3. 1Андреева Р. В. 1971. До вивчения патогенно! микофлоры гед-
3iB (Diptera, Tabanidae) на Украшъ Доп. АН УРСР, сер. Б, 11 : 1042—1044.
Колесова О. Ф. 1966. К экологии и фенологии преимагинальных стадий развития
слепней (Diptera, Tabanidae). В кн.: Вопросы краевой зоологии и паразитологии,
2. Иваново : 47—52. \
Колесова О. Ф. 1968. Об экологии и фенологии Heptatoma pellucensF.
(Tabanidae, Diptera). Мед. паразитол. и паразит, болезни, 37, 5 : 585—588.
Колонии Г. В. 1969. Значение кровососущих двукрылых в эпизоотологии и
эпидемиологии сибирской язвы (обзор литературы). Мед. паразитол. и паразит,
болезни, 38, 3 : 354—357.
Колосов Ю. М. и Попов Л. В. 1932. Каталог двукрылых Среднего Урала.
В сб.: Работы энтомологического отделения паразитологического отдела,
Свердловск, 1 : 3—15.
Колычева Р. В.и Паонко Н.К. 1964. О сезонности лёта и физиологическом
возрасте массовых видов слепней (Diptera, Tabanidae) в речных поймах на
востоке Воронежской области. В кн.: Охрана природы Центральночерноземной
полосы, 5.Воронеж : 232—236.
Коренчевская Г. А. 1960. Материалы по фауне слепней южноукраинских
степей, включая прилегающие части лесостепи. Научн. ежегодн. Одесск. гос.
унив., биолог, фак., 2 : 162—164.
Крыжановский О.Л. 1965. Состав и происхождение наземной фауны Средней
Азии. М.'—Л.
Куандыкова У. С. 1966. Фаунистический состав и распространение слепней
в бассейне р. Или. Тр. Инст. зоол. АН Каз. ССР, 25 : 96—101.
Куандыкова У. С. 1968. Слепни бассейна р. Или. Автореф. канд. дисс. Алма-Ата.
Кузякин А. П. 1962. Зоогеография СССР. Уч. зап. Моск. обл. Педагог, инст.
им. Н. К. Крупской, Биогеография, 1 : 3—182.
Купрессова В. В., Савельева И. П., Романенко В. Н. и
Орлов В. М. 1972а. Некоторые вопросы экологии слепней бассейна р. Подка-
менная Тунгуска. В кн.: Зоологические проблемы Сибири. Томск : 124—125.
Купрессова В. Б., Савельева И. П., Романенко В. Н. и
Орлов В. М. 19726. Видовой состав и экология слепней бассейна р. Подкамен-
ной Тунгуски. Мед. паразитол. и паразит, болезни, 41, 3 : 314—318.
ЛИТЕРАТУРА i Ш
Кучеров Е. В. 1959. Материалы по фауне слепней Южного Урала и
Башкирского Зауралья. Зоол. журн., 38, 9 : 1383—1395.
Кучеров Е. В. 1964. О фауне слепней Уфимского плато и Месягутовской лесостепи
Башкирии. Зоол. журн., 43, 4 : 614—616.
Кучеров Е. В. и Еникеев Р. X. 1965. Слепни и борьба с ними. Уфа.
Лейкина Г. А. 1964. К изучению кровососущих двукрылых в нижнем течении
Терека и Кумы и их роль в туляремийном очаге. Мед. паразитол. и паразит,
болезни, 33, 1 : 40—44.
Лобанов А. М. 1959. К биологии Tabanus bovinus L. Мед. паразитол. и паразит,
болезни, 28, 2 : 234—235.
Л утт а А. С. 1950. Слепни Кара-Калпакии. В кн. : Кара-Калпакская АССР.
Материалы по производительным силам Узбекистана, I. Ташкент : 311—360.
Л у т т а А. С. 1959. Материалы по видовому составу и биологии слепней (Tabanidae)
Карельской АССР. Тр. Карельск. фил. АН СССР, 14 : 84—109.
Л у т т а А. С. 1960. О распространении слепней в Карелии. Паразитол. сб. 19 : 291 —
. 307.
Л у т т а А. С. 1964а. Болота и заболоченные луга — места выплода слепней в
Карелии. В кн.: К природной очаговости паразитарных и трансмиссивных
заболеваний в Карелии. М.—Л. : 120—130.
Л у т т а А. С. 19646. Материалы по гонотрофическому циклу слепней Карелии.
В кн.: К природной очаговости паразитарных и трансмиссивных заболеваний
в Карелии. М.—Л. : 131—153.
Л у т т а А. С. 1964в. О зараженности кладок слепней яйцеедом Telenomus tabani
Mayr. (Hymenoptera, Scelionidae). В кн.: К природной очаговости
паразитарных и трансмиссивных заболеваний в Карелии. М.—Л. : 176—180.
Л у т т а А. С. 1964г. Личинки слепней в дельте Аму-Дарьи и в бассейне Сыр-
Дарьи. Тр. Инст. зоол. АН Каз. ССР, 22 : 184—191.
Л утт а А. С. 1965. Экологические особенности распространения слепней (Diptera,
Tabanidae) в Узбекистане. Энтомол. обозр., 44, 3 : 595—604.
Л утт а А. С. 1966а. К биологии Atylotus karybenthinus Sz. (Diptera, Tabanidae).
Тр. Инст. зоол. АН Каз.ССР, 25 : 102—115.
Л]у т т а А. С. 19666. О влиянии слепневых яйцеедов на численность слепней. Тр.
Инст. зоол. АН Каз: ССР, 25 : 116—120.
Л]у тт а А. С. 1970. Слепни Карелии. Л.
Л у т т а А. С. 1972а. Ландшафтно-географические особенности сезонной активности
слепней (сем. Tabanidae). В кн.: Биологические основы борьбы с
трансмиссивными и паразитарными заболеваниями на севере. Петрозаводск : 154—160.
Л у т т а А. С. 19726. Зимовка и холодостойкость личинок слепней (сем. Tabanidae)
Карелии. В кн.: Биологические основы борьбы с трансмиссивными и
паразитарными заболеваниями на севере. Петрозаводск : 161—183.
Ма'евский А. Г. 1956. Некоторые данные о фауне слепней (Tabanidae) БССР.
Вестн. АН БССР, сер. биол. наук, 3 : 113—117.
Маевский А. Г. 1960. Сезонная динамика численности слепней Белорусской ССР.
Зоол. журн., 39, И : 1644—1651.
Майр Э. 1947. Систематика и происхождение видов с точки зрения зоолога. М.
(Перевод с английского).
Майр Э., Линсли Э., Юзингер Р. 1956. Методы и принципы зоологической
систематики. М. (Перевод с английского).
Мамедниязов О. 1971. Кровососущие двукрылые насекомые (гнус) в районах
4-й очереди Каракумского канала и мероприятия по борьбе с ними. Автореф.
канд. дисс. Ашхабад..
Маршенкулов 3. М. 1967. Слепни (Diptera, Tabanidae) Кабардино-Балкарии.
Автореф. канд. дисс. Нальчик.
М а с л о в А. В. 1960. Кровососущие двукрылые насекомые Дальнего Востока.
Вопросы геогр. Дальн. Востока, 4 : 269—299.
Мезенев Н. П. 1972. Комплекс гнуса в северных районах Таймыра. В кн.:
Зоологические проблемы Сибири : 140—142.
Мельникова Т. Г. 1965. О слепнях (Tabanidae) Таджикистана. Зоол. журн.,
44, 7 : 1028—1034.
М е н з б и р М. А. 1914. Зоологические участки Туркестанского края и вероятное
происхождение фауны последнего. М.
Митрофанова Ю. Г. 1951. Материалы к изучению слепней Предуралья. Уч.
зап. Пермск. унив., 6, 2 : 3—26.
Митрофанова Ю.Г. 1955. Фауна и экология слепней (Tabanidae) Пермской
области (Кунгурский и Кишертский районы) и меры борьбы с ними. Уч. зап.
Пермск. унив., 7, 3 : 3—22.
Мончадский А. С. 1939. Возможные переносчики весенне-летнего энцефалита.
Тр. Военно-мед. акад. им. С. М. Кирова, 18 : 189—197.
1J° ВВЕДЕНИЕ
(Мочульский В.) Motschulsky V. 1859. Catalogue des Insectes rappor-
tes des environs du fl. Amour, depuis la Schilka jusqu'.a Nikolaevsk, examines
et enumeres. Bull. Soc. Natur. Moscou, 32, 2 : 487—507.
Мяло И. И. 1962. Слепни Зейско-Буреинской низменности и защита от них скота.
Тр. Дальневосточн. н.-иссл. ветер, инст., 4 : 147—152.
Нагорский В. 1902. Опыт эпизоотологии как учение о причинах и процессе
массового развития заразных болезней домашних животных. Сибирская язва.
Архив ветерин. наук, 7, приложение : 1—48.
Никольская М. Н. 1948. Виды рода Telenomus (Hymenoptera, Scelionidae)
паразиты яиц слепней. ДАН СССР, 62, 5 : 729—732.
О л и г е р И. М. 1960. Фауна слепней Чувашской АССР. Зоол. журн., 39, 7 : 1099—
1101.
О л и г о р И. М. 1967. Фауна слепней зоны южной тайги Среднего Поволжья. Зоол.
журн., 46, 5 : 721—729.
Олсуфьев Н. Г. 1933. Tabanus L. Слепень. В кн.: Штакельберг А. А.
Определитель мух Европейской части СССР. Изд. АН СССР, Л. : 69—88.
Олсуфьев Н. Г. 1934. Материалы по фауне слепней Ленинградской области. Па-
разитол. сб. 4:-111—201.
Олсуфьев Н. Г. 1935а. Слепни. В кн.: Практикум медицинской паразитологии,
под ред. Е. Н. Павловского. Л. : 159—180.
Олсуфьев Н. Г. 19356. Материалы по изучению слепней Ленинградской области.
В кн.: Вредители животноводства. Л. : 251—316.
Олсуфьев Н. Г. 1935в. Материалы по фауне слепней (Tabanidae) Урала. Пара-
зитол. сб. 5 : 205—215.
Олсуфьев Н. Г. 1936а. Материалы по фауне слепней Западной Сибири. Парази-
тол. сб. 6 : 201—245.
Олсуфьев Н. Г. 19366. К микроскопической анатомии головы и
пищеварительного тракта личинок слепней. Паразитол. сб. 6 : 247—278.
Олсуфьев Н. Г. 1937а. Слепни (Tabanidae). Фауна СССР. Двукрылые насекомые,
VII, 2. Изд. АН СССР. Л.
Олсуфьев Н. Г. 19376. Материалы по фауне слепней Казахстана. Тр. Казахстан.
фил. АН СССР, 2 : 123—180.
Олсуфьев Н. Г. 1938. Слепни СССР. Дисс. докт. М.
Олсуфьев Н. Г. 1939а. Видовой состав и сезонная динамика численности
кровососущих двукрылых в дельте Волги и их возможная роль в эпидемиологии
туляремии. Зоол. журн., 18, 5 : 786—798.
Олсуфьев П. Г. 19396. Материалы по фауне слепней (Diptera, Tabanidae)
Таджикистана. Тр. Таджикск. базы АН СССР, 11 : 93—112.
Олсуфьев Н. Г. 1940. Двойственный характер питания и половой цикл у самок
слепней (Diptera, Tabanidae). Зоол. журн., 19, 3 : 445—455.
Олсуфьев Н. Г. 1941. Материалы по фауне слепней Кавказа. Тр. зоол. сектора
Груз. фил. АН СССР, 3 : 45—90.
Олсуфьев Н. Г. 1949а. К изучению фауны слепней (Tabanidae) южной части
Московской области. Бюлл. Моск. общ/испыт. природы, отд. биологии, 54, 1 : 35—
46.
Олсуфьев Н. Г. 19496. О слепнях северной части Барабинской лесостепи и
некоторых способах борьбы с ними. В кн.: Вопросы краевой,, общей и
экспериментальной паразитологии, 6. М. : 34—58.
Олсуфьев Н. Г. 1952. Новые виды слепней (Diptera, Tabanidae) фауны СССР.
Энтомол. обозр., 32 : 311—315.
Олсуфьев Н. Г. 1958. Семейство Tabanidae — слепни. В кн.: В. Н. Б е к л е м и-
ш е в (ред.). Определитель членистоногих, вредящих здоровью человека.
М. : 109—125.
Олсуфьев Н. Г. 1962а. О слепнях (Tabanidae) Алтайского края. Зоол. журн.,
41, 6 : 882—892.
Олсуфьев Н. Г. 19626. О диагностическом значении строения гениталий самок .
в группе Tabanus (Tylostypia) tropicus Panz. (Diptera, Tabanidae). В кн. : Вопросы
общей зоологии и медицинской паразитологии. М. : 524—528.
Олсуфьев Н. Г. 1962в. Слепни (Diptera, Tabanidae). В кн.: Переносчики
возбудителей природноочаговых болезней. М. : 144—178.
Олсуфьев Н. Г. 1964. К изучению фауны слепней (Tabanidae) западной части
Большого Кавказа, с описанием нового рода. Бюлл. Моск. общ. испыт. природы,
отд. биологии, 69, 3 : 73—76.
Олсуфьев Н. Г. 1966. О древних атлантических связях фауны южной части
Северной Америки и Средней Азии на примере слепней (Tabanidae). Бюлл. Моск.
общ. испыт. природы, отд. биологии, 71, 6 : 52—58.
Олсуфьев Н. Г. 1967. Новые виды слепней (Diptera, Tabanidae) Палеарктики.
Энтомол. обозр., 46, 2 : 379—390.
ЛИТЕРАТУРА
111
(Олсуфьев Н. Г.) О 1 s u f j e v N. G. 1969. То the taxonomy and distribution
of Tabanidae (Diptera) of the Palaearctic Region. Acta entomol. bohemoslov.,
66, 2 : 115—121.
Олсуфьев Н. Г. 1970. Новые и малоизвестные слепни (Diptera, Tabanidae)
фауны СССР и сопредельных стран. Энтомол. обозр., 49, 3 : 683—687.
Олсуфьев Н. Г. 1971. Некоторые вопросы номенклатуры и таксономии палеаркти-
ческих Tabanidae (Diptera). Бюлл. Моск. общ. испыт. природы, отд. биологии,
76, 6 : 138—139.
Олсуфьев Н. Г. 1972. Новые и малоизвестные слепни (Diptera, Tabanidae)
фауны СССР и сопредельных стран. Сообщение II. Энтомол. обозр., 51, 2 : 446—
453.
Олсуфьев Н. Г. 1973. Ревизия типовых экземпляров палеарктических слепней
(Diptera, Tabanidae) Зоологического музея Университета им. Гумбольдта в
Берлине. Энтомол. обозр., 52, 1 : 195—200.
Олсуфьев Н. Г. и Боженко В. П. 1950. Материалы по изучению слепней
Восточного Казахстана. Изв. АН Каз. ССР, серия паразитолог., 8 : 54—61.
Олсуфьев Н.Г. и Голов Д. А. 1936. Роль слепней в передаче и хранении
туляремии. В кн.: Патогенные животные. М. : 187—224.
(Олсуфьев Н. Г. и Лек л.е р к М.) Olsufjev N. G. and Leclercq M.
1971. Some problems of the nomenclature of Palaearctic Tabanidae (Diptera).
Bull. Ann. Soc. R. Ent. Belg., 107 : 325—326.
Олсуфьев Н. Г. и Лелеп П. П. 1935. О значении слепней в распространении
сибирской язвы. В кн.: Паразиты, переносчики и ядовитые животные. М. :
145—197.
Олсуфьев Н.Г. и Мельникова Т. Г. 1962. К фауне слепней (Diptera,
Tabanidae) Крыма. Энтомол. обозр., 41, 3 : 576—578.
Олсуфьев Н.Г. и Мосолов Л. П. 1964. О слепнях Московской области.
Зоол. журн., 43, 10 : 1480—1487.
(Олсуфьев Н., Моуха Й. и Хвала М.) Olsufjev N., M ouch a J.
und Cli v a j. a M. 1964. Chrysozona scutellata sp. n. aus Mitteleuropa (Diptera,
Tabanidae). Casopis Ceskoslovenske Spolecnosti Entomologicke, 61, 3 : 284—286.
(Олсуфьев Н., Моуха И. и Хвала M.) Olsufjev N., Moucha J.
und С h v a 1 a M. 1967. Zur Taxonomie und Verbreitung dei europaischen und
kleinasiatischen Arten der Tabanus bovinus-Grwp'pe (Diptera, Tabanidae). Acta
entomol. bohemoslov., 64, 4 : 303—313.
Олсуфьев Н. Г., Петров В. Г. и Хлюстова А. И. 1958. Фауна,
распространение и сезонная численность кровососущих двукрылых в северной
части Волро-Ахтубинской поймы и в долине Волги в пределах Волгоградской
области. В кн.: Вопросы эпидемиологии и профилактики туляремии. М. : 91—
104.
Олсуфьев Н. Г. и Формозов А. Н. 1953. О фауне слепней Центрального
Казахстана. Тр. Инст. географии, 54, Материалы по биогеографии СССР,
М. : 12-32.
Олсуфьев Н.Г. и Шевченко В. В. 1964. Обнаружение нового для фауны
СССР вида слепня Tabanus ansarii badhysi subsp. nova. Тр. Инст. зоол. АН Ка-
захск. ССР, 22 : 197—198.
Остен-Сакен Р. 1858. Очерк современного состояния познания
энтомологической фауны окрестностей Санкт-Петербурга. Журн. Мин. народн. просвещения,
12 (1857) : 1—166 (отд. оттиск).
Павлов С. Д. 1965. О перспективах борьбы с гнусом в зоне Сибири и мерах по
массовой защите крупного рогатого скота. В кн.: Проблемы ветеринарной
санитарии, Тр. Всесоюзн. н.-иссл. инст. ветеринарн. санитар. 26 : 228—240.
Павлов С. Д. 1973. Химические методы борьбы с кровососущими двукрылыми
насекомыми в ветеринарии. В кн.: Одиннадцатое совещание по паразитологическим
проблемам. Тезисы докладов. Л. : 96—101.
Павлова Р. П. 1965. Влияние массовых обработок крупного рогатого скота поли-
хлорпиненом на численность и возрастной состав слепней. В кн.: Проблемы ве-
тери арной санитарии. Тр. Всесоюзн. н.-иссл. инст. ветеринарн. санитар.,
26 : 66—275.
Павлова Р. П. 1972. Слепни (Diptera, Tabanidae) Тюменской области. Автореф.
канд. дисс. М.
Павловский Е. Н. 1948. Руководство по паразитологии человека, 2. Л.
Павловский Е. Н., Благовещенский Л. И. и Алфеев Н. И.
1935. К фауне наружных паразитов животных в Кустанайском районе
(Северный Казахстан). В кн.: Вредители сельскохозяйственных животных. Л. : 229—
241.
Павловский Е. Н., Штейн А. К. и Олсуфьев Н. Г. 1935.
Экспериментальное исследование над действием слюны слепней на кожные покровы
человека. В кн.: Паразиты, переносчики и ядовитые животные. М. : 426—447.
112
ВВЕДЕНИЕ
П а е н к о Н. К. 1966. О методике определения физиологического возраста слепней
(Tabanidae, Diptera). Сборн. зоол. и паразитол. работ Воронежск. гос. унив.,
Воронеж : 91—97.
П а е н к о Н. К. 1968. Гонотрофический цикл и плодовитость слепней (Diptera,
Tabanidae) в условиях Средне-Русской лесостепи. Автореф. канд. дисс. Воронеж.
(П а л л а с М.) Pallas М. 1771—1776. Reise durch verschiedeae Provinzen des
Russischen Reiches. St.-Petersburg.
(Парамонов С.) Paramonov S. 1925. Zur Kenntnis der Insectenfauna von
Bessarabien und Ukraine. Soc. Entomologica, 6 : 21—23.
(Парамонове.) Paramonov S. 1926. Fragmente zur Kenntnis der Dipte-
renfauna Armeniens. Soc. Entomologica : 33—34, 38—39, 44, 46—47.
Парамонов С. Я. 1929. Диптеролопчт фрагменти. Тр. ф1зичн.-мат. Ввддшу
ВУАН, 13, 1 : 177-183.
Парамонов С. Я. 1933. Про деяш щкав! знахвдки двукрилыцв в Арменп.
Журн. Bio-золог. циклу ВУАН, 4 : 31—39.
П е г а с о в а Д. П. и Н о с к о в а В. А. 1961. О содержании гиалуронидазы
в слюнных железах и теле кровососущих двукрылых. Уч. зап. Горьковск. унив.,
21 : 157—163.
Петрова Е. Ф. 1953. К фауне слепней низовий реки Чу. Тр. Инст. зоол. АН
КазССР, 1 : 108-111.
Петрова Р. Г.11955. К изучению Слепней Московской и Астраханской областей
и влияние их паразитирования на организм животных. Автореф. канд. дисс. Л.
Петрова Р. Г. 1956. К изучению видового состава, сезонной и суточной активности
слепней в Астраханской области. Энтомол. обозр., 35, 2 : 359—370.
(Плеске Ф.) Р 1 е s k e Th. 1910a. Bescbreibung des noch unbekannten Mannchens
der Chrysops divaricatus Loew (Diptera, Tabanidae). Ann. Zool. Mus. Akad. Sci.,
15 : 430—435.
(Плеске Ф.) P 1 e s k e Th. 19106. Uber einige der genaueren Definition bediirftige
Chrysops-Arten aus dem palaearktischen Faungebiete (Diptera, Tabanidae). Ann.
Zool. Mus. Akad. Sci., 15 : 436—456.
(Плеске Ф.) Pleske Th. 1910b. Beschreibung einiger unbekannter palaearkti-
scher Chrysops-Arten (Diptera, Tabanidae). Ann. Zool. Mus. Akad. Sci., 15 : 457—
473.
Поляков В. А. 1965. Некоторые особенности биологии слепней Дудинского
района Таймырского национального округа. В кн.: Проблемы ветеринарной
санитарии. Тр. Всесоюзн. н-иссл. инст. ветеринарн. санитар., 26 : 351—357.
Поляков В. А. 1967. Слепни Таймыра и меры защиты от них северных оленей.
Автореф. канд. дисс. Омск.
Поляков В. А. 1968. Суточный ритм численности слепней, нападающих на
северного оленя. Докл. ВАСХНИЛ, 6 : 37—40.
Поляков В. А. 1971а. Фауна и фенология слепней центральной части Эвенкии-,
ского национального округа. В кн.: Проблемы ветеринарной санитарии. Тр.
Всесоюзн. н.-иссл. инст. ветеринарн. санитар., 39 : 320—325.
Поляков В. А: 19716. Результаты поисков кладок яиц слепней из рода Chrysops
на Таймырском полуострове. В кн.: Проблемы ветеринарной санитарии. Тр.
Всесоюзн. н.-иссл. инст. ветеринарн. санитар., 39 : 326—329.
Поляков В. А. 1971в. Видовой состав и численность слепней, нападающих на
северных оленей Таймырской тундры. Сиб. вестн. с.-х. науки, 3 : 77—81.
Поляков В. А. 1972. Слепни лесотундры как возможные носители
микроорганизмов. Ветеринария, 6 : 51—53.
Поляков В. А. 1974. Слепни севера Азиатской части СССР и защита от них
северных оленей. Магаданский зональный научно-исследовательский институт
сельского хозяйства северо-востока.
Попов В. В. 1962. О фауне слепней (Tabanidae) Тюменской области. Зоол. журн.,
41, 1 : 100—109.
(Порчинский И.) Portschinsky I. 1881. Diptera europaea et asiatica
nova aut minus cognita. II. Horae Soc. Ent. Ross., 16 : 273—284.
(Порчинский И.) Portschinsky I. 1887. Diptera europaea et asiatica
nova aut minus cognita. VI. Horae Soc. Ent. Ross., 21 : 176—200.
(Порчинский И. )Portschinsky I. 1892. D iptera europaea et asiatica nova
aut minus cognita. VII. Horae Soc. Ent. Ross., 26 : 201—227.
Порчинский И. А. 1899. Слепни (Tabanidae) и простейшие способы их
уничтожения. Тр. Бюро по энтомол., 2, 8. (6-е издание, 1915).
Порчинский И. 1901. Очерк фауны Валуйского уезда Воронежской губернии.
Харьков : 1—-18.
Потапов А. А. 1961. К методике определения численности слепней и дальности
их полета. Мед. паразитол. и паразит, болезни, 30, 1 : 51—54.
1 Она же Соболева Р. Г., см. дальше.
ЛИТЕРАТУРА
ИЗ
Потапов А. А. 1964. Автоматическая ловушка (АЛ-1) для определения суточного
хода численности слепней (Tabanidae) и мошек (Simuliidae). Мед. паразитол. и
паразит, болезни, 33, 2 : 201—203.
Потапов А. А. и В л адимиров Н. Н. 1962. Приборы для автоматического
включения ловушек и для фракционированного учета численности слепней.
Мед. паразитол. и паразит, болезни, 31, 1 : 109—114.
Потапов А. А. и Владимирова В. В. 1963. Численность и видовой состав
слепней (Tabanidae) на некоторых водохранилищах лесной зоны европейской
части СССР. Вопросы мед. паразитол., М. : 332—341.
Расницын СП. 1963. Обзор методов борьбы со слепнями. Мед. паразитол. и
паразит, болезни, 32, 5 : 611—616.
Растет'а ев Ю. М. 1965. Защита животных от гнуса в хозяйствах треста «Ското-
откорм» Тюменской области. В кн.: Проблемы ветеринарной санитарии. Тр.
Всесоюзн. н.-иссл. инст. ветеринарн. санитар., 26 : 332—334.
Растегаева К. С. 1956. Слепни Омской области и новый способ защиты скота от
их нападений. Тр. Томск, гос. унив. им. Куйбышева, 42 : 243—246.
Растегаева К. С. 1958. Слепни, меры борьбы с ними и опыт защиты
сельскохозяйственных животных от их нападения в Омской области. Тр. Омск. с.-х. инст.
им. С. М. Кирова, 23,-2 : 34—49.
Растегаева К. С. I960: Слепни Омской области и защита сельскохозяйственных
животных от их нападения. Зоол. журн., 39, 1 : 97—110.
Растегаева К. С. 1962. Слепни Омской области. Автореф. канд. дисс. Омск.
(Р е м м X.) Remm H. 1959. Eesti parmlaste okoloogiast. Entomoloogiline Kogumik,
1 : 181—188.
Родендорф Б. Б. 1964. Историческое развитие двукрылых насекомых. М.
Родионов И. М. 1936. Опыт выяснения роли, двукрылых насекомых в переносе
инфекционной анемии лошадей. Сов. ветеринария, 12 : 11—18.
Сазонова О. Н. 1949. О слепнях (Tabanidae) низовьев рек Оби и Иртыша. В кн.:
Вопросы краевой, общей и экспериментальной паразитологии, 6. М. : 52—58.
С а м к о К. 1929. О массовом появлении летом 1929 года в Тобольском округе
слепней. Бюлл. общества изучения края при Музее Тобольского севера. Тобольск,
4 : 31-34.
Сахибзадаев К. С. 1955. Слепни Северного Прикаспия и меры борьбы с ними.
Автореф. канд. дисс. Алма-Ата.
Сахибзадаев К. С. 1957. Слепни (Tabanidae) Северного Прикаспия. Тр. Инст.
зоол. АН Казахск. ССР, 7 : 95—120.
Сахибзадаев. К. С. 1958а. Материалы по биологии слепней (Tabanidae) в
Северном Прикаспии. Тр. Инст. зоол. АН Казахск. ССР, 8 : 215—221.
Сахибзадаев К. С. 19586. Численность слепней (Tabanidae) и распределение
их по биотопам в Северном Прикаспии. Тр. Инст! зоол. АН Казахск. ССР, 9 :
158-163.
Северцов Н. А. 1877. О зоологических (преимущественно орнитологических)
областях внетропических частей нашего материка. Изв. Русск. географ, общ.,
13, 3 : 125—155.
Семенов-Тян-ШанскийА. П. 1936. Пределы и зоогеографические
подразделения Палеарктической области для наземных сухопутных животных на
основании географического распределения жесткокрылых насекомых. М.—Л.
Сергеев А. Ф. 1962. Фауна слепней (Diptera, Tabanidae) Адыгеи. Зоол. журн.,
41, i ; 143—146.
Симбирцев Н. Л. 1965. Передвижные лагери как способ защиты животных.
В кн.: Проблемы ветеринарной санитарии. Тр. Всесоюзн. н.-иссл. инст.
ветеринарн. санитар., 26 : 287—296.
Синельщиков В. А. 1962. О слепнях среднего течения реки Иртыша. Тр. Инст.
зоол. АН Казахск. ССР, 18 : 241—253.
Синельщиков В. А. и Моторный И. А. 1970. О слепнях (Diptera,
Tabanidae) Молдавской ССР. В кн.: Паразиты животных и растений. Кишинев,
5 : 105—110.
С.иницын В. М. 1962. Палеогеография Азии. М.—Л.
Синтенис Ф. 1904. Diptera и Hymenoptera, собранные Канинской экспедицией.
Зап. Русск. геогр. общ. по общей географии, 41, 1 : 279—284,
Скуфьин К. В. 1938. К изучению фауны слегшей Карачаевской области. Изв.
Воронежск. гос. пед. инст., 2 : 73—80.
Скуфьин К. В. 1949а. К экологии слепней Воронежской области. Зоол. журн.,
28, 2 : 145—156.
Скуфьин К. В. 19496. Материалы по исследованию мест выплода слепней
(Tabanidae) в окрестностях Воронежа. Зоол. журн., 28, 6 : 527—534.
Скуфьин К. В. 1951а. Использование занефтеванных водоемов как метод
экологического исследования слепней и их истребления, Зоол. журн., 30, 2 : 130—139.
8 Н. Г. Олсуфьев
114 ВВЕДЕНИЕ
Скуфьин К. В. 19516. Опыт применения чучелообразной ловушки для слепней.
Зоол. журн., 30, 4 : 378—380.
Скуфьин К. В. 1952а. Слепни Воронежской области. Дисс. докт. Воронежский гос.
унив.
Скуфьин К. В. 19526. Экология пестряка реликтового Chrysops relictus Mg. Зоол.
журн., 31, 5 : 664-668.
Скуфьин К. В. 1953. О фауне слепней (Tabanidae) поймы Хопра в пределах
Воронежской области. Бюлл. Общ. естествоиспыт. при Воронежск. гос. унив.,
8 : 47-52. '
С к у ф ь и н^ К. В. 1956. Заметки по морфологии яиц и яйцекладок некоторых видов
слепней (Tabanidae) Воронежской области. Бюлл. Общ. естествоиспыт. при
Воронежск. гос. унив., 10 : 57—63.
Скуфьин К. В. 1958а. Экология нападения слепней на добычу. Тр. Воронежск.
гос. унив., 45, 1 : 55—64.
Скуфьин К. В. 19586. Качественная и количественная характеристика
распределения слепней по Воронежской области. Тр. Воронежск. гос. унив., 56, 2 : 3—18.
Скуфьин. К. В. 1959. К вопросу о гонотрофическом цикле слепней Tabanidae
(Diptera) в условиях окрестностей Воронежа. Бюлл. Воронежск. общ.
естествоиспыт., 9 : 85—88.
Скуфьин К. В. 1960а. О распределении слепней в Воронежской области и
химическом методе борьбы с ними. В кн.: Охрана природы центральной черноземной
полосы. Воронеж, 3 : 179—205.
Скуфьин К. В. 19606. К изучению фауны слепней (Tabanidae) северо-восточной
части Румынских Карпат и Прикарпатья. В кн.: Конференция по вивченню флори
i фауни Карпат та прилеглих територш. Киев : 32—36.
Скуфьин К. В. 1966а. Материалы по экологии слепней (Tabanidae, Diptera)
Воронежской области. Сб. зоол. и паразитол. работ Воронежск. гос. унив.:
2iZ—83.
Скуфьин К. В. 19666. О местах выплода слепней (Diptera, Tabanidae) в западной
части Московской области. Сб. зоол. и паразитол. работ Воронежск. гос. унив.:
Скуфьин К. В. 1967. Заметки по морфологии личинок некоторых видов слепней
(Tabanidae, Diptera). В кн.: Вредные и полезные насекомые. Воронеж : 168—206.
Скуфьин К. В. 1968. Стациальная и сезонная характеристика личинок слепней
(Diptera, Tabanidae) Воронежской области. Паразитол., 2, 2 : 97—104.
Скуфьин К. В. 1973. Методы сбора и изучения слепней. Методы паразитологи-
ческих исследований, 8. Л.
Скуфьин К. В. и Калита СР. 1959. К изучению фауны слепней
Кавказского государственного, заповедника. Тр. Кавказск. гос. заповеди., 5-: 207—
209.
Скуфьин К. В. и Паенко Н. К. 1967. О плодовитости и гонотрофическом
цикле дождевки Haematopota pluvialis L. (Diptera, Tabanidae). Паразитол.,
1, 6 : 512-518.
Скуфьин К. В.иПаенкоН. К. 1971. О плодовитости и гонотрофическом цикле
пестряка Chrysops relictus Mg. (Diptera, Tabanidae). В кн.: Вопросы.зоологии и
физиологии. Тр. Воронежск. гос. унив., 93 : 3—13.
Смирнов Е. С. 1923. К строению мужской половой системы двукрылых
насекомых. Биол. известия, 1 : 39—56.
Соболева Р. Г. 1962. Видовой состав и фенология слепней на юге Приморского
края. В кн.: Проблемы ветеринарной санитарии. Тр. Всесоюзн. н.-иссл. инст.
ветеринарн. санитар., 20 : 138—146.
Соболева Р. Г. 1965а. Токсичность слюны слепней для организма животных.
Зоол. журн., 44, 3 : 396—402.
Соболева Р. Г. 19656. Изучение экологии наиболее массовых видов слепней
юга Приморского края. Тезисы докл. на сессии Совета Дальневосточного
филиала Сибирск. отделения АН СССР по итогам научных исследований 1964 года.
Владивосток : 18—21.
Соболева Р. Г. 1965в. Слепни юга Приморья — эктопаразиты крупного рогатого
скота и борьба с ними. В кн.: Ученые — сельскому хозяйству Дальнего Востока.
Владивосток : 99—107.
Соболева Р. Г. 1968а. Гонотрофический цикл, плодовитость и яйцекладка у
некоторых видов слепней Tabanidae на юге Приморского края. В кн.: Фауна и эко^
логия насекомых Дальнего Востока. Владивосток : 126—146.
Соболева Р. Г. 19686. О морфологии и биологии некоторых редких и
малоизвестных видов слепней (сем. Tabanidae) Приморского края. В кн.: Фауна и экология
насекомых Дальнего Востока. Владивосток : 161—169.
Соболева Р. Г. 1968в. Описание самца Hybomitra stigmoptera N. Ols. (Tabanidae)
из Приморского края. В кн.: Фауна и экология насекомых Дальнего Востока.
Владивосток : 170—171.
ЛИТЕРАТУРА
115
Соболева Р. Г. 1970а. Ранее неизвестные личинки и куколки некоторых видов
слепней рода Tabanus (Diptera, Tabanidae) из Приморского края. Паразитол., 4,
3 : 250-260.
Соболева Р. Г. 19706. Морфология личинок и куколок слепней рода Hybomitra
(Diptera, Tabanidae) из Приморского края. В кн.: Энтомологические
исследования на Дальнем Востоке, Тр. Биолого-почв. инст. Дальневосточ. фил. АН СССР,
2 : 213—226.
Соболева Р. Г. 1970в. Морфология личинки и куколки Chrysops suavis Lw.
(Diptera, Tabanidae) из Приморского края. В кн.: Энтомологические исследования
на Дальнем Востоке. Тр. Биолого-почв. инст. Дальневосточ. фил. АН СССР,
2 : 227—230.
Соболева Р. Г. 1970г. Биология слепня Hybomitra brevis Lw. (Diptera, Tabanidae)
на юге Приморского края'. Энтомол. обозр., 49, 3 : 545—554.
Соболева Р. Г. 1971. Описание личинок пяти видов слепней (Diptera, Tabanidae)
из Приморского края. Паразитол., 5, 2 : 140—146.
Соболева Р. Г. 1974. Слепни юга Приморского края. Новосибирск.
Соболева Р. Г. и Кадырова М. К. 1972. Морфология ранее неизвестных
куколок некоторых видов слепней (Diptera, Tabanidae) Узбекистана. В кн.:
Паразитические членистоногие Ферганской долины. Ташкент : 42—51.
Сомов П. В., Романова В. П. и Данилова М.И. 1939. Роль
кровососущих двукрылых в эпидемиологии туляремии. Тр. Всесоюзн. конфер. микробиол.,
эпидемиол. и инфекционистов, М.—Л. (1940) : 272—274.
Сперанская В. Н., Тальдрик А. А., Гусев В. Ф., Озеров В. Г.,
Хисматуллина Р. Н., Костенко Л. А., Малышев М. А.,
Маслий Я. К. и Качанова А. П. 1965. Опыт защиты крупного
рогатого скота от гнуса в условиях Ленинградской области. В кн.: Проблемы
ветеринарной санитарии. Тр. Всесоюзн. н.-иссл. инст. ветеринарн. санитар., 26 :
322—327.
Старобогатов Я. И. 1970. Фауна моллюсков и зоогеографическое
районирование континентальных водоемов земного шара. Л.
Страхов Н. М. 1938. Историческая геология. М.
Сурков B.C., Тимофеева Н.С. и С у ч к о в а Н. Г. 1972. К вопросу о
природной очаговости эризипелоида на Сахалине. Журн- микробиол., эпидемиол.
и иммунобиол., 7 : 3—5.
С у ш к и н П. П. 1925. Зоологические области средней Сибири и ближайших частей
нагорной Азии и опыт истории современной фауны Палеарктической Азии.
Бюлл. Моск. общ. испыт. природы, отд. биол., нов. серия, 34, вып. (поев. М. А.
Мензбиру), М.
Тальдрик А. А. 1965. Результаты испытания карбоксита и гексамида для
защиты крупного скота от слепней, мокрецов, мух-жигалок в хозяйствах
Тихвинского района Ленинградской области. В кн.: Проблемы ветеринарной санитарии,
Всесоюзн. н.-иссл. инст. ветеринарн. санитар., 26 : 319—321.
Тамарина Н. А. 1951. К биологии слепня-златоглазика Chrysops relictus Mg.
Вестн. Московск. гос. унив., 6 : 101—108.
Тамарина Н. А. 1956. Морфология личинок и куколок слепней-златоглазиков
Chrysops relictus Mg. и Chrysops rufipes Mg. Тр. Всесоюзн. энтомол. общ., 45 :
167—192.
Таскаева Е. 3. 1951. К фауне слепней Троицкого лесостепного заповедника.
Изв. ест.-науч. инст. при Пермском гос. унив. им. А. М. Горького, 13, 2—3 :
157—167.
Тертерян А. Е. 1954. Сравнительная эффективность метода непосредственного
сбора кровососущих слепней (Diptera, Tabanidae) с разных участков тела
животного. Изв. АН Армянск. ССР, 7, 7 : 71—78.
(Тертерян А. Е.) Terterjan А. Е. 1960. Zur Okologie der Larven und Pup-
pen einiger in Armenien verbreiteter Tabaniden. XI. Intern. Kongress f. Ento-
mologie, B, 1 : 192—193.
Тертерян А. Е. 1969. Chrysops buxtoni Aust., новый вид слепня (Diptera,
Tabanidae) для фауны СССР из Армянской ССР. Биолог, журн. Армении, АН Армянск.
ССР, 22, 3 : 77—79.
Тертерян А. Е. 1972. К морфологии гениталий самцов слепней (Diptera,
Tabanidae) Армянской ССР. Энтомол, обозр., 51, 3 : 573—560.
Тупикова Н. В. 1969. Зоологическое картографирование. М.
Туров И. С. 1953. О роли слепней в биологии лося. Зоол. журн., 32, 5 : 886—892.
Федоров В. Г. 1971. О слепнях лесостепных предгорий и низкогорья Салаира,
В кн. : Вопросы инфекционной патологии, природноочаговые болезни. Омск ;
280—284.
Федченко Б. 1892. Diptera. В : I. Dwigubsky. Primitiae Faunae Mosquensis,
2-е изд. М. : 83—99 (Tabanidae : 88).
8*
116
ВВЕДЕНИЕ
Фоминых В. Г., Лужкова А. Г., Панков а Т. Ф. и Вечер Л. Ф.
1972. Некоторые вопросы фауны и биологии слепней Томской области. В кн.:
Зоологические проблемы Сибири, Новосибирск: 199—200.
Ф р и д о л и н В. Ю. 1936. Животно-растительное сообщество горной страны
Хибин. Тр. Кольск. базы им. С. М. Кирова, 3 : 1—295.
Харитонова Н. Я. 1939. К вопросу об эктопаразитах домашнего скота в районе
Оханска и Перми. Уч. зап. Пермск. гос. пед. инст., Пермь, 4 : 1—17.
Чарыкулиев Д. 1967. К изучению слепней [Diptera, Tabanidae) Туркмении.
Изв. АН Туркмен. ССР, серия биолог, наук, 4 : 73—79.
Черепанов А. И. 1966. Система мероприятий по борьбе с гнусом. В кн.:
Биологические основы борьбы с гнусом в бассейне Оби. Новосибирск : 238—268.
Чиров П. А. 1968а. Слепни (Diptera, Tabanidae) Чуйской долины и северных
склонов Киргизского Ала-тоо. Паразитол., 2, 1 : 27—32.
Чиров П. А. 19686. Слепни (Diptera, Tabanidae) Киргизии. Автореф. канд.
дисс. Фрунзе.
Ч у п и с Т. С. 1947. До вивчення гедз!в в л1вобережно! Украши. Зб1рник праць з
паразитол., 1 : 143—144.
Ч у п и с Т. С. 1954. Бюлоия личинок i лялячек гедз!в Л1вобережжя УРСР. 36ip-
ник праць зоолопчного музею, 26 : 30—39.
Шавкунова Т. Г. 1953. Новые для окрестностей Томска виды слепней. Заметки
по фауне и флоре Сибири, 17, Томск : 53—54.
Шакирзянова М. С. 1958. Материалы по фауне кровососущих двукрылых
Северного Прикаспия. Тр. Инст. зоол. АН Каз. ССР, 8 : 197—208.
Шаров А. Г. 1956. Северные представители двукрылых в энтомофауне
высокогорного Кавказа. Зоол. журн., 35, 10 : 1514—1516.
Шатько П. Д. 1952. Роль слепней как переносчиков эмфизематозного карбункула.
Ветеринария, 7 : 28..
Шевченко В. В. 1953а. Слепни бассейна реки Джабаглы (государственный
заповедник Аксу-Джабаглы в Западном Тянь-Шане). Тр. Инст. зоол. АН Ка-
захск. ССР, 2 : 145—156.
Шевченко В. В. 19536. Материалы по фауне и экологии слепней Таласского
Ала-Тау и Кара-Тау. Бюлл. Моск. общ. испыт. природы, отдел биологии, 58,
1 : 18—34.
Шевченко В. В. 1955. Новые данные о слепнях (Tabanidae) Западного Тянь-Шаня.
Тр. Инст. зоол. АН Казахск. ССР, 3 : 47—53.
Шевченко В. В. 1960. О таксономическом значении особенностей строения
гениталий некоторых палеарктических видов слепней подсемейства Chrysopinae
(Diptera, Tabanidae). Тр. Инст. зоол. АН Казахск. ССР, 11 : 157—172.
Шевченко В. В. 1961. Слепни Казахстана. Алма-Ата.
. Ш (в ч е н к о В. В. 1962. О гениталиях некоторых палеарктических дождевок
(Chrysozona, Tabanidae, Diptera). Тр. Инст. зоол. АН Казахск. ССР, 18 : 224—
234.
Штакельберг А. А. 1916. К диптерофауне окрестностей Нижней Бронной
Петергофского уезда. Русск. энтомол. обозр., 16, 3—4 : 299—307.
Штакельберг А. А. 1922. К диптерофауне Черниговской губернии. Ежегодн.
Зоол. музея Российск. АН, 23 : 404—410.
Штакельберг А. А. 1926а. Материалы для определения кровососущих
двукрылых. 2.Определитель семейств и родов по взрослой фазе. Вестн. микробиол. и
эпидемиол., 5, 1—2 : 43—56. •
Штакельберг А. А. 19266. К диптерофауне Армении. Русск. энтомол. обозр.,
20 : 65—68.
(Штакельберг A. )Stackelberg A. 1926b. Notes on the genus Chrysops.
Bull. Ent, Res., 16, 4 : 325—328.
Штакельберг A. A. 1951. Двукрылые. В кн.: Ущелье Кондара. М.—Л. : 129—
148.
Штакельберг А. А. 1954. Материалы по фауне двукрылых Ленинградской
области. Т. II, Diptera, Brachycera. Тр. Зоол. инст. АН СССР, 15 : 199—228.
Штакельберг А. А. и Тертерян А. Е. 1953. О морфологическом
строении придатков полового аппарата самок слепней. Докл. АН Армянской ССР,
16, 2 : 53-64.
Штегман Б. К. 1938. Основы орнитологического деления Палеарктики. Фауна
СССР, Птицы, I, вып. 2, Изд. АН СССР, М.—Л.
Штейнгель Ф. 1897. О принятии пищи у Tabanus. Работы лаборатории Зоолог,
кабинета Варшавск. унив. 1896 года. Варшава (отд. оттиск : 1—8).
Щербина В. П. 1972. Слепни (Diptera, Tabanidae) Северного Кавказа. В кн.:
Чтения памяти Николая Александровича Холодковского 1970 г. Л. : 93—118.
Ягодинский В. Н. 1962. Случай выделения вируса из группы клещевого
энцефалита от слепней. Мед. паразитол. и паразит, болезни, 31, 2 : 19.
ЛИТЕРАТУРА ' 117
Янович Г. И. 1958. Опыт борьбы с двукрылыми кровососущими насекомыми.
Ветеринария, 6 : 49—50.
Ярошевский В. 1876. Список двукрылых насекомых, собранных
преимущественно в Харькове и его окрестностях. Тр. Общ. испыт. природы при Харьков,
унив., 10 : 1—150.
Ярошевский В. 1877. Дополнение к списку двукрылых насекомых Харькова и его
окрестностей с указанием распространения их в пределах Европейской России.
Тр. Общ. испыт. природы при Харьков, унив., 11 (отд. оттиск : 1—138). ■
Abbassian-LintzenR. 1960a. Tabanidae (Diptera) of Iran. II. Records of
horseflies in the Kazeroon area (province of Fars, Southern Iran). Arch. Inst. Razi,
12 : 25—37.
Abbassian-LintzenR. 1960b. Tabanidae (Diptera) of Iran. V. Description of
three new subspecies of the genus Haematopota. Bull. Soc. Path. Exotique, 53, 5 :
824—827.
Abbassian-LintzenR. 1961. Tabanidae (Diptera) of Iran. VI. Records of
horseflies from Southeast Iran (Iranian Baluchistan and the Jiroft area). Bull. Soc. Path.
Exotique, 54, 1 : 128—147.
Abbassian-LintzenR. 1963. Tabanidae (Diptera) of Iran. XI. On the horsefly
fauna of Khuzistan (Southwest Iran). Bull. Soc. Path. Exotique, 56, 1 : 77—88.
Abbassian-Lintzen R. 1964a. Tabanidae (Diptera) of Iran. X. List, keys and
distribution of species occuring in Iran. Annales Parasitol. Hum. compar., 39, 3 :
285—327.
Abbassian-Lintzen R. 1964b. Tabanidae (Diptera) of Iran. IX. Horseflies
from Azerbaijan (N. W. Iran). Bull. Soc. Path. Exotique, 57, 1 : 164—173.
Austen E. 1919—1920. A contribution to knowledge of the Tabanidae of Palestina.
Bull. Ent. Res., 10 : 277—321.
Becker Th. 1900. Beitrage zur Dipteren-Fauna Sibiriens. Acta Soc. Sc. Fenn., 26,
9 : 1—66.
Becker Th. 1912. Persische Dipteren. Ann. Mus. Zool. St. Petersb., 17 : 503—654.
Bequaert J. 1924. Notes upon Surcouf's treatment of the Tabanidae in the genera
insectorum and upon Enderlein's proposed new classification on this family.
Psyche, Boston : 24—40.
Bequaert J. 1930. Entomology. Tabanidae. The African Republic of Liberia and
the Belgian Congo, 2 : 858—971 (Harvard Univ. Press).
Bigot J. 1880. Dipteres nouveaux ou peu connus. XX. Quelques Dipteres de Perse
et du Caucase. Ann. Soc. Ent. France, 10 : 139—154.
Bigot J. 1892. Description de Dipteres nouveaux. Mem. Soc. Zool. France, 5 : 602—
691.
В r a u e r F. 1880. Die Tab anus-Aiten der Europaischen, Mediterranen und Sibirischen
Subregionen. Denkschr. d. Mat.-Naturwissensch. Classe d. Kais. Akad. d. Wissen-
schaften, Wien, 42 : 119—216.
Bruce W. and Decker G. 1951. Tabanid control on dairy and beef catties with
synergized pyrethrins. J. Econ. Entpm., 44, 2. .
Cameron A. 1926. Bionomics of the Tabanidae (Diptera) of the Canadian prairie.
Bull. Entomol. Research, 17 : 1—42.
Cameron A. 1934. The life-history and structure of Haematopota pluvialis L. Trans.
Roy. Soc. Edinb., 58, 10 : 211—250.
С a t t s E. P. 1970. A canopy trap for collecting Tabanidae. Mosquito News, 30, 3:472 —
474. '
Cederhielm I. 1798. Faunae Ingricae prodromus.
Chen S. H. and Q u о F. 1949. On the opistacanthous Tabanidae of China. Chinese
Journ. Zool., 3 : 1—10.
С h e n f u F. a. W u M. 1940. Catalog insectorum Sinensinium, order Diptera,
Tabanidae, V. Peiping : 173—192.
С h v a,l a M. „1964. Some new or little known Tabanidae (Diptera) from Szechoslovakia.
Casopis Ceskoslovenske Spolecnosti Entomologicke, ©1, 4 : 374—383.
С h v a 1 a M. and Jezek J. 1969. Immature stages of 5 European Hybomitra
species of the bimaculata- and montana-gTowps (Diptera, Tabanidae). Folia Para-
sitologia (Praha), 16 : 329—347.
С h v a 1 a M. and Lyneborg L. 1970. A revision of Palearctic Tabanidae (Diptera)
described by J. С Fabric.ius. J. Med. Entom., 7 (5) : 543—555.
С h v a 1 a M., Lyneborg L. and M о u с h a J. 1972. The horse flies of Europe
(Diptera, Tabanidae). Copenhagen.
Chvala M. und M ouch a J. 1971. Zur Taxohomie • von Hybomitra nitidifrons
(Szilady, 1914) (Diptera, Tabanidae). Notulae entomolog., 51 : 109—112.
Ciurea I., Seguy E. et Stefanescu T. 1937. Sur une espece de Taon
. (Sziladynus solstitialis var. ciureai, nov. Seguy 1937) qui attaque impetueusement
118 ВВЕДЕНИЕ
l'homme et les animaux dans le delta du Danube. Arch. Roum. Pathol. Exper.,
10, 3 : 205-209.
Collin J. 1940. A note on certain structure distinction between the females of Theri-
oplectes tropicus L., solstitialis Mg. and distinguendus Verr. Entomol. Monthly Ma-
gaz., 76, 4 ser., 8 (915) : 178—179.
С о n n a 1 A. and С о n n a 1 S. 1922. The development of Loa loa (Guyot) in Chrysops
silacea (Aust.) and in Chrysops dimidiata (van der Wulp). Trans. Roy. Soc. Trop.
Med. Hyg., 16, 1—2 : 64-89.
D a v i e s D. M., Golini V. I. and Raastad J. E. 1971. Observations on
some Scandinavian Tabanidae (Diptera). Norsk Entomol. Tidsskrift, 18 : 113—
117.
Dinulescu Gh. 1958. Diptera Tabanidae. Fauna Republicii Populare Romine, In-
secta, 11, 2.
Efflatoun-Bey H. 1930. A monograph of Egyptian Diptera. Part III. Family
Tabanidae. Mem. Soc. Ent. d'Egypte, 4, 1.
Enderlein G. 1922. Ein neues Tabanidensystem. Mitt. Zool. Mus. Berlin, 10, 2 :
335—351.
Enderlein G. 1925. Studien an blutsaugenden Insekten. 1. Grundlagen eines neuen
Systems der Tabaniden. Mitt. Zool. Mus. Berlin, 11, 2 : 255—409.
Enderlein G. 1933. Entomologische Ergebnisse der Deutsch-Russischen Alai-
Pamir-Expedition, 1928. I. Diptera. Deutsche Entomol. Zeitschr., 2/3 : 144—145.
Erichson 1851. Tabanus astur in Middendorff's «Sibirische Reise», 2, 1 : 66.
Eversmann E. 1834. Diptera Wolgam fluvium inter et montes Uralenses obser-
vata. Bull. Soc. Natur, Moscou, 7 : 420—432.
Fairchild G. B. 1961. The Adolpho Lutz collection of Tabanidae (Diptera). Mem.
Inst. Oswaldo Cruz, 59, 2 : 185—249.
Fairchild G. B. 1969. Climate and the phylogeny and distribution of Tabanidae.
Bull. Entom. Soc. America, 15, 1 : 7—11.
Frey R. 1911. Zur Kenntnis der Dipterenfauna Finnlands. Acta Soc. pro Fauna et
Flora Fennica, 34, 6 : 1—57.
Frey R. 1915. Diptera Brachycera aus der arktischen Kiistengegenden Sibiriens. Mem.
l'Acad. Sc. Russ., ser. VIII, 29, 10 : 1—35.
Gimmerthal R. 1847. Dritter Beitrag zur Dipterologie Russlands. Bull. Soc.
Nat. Moscou : 175—223.
Hoare C. et Coutelen F. 1933. Essai de classification des trypanosomes des
mammiferes et de l'homme basee sur leurs caracteres morphologiques. Ann. Para-
sit. Hum. сотр., 11 : 196—200.
Hummel A. 1821—1829. Essais Entomologiques. I—VII.
Isaac P. 1924. Papers on Indian Tabanidae. Mem. Depart. Agric. India, Entom. Series,
8, 5 : 53—62.
Isaac P. 1925. Papers on Indian Tabanidae. Mem. Depart. Agric. India, Entom.
Series, 9, 10 : 93—109.
Jenkins D. W. 1960. Pathogens, parasites and predators of medically important
insects. Biological control of insects of medical importance. Technical Report,
Washington : 6—16.
I ezek J. 1970. Larvae and pupae of four European Chrysops species (Diptera,
Tabanidae). Acta entom. bohemosl., 67, 6 : 375—383.
I ezek J. 1971. Larven und Puppen der Art Heptatoma pellucens (Fabr.) und vier euro-
paischer Arten der Gattung Haemaiopota Meig. (Diptera, Tabanidae). Acta entom.
bohemosl., 68, 5 : 341—351.
Kauri H. 1951. Bemerkungen iiber schwedische Tabaniden. Opuscula Entomologica,
16 : 97—109.
Kauri H. 1954. Bemerkungen iiber schwedische Tabaniden. II. Opuscula Entomologica,
19 : 239—244. ..
Kauri H. 1958. Uber zwei fur Europa und Nord-America gemeinsame Tabaniden-
Arten. Opuscula Entomologica, 23 : 95—104.
Kauri H. 1968. Uber die norwegischen Tabaniden (Diptera). Norsk Entomol.
Tidsskrift, 15, 1 : 63—64.
Kertesz С 1908. Catalogue Dipterorum. Budapest, 3 (Tabanidae : 146—293).
К r 6 b e r O. 1920. Die Chrysops-Aiten der palaearktischen Region nebst den Arten der
angrenzenden Gebiete. Zool. Jahrbiicher, Abt. Syst., 43 : 41—160.
К r 6 b e r O. 1921. Die palaearktischen Arten der Gattung Pangonia Latr. Arch. f. Na-
turg., 87, A, 1 : 1—67.
К r 6 b e r O. 1922. Beitrage zur Kenntnis palaearktischer Tabanidae. (Teil I: Surcoufia,
Heptatoma, Silviusu. Chrysozona). Arch. f. Naturg., 88, A, 8 : 114—164.
К r 6 b e r O. 1923. Beitrage zur Kenntnis palaearktischer Tabaniden. II Teil: die
Untergattung Therioplectes Zell. nebst Bemerkungen zu den mir bekannt geworde-
nen Bigotschen Tabaniden Typen der Collectionen Mr Collin's in Neumarket. Arch.
f. Naturg., 89, A, 12 : 55—118.
ЛИТЕРАТУРА П9
К r 6 b e r O.. 1924. Beitrage zur Kenntnis palaearktischer Tabaniden. (Teil III: Ochrops,
Atylotus, Tabanus s. str., Baikalia, Isshikia). Arch. f. Naturg., 90, A, 9 : 1 —195.
К r 6 b e r O. 1925. Tabanidae in Lindner: Die Fliegen der Palaearktischen Region.
Lief. 5-8.
К r 6 b e r O. 1928. Neue palaearktische Tabaniden. Zool. Anzeiger, 76 : 261—272.
К r 6 b e r O. 1938. Katalog der palaearktischen Tabaniden nebst Bestimmungstabellen
und Zusatzen zu einze'lnen Arten sowie Neubeschreibungen. Acta Inst, et Musei
Zool. Univers. Atheniensis, 2, 3/4 : 57—245.
Lecaillon A. 1905. Sur la ponte des oeufs et la vie larvaire des Tabanides, particu-
- lierement du taon a quatre tSches (Tabanus quatuornotatus Mg.). Ann. Soc. Ent.
France: 20—28.
Leclercq M. 1952. Introduction a l'etude des Tabanides et revision des especes
de Belgique. Memoires de l'lnstitut Royal des sciences naturelles de Belgique,
123:1—80.
Leclercq M. 1957. Revision systematique et biogeographique des Tabanidae
(Dipt.) de France. Annales Parasitol. Hum. compar., 32, 4 : 303—425.
Leclercq M. 1960a. Revision des Nemorius Rondani (Dipt., Tabanidae). Nemorius
abbassianae n. sp. Bull. Ann. Soc. Ent. Belg., 96, 1—2 : 32—36.
Leclercq M. 1960b. Revision systematique et biogeographique des Tabanidae
(Diptera) Palearctiques. I. Pangoniinae et Chrysopinae. Mem. Inst. Roy. Sci. Nat.
Belgique, Deuxieme ser. Fasc, 63.
Leclercq M. 1966. Revision systematique et biogeographique des Tabanidae
(Diptera) Palearctiques. II. Tabaninae. Mem. Inst. Roy. Sci. Nat. Belgique, Deuxieme
ser., Fasc, 80.
Leclercq M. 1967. Tabanidae (Diptera) des Pays-Bas. Zoolog. Bijdragen, 9 : 3—34.
Leclercq M. et Olsufjev N. G. 1975. Catalogue des Tabanidae (Diptera)
Palearctiques. Bull.. Ann. Soc. r. beige Ent., Ill : 25—36.
Liu Wi-teh 1958. Uber die Chrysozona Bremsen aus China. Acta Zool. Sinica, 10,
2 : 151—160.
Liu Wi-teh 1959a. Preliminary notes on the Tabanid flies from North-West China.
Acta Zool. Sinica, 11, 2 : 158—170.
Liu Wi-teh 1959b. On the subgenus Ochrops (gen. Tabanus, Tabanidae, Diptera) from
China. Acta Entomol. Sinica, 9 : 388—391.
L о e w H. 1858a. Zur Kenntnis der europaischen Tabanus-Arten. Verh. Zool.-bot. Ge-
sell. Wien, 8 : 573—612.
L о e w H. 1858b. Versuch einer Aneinandersetzung der europaischen Chrysops-Arten.
Verh. Zool.-bot. Gesell. Wien, 8 : 613—634.
L о e w H. 1860. Dipterenfaunas Siid-Afrikas : 14, 31 (Berlin).
L о e w H. 1871. Beschreibungen europaischer Dipteren. Halle, 2 : 61.
L о e w H. 1874. Diptera nova a Hug. Theod. Christopho collecta. Zeitschr. Ges.
Naturw., 43 : 413—420.
Lundbeck W. 1907. Diptera Danica, 1. Copenhagen.
Lutz A. 1909. Tabaniden Brasiliens und einiger Nachbarstaaten. Zool. Jb:, Suppl.,
10 : 619—692.
Lyneborg L. 1959. A revision of the Danish species of Hybomitra End. (Dipt.,
Tabanidae). Entom. Medd., 29 : 78—150.
Lyneborg L. 1961. On Tabanus tropicus and other Linnean species of Palaearctic
Tabanidae (Diptera). Entom. Medd., 31 : 97—103.
Lyneborg L. and Chvala M. 1970. Revision of Haematopota Meig. in North
Europe (Dipt., Tabanidae). Ent. Scand., 1 : 30—40.
Mackerras I. 1954. The classification and distribution of Tabanidae (Diptera).
I. General review. Austral. J. Zool., 2, 3 : 431—454.
Mackerras I. 1955. The classification and distribution of Tabanidae (Diptera).
II. History; Morphology; Classification; Subfamily Pangoniinae. Austral. J. Zool.,
3, 3 : 439—511. • .
Mackerras I. 1959. Distributional patterns of the Tabanidae (Diptera). XV Intern.
Congress of Zool., 48 : 1—4.
Mackerras I. and Fuller M. 1942. The genus Pelecorhynchus (Diptera, Tabanoi-
dea). Proceed. Linnean Soc. N. S. Wales, 67, 1—2 : 9—76.
Marchand W. 1920. The early stages of Tabanidae. Monogr. Rockefeller Inst. Med.
Res., 13, New York.
McAlpine J. 1961. Variation, distribution and evolution of the Tabanus (Hybomitra)
frontalis complex of horse flies (Diptera, Tabanidae). Canad. Entomol., 93 : 894—
924.
Meigen J.W. 1820. Systematische Beschreibung der bekannten europaischen zwei-
fliigeligen Insekten, 2. Aachen.
Mitzmain M. 1913. The mechanical transmission of surra by Tabanus striatus
F. Philipp. J. Sci., Ser. В., 8, 3 : 223—229.
120 ВВЕДЕНИЕ
М i t z m a i n M. 1914. Insect transmission of anthrax. J. Trop. Med. Hyg., 17, 4 : 61
(abstract).
Morris H. 1918. Blood-sucking insects as transmitters of anthrax or charbon.
Louisiana St. Univ. Agric. St., Bull. 163 : 1—15.
M о u с h a J. 1959. Zur Kenntnis der Tabaniden (Diptera) der Kaukasuslander. Acta
Soc. ent. Ceschosloveniae, 56 : 129—136.
M о u с h a J. 1969. Die Gattungen und Untergattungen der Tabanidae. Angew. Parasi-
tol., 10, 3 : 170—184.
M о u с h a J. 1971. Type-specimens of Palaearctic Tabanidae (Diptera) in the British
Museum (Natural History). Acta faun, entom. mus. nat. Pragae, 14 : 21—31.
Moucha J. a Chvala M. 1955. Revise ovadovitych (Dipt., Tabanidae) Ces-
koslovenska. I. cast: Chrysopinae. Zool. a. Entomolog. Listy, 4 (18), 3 : 227—238.
Moucha J. a Chvala M. 1956. Revise ovadovitych (Dipt., tabanidae) Ces-
koslovenska. II. cast. Haematopotinae. Zoolog. Listy, 5 (19), 3 : 259—270.
Moucha J. und Chvala M. 1957. Beitrag zur Kenntnis der Gattung Chrysops.
nebst Beschreibung einer neuen Untergattung Pseudochrysops n. subgen. (Diptera,
Tabanidae): Acta entom. mus. nat. Pragae, 31 : 159—162.
Moucha J. a Chvala M. 1958a. Revise ovadovitych (Dipt., Tabanidae) Cesko-
slovenska. Ill, cast: Tabaninae: Tabanus s. str. Zoolog. Listy, 7 (21), 2 : 182—198.
Moucha J. und Chvala M. 1958b. Zur Kenntnis der Gattung Theriorlectes Zel-
ler 1842 (Diptera, Tabanidae). Acta entom. mus. nat. Prage, 32 : 571—582.
Mouc.lia J. а С h v a 1 a M. 1959a. Revise ovadovitych (Dipt., Tabanidae}
Ceskoslovenska. IV. cast: Tabaninae 2. Zoolog. Listy, 8 (22), 2 : 161—174.
Moucha J. und Chvala M. 1959b. Die Tabaniden der Afghanistan-Expedition
(1952—1953) J. Klapperich (Diptera, Tabanidae). Acta entom. mus. nat. Pragae,
33 : 273-286.
Moucha J. und Chvala vM. 1959c. Revision der Gattung Nemorius Rond.
(Diptera, Tabanidae). Casopis Ceskoslovenske Spolecnosti Entomologicke, 56, 2 : 137—
141.
Moucha J. and Chvala M. 1963. Tabanidae (Diptera) collected by Dr. K. Lind-
berg in Afghanistan. Acta entom. mus. nat. Pragae, 35 : 521—525.
Moucha J. and Chvala M. 19v64. Notes on the genus Therioplectes Zeller, 1842
(Diptera, Tabanidae). Casopis Ceskoslovenske Spolecnosti Entomologicke, 61,.
2 : 100-105.
Moucha J. und Chvala M. 1965. Bemerkungen zur Kenntnis der Asiatischen
Hybomitra-Arlen (Diptera, Tabanidae). Acta entom. mus. nat. Pragae, 36 : 335—338.
Moucha J. und Chvala M. 1967a. Beschreibung des Mannchens von Haematopota
scutellata nebst Bemerkungen uber die Gattung Haematopota Meig. in der Tsche-
choslowakei (Diptera, Tabanidae). Acta entomol. bohemoslov. 64, 3 : 224—231.
Moucha J. und Chvala M. 1967b. Beitrage zur Kenntnis der Fauna Afganistans..
Tabanidae, Diptera. Casopis Moravskeho Musea, 52, Suppl. : 183—186.
Moucha J. und Chvala M. 1968'. Die Gattung Hybomitra Enderlein, 1922 in
der Tschechoslowakei. Acta faun, entom. mus. nat. Pragae, 12 : 263—294.
Moucha J. und Chvala M. 1969a. Zur Kenntnis der Bremsengattung A tylotus
Osten-Sacken in der Tschechoslowakei (Diptera, Tabanidae). Acta entom.
bohemoslov. 66, 5 : 321—329.
Moucha J. und Chvala M. 1969b. Ergebnisse der zoologischen Forschungen-
von Dr Z. Kaszab in der Mongolei Nr. 156 Diptera: Tabanidae. Acta entom. mus.
nat. Pragae, 38 : 199—205.
Murdoch W. and Takahasi H. 1969. The female Tabanidae of Japan, Korea
and Manchuria. Mem. Entom. Soc. Washington, 6.
N e a v e S. 1912. Notes on the blood-sucking insects of eastern tropical Africa. Bull..
Ent. Res., 3 : 275—323.
Nieschulz O. 1927. Uber die Lebensdauer der Tabaniden. Zbl. Bakt. I. Orig.,
103 : 6-8. .. .. . ■ .
Nieschulz O. 1929. Uber die mechanische Ubertragung voneinigen Bakterienkrank-
heiten durch blutsaugende Insecten. Arch. Schiffs-Tropenhyg., 33, 3 : 282—287.
Nieschulz O. 1935. Uber die Larvenstadien von Tabanus rubidus Wd. und Tabanus-
striatus F. Ztschr. Parasitenk., 7, 8 : 639—656.
Nieschulz O. und Kraneveld F. 1929. Experimented Untersuchungen iiber
die Ubertragung der Biiffelseuche durch Insekten. Zbl. Bakt., I. Orig., 113 :
403-417.
N 6 11 e r W. 1925. Der Nachweis des Ubertragers des gemeinen Rindertrypanosoma
Trypanosoma theileri, mit Hilfe des Kulturverfahrens. Zbl. Bakt. Infect. Paras. I»
Abt. Ref., 79 : 133—142.
OldroydH. 1957. The horse flies of the Ethiopian Region, 3. London.
О u с h i У. 1943. Diptera Sinica. Tabanidae IV. Notes on some Tabanid flies belonging
to the subfamilies Tabaninae and Bellardiinae from East China. Shanghai
Sizenkagaku Kenkyusyo Iho, 13 (16) : 505—552.
ЛИТЕРАТУРА 121
О v a z z a M., Camicas J.etPichonG. 1968. Notes pour une revision systemati-
que de l'espece Atylotus agrestis (Wiedemann, 1828) Diptera: Tabanidae. O. R. S.
Т. О. М., ser. Ent. med., 6, 1 : 3—14.
Pandelle L. 1883. Synopsis des Tabanides de Prance. Rev. Entomol. Fr.,.
2 : 165—228.
P a r m a n D. 1928. Experimental dissemination of the Tabanid egg parasite, Phanurus
emersoni Girault, and, biological notes on the species. Circ. U. S. Dept., 18 : 1—6.
Pechuman L. 1957.. The Tabanidae of New York. Proc. Rochester Acad. Sci., 10,-
3 : 121—179.
Pechuman L. and Stone A. 1968. A new synonymy in Hybomitra. Proc. Entomol.
Soc. Washington, 70, 4 : 302.
Pechuman L., Teskey H. and D a v i e s D. 1961. The Tabanidae (Diptera) of
Ontario. Proc. Ent. Soc. Ont., 91 (1960) : 77—121.
P h i 1 i p C. 1928. Methods of collecting and rearing the immature stages of Tabanidae.
J. Parasitol., 14, 4:243—253.
Philip С 1941. Comments on the supra-specific categories of Nearctic Tabanidae
(Diptera). Canad. Entomol., 73, 1 : 2—14.
Philip C. 1947. A catalog of the blood-sucking fly family Tabanidae (horseflies and
deerflies) of the Nearctic Region North of Mexico. Amer. Midland Natur., 37,.
2:257-324. '
P h i 1 i p C. 1956. Records of horseflies in Northeast Asia (Diptera, Tabanidae). Jap. J..
Sanit. Zoology, 7, 3—4 : 221—230.
Philip С 1960. Another, holarctic species of Tabanidae (Diptera). Canad. Entomol.,
92, 9 : 697—699.
P h i 1 i p С 1961. Notes on Palaearctic Nemorius (Diptera : Tabanidae) with description-
of one new species. BulL et Annales Soc. Roy. Ent. Belgique, 97, 7—8 : 225—236.
R а о N. and SyedM. 1958. Tabanus flies as transmitters of anthrax, a field
experience. Indian Veter. J.', 35, 7 : 348—353.
Ricardo G. 1911. A revision of the species of Tabanus from the Oriental region,
including notes of species from surrounding countries. Rec. Ind. Mus., 4 (6) : 111—258.
R о n d a n i С 1856. Dipterologicae Italicae prodromus. 1. Parmae : 1—226.
S a i t о Y. 1967. The life-history of Tabanus iyoensis Shiraki, 1918, with particular
reference to Its ability of autogeny. Acta Medica et Biologica, 14 : 181—195.
SchinerJ. 1862. Fauna austriaca. Die Fliegen. Wien.
Schwetz J. 1919. Quelques observations preliminaires sur les moeurs de la Pangonia,
zonata Walk. Ann. Trop. Med. Parasitol., 12, 3—4 : 281—288.
Scopoli J. A. 1763. Entomologia Carniolica exhibens Insecta Carnioliae indigena
et distributa in ordinis, genera, species, varietates methodo Linneana. 36+ •
+420 pp. Vindobonae (-Wien.)
Scott J.. 1922. Insect transmission of swamp fever or infectious anemia of horses.
. Wyoming Agric. Exp. Sta., Bull.: 1—133.
S h i г а к i T. 1918. Blood-sucking insects of Formosa. I. Tabanidae. Formosa.
Stammer H. 1924. Die Larven der Tabaniden. Zschr. wissensch. Biol., Abt. A, 1 :
121—170.
Stone A. 1938. The horseflies of the subfamily Tabaninae of the Nearctic Region.
U. S. Dept. Agr. Miscel. Pub., 305, Washington : 1—171.
SurcoufJ. 1912. Note sur les Dupteres piqueurs. Annales d'Historie Naturelle, II,.
1 : 71—73.
SurcoufJ. 1921. Tabanidae (avec Suppl.). In Wytsman's Genera Insectorum, Fasc,
1 5 BttixgIIgs* ■•
SurcoufJ. 1924. Les Tabanides de France et des pays limitrophes. Encyclop. Ento-
Szilady Z. 1914. Neue. oder wenig bekannte palaarktische Tabaniden. Ann. Mus.
Nat. Hungar., 12 : 661—673.
S z i 1 a d у Z. 1915. Subgenus Ochrops, eine neue Untergattung der Gattung Tabanus L.
1761 (Diptera). Ent. Mitt., 4 : 93.
Szilady Z. 1917. Revision der Palaarktischen Chrysops-Arten (Diptera) nebst anato-
mischen und phylogenetischen Bemerkungen. Arch. f. Naturg., 83, A, 4 : 85—135..
Szilady Z. 1923. New or little known horseflies (Tabanidae).Biolog. Hungar, 1,1:1—39..
S z i 1 a d у Z. 1926. Dipterenstudien. Ann. Mus. Nat. Hungar., 24 : 586—611.
T а к a g i S. 1941. Tabanidae of the North Manchuria. Report Inst, for horse-diseases,.
Manchoukuo, 2.
Takahasi H. 1962. Fauna Japonica: Tabanidae. Tokyo.
T e s к е у Н. 1962. A method and apparatus for collecting larvae of Tabanidae (Diptera)-
and other invertebrate inhabitants of wetlands. Proc. Entomol. Soc. Ont. 92
(1961) : 204—206.
T e s к е у Н. 1969. Larvae and pupae of some Eastern North American Tabanidae
(Diptera). Memoirs Entomol. Soc. Canada, 63 : 3—147.
TheodorO. 1965. Tabanidae of Israel. Israel J. Zool., 14 : 241—257.
.122
ВВЕДЕНИЕ
Trojan P 1959. Klucze do oznaczania owadow Polski, Diptera, Tabanidae. Polski
Zwiazek Entomologiczny, 28 : 1—69.
V e г г a 11 G. 1909. Tabanidae. Britishv Flies, 5 : 320—439
W a h I b fl8 Pl Fl 1848- МУа svenska Diptera. Of v. K. Svenska Vetensk. Akad. Forh.
о : 198—201.
W a t a n a b e M. and К a m i m u r a K. 1971. Observations on the autogeny of
horsefly, Tabanus iyoensis Shiraki. Jap. J. Sani. Zoo., 22 : 17—176
Z e 11 e r P. C. 1842. Beitrag zur Kenntnis der Dipteren aus den Familien der Bombvlier
Anthracier u. Asiliden. Isis, 2 : 807—848. . '
-Zetterstedt J. W. 1842. Diptera Scandinaviae disposita et descripta. I. Lund.
И. СПЕ ЦИ А Л Ь Н А Я ЧАСТЬ
ДИАГНОЗ СЕМЕЙСТВА TABANIDAE
Слепни — коренастые, средних или крупных размеров мухи, от 10 до 30 мм, редко
мельче (до 6 мм). Тело покрыто мягкими волосками, без грубых щетинок. Голова
поперечная, по ширине б. м. равная груди. Усики 3-члениковые; 3-й членик усиков, с
палочковидным концом, поделенным на несколько вторичных члеников (3—7). Щиток
по заднему краю без шипов. Брюшко состоит из 7 видимых сегментов, очень редко из 6.
Закрыловые чешуйки хорошо развиты. Крылья с полным жилкованием; число задне-
крайних ячеек составляет 5. Костальная жилка обходит все крыло; радиальные
жилки не сдвинуты к переднему краю крыла. 4-я заднекрайняя ячейка, как правило,
открытая, анальная ячейка чаще замкнутая. Лапки ног с хорошо развитым эмподием,
равным по величине пульвиллам. Средние и часто задние голени на конце со
шпорами.
Самки (за немногим исключением) нападают на теплокровных животных и сосут
кровь; самцы питаются соками растений. Личинки б. м. сигаровидной формы, состоят
из 12 сегментов (не считая головы), амфипнейстичные, с соединенными вместе обеими
задними стигмами, расположенными в общей щели; передние стигмы в числе 2 в виде
ничтожных рудиментов на границе I и II сегмента. Личинки по преимуществу хищные
обитают в воде или в почве. Куколка свободная.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ И РОДОВ
СЕМЕЙСТВА TABANIDAE
4 (8). На темени имеются'З глазка. Задние голени на вершине с концевыми шпорами
(рис. 6). У с? гоностиль с конической вершиной, одинарный или двойной
(рис. 54, 1,11, III, А). х
Рис. 53. Строение усиков слепней: 1 -*- Heptatoma;
2 — Silvius; 3 — Pangonius; 4 —■ Chrysops; 5 — Hae-
matopota; 6 — Tabanus.
.2 (5). 3-й членик состоит из 8 (иногда неясно поделенных) сегментов (рис. 53, 3).
Хоботок по большей части длинный, на большое расстояние выдается из ротовой
124 ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМ. И РОДОВ
полости, с узкими сосательными лопастями (рис. 56). У 3 гоностиль двойной,
(рис. 54, /, А). (Подсем. Pangoniinae).
3 (4). 1-я заднекрайняя ячейка на конце замкнутая
1. Pangonius Latr. (стр. 127)\.
4 (3). 1-я заднекрайняя ячейка на конце открытая . . . ...»
2. Stonemyia Bren. (стр. 128).
5 (2). 3-й членик усиков состоит из 5 сегментов (рис. 53, 2, 4). Хоботок относительно
короткий, с довольно широкими сосательными лопастями (рис. 62, 70). У 6
гоностиль одинарный, с б. м. конической вершиной (рис. 54, //, ///, А). (Подсем..
Chrysopsinae).
6 (7). 2-й членик усиков удлиненный, обычно почти равен по длине первому
(рис. 53, 4). Лицевые мозоли имеются (рис. 70), крайне редко едва выражены:
(подрод Peter sent si Ivius, рис. 111). Крылья с буро-черным рисунком в виде-
поперечных перевязей и пятен, крайне редко целиком бесцветные (подрод.
Turanochrysops) 4. Chrysops Mg. (стр. 139).
7 (6). 2-й членик усиков короткий, значительно короче 2-го (рис. 53, 2), очень редко-
оба членика равны. Лицевые мозоли отсутствуют, реже имеются (подрод Пе-
terosilvius). Крылья бесцветные (без темного рисунка) (рис. 57) -
3. Silvius Mg. (стр. 130).
8 (1). Темя без глазков, иногда имеется небольшой глазковый бугорок. Задние голени:
на вершине без шпор. У £ гоностиль с усеченной вершиной (рис. 54, IV, V, VI, А)..
(Подсем. Tabaninae).
9 (24). Крылья бесцветные или слегка затемненные, но без сетчатого рисунка
(рис. 169, 195, 212, 234, 261). По большей части крупные, коренастые виды,,
10—26 мм, очень редко мельче.
10 (23). Усики короткие, неправильной формы. Их 3-й членик сдавлен с боков, с б. м..
ясно выраженным дорсальным углом, концевая часть членика («палочка»)
обычно состоит из 4 сегментов (рис. 53, 6).
11 (16). Базикоста голая (имеются лишь микротрихии, видимые при сильном
увеличении, крайне редко имеются также единичные короткие волоски) (рис. 5,.
слева).
12 (15). Хоботок хорошо развит, массивный, у самки его длина составляет не менее-
половины высоты головы (рис. 2). Пульвиллы и эмподий нормально развиты..
13 (14). Крылья затемненные по крайней мере ниже птеростигмы в основании ячеек
i?i, R2 и Pv Щупальца, ноги и брюшко черные
5. Dasyrhamphis End. (стр. 185).
14 (13). Крылья без заметного затемнения ниже птеростигмы. Щупальца и голени ног
желто-коричневые, брюшко с большими коричневыми пятнами по бокам . ..
6. Philipomyia Ols. (стр. 186)..
15 (12). Хоботок рудиментарный, составляет приблизительно V3 высоты головы
(рис. 123). Пульвиллы и эмподий сильно укорочены (рис. 123, В), лишь на
передних лапках самца нормально развиты
7. Nanorhynchus Ols. (стр. 190).
16 (11). Базикоста покрыта короткими волосками (щетинками), такими же, как
основание костальной жилки (рис. 5, справа).
17 (22). Глаза яркие — зеленые, синие, коричневые, с полосками или без них; у сухих
коллекционных экземпляров глаза принимают черную окраску, у самцов иногда
двуцветную: верхняя часть глаз с крупными фасетками остается светлой,
нижняя с мелкими фасетками становится черной. У самки мозоли на лбу, как,
правило, хорошо развиты. Темные или светлые виды различных размеров: 10—
26 мм.
18 (19). Темя с б. м. блестящим глазковым бугорком * (рис. 2) (у самок иногда
последний может быть рудиментарным); глаза покрыты волосками ** (в редких
случаях у самок волоски плохо заметны) и, как правило, с тремя поперечными
полосками *** - 11. Hybomitra End. (стр. 308).
19 (18). Глазковый бугорок на темени отсутствует; у самца, если имеется, то матовый,
так как целиком покрыт налетом; глаза голые или в волосках; число полосок:
на глазах варьирует от 1 до 4 или последние отсутствуют совершенно.
20 (21). Средние и задние голени утолщены, в густых беловатых волосках. Крупные,
мохнатые, пестроокрашенные слепни, напоминающие шмелей (рис. 234) . . -
10. Therioplectes Zell. (стр. 304).
* Не смешивать с темными пятнами обнаженной кутикулы, которые иногда*
бывают развиты на темени у видов рода Tabanus.
** Волоски заметны лишь при рассматривании глаз сбоку.
*** Полоски на глазах видны у живых или свежеумерщвленных экземпляров;
и исчезают при сушке последних; полоски могут быть восстановлены, если поместить,
слепня на несколько часов во влажную камеру.
Рис" 54. Детали строения терминалий 6г .<£ I -^геГм?;Т- Й^^^ЙГЙ j
у// _ Chrysops caecutiens L.; IV - Dasyrhamphis urn annusi mg , „ (IX , x тергиты) 6\
•° F приемника ?.
126 ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ЛОДСЕМ. И РОДОВ
21 (20). Средние и задние голени не утолщены, в коротких черных или серых
прилегающих волосках. Мелкие или крупные виды, обычно с б. м. монотонной
окраской, реже окрашены ярко, но никогда не имеют вида шмелей (рис. 169, 195);
8. Tabanus L. (стр. 192).
22 (17). Глаза бледные охряно-желтые или зеленоватые, с тонкой, не достигающей
внешнего края срединной полоской или без нее; у сухих экземпляров окраска
глаз обычно сохраняется или темнеет до красно-коричневых тонов. У самки
мозоли на лбу лишь в виде рудиментарных округлых точек или отсутствуют
совершенно. Светло-серые или охряно-желтые, небольшие (9—16 мм) виды
(рис. 212) 9. Atylotus О. S. (стр. 281).
23(10). Усики длинные, шнуровидные (рис. 53, 1). Деление 3-го членика усиков на
сегменты выражено настолько резко, что усики выглядят 6-члениковыми . . .
12. Heptatoma Mg. (стр. 396).
24-(9). Крылья с сетчатым серым рисунком («мраморные» — на сером фоне имеются
явственные мелкие светлые пятна) (рис. 330). Палочка 3-го членика усиков
трехчлениковая (рис. 53, 5). Мелкие виды с узким телом: 8—11 мм . . . .
13. Haematopota Mg. (стр. 399).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ЗРЕЛЫХ (ВЗРОСЛЫХ) ЛИЧИНОК
СЕМЕЙСТВА TABANIDAE ДО РОДА (ПО CHVALA, JEZEK, 1969)
1 (2). 3-й членик усиков такой же длины или длиннее 2-го. Дыхательная трубка
длинная (рис. 55, А), на конце с остроконечным выступом. Брюшные сегменты
с 3 парами псевдоподий. Центральные трахейные стволы не расширены . _
Chrysops Mg.
2 (1). 3-й членик усиков короче 2-го. Дыхательная трубка умеренной длины или
короткая, на конце тупая. Брюшные сегменты с 4 тарами псевдоподий.
Центральные трахейные стволы расширены.
Рис. 55. Детали строения личинок слепней. А —Д — дорсальная сторона задней части
тела: А — Chrysops caecutiens L., Б — Heptatoma pellucens F., В — Haematopota plu-
vialis L., Г— Tabanus bromius L., Д — Hybomitra muehlfsldi Br., E — мандибула
H. muehlfeldi Вт. А, Г — по К. В. 'Скуфьину, 1967; Б, В — no Jezek, 1971; Д, Е —
по Chvala a. Jezek, 1969.
3 (4). Дорсальная сторона личинки с темно-коричневым рисунком из густых, тонких
и маленьких шипиков (рис. 55, Б). Верхняя челюсть примерно с 17
поперечными бороздками по всему периметру. По бокам преанальной складки по
небольшому утолщению с очень длинными шипиками Heptatoma Mg.
4 (3). Дорсальная сторона личинки без темно-коричневого рисунка. Верхняя челюсть
зазубрена лишь по нижнему краю. По бокам преанальной складки утолщения
с длинными шипиками отсутствуют.
5 (6). Анальный сегмент округлый, дыхательная трубка короткая и почти
полусферическая, составляет около V3 Длины анального сегмента (рис. 55, В). Взрослая
личинка небольшой величины, до 20 мм Haematopota Mg.
6 (5). Анальный сегмент удлиненный, дыхательная трубка длиннее х/з длины
анального сегмента. Взрослая личинка обычно больше.
7 (8). Личинка белая или желтовато-белая. Анальный сегмент с дорсо-латеральными
1. PANGONIUS
12Т
пятнами (рис. 55, Г). Нижний край верхней челюсти с 13—23 зубцами по всей
длине. 1-й членик щупалец в 21/2—3 раза длиннее 2-го Tabanus L.
8 (7). Личинка оранжетзая, красноватая, зеленоватая или коричневатая. Анальный
сегмент без дорсо-латеральных пятен (рис. 55, Д). Нижний край верхних
челюстей с 7—9 зубцами лишь в апикальной части (рис. 55, Е). 1-й членик
щупалец приблизительно в два раза длиннее 2-го Hybomitra End.
I. Подсем. PANGONIINAE Loew, 1860
Задние голени на вершине с концевыми шпорами (шипами). На темени имеются
глазки в числе 3. 3-й членик усиков поделен на 8 сегментов. Хоботок длиннее высоты
головы, с узкими сосательными лопастями. Базикоста голая. IX тергит брюшка с??
в виде цельной пластинки. Муфта протока семеприемника $ цилиндрическая, без
грибовидного расширения в дистальной части (рис. 54, /, Е).
1. Триба PANGONIINI (Loew, 1860) Enderlein, 1922
Гоностиль гоноподита в двойной (рис. 54, I, А).
1. Род PANGONIUS Latr. — ПАНГОНИИ *
Latreille, 1802, Hist. nat. crust, ins., Ill : 437 (Pangonius); К г б b e r,.
1921 :16 (Pangonia); S z i la d y, 1923 : 21 (Pangonia); S u г с о u f, 1924 : 197 (Pangonia);
Krober in Lindner, 1925 : 25 {Pangonia); Олсуфьев, 1937a : 57 (Pangonia);.
Leclercq, 1960b : 7, 18 (Pangonius). — Pangonia R о n d a n i, 1863 : 84, цит.
no: Leclercq, 1960b : 17. —- Melanopangonia S z i 1 a d y, 1923 : 30. — Taeniopan-
gonia Szilady, 1923 : 30.
- Тип рода Tabanus mauritanus Linne, 1767.
Коренастые, по б. ч. крупные мухи 15—20 мм с широко расставленными крыльями:
и б. м. длинным хоботком (рис. 56). Усики длиною с голову или несколько короче;
1-й и 2-й членики маленькие, 3-й длинный, узкий, поделен на 8 сегментов, из них 1-й
сегмент лишь немногим больше последующих. Лоб (у самок) относительно неширокий,,
слегка сужающийся кверху. Лобные мозоли чаще отсутствуют. Нижняя часть головы
(лицо) обычно выдается вперед, блестящая или в налете. Хоботок длинный, тонкий,
часто значительно превосходит длину головы, с узкими сосательными лопастями (1а~
bellae). Крылья широкие, бесцветные или затемненные, иногда с явственными пятнами
на поперечных жилках; 1-я заднекрайняя ячейка замкнута на конце. В Палеарктике-
известны 30 видов, распространенных в Средиземноморской подобласти. В СССР
известен 1 вид.
Самки Pangonius сосут кровь домашних и диких животных, но известны
некровососущие формы, встречающиеся только на цветах. ч
Куколки Pangonius с резкими выростами на голове.
1. Pangonius pyritosus Lw. (рис. 56) — Пангоний восточный.
Loew, 1859, Neue Beitr., VI : 27; Krober, 1921 : 54; Szilady, 1923 : 25r.
Krober in Lindner, 1925 : 33; О л с у ф ь е в, 1937а : 58; L е с 1 е г с q, 1960b :
22; С h v a 1 а, Lyneb.org' a. Moucha, 1972 : 82.
9. Лоб серый, мозоли отсутствуют. Лицо блестяще-черное или черно-коричневое,.
выпуклое. Щеки в сером налете, снизу с блестящей полоской. Усики: 1-й и 2-й членики
желтые, 3-й членик красно-желтый, на конце черный (последние 2—3 сегмента).
1-й сегмент концевого членика не шире 2-го членика. Щупальца узкие, коричневые,
небольшие, остроконечные. Грудь буровато-серая, в коротких серебристых волосках.
Спинка без продольных полосок. Крылья слегка дымчатые, с намечающимися
буроватыми пятнышками на поперечных жилках; г4 с придатком. Ноги: бедра
черно-коричневые, голени коричнево-желтые. Брюшко сверху темное, в большей своей части покрыто-
прилегающими густыми, но короткими желтовато-серыми волосками. I и II тергиты
и частью III по бокам с небольшими желтыми пятнами. Снизу брюшко коричнево-
черное, в коротких густых, серых волосках. VIII стернит крупный, слабохитинизиро-
ванный, с округлыми боковыми сторонами; гононофиз небольшой, с апикальной
выемкой (рис. 54, /); церки треугольной формы, темно-коричневые; муфта
семеприемника бесцветная. 17—19 мм.
cJ. Лобный треугольник в желтоватом налете. Лицо блестяще-черное или черно-
коричневое. Щеки в сером налете и в длинных желтовато-серых волосках; снизу
каждая щека с небольшой блестящей поперечной полоской. Усики и щупальца как у ?;.
* Здесь и далее характеристика родов дается применительно к фауне Палеарктики..
128 I. ПОДСЕМ. PANGONIINAE
концевой членик щупалец несколько короче. Грудь черная, спинка и щиток в буроватом
налете и тонких, желтоватых волосках. Крылья немного длиннее, чем у ?, с
коричневатым оттенком. Ноги как у $>. Брюшко: I—IV тергиты красновато-желтые,
последующие кольца черные, с узкими желтыми каемками. II—III тергиты каждый посредине
Рис. 56. Pangonius pyritosus Lw. 9.
имеет по темному почти квадратному пятну, IV тергит с поперечным пятном. Снизу
брюшко в основной половине желто-коричневое, IV и последующие кольца темные.
II стернит брюшка с темным пятном посредине. Брюшко в коротких, густых,
желтоватых волосках, с примесью черных волосков на темных частях. 18.5 мм.
Распространение: восточносредиземноморский (степной?) вид. Украина
(Львовская, Волынская, Хмельницкая, Житомирская, Одесская, Киевская,
Черниговская, Сумская, Харьковская, Запорожская области; Ростовская обл. (хутор
Коньков Милютинского р-на). Румыния, Болгария, Югославия, Турция. Июнь—июль.
Не часто.
Самцы и самки на цветах различных растений; имеется указание о нападении
самок пангоний на скот с целью кровососания (Бошко, 1950).
2. Род STONEMYIA Вгеп,
V
Brennan, 1935, Bull. Univ. Kansas, 22:360; L e с 1 e г с q, 1960b : 6, 17;
Takahasi, 1962 : 10. — Corizoneura auct. nee Rondani, 1863.
Тип рода Pangonia tranquilla 0. S., 1875.
Относительно небольшие, 11—15 мм, реже крупные (до 18 мм) мухи с умеренно
развитым хоботком. 3-й членик усиков узкий, поделен на 8 сегментов. 1-я заднекрай-
няя ячейка открытая.
2. STONEMYIA 139
В Палеарктике известно 5 видов, в том числе 3 вида в Восточноазиатской
подобласти; несколько видов в Северной Америке.
2 вида описаны с Кавказа или прилегающих к нему частей Ирана и Турции. Нам
в натуре неизвестны.
В первое издание «Фауна СССР» нами был включен описанный «с Кавказа» Tabanus
oritensis Bigot, 1892, позднее отнесенный Кребером (КгбЬег, 1921) к Corizoneura. Moyxa
(Moucha, Прага), изучив коллекцию Биго (Bigot), хранящуюся в Британском музее,
сообщил нам, что «Г. oritensis Bigot, 1892» — это распространенная в Африке
Mesomyia и указание ее'для Кавказа ошибочно.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА
STONEMYIA BREN. (<5<5?9)
1 (2). Усики коричнево-красные 1. S. tigris Big.
2 (1). Усики целиком черные , . , . , , » * «».»»»• 2. S. caucasica Krob.
1. Stonemyia tigris Big.
Bigot, 1880, Annal. Soc. Ent. France, 5, 10 : 143 (Pangonia); Krober, 1921 :
11 (Pangonia); Krober in Lindner, 1925 : 24 (Pangonia); Олсуфьев, 1937а :
59 (Pangonia); L e с 1 e г с q, 1960b : 18.
cJ (no Bigot) черно-коричневый, края сегментов красноватые. Задние голени
с 2 длинными шпорами на конце. Усики красно-коричневые. Хоботок значительно
короче высоты головы, сосательные лопасти расширены, черные. Щупальца беловатые,
усеяны маленькими черными волосками. Лицо снизу в желтовато-белых волосках.
Глаза голые, коричневые, нижняя половина и задние края черноватые. Спинка с
темными продольными полосками сероватого цвета; возле основания крыльев рыжеватая
мозоль (нотоплевры?). Щиток черноватый. Бочки груди в рыжеватых волосках; брюшко
возле основания несколько рыжеватое с каждой стороны, снизу рыжее, коричневатое
посредине, конец коричневый. Жужжальца коричневые с беловатой головкой. Коксы
и бедра черноватые, в серых волосках, колени и голени буро-желтые. Передние голени
по задней стороне в большей своей части, как и лапки, черные; средние и задние лапки
с коричневыми концами, присоски беловатые. Крылья бледно-серые, г4 без придатка.
Все заднекраиние ячейки широко открытые, жилки окаймлены очень
бледно-рыжеватым, с маленьким расплывчатым рыжеватым пятном; птеростигма рыжеватая.
?. Похожа на S, за исключением: основания усиков более красноватые, лицо
и волоски на щеках беловатые; щупальца без черных волосков. Лоб сероватый, мозоль
черноватая, снизу овальная, кверху линейная, узкая, слегка расширена на конце.
Глаза целиком коричневые.. Грудь и щиток целиком покрыты желтовато-серым налетом,
бочки груди менее' оволосены. Края брюшных сегментов с более бледным оттенком.
18 ым.
Распространение: Иран или Кавказ.
*2. Stonemyia caucasica Krob.
Krober, 1921, Arch. f. Naturg., 1 : 12 (Pangonia); Krober in Lindner,
1925 : 23 (Pangonia); Олсуфьев, 1937a : 61 (Pangonia); L e с 1 e г с q, 1960b : 17.
cJ (no Krober). Голова матово-беловато-серая, лицо в длинных светло-желтых
волосках; по краю глаз черные волоски. Щупальца узкие; 1-й членик серый, 2-й светло-
коричневый, тупоконечный, в длинных белых волосках. Хоботок короткий. Глаза
соприкасаются; верхние 2/3 глаза в крупных* фасетках, светло-коричневые, нижняя V3
черно-коричневая. Глазковый бугорок выступающий, с пучком черных волосков; 3
желтых глазка. Усики черные; 1-й членик утолщен, его длина превосходит ширину в
2 раза; 2-й членик почти кубический, оба в сером.налете и в черных волосках. 3-й
членик длиннее обоих предыдущих. Спинка черная, слегка блестящая, покрыта почти
войлоковидными желто-коричневыми волосками. Крыловые пластинки и жужжальца
светло-коричневые, головка черная. Брюшко короткое, не длиннее спинки и щитка,
черное; I и II тергиты с желто-красными боковыми пятнами; каемки желтоватые; II—
VI тергиты с желтыми срединными треугольниками. Снизу брюшко окрашено так же,
в светлом налете. Коксы, бедра и лапки черные, голени в большей своей части желто-
коричневые и в беловатых волосках. Крылья почти прозрачные. 1-я и 4-я заднекраиние
ячейки широко открытые, г4 без придатка.
$. Голова очень широкая, беловато-серая, щупальца охряно-желтые, концевой
членик имеет форму клинка. Лоб очень широкий, шире, нежели глаз; ширина верхней
части лба составляет приблизительно 3/4* ширины» основания. На лбу имеется крупная,
9 H. Г. Олсуфьев
130 II. ПОДСEM. CHRYSOPSINAE
блестяще-черная мозоль. Спинка пепельно-черная, с 3 оливково-коричневыми
полосками. 12.5—13 мм.
Распространение: Турция—Курдистан (Казикопоран).
II. Подсем. CHRYSOPSINAE Lutz, 1905
Задние голени на вершине с концевыми шпорами. На темени имеются глазки
в числе 3. 3-й членик усиков поделен на 5 сегментов. Хоботок немного короче высоты
головы с довольно широкими сосательными лопастями. Базикоста голая. IX тергит
брюшка у вЧ разделен на две пластинки (рис. 54, II, III, Б, Г). Гоностиль у $
одинарный, с б. м. конической вершиной (рис. 54, //, III, А). Муфта протока семеприемника <j>
цилиндрическая, без грибообразного расширения в дистальной части (рис. 54, //,
III, E).
2. Триба CHRYSOPSINI (Lutz, 1905) Enderlein, 1922
Усики длиннее, чем передне-задняя .толщина.головы. 1-й членик усиков явственно
длиннее ширины. Лоб относительно широкий, его высота превосходит ширину не более
чем в 2 раза. Тело относительно узкое.
3. Род SILVIUS Mg. — СИЛЬВИИ
М е i g е п, 1820, Syst. Beschreib., 11:27; К г 6 b е г, 1922:117; Surcouf,
1924 : 204; Krober in Lindner, 1925 : 37; Олсуфьев, 1937а : 112; M о-
ucha et Chvala, 1955:237; Leclercq, 1960b : 26, 40; Шевченко,
1961 : 55; Takahasi, 1962 : 16. — Nemorius R on dan i, 1856 :171. — Heterosilvius
Olsufjev, 1970:683.
Тип рода Tabanus vituli Fabricius, 1805.
Средних размеров или маленькие мухи с прозрачными крыльями. Усики длиной
с голову или слегка длиннее. Первые 2 членика б. м. цилиндрические, 2-й членик
короткий (длина равна ширине), реже удлиненный, 3-й членик узкий и длинный. Палочка
4-члениковая. Глаза голые, очень редко в волосках. У $ фасетки верхних частей глаз
значительно крупнее нижних, с резкой границей между теми и другими. Лоб (у $>)
б. м. широкий, в нижней части обычно имеется лобная мозоль. На темени расположены
глазки в числе 3. Лицо целиком в налете, без блестящих мозолей, реже мозоли имеются
(подрод Heterosilvius Ols.); щеки иногда в заметной черной пунктировке. Хоботок
небольшой, не превосходит высоту головы. Щупальца у ? ножевидные, немного короче
хоботка, у в маленькие, притуплённые на конце и часто изогнутые внутрь. Крылья
обычно бесцветные, г4 с небольшим придатком или без него. Иногда бывает хорошо
развита птеростигма — хитиновое утолщение мембраны крыла возле впадения гг
в костальную жилку. Задние голени с концевыми шпорами (иногда слабо развиты).
Терминалии <j> (рис. 54, //, В, Г, Д, Е): VIII стернит шестигранной или трапециевидной
(грушевидной) формы, гонопофиз обычно крупный, с небольшой апикальной выемкой;
церки с округлой или уплощенной вершиной; семеприемники перед сгибом с
небольшим расширением, сперматека цилиндрическая с тупым концом или сигаровидная,
обычно окрашена в черный цвет. Терминалии S (рис. 54, //, А, Б): гоностиль б. м.
остроконечный, имеет наибольшую ширину в передней или средней части; эпандрий с
четкой границей между IX и X тергитами.
Небогатый видами род, насчитывающий около 40 видов, распространенных в Па-
леарктической, Неарктической, Неотропической и Ориентальной областях; отсутствует
в Австралии и в Африке к югу от Сахары. Виды Silvius, ранее указывавшиеся для этих
континентов, ныне отнесены к другим родам. В Палеарктике известно 20 видов,
распространенных в Средиземноморской и Восточноазиатской подобластях; 1 вид
(Silvius (s. str.) vituli) частично проникает в Боревразийскую подобласть. В СССР 6 видов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ И ВИДОВ РОДА
' SILVIUS MG. (?? S3)
1 (10). Лицевые мозоли отсутствуют: лицо целиком в желтом или сером налете.
2 (5). Относительно коренастые охряно-желтые или черные виды, в последнем случае
с резкими желто-серыми каемками по заднему краю тергитов брюшка. По бокам
основания усиков хитин в сером-налете. Концевой членик щупалец ? на внешней
стороне без бороздки. Крупнее: 10—13, редко 9 мм. (Подрод Silvius Mg.).
3 (4). Брюшко охряно-желтое, без темного рисунка. У J лоб относительно узкий,
продольный (рис. 58) 1. S. (s. str.) vituli F.
4 (3). Брюшко черное, с резкими желто-серыми каемками по заднему краю тергитов
и желтыми боковыми пятнами на I—II тергитах. У 5 лоб широкий, поперечный
(рис. 61) • 2. S. (s. str.) latifrons Ols.
3. SILVIUS
131
5 (2). Пепельно-серые или светло-серые с относительно узким телом виды. По бокам
основания усиков .'часто небольшие участки блестяще-черного хитина (рис. 62).
Концевой членик щупалец <j> на внешней стороне с продольной бороздкой.
Мельче: 7—9, редко 10 мм. (Подрод Nemorius Rond.).
6 (9). Бедра ног черные в сером налете. Лобная мозоль у $> занимает 1/3—V« ширины
лба (рис. 63, 65). Жилки крыла коричневые.
7 (8). 2-й членик усиков короткий, его длина б. м. равна ширине (рис. 65) . . .
4. S. (Nemorius) caucasicus Ols.
8 (7). 2-й членик усиков удлиненный, его длина превосходит ширину не менее чем
в 2 раза (рис. 63) 3. S. (Nemorius) vitripennis Mg.
9 (6). Бедра ног желтые. Лобная мозоль у J занимает г/ъ—V? ширины лба (рис. 66).
Жилки крыла бледно-желтые 5. S. (Nemorius) irritans Ric,
10 (1). Лицевые мозоли имеются (рис. 67). (Подрод Heterosilvius Ols.) ,
6. S. (Heterosilvius) zaitzevi Ols.
1. Подрод SILVIUS s. str.
M e i g e n, 1820, Syst. Beschreib., II : 27.
Тип подрода Tabanus vituli Fabricius, 1805.
Лицевые мозоли и мозоли по бокам основания усиков отсутствуют. Средней
величины или небольшие б. м. коренастые охряно-желтые или темные с черно-желтым
рисунком брюшка виды. У J глаза при жизни с темным рисунком из мелких^пятнышек;
концевой членик щупалец на внешней стороне без бороздки.
1. Silvius (s. str.) vituli F. (рис. 57) — Сильвий золотистый,
Fabricius, 1805, Syst. Antl. : 97 (Tabanus); Krober, 1922:130; S u r"
с о u f, 1924 : 206; Krober in Lindner, 1925 : 41; Олсуфьев, 1937a : 118;
Mouchaet Chvala, 1955 : 237; Leclercq, 1960b : 42. — alpinus Sc о р о 1 i,
1763 : 372 (Tabanus) (nomen oblitum, Olsufjev a. Leclercq, 1971 : 325; О л-
c у ф ь е в, 1971 : 138).
$. Лоб и лицо в охряно-желтом налете и коротких желтых волосках. Высота лба
превосходит ширину в 2 раза (рис. 58). Лобная мозоль небольшая, блестяще-черная.
Рис. 57. Silvius (s. str.) vituli F. ?.
щитовидной формы. Усики красновато-желтые, палочка коричнево-черная. ЧЦупальца
красновато-желтые. Крылья слегка дымчатые, с желто-коричневыми жилками. Ноги
желтые, передние лапки, концы передних голеней и лапок последующих ног чернова-
9*
132 П. ПОДСЕМ. CHRYSOPSINAE
тые. Брюшко охряно-желтое, в коротких черных волосках; по заднему краю колец
преобладают желтые волоски. VIII стернит широкий, с умеренно развитыми
латеральными лопастями, слабо окрашен (желтый); перки с округлой или слегка уплощенной
вершиной (рис. 54, //, В, Г, Д, Е). 10.5—12 мм.
Рис. 58. Silvius (s. str.) vituli F. Лобная полоска,
VIII стернит, перки и сперматека (варианты) $>.
cJ. Очень похож на $. Фасетки верхних 2/3 глаз значительно крупнее нижних,
с резкой границей между теми и другими. Затылок за глазами в длинных,
выступающих желтых волосках, расположенных в ряд. Усики как у 5; 1-й членик несколько
короче и утолщен, вместе с последующим члеником покрыт длинными черными и жел-
Рис. 59. Ареалы: 1 — Silvius (s. str.) vituli; 2 ~ Chrysops (s. str.) suavis sua-
vis; 3 — Ch. (s. str.) suavis sakhalinensis. Значками (точки, кружки) отмечены
места обнаружения вида, причем близрасположенные местонахождения
объединены одним значком. Сплощная линия — выявленные контуры ареалов;
прерывистая линия — провизорные границы. Аналогичные обозначения
приняты на последующих картосхемах с изображением ареалов других
видов слепней. .
тыми волосками, с преобладанием первых. Щупальца сигаровидные, желтые. Лицо
в желтом налете и коротких желтых волосках. Крылья, брюшко и ноги как у $. г4
иногда с коротким придатком. 13 мм.
Распространение: средиземноморский (южноевропейский) лесной вид,
•часто в горах (рис. 59, 1). В СССР от запада Украины и Литвы на восток до Урала
;(Кунгур Пермской обл.; Губерлинские горы в Оренбургской обл.), на север до
Даниловского р-на Ярославской обл1 (58° с. ш.) й Кирова, на юг до Молдавии,
Черновицкой, Киевской, Тульской, Воронежской (Хоперский заповедник) и Куйбышевской об-
3. SILVIUS 133
ластей и далее, после перерыва в распространении в южнорусских степях, обитает
.'на Сев. Кавказе и в Закавказье (преимущественно в предгорьях и горах). Средн. и
Ю. Европа, Сев. Африка (Марокко, Алжир), Турция. Обычен местами на Кавказе-,
в других частях ареала (в пределах СССР) не часто.
33 из горных частей Кавказа (район г. Адлера) отличаются чисто-черными
волосками на затылке и более темными крыльями — var. alpinus Drap.
Июль—август, иногда раньше. На зонтичных. Нами §§ этого вида собраны во
время нападения и кровососания на волах, что подтверждает наблюдения Сюркуфа
(Surcouf, 1924) о гематофагии этого вида.
Хвала и Линеборг (Chvala a. Lyneborg, 1970) предложили для Silvius (s. str.)
vituli Fabricius, 1805, восстановить более старое название S. alpinus Scopoli, 1763,
но оно является nomen oblitum, так как не применялось в основной зоологической
литературе свыше 50 лет.
2. Silvius (s. str.) latifrons Ols. (рис. 60, 61) — Сильвий широколобый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 116; L е с 1 е г с q,
1960b : 41.
5. Лоб шириной явственно превосходит высоту, в сером налете; кверху лоб
заметно сужен. Лобная мозоль большая, поперечная, боковыми сторонами не достигает
края глаз, блестяще-черная. Лицо в сером налете п светлых волосках. Усики длинные,
черные, 3-й членик иногда слегка буроватый в основании. Первые два членика в
легком сером налете. Щупальца желтые или светло-коричневые. Крылья слегка
дымчатые, жилки коричневые; г4 иногда с коротким придатком. Ноги: бедра черно-серые,
Рис. 60. Silvius (s. str.) latifrons Ols. $.
голени желтые, с черноватыми концами. Лапки черные, последние две пары в
основании коричневые. Брюшко: I и II тергиты буровато-желтые, I тергит с черным срединным
пятном, более широким, чем щиток, II тергит в передней половине с двойным черным
пятном, позади которого лежит серый треугольник. Последующие тергиты черные,
с резкими желто-серыми каемками, расширенными посредине в небольшие
треугольники. Снизу брюшко черное, в густом сером налете. Стерниты по заднему краю с
желтоватыми каемками. II (иногда и III) стернит по бокам желтоватый. VIII стернит
грушевидной формы, его боковые стороны коричневые, основание с небольшой выемкой,
церки с округлой вершиной; проток семеприемника перед сгибом заметно расширен,
сперматека цилиндрическая, с тупой вершиной, черная. 9—12 мм.
<5. Очень похож на <j>. Фасетки верхних 2/3 глаз значительно крупнее нижних,
граница между теми и другими резкая. Затылок в коротких, глазковый бугорок в длинных
134 II. ПОДСЕМ. CHRYSOPSINAE
черных волосках. Усики черные, 1-й и 2-й членики в легком сером налете и в длинных
черных волосках. Щупальца: концевой членик удлиненный, тупой на конце,
коричневый. Ноги и крылья как у <j>. Лапки целиком темные. Брюшко как у 5; двойное черное
пятно на II тергите крупнее, с едва намечающимся треугольником позади. 10—12 мм.
Рис. 61. Silvius (s. str.) latifrons Ols. Лобная .голоска, усик.
VIII стернжг, церки и сперматека <j>.
Распространение: горный балкано-кавказский вид. Кавказ: Краснодар,
ский край (Майкоп; Гузерипль; Красная Поляна), Грузия (Хочал-даг; Пасанаури),
Азербайджан (Закатальский заповедник; Гасанриз). Вост. Турция (б. Турецкая
Армения), Греция (подвид S. (s. str.) latifrons graecus Chv., Lyn. et M.). H,e' часто. Июль.
Голотип: g, Хочал-даг, субальпийский луг, Дагестан, 29 VI 1913 (ст. ст.),
Млокосевич. В коллекции Зоологического института Академии наук СССР, Ленинград.
2. Подрод NEMORIUS Rond.
R о n d a n i, 1856, Dipterol. Ital. prodr., I : 171; Moucha et Chvala, 1959c :
137 (genus); Leclercq, 1960b : 28, 42 (genus); Philip, 1961 : 225 (genus); О 1-
8 u f j e v, 1969:116 (subgenus).
Тип подрода Chrysops vitripennis Meigen, 1820.
Лицевые мозоли отсутствуют; по бокам основания усиков часто небольшие паран-
тенальные мозоли в виде участков блестяще-черного хитина. У $ концевой членик
щупалец по внешнему краю с продольной бороздкой. Пепельно-серые или
светло-серые небольшие виды с относительно узким телом. У $> глаза при жизни с темным
рисунком в виде поперечной срединной полоски или пятна и неширокой каймы по внешнему
краю глаза. Задние голени с очень маленькими шпорами.
Всего известно 5 видов, распространены 1 вид в Гималаях и 4 вида в
Средиземноморской подобласти, из них 3 вида в СССР. У S. (Nemorius) vitripennis и S. (Nemorius)
caucasicus строение лобных мозолей и степень развития парантенальных мозолей у
основания усиков изменчивы, особенно у первого вида, вследствие чего его неоднократно
описывали под разными названиями.
Положение Nemorius в системе дискуссионно. Кребер (КгбЬег, 1921, 1925, 1938),
Сюркуф (Surcouf, 1924), Маккерас (Mackerras, 1955) считали Nemorius подродом в роде
Chrysops. Moyxa и Хвала (Moucha и Chvala, 1959c), Леклерк (Leclercq, 1960b) и Филип
(Philip, 1961) выделили Nemorius в самостоятельный род. Нами (Олсуфьев, 1937а,
1969; Leclercq et Olsufjev, 1975) представители Nemorius включены в род Silvius.
Moyxa и Хвала обосновывают род Nemorius наличием у него удлиненного 2-го членика
усиков в сочетании с прозрачными крыльями и отсутствием лицевых мозолей; два
последних признака свойственны также Silvius, но у последнего 2-й членик короткий.
Леклерк указывает для Nemorius удлиненные усики, прозрачные крылья и отсутствие
лицевых мозолей и дополняет характеристику окраской глаз: у Nemorius имеется
срединная поперечная полоска, у Silvius— многочисленные мелкие округлые пятна.
Филип, сближая Nemorius и Silvius, главное значение для родового обоснования
Nemorius придает рисунку глаз, а также ширине лба $>, дополнительным мозолям на лбу
и по бокам оснований усиков, серой окраске тела, наличию бороздки на щупальцах
и т. д. Однако все эти признаки не могут иметь родового значения. Из входящих в
состав Nemorius 4 видов только один вид — S. (Nemorius) vitripennis — имеет удлиненные
усики с относительно длинным 2-м члеником (длина которого изменчива в известных
пределах!). Остальные 3 вида — S. (Nemorius) caucasicus и особенно S. (Nemorius)
3. SILVIUS 135
shapuricus и S. (Nemorius) irritans — имеют короткие усики с коротким 2-м члеником,
как у Silvius. По строению головы и ее придатков, а также тела и его окраске Nemorius
очень близок североамериканскому Griseosilvius, включаемому Филипом в качестве
подрода в род Silvius. Этот автор отметил близость Griseosilvius и Nemorius. Окраска
глаз различна у $5'Nemorius и Silvius, но у cJcJ она не изучена. Этот признак, однако,
не может считаться достаточным для обоснования рода, если учесть разнообразие в
окраске глаз в других группах Tabanidae, например в роде Tabanus. Дополнительные
мозоли на лбу и по бокам основания усиков у $> могут отсутствовать у Nemorius,
например у S. (Nemorius) caucasicus molitor или у S. (Nemorius) vitripennis abbassianae.
Следовательно, этот признак нестоек даже в пределах вида и не может служить
критерием для родовой дифференциации. Наконец, ширина лба у 5 может быть
значительной и у Silvius, например у S. (s. str.) latifrons.
Все изложенное дает нам основание считать Nemorius лишь подродом в роде
Silvius.
3. Silvius (Nemorius) vitripennis Mg. (рис. 62, 63) — Сильвий бесцветнокрылый.
Meigen, 1820, Syst. Beschreib., II : 74 (Chrysops); R о n d a n i, 1856, I. c. : 171
(Nemorius); КгбЬег, 1921 : 61 (Chrysops); Surcouf, 1924 : 212 (Chrysops);
КгбЬег in Lindner, 1925 : 21 (Chrysops); Олсуфьев, 1937a : 113 (Silvius); M o-
uchaet Chvala, 1959c : 138; 1963 : 522 (Nemorius); Leclercq, 1960b : 43
(Nemorius); Шевченко, 1961 : 56 (Silvius); Philip, 1961 : 230 (Nemorius); Chvala,
Lyneborg a. Moucha, 1972 : 165 (Nemorius). — ranzonii S с h i n e r, 1858 :
34 (Chrysops),, цит. no: Leclercq, 1960b : 43. — horvathi S z i 1 a d y, 1926 : 597
(Nemorius). — klapperichi Moucha et Chvala, 1959c : 141 (Nemorius). — bou-
vieri Philip, 1961 : 232 (Nemorius). — abbassianae Leclercq, 1960b : 32
(Nemorius). — kondarensis В a r a t о v, 1961 : 186 (Chrysops, <$).
Небольшой пепельно-серый вид, похожий на дождевку.
д. Лоб широкий, слегка превосходит ширину глаза, в сером налете. Лобная
мозоль небольшая, б. м. поперечная, блестяще-черная; по бокам мозоли на лбу имеется
по небольшой блестящей косой черточке или треугольничку (боковые лобные мозоли).
Сбоку у основания усиков с каждой стороны имеется по треугольному
блестяще-черному пятну с раздвоенной вершиной, достигающей обычно концом края глаза (паран-
тенальные мозоли).. Лицо голубовато-серое, в
коротких беловатых волосках; щеки в заметной
черной пунктировке. Щупальца желтоватые. Усики
длинные, тонкие, черные; 1-й и 2-й членики в сером
налете, 2-й членик слегка удлинен, его длина
приблизительно в два раза превосходит ширину. 1-й
членик длинный, тонкий, его длина превосходит ширину
в 4—5 раз; перед вершиной членик с явственной
перетяжкой. Крылья бесцветные; г4 часто с коротким
придатком. Жилки в внешней половине крыла
коричневые, к основанию желтоватые. Птеростигма
небольшая, желтая. Жужжальца коричневые. Ноги:
Рис. 62. Silvius (Nemorius) vitripennis Mg. 5.
Голова спереди.
бедра черно-серые, основания передних голеней, средние и задние голени (за
исключением черных кончиков) и основания двух последних пар лапок желтые.
Концы передних голеней и лапки черные. Брюшко темное, в сером налете, тергиты
•по заднему краю с узкой серой каемкой; II тергит по бокам с небольшим коричневато-
желтым пятном, слабо заметным из-за прикрывающего его серого налета. Последний
образует посредине тергита неясный серый треугольник, а по бокам большие серые
пятна, занимающие всю боковую сторону полукольца; на последующих тергитах этот
рисунок постепенно теряется, сливаясь в монотонный серый цвет. Снизу брюшко
темное, в густом сером налете. VIII стернит грушевидный, затемнен по бокам (окраска
изменчива), гонопофиз с значительной апикальной выемкой; церки^с уплощенной
вершиной; семенной проток значительно расширен перед сгибом, сперматека с тупой
вершиной, черная. 7—10, редко 6 мм.
<5. Глаза голые; фасетки верхних 2/3 глаза значительно крупнее нижних, граница
между теми и другими резкая. Лобный треугольник серый. Усики черные, 1-й и 2-й
членики в сером налете и длинных волосках, 1-й членик длинный, цилиндрический,
•перед вершиной с явственной перетяжкой; 2-й членик по длине приблизительно в два
136 П. ПОДСЕМ. CHRlfSOPSlNAE
раза прэвосходит ширину. Лицо в сером налете и беловатых волосках; щеки в черной
пунктировке. У основания усиков сбоку имеется блестяще-черная парантенальная
мозоль. Щупальца маленькие, темные, в сером налете и в беловатых волосках.
Концевой членик слегка изогнут, тупоконечный. Крылья, ноги и жужжальца как
у $. Брюшко черное, тергиты по заднему краю с узкими беловатыми каемками. III и
последующие кольца в большей части в тонком сероватом налете. II—III тергиты по
91
Рис. 63. Silvius (Nemorius) vitripennis Mg. Лобная полоска,
усик, VIII стернит, церки и сперматека <j>. А — гоностиль в.
бокам просвечивают желтым. Снизу брюшко в сером налете, с просвечивающей
желтизной по бокам II—III стернитов. 7.5—9 мм.
Распространение: средиземноморско-среднеазиатский предгорно(горно)-
степной вид (рис. 64, 1). Западная Украина, юг Туркмении, Таджикистан,
Узбекистан (Ташкентская, Андижанская, Сурхандарьинская области), Чимкентская обл.
Рис. 64. Ареалы: 1 — Silvius (Nemorius) vitripennis; 2 —■ S. (Nemorius)
caucasicus; 3 — Chrysops (s. str.) makerovi.
(хребты Каратау, Таласский Алатау), Киргизия (хребты Чаткальский и Ферганский).
Южная и частью средняя Европа (Австрия), Турция, Иран, Афганистан. Обычен
местами в Узбекистане ж Таджикистане. Май—июль, иногда позднее. Личинки найдены
в родниках (Кадырова, 19726).
Изменчивый вид. Лобная мозоль округлой, пятигранной, шестигранной,
треугольной или ромбической формы, иногда соединена внешней стороной с боковыми лобными
мозолями. У многих самок с Гиссарского хребта (Таджикистан) и всех самок с
Западного Тянь-Шаня и хр. Каратау отсутствуют боковые лобные'мозоли (var. horvathi Sz.)r
а в средней части наличника часто имеется небольшое блестяще-черное пятно
неправильной формы. У самок из Южной Туркмении желтые боковые пятна на брюшке
захватывают I и II и даже III и IV тергиты, соответственно I и II стерниты по бокам
3. SILVIUS 137
желтые (var. bouvieri Phil.). Парантенальные мозоли к внешнему краю могут не быть
раздвоенными или, наоборот, имеют глубокую выемку, достигают или не достигают
края глаз. Пунктировка лица варьирует от хорошо выраженной до очень тонкой. У
самок из южного Ирана (Шираз) парантенальные мозоли отсутствуют (полностью
прикрыты налетом); эта форма описана Леклерком (Leclercq) как самостоятельный вид
N. abbassianae. .Она представляет собой интересный пример изменчивости,
гомологичной таковой S. (Nemorius) caucasicus molitor Bod. et- Sam. Если эта форма
географически б. м. обособлена, то она может быть обозначена как S. (Nemorius) vitripennis
abbassianae Led.
4. Silvius (Nemorius) caucasicus Ols. (рис. 65) — Сильвий кавказский.
Диморфный вид, представлен двумя подвидами : S, (Nemorius) caucasicus cauca- ■
sicus Ols. и S. (Nemorius) caucasicus molitor Bog. et Sam. Аллопатричен S. vitripennis Mg.
4a. Silvius (Nemorius) caucasicus caucasicus Ols. — Сильвий кавказский
номинативный.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 115; Г а у з е р, 1939 :
153 (личинка, куколка); Leclercq, 1960b : 43 (Nemorius).
?. Глаза голые. Лоб широкий, равен по ширине глазу, в сером налете. Над усиками
небольшая, продолговатая, блестяще-черная лобная мозоль; боковые полоски на лбу
N отсутствуют. Усики: 1-й и 2-й членики коричневые или черные, в сером налете, 3-й
членик черный, 1-й членик приблизительно в 2х/2—3 раза длиннее своей толщины, перед
вершиной с нерезкой перетяжкой, 2-й членик усиков лишь немногим превосходит
ширину (не более чем. в 1г/2 раза). Возле основания усиков по бокам по' блестящему
черному конусовидному пятну (парантенальная мозоль), достигающему концом края
глаз. Лицо в сером налете и в коротких серых волосках; щеки в тонком, черном
пунктире. Щупальца желтоватые. Крылья бесцветные; жилки в вершинной части крыла
коричневые, к основанию желтоватые. Птеростигма хорошо развита, коричневая;
г4 иногда с коротким придатком. Жужжальца коричневые.
Ноги: бедра серые, голени средней и задней пар ног
желтые, с черными кончиками, передние голени в основной
части желтые, к вершине черноватые. Передние лапки чер-
ные целиком, последующие с желтыми основаниями.
Брюшко темное, в сером налете; тергиты по заднему краю
с узкой, серой каемкой; II тергит с неясным серым
треугольником посредине, вершиной простирающимся до
переднего края кольца, тогда как по бокам с серыми
пятнами; последние занимают всю боковую часть тергита. На
Рис. 65. Silvius (Nemorius) caucasicus caucasicus Ols.
Лобная полоска и усик $.
последующих тергитах этот рисунок постепенно теряется, переходя в однообразно-
серый тон. У потертых экземпляров брюшко монотонно-серое. 6—8 мм.
cJ. Фасетки верхних 2/3 глаз значительно крз'пнее нижних, граница между теми
и другими резкая. Глаза голые, лобный треугольник серый. Усики черные, 1-й и 2-й
членики в тонком сером налете и длинных, черных волосках. 1-й членик слегка утолщен,
за серединой с легкой перетяжкой, его длина превосходит толщину в 2—21/., раза.
2-й членик короткий, длиной равен или лишь едва превосходит собственную ширину
(не более чем в 1V2 раза). Лицо в сером налете и такого же цвета волосках; щеки в
тонкой пунктировке, в верхних частях в длинных черных волосках. Сбоку основания
усиков небольшая блестяще-черная парантенальная мозоль. Щупальца коричневые,
в светлых волосках, концевой членик слегка изогнут, с притуплённым кончиком.
Крылья, жужжальца и ноги как у '$. Брюшко черное, с очень узкими серыми каемками
по заднему краю тергитов. II—III тергиты по бокам грязно-желтые. Снизу брюшко
в основании желтое, IV и последующие стерниты черные, в сером налете. I стернит
имеет посредине темное пятно, которое слегка заходит на II кольцо. 7.5—9 мм.
Очень похож на 5. (Nemorius) vitripennis, от которого, однако, легко отличается
коротким 1-м, и в особенности 2-м члеником усиков; кроме того, у $ отсутствуют
боковые лобные мозоли, а II тергит без боковых желтоватых пятен; форма лобной мозоли
изменчива. От S. (Nemorius) irritans отличается коричневыми жужжальцами и жилками
(у S. (Nemorius) irritans жужжальца белые, а жилки на крыльях бледно-желтые).
Распространение: Кавказский предгорно(горно)-степной подвид
(рис. 64, 2). Краснодарский край (Майкоп), Северная Осетия (Камгарон), Дагестан,
4 Азербайджан, Армения, Грузия. Июнь—июль, иногда позднее. Не часто. Личинки об-
138 II. ПОДСЕМ. CHRYSOPSINAE
наружены в верхнем слое береговых песчаных наносов поймы небольшой речки в зоне
предгорной степи (Гаузер, 1939).
Голотип: 5, б. Еленендорф Ганджинского у. (ныне Кировабад, Азербайджан),
23 VIII 1925, С. Тарбинский. В коллекции Зоологического института Академии наук
СССР, Ленинград.
4b. Silvius (Nemorius) caucasicus molitor Bog. et Sam. — Сильвий кавказский
подобный.
Богачев и Самедов, 1949, Изв. Акад. наук Азерб. ССР, 5 : 67.
?. От номинативной формы отличается отсутствием парантенальных мозолей
■(полностью прикрыты серым налётом). Встречаются промежуточные экземпляры,
у которых налет лишь частично прикрывает парантенальные мозоли.
cJ неизвестен.
Распространение: Нахичеванская АССР, Армения. Не часто.
Описанный Леклерком (Leclercq, 1960b) из Южного Ирана S. (Nemorius) abbassia-
пае, позднее идентифицированный нами с S. (Nemorius) caucasicus molitor Bog. et Sam.
(Abbassian-Lintzen, 1964), представляет собой вариант S. (Nemorius) vitripennis с
прикрытыми налетом мозолями по бокам оснований усиков.
5. Silvius (Nemorius) irritans Ric. (рис. 66) — Сильвий бледный.
R i с а г d о, 1901, Ann. Mag. Nat. Hist., 8 : 292; К г 6 b e г, 1922 : 122; К г 6-
ber in Lindner, 1925 : 40; Leclercq, 1960b : 41; — unicolor Becker,
1912 : 588.
$. Лоб шире поперечника глаза, в беловато-сером налете. Лобная мозоль
блестяще-черная или коричневатая, маленькая, составляет 1/6—1/7 ширины лба; боковые
лобные мозоли отсутствуют. Усики короткие, желто-коричневые, концевая часть
3-го членика черная. 1-й членик в 2—3 раза длиннее своей толщины, 2-й членик по
,,,,,„, и1 ,,,„,,,, длине'равен или едва превышает собственную тол-
>^»*ий^:{:,"';;Jt^l^f"^''!У*';'.;^^*<л. щину. Парантенальные мозоли едва развиты: имеет-
Ч>--'' .'!)'!';'Г ся небольшая черная поперечная черточка возле гла-
■''"■-•' за и ниже ее возле границы с щекой небольшое
черное пятно. Лицо в светло-сером налете, щеки в
грубоватой, наличник в более мелкой черной
пунктировке. Щупальца желтоватые. Крылья бесцветные,
жилки бледно-желтые. Жужжальца с белой голов-
Рис. 66. Silvius (Nemorius) irritans Ric. Лобная
полоска и усик $>.
кой. Ноги желтые, большая часть передних голеней, кончики средних и задних голе-,
ней, передние лапки и конец средних и задних лапок черноватые; средние и задние
бедра к вершине иногда затемнены. Брюшко светло-серое, тергиты по заднему краю
с желтоватой каймой, I и II, иногда также III и IV тергиты по бокам с желтоватым
пятном. Снизу брюшко сероватое, обычно с желтоватыми I и II стернитами,
последующие стерниты с желтоватой каймой. 6—8.5 мм.
(J неизвестен.
Распространение: переднеазиатский степной вид. Юг Туркмении (Се-
рахс; Пуль-и-Хатум на р. Теджен, Бадхыз). Турция, Ирак, Иран, Афганистан. Редок.
Июнь.
3. Подрод HETEROSILVIUS Ols.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 683.
Средней величины виды с оранжево-желтым или коричневым брюшком. Лицевые
мозоли имеются. Лоб уже поперечника глаза.
Тип подрода Silvius zaitzevi Olsufjev, 1941.
В Палеарктике 6 видов: 1 вид в СССР, 3 вида в Японии, 1 вид в Китае и 1 вид
в Турции.
6. Silvius (Heterosilvius) zaitzevi Ols. (рис. 67) — Сильвий Зайцева.
Олсуфьев, 1941, Тр. Зоол. сектора Груз. фил. АН СССР, III : 59 (?);
L е с 1 е г с q, 1960b : 42; Гургенидзе, 1972а : 201 ($)•
4. CHRYSOPS 139
д. Глаза голые. Лоб слегка расширен книзу, покрыт желтым налетом, высота лба
немного превосходит ширину основания. Лобная мозоль крупная, выпуклая, округлая
или неправильной формы, блестяще-коричневая, иногда черноватая по краям.
Боковыми сторонами мозоль не достигает края глаз. На темени возле глазков обычно имеются
небольшие блестяще-черные участки. Лицо в желтом налете и с хорошо развитыми
блестяще-желтыми лицевыми мозолями. Последние занимают верхнюю часть щек и
боковые части наличника. Усики красновато-желтые, 3-й членик на конце черноватый.
1-й членик короткий, утолщенный; 2-Й членик тоже короткий, его длина лишь немного
превосходит собственную толщину. Щупальца узкие, красновато-желтые. Крылья
слегка дымчатые, жилки желто-коричневые. Ноги красновато-желтые, концы лапок
коричневатые. Брюшко красновато- или оранжево-
желтое, в коротких черных и желтых волосках.
Тергиты II—V (или VI) каждый с небольшим
продолговатым черным пятном посредине; в совокупности эти
пятна составляют узкую срединную полоску или
цепочку. Снизу брюшко оранжево-желтое, с
небольшим продолговатым темным пятном в основании.
11—12 мм.
$. Глаза голые. Фасетки верхних 2/;ч глаза
крупнее нижних приблизительно в 4 раза, граница
с мелкими фасетками довольно резкая. Затылок за
глазами в длинных, тонких, желтоватых волосках.
Усики: 1-й членик коричневато-желтый, блестящий,
удлиненно-яйцевидный, приблизительно в 13/4 раза
длиннее толщины, 2-й членик того же цвета,
короткий, его длина примерно равна толщине, 3-й членик
очень узкий, в основании желтовато-коричневый,
конец площадки и палочка, черные. Лицо в желтом
Рис. 67. Silvius (Heterosilvius) zaitzevi Ols. Голова
спереди, усик и щупальце §.
налете, лицевые мозоли имеются, приблизительно как у $, блестяще-желтые. Концевой
членик щупалец желтый, маленький, конический, в длинных желтых волосках. Ноги
и крылья как у 9; кончики задних бедер блестяще-черные. Брюшко ярко-оранжево-
желтое (как у ?). I тергит с крупным черным пятном, захватывающим
передне-боковую его часть. II—IV тергиты каждый с небольшим продольным черным пятном
посредине. Пятна не достигают заднего края тергитов и образуют цепочку, в которой каждое
следующее пятно меньше предыдущего. Последующие тергиты без черного пятна,
желтовато-оранжево-коричневые. Снизу брюшко оранжево-желтое, с крупным
продолговатым черным срединным пятном на II стерните, почти достигающим заднего края
полукольца; III—IV стерниты каждый с небольшим черным срединным пятном,
последующие стерниты коричневатые в передней части. 10 мм.
Распространени е: Большой Кавказ — Красная Поляна Краснодарского
края, Лентехский, Амбролаурский и Онский р-ны Грузии; Малый Кавказ — Баку-
риани и Ломис-Мта Боржомского р-на Грузии. В пределах 1500—2300 м в лесной и
субальпийской зонах. Не часто. Указание Ш. Джафарова (1960) для Муганской степи,
Азербайджан, видимо, ошибочно.
Голотип: 5, Бакуриани (Грузия), 14 VIII 1934, Я. Киршенблат. В коллекции
Зоологического института Академии наук СССР, Ленинград.
4. Род CHRYSOPS Mg. — ЗЛАТОГЛАЗКИ, или ПЕСТРЯКИ
М е i g е п, 1800, Nouv. classif. : 23; S z i 1 a d у, 1917 : 85; К г 6 b e г, 1920 :
41; S ur со uf, 1924 : 208; Krober in Lindner, 1925 : 2; О л с у ф ь е в, 1937а :
61; М ouch a et Chvala, 1955 : 228; L е с 1 е г с q, 1960b : 28, 43; Ш е в ч е н к о,
1961 : 30; Takahasi, 1962 : 20; В и о л о в и ч, 1968 : 31. — Heterochrysops
Krober, 1920 : 50. — Turanochrysops Stackelberg, 1926b : 327. — Pseudochrysops
Moucha et Chvala, 1957 : 160; Petersenichrysops Moucha et Chvala, in
M о u с h a, 1969 : 173.
Тип рода Tabanus caecutiens Linne, 1758.
Стройные, средних размеров, пестроокрашенные мухи (рис. 68, 69). Усики
длиннее головы; 1-й и 2-й членики цилиндрические (очень редко 1-й членик утолщен),
длинные, 2-й членик лишь едва короче первого или равен ему, 3-й членик узкий, почти без
«дорсального угла, лишь слегка расширен в основании; палочка 4-члениковая. Глаза с
140
II. ПОДСEM. CHRYSOPSINAE
пятнами или без них. У 3 фасетки верхних частей глаз обычно крупнее, нижних, реже-
б. м. одинаковой величины. Глазки на темени имеются в количестве 3. Лоб широкий,
его высота лишь едва превосходит ширину. В основании лба имеется довольно крупная,
блестящая лобная мозоль. Нижняя часть головы с хорошо развитыми лицевыми
мозолями (рис. 70, 71). Из них различают: 1) парные лицевые мозоли, имеющие обычно'
округлые очертания; они захватывают верхнюю часть щек и прилегающие части
наличника; 2) парные щёчные мозоли в виде блестящих пятен на нижней стороне щек и 3)
непарную наличниковую мозоль, которая занимает нижнюю (концевую) часть наличника;
ее верхние концы в виде перемычек присоединены к лицевым мозолям, разделяясь >
неглубокой бороздкой. Хоботок тонкий, умеренной длины, не превосходит высоту
головы.. Крылья большей частью с резким рисунком из темных поперечных пятен и пе-
Рис. 68. Златоглазик Chrysops (s. str.) caecutiens
caecutiens L. J.
ревязей, очень редко бесцветные. Хорошо развита птеростигма — хитиновое
утолщение мембраны крыла возле впадения жилки Г[ в костальную жилку. В покое крылья
расставлены в стороны б. м. под прямым углом друг к другу; г4 без придатка. Рисунок
крыла (рис. 72) в основном состоит из узкой темной полосы вдоль переднего края, от
которой за серединой крыла отходит поперечная перевязь, шириной слегка
превосходящая длину дискоидальной ячейки. В вершинной части крыла вдоль костальной жилки
расположено вершинное пятно. У S6 темный рисунок крыла развит сильнее, нежели
У ??• Задние голени с 2 концевыми шпорами. Брюшко обычно с резким, темным
рисунком на желтом фоне, реже целиком черное; у $ брюшко чаще окрашено темнее, чем'
у ?. Терминалии 5 (рис. 54, III, В, Г, Д, Е): VIII стернит трапециевидной или
грушевидной формы, обычно темно окрашен, с выделяющимся более светлым треугольным
пятном, реже боковые стороны светлее срединного треугольного пятна, гонопофиз
небольшой, обычно с апикальной выемкой; церки с округлой вершиной; семеприемники
короткие, сперматека большая, сигаровидная или цилиндрическая, ее базальная часть
полупрозрачная, а апикальная — коричневая или черноватая. Терминалии 3 (рис. 54,
III, А, Б): гоностиль удлиненный, приострен на конце; эпандрий неясно поделен на
IX и X тергиты; церки со слегка уплощенной вершиной.
При определении златоглазиков наибольшее значение имеет окраска крыльев
и брюшка, которая обычно довольно постоянна. Известное значение имеют также
окраска ног, форма и окраска лицевых мозолей и отчасти окраска лба и усиков. Окраска
усиков подвержена некоторой изменчивости. Диагностическое значение терминалии
ограничено вследствие значительного однообразия строения, в том числе VIII стер-
Рис. &9. Златоглазик Chrysops (s. str.) caecutiens
caecutiens L. S.
Рис. 70. Chrysops (s. str.) caecutiens
caecutiens L. Голова ? спереди, г — сложный
фасетчатый глаз; гл — глазки; у — уси-
ковая ямка (усики удалены); лбм —
лобная мозоль; лм —'лицевая мозоль; щм —
щечная мозоль; нм — наличниковая
мозоль; х — хоботок; щ — щупальце.
Рис. 71. Chrysops (s. str.) caecutiens
caecutiens L. Голова S спереди.
142 П. ПОДСЕМ. CHRYSOPSfNAE
нита $. У близких видов, например Ch. (s. str.) caecutiens и Ch. (s. str.)
pictus,индивидуальные различия- VIII стернита <j> перекрывают видовые различия.
д
Рис. 72. Крыло J Chrysops. n — перевязь; вк —
внешний край перевязи; вп — вершинное пятно; пр —
перемычка между перевязью и вершинным пятном; д —
дискоидальная ячейка; пт — птеростигма.
Род Chrysops насчитывает в своем составе около 250 видов. Распространен во всех:
5 частях света, но наиболее многочислен в Америке (Северной и Южной). В Австралии
всего 1 виг. В Палеарктике известно 46 видов, в том числе в СССР 27 видов.
Питавшиеся детритом или хищные личинки обитают по преимуществу в ручьях
или в прибрежных частях рек и озер.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА 110ДРОДОВ
РОДА CHRYSOPS MG.
1 (6). Крылья с пестрым рисунком, состоящим из темной поперечной перевязи и пятен
(рис. 68). Лицевые мозоли нормально выпуклые (рис. 70).
2 (5). На крыле вершинное пятно или узкая темная полоска на его месте вдоль
костальной жилки имеется (рис. 73, 80, 81). Наличниковая мозоль или мозольки
по бокам дистальной части наличника имеются.
3 (4). Дискоидальная ячейка одноцветно темная, не выделяется окраской от прочих
участков перевязи (изредка слегка просветлена) (рис. 73, 81, 94)
1. Chrysops Mg. s. str. (стр. 142).
4 (3). Дискоидальная ячейка в средней части явственно просветлена (с «окошечком»)»
или целиком прозрачная (рис. 102, 103, 108)
2. Heterochrysops Krob. (стр. 172).
5 (2). На крыле вершинное пятно или узкая темная полоска на его месте вдоль
костальной жилки отсутствует (рис. 111). Наличник в дистальной части в налете,,
без мозоли 3. Petersenichrysops M. et Chv. (стр. 182).
6 (1). Крылья бесцветные (рис. 112). Лицевые мозоли конусовидно выступают вперед
(рис. 113, 114) 4. Turanochrysops Stack, (стр. 183).
1. Подрод CHRYSOPS Mg. s. str.
Тип подрода Tabanus caecutiens Linne, 1758.
Крылья с рисунком из темных поперечных перевязей и пятен. Лицевые мозоли
нормально выпуклые. Дискоидальная ячейка одноцветно темная или же слегка
просветлена в средней части (но без явственного «окошечка»).
Подрод наиболее богато представлен в таежно-лесном и степном поясе Голарк-
тики, и лишь немногие виды заходят в пустынно-степную зону.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА
CHRYSOPS MG. S. STR.
А. Глаза разделены лбом: $ $
1 (4). Лицевые и щечные мозоли слиты вместе, лицо блестяще-черное. Грудь, брюшко
и ноги целиком черные.
2 (3). Передняя базальная ячейка целиком темная (рис. 73). Вершинное пятно крыла
соединено с перевязью очень узкой перемычкой, не превосходящей V2 ширины
ячейки R± 1. ch. (s. str.) japonicus Wied.
3 (2). Передняя базальная ячейка в вершинной V3 с прозрачным пятном (рис. 75).
Вершинное пятно крыла соединено с перевязью по всей ширине ячейки Rt
2. Ch. (s. str.) sepulcralis F,.
4. CHRYSOPS Ш
4 (1). Лицевые и щечные мозоли явственно обособлены между собой окружающим
их желтым или серым налетом (рис. 70). По крайней мере 2-й тергит брюшка по-
бокам желтоватый.
5 (6). Вершинное пятно крыла не достигает г4 и очень узкое, не шире половины.
ячейки Rt (рис. 80) 5. Ch. (s. str.) anthrax Ols..
6 (5). Вершинное пятно достигает г4 и обычно не уже ширины ячейки R± (рис. 76, 78,
81 и др.).
7 (18). Голени черные, лишь иногда средние голени в основании коричневатые.
8 (11). Вершинное пятно крыла узкое, его вершина налегает на г4 не более чем на V2
ее длины. Внешний край темной перевязи крыла с резким выступом в ячейке R2.
(рис. 76, 78).
9 (10). 2-й тергит брюшка преимущественно желтый, с черным пятном в виде
перевернутой латинской буквы V (рис. 78). 3-й стернит брюшка желтый, с черным
поперечным пятном посредине. Крупнее: 9—12 мм
4. Ch. (s. str.) divaricatus Lw.
10 (9). 2-й тергит брюшка преимущественно черный, с незначительными желтыми.
пятнами по бокам и небольшим серым треугольником посредине (рис. 76).
3-й стернит брюшка черный. Мельче: 7—11 мм
3. Ch. (s. str.) nigripes Ztt.
11 (8). Вершинное пятно крыла широкое, налегает на г4 явственно более чем на 1/2 ее-
длины (рис. 81, 84). Внешний край темной перевязи крыла ровный или лишь
с незначительным уголком в ячейке R2.
12 (17). Вершинное пятно крыла соединено с перевязью перемычкой по всей ширине-
ячейки Дг (рис. 81, 84).
13 (14). Брюшко с резкими светлыми каемками по заднему краю тергитов (рис. 81)
6. Ch. (s. str.) makerovi PL
14 (13). Тергиты брюшка без светлых каемок (рис. 68, 83).
15 (16). II тергит брюшка желтый, посредине с черным пятном в виде перевернутой
буквы V; III тергит целиком черный (рис. 68)
7а. Ch. (s. str.) caecutiens caecutiens L.
16(15). IГ тергит брюшка целиком желтый или же с двумя едва намечающимися
расходящимися кзади полосками (редукция V-образного пятна). III тергит
черный, посредине с небольшим желтым треугольным пятном (рис. 83) . .
7b. Ch. (s. str.) caecutiens ludens Lw.
17 (12). Вершинное пятно крыла явственно обособлено от перевязи просветом в ячейке
R± (рис. 87). II тергит брюшка желтый с двойным черным пятном посредине-
, 10. Ch. (s. str.) validus Lw.
18 (7). По крайней мере голени средней пары ног коричневые или желтые.
19 (36). II тергит брюшка желтый, в средней части с одним или двумя черными
пятнами (рис. 84, 89 и др.). Анальная ячейка на конце замкнутая, очень редко-
открытая.
20 (25). Голени задних ног черные.
21 (24). Вершинное пятно крыла в виде узкой полоски, отходящей от перевязи
пошире ячейки Лг и далее постепенно сходящей на нет; пятно вершиной лишь
незначительно налегает на г4 (рис. 91, 95).
22 (23). Уже и мельче: 7—9 мм. II тергит брюшка с продолговатым двойным черным
пятном, не доходящим концами до заднего края тергита. Последующие тергиты
черные, с желтыми каемками (рис. 91) . . . 12. Ch. (s. str.,) angaricus Ols.
23 (22). Шире и крупнее: 9—10.5 мм. II тергит брюшках 2 черными пятнами,
продолженными на последующие тергиты в виде 2 черных полосок. Последние
поделены резкой продольной желтой полоской (рис. 95)
15. Ch. (s. str.) suavis Lw.
24 (21). Вершинное пятно крыла широкое, налегает на г4 на г/2 ее длины, соединено
с перевязью очень узкой перемычкой; последняя занимает не более V2 ширины
ячейки i?! (рис. 86). II тергит брюшка желтый, в передней части с
незначительной черной точкой или черточкой посредине
9. Ch. (s. str.) turanicus Ols.
25 (20). Голени задних ног коричневые или желтые.
26 (33). Перемычка, соединяющая вершинное пятно крыла с перевязью, занимает
ячейку R1 целиком (рис. 84, 92).
27 (28). II тергит брюшка с небольшим черным пятном посредине; последнее имеет
форму лунки, точки или прямоугольника (рис. 84) .
8. Ch. (s. str.) pictus Mg.
28 (27). II тергит брюшка с 2 черными пятнами, имеющими б. м. треугольную форму
(рис. 89, 94).
29 (30). Черные пятна на II тергите соединены между собой в его передней части
(рис. 88, 89). Ячейка Р4 не сужена к вершине . . 11. Ch. (s. зЦ.) relictus Mg.
30 (29). Черные пятна на II тергите обособлены друг от друга (рис. 92, 94). Ячейка.
Р4, как правило, сужена к вершине.
144 11. ПОДСЕМ. CHRYSOPSINAE
31 (32). Бедра черные. Пятна на II тергите б. м. сближены (рис. 92). Лобная мозоль
крупная, обычно почти достигает края глаз. Темя вокруг глазков блестяще-
черное 13. Ch. (s. str.) parallelogrammus Zell.
32 (31). Бедра желтые. Пятна на II тергите широко расставлены друг от друга (рис. 94).
Лобная мозоль умеренной величины, далеко не достигает края глаз. Темя
вокруг глазков обычно покрыто серым налетом
14. Ch. (s. str.) concavus Lw.
33 (26). Перемычка, соединяющая вершинное пятно крыла с перевязью, занимает
не более г/2 ширины ячейки R± (иногда перемычка отсутствует совершенно)
(рис. 96, 98, 99).
34 (35). Вершинное пятно крыла налегает на г4 обычно не менее чем на х/з ее длины.
Темное пятно на I тергите брюшка обычно не превышает ширину щитка (рис. 96).
В типичном случае II тергит брюшка с 2 относительно круппыми овальными
черными пятнами, расположенными возле заднего края тергита; иногда пятна
в паре соединены между собой (рис. 98, Б) или отсутствуют полностью . .
16. Ch. (s. str.) ricardoae PI.
35 (34). Вершинное пятно крыла налегает на г4 обычно не более чем на V4 ее длины.
Темное пятно на I тергите брюшка захватывает всю его переднюю часть
(рис. 99). В типичном случае II тергит брюшка с двумя маленькими круглыми
черными пятнами, расположенными отступя от заднего края тергита; иногда
пятна в паре соединены между собой или отсутствуют полностью .....
17. Ch. (s. str.) dissectus Lw.
36 (19). II тергит брюшка черный, с небольшим серым треугольничком посредине
(рис. 100). Анальная ячейка на конце открытая . . .
18. Ch. (s. str.) rufipes Mg.
Б. Глаза соприкасаются (или почти соприкасаются):^
1 (6). Лицо блестяще-черное, без пятен налета.
2 (3). Передняя базальная ячейка затемнена целиком
1. Ch. (s. str.) japonicus Wied.
3 (2). Передняя базальная ячейка в вершинной части с прозрачным пятном.
4 (5). Вершинное пятно крыла соединено с перевязью перемычкой j .
. . . 2. Ch. (s. str.) sepulcralis F.
5(4). Вершинное пятно крыла обособлено ..... 10. Ch. (s. str.) validus Lw.
6 (1). Лицевые и щечные мозоли обособлены окружающим их серым налетом
(рис. 71).
7 (14). Голени средних ног черные.
8 (11). Внешний край темной перевязи крыла с резким выступом в ячейке R2.
9 (10). Вершинное пятно крыла узкое, не шире 1-й радиальной ячейки. III стернит
брюшка желтый, с черным поперечным пятном посредине
4. Ch. (s. str.) divaricatus Lw.
10 (9). Вершинное пятно значительно шире 1-й радиальной ячейки. III стернит брюшка
черный 3. Ch. (s. str.) nigripes Ztt.
11 (8). Внешний край перевязи крыла в ячейке R2 без выступа.
12 (13). Бочки груди с пучком золотисто-желтых волосков
7а. Ch. (s. str.) caecutiens caecutiens L.
13(12). Бочки груди в черных волосках. . . 7b. Ch. (s. str.) caecutiens ludens Lw.
14 (7). Голени средних ног коричневые или желтые.
15 (32). Глаза соприкасаются на некотором протяжении. II тергит брюшка по бокам
желтый.
16 (19). Притуплённые на вершине щупальца короче 1/2 длины хоботка (рис. 89, 91).
17 (18). Голова относительно большая, явственно шире груди. Задние голени черные.
Вершинное пятно крыла узкое (рис. 91) . . . 12. Ch. (s. str.) angaricus Ols.
18 (17). Голова нормальной величины, не шире груди. Задние голени коричнево-
желтые. Вершинное пятно крыла широкое (рис. 89)
11. Ch. (s. str.) relictus Mg.
19 (16). Заостренные на конце Щупальца длиннее V2 хоботка (рис. 84).
20 (21). 4-я заднекрайняя ячейка не сужена к вершине. Назальные ячейки лишь
слегка просветлены у вершины 8. Ch. (s. str.) pictus Mg.
21 (20). 4-я заднекрайняя ячейка сужена к вершине. Базальные ячейки с хорошо
выраженным просветом в вершинной части.
22 (25). Вершинное пятно крыла в виде равномерно сужающейся полоски, без
расширения концевой части.
23 (24). Фасетки верхних 2/з гпаз лишь едва крупнее нижних; голова не шире груди.
Внешний край перевязи крыла прямой 15. Ch. (s. str.) suavis Lw.
24 (23). Фасетки верхних 2/з глаз намного крупнее нижних, приблизительно в 5—
6 раз; голова большая, заметно шире груди. Внешний край перевязи с резким
выступом в ячейке R2 5. Ch. (s. str.) anthrax Ols.
4. CHRYSOPS 145
25 (22). Вершинное пятно крыла расширено в концевой части, соединено более узкой,
нежели пятно, перемычкой с перевязью.
26 (31). Перемычка, соединяющая вершинное пятно крыла с перевязью, не шире г/2
радиальной ячейки.
27 (30). II тергит брюшка с. крупным б. м. прямоугольным черным пятном посредине,
занимающим всю среднюю часть тергита, по бокам желтый. Вершинное пятно
крыла налегает на г4 примерно на г/2 ее длины.
28 (29). III и IV тергиты брюшка желтые, по переднему краю с поперечной черной
полоской, расширенной посредине в два направленных вершинами назад
треугольника (рис. 86) 9. Ch. (s. str.) turanicus Ols.
29 (28). Ill и IV тергиты в большей своей части черные, III тергит иногда с небольшим
желтоватым треугольником по заднему краю (рис. 96)
16. Ch. (s. str.) ricardoae PI.
30 (27). II тергит брюшка желтый, с небольшим, обратносердцевидным пятном
посредине (рис. 99). Вершинное пятно узкое, налегает вершиной на г4 не более
чем на 11в ее длины 17. Ch. (s. str.) dissectus Lw.
31 (26). Перемычка, соединяющая вершинное пятно с перевязью, равна или слегка
превосходит ширину Rx 13. Ch. (s. str.) parallelogrammus Zell.
32 (15). Глаза на лбу не соприкасаются (между ними имеется очень узкий, но
явственный промежуток; рис. 100). II тергит брюшка черный, с очень
незначительными буроватыми пятнами по бокам. Анальная ячейка открытая на конце
18. Ch. (s. str.) rufipes Mg.
1. Chrysops (s. str.) japonicus Wied. (рис. 73) —Пестряк японский.
Wiedemann, 1828, Aussereur. zweifl. Ins., 1 : 208; Szilady, 1917 : 114;
S h i r a k i, 1918 : 83; К r 6 b e r, 1920 : 103; Krober in Lindner, 1925 : 13;
Олсуфьев, 1937a : 68; L e с 1 e г с q, 1960b : 47; Takahasi, 1962 : 27; В и о-
л о в и ч, 1968 : 36.
§. Лоб и лицо лишены налета, блестяще-черные. Усики и щупальца черные. Грудь
черная, спинка блестящая, без полосок, в коротких черных волосках. Бочки груди
в черных волосках. Ноги целиком черные. Крылья с резко очерченным
черно-коричневым рисунком. От основания крыла вдоль его внешнего края идет темная полоса,
которая за серединой треугольно расширяется, образуя перевязь. Последняя обычно
достигает заднего края крыла в пределах 4-й заднекрайней ячейки. Задняя базальная
ячейка прозрачная, передняя целиком темная. Внешний край перевязи во 2-й радиаль-
Рис. 73. Chrysops (s. str.) japonicus Wied. Крыло J.
ной ячейке с небольшим углевидным выступом, достигающим основания rt.
Вершинное пятно узкое, резко очерченное, соединено узкой полоской с перевязью. Брюшко
целиком черное, сильно блестящее, покрыто короткими черными волосками. 7—10 мм.
S. Похож на $. Глаза соприкасаются в средней трети. Лицо блестяще-черное.
Щупальца составляют 1/2 длины хоботка. Усики и ноги одноцветно-черные. Спинка
и брюшко блестяще-черные, в черных волосках. Крылья как у $>, но задняя базальная
ячейка темная, как передняя.
Распространение: представитель восточноазиатского типа фауны;
лесной вид, частично в горах (рис. 74, 2). Приморский край и юг Хабаровского края (на
север до Хабаровска и р. Сулук притока Амгуни). Сев.-Вост., Вост. и Ю.-Вост. Китай,
Корейский п-ов, Япония. Местами обычен. Конец мая—июнь.
2. Chrysops (s. str.) sepulcralis F, (рис. 75) — Пестряк траурный.
F a b r i с i u s, 1794, Entom. syst., 4 : 374 (Tabanus); Lundbeck, 1907 : 94;
V e r r a 11, 1909 : 437; P 1 e s k e, 1910b : 436; Szilady, 1917 : 127; Krober
1920 : 98; S urcouf, 1924:218; Krober in Lindner, 1925:20; Олсуфьев,
1937a : 68; M о u с h a et С h v a 1 a, 1955 : 230; Шевченко, 1961 : 33; L e c-
10 H. Г. Олсуфьев
146 II. ПОДСЕМ. CHRYSOPSINAE
1 е г с q, 1960b : 50; В и о л о в и ч, 196.8 :37;Chvala, Lyneborg а. М о и с h a,
1972 : 112. — maurus S i e b к е, 1864 : 108, цит. по: К г 6 Ь е г, 1920 : 98.
$. Лоб и лицо блестяще-черные; лишь от основания усиков на наличник
спускается короткая полоска коричневого налета. Усики черные, относительно короткие.
Щупальца черные, остроконечные, превосходят V2 длины хоботка. Грудь черная;
спинка блестящая, в легком буроватом налете, без явственных полосок, в коричнево-
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° «0° 180° 160°
Рис. 74. Ареалы: 1 — Chrysops (s. str.) sepulcralis; 2 — Ch. (s. stf.) japonicus.
желтых волосках. Бочки груди в желтых, коричневых, реже черных волосках. Ногв
целиком черные. Крылья с нерезким темным рисунком, так как прозрачные части слегка
буровато затемнены. Внешний край перевязи во 2-й радиальной ячейке с углевидным
выступом в сторону основания г4. Задняя базальная ячейка затемнена от основания
на 1/31 передняя на 2/3. Вершинное пятно б. м. узкое, с неясными очертаниями на конце.
Брюшко целиком черное, сверху слабо блестящее, задние края тергитов в желто-серых
или коричневых волосках, тогда как прочие части — в черных. Иногда брюшко
целиком в черных волосках. 7—9.5 мм.
6. Глаза на лбу соприкасаются на
некотором протяжении. Лицо почти
целиком блестяще-черное; лишь от основания
усиков на наличник спускается корот-
Рис. 75. Chrysops (s. str.) sepulcralis F.
Крыло $.
кая, узкая полоска буроватого налета. Усики относительно короткие, черные. Щупальца:
черные, очень узкие, составляют 1/2 длины хоботка. Грудь черная, спинка без полосок,
бочки груди в черных, реже желтых волосках! Ноги целиком черные. Крылья с нерезким
темным рисунком, так как их прозрачные части слегка буровато затемнены. Основание,
большая часть базальных ячеек и передний край крыла коричневые, за серединой от
последнего отходит такого же цвета широкая, поперечная перевязь, б. м. резко
ограниченная в начале, но далее возле заднего края крыла мягко расплывающаяся. Ее
внешний край как у ?. Вершинное пятно к концу несколько шире, нежели у $, кзади с
размытыми очертаниями. Прочее как у ?. 6—8 мм.
Экземпляры с черными волосками на бочках груди и на брюшке, описанные в свое
время Зибке (Siebke) в качестве самостоятельного вида Ch. (s. str.) maurus,
представляют собой темную форму Ch. (s. str.) sepulcralis var. maurus, причем среди самок эта
форма встречается в меньшем проценте, тогда как среди самцов преобладает. Между
основной формой и var. maurus можно найти все переходы. Географическое
распространение у обеих форм б. м. одинаковое.
Распространение: европейско-западносибирский лесной вид (рис. 74, 1).
От Карелии, Эстонии, Литвы и запада Украины на восток до Павлодарской обл.,.
4. CHRYSOPS
147
на север до Хибин, Архангельска, низовий р. Пеша, Печоро-Илычского заповедника
и Тюменской обл. (Нйжнетавдинский р-н), на юг до Львовской, Черниговской, Курской,
Воронежской областей, Южного Урала и Целиноградской обл. Сев. и Средн. Европа.
Не часто. Связан обитанием с болотами.
3. Chrysops (s. str.) nigripes Ztt. (рис. 76) — Пестряк черноногий.
Zetterstedt, 1840, Ins. lap. : 519; Lundbeck, 1907 : 98; Szilady,
1917 : 120; К r 6 b e r, 1920 : 104; S u г с о u f, 1924 : 217; Kroberin Lindner,
1925 : 16; Олсуфьев, 1937a : 70; Moucha et Chvala, 1955 : 230; L e c-
1 er с q, 1960b : 47; Ш е в ч е н к о, 1961 : 34; Т a ka h as i, 1962 : 30; В и о л о в и ч,
1968 :38; Chvala, Lyneborg a. Moucha, 1972 : 114. — lapponicus L о е w,
1858b : 624. — nigricomis (Matsumura) apud S h i r a k i, 1918 : 77.
$. Лоб и лицо покрыты пепельно-серым налетом, иногда с желтоватым оттенком.
Лобная мозоль черная, боковыми сторонами явственно не достигает края глаз. Темя
возле глазков блестяще-черное, возле глаз обычно в сером налете. Лицевые мозоли
Рис. 76. Chrysops (s. str.) nigripes Ztt. Брюшко, крыло, VIII стернит,
церки и, сперматека $>.
округлые, блестяще-черные, наличниковая мозоль такого же цвета, верхние концы
в месте присоединения к лицевым мозолям с легкой перетяжкой. Щечные мозоли
черные, занимают всю нижнюю часть щек. Усики черные; иногда 1-й членик изнутри
коричнево-красный. Щупальца узкие, б. м. остроконечные, коричнево-черные, по длине
равны или слегка превосходят 1/2 длины хоботка. Грудь черная, спинка блестящая,
с двумя матовыми голубовато-серыми полосками, почти достигающими основания
щитка. Боковые стороны спинки и груди в желтовато-сером налете и в
золотисто-желтых волосках. Ноги черные; иногда основания двух последних пар лапок коричневатые.
Крылья прозрачные с резким темным рисунком. В основании и далее по переднему краю
крыло коричнево-черное, за серединой с такого же цвета широкой поперечной
перевязью, достигающей заднего края лишь узким языком вдоль анальной ячейки;
вершинное пятно узкое. Внешний край перевязи во 2-й радиальной ячейке с угловидным
выступом в сторону основания г4. Брюшко широкоовальное, блестяще-черное. II тергит
узкопоперечный, приблизительно в 3—31/2 раза шире своей длины; желтая окраска
кольца почти целиком вытеснена черным рисунком, состоящим из двух б. м.
ромбических пятен, в паре широко сливающихся вершинами. II и последующие тергиты по
заднему краю с желто-серыми каемками, посредине расширенными в небольшие серые
треугольники. Снизу брюшко преимущественно черное, боковые стороны II стернита
часто с грязно-желтыми пятнами; все стерниты по заднему краю с неширокими
желтыми каемками. VIII стернит шире своей длины, гонопофиз с апикальной выемкой,
треугольник посредине обычно темнее боковых сторон; церки обычной формы.
7—11 мм.
в. Глаза на лбу соприкасаются на некотором протяжении. Лицо в желтовато-
сером налете, с очень крупными, блестяще-черными мозолями, слитыми между собой
вдоль наличника. Щупальца черные, узкие, на конце б. м. остроконечные, составляют
около V2 длины хоботка. Усики черные, иногда 1-й членик коричневый. Грудь черная,
спинка с двумя едва заметными узкими желто-серыми полосками; нотоплевры и мезо-
плевры преимущественно в черных волосках, реже целиком в черных или же только
10*
148
П. ПОДСЕМ. CHRYSOPSINAE
в желтых. Ноги черные, основания двух последних пар лапок иногда коричневые.
Крылья в основании и далее по переднему краю темно-коричневые, за серединой с
такого же цвета широкой поперечной перевязью, резко очерченной в начале, но далее
возле заднего края крыла мягко расплывающейся. Базальные ячейки лишь в концевой
трети с прозрачным пятном. Внешний край перевязи как у $. Вершинное пятно к концу
шире, чем у $,'кзади с размытыми очертаниями. Окраска брюшка как у $, .тергиты
с более короткими треугольниками посредине. 8—9 мм.
0° К ВОСТОКу ОТ ГриНВИЧа 90° <20° 150° 180° 150°
Рис. 77. Ареал Chrysops (s. str.) nigripes.
Распространение: голарктический таежный вид. В СССР распространен
от Мурманской и Ленинградской областей на восток до Камчатки и Сахалина (рис. 77).
На север известен до Мурманска, низовий Печоры, Оби, Енисея, Жиганска на Лене
и Анадыря, на юг до Витебской обл., окр. Москвы (Мытищи), Горького, Южного Урала,
Алтая, Забайкалья и юга Приморской обл. Сев. Европа, Вост. Монголия, Сев.-Вост.
Китай, Япония (о. Хоккайдо), Сев. Америка. Часто на болотах. Обычен на севере,
в южной части ареала редок. Летний вид — середина июня—июль. Кладки яиц
обнаружены на нижней стороне листьев осоки по берегам озер (Поляков, 19716).
4. Chrysops (s. str.) divaricatus Lw. (рис. 78) — Пестряк болотный.
Loew, 1858b, Verh. Zool.-bot. Gesell. Wien, 8 : 624; Pleske, 1910a : 431 (<});
S z i 1 a d y, 1917 : 110; К г 6 b e r, 1920 : 106; Krober in Lindner, 1925 : 11;
Олсуфьев, 1937a : 71; M о u с h a et С h v a 1 a, 1955 : 231; L e с 1 e г с q, 1960b :
46- Шевченко, 1961 : 36; В и о л о в и ч, 1968 :40;Chvala, Lyneborg
a. Mouch a, 1972 : 116.
$. Лоб и лицо покрыты желтым налетом. Лобная мозоль крупная, черная,
боковыми сторонами почти соприкасается с краями глаз. Темя возле глазков на
значительном протяжении (до края глаз) блестяще-черное. Лицевые мозоли блестяще-
черные, крупные, округлые, почти соприкасаются с краями глаз, узко соединены
с верхними концами наличниковой мозоли. Щечные мозоли крупные, занимают всю
нижнюю часть лица, узкой блестящей полоской .продолжены к наличнику, обычно
достигая последнего. Усики черные. 1-й членик по крайней мере с внутренней стороны
и часто основание 3-го коричнево-красные. Щупальца коричневые, равны или едва
превосходят Ч2 длины хоботка. Грудь черная, спинка блестящая, с двумя узкими
Желто-серыми полосками, почти достигающими основания щитка. Боковые стороны
спинки и груди в голубовато-желтом налете и в густых золотисто-желтых волосках.
Ноги черные, основание первых двух пар голеней и последних двух пар лапок
коричневые. Крылья в основании и далее по переднему краю черно-коричневые, за серединой
с широкой коричневой поперечной перевязью, достигающей заднего края лишь узким
языком вдоль анальной ячейки. Вершинное пятно узкое, не шире 1-й радиальной
ячейки. Внешний край перевязи во 2-й радиальной ячейке с углевидным выступом
в сторону основания жилки г4. Брюшко широкое, б. м. выпуклое, блестяще-черное,
■с резким желтым рисунком. Боковые стороны I и большая часть II тергита желтые.
Последний посредине с черным пятном в виде широко расставленной концами пере-
4. CHRYSOPS
149
вернутой буквы V.Ee концы слегка изогнуты наружу, не достигают заднего края
тергита, отделяясь от последнего желтой каемкой. Иногда концы пятна сильно укорот
чены и в таком случае рисунок напоминает таковой Ch. (s. str.) relictus. Последующие
тергиты по заднему краю с резкими желтыми каемками на III—V тергитах,
расширенными посредине в желтые треугольники. Снизу брюшко преимущественно желтое.
Первые 2 кольца посредине с небольшим, общим округлым темным пятном. III стер-
нит по переднему краю с полуовальной, далеко не достигающей боковых краев
поперечной полоской. Последующие кольца черные, по заднему краю с очень широкими
желтыми каемками, занимающими до V2 стернита. VIII стернит шире своей длины,
гонопофиз с очень слабой апикальной выемкой; церки удлиненные, с округлой,
вершиной; хитинизированная часть сперматеки с слегка неровной (мелкобугорчатой)
поверхностью. 9—12 .мм.
Рис. 78. Chrysops (s. str.) divaricatus Lw. Брюшко, крыло, VIII стернит,
церки (А — асимметричные) и сперматека $.
в- Глаза на лбу соприкасаются на коротком протяжении. Лицо в желтоватом
налете. Лицевые мозоли блестяще-черные, крупные, округлые, широко прилегают
к нижнему краю глаз, узкими полосками продолжены вниз по бокам наличника.
Щечные мозоли широкие. Щупальца длинные, черно-коричневые. Усики черные,
1-й членик иногда в основании красноватый. Грудь черная, спинка блестяще-черная,
с двумя явственными беловато-серыми полосками, исчезающими возле щитка. Боковые
края спинки и бочки груди в густых, золотисто-коричневых волосках; от основания
крыла к плечевому углу проходит полоска черных волосков. Ноги целиком черные;
иногда основания последних двух пар лапок и основания передних и средних голеней
коричневые. Крылья: основание, большая часть базальных ячеек и передний край
крыла коричневые; от последнего за серединой отходит широкая поперечная перевязь,
достигающая заднего края крыла лишь узким языком вдоль анальной ячейки.
Прозрачное пятно в нижней базальной ячейке больше, нежели в верхней. Вершинное
пятно узкое, не шире 1-й радиальной ячейки, едва заходит за г4. Внешний край
перевязи как у §. Брюшко черное, с желтым рисунком. II тергит желтый, с крупным
черным пятном, занимающим его среднюю часть. Боковые края пятна округло вырезаны,
тогда как по заднему краю вклинивается желтоватый срединный треугольник. III
тергит по переднему краю с поперечной черной полоской, не достигающей внешних
сторон, с небольшим вклинивающимся в нее сзади срединным треугольником.
Последующие тергиты черные, по заднему и боковым краям с желтой каймой, треугольно
расширенной на IV тергите. Снизу брюшко преимущественно желтое. Два первых
стернита с небольшим, темным продолговатым пятном посредине, III стернит с
поперечным пятном по переднему краю; последующие стерниты черные, по заднему краю
с желтыми каемками. 9.5—10.5 мм.
Распространение: таежный вид (рис. 79). От Карелии, Эстонии,
Белоруссии и запада Украины на восток до Магаданской обл. (Верхний Сеймчан
на р. Колыме), на север до северной Карелии (66° с. ш.), ст. Обозерской
Архангельской обл., Лабытнанги в низовье Оби, Снежногорска в низовье Енисея (68° с. ш.)
и Жиганска на Лене, на юг до Воронежской обл., Среднего Урала (Кыштым,
Ильменский хребет), Кокчетавской обл. (Боровое), Алтайского края и Ксеньевки Читинской
ISO II. ПОДСEM. CHRYS0PS1NAE
обл. Сев. и Сред. Европа. Обнаружен в заметном количестве в Ногинске и Учинском
водохранилище Московской обл. (Л. П. Мосолов), в Хреновском бору Воронежской
обл. (Скуфьин, 1952а), в лесах левобережья Волги в Татарской АССР (Волкова, 1950),
Рис. 79. Ареал Chrysops (s. str.) divaricatus.
в некоторых местах лесостепной и таежной зон Сибири и низовий р. Оби (Виолович,
1968; Федоров, 1971), а за рубежом — на юго-востоке ГДР и в южной Баварии, ФРГ
(И. Моуха, личное сообщение); в других частях ареала не часто. Летний вид: июнь—
июль, редко позднее. Личинки найдены во мху по краю гипнового болота (Н. Г.
Олсуфьев).
5. Chrysops (s. str.) anthrax Ols. (рис. 80) — Пестряк стройный.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 74; L е с 1 е г с q,
1960b : 44; Виолович, 1968 : 40.
<j>. Лоб в сероватом, лицо в желтом налете. Лобная мозоль крупная, выпуклая,
блестяще-черная, боковыми сторонами не достигает края глаз, отделяясь полосками
налета. Темя на значительном протяжении (до края глаз) блестяще-черное, покрыто
короткими светлыми волосками. Лицевые мозоли двуцветные: расположенные на
щеках — блестяще-черные или черно-коричневые, на наличнике — коричневые или"
желтоватые. Наличниковая мозоль коричневая, соединена широкими перемычками
с лицевыми мозолями. Щечные мозоли крупные, блестяще-черные, занимают всю
нижнюю часть лица и достигают наличника. Усики умеренной длины. 1-й и 2-й членики
желтовато-коричневые, в черных волосках, 3-й членик черный, в основании
коричневый. Щупальца черные или черно-коричневые, остроконечные, превосходят 1/2 длины
хоботка. Грудь черная, спинка слабо блестящая, с двумя довольно широкими
голубовато-серыми полосками посредине, почти достигающими основания щитка'. Боковые
стороны спинки и груди в желто-сером налете и в золотисто-желтых волосках; от
основания крыла к плечевому углу проходит коричневая полоска, покрытая более
темными волосками. Ноги черные, средние голени темно-коричневые, основания лапок
последних двух пар ног коричневые. Крылья прозрачные, с интенсивным темным
рисунком. Внешний край перевязи с резким углевидным выступом в пределах 2-й
радиальной ячейки; позади г4+5 перевязь не превышает шириной дискоидальную ячейку.
Вершинное пятно в виде очень узкой полоски, идущей от перевязи вдоль внешнего
края крыла и исчезающей, не достигая г4. Передняя базальная ячейка затемнена в
основной половине, задняя в основной четверти. 4-я заднекрайняя ячейка б. м. сужена
к вершине. Брюшко слегка трапециевидное, б. м. узкое. I тергит по бокам коричнево-
желтый, посредине черный, II тергит грязно-желтый, посредине с черным пятном
в виде перевернутой буквы V с широко расставленными концами (как у Ch. (s. str.)
divaricatus). Последующие кольца черные, с желтыми каемками по заднему краю,
расширенными посредине в небольшие треугольники. Эти каемки, так же как и
желтые участки на I и II тергитах, покрыты короткими желтыми волосками, прочее в
черных волосках. Снизу брюшка первые два кольца желтые, последующие черные. 8—
10.5 мм.
4. CHRYSOPS 151
cJ. Голова относительно большая, в поперечнике немного шире спинки. Глаза
соприкасаются на значительном протяжении. Фасетки верхних 2/3 глаз значительно
.крупнее нижних, приблизительно в 5—6 раз, граница между теми и другими резкая.
Усики: 1-й членик желтый, 2-й желто-коричневый, оба в довольно длинных черных
волосках, 3-й членик узкий,, черный, площадка в базальной половине коричневая.
Лицевые мозоли двуцветные: расположенные на щеках черные, на наличнике —
светло-коричневые; щечные мозоли большие, блестяще-черные. Щупальца коричневые,
длинные, остроконечные, составляют не менее 1/2 длины хоботка. Темный рисунок
крыла сходен с таковым самки, но вершинное пятно немного шире ячейки Rx и концом
достигает г4, перевязь по внешнему краю немного шире дискоидальнои ячейки и кзади
достигает края крыла в пределах ячеек Pi и А, обходя среднюю часть Р5, передняя
базальная ячейка затемнена от основания на 2/3 и задняя — на 1/3. 4-я заднекрайняя
ячейка сужена к вершине. Ноги черные, средние голени и основание средних и задних?
лапок темно-коричневые. Брюшко сверху преимущественно коричневое, V—VI тер-
гиты и небольшой участок на I тергите кзади от щитка черные. I тергит по бокам и
II тергит беловато-желтые, срединное коричневое V-образное пятно на последнем
с широко расставленными концами, кзади от пятна имеется небольшой светло-желтый
треугольник. Последующие тергиты каждый по заднему, краю с неширокой желтоватой
или сероватой каймой, посредине расширенной в маленький треугольник. Светлые
части брюшка в светлых, темные в коротких черных прилегающих волосках. Снизу
брюшко в основной половине желтое, IV—VII стерниты черноватые. 9 мм.
5 имеет сходство с Ch. (s. str.) divaricatus, но хорошо отличается от последнего-
двуцветными лицевыми мозолями, рисунком крыла и брюшка. Более светлая окраска
брюшка у с? в сравнении с 5, а также относительно большая голова у $ с очень
крупными фасетками верхних частей глаз выделяют этот вид от других палеарктических
видов подрода Chrysops. В подроде Heterochrysops большую голову и укрупненные-
фасетки верхних частей глаз имеет S Ch. (Heterochrysops) vanderwulpi.
Распространение: юг Приморского края (оз. Ханка; Новокиевское
в р-не Посьета; оз. Хасан; Спасск). Сев.-Вост. Китай. Редок. Июль—август.
Голотип: 5, берег оз. Ханка, Приморский край, 7 VII 1932, А. Булдовский. В
коллекции Зоологического института Академии наук СССР, Ленинград. S описывается
нами впервые по экземпляру из окр. Спасска Приморского края, пойманному 19 VIII
1963 Э. П. Нарчук.
6. Chrysops (s. str.) makerovi PL (рис. 81) — Пестряк Макерова.
Р 1 е s k e, 1910b, Ann. Mus. Zool. St. Petersb., 15 : 469; S z i1 a d y, 1917 : 116;
К г о b e r, 1920 : 96; К r 6 b e r in Lindner, 1925 : 13; О л с у ф ь е в, 1937а : 75;
Leclercq, 1960b : 47; takahasi, 1962:29; Виолович, 1968:41.—
loewi К г 6 b e r, 1920 : 106. — nigricornis Matsumura, 1911 : 66, цит. по:
Leclercq, 1960b : 47.
<j>. Лицо и лоб в серовато-желтом налете. Лобная мозоль крупная, блестяще-
черная. Темя на значительном протяжении блестяще-черное. Лицевые мозоли
крупные, округлые, черные, наличниковая мозоль такого же цвета, ее верхние концы
в месте присоединения к лицевым мозолям с легкой перетяжкой. Щечные мозоли
небольшие, черные. Усики относительно короткие, черные. Щупальца узкие,
коричнево-черные, остроконечные. Грудь черная; спинка с 2 серыми полосками,
исчезающими за серединой. Боковые края спинки и бочки груди в сером налете и серовато-
152 П. ПОДСЕМ. CHRYSOPSINAE
желтых волосках. Крылья: внешний край перевязи с небольшим уголком возле г4+5,
далее прямой. Вершинное пятно б. м. крупное, соединено с перевязью по ширине
ячейки i?j. Дискоидальная ячейка часто слегка (у облетанных экземпляров сильно!)
просветлена в средней части. Ноги одноцветно-черные, изредка средние голени в
основании коричневые. Брюшко: II тергит желтый, с V-образным черным пятном посредине;
концы пятна не достигают заднего края тергита. Иногда это пятно имеет форму двух
треугольников, соединенных в паре, напоминая рисунок брюшка Ch. (s. str.) relictus.
Треугольник позади пятна беловатый. Последующие кольца блестяще-черные, с
неширокими, но резкими желтоватыми или беловатыми задними каемками,
расширенными посредине в небольшие треугольники. Снизу брюшко в основании желтое (I—
II стерниты) с темным пятном посредине, III и последующие стерниты черные, с
желтовато-серыми широкими каемками по заднему краю. 10—12.5 мм.
3 неизвестен.
Наиболее близок к Ch. (s. str.) divaricatus и Ch. (s. str.) nigripes, от которых легко
отличается крупным вершинным пятном на крыльях, незначительным выступом по
внешнему -краю перевязи возле г4+5, а также рисунком брюшка.
Распространение: восточносибирский таежный вид, преимущественно
в горах (рис. 64, 3). От Алтая и Красноярска на восток включая Камчатку, Сахалин
и о. Кунашир; на север до Северо-Енисейска Красноярского края, устья р. Алдан
севернее Якутска, Верхнего Сеймчана на Колыме и Маркова на р. Анадырь, на юг
до Забайкалья, Амурской обл. и юга Приморского края (Супутинский заповедник).
Сев.-Вост. Китай, Япония (о. Хоккайдо). Обычен местами в Восточной Сибири и на
Сахалине. Июнь—август.
7. Chrysops (s. str.) caecutiens L. — Пестряк лесной.
Представлен двумя формами: северным номинативным подвидом Ch. (s. str.)
caecutiens caecutiens L. и южным подвидом Ch. (s. str.) caecutiens ludens Lw.
7a. Chrysops (s. str.) caecutiens caecutiens L. (рис. 68, 69, 70, 71) — Пестряк лесной
номинативный.
L i n n ё, 1758, Syst. Nat. : 602 (Tabanus); Lundbeck, 1907 : 97; V e г г a 1 1,
1909:427; S z i 1 a d у, 1917:106; К г 6 b e г, 1920:118; S u г с о u f, 1924:221;
Krober in Lindner, 1925 : 8; Олсуфьев, 1937a : 76; M о и с h a et
Chvala, 1955 : 231; L e с 1 e г с q, 1960b : 44; Шевченко, 1961 : 37; Виол о-
вич и Евстигнеева, 1964 : 106 (экология); С к у ф ь и н, 1967 : 184
(личинка); Виолович, 1968 : 43; J e z e к, 1970 : 376 (личинка, куколка); Chvala,
Lyneborg a. Moucha, 1972:118.
$>. Лоб и лицо покрыты желтовато-серым налетом. Лобная мозоль черная,
крупная, боковыми сторонами почти соприкасается с краями глаз. Темя возле глазков
на значительном протяжении блестяще-черное (до края глаз). Лицевые мозоли
крупные, блестяще-черные, почти соприкасаются с краями глаз, широко соединены с на-
личниковой мозолью (рис. 70). Щечные мозоли черные, небольшие. Усики целиком
черные, реже 1-й членик коричнево-желтый. Щупальца черные. Грудь черная, спинка
блестящая, с двойной, широкой, желто-серой полосой; последняя исчезает за
серединой спинки. Боковые стороны спинки и бочки груди в голубовато-желтом налете и
густых золотисто-желтых волосках. Ноги черные; основания средних голеней и по-
4. CHRYSOPS
153
следних двух пар лапок слегка коричневатые. Крылья с широкой черно-коричневой
поперечной перевязью, достигающей заднего крыла и с крупным вершинным пятном;
последнее соединяется с перевязью по ширине радиальной ячейки. Внешний край
перевязи кзади от г2+3 выпуклый или же образует легкий угол, вершина которого
лежит возле г4+5. Брюшко б. м. трапециевидное; боковые стороны I и большая часть
II тергита желтые. Последний посредине с черным пятном в виде перевернутой буквы V".
Пятно иногда настолько развито, что лежащий за ним светлый срединный треугольник
почти вытеснен, иногда же пятно в виде тонких, почти исчезающих черточек. Концы
пятна достигают заднего края кольца, образуя на некотором протяжении черную
каемку. Последующие тергиты целиком черные, иногда с едва намечающимися
маленькими буроватыми треугольниками посредине. Первые два стернита желтые, из них
I стернит посредине с большим прямоугольным, II с треугольным или также
прямоугольным темным пятном, часто слиты вершиной с предыдущим. Желтая окраска
иногда заходит по бокам на III стернит, который вместе с последующими кольцами
Рис. 82. Ареалы: 1 — Chrysops (s. str.) caecutiens caecutiens; 2 — Ch. (s. str.)
caecutiens ludens.
черный, в сероватом налете. VIII стернит шире длины, часто по бокам угловатый,
гонопофиз с небольшой апикальной выемкой или без нее; церки обычной формы;
вершина сперматеки коническая (рис. 54, ///, В, Г, Д, Е). 7.5—11 мм.
cJ. Глаза на лбу соприкасаются на некотором протяжении. Лицо в тонком,
желтовато-сером налете, с очень крупными, блестяще-черными лицевыми мозолями (рис. 71).
Щупальца черные, б. м. остроконечные, составляют приблизительно V2 длины
хоботка. Усики черные. Грудь черная; нотоплевры в черных, мезоплевры в густых
золотисто-желтых волосках. Ноги черные; основания двух последних пар лапок
коричневатые. Крылья в основной части целиком затемнены, так что перевязь и
затемненное основание крыла слиты. Внешний край перевязи кзади от г2+3в виде слегка
выпуклой линии. Вершинное пятно крупное, широко соединено с перевязью. На
медиальной жилке возле дискоидальной ячейки лежит маленькое беловатое пятнышко.
Брюшко сверху матово-черное, в черных волосках, на последних кольцах с примесью
желтых. Боковые края II тергита коричневые. Снизу брюшко черное; II стернит по
бокам часто коричневый. 8—10 мм.
Распространение: европейско-сибирский лесной подвид, широко
проникает в таежную зону (рис. 82, 1). В СССР от Мурманской обл. (р. Лотта),
Прибалтики, запада Украины и Молдавии на восток включая Якутию и север Хабаровского
края (Нелькан), на север до низовий Печоры, Енисея, Хатанги (72° с. ш.) и Жиганска
на Лене, на юг включая Кавказ (горы), Сев. Казахстан (Боровое, Павлодар), Алтай,
Джунгарский Алатау (на юг до Панфиловского р-на), Туву и Забайкалье. Европа,
Зап. Китай (Синьцзян), Сев. Монголия. Обычен. С конца мая по август. Держится
преимущественно близ ручьев и речек. Откладка яиц над водой, на нижней
поверхности листьев травянистой и кустарниковой растительности. Нехищные личинки
обитают на дне и в берегах ручьев, речек, реже озер, канав и прудов; куколки в тех же
местах, но над урезом воды.
Экспериментально доказанный переносчик туляремии (Боженко, 1941),
154 II. ПОДСЕМ. CHRYSOPSINAE
7b. Chrysops (s. str.) caecutiens ludens Lw. (рис. 83) — Пестряк лесной южный.
L о е w, 1858b, Verh. Zool.-bot. Gesell. Wien, 8 : 628 (Chrysops ludens); S z i 1 a d y,
1917 : 107 (Ch. caecutiens var. ludens); Krober in Lindner, 1927 : 13. — me-
ridionalis Strobl, 1906 : 277 ($) (пит. по: L e с 1 e г с q, 1960b : 45); S u г с о u f,
1924 : 223. — trifenenstratus Krober, 1920:119 (<J); S u г с о u f, 1924:223.—
hermanni Krober, 1920 : 117 ($).
Отличается от основной формы следующим. §: V-образный рисунок на II тергите
■брюшка отсутствует целиком или же едва намечен; на III—-IV тергитах имеются не-
Рис. 83. Chrysops (s. str.) caecutiens caecutiens L. VIII стернит, церки
и семеприемники j. A — Ch. (s. str.) caecutiens ludens Lw. Брюшко J.
•большие срединные желтые пятна, в'- волоски на бочках груди чисто-черные и
вершинная часть базальных ячеек светлая. Кроме того, у обоих полов вершинное пятно
крыла более широкое.
Распространение: Ю.-З. Украина, Крым, Кавказ. Ю. Европа, Турция,
Сев. Иран (рис. 82, 2).
8. Chrysops (s. str.) pictus Mg. (рис. 84) — Пестряк украшенный.
М е i g e n, 1820, Syst. Beschreib., II : 70; L u n d b e с k, 1907 : 95; V e r r a 11,
1909:429; S z i 1 a d y, 1917:122; Krober, 1920:125; S u г с о u f, 1924:225;
Krober in Lindner, 1925 : 18; Олсуфьев, 1937a : 78; M о и с h a et
Ch v a 1 a, 1955 : 233; L e с 1 e г с q, 1960b : 48; Ш е в ч е н к о, 1961 : 39; Скуфьин,
1967:181 (личинка); Виолович, 1968:44; J e z e к, 1970:378 (личинка,
куколка). — quadratus M e i g e n, 1820, 1. с. : 70. — viduatus Fabricius, 1794,
1. с. : 374 (nomen oblitum, Olsufjev a. Leclercq, 1971 : 325; Олсуфьев,
1971 : 138).
J. Лоб и лицо покрыты желтым налетом. Лобная мозоль черная, крупная, почти
соприкасается с краями глаз. Темя возле глазков на значительном протяжении (до
края глаз) блестяще-черное. Лицевые мозоли блестяще-черные, крупные, округлые,
б. м. широко соединены с наличниковой мозолью. Щечные мозоли маленькие, черные,
кругом изолированы налетом. Усики черные; 1-й и внутренняя сторона 2-го членика
коричнево-красные. Щупальца длинные, с слегка притуплённой вершиной,
коричневые, превосходят 1/2 длины хоботка. Грудь черная, спинка блестящая, с двойной,
широкой, за серединой исчезающей, голубовато-желтой полоской. Боковые стороны
спинки и бочки груди в желтом налете и густых золотисто-желтых волосках. Ноги:
основания передних, целиком средние и задние голени и основания двух последних
пар лапок коричнево-желтые; прочие части ног черные. Изредка средние и задние
бедра коричневатые. Крылья: перевязь широкая, достигает заднего края '.крыла,
ее внешний край позади г2+3 выпуклый. Вершинное пятно широкое, захватывает
2/3 длины жилки г4; пятно соединено с перевязью широкой полоской, слегка
превосходящей ширину 1-й радиальной ячейки. Брюшко трапециевидное, с сильно развитым
желтым рисунком. II тергит желтый, у переднего края с незначительным лунковид-
ным, квадратным или прямоугольным черным пятном (изредка может отсутствовать).
Последующие тергиты черные, III с широкой, остальные с более узкими желтыми
каемками по заднему краю; на III—IV кольцах каемки расширены посредине в
короткие треугольники, не достигающие переднего края тергита. Брюшко снизу в основ-
4. CHRYSOPS H5
ной половине желтое; I и II стерниты с незначительным, III с б. м. крупным поперечно-
овальным темным пятном посредине. Последующие стерниты черные, с желтыми
каемками по заднему краю. VIII стернит шириной превосходит длину, боковые стороны
угловатые, гонопофиз с значительной апикальной выемкой; церки обычной формы;
сперматека с конической вершиной. 8—12 мм.
3. Глаза на лбу на некотором протяжении соприкасаются. Лицо в желто-сером
налете. Лицевые мозоли крупные, округлые, блестяще-черные, продолжены вниз
по бокам наличника сначала узкими, затем резко расширенными блестящими
полосками. Щечные мозоли блестяще-черные, поперечные, внутренним концом сливаются
с блестящим наличником. Концевой членик щупалец черный, узко заостренный,
длинный, явственно превосходит 1/2 длины хоботка (рис. 84, Г). Усики черные, иногда
Рис. 84. Chrysops (s. str.) pictus Mg. A—E — брюшко, крыло, VIII стернит
(варианты) и церки (варианты) $; Ж, 3 — брюшко и голова 6.
в основании коричневатые. Грудь черная; спинка без полосок, блестящая. Мезоплевры
в черных, нотоплевры в желтых, реже также в черных волосках. Ноги
преимущественно черные; передние и задние голени в основании, средние голени целиком и
основания последних двух пар лапок коричневые. Крылья: перевязь как у <j>,
вершинное пятно очень широкое, почти целиком налегает на г4. Базальные ячейки затемнены
почти нацело; исключение составляет небольшой просвет в вершинной \/3 передней
базальной ячейки, который заходит на незначительном протяжении в заднюю ячейку.
Прозрачные участки крыла в вершинной и задней его частях слегка буровато
затемнены. Прочее как у <j>. Брюшко черное, с желтым рисунком. II тергит желтый,
посредине с большим, трапециевидным черным пятном, занимающим всю среднюю часть
тергита. Последующие тергиты черные, по заднему краю с буро-желтыми каемками;
на III тергите желтые каемки по бокам резко расширены и по самому краю вытесняют
черный рисунок. Снизу брюшко черное; I—II (или I—III) стерниты по бокам желтые,
последующие с узкими желтыми каемками по заднему краю. 8—10 мм.
Распространение: европейско-западносибирский лесной вид, широко
распространен в европейской части СССР в лесной и лесостепной полосе, частично
•заходит в таежную зону (рис. 85,1). На север известен до южных частей Карелии (Мед-
вежьегорск), Архангельской обл. (Котлас), Среднего Урала (Никито-Ивдель), на юг
включая Одесскую обл., Молдавию, Крым, Северный Кавказ (юг Ставропольского
края; Северо-Осетинская, Кабардино-Балкарская, Чечено-Ингушская и
Дагестанская АССР) и Закавказье (Закатальский заповедник, Азербайджан; Махарадзевский р-н,
Грузия); по ту сторону Урала спорадические находки в Челябинской (Троицк),
Курганской (Шадринск), Кокчетавской (Боровое) и Томской областях (окр. Томска и
156 II. ПОДСЕМ. CHRYSOPSINAE
Асиновский р-н). Европа (кроме крайнего севера), Турция. Обычен в южных частях
лесной зоны. Летний вид — с середины июня по середину августа. Личинки найдены
в небольших водоемах в черноольховой топи (Скуфьин, 1952а).
Рис. 85. Ареалы: 1 — Chrysops (s. str.) pictus; 2 — Hybomitra (s. str.) ussu-
riensis.
Хвала и Линеборг (Ghvala a. Lyneborg, 1970) предложили для Chrysops (s. str.)
pictus Meigen, 1820, восстановить более старое название Ch. (s. str.) viduatus Farbicius,
1794, но оно является nomen oblitum, так как не применялось в основной
зоологической литературе свыше 50 лет.
9. Chrysops (s. str.) turanicus Ols. (рис. 86) — Пестряк туранский.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 80; L е с 1 е г с сг,
1960b : 51; Шевченко, 1961 : 40.
$>. Лоб в желтом налете. Лобная мозоль умеренной величины, блестяще-черная,
не достигает края глаз. Темя возле глазков в тонком налете, не блестящее. Лицевые
мозоли коричневые, небольшие, соединены с наличниковой мозолью б. м. широкой
перемычкой. Щечные мозоли блестяще-черные, крупные, достигают наличника. Усики
умеренной длины, 1-й членик коричнево-желтый, 2-й членик коричневатый изнутри,
3-й целиком черный. Щупальца б. м. остроконечные, коричнево-черные. Грудь
черная; спинка с двумя сравнительно узкими желтовато-серыми полосками. Боковые
стороны^ спинки и бочки груди в желтом налете и коротких желтых волосках. Крылья:
внешний край позади г2+3 слегка выпуклый, далее, перейдя т^, вогнутый. Вершинное
пятпо б. м. широкое, налегает на г4 на V2 ее длины; пятно соединено с перевязью
очень узкой полоской вдоль переднего края крыла, не превышающей V2 ширины 1-й
радиальной ячейки. Назальные ячейки затемнены в основании, передняя на 1/3, задняя
на VB длины. 5-я заднекраиняя ячейка просветлена по средней линии. 4-я
заднекраиняя ячейка сильно сужена в вершине. Ноги черные; средние голени и основания
средних и задних лапок светло-коричневые. Брюшко: I и II тергиты желтые; I тергит
с черным прямоугольным пятном посредине, пятно несколько ^же щитка. II тергит
с короткой, черной продольной черточкой (или точкой или без нее) в передней части
кольца. III и IV тергиты черные, с широкой желтой каемкой по заднему краю; эти
каемки расширены посредине в треугольники, достигающие вершиной переднего края
тергитов. Последующие тергиты желто-серые. Снизу брюшко в основной половине
желтое, IV и последующие стерниты темные. 9—9.5 мм.
в- Глаза соприкасаются на некотором протяжении. Лицо в желтом налете.
Лицевые мозоли округлые, блестяще-черные, соединены б. м. узкими перемычками с
наличниковой мозолью. Щечные мозоли б. м. крупные, достигают наличника. Усики
умеренной длины, черные. 1-й членик и внутренняя сторона 2-го
желтовато-коричневые. Щупальца коричнево-черные, остроконечные, превышают V2 длины хоботка.
Грудь черная; спинка в передней части с двумя неясными серыми полосками. Боковые
стороны груди и спинки в желтоватом налете и желтых волосках. Крылья: перевязь
и вершинное пятно как у д. Передняя базальная ячейка затемнена на половину от
4. CHRYSOPS 157
основания, задняя на две трети, причем затемнение идет вдоль всей си2 и достигает
заднего края крыла; 5-я заднекраиняя- ячейка просветлена по средней линии; прочее
как у $. Ноги как у $. Брюшко желтое, с черным рисунком; I тергит желтый по бокам,
тогда как в средней части и по переднему краю черный. II тергит с крупным
прямоугольным черным пятном посредине, занимающим V3 ширины брюшка. III и IV
тергиты по переднему краю с черными каемками, расширенными в средней части в два
треугольничка, не достигающих вершинами заднего края тергита. Последующие
Рис. 86. Chrysops (s. str.) turanicus Ols. А, В — брюшко и крыло ?; Б и Г — брюшко
и крыло 6.
тергиты желтые. Снизу брюшко в основной половине желтое, с небольшим темным
пятном в основании, IV и последующие стерниты черные. 9 мм.
Близок к Ch. (а. str.) pictus, отличаясь в основном следующим: 1) менее широким
вершинным пятном, узко соединенным с перевязью; 2) суженной в вершине 4-й задне-
крайней ячейкой; 3) брюшко с более развитым желтым рисунком, в частности, у $
темное пятно на II тергите в виде незначительной черточки.
Распространение: Узбекистан (Ташкентский р-н; Куропаткино близ
Самарканда), Киргизия (Джалалабад; Чаткальский хребет), Чимкентская обл. КазССР
(хр. Каржантау; долина Угама притока Чирчика). Редок.
Голотип: ?, Джалалабад б. Андижанского у., 10—13 VI 1909 (ст. ст.), Рыжикова.
В коллекции Зоологического института Академии наук СССР, Ленинград.
10. Chrysops (s. str.) validus Lw. (рис. 87) — Пестряк большой.
L о е w, 1858b, Verh. Zool.-bot. Gesell. Wien, 8 : 619; S z i 1 a d y, 1917 : 129;
К r 6 b e r, 1920 : 72; К r 6 b e r in Lindner, 1925 : 21; S z i 1 a d y, 1926 : 598
(6); Олсуфьев, 1937a : 82; L e с 1 e г с q, 1960b : 51; Виолович, 1968 : 45.
Крупный, коренастый вид с резко выраженным половым диморфизмом в окраске.
<j>. Лицо и лоб в желтом налете. Лобная мозоль очень крупная, блестяще-черная.
Темя возле глазков блестящее. Лицевые мозоли черные, широко слиты с концами
наличниковой мозоли. Щечные мозоли маленькие. Усики умеренной длины, 1-й и
частью 2-й членик коричнево-желтые, 3-й членик черный. Щупальца черные. Грудь
черная; спинка в буроватом налете с двумя желтыми нерезкими полосками
посредине. Боковые стороны спинки и бочки груди в желтом налете и густых золотисто-
желтых волосках. Ноги черные. Основания средних голеней и лапок двух последних
пар ног слегка коричневые. Крылья: внешний край перевязи изогнут под тупым углом,
вершина которого лежит возле г4+5. Вершинное пятно широко обособлено от
перевязи просветом. Перевязь целиком простирается до заднего края крыла. Базальные
ячейки затемнены от основания наполовину. Брюшко: II тергит желтый, посредине
с двумя черными б. м. треугольными пятнами, соединенными в паре (как у Ch. (s. str.)
' relictus). Последующие кольца черные, с неясными задними каемками и
треугольниками посредине, образованными налетом и желтоватыми волосками. Снизу брюшко
в основании желтое, с темным пятном посредине, III и последующие стерниты темные.
Иногда III стернит по бокам желтый. 10—13 мм.
S. Лицо блестяще-черное; имеется лишь узкая полоска желтоватого налета на
наличнике. Усики целиком черные. Концевой членик щупалец маленький, черный,
тупой на конце, составляет 1/3 длины хоботка. Грудь и брюшко целиком черные, в чер-
158
II. ПОДСЕМ. CHRYSOPSINAE
ных волосках. Ноги черные, лишь основания лапок двух последних пар ног слегка,
коричневатые. Крылья: внешний край перевязи и вершинное пятно как у $
(последнее несколько шире, нежели у 5); основная половина крыла почти целиком затемнена,
за исключением небольшого просвета («окошечка»), расположенного в концевой трети,
обеих базальных ячеек. 9—11.5 мм.
Рис. 87. Chrysops (s. str.) validus Lw. Брюшко и
крыло §.
6 похож на Ch. (s. str.) caecutiens ludens, но легко отличается от последнего более-
крупными размерами, одноцветно-черным брюшком и обособленным от перевязи
вершинным пятном, g напоминает Ch. (s. str.) relictus, легко отличаясь
обособленным вершинным пятном крыла.
Распространение: восточносибирский таежный вид (рис. 101, 2).
От Братска и Иркутска на восток до низовий Амура и юга Приморского края, на
север до Жиганска в Якутии и низовий Алдана. Вост. Китай, Вост. Монголия,
Корейский п-ов. Не часто. Июль. Указание В.|Г. Фоминых с соавт. (1972) для Томской обл.
ошибочно. 4
»
11. Chrysops (s. str.) relictus Mg. (рис. 88, 89) — Пестряк обыкновенный.
М е i g е п, 1820, Syst. Beschreib., II : 69; Lundb.eck, 1907 : 96; V е г г а 1 1,.
1909 : 434; S z i 1 a d у, 1917 : 124; К г 6 Ь е г, 1920 : 124; S u г с о и f, 1924 : 224;.
Krober in Lindner, 1925 : 18; Олсуфьев, 1937а : 80; Т а м а р и н а,
1951 : 101 (биология); С к у ф ь и н, 19526 : 664 (экология); MouchaetChvala,
1955 : 234; Тамарина, 1956 : 172 (личинка, куколка);' Leclercq, 1960b :
48; Шевченко, 1961 : 40; С к у ф ь и н, 1967 : 185 (личинка); В и о л о в и ч,.
1968:45; J e z e к, 1970:378 (личинка, куколка); Chvala, Lyneborg a.
Moucha, 1972 : 124. — melanopleurus W a h 1 Ь е г g, 1848 : 200.
$. Лоб в сером, лицо в желтоватом налете. Лобная мозоль умеренной величины,
черная, поперечно-овальная, боковыми сторонами не достигает края глаз. Темя
вокруг глазков блестяще-черное, возле глаз в сером налете. Лицевые мозоли блестяще-
черные, умеренных размеров, не соприкасаются с краями глаз. Наличниковая мозоль-
посредине с тонкой полоской налета, делящей ее на две половинки. Эти последние
соединены узкой перемычкой с лицевыми мозолями. Щечные мозоли незначительной
величины, блестяще-черные, со всех сторон изолированы налетом. Усики черные;
1-й, 2-й (изнутри) и основание 3-го членика коричнево-красные, но иногда усики
целиком черные. Концевой членик щупалец коричневый, б. м. остроконечный. Грудь-
черная, сверху с широкой, двойной, матово-серой полосой посредине, достигающей
основания щитка. Боковые стороны спинки и груди в желтовато-сером налете и
в серо-желтых волосках. Щиток в матово-сером налете. Ноги: основания V2
передних голеней, целиком средние и задние голени и основания последних двух пар лапок
желто-коричневые; прочие части ног черные. Крылья: перевязь широкая, достигает
заднего края крыла. Дискоидальная, 4-я и 5-я заднекрайние ячейки и 2-я
радиальная ячейка в средней части слегка просветлены. Внешний край перевязи кзади от г2+3:
в виде выпуклой линии. Вершинное пятно большое, его соединение с перевязью слегка
превосходит ширину 1-й радиальной ячейки. На конце пятно расплывчато сливается
с слегка буровато затемненной прозрачной частью крыла, тогда как возле основания
г4 имеет с перевязью неясную перемычку, обособляя таким образом во 2-й
радиальной ячейке округлое светлое пятно. 4-я заднекрайняя ячейка к концу иногда слегка.
4. CHRYSOPS J5y
сужена. Анальная ячейка часто (у 1/3 особей) не замкнута на конце. Брюшко черное,
с сильно развитым желтым рисунком; II тергит желтый, с двумя обычно соединенными
в паре че-рными треугольными пятнами. Последующие тергиты блестяще-черные,
по заднему краю с желтыми каемками, посредине расширяющимися в желто-серые
треугольники.' I и II стерниты желтые, с темным срединным пятном, последующие
черные, с желтыми каемками по заднему краю; иногда желтые каемки на III—IV
стернитах почти целиком вытесняют черный цвет. VIII стернит имеет ширину, б. м.
равную длине, боковые стороны округлены, гонопофиз с небольшой апикальной
выемкой; церки иногда слегка удлинены, X тергит крупнее, чем обычно; вершина спер-
матеки приострена или тупая. 7.5—12 мм.
3- Глаза на лбу соприкасаются на некотором протяжении. Лицо в желто-сером
налете. Лицевые мозоли крупные, округлые, блестяще-черные, узкими полосками:
Рис. 88. Chrysops (s. str.) relictus Mg. ?.
продолжены вниз по бокам наличника и здесь между собой не слиты, широко
разделяясь налетом. Щечные мозоли блестяще-черные, узкопоперечные, внутренним
концом достигают блестящих полосок наличника, сливаясь с последними. Концевой
членик щупалец черный, маленький, тупоконечный, короче V2 длины хоботка. Усики,
черные; 1-й и 2-й членики иногда изнутри коричневатые. Грудь черная; спинка с
неясной, двойной, б. м. широкой серой полоской, исчезающей за серединой. Боковые
стороны спинки и груди в тонком сером налете, покрыты черными и желтыми
волосками. Ноги и крылья как у $; базальные ячейки в основной половине затемнены.
Брюшко желтое, с черным рисунком. II тергит с двумя широко соединенными в паре-
черными треугольными пятнами, занимающими всю среднюю часть тергита. Два
последующих тергита спереди с черной поперечной полоской, не достигающей боковых
. краев кольца; полоска в средней части расширена в два полукруглых или
треугольных выступа. Последующие тергиты черные. Снизу брюшко в основной половине-
.желтое. I—II стерниты с общим пятном, два последующих с изолированными
небольшими темными пятнами посредине. V и последующие стерниты черные, с желтыми
каемками по заднему краю. 7.5—11 мм.
Распространение: лесостепной вид, в СССР широко распространен
в степной, лесостепной и лесной полосе, на севере частично в таежной и тундровой,
зоне, а на юге — в полупустынной зоне (рис. 90). На север известен до Мурманска,
Канина п-ова (оз. Парусное), южной части о. Колгуева, Усть-Цильмы на Печоре,
низовий Оби, Игарки на Енисее, Сев. Приангарья и Жиганска на Лене, на восток
до Якутска, Сев. Прибайкалья и Иркутска, на юг включая Одесскую обл.,
Молдавию, Крым, Калмыцкую АО (Элиста), дельту Дона, Волги и Урала, Центральный
Казахстан (до 48° с. ш.), Аягуз (в 50 км от устья) и оз. Зайсан. Европа (кроме
крайнего юга), Зап. Китай (Синьцзян), Сев. Монголия (Улясутай). Распространение-
МО П. ПОДСЕМ. CHRYSOPSINAE
в|Якутии и Сев. Монголии, возможно, изолировано от основного ареала. Обычен
по берегам водоемов степной и лесостепной полосы Европы и Зап. Сибири, в особен-
Рис. 89. Chrysops (s. str.) relictus Mg. А— Д — брюшко, крыло, VIII стернит,
церки и сперматека $; Е, Ж — брюшко и голова 6.
ности многочислен в поймах больших рек, по которым распространен далеко на север.
Июль—август, иногда раньше или позднее. Кладка яиц и личинки по берегам
пресных озер, рек с замедленным течением, стариц, каналов и прудов.
Рис. 90. Ареал Chrysops (s. str.) relictus.
Часто нападает на человека. Доказанный переносчик туляремии (Сомов с соавт
1940: Боженко, 1941),
4. CHRYSOPS 161
12. Chrysops (s. str.) angaricus Ols. (рис. 91) — Пестряк ангарский.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2:85; L е с 1 е г с q,
1960Ь : 44; В и о л о в и ч, 1968 : 48.
$. Лоб и' лицо в желтоватом налете. Лобная мозоль крупная, матово-черная.
Темя возле глазков слегка блестящее. Лицевые мозоли округлые, блестяще-черные;
наличниковая мозоль небольшая, черная, соединена с лицевыми мозолями очень
узкими перемычками. Щечные мозоли умеренных размеров, далеко не достигают
наличника. Усики длинные, 1-й членик желто-коричневый, последующие черные. Щупальца
коричневые, остроконечные. Боковые стороны спинки и бочков в груди в желтом
налете и коротких желтых волосках. Крылья: внешний край перевязи позади г2+3 прямой
или слегка выпуклый. Вершинное пятно в виде узкой полоски, последняя отходит
от перевязи по ширине радиальной ячейки и, постепенно сужаясь, доходит до г4. Ноги
черные; основания средних голеней и лапок 2-й и 3-й пары ног коричневые; иногда
Рис. 91. Chrysops (s. str.) angancus Ols А, Б — брюшко и..
->- -крыло $. В, Г — брюшко и голова <$.'
средние голени целийом коричневые. Брюшко: I и II тергиты желтые, с небольшими
черными пятнами посредине. Пятно на I тергите по ширине соответствует щитку,
имеет небольшую выемку по заднему краю и округлые боковые стороны. Пятнб на
II тергите состоит из двух узких треугольников, слитых в паре в' передней части
тергита. Последующие тергиты черные, с желтыми каемками по заднему краю; III и
IV тергиты посредине имеют по узкому, желтому, треугольному пятну, достигающему
переднего края тергита. Снизу брюшко в основании желтое, II стернит с маленьким
продольным, III с более крупным темно-коричневым пятном посредине. IV и
последующие кольца темные, с желтыми каемками. ,7—9 мм.
в. Глаза соприкасаются на некотором протяжении. Лицо в желтом налете.
Лицевые мозоли округлые, блестяще-черные, почти обособлены от наличниковой мозоли.
Щечные мозоли крупные, соединены с наличником. Усики умеренной длины, 1-й
членик желто-коричневый, последующие черные. Концевой членик щупалец коричнево-
черный, на конце притуплённый, короче половины длины хоботка. Боковые стороны
спинки и бочки груди в редких, длинных, желтых волосках. Крылья: перевязь как
у 5, вершинное пятно слегка шире 1-й радиальной ячейки. Передняя базальная ячейка
затемнена на V2— 2/3i задняя на V3 от основания. Остальное как у §. Ноги черные;
средние голени в большей своей части и.основания средних и задних лапок коричнево-
желтые. Брюшко: II и III тергиты и боковые стороны I оранжево-желтые. II тергит
посредине с большим черным трапециевидным пятном, III тергит с двумя небольшими
треугольными пятнами. Последующие кольца черные, с узкими желтыми каемкаци.
11 Н. Г. Олсуфьев
162 II. ПОДСЕМ. CHRYSOPSINAE
Иногда IV тергит желтоватый, с двумя черными треугольными пятнами посредине
(как на III кольце). Снизу брюшко в основной половине желтое. 3 (или 4) концевых
стернита черные, с узкими желтыми каемками. Светлые стерниты имеют посредине
по темному пятну. 7—7.5 мм.
Распространение: восточносибирский (степной?) вид. Иркутская,
Читинская, Амурская области, Бурятская АССР. Сев. Монголия, Сев.-Зап. Китай. Редок.
Июнь—июль.
Голотип: 9, окр. Толбузино, верх. Амура, 13 VI—10 VII 1929, Принада. В
коллекции Зоологического института Академии наук СССР, Ленинград.
13. Chrysops (s. str.) parallelogrammus Zell. (рис. 92) — Пестряк западный.
Z е 11 е г, 1842, Isis : 823; Р 1 е s k е, 19106 : 449, S г i 1 a d у, 1917 : 122; К г б-
Ь е г, 1920 : 88; S u г с о и f, 1924 : 216; КгбЬег in Lindner, 1925 : 17; О л-
с у ф ь е в, 1937а : 87; М о и с h a et С h v а 1 а, 1955 : 235; L е с 1-е г с q, 1960b :
48; Chvala, Lyneborg a. Moucha, 1972 : 128.
5. Лоб в сероватом, лицо в желтоватом налете. Лобная мозоль крупная, обычно
рточти достигает края глаз, блестяще-черная. Темя вокруг глазков блестящее.
Лицевые мозоли овальные, блестяще-черные, соединены б. м. широкими перемычками
с черной наличниковой мозолью, соответственно разделяющая их срединная полоска
налета значительно -уже наличника. Щечные мозоли небольшие, блестяще-черные.
Усики: 1-й и внутренние стороны 2-го членика желто-коричневые, 3-й членик черный.
Рис. 92. Chrysops (s. str.) parallelogrammus Zell. A—В —•
брюшко, крыло и лобная полоска <j>; Г — брюшко 3.
Щупальца остроконечные, коричневые. Грудь черная, в сером налете, спинка
посредине с двумя серыми полосками, достигающими щитка. Последний в сером налете.
Бочки груди и боковые стороны спинки в желтоватых волосках. Крылья: перевязь
достигает заднего края крыла, ее внешний край перед г4+5 образует легкий угол, далее
между т1 и задним краем крыла вогнутый. Вершинное пятно ланцетовидное, налегает
на г4, на ее вершинную треть. Пятно соединено с перевязью по ширине 1-й радиальной
ячейки. 4-я заднекрайняя ячейка сужена к вершине, анальная ячейка иногда не
замкнута на конце. Ноги коричнево-желтые; бедра, вершины передних голеней,
передние лапки и концы последующих черные, иногда передние и средние бедра
коричневые. Передние коксы черные, в сером налете. Брюшко: I и II тергиты желтые. I тергит
посредине с черным, кзади слегка трапециевидно сужающимся пятном. Его вершина
не превосходит ширину щитка. Задний край пятна с небольшой* выемкой. II тергит
с двумя узкими, запятовидными или треугольными черными пятнами. Эти пятна б. м.
сближены между собой. III и IV тергиты желто-серые, по их переднему краю с каЖ-
4. CHRYSOPS 163
дой стороны имеется по паре направленных вершинами назад черных треугольников,
из них внешние треугольники короче внутренних. Последующие тергиты темные, в
сером налете и волосках. Снизу брюшко в основной половине желтое, с небольшим
темным пятном посредине. IV (или III) и последующие стерниты черные, в сером налете.
9—10 мм.
6. Лицевые мозоли крупные, округлые, блестяще-черные. Наличниковая мозоль
черная, соединена с лицевыми мозолями б. м. пшрокими перемычками. Щечные мозоли
большие, достигают наличника. Усики черные. Щупальца остроконечные, коричнево-
Рис. 93. Ареалы: 1 — Chrysops (s. str.) parallelogrammus; 2 — Ch. (s. str.)
concavus; 3 — Ch. (s. str.) dissectus.
черные, превосходят V2 длины хоботка. Грудь черная, спинка с двумя узкими серыми
полосками, достигающими основания щитка. Боковые стороны спинки и бочки груди
в густых желтых волосках. Крылья: перевязь как у $, вершинное пятно несколько
шире, налегает на rt на 2/3 ее длины; Соединение пятна с перевязью превосходит
ширину радиальной ячейки. Базальные ячейки затемнены от основания на две трети.
Ноги: основная часть передних голеней, целиком средние и задние голени и основания
лапок последних двух пар ног светло-коричневые. Прочие части ног черные. Брюшко:
I—IV тергиты желтые, с черным рисунком. I тергит с крупным черным пятном,
занимающим всю его среднюю часть. II—IV тергиты имеют по паре черных треугольных
пятен, направленных вершинами назад. На II тергите эти треугольники слиты в
основании между собой и с впереди лежащим пятном на I кольце. V и последующие
кольца черные, с желтыми каемками по заднему краю. Снизу брюшко в основной
половине желтое, с небольшим темным пятном на I—II стернитах, IV и последующие
стерниты черные, с узкими желтыми каемками. 9.5 мм.
Распространение: западноевропейский лесостепной вид, аллопатричен
Ch. (s. str.) concavus (рис. 93, 1). В СССР известны единичные находки: Ржев
Калининской обл.; пос. Оболь Витебской обл.; Столинский р-н Брестской обл.; Даугавпилс-
ский р-н Латвийской ССР (К: Эльберг); р. Тетерев Иванковского р-на Киевской обл.;
Цюрупинск на Днепре Херсонской обл. Ср. и Ю. Европа. Не часто. 3-я декада июня—
Июль.
14. Chrysops (s. str.) concavus Lw. (рис. 94) — Пестряк пойменный.
Loew, 1858b, Verh. Zool.-bot. Gesell. Wien, 8:622; P 1 e s k e, 19106:454;
Szilady, 1917 : 107; Krober, 1920 : 85; К г о Ь е г in Lindner, 1925 : 9;
Олсуфьев, 1937а : 89; L е с 1 е г с q, 1960b : 46; Ш е в ч е н к о, 1961 : 44; В и о-
л о в и ч, 1968 :49; Chvala, Lyneborg a. Moucha, 1972 : 130.
9. Лоб в голубовато-сером, лицо в желтоватом налете. Лобная мозоль
умеренной величины, овальная, не достигает края глаз, слабо блестящая, черная. Темя между
глазками покрыто налетом, реже слегка блестящее. Лицевые мозоли небольшие,
вытянутые, блестяще-черные, не достигают края глаз. Наличниковая мозоль коричневая,
соединяется узкими перемычками с лицевыми мозолями, соответственно
-разделяющая их срединная полоска налета б. м. равна ширине наличника. Иногда наличнико-
11*
164
II. ПОДСЕМ. CHRYSOPSINAE
вая мозоль посредине прикрыта полоской налета, делящей ее на две половинки.
Щечные мозоли небольшие, черные, обособлены от наличника налетом. Усики: 1-й членик
красновато-желтый, 2-й темно-коричневый, 3-й черный, в основании коричневый.
Иногда оба последних членика целиком черные. Щупальца б. м. остроконечные, желто-
коричневые. Грудь черная, в сероватом налете; спинка с двумя голубовато-серыми
срединными полосками, достигающими щитка. Щиток в сером налете. Боковые стороны
спинки и бочки груди в серовато-желтых волосках. Крылья: перевязь достигает
заднего края крыла, слегка просветляясь в.4-й и 5-й заднекрайних ячейках. Дискоидаль-
ная ячейка также слегка просветлена в средней части (у облетанных экземпляров
иногда просветлена значительно!). Внешний край перевязи перед г4+5 образует легкий
угол, далее край вогнутый. Вершинное пятно умеренной ширины, захватывает г4
не более чем на V2 ее длины. Средняя часть ячейки Л2 в пределах вершинного пятна
слегка продольно просветлена. Пятно соединено с перевязью по ширине 1-й радиаль-
Рис. 94. Chrysops (s. str.) concavus Lw. Бркнпко,'крыло, лобная
полоска, VIII стернит, церки и сперматеки <j>.
ной ячейки. 4-я заднекрайняя ячейка заметно сужена к вершине. Ноги коричневато-
жеятые, колени и концы лапок черноватые. Коксы первой, пары ног желтые, вершина
затемнена. Брюшко: I и II тергиты желто-серые; I тергит.е черным пятном посредине,
соответствующим или слегка превосходящим ширину щитка. Углы этого пятна
закруглены, тогда как по заднему.краю имеется небольшая выемка. Птергит с двумя
небольшими треугольными пятнами,, широко ^доставленными между собой. Между
ними лежит крупный серый треугольник. Последующие тергиты черные, в сером
налете и волосках. На III и IV тергитах по переднему краю имеется с каждой стороны
по черной поперечной полоске,, слегка расширяющейся в небольшие треугольнички.
Снизу брюшко в основании жёлтое, с небольшим темным пятном посредине, III и
последующие стерниты черные, в густом сером налете. VIII стернит шире длины,
б. м. треугольной, формы, говюпофиз с незначительной' апикальной выемкой; церки
обычной формы; сдерматека тупоконечная. 8.5—11 мм,
в неизвестен.
Очень близок к Ch. (s. str.) parallelogrammus (возможно, его подвид), отличаясь
от него следующим: 1) целиком светлыми бедрами и, передними тазиками; 2) широко
расставленными черными треугольными пяхнами на II тергите брюшка; 3) меньшей
величиной лобной мозоли; 4) матово-серым.,теменем; 5), .коричневой наличниковой
мозолью, соединенной, с лицевыми мозолями более узкими перемычками.
Распространение: лесостепной (лугавой) вид.,;:,мес,тами,:ДОвольно далеко
проникает в .таежн.Ог.ле.сную^зону.. Ал:ша№Триче,н Ск-..р^г1а},1ф^гащгпц8,{^жс.,%^г 2).
4. CHRYSOPS 165
От Киева на восток до Иркутска, на север до Иловны б. Мологского у. Ярославской
губ-., Сыктывкара, Усть-Кулома Коми АССР, Соликамска Пермской обл., с.-з.
Томской обл. (пос. Назино на Оби) и устья Подкаменной Тунгуски, на юг до
Воронежской обл. (Богучарский р-н), Волгоградской обл. (Котельниковский р-н), низовий
Урала, юго-востока Павлодарской обл. (пос. Канонерка), окр.
Семипалатинска и Барнаула. Указание Силади (Szilady, 1917) и Крёбера (КгбЬег, 1925)
на распространение этого вида в Австрии ошибочно, в музее Вены 1 5 с определением
«.Ch. concavus» на самом деле Ch. divaricatus (Moyxa, личное сообщение). Неправильно
также указание Н. А. Виоловича (1968) на распространение Ch. concavus в ФРГ и ГДР.
Преимущественно по долинам крупных рек, особенно в степной полосе. Не часто.'
15. Chrysops (s. str.) suavis Lw. (рис. 95) — Пестряк привлекательный.
Изменчив. В пределах СССР известны два подвида: номинативный Ch. (s.'str.)
suavis suavis Lw. и сахалинский Ch. (s. str.) suavis sakhalinensis Pleske.
Рис. 95. Chrysops (s. str.) suavis suavis Lw. A—Г — брюшко,
крыло, VIII стернит и церки $; Д — брюшко 3.
15а. Chrysops (s. str.) suavis suavis Lw. — Пестряк привлекательный
номинативный.
L о e w, 1858b, Verb. Zool.-bot. Gesell. Wien, 8 : 620; Szilady, 1917 : 128;
S h i r a k i, 1918 : 53; КгбЬег, 1920 : 79; КгбЬег in Lindner, 1925 : 20;
Олсуфьев, 1937a : 90; L e с 1 e г с q, 1960b : 50; T a k a h a s i, 1962 : 32; Ш е в-
ч е н к о, 1961 : 43; В и о л о в и ч, 1968 : 49; Соболева, 1970в : 227 (личинка,
куколка); —sakhalinensis Pleske, 1910в : 472.
$. Лоб и лицо в желтоватом налете. Лобная мозоль крупная, блестяще-черная.
Темя вокруг глазков блестящее. Лицевые мозоли черные, не достигают наличника.
Усики умеренной длины, черные, 1-й членик и внутренние стороны 2-го желтовато-
коричневые. Щупальца коричнево-черные. Грудь черная, спинка с 2 ясными серыми
или желтоватыми полосками, исчезающими возле щитка. Боковые стороны спинки
и бочки груди в желтом налете и в желтых волосках. Крылья: внешний край
перевязи прямой от г2+3 до заднего края крыла. Вершинное пятно в виде узкой полоски,
идущей от перевязи вдоль внешнего края крыла и сходящей на нет за жилкой г4.
Ширина полоски равна или едва превышает ширину 1-й радиальной ячейки.
Анальная ячейка иногда на конце открытая. 4-я заднекрайняя ячейка сужена к вершине.
Ноги черные; голени средних ног и основания средних и задних лапок коричневые.
Брюшко слегка трапециевидно сужено кзади; II тергит желтый, с двумя черными
параллельными полосками, не достигающими переднего края тергита. Последующие
тергиты черные, с желтой полоской посредине, сходящей на нет к концу брюшка.
166 II. ПОДСЕМ, CHRYSOPSINAE
Рисунок варьирует: иногда желтая окраска появляется по бокам на III и
последующих тергитах, выделяя две резких черных продольных полоски, начинающихся на
II тергите; наиболее светлая окраска брюшка у экземпляров из Тувы. Иногда
срединная желтая полоска, лежащая между черными продольными, слегка треугольно
расширена кзади на каждом кольце. Снизу брюшко в основной части желтое, IV или III
и последующие кольца черные. II стернит иногда с небольшим темным пятном
посредине. VIII стернит шире длины, слегка угловатый по бокам, гонопофиз с
незначительной апикальной выемкой; церки обычной формы. 7—11 мм.
6. Лицевые мозоли б. м. крупные, блестяще-черные. Щечные мозоли хорошо
развиты. Усики умеренной длины; 1-й и 2-й членики коричневые, 3-й черный. Щупальца
черные, остроконечные, равны 3/4 длины хоботка. Боковые стороны спинки и бочки
груди в желтом налете и такого же цвета волосках. Крылья как у §; базальные ячейки
затемнены почти целиком, за исключением небольшого прозрачного участка в
вершинной части. Окраска ног как у §. Брюшко сверху желтое, с двумя черными
продольными полосками посредине, простирающимися на II—IV тергиты. Эти полоски слиты
вместе в передней части II тергита. III—IV тергиты по бокам, последующие тергиты
целиком черные. Снизу брюшко в основной половине желтое, с небольшим темным
пятном на II стерните, IV (или III) и последующие кольца черные. 8—9 мм.
Распространение: восточносибирско-дальневосточный лесной вид
(рис. 59, 2). В СССР распространен от окр. Семипалатинска, Кемерова, Тогучина
Новосибирской обл. и Томска на восток до Хабаровского и Приморского краев, к
северу известен до Сев. Приангарья Красноярского края, Усть-Кута в Иркутской обл.,
Центральной Якутии и севера Амурской обл., к югу до Тувы, южного Забайкалья и
Посьета Приморского края. Сев. и Вост. Монголия, Сев.-Вост.. Китай, Корейский
п-ов, Япония. Местами обычен. Середина июня—июль, иногда позднее. Указание
Ченфу и By (Chenfu a. Wu, 1940) на нахождение этого вида в Китае (о. Тайвань),
видимо, ошибочно, так как он не упоминается для этого острова в известных
монографиях Шираки (Shiraki, 1918) и Такахази (Takahasi, 1962).
15b. Chrysops (s. str.) suavis sakhalinensis PI.
P 1 e s k e, 1910b, Ann. Mus. Zool. St. Petersb., 15, 4 : 472 {Chrysops sakhalinensis);
Stackelberg, 1926b : 325.
Представляет собой меланистическую форму.
$ отличается от номинативного подвида следующим: на II тергите брюшка
черные полоски достигают его переднего края, сливаясь с черным пятном на I тергите;
III и последующие тергиты по бокам черные; снизу брюшко черное начиная с III стер-
нжта, I и II стерниты обычно с черным срединным пятном; иногда средние голени
целиком черные. 9—10 мм.
6 также отличается от номинативной формы более темной окраской брюшка:
на II тергите вместо двух черных полос имеется образованное их слиянием крупное
срединное прямоугольное черное пятно с маленьким желтым треугольником по
заднему краю, боковые стороны тергита желтые; III тергит черный с 3 небольшими
желтыми продольными полосками — срединной и боковыми, IV и последующие тергиты
черные, с едва намечающейся желтоватой срединной полоской; снизу брюшко черное,
боковые стороны I—II стернитов желтые; средние голени темно-коричневые. 8.5 мм.
Распространение: о. Сахалин, о. Кунашир (рис. 59, 3). Обычен. Июнь—
сентябрь.
Отмечаются переходные экземпляры к номинативной форме — у ? черные
продольные полоски на II тергите немного не достигают его переднего края или на III
тергите по бокам имеется по небольшому желтому пятну. С этими переходными
экземплярами сходны экземпляры из Центральной Якутии.
1 $ с о. Кунашир от 1 IX 1953 (Н. А. Виолович) имеет резко отличающийся
темный рисунок на II тергите брюшка: вместо двух продольных черных пятен имеется
одно черно-коричневое поперечное полукруглое пятно по заднему краю тергита, III
тергит целиком черно-коричневый, IV тергит черный, с едва заметной желтоватой
полоской посредине. Строение головы, окраска крыльев, ног, низа брюшка и так далее,
как у Ch. (s. str.) suavis sakhalinensis. Мы обозначаем эту уклоняющуются форму как
Ch. (s. str.) suavis var. kunashiri Ols., var. nova.
16. Chysops (s. str.) ricardoae PI. — Пестряк Рикардо.
Изменчив. В СССР представлен тремя подвидами: номинативным Ch. (s. str.)
ricardoae ricardoae PL, Ch. (s. str.) ricardoae volgensis Ols. и Ch. (s. str.) ricardoae jaku-
lensis Ols.
16a. Chrysops (s. str.) racardoae' ricardoae Pi. (рис. 96) — Пестряк Рикардо
номинативный.
Р 1 е s k е, 1910в, Ann. Mus. Zool. St. Petersb., 15 : 461; Szila'dy, 1917 : 124;
Krober, 1920 : 76; К r 6 b e r in L i n d n e r, 1925 : 19; Олсуфьев, 1937а :
4. CHRYSOPS 167
91; Шевченко, 1961.: 45; Виолович, 1968 : 50. — wagneri P 1 e s k e, 1910b :
466. — przewalskii Pl.eske, 1910b : 464.— mongolicus S z i l a d y, 1917 : 109
(Ch. dissectus Lw., var.). -— pseudoricardoae Krober, 1920 : 78 (<J).
$. Лоб и лицо в желтоватом налете. Темя возле глазков слегка блестящее.
Лобная мозоль крупная, блестяще-черная. Лицевые и наличниковые мозоли черные или
коричнево-черные (или желто-коричневые). Щечные мозоли довольно крупные,
черные, не достигают наличника. Щупальца коричневые. Усики длинные, черные, 1-й
членик в основании (и в особенности с внутренней стороны) темно-коричневый. Крылья:
внешний край перевязи образует легкий угол, вершина которого лежит немного выше
г4+5; позади этой жилки край перевязи б. м. прямой. Вершинное пятно довольно
крупное, б. м. обособлено от перевязи светлым перерывом, захватывает приблизительно
Рис. 96. Chrysops (s. str.) ricardoae ricardoae PI. A—Г — брюшко,
крыло, VIII стернит и церки $; Д — брюшко 3.
1/3—1/2 длины г4. 5-я заднекрайняя ячейка просветлена на значительном протяжении
к внешнему краю крыла. 4-я заднекрайняя ячейка резко сужена к вершине. Ноги:
бедра черные, передние голени в основании коричневые, далее черные, средние
голени желтые с коричневым концом, задние целиком коричневые. Передние лапки
черные, средние и задние в основной части коричневые, далее также черные. Брюшко:
I тергит с черным прямоугольным пятном посредине, по бокам желтый. Пятно не
превосходит ширину щитка и задней стороной достигает края тергита. Задние углы пятна
слегка закруглены. II тергит желтый, возле заднего края с двумя сближенными
черными пятнами б. м. овальной формы. Величина пятен варьирует, иногда они не
выражены. Последующие тергиты черные, по заднему краю часто коричневые. III
тергит с резким, последующие с неясными желтыми срединными треугольниками из
желтого налета и такого же цвета волосков, сливающихся с общим налетом и волосяным
покровом колец. II или III стерниты желтые, первый иногда с небольшим темным
пятном посредине, последующие черные, в желтоватом налете и волосках. VIII стернит
шире длины, с округлыми боковыми сторонами, иногда целиком темный, гонопофиз
с слабой апикальной выемкой или без нее; церки обычной формы. 7—Ю мм.
3- Глаза на лбу соприкасаются. Лицо в желтоватом налете. Лицевые мозоли
блестяще-черные, соприкасаются с краем глаз. Щечные мозоли большие, соединены
с наличниковой мозолью. Щупальца черные, концевой членик остроконечный, слегка
превосходит V2 длины хоботка. Усики черные. Боковые края спинки и бочки груди
в желтых волосках. Крылья как у $; основная 1/2 базальных ячеек затемнена. Ноги:
средние голени и основания двух последних пар лапок коричневые, прочие части
Ш п. ПОДСЕМ. CHRYSOPSINAE
черные. Брюшко: II тергит желтый, с большим черным пятном посредине, занимающим
примерно 1/3 ширины тергита; пятно б. м. прямоугольной формы. Последующие тер-
гиты черные, с неясным сероватым налетом по заднему краю колец, образующим
посредине треугольнички (заметны лишь у непотертых экземпляров). Снизу брюшко
в основании желтое, III стернит по заднему краю и частью посредине черный,
последующие стерниты целиком черные. II стернит с небольшим черным пятном посредине.
7.5—9 мм.
Небольшой вид, с б. м. обособленным вершинным пятном на крыле и двумя
небольшими овальными- черными пятнами на II тергите брюшка. Наиболее близок
к Ch. (s. str.) dissectus.
Распространение: казахстано-монгольский пустынно-степной подвид
(рис. 97, 1). Казахстан (от восточной части хр. Каржантау и Джабаглы-су по
предгорьям и низкогорьям Тянь-Шаня на восток до Нарынкола и к северу до Уч-Арала
Талды-Курганской обл.), Киргизия (дол. Таласа и Чу; Иссык-Кульская котловина),
Хакасская АО, Тувинская АССР и Забайкалье. Монголия, Китай. Июнь—июль,
иногда позднее. Местами обычен.
Рис. 97. Ареалы: 1 — Chrysops (s. str.) ricardoae ricardoae; 2 — Ch. (s. str.)
ricardoae volgensis; 3 — Ch. (s. str.) ricardoae jakutensis.
16b. Chrysops (s. str.) ricardoae volgensis Ols. (рис. 98, A) — Пестряк Рикардо
ВОЛЖСКИЙ.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 446.
§. От номинативной формы отличается светлой окраской: 1-й и 2-й членики
усиков, бедра ног и лицевые мозоли целиком желтые или коричневатые; черные пятна
на II тергите брюшка небольшие, иногда целиком отсутствуют; снизу брюшка желтая
окраска охватывает I—III или I—IV стерниты, черные пятна посредине отсутствуют;
темный рисунок крыла более светлый, коричневатый, вершинное пятно налегает
на г4 более чем на х/2 ее длины; лобная мозоль в срединной части коричневатая. 9—
10 мм. :'
3 неизвестен.
Распространение: долина Волги от окр. Волгограда до Астрахани,
долина Урала от Уральска до Гурьева и местами в Волжско-Уральском междуречье
(рис. 97, 2). Ареал подвида изолирован от ареала номинативного подвида
значительными пространствами.
16с. Chrysops (s. str.) ricardoae jakutensis Ols, (рис. 98, В) — Пестряк Рикардо
якутский. .
О л с у ф ь е в, 1972, Энтомол. обозр., 51, 2:447.
$. От номинативной формы отличается более темной окраской. 1-й и 2-й членики
усиков черные, иногда лишь основание 1-го членика коричневое; лицевые мозоли
черные или черно-коричневые; бедра ног черные; черное пятно на I тергите брюшка
захватывает также переднебоковую часть полукольца; черные пятна на II тергите
крупные, впереди часто соединены, между собой и слиты с черным пятном на I тергите;
4. CHRYSOPS 169
снизу брюшка желтая окраска охватывает I—II стерниты, на обоих часто по
срединному черному пятну; лобная мозоль черная, крупнее, чем у номинативного, подвида;
темный рисунок крыла черноватый, вершинное пятно налегает на жилку г4 не более
чем на V3 ее длины.
3 неизвестен.
Распространение: Центральная Якутия, по-видимому, изолированно
от ареала номинативного подвида (рис. 97, 3).
Рис. 98. А — Chrysops (s. str.) ricardoae volgensis Ols.,
крыло и брюшко $; Б — Ch. ricardoae jakutensis Ols.,
крыло и брюшко §.
17. Chrysops (s. str.) dissectus Lw. (рис. 99) — Пестряк дальневосточный.
L о e w, 1858b, Verh. Zool.-bot. Gesell. Wien, 8 : 618; S z i 1 a d y, 1917 : 109;
К r 6 b e r, 1920 : 70; Krober in Lindner, 1925 : 10; S t а с k e 1 Ь е г g,
1926b : 325 (3); Олсуфьев, 1937a : 93; L e с 1 e г с q, 1960b : 46; В и о л о в и ч,
1968 : 51. — bipunctatus М о t s с h ul sky, 1859 : 487. — amurensis P 1 e s k e, 1910b:
459. — binoculatus S z i l a d y, 1917 : 105.
$. Лоб и лицо в желто-сером налете. Лобная мозоль черная, крупная, боковыми
сторонами не достигает края глаз, отделяясь полоской налета. Темя возле' глазков
на небольшом протяжении блестящее. Лицевые мозоли коричневые или двуцветные;
часть, расположенная на наличнике, — коричневая, на щеках — черная. Налични-
ковая мозоль желто-коричневая, верхними концами б. м. широко соединена с
лицевыми мозолями. Щечные мозоли блестяще-черные, обычно хорошо развиты. Усики
б. м. длинные, 1-й и 2-й членики желтые или желто-коричневые, 3-й членик черный.
Щупальца темно-коричневые, б. м. остроконечные. Ноги в основном
коричнево-желтые; основание бедер, большая часть передних голеней, передние лапки и концевые -
членики лапок последних двух пар ног черноватые. Иногда бёдра всех ног целиком
черные. Крылья: внешний край перевязи в виде слегка выпуклой линии возле г4, далее
край перевязи прямой или вогнутый. Вершинное пятно очень узкое, чаще обособлено
от перевязи светлым перерывом. Ширина пятна едва составляет ширину 1-й
радиальной ячейки или же значительно уже ее. 4-я заднекрайняя ячейка к вершине сильно
сужена, иногда замкнута. Брюшко слегка сужающееся кзади. I тергит с черным
пятном, захватывающим всю среднюю и переднебоковую часть тергита; пятно, таким
образом, явственно шире щитка. Заднебоковые стороны тергита и узкая каемка пятна
желтые. II тергит желтый, с двумя маленькими круглыми черными пятнами,
расположенными отступя от заднего края тергита. Пятна могут варьировать от
незначительных точек до овальных или треугольных пятен и даже сливаться в общее пятно
б. м. сердцевидной формы или же отсутствовать нацело (var. apuncta Phil.). Ill и
IV тергиты блестяще-черные, с неясными, желтоватыми, узкими каемками по заднему
краю и такого же цвета треугольниками посредине. Последующие тергиты темные.
в сероватом налете и волосках. I—II стерниты желтые, последующие черные. Иногда
желтый цвет простирается на III стернит, в той или иной мере вытесняя черный цвет.
9.5—11 мм.
tJ. Глаза на лбу соприкасаются на некотором протяжении. Лицо в желтовато-
сером налете; наличниковые мозоли желтые, лицевые мозоли двуцветные: на щеках
черные, на наличнике коричневые. Щечные мозоли сравнительно небольшие, блестяще-
черные, достигают наличника. Щупальца черные, слегка притуплены на конце, не-
170 II. ПОДСЕМ. CHRYSOPSINAE
сколько превышают х/2 длины хоботка. Усики б. м. тонкие. Первые 2 членика желто-
коричневые, 3-й черный; иногда 2-й членик и вершина первого сильно затемнены,
почти черные. Боковые края спинки и бочки груди в желтовато-сером налете и в густых
золотисто-желтых волосках. Крылья как у g; основная половина базальных ячеек
затемнена. Передние ноги: тазики желто-коричневые, бедра и основание голеней
темно-коричневые, далее ноги черные. Бедра средних и задних ног черные, средние
голени желто-коричневые, задние темно-коричневые; лапки в основной части
коричневые, концы черные. Брюшко: II тергит желтый, в передней части с небольшим черным
Рис. 99. Chrysops (s. str.) dissectus Lw. А, В — брюшко и крыло
$; В — брюшко с?; Г — изменчивость темных пятен на II тергите брюшка §,
сердцевидным пятном посредине. III тергит черный, с широкой желтой каемкой по
заднему краю, треугольно расширяющейся посредине и по бокам. IV тергит черный,
с легким желтоватым налетом по заднему краю, последующие тергиты целиком в
налете. Снизу брюшко в основной половине желтое, IV и последующие стерниты черные.
9—10.5 мм.
Близок к Ch. (s.str.) ricardoae, отличаясь от него большей величиной, отсутствием
на крыльях ясного угла по внешнему краю перевязи, более узкому вершинному пятну
и рисунку брюшка.
Распространение: дальневосточный лесной или луго-лесной вид (рис. 93,
3). Амурская обл. (окр. г. Зеи; Тыгда приток Зеи), Хабаровский край (на север до
Нижнетамбовского на Амуре, 51° с. ш.), Приморский край (на юг до Хасанского р-на).
Сев.-Вост. Китай, Корейский п-ов, Япония. Не часто. Июль—август.
18. Chrysops (s. str.) rufipes Mg. (рис. 100) — Пестряк красноногий.
М е i g e n, 1820, Syst. Beschreib., II : 71; L u n d Ь е с k, 1907 : 93; P 1 e s k e,
19106 : 445; S z i 1 a d y, 1917 : 125; К г 6 b e r, 1920 : 115; S u г с о u f, 1924 : 220;
Krober in Lindner, 1925 : 19; Олсуфьев, 1937a : 95; Mo u с h a
et С h v a 1 a, 1955 : 236; Тамарина, 1956 : 182 (личинка, куколка); L e c-
1 e г с q, 1960b : 49; Шевченко, 1961 : 47; С к у ф ь и н, 1967 : 187 (личинка);
Виолович, 1968:51; J e z е к, 1970:381 (личинка, куколка); С h v a 1 а,
Lyneborg a. Moucha, 1972 : 132.
4. CHRYSOPS 171
$. Лоб в тонком буроватом налете, темя блестяще-черное, лицо в желто-сером
налете. Верхние лицевые мозоли небольшие, блестяще-черные, наличниковая мозоль
соединена с лицевыми мозолями узкими перемычками, блестяще-желтая. Щечные
мозоли черные, занимают почти целиком нижнюю часть щек. Усики короткие, их
первые два членика коричнево-желтые, 3-й членик черный. 1-й членик явственно
утолщен. Щупальца коричневые, остроконечные, едва короче хоботка. Ноги коричнево-
желтые, концы лапок темные. Иногда средние и задние голени в основной части
затемнены. Крылья, в особенности вдоль жилок, буровато затемнены, с более темным
нерезким рисунком. Основание, передний край и перевязь за серединой крыла
коричневые. Последняя б. м. широкая, достигает края крыла; дискоидальная, 4-я и 5-я зад-
некрайние и 2-я радиальная ячейки в средней части слегка просветлены. Внешний
край перевязи кзади от г2+3 в виде выпуклой линии. Вершинное пятно расширено,
перемычка с перевязью занимает ширину радиальной ячейки. Анальная ячейка на
Рис. 100. Chrysops (s. str.) rufipes Mg. Брюшко, крыло, VIII стер-
нит (варианты), церки (варианты) и сперматека §. А — голова <?.
конце открытая. Брюшко продолговато-овальное, слабо блестящее, черное. I и II
тергиты по боковым сторонам с незначительными грязно-желтыми пятнами. Все тергиты
по заднему краю с узкой желтой каемкой, II—IV кольца посредине с маленькими
желтовато-серыми треугольниками. Снизу брюшко черное, стерниты по заднему краю
с коричневатыми каемками. II стернит иногда по бокам желтоватый. Терминалии:
VIII стернит не шире длины или слегка шире; боковые стороны округлые или слегка
угловатые, гонопофиз без апикальной выемки; церки обычной формы; вершина спер-
матеки приострена. 6.5—8.5 мм.
(J. Глаза не соприкасаются (между ними имеется очень узкий, но явственный
промежуток). Фасетки глаз б. м. одинаковой величины. Лицо в желто-сером налете.
Верхние лицевые мозоли округлые, коричнево-черные, блестящие, продолжены вниз
вдоль наличника блестяще-желтыми полосками. Щечные мозоли черные, занимают
почти целиком нижнюю часть щек. Усики короткие,' черные, 1-й членик явственно
утолщен. Щупальца черные, тупоконечные, превосходят половину длины хоботка.
Ноги желто-^коричневые. Передние и средние бедра в основной половине, задние
на 2/3 своей длины, а также концы всех лапок буро-черные; иногда 1-я пара ног
целиком черная, за исключением колен. Крылья: обе базальных ячейки от основания
на 2/з коричневые; остальное как у $. Брюшко черное, узкое. I—III тергиты по
боковому краю с узкой желтоватой полоской. Все кольца по заднему краю с очень узкими
желто-коричневыми каемками; II—IV тергиты посредине с маленькими серыми
треугольниками. Снизу брюшко как у <j>. 6.5—8.5 мм.
Распространение: европейско-западносибирский лесостепной вид
(рис. 101, 1). В СССР известен от Эстонии, Ленинградской обл. и запада Украины
на восток до Томска и Тогучинского р-на Новосибирской обл., на север до юга Каре-
172
П. ПОДСЁМ. CHRYSOPSINAE
лии (Сортавала), Ярославского р-на, Кинешмы, Тобольска и Верхне-Имбатского на
Енисее (63° с. ш.), на юг до низовий Днепра (Цюрупинск Херсонской обл.), Крыма,
Ростовской обл. (пос. Краснодвор), дельты Волги, Центрального Казахстана (р. Бала-
Джезде к северу от Карсакпая Карагандинской обл.), Новосибирска, Павлодара и
Локтевского р-на Алтайского края; северная и восточная граница распространения
изучена недостаточно. Средн. и частью Сев. Европа.
Не кровосос. Июнь—июль. Самцы и самки на растениях по краю болот или в пойме
рек, иногда в больших количествах; личинки в воде по краю гипнового болота, в
прибрежных частях озер или речек с медленным течением.
Рис. 101. Ареалы: 1 - Chrysops (s. str.) rufipes; 2 - Ch. (s. str.) valtdus.
К г б Ь е г, 1920,
L е с 1 е г с q, 1960b :
2. Подрод HETEROCHRYSOPS КгбЬ.
Олсуфьев, 1937а : 87;
Zool. JahrMcher, Syst., 43 : 50;
34; Шевченко, 1961 : 48.
Тип подрода Chrysops mlokosiewiczi Bigot, 1880.
Крылья с рисунком из темных поперечных перевязей и пятен. Дискоидальная
ячейка в средней части просветлена (с «окошечком») или целиком прозрачная.
Виды, относящиеся к этому подроду, распространены главным образом на юге
и лишь один вид заходит в таежную зону (Вост. Сибирь).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА
HETEROCHRYSOPS KR0B.
А. Глаза разделены лбом:
??
2(3).
3(2).
4(7).
1 (10). Крылья с хорошо выраженным темным рисунком; перевязь имеется;
дискоидальная ячейка прозрачная лишь в средней части, по краям затемнена (рис. 102
103 и др.). '
Задние голени черные. Вершинное пятно крыла широкое, обособлено от
перевязи просветом в i?! (рис. 102) ... 19. Ch. (Heterochrysops) sejunctus Szil.
Задние голени желтые. Вершинное пятно крыла узкое или широкое, всегда
соединено с перевязью перемычкой.
Лобная мозоль черная. Брюшко желтое, с темными пятнами, не образующими
продольных полосок (рис. 103, 104).
, 5 (6). Вершинное пятно крыла широкое, налегает на г4 на 3/4 ее длины;
дискоидальная ячейка расширена (рис. 103) . . .20. Ch. (Heterochrysops) italicus Mg..
6 (5). Вершинное пятно крыла умеренной ширины или узкое, не превышает половины
длины г4; дискоидальная ячейка нормальная (рис. 104)
21. Ch. (Heterochrysops) Havipes Mg.
7 (4). Лобная мозоль по крайней мере в нижней части коричневая. Брюшко желтое,
с 4 узкими черными продольными полосками (рис. 108, 109).
8 (9). 3-й членик усиков красно-желтый, с черной палочкой; 2-й членик желтый,
слегка утолщен, его длина превосходит ширину не более чем в 2 раза. Внешний
4. CHRYSOPS 173
край перевязи позади г4+5 образует уступ (рис. 108)
22. Ch. (Heterochrysops) vanderwulpi Krob.
9 (8). 3-й членик усиков целиком черный; 2-й членик коричнево-черный, его длина
превосходит ширину в 3 раза. Внешний край перевязи прямой (рис. 109) . ,
. 23. Ch. (Heterochrysops) mlokosiewiczi. Big.
10 (1). Крылья почти прозрачные; на месте перевязи имеются лишь 2 узких
поперечных полоски; дискоидальная ячейка почти целиком прозрачная (рис. 110)
24. Ch. (Heterochrysops) oxianus PI.
Б. Глаза соприкасаются или сильно сближены: S3
1 (4). Глаза сильно сближены, но не соприкасаются (между глазами имеется узкий,
но явственный промежуток) (рис. 102, 106).
2 (3). Вершинное пятно крыла широкое, обособлено от перевязи просветом в Яц.
Брюшко сверху черное 19. Ch. (Heterochrysops) sejunctus Szil.
3 (2). Вершинное пятно крыла соединено с перевязью перемычкой по ширине Rt.
Брюшко, по крайней мере на II тергите, с желтыми пятнами
21. Ch. (Heterochrysops) flavipes Mg.
4 (1). Глаза соприкасаются на некотором протяжении (рис. 103).
5 (6). Брюшко черное, с незначительными желтыми пятнами по бокам II тергита.
Вершинное пятно крыла очень широкое, налегает да г4 на 3/4 ее длины . . .
: 20. Ch. (Heterochrysops) italicus Mg.
6 (5). Брюшко желтое, сверху с двумя резкими черными продольными полосками
посредине и незначительными полосками по бокам. Вершинное пятно крыла
узкое, налегает на г4 не более чем на г/3 ее длины.
7 (10). Перевязь в задней части нормально широкая, просветлена в дискоидальной
ячейке и слегка в основании R2. Основание 1-й заднекрайней ячейки темное.
8 (9). 2-й членик усиков желтый, б. м. овальный, короткий, его длина превосходит
ширину не более чем в 2 раза. Бедра желтые
22. Ch. (Heterochrysops) vanderwulpi Krob.
9 (8). 2-й членик усиков коричнево-черный, цилиндрический, его длина превосходит
ширину в- 3 раза. Бедра черные
23. Ch. (Heterochrysops) mlokosiewiczi Big.
10(7). Перевязь треугольная, сужающаяся кзади (рис. 110). Центральная часть
перевязи просветлена (основание 2-й радиальной, 1-й заднекрайней и
центральная часть дискоидальной ячеек) .... 24. Ch. (Heterochrysops) oxianus PI.
19. Chrysops (Heterochrysops) sejunctus Szil. (рис. 102) — Пестряк горный.
S z i 1 a d у, 1917, Arch. f. Naturgv 83, 4 : 126; Krober in Lindner,
1925 : 19; Олсуфьев, 1937a : 98; Leclercq, 1960b : 54; — interruptus К г 6-
Ь е r, 1920 : 134.
Рис. 102. Chrysops (Heterochrysops) sejunctus Szil. Брюшко и крыло $.
A — голова 6.
g. Лоб в сероватом, лицо в желтом налете. Лобная мозоль крупная, блестяще-
черная, боковыми сторонами почти достигает края глаз. Темя на значительном
протяжении блестяще-черное. • Лицевые мозоли черные или черно-коричневые, слегка
продолговатые. Наличниковая мозоль поделена на две половины полоской налета,
174 П. ПОДСЕМ. CHRYSOPSINAE
спускающейся сверху острым мысом. Эти половинки соединены б. м. широко с
лицевыми мозолями. Щечные мозоли небольшие, блестяще-черные. Усики короткие,
черные; 1-й членик слегка утолщен, часто в основной части коричневый. Щупальца
остроконечные, черные. Бочки груди и боковые стороны спинки в густых золотисто-желтых
волосках. Крылья: перевязь резкая, темная, достигает края крыла лишь внутренней
стороной вдоль жилки си2. Дискоидальная ячейка в средней части просветлена.
Внешний край перевязи кзади от г2+3 резко выпуклый. Вершинное пятно широкое, налегает
на жилку г4 на 3/4 ее длины, обособлено от перевязи просветом в ячейке Rx. Ноги
черные; средние голени, основания передних голеней и основания лапок последних
двух пар ног коричневые. Брюшко: I и II тергиты желтые; I тергит с крупным, черным
трапециевидным пятном, сужающимся кзади. Его ширина превосходит ширину щитка.
II тергит посредине с черным F-образным пятном, концами упирающимся в задний
край тергита (как у Ch. (s. str.) caecutiens). Последующие тергиты черные, из них III—
IV или III—V тергиты имеют по заднему краю по небольшому срединному
треугольнику желтовато-серого цвета. Снизу брюшко в основной половине желтое; II стернит
с небольшим темным пятном посредине, следующий за ним стернит с более крупным
поперечным пятном; последующие стерниты целиком черные, лишь иногда IV стернит
по бокам желтоватый. 9—9.5 мм.
3- Глаза не соприкасаются, разделяясь узким, но явственным промежутком.
Фасетки глаз б. м. одинаковой величины. Лицевые мозоли блестяще-черные, крупные,
соединены с наличниковой мозолью широкими перемычками. Кроме того, лицевые
мозоли узкими полосками вдоль края глаз соединены с хорошо развитыми щечными
мозолями. Последние достигают наличника. Усики короткие, целиком черные; 1-й
членик явственно утолщен. Щупальца черные, небольшие, короче J/2 длины хоботка;
концевой членик притуплён. Грудь одноцветно-черная, в черных волосках. Ноги
целиком черные. Крылья: внешний край перевязи и вершинное пятно как у §;
последнее также обособлено. Дискоидальная ячейка просветлена, но менее резко,
нежели у $• Основная половина крыла буро затемнена, просветляясь в нижней части.
Назальные ячейки в вершинной трети имеют по светлому округлому пятну, от которого
к основанию направляется легкий луч. На медиальной жилке, возле места ее
ветвления, лежит маленькое беловатое пятнышко. Жилки темно-коричневые. Брюшко
черное, в черных волосках. II—HI стерниты по бокам слегка буроватые. 6.5—8 мм.
Похож на Ch. (s. str.) caecutiens (3 на Ch. (s. str.) caecutiens ludens), но легко
отличается обособленным вершинным пятном на крыльях и просветленной дискоидальной
ячейкой.
Распространение: горный вид, известен с М. Кавказа. Грузия (включая
Аджарскую АССР), Армения, Нахичеванская АССР. Вост. Турция (б. Турецкая
Армения). Не часто. 21 VI—15 VIII. Указание Крёбера (КгоЬег, 1921) на нахождение
этого вида в Крыму и Астрахани ошибочно.
20. Chrysops (Heterochrysops) italicus Mg. (рис. 103) — Пестряк окончатьш.
М е i g e n, 1804, Klass., I : 158; S z i 1 a d у, 1917 : 113; К г б Ь е г, 1920 : 143;
Surcouf, 1924:233; КгоЬег in Lindner, 1925:12; Олсуфьев,
1937а : 100; L е с 1 е г с q, 1960Ь : 53; Б о ш к о, 1968 : 37 (экология); С h v a 1 а,
Lyneborg a. Moucha, 1972 : 139. — marmoratus, Rossi, 1790 : 322, цит.
по: К г 6 Ь е г, 1938 : 59. — fenestratus Latreille, 1809 : 285, цит. по: К г 6 Ь е г,
1938 : 59.
§ Лоб и лицо в сером налете. Лобная мозоль небольшая, поперечно-овальная,
черная, далеко не достигает края глаз. Темя возле глазков блестяще-черное. Лицевые
мозоли небольшие, продолговатые, коричневые. Наличниковая мозоль такого же
цвета, разделена на две половинки б. м. широкой полосой налета. Последние
соединены с лицевыми мозолями б. м. узкими перемычками. Щечные мозоли умеренных
размеров, блестяще-черные. Усики длинные, тонкие; 1-й членик желтый, к концу
коричневый, 2-й темно-коричневый, 3-й черный с коричневым основанием. Щупальца
желтые. Крылья: перевязь просветлена в дискоидальном поле («окошечко»), основной
части 2-й радиальной ячейки и концевой части 5-й заднекрайней ячейки. Внешний
край перевязи с небольшой выпуклостью между г2+3 и г4+5 и затем далее
широковыпуклый между г4+б и задним краем крыла. Вершинное пятно широкое, налегает на г4
на 2/з ее длины; пятно соединено с перевязью узкой полоской, не превышающей 1/2
ширины 1-й радиальной ячейки. Анальная ячейка не замкнутая. Дискоидальная
ячейка очень широкая. Ноги желтые; колени, вершины передних голеней, передние
лапки и концы последующих лапок черные. Брюшко: I и II тергиты желтые, I тергит
с крупным поперечным черным пятном, значительно превосходящим ширину щитка.
II тергит с 2 косыми треугольными пятнами; эти пятна в передней части тергита обычно
соединены концами, обособляя позади себя крупный желтый треугольник.
Последующие тергиты грязно-желтые или серые, с четырьмя продольными рядами небольших
черных треугольных пятен, убывающих в размерах спереди назад. Пятна
расположены по переднему краю тергитов, причем в средних рядах пятна крупнее, нежели
4. CHRYSOPS
175
в боковых. Снизу брюшко желтоватое, III и последующие кольца с широкой темной
срединной полосой, составляющей около V2 ширины стернитов; последние стерниты
затемнены целиком. VIII стернит б. м. треугольной формы, шире длины, с темным
срединным треугольником, боковые стороны округлые, гонопофиз с незначительной
апикальной выемкой или без нее; церки обычного строения; сперматека
остроконечная. 9—10.5 мм.
3- Глаза соприкасаются на очень незначительном протяжении. Лицевые мозоли
черные, соединены с наличниковой мозолью б. м. широкой перемычкой. Щечные
мозоли черные, соединены с наличником. Усики длинные, черные или черно-коричневые.
Рис! 103. Chrysops (Heterochrysops) italicus Mg. Брюшко, крыло, VIII стернит
(варианты) и церки $. А—голова <J.
Щупальца коричневые, остроконечные, явственно длиннее 1/г длины хоботка. Крылья:
перевязь достигает заднего края крыла, ее внешний край как у §. Дискоидальная
ячейка с округлым светлым пятном. Вершинное пятно как у $• Обе базальные ячейки
затемнены на 2/3 от основания. Ноги черные; средние голени . желтые, с коричневой
вершиной, задние голени и основания лапок последних двух пар ног коричневые.
Брюшко сверху черное, II (или I—II) тергит по бокам с небольшими желтыми
пятнами; II—V тергиты по заднему краю с маленькими серыми срединными
треугольничками. II (или I—IV)'cTepHHT грязно-желтый, с темной полоской посредине,
последующие черные. 9 мм;
Близок к Ch. (Heterochrysops) flavipes, отличаясь: 1) большей величиной, 2) более
широким вершинным пятном и 3) более крупными и широко расходящимися кзади
черными пятнами на II тергите; у 3 глаза соприкасаются.
Распространение: средиземноморский вид, преимущественно близ
морского побережья. Юг Украины (от дельты Дуная до Бердянска, включая Крымский
п-ов); Краснодарский край (Темрюк, Анапа). Юг Европы, Сев. Африка, М. Азия,
РИран. Местами обычен. Май—июль, иногда позднее.
Кладки яиц на растениях, хищные личинки в берегах морских заливов и соленых
озер (Бошко, 1968, 1973).
Обнаружен спонтанно зараженным возбудителем туляремии.
21. Chrysops (Heterochrysops) flavipes Mg. — Пестряк желтоногий.
Политипичный вид. В СССР распространены две географических расы —
номинативная Ch. (Heterochrysops) flavipes flavipes Mg. и южная Ch. (Heterochrysops)
flavipes punctifer Lw., а также закавказский варьетет для обеих рас — var. abdomi-
nalis Krob., определяемый только по самцам.
21а. Chrysops (Heterochrysops) flavipes flavipes Mg. (рис. 104, 105, 106) —
Пестряк желтоногий номинативный.
М е i g e n, 1804, Klassific, 1 : 159; S z i 1 a d у, 1917 : 110; КгоЬег, 1920 :
137; Surcouf, 1924 : 231; КгоЬег in Lindner, 1925 : 11; О л с у ф ь е в,
1937а : 102; Moucha et Chvala, 1955 : 229; L е с 1 е г с q, 1960Ь : 52; Ш е в-
ч е н к о, 1961:51; Chvala, Lyneborg a. Moucha, 1972:137.
176
П. ЛОДСЕМ. CHRYSOPSINAE
$. Лоб и лицо в желто-сером налете. Лобная мозоль небольшая, блестяще-черная,
полуовальной формы; нижний край мозоли посредине с небольшим мысиком,
направленным вниз. Темя возле глазков блестяще-черное. Лицевые мозоли небольшие,
далеко не соприкасаются с краем глаз, черные или коричневые. Наличниковая мозоль
одноцветная с лицевыми мозолями, поделена узкой полоской налета на две половинки;
последние соединены с лицевыми мозолями б. м. узкими перемычками. Щечные
мозоли небольшие, блестяще-черные. Усики длинные, тонкие. 1-й членик желтый, с
коричневой вершиной, 2-й темно-коричневый, 3-й черный с коричневым основанием.
Щупальца желтые. Крылья: перевязь достигает заднего края крыла, слегка
просветляясь в задней части 5-й заднекрайней ячейки. Дискоидальная ячейка с овальным
светлым пятном. Внешний край перевязи слегка выпуклый. Вершинное пятно
относительно узкое, налегает на г4 на 1j3 ее длины. Пятно соединено с перевязью пере-
Рис. 104. Chrysops (Heterochrysops) flavipes Рис. 105. Chrysops (Heterochrysops) fla-
flavipes Mg. $. vipes flavipes Mg. <$■
мычкой, занимающей У2 ширины ячейки В.г. Ноги желтые, колени черные; концы
передних голеней, передние лапки и концы последующих лапок черные. Задние бедра
часто в основании коричнево-затемнены, реже целиком черные. Брюшко сверху
желтое; I тергит посредине с небольшим темным пятном, не превосходящим ширину щитка.
Пятно сзади с небольшой, желтой б. м. треугольной выемкой. II тергит с двумя
узкими, черными пятнами в виде косых черточек или треугольников, слегка
расходящихся концами в заднем направлении. Последующие тергиты каждый имеет спереди
поперечную узкую черную полоску, прерванную посредине. Возле перерыва полоска
с каждой стороны слегка треугольно расширена кзади. Два концевых тергита
целиком желтые. Снизу брюшко в основной половине желтое, II стернит с небольшим,
III (и часто IV) с более крупным темным пятном посредине. Последующие стерниты
целиком темные. VIII стернит грушевидный, б. м. равномерно темный, ширина
превосходит длину, боковые стороны слегка угловатые, гонопофйз без заметной
апикальной выемки; церки обычного строения; базальная часть IX стернита изменчива; спер-
матека с приостренной вершиной. 6—9 мм.
$. Глаза не соприкасаются, разделяясь узкой, но явственной, блестящей
полоской лба. Фасетки верхних 3/3 глаз крупнее нижних. Лицевые мозоли округлые,
блестяще-черные или темно-коричневые, соединены с наличниковой мозолью относительно
широкими перемычками. Щечные мозоли умеренной величины, соединены с
наличником. Усики умеренно длинные, черные, 1-й членик часто коричневый изнутри.
Щупальца черные, остроконечные, превосходят */2 длины хоботка. Крылья: перевязь
достигает заднего края; дискоидальная ячейка с небольшим, округлым просветом.
Внешний край перевязи выпуклый. Вершинное пятно б. м. широкое, налегает на
г4нал1г ее длины. Пятно соединено с перевязью по ширине ячейки Rt. Базальные ячейки
затемнены от основания на половину. Ноги в основном черные; средние голени и
основания средних и задних лапок коричнево-желтые. Брюшко: I тергит черный е
незначительными желтыми пятнами по бокам; II тергит желтый с, большим черным пятном,
4. CHRYSOPS 177
занимающим всю среднюю часть тергита; позади пятна посредине имеется небольшой
желтый треугольник. От темного пятна в обе стороны отходит по узкой черной
полоске; последние идут вдоль заднего края кольца. Последующие тергиты черные,
из них III и IV тергиты имеют каждый по заднему краю по три желтых треугольных
пятна. Снизу брюшко в основной половине желтое, I—VI стерниты посредине с
темными пятнами, постепенно увеличивающимися кзади. V и последующие тергиты
целиком черные. 7.5—8 мм.
Распространение: южноевропейский (средиземноморский) лесостепной
подвид (рис. 107, 1). В СССР широко распространен на юге европейской части страны,
на север до Киева, Сум, Воронежа, Казани, Оренбурга, на юг включая дельту Дуная,
Рис. 106. Chrysops (Heterochrysops) flavipes Mg. A — брюшко З
Ch. (Heterochrysops) flavipes var. abdominalis Krob.; Б — голова 6
I. typica; В — вершина крыла § Ch. (Heterochrysops) flavipes pun-
ctifer Lw.; Г —. вершина крыла 5 номинативной формы Ch.
(Heterochrysops) flavipes flavipes Mg.; Д, E — VIII стернит и церки
Ch. (tleterochrysops) flavipes flavipes Mg.
Крым, Сев. Кавказ, Закавказье (Грузия, Армения, Азербайджан), дельту Волги и
Урала; по ту сторону Урала известна находка в Актюбинской обл. (Кара-Сай, 75 км.
с.-з. Иргиза). Ю. и частью Средн. Европа, Сев. Африка (Марокко, Алжир), Турция.
Обычен. Середина июня—август. Преимущественно в поймах рек; многочисленные
кладки яиц на растениях по песчаным берегам, там же обитают личинки.
Экспериментально доказанный переносчик туляремии (Боженко, 1941).
Примечание: в Закавказье 36 чаще представлены темным варьететом — var.
abdominalis Krob. Он отличается почти целиком черным брюшком, лишь по бокам
II тергита имеются незначительные желтые пятна (рис. 106, А). II—V тергиты с
незначительными сероватыми треугольниками посредине; III тергит иногда
дополнительно с легкими боковыми треугольничками серого цвета. Снизу брюшко черное,
II—III стерниты по бокам с довольно крупными желтыми пятнами.-§ не отличается
от основной формы. Вершинное пятно на крыльях варьирует у обоих полов в размерах
от Ch. (Heterochrysops) flavipes flavipes к Ch. (Heterochrysops) flavipes punctifer
(Закавказье. Сев.-Зап. Иран, Сев.-Вост. Турция, Сирия).
21Ь. Chrysops (Heterochrysops) flavipes punctifer Lw. — Пестряк желтоногий
средиземноморский.
L о е w, 1856, Neue Beitr., 4 : 24 (Ch. punctifer); S u г с о u f, 1924 : 232 (Ch.
punctifer); Олсуфьев, 1937a : 104; L e с 1 e г с q, 1960b : 52; Шевченко,
1961 : 51. — maculiventris Becker, 1912 : 587 (3). — beckeri К г б b e r, 1920 :
135 ($). — askahabadensis S z i 1 a d y, 1917 : 111 (Ch. flavipes Mg., var.).
Южный подвид. Отличается от основной формы более широким вершинным
пятном, налегающим у $ на 1/2, у 3 на 2/8 длины жилки г4, и более широкой перемычкой
между пятном и перевязью (рис. 106, В). Перемычка занимает почти целиком ширину
ячейки jRj.
Распространение: средиземноморско-среднеазиатский степной подвид
(рис. 107, 2). Восточное Предкавказье (Каякент), Закавказье (Грузия; Азербайджан,
включая Нахичеванскую АССР); юг Туркмении; Таджикистан; Узбекистан; Киргизия
(Ферганская долина); Казахстан (Чимкентская и Джамбулская области). Ю. Европа,
Сев. Африка, Передняя Азия, Иран, Афганистан, Зап. Индия. Местами обычен.
Июнь—июль, иногда раньше или позднее. На Кавказе рассматриваемый подвид рас-
12 Н. Г. Олсуфьев
178 II. ПОДСЕЛ. CHRYSOPSINAE
пространен наряду с номинативным подвидом и переходными формами. Один из
вариантов этого изменчивого вида с почти прозрачными в основании базальными
ячейками описан Крёбером из Туркестана как Ch. beckeri. Подобные экземпляры имеются
в коллекциях Зоологического института Академии наук из Туркмении; мы их
относим к подвиду Ch. (Heterochrysops) flavipes punctifer Lw. Тип Беккера Ch. maculiventris
идентичен экземплярам из Ср. Азии.
0° К 60CTOKV ОТ ГриНВИЧа 90° W 150° 180° 150°
0° 30° 60° 90° 120°
Рис. 107. Ареалы: 1 — Chrysops (Heterochrysops) flavipes flavipes; 2 — Ch.
(Heterochrysops) flavipes punctifer; 3 — Ch. (Heterochrysops) flavipes simil-
limus; 4 — Ch. (Heterochrysops) flavipes gedrosiana; 5 — Ch.^ (Heterochrysops)
vanderwulpi.
22. Chrysops (Heterochrysops) vanderwulpi Krob. (рис. 108). — Пестряк Вандер-
вульпа.
К г 6 Ь е г, 1929, Zoolog. Jahrb. (Ch. (Heterochrysops) mlokosiewiczi Bigot,4 var.),
56 : 467; Олсуфьев, 1937a : 105; L e с 1 e г с q, 1960b : 54; Takahas i,
1962 : 23; В и о л о в и ч, 1968 : 54. — striatus van-der-Wulp, 1885 : 79,
цит. по: К г 6 b е г, 1929 : 467. — mlokosiewiczi (Big.) apud S h i г a k i, 1918 : 25
et auct.
5. Лоб в сером, лицо в желтом налете., Лобная мозоль умеренно крупная, не
достигает края глаз, светло-коричневая, с черноватыми верхними краями. Темя возле
глазков в сером налете. Лицевые мозоли вместе с наличниковой желтые. Щечные
мозоли в виде рудиментарных черных точек возле края глаз. Усики короткие, толстые;
1-й и 2-й членики желтые, 3-й членик красно-желтый, с черной палочкой. Длина
2-го членика превосходит ширину в 1 V2 раза. Щупальца оранжево-желтые, б. м.
остроконечные. Ноги оранжево-желтые, концы лапок коричнево-черные. Крылья
с желто-бурым рисунком. Перевязь достигает заднего края лишь узким языком вдоль
сиг. Птеростигма желтая. Дискоидальная ячейка просветлена. Внешний край пере-
■ вязи возле ?4+5 с небольшим угловидным выступом, далее б. м. прямой. Вершинное
пятно в виде полоски, отходящей от перевязи и постепенно сужающейся к концу;
перейдя rt пятно исчезает, ширина полоски в начале превосходит ширину 1-й
радиальной ячейки. Брюшко светло-желтое, с 4 продольными полосками; каждая
полоска состоит из ряда узкоклиновидных черных пятен, направленных острием назад.
Снизу брюшко желтое, с серединным рядом небольших черных пятен, по пятну на
каждом стерните. V и последующие стерниты иногда целиком темные. VIII стернит
б. м. грушевидной формы, его длина почти равна ширине, боковые стороны округлые,
гонопофиз с слабой апикальной выемкой; церки слегка удлиненные; сперматека с
тупой вершиной. 7—9.5 мм.
3- Глаза соприкасаются на некотором протяжении. Фасетки верхних 2/3 глаз
значительно крупнее нижних, граница между теми и другими явственная. Лицевые
мозоли желто-коричневые, блестящие, наличниковая мозоль такого же цвета,
соединена с лицевыми мозолями широкими перемычками. Щечные мозоли в виде маленьких
4. CHRYSOPS 179
черных треугольников возле края глаз. Усики короткие: первые два членика сильно
утолщены, желтые, 3-й членик коричневый, палочка черная; 2-й членик овальный,
его длина лишь в 1 1/2—2 раза превосходит ширину. Щупальца желтые, явственно
превосходят V2 длины хоботка. Крылья как у §■; вершинное пятно с более резкими
очертаниями. Ноги желто-коричневые, концы лапок черные; иногда также концы
передних голеней черные (в таком случае передние лапки целиком черные). Брюшко
красновато-желтое, с двумя черными продольными полосками посредине, состоящими,
как У ?, из узкотреугольных пятен, направленных вершинами назад. III—V тергиты
по бокам с незначительными черными полосками или узкими треугольничками. Снизу
брюшко желтое, I—IV стерниты с небольшими черными пятнами посредине,
последующие кольца черные. 9—10 мм.
Рис. 108. Chrysops (Heterochrysops) vanderwulpi Krob. Брюшко, крыло,
усик, VIII стернит и церки $.
Похож на Ch. (Heterochrysops) mlokosiewiczi и ранее неправильно считался его
синонимом (или варьететом). Резко отличается от последнего короткими и более
светлыми усиками, более широким вершинным пятном и угловидным выступом по внешнему
краю перевязи возле г4+5.
Распространение: представитель восточноазиатского типа фауны
(рис. 107, 5). В СССР распространен отдельными участками: 1) север Омской обл.
(единственная находка в Усть-Ишимском р-не); 2) Алтайский край — пойма р. Оби
(от Фоминского до Обского); 3) Красноярский край — Сев. Приангарье, Столбы близ
Красноярска и Хакассия; 4) Прибайкалье; 5) Центральная Якутия (на север
до 63°с. гл.); 6) Амурская обл., Приморский и Хабаровский края (на север до устья Зеи
и низовий Амура) и 7) о. Кунашир. Два последних участка являются северной частью
основного ареала, охватывающего Сев.-Вост., Вост. и Ю.-Вост. Китай (включая о.
Тайвань и о. Хайнань), Корейский п-ов и Японию. Популяции вышеупомянутых 1—5
участков, видимо, обитают изолированно. Обычен на Дальнем Востоке, в западных
частях ареала редок. Июль, иногда раньше или позднее.
23. Chrysops (Heterochrysops) mlokosiewiczi Big. (рис. 109) — Пестряк Млоко-
севича.
Bigot, 1880, Ann. Soc. Entom. France, X : 146; Олсуфьев, 1937а : 107;
L.eclercq, 1960b : 53; Шевченко, 1961 : 52. — iranensis Bigot, 1892 :
602.
9- Лоб в желтовато-сером, лицо в желтом налете. Лобная мозоль крупная, слабо
блестящая, двуцветная: в нижнецентральной части коричневая, по краям черная.
Лицевые и наличниковые мозоли желтые или желто-коричневые. Щечные мозоли
отсутствуют. Щупальца желтые или коричневатые. Усики длинные и тонкие. 1-й
членик желтый или желто-коричневый, с темным концом, 2-й членик коричневый, часто
снаружи черноватый, 3-й целиком черный; 2-й членик цилиндрический, его'длина
превышает ширину в три раза. Крылья с черноватым рисунком. Перевязь достигает
заднего края лишь узким языкам вдоль си2; внешний край перевязи прямой.
Птеростигма черноватая. Дискоидальное поле сильно просветлено в центральной части;
2-я радиальная ячейка в основании слегка просветлена. Вершинное пятно очень
узкое, в виде незначительной полоски вдоль костальной жилки, занимает
примерно г/2 ширины ячейки Rx; вершинное пятно исчезает не доходя до г4. Ноги желтые;
12*
180 II. ПОДСЕМ. CHRYSOPSINAE
передние голени и лапки и концы средних и задних лапок черные; большая часть
задних голеней и колени средней пары ног коричневые. Брюшко желтое, сверху
с четырьмя продольными черными полосками. Последние состоят из узких
клиновидных пятен. Снизу брюшко желтое, с слегка затемненным кончиком. VIII стернит
грушевидный, его ширина превосходит длину, боковые стороны округлые, гонопо-
физ с апикальной выемкой; перки обычного строения; сперматека с тупым концом.
7—10.8 мм.
<J. Глаза соприкасаются на некотором протяжении; фасетки верхних 2/3 глаз
значительно крупнее нижних; граница между теми и другими б. м. резкая. Лицевые
мозоли небольшие, округлые, грязно-желтые; наличниковая мозоль разбита
срединной полоской налета на две половинки; последние узкими перемычками соединены
с лицевыми мозолями. Щечные мозоли отсутствуют. Усики б. м. длинные; 1-й членик
слегка утолщен, желто-коричневый, 2-й членик коричнево-черный, цилиндрический,
его длина превышает ширину в 3 раза, 3-й членик узкий, целиком черный. Щупальца
Рис. 109. Chrysops (Heterochrysops) mlokosiewiczi Big. Брюшко,
крыло, усик, VIII стернит, церки и сперматека §.
коричневато-желтые, слегка притуплённые на конце; их длина не превышает *72 длины
хоботка. Крылья: перевязь и вершинное пятно как у $; передняя базальная ячейка
затемнена от основания на половину, задняя ячейка затемнена в основании и далее
вдоль нижнего края до конца ячейки. Ноги: средние голени и основания лапок двух
последних пар ног желто-коричневые; задние голени и основания передних голеней
коричневые или также желто-коричневые; бедра, вершины передних голеней, передние
лапки и концы последующих лапок черные. Врющко серовато-желтое; сверху рисунок
как у ?, но обе срединных полоски состоят из несколько более широких треугольных
пятен, в особенности на II тергите. Снизу брюшко в основной половине
грязно-желтое, с рядом черных пятен посредине, IV и последующие стерниты черные. 7—10 мм.
Распространение: турано-монгольский пустынный вид. В СССР
известен из Туркмении (окр. Мерва; окр. Керков-Келифский Узбой), Таджикистана,
Узбекистана (Сурхандарьинская, Ташкентская, Андижанская области), Киргизия
(Ферганская, Чуйская, Таласская долины) и юга Казахстана (Кзыл-Ординская,
Чимкентская, Джамбулская и Алма-Атинская области), на север до-Кзыл-Орды и устья Или,
на восток до Нарынкола и Панфиловского р-на Талды-Курганской обл. Китай:
Внутренняя Монголия (Ордос, Тунляо); Афганистан. Местами обычен, обитает в поймах
рек, по берегам озер, прудов, на посевах риса (Гребенюк и Чиров, 1971).
Биго (Bigot, 1880) описал этот вид по $, добытой Млокосевичем в Сев. Иране или
на Ю. Кавказе. Однако в сводной публикации (Abbassian-Lintzen, 1964) о слепнях
Ирана этот вид не упоминается, не найден он и исследователями фауны слепней
Восточного Закавказья (Гаузер, 1939, 1953, 1960; Джафаров, 1960), хотя вероятность
его нахождения в низовьях Куры весьма велика. Указания на распространение этого
вида в Японии (Shiraki, 1918), во многих местах Восточного Китая, включая о.
Тайвань (Chen a. Quo, 1949), повторенные затем Леклерком (Leclercq, 1960a), относятся.
к Ch. (Heterochrysops) vanderwulpi.
24. Chrysops (Heterochrysops) oxianus PL (рис. 110) — Пестряк тугайный.
P 1 e s k e, 1910b, Ann. Mus. Zool. St. Petersb., 15 : 458; Szilady, 1917 : 121;
К r 6 b e r, 1920 : 69; К r 6 b e r in Lindner, 1925 : 16; S t а с k e 1 b e r g,
1926b : 326 (<J); Олсуфьев, 1937a : 108; Leclercq, 1960b : 54; 1П e в-
ч е н к о, 1961 : 54.
4. CHRYSOPS 181
$. Лоб и лицо в голубовато-сером налете. Лобная мозоль маленькая, черная.
Темя возле глазков в густом налете. Лицевые и наличниковая мозоли желтые,
соединены между собой б. м. широко. Щечные мозоли в виде узких черных полосок. Усики
тонкие, длинные, 1-й и 2-й членики желтые или коричневые, 3-й членик черный,
в основании коричневый. Щупальца желтые. Крылья: темный рисунок почти
отсутствует; перевязь редуцирована, вместо нее имеются две бледных серых поперечных
полоски: более длинная, почти достигающая заднего края крыла, внешняя полоска
и более короткая внутренняя полоска; эти полоски соответствуют внешней и
внутренней сторонам перевязи. Вершинное пятно в виде очень узкой полоски, идущей от
перевязи вдоль края крыла и сходящей на нет возле г4. Ширина полоски не превосходит
1/2 ширины 1-й радиальной ячейки. Ноги желтые; вершина передних голеней,
передние лапки и концы последующих лапок черные. Задние бедра в средней части
коричневые. У темных экземпляров задние голени черные, средние и передние голени в той
или иной мере коричнево затемнены. Брюшко желтое, с четырьмя продольными
черными полосками. Срединные полоски состоят из узких клиновидных пятен,
направленных остриями назад, боковые полоски — из черточек. Снизу брюшко желтое,^с не-
Рис. 110. Chrysops (Heterochrysops) oxianus PI. Брюшко
и крыло §. А — крыло 6.
большими сероватыми пятнами посредине II—IV стернитов; последующие стерниты
серые. У темных экземпляров брюшко снизу целиком серое. 8—10 мм.
<J. Глаза соприкасаются на некотором протяжении. Лицевые мозоли коричнево-
желтые, небольшие, соединены б. м. широкими перемычками с наличниковой мозолью.
Щечные мозоли в виде ничтожных черных точек возле края глаз. Усики умеренной
длины, 1-й и 2-й членики коричневые, 3-й черный. Щупальца коричневые,
остроконечные, превосходят 1/2 длины хоботка. Крылья с более развитым, нежели у $,
темным рисунком. Перевязь имеет форму треугольника, упирающегося вершиной в
задний край крыла. Центральная часть перевязи (основание ячейки R2, 1-й заднекрайней
и центральная часть дискоидальной ячейки) просветлена. Внешний край перевязи
прямой или слегка выпуклый. Вершинное пятно как у $, но шире, соответствует
ширине 1-й радиальной ячейки. Вдоль сщ имеется легкое затемнение. Верхняя
базальная ячейка затемнена на V3 от основания, нижняя лишь слегка. Ноги в основном
черные; средние голени, основания передних голеней и лапок последних двух пар
ног коричневато-желтые. Вершина средних и задних бедер коричневая: Брюшко
сверху желтое, с четырьмя продольными черными полосками; срединные полоски
сближены между собой, частично сливаясь в передней части брюшка. Снизу брюшко
желтое, с небольшими черными пятнами, образующими срединную полоску, и
короткими боковыми черточками на II и последующих кольцах. Последние два стернита
темные. 7—9 мм'. •
Распространение: турано-закавказский вид, долины рек пустынной
зоны, известен только из СССР. Азербайджан (пойма Куры близ Евлаха); Туркмения
(Чарджоу); Каракалпакия (Нукус, Кипчак, Кунград, Чимбай); Узбекистан (близ
Термеза; долина Сырдарьи между впадением Ангрена и Келеса; Джиниз-Кетык близ
Байраккума в дол. Сырдарьи; Хива); Казахстан (в р-не пос. Ильич р-н
Чимкентской обл.; Жулек, 100 км на ю.-в. от Кзыл-Орды; Казалинск). В пределах Средней
Азии все находки приурочены к долинам рек Амударьи и Сырдарьи. Июнь—июль,
иногда позднее.
Личинки найдены у берега реки (в воде ?) в зарослях вейника (Лутта, 1964г).
182 П. ПОДСЕМ. CHRYSOPSINAE
3. Подрод PETERSENICHRYSOPS M. et Chv.
Angew. Parasitol., 10,
159 nee Pseudochrysops
Moucha et Chvala, 1969, in M о и с h a, 1969,
3 : 173. — Pseudochrysops Moucha et Chvala, 1957 :
Nabokov, 1945 (Lepidoptera, Lycaenidae).
Тип подрода Chrysops hamatus Loew, 1858.
Мозоли на лице у $ небольшие, у # отсутствуют или очень маленькие. Дисталь-
ная часть наличника в налете. Перевязь на крыле умеренно развита, вершинное пятно
отсутствует. У S светлый рисунок брюшка развит сильнее, чем у $•
Известны 3 вида, все из Восточного Средиземноморья. В СССР обнаружен 1 вид.
Рис. 111. Chrysops (Petersenichrysops) buxtoni armeniensis Ols. Крыло, брюшко,
голова спереди и усик $>•
25. Chrysops (Petersenichrysops) buxtoni Aust. — Пестряк Бакстона.
Austen, 1922, Bull. Ent. Res., 13 : 151 (Ch. buxtoni); К г о Ь е r in
Lindner, 1925 : 8; L e с 1 e г с q, 1960b : 44; T e p т е р я н, 1969 : 77.
Описан из Израиля. Изученный нами экземпляр $ из Армении имеет некоторые
отличия от $, описанной Остином (Austen), но сходен с $, описанной А. Е. Тертеря-
ном из Армении. Это дает нам
основание считать экземпляры из СССР
относящимися к отдельному подвиду.
25а. Chrysops (Petersenichr ysops)
buxtoni armeniensis subsp. nova (рис.
Ill) — Пестряк Бакстона армянский.
?. Лоб и лицо в светлом
желтовато-сером налете. На темени между
глазками блестяще-черное треугольное
пятно. Лобная мозоль
блестяще-черная, поперечная, ее боковые края
далеко не достигают края глаз. На лице
по бокам наличника по
блестяще-коричневой продолговатой мозоли. Усики
длинные, тонкие, черные; 1-й членик
на 3/4 от основания коричневато-желтый.
Концевой членик щупалец очень узкий,
темно-коричневый. Крылья: перевязь
широкая, светло-коричневая, не дости-
Рис. 112. Chrysops (Turanochrysops) hya-
_ lipennis Stack. $.
гает заднего края крыла, шире дискоидальной ячейки, последняя целиком темная. Мезо-
плевры в верхней части коричневые. Ноги желто-коричневые, концы голеней, передние
лапки целиком, последующие лапки черные кроме основания, передние и задние бедра
4. CHRYSOPS 183
затемнены к вершине. Брюшко преимущественно желтое, с черным рисунком. I тер-
гит в передней части с черным поперечным пятном. II тергит посредине с небольшим
черным пятном в виде перевернутой буквы V, концы которого не достигают заднего
края тергита. III—IV тергиты коричневатые, в передней части каждый с 4
небольшими б. м. треугольными черными пятнами, из них средняя пара крупнее боковых.
Три последних тергита черные, в сером налете, с коричневато-желтой каймой. Брюшко
снизу преимущественно желтое, I—II стерниты посредине с узким сероватым пятном,
последующие два стернита каждый с крупным коричневым пятном посредине, V—
VII стерниты черные, в сером налете, с желтоватой каймой. 8.5 мм.
Голотип: 9> 03- Айгер-лич Эчмиадзинского р-на, Армения, 4 VII 1963, Амбарцу-
мян. В коллекции Зоологического института Академии наук СССР, Ленинград.
А. Е. Тертерян (1969) имел ? этого подвида из окр. с. Агарак Аштаракского
района, Армения. -
От номинативной формы описываемый подвид отличается следующим: перевязь
на крыле менее развита, она не достигает его заднего края, на II—IV тергитах брюшка
преобладает желтая окраска, тогда как черный рисунок развит слабее, лицевые мозоли
коричневые, бедра по крайней мере средние коричневые.
3 неизвестен.
4. Подрод TURANOCHRYSOPS Stack.
Stackelherg, 1926в, Bull. Ent. Res., 16 (4) : 326; Олсуфьев,
1937a : 110; Leclercq, 1960b : 43.
Тип подрода Chrysops (Turanochrysops) hyalipennis Stack., 1926.
Крылья на всем протяжении бесцветные (рис. 112). Лицевые мозоли конусовидно
выступают вперед, блестяще-желтые (рис. 113, 114).
Известны 2 вида, оба из Средней Азии.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА
TURANOCHRYSOPS STACK. (<}<Z)
1 (2). 1-й членик усиков сильно утолщен (рис. 114); лобная мозоль крупная, желтая
27. Ch. (Turanochrysops) hyalipennis Stack.
2 (1). 1-й членик усиков тонкий, цилиндрический (рис. 113); лобная мозоль
маленькая, черная 26. Ch. (Turanochrysops) stackelbergi Ols.
26. Chrysops (Turanochrysops) stackelbergi Ols. (рис. 113) — Пестряк Штакель-
берга.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 110; Leclercq,
1960Ь : 44; Шевченко, 1961 : 55.
$. Лоб в голубовато-сером, лицо в желтоватом налете. Лобная мозоль маленькая,
плоская, блестяще-черная. Темя между глазками в налете. Лицевые мозоли блестяще-
желтые, конусовидно выдаются вперед. Наличниковая мозоль одноцветная с лицевыми
мозолями, соединена с последними б. м. широкими перемычками. Щечные мозоли
незначительных размеров, коричневые. Щупальца желтые, тупоконечные. Усики очень
тонкие, небольшие. 1-й и 2-й членики желтые, 3-й коричневый, палочка черная. Ноги
желтые, кончики лапок коричневатые. Крылья бесцветные. Анальная ячейка открытая.
Брюшко желтое, сверху с четырьмя узкими, продольными, черными полосками, снизу
стремя неясными темными полосками: более широкой посредине и узкими близ края
брюшка. VIII стернит грушевидный, его длина равна ширине, гонопофиз с
апикальной выемкой; церки обычного строения; сперматека остроконечная. 6—8 мм.
с? неизвестен.
Похож на Ch. (Heterochrysops) oxianus и Ch. (Turanochrysops) hyalipennis.
От первого легко отличается совершенно бесцветными крыльями и сильно выпуклыми
лицевыми мозолями, от второго — тонким 1-м члеником усиков и маленькой черной
лобной мозольной.
Распространение: среднеазиатский (туранский) вид, обитатель долин
рек пустынной зоны. Узбекистан (Термезский р-н: о. Арал-Пайчамбар на Амударье);
Таджикистан (Старая- Пристань на Вахше; окр. Ленинабада: тугай Сары-Камыш на
Сырдарье); Казахстан, Кзыл-Ординская обл. (Тюмень-Арык; Казалинск). Не часто.
Июнь—сентябрь. Все находки приурочены к долинам Амударьи (включая приток Вахш)
и Сырдарьи.
Голотип: 9, Казалинск, Казахстан, 29 VII 1934, Д. Благовещенский. В коллекции
Зоологического института Академии наук СССР, Ленинград.
184
III. ДОДСЕМ. TABANINAE
27. Chrysops (Turanochrysops) hyalipennis Stack, (рис. 112, 114). — Пестряк про
зрачнокрыпый.
Stackelberg, 1926в, Bull. Ent. Res.,
1937a : 111; L e с ] e г с q, 1960b : 44; Ш е в ч е н к о,
1965 : 1029.
16 (4) 327; Олсуфьев,
1961 : 55; Мельникова,
$ Лобная мозоль крупная, блестяще-желтая. Темя между глазками лишь слегка
блестящее, в остальных частях в сером налете. Все мозоли нижней части головы
желтые. Лицевые мозоли конусовидно выдаются вперед. Наличниковая мозоль соединена
с лицевыми мозолями широкими перемычками. Шечные мозоли в виде узкой полоски,
достигающей наличника. Щупальца желтые,
толстые, тупоконечные. Усики длинные,
целиком желтые; 1-й членик сильно утолщен,
блестящий, >в коротких черных волосках,
Рис. 113. Chrysops (Turanochrysops)
stackelbergi Ols. Голова, VIII стернит
и церки $.
Рис. 114. Chrysops (Turanochrysops)
hyalipennis Stack. Голова $.
2-й членик тонкий, цилиндрический, 3-й членик очень узкий, по длине равен первым
двум членикам, взятым вместе. Крылья бесцветные; анальная ячейка на конце не
замкнута на небольшом протяжении. Ноги целиком желтые. Брюшко желтое, сверху
С четырьмя узкими продольными черными полосками: двумя средними и двумя
боковыми, идущими возле края брюшка. V—VII тергиты блестящие, предыдущие тергиты
матовые. Снизу брюшко с 3 узкими черными продольными полосками: срединной и
двумя боковыми, расположенными возле края брюшка. 8.5 мм.
S неизвестен.
Распространение: среднеазиатский (туранский) вид, обитатель долин рек
пустынной зоны. Известен из немногих точек: Туркмения (Теджен; Серахс; Иолотань
на Мургабе); Каракалпакия (Чимбай); Таджикистан (Дагана на Пяндже Московского
р-на; Гусар на Зеравшане выше Пенджикента). Редок. Август—сентябрь, иногда раньше.
Все находки относятся к современному или древнему бассейну Амударьи. Ближайшие
родственные виды с конусовидными лицевыми мозолями и утолщенным 1-м члеником
усиков обитают в пустынных частях Сев. Америки.
III. Подсем. TABANINAE (Latreille, 1802) Loew, 1860
Задние голени на вершине без шпор. Темя без глазков, иногда имеется небольшой
глазковый бугорок. 3-й членик усиков поделен на 5 или 4 сегмента (редко на. меньшее
число). Хоботок обычно не длиннее половины высоты головы, с довольно широкими
сосательными лопастями. Базикоста в коротких волосках (щетинках), реже голая
(триба Diachlorini). IX тергит брюшка у <Jg разделен на две пластинки. У в гоностиль
с усеченной вершиной, имеющей обычно на внешней стороне зубчик; муфта семеприем-
ника $ с грибообразным расширением в дистальной части (рис. 54, IV, V, VI, Е).
3. Триба DIACHLORINI Enderlein, 1922
Базикоста голая (рис. 5, слева). Усики: первые 2 членика короткие, 3-й членик
удлинен, поделен на 5 сегментов, соответственно палочка 4-члениковая. Лобная
полоска у $ умеренной ширины или узкая. Крылья бесцветные или затемнены, но без
сетчатого рисунка. Тело обычно б. м. коренастое.
5. DASYRHAMPHIS 185
5. Род DASYRHAMPHIS End.
Enderlein, 1922, Mitt. Zool. Mus. Berlin, 10 : 346; L e cl er'c q, 1966 : 96. —
Efflatounanus Philip, 1948; 79, цит. по: L e с 1 e г с q, 1966 : 96.
Тип рода Tabanus ater Rossi, 1790.
Коренастые, средних размеров темноокрашенные мухи облика Tabanus. Глаза
в волосках или голые, без полосок. Крылья обычно в той или иной мере затемнены;
г4 без придатка. У 5 лоб умеренной ширины, глазковый бугорок отсутствует, обе
лобных мозоли хорошо развиты, соединены между собой. Брюшко обычно темно окрашено.
Средиземноморский род, насчитывающий в своем составе 12 видов. В СССР 1 вид.
1. Dasyrhamphis umbrinus Mg. (рис. 115) — Слепень темный.
М е i g е п, 1820, Syst. Beschreib., II : 35 {Tabanus); В г а и е г, 1880 : 162
(Tabanus); Surcouf, 1924 : 126 (Tabanus); КгоЬег in Lindner, 1925 : 97
(Tabanus); Олсуфьев, 1937а : 289
(Tabanus); Leclercq, 1966 : 98; С h v a 1 a, L y-
neborg a. Moucha, 1972:461.
$. Глаза голые (при 16-кратном
увеличении видны редкие очень короткие волоски),
без полосок. Лоб широкий, его высота
приблизительно в 3 раза превосходит ширину,
параллельный или слегка расширяющийся
книзу, в голубовато-сером налете и
коротких серебристых волосках. Лобные мозоли
блестяще-черные; слиты между собой в про-
Рис. 115. Dasyrhamphis umbrinus Mg. Лобная
полоска, усик и щупальце д.
долговатую полосу, имеющую перетяжку в
сторонами мозоли далеко не достигают края глаз. Лобный треугольник
блестяще-черный, реже в тонком сером налете. Усики черные; 3-й членик с незначительным закруг-
Рис. 116. Ареал Dasyrhamphis umbrinus.
ленным дорсальным углом. Щупальца и ноги черные. Крылья с коричневым
расплывчатым пятном ниже птеростигмы. Брюшко черное, сверху в коротких черных и серых
волосках. Последние сгруппированы главным образом по заднему краю тергитов,
образуя каемки, расширенные по бокам; концевые тергиты почти целиком в серых
волосках. Снизу брюшко черное, в коротких серых волосках; стерниты ц цередцеи части
месте их соединения. Боковыми
186 III. ПОДСЕМ. TABANINAE
обычно в черных волосках. VIII стернит (рис. 54, IV, В) темно окрашен, с узким гоно-
пофизом, имеющим небольшую апикальную выемку; латеральные лопасти большие;
базальная лопасть короткая, шире гонопофиза. Церки округлые. 12—15 мм.
с?. Голова небольшая, слабо выпуклая. Глаза голые (при 16-кратном увеличении
видны редкие очень короткие волоски), без полосок. Фасетки верхних частей глаз
заметно крупнее нижних. Глазковый бугорок и возле него затылок в черных или серых
слегка выступающих волосках. Вдоль глазного шва расположен ряд заметных серых
волосков. Лобный треугольник в сером налете. Усики как у $. Щупальца черные. Ноги
и крылья как у $• Брюшко короткоконическое, черное, II—III тергиты по бокам
неясно каштаново-коричневые. Волоски короткие, прилегающие, черные и серые.
Последние сгруппированы по заднему краю тергитов, образуя неясные каемки. Снизу
брюшко черное, в черных и по заднему краю тергитов в светлых волосках. Гоностиль
(рис. 54, IV, А) немного утолщен в основании; церки округлые. 12.5—16 мм.
Распространение: восточносредиземноморский вид (рис. 116). В СССР
известен из восточной Грузии (Эльдар), Армении (Чиманкенд), Азербайджана
(низовья Самура; Куба — Хачмасская низменность; Боз-даг; Усунли на Иори) и
Туркмении (Каракала; Чандыр приток Сумбара; Кизыл-Арват; Ашхабад). Италия, Ю.
Австрия, Балканы, Турция, Ливан, Израиль, Иран. Не часто. Ранний вид — с середины
апреля по июнь.
6. Род PHILIPOMYIA Ols.
Олсуфьев, 1964, Бюлл. Моск. об-ва исп. природы, отд. биологии. LXIX,
3 : 74.
Тип рода Tabanus graecus Fabricius, 1794.
Коренастые средних размеров виды, сходные с Tabanus (рис. 117). Глаза голые,
без полосок. Крылья почти прозрачные, яйчека Р1 открытая, г4 без придатка. У g лоб
умеренной ширины, глазковый бугорок
отсутствует, обе лобных мозоли слиты
в узкую продольную килевидную
полоску. У 3 голова небольшая, не больше,
чем у $. Брюшко короткое, широкое, с
коричнево-желтыми пятнами по бокам.
У § необычайно длинные протоки семе-
приемника (рис. 118), последний кпереди
достигает II—III кольца брюшка.
Средиземноморский род,
насчитывающий 4 вида; из них 2 вида в СССР.
Преимущественно в горах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
ВИДОВ РОДА PHILIPOMYIA OLS.
(??<?(?)
1 (4). 3-й членик усиков желто- или
красно-коричневый, с черной
палочкой или целиком оранжевый.
У $ брюшко снизу по крайней
мере по бокам II стернита желто-
коричневое.
2 (3). 3-й членик усиков с резким, в
виде зубца, остроугольным
дорсальным выступом. У § лобная
килевидная мозоль нижним концом
обычно значительно не дости-
Рис. 117. Philipomyia rohdendorfi Ols. <j>.
гает границы лобного треугольника (рис. 119). II стернит брюшка обычно
с крупным, черным квадратным пятном посредине, лишь немного не
достигающим заднего края стернита. У $ фасетки верхних 2/з глаз в 5—6 раз крупнее
нижних, граница между теми и другими резкая. У обоих полов жилки переднего
края крыла и мембрана между ними желтые; передние лапки черные . . .
t ■• ■ ••■ • t 1 ...... . 1. ph. graeca F.
6. PHILIPOMYIA
187
3(2).
4(1).
3-й членик усиков обычно с прямоугольным дорсальным выступом, реже
последний зубцевидный. У $ лобная килевидная мозоль нижним концом
достигает или почти достигает границы лобного треугольника (рис. 120). II стернит
обычно без темного пятна посредине; реже имеется небольшое пятно, не
простирающееся далее половины стернита. У 6 фасетки средней Vs глаз в 2—3 Раза
крупнее нижних, переход постепенный. У обоих полов жилки переднего края
ксыла и мембрана между ними коричневатые; передние лапки
темно-коричневое 2. Ph. apnea Mg.
3-й члёник'усиков черный. У Р брюшко снизу черное. Высокогорья Большого и
Малого Кавказа 3. Ph. rohdendorfi Ols.
Рис. 118. А — семеприемники $ Philipomyia aprica Mg.-; В — то же
Tabanus bovinus L.
*1. Philipomyia graeca F. (рис. 119) — Слепень греческий.
Fabricius, 1794, Ent. Syst., 4 : 38 {Tabanus); В r a u e r, 1880 : 178
(Tabanus); S u г с о u f, 1924 : 166 (Tabanus), Krober in Lindner, 1925 : 115
(Tabanus); Moucha et Chvala, 1958 : 185 (Tabanus); L e с 1 e г с q, 1966 : 151
(Tabanus); Chvala, Lyneborg a. Moucha, 1972:448.
$. Лобная полоска узкая, с параллельными сторонами, ее высота превосходит
ширину основания в 41/2—5 раз, покрыта желтоватым налетом. Килевидная полоска
на лбу (нижняя+средняя мозоль) нижним концом обычно значительно не достигает
границы лобного треугольника. Лобный треугольник в желтоватом налете. Усики
желто- или красно-коричневые, палочка обычно черноватая; 3-й членик^ широкий,
дорсальный выступ остроугольный, резкий. Концевой членик щупалец узкий,
желтоватый, в желтоватых волосках, иногда с примесью черных. Жилки переднего края крыла
и мембрана между ними желтые. Бедра черно-серые, средние и задние голени и базаль -
ная часть передних голеней желто-коричневые. Концевая часть передних голеней и
передние лапки черные, лапки последующих ног темно-коричневые. Брюшко черное,
188
III. ПОДСЕМ. TABANINAE
с большими желто-коричневыми пятнами по бокам I—III тергитов. Тергиты по
заднему краю в густых, желтовато-серых волосках, образующих явственные каемки;
последние в средней части слегка треугольно расширены; передне-боковая часть II тер-
гита обычно в золотисто-желтых волосках. Снизу брюшко преимущественно черное,
с широкими сероватыми каемками по заднему
краю стернитов; II стернит по бокам желто-
коричневый, посредине обычно с крупным
черным пятном, лишь немного не достигающим
заднего края полукольца, III стернит по
бокам желтоватый. 16—18 мм.
(J. Фасетки верхних 2/3 глаз значительно
крупнее нижних, приблизительно в 5—6 раз.
Лобный треугольник в сером налете. Усики
как у $, но 3-й членик более узкий. Концевой
членик щупалец коричневато-желтоватый,
продолговато-овальный. Ноги и крылья как у $.
Брюшко с большими желто-коричневатыми
пятнами по бокам I—III тергитов, разделяющая
их узкая срединная полоска и конец брюшка
черные. Снизу брюшко в основной половине
желто-коричневатое, II стернит с темным пят-
Рис. 119. Philipomyia graeca F. Лобная
полоска, усик и щупальце $.
ном посредине, IV и последующие стерниты черные, с серовато-желтоватой каймой.
16—17 мм.
Распространение: средиземноморский (южноевропейский) вид. В СССР
не известен, но весьма вероятно его нахождение в Молдавии, поскольку он довольно
обычен близ Ясс в Румынии (Скуфьин, 19606). Ю. и частично Средн. Европа. Май-
июль.
2. Philipomyia aprica Mg. (рис. 120) — Слепень теплолюбивый.
М е i g е п, 1820, Syst. Beschreib., II : 37 (Tabanus); В г а и е г, 1880 : 176 (Та-
banus); Surcouf, 1924 : 164 (Tabanus); К г б Ь е г, 1924 : 119 (Tabanus);
КгоЬег in Lindner, 1925 : 105 (Tabanus); О л с у ф ь е в, 1937а : 290
(Tabanus); Leclercq, 1966 : 15 (Т. graecus apricus Meig.); Chvala, Lyneborg
а. M о u с h a, 1972 : 946.
$. Лобная полоска узкая, с параллельными сторонами, ее высота превосходит
ширину основания в 41/2—5 раз, покрыта желтоватым налетом. Килевидная полоска
на лбу (нижняя+средняя мозоль) нижним концом достигает или почти достигает
границы с лобным треугольником. Лобный треугольник в желтоватом налете. Усики
красновато-коричневые с черной палочкой; 3-й членик широкий, дорсальный угол б. м.
прямоугольный. Концевой членик щупалец узкий, желтоватый, в светлых и частью
черных волосках. Крылья в передней части с легким буроватым оттенком, жилки
коричневые. Бедра черные в сером налете, голени коричнево-желтые. Концевая часть
передних голеней и передние лапки темно-коричневые, лапки последующих ног коричневые
с более светлым основанием. Брюшко блестяще-черное с большими
желтовато-коричневыми пятнами по бокам, захватывающими I—III тергиты. Тергиты по заднему краю
в густых, желтовато-серых волосках, образующих явственные каемки; последние
слегка расширены посредине в небольшие треугольники. Прочие части брюшка в
черных волосках. Переднебоковая часть II тергита в черных волосках. Снизу брюшко
желтовато-коричневое, концевые 3 или 4 стернита черно-серые. В основании брюшка
часто имеется небольшое темное пятно. VIII стернит с крупным гонопофизом,
апикальная выемка умеренная; латеральные лопасти небольшие, боковые коричневые пятна
крупные, иногда сливающиеся посредине стернита; базальная лопасть короткая,
одинаковой ширины с гонопофизом, основной край прямой или слегка выпуклый. Церки
б. м. треугольные. Сперматека узковеретеновидная. 14.5—19 мм.
(J. Фасетки средней трети глаз немного (в 2—3 раза) крупнее нижних, переход
постепенный. Лобный треугольник в сером налете. Усики как у $; 3-й членик более
узкий, его дорсальный угол б. м. тупоугольный. Концевой членик щупалец
светло-коричневато-желтоватый, продолговато-овальный. Ноги и крылья как у $• Брюшко
с большими желтовато-коричневыми пятнами по бокам, захватывающими I—IV или
I—III тергиты. Разделяющая их узкая срединная полоска и конец брюшка черные.
6. PHILIPOMYIA
189
Снизу брюшко в основной половине желто-коричневое, концевые 3 или, 4 стернита
черно-серые. 14—17.5 мм.
Распространение: средиземноморский (южноевропейский)
горно-лесной вид. В СССР широко распространен на Кавказе: Большой Кавказский хребет
(включая северные и южные склоны и частично предгорья), от Варениковской и Анапы Крас-
Рис.' 120. Philipomyia aprica Mg. Лобная полоска, усик, щупальце,
VIII стернит, церки, сперматеки и муфта семеприемника $.
нодарского края на западе, до Кубы в Азербайджане на востоке; Малый Кавказский
хребет в пределах Грузии (на запад включая Аджарскую АССР), Армении и
Азербайджана (включая Нахичеванскуго АССР); Талышский хребет. Средн. и Ю. Европа
(на запад до Португалии), Турция, Иран. В горах преимущественно в лесной
и частично в альпийской зоне. Обычен. Июнь—август.
Леклерк считает Ph. aprica лишь подвидом Ph. graeca, хотя большинство
энтомологов признает их самостоятельными видами. Мы хорошо различаем оба вида по
признакам, указанным в определительной таблице.
3. Philipomyia rohdendorfi Ols. (рис
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII
1 е г с q, 1966 : 166 (Tabanus); Chvala, Lyneborg
$. Лобная полоска с параллельными
сторонами, ее высота превосходит основание
в 4—41/2 раза, покрыта пепельно-серым или
слегка желтоватым налетом. Килевидная
полоска на лбу (нижняя+средняя мозоли)
нижним концом обычно достигает ■ границы
лобного треугольника. Лобный
треугольник в сером налете. 3-й членик усиков
целиком черный, реже в основании слегка
коричневый, широкий, с б. м. прямоугольным
дорсальным выступом. Концевой членик
щупалец свётло-желтовато-коричневатый. Крылья
в передней части с легким буроватым
оттенком, жилки коричневые. Бедра черные
Рис. 121. Philipomyia rohdendorfi Ols. Лобиая
117, 121) — Слепень Родендорфа
2 : 291 (Tabanus)
L е с-
а. М о и с h а, 1972 : 450.
4 полоска, усик и щупальце $.
в сером налете, голени желтовато-коричневые. Концевая половина передних
голеней и передние лапки черные, лапки последующих ног темно-коричневые с более
светлым основанием. Брюшко блестяще-черное с красновато-коричневыми нерезко
отграниченными пятнами по бокам, захватывающими I—III тергиты. Задний край тер-
190
III. ПОДСЕМ. TABANINAE
гитов в густых, блестяще-серых или желтоватых волосках, образующих довольно
резкие каемки. Эти последние посредине немного расширены. Черные части брюшка в
коротких черных волосках, коричневые боковые пятна — в золотисто-желтых волосках.
Снизу брюшко черное, в тонком буровато-сером налете и желтовато-серых волосках.
Задний край стернитов с неясной коричневатой каймой. 16—19 мм.
tJ. Фасетки средней трети глаза слегка крупнее (в 2—3 раза) нижней части,
переход нерезкий. Лобный треугольник в сером налете. Усики как у $; 3-й членик более
узкий. Щупальца коричневые, концевой членик продолговато-овальный. Ноги и
крылья как у $. Брюшко с большими красновато-коричневыми пятнами по бокам,
захватывающими I— III или I—1Утергиты. Разделяющая их срединная узкая полоска
и конец брюшка черные. Снизу брюшко в основной половине желто-коричневое,
концевые 3 или 4 стернита черные. II стернит посредине обычно с крупным черно-серым
пятном. 16—18 мм.
Распространение: кавказский горно-альпийский вид. Большой
Кавказский хребет (включая северные и южные склоны) от Кавказского заповедника на западе
до юга Дагестана (Каякентский р-н) и северо-запада Азербайджана (Белоканы) на
востоке; северо-запад Малого Кавказского хребта (Хуло Аджарской АССР; Бакуриани
и Цхрацкаро Грузинской ССР). Обычен. Июль—август.
Голотип: 9, Салги Чечено-Ингушской АССР, 5462 фут., 3 VIII 1927, А. Кириченко.
В коллекции Зоологического института Академии наук СССР, Ленинград.
7. Род NANORHYNCHUS Ols. — КОРОТКОХОБОТНИКИ
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 325 {Nanorhynchus);
Leclercq, 1966 : 99; Олсуфьев, 1966 : 52 {Nanorhynchus, emend).
Тип рода Tabanus crassinervis Villeneuve, 1924.
Средней величины мухи облика Tabanus. Хоботок у обоих полов очень маленький,
его длина составляет не более 1/3—V4 высоты головы. Затылок и щеки сильно
утолщены. Глаза в коротких волосках, без полосок. Глазки или глазковый бугорок
отсутствуют. Усики: 1-й и 2-й членики небольшие, б. м. бокаловидные, 3-й членик длинный, без
выраженного дорсального угла, палочка 4-члениковая. Крылья без темных пятен или
перевязей; все заднекрайние ячейки открытые. Эмподий и пульвиллы сильно
укорочены, у с? на передних лапках нормально
развиты. Брюшко сверху в сером налете,
без темного рисунка.
Единственный вид этого рода известен
из Средней Азии. Родственные виды рода
Aegialomyia Phil, обитают на юге Сев.
Америки, преимущественно на морских
побережьях (Олсуфьев, 1966).
1. Nanorhynchus crassinervis Villen.
(рис. 122, 123) — Короткохоботник туран-
ский.
Villeneuve in К г 6 Ь е г, 1924,
Arch. i. Naturg., 90 : 61 {Tabanus); К г о-
ber in Lindner, 1925 : 91 {Tabanus);
Олсуфьев, 1937а : 325; Leclercq,
1966 : 99.
'$. Глаза в очень коротких едва
заметных серых волосках. Лобная полоска
широкая, параллельная или слегка расширена
кверху, ее высота в 2—2х/4 раза превосхо-
Рис. 122. Nanorhynchus crassinervis
Villen. §.
дит ширину, покрыта тонким желтоватым налетом. Нижняя лобная мозоль бледно-
желтая или коричневатая, боковыми сторонами достигает края глаз. Средняя лобная
мозоль отсутствует. Лобный треугольник в желтоватом налете. Усики: 1-й и 2-й
членики светло-коричнево-желтые, в б. м. коротких волосках, 3-й членик одноцветно-
черный, узкий, дорсальный угол не выражен. Концевой членик щупалец беловатый,
тонкий, заостренный, в беловатых волосках. Нотоплевры желтоватые. Крылья с лег-
7. NANORHYNCHUS 191
ким молочным оттенком, жилки коричневые, вдоль жилок мембрана крыла слегка
затемнена; г4 с придатком или без него. Жужжальца желтоватые или беловатые.|Ноги
желтые, бедра иногда сероватые, конец передних голеней, средних и задних лапок
коричневый, передние лапки черноватые. Брюшко длинное, в густом светло-сером
налете и коротких серых волосках; боковые стороны II тергита и все кольца по
заднему краю просвечивают желтоватым. VIII стернит широкий, гонопофиз крупный,
в длинных и густых черных волосках, вершина без апикальной выемки, латеральные
лопасти остроугольные, светлые, базальная лопасть короткая, незначительно затем-
Рис. 123. Nanorhynchus crassinervis Villen. Голова, усик, VIII стернит, церки,
сперматека и муфта семеприемника J. А — гоноподит; Б — церки и эпандрий;
В — концевой членик лапки задней ноги 3 с вентральной стороны: эмподий
и пульвиллы резко укорочены.
нена по бокам, нижний край округлый. Церки с округлой вершиной, IX тергит в
желтых и длинных черных волосках, с округлым черноватым расширением по внешнему
краю. Семеприемники умеренной длины с небольшой, удлиненной сперматекой; муфта
семеприемника короткая, с очень маленьким грибовидным расширением. 13—14.5 мм.
в- Голова небольшая, слабо выпуклая. Фасетки верхних 3/4 глаза крупнее
нижних (приблизительно в 3—4 раза). Глаза в короких серых волосках. Затылок
сильно утолщен, в длинных светлых волосках, выступающих над поверхностью глаз.
Лобный треугольник в сером налете. Усики как у $, но более узкие. Концевой членик
щупалец узкий, беловатый, в светлых волосках. Крылья и жужжальца как у $. Ноги:
бедра черные в сером налете, голени коричнево-желтые. Конец передних голеней,
лайки средней и задней пары ног темно-коричневые, передние лапки черные. Брюшко
серое, боковые стороны II тергита и задний край всех колец желтоватые. Снизу
брюшко серое, стерниты по заднему краю желтоватые. У светлого экземпляра из
Бадхыаа желтая окраска распространяется на боковые стороны I—V тергитов, а снизу
соответственно I—V стерниты желтые. Терминалии обычного для Tabaninae строения.
12—14 мм.
192
III. ПОДСЕМ. TABANINAE
Распространение:, среднеазиатский (туранский) пустынный вид.
Туркмения: Красноводск, окр. Джебела, Большой Балхан, Кизыл-Арват, Арман-Саад
близ Кизыл-Атрека, Имам-баба, Бадхыз; Узбекистан: близ хр. Нуратау; Казахстан:
низовья Чу. Редок. Май—начало июня. Не кровосос.
4. Триба TABANINI (Latreille, 1802) Enderlein, 1922
Базикоста в волосках (рис. 5, справа). Усики: первые 2 членика короткие, б. м.
бокаловидные, 3-й членик удлинен, неправильной формы, обычно имеет дорсальный
угол, поделен на 5 сегментов, соответственно палочка 4-члениковая, крайне редко
3-члениковая (подрод Surcoufiella в роде Atylotus, Glaucops в роде Tabanus, Tibeto-
myia в роде Hybomitra и др.). Лобная полоска у g относительно узкая, ее высота
превосходит ширину основания более чем в 2 раза. Крылья бесцветные или затемнены,
но без сетчатого («мраморного») рисунка. Тело обычно б. м. коренастое.
8. Род TABANUS L. — СЛЕПНИ
Li ппе, 1758, Syst. Nat., X : 601; В г a u e г, 1880:137; S h i г a k i, 1918:
119; Surcouf, 1924:158; К г 6 Ь е г, 1924:79; КгбЬег in Lindner,
1925 : 54; Олсуфьев, 1937а : 118; Moucha et Chvala, 1958 : 182;
Шевченко, 1961 : 58; T a k a h a s i, 1962 : 65; L e с 1 e г с q, 1966 : 136; Виолович,
1968 : 83; Murdoch et Takahasi, 1969 : 56. — Phyrta Enderlein, 1922 :
344. — Straba Enderlein, 1923 : 544, цит. no: L e с 1 e г с q, 1966 : 136.
Тип рода Tabanus bovinus Linrie, 1758.
Коренастые виды различных размеров, от 10 до 30 мм, по преимуществу темной
окраски (светло окрашены пустынные виды). Глазковый бугорок у 9? отсутствует
(иногда на темени бывает развито небольшое блестящее пятно — мозоль), у 33
имеется, глубоко посажен между глазами и покрыт налетом или волосками. Глаза по
преимуществу голые, реже покрыты волосками. У 33 фасетки верхних частей глаз
обычно значительно крупнее нижних. При жизни глаза ярко окрашены — коричневые,
зеленые, синие, с полосками или без них. У сухих экземпляров глаза принимают
черную окраску, причем у 33 крупнофасетчатое поле иногда остается светлым,
вследствие чего глаза резко двуцветные. Лобные мозоли у 99 обычно хорошо развиты,
нижняя прямоугольная, треугольная или овальная, средняя веретеновидная или в виде
обособленного пятна. Лобный треугольник без волосков по бокам. Крылья бесцветные,
реже затемнены; г4 иногда с развитым придатком. Средние и задние голени не
утолщены, в коротких черных или светлых прилегающих волосках. VIII стернит у 99
обычно с развитыми латеральными лопастями. У ряда видов терминалии 99 имеют
различия, что, несмотря на индивидуальную изменчивость, позволяет использовать
строение терминалии для диагностики как дополнительный признак.
Гетероморфная группа. Обширный род, насчитывающий в своем составе около
1000 видов (Moucha, личное сообщение) и распространенный во всех частях Света.
В Палеарктике известно 182 вида. В СССР 64 вида, встречающиеся главным образом
в южных частях страны.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА TABANUS Ъ.
А. Глаза разделены лбом: 99
1 (142). 1-я заднекрайняя ячейка на конце не замкнутая (иногда лишь сужена к
вершине).
2 (91). Небольшие, менее 18 мм, виды. Глаза с полосками или без таковых.*
3 (50). Средняя лобная мозоль в виде овального, сердцевидного или поперечного
пятна, вполне обособлена от нижней мозоли (рис. 127, 136 и др.).; иногда
средняя мозоль нацело скрыта налетом (у потертых экземпляров обе мозоли иногда
соединяются тонкой линией).
4(17). Нижняя лобная мозоль расположена отступя от границы лобной полоски
с лобным треугольником (рис. 127). (I группа: Т. bifarius Lw.).
5 (6). Темя, как правило, с блестяще-черным, часто двойным пятном. Глаза в хорошо
заметных довольно длинных волосках. 3-й членик усиков черный, реже темно-
или красно-коричневый 5. Т. quatuornotatus Mg.
* У видов рода Tabanus наличие полосок на глазах, их число или отсутствие
таковых имеет большое значение для определения и должно быть отмечено либо у све-
жеумерщвленных экземпляров, либо после помещения сухих экземпляров во влажную
камеру.
8. TABANUS 193
6 (5). Темя целиком в налете, крайне редко с незначительным блестяще-черным
пятном. Глаза голые или в малозаметных коротких волосках. 3-й членик усиков
обычно оранжево-желтый или коричневый, иногда с более темной палочкой.
7 (8). Нижняя лобная мозоль желтая или грязно-желтая . . . . 1. Т. bifarius Lw.
8 (7). Нижняя лобная мозоль коричневая или черная.
9 (10). Брюшко сверху черно-серое, без заметных коричневых пятен по бокам; серые
пятна налета обычно имеют оливковый оттенок .... 6. Т. caucasius КгбЬ.
10 (9). По крайней.мере II—III тергиты брюшка по бокам коричневые; серые пятна
налета без оливкового оттенка.
11 (12). Лобный треугольник в верхней части блестяще-коричневый
7. Т. glaucopis Mg.
12 (11). Лобный треугольник целиком в сером налете (может быть частично оголен
у потертых экземпляров!).
13 (14). Глаза в коротких волосках 2. Т. lunatus F.
14 (13). Глаза голые.
15 (16). Бедра ног черные, в сером налете. Крупнее: 14—17.5 мм. Ср. Азия . . .
3. Т. bactrianus Ols.
-г-""?'гД\
Рис. 124. Голова Tabanus filipjevi Ols. 9 {.слева) и
Т. zimini Ols. 9 (справа).
16 (15). Бедра ног коричневые, в сером налете. Мельче: 11—14 мм. Талыш . . .
4. Т. talyshi Ols.
17 (4). Нижняя лобная мозоль расположена на уровне границы лобной полоски
с лобным треугольником (рис. 136). (II группа: Т. eordiger Mg.).
18 (35). rt с придатком (рис. 4). Крылья обычно с легким молочным оттенком.
19 (32). Глаза с одной или несколькими полосками.
20 (21). Лобная полоска узкая, ее высота превосходит ширину основания в 4—5 раз
(рис. 136) 8. Т. accipiter Szil.
21 (20). Лобная полоска более широкая, ее высота превосходит ширину в 21/«—
З1/* раза (рис. 138, 139 и др.).
22 (29). Глаза с 3—4 полосками (рис. 138, 139, 143 ж др.).
23 (26). На уровне основания усиков имеется кофейно-коричневая полоска,
отделяющая верхнюю пепельно-серую часть головы от белоснежного лица.
24(25). Глаза с 3 полосками 14. Т. ansarii Abbas.-L.
25 (24). Глаза с 4 полосками . 13. Т. freyi Szil.
26 (23). Темная полоска, возле основания усиков отсутствует.
27 (28). Бедра ног черные в сером налете. Лобная полоска широкая, ее высота в 21/i—
3 раза превосходит ширину основания (рис. 139). Брюшко сверху с довольно
резким рисунком из продольных рядов беловато-серых пятен, чередующихся
с рядами черноватых пятен; по бокам брюшка часто небольшие коричневые
пятна, но обычно не более как на II—III тергитах. Если рисунок брюшка
нерезкий, то по бокам брюшка коричневые пятна отсутствуют или едва развиты
f 10. Т. sabuletotum Lw.
28 (27). Бедра ног красновато-желтые, крайне редко сероватые. Лобная полоска
умеренно широкая, ее высота в З1/*—3V2 раза превосходит ширину основания
(рис. 138). Брюшко сверху с неясным рисунком из продольных рядов беловато-
серых пятен, чередующихся с рядами коричневых пятен; боковые стороны
по крайней мере II—IV тергитов желто-коричневые
9. Т. subsabuletorum Ols.
29 (22). Глаза с 1 полоской посредине и иногда дополнительной второй полоской по
нижнему краю (рис. 124).
30 (31). Глаза с 1 полоской. Ширина глаза в поперечнике б. м. равна его наибольшей
высоте, вследствие чего голова слабо поперечная (рис. 124, слева) ....
....!• 16. Т. filipjevi Ols.
З1'(30). Глаза с 1 полоской посредине и, кроме того, дополнительной полоской по
нижнему краю. Ширина глаза в поперечнике превосходит его наибольшую-
13 Н. Г. Олсуфьев
. 194 III. ПОДСЕМ. TABANWAE
высоту, вследствие чего голова сильно поперечная (рис. 124, справа) . . .
15. Т. zimini Ols.
32 (19). Глаза без полосок.
33 (34). Лобный треугольник по бокам блестяще-коричневый. Высота лба превосходит
ширину основания в 2—2х/2 раза (рис. 147). Мельче: 10—13.5 мм
17. Т. brunneocallosus Ols.
34 (33). Лобный треугольник целиком в желтовато-сером налете. Высота лба
превосходит ширину основания в 4—4V2 раза (рис. 157). Крупнее: 14—16 мм . .
23. Т. fumidus Aust.
35 (18). г4 без придатка (лишь в редких случаях с незначительным рудиментом).
Крылья обычно с сероватым оттенком.
36 (43). Глаза с полосками.
37 (40). Глаза с4 полосками (рис. 141, 142). Темная полоска возле основания усиков-
отсутствует.
38 (39). Бедра ног черные в сером налете. Нижняя лобная мозоль черная. Глаза
голые. Таджикистан, Иран 12. Т. mistshenkoi Ols.
39 (38). Бедра ног желтые. Нижняя лобная мозоль коричневая. Глаза в коротких
волосках. Монголия, КНР 11. Т. stackelbergiellus Ols.
40 (37). Глаза с 1 полоской (рис. 149, 151). На уровне основания усиков имеется
коричневая поперечная полоска.
41 (42). Лобная полоска умеренно широкая, ее высота превосходит ширину
основания в 3V4—4 раза; кверху полоска расширяется лишь слегка (рис. 151). Затылок
за глазами обычно в черных волосках или в светлых с примесью черных. Ното-
плевры, как правило, черные 19. Т. unifasciatus Lw.
42 (41). Лобная полоска более широкая, ее высота превосходит ширину основания
приблизительно в 3 раза; кверху полоска расширена довольно значительно
(рис. 149). Затылок за глазами в светлых волосках. Нотоплевры, как правило,
желто-коричневые 18. Т. leleani Aust.
43 (36). Глаза без полосок.
44 (45). Лобный треугольник блестяще-черный (рис. 158)
24. Т. kinoshitai Kono et Так.
45 (44). Лобный треугольник в сером налете.
46 (49). На уровне основания усиков имеется кофейно-коричневая полоска. Средняя
. . . лобная мозоль крупная, черная (рис. 153).
47 (48). Лобная полоска очень широкая, ее высота превосходит ширину основания
в 3V2—3V4 раза, заметно расширена кверху (рис. 153). Нотоплевры желто-
коричневые. Крупнее и коренастее: 13—17 мм. Европейская часть СССР,
Кавказ • . . 21. Т. cordiger Mg.
48 (47). Лобная полоска менее широкая, ее высота превосходит ширину основания
приблизительно в 4 раза, едва расширена кверху (рис. 152). Нотоплевры черные.
Мельче и у^ке: 11—15 мм. Горный Крым . 20. Т. smirnovi Ols.
49(46). На уровне основания усиков имеется светло-коричневая полоска. Средняя
лобная мозоль небольшая, черная, часто целиком покрыта налетом (рйс. 155).
Ср. Азия 22. Т. golovi Ols.
50 (3). Средняя лобная мозоль б. м. веретеновидная, обычно достигает нижним
концом до нижней лобной мозоли (рис. 159, 160 и др.).
■51 (82). 1-я заднекрайняя ячейка не сужена или едва сужена к вершине (III группа:
Т. bromius . L,; рис.. 169).
52 (53). Глаза явственно покрыты волосками 25. Т. rupium Br.
53 (52). Глаза голые.
54 (61). Глаза с 1—3 полосками.
55 (56). Глаза с 3 полосками (рис. 176) 38. Т. tergestinus Egg.
56 (55). Глаза с 1 полоской.
57 (58). Концевой членик щупалец узкий, коричневый (рис. 172). Брюшко сверху
без боковых светлых рядов пятен. г4 с длинным придатком. Таджикистан . .
. 34.. Т.. hissaricus Bar.
58 (57). Концевой членик щупалец беловатый или желтовато-серый, в основании
умеренно утолщен. Брюшко сверху помимо срединного ряда серых
треугольников имеет по боковому ряду серых пятен. г4 без придатка (изредка с его
рудиментом).
59 (60). Затылок в виде относительно утолщенной полоски (рис. 125, слева). Нижняя
лобная мозоль треугольно-щитовидная, -мелкоморщинистая. Брюшко сверху
серо-черное, со слабо выраженным серым рисунком у. . ;.
26. Т. maculicornis Ztt.
60(59). Затылок в виде. относительно узкой полоски (рис. 125, справа). Нижняя
, лобная мозоль б. м. прямоугольная, блестящая. Брюшко,-сверху с хорошо
развитым светло-серым рисунком, иногда с развитой желто-коричневой окраской
по бокам (южный подвид Т. Ъ. flavofemoratus Strobl.) . . . 33. Т. bromius L.
%i (54). Глаза без полосок. .....
^8?.tab-anust~
Ш5
62 (63). Концевой членик шупалец узкий, коричневый. Брюшко с одним срединным
■ . .рядом небольших, белых треугольников . . ',. . . ; . 39. Т. armenicus Szil.
63 (62). Концевой членик щупалец утолщен, беловатый или бледно-ко,ричневатый.
Брюшко с 3 рядами неясных, серых пятен или целиком оранжево-желтое.
-64 (67).. По.крайней мере средние и задние бедра> желтые или оранжево-желтые.
65 (66). Мельче 10—14 мм .(редко 15 мм). Передние, бедра, как .правило, .желтые.
VIII стернит с небольшой апикальной выемкой, церки с округлой вершиной
(рис. 174) i 36. Т. laetetinctus Beck.
66 (65). Крупнее:'14—16 мм. Передние бедра черные или черно-коричневые, в сером
налете. VIII стернит без апикальной выемки, церки с слегка уплощенной
вершиной (рис. 175) .. . .. . .37,. T..hauseri Ols.
67 (64). Бедра черно-серые. . •:
68 (69). Брюшко сверху по бокам в густых золотисто-оранжевых волосках. Горы
Средней Азии •. . . 35. Т. semenovi Ols.
69 (68). Брюшко сверху по бокам в серых или желтоватых волосках, обычно
чередующихся с пятнами черных волосков.
Рис. 125. Затылочная полоска Tabanus maculicornis Ztt. $
(слева) и Т. bromius L. g (справа).
70 (71). Высота лобной полоски превосходит ширину основания в 5—6 раз (рис. 168).
Лобный треугольник в коричневатом налете, резко отличающемся окраской от
белоснежной нижней части лица . . . . 32. Т. regularis Jaenn.
71 (70). Высота лобной полоски превосходит ширину основания в 3V2—5 раз (рис. 162,
163 и др.). Лобный треугольник в серовато-желтоватом налете, лишь слегка
отличающемся от беловато-серой окраски нижней части лица.
72 (73). Нижняя лобная мозоль расположена несколько выше основания лобной
' полоски (рис. 162). Небольшой, 10—14 мм, вид серого цвета. Алтай, Восточная
Сибирь, Дальний Восток . . . . . . .".'.". ....'. 27. Т; geminus Szil.
73 (72). Нижняя лобная мозоль расположена б. м. на уровне основания лобной
полоски (рис. 163, 165 и др.). Крупнее: 14—18 мм, редко 13 мм. Различные места
СССР, на восток до Байкала.
,74,(77). Нижняя лобная мозоль треугольно-щитовидная (рис. 163, 208).
75 (76). Концевой членик щупалец преимущественно в черных волосах. Затылок
в выступающих над галазами черных или светлых волосках t .
' ' .:............ 28. Т. miki Br.
76 (75). Концевой членик щупалец' в" белых волосках, 'изредка- с незначительной
примесью черных. Затылок в относительно коротких светлых или черных
волосках (из группы Т. autumnalis L.) ...... 62. Т.' atropathenicus , QJs.
77 (74). Нижняя лобная мозоль б. м. прямоугольная (рис.'165, 166, 167).
78 (81). 3-й членик усиков по крайней мере в основании коричневый или
черно-коричневый (иногда целиком коричневый или оранжевый). Лобная полоска йверху
обычно слегка расширена, ее высота превосходит ширину основания в 4—5 раз
(рис. 165, 166). . , , „
79 (80). Нижняя лобная мозоль, как правило, в базальной части коричневая
. (рис. 165), если целиком черная, то концевой членик щупалец преимущественно
в черных волосках. Лобная полоска'обычно в желтовато-сером налете. Брюшко
черно-серое, чаще с }эа'звить1ми" коричневыми пятнами по бокам II—III или
II—IV тергитов. Карпаты, горы Крыма, Кавказа, Ср. Азли . , . . .
, . . , . . , ,..:....,... 29. Т. indrae Haus.
80 (79). Нижняя лобная мозоль целиком черная (рйс. 166). Концевой членик щупалец
в беловатых и единичных черных волосках. Лобная полоска в пепельнб-'с'ёзйм
196 III. ПОДСЕМ. TABANINAE
налете. Брюшко черно-серое, иногда лишь слегка коричневатое по бокам II
(или II—III) тергита. Малый Кавказ (Нахичеванская АССР) ......
30. Т. infestus Bog. et Sam.
81 (78). Усики целиком черные. Лобная полоска б. м. параллельная, ее высота
превосходит ширину основания в Зг/2—4 раза (рис. 167)
31. Т. armeniacus КгбЬ.
82 (51). 1-я заднекрайняя ячейка б. м. резко сужена к вершине. (VII группа: Т. autum-
nalis L.; рис. 205).
83 (84). Брюшко сверху с 2 боковыми резкими, широкими, розовато-серыми
продольными полосками, разделенными посредине черной полосой (рис. 202)
60. Т. spectabilis Lw.
84 (83). Брюшко сверху с иным рисунком: имеется срединный ряд серых или беловатых
треугольников (рис. 205).
85 (90). Брюшко черное, с б. м. резким серым рисунком.
86 (87). Боковые серые пятна имеются на I—II или I—III тергитах брюшка, на
последующих тергитах вместо них серые заднекрайние каемки. Высота лобной
полоски превосходит ширину основания в 6 раз (рис. 210)
64. Т. trigeminus Coq.
87 (86). Боковые серые пятна имеются по крайней мере на II—V тергитах брюшка.
Высота лобной полоски превосходит ширину основания в 4—5 раз (рис. 206, 209).
88 (89). Нижняя лобная мозоль продолговато-овальная (рис. 206). На III тергите
срединный серый треугольник вершиной обычно не достигает заднего края
предыдущего тергита. Коренастее и крупнее: 16 мм и более (очень редко 14 мм).
Европа, Зап. Сибирь, на восток до Енисея 61. Т. autumnalis L.
89 (88). Нижняя лобная мозоль вытянуто-треугольная (рис. 209). На III тергите
срединный серый треугольник вершиной достигает заднего края предыдущего
тергита. Стройнее и мельче: 15—18 мм. Дальний Восток
63. Т. signatipennis Portsch
90 (85). Брюшко с коричневой окраской по бокам или целиком коричнево-желтое
Рисунок из желтовато-серых пятен менее резкий
. 61Ь. Т. autumnalis brunnescens Szil
91 (2). Крупные коренастые виды: 18—28 мм.* Глаза без полосок.
92 (99). г4 с придатком. Лобная полоска относительно широкая, ее высота превосходит
ширину основания в 3—3V2 раза (рис. 178, 179 и др.). (IV группа: Т. argenteo-
maculatus Krob.).
93 (94). Глаза явственно покрыты волосками. Крылья коричнево затемнены . . .
40. Т. argenteomaculatus КгбЬ.
94 (93). Глаза голые. Крылья бесцветные или слегка затемнены.
95 (96). Брюшко резко двуцветное: первые 3 сегмента в густом сером налете и белых
волосках, последующие сегменты черные, в черных волосках. Армянская ССР,
Нахичеванская АССР 43. Т. shelkovnikovi Param.
96 (95). Брюшко одноцветно-серое или черное. Горы Средней Азии.
97 (98). Брюшко сверху в светло-сером налете, боковые стороны I—IV тергитов
преимущественно в коротких беловатых волосках
41. Т. montiasiaticus Ols. sp. nova.
98 (97). Брюшко сверху в темно-сером налете, боковые стороны I—IV тергитов в
коротких черных волосках 42. Т. sehiva M. et Chv.
99 (92). г4 без придатка. Лобная полоска относительно узкая, ее высота превосходит
ширину основания в 4—6 раз.
.100 (103). Черные тёргиты брюшка с резкими золотисто-желтыми каемками по
заднему краю (типа Hybomitra tarandina L.) (VI группа: Т. chrysuru,s Lw,).
101 (102). Передние лапки желто-коричневые. Ведра ног коричневые
. . . ... . . . . ................ 59. Т. buddha Portsch.
102 (101). Передние лапки черные. Бедра ног в основной части черные
58. Т. chrysurus Lw.
103 (100). Брюшко без бросающихся в глазах резких, золотистых каемок по заднему
краю тергитов.
104 (131). Брюшко сверху с 1 срединным рядом беловатых или серых треугольников,
если целиком черное, то треугольники могут отсутствовать (V группа: Т. Ъо-
vinus L.; рис. 195).
105 (112). Брюшко черное, в черных волосках, иногда 1—3 тергиты по бокам темно-
коричневые. Средние и задние голени обычно целиком черные.
106 (107). Крылья в основной половине коричнево затемнены. Лобный треугольник
и лицо в коричневом налете 47. Т. anthrax Ols.
* Виды группы Т. autumnalis L. По размерам тела занимают промежуточное
положение: от 14 до 22 мм, экземпляры этих видов, имеющие размеры тела менее 18 мм
определяются по таблице после тезы 2.
8. TABANUS 197
107 (106). Крылья б. м. бесцветные, без бросающегося в глаза затемнения в основной
половине. Лобный треугольник и лицо в сером или светло-коричневатом налете.
108 (109). Верхний край усиковых ямок широкий, занимает более V3 высоты лобного
треугольника (рис. 186). Лобная полоска и лобный треугольник в
светло-коричневатом налете. Нижняя часть лица в светлых волосках с примесью
коричневатых . . 48. Т. subparadoxus Ols.
109 (108). Верхний край усиковых ямок узкий, занимает не более 1/i—V5 высоты'
лобного треугольника (рис. 187, 188). Лобная полоска и лобный треугольник
в беловато-сером налете. Нижняя часть лица в белых волосках.
110 (111). По крайней мере II тергит брюшка по бокам темно-коричневый. Спинка
в умеренно густых серых волосках 49. Т. swiridowi Portsch.
111 (110). II тергит брюшка по бокам черный. Спинка в густых беловато-серых
волосках 50. Т. perelegans Ols.
Примечание: достоверное определение Т. swiridowi и Т. perelegans может быть
сделано лишь по самцам.
112 (105). Брюшко с коричневыми или желто-коричневыми пятнами по бокам,
захватывающими по крайней мере I—III тергиты, покрыто черными и (или) светлыми
(желтыми, серыми или белыми) волосками. Средние и задние голени коричневые'
или желто-коричневые.
113 (118); Боковые стороны II—III тергитов брюшка в одноцветных белых или желтых
волосках, без примеси черных.
114 (115). Боковые стороны брюшка на II—III тергитах в золотисто-желтых волосках.
3-й членик усиков с резким, острым дорсальным углом (рис. 199). О-ва Сахалин,
Кунашир; Япония 57. Т. sapporoenus Shir.
115 (114). Боковые стороны брюшка на II—III тергитах в беловатых волосках. 3-й
членик усиков с прямым дорсальным углом. Кавказ.
116 (117). Конец брюшка сероватый. Брюшко сверху с срединным рядом относительно
узких, неясно отграниченных желтовато-серых треугольников. По крайней
мере 1-й и 2-й членики усиков коричневые . . . ■. . . 46. Т. olsufjevi Haus.
117 (116). Конец брюшка черноватый. Брюшко сверху с срединным рядом
относительно широких, б. м. ясно отграниченных беловатых треугольников. 1-й и
2-й членики усиков черные 45. Т. semiargenteus Ols.
118(113). Боковые стороны II—III тергитов' брюшка кпереди преимущественно
в черных, кзади в светлых (белых или желтых) волосках.
119 (124). Концевой членик щупалец серый или коричневый, в густых коротких
черных или серых волосках. Усики целиком черные или черно-коричневые
с черным концом.
120 (123). Беловатые срединные треугольники сверху брюшка четкие, с прямыми
сторонами. Брюшко снизу с резко отграниченной черно-коричневой срединной
полосой. Бедра черные, в сером налете-.
121 (122). Брюшко с желто-коричневыми пятнами на бокам. Европа, запад Украины,
Молдавская ССР 53. Т. spodopterus Mg.
122 (121). Брюшко с красно-коричневыми или каштановыми пятнами по бокам. Кавказ
52. Т. portschinskii Ols.
Примечание: достоверное определение Т. spodopterus и Т. portschinskii может
быть сделано лишь по самцам.
123 (120). Сероватые срединные треугольники сверху брюшка менее четкие, с
вогнутыми сторонами. Брюшко снизу с неясной срединной полосой или последняя
отсутствует. По крайней мере средние и задние бедра в большей своей части
темно-коричневые, реже черные, в сером налете ... 44. Т. prometheus Szil.
124 (119)." Концевой членик щупалец беловато-желтоватый или светло-коричневый,
в светлых и черных волосках. По крайней мере 1-й и 2-й членики усиков
коричневые.
125 (130). Нижняя лобная мозоль б. м. треугольная, ее наибольшая ширина
приходится на основание'(рис. 189, 196). 3-й членик усиков с резким б. м.
остроугольным дорсальным выступом.- Брюшко снизу с буро-черной срединной полосой.
126(127). Брюшко снизу коричнево-черное или черное, с широкими беловатыми
каемками по заднему краю стернитов; сверху срединные треугольные пятна
обычно с б. м. прямыми сторонами 51. Т. sudeticus Zell.
127 (126). Брюшко снизу коричневато- или красновато-желтое в сером налете, с
коричнево-черной срединной полосой; беловатые каемки по заднему краю
стернитов отсутствуют. Сверху срединные треугольные пятна обычно с вогнутыми
сторонами.
128 (129). 3-й членик усиков черный, лишь основание иногда коричневатое (рис. 196).
Срединные треугольники на тергитах брюшка б. м. резкие, беловато-серые.
Европа, Зап. и Средн. Сибирь ............. .55. Т. bovinuS L.
129 (128). 3-й членик усиков коричневый йлй коричнево-желтый с/черным концом
(палочка и часть площадки) (рис. 198). Срединные треугольники на тергитах
198 III. ПОДСЕМ. TABANINAE
: брюшка нерезкие, сероватые. Средн. и Вост. Сибирь, Дальний Восток . .
56. Т. pleskei КгбЬ.
130 (125). Нижняя лобная мозоль овальная или продолговатая, ее наибольшая
ширина приходится на середину (рис. 194). 3-й членик усиков с менее резким
б. м. прямоугольным дорсальным выступом. Брюшко снизу с красноватой
срединной полосой 54. Т. tinctus Wlk.
131 (104). Брюшко сверху с 3 рядами беловатых или серых пятен из налета и светлых
волосков (если брюшко коричневое, то боковые пятна менее заметны и видны,
если рассматривать брюшко сзади!) или по крайней мере боковые серые пятна
имеются на II тергите (помимо срединного ряда треугольников), или брюшко
с 2 боковыми резкими, широкими розовато-серыми продольными полосами,
разделенными посредине черной полосой. (VII группа: Т. autumnalis L.;
рис. 202, 205).
132 (141). 1-я заднекрайняя ячейка сужена к вершине.
133 (134). Брюшко сверху без срединного ряда светлых треугольников, но с 2
резкими, широкими, розовато-серыми продольными полосами, разделенными
посредине черной полосой (рис. 202) 60. Т. spectabilis Lw.
134 (133). Брюшко с иным рисунком: имеется срединный ряд серых или беловатых
треугольников (рис. 205).
Рис. 126. Вершина крыла Tabanus amaenus
Wlk. 1-я заднекрайняя ячейка замкнутая.
135 (140). Брюшко черное, с б. м. резким серым рисунком.
136 (137). Боковые серые пятна имеются на I—II или I—III тергитах брюшка, на
последующих тергитах вместо них серые заднекрайние каемки. Высота лобной
полоски превосходит ширину основания в 6 раз (рис. 210)
64. Т. trigeminus Coq.
137 (136). Боковые серые пятна имеются по крайней мере на II—V тергитах брюшка.
Высота лобной полоски превосходит ширину основания в 4—5 раз (рис. 206, 209).
138 (139). Нижняя лобная мозоль продолговато-овальная (рис. 206). На III тергите
срединный серый треугольник вершиной обычно не достигает заднего края
предыдущего тергита. Коренастее и крупнее : до 22 мм. Европа, Зап. Сибирь,
на восток до Енисея 61. Т. autumnalis L.
139 (138). Нижняя лобная мозоль вытянуто-треугольная (рис. 209). На III тергите
срединный треугольник достигает заднего края предыдущего тергита. Стройнее
и мельче: до 18 мм. Дальний Восток .... 63. Т. signatipennis Portsch.
140 (135). Брюшко с коричневой окраской по бокам или целиком коричнево-желтое.
Рисунок из желтовато-серых пятен, менее резкий
61Ь. Т. autumnalis brunnescens Szil.
141 (132). 1-я заднекрайняя ячейка не сужена к вершине
62. Т. atropathenicus Ols.
142 (1). 1-я заднекрайняя ячейка замкнута и на конце с коротким стебельком
(рис. 126) * Дальний Восток 65. Т. amaenus Wlk.
( Б. Глаза соприкасаются:^
1 (НО). 1-я заднекрайняя ячейка на конце не замкнутая.
2 (15). Глаза явственно покрыты волосками.
3(4). Голова очень большая, сильно выпуклая. Крупный, черный вид с темными
крыльями и черными ногами ........ 40. Т. argenteomaculatus Krob.
4 (3). Голова нормально выпуклая. Средних размеров виды с прозрачными крыльями.
* В редких случаях замкнутая ячейка с коротким стебельком может быть у
Т. signatipennis и Т. trigeminus, которые отличаются от Т. amaenus более резким серым
рисунком сверху брюшка; для их определения см. тезу 82 или 132. Оба вида
распространены на Дальнем Востоке.
8. TABANUS
199
5 (14). Средние и задние голени желто-коричневые.
6 (13). Концевой членик щупалец б. м. тупоконечный.
7(10). Брюшко черно-серое, с незначительными коричневыми пятнами по. бокам
II тергита. Усики черные, реже красно-коричневые.
8 (9). 1-й членик усиков сверху в длинных волосках, превосходящих длину 1-го
и 2-го члеников вместе взятых. Концевой членик щупалец с клювообразной
вершиной. Европа, Передняя Азия, Кавказ, Туркмения
5. Т. quatuomotatus Mg.
9 (8). 1-й членик усиков сверху в коротких волосках, не превосходящих длину
членика. Концевой членик щупалец с конусообразной притуплённой вершиной.
Монголия, Сев.-Зап. Китай 11. Т. stackelbergiellus Ols.
10 (7). Брюшко серое, с желто-коричневыми пятнами по бокам, простирающимися
на I—III тергиты. Усики оранжевые, палочка черная.
11 (12). Затылок за глазами с рядом длинных, выступающих, коричневых волосков.
Глаза в длинных волосках 2. Т; lunatus F.
12 (И). Затылок за глазами в коротких волосках. Глаза в коротких, слабо заметных
волосках 3. Т. bactrianus Ols.
13 (6). Концевой членик щупалец с оттянутым вперед приостренным кончиком
(рис. 127) 1. Т. bifarius Lw.
14(5). Средние и задние голени черные, лишь слегка'коричневые в основании . .
25. Т. rupium Br.
15 (2). Глаза голые (лишь при увеличении Х16 и более удается иногда заметить ред-
■ кие, короткие волоски).
16 (19). Лобный треугольник в верхней половине блестяще-коричневый или целиком
блестяще-черный.
17 (18). Лобный треугольник в верхней половине блестяще-коричневый
7. Т. glaucopis Mg.
18 (17). Лобный треугольник целиком блестяще-черный . .
j 24. Т. kinoshitai Kono et Так.
19 (16). Лобный треугольник целиком в сером налете.
20 (37). г4 с придатком.
21 (24). Крупные виды: 19—20 мм. Глаза без полосок.
22 (23). Брюшко резко двуцветное: первые 3 сегмента в густом сером налете и белых
волосках, последующие сегменты черные, в черных волосках
43. Т. shelkovnikovi Param.
23 (22). Брюшко одноцветное серое 41. Т. montiasiaticus Ols., sp. nova
24 (21). Небольшие виды: 12—15 мм. Глаза с полосками.
25 (32). Глаза с 2—3 полосками.
26 (27). Затылок в коротких волосках 8. Т. accipiter Szil.
27 (26). Затылок в длинных, выступающих над поверхностью глаз волосках.
28 (29). Голова большая, б. м. полушаровидная. Возле основания усиков поперечная
темная полоска («уздечка») отсутствует 10. Т. sabuletorum Lw.
29 (28). Голова умеренной величины, не больше, чем у $. На уровне основания усиков
имеется поперечная кофейно-коричневая полоска («уздечка»).
30(31). Мелкие фасетки глаз резко отличаются от крупных; затылочная полоска
широкая, с длинными белыми волосками; голова широкая
15. Т. zimini Ols.
31 (30). Мелкие фасетки глаз постепенно переходят в большие; затылочная полоска
узкая, с черными, более короткими волосками; голова нормальной ширины *
14. Т. ansarii Abbas.-L.
32 (25). Глаза с 1 полоской или без нее.
33 (36). Голова сильно выпуклая, полушаровидная. Фасетки верхних 3/4 глаз
приблизительно в 5—6 раз крупнее нижних. Затылочная полоска с рядом слегка
выдающихся над поверхностью глаз волосков.
34 (35). Усики черные 16: Т. filipjevi Ols.
35 (34). Усики красновато-желтые 23. Т. fumidus Aust.
36 (33). Голова умеренно выпуклая. Фасетки верхних 2/з глаз приблизительно в 4 раза
крупнее нижних. Затылочная полоска с рядом относительно длинных волосков,
значительно выступающих над поверхностью глаз
17. Т. brunneocallosus Ols.
37 (20). г4 без придатка (лишь иногда имеется незначительный рудимент).
38 (95). Фасетки верхних 2/з или 3U глаз значительно крупнее нижних, обычно
с б. м. резкой границей между теми и другими. Голова крупнее, чем у $, часто
полушаровидная. <
39 (68). Небольшие или средних размеров виды: 12—17 мм.
40 (51). Затылочная полоска с рядом заметно выступающих над поверхностью глаз
волосков.
* Различия 66 Т. zimini и Т. ansarii нами указаны по: Abbassian-Lintzen, 1964.
200 HI. ПОДСЕМ. TABANINAE
41 (42). Мелкофасетчатое поле глаз с 3 полосками, крупнофасетчатое поле с 1
довольно широкой темной полоской, заметной и у сухих экземпляров . . . .
* . . 12. Т. mistshenkoi Ols.
42 (41). Глаза с 1 полоской, расположенной на границе крупных и мелких фасеток,
или таковая отсутствует.
43 (46). На уровне основания усиков имеется явственная, коричневая, поперечная
полоска. Концевой членик щупалец сильно утолщен (рис. 149). Голова очень
большая, полушаровидная; крупные фасетки занимают 3/i глаза.
44 (45). Волоски на затылке длинные, далеко выдаются над поверхностью глаз. Глаза
с узкой, нерезкой полоской на границе крупных и мелких фасеток или совсем
без полоски 21. Т. cordiger Mg.
45 (44). Волоски на затылке лишь немного выдаются над поверхностью глаз. Глаза
с резкой, широкой полоской на границе крупных и мелких фасеток ....
: 18. Т. leleani Aust.
46 (43). На уровне основания усиков коричневая полоска отсутствует. Концевой
членик щупалец продолговато-овальный. Голова умеренно большая. Крупные
фасетки занимают 2/з глаза.
47 (48). Брюшко снизу в основной половине коричнево-желтое. Крупнее: 15—16 мм
28. Т. miki Br.
48 (47). Брюшко снизу целиком черное или черно-серое. Мельче: 11.5—14.5 мм.
49 (50). Усики коричнево-желтые, палочка обычно темнее площадки. Затылок в
светлых, коричневато-желтых волосках 26. Т. maculicomis Ztt.
50 (49). Усики целиком оранжево-желтые. Затылок в темных, преимущественно черных
волосках 27. Т. geminus Szil.
51 (40). Затылочная полоска в коротких волосках, не выдающихся заметно над
поверхностью глаз.
52(53). Брюшко сверху по бокам в густых золотисто-оранжевых. волосках . . .
х , 35. Т. semenovi Ols.
53 (52). Брюшко сверху по бокам в серых или желтоватых волосках, чередующихся
с пятнами черных волосков.
54 (65). Голова пропорционально туловищу очень большая, полушаровидная
(рис. 156), крупные фасетки занимают около sli глаза.
55 (58). Концевой членик щупалец сильно утолщен (рис. 149).
56 (57). На уровне основания усиков имеется поперечная коричневая полоска,
отграничивающая белоснежное лицо от сероватого лобного треугольника. Глаза
с резкой полоской на границе крупных и мелких фасеток. . 18. Т. leleani Aust.
57 (56). На уровне основания усиков коричневая полоска отсутствует (имеется лишь
легкая желтизна). Глаза с нерезкой полоской на границе крупных и мелких
фасеток ._ ; 22. Т. golovi Ols.
58 (55). Концевой членик щупалец продолговато-овальный, часто на конце с
оттянутым вперед кончиком.
59(62). Средние и задние бедра желтые.
60 (61). Передние бедра желтые ' 36. Т. laetetinctus Beck.
61 (60). Передние бедра черные, в сером налете 37. Т. hauseri Ols.
62 (59). Средние и задние бедра черные, в сером налете.
63 (64). Крупнее: 16—17 мм. Лобный треугольник в нижней части и по бокам усиков
в светло-сером налете . 62. Т. atropathenicus Ols.
64 (63). Мельче: 14 мм. Лобный треугольник в нижней части и особенно по бокам
усиков в коричневатом налете 32. Т. regularis Jaenn.
65 (54). Голова умеренных размеров; крупные фасетки занимают' не более 2/3 глаза.
66 (67). Голова менее выпуклая. Глаза с резкой полоской на границе крупных й
мелких фасеток 33. Т. bromius L.
67 (66). Голова более выпуклая. Глаза с нерезкой полоской на границе крупных
и мелких фасетов.,,,...,., 29. Т. indrae Haus.
Примечание: достоверное определение Т. bromius и Т. Indrae может быть
сделано только по самкам.
68 (39). Крупные, коренастые виды: 17—25 мм.
69. (70). Черные тергиты брюшка с резкими, золотисто-желтыми каемками по заднему
краю (типа Hybomitra tarandina L.) 58. Т. chrysurus Lw.
Т. buddha Portsch.
70 (69). Брюшко без бросающихся в глаза резких, золотисто-желтых каемок по
заднему краю тергитов.
71 (74). Брюшко и ноги черные.
72(73). Крылья в основной половине коричнево затемнены . . . 47. Т. anthrax Ols.
73 (72). Крылья б. м. бесцветные • 50. Т. perelegans Ols.
74 (71). Брюшко с развитой коричневой окраской по бокам или черное, сверху с
продольными рядами серых пятен. По крайней мере базальная часть голеней
коричневая.
8. TAB ANUS 201.
75 (76). Брюшко сверху с 2 резкими розовато-серыми продольными полосами,
разделенными черной срединной полосой; срединный ряд светлых треугольных
пятен отсутствует 60. Т. spectabilis Lw.
76 (75). Брюшко с иным рисунком; срединный ряд светлых треугольных пятен
имеется.
77 (84). Брюшко сверху с 3 рядами беловатых или серых пятен из налета и светлых
волосков (если брюшко коричневое, то боковые пятна менее заметны и видны,
если рассматривать брюшко сзади!) или по крайней мере боковые серые пятна
имеются на II тергите (помимо срединного ряда треугольников),
78 (83). 1-я заднекрайняя ячейка сужена к вершине.
79 (82). По меньшей мере II—III тергиты брюшка с боковыми коричневыми
пятнами.
80 (81). Коренастее и крупнее: 17—21 мм. Европа, Ср. Азия, Зап. Сибирь, на восток
до Енисея 61. Т. autumnalls L.
81 (80). Стройнее и мельче: 17 мм. Дальний Восток . . 63. Т. signatipennis Portsch.
82 (79). Коричневая окраска по бокам II тергита неясная . . 64. Т. trigeminus Coq.
83 (78). 1-я заднекрайняя ячейка не сужена к вершине . . 62. Т. atropathenicus Ols.
84 (77). Брюшко сверху с 1 срединным рядом беловатых или серых треугольников.
85 (88). Боковые стороны II—III тергитов брюшка преимущественно в белых или
желтых волосках.
86 (87). Боковые стороны брюшка на II—III тергитах в золотисто-желтых волосках.
3-й членик усиков с острым дорсальным углом. О-ва Сахалин, Кунашир; Япония
57. Т. sapporoenus Shir.
87 (86). Боковые стороны брюшка на II—III тергитах в беловатых волосках. 3-й
членик усиков с прямым дорсальным углом. Кавказ . . . 46. Т. olsufjevi Haus.
88 (85). Боковые стороны II—III тергитов брюшка преимущественно в черных
волосках.
89 (94). Крупнофасетчатая часть глаз резко отграничена от мелкофасетчатой части.
Крупные фасетки в 5—6 раз больше мелких. Брюшко сверху по бокам кашта-
ново-коричневое.
90 (91). Брюшко снизу черно-коричневое, с б. м. резкими желтоватыми или
беловатыми каемками по заднему краю стернитов 51. Т. sudeticus Zell.
91 (90). Брюшко снизу коричневое, без резких светлых каемок по заднему краю
стернитов.
92 (93). Брюшко снизу с хорошо выделяющейся коричневой продольной полосой
посредине. Сверху брюшка срединный ряд состоит из беловатых средней
величины треугольников. Бедра средних и задних ног черные в сером налете
. 52. Т. portschinskii Ols.
93 (92). Брюшко снизу без бросающейся в глаза темной полосы посредине (вдоль
средней линии имеется лишь небольшое затемнение). Сверху брюшка срединный
ряд состоит из сероватых небольших треугольников. Бедра средних и задних
ног преимущественно темно-коричневые, в сером налете •
..'.....■ 44. Т. prometheus Szil.
94 (89). Крупнофасетчатая часть глаз нерезко отграничена от мелкофасетчатой части,
переход постепенный. Крупные фасетки примерно в 3—4 раза крупнее мелких.
Брюшко сверху по бокам красно-коричневое 54. Т. tinctus Walk.
95 (38). Фасетки верхних частей глаз мало отличаются по величине от нижних, если же
несколько крупнее, то они занимают лишь. небольшое пространство возле
глазного шва и постепенно переходят в мелкие фасетки, без заметной границы
между теми и другими. Голова не крупнее,, чем у ?.
96 (97). Щупальца и ноги черные 49. Т. swiridowi Portsch.
97 (96). Щупальца белые, желтоватые или коричневатые; голени по крайней мере
средней пары ног коричневые или коричнево-желтые.
98 (101). Глаза с 1—3 полосками.
99 (100). Глаза с 1'полоской. Брюшко снизу черно-серое. Затылок в заметно
выступающих над поверхностью глаз преимущественно черных волосках ....
19. Т. unifasciatus Lw.
100 (99). Глаза* с 2—3 полосками. Брюшко снизу желто-коричневое, с темным
концом. Затылок в коротких волосках . . . . . - .. . 38. Т. tergestinus Egg.
101 (98). Глаза без полосок.
102 (105). Небольшие виды: 13—15 мм. Брюшко черное, с едва заметными
коричневыми пятнами по бокам II тергита или без них.
103 (104). На уровне основания усиков имеется коричневая поперечная полоска.
Затылок с рядом выступающих над поверхностью глаз волосков. Крым . .
_ _ ......... .', 20. Т. smirnovi Ols.
104 (103)! На уровне основания усиков коричневая поперечная полоска отсутствует.
Затылок в коротких волосках. Кавказ 31. Т. armeniacus Krob.
105 (102). Крупные виды: 17.5—24 мм. Брюшко с "развитыми боковыми коричневыми
■ пятнами на 1—3 тергитах.
202
III. ПОДСЕМ. TABANINAE
106 (107). 3-й членик усиков желтовато-коричневый, с черноватой палочкой.
Срединные треугольники по заднему краю тергитов брюшка слабо намечены,
сероватые. Вост. Сибирь, Дальний Восток 56. Т. pleskei Krob.
107 (106). 3-й членик усиков черный, иногда лишь в основании слегка коричневатый,
по /<тРеДИШЫе треугольники по заднему краю тергитов брюшка резкие, беловатые.
108 (109). Брюшко снизу красно-коричневое, по бокам в серебристо-сером налете,
выделяющем резкую тешую среданную полосу . . . . 53. Т. spodopterus Mg.
109 (108). Брюшко снизу желто-коричневое, с буро-черной полосой посредине . .
..п ,. • ; i 55. Т. bovinus L.
ни (1). 1-я заднекраиняя ячейка замкнута и на конце с коротким стебельком
(рис. 126). Дальний Восток 65. Т. amaenus Wlk.
I. Группа Tabanus bifarius Lw.
Нижняя лобная мозоль расположена отступя от нижней границы лобной полоски.
Средняя лобная мозоль овальная, вполне обособлена от нижней мозоли (иногда
целиком прикрыта налетом). Лоб относительно узкий, с б. м. параллельными сторонами.
Глаза покрыты волосками или голые. г4 часто с коротким придатком. Глаза с 3
полосками. Небольшие или средних размеров виды, распространенные преимущественно
в Средиземноморской подобласти.
1. Tabanus bifarius Lw. — Слепень южный.
Представлен 3 формами: номинативным подвидом Т. bifarius bifarius Lw. и
распространенными в Закавказье подвидами: Т. bifarius tarjukini Haus. и Т. bifarius kurensis
Рис. 127. Tabanus bifarius Lw. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека ?. Л — щупальце в.
la. Tabanus bifarius bifarius Lw. (рис. 127) — Слепень южный номинативный.
с hoews< А8п5,8а', У?ТЪ Zo?L-bot- Geedl. Wien, 8: 595; В г а и е г, 1880:172;
S u г с о u f, 1924 : 144; КгоЬег, 1924 : 74; К г о Ь е г in L i n d n er, 1925 : 90
?Qfift° \%b er\ i93?a : 24т4; M °i" c h a et C h v a L a' i958 : i95; L ё с 1 ет с q
1966 142 С h v а 1 a, L у n e Ь о г g а. М о и с h а, 1972 : 316. - tenuicornis
Е n d е г I е 1 п, 1932 : 63, цит. по: С h v а 1 a, L у п е Ь or g а. М о и с h a, 1972:
?. Глаза в коротких, белесых волосках, с 2 или 3 тонкими полосками. Лоб узкий
его высота превосходит основание в 5-5 V2 раза, слегка расширяющийся кверху
в желтоватом налете. Затылок за глазами в б. м. коротких черных или желтовато-
серых волосках. Нижняя лобная мозоль желтая, прямоугольная или овальная,
расположена отступя от нижней границы лба, боковыми сторонами не достигает края
глаз, средняя мозоль черная, овальная, обособленная от нижней. Иногда средняя
мозоль в той или иной мере (реже целиком) прикрыта налетом; как исключение обе мо-
\
8. TABANUS '20B'
золи скрыты налетом. Темя без темных пятен. Лобный треугольник в густом,
желтоватом налете. Усики желтые, узкие, 3-й членик слегка коричневый, с темной палочкой.
.Дорсальный угол слабый, часто с притуплённым уголком. Щупальца бледно-желтые,
-концевой членик тонкий, приблизительно в 4 раза длиннее ширины, слабо изогнут,
покрыт тонкими, короткими, светлыми волосками. Нотоплевры темные. Крылья
прозрачные, жилки коричневые; г4 с небольшим придатком. Ноги: бедра серые; основная
половина передних голеней и большая часть средних и задних голеней желтые. Концы ■
передних голеней и передние лапки черные, последующие лапки и кончики голеней
.коричнево-черные. Брюшко черное, в б. м. густом оливково-сером налете, с 3
продольными рядами светлых пятен (налет-(-волоски): треугольных или трапециевидных
посредине, образующих общую продольную полосу, и б. м. округлых по бокам.
Промежутки между пятнами преимущественно в черных волосках; у потертых
экземпляров благодаря отсутствию волосков светлые пятна б. м. сливаются с общим фоном
'брюшка. Снизу брюшко в густом оливково-сером налете. VIII стернит: гонопофиз
небольшой, без апикальной выемки; латеральные лопасти слабо выпуклые, темно-
коричневые; базальная лопасть шире гонопофиза, с вогнутым основным краем. Церки
округло-треугольные. Сперматека игловидная. 14—17 мм.
6. Глаза в умеренно длинных, белесых волосках, с 2 полосками. Голова
относительно небольшая. Фасетки верхних 2/3 глаз приблизительно в 4 раза крупнее
нижних. Затылок за глазами в средней 1/3 в длинных, коричневатых или серых волосках.
Лобный треугольник и лицо в светло-сером налете, последнее в тонких, такого же
цвета волосках. Усики желтые, палочка темная; 3-й членик очень узкий, площадка
равномерно переходит в палочку, дорсальный угол незначительный. Концевой
членик щупалец бледно-желтый, узкий, с оттянутым вперед и приостренным кончиком,
покрыт тонкими, длинными, светло-серыми волосками. Нотоплевры одноцветные
■с грудью. Крылья и ноги как у самки. Брюшко черное, в тонком оливково-сером
налете; II—III тергиты по бокам с небольшими коричневыми пятнами. 3 продольных
ряда светлых пятен из налета+волосков: треугольных посредине и косо-овальных
по бокам. Срединный ряд треугольников б. м. сливается в общую продольную полоску.
Снизу брюшко одноцветно-оливково-серое. 15—16 мм.
От всех видов Tabanus с оволосенными глазами 5 легко отличается желтой
нижней лобной мозолью, а с? — остроконечными щупальцами.
Распространение: южноевропейский (средиземноморский) степной вид.
Юг европ. части СССР (на север известен до Львова, Умани Киевской обл., Воро-
шиловградской обл., Красноармейска близ Волгограда), Молдавия, Крым, Сев.
Кавказ (Краснодарский, Ставропольский края, Кабардино-Балкарская, Дагестанская
АССР), Закавказье — Азербайджан (включая Нахичеванскую АССР), Грузия,
Армения. Средн. и Ю. Европа (на запад до Испании), Сев. Африка (Тунис, Марокко),
Турция, Иран.
На Кавказе и в Крыму по предгорьям и низкогорьям местами обычен. Часто
на цветах. Личинки найдены в почве вдали от воды (Гургенидзе, 19726).
lb. Tabanus bifarius tarjukini Haus. — Слепень южный Тарьюкина.
Г а у з е р, 1941, Изв. Азерб. фил. АН СССР, 2 : 73; L е с 1 е г с q, 1966 : 142.
5. Брюшко в сером налете, сверху в густых коротких черных и серебристо-серых
волосках; 3-й членик усиков темно-бурый. 15—16 мм.
(У неизвестен.
Распространение: ю.-з. Азербайджан: Нагорный Карабах (г. Кире),
Кельбаджарский р-н; Нахичеванская АССР (Шахбуз); Армения: Еленовка. Местами
обычен.
1с. Tabanus bifarius kurensis Djaf. — Слепень южный куринский.
Джафаров, 1960, Зоол. жури., 39, 5 : 714; L е с 1 е г с q, 1966 : 142.
5. Брюшко сверху с боковыми розовато-коричневыми пятнами на I—III или I—
IV тергитах. 16—17 мм.
(У неизвестен.
Распространение: Азербайджан — Прикуринская низменность (Шам-
.хорский и Акстафинский р-ны).
2. Tabanus lunatus F. (рис. 128). — Слепень средиземноморский.
F.abricius, 1794, Ent. Syst., 6:370; В г a u e r, 1880:171; S иг сои f,
1924 : 145; Krober, 1924 : 76; К г б Ь е г in L i n d n e r, 1925 : 93; О л с у ф ь е в,
1937а: 246; L е с 1 е г с q, 1966 : 156; Chvala, Lyneborg a. Moucha,
1972 : 312.
'* ?. Глаза в коротких, серых волосках, с 3 полосками. Лоб слегка расширен кверху,
узкий, его высота превосходит основание в 4—5 раз, в желтовато-сером налете. За-
204 III. ПОДСЕМ.. TABANINAR
тылок в коротких, беловатых волосках. Нижняя лобная мозоль темно-коричневая,
округлая или прямоугольная, расположена отступя от нижней границы лба и
боковыми сторонами далеко не достигает края глаз. Средняя мозоль широкоовальная,
черная, обособленная от нижней. На темени часто небольшое темное пятно. Лобный
треугольник целиком в желтоватом налете. Усики, как правило, оранжевые, с темной
палочкой, реже черные; дорсальный угол незначительный. Концевой членик щупалец
бледно-желтый или беловатый, утолщен, его длина превосходит ширину не более чем
в 3 раза, покрыт короткими черными и светлыми волосками. Нотоплевры серые, реже
желтоватые. Крылья прозрачные, жилки коричневые; г4 без придатка. Ноги: бедра
серые, основная половина передних голеней и почти целиком средние и.задние голени
коричневато-желтые; концы передних голеней и передние лапки черные, лапки
последующих ног и кончики голеней коричнево-черные. Брюшко черно-серое,
боковые стороны II (или I—IV) тергита
светло-коричневые. Посредине брюшка продольный ряд
неясных треугольников из налета и серых
волосков, по бокам их черные волоски. Боковые
стороны тергитов преимущественно в светлых
волосках. Снизу брюшко одноцветно-серое,
реже в основании желтоватое. 12—16 мм.
<?. Голова относительно небольшая,
примерно как у 5- Глаза в длинных, серых
волосках, с двумя полосками; фасетки верхних
2/3 глаз крупнее нижних приблизительно
|Ш1ШЦМ11Ш1пп{
Рис. 128. Tabanus lunatus F. Лобная полоска,
усик и щупальце $.
в 3—4 раза. Затылок за глазами с рядом длинных, выступающих над поверхностью
глаз, коричневых волосков. Лобный треугольник в желтовато-сером налете, перед
вершиной с небольшой коричневой, блестящей, поперечной полоской. Усики оранжевые,
палочка на конце черная, 3-й членик очень узкий, площадка равномерно переходит
в палочку, дорсальный угол едва выражен. Концевой членик щупалец
бледно-желтый, овальный, на конце тупой, покрыт длинными черными и светлыми волосками.
Крылья и ноги как у самки. Брюшко черно-серое с большими желто-коричневыми
пятнами по бокам I—III (или I—IV) тергитов. 3 ряда продольных, весьма неясных, олив-
ково-серых пятен из налета и светлых волосков, из них срединные пятна б. м.
треугольной формы. Снизу брюшко в основной половине желтое, V и последующие стерниты
черно-серые. II стернит посредине с небольшим черным пятном, иногда продолженным
на III стернит узкой полоской. 12—14 мм.
Близок к Т. bifarius, но, помимо меньшей величины, легко отличается'строением
концевого членика щупалец (у <£ с тупым концом, а у 5 — короткий, утолщенный).
Распространение: южноевропейский (средиземноморский) вид. Нахи-
чеванская АССР — Дисар и Парага близ Ордубада. Редок. Ю. Европа (на запад до
Португалии), Сев. Африка, Передняя Азия.
3. Tabanus bactrianus Ols. (рис. 129) — Слепень бактрийский.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 247 (Т. lunatus
bactrianus); 1939b : 106; Шевченко, 1961:136; L е с 1 е г с q, 1966:157 (Т. lunatus
bactrianus Ols.).
5. Глаза голые, с 3 полосками. Лобная полоска слегка расширена кверху, ее
высота превосходит ширину основания в 41/2—5 раз; полоска в желтовато-сером налете.
Нижняя лобная мозоль б. м. прямоугольная, темно-коричневая. Средняя мозоль,
продолговато-овальная, черная. Темя иногда с более темным пятном. Лобный
треугольник в желтовато-сером налете. Усики красновато-коричневые, палочка
черноватая, 3-й членик относительно узкий, с тупым дорсальным углом. Концевой членик
щупалец желтовато-белый, преимущественно в светлых волосках, его длина превосходит
наибольшую толщину приблизительно в 3 раза. Нотоплевры. коричневые. Крылья
бесцветные; г4 без придатка. Ноги: бедра черно-серые, голени в большей своей части светло
желтовато-коричневые, концевая половина передних голеней и лапки всех ног
черноватые. Брюшко черно-серое, II—III (или II—IV) тергиты по бокам коричневые. Сверху
брюшка 3 продольных ряда нерезких серых пятен (налет+волоски); треугольных
посредине и б. м. ромбических по бокам. Снизу брюшко черно-серое, в основании иногда
светло-коричневатое. VIII стернит: гонопофиз широкий и короткий, с небольшой
апикальной, выемкой; латеральные лопасти большие, коричневые; базальная лопасть ко-
8. TABANUS 205
роткая, с слабо вогнутым основным краем. Церки с уплощенной вершиной, волоски на
X тергите очень длинные. Сперматека узковеретеновидная. 14—17.5 мм.
<?. Голова относительно небольшая, лишь немногим больше, чем у 5- Глаза в
коротких серых волосках, с 2 полосками; фасетки верхних г/3 глаз крупнее нижних
приблизительно в 4 раза. Затылок за глазами в коротких светлых волосках, немного
выступающих над поверхностью глаз лишь возле места соприкосновения глаз. Лобный
треугольник в сером налете, в верхней части слегка коричневатый. Усики как у 5, но
■более узкие. Концевой членик щупалец продолговато-овальный, тупоконечный,~жел-
Рис. 129. Tabanus bactrianus Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека §.
товато-белый, в светлых волосках. Грудь и ее придатки как у 5- Окраска брюшка как
у 5, коричневые пятна по бокам II—IV тергитов выражены более четко. 14.5—16.5 мм.
Близок к Т. lunatus и первоначально описан нами в качестве подвида этого
последнего. Однако изучение сУсУ показало, что Т. bactrianus следует рассматривать в качестве
самостоятельного вида. Он отличается от Т. lunatus более крупными размерами тела,
а также слабым опушением глаз и затылка.
Распространение: бактрийско-иранский горно-лесной вид. Узбекистан
(Сурхандарьинская, Самаркандская, Ташкентская, Ферганская, Андижанская
области), Таджикистан (сев.-зап. Гиссарского хребта и долина Зеравшана), Киргизия
(долина р. Чаткал, Чаткальский, Ферганский и Алайский хребты), Казахстан (хребты
Каржантау и западная часть Таласского Алатау). Встречается в пределах 1300—3000 м,
в основном приурочен распространением к лесам грецкого ореха. Местами обычен.
Афганистан, Иран. Июль—август. Личинки найдены в реках, родниках, арыках и
болотах (Кадырова, 19726).
Голотип: 5, урочище, Хан-тахта сев.-зап. Гиссарского хребта, 24 VII 1933, Вель-
тищев. В коллекции Зоологического института Академии наук СССР, Ленинград.
4. Tabanus talyshi Ols. (рис. 130) — Слепень талышский.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 447.
5. Глаза голые, с 3 полосками. Лобная полоска слегка расширена кверху, ее
высота превосходит ширину основания в 41/2—5 раз; полоска, включая темя, в сером
налете. Нижняя лобная мозоль б. м. прямоугольная или неправильной формы, темно-
коричневая, расположена отступя от границы лобного треугольники. Средняя мозоль
продолговато-овальная, черная. Лобный треугольник в желтовато-сером налете
(у потертых экземпляров может быть частично обнажен!). Усики
красновато-коричневые, 3-й членик относительно узкий, с тупым дорсальным углом. Концевой членик
щупалец светлый, слегка желтовато-сероватый, преимущественно в черних
волосках, его длина превосходит наибольшую толщину приблизительно в 3 раза.
Нотоплевры коричневые. Крылья бесцветные, жилки желтые; г4 обычно без придатка. Ноги:
бедра светло-коричневые, в сером налете, голени желто-коричневые, концы передних
голеней и передние лапки черноватые, последующие лапки желто-коричневые. Брюшко
черно-серое, II—III (или II—IV) тергиты по бокам коричневатые. Сверху брюшка 3
■206 т. подсем: tabaninae
продольных ряда нерезких серых пятен (налет+волоски), треугольных посредине и
неправильной формы по бокам. Снизу брюшко черно-серое, в основании неясно
коричневатое. 11—14.5 мм.
Похож на Т. lunatus и Т. bactrianus, но
хорошо отличается от' обоих видов
коричневыми бедрами и светлыми лапками средних и
задних ног; у первых двух видов бедра черно-
серые, а средние и задние лапки черноватые.
cJ неизвестен.
Распространение:
Азербайджанская ССР, Массалинский и Джалилабадский
р-ны. Июль—август.
Рис. 130. Tabanus talyshi Ols. Лобная полоска,,
усик и щупальце $.
5. Tabanus quatuornotatus Mg. — Слепень-
четырехточечный.
Представлен двумя формами: номинативным подвидом Т. quatuornotatus quatuor-
. notatus Mg. и закавказским подвидом Т. quatuornotatus araxis Ols.
5а. Tabanus quafuornotafus quatuornotatus Mg. (рис. d 41) — Слепрнь
четырехточечный номинативный.
M е i g e n, 1820, Syst. Beschreib., II : 51; В г a u e r, 1880 : 173; Surcouf,
1924 : 140; К г о Ь е г, 1924 : 67; К г 6 Ь е г in L i n d n e г, А925 : 95; О л с у ф ь е вг
1937а : 248; Г а у з е р, 1939 : 141 (<J); Moucha et С h ч а 1 а, 1958 : 194; L e c-
lercq, 1966 ^164; С hvala, L у п е_Ь о г g а. М о и с h a, 1972:301.
$. Глаза в б. м. длинных серых волосках, с 3 полосками. Лобная полоска
параллельная или слегка^расширена кверху, в сером налете, высотг> ее превосходит ширину
Рис. 131. Tabanus quatuornotatus quatuornotatus Mg. Лобная
полоска, усик, щупальце, VIII стернит., церки и сперматека $.
основания в 3V2—4 раза. Нижняя лобная мозоль черная, блестящая, б. м. квадратная,
расположена отступя от границы лба с лобным треугольником, по б.окам обычно
недостигает края глаз. Средняя мозоль большая, овальная, черг-*я, обособлена от
нижней пространством налета. На темени небольшое (чаще двойное), блестяще-черное-
пятно. Лобный треугольник низкий, возле границы с лбом блестяще-черный или ко^
ричневый, в прочих частях в желтовато-сером налете. Затылок в виде относительно
широкой полоски, покрыт длинными черными иди серыми волосками. Усики
одноцветно-черные, реже площадка темно-коричневая; 1-й членик в тонких, б. м. длинных
.волосках; 3-й членик узкий, с слабым дорсальным-углом. Концевой членик щупалец,
желтовато-белый, б. м. узкий-, покрыт тонкими и б. м. длинными светлыми волосками
и единичными, короткими черными волосками. Дотоплевры темные. Крылья
бесцветные; гй с придатков .или без него: Ноги;,.бедра-.черно-серые, голени,яордчневато-жел-
8. TABANUS 207
тые; концы передних голеней и лапки черные; последующие лапки и частью кончики
голеней коричнево-черные. Брюшко черное, сверху с 3 рядами серых пятен (налет+
волоски), треугольных посредине и б. м. ромбических или округлых по бокам. Задние
каемки тергитов <!ерые. Брюшко снизу одноцветно-темно-серое. VIII стернит: гонопофиз
короткий и широкий, с небольшой апикальной выемкой; латеральные лопасти большие,
темно-коричневые; базальяая лопасть короткая, широкая, с б. м. прямым основным
краем. Церкй с округлой вершиной. Сперматека игловидная. 13—15 мм.
<J. Голова не больше и не выпуклее, нежели у $. Глаза в густых и длинных серых
волосках, с 2 полосками, фасетки верхних частей глаз в 2—3 раза крупнее нижних,
переход крупных фасеток в мелкие постепенный. Затылок в длинных, изогнутых вперед,
черных волосках. Лобный треугольник в сером налете, перед вершиной с поперечной
блестяще-черной полоской. Усики: 1-й членик сверху в длинных черноватых
волосках, 3-й членик черный или коричневый, узкий, с тупым углом. Концевой членик
щупалец светло-желтовато-коричневый, в основании толстый, на конце клювовидный
или тупоконический, в черных и светлых волосках. Грудь и ее придатки как у $.
Брюшко черное, II или И—III тергиты по бокам каштаново-коричневые, сверху с 3
рядами буроватых пятен: треугольных пе/средине неокруглых по бокам. Снизу брюшко
черно-серое, задний край стернитов коричневатый. 12—14 мм.
Распространение: южноевропейский (средиземноморский)
горно-лесостепной подвид. В СССР известен из ю.-з. Украины (Кодымский р-н Одесской обл.),
Молдавии, Краснодарского края (Геленджик, Майкоп, Кавказский заповедник),
Ставропольского края (Пятигорск), Кабардино-Балкарской АССР, Дагестанской АССР,
Азербайджана, Грузии, Армении и Туркмении (Копет-Даг). Местами обычен. Средн.
и Ю. Европа (на запад до Испании), Марокко, Тунис, Иран. Май—июль. Яйца
откладывают на растения на лугах; личинки развиваются в почве (Lecaillon,^1905).
5b. Tabanus quatuornotatus araxis Ols. (рис. 132) — Слепень четырехточечный
араксинский.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 448.
От номинативной формы 5 отличается отсутствием блестящей черной мозоли в
верхней части лобного треугольника (иногда имеется лишь незначительное коричневое по-
Рис. 132. Tabanus quatuornotatus Рис. 133. Tabanus caucasius
araxis Ols. Лобная полоска, усик и Krob. Лобная полоска, усик
щупальце $. и щупальце $.
перечное пятнышко!) и коричневой нижней лобной мозолью, <J отличается несколько
более крупными фасетками .верхних 2/3 глаз, более четко отграниченными от нижних
мелких фасеток. Кроме того, оба пола отличаются от номинативной формы:
преимущественно красно-коричневым и более узким 3-м члеником усиков, светлыми волосками
на затылке и 1-м членике усиков и более светлой-окраской тела; брюшко сверху с
более четким срединным рядом серых треугольных пятен. 5 14—17 мм, 3 14—17 мм.
Распространение: Армения (Азии близ Араздаяна; Мегри). Середина
июня—начало июля. Не часто.
9$ с частично или полностью прикрытой налетом мозолью в верхней части лобного
треугольника встречаются в некоторых аридных местах Азербайджана и Грузии, но
они имеют обычно черную нижнюю лобную мозоль, преимущественно черные усики
и черные волоски на затылке, т. е; по этим признакам они соответствуют номинативной
форме; серый рисунок брюшка у них светлее, чем у номинативной формы. Эти
экземпляры можно рассматривать как переходные между номинативной и араксинской
формами. , ' *
208 III. ПОДСЕМ. TABANINAE
6. Tabanus caucasius Krob. (рис. 133) — Слепень горно-лесной.
К г б Ь е г, 1924, Arch. i. Naturg., A, 90, 9 : 183; Krober in Lindner,
1925 : 110; Олсуфьев, 1937а : 250; L е с 1 е г с q, 1966 : 145; С h v a 1 a, L у-
neborg a. Moucha, 1972 : 328.
5. Глаза голые или в очень коротких редких волосках (видны при увеличении
Х16), с 3 полосками. Лобная полоска б. м. параллельная, в желтовато-сером налете,
высота превосходит ширину в Зх/2—4 раза. Нижняя лобная мозоль б. м.
прямоугольная, блестяще-черная или темно-коричневая, расположена отступя от нижней границы
лба, боковыми сторонами немного не достигает края глаз; ее нижний край часто с 3
выступами. Средняя мозоль большая, овальная, черная, вполне обособлена от нижней.
Лобный треугольник низкий, выпуклый, в верхней части с двойным,
блестяще-черным или коричневым пятном (иногда пятно целиком прикрыто налетом).' Затылочная
полоска утолщена, с рядом слегка выступающих желтоватых волосков. Усики узкие,
1-й и 2-й членики желто-коричневые, 3-й членик черный, с желто-коричневым
основанием или целиком желто-коричневый. Дорсальный угол едва выражен. Концевой
членик щупалец узкий, бледно-желтовато-коричневый, покрыт короткими серыми и
черными волосками. Нотоплевры темные. Крылья бесцветные, жилки желто-коричневые;
г4 без придатка. Ноги: бедра черно-серые, голени желтые, передние голени в
вершинной 1/3 черные, последующие голени с коричневатыми кончиками, передние лапки
коричневые. Брюшко пропорционально груди большое, широкое, черно-серое, обычно
с хорошо заметным оливковым оттенком. Дорсальная сторона с 3 рядами серых пятен:
б. м. треугольных посредине и округлых по бокам. Снизу брюшко черное, в густом
оливково-сером налете. 9.5—14.5 мм.
3 неизвестен.
Похож на Т. glaucopis, но значительно мельче, и по бокам брюшка отсутствуют
коричневые пятна.
Распространение: кавказский горно-лесной вид. Большой Кавказский
хребет — по северным и южным склонам от Майкопа, Кавказского заповедника и
Хосты Краснодарского края на восток до Кусар и Варташена в Сев. Азербайджане;
горный Дагестан; Армения. Обычен. Июль—август.
Рис. 134. Tabanus glaucopis Mg. Лобная полоска, усик, щупальце, VIII стер-
нит, церки и сперматека $.
7. Tabanus glaucopis Mg. (рис. 134) — Слепень поздний.
М е i g e n, 1820, Syst. Beschreib., II : 48; В г а и е г, 1880 : 198; V е г г а 11,
1909 : 417; Sur со u f, 1924 : 193; К г б Ь е г, 1924 : 185; К г б Ь е г ia
Lindner, 1925 : 114; Олсуфьев, 1937а : 250; Moucha etChvala, 1958 : 186;
Leclercq, 1966 : 149; В и о л о в и ч, 1968 : 87; С h v а 1 a, Lyneborg a.
Moucha, 1972 : 322.
5. Глаза голые, с 3 полосками. Лоб почти параллельный, в жёлтовато-сером налете,
его высота в 4—5 раз превосходит ширину основания. Нижняя лобная мозоль
расположена отступя от нижнего края лобной полосы, прямоугольная, блестяще-черная или
темно-коричневая, боковыми сторонами немного не достигает края глаз. Средняя мо-
8. TABANUS 209
золь большая, овальная, черная, целиком обособлена от нижней мозоли. Иногда
средняя мозоль почти целиком прикрыта налетом. Затылочная полоска слегка утолщена,
с рядом коротких, светлых волосков. Лобный треугольник в верхней части блестящий,
коричневый или черный, тогда как ниже, возле основания усиков, в желтовато-сером
налете. Усики коричневые, 3-й членик узкий, с небольшим дорсальным углом и
короткой палочкой. Концевой членик щупалец утолщен, светлый, желтоватый, покрыт
короткими желтовато-серыми и черными волосками. Нотоплевры желто-коричневые,
реже темные. Крылья бесцветные, жилки коричневые; г4 иногда с коротким придатком.
Ноги: бедра черно-серые, голени коричневато-желтые, передние голени в концевой
половине черные, последующие голени с коричневыми кончиками. Передние лапки
черные, последующие коричневые. Брюшко удлиненное, черно-серое, II—III тергиты
по бокам коричневатые. Тергиты с узкими коричневато-серыми каемками и, кроме
того, боковые стороны тергитов II—V или II—VI с небольшим, овальным, серым
пятном. Посредине обычно имеется ряд сероватых треугольников. Брюшко снизу черно-
серое, с желтовато-коричневым основанием, реже целиком темное. VIII стеряит: го-
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180° 160°
Рис. 135. Ареал Tabanus glaucopis.
нопофиз широкий, с небольшой апикальной выемкой; латеральные лопасти большие,
коричневые; базальная лопасть короткая, с слегка вогнутым основным краем. Церки
с округлой вершиной. Сперматека игловидная. 14—18 мм.
в. Голова очень большая, полушаровидная. Глаза голые. Фасетки верхних
3/4 глаз значительно крупнее нижних, с резкой границей между теми и«другими.
Затылок в длинных, выступающих над поверхностью глаз коричнево-черных волосках.
Лобный треугольник в верхней части с двойной блестяще-черной или
темно-коричневой мозолью, возле основания усиков в желтоватом налете. Усики как у 5i но более
узкие. Концевой членик щупалец продолговато-овальный, с тупым концом,
желтоватый, покрыт серыми и единичными черными волосками. Грудь и ее придатки как
у 5- Брюшко коническое, его окраска как у §, коричневая окраска по бокам часто
простирается на IV тергит. 14—17 мм.
Распространение: европейско-сибирский лесной вид (рис. 135). В СССР
от юга Карелии (на север до Медвежьегорска), Ленинградской обл., Эстонии, Литвы
и запада Украины, на восток до Забайкалья (Хаимская, Баргузинский тракт), на
север до юга Коми АССР (с. Летка), Сельярова на Оби (250 км к востоку от
Ханты-Мансийска, 61°30' с. ш.) и Южно-Енисейска Красноярского края, на юг до
Кагула Молдавской ССР, Харьковской, Воронежской, Саратовской областей,
Южного Урала (Янтышево), юга Тюменской обл., Королевки близ Омска, Майны к
югу от Минусинска; Закавказье: Азербайджан (Закаталы, Карабахская АО, Та-
лыш, Нахичеванская АССР), Армения (Красное село), Грузия (Бакуриани). Европа
(на запад до Испании), Турция, Иран, Вост. Монголия, Сев.-Вост. Китай (Наи-
лумту). Местами обычен. Поздний вид: июль—август, редко раньше.
Указание Н. А. Виоловича (1968) на распространение этого вида в Приморье
сомнительно. На Сев. Кавказе не обнаружен, но вероятность нахождения его здесь
велика. В Закавказье местами поднимается в горы.
14 Н. Г. Олсуфьев
UJO III. ПОДСЕМ. TABANINAE
Кладки яиц находили вдали от воды на листьях подорожника, клевера и других
растений (Richards, 1930; Wyniger, 1953, цит. по: Лутта, 1970 : 225), личинки
развиваются в почве (Wyniger, 1953, цит. по: Лутта, 1970 : 225). Переносчик Trypanosoma
theileri Laveran (Noller, 1925).
II. Группа Tabanus cordiger Mg.
Средняя лобная мозоль б. м. поперечная или округлая, обособлена от нижней
(иногда целиком прикрыта налетом). Нижняя лобная мозоль расположена на уровне
границы лба и лобного треугольника. Лоб обычно широкий, расширенный кверху.
г4 с придатком или без него. Крылья часто с легким молочным оттенком. Глаза с 1 —
3 полосками, реже без полосок, голые или (редко) в волосках.
Небольшие или средней величины, преимущественно светлые виды,
свойственные главным образом побережьям водоемов аридных ландшафтов Евразии и Африки;
1 вид в Сев. Америке.
Рис. 136. Tabanus accipiter Szil. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека §.
8. Tabanus accipiter Szil. (рис. 136) — Слепень бледнолицый.
S z i 1 a d у, 1923, Biolog. Hungar., 1, 1 : 3; К г б b е г in Lindner, 1925 :
105; Олсуфьев, 1937а : 252; Шевченко, 1961 : 138; L е с 1 е г с q, 1966 : 137.
5. Глаза голые, с 3 полосками. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 4 раза, кверху слегка расширяется, покрыта
серым налетом. Нижняя лобная мозоль блестяще-черная или темно-коричневая, б. м.
прямоугольная, боковыми сторонами почти достигает края глаз. Средняя мозоль
черная, овальная, вполне обособлена от нижней. Затылочная полоска относительно
узкая, с рядом светлых, слабо выступающих волосков. Лобный треугольник в
желтовато-сером налете. Усики оранжево-желтые или коричневые. 3-й членик усиков
узкий, с тупым дорсальным углом. Концевой членик щупалец беловатый, утолщен,
с острым кончиком, покрыт преимущественно беловатыми волосками. Нотоплевры
желтоватые. Крылья бесцветные; г4 с придатком. Ноги: бедра серые, голени бледно-
желтые, концы передних голеней и передние лапки черные, лапки последующих ног
коричневые. Иногда бедра сквозь серый налет просвечивают желтовато-коричневым.
Брюшко удлиненное, буровато-серое, с развитой коричневой окраской по бокам I—
III или I—V тергитов, просвечивающей сквозь серый налет. Последний образует вместе
с светлыми волосками 3 ряда пятен: треугольных посредине и косых или ромбических
по бокам. Тергиты по заднему краю с желтовато-серыми каемками. Снизу брюшко
в густом, сером налете, сквозь который в базальной части просвечивает желтовато-
коричневая основная окраска. У светлых экземпляров брюшко снизу целиком
коричневато-желтое. VIII стернит желтый, много шире длины, гонопофиз с
незначительной апикальной выемкой; латеральные лопасти большие, слабо хитизированы; базаль-
ная лопасть короткая, с прямым основным краем. Церки с округлой вершиной.
Сперматека веретеновидная. 13—16 мм.
8. Голова не крупнее или же лишь немногим крупнее, нежели у §. Глаза голые,
фасетки верхних 2/3 глаза значительно крупнее нижних, с резкой границей между
8. TABANUS ' 211
теми и другими. ЭДелкофасетчатое поле с 2 полосками. Затылочная полоска очень
узкая, в коротких светлых волосках. Лобный треугольник в сером налете. Усики очень
узкие, светло-коричневые. Концевой членик щупалец беловатый,
продолговато-овальный, в белых волосках. Грудь и ее придатки как у $. Брюшко коническое, его рисунок
как у $. 13.5—15 мм.
Распространение: бактрийско-иранский предгорно-низкогорный вид
(рис. 137). Юг Туркмении (от Казанджика и Кара-Калы на западе до окр. Кушки и
р. Кашан на востоке), Узбекистан (Сурхандарьинская, Ферганская области),
Таджикистан (долина Амударьи и Пянджа от Айваджа до Московского и долина Вахша
вверх до Курган-Тюбе; окр. Ленинабада в Ферганской долине); юг Казахстана (Жулек
на Сырдарье). Иран, Афганистан. Обычен на юге Туркмении, в других местах не часто.
Июнь—июль.
Рис. 137. Ареал Tabanus accipiter.
9. Tabanus subsabuletorum Ols. (рис. 138) — Слепень тугайный.
Олсуфьев, 19376, Труды Казахстан, фил. АН СССР, 2 : 169; 1937а : 254;
Шевченко, 1961 : 139; L е с 1 е г с q, 1966 : 171.
5. Глаза голые, зеленовато-серые, с 4 узкими медными, полосками (иногда
верхняя и нижняя отсутствуют). Лобная полоска умеренной ширины, ее высота
превосходит ширину основания в З1/,,—3V2 раза, покрыта серым налетом. Нижняя лобная
мозоль блестяще-черная, б. м. квадратная, боковыми сторонами достигает края глаз,
средняя мозоль черная, небольшая, чаще сердцевидная. Обычно обе мозоли вполне
изолированы между собой, реже их соединяет тонкая полоска; иногда средняя мозоль
целиком прикрыта налетом. Затылок в виде умеренно широкой полоски, с рядом
выступающих серых волосков. Усики узкие, целиком желто-коричневые, иногда с темно-
коричневой палочкой. Лобный треугольник в желтовато-сером налете, темная полоска
возле основания усиков отсутствует. Концевой членик щупалец утолщен, беловатый
или слегка желтоватый, покрыт короткими, преимущественно черными волосками.
Нотоплевры коричнево-желтые. Крылья бесцветные, жилки коричневые; г4 с
придатком. Ноги: бедра красновато-желтые, крайне редко сероватые, голени желтые,
концы передних голеней и передние лапки черноватые, средние и задние лапки
коричневые, в основании желтоватые. Брюшко сероватое, с нерезко отграниченными
коричневато-желтыми пятнами по бокам II—IV или II—V тергитов. Сверху 3 ряда нерезких
беловато-серых пятен: треугольных посредине и б. м. ромбических по бокам,
разделенных двумя рядами коричневатых, слегка косо расположенных пятен. Снизу брюшко
розовато-желтое, 3 последних стернита серые. VIII стернит желтый, без темного
рисунка, гонопофиз конический, апикальная выемка не выражена; латеральные лопасти
большие, слабо хитинизированные; базальная лопасть с вогнутым основным краем,
уже гонопофиза. Церки угловатые. Сперматека б. м. веретеновидная. 9.5—15 мм.
8 неизвестен.
14*
212
III. ПОДСЕШ. TABANINAE
Похож на Т. sabuletorum, § отличается: 1) более узким лбом и менее развитой
средней лобной мозолью; 2) красновато-желтыми бедрами; 3) более развитой
коричневато-желтой окраской брюшка, обычно захватывающей боковые стороны II—IV
тергитов; 4) менее резким рисунком брюшка — косые пятна по бокам серых срединных
треугольников коричневатые и 5) строением терминалий. Ранее указывавшаяся нами
в качестве одного из дифференциальных признаков — ширина затылочной полоски —
оказалась варьирующей у Т. sabuletorum и непригодной для различения его от Т. sub-
sabuletorum. Наше описание в Т. subsabuletorum следует отнести к Т. sabuletorum.
Распространение: среднеазиатский пустынный вид, преимущественно
по долинам рек. Восточная Туркмения (Чарджоу, Фараб, Дарганата, Келифский
Узбой близ Керков), Таджикистан (Куляб, Айвадж, низовья Пянджа, Ленинабад),
Рис. 138. Tabanus subsabuletorum Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека §■
Узбекистан (Термез, Бухара, Самарканд, Каттакурган, Ташкент, Ферганская
долина, Хива, Каракалпакия), Казахстан (Мангышлак; долина Сырдарьи: от Кзыл-
Орды на севере до Чардара на юге). Конец мая—начало августа. Личинки найдены
на рисовых полях и в арыке (Кадырова, 19726). Наши указания (Олсуфьев, 1937а)
о распространении этого вида в бассейне Балхаша, а также в Зап. Китае и Монголии
должны быть отнесены к Т. sabuletorum. Видимо, неточно и наше указание для
Азербайджана. Указание Такаги (Takagi, 1941) на обнаружение Т. subsabuletorum во
Внутренней Монголии КНР (оз. Далайнор), вероятно,- относится к Т. stackelbergiellus:
на табл. VII, рис. 5 видно, что г4 без придатка, что характерно для последнего вида.
Наше указание Т. subsabuletorum для Палестины (Иерусалим) мы относим к близкому
виду, идентификация которого пока неясна.
Голотип: 5> Сырдарья, Жулек б, Перовского у., 9 VI 1912 (ст. ст.), Вольман.
В коллекции Зоологического института Академии наук СССР, Ленинград.
10. Tabanus sabuletorum Lw. — Слепень песчаный.
Широко распространенный изменчивый вид. В СССР представлен 2 подвидами:
южным номинативным Т. sabuletorum sabuletorum Lw. и северным Т. sabuletorum
gerkei Br.
10а. Tabanus sabuletorum sabuletorum Lw. (рис. 139) — Слепень песчаный
номинативный.
L e e w, 1874, Zeitschr. Ges. Naturw., 9 : 414; S z i 1 a d у, 1914 : 670; К r 6 b e г,
1924 : 66; К r 6 b e r in Lindner, 1925 : 96; Олсуфьев, 1937а : 255;
Шевченко, 1961 : 140; L e с 1 e г с q, 1966 : 167; Виолович, 1968 : 88; С h v a 1 a,
Lyneborg a. Moucha, 1972 : 344. — lama Portschinsky, 1892:201.
5- Глаза голые (при увеличении Х16 иногда удается заметить короткие волоски),
с 4 полосками, из них 2 синего цвета расположены в средней части глаза, а 2 других
медного цвета — по верхнему и нижнему краям глаза. Лобная полоска широкая,
расширяется кверху, ее высота превосходит ширину основания в 21/4—3 раза, покрыта
светло-серым налетом. Нижняя лобная мозоль блестяще-черная или
темно-коричневая, прямоугольная, достигает боковыми сторонами края глаз. Средняя лобная мозоль
8. TABANUS 213
в виде двойного, черного, поперечного пятна, вполне обособлена от нижней мозоли
(иногда целиком скрыта налетом). Затылочная полоска широкая, в особенности возле
темени, с рядом длинных, заметно выдающихся серых волосков. Лобный треугольник
низкий, в сером налете. Возле основания усиков темная полоска отсутствует. Усики
узкие, 1-й и 2-й членики коричневые, 3-й членик черный, с коричневым основанием,
с едва выраженным дорсальным углом. Окраска усиков варьирует и встречаются
экземпляры с целиком желто-коричневыми усиками. Концевой членик щупалец белый,
утолщен, покрыт, как правило, только белыми волосками; у экземпляров из
Монголии и Вост. Казахстана иногда на щупальцах единичные черные волоски. Крылья
с легким молочным оттенком, жилки коричневые, в основной части крыла часто
желтоватые; г4 с придатком. Ноги: бедра серые, голени бледно-желтые, передние голени
в вершинной 1/3 черные, последующие голени с коричневатым кончиком. Передние
лапки черные, последующие лапки коричневые, с желтым осповаписм. Окраска бедер
варьирует и изредка встречаются экземпляры с целиком серовато-желтыми бедрами
и передними коксами. Брюшко черновато-серое, с четким беловатым рисунком (налет+
Рис. 139. Tabanus sabuletorum sabuletorum Lw. Лобная полоска,
усик, щупальце, VIII стернит и сперматека §. .4 — Т. sabuletorum
gerkei Br., VIII стернит и церки.
волоски) в виде 3 продольных рядов пятен: треугольных посредине и б. м.
ромбических по бокам. Тергиты по заднему краю с беловато-серыми каемками. По бокам брюшка
на I—III тергитах обычно бывает развита коричневато-желтая окраска, которая
просвечивает сквозь налет розоватым. Снизу брюшко пепельно-серое, в тонком
беловатом налете, в базальной части иногда просвечивает розоватым. VIII стернит: гоно-
пофиз с апикальной выемкой; латеральные лопасти небольшие, коричневатые; базаль-
ная лопасть с прямым основным краем. Церки с округлой вершиной. Сперматека б. м.
веретеновидная. 8.5—15 мм.
6. Голова большая, сильно выпуклая. Глаза голые (возле глазного шва в очень
коротких волосках), фасетки верхних 2/3 глаз значительно крупнее нижних,
приблизительно в 5—6 раз. Мелкофасетчатое поле с 3 полосками, кроме того
крупнофасетчатое поле с неясной, темноватой поперечной полоской. Затылочная полоска
утолщена, снабжена рядом длинных, изогнутых вперед концами серых волосков. Лобный
треугольник в сером налете, в верхней части с коричневатой матовой поперечной
полоской. Усики как у $, но более узкие. Возле основания усиков темная полоска
отсутствует. Концевой членик щупалец утолщен, белый, в белоснежных волосках. Грудь
и ее придатки как у §. Брюшко коническое, рисунок как у $. Боковые стороны II
или I—IV тергитов и соответственных стернитов с коричневато-желтоиили розоватой
окраской. 11.5—13 мм.
Распространение: пустынная зона, преимущественно около
солоноватых водоемов (рис. 140, 1). В СССР известен из Закавказья (Восточная Грузия,
Азербайджан), Туркмении, Таджикистана, Узбекистана, юга Казахстана (на север до 46—
48° с. ш.), Тувы и Забайкалья. Иран, Зап. и Центр. Китай, Монголия. Местами обычен.
Июнь—июль (в Туркмении — со второй половины мая). Личинки найдены на
рисовых полях и в арыке (Кадырова, 19726).
У $5 ширина лобной и затылочной полоски варьирует — наряду с типичными
широколобыми экземплярами встречаются особи, у которых лобная полоска сужена
2U III. ПОДСЕМ. TABANINAE
(отношение ширины основания к высоте 1 : 3) и соответственно затылочная полоска
утолщена умеренно. Эти экземпляры ранее ошибочно мы принимали за Т. subsabu-
letorum. Экземпляры 5$ из Восточно-Казахстансткой обл. (хр. Саур), Тувинской АССР,
некоторых мест Монголии (Кобдо, оз. Убса) и Китая (Алашань, Цайдам) обращают
на себя внимание крупными размерами тела — до 15 мм.
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 130° -150° 180° К0°
Рис. 140. Ареалы: 1 — Tabanus sabuletorum sabuletorum; 2 — Т. sabuletorum
gerkei.
10b. Tabanus sabuletorum gerkei Br. — Слепень песчаный северный.
В r a u e г, 1880, Denkschr. Akad. Wiss. Wien, 42 : 205 (T. gerkei).
$c? отличаются от номинативной формы менее четким пепельно-серым рисунком
брюшка, у 5 коричневые пятна по его бокам обычно отсутствуют, лоб очень широкий,
иногда отношение ширины основания к высоте 1 : 21/4, соответственно сильно утолщен
затылок возле темени. Концевой членик щупалец обычно только в белых волосках.
9—13 мм.
Распространение: экземпляры, послужившие Брауэру (Brauer, 1880)
для описания Т. gerkei, сведенного затем нами (Олсуфьев, 1937а) в синонимы Т.
sabuletorum, были из южной России и Кавказа; номинативную форму Лёв (Loew, 1864)
описал из Ирана (Шахруд), т. е. из южной части ареала вида. Подвид охватывает
(рис. 140, 2) распространением юг Украины (на западе до устья Дуная, на юге
включая Крым), Сев. Кавказ, Сев. Прикаспий, Сев. Казахстан и Зап. Сибирь, на север
известен до Самарского леса близ Днепропетровска, окр. Оренбурга, Кустанайской
(Семиозерный), Курганской (Половинное), юга Омской (Нововаршавка) и
Новосибирской (Карасукский р-н) областей, на юг примерно до 48—46° с. ш., где смешивается
с популяцией номинативного подвида. В северных частях ареала не часто, на юге
обычен.
*11. Tabanus stackelbergiellus Ols. (рис. 141)—Слепень Штакельберга.
Олсуфьев, 1967, Энтом. обозр., 46, 2:385. —subsabuletorum (Ols.) apud
Т a k a g i, 1941 : 74. — kimbarai Murdoch etTakahasi, 1969 : 77.
5. Глаза в коротких сероватых, волосках, с 4 полосками. Лобная полоска
широкая, слегка расширена кверху, ее высота превосходит ширину основания в 2V4—2V2
раза, покрыта светло-серым налетом. Нижняя лобная мозоль темно-коричневая или
черная, блестящая, б. м. прямоугольная, боковыми сторонами достигает края глаз.
Средняя лобная мозоль в виде двойного, черного поперечного пятна, обособлена
от нижней мозоли. Затылочная полоска широкая, с рядом выступающих серых
волосков. Лобный треугольник низкий, в сером налете. Возле основания усиков темная
полоска отсутствует. Усики узкие, 1-й и 2-й членики желто-коричневые, 3-й членик
черный, обычно с коричневым основанием, со слабым дорсальным углом. Концевой
членик щупалец белый, слабо утолщен, покрыт беловатыми и единичными черными
волосками. Крылья бесцветные, жилки желто-коричневые; г4 без придатка (или с руди-
5. TABANUS
215
ментом). Бедра ног, голени и основание средних и задних лапок бледно-желтые, конец
передних голеней, передние лапки, конец средних и задних лапок коричнево-черные.
Брюшко серое, II—IV тергиты по бокам просвечивают желто-розовым цветом.
Беловато-серый налет и волоски образуют сверху брюшка 3 продольных ряда пятен,
треугольных посредине (могут отсутствовать) и б. м. ромбических по бокам, а также
каемки по заднему краю тергитов. Снизу брюшко желтовато-розовое или серое,
покрыто беловато-серым налетом и светлыми волосками. 11—13 мм.
8. Голова крупнее и более выпуклая, чем у $. Глаза в довольно длинных,
сероватых волосках. Фасетки верхних 2/3 глаз в 3—4 раза крупнее нижних, резко отделяясь
от них. Мелкофасетчатое поле с 3 полосками. Затылочная полоска узкая, снабжена
рядом длинных коричневых и сероватых волосков. Лобный треугольник в сером
налете, в верхней половине с черноватой
слабо блестящей поперечной полосой. Усики
ШШШШШШ////// _-j^ как у ?'н0 3-й членик более узкий; 1_й чле*
ник серый. Возле основания усиков темная
Рис. 141. Tabanus stackelbergiellus Ols.
Лобная полоска, усик и щупальце $.
Рис. 142. Tabanus mistshenkoi Ols.
Лобная полоска, усик и щупальце $.
полоска («уздечка») отсутствует. Концевой членик щупалец беловатый, продолговато-
овальный, к вершине слегка конический, в беловатых волосках. Крылья бесцветные,
г4 без придатка, жужжальца черноватые. Ноги: бедра черные в сером налете, голени
желтоватые с темным кончиком, лапки черноватые, средние и задние лапки в
основании желтоватые. Брюшко сверху, с широкой черной срединной полосой, в ее пределах
на II тергите неясный сероватый треугольник, на последующих едва заметный
буроватый налет посредине; по бокам брюшка по ряду резких косо расположенных
беловатых пятен; боковые стороны II тергита и краевая часть последующих
розовато-желтоватые. Снизу брюшко серое, в беловатых волосках, задний край стернитов
желтоватый. 12.5—14 мм.
Сходен с Т. sabuletorum, но легко отличается от последнего глазами, покрытыми
волосками, и отсутствием придатка у жилки г4. „' „ .
Распространение: Монголия (Центральный и Восточный аймаки),
Китай — Внутренняя Монголия (Тунляо; Хинган). По-видимому, к этому же виду
относятся 2 $$ с оз. Далайнор, Внутренняя Монголия, отмеченные Такаги (Takagi,
1941) как Т. subsabuletorum. Т. kimbarai Murdoch et Takahasi, 1969, $, судя по
описанию и рисунку идентичен Т. stackelbergiellus Ols., 1967. В СССР возможно
нахождение Т. stackelbergiellus в даурских степях, расположенных к северу от упомянутого
озера.
12. Tabanus mistshenkoi Ols. (рис. 142) — Слепень Мищенко.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 257; Ш е в ч е н к о,
1961 : 142; L е с 1 е г с q, 1966 : 159. — fasciatus Belanovsky, 1950 : 137,
syn. nov.
5. Глаза голые, с 4 'полосками (типа Т. sabuletorum). Лобная полоска умеренно
широкая, слегка расширена кверху, ее высота превосходит ширину основания
приблизительно в 3V2 раза, покрыта серым налетом. Нижняя мозоль б. м. квадратная,
блестяще-черная, боковыми сторонами достигает края глаз. Средняя мозоль черная,
в виде большого поперечного пятна, вполне обособлена от нижней; боковыми
сторонами средняя мозоль лишь немного не достигает края глаз, отделяясь узкими
полосками серого налета. Затылочная полоска умеренно утолщена, снабжена несколько
выдающимися вверх белыми волосками. Лобный треугольник низкий, в сером налете.
216
III. ПОДСЕМ. TABANINAE
Усики узкие, черные, 3-й членик в основании коричневатый. 1-й членик в сером
налете. На уровне основания усиков темная поперечная полоска отсутствует. Концевой
членик щупалец белый, сильно утолщен, с острым концом, покрыт белыми и
немногочисленными черными волосками. Нотоплевры коричневато-желтые. Крылья
бесцветные, жилки коричневатые, более темные в вершинной части; г4 без придатка.
Ноги: бедра черно-серые, голени желтые; вершинная половина передних голеней,
кончики последующих голеней и лапки всех ног черные. Средние и задние лапки
в основании желтоватые. Брюшко черное, с 3 довольно резкими рядами серых пятен
(налет+волоски): треугольных посредине и б. м. ромбических по бокам. Задние края
тергитов с узкими желтоватыми каемками. Иногда срединный ряд треугольников
выражен менее отчетливо, нежели боковые пятна. Снизу брюшко черно-серое, в беловато-
сером налете. 9.5—11.5 мм.
8. Голова крупнее и более выпуклая, нежели у $. Глаза (верхняя часть) в
коротких, малозаметных, светлых волосках. Фасетки верхних 2/3 глаз значительно крупнее
нижних, приблизительно в 4 раза, резко отделяясь от нижних. Мелкофасетчатое поле
с 3 полосками, из них одна полоска на границе с крупными фасетками,
крупнофасетчатое поле с одной довольно резкой, темной, широкой, поперечной полосой.
Затылочная полоска узкая, снабжена рядом умеренно длинных серых волосков. Лобный
треугольник в сером налете, в верхней трети с довольно резкой, черноватой поперечной
полосой. Усики как у $, 3-й членик очень узкий. Концевой членик щупалец сильно
утолщен, лимоновидный, покрыт белыми и немногочисленными черными волосками.
Грудь, ее придатки и брюшко как у 5- Последнее с очень незначительной
коричневатой окраской по боковому краю II тергита, отчасти прикрытой серыми боковыми
пятнами — налетом. 10—11.5 мм.
Похож на Т. sabuletorum, но легко отличается от него по отсутствию придатка
у г4. 36 отличаются более короткими волосками на затылке (короче почти вдвое).
Распространение: юг Таджикистана — долина Пянджа и его
притоков, предгорья и низкогорья Гиссарского хребта и его южных отрогов. Иран. Май—
июнь. Не часто.
В. В. Шевченко (1961) указал этот вид по 1 5 Для пос. Ильич Чимкентской обл.;
находка нуждается в подтверждении.
Голотип: 5, окр. Душанбе, Таджикистан, 9 VI 1935, Л. Мищенко. В коллекции
Зоологического института Академии наук СССР, Ленинград.
13. Tabanus freyi Szil.
S z i 1 a d у, 1926, Ann. Mus. Nat. Hungar., 24 : 600 (Q); Олсуфьев, 1937а :
258; Lee 1 ere q, 1966 : 148.
Нам в натуре неизвестен. Силади (Szilady) дает следующее краткое описание $:
«Близок к Т. gerkei Br., но отличается от него следующими признаками: лицо резко
разделяется посредством кофейно-коричневой поперечной полоски возле основания
усиков на пепельно-серую верхнюю часть головы и белоснежно-белую, покрытую
такого же цвета волосками, нижнюю часть головы (напоминая Т. cordiger Mg.). Усики
целиком черные. Среднее лобное пятно разделено на два боковых пятна. Глаза с
четырьмя поперечными полосками».
Распространение: «Туркмения — Перевалье 24 V 1898».
Идентификация этого вида представляется затруднительной ввиду неполноты
оригинального описания, а также утери типового экземпляра (см. замечание в конце
описания Т. ansarii Abbas.-L.).
14. Tabanus ansarii Abbas.-L. — Слепень Анзари.
Abbas sian-Lintzen, 1960, Arch. Inst. Razi, 12 : 30; L e с 1 e г с q,
1966 : 138. . .
Описан по сборам из Южного Ирана (провинция Фарс). Изменчив, в частности,
популяции, обитающие в юго-восточной части Ирана, образуют подвиды Т. ansarii
nigrinervis Abbas.-L. и Т. ansarii gedrosiae Abbas.-L. В СССР распространен подвид
Т. ansarii badhysi Ols. et Shevtsh. Так как номинативный подвид Т: ansarii ansarii
Abbas.-L. у нас отсутствует, его описание мы не даем. Но для подвида Т. ansarii badhysi
Ols. et Shevtsh. неизвестен 3, поэтому ниже дано описание 6 номинативного подвида
(по Abbassian-Lintzen).
14а. Tabanus ansarii badhysi Ols. et Shevtsh. (рис. 143) — Слепень Анзари бадхыз-
ский.
Олсуфьев и Шевченко, 1964, Тр. Инст. зоол. АН Казахск. ССР,
XXII : 197; L е с 1 е г с q, 1966 : 138.
5. Глаза голые, с 3 полосками. Высота лобной полоски превосходит ширину
основания в 2V2 раза, кверху полоска значительно расширена, в сером налете. Ниж-
8. TABANUS
217
няя лобная мозоль блестяще-черная, поперечная, боковыми сторонами достигает
или почти достигает края глаз. Средняя мозоль черная, сердцевидная или округлая.
В верхней части лба выше средней мозоли обычно имеется по боковой короткой
блестящей черной черточке, параллельно краю глаз. Усики черные, узкие, дорсальный угол
едва выражен, площадка 3-го членика приблизительно в 2—21/2 раза длиннее ширины.
Лобный треугольник серый, на уровне основания усиков имеется поперечная
коричневая полоска («уздечка»). Концевой членик щупалец в основании слабо утолщен,
беловатый, в беловатых волосках и единичных черных. Крылья с заметным молочным
оттенком, жилки коричневые, птеростигма бледно-желтая или светло-коричневая;
г4 с придатком. Бедра целиком или на большем протяжении черно-серые,__реже
коричневые. Средние и задние голени и базальная половина передних голеней коричнево-
желтые, концевая часть передних голеней и передние лапки черные, последующие
лапки черные с коричневым основанием. Брюшко серое, боковые стороны I—IV или
I—V тергитов желтовато-коричневые,
задний край всех тергитов также желтовато-
коричневый. По бокам тергитов имеются
округлые нерезкие пятна беловатого налета.
Срединный ряд светлых треугольников
слабо выражен. Снизу брюшко в основной части
желтовато-коричневое ,i концевые 2 или
3 стернита черно-серые с желтоватой
каймой по заднему краю. 9—12 мм.
Рис. 143. Tabanus ansarii badhysi Ols.
et Shevtsh. Лобная полоска, усик и
щупальце $•
S. Т. ansarii ansarii Abbas.-L(. no
Abbassian-Lintzen, 1963, сокращенно). Голова
не увеличена. Глаза голые, с 2 полосками, область постепенно увеличивающихся
фасеток составляет 2/3 глаза и отличается от мелких фасеток. Затылок узкий,
преимущественно в черных длинных волосках и немногих белых. Лобный треугольник в сером
налете, в верхней части с более темной поперечной полоской. На уровне основания
усиков имеется черно-коричневая поперечная полоска. Усики целиком черные, форма
их как у $. Концевой членик щупалец грязновато-желтый, преимущественно в
черных волосках, приблизительно в 2. 3 раза длиннее толщины, с вытянутым острым
кончиком, направленным слегка вниз. Крылья слегка сероватые. Ноги как у $ Г. ansarii
gedrosiae (т. е. бедра черные, в сером налете; передние голени в основной 1/3 желтые,
остальное черное; средние и задние голени грязно-желтые с черным концом, лапки
черные). Брюшко: рисунок и окраска сходны с таковыми $ последнего подвида (т. е.
брюшко сверху в основном черное, с 3 рядами пятен: серых треугольных посредине
и косых серебристо-серых по бокам), но II тергит по бокам с крупным
красно-коричневым пятном. 12 мм.
Распространение: Туркмения (Бадхыз; Кара-кала); Казахстан (Джу-
салы Кзыл-Ординской обл.). Последняя декада мая—начало июня. Редок.
Т. ansarii похож на Т. sabuletorum, но отличается от него наличием ясной
коричневой поперечной полоски («уздечки») на уровне основания усиков и 3 полосками на
глазах вместо 4. Похож также на Т. zimini, но у последнего всего 2 полоски на глазах,
средняя лобная мозоль часто" поделена на 2 мозольки. У Т. ansarii средняя лобная
мозоль б. м. сердцевидной формы. Т. ansarii мог бы быть принят за Т. freyi, описанного
Силади (Szilady) в 1926 г. из Перевалья (Туркмения), но, судя по оригинальному
описанию автора, у Т. freyi глаза с 4 поперечными полосками, а средняя лобная мозоль
разделена на 2 боковых пятна. По сообщению Аббассиан-Линтцен
(Abbassian-Lintzen,1961), типовой экземпляр, хранившийся в Венгерском национальном музее в
Будапеште, погиб, и поэтому вопрос об этом виде трудно разрешим.
15. Tabanus zimini Ols. (рис. 144) — Слепень Зимина.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 258; Шевченко,
1961 : 143; L е с 1 е г с q, 1966 : 176; Соболева и Кадырова, 1972 : 42
(куколка). — luppovae В а г a t о v, 1961 : 184.
5. Глаза голые, с 1 широкой срединной полоской и, кроме того, 1 полоской по
нижнему краю. Голова сильно поперечная. Лоб широкий, сильно расширяющийся кверху,
приблизительно на -Vsi покрыт светло-серым налетом. Высота лба превосходит ширину
основания приблизительно в 21/2—23/i раза. Нижняя лобная мозоль поперечно-прямо-
^W HI. ПОДСЕШ. TABANINAE
угольная, блестяще-черная или темно-коричневая, боковыми сторонами достигает
края глаз. Средняя мозоль вполне обособлена от нижней, обычно представлена 2
изолированными, небольшими, блестяще-черными пятнами; иногда оба пятна слиты в
небольшую, поперечную, сильно блестящую мозоль и, наоборот, могут быть целиком
покрыты налетом. Затылочная полоска очень сильно утолщена, с рядом длинных,
беловатых волосков. Лобный треугольник низкий, в светло-сером налете. На уровне
основания усиков имеется поперечная коричневая полоска («уздечка»). Усики узкие,
черные, 3-й членик с едва выраженным дорсальным углом. Концевой членик щупалец
белый, в коротких, белых волосках, в основании умеренно утолщен, с острым
кончиком. Боковые края спинки, включая нотоплевры и плечи, бледно-розовато-желтые.
Мезоплевры в верхней половине просвечивают бледно-розовато-желтым. Крылья с
легким молочным оттенком; >4 с придатком. Ноги: бедра светло-серые, голени бледно-
желтые, концы передних голеней и передние лапки черные, лапки последующих ног
коричневые. Брюшко узкое, черно-серое, боковые стороны I—III или I—IV тергитов
бледно-розово-желтые. Сверху брюшко с 3 рядами б. м. резких светло-серых пятен:
треугольных посредине и б. м. ромбических по бокам; светлые пятна разделяют косые
коричнево-черные черточки. Снизу брюшко розово-желтое, последние 4 или 3 стернита
Рис. 144. Tabanus zimini Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $.
серые. У темных экземпляров розово-желтая окраска на брюшке может целиком
отсутствовать; у наиболее светлых особей, наоборот, занимать его большую часть. VIII
стернит темно окрашен, гонопофиз широкий, двулопастной, с глубокой выемкой;
латеральные лопасти маленькие, слабо хитинизированы; базальная лопасть с прямым основным
краем. Церки с округлой вершиной. Сперматека б. м. веретеновидная. 10—14 мм.
в. Голова немного шире груди, поперечная, умеренно выпуклая. Глаза голые,
фасетки верхних 2/3 крупнее нижних в 4—5 раз, граница между теми и другими резкая.
Мелкофасетчатое поле с двумя резкими широкими темно-лиловыми полосками: одна
полоска на границе крупных и мелких фасеток и вторая по нижнему краю глаза. У
сухих экземпляров глаза резко двуцветные: крупнофасетчатое поле желтовато-палевое,
мелкофасетчатое поле — черное. Затылочная полоска сильно утолщена, в довольно
длинных изогнутых белых волосках. Лобный треугольник в сером налете, в верхней
части с черной матовой поперечной полоской. Усики целиком черные, 3-й членик
немного у^ке, чем у $. На уровне основания усиков имеется поперечная кофейно-корич-
невая полоска («уздечка»). Лицо в беловатом налете и в белоснежных волосках.
Концевой членик щупалец белый, продолговато-овальный, с оттянутым вперед кончиком,
в белых волосках. Грудь и ее придатки как у $. Брюшко коническое, сверху рисунок
как у 5; боковые стороны II—III тергитов желтовато-розоватые. Снизу брюшко
светлосерое, задний край стернитов желтоватый. 9—11.5 мм.
Распространение: переднеазиатско-кавказско-тураяский вид, побережья
водоемов пустынной зоны. В СССР обнаружен в Азербайджане (включая Нахичеван-
скую АССР), Туркмении, Узбекистане (включая Хиву и Каракалпакию), Таджикистане
и Казахстане (на север до оз. Индер Гурьевской обл., оз. Ащи-Алакуля в 135 км
к востоку от Иргиза, 48°30' с. ш., среднего течения р. Сары-Су, 47° с. ш. и дельты
Или). Афганистан, Иран, Ирак. Не часто. Июнь—июль, иногда раньше или позднее.
Предпочитает солоноватые водоемы, а также обитает возле колодцев в пустыне.
Личинки найдены на соленом озере (Соболева и Кадырова, 1972).
Голотип: 5, Хива, Рават, 24 VI 1927, Л. Зимин. В коллекции Зоологического
института Академии наук СССР, Ленинград.
8. TABANUS 219
16. Tabanus filipjevi Ols. (рис. 145) — Слепень Филипьева.
Олсуфьев, 19376, Тр. Казахстан, фил. АН СССР, 2 : 165; 1937а : 260;
Шевченко, 1961:144; Leclercq, 1966:148; Соболева и Кадырова,
1972 : 44 (куколка).
$. Глаза голые, с 1 резкой полоской. Лобная полоска широкая, кверху сильно
расширена, ее высота превосходит ширину основания в 21/2—23/4 раза, покрыта серым
налетом. Нижняя лобная мозоль б. м. прямоугольная, почти достигает края глаз,
блестяще-черная, реже темно-коричневая. Средняя мозоль в виде небольшого,
черного двойного (или одного округлого) пятна, вполне обособленного от нижней мозоли.
Часто средняя мозоль бывает целиком покрыта налетом. Затылок в виде относительно
широкой полоски, снабжен довольно длинными серыми волосками, выступающими
над поверхностью глаз. Лобный треугольник в сером налете. Усики узкие, черные,
3-й членик с тупым дорсальным углом. На уровне основания усиков имеется легкая,
коричневая, поперечная полоска («уздечка»). Концевой членик щупалец белый, в
основании сильно утолщен, на конце заострен, покрыт нежными белыми волосками.
Рис. 145. Tabanus filipjevi Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $■
Нотоплевры желтоватые. Крылья с легким молочным оттенком, жилки коричневые;
г4 с придатком. Ноги: бедра серые, голени желтые; вершина передних голеней черная.
Передние лапки черные, последующие коричнево-черные, с желтоватым основанием.
Брюшко пепельно-серое, с тремя довольно резкими продольными рядами
светлосерых пятен: треугольных посредине и косых по бокам; II—III тергиты (реже I—IV)
потокам просвечивают розово-желтым. Снизу брюшко серое, реже в основании бледно-
розовато-желтое. VIII стернит: гонопофиз с небольшой апикальной выемкой;
латеральные лопасти маленькие, слабо хитинизированы, боковые стороны склерита
коричневые; базальная лопасть с вогнутым основным краем. Церки с о. м. округлой
вершиной. Сперматека миндалевидная. 12—15 мм.
S. Голова сильно выпуклая, полушаровидная. Глаза голые, с ,1 полоской;
фасетки верхних 3/4 глаз приблизительно в 5—6 раз крупнее нижних. Затылочная
полоска относительно широкая, снабжена рядом слегка выдающихся над поверхностью
глаз серых волосков. Лобный треугольник серый. Усики как у $, но более узкие. На
уровне основания усиков имеется коричневая полоска. Лицо в белоснежных волосках.
Концевой членик щупалец яйцевидный, белый, покрыт белыми волосками. Грудь,
ее придатки и окраска брюшка как у $• 12 мм.
Похож на Т. sabuletorum, но легко отличается наличием одной полоски на глазах.
Распространение: кавказско-турано-монгольский вид, побережья
водоемов пустынной зоны. Азербайджан, Туркмения, Узбекистан (Хива, Каракалпакия),
Казахстан (на север до Мангышлака, Иргиза, Тургая, среднего течения Сары-су,
47° с. ш., Прибалхашья и хребта Саур). Китай: Синьцзян, Внутренняя Монголия
(Алашань). Не часто. Конец мая—конец августа. Личинки найдены на соленом озере,
(Соболева и Кадырова, 1972).
220 III. ПОДСЕМ. TABANINAE
Голотип $: окр. Иргиза Актюбинской обл. Казахской ССР, 14 VI 1928, Оленев
и Попов.
17. Tabanus brunneocallosus Ols. (рис. 146, 147) — Слепень такырный.
Олсуфьев, 19376, Тр. Казахстан, фил. АН СССР, 2 : 167; 1937а : 261;
Шевченко, 1961 : 145; L е с 1 е г с q, 1966 : 145; Chvala, Lyneborg a. М о-
u с h а, 1972 : 342. — gerkei (Br.) apud К г б b e r, 1924 : 174.
$. Глаза голые или же в едва заметных волосках, без полосок. Лобная полоска
очень широкая, ее высота превосходит ширину в 2—2 V2 раза, покрыта серым
налетом. Лобные мозоли коричневые, нижняя поперечная, достигает края глаз, средняя
также в виде большого, поперечного пятна, сильно сближенного с нижней мозолью.
Иногда между обеими мозолями посредине
имеется узкая перемычка, чаще же они
вполне обособлены. В редких случаях
средняя мозоль целиком прикрыта налетом.
Лобный треугольник низкий, по бокам
блестяще-коричневый, в средней части
прикрыт желтовато-серым налетом. Усики
узкие, черные; 3-й членик с незначительным
Рис. 146. Tabanus brunneocallosus Ols 5. Рис. 147. Tabanus brunneocallosus Ols.
Лобная полоска, усик и щупальце $■
дорсальным углом. Щупальца белые, концевой членик слабо утолщен, остроконечный,
покрыт тонкими белыми волосками. Затылочная полоска широкая, с рядом
выступающих над поверхностью глаз светлых волосков. Нотоплевры часто желтоватые.
Крылья с хорошо заметным молочным оттенком, жилки коричневые; г4 с придатком.
Ноги: бедра серые, голени желтые; передние голени на конце черные. Лапки черные,
средние и задние лапки в основании темно-коричневые. Брюшко черное, посредине
с продольным рядом небольших серых треугольников и по бокам с довольно широкой
полоской из крупных серых пятен. Тергиты по заднему краю с узкой желтоватой
каемкой. Снизу брюшко серое. 10—13.5 мм.
в- Голова умеренно выпуклая, лишь немногим больше, чем у $. Глаза голые,
фасетки верхних 2/3 глаз приблизительно в 4 раза крупнее нижних, граница между
крупными и мелкими фасетками резкая, вдоль нее имеется полоска. Затылок в
относительно длинных, серых волосках, значительно выступающих над поверхностью
глаз. Лобный треугольник в сероватом налете, в верхней части желтоватый. Усики
как у $• Концевой членик щупалец продолговато-овальный, беловатый, в светлых
волосках. Грудь, ее придатки и рисунок брюшка как у $• II тергит брюшка по бокам
коричневый. 13 мм.
Похож на Т. sabuletorum, но 5 легко отличается от 55 этого и родственных видов
блестяще-коричневым по бокам лобным треугольником, коричневыми, сильно
сближенными между собой лобными мозолями и глазами без полосок. Сделанное Крёбе-
ром (Krober, 1924) описание самки Т. gerkei явно относится к Т. brunneocallosus.
Распространение: монголо-казахстанский пустынно-степной вид, пре-
8. TABANUS
221
имущественно около солоноватых водоемов (рис. 148,1). Ставропольский край (Маныч.
40 км восточнее Дивное), Волгоградская обл. (40 км южнее оз. Эльтон), Алтайский
край (с. Угловское), Казахстан: на западе от Чижинских разливов 60 км севернее
Фурманово Уральской обл. и залива Кайдак Гурьевской обл., на,востоке до среднего
течения р. Или Талды-Курганской обл., на север до Кустаная и Ермен-тау 150 км
северо-восточнее Целинограда, на юг до Джусалов Кзыл-Ординской обл. и Бетпа-
кдалы. Монголия (Южно-Гобийский и Восточный аймаки), Китай: запад Внутренней
Монголии (Эдзин-гол). Не часто. Конец мая—середина августа.
Голотип: $! Кустанай, Карагай 26 VI 1923. В коллекции Зоологического
института Академии наук СССР, Ленинград.
Рис. 148. Ареалы: 1 — Tabanus brunneocallosus; 2 — Т. indrae (все подвиды).
18. Tabanus leleani Aust. — Слепень Лелеана.
Представлен двумя формами: более темным широко распространенным
номинативным подвидом Т. leleani leleani Aust. и более светлым, свойственным некоторым местам
Средней Азии, подвидом Т. leleani turkestanicus Ols.
18а. Tabanus leleani leleani Aust. (рис. 149) — Слепень Лелеана номинативный.
Austen, 1920, Bull. Ent. Res., X : 312; К г 6 b e r, 1924 : 172; К г б b e г
in Lindner, 1925 : 118; О л с у ф ь е в, 1937а : 262; М о и с h a et Chvala,
1959b : 281; Шевченко, 1961:147; L е с 1 е г с q, 1966:155; Виолович,
1968 :89; Chvala,- Lyneborg a. Moucha, 1972 : 338; К а д ы р. о в а,
1972а : 214 (биология); Соболева и Кадырова, 1972 : 46 (куколка).
$. Глаза голые, с 1 довольно широкой полоской посредине. Лобная полоска
широкая, расширяющаяся кверху, в пепельно-сером налете. Ее высота превосходит
ширину основания в 2 1/2—3 V2 раза. Нижняя лобная мозоль б. м. квадратная,
блестяще-черная, боковыми сторонами достигает края глаз. Средняя мозоль вполне
обособлена от нижней, имеет вид двойного черного пятна. Затылок умеренно утолщен,
с рядом слегка выдающихся светлых волосков. Лобный треугольник в пепельно-сером
налете. На уровне основания усиков имеется кофейно-коричневая полоска («уздечка»),
резко отделяющая серый лобный треугольник от белоснежного лица. Усики черные,
реже площадка 3-го членика в той или иной мере коричневая. 3-й членик умеренной
ширины, с б. м. тупоугольным дорсальным углом. Концевой членик щупалец
белоснежный, сильно утолщен, с острым кончиком. Нотоплевры желто-коричневые.
Крылья с легким сероватым оттенком, жилки коричневые; г4 без придатка. Ноги:
бедра черно-серые, основная часть передних голеней, средние и задние голени
коричнево-желтые; концевая часть передних голеней и лапки всех ног черные. Голени
задних ног иногда темные. Брюшко черно-серое, с боковыми, хорошо заметными рядами
серых косых пятен, по одному с каждой стороны тергита, и менее ясным срединным
рядом серых треугольничков. Тергиты по заднему краю с узкими желтовато-серыми
каемками. Брюшко снизу черно-серое, часто посредине слегка намечена более темная,
чем боковые стороны брюшка, продольная полоска. VIII стернит коричневый, с
темным гонопофизом, последний конический, с небольшой апикальной выемкой; латераль-
222
III. ПОДСЕМ. TABANINAE
ные лопасти небольшие; базальная лопасть с слабо вогнутым основным краем Цевки
с округлой вершиной. Сперматека миндалевидная. 11.5—15.5 мм.
3. Голова большая, полушаровидная. Глаза голые. Фасетки верхних 3/„ глаз
резко крупнее нижних. На границе крупных и мелких фасеток имеется широкая
резкая полоска. Затылок в слегка выдающихся над поверхностью глаз беловатых
Рис. 149. Tabanus Meant Meant Aust. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $. А — щупальце <J,
волосках. Лобный треугольник в нижней части серый, в верхней —коричневатый
На уровне основания усиков имеется б. м. резкая, кофейно-коричневая поперечная
полоска. Усики как у $, 3-й членик несколько более узкий. Концевой членик
щупалец белоснежный, сильно утолщен. Грудь и ее придатки как у $. Брюшко короткое
коническое, рисунок как у $. II тергит по бокам коричневый. 12—15 мм
Рис. 150. Ареалы: 1 — Tabanus Meant Meant; 2 — Т. Meant turkestanicus.
Очень похож на Т. cordiger, но $9 легко отличаются от этого последнего
благодаря наличию полоски на глазах. 33 отличаются с большим трудом, главным образом
более короткими и более светлыми волосками на затылке и более резкой и широкой
полоской на глазах. Т. Meant весьма близок также к Т. unifasciatus, настолько, что
их $5 иногда внешне почти не отличимы (терминалии различны !), в то время как 36
легко различаются по величине головы и размерам фасеток (см. указание при
описании Т. unifasciatus).
Распростран-ение: средиземноморско-азиатский предгорно (горно)-степ-
ной подвид (рис. 150, 1). В СССР распространен в восточном Предкавказье (Севере
8. TABANUS 223
Осетинская и Дагестанская АССР), Закавказье (Азербайджан, включая Нахичеван-
скую АССР, Грузия, Армения), Туркмении, Узбекистане (включая Каракалпакию),
Киргизии, юге и востоке Казахстана (включая Прибалхашье, Зайсан и р-н Зыряновска
Восточно-Казахстанской обл. и Жанаарка Карагандинской обл.), Алтайском крае
(окр. Змеиногорска). Балканы, Сев. Африка, Передняя Азия, Иран, Афганистан,
сев.-зап, Пакистан, Китай, Монголия. Местами весьма обычен. Конец
апреля—август, иногда сентябрь и позднее.
Личинки в берегах рек, оросительных каналах (арыках), ямах с водой, рисовых
полях и т. д., кладка яиц на листьях ивы, тутовых и других деревьев на высоте от 1.5
до 3 м и злаковых растений на берегу водоемов (Кадырова и Ган, 1967; Кадырова,
19726).
В горах Средней Азии распространен по степным участкам до 2600 м. Экземпляры
из Закавказья отличаются коричневым 3-м члеником усиков; у 66 волоски на затылке
более заметно выступают над поверхностью глаз. Указание Г. В. Бошко на
нахождение Т. Meant близ г. Остер Черниговской обл. нуждается в подтверждении. Вероятно,
распространен на юге Зап. Европы, но ошибочно принимался за какой-либо другой
вид, например, Т. unifasciatus или Т. cordiger.
18Ь. Tabanus leleani turkestanicus Ols. — Слепень Лелеана туркестанский.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 684. — leleani pallidus О 1 s u f j e v,
1937a : 264, пес Т. pallidus Palisot de Beauvois, 1809.
Отличается от основной формы более светлой окраской спинки и брюшка. У обоих
полов брюшко по бокам с коричневой окраской, простирающейся на I—III или I—
IV тергиты. Более крупные и резкие боковые светлые пятна из налета и волосков имеют
слегка желтоватую окраску и достигают заднего края тергитов. Срединные
треугольники более резкие, достигают вершиной переднего края тергитов. Заднекраиние
каемки более широкие. Брюшко снизу в основной половине просвечивает сквозь
густой беловатый налет розовато-коричневым. Ноги светлее: средние и задние голени
бледно-желтые, соответствующие им лапки коричневые. 5 13—15 мм, 6 13.5—15 мм.
Распространение: среднеазиатский подвид, свойственный долинам рек
пустынь южного типа (рис. 150, 2). Туркмения (Байрам-али, Теджен, Ашхабад),
Таджикистан (Гиссарская долина; Московский р-н и др.), Узбекистан (Бухара).
Май—август.
Голотип: $, Байрам-али, Туркмения, 1 V 1933, Богуш. В коллекции
Зоологического института Академии наук СССР, Ленинград.
19. Tabanus unifasciatus Lw. (рис. 151) — Слепень однополосый.
L о е w, 1858a, Verb. Zool.-bot. Gessell. Wien, 8 : 600; В г a u e г, 1880 : 202;
Surcouf, 1924:187; К г 6 b e г, 1924:171; Krober in Lindner, 1925:
131; Олсуфьев, 1937a : 264; С h v a 1 a, 1964 : 349; L e с 1 e г с q, 1966 : 175;
Chvala, Lyneborg a. Moucha, 1972:340.
$. Глаза голые, с 1 полоской. Лобная полоска умеренно широкая, кверху
немного расширена, покрыта серым налетом. Ее высота превосходит ширину основания
в 3 1/2—4 раза. Нижняя лобная мозоль б. м. квадратная, блестяще-черная, боковыми
сторонами достигает края глаз, средняя мозоль вполне обособлена от нижней, имеет
вид черного, округлого двойного пятна. Затылочная полоска умеренной ширины,
снабжена рядом выдающихся над глазами черных и частью светлых волосков. Лобный
треугольник серый. На уровне основания усиков имеется темно-коричневая
поперечная полоска («уздечка»). Усики целиком черные, 3-й членик с умеренно широкой
площадкой и тупоугольным дорсальным углом. Концевой членик щупалец белоснежный,
сильно утолщен, с острым кончиком, покрыт белыми и единичными черными
волосками. Нотоплевры черно-серые, реже желтовато-коричневые. Крылья прозрачные,
с легким сероватым оттенком; г4 без придатка. Ноги: бедра черно-серые, голени светло-
коричневато-желтые, передние голени в вершинной J/3 или V2 черные, последующие
голени с черноватыми кончиками. Лапки черные, иногда средние и задние лапки
в основании коричневатые. Окраска ног варьирует и встречаются экземпляры, у
которых задние и передние голени целиком черно-серые. Брюшко черное или черно-серое,
с боковыми хорошо заметными рядами небольших косых серых пятен, по одному с
каждой стороны. Вдоль средней линии брюшка намечается срединный, ряд неясных
сероватых треугольников. Тергиты по заднему краю с узкими желтовато-серыми каемками.
Брюшко снизу целиком черно-серое. Часто вдоль средней линии намечается неясная
более темная продольная полоска. VIII стернит: гонопофиз б. м. конический с слабой
апикальной выемкой; латеральные лопасти умеренные, темные; базальная лопасть
с слегка вогнутым основным краем. Церки с уплощенной вершиной. Сперматека
миндалевидная. 12—16 мм.
224
III. ПОДСЕМ. TABANINAE
в. Голова небольшая, не больше, чем у £. Глаза голые или в коротких,
малозаметных волосках, с 1 полоской. Фасетки глаз б. м. одинаковой величины или верхние
слегка крупнее нижних. Затылочная полоска слегка утолщена, в особенности возле
темени, и снабжена рядом довольно длинных, выдающихся над глазами, черных и
серых волосков. Лобный треугольник серый. Усики как у 5, но 3-й членик несколько
уже. На уровне основания усиков имеется темно-коричневая поперечная полоска.
Концевой членик щупалец белый, сильно утолщен, покрыт белыми и единичными
черными волосками. Грудь и ее придатки как у §. Брюшко коническое, его рисунок
как у 5- Боковые стороны II или II—III тергитов коричневатые. 14—15 мм.
66 этого вида резко отличаются от всех прочих видов группы Т. cordiger (кроме
Т. smirnovi) маленькой головой, с б. м. одинаковыми фасетками глаз. 59 весьма по-
Рис. 151. Tabanus unifasciatus Lw. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
хожи на Т. leleani и различаются лишь с трудом, главным образом по ширине лба.
Однако этот признак не всегда надежен и в сомнительных случаях точное определение
может быть произведено только по 66-
Распространение: южноевропейский (средиземноморский) лесной вид,
часто в горах. В СССР известен из западных и южных частей УССР (Великоберезнян-
ский р-н Закарпатской обл.; Жданов Донецкой обл.; Керчь Крымской обл.),
Молдавии, с Северного Кавказа (от Абрау-Дюрсо близ Новороссийска, на восток включая
Дагестанскую АССР), из Закавказья (Азербайджан, включая Нахичеванскую АССР,
Армения, Грузия) и Туркмении (Копет-Даг: Бахарденский и Кара-Калинский р-ны).
Средн. и 10. Европа (на запад до Португалии), Передняя Азия, Иран. Редок на
Украине, но на Кавказе местами обычен. Июнь—июль.
Указание Крёбера (Krober, 1924) на нахождение этого вида в окр. Кушки, по всей
вероятности, должно быть отнесено к Т. leleani.
20. Tabanus smirnovi Ols. (рис. 152). — Слепень Смирнова.
Олсуфьев, 1962, Энтом. обозр., 41, 3 : 577; L е с 1 е г с q, 1966 : 169;
Chvala, Lyneborg a. Moucha, 1972 : 336. — tauricus 0 1 s u f j e v,
1952 : 312; nee T. tauricus Knderlein, 1932; Б о ш к о, 1962 : 1251 (6).
$. Глаза голые (или в очень коротких волосках), без полоски. Лобная полоска
умеренно широкая, кверху немного расширена, покрыта серым налетом. Ее высота
превосходит ширину основания в 3 3/4—4 раза. Нижняя лобная мозоль б. м.
квадратная, блестяще-черная, боковыми сторонами достигает края глаз; средняя мозоль
в виде двойного черного пятна, обособлена от нижней мозоли. Затылочная полоска
умеренной ширины, с рядом выступающих над глазами черных и серых волосков.
Лобный треугольник серый. На уровне основания усиков имеется темно-коричневая
попеперечная полоска («уздечка»). Усики целиком черные, 3-й членик б. м. узкий,
с тупым дорсальным углом. Концевой членик щупалец беловатый, умеренно
утолщен, покрыт белыми и немногочисленными черными волосками. Нотоплевры черно-
серые. Крылья с незначительным сероватым оттенком, жилки черно-коричневые; г4
без придатка. Ноги: бедра черные, в сером налете, передние голени в базальной части,
последующие голени в большей своей части желто-коричневатые, но могут быть почти
целиком черноватыми. Брюшко черно-серое, II—V тергиты с хорошо заметными
небольшими серыми боковыми косыми пятнами. Средний ряд светлых треугольных
8. TABANUS 225
пятен отсутствует или имеются его следы. Брюшко снизу целиком черно-серое,
посредине имеется неясная, более темная продольная полоска. 11—15 мм.
3- Голова не больше, чем у J. Глаза голые, без полосок, фасетки глаз б. м.
одинаковой величины. Затылочная полоска слегка утолщена, в особенности возле темени,
и снабжена рядом довольно длинных, выступающих над глазами, черных волосков.
Лобный треугольник серый. Усики как у $, но 3-й членик немного уже. На уровне
основания усиков имеется темно-коричневая поперечная полоска. Концевой членик
щупалец яйцевидный, беловатый, покрыт белыми и единичными черными волосками.
Грудь и ее придатки как у $. Брюшко коническое, черное, с серым рисунком как у $;
II тергит по бокам иногда слегка коричневатый. 13.5—14 мм.
Похож на Т. unifasciatus, но легко отличается от последнего отсутствием полоски
на глазах и более узкими щупальцами.
, Распространение: горный Крым, где этот вид эндемичен. Июль—август,
иногда позднее.
Голотип: 5, долина Альмы, 30 км северо-западнее Алушты, 3 IX 1950, Н. Г.
Олсуфьев. В коллекции Зоологического института Академии наук СССР, Ленинград.
Рис. 152. Tabanus smirnovi Ols. Лоб- Рис. 153. Tabanus cordiger Mg. Лобная по-
ная полоска, усик и щупальце $. лоска, усик и щупальце $.
21. Tabanus cordiger Mg. (рис. 153) — Слепень широколобый.
М е i g e n, 1820, Syst. Beschreib., II : 47; В г а и е г, 1880 : 201; V е г г а 11,
1909:413; Surcouf, 1924:185; Krober, 1924:162; Krober in
Lindner, 1925 : 111; Олсуфьев, 1937а : 266; Moucha et Chvala, 1958 : 187;
L eel arc q, 1966:146; Chvala, Lyneborg a. Moucha, 1972:334.
5- Глаза голые, без полосок. Лобная полоска широкая, расширяющаяся кверху,
в сером или желтовато-сером налете; ее высота превосходит ширину основания в 2 V2—
3 V* раза. Нижняя лобная мозоль блестяще-черная, б. м. квадратная, боковыми
сторонами обычно достигает края глаз. Средняя мозоль в виде большого, черного,
округлого или двойного пятна, вполне обособлена от нижней; крайне редко мозоль
полностью прикрыта налетом. Затылочная полоска умеренно утолщена, с рядом
светлых и черных выступающих над глазами волосков. Лобный треугольник низкий,
в сером налете. На уровне основания усиков имеется коричневая поперечная полоска
(«уздечка»). Усики целиком черные; 3-й членик с довольно широкой площадкой,
дорсальный угол" б. м. прямоугольный. Иногда 3-й членик в основании
темно-коричневый. Концевой членик щупалец желтовато-белый, сильно утолщен, с острым концом,
покрыт беловатыми волосками, с примесью коротких черных волосков. Нотоплевры
желто-коричневые. Крылья бесцветные; г4 без придатка. Ноги: бедра черно-серые,
голени желто-коричневые; вершинная половина передних голеней черная, кончики
последующих голеней черноватые; лапки всех ног черные, лишь основания средних
и задних лапок коричневые. Брюшко черное, с двумя боковыми рядами небольших,
но хорошо заметных, косо расположенных серых пятен и срединным рядом неясных
сероватых треугольников. Тергиты по заднему краю с узкими желтовато-серыми
каемками. II тергит по бокам иногда слегка буроватый. Брюшко снизу черно-серое,
посредине часто с неясной темной полоской. VIII стернит (рис. 54, V, В) удлиненный,
15 Н. Г. Олсуфьев
226 III. ПОДСЕМ. TABANINAE
гонопофиз с небольшой апикальной выемкой, латеральные лопасти небольшие,
коричневые; базальная лопасть с вогнутым основным краем. Церки с округлой
вершиной. Сперматека миндалевидная. 13—17 мм, редко 10.5 мм.
в- Голова большая, полушаровидная. Глаза голые. Фасетки верхних 3Д, глаза
резко крупнее нижних, на границе тех и других обычно имеется узкая полоска.
Затылок в длинных, далеко выступающих над глазами коричневых или желтоватых
волосках. Лобный треугольник серый. На уровне основания усиков имеется
коричневая поперечная полоска. Усики как у $. Концевой членик щупалец желтовато-белый,
сильно утолщен, покрыт беловатыми волосками и единичными черными. Грудь и ее
придатки как у $. Брюшко коническое, его рисунок как у 5, II—III тергиты по бокам
коричневатые. Снизу брюшко черно-серое. 12—15 мм.
Рис. 154. Ареал Tabanus cordiger.
Распространение: европейский лесной вид, часто в горах (рис. 154).
Европ. часть СССР, на север до юга Карельской АССР (Медвежьегорск), Вельска
Архангельской обл., Удмуртской АССР, Ухты и Инты (66° с. ш.) Коми АССР, на восток
до Никито-Ивделя и Свердловска, на юг включая Молдавию, Закарпатскую,
Киевскую, Московскую обл., Татарскую, Башкирскую АССР и Оренбургскую обл., а после
перерыва в степной зоне распространен на Северном Кавказе в Краснодарском крае
(Туапсе,- Сочи, Майкоп, Кавказский заповедник), Кабардино-Балкарской, Северо-
Осетинской, Дагестанской АССР и в Закавказье — в Азербайджане (включая Нахи-
- чеванскую АССР), Грузии и Армении. Европа (кроме крайнего севера), Марокко,
Турция, Иран. Указания на распространение этого вида в Китае (Ouchi, 1943), на
п-ове Корея (Philip, 1956) и в Японии (Ricardo, 1911; Shiraki, 1918) ошибочны.
Редок, в горах Кавказа и в Ю. Урале местами обычен. Июнь—июль.
22. Tabanus golovi Ols. — Слепень Голова.
Представлен двумя формами: более темным, встречающимся в горах Средней Азии
номинативным подвидом Т. golovi golovi Ols. и более светлым, распространенным на
равнинах и низкогорьях среднеазиатским подвидом Т. golovi mediaasiaticus Ols.
22а. Tabanus golovi golovi 01.=. (рис. 155) — Слепень Голова номинативный.
Олсуфьев, 19376, Тр. Казахстан, фил. АН СССР, 2 : 163; 1937а : 267; М о-
ucha et Chvala, 1959b : 281; Шевченко, 1961 : 149; L e с 1 e г с q, 1966 :
150; Б а р а т о в, 1966 : 76 (8).
$. Глаза голые, темно-зеленые, без полосок. Лобная полоска широкая, ее высота
в 3—3 1/2 раза превосходит ширину, в желтовато-сером налете. Нижняя лобная
мозоль б. м. квадратная, блестяще-черная, не достигает края глаз, отделяясь узкими
серыми полосками. Средняя мозоль отсутствует (прикрыта налетом) или имеется,
округлая, черная, вполне обособлена от нижней. Усики черные, узкие, 2-й членик
. и немного основание 3-го членика коричневые. Затылок в виде относительно узкой
полоски и в коротких волосках. Лобный ■ треугольник желтовато-серый. Концевой
8. TABANUS 227
членик щупалец утолщен, белый, покрыт белыми и черными волосками. На уровне
основания -усиков имеется желтоватая или коричневатая полоска. Нотоплевры
коричневые. Крылья бесцветные, жилки черно-коричневые; г4 без придатка. Ноги:
бедра черно-серые, голени желтые, с черноватыми кончиками, лапки черные. Брюшко
черное, сверху с 3 рядами серых пятен: треугольных, достигающих вершиной
переднего края тергитов, посредине и косо-овальных по бокам. Тергиты по заднему краю
с узкими, желтовато-серыми каемками. Снизу брюшко пепельно-серое. VIII стернит
короткий, гонопофиз крупный с апикальной выемкой; латеральные лопасти умеренно
развиты, слабо хитинизированы; базальная лопасть с прямым основным краем. Церки
с округлой вершиной. Сперматека миндалевидная. 15—18 мм.
Рис. 155. Tabanus golovi golovi Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека 2-
6 (по Баратову, 1966, сокращенно). Голова большая, но меньше, чем у Т. golovi
mediaasiaticus, выпуклая. Глаза голые. Фасетки верхних 3/4 глаз приблизительно в 5 раз
крупнее нижних. Затылок за глазами в коротких серых волосках. Возле темени
волоски слегка выдаются над поверхностью глаз. Усики: 1-й и 2-й членики желтые,
3-й членик узкий, желтовато-коричневый. Щупальца белые, концевой членик сильно
утолщен, покрыт длинными белыми и единичными короткими черными волосками.
На уровне основания усиков имеется легкая желтизна. Нотоплевры коричневато-
желтые. Крылья прозрачные; г4 без придатка. Ноги: бедра черно-серые, голени
желтые, вершина передних голеней и лапки черные, лапки последующих ног коричневые.
Брюшко темно-серое, с 3 рядами светло-серых пятен, треугольных.посредине и косо-
овальных по бокам. По бокам II тергита имеются светло-коричневые пятна. Тергиты
по заднему краю с желтовато-серыми каемками. Брюшко снизу: I—III стерниты
розовато-желтые и в сером налете, остальные темно-серые. 16 мм.
Напоминает Т. cordiger, но 5 отличается от последнего несколько более узким
лбом, менее резкой коричневой полоской возле основания усиков, отсутствием, как
правило, средней мозоли и строением терминалий. в легко отличается короткими
волосками на затылке.
Распространение: среднеазиатский горный подвид. Таджикистан
(хребты Гиссарский, Каратегинский, Дарвазский на юг до Московского р-на),
Узбекистан (Сурхандарьинская, Ташкентская, Андижанская области, Бируни
Каракалпакской АССР), Киргизия (Чаткальская, Ферганская долины, долина р. Кекемерен),
Казахстан (Заилийский и Джунгарский Алатау). Зап. Китай (Кульджа), Афганистан.
Обычен в южных частях ареала. Май—август, редко раньше или позже.
Голотип: 5> 0КР- Алма-Ата, 8 VII 1934, Н. Олсуфьев. В, коллекции
Зоологического института Академии наук СССР, Ленинград. <
15*
228 HI. ПОДСЕМ. TABANINAE
22b. Tabanus golovi mediaasiaticus 01s. (pnc. 156) — Слепень Голова
среднеазиатский.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 684. — golovi pallidas Олсуфьев.
19376 : 164. *
$. Отличается от основной формы более светлой окраской и несколько меньшими
размерами. Лобная мозоль и усики коричневые. Брюшко сверху с нерезко
отграниченными желтовато-коричневыми боковыми пятнами, снизу брюшко в основной половине
желтовато-коричневое.
Экземпляры из Туркмении (Тахта-Базар, Имам-Баба и др.) отличаются наиболее
светлой окраской: бедра желтые, усики коричнево-желтые, щупальца целиком в
белых волосках. Брюшко
светло-коричневое, с более темной срединной полосой,
посредине которой расположен
продольный ряд серых треугольников. Крылья
с коричневыми жилками. Жужжальца
с почти целиком беловатой головкой.
14—17 мм.
<?. Голова большая, сильно
выпуклая. Глаза голые. Фасетки верхних 3/i
глаз значительно крупнее нижних,
приблизительно в 5—6 раз. Затылок в
коротких светлых волосках. Усики узкие,
коричнево-желтые. Щупальца белые,
концевой членик утолщен, в белых
волосках. Лобный треугольник серый.
Нотоплевры желто-коричневые. Крылья,
ноги, жужжальца и брюшко как у §•
14—16 мм.
Распространение:
предгорья и низкогорья гор Средней Азии,
местами до 2000 м, встречаясь вместе
с номинативным подвидом; по долинам
крупных рек южных частей
пустынной зоны. Южная Туркмения,
Таджикистан, Узбекистан (на север до
Рис. 156. Tabanus golovi mediaasiaticus
Ols. <J.
Шаббаза Каракалпакской АССР), Киргизия (Чуйская, Таласская, Ферганская
долины, пойма р. Нарына в окр. Казармана), Южный Казахстан (на север до Каза-
линска и дельты р. Или). Зап. Китай (Синьцзян). Обычен. Май—август, иногда
сентябрь. Места выплода: реки, каналы (арыки), рисовые чеки (Кадырова и Ган, 1967).
Г о л о т и п: 5. пос- Илийский Алма-Атинской обл., 4 VIII 1934, Н. Олсуфьев.
В коллекции Зоологического института Академии наук СССР, Ленинград.
23. Tabanus fumidus Aust. (рис. 157) — Слепень дымчатый.
Austen, 1923, Bull. ent. Res., 13 : 281; L e с 1 e г с q, 1966 : 149. — turpis
Bogatshev et Samedov, 1949 : 70; — sordes (Bog. et Sam.) apud H a u s e r,
1953 : 250. *
?. Глаза голые, без полосок) Лобная полоска умеренно широкая, ее высота
в 3 1/2—4 раза превосходит ширину основания, с почти параллельными сторонами,
в светлом желтовато-сером налете. Нижняя лобная мозоль б. м. квадратная, блестяще-
черная или темно-коричневая, отделена от края глаз очень узкой полоской налета;
средняя лобная мозоль черная, овальная, иногда с выемкой вверху, отделена от
нижней. Лобный треугольник в желтовато-сером налете, у основания усиков желтизна
выражена сильнее. Усики узкие, оранжево-желтые, 3-й членик с незначительным
дорсальным углом. Концевой членик щупалец беловатый, в основании значительно
утолщен, в белых волосках. Затылочная полоска умеренно утолщена, в относительно
коротких, едва выступающих над поверхностью глаз, серых волосках. Нотоплевры
коричневато-желтые. Крылья бесцветные, жилки желтые; г4 с придатком. Ноги:
бедра желто-коричневые или черноватые, в сером налете, голени желтые, вершинная
1/3 передних голеней и лапки черные, последующие лапки коричневые. Брюшко сверху
.посредине с широкой темной полосой, по бокам I—V или I—VI тергитов с желтовато-
5. TABANUS
229
розовыми пятнами, а также с 3 рядами нерезких сероватых пятен налета и светлых
волосков: треугольных посредине и б. м. ромбических по бокам. Снизу брюшко
желтовато-розовое, концевая часть сероватая. 13.5—16 мм.
8. Голова значительно крупнее, чем у $, полушаровидная. Глаза голые,
фасетки верхних 3/4 глаза приблизительно в 5—6 раз крупнее нижних. Затылок в
коротких, серых волосках, едва выступающих над поверхностью глаз лишь возле^ темени.
Лобный треугольник в желтовато-сером налете, в верхней части коричневатый. Усики
как у 5, но 3-й членик более узкий. Концевой членик щупалец белый, продолговато-
овальный, в белых волосках. Грудь и ее придатки как у $. Брюшко коническое,
рисунок как у $, но сверху продольные ряды серых пятен более четкие. 13.5 мм.
Распространение: переднеазиатско-туранский вид долин пустынно-
степных рек. Азербайджан (долина р. Алазани; Самух на Куре; Ордубад и Дасты
Нахичеванской АССР); Армения (Мег-
■^^m^m^ii^^ ^?. ри); Туркмения (Серахс);
Каракалпакия (Бадайтугай Бирунийского р-на).
Иран, Афганистан, Ирак. Редок. Июнь-
август.
^ШШШ|1ИЩ!;Шш.
Рис. - 157. Tabanus fumidus Aust.
Лобная полоска, усик и щупальце $.
Рис. 158. Tabanus kinoshitai Kono
et Так. Лобная полоска и щупальце $.
24. Tabanus kinoshitai Kono et Так. (рис. 158) — Слепень Киношита.
Kono et Takahasi, 1939, Zool. Mag. Tokyo, 51:759; L e с 1 e г с q,
1966 : 154; Виолович, 1968 : 90. — cordiger (Mg.) apud Ricardo, 1911 : 239
(partim).
5. Глаза голые, без полосок. Лобная полоска широкая, ее высота превосходит
ширину основания в 3—3 Va раза, с почти параллельными сторонами, в сером налете.
Нижняя лобная мозоль квадратная или поперечная, блестяще-черная, боковыми
сторонами достигает края глаз. Средняя лобная мозоль черная, большая,
продолговато-овальная, б. м. изолирована от нижней мозоли. Лобный треугольник блестяще-
черный. Усики черные или темно-коричневые, узкие, 3-й членик с тупым дорсальным
углом. На уровне основания усиков по верхнему краю щек имеется кофейно-коричне-
вая поперечная полоска («уздечка»). Концевой членик щупалец желтовато-белый,
в основании значительно утолщен, в беловатых и черных волосках. Нотоплевры желто-
коричневые. Крылья бесцветные; vt без придатка. Бедра черно-серые, голени желто-
коричневые, концевая 73 передних, конец средних и задних голеней, а также лапки
черные. Брюшко черное, сверху с 3 рядами серых пятен: треугольных посредине
и округлых по бокам. Снизу брюшко черно-серое с неясной черноватой срединной
полосой. 14—16 мм.
3 (по Takahasi, 1962, сокращенно). Голова большая, крупные фасетки занимают
2/3 глаза, граница с мелкими фасетками явственная. Лобный треугольник блестяще-
черный, выпуклый. Концевой членик щупалец овальный, белый, в длинных белых
волосках. Брюшко как у $; И—III (иногда также IV) тергиты по бокам с
коричневыми пятнами. Брюшко снизу серое. 13—14 мм.
От всех видов группы Т. cordiger $c? рассматриваемого вида легко отличаются
блестяще-черным лобным треугольником.
Распрост р а н е н и е: представитель фауны Восточноазиатской подобласти.
В СССР найден на о. Кунашир, вулкан Раусуяма (Конаков). Япония (о-ва Хоккайдо,
Хонсю, Кюсю). Редок. Июнь—июль.
III. Группа Tabanus bromiusL.
Средняя лобная мозоль б. м. веретеновидная и обычно соединена с нижней
мозолью; последняя б. м. прямоугольная или щитовидно-треугольная. Лобная полоска
относительно узкая, кверху слабо расширена. Глаза голые, реже в волосках, с по-
230
III. ПОДСЕМ. TABANINAE
лоскаыи или без таковых. Крылья бесцветные; г4 без придатка (как исключение —
иногда с небольшим рудиментом и очень редко придаток хорошо развит — Т. hissa-
ricus Bar.). Средних размеров или небольшие виды, распространенные в Евразии
преимущественно в лесном (горно-лесном) или степном ландшафтах; ряд видов в Сев.
Америке.
25. Tabanus rupium Br. (рис. 159) — Слепень скалистый.
В г а и е г, 1880, Denkschr. Akad. Wissensch. Wien, 42 : 163; К г б Ь е г, 1923:
93; S u г с о u f, 1924 : 105; Krober in Lindner, 1925 : 69; Олсуфьев,
1937a : 269; L e с 1 e г с q, 1968 : 167; Chvala, Lyneborg a. Moucha,
1972 : 307. — abazus Bigot, 1880 : 146. — alazinus Bigot, 1892 : 647.
$. Глаза в коротких, но явственных серых волосках, с рудиментарной полоской.
Лобная полоска умеренно широкая, высота приблизительно в 3—3 1/2 раза
превосходит ширину, параллельная или слегка расширяющаяся кверху, в сером налете.
Затылок за глазами в виде относительно
широкой полосы, в длинных, выступающих над
поверхностью глаз, черных и светлых
волосках. Нижняя лобная мозоль черная,
слабо морщинистая, плоская, б. м.
прямоугольная, боковыми сторонами почти
достигает края глаз. Нижний край мозоли
обычно с 3 выступами. Средняя мозоль узко-
веретеновидная, черная, упирается в
нижнюю мозоль. Лобный треугольник и верхний
край щек в буровато-сером налете. Усики
черные; 3-й членик узкий, с небольшим
Рис. 159. Tabanus rupium Br. Лобная
полоска, усик и щупальце §.
дорсальным углом. Концевой членик щупалец светло-коричневый, в основании
утолщен, б. м. остроконечный, с внешней стороны равномерно покрыт короткими черными
волосками. Нотоплевры черные. Крылья бесцветные, жилки черно-коричневые; г4
без придатка. Ноги черные, большая часть средних и задних голеней и основание
передних голеней темно-коричневые. Иногда задние голени целиком черные. Брюшко
черное, сверху с срединным рядом небольших серых пятен (налет-|-волоски), по
бокам I—II (или I—IV) тергитов по небольшому серому пятну. Изредка II тергит по
бокам слегка коричневатый. Снизу брюшко черное, в голубовато-сером налете. 12—
14.5 мм.
<?. Голова немного больше, чем у $. Глаза в густых и длинных коричневато-серых
волосках. Фасетки верхних 2/3 глаз немного крупнее нижних, переход постепенный,
на границе одна полоска. Линия соприкосновения глаз превосходит высоту лобного
треугольника не более чем в 1.5 раза. Последний в сером налете. Затылок за глазами
в длинных, черных и серых, вперед изогнутых волосках. Концевой членик щупалец
желтоватый, в основании серый, умеренно утолщен, приблизительно в 1.5 раза
длиннее наибольшей толщины, тупоконечный, покрыт сероватыми волосками с примесью
черных. Усики как у §, отличаясь более узким 3-м члеником. Волоски на 1-м членике
длиннее его самого. Грудь и ее придатки как у $• Средние и задние голени
темно-коричневые, с черноватым концом, благодаря опушению из черных и серых волосков выглядят
темнее, передние голени черные, в основании темно-коричневые. Брюшко
черное, сверху с нерезким беловато-серым рисунком как у §. II и частично III тер-
гиты по бокам коричневатые. Снизу брюшко в сером налете, с неясной темной
срединной полоской, задний край стернитов с узкой коричневато-сероватой полоской. 12—
12.5 мм (по Крёберу, 15 мм).
От всех видов группы Т. bromius L. легко отличается наличием волосков на глазах.
Распространение: европейско-кавказский горно-лесной вид. В СССР
известен с Большого Кавказа (от сев. и ю. отделов Кавказского заповедника и
Абхазии на восток до Дагестана и Закатал), Малого Кавказа — в пределах Грузии (Ба-
куриани, Аспиндза, окр. Батуми), Армении (Дилижан, оз. Севан, Мегринский р-н) и
Азербайджана (Нахичеванская АССР: Ордубад); Талыш. Горы Средн. и Ю. Европы,
Турция. Местами обычен. Июнь—начало августа.
26. Tabanus maculicornis Ztt. (рис. 160) — Слепень черносерый.
Zetterstedt, 1842, Dipt. Scand.; 1 : 117; В г a u e r, 1880 : 197; Lund-
beck, 1907:131; V е г г а 1 1, 1909:410; Surcouf, 1924:180; Krober,
8. TABANUS 231
1924 : 152; Krober in Lindner, 1925 : 118; Олсуфьев, 1937а : 271;
Moucha et Chvala, 1958:188; Шевченко, 1961:151; Leclercq,
1966 : 157; Виолович, 1968 : 91; И в а н и щ у к, 1970 : 142 (личинка);
Chvala, Lyneborg a. Moucha, 1972 : 360.
$. Глаза голые, с 1 полоской. Лобная полоска узкая, ее высота превосходит
ширину основания в 4—4 V2 раза, покрыта желтовато-серым налетом. Нижняя лобная
мозоль черная, треугольно-щитовидная, мелкоморщинистая. Ее нижний край обычно
с 3 выступами: небольшими по бокам и более широким посредине. Средняя мозоль
веретеновидная, соединена с нижней мозолью. Лобный треугольник желтовато-серый.
Затылочная полоска утолщена, снабжена слегка выдающимися над глазами
желтовато-серыми волосками. Усики коричневые или желто-коричневые, палочка и 1-й
членик часто черноватые. 3-й членик с довольно широкой площадкой и прямым или
тупым дорсальным углом. Концевой членик щупалец умеренно утолщен, светло
желтовато-серый, в коротких, преимущественно черных волосках. Нотоплевры черные,
иногда коричневатые. Ноги черные, бедра в сером налете, основание передних и боль-
Рис. 160. Tabanus maculicornis Ztt. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $.
шая часть средних и задних голеней коричнево-желтые. Крылья прозрачные, с легким
сероватым оттенком; г4 без придатка. Брюшко черное, с неясным серым рисунком
из 3 продольных рядов небольших пятен: треугольных посредине и неправильно
прямоугольных по бокам. Лишь иногда рисунок бывает выражен б. м. резко (у
совершенно непотертых экземпляров). Светлые пятна в беловато-серых, прочие части
•брюшка в черных, прилегающих волосках. Снизу брюшко одноцветно-черно-серое:
VIII стернит преимущественно темный, гонопофиз крупный, с небольшой апикальной
выемкой; латеральные лопасти небольшие; базальная лопасть с прямым основным
краем. Церки с слегка уплощенной вершиной. Сперматека миндалевидная. 10—14.5 мм.
$. Голова больше, чем у $, полушаровидная. Глаза голые, с одной полоской
на границе мелких и крупных фасеток. Крупные фасетки резко отличаются по
величине от мелких и занимают верхние 2/3 глаза. Затылок за глазами утолщен и снабжен
рядом длинных, выдающихся над поверхностью глаз, желтовато-серых волосков.
Лобный треугольник у вершины темно-серый, в остальной части в коричневато-сером
налете. Концевой членик щупалец узкоовальный или ланцетовидный,
светло-желтовато-серый, в длинных серых и черных волосках. Усики как у $, отличаясь более
узким 3-м члеником. Грудь и ее придатки как у $. Брюшко черное, Н—III (илнтолько
II) тергиты по бокам с незначительными коричневыми пятнами. Задние каемки
тергитов по боковым сторонам и небольшие срединные треугольники в серых, прочие части
<брюшка в черных, б. м. длинных волосках. Снизу брюшко черное, в серых волосках;
задние края стернитов с узкими, желтовато-серыми каемками. 11.5—14.5 мм.
Небольшой черно-серый вид. Похож на Т. bromius, но 5 легко отличается
утолщенной, затылочной полоской (рис. 125, слева) и более темным брюшком, а $•—
наличием на затылке длинных волосков.
Распространение: европейско-западносибирский лесной вид (рис. 161,1);
ранее мы неправильно считали его таежным видом (Олсуфьев, 1937а). В СССР от
Карелии и запада Украины на восток до Хакасской АО и Алтая, на север до Мурманской
232
III. ПОДСЕМ. TABANINAE
™- (£p Л? }' ^? ACCP (УхТ^' севеРа 0мской обл- и Томска, на юг до
Молдавской ССР Кагул, Котовск и др.) Воронежской, Оренбургской областей (Бузулук),
Ю. Урала (Иргизла), Тары Омской обл., Семипалатинска, хр. Саура и Джунгарского
Алатау (Саркандскии р-н); изолированно обитает в Закавказье: окрест, г Батуми
Ьакуриани, Талыш. Европа (на запад до Испании). Указание о нахождении этого
Рис. 161. Ареалы: 1 _ Tabanus maculicornis; 2 — Т. geminus.
™£LHhP" ЛЯПИН' СеВ- УраЛ (64> ^ и на юге Одесской обл. нуждаются в
подтверждении. Несомненно неправильно Н. А. Виолович (1968) указал Г. maculicornis для
Супутинского заповедника Приморского края. "<-<"«« для
Весьма обычен на северо-западе,, в средней полосе европейской части СССР местами
на Урале и Алтае. Июнь-июль. Личинок обнаруживали по берегам речек ручьев
т^ИТНЫХйЧП^еХ0ДНЫХ б£ЛОтах (Лутта' 1970>' заболоченных низинах на пастбище
(Олсуфьев, 19356) и т. д Куколок находили в заболоченных берегах ручья "реки
и на заболоченном лугу (Лутта, 1970). уу'«•игреки
ШШуьШШщжшшш
I'lllll'/,'
щ
I'll'
c^^ko
Рис. 162. Tabanus geminus Szil. Лобная "полоска, усик,
щупальце, VIII стернит, церки и сперматека $.
27. Tabanus geminus Szil. (рис. 162) — Слепень парный.
<н ч. Sn * * а dJ' im\ J£0l08A Нш12аг" 1, 1 : 3; К г 6 Ь е г in Lindner, 1925
113; Олсуфьев, 1937а : 272; Шевченко, 1961:152; L е с 1 е г с q 1966
149; Виолович, 1968:92; Murdoch et Т a k a h a s i, 1969 • 79
8. TABANUS 233
§. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания в 3 */2—4 раза, в сером налете. Нижняя лобная мозоль блестяще-
черная, расположена отступя от границы лба и лобного треугольника, боковыми
сторонами не достигает края глаз. Форма мозоли б. м. прямоугольная или щитовидно-
треугольная, ее нижний край часто с 3 выступами. Средняя мозоль блестяще-черная,
короткая, веретеновидная, часто не соединена концом с вершиной нижней мозоли.
Лобный треугольник в буровато-сером налете. Затылочная полоска утолщена, в слегка
выдающихся над глазами серых и черных волосках. Усики оранжево-желтые, 3-й чле
ник с умеренно широкой площадкой и прямо- или тупоугольным дорсальным углом.
Концевой членик щупалец слабо утолщен, бледно-желтовато-серый, в коротких,
преимущественно черных волосках. Нотоплевры темные. Крылья бесцветные; г4 обычно
без придатка. Ноги: бедра черно-серые, голени коричневато-желтые, вершинная
половина передних голеней, кончики последующих голеней и лапки всех ног черные.
Брюшко черное, с серыми каемками (налет-j-волоеки) по заднему краю тергитов,
расширенными в небольшие срединные треугольники; боковые продольные ряды
серых пятен слабо выражены и часто последние заметны лишь на II тергите. Снизу
брюшко целиком черно-серое, стерниты по заднему краю с узкой желтоватой каемкой.
11—12 мм.
cj. Голова немного больше, чем у $, полушаровидная. Глаза голые. Фасетки
верхних 2/3 глаз значительно крупнее нижних, с резкой границей между теми и
другими, вдоль которой расположена неширокая полоска. Затылок слегка утолщен,
снабжен рядом длинных, выдающихся над поверхностью глаз черных и частью серых
волосков. Лобный треугольник коричневый, в нижней части в сером налете. Усики
оранжево-желтые, 3-й членик более узкий, нежели у $• Концевой членик щупалец
бледно-желтоватый, овальный, покрыт длинными серыми и черными волосками.
Грудь и ее придатки как у $. Брюшко коническое, окрашено как у $, но II или II—
III тергиты по бокам коричневатые. 12 мм.
Похож на Т. maculicornis, но отличается меньшей величиной и целиком оранжево-
желтыми усиками; кроме того, 5$ легко отличаются по отсутствию полоски на глазах
и по строению лба, а 36 — несколько более короткими, преимущественно черными
волосками на затылке.
Распространение: восточносибирский лесной вид, аллопатричен
Т. maculicornis (рис. 161, 2). От Алтая, Томска и Тогучина Новосибирской обл. на
восток до Приморского края, на север до окр. Красноярска, Сев. Прпангарья (Усоль-
цево, 59° с. ш.), Кумара на Амуре и окр. Хабаровска, на юг до Тувы, Иркутска и Ха-
санского р-на Приморского края. Монголия (Булганский аймак), Сев.-Вост. Китай.
Обычен в Приморском крае, в западных частях ареала не часто. Июль—август.
28. Tabanus miki Br. — Слепень Мика.
Изменчивый вид, представлен 3 формами: номинативным широко.
распространенным подвидом Т. miki miki Br. и двумя горными подвидами: Т. miki colchidicus
Ols. и Т. miki australis Haus.
28a. Tabanus miki miki Br. (рис. 163) — Слепень Мика номинативный.
В г a u e r, 1880, Denkschr. Akad. Wissensch. Wien, 42 : 195; Lundbeck,
1907 : 130; S u г с о u f, 1924 : 179; Krobet, 1924 : 152; Krober in
Lindner, 1925:119; О л с у ф ь е в, 1937а : 274; Moucha etChvala, 1958:190;
Шевченко, 1961 : 153; L е с 1 е г с q, 1966 : 158; Виолович, 1968 : 92;
Иваяищук, 1970 : 143 (личинка); Chvala, Lyneborg a. Moucha,
1972 : 349.
5. Глаза голые, без полосок. Лобная полоска узкая, ее ^высота превосходит
ширину основания в 4—5 раз, покрыта желтовато-серым налетом. Нижняя лобная
мозоль треугольно-щитовидная, черная или черно-коричневая, часто в тонких
морщинках. Боковыми сторонами мозоль не достигает края глаз. Средняя мозоль веретено-
видная или копьевидная, соединена тонкой линией с вершиной нижней мозоли.
Затылочная полоска слегка утолщена, с рядом преимущественно черных, волосков (реже
целиком желто-серых), слегка выдающихся над поверхностью глаз (смотреть спереди!).
Лобный треугольник высокий, в желтовато-сером налете. Концевой членик щупалец
палево-желтый, в основании утолщен, к концу равномерно сужен, б. м. остроконечный,
покрыт короткими, преимущественно черными волосками. Усики черные; 1-й, 2-й
и основание 3-го членика часто коричневые. 3-й членик в основании умеренно
расширен, с тупоугольным дорсальным выступом. Нотоплевры черные. Крылья бесцветные;
г4- без придатка. Ноги : бедра черные, в сером налете, их кончики, основная половина
передних и большая часть средних и задних голеней желто-коричневые, кончики
голеней и основание лапок последних двух пар ног коричневые, прочее черное.' Брюшко
черно-бурое, сверху с коричневыми боковыми пятнами на I—III (или I—V) тергитах
и с темной срединной полосой, занимающей х/з ширины брюшка. Боковые стороны
234
III. ПОДСЕМ. TABANINAE
полосы, в пределах каждого кольца, полукругло вырезаны. II—V тергиты с 3 неясными
желтовато-серыми рядами пятен: треугольными посредине и округлыми или косыми
по бокам. Пятна и задние каемки тергитов в желтовато-серых, прочие части брюшка
в черных коротко прилегающих волосках. Снизу брюшко в основной половине
желтовато-коричневое, последние 3—4 стернита темные. VIII стернит: гонопофиз широкий
Рис. 163. Tabanus miki miki Br. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
с апикальной выемкой; латеральные лопасти умеренно развиты, коричневые;
базальная лопасть с слегка вогнутым основным краем. Церки с округлой вершиной.
Сперматека миндалевидная. 14—16 мм.
cJ. Голова немного крупнее, нежели у 5- Глаза коричневые, с одной полоской,
расположенной на границе крупных и мелких фасеток. Фасетки верхних 2/3 глаз
0° К ВОСТОКУ ОТ ГрИНВИЧа 90° 120= 150° ' 130° 150°
Рис. 164. Ареалы: 1 — Tabanus miki (все подвиды); 2 — Т. baddha.
значительно крупнее нижних (примерно в 4 раза), граница между теми и другими
резкая. Затылок за глазами с рядом слегка выступающих над поверхностью глаз
черных и светлых волосков. Лобный треугольник в желтовато-сером налете, возле
вершины буроватый. Концевой членик щупалец палево-желтый, овальный, покрыт
б. м. длинными, преимущественно черными волосками. Усики как у §, отличаясь
меньшей шириной 3-го членика. Грудь и ее придатки как у 5- Брюшко коническое, корич-
8. TABANUS
235
невые пятна захватывают I—V тергиты, срединная черная полоса на II тергите
занимает приблизительно 1/7 его ширины, на последующих тергитах расширяется.
Остальное как у 5- 15—16 мм.
Похож на Т. bromius, но отличается: $ по отсутствию полоски на глазах, 3 —
по наличию за глазами черных выдающихся волосков.
Распространение: европейско-спбирский лесной подвид (рис. 164, 1).
В СССР от Прибалтики и Молдавии на восток до Иркутской обл. (Братск, Зима),
на север до окрест. Сортавала Карельской АССР, Иванова, Перми, севера Омской обл.
и Енисейска Красноярского края, на юг до Николаева УССР, Крыма, Сев. Кавказа
и Закавказья (Азербайджан, включая Нахичеванекую АССР; Армения; Грузия),
10. Урала (Стерлитамак, Салават), Челябинской обл. (Троицк) и Семипалатинска.
Средн. и 10. Европа, Турция. Указание Г. А. Бабаянца (1962) на нахождение в
Туркмении ошибочно. Многочислен в подзоне южной тайги и лесной зоне Западной Сибири,
в других частях ареала не часто. Июнь—июль.
28b. Tabanus miki colchidicus Ols. — Слепень Мика колхидский.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 684. — miki niger О 1 s u f j e v,
1937a : 275.
$ этого вида отличается от номинативной формы целиком черным брюшком и
слегка уплощенными церками, у 3 боковые коричневые пятна на брюшке расположены
на II—III тергитах и лишь слегка захватывают I тергит, не заходя на 4-е кольцо.
Затылочная полоска несколько более утолщена и снабжена несколько более длинными,
чаще только черными волосками. Усики целиком черные, реже 3-й членик в основании
слегка коричневый. Передние и задние голени иногда целиком черно-серые. Пятна
сверху брюшка (волоскиЦ-налет) обычно серые. 5 14—16 мм, 3 15—15.5 мм.
2 похожа на Т. maculicornis, отличаясь,отсутствием полоски на глазах и^черными
волосками на затылке. 3 мало отличается по внешнему виду от номинативной формы.
Распространение: горно-лесной подвид. Горный Крым; Карпаты;
Большой Кавказ: Кавказский заповедник (сев. и ю. отделы), Теберда, Абхазия,
Казбек, Кусарский р-н Азербайджана; Малый Кавказ: Грузия — Бакуриани; Аджарская
АССР; Талыш. Не часто. Июль—август.
Голотип: 5, Кавказ — Казбек, из коллекции Порчинского. В коллекции
Зоологического института Академии наук СССР, Ленинград.
28с. Tabanus miki australis Haus. — Слепень Мика южный.
Г а у з е р, 1960, Энтомол. обозр., 39, 3 : 657; L е с 1 е г с q, 1966 : 159.
5. От номинативной формы отличается меньшими размерами тела — 12—14 мм,
! преимущественно черной окраской брюшка — коричневые пятна по бокам имеются
I только на II—III (или II—IV) тергитах, светлые волоски сверху брюшка серые.
Распространение: Азербайджан (Ханларский р-н; Нахичеванская
АССР — Шахбузский р-н); Армения (Мегринский р-н); Грузия (Бакуриани). Иран.
Не часто. Июль.
29. Tabanus indrae Haus. — Слепень бесполосый.
Изменчивый вид, известны 3 географических формы: Т. indrae indrae Haus.,
Т. indrae vappa Bog. et Sam. и Т. indrae montivagus Ols.
1 29a. Tabanus indrae indrae Haus. (рис. 165) — Слепень бесполосый номинативный.
Г а у з е р, 1939, Тр. Зоол. инст. Азерб. фил. АН СССР, X : 142; Leclercq,
1966 : 153; Олсуфьев, 1970: 683; Chvala, Lyneborga. Moucha,
1972 : 351.
5. Глаза голые, без полосок. Лобная полоска относительно узкая, ее высота
превосходит ширину основания приблизительно в 4 1/2—5 раз, кверху слегка
расширена, покрыта желтовато-серым налетом. Нижняя лобная мозоль б. м. прямоугольная,
блестяще-черная или темно-коричневая, боковыми сторонами обычно немного не
достигает края глаз. Средняя мозоль черная, веретеновидная, концом обычно достигает
нижней мозоли. Лобный треугольник в желтовато-сером налете. Усики коричневые,
палочка черноватая, дорсальный угол 3-го членика б. м. прямой или тупой.
Затылочная полоска узкая, в коротких, едва выступающих над поверхностью глаз,
преимущественно светлых волосках. Концевой членик щупалец палевый, в основании
утолщен, покрыт преимущественно черными волосками. Нотоплевры черные или
коричневые; Крылья бесцветные; г4 без придатка (или с его рудиментом). Ноги: бедра черно-
серые, базальная часть передних, средние и задние голени желто-коричневые, конец
передних голеней и передние лапки черные, последующие лапки коричневые. Брюшко
черно-серое, боковые стороны II—III тергитов коричневые. Брюшко сверху с 3 ря-
236
III. ПОДСЕМ. TABANINAE
дами сероватых пятен (налет +волоски): треугольных посредине и овальных или
ромбических по бокам. Снизу брюшко серое, в основании желтовато-коричневое. VIII стер-
нит желтый, гонопофиз б. м. конический, с апикальной выемкой; латеральные
лопасти умеренно развиты, слабо хитинизированы к вершине; базальная лопасть с
вогнутым основным краем. Церки с округлой вершиной. Сперматека веретеновидная.
11.5—17 мм.
сУ. Голова немного больше и выпуклее, чем у $. Фасетки верхних 2/3 значительно
крупнее нижних, приблизительно в 4—5 раз, граница между крупными и мелкими
фасетками резкая. Имеется одна полоска. Затылок узкий, в коротких серых волосках.
Лобный треугольник в желтовато-сером налете, в верхней части более темный. Усики:
1-й членик желтоватый или серый, сверху, особенно кончик, в коротких черных
волосках, 3-й членик черноватый, с коричневым основанием, узкий, дорсальный выступ
тупоугольный. Концевой членик щупалец слегка желтоватый, с тупым концом, в.2 раза
Рис. 165. Tabanus indrae indrae Hans. Лобная полоска, усик, щупальце, VIII стернит
и сперматека $. А — Т. indrae montivagus Ols. VIII стернит, церки и сперматека $
длиннее наибольшей толщины, покрыт умеренно длинными светлыми и черными
волосками. Грудь и ее придатки как у $. Брюшко черное, с темно-коричневыми
боковыми пятнами на II—III тергитах, частично, захватывающими IV тергит. Эти пятна
разделяет черная продольная полоса, занимающая около 1/s ширины брюшка, на
каждом тергите она кзади немного расширена. Тергиты по заднему краю с узкой
коричневой каймой. Брюшко сверху с 3 рядами небольших сероватых пятен (налет+волоски)г
треугольных посредине и округлых по бокам. Снизу брюшко серое, его базальная
часть и задний край стернитов желтоватые. 14 мм.
Похож на Т. bromius, но крупнее, 5 легко отличается по отсутствию полоски
па глазах. Похож также на Т. miki, но в легко отличается короткими волосками на
затылке, а 5 иным строением нижней лобной мозоли.
Распространение: понтийско-гирканский предгорно-горный подвид
(рис. 148, 2). В СССР известен из Вост. Предкавказья (Гудермес, Дагестан),
Азербайджана (Кусары, Закаталы, Ханлар, Талыш, Нахичеваяская ССР), Грузии (окрест.
Батуми), Армении (Мегри, Бюракан) и Туркмении (Копет-Даг: Бахарденский р-н).
Югославия, Болгария, Иран. Не часто. Июнь—август.
5$ из Туркмении (Копет-Даг) часто с целиком черным 3-м члеником усиков;
по окраске нижней лобной мозоли некоторые 99 переходные к Т. indrae vappa.
Описание 6 сделано по экземпляру из Копет-Дага.
29b. Tabanus indrae vappa Bog. et Sam. — Слепень бесполосый светлый.
Богачев и Самедов, 1949, Изв. АН Азерб. ССР, 5:71 (Т. vappa);
Г а у з е р, 1960 : 656 (Т. vappa); Leclercq, 1966 : 175 (Т. vappa); Олсуфьев,
1970 : 684.
5<У. От номинативной формы отличаются более светлой окраской: 3-й членик
усиков оранжевый или красновато-коричневый, иногда с более темной палочкой,
боковые коричневые пятна имеются на II—IV или I—V тергитах брюшка, у 5 нижняя
часть нижней лобной мозоли часто желто-коричневая. 13—16 мм.
8. TABANUS
237
Распространение: Закавказско-среднеазиатский предгорно-горный под-
1шд. Азербайджан (включая Нахичеванскую АССР), Таджикистан (хребты Гиссар-
ский, Каратегинский, Зеравшанский, 1100—2000 м), Узбекистан (Сурхандарьинская
и Самаркандская области). Не'часто. Июнь—август.
29с. Tabanus indrae montivagus Ols. — Слепень бесполосый горный.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 684.
$. От номинативной формы отличается более темной окраской: 3-й членик усиков
черный с темно-коричневым основанием или целиком темно-коричневый, нижняя
лобная мозоль часто целиком черная, брюшко черно-серое, с едва заметными боковыми
коричневыми пятнами на II—III тергитах, средние и задние голени коричневые с
черноватой вершиной. VIII стернит с боковыми коричневыми пятнами. 13—16 мм.
в неизвестен.
Рассматриваемый подвид имеет большое сходство с Т. miki colchidicus, отличаясь
от последнего более крупной, прямоугольной нижней мозолью, почти достигающей
боковыми сторонами края глаз, и более короткими волосками на затылочной полоске.
Распространение: горный подвид. Горный Крым; Большой Кавказский
хребет (в р-не Адлера Краснодарского края; Абхазия; Сванетия); УССР: Йвано-Фран-
ковская обл. (Тлумачский р-н). Не часто. Июль—начало августа.
Между тремя подвидами Т. indrae имеются переходы.
Рис. 166. Tabanus infestus Bog. Рис. 167. Tabanus armeniacus Krob. Лоб-
et Sam. Лобная полоска, усик ная полоска, усик и щупальце $.
и щупальце £.
30. Tabanus infestus Bog. et Sam. (рис. 166) —Слепень опасный.
Богачев иСамедов, Изв. АН Азерб. ССР, 5 : 71; Г а у з е р, 1960 :
656; Leclercq, 1966 : 153.
5. Глаза голые, без полосок. Лобная полоска в сером налете, ее высота
превосходит ширину основания приблизительно в 4 V2 раза, кверху слегка расширена.
Нижняя лобная мозоль блестяще-черная, б. м. прямоугольная, боковыми сторонами
немного не достигает края глаз. Средняя мозоль черная, веретеновидная, концом
достигает нижней мозоли. Лобный треугольник в сером налете. Усики: 1-й и 2-й членики
черные, 3-й членик темно-коричневый или черноватый, дорсальный угол б. м. прямой
или тупой. Затылочная палочка узкая, в коротких светлых волосках. Концевой членик
щупалец желтовато-беловатый, в беловатых и единичных черных волосках, в
основании умеренно утолщен, в 2 1/2—3 раза длиннее наибольшей толщины. Нотоплевры
черно-серые. Крылья бесцветные; г4 без придатка. Ноги: бедра черно-серые,
базальная часть передних, средние и задние голени на большем протяжении светло-желто-
коричневые, остальное черное. Брюшко черно-серое, сверху с 3 рядами серых пятен
(налет-(-волоски): треугольных посредине и б. м. ромбических по бокам. На II—
III тергитах боковые серые пятна маскируют незначительную коричневую окраску
хитина полуколец. Снизу брюшко черное, в густом сером налете, стерниты по заднему
краю с узкой желтоватой каемкой. 13—14 мм.
6 неизвестен.
Похож на Т. bromius, но легко отличается от него отсутствием полоски на глазах.
От Т,..indrae, с которым также сходен, отличается меньшей величиной и более
светлым рерым рисунком сверху брюшка.
238
III. ПОДСЕМ. TABANINAE
Распространение: Нахичеваиская АССР (Шахбузский р-н). Редок.
Август.
31. Tabanus armeniacus Krob. (рис. 167) — Слепень армянский.
К г б Ь е г, 1928, Zool. Anzeiger., 76 : 267;. Олсуфьев, 1937а : 275;
Скуфьи и, 1938 : 80 •(<?); L е с 1 о г с q, 1966 : 139; Clival a, Lyneborg
a. Mouch a, 1972 : 348.
$. Глаза голые, без полосок. Лобная полоска умеренной ширины, с почти
параллельными сторонами, ее высота превосходит ширину основания в 3 1/1—4 раза,
покрыта серым или слегка желтоватым налетом. Нижняя лобная мозоль б. м.
квадратная, блестяще-черная, боковыми сторонами не достигает края глаз. Средняя мозоль
черная, веретеновидная, концом достигает до нижней мозоли. На темени маленькое
блестяще-черное или коричневое пятнышко — мозоль. Затылочная полоска узкая,
в коротких серых и частью черных волосках. Усики черные, 1-й членик сверху сильно
клювовидно выдается вперед, в сером налете, 3-й членик с широкой площадкой,
дорсальный угол б. м. прямоугольный. Иногда площадка темно-коричневая. Концевой
членик щупалец желтовато-белый, утолщен, покрыт довольно грубыми черными и
и частью беловатыми волосками. Нотоплевры черно-серые. Ноги черно-серые, средние
голени и частью основания передних и задних голеней желтовато-коричневые. Окраска
голеней маскируется серым налетом и густыми белыми и черными волосками. Крылья
бесцветные; ?-4 без придатка. Брюшко черное, с двумя боковыми рядами серых
ромбических пятен (налет+волоски). Кроме того, имеется срединный ряд из небольших
треугольников. Задние каемки тергитов сероватые. Снизу брюшко черное, в тонком
голубовато-сером налете; посредине часто имеется темная продольная полоса. 13—
16 мм.
в. Голова не больше, чем у J. Глаза голые без полосок, фасетки верхних частей
глаз лишь немногим крупнее нижних, переход постепенный. Затылок в коротких,
светлых волосках. Лобный треугольник серый, более темный в верхней части. Усики
как у $, но 3-й членик более узкий. Концевой членик щупалец яйцевидный,
желтовато-беловатый, в основании серый, покрыт преимущественно черными волосками.
Грудь и ее придатки как у $. Брюшко черное, боковые стороны II (или II—III) тер—
гита слегка коричневые. Сверху брюшка 3 неясных продольных ряда серых пятен
(налет+волоски): треугольных посредине и б. м. округлых по бокам. Снизу брюшко
как у $. 15—16 мм.
Похож на Т. miki colchidicus и Т. indrae montivagus: $ отличается более широкой
лобной полоской с б. м. параллельными сторонами, квадратной нижней лобной мозолью
и клювовидно вытянутой верхнёпередней частью 1-го членика усиков; в легко
отличается небольшой головой и б. м. одинаковыми фасетками глаз.
Распространение: кавказский горно-лесной вид. Большой
Кавказский хребет (от Карачаево-Черкесской А. О. и Абхазской АССР на восток до гор Шах-
даг и Тфан с.-в. Азербайджанской ССР), Малый Кавказский хребет в'пределах
Грузии (окр. Батуми), Армении (оз. Севан, Памбак) и Нахичеванской АССР; Талыш.
Не часто. Июль—середина августа.
Рис. 168. Tabanus regularis Jaenn. Лобная полоска
усик и щупальце ?. А - щупальце в-
32. Tabanus regularis Jaenn. (рис. 168) — Слепень стройный.
Jaennicke, 1866, Berl. Ent. Zei'tschr., X : 85 (9); Brauer, 1880 : 194;
Surcouf, 1924:178; S z i 1 a d y, 1923 : 18 (<?); К г б b e r, 1924 : 141; К г 6-
ber in Lindner, 1925 : 124; Олсуфьев, 1937а : 277; Leclercq, 1966:
165; Chvala, Lyneborg a. Mouch a,. 1972:354.
$. Глаза голые, без полосок. Лобная полоска очень узкая, ее высота превосходит
ширину основания в 5—6 раз, покрыта желтовато- или коричневато-серым налетом.
8. TABANUS 239
Нижняя лобная мозоль посредине с тонкой продольной бороздкой, коричневая,
продолговатая, верхним концом переходит в узковеретеновидную черную среднюю
мозоль. Лобный треугольник высокий, покрыт коричневато-серым налетом. Его окраска,
равно как и коричневая окраска верхних уголков щек, представляет собой контраст
в сравнении с серебристо-серым лицом, покрытым беловатыми волосками. Усики
коричневые, 3-й членик умеренно широкий, дорсальный угол тупоугольный.
Затылочная полоска относительно узкая, снабжена короткими, серыми волосками.
Концевой членик щупалец беловатый, в основании умеренно утолщен, покрыт белыми и
черными волосками. Нотоплевры желтовато-коричневые, реже черно-серые. Крылья
бесцветные; г4 .без придатка. Ноги: бедра серые, голени желтые, вершинная треть
или половина цередних голеней и передние лапки черные, последующие лапки темно-
коричневые. Брюшко черное, с явственными 3 рядами серых пятен (налет+волоски):
треугольных посредине и б. м. ромбических по бокам. Эти пятна достигают переднего
края тергитов, так что в общей сложности часто создают впечатление 3 серых
продольных полос. Боковые стороны II или II—III тергитов обычно слегка просвечивают
сквозь налет коричневым. Снизу брюшко черно-серое, в основании иногда
коричневое, в густом сером налете, посредине часто намечается более темная продольная
полоса. 12.5—15 мм.
6. Голова больше, чем у $, полушаровидная. Крупные фасетки занимают
верхние 2/3—3/4 головы, в 4—5 раз крупнее мелких, на границе крупных и мелких
фасеток имеется довольно широкая медно-лиловая полоска. Затылок в очень коротких
волосках. Лобный треугольник в коричневато- или желтовато-сером налете, в
верхней части, перед вершиной имеется коричневатая поперечная полоска. Усики как у $,
но более узкие. Концевой членик щупалец беловатый, продолговато-овальный, обычно
с оттянутым кончиком, покрыт белыми волосками. Грудь и ее придатки как у $.
Брюшко коническое, черно-серое, с 3 рядами серых пятен (как у $). II—III тергиты
по бокам коричневые. Снизу брюшко черно-серое, в основной половине слегка
коричневатое. 13—1-4.5 мм.
Небольшой, черно-серый вид. Похож на Т. bromius, $ отличается значительно
более узким лбом и глазами без полоски, в — более крупной головой.
Распространение: южноевропейский (средиземноморский) степной?
вид, преимущественно по предгорьям и низкогорьям. В СССР известен из
Азербайджана (Закаталы, Самур, Геок-Тепе Агдашского р-на, Талыш, Нахичеваиская АССР —
Ордубад, Шахбуз), Грузии, Армении (Дилижан). Ю. Европа, Сев. Африка, Передняя
Азия, Иран, Ирак. Местами обычен. Июль—по 2-ю декаду августа. Указание Щербины
(1972) для Приэльбрусья, 2200 м,
нуждается в подтверждении.
33. Tabanus bromius L. (рис.
169) — Слепень серый.
Широко распространенный
изменчивый вид, представлен 2
формами: номинативным подвидом Т.
bromius bromius L. и южным более
светлым подвидом Т. bromius flavofemo-
ratus Strobl.
33a. Tabanus-bromius bromius L.
(рис. 170) — Слепень серый
номинативный.
L i n n ё, 1761, Fauna Suec. : 463;
Brauer, 1880 : 187; Lundbeck,
1907:127; V e r r a 11, 1909:406;
Surcouf, 1924 : 182; К г б b e r
in Lindner, 1925 : 109;
Олсуфьев, 1937a : 278; M о и с h a
et Chvala, 1958a : 189; 1959b :
282; Шевченко, 1961 : 154;
Leclercq, 1966 : 143; С к у ф ь-
и н, 1967 : 194 (личинка); В и о л о-
в и ч, 1968 : 93; Chvala, L у-
neborga. Mouch a, 1972 : 357.
— nigricans S z i 1 a d у, 1914 : 668.
,Рис. 169. Tabanus bromius bromius L. J.
840
III. ПОДСЕМ. TABANINAE
<j>. Глаза голые, с одной относительно широкой, не достигающей внешнего края
глаза полоской. Лобная полоска узкая, слегка сужающаяся к основанию, ее высота
в 4—5 раз превосходит ширину основания, покрыта желтовато-серым налетом.
Затылочная полоска узкая (рис. 125, справа), с рядом коротких, желто-серых волосков. Нижняя
лобная мозоль слабо выпуклая, блестяще-черная, б. м. прямоугольная. Боковыми
сторонами мозоль не достигает края глаз. Средняя мозоль узковсретеповидпая или
клиновидная, топкой линией соединена с нижней мозолью. Лобный треугольник высокий, вместе
с прилегающими уголками щек в желтоватом налете. Концевой членик щупалец палевый
или беловатый, в основании утолщен, к концу равномерно сужоя, с слегка
притуплённым кончиком, покрыт короткими черными и светлыми волосками. Усики
коричнево-черные; 3-й членик в основной 1/2 коричневый или красноватый, б. м. расширен, с
тупоугольным дорсальным выступом. Палочка тонкая, остроконечная, по длине б. м.
равна площадке; 1-й членик часто коричнево-красный. Нотоплевры чаще коричневые.
Крылья бесцветные, с темно-коричневыми жилками; rt обычно без придатка. Ноги:
бедра черно-серые, основная часть (или целиком) средних и задних голеней грязно-
Рис. 170. Tabanus bromius bromius L. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $. А — двувершинная
сперматека (уродство).
желтая или коричневая. Концы передних голеней и лапки Черные, часто средние
и задние лапки в основании коричневые. Брюшко черно-серое, сверху с явственным
более светлым серым рисунком, состоящим из 3 рядов пятен (налет+волоски):
треугольных посредине и ромбических по бокам. Эти последние варьируют по форме
и величине и могут быть представлены косыми черточками. II—III тергиты по бокам
часто неясно буроватые. Снизу брюшко темно-серое, с неясной, более темной
срединной полоской. I—III стерниты часто слегка буровато-желтые. VIII стернит
удлиненный, гонопофиз крупный, б. м. конический, обычно с резкой апикальной выемкой;
латеральные лопасти небольшие, с темным пятном; базальная лопасть с слегка
вогнутым, реже прямым основным краем. Церки с округлой вершиной. Сперматека
миндалевидная. 11—16 мм.
в. Голова не больше, чем у $. Глаза голые, с одной узкой полоской на границе
крупных и мелких фасеток. Иногда удается заметить на глазах очень короткие
волоски. Фасетки верхних 2/3 глаз / значительно крупнее нижних, граница между теми
и другими б. м. резкая. Величина фасеток верхней части глаз варьирует: они могут
быть крупнее нижних в 3—4 раза и в 5—6 раз. Затылок за глазами лишь возле глаз-
кового бугорка в очень коротких, светлых волосках. Лобный треугольник перед
вершиной буроватый, тогда как в нижней части в сером налете. Концевой членик
щупалец палево-желтый, продолговато-овальный, покрыт б. м. длинными светлыми
и черными волосками. Усики как у $, но 3-й членик более узкий. Грудь и ее придатки
как у $. Брюшко буро- или серо-черное, сверху с коричневыми боковыми пятнами
на II—III (или II—IV) тергитах. Разделяющая их темная срединная полоска
занимает 1/3 ширины брюшка; боковые стороны полоски в пределах каждого тергита
полукругло вырезаны. Окраска дополняется 3 желтовато-серыми продольными рядами
пятен: треугольных посредине и косых по бокам. Брюшко снизу буро-серое, сб. м.
8. TABANUS 241
явственной темной срединной полоской. Базальные стерниты желто-коричневые.
14—16 мм.
Распространение: европейско-западносибирский лесостепной подвид,
широко проникает в лесную зону и в горы (рис. 171, 1). В СССР распространен
от Карелии, Прибалтики, запада Украины и Молдавии на восток до Томска и
предгорий Алтая, на север известен до Карельской АССР, Котласа Архангельской обл.,
Ухты Коми АССР, севера Омской и Новосибирской областей, на юг до устья Дуная,
Крыма, Сев. Кавказа, Закавказья (включая Азербайджан, Грузию, Армению),
низовий Урала (80 км сев. Гурьева), Целинограда, Каркаралинска и далее, огибая пустыни
Казахстана и Средней Азии, до Алма-Аты, Киргизии, Узбекистана и Таджикистана
(Гиссарский хребет, 1500—3500 м). Европа (кроме крайнего севера), Сев. Африка,
Передняя Азия, Иран, Афганистан. Обычен. Июнь—август. В южных частях ареала
на значительном пространстве наряду с типичной формой встречаются промежуточные
экземпляры между Т. bromius bromius и Т. Ъ. flavofemoratus.
Рис. 171. Ареалы: I — Tabanus bromius bromius; 2 — Т. bromius flavofemoratus.
Личинки нами найдены в слабопесчаном грунте в мокрой осоковой низине по дну
оврага (Ленинградская обл.) и в прибрежных частях озера (Зап. Сибирь). В Средней
Азии (Ферганская долина) личинок обнаруживали в арыках, родниках и реках
(Кадырова, 19726).
Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 1936)
и сибирской язвы (Олсуфьев и Лелеп, 1935).
33b. Tabanus bromius flavofemoratus Strobl. — Слепень серый желтоногий.
Strobl, 1908, Mem. Real Soc. Esp. Hist. Nat., Ill : 292; S u г с о u f, 1924 :
184; Rrober, 1924 : 156; Krober in Lindner, 1925 : 109; Олсуфьев,
1937a : 280; Leclercq, 1966 : 144. — anthophilus L о e w, 1858a : 594. — bro-
miolus S z i 1 a d y, 1923 : 4. — ? appendiculatus S z i 1 a d y, 1923 : 3.
Отличается в основном более стройным телом и более светлой окраской главным
образом брюшка.
$. Глаза с одной узкой, иногда почти исчезающей золотисто-медной полоской,
которая у коллекционных экземпляров при их последующей размочке во влажной
камере иногда может не восстановиться. Лобная полоска с почти параллельными
сторонами, ее высота превосходит ширину в 3-3/4 —4 раза. Нижняя лобная мозоль
б. м. квадратная, темно-коричневая или черная. Концевой членик щупалец белый,
часто целиком в белых волосках. Усики желто-коричневые, с более темной палочкой.
Крылья с желто-коричневыми жилками; ri иногда с рудиментарным придатком. Ноги:
бедра светло-серые, иногда сквозь серый налет просвечивают желтоватым, голени
желтые, кончики передних голеней и передние лапки черные, лапки последующих ног
коричневые. Брюшко с сильно развитой желто-коричневой окраской по бокам,
захватывающей I—IV (или I—VI) тергиты. Продольные ряды пятен из налета+волос-
ков, желтоватые, поэтому на желтоватом фоне брюшка выделяются менее резко.
16 н. г. Олсуфьев
242 • HI. ПОДСЕИ. TABANINAE
Снизу брюшко почти целиком коричневато-желтое. Терминалии как у номинативного
подвида. 11—16 мм.
в. Голова не больше, чем у <j>. Иногда на глазах довольно хорошо заметны
короткие волоски. На границе крупнофасетчатого и нижнего мелкофасетчатого поля
имеется узкая, медно-коричневая полоска. Усики как у $, но более узкие; концевой
членик щупалец беловатый, овальный, в тонких белых и единичных черных волосках
Грудь и ее придатки как у $. Брюшко по бокам с желто-коричневой окраской,
простирающейся на II—IV или I—V тергиты. Продольные ряды желтоватых пятен (на-
лет+волоски) слабо выражены. Снизу брюшко в основной половине коричнево-желтое,
последние 3 или 2 стернита серые. 11—16 мм.
Распространение: средиземноморско-среднеазиатский степной подвид,
обычно по предгорьям и низкогорьям (рис. 171, 2). Низовья Дона, Краснодарский
и Ставропольский край (Александровское), Кабардино-Балкарская АССР
(Урожайное), Северо-Осетинская и Дагестанская АССР; пустынно-степные районы
Закавказья, включая Азербайджан, Грузию и Армению; Таджикистан, Узбекистан,
Киргизия, Ю.-В. Казахстан (в последних двух
республиках чаще переходные особи к Т.
bromius bromius). Ю. Европа, Сев. Африка,
Турция, Иран. Местами обычен. Июнь—август.
34. Tabanus hissarieus Bar. (рис.
172) — Слепень гиссарекий.
Б а р а т о в, 19626, Докл. АН Тадж.
ССР, V, 6 : 44.
$. Глаза голые, с одной полоской.
Лобная полоска умеренной ширины, ее высота
превосходит ширину основания
приблизительно в 3 1/2—4 раза, кверху слегка рас-
Рис. 172. Tabanus hissarieus Ваг. Лобная
полоска, усик и щупальце $.
ширена, покрыта серым налетом. Нижняя лобная мозоль крупная, блестяще-черная,
боковыми сторонами почти достигает края глаз, верх округлый, соединена с средней
черной веретеновидной мозолью. На темени черное пятно, частично прикрытое
налетом. Затылок узкий, в слегка выступающих над глазами светлых волосках. Лобный
треугольник в сером налете. Верхний край усиковых ямок широкий. Усики: 1-й
членик коричневый или черноватый в сером налете, 3-й членик коричневый, палочка
черноватая, почти в 2 раза короче длины площадки, последняя с тупым дорсальным
углом. Концевой членик щупалец коричневый, в сером налете, узкий, его длина в
31/2 раза превосходит наибольшую ширину, покрыт преимущественно светлыми
волосками. Нотоплевры черно-серые. Крылья б. м. бесцветные, жилки коричневые;
г4 с длинным придатком. Ноги: бедра черно-серые, основание передних и большая часть
средних и задних голеней желтовато-белые, остальное черное. Брюшко черно-серое,
II тергит посредине с неясным серым треугольным пятном (по Баратову, II—III
тергиты с срединными серыми треугольниками, покрытыми золотисто-желтыми волосками),
тергиты по заднему краю в серых волосках и с узкой серой каемкой. Снизу
брюшко черно-серое, посредине имеется неясная темная продольная полоса. 14—16 мм.
в неизвестен.
Своеобразный вид, легко отличающийся от остальных представителей группы
Т. bromius относительно узкими, темными щупальцами, отсутствием сверху брюшка
боковых продольных рядов светлых пятен и длинным придатком у г4.
Распространение: Таджикистан: Гиссарекий хребет, 1200—2000 м.
Редок. Конец июня—начало июля.
35. Tabanus semenovi Ols. (рис. 173) — Слепень Семенова.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 281; М о и с h a et
Clival a, 1959b : 283; Шевченко, 1961:159; Б аратов, 1962в : 97 (6);
L е с 1 е г с q, 1966 : 168.
$. Глаза голые, без полосок. Лобная полоска довольно узкая, слегка расширена
кверху, покрыта желтым налетом. Высота полоски превосходит ширину основания
в 4—4 !/2 раза. Нижняя лобная мозоль б. м. квадратная, блестящая, черная или
темно-коричневая, средняя мозоль в виде тонкого киля, соединена с нижней. Лобный
8. TABANUS 343
треугольник в желтоватом налете. Усики желто-коричневые, с коричневой палочкой.
3-й членик довольно узкий, с б. м. прямоугольным дорсальным выступом. Концевой
членик щупалец желтовато-белый, в основании утолщен, к концу равномерно сужен,
с острым кончиком. Членик покрыт смесью черных и беловатых волосков. Нотоплевры
черные, в сером налете. Ноги: бедра черно-серые, голени желтые. Концы передних
голеней и передние лапки черные, средние и задние лапки желтые, с коричневым
концом. Крылья б. м. бесцветные; г4 без придатка, крайне редко он имеется. Брюшко
оранжево-желтое, в коротких, такого же цвета волосках. Посредине имеется
неширокая, черноватая продольная полоса, отчасти прикрытая срединным рядом желтых
треугольных пятен (налет+волоски). Темные участки полосы в немногочисленных
черных волосках. Боковые пятна из желтоватого налета имеются, но они не
выделяются на общем оранжево-желтом фоне брюшка. Снизу брюшко целиком оранжево-
желтое. VIII стерыит: гонопофиз б. м. конический, с едва заметной апикальной
выемкой, в тонких светлых коричневых волосках; латеральные лопасти небольшие,
Рис. 173. Tabanus semenovi Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
коричневые, с слабо хитинизированной вершиной; базальная лопасть с вогнутым
основным краем. Церки с округлой вершиной. Сперматека миндалевидная. 13.5—
17 мм.
в. Голова крупнее, чем у $, полушаровидная. Глаза голые, фасетки верхних
3/4 глаз крупнее нижних в 5—6 раз, граница между теми и другими резкая. Затылок
в коротких светлых волосках. Лобный треугольник в желтовато-сером налете, в
верхней части коричневый. Усики как у $, но более узкие. Концевой членик щупалец
утолщен к вершине, беловатый, в светлых волосках. Грудь и ее придатки как у $.
Брюшко коническое, его окраска как у $. 16 мм.
От всех видов группы Т. bromius хорошо отличается ярко-оранжевой окраской
боковых сторон брюшка, покрытых золотисто-желтыми волосками.^
Распространение: горно-лесной бактрийско-иранский вид. От гор
Байсунтау, Гиссарского и Зеравшанского хребтов и зап. Памира (Хорог), на восток
до Таласского Алатау и Каратау, на юг включая хребты Каржантау, Угамский,
Чаткальский, Кураминский, Ферганский и Алайский: от 1200 до 3000 м. Афганистан,
Иран. Местами обычен. Июнь—август. Личинки найдены в родниках, арыках и речке
(Кадырова, 19726).
Голотип: ?, Арсланбобский горный массив, ур. Ак-Терек 1500 м, Ферганский
хребет, 28 VII1935, И. Корженевская. В коллеции Зоологического института Академии
наук СССР, Ленинград.
(
36. Tabanus laetetinctus Beck. (рис. 174) — Слепень светлый.
Изменчивый вид, представлен двумя формами: номинативным светлым подвидом
Т. laetetinctus laetetinctus Beck, и более темноокрашенным подвидом Т. laetetinctus
sordes Bog. et Sam.
16*
244
III. ПОДСЕМ. TABANINAE
36a. Tabanus laetetinctus laetetinctus Beck. — Слепень светлый номинативный.
Becker, 1912, Ann. Mus. Zool. St. Petersb., XVII : 595; К г б b e r, 1924 •
164; К rober in Lindner, 1925 : 118; Олсуфьев, 1937а : 282; 19396 : 99
$LM °ur c h a et С h v a 1 a, 1959b : 283; Шевченко, 1961 : 158; Leclercq,
1966 : 155. — cuculus S z i 1 a d y, 1923 : 2.
?. Глаза голые, без полосок. Лобная полоска узкая, с почти параллельными
сторонами, ее высота превосходит ширину основания в 5—5 V2 раза, покрыта желтовато-
серым налетом. Нижняя лобная мозоль б. м. прямоугольная, блестящая, темно-
коричневая или черная, боковыми сторонами не достигает края глаз. Средняя лобная
мозоль черная, б. м. веретеновидная, тонкой линией соединена с нижней мозолью;
иногда соединение прервано налетом. Лобный треугольник высокий, в легком
желтоватом налете, заметно отличаясь окраской от беловато-серого лица. Усики целиком
оранжево-желтые. 1-й членик довольно резко бокаловидный, с сильно оттянутым
вперед кончиком, 3-й членик относительно узкий, с тупоугольным дорсальным углом. За-
Рис. 174. Tabanus laetetinctus laetetinctus Beck.
Лобная полоска, усик, щупальце, VIII стернит
и церки 5-
тылочная ?полоска узкая.гв коротких серых волосках. Концевой членик-щупалец белый
или слегка желтоватый, в основной половине утолщен, покрыт'белыми и отчасти черными
волосками, реже целиком в белых волосках. Нотоплевры бледно-коричневато-желтые.
Крылья бесцветные; г4 без придатка или же с небольшим его рудиментом. Ноги светло-
коричневато-желтые, лишь передние лапки коричнево-черные. Брюшко коричнево-
желтое, II тергит иногда посредине с короткой темной черточкой возле переднего
края (иногда продолжена на последующие тергиты) и, кроме того, последние 2 или
3 тергита сероватые. Брюшко сверху с 3 рядами желтовато-серых пятен: треугольных
посредине и б. м. округлых или неправильной формы по бокам, покрыто короткими
желтыми и черными волосками; первые занимают боковые стороны тергитов и
срединный ряд треугольных пятен, слитых иногда в продольную полоску, тогда как
черные волоски расположены по бокам срединного ряда, группируясь в 2 боковые
продольные полоски. Рисунок из волосков обычно слабо выделяется на светлом
фоне брюшка и заметен лишь у экземпляров хорошей сохранности. Снизу брюшко
коричневато-желтое, последние 3—4 тергита часто сероватые. VIII стернит желтый,
гонопофиз б. м. конический, с небольшой апикальной выемкой; латеральные лопасти
небольшие; базальная лопасть с слегка вогнутым основным краем. Перки с округлой
вершиной. 10.5—15 мм.
6. Голова больше, чем у $, полушаровидная. Фасетки верхних 3/4 глаз крупнее
нижних приблизительно в 5—6 раз, граница между теми и другими резкая, полоска
отсутствует. Затылок в коротких волосках, не выступающих над поверхностью глаз.
Лобный треугольник в желтовато-сером налете, в верхней части иногда
коричневатый. Усики как у ?, но 3-й членик более узкий. Концевой членик щупалец беловатый,
продолговато-овальный, в светлых и единичных черных волосках. Грудь и ее при-
8. TABANUS 245
датки как у $. Брюшко: желто-коричневая окраска захватывает боковые стороны
I—IV тергитов, посредине продольная темная полоса занимает примерно 1/4—1/3
ширины брюшка. Задний край тергитов с умеренно широкой желтоватой каймой.
Имеются 3 продольных ряда желтовато-серых пятен (налет -j-волоски), их форма как у $.
Снизу брюшко как у $. 10—13.5 мм.
От всех видов группы Т. bromius (кроме Т. hauseri) отличается светлыми коричне-.
вато-желтыми бедрами; по окраске брюшка похож на Т. bromius flavofemoratus, но J
легко отличается отсутствием полоски на глазах, а в более крупной головой.
Распространение: понтийско-гирканский предгорно(горно)-степной
подвид. В СССР известен в Азербайджане (включая Нахичеванскую АССР), на юге
Туркмении, в Таджикистане (хребты Гиссарский, Дарвазский и их предгорья),
Узбекистане (Сурхандарьинская, Ташкентская, Андижанская, Ферганская области)
и Киргизии (Ферганский хребет, 1000—1700 м). Афганистан, Иран, Ирак, Турция.
Местами обычен. Июнь—август, иногда позднее. Личинки в арыках, родниках и
реках (Кадырова, 19726).
36b. Tabanus laetetinctus sordes Bog. et Sam. — Слепень светлый серобрюхий.
Богачев и Самедов, 1949, Изв. АН Азерб. ССР, 5 : 70 (Т. sordes);
Leclercq, 1966 : 169 (Т: sordes); — sordes assuetus (Hauser) apud Leclercq, 1966 :
140. — assuetus Hauser, 1960 : 654. — assuetus cinereus Hauser, 1960 : 655.
От номинативной формы $<? отличаются преобладанием черно-серой окраски
брюшка: желто-коричневые пятна по его бокам небольшие, захватывают II—III
(или I—IV) тергиты, но могут быть лишь на II тергите. Соответственно на темном
фоне брюшка серые продольные ряды пятен из волосков и налета выступают более
четко. Снизу брюшко в базальной части розовато-желтоватое, но изредка целиком
серое, стерниты по заднему краю желтоватые. Ноги как у номинативной формы,
изредка бедра сероватые. $ 10—14 мм, 6 10—13.5 мм.
Описан А. В. Богачевым и Н. Г. Самедовым (1949) в качестве самостоятельного
вида Т. sordes по сборам Гаузера в 1939 г. в Нахичеванской АССР. Судя по описанию,
в распоряжении авторов наряду с экземплярами с темным рисунком брюшка были
особи номинативной формы и переходные. Позднее Т. sordes был описан Е. Г. Гаузером
(1960) как Т. assuetus по тем же сборам 1939 г. в Нахичеванской АССР.
Изучив коллекционные материалы вышеуказанных авторов, а также
опубликованные ими описания, мы (Олсуфьев, 1969) таксономировали Т. sordes Bog. et Sam.
как темноокрашенный подвид Т. laetetinctus Beck., a T. assuetus Haus. вместе с
описанным Гаузером подвидом Т. assuetus cinereus Haus. отнесли в синонимы к Т. laetetinctus
sordes Bog. et Sam.
Распространение: преимущественно горный подвид. Азербайджан
(включая Нахичеванскую АССР), Армения (Мегринский р-н), Грузия, Туркмения
(Копет-Даг), Таджикистан. Иран, Турция. Не часто. Конец июня—август.
37. Tabanus hauseri Ols. (рис. 175) — Слепень Гаузера.
Олсуфьев, 1967, Энтомол. обозр., 46, 2 : 387. — turpis (Bog. et Sam.) apud
Hauser, 1953 : 251.
<j>. Глаза голые, без полосок. Лобная полоска слегка расширена кверху, ее высота
приблизительно в 5 раз превосходит ширину основания, в желтовато-сером налете.
Нижняя лобная мозоль коричневая, блестящая, прямоугольная, ее высота обычно
превосходит ширину, боковыми сторонами почти достигает края глаз. Средняя
лобная мозоль черная или темно-коричневая, веретеновидная, обычно не соединена с
нижней лобной мозолью, реже соединена с ней тонкой линией, еще реже — полностью
прикрыта налетом. Затылок узкий, в коротких серых волосках. Лобный треугольник
коричневый, в желтовато-сером налете, последний часто частично стерт с лобного
треугольника. Усики целиком оранжево-желтые, 3-й членик с тупоугольным
дорсальным углом. Концевой членик щупалец белый или слегка желтоватый, умеренно
утолщен, в белых и единичных черных волосках. Нотоплевры коричнево-желтые. Крылья
бесцветные, жилки коричневые; rt без придатка. Ноги: передние бедра черные или
черно-коричневые, в сером налете, средние и задние бедра оранжево-желтые, голени
желтые, концы передних голеней и передние лапки темно-коричневые или черноватые,
средние и задние лапки желто-коричневые.-Брюшко коричневое с рыжеватым
оттенком. Сверху посредине брюшка имеется продольная узкая серая полоска, занимающая
на II—III тергитах V8—х/10 ширины брюшка, на последующих тергитах она
расширяется и VI—VII тергиты целиком темные; иногда полоска отсутствует на III —
IV тергитах. 3 продольных ряда желтовато-серых пятен (налет+волоски):
треугольных посредине и неправильной формы по бокам; по бокам среднего ряда
треугольников по ряду косых пятен из черноватых волосков, оттеняющих светлый рисунок
брюшка. Снизу брюшко розовато-желтое, в сероватом налете, посредине неясная
более темная полоска, VII или VI—VII стерниты сероватые. VIII стернит: гонопофиз
246 III. ПОДСЕМ. TABANINAE
лопатообразный, без выраженной апикальной выемки; латеральные лопасти
небольшие, коричневатые; базальная лопасть удлиненная, ее основной край прямой. Церки
со слегка уплощенной вершиной. 14—16 мм.
в (по Гаузеру, 1953, сокращенно). Голова большая. Глаза голые, фасетки верхних
частей глаз значительно крупнее нижних, граница между теми и другими резкая.
Затылок в коротких светлых волосках. Лобный треугольник перед вершиной темно-
бурый, в остальной части в сероватом налете. Концевой членик щупалец
светлосерый, со слегка желтоватым оттенком, продолговато-овальный, в беловатых и
отчасти черных волосках. Усики как у J, но 1-й членик с более вытянутым вперед дор-
Рис. 175. Tabanus hauseri Ols. Лобная полоска, усик,
щупальце, VIII стернит и церки $>•
сальным углом. Нотоплевры темные. Ноги как у $. Брюшко рыжевато-бурое; I —
IV тергиты со светло-коричневыми боковыми пятнами. Сверху (снизу?) брюшко
охристо-рыжее, с неясной темной полоской посредине. Последние 2—3 тергита (стернита?)
темно-серые. 14 мм.
Близок к Т. laetetinctus, но отличается от него большей величиной, темными
передними бедрами и более продолговатой нижней мозолью обычно коричневого цвета ($).
Имеются также различия в строении терминалий J.
Т. hauseri Ols. был ошибочно определен А. В. Богачевым и Н. Г. Самедовым как
Т. laetetinctus Beck, и под этим названием опубликован в их работе 1949 г. Е. Г. Гаузер
(1953) описал $ и с? этого вида, неправильно обозначив его как Т. turpis Bog. et
Sam., 1949.
Распространение: Азербайджан — Нахичеванская АССР (Ордубад-
ский р-н) и окр. Закатал. Редок.
38. Tabanus tergestinus Egg. (рис. 176) — Слепень трехполосый.
Е g g е г, 1859, Verh. Zool.-bot. Gesell. Wien, 9 : 391; В r a u e г, 1880 : 189;
Surcouf, 1924:185; К r 6 b e г, 1924:158; Kroberin Lindner, 1925:
129; Олсуфьев, 1937a : 283; Moucha et Chvala, 1958a : 188; L e c-
lercq, 1966 : 173; С h v a 1 a, Lyneborg a. Moucha, 1972:362.
$. Глаза голые, с 3 полосками. Лобная полоска очень узкая, кверху слегка
расширена, ее высота в 5 1/2—6 раз превосходит ширину основания, покрыта серым
налетом. Нижняя лобная мозоль треугольно-щитовидная, иногда овальная, черная или
темно-коричневая, средняя мозоль в виде узкой, веретеновидной, черной полоски,
соединена концом с вершиной нижней мозоли. Затылочная полоска узкая, в коротких
серых волосках. Лобный -треугольник высокий, в сером или слегка желтоватом налете.
Лицо в сером налете и такого же цвета волосках. Усики коричневые или красновато-
8. TABANUS
247
коричневые, концевая половина (часть площадки и палочка) черная. 3-й членик
умеренной ширины, дорсальный угол прямо- или тупоугольный. Концевой членик
щупалец беловатый или бледно-желтовато-коричневый, в основной половине утолщен,
LIT ч.'\Ч / ('!'■ .
Рис. 176. Tabanus tergestinus Egg. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
равномерно покрыт короткими черными волосками, с незначительной примесью серых.
Нотоплевры черно-серые. Крылья с легким буроватым оттенком; г4 без придатка.
1-я заднекрайняя ячейка не сужена к вершине. Ноги: бедра черно-серые, голени
Рис. 177. Ареалы: 1 — Tabanus tergestinus; 2 — Т. chrysurus.
коричнево-желтые; передние голени в вершинной половине черные, кончики
последующих голеней черноватые. Лапки всех ног черные, реже средние и задние лапки
коричневые. Брюшко удлиненное, черно-серое, по бокам с ржаво-коричневыми
пятнами, простирающимися на I—IV тергиты. Разделяющая пятна темная срединная
полоска занимает приблизительно L/i—V6 ширины брюшка. Снизу брюшко
желтовато-коричневое, последние два стернита сероватые. VIII стернит преимущественно
248 ' III. ПОДСЕМ. TABANINAE
темно окрашен и весьма своеобразной формы: гонопофиз двулопастной, относительно
узкий, с апикальной выемкой; латеральные лопасти большие; базальная лопасть
короткая, значительно шире гонопофиза, с прямым основным краем. Церки с округлой
вершиной. Сперматека миндалевидная. 15—18 мм.
6. Голова не больше, чем у $. Глаза голые, в нижней 1/3 с 2 полосками. Фасетки
верхних частей глаз, в особенности возле глазного шва, заметно крупнее нижних;
крупные фасетки постепенно переходят в мелкие, без резкой границы. Затылок
в коротких^ желтовато-серых волосках. Лобный треугольник в сером налете. Усики
как у $, 3-й членик более узкий. Концевой членик щупалец желтовато-белый,
продолговато-овальный, с оттянутым кончиком, покрыт длинными черными и серыми
волосками. Грудь и ее придатки как у $. Брюшко с большими ржаво- или желто-
коричневыми пятнами по бокам, захватывающими I—IV тергиты. Пятна разделяет
очень узкая черно-серая полоска, которая постепенно расширяется в направлении
кзади. Последние 3 тергита-черно-серые. Снизу брюшко желто-коричневое,
последние 2—3 стернита сероватые. 15—18 мм.
Легко отличается от прочих видов группы Т. bromius наличием нескольких
полосок на глазах (3 у $ и 2 у .#).
Распространение: средиземноморский (южноевропейский) лесной вид,
преимущественно в горах (рис. 177, 1). В СССР распространен в западной и северной
Украине (на север до Киева и Сум, но не найден в Крыму), в Молдавии, в
Воронежской (Вогучар), Куйбышевской (Жигули) и Саратовской (с. Воскресенское) областях,
далее, после перерыва в южнорусских степях, на Сев. Кавказе (от Новороссийска
Краснодарского края до Дагестана) и в Закавказье — в Азербайджане, Армении
и Грузии. Ю. и отчасти Средн. Европа, Турция, Иран. Местами обычен. Июнь-
август.. Личинки найдены в почве вдали от воды (Гургенидзе, 19726).
39. Tabanus armenieus Szil.
S z i 1 a d у, 1926, Ann. Mus. Nat. Hungar., 24 : 599; Олсуфьев, 1937а : 284;
Leclercq, 1966 : 173.
Нам в натуре этот вид неизвестен. Силади (Szilady) дает следующее описание $:
«Длина 14 мм. Темно-коричневый Tabanus с бесцветными крыльями, голыми глазами,
без полосок; лобная полоска напоминает bovinus. Глаза темно-коричнево-зеленые,
голые, без полосок. Голова спереди серо-коричневая, сзади серая, в скудных белых
волосках. Усики темно-коричневые, базальная часть вплоть до дорсального угла
3-го членика красноватая, 1-й членик сверху с оттянутым кончиком. Щупальца
коричневые, концевой членик в основании утолщен, в черных волосках. Лобная полоска
узкая, книзу сужена, нижняя лобная мозоль блестящая, ее веретеновидное
продолжение (средняя мозоль) составляет более V3 длины лба, глазковая мозоль отсутствует,
место последней одинаково окрашено с лбом. Грудь темно-коричнево-серая, на
матовой спинке спереди заметны следы 3 беловатых продольных полосок. Нотоплевры
красно-коричневые. Ноги коричнево-желтые, коксы, лапки, передние бедра и концы
передних^ голеней черно-коричневые. Крылья бесцветные, с черными жилками; 1-я
заднекрайняя ячейка параллельная; г4 без придатка. Брюшко темно-коричневое,
блестящее, в очень скудных волосках, возле задних каемок сегментов несколько
светлее. Основной сегмент с неясным, черным срединным пятном, на котором и
последующем сегментах ряд маленьких, равносторонних белых треугольников.
«1 $ из. Талышана Каспийском море в Мюнхенском музее. Этот новый вид югохож
на ориентальных Т. rubicundus Macq. или Т. fuscicauda Big. От обоих отличается
меньшей величиной, иначе окрашенным брюшком и ногами и иначе устроенной лобной
мозолью».
Типовой экземпляр в Мюнхенском музее не сохранился.
IV. Группа Tabanus argenteomaculatus Krob.
Крупные, черные или черно-серые виды: 21—24 мм. Глаза без полосок, голые
или в волосках, г4 с придатком. 1-я заднекрайняя ячейка к вершине сужена. Лоб
широкий, его высота превосходит ширину в 2 V2—3 Ч2 раза. Нижняя лобная мозоль
большая, ее нижний край б. м. на границе с лобным треугольником. Средняя лобная
мозоль б. м. веретеновидная или клиновидная, часто целиком скрыта налетом.
Концевой членик щупалец утолщен. Горные виды.
По-видимому, группа Т. argenteomaculatus родственна группе североамериканских
крупных темноокрашенных широколобых видов, например, Т. atratus F., Т. punctifer
О. S., Т. stygius Say и др.
40. Tabanus argenteomaculatus Krob. (рис. 178) — Слепень серебристый.
К г 6 Ь е г, 1928, Zool. Anzeiger, 76 : 263; О лсуфьев, 1937а : 285;
Шевченко, 1961:160; Leclercq, 1966:139.
8. TABANUS 249
$. Глаза в коротких, серых волосках, без полосок. Лоб широкий, его высота
превосходит ширину приблизительно в 2 V2—3 раза, слегка расширяется кверху,
покрыт темно-серым налетом. Нижняя лобная мозоль черная, блестящая, большая,
б. м. треугольная или прямоугольная, не достигает края глаз, средняя мозоль в виде
продолговатого черного блестящего пятна с продольной бороздкой посредине,
широко соединена с. нижней мозолью. Обе мозоли частично, реже целиком прикрыты
налетом. Затылок в коротких волосках. Лобный треугольник в сером налете. Усики
черные; 3-й членик усиков узкий, дорсальный угол тупоугольный. Концевой членик
щупалец желтовато-серый, в основании утолщен, покрыт короткими черными
волосками. Ноги одноцветно-черные, в черных волосках, лишь в основании средних и
задних голеней имеется ничтожная примесь коротких серебристых волосков. Крылья
интенсивно коричнево затемнены; вершина и задний край слегка просветлены; г4 с
длинным придатком. Брюшко черное, в черных волосках, боковые стороны III и частью
II тергитов брюшка с большими пятнами сероватого налета и серебристых волосков.
Снизу брюшко блестяще-черное. 19—22 мм.
Рис. 178. Tabanus argenteomaculatus Krob. Лобная полоска, усик
и щупальце $. А — вариант, лобные мозоли целиком прикрыты
налетом.
в. Голова большая, сильно выпуклая. Глаза в длинных, серых волосках.
Фасетки верхних частей глаз значительно крупнее нижних (приблизительно в 4—5 раз),
граница между теми.и другими резкая. Затылок за глазами в коротких черных
волосках. Усики как у §,• с более узким 3-м члеником. Лобный треугольник в сером налете.
Концевой членик щупалец коричневый, овальный, с острооттянутым вниз маленьким
кончиком, покрыт длинными, черными волосками. Придатки груди и брюшко как
у $; серебристые пятна на II—III тергитах меньшей величины, нежели у $ . 22 мм.
Распространение: среднеазиатский г'орный вид. Таджикистан (Гиссар-
ский хребет), Узбекистан (Сурхандарьинская, Самаркандская, Ферганская области),
Казахстан (Джуигарский Алатау, дол. р. Теректы, яблоневая зона, 29 VII 1936 1 ?
В. Н. Шнитников). Редок. Июнь—август.
41. Tabanus montiasiaticus Ols. sp. nova (рис. 179) — Слепень горноазиатский.
— grandis (Szil.) apud О 1 s u f j e v, 1937, et auct.
$. Крупный, черно-серый вид с продолговатым телом. Глаза голые, без полосок.
Лобная полоска широкая, слегка расширена кверху, ее высота превосходит ширину
основания приблизительно в 3 1/2 раза, покрыта пепельно-серым налетом. Нижняя
лобная мозоль блестяще-черная, б. м. квадратная, с закругленными углами,
боковыми сторонами не достигает края глаз. Средняя мозоль б. м. веретеновидная или
клиновидная, обычно тонкой полоской соединена с нижней; иногда мозоль целиком
250 IIL ПОДСЕМ. TABANINAE
прикрыта налетом. Лобный треугольник в сером налете. Затылочная полоска узкая,
в коротких черных волосках. Усики целиком черные, 1-й и 2-й членики в сероватом
налете. 3-й членик относительно узкий, дорсальный угол тупоугольный. Концевой
членик щупалец беловатый или сероватый, в основной половине довольно сильно
утолщен, покрыт короткими черными и белыми волосками. Нотоплевры коричневые,
в сером налете. Ноги целиком черные или же голени в той или иной мере коричневатые.
Крылья бесцветные; г4 с придатком. 1-я заднекрайняя ячейка к вершине заметно
сужена. Брюшко продолговатое, черное, в тонком голубовато-сером налете и коротких
беловатых и черных волосках. Светлые волоски преобладают на I—III или I—IV тер-
гитах, особенно по бокам, черные — на последующих тергитах; переход не резкий.
Снизу брюшко черно-серое, боковые стороны в беловатом налете и светлых волосках,
выделяющих темную срединную полоску. VIII стерпит: гонопофиз б. >м. конический,
с небольшой апикальной выемкой; латеральные лопасти умеренно развиты, с
коричневым пятном; базальная лопасть со слабо вогнутым основным краем. Вазальная пла-
Рис. 179. Taba.nus montiasiaticus Ols. Лобная полоска, усик, щупальце, VIII и
IX стерниты, церки и сперматека $.
стинка IX стернита с небольшой, но глубокой выемкой по основному и внутреннему
краям. Церки с уплощенной вершиной. Сперматека бомбовидная. 17—24 мм.
S. Голова значительно крупнее, чем у $, полушаровидная. Глаза голые.
Фасетки верхних 3/4 глаз приблизительно в 5—6 раз крупнее нижних, граница между
ними резкая. Затылок узкий, в очень коротких волосках, едва заметных лишь возле
темени. Лобный треугольник в сером налете, более темный в верхней части. Усики
как у $, но 3-й членик более узкий. Концевой членик щупалец желтовато^палевый,
продолговато-овальный, в черных и светлых волосках. Грудь и ее придатки как у $.
Брюшко коническое, в сером налете, короткие серые волоски с примесью черных
имеются главным образом по бокам II—III тергитов, остальное преимущественно в черных
волосках. Снизу брюшко как у $. 20—21 мм.
Описываемый вид был нами (Олсуфьев, 1937а) без изучения типа идентифицирован
с Т. grandis, описанным Силади (Szilady) в 1923 г. по 1 $ из Малой Азии (Гамидие),
хотя имевшиеся в нашем распоряжении экземпляры имели происхождение из горных
частей Средней Азии. Изучение накопившегося за ряд лет коллекционного материала,
а также данных о распространении среднеазиатской формы ставят под сомнение нашу
идентификацию этой формы с Т. grandis Szil. В описании последнего Силади
указывает: «. . .брюшко черное, с каймами из тонких серебристо-белых волосков и
серебристо-серого покрытия (tomentum), которое образовано короткими прилегающими
волосками и налетом того же цвета, покрытие прервано лишь широкой черной
полосой на вентральной стороне». У $$ среднеазиатской формы беловато-серые волоски
на брюшке не образуют каймы по заднему краю колец. Голубовато-серый налет
сверху покрывает все брюшко, а короткие беловато-серые волоски расположены по
бокам I—III тергитов (иногда также в средней части), на IV тергите светлые волоски
имеются в небольшом количестве (могут отсутствовать), остальное в черных волосках.
8. TABANUS
251
Поскольку в группе Т. argenteomaculatus окраска брюшка имеет особо важное
таксономическое значение, следует признать, что среднеазиатская форма, принятая
нами ранее за Т. grandis Szil. и ныне описываемая как Т. montiasiaticus sp. nova,
и истинный Т. grandis Szil. не идентичны. Данные о распространении обоих видов
подтверждают их самостоятельность. Т. montiasiaticus распространен в горах Средней
Азии и западнее Гиссарского и Зеравшанского хребтов не известен. Т. grandis,
очевидно, свойствен только Передней Азии, поскольку он не найден в Иране, где обитает
близкий вид Т. 'shelkovnikovi. В Афганистане обитает другой близкий вид ■— Т. schiva
М. et Chv.
Голотип: ?, урочище Квак Гиссарского хребта, 2000 м, 35 км к сев. от Душанбе,
'29 VII 1937, Гуссаковский. В коллекции Зоологического института Академии наук
СССР, Ленинград.
Паратипы: 32 J 4 в из Таджикистана и других мест Средней Азии, в коллекции
Н. Г. Олсуфьева.
Распространение: горы Средней Азии, преимущественно средний пояс
и низкогорья. Таджикистан (хребты Гиссарский, Зеравшанский, Каратегинскии,
1000—2500 м), Узбекистан (Ташкентская обл.; Наманганский р-н; Фергана),
Киргизия (Чаткальский и Ферганский хребты), Казахстан (хребты Каржантау, Каратау,
Таласский, Заилийский, Джунгарский, Саур). Зап. Китай (Синьцзян). Обычен в
горах Таджикистана, в других местах
не часто. Июнь—июль, иногда рань- 1ШЩш1шп
ше или позднее. Личинки найдены ^
в речках, родниках и арыках
(Кадырова, 19726).
42. Tabanus schiva M. et Chv.
(рис. 180) — Слепень шива.
Moucha etChvala, 1959,
Acta Entom. Mus. Nat. Pragae,
XXXIII : 284 (T. grandis schiva);
L e с 1 e г с q, 1966 : 152 (Т. grandis
schiva).
J. Крупный черный вид. Глаза
голые, без полосок. Лобная полоска
широкая, слегка расширена кверху,
ее высота превосходит ширину
основания приблизительно в 3 х/2 раза,
покрыта серым налетом. Нижняя
лобная мозоль блестяще-черная, б. м.
Рис. 180. Tabanus schiva M. et Chv.
Лобная полоска, усик и щупальце.
Щ0Ш
w
ж
Шш
квадратная, с слегка закругленными верхними углами, боковыми сторонами немного
не достигает края глаз. Средняя мозоль черная, продолговатая или клиновидная,
соединена более узкой полоской с нижней. Лобный треугольник в сером налете.
Затылочная полоска узкая, в коротких черных волосках. Усики черные, 3-й членик
■относительно узкий, дорсальный угол б. м. тупой. Концевой членик щупалец
желтовато-палевый, в основании утолщен, в черных волосках. Нотоплевры коричневые
или черно-коричневые, в сером налете. Крылья дымчатые, слегка коричневатые вдоль
жилок, последние темно-коричневые; г4 с придатком. Ноги черные. Брюшко черное,
в коротких черных волосках, снизу б. м. блестящее. Терминалии как у Т.
montiasiaticus, лишь гонопофиз почти без апикальной выемки. 17—22.5 мм.
в неизвестен.
Распространение: горный афгано-таджикский вид, обитает в лесном
и субальпийском поясах (1500—2800 м). Таджикистан: Гиссарский и Каратегинскии
хребты. Афганистан. Местами многочислен. Июнь—июль.
43. Tabanus shelkovnikovi Param. (рис. 181) — Слепень Шелковникова.
Парамонов, 1933, Журнал Bio-Зоолог, циклу ВУАН, 4 : 34 (38); О л-
суфьев, 1937а : 288; 1941:75 {$); Abbassian-Lintzen, 1960b : 821;
L е с 1 е г с q, 1966 : 168.
9. Глаза голые, без полосок. Лобная полоска широкая, слегка расширена кверху,
■ее высота превосходит ширину основания в 3—3 V2 раза, покрыта серым налетом.
252
III. ПОДСЕШ. TABANINAE
Нижняя лобная мозоль блестяще-черная, б. м. квадратная, с закругленными
верхними углами, боковыми сторонами немного не достигает края глаз. Средняя мозоль
черная, продолговатая или клиновидная, соединена более узкой полоской с нижней
мозолью; средняя'мозоль часто целиком прикрыта налетом. Лобный треугольник в
сером налете. Затылочная полоска узкая, в коротких светлых и частью черных волосках.
Усики черные, иногда 1-й членик и основание 3-го коричневатые. Концевой членик
щупалец желтовато-белый, в ос-
новации утолщен, покрыт
преимущественно черными
волосками. Нотоплевры коричневые.
Крылья бесцветные; г4 с
придатком. Ноги черные, основания
голеней коричневатые. Брюшко
резко двуцветное: I—III тергиты
в густом голубовато-сером налете
и прилегающих беловатых
волосках, последующие тергиты
черные, в черных волосках. Снизу
брюшка боковые стороны I—III
или I—IV стернитов в
голубовато-сером налете и беловатых
волосках, посредине стернитов
широкая черная полоса, IV—VII
стерниты черные, в черных во-
тосках. 20—23 мм.
Рис. 181. Tabanus shelkovnikovi
Param. Лобная полоска, усик и
щупальце $.
в. Голова большая, сильно выпуклая. Глаза голые, фасетки верхних 8/* глаз
значительно крупнее нижних, приблизительно в 4—5 раз, граница между теми и
другими резкая. Затылок в коротких волосках. Лобный треугольник в сером налете,
вершинная часть затемнена. Усики как у $, но более узкие. Концевой членик щупалец
желтовато-белый, умеренно утолщен, в беловатых и черных волосках. Крылья и ноги
как у $. Брюшко окрашено как у J. 20 мм.
Распространение: горный закавказско-иранский вид. Нахичеванская
АССР (Ордубадский р-н); Армения (Мегринский р-н; Джермук). Сев. Иран. Редок.
Июнь—середина августа.
Аббассиан-Линтцен (Abbassian-Lintzen, 1961) описала из юго-восточного Ирана
(область Жирофт) подвид Т. shelkovnikovi meridionalis, отличающийся от
номинативной формы более темной мышино-серой окраской светлоокрашенных частей брюшка.
Однако название meridionalis является младшим омонимом Tabanus meridionalus Thun-
berg, 1827 : 58 и согласно Международным правилам номенклатуры должно быть
заменено другим названием. Мы предлагаем его изменить на Tabanus shelkovnikovi abbassi-
anae nom. novum.
V. Группа Tabanus bovinusL.
Крупные коренастые виды: 18—28 мм. Глаза голые, без полосок. Лобная полоска
относительно узкая, нижняя лобная мозоль треугольная или овальная, средняя
мозоль в виде узкой килевидной или линейной полоски, соединенной с нижней мозолью.
Усики чаще с умеренным дорсальным выступом. Концевой членик щупалец б. м.
узкий, у 99 обычно плоский, тупоконечный. Крылья бесцветные, реже затемнены
в основной половине; г4 без придатка. 1-я заднекрайняя ячейка чаще заметно сужена
к вершине. Сверху брюшко с одним срединным рядом светлых треугольных пятен
(у чериоокрашенных видов они могут отсутствовать).
Многочисленная группа, в Палеарктике свойственная в основном
Средиземноморской и Восточноазиатской подобластям; преимущественно в лесных ландшафтах,
ряд видов в горах. Некоторые виды обитают в Сев. Америке.
44. Tabanus prometheus Szil. (рис. 182) — Слепень Прометея.
S z i I a d у, 1923, Biolog. Hungar., 1.1 : 12 (Т. eggeri Schin. var. prometheus);
Krober in Lindner, 1925 : 117 (T. eggeri var. prometheus Szil.); Олсуфьев,
8. TABANUS 253
1937a : 314 (T. eggeri var. prometheus Szil.); Leclercq, 1966 : 174 (T. tinctus
prometheus Szil.); Chvala, Lyneborg a. Moucha, 1972 : 383. — capita О 1-
s u f j e v, 1937a : 299.
9. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 5 V2 раз, покрыта желтовато-серым налетом.
Нижняя лобная мозоль черная, реже темно-коричневая, овальная, слегка угловатая,
ее высота превосходит ширину в 1 1/2—2 раза. Средняя мозоль в виде килевидной
или веретеновидной- полоски, соединенной с нижней мозолью. Лобный треугольник
в желтовато-сером налете. Усики черные, иногда площадка темно-коричневая, палочка
черная. 3-й членик умеренно широкий, дорсальный угол прямоугольный или
тупоугольный. Верхний край усиковых ямок широкий, его ширина составляет 1/i—х/3
высоты лобного треугольника. Концевой членик щупалец коричневато-серый, длинный,
плоский, со слегка притуплённым кончиком, густо покрыт короткими черными
волосками, придающими щупальцам
черноватый оттенок; иногда
имеется примесь желто-серых
волосков. Нотоплевры коричневые.
Крылья с очень слабым
дымчатым оттенком; г4 без придатка.
1-я заднекрайняя ячейка сильно
сужена к вершине. Ноги:
передние бедра черные или темно-
коричневые, средние и задние
■бедра темно-коричневые, в
сероватом налете и светлых волосках,
передние голени в, основании,
средние и задние голени на
большом протяжении
желтовато-коричневые, остальное черное.
Брюшко сверху, шоколадно-ко-
ричневое, вдоль средней линии и
на конце черноватое. Срединные
т
Рис. 182. Tabanus prometheus
Szil. Лобная полоска, усик и
щупальце 9-
"**
треугольники серые, с вогнутыми боковыми сторонами, вершиной почти достигают
заднего края предыдущего кольца. Заднебоковые края тергитов покрыты серыми
волосками, образующими боковые нерезкие полукаймы, прочее в коротких черных
волосках. Снизу брюшко шоколадно-коричневое, последние 2—3 стернита черновато-
серые. Срединная темная полоска отсутствует нацело или же имеется легкое
продольное затемнение, не имеющее, однако, резких контуров. 20—24 мм.
3- Голова очень большая, сильно выпуклая, много крупнее, нежели у 9- Глаза
голые, без полосок. Верхние фасетки значительно крупнее нижних, приблизительно
в 5—6 раз. Граница между теми и другими резкая. Нижнее мелкофасетчатое поле
сильно развито, занимая явственно более 1/3 всей поверхности глаз. Затылок в очень
коротких волосках. Лобный треугольник серый, с темной вершиной. Усики как у 9»
но значительно уже. Концевой членик щупалец бледно-коричневато-серый,
продолговатый, на конце тупой или слегка конический, покрыт преимущественно черными
волосками. Грудь и ее придатки как у 9- Брюшко окрашено как у 9; срединные треуголь- •
ники более узкие и вершиной достигают лишь до половины тергитов, заднебоковые
полукаемки из серых волосков очень небольшие, расположены в задних углах
тергитов. Снизу брюшко шоколадно-коричневое, последние 2 или 3 стернита черноватые;
темная срединная полоса отсутствует, имеется лишь легкое затемнение без
обозначенных границ. 22—24 мм.
Крупный, шоколадно-коричневый вид. Из нашей фауны может быть смешан
с Т. tinctus, но легко отличается от него темно-коричневыми бедрами и темноокрашен-
ным концевым члеником щупалец. Т. prometheus описан Силади (Szilady) как подвид
Т. eggeri по экземплярам, собранным в долине Аракса. Произведенное нами изучение
типового экземпляра 9. полученного при содействии д-ра И. Моухи (Прага) из Музея
Вены, выяснило, что описанный нами Т. capita является синонимом Т. prometheus
Szil. Этот вид значительно отличается от Т. eggeri.
254 III. ПОДСЕМ. TABANINAE
Распространение: понтийско-гирканский вид. Азербайджан (низовья
Самура; Кусары; Нахичеванская АССР), Армения (Герусы, Мегри, Кафан).
Болгария, Иран. Не часто. Июль—август.
Нападает на животных преимущественно в сумерки (Гаузер, 1960).
45. Tabanus semiargenteus Ols. (рис. 183) — Слепень красивый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 301; L е с 1 е г с q,
1966 : 168.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит ширину
основания в А1/^—5 раз, покрыта серым, реже желтоватым налетом. Нижняя лобная
мозоль черная, продолговато-овальная, кверху слегка сужена; ее нижний край
округлый или с едва намечающимися двумя выступами. Средняя мозоль в виде узкой чер-
^-. ной, килевидной полоски, соединен-
^^г^у^Ш&яп нои с вершиной нижней мозоли. Лоб-
«1«.>.1цш»..-^!'двдр;^;^.'""'''-- ^^^ЩщЩЙйЩч. ный треугольник в светло-сером на-j
«-■ли,-. «/,»-^^^ss;.o«Vv лете. Усики целиком черные, 3-й
членик довольно узкий, с прямым
или тупым дорсальным углом.
Концевой членик щупалец желтовато-
серый, плоский, узкий, с тупым
кончиком, равномерно покрыт черными
волосками, придающими щупальцам
черноватую окраску. Реже членик
расширен или же покрыт смесью
черных и желтовато-серых волосков.
Нотоплевры коричневые или черно-
серые. Крылья бесцветные, лишь
костальное поле слегка буроватое;
г4 без придатка. 1-я заднекрай-
няя ячейка к вершине сильно сужена.
Ноги в большей своей части черные,
Рис. 183. Tabanus semiargenteus
Ols. Лобная полоска, усик и
щупальце J.
лишь основная 1/3 голеней желтовато-коричневая; иногда средние и задние голени
светлые на 2/3 своей длины. Брюшко производит впечатление покрытого плесенью, так как
I—III тергиты и отчасти боковые стороны IV тергита покрыты тонким беловатым нале-
■томи густыми, короткими, серебристо-белыми волосками. Сквозь волоски и налет
просвечивает основной коричнево-желтый цвет брюшка, принимающий благодаря налету
розоватый оттенок. IV—VII или V—VII тергиты черные, в черных волосках, II—III
или II—V тергиты имеют по срединному беловатому треугольнику из налета, но на
фоне белых волосков эти треугольники слабо выделяются и бывают резкими лишь на
черных IV и V тергитах. Снизу брюшко в основной половине желто-коричневое, V—
VII стерниты черные. Боковые стороны брюшка в густом беловатом налете,
выделяющем посредине широкую темную продольную полосу. 20—22 мм.
(J неизвестен.
Красивый, крупный вид, выделяющийся из всех известных нам видов группы
Т. bovinus по наличию густых беловатых волосков на брюшке, прикрывающих
большую часть I—III или I—IV тергитов.
Голотип: $, Кущи-беляк Доралагезского р-на, Армения, 16 VII 1934, Тер-Мина-
сян. В коллекции Зоологического института Академии наук СССР, Ленинград.
Распространение: ирано-закавказский горный вид. Армения;
Нахичеванская АССР. Сев. Иран. Июль—середина августа. Местами многочислен.
46. Tabanus olsufjevi Haus. (рис. 184) — Слепень Олсуфьева.
Гаузер, 1960, Энтомол. обозр., 39, 3:658; L е с 1 е г с q, 1966:162.—
olsufjevi Bogatshev etSamedov, 1949 : 72 (Т. semiargenteus Ols., ab.
olsufjevi).
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит ши
рину основания приблизительно в 5 раз, кверху слегка расширена, покрыта серовато"
желтым налетом. Нижняя лобная мозоль б. м. овальная или треугольно-щитовидная,
черно-коричневая, слабо блестящая, вверх переходит в линейную черную среднюю
т
Ш
8. TABANUS
255
мозоль. Лобный треугольник в желтовато-сером налете. Усики: 1-й и 2-й членики
темно-коричневые, 3-й черный, в основании слегка коричневый; площадка с прямым
или слегка тупым дорсальным углом. Концевой членик щупалец удлиненный, палево-
коричневатый, в'черных и светлых волосках. Нотоплевры коричневые или черные,
в сером налете. Крылья бесцветные; г4 без придатка; 1-я заднекрайняя ячейка сильно
сужена к вершине. Ноги: бедра черные, в сером налете, голени желтые с черноватой
вершиной, лапки черные. Брюшко сверху как бы припудрено, в желтовато-сером
налете и преимущественно желтовато-
серых коротких волосках. Сквозь
волоски и налет просвечивает
основной коричневато-желтый цвет
брюшка, захватывающий боковые стороны
I—IV (или I—V) тергитов,
посредине имеется неясная темная
продольная полоска и ряд сероватых,
нерезких, вытянутых треугольников.
Последние 2 или 3 тергита черновато-
серые, преимущественно в черных
волосках. Снизу брюшко в большей
своей части желтовато-коричневое,
в желтовато-сером налете и светлых
волосках, V—VII стерниты
черноватые, преимущественно в черных
волосках. Посредине имеется
нерезкая широкая темная полоса. 22—
25 мм.
(J. Голова больше, чем у ?,
полушаровидная. Глаза голые.
Фасетки верхних 3/4 глаз крупнее
нижних приблизительно в 5—6 раз,
граница между теми и другими резкая.
Лобный треугольник в сером налете.
Усики как у ?, но 3-й членик немного
Рис. 184. Tabanus olsufjevi Haus.
Лобная полоска, усик и щупальце $.
^1Ц
уже. Концевой членик щупалец продолговатый, почти в 2 раза длиннее наибольшей
толщины, светло-коричневый, в длинных светлых и черных волосках. Ноги, крылья,
брюшко как у $. 23 мм.
Похож на Т. semiargenteus, отличаяясь главным образом желтоватым оттенком
окраски светлых волосков брюшка и коричневыми 1-м и 2-м члениками усиков.
Этот вид был описан А. В. Богачевым и Н. Г. Самедовьгм (1949) в качестве абер-
рации Tabanus semiargenteus Ols., тогда как Е. Г. Гаузер (1960) выделил его в
самостоятельный вид; в- соответствии со статьей 10 (Ь) Международного кодекса
зоологической номенклатуры автором названия вида Т. olsufjevi является Е. Г. Гаузер.
Распространение: ирано-закавказский горный вид. Нахичеванская
АССР (Ордубадский р-н); Армения (Азизбеков; Личк-Мегри). Иран. Июль—начало
августа. Местами многочислен.
47. Tabanus anthrax Ols. (рис. 185) — Слепень угольный.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2:302; L е с 1 е г с q,
1966 : 138. — pfeifferi К г б b e r, 1938 : 239.
$. Глаза голые, без полосок. Лобная полоска узкая, высота превосходит ширину
основания в 5 раз, покрыта коричневым налетом. Нижняя лобная мозоль блестяще-
черная, крупная, продолговато-овальная с продольной бороздкой посредине.
Средняя мозоль узковеретеновидная, соединена с нижней мозолью. Лобный треугольник
и лицо в коричневом налете, последнее в коричнево-чериых волосках. Усики целиком
черные, 3-й членик умеренной ширины, с тупоугольным дорсальным углом.
Концевой членик щупалец черный, узкий, плоский, в черных волосках. Грудь, ноги и
жужжальца целиком черные. Бочки груди в черных волосках. Базальные 2/3 крыла сильно
коричнево затемнены, вершина прозрачная, слегка дымчатая; переход не резкий;
г4 без придатка. 1-я заднекрайняя ячейка к вершине слегка сужена. Брюшко широ-
256 III. ПОДСЕМ. TABANINAE
кое, с обеих сторон блестяще-черное, только в черных волосках. Светлые срединные
треугольники отсутствуют. 22—23 .мм.
3- Голова очень большая, резко выпуклая. Глаза голые, без полосок. Фасетки
верхних 3/4 глаз значительно крупнее нижних, приблизительно в 5—6 раз, граница
между теми и другими резкая. Затылок в очень коротких волосках. Лобный треуголь.
ник в коричневом налете, сверху черноватый. Усики целиком черные, 3-й членик
узкий, с б. м. прямым дорсальным углом. Лицо в коричневом налете и черных волос,
ках. Концевой членик щупалец коричнево-черный, продолговатый, в черных волосках#
Рис. 185. Tabanus anthrax Ols. Лобная полоска, усик
и щупальце $.
Ноги и брюшко целиком черные, в черных волосках. Крылья как у $, с сильным
коричневым затемнением, захватывающим базальные 2/3 крыла. 22 мм.
Характерный крупный, черный вид, с сильно затемненными в основной части
крыльями.
Голотип: $, Тылляк на р. Гилянчай Нахичеванской АССР, 1 VIII 1933, Знойно.
В коллекции Зоологического института Академии наук СССР, Ленинград.
Распространение: закавказско-ирано-турецкий горный вид. Нахиче-
ванская АССР (Ордубадский р-н); Армения (Гарнинский заповедник; Кавушуг). Иран,
Турция. Июль—начало августа. Редок.
48. Tabanus subparadoxus Ols. (рис. 186) — Слепень сумеречный.
Олсуфьев, 1941, Тр. зоол. сектора Груз. фил. АН СССР, III : 79; L е с-
lercq, 1966:171; Chvala, Lyneborg a. Moucha, 1972 : 368. — morio
Olsufjev, 1937a : 304 nee T. morio Fabricius, 1794, 1. с 363.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота приблизительно
в 5 раз превосходит ширину основания, покрыта светло-коричневатым налетом.
Нижняя лобная мозоль блестяще-черная или темно-коричневая, продолговато-овальная
или треугольно-щитовидная. Нижний край мозоли округлый или слегка раздвоен.
Средняя мозоль в виде узкой, черной, килевидной полоски, соединена с вершиной
нижней мозоли. Лобиый треугольник в светло-коричневатом налете. Верхний край усико-
вых ямок очень широкий, занимает от V3 до 1/2 высоты лобного треугольника,
желтоватый, плоский. Усики целиком черные. 3-й членик умеренной ширины, с бч м.
пр'ямым дорсальным углом. Концевой членик щупалец черно-серый, изнутри
коричневый, длинный, плоский, с тупым кончиком, густо покрыт короткими черными
волосками, придающими ему черноватую окраску. Нотоплевры черные. Крылья слегка
дымчатые; г4 без придатка. 1-я заднекрайняя ячейка к вершине несколько сужена. Ноги
8. TABANUS 257
целиком черные и в черных волосках, лишь передние голени в основании слегка
коричневые. Брюшко черное, слабо блестящее, в черных волосках; II тергит с
маленьким сероватым срединным треугольничком, последующие 2 или 3 тергита обычно
по заднему краю с крошечными сероватыми пятнышками — рудиментами срединных
треугольников; иногда брюшко целиком черное. Снизу брюшко блестяще-черное,
в черных волосках, без серого налета по бокам. Задний край стернитов, как правило,
целиком в черных волосках, реже его боковые стороны в немногочисленных,
малозаметных серых волосках. VIII стернит преимущественно темный, гонопофиз крупный,
Рис. 186.. Tabanus subparadoxus Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
с небольшой апикальной выемкой; латеральные лопасти умеренно развиты; базаль-
ная лопасть со слегка вогнутым основным краем. Церки со слабо уплощенной
вершиной. Сперматека миндалевидная. 21—24.5 мм.
S неизвестен.
Близок к Т. paradoxus Jaenn., отличаясь более темной окраской брюшка.
От Т. swiridowi и Т. perelegans, с которыми Т. subparadoxus также имеет много
сходства, последний легко отличается признаками, указанными в определительной
таблице.
Голотип: $, Нальчик Кабардино-Балкарской АССР, Ю. Вагнер. В коллекции
Зоологического института Академии наук СССР, Ленинград.
Распространение: предгорно-горный вид, эндемик Кавказа.
Кабардино-Балкарская, Чечено-Ингушская (Салги), Дагестанская АССР, Грузия (Тбилиси,
Лагодехи, Боржоми). Июль—август. Не часто.
Нападает на животных преимущественно рано утром или в сумерки (Исмаилов,
1970).
49. Tabanus swiridowi Portsch. (рис. 187) — Слепень Свиридова.
Portschinsky, 1881, Ногае Soc. Ent. Ross., XVI : 273; S z i 1 a d у, 1923 :
17; К г 6 b e г, 1924 : 107; Krober in Lindner, 1925 : 128; К г о b e r, 1928 :
268; Парамонов, 1929:182 (в); Олсуфьев, 1937а : 306; L e с 1 e г с q,
1966 : 172.
?. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 5 раз, покрыта серым налетом. Нижняя лобная
мозоль черная, тусклая, б. м. овальная, кверху слегка сужена. Нижний край мозоли
с 2 или 4 небольшими выступами. Средняя мозоль в виде тонкой, килевидной, черной
полоски, соединенной с вершиной нижней мозоли. Лобный треугольник светло-серый,
17 Н. Г. Олсуфьев
258 HI. ПОДСЕМ. TABANINAE
верхний край усиковых ямок узкий, составляет не более 1/в высоты лобного
треугольника. Усики целиком черные. 3-й членик довольно узкий, дорсальный угол б. м.
прямой. Концевой членик щупалец черно-серый, изнутри коричневый, плоский, со слегка
притуплённым кончиком, снаружи покрыт густыми, короткими черными волосками.
Нотоплевры черные или коричневые. Крылья почти бесцветные, лишь с очень слабым
дымчатым оттенком, жилки коричневые; ri без придатка. 1-я заднекрайняя ячейка
довольно сильно сужена к вершине. Ноги черные, базальная часть голеней темно-
коричневая. Иногда средние и задние голени в большей своей части коричневые.
Брюшко черное, в черных волосках. II или II—III тергиты по бокам
темно-коричневые. На II—V тергитах иногда бывают выражены крошечные, сероватые, срединные
треугольнички. Снизу брюшко блестяще-черное, в черных волосках. Задние края
стернитов по бокам в беловатых волосках, образующих хорошо заметные полукаемки.
Иногда брюшко в основной половине слегка коричневое. 19—24 мм.
Рис. 187. Tabanus swiridowi Portsch. Рис. 188. Tabanus perelegans Ols. Лоб-
Лобная полоска, усик и щупальце $. ная полоска и щупальце $; усик с?.
<J. Голова не больше, чем у $. Глаза голые, без полосок. Фасетки глаз б. м.
одинаковые. Лобный треугольник светло-серый. Усики целиком черные, узкие;
дорсальный угол 3-го членика почти прямой. Концевой членик щупалец черный или
коричневый, овальный, со слегка оттянутым вперед кончиком, покрыт черными волосками.
Грудь и ее придатки как у $. Брюшко черное, в черных волосках. II или II—III
тергиты с неясными темно-коричневыми боковыми пятнами. Снизу брюшко как у $. 17—
20 мм. х
Похож на Т. subparadoxus; $ отличается наличием темио-коричневых пятен по
бокам II—III тергитрв, чисто-белыми волосками лица и более узкими краями усиковых
ямок; мельче.
Распространение: горный вид, система Малого Кавказа: Армения,
Нахичеванская АССР. Восточная Турция. Июль. Редок.
Указание А. В. Богачева и Н. Г. Самедова (1949) этого вида для Боржоми и Баку-
риани, видимо, должно быть отнесено к Т. subparadoxus.
50. Tabanus perelegans Ols. (рис. 188) — Слепень видный.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 448.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 5 раз, кверху слегка расширена, покрыта серым
налетом. Нижняя лобная мозоль блестяще-черная, продолговато-овальная, в
основании с 2 небольшими выступами. Средняя мозоль черная, линейная, соединена с
нижней. Лобный треугольник и лицо в светло-сером налете, последнее в белых волосках.
Верхний край усиковых ямок узкий, составляет приблизительно V4 высоты лобного
треугольника. Усики: 1-й и 2-й членики черные, 3-й членик обломан. Концевой членик
8. TABANUS 259
щупалец черно-коричневый, удлиненный, в черных волосках. Нотоплевры черные,
в сером налете. Ноги черные. Крылья б. м. бесцветные; жилки темно-коричневые;
г4 без придатка; 1-я заднекрайняя ячейка резко сужена к вершине. Брюшко черное,
в черных волосках; задние каемки стернитов по бокам с небольшим количеством
светлых волосков. 23 мм.
$. Голова не крупнее, чем у $. Фасетки верхних 2/3 глаз крупнее нижних
приблизительно в 4 раза, переход нерезкий. Лобный треугольник в сером налете. Усики
черные, 3-й членик узкий, дорсальный угол б. м. прямой. Концевой членик щупалец
коричневый, продолговато-овальный, со слегка оттянутым вперед кончиком,
преимущественно в черных волосках. Грудь и ее придатки как у $. Брюшко черное. II
(или II—III) тергит по бокам с темно-коричневым оттенком. Снизу брюшко как у $.
21 мм.
Похож на Т. subparadoxus и Т. swiridowi. От первого,отличается более узким
верхним краем усиковых ямок и белыми волосками нижней части лица, от второго — $
отличается отсутствием коричневых пятен по бокам брюшка, 3 — различной
величиной фасеток верхних и нижних частей глаз.
Распространение: Армения, Саданахач, 2 VIII 1927, 2 <$ 1 $.
Рис. 189. Tabanus sudeticus sudeticus Zell. Лобная полоска, усик, щупальце,
VIII стернит и церки $.
51. Tabanus sudeticus Zell. (рис. 189) — Слепень судетский.
Изменчивый вид, представлен 2 подвидами: северным темноокрашенным Т.
sudeticus sudeticus Zell. и южным светлоокрашенным Т. sudeticus verralli Oldr.
51а. Tabanus sudeticus sudeticus Zell. — Слепень судетский номинативный.
Z e 11 e г, 1842, Isis, II : 815; Brauer, 1880 : 184; Lundbeck, 1907 : 126;'
V e г г a 11, 1909 : 397; S u г с о u f, 1924 : 169; К г б b e r, 1924 : 122; К г б Ь е г
in Lindner, 1925:127; Олсуфьев, 1937а : 307; Moucha etChvala,
1958а : 192; L е с 1 е г с q, 1966 : 171; Chvala, Lyneborg a.Moucha, 1972:
385.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину приблизительно в 4 раза, в желтом налете и в коротких преимущественно
черных волосках. Нижняя лобная мозоль матово-черная или черно-коричневая,
узкотреугольная, с б. м. выпуклыми сторонами. Нижний край мозоли с 4 небольшими вы-
17*
260 III. ПОДСЕШ. TABANINAE
ступами; последние варьируют в зависимости от налегания на мозоль серого налета
и могут не быть выражены. Вершина мозоли б. м. широко сливается с короткой,
грубой, килевидной полоской (средней мозолью). Лобный треугольник в желто-сером
налете. Верхний край усиковых ямок широкий, занимает 1/i—V3 высоты лобного
треугольника. Концевой членик щупалец желтовато-коричневый, плоский, длинный,
на конце тупой, покрыт короткими, главным образом черными волосками. Усики:
1—2-й членики и примерно базальная 1/i площадки 3-го членика красно-коричневые;
далее площадка коричнево-черная. Палочка черная. 3-й членик очень широкий, его
нижний край выпуклый, верхний с резким б. м. остроугольным дорсальным выступом,
за которым следует вырезка. Нотоплевры коричневые. Ноги: бедра черные, в сером
налете, большая часть передних голеней, средние и задние голени желтые. Концы
передних голеней и все лапки черные, лишь основания последних двух пар лапок
коричневые. Крылья с легким буроватым оттенком, хорошо заметным в костальном
поле. Брюшко широкое, черное, сверху с срединным рядом резких беловатых
треугольных пятен; последние короткие, с б. м. прямыми боковыми сторонами; вершина
0° К ВОСТОКУ ОТ Гринвича 90° КО" 150° Ш° 150°
0° 30° 60° 90° «0°
Рис. 190. Ареалы: 1 — Tabanus sudeticus (оба подвида); 2 — Т. amaenus
(оба подвида).
треугольников не достигает заднего края предыдущего тергита. Боковые стороны
I—III тергитов с крупными, но неясно отграниченными каштаново-коричневыми
пятнами. Задние края всех тергитов по бокам с широкими желтыми каемками,
расширяющимися к внешнему краю. Брюшко сверху в коротких прилегающих волосках,
усиливающих основной рисунок; треугольные пятна в беловатых, боковые стороны
каемок в золотисто-желтых, прочие темные части в черных волосках. Брюшко снизу
черное, в сером налете, с черной срединной полосой и широкими беловато-серыми
каемками по заднему краю стернитов, суженными в средней части колец. Иногда I—III
стерниты по бокам красновато-коричневые. Срединная полоса покрыта короткими,
черными, прочие части брюшка и каемки серыми волосками. VIII стернит: гонопофиз
крупный, с небольшой апикальной выемкой; латеральные лопасти умеренно
развиты, с коричневым пятном; базальная лопасть с прямыми или слегка вогнутыми
боковыми сторонами и основным краем. Церки с б. м. округлой вершиной. Сперматека
миндалевидная. 23—28 мм.
в- Голова ^умеренно большая, несколько выпуклее, нежели у 5- Глаза голые,
без полосок. Фасетки верхних 2f3 глаз крупнее нижних приблизительно в 5 раз,
граница между теми и другими довольно резкая. Лобный треугольник в желтовато-сером
налете, вершина коричневая. Концевой членик щупалец коричнево-желтый, оваль
ный, с оттянутым вперед кончиком, в умеренно длинных черных волосках. Усики
как у $; 3-й членик в основной части менее расширен. Нотоплевры коричневые. Ноги:
бедра и лапки черные, голени коричневые, их концы черно-коричневые. Крылья как
у $. Брюшко сверху окрашено как у ?, отличаясь несколько меньшими размерами
срединных треугольных пятен. Снизу брюшко черное, задние края стернитов с
резкими желтовато-серыми каемками, покрытыми светлыми волосками; прочие части
в черных волосках. II—IV стерниты по бокам часто темно-коричневые. 20—24-мм.
8. TABANUS 261
Распространение: европейский лесной подвид, часто в горах (рис. 190,
1). В СССР от Эстонии, Ленинградской обл., Белоруссии, Молдавии и запада Украины
на восток до Урала (крайнее восточное местонахождение — Тургояк Челябинской
обл., восточный склон Урала), на север до ст. Ревсельга Мурманской ж. д. (близ 61°
с. ш.), юга Архангельской обл. (Черевково) и Перми, на юг до средней полосы
Украины, Воронежской, Саратовской области и Ю. Урала (Залаир); Кавказ (Кавказский
заповедник; Северо-Осетинская АССР). Европа (кроме севера), ? Марокко. Июнь-
июль, редко позднее.
Довольно обычен в горах Ю. и частью Средн. Урала, а также в горах Европы,
в прочих частях ареала не часто. Распространение на Кавказе изолировано от
основного ареала, причем преобладает подвид Т. sudeticus verralli Oldr. Указание Т.
sudeticus для Омска (Zychy, 1901, цит. по: Виолович, 1968 : 97) следует считать ошибочным.
51b. Tabanus sudeticus verralli Oldr. — Слепень судетский южный.
О 1 d г о у d, 1939, Brit. Bloodsuck. Flies : 103 (Т. verralli). — perplexus V e r-
r a 11, 1909 : 399 (T. sudeticus Zell. var. perplexus).
От номинативной формы отличается более светлой окраской и меньшими
размерами тела. У 5 коричневые боковые пятна сверху брюшка захватывают I—IV тергиты,
а снизу соответственно I—IV стерниты, заднебоковые каемки тергитов сероватые.
Концевой членик щупалец преимущественно в светлых волосках. Средние бедра,
а также конец средних и задних голеней коричневые. 19—23 мм.
Распространение: в СССР изучено недостаточно. Достоверные находки
этого подвида известны в низовьях Днепра (Цюрупинск), в Белоруссии
(Борисовский р-н), в окр. Волгограда (Средняя Ахтуба) и на Кавказе: от Сочи на восток по
южным склонам Главного Кавказского хребта до Нухи: Аджарская АССР (окр. Батуми
и Кобулети). Средн. и Ю. Европа. Июль. Не часто.
52. Tabanus portschinskii Ols. (рис. 191) — Слепень Порчинского.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2:309; L е с 1 е г с а,
1966 : 163.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 5 раз, покрыта серым или желтовато-серым налетом.
Нижняя лобная мозоль черная или черно-коричневая, продолговатая, кверху слегка
сужена, ее нижний край с небольшим двойным выступом. Средняя мозоль в виде
тонкой килевидной полоски, соединенной с вершиной нижней мозоли. Лобный
треугольник и лицо в светло-сером налете, последнее в белых волосках. Усики целиком
черные, 3-й членик умеренной ширины, дорсальный угол б. м. прямой. Концевой
членик щупалец серый или слегка коричневатый, узкий, плоский, с острым или слегка
притуплённым кончиком, равномерно покрыт короткими черными волосками,
придающими щупальцам более темный оттенок. Нотоплевры темно-коричневые. Крылья
с легким'дымчатым оттенком, жилки черно-коричневые; г4 без придатка. 1-я заднекрай-
няя ячейка б. м. резко сужена к вершине. Ноги: бедра черно-серые, голени
коричневые; вершинная 1/2 или 2/3 передних голеней, кончики последующих голеней и лапки
всех ног черные. Брюшко черное с темными красно-коричневыми боковыми пятнами,
простирающимися на I—III, реже I—IV тергиты. Граница пятен неясная. Посредине
брюшка имеется продольный ряд белых б. м. равносторонних треугольников;
последние не достигают вершиной до заднего края предыдущего тергита. По заднебоковому
краю тергитов имеются довольно широкие светлые каемки. Эти последние покрыты
золотисто-желтыми волосками, а на задневнешних углах — серебристо-белыми.
Каемки посредине прерваны и не достигают белых треугольников, разделяясь темными
промежутками, покрытыми короткими черными волосками. Черные волоски
покрывают также всю переднюю часть тергитов. Снизу брюшко с широкой
черно-коричневой полосой посредине, а по бокам в густом беловатом налете. В основной половине
сквозь налет просвечивает основной коричневый цвет брюшка, благодаря чему налет
имеет розоватый оттенок. 20—21 мм.
с?. \Голова крупнее и значительно выпуклее, нежели у $, полушаровидная (рис. 192,
внизу). Глаза голые, без полосок. Фасетки верхних 2/3 глаз значительно крупнее
нижних, приблизительно в 5—6 раз. Граница между крупными и мелкими фасетками
резкая. Лобный треугольник в желтовато-белом налете, возле вершины темно-серый.
Усики как у 5, но более узкие. Концевой членик щупалец
бледно-желтовато-коричневый, продолговатый, со слегка оттянутым вперед кончиком, покрыт довольно
длинными черными волосками. Нотоплевры коричневые. Придатки груди как у $. Брюшко
сверху окрашено в общем' как у 5, белые срединные треугольники значительно меньше,
а заднебоковые светлые каемки несколько уже. Снизу брюшко
красновато-коричневое, последние 3 стернита черные. Посредине имеется коричневая продольная полоса.
Боковые стороны брюшка в тонком беловатом налете. 18.5—20 мм.
262
III. ПОДСЕЙ, TABANINAE
Очень похож на Т. spodopterus, отличие заключается главным образом в несколько
более темной окраске брюшка. S6 различаются по строению глаз и размерам головы
настолько резко, что это дало нам основание рассматривать их как самостоятельные
виды.
Распространение: Закавказье, в предгорьях и горах Большого и Малого
Кавказа; Грузия (окр. Тбилиси; Коджоры; Бакуриани; Омало, Большой
Кавказский хребет); Азербайджан (Закатаны; Аджикент; хр. Пант-даг; Ленкорань);
Армения (Дилижан; Красносельск). Не часто. Июнь—август.
Рис. 191. Tabanus portschinskii Ols. Лобная по- Рис. 192. Голова Tabanus spo-
лоска, усик и щупальце $. dopterus Mg. в {вверху) и Т.
portschinskii Ols. (J (внизу).
Голотип: $, Аджикент, VII 1909, Бабаджаниди. В коллекции Зоологического
института Академии наук СССР, Ленинград.
53. Tabanus spodopterus Mg. — Слепень пестрый.
Изменчив. В пределах своего обширного ареала представлен 3 подвидами:
номинативным Т. spodopterus spodopterus Mg., иберийским Т. spodopterus ibericus Ols., M.
et Chv. и понтийским Т. spodopterus ponticus Ols., M. et Chv. В СССР найден только
номинативный подвид Т. spodopterus spodopterus Mg.
53а. Tabanus spodopterus spodopterus Mg. (рис. 193) — Слепень пестрый
номинативный.
Meigen, 1820, Syst. Beschreib., 11:46; В г a u e r, 1880:181; S u г с о u i,
1924:170; К г б Ь е г, 1924:131; Krober in Lindner, 1925:126;
Олсуфьев, 1937а : 311; Moucha etChvala, 1958a : 191; L е с 1 е г с q, 1966 : 170;
Olsufjev, Moucha et Chvala, 1967 : 304; Chvala, Lyneborg a.
Moucha, 1972 : 379.
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 5 раз, покрыта желтовато-серым налетом. Нижняя
лобная мозоль тускло-черная, продолговатая, слегка суженная кверху. Нижний край
округлый или в виде 2 тупых выступов. Высота мозоли в 1V2—2 раза превосходит
ширину. Средняя мозоль в виде черной килевидной полоски, соединенной с вершиной
нижней мозоли. Лобный треугольник в светло-сером налете. Усики целиком черные.
3-й членик сравнительно узкий, с прямо- или тупоугольным дорсальным углом.
Концевой членик щупалец коричневато-серый, узкий, плоский, со слегка притуплённым
кончиком, равномерно покрыт короткими черными волосками, придающими
щупальцам более темную окраску. Нотоплевры коричневые. Крылья с легким дымчатым
оттенком, костальное поле буроватое. Жилки коричневые; г4 без придатка. 1-я задне-
8. TABANUS
263
крайняя ячейка довольно резко сужена к вершине, иногда почти замкнутая. Ноги:
бедра черно-серые, голени коричневые; вершинная 1/2 передних голеней и передние
лапки черные, кончики последующих голеней темно-коричневые. Средние и задние
лапки черные, с темно-коричневым основанием. Брюшко сверху весьма напоминает
таковое Т. bovinus, но окраска несколько более сочная, коричневая, с широкими задне-
боковыми каемками из белых и желтых волосков. Последние узкой полоской вдоль
заднего края немного не достигают срединных треугольников. Эти последние более
короткие, нежели у Т. bovinus, белые, с прямыми боковыми сторонами. Снизу брюшко
желтовато- или красновато-коричневое, боковые стороны в тонком беловатом налете,
резко выделяющем широкую, срединную красновато-коричневую продольную полосу.
19—23 мм.
S. Голова не крупнее, нежели у £ (рис. 192, вверху). Глаза голые, без полосок,
фасетки верхних частей глаз, в особенности возле шва, лишь немного крупнее ниж-
Рис. 193. Tabanus spodopterus spodopterus Mg.
Лобная полоска, усик и щупальце $.
них. Лобный треугольник серый, возле вершины черноватый. Усики как у $, но
более узкие. Концевой членик щупалец бледно-желтовато-коричневый, продолговатый,
на переднем конце конический, покрыт черными и частью желтовато-серыми
волосками. Нотоплевры коричневые. Придатки груди как у $, лишь голени несколько
более темные, коричневые. Брюшко в общем окрашено как у $, но более темное;
срединные треугольники меньшей величины, заднебоковые каемки из желтых волосков
несколько уже, примерно на 1/3. 17.5—21.5 мм.
Очень похож на Т. portschinskii, распространенного в Закавказье; различаются
главным образом t?t? по строению головы. Похож также на Т. bovinus, отличаясь
более узкими, чисто-черными усиками, более узкой лобной мозолью (5), более короткими,
резкими, белыми срединными треугольниками на дорсальной стороне и более резкой
красно-коричневой полосой на вентральной стороне брюшка (у Т. bovinus полоса
черно-бурая, часто распадается на отдельные пятна, по одному на каждом стерните).
Распространение: южноевропейский (средиземноморский) лесной
подвид, часто в горах. В СССР известен из Молдавии (Котовск, Унгены и др.),
Винницкой (Могилев-Подольский) и Одесской областей (Кодыма). Средн. и Ю. Европа. В СССР
редок, на западе местами обычен. Июль.
54. Tabanus tinctus Wlk. (рис. 194) — Слепень меченый.
Walker, 1850, Ins. Saunders. Dipt., 1 : 29; M о и с h a, 1971 : 30; Chvala,
Lyneborg a. Moucha, 1972 : 390. — mixtus S z i 1 a d y, 1914, 12 : 672 et auct.
J. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания в 5—5Va раза, покрыта серым налетом. Нижняя лобная мозоль тускло-
черная или темно-коричневая, продолговатая, с округлой вершиной и б. м. прямым
основанием или овальная. Высота мозоли превосходит ширину в 1V2 раза. Средняя
мозоль в виде тонкой, черной килевидной полоски, отходящей от вершины нижней
264
т. подсей, tabaninae
мозоли. Лобный треугольник в светло-сером налете. Усики красновато-коричневые,
палочка черноватая. 1-й членик усиков желтоватый, 3-й членик умеренной ширины,
с б. м. прямоугольным дорсальным углом. Концевой учленик щупалец желтовато-
белый, узкий, плоский, с б. м. острым кончиком, покрыт короткими беловатыми
волосками. Иногда имеется незначительная примесь черных волосков, в особенности
на кончике щупалец. Нотоплевры коричневые или желто-коричневые. Крылья
бесцветные, жилки коричневые; костальное поле слегка буроватое; г4 без придатка. 1-я
заднекрайняя ячейка заметно сужена к вершине. Ноги: бедра черно-серые, в
беловатых волосках, голени коричневые или желтые; вершинная половина передних
голеней и передние лапки черные, кончики средних и задних голеней коричневые или
черноватые. Средние и задние лапки черные, с коричневым основанием. Брюшко
красновато-коричневое, сверху с узкой черной срединной полоской; последняя на
II—Ш тергитах прервана возле заднего края, на V тергите расширена кзади и
захватывает последующие VI и VII тергиты 'целиком. Вдоль средней линии полоска
Рис. 194. Tabanus tinctus Wlk. Лобная'полоска,'
усик и щупальце $.
в значительной мере или почти целиком скрыта крупными, беловато-серыми
срединными треугольниками. Последние имеют почти прямые боковые стороны; их вершина
немного не достигает до заднего края предыдущего тергита. Задний край тергитов
с узкой беловато-серой каемкой, расширенной по боковым сторонам. Каемки в
желтоватых и беловато-серых, треугольники в беловатых волосках, прочие части брюшка
в черных коротких волосках. Снизу брюшко красновато-желтое, последние 2 стер-
нита черно-серые. Боковые стороны брюшка в нежном беловатом налете,
выделяющем посредине нерезкую красновато-желтую или коричневатую продольную полосу.
21—25 мм.
в. Голова не шире груди. Фасетки верхних 2/3 глаз крупнее нижних
приблизительно в 3—4 раза, граница между ними нерезкая. Усики красно-коричневые, с
коричневым концом, немного уже, чем у J. Концевой членик щупалец желтовато-белый,
продолговато-овальный, преимущественно в черных волосках. Ноги как у J. Брюшко
сверху с красновато-коричневыми боковыми пятнами на I—III, частично IV
тергитах. Темная срединная полоса, разделяющая эти пятна, занимает на II тергите
приблизительно 1/5 его ширины, на III тергите — Vg и на IV тергите — почти всю его
ширину. Последующие тергиты целиком черные. Срединные треугольники слабо раз
виты. Брюшко снизу коричневое, V—VII стерниты черно-коричневые. Темная
срединная полоса неясная. Стерниты по заднему краю с узкой беловатой каймой, которая
по бокам лишь в скудных беловатых волосках; другие части стернитов в черных
волосках. 21.5 мм.
Описание в нами сделано по экземплярам из Болгарии; ранее опубликованное
нами (Олсуфьев, 1937а) описание в, заимствованное у Крёбера (КгбЬег, 1925), видимо,
относится к какому-то другому виду.
8. TABANUS
265
Близок к Т. bovinus, отличаясь более светлыми усиками, иным строением
нижней лобной мозоли (J) и различием в величине фасеток верхней и нижней части
глаз (в).
Распространение: южноевропейский (средиземноморский)? степной вид.
В СССР известен из Закавказья: окр. Тбилиси, Ханлар, Варташен, дол. Аракса
РКусары. Ю. Европа, Сев. Африка, Турция. Июль. Редок.
Экземпляры из Закавказья ($$) отличаются от южноевропейских немного более
узким 3-м члеником усиков, желтоватым оттенком окраски брюшка и более светлыми
голенями; эту форму мы обозначаем как Т. tinctus var. transcaucasiensis Ols. var. nova.
55. Tabanus bovinus L. (рис. 195, 196) — Слепень бычий.
Linne, Syst. Nat., 1758:601; L о e w, 1858a : 606; В г а и е r, 1880:185;
Lundbeck, 1907 : 125; V e г r a 1 1, 1909 : 393; Surconf, 1924 : 167; К г б Ь е r,
1924 : 123; Krober in Lindner, 1925:108; Олсуфьев, 1937a : 316; Мои
cha et Chvala, 1958a : 193; Шевченко, 1961 : 168; L e с 1 e г с q, 1966 :
142; С к у ф ь и н, 1967 : 189 (личинка); В и о л о в и ч, 1968 : 97; С h v a 1 a, L у-
neborg а. Мои cha, 1972 : 388.
Рис. 195. Tabanus bovinus L. J.
'$. Глаза голые, без полосок. Лобная полоска узкая, слегка сужена к основанию,
ее высота превосходит ширину в 4—41/2 раза, в желтом налете. Нижняя лобная мозоль
слабо выпуклая, черная или черно-коричневая, треугольная, тонко
поперечно-морщинистая. Мозоль по нижнему краю с 4 небольшими выступами, почти
достигающими основания лба, тогда как ее вершина б. м. тонко соединена с,узкой и длинной
килевидной средней мозолью. Лобный треугольник в желто-сером налете. Концевой
членик щупалец палево-желтый, плоский, длинный, в основной половине умеренно
расширен, на конце тупой, покрыт короткими преимущественно светлыми волосками.
Усики в большей своей части черные: 1-й и 2-й членики, а иногда незначительная часть
основания 3-го членика коричневые. 3-й членик широкий, с резким остро- или
прямоугольным дорсальным выступом. Нотоплевры обычно коричнево-желтые, редко черно-
серые. Ноги: бедра черные, в сером налете; бблыпая часть передних, средние и
задние голени желтые. Средние и задние лапки в основной половине коричневые. Концы
передних голеней, передние лапки и концы последующих лапок черные. Крылья
прозрачные, с легким буроватым оттенком; г4 без придатка. 1-я заднекрайняя ячейка
266
III. ПОДСЕМ. TABANINAE
к вершине слегка сужена. Брюшко буро-черное, сверху с срединным рядом резких
серых или желтоватых треугольных пятен; последние длинные, с вогнутыми боковыми
сторонами, вершиной достигают или почти достигают заднего края предыдущего тер-
гита. Боковые стороны I—III тергитов с крупными, нерезко отграниченными желто-
коричневыми пятнами, тогда как последующие тергиты по бокам с такого же цвета
каемками. Каемки в желтых, срединные треугольные пятна обычно в беловатых,
прочие части брюшка в черных коротко прилегающих волосках. Снизу брюшко в
основной половине коричнево- или розово-желтое, к концу темное; посредине имеется
коричневая или черноватая продольная полоса. Концевые 3 стернита по заднему краю
с узкими желтыми каемками. Срединная полоса в черных, тогда как боковые стороны,
покрытые тонким серым налетом, б. м. скрывающим основную окраску, — в серых,
коротко прилегающих волосках. VIII стернит: гонопофиз довольно' широкий, с
небольшой апикальной выемкой; латеральные лопасти умеренно развиты, с коричне-
Рис. 196. Tabanus bovinus L. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
вым пятном; базальная лопасть вогнута по бокам и основному краю. Церки с
округлой или слегка уплощенной вершиной. Сперматека миндалевидная. Терминалии сходны
с таковыми Т. sudeticus и не всегда различимы между собой. 19—24 мм.
в. Голова по своим размерам не превосходит таковую у $. Глаза голые, без
полосок. Фасетки глаз б. м. одинаковой величины, лишь возле глазного шва слегка
крупнее нижних. Лобный треугольник в желтовато-сером налете, его вершина коричневая.
Концевой членик щупалец продолговато-овальный, бледно-желтовато-коричневый,
покрыт умеренно длинными черными и светлыми волосками. Усики как у $; 3-й членик
в основании менее расширен, 2-й членик иногда черный. Нотоплевры коричневые.
Прочее как у $, за исключением треугольных пятен на брюшке, которые у в несколько
меньших размеров. 20—24 мм.
Распространение: европейско-сибирский лесной вид (рис. 197, 1).
В СССР распространен от Прибалтики и запада Украины на восток до Тайшета
Иркутской обл. и Минусинска, на север до Суоярвского и Кондопожского р-нов
Карельской АССР, Усть-Вымского р-на Коми АССР (62° с. ш.), Ханты-Мансийска Тюменской
обл., Назина с.-з. Томской обл. (60° с. ш.) и Томска, на юг до дельты Дуная, Крыма,
Предкавказья, Закавказья (включая Грузию, Армению, Азербайджан), Кулагине
на р. Урале, Оренбурга, Павлодара и Зайсана. Европа (на запад до Испании), Сев.
Африка. Обычен в лесной зоне и лесостепи. Июнь—июль, редко позднее. Обычен.
Сообщение Г. А. Бабаянца (1962) о нахождении Т. bovinus в Туркмении ошибочно.
Яйца откладывают по берегам озер, болот, стариц, затонов; личинки в берегах
речек, в черно-ольховых топях, полузаболоченных пойменных озерах, канавах, бере-
8. TABANUS Ж7
гах пруда (Скуфьин, 19526), взрослые личинки иногда в почве вдали от воды
(Олсуфьев, 1937а). Куколок находили в моховых кочках небольших лесных осоково-мохо-
вых болот (Виолович, 1968).
Экспериментально доказанный переносчик сибирской язвы (Олсуфьев и Лелеп,
1935).
0° К ВОСТОКу ОТ Гринвича 90° 120° 150° 180° «0°
0° 30° 60° 30° 120°_
Рис. 197. Ареалы: 1 — Tabanus bovinus^ 2 — Т. pleskei.
Рис. 198. Tabanus pleskei Krob. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
56. Tabanus pleskei Krob. (рис. 198) — Слепень Плеске.
К г б Ь е г, 1924, Arch. Naturg., 90 А, 9 :124; КгбЬег in Lindner, 1925 :
122; Олсуфьев, 1937а : 318; L е с 1 е г с q, 1966 : 163; Murdoch etTaka-
h a s i, 1969 : 61; Соболева, 1970а : 250 (личинка, куколка).
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота в 4—41/2 раза
превосходит ширину основания, покрыта голубовато- или желтовато-серым налетом.
-1 . Я/. ПОДСЕМ. TABANINAE
Нижняя лобная мозоль черная или черно-коричневая, треугольная, с слегка
выпуклыми боковыми сторонами. Ее ■ поверхность в тонких морщинках. Нижний край
мозоли с 4 б. м. явственными выступами. Средняя мозоль узкая, килевидная, черная,
соединена с вершиной нижней мозоли. Лобный треугольник в желтовато-сером
налете. Усики коричнево- или оранжево-желтые, 3-й членик в вершинной половине
коричнево-черный. Последний в основании довольно широкий, с резким, прямо- или
остроугольным дорсальным выступом. Концевой членик щупалец желтовато-серый,
светлый, покрыт короткими черными и.желтовато-серыми волосками; обычно
преобладают черные волоски. Форма членика как у Т. bovinus. Нотоплевры черно-серые,
реже коричневатые. Крылья с легким буроватым оттенком; г4 без придатка. 1-я задне-
крайняя ячейка слегка сужена к вершине. Ноги: бедра черно-серые, кончики
средних и задних бедер, базальные 2/3 или V2 передних голеней и большая часть средних
и задних голеней желтые. Вершинная V3 передних голеней и передние лапки черные,
последующие лапки черно-коричневые, с более светлым основанием. Кончик средних
и задних голеней часто коричневый. Брюшко широкое, черное, I—III тер гиты по
бокам с крупными, неясно отграниченными коричневыми пятнами. Посредине имеется
ряд узких, серых треугольников, вершиной почти достигающих до заднего края
предыдущего тергита. Тергиты по задпему краю с коричнево-желтыми каемками;
последние в средней части узкие, тогда как по бокам довольно сильно расширены. Каемки
покрыты сероватыми или желтоватыми волосками, треугольники — серыми, прочие
части брюшка — короткими черными волосками. Снизу брюшко в основной половине
светло-желтоватое или розовато-коричневое, далее черно-серое. Посредине имеется
темная, черно-бурая продольная полоса. Терминалии весьма сходны с таковыми
Т. bovinus. Гонопофиз может иметь более глубокую апикальную выемку, а базальная
лопасть VIII стернита — значительно более вогнутые боковые стороны и основной
край. 20—26 мм.
6. Голова не крупнее, нежели у $. Фасетки глаз б. м. одинаковые. Глаза голые,
без полосок. Лобный треугольник в сером налете. Усики как у $?, но менее широкие.
Концевой членик щупалец бледно-желтовато-коричневатый,. продолговато-овальный,
на переднем конце с слегка оттянутым вниз кончиком, покрыт преимущественно
черными волосками. Крылья как у $. Ноги несколько темнее окрашены, нежели у ?,
а именно: голени коричневые, средние и задние лапки черноватые. Брюшко как у $;
желтоватые каймы по заднему краю тергитов более узкие, серые срединные
треугольники маленькие, слабо выделяются. 21—22 мм.
Весьма близок к Т. bovinus, отличаясь от последнего более светлыми усиками,
темными нотоплеврами, менее ясными срединными треугольниками на брюшке и
общим свинцово-серым оттенком в окраске тела.
Распространение: восточносибирский лесной вид, аллопатричен Т.
bovinus (рис. 197, 2). Тогучин Новосибирской обл., Красноярский край (от Минусинска
на север до Енисейска и Среднего Приангарья), Тувинская АССР, Прибайкалье и
Забайкалье, Амурская обл., Хабаровский край (на север до Нижнетамбовского на
Амуре), Приморский край. Сев. Монголия, Сев.-Вост. Китай, север КНДР. Июль.
Местами обычен.
Указание Н. А. Виоловича (1968) об обнаружении Т. pleskei на Алтае (Усть-Кокса)-
нуждается в подтверждении.
57-. Tabanus sapporoenus Shir. (рис. 199) — Слепень японский.
S h i г а к i, 1918, Bloodsuck. Ins. Formosa, 1 : 295; Krober in Lindner,
1925 : 125; Takahasi, 1962 : 74; Виолович, 1968 : 102; Murdoch et
Takahasi, 1969 : 60.
$. Глаза голые, без полосок. Лобная полоска умеренной ширины, ее высота
превосходит ширину основания приблизительно в 4 раза, с б. м. параллельными сторонами,
покрыта желтовато-сероватым налетом. Нижняя лобная мозоль черная или темно-
коричневая, продолговато-треугольная или щитовидная, кверху продолжена в
короткую черную среднюю мозоль в виде клиновидно сужающейся к вершине полоски.
Лобный треугольник в желтоватом налете. Верхний край усиковых ямок широкий. Усики:
1-й и 2-й членики желто-коричневые, 3-й членик в основании красно-коричневый,
остальное черное. Дорсальный угол острый, в виде направленного вперед зубца.
Концевой членик щупалец узкий, желто-коричневый, в коротких черных волосках.
Нотоплевры черные, в сером налете, иногда коричневатые. Крылья слегка коричневато
затемнены, жилки коричневые; г4 без придатка. Ноги: бедра черные, в сером налете,
вершинная 1/3 или % средних и задних бедер желто-коричневая, голени желтые, конец
передних голеней и лапки черные, средние и задние лапки коричневые с черноватым
концом. Брюшко: боковые стороны II—III и частично IV тергитов
рыжевато-коричневые, посредине имеется неясно отграниченная темная полоса, занимающая на II—
III тергитах 115—111 их ширины; IV—VII тергиты черноватые, в желтовато-буром
налете, задний край тергитов с желтой каймой. Вдоль средней линии• имеются неясные
серые треугольники, немного не достигающие вершиной края предыдущего тергита.
8. TABANUS 269
Брюшко сверху преимущественно в коротких золотисто-желтых волосках, лишь на
III—IV (или II—IV) тергитах кпереди от срединных треугольников небольшое пятно
черно-коричневых волосков. Снизу брюшко черное, с широкими жёлтыми каймами
по заднему краю стернитов; боковые стороны брюшка в буроватом налете и
желтоватых волосках, посредине неясная черноватая продольная полоса преимущественно
в черных волосках. 22—25 мм (по Виоловичу, 1968, 16—20 мм).
в (по Takahasi, 1962). Голова шире груди, полушаровидная. Глаза голые.
Крупные фасетки занимают верхние 2/3 глаз, граница с мелкими фасетками нерезкая. Лобный
Рис. 199. Tabanus sapporoenus Shir. Лобная
полоска, усик и щупальце $.
треугольник, лицо и щеки в серовато-желтом налете. Усики уже, чем у $,
дорсальный выступ резкий, направленный вперед. Брюшко более коричневое, чем у $,
срединная черная полоса занимает около 1/4 ширины тергитов, с серовато-желтыми
срединными треугольниками. 21—24 мм.
Своеобразный крупный вид с зубцовидным дорсальным выступом на 3-м членике
усиков и рыжевато-коричневым брюшком (сверху), преимущественно покрытым
золотисто-желтыми короткими волосками.
Распространение: представитель восточноазиатского типа фауны.
О-ва Сахалин и Кунашир. Япония (о-ва Хоккайдо и Хонсю). Местами многочислен.
Июль—август.
VI. Группа Tabanus chrysurus Lw.
Крупные виды с резкими золотисто-желтыми каемками на брюшке по. заднему
краю тергитов (типа Hybomitra tarandina L.). Глаза голые, без полосок. Усики с
острым дорсальным выступом. Лобная полоска умеренно широкая, нижняя мозоль б. м.
треугольная, средняя линейная соединена с нижней. Концевой членик щупалец $
плоский, б. м. узкий; г4 без придатка. 1-я заднекрайняя ячейка почти не сужена
к вершине.
58. Tabanus chrysurus Lw. (рис. 200) — Слепень золотистый.
L о е w, 1858, Wien Ent. Monatschr., II : 103; S h i r a k i, 1918 : 407; Krober,
1924 : 116; Krober in Lindner, 1925 : 110; Олсуфьев, 1937a : 320; T a-
k a h a s i, 1962 : 70; В и о л о в и ч, 1968 : 104; Murdoch et Takahasi, 1969 :
59.
270 III. ПОДСЕМ. TABANINAE
$. Глаза голые, без полосок. Лобная полоска умеренной ширины, ее высота
превосходит ширину основания в 4—4Уа раза, покрыта желтым налетом. Нижняя
лобная мозоль треугольная, коричневая, ее вершина переходит в короткую килевидную
или линейную среднюю мозоль. Лобный треугольник в желтом налете. Усики
целиком оранжево-желтые, 3-й членик широкий, с резким дорсальным выступом.
Концевой членик щупалец оранжевый, узкий, с притуплённым концом, покрыт желтыми
и черными волосками. Нотоплевры желто-коричневые. Бочки груди в верхней части
Рис. 200. Tabanus chrysurus Lw. Лобная полоска, усик и ]
щупальце $.
в желтых волосках, мезоплевры и нижняя часть груди в черных волосках. Крылья
« буроватым оттенком, в основной половине возле костального поля — желтоватые.
Жилки желто-коричневые; г4 без придатка. Ноги: бедра черные или черно-коричневые,
с желтоватыми вершинами, голени желтые, передние лапки черные, лапки
последующих ног коричневые, с желтоватым основанием. Брюшко продолговатое, черное, с
широкими, резко выделяющимися золотисто-желтыми каемками (налет + волоски) по
заднему краю тергитов и стернитов. 23—30 мм.
6 (по Takahasi, 1962). Окраска как у $, желтые заднекрайние каймы на брюшке
■> уже, чем у $, срединные направленные вперед пятна неясные. Крупнофасетчатое поле
глаз занимает верхние 3/4 глаз, весьма резко отграничено от мелкофасетчатого поля.
Линия соприкосновения глаз длинная, примерно в 5 раз превосходит высоту лобного
треугольника. 23—25 мм.
В нашей фауне самый крупный вид — $$ до 30 мм.
Распространение: представитель восточноазиатского типа фауны
(рис. 177, 2). Приморский край (п. Дальний Кут в среднем течении Имана, Спасский,
Чугуевский р-ны, Супутинский заповедник). Сев.-Вост. Китай, Корейский п-ов,
Япония. Редок в Приморском крае и обычен в Японии. Июль—август. Указание
Н. А. Виоловича (1968) о распространении этого вида «на большей территории Китая,
включая Тайвань», неточно.
8. TABANUS
271
59. Tabanus buddha Portsch. (рис. 201) — Слепень Будды.
Portschinsky, 1887, Ногае Soc. Ent. Ross., 21 : 181; К г 6 b е г in L i n-
d n e г, 1925 : 110; S z i 1 a d у, 1926 : 604; Олсуфьев, 1937а : 322; L e с г е с q,
1966 : 145; Виолович, 1968 : 106; Соболева, 1968а : 165 (биология);
Murdoch et Takahasi, 1969 : 59; Соболева, 1970а : 254 (личинка, куколка).
$. Глаза голые, без полосок. Лобная полоска довольно широкая, с
параллельными сторонами, ее высота превосходит ширину основания в 3—3*/4 раза, покрыта
желтым налетом. Нижняя лобная мозоль блестяще-коричневая, широкотреугольная,
Рис. 201. Tabanus buddha Portsch. Лобная полоска, усик, щупальце, VIII стер-
Г нит, церки и сперматека $.
Ш'
с выпуклыми боковыми сторонами. Ее нижний край с 2 слегка выдающимися
закруглениями. Средняя мозоль отходит от нижней в виде тонкой, короткой, черной линии,
длиной не превосходящей высоту нижней мозоли. Лобный треугольник в желтом
налете. Усики целиком оранжевые, 3-й членик очень широкий, с резким, в виде зубца,
дорсальным выступом, за которым следует глубокая вырезка. Концевой членик
щупалец оранжево-желтый, плоский, довольно узкий, с тупым концом, покрыт короткими
желтыми волосками. Нотоплевры желто-коричневые. Бочки груди в черных волосках,
мезоплевры с пятном желто-коричневых волосков. Крылья буровато затемнены,
костальное поле и прилегающие к нему части крыла желтые. Жилки желто-коричневые,
к вершине и по заднему краю крыла — коричневые; г4 без придатка. Ноги целиком,
включая передние лапки, желто-коричневые; бедра несколько темнее, нежели голени.
Брюшко черное, с резкими желтыми каемками (налет-j-волоски) по заднему краю
тергитов. На I—II тергитах эти каемки сравнительно узкие, на двух последующих
кольцах значительно шире, тогда как 3 концевых тергита обычно целиком желтые. Снизу
брюшко повторяет рисунок верха, однако желтые каемки, начинаясь на II стерпите,
расширяются на последующих кольцах более равномерно. VIII стерпит желтый*
с боковыми коричневыми пятнами; гонопофиз конический, без апикальной выемки,
в тонких желтых и единичных черных волосках; латеральные лопасти небольшие;
272 III. ПОДСЕМ. TABANINAE
базальная лопасть удлиненная, с вогнутыми боковыми и основным краями. Церки
с слегка уплощенной вершиной. Сперматека миндалевидная. 22—26 мм.
в. Голова умеренной величины. Линия соприкосновения глаз в 21/2 раза больше
высоты лобного треугольника. Глаза голые, без полосок. Фасетки верхних 2/3 глаз
немного крупнее нижних, приблизительно в 2—3 раза. Затылок в очень коротких
волосках. Усики целиком оранжевые. 3-й членик усиков немного уже, чем у $, с
резким дорсальным зубцом. Лобный треугольник и лицо в оранжево-желтом налете,
последнее в золотистых волосках. Концевой членик щупалец оранжевый,
приблизительно в lV2 раза длиннее наибольшей ширины, с конической вершиной, в оранжевых
волосках. Грудь и ее придатки, а также брюшко как у $. 21 мм.
Похож на Т. chrysurus, легко отличаясь желто-коричневыми передними лапками,
более широким лбом, нижней лобной мозолью и черными волосками бочков груди.
Распространение: представитель восточноазиатского типа фауны
(рис. 164, '2). Юг и запад Приморского края, юг Хабаровского края (с. Бичевая и
с. Ходы на р. Хор). Сев.-Вост. и Вост. Китай (на юг до Сычуани), Корейский п-ов.
Не часто. Июнь—август. Указание Н. А. Виоловича (1968) о распространении этого
вида в Китае (о. Тайвань) неправильно.
Личинки обитают во влажной почве по берегам непроточных водоемов, в
заболоченных низинах. Кладки яиц белого цвета, на нижней стороне листьев осоки, ивы,
ольхи и других растений (Соболева, 1968а).
VII. Группа Tabanus autumnalisL.
Средней величины или крупные виды: 15—22 мм, крайне редко 14 мм. Глаза
голые, без полосок. Лобная полоска относительно узкая, нижняя лобная мозоль
треугольная или овальная, средняя мозоль в виде веретеновидной или линейной полоски,
соединенной с нижней мозолью. Усики чаще с умеренным дорсальным выступом.
Концевой членик щупалец узкий или умеренно утолщен, б. м. остроконечный. Крылья
бесцветные; г4 без придатка. 1-я заднекрайняя ячейка резко сужена к вершине, иногда
замкнута (исключение Т. atropathenicus — ячейка не сужена к вершине). Сверху
брюшко с 3 рядами светлых пятен из налета-f-волосков (иногда срединный ряд
отсутствует или боковые ряды ограничены I—II тергитами).
Численно небольшая группа, в Палеарктике свойственная в основном
Средиземноморской и Восточноазиатской подобластям. В отличие от группы Т. bovinus обитают
преимущественно в открытых ландшафтах.
60. Tabanus spectabilis Lw. (рис. 202, 203) — Слепень двуполоеый.
L о е w, 1858a, Verb. Zool.-bot. Gesell. Wien, 8 : 605; В г а, и е г, 1880 : 190;
Surcoui, 1924 : 173; К г б Ь е г, 1924 : 137; Krober in Lindner, 1925 :
126; Олсуфьев, 1937а : 294; Шевченко, 1961:183; L е с 1 е г с q, 1966:
169; Chvala, Lyneborg a. Moucha, 1972:370.
$?. Глаза голые', без полосок. Лобная полоска умеренно широкая, ее высота
в 3!/2—4 раза превосходит ширину основания, слегка расширена кверху, покрыта
желтовато-серым налетом. Нижняя лобная мозоль округлая или короткоовальная,
блестяще-черная. Средняя мозоль черная, веретеновидная, тонкой линией соединена
с вершиной нижней мозоли. Лобный треугольник в сером налете. Усики: 1-й и 2-й
членики бледно-коричневато-желтые, 3-й членик черный или коричневый с черной
палочкой, узкий, дорсальный угол тупоугольный. Концевой членик щупалец белый,
в основании умеренно утолщен, покрыт короткими, нежными, белыми волосками,
иногда на кончике имеются единичные черные волоски. Нотоплевры
светло-коричневые. Крылья бесцветные; г4 без придатка. 1-я заднекрайняя ячейка заметно сужена
к вершине. Ноги: бедра черно-серые, голени желтые. Вершинная 1/3 передних
голеней, кончики последующих голеней и лапки всех ног черные. Брюшко черное, по
бокам с двумя продольными, резкими, широкими розовато-белыми полосами. Эти
последние образованы соединенными между собой в непрерывный ряд пятнами, имеющими
слегка округлые очертания. Снизу брюшко розовато-серое, с более темной
коричневатой или черноватой нерезкой продольной полосой посредине. VIII стерпит
преимущественно темно окрашен; гонопофиз короткий и широкий, без выраженной
апикальной выемки; латеральные лопасти небольшие; базальная лопасть с вогнутым
основным краем. Церки б. м. треугольные. Сперматека бомбовидная. 16—21 мм.
<3. Голова несколько более выпуклая, нежели у $. Глаза голые, без полосок.
Фасетки верхней V2 глаз значительно крупнее нижних, приблизительно в 4—5 раз.
Граница между теми и другими резкая. Лобный треугольник в желтовато-сером
налете; его вершина коричневатая. Усики как у $,. но более узкие. Концевой чденик
белый, продолговато-овальный, на конце тупой, покрыт белыми и единичными черными
волосками. Грудь, ее придатки и брюшко как у ?. 16.5—20 мм.
8. TABANUS
273
Красивый, черно-серый вид, легко отличающийся от всех прочих видов группы
Т. autumnalis наличием на брюшке двух широких, розовато-белых полос, резко
выделяющихся на черном фоне.
\
V
Рис, 202. Tabanus spectabilis Lw. $.
та
штш
Рис. 203. Tabanus spectabilis Lw. Лобная полоска, усик, щупальце,
VIII стерпит, церки и сперматека $.
Распространение: средиземноморско-среднеазиатский степной вид,
частично в горах (рис. 204). Молдавская ССР (на север до Унген), Одесская обл. (Ко-
дым, Ананьев), Дагестан (на север до Буйнакска), Грузия, Армения, южная
Туркмения, Таджикистан, Узбекистан (включая Ферганскую долину), Казахстан —
Таласский Алатау и Каратау. Ю. Европа (па запад до Испании), Марокко, Передняя Азия,
Иран. Обычен. Июнь—август. '
18 Н. Г. Олсуфьев
274 Ш. ПОДСЕМ. TABANINAE
36 и $9 часто на стволах деревьев, в особенности на тополях. Личинок
обнаруживали в теплых проточных водоемах (Армения; Тертерян, 1960 и его личное
сообщение).
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180° 150°
Рис. 204. Ареал Tabanus spectabilis.
61. Tabanus autumnalis L. (рис. 205) — Слепень большой.
Политипический вид, в пределах своего огромного ареала представлен 2 формами:
темноокрашенным номинативным подвидом Т. autumnalis autumnalis L. и
светлоокрашенным южным подвидом Т. a. brunnescens Szil.
Рис.205. Tabanus autumnalis autumnalisL. J.
8. TABANUS 275
61a. Tabanus autumnalis autumnalis L. (рис. 206) — Слепень большой
номинативный.
L i n n ё, 1761, Fauna Suec. : 462; В г a u e r, 1880 : 192; Lundbeck, 1907 :
129; V err a 11, 1909:401; S u г с о u f, 1924:175; К r 6 b e r, 1924:139;
Kroner m Lindner, 1925 :107; Олсуфьев, 1937а : 295; Moucha et Chvala,
1958a : 191; Ш е в ч е н к o, 1961 : 165; L e с 1 e г с q, 1966 : 140; С к у ф ь и н, 1967:
193 (личинка); Виолович, 1968 :95; Chvala, Lyneborga. Moucha
$. Глаза голые, без ^полосок. Лобная полоска узкая, ее высота превосходит
ширину основания в 4—41/4 раза, кверху слегка расширена, покрыта пепельно-серым
налетом и короткими, преимущественно черными волосками. Нижняя лобная мозоль
плоская, овальная, коричнево-черная, слабо блестящая. Высота мозоли менее чем
Рис. 206. Tabanus autumnalis autumnalis L. Лобная полоска, усик, щупальце, VIII стер-
нит (варианты), перки и сперматека $.
■ в 2 раза превосходит ширину. Средняя мозоль в виде узкой килевидной черной полоски,
соединенной с вершиной нижней мозоли. Лобный треугольник в сером налете.
Концевой членик щупалец белый или желтовато-белый, тонкий, длинный, в оснований
слегка расширен, покрыт короткими белыми и частью черными волосками. Усики
узкие, чернью; 1-й членик и незначительная часть основания 3-го членика часто
коричневатые. 3-й членик в основании умеренно расширен, с б. м. тупоугольным
дорсальным углом. Нотоплевры коричневые или коричнево-желтые. Ноги: бедра черно-серые,
основная половина передних голеней и большая часть средних и задних голеней
палево-желтые, их концы, а также лапки черные. Крылья бесцветные; г4 без придатка.
1-я заднекрайняя ячейка заметно сужена к вершине. Брюшко продолговатое, черное,
сверху с резким серым рисунком, состоящим из 3 рядов крупных пятен из серебристо-
серого налета, треугольных посредине и б. м. ромбических по бокам. Треугольные
пятна б. м. длинные, с вогнутыми боковыми сторонами, вершиной обычно достигают
заднего края предыдущего тергита. Серые пятна в светлых, прочие части брюшка
в черных, коротких, прилегающих волосках. Иногда II—III тергиты по бокам
коричневатые (переходная форма к Т. a. brunnescensl). Снизу брюшко черно-серое, по бокам
в голубовато-сером налете и в беловатых волосках, посредине с б. м. широкой буро-
черной полосой, покрытой черными коротко прилегающими волосками. VIII стерпит
своеобразной формы: гонопофиз широкий, лопатообразный, с незначительной
апикальной выемкой; латеральные лопасти маленькие, слабо хитинизированы; по бокам
склерита по крупному коричневому пятну; базальная лопасть с вогнутыми боковыми
краями, основной край обычно двулопастной благодаря угловидной выемке. Церки с
уплощенной вершиной. Сперматека веретеновидная, с вытянутой вершиной. 16—22 мм
крайне редко 14 мм. '
в- Голова несколько более выпуклая, нежели у ?. Глаза без полосок. Фасетки
верхних 2/j глаз значительно крупнее нижних, примерно в 5—6 раз. Граница между
теми и другими резкая. Лобный треугольник в пепельно-сером налете с коричнево-
18*
376 Ш. ПОДСЕМ. TABANINAE
желтой вершиной. Концевой членик щупалец ланцетовидный или
продолговато-овальный в умеренно длинных, преимущественно черных волосках. Усики как у ?, но 3-й
членик в основании менее расширен. Грудь и ее придатки как у ?; иногда передние
голени целиком черные. Брюшко черное, с коричневыми боковыми пятнами,
захватывающими I—IV тергиты, и с 3 рядами серых пятен (налет+волоски). Последние имеют
ту же форму, что у ?, но боковые пятна выражены менее резко. Разделяющая
коричневые боковые'пятна черная срединная полоса занимает 1/3—1U ширины брюшка. Снизу
брюшко черное, с коричневым основанием, по^ бокам в голубовато-сером налете,
посредине с б. м. широкой буро-черной полосой. 17.5—20 мм.
Распространение: европейско-западносибирский лесостепной подвид
(рис 207 1). В СССР от Молдавии, запада Украины и Белоруссии на восток до
Енисея (устье Ангары, Красноярск, Минусинск), на север до Пскова, Ивановской обл.,
Кирова Никито-Ивделя на Урале и верховий р. Полуя Тюменской обл., на юг
до дельты Дуная, Крыма, Кавказа (преимущественно его западная часть), низовий
Рис. 207. Ареалы: 1 — Tabanus autumnalis autumnalis; 2 — Т. autumnalis
brunnescens.
Урала, Тургая, Бетпак-Далы (46° с. ш.), Алма-Аты, Фрунзе, Таласса и Чимкента.
Средн. и Ю. Европа, Турция, Зап. Монголия, Зап, Китай (Синьцзян), Обычен в
лесостепной и редок в лесной зоне. Июнь—август.
В южных частях ареала на значительном пространстве наряду с типичной формой
встречаются промежуточные экземпляры между Т. a. autumnalis и Т. a. brunnescens.
Указание А. В. Гуцевича (1940) на основе нашего определения иа нахождение
этого вида в Хабаровском крае (Обор) ошибочно, его следует отнести к Т. signatipen-
nis Portsch. .
Кладки яиц нами найдены в большом количестве в июле на стеолях bcirpus mari-
timus и соцветиях Butomus umbellatus вдоль берегов медленнотекущих речек, стариц
и пресных озер, личинки осенью и весной в слабо песчаных берегах упомянутых
водоемов, главным образом выше ватерлинии (Зап. Сибирь),
Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 193Ь)
и сибирской язвы (Олсуфьев и Лелеп, 1935).
61b. Tabanus autumnalis brunnescens Szil, — Слепень большой южный.
S z i 1 a d у, 1914, Ann. Mus. Nat. Hungar., 1'2 : 671 (T. brunnescens); S'uicouf,
1924 • 177 (T. brunnescens); К г 6 b e r, 1924 : 139 (T. brunnescens); Krober in
Lindner, 1925:107; Олсуфьев, 1937а : 297; Moucha etChvala, 1959b:
285; Шевченко, 1961 : 166; L e с 1 e г с q, 1966 : 141. — ? polygonus Walker,
1854 : 237, цит. no: L e с 1 e г с q, 1966 : 163. — ? polyzonatus Bigot, 1892 : 648.
Южный подвид. Отличается от основной формы значительно более светлой
окраской, главным образом желто-коричневым брюшком.
$. Нижняя лобная мозоль коричневая, ее высота часто в 2 раза превосходит
ширину. Лобная полоска в желтовато-сером налете. Усики: 1-й и 2-й членики желто-
коричневые, 3-й членик коричневый, с черной палочкой. Окраска сильно варьирует —
8. TABANUS 277
встречаются экземпляры с целиком оранжево-желтыми усиками (Туркмения).
Концевой членик щупалец белый, покрыт обычно только белыми волосками. Ноги:
средние и задние голени целиком коричневато-желтые, без затемненных кончиков,
средние и задние лапки коричневые. Встречаются экземпляры, у которых бедра средних
и задних ног целиком коричневато-желтые,, одноцветные с голенями, а передние
голени лишь на'самом конце черноватые. Брюшко коричневое или желто-коричневое,
лишь концевые -2—3 тергита и узкая продольная полоска посредине черноватые.
Светлые пятна из налета и волосков имеют легкий желтоватый оттенок; из них срединный
ряд'треугольников выражен довольно резко, тогда как боковые пятна часто несколько
стушевываются на основном желто-коричневом фоне и заметны лишь при
рассматривании брюшка сзади (у сильно потертых экземпляров пятна могут быть незаметны).
Снизу брюшко светло-коричневато-желтое, без выделяющейся темной полосы
посредине или лишь со слегка заметной полосой. Терминалии как у номинативного
подвида, лишь VIII стернит окрашен светлее, преимущественно желтый, боковые
коричневые пятна небольшие, светлые. 16—21 мм.
$. Очень похож на $. Крупнофасетчатое поле глаз резко выделяется более
светлой, песочной окраской от мелкофасетчатого поля, которое при жизни ярко-зеленое.
Усики, ноги и крылья как у $. Брюшко как у $, боковые пятна из желтоватого налета
и волосков часто менее явственны. 17—21 мм.
Распространение: средиземноморско-азиатский пустынно-степной
подвид (рис. 207, 2). Одесская (Балта), Николаевская области (Вознесенск, Снегиревка),
Кавказ (преимущественно восточная его часть), Туркмения, Узбекистан, Таджикистан,
Казахстан (на север до 48° с. ш.). Ю. Европа, Сев. Африка (включая Египет),
Передняя Азия, Иран, Афганистан. Обычен. Май—сентябрь. Хвала, Линеборг и Моуха
(Chvala, Lyneborg a. Moucha, 1972) считают наиболее светлые экземпляры Т.
autumnalis brunnescens Szil. идентичными Т. polygonus Walker, 1854= Г. polyzonatus Big.,
1892. Вопрос этот нуждается в дальнейшем изучении.
9? и SS часто на стволах тополей по краю дороги. Личинок обнаруживали в
Армении в теплых'проточных водоемах (Тертерян, 1960 и его личное сообщение), а в
Средней Азии (Ферганская долина) в рисовых
полях, болотах, арыках, речках и на озере -^^^^^ISiSMi^^^^
(Кадырова, 19726).
62. Tabanus atropathenicus Ols. (рис.
208) — Слепень привлекательный.
Олсуфьев, 1937а, Фауиа СССР,
Двукрылые, VII, 2 : 298; Leclercq,
1966 : 140.
$. Глаза голые, без полосок. Лобная
полоска умеренно широкая, кверху слегка
расширяется, ее высота превосходит
ширину основания в 33/4—4 раза, покрыта серым
налетом. Нижняя лобная мозоль выпуклая,
треугольно-щитовидная, слабо блестящая,
черная или черно-коричневая. Ее нижний
край без заметных выступов. Средняя мо-
Рис. 208. Tabanus atropathenicus Ols.
Лобная полоска, усик и щупальце J.
золь короткая, б. м. веретеновидная, черная, нижним концом соединена с вершиной
нижней мозоли. Затылок узкий, с рядом б. м. коротких черных и серых волосков.
Лобный треугольник в сером налете. Усики желто-коричневые, базальные членики
несколько светлее концевого. Палочка темно-коричневая.4 3-й членик усиков узкий,
его дорсальный выступ тупоугольный. Концевой членик щупалец беловатый, довольно
сильно утолщен, покрыт короткими белыми и единичными черными волосками. Ното-
плевры черно-серые. Крылья бесцветные; г4 без придатка. 1-я заднекрайняя ячейка
к концу не сужена. Ноги: бедра черно-серые, голени коричнево-желтые. Вершина
передних голеней и передние лапки черные, лапки последующих ног коричневые,
с черноватой вершиной. Кончики задних голеней коричневые. Брюшко черно-серое,
матовое, II—IV (или II) тергиты по бокам шоколадно-коричневые. Коричневая окраска
слегка заходит на I тергит. Продольно посредине имеется ряд хорошо заметных,
узких, серых треугольников, вершиной достигающих до переднего края тергита.
Боковые стороны брюшка с легким сероватым или желтоватым налетом, образующим
неясный ряд боковых пятен. Тергиты по заднему краю с узкой желтовато-серой каем-
278 III. ПОДСЕМ. TABANINAE
кой, покрытой серыми волосками. Снизу брюшко шоколадно- или желто-коричневое,
в нежном беловато-сером налете, 3 концевых стернита серые. Темная продольная
полоса посредине отсутствует. 16—20 мм.
в- Голова крупнее, чем у $, полушаровидная. Глаза голые. Фасетки верхних
3/4 глаз крупнее нижних в 5—6 раз, граница между ними резкая. Лобный
треугольник в сером налете, к вершине коричневый. Усики как у $, но более узкие. Концевой
членик щупалец беловатый, продолговато-овальный, приблизительно в 2 раза
длиннее наибольшей толщины, в светлых волосках. Ноги, крылья и брюшко окрашены
как у $. 16—19 мм.
Своеобразный вид, напоминающий по окраске переходную форму от Т. a. autu-
mnalis к Т. a. brunnescens, но легко отличающийся от нее несуженной к вершине 1-й
заднекрайней ячейкой и темными нотоплеврами; $ отличается еще утолщенными
щупальцами и иным строением лба.
Распространение: Понтийско-Гирканский (восточно-средиземноморский)
степной вид. Азербайджан: Ханларский р-н (Аджикенд), Нагорный Карабах, Нахи-
чеванская АССР; Армения (Армаж в дол. Аракса; Джермук); Туркмения (Геок-Тепин-
ский р-н). Иран. Редок. Июнь—июль.
Голотип: $, Белев на р. Гилян-чай Нахичеванской АССР, 27 VI 1933, Знойко.
В коллекции Зоологического института Академии наук СССР, Ленинград.
Рис. 209. Tabanus signatipennis Portsch. Лобная полоска,
усик, щупальце, VIII стернит, церки и семеприемник $.
63. Tabanus signatipennis Portsch. (рис. 209) — Слепень отличный.
Portschinsky, 1887, Ногае Soc. Ent. Ross., XXI : 180; О 1 s u f j e v,
1969 : 118; Соболева, 1971 : 144 (личинка, куколка). — tahasagoensis Shiraki,
1918 : 323 et auct. — mandarinus (Schiner) apud О u с h i, 1943 : 531 (Phyrta).
$. Глаза голые, без полосок. Высота лобной полоски приблизительно в 4—5 раз
превосходит ширину основания, кверху слегка расширена, покрыта желтовато-серым
налетом. Нижняя лобная мозоль вытянуто-треугольная, слабо блестящая, в нижней
части светло-коричневая, кверху темнее (иногда целиком черная), боковыми
сторонами немного не достигает края глаз, вершиной переходит в короткую, черную киле-
видную среднюю мозоль. Лобный треугольник в сером или слегка желтоватом налете.
Усики: J-й и 2-й членики темно-коричневые, 3-й членик черный, иногда коричневый
в основании, узкий, дорсальный угол б. м. прямой. Концевой членик палево-белый,'
узкий, в черных волосках с примесью светлых. Нотоплевры коричневые, в сером
налете. Крылья слегка дымчатые, 1-я заднекрайняя ячейка резко сужена к вершине,
иногда замкнутая с коротким стебельком (как у Т. amaenus); r4 без придатка (иногда
имеется незначительный рудимент). Ноги: бедра черные, в сером налете, средние и
задние бедра иногда просвечивают коричневым, голени желтые, концевая V3
передних голеней и передние лапки чёрные, кончики средних и задних голеней, а также
средние и задние лапки темно-коричневые или черноватые. Брюшко черное, сверху
с 3 рядами резких светло-серых пятен: треугольных посредине и ромбических или
овальных по бокам. Последние имеются по крайней мере на I—V ('или I—IV) тергитах.
Срединные треугольники с вогнутыми боковыми сторонами, вершиной достигают
8. TABANUS 379
заднего края тергитов. Иногда II тергит по бокам неясно коричневатый. Снизу брюшко
черно-серое, с неясной более темной срединной полосой. VIII стернит: гонопофиз
лопатообразный, без заметной апикальной выемки; латеральные лопасти небольшие,
коричневые; базальная лопасть с б. м. прямыми боковыми сторонами и основным краем.
Церки с уплощенной вершиной. Сперматека веретеновидная. 15—-18 мм.
3 (по Takahasi, 1962). Глаза голые, фасетки верхних 2/3 глаз крупнее нижних.
Линия соприкосновения глаз в 7 (?) раз превосходит высоту лобного треугольника.
Усики уже, чем у $. Концевой членик щупалец темно-коричневый, в сером налете,
цилиндрический, равномерно утолщен к концу. Брюшко как у $ или иногда его ба-
зальные 2 тергита с боковыми коричневыми пятнами. 17 мм.
Очень похож на Т. autumnalis autumnalis, но мельче и стройнее, 1-я
заднекрайняя ячейка к вершине сужена сильнее (иногда замкнутая), нижняя лобная мозоль
у $ вытянуто-треугольная.
Распространение: представитель восточноазиатского типа фауны. Юг
Приморского края (Хасанский р-н); Хабаровский край (Обор); Амурская обл. (Тыгда,
бассейн Зеи, 7 VIII 1959 1 ? Грунин).
Китай, Корейский п-ов, Япония. В СССР
редок, в Китае местами обычен. Июль—
август.
64. Tabanus trigeminus Coq. (рис. 210)
— Слепень дальневосточный.
Coquillet, 1898, Proc. U. S. Nat.
Mus., XXI : 310; Murdoch et
Takahasi, 1969 : 81; Соболева, 1968а : 162
(биология). —: mandarinus (Schiner) apud
Shiraki, 1918 : 301 et auct. — yama-
sakii О u с h i, 1943 : 532 (Phyrta).
$. Глаза голые, без полосок. Лобная
полоска узкая, ее высота- приблизительно
в 6 раз превосходит ширину основания,
кверху слегка расширена, покрыта серым
Рис. 210. Tabanus trigeminus Coq. Лобная
полоска, усик и щупальце $.
налетом. Нижняя лобная Мозоль тускло-черная, реже коричневая, вытянуто-тре
угольная, боковыми сторонами не достигает края глаз, кверху продолжена в килевид-
ную черную полоску. Лобный треугольник в сером налете. Усики: 1-й и 2-й членики
черные, в сером налете или коричневые, 3-й членик усиков черный, в основании
красновато-коричневый (иногда коричневая вся площадка), относительно узкий,
дорсальный угол прямой или немного острый. Концевой членик щупалец узкий,
светло-коричневый, в коротких черных волосках. Нотоплевры черные или коричневые, в сером
налете. Крылья слегка дымчатые, 1-я заднекрайняя ячейка резко сужена к вершине,
иногда замкнутая, с небольшим стебельком (как у Т. amaenus); r4 без придатка. Ноги:
бедра черные, в сером налете, голени желтые, их вершина, а также лапки черные.
Брюшко черное, сверху с резким беловато-серым рисунком из налета+волосков.
Посредине имеется продольный ряд крупных беловато-серых треугольников с
прямыми боковыми сторонами, вершиной достигающих заднего края предыдущих
тергитов; I —-II или I—III тергиты по бокам с неправильной формы светлыми пятнами,
образованными расширенными заднекрайними каемками, последующие тергиты с узкими
заднекрайними каемками из серого налета и беловатых волосков. Снизу брюшко черно-
серое, заднекрайние каемки желтоватые, посредине имеется резкая продольная
черноватая полоса. 15—20 мм.
в- Голова крупнее, чем у $, полушаровидная. Глаза голые. Фасетки верхних
3/4 глаз крупнее нижних приблизительно в 6—7 раз, граница между теми и другими
резкая. Лобный треугольник в желтовато-сером налете, вершина коричневая. Усики как
у $, но более узкие. Концевой членик щупалец палевый, продолговатый, к концу
немного утолщен, покрыт черными и светлыми волосками. Крылья и ноги как у $,
передние голени черноватые на 2/3 от вершины. Брюшко коническое, черное, I—III тергиты
по бокам иногда неясно коричневые, светлый рисунок из налета+волосков как у $.
Снизу брюшко как у $. 18—19 мм.
Близок к Т.. signatipennis, но в среднем крупнее и коренастее, светлые пятна
по бокам брюшка кзади не распространяются далее III тергита, у $ лоб более узкий.
280 т. ПОДСЕМ. TABANINAE
По заключению Филипа (Philip), Т. mandarinus Schin., синонимом которого
долгое время считался Т. trigeminus Coq., не идентичен последнему (Murdoch et
Takahasi, 1969).
Распространение: представитель восточноазиатского типа фауны. Юг
Приморского края (Кедровая падь и ст. Приморская Хасанского р-на). Китай (на юг
включая Тайвань), Япония. В СССР редок, в Китае и Японии местами обычен. Июль-
август.
Личинки в илистых берегах ручьев на пастбищах (Takahasi, 1962).
65. Tabaniis amaenus Wlk. — Слепень бурый.
Широко распространенный на юго-востоке Азии, изменчивый вид. Представлен
2 подвидами: южным номинативным Т. amaenus amaenus Walk, и северным Т.
amaenus griseinus Phil.
Рис. 211. Tabanus amaenus Wlk. Лобная полоска,
усик, щупальце, VIII стернит, перки и сперматека $.
65а. Tabanus amaenus amaenus Wlk. (рис. 211) — Слепень бурый номинативный.
Walker, 1848, List Dipt. Brit. Mus. : 103; Takahasi, 1962 : 104; Lec-
1 e г с q, 1966 : 138; Виолович, 1968 : 107; — clausacella Macquart, 1855 :
25, цит. no: L'e с 1 e г с q, 1966 : 138. — sinicus Bigot, 1892 : 629. — lateralis S h i-
r a k i, 1918 : 322 (T. amaenus Wlk., var.) — pallidiventris Olsufjev, 1937a : 324
(T. griseus Krob., subsp.).
$. Глаза голые, без полосок. Лобная полоска узкая, ее высота превосходит
ширину основания приблизительно в 51/2—6 раз, покрыта желтовато-серым налетом.
Нижняя лобная мозоль вытянуто-треугольная, темно-коричневая, блестящая, в
верхней части черноватая. Средняя мозоль не длиннее нижней, веретеновидная, черная,
соединена с вершиной последней; в этом месте обычно имеется небольшая перетяжка.
Лобный треугольник в беловато-сером налете. Усики оранжево-желтые или
коричневатые, палочка черная. 3-й членик усиков узкий, дорсальный угол с небольшим
зубчиком, снабженным черными волосками. Концевой членик щупалец
беловато-палевый, тонкий, покрыт короткими черными и белыми волосками. Нотоплевры желто-
коричневые. Крылья бесцветные, жилки темно-коричневые; г4 без придатка. 1-я задне-
крайняя ячейка замкнута и на конце с коротким стебельком. Ноги: бедра серые,
с желтыми кончиками, у средних бедер концевая 1/3 желтоватая, голени
бледно-коричневато-желтые, в белых и частью черных волосках; концы передних голеней и
передние лапки черные, лапки последующих ног черно-коричневые. Брюшко продолговатое,
черно-серое, с буроватым оттенком, II—III или I—IV тергиты по бокам коричневые.
Имеются 3 ряда неясных серых пятен (налет-|-волоски): срединный ряд узких пятен,
б. м. сливающихся в общую узкую продольную полоску и боковые ряды из
неправильной формы более крупных пятен; срединный ряд иногда не выражен или представлен
узкими неясными треугольниками. Снизу брюшко буровато-серое, в основной части
коричневато-желтое, посредине с нерезкой темной продольной полосой. Задние каемки
стернитов желтоватые. VIII стернит: гонопофиз конический, с маленькой
апикальной выемкой; латеральные лопасти небольшие, коричневые; базальная лопасть
удлиненная, с вогнутыми боковыми сторонами и основным краем. Церки с округлой
вершиной. Сперматека веретеновидная. 15—17 мм.
9. ATYLOTUS 381
3- Голова крупнее, нежели у $, полушаровидная. Глаза голые. Фасетки
верхних 3/4 глаз крупнее нижних приблизительно в 6—7 раз, граница между теми и
другими резкая. Лобный треугольник серый, его вершина коричневая. Усики как у ?,
но более узкие. Концевой членик щупалец продолговатый, к концу слегка утолщен,
желтоватый, покрыт беловатыми волосками. Ноги и крылья как у ?. Брюшко
коричнево-желтое с 2 боковыми рядами слегка косых желтоватых пятен (налет+волоски)
и с срединной черновато-серой полоской. Последняя состоит из продолговатых пятен,
не достигающих задним концом до края тергита. На II тергите полоска составляет
1/10—1/12 ширины кольца, на последующих тергитах постепенно расширяется, так что
последние 2 тергита целиком черно-серые. Средний ряд серых пятен не выражен или
едва заметен. Снизу брюшко коричневато-желтое, последние 3 стернита темно-серые.
16—16.5 мм.
Распространение: представитель восточноазиатского типа фауны
(рис. 190, 2). Китай (включая Тайвань), Корейский п-ов, Япония. Обычен. В СССР,
по-видимому, отсутствует.
65b. Tabanus amaenus griseinus Phil. — Слепень бурый северный.
Philip, 1960, Malaysian Parasites, XXXV : 31 (Т. griseinus); Takahasi,
1962 : 108 (Т. griseinus Phil.); Соболева, 1968а : 163 (биология); 1970а : 257
(личинка, куколка). —griseus К г б b e r, 1928:270, пес Tabanus (Therioplectes)
griseus Е п d e r 1 е i n, 1925 : 367.
$. От номинативной формы отличается более темной окраской брюшка: оно
целиком черно-серое, лишь иногда II или II—III тергиты по бокам слегка буроватые.
Нижняя лобная мозоль иногда целиком черная. Средние голени черно-серые, лишь
кончик желтый. 15—16 мм.
Распространение: Амурская обл. (близ устья Зеи), Хабаровский край
(на север до Хабаровска), Приморский край. Сев.-Вост. Китай, Корейский п-ов,
Япония. В СССР редок, в Китае обычен.
Примечание. Tabanus amaenus Wlk. был описан из Гонконга, поэтому
номинативной формой следует считать популяцию этого вида из Китая, по крайней мере из его
южных частей. В просмотренной нами коллекции Энтомологического института
Академии наук КНР (Шанхай), определение которой произведено Лю-Ве-де (Liu-Wi-teh),
Т. amaenus представлен большой серией экземпляров и с ними мы произвели
идентификацию особей Т., griseinus Phil, из СССР и Сев.-Вост. Китая. В наших сборах
из Сев.-Вост. Китая (Чанчунь, Гирин, Пекин) оказались как номинативная форма
Т. amaenus amaenus Wlk., так и северный подвид Т. amaenus griseinus Phil., а также
переходные экземпляры.
В монографии Шираки (Shiraki, 1918) описание и рисунки Т. amaenus Wlk.
довольно хорошо соответствуют северной форме Т. amaenus griseinus, тогда как
описываемый Шираки вновь Т. amaenus var. lateralis соответствует номинативной форме
Т. amaenus amaenus Wlk. В противоположность этому описание и рисунок Т.
amaenus Wlk. в недавно опубликованной монографии Мардок и Такахаси (Murdoch et
Takahasi, 1969) явно не соответствуют виду, который китайские энтомологи и вслед
за ними мы считаем Т. amaenus Wlk. Мардок и Такахаси отмечают у $ красно-желтые
щупальцы, коричнево-черные усики, срединный ряд четких серых треугольников
сверху брюшка и т. д., что не совпадает с признаками Т. amaenus Wlk. Но в той же
монографии этому последнему виду хорошо соответствуют рисунки и описание Т.
pallidiventris Ols. (=T. amaenus amaenus Wlk.), которого авторы считают
самостоятельным видом. В монографии Такахаси (Takahasi, 1962) Т. amaenus Wlk. и Т. griseinus
Phil, рассматриваются как самостоятельные виды, причем описание и рисунки Т.
griseinus соответствуют тому, что мы считаем Т. amaenus griseinus Phil', (северная форма),
тогда как описание и рисунки Т. amaenus Wlk. имеют некоторые отличия.
9. Род ATYLOTUS О. S. — СВЕТЛОГЛАЗЫЕ СЛЕПНИ
Osten-Sacken, 1876, Mem. Boston Soc. Nat. Hist., 2 : 426 (Tabanus,
subgenus); Stone, 1938 : 19 (Tabanus, subgenus); Philip, 1941 : 11; 1947 : 287; M о u-
cha etChvala, 1959a : 170; 1969a : 321; Виолович, 1968 : 74. — Ochrops
S z i 1 a d y, 1915:93 (Tabanus, subgenus). — Вaikalia S u г с о u f, 1921:39.—
Dasystypia Enderlein, 1922 : 347. — Surcoufiella Bequaert, 1924 : 26. —
Baihalomyia Stackelberg, 1926a : 53.
Тип рода Tabanus bicolor Wiedemann, 1821.
Светло-серые или охряно-желтые, небольшие (8—16 мм) виды, обычно с сильно
выпуклой головой (в особенности у 33) (рис. 212, 213). Глаза у живых насекомых
бледные, зеленоватые, желтые или серые, с одной тонкой, не достигающей внешнего
края глаз, темной полоской или без нее. У сухих экземпляров окраска глаз б. м.
сохраняется или темнеет до красно-коричневых тонов. Глаза голые или в волосках.
282 т. ПОДСЕМ. TABANINAE
Глазковый бугорок у $$ отсутствует, у SS имеется, расположен ниже уровня глаз,
в сером налете. Лобные мозоли у $ небольшие, округлые, плоские, далеко не
достигают края глаз и обычно обособленные; иногда мозоли отсутствуют (прикрыты
налетом). Лобный треугольник без волосков по бокам. Крылья бесцветные; г4 с придатком
или без него. Терминалии обычного для трибы Tabanini строения, у $$ VIII стер-
нит с б. м. развитыми латеральными лопастями и боковыми коричневыми пятнами.
Рис. 212. Atylotus (s. str.) rusticus L. 5. Рис. 213. Atylotus (s. str.) rusticus L. (J.
У некоторых видов терминалии $$ имеют различия, что может быть использовано для
диагностических целей.
Относительно немногочисленный род, насчитывающий в своем составе около
60 видов, распространенных в Евразии, Сев. Америке и Африке. В СССР известно
14 видов, из них большинство свойственно югу (степная и пустынная зоны), немногие
виды распространены в таежно-лесной полосе, заходя довольно далеко на север
{A. (Surcoufiella) sublunaticornis, A. (s. str.) fulvus).
Трудная для определения группа вследствие однообразия в строении и
изменчивости в окраске; для определения необходимы экземпляры хорошей сохранности и
сериями по нескольку штук.
В первое издание «Фауны СССР» нами ошибочно был включен описанный Силади
(Szilady) из «Сарепты» Tabanus (Ochrops) kerteszi: выяснилось, что это Сарепта,
находящаяся в Сирии, но не близ Волгограда (Сарепта — ныне Красноармейск).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА ATYLOTUS О. S.
1 (2). г4 с придатком, обычно хорошо развитым. Палочка усиков состоит из 4 члеников.
Глаза у $ голые, реже прикрыты заметными волосками (A. (s. str.) latistria-
tus Br.). Средней величины мухи, 11—16 мм, очень редко 8—10 мм. $$
кровососы 1. Подрод Atylotus О. S.
2 (1). г4 без придатка (иногда имеется его рудимент). Палочка усиков с варьирующим
числом члеников от 2 до 4. Глаза у § покрыты короткими волосками.
Небольшие мухи, 8—10 мм, редко до 12 мм. $$ не кровососы
2. Подрод Surcoufiella Bequ.
1. Подрод ATYLOTUS О. S. s. str.
Osten-Sacken, 1876, Mem. Boston Soc. Nat. Hist., 2 :'425.
Тип подрода Tabanus (Atylotus) bicolor Wiedemann, 1821.
Глаза у $ голые, реже покрыты заметными волосками; у $ глаза покрыты
волосками, очень редко голые. г4 с придатком, обычно хорошо развитым. Палочка усиков
9. ATYLOTUS 383
состоит из 4 члеников. Средней величины мухи, 11—16 мм, очень редко 8—10 мм.
$$ кровососы.
Подрод насчитывает свыше 50 видов, распространенных в Евразии, Сев. Америке
и Африке; в СССР известно 12 видов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА ATYLOTUS О. S.
А. Глаза разделены л б о м: $>$>
1 (2). Глаза явственно покрыты волосками. Южное побережье Крыма
1. A. (s. str.) latistriatus Br.
2 (1). Глаза голые (или в едва заметных волосках).
3 (12). Желто-бурые виды. Светлые волоски брюшка сверху охряно-желтые.
4 (5). Усики очень широкие, длина площадки превосходит ширину менее чем
в 1V2 раза (рис. 215). Нотоплевры обычно темные ... 2. A. (s. str.) fulvus Mg.
5 (4). Усики менее широкие, длина площадки превосходит ширину в 1V2 раза и более
(рис. 217, 218 и др.). Нотоплевры обычно желтоватые.
6 (7). Лобные мозоли относительно крупные (рис. 218). Сверху брюшка срединная
темная полоска очень узкая, она занимает на II—III тергитах 1/7—V» ширины
кольца 4. A. (s. str). loewianus Villen.
7 (6). Лобные мозоли небольшие (иногда средняя мозоль целиком прикрыта налетом).
Сверху брюшка срединная темная полоска обычно широкая, она занимает
на II—III тергитах не менее 1/t—V5 ширины кольца.
8 (11). Желто-коричневые пятна по бокам брюшка захватывают по меньшей мере
I—III тергиты. Бедра оранжево-желтые целиком или немного затемнены в
основании.
9 (10). Горы Средней Азии. Площадка усиков с тупоугольным дорсальным выступом
(рис. 217) 3. A. (s. str.) chodukini Ols.
10 (9). Дальний Восток. Площадка усиков с б. м. прямоугольным дорсальным
выступом (рис. 219) 5. A. (s. str.) horvathi Szil.
11 (8). Желто-коричневые пятна по бокам брюшка захватывают I—II тергиты и лишь
незначительно заходят на переднюю часть III тергита. Бедра обычно не менее
чем на Vg от основания черно-серые, вершина желто-оранжевая .....
8. A, (s. str.) rusticus L. var. ochraceus Ols. et Meln.
12 (3). Голубовато-серые или черно-серые виды. Светлые волоски брюшка беловато-
серые (редко слегка желтоватые).
13 (22). Затылок за глазами в коротких волосках, лишь возле темени едва
выступающих над поверхностью глаз (смотреть спереди!).
14 (17). Бедра ног (по крайней мере средних и задних) в основной */з или почти целиком
черно-серые, с желтоватыми концами.*
15 (16). Дорсальный угол 3-го членика усиков расположен б. м. посредине площадки
(рис. 223). Бедра ног обычно черно-серые на 2/з от основания. Средние и задние
лапки коричнево-черные. Коренастее и крупнее. Европейская часть СССР и
Зап. Сибирь, на восток до Енисея 8. A. (s. str.) rusticus L.
16 (15). Дорсальный угол 3-го членика усиков расположен в базальной г/3 площадки
(рис. 220). Бедра ног обычно черно-серые на Vs или Va 0T основания. Средние
и задние лапки желтые, с коричневыми кончиками. Уже и мельче. Восточная
Сибирь, Дальний Восток 6. A. (s. str.) miser Szil.
17 (14). Бедра ног желтые, иногда в основании слегка серые.**
18 (19). Усики широкие, длина площадки превосходит ширину менее чем в 1V2 раза
(рис. 225). Горный вид. Малый Кавказ (Нахичеванская АССР и Армения)
9. A. (s. str.) proditor Bog. et Sam.
19 (18). Усики менее широкие, длина площадки обычно превосходит ширину в 1V2 раза
и более (рис. 221, 226). Равнинные степные и пустынные виды.
20 (21). Передние лапки черные, в основании часто темно-коричневые. Брюшко сверху
на фоне светлых волосков с 4 продольными рядами пятен из черных волосков,
из них средние пятна образуют 2 довольно резкие параллельные полоски
(рис. 226); боковые ряды пятен вдоль внешнего края брюшка иногда неясные
или могут отсутствовать. Снизу брюшко преимущественно розово- или
оранжево-желтое. Крупнее и коренастее .... 10. A. (s. str.) quadrifarius Lw.
21 (20). Передние лапки светло-коричневые или желтоватые. Брюшко
преимущественно черно-серое, сверху с широкой срединной черноватой полосой,
покрытой главным образом черными волосками (рис. 221). Снизу брюппсо
преимущественно черно-Серое. Мельче и уже .... 7. A. (s. str.) pallitarsis Ols.
* Изредка встречаются экземпляры A. (s. str.) miser Szil. с целиком желтыми
бедрами!
** Изредка встречаются экземпляры A. (s. str.) quadrifarius Lw. с
затемненными в основной 1/3—1/2 средними и задними бедрами!
884 Ш. ПОДСЕМ. TABANINAE
22 (13). Затылок за глазами в длинных, светлых волосках, заметно выступающих над
поверхностью глаз (рис. 228, 230) (смотреть спереди!).
23 (24). Бедра в большей своей части черно-серые. Брюшко сверху с широкой, темной
срединной полосой, покрытой черными волосками; по бокам брюшко в серых
волосках (рис. 228). Палочка слегка темнее площадки усиков
11. A. (s. str.) flayoguttatus Szil.
24 (23). Бедра желтые, лишь иногда в основании слегка серые. Брюшко сверху
преимущественно серое или желтовато-розовое, с 4 продольными рядами узких
темных пятен из черных волосков (рис. 230). Палочка одноцветная с площадкой
усиков 12. A. (s. str.) pulchellus Lw.
Б. Глаза соприкасаются: 36
1 (20). Затылок за глазами в коротких волосках.
2 (11). Желто-бурые виды. Светлые волоски брюшка сверху охряно-желтые.
3 (6). Глаза в длинных волосках. Голова умеренно выпуклая.
' 4 (5). II стернит с крупным черным б. м. прямоугольным пятном посредине . .
1. A. (s. str.) latistriatus Br.
5 (4). II стернит без темного пятна посредине . . 4. A. (s. str.) loewianus Villen.
6 (3). Глаза в умеренно длинных или коротких волосках. Голова сильно выпуклая,
полушаровидная.
7 (8). Усики относительно широкие, длина площадки превосходит ширину не более
чем в 1V2 раза. Нотоплевры темные 2. A. (s. str.) lulvus Mg.
8 (7). Усики относительно узкие, длина площадки превосходит ширину более чем
в 1V2 раза. Нотоплевры чаще желтые.
9 (10). Горы Средней Азии. Площадка усиков с тупоугольным дорсальным углом
3. A. (s. str.) chodukini Ols.
10 (9). Дальный Восток. Площадка усиков с б. м. прямоугольным дорсальным углом
5. A. (s. str.) horvathi Szil.
11 (2). Серые или желтовато-серые виды. Светлые волоски брюшка сверху беловато-
серые.
12 (15). Глаза в длинных волосках.
13 (14). Бедра не менее чем на Vs от основания черно-серые. Европа, Зап. Сибирь
8. A. (s. str.) rusticus L.
14 (13). Бедра желтые, лишь в основании слегка сероватые. Горный вид — Малый
Кавказ (Нахичеванская АССР, Армения)
9. A. (s. str.) proditor Bog. et Sam.
15 (12). Глаза в умеренно длинных или коротких волосках.
16 (17). Передние лапки желто-коричневые .... 7. A. (s. str.) pallitarsis Ols.
17 (16). Передние лапки черные или черно-коричневые.
18 (19). Брюшко сверху на фоне светлых волосков с 4 продольными рядами пятен
из черных волосков, из них средние пятна образуют 2 довольно резкие
параллельные полоски 10. A. (s. str.) quadrifarius Lw.
19 (18). Брюшко сверху в светлых волосках, без пятен или полосок из черных
волосков 6. A. (s. str.) miser Szil.
20 (1). Затылок в длинных, выступающих над глазами волосках.
21 (22). Глаза в хорошо заметных волосках. Бедра ног черно-серые с
желто-коричневым кончиком 11. A. (s. str.) flavoguttatus Szil.
22 (21). Глаза голые (при сильном увеличении видны очень короткие волоски). Бедра
желтые, в основании сероватые 12. A. (s. str.) pulchellus Lw.
1. Atylotus (s. str.) latistriatus Br. (рис. 214) — Слепень южный.
Biauer, 1880, Denkschr. Akad. Wissensch. Wien, 42:170 (Tabanus); V e r-
rall, 1909:388 (Tabanus); К г б b e r, 1924:15 (Tabanus); Chvala, Lyne-
borg a. Moucha, 1972 : 276. — nigrifacies G о b e г t, 1897 : 303 (Tabanus), пит.
no: Chvala, Lyneborg a. Moucha, 1972:267.
$. Глаза в умеренно длинных серых волосках. Лобная полоска широкая, ее
высота в 3—Зх/2 раза превосходит ширину, в сером налете. Нижняя лобная мозоль
крошечная, коричневая или черная, часто целиком прикрыта налетом, средняя мозоль
имеется лишь изредка, небольшая, черная. Затылок за глазами в коротких
желтоватых волосках. Усики оранжево-желтые, 3-й членик широкий, площадка
приблизительно в 1V2 раза длиннее ширины, дорсальный угол расположен в базальной 1/3
площадки, с явственным зубчиком. Щупальца желтовато-белые, концевой членик сильно
утолщен, за серединой сужен и б. м. заострен на конце, покрыт довольно грубыми
черными и светлыми волосками. Нотоплевры коричневато-желтые. Крылья
бесцветные; г4 с длинным придатком. Ноги коричневато-желтые; передние лапки и концы
передних голеней черные: средние и задние лапки на конце коричневые; средние и
9. ATYLOTUS 285
illlH/lfl/l,(l^
задние бедра (частью Передние) в основной трети серые. Брюшко черное, в буроватом
налете, по бокам с красновато-желтыми пятнами, простирающимися на I—III тер-
гиты. Разделяющая их темная срединная полоса занимает приблизительно 1/3—1/5
ширины брюшка. Брюшко сверху покрыто преимущественно золотисто-желтыми
волосками с примесью черных, особенно
на темных частях те.ргитов. Снизу брюшко
коричневато-желтое, в светлых волосках, ,#swf>W
в основании имеется темное пятно (про- """'•""■""'
стирается иногда до 3 кольца); конец
брюшка темно-серый.- 11—15.5 мм.
(J. Голова выпуклее, чему?. Глаза в .<.',",,...'.•] Will Ч
густых, длинных желтовато-серых волосках. У'!•'.'•'•';"'^;l wif/ II
Волоски на затылке лишь возле глазкового ''','•'•''■'':'- '(/II/ '
бугорка слегка длиннее, нежели волося- :■.'.';. •■;'.•; {//// 'I1
ной покров глаз. Крупнофасетчатая часть '•:■ L Ч
глаза резко отграничена от мелкофасет- 1_:Л'Д v' '\U
7/'Г 'V
Рис. 214. Atylotus (s. str.) latistriatus Br.
Лобная полоска, усик и щупальце $.
чатой. Усики как у $, но более узкие. Концевой членик щупалец слегка желтоватый,
продолговато-овальный, преимущественно в темных волосках. Грудь, крылья и ноги
как у ?. Сверху брюшка боковые красновато-коричневые пятна захватывают
I—IV тергиты, средняя темная полоса на II тергите в виде б. м. широкого пятна,
занимающего 1/5 ширины тергита, на III тергите вдвое уже, чем на предыдущем тергите,
на IV тергите несколько шире предыдущего. Брюшко сверху покрыто смесью черных
и золотисто-желтых волосков, на темных частях преобладают черные, на светлых —
желтые. Снизу брюшко окрашено как у $. 14—16 мм.
Распространение: южноевропейский (средиземноморский) вид. Крым:
Карадаг, Судак (Новый Свет), Соколиное у р. Сухой, окр. Евпатории. Указание
Силади (Szilady, 1915) этого вида для Валуек Воронежской обл. явно ошибочно. Ю. и
частью Средн. Европа, Сев. Африка. Июнь—июль. Не часто.
2. Atylotus (s. str.) fulvus Mg. (рис. 215) — Слепень рыжий. .
Изменчивый вид, в пределах своего обширного ареала представлен подвидами:
номинативным A. (s. str.) fulvus fulvus Mg., кавказским A. (s. str.) fulvus aureus Haus.
и средиземноморским A. (s. str.) fulvus rufipes Mg. Последняя форма значительно
отличается от номинативной и, возможно, представляет собой самостоятельный вид;
в СССР она не встречается.
2а. Atylotus (s. str.) fulvus fulvus Mg. — Слепень рыжий номинативный
М е i g e n, 1820, Syst. Beschreib., II : 61 (Tabanus); В r a u e г, 1880 : 169
(Tabanus); Lundbeck, 1907:124 (Tabanus); V e r r a 11, 1909:381 (Tabanus); S u r-
c о u f, 1924 : 154 (Tabanus); Krober, 1924 : 21 (Tabanus); К г б b e г in
Lindner, 1925:79 (Tabanus); Олсуфьев, 1937а : 211 (Tabanus); Moucha et
Chvala, 1959a : 171 (Tabanus); Шевченко, 1961 : 120 (Tabanus); Leclercq,
1966 : 128; Виолович, 1968 : 78; И в а н и щ у к, 1970 :148 (личинка); Chvala,
Lyneborg a. Moucha, 1972 : 270.
9. Глаза голые (лишь при увеличении X 16 удается заметить редкие, очень
короткие волоски). Лобная полоска умеренной ширины (ее высота превосходит в Зх/4—
4 раза), с параллельными сторонами, в желтом налете, покрыта короткими,
преимущественно черными волосками. Нижняя и средняя лобные мозоли в виде плоских,
черно-блестящих мозолей, из них верхняя б. м. клиновидная, обращена острием вниз,
тогда как нижняя округлая, часто разделена пополам тонкой полоской налета. Степень
покрытия налетом варьирует, и верхняя мозоль (редко нижняя) может быть целиком
прикрыта. Затылок за глазами в коротких, светлых волосках. Концевой членик
щупалец палево-желтый, в основании сильно утолщен, за серединой резко сужен,
остроконечный, покрыт короткими, преимущественно черными волосками. Усики
целиком оранжево-желтые. 3-й членик очень широкий, с почти прямоугольным
дорсальным выступом, вершина которого расположена в основной 1/3 площадки.
Нотоплевры черные, в желто-сером налете. Ноги в большей своей части оранжево-желтые;
бедра в основной 1/3 черно-серые, концевая 1/3 передних голеней, передние лапки и
концевые членики последующих лапок черные. Крылья бесцветные;- г4 с придатком.
286
III. ПОДСЕК. TABANINAE
Брюшко в густом желто-буром налете, сверху равномерно покрыто желтыми и
черными, короткими, прилегающими волосками. I—III тергиты и соответствующие им
стерниты по бокам с коричневато-желтыми пятнами. Разделяющая их темная полоса
занимает около l/3—V4 ширины брюшка. VIII стернит: гонопофиз обычной величины
^аМИЫ """");; jj'l'Д to i it Цц^
Рис. 215. Atylotus (s. str.) fulvus fulvus Mg. Лобная полоска, усик, щупальце,
VIII стернит (варианты), церки и сперматека ?.
в черных волосках, с выраженной апикальной выемкой; латеральные лопасти умеренно
развиты, боковые коричневые пятна крупные, между ними неширокая светлая полоса-
базальная лопасть короткая, слегка сужена к основному краю, последний немного
вогнутый. Церки с округлой вершиной. Сперматека узковеретеновидная. 13—16 мм.
Рис. 216. Ареалы: 1 — Atylotus (s. str.) fulvus fulvus; 2 — A. (s. str.) fulvus
aureus; 3 — A. (s. str.) chodukini.
в- Голова значительно больше, чем у $, полушаровидная. Фасетки верхних 2/ч
глаз значительно крупнее нижних (в 4—5 раз); граница между теми и другими резко
выражена. Крупнофасетчатая часть глаз в коротких, прочие части в едва заметных
серых волосках. Затылок за глазами в очень коротких, серых волосках. Усики
целиком оранжево-желтые; 1-й членик сверху в коротких черных волосках; 3-й членик зна-
9. ATYLOTUS 287
чительно уже, нежели у ?, с тупоугольным дорсальным выступом, вершина которого
расположена в оеновной 1/3 площадки. Концевой членик щупалец
вытянуто-овальный, покрыт б. м. короткими, преимущественно черными волосками. Нотоплевры,
крылья и ноги как у $. Брюшко матовое, в густом желтовато-буром налете, сверху
равномерно покрыто желтыми и черными коротко прилегающими волосками.
1—4 кольца брюшка по бокам с желтыми пятнами; разделяющая их полоска б. м.
узкая, занимает х/4—1/5 ширины брюшка. 13—16 мм.
Распространение: европейско-сибирский лесной подвид (рис. 216, 1),
в СССР от Карелии и Западной Украины на восток до Забайкалья. На север известен
до Хибин, ст. Обоверской Архангельской обл., Усть-Цыльмы Коми АССР, Березова
на Оби, Верхне-Имбатского на Енисее и Ванавары на Подкаменной Тунгуске, на юг
до Херсонской обл. (север), р. Кушугум близ Запорожья, Боброва и Хреновского
бора Воронежской обл., Куйбышевской обл., а после перерыва распространения
в степях юго-востока — на Кавказе (подвид A. (s. str.) fulvus aureus Haus.); в Сибири
на юг распространен до Кустаная, Борового Кокчетавской обл., Алтая, Тувы,
южного и северного Прибайкалья, по предгорьям и горам востока Казахстана (Саур,
Тарбагатай, Джунгарский и Заилийский Алатау) до севера Киргизии (Киргизский
хребет). Европа, Турция. Июнь—июль. Не часто. Возможно, что часть указаний этого
вида для юга Европы относится к A. (s. str.) loewianus, которого ранее смешивали
с A. (s. str.) fulvus; в Сев. Африке (Марокко) распространен A. (s. str.) fulvus rufipes
Mg. Указание Шираки (Shiraki, 1918) на распространение A. (s. str.) fulvus в Японии
ошибочно.
В Карелии личинки найдены в низинных и переходных болотах, редко в сфагно-
.вых болотах (Лутта, 1970).
2Ь. Atylotus (s. str.) fulvus aureus Haus. — Слепень рыжий кавказский.
Г а у з е р, 1941, Изв. Азерб. фил. АН СССР, 2 : 70 {Tabanus); 1939 : 140 (Taba-
nus). — transcaucasicus Bogatshev et Samedov, , 1949 : 68 (Tabanus fulvus
transcaucasicus).
От номинативной формы 5 $ отличаются более выраженной золотистой
окраской волосков тела, в особенности брюшка, почти без примеси черных волосков, и более
светлыми ногами: передние бедра целиком желтые, средние и задние бедра затемнены
в основании примерно на V4 своей длины; у $ лобные мозоли обычно целиком прикрыты
налетом. 13—15 мм.
Распространение: кавказский горно-лесной подвид (рис. 216, 2).
Большой Кавказский хребет (от Майкопа на восток до Дагестана и Омало в Грузии),
Малый Кавказский хребет в пределах Грузии (Бакуриани), Армении и Азербайджана
(включая Нахичеванскую АССР). Сев.-Вост. Турция. Июль—август. Не часто.
Подвид кратко описан Гаузером (1939) по сборам из Ханларского р-на, Азербайджан,
но без латинского названия; последнее дано Гаузером как Tab anus fulvus aureus subsp. n.
в его публикации 1941 г. Позднее предложенное Богачевым и Самедовым (1949) для
этого подвида название Т. fulvus subsp. transcaucasicus n. оказывается синонимом.
Леклерк (Leclercq, 1966) неправильно отнес
рассматриваемый подвид в синонимы A. (s. str.)
fulvus rufipes M*.
3. Atylotus (s. str.) chodukini Ols. (рис. 217)
— Слепень. Ходукина.
Олсуфьев, 1952, Энтомол. обозр.,
32 : 314 (Tabanus); Шевченко, 1961 :122
(Tabanus); Leclercq, 1966 : 128.
$. Глаза голые. Лобная полоска слегка
расширена кверху, ее высота превосходит
Рис. 217. Atylotus (s. str.) chodukini
Лобная полоска, усик и щупальце $.
Ols.
ширину основания в 4—41/2 раза, в желтом налете. Нижняя лобная мозоль
в виде маленького поперечного или округлого черного пятнышка, средняя мозоль
отсутствует или имеется в виде маленького неправильной формы черного пятнышка.
Затылок за глазами в коротких волосках. Усики оранжево-желтые; 3-й членик
относительно узкий, дорсальный выступ образует тупой угол, расположен в базальной
трети площадки. Концевой членик щупалец в основании заметно утолщен, к концу
2SS
III. ПОДСЕМ. TABANINAE
б. м. равномерно сужен, беловатого цвета, покрыт светлыми и черными волосками.
Нотоплевры желтые. Ноги оранжево-желтые, передние лапки и концевая часть
последующих лапок коричневые. Средние и задние бедра в основании слегка затемнены.
Крылья бесцветные, жилки желтые; г4 с придатком. Брюшко сверху с крупными
рыжевато-желтыми боковыми пятнами, захватывающими I—IV или I—V (иногда I—III)
тергиты, покрыто желтоватым налетом и короткими золотисто-жёлтыми и черными
волосками; первые преобладают на светлых частях брюшка, вторые — на темных.
Снизу брюшко рыжевато-желтое, с небольшим темным пятном в основании, V—VI
(или IV—VI) стерниты часто сероватые. 11—16 мм.
в- Голова крупнее, чем у $, полушаровидная. Глаза в относительно коротких
волосках. Верхние фасетки крупнее нижних в 4—5 раз, граница между ними резкая.
Затылок в коротких волосках. Усики как у ?, но 3-й членик более узкий. Концевой
членик щупалец беловатый, в средней части немного утолщен, покрыт черными и
беловатыми волосками. Нотоплевры, крылья и ноги как у $. Брюшко сверху с рыжевато-
коричневыми пятнами, захватывающими боковые стороны I—IV тергитов;
разделяющая их темная срединная полоса занимает почти 1/3 ширины брюшка. Окраска
волосков сверху брюшка как у $. Снизу брюшко как у 9, но темное пятно на II
стершие более крупное. 14—15 мм.
Похож на A. fulvus, но отличается от него более узкой площадкой 3-го членика
усиков, светлыми нотоплеврами и преобладающей желтой окраской низа брюшка.
Распространение: горы Средней Азии (рис. 216, 3). Западный Тянь-
Шань в пределах Казахстана и Киргизии (хребты Каржантау, Угамский, Пскемский,
Чаткальский и Таласский); Памиро-Алай в пределах Таджикистана (хребты Гиссар-
ский, Вахшский, Петра Первого, Дарвазский). Лесная зона и альпийские луга от 1300
до 3000 м. Июль—август. Местами обычен.
Голотип: 5,- хр. Каржантау, берег оз. Су-Синган, 27 VII 1939,. Обухова. В
коллекции Зоологического института Академии наук СССР, Ленинград.
У экземпляров $$ с Тянь-Шаня обычно средняя мозоль отсутствует, тогда как
у экземпляров с Гиссаро-Дарваза она, как правило, имеется. Эту форму мы
обозначаем как A. (s. str.) chodukini var. callosus Ols., var. nova.
Рис. 218. Atylotus (s. str.) loewianus Villen. Лобная полоска,
усик, щупальце, VIII стернит, церки и сперматека $.
4. Atylotus (s. str.) loewianus Villen. (рис. 218) — Слепень Лева.
V i 11 е n e u v e, 1920, Ann. Soc. ent. Belg., 60 : 65 (Tabanus); S u г с о u f, 1924:
155 (Tabanus); Krober in Lindner, 1925 : 79 (T. fulvus loewianus Villen.):
С h v a 1 a, 1964 : 376. - znojkoi О 1 s u f j e v, 1937a : 212 (Tabanus).
?. Глаза голые (лишь при увеличении Х16 удается заметить очень короткие,
редкие волоски), без полоски. Лоб узкий, его высота в 4—5 раз превосходит ширину
основания, слегка расширен кверху, в желтоватом налете. Лобные мозоли крупные,
слегка выпуклые, блестяще-черные, вполне изолированы одна от другой. Нижняя
мозоль округло-поперечная, средняя б. м. сердцевидная, обычно крупнее нижней.
Усики оранжево-желтые. 3-й членик б. м. узкий, с тупым дорсальным углом;
последний расположен в базальной V3 площадки. Длина площадки превосходит ширину почти
9. ATYLOTUS 289
в 2 раза. Концевой членик щупалец желтоватый, б. м. узкий, в основной части слегка
утолщен, беспорядочно покрыт довольно грубыми светлыми и черными волосками.
Нотоплевры желтые. Крылья бесцветные; г4 с придатком. Ноги желтые; средние и
задние бедра часто в основной 1/3 сероватые, передние лапки и концы передних голеней
черные, концы последующих лапок коричнево-черные. Брюшко черное в буроватом
налете, с большими рыжевато-желтыми пятнами по бокам, простирающимися на I—
III (или I—IV) тергиты. Их разделяет очень узкая темная срединная полоска,
занимающая приблизительно 1/s—1/9 ширины брюшка. Волосяной покров состоит из смеси
коротких желтых и черных волосков. Снизу брюшко рыжевато-желтое, последние
стерниты сероватые. VIII стернит: гонопофиз короткий и широкий, в черных волосках,
с незначительной апикальной выемкой; латеральные лопасти крупные, боковые
коричневые пятна в виде редуцированного контура; базальная лопасть короткая,
суженная к основному краю, последний б. м. прямой. Церки с округлой вершиной.
Сперматека веретеновидная. 13—15 мм.
в- Голова лишь немногим больше, чем у $. Глаза в длинных, светлых волосках.
Фасетки верхних частей глаз приблизительно в 4 раза крупнее нижних, граница между
теми и другими резкая. Затылок за глазами в коротких волосках. Усики как у $,
но 3-й членик более узкий. Концевой членик щупалец желтоватый, продолговато-
овальный, в черных и светлых волосках. Нотоплевры, ноги и крылья как у $.
I—IV тергиты брюшка рыжевато-желтые, с срединным рядом очень узких сероватых
небольших пятен, V—VII тергиты черно-серые. Сверху брюшко покрыто смесью
черных и желтых волосков. Снизу брюшко как у $>. 15 мм.
Буро-желтый вид, напоминающий A. (s. str.) fulvus. $? легко отличаются более
узким лбом, крупными лобными мозолями, более узкими щупальцами и усиками и,
наконец, сильно развитой желтой окраской брюшка. 3<$ отличаются более' длинными
волосками на глазах и светлой окраской брюшка.
Распространение: южноевропейский (средиземноморский)
лесостепной вид; проникает в горы. В СССР распространен в западных частях УССР
(Закарпатская, Ивано-Франковская, Житомирская, Одесская области), в Молдавии,
Восточном Предкавказье (Каякентский р-н) и в Закавказье (Азербайджан: Куба,
Ленкорань, Алазань, Закаталы). Средн. и Ю. Европа, Сев. Африка (Марокко),
Ливан. Июль—август. Не часто.
5. Atylotus (s. str.) horvathi Szil.
(рис. 219) — Слепень желтый.
S z i 1 a d у, 1926, Ann. Mus. Nat.
Hungar, 24 : 601 (Tabanus); Takahasi,
1962 : 63 (Tabanus); Murdoch et
Takahasi, 1969 : 55. — bivittatus (Matsu-
mura) apud S h i r a k i, 1918 : 180
(Tabanus) et auct. — pyrrhus (Wlk.) apud С о-
q u i 1 e t, 1899:311 (Tabanus), цит. по:
Murdoch et Takahasi, 1969:55.
— fulvus (Mg.) apud S h i r a k i, 1918 : 163
(Tabanus). — bivittateinus (Takahasi) no
Виоловичу, 1968 : 79.
$. Глаза голые, стойкой полоской или
без нее. Лобная полоска узкая, ее высота
Рис. 219. Atylotus (s. str.) horvathi Szil.
Лобная полоска, усик и щупальце $.
в 41/2—5 раз превосходит ширину основания, с почти параллельными сторонами,
покрыта желтоватым налетом. Лобные мозоли небольшие, нижняя мозоль коричневая
или черная, б. м. округлая или поперечная, средняя черная, часто сердцевидная,
иногда целиком скрыта налетом. Затылок в коротких волосках. Усики оранжево-
желтые; 3-й членик умеренной Ширины, дорсальный угол б. м. прямой, расположен
в базальной 1/3 площадки. Длина площадки превосходит ширину приблизительно
в 11/2—2 раза. Концевой членик щупалец желтый, утолщен, б. м. равномерно сужен
к концу, покрыт короткими черными и светлыми волосками; г4 с придатком.
Нотоплевры желтые. Ноги оранжево-желтые; средние и задние бедра в основной 1/3 серые;
передние лапки и концы передних голеней черные; последующие лапки с
черно-коричневыми концами. Окраска бедер варьирует: встречаются экземпляры с целиком желтыми
бедрами, с другой стороны — серая окраска иногда 'занимает более 1/2 их длины, в том
числе и на передних бедрах. Брюшко темное, в желтовато-сером налете, с большими
желтыми пятнами по бокам. Пятна простираются на I—III (реже I—IV) тергиты. Разде-
19 Н. Г. Олсуфьев
290
III. ПОДСЕМ. TABANINAE
ляющая их срединная темная полоса занимает 1/i~1/3 ширины брюшка. Брюшко
покрыто короткими, густыми, желтыми волосками, на конце с незначительной примесью
черных. Последние иногда образуют 2 неясных продольных полоски по краям
срединной темной полосы. Снизу брюшко серое, I—II или I—III стерниты по бокам
оранжево-желтые. Иногда брюшко снизу в большей своей части желтое, крайне редко —
целиком черно-серое. 12.5—16 мм.
(J. Голова значительно больше, чем у ?, полушаровидная. Фасетки верхних
частей глаз в 4—5 раз крупнее нижних, граница между теми и другими резкая. Глаза
в коротких, серых волосках. Затылок в коротких волосках. Усики как у $, но несколько
у^ке. Концевой членик щупалец бледно-желтый, продолговато-овальный,
в тонких, светлых и черных волосках. Нотоплевры, крылья и ноги как у $. Брюшко
в буровато-желтом налете, с большими желтыми пятнами по бокам,
простирающимися на I—IV (редко I—III) тергиты. Разделяющая их темная срединная полоса
занимает приблизительно V7 ширины брюшка (редко шире). Брюшко покрыто
прилегающими желтыми волосками, с примесью черных (последние хорошо заметны на фоне
желтых боковых пятен). Снизу брюшко желтое, с узкой серой полоской посредине
(иногда имеется только темное пятно на II стерните), последние 3 (или 2) стернита
сероватые. 12—14.5 мм.
Близок к А. (s. str.) fulvus, отличаясь более стройным телом и менее широким 3-м
члеником усиков. Сходен с A. (s. str.) chodukini, отличаясь несколько иным строением
усиков и более развитыми лобными мозолями ($).
Распространение: дальневосточный лесной вид. Приморский край
(на юг включая Хасанский р-н); Амурская обл. близ Свободного ($>с? темноокрашенные
недостаточной сохранности, определение условное). Сев.-Вост. Китай, Япония. Июнь— s
август. Местами обычен.
Рис. 220. Atylotus (s. str.) miser Szil. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $.
6. Atylotus (s. str.) miser Szil. (рис. 220) — Слепень изменчивый.
S z i 1 a d у, 1915, Ent. Mitt., 4 : 103 (Tabanus); Krober, 1924 : 20 {Tabanus);
Krober in Lindner, 1925:81 {Tabanus); Олсуфьев, 1937а : 215
{Tabanus); Шевченко, 1961 : 123 {Tabanus); Leclercq, 1966 : 131; Виолович,
1968 : 79; Murdoch et T a k a h a s i, 1969 : 53; Соболева, 1971 : 142
(личинка, куколка). — bivittateinus Takahasi, 1962 : 62 {Tabanus). — rusticus (L.)
apud S h i r a k i, 1918 : 173 {Tabanus).
$. Глаза голые (лишь при сильном увеличении заметны короткие волоски), с 1
узкой полоской.. -Затылок за глазами в коротких, светлых волосках. Лобная полоска
умеренной ширины, ее высота в 4—5 раз превосходит ширину основания, с
параллельными сторонами, покрыта серым или желтоватым налетом. Лобные мозоли небольшие,
черные, округлые, нижняя иногда несколько крупнее средней; изредка нижняя
мозоль коричневатая. Усики желтые, 3-й членик б. м. узкий, с прямым или тупым
дорсальным углом (иногда острым); угол расположен в базальной 1/3 площадки.
Концевой членик щупалец желтоватый, в основной части утолщен, покрыт б. м. короткими
9. ATYLOTUS 291
светлыми и черными волосками. Нотоплевры желтые, реже серые, одноцветные со
спинкой. Крылья бесцветные; г4 с придатком. Ноги желтые. Базальная половина бедер
серая, передние лапки и концы передних голеней черные,"концы последующих лапок
коричневые. В редких случаях бедра целиком желтые или же, наоборот, серые. Брюшко
черно-серое, в сером или буроватом налете, I—II или I—III (редко I—IV) тергиты
по бокам грязно-желтые, разделяющая их срединная темная полоса занимает обычно
V3 ширины тергитов, редко больше или меньше. Брюшко сверху покрыто
маскирующими основную окраску тергитов короткими серыми или желтоватыми волосками с той
или иной примесью черных волосков, последние иногда образуют 2 довольно
явственных продольных параллельных полоски по краям срединной темной полосы; изредка
помимо срединных полосок имеются еще по боковому ряду пятен из черных волосков
с каждой стороны брюшка, в таком случае четырехполосый рисунок напоминает
таковой A. (s. str.) quadrifarius. Снизу брюшко черно-серое, в сером или буроватом налете.
II стернит по бокам часто желтоватый, но иногда желтая окраска распространяется
на боковые стороны I—IV стернитов и в редких случаях брюшко снизу почти целиком
желтое. VIII стернит: гонопофиз слегка конусовидный, в черных волосках, с
незначительной апикальной выемкой; латеральные лопасти умеренно развиты, боковые
коричневые пятна крупные, между ними небольшое светлое пространство; базальная лопасть
со слабо вогнутым основным краем. Церки обычно со слегка уплощенной вершиной.
Сперматека узковеретеновидная или игловидная. 11—16 мм.
<?. Голова крупнее, чем у $. Глаза в коротких серых волосках; фасетки верхних
частей глаз значительно крупнее нижних. Затылок за глазами-в коротких волосках.
Усики как у $, но несколько уже. Концевой членик щупалец желтоватый, удлиненно-
овальный, в тонких, светлых и черных волосках. Нотоплевры желтые или серые,
одноцветные со спинкой. Крылья и ноги как у $>. Брюшко серое, с большими желтыми
пятнами по бокам, простирающимися на I—III или I—IV тергиты. Разделяющая их
срединная темная полоска занимает 1/4—1/3 ширины брюшка. Брюшко покрыто серыми
или желтоватыми волосками, к которым лишь на конце брюшка примешиваются
черные волоски. Снизу брюшко по бокам желтое, посредине с серой полосой; последние
3 или 4 стернита серые. 11 — 15 мм.
Похож на A. (s. str.) rusticus, но мельче и уже. В типичном случае бедра затемнены
на V2 от основания, в концевой части желтые, дорсальный угол 3-го членика усиков
расположен в'базальной 1/3, лоб у 5 уже, нежели у A. (s. str.) rusticus, мозоли мельче,
средние и задние лапки светлее. У 3 глаза покрыты значительно более короткими
волосками, голова относительно груди шире.
Распространение: восточносибирский степной (или луголесной) вид,
аллопатричен A. (s. str.) rusticus L. (рис. 224, 2). В СССР от Кемеровской обл. (Тисуль
б. Марьинского у.) на восток до низовий Амура и юга Приморской обл. (Хасанский
р-н); юг Красноярского края, Хакасия, южное Прибайкалье, Забайкалье,
Приамурье. Китай, Монголия, Корейский п-ов, ? Япония. Июль—август. Местами обычен.
- 7. Atylotus (s. str.) pallitarsis Ols. (рис. 221) — Слепень бледноногий.
Олсуфьев, 19376, Паразитол. сб. 6 : 236 {Tabanus); 1937a : 216 (Tabanus);
Шевченко, 1961 : 124 (Tabanus); Leclercq, 1966 : 131; Виолович, 1968 :
80; Murdoch et Takahasi, 1969 : 54; С h v a.l a, L у n e b о r g a. M о и с h a,
1972 : 279.
$. Глаза голые или в очень коротких волосках, видимых лишь возле лба. При
жизни глаза бледно-серые или беловатые, с одной узкой полоской или без нее. Лоб
б. м. широкий, высота превосходит ширину приблизительно в 3—4 раза, с
параллельными сторонами или слегка расширенный книзу, покрыт светло-серым налетом.
Затылок в коротких волосках. Лобные мозоли маленькие, нижняя б. м. поперечная,
коричневая или черная, средняя округлая, черная, часто совершенно прикрыта
налетом. Усики желтые. 1-й членик бледно-желтый, сверху в коротких, черных
волосках. 3-й членик узкий, дорсальный угол прямой или тупой, расположен в базальной
г/3 площадки. Концевой членик щупалец белый, в основании слегка утолщен, далее
постепенно сужается, покрыт короткими белыми и черными волосками. Нотоплевры
желтые, реже серые, одноцветные со спинкой. Крылья прозрачные; г4 с придатком.
Ноги целиком желтые, лишь передние лапки светло-коричневые. Средние и задние
бедра иногда в основании слегка сероватые. Брюшко темно-серое, посредине с
черноватой продольной полоской, занимающей приблизительно V3—1/4 ширины брюшка.
Полоска покрыта короткими черными, боковые стороны брюшка серыми волосками,
выделяющими полоску. Иногда посредине полоски к черным волоскам
примешиваются серые волоски, и в таком случае полоска менее резкая. II тергит (или I—III)
по бокам буроватый или желтоватый. Снизу брюшко темно-серое, II—III стерниты
по бокам слегка просвечивают желтоватым, но иногда желтая окраска
распространяется на 'I—V стерниты. VIII стернит бледно-желтый, гонопофиз небольшой, в черных
волосках, с выраженной апикальной выемкой; латеральные лопасти умеренно
развиты, слабо хитинизированы, боковые коричневые пятна отсутствуют; базальная ло-
19*
292
III. ПОДСЕМ. TABANINAE
пасть с прямым или слегка вогнутым основным краем. Церки продолговатые, с
округлой вершиной. Сперматека игловидная. 10—13 мм.
$. Голова немного крупнее, чем у $. Зона крупных фасеток, занимающих
верхние 2/3 глаз, в коротких светлых волосках. Затылок в коротких волосках. Усики как
у $, но более узкие. Концевой членик щупалец беловатый, удлиненно-овальный,
Рис. 221. Atylotus (s. str.) pallitarsis Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки, сперматека и брюшко сверху $.
в тонких беловатых и единичных черных волосках. Грудь и ее придатки как у $. Ноги
желтые, основание средних и задних бедер серое, концы передних голеней и передние
лапки коричневые, средние и задние лапки целиком желтые. Брюшко черно-серое,
I_IV тергиты по бокам желтовато-коричневые, покрыты сероватыми волосками,
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180" 150°
Рис. 222. Ареал Atylotus (s. str.) pallitarsis.
срединная темная полоса, покрытая преимущественно черными волосками, занимает
1/3 ширины брюшка. Снизу брюшко черно-серое, с желтовато-коричневатыми пятнами
по бокам I—IV стернитов. 11—11.5 мм.
Небольшой вид, с узким телом, близкий к A. (s. str.) miser, но отличающийся
главным образом следующим: 1) более светлыми лапками, 2) у $ более широким лбом,
часто слегка расширяющимся книзу.
Распространение: сибирско-монгольский степной вид (рис. 222).
Челябинская (Верхне-Уральск, Магнитогорск, Троицк), юг Тюменской (Казанское), Ку-
9. ATYLOTUS 293
станайская, Омская, Новосибирская обл., Алтайский край, Хакасия, Тува,
Приморский край (Хасанский, Ивановский р-ны). Монголия, Сев.-Вост. и Вост. Китай. Июль-
август. Не часто.
Голотип: 2, ст. Чаны Новосибирской обл., 14 VII 1932, Н. Олсуфьев. В
коллекции Зоологического института Академии наук СССР, Ленинград.
8. Atylotus (s. str.) rusticus L. (рис. 212, 213, 223) — Слепень полевой.
L i n n e, 1767, Syst. Nat. ed. 12, 2 : 100 (Tabanus); Bra'uer, 1880 : 168
(Tabanus); L un d b ec k/1907 : 122 (Tabanus); Verrall, 1909 : 384 (Tabanu*); Surcouf,
1924:156 (Tabanus); К г 6 be r, 1924:32 (Tabanus); Krober in Lindner,
1925 : 83 (Tabanus); Олсуфьев, 1937а : 219 (Tabanus); Moucha etChvala,
1959a : 171 (Tabanus); Шевченко, 1962 : 124 (Tabanus); L e с 1 e г с q, 1966 : 133;
Виолович, 1968 : 81; Clivala, Lyneborg a. Moucha, 1972 : 274.
Рис. 223. Atylotus (s. str.) rusticus L. Лобная полоска, усик, щупальце,
VIII стернит, церки $ и сперматека. А — гоностиль; Б — церки и эпан-
дрий <$■
2- Глаза с тонкой срединной полоской, голые (лишь при увеличении Х16
удается заметить редкие, очень короткие светлые волоски). Лобная полоска б. м. узкая
(ее высота в З1^—4 раза превосходит основание), с параллельными сторонами, в желто-
сером налете, покрыта короткими, преимущественно серыми волосками. Нижняя и
средняя лобные мозоли в виде блестяще-черных точек: нижней б. м. округлой и
средней неправильной формы. Затылок за глазами в коротких, светлых волосках.
Концевой членик щупалец желтовато-белый, в основной половине слабо утолщен, на конце
тупой, покрыт тонкими серыми волосками, с небольшой примесью черных. Усики
желтые, палочкэ часто слегка буроватая. 3-й членик умеренно широкий, с тупоугольным
дорсальным выступом, вершина которого расположена б. м. посредине площадки.
Нотоплевры серые, одноцветные со спинкой, реже желтоватые. Ноги в большей своей
части желтые; тазики и бедра по крайней мере на 3li или 1/2 своей длины от основания
серые. Концевая 1/3 передних голеней и целиком лапки черные; лапки 2-й и 3-й пары
ног на конце коричневые. Крылья бесцветные; г4 с придатком. Брюшко черно-серое,
в густом сером налете, б. м. равномерно покрыто прилегающими серыми волосками,
сверху иногда с 2 продольными полосками из черных волосков и небольшими пятнами
таких же волосков по бокам I—V тергитов (рисунок напоминает таковой A. (s. str.)
quadrifarius\). II (или I—III) тергит по бокам с небольшим желто-коричневым
пятном. Такие же пятна имеются по бокам соответствующих им стернитов. Эти пятна
иногда мало заметны благодаря покрывающему их налету и волоскам, но могут
отсутствовать целиком. VIII стернит: гонопофиз короткий и широкий, в черных волосках,
с нерезкой апикальной выемкой; латеральные лопасти развиты, боковые коричневые
пятна умеренные, разделены широкой светлой полосой; базальная лопасть короткая,
1
294 III. ПОДСЕМ. TABANINAE
основной край прямой или слегка вогнутый. Церки с округлой вершиной. Сперматека
веретеновидная. 12—16 мм.
cJ. Голова немного крупнее, чем у 2i полушаровидная. Крупнофасетчатая часть
глаз в длинных, прочие части в коротких серых волосках. Затылок за глазами в
коротких серых волосках. Усики как у 9, но 3-й членик более узкий. Концевой членик
щупалец желтовато-белый, эллиптический, с прямой верхней и слегка выпуклой
нижней сторонами, в длинных, серых волосках, с незначительной примесью более коротких,
темных волосков. Нотоплевры серые, одноцветные со спинкой. Ноги и крылья как у 2-
Брюшко в сером налете и в густых, коротко прилегающих серых волосках. Волоски
б. м. равномерно прикрывают кольца и затушевывают их основную окраску. Брюшко
по бокам I—III (реже I—IV) тергитов с желтыми пятнами и с темной полоской
посредине, составляющей 1/3—1/& ширины брюшка. Снизу брюшко в основной половине
0° К ВОСТОКУ ОТ Гринвича 90° 120° 150° 180° 150°
Рис. 224. Ареалы: 1 — Atylotus (s. str.) rusticus; 2 — A- (s. str.) miser.
желтое, I стернит, затем прямоугольное пятно на II стерните (часто продолженное
на последующие стерниты в виде б. м. широкой полоски) и последние 3—4 стернита
темные. 11—15 мм.
Распространение: европейско-западносибирский лесостепной вид
(рис. 224,1). В СССР распространен от Прибалтики и запада УССР на восток до
Красноярского края (Сорокино). На север известен до юга Карелии (Повенец, Сортавала),
Кандалакшского залива (о. Великий — 66°35' с. ш.), Котласа Архангельской обл.,
Печоро-Илычского заповедника в Коми АССР и Копыловки Томской обл., на юг
включая Молдавию, Крым, Предкавказье (от Краснодарского края по Дагестан, местами
поднимается в горы!), Азербайджан (Кусары и отроги Шах-Дага), Саратовской обл.,
Оренбурга, Борового Кокчетавской обл., Павлодарской обл. и Малокрасноярска на
Иртыше Восточно-Казахстанской обл. Европа. Указания, на распространение
в Марокко и Турции нуждаются в подтверждении, а данные о распространении в
Туркмении (Бабаянц, 1962) и Вост. Китае ошибочны.
Обычен в степной и лесостепной части ареала, в лесной зоне, куда он проникает
вслед за вырубкой лесов, не часто. Июль—август.
Некоторые экземпляры из Крыма и Запорожской обл. имеют желтую окраску
волосков брюшка — A. (s. str.) rusticus L. var. ochraceus Ols. et Meln.
9. Atylotus (s. str.) proditor Bog. et Sam. (рис. 225) — Слепень закавказский.
Богачев и Самедов, 1949, Изв. АН Азерб. ССР, 5 : 68 (Tabanus); L е с-
lercq, 1966: 132. .
$. Глаза голые, без полоски. Лобная полоска слегка расширена кверху, ее вы-"
сота превосходит ширину основания приблизительно в 4 раза, прикрыта желтовато-
серым налетом. Лобные мозоли отсутствуют, реже имеются в виде одной или двух
небольших черных мозолек. Затылок в относительно коротких светлых волосках, .слегка
выступающих над поверхностью глаз возле темени. Усики желтые, 3-й членик слегка
оранжевый, широкий, дорсальный угол б. м. прямой или слегка тупой, расположен
9. ATYLOTUS
295
в базальной 1/3 площадки. Концевой членик щупалец беловатый, умеренно утолщен
в основании^ покрыт беловатыми и единичными черными волосками. Нотоплевры
серые, одноцветные со спинкой или желтые. Крылья бесцветные; г4 с придатком. Ноги
желтые, средние и задние бедра в основании сероватые, вершина передних голеней
коричневая, передние лапки черные, последующие с коричневатым концом. Брюшко
черно-серое, сверху в серых и черных волосках, последние преимущественно собраны
в 2.довольно широких продольных полоски
в средней части брюшка. II или I—II тергиты
по бокам имеют по небольшому желтовато-
коричневому округлому пятну. I—II или
I—III стерниты по бокам
желтовато-коричневатые. 14—15 мм.
А. Голова значительно крупнее, чем у 2,
полушаровидная. Крупнофасетчатая часть
занимает верхние 3/4 глаз, покрыта густыми,
длинными серыми волосками. Затылок за
глазами в коротких волосках, лишь возле
глазкового бугорка имеется пучок более
длинных волосков. Концевой членик
щупалец беловато-желтоватый, удлиненно-оваль-
Рис. 225. Atylotus (s. str.) proditor Bog. et Sam.
Лобная полоска, усик и щупальце $>. -
ный, в тонких светлых и единичных черных волосках. Усики: 1-й и 2-й членики
желтые, 3 й обломан. Нотоплевры серые или желтые. Ноги и крылья как у ?. Брюшко
коническое, окрашено в основном как у 2. но желтые пятна по бокам более развиты —
они захватывают I—III тергиты и соответствующие им стерниты, разделяющая их
темная полоса занимает около 1/3 ширины тергитов. 16 мм.
Похож на A. (s. str.) rusticus, оба пола отличаются желтыми бедрами, SS более
крупной головой, $2 слегка расширяющейся кверху лобной полоской, слабым
развитием мозолей (чаще отсутствуют) и более широкой площадкой 3-го членика усиков.
А. В. Богачев и Н. Г. Самедов при описании 22 этого вида отметили у него
длинные волоски на затылочной полоске. На самом деле эти волоски умеренно развиты,
и как показало изучение $3, описываемых нами впервые, А. (s. str.) proditor
принадлежит к группе видов с короткими волосками на затылке.
Распространение: горный вид, обитатель Малого Кавказа. Нахиче-
ванская АССР, Шахбузский р-н, верхняя граница леса и субальпийские луга;
Армения, Эчмиадзинский р-н. Конец июля—август. Местами обычен.
10. Atylotus (s. str.) quadrifarius Lw. (рис. 226) — Слепень четырехполосый.
Loew, 1874, Zeitschr. ges. Naturw., 9:414 {Tabanus); К г о b e r, 1924:12
{Tabanus); Krober in Lindner, 1925 : 83 (Tabanus); Олсуфьев, 1937а :
222 {Tabanus); L e с 1 e г с q, 1966 : 132; Chvala, Lyneborga. Mbucha,
1972 : 276; Олсуфьев, 1973 : 195. — agrestis auct., nee Wiedemann, 1828 :
55 (Tabanus), цит. по: Chvala, Lyneborg a. M ouch a, 1972 : 276. — afgha-
nistanicus Moucha et Chvala, 1959b : 279 (Atylotus agrestis afghanistanicus).
2. Глаза голые, с одной полоской или без нее. Лобная полоска б. м. узкая, ее
высота в 4—5 раз превосходит ширину основания, с параллельными сторонами или
слегка расширяющаяся кверху, покрыта светло-серым налетом. Лобные мозоли
небольшие, средняя б. м. сердцевидная или клиновидная, черная, нижняя — округлая,
коричневая; реже обе мозоли одновременно черные или же коричневые. Затылок в
коротких волосках. Усики желтые, 3-й членик красноватый, узкий, с тупым
дорсальным углом; палочка иногда слегка затемнена. Щупальца белые, концевой членик
узкий, покрыт короткими беловатыми и черными волосками. Нотоплевры желтые.
Крылья бесцветные; г4 с б. м. длинным придатком. Ноги желтые; передние лапки и
концы передних голеней коричнево-черные, концы последующих лапок коричневые.
Брюшко розово-желтое, к концу иногда сероватое, в светлых волосках, в средней части
с 2 узкими, довольно резкими продольными полосками из черных волосков; кроме
того, по бокам тергитов обычно имеются небольшие пятна из черных волосков,
образующие боковые ряды или полоски. Снизу брюшко одноцветно-розово-желтое.
Иногда полоски на брюшке едва намечены. VIII стернит светло-желтый, иногда (у
темных экземпляров) слегка коричневый по бокам; гонопофиз в черных волосках, с
незначительной апикальной выемкой; латеральные лопасти небольшие; базальная ло-
296 III. ПОДСЕМ. TABANINAE
пасть с прямыми боковыми сторонами и почти прямым основным краем. Церки с
округлой или слегка уплощенной вершиной. Сперматека узковеретеновидная. 11—16 мм.
3. Голова лишь немногим крупнее, чем у 2- Крупнофасетчатая часть глаз в
умеренно длинных или коротких, серых волосках. Затылок в коротких, светлых волосках,
лишь незначительно выступающих над поверхностью глаз возле темени. Усики как
у J, но более узкие. Концевой членик щупалец беловатый, удлиненно-овальный,
в тонких беловатых и единичных черных волосках. В остальном целиком повторяет ?.
Брюшко снизу розово-желтое, с небольшим серым пятном в основании. 12—15 мм.
Изменчивый вид. Окраска ног и брюшка у § варьирует — у некоторых
экземпляров из Восточно-Казахстанской обл. (оз. Зайсан) и Тувинской АССР бедра ног на
значительном протяжении от основания серые, а брюшко сверху почти целиком черно-
серое, за исключением небольших желтоватых пятен по бокам II тергита. Эти
экземпляры похожи на A. (s. str.) rusticus, но отличаются тем, что снизу брюшко у них
преимущественно или целиком розовато-желтое, а сверху продольные полоски из черных
волосков относительно резкие. Варьирует также ширина и густота покрытия черными
волосками продольных полосок сверху брюшка.
Рис. 226. Atylotus (s. str.) quadrifarius Lw. Лобная полоска, усик, щупальце, брюшко
сверху, VIII стернит, церки и сперматека 2-
В литературе по палеарктическим Tabanidae рассматриваемый вид длительное
время публиковался как Atylotus (s. str.) agrestis Wd., а темные экземпляры относили
к A. (s. str.) quadrifarius Lw. Недавно выяснили (Ovazza, Camicas и Pichon, 1968),
что истинный A. (s. str.) agrestis Wd. распространен в Африке, тогда как в Европе
обитает близкий к нему вид, отличающийся по тсрмипалиям самцов; для последнего
авторы использовали название A. (s. str.) afghanistanicus M. et Chv., 1959, описанного
из Афганистана, как подвид (мелкая форма) A. (s. str.) agrestis Wd. Значительная
индивидуальная изменчивость A. (s. str.) quadrifarius Lw. не позволяет обосновать под-
видовые различия между афганской популяцией этого вида и популяциями,
обитающими в других местах СССР. На основании изучения типового экземпляра 2 A. (s. str.)
quadrifarius Lw. нами установлена идентичность этого вида с A. (s. str.) agrestis (Wd.)
auct.=/l. (s. str.) afghanistanicus M. et Chv. (Олсуфьев, 1973).
Очень возможно, что A. (s. str.) agrestis Wd. и A. (s. str.) quadrifarius Lw.
представляют собой лишь подвиды одного вида A. (s. str.) agrestis Wd.: различия между 33
обеих форм незначительны, а 22 неразличимы между собой.
Следует отметить, что у 33 A. (s. str.) quadrifarius из Волгоградской
и'Воронежской областей крупнофасетчатая часть глаза покрыта относительно длинными
волосками, в 3—4 раза превышающими поперечник фасетки. У 33 из Ставропольского края
(оз. Тамбукан близ Пятигорска), Казахстана (Карагандинская, Джамбулская обл.)
и Афганистана (паратип A. (s. str.) agrestis afghanistanicus) волоски на
крупнофасетчатой части глаз более короткие, приблизительно равные поперечнику фасетки.
Типовой экземпляр (?) Лёва (Loew) происходит из Волгоградской обл. (б. Сарепты).
Таксономическое положение «A. agrestis», распространенного в Ориентальной области
(Гонконг, Шри Ланка и др.), остается неясным.
9. ATYLOTUS 297
Распространение: побережья водоемов пустынной и степной зон
(рис. 227). Дельта Дуная, Днепра, Одесса, Крым, северное Приазовье, низовье Дона,
Волгоградская (Волго-Ахтубинская пойма), Саратовская (пойма Волги и Хопра),
Воронежская области (с. Рождественская Хава близ Воронежа), Сев. Кавказ
(Майкоп; близ Пятигорска; Дагестан); Азербайджан (включая Нахичеванскую АССР),
Армения (Араратский и Эчмиадзинский р-ны), Грузия (окр. Тбилиси), окр. Оренбурга,
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180° 150°
Рис. 227. Ареал Atylotus (s. str.) quadrifarius.
Челябинск, Алтайский край, Тува, большая часть Казахстана (на запад включая
Уральскую и Гурьевскую области, на север Кустанайскую, Целиноградскую и
Павлодарскую области, на восток — до оз. Зайсан, на юг включая Талды-Курганскую,
Алма-Атинскую и Чимкентскую области!, Узбекистан, Туркмения. Ю. Европа,
Израиль (Элат), Ирак, Иран, Афганистан, Китай, Монголия.
Поздний вид, встречается в августе—сентябре, реже раньше, Предпочитает для
выплода^непроточные солоноватые водоемы (в том числе колодцы в пустыне), возле
которых местами обычен.
Экспериментально доказанный переносчик туляремии (Олсуфьев, 1938).
11. Atylotus (s. str.) flavoguttatus Szil. (рис. 228) — Слепень украшенный.
• S z i 1 a d у, 1915, Ent. Mitt., 4 : 98 (Tabanus); К r 6 Ь е r, 1924 : 33 (Tabanus);
Krober in Lindner, 1925 : 84 {Tabanus); Олсуфьев, 1937а : 223 (Tabanus);
Шевченко, 1961 : 128 (Tabanus); L e с 1 e г с q, 1966 : 128; Chvala, Lyne-
borg a. Moucha, 1972 : 280.
2- Глаза голые (при увеличении X 16 и более видны очень короткие серые волоски),
с одной узкой полоской или без нее. Затылок с рядом длинных, выступающих над
поверхностью глаз серых волосков. Лоб умеренна широкий, его высота превосходит
ширину приблизительно в 3—Зх/3 раза, с параллельными сторонами, в беловато-сером
налете. Лобные мозоли небольшие, черные; нижняя несколько крупнее, б. м.
поперечная, часто коричневатая, средняя округлая, иногда целиком прикрыта налетом. Усики:
1-й членик бледно-желтый, 2-й и 3-й коричневато-желтые, палочка коричневатая или
черноватая. 3-й членик б. м. узкий, с тупым дорсальным углом. Щупальца: концевой
членик белый, несколько утолщен в основной части, покрыт нежными, белыми и
единичными черными волосками. Нотоплевры желтые. Крылья бесцветные; г4 с придатком.
Ноги: бедра серые, кончики желтые. Средние и задние голени желтые, с темными
кончиками, передние голени черные, с желтым основанием. Лапки черные, основания
средних и задних лапок коричневые. Брюшко снизу черно-серое, с небольшими
грязно-желтыми пятнами по. бокам I—II тергитов и, кроме того, иногда с парой маленьких
изолированных желтоватых пятен на III тергите. Посредине брюшка имеется широкая,
продольная, темная полоса, покрытая короткими, преимущественно черными
волосками; боковые стороны тергитов в серых волосках. Снизу брюшко серое, I и II стерниты
по бокам слегка желтоватые. VIII стернит: гонопофиз короткий, в тонких желтоватых
волосках, с небольшой апикальной выемкой; латеральные лопасти и боковые
коричневые пятна крупные, последние разделены неширокой светлой полосой; базальная
298
III. ПОДСЕМ. TABANMAE
лопасть с б. м. прямым основным краем. Церки с округлой вершиной. Сперматека узко-
воретеновидная. 11.5—16 мм.
6- Голова немного шире, чем у 2i полушаровидная. Крупнофасетчатая часть
' глаз в коротких, серых волосках. Затылок с рядом длинных, серых волосков. Усики
как у 2, по несколько уже. Концевой членик щупалец белый, вытянуто-овальный,
Рис. 228. Atylotus (s. str.) flavoguttatus Szil. Лобная полоска, усик, щупальце, брюшко
сверху, VIII стернит, церки и сперматека 2-
в нежных белых волосках. Нотоплевры желтые. Придатки груди как у $. Брюшко
сверху с резкой, продольной, блестящей, черной полосой, занимающей приблизительно
Vs ширины брюшка. Боковые стороны брюшка на I—III и частью IV тергитах корич-
Рис. 229. Ареал Atylotus (s. str.) flavoguttatus.
нево-желтые, далее черные. Темная срединная полоса покрыта преимущественно
черными, боковые стороны брюшка серыми волосками, выделяющими полосу. Снизу
брюшко в основной части коричневато-желтое; посредине проходит серая продольная
полоса; последние 3—4 кольца целиком серые. 13—14 мм. " "
9. ATYLOTUS
299
Своеобразный вид, несколько похожий на A. (s. str.) rusticus, но легко
отличающийся от него'длинными волосками на затылке и широкой, темной полосой на брюшке
сверху.
Распространение: побережья водоемов пустынно-степных ландшафтов
(рис. 229). Одесская обл. (дельта Дуная, устье Днестра), Молдавская ССР (низовье
Прута в Кагульском р-не; Котовск; Оргеев), Николаевская обл. (Вознесенск; Сниги-
ревка), Днепропетровская (р. Самара), Ростовская (низовье Дона), Волгоградская
обл. (Дон у Калача; Волго-Ахтубинская пойма), дельта Волги, Сев. Кавказ (от
Краснодарского края на восток включая равнинный Дагестан, к югу до Дербента),
Азербайджан (включая Нахичеванскую АССР), Грузия (Цители-Цкаро), Казахстан (на
запад включая Уральскую и Гурьевскую области, на север.до Семиозерного р-на Ку-
станайской обл., на восток до Капала Талды-Курганской обл. и на юг включая
Чимкентскую обл.), Киргизия (Чуйская и Ферганская долины), Узбекистан
(включая Каракалпакскую АССР), Таджикистан. Спорадические находки известны на юге
Франции, в Венгрии, Турции (Эдирне), Румынии и Алжире. На западе некоторые
энтомологи не различали A. (s. str.) flavoguttatus от A. (s. str.) rusticus, вследствие чего
распространение A. (s. str.) flavoguttatus вне СССР изучено недостаточно. В
Народном музее Праги мы видели вполне типичную $ А. (&. str.) flavoguttatus, пойманную
в Бугаце, Венгрия, и снабженную определительной этикеткой: «Tabanus rusticus
bugacicus Sz. det. Szilady, 1925», экземпляр получен из коллекции Будапештского
музея.
С конца мая по конец июля. Обычен по побережьям рек и озер в подзоне северной
пустыни.
Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 1936).
12. Atylotus (s. str.) pulehellus Lw. (рис. 230) — Слепень изящный.
Представлен 2 подвидами: более светлым A. (s. str.) pulehellus pulehellus Lw. и
темнее окрашенным A. (s. str.) pulehellus karybenthinus Szil.
Рис. 230. Atylotus (s. str.) pulehellus pulehellus Lw. Лобная полоска, усик,
щупальце, брюшко сверху, VIII стернит, це^жи и сперматека <j>. A —
A. (s. str.) pulehellus karybenthinus Szil., брюшко сверху 2-
12a. Atylotus (s. str.) pulehellus pulehellus Lw. — Слепень изящный номинативный.
Loew, 1858a, Verh. Zool.-bot. Gesell. Wien, 8:597 (Tabanus); Szilady,
1915 : 98 (Tabanus); К г б b e r, 1924 : 19 (Tabanus); Krober in Lindner, 1925 :
83 (Tabanus); Олсуфьев, 1937а : 226 (Tabanus); Шевченко, 1961 : 130
(Tabanus); L e с 1 e г с q, 1966 : 132; Chvala, Lyneborg a. Mouch a, 1972 : 282.—
cyprianus R i с a r d o, 1911 : 248 (Tabanus).
2- Глаза голые, с тонкой полоской или без нее. Лоб умеренно широкий, его
высота в 3 3/4—4 раза превосходит основание, с параллельными сторонами или слегка
расширяющийся кверху, покрыт тонким, беловатым налетом. Затылок за глазами
300 III. ПОДСЕМ. TABANINAE
слегка утолщен, с рядом длинных, тонких, беловатых волосков. Лобные мозоли
небольшие, округлые, обе коричневые; средняя часто в виде двойной мозольки, иногда
черная. Усики желтые, 1-й членик бледно-желтый, 3-й членик б. м. узкий,
дорсальный угол притуплён, площадка приблизительно в 2 раза длиннее ширины. Палочка
короткая, одноцветная с площадкой. Концевой членик щупалец белый, утолщен,
покрыт нежными белыми волосками. Нотоплевры бледно-желтые. Крылья бесцветные;
г4 с длинным придатком. Ноги бледно-желтые, передние лапки и концы передних
голеней черные или темно-коричневые, лапки последующих ног на конце
коричневатые. Брюшко серое, боковые стороны I—IV тергитов бледно-розово-желтые с
широкой серой полоской посредине. Последняя занимает приблизительно 1/4—1/3 ширины
брюшка. Волосяной покров состоит из коротко прилегающих серых и черных
волосков. Последние сгруппированы в 4 продольных ряда небольших, б. м. треугольных
пятен (видны только у экземпляров хорошей сохранности!). Пятна расположены
по переднему краю тергитов и задними концами достигают не более чем до их середины.
Каемки тергитов желтые. Снизу брюшко бледно-розово-желтое, со слегка сероватым
Рис. 231. Ареалы: 1 — Atylotus (s. str.) pulchellus pulchellus; 2 — A. (s. str.)
pulchellus karybenthinus.
концом. Розоватые пятна по бокам брюшка варьируют в размерах и иногда
отсутствуют совершенно или же, наоборот, брюшко целиком желтовато-розовое. VIII
стерши бледно-желтый, без боковых коричневых пятен; гонопофиз в тонких желтоватых
волосках, с небольшой апикальной выемкой; латеральные лопасти небольшие, слабо
хитинизированы; базальная лопасть со слегка вогнутым основным краем. Церки
с округлой вершиной. Сперматека узковеретеновидная или игловидная. 8—14 мм.
(J. Голова сильно выпуклая, большая. Глаза голые, фасетки верхних 2/3
значительно крупнее нижних. Затылок за глазами с рядом длинных, белых волосков. Усики
как у 2> но более узкие. Концевой членик щупалец белый, овальный, в нежных белых
волосках. Нотоплевры бледно-желтые. Крылья как у 2- Ноги как у 2. бедра иногда
почти целиком серые. Брюшко розовато-желтое, I тергит возле щитка, небольшая
узкая полоска посредине и последние 3 тергита серые. 4 ряда узкотреугольных пятен
из черных волосков; пятна достигают концами до заднего края тергитов; прочее в
серых волосках. Снизу брюшко бледно-розово-желтое, II стернит с небольшим серым
пятном посредине. 12 мм.
Распространение: побережья водоёмов пустынной зоны (рис. 231, 1).
Азербайджан (Прикуринско-Араксинская низменность; Нахичеванская АССР —
долина Аракса), юг Туркмении, Узбекистана, Таджикистана. Алжир, Египет, Турция,
Кипр, Израиль, Ирак, Иран, Центральный Китай (провинция Шенси); ? о. Сардиния.
Обычен возле водоемов в подзоне южных пустынь.
Экспериментально доказанный переносчик трипаносомоза верблюдов в Сев.
Африке.
12b. Atylotus (s. str.) pulchellus karybenthinus Szil. (рис. 230, A) — Слепень
изящный северный.
S z i 1 a d у, 1915, Ent. Mitt., 4 : 103 (Tabanus karybenthinus); К г б b e г, 1924 :
18 (Tabanus); Krober in Lindner, 1925 : 80 (Tabanus); Олсуфьев, 1937а :
9. ATYLOTUS
301
224 (Tabanus); 19396 : 98 (T. pulchellus karybenthinus Szil.); L e с 1 e г с q, 1966 : 129
(Atylotus karybenthinus Szil.); Л у т т а, 1966а : 102 (биология).
Отличие 22 следующее: у A. (s. str.) pulchellus karybenthinus пятна из черных
волосков сверху брюшка, образующих 2 срединных ряда, имеют
вытянуто-треугольную форму и б. м. достигают заднего края тергитов, тогда как у A. (s. str.)
pulchellus pulchellus эти пятна в виде небольших треугольников и далеко не достигают
заднего края тергитов. Известны переходные формы. Терминалии как у номинативного
подвида. 8—15 мм.
У 3 A. (s. str.) pulchellus karybenthinus серая окраска брюшка более развита, чем
у номинативного подвида: сверху брюшка срединная темная полоса, разделяющая
боковые желто-розовые пятна, занимает приблизительно 113 ширины тергитов, а сами
пятна захватывают боковые стороны I—III или I—IV тергитов. Снизу брюшко в ба-
зальной части желтоватое, со срединным крупным серым пятном на I—II стернитах,
IV—VII стерниты серые; иногда брюшко снизу целиком серое, лишь задняя кайма
стернитов желтоватая. 13—14 мм.
Распространение: (рис. 231, 2). Азербайджан
(Прикуринско-Араксинская низменность), Казахстан (на север до Казалинска и Прибалхашья), Киргизия
(Ферганская долина), Узбекистан (включая Каракалпакскую АССР), Таджикистан,
Туркмения. Иран, Китай (Синьцзян, Алашань). Сообщение М. С. Шакирзяновой (1958)
об обнаружении A. (s. str.) karybenthinus Szil. в окр. Гурьева относится к A. (s. str.)
flavoguttatus Szil. (В. В. Шевченко, личное сообщение!). Указание Такаги (Takagi,
1941) на распространение A. (s. str.) pulchellus karybenthinus в Сев.-Вост. Китае, по-
видимому, ошибочно.
Обычен возле водоемов в северных частях подзоны южных пустынь; в южных
частях ареала часто встречается вместе с номинативным подвидом и переходными
формами. Кладки яиц на растениях в прибрежных частях стоячих водоемов речных
долин (Лутта, 1966а). Личинок находили в рисовых полях, болотах, реках и озере
(Кадырова, 19726).
Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 1936).
2. Подрод SURCOUFIELLA Bequ.
Bequaert, 1924, Psyche, 31, 1 : 26. — Baikalia S u г с о u i, 1921:39 nee
Baicalia Martens, 1886 : 182 (Mollusca), пит. по: Штакельберг, 1926a : 53.—
Baikalomyia Штакельберг, 1926а : 53.
Тип подрода Baikalia vaillanti Surcouf, 1921= Tabanus sublunaticornis Zetterstedti
1842.
Глаза у 2с? в заметных волосках. г4 без придатка, иногда лишь с его рудиментом.
Палочка усиков с варьирующим числом члеников от 2 до 4. Небольшие мухи, 8—10 мм,
редко до 12 мм. 22 не кровососы, обитают на болотах.
Подрод насчитывает 6 видов, из них 2 обитают в Евразии, в том числе на
территории СССР, и 4 вида — в Сев. Америке.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА
SURCOUFIELLA BEQU. (??сТсТ)
1 (2). Затылок за глазами в коротких, желтовато-серых волосках. У 2 1-й (или 1—3-й)
тергит по бокам с небольшими грязно-желтыми пятнами
13. А. (Surcoufiella) plebejus Flln.
2 (1). Затылок с рядом заметно выступающих над поверхностью глаз черных волосков.
У 2 брюшко одноцветное черно-серое
.,,,.,... 14. A. (Surcoufiella) sublunaticornis Ztt.
13. Atylotus (Surcoufiella) plebejus Flln. — Слепень невзрачный.
Изменчивый вид. Представлен 2 подвидами: номинативным A. (Surcoufiella)
plebejus plebejus Flln. и сибирским A. (Surcoufiella) plebejus sibiricus Ols.
13a. Atylotus (Surcoufiella) plebejus plebejus Flln. (рис. 232) — Слепень
невзрачный номинативный.
Fallen, 1817, Dipt. Suec. : 8 (Tabanus); Kauri, 1951 : 107; 1954 : 243; Lec-
lercq, 1966:131; Chvala, Lyneborg a. M о u с h a, 1972 : 264. — aethe-
reus Bigot, 1892 : 637 (Tabanus) et auct. — calvus S z i 1 a d y, 1915 : 94 (T. plebejus
var. calvus).
2- Голова небольших размеров. Глаза с срединной полоской, в коротких серых
волосках. Лобная полоска относительно узкая, ее высота приблизительно в З1/? раза
превосходит ширину, с параллельными сторонами, в сером налете, покрыта б. м.
302 III. ПОДСЕМ. TABANINAE
редкими, короткими, светлыми и темными волосками. Нижняя лобная мозоль
маленькая, округлая, блестяще-черная, иногда разделена вертикально пополам или почти
целиком прикрыта налетом; средняя мозоль в виде небольшой тонкой линии. Затылок
аа глазами в виде относительно узкой полоски, с рядом коротких серых волосков.
Концевой членик щупалец относительно узкий, желтоватый, в основании слегка
утолщенный, остроконечный, покрыт длинными черными и серыми волосками. Усики
целиком желтые; 1-й членик сверху в б. м. длинных, черных волосках. 3-й членик
короткий, в основании слегка расширен, с небольшим дорсальным углом. Палочка
короткая, толстая, обычно короче площадки; ее членики иногда неясно разграничены между
собой. Нотоплевры черно-серые. Ноги коричнево-желтые; тазики и основания
средних и задних бедер темно-серые; концы передних голеней и лапки черные. Средние и
задние лапки на конце коричневые. Крылья бесцветные; г4, как правило, без придатка.
Брюшко черное, в пепельно-сером налете, покрыто б. м. беспорядочно короткими
светло-серыми и черными волосками. Задние каемки тергитов желтоватые, в светлых
волосках. I—II тергиты по бокам с небольшими грязно-желтыми пятнами. 9—И мм.
Рис. 232. Atylotus (Surcoufiella) plebejus
plebejus Flirt. Лобная полоска, усик и щупальце $.
A —A. (Surcoufiella) plebejus sibiricus Ols.,
лобная полоска и щупальце $.
cJ. Голова значительно крупнее, чем у 2> помушаровидная. Глаза в длинных,
серых волосках. Фасетки верхних 2/3 глаз заметно крупнее нижних (приблизительно
в 4 раза), граница между теми и другими б. м. резко выражена. Лобный треугольник
желтый, в пепельно-сером налете. Концевой членик щупалец овальный, желтовато-
белый, покрыт б. м. длинными, преимущественно черными волосками. Затылок за
глазами с рядом коротких, светлых волосков, не превосходящих длину волосяного покрова
глаз. Усики целиком желтые, 1-й членик сверху в умеренно длинных черных волосках,
по длине б. м. равных 1-му членику. 3-й членик как у §. Нотоплевры и крылья как у 2-
Ведра ног в большей своей части серые; остальное как у §. Брюшко черное,
в сером налёте и в длинных, тонких, серых и черных волосках; I—III тергиты по
бокам с небольшими грязно-желтыми пятнами. Снизу брюшко одноцветно-темное.
11—12 мм.
Очень похож на A. sublunaticomis, но легко отличается от него отсутствием
длинных черных волосков на затылке.
Распространение: лесная и частью таежная зона. Ленинградская обл.
(Луга); юг Карельской АССР; Эстония; Калининская обл. (Нелидовский р-н); окр.
Москвы; Ивановская обл.; Украина (Хмельницкий). Сев. и Средн. Европа. Редок.
Конец июня—июль. Обитает по краю болот; личинки найдены в низинном болоте и
сильно заболоченной низине близ сфагнового болота (Карелия, Лутта, 1970).
13b. Atylotus (Surcoufiella) plebejus sibiricus Ols. — (рис. 232, A) — Слепень
невзрачный сибирский.
Олсуфьев, 1937а, Паразитол. сб. 6:236 (Tabanus); Leclercq, 1966:
132; Виолович, 1968 : 77; Murdoch et T a k a h a s i, 1969 : 52.
От номинативной формы отличается желтоватым оттенком окраски налета и
волосков тела, особенно брюшка, нотоплевры часто красновато-коричневые; кроме того,
$ отличается несколько более крупной головой, а 2 — более широкой лобной
полоской: ее высота превосходит ширину основания приблизительно в 21/2—3 раза. 9—
12.5 мм.
9. ATYLOTUS 303
Распространение: Новосибирская обл. (Венгеровский р-н); Тувинская
АССР (хр. Танну-Ола); Иркутск; Якутская АССР (Качикатский наслег); Приморский
край (Супутинский и Судзухинский заповедники; оз. Ханка; Дальнереченский р-н);
Камчатка. Сев.-Вост. Китай. На цветах зонтичных по краю болот. Редок.
Июль—август.
Голотип: 2, о. Венгерово (б. Спасск) Новосибирской обл., 17 VII 1932, Н.
Олсуфьев. В коллекции Зоологического института Академии наук СССР, Ленинград.
14. Atylotus (Surcoufiella) sublunaticomis Ztt. (рис. 233) — Слепень карликовый.
Zetterstedt, 1842, Dipt. Scand., 1:118 (Tabanus); Kauri, 1951:106;
1954 : 243 (Tabanus); Leclercq, 1966 : 133; Виолович, 1968 : 77; M u r d о с h
et Takahasi, 1969 : 51; С h v a 1 a, Lyneborga. Mo.ucha, 1972 : 262. —
plebejus (Fallen) apud В r a u e r, 1880 : 167 (Tabanus) et auct. — vaillanti S u г с о u f,
1921 : 38 (Balkalla).
?. Голова маленькая. Глаза с срединной полоской, в коротких серых волосках.
Лобная полоска широкая, ее высота превосходит ширину в 21/i—21/2 раза, с
параллельными сторонами или слегка расширяющаяся
к основанию. Полоска в голубовато-сером или
желтоватом налете, покрыта редкими,
короткими, светлыми и темными волосками. Лобные
мозоли отсутствуют. Затылрк за глазами в виде
относительно широкой полоски, с рядом
выступающих черных волосков. Концевой членик
щупалец желтовато-белый, короткий, в
основании утолщен, за серединой сужен, покрыт от-
Рис. 233. Atylotus (Surcoufiella) sublunaticor-
nis Ztt. Лобная полоска, усик и щупальце 2-
носительно длинными черными и светлыми волосками. Усики целиком желтые; 1-й
членик сверху в.б. м. длинных черных волосках; 3-й членик короткий, слабо расширен,
с едва выраженным дорсальным углом; палочка толстая и очень короткая, обычно
значительно короче площадки; ее членики часто между собой б. м. слиты, так что
палочка становится трех- и даже двучлениковой. Нотоплевры черно-серые. Ноги
коричнево-желтые, тазики, основания передних и большая часть средних и задних бедер
темно-серые; концы передних голеней и передние лапки черные, концы средних и
задних лапок коричневые. Окраска ног варьирует, и у наиболее темных экземпляров
желтые лишь колени, средние голени, основания передних голеней и 1-й
членик'средних и задних лапок. Крылья бесцветные; г4, как правило, без придатка. Брюшко
овальное, черное, в сером налете, покрыто б. м. беспорядочно короткими серыми и
черными волосками. 8—10 мм.
cJ. Голова немного больше, чем у <}, овально-выпуклая. Глаза в длинных, серых
волосках. Фасетки верхних 2/3 глаз заметно крупнее нижних (приблизительно в 4
раза); граница между теми и другими явственная, но не резкая. Лобный треугольник
желтовато-серый. Концевой.членик щупалец продолговато-овальный, желтовато-белый,
покрыт длинными темными и светлыми волосками. Затылок за глазами с рядом длинных
черных волосков. Усики целиком желтые, 1-й членик сверху в длинных черных
волосках, по длине часто превосходящих 1-й и 2-й членики, вместе взятые, 3-й членик
как у §. Нотоплевры и крылья как у 2- Ноги как у 2, за исключением передних бедер,
которые в большей своей части серые. Брюшко черное, в сером налете и в длинных
тонких серых волосках, сверху в средней части с примесью черных. I—III тергиты
(иногда только .II) по бокам с небольшими грязно-желтыми пятнами. Снизу брюшко
одноцветно-черно-серое. 9.5—11 мм.
Распространение: таежная и лесная зоны. Известен из немногих мест:
Хибины; Соловецкие о-ва; юг Карельской АССР; Эстония; Ленинградская,
Псковская области (Новоржевский р-н); Коми АССР (Ухта); Сев. Урал (басе. р. Войкар);
Омская обл. (Тара); Новосибирская обл. (Куйбышевский р-н); Алтай; Тувинская
АССР (хр. Танну-Ола); окр. Томска; Байкал; Яблоновый хребет; Амурская обл.
(Сосновая-Джелтулак); Якутская АССР (Нагорный-Цыганка); Магаданская обл. (Атка);
Камчатка; Приморский край (Б. Уллис; Супутинский заповедник); Сахалин. Сев. и
Средн. Европа, Сев.-Вост. Китай, Монголия. Редок. Конец июня—август.
Приурочен обитанием к болотам, особенно сфагновым. Личинки обнаружены в Карелии
в низинном болоте и сильно заболоченном лугу близ сфагнового болота (Лутта, 1970).
304 HI. ПОДСЕМ. TABANINAE
10. Род THERIOPLECTES Zell. — ШМЕЛЕВИДНЫЕ СЛЕПНИ
Z e 1 1 e г, 1842, Isis, 2 : 819; Moucha etChvala, 1958b : 571; 1964 : 100. —
Tabanus (L.) auct. (partim). — Sziladya Enderlein, 1923 : 545, цит. по: L e с 1 e-
rcq, 1966 : 134.
Тип рода Therioplectes tricolor Zell., 1842.
Крупные (19—26 мм, очень редко мельче!), коренастые виды, несколько
напоминающие шмелей по густому волосяному покрову тела и пестрой окраске (рис. 234).
Голова у обоих полов пропорционально
телу небольшая, сильно поперечная,
спереди почти плоская. Глаза в густых
и относительно длинных волосках, при
жизни черные или синие, без полосок,
у сухих экземпляров
интенсивно-черные. У 36 фасетки глаз одинаковой
величины. Усики. узкие, с
незначительным дорсальным углом. Глазковый
бугорок у 9 отсутствует, у 6 расположен
ниже уровня глаз, в налете и в
волосках. Лобный треугольник по бокам
с небольшим участком коротких
волосков. Лобные мозоли у 2 неясно
отграничены. Волоски на теле, в особенности
на бочках груди, длинные и очень
густые. Средние и задние бедра утолщены
и равномерно покрыты густыми,
длинными волосками, делающими их еще
толще. Крылья в средней части более
или менее буровато затемнены; г4 без
придатка (иногда имеется его рудимент).
Рис. 234. Therioplectes tricolor Zell. 2»
Род распространен только в Палеарктике, главным образом в Средиземноморской
подобласти и насчитывает всего 7 видов. Последние обитают преимущественно в
предгорьях и горах. Therioplectes однообразны по размерам и строению, но хорошо
отличаются окраской волосков тела. Олдройд (Oldroyd, 1957) указывает 2 вида
Therioplectes для горных частей экваториальной Африки, но, по-видимому, они принадлежат
к роду Hybomitra. В СССР известны 4 вида, распространенных на Кавказе, из них
один вид заходит в Крым, а другой, огибая с юга Каспийское море, в горы Туркмении
(Копет-Даг).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ПОДВИДОВ РОДА
THERIOPLECTES ZELL. {36 99)
1 (8). I тергит брюшка в густых, белых или желтовато-серых волосках.
2 (7). Лицо в черных или черно-коричневых волосках. Светлые волоски
покрывают только I тергит брюшка.
3 (6). Светлые волоски на бочках груди, задней части спинки, на щитке, а также
на I тергите брюшка белого цвета. Рыжие или серовато-желтые волоски на конце
брюшка расположены на IV—VII тергитах (включая переднюю часть IV тер-
гита). Кавказ.
4 (5). IV—VII тергиты брюшка покрыты ярко-рыжими волосками
la. Th. tricolor tricolor Zell.
5 (4). IV—VII тергиты брюшка покрыты серовато-желтыми волосками ....
lb. Th. tricolor luminosus Ols. subsp. nova.
6 (3). Светлые волоски на бочках груди, задней части спинки, на щитке, а также
на I тергите брюшка желтоватые. Желтовато-серые или рыжие волоски на
конце брюшка расположены на V—VII тергитах и лишь незначительно
захватывают заднюю часть IV тергита. Крым
1с. Th. tricolor pallidicauda Ols.
7 (2). Лицо в желтовато-серых волосках, с примесью единичных черных волосков.
Светлые волоски покрывают I тергит и заднюю половину II тергита брюшка
2. Th. tunicatus Szil.
8(1). I тзр гит брюшка в черных волосках.
10. THERIOPLECTES
305
9 (10). Конец'-брюшка в интенсивно-рыжих волосках. Крылья обычно с сильным
коричневым затемнением в средней или большей их части
.....' 3. Th.. carabaghensis Portsch.
10 (9). Конец брюшка в белых волосках. Крылья с легким затемнением по переднему
краю и возле птеростигмы 4. Th. albicauda Ols.
1. Therioplectes tricolor Zell. (рис. 234) — Слепень трехцветный.
Представлен 4 подвидами, из них 3 подвида обитают на территории СССР:
номинативный: Th. tricolor tricolor Zell. — на Кавказе, Th. tricolor luminosus Ols. subsp.
nova — в западных частях Большого Кавказа, Th. tricolor pallidicauda Ols. — в Крыму.
Четвертый подвид — Th. tricolor kirchbergi M. et Chv. — описан с о. Самос Греческого
архипелага.
la. Theribplectes tricolor tricolor Zell. — Слепень трехцветный номинативный.
Z е 11 е г, 1842, Isis : 819 (Therioplectes); В г а и е г, 1880 : 165 (Tabanus); К г
fiber, 1924 : 54 (Tabanus); Krober in Lindner, 1925:97 (Tabanus);
Олсуфьев, 1937a : 228 (Tabanus); Moucha et Clival a, 1958b : 576; L e c-
1 e г с q, 1966 : 135; Chvala, Lyneborg a. Moucha, 1972 : 289.
2- Глаза в густых, коричневых волосках. Лобная полоска широкая, с
параллельными сторонами или слегка расширена кверху, ее высота превосходит ширину
основания приблизительно в 2—21/2 раза, черная в коричневатом налете и густых желто-
коричневых волосках. Нижняя лобная мозоль б. м. треугольная или овальная, черная,
с неровной поверхностью, слабо выделяется на лбу. Мозоль продолжена кверху
короткой полоской. Лобный треугольник низкий, сильно выпуклый, черный, в
коричневатом налете. Усики черные или черно-коричневые. 3-й членик узкий, дорсальный угол
тупой; палочка короткая, толстая. Лицо черное,, в густых черных волосках.
Концевой членик щупалец черный, слегка отсвечивает коричневатым, широкий, тупой на
конце, покрыт короткими, черными волосками. Грудь: спинка двуцветная,
передняя половина в черных, задняя часть и щиток в густых, белых волосках. Нотонлевры
и бочки груди с общим продолговатым пятном в густых и длинных белых волосках.
Нижняя часть груди в черных волосках. Крылья прозрачные, слегка серые, с
расплывчатым, буроватым пятном ниже птеростигмы. Ноги: бедра черные, в черных
волосках, средние и (задние голени и основание передних бледно-желтые, в густых белых
волосках; передние лапки и большая часть передних голеней черные, лапки
последующих ног и кончики голеней коричнево-черные. Брюшко широкое, лопатообразное,
черное. I тергит в густых белых волосках, II—III в черных, IV—VII тергиты в
интенсивно-рыжих волосках. Цветовые границы резкие. Снизу брюшко черное, в черных
волосках, конец в тонких, рыжих волосках. 20—25 мм.
3- Глаза в длинных, коричневых волосках. Лобный треугольник выпуклый,
в коричневом налете. Затылок в б. м. длинных, желтовато-серых волосках, в
особенности возле глазка. Усики как у 9-3-й членик немного уже. Концевой членик щупалец
темно-коричневый, яйцевидный, покрыт черными, длинными волосками. Лицо черное,
в густых черных волосках. Грудь и ее придатки как у 2- Брюшко короткоконическое,
раскраска волосков как у 2- 19—24 мм (очень редко 16 мм).
&** Распространение: балкано-кавказский вид. В СССР обитает в
Предкавказье (от Майкопа на западе до Дагестана и низовий Самура на востоке) и в
Закавказье (Азербайджан, Грузия, Армения). Турция, Болгария, Румыния, Греция.
РСицилия. Указание Г. А. Бабаянца (1962) о распространении этого подвида в
Туркмении ошибочно.
Преимущественно в горах. Июнь—июль. Не часто.
22 нападают на домашних животных; Порчинский наблюдал в горах 36 и 22 на
цветах зонтичных (Heracleum).
lb. Therioplectes tricolor luminosus Ols. subsp. nova. — Слепень трехцветный
светлый.
2cJ. От номинативной формы отличается серовато-желтыми волосками брюшка
на IV—VII тергитах. От Th. tricolor pallidicauda, с которым описываемый подвид
сходен по окраске волосков конца брюшка, отличается белым цветом светлых волосков
груди и I тергита- брюшка (как у номинативной формы). 20—22 мм.
Ы Голотип: 2' Майкоп Краснодарского края, 1951 г. В коллекции Зоологического
института Академии наук СССР, Ленинград.
Паратипы: 5 2 оттуда же; 1 2 верховья Белой Краснодарского края, Филипченко;
3 3 окр. Пятигорска. В коллекции Зоологического института Академии наук СССР,
Ленинград и в нашей коллекции.
Экземпляры этого подвида нами (Олсуфьев, 1937а) были ошибочно отнесены
"к Th. tricolor pallidicauda Ols.
20 H. Г. Олсуфьев
306 HI. ПОДСЕМ. TABANINAE
lc. Therioplectes tricolor pallidicauda 01s. — Слепень трехцветный бледнохвостый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII : 230 (Tabanus); М о и с h a
et С h v а 1 а, 1958Ь : 578; L е с 1 е г с q, 1966 : 136; Chvala, Lybeborg a.
М о и с h a, 1972 : 290.
$(J. От номинативной формы отличается желтоватым или буроватым оттенком
окраски светлых волосков груди и I тергита брюшка и желтовато-серыми волосками
на конце брюшка, занимающими V—VII тергиты (и лишь немного заднюю часть IV
тергита); эти волоски изредка бывают рыжего цвета, как у номинативной формы. 20—
22 мм.
Распространение: Крым, предгорья и горы. Конец мая—июнь. Не часто.
Голотип: <$, Чумакарка Симферопольского р-на, Крым, 5 VI 1923, Олсуфьев.
В коллекции Зоологического института Академии наук СССР, Ленинград.
Указание Е. Г. Гаузера (1960) о нахождении этого вида в Нахичеванской АССР
(1 экз. с этикеткой «Джульфа» в Музее Грузии) следует считать сомнительным, равно
как указание Леклерка (Leclercq, 1966) для Ирана.
2. Therioplectes tunicatus Szil. — Слепень светлобородый.
S z i 1 a d у, 1927, Zool. Anzeiger, 74 : 204 (Tabanus); Moucha et Chvala,
1958b : 574; 1964 : 101; L e с 1 e г с q, 1966 : 136; Chvala, Lyneborg a.
Moucha, 1972 : 287. — alazanicus Hauser, 1953 : 247.
$>. Глаза в серых волосках. Лобная полоска с параллельными сторонами или
слегка расширяется кверху, ее высота превосходит ширину основания в 2—21/2 раза,
в сером налете и желтоватых волосках. Нижняя лобная мозоль б. м. треугольная или
овальная, черная, слабо блестящая. Верхним концом мозоль продолжена в короткую,
черную полоску. Затылок в желтых волосках. Лобный треугольник черный, иногда
в центре коричневатый., покрыт серым налетом. Усики черные; 1-й членик в сером
налете, иногда коричневый, 3-й членик в основании коричневатый, дорсальный угол
тупой. Лицо в серо-желтых волосках, с примесью единичных черных волосков.
Концевой членик щупалец коричневый или черноватый, в черных волосках, в основании
утолщен, тупоконечный. Грудь в густых серовато-желтых волосках, па спинке и щитке
волоски иногда золотистого оттенка. Крылья с расплывчатым коричневым пятном
ниже птеростигмы. Ноги: бедра черно-серые, в желтовато-серых волосках, иногда
с примесью черных, средние и задние голени и основание передних бледно-желтые,
в густых такого же цвета волосках; передние лапки и концевая часть передних голеней
черно-коричневые, лапки последующих ног и кончики голеней коричневые. Брюшко
черное, I тергит и широкая кайма по заднему краю II тергита в беловато-сером
налете, оба тергита покрыты желтовато-серыми волосками; последующие тергиты в
черных волосках, VI—VII или V—VII тергиты в золотисто-рыжих волосках, иногда
такие же волоски в небольшом количестве имеются на III—IV тергитах. Снизу брюшко
черное, преимущественно в черных волосках, II стернит с широкой беловатой
каймой из налета и волосков, VI—VII стерниты в золотисто-рыжих волосках. 19—21 мм.
cJ. Глаза в длинных, серых волосках. Лобный треугольник в сером налете.
Затылок в относительно длинных желтоватых волосках. Усики как у $, но более узкие.
Концевой членик щупалец коричневый, яйцевидный, покрыт преимущественно
черными волосками. Лицо в густых желтовато-серых волосках, в нижней части с примесью
черных. Грудь и ее придатки как у §. Брюшко коническое, окраска его как у $. 18 мм.
Распространение: балкано-кавказский вид. В СССР известен из
немногих мест Закавказья: Муганлы, Закатальского р-на, Сев.-Зап. Азербайджан; Лаго-
дехи, Сев.-Вост. Грузия. Турция, Сирия, Израиль, Греция, Албания, Венгрия,
Югославия, Италия (Сицилия). Июнь. Не часто.
3„ Therioplectes carabaghensis Portsch, (рис. 235) — Слепень карабахский.
Изменчивый вид. В пределах СССР представлен 2 формами: распространенным
в горах Закавказья номинативным подвидом Th. carabaghensis carabaghensis Portsch.
и обитающим в Копет-Даге, Туркмения, Th. carabaghensis orientalis Ols., subsp. nova.
3a. Therioplectes carabaghensis carabaghensis Portsch. — Слепень карабахский
номинативный.
Порчинский, 1877, Тр. Русск. энтом. общ., X : 154 (Tabanus); КгоЬег
in L i n d n e r, 1925 : 90 (Tabanus); Олсуфьев, 1937а : 230 (Tabanus); Moucha
et Chvala, 1958b : 576; Leclercq, 1966 : 134.
9. Глаза в коричневых волосках. Лоб широкий, его высота превосходит ширину
приблизительно в 2—2х/2 раза, с параллельными сторонами или слегка
расширяющийся кверху, коричнево-черный, в б. м. длинных, коричневых волосках. Нижняя
лобная мозоль б. м. треугольная, плоская, черная, слабо блестящая. Верхним кон-
10. THERIOPLECTES 307'
цом мозоль продолжена в короткую, черную полоску. Затылок в коричнево-черных
волосках. Лобный треугольник черный или темно-коричневый, в коричневом налете.
Усики: базальные членики темно-коричневые, 3-й членик черный, часто с коричневым
основанием со слабо выраженным дорсальным углом; палочка короткая, толстая. Лицо
черное, в черных волосках. Концевой членик щупалец черный, широкий, на конце
тупой, покрыт короткими, черными волосками. Грудь черная, в черных волосках,
спинка и щиток в густых, желтовато-серых волосках; последние обычно образуют
пятно на бочках груди. Крылья слегка дымчатые, с большим коричневым пятном ниже
птеростигмы. Ноги: бедра черные, в черных волосках, средние и задние голени и
основания передних голеней бледно-желтые, в густых белых волосках; передние лапки
и большая часть передних голеней черные, лапки последующих ног и кончики
голеней коричнево-черные. Брюшко широкое, лопатообразное, блестяще-черное, в черных
волосках, последние 3 кольца в рыжих волосках. VIII стернит: гонопофиз широкий,
в густых светлых, желтовато-коричневых волосках, со слабой апикальной выемкой;
Рис. 235. Therioplectes carabaghensis carabaghensis Portsch. Лобная
полоска, усик, щупальце, VIII стернит, церки и сперматека $.
лате ральные лопасти небольшие, боковые коричневые пятна соприкасаются по сред»
ней линии стернита; базальная лопасть короткая, со слегка выпуклым основным краем.
Церки небольшие, округло-треугольные, X тергит в густых, длинных волосках.
Сперматека миндалевидная. 19—23 мм.
в- Глаза в длинных, коричневых волосках. Затылок за глазами в б. м. длинных,
коричнево-черных волосках, в особенности возле глазка. Лобный треугольник
выпуклый, коричнево-черный. Усики черные или темно-коричневые, их форма как у 9;
3-й членик несколько уже. Концевой членик щупалец темно-коричневый,
яйцевидный, в длинных, черных волосках. Грудь и ее придатки как у $. Брюшко короткокони-
ческое, блестяще-черное; расцветка волосков как у 2- 21—23 мм.
Окраска волосков груди несколько варьирует: в передней части спинки, возле
головы, часто имеется значительная примесь черных волосков; у 1 $ из Сев.-Зап. Ирана
(Савалян) спинка и бочки груди в коричнево-желтых волосках. Некоторые
экземпляры, в том числе 1 с? 1 2 ив типовой серии Порчинского, не имеют пятна светлых волос-,
ков на бочках груди. 3 29 из Лерикского (Талыш) имеют почти бесцветные крылья,
с едва заметным коричневым затемнением в базальной части радиальных ячеек, но у $
оттуда же крылья с нормально развитым коричневым пятном. ^
Распространение: восточная часть Большого Кавказа в пределах
Азербайджана (Тубикенд и Чухуръюрт); Малый Кавказ в пределах Армении (Арзни; Су-
сиан; ю.-в. берег Севана; Мегринский р-н) и Азербайджана (Аджикенд; Ханлар5
г. Кире близ Шуши; Биченак Шахбузского р-на Нахичеванской АССР); Талыш. Иран.
Июнь—июль. Не часто.
20*
308 III. ПОДСЕМ. TABANINAE
3b. Therioplectes carabaghensis orientalis 01s. subsp. nova — Слепень карабахский
восточный.
§. От номинативного подвида отличается очень темными крыльями — они имеют
интенсивную коричневую окраску, за исключением более светлых вершины, заднего
края крыла, средней части дискоидальпой ячейки, иногда также нижней базальной
ячейки. Бочки груди только в черных волосках. Рыжие волоски расположены на
VI—VII тергитах, тогда как V тергит в черных волосках. 20—23 мм.
Голотип: 2. Ипай-кала Бахарденского р-на, Копет-Даг, Туркмения И VI 1972,
Д. Чарыкулиев. В коллекции Зоологического института Академии наук СССР,
Ленинград.
Паратипы: 6 2 там же и того же числа; 1 2 Кара-кала, Арпакленское ущелье,
Туркмения, 7 V 1932, П. Петрищева; 1 5 Иол-Дере, близ Кара-кала, Туркмения,
20 VI 1952, Борисова; 12 г. Сюит, Зап. Копет-Даг, 20—22 V 1953, Пономарева. В
коллекции Зоологического института Академии наук СССР, Ленинград, и в коллекции
Н. Г. Олсуфьева, Москва.
К описываемому подвиду, видимо, принадлежит 1 2 из Ашхабада, описание
которой дает Крёбер (КгбЬег, 1924 : 53), неправильно отнеся ее в варьететы без латинского
названия к Th. gigas.
4. Therioplectes albicauda Ols. — Слепень белохвостый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 231 (Tabanus); M о u-
cha etChvala, 1958b : 575; L e с 1 e г с q, 1966 : 134.
2- Глаза в коричневых волосках. Затылок за глазами в длинных, желтых
волосках. Лоб широкий, его высота приблизительно в 2—21/i раза превосходит ширину,
слегка расширяется кверху, черный, в буроватом налете, покрыт б. м. длинными
желтыми волосками. Нижняя лобная мозоль плоская, черная, б. м. треугольная, неясно
отграничена от лба. Верхним концом мозоль переходит в короткую, черную полоску.
Лобный треугольник черный или коричневатый, в коричневом налете. Усики темно-
коричневые, с черным концом. 3-й членик узкий, с небольшим дорсальным углом,
палочка короткая, толстая. Лицо черное, в черных волосках, наличник в желтых
волосках, нижняя часть лица с примесью желтовато-серых волосков. Концевой членик
щупалец черный или черно-коричневый, широкий, с тупым концом, покрыт короткими
черными волосками. Грудь черная, в густых черных волосках; спинка и щиток в
коричневато-желтых или сероватых волосках. Крылья с легким затемнением по переднему
краю и ниже птеростигмы, в прочих частях б. м. прозрачные. Ноги: бедра черные,
в черных волосках, средние и задние голени бледно-желтые, в густых беловатых
волосках. Передние лапки и голени черные, последние снаружи в основании слегка
коричневые и на небольшом протяжении в желтовато-серых волосках. Лапки
последующих ног и кончик голеней коричневые. Брюшко широкое, лопатообразное,
блестяще-черное, в черных волосках, лишь последние 3 тергита (и частью
предыдущий) в густых белых волосках. Снизу брюшко черное, в основании в черных, на конце
в тонких белых волосках. 22—23 мм.
3- Глаза в длинных коричневых.волосках. На темени пучок желтоватых
волосков. Лобный треугольник коричнево-черный. Усики: базальные членики
темно-коричневые, 3-й членик черный, с темно-коричневым основанием, дорсальный угол тупой.
Концевой членик щупалец черно-коричневый, яйцевидный, в длинных черных
волосках. Грудь и ее придатки как у 2- Брюшко коническое, блестяще-черное; расцветка
волосков.как у 2- 21 мм.
3 описывается впервые.
Распространение: в СССР известен только из юго-восточного
Азербайджана: Джалилабадский (Астрахан-Базар), Ленкоранский и Лерикский р-ны. Иран
(побережье Каспийского моря). Редок. Конец мая—июнь.
Голотип: $, Ленкорань, 8 V 1909 (ст. ст), А. Кириченко. В коллекции
Зоологического института Академии наук СССР, Ленинград.
11. Род HYBOMITRA End.
Enderlein, 1922, Mitt. Zool. Mus. Berlin, 10 : 347; Philip, 1941 : 7; 1947 :
291; Lyneborg, 1959 : 78; T a k a h a s i, 1962 : 35; L e с 1 e г с q, 1966 : 59; В и о-
л о в и ч, 1968 : 112. — Tabanus (L.) aucts- (partim.). — Tylostypia Enderlein,
1922 : 347. — Sipala Enderlein, 1923, 1. с : 545. — Tylostypina Enderlein,
1923, 1. с : 545. — Sziladynus Enderlein, 1925:181, пит. no: L e с 1 e г с q,
196б': 99. — Mouchaemyia 0 1,'sufjev, 1972:450.
Тип рода Hybomitra so-lox Enderlein, 1925= Tabanus rhombicus Osten-Sacken, 1876.
Средних размеров или крупные, по преимуществу темноокрашенные виды (рис. 261,
272, 311, 312). Глазковый бугорок имеется у обоих полов! блестящий, реже у?в виде
11. HYBOMITRA 309
незначительного рудимента, тогда как у 3 в легком налете. Голова у 3 обычно лишь
немногим больше, чем у 2. редко значительно крупнее, полушаровидная. Глаза
покрыты волосками; в очень редких случаях (группа Н. (s. str.) erberi Br.) глаза у 2
в едва заметных волосках. Глаза у живых особей яркие: зеленые, пурпурные, синие,
У 29' как правило, с 3 полосками, у 33 с 2 полосками в зоне мелких фасеток. У сухих
экземпляров глаза принимают буро-черную окраску. У 33 фасетки верхних 2/3 глаз
крупнее нижних или относительно одинаковые. Лобные мозоли у 22 обычно хорошо
развиты, нижняя б. м. прямоугольная или треугольно-щитовидная, средняя веретено-
видная или линейная, обычно соединена с нижней. Лобный треугольник без волосков
по бокам, в редких случаях волоски имеются (подрод Mouchaemyia Ols.). Палочка 3-го
членика усиков явственно 4-члениковая, очень редко членики частично редуцированы
(некоторые Hybomitra s. str., подрод Tibetomyia Ols., Ц. Азия). Крылья бесцветные или
слегка затемнены, иногда вокруг поперечных жилок имеются буроватые пятна; г4
иногда с коротким придатком. Брюшко у 33 почти всегда, у 22 чаще, с коричнево-
желтыми пятнами по бокам. Терминалии обычного для трибы. Tabanini строения,
у 2? VIII стернит с б. м. развитыми латеральными лопастями и боковыми коричневыми
пятнами. У некоторых видов терминалии 22 различаются, что позволяет
использовать их строение для уточнения диагностики.
Обширный род, насчитывающий около 150 видов и распространенный
преимущественно в Голарктике; немногие виды .известны из Эфиопской и Ориентальной
областей. В Палеарктике 96 видов. Основная масса видов приурочена обитанием к
равнинной таежно-лесной полосе или горам, лишь немногие виды свойственны водоемам
степной или пустынной зон. В СССР известны 50 видов.
Некоторые группы видов трудны для определения, вследствие чего необходимо
сличение с точно определенными экземплярами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА
HYBOMITRA END.
1 (2). Глаза покрыты очень длинными волосками. Лобный треугольник по бокам
в волосках (рис. 236) 1. Mouchaemyia Ols.
2 (1). Глаза в умеренно длинных или коротких волосках (у 2? иногда почти голые!).
Лобный треугольник по бокам без волосков, крайне редко в единичных волосках.
3(4). Концевые сегменты брюшка не сжаты с боков 2. Hybomitra End.
4 (3). Концевые сегменты брюшка явственно сжаты с боков (рис. 324)
• . • 3. Sipala End.
1. Подрод MOUCHAEMYIA Ols.
Олсуфьев, 1972, Энтомол. обозр.," 51,|2 : 450.
У обоих полов лобный треугольник по бокам в волосках (рис. 236). Глаза с 1—3
полосками (иногда без них), покрыты длинными волосками, особенно у 3- Палочка
3-го членика усиков явственно 4-члениковая. Конец
брюшка не сжат с боков.
Тип подрода Hybomitra (Mouchaemyia) caucasi Szilady,
1923.
Средиземноморский подрод. Кроме распространенного
на Кавказе Н. (Mouchaemyia) caucasi Szil., мы включаем
в него Н. (Mouchaemyia) lateralis Mg. (Ю. Европа),
Н. (Mouchaemyia) decora Lw. (Балканы, Передняя Азия)
и Н. (Mouchaemyia) mendica Villen. (Передняя Азия).
Наличие у Mouchaemyia волосков по бокам лобного
треугольника хорошо отличает этот подрод от других
Hybomitra и сближает с Therioplectes. Однако по другим приз-
Рис. 236. Hybomitra (Mouchaemyia) caucasi Szil. Лобный
треугольник 2i покрытый по бокам тонкими волосками.
накам, в том числе наличию глазкового бугорка, Mouchaemyia, несомненно, относятся
к Hybomitra. Любопытно, что на основании внешнего сходства Теодор (Theodor, 1965)
отнес Н. (Mouchaemyia) decora Lw. и Н. (Mouchaemyia) mendica Villen. к
Therioplectes.
910 III. ПОДСЕМ. TABANINAE
1. Hybomitra (Mouchaemyia) caucasi Szil. (рис. 237) — Слепень кавказский.
S z i 1 a d у, 1923, Biolog. Hungar., 1,1:5 (Tabanus); Krober in Lindner,
1925 : 62 (Tabanus); Олсуфьев, 1937a : 158 (Tabanus); L e с 1 e г с q, 1966 : 104;
Chvala, Lyneborg a. Moucha, 1972 : 192.
?. Глаза в длинных, серых волосках, с 1 полоской или без нее. Лобная полоска
широкая, с параллельными сторонами, ее высота в 21/%—3 раза превосходит ширину,
покрыта длинными, преимущественно светлыми волосками. Темя с
блестяще-коричневым треугольником, в передней части которого расположен овальный, коричневый,
глазковый бугорок. Нижняя лобная мозоль блестяще-черная, прямоугольная или
щитовидная, обычно достигает края глаз. Средняя мозоль широковеретеновидная,
чаще достигает нижней мозоли. Лобный треугольник низкий, в густом сером налете.
Усики черные: 1-й членик в сером налете и в длинных светлых волосках, 3-й членик
иногда в основании коричневатый, узкий, дорсальный угол почти совсем сглажен.
Палочка короткая, приблизительно в 2 раза короче площадки. Щупальца беловато-
желтые, концевой членик слабо утолщен или узкий, покрыт относительно длинными
Рис. 237. Hybomitra (Mouchaemyia) caucasi Szil. Лобная
полоска, усик, щупальце, VIII стернит, церки и сперматека ?•
светлыми волосками. Нотоплевры темные. Крылья бесцветные, жилки
темно-коричневые. Жужжальца черно-коричневые. Ноги: бедра черно-серые, голени желтые,
с черноватыми кончиками, лапки черные. Брюшко широкое, чёрное. II тергит с
крупными, беловатыми пятнами по бокам (налет+волоски) и небольшим треугольником
посредине, последующие тергиты со срединными треугольниками и каемками из
светлых волосков, без боковых пятен. Снизу брюшко в густом сером налете. VIII стернит:
гонопофиз крупный, в тонких желтых волосках, с выраженной апикальной выемкой;
латеральные лопасти небольшие, остроугольные; боковые коричневые пятна крупные;
базальная лопасть со слабо вогнутым основным краем. Церки с уплощенной
вершиной. Сперматека веретеновидная. 13—16 мм.
в- Глаза в очень длинных серых волосках. Фасетки глаз б. м. одинаковой
величины (или верхние фасетки слегка крупнее нижних). Лобный треугольник серый. Усики
как у 2, но 3-й членик немного уже; 1-й членик сверху в очень длинных желтовато-
серых волосках. Концевой членик щупалец вытянуто-овальный, желтовато-серый,
в светлых волосках. Грудь и ее придатки как у £. Брюшко короткое, черное; II—III
(иногда также IV) тергиты по бокам коричневые. Кроме того, II тергит по бокам
с крупными светлыми пятнами из серых волосков и налета, посредине с небольшим
треугольником, последующие тергиты со срединными треугольниками и каемками
из серых волосков, без боковых пятен. Снизу брюшко в густом сером налете и
светлых волосках. 15 мм.
Распространение: кавказский предгорно(горно)-степной вид.
Дагестан (Хасавюртовский и Каякентский р-ны); Азербайджан (Кубинский р-н; Карабах);
Грузия (Манглиси; Тетри-Цкаро; Ахалкалаки; Цалка); Армения (Степанаван; Дили-
жан; Нор-Баязет и другие пункты). Сев.-Вост. Турция. Май—июль. Не часто.
11. HYBOMITRA 311
2. Подрод HYBOMITRA End. s. str.
E n d e г 1 e i n, 1922, Mitt. Zool. Mus. Berlin, 10, 2 : 347.
Тип подрода Hybomitra solox Enderlein, 1925= Tabanus rhombicus Osten-Sacken,
1876.
У обоих полов лобный треугольник по бокам без волосков.* Глаза обычно с 3
полосками, крайне редко без них (Н. expollicata expollicata Pand.), покрыты умеренно
длинными или короткими волосками (иногда у самок едва заметными — группа
Н. (s. str.) erberi Br.). Палочка 3-го членика усиков обычно явственно 4-члениковая.
Конец брюшка не сжат с боков.
В Палеарктике около 70 видов, в СССР — 48 видов. В равнинных таежно-лес-
ных, лесостепных ландшафтах и в горах господствующая группа по обилию видов и
особей.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА
HYBOMITRA END.
А. Глаза разделены лбом: 2?
1 (98). Жужжальца коричневые или черные (иногда вершина головки желтоватая).
Глаза покрыты относительно хорошо развитыми волосками.
2 (51). Концевой членик щупалец узкий, его длина приблизительно в З1^—4 раза
превосходит толщину (рис. 239, 241 и др.). Щупальца коричневые, оранжевые
или черные, в последнем случае иногда в основании утолщены.
3,(6). Лобный треугольник частично оголен, блестяще-желтый. Черные тергиты
брюшка с резкими широкими желтыми каемками, покрытыми
золотисто-желтыми волосками. Крупные виды (17—23 мм), напоминающие по окраске ос
(рис. 261). (II группа: Н. (s. str.) tarandina L.).
4 (5). Бочки груди в желтоватых волосках 16. Н. (s. str.) tarandina L.
5 (4). Бочки груди в одноцветно-черных волосках
17. Н. (s. str.) tarandinoides Ols.
6 (3). Лобный треугольник в сероватом или желтоватом налете, если же оголен, то
блестяще-черный.
7 (20). Лобный треугольник на значительном протяжении блестяще-черный (рис. 267)
или же крылья с хорошо заметными коричневатыми пятнами вокруг
поперечных жилок, в особенности в основании жилки г4 (рис. 272). (III группа:
Н. (s. str.) astur Erichs.).
8 (9). Лобный треугольник целиком в сером налете (рис. 273) .
23. Н. (s. str.) astur Erichs.
9 (8). Лобный треугольник на значительном протяжении блестяще-черный (рис. 267,
269 и др.).
10 (11). Вершинная Vs бедер оранжевая .... 18. Н. (s. str.) aequetincta Beck.
11 (10). Бедра целиком черные.
12 (15). Лобная полоска в относительно коротких, -черных и серых волосках. II тергит
брюшка без красноватых пятен по бокам.
13 (14). Средние и задние голени черные. Брюшко черное, в черных волосках, II тергит
по бокам с беловато-серыми пятнами из налета и волосков, II и последующие
тергиты по заднему краю с небольшим срединным треугольничком из беловатых
волосков. Западная Европа 26. Н. (s. str.) micans Mg.
14 (13). Средние и задние голени светло-коричневые, с черными кончиками. Брюшко
сверху в буроватом налете и в коротких прилегающих золотисто-желтых или
серо-желтых волосках. Дальний Восток . . 19. H.,(s. str.) stigmoptera Ols.
15 (12). Лобная полоска в длинных, густых желтых волосках. II тергит брюшка по
бокам с красноватыми пятнами. Горы Средней Азии.
16 (19). Брюшко сверху целиком в красновато-желтых или золотистых волосках.
17 (18). Бочки груди и низ лица в желтых волосках . .'
20. Н. (s. str) tatarica Portsch.
18 (17). Бочки груди и низ лица в черных волосках
21. Н. (s. str.) shevtshenkoi Ols.
19 (16). Ill—IV тергиты в черных, прочие тергиты в золотисто-желтых волосках
- 22. Н. (s. str.) zonata Szil.
20 (7). Лобный треугольник в желтоватом или сером налете. Крылья не имеют хорошо
заметных темных пятен вокруг поперечных жилок (иногда имеются лишь
следы). (I группа: Н. (s. str.) kaurii Chv. et Lyn.).
* Иногда единичные волоски отмечаются у 2? Н. (s. str.) aterrima и Н. (s. str.)
auripila.
312 III. ПОДСЕМ. TABANINAE
21 (40). Брюшко сверху с незначительными темно-коричневыми пятнами по бокам II
(реже I—III) тергита или целиком черное. Снизу брюшко, как правило,
одноцветно-черное, лишь изредка (светлые Н. (s. str.) lapponica !) II (или I—III)
стернит по бокам коричневатый.
22 (33). Лобная полоска широкая, ее высота превосходит ширину основания в 21/i—
3 раза (рис. 239, 241, 275 и др.).
23 (24). Бочки груди в черных волосках; нив лица обычно со значительной примесью
черных волосков 25. Н. (s. str.) aterrima Mg.
(из группы Н. (s. str.) astur Erichs.).
24 (23). Бочки груди в серых или желтоватых волосках, на мезоплеврах часто примесь
черных волосков.
25 (26). Брюшко с каймами из золотисто-желтых волосков по заднему краю тергитов.
Коричневые пятна по бокам II тергита отсутствуют
, 24. Н. (s. str.) auripila Mg.
(из группы И. (s. str.) astur Erichs.).
26 (25). Брюшко с каймами из беловатых или желтовато-сероватых волосков по
заднему краю тергитов. Коричневые пятна по бокам II тергита имеются, если
отсутствуют, то брюшко сверху с рисунком из 3 продольных рядов серых пятен.
27 (32). Брюшко с небольшими темно-коричневыми пятнами по бокам II тергита.
Щупальца коричнево-черные.
28 (29). Голени одноцветно-черные 6. Н. (s. str.) altaica Ols.
29 (28). Голени по крайней мере средней пары ног в основной части коричневые.
30 (31). 1-й членик усиков черный, веером налете. 3-й членик усиков короткий,
площадка обычно преимущественно черная с коричневой базальной частью
(рис. 241). Тергиты брюшка по заднему краю с нерезкой сероватой или
желтоватой каймой. Мельче: 11—13 мм, редко до 15 мм. ^
3. Н. (s. str.) sexfasciata Hine.
31 (30). 1-й членик усиков желто-красный или частично сероватый. 3-й членик усиков
удлиненный, площадка обычно целиком или преимущественно желто-красная
(рис. 239). Тергиты брюшка по заднему краю с резкой беловатой каймой. 14—
16 мм, редко 13 мм 2. Н. (s. str.) kaurii Chv. et Lyn.
32 (27). Брюшко одноцветно-черное с серым рисунком из 3 продольных рядов пятен.
Щупальца бледно-коричневые 8. Н. (s. str.) astuta 0. S.
33 (22). Лобная полоска менее широкая, ее высота превосходит ширину основания
в 3V2—4 раза (рис. 243, 244, 247 и др.).
34 (37). Брюшко черно-серое, без оливкового налета; светлые волоски на тергитах
беловато-серые.
35 (36). Мельче: 10—14 мм. Глаза в коротких, но хорошо заметных волосках. Брюшко-
с неясными узкими сероватыми каемками по заднему краю тергитов. Нижняя
лобная мозоль крупная, б. м. прямоугольная, достигает или почти достигает
края глаз (рис. 247). II тергит брюшка одноцветно-черный или с небольшими
буроватыми пятнами по бокам. Север европейской части СССР, Сибирь,
Дальний Восток 7. Н. (s. str.) lapponica Wahlbg,
36 (35). Крупнее: 13—16 мм. Глаза голые (при увеличении Х16 едва заметны очень
короткие волоски). Брюшко с резкими умеренно широкими беловатыми
каемками по заднему краю тергитов. Нижняя лобная мозоль небольшая, чаще
треугольная, не достигает края глаз (рис. 243). II тергит брюшка с небольшими
каштаново-коричневыми пятнами по бокам. Дальний Восток
.... i 4. Н. (s. str.) stenopselapha Ols.
37 (34). Брюшко черное, в оливковом налете; светлые ,волоски на тергитах золотисто-
желтые.
38 (39). Брюшко сверху в интенсивном оливковом налете и в коротких, густых
золотисто-желтых волосках. Концевой' членик щупалец оранжево-желтый, в
такого же цветка волосках 15. Н. (s. str.) bagheera Viol.
39 (38). Брюшко сверху в умеренном оливковом налете. Тергиты по заднему краю
в желтых волосках, тогда как в передней части в черных волосках. Концевой
членик щупалец коричневатый, в черных волосках
14. Н. (s. str.) obscurinervis Viol.
40 (21). Брюшко сверху с хорошо развитыми желто-коричневыми или золотисто-
оранжевыми пятнами по меньшей мере по бокам I—III тергитов. Снизу брюшко,
как правило, в основании желто-коричневое, лишь изредка (темные И. brevis)
черное, с желтыми каймами по заднему краю стернитов.
41 (42). Брюшко резко двуцветное: основная часть брюшка оранжево-желтая, в
золотисто-желтых волосках, IV и последующие кольца одноцветно-черные, в черных
волосках. Дальний Восток 12. Н. (s. str) nigella Szil.
42 (41). Брюшко иной окраски; IV и последующие кольца всегда с б. м. выраженной
светлой каемкой или в светлых волосках.
43 (48). Усики оранжевые или желто-коричневые, палочка одноцветная с площадкой.
44 (47). Светлые волоски брюшка рыжие или золотисто-желтые.
11. HYBOMITRA
313
45 (46). Щупальца желто-оранжевые. 3-й членик усиков с резким дорсальным
выступом (рис. 257). Крупнее: 16—21 мм 13. Н. (s. str.) pavlovskii Ols.
46 (45). Щупальца черные; 3-й членик усиков с б. м. прямым дорсальным углом
(рис. 251). Мельче: 13—16 мм 9. Н. (s. str.) brevis Lw.
47 (44). Светлые волоски брюшка серебристо-серые
10. Н. (s. str.) ussuriensis Ols.
48 (43). Усики коричневые, с черной палочкой.
49 (50). 3-й членик усиков с резким дорсальным выступом (рис. 254). Нижняя лобная
мозоль не достигает края глаз, слегка морщинистая. Крупнее: 13—18.5 мм
11. Н. (s. str.) arpadi Szil.
50 (49). 3-й членик усиков с б. м. прямым дорсальным углом (рис. 244). Нижняя
лобная мозоль выпуклая, блестящая, достигает края глаз. Мельче: 12.5—
16.5 мм 5. Н. (s. str.) olsoi Takah.
51 (2). Концевой членик щупалец утолщен, его длина превосходит толщину не более
чем в 3 раза (рис. 279, 281, 292 и др.). Щупальца светлые желтовато-белые,
бледно-желто-коричневые или сероватые, но никогда не черные или темно-
коричневые. (IV группа: Н. (s. str.) bimaculata Macq.).
52 (55). Лобный треугольник на б. м. значительном протяжении блестяще-черный
или черно-коричневый (рис. 279, 281).
53 (54). Сильно выпуклый лобный треугольник целиком блестяще-черный (рис. 279).
Нижняя лобная мозоль, как правило, черная. Мельче: 11.5—15 мм ....
28. Н. (s. str.) lurida Plln.
54 (53). Умеренно выпуклый лобный треугольник лишь в центральной части
блестящий, коричнево-черный, по краям в сером налете (рис. 281), крайне редко
целиком голый. Нижняя лобная мозоль чаще коричневая, тонкоморщинистая.
Крупнее: 14—18 мм 29. Н. (s. str.) nitidifrons Szil.
55 (52). Лобный треугольник целиком прикрыт налетом, матово-серый или
желтоватый.
56 (77). Брюшко сверху одноцветно-черное или черно-серое, крайне редко (Я. (s. str.)
sogdiana, H. (s. str.) nigricorpa) II тергит по бокам просвечивает коричневатым,
но в таком случае брюшко снизу целиком одноцветно-черное.
57 (58). Лобная полоска сильно сужена к основанию, ее высота превосходит ширину
основания приблизительно в 4V2—5 раз (рис. 297). 3-й членик усиков коричнево-
красный, палочка обычно черная
37. Н. (s. str.) bimaculata Macq. var. bisignata Jaenn.
58 (57). Лобная полоска лишь слегка сужена к основанию и более широкая: ее высота
превосходит ширину основания приблизительно в 3—4 раза (рис. 299, 300 и др.).
3-й членик усиков целиком черный, крайне редко в основании коричневый.
59 (68). Брюшко снизу блестяще-черное, лишь иногда в легком голубоватом, налете.
60 (61). Нижняя лобная мозоль, как правило, достигает края глаз, блестяще-черная
(рис. 299).. Бочки груди в черных волосках, реже в серых. Горы Кавказа,
Средн. Европы 38. Н. (s. str.) caucasica Enderl.
61 (60). Нижняя лобная мозоль не достигает края глаз, обычно в тонких поперечных
морщинках (рис. 300, 302 и др.). Бочки груди в серых, реже в черных волосках.
Горы Средней Азии.
62 (63). I и II тергиты в легком сером налете и в коротких, густых беловато-серых
волосках, последующие тергиты черные, в черных волосках, часто с небольшой
примесью белых волосков посредине и по заднему краю. II тергит обычно
с неясно просвечивающими коричневыми пятнами по бокам
41. Н. (s. str.) sogdiana Ols.
63 (62). Брюшко окрашено иначе.
64 (65). II и последующие стерниты брюшка по бокам в беловато-серых волосках,
вследствие чего посредине неясно выделяется более темная широкая полоса,
покрытая черными волосками. I—II тергиты брюшка в легком голубоватом
налете и преимущественно в серых волосках, II тергит в средней части обычно
с примесью черных волосков, последующие тергиты черные, в черных волосках,
с незначительной примесью серых волосков посредине и по заднебоковому
краю 42. Н. (s. str.) semipollinosa Ols.
65 (64). II и последующие стерниты брюшка по бокам в черных волосках, вследствие
чего брюшко равномерно блестяще-черное, иногда по заднему краю стернитов
незначительная примесь светлых волосков. II тергит блестяще-черный, покрыт
черными волосками, иногда с примесью серых.
66 (67). I тергит в легком сероватом налете и преимущественно в беловато-серых
волосках. Последующие тергиты блестяще-черные, в коротких черных
волосках, последние 2—3 тергита по заднему краю в серых волосках. Нижняя
лобная мозоль обычно в центральной части коричневая. Коренастее и крупнее:
16—19 мм 40. Н (s. str.) nigricorpa Krob.
67 (66). I тергит, как и последующие, блестяще-черный; тергиты брюшка в черных
волосках, часто со значительной примесью серых по заднему краю и по бокам.
314 1П. ПОДСЕМ. TABANINAE
Нижняя лобная мозоль черная. Уже и мельче: 13—17 мм
39. Н. (s. str.) hunnorum Szil.
68 (59). Брюшко снизу матовое, в сером налете.
69 (70). Средние голени черные (окраска маскирована желтовато-серыми волосками!).
Кавказ (из группы Н. (s. str.) astuA) 27. Н. (s. str.) popovi Ols.
70 (69). Средние голени по крайней мере в основной части коричневые.
71 (72). Брюшко сверху с 2 боковыми рядами беловато-серых пятен (налет+волоски),
разделенных посредине широкой черной полосой; срединный ряд серых
треугольников отсутствует. Дальний Восток ... 32. Н. (s. str.) adachii Takag.
72 (71). Брюшко сверху с 3 рядами серых пятен (налет+волоски): треугольных
посредине и б. м. ромбических по бокам.
73 (74). Нижняя лобная мозоль б. м. треугольная или пятиугольная, не достигает
края глаз, в тонких поперечных морщинках (рис. 306). 3-й членик усиков
одноцветно-черный. Горы Средней Азии
43. Н. (s. str.) shnitnikovi Ols.
74 (73). Нижняя лобная мозоль б. м. прямоугольная, обычно достигает края глаз,
гладкая, сильно блестящая (рис. 309, 315). 3-й членик усиков в основании
коричневый, реже целиком черный. Таежно-лесная полоса европейской части
СССР и Сибири.
75 (76). Лобный треугольник низкий (рис. 309). 3-й членик усиков очень узкий, с
прямым или слегка изогнутым под углом нижнем краем; дорсальный угол слабо
выражен. Лобная полоска относительно широкая, ее высота в 3—3V2 раза
превосходит ширину основания 45. Н. (s. str.) nigricornis Ztt.
76 (75). Лобный треугольник относительно высокий (рис. 315). 3-й членик усиков
менее узкий, с б. м. равномерно выпуклым нижним краем; дорсальный угол
явственный. Лобная полоска относительно узкая, ее высота приблизительно
в 4 раза превосходит ширину основания
47. Н. (s. str.) montana Mg. var. flaviceps Zett.
77 (56). Брюшко сверху с коричнево-желтыми пятнами по бокам (иногда только
на II тергите!); снизу брюшко по крайней мере в основании коричневое (иногда
лишь по бокам II стернита).
78 (85). Площадка 3-го членика усиков черная, лишь в основании иногда
коричневатая; дорсальный угол площадки слабо развит, тупоугольный (рис. 283, 285,
315 и др.).
.79 (80). Концевой членик щупалец сильно утолщен (рис. 283),. его длина обычно
превосходит толщину не более чем в 2V2 раза (смотреть сбоку!), в светлых
волосках, иногда с незначительной примесью черных. Высота лобной полоски
превосходит ширину основания в 3V2—4 раза. Площадка 3-го членика усиков
относительно широкая, ее длина превосходит наибольшую ширину
приблизительно в IV2 раза 30. Н. (s. str.) sareptana Szil.
■80 (79). Концевой членик щупалец умеренно утолщен (рис. 285, 315 и др.), его длина
обычно превосходит толщину в 3 раза, преимущественно в черных волосках.
Высота лобной полоски превосходит ширину основания в 4—4V2 раза.
Площадка 3-го членика усиков менее широкая, ее длина обычно превосходит
наибольшую ширину приблизительно в 13/4—2 раза.
•81 (82). Лобная мозоль плоская, тонкоморщинистая, б. м. треугольная, обычно
в центре коричневатая (рис. 307) 44. Н. (s. str.) turkestana Szil.
82 (81). Лобная мозоль выпуклая, блестяще-черная, б. м. прямоугольная (рис. 285,
315).
83 (84). г4 без придатка (иногда имеется незначительный рудимент). Светлые волоски
по бокам тергитов брюшка, как правило, желтоватые. Таежно-лесная полоса
европейской части СССР и Сибири . . . 47а. Н. (s. str.) montana montana Mg.
84 (83). r4 с придатком. Светлые волоски по бокам тергитов брюшка беловатые. Горный
Алтай (Чуйская степь) 31. Н. (s. str.) tschuensis Ols.
■85 (78). Площадка 3-го членика усиков желтая или красно-коричневая, палочка
обычно черная; дорсальный угол площадки хорошо развит, прямоугольный
или слегка остроугольный (рис. 287, 292, 294 и др.).
■86 (95). Лобная полоска узкая, ее высота превосходит ширину основания в 4V2—
6 раз (рис. 287, 292, 294 и др.).'
Примечание: последующие виды (33—37) трудны для определения, в
сомнительных случаях. необходимо исследование терминалий.
87 (88). Крупнее: 17—19 мм. Нижняя лобная мозоль слабо выпуклая, черная, в
незначительных морщинках, умеренно блестящая. Концевой членик щупалец
в светлых волосках или с примесью единичных черных волосков
35. Н. (s. str.) ukrainica Ols.
88 (87). Мельче: 14—17 мм. Нижняя лобная мозоль плоская, черная или
темно-коричневая, морщинистая, не блестящая. Концевой членик щупалец, как правило,
преимущественно в черных волосках.
11. HYBOMITRA
315
■89 (90). Общий тон окраски тела свинцово-серый. Задние голени коричневые,
вершинная, половина черноватая. Нотоплевры черные. Темная срединная полоска
сверху брюшка, разделяющая боковые коричневые пятна, занимает не менее
Vs ширины брюшка. Церки короткие, не длиннее или едва длиннее X тергита.
Гонопофиз и латеральные лопасти VIII стернита темно-коричневые, первый
лишь немного светлее последних; вместе они образуют б. м. ровный внешний
край в виде полукруга (рис. 288, А) .... 37. Н. (s.str.) bimaculata Macq.
90(89). Общий тон окраски тела буроватый. Задние голени желтые, с коричневой
вершиной. Нотоплевры желто-коричневые или черные. Темная срединная
полоска сверху брюшка, разделяющая боковые желто-коричневые пятна,
занимает обычно 1/4—Ve ширины брюшка, реже Vs ег0- Церки заметно, обычно
не менее чем в 2 раза, длиннее X тергита. Гонопофиз у VIII стернита обычно
с апикальной выемкой, коричневато-желтый, хорошо отличается своим цветом
от темно-коричневых латеральных лопастей.
91 (94). II тергит брюшка по бокам со значительной примесью черных волосков
(у экземпляров II. (s. str.) ciureai с юга они могут отсутствовать!). II—IV тер-
гиты брюшка по заднему краю со слабо намеченными треугольными пятнами
серых волосков (у потертых экземпляров отсутствуют!). Назальная лопасть
VIII стернита с вогнутым основным краем (рис. 288, Б, В).
92 (93). Коричневые пятна по бокам брюшка захватывают лишь I—III тергиты.
Гонопофиз у VIII стернита двулопастной вследствие наличия хорошо
выраженной апикальной выемки. Церки удлиненные, с округлой вершиной (рис. 288, В)
36. Н. (s. str.) muehlfeldi Br.
93 (92). Коричневые пятна по бокам брюшка обычно захватывают I—IV тергиты.
Гонопофиз у VIII стернита с едва заметной апикальной выемкой. Церки не
удлиненные, с уплощенной вершиной (рис. 288, В)
34. Н. (s. str.) ciureai Seg.
94 (91). II тергит брюшка по бокам в золотисто-желтых волосках, лишь иногда с
незначительной примесью черных,(экземпляры из Восточной Сибири и Дальнего
Востока!). II—IV тергиты брюшка по заднему краю с короткими беловатыми
срединными треугольниками (налет+белые волоски). Базальная лопасть
VIII стернита с прямым или слегка вогнутым основным краем (рис. 288, Г и 291)
33. Н. (s. str.) distinguenda Verr.
95 (86). Лобная полоска умеренно широкая, ее высота превосходит ширину основания
приблизительно в 3—4 раза (рис. 281, 313).
96 (97). Концевой членик щупалец сильно утолщен, его длина превосходит толщину
не более чем в 2V2 раза (смотреть сбоку!) (рис. 281). Нотоплевры
красно-коричневые. Нижняя лобная мозоль плоская, морщинистая, обычно в центральной
части коричневая 29. Н. (s. str.) nitidifrons Szil.
97 (96). Концевой членик щупалец умеренно утолщен, его длина превосходит толщину
почти в 3- раза (рис. 313). Нотоплевры черные, редко коричневые. Нижняя
лобная мозоль выпуклая, гладкая, блестяще-черная
46. Н. (s. str.) lundbecki Lyn.
98 (1). Жужжальца (по крайней мере головка) белые или желтоватые (у зажиревших
экземпляров грязно-желтые или коричневатые). Примечание: последующие
виды (47—;51) исключительно однообразны и очень трудны для определения,
вследствие чего необходимо сличение с точно определенными коллекционными
экземплярами. (V группа: Н. (s. str.) erberi Br.).
99 (104). Темя с явственным глазковым бугорком обычно в виде коричневого
треугольничка.
100 (101). Концевой членик щупалец сильно утолщен, его длина превосходит толщину
приблизительно в 2V2 раза (рис. 323). Брюшко светло-шоколадно-коричневое,
сверху без темной срединной полоски на II—IV тергитах (или с ее
незначительным рудиментом). Крупнее: 18—20 мм (редко 16 мм)
51. Н. (s. str) turanica Ols.
101 (100). Концевой членик щупалец умеренно утолщен, его длина превосходит
толщину приблизительно в 3 раза (рис. 317, 319 и др.). Брюшко сверху с б. м.
развитыми коричнево-желтыми пятнами по бокам, разделенными черно-серой
срединной полоской. Мельче: 13—18 мм.
102 (103). Коричневые пятна сверху брюшка обычно захватывают боковые стороны
I—IV тергитов. Глаза в коротких волосках, иногда почти голые .....
47Ь. Н. (s. str.) montana morgani Sure.
103 (102). Коричневые пятна сверху брюшка обычно захватывают боковые стороны
I—III тергитов (иногда только II тергита). Глаза в умеренно длинных волосках
47с. Н. (s. str.) montana reinigiana End.
104 (99). Темя с глазковым рудиментом в виде крошечных, обычно разобщенных между
собой, бугорков или плоских мозолек в числе 1—3 (иногда почти целиком при>
крыты налетом). (V группа: Я. (s. str.) erberi Br.).
316 III. ПОДСЕМ. TABANINAE
105 (106). Брюшко снизу с широкой, резко выделяющейся черной срединной полосой
48. Н. (s. str.) expollicata Pand.
106 (105). Брюшко снизу без широкой черной полосы, но на II стерните посредине
может быть темное пятно.
107 (108). На темени 1—3 черных или коричнево-черных бугорочка (рис. 319, А).
Брюшко сверху с относительно широкой черной срединной полосой, по которой
продольно проходит б. м. резкая, серая полоска налета. Темная полоса занимает
приблизительно %—V6 ширины брюшка. II стернит обычно с темным пятном*
посредине 49. Н. (s. str.) erberi Br.
108 (107). На темени 2 продолговатых коричневых мозольки, впереди которых лежит-
небольшая точка или бугорочек (рис. 321, А). Брюшко сверху с узкой, темной
срединной полоской, прикрытой тонким сероватым налетом. Полоска
составляет приблизительно 1/8—Vio ширины брюшка. II стернит без темного пятна
посредине 50. Н. (s. str.) peculiaris Szil.
Б- Глаза соприкасаются: 6S
1 (78). Жужжальца темно-коричневые или черные.
2 (7). 1—3-й или 1—4-й членики передних лапок с длинными, почти вертикально-
стоящими щетинками (рис. 266).
3 (4). Лобный треугольник на небольшом протяжении блестяще-черный. Западная
Европа ^ 26. Н. (s. str.) micans Mg,
4 (3). Лобный треугольник целиком в сером налете.
5 (6). Крылья с интенсивными коричневыми пятнами вокруг поперечных жилок.-
Средние голени коричневые?. Восточная Сибирь ,
23. Н. (s. str.) astur Erichs.
6 (5). Крылья без коричневых пятен вокруг поперечных жилок (имеются лишь-
следы). Средние голени черные. Малый Кавказ
. . .n 27. Н. (s. str.) popovi Ols,
7 (2). Передние лапки без длинных щетинок.
8 (15). Лобный треугольник целиком или по крайней мере по бокам блестяще-черный.
Крылья часто с явственными темными пятнами у поперечных жилок.
9 (10). Вершина бедер, средние и задние лапки оранжевые
18. Н. (s. str.) aequetincta Beck.
10 (9). Бедра целиком черные, средние и задние лапки коричневые, крылья с легкими-
пятнами у поперечных жилок.
11 (12). Лобный треугольник блестящий по бокам, в средней части в сером налете,
. Дальний Восток 19. Н. (s. str.) stigmoptera Ols,
12 (11). Лобный треугольник целиком блестяще-черный. Горы Средней Азии.
13 (14). Брюшко сверху в золотисто-желтых волосках
20. Н. (s. str.) tatarica Portsch.
21. H. (s. str.) shevtshenkoi Ols.
14 (13). Брюшко сверху в черных волосках 22. Н. (s. str.) zonata Szil.
15 (8). Лобный треугольник в сером или желтом налете. Крылья без пятен у
поперечных жилок (иногда бывают следы!).
16 (19). Тергиты брюшка с резкими, бросающимися в глаза,, золотисто-желтыми
каемками. Крупные виды, напоминающие по окраске ос.
17(18). Бочки груди в желтоватых волосках 16. Н. (s. str.) tarandina L,
18 (17). Бочки груди в черных волосках .... 17. Н. (я. str.) tarandinoides Ols,
19 (16). Тергиты брюшка без бросающихся в глаза желтых каемок.
20 (29). Голени ног черные (иногда средние голени в базальной части
черно-коричневые).
21 (28). Брюшко снизу одноцветно-черное.
22 (23). Бочки груди в серых волосках. Крупнее: 17—19 мм ,
42. Н. (s. str.) semipollinosa Ols.
23 (22). Бочки груди в черных волосках. Мельче: 14.5—16 мм.
24 (25). Линия соприкосновения глаз не более 1V2 высоты лобного треугольника.
Кавказ; Средн. Европа 38. Н. (s. str.) caucasica Enderl.
25 (24). Линия соприкосновения глаз вдвое больше высоты лобного треугольника.
26 (27). Вокруг основания г4 имеется небольшое коричневое пятнышко. Брюшко»
обычно целиком черное. Эстония; Латвия; Карпаты; Сев. и Средн. Европа
24. Н. (s. str.) auripila Mg.
Западная Европа 25. Н. (s. str.) aterrima Mg.
27 (26). Вокруг основания г4 пятнышка нет. II тергит брюшка по бокам неясно
коричневатый. Горы Ср. Азии 39. Н. (s. str.) hunnorum Szil.
28(21). Брюшко снизу в основании темно-коричневое
6. Н. (s. str.) altaica Ols.
29 (20). По крайней мере средние и задние голени ног коричневые или
желто-коричневые.
П. HYBOMITRA 31?
-30 (49). Концевой членик щупалец узкий, б. м. цилиндрический, коричневый или
черный.
31 (48). Брюшко сверху с коричневыми или желто-коричневыми пятнами по бокам,
разделенными срединной черной полосой.
-32 (45). Мельче: 11—15 мм. Черная срединная полоска на II тергите брюшка в б. м.
стоячих, на последующих тергитах в прилегающих черных волосках.
33 (36). Голова сильно выпуклая, полушаровидная. Фасетки верхних % значительно
крупнее нижних (приблизительно в 4—5 раз). Граница между теми и другими
довольно резкая.
-34 (35). Брюшко с беловатыми волосками по заднему краю тергитов, снизу черное.
Таежно-лесная полоса европейской части СССР и Сибири
2. Н. (s. str.) kaurii Chv. et Lyn.
35 (34). Брюшко с желтоватыми волосками по заднему краю тергитов, снизу в
основании темно-коричневое (III и частью II стерниты). Дальний Восток . . .
. : 4. Н. (s. str.) stenopselapha Ols.
36 (33). Голова умеренно выпуклая. Фасетки верхних частей глаз лишь незначительно
отличаются от нижних (в 2—3 раза) или б. м. одинаковые.
-37 (38). Спинка преимущественно в серых волосках. Светлый налет на спинке и по
заднему краю тергитов с оливковым оттенком .
14. Н. (s. str.) obscurinervis Viol.
-38 (37). Спинка преимущественно в черных волосках. Светлый налет,на спинке и по
заднему краю тергитов сероватый или желтоватый.
.39 (40). Брюшко снизу черное, стерниты по заднему краю с желтоватой каймой . .
3. Н. (s. str.) sexfasciata Hine.
40 (39). Брюшко снизу в основании коричневое (III и частью II стерниты).
-41(44). 3-й членик усиков черно-коричневый или желто-коричневый с черной
палочкой.
•42 (43). II—III тергиты брюшка по бокам (в области коричневых пятен) с
треугольными пятнами беловатого налета (смотреть сзади!). Площадка 3-го членика
усиков желто-коричневая 7. Н. (s. str.), lapponica Wahlbg.
•43 (42). II—III тергиты брюшка по бокам без пятен беловатого налета. Площадка
3-го членика усиков черная, с коричневым основанием
5. Н. (s. str.) olsoi Takah.
44 (41). 3-й членик усиков оранжевый, лишь концевой членик палочки слегка
затемнен- . . i 9. ,Н. (s. str.) brevis Lw.
45 (32). Крулнее: 16—18 мм. Черная срединная полоска по крайней мере на II—III
тергитах в стоячих густых, черных волосках.
46 (47). Глаза в коричневых волосках; коричневые пятна по бокам II—III тергитов
без серого налета М. Н. (s. str.) arpadi Szil.
47 (46). Глаза в серых волосках; коричневые пятна по бокам II—III тергитов в
тонком сером налете, обособленном в пятна (смотреть сзади1)
: 10. Н. (s. str.) ussuriensis Ols.
48 (31). Брюшко сверху в основной части оранжево-желтое (окраска частично
маскируется многочисленными черными волосками), IV и последующие кольца
одноцветно-черные, в черных волосках 12. Н. (s. str.) nigella Szil.
49 (30). Концевой членик щупалец сильно утолщен или продолговато-овальный,
в последнем случае всегда светлый, беловато-серый или коричневато-желтый.
-50 (63). 3-й членик усиков черный, лишь иногда слегка коричневатый в основании.
Дорсальный угол б. м. тупой, иногда 3-й членик очень узкий.
51 (02). Фасетки верхних частей глаз заметно крупнее пижних (приблизительно
в 3—4 раза).
52 (61). 1-й членик усиков сверху в длинных волосках, б. м. равных по длине членику
или длиннее его (рис. 306, 307, 309).
53 (58). Брюшко снизу черное (или черно-серое).
54 (57). Брюшко снизу блестяще-черное.
55 (56). Коричневые пятна по бокам I—II тергитов брюшка преимущественно в белых
волосках 41. Н. (s. str.) sogdiana Ols.
56 (55). Коричневые пятна по бокам I—II тергитов брюшка в черных волосках . .
40. Н. (s. str.) nigricorpa Krob.
57 (54). Брюшко снизу матово-серое ....... 43. Н. (s. str.) shnitnikovi Ols.
58 (53). Брюшко снизу в основании желто-коричневое, конец брюшка черный.
59 (60). Глаза в коричневых волосках. 3-й членик усиков очень узкий
45. Н. (s.. str.) nigricornis Ztt.
60 (59). Глаза в серых волосках. 3-й членик усиков менее узкий
44. Н. (s. str.) turkestana Szil.
61 (52). 1-й членник усиков сверху в коротких волосках, длина которых меньше
длины членика (рис. 315, Б) .
47а. Н. (s. str.) montana montana Mg.
■62 (51). Фасетки глаз б. м. одинаковые 30. Н. (s. str,) sareptana Szil.
318 III. ПОДСЕМ. TABANINAE
63 (50). 3-й членик усиков желто-коричневый, палочка часто коричнево-черная..
Дорсальный угол б. м. прямой.
64 (67). Линия соприкосновения глаз не более I1/» высоты лобного треугольника
(рис. 279).
65 (66). Мельче: 12—14 мм. Черная срединная полоскана III терпите брюшка кзади
слегка расширена или параллельная, занимает 1/s—г/ь ширины брюшка.
Вершина щитка в темных волосках. На передних лапках коготки равной величины
28. Н. (s. str.) lurida Flln.
66 (65). Крупнее: 14—17 мм. Черная срединная полоска на III терпите кзади заметно'
сужена, занимает г/6—78 ширины брюшка. Вершина щитка в светлых волосках.
На передних лапках внутренний коготок немного короче внешнего
29. Н. (s. str.) nitidifrons Szil..
67 (64). Линия соприкосновения глаз вдвое больше высоты лобного треугольника
(рис. 297).
68 (75). Фасетки глаз б. м. одинаковой величины или же верхние заметно (но не резко)-
отличаются от нижних (крупнее не более чем в 2—4 раза) (рис. 289, А).
69 (74). Концевой членик щупалец сильно утолщен, как бы вздут (рис. 287, 295, 297).
1-й членик усиков сверху в длинных волосках, равных по длине 1-му и 2-му
членикам вместе взятым (рис. 297). Глаза в коричневых волосках.
70 (71). Спинка в сероватом налете. Фасетки глаз б. м. одинаковой величины . ..
37. Н. (s. str.) bimaculata Macq.
71 (70). Спинка в буроватом налете. Фасетки глаз заметно крупнее нижних (в 2—
4 раза).
72 (73). Мельче: 13—15 мм. Фасетки в верхней части глаз крупнее нижних в 3—
4 раза 36. Н. (s. str.) muehlfeldi Br..
73 (72). Крупнее: 15—17.5 мм. Фасетки в верхней части глаз крупнее нижних
приблизительно в 2 раза (рис. 289, А) 33. Н. (s. str.) distinguenda Verr.
74 (69). Концевой членик щупалец слабо утолщен (рис. 313). 1-й членик усиков сверху-
в коротких волосках, не превосходящих длину членика. Глаза в белесых
волосках 46. Н. (s. str.) lundbecki Lyn.
75 (68). Фасетки верхних частей глаз в 5—6 раз крупнее нижних, причем граница
между крупными и мелкими фасетками б. м. резкая (рис. 289, Б).
76 (77). 1-й членик усиков в длинных волосках, равных по длине 1-му и 2-му членикам.
вместе взятым. Мельче: 14—17 мм 34. Н. (s. str.) ciureai Seg.
77 (76). 1-й членик усиков в коротких волосках; не превосходящих длину членика.
Крупнее: 18 мм 35. Н. (s. str.) ukrainica Ols.
78 (1). Жужжальца белые или желтоватые.
79 (84). II стернит брюшка с темным пятном посредине или же брюшко снизу с черной:
срединной полоской.
80 (83). Брюшко снизу с темным пятном на II стерните.
81 (82). Бедра почти целиком черно-серые. Фасетки верхних'2/з глаз лишь немного
крупнее нижних (приблизительно в 2—3 раза)
47b. Н. (s. str.) montana morgani Sure.
82 (81). Вершинная 1/s бедер коричнево-желтая. Фасетки верхних 2/3 резко крупнее-
нижних (приблизительно в 5—6 раз). 49. Н. (s. str.) erberi Br.
83 (80). Брюшко снизу с черной срединной полоской
48. Н. (s. str.) expollicata Pand..
84 (79). II стернит брюшка без темного пятна или полоски посредине .;....
50. Н. (s. str.) peculiaris Szil.
I. Группа Hybomitra (s. str.) k a u r i i Chv. et Lyn.
Щупальца узкие, черные, коричневые или оранжевые. Лобный треугольник
целиком в сером налете. Крылья без заметных пятен на поперечных жилках. Брюшко-
без резких золотисто-желтых каемок по заднему краю тергитов. Жужжальца
коричневые. У 33 многих видов этой группы на передних лапках внутренние коготок и пуль-
вилла значительно короче внешних (рис. 238).
2. Hybomitra (s. str.) kaurii Chv. et Lyn. (рис. 239). — Слепень Каури.
Chvala a. Lyneborg, 1970, J. Med. Entom., 7, 5 : 546; С h v a 1 a, L y-
neborg a. Moucha, 1972 : 202. — borealis M e i g e n, 1820, 1. с : 37 (Ta-
banus); L о e w, 1858a : 586 (Tabanus) et auct. пес Т. borealis Fabricius, 1791.
$. Глаза в коротких серых волосках, с 3 широкими полосками. Лобная полоска
широкая, ее высота превосходит ширину в 2V2—3 раза, с параллельными сторонами
или слегка сужающаяся к основанию, в пепельно-сером или желто-сером налете.
Нижняя лобная мозоль широкая, почти достигает до края глаз, б. м. прямоугольная,
слегка выпуклая, блестяще-черная. Средняя мозоль узковеретеновидная, блестяще—
11. HYBOMITRA
зт
черная; чаще она бывает обособлена от нижней лобной мозоли. Глазковыи бугорок
выпукло-овальный, красно-коричневый. Лобный треугольник б. м. низкий, пепельно-
серый. Щеки и лицо серые, покрыты желтовато-серыми волосками. Усики
красно-коричневые, палочка черная; 3-й членик умеренно широкий, с б. м. прямым дорсальным
углом, верхний край площадки часто черноватый. Концевой членик узкий, темно-
коричневый или черный, покрыт короткими темными волосками. Нотоплевры черные,.
Рис. 238. Левая передняя лапка с дорсальной
стороны $ Hybomitra sexfasciata Hine.
Внутренние коготок и пульвилла значительно короче
внешних.
редко коричневые. Жужжальца коричнево-черные. Ноги черные, основание голеней,
передних ног и' значительная часть средних и задних голеней коричневые. Крылья
слегка затемнены к переднему краю, жилки темно-коричневые; г4 без придатка.
Брюшко черное, сильно блестящее, с небольшими каштаново-коричневыми пятнами по
бокам II реже III тергитов. Задние края тергитов с узкими, резко выраженными
беловато-серыми каемками, покрытыми серебристо-серыми волосками. Иногда эти каемки
посредине (особенно на II тергите) расширены в виде направленных вперед неясных
беловатых треугольных пятен. Брюшко снизу целиком черное, с беловатыми каемками
по заднему краю стернитов. VIII стернит: гонопофиз, умеренной величины, в черных.
Рис. 239. Hybomitra (s. str.) kaurii Chv. et Lyn. Лобная
полоска, усик, щупальце, VIII стернит, церки и сперматека $.
А — щупальце <?.
волосках, с небольшой апикальной выемкой; латеральные лопасти небольшие; боковые
коричневые пятна крупные; базальная лопасть с прямым или слегка вогнутым
основным краем. Церки короткие, с уплощенной вершиной. Сперматека узковеретеновидная.
13—15.5 мм. . .,-
S Голова сильно выпуклая, полушаровидная. Глаза в длинных белесых или светло-
коричневатых волосках, фасетки верхних 3/4 глаз значительно крупнее нижних,
граница между теми и другими выражена б. м. резко. Глазковыи бугорок овальный,
коричнево-красный. Лобный треугольник серебристо-серый, линия соприкосновения глаз
в 11/ _2 раза больше его высоты. Усики красно-коричневые, большая часть 1-го
членика^ концевая часть 3-го темные. 1-й членик сверху в коротких, не превосходящих
его длину черных волосках. 3-й членик узкий, с небольшим дорсальным выступом.
Концевой членик щупалец коричнево-черный, узкий, цилиндрический, на конце ту-
>
320 III. ПОДСЕМ. TABANINAE
пой, покрыт длинными, черными волосками. Нотоплевры черные. Жужжальца, ноги
и крылья как у $. Внутренние коготок и пульвилла на передних лапках укорочены.
Брюшко, как у §, с более крупными каштаново-коричневыми пятнами по бокам,
которые, как правило, продолжены до заднего края III тергита. 14—16 мм.
Распространение: европейско-сибирский лесной и частью таежный вид.
(рис. 240). Ранее мы не отличали его от Н. (s. str.) sexfasciata Hine (см. стр. 322),
поэтому ареал мы можем указать лишь фрагментарно. Достоверно известен из еледую-
Рис. 240. Ареал НуЪотЫга (s. str.) kaurii.
щих мест: Кольский п-ов (Хибины), Эстония, Ленинградская, Псковская, Московская
области, Иваново, Карельская и Коми АССР, Ю. Урал, Томская обл. (Томск; Колпа-
шово; Назино), Красноярский край (Шумиха; Северо-Енисейск; Абакан), Иркутск.
Сев., Средн. и частью Ю. Европа. Не часто. Июнь—июль. Личинка найдена в
береговой моховой кочке лесного озерка (Бей-Биенко, 1963).
Рассматриваемый вид известен как Hybomitra (s. str.) borealis Meigen (или Я. (s str.)
borealis Loew), но это название является младшим омонимом другого вида —
Н. (s. str.) borealis Fabrieius, 1781 (=#. (s. str.) lapponica Wahlb., 1848), поэтому оно
упразднено и заменено на Н. (s. str.) kaurii Chv. et Lyn.
3. Hybomitra (s. str.) sexfasciata Hine (рис. 241) — Слепень северный.
Hine, 1923, Canad. Ent., 55 : 144 (Tabanus); Philip, 1956:229; Kauri,
1958:96; Philip, 1960:697; Chvala, Lyneborg a. Moucha, 1972:
200; Олсуфьев, 1972 : 451. — anderi Kauri, 1951 : 101 (T. borealis anderi).
?. Глаза в коротких серых волосках с 3 полосками. Лобная полоска с б. м.
параллельными сторонами, ее высота превосходит ширину основания приблизительно
в 21/2 раза, в сером налете. Нижняя лобная мозоль блестяще-черная, б. м.
прямоугольная или сверху округлая, почти достигает края глаз. Средняя мозоль черная, верете-
новидная, обычно достигает нижней мозоли. Глазковый бугорок выпуклый, блестящий,
красно-коричневый. Лобный треугольник низкий, в сером налете. Усики черные, 1-й
и 2-й членики в сером налете, 3-й членик относительно короткий, в основании
коричневый, с прямым или тупым дорсальным углом. Концевой членик щупалец темно-
коричневый, узкий, преимущественно в черных волосках. Нотоплевры черные.
Жужжальца коричнево-черные. Крылья с легким буроватым затемнением вдоль жилок,
особенно в передней половине; г4 без придатка. Ноги черные, основание голеней
передней пары и значительная часть средних1 и задних голеней коричневые. Брюшко черное,
блестящее, с небольшими каштаново-коричневыми пятнами по бокам II и частично I—
III тергитов. Задний край тергитов с узкой сероватой или желтоватой каемкой и в
такого же цвета коротких волосках, остальное в черных волосках; каемки на II—
IV тергитах слегка расширены посредине в небольшие треугольники (часто мало
заметны). Снизу брюшко черное, преимущественно в желтовато-серых волосках, стер-
ниты по заднему краю с желтовато-серыми каемками. Терминалии сходны с таковыми
Н. (s. str.) kaurii, лишь церки немного длиннее. 11—15 мм.
11. HYBOMITRA
3S1
$. Голова умеренно выпуклая. Глаза в длинных светло-коричневых волосках.
Фасетки верхние частей глаз лишь незначительно (в 2—3 раза) крупнее нижних,
граница между теми и другими нерезкая. Линия соприкосновения глаз в 1V2—134 раза
превосходит высоту лобного треугольника; последний в пепельно-сером налете. Усики
черные, 3-й членик в базалыгой части коричневый; дорсальный угол б. м. прямой.
Рис. 241. Hybomitra (s. str.) sexfasciata Hine. Лобная
полоска, усик, щупальце, VIII стернит и церки $.
1-й членик сверху в густых, черных волосках, их длина заметно превосходит длину
членика. Концевой членик щупалец черно-коричневый, узкий, цилиндрический, со
слегка суженным кончиком, в черных волосках. Лицо и бочки груди в серых волосках
с примесью черных. Ноги и крылья как у ?. Внутренние коготок и пульвилла передних
Рис. 242. Ареал Hybomitra (s. str.) sexfasciata.
лапок укорочены (рис. 238). Брюшко черное, по бокам с небольшими
каштаново-коричневыми пятнами на II и частично на III тергитах. Тергиты по заднему краю с узкой
желтоватой каемкой и такого же цвета волосками. На II терпите каемка слегка
расширена в небольшой срединный треугольник. Снизу брюшко черное, в черных волосках,
стерниты по заднему краю с желтоватой каемкой и в желтоватых волосках. 13 мм.
Примечание: описание 5 сделано по многочисленным экземплярам из разных мест
севера Евразии, описание 6 — по экземпляру, пойманному в бассейне р. Маньи,
Лесной Урал, Ляпин, сбор Флерова 22—23 VI 1927, там же поймана $ 15—23 VII 1927.
Дополнительно для описания в использован экземпляр, добытый А. Н. Желоховце-
вым 10 VII 1947 на р. Джила, притоке р. Ингоды, Читинской обл.
21 Н. Г. Олсуфьев
38S т. ПОДСЕМ. TABANINAE
Этот вид описан из Северной Америки (Аляска, Лабрадор), но экземпляры его
из СССР нами ошибочно были приняты за Н. (s. str.) borealis Lw. (=#. (s. str.) kaurii
Chv. et Lyn.), с которым он очень близок. Каури (Kauri, 1951) описал из северной
Скандинавии Н. (s. str.) borealis anderi, которого впоследствии он идентифицировал с Н.
(s. str.) sexfasciata Hine (Kauri, 1958). H. (s. str.) sexfasciata отличается от H. (s. str.)
kaurii меньшими размерами тепа, более темными и короткими усиками и менее резкими
(иногда с желтоватым оттенком) каемками колец брюшка.
Распространение: голарктический тундрово-таежный вид (рис. 242).
В СССР ареал изучен недостаточно ввиду смешения с Н. (s. str.) kaurii. Достоверно
известен из следующих мест: Кольский п-ов, Архангельская обл., Коми АССР, Сев.
Урал, север Тюменской обл., Томская обл., Алтай, Красноярский край, Тува,
Иркутская обл., Прибайкалье и Забайкалье, Якутия, Амурская обл., Приморский и
Хабаровский края. Магаданская (включая Анадырь и Чукотский п-ов), Камчатская и
Сахалинская области. Сев. Скандинавия, Сев. и Вост. Монголия, Сев. Америка. Обычен
на севере и востоке Сибири. Июль.
Рис. 243. Hybomitra (s. str.) stenopselapha Ols. Лобная полоска,
усик, щупальце, VIII стернит, церки и сперматека J.
4. Hybomitra (s. str.) stenopselapha Ols. (рис. 243) — Слепень узкощупальцевый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 138 (Tabanus); L е с-
1 е г с q, 1966 : 120; Виолович, 1968 : 126; Murdoch et Takahasi
1969 : 38.
?. Глаза в очень коротких (видимых лишь при увеличенииХ 16) волосках, с 3
полосками. Лобная полоска заметно сужена к основанию, ее высота превосходит ширину
приблизительно в 4 раза, в сером или желтоватом налете. Глазковый бугорок овальный,
блестяще-коричневый. Нижняя лобная мозоль б. м. треугольная или щитовидная,
слабо блестящая или темно-коричневая, боковыми сторонами не достигает края глаз.
Средняя мозоль в виде узкой черной полоски, иногда целиком прикрыта налетом.
Лобный треугольник в сером или желтоватом налете. Усики коричневые, с черноватым
концом. 1-й членик в коротких, черных волосках. 3-й членик умеренно широкий, с
резким дорсальным выступом, палочка короткая, толстая, в 2 раза короче площадки.
Концевой членик щупалец узкий, коричневатый, в коротких черных волосках. Нотоплевры-
черные. Крылья бесцветные. Жужжальца коричневые. Ноги черные, средние и
задние голени темно-коричневые. Брюшко черное, с узкими, серыми, реже желтоватыми
каемками по заднему краю тергитов; I и II тергиты по бокам с небольшими каштаново-
коричневыми пятнами. II—IV тергиты посредине с серыми треугольными пятнами из
налета и волосков. Снизу брюшко черное, в сером налете, стерниты по заднему краю
с беловатыми каемками. II стернит иногда по бокам слегка бурый. VIII стернит: гоно-
пофиз умеренной величины, в черных волосках, с небольшой апикальной выемкой;
латеральные лопасти обычные, боковые коричневые пятна большие; базальная лопасть
короткая, с прямым основным краем. Церки с округлой вершиной. Сперматека узко-
веретеновидная. 13—16 мм.
(?. Голова крупнее, чем у ?, полушаровидная. Глаза в умеренно длинных
сероватых волосках. Фасетки верхних 3/4 глаз в 4—5 раз крупнее нижних, граница между
теми и другими довольно резкая. Лобный треугольник серебристо-серый. Усики как
у $, но 3-й членик более узкий. 1-й членик сверху в коротких черных волосках, не
превышающих длину членика. Концевой членик щупалец узкий, коричневый, в черных
11. HYBOMITRA
323
волосках. Грудь и ее придатки как у 5- Внутренние коготок и пульвилла передних
лапок укорочены. Брюшко в основном окрашено как у ?, но темно-коричневые пятна
занимают боковые стороны I—III тергитов, соответственно II—III стерниты по бокам
также коричневые. Каемки по заднему краю тергитов и срединные треугольники
в слегка желтоватых волосках. 14.5 мм.
Похож на Н. (s. str.) kaurii, $ хорошо отличается более узким лбом и почти
голыми глазами, 3 отличается желтоватой окраской волосков по заднему краю тергитов
брюшка и развитой коричневой окраской на II—III стернитах.
Распространение: дальневосточный лесной вид. Известен из Приморского
края (на север до среднего течения Имана) и Амурской обл. (Архара), не часто. Сев.-
Вост. Китай (провинция Хейлунцзян). Июль.
- Голотип: 5, Виноградовка Приморского края, 20 VII 1929 Дьяконов-Филипьев,
В коллекции Зоологического института Академии наук СССР, Ленинград.
Рис. 244. Hybomitra (s. str.) olsoi Takah. Лобная полоска,
усик, щупальце, VIII стернит, церки и сперматека <j>.
5. Hybomitra (s. str.) olsoi Takah. (рис. 244) — Слепень восточный.
Takahasi, 1962, Fauna Japonica, Tabanidae : 48; L e с 1 e г с q, 1966 : 115j
Виолович, 1968 : 126; Murdoch et Takahasi, 1969 : 41. — angustipal-
pis О 1 s u f j e v, 1936a : 228 (Tabanus) et auct., пес Т. angustipalpisS chuurmans
Stekhoven, 1926.
$. Глаза в коротких, серых волосках, с 3 полосками. Лобная полоска умеренно
широкая, ее высота превосходит основание в Зг/2—4 раза, слегка расширяется кверху,
покрыта серым или буроватым налетом и короткими преимущественно темными воло-
сками. Нижняя лобная мозоль блестяще-черная, слегка выпуклая, б. м.
прямоугольная, боковыми сторонами обычно соприкасается с краями глаз. Средняя лобная мозоль
представлена узкой, блестяще-черной веретеновидной полоской; часто она бывает
обособлена от нижней мозоли. Глазковый бугорок выпукло-овальный,
блестяще-коричневый или красноватый. Лобный треугольник в пепельно-сером налете. Усики
преобладающе черные, 3-й членик широкий, в основной половине коричневый; дорсальный
угол б. м. прямой. Концевой членик щупалец узкий, коричневый, покрыт короткими
черными волосками. Нотоплевры черные. Жужжальца коричнево-черные. Ноги преиму-
щественно черные, основание голеней передних ног и значительная часть средних и
задних голеней коричневые. Крылья с легким буроватым затемнением к переднему краю;
жилки тёмно-коричневые; г4 без придатка. Брюшко овальное, слабо блестящее, с б. м.
крупными коричневыми пятнами по бокам, захватывающими I—III (или I—IV) тер»
гиты и разделенными посредине черной полоской. Ее ширина составляет */3—1/4
ширины брюшка. Брюшко сверху покрыто короткими, прилегающими, светлыми и
темными волосками; преобладают последние. Светлые волоски расположены по бокам и
затем по заднему краю тергитов. Последние со светлыми каемками. Снизу брюшко
в основной части красноватое или коричневое, блестящее, последние 3 стернита (реже
4) черные. II стернит иногда с темным прямоугольным пятном посредине. VIII стернит:
гонопофиз довольно крупный, в черных волосках, с выраженной апикальной выемкой;
латеральные лопасти и боковые коричневые пятна умеренной величины; базальная
лопасть с прямым основным краем. Церки с уплощенной вершиной. Сперматека
игловидная. 12—16.5 мм.
(J. Голова умеренно выпуклая, но довольно широкая. Глаза в умеренно длинных
серых волосках. Фасетки верхней части глаз лишь незначительно (в 2—3 раза) круп-
I
21*
324 III. ПОДСЕМ. TABANINAE
нее нижних, переход постепенный. Линия соприкосновения глаз в 13/4 раза больше-
высоты лобного треугольника, последний в пепельно-сером налете. Усики как у $,
3-й членик более узкий. 1-й членик сверху в умеренно длинных черных волосках,
обычно не превосходящих длину членика. Концевой членик щупалец
темно-коричневый, узкий, цилиндрический, со слегка суженным концом, покрыт черными волосками.
Грудь и ее придатки как у $. Внутренние коготок и пульвилла передних лапок
укорочены. Брюшко блестящее, с большими красновато-коричневыми пятнами по бокам I—
III или I—IV тергитов, разделяющая их черная срединная полоска обычно наиболее
узкая на III тергите, где она составляет 1/4—1/0 ширины полукольца. IV"—VI или V—
VI тергиты черные. Тергиты по заднему краю с узкой желтовато-серой каймой и
такого же цвета короткими волосками, остальные части тергитов в черных волосках.
Последние более длинные и стоячие в средней части II тергита, на последующих тер-
гитах они короче и более прилегающие. Снизу брюшко блестящее, в основании красно-
Рис. 245. Ареал Hybomitra (s. str.) olsoi.
коричневое, крупное срединное пятно на II стерните и V—VII или IV—VII стерниты
черные. По заднему краю стернитов узкая желтовато-серая каемка и желтоватые
волоски, остальное преимущественно в черных волосках. 13—14.5 мм.
В сравнении с ранее опубликованным (Олсуфьев, 1937а) описанием 3, сделанным
по несколько дефектному экземпляру, нами внесены некоторые изменения, основанные
на изучении дополнительного коллекционного материала (11 3).
Занимает промежуточное положение между Н. kaurii и Н. arpadi. От первого легко
отличается коричневым основанием брюшка и отсутствием резких беловатых каемок
по заднему краю тергитов, от второго вида — меньшей величиной, блестящей, более
крупной нижней лобной мозолью, менее резким дорсальным углом у 3-го членика
усиков (5) и отсутствием густых стоячих волосков в средней части III—IV тергитов (3).
Распространение: восточносибирский горно-таежный вид (рис. 245).
От Нижне-Вартовского (на Оби), Новосибирской обл. (Тогучин) и горного Алтая
(включая его восточноказахстанскую часть) на восток до Колымы, Охотского побережья
(Тауй) и Сахалина, на север до Верхне-Имбатского на Енисее, Нижней Тунгуски (Учами),
Жиганска на Лене, Верхоянска и Средне-Колымска, на юг до Тувы, южного
Прибайкалья, Приамурья и района Хабаровска (Обор). Сев. Монголия, Сев.-Вост. Китай
(Б. Хинган), Япония (о. Хонсю). Местами обычен. Середина июня—июль. Указание
Н. А. Виоловича (1968) о распространении этого вида наряду с зоной тайги «в
широколиственных лесах субтропического облика в Приморье» основано на недоразумении.
6. Hybomitra (s. str.) altaica 01s. (рис. 246) — Слепень алтайский.
Олсуфьев, 1936а, Паразитол. сб. 6 : 223 (Tabanus); 1937a : 133 (Tabanus);
Шевченко, 1961 : 66 (Tabanus); L е с 1 е г с q, 1966:100; Виолович,
1968 : 121.
9. Глаза в коротких, коричневатых волосках, с 3 полосками. Лобная полоска
Широкая, ее высота превосходит ширину в 2V4—2V2 раза, с б. м. параллельными
сторонами, в сером налете. Нижняя лобная мозоль блестяще-черная, широкая, обычно
11. HYBOMITRA 325
достигает боковыми сторонами края глаз, квадратная или поперечная. Средняя
лобная мозоль в виде короткой, довольно широкой черной полоски, последняя нижним
концом соединена с нижней мозолью. Глазковый бугорок овальный, выпуклый,
блестяще-черный. Лобный треугольник очень низкий, в пепельно-сером налете. Усики
черные; 3-й членик в основании слабо расширен, с тупым дорсальным углом.
Концевой членик щупалец узкий, темно-коричневый, покрыт длинными темными и частью
светлыми волосками. Нотоплевры черные. Бочки груди в коричневых и черных
волосках с примесью светлых. Жужжальца коричнево-черные. Ноги одноцветно-черные.
Крылья бесцветные, жилки коричневые; изредка г4 с небольшим придатком. Брюшко
черное, слабо блестящее; II и частью I
тергиты по бокам коричневые. Тергиты по
заднему краю с незначительными желтоватыми
каемками налета и со светлыми волосками.
У некоторых экземпляров светлые волоски
образуют срединный ряд треугольных
пятен. Снизу брюшко одноцветно-черное.
11—15 мм.
3- Голова умеренно выпуклая. Глаза
в длинных коричневых волосках. Фасетки
верхних 2/3 глаз несколько крупнее нижних,
но без резкой границы между теми и дру-
Рис. 246. Hybomitra (s. str.) altaica Ols.
Лобная полоска, усик и щупальце J.
гими. Глазковый бугорок овальный, коричневый, блестящий. Лобный треугольник
пепельно-серый; линия соприкосновения лба в 1V2—13/4 раза превосходит высоту
треугольника; щеки и лицо серые, покрыты преимущественно темными волосками.
Усики как у 9; 1-й членик сверху в б. м. длинных, черных волосках, превосходящих длину
членика. Концевой членик щупалец темно-коричневый, узкий, на конце слегка
заострен, покрыт темными волосками. Нотоплевры черные. Бочки груди в черных волосках.
Жужжальца коричнево-черные. Ноги и крылья как у 9- Внутренние коготок и
пульвилла передних лапок укорочены. Брюшко черное, слабо блестящее. I—III тергиты
по бокам с коричневыми пятнами. Брюшко покрыто черными волосками, с ничтожной
примесью светлых по заднему краю тергитов. Снизу брюшко черное, III и частью
боковые стороны II и IV стернитов (не всегда) коричневые. 13.5—15 мм.
Близок к Н. kaurii и особенно к Н. sexfasciata, но легко отличается от обоих видов
одноцветно-черными ногами и усиками.
Распространение: высокогорный вид, найден на Алтае, на западе
включая Катон-Карагайский р-н Восточно-Казахстанской обл., на востоке заходит в Туву
по хребту Западный Танну-Ола. Монгольский Алтай (р. Олонур 48°19' с. ш. и 88°3' в. д.).
Указание этого вида для ущелья Нухт в окр. Улан-Батора (Moueha et Chvala, 1968)
нуждается в подтверждении. В основном обитает в альпийском поясе, где местами весьма
обычен. С конца июня по середину августа.
Голотип: 9, Катон-Карагай, р. Сарымсакты Восточно-Казахстанской обл. 8 VIII
1926, Верещагин. В коллекции Зоологического института Академии наук СССР,
Ленинград.
7. Hybomitra (s. str.) lapponica Wahlbg. рис. 247 — Слепень болотный.
W a hi berg, 1848, Oefv. Vet. Akad. Forh., IX : 199 (Tabanus); Braue'r,
1880 : 138 (Tabanus); К г б b e r, 1923 : 74 (Tabanus); Krober in Lindner,
1925 : 65 (Tabanus); Олсуфьев, 1937a : 134 (Tabanus); Moueha et Chvala,
1959a : 165 (Tylostypia); Takahasi, 1962 : 44; L e с 1 e г с q, 1966 : 109; В и-
o л о в и ч, 1968 : 122; Murdoch et Takahasi, 1969 : 36; Соболева,
1971:140 (личинка, куколка). — borealis Fabricius, 1781 (Tabagnus) (nomen
oblitum, О 1 su f j e v et L e с 1 e г с q, 1971:325; Олсуфьев, 1971:138).—
typha Whitney, 1904 : 206 (Tabanus), цит. по: Stone, 1938 : 148. — hirticeps
(Lw.) apud S h i г a k i, 1918 : 131 (Tabanus).
9. Глаза в коротких серых волосках, с 3 полосками. Лобная полоска умеренно
широкая, слегка сужена к основанию, ее высота превосходит ширину в 31/i—33/4 раза,
в желто-сером налете. Нижняя лобная мозоль слегка выпуклая, блестяще-черная,
б. м. прямоугольная. Боковые стороны мозоли достигают или почти достигают края
глаз. Средняя лобная мозоль в виде узкой, веретеновидной блестяще-черной полоски,
слегка расширенной в нижней трети; полоска обычно соединяется с нижней мозолью.
Глазковый бугорок выпуклый, овальный, красно-коричневый. Лобный треугольник
326 111. ПОДСЕМ. TABANINAE
низкий, пепельно-серый. Усики красно-коричневые, концевая часть 3-го членика
черная, дорсальный угол б. м. прямой. Концевой членик щупалец темно-коричневый,
узкий, покрыт короткими черными волосками. Нотоплевры черные, редко коричневые.
Жужжальца коричнево-черные. Ноги черные, основание передних и задних голеней
и значительная часть средних голеней коричневые. Крылья слегка затемнены к
переднему краю, жилки темно-коричневые. Брюшко черное, слабо блестящее с серым
рисунком, иногда с небольшими коричневатыми пятнами по бокам II тергита, крайне редко
также III тергита. Задние края тергитов с узкой, неясно выраженной, сероватой
каемкой, расширенной на II тергите посредине и по бокам, на III только по бокам, в
сероватые треугольные пятна (налет-|-волоски). Брюшко снизу черное, с узкими светлыми
каемками по заднему краю. II—III, реже II—-IV стерниты иногда с небольшими
красновато-коричневыми пятнами по бокам. VIII стернит: гонопофиз небольшой, в черных
волосках, обычно без апикальной выемки; латеральные лопасти небольшие; боковые
коричневые пятна хорошо развиты; базальная лонасть короткая, с прямым или слегка
вогнутым основным краем. Церки с округлой вершиной. Сперматека узковеретеновид-
ная. 10—14 мм.
Рис. 247. Hybomitra (s. str.) lapponica Wahlbg.
Лобная полоска, усик, щупальце, VIII стернит
и церки §•
6- Голова умеренно выпуклая. Глаза в длинных светло-коричневых волосках.
Фасетки верхних 2/3 глаз заметно (но не резко) крупнее нижних. Глазок овальный,
коричнево-красный. Лобный треугольник серебристо-серый. Усики
желто-коричневые, концевая часть 3-го членика темная. 1-й членик сверху в длинных черных
волосках, по длине равных или слегка превосходящих 1-й и 2-й членики вместе взятые.
3-й членик узкий, с умеренным дорсальным углом. Концевой членик щупалец
коричнево-черный, узкоцилиндрический, покрыт черными волосками. Грудь и ее придатки
как у J. Внутренние коготок и пульвилла передних лапок укорочены. Брюшко черное,
слабо блестящее, с коричневыми пятнами по бокам II—III тергитов; разделяющая их
срединная черная полоска занимает приблизительно 1/3 ширины брюшка. Задние каемки
тергитов сероватые, II—III тергиты по бокам на коричневом фоне с крупными
треугольными беловатыми пятнами (налет-j-волоски). II тергит посредине с коротким светлым
треугольником. Снизу брюшко в основной половине желто-коричневое, I и IV—VII
(или V—VII) стерниты черные, с серыми каемками; II стернит посредине с крупным
черным пятном. 11—13 мм. •
Распространение: голарктический таежный вид (рис. 248). От
Эстонской и Литовской ССР на восток до Камчатки, Сахалина и о. Кунашир, на север до
сев. побережья Кольского п-ова (Кольский залив), Архангельска, Малоземельской
тундры, низовьев Оби, Енисея Жиганска на Лене и Маркова на Анадыре, на юг до
Глусского р-на Могилевской обл., Нелидова Калининской обл., Иванова, Горького,
Чебоксар, Свердловска, Омска, Кемерова, Алтая (на юг до Телецкого оз.), Тувы,
южного Забайкалья, Приамурья и Приморского края (включая Хасанский р-н на юге).
Сев. и Центр. Европа, Сев.-Вост. Китай, Сев. Япония (Хоккайдо), Сев. Америка.
Многочислен в северной и средней тайге и в лесах Приморского края, в прочих частях ареала
не часто. Июнь—август.
По-видимому, распространение в Центральной Европе оторвано от основного ареала.
Каури (Kauri, 1958) по внешнему строению установил идентичность североамериканской
Н. (s. str.) typha Whitney и H. (s. str.) lapponica Wahlbg. Мы не нашли различий в
строении терминалий североамериканской популяции от таковых евразиатской популяции.
Но американские Н. (s. str.) typha имеют коричневые нотоплевры, что не характерно
11. HYBOMITRA
327
для наших особей этого вида; североамериканскую популяцию следует рассматривать
как подвид Н. (s. str.) lapponica Wahlbg. и обозначать Н. (s. str.) lapponica typha Whitney.
Хвала и Линеборг (Chvala a. Lyneborg, 1970) предложили для H. (s. str.) lapponica
Wahlberg, 1848, восстановить более старое название Н. (s. str.) borealis Fabricius,
1781, но оно является nomen oblitum, так как не применялось в основной
зоологической литературе свыше 50 лет. Название «borealis F.» неприемлемо и потому, что оно
дублирует «borealis Mg., Lw.», длительное время применявшееся для другого вида
Hybomitra [Н. (s. str.) kaurii Chv. et Lyn.).
0° К ВОСТОКУ ОТ ГрИНВИЧа 90° 120° «0° 180° 150°
Рис. 248. Ареал Hybomitra (s. str.) lapponica.
8'. Hybomitra (s. str.) astuta O. S. (рис. 249) — Слепень полярный.
Osten Sacken, 1876, Mem. Boston Soc. Nat. Hist., 2 : 471 (Tabanus); Phi-
1 i?p, 1947 : 292. — polaris F r e y, 1915 : 8 (Tabanus) et auct. — boreus Stone,
1938 : 147 (Tabanus).
$. Голова по отношению к туловищу небольшая. Глаза в коротких, серых воло-
сках,*с 3 полосками. Лобная полоска широкая, с параллельными сторонами, высота
Рис. 249. Hybomitra (s. str.) astuta О. S. Лобная полоска, усик и щупальце $. А —
VIII стернит и церки $ из СССР (Таймыр); Б — то же § из США (Аляска).
превосходит ширину в 21/4—3 раза, в голубовато-сером налете. Нижняя лобная мозоль
слегка выпуклая, блестяще-черная, поперечно-прямоугольная или квадратная.
Ее боковые стороны почти достигают края глаз, отделяясь лишь узкими серыми
полосками. Средняя мозоль веретеновидная, блестяще-черная, обычно достигает до нижней
мозоли. Лобный треугольник низкий, в голубовато-сером налете. Глазковый бугорок
относительно крупный, выпукло-овальный, коричнево-красный. Усики короткие,
328 III. ПОДСЕМ. TABANINAE
черные; иногда 2-й членик, внутренняя сторона 1-го и основание 3-го членика
коричневатые. Спинная сторона 1-го членика в относительно коротких темных волосках,
обычно не превосходящих длину членика, 3-й членик короткий, в основании б. м.
сильно расширен, с тупым дорсальным углом; палочка короткая, толстая. Концевой
членик щупалец коричневатый, иногда палевый, узкий, покрыт б. м. короткими
темными и частью светлыми волосками. Нотоплевры черные. Жужжальца
черно-коричневые. Ноги черные, кончик бедер, основание передних и большая часть средних и
задних голеней коричневые. Иногда ноги почти целиком черные. Крылья бесцветные,
жилки коричневые; г4 без придатка. Брюшко пропорционально груди большое, черное,
сверху с явственным серым рисунком (налет+волоски). Последний состоит из серых
треугольных пятен посредине и б. м. ромбических по бокам, образующих в общей
сложности 3 продольных ряда небольших пятен. Снизу брюшко черно-серое, в коротких
серых волосках, с узкими желтовато-серыми каемками по заднему краю стернитов.
VIII стернит широкий, гонопофиз короткий и широкий, в черных волосках, с
неглубокой выемкой; латеральные лопасти небольшие, боковые коричневые пятна крупные;
базальная лопасть короткая, с б. м. прямым основным краем. Церки с округлой или
слегка уплощенной вершиной. Сперматека узковеретеновидная. 12—15 мм.
Рис. 250. Ареал Hybomitra (s. str.) astuta.
' 3 (no Kauri, 1954). Глаза покрыты длинными коричневыми волосками. Фасетки
глаз верхних 3/4 немного крупнее, чем внизу, без резкой границы между теми и
другими. Лобный треугольник серый, блестящий, возле основания усиков немного
желтоватый. Линия соприкосновения глаз приблизительно в 2 раза превосходит высоту
лобного треугольника. 1-й и 2-й членики усиков темно-красно-коричневые и сверху
в длинных волосках, 3-й членик черный и очень узкий, с тупым, очень плоским
дорсальным углом. Палочка короткая и черная, щупальце черно-коричневое, в длинных
черных волосках, концевой членик короткий и тупой, толще, чему borealis (=kaurii) и
lapponicus. Спинка груди черная, блестящая и, как плевры, покрыта обильными
длинными черными волосками. Нотоплевры черные. Бедра ног черные, голени
темно-коричневые, лишь концевая часть передних голеней черная, лапки черные. Брюшко блестяще-
черное. По бокам II и III тергитов коричневые пятна, также задний край I тергита
коричневый. Маленькое коричневое пятно имеется еще на IV тергите. Следующие друг
за другом серые боковые пятна слабо заметны. Задний край тергитов серый и в черных
волосках. Лишь в средней части (места светлых треугольников) и на коротком
пространстве по бокам имеются также светлые волоски. Лишь на II сегменте имеется слабо
заметный светлый срединный треугольник. Черная срединная полоса шире, чем 1/3
брюшка. Снизу брюшко коричневое. I и 3 последних стернита целиком черные.
Посредине II и IV и возле заднего края III стернита расположены черные пятна. Задний
край стернитов с рядом светлых волосков. 14.3 мм.
Похож на Н. (s. str.) nigricornis. 22 отличаются: 1) меньшей величиной, 2) более
широкими и короткими усиками, 3) тонкими коричневатыми щупальцами, 4) более
широким лбом, 5) более темными ногами и 6) строением терминалий. От Н. (s. str.)
lapponica 25 легко отличаются более широким лбом, черными усиками и более резкими
серыми пятнами на брюшке. Описание 3, сделанное Каури (Kauri), не обосновывает
11. HYBOMITRA
329
различия с 3 Н- (s. str.) nigricornis, вследствие чего мы не включили 3 Я. (s. str.) astuta,
который нам в натуре неизвестен, в определительную таблицу.
Распространение: голарктический тундровый вид, по горам проникает
далеко на юг (рис! 250). В СССР основной ареал охватывает крайний север от Хибин и
Сев. Карелии, на восток включая Камчатку. На севере крайние местонахождения —
низовья Печоры, Енисея (Дудинка), Хатанги (72° с. ш.), Лены (Тикси), Яны (Казачье)
и Колымы (Нижние Кресты), на юг до Байкита на Подкаменнои Тунгуске, Северо-
Енисейска Красноярского края и окр. Петропавловска-на-Камчатке. Южнее основного
ареала спорадические находки в горах Тувы (хр. Танну-Ола) и прилегающей части
Алтая, в южном Прибайкалье (р. Саган-Хара, приток р. Китоя, Вост. Саян). Сев.
Монголия (Архангайский аймак) и в горах Средней Швеции (Херьехогна в обл. Даларна_,
61°40' с. ш.), Сев. Америка. Не часто. Июль—август. По-видимому, в горах (горной
тундре) Восточной Сибири II. (s. str.) astuta распространена значительно шире, чем это
известно в настоящее время.
Мы не нашли различий во внешнем строении 4 22 Я. (s- str-) astuta О. Ь. из Оев.
Америки и серии 22 Я. (s. str.1! polaris Frey из разных мест СССР, терминалий их также
оказались одинаковыми, что дает нам основание считать Н. (s. str.) polaris Frey
синонимом H. (s. str.) astuta О. S.
Рис. 251. Hybomitra (s. str.) brevis Lw. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека 2-
9. Hybomitra (s. str.) brevis Lw. (рис. 251) — Слепень короткобрюхий.
Loew, 1858a, Verh. Zool.-bot. Gesell. Wien, 8:584 (Tabanus); Brauer,
1880'.: 145 (Tabanus); К г о b e r, 1923 : 81 (Tabanus); Szilady, 1923 : 14
(Tabanus); Krober in. Lindner, 1925 : 62 (Tabanus); Олсуфьев, 1937а : 141
(Tabanus); Шевченко, 1961:66 (Tabanus); Takahasi, 1962:42; L e c-
1 e г с q,' 1966 : 103; Виолович, 1968:127; Murdoch et Takahasi,
1969 : 39; Соболева, 19706 : 217 (личинка, куколка); 1970г : 545 (биология).
2- Глаза в- коротких, серых волосках с 3 полосками. Лобная полоска умеренно
широкая, ее высота превосходит основание в З1^ раза, в желтом налете. Глазковый
бугорок красновато-коричневый, овальный, блестящий. Нижняя лобная мозоль б. м.
квадратная, блестяще-коричневая. Средняя мозоль черная, узковеретеновидная,
концом б. м. достигает нижней мозоли. Лобный треугольник серый. Усики целиком,
оранжевые, иногда кончик палочки черноватый; 3-й членик умеренно широкий, с б. м.
прямоугольным дорсальным выступом. Концевой членик щупалец черный или темно-
коричневый, узкий, в сером налете, покрыт светлыми и темными волосками. Нотоплевры
темные. Крылья слегка буровато-дымчатые. Жилки коричневые, к основанию крыла
желтые; г4 без придатка. Жужжальца черно-коричневые. Ноги: бедра черные, средние
и задние голени желто-коричневые, их кончик и лапки черноватые. Передние голени
черные, в основании коричневые, лапки черные. Брюшко короткое, широкое, черное,
с большими желтыми боковыми пятнами на I—III тергитах. Тергиты по заднему краю
с желтыми каемками, покрыты золотисто-желтыми или сероватыми волосками; на II—
IV тергитах каемки расширены посредине в неясные треугольники. Снизу брюшко
черное, в сером налете, стерниты по заднему краю с широкой желтой каймой; часто II
стернит по бокам и иногда почти весь III стернит желтые. VIII стернит: гонопофиз
умеренной величины, в черных волосках, со слабой апикальной выемкой; латеральные лопасти
-330 III. ПОДСЕМ. TABANINAE
умеренно развиты, боковые коричневые пятна большие; базальная лопасть короткая,
с б. м. прямым основным краем. Церки короткие, с б. м. округлой вершиной.
Сперматека узковеретеновидная, с притуплённой вершиной. 12—16 мм.
(J. Глаза в длинных, светло-коричневых волосках. Фасетки верхних частей глаз
лишь незначительно отличаются по величине от нижних. Глазковый бугорок округлый,
коричнево-красный. Усики целиком оранжевые; 1-й членик в длинных, черных
волосках, 3-й членик с явственным дорсальным углом. Лобный треугольник серый.
Концевой членик щупалец черный, узкий, цилиндрический, со слегка суженным кончиком,
покрыт желтоватыми и черными волосками. Нотоплевры черные. Жужжальца и крылья
как у $. Ноги: средние и задние голени и основание передних голеней коричневые,
лрочие части ног черные. Внутренние коготок и пульвилла передних лапок укоро-
Рис. 252. Ареал Hybomitra (s. str.) brevis.
■чены. Брюшко блестяще-черное, с коричневыми пятнами-по бокам I—III тергитов.
Тергиты по заднему краю с неширокими желтыми каемками и в желтых волосках; каемки
посредине слегка треугольно расширены на II и отчасти последующих тергитах. Снизу
брюшко черное, стерниты с широкими желтыми каемками; II стернит (и частью III)
по бокам желто-коричневый. 14—15 мм.
Распространение: восточносибирский лесной вид (рис. 252). От Алтая
(включая его восточноказахстанскую часть) и Томска на восток до низовий Амура (Ниж-
детамбовское) и Приморского края, на север до Байкита на Подкаменной Тунгуске,
•окр. Якутска и р. Деп, притока Зеи (53°45' с. ш.). Сев.-Вост. и Вост. (Ханькоу) Китай,
Корейский п-ов, Япония (о. Хонсю). Местами обычен. Раннелетний вид: июнь—сере-
.дина июля, редко позднее. Личинки обитают во влажной почве берегов непроточных
водоемов и заболоченных низин (Виолович, 1968).
10. Hybomitra (s. str.) ussuriensis'Ols. (рис. 253) — Слепень уссурийский.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 142 (Tabanus); T а-
k a h a s i, 1962 : 43; L е с 1 е г с q, 1966 : 123; Виолович, 1968 : 128;
Murdoch et Takahasi, 1969 : 40.
$. Глаза в коротких, серых волосках, с 3 полосками. Лобная полоска б. м. узкая,
-явственно сужена к основанию, ее высота превосходит ширину приблизительно в 4 раза,
в сером налете. Глазковый бугорок блестяще-коричневый. Нижняя лобная мозоль
коричнево-черная, блестящая (не морщинистая), с тонкой бороздкой посредине. Краями
мозоль часто достигает глаз. Средняя лобная мозоль черная, коротковеретеновидная,
на свободном конце вильчато расщеплена, нижним концом б. м. достигает нижней
мозоли. Лобный треугольник высокий, серый. Усики целиком коричневато-желтые,
-лишь последний членик палочки слегка затемнен. 1-й членик в коротких, черных
волосках. 3-й членик широкий, с прямым или острым дорсальным утлом (без резкого
выступа). Щупальца бледно-коричневые, концевой членик б. м. узкий, коленчато
изогнут, покрыт короткими светлыми и черными волосками. Нотоплевры
красно-коричневые. Жужжальца коричнево-черные. Крылья прозрачные, с легким затемнением в
передней части. Жилки коричневые; г4 без придатка. Ноги: бедра черные, с коричневым
11. HYBOMITRA 331
кончиком, средние и задние голени и основание лапок желтовато-коричневые, передние
голени черные; С коричневым основанием; лапки черные. Брюшко черное, с
коричневыми пятнами по бокам, захватывающими I—III (или I—IV) тергиты. Разделяющая
пятна черная полоска составляет 1/3 или V4 ширины брюшка. Тергиты по заднему краю
•с узкими каемками из серых волосков, II и частью III тергиты посредине с небольшими
беловатыми треугольниками, тогда как по бокам, на коричневом фоне, с крупными,
косо-треугольными, желтовато-серыми пятнами (налет+волоски). На последующих
тергитах боковые пятна слабо выражены. Снизу брюшко желто- или
красновато-коричневое, небольшое треугольное пятно в основании и последние 3 кольца темные. VIII
стернит: гонопофиз короткий, в черных волосках, со слабой апикальной выемкой;
латеральные лопасти и боковые коричневые пятна большие; базальная лопасть очень
короткая, с прямым или слегка вогнутым основным краем. Церки с уплощенной
вершиной. Сперматека веретеновидная. 15—18 мм.
сь о
Рис. 253. Hybomitra (s. str.) ussuriensis Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $. А — щупальце сГ.
3. Глаза в длинных, серых волосках. Фасетки верхних 2/3 глаз слегка крупнее
■нижних. Голова сильно выпуклая. Глазковый бугорок крупный, темно-коричневый.
Лобный треугольник серый. Усики: 1-й и 2-й членики желто-коричневые, в б. м.
коротких черных волосках (3-й членик обломан). Концевой членик щупалец узкий,
коричневый, в длинных, преимущественно черных волосках. Грудь и ее придатки как
у 9; нотоплевры темно-коричневые. Брюшко черное, I—III тергиты по бокам с
коричневыми пятнами. Разделяющая их черная полоска занимает приблизительно V4 ширины
брюшка, на II—III тергитах эта полоска в стоячих черных волосках. Последующие
кольца черные, в прилегающих черных волосках. Тергиты по заднему краю с узкими
светлыми каемками и в серых волосках; II—III тергиты по бокам на коричневом фоне
■с косо-треугольными пятнами сероватого налета (смотреть сзади!). Снизу брюшко
желтовато-коричневое, V—VII стерниты темные. II стернит с темным пятном посредине.
17 мм.
Близок к Н. (s. str.) arpadi. 9 отличается: 1) целиком желтыми усиками с менее
резким дорсальным углом, 2) более крупной и гладкой нижней лобной мозолью, 3) более
короткой и вильчато расщепленной средней мозолью, 4) серыми волосками глаз,
5) красноватыми нотоплеврами, 6) крупными пятнами из желтовато-белого налета и
волосков по бокам тергитов и 7) строением терминалий. в отличается: 1) серыми волосками
глаз, 2) более выпуклой головой, 3) наличием стоячих волосков на брюшке лишь на Н
и III тергитах (у Н. (s. str.) arpadi на II—IV тергитах), 4) пятнами беловатого налета
яо бокам II—III тергитов.
Распространение: дальневосточный лесной вид, аллопатричен Н. (s. str.)
arpadi (рис. 85, 2). Амурская обл. (близ Кумара на Амуре, Шимановский р-н);
Хабаровский край, на север до низовий Амура (Нижнетамбовское; протока Малышевская;
3j^_ Ш. ПОДСЕШ. TABANINAE
р. Сулук приток Амгуни); Приморский край (на юг до Хасанского р-на). Сев.-Вост-
Китай; Япония (о. Хоккайдо). Местами обычен. Июнь—июль.
Г о л о т и п: 2, протока Малышевская, низ. Амура, 23 VI 1910 (ст. стиль), Сол-
датов. В коллекции Зоологического института Академии наук СССР, Ленинград.
11. Hybomitra (s. str.) arpadi Szil. (рис. 254) — Слепень Арпада.
S z i 1 a d у, 1923, Biolog. Hungar., 1, 1 : 7 (?) (Tabanus); Kroberin
Lindner, 1925 : 60 (Tabanus): Олсуфьев, 1934 : 169 (3) (Tabanus); 1937a : 144
(Tabanus); Philip, 1960:697; Takahasi, 1962:46; L e с 1 e г с q, 1966:100;
Виолович, 1968:128; Murdoch et Takahasi, 1969:40; Teskey,
1969 : 97 (личинка, куколка); Chvala, Lyneborg a. Moucha, 1972 : 204.—
gracilipalpis Hine, 1923, 1. с : 143 (Tabanus) et auct. — stackelbergi О 1 s u f j e v,
1933 : 75 (Tabanus). — ochotscana Enderlein, 1934 : 184 (Stypommia) (partim)„
цит. по: К г о b e r, 1938 : 240.
Рис. 254. Hybomitra (s. str.) arpadi Szil. Лобная полоска, усик
щупальце, VIII стернит, церки и сперматека $. '
д. Глаза в коротких, коричневых волосках, с 3 полосками. Лобная полоска б. м.
узкая, заметно сужена к основанию, ее высота превосходит основание в 4—41/2 раза,
в сером или желтоватом налете. Нижняя лобная мозоль б. м. плоская, морщинистая,,
черная, с небольшой продольной полоской посредине. Боковые стороны мозоли не
достигают края глаз, вершина ее, как правило, слита со средней мозолью. Последняя в виде-
длинной, плоской (реже слегка выпуклой) тускло-черной полоски. Верхний конец
мозоли иногда вильчатый. Глазковый бугорок крупный, выпуклый, блестяще-коричнево-
красный. Лобный треугольник плоский, высокий, желтовато-серый. Усики
коричневые; палочка, верхний край 3-го и основание 1-го членика (или целиком 1—2-й членики)
черные. 1-й членик в сером налете, сверху в коротких, прилегающих волосках. 3-й
членик в основании сильно расширен, с резким, б. м. остроугольным дорсальным выступом,
благодаря чему его верхний край с вырезкой. Палочка короткая, толстая. Концевой
членик щупалец светло-коричневый, узкий, покрыт короткими светлыми и черными
волосками. Нотоплевры черные, очень редко буроватые. Жужжальца темно-коричневыег
иногда с более светлым кончиком. Ноги: кончик бедер, средние и задние голени,
основание средних и задних лапок и передних голеней коричнево-желтые, прочие части ног
черные. Крылья слегка буро затемнены к переднему краю, жилки коричневые; г4 без
придатка. Брюшко широкоовальное, черное, слабо блестящее, с коричневыми пятнами
по бокам, захватывающими I—III тергиты. Разделяющая их черная серединная
полоска занимает 1/3 ширины брюшка. Задние края тергитов с узкими желтыми каемками
и в коротких золотистых или сероватых волосках. Такого же цвета волоски образуют
посредине брюшка ряд небольших, большей частью явственных треугольных пятен.
Снизу брюшко коричнево-желтое; небольшое треугольное пятно в основании и конец
брюшка темные. VIII стернит: гонопофиз до-вольно крупный, в черных волосках, со
слабой апикальной выемкой; латеральные лопасти и.боковые коричневые пятна
умеренной величины; базальная лопасть короткая, с б. м. прямым основным краем. Церки
с округлой вершиной. Сперматека узкая, с притуплённой вершиной, иногда веретено-
видная. 13—18.5 мм.
11. HYBOMITRA
333
в-. Голова умеренно выпуклая. Глаза в длинных, коричневых волосках. Фасетки
в верхней части глаз несколько крупнее нижних (но не резко). Глазковый бугорок б. м.
крупный, красновато-коричневый, блестящий. Лобный треугольник серый, линия
соприкосновения глаз в 2 раза больше его высоты. Усики как у 5, 3-й членик более
узкий и с менее резким дорсальным выступом. 1-й членик сверху в относительно
коротких черных волосках, обычно не превосходящих длину членика. Концевой членик
щупалец бледно-коричневый, узкоовальный, на конце тупой, покрыт длинными
преимущественно темными волосками. Нотоплевры, как правило, черные, редко буроватые.
Жужжальца коричневые, ноги и крылья как у $. Брюшко по бокам с коричневыми
пятнами, простирающимися на I—III тергиты; разделяющая их черная срединная полоска
занимает около 1/3—1/4 ширины брюшка. Большая часть I и IV—VII тергиты черные;
иногда коричневые боковые пятна заходят на V тергит. Тергиты в черных, их каемки
в желтых волосках. Черная срединная полоска на II—IV тергитах покрыта густыми,
■стоячими, черными волосками. Снизу брюшко коричнево-желтое; его основание,
прямоугольное пятно на II стерните и 3 последних кольца — черные. 16—18 мм.
Рис. 255. Ареал Hybomitra (s. str.) arpadi.
Распространение: голарктический таежный вид (рис. 255). В СССР от
Кольского п-ова, Карелии, Эстонии и Белоруссии на восток до Чукотского нац. округа,
Камчатки и Сахалина. На север известен до Мурманска, низовий Печоры, Оби, Енисея,
Лены (пос. Кгоеюр, 70с40' с. ш.), Колымы (Походск) и Анадыра, на юг до Глусского
р-на Могилевской обл., Брянска, юга Московской обл. (с. Семеновское), Ивановской
обл., Чебоксар, Ю. Урала (Бакал, 55° с. ш.), Тюмени, Куйбышевского (Зоново) и То-
гучинского р-нов Новосибирской обл., Алтая, Тувы, южного Прибайкалья, Приамурья,
района Хабаровска (Сарапульское на Амуре, 75 км ниже Хабаровска) и
Приморского края (п. Дальний Кут в среднем течений Имана); М. Шантарские острова. Сев. и
Средн. Европа (Бельгия, ФРГ), Сев.-Вост. Китай (Б. Хинган), Япония, Сев. Америка.
Многочислен в северных и средних частях ареала. Июнь—июль. -
12. Hybomitra (s. str.) nigella Szil. (рис. 256) — Слепень чернохвостый.
S z i 1 a d у, 1914, Ann. Mus. Nat. Hungar., 12 : 664 (Tabanus); Олсуфьев,
1973 : 196. — nigricauda О 1 s u f ] e v, 1937a : 146 (Tabanus) et auct.
§. Глаза в очень коротких коричневых волосках, с 3 полосками. Лобная полоска
б. м. узкая, сужена к основанию, ее высота превосходит основание в 4 раза, покрыта
желтовато-серым налетом. Глазковый бугорок выпукло-овальный,
блестяще-коричневый. Нижняя лобная мозоль б. м. треугольная или щитовидная, блестяще-черная или
коричневая, не достигает края глаз. Средняя мозоль узковеретеновидная или
линейная, упирается в нижнюю. Лобный треугольник в желтовато-сером налете. Усики:
1—2-й членики красноватые, в сером налете, 3-й членик красновато-коричневый,
с черноватым концом. Дорсальный выступ резкий, палочка короткая, в 2 раза короче
площадки. 1-й членик сверху в коротких черных волосках. Концевой членик щупалец
светло-коричневый, узкий, в коротких, преимущественно черных волосках. Ното-
ллевры коричневые. Крылья слегка дымчатые; жилки коричневые; ri без придатка,
334 IH. ПОДСЕМ. TABANINAE
Жужжальца коричневые. Ноги черные, средние и частью задние голени коричневые-
Брюшко резко двуцветное: II и III тергиты оранжево-желтые, каждый с темным
пятном посредине, покрыты коротко прилегающими золотисто-желтыми волосками, IV и
последующие тергиты черные, в одноцветно-черных волосках. Снизу брюшко также
двуцветное; основная часть грязновато-желтая, IV и последующие стерниты черные.
II (иногда также^Ш) стернит посредине с неясным темным пятном. VIII стернит: го-
нопофиз крупный^ в черных волосках, со слабой апикальной выемкой; латеральные
лопасти умеренной величины, остроугольные; боковые коричневые пятна средних
размеров; базальная лопасть со слегка вогнутым основным краем. Церки с уплощенной,
вершиной. Сперматека веретеновидная. 17—22 мм.
3- Голова небольшая, не шире груди. Глаза в умеренно длинных
темно-коричневых волосках. Фасетки верхних частей глаз лишь едва крупнее нижних. Лобный
треугольник в желтовато-сером налете. 1-й членик усиков в базальной части черно-серый,,
вершина его красновато-коричневая, сверху покрыт короткими, черными волосками.
Рис. 256. Hybomitra (s. str.) nigella Szil. Лобная полоска, усик,
щупальце, VIII стернит и церки J.
3-й членик как у §, но более узкий. Концевой членик щупалец коричневый,
приблизительно в 1V2 раза длиннее толщины, в черных волосках. Грудь и ее придатки как у Р.
Брюшко окрашено сходно с 5, но II и III тергиты и соответствующие им стерниты в зоне
оранжевой окраски покрыты преимущественно короткими черными волосками, а
золотистые волоски расположены лишь по заднему краю тергитов. 19 мм.
8 описывается нами впервые.
Этот крупный, красивый вид легко отличается от прочих известных палеарктиче-
ских видов Hybomitra резко двуцветной окраской брюшка: желтой в основной части
и черной на конце. Любопытно, что аналогичную окраску брюшка имеют
североамериканские Н. (s. str.) criddlei Brooks и Н. (s. str.) cinctaF., которым несомненно Н. (s. str.)
nigella Szil. близкородствен по ряду признаков.
Распространение: дальневосточный лесной вид. Приморский (на юг
до Хасанского р-на) и Хабаровский края (близ впадения р. Уссури в Амур; с. Обор).
Сев.-Вост. Китай. Не часто. Июль—начало августа.
13. Hybomitra (s. str.) pavlovskii Ols. (рис. 257) — Слепень Павловского.
Олсуфьев, 1936а, Паразитол. сб. 6 : 227 (Tabanus); 1937а : 147 (ТаЬапш)-
L е с 1 е г с q, 1966 : 116; Виолович, 1968 : 130; Murdoch et Takahasi,
1969 : 39.
?. Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска б. м. узкая,
ее высота превосходит ширину основания в 4—4V2 раза. К основанию лобная полоса
слегка сужена, покрыта желтовато-серым налетом. Нижняя лобная мозоль коричнево-
черная, блестящая, б. м. прямоугольная, боковыми сторонами не достигает края глаз.
Средняя мозоль черная, б. м. веретеновидная, обычно соединяется концом с нижней,
мозолью. Лобный треугольник в желтовато-сером налете. Усики оранжево-красные,
3-й членик в основании сильно расширен, с б. м. резким дорсальным выступом. Палочка
11. HYBOMITRA 335
Рис. 257. Hybomitra (s. str.) pavlovskii Ols. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека J.
коричневые. Бедра ног черные, в сером налете; их вершина, затем голени и лапки по-,
следних 2 пар ног коричнево-оранжевые. Конец передних голеней и передние лапки,
черные. Крылья слегка буровато затемнены, особенно к переднему краю, жилки корич-.
невые. Брюшко широкое, лопатообразное, сверху с большими оранжево-красными пят-_
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180° 150°
0° 30° 60° 90° 120°
Рис. 258. Ареал Hybomitra (s. str.) pavlovskii.
нами по бокам, захватывающими I—IV тергиты и разделенными черной срединной
полоской. Ширина этой полоски составляет 1li—1U ширины брюшка. Сверху брюшко.
1 равномерно покрыто густыми, золотисто-рыжими волосками, без примеси черных.
Снизу брюшко оранжево-красное, в золотистых волосках, 3 концевых стернита томныо;
II стернит посредине с темным прмоугольным пятном, реже последнее отсутствует.
VIII стернит: гонопофиз короткий и широкий, в коричневато-желтых, в средней чисти,
336
III. ПОДСЕМ. TABANINAE
в черных волосках, со слабой апикальной выемкой; латеральные лопасти б. м.
остроугольные, боковые коричневые пятна умеренной величины; базальная лонасть по
■бокам и основному краю вогнутая. Церки короткие, их вершина плоская. Сперматека
веретеновидная. 16—21 мм.
в неизвестен.
Распространение: восточносибирский таежно-лесной вид (рис. 258).
От Томской обл. (Колпашево) и Алтая на восток до Колымы и низовьев Амура (с. Удин-
•екое на Амгуни). На север известен до Верхне-Имбатского на Енисее, Жиганска на
Лене и Верхнего Сеймчана на Колыме, на юг до Тувы, Южного Прибайкалья и юга
Приморского края (к сев.-востоку от Владивостока). Сев. Монголия, Сев.-Вост. Китай.
Местами обычен в южных частях ареала, в северных его частях редок. Июль—начало
августа. Откладку яиц наблюдали на листе тростника (Виолович, 1968).
Г о л о т и п: 2, Бунбуй Канского р-на Красноярского края, 22 VI1915 (ст. стиль),
Вараксина. В коллекции Зоологического института Академии наук СССР, Ленинград.
14. Hybomitra (s. str.) obscurinervis Viol. (рис. 259) — Слепень темнокрылый.
Виолович, 1956а, Паразитол. сб. 16 : 152 (Tabanus); 1968 : 136;
Murdoch et T-a k a ha si, 1969:38.
2- Глаза в коротких сероватых волосках, с 3 полосками. Лобная полоска с почти
параллельными сторонами, ее высота превосходит ширину основания в 4 раза, в
сероватом налете. Нижняя лобная мозоль
блестяще-черная, б. м. прямоугольная или щитовидная,
боковыми сторонами не достигает края глаз. Средняя
лобная мозоль черная, узковеретеновидная, б. м.
достигает нижнюю лобную мозоль. Глазковый бугорок
выпуклый, коричнево-красный. Лобный
треугольник в желтовато-сером налете. Усики: 1-й и 2-й
членики черные, в сером налете, 3-й членик
узкий, короткий, темно-коричневый с черной- па-
Рис. 259. Hybomitra (s. str.) obscurinervis Viol.
Лобная полоска, усик и щупальце §.
лочкой; дорсальный угол слегка тупой. Концевой членик щупалец узкий,
коричневый, в коротких черных. волосках. Нотоплевры коричневые. Жужжальца
темно-коричневые. Крылья слегка буровато затемнены возле жилок, особенно в передней
части крыла; г4 с небольшим придатком. Ноги: бедра черно-серые, голени
желто-коричневые, конец передних голеней и передние лапки черноватые, средние и задние лапки
коричневые, с желтоватым основанием. Брюшко черное, в буровато-оливковом налете.
Тергиты по заднему краю с неширокими желтыми каймами, покрытыми короткими
золотисто-желтыми волосками, остальное в коротких черных волосках. Каемки посредине
расширены в небольшие треугольники. Снизу брюшко в коротких желтоватых и
единичных черных волосках, стерниты по заднему краю с неширокими желтыми каймами.
14—15 мм.
3- Голова не крупнее, чем у 2- Глаза в умеренно длинных серых волосках.
Фасетки верхних и нижних частей глаз б. м. одинаковой величины. Глазковый бугорок
маленький, коричневый. Лобный треугольник в желтовато-сером налете. Усики по
форме и окраске как у 5, но 3-й членик более короткий и узкий. 1-й членик сверху
в черных волосках, по длине равных членику. Концевой членик щупалец
светло-коричневый, продолговато-овальный, преимущественно в черных волосках. Спинка в легком
серовато-оливковом налете и в коротких и длинных серых волосках. Нотоплевры черно-
серые. Крылья окрашены как у 2; гл без придатка. Ноги и жужжальца как у $. Брюшко
черное, в легком буровато-оливковом налете, II—III тергиты по бокам с небольшими
коричневыми пятнами. Тергиты по заднему краю с желтовато-оливковой каймой и в
золотистых волосках, передняя часть тергитов в довольно длинных черных волосках;
II—III тергиты каждый посредине с нерезким треугольником оливкового налета.
Снизу брюшко коричневое, с широкой черноватой полосой посредине, IV и
последующие стерниты преимущественно черноватые, с желтой каймой по заднему краю.
Черные участки в черных, коричневые,участки и задний край стернитов — в золотисто-
желтых волосках. 12 мм.
Близок к Н. (s. str.) bagheera, но легко отличается коричневым концевым члеником
щуналец, черными 1-м и 2-м члениками усиков и наличием сверху брюшка наряду с
золотистыми волосками, образующими каймы по заднему краю тергитов, черных
волосков, прикрывающих большую часть тергитов кпереди от золотистых каемок.
11. HYBOMITRA
337
3 описывается нами впервые.
Распрос.т ранение: о. Кунашир Сахалинской обл. Август. Редок.
15. Hybomitra (s. str.) bagheera Viol. (рис. 260) — Слепень сахалинский.
Виолович, 1968, Слепни Сибири : 137; Murdoch et Takahasi
1969 : 37. — olivaceus Violovitsh, 1956a : 153 (Tabanus), nee T. olivaceus
D e G e e r, 1776 et T. olivaceus Schuurmans Stekhoven, 1926.
2- Глаза в коротких, желтоватых
волосках, с 3 полосками. Лобная полоска с б. м.
параллельными сторонами, ее высота
превосходит ширину основания в 3 раза, в
желтоватом налете. Нижняя лобная мозоль
темно-коричневая, блестящая, щитовидная,
боковыми сторонами не достигает края глаз.
Средняя мозоль черная, узковеретеновидная,
почти достигает нижнюю мозоль. Глазковый
бугорок слегка выпуклый,
блестяще-коричневый. Лобный треугольник в
желтоватом налете. Усики: 1-й и 2-й членики оран-
Рис. 260. Hybomitra (s. str.) bagheera Viol.
Лобная полоска, усик и щупальце $.
жево-желтые, 3-й членик узкий, черный, площадка темно-коричневая, дорсальный
угол б. м. прямой. Концевой членик щупалец узкий, оранжево-желтый, в такого же
цвета волосках. Нотоплевры коричневые. Жужжальца темно-коричневые. Крылья
б. м. бесцветные; г4 без придатка. Бедра черно-серые, их конец, а также голени и
основание лапок коричнево-желтые, конец передних лапок черноватый, средних и задних
лапок коричневый. Брюшко черное, в густом оливковом налете, тергиты и стерниты по
заднему краю с неширокой желтой каймой. Брюшко покрыто короткими золотисто-
желтыми волосками, более густыми сверху. 16 мм.
$ неизвестен.
Распространение: описан по единственной 2 из Анивского р-на
Сахалинской обл. 1 августа 1950 г.
II. Группа Hybomitra (s. str.) tarandina L.
Щупальца узкие, оранжевые. Тергиты брюшка с резкими желтыми каемками по
заднему краю. Нижняя лобная мозоль грязно-желтая. Лобный треугольник часто на
некотором протяжении коричнево-желтый. Жужжальца коричневые.
16. Hybomitra tarandina L. (рис. 261, 262) — Слепень олений.
Linne, 1761, Fauna Suec. : 462 (Tabanus); Braue'r, 1880:139 (Tabanus);
Shiraki, 1918 : 124 (Tabanus); Krober in Lindner, 1925 : 71 (Tabanus);
Олсуфьев, 1937а : 149 (Tabanus); Шевченко, 1961 : 71 (Tabanus);
Takahasi, 1962:39; L e с 1 e г с q, 1966:120; Виолович, 1968:131;
Murdoch et Takahasi, 1969 : 35; С h v a 1 a, L у n e Ь о г g a. M о u с h a, 1972 :
178. — karafutonis M a t s u m u r a, 1911 : 64, цит. no: L e с 1 e г с q, 1966 : 120
(Tabanus).
2- Глаза в коротких, желто-коричневых волосках, с 3 тонкими полосками.
Лобная полоска широкая, ее высота превосходит ширину основания в 3—З1^ раза, с
параллельными или слегка расширяющимися кверху сторонами, в желтом налете. Нижняя
лобная мозоль и частично оголенный лобный треугольник грязно-желтые.. Средняя
лобная мозоль блестяще-черная, узковеретеновидная, с хорошо выраженным килем
посредине. Конец мозоли чаще соединен с нижней мозолью тонкой линией. Глазковый
бугорок овальный, коричнево-желтый, блестящий. Усики оранжево-красные, их 3-й
членик широкий, с резким б. м. остроугольным дорсальным выступом. Щупальца
оранжево-желтые; концевой членик узкий и длинный, коленчатый, покрыт короткими,
прилегающими желтыми волосками. Грудь черная, в буроватом налете. Нотоплевры и
небольшие участки спинки позади них коричнево-желтые, густо покрыты золотисто-желтыми
волосками. Бочки груди в густых, длинных, желтых волосках. Жужжальца
желто-коричневые. Ноги коричнево-желтые, лишь тазики и основание бедер черные. Крылья
прозрачные, буровато-желтые, с коричнево-желтыми жилками. Брюшко черное, I—II или
22 Н. Г. Олсуфьев
338 III. ПОДСЕМ. TABANINAE
III тергиты по бокам слегка коричневатые; все кольца брюшка по заднему краю с
широкими желтыми каемками, покрытыми золотисто-желтыми волосками. В основной
части кольца в черных волосках. VIII стернит своеобразной формы: гонопофиз очень
широкий и короткий, в тонких желто-коричневых волосках, без апикальной выемки;
латеральные лопасти и боковые коричневые пятна небольшие; основной край базаль-
ной лопасти резко вогнутый. Церки с уплощенной вершиной. Сперматека
миндалевидная, черноватая. 17—22 мм.
3. Голова умеренно выпуклая. Глаза покрыты густыми, светло-коричневыми
волосками; фасетки глаз б. м. одинаковой величины. Лобный треугольник в желтовато.
Рис. 261. Hybomitra (s. str.) tarandina L. 5.
сером налете. Щеки и лицо желтые, в нижней части в длинных золотисто-желтых
волосках, в верхней с примесью черных. Усики оранжево-красные, их 3-й членик с
резким дорсальным выступом. Концевой членик щупалец узкий, оранжево-желтый, покрыт
длинными светлыми волосками, с примесью темных. Грудь черная, в буроватом
налете. Нотоплевры и небольшие участки спинки позади них коричнево-желтые, густо
покрыты золотисто-желтыми волосками. Спинка в густых, стоячих, буро-желтых и
черных волосках, щиток в тонких жёлтых волосках. Бочки груди в желто-бурых
волосках, иногда с примесью черных. Жужжальца коричневые. Ноги, крылья и брюшко
как у §. 17—20 мм.
Распространение: таежный вид (рис. 263). В СССР от Кольского п-ова,
Карелии, Эстонии и северо-запада Украины на восток до Охотского побережья (р. Тауй),
Сахалина и о. Кунашир. На север известен до Кандалакши Мурманской обл.,
Архангельска, низовий Печоры, Оби, Туруханска на Енисее и Индигирки (Якутия), на юг
до Дубровицкого р-на Ровенской обл. (51°30' с. ш.), Глусского р-на Могилевской обл.,
окр. Москвы, Ивановской обл., Чебоксар, Казани, Ю. Урала (Белорецк), Тары на
Иртыше, Тогучинского р-на Новосибирской обл., Алтая, южного Прибайкалья,
Приамурья и юга Приморского края (до Хасанского р-на включительно). Сев. и Средн.
Европа (спорадические находки в ФРГ и ГДР; Тироль в Австрии — 1 3 в коллекции
ЗИН АН СССР), Сев. Монголия, Китай (Сев.-Вост. и Тайвань) (в горах на высоте
7800 футов!), Корейский п-ов, Япония (острова Хоккайдо и Хонсю). Обычен в
северной и средней тайге, редок в южных и западных частях ареала. Июнь—июль, иногда
позднее. Связан с хвойными лесами и моховыми болотами. В Карелии личинки най-
11. HYBOMITRA
339
дены в низинных и переходных болотах, а также по берегу ручья и в заболоченных
местах в лесу (Лутта, 1970). Куколок находили на сфагновом (И. А. Порчинский) и
низинном болоте (Лутта, 1970), а также в моховой кочке небольшого таежного болота
(Виолович, 1968).
Рис. 262. Hybomitra (s. str.) tarandina L. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $.
Рис. 263. Ареал Hybomitra (s. str.) tarandina.
17. Hybomitra (s. str.) tarandinoid.es Ols. (рис. 264) — Слепень чернобокий.
Олсуфьев, 1936а, Паразитол. сб. 6 : 224 (Tabanus); 1937a : 150 (Tabanus);
Шевченко, 1961 : 71 (Tabanus); L е с 1 е г с q, 1966 : 121; Виолович, 1968 :
133; Соболева, 1971 : 141 (личинка, куколка).
§. Глаза в коротких, коричневых волосках. Лобная полоска широкая, ее высота
превосходит основание в 3—3V4 раза, слегка расширяется кверху, покрыта желтым
налетом и такого же цвета волосками. Обе лобные мозоли коричнево-желтые,
блестящие, нижняя щитовидная, верхним концом широко соединена с средней мозолью.
Последняя в виде короткой, широкой полоски, с небольшим вдавлением посредине и
вильчато расщепленным верхним концом. Глазковый бугорок овальный, коричнево-
22*
340 III. ПОДСЕМ. TABANINAE
желтый, блестящий. Лобный треугольник в желтом налете, в средней части имеются
участки оголенного блестяще-желтого хитина. Усики оранжево-красные, их 3-й членик
б. м. широкий, с резким дорсальным выступом. Щупальца оранжево-желтые, в желтых
Рис. 264. Hybomitra (s: str.) tardndinoides 01s. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека J.
волосках; концевой членик слегка утолщен в основании, далее коленчато изогнут,
на конце притуплён. Грудь черная, нотоплевры коричневые. Спинка в коротких
желтых и коричневых, щиток в черных волосках. Бочки груди в густых черных волосках,
без примеси светлых. Жужжальца темно-коричневые. Ноги коричнево-желтые, тазики
Рис. 265. Ареал Hybomitra (s. str.) tarandinoides.
и основание бедер черные. Крылья слегка буровато-желтые, с коричнево-желтыми
жилками. Брюшко черное, I—II или I—III тергиты по бокам слегка коричневые; все
кольца брюшка по заднему краю с широкими желтыми каемками, покрытыми
золотисто-оранжевыми волосками. VIII стернит: гонопофиз б. м. конический, в тонких,
желтых волосках, с небольшой апикальной выемкой; латеральные лопасти и коричневые
11. HYBOMITRA
341
боковые шйнд.умеренной величины; основной край базальной лопасти слабо вогнутый.
Церки с упл'ощенной вершиной. Сперматека веретеновидная, черноватая. 21—23 мм.
3- Голова умеренно выпуклая. Глаза в длинных, желтоватых волосках. Фасетки
глаз б. м. одинаковой величины. Лобный треугольник в желтоватом налете. Щеки и
лицо в коричнево-черных волосках. Усики оранжево-красные, 3-й членик с б. м.
резким дорсальным выступом. Концевой членик щупалец узкий, грязно-оранжевый,
в темных, длинных волосках. Грудь черная, нотоплевры коричневые. Спинка покрыта
черными волосками, к которым в передней ч:асти примешиваются желтые. Бочки груди
в черных волосках. Жужжальца, ноги, крылья и брюшко как у §. 19 мм.
Описываемый вид близок к Н. (s. str.) tarandina. Хорошо отличается от последнего
строением лобных мозолей и терминалий (§) и черной окраской волосков бочков груди
(оба пола). Вызывает удивление, что некоторые авторы (Takahasi, 1962; Murdoch et
Takahasi, 1969) не различают эти 2 вполне самостоятельных вида.
Распространение: восточносибирский лесной вид, преимущественно
в горах (рис. 265). От Алтая (включая его восточноказахстанскую часть) и Кемеровской
обл. (Сарбалы, 35 км к югу от Новокузнецка) на восток до низовий Амура и Приморского
края. На севере известен в Центральной Якутии (вероятно, изолированное от
основного ареала местонахождение), на юг до южного Прибалхашья, Приамурья и Хасан-
ского р-на Приморского края. Сев. Монголия, Сев.-Вост. Китай. Не часто. Июнь—
июль.
Голотип: §, дорога Спасск—Яковлевка Приморского края, 4 VII 1927, А. Шта-
кельберг. В коллекции Зоологического института Академии наук СССР, Ленинград.
i III. Группа Н у Ь omitra (s. str.) astur Eriehs.
Щупальца узкие, реже утолщены, черные или темно-коричневые, редко желтые.
Лобный треугольник блестяще-черный или же крылья с небольшими коричневыми
пятнами вокруг поперечных жилок. Тергиты по заднему краю часто с каемками желтых
волосков. Жужжальца коричневые. У 33 некоторых видов на передних лапках
длинные, стоячие щетинки (рис. 266).
Рис. 266. Передняя лапка: А — 3 Hybomitra (s. str.) astur Eriehs.;
Б — 3 H. (s. str.) popoui 01s. I
18. Hybomitra (s. str.) aequetincta Beck. (рис. 267) --*- Слепень желтоногий.
Becker, 1900, Acta Soc. Sci. Fenn., 26 : 8 (Tabanus); Philip, 1947 : 281;
Chvala, Lyneborg a. Moucha, 1972 : 180. — flavipes Wiedemann,
1828 : 137, пит. по: Chvala, Lyneborg a. Moucha, 1972 : 180 (Tabanus)
et auct. пес Т. jlavipes Gravenhorst, 1807 : 363, цит. по: Chvala,
Lyneborg a. Moucha, 1972 : 180.
§. Глаза в коротких коричневых волосках, с 3 полосками. Лоб широкий, его
высота превосходит основание в 2г/2—3 раза. Глазковый бугорок овальный, блестяще-
черно-коричневый. Нижняя лобная мозоль большая, блестяще-черная, соединена с
короткой, широкой средней мозолью. Лобный треугольник блестяще-черный, по краям
слегка в налете. Усики красно-коричневые, с черным концом. 3-й членик широкий,
палочка короткая, толстая. Лицо серое, в длинных, желтоватых волосках. Щупальца
черные или темно-коричневые, концевой членик тонкий, заостренный на конце,
покрыт короткими черными волосками. Грудь черная; бочки груди в густых желтоватых
волосках, спинка блестящая, без полосок. Нотоплевры черные. Крылья слегка
буроватые, с коричневыми жилками; г4 без придатка. Ноги: бедра в основной части черные,
вершинная х/3 бедер, голени и лапки последних двух пар ног оранжевые. Конец
передних голеней и передние лапки черные. Брюшко широкое, черное, тергиты по заднему
краю с широкими каемками густых, золотисто-желтых волосков. II тергит по бокам
с небольшими коричневыми пятнами. Снизу брюшко черное, с желтыми каемками по
заднему краю стернатов. VIII стернит: гонопофиз средней величины в тонких желтых
волосках, с выраженной апикальной выемкой; латеральные лопасти и боковые
коричневые пятна умеренно развиты; базальная лопасть со слегка вогнутым или прямым краем.
Церки короткие, с уплощенной вершиной. Сперматека веретеновидная. 15—19 мм.
342 Ш. ПОДСЕМ. TABANINAE
S (noKrober) весьма похож на J. Фасетки в верхней части глаз явственно крупнее
нижних. Брюшко черное, I и II тергиты красноватые.
Распространение: голарктический тундровый вид (рис. 268). От Индиги
(северо-восток Архангельской обл.) и Усть-Цильмы (Коми АССР) на восток включая
Рис. 267. Hybomitra (s. str.) aequetincta Beck. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека J.
Чукотский нац. округ (Марково на Анадыре; р. Великая-Большая) и Камчатку. На
север известен до низовий Печоры, Оби (Лабытнанги), Енисея (Норильск), Лены (Кю-
сюр) и Колымы (Походск), в Средней, Южной и Вост. Сибири обнаружен в ряде
пунктов, на юге до горного Алтая, Тувы, Хакасии, южного Прибайкалья, Забайкалья,
Рис. 268. Ареал Hybomitra (s. str.) aequetincta.
Камчатки и Приморского края (Сихртэ-Алинь), где в основном обитает в горной тундре.
Сев.-Вост. Китай (хр. Б. Хинган), Сев. Америка. Любопытно, что в Америке Н. (s. str.)
aequetincta распространена в сев.-восточной ее части (в основном в Лабрадоре) и не
найдена на Аляске. На севере местами массовый вид. Июль, иногда позднее. Указание
В. Г. Фоминых с соавт. (1972) об обнаружении этого вида в Томской обл. нуждается
в подтверждении.
11. HYBOMITRA 343
19. Hybomitra (s. str.) stigmoptera Ols. (рис. 269) — Слепень пятнистокрылый.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 153 (Tabanus); T a k a-
hasi, 1962:41; L е с 1 е г с q, 1966:120; Виолович, 1968:134;
Соболева, 19686 : 170 (ЗУ, Murdoch et Takahasi, 1969 : 34; Соболева,
19706 : 219 (личинка, куколка).
§. Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска с 6. м.
параллельными сторонами, широкая, ее высота превосходит ширину приблизительно
в 3 раза, в сером налете. Темя с небольшим, блестящим, коричнево-черным
треугольником, в передней части которого расположен глазковый бугорок. Нижняя лобная
мозоль большая, квадратная, блестяще-черная, соприкасается с краями глаз. Средняя
мозоль черная, узковеретеновидная, концом достигает до нижней мозоли. Лобный
треугольник выпуклый, в средней части в налете, по краям блестяще-черный; иногда
треугольник целиком без налета. Лицо серое, в тонких, длинных, серых волосках;
Рис. 269. Hybomitra (s. str.) stigmoptera Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека §•
щеки обычно на некотором протяжении блестяще-черные. Усики: 1-й членик черный,
сверху в б. м. длинных волосках, 2-й членик черный или коричневый, 3-й членик
умеренно широкий, коричнево-красный, с черной палочкой. Дорсальный угол тупой,
иногда закруглен, палочка короткая, толстая, приблизительно в 2 раза короче
площадки. Щупальца черные или коричневые, концевой членик в основании слегка
утолщен, далее коленчато изогнут и к концу утончается; членик покрыт тонкими, золотисто-
желтыми и на конце черными волосками. Жужжальца темно-коричневые. Ноги:, бедра
черные, средние и задние голени светло-коричневые, с черным кончиком, передние
голени черные, с коричневатым основанием. Лапки всех ног черные. Крылья с довольно
явственными коричневатыми пятнами вокруг поперечных жилок. Жилки коричневые;
г4 без придатка. Брюшко широкое, округлое, черное, сверху в буроватом налете и в
коротко прилегающих золотисто-желтых или серо-желтых волосках. Снизу брюшко
черное, в сероватых или желтоватых волосках. Коричневые пятна по бокам брюшка
отсутствуют. VIII стернит темно окрашен, гонопофиз умеренно развит, в
темно-коричневых волосках, с выраженной апикальной выемкой; латеральные лопасти
остроугольные, боковые коричневые пятна крупные; базальная лопасть со слегка вогнутыми
боковыми сторонами, основной край прямой или слегка вогнутый. Церки со слегка
уплощенной вершиной. Сперматека лимонообразная. 15—18 мм.
(J. Голова небольшая, не шире груди. Глаза в длинных темно-коричневых
волосках. Фасетки верхних частей глаз лишь незначительно крупнее нижних (в 2—3 раза),
переход нерезкий. Лобный треугольник по бокам блестяще-черный, в средней части
в сером налете; линия соприкосновения глаз приблизительно в 2 раза превосходит его
высоту. Усики как у §, но 3-й членик более узкий; 1-й членик сверху в густых и:
длинных черных волосках, превосходящих длину 1-го и 2-го члеников вместе взятых.
Концевой членик щупалец черный, овальный, к вершине немного сужен, в черных волосках.
Щеки в сером налете, их верхняя часть преимущественно в черных волосках. Грудь и
ее придатки как у $. Брюшко блестяще-черное, II тергит по бокам неясно коричневый.
344 III. ПОДСЕМ. TABANINAE
Тергиты покрыты черными волосками, по заднему краю в сероватых или желтоватых
волосках. Снизу брюшко блестяще-черное, стерниты в черных, по заднему краю в
серых волосках. 15—16 мм.
Распространение: дальневосточный лесной вид. Юг Приморского края
и восточные склоны Сихотэ-Алиня. Сев.-Вост. Китай; Корейский п-ов; Япония
(о. Хонсю). Июнь, начало июля. Местами обычен.
Г о л о т и п: 9, Кедровая Падь близ Барабаша, Приморский край, 15 VI 1926,
Мордвилко. В коллекции Зоологического института Академии наук СССР,
Ленинград.
20. Hybomitra (s. str.) tatarica Portsch. (рис. 270) — Слепень татарский.
Portschinsky, 1887, Ног. Soc. Ent. Ross., 21 : 178 (Tabanus); К г о Ь е г,
1923 : 77 {Tabanus); S z i 1 a d у, 1923 : 10 {Tabanus); Krober in Lindner,
Рис. 270. Hybomitra (s. str.) tatarica Portsch. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека §.
1925 : 71 {Tabanus); Олсуфьев, 1937а : 154 {Tabanus); Шевченко, 1961:
71 {Tabanus); L е с 1 е г с q, 1966 : 121. — astur (Erichs.) apud В r a u e г, 1880 : 140
(в). — bicolor S z i 1 a d у, 1923 : 11 (<J) {T. tataricus var. bicolor). — aurantiacus S z i-
1 a d y, 1923 : 11 (J) {T. tataricus var. aurantiacus).
§. Голова сильно поперечная. Глаза в коротких, коричневатых волосках, грязно-
зеленые, с 3 широкими фиолетово-синими полосками. Лоб широкий, его высота в 2—
2х/2 раза превосходит ширину, в коричневом налете, покрыт густыми и длинными
золотисто-желтыми волосками. Глазковый бугорок маленький, коричневатый, неясно
выделяется на темени. Нижняя лобная мозоль большая, б. м. треугольная, блестяще-
черная, вершиной сливается с короткой средней мозолью. Лобный треугольник
низкий, блестяще-черный. Усики черные, часто 2-й членик и основание 3-го красноватые.
1-й членик сверху в длинных черных волосках. 3-й членик узкий, дорсальный угол
сглажен, палочка короткая, толстая. Щеки на значительном протяжении блестяще-
черные, средняя часть лица в желтом налете, и то, и другое покрыто густыми и длинными
желтыми волосками. Концевой членик щупалец коричнево-черный, узкий, сильно колен-
чато изогнут, покрыт черными и более длинными желтыми волосками. 1-й членик
щупалец в длинных, желтых волосках. Грудь черная, в густых и длинных желтых волосках.
Крылья прозрачные, с легкими коричневатыми пятнышками вокруг поперечных
жилок. Жилки коричневые, в основной половине крыла желтоватые; г4 иногда с
придатком. Жужжальца темно-коричневые. Ноги: бедра черные, голени последних 2 пар ног
красновато-желтые, лапки коричневые; передние голени черные с коричневым
основанием, передние лапки черные. Брюшко короткое, широкое, сверху черное, с
красно-бурыми пятнами по бокам.II—III тергитов, покрыто густыми, длинными, золотисто-
желтыми волосками, маскирующими основную окраску брюшка. Снизу брюшко
черное, покрыто тонкими желтыми и черными волосками. VIII стернит: гонопофиз
умеренной величины, в тонких коричнево-желтых волосках, апикальная выемка едва
заметна; латеральные лопасти средней величины, боковые коричневые пятна довольно
большие; базальная лопасть короткая, основной, край прямой или слегка выпуклый.
Церки со слегка уплощенной вершиной. Сперматека миндалевидная. 10—16 мм.
11. HYBOMITRA 345
3. Глаза в длинных коричневых волосках; фасетки верхних и нижних частей глаз
мало разнятся между собой по величине. Глазковый бугорок маленький, коричневый.
Лобный треугольник блестяще-черный. Усики как у §; 1-й членик в густых и очень
длинных черных волосках. Лицо черно-бурое, в черных и бурых волосках, щеки возле
края глаз слегка блестящие. Щупальца черные, концевой членик овальный. Грудь
черная, бочки груди и боковые стороны спинки в черных, средняя часть спинки и
щиток в желтоватых волосках. Крылья, ноги и жужжальца как у <}. Брюшко короткое,
сверху в густых красновато-желтых волосках, более длинных возле заднего края
тергитов. II — III тергиты по бокам с красноватыми пятнами. Снизу брюшко
блестяще-черное, в черных волосках. 15—16 мм.
Своеобразный слепень, напоминающий по внешнему виду пчелу.
Распространение: среднеазиатский горный вид (рис. 271). Обитает на
высоте 1400—4000 м, чаще в субальпийском и альпийском поясе, реже в горных лесах,
по всему Памиро-Алаю (кроме восточного Памира) и Тянь-Шаню, на восток включая
Джунгарский Алатау и на север — Каратау. Зап. Китай (Каш, приток Или). Обычен.
Июнь—август. Куколки найдены в моховом покрове близ родника (Шевченко, 1961).
В нижних и средних поясах гор преобладают более крупные, хорошо опушенные §2,
в альпийском поясе чаще встречаются
мелкие (10—12 мм), слабее опушенные,
светлоглазые особи (Шевченко, 1961). Мы
предлагаем их обозначать как Н. (s. str.) tatarica
var. nana Ols., var. nova.
21. Hybomitra shevtshenkoi Ols. —
Слепень Шевченко.
О л, с у ф ь е в, 1952, Энтомол. обозр.,
32, 2:314 {Tabanus); Шевченко,
1961 : 76 {Tabanus): Leclercq, 1966:119.
9- Глаза в коротких коричневатых
волосках, с 3 полосками. Лобная полоска
широкая , е"е высота превосходит ширину
основания в 2г—21/2 раза, покрыта густыми,
длинными, золотисто-желтыми волосками.
Нижняя лобная мозоль блестяще-черная, пря-
Рис. 271. Ареал Hybomitra (s. str.) tatarica.
моугольная, достигает боковыми сторонами края глаз, вверх переходит в короткую,
черную среднюю мозоль. Глазковый бугорок небольшой, коричневый. Лобный
треугольник низкий, блестяще-черный. Щеки на значительном протяжении
блестяще-черные. Средняя часть лица в желтоватых волосках, нижняя — в черных. Усики черные,
2-й и основание 3-го членика коричневатые. 1-й членик усиков сверху в длинных
черных волосках, 3-й членик узкий, дорсальный угол сглажен. Концевой членик щупалец
узкий, черный, покрыт черными волосками. Грудь черная; среднеспинка и щиток
покрыты густыми, довольно длинными желтовато-бурыми волосками. У основания крыла
сверху имеется небольшое, но заметное пятно черных волосков. Бочки груди в густых
длинных черных волосках; иногда на мезоплеврах имеется незначительная примесь
желтоватых волосков. Крылья бесцветные, с незначительными коричневыми
пятнышками вокруг поперечных жилок; г4 часто с придатком. Жужжальца темно-коричневые.
Бедра ног черные, средние и задние колени красновато-желтые, передние голени
черные с коричневым основанием; передние лапки черные, средние и задние коричневые.
Брюшко короткое, широкое, сверху черное, с красно-бурыми пятнами по бокам II—
III тергитов, покрыто густыми, длинными золотисто-желтыми волосками,
маскирующими основную окраску брюшка. Снизу брюшко черное, покрыто черными волосками,
лишь на 3 последних стернитах примешиваются золотистые волоски. 13—17 мм.
S (по Шевченко, 1961) «Глаза в длинных коричневых волосках с 2 или 3 полосками;
фасетки верхних частей глаз несколько крупнее нижних. Глазковый бугорок очень
маленький, темно-бурый. Лобный треугольник с узким основанием, блестяще-черный,
Щеки на значительном протяжении блестяще-черные, все лицо в одноцветных черных
волосках, без примеси желтых. Усики: 1—3-й членики черные, 2-й членик
коричневый; первые 2 членика в длинных черных волосках; концевой членик узкий,
дорсальный угол, как и у самок, сглажен. Концевой членик щупалец овальный, покрыт
черными волосками. Грудь черная, среднеспинка и щиток покрыты густыми и длинными
золотисто-желтыми волосками. Бочки груди в густых и длинных черных волосках.
Крылья прозрачные, с небольшими размытыми коричневыми пятнами вокруг попереч-
50° 60° 70° 80°
60° 70° 80°
346 III. ПОДСЕМ. TABANINAE
ных жилок и слегка коричнево затемненные у оснований; жилка г4 без придатка.
Жужжальца темно-коричневые. Бедра черные, средние и задние голени коричнево-желтые,
передние голени черные, с коричневым основанием. Брюшко как у§, но несколько
уже». 11.5—13.3 мм.
9 В. (s. str.) shevtshenkoi хорошо отличается от 2 В. (s. str.) tatarica черными
волосками на бочках груди и нижней части лица; те же различия В. В. Шевченко (1961)
указывает и для cJcJ этих видов. Однако S6 В. (s. str.) tatarica из Сарыджаса
(Восточная Киргизия), использованные нами для описания этого вида в Фауне СССР
(Олсуфьев, 1937а, стр. 155), также имеют чисто-черные волоски на бочках груди
и на лице, по остальным признакам они также соответствуют описанию В. В. Шевченко
S В. (s. str.) shevtshenkoi. Но в Восточной Киргизии обитает только Н. (s. str.)
tatarica. На основании изложенного вопрос о различиях между SS В. (§. str.) tatarica
Portseh. и В. (s. str.) shevtshenkoi Ols. следует считать открытым, необходимо
дальнейшее их более детальное изучение.
Распространение: среднеазиатский горный вид, обитающий на высоте
1300—3000 м, преимущественно в субальпийском поясе Западного Тянь-Шаня: хребты
Таласский и Пскемский. Указан для гор Таджикистана без более точной локализации.
Не часто. С июня по первую декаду августа. Среди субальпийских и особенно
альпийских популяций преобладают мелкие, светлоглазые экземпляры самок (Шевченко,
1961). Их можно обозначить как В. (s. str.) shevtshenkoi var. alpina Ols., var. nova.
Кладки яиц на листьях ивы близ русла горной речки; личинка в увлажненном
моховом покрове (Шевченко, 1961).
Голотип: 2, заповедник Аксу-Джабаглы, Талды-Булак 13 VI 1949, Шевченко.
В коллекции Зоологического института Академии наук СССР, Ленинград.
22. Hybomitra (s. str.) zonata Szil. — Слепень пестрый.
S z i 1 a d у, 1923, Biolog. Hungar., 1 : 11 (Tabanus tataricus var. zonatus); К г o-
ber in Lindner, 1925 : 71 (T. tataricus zonatus); Олсуфьев, 1937а : 155
(Tabanus); Moucha et Chvala, 1963 : 523 (Tylostypia); L e с 1 e г с q, 1966:
124. — nigrocincta Enderlein, 1925 : 344 (Tylostypina).
2- Голова сильно поперечная; глаза в коротких серых волосках. Лоб широкий,
его высота в 2—21/2 раза превосходит ширину, в буром налете и густых, длинных,
желтых волосках. Глазок маленький, округлый, неясно выделяется на темени. Нижняя
лобная мозоль большая, блестяще-черная. Лобный треугольник низкий, блестяще-
черный. Усики черные; 2-й членик и основание 3-го коричневатые. 1-й членик сверху
в б. м. длинных черных и желтоватых волосках. 3-й членик узкий, дорсальный угол
совершенно сглажен. Щеки на значительном протяжении блестяще-черные. Лицо
в длинных желтых волосках. Щупальца коричнево-черные, концевой членик б. м.
узкий, коленчато изогнутый, покрыт черными и более длинными желтыми волосками.
Бочки груди и спинка в густых золотисто- или красновато-желтых волосках. Крылья
бесцветные, возле поперечных жилок с легкими коричневыми пятнышками. Жилки
коричневые, в основной половине крыла желтоватые; г4 иногда с придатком.
Жужжальца темно-коричневые. Ноги: бедра черные, средние и задние голени красновато-
желтые, лапки коричневые; передние лапки и голени черные, последние с коричневым
основанием. Спинка и бочки груди в желтых волосках. Брюшко короткое, широкое,
черное, II тергит по бокам с небольшим красновато-коричневым пятном, иногда
продолженным на III тергит. I и II тергиты в густых золотисто-желтых волосках, III и
IV тергиты в черных волосках, в средней части IV тергита с незначительной примесью
желтых, последующие тергиты в желтых волосках. Снизу брюшко блестяще-черное,
в черных волосках, иногда на II и V—VII стернитах волоски желтые. 12—15 мм.
$. Глаза в длинных, коричневых волосках; фасетки верхних и нижних частей
глаз мало разнятся между собой по величине. Глазковый бугорок небольшой,
коричневый, плохо выделяется на темени. Лобный треугольник блестяще-черный. Усики
черные; их форма как у 2; 1-й членик сверху в длинных, черных волосках. Лицо в черных
волосках, края щек блестяще-черные, в черных волосках, концевой членик яйцевидно
утолщен. Грудь черная; бочки груди и спинка в черных волосках, лишь возле
основания щитка единичные светлые волоски. Ноги, крылья и жужжальца как у 2- Брюшко
сверху блестяще-черное, в черных волосках, с единичными светлыми волосками на
кончике брюшка. II—III тергиты по бокам с небольшими каштаново-коричневыми
пятнами. Снизу брюшко блестяще-черное, в черных волосках. 14 мм.
Очень похож на В. (s. str.) tatarica : 2 отличается двуцветной окраской верхней
стороны брюшка (черные волоски на III и IV тергитах) и частью черными волосками
на его нижней поверхности; g отличается чисто-черными волосками брюшка и спинки
и более темными каштановыми боковыми пятнами на II—III тергитах.
Распространение: среднеазиатский горный вид. Таджикистан — хребты
Каратегинский, Петра Первого и Дарвазский. Силади (Szilady, 1923) описал этот вид
по экземпляру (2) из Самарканда, но это указание, видимо, неточно (Олсуфьев, 1973).
Афганистан. Редок.
11. HYBOMITRA
347
23. Hybomitra (s. str.) astur Erichs. (рис. 272, 273) — Слепень сибирский.
E r i с h s о n, 1851, in Middendorfs Sibirische Reise, II : 66 [Tabanus) : В r a u e r,
1880 : 140 {Tabanus) (S=H. tatarica Portseh.); Krober, 1923 : 78 (Tabanus); К. г o-
ber in Lindner, 1925:60 (Tabanus); Олсуфьев, 1937а : 156 (Tabanus);
Moucha et Chvala, 1965 : 335 (<?); L e с 1 e г с q, 1966 : 101; В и о л о в и ч,
1968 : 135; Murdoch et Takahasi, 1969 : 34..— guttipennis Enderlein,
1925 : 360 (Therioplectes).
Рис. 272. Hybomitra (s. str.) astur Erichs. 2-
О Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска
умеренно широкая, ее высота в 3-3V2 раза превосходит ширину, с параллельными
сторонами в сером налете. Темя с небольшим блестяще-коричневым треугольником, в
передней части которого расположен овальный глазковый бугорок. Нижняя лобная
мозоль блестяще-черная, прямоугольная, соприкасается с краем глаз. Средняя мозоль
черная веретеновидная или линейная, упирается в нижнюю мозоль. Лобный
треугольник низкий серый. Усики: 1-й и 2-й членики черные, 3-й членик
красновато-коричневый с черным концом. Палочка короткая, толстая, приблизительно в 2 раза короче
площадки. Последняя с тупым дорсальным углом. 1-й членик сверху в б. м. длинных
черных волосках. Щупальца черные, концевой членик узкий, коленчато изогнут, на
конце приострен, покрыт тонкими светлыми волосками. Нотоплевры черные. Ноги:
бедра черные средние и задние голени желто-коричневые, с черноватым кончиком,
передние голени черные, в основании коричневые, лапки всех ног черные. Жужжальца
темно-коричневые. Крылья с явственными коричневатыми пятнышками вокруг
поперечных жилок. Жилки коричневые; г4 часто с придатком. Брюшко черное, блестящее,
тергиты по заднему краю с б. м. широкими каемками серых или желтоватых волосков.
II тергит часто по бокам с небольшими красновато-коричневыми пятнами. Снизу
брюшко черное, в сероватом налете и серых волосках. VIII стернит темно окрашен,
гонопофиз немного удлиненный, в черных волосках, со слабой апикальной выемкой;
348 III. ПОДСЕМ. TABANINAE
латеральные лопасти б. м. остроугольные, боковые коричневые пятна крупные; ба-
зальная лопасть с прямым основным краем. Церки с уплощенной вершиной. Сперма-
тека веретеновидная. 13—16 мм.
Рис. 273. Hybomitra (s. str.) astur Erichs. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека §■
3- Голова не больше, чем у $. Глаза в умеренно длинных коричневых волосках.
Фасетки верхних частей глаз едва крупнее нижних. Лобный треугольник в сером
налете. Усики как у 9, но 3-й членик более узкий. 1-й членик сверху в густых длинных
черных волосках, превосходящих длину 1-го и 2-го члеников вместе взятых. Концевой
членик щупалец черный, продолговато-овальный, в черных волосках. Грудь и ее
придатки как у $, коричневые пятна вокруг поперечных жилок довольно резкие. Перед-
Рис. 274. Ареал Hybomitra (s. str.) astur.
ние лапки с длинными почти вертикальными щетинками, расположенными дорсально
в передней части 1—3-го члеников (рис. 266, А). Брюшко черное, блестящее, I и II
тергиты по бокам с неясными темно-коричневыми пятнами. Тергиты покрыты черными
волосками, к которым в конце брюшка примешиваются серые волоски, кольца по
заднему краю с узкой, неясной сероватой каемкой. Снизу брюшко черное, в черных и
серых волосках. 13 мм.
3 впервые был описан Моуха и Хвала (Moucha u. Chvala, 1965), но авторами не
упомянут очень важный признак — наличие на передних лапках длинных, почти верти-
11. HYBOMITRA 349
кальных щетинок. Это интересное «вооружение», назначение которого неясно, до сих
пор было известно среди палеарктических видов только у 3 Н. (s. str.) micans Mg.,
обитающего в Европе. Мы отмечаем эту особенность для 33 Н. (s. str.) astur Erichs.
и H. (s. str.) popovi 01s.
Распространение: восточносибирский лесной вид (рис. 274). От Алтая
и окр. Томска на восток до Сахалина, на север известен до Подкаменной Тунгуски (Тэ-
тэрэ), Центральной Якутии, Охотского побережья (Аян), на юг до Тувы, южного
Прибайкалья, Сихотэ-Алиня и юга Приморского края (Седанка близ Владивостока). Сев.
Монголия, Корейская Народная Демократическая Республика (бассейн р. Ялу), Сев.-
Вост. Китай. Местами обычен. Конец июня—июль.
24. Hybomitra (s. str.) auripila Mg. (рис. 275) — Слепень золотистоволосый.
М е i g e n, 1820, Syst. Beschreib., 11:41 (Tabanus aterrimus var. auripilus);
Moucha et Chvala, 1959a : 164 (Tylostypia); Б о ш к о, 19586 : 351 (Tabanus);
Chvala, Lyneborg a. Moucha, 1972:184.
§. Глаза в умеренно длинных коричневато-серых волосках, с 3 полосками.
Лобная полоска широкая, слегка расширена кверху, ее высота превосходит ширину
основания приблизительно в 3 раза, в сером или
желтоватом налете. Нижняя лобная мозоль
блестяще-черная, б. м. прямоугольная, бо- \\\\\\ft\\\№ |Щ(fflflj.fe^fea, ^яг-г^й^
ковыми сторонами достигает края глаз, час- "ч" "*г """""
то посредине с тонкой штриховкой,
сходящейся к центру «елочкой». Средняя мозоль
черная, веретеновидная, нижним концом
достигает ни жней мозоли. Глазковый бугорок
выпуклый, блестящий, черно-коричневый.
Лобный треугольник низкий, в сером налете,
иногда по бокам с единичными волосками.
Усики черные, 3-й членик с тупым
дорсальным углом, в основании
коричневатый; палочка обычно 3-члениковая.
Концерне. 275. Hybomitra (s. str.) auripila Mg.
Лобная полоска, усик и щупальце 2.
вой членик щупалец относительно узкий, черно-коричневый, преимущественно в
черных волосках. Нотоплевры черные. Бочки груди в желтоватых волосках с примосыо
черных. Жужжальца коричнево-черные. Крылья с легким коричневым затемненном
вдоль жилок в передней части крыла, в основании г4 небольшое коричневое пятнышко;
г4 без придатка. Ноги черные, в черных волосках, базальная часть голеней в жо'лто-
ватых волосках. Брюшко черное, задний край и боковые стороны тергитов в золотисто-
желтых волосках, остальное в черных волосках. Снизу брюшко черное, в золотисто-
желтых волосках. 14—17 мм.
3- Голова не крупнее, чем у $, но слегка более выпуклая. Глаза в умеренно
длинных коричневато-серых волосках. Фасетки верхних 2/з глаз в 2—3 раза крушюо
нижних. Линия соприкосновения глаз в 2 раза превосходит высоту лобного треугольники,
последний в сером налете. Усики как у $, но более узкие. Лицо в черных волосках.
Концевой членик щупалец черный, умеренно утолщен, в черных волосках. Бочки груди
в черных волосках. Ноги, крылья и жужжальца как у 2- Брюшко черное, боз
коричневых пятен по бокам. Тергиты брюшка в черных волосках, II тергит по задшшу крню
в средней части, а последующие дополнительно и по бокам с небольшим количпитном
серых волосков. 15 мм.
Примечание: описание нами сделано по серии 22 и 1 3 из Чехословакии (Ноги-
мия, сбор И. Моухи), 3 определен М. Хвалой. Н. (s. str.) auripila шшболои Гшшкн
к Н. (s. str.) aterrima и ранее считалась варьететом последнего вида. 22 II. (я. hIp.) rt»W-
pila хорошо отличаются от Н. (s. str.) aterrima золотисто-желтыми волосками Гционим
и преимущественно желтовато-серыми волосками низа лица и бочков груди. Рииличин
33 неясны из-за недостаточного количества исследованных экземпляре]).
P а сп ростране ни е: европейский горно-лесной вид. В СССР ппйдои и Пи-
карпатской (гора Говерла) обл., Дрогобычском р-не Львовской обл., н окр. .Пыюии,
в Латвии (близ г. Валка; К. Эльберг) и в Эстонии (Выруский р-п), п последнем случим
указан X. Реммом (1959) под неправильным определением II. (я. Hlr.) iilvrrlnia и
Н. (s. str.) micans. Сев. и Средн. Европа. Местами обычен. Июнь—июль.
350 III. ПОДСЕМ. TABANINAE
*25. Hybomitra (s. str.) aterrima Mg. (рис. 276) — Слепень черный.
Meigen, 1820, Syst. Beschreib., II : 33 (Tabanus); Brauer, 1880 : 142 (Ta-
banus); S u г с о u f, 1924:102 (Tabanus); Krober in Lindner, 1925:61
(Tabanus); Leclercq, 1966 : 101; С h v a 1 a, Lyneborga. Moucha, 1972 :
186. — lugubris Zetterstedt, 1840, 1. с : 515 (Tabanus).
J. Глаза в умеренно длинных серых волосках, с 3 полосками. Лобная полоска
широкая, с б. м. параллельными сторонами, ее высота превосходит ширину основания
приблизительно в 3 раза, в сером налете. Нижняя лобная мозоль треугольная или
прямоугольная, черная, слабо блестящая, в тонких бороздках, сходящихся под углом
(«елочкой») к срединной вертикальной бороздке, боковыми сторонами достигает края
глаз. Средняя мозоль черная, веретеновидная, концом обычно достигает нижней
мозоли. Глазковый бугорок выпуклый, черно-коричневый. Лобный треугольник низкий,
в сером налете, по бокам с единичными волосками. Верхний край усиковых ямок
относительно широкий, составляет 1/3 высоты лобного треугольника. Усики черные, 3-й чле-
Рис. 276. Hybomitra (s. str.) aterrima Mg. Рис. 277. Hybomitra (s. str.) micans Mg.
Лобная полоска, усик и щупальце J. Лобная полоска, усик и щупальце $.
ник в основании коричневатый, с б. м. тупым дорсальным углом; 1-й членик
сверху в длинных черных волосках, немного превосходящих длину членика.
Концевой членик щупалец узкий, черно-коричневый, преимущественно в черных
волосках. Низ лица преимущественно в черных волосках. Нотоплевры черные. Бочки
гРУйи в черных волосках. Жужжальца черно-коричневые. Крылья с легким
коричневым затемнением вдоль жилок в передней части крыла, в основании жилки г4
небольшое коричневое пятнышко; г4 без придатка. Ноги черные, в черных волосках. Брюшко
черное, слабо блестящее, в черных волосках, II тергит по бокам с небольшими, неясно
отграниченными темно-коричневыми пятнами. 14—16.5 мм.
<?. Голова не крупнее, чем у $. Глаза в умеренно длинных серых волосках.
Фасетки верхних 2/3 глаз лишь немного крупнее нижних. Линия соприкосновения глаз
в 2 раза превосходит высоту лобного треугольника, последний в сером налете. Лицо
и бочки груди в черных волосках. Усики как у 2, 1-й членик сверху в длинных волосках.
Концевой членик щупалец черный, узкий, в черных волосках. Ноги, крылья и
жужжальца как у 2- Брюшко блестяще-черное, в черных волосках. II (или II—III)
тергиты по бокам неясно коричневатые. 13.5—16 мм.
Известен варьетет Н. (s. str.) aterrima var. lugubris Ztt. (2), у которого на теле,
кроме черных, имеются серые волоски.
Распространение: Сев. и Зап. Европа, преимущественно в горах. Редок.
Июнь—июль. Указание X. Ремма (1959) для Эстонии относится к Н. (s. str.) auri-
pila Mg.
*26. Hybomitra (s. str.) micans Mg. (рис. 277) — Слепень западный.
Meigen, 1804, Klassific, 1 : 167 (Tabanus); Brauer, 1880 : 137 (Tabanus);
Surcouf, 1924 : 100 (Tabanus); Krober in Lindner, 1925 : 67 (Tabanus);
Moucha et Clival a, 1959a : 163 (Tylostypia); Leclercq, 1966:111;
Chvala, Lyneborg a. Moucha, 1972 : 182.
.$. Глаза в умеренно длинных серых волосках, с 3 полосками. Лобная полоска
относительно широкая, ее высота превосходит ширину основания приблизительно
11. HYBOMITRA 351
в 3 раза, с параллельными сторонами или едва сужена книзу, в сером налете. Нижняя
лобная мозоль черная, б. м. прямоугольная или щитовидная, боковыми сторонами
достигает края глаз, блестяще-черная. Средняя мозоль веретеновидная, черная, концом
обычно достигает нижней мозоли. Глазковый бугорок коричневый или черный, плоский,
окружен небольшим блестящим пятном. Лобный треугольник низкий,
блестяще-черный, иногда в той или иной мере прикрыт серым налетом. Низ лица в серых волосках.
Усики черные. 1-й членик сверху в умеренно длинных черных волосках. 3-й членик
умеренной ширины, дорсальный угол прямой или слегка тупой. Концевой членик
щупалец черноватый, покрыт преимущественно черными волосками, его длина превосходит
наибольшую толщину приблизительно в 3—4 раза. Нотоплевры черные. Жужжальца
черно-коричневые. Крылья б. м. бесцветные, жилки коричневые, вокруг основания
жилки г4 имеется небольшое коричневое пятнышко. Ноги черные. Брюшко блестяще-
черное, широкое. II тергит по бокам с беловато-серыми пятнами из налета и волосков,
II и последующие тергиты по заднему краю с небольшим срединным треугольником
из беловатых волосков. Снизу брюшко черное, в легком сероватом налете, в черных
и беловатых волосках, последние расположены преимущественно по заднему краю стер-
нитов. 14—16 мм.
S- Голова не больше, чем у $. Глаза в серых волосках. Фасетки верхних 2/3 глаз
лишь немного крупнее нижних. Лобный треугольник в средней части голый, блестяще-
черный. >сики как у 2, но 3-й членик немного уже, а 1-й членик более вздут, сверху
в длинных черных волосках. Концевой членик щупалец черноватый, продолговато-
яйцевидный, на конце приострен. Крылья и грудь как у 2- Ноги черные. На дорсальной
поверхности первых 4 члеников передних лапок имеются очень длинные, стоячие,
черные щетинки. Брюшко черное, боковые стороны II тергита неясно темно-коричневые.
II—IV тергиты по заднему краю посредине с небольшим количеством беловатых
волосков. Снизу брюшко черное. 16 мм.
Распространение: европейский лесной вид. Зап. Европа. Редок. Июнь—
июль. Указание X. Ремма (1959) для Эстонии относится к Н. (а. str.) auripila Mg.
27. Hybomitra' (s. str.) popovi
Ols. (рис. 278) — Слепень Попова.
Олсуфьев, 1937а, Фауна
СССР, Двукрылые, VII, 2 : 174
(Tabanus); Leclercq, 1966 : 117.
9- Глаза в умеренно длинных
серых волосках, с 1—3 полосками
(не всегда восстанавливаются при
размачивании). Лобная полоска
умеренной ширины, ее высота прево-
Рис. 278. Hybomitra (s. str.) popovi
Ols: Лобная полоска, усик и
щупальце $.
сходит ширину основания в 3—3V4 раза, слегка расширена кверху, в сером или желто-
ватом налете. Нижняя лобная мозоль б. м. прямоугольная или треугольная, блостшце-
черная, обычно достигает края глаз. Средняя мозоль узковеретеновидная, б.
и.'соприкоснется с нижней. Темя иногда слегка вдавлено. Глазковый бугорок уплощен, коричиепыИ
или черноватый, обычно вокруг него имеется небольшое блестяще-коричневое прострой
ство. Лобный треугольник низкий, в сером налете. Усики чёрные, 1-й членик и сиром
налете, сверху в светлых, умеренной длины волосках, 3-й членик со слабо расширен
ной площадкой, дорсальный угол обычно тупо закруглен. Концевой членик щунилец
желтовато-палевый, умеренно утолщен, покрыт относительно длинными светлыми
полосками, иногда лишь с незначительной примесью черных волосков..Бочки груди и
серых или желтоватых волосках. Нотоплевры черно-серые. Ноги целиком чорпые;
голени, особенно средние и задние, в светлых волосках. Крылья бесцветные, жилки темно
коричневые; г4 без придатка. Вокруг основания г4 обычно имеется небольшое кормчие
вое пятнышко. Жужжальца черно-коричневые. Брюшко широкое, черное. 11 тергит
по бокам с заметными, но не резкими пятнами серого налета и беловатых или желтопн-
тых волосков. Последние образуют также срединный ряд небольших треугольником
и располагаются по заднебоковой стороне III и последующих тергитов, а также ни
I тергите. Остальное в черных волосках. Снизу брюшко черное, в сером налете и
желтовато-серых волосках. 14—17.мм.
сТ. Голова не больше, чем у 2- Глаза в длинных, серых волосках. Фасетки иерхпих
частей глаз по величине почти не отличаются от нижних.,Глазковый бугорок темно-
352 П1. ПОДСЕМ. TABANINAE
коричневый, часто в тонком налете. Лобный треугольник в сером налете, линия
соприкосновения глаз превосходит высоту треугольника приблизительно в 1V2 раза. Усики
как у J, но 3-й членик более узкий. 1-й членик усиков черно-серый, сверху в густых
длинных черных волосках, по длине превосходящих 1-й и 2-й членики вместе взятые.
Концевой членик щупалец светло-коричневатый, яйцевидный, покрыт желтовато-
серыми и черными волосками. Грудь и ее придатки как у §; бочки груди в желтрвато-
серых волосках с значительной примесью черных. Передние лапки с длинными, почти
вертикальными щетинками, расположенными дорсально в передней части 1 —4-го
члеников (рис. 266, Б). Костальная и субкостальная ячейки коричнево затемнены, более
интенсивно, чему J. Брюшко черное, блестящее, в черных волосках, слегка
коричневатое по бокам II тергита и здесь же по бокам расположены едва заметные небольшие
сероватые пятна налета и светлых волосков; небольшие группы серых волосков
расположены по заднему краю II—IV тергигов в их средней части, а также на концевых терги-
тах и по их заднебоковому краю. Снизу брюшко черное, слабо блестящее, в тонком
сероватом налете, покрыто преимущественно сероватыми волосками, с небольшой примесью
черных волосков. 14—16 мм.
Своеобразный черноокрашенный вид с редуцированным количеством полосок
на глазах. Среди палеарктических Hybomitra редукция полосок на глазах встречается
редко — она известна у Н. (s. str.) expollicata, Н. (Mouchaemyia) caucasi, H. (Mouchae-
myia) decora и немногих других видов. У 3 любопытно наличие на передних лапках
длинных почти вертикальных щетинок, что известно у в Н. (s. str.) micans и И. (s. str.)
astur. По этому признаку, а также наличию коричневого пятнышка вокруг основания г4
Н. (s. str.) popovi следует отнести в группу Н. (s. str.) astur, несмотря на относительно
светлые щупальца, особенно у <J.
Нами этот вид был описан по 3 99 из Армении и, как выясняется, недостаточно
полно. Это описание мы дополняем данными изучения 11 99 из Бакуриани, Грузия.
8 33 собраны там же, описание их дается нами впервые.
Распространение: закавказский горный вид. Грузия (Бакуриани;
Цхра-Цхаро); Армения (Дилижан; Семеновка; Воронцовка; Степанаван). Не часто.
Июль.
Голотип: 9. Дилижан—Семеновка, Армения, 4 VII 1931, В. Попов. В коллекции
Зоологического института Академии наук СССР, Ленинград.
* IV. Группа Hybomitra (s. str.) bimaculata Macq.
Щупальца утолщены, желтовато-белые или сероватые. Лобный треугольник в
налете, очень редко (у 99) блестяще-черный. Жужжальца, как правило, коричневые.
Волоски на глазах и глазковый бугорок на темени хорошо развиты.
28. Hybomitra (s. str.) lurida FUn. (рис. 279) — Слепень ранний.
Fallen, 1817, Dipt. Suec, Taban! : 5 (Tabanus); В г a u e r, 1880 : 148 (Ta-
banus);. Lundbeck, 1907 : 116 {Tabanus); V e r r a 11, 1909 : 367 (Tabanus);
К г о b e г, 1923 : 90 (Tabanus); КгбЬег in Lindner, 1925 : 66 (Tabanus);
Олсуфьев, 1937a : 159 (Tabanus); Moucha et Chvala, 1959a : 166 (Tylo-
stypia); Lyneborg, 1959 : 91; Ш е в ч е н к o, 1961 : 79 (Tabanus); L e с 1 e г с q,
1966 : ПО; В и о л о в и ч, 1968 : 137; Pechuman a. Stone, 1968 : 302;
Murdoch et Takahasi, 1969 : 43. — hirticeps L о е w, 1858 : 105 (Tabanus),
цит. no: Chvala, Lyneborg a. Moucha, 1972 : 211. — metabolus
McDunnough, 1922 : 239 (Tabanus), цит. по: Stone, 1938 : 151.
9. Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска
умеренной ширины, слегка сужающаяся к основанию, ее высота в Зх14—33/4 раза
превосходит основание, в желтовато-сером налете. Нижняя лобная мозоль б. м. прямоугольная,
морщинистая, блестящая, коричнево-черная. Боковые стороны мозоли не достигают
края глаз, вершина часто сливается с средней мозолью; последняя блестяще-черная,
.веретеновидная, с узким килем посредине. Глазковый бугорок выпуклый, блестящий,
коричнево-красный. Лобный треугольник сильно выпуклый, широкопоперечный, почти
целиком лаково-блестящий, коричнево-черный. Усики желто-коричневые; 1-й членик,
верхний край 2-го членика и палочка темные. 1-й членик сверху в б. М. длинных,
преимущественно светлых волосках. 3-й членик в основании широкий, с явственным б. м.
прямоугольным дорсальным выступом; палочка короткая, толстая. Концевой членик щупалец
короткий, беловато-желтый, в основании очень сильно утолщен, остроконечный,
покрыт короткими, светлыми и темными волосками. Нотоплевры коричневые или черные.
Жужжальца темно-коричневые. Ноги: большая часть передних, целиком средние и
задние голени и основание средних и задних лапок коричневые, прочие части ног
черные. Крылья слегка буровато затемнены по переднему краю и возле поперечных
жилок (особенно в основании г4); жилки темно-коричневые; г4 иногда с небольшим
придатком. Брюшко широкоовальное, короткое, слабо блестящее, черное, по бокам с гря-
11. HYBOMITRA
353
зно-коричневыми пятнами, простирающимися на I—III тергиты. Разделяющая их
срединная темная полоса занимает около 1/2—1/8 ширины брюшка. Тергиты по заднему
краю с небольшими срединными пятнами из сероватых волосков (иногда отсутствуют).
Боковые стороны тергигов, в особенности II—III, с сероватыми пятнами из налета
Рис. 279. Hybomitra (s. str.) lurida Flln. Лобная полоска, усик,
щупальце, VIII стернит и церки 9- А — линия соприкосновения
' глаз $.
и серых волосков. Снизу брюшко в базальной части желто-коричневое; I кольцо,
срединное пятно на II стерните и 4 конечных кольца темные. VIII стернит темно окрашен,
гонопофиз короткий и широкий, в черных волосках, без апикальной выемки;
латеральные лопасти умеренной величины, боковые коричневые пятна большие; базальная ло-
0° К ВОСТОКУ ОТ ГриНВИча 90° 120° 150° 180° 150°
0° 30° ' 60° 90° 120°
Рис. 280. Ареал'Hybomitra (s. str.) lurida.
пасть короткая, с б. м. прямым основным краем. Церки округлые или со слегка
уплощенной вершиной. Сперматека веретеновидная. 11.5—15 мм.
Экземпляры из северных частей Вост. Сибири (Якутская АССР, Магаданская обл.)
часто с целиком черным брюшком и темными усиками, мы обозначаем ее как Н. (s. str.)
lurida var. sordida Ols., var. nova.
<J. Глаза в длинных, темно-коричневых волосках;, фасетки глаз б. м. одинаковой
величины, лишь возле лобного шва несколько крупнее. Глазковый бугорок округлый,
красно-коричневый, блестящий. Лобный треугольник в нижней части сильно
выпуклый, в серебристо-сером налете; линия соприкосновения глаз в l1^ раза больше его
высоты (рис. 279, А). Усики как у 9; 3-й членик более узкий. 1-й членик на спинной
23 Н. Г. Олсуфьев
354 П1. ПОДСЕМ. TABANINAE
стороне в длинных черных волосках, равных по длине 1-му и 2-му членикам, вместе
взятым. Концевой членик щупалец беловато-желтый, в основной половине лишь
иногда в сероватом налете, короткоовальный, покрыт длинными, преимущественно
темными волосками. Нотоплевры коричневые или черные. Крылья и ноги как у 2-
Брюшко черное, блестящее, по бокам с коричневыми пятнами, простирающимися на
I—III тергиты (реже незначительно заходят на IV терпит). Разделяющая их черная
полоска относительно широкая. На III тергите она в виде слегка расширяющейся
кзади, реже параллельной, полоски, занимающей 1/я—1/& его ширины. Брюшко сверху
в черных волосках, лишь задние каемки стернитов по боковым сторонам и в центре
в желто-серых волосках. Снизу брюшко желто-коричневое, I стернит, квадратное
пятно (иногда отсутствует) на II стерните и концевые 3—5 колец темные. 12—14 мм.
Распространение: голарктический таежно-лесной вид (рис. 280).
В СССР известен от Кольского п-ова, Эстонии, Литвы и Сев.-Зап. Украины (Шацк
Волынской обл.) на восток до Камчатки и Сахалина, на север до Хибин, Низовий Печоры,
Оби, Енисея, Лены (Жиганск), Индигирки и Колымы, на юг до Киева, Воронежа,
Оренбургской обл., Борового, юго-востока Павлодарской обл., Алтая, Тувы, южного
Прибайкалья, Приамурья и юга Приморского края. Изолированное местонахождение в
Закавказье (Бакуриани). Сев. и Средн. Европа (на запад до Пиренеев), Сев. Монголия,
Сев.-Зап. Китай, Япония. Ранний вид — май—июнь, на севере июль—август. Обычен
в лесотундре, северной и средней тайге, многочислен местами в Туве и Сев. Монголии,
в прочих частях ареала не часто.
По нашим исследованиям, терминалии 22 североамериканской популяции не
имеют отличий от таковых палеарктической популяции.
29. Hybomitra (s. str.) nitidifrons Szil. — Слепень гололобый.
Голарктический вид, в пределах своего обширного ареала представлен 3 подвидами:
восточносибирским номинативным Н. nitidifrons nitidifrons Szil., европейско-западно-
сибирским Н. nitidifrons confiformis М. et Chv. и североамериканским Н. nitidifrons
nuda McDunn.
29а. Hybomitra (s. str.) nitidifrons nitidifrons Szil. (рис. 281) — Слепень гололобый
номинативный.
S z i 1 a d у, 1914, Ann. Mus. Nat. Himgar. XII: 664 (Tabanus); Олсуфьев,
1937a : 163 (T. confinis nitidifrons Szil.): Leclercq, 1966 : 194 (Hybomitra canfor-
mis nitidifrons Szil.); В и о л о в и ч, 196Ь : 141 (Н. confinis nitidifrons Szil.).
2- Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска
умеренно широкая, слегка сужена к основанию (высота полоски в Зх/4—4 раза превосходит
основание), в желтовато-сером налете. Нижняя лобная мозоль почти плоская, слабо
блестящая, черная, с коричневатой серединой и с небольшой продольной бороздкой,
к которой сходятся морщинки; боковые стороны мозоли не достигают края глаз.
Средняя мозоль узковеретеновидная, черная, слабо блестящая, тонкой линией соединена
с нижней мозолью. Глазковый бугорок овальный, выпуклый, блестяще-коричневый.
Лобный треугольник слегка выпуклый, широкопоперечный; его верхне-центральная
часть без налета, слабо блестящая, коричнево-черная, тогда как прочие части в
желтовато-сером налете. Величина оголенных участков варьирует, и лобный треугольник
может быть весь покрыт налетом или целиком блестящим. Усики коричнево-красные,
палочка, верхний край 3-го и 1-й членик темные, 1-й членик сверху с б. м. длинными
светлыми волосками, 3-й членик в основании широкий, с б. м. прямоугольным
дорсальным выступом. ГГалочка короткая, толстая. Концевой членик щупалец светлый, палево-
желтый, в основании очень сильно утолщен, на конце слегка притуплён, покрыт
короткими светлыми и темными волосками; последние сгруппированы преимущественно
в концевой части членика. Нотоплевры желто-коричневые, редко темные. Жужжальца
темно-коричневые. Ноги: большая часть передних, целиком средние и задние голени
и большая часть средних и задних лапок коричнево-желтые; остальное черное. Крылья
слегка буро затемнены к переднему краю, жилки коричневые. Брюшко
широкоовальное, слабо блестящее, черное, по бокам с коричнево-желтыми пятнами,
простирающимися на I—III (или I—IV) тергиты. Разделяющая их срединная темная полоска
занимает около 1/3—XU ширины брюшка. II и последующие тергиты по заднему краю с
коротким, неясным, беловато-серым срединным треугольным пятном (налет+волоски).
Боковые стороны II—III тергитов с крупными, но не резкими, беловато-серыми
пятнами, покрытыми преимущественно белыми волосками. Каемки всех тергитов
по бокам в коротких беловатых волосках. Снизу брюшко в большей части
светло-коричнево-желтое, в беловато-серых волосках, треугольное или квадратное
пятно на II стерните и 3—4 последних кольца черно-бурые. Окраска брюшка варьирует
за счет вытеснения светлых пятен, в редких случаях вплоть до почти полного их
исчезновения; в таком случае брюшко одноцветное, буро-черное, с неясным серым рисунком.
Эту темную форму мы обозначаем как Н. (s. str.) nitidifrons var. fusca Ols., var. nova.
11. HYBOMITRA
355
VIII стернит: гонопофиз широкий и короткий, в черных волосках, с маленькой
апикальной выемкой, внешний край гонопофиза равномерно переходит в латеральные
лопасти; последние крупные, б. м. остроугольные; боковые коричневые пятна большие;
базальная лопасть удлиненная, с прямым основным краем. Церки с уплощенной
вершиной. Сперматека миндалевидная или веретеновидная. 16—18 мм.
$. Глаза в длинных, серовато-коричневых волосках. Фасетки глаз б. м.
одинаковой величины, лишь возле лобного шва несколько крупнее. Глазковый бугорок
округлый, красно-коричневый, блестящий. Лобный треугольник умеренно выпуклый, в
серебристо-сером налете; линия соприкосновения глаз в V-l2 раза больше его высоты.
Усики как у 2; 3-й членик более узкий. 1-й членик сверху в длинных, черных волосках,
по длине равных 1-му и 2-му членикам вместе взятым. Концевой членик щупалец
палево-желтый, в основной половине в сером налете, сильно тыквовидно утолщен,
покрыт длинными, преимущественно темными волосками. Грудь и ее придатки как у $.
Рис. 281. Hybomitra (s. str.) nitidifrons confiformis M. et Chv. Лобная
полоска, усик, щупальце, VIII стернит, церки и сперматека 2-
Коготки на передних лапках асимметричны: внутренний коготок немного короче
внешнего. Брюшко черное, блестящее, по бокам с красновато- или желто-коричневыми
пятнами, простирающимися на I—III тергиты (реже незначительно заходят на IV тергит).
Разделяющая их срединная полоска на III тергите сильно сужена и здесь занимает
1/5—1/ц его ширины. Брюшко сверху в черных волосках, лишь задние каемки тергитов
по боками в середине в беловато^ерых волосках. II и частью III тергиты по бокам
с нерезкими беловато-серыми пятнами налета. Снизу брюшко светло-коричнево-жол-
тое, треугольное пятно на II стерните и 4 последних кольца темные. 17 мм.
Распространение: восточносибирский таежно-лесной подвид (рис. 282,
2). От Байкала и Якутии до Дальнего Востока, на север до Жиганска на Лене и
Охотского побережья, на юг до Южного Прибайкалья, Приамурья и юга Приморского края.
Сев. Монголия, Сев.-Вост. Китай, Корейский п-ов, Япония. Июнь—июль. Не часто.
Принадлежность к этому подвиду популяции, обитающей в Японии (о. Хопсю),
остается неясной. Такахаси (Takahasi, 1962, р. 54), описывая Н. (s. str.) confinis
(=Н. (s. str.) nitidifrons) по единственному экземпляру (2) из Никко, префектура То-'
чиги, указывает на брюшке (сверху) «срединный ряд маленьких желтых треугольпикои
и весьма ясные косые желтые боковые пятна на II и III тергитах; IV—VI тергиты
черные с боковыми пятнами золотисто-желтых волосков». Для номинативного подпидн
Н. (а. str.) nitidifrons характерно наличие на дорсальекой стороне брюшка беловатых ио-
лосков в отличие от западного подвида Н. (s. str.) nitidifrons confiformis, имеющого iio-
лотисто-желтые волоски.
29b. Hybomitra (s. str.) nitidifrons confiformis Chv. et M. — Слепень гололобый
западный.
Chvala et Moucha, 1971, Notulae entomolog., 51 : 109. — conforml»
F r e y, 1911 : 27-(Tabanus) пес Т. conformist alker, 1848: 150, цит. по: С 1) V n 1 I),
Lyneborg a. Moucha, 1972 : 214. — confinis auct. nee T. confinis Z с I, I; о r-
s t e d t, 1840 : 516, цит. по: Leclercq, 1966 : 104.
2M*
356 III. ПОДСЕМ. TABANINAE
9cJ. От номинативной формы отличается красновато-коричневой окраской
боковых пятен брюшка и наличием на них, а также по заднему краю тергитов золотисто-
желтых волосков; серовато-белые пятна налета по бокам II—III тергитов отсутствуют;
снизу брюшко в основной части красновато-коричневое. 2 14—18 мм, 3 14—17 мм.
Распространение: европейско-западносибирский таежно-лесной
подвид (рис. 282,1). В СССР от Кольского п-ова, Эстонии, Литвы и запада Украины (Ивано-
Франковская обл.) на восток до Енисейско-Ленского водораздела, на север до Пул-
озера Мурманской обл., Индиги Архангельской обл., Усть-Цильмы Коми АССР,
низовий Оби и Енисея, на юг до Днепропетровской и Ворошиловградской областей УССР,
Пензы, Ю. Урала, севера Карагандинской обл. (урочище Карагаш), Павлодарской
обл., Семипалатинска и Алтая; изолированно распространен в Закавказье (Бакуриани).
Сев. и Средн. Европа (на запад до Бельгии). Обычен в зоне тайги и местами лесотундры,
а также в западносибирской лесостепи, в других частях ареала не часто. Ранний
слепень: май—июнь, на севере июль—агуст. В Ленинградской и Московской областях
появление первых 22 обычно совпадает с началом цветения черемухи. Откладка яиц
Рис. 282. Ареалы: 1 — Hybomitra (s.'str.) nitidifrons confiformis; 2 — H. (s. str.)
nitidifrons nitidifrons.
нами отмечена на стебле злака по краю запущенной мелиорационной канавы с водой
(окр. Ленинграда). Личинки найдены в черноольховой топи (Воронежская обл.; Ску-
фьин, 1952а).
Распространенный в Сев. Америке Н. (s. str.) nuda McDurm., которого на основании
изучения 6 22 и 1 S мы считаем подвидом В- (s. str.) nitidifrons Szil., отличается
главным образом тем, что у 2 лобный треугольник всегда голый, а коричневые пятна по
бокам брюшка обычно захватывают I—IV тергиты, тогда как у номинативной формы
лобный треугольник чаще оголен лишь частично, а коричневые пятна обычно расположены
по бокам I—III тергитов; строение терминалий 22 б. м. одинаковы. У Н. (s. str.)
nitidifrons nuda, как и у номинативного подвида, на II—III тергитах брюшка имеются
ясные боковые беловато-серые пятна налета, а светлые волоски сверху брюшка чаще
беловатые. У 33 Н. (s. str.) nitidifrons nuda, так же как у 33 других подвидов
Н. (s. str.) nitidifrons, отмечается небольшая асимметричность коготков передних
лапок, на что наше внимание обратил Филиц (Philip). Подобная асимметрия коготков
(и пульвилл) нами обнаружена у 33 некоторых Hybomitra в группе Я. (s. str.) kaurii.
30. Hybomitra (s. str.) sareptana Szil. (рис. 283) — Слепень еарептский.
S z i 1 a d у, 1914, Ann. Mus. Nat. Hungar., 12 : 662 (2) {Tabanus); К г о b e r,
1923 : 95 (Tabanus); Krober in Lindner, 1925 : 70 (Tabanus); Олсуфьев,
1936a : 234 (3) (Tabanus); 1937a : 164 (Tabanus); Ш евченко, 1961 : 83 (Tabanus);
Leclercq, 1966 : 117; Виолович, 1968 : 141; C/hvala, Lyneborga.
M ouch a, 1972:218. '
2. Глаза в коротких, серых волосках, с 3 полосками. Лобная полоска умеренно
широкая, слегка сужена к основанию (высота полоски приблизительно в 31/2—4 раза
превосходит основание), в сером'налете. Глазковый бугорок овальный, блестяще-
11. HYBOMITRA
357
коричневый, темя вокруг бугорка часто слегка блестящее. Нижняя лобная мозоль
гладкая, б. м. плоская, слегка выпуклая, блестяще-черная, реже морщинистая или
коричневая. Боковыми сторонами мозоль чаще достигает края глаз. Средняя мозоль узко-
веретеновидная, нижним концом достигает нижней мозоли. Лобный треугольник б. м.
высокий, всегда в сером налете. Усики черные; 3-й членик часто в основании слегка
коричневый. 1-й и 2-й членики в густом сером налете, сверху в коротких,
преимущественно светлых волосках. 3-й членик умеренно широкий, с тупым дорсальным углом.
Щупальца белые или желтоватые, концевой членик сильно утолщен, покрыт очень
короткими и нежными светлыми волосками, иногда с незначительной примесью черных.
Нотоплевры желто-коричневые. Крылья бесцветные; жилки коричневые; г4 без
придатка. Жужжальца темно-коричневые. Ноги: бедра черные, в сером налете, основание
передних голеней, средние и задние голени целиком и основание лапок
желто-коричневые; конец передних голеней и передние лапки черные. Брюшко черное, по бокам
со светло-коричневыми пятнами, захватывающими I—III тергиты. Разделяющая
пятна черная полоса составляет приблизительно 1/3 брюшка. Посредине брюшко с ря-
Рис. 283. Hybomitra (s. str.) sareptana Szil. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека 2-
дом небольших, серых треугольных пятен (налет4-,волоркй), по бокам с более крупными
пятнами б. м. ромбической формы. Снизу брюшко розово-желтое; I стернит, квадратное
. или прямоугольное пятно на II стерните и 3—4 концевых стернита темные. Окраска
брюшка: варьирует, и иногда оно целиком черное, без боковых коричневых пятен
(Н. (s; str.) sareptana var. melas Szil.). VIII стернит своеобразной формы: гонопофий
широкий и короткий, в черных волосках, с небольшой апикальной выемкой; латераль1-
ные лопасти удлиненные, остроугольные, боковые коричневые пятна крупные; базаль-
ная лопасть с прямым основным краем. Церки с уплощенной вершиной. Сперматека
миндалевидной формы. 14—19 мм.
cJ. Глаза в довольно длинных серовато-желтых волосках, фасетки глаз б. м.
одинаковой величины. Глазковый бугорок овальный, красно-коричневый, блестящий.
Лобный треугольник слабо выпуклый, в серебристо-сером налете; линия соприкосноио-
ния глаз в 13/4 раза больше его высоты. Усики одноцветно-черные, 3-й членик с б. м.
тупым дорсальным углом. 1-й членик сверху в длинных, светлых и черных волосках,
по длине равных 1-му и 2-му членикам вместе взятым: Концевой членик щупалец
желтоватый или серый, утолщенный, покрыт б. м: длинными светлыми.волосками, с ноани-
чительной примесью черных. Грудь и ее придатки как у 2- Брюшко блестящее, по
бокам с крупными желто-коричневыми пятнами, простирающимися на I—IV тергиты и
разделенными черной срединной полоской. Последняя немного сужена на III тергито,
и здесь ее ширина составляет 1/4—1/5 ширины кольца. Окраска брюшка дополняете)!
небольшими пятнами тонкого, беловатого налета (видны при рассматривании брюшки
сзади), расположенными по бокам II—VI тергитов. Брюшко покрыто черными
полосками, с небольшой примесью светлых волосков на месте светлых пятен из налотп п
кое-где по заднему краю тергитов. Снизу брюшко желто-коричневое; I стернит, кннД"'
ратное пятно на II стерните и 3 последних кольца черные. 15.5—17 мм.
358 III. ПОДСЕМ. TABANINAE
Весьма близок к Н. (s. str.) nitidifrons, отличаясь от него более светлыми волосками
глаз и черными усиками с тупоугольным дорсальным выступом. Кроме того, у §
лобный треугольник целиком в сером налете, щупальца чаще только в светлых волосках.
Распространение: (рис. 284, 1) преимущественно по долинам рек
степной зоны юго-востока европейской части СССР (Сев. Донец Ворошиловградской обл.;
Дон ниже Калача; Волга от Воскресенского выше Саратова до Астрахани; среднее и
нижнее течение Урала), Западной Сибири (Челябинская обл. — близ Троицка;
Омская, Новосибирская, юг Томской обл., пойма Оби в Алтайском крае) и Сев.
Казахстана (Кустанайская обл. — на юг до б. Наурзумского заповедника Семиозерного р-на;
Павлодар). Указание для Дербента (Олсуфьев, 1941) нуждается в подтверждении.
Многочислен в Волго-Ахтубинской пойме близ Волгограда, в пойме Оби около Барнаула и
в некоторых других местах. Ранний вид — обычно в мае, реже в июне и позднее.
По нашим наблюдениям, в Волго-Ахтубинской пойме $§ откладывают яйца на тростник
и другие растения на сухих участках до затопления почвы водой.
Рис. 284. Ареалы: 1 — Hybomitra (s. str.) sareptana; 2 — H. (s. str.) tschuensis;
3— H. (s. str.) adachii. ч
31. Hybomitra (s. str.) tschuensis Ols. (рис. 285) — Слепень чуйский.
• -Олсуфьев; 1962( Зоол. журн.; 41, <6 : 886 (Tabanus sareptanus tschuensis);
L е с 1 е г с q, 1966 : 118 {Hybomitra sareptana tschuensis Ols.); Виолович, 1968 :
142 (Я. sareptana tschuensis Ols.); Олсуфьев, 1969:117.
5. Глаза в коротких серых волосках, с 3 полосками. Лобная полоска слегка
расширена кверху, ее высота превосходит ширину осповапия приблизительно в 4 раза,
в сером налете. Нижняя лобная мозоль блестяще-черная, б. м. прямоугольная,
боковыми сторонами почти достигает края глаз. Средняя лобная мозоль черная, веретено-
видная, обычно не достигает нижней мозоли, иногда целиком прикрыта налетом.
Глазковый бугорок небольшой, коричнево-красный, темя вокруг и сзади обычно
блестящее. Лобный треугольник в сером налете. Усйки черные, 3-й членик иногда в
основании слегка коричневый, относительно узкий, с тупым дорсальным углом; 1-й
членик сверху в коротких и умеренно длинных, преимущественно черных волосках.
Концевой членик щупалец светло-коричневато-желтый, умеренно утолщен, его длина
превосходит наибольшую толщину приблизительно в 3 раза, покрыт преимущественно
черными волосками. Нотоплевры коричневые, реже черно-серые. Крылья
бесцветные, жилки коричневые; г4 обычно с хорошо развитым придатком. Жужжальца темно
коричневые. Бедра черно-серые, голени желто-коричневые, более темные к вершине,
лапки черные. Брюшко черное, боковые стороны II или I—III тергитов по бокам
коричневые. Сверху брюшка 3 ряда серых пятен из налета и волосков; треугольных
посредине и б. м. ромбических по бокам. Снизу брюшко в основной части
желтовато-коричневое, концевые 3—4 стернита черные, волоски снизу преимущественно беловато-
серые, к концу брюшка примешиваются черные волоски. Иногда брюшко с обеих
сторон целиком чёрное, сверху с типичным серым рисунком, снизу в сероватом налете
и с желтоватыми каймами по заднему краю стернитов. Мы обозначаем эту темнуюформу
как Н. (s. str.) tschuensis var. nigrescens Ols., var. nova. VIII стерши: гонопофиз удли-
11. HYBOMITRA 359-
ненный, в черных волосках, с глубокой апикальной выемкой; латеральные лопасти
небольшие, боковые коричневые пятна довольно крупные; базалъная лопасть
короткая, с вогнутым основным краем. Церки с б. м. округлой вершиной. 14—17.5 мм.
(J неизвестен.
Близок к Н. (s. str.) sareptana и первоначально нами описан как его подвид.
Отличается от Н. (s. str.) sareptana более узким лбом и щупальцами, наличием придатка
у г4 и в особенности строением терминалий.
Распространение: (рис. 284, 2) горный Алтай, в Чуйской долине и Ку-
райской степи, на запад до Чибита.
Рис. 285. Hybomitra (s. str.). tschuensis Ols. Лобная полоска,
у^ик, щупальце, VIII,стерниг и церки $.
Голотип: 5. Красная Горка,'Чаган-Узун, долина Чуй, 15 VII 1957, Олсуфьев.
В коллекции Зоологического института Академии наук СССР, Ленинград.
32. Hybomitra (s. str.) adachii Takag.. (рис. 286) —Слепень Адачи. ,
Т a k a g i, 1941, Report Inst. Horse-diseases, 2:49 (Tabanus); L e с 1 e г с q,
1966 : 100; Murdoch et Takahasi, 1969 : 44. — sareptana adachii (Так.)
apud 0.1 $ u f j e v, 1969 : 117.
5. Глаза в коротких серых
волосках, с 3 полосками. Лобная
полоска слегка расширена кверху, ее
высота превосходит ширину
основания приблизительно в 3 раза, в
желтовато-сером налете. Нижняя
лобная мозоль черная, с коричневым
центром, блестящая, б. м.
прямоугольная, боковыми сторонами почти
достигает края глаз. Средняя
мозоль черная, продолговато-овальная,
иногда не соединена с нижней мо-
Рис. 286. Hybomitra (s. str.) adachii
Takag. Лобная полоска, усик и
щупальце 2- ■
золью. Глазковый бугорок блестяще-коричневый, темя вокруг и сзади блестящее.
Лобный треугольник в сером налете. Усики: 1-й и 2-й членики черные, в сером налете,
1-й членик сверху в умеренно длинных светлых волосках; 3-й членик коричневый,
палочка и верхний край площадки черные; членик умеренно широкий, дорсальный
угол б. м. прямой. Концевой членик щупалец беловато-желтоватый, утолщен, его
360 III. ПОДСЕМ. TABANINAE
длина превосходит наибольшую толщину приблизительно в 21/2 раза, в светлых
волосках, иногда с примесью единичных черных волосков. Нотоплевры коричневые,
жужжальца черно-коричневые. Крылья бесцветные, жилки коричневые; г4 без придатка.
Ноги: бедра черно-серые, голени желто-коричневые, с более темным концом, лапки
черные. Брюшко черное, с 2 боковыми рядами крупных беловато-серых пятен из
налета и волосков; разделяющая,их широкая черная полоса в черных волосках. Снизу
брюшко черное, в сером налете и коротких беловато-серых волосках, стерниты по
заднему краю желтоватые. 16.5—17 мм.
с} неизвестен.
Близок к Н. (s. str.) sareptana и особенно похож на темный варьетет этого вида —
Н. (s. str.) sareptana var. melas Szil., отличаясь более широким лбом, отсутствием сверху
брюшка срединнрго ряда серых треугольников и некоторыми деталями в строении
терминалий.
Распространение: (рис. 284, 3) Читинская обл., Онон между Усть-Иля
и Акша; юг Приморского края — Шкотовский р-н (Р. Г. Соболева, 1974). Сев.-Вост.
Китай. Редок.
33. Hybomitra (s. str.) distinguenda Verr. — Слепень лесной.
В пределах своего обширного ареала представлен 2 подвидами: европейско-си-
бирским номинативным Н. (s. str.) distinguenda distinguenda Verr. и дальневосточным
H. (s. str.) distinguenda contigua Ols.
Рис. 287. Hybomitra (s. str.) distinguenda distinguenda Verr. Лобная
полоска, усик и щупальце %. А — щупальце £.
33а. Hybomitra (s. str.) distinguenda distinguenda Verr. (рис. 287) — Слепень
лесной номинативный.
V е г г а 11, 1909, Brit. Flies, 5 : 371 (Tabanus); S h i r a k i, 1918 : 150
{Tabanus); К r 6 b e r, 1923 : 107 (Tabanus); S и г с о и f, 1924 : 119 (Tabanus); Krober
in Lindner, 1925:64 (Tabanus); Олсуфьев, 1937а : 165 (Tabanus);
Collin, 1940 : 178 (Tabanus); M о и с ha et С h v a 1 a, 1959a : 168 (Tylostypia); L y-
n e b о r g, 1959 : 115; Шевченко, 1961 : 84 (Tabanus); Takahasi, 1962 : 56;
Олсуфьев, 19626 : 526 ; L e с 1 e г с q, 1966 : 105; Виолович, 1968 : 142;
Murdoch et Takahasi, 1969 : 47; И в а н и щ у к, 1970 : 145 (личинка);
Chvala, Lyneborg a. М о и с h a\ 1972 : 235.
5. Глаза в коротких, коричневых волосках, с 3 полосками. Лобная полоска узкая,
заметно сужающаяся к основанию, ее высота в 41/2—5х/4 раза превосходит основание,
в серовато-желтом налете. Нижняя лобная мозоль почти плоская, мелкоморщинистая,
тускло-черная (иногда коричневатая), вытянуто-прямоугольная. Боковые стороны
мозоли не достигают края глаз, отделяясь от последних узкими, серыми полосками;
вершина сливается с средней мозолью, представленной узким, черным, слабо
блестящим валиком. Глазковый бугорок овальный, выпуклый, блестяще-коричневый.
Лобный треугольник высокий, слегка выпуклый, желтовато-серый. Усики коричнево-
желтые, 1-й членик, верхний край 3-го членика и палочка темные. 1-й членик в сером
налете, сверху в б. м. длинных черных волосках, 3-й членик в основании сильно
расширен, с б. м. прямоугольным дорсальным выступом. Палочка короткая, толстая.
Концевой членик щупалец светлый, палево-желтый, в основании сильно утолщен,
со слегка притуплённым кончиком, покрыт короткими, преимущественно черными
волосками. Нотоплевры черные, реже коричневые. Жужжальца темно-коричневые.
11. HYBOMITRA ш
Ноги: кончик бедер, средние и задние голени, основание средних и задних лапок,
а также базальная часть передних голеней коричневато-желтые, остальное черное.
Крылья слегка буровато затемнены к переднему краю, жилки темно-коричневые.
Брюшко широкоовальное; общий тон окраски желто-бурый, по бокам с крупными,
оранжево-желтыми пятнами, простирающимися на I—IV тергиты (реже I—III).
Разделяющая их срединная черная полоска составляет около х/5—1/t ширины брюшка,
II—IV тергиты по заднему краю с короткими, беловатыми срединными треугольнич-
Рис. 288. VIII стернит, церки и сперматека ?$• А — Hybomitra
(s. str.) bimaculata Macq.; Б — H. (s. str.) muehlfeldi Br.; В —
H. (s. str.) ciureai Seg.; Г— H. (s. str.) distinguenda distinguenda
Verr.
ками (налет+беловатые волоски). Боковые стороны II и отчасти III тергитов в
коротких золотисто-желтых волосках, без примеси черных волосков. Снизу брюшко в
большей части желтое, основание I стернита, небольшое треугольное пятно на II стерните
и 3 последних кольца буровато-черные. VIII стернит (рис.. 288, Г): гонопофиз средней
величины, в коричневых и черных волосках, двулопастной благодаря хорошо
выраженной апикальной выемке; латеральные лопасти и боковые коричневые пятна
средней величины; базальная лопасть с б. м. прямым нижним краем. Ширина церок слегка
превосходит их длину, вершина у них округлая или слегка уплощенная. Сперматека
миндалевидная. 13—17 мм.
3. Глаза в длинных коричневых волосках. Фасетки верхних 2/3 глаз заметно
(но не резко) крупнее нижних (рис. 289, А). Глазковый бугорок округлый, коричпопо-
красный, блестящий, сзади обычно с пучком черных волосков. Лобный треугольник
пепельно-серый, линия соприкосновения глаз в 2 раза больше его высоты. Усики кпк
562 HI. ПОДСЕМ. TABANINAE
у 2; 3-й членик более узкий. 1-й членик сверху в длинных черных волосках, по длине
равных 1-му и 2-му членикам вместе взятым. Концевой членик щупалец светлый,
палево-желтый, в основной половине в сером налете; он сильно утолщен, почти
шаровидный, покрыт длинными, преимущественно темными волосками. Нотоплевры чер-
Рис. 289. Фрагмент сложного фасетчатого глаза $: А —
Hybomitra (s. str.) distinguenda distinguenda Yen.; Б — H. (s. str.)
ciureai Seg.
ные, реже буроватые. Жужжальца коричневые. Крылья и окраска ног как у 5;
средние голени в умеренно длинных, тонких, темных волосках. Общий тон окраски брюшка
желто-бурый; боковые стороны брюшка с большими, желто-коричневыми пятнами,
разделенными узкой, черной срединной полоской. Последняя сужена на III тергите,
и здесь ширина ее составляет приблизительно 1/е—V7 ширины брюшка. Основание I,
Рис. 290. Ареалы: 1 - Hybomitra (s. str.) distinguenda distinguenda; 2 -
H. (s. str.) distinguenda contigua.
боковые края IV и последующие тергиты целиком черные: Брюшко сверху в черных
волосках, лишь задние каемки II—IV тергитов по бокам и в центре в желтовато-серых
волосках. Снизу брюшко коричневато-желтое; I стернит, небольшое треугольное пятно
на II стерните и последние 3 кольца темные., 17—17.5 мм.
Распространение: европейско-сибирский лесной подвид (рис. 290, 1).
В СССР известен от Ленинградской обл., Литвы, Молдавии и запада Украины (Закар
патская обл.), на восток до Байкала, на север до южной части Карелии (Медвежье-
горский р-н), Онеги Архангельской обл. Ухты в Коми АССР, севера Свердловской обл.
11. HYBOMITRA
363
(Никито-Ивдель), севера Омской обл., Томска и Подкаменной Тунгуски. На юге
европейской части СССР вид известен до Полтавской (Миргородский р-н),
Воронежской областей, Пензы, Ю. Урала (Янтышево), а после перерыва в
распространении в степях юго-востока — на Северном Кавказе (Майкоп, Нальчик, Орджоникидзе,
Нечаевка в Дагестане) и в Закавказье (Гагра в Абхазии; Манглиси и Бакуриани в
Грузии; Дилижан в Армении; Талыш в Азербайджане). В Азии на юг распространен до Кок-
четавской обл. (Боровое), Павлодарской обл., Семипалатинска,
Восточно-Казахстанской обл., Саура, Джунгарского Алатау (на юг до Панфиловского р-на), Алтая,
Хакасской АО и Иркутска. Европа, Турция. Не часто. Июнь—июль, редко позднее.
Указание Н. (s. str.) distinguenda для степного юга УССР (Гладенко и Фортуш-
ный, 1951; Коречевская, 1960; Бошко, 1964) нуждается в подтверждении, так как
при определении самок возможно смешение с южными экземплярами Н. (s: str.)
ciureai, не имеющими примеси черных
волосков на II тергите брюшка.
ЗЗЬ. Hybomitra (s. str.) distinguenda
contigua Ols. (рис. 291) — Слепень
лесной дальневосточный.
Олсуфьев, 1972, Энтомол. обозр.,
51, 2 : 452.
2. Б оковые желто-коричневые пятна
сверху брюшка в желтовато-серых волос-
Рис. 291. Hybomitra (s. str.) distinguenda
contigua Ols. VIII стернит и церки 5
(варианты).
О О с^> *о
ках, на II тергите в его переднебоковой части часто имеется примесь черных волосков.
Нотоплевры, как правило, коричневые. Глаза обычно в сероватых волосках. Терми-
налии имеют небольшие отличия от таковых номинативной формы: гонопофиз VIII
стернита часто с незначительной апикальной выемкой, основной край базальной
лопасти слегка вогнутый, церки иногда с более уплощенной вершиной. 13—18 мм. При
определении этот подвид может быть легко принят за Н. (s. str.) ciureai; от последнего
вида отличается более коренастым сложением, более широким брюшком, беловатыми
срединными треугольничками по заднему краю тергитов (хорошо заметны только
у непотертых экземпляров!) и более короткой базальной лопастью VIII стернита.
33 хорошо отличаются от 36 Н. (s. str.) ciureai менее крупными фасетками глаз, но
отличия от номинативной формы неясны.
Распространение: восточносибирский лесной подвид. Забайкалье,
Якутия, Амурская обл., Хабаровский и Приморский края,, Сахалин. Сев-Воет. Китай,
Япония. Экземпляры из Якутии (Намский р-н) имеют темные нотоплевры и более
яркие золотисто-желтые волоски по бокам II тергита, но с примесью черных волосков
в его переднебоковой части.
34. Hybomitra (s. str.) ciureai Seg. (рис. 292) — Слепень узколобый.
S ё g u у, 1937 in: Ciurea, Seguy et Stefanescu, Arch. Roum.
Pathol. Exper., 10, 3:207 {Sziladynus solstitialis Schiner, var. ciureai); Chvala
et J e z e k, 1969 : 337 (личинка, куколка); Иванищук, 1970 : 147 (личинка);
Chvala, Lyneborg a. Moucha, 1972 : 237. — solstitialis Schiner,
1862 (Tabanus) et auct. nee T. solstitialis M e i g e n, 1820. — schineri Lyneborg,
1959 (Hybomitra).
$. Глаза в коротких, буровато-серых волосках, с 3 нерезкими полосками. Лобная
полоска очень узкая, сильно сужена к основанию, высота полоски в 4!/3—51/2 раза
превосходит основание, в желтовато-сером налете. Нижняя лобная мозоль
мелкоморщинистая, коричневая, реже черная, овальная или щитовидная. Боковые стороны
мозоли немного не достигают края глаз. Средняя мозоль представлена узким, черным,
слабо блестящим валиком, соединенным,с нижней мозолью. Глазковый бугорок
овальный, выпуклый, блестяще-коричневый. Лобный треугольник плоский, высокий, и
сером налете. Усики красно-желтые, палочка и иногда верхний край площадки черноии-
тые; 1-й членик сверху в коротких и отдельных относительно длинных, черных
волосках; 3-й членик в основании сильно расширен, с явственным б. м. прямоугольным
дорсальным выступом; палочка короткая, толстая. Концевой членик щупалец палишь
364
III. ПОДСЕМ. TABANINAE
желтый, в основании утолщен, остроконечный, покрыт короткими, преимущественно
черными волосками. Нотоплевры желто-коричневые. Жужжальца коричневые, головка
часто со светлым кончиком. Ноги: кончик бедер, средние и задние голени, основание
средних и задних лапок, а также базальная половина передних голеней коричнево-
желтые, прочее черное. Крылья бесцветные, жилки темно-коричневые. Брюшко
узкоовальное или слегка коническое, общий тон окраски тусклый, черно-серый, по бокам
с коричнево-желтыми пятнами, простирающимися на I—IV (редко на I—III) тергиты.
Пятна разделены умеренно широкой срединной черной полоской, составляющей около
1/5—V7 ширины брюшка. II—IV тергиты по заднему краю со слабо намеченными
треугольными пятнами из налета я серых волосков (у потертых экземпляров
отсутствуют). Боковые стороны II и III тергитов покрыты короткими, желтыми волосками,
с незначительной примесью черных (у экземпляров с юга могут отсутствовать!). Снизу
брюшко в большей части желтое, небольшое пятно на II стерните и 3 последних кольца
черно-серые. VIII стернит: (рис. 288, В): гонопофиз средней величины, в густых черных
волосках, без апикальной выемки или с ее следами; латеральные лопасти и боковые
pit
Рис. 292. Hybomitra (s. str.) ciureai Seg. Лобная
полоска, усик и щупальце ?. А — щупальце $.
коричневые пятна умеренно развиты; базальная лопасть с вогнутым основным краем.
Ширина церок слегка превосходит их длину, вершина их значительно уплощена.
Сперматека миндалевидная. 14—17 мм.
$. Глаза в длинных буровато-серых волосках. Фасетки верхних 2/3 глаз
значительно крупнее нижних (приблизительно в 5—6 раз), граница между теми и другими
резко выражена (рис. 289, Б). Лобный треугольник пепельно-серый, линия
соприкосновения глаз в 2—2V2 раза больше его высоты. Усики как у $; 3-й членик более узкий.
1-й членик сверху в длинных черных волосках, по длине равных 1-му и 2-му членикам
вместе взятым. Концевой членик щупалец светлый, палево-желтый, в основной
половине в сером налете. Членик сильно утолщен, яйцевидный или тыквовидный, покрыт
длинными, преимущественно темными волосками. Грудь и ее придатки как у ?. Общий
тон окраски брюшка желто-бурый; брюшко по бокам с коричневато-желтыми пятнами,
простирающимися на I—IV (реже на I—III) тергиты и разделенными срединной
черной полоской. Последняя сильно сужена на III тергите, и здесь ее ширина составляет
около 1/в—Vs ширины брюшка. Основание I и V—VII тергиты целиком черные. Брюшко
сверху в черных волосках; задние каемки II—IV (или II—V) тергитов по бокам и
в середине в-серых волосках. Снизу брюшко желтое, небольшое пятно на II стерните
и последние 3 кольца темные. 14—17 мм.
Распространение: европейско-сибирский лесостепной вид (рис. 293).
В СССР от Эстонии, Ленинградской, Калининградской, Одесской областей и Молдавии
на восток до Якутска и Забайкалья, на север до Карелии (Лоухский р-н), Сыктывкара,
низовий Оби (Питляр), Енисея (Снежногорск на Хантайке) и среднего течения Вилюя
(Мирный), на юг до дельты Дуная, Днепра, Крыма, Сев. Кавказа и Закавказья
(Азербайджан: Закаталы, Самур, Акстафа, Ленкорань; Грузия: Тбилиси, Касрис-цкали),
низовий Урала (49° с. ш.), оз. Аксуат Кустанайской обл., Целинограда,
Павлодарской обл., Семипалатинска, Саура, предгорий Дшунгарского Алатау, юга
Алма-Атинской обл., Тувы и южного Прибайкалья. Европа (кроме севера Скандинавии), Турция,
11. HYBOMITRA
365
Сев. Иран, Сев.-Зап. Китай (окр. г. Бурчум на Черном Иртыше). Весьма обычен в
лесостепной и частью степной зоне, особенно в поймах крупных рек, в Сибири проникая
по ним далеко на север; в других местах не часто. Июнь—август. Имеющиеся в
литературе указания о распространении этого вида на Дальнем Востоке ошибочны.
Рассматриваемый вид длительное время публиковался как Hybomitra (s. str.)
solstitialis Schiner,,1862, но это название является младшим омонимом другого вида —
Н. (s. str.) solstitialis Meigen, 1820 ( = #. (s. str.) bimaculata Macquart), и поэтому оно
отвергнуто. Взамен предложено название Н. (s. str.) schineri Lyneborg, 1959, но
позднее выяснилось, что для этого вида уже имеется старший синоним Н. (s. str.) ciureai
Seguy, 1937 (описан как варьетет И. (s. str.) solstitialis Schin.).
Под названием Н. (s. str.) solstitialis Schin. ранее определялись II. (s. str.) ciureai
Seg. и H. (s. str.) muehlfeldi Br. (apud Lyneborg), поэтому старые указания (до 1962 г.)
относятся к обоим видам и нуждаются в пересмотре.
Кладки яиц мы находили на нижней поверхности осок и других растений по краям
озер и болот, личинки обнаруживали в мелких болотцах и в берегах рек.
Экспериментально доказанный переносчик сибирской язвы (Олсуфьев и Лелеп,
1935) и туляремии (Олсуфьев и Голов, 1936).
Рис. 293. Ареал Hybomitra (s. str.) ciure ai.
35. Hybomitra (s. str.) ukrainica Ols. (рис. 294) — Слепень украинский.
Олсуфьев, 1952, Энтомол. обозр., 32 : 311 (Tabanus); Leclercq, 1966 :
123; Chvala, Lyneborg a. Moucha, 1972:240.
5. Глаза в относительно коротких серых волосках, с 3 полосками. Лобная полоска
узкая, ее высота превосходит ширину основания в 41/2—5 раз, кверху заметно
расширена, в желтовато-сером налете. Глазковый бугорок выступающий,
блестяще-коричневый. Нижняя лобная мозоль б. м. прямоугольная или щитовидная, черная, умеренно
блестящая, боковыми сторонами почти достигает края глаз. Средняя лобная мозоль
черная, в виде узкого киля, соединена с нижней мозолью. Лобный треугольник в
желтовато-сером налете. Усики коричнево-жвлтыз, палочка черная; 1-й членик сверху
в относительно коротких, черных волосках, площадка 3-го членика умеренной
ширины, дорсальный угол тупой или прямой. Концевой членик щупалец беловатый
с кремовым оттенком, слабо утолщзн, покрыт светлыми и единичными черными
волосками. Нотоплевры желто-коричневые. Бедра черные, в сером налете. Передние
голени в основной 1/3 , средние и задние голени целиком жзлто-коричневые. Конец
передних голеней и лапки черные, средние и задние лапки коричневые, с черным
концом. Крылья бесцветные, жилки коричневые; г4 без придатка. Жужжальца темно-
коричневые. Брюшко широкое, оранжево-жзлтое по бокам I—IV тергитов,
черно-серая полоска посредине на II—III тергитах занимает 1/8—У10 ширины колец, V—
VII тергиты черно-серые. Коричнево-жзлтая часть брюшка по бокам покрыта
короткими оранжево-желтыми волосками, черно-серые его части — черными волосками,
на I—IV тергитах по заднему краю имеются небольшие срединные треугольнички
из беловатых волосков. Снизу брюшко оранжево-желтое, V—VII стерниты черно-
серые, с желтоватой каймой, волоска снизу брюшка преимущественно желтовато-
серые. VIII стернит: гонопофиз небольшой, в черных и коричневых волосках, с едва
366 III. ПОДСЕМ. TABANINAE
заметной апикальной выемкой; латеральные лопасти небольшие, боковые
коричневые пятна умеренной величины; базальная лопасть удлиненная, с вогнутым
основным краем. Церки с сильно уплощенной вершиной. Сперматека булавовидная или
миндалевидная. 17—19 мм.
с?. Голова не больше, чем у 2- Глаза в умеренно длинных коричневато-серых
волосках. Фасетки верхних частей глаз значительно крупнее нижних, приблизительно
в 5—6 раз, граница между теми и другими довольно резкая. Лобный треугольник
в сером налете, линия соприкосновения глаз в 2 раза больше его высоты. Усики как
у 2. но 3-й членик более узкий; 1-й членик сверху в коротких черных волосках.
Концевой членик щупалец сильно утолщен, как бы вздут, светло-желтовато-кремовый,
в основании сероватый, покрыт преимущественно черными волосками. Грудь и ее
придатки как у 2- Брюшко окрашено как у 2, но оранжево-желтые пятна по бокам I—IV
тергитов преимущественно в черных волосках. 18 мм.
Рис. 294. Hybomitra (s. str.) ukrainica Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека 2-
Крупный, эффектный вид, напоминающий окраской Н. (s. str.) ciureai. От
последнего, кроме величины, 5 отличается более крупной и гладкой нижней лобной мозолью,
а в — короткими волосками сверху 1-го членика усиков.
Распространение: дельта Дуная (Измаил, Килия, Вилково);
Молдавская ССР (юг, средняя часть); Маяк близ устья Днестра и Кодыма, с.-з. Одесской
обл.; Днепровский лиман (у Ивано-Франковска). Румыния. Обычен в дельте Дуная.
Конец мая—июнь.
Г о л о т и п: 5' Вилково-Килия б. Измаильской обл., по Дунаю, 1947, И. В. Ку-
цын. В коллекции Зоологического института Академии наук СССР, Ленинград.
36. Hybomitra (s. str.) muehlfeldi Br. (рис. 295) — Слепень Мюльфельда.
В г a u e r, 1880, Denks. Nat. Akad. Wien, 42 : 149 (Tabanus muhlfeldi, (J); L у n e-
b о г g, 1959 : 119; Шевченко, 1961 : 87 (Tabanus); Олсуфьев, 19626 : 526
(Tabanus); L e с 1 e г с q, 1966:114; Виолович, 1968:146; Murdoch
et Takahasi, 1969 : 46; С h v a 1 a et J e z e k, 1969 : 331 (личинка, куколка);
Chvala, Lyneborg a. Moucha, 1972 : 228. — tropicus (L.) apud
Collin, 1940:178 (Therioplectes). — solstitialis (Schiner) apud auct. (partim).
2- Глаза в коротких, коричневато-серых волосках, с 3 полосками. Лобная полоска
очень узкая, сильно сужена к основанию, ее высота в 5—6 раз превосходит ширину
основания, обычно в сером, реже желтоватом налете. Нижняя лобная мозоль б. м.
прямоугольная, обычно тускло-черная, реже коричневая, плоская, мелкоморщинистая.
Боковыми сторонами мозоль немного не достигает края глаз. Средняя мозоль в видя
узкой килевиднои черной полоски, концом достигает нижней мозоли. Глазковыи
бугорок выпукло-овальный, блестяще-коричневый. Лобный треугольник в сером
налете. Усики: 1-й членик обычно черно-серый, значительно реже коричневый, сверху
преимущественно в черных волосках, 3-й членик красновато-коричневый, площадка
П. HYBOMITRA
367
к концу затемнена, палочка черная; членик умеренно широкий, с б. м. прямоугольным
дорсальным выступом. Концевой членик шупалец желтовато-палевый, умеренно
утолщен, покрыт преимущественно черными волосками. Нотоплевры желто-коричневые,
значительно реже черно-серые. Крылья бесцветные, жилки коричневые; г4 без
придатка. Жужжальца темно-коричневые. Ноги: бедра черно-серые, голени желто-корич-
Рис. 295. Hybomitra (s. str.) muehlfeldi Br. Лобная полоска, усик
и щупальце 2-^4 — щупальце (J.
невые, конец передних голеней и лапки черные, средние и задние лапки коричневые,
с черноватым концом. Брюшко тускло-черно-серое, с грязно-желто-коричневыми
пятнами по бокам I—III (иногда I—II) тергитов; разделяющая их темная полоса
составляет около V3—1U ширины брюшка. II—V тергиты по заднему краю со слабо
намеченными треугольными пятнами из налета и серых волосков (у потертых экземпляров
отсутствуют!). Боковые стороны II и III тергитов покрыты короткими, преимуще-
Рис. 296. Ареал Hybomitra (s. str.) muehlfeldi.
ственно черными волосками с примесью желтых. Снизу брюшко в большей части
желто-коричневое, пятно посредине II стернита и 3 (или 4) последних кольца черно-
серые. VIII стернит (рис. 288, Б): гонопофиз удлиненный, конусовидный, в тонких
желтовато-коричневых волосках, с довольно резкой апикальной выемкой;
латеральные лопасти и боковые коричневые пятна умеренно развиты; базальная лопасть
короткая, основной край довольно резко вогнутый. Длина церок обычно немного
превосходит их ширину. Сперматека миндалевидная. 11—16.5 мм.
3. Глаза в длинных коричневых волосках. Фасетки верхних частей глаз в 3—
4 раза крупнее нижних, переход между ними нерезкий. Лобный треугольник
серебристо-серый, линия соприкосновения глаз в 2 раза больше его высоты. Усики как у 2,
3-й членик более узкий, 1-й членик сверху в черных волосках, превосходящих длину
368 III. ПОДСЕМ. TABANINAE
членика. Концевой членик шупалец сильно утолщен, светло-желтовато-коричневатый,
в основании обычно сероватый, покрыт преимущественно черными волосками. Грудь
и ее придатки как у $■ Брюшко в основном окрашено как у $, срединная темная полоска
значительно сужена на III тергите, где она занимает приблизительно 1/в—1/7 ширины
кольца. 13—15 мм.
От ближайших видов, особенно от Н. (s. str.) ciureai, 92 В. (s. str.) muehljeldi
достоверно отличимы лишь по строению терминалий. $в различимы с трудом по
величине фасеток глаз и другим признакам.
Распространение: таежно-лесной вид (рис. 296). Ареал изучен
недостаточно, так как ранее смешивался с Н. (s. str.) ciureai. В СССР известен от Карелии,
запада Белоруссии (Беловежская пуща) и Могилев-Подольского, на восток до
Чукотского нац. округа и Камчатки, на север до северных частей Карелии, Онеги Архан-
Рис. 297. Hybomitra (s. str.) bimaculata Macq. Лобная полоска,
усик и щупальце <%. А — щупальце; Б — усик; В — линия
соприкосновения глаз 3-
гельской обл., Сыктывкара, Печоро-Илычского заповедника, Обской губы, Верхне-
Имбатского на Енисее, Якутска, Средне-Колымска и Маркова на р. Анадырь, на юг
до Полтавской (Миргородский р-н), Воронежской областей, низовий Дона, Ю. Урала,
севера Курганской обл., Семипалатинска, предгорий Дшунгарского и Заилийского
Алатау (на запад до Чу-Илийских гор), Алтая, Тувы и Иркутска. Обнаружен также
на Северном Кавказе (от Краснодара до Дагестана) и в Закавказье (Бакуриани). Сев.
и Средн. Европа (на запад включая Францию и Англию). Указание Бабаянца (1962)
о распространении этого вида в Туркмении ошибочно. Нуждается в подтверждении
указание (Murdoch et Takahasi, 1969) о распространении этого вида на Дальнем
Востоке, включая Японию. Обычен в полосе смешанных лесов, но в южных частях ареала
и в Вост. Сибири не часто. Июнь—июль. Личинки найдены в переходном болоте (Лутта,
1970).
37. Hybomitra (s. str.) bimaculata Macq. (рис. 297) — Слепень полуденный.
Macquart, 1826, Recueil Soc. Agric. Lille : 163 (Tabanus); Lyneborg,
1959 : 101 (partim); L e с 1 e г с q, 1966 : 122; С к у ф ь и н, 1967 : 196 (личинка);
Chvala et Jezek, 1969 : 335 (личинка, куколка); Иванищук, 1970 : 148
(личинка); Chvala, Lyneborg a. Moucha, 1972 : 231. — tropicus
Panzer, 1794 : 22 (Tabanus), цит. по: L e с 1 ere q, 1966:121; В г а и е r, 1880:146
(Tabanus) et auct. — solstitialis M e i g e 11, 1820, 1. c. : 561 (Tabanus) nomen obli-
tum, nee T. solstitialis Schiner, 1862 (=Hybomitra ciureai Seguy, 1937). — bisignatus
Jaennicke, 1866, 1. с :74 (Tabanus). — collini Lyneborg, 1959 : 94
(Hybomitra).
9. Глаза в коротких коричневых волосках, с 3 полосками. Лобная полоска узкая,
обычно сужена к основанию, ее высота в 4х/2—5 раз превосходит основание, в
желтовато-сером или пепельном налете. Нижняя лобная мозоль вытянуто-прямоугольная,
11. HYBOMITRA
369
плоская, мелкоморщинистая, тускло-черная. Боковые стороны мозоли не достигают
края глаз, отделяясь от последних узкими серыми полосками. Средняя мозоль в виде
узкой, слабо блестящей, черной, килевидной полоски: последняя соединена с нижней
мозолью. Глазковый бугорок овальный, выпуклый, блестяще-коричневый. Лобный
треугольник плоский, высокий,- в голубовато-сером налете. Усики коричневые или
желто-красные, палочка, верхний край 3-го и первые 2 членика черные. 1-й членик
в сером налете, сверху в б. м. длинных черных волосках. 3-й членик в основании сильно
расширен, с прямоугольным дорсальным выступом. Палочка короткая, толстая.
У экземпляров с Дальнего Востока усики обычно целиком красновато-оранжевые.
Концевой членик щупалец светлый, обычно в сероватом налете. Он в основании сильно
утолщен, остроконечный, прикрыт короткими, преимущественно черными волосками.
Нотоплевры, как правило, черные, лишь очень редко коричневатые. Жужжальца
темно-коричневые. Ноги: кончик бедер, основание передних и большая часть средних
и задних голеней коричневые, прочие части ног черные. Крылья прозрачные, почти
не затемненные к переднему краю, жилки черно-коричневые. Брюшко: общий тон
окраски тусклый, свинцово-черный; по бокам брюшка красновато-коричневые пятна,
простирающиеся на I—III тергиты; их разделяет широкая черная полоска,
занимающая около 1/3 ширины брюшка. Тергиты по заднему краю с небольшими срединными
треугольниками из серебристо-серых волосков (у потертых экземпляров треугольники
неясные). Каемки тергитов в такого же цвета волосках. II тергит по бокам
преимущественно в черных волосках. Снизу брюшко в основной половине красноватое; I стер-
нит, треугольное пятно на II и 3—4 концевых стернита темные. Окраска брюшка
сильно варьирует за счет вытеснения коричневой окраски, вплоть до полного ее
исчезновения. В последнем случае брюшко целиком одноцветно-черное, с серым рисунком
(Н. (s. str.) bimaculata var. bisignata Jaenn.). VIII стернит (рис. 288, А): гонопофиз и
латеральные лопасти темно-коричневые, первый лишь немного светлее последних и
вместе они образуют ровный внешний край в виде полукруга; гонопофиз широкий и
короткий, в тонких коричневых волосках; базальная лопасть оо слегка вогнутым
основным краем. Церки очень короткие, едва длиннее X тергита, вершина их сильно
уплощена. Сперматека миндалевидная. 14—17 мм.
<J. Глаза в длинных, коричневых волосках. Фасетки глаз б. м. одинаковой
величины, лишь возле лобного шва несколько крупнее. Глазковый бугорок красновато-
коричневый, блестящий- Лобный треугольник серебристо-серый, линий
соприкосновения глаз в 2 раза больше его высоты. Усики как у 9. 3-й членик более узкий. 1-й чло-
ник сверху в длинных черных волосках, по длине равных 1-му и 2-му членикам вместе
взятым. Концевой членик щупалец желто-белый, в основной половине серый,, сильно
тыквовидно утолщен, покрыт длинными, преимущественно темными волосками.
Нотоплевры черные. Жужжальца коричневые. Крылья и окраска ног как у 9; средние голени
в густых, длинных, тонких темных волосках. Общий тон окраски брюшка свинцово-чор-
ный, по бокам с красновато-коричневыми пятнами, простирающимися на I—III тор-
гиты (реже незначительно заходят на IV тергит). Разделяющая их срединная чернил
полоска составляет около 1/л—х/5 ширины брюшка, причем на III тергите полоска
несколько сужена. II тергит посредине с неявственным серым треугольным пятном.
Брюшко сверху в черных волосках, лишь задние каемки тергитов по бокам и в сродной
части в серебристо-серых волосках. Снизу брюшко красновато-желтое; I стернит,
небольшое треугольное пятно на II стерните и 3 последних кольца черные. 13.5—16 мм.
После опубликования монографии Брауэра (Вгаиег, 1880) длительное время л
основной зоологической литературе рассматриваемый вид публиковался под названном
Tabanus tropicus Panzer, 1794. Однако оно является младшим омонимом Т. tropicus
Linne, 1758, и от него отказались, хотя последнее также отвергнуто как потоп obli-
tum. Для Н. (s. str.) bimaculata. Macq., 1826, имеется старший синоним Н. (s. str.)
solstitialis Mg., 1820, но так как это название является nomen oblitum и оно длительно
применялось для другого вида — Tabanus solstitialis Schiner, 1862 (ныне Н. (s. str.) ciunml
Seguy, 1937), от него также следует отказаться во избежание недоразумений при
использовании старой литературы.
Распространение: таежно-лесной вид (рис. 298). В СССР изноотон
от Кольского п-ова, Прибалтийских республик и запада Украины (Закарпатская обл.)
на восток до Камчатки и Сахалина, на север до Хибин Мурманской обл.,
Архангельска, Усть-Цильмы Коми АССР, Низовий Оби (Лабытнанги),
Енисея (Снежногорск на Хантайке, притоке Енисея), Центральной Якутии и Ворхиого
Сеймчана на Колыме, на юг до Запорожья, Воронежской обл., Пензы, 10. Урала,
Кокчетавской обл. (Боровое), Павлодарской обл. (Баянаульский р-н), хр. Саурм,
Дшунгарского Алатау (Саркандский р-н), Тувы, южного Прибайкалья, Приаму рьк
и Владивостока. Оторванно от основного ареала локальные находки.па Капкано:
Кабардино-Балкарская и Северо-Осетинская АССР, Азербайджан (Ленкорань; Л pout),
Грузия (Бакуриани; Аспиндза). Большая часть Европы, Вост. Монголия', Con.-Пост,
Китай, Япония (острова Хоккайдо, Хонсю). Весьма обычен в средних и.дожнмх частнх
тайги и в лесной зоне. Июнь—июль, редко раньше или позднее.
24 Н. Г. Олсуфьев
370 _ III. ПОДСЕМ. TABANINAE
Экземпляры из Приморского края, имеющиеся в коллекции Зоологического
института АН СССР, относятся к Н. (s. str.) bimaculata var. bisignata, которая, очевидно,
здесь преобладает. В западных частях своего ареала эта форма встречается обычно
в единичных экземплярах как примесь к основной форме. Откладка яиц на нижнюю
поверхность листьев осок, Scirpus, Glyceria и других прибрежных растений. Личинки
преимущественно в мелких заболоченных водоемах и болотах разных типов, включая
сфагновые (Олсуфьев, 19356; Скуфьин, 19666; Лутта, 1970); куколок находили в
низинном болоте (Лутта, 1970).
0° к востоку от Гринвича 90° izo° «о° 180° 150°
Рис. 298. Ареал Hybomitra (s. str.) bimaculata.
38. Hybomitra (s. str.) caucasica Enderl. (рис. 299) — Слепень альпийский.
Enderlein, 1925, Mitt. Zool. Mus. Berlin, 11, 2 : 358 (2) (Therioplectes). —
tetricus S z i 1 a d y, 1914 : 661 (Tabanus) пес T. tetricus Marten, 1883 : 111, пит. по:
Stone, 1938 : 159. — bimaculata Enderlein, 1925 : 358 (3) (Therioplectes) nee
Tabanus bimaculatus Macquart, 1826, 1. с : 163. — olsoufievi Philip, 1956 :
230. — olsufievina Philip, 1961 : 479, цит. по: Chvala, Lyneborg a.
M о u с h a, 1972 : 189.
5. Глаза в умеренно длинных, серых волосках. Лобная полоска умеренной ширины,
ее высота превосходит основание в 3V4—4 раза, слегка сужается к основанию. Нижняя
лобная мозоль треугольная или прямоугольная, сверху часто округлая, блестяще-
черная, в мелких морщинках, обычно достигает края глаз. Средняя мозоль узко-
веретеновидная, нижним концом б. м. достигает нижней мозоли. Глазковый бугорок
овальный, блестяще-коричневый. Лобный треугольник серый. Верхний край усиковых
ямок узкий, не более 1/i высоты лобного треугольника. Усики черные, иногда 1—2-й
членики и основание 3-го коричневатые. 1-й членик сверху в длинных черных волосках;
3-й членик узкий, с тупым дорсальным углом. Последний иногда закруглен. Лицо
в длинных серых волосках. Щупальца бледно-коричневато-желтые; концевой членик
утолщен, на конце приострен, покрыт короткими черными и светлыми волосками.
Грудь черная, бочки в черных волосках. Крылья бесцветные, жилки коричневые;
основание крыла, костальная и субкостальная ячейки желтые. Ноги и брюшко
одноцветно-черные, в черных волосках. Иногда средние голени в основании слегка
коричневатые. На брюшке сверху в базальной части легкий „синеватый налет, хорошо
заметный при рассматривании брюшка сзади. Снизу брюшко черное, слабо блестящее,
в легком сизом налете и в черных волосках, на концевых стернитах небольшая примесь
серых волосков. Изредка бочки груди с значительной примесью серых волосков, тер-
гиты по заднему краю с каемкой из серых или желтовато-серых волосков, слегка
расширенной в срединные треугольники. Соответственно стерниты по заднему краю
с узкой каемкой серых волосков. VIII стернит: гонопофиз крупный, в черных волосках,
с едва заметной апикальной выемкой; латеральные лопасти небольшие, боковые
коричневые пятна умеренной величины; базальная лопасть с вогнутыми боковыми сторонами
и основным краем. Церки небольшие, со слегка уплощенной вершиной. Сперматека
миндалевидная. 15—17 мм.
11. HYBOMITRA
371
3 Фасетки глаз б. м. одинаковой величины, глаза в длинных, серых волосках.
Лобный треугольник серый, линия соприкосновения глаз приблизительно в 1V2 раза
длиннее его высоты. Усики как у 5, 3-й членик более узкий; 1-й членик в длинных
волосках по длине превосходящих 1-й и 2-й членики вместе взятые. Концевой членик
щупалец б. м. яйцевидно утолщен, коричневато-желтый или желто-серый, в длинных
черных волосках. Лицо темно-серое, в черных и серых волосках. Грудь, ноги и крылья
как у 2 Брюшко блестяще-черное, в черных волосках, с неясными темно-коричневыми
пятнами по бокам II—III тергитов; снизу брюшко одноцветно-черное. 14.5—15 мм.
Распространение: высокогорные части Б. и М. Кавказа, где обычен.
Горы Средн. Европы и Сев. Ирана. Июль—август. Указание Леклерка (Leclercq,
1966) на распространение этого вида в Афганистане и Сибири ошибочно. Н. (s. str.)
£^^)
Рис. 299. Hybomitra (s. str.) caucasica Enderl. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека ?.
caucasica наиболее близка к Н. (s. str.) hunnorum, связываясь через нее с группой
nigricorpa—semipollinosa—shnitnikovi—nigricornis. Все они характеризуются темной
окраской тела, светлыми и б. м. утолщенными щупальцами и относительно узкими,
преимущественно черными усиками с довольно длинными волосками на 1 -м членике.
Форма лба, строение мозолей у 2, а также глаз у 3 весьма близки. Благодаря темной
окраске тела, его размерам и форме, Н. (s. str.) caucasica имеет значительное внешноо
сходство с Н. (s. str.) aterrima настолько, что западные энтомологи до нашего
указания (Олсуфьев, 1937а) эти виды не различали. Н. (s. str.) caucasica отличается от
Н. (s. str.) aterrima более светлыми и утолщенными щупальцами и отсутствием
коричневатого пятнышка вокруг основания жилки г4; у 2 Н. (s.str.) caucasica лобная
полоска менее широкая, верхний край усиковых ямок узкий, не шире V4 высоты
лобного треугольника. У в Н. (s. str.) caucasica линия соприкосновения глаз в 11/2 ра:ш
превосходит высоту лобного треугольника, у Н. (s. str.) aterrima — в 2 раза.
39. Hybomitra (s. str.) hunnorum Szil. (рис. 300) — Слепень тяныпанский.
S z i 1 a d у, 1923, Biol. Hungar., 1, 1 : 8 (Tabanus); Олсуфьев, 1937а : 175
(Tabanus); Moucha et Chvala, 1959b : 278 (Tylostypia); Шевченко, 1901 :
90 (Tabanus); Leclercq, 1966:108 (Hybomitra). — pamirensis Enderlein,
1933 : 145 (Aplococera).
9. Глаза в коротких, серых волосках, с 3 узкими, неясными, медными полосками
(нижняя и верхняя иногда отсутствуют). Лобная полоска умеренной ширины, оо иы<
сота превосходит основание в 3%—З1^ раза, слабо расширяющаяся кверху. Глазкопый
бугорок овальный, коричневый, обычно уплощен. Нижняя лобная мозоль небольшая,
овальная, черная, редко коричневатая, морщинистая, не достигает края гЛаз. Ипогдп
мозоль почти целиком прикрыта налетом или же наоборот часть налета вокруг моаоли
стерта и тогда последняя имеет б. м. пятиугольную форму. Средняя мозоль малопькин,
овальная или линейная, с неясными границами, разобщена с нижней. Лобный
треугольник низкий, в сером налете. Концевой членик щупалец желтовато-серый, уморошюи
толщины к концу приострен, покрыт короткими, темными и светлыми полоскаМИ|
24*
372 III. П'ОДСЁМ. TABANINAE
Предыдущий членик в длинных светлых волосках. Усики целиком черные. 1-й членик
сверху в длинных, темных волосках. 3-й членик узкий, с тупым дорсальным углом;
последний часто закруглен. Лицо серое, в длинных серых волосках. Грудь черная;
бочки в б. м. длинных желтовато-серых волосках, с небольшой примесью темных,
иногда только в черных волосках. Нотоплевры черные. Крылья бесцветные, жилки
Рис. 300. Hybomitra (s. str.) hunnorum Szil. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $•
темно-коричневые. Жужжальца коричнево-черные. Ноги целиком черные; иногда
основная часть средних голеней, реже задних и передних, б. м. коричневая. Брюшко
сверху блестяще-чёрное, покрыто черными и желтовато-серыми волосками.
Последние образуют на брюшке нерезкие пятна — треугольные посредине и ромбические или
Рис. 301. Ареал Hybomitra (s. str.) hunnorum.
неправильно треугольные по бокам, а также каемки' по заднему краю тергитов. Снизу
брюшко блестяще-черное, в черных и серых волосках.! Иногда брюшко с обеих сторон
целиком в черных волосках. VIII стернит: гонопофиз! довольно крупный, в черных
волосках,, со слабой апикальной выемкой; латеральные лопасти и боковые
коричневые пятни большие; ба'зальная лойасть короткая, - со слабо1 вогнутым Основным краем.
11. HYBOMITRA 373
Церки крупные, с округлой или слабо уплощенной вершиной. Сперматека
миндалевидная. 15—16.5 мм.
cJ. Глаза в длинных серых волосках; фасетки верхней части глаз значительно
крупнее нижних, граница между теми и другими хорошо выражена. Глазок овальный,
коричневый. Лобный треугольник в сером налете, линия соприкосновения глаз в 2 раза
длиннее высоты треугольника. Усики целиком черные. 1-й членик к концу сильно
расширен, сверху клювовидно вытянут вперед (смотреть сбоку) и покрыт длинными,
черными волосками, по длине превосходящими 1-й и 2-й членики вместе взятые. 3-й
членик узкий, с тупым дорсальным углом. Концевой членик шупалец овальный,
коричнево-желтый, покрыт б. м. длинными светлыми и темными волосками. Лицо серое,
в серых волосках, на щеках со значительной примесью темных. Грудь черная; бочки
груди в черных волосках, с незначительной примесью серых, спинка и щиток в черных
волосках. Нотоплевры черные. Крылья и жужжальца как у $• Ноги
одноцветно-черные, реже средние и задние голени коричневые. Брюшко черное, слабо блестящее,
с каштаново-коричневыми пятнами по бокам II и частью III тергитов. Брюшко покрыто
черными волосками, с незначительной примесью желтоватых или серых по заднему
краю тергитов. Снизу брюшко одноцветно-черное. 15.5—16 мм.
Распространение: среднеазиатский горный вид (рис. 301). Обитает
на высоте 1200—3500 м и выше (до 4200 м), преимущественно в альпийском и
субальпийском поясах, реже в лесах, на Памиро-Алае, Тянь-Шане (включая Джунгарский
Алатау) и Каратау. Афганистан (Гиндукуш), Зап. Китай (хр. Кучур-Таг и Кашгария).
Местами массовый вид. Июнь—август.
Рис. 302. Hybomitra (s. str.) nigricorpa Krob.
Лобная полоска, усик и щупальце §•
40. Hybomitra (s. str.) nigricorpa Krob. (рис. 302) — Слепень чернотелый.
К г б b е г, 1923, Arch. f. Naturg., 89, А, 12 : 72 (Tabanus); Олсуфьев, 1937а :
176 (Tabanus); Шевченко, 1961 : 92 (Tabanus); Leclercq, 1966 : 115. — hunnorum
(Szil.) apud К r 6 b e r in Lindner, 1925:65.
5. Глаза с З полосками, покрыты умеренно длинными серыми волосками. Лобная
полоска б. м. широкая, ее высота превосходит ширину в 3—З1^ раза. Глазковый
бугорок продолговатый, слабо выпуклый, коричневый. Нижняя лобная мозоль плоская,
б. м. пятиугольная; ее средняя часть темно-коричневая, мелкоморщинистая, тогда как
боковые стороны блестяще-черные. Средняя мозоль представлена короткой веретено-
видной черной полоской, обособленной или достигающей концом нижней мозоли.
Лобный треугольник в сером налете. Усики целиком черные; 1-й членик сильно
бокаловидно расширен, сверху в длинных черных и серых волосках, 3-й членик узкий, с
тупым дорсальным углом; вершина угла слегка закруглена. Лицо в сером налете и тонких,
но довольно густых и длинных серых волосках. Щупальца желтовато-белые, концевой
членик в основании утолщен, в вершинной части густо покрыт короткими черными
волосками, к основанию членика, сменяющимися более тонкими и длинными белова-
374 III. ПОДСЕМ. TABANINAE
тыми волосками. Бочки груди покрыты преимущественно серыми волосками. Ното-
плевры черные. Крылья бесцветные, жилки коричневые; г4 без придатка. Жужжальца
коричнево-черные. Ноги черные; основная часть голеней, в особенности средней пары
ног, коричневая и покрыта короткими серыми волосками, делающими ее светлее,
тогда как прочие части ног в черных волосках. Брюшко широкое, блестяще-черное,
II тергит в переднебоковой части с небольшим, темно-коричневым пятном. I и задние
края V—VII тергитов в густых, но коротких, прилегающих, беловато-серых волосках;
прочие тергиты в коротких черных волосках. Светлые каемки по заднему краю тергитов
отсутствуют нацело. Снизу брюшко блестяще-черное, в черных волосках; 3 последних
стернита по заднему краю в беловато-серых волосках. 15—19 мм.
в (по Krober). Похож на §. Глаза большие, различие фасеток минимальное,
граница не резкая, крупные фасетки занимают приблизительно 3/4 глаз. Волосяной покров
очень густой, короткий, беловатый. На глазах имеются 2 полоски. Лобный
треугольник менее выпуклый, нежели у 2> в беловатом налете. Затылок в коротких, беловатых
волосках, не превосходящих длину волосяного покрова глаз. Усики черные; 3-й
членик уже, нежели у $.' 1-й и 2-й членики в длинных, стоячих, серых волосках. Лицо
черное, благодаря налету беловато-серое. Волоски длинные, беловатые, на щеках
с многочисленными черными волосками. Щупальца желто-коричневые, снизу черные,
в длинных отстоящих черных и белых волосках. Концевой членик удлиненно-овальный,"
довольно толстый. Грудь и щиток блестяще-сине-черные, лишь задний край спинки
между основанием крыльев и щитком с коричневым оттенком. Полоски на спинке
отсутствуют. Волосяной покров нежно-серый, бочки груди в многочисленных черных
волосках. Брюшко одноцветно-черное, с каштаново-коричневыми пятнами по бокам I
и II колец, покрыто шелковистыми черными волосками. Лишь I кольцо с маленьким
треугольным пятном из густых беловатых волосков, IV со следами, V с треугольником,
который продолжен к переднему краю, и с каемкой по заднему краю из беловатых
волосков; VI и VII кольца в желтовато-белых волосках. Светлые заднекрайние каемки
отсутствуют. Жужжальца темно-коричневые, с желтоватой головкой. Закрыловые
пластинки коричнево-прозрачные, с нежными желтыми и коричневыми жилками.
Птеростигма и придаток у г4 отсутствуют. Ноги темно-коричневые, в черных волосках,
лишь голени светлогкоричневые и в коротких беловатых волосках. Передние голени
почти целиком, последующие б. м. сильно затемнены. 15 мм.
Распространение: среднеазиатский горный вид с относительно
локальным ареалом. Обнаружен на высоте 1800—3000 м в западной части Таласского Алатау
и на Угамском хребте, а на юге — на северных склонах Алайского и Туркестанского
хребтов. Обитает преимущественно в субальпийском и альпийском поясах. Не часто.
Июль—часть августа.
Описан Крёбером (КгоЬег, 1923) из окр. Гульчи (Алайский хр.) и позднее (1925)
им же самим сведен в синонимы к Н. (s. str.) hunnorum со следующим примечанием:
«Моя 2 nigricorpus, по разъяснению Szilady, идентична его hunnorum; 3 относится
к новому виду».
41. Hybomitra (s. str.) sogdiana Ols. (рис. 303) — Слепень согдианский.
Олсуфьев, 1952, Энтомол. обозр., 32 : 313 (Tabanus); Шевченко, 1961 :
93 (Tabanus); L е с 1 е г с q, 1966 : 119.
5- Глаза в умеренно длинных серых волосках, с 3 полосками. Лоб умеренной
ширины, заметно расширяется кверху, его высота превосходит ширину основания в 3—
З1/^ раза, в сером налете. Нижняя лобная мозоль обычно в виде небольшой треугольной
или щитовидной морщинистой мозольки темно-коричневого или черного цвета, далеко
не достигает сторонами края глаз. Средняя мозоль либо отсутствует, либо представлена
небольшой черной веретеновидной полоской, часто не соединенной с нижней мозолью.
Глазковый бугорок продолговато-овальный, блестяще-коричневый. Лобный
треугольник в сером налете. Усики черные, 1-й членик в сером налете и в относительно длинных,
преимущественно серых волосках, 3-й членик узкий, его дорсальный выступ
тупоугольный, с закругленной вершиной. Лицо в пепельно-сером налете и в довольно густых и
длинных белых волосках. Концевой членик щупалец умеренно утолщен,,
беловато-кремовый, покрыт белыми и немногочисленными черными волосками. Нотоплевры
черные. Крылья бесцветные, жилки желтые или желто-коричневые, более темные к задне-
внешнему краю крыла; г4 изредка с небольшим придатком. Жужжальца коричнево-
черные. Бедра черно-серые, передние голени в основании, средние и задние голени
на большем протяжении желто-коричневые, конец их черноватый. Лапки черные.
Брюшко широкое, блестяще-черное, II тергит обычно с неясно просвечивающими
коричневыми пятнами по бокам. I и II тергиты в легком сером налете и в коротких густых
беловато-серых волосках. Последующие тергиты черные, в черных волосках, задний
край V—VII тергитов в густых, прилегающих, серебристо-белых волосках. Иногда
белые волоски имеются по заднему краю III—IV тергитов, образуя » средней части
небольшие треугольники. Снизу брюшко блестяще-черное, II стернит по бокам и 3
noil. HYBOMITRA . 375
следних стернита (иногда также предыдущие) по заднему краю в беловатых волосках,
остальное в черных волосках. 15.5—20 мм.
$. Глаза в длинных, коричневато-серых волосках. Фасетки верхних 2/3 глаз
крупнее нижних приблизительно в 4-5 раз. Лобный треугольник в сером налете. Усики как
у 2, но 3-й членик более узкий. 1-й членик сверху в б. м. длинных черных волосках.
Концевой членик щупалец светло-коричневато-палевый, овальный, преимущественно
в черных волосках. Грудь и ее придатки как у §• Брюшко окрашено в основном как у 5,
но на I—II тергитах белые волоски располагаются более крупным пятном по бокам и
маленьким пятном по их заднему краю. 17 мм.
Крупный, эффектный вид, хорошо отличающийся от всех черноокрашенных горных
среднеазиатских видов довольно четкой двуцветной окраской верхней стороны брюшка:
серовато-белой матовой в базальной части и блестяще-черной — в апикальной.
Распространение: среднеазиатский горный вид, с локальным ареалом
(рис. 305, 2). Обитает на высоте 1500—3500 м, преимущественно в альпийском поясе,
в западной части Таласского Алатау, включая хребты Угамский и Каржантау. Указан
для гор Таджикистана без более точной локализации. Июнь—август. Не часто.
Голотип: 2, Каржантау, Мын-Булак, 10 VII 1939, Обухова. В коллекции
Зоологического института Академии наук
СССР, Ленинград.
42. Hybomitra (s. str.) semipollinosa
Ols. (рис. 304) — Слепень гладколобый.
Олсуфьев, 1937а, Фауна СССР,
ч Двукрылые, VII, 2 : 178 (Tabanus);
Moucha et Chvala, 1959b : 278
(T ylostypia); Шевченко, 1961 : 95
(Tabanus); L e с 1 e г с q, 1966:119.
5- Глаза в умеренно длинных серых
волосках, с 3 полосками. Лобная полоска
умеренно широкая, ее высота в 3—3V2
раза превосходит ширину, кверху слегка
Рис. 303. Hybomitra (s. str.) sogdiana Ols.
Лобная полоска, усик и щупальце $•
расширяется, в сером налете. Глазковый
плоский, коричневый, за ним имеется небольшое, блестяще-черное пространство.
Нижняя лобная мозоль б. м. пятиугольная или округлая, черная, слабо блестящая,
тонкоморщинистая. Средняя мозоль в виде короткой, черной полоски, обычно не
соединенной с нижней мозолью, иногда целиком прикрыта налетом. Лобный треугольник
в сером налете. Щупальца серые или желтоватые, концевой членик в основании
утолщен, густо покрыт короткими, преимущественно черными волосками. Усики целиком
черные. 1-й членик бокаловидно расширен, сверху в б. м. длинных черных и светлых
волосках. 3-й членик узкий, с тупым дорсальным углом. Лицо серое, в такого же
цвета волосках. Бочки груди в серых волосках, с незначительной примесью темных.
Нотоплевры черные. Крылья бесцветные, жилки темно-коричневые; г4 без придатка.
Жужжальца темно-коричневые. Ноги черные, часто значительная часть средних и
задних и основание передних голеней темно-коричневые. Брюшко черное, слабо
блестящее. I и II тергиты в тонком голубовато-сером налете (припудрены). Волосяной
покров брюшка короткий, прилегающий. I тергит, боковые стороны II тергита и затем
боковой край всех последующих тергитов в беловатых волосках. Кроме того,
незначительное количество белых волосков имеется по заднему краю II—V тергитов, образуя
в средней части небольшие треугольнички (заметны лишь у непотертых экземпляров).
Прочие части брюшка в коротких черных волосках. Снизу брюшко черное, по бокам
в легком голубовато-сером налете и коротких прилегающих беловато-серых волосках,
посредине с широкой полосой черных волосков. VIII стернит: гонопофиз небольшой,'
преимущественно в черных волосках, со слабой апикальной выемкой; латеральные
лопасти и боковые коричневые пятна небольшие; базальная лопасть удлиненная,
затемнена, со слабо вогнутым основным краем. Церки короткие, с уплощенной
вершиной. Сперматека лимоновидная или миндалевидная. 15—20 мм.
S- Голова дашпь немного больше, чем у 2, умеренно выпуклая. Глаза в длинных
серых волосках. Фасетки верхних частей глаз в 3—4 раза крупнее нижних, переход
горок небольшой, продолговато-овальный,
376
III. ПОДСЕМ. TABANINAE
не резкий. Лобный треугольник в серебристо-сером налете, линия соприкосновения
глаз приблизительно в 13/4 раза превосходит высоту треугольника. Усики как у 9,
но 3-й членик более узкий. 1-й членик сверху в длинных черных и серых волосках, их
длина превосходит длину членика. Концевой членик щупалец овальный, серовато-
Рис. 304. Hybomitra (s. str.) semipollinosa Ols. Лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека $.
палевый, в черных и серых волосках. Грудь и ее придатки как у ?. Ноги целиком
черные, реже средние голени в базальной части черно-коричневые. Брюшко сверху
черное, слабо блестящее, II или II—III тергиты по бокам с темно-коричневыми
пятнами, разделяющая их темная срединная полоса занимает приблизительно 1/3 ширины
полуколец. II тергит по бокам в зоне
коричневой окраски в тонком
голубовато-сером налете и здесь же иногда
небольшие группы беловатых волосков.
На II—V тергитах белые волоски
образуют небольшие заднекрайние
треугольники и в небольшом количестве
располагаются по заднебоковому краю терги-
тов, остальное в черных волосках. Снизу
брюшко черное, умеренно блестящее,
в тонком голубовато-сером налете,
покрыто черными волосками и по
заднебоковому краю стернитов — серыми
волосками. 16—19 мм.
Распространение:
среднеазиатский горный вид (рис. 305,1). Оби-
Рис. 305. Ареалы: 1 — Hybomitra
(s. str.) semipollinosa; 2 —• II. (s. str.)
sogdiana.
тает на высоте 800—3000 м, преимущественно в лесном, реже в субальпийском и
альпийском поясах, на Зап. Памиро-Алае (хребты Гиссарский, Зеравшанский,
Туркестанский), Зап. и'Сев. Тянь-Шане (хребты Ферганский, Таласский, Пскемский, Угамский,
Чаткальский, Каржантау, Заилийский, Джунгарский), Тарбагатае<и Сауре.
Афганистан (Бадахшан). Местами обычен. С начала июня до середины августа.
Голотип: 5, Алма-Атинское ущелье, Заилийский Алатау, 24 VII1934, Н. Олсуфьев.
В коллекции Зоологического института Академии наук СССР, Ленинград.
П. HYBOMITRA
377
43. Hybomitra (s. str.) shnitnikovi Ols. (рис. 306) — Слепень Шнитникова.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII. 2 : 180 (Tabanus); М о и с h a
et Chvala, 1959b; 279 (Tylostypia); Шевченко, 1961:97 (Tabanus); L e
frier с q, 1966 : 119. — bouvieH Philip, 1958 : 333, цит. по: L e с 1 e г с q, 1966 :
119.
§. Глаза в умеренно длинных серых волосках, зеленые, с 1—3 узкими полосками.
Лоб умеренной ширины, его высота превосходит ширину в 31/2—4 раза, слегка
расширяющийся кверху. Глазковый бугорок в виде небольшого, темно-коричневого
треугольника или овальной мозольки, слабо блестящий. Нижняя лобная мозоль
треугольная или округлая, блестяще-черная, тонкоморщинистая. Средняя мозоль небольшая,
Рис. 306. Hybomitra (s. str.) shnitnikovi Ols. Лобная полоска, усик и
щупальце $. А ~ щупальце; В — усик $.
широковеретеновидная, чаще соприкасается с нижней. Лобный треугольник б. м.
низкий, в сером налете. Усики целиком черные. 1-й членик в сером налете, бокаловидно
расширен, сверху в б. м. длинных, светлых и черных волосках; 3-й членик узкий,
с б. м. тупым дорсальным углом. Шупальца слегка утолщены, на конце приострены,
желтовато-белые, покрыты короткими, преимущественно черными волосками. Ното-
плевры черные или коричневые. Крылья бесцветные, жилки темно-коричневые; г4
иногда с небольшим придатком. Жужжальца темно-коричневые. Ноги черно-серые;
основная часть средних и частью передних голеней коричневая, реже голени целиком
черные. Брюшко черное, сверху с 3 рядами пятен из серого налета и светлых волосков.
Срединный ряд состоит из узких, треугольных пятен, боковые — из ромбических или
неправильно треугольных. Снизу брюшко одноцветно-черно-серое, в густом сером
налете. 15—20 мм.
с?. Глаза в длинных, серых волосках; фасетки верхних 2/3 глаз значительно
крупнее нижних. Глазок маленький, коричневый. Лобный треугольник серый. Усики
черные. 1-й членик резко бокаловидно расширен, сверху клювовидно вытянут вперед
(смотреть сбоку), покрыт умеренно длинными, преимущественно светлыми волосками.
Их длина не превосходит 1-й членик. 3-й членик как у $, но более узкий. Щупальца
желтовато-белые или сероватые, концевой членик яйцевидно утолщен, со слегка
оттянутым кончиком, покрыт длинными светлыми и черными волосками. Ноги, крылья и
жужжальца как у $• Брюшко матово-черное, с 3 рядами неясных сероватых пятен
(налет+волоски): треугольных посредине и косых по бокам. II—III тергиты по
бокам буроватые. Снизу брюшко черное, в сером налете и серых волосках. 16—19 мм.
Распространение: горный среднеазиатский вид. Обитает на высоте
900—3000 м, преимущественно в лесном и реже в альпийском поясе, на Гиссаро-Алае
(хребты Зеравшанский, Гиссарский, Каратегинский), Тянь-Шане (включая
Джунгарский Алатау), Каратау и Сауре. Афганистан. Обычен. С конца мая по начало
августа. Личинки найдены в реках, родниках, арыках и болотах (Кадырова, 19726).
378
III. ПОДСЕМ. TABANINAE
Голотип: 5, Алма-Атинское ущелье близ Алма-Аты, 1500 м, 24 VII 1934, Н.
Олсуфьев. В коллекции Зоологического института Академии наук СССР, Ленинград.
44. Hybomitra (s. str.) turkestana Szil. (рис. 307) — Слепень туркестанский.
S z i 1 a d у, 1923, Biol. Hungar., 1 : 8 (Tabanus); Krober in Lindner,
1925 : 72 (Tabanus); Олсуфьев, 1937а : 182 (Tabanus); Шевченко, 1961 : 99
(Tabanus); Leclercq, 1966 : 123. — immaculiventris Krober, 1922 : 103
(T. montanus var. immaculiventris), цит. по: Leclercq, 1966 : 123. — ater S z i-
lady, 1923 : 9 (T. turkestanus var. ater). — minor S z i 1 a d y, 1923 : 9 (T. turkesta-
nus var. minor).
<j>. Глаза в умеренно длинных, светлых волосках, изумрудно-зеленые, с 1—3
узкими, медными или золотистыми полосками. Лоб узкий, его высота превосходит
основание приблизительно в 4 раза, слегка расширяется кверху. Глазковый бугорок в виде
Рис. 307. Hybomitra (s. str.) turkestana Szil. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека $. А — усик 3.
темного треугольничка или продолговатой плоской мозольки; иногда бугорок почти
целиком покрыт налетом. Нижняя лобная мозоль треугольная, тонкоморщинистая,
матово-черная, обычно с коричневой серединой. Средняя мозоль в виде короткой,
тонкой линии, обычно соединенной с вершиной нижней мозоли, или же отсутствует
совершенно. Лобный треугольник в желтоватом налете. Усики: 1-й членик бокаловидно
расширен, темно-серый, в б. м. длинных черных и светлых волосках; 2-й членик в
основании красноватый; 3-й членик черный, узкий, с тупым дорсальным углом; основание
членика иногда коричневое. Щупальца желтовато-белые, концевой членик умеренной
толщины, покрыт светлыми и в концевой части черными волосками. Нотоплевры
темные, реже коричневатые. Крылья бесцветные, жилки коричневые; г4 часто с коротким
придатком. Жужжальца темно-коричневые. Ноги: бедра черно-серые, средние и
задние голени и основная часть передних голеней коричневато-желтые. Кончик средних
и задних голеней черноватый. Конец передних голеней и лапки всех ног черные.
Брюшко широкое, темное, с коричнево-желтыми пятнами по бокам I—IV тергитов,
покрыто довольно густыми черными и желтовато-серыми волосками. Светлые волоски
преобладают по заднему краю тергитов и образуют продольный ряд треугольных
пятен посредине брюшка. Снизу брюшко коричневато-желтое. II стернит без темного
пятна посредине. V и последующие стерниты темные. VIII стернит: гонопофиз
крупный, преимущественно в черных волосках, с выраженной апикальной выемкой;
латеральные лопасти и боковые коричневые пятна небольшие; базальная лопасть со слегка
11. HYBOMITRA
379
вогнутым основным краем. Церки крупные, со слегка уплощенной вершиной.
Сперматека миндалевидная. 15—20 мм.
3. Глаза в длинных, светлых волосках. Фасетки верхней части глаз заметно
крупнее нижних. Глазковый бугорок маленький, коричневый. Лобный треугольник в сером
налете. Усики черные; 1-й членик сильно бокаловидно расширен, сверху клювовидно
выдается вперед, покрыт длинными светлыми и темными волосками. 3-й членик узкий,
с тупым дорсальным углом. Основание
членика иногда коричневое. Лицо серое,
в густых серых волосках, с примесью
темных в верхней части щек. Концевой
членик щупалец желтовато-белый или
сероватый, овальный, со слегка
оттянутым кончиком. Членик покрыт
длинными, светлыми и темными волосками.
Крылья, ноги и жужжальца как у 3-
Брюшко черное, с желто-коричневыми
пятнами по бокам, захватывающими
I—IV тергиты. Разделяющая их
темная срединная полоска составляет
приблизительно l/5—Ve ширины брюшка,
покрыта б. м. короткими,
прилегающими черными волосками. Каемки
тергитов в желтовато-серых волосках,
образующих посредине небольшие тре-
Рис. 308. Ареал Hybomitra (s. Str.)
turkestana.
угольнички. Снизу брюшко желто-коричневое, II стернит без темного пятна посредине;
V и последующие стерниты темные. 13—18 мм.
Похож на Н. (s. str.) montana (включая терминалии з), но коренастее и крупнее;
3 легко отличается треугольной морщинистой нижней лобной мозолью, $ —
наличием длинных волосков на 1-м членике усиков.
Распространение: горный среднеазиатский вид (рис. 308). Обитает
на высоте 1200—3800 м, в лесном и альпийском поясах, на Гиссаро-Алае, Тянь-Шане
(включая Джунгарский Алатау), Каратау, Тарбагатае, Сауре и Южном Алтае
(Восточный Казахстан). Зап. Китай (Синьцзян; Кашгария); Ганьсу. На Тянь-Шане местами
массовый вид. Конец мая—середина августа. Кладки яиц на нижней стороне листьев
ивы у русла горных речек, личинки в моховом покрове и влажной почве (Шевченко,
1961). Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 1936).
45. Hybomitra (s. str.) nigricornis Ztt. (рис. 309) — Слепень черноусый.
Zetterstedt, 1842, Dipt. Scand., 1 : 112 (Tabanus); В г a u e r, 1880 : 155;
Krober, 1923:92 (Tabanus); S u г с о u f, 1924:107 (Tabanus); Krober in
Lindner, 1925 : 69 (Tabanus); Олсуфьев, 1937a : 183 (Tabanus); Ш о i>-
4 е н к o, 1961 : 102 (Tabanus); С h v a 1 a, 1964 : 375; Leclercq, 1966 : 114;
Виолович, 1968:151; Murdoch et Takahasi, 1969:45; С h v a 1 n,
Lyneborg a. Moucha, 1972 : 209. — ketonensis К о n о et Takahasi,
1939, 1. с : 21 (Tabanus). — altainus S z i 1 a d y, 1926 : 20 (Tabanus), цит. по:
Chvala, Lyneborg a. Moucha, 1972:209.
3. Глаза в коротких серых волосках, с 3 полосками. Лобная полоска б. м.
широкая, ее высота в 3—31/2 раза превосходит ширину, с параллельными сторонами или
слегка сужающаяся к основанию, в голубовато-сером налете. Нижняя лобная моноль
слегка выпуклая, блестяще-черная, б. м. квадратная. Ее боковые стороны обычно
достигают края глаз. Средняя мозоль блестяще-черная, короткая, обычно соодилони
с нижней мозолью. Глазковый бугорок слабо выпуклый, овальный, коричнопо-крш)-
ный, умеренно блестящий или матовый. Лобный треугольник выпуклый, шшкий,
в сером налете. Усики красно-коричневые, концевая часть 3-го членика черпая; иногда
усики целиком черные, за исключением основания 3-го членика. 1-й членик (яюрху
в тонких, б. м. длинных волосках. 3-й членик длинный, узкий, дорсальный угол тупо*
угольный, палочка длинная, тонкая. Концевой членик щупалец желтоипто-болЫЙ,
в базальной части утолщен, покрыт преимущественно черными волосками. НотоплшфЫ
черные, редко буроватые. Жужжальца темно-коричневые. Ноги: кончик бодор, «|ШД«
ние и задние голени, основание средних и задних лапок и передних голоной Ж0ЛТ0-
коричневые; прочие части ног черные. Крылья прозрачные, жилки коричионыо; 1\ ЧПОФО
с небольшим придатком. Брюшко пропорционально груди большое, широкой, чкрнов,
50° 60° 70° 80°
60° 70° 80°'
380_ III. ПОДСЕМ. TABANINAE
сверху е явственным еерым рисунком из треугольных пятен посредине и ромбических
по бокам (налет-|-волоски), образующих в общей сложности 3 продольных ряда.
Снизу брюшко черно-серое, с узкими, желтоватыми каемками и светлыми волосками
по заднему краю стернитов. Как редкое исключение II и III тергиты по бокам и соот-
Рис. 309. Hybomitra (s. str.) nigricornis Ztt. Лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека <j>. А — щупальце; В — усик <J.
ветствующие им стерниты красновато-коричневые. VIII стернит: гонопофиз крупный,
в черных и коричневых волосках, с б. м. глубокой апикальной выемкой; латеральные
лопасти умеренно развиты, боковые коричневые пятна крупные; базальная лопасть
Рис. 310. Ареал Hybomitra (s. str.) nigricornis.
с прямым или слегка вогнутым основным краем. Церки со слегка уплощенной
вершиной. Сперматека веретеновидная. 13—18 мм.
в. Глаза в длинных коричневых волосках. Фасетки верхних 2/3 глаз заметно
(но не резко) крупнее нижних. Глазок овальный, коричневый, слабо блестящий. Лоб-
11. HYBOMITRA
381
ный треугольник темно-серый, линия соприкосновения глаз в 11/2 раза больше его
высоты. Усики черные, основание 3-го членика и иногда 2-й членик красно-коричневые.
1-й членик сверху в длинных, черных волосках, по длине равных или слегка
превосходящих 1-й и 2-й членики вместе взятые. 3-й членик очень узкий, с незначительным
дорсальным углом. Концевой членик щупалец желтоватый или светло-коричневый,
б. м. утолщенный, на конце обычно клювовидно вытянут, покрыт длинными,
преимущественно черными волосками. Нотоплевры черные. Жужжальца, крылья и ноги как
у $. Брюшко черное, II—III тергиты с коричневыми пятнами по бокам. Разделяющая
Рис. 311. Hybomitra (s. str.) lundbecki Рис. 312. Hybomitra (s. str.) lundbecki
lundbecki Lyn. $. lundbecki Lyn. (J.
их черная полоска занимает приблизительно 1/3 ширины брюшка. Задние края терги-
тов по бокам с каемками, посредине с небольшими треугольниками из серых волосков.
Снизу брюшко в основании коричневое, I и V—VII стерниты черные. II стернит с
черным пятном посредине. 13.5—16 мм.
Распространение: таежный вид (рис. 310). В СССР известен от
Кольского п-ова, Ленинградской обл. и севера Украины на восток до Чукотского
нац. округа, Камчатки и Сахалина, на север до Имандры Мурманской обл., низовий
Оби (Лабытн^нги), Верхне-Имбатского на Енисее, Жиганска на Лене и Нижних
Крестов на р. Колыме, на юг до Чигиринского р-на Киевской обл., Воронежа,
Пензенской обл. (Сердобский р-н), Ю. Урала, Кокчетавской обл. (Боровое), Семипалатинска,
Джунгарского Алатау (Панфиловский р-н), Алтая, Тувы, южного Прибайкалья,
Приамурья и Приморского края (Яковлевка). Сев. и Средн. Европа, Сев. Монголия,
Сев.-Вост. Китай. Обычен местами в Ю. Урале, на Алтае, в Туве, Вост. Сибири и
на Сахалине, в западных частях ареала редок. Июль—август, редко раньше или
позднее. Кладки яиц на листьях тростника и других растений по прирусловым заболочои-
ностям.
46. Hybomitra (s. str.) lundbecki Lyn. (рис. 311, 312) — Слепень Лундбека.
Изменчивый вид, представлен 2 подвидами: номинативным, распространенным
в Европе и Зап. Сибири, — Н. (s. str.) lundbecki lundbecki Lyn. и восточным,
обитающим в Восточной Сибири и на Дальнем Востоке — Н. (s. str.) lundbecki sibiriensis Ola.
382
III. ПОДСЕМ. TABANINAE
46a. Hybomitra (s. str.) lundbecki lundbecki Lyn. (рис. 313) — Слепень Лундбека
номинативный.
L у n e Ь о г g, 1959, Ent. Medd., 29 : 127; L е с 1 е г с q, 1966 : 110; В и о л о-
в и ч, 1968 : 153; Murdoch et Takahasi, 1969 : 48; С h v a 1 a a. J e z e k,
1969:340 (личинка, куколка); И в а н и щ у к, 1970:146 (личинка); С h v a 1 а,
Lyneborg a. Moucha, 1972 : 220. — fulvicornis (Meigen) apud S u г с о u f,
1924 : 116 (Tabanus) et auct. — montanus (Meigen) apud В г a u e r, 1880 : 144 (Taba-
nus) (partim) et auct.
5. Глаза в коротких, серых волосках, с 3 резкими, широкими полосками.
Лобная полоска умеренно широкая, заметно сужена к основанию, ее высота превосходит
основание приблизительно в 3V2 раза, в серо-желтом налете. Нижняя лобная мозоль
слегка выпуклая, гладкая, блестяще-черная, щитовидная или округлая. Ее боковые
стороны не достигают края глаз, верхняя сторона округлая, как правило, вершиной
не соединяется с средней мозолью. Последняя короткая, черная, слабо блестящая.
Глазковый бугорок слабо выпуклый, овальный, его передняя половина в сером налете,
задняя блестяще-коричневая, с небольшим в давлением. Лобный треугольник в
желтовато-сером налете. Усики коричневато-желтые, палочка и основание 1-го членика
темные. 1-й членик сверху в коротких, прилегающих, черных волосках. 3-й членик
короткий, в основной части широкий, с прямоугольным, реже остроугольным
дорсальным выступом, палочка короткая, толстая. Концевой членик щупалец желтовато-бе-
Рис. 313. Hybomitra (s. str.) lundbecki lundbecki Lyn. Лобная полоска, усик, щупальце,
VIII стернит (варианты), церки и сперматека $. А — щупальце; Б — усик (J.
лый, в базальной части умеренно утолщен, тупоконечный, покрыт короткими,
преимущественно черными волосками. Нотоплевры, как правило, черные, реже коричневые.
Жужжальца коричневые, головка иногда с более светлым кончиком. Ноги: бедра
черные, их кончик, большая часть средних и задних голеней и основание передних
голеней коричневато-желтые; передние лапки черные, последующие коричневые, с
черным концом. Крылья бесцветные или слегка затемненные к переднему краю. Жилки
коричневые; г4 иногда с небольшим придатком. Брюшко широкоовальное, сверху
туекдо-буровато-серое, с коричнево-желтыми пятнами по бокам. Последние
захватывают I—III тергиты (реже I—IV) и разделяются широкой, черно-серой срединной
полосой. Тергиты по заднему краю и по бокам в коротко прилегающих б. м.
многочисленных серо-желтых волосках; последние образуют посредине брюшка ряд б. м.
явственных треугольных пятен. Снизу брюшко коричнево-желтое. Квадратное пятно
на II стерните и конец брюшка темные. VIII стернит: гонопофиз широкий и короткий,
в тонких желто-коричневых волосках, иногда с примесью черноватых, без апикальной
выемки или с ее следами; латеральные лопасти и боковые коричневые пятна небольшие;
базальная лопасть удлиненная, с вогнутым основным краем. Церки короткие, с сильно
уплощенной вершиной. Сперматека миндалевидная. 12.5—16.5 мм.
(J. Глаза в длинных, серых или белесых волосках. Фасетки глаз б. м. одинаковой
величины. Глазковый бугорок небольшой, овальный, красновато-коричневый. Лобный
треугольник пепельно-серый, линия соприкосновения глаз приблизительно в 1х/2 раза
больше его высоты. Усики коричневато-желтые, палочка и основание 1-го членика
темные, реже усики целиком темные, за исключением 3-го членика. 1-й членик сверху
в коротких черных волосках, не превышающих длину членика, 3-й членик узкий,
П. HYBOMITRA
383
с явственным прямоугольным (реже остроугольным) дорсальным выступом. Концевой
членик щупалец светлый, желтовато-палевый, овальный, покрыт длинными,
преимущественно темными волосками. Нотоплевры, как правило, черные, редко коричневые.
Жужжальца, ноги и крылья как у ?. Брюшко по бокам с большими
желто-коричневыми пятнами, простирающимися на I—IV тергиты. Разделяющая пятна черная
срединная полоска сужена на III тергите и занимает здесь около 1/5—У6 ширины брюшка.
Основание I и целиком V—VII тергиты черные. Коричнево-желтая окраска по бокам
IV тергита часто в той или иной мере вытеснена черной, реже тергит целиком черный.
Брюшко сверху в черных волосках. II тергит посредине, на фоне полоски, с неясным
узким серым треугольным пятном; сама полоска на этом тергите покрыта б. м.
длинными стоячими черными волосками, тогда как на последующих тергитах — коротко
прилегающими. Снизу брюшко коричнево-желтое; основание, квадратное пятно на II
стерните и 3 копцевых стернита темные. 14—16 мм.
Рис. 314. Ареалы: 1 — Hybomitra (s. str.) lundbecki lundbecki; 2 — H. (s. str.)
lundbecki sibiriensis.
Наиболее близок к Н. (s. str.) montana, легко отличаясь от него более широкими,
коричневато-желтыми усиками и темными нотоплеврами (не всегда!). Насколько
позволяют судить описания, большинство старых авторов смешивало Н. (s. str.)
lundbecki с Н. (s. str.) montana и описания этого последнего на самом деле должны быть
отнесены к Н. (s. str.) lundbecki.
Распространение: европейско-западносибирскйй таежно-лесной подвид
(рис. 314, 1). В СССР обитает от Кольского п-ова, Эстонии, Литвы и запада Украины
(Львов), на восток до Енисейско-Ленского водораздела. На север известен до Кольского
залива, низовий Печоры, Оби и Енисея, на юг до Могилев-Подольского, Полтавской обл.
(Миргородский р-н), Воронежской обл., Пензы,-Ю. Урала, Кокчетавской обл.
(Боровое), Семипалатинска и хр. Саура. Сев. и Средн. Европа. Обычен, особенно в таежных
частях ареала. Июнь—июль, редко позднее. Откладка яиц нами отмечена на осоко и
листьях серой ольхи по краю заболоченной ямы на пастбище (Ленинградская обл.).
Личинки преимущественно в болотах разных типов, включая сфагновые, а такжо
на заболоченных участках на лугу и в лесу (Скуфьин, 19666; Лутта, 1970). Куколок
находили на заболоченном лугу и низинном болоте (Лутта, 1970).
46b. Hybomitra lundbeeki sibiriensis Ols. — Слепень Лундбека сибирский.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 452. — siUrica О 1 я u i j e v, 1070 :
684, пес Therioplectes (=Hybomitra) sibiricus Enderlein, 1925 : 358.
$c? отличаются от номинативной формы целиком одноцветными оранжоно-жол-
тыми усиками, включая палочку; у последней иногда концевое колечко с коричшши-
тым кончиком.
Распространение: восточносибирский таежный подвид (рис. 31 Л, У),
От горного Алтая (Чуйская степь), Тувы и Енисейско-Ленского водораздела па иоотоК
до Камчатки, на север включая Центральную Якутию и Средне-Колымск, на юг до
южного Прибайкалья, Приамурья и юга Приморского края (Хасанский р-н). Сев.
Монголия, Сев.-Вост. Китай. Местами обычен. Июнь—июль, редко позднее.
У д$ из горного Алтая и Тувы концевая V4 или 1/3 средних и частью задиих
голеней желто-коричневая.
384 III. ПОДСЕМ. TABANINAE
47. Hybomitra (s. str.) montana Mg. — Слепень обычный.*
Голарктический весьма изменчивый вид. В СССР распространены 4 подвида:
номинативный европейско-сибирский таежно-лесной подвид — Н. (s. str.) montana
montana Mg., степной — H. (s. str.) montana morgani Sure, горно-кавказский — H. (s. str.)
montana karatschajensis Skuf. и горно-среднеазиатский Н. (s. str.) montana reinigiana End.
В южной Монголии обитает Н (s. str.) montana zaitzevi Ols., в Сев. Америке — H.
(s. str.) montana frontalis Wlk. Линеборг (Lyneborg, 1959) описал в Дании Н. (s. str.)
tuxeni, представляющий собой крупные экземпляры (16—18 мм) Н. (s. str.) montana
montana, встречающиеся среди основной популяции, но не имеющие определенной
географической локализации. Двумя годами позже он же (Lyneborg, 1961) предложил
для этой формы применить название Н. tropica Linne, 1758, изучив типовой экземпляр
последнего. Но Н. (s. str.) tropica Linne является nomen oblitum, поскольку оно
длительно не применялось в основной зоологической литературе. Название «tropica L.»
неприемлемо и потому, что оно дублирует «tropica Panz.», длительное время
применявшееся для другого вида Hybomitra (Н. (s. str.) bimaculata Macq.).
; 47a. Hybomitra (s. str.) montana montana Mg. (рис. 315) — Слепень обычный
номинативный.
М е i g е n, 1820, Syst. Beschreib., II : 55 (Tabanus); К г о b e г, 1923 : 99 (Ta-
banus); S u г с о u f, 1924 : 113 (Tabanus); Krober in Lindner, 1925 : 67
(Tabanus); 0,ta суфьев, 1937а : 187 (Tabanus); Moucha et Chvala, 1959a :
167 (Tylostypia); Lyneborg, 1959:139; Шевченко, 1961:104 (Tabanus);
Takahasi, 1962:54; L e с 1 e г с q, 1966:112; Виолович, 1968:155;
Murdoch, et Takahasi, 1969 : 47; С h v a 1 a a. Jezek, 1969 : 341
(личинка, куколка); Соболева, 19706 : 223 (личинка, куколка); Иванищук,
1970:147 (личинка); Chvala, Lyneborg a. Moucha, 1972:224.—
flaviceps Zetterstedt, 1842 : 111 (Tabanus). — sachalinensis Enderlein,
1925 : 359 (Therioplectes). — bimaculatus Enderlein, 1925 : 360 (Th. borealis Mg. var.
bimaculatus). — ochotscana Enderlein, 1934, 1. c. : 184 (Stypommia) (partim). —
montana manshurensis Philip, 1956 : 227 (Hybomitra). — tuxeni Lyneborg,
1959 : 133 (Hybomitra). — ? calluneticola Krober, 1935 : 159 (Sziladynus), цит. по:
Chvala, Lyneborg a. Moucha, 1972 : 224. — tropicus Linne, 1758
(nomen oblitum, Olsufjev et Leclercq, 1971 : 325; Олсуфьев, 1971 :
138). •
$>. Глаза в коротких, серых волосках, ярко-зеленые, с 3 тонкими полосками; у
некоторых экземпляров как исключение глаза лишь с 1 средней полоской или последние
отсутствуют целиком. Лобная полоска неширокая, с параллельными сторонами или
слегка сужающаяся к основанию, ее высота в 4—4х/4 раза превосходит ширину, в
желтовато-сером налете. Нижняя лобная мозоль б. м. прямоугольная, слегка выпуклая,
блестяще-черная. Ее боковые стороны обычно достигают края глаз. Средняя мозоль
короткая, веретеновидная, черная, чаще соединена тонкой полоской с нижней
мозолью. Глазковый бугорок в виде слабо выпуклой, продолговатой, коричневой мо-
зольки, иногда с легкой продольной бороздкой посредине. Лобный треугольник б. м.
плоский, высокий, желтовато-серый. Усики преимущественно черные, основная
часть 3-го членика, иногда 2-й и редко 1-й членики коричнево-красные. 1-й членик
в сером налете, сверху в коротких, прилегающих, черных волосках. 3-й членик
длинный, б. м. узкий, с тупым дорсальным углом, палочка тонкая. Концевой членик
щупалец светлый, желтовато-белый, в основании слегка утолщен, покрыт короткими,
преимущественно черными волосками. Нотоплевры желто-коричневые, очень редко
темные. Жужжальца коричневые, головка иногда с желтовато-белым кончиком. Ноги:
бедра черно-серые, их кончик, средние и задние голени, основание средних и задних
лапок, а также основная половина передних голеней желто-коричневые; прочие части
черные. Крылья слегка затемнены к переднему краю или бесцветные, жилки
коричневые; г4 иногда с небольшим придатком. Брюшко широкое, сверху тускло-буровато-
серое или черноватое, с грязно-желтыми или коричневыми пятнами по бокам,
захватывающими I—III или I—IV тергиты. Эти пятна покрыты желтыми или сероватыми
волосками с примесью черных, располагающихся в переднебоковой части тергитов.
Количество черных волосков варьирует — они могут отсутствовать почти целиком
или значительно преобладать над светлыми волосками. Разделяющая коричневые пятна
черно-серая срединная полоска составляет приблизительно г/3—1U ширины брюшка,
на каждом тергите имеет по срединному треугольному пятну, образованному сероватым
налетом и желтыми или серыми волосками (часто бывают стерты!); треугольники верши-
* Ранее нами (Олсуфьев, 1958) и В. В. Шевченко (1961) этот слепень по его
латинскому названию был обозначен горным," но это мало согласуется с его
распространением, и поэтому русское название целесообразно изменить во избежание
недоразумений.
11. HYBOMITRA
385
ной достигают или почти достигают заднего края предыдущего тергита. Снизу брюшко
коричнево-желтое, его основание, а также V—VII стерниты черно-серые. II стернит
часто без темного пятна посредине. VIII стернит: гонопофиз средней величины, в
черных волосках, с незначительной апикальной выемкой; латеральные лопасти небольшие,
боковые коричневые пятна обычно умеренно развиты; базальная лопасть с прямым
или слегка вогнутым основным краем. Церки со слегка уплощенной или округлой
вершиной. Сперматека миндалевидная. 12—18.5 мм.
Окраска брюшка варьирует за счет вытеснения коричневых пятен, вплоть до
полного их исчезновения. В последнем случае брюшко целиком черно-серое, с неясным
серым рисунком — Н. (s. str.) montana var. flaviceps Zett. Экземпляры с подобной ок-
Рис. 315. Hybomitra (s. str.) montana montana Mg. Лобная полоска, усик, щупальце,
VIII стернит и церки <j>. А — щупальце; В — усик 3; В — VIII стернит и церки <j
Н. montana morgani Sure; Г — то же Н. montana frontalis Wlk. (Канада).
раской по преимуществу северного происхождения очень похожи на Н. (s. str.) nigri-
cornis, но легко отличаются от этого вида более узким лбом, высоким лобным
треугольником, несколько более широкими усиками, желто-коричневыми нотоплеврами
(не всегда), менее резким серым рисунком на брюшке и строением терминалий.
с?. Глаза в умеренно длинных серых волосках. Фасетки верхних 2/3 глаз заметно
(но не резко) крупнее нижних. Глазковый бугорок небольшой, коричневый, тусклый.
Лобный треугольник темно-серый, линия соприкосновения глаз в 2 раза больше ого
высоты. Усики как у 2, 3-й членик более узкий. 1-й членик сверху в относительно
коротких черных волосках, не превосходящих длину членика. Концевой членик
щупалец светлый, желтовато-палевый, вытянуто-овальный, на конце б. м. тупой, покрыт
длинными, преимущественно темными волосками. Нотоплевры желто-коричиепыо.
Жужжальца, ноги и крылья как у ?. Брюшко: I—IV тергиты красно-коричнепыо,
с б. м. узкой черной серединной полоской, V—VII тергиты черные. Полоска на III
тергите сужена и здесь занимает около х/5—111 ширины брюшка. Брюшко сверху в чор»
ных волосках, лишь задние каемки тергитов по бокам и в середине в желтых или
сероватых волосках. Срединная полоска на II тергите в б. м. длинных, стоячих, черных
волосках, тогда как на последующих — в коротко прилегающих. Брюшко спину ко»
ричневое, V—VII стерниты и срединное прямоугольное пятно на II кольце томный.'
13—18 мм.
25 Н. Г. Олсуфьев
386 III. ПОДСЕМ. TABANINAE
Распространение: палеарктический таежно-лесной подвид (рис. 316, 1).
В СССР от Мурманской обл., Эстонии, Литвы и Волынской обл. (Шацк) на восток до
Чукотского нац. округа, Камчатки, Сахалина и Курильских о-вов. На северу известен
до сев. побережья Кольского п-ова, низовий Печоры, Оби, Енисея (Дудинка), Лены
(Жиганск), Яны (Верхоянск), Колымы (Нижние Кресты) и Анадыря (Марково), на юг
до Киевской, Воронежской, Тамбовской (Моршанск), Куйбышевской областей
Ю. Урала, Центрального Казахстана (Жаман-Сарысу и устье Атасу), Павлодарской,
обл., Алтая, южного Прибайкалья, Приамурья и Приморского края (Владивосток).
Сев. и Средн. Европа, Сев. Монголия, Сев.-Вост. Китай, Япония (острова Хоккайдо
и Хонсю).
Неправильно указан Н. А. Виоловичем (1968) для Китая (о. Тайвань), а также
для южных японских островов Сикоку и Кюсю (см. Takahasi, 1962, стр. 56). Обычен
в таежной полосе и особенно на Алтае, в Туве, Восточной Сибири и на Сахалине, в
европейской части ареала в лесной зоне не часто. Экземпляры с Крайнего Севера в
большинстве относятся к Н (s. str.) montana var. flaviceps Zett.Hionb—август, иногда раньше.
0° К вОСТОКу ОТ ГрИНВИЧа 90° 120° 150° 180° «0°
Рис. 316. Ареалы: 1 — Hybomitra (s. str.) montana montana; 2 —■ H. (s. str.)
montana morgani; 3 — H. (s. str.) montana karatschajensis; 4 — H. (s. str.)
montana reinigiana; 5 — H. (s. str.) montana zaitzevi.
Личинки найдены в Ленинградской обл. на заболоченных участках луга (Олсуфьев,
19356), в Карелии — в низинных и переходных болотах, в берегах речек и небольших
водоемах типа пруда (Лутта, 1970); куколка найдена в заросшем карьере (Лутта,
1970).
Североамериканский подвид Н. (s. str.) montana frontalis Walk, наиболее близок
к Н. (s. str.) montana montana Mg. До сих пор H. (s. sir.) frontalis считался
самостоятельным видом, но мы, изучив серию экземпляров (12$ 46) из США и Канады,
пришли к заключению, что это не более как подвид Н. (s. str.) montana. От
номинативного подвида Н. (s. str.) montana frontalis отличается преимущественно беловато-
серыми волосками сверху брюшка, немного более темными волосками глаз и несколько
меньшими размерами тела. Мы не нашли различий в строении терминалий у 22 обоих
подвидов (рис. 315, Г).
47b. Hybomitra (s. str.) montana morgani Sure. — Слепень обычный Моргана.
S u г с о u f, 1912, Annales d'Historie Naturelle, II, 1:71 (Tabanus morgani);
С h v a 1 a, Lyneborg a. M о u с h a, 1972:245 (Hybomitra morgani Sure);
О'л с у ф ь е в, 1973 : 199. — sibiricus Enderlein, 1925 : 358 (Therioplectes). —
staegeri Lyneborg, 1959:145 (Hybomitra). — miilefeldi (Br.) apud Krober,
1923 : 97 (Tabanus) et auet.
5(J отличаются от номинативного подвида белой или желтоватой головкой
жужжалец, беловато-серой окраской светлых волосков брюшка (не всегда!) и более короткими
волосками на глазах; у 2 нижняя лобная мозоль часто коричневая, желто-коричневые
пятна сверху брюшка обычно захватывают боковые стороны I—IV тергитов (изредка
11. HYBOMITRA
387
также V тергита), разделяющая их срединная темная полоса занимает 1/t—г/7 ширины
колец. VIII стернит 2 более светлый, базальная лопасть с сильнее вогнутым
основным краем; церки с более уплощенной вершиной (рис. 315, В). 2 14—18 мм, 6 15—16 мм.
Распространение: южный степной подвид (рис. 316, 2). В СССР известен
в Ю-.Зап. Украине (Винницкая, Одесская, Николаевская области), на юге Ростовской
(низовье Дона), Оренбургской, Челябинской, Тюменской, Омской и Новосибирской
областей, в Зап., Сев., Центр, и 10.-Вост. Казахстане, в Киргизии (оз. Иссык-Куль),
степных частях горного Алтая, в Туве, южном Прибайкалье и Забайкалье и на юге
Приморского края. Обнаружен также в Предкавказье (Майкоп) и в Закавказье:
Армения (Еленино, Шордж, Гилли на Севане, Загалу и другие пункты) и Грузия (Ахал-
калаки). Дания, Румыния, Иран, Монголия, Китай. Обычен в Казахстане, на Алтае,
в Туве и Монголии, в других частях ареала не часто. Переходные формы к
номинативному подвиду обычны в местах стыка ареалов обоих подвидов. Июнь—июль, иногда
позднее. Откладку яиц наблюдали на лист тростника и лист сусака на заболоченном
берегу озера (Виолович, 1968).
47с. Hybomitra (s. str.) montana reinigiana End. — Слепень обычный Рейнига.
Enderlein, 1933, Deutsch. Ent. Zeitschr., 2/3 : 145 (Tylostypia); Олсуфьев,
1937 : 19 (Tabanus); Шевченко, 1961 : 108 (Т. staegeri reinigianus End.); Б а р a-
т о в, 1962b : 96 (Tabanus); Leclercq, 1966:113.
2 (J от номинативного подвида отличаются беловатой головкой жужжалец,
серовато-белыми волосками брюшка и обычно целиком черными усиками; снизу брюшко
на II стерните с крупным черно-серым пятном посредине, которое бывает часто
продолжено на последующие стерниты, образуя нерезкую темную полосу. От Н. (s. str.)
montana morgani Sure, отличается преимущественно черными усиками, 2 — наличием
коричневых пятен на брюшке по бокам I—III тергитов (иногда пятна имеются лишь
на II тергите или незначительно захватывают IV тергит), более темной окраской низа
брюшка (см. выше) и более длинными волосками на глазах. Окраска тела пепельно-
серая. 13—15 мм.
Различия Н. (s. str.) т. reinigiana и Н. (s. str.) m. morgani нечеткие и, возможно,
что обе формы должны рассматриваться как один подвид.
Распространение: высокогорный среднеазиатский подвид (рис. 316, 4).
Степные нагорья Тянь-Шаня и Памиро-Алая, на высоте 1800—3000 и до 3700 м (Памир).
Китай (Зап. Кашгария). Местами обычен. Конец июня—июль. По-видимому, к этому
подвиду надо отнести указание Леклерка (Leclercq, 1966) о нахождении II. (s. str.)
montana montana в горах западнее Кабула, Афганистан.
47d. Hybomitra (s. str.) montana karatschajensis Skuf. — Слепень обычный карачай-
ский.
С к у ф ь и н, 1938, Изв. Воронежск. гос. пед. инст., 2:79 (Tabanus).
2- От номинативного подвида отличается главным образом целиком черными
усиками, длинными неприлегающими волосками на 1-м членике усиков, темными ното-
плеврами и черным низом брюшка. От Н. (s. str.) montana var. flaviceps, встречающегося
преимущественно на севере Евразии, подвид отличается, помимо первых двух
признаков, наличием бурых пятен на I—III тергитах брюшка. 15—18 мм.
Р аспространение: горный (? степной) кавказский подвид (рис. 316, 3):
Карачаево-Черкесская А. О. (долина р. Доут и Учкулан, 1500—2400 м) и Кабардино-
Балкарская АССР. Конец июля—начало августа. Редок.
V. Группа Hybomitra (s. str.) е г b e r i Br.
Жужжальца белые или желтоватые. Щупальца узкие или утолщены, желтовато-
белые. У 2 волоски на глазах и глазковый бугорок плохо развиты (рудиментарные)).
Терминалий 22 видов этой группы плохо различимы (Н. (s. str.) turanica не изучена),
48. Hybomitra (s. str.) expollicata Pand. — Слепень чернополосый.
Изменчивый вид, представлен 2 подвидами: номинативным европейско-западно-
сибирским Н. (s. str.) expollicata expollicata Pand. и восточносибирским Н. (s. sir.)
expollicata orientalis Ols.
48a. Hybomitra (s. str.) expollicata expollicata Pand. (рис, 317).
Pandelle, 1883, Rev. Ent. Fr., 2:218 (Tabanus); S u г с о u f, 1924 : ИМ
(Tabanus); Leclercq, 1966:107; С h v a 1 a, Lyneborg a. M о и о hi»,
1972 : 243. — nigrivitta Olsufjev, 1936a : 231 (Tabanus). — pseuderberi P h i 1 I J»
et Ait ken, 1958:88 (Hybomitra), пит. по: Leclercq, 1966 : 107. — «rlifiH
В г a u e r (5) (Tabanus).
388 III. ПОДСЕМ. TABANINAE
5. Глаза в очень коротких серых волосках, при жизни ярко-изумрудно-зеленые,
с 1—3 узкими, золотисто-медными полосками (обычно не восстанавливаются во влажной
камере) или без них. Лобная полоска б. м. узкая с параллельными сторонами или слегка
суженная к основанию, ее высота в 4—5 раз превосходит ширину, в сером налете.
Глазковый бугорок рудиментарный, в виде 3 крошечных коричнево-черных бугорков
или мозолек, разобщенных между собой налетом, реже слитых в общую мозоль.
Нижняя лобная мозоль блестяще-черная, прямоугольная или треугольная, не достигает
края глаз. Ее нижний край часто с 3 выступами. Средняя мозоль узковеретеновидная,
часто достигает концом до нижней мозоли. Иногда средняя мозоль целиком скрыта
налетом. Лобный треугольник высокий, в сером налете. Усики одноцветно-черные;
1-й членик в сером налете и коротких волосках. 3-й членик узкий, его дорсальный угол
тупой; палочка длинная, тонкая. 'Щупальца беловатые, концевой членик тонкий,
заостренный на конце, покрыт короткими светлыми волосками; изредка
незначительная примесь черных. Нотоплевры желто-коричневые. Крылья слегка
буровато-дымчатые; г4 без придатка. Жужжальца белые или желтоватые. Ноги: бедра черные, в сером
Рис. 317. Hybomitra (s. str.) expollicata expollicata Panrl. Лобная полоска, усик, щупальце,
VIII стернит, церки, сперматека и брюшко снизу 2-
налете, вершинная 1/3 бедер коричнево-желтая; средние и задние голени и основание
передних голеней коричнево-желтые, прочие части ног черные. Брюшко темное, с
небольшими коричневыми пятнами по бокам I—III или I—IV тергитов и с широкой
блестяще-черной продольной полосой посредине. Последняя занимает^ V3—1/1 ширины
брюшка и покрыта короткими черными волосками. Посредине черной полосы обычно
имеется линейная полоска из легкого налета и желтоватых волосков. Боковые стороны
тергитов преимущественно в желтовато-серых, коротких волосках. Снизу брюшко
повторяет рисунок верха. I—III стерниты светло-коричневые, с широкой черной
полосой посредине, последующие стерниты целиком черные. VIII стернит заметно длиннее
ширины; гонрпофиз удлиненный, округлый или слегка конический, в черных волосках,
без апикальной выемки; латеральные лопасти и боковые коричневые пятна небольшие;
базальная лопасть с вогнутым основным краем. Церки крупные, с сильно уплощенной
вершиной. Сперматека булавовидная или миндалевидная. 12—18 мм.
$. Глаза в длинных, серых волосках. Фасетки верхних 2/3 глаз крупнее нижних
приблизительно в 2—3 раза. Глазковый бугорок маленький, коричневый, в тонком
сером налете. Лобный треугольник серый. Усики одноцветно-черные; 1-й членик в сером
налете и в коротких волосках. 3-й членик узкий, с тупым дорсальным углом, палочка
тонкая. Концевой членик щупалец узкоовальный, желтовато-белый, покрыт тонкими,
светлыми и черными волосками; иногда членик со слегка приостренным концом. Грудь
и ее придатки как у 2* Брюшко с коричневыми пятнами по бокам, простирающимися
на I—IV тергиты. Их разделяет блестящая черная полоска, ширина которой
составляет приблизительно 1/3—V, ширины брюшка. Налет на срединной полоске отсутствует.
V—VII кольца черные. Рисунок дополняется волосками: по бокам II—V тергитов
расположены довольно крупные (но не резкие) пятна из коротких желтых волосков, тогда
как вся средняя часть брюшка покрыта черными волосками. Снизу брюшко коричне-
11. HYBOMITRA 389
вое, с б. м. резкой черной срединной полосой; последние 3 (или 4) стернита черные.
Иногда черная полоса на III—IV стернитах слабо выражена. 13—16 мм.
Очень похож па Н. (s. str.) montana morgani и II. (s. str.) erberi, но легко отличается
от обоих видов черной срединной полосой на нижней стороне брюшка и слегка
дымчато-бурыми крыльями. Кроме того, от первого вида отличается более тонкими
щупальцами, одноцветно-черными усиками и строением терминалий 2-
Описанная Брауэром (Вгаиег, 1880) 2 Tabanus erberi на самом деле представляет
собой Н. expollicata expollicata Pand., что нами выяснено путем изучения типового
экземпляра, сохраняемого в Венском музее и любезно присланного нам на проверку
Линеборгом (Lyneborg).
Рис. 318. Ареалы: 1 — Hybomitra (s. str.) expollicata expollicata; 2 — H. (s. str.)
expollicata orientalis.
Распространение: побережья водоемов степной зоны (рис. 318, 1).
В СССР от Одесской обл. (Килия) на восток до Енисейско-Ленского водораздела,
на север до Черниговской обл. (Остер), Воронежской обл. (Анна), юга Удмуртской
АССР (Каракулинский р-н), юга Тюменской обл., Подкаменной Тунгуски (возможно,
изолированное местонахождение), на юг до сев. побережья Черного и Азовского морей,
низовья Дона, Калмыцкой АССР (оз. Сарпа; Ики-Багуты), дельты Волги и Урала,
Аральского моря (о. Барса Кельмес), Бетпак-Далы, Джунгарского Алатау
(Панфиловский р-н) и Восточно-Казахстанской обл. (Тарбагатайский р-н).; Грузия (Ахалкала-
ки). Средн. и 10. Европа (преимущественно в зоне морского побережья), Монголия.
Предпочитает солоноватые водоемы. Обычен в Зап. Сибири и Сев. Казахстане, в
западных частях ареала не часто. Июнь—июль, иногда позднее. Личинки нами найдены
в кочке на берегу озера (Зап. Сибирь).
48b. Hybomitra (s. str.) expollicata orientalis Ols.
Олсуфьев, 1970, Энтомол. обозр., 49, 3 : 686.
I 2- Отличается от номинативного подвида более развитой черно-серой окраской
средних и задних бедер, благодаря чему желто-коричневым остается только их кончи к
на колене; глаза с 3 полосками, у сухих экземпляров, восстанавливающимися при
содержании во влажной камере; глазковый бугорок часто более выражен. 15—18 мм.
Распространение: (рис. 318, 2), от Тувы, Забайкалья и Приамурьн
на восток включая юг Приморского края. Монголия, Сев.-Вост. Китай. Местами
обычен. Июнь—июль, иногда позднее.
49. Hybomitra (s. str.) erberi Br. (рис. 319) — Слепень Эрбера.
Вгаиег, 1880, Denkschr. Akad. Wien, 42 : 151 6 (Tabanus); К г 6 Ь е г, 11)2(1 :
104 (Tabanus); S и г с о и f, 1924 : 132 (Tabanus); Krober in Lindner, 1025 !
91 (Tabanus); Олсуфьев, 1937a : 196 (Tabanus); Шевченко, 1961 : 11 !1
(Tabanus); L e с 1 e г с q, 1966 : 106; Murdoch et Takahasi, 1969 : f>();
С h v a 1 a, Lyneborg a. Moucha, 1972 : 248.
390 III. ПОДСЕМ. TABANINAE
5. Глаза в очень коротких светлых волосках (видны при увеличении X16), с 3 узкими
медно-коричневыми полосками. Лоб узкий, его высота в 4—4V2 раза превосходит
основание, заметно расширяется кверху. Нижняя лобная мозоль прямоугольная или
треугольная, блестяще-черная, ее нижний край часто с 3 выступами. Средняя мозоль
б. м. веретеновидная, нижним концом чаще не достигает нижней мозоли; в редких
случаях срединная мозоль почти целиком скрыта налетом. Глазковый бугорок в виде
рудимента, состоящего из 1—3 незначительных черных бугорков, часто почти целиком
прикрытых налетом. Лобный треугольник высокий, в сером налете. Усики: 1-й и 2-й
членики черные или коричневые, в сером налете, 3-й членик узкий, с тупым
дорсальным углом, черный, в основании часто слегка красновато-коричневый. Концевой
членик щупалец белый, узкий, с приостренным концом, покрыт короткими, светлыми
и частью черными волосками. Нотоплевры красновато-желтые. Крылья бесцветные,
Рис. 319. Hybomltra (s. str.) erberi erberi Br. Лобная полоска, усик, щупальце,
VIII стернит, церки, сперматека и брюшко снизу <j>. A — рудимент глазкового
бугорка $.
жилки светло-коричневые. Жужжальца белые. Ноги: бедра в основной части серые,
их вершинная 1/3) основание передних голеней и целиком голени последующих 2 пар
ног коричневато-желтые. Концы передних голеней и передние лапки черные,
последующие лапки коричневые, с черными концами. Брюшко коричневато-желтое, в
коротких волосках, сверху с умеренно широкой темной срединной полоской, слегка
расширяющейся спереди назад. • Ее ширина составляет приблизительно 1IS—1U ширины
брюшка. Вдоль средней линии по темной полоске проходит б. м. резкая серая полоска
(налет-(-светлые волоски). V и последующие тергиты темные. II стернит обычно с
небольшим темным пятном посредине. V и последующие стерниты темные. Терминалии
сходны с таковыми Н. (s. str.) expollicata, но у VIII стернита основной край базальной
лопасти обычно прямой, а гонопофиз иногда со следами апикальной выемки. 12—17 мм.
(J. Глаза в умеренно длинных серых волосках. Фасетки верхних 2/3 глаз
значительно крупнее нижних (приблизительно в 5—6 раз), с резкой границей между теми и
другими. У сухих экземпляров верхние фасетки светлее нижних, вследствие чего
глаза двуцветные. Глазковый бугорок небольшой, в тонком сером налете за
исключением 2 (или 3) крошечных блестяще-коричневых бугорочков, расположенных на нем.
Лобный треугольник в сером налете. Усики как у 2, но с более узким 3-м члеником;
1-й членик, в коротких волосках. Щупальца беловатые, концевой членик вытянуто-
овальный, со слегка оттянутым кончиком, покрыт длинными, серыми и черными
волосками. Грудь и ее придатки как у §. Брюшко коричневато-желтое, сверху посредине
имеется очень узкая черная полоска, прикрытая серебристо-серым налетом (полоска
налета хорошо видна при рассматривании брюшка сзади). Ширина темной полоски
составляет приблизительно 1/10—Vi2 ширины брюшка. V и последующие кольца
темные. II стернит с темным пятном посредине. 14—16 мм.
11. HYBOMITRA
391
Благодаря любезности Линеборга (Lyneborg) мы имели возможность изучить
типовые экземпляры J и 2, пойманные на Корфу и послужившие Брауэру (Вгаиег, 1880)
для описания этого вида. Как мы и предполагали (Олсуфьев, 1937а, стр. 198), 3 вполне
идентичен S3, определенным нами как It. (s. str.) erberi Br., тогда как 2 оказалась
Н (s. str.) expollicata Pand.
H. (s. str.) erberi близок к II. (s. str.) montana morgani, H. (s. str.) peculiaris и Н.
(s. str.) expollicata. <$$ H. (s. str.) erberi легко отличаются от всех перечисленных видов
•очень крупными фасетками верхних 2/3 глаз, резко отделяющимися от
мелкофасетчатого поля. Различия 22 более тонки: от Н. montana morgani отличается: 1) рудиментом
черного глазкового бугорка и 2) желто-коричневой вершиной бедер (у Н. (s. str.)
montana morgani глазковый бугорок в виде треугольной коричневой мозольки, а у бедер
желтый лишь самый кончик). От Н. (s. sir.) peculiaris рассматриваемый вид отличается:
1) строением рудимента глазкового бугорка в виде маленьких черных бугорочков
(у Я. (s. str.) peculiaris 2 коричневых мозольки), 2) более широкой темной полосой сверху
Рис. 320. Ареалы: 1 — Hybomltra (s. str.) erberi erberi; 2 — H. (s. str.) erberi
fuscipennis.
брюшка, с резкой серой продольной полоской посредине и 3) наличием темного пятна
на II стерните (не всегда!). От Н. (s. str.) expollicata отличается: 1) отсутствием черной
срединной полосы на нижней стороне брюшка (имеется лишь пятно на II стерните),
2) более резкой и широкой серой продольной полоской на дорсальной стороне брюшка,
на фоне черной срединной полосы и 3) более светлыми крыльями.
Распространение: побережья водоемов пустынной и частью степной зон
{рис. 320, 1). Дельта Дуная, Днепра, Крым, Азовское побережье УССР, низовья Дона,
Кубани, Волгоградская обл. (оз. Цаца), Калмыцкая АССР, Вост. Предкавказье (от Ма-
ныча близ Дивное до Судака), Азербайджан (Апшерон, Прикуринско-Араксинская
низменность, Кызыл-Агачский заповедник, Нахичеванская АССР), Армения
(Араратский, Эчмиадзинский, Октемберянский р-ны), низовья Волги, большая часть
Казахстана (к северу до с. Круглое на Урале, 49°30', Тургая и к востоку до Зайсана),
Киргизия,' Узбекистан, Таджикистан, Туркмения. Ю. Европа (локальные
местонахождения), Иран, Монголия, Китай. В Вост. Китае представлен подвидом Н. erberi
fuscipennis Szil. (рис. 320, 2). Обычен по побережьям водоемов в подзоне северных пустынь,
предпочитает непроточные солоноватые водоемы. Июнь—август. Личинки найдены
на рисовых полях и болотах (Кадырова, 19726). Экспериментально доказанный
переносчик туляремии (Олсуфьев и Голов, 1936).
Экземпляры из Центрального Казахстана (Тургай) отличаются более темной
окраской брюшка — снизу последнее иногда с темной широкой полосой вместо обычного
пятна на II стерните, сверху боковые желтые пятна достигают лишь III тергита (27.
(s. str.) erberi Br. var. obscuriventris Ols., var. nova). Эти экземпляры очень похожи
на Н. (s. str.) expollicata, с достоверностью отличаясь лишь по II и III признакам.
Наоборот, экземпляры из Туркмении наиболее светлые: желтые боковые пятна
простираются на IV тергит, II стернит лишь с незначительным намёком на темное пятио или >ко
совершенно без него. Такие экземпляры отличаются от Н. (s. str.) peculiaris, па
которого они очень похожи, лишь по глазков'ому рудименту и более резкой серой
срединной полоске сверху брюшка.
392 III. ПОДСЁМ. TABANINAE
50. Hybomitra (s. str.) peculiaris Szil. (рис. 321) — Слепень тугайный.
S z i 1 a d у, 1914, Ann. Mus. Nat. Hungar., 12:665 (Tabanus); Surcouf,
1924 : 136 (Tabanus); Krober in Lindner, 1925 : 95 (Tabanus); Олсуфьев,
1937a : 198 (Tabanus); Шевченко, 1961 : 115 (Tabanus); L e с 1 e г с q, 1966 : 117;
Chvala, L у n e Ь о г g' a. Moucha, 1972 : 250. — ?strix S z i 1 a d y, 1923 :
1 (Tabanus).
9. Глаза голые или в очень коротких, серых волосках (едва заметны при
увеличении Х16), с 3 узкими полосками. Лоб узкий, его высота превосходит основание в 4—
41/2 раза, чуть расширяющийся кверху. Нижняя лобиая мозоль б. м. прямоугольная,
блестяще-черная или коричневая, средняя веретеновидная, черная, нижним концом
часто достигает нижней мозоли. Глазковый бугорок в виде рудимента, состоящего
из 2 плоских, продолговатых, коричневатых мозолек и ниже расположенной
небольшой черной точки (или бугорочка). Лобный треугольник высокий, в сером налете.
Усики: 1-й и 2-й членики красио-коричпевые, 3-й членик узкий, черный, в основании
красноватый; дорсальный угол тупой. 1-й членик сверху в коротких, черных волосках.
Рис. 321. Hybomitra (s. str.) peculiaris Szil. Лобная полоска, усик, щупальце, VIII стер-
нит, церки, сперматека и брюшко снизу $. А — рудимент глазкового бугорка $.
Щупальца беловатые, узкие, б. м. заостренные к концу, в светлых и кое-где в
коротких черных волосках. Нотоплевры желто-коричневые. Крылья бесцветные, жилки
светло-коричневые. Жужжальца белые. Ноги: бедра в основной части серые, их конец
и затем голени последних двух пар ног коричневато-желтые, иногда бедра целиком
желтые. Передние голени черные, в основании коричневые. Передние лапки черные,
последующие коричневые, с черным концом. Брюшко коричневато-желтое, в коротких
волосках, сверху с узкой темной полоской посредине, занимающей примерно 1/в—
1/10 ширины брюшка. Эта полоска иногда распадается на отдельные пятна — черточки,
по одному на каждом тергите, в редких случаях полоска отсутствует совершенно;
V и последующие кольца брюшка темные. Иногда темная окраска заходит на
предыдущий тергит или, наоборот, желтая окраска частично вытесняет темную на V кольце.
По темной срединной полоске намечена узкая неясная продольная сероватая полоска
(налет+волоски). II стерпит без темного пятна посредине; VI (реже V) и последующие
стерниты темные. Терминалии сходны с таковыми Н. (s. str.) erberi и Н. (s. str.) expolli-
cata; гонопофиз с небольшой апикальной выемкой, основной край базальной лопасти
слегка вогнутый. 11—17 мм.
S- Глаза в умеренно длинных, белесых волосках. Фасетки верхней части глаз-
крупнее нижних приблизительно в 3 раза. Глазковый бугорок маленький,
коричневатый, обычно покрыт серым налетом; 2—3 крошечных коричневых точки или бугорочка
видны на нем при сильном увеличении. Лобный треугольник серый. Усики как у $,
3-й членик очень узкий; 1-й членик в коротких волосках. Щупальца беловатые,
концевой членик вытянуто-овальный, со слегка оттянутым кончиком, покрыг тонкими
светлыми волосками, иногда с незначительной примесью темных. Грудь и ее придатки
как у $. Брюшко коричневато-желтое, сверху с очень узкой (почти линейной)
черновато-серой полоской посредине. Полоска иногда почти исчезает на III тергите. V и
последующие кольца брюшка черно-серые. II стернит без темного пятна посредине.
14.5—16 мм.
11. HYBOMITRA 393
Распространение: побережья водоемов пустынной и частью степной зон
(рис. 322). Дельта Дуная, Бердянск, низовья Дона, Сев. Кавказ (от Краснодарского
края до Дагестана), Азербайджан (Куба-Хачмас; Прикуринско-Араксинская
низменность; Кызыл-Агачский заповедник), Армения (Араздаян; Давалух на Араксе; Веди;
Басаргечар на оз. Севан), Нижняя Волга (Волгоград; оз. Цаца; дельта Волги),
Казахстан (на север до с. Круглое на Урале, 49°30', северного Приаралья, Бетпакдалы,
г. Зыряновска Восточно-Казахстанской обл. и хр. Саура), Узбекистан, Киргизия,
Таджикистан, Туркмения. Греция, Румыния, Ирак, Иран, Афганистан, Монголия,
Китай. Обычен, особенно в плавнях Сырдарьи, Амударьи, Чу и Или; в западном Тянь-
Шане местами поднимается в горы до 1500 м (Чаткальский хр.). Июнь—август, иногда
раньше или позднее. Экспериментально доказанный переносчик туляремии (Олсуфьев
и Голов, 1936). Личинки найдены при обследовании озера в плавающей
растительности, в воде у самого уреза и в почве у берега (Лутта, 1964г), а также на рисовых
полях, болотах и речках (Кадырова, 19726).
Рис. 322. Ареал Hybomitra (s. str.) peculiaris.
Tabanus strix Szilady, 1923, описанный по экземплярам (2$) из Туркмении (Кари-
бент=Теджен) и Ирака (Багдад), по-видимому, представляет собой светлую форму
Н. (s. str.) peculiaris Szil. с желтыми бедрами и частично редуцированной срединной
темной полоской сверху брюшка (Олсуфьев, 1970).
51. Hybomitra (s. str.) turanica Ols. (рис. 323) — Слепень туранский.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII,' 2 : 200 (Tabanus); HI e в-
чен'ко, 1961:117 (Tabanus); L е с 1 е г с q, 1966:123.
2- Глаза в очень коротких, серых волосках, с 3 полосками. Лобная полоска б. м.
узкая, слегка сужена к основанию, ее высота в 41/2 раза превосходит ширину, в сером
налете. Глазковый бугорок в виде маленькой, плоской, овальной,
красновато-коричневой мозольки. Нижняя лобная мозоль б. м. прямоугольная, коричнево-черная, слабо
блестящая. Средняя мозоль б. м. веретеновидная, обособлена от нижней. Лобный
треугольник высокий, в сером налете. 1-й членик в сером налете и в коротких волосках,
изутри красноватый, 2-й членик желто-красный, 3-й узкий, с тупым дорсальным углом,
в основной половине красновато-коричневый, к концу черный. Щупальца желтовато-
белые, концевой членик сильно утолщен, покрыт очень короткими светлыми волосками.
Нотоплевры и частью мезоплевры желто-коричневые. Крылья бесцветные, жилки
светло-коричневые. Ноги: бедра темные, в сером налете, их кончик желтый. Голом и
желто-коричневые, передние голени с черной вершиной. Передние лапки черные,
последующие коричневые, с черным концом. Задневнешний край задних голеней с
плотным рядом серебристых волосков. Жужжальца белые, головка в основании
коричневая. Брюшко сверху коричневое, с шоколадным оттенком, покрыто очень коротким»
черными и светлыми волосками. Последние сгруппированы в небольшие пятна по
бокам тергитов (по 1 с каждой стороны) и затем посредине, образуя па II—III торгнтпх
неясные треугольники. Срединная темная полоска на II—IV тергитах отсутстиуит
394 III. ПОДСЕМ. TABANINAE
(реже имеются небольшие узкие темные пятна), на V—VI тергитах посредине по черно-
серому трапециевидному пятну и по бокам по небольшому косому пятну; VII (иногда
также VI) тергит черно-серый. Снизу брюшко желтовато-коричневое, в тонком сером
налете. II стернит без темного пятна.
16—20 мм.
(J неизвестен.
Своеобразный вид, выделяющийся
от прочих видов с белыми
жужжальцами 1) более крупными размерами, 2)
утолщенными щупальцами и 3) светло-
шоколадной окраской брюшка, без
черной срединной полоски (или с ее
рудиментом).
Распространение:
известен с Амударьи — Туркмения (Фараб
близ Чарджоу); Каракалпакия
(Кипчак; Кок-Дарья Ак-Тау). Редок. Май—
июнь, иногда позднее.
Рис. 323. Hybomitra (s. str.) turanica Ols.
Лобная полоска, усик и щупальце $.
Голотип: $, Фараб близ Чарджоу, 15 V 1911 (ст. стиль), А. Гольбек. В коллекции
Зоологического института Академии наук СССР, Ленинград.
3. Подрод SIPALA End.
Enderlein, 1923, Deutsche Entom. Zeitschr., 5 : 545 (genus); Leclercq,
1966 : 124 (subgenus).
Тип подрода Tabanus acuminatus Loew, 1858.
Конец брюшка сжат с боков. Глаза с 3 полосками. Остальное как у подрода
Hybomitra.
Подрод насчитывает 3 вида, распространенных в Палеарктике, из них 1 вид в СССР,
52. Hybomitra (Sipala) acuminata Lw. (рис. 324, 325) — Слепень остробрюхий.
Loew, 1858a, Verh. Zool.-bot. Gesell. Wien, 8 : 590 (Tabanus); В г a u e r, 1880:
157 (Tabanus); Krober, 1923 : 82 (Tabanus); Krober in Lindner, 1925 : 60
(Tabanus); Олсуфьев, 1937a : 194 (Tabanus); Шевченко, 1961 : 111
(Tabanus); Leclercq, 1966 : 124; Chvala, Lyneborg a. Mouch a, 1972:253.
5. Глаза в коротких, серых волосках, с 3 полосками. Лобная полоска сильно
сужена к основанию, ее высота превосходит ширину последнего приблизительно в 4 раза,
в сером налете. Нижняя лобная мозоль б. м. прямоугольная, блестяще-черная, почти
достигает края глаз. Средняя мозоль вытянуто-овальная, черная, обособлена от
нижней. Глазковый бугорок маленький, круглый, красновато-коричневый. Затылок за
глазами в виде относительно широкой полоски. Усики черные; 2-й и основание 3-го членика
часто красноватые. 1-й членик в коротких волосках, 3-й членик узкий, со слабым
дорсальным углом. Лобный треугольник в сером налете. Концевой членик щупалец белый,
узкий, покрыт тонкими белыми и черными волосками. Нотоплевры желто-коричневые.
Крылья бесцветные; г4 без придатка. Жужжальца белые или желтоватые. Ноги: бедра
черные, в сером налете, с желтоватым кончиком; голени и основание лапок последних
2 пар йог желто-коричневые; к концу лапки коричнево-черные. Вершина передних
голеней и передние лапки черные. Брюшко узкое, сверху светло-коричневое, с
продольной черной полоской посредине, занимающей 1/4,—1/а ширины брюшка. IV (или III) и
последующие тергиты целиком темные. Вдоль средней линии полоска в тонком сером
налете и серых волосках. Снизу брюшко в основной части красновато-желтое или
коричневое, II стернит с крупным темным пятном посредине, IV и последующие стерниты
темные. Терминалиисходны с таковыми Н. (s. str.) peculiaris, но боковые коричневые
пятна на VIII стерните значительно крупнее. 10.5—16 мм.
с?. Голова сильно выпуклая. Глаза в длинных, серых волосках. Фасетки верхних
2/3 глаз крупнее нижних приблизительно в 2—3 раза. Глазковый бугорок маленький,
круглый, коричневый. Лобный треугольник серый. Усики как у ?, 3-й членик очень
узкий, 1-й членик сверху в коротких волосках. Концевой членик щупалец беловатый,
11. HYBOMITRA
395
вытянуто-овальный, в длинных светло-серых и единичных черных волосках. Грудь и
ее придатки как у §. Брюшко коническое, рисунок как у ?, кончик иногда сдавлен
с боков. 12—13 мм.
Рис. 324. Hybomitra (Sipala) acuminata Lw. 9.
Близок к Н. (s. str.) erberi и Н. (s. str.) peculiaris, но меньших размеров. 5_хорошо
отличается круглым глазковым бугорком, более широкой затылочной полоской и ежа-
Рис. 325. Hybomitra (Sipala) acuminata Lw. Лобная полоска,
усик, щупальце, VIII стернит, перки и сперматека (варианты) ?.
тым с боков кончиком брюшка; 6, помимо меньших размеров, выделяется относительно
более выпуклой головой и более широкой темной продольной полоской сверху брюшка,
его кончик иногда сжат с боков.
396
III. ПОДСЕМ. TABANMAE
Распространение: побережья водоемов пустынной и частью степной зон
(рис. 326). В СССР известен на юге УССР (дельта Дуная, устье Днестра, дельта Днепра,
Бердянск), в Воронежской (Елань-Колено, видимо, изолированное местонахождение),
Волгоградской областях (Волго-Ахтубииская пойма), Краснодарском крае (запад),
Кабардино-Балкарской АССР, Сев.-Зап. Прикаспии (от Дагестана до дельты Волги ж
Рис. 326. Ареал Hybomitra (Sipala) acuminata.
восточнее), Азербайджане (Пушкино, Акстафа, Кюрдамир), Армении (Эчмиадзин,
Зейва), большей части Казахстана (на север до с. Круглое на р. Урал, 49°30', Семиозер-
ного р-на Кустанайской обл., на восток до Зайсана), в Киргизии, Узбекистане (включая
Ферганскую долину), Таджикистане и Туркмении. Спорадически распространен в 10.
и частью Средн. Европе (бассейн Дуная), Иране и Китае. Многочислен в подзоне
северных пустынь, в остальных частях ареала не часто. Май—август.
5. Триба HAEMATOPOTINI (End«rlein, 1922) Bequaert, 1930
Базикоста в волосках, иногда разреженных. Усики: 1-й членик удлиненный,
3-й членик поделен на 4 сегмента, соответственно палочка 3-члениковая. Лобная
полоска у 5 широкая. Терминаляи сходны с таковыми Tabanini; у 22 VIII стернит с б. м.
развитыми латеральными лопастями.
Небольшие мухи с узким телом, крылья обычно с характерным сетчатым
(«мраморным») рисунком. Глазки и глазковый бугорок на темени отсутствуют.
12. Род НЕРТАТОМА Mg. — ДЛИННОУСКИ
Meigen, 1803, Illig. Magaz., 11:267; К г 6 b е г, 1923:133; S u г с о и i,
1924 : 66; Олсуфьев, 1937а : 293; Moucha etChvala, 1956 : 262;. L e c-
lercq, 1966:176; Виолович, 1968 : 72. — Hexatoma M e i g e n, 1820, 1. с:
83.
Тип рода Tabanus pellucens Fabricius, 1776.
Небольшие, коренастые мухи (рис. 327). Крылья бесцветные, в покое
складываются вдоль тела слегка кровлеобразно. Усики длинные, шнуровидные. 3-й членик
резко поделен на 4 сегмента, настолько резко, что усики могут быть приняты за 6-чле-
никовые. Глаза у 3 соприкасаются, у J разделены широким лбом, имеют поперечные
полоски и в коротких волосках. Хоботок немного короче высоты головы. У 3 брюшко
состоит из 6 видимых колец. Известен всего 1 вид, распространенный в Палеарктике.
1. Heptatoma pellucens F. — Длинноуска обыкновенная.
Представлена 2 подвидами: номинативным европейско-западносибирским Н.
pellucens pellucens F. и восточным или среднесибирским Н. pellucens orientalis Ols
12. НЕРТАТОМА
397
la. Heptatoma pellucens pellucens F. —Длинноуска обыкновенная номинативная.
Fabricius, 1776, Gen. Ins. Mant. : 307 (Tabanus); Lundbeck, 1907:
100; К г 6 b e r, 1922 : 133; S u г с о u f, 1924 : 66; Stammer, 1924 : 121
(личинка); Krober in Lindner, 1925 : 41; О л с у ф ь е в, 1937а : 293; Moucha
et Chvala, 1956:262; Leclercq, 1966:176; Виолович, 1968:72;
И в а н ищ у к, 1970 : 137 (личинка); J e z e к, 1971 : 342 (личинка, куколка);
Chvala, Lyneborg a. Moucha, 1972 : 399.
2- Глаза в очень коротких волосках, видимых лишь при сильном увеличении
(Х16). Лобная полоска широкая, блестяще-черная, покрыта б. м. длинными желтыми
Рис. 327. Heptatoma pellucens pellucens F. $.
волосками.) Усики целиком черные, шнуровидные; 1-й членик цилиндрический, 2-й
короткий, составляет 1/2 длины первого, оба в тонком сером налете; 3-й членик вдвое
длиннее обоих предыдущих члеников вместе взятых, резко поделен на 4 вторичных
членика. Лицо плоское, покрыто голубовато-серым налетом и б. м. густыми желтоватыми
волосками. Концевой членик щупалец черный, небольшой, покрыт б. м. короткими,
снаружи черными, изнутри серыми волосками; членик в основании утолщен, за
серединой сужен, остроконечный. Бочки груди в светло-коричневато-желтых
волосках. Нотоплевры черные, в густых коричневато-желтых волосках. Жужжальца
коричневые. Ноги в большей своей части черные; бедра в сером налете,
основная половина передних, большая часть средних и задних голеней беловатые.
Основание лапок 2 последних пар ног желтые. Крылья бесцветные, жилки
коричневые; г4 без придатка. Брюшко блестяще-черное, в коротких черных волосках. II тер-*
гит с крупными боковыми пятнами беловатого налета. I—II и VI—VII тергиты в
коротких желтоватых волосках. Снизу брюшко черное; боковые стороны стернитов в
беловатом налете+светлых волосках, благодаря чему посредине брюшка выделяется резкая
черная продольная полоса. Часто светлый налет образует лишь боковые полукаомки
по заднему краю стернитов. Терминалии своеобразного строения (рис. 328, А—Г).
VIII стернит широкий, гонопофиз короткий, с апикальной выемкой; латералышо ло-
398 III. ПОДСЕМ. TABANINAE
пасти небольшие, боковые коричневые пятна крупные, почти достигают центра стер-
нита; базальная лопасть короткая, с прямым основным краем. Дистальная часть IX
стернита с пластинками, вооруженными шипиками. Семеприемники небольшие, муфта
умеренной длины, бесцветная, дистальная часть с очень узким слегка коричневоокра-
шенным бортиком; сперматека сигаровидиая, черная, без сужения переходит в проток
семеприемника. Церки треугольные', склериты X тергита широкие, IX тергита — очень
узкие. 9.5—13.5 мм.
3- Голова по размерам не больше, нежели у $. Глаза в коротких светлых волосках.
Фасетки верхних а/3 глаз крупнее нижних, граница между теми и другими нерезкая.
Глазковый бугорок отсутствует. Затылок за глазами с рядом заметно выступающих,
длинных, черных волосков. Лобный треугольник блестяще-черный, его вершина в
сером налете, тогда как боковые стороны возле щек в желтовато-серых волосках. Усики
как у ?. 1-й членик и вершина 2-го в длинных черных волосках. Концевой членик
щупалец черный, б. м. яйцевидный, покрыт длинными черными и частью серыми
волосками. Щеки очень толстые, блестяще-черные, вместе с лицом покрыты желтовато-
серыми волосками; верхний край щек возле лобного треугольника в сером налете и
покрыт черными волосками. Грудь и ее придатки как у ?. Брюшко черное, сверху
блестящее, I и II тергиты в густых желтоватых волосках, последующие в более коротких
и редких черных волосках. Последнее кольцо брюшка и терминалии в светлых
волосках. II—III тергиты иногда по бокам неясно коричневатые. Снизу брюшко черное,
I—II стерниты в сероватом налете и желтых волосках. Последующие стерниты черные,
с узкими каемками из желтовато-серых волосков, широко разделенными посредине.
Терминалии (рис. 328, А—В): гоностиль как у Tabanini; вершина двузубчатая; эпанд-
рий сросшийся дистальными концами, граница между IX и X тергитами неясная;
субанальный склерит цельный, не поделен на 2, как обычно. 13—14.5 мм.
Распространение: европейско-западносибирский лесной подвид
(рис. 329, 1). От Эстонии и запада Украины на восток до Новосибирской обл., на север
до Сегежского р-на Карельской АССР, Инты Коми АССР (66° с. ш.) и севера Омской обл.,
на юг до Ужгородского р-на Закарпатской обл., Шепетовки Хмельницкой обл.,
Брянска, окр. Воронежа, Ю. Урала (Атиково, 53° с. ш.) и Зонова Куйбышевского р-на
Новосибирской обл. Н. А. Виоловичем (1968) номинативный подвид указан еще для
Среднего Приобья (Копыловка, Белый Яр и др.) и Нижнего Енисея (Верхне-Имбатское,
Туруханск), но, возможно, обитающие здесь популяции являются промежуточными
к восточному подвиду; исследованный нами экземпляр $ из окр. Томска от В. М.
Попова и серия 5J из Колпашева на Оби от В. Г. Фоминых относятся к Н. pellucens
orientalis. Средн. Европа (на запад включая Францию), юг Скандинавии, Финляндия.
Не часто (в Тульской обл. местами до 3% обилия). Июнь—август.
13. НАЕМАТОРОТА
399
Личинок находили в загрязненных водоемах (включая цементированные), особенно
возле свинарников, а также под корой упавших в воду стволов деревьев.
lb. Heptatoiria pellucens orientalis 01s. — Длинноуска обыкновенная восточная.
Олсуфьев, 1962а, Зоол. жури., 41, 6 : 889; L е с 1 е г с q, 1966 : 177; В и о-
лович, 1968 : 74.
0° К ВОСТОКУ ОТ ГриНВИЧа 90° 120° 150° 180° 150°
Рис. 329. Ареалы: 1 — Heptatoma pellucens pellucens; 2 — Н. pellucens
orientalis.
$<J. От номинативной формы отличается беловато-серой окраской волосков на
бочках груди и в нижней части лица. 5 Ю—14 мм, 3 14—15 мм.
Распространение: среднесибирский лесной подвид (рис. 329, 2).
Алтайский край (Красногорский р-н; Чойский аймак; Сайдыс), Томская обл. (Корнилово,
Колпашево), Кемерово, Красноярский край (от Хакасской А. О. на юге до Теи близ
Северо-Енисейска на севере), Иркутская обл. (Тайшет; Усольцево и Братск в среднем
течении Ангары; Усть-Кут). Июнь—август. Не часто.
Голотип: 5> Еронда Старо-Бардинского р-на Алтайского края, 20 VI 1957,
Н. Г. Олсуфьев. В коллекции Зоологического института Академии наук СССР,
Ленинград.
13. Род НАЕМАТОРОТА Mg. — ДОЖДЕВКИ
М е i g e п, 1803, Illig. Magaz. Ins., II : 267; L е с 1 е г с q, [1966 : 177; В и о л о-
в и ч, 1968 : 55. — Chrysozona Moigen, 1800 : 23; название отвергнуто
Международной комиссией Зоологической номенклатуры, 1963.
Тип рода Tabanus pluvialis Linne, 1758. v
Небольшие виды, с узким черно-серым телом (рис. 330, 331). Крылья серые,
со светлым сетчатым рисунком («мраморные»), в покое складываются кровлеобразно
вдоль тела; г4 с придатком. Глаза с волнистыми поперечными полосками, обычно в
заметных волосках. У 33 фасетки верхних 2/3 или 3/4 глаз крупнее нижних. Усики
длиннее головы: 1-й членик довольно крупный, цилиндрический или яйцевидный, часто
сильно блестящий, 2-й членик маленький, бокаловидный, 3-й членик б. м. длинный,
с 3-члениковой палочкой. Площадка в основной части слабо расширена, сверху без
выраженного дорсального угла. Лоб 5 широкий (рис. 332), в основании с поперечной
лобной мозолью и в средней части с 3 бархатистыми темными пятнами, расположенными
по треугольнику: 2 более крупными боковыми и маленьким средним (может иногда
отсутствовать). Между основаниями усиков и над ними, как правило, имеется крупноо,'
черное, бархатистое пятно, соприкасающееся с нижней лобной мозолью. У 3 лобный
треугольник с черным бархатистым пятном. Щеки в заметной темной пунктировке
Хоботок короткий, не более высоты головы. Терминалии <J (рис. 54, VI, А, Б):
кондовая часть гоностиля без внешнего (верхнего) зубчика. Терминалии 5 (рис. 54, VI, В—К)\
VIII стернит обычно в большей своей части темно окрашен, гонопофиз уморошшй
400 III. ПОДСЕМ. TABANINAE
величины, с апикальной выемкой, латеральные лопасти чаще небольшие; семеприем-
ники у большинства видов удлиненные, обычного для Tabaninae строения, с
коричневой веретеновидпой сперматекой, у немногих видов маленькие, сперматека в виде б. м.
прозрачной трубки.
Рис. 330. Haematopota pluvialis
pluvialis L. 5.
Рис. 331. Haematopota pluvialis
pluvialis L. <£.
Обширный род, насчитывающий около 390 видов (Moucha, 1969). Свойствен глав-
нымрбразом Африке, частью Индии и Палеарктике, совершенно отсутствует в Австра-
лии!и представлен всего несколькими видами в Сев. Америке. На долю Палеарктики
приходится 58 видов, распространенных главным образом в Средиземноморской
подобласти. В СССР известно 17 видов дождевок,
которые весьма однообразны по внешности и различаются
с большим трудом. Мы исключаем из рассмотрения
Haematopota obscurata Bigot, описанную с Кавказа (или
Сев. Ирана) и упомянутую нами в 1-м издании «Фауна
СССР». Оригинальное описание Виго (Bigot)
недостаточно для распознания экземпляров этого вида, а тип
утерян.
^^ШтШ
Рис. 332. Лобная полоска и усик § Haematopota: A —
среднее; Б — боковые бархатистые пятна; В —
лобная мозоль; Г — перетяжка перед вершиной 1-го
членика усиков.
При определении дождевок наибольшее значение имеет строение и окраска
усиков, затем строение лба и рисунок крыльев. Последний, впрочем, изменчив и целиком
на него полагаться нельзя. Известную помощь оказывает также окраска брюшка —
главным образом степень развития боковых серых пятен, но у потертых или
зажиревших экземпляров пятна могут в той или иной мере исчезнуть. Терминалии 22
большинства видов слабо дифференцированы и подвержены значительной изменчивости,
что затрудняет их использование для диагностики.
22 сосут кровь, нападают почти бесшумно; в отличие от пестряков и слепней часто
летают и в пасмурную погоду и особенно назойливы перед дождем, за что и получили
13. HAEMATOPOTA 401
название «дождевки». Охотно нападают на человека. Хипшых личинок находили
в прибрежных частях водоемов или в почве, вплоть до огородных грядок.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА
HAEMATOPOTA MG.
А. Глаза разделены лбом: 2?
1 (2). Крылья с интенсивным коричневым рисунком. Лобная мозоль
темно-коричневая (рис. 344). Мелкий вид, 6—8 мм, с почти целиком желтыми усиками.
Приморский край 10. Н. koryoensis Shir.
2 (1). Крылья с серым или буровато-серым рисунком. Лобная мозоль черная. Обычно
более крупные виды с черными, серыми, реже красновато-желтыми усиками.
3 (20). 1-й членик усиков относительно узкий, б. м. цилиндрический, иногда
вследствие перетяжек узловатый, в сером налете, его длина обычно превосходит
собственную толщину более чем в 3 раза (рис. 333, 335, 340 и др.), если менее
(1 : 2^1^), то членик по крайней мере изнутри коричневый.
4 (5). Бархатистое черное пятно между усиками отсутствует. Ширина лба равна 1/%
ширины головы. 1-й членик усиков в густых волосках, на конце без перетяжки.
Крылья беловатые, с серыми пятнами. Задний край крыла с широкой,
непрерывной, белой каймой. 7 мм 7. Н. eaenofrons Krob.
5 (4). Бархатистое черное пятно между усиками имеется. Ширина лба
приблизительно равна х/3 ширины головы.
6 (9). Задний край крыла с широкой беловатой каймой (у экземпляров Н. turkesta-
nica из Сибири и Сев.-Вост. Китая может отсутствовать!). 1-й членик усиков
часто с несколькими перетяжками или слегка утолщен в средней части. 3-й
членик усиков узкий, обычно явно уже, нежели 1-й (смотреть сбоку!).
7 (8). Длина 1-го членика усиков превосходит толщину приблизительно в 3V2—
4 раза (рис. 340). Членик целиком черно-серый, изредка в основании
коричневый 8. Н. pallens Lw.
8 (7). Длина 1-го членика усиков превосходит толщину приблизительно в 2У2—
3 раза (рис. 342). Членик изнутри (иногда целиком) коричневый или красно-
коричневый, значительно реже целиком черно-серый
. 9. Н. turkestanica Krob.
9 (6). Задний край крыла без широкой беловатой каймы (иногда лишь с узкой
каемкой). 1-й членик усиков б. м. цилиндрический и, как правило, не более чем
с 1 перетяжкой перед вершиной. 3-й членик усиков равен по ширине 1-му или
даже шире его (рис. 333, 334, 335 и др.).
10 (15). 1-й членик усиков коричнево-желтый или красноватый, с темной вершиной,
реже целиком черно-серый, но в таком случае светлые боковые пятна сверху
брюшка резкие начиная с I тергита.
11 (14). Концевой членик щупалец утолщен, его длина превосходит наибольшую
толщину в 2—2V2 раза (рис. 333). Боковые сероватые округлые пятна на брюшке
со II. (или с I) тергита.
12 (13). Крупнее: 11.5—13 мм. Светлые пятна сверху брюшка оливково-серые;
боковые округлые пятна имеются на II и последующих тергитах. 3-й членик усиков
слегка шире 1-го (смотреть сбоку!) 1. Н. grandis Macq.
13 (12). Мельче: 10—11 мм. Светлые пятна сверху брюшка беловато-серые; боковые
округлые пятна имеются на I и последующих тергитах. 3-й членик усиков не
шире 1-го 2. Н. pavlovskii Haus.
14 (11). Концевой членик щупалец узкий, его длина превосходит наибольшую
толщину в 3—Зх/2 раза (рис. 335). Боковые сероватые округлые пятна на брюшке
с IV (или с III) тергита 3. Н. easpiea Abbas.-L.
15 (10). 1-й членик усиков черно-серый, иногда в основании слегка коричневатый;
боковые светлые пятна сверху брюшка на концевых тергитах, иногда неясные.
16 (19). Ширина лба явственно превосходит высоту (рис. 338, 339), 3-й членик усиков
черный. Бедра черные, в сером налете.
17 (18). 1-й членик усиков умеренно длинный, его длина превосходит толщину
приблизительно в 3—3V2 Раза (рис. 338). Горы Средней Азии . . 5. Н. baetriana Ols.
18 (17). 1-й членик усиков очень длинный, его длина превосходит толщину в 4—5 раз
(рис. 339). Кавказ 6. Н. longeantennata Ols.
19 (16). Высота лба слегка превосходит ширину (рис. 336). По крайней мере средние
бедра на некотором протяжении желтоватые. Усики часто в основании
коричневые 4. Н. italica Mg.
20 (3). 1-й членик усиков б. м. утолщен, его длина превосходит толщину не более чем
в 2х/г Раза (рис. 345, 347, 348 и др.). Членик черный, обычно блестящий, иногда
сверху прикрыт серым налетом.
1U 26 Н. Г. Олсуфьев
402 III. ПОДСЕМ. TABANINAE
21 (30). 3-й членик усиков по крайней мере в основании коричневый или красновато-
желтый (рис. 345, 347, 348 и др.).
22 (29). Вершина крыла с узкой запятовидной беловатой полоской. По краю крыла
в заднекрайних ячейках иногда имеется ряд светлых парных пятен или
колечек (рис. 345, 348, 351).
23 (28). Высота лба слегка превосходит ширину, реже равна ей (рис. 348, 351).
Боковые бархатистые пятна на лбу хорошо развиты, округлые или в виде запятой,
среднее пятно обычно имеется. Сверху брюшка боковые светлые пятна налета
небольшие, обычно начинаются со II или с последующих тергитов.
24 (25). 1-й членик усиков на спинной стороне перед вершиной с б. м. резкой
перетяжкой (смотреть сбоку!) (рис. 348). Обычно общий оттенок окраски тела и
особенно светлого рисунка сверху брюшка буровато- или оливково-серый,
реже мышино-серый 13. Н. pluvialis L.
25 (24). 1-й членик усиков на спинной стороне перед вершиной с неясной перетяжкой
или без нее (рис. 351). Общий оттенок окраски тела и особенно светлого рисунка
сверху брюшка голубовато-серый.
26 (27). 1-й членик усиков сверху в светло-сером налете, прикрывающем почти весь
членик. Щиток, как правило, в голубовато-сером налете, иногда в средней
части имеется небольшая примесь коричневатого налета. Брюшко сверху
с светлыми боковыми пятнами обычно начиная со II тергита. Крылья бледно-
серые 14. Н. subcylindrica Pand.
27 (26). 1-й членик усиков сверху лишь в основании в темно-сером или буроватом
налете. Щиток посредине с округлым пятном серого налета, тогда как по бокам
в коричневатом налете (рис. 353). Брюшко сверху со светлыми боковыми
пятнами начиная с V или IV тергита (у кавказского подвида Н. scutellata pontica
Ols. с III тергита). Крылья серые или слегка буроватые
15. Н. scutellata Ols., M. et Chv.
28 (23). Ширина лба обычно слегка превосходит высоту (рис. 345). Боковые
бархатистые пятна на лбу небольшие, треугольные или округлые, среднее пятно
отсутствует или в виде ничтожной точки. Сверху брюшка боковые светлые пятна
налета сильно развиты и имеются начиная с I (реже со II) тергита ....
11. Н. pallidula Krob.
29 (22). Вершина крыла с широкой поперечной беловатой полосой, продолженной
по его заднему краю в непрерывную светлую широкую кайму (рис. 347) . .
12. Н. desertorum Szil.
30 (21). 3-й членик усиков целиком черный (рис. 354, 355).
31 (34). Светлые пятна на крыльях расположены в беспорядке или собраны б. м.
кольцеобразно (рис. 354). Заднекрайние ячейки с косыми пятнами,
расположенными отступя от края крыла. 1-й членик усиков относительно небольшой,
заметно короче высоты лба (рис. 353, 354). Европ. часть СССР, Кавказ,
Зап. Сибирь.
32 (33). 1-й членик усиков приблизительно в 2—2х/2 раза длиннее своей наибольшей
толщины (рис. 353). Высота лба обычно слегка превосходит его ширину.
Мембрана крыла вокруг поперечных жилок без коричневатого затемнения . .
15. Н. scutellata Ols., M. et Chv.
33 (32). 1-й членик усиков приблизительно в 13/4—2 раза длиннее своей наибольшей
толщины (рис. 354). Ширина лба в основании обычно слегка превосходит его
высоту. Мембрана крыла вокруг поперечных жилок с легким коричневатым
затемнением 16. Н. crassicornis Wahlbg.
34 (31). Светлые пятна на крыльях сгруппированы в резкие розетки (рис. 356).
Заднекрайние ячейки обычно со светлыми пятнами — колечками, образующими
кайму. 1-й членик усиков большой, муфтообразный, почти равен высоте лба
(рис. 356, 357). Восточная Сибирь, Дальний Восток.
35 (36). 3-й членик немного длиннее 1-го (рис. 356). II тергит брюшка посредине
с серой полоской или треугольником, последующие тергиты часто с срединной
серой полоской; V—VI или IV—VI тергиты по бокам с округлыми серыми
пятнами (рис. 357, В) 17. Н, tamerlani Szil.
36 (35). 3-й членик равен или слегка короче 1-го (рис. 357). II тергит брюшка посредине
без серой полоски или треугольника, последующие тергиты без серых пятен,
лишь иногда VI—VII тергиты с неясными боковыми сероватыми пятнами
18. Н. stackelbergi Ols.
Б. Глаза соприкасаются: SS
1 (4). Бархатистое черное пятно на лобном треугольнике верхними углами не
достигает края глаз (рис. 340).
2(3). 1-й членик усиков короткоовальный, его длина превосходит наибольшую
толщину приблизительно в lVg раза (рис. 342) .... 9. Н. turkestanica Krob.
13. НАЕШАТОРОТА 4°8
3 (2). 1-й членик усиков удлиненно-овальный, его длина превосходит наибольшую
толщину приблизительно в 2 раза (рис. 340) 8. Н. pallens Lw.
4 (1). Бархатистое черное пятно на лобном треугольнике верхними углами достигает
или почти достигает края глаз.
5 (16). 3-й членик усиков в основании коричневый.
€ (9). 1-й членик усиков большой, эллиптический, его длина превосходит толщину
явственно более чем в 2 раза (рис. 336).
7 (8). 1-й членик усиков сверху целиком в сером налете. Коричневая окраска по
бокам брюшка сверху хорошо развита, на III тергите она может занимать
с каждой стороны кольца до V3 его ширины. I—III стерниты по бокам
коричневые. Боковые серые пятна начиная с IV тергита брюшка хорошо развиты . .
1. Н. grandis Macq.
8 (7). 1-й членик усиков сверху по крайней мере в концевой части не прикрыт
налетом, блестяще-черный. Коричневая окраска по бокам брюшка сверху слабо
развита, снизу стерниты целиком черно-серые. Боковые серые пятна налета
начиная с V тергита брюшка слабо развиты 4. Н. italica Mg.
9 (6). 1-й членик усиков б. м. яйцевидный, его длина превосходит толщину не более
чем в 2 раза (рис. 347, 348).
10 (15). Крылья без широкой беловатой каймы по заднему краю (иногда лишь с узкой
каемкой).
11 (12). Щиток посредине с округлым пятном серого налета, тогда как по бокам в
коричневом налете 15. Н. scutellata 01s.,M. et Chv.
12 (11). Щиток равномерно покрыт темно-серым или буроватым налетом.
13 (14). 1-й членик усиков сверху почти'на всем протяжении (или целиком) в сером
налете. Светлый рисунок брюшка голубовато-серый. Крылья с бледным серым
рисунком 14. Н. subcylindrica Pand.
14 (13). 1-й членик усиков сверху по крайней мере в концевой г/2 блестяще-черный.
Светлый рисунок брюшка буровато-оливковый. Крылья с хорошо выраженным
буроватым рисунком 13. Н. pluvialis L.
15 (10). Крылья с1 широкой беловатой каймой по заднему краю (рис. 347) ....
12. Н. desertorum Szil.
16 (5). 3-й членик усиков целиком черный (рис. 354, 356).
17 (18). 1-й членик усиков эллиптический, кпереди слегка сужен, целиком в сером
налете. Горы Средн. Азии 5. Н. bactriana Ols.
18 (17). 1-й членик усиков яйцевидный, блестяще-черный. Распространение иное.
19 (20). Брюшко целиком черное, без коричневых боковых пятен в основании. 1-й
членик усиков сверху в черных волосках 16. Н. crassicornis Wahlbg.
20 (19). II тергит брюшка с небольшими коричневыми пятнами по бокам. 1-й членик
усиков сверху в желтых волосках 17. Н. tamerlani. Szil.
1. Haematopota grandis Macq. (рис. 333) — Дождевка большая.
. Macquart, 1834, Suit, a Buff., 1 : 211; Krober, 1922 : 147 (Chrysozona
italica var. grandis Macq.); S u г с о u f, 1924 : 80 (Chrysozona); Krober in Lin d-
n e r, 1925 : 48 (Ch. italica grande Macq.); Олсуфьев, 1937а : 333 (Chrysozona);
Moucha et Chvala, 1956 : 262; L e с 1 e г с q, 1966 : 181; Chvala, Lyno-
borg a. Moucha, 1972 : 409. — fraseri Austen, 1925 : 9, пит. по: С h v a 1 a,
L у n e b о r g a, Moucha, 1972 : 409. — iranica Abbassian-Lintzen,
1960b : 824 (H. grandis iranica).
2- Глаза в коротких серых волосках. Лоб широкий, с параллельными сторонами,
его высота и ширина б. м. равны, покрыт серым надетом. Лобная мозоль черная, бло-
стящая, приблизительно в Зх/а раза шире высоты, боковыми сторонами достигает крик
глаз. Боковые бархатистые пятна небольшие, округлые или лунковидные, сродное
пятно маленькое, овальное. Усики длинные, коричневые или красноватые, палочки И
концевая часть 1-го членика черные. Иногда также вся наружная сторона 1-го члепикн
черно-серая. Последний длинный, цилиндрический, его длина превосходит ширину
приблизительно в 4 раза, перед вершиной с резкой перетяжкой, отделяющей почти
сферический кончик; членик матовый, в тонком сером налете. 3-й членик слегка широ
1-го,, с толстой палочкой. Концевой членик щупалец желтоватый, довольно силыш
утолщен, его длина превосходит толщину приблизительно в 2—21/2 раза, покрыт
короткими беловатыми и черными волосками. Крылья бледно-серые, жилки
коричневые. Светлые пятна мелкие, обычно не образуют явственных розеток; задний нрпЙ
крыла целиком серый, без светлой каймы. Вершинная запятовидная поперечна)! 11(1"
лоска темная, чаще не достигает заднего края крыла. Ноги: бедра и голени жолтЫН|
концевая 1/3 (или 3/3) передних голеней черная, последующие голени с серыми
колонками. Лапки черные, основание средних и задних лапок желтое. Брюшко чорноо, об»
щий оттенок светлого рисунка оливково-серый. Тергиты по заднему краю с уакими
серыми каемками, расширенными посредине в узкий треугольник; последний достигши1
вершиной до заднего края предыдущего тергита. II и последующие тергиты но (1оКЙМ
26 н. Г. Олсуфьев
404 ■ III. ПОДСЕМ. TABANINAE
с хорошо заметными, оливково-серыми, округлыми пятнами. Снизу брюшко черно-
серое. 11.5—13 мм.
3. Глаза в густых, длинных, сероватых волосках. Затылок в длинных черных
волосках, иногда светлых в центральной части темени. 1-й членик усиков удлиненно-
эллиптический, приблизительно в 2V2 раза длиннее наибольшей толщины (смотреть
сбоку!), черный, в сером налете, иногда к вершине бока членика блестяще-черные.
3-й членик усиков красновато-коричневый, палочка черпая, площадка менее широкая,
чем у $>. Концевой членик щупалец конический, в основании сильно утолщен, его длина
превосходит наибольшую толщину в 2—21/2 раза, беловато-кремовый, в длинных белых
и черных волосках. Бедра ног красно-желтые, иногда передние и задние бедра на
значительном протяжении серые. Крылья как у $>. Брюшко: II—III (или I—IV) тергиты
Рис. 333. Haematopota grandis Macq. Крыло, лобная полоска,
усик и щупальце §.
по бокам с крупными коричневыми пятнами, на III тергите они занимают около 1/3
(и более) ширины кольца. Серые боковые пятна налета начиная с IV (или III) тергита
хорошо развиты. I—III стерниты брюшка по бокам желто-коричневые, остальное
черно-серое. 11—12 мм.
Похожа на И. italica, но 5 легко отличается желто-коричневым 1-м члеником
усиков с резкой перетяжкой на конце, более толстыми щупальцами и более широким
лбом; общий тон окраски брюшка сверху буровато-оливковый, боковые светлые пятна
начиная с II тергита. 8 отличается главным образом более развитой коричневой
окраской по бокам брюшка и более значительным покрытием серым налетом 1--го членика
усиков; боковые серые пятна с IV (или III) тергита.
Распространение: южноевропейский (средиземноморский) вид. В СССР
известен из немногих мест: Полтавская (Миргородский р-н), Одесская области (Ново-
Ивановка), Молдавия (Страшены, Котовск), окр. Ставрополя, Абхазия (Новый Афон).
Ю. и частью Средн. Европа (на запад до Испании), Марокко, Турция. В СССР редок.
Июль.
Аббассиан-Линцеи (Abbassian-Lintzen, 1960b) описала из Ирана подвид Я.
grandis iranica, но он заметно отличается от номинативной формы и, возможно,
представляет собой самостоятельный вид. Но Н. grandis iranica близок, если не идентичен,
Н. pavlovskii.
2. Haematopota pavlovskii Haus. (рис. 334) — Дождевка Павловского.
Г а у з е р, 1960, Энтом. обозр., 39, 3:659 (Chrysozona); L е с 1 е г с q,
1966 : 186.
$. Глаза в очень коротких, беловатых волосках. Лоб с параллельными сторонами,
его высота и ширина приблизительно равны, покрыт светло-серым налетом. Боковые
бархатистые пятна черные, умеренной величины, овальные, среднее пятно небольшое.
Лобная мозоль черная, блестящая, ее ширина в 4 раза превосходит высоту, боковыми
сторонами достигает края глаз. Усики: 1-й членик б. м. цилиндрический, его длина
13. HAEMATOPOTA
405
превосходит толщину в 3V2 раза, с едва заметной перетяжкой перед вершиной,
красновато-коричневый, к вершине сероватый (иногда целиком серый); 3-й членик в базаль-
ной части равен ширине 1-го, красновато-коричневый, к концу темнее, палочка черная.
Концевой членик щупалец палевый, утолщен, его длина превосходит наибольшую
толщину в 2 раза, покрыт беловатыми и единичными черными волосками. Спинка с 5
беловато-серыми, хорошо заметными продольными полосками. Крылья бледно-серые,
светлый рисунок из умеренной величины пятен, не образующих явственных розеток.
Ш£Ш 4-...-^..v;«,:
.,.,мт I m^
Рис. 334. Haematopota pavlovskii Haus. Крыло,
лоска, усик и щупальце ?.
лобная по-
Вершинная поперечная полоска узкая, достигает заднего края крыла, последний с
узкой светлой каймой. Ноги: бедра и голени желтые, бедра с сероватой вершиной
(иногда основание также сероватое), концевые 2/3 передних голеней черноватые,
последующие голени с коричневато-серыми колечками; лапки черные, основание средних и
задних лапок желто-коричневое. Брюшко черно-серое, сверху с четким светло-серым
рисунком из налета и волосков: среднего ряда узких треугольников (или продольных
полосок), вершиной достигающих края предыдущего тергита, Заднекрайних умеренно
широких каемок, переходящих на боковой край тергитов, и крупных боковых
округлых пятен начиная с I тергита. Снизу брюшко светло-серое. 10—11 мм.
(J неизвестен.
Распространение: Нахичеванская АССР (долина Аракса). Редок.
Июнь—начало августа.
Н. pavlovskii весьма близок к Я. grandis iranica Abbas.-L., 1960, возможно, что они
идентичны, но для решения этого вопроса необходим дополнительный материал.
Взаимоотношения этих форм с Я. grandis не совсем ясны.
3. Haematopota caspica Abbas.-L.—Дождевка каспийская.
Представлена 2 подвидами: номинативным, распространенным в южном Приказ
пии Н. caspica caspica Abbas.-L. и кавказским Н. caspica transcaucasica Ols.
За. Haematopota caspica caspica Abbas.-L. (рис. 335, A) — Дождевка каспийская
номинативная.
Abbassian-Lintzen, 1960b, Bull. Soc. Path. Exotique, 53, 5 : 825
(Я. pluvialis caspica); 1964 : 300; L e с 1 e г с q, 1966 : 179.
$. Глаза в очень коротких, едва заметных волосках. Лоб с параллельными
сторонами, его высота заметно превосходит ширину. Лобная мозоль блестяще-черная, оо
ширина превосходит высоту в 3 раза. Боковые черные бархатистые пятна округлые
или слегка продолговатые, небольшие, среднее пятно крошечное. 1-й членик усикоп
в основании красно-коричневый, к вершине черноватый, в 21/а раза длиннее
наибольшей толщины, в дистальной части слегка утолщен и перед вершиной сверху с
небольшой перетяжкой (смотреть сбоку!). 3-й членик в базальной части равен шириио 1-го
26*
406 III. ПОДСЕМ. TABANINAE
или слегка уже последнего, площадка коричневая, в основании красноватая, палочка
черная. Концевой членик щупалец беловато-палевый, его длина превосходит
наибольшую толщину в 3—Зх/2 раза, покрыт светлыми и черными волосками. Ноги: бедра
желтые, с сероватой вершиной, передние бедра иногда сероватые; остальное как обычно.
Крылья слегка буроватые, с мелкими светлыми пятнами. Вершинная поперечная
полоска редуцирована до небольшого продолговатого пятнышка. Задний край крыла
без светлой каймы. Брюшко сверху черно-коричневое, тергиты по заднему краю с узкой
буровато-серой каймой, II тергит посредине с такого же цвета узким треугольником,
на последующих тергитах узкая слабо заметная сероватая срединная полоска.
Боковые буровато-оливковые пятна небольшие, расположены начиная с IV или III тергита.
Снизу брюшко черно-серое. 9—11 мм.
в неизвестен.
Близка к Н. variegata F., отличаясь главным образом более коротким и слегка
утолщенным в дистальной части 1-м члеником усиков.
Распространение: Ленкоранская низменность в Азербайджане. Иран.
JB СССР не часто. Июль—начало сентября.
Рис. 335. А — лобная полоска, усик и щупальце Haematopota
caspica caspica Abbas.-L.; Б — то же 2 Н. с. transcaucasica Ols.
3b. Haematopota caspica transcaucasica Ols. (рис. 335, Б) — Дождевка каспийская
закавказская.
Олсуфьев, 1970, Энтомол. обозр., 49, 3:687.
2. От номинативной формы отличается более округлыми боковыми бархатистыми
пятнами на лбу, менее узким лбом и несколько более длинным 1-м члеником усиков —
его длина превосходит толщину в 3 раза; светлые пятна на крыльях немного крупнее,
вершинное поперечное пятно нормально развито в виде запятой. 9—10 мм.
Распространение: Грузия (Тбилиси); Азербайджан (Белоканы,
Закатали, Куткашен, низовья р. Самур). Июнь—сентябрь. Местами обычен, приурочен
обитанием к лесным участкам.
Ранее эту форму мы (Олсуфьев, 1937а, 1941) и другие авторы (Гаузер, 1953; Джафа-
ров, 1960) определяли неправильно как Н. variegata F. Наше указание (1937а) Н.
variegata для Нальчика, Кавказ, основано на ошибочной этикетке.
4. Haematopota italica Mg. (рис. 336) — Дождевка западная.
Meigen, 1804, Klassific, 1:163; Lundbeck, 1907:106; Verra II,
1909 : 341; К г о Ь е г, 1922 : 145 (Chrysozona); Surcouf, 1924 : 79 (Chrysozona);
Krober in Lindner, 1925:48 (Chrysozona); Олсуфьев, 1937а : 337
(Chrysozona); Moucha et Chvala, 1956 : 263 (Chrysozona); Leclercq, 1966 :
182; Chvala, Lyneborg a. Moucha, 1972 : 414.
5. Глаза в очень коротких серых волосках. Лоб с параллельными сторонами, его
высота слегка превосходит ширину, реже равняется ей, покрыт серым налетом.
Боковые бархатистые пятна крупные, округлые, среднее пятно маленькое, овальное.
Лобная мозоль блестяще-черная, ее ширина в З1^—4 раза превосходит высоту, боковыми
сторонами достигает края глаз. Усики черные, 3-й членик в основании обычно
красновато-коричневый. 1-й членик черный, в сером налете, цилиндрический, его длина
превосходит толщину приблизительно в 4 раза. Перед вершиной членик с легкой перетяжкой.
Концевой членик щупалец желтоватый, серый, узкий, его длина превосходит толщину
в 3—3Vg раза, покрыт белыми и черными волосками. Крылья бледно-буровато-серые,
13. HAEMATOPOTA 407
жилки коричневые. Светлые пятна довольно мелкие, расположены б. м. беспорядочно.
Вершинная поперечная полоска узкая, запятовидная, часто не достигает заднего края
крыла. Этот последний с узкой беловатой каемкой или целиком серый. Ноги: бедра
серые, средние и задние бедра часто на некотором протяжении желтые; голени желтые,
вершинные 2/3 передних голеней черные, последующие голени с коричневыми колеч-
Рис. 336. Haematopota italica Mg. Крыло, лобная полоска, усик, щупальце, VIII стер-
нит И' церки д. А — усик #.
/
ками. Лапки черные, основание средних и задних лапок желтое. Брюшко черное,
тергиты с резкими, светло-серыми (голубоватыми) узкими каемками по заднему краю.
Посредине имеется узкая серая продольная полоса (последняя часто на II тергите
кзади слегка расширена). Серые боковые пятна небольшие, малозаметные, на V—VI
Рис. 337. Ареал Haematopota italica.
или IV—VI тергитах. Снизу брюшко черно-серое, в густом беловатом налете. УШ,стер-
нит широкий, коричнево затемнен, гонопофиз с умеренной апикальной выемкой;
латеральные лопасти небольшие, не отграничены от гонопофиза, основной край базалыюи
лопасти прямой. Церки и семеприемники как у Н. pluvialis. 9.5—12.5 мм.
3- Глаза в длинных, желтовато-серых волосках. Затылок за глазами с рядом
длинных, желтовато-серых и более темных коричнево-черных волосков. Лобный
треугольник с черным пятном, почти достигающим верхними углами края глаз. Усики
408 III. ПОДСЕМ. TABANINAE
в большей своей части черные; 3-й членик в основной половине коричневатый. 1-й
членик крупный, блестяще-черный, длинноовальный; его длина превосходит толщину
в 21/2—3 раза, покрыт длинными черными волосками. Спинная сторона членика в
большей части в сером налете. У лекоторых экземпляров в коллекции 1-й членик темно-
коричневый (выцветание). Концевой членик щупалец слегка желтоватый, довольно
толстый, конический, покрыт белыми волосками. Крылья и ноги как у $>. Брюшко
черное, II терпит по бокам коричневатый. Все тергиты по заднему краю с узкими
желтовато-серыми каемками. II тергит посредине с узким сероватым треугольником,
последующие с узкой продольной полоской. V—VI тергиты с неясными, округлыми,
серыми боковыми пятнами. Снизу брюшко одноцветно-серое. 8—11 мм.
Распространение: европейский лесной вид (рис. 337). В СССР
распространена от Прибалтики и запада Украины на восток до Урала (Свердловск, разъезд
Укагач), на север до оз. Яписярви Карельской АССР (61°55' с. ш.), Вельска
Архангельской обл. и Печоро-Илычского заповедника Коми АССР (62° с. ш.), на юг до
Молдавской ССР (Унгены, Страшены), Киевской обл. (Канев), Сум, Пензы, Татарской
АССР (Бугульма) и Башкирской АССР (Караидель).. Европа (кроме севера), Сев.
Африка. Не часто. Июнь—август.
Рис. 338. Haematopota bactriana Ols. Лобная полоска, усик, VIII стер-
нит и церки (варианты) §.
5. Haematopota bactriana Ols. (рис. 338) — Дождевка бактрийская.
Олсуфьев, 1939, Тр. Таджикск. базы АН СССР, И : 100 (Chrysozona); Б а р а-
т о в, 1961 : 187 (<J); L е с 1 е г с q, 1966 : 178.
$>. Глаза в очень коротких светлых волосках. Лоб с параллельными сторонами
или слегка расширен книзу, его ширина намного превышает высоту., Лобная мозоль
блестяще-черная, узкопоперечная, ее ширина в 5—6 раз превосходит высоту.
Боковые черные бархатные пятна небольшие, б. м. троугольпой формы, реже круглые,
среднее пятно небольшое, черное. 1-й членик усиков б. м. цилиндрический, перед
вершиной обычно с незначительной перетяжкой, в 2V2—3 раза длиннее наибольшей
толщины, черный, целиком в сером налете. 3-й членик не шире 1-го, целиком черный.
Концевой членик щупалец беловато-палевый, иногда сероватый по внутренней
стороне, в 2х/2—3 раза длиннее наибольшей толщины, в светлых волосках, иногда с
незначительной примесью черных. Ноги: бедра черные, в сером налете, остальное как обычно.
Крылья бледно-серые с обычным мелким светлым рисунком. Брюшко сверху
черное с рисунком из голубовато-серого налета: узких каемок по заднему краю тергитов,
срединной продольной полоски, на II тергите расширенной в узкий треугольник и
небольших боковых округлых пятен начиная с V, реже IV тергита. Снизу брюшко
черно-серое. VIII стернит: длина немного превосходит ширину, гонопофиз с умеренной
апикальной выемкой, латеральные лопасти небольшие, боковые коричневые пятна
средней величины или большие, основной край базальной лопасти б. м. прямой. Церки
и семеприемники как у Н. pluvialis. 8—11 мм.
в- Глаза в длинных светло-серых волосках. Затылок в длинных, преимущественно
светлых волосках. Лобный треугольник с черным пятном, достигающим верхними
углами края глаз. Усики: 1-й членик черный, в густом сером налете, в 21/2 раза
длиннее наибольшей толщины, сверху в длинных, преимущественно черных волосках.
3-й членик черный. Концевой членик щупалец сероватый, в 2 раза длиннее
наибольшей толщины, в беловато-серых волосках. Крылья и ноги как у ?. Брюшко черное,
V.
13. HAEMATOPOTA
409
без коричневых пятен по бокам (по Баратову, 2—3-й тергиты по краям желтые).
Тергиты по заднему краю с узкими желтовато-серыми каемками, V—VI (или IV—VI)
тергиты по бокам с небольшими голубовато-серыми округлыми пятнами и посредине
с небольшой продольной полоской такого же цвета. Снизу брюшко черно-серое. 9—
11 мм.
Близка к Н. italica, 5 отличается широким лбом и более коротким 1-м члеником
усиков, в — целиком покрытым серым налетом 1-м члеником усиков и отсутствием
коричневых пятен по бокам брюшка.
Распространение: среднеазиатский горно-лесной вид. В СССР известен
из Узбекистана — горы Байсунтау, западный отрог Гиссарского хр. и Таджикистана —
хребты Гиссарскип, Зеравшанскнй, Каратегинский, на высотах от 1300 до 3600 м.
Шугпан, Афганистан. Местами обычен. Июнь—август.
Голотип: 2, урочище Квак, Гиссарский хр., 2000 м, 35 км сев. Душанбе, 20 VII1937.
Гуссаковский. В коллекции Зоологического института Академии паук СССР,
Ленинград.
Рис. 339. Haematopota longeantennata Ols. Крыло, лобная
полоска, усик и щупальце §.
6: Haematopota longeantennata Ols. (рис. 339) — Дождевка длинноусая.
Олсуфьев, 1937а, Фауна СССР, Двукрылые, VII, 2 : 338 (Chrysozona); L е с-
1 е г с q, 1966 : 184; Chvala, Lyneborg a. Moucha, 1972 : 416.
§. Глаза в коротких, скудных волосках. Лоб широкий, с б. м. параллельными
сторонами, его ширина слегка превосходит высоту, покрыт серым налетом. Боковые
бархатистые пятна небольшие, черные, б. м. округлые. Среднее пятно небольшое, округлое,
коричневое. Лобная мозоль блестяще-черная, ее ширина приблизительно в 5—6 раз
превосходит высоту, боковыми сторонами немного не достигает края глаз, отделяясь
от него узкими полосками серого налета. Усики очень длинные, целиком черные.
1-й членик в сером налете и Коротких черных волосках, цилиндрический, его длина
в 4—5 раз превосходит толщину. Перетяжка перед вершиной слабо выражена. 3-й
членик часто слегка шире 1-го. Концевой членик щупалец желтоватый, в белых и черных
волосках; его длина превосходит толщину в 2V2—3 раза. Крылья
бледно-буровато-серые, жилки коричневые, светлые пятна небольшие, расположены беспорядочно,
не образуя розеток. Вершинная поперечная полоска узкая, запятовидная, не достигав*
заднего края крыла. Последний целиком серый, без беловатой каемки или с узкой
каемкой. Ноги: бедра черные в сером налете, передние голени в основной 1/3 желтые,
далее черные, средние и задние голени желтые, с 3 черноватыми колечками; лапки
черные, основание средних и задних лапок желтоватое. Брюшко черное, тергиты по
заднему краю с узкой светло-серой каемкой. II тергит с узким светло-серым срединным
треугольником, последующие с продольной полоской того же цвета. V—VI тергиты
по бокам с округлыми, неясными, сероватыми пятнами. Снизу брюшко черно-серое,
в беловатом налете. 8—11 мм.
3 неизвестен.
Похожа на Н. italica, отличаясь более широким лбом, узкой лобной мозолью и
длинными, целиком черными усиками.
410 III. ПОДСЕМ: TABANINAE
Распространение: балкано-кавказский (восточносредиземноморский)
вид. В СССР известен в Закавказье — Грузия (Гагра, Кутаиси), Азербайджан (Геок-
тапа Агдашского р-на; Кусары; низовья Самура; Белоканы; Закаталы). Балканы.
Редок. Июнь.
Голотип: g, Кутаиси, Грузия, 1875, А. Ильин. В коллекции Зоологического
института Академии наук СССР, Ленинград.
*7. Haematopota caenofrons Krab.
К г 6 b е г, 1922, Arch. f. Naturg., 88, А, 8 :141 (Chrysozona); Krober in
Lindner, 1925 : 45 (Chrysozona); Олсуфьев, 1937а : 339 (Chrysozona); Leclercq,
1966 : 179.
В коллекциях Зоологического института Академии наук СССР этот вид
отсутствует, поэтому мы приводим оригинальное описание (Krober, 1922).
£. «Лоб составляет около 1/2 ширины головы, кверху слегка сужен, темно-пепельно-
серый. Посредине протянута вниз одна бело-серая полоска, которая на конце пуговице-
образно расширена, именно там, где обычно расположено бархатистое пятно. Оба
боковых бархатистых пятна очень маленькие и расположены в бело-сером квадрате.
Границу с лицом составляет узкая, черная, умеренно блестящая, поперечная полоска.
Черное бархатистое пятно между усиками отсутствует. Лицо целиком бело-серое,
с незначительными слабо заметными темными точечками. Наличник без пятен. Волоски
длинные, беловатые, на лбу такие же, но с вкрапленными черными волосками.
Концевой членик щупалец узкий, бело-желтый, лишь в белых волосках. Затылок серый,
в коротких, желтоватых волосках. Усики черно-коричневые. 1-й членик
длинно-цилиндрический (1 мм), по крайней мере в 4 раза длиннее толщины. 2-й очень короткий,
3-й отсутствует. Грудь черно-коричневая, с 3 беловатыми продольными линиями.
Боковые волоски и бочки груди светло-серые. Черные части в шоколадно-коричневом
налете. Волосяной покров нежный, беловатый. Закрыловые чешуйки коричневатые.
Жужжальца беловатые, основание головки коричневое. Брюшко белогсерое. Боковые
пятна окаймлены чисто-коричневым, до III кольца включительно. Задние края
светлые, в беловатых волосках. Брюшко снизу светло-пепельно-серое, в белых волосках.
Тазики и бедра черно-коричневые, благодаря налету серые, передние голени с светлой
базальной третью, остальные с 2 светлыми колечками. Лапки черные, основание
средних и задних лапок желтоватое. Крылья молочно-белые с коричневыми, грубыми
пятнами, которые скоплены к переднему краю. Передняя базальная ячейка и весь задний
край крыла почти без пятен. Придаток г4 очень длинный до дискоидальной ячейки.
Длина 7 мм, длина крыла 8, ширина 2.7 мм».
Распространение: Казикопоран, Сев.-Вост. Турция.
8. Haematopota pallens Lw. (рис. 340) — Дождевка бледная.
Loew, 1870, Schrift. Ges. Freund. Nat. Moscou : 54; 1871, 2:61; В е с k e r,
1912:587; Олсуфьев, 1937а : 340 (Chrysozona); Шевченко, 1961:171
(Chrysozona); Leclercq, 1966 :485; Chvala, Lyneborg a. Moucha,
1972 : 407. — caucasica Krober, 1922 : 142 (Chrysozona). — ? araxis S z i 1 a d y,
1923 : 33. i
g. Глаза в коротких волосках. Лоб широкий, с параллельными сторонами, его
ширина заметно превосходит высоту, покрыт светло-серым налетом. Боковые
бархатистые пятна черные, небольшие, округлые. Среднее пятно маленькое, черное. Лобная
мозоль блестяще-черная, узкопоперечная, ее ширина в 4—5 раз превосходит высоту.
Боковыми сторонами мозоль немного не достигает края глаз, отделяясь узкой,
клиновидной полоской налета. Усики длинные, черные, 3-й членик очень узкий, уже, чем 1-й,
в основании коричневый. 1-й членик в сером налете, б. м. цилиндрический, его длина
превосходит толщину приблизительно в 31/2—4 раза. Членик в профиль неровный,
имеет довольно резкую перетяжку перед вершиной и, кроме того, часто имеется легкий
перехват в средней части. Здесь членик иногда бывает заметно утолщен и тогда
перехват выделяется более резко. В отдельных случаях на переднем конце имещтся еще
дополнительно мелкие перетяжки, придающие членику узловатый вид. Членик иногда
в основной части, в особенности изнутри, коричневый. Концевой членик щупалец
слегка желтоватый или белый, узкий, покрыт белыми и единичными черными волос--
ками. Крылья светло-серые, жилки желто-коричневые, к вершине более темные.
Светлые пятна многочисленны и расположены беспорядочно, не образуя замкнутых
розеток. Вершинная поперечная полоска двойная, разделена на 2 полоски: внешняя
полоска более широкая, достигнув заднего конца крыла переходит затем в широкую
беловатую кайму, внутренняя полоска узкая, волнисто изогнута. Эти полоски
варьируют: они могут быть слиты в одну общую широкую полоску и, наоборот, внутренняя
полоска может почти совсем отсутствовать. Ноги: бедра черные, *в сером налете,
передние голени черные, в основной 1/3 желтые, последующие голени желтые, с 3 черно-
13. HAEMATOPOTA
411
ватыми колечками. Лапки черные, основание средних и задних лапок желтое.
Встречаются экзем^ляртЕ с желтоватыми средними и задними бедрами. Брюшко черное, тер-
ги^ы по зад™Ркраю с узкими светло-серыми каемками. Посредине имеется узкая,
Рис. 340. Haematopota pallens Lw. Крыло, лобная полоска, усик VIII этернит
(варианты), церки и сперматека ?. А — усик; Ь — лоо в.
серая, продольная полоска. II—VI тергиты с большими, светло-серыми б°,к°г™
пятнами, отчасти сливающимися с боковым светло-серым рантом брюшка.. Снизу
брюшко черно-серое, в светло-сером налете. VIII стернит: гонопофиз со слабо^выем-
о° к востоку от Гринвича 90° <го° ^о°
150°
Рис. 341. Ареал Haematopota pallens.
кой, латеральные лопасти умеренно развиты, ясно отграничены от гонопофиза
боковые коричневые пятна крупные, базальная лопасть со слегка вогнутым основным
краем. Церки и семеприемники как у Н. pluviahs. 7.5—11 мм. „„„TTV „,,„
Л Глаза в длинных светло-серых волосках. Затылок в длинных, светло-серых или
желтоватых волосках. Лобный треугольник в серебристо-сером налете; черное Oupxu-
тистое пятно небольшое, далеко не достигает верхними углами края wiuj. Усики.
412
III. ПОДСЕМ. TABANINAE
1-й членик блестяще-черный, продолговато утолщенный, его длина превосходит
толщину приблизительно в 2 раза. Спинная сторона членика в основной половине в сером
налете- и в довольно длинных светло-серых волосках, тогда как в концевой части в
черных волосках. 3-й членик черный, в основании коричневатый, узкий. Концевой членик
щупалец желтоватый, тупоконический, относительно узкий, покрыт белыми волосками.
Бочки груди в сером налете и в светло-серых волосках, иногда с значительной
примесью черных волосков на мезоплеврах. Придатки груди и брюшко как у $, II тергит
с желто-коричневым пятном по бокам. Последнее часто немного заходит на III тергит
9—12 мм.
Характерный вид с широким лбом, длинным 1-м члеником усиков и широкой
беловатой каймой по заднему краю крыла:
Распространение: берега водоемов пустынной зоны, преимущественно
проточных, частично проникает в степную зону (рис. 341). Аллопатричеп И.
turkestanica. Юг Украины (дельта Дуная, устье Днепра, Крым, Приазовье), Молдавия, восток
Воронежской (Хоперский заповедник), Ростовская (Цимлянская, низовья Дона),
Волгоградская области (Ильмень-Суворовский на Дону, 48° с. ш.), север
Калмыцкой АССР (Обильное), дельта Волги, Урала, Сев. Кавказ (от Ейска и Анапы па восток
до 10. Дагестана), Азербайджан (Курино-Араксинская и Ленкоранская низменности,
Нахичевань), Армения (Степапавап, дол. Аракса), Грузия (окр. Тбилиси; Шираки-
Эльдари), юг Туркмении, Таджикистан, Узбекистан, Киргизия (Таласская, Чуйская
и Ферганская долины; Дараут-Курган в Алайской долине, 2500 м); Казахстан
(предгорья Зап.^Тянь-Шаня и Каратау, по Сырдарье до Казалипска). Турция, Ирак, Иран,
Зап. Китай (дол. Кызылсу, Зап. Кашгария). Местами обычна. Июнь—август, иногда
раньше или позднее.
Указания на распространение Н. pollens в Прибалхашье относятся к Н.
turkestanica (В. В. Шевченко, личное сообщение). Сообщение Г. В. Бошко (1950) об
обнаружении этого вида близ г. Остер Черниговской обл. нуждается в подтверждении.
9. Haematopota turkestanica Krob. — Дождевка туркестанская.
Изменчивый вид, представлен 2 подвидами: западным наминативным Н.
turkestanica turkestanica Krob. и восточным И. turkestanica orientalis Ols. Описанная Е. Г. Гау-
зером (1960) из Нахичеванской АССР Н. nubilis, видимо, является локальной формой
Н. turkestanica; в публикации А. В. Богачева и Н. Г. Самедова (1949),она определена
как Н. sobrina Krob. С Н. nubilis Haus. большое сходство обнаруживает Н. atropathe-
nica, описанная Аббассиан-Линцен (Abbassian-Lintzen, 1964b) из Ирана. Недостаток
коллекционного материала (мы видели только 2 22 Н- nubilis и не имели Н. atropathe-
nica) не позволяет высказаться более определенно.
9а. Haematopota turkestanica turkestanica Krob. (рис. 342) — Дождевка
туркестанская номинативная.
К г б Ь е г, 1922, Arch. i. Naturg., 88, А, 8 :151 (Chrysozona); Krober in Lind-
nе г, 1925 : 53 (Chrysozona); Олсуфьев, 1936а : 218 (3) {Chrysozona); 1937a : 342
(Chrysozona); Шевченко, 1961:172 (Chrysozona); L e с 1 e г с q, 1966-189-
Виолович, 1968 :57; Chvala, Lyneborg a. Moucha, 1972 : 424. —
■ mmuscula var. turkestanica S z i 1 a d y, 1922 : 21, цит. по: L e с 1 ет с q, 1966 : 189 —
?nubilis Hauser, 1960 : 659 (Chrysozona). — ? atropathenica
Abbassian-Lintzen, 1964b : 170.
?. Глаза в очень коротких волосках. Лоб широкий, с параллельными сторонами,
его ширина значительно превосходит высоту, покрыт светло-серым налетом. Боковые
•бархатистые пятна небольшие, треугольные или округлые, черные. Среднее пятно
лишь немного меньше боковых, округлое, черное. Лобная мозоль блестяще-черная
узкопоперечная, ее ширина превосходит высоту в 5 раз, в средней части выдается вверх
узким мысом. Боковыми сторонами мозоль не достигает края глаз, отделяясь полоской
налета. Усики, умеренной длины, 1-й членик черный, изнутри коричнево-желтый
целиком покрыт серым налетом; в средней части членик муфтообразно утолщен перед
вершиной имеется резкая перетяжка. Длина членика превосходит наибольшую
толщину в 2 /2—3 раза. Его окраска варьирует, и встречаются экземпляры, у которых
членик почти целиком коричнево-желтый, с сероватым кончиком. 2-й членик коричневый
d-и членик черный, в основной части желтовФго-коричневый, узкий, заметно уже 1-го
Концевой членик щупалец беловатый или слегка желтоватый, узкий, покрыт
преимущественно белыми волосками. Крылья светло-серые, жилки коричневые. Светлые пятна
многочисленные, сливающиеся между собой, часто образуют розетки. Вершинная
поперечная полоска состоит из 2 полосок: внешняя широкая, оставляет у крыла лишь
ничтожный сероватый кончик, внутренняя узкая, изогнутая кнаружи. Задний край
крыла с широкой беловатой каймой. Ноги: бедра желто-коричневые, более темные
к вершине, голени желтые. Передняя пара голеней в концевой V» или 2Д< черная
последующие пары с 3 черноватыми колечками. Лапки черные, основание средних и
13. HAEMATOPOTA
413
задних лапок желтое. Брюшко черное, с узкой, голубовато-серой продольной
полоской посредине и начиная с II тергита с довольно резкими боковыми,
голубовато-серыми пятнами. Последние иногда настолько сильно развиты, что сливаются в
боковые, довольно широкие, продольные полосы. Задние края тергитов с узкими серыми
Рис. 342. Haematopota turkestanica turkestanica Krob. Крыло, лобная полоска, усик,
щупальце, VIII стернит, церки и сперматека (варианты) $.4 — усик $.
каемками. Снизу брюшко черно-серое, в светлом,голубовато-сером налете. Терми
налии сходны с таковыми Н. pollens, базальная лопасть немного длиннее. 8—11мм.
S- Глаза в довольно длинных, светло-серых волосках. Затылок за глазами с рядом
умеренно длинных светло-серых волосков. Лобный треугольник в сером налете, посро-
Рис. 343. Ареалы: 1 — Haematopota turkestanica turkestanica; 2 — Н.
turkestanica orientalis.
дине с небольшим бархатисто-черным треугольным пятном. Верхние углы этого шпиц
закруглены и далеко не достигают края глаз, отделяясь серым налетом. Усики и Cwjiir
шей своей части черные; 3-й членик в основной половине коричневатый. 1-й члшшк
сильно утолщен, короткоовальный (его длина превосходит толщину приблинитолыш
в lVj раза), покрыт умеренно длинными черными волосками, тогда как и ог.моииои
части — серыми волосками. Концевая 1/3 членика блестящо-чориая, остальное н том
ком сером налете. Концевой членик щупалец желтовато-белый, в основании утолщим,
далее равномерно сужен, покрыт светлыми, умеренно длинными, болоиатыми полис-
414 ' III. ПОДСЕМ. TABANINAE
ками. Ноги и крылья как у g.- Брюшко черное, с небольшими коричневыми пятнами
по бокам II и частью I и III тергитов. II тергит с небольшими, III и последующие
с довольно крупными, хорошо заметными серебристо-серыми пятнами по бокам. Эти же
тергиты по заднему краю с узкой серой каемкой. IV—VI тергиты посредине с серой
продольной полоской. Снизу брюшко черно-серое, в густом светлом налете. 10—11 мм.
Очень похожа на Н. pallens, главным образом отличаясь более коротким 1-м
члеником усиков.
Распространение: берега водоемов, в том числе соленых, пустынной и
частью степной зон (рис. 343, 1). Аллопатричен Н. pallens. В СССР подвид
распространен от устья Днепра на восток до Обско-Енисейского водораздела, на север до Канева
Киевской обл., востока Воронежской обл. (Хопер), окр. Оренбурга, юго-востока
Башкирской АССР (Новозирганово, Баилово), Челябинской обл. (Троицк), юга Тюменской
(Казанское) и Омской (Горьковское) областей, на юг до Крыма,
Кабардино-Балкарской АССР (степная часть), дельты Волги (Камызякский р-н), окр. Гурьева, низовий
Амударьи, Термезского р-на Сурхандарьинской обл., Туркестанского хребта
(заповедник Гуралаж), Папского р-на Наманганской обл. и Тянь-Шаня (Киргизский хр.^
Казарман; Нарын; Иссык-Кульская котловина; Нарынкол). Монголия (юг и восток).
Обычен в Казахстане, в западных частях ареала не часто. Май—сентябрь.
У экземпляров из северных частей ареала распространения (Зап. Сибирь,
Оренбургская обл.) светлый рисунок на брюшке, образованный налетом, не имеет
голубоватого оттенка, а слегка оливковый.
Личинки нами найдены в прибрежных частях речки (окр. Венгерова Зап. Сибирь).
Экспериментально доказанный переносчик туляремии (Олсуфьев и Голов, 1936).
9b. Haematopota turkestanica orientalis Ols. — Дождевка туркестанская
восточная.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 452.
2. От номинативного подвида отличается менее широким лбом, более крупными
боковыми бархатистыми пятнами и более узкой светлой каймой по заднему краю
крыльев, последняя изредка почти не выражена; 1-й членик усиков иногда целиком
черно-серый. 8.5—11 мм.
Распространение: (рис. 343, 2) Тувинская АССР, окр. Минусинска,
Иркутская обл( (Балаганск), Забайкалье, Приамурье и Приморский край. Сев.
Монголия, Сев.-Вост. Китай. Местами обычен. Июнь—июль, иногда позднее.
Рис. 344. Haematopota koryoensis Shir. Крыло,
лобная полоска, усик и щупальце ?.
10. Haematopota koryoensis Shir. (рис. 344) — Дождевка карликовая.
S h i r a k i, 1932, Trans. Nat. Hist. Soc. Formosa, 22 : 268 (Chrysozona);
Murdoch et Takahasi, 1969 : 88. — nana Olsufjev, 1937a : 345.
g. Глаза голые или в ничтожных волосках. Лоб с параллельными сторонами или
слегка расширен книзу, его ширина слегка превосходит высоту, покрыт серым налетом.
Боковые бархатистые пятна довольно крупные, овальные или округлые, черные.
Среднее пятно небольшое, округлое, черное. Нижняя лобная мозоль темно-коричневая,
блестящая, ее ширина превосходит высоту в 4 раза, боковыми сторонами достигает
края глаз. Усики короткие, желтые. 1-й членик усиков блестящий, б. м.
цилиндрический, его длина превосходит ширину в 3 раза, без перетяжки на конце ибез серого
13. HAEMATOPOTA
415
налета. 3-й членик с коррткой, толстой, черноватой палочкой. Концевой членик
щупалец бледно-коричневый, с сероватым оттенком, узкий, его длина превосходит толщину
приблизительно в 4 раза, покрыт преимущественно черными волосками. Крылья
коричневые, с мелкими беловатыми пятнами: последние разбросаны б. м. беспорядочно или
■образуют нечеткие розетки. Вершинная перевязь резкая, довольно широкая, часто
двойная, достигает заднего края крыла. Кончик крыла коричневый. Задний край
крыла начиная с 1-й заднекрайней ячейки окаймлен цепочкой светлых пятен. Жилки
коричневые. Ноги: бедра коричневые, передние голени преобладающе
черно-коричневые, с желтым колечком возле основания, средние и задние голени желтые, с
коричневыми колечками. Передние лапки черные, последующие темно-коричневые, с желтым
•основанием. Брюшко темно-коричневое, тергиты по заднему краю с узкой сероватой
или коричневатой каемкой. Посредине едва намечается серая продольная полоска.
Боковые серые пятна не выражены. Снизу брюшко коричневое, в сероватом налете.
6—8 мм.
3 неизвестен.
Распространение: юг Приморского края — р. Тумень-ула, граница
с КНДР; оз. Тальми и оз. Хасан; Славянский залив; Барабаш близ Сидими;
заповедник Кедровая Падь; окр. Приморского; п. Мирный Надеждинского р-на. Корейский п-ов,
-Сев.-Вост. Китай. Не часто. Июль—начало августа.
Рис. 345. Haematopota pallidula Krob. Крыло, лобная
полоска, усик и щупальце g.
11. Haematopota pallidula Krob. (рис. 345) — Дождевка ранняя.
К г о Ь е г, 1922, Archiv f. Naturg., 88, А, 8 : 152 (Chrysozona); К г б b e r in
Lindner, 1925 : 51 (Chrysozona);. Олсуфьев, 1937а : 346 (Chrysozona);
Шевченко, 1961 : 174 (Chrysozona); Leclercq, 1966 : 186; Chvala, Lyno-
borga. Moucha, 1972 : 443.
g. Глаза в очень коротких волосках. Лоб широкий, его ширина в основании слегка
превосходит высоту, покрыт желтовато-серым налетом. Боковые бархатистые пятпа
черные, небольшие, б. м. треугольные. Среднее пятно отсутствует или в виде
ничтожной точки. Лобная мозоль блестяще-черная, ее ширина превосходит высоту
приблизительно в 4 раза. Усики черные, лишь базальная половина 3-го членика красновато-
коричневая. 1-й членик блестяще-черный, утолщен, его длина превосходит толщину
приблизительно в 2 раза, перетяжка перед вершиной отсутствует. Концевой члоник
щупалец желтоватый, довольно узкий, покрыт преимущественно светлыми волосками.
Крылья светло-серые, с сильно развитым светлым рисунком. В средней части крыли
светлые пятна б. м. соединены между собой и образуют явственные розетки. Вершинная
поперечная полоска относительно неширокая, изогнутая, не достигает заднего кран
крыла. Последнее в пределах заднекрайних ячеек с широкой беловатой каймой,
состоящей из ряда пятен-колечек, по одному в каждой ячейке. Анальное поле почти
целиком светлое. Жилки коричневые, вдоль жилок мембрана крыла слегка коричиоватсш.
Ноги: бедра черно-серые, в густом сером налете, средние и задние бедра иногда на
некотором протяжении желтоватые, передние голени в основной 1/3 желтые, дилоо чор-
шыв, средние и задние голени желтые, с 3 черно-коричневыми колечками. Лапки чор-
416 III. ПОДСЕМ. TABANMAE
ные, основание средних и задних лапок желтое. Брюшко продолговатое, общий тоа
окраски оливково-серый. Тергиты по бокам с крупными, но не резкими, серыми
пятнами начиная с I кольца (реже со II). Эти пятна б. м. сливаются в 2 продольных
боковых полосы. Срединная серая полоска выражена на II тергите, далее обычно-исчезает.
Заднекрайние каемки неясные. Снизу брюшко одноцветно-серое, с оливковым
оттенком. 9—11 мм.
3 неизвестен.
Похожа на Н. desertorum, отличаясь меньшей величиной боковых бархатистых
пятен на лбу, отсутствием, как правило, срединного пятна, меньшей толщиной 1-го
членика усиков и иным рисунком крыла.
Распространение: восточноевропейско-североказахстанский степной
вид (рис. 346, 1). Восточная Украина, юг (ст. Кантемировка) и восток (Хоперский
заповедник) Воронежской обл., Волгоградская обл. (по Дону к югу от Калача; Тунгу-
тово), Чувашская АССР (Порецкое), Удмуртская АССР, юг Пермской обл. (близ Кун-
гура), Оренбургская обл.; юго-восток Башкирской АССР (Новозирганово), Зап. и
Сев. Казахстан, на восток до Павлодарской обл. (Баянаульский р-н). Редок. Конец
апреля—июнь, иногда июль.
Рис. 346. Ареалы: 1 — Haematopota pallidula; 2 — Н. desertorum.
12. Haematopota desertorum Szil. (рис. 347) — Дождевка степная.
S z i 1 a d у, 1923, Biolog. Hungar., 1, 1 : 35; К г б b e r in Lindner, 1925 :
47 (Chrysozona); Олсуфьев, 1936a : 221 (#) (Chrysozona); 1937a : 348 (Chrysozona);
L e с 1 e г с q, 1966 : 180; Виолович, 1968 :60; Murdoch et Takahasi,
1969 : 87. — pallens (Lw.) apud К г б b e r, 1922 : 151; 1925 : 51 (Chrysozona).
$>. Глаза в очень коротких волосках. Лоб обычно книзу слегка расширен. Ширина
лба в основании слегка превосходит высоту или равна ей. Боковые черные бархатистые
пятна на лбу довольно крупные, округлые, среднее пятно маленькое. Нижняя лобная
мозоль блестяще-черная, поперечная, приблизительно в 3—4 раза шире своей высоты,
боковыми сторонами б. м. достигает края глаз. Посредине мозоль обычно расширена'
вверх в небольшой угол. Усики в большей части черные; 2-й членик целиком, 3-й
членик в основной половине коричневые. Граница слияния обоих вторичных члеников,
образующих площадку, обычно хорошо заметна. 1-й членик блестяще-черный, овальный г
его длина превосходит наибольшую ширину приблизительно в l1^—2 раза; сверху
в основной половине членик в тонком сероватом налете, перетяжка перед вершиной
отсутствует. Концевой членик щупалец палево-желтый или коричневый, узкий, со-
слегка притуплённым кончиком, покрыт черными и серыми волосками. Ноги в
большей части черные, бедра в сером налете, иногда в средней части коричневатые.
Основание передних, 2 кольца на средних и задних голенях и основание последних 2 пар
лапок желтые. Иногда желтые кольца голеней между собой почти слиты. Крылья
буровато-серые, с мелким сетчатым рисунком, частично образующим розетки. Вершинная
поперечная полоска широкая, оставляет затемненным лишь самый кончик вершины
крыла, далее переходит в широкую светлую кайму, огибающую весь задний край
крыла. Жилки коричневые. Брюшко черное, с нерезким рисунком; общий тон окраски
13. HAEMATOPOTA
417
оливково-серый. Тергиты по заднему краю с узкими каемками серого налета; посредине
от каемки отходит вперед б. м. широкая полоска, достигающая края предыдущего
кольца, так что в совокупности эти полоски образуют хорошо заметную общую
срединную полоску, идущую продольно по брюшку. Боковые стороны тергитов начиная с II
кольца с крупными, но не резкими светлыми пятнами. Снизу брюшко одноцветно-серое.
VIII стернит коричнево затемнен, длиннее ширины, гонопофиз с умеренной
апикальной выемкой, латеральные лопасти маленькие, базальная лопасть с прямым или слегка
выпуклым основным краем. Церки и семеприемпики как у И. pluvialis, сперматека
игловидная. 8—10 мм.
$. Глаза в умеренно длинных светлых волосках. Затылок за глазами в длинных
темных волосках, с небольшой примесью светлых. Лобный треугольник в сером на-
Рис. 347. Haematopota desertorum Szil. Крыло, лобная полоска, усик, щупальце,
VIII стернит, церки и сперматека §. А — усик $.
лете, посредине с бархатисто-черным -треугольником, достигающим верхними углами
нижнего края глаз. 1-й членик усиков сильно вздут, его длина превосходит
наибольшую ширину едва в 11/2 раза, блестяще-черный, покрыт длинными черными волосками;
сверху в основании членик в легком сером налете. 2-й членик коричневый, 3-й членик
также коричневый, палочка черная. Концевой членик щупалец коричневый, на конце
слегка притуплён, покрыт довольно длинными светлыми и темными волосками. Крылья
и ноги как у 5- Брюшко черное, II—III и частью IV тергиты по бокам коричневые.
Посредине брюшка проходит нерезкая полоска серого налета, начинающаяся на II
кольце, III и последующие тергиты по бокам с крупными, но неясными серыми пятнами.
Снизу брюшко черно-серое, II—III стерниты по бокам коричневые. 8.5—9.5 мм.
Похожа на Н. pluvialis, но легко отличается от последней по отсутствию перетяжки
на конце 1-го членика усиков и наличию на крыльях широкой беловатой каймы.
Крёбер (Krober, 1922, 1925) неправильно отнес этот вид к Н. pallens, описанный Левом
(Loew) из Узбекистана (Зеравшан).
Распространение: восточносибирский степной вид (рис. 346, 2), аллопат-
ричен Н. pallidula. От Омской (Горьковский р-н) обл. на восток до Приморского края,
на север до Зонова Куйбышевского р-на и Тогучина Новосибирской обл., низовий
Ангары (Мурский порог), Братска Иркутской обл., Амурской обл. (близ Кумара) и
окр. Хабаровска, на юг до Тувы, южного Прибайкалья и юга Приморского края (оз. Ха-
сан). Сев. и Вост. Монголия, Сев.-Вост. и Вост. Китай. Обычен в Туве и на Суйфуно-
Приханкайской равнине Приморского края, в остальных частях ареала не часто.
Июль—август.
13. Haematopota pluvialis L. (рис. 330, 331) — Дождевка обыкновенная.
Изменчивый вид, в пределах своего огромного ареала представлен рядом форм;
на территории СССР обитают 3 подвида: номинативный европейско-азиатский Н.
pluvialis pluvialis L., дальневосточный Н. pluvialis tristis Big. и кавказский горный //.
pluvialis ioffi Ols.
13a. Haematopota pluvialis pluvialis L. (рис. 348) — Дождевка обыкновенная
номинативная.
Li n n ё, 1758, Syst. Nat. : 602 (Tabanus); Lundbeck, 1907 : 104; V о r-
rall, 1909 : 331; Krober, 1922 : 156 {Chrysozona); S u г с о u f, 1924 : 77 (Chry-
418 III. ПОДСЕМ. TABANINAE
sozona); Kroner in Lindner, 1925:51 (Chrysozona); Олсуфьев, 1937а:
349 {Chrysozona); Moucha et Chvala, 1956 : 266; Шевченко, 1961 : 174
(Chrysozona); Leclercq, 1966:186; В и о л о в и ч, 1968:61; Cameron,
1934: 211 (биология, личинка, куколка); С к у ф ь и н, 1967 : 200 (личинка); J e z e к,
1971 : 345 (личинка, куколка); Chvala, Lyneborg a. Moucha, 1972 : 43.
g. Глаза в очень коротких светлых волосках. Лоб умеренно широкий, с б. м.
параллельными сторонами, его высота слегка превосходит ширину, реже равняется ей,
покрыт желтовато-серым налетом. Черные боковые пятна на лбу крупные, круглые,
отделены от нижней мозоли очень узкой полоской налета. Среднее пятно маленькое,
Рис. 34. Haematopota 'pluvialis pluvialis L. Крыло, лобная полоска, усик,
щупальце, VIII стернит (варианты), церки и семеприемник g. A — усик g.
округлое, иногда в виде незначительной черной точки. Нижняя лобная мозоль
блестяще-черная, поперечная, приблизительно в 3—4 раза шире своей высоты, боковыми
сторонами достигает края глаз. Усики в большей части черные; 3-й членик в основании
красно-коричневый. 1-й членик блестяще-черный, утолщенный (его длина превосходит
наибольшую ширину в 2—2V2 раза), сверху возле вершины с резкой перетяжкой
(смотреть сбоку!), тогда как ближе к основанию членик в сероватом налете. Концевой
членик щупалец палево-желтый или коричневато-серый, приблизительно в 2х/2—3 раза
длиннее наибольшей толщины, покрыт черными и серыми волосками. Щиток в
буровато-сером налете. Ноги в большей своей части черные; бедра в сером налете,
основание передних, 2 кольца на средних и задних голенях и основание последних двух пар
лапок темно-желтые. Иногда желтые кольца голеней между собой почти слиты. Крылья
буровато-серые, с явственным сетчатым рисунком; жилки коричневые. Светлые пятна
относительно мелкие, довольно беспорядочно расположены по крылу, реже образуют
розетки. Вершинная поперечная полоска обычно довольно широкая, запятовидная,
не достигает заднего края крыла. Последнее, почти как правило, с узкой беловатой
каймой. Иногда светлый рисунок на крыльях сильно развит. Брюшко черное с
буроватым оттенком и с нерезким рисунком; общий тон его окраски оливково-серый. Тергиты
по заднему краю о узкой каемкой серого налета; каемки посредине расширены в виде
направленной вперед узкой полоски, достигающей заднего края предыдущего кольца.
II тергит с маленькими, последующие с более крупными округлыми оливково-серыми
пятнами по бокам. Снизу брюшко одноцветно-серое. VIII стернит по бокам коричнево
затемнен, реже целиком темный,' гонопофиз с выраженной апикальной выемкой,
латеральные лопасти небольшие, базальная лопасть короткая, часто со слегка вогну-
13. HAEMATOPOTA
419
тыми боковыми сторонами, основной край лопасти прямой или слегка вогнутый, реже
выпуклый. Церки округло-треугольные. Семеприемник с длинным семепротоком,
приблизительно в 15—17 раз длиннее муфты, сперматека веретеновидная, коричневатая.
8—12, редко 6.5 мм.
(J. Глаза в б. м. длинных желтовато-серых или коричневатых волосках. Затылок
за глазами с рядом выступающих, длинных, черных волосков. Иногда в средней части
к черным примешиваются желтые волоски. Лобный треугольник в сером налете,
посредине с черным треугольным пятном, верхними углами достигающим края глаз. Усики
в большей своей части черные; 3-й членик в основной половине коричневый. 1-й членик
блестяще-черный, сильно яйцевидно утолщен (его наибольшая ширина относится
к длине как 1:2), покрыт длинными черными волосками. Спинная сторона членика
у основания в сером налете. Концевой членик щупалец желто-серый или коричневатый,
в основании утоящен5 далее сужен; на конце б. м. притуплён. Ноги и крылья как
у $. Брюшко черное, с небольшими коричневатыми пятнами по бокам II—III (или
Рис. 349. Ареалы: 1 — Haematopota pluvialis pluvialis; 2 — Н. plavialis
tristis.
I—III) тергитов и как у g с неясным серым рисунком. Серые боковые пятна начиная
с III тергита. Снизу брюшко черно-серое, боковые стороны II—III стернитов желто-
коричневые; реже брюшко снизу одноцветно-черно-серое. 8—11 мм.
Распространение: европейско-сибирский лесной подвид, широко
проникает на север в тайгу и на юг в степную зону (рис. 349,1). В СССР распространен от
западных границ на восток до Якутска и Забайкалья, на север до Мурманска, дельты
Печоры, Обской губы, низовий Енисея (Снежногорск, 68° с. ш.; Тэтэрэ, приток Под-
каменной Тунгуски) и севера Иркутской обл. (Среднее Приангарье; Усть-Кут), па юг
ЯО низовий Днепра (Цюрупинск), Дона, Крыма, Предкавказья, Закавказья, включая
Грузию и Азербайджан (на Кавказе часто в горах), нижнего течения Урала (нос. Круглое),
Темира, Джезказгана (48° с. ш.) Павлодарской обл., Джунгарского Алатау, Алтая,
Тувы и южного Прибайкалья. Вся Европа, Турция, Зап. Китай (Синьцзян).
Распространение в Якутии, видимо, оторвано от основного ареала. Массовый вид в лосной
эоне и подзоне южной тайги, а также местами в лесных предгорьях Северного
Кавказа. Конец июня—август, иногда раньше.
Личинки найдены в берегах водоемов, а также при почвенных раскопках, вплоть
ЯО огородных грядок. Экспериментально доказанный переносчик сибирской яипи
(Олсуфьев и Лелеп, 1935); в природе обнаруживали зараженным возбудителем
туляремии (Сомов с .соавт., 1939).
13Ь. Haematopota pluvialis tristis Big. (рис. 350) — Дождевка обыкновенная
дальневосточная.
Bigot, 1891, Bull. Soc. Zool. France, 16 : 77 (H. tristis); Rrobor in
Lindner, 1925 : 53 (Chrysozona); Takahasi, 1962 : 121 (Chrysozona); L о С 1 О г о п,
1966 : 189; Виолович, 1968 :65; Murdoch et Takahasi, 1000 : 00;
Олсуфьев, 1969 : 118. — sakhalinensis S h i r a k i, 1918 : 104 (Haematopota).
420 III. ПОДСЕМ. TABANINAE
$(J. От номинативной формы отличается главным образом рисунком крыла: в его
внешней половине светлые пятна образуют 2 широких кольца, внутри которых более
мелкие светлые пятна обычно отсутствуют. Боковые светлые пятна на брюшке с IV
или III тергита, малозаметные. 2 7—12 мм, 3 8—11 мм.
Мы не обнаружили различий в строении терминалий 52 номинативной формы и
Н. pluvialis tristis.
Распространение: дальневосточный лесной подвид, ареал оторван
от ареала номинативного подвида (рис. 349, 2). Низовья Амура (Нижнетамбовка,
Сусанине), Сахалин, о. Кунашир, острова Малой Курильской гряды. Япония (о-ва
Хоккайдо, Хонсю). Обычен.
Рис. 350. Haematopota pluvialis tristis Big. Вершинная часть
крыла, VIII стернит и церки g.
13с. Haematopota pluvialis ioffi Ols. — Дождевка обыкновенная Иоффа.
Олсуфьев, 1972, Энтомол. обозр., 51, 2 : 453.
$. От номинативного подвида отличается черной основной окраской брюшка и
голубовато-серым налетом, образующим на нем светлый рисунок, менее развитыми
боковыми пятнами сверху брюшка, начинающимися с IV тергита, более бледной
окраской крыла и несколько иным его рисунком: светлые пятна более мелкие, задний край
крыла без светлой каемки, вершинная поперечная запятовидная полоска слабо
выражена или отсутствует. 9—10.5 мм.
Распространение: Шови близ Мамисонского перевала, верховья Риона,
Грузия, 1600—2500 м VI—VIII 1937, И. Г. Иофф. Серия экземпляров.
14. Haematopota subcylindrica Pand. (рис. 351) — Дождевка светлая.
Pandelle, 1883, Rev. Ent. Fr., 2 : 196 (Haematopota pluvialis var.
subcylindrica); S u г с о u i, 1924 : 78 (Chrysozona pluvialis subcylindrica); Krober in
Lindner, 1925 : 52 (Ch. pluvialis var. subcylindrica); Leclercq, 1966 : 187; Moueha
et С h v a 1 a, 1967a : 225; J e z e k, 1971 : 349 (личинка, куколка); С h v a 1 a,
Lyneborg a. Moueha, 1972 : 436. — hispanica (Szil.) apud О 1 s u f j e v,
1937a : 351 (Chrysozona) et auct. — elbursiensis Abbassian-Li n't z e n, 1960b :
826 (H. pluvialis elbursiensis).
$• Глаза в коротких светлых волосках. Лоб умеренно широкий с параллельными
сторонами, высота лба слегка превосходит ширину, реже равна ей, покрыт голубовато-
серым налетом. Черные боковые пятна на лбу чаще небольшие, округлые или в виде
запятой. Среднее пятно маленькое, черное, иногда почти исчезающее. Нижняя лобная
мозоль блестяще-черная, поперечная, приблизительно в 3—4 раза шире своей высоты,
боковыми сторонами достигает края глаз. Усики черные, лишь 3-й членик в основании
слегка коричневатый. 1-й членик утолщен, его длина превосходит наибольшую
ширину в Z1/^—3 раза, без заметной перетяжки возле вершины, сверху почти целиком
в голубовато-сером налете. Концевой членик щупалец палево-серый, довольно узкий,
его длина превосходит наибольшую толщину приблизительно в 3 раза, покрыт черными
и серыми волосками. Щиток в голубовато-сером налете, иногда в средней части имеется
небольшая примесь коричневого налета. Ноги в большей своей части черные; бедра
в сером налете, основание передних, 2 кольца на средних и задних голенях и основание
последних 2 пар лапок желтые. Крылья бледно-серые, с неясным светлым рисунком.
Светлые пятна б. м. соединены между собой и часто образуют розетки. Заднекрайние
ячейки с поперечными светлыми черточками, расположенными далеко отступя от края
крыла. Эти черточки образуют параллельный заднему краю ряд, который в вершинной
13. HAEMATOPOTA 431
части заканчивается вершинной поперечной полоской. Последняя узкая, изогнутая,
направляется от г5 вверх и далее упирается в передний край чкрыла возле г2. Задний
край крыла без светлой каемки, реже таковая имеется, узкая и расположена в
пределах заднекрайних ячеек. Брюшко черное, с нерезким рисунком; общий тон окраски
последнего голубовато-серый. Тергиты по заднему краю с узкой каемкой серого
налета; каемки посредине расширены в направленную вперед узкую полоску,
достигающую заднего края предыдущего кольца. II тергит с маленькими, последующие с более
крупными округлыми голубовато-серыми пятнами по бокам. Снизу брюшко одноцветно-
серое. Терминалий сходны с таковыми Н. pluvialis. 8—13 мм.
3- Глаза в б. м. длинных светлых волосках. Затылок за глазами с рядом длинных
темных волосков, с небольшой примесью светлых. Лобный треугольник в сером
налете, посредине с черным треугольным пятном, достигающим верхними углами края
глаз. Усики черные, лишь 3-й членик в основании коричневый. 1-й членик утолщен,
его длина превосходит ширину приблизительно в 2 раза, покрыт б. м. длинными чер-
Рис. 351. Haematopota subcylindrica Pand. Крыло, лобная полоска, усик, щупальце,
VIII стернит (варианты) и церки g. А — усик 3-
ными и частью светлыми волосками. Спинная сторона членика почти целиком в сером
налете. Концевой членик щупалец палево-серый, утолщен, на конце слегка притуплён,
покрыт довольно длинными черными и серыми волосками. Ноги как у g; крылья слегка
темнее, но рисунок тот же. Брюшко черное, I—III тергиты по бокам коричневые.
Рисунок из серого налета как у g, но боковые серые пятна начинаются с III кольца и хорошо
заметны на IV—VI тергитах. Брюшко снизу черное, II стернит по бокам
коричневатый. 10.5 мм.
От Н. pluvialis, с которой рассматриваемый вид имеет много сходства, g легко
отличается общим голубовато-серым тоном окраски тела, отсутствием на 1-м членике
усиков резкой перетяжки перед вершиной, хорошо развитым налетом на нем же,
меньшей величиной (не всегда) боковых бархатистых пятен на лбу и более бледной окраской
крыльев. 3 отличается с большим трудом — по налету на спинной стороне 1-го членика
усиков, более бледным крыльям и по общему голубовато-серому тону окраски тела,
в особенности груди. В общей массе gg H. subcylindrica несколько крупнее gg H.
pluvialis. Лёт Н. subcylindrica происходит несколько раньше, нежели Н. pluvialis, а именно
в июне и частью июле, тогда как обыкновенная дождевка в массе летает в июле и августе.
Распространение: европейско-западносибирский лесостепной вид
(рис. 352). От Прибалтики, запада Украины и от Молдавии на восток до Енисея
(Хакасская АО) и Подкаменной Тунгуски (Тэтэрэ) (вероятно, изолированные
местонахождения), на север до Витебской обл., БССР (Ушачи), Московской обл. (на север до
Дмитрова), Ивановской обл., окр. Сыктывкара, Пермской обл. (Пашня), Березовского р-на
(Полноват) и Назина на Оби, на юг до дельты Дуная и Днепра, Крыма, низовий Дона,
Предкавказья, Закавказья (Грузия, Армения и Азербайджан, включая Нахичеван-
скую АССР), дельты Волги, низовий Урала, Кустанайской обл. (Сёмиозерный р-н),
Карагандинской обл. (урочище Карагаш), Семипалатинска и Джунгарского Алатау.
27 Н. Г. Олсуфьев
422
III. ПОДСЕМ. TABANINAE
Изолированно отмечен для Ю. Карелии (ст. Хиитола). Средн. и Ю. Европа, Вост.
Турция, Сев. Иран. Обычен возле водоемов степной зоны; на Кавказе проникает довольно
высоко в горы.
Личинки нами найдены в кочках по берегу небольшого озера вместе с личинками
Tabanus autumnalis и др. (Венгерово, Зап. Сибирь).
Рис. 352. Ареал Haematopota subcylindrica.
15. Haematopota scutellata Ols., M. et Chv. — Дождевка горная.
В пределах своего ареала представлена 2 подвидами: европейским номинативным
Н. scutellata scutellata Ols., M. et Chv. и кавказским Н. scutellata pontica Ols.
15a. Haematopota scutellata scutellata Ols., M. et Chv. (рис. 353) — Дождевка
горная номинативная.
Moucha u. Chvala, 1964, Casopic Ceskoslovenske Spol.
3 : 284 (Chrysozona); L e с 1 e г с q, 1966 : 188; Moucha a.
: 227 ((J); J e z e k, 1971 : 346 (личинка, куколка); Chvala,
Moucha, 1972 : 434.
редкие волоски).
О 1 s u i j e v,
Entomologicke, 61
Chvala, 1967a
Lyneborg a.
g. Глаза голые (лишь при сильном увеличении видны короткие
Лоб с параллельными сторонами или слегка расширен книзу, его высота немного
превосходит ширину или они равны; лоб в
голубовато-сером налете. Боковые черные
бархатистые пятна большие, неправильной
формы, в верхней части с выступом, реже
округлой формы. Среднее бархатистое пятно часто
довольно крупное, округло-треугольной
Рис. 353. Haematopota scutellata scutellata
Ols., M. et Chv. Лобная полоска, усик и
щиток g.
формы. Нижняя лобная мозоль блестяще-черная, ее ширина превосходит высоту
в 3—4 раза и боковыми сторонами достигает края глаз. Усики черйые, 3-й членик
в основании слегка коричневатый или целиком черный. 1-й членик утолщен, его длина
превосходит наибольшую толщину в 2—21/2 раза, блестяще-черный, сверху перед
вершиной с неясной перетяжкой или без нее; в базальной части на небольшом протяжении
в коричневатом или сероватом налете. 3-й членик усиков очень узкий, площадка к
вершине лишь немного уже базальной части. Концевой членик щупалец узкий,
темно-серый, в светлых и черных волосках. Щиток черный, с явственным округлым пятном се-
13. HAEMATOPOTA
423
рого налета, занимающим всю его среднюю часть; боковые стороны щитка в
коричневом налете. Ноги в большей своей части черные; бедра в сером налете, основание
передних, 2 кольца на средних и задних голенях и основание последних 2 пар лапок желтые.
Средние и задние бедра иногда желтоватые. Крылья коричневато-серые со светлым
рисунком из относительно мелких пятен, которые в средних частях крыла обычно обра-,
зуют нерезкие розетки. Поперечная запятовидная полоска перед вершиной крыла
узкая, не достигает его заднего края. Последний без светлой каймы или с узкими
пятнами, по 1 в каждой заднекрайней ячейке. Брюшко черное, сверху с
голубовато-серым рисунком: тергиты по заднему краю с узкими каемками, II тергит посредине с
длинным узким треугольником, на последующих тергитах иногда едва заметная срединная
полоска. Боковые светлые пятна маленькие обычно начиная с V тергита, реже с IV.
Снизу брюшко черное, в сером налете. 9.5—11 мм.
3 (по Moucha a. Chvala, 1967a, сокращенно). Глаза в коричневых волосках.
Затылок в длинных черных волосках. Лобный треугольник с крупным черным бархатистым
треугольным пятном. 1-й членик усиков блестяще-черный, эллиптической формы, его
длина превосходит наибольшую толщину приблизительно в 2 раза, в длинных черных
волосках, в основании в сером налете, 3-й членик узкий, черно-коричневый, в
основании более коричневый. Концевой членик щупалец серый, конусовидный, в 2 раза
длиннее своей наибольшей толщины, в светло-серых волосках. Щиток как у g с крупным
округлым пятном серого цвета. Ноги как у g, но светлые части более коричневые.
Крылья как у g. Брюшко черное, тергиты по заднему краю с светло-серой каймой,
2 предпоследних сегмента с серыми боковыми пятнами. II тергит по бокам иногда
коричневатый. Снизу брюшко в сером налете. 9—10 мм.
Распространение: европейский горно-лесной подвид. В СССР
обнаружен в Ивано-Франковской обл. УССР (Косов). Средн. Европа, от Румынии на запад
до Франции. Июль—начало сентября. Не часто.
15b. Haematopota scutellata pontica Ols. — Дождевка горная кавказская.
Олсуфьев, 1964, Бюлл. Московск. общества испытателей природы, отд.
биологический, 69, 3:75 (Chrysozona); L е с 1 е г с q, 1966:188.
g. От номинативного подвида отличается меньшим развитием бархатистых пятен
на лбу, наличием серых боковых пятен начиная с III тергита брюшка и более узким
1-м члеником усиков. 10—10.5 мм.
Распространение: кавказский горно-лесной подвид. Кавказский
заповедник (сев. и ю. отделы); близ Красной Поляны; Кабардино-Балкарская АССР;
Дагестан; Абхазия (Ажары, Латы, Квезани); Сев.-Вост. Грузия (Лагодехи);
Аджарская АССР (р-н Батуми и Кедский р-н). Не часто. Июль.
16. Haematopota crassicornis Wahlbg. (рис. 354) — Дождевка черноусая.
W a h I b е г g, 1848, Oefv. Svenska Vet.-Akad. Foerh, 5:9; Lundbeck,
11907 : 105; V e r r a 11, 1909 : 337; К r 6 b e r, 1922 : 155 (Chrysozona); S u г с о u i,
1924 : 76 (Chrysozona); Krober in Lindner, 1925 : 46 (Chrysozona); Олсуфьев,
1937a : 354 (Chrysozona); Moucha et Chvala, 1956 : 264; L e с 1 e г с q, 1966 :
180; Виолович, 1968 : 68; И в а н и щ у к, 1970 : 149 (личинка); J e z e к,
1971 : 347 (личинка, куколка); Chvala, Lyneborg a. Moucha, 1972 :
441. —flavopilosa К г б b e r, 1922:155 (3).
g. Глаза в очень коротких светлых волосках. Лоб обычно слегка расширен книзу
и здесь его ширина слегка превосходит высоту. Черные боковые пятна на лбу
бобовидной или лунковиднои формы, среднее маленькое, округлое. Нижняя лобная мозоль,
блестяще-черная, приблизительно в 4 раза шире своей высоты, боковыми сторонами
достигает края глаз. Усики короткие, целиком черные. 1-й членик утолщен, овальный,
его наибольшая, ширина обычно укладывается в длине 13/4—2 раза, целиком
блестящий. Очень редко 3-й членик в основании слегка коричневатый. 1-й членик на конце
без перетяжки. Концевой членик щупалец палево-серый или желтоватый, узкий, его,
длина приблизительно в 3—ЗЧ2 раза превосходит толщину. Членик покрыт серыми и
черными волосками. Щиток черный, в буроватом налете. Ноги в большей своей части
черные. Бедра в сером налете, основание передних, 2 кольца на средних и задних
голенях и основание последних 2 пар лапок желтые. Крылья буровато-серые, с легким,
коричневатым затемнением вокруг поперечных жилок. Жилки коричневые. Светлые
пятна довольно мелкие, б. м. сгруппированы и частью соединены в кольца, реже
образуют розетки. Вершинная поперечная полоска узкая, запятовидная, не достигает
заднего края крыла. Заднекрайние ячейки с поперечными черточками (или двойными
пятнами), расположенными отступя от заднего края крыла и образующими параллельную,
ему цепочку. Задний край крыла без светлой каемки. Брюшко черное, задние края
тергитов с узкими серыми каемками. На II тергите каемка расширена вперед в
короткий срединный треугольник. IV (иногда III) и последующие тергиты с легкой сероватой
27*
48'4 III. ПОДСЕМ. TABANINAE
срединной полоской. Ill тергит по бокам с маленькими, последующие с более крупными
округлыми серыми пятнами. Снизу брюшко одноцветно-темно-серое. VIII стернит
широкий, коричневый, базальная лопасть со светлым треугольником, гонопофиз со
слабой апикальной выемкой, латеральные лопасти средней величины, базальная лопасть
чаще короткая, с б. м. вогнутыми боковыми сторонами и прямым основным краем.
Церки б. м. треугольные. Семеприемники маленькие, лишь в 4—5 раз длиннее муфты,
сперматека в виде прозрачной трубки, немного более короткой, чем семепровод. 9—
10 мм.
в- Глаза в б. м. длинных, светло-коричневых волосках. Затылок за глазами с
рядом выступающих, длинных, темных и светлых волосков. Усики целиком черные,
Рис. 354. Haematopota crassicornis Wahlbg. Крыло, лобная полоска, усик, щупальце,
VIII стернит (варианты), церкй (варианты) и семеприемники g. А — усик S-
1-й членик сильно утолщен, яйцевидный, блестяще-черный, покрыт длинными черными
волосками. Концевой членик щупалец палево-желтый, короткий, в основании
утолщен, далее слегка сужен, на конце тупой. Членик покрыт длинными желто-серыми
волосками. Лобный треугольник в желтовато-сером налете, посредине с черным округло-
треугольным пятном, достигающим верхними углами края глаз. Ноги и крылья как
У g. Брюшко черное. Задние края тергитов с узкими каемками серого налета; IV—VI
кольца по бокам с небольшими, округлыми, серыми пятнами. Снизу брюшко
черно-серое. 8—11 мм.
Распространение: европейско-западносибирский лесной вид, аллопатри-
Чен Н. tamerlani (рис. 355, 1). Широко распространен в европейской части СССР, на
север известен до средней Карелии (Беломорский р-н, 64°40'), Архангельска и устья
Печоры, на юг до Николаевской обл. УССР (Вознесенск), Харькова, Волгоградской обл.
(пойма Дона к югу от Калача) и Ю. Урала, а после перерыва в южнорусских степях
распространен на Сев. Кавказе и в Закавказье (включая Грузию, Армению и
Азербайджан). По ту сторону Урала известны спорадические находки близ Свердловска (Уктус),
Тюмени, севера Омской обл. и, по-видимому, изолированно в Алтайском крае (окр. Бар-
Паула и Кытмановскийр-н). Европа, кроме крайнего севера и юга, Турция.
Многочислен в горах Кавказа, в остальных частях ареала (в пределах СССР) не часто. Июнь—
июль, редко позднее. Указание В. Г. Фоминых с соавт. (1972) для Томской обл.
нуждается в подтверждении.
Изменчивый вид. gg с Большого и Малого Кавказа отличаются от
среднеевропейских наличием на щитке в его апикальной части небольшого округлого серого пятна
налета и слегка более утолщенным 1-м члеником усиков, иногда покрытым сверху,
13. HAEMATOPOTA 425
особенно в базальной части, серым налетом. Мы обозначаем ее как Н. crassicornis var.
griseopunctata Ols., var. nova.. У gg из Алтайского края боковые серые пятна на брюшке
расположены начиная с II тергита.
Личинки нами найдены в песчаных берегах лесного ручья и небольшой речки
(Ленинградская обл.).
о° к востоку от Гринвича 90° ш° т° т° <50°
0° 30° 60° 90° 120°
Рис. 355. Ареалы: 1 — Haematopota crassicornis; 2 — Н. tamerlani.
17. Haematopota tamerlani Szil. (рис. 356) — Дождевка Тамерлана.
S z i 1 a d у, 1923, Biolog. Hungar., 1, 1 : 34 (H. crassicornis tamerlani); Kriiber
in Lindner, 1925:46 {Chrysozona crassicornis tamerlani Szil.); Олсуфьев,
1937a : 356 {Chrysozona) (S); Takahasi, 1962 : 119 {Chrysozona); L e с 1 e г с q,
1966 : 189; В и о л о в и ч. 1968:69; Murdoch et Takahasi, 1969:87;
Соболева, 1971 : 144 (личинка, куколка).
Рис. 356. Haematopota tamerlani Szil. Крыло, лобная полоска,
усик и щупальце g. А — усик S-
g. Глаза в очень коротких, светлых волосках. Лоб широкий, обычно несколько
расширен книзу, ширина основания лба явственно превосходит высоту. Боковые
бархатистые пятна крупные, черные, б. м. треугольные. Среднее пятно небольшое, черное.
Лобная мозоль блестяще-черная, ее ширина превосходит высоту приблизительно
в 4—5 раз, боковыми сторонами достигает края глаз. Усики целиком черные. 1-й чле-
426 HI. ПОДСЕМ. TABANINAE
ник большой, удлиненно-овальный, часто муфтообразный, блестяще-черный, сверху
в базальной 1/3 часто в сероватом налете; на конце членика перетяжка отсутствует.
3-й членик немного длиннее 1-го. Концевой членик щупалец желтоватый, в основной
половине утолщен, далее узкий, с тупым кончиком, покрыт серыми и черными
волосками. Спинка с явственными серыми продольными полосками, из них срединная не
имеет перерыва в средней 1/3 спинки. Щиток преимущественно или целиком в сером
налете. Крылья коричневато-серые, с резким рисунком: светлые пятна сгруппированы,
как правило, в бросающиеся в глаза розетки. Вершинная поперечная полоска довольно
резкая, запятовидная, обычно не достигает заднего края крыла. Последнее со светлой
Рис. 357. Haematopota stackelbergi Ols. Лобная полоска, усик. А — распределение
серого рисунка (пунктир) на спинке и брюшке сверху $; Б — то же у О. Н.
tamerlani Szil.
каймой^ образованной цепочкой пятен-колечек, расположенных по 1 в каждой задне-
крайней ячейке. Эти колечки могут распасться на самостоятельные пятна, образующие
2 параллельных цепочки, из них задняя располагается по краю крыла. Ноги в большей
своей части черные, основание передних голеней, 2 кольца на средних и задних
голенях и основание средних и задних лапок желтые. Брюшко черное, тергиты с узкими
заднекрайними серыми каемками. Посредине обычно имеется узкая серая продольная
полоска, начинающаяся на II тергите. V—VI или IV—VI тергиты с небольшими
сероватыми боковыми пятнами. Снизу брюшко одноцветно-черно-серое. VIII стернит (рис. 358,
А,. Б) широкий, коричневый, базальная лопасть со светлым треугольником, гонопо-
физ с б. м. умеренной выемкой, латеральные лопасти обычно хорошо развиты, базальная
лопасть короткая, с прямым основным краем. Церки с округлой вершиной. Семе-
приемники маленькие, лишь в 4—5 раз длиннее муфты, сперматека в виде слегка
затемненной трубки, немного длиннее семепровода. 8.5—13 мм.
<$. Глаза в довольно длинных светло-коричневых волосках. Затылок в длинных
волосках: в средней части в желтых, по бокам в коричневых. Усики целиком черные.
1-й членик крупный, сильно яйцевидно утолщен; блестяще-черный, покрыт длинными
желтыми волосками, к которым в концевой части примешиваются в небольшом
количестве черные волоски. 3-й членик усиков слегка короче 1-го. Грудь и ее придатки как
13. HAEMATOPOTA
427
у Р. Брюшко черное, I (или II—III) тергит по бокам с небольшим коричневым пятном.
Тергиты с узкими сероватыми заднекрайними каемками, V—VI тергиты по бокам
с небольшими сероватыми пятнами. Снизу брюшко черно-серое. 9—10 мм.
Распространение: восточносибирский лесной вид, часто в горах; алло-
патричен Н. crassicomis (рис. 355, 2). В СССР от Томской, Новосибирской областей (Тогу-
чинский р-н) и Алтая (Шебалино), на восток до Сахалина и о. Кунашир, на север до Под-
каменной Тунгуски, Якутска, Нелькана (север Хабаровского края) и Усть-Омчука
(юг Магаданской обл.), на юг до Тувы, южного Забайкалья, Приамурья и юга
Приморского края. Сев. Монголия, Сев.-Вост. и Вост. Китай, Корейский п-ов, Япония
(о. Хоккайдо). Обычен в Приморском крае, в других частях ареала не часто. Конец
июня—середина августа, иногда позднее.
На Сахалине обнаружен спонтанно зараженным возбудителем эризипелоида
(Евсеева и Тимофеева, 1971), но, возможно, это указание следует отнести к II. yluuialis
tristis, значительно более обильной на Сахалине, нежели Н. tamerlani.
Рис. 358. Терминалии $$: А, Б — Haematopota tamerlani Szil.; В, Г -
Н. stackelbergi Ols. VIII стернит (варианты), церки (варианты) и сомонриомпики.
18. Haematopota stackelbergi Ols. (рис. 357) — Дождевка Штаксльбергн.
Олсуфьев, 1967, Энтомол. обозр., 46, 2 : 388.
5- Глаза в очень коротких светлых волосках. Лоб обычно носколт.ко ршчпирви
книзу, ширина основания лба явственно превосходит высоту. Вокоиыо бирхн'ПШ'ШВ
пятна крупные, черные, б. м. треугольной формы. Среднее пятно небольшое, чорши».
Лобная мозоль блестяще-черная, ее ширина превосходит высоту нриблииитольно
в 4—4!/2 раза, боковыми сторонами достигает края глаз. 1-й члоиик усиком больший,
удлиненно-овальный или муфтообразный, блестяще-черпый, на коицо норотнЖМЙ
отсутствует. 3-й членик равен или слегка короче 1-го. Концопой члоиик щуинлиц
сероватый, умеренно утолщен. Спинка в передней части со слабо ршпнмыми оормми
продольными полосками, из них средшшая полоска поело инпчитолытго uopollMHtl
в средней х/з спинки, в задней ее части представлена короткой уикой полоской. Щитои
преимущественно или целиком в темно-буром налото. Крылья длинный, кормчноингмм,
428 III. ПОДСЕМ. TABANINAE
с резким светлым рисунком; светлые пятна вокруг поперечных жилок сгруппированы,
как правило, в бросающиеся в глаза розетки. Вершинная поперечная полоска
относительно узкая, занято видная, не достигает заднего края крыла. Отступя от заднего
края крыла имеется цепочка светлых небольших двойных пятен, по паре в каждой
заднекрайней ячейке; обычно в передневнешнем углу 2-й, 3-й и 5-й (или всех) задне-
крайних ячеек по светлому угловидному пятну. Бедра ног черные, в сером налете,
остальное как обычно. Брюшко черное, тергиты по заднему краю с очень узкой серой
каемкой, не расширяющейся в средней части кольца в заметный треугольник или
полоску. VI тергит с незначительными боковыми округлыми сероватыми пятнами.
VII тергит с такими же пятнами или целиком черно-бурый. Снизу брюшко одноцветно-
черно-серое. Терминалии (рис. 358, В, Г) сходны с таковыми Н. tamerlani. 10—12 мм.
S неизвестен.
Похожа на Н. tamerlani, отличаясь более массивным 1-м члеником усиков,
удлиненными более темными крыльями и менее развитым серым рисунком на спинке и
брюшке.
Распространение: дальневосточный (? лесной) вид. Амурская обл.
(Корсаково к югу от Кумара); Приморская обл. Сев.-Вост. Китай. Не часто. Конец
июня—начало августа.
Добавление
В статье Т. П. Худавердыева и Ш. М. Джафарова «Описание новых видов
слепней (Diptera, Tabanidae) из низменных районов Нах. АССР» (Ученые записки
Министерства высшего и среднего специального образования Азербайджанской ССР,
серия биологических наук, 1974, 3 : 42—46) опубликованы два новых вида: Chry-
sops nachitschevanicus sp. п., $>$ собраны около р. Нахичеванчай (с. Бадашхан)
в июле 1973 г. и Taban'us dzhafarovi sp. п., jj собраны там же в октябре—ноябре
(год не указан).
В статье Н. Г. Олсуфьева и П. П. Иванищука «Обнаружение в средней
полосе европейской части СССР мухи-дождевки Haematopota scutellata Ols., M. et
Chv. (Diptera, Tabanidae) и описание нового подвида» (Энтомологическое обозрение,
1975, 54, 4 : 890—895) опубликован новый подвид: Haematopota scutellata rossica
Ols. et Ivan., $g<Jc? выведены из окуклившихся личинок, собранных на р. Моча
Сокольского р-она Ивановской обл. в 1972—1974 гг.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ *
abazus Big., Tabamis 230
abbassianae Lecl., Silvius vitripennis var.
135, 137
abbassianae Ols., Tabanus shelkovnikovi
subsp. 252
abdominalis Krob., Chrysops flavipes var.
175, 177
accipiter Szil., Tabanus 193, 199, 210
acuminata Lw., Hybomitra 394
adachii Talc., Hybomitra 314, 359
aequetincta Beck., Hybomitra 311, 316,
347
aethereus Big., Atylotus 301
afghanistanicus M. et Chv., Atylotus 295
agrestis auct., Atylotus 295
alazanicus Hauser, Therioplectes 306
alazinus Big., Tabanus 230
albicauda Ols., Therioplectes 305, 308
alpina Ols., Hybomitra shevtshenkoi var.
346
alpinus Drap., Silvius vituli var. 133
alpinus Scop., Silvius 131, 133
altaica Ols., Hybomitra 312, 316, 324
altainus Szil., Hybomitra 379
amaenus Wlk., Tabanus 198, 202, 280
amurensis PIT, Chrysops 169
anderi Kauri, Hybomitra 320
angaricus Ols., Chrysops 143, 144, 161
angustipalpis Ols., Hybomitra 323
ansarii Abbas.-L., Tabanus 193, 199, 216
anthophilus Lw., Tabanus 241
anthrax Ols., Chrysops 143, 144, 150
anthrax Ols., Tabanus 196, 200, 255
Aplococera Enderl. 371
appendiculatus Szil., Tabanus 241
aprica Mg., Philipomyia 187, 188
apuncta Phil., Chrysops dissectus var. 169
araxis Ols., Tabanus quatuornotatus subsp.
206, 207
araxis Szil., Haematopota 410
arg'enteomaculatus Krob., Tabanus 196,
198, 248
armeniacus Krob., Tabanus 196, 201, 238
armenicus Szil., Tabanus 195, 248
armeniensis Ols., Chrysops buxtoni subsp.
182
arpadi Szil., Hybomitra 313, 317, 332
askahabadensis Szil., Chrysops flavipes var.
177
assuetus Haus., Tabanus 245
astur Erichs., Hybomitra 311, 316, 341,
347
astur (Erichs.) apud Brauer (<J),
Hybomitra 344
astuta O. S., Hybomitra 312, 327
ater Rossi, Dasyrhamphis 185
ater Szil., Hybomitra 378
aterrima Mg., Hybomitra 311, 312, 316,
350
atropathenica Abbas.-L., Haematopota 412
atropathenicus Ols., Tabanus 195, 200,
201, 277
Atylotus O. S. 126, 281, 282
aurantiacus Szil., Hybomitra 344
aureus Haus., Atylotus fulvus subsp. 285,
287
auripila Mg., Hybomitra 311, 312, 316,
349
australis Haus., Tabanus miki subsp. 235
autumnalis L., Tabanus 196, 198, 201,
272, 274
bactriana Ols., Haematopota 401, 403,
408
bactrianus Ols., Tabanus 193, 199, 204
badhysi Ols. et Shev., Tabanus ansarii
subsp. 216
bagheera Viol., Hybomitra 312, 337
Baikalia Sure. 281, 301
Baikalomyia, Stack. 281, 301
beckeri Krob., Chrysops 177
bicolor Szil., Hybomitra 344
bicolor Wd., Atylotus 281, 282
bifarius Lw., Tabanus 192, 193, 199, 202
bimaculata Macq., Hybomitra 315, 318,
352, 368
bimaculata Enderl., Hybomitra 370
bimaculata Enderl., Hybomitra borealis
var. 384
binoculatus Szil., Chrysops 169
bipunctatus Motsch., Chrysops 169
bisignata Jaenn., Hybomitra bimaculata
var. 313, 368, 369
bivittateinus Takah., Atylotus 290
bivittateinus (Takah.) apud Violovitsh,
Atylotus 289
bivittatus Mats., apud Shiraki et auct.,
Atylotus 289
borealis F., Hybomitra 325, 327
1 Курсив — синонимы; полужирный — номера страниц, на которых прино-
дено описание таксона. Названия видов приведены в соответствии с грамматичоским
родом их современного родового названия.
430 УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ
borealis Lw., Hybomitra 318, 327
■borealis Mg., Hybomitra 318, 327
boreus Stone, Hybomitra 327
bouvieri Phil., Hybomitra 377
bouvieri Phil., Silvius vitripennis var.
135, 137
bovinus L., Tabanus 192, 197, 202, 265
brevis Lw., Hybomitra 313, 317, 329
bromiolus Szil., Tabanus 241
bromius L., Tabanus 194, 200, 229, 239
brunneocallosus Ols., Tabanus 194, 199,
220
brunnescens Szil., Tabanus autumnalis
subsp. 196, 198, 276
buddha Portsch., Tabanus 196, 200, 271
buxtoni Aust., Chrysops 182
caecutiens L., Chrysops 139, 142, 143,
144, 152
caenofrons Krob., Haematopota 401, 410
callosus Ols., Atylotus chodukini var. 288
■calluneticola Krob., Hybomitra 384
calvus Szil., Atylotus 301
■capito Ols., Tabanus 253
carabaghensis Portsch., Therioplectes 305,
306
caspica Abbas.-L., Haematopota 401, 405
caucasi Szil., Hybomitra 309
caucasici Enderl., Hybomitra 313, 316,
370
caucasica Krob., Haematopota 410
caucasica Krob., Stonomyia 129
caucasicus Ols., Silvius 131, 134, 136, 137
caucasius Krob., Tabanus 193, 208
chodukini Ols., Atylotus 283, 284, 287
Chrysops Mg. 124, 139, 142
Chrysopsinae Lutz 124, 130
Chrysopsini Lutz 130
Chrysozona Mg. 399
chrysurus Lw., Tabanus 196, 200, 269
■cinereus Haus., Tabanus laetetinctus subsp.
245
•ciureai Seg., Hybomitra 315, 318, 363
clausacella Macq., Tabanus 280
colchidicus Ols., Tabanus miki subsp. 235
collini Lyn., Hybomitra 368
concavus Lw., Chrysops 144, 163
■confiformis M. et Chv., Hybomitra niti-
Mifrons subsp. 355
•confini's Ztt., apud auct., Hybomitra 355
■conformis Frey, Hybomitra 355
contigua Ols., Hybomitra distinguenda
subsp. 360, 363
cordiger Mg., Tabanus 193, 194, 200, 210,
225
cordiger (Mg.) apud Ricardo, Tabanus 229
•Corizoneura auct. 128, 129
crassicornis Wahlb., Haematopota 402,
403, 423
■crassinervis Villen., Nanorhynchus 190
cuculus Szil., Tabanus 244
•cyprianus Ric, Atylotus 299
Dasyrhamphis Enderl. 124, 185
Oasystypia Enderl. 281
decora Lw., Therioplectes 309
desertorum Szil., Haematopota 402, 403,
416
Diachlorini Enderl. 184
dissectus Lw., Chrysops 144, 145, 169
distinguenda Verr., Hybomitra 315, 318,
360
divaricatus Lw., Chrysops 143, 144, 148
Efflatounanus Phil. 185
elbursiensis Abbas.-L., Haematopota plu-
vialis subsp. 420
erberi Br., Hybomitra 311, 316, 318, 375,
387, 389, 392
erberi Br. (5), Hybomitra 387
expollicata Pand., Hybomitra 316, 318,
387, 391, 392
fasciatus Belan., Tabanus 215
fenestratus Latr., Chrysops 174
filipjevi Ols., Tabanus 193, 199, 219
flaviceps Zett., Hybomitra montana var.
314, 385
flavipes Mg., Chrysops 172, 173, 175
flavipes Wd., Hybomitra 341
flavoferomatus Strobl., Tabanus bromius
subsp. 241
flavoguttatus Szil., Atylotus 284, 297
flavopilosa Krob., Haematopota 423
fraseri Aust., Haematopota 403
freyi Szil., Tabanus 193, 216
frontalis Walk., Hybomitra montana subsp.
385, 386
fulvicornis (Mg.) apud Surcouf et auct.,
Hybomitra 382
fulvus Mg., Atylotus 283, 284,. 285
fulvus (Mg.) apud Shiraki, Atylotus 289
fumidus Aust., Tabanus 194, 199, 228
fusca Ols., Hybomitra nitidifrons var. 354
fuscipennis Szil., Hybomitra erberi subsp.
391
gedrosiana Abbas.-L., Chrysops flavipes
subsp. 178
geminus Szil., Tabanus 195, 200, 232
gerkei Br., Tabanus sabuletorum subsp.
214
gerkei (Br.) apud Krober, Tabanus 220
glaucopis Mg., Tabanus 193, 199, 208
golovi Ols., Tabanus 194, 200, 226
gracilipalpis Hine, Hybomitra 332
graeca F., Philipomyia 186, 187
graecus Chv., Lyn. et M., Silvius lati-
frons subsp. 134
• grandis Macq., Haematopota 401, 403
grandis (Szil.) apud Olsufjev et auct.,
Tabanus 249
grandis Szil., Tabanus 250
griseinus Phil., Tabanus amaenus subsp.
281
griseopunctata Ols., Haematopota
crassicornis var. 425
Griseosilvius Phil. 135
griseus Krob., Tabanus 281
guttipennis Enderl., Hybomitra 347
Haematopota Mg. 126, 399, 401
Haematopotini Enderlein 396
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ
431
hauseri Ols., Tabanus 195, 200, 245
Heptatoma Mg. 126, 396
hermanni Krob., Chrysops 154
Heterochrysops Krob. 139, 142, 172
Heterosilvius Ols. 131, 138
Hexatoma Mg. 396
hirticeps (Lw.) apud Shiraki, Hybomitra
325
hirticeps Lw., Hybomitra 352
hissaricus Bar., Tabanus 194, 242
hispanica (Szil.) apud Olsufjev et auct.,
Haematopota 420
horvathi Szil., Atylotus 283, 284, 289
horvathi Szil., Silvius vitripennis var.
135, 136
hunnorum Szil., Hybomitra 314, 316, 371
hunnorum (Szil.) apud Krober,
Hybomitra 373 v
hyalipennis Stack., Chrysops 183, 184
Hybomitra Enderl. 124, 308, 309, 311
immaculiventris Krob., Hybomitra 378
indrae Haus., Tabanus 195, 200, 235
infestus B. et Sam., Tabanus 196, 237
interruptus Krob., Chrysops 173
ioffi Ols., Hadmatopota pluvialis subsp.
417, 420 '.
iranensis Big., Chrysops 179
iranica Abbas.-L., Haematopota grandis
subsp. 403, 405
irritans Ric, Silvius 131, 135, 138
italica Mg., Haematopota 401, 403, 406
italicus Mg., Chrysops 172, 173, 174
jakutensis Ols., Chrysops ricardoae subsp.
166, 168
japonicus Wd., Chrysops 142, 144, 145
Jearafutnnis Mats., Hybomitra 337
karatschajensis Scuf., Hybomitra
montana subsp. 384, 387
karybenthinus Szil., Atylotus pulchellus
subsp. 299, 301
kaurii Chv. et Lyn., Hybomitra 312,
317, 318
kerteszi Szil., Atylotus 282
ketanensis Kono et Так., Hybomitra' 379
kimbarai Murd. et Так., Tabanus 214
kinoshitai Kono et Так., Tabanus 194,
199, 229
kirchbergi M. et Chv., Therioplectes
tricolor subsp. 305
klapperichi M. et Chv., Silvius 135
kondarensis Bar., Silvius 135
koryoensis Shir., Haematopota 401, 414
kunashiri Ols., Chrysops suavis var. 166
kurensis Djaf., Tabanus bifarius subsp.
202, 203
laetetinctus Beck., Tabanus 195, 200, 244
lama Portsch., Tabanus 212
lapponica Wahlb., Hybomitra 312, 317,
325
lapponicus Lw., Chrysops 147
lateralis Mg., Therioplectes 309
lateralis Shir., Tabanus amaenus var. 281
latifrons Ols., Silvius 130, 133
latistriatus Br., Atylotus 283, 284
leleani Aust., Tabanus 194, 200, 221
loewi Krob., Chrysops 151
loewianus Villen., Atylotus 283, 284,
288
longeantennata Ols., Haematopota 401,
409
ludens Lw., Chrysops caecutiens subsp.
143, 144, 154
lugubris Zett., Tabanus 350
luminosus Ols., Therioplectes tricolor
subsp. 305
lunatus F., Tabanus 193, 199, 203
lundbecki Lyn., Hybomitra 315, 318, 381
luppovae Bar., Tabanus 217
lurida Flln., Hybomitra 313, 318, 352
maculicornis Ztt., Tabanus 194, 200, 230
maculiventris Beck., Chrysops 177
makerovi PL, Chrysops 143, 151
mandarinus (Schin.) apud Ouchi, Tabanus
278
mandarinus (Schin.) apud Shiraki,
Tabanus 279
manshurensis Phil., Hybomitra montana
subsp. 384
marmoratus Rossi, Chrysops 174
mauritanus L., Pangonius 127
maurus Siebke, Chrysops sepulcralis var.
146
mediaasiaticus Ols., Tabanus golovi subsp.
228
Melanopangonia Szil. 127
melanopleurus Wahlb., Chrysops 158
melas Szil., Hybomitra sareptana var. 357
mendica Villen., Therioplectes 309
meridionalis Abbas.-L., Tabanus 252
meridionalis Strobl., Chrysops 154
Mesomyia 129
metabola McDun., Hybomitra 352
micans Mg., Hybomitra 311, 316, 350
miki Br., Tabanus 195, 200, 233
minor Szil., Tabanus turkestanus var. 378
miser Szil., Atylotus 283, 284, 290
mistshenkoi Ols., Tabanus 194, 200, 215
mixtus Szil., Tabanus 263
mlokosiewiczi Big., Chrysops 173, 179
mlokosiewiczi (Big.) apud Shiraki et auct.,
Chrysops 178
molitor B. ot Sam., Silvius caucasicus
subsp. 135, 138
mongolicus Szil., Chrysops dissectus var..
167
montana Mg., Hybomitra 314, 317, 384
montanus (Mg.) apud Brauer et auct.,
Hybomitra 382
montiasiaticus Ols., Tabanus 196, 199, 249
montivagus Ols., Tabanus indrae subsp.
235, 237
morgani Sure, Hybomitra montana subsp.
315, 386, 391
morio Ols., Tabanus 256
'Mouchaemyia Ols. 308, 309
muehlfeldi Br., Hybomitra 315, 318, 366
mtthlfeldi (Br.) apud Krober et auct.,
Hybomitra 386
nana Ols., Haematopota 414
nana Ols., Hybomitra tatarica var. 345
432
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ
Nanorhynchus Ols. 124, 190
Nemorius Rond. 131, 134
nigella Szil., Hybomitra 312, 317, 333
niger Ols., Tabanus 235
nigrescens Ols., Hybomitra tschuensis var.
358
nigricans Szil., Tabanus 239
nigricauda Ols., Hybomitra 333
nigricomis Mats., Chrysops . 151
nigricornis (Mats.) apud Sniraki, Chrysops
147
nigricornis Ztt., Hybomitra 314, 317, 379
nigricorpa Krob., Hybomitra 313, 317,
373
nigrifacies Gob., Atylotus 284
nigripes Ztt., Chrysops 143, 144, 147
nigrivitta Ols., Hybomitra 387
nigrocincta Enderl., Hybomitra 346
nitidifrons Szil., Hybomitra 313, 315,
318, 354
nubilis Haus., Haematopota 412
nuda McDunn., Hybomitra nitidifrons sub-
sp. 354
obscurata Big., Haematopota 400
obscurinervis Viol., Hybomtira 312, 317,
336
obscuriventris Ols., Hybomitra erberi var.
391
ochotscana Enderl., Stypommia 332, 384
ochraceus Ols. et Meln., Atylotus rusti-
cus var. 283, 294
Ochrops Szil. 281
clivacea Viol., Hybomitra 337
olivaceus De Geer, Tabanus 337
olivaceus Schuurmans Stekhoven,
Tabanus 337
olsoi Takah., Hybomitra 313, 317, 323
olsoufievi Phil., Hybomitra 370
olsufievina Phil., Hybomitra 370
olsufjevi Haus., Tabanus 197, 201, 254
olsufjevi B. et Sam., Tabanus semiargen-
teus ab. 254
orientalis Ols., Haematopota
turkestanica subsp. 414
orientalis Ols., Heptatoma pellucens subsp.
396, 399
orientalis Ols., Hybomitra expollicata
subsp. 387, 389
orientalis Ols., Therioplectes carabaghen-
sis subsp. 308
oritensis Big., Mesomyia 129
oxianus PL, Chrysops 173, 180
pallens Lw., Haematopota 401, 403, 410
pallens (Lw.) apud Krober, Haematopota
416
pallidicauda Ols., Therioplectes tricolor
subsp. 305, 306
pallidiventris Ols., Tabanus griseus subsp.
280
pallidula Krob., Haematopota 402, 415
pallidus Ols., Tabanus golovi subsp. 228
pallidus Ols., Tabanus leleani subsp. 223
pallitarsis Ols., Atylotus 283, 284, 291
pamirensis Enderl., Hybomitra 371
Pangoniinae Loew 123, 127 .
Pangoniini Loew 127
Pangonius Latr. 123, 127
paradoxus Jaenn., Tabanus 257
parallelogrammus Zell., Chrysops 144, 162
pavlovskii Haus., Haematopota 401, 404
pavlovskii Ols., Hybomitra 313, 334
peculiaris Szil., Hybomitra 316, 318,
391, 392
pellucens F., Heptatoma 396, 397
Petersenichrysops M. et Chv., Chrysops
subgen. 139, 142, 182
perelegans Ols., Tabanus 197, 200, 258
perplexus Verr., Tabanus sudeticus var.
261
pfeifferi Krob., Tabanus 255
Philipomyia Ols. 124, 186
Phyrta Enderl. 192
pictus Mg., Chrysops 143, 144, 154
plebejus Flln., Atylotus 301
plebejus (Flln.) apud Brauer, Atylotus 303
pleskei Krob., Tabanus 198, 202, 267
pluvialis L., Haematopota 399, 400, 402,
403, 417
polaris Frey, Hybomitra 327, 329
polygonus Walk., Tabanus 276, 277
polyzonatus Big., Tabanus 276, 277
pontica • Ols., Haematopota scutellata
subsp. 422, 423
popovi Ols., Hybomitra 314, 316, 341,
351
portschinskii Ols., Tabanus 197, 201, 261
proditor B. et Sam., Atylotus 283, 284,
294
prometheus Szil., Tabanus 197, 201, 252:
prometheus Szil., Tabanus eggeri var. 252.
przewalskii PL, Chrysops 167
pseuderberi Phil, et Aitk., Hybomitra 387
Pseudochrysops M. et Chv. 139, 182
pseudoricardoae Krob. (8), Chrysops 167
pulchellus Lw., Atylotus 284, 299
punctifer Lw., Chrysops flavipes subsp.
175, 177
pyritosus Lw., Pangonius 127
pyrrhus (Wlk.) apud Coquil., Atylotus 289
quadratus Mg., Chrysops 154
quadrifarius Lw., Atylotus 283, 284, 295
quatuornotatus Mg., Tabanus 192, 199,
206
regularis Jaenn., Tabanus 195, 200, 238
reinigiana Enderl., Hybomitra montana
subsp. 315, 387
relictus Mg., Chrysops 143, 144, 158
rhombica O. S., Hybomitra 308, ,311
ricardoae PL, Chrysops 144, 145, 166
rohdendorfi Ols., Philipomyia 187, 189,
rufipes Mg., Atylotus fulvus subsp. 285
rufipes Mg., Chrysops 144, 145, 170
rupium Br., Tabanus 194, 199, 230
rusticus L., Atylotus 283, 284, 293
rusticus (L.) apud Shiraki, Atylotus 2901
sabuletorum Lw., Tabanus 193, 199, 212
sachalinensis Enderl., Hybomitra 384
sakhalinensis PL, Chrysops suavis subsp.
165, 166
sakhalinensis Shir., Haematopota 419
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ
433
eapporoenus Shir., Tabanus 197, 201, 268
sareptana Szil., Hybomitra 314, 317, 356
schineri Lyn., Hybomitra 363, 365
schiva M. et Chv., Tabanus 196, 251
scutellata Ols., M. et Chv., Haematopota
402, 403, 422
sejunctus Szil., Chrysops 172, 173
semenovi Ols., Tabanus 195, 200, 242
semiargenteus Ols., Tabanus 197, 254
semipollinosa Ols., Hybomitra 313, 316,
375
sepulcralis F., Chrysops 142, 144, 145
sexfasciata Hine, Hybomitra 312, 317,
319, 320
shapuricus Abbas.-L., Silvius 135
shelkovnikovi Param., Tabanus 196, 199,
251
shevtshenkoi Ols., Hybomitra 311, 316,
341
shnitnikovi Ols., Hybomitra 314, 317,
377
sibirica Ols., Hybomitra 383
sibirica Enderl., Hybomitra 386
sibiricus Ols., Atylotus plebejus subsp.
301, 302
sibiriensis Ols., Hybomitra lundbecki
subsp. 381, 383
■signatipennis Portsch., Tabanus 196, 198,
201, 278
Silvius Mg. 124, 130, 131
simillimus Aust., Chrysops flavipes subsp.
178
sinicus Big., Tabanus 280
Sipala Enderl. 308, 309, 394
smirnov'i Ols., Tabanus 194, 201, 224
sogdiana Ols., Hybomitra 313, 317, 374
sobrina (Krob.) apud B. et Sam.,
Haematopota 412
solox Enderl., Hybomitra 308, 311
solstitialis Mg., Hybomitra 363, 368, 369
solstitialis Schin., Hybomitra 363, 365
solstitialis (Schin.) apud auct. (partim),
Hybomitra 366
sordes B. et Sam., Tabanus laetetinctus
subsp. 245
sordes (B. et Sam.) apud Hauser, Tabanus
228
sordida Ols., Hybomitra lurida var. 353
spectabilis Lw., Tabanus 196, 198, 201,272
spodopteras Mg., Tabanus 197, 202, 262
stackelbergi Ols., Chrysops 183
stackelbergi Ols., Haematopota 402, 427
stackelbergi Ols., Hybomitra 332
stackelbergiellus Ols., Tabanus 194, 199,
214
staegeri Lyneb., Hybomitra 386
stenopselapha Ols., Hybomitra 312, 317,
322
stigmoptera Ols., Hybomitra 311, 316, 343
Stonemyia Bren. 124, 128
Straba Enderl. 192
striatus Wulp, Chrysops 178
strix Szil., Hybomitra 392, 393
suavis Lw., Chrysops 143, 144, 165
subcylindrica Pand., Haematopota 402,
403, 420
sublunaticornis Ztt., Atylotus 301, 303
subparadoxus Ols., Tabanus 197, 256, 258
subsabuletorum Ols., Tabanus 193, 211
subsabuletorum (Ols.) apud Takagi,
Tabanus 214
sudeticus Zell., Tabanus 197, 201, 259
Surcoufiella Bequ., Atylotus subgen. 281,
282, 301
swiridowi Portsch., Tabanus 197, 201,
257
Sziladya Enderl. 304
Sziladynus Enderl. 308
Tabaninae Latreille 124, 184
Tabanini Latreille 192
Tabanus L. 126, 192
Taeniopangonia Szil. 127
takasagoensis Shir., Tabanus 278
talyshi Ols., Tabanus 193, 205
tamerlani Szil., Haematopota 402, 403,
425
tarandina L., Hybomitra 311, 316, 337
tarandinoides Ols., Hybomitra 311, 316,
339
-tarjukmi Haus., Tabanus bifarius subsp.
202, 203
tatarica Portsch., Hybomitra 311, 316, 344
tauricus Ols., Tabanus 224
tenuicornis Enderl., Tabanus 202
tergestinus Egg., Tabanus 194, 201, 246
tetricus Szil., Hybomitra 370
Therioplectes Zell. 126, 304
tigris Big., Stonemyia 129
tinctus Walk., Tabanus 198, 201, 263
transcaucasica Ols., Haematopota caspicn
subsp. 405, 406
transcaucasicus B. et Sam., Atylotus 287
transcaucasiensis Ols., Tabanus tinclus
var. 265
tranquilla O. S., Stonemyia 128
tricolor Zell., Therioplectes 304, 305
trifenestratus Krob. (<}), Chrysops 154
trigeminus Coq., Tabanus 196, 198, 201,
279
tristis Big., Haematopota pluvialis subsp,
417, 419
tropica (L.) apud Collin, Hybomitra 3(1(1
tropica L., Hybomitra 384
tropicus Pz., Hybomitra 368
tschuensis Ols., Hybomitra 314, 358
tunicatus Szil., Theorioplectes 304, 30(1
turanica Ols., Hybomitra 315, 393
turanicus Ols., Chrysops 143, 145, 150
Turanochrysops Stack. 139, 142, 183
turkestaria Szil., Hybomitra 314, 317, 378
turkestanica Krob., Haematopota 401, 402,
412
turkestanica Szil., Haematopota minuscula
var. 412
turkestanicus Ols., Tabanus leleani subsp,
221, 223
turpis B. et Sam., Tabanus 228
turpis (B. et Sam.) apud Hauser, Tabanus
245
tuxeni Lyneb., Hybomitra 384
Tylostypia Enderl. 308
Tylostypina Enderl. 308
typha Whit., Hybomitra lapponica subsp.
325, 327
ukrainioa Ols., Hybomitra 314, 318, 3(15
434
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЛЕПНЕЙ
umbrinus Mg., Dasyrhamphis 185
unicolor Beck., Silvius 138
unifasciatus Lw., Tabanus 194, 201, 223
ussuriensis Ols., Hybomitra 313, 317, 330
vallanti Sure, Atylotus 303
validus Lw., Chrysops 143, 144, 167
vanderwulpi Krob., Chrysops 173, 178
vappa B. et Sam., Tabanus indrae subsp.
235, 236
variegata F., Haematopota 406
verralli Oldr., Tabanus sudeticus subsp.
261
viduatus F., Chrysops 154, 156
vitripennis Mg., Silvius 131, 134, 135
vituli F., Silvius 130, 131
volgensis Ols., Chrysops ricardoae subsp -
166, 168
wagneri PI., Chrysops 167
yamasakii Ouchi, Tabanus 279
zaitzevi Ols., Hybomitra montana subsp»
384
zaitzevi Ols., Silvius 138
zimini Ols., Tabanus 194, 199, 217
znojkoi Ols., Atylotus 288
zonata Szil., Hybomitra 311, 316, 346
СОДЕРЖАНИЕ
Предисловие
Систематический указатель видов
I. Введение
1. Краткая история изучения слепней в СССР
2. Общая характеристика семейства
3. Строение слепней
Взрослое насекомое
Фазы развития
4. Филогения и классификация
5. Изменчивость слепней
Индивидуальная изменчивость
Географическая изменчивость '.
6. Географическое распространение и вопросы палеогенеза фауны слепней
Общие данные о фауне слепней Палеарктики и СССР
Типология (классификация) ареалов слепней
Боревразийский тип фауны . . ■
Средиземноморский тип фауны
Афроевразийский аридный тип фауны ;
Восточнразиатский тип фауны
Количественные аспекты фауны слепней ландшафтных зон и горных
систем СССР
Аспекты фаун равнинных ландшафтных зон ............
Аспекты фаун горных систем
Зоогеографическое деление Палеарктики . . . .'
Некоторые вопросы палеогенеза фауны Палеарктики
7. Биология слепней
8. Вредоносное значение слепней
Пораженность различных ландшафтных зон СССР
Сезонная численность слепней
Патогенное значение слепней '
9. Сбор, хранение и определение слепней
10. Борьба со слепнями
11. Литература ^
II. Специальная часть . . .
Диагноз.семейства Tabanidae . .
Определительная таблица подсемейств и родов
I. Подсемейство Pangoniinae . . . .■
1. Триба Pangoniini
1. Род Pangonius
2. Род Stonemyia
II. Подсемейство Chrysopsinae . . ,
2. Триба Chrysopsini
3. Род Silvius
• 4. Род Chrysops
III. Подсемейство Tabaninae
3. Триба Diachlorini
5. Род Dasyrhamphis
6. Род Philipomyia
7. Род Nanorhynchus :.
4. Триба Tabanini .
8. Род Tabanus
9. Роя Atylotus 281
10. Род Therioplectes ; 304
11. Род Hybomitra 308
5. Триба Haematopotini 396
12. Род Heptatoma 396
13. Род Haematopota 399
Добавление 428
Указатель латинских названий слепней : 429
СЛЕПНИ.
Сем. TABANIDAE
(Фауна СССР.
Насекомые двукрылые. Т. VII. Вып. 2)
Утверждено к печати
Зоологическим институтом Академии наук СССР
Редактор издательства А. А. Ройтман
Технический редактор М. 9. Нарлайтис
Корректоры Г. Я. Атлас и Л. Б. Наместникова
Сдано в набор 20/IV 1976 г. Подписано к печати 14/IV 1977 г.
Формат 70xl08Vie- Бумага М2. Печ. л. 27'/4=38.15 усл. печ. л.
Уч.-изд. л. 47.41. Изд. № 5890/6115. Тип. зак. N5 1159. М-17243.
Тираж 1900. Цена 5 р. 27 к.
Ленинградское отделение издательства «Наука»
199164, Ленинград, В-164, Менделеевская линия, д. 1
1-я тип. издательства «Наука»
199034, Ленинград, В-34, 9 линия, Д. 12