/


















































































































































































































































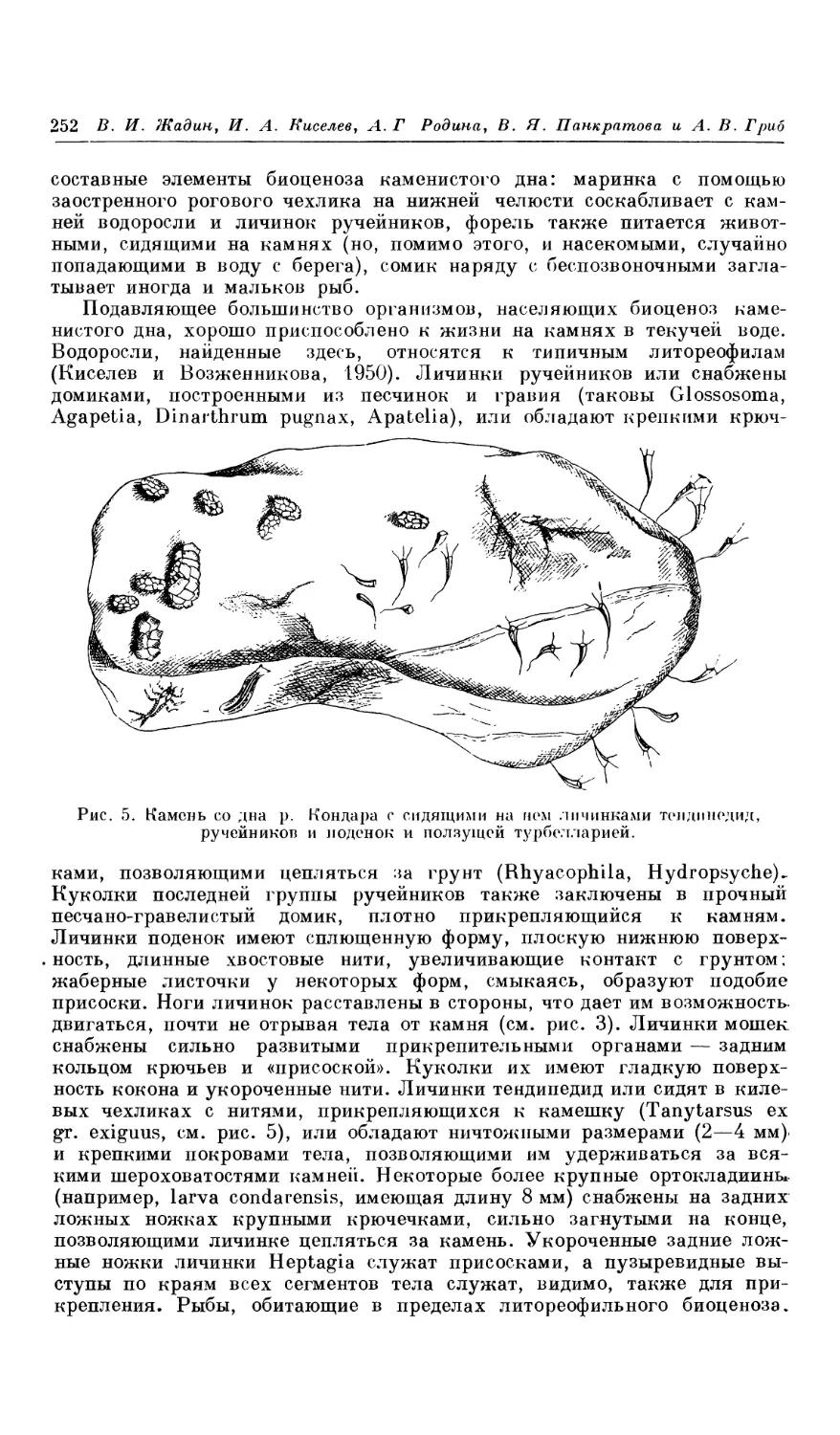























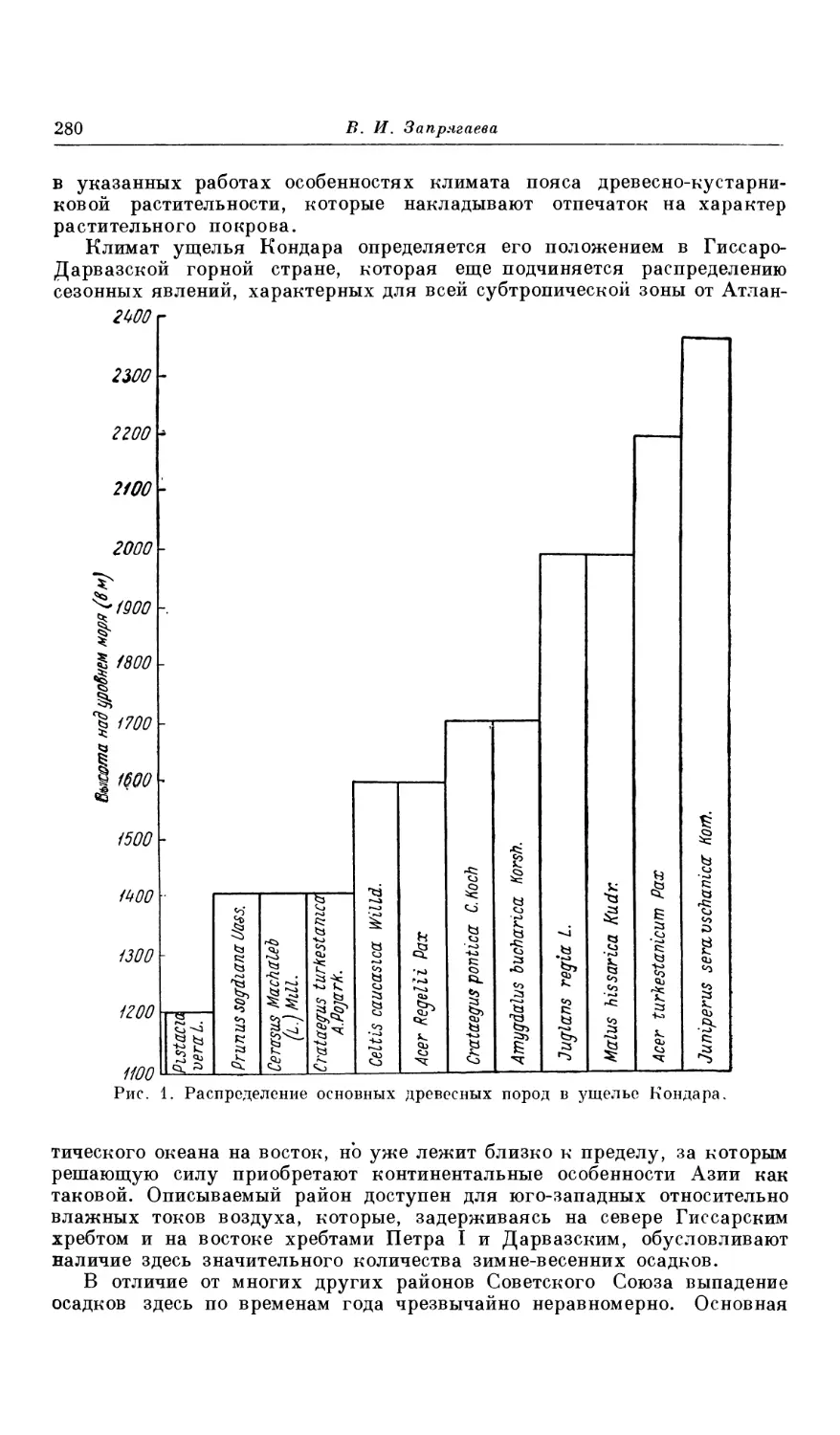






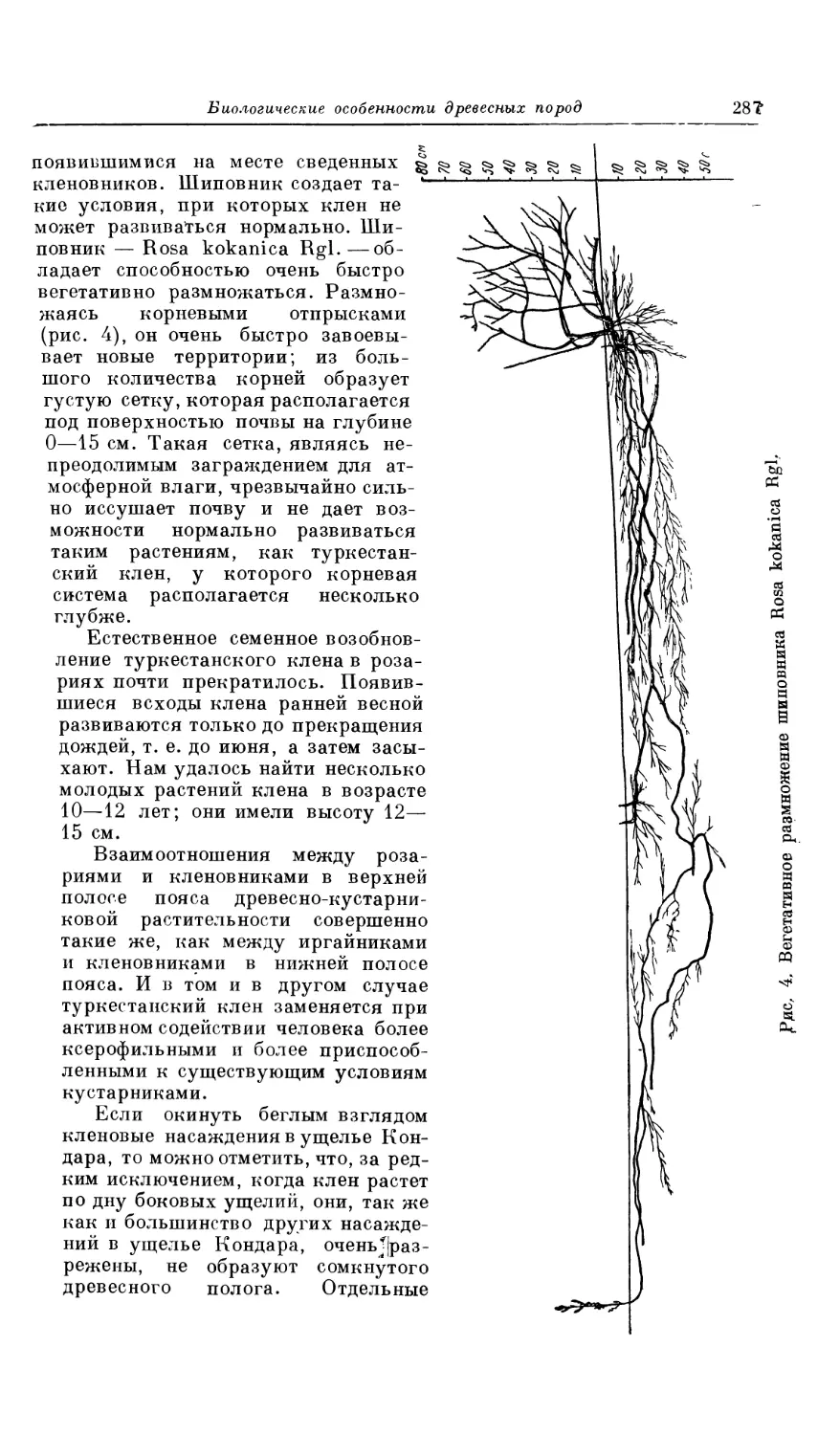
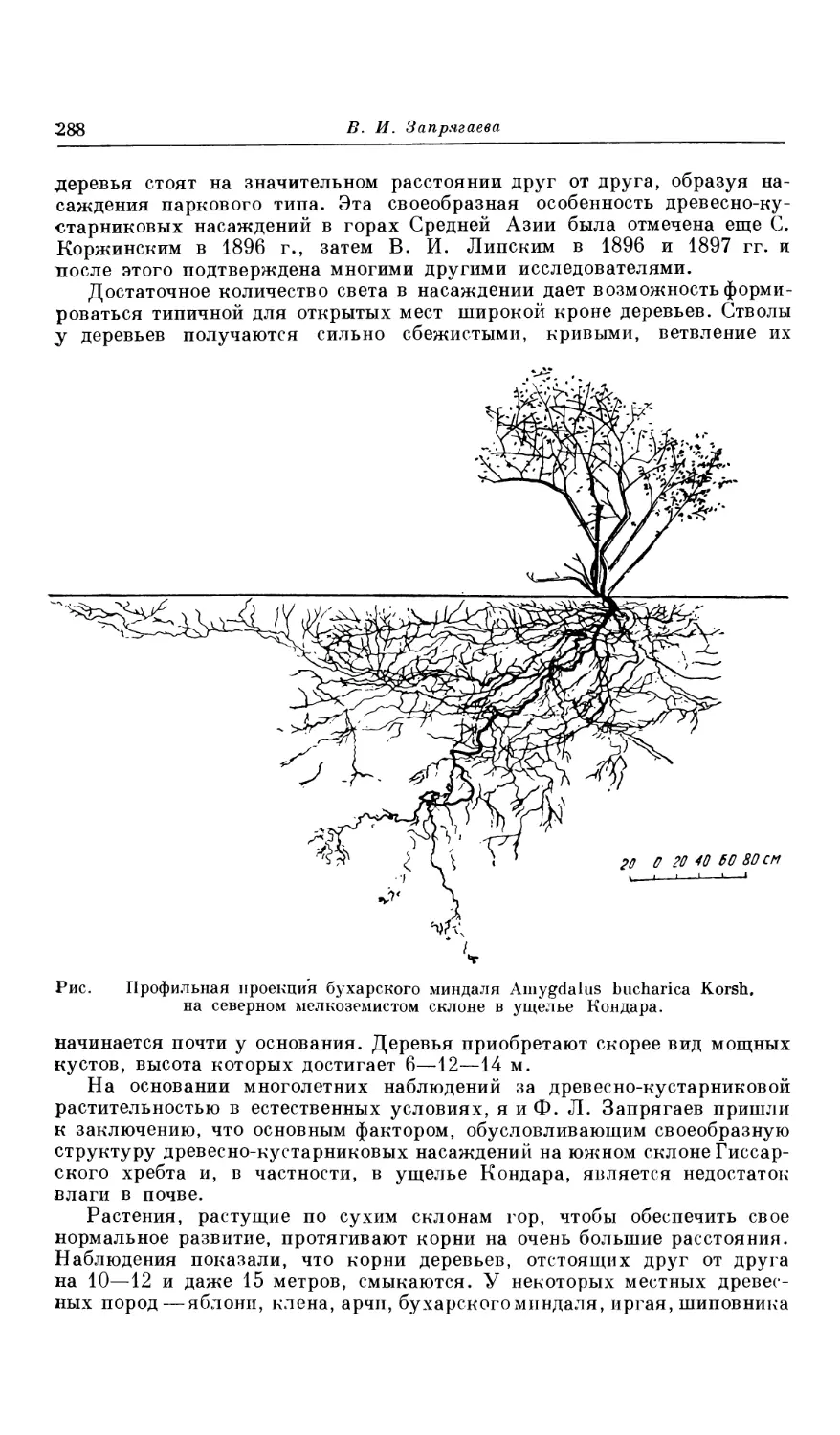


































































































































Текст
АКАДЕМИЯ
НАУК
СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
УЩЕЛЬЕ БОНДАРА
(ОПЫТ БИОЛОГИЧЕСКОЙ МОНОГРАФИИ)
Под редакцией
акад. Е. Н. ПАВЛОВСКОГО и проф. В. И. ЖАДИНА
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА
1951
ЛЕНИНГРАД
Ответственный редактор акад. Е. Н. Павловский
Редактор проф. В, И. Жадин
Посвящается
Академии Наук Таджикской ССР
в связи с ее организацией на основе
Таджикского филиала
Академии Наук СССР
ВВЕДЕНИЕ
Ущелье Кондара — одно из многих подобных ущелий в горах Таджи-
кистана — расположено на южном склоне Гиссарского хребта близ
Сталинабада. На коротком протяжении ущелья сменяются различные
высоты (от 1100 до 2600 м над ур. м.) со своими климатическими особен-
ностями, своеобразным сочетанием условий существования для растений
и животных, зависящих не только от высоты местности, но и от распо-
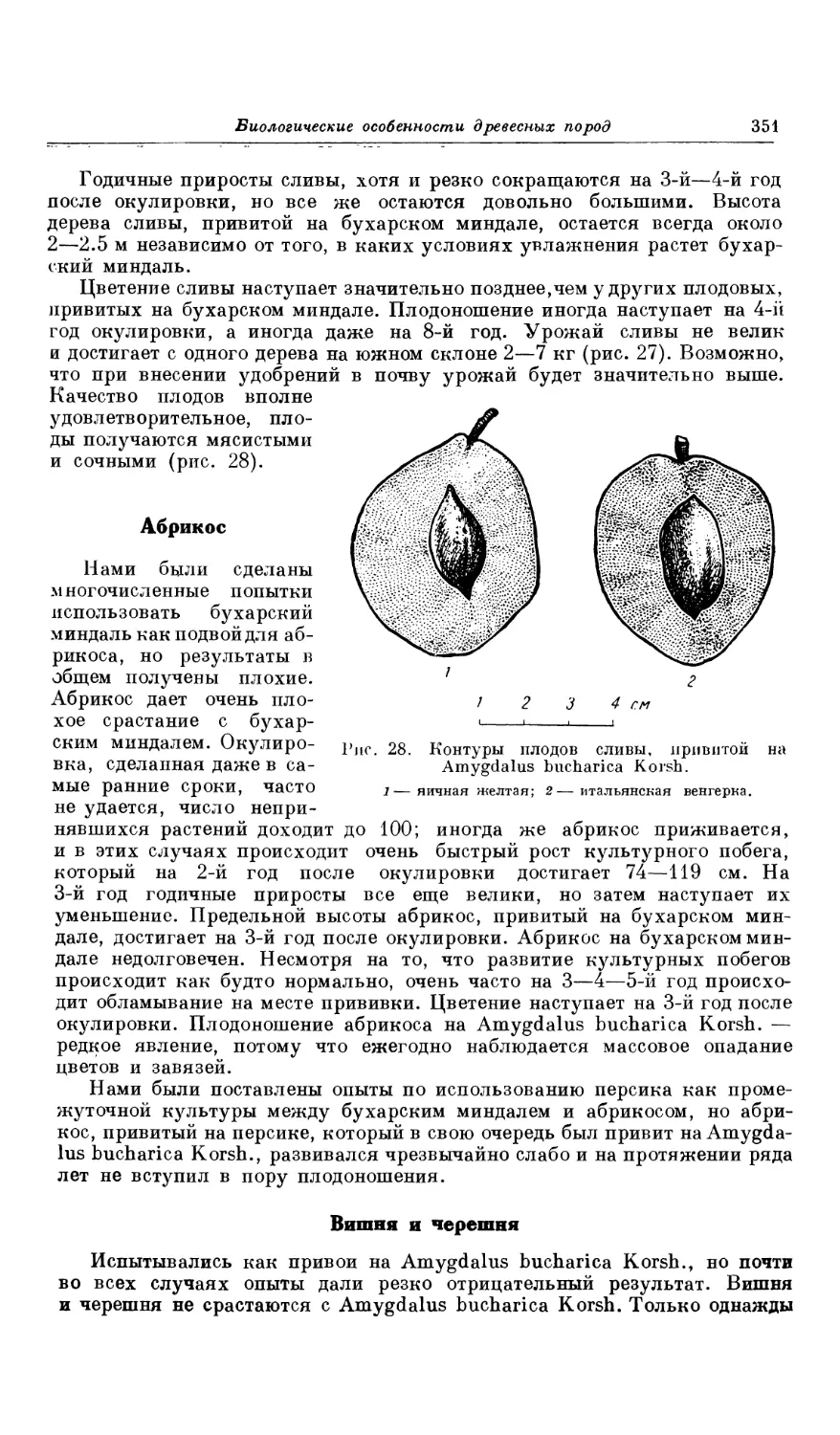
ложения склонов в отношении к странам света, близости или отдален-
ности от водных источников и др.
Склоны ущелья поросли многочисленными древесными, кустарнико-
выми и травянистыми растениями, среди которых многие представляют
ту или иную хозяйственную ценность. Однако использование раститель-
ности в ее естественном состоянии носит ограниченный характер: дре-
весные породы отличаются большей частью медленным ростом и вос-
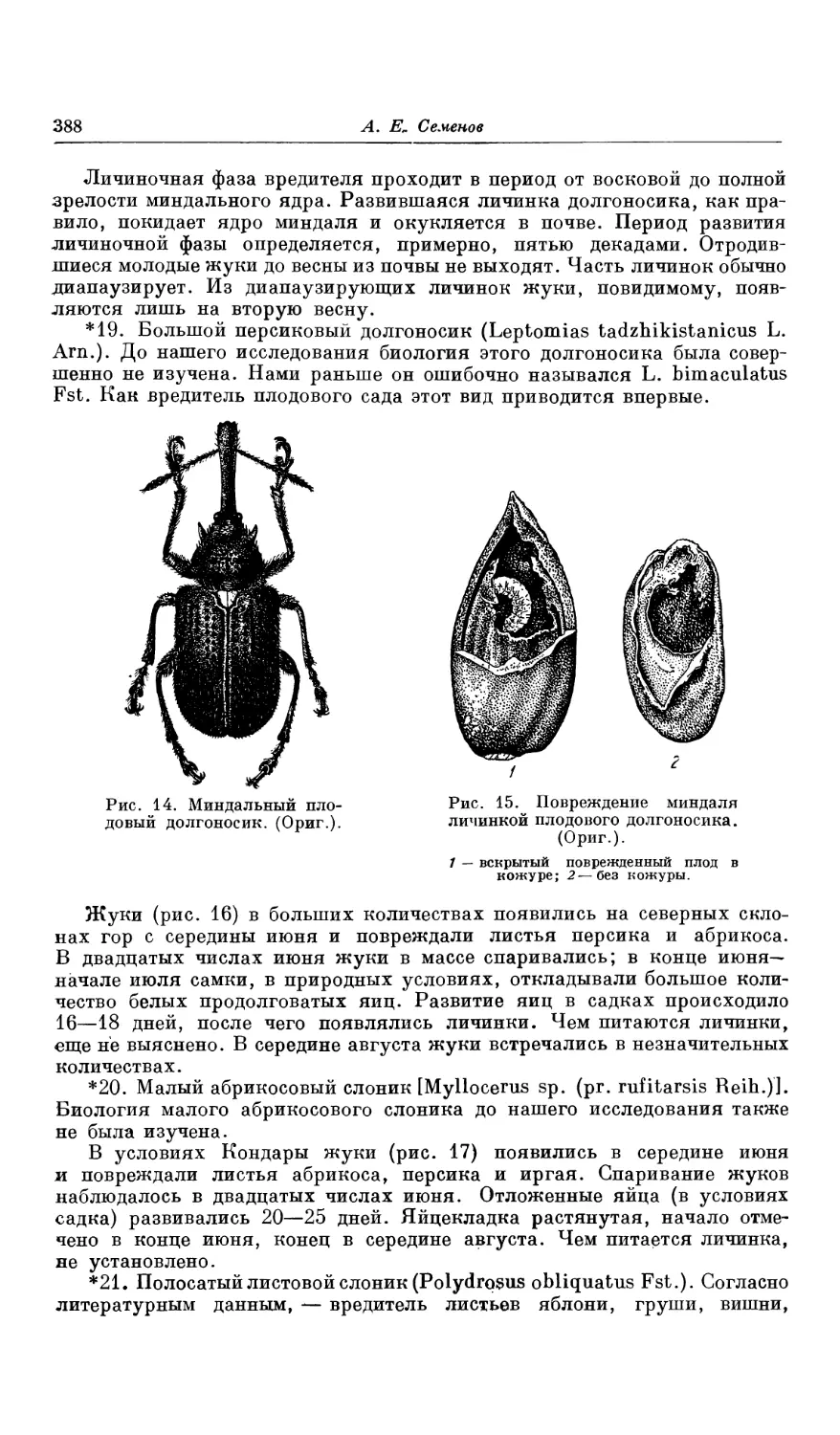
производством, плоды, фрукты и ягоды дикорастущих плодовых обычно
не удовлетворительны по качеству. Примитивное земледелие нередко
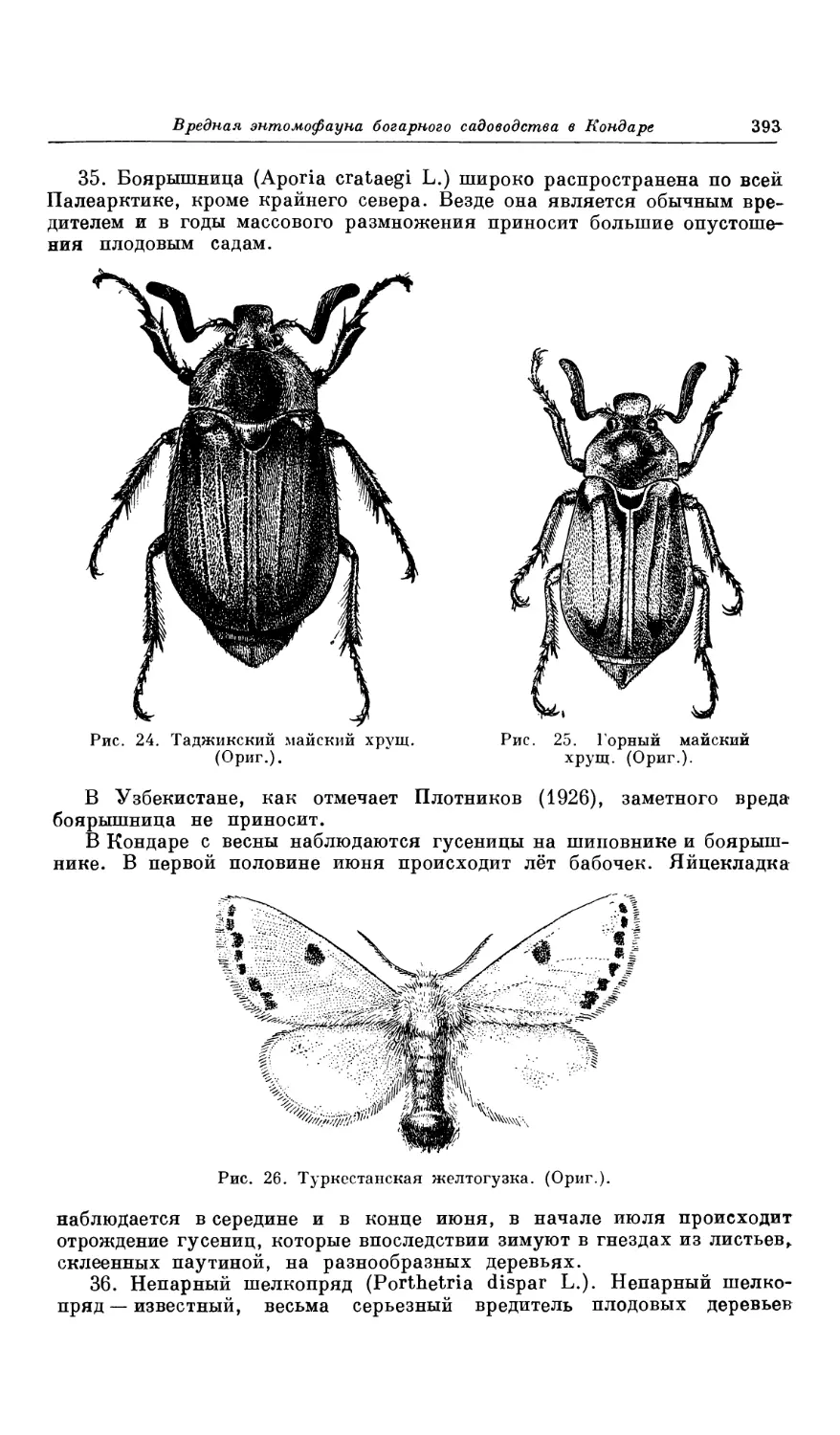
влекло за собою эрозию горных склонов и полное уничтожение почвен-
ного покрова.
При умелом хозяйствовании, при проведении необходимых меро-
приятий по улучшению растительности, переделке природы растений
мичуринскими методами, коренном улучшении водного режима почв
гидротехническими приемами можно добиться превращения горных
ущелий в богатые урожаями сады и культурные полевые угодья.
Мысль о возможности «облагораживания» естественной раститель-
ности в горах Таджикистана впервые высказал Владимир Леонтьевич
Комаров (впоследствии академик и президент Академии Наук СССР),
когда он, еще студентом, посетил отсталую феодальную Горную
Бухару.
Практическая задача культурного освоения и преобразования при-
роды горных ущелий выдвигает перед исследователями громадное коли-
чество вопросов первостепенного научно-теоретического значения.
Ставя вопросы хозяйственного освоения биологических ресурсов гор.
исследователь неминуемо подходит к решению многих основных теоре-
тических проблем современной биологии. Единство теории и практики
становится ведущей линией работы.
Таджикский филиал (прежде Таджикская база) Академии Наук
СССР с самого своего возникновения предпринял работы по ботаническому
исследованию Гиссарского хребта и организовал в 1935 г. в ущелье Кон-
дара сначала опорный пункт, а затем опытную станцию, получившую
наименование Варзобской горной ботанической станции. Эта Станция,
руководимая ее основателем Ф. Л. Запрягаевым (погибшим во время
Великой Отечественной войны), наряду с описательными полевыми
6
Введение
работами заложила многолетние опыты в целях наилучшего плодовод-
ческого, лесоводческого и сельскохозяйственного освоения ущелья:
его склонов — методами горного бесполивного (богарного) земледелия
и его дна — методами обычного для Средней Азии поливного хозяй-
ства.
В своей плодотворной работе Варзобская горная ботаническая стан-
ция не была одинока. В ее работах непосредственное участие принимали
ученые различных институтов Академии Наук СССР (Ботанического,
Института почвоведения и др.) и ее Таджикского филиала (институтов
ботаники, зоологии и паразитологии). Во время Великой Отечественной
войны в Сталинабад из блокированного Ленинграда был эвакуирован
Зоологический институт Академии Наук СССР, также включившийся
в научно-исследовательские работы по ущелью Кондара.
В итоге длительных работ большого научного коллектива были изу-
чены многие существенные вопросы, касающиеся биологических ресур-
сов ущелья — растительного и животного мира. К этому же вре-
мени одним из основателей Варзобской ботанической станции —
непрерывно работавшей на ней В. И. Запрягаевой — была подытожена
десятилетняя работа Станции по освоению и мелиорации древесной
растительности Кондары (диссертация на соискание ученой степени кан-
дидата биологических наук).
Новизна подхода к развитию научных исследований, непосред-
ственно слитых с практическими требованиями народного хозяйства,
их глубокий теоретический интерес, обилие ботаников, зоологов и дру-
гих специалистов, работавших в Кондаре или над материалами из Кон-
дары, породили довольно большую литературу, которая требовала сводки
и опубликования таковой. Отсылая за подробностями к статье
Е. Н. Павловского «Возникновение и развитие деятельности Варзобской
горной ботанической станции Таджикского филиала Академии Наук»,
отметим, что в настоящее время эта работа закончена и представляется
вниманию читателей. В ней помещены статьи только о биологических
ресурсах ущелья, так как сведения о геоморфологии, климате и почвах
Таджикистана можно почерпнуть из других литературных источников,
включая фонды Филиала.
Сборник-монография начинается статьей академика Е. Н. Павлов-
ского, освещающей историю возникновения и деятельности Варзобской
горной ботанической станции; далее следуют статьи В. В. Письяуковой и
А. С. Королевой о флоре и растительном покрове Кондары; ряд статей —
Б. С. Виноградова, А. Я. Тугаринова, С. А. Чернова, Г. X. Шапошни-
ковой, Е. П. Голубцовой, А. А. Штакельберга, И. А. Рубцова, С. Г. Леп-
невой, В. В. Попова, М. Н. Никольской, А. Н. Кириченко, Л. Л. Ми-
щенко, А. Н. Поповой, Д. Е. Харитонова, Д. И. Благовещенского,
Е. С. Кирьяновой, Б. В. Лотоцкого и Е. Ф. Сосниной — дает сведе-
ния о животном мире ущелья.
Следует отметить, что, несмотря на большой коллектив зоологов,
принимавших участие в изучении животного мира Кондары, не все группы
фауны оказались достаточно исследованными, некоторые же остались
вовсе не затронутыми; вследствие этого в настоящем сборнике не нашли
себе места статьи о таких группах животных, как бабочки, жуки, скор-
пионы, фаланги и некоторые другие. Одной из причин неполноты охвата
фауны насекомых является преждевременная кончина одного из ста-
рейших исследователей фауны Кондары, б. научного сотрудника Тад-
жикского филиала АН СССР — В. В. Гуссаковского.
Введение
7
Статья В. И. Жадина, И. А. Киселева, А. Г. Родиной, В. Я. Панкра-
товой и А. В. Гриб посвящается краткому очерку жизни (бактериальной,
растительной и животной) в водах Кондары.
Описательная часть монографии завершается статьей скончавшегося
в 1948 г. замечательного натуралиста проф. А. Я. Тугаринова о кален-
даре жизни в ущелье Кондара.
Большой раздел книги посвящен результатам опытов хозяйствен-
ного освоения растительных ресурсов ущелья: так, статья В. И. Запря-
гаевой говорит о мелиорации древесной растительности; А. С. Коро-
лева написала о культуре кормовых трав. Здесь же приводятся данные
о вредителях богарного садоводства (статья А. Е. Семенова) и о болез-
нях растений в ущелье Кондара (статья В. Ф. Купревича).
Редакция и составители этой книги выражают надежду, что научно-
исследовательские работы по освоению богатейших возможностей при-
роды горного Таджикистана будут продолжаться в возрастающих по глу-
бине и охвату темпах и что настоящая книга окажется полезной при
дальнейших исследованиях Таджикистана и всей Средней Азии. Закон-
чена она была незадолго до реорганизации Таджикского филиала Ака-
демии Наук СССР в Академию наук Таджикской ССР. Этот коллектив-
ный труд, вместе со всей научной продукцией филиала и некоторых
центральных институтов Академии Наук СССР, послужит основой для
дальнейшего мощного развития науки в Таджикистане, где она выросла
на «невспаханной почве». Так же как и развитие Советского Таджики-
стана в целом, организация Академии является блестящим образцом
претворения в жизнь ленинско-сталинской национальной политики, j
Акад. Е. И. Павловский х
Проф. В. И. }Кадин. \
Акад. Е. Н. ПАВЛОВСКИЙ
Председатель президиума Таджикского филиала и директор Зоологического
института Академии Наук СССР
ВОЗНИКНОВЕНИЕ И РАЗВИТИЕ ДЕЯТЕЛЬНОСТИ ВАРЗОБСКОЙ
ГОРНОЙ БОТАНИЧЕСКОЙ СТАНЦИИ ТАДЖИКСКОГО ФИЛИАЛА
АКАДЕМИИ НАУК СССР
Варзобская горная ботаническая станция, входящая в состав учре-
ждений, совокупность которых образует Ботанический институт Таджик-
ского филиала Академии Наук СССР, является одной из старейших ча-
стей этого филиала. Начало ее организации восходит к 1935 г.; сам же
филиал возник, сначала на положении базы Академии Наук СССР,
в 1933 г. Осенью 1932 г. в Сталинабад, по приглашению Таджикского
правительства, прибыла бригада академиков и профессоров для озна-
комления на месте с работами экспедиции 1932 г., отряды которой дей-
ствовали во многих местах Таджикистана. В состав делегации входили
академики В. Л. Комаров, С. Ф. Ольденбург, А. Ф. Иоффе, Д. Н. Пря-
нишников, профессора Д. В. Наливкин, Е. Н. Павловский. В результате
совещаний с правительством Таджикской ССР возникла идея об орга-
низации в Сталинабаде постоянной базы Академии Наук СССР для
стационарных и экспедиционных исследований Таджикистана, каковые
исследования при такой форме организации могли быть осуществляемы
планово и более результативно. Таджикская база Академии Наук была
открыта с начала 1933 г. Она создавалась абсолютно заново, на «невспа-
ханной почве», не имея от прошлого никакого наследия в отношении кад-
ров, лабораторий, оборудования, библиотеки, зданий, средств, не го-
воря уже о научной тематике.
Потребность в учреждении такого рода была очевидна особенно для
Таджикистана, который в дореволюционном прошлом являлся Восточ-
ной (или Горной) Бухарой; долицы Зеравшана и Ферганы входили в со-
став Самаркандской области. Восточная Бухара далеко отстояла от
основных путей сообщения и по географическому положению своему была
трудно доступна. Не приходится поэтому удивляться, что в прежнее
время в нее пробирались лишь редкие путешественники (Леман, Север-
цов, Мушкетов, Липский, Богоявленский и др.). Вошедший в состав
Таджикской ССР Памир — Автономная Горно-Бадахшанская область —
был еще менее доступен для сколько-нибудь регулярной научно-иссле-
довательской работы.
Народное хозяйство молодой республики должно было строиться на
основе использования местных природных ресурсов, развития произ-
водительных сил и перестройки прежнего хозяйственного модуса в связи
с новой специальностью Таджикистана, по общегосударственному
10
Е. Н. Павловский
плану. Такой специальностью явилось хлопководство, для развития
которого потребовалось строительство Вахшской ГЭС в целях орошения
долины Вахша.
Ясно, что с первых же лет жизни Таджикистана в семье братских
советских республик ему потребовалась научная основа для социали-
стического строительства.
Практика экспедиционных исследований, при всей важности добытых
ими результатов, все же оказалась недостаточной, хотя бы по одному тому,
что по множеству вопросов, касавшихся живой природы, требовались
круглогодичные и, во всяком случае, продолжительные работы на ме-
стах; это было особенно очевидно для исследований эксперименталь-
ного характера. По указанной причине и возникла Варзобская горная
ботаническая станция при ближайшем участии, тогда — вице-президента
Академии Наук СССР, академика В. Л. Комарова. Варзобское ущелье —
Кондара, — расположенное по южному склону Гиссарского хребта,
было известно как местность, представляющая особую ценность для
научных исследований. Первые же годы деятельности Станции подтвер-
дили удачность выбора места для ее основания.
Организации Варзобской станции предшествовали двухгодичные ста-
ционарные работы по изучению главнейших растительных ассоциаций
всего вертикального профиля южных склонов центральной части Гиссар-
ского хребта. Основной целью являлось выяснение особенностей посезон-
ного развития растительного покрова и изучение основных биологи-
ческих свойств различных видов растений и растительных ассоциа-
ций в целом. Данных такого рода по Таджикистану до сих пор не было.
Оказалось, что урочище Кондара вмещает в себя особенности сколь
оригинальные, столь и типичные для гор центрального Таджикистана.
Уже первые ориентировочные обследования Ф. Л. Запрягаева поз-
волили выделить на южном склоне Гиссарского хребта следующие зоны
растительности:
Над ур. м.
I. Полупустыня . до 800 м
II. Полу степь от 700 » 1250 »
III. Нижняя лесная полоса » 1150 » 2050 »
IV. Верхняя » » » 1900 » 2550 »
V. Субальпийская зона » 2400 » 3500 »
VI. Альпийская полоса . » 3300 » 3950 »
VII. Снег и ледники » 3600 » 3950 » и выше
В непосредственной близости Станции, лежащей в нижней полосе
древесно-кустарникового пояса (около 1100 м над ур. м.), имеются опор-
ные пункты: Квак — в верхней подзоне этого пояса (на высоте около
1800 м) и урочище Руидашт — в субальпийской полосе (2600 м). На-
блюдения и работы производились и в альпийском поясе (на высоте
3300—4000 м).
Полевые, стационарные и экспериментальные исследования ботани-
ков Таджикского филиала (базы) Академии Наук СССР имели конечной
целью преобразование естественной растительности на основе геобота-
нических и эксперементальных работ, отвечающих производственным
требованиям народного хозяйства республики, которые были столь же
четки, как и разнообразны. Предстояло повышение производительности
естественных кормовых угодий, облесение горных склонов для борьбы
с эрозией, создание богарных лесосадов и садов, получение лесных строи-
тельных материалов и др. Должны были быть разработаны и проверены
Возникновение и развитие деятельности Варзобской станции
11
на практике методы всех этих работ в такой форме, чтобы они могли
быть использованы и для других районов Таджикистана, где могли быть
организованы, смотря по потребности, опытные пункты. Урочище
Кондара оказалось вполне подходящим местом для указанных работ.
Необходимость развития ботанических исследований именно в на-
званных направлениях властно диктовалась многими побудительными
причинами. Они были подробно изложены в докладной записке директора
Ущелье Кондара.
(Фот. А. И. Иванова).
и одного из основателей Варзобской горной ботанической станции —
Ф. Л. Запрягаева, представленной в ЦК КП(б) Таджикистана. В ней
говорилось следующее.
Несмотря на наличие больших площадей, покрытых древесной расти-
тельностью (около 170 000 га), Таджикистан не имеет своих лесных
строительных материалов и достаточного количества дров. Все это при-
ходилось завозить из отдаленных частей СССР, что весьма удорожало
стоимость доставленных материалов. Кроме того, в такой горной стране,
как Таджикистан, есть районные центры, куда' не было физической воз-
можности доставлять привозные лесные материалы, что препятствовало
развитию строительства.
12
Е. Н. Павловский
Выход из положения мог быть найден созданием высококачественных
горных лесных насаждений. Станцией были поставлены исследования
по следующим направлениям: а) разработка методов разведения леса на
горной богаре; б) мелиорация уже существующих насаждений (лесо-
сады) и в) интродукция ценных иноземных древесных пород. Эти задачи
могли быть разрешаемы на основе изучения биологии и физиологии
отдельных деревьев разных видов и цельных насаждений.
Значение теоретической основы для практики лесоразведения на бо-
гаре явствовало из неоднократных попыток местных лесхозов заклады-
вать на богаре леса производственного значения. Так как никакой мето-
дики лесоразведения на богаре в Таджикистане не было разработано,
то посадки производились «вслепую» с затратой больших средств; обыч-
ным результатом была неудача, ибо посадки погибали от летней
засухи на 80—100% ; оставшиеся в живых давали прирост от 3 до 40 мм
в год.
Применение разработанной на Станции методики посадки и ухода
за деревьями дало отличные результаты: например — 20-сантиметровый
айлант осенней посадки 1935 г. через год достиг высоты 90 см, а к весне
1938 г. он вырос до 3 м; посаженная одновременно с ним белая ака-
ция за тот же срок выросла с 65 см до 240—270 см; и эти примеры
не являлись исключением. Ясень и шелковица давали в год метровый
прирост.
На опытных участках Станции хороший рост дали и чуждые для
флоры Таджикистана дубы, софора, гледичия и др.
Параллельно производилось изучение распределения корневой
системы деревьев на богаре и важнейшей проблемы водного баланса ксе-
рофитов из древесных пород в целях разработки способов ускорения их
медленного в естественных условиях роста.
Второй задачей, поставленной перед собою Станцией, явилось созда-
ние лесосадов и культурных горных садов. Лесосады возникают на базе
дико произрастающих лесов через перепрививку составляющих их
деревьев и кустарников различными сортами культурных плодовых,
другими словами — в результате преобразования диких, ни на что
не используемых в пищевом отношении деревьев и кустарников.
Сады же закладываются на расчищенных участках с подготовленной
почвой — пересадкой культурных растений, выращенных в питом-
никах.
Идея организации лесосадов не совсем нова, ибо в литературе встре-
чаются указания по этому вопросу, в большинстве — предположитель-
ного характера. Значение работ Станции в рассматриваемом отношении
определяется, во-первых, полнейшей новизной их для Таджикистана
и массовостью.
Успех работ зависел от изучения биологии естественно произрастаю-
щих подвоев, подбора привоев, исследования их физиологии в условиях
роста на ином виде подвоя. Необходимо было разработать и самые методы
перепрививки.
Работы Станции по организации лесосадов увенчались отличными
успехами.
Для облагораживания естественно произрастающих деревьев и ку-
старников рекомендуются следующие ^комбинации:1
1 Подробности см. в статье В. И. Запрягаевой— одного из организаторов Вар-
зобской горной ботанической станции (стр. 279 настоящего сборника).
Возникновение и развитие деятельности Варзобской станции 13
Подвой
Привой
1. Бухарский миндаль, плоды которого
почти никак не используются
2. Иргай
3. Яблоня дикая
4. Груша Коржинского
5. Груша разнолистная
6. Вишня махалебская
7. Вишня карликовая
8, Алыча
( Сладкий миндаль
t Персик
Слива
Абрикос
Айва
Груша
Яблоня
Яблоня
Груша
Груша
Г Вишня
( Черешня
f Вишня
I Черешня
Слива
Отметим важность широкого продвижения в колхозную практику
технически несложной закладки лесосадов, отличающейся дешевизной;
начало плодоношения наступает на третьем году. Негодные под другие
культуры угодья (а их много) вполне могут быть использованы под лесо-
сады.
Работы в этом направлении превращаются в подлинно мичуринские:
необходимо выяснить, происходит ли изменение наследственных особен-
ностей культурного привоя, произрастающего на диком подвое другого
вида. Вопрос этот разрешается высевом семян привоя и определением
того, приобретена ли сеянцами из этих семян большая стойкость к су-
хости. Возможно, что потребуются испытания сеянцев ряда поколений,
повторно используемых для перепрививки.
Во вкусовом отношении плоды привоев обладали сладким вкусом
и сильным ароматом.
Устройство лесосадов ограничивается участками с наличием диких
деревьев и кустарников в ближайшем окружении колхоза. Трудность
разведения садов на богаре создается необходимостью хорошего оро-
шения, практически неосуществимого на богаре. Поэтому мысль иссле-
дователя останавливается над вопросом, как можно лучше задержать
весеннюю влагу. На Станции был применен метод террасирования, от-
крывший возможности хорошего роста на богаре персика, абрикоса,
яблони, айвы и шелковицы. Прирост в первый год пересадки доходил*
до 36 см, а на второй — до 60—100 см. Возможность разведения шелко-
вицы на богаре открывает новые и весьма широкие перспективы для раз-
вития шелководства.
Устройство лесосадов и насаждение садов способствуют укреплению
склонов от размыва и являются, со своей стороны, способом борьбы
с эрозией. В целях рационализации работы с такой направленностью
Варзобской станцией поставлен вопрос об организации садово-сенокос-
ного или садово-сенокосно-зернового хозяйства. Суть дела заключается
в том, что одновременно с закладкой плодового или тутового сада дол-
жен производиться посев зерновых культур или создание богарных
сенокосов — и то и другое в широких междурядьях между посаженными
деревьями. Таким способом достигается более надежное закрепление
склонов от размывов, при которых легко может быть нацело снесен плодо-
родный слой почвы, и соответственный участок в хозяйственном отно-
шении погибнет.
Богарное травосеяние имеет и самодовлеющее народнохозяйственное
значение. Задачей опытников-исследователей является подбор и испы-
14
Е. Н. Павловский
тание различных диких местных и культурных кормовых трав для поды-
скания такого видового состава смесей, при котором обеспечиваются
урожайные и стойкие сенокосы. Работы эти усложняются тем, что их
приходится вести на разных высотах. Уже первые опыты дали хорошие
результаты: оказалось, что ряд сортов люцерны вполне благоприятно
развивается даже на высоте 2600 м над ур. м., где в естественных усло-
виях она никогда не встречается.
Эти работы по травосеянию способствуют кормовому улучшению
пастбищ. Летние пастбища с давних пор оказались сосредоточенными
в высокогорной части Таджикистана. Использовались они без всякой
системы; неправильный искони выпас привел к засорению пастбищных
угодий несъедобными колючими и ядовитыми травами. В некоторых слу-
чаях непоедаемая масса составляла три четверти всего урожая травы
соответственного угодья. Так обстояло дело летом; зимой же все пого-
‘ловье скота питалось скудным подножным кормом; в холодные и снеж-
ные периоды скот получал подкормку из мягкой соломы или из жестких,
малопитательных трав. Да и их заготовлять было нелегко, ибо вслед-
ствие разобщенности мест их произрастания сенокошение производилось
серпом и выборочно.
Развитие животноводства в Таджикистане требовало коренной реор-
ганизации естественной растительности в целях создания всеми необхо-
димыми способами высококачественных горных пастбищ и сенокосов.
Эта задача являлась третьей основной проблемой работ Станции. Важ-
ной ее задачей являлась борьба с пастбищными сорняками, о которых
говорилось выше. Многочисленные опыты в этом направлении дали отлич-
ные результаты. Подрезка растений кетменем на определенном уровне
основания стебля полностью уничтожала растения, которые больше
не возобновлялись. Освобождавшиеся места засевались кормовыми тра-
вами (пажитником).
Итак, Станция с первых же месяцев своих работ взяла напра-
вления, отвечающие потребностям народного хозяйства Таджикистана
и вместе с тем полные глубокого теоретического интереса.
Нельзя не отметить характерную общую черту всех работ Станции —
это их комплексность, что было весьма компетентно засвидетельствовано
заключением Стационарной комиссии Государственного Ботанического
общества по докладу о геоботанических исследованиях Варзобской гор-
ной ботанической станции.
Это заключение гласило:
«Заслушав и обсудив на собрании 14 марта 1937 г. доклад
Ф. Л. Запрягаева о ботанических исследованиях Варзобской гор-
ной станции Таджикистанок ой базы Академии Наук СССР, Ко-
миссия сочла необходимым отметить следующее:
«1. На основании неоднократных в течение последних трех лет
заслушанных сообщений о работе упомянутой Станции, Комиссия
констатирует крупные успехи ее работы. Станция Таджикистан-
ской базы опередила многие другие станции, поскольку от из-
учения природных закономерностей растительного покрова она
перешла уже к преобразованию растительности и растений. Сделано
это с таким успехом, что уже в настоящее время результаты иссле-
дований Станции могут быть использованы местными производ-
ственными организациями.
«2. Комиссия отмечает, как пример для аналогичных станций,
правильную организацию исследовательской работы Станции Тад-
Возникновение и развитие деятельности Варзобской станции
15
жикистанской базы, выразившуюся в проведении принципа ком-
плексности исследований с общим направлением в сторону рацио-
нального освоения горных территорий.
«3. Комиссия находит целесообразным при дальнейшем раз-
вертывании работы Станции обратить больше внимания на углу-
бление эколого-физиологических исследований.
Председатель Государственного Ботанического
общества академик — В. Л. Комаров. Ученый
секретарь Государственного Ботанического об-
щества, председатель Стационарной комиссии
д-р биол. наук — А. П. Шенников».
Работы Станции вообще шли в связи с Ботаническим институтом,
ныне имени В. Л. Комарова, Академии Наук СССР. Ежегодно на Уче-
ном совете этого Института заслушивались и обсуждались доклады
директора Станции о произведенных работах, что обеспечивало необхо-
димое коррегирование исследований. Некоторые материалы оканчива-
лись обработкой в стенах Ботанического института Академии Наук во
время командировок в Ленинград и отпусков сотрудников Станции.
В самом Таджикском филиале Академии Наук Варзобская горная
ботаническая станция входит в состав Ботанического института
филиала, через Ученый совет которого проходят научные доклады,
отчеты и планы работ Станции.
Из всего сказанного очевидно, что все начальные работы Станции
являлись ботаническими; но в ближайшие же годы определилась необ-
ходимость организации на территории Станции энтомологических иссле-
дований. Работы по выявлению общей фауны велись ныне покойным
энтомологом, старшим научным сотрудником Института зоологии и па-
разитологии филиала В. В. Гуссаковским, которым были накоплены
обширные и весьма ценные материалы.
По мере развития основных работ Станции потребовалось проведение
специальных исследований по фауне и биологии вредных насекомых,
прежде всего для привоев в опытном лесосаде на территории самой Стан-
ции. Эти работы проводились кандидатом сельскохозяйственных наук
А. Е. Семеновым. Как и следовало ожидать, в составе энтомофауны Кон-
дары было обнаружено большое число (40 видов) вредных в различном
отношении насекомых. Следовательно, природа Кондары являлась ре-
зервуаром потенциально вредной энтомофауны, существующей за счет
использования естественной флоры Кондары.
При появлении такого нового элемента флоры, как плодовые деревья,
развившиеся из привоев к диким подвоям, можно было ожидать перехода
различных вредных насекомых на новый, заманчивый для них, источник
питания. Так оно и оказалось в действительности. 29 видов вредных
насекомых Кондары перешли на культурные растения. В связи с этим
зоологами проводились работы по методам борьбы с важнейшими вреди-
телями из мира насекомых.
В сходном положении оказался вопрос о вредителях люцерны, кото-
рая в опытных условиях хорошо произрастала на такой высоте, где до
сей поры ее не было. Исследования А. И. Карповой показали, что на
люцерну, как на новый привлекательный источник питания, переходит
значительное число видов вредных насекомых из окружающей природы.
Зоологические исследования на Станции охватили и другие группы
животных — млекопитающих, птиц и проч.
16
Е. Н. Павловский
В годы Великой Отечественной войны, когда Зоологический институт
Академии Наук СССР был перебазирован в Сталинабад, ряд специалистов
Института работали в Кондаре, и их статьи нашли место в предлагаемой
книге. Усиление зоологических исследований отразилось прежде всего
на выявлении фауны Кондары. Она оказалась чрезвычайно обильной,
но выясненной далеко не равномерно.
Количественное разнообразие выявленной доселе фауны Кондары
и ее окрестностей выражается следующим числом наземных видов, кото-
рое является, без сомнения, далеко не исчерпывающим:
Млекопитающие 16 видов
Птицы .... 135 видов и подвидов
Пресмыкающиеся И видов
Амфибии 1 »
Рыбы 3 »
Моллюски наземные 26 »
Насекомые:
Двукрылые около 250 видов
Мошки 16 »
Ручейники. 10 »
Пчелиные 203 »
Осы блестянки 70 »
Настоящие полужесткокрылые 256 »
Уховертки 2 »
Таракановые 13
Палочники 1 »
Кузнечиковые 27 »
Саранчевые 53 »
Стрекозы 9 »
Пауки 36 »
Сенокосцы . . . 1 »
Иксодовые клещи 13 »
Пухоеды с птиц 20 »
Паразиты мышевидных грызунов 48 »
Волосатики и почвенные нематоды до 10 »
Кроме того обнаружено много видов в источниках Кондары.
В целом фауна Кондары весьма богата; число ее видов приближается
к 1800. Некоторые группы остались необработанными или обработан-
ными в разной степени, что неизбежно привело к пестроте изложения
данных по фауне Кондары.
Все же, в целом сделано много; если добавить, что число видов од-
них цветковых растений Кондары доходит до 756, то очевидным станет,
сколь привлекательной и ценной для разнообразных биологических
исследований является Кондара и ее окружение.
Это отразилось на разнообразии основных исследований, выполняв-
шихся на Варзобской горной ботанической станции, что явствует из
приводимого ниже перечня, составленного Ю. С. Григорьевым.
I. Изучение физико-географических условий
Станции
Исследование климатических условий и их дифференциации в зави-
симости от рельефа (проф. Сапожникова).
Динамика влажности почв (ст. научн. сотр. Запрягаев).
Характер и закономерности распределения почвенного покрова (проф.
Антипов-Каратаев).
Возникновение и развитие деятельности Варзобской станции
17
II. Ботанические и растениеводческие
исследования
Общее изучение флоры высших растений (канд. биол. наук
Письяукова, ст. научи, сотр. Запрягаев, Королева и др.).
Закономерности распределения раститель ного покрова (ст. научи,
сотр. Запрягаев, Королева).
Ритмика растительного покрова (ст. на учн. сотр. Запрягаев).
Общее изучение микрофлоры и, в ча стности, паразитных грибов
(проф. Купревич).
Биология дикорастущих древесных пород (канд. биол. наук Запря-
гаева).
Водный режим некоторых древесных пород (канд. биол. наук Матвеев
и проф. Купревич).
Опытная культура плодовых на богаре (ст. научн. сотр. Запрягаев).
Опыты по прививкам культурных плодовых на дикорастущих кустар-
никах (канд. биол. наук Запрягаева).
Опыты по богарному лесоразведению (ст. научн. сотр. Запрягаев)^
Богарное травосеяние и, в частности, культура люцерны на богаре
(ст. научн. сотр. Королева).
Изучение разнообразия форм грецкого ореха (ст. научн. сотри-
Печникова).
Опытные работы по улучшению пастбищ (ст. научн. сотр. Запрягаев).
Изучение дикорастущих лекарственных и витаминосодержащих
растений (ст. научн. сотр. Варивцева и Королева).
Изучение дикорастущих каучуконосов (ст. научн. сотр. Королева)..
Выявление и изучение растений-красителей (ст. научн. сотр. Варив-
цева и Пояркова).
III. Зоологические и паразитологические
исследования
Общее изучение фауны насекомых (канд. биол. наук Гуссаковский,
проф. Штакельберг и др.).
Исследование фауны млекопитающих (проф. Виноградов).
Исследование фауны пресмыкающихся (доктор биол. наук Чернов).
Фауна птиц и их биология (доктор биол. наук Иванов, проф. Туга-
ринов).
Гидробиологические исследования (проф. Жадин, Киселев, Родина).
Насекомые — вредители плодовых (канд. биол. наук Семенов).
Насекомые — вредители люцерны (канд. биол. наук Карпова).
Наземные моллюски как вредители сельского хозяйства (проф.
Жадин).
Биология малярийных комаров (канд. биол. наук Луппова, проф.
Лепнева).
Распространение и биология клещей-переносчиков возвратного тифа
Crnithodorus papillipes (канд. биол. наук: Благовещенский, Сердюкова,
Соснина).
Биоценозы гнезд птиц (канд. биол. наук Мулярская).
Экологические наблюдения над Crnithodorus papillipes в постройках;
меры борьбы с ним (канд. Сиол. наук Соснина, Теравский, Шустров).
Анатомия насекомых (акад. Павловский, Теравский и др.).
Из этого, неисчерпывающего, перечня очевидно, что Варзобская гор-
ная ботаническая станция фактически превратилась в базу для иссле-
2 Ущелье Кондара
18
Е. Н. Павловский
дований по самым разнообразным отраслям биологических наук, в ком-
плексную биологическую станцию.
Многосторонняя изученность природы Станции и в особенности ее
весьма богатых растительного и животного мира дают такую полную
картину флоры и фауны территорйи Варзобской станции, какую имеют,
пожалуй, лишь единичные участки нашей страны, которые исследова-
лись с давнего времени. Во всяком случае, в других районах Средней
Азии этого нет.
Естественно, что Варзобская станция открывает совершенно исклю-
чительные возможности для самых разнообразных ботанических, зооло-
гических, сельскохозяйственных и других работ, в частности, по вопро-
сам биоценологии. Именно в условиях Кондары и Варзобской станции
открылись особенно благоприятные предпосылки для исследований
в направлении мичуринской генетики и советского творческого дарви-
низма, что явилось новой струей в деятельности Станции и что, несо-
мненно, привлечет к себе и других специалистов сверх штатного персо-
нала Станции.
Но этому должна быть оказана особая помощь. По Кондаре уже на-
копилась научная литература, немало имеется неопубликованных данных,
и многие материалы еще ждут обработки; однако и то, что уже есть,
представляет собой большую ценность, которая должна стать широко
доступной. Первый опыт такого рода и предлагается в виде настоящей
книги, долженствующей стать как бы научным путеводителем по Кон-
даре. Идея составления книги такого рода зародилась у автора еще
в 1928 г., во время работы в Среднеазиатской паразитологической экспе-
диции на станции Репетек Ашхабадской железной дороги. Эта станция,
расположенная в Каракумах, широко известна по многим исследованиям
природы песчаной жаркой пустыни. Казалось бы, вполне целесообразно
составить сводку важнейших научных работ разных направлений, чтобы
дать своего рода «введение в изучение природы Каракумов», которое
одновременно явилось бы научным путеводителем-справочником по
Репетеку, где работала Репетекская станция по изучению песчаной
пустыни. Пользуясь такой книгой, натуралист, намеревающийся при-
ехать для научной работы в Репетек, мог бы заранее ориентироваться
в составе и особенностях местной флоры и фауны и выбрать объект для
исследования. Предложение о составлении книги о природе Репетека
не встретило отклика со стороны руководства станции, и дело осталось
неосуществленным.
Продолжительные работы автора в Таджикистане во многом были
связаны с Кондарой. Близкое знакомство с Варзобской горной ботани-
ческой станцией на протяжении 15 лет воскресило у автора идею состав-
ления монографической сводки — научного путеводителя, или «введения
в изучение Кондары». Приезд и двухлетнее пребывание в Сталинабаде
в годы Великой Отечественной войны большинства научных сотруд-
ников Зоологического института Академии Наук СССР обеспечили
основательное изучение фауны Кондары. Профессор В. И. Жадин, неза-
висимо от автора, пришел к мысли о желательности составления моно-
графии о природе Кондары.
Идея составления монографии по Кондаре встретила сочувствие со
стороны специалистов Зоологического института Академии Наук СССР
и научных сотрудников Таджикского филиала Академии Наук СССР.
Работы по составлению монографии были начаты еще в Сталинабаде, но
закончились после возвращения Зоологического института в Ленинград.
Возникновение и развитие деятельности Варзобской станции 19
Предлагаемая книга является первым опытом составления монографии
о природе типовой местности, в частности, в Таджикистане; несмотря
на, казалось бы, сугубую локальность использованных материалов,
законченная монография будет полезна для научно-исследовательских
и экспериментальных работ и в других местах Средней Азии, близ-
ких по характеру природы к Кондаре.
К сожалению, не удалось избежать недостатков, прежде всего, в
отношении пробелов по целым группам насекомых и других беспозвоноч-
ных; вторым недостатком является неоднородность составления статей
по отдельным группам животных; но и с этими недостатками, как можно
на то надеяться, предлагаемая монография окажется полезной. Допол-
нительные материалы, необходимые приезжающим работникам, могут
быть использованы из фондов Таджикского ({илиала Академии Наук
СССР.
Коллектив авторов этой книги в дальнейшем будет работать над ее
усовершенствованием и пополнением. Поэтому просьба ко всем исследо-
вателям, в какой-либо степени работавшим в Кондаре или над материа-
лами из Кондары, не отказать в присылке оттисков напечатанных работ,
информаций и других сообщений по адресу: Ленинград, 164, Зоологи-
ческий институт Академии Наук СССР. Директору Института.
2
растительный мир
В. В. ПИСЬЯУКОВА
ФЛОРА УЩЕЛЬЯ КОНДАРА
Ущелье небольшой горной речки Кондара находится в отрогах Гис-
сарского хребта в 30 км севернее Сталинабада. Кондара является право-
бережным притоком реки Варзоб и впадает в нее на высоте около
1100 м (здесь и в дальнейшем высота дана в метрах над уровнем моря).
Свое начало Кондара берет от целой системы родников, вытекающих на
высотах Д500—1700 м. Ущелье, образованное Кондарой, имеет в длину
несколько километров при средней ширине (до вершин прибрежных
хребтов) около 2 км и простирается с запада на восток. В результате
такого направления ущелья хребты, ограничивающие реку, обращены
к ней северным (правобережный) и южным (левобережный) склонами.
Склоны хребтов то пологи, мелкоземисты и местами распаханы, то очень
круты и скалисты. Хребты поднимаются над уровнем Кондары на высоту
от 200 до 1000 м, все время повышаясь к верховьям ее. В верховьях ущелье
Кондары резко ограждено мощной крутой стеной водораздела рек Лю-
чоб—Варзоб, носящего название Руидашта. Это урочище представляет
собой полого всхолмленное плато 10—12 км длиной и 6—7 км шириной,
покрытое толстым чехлом мелкозема. Восточный край плато, обращенный
к ущелью р. Кондара, очень крут и скалист. Высоты Руидашта колеб-
лются от 2400 до 2850 м.
Территория, флора которой включена в список ущелья Кондара,
захватывает все ущелье ее от устья (берег р. Варзоб) до верховий (уро-
чища Квак и Тереакба), восточный склон Руидашта и восточный сектор
самого плато. Эти пределы ограничивают площадь около 7—8 тыс. га
с вертикальным диапазоном от 1100 до 2850 м.
Основная часть этой территории лежит в поясе древесно-кустарни-
ковой растительности, которая на южных склонах Гиссарского хребта
начинается от 1000—1200 м и исчезает на высотах около 2500 м.1 В этом
поясе Гиссарского хребта реализуется самое большое количество осад-
ков — свыше 1000 мм в год. Только в нижней части ущелья до высоты
1300—1400 м на щебнистых открытых склонах можно встретить фраг-
менты горной полустепи (по терминологии М. Г. Попова, 1914 г.) с Agro-
pyrum trichophorum, Inula grandis, Scabiosa songorica, Alcea nudi-
flora, Rhaponticum integrifolium, Agropyrum repens, Scaligeria allio-
ides, Grupina vulgaris. Субальпийские флористические элементы также
появляются на Руидаште в небольшом количестве, так как это плато
лежит скорее в зоне переходной от древесно-кустарниковой к субаль-
пийской. И действительно, из 250 видов, населяющих Руидашт, едва
1 Имеется в виду высотное распределение древесно-кустарниковых формаций,
отдельные же виды этого пояса идут и выше.
В. В. Письяукоеа
50 видов могут быть отнесены к субальпийцам, а остальные являются
обитателями пояса древесно-кустарниковой растительности (хотя ча-
стично только верхних его пределов).
На площади в 7—8 тыс. га и в пределах от 1100 до 2850 м высоты
нами зарегистрировано и собрано 756 видов высших папоротникообраз-
ных и семенных растений.1 Такая флора даже для горной Средней Азии
может быть признана богатейшей. К сожалению, в литературе имеется
не много материала для сравнения, так как полные исчисления видового
состава флор по районам нашей страны встречаются до сих пор редко.
Однако некоторые параллели провести можно.
М. Г. Попов (1941) флору бассейна Малой Алмаатинки (Заилийский
Ала-тау) исчисляет приблизительно в 800 видов. Однако эта флора заре-
гистрирована им, во-первых, на площади, в два раза превышающей нашу
(16 000га), и, во-вторых, по высотному профилю, поднимающемуся от 1200
до 4000 м. В списке М. Г. Попова фигурируют элементы флоры полу-
пустыни, полустепи, степи, леса лиственного и леса хвойного, субаль-
пийского и альпийского поясов. Имеющиеся у меня материалы позволяют
утверждать, что на южных склонах Гиссарского хребта, по высотному
профилю от 1200 м до снежных полей (4500—4700 м), на площади в 16—
20 тыс. га обитает около 1200 видов.
Флору Кугитанга (крайний юго-зап. отрог Памиро-Алая) С. А. Нев-
ский (1937) исчисляет в 800 видов. На Кара-тау (зап. Тянь-шань),
по его мнению, не больше 1000 видов, и «немногим богаче, — говорит
он, — вероятно, и Копет-даг».
Какую же сокровищницу флоры представляют южные склоны Гис-
сарского хребта, если здесь, в маленьком горном ущелье, почти не выхо-
дящем за пределы древесно-кустарникового пояса, обитает свыше 756
видов!
Для того, чтобы понять характер этой флоры и разобраться в элемен-
тах, ее слагающих, необходимо произвести, прежде всего, некоторые
статистические сопоставления.
Семьсот пятьдесят шесть видов флоры Кондары относятся к 68 семей-
ствам и 348 родам. Из этого числа: папоротникообразных — 6 видов,,
голосеменных — 2, однодольных — 159 и двудольных — 588. По своему
составу это флора умеренных широт, в которой господствующими па
числу видов являются следующие семейства:
Число о/л
видов /0
Сложноцветные (Compositae) 102 13.64
Злаки (Gramineae) . . 83 11
Бобовые (Leguminosae) 61 8
Лилейные (Liliaceae) . . . 40 5.3
Гвоздичные (Caryophyllaceae) 40 5.3
Зонтичные (Umbelliferae) 38 5
Губоцветные (Labiatae) 38 5
Розоцветные (Rosaceae) . 37 4.9
Крестоцветные (Cruciferae) 35 4.6
Лютиковые (Ranunculaceae) 29 3.84
Норичниковые (Scrophulariaceae) 23 3.04
Бурачниковые (Boraginасеае) 21 2.8
Осоковые (Сурегасеае) 19 2.5
Гречишные (Polygonaceae) 19 2.5
Мареновые (Rubiaceae) 16 2.1
Список их см. в конце этой статьи.
Флора ущелья Кондара
2^
Заслуживают внимания следующие особенности флоры Кондары.
1) Большой удельный вес злаков в ее составе (11%). Это зависит
как от родового разнообразия их (35 родов), так и от богатства видами
некоторых из них (мятлик — 13 видов, костер — 8, рисовидка, поле-
вица, вейник, ячмень — по 5 видов). Второе место по богатству видами
злаки занимают также и во флоре Алмаатинского заповедника
(М. Г. Попов, 1940, 1941), где их 9%, и во флоре Кугитанга (С. А. Нев-
ский, 1937), где их 11.7%.
2) Бобовые занимают третье место, однако род Astragalus во флоре
ущелья Кондара самый большой, он содержит 21 вид. Богатыми также
оказались роды люцерна (7 видов) и вика (6 видов).
3) Необычайно богатыми во флоре Кондары оказались семейства
лилейных и гвоздичных. Вместо обычного для Средней Азии 6—7-го места,
лилейные по числу видов в Кондаре заняли 4-е место, оттеснив зонтич-
ные, губоцветные и крестоцветные.1
В Средиземноморской флористической области лилейные выдвигаются
обычно богатейшим родом Allium (лук). Это нашло свое отражение и во
флоре Кондары, где произрастает 16 видов лука. Кроме того, в ущелье
обитают 8 видов рода Gagea и 6 видов рода Eremurus.
Гвоздичные выдвинулись также благодаря наличию молодых энде-
мичных циклов в составе некоторых родов, например Silene и Meland-
ryum. Род Silene в Кондаре имеет 12 видов, из них 5 видов — обитатели
скал, широко представленные на Руидаште: S. samarkandensis, S. con-
formifolia, S. scabrifolia, S. tachtensis, S. guntensis. Следует заметить,
что первые три вида — эндемы Памиро-Алая, a S. tachtensis кроме Па-
миро-Алая обитает и в Тянь-шане. В кленово-ореховых рощах обитают
S. Komarovii, S. baldshuanica и S. bucharica, первые две — виды памиро-
алайско-тяныпанские, a S. bucharica известна пока с Гиссарского хребта.
Кондаринские виды рода Melandryum почти все относятся к ряду
Elata Scbischk., подрода Elisanthae. Все виды этого ряда — памиро-
алайско-тяньшанские эндемы (у нас — М. ovalifolium и М. ferganicum),
а М. erubescens пока известна только с Гиссарского хребта.
4) Необычайно много оказалось во флоре Кондары семенных расте-
ний-паразитов. Род Cuscuta в ущелье имеет 7 видов, a Orobanche — 6.
Такие из них, как С. Lehmanniana, О. gigantea, О. Kotschyi, О. pamiro-
alaica, играют даже большую ландшафтную роль в Кондаре, резко бро-
саясь в глаза огромной своей величиной, а в иные годы и обилием.
5) Следует отметить также богатство рода Euphorbia во флоре Кон-
дары.2 Молочай в Кондаре имеет 9 видов, что составляет 1.2% ее флоры.
В остальном состав и соотношение господствующих во флоре ущелья
семейств дает обычную для Средней Азии картину. В Кондаре также
первое место занимают семейства с циклами мощных ирано-переднеази-
атских или горных центральноазиатских форм в своем составе:
Сложноцветные (Cousinia) 14 видов Гвоздичные (Silene) 12 видов
(Erigeron) 5 » Зонтичные (Ferula) . 6
Бобовые (Astragalus) 21 » (Scaligeria) 4 »
Лилейные (Allium) 16 » Губоцветные (Eremostachys) 9 »
(Gagea). . 8 » (Nepeta) . . . 4 »
(Eremurus) 6 » (Scutellaria). 4—5
1 Так же обильны лилейные, гвоздичные, крестоцветные и во флоре Кара-тау,.
судя по данным Н. В. Павлова и С. Ю. Липшица (1934).
2 Как, по С. А. Невскому, и во флоре Кугитанга.
26
В. В. Письяукова
Также и бурачниковые, лютиковые, некоторые роды касатиковых,
злаков, крестоцветных, розоцветных и других семейств.
6) При сравнении удельного веса основных семейств во флорах Кон-
дары, Кугитанга и Алмаатинского заповедника интересно отметить также
следующее. Сем. лебедовые (Chenopodiaceae) в Кондаре никакого зна-
чения не имеет, оно представлено 3 видами — сорняками. На Куги-
танге же маревых 25 видов, что составляет 4% его флоры. Там маревые
по числу видов превышают розоцветные и крестоцветные. В Алмаатин-
ском заповеднике, наоборот, два последние семейства занимают 4-е и
5-е места,1 уступая только злакам, сложноцветным и бобовым, а маревые
стоят на 11-м месте. Интересно также соотношение осоковых: в заповед-
нике их 3%, на Кугитанге — 1.5, а в Кондаре — 2.5.
Следует особо подчеркнуть, что приведенный на стр. 24 список
господствующих семейств не исчерпывает флоры ущелья. Больше
того, представители этих мощных семейств далеко не отражают свое-
образия этой флоры и даже не они одни создают ландшафты в Кондаре.
Господство свое эти растения делят с растениями совсем иной группы,
большая часть которых относится к малым в нашей флоре семействам.
Однако значение этих малых семейств исключительно велико, не-
смотря на то, что статистический анализ маскирует их бесконечно малыми
дробями процентных отношений. Представители этих малых семейств
главным образом и дают нам понятие о характере растительного покрова
Кондары и его строе, о богатстве, своеобразии и родственных связях
его флоры, а следовательно, и об истории ее развития.
В составе этих семейств скрываются настоящие шедевры среднеази-
атской лесной флоры, такие, например, как: грецкий орех, образующий
рощи на северных и северо-западных склонах хребтов Кондары; 2 вида
клена, из которых клен туркестанский образует лесные формации; кар-
кас — обычный член лесов, фисташка, ясень, крушина, иргай, виноград
и др. Под покровом этого древесного полога располагаются столь же за-
мечательные представители травянистых растений, фитоценологические
и генетические связи которых уводят нас далеко за пределы нашей
страны и далеко в глубь веков.
Список растений Кондары поражает также обилием древесных и ку-
старниковых форм в его составе. Их оказывается 56 видов, в числе кото-
рых 2 лианы. По отношению к общему числу видов это составляет~7.4%.
Для современных широколиственных лесных формаций умеренной зоны
это количество деревьев и кустарников довольно велико.1 2 Это несоответ-
ствие лесов Кондары современному строю их в большей части Европы
и Азии свидетельствует о большой древности основного ядра древесно-
кустарниковой растительности ущелья. Заслуживает большого внима-
ния также и то, что из этих деревьев и кустарников 20 видов являются
плодово-ягодными, а склоны ущелья, даже при беглом взгляде на них,
производят впечатление коллекционного питомника.
Попытаемся выяснить, в этой статье — лишь в общих чертах, на чем
основано такое необычайное богатство флоры Кондары, т. е. посмотрим,
какие элементы ее слагают.
1 Особенно богаты в заповеднике роды Potentilla (21 вид), Geum — (5) и др.,
а из крестоцветных: Draba, Lepidium, Erysium, Sisymbrium и др.
2 Во флоре Кавказа (Гроссгейм, 1949) дикорастущих древесно-кустарниковых
форм — 6.5%, во флоре Талыша (Гроссгейм, 1926) их около 8%, во флоре Дальнего
Востока (Комаров и Клобукова, 1931—1932) — около 11.5%.
Флора ущелья Кондара
27
Вопрос о географо-генетической номенклатуре здесь обсуждать мы
не можем.
Для целей этой статьи мы воспользуемся наиболее общепринятыми
для Средней Азии градациями.
М. Г. Попов (1941) при анализе флоры бассейна р. Малой Алмаатинки
устанавливает три ее составных географо-генетических элемента: боре-
альный, восточно-древне-средиземноморский 1 и восточно- и центрально-
азиатский (горные элементы флоры Гинкго).
Во флоре Кугитанга, более близкой к нашей,* 2 С. А. Невский уста-
навливает 8 генетических элементов: средиземноморский в широком
смысле (включающий 6 особых генетических групп), реликтовый
арктотретичный, бореальный и сорняки — мигранты тропической
флоры.
I. К группе бореальных видов в Кондаре относятся (выборочно)
следующие: Digraphis arundinacea, Роа pratensis, Р. annua, Р. nemo-
ralis, Agrostis canina, A. alba, Calamagrostis epigejos, C. pseudophrag-
nrites, Milium vernale, Festuca pratensis, F. gigantea, Dactylis glomerate,
Glyceria plicata, Bromus inermis, Brachypodium silvaticum, Agropyrum
repens, Carex compacta, C. polyphylla, Trichophorum, Heleocharis, Jun-
cus lampocarpus, Rumex crispus, R. conglomeratus, R. syriacus, Polygo-
num persicaria, P. hydropiper, P. convolvulus, Stellaria media, Herniaria,
Ranunculus laetus, Rubus caesius, Geum urbanum, Rosa canina, R. corym-
bifera, Medicago lupulina, M. falcate, M. minima, Melilotus officinalis,
Trifolium repens, T. pratense, Lathyrus pratensis, L. tuberosus, Geranium
Robertianum, G. Regelii, Hypericum perforatum, Epilobium hirsutum,
Ghamaenerium angustifolium, Aegopodium podagraria, Lycopus europeus,
Scrophularia alata, Veronica beccabunga, V. anagallis, Asperula arvensis,
A. aparine, Galium verum, G. aparine, Plantago, Valeriana, Solidago
virga aurea, Achillea millefolium, Artemisia absinthium, A. vulgaris,
Tussilago farfara, Cirsium lanceolatum, Cichorium intybus, Crepis sibirica
и многие другие виды, так резко бросающиеся в глаза на фоне свое-
образной основной флоры ущелья. В нашем списке бореальных видов
всего около 135, что дает почти 18% флоры Кондары. Такое количество
бореальных элементов является довольно значительным. Правда,
Кондара уступает бореальной группе бассейна р. Малой Алмаатинки,
где, по данным М. Г. Попова, их имеется свыше 25% (и даже
ДО 40%).
Во флоре Кугитанга бореальные элементы дают около 13%.3 Все
виды этой группы в Кондаре приурочены к влажным тенистым местам.
Они обитают по берегам р. Кондара и ключей, в нее впадающих, в те-
нистых кленовниках и ореховых рощах на северных и северо-западных
склонах, на влажных лужайках Руидашта. Многие из этих видов обра-
зуют в Кондаре (как и вообще на южных склонах Гиссарского хребта)
группировки ландшафтного порядка, соучаствуя в них с аркто-третич-
ными и горными азиатскими мезофильными видами или чередуясь с ними.
К числу таких группировок относятся, например, развитые в Кондаре
L Ирано-переднеазиатский.
2 Из 588 видов флоры Кугитанга (Невский, 1937) общими с Кондарой являются
около 280 видов, а из 1264 видов флоры Алмаатинского заповедника (Попов, 1940)
общими с Кондарой оказались 264 вида.
3 Из 14.7%, для групп аркто-третичной и бореальной на первую приходится
28
В. В. Писъяукова
(и вообще на Гиссарском хребте) ежово-разнотравные и эгоподиевые
кленовники или кленово-ореховые рощи, кленовники с кипреем, яблоне-
вые рощи с почти чистым покровом из Роа nemoralis и Melissa officina-
lis, обычные на северных склонах холмов, и в ложбинках на Руидаште
ассоциации Geranium saxatile с Ligularia persica.
Большинство растений, относящихся к группе бореальных, в дре-
весно-кустарниковых формациях Кондары и на Руидаште распростра-
нены довольно широко и не обнаруживают заметных' признаков угне-
тения.
Однако имеются виды очень ограниченные в своем распространении.
Lavatera, caschemiriana Chamaenerium angustifolium, Digraphis arun-
dinacea, Festuca gigantea встречаются только в пределах от 1600
до 2400 м и только в очень тенистых кленовниках северных скло-
нов. М. Г. Попов (1938) такие растения относит к бореальным релик-
товым элементам во флоре Средней Азии.
К видам бореальным относят обычно и Geranium Robertianum, что
мне кажется не совсем верным. Правда, это растение связано в своем
современном распространении с лесной зоной Европы (вплоть до При-
балтики), но генетические связи его, вероятно, не бореальные. Воронов
(«Материалы для флоры Кавказа») пишет, что «Geranium Robertianum L.,
по всей вероятности, древняя третичная форма Средиземноморской
области, в последние геологические эпохи расширившая свой ареал за
пределы отечества». Заслуживает внимания в связи с этим то, что
G. Robertianum морфологически сходна с макронезийской G. anemonifo-
lium, обитающей в лавровых лесах Мадейры и Канарских островов
в полосе постоянных морских туманов.
Во всяком случае, в условиях лесного пояса Гиссарского хребта
G. Robertianum является растением безусловно реликтовым. Она отно-
сится к той группе реликтов, которую М. Г. Попов (1938) называет
реликтами древнесредиземноморской флоры. Будучи растением глубоко
теневым и очень чувствительным не только к влажности почвы, но и,
особенно, воздуха, G. Robertianum оказалась неспособной к извест-
ной ксерофилизации, необходимой для жизни на ее родине в послелед-
никовое время (как это случилось, например, с Onobrychis grandis).
Поэтому она распространилась в более влажные районы умеренных
широт Европы.
Встречается G. Robertianum в ущельях Гиссарского хребта очень
редко и всегда только у самой воды под защитой кроны ореховых
деревьев или в тени под карнизами скал.1
В бореальной группе флоры Кондары имеется большое количество
полезнейших кормовЬтх растений, таких, как ежа, мятлики, овсяница,
виды костра, чины, люцерны, клевера, горошков и др. В светлых кле-
новых рощах они все образуют мощные заросли. Плодовых и ягодных
форм, кроме ежевики, в этой группе нет.
II. Аркто-третичная реликтовая группа во флоре Кондары предста-
влена почти 30 видами. В числе их обитают в ущелье: Muehlenbergia
Hiigelii, Fritillaria Eduardii (рис. 1), Epipactis Royleana (рис. 2), Codo-
nopsis clematidea, Juglans regia, Corydalis nudicaulis, C. sp. (близкая
1 Весьма вероятно, что к этой группе относятся и такие тенелюбы-средиземно-
морцы, как Buchingera axillaris из крестоцветных и Physocaulis nodosus из зонтичных,
ютящиеся в массе под защитой кроны Juglans regia. Здесь они образуют такие же
чистые зарослп, как и Impatiens parviflora.
Гис. 1. Рябчик Эдуарда — Fritillaria Eduardii Rgl.
30
В. В, Письяукова
Рис. 2. Дремлик Ройля — Epipact is Royleana Lindl.
к предыдущей), Plata-
nus, Cotoneaster insignis,
Euonymus Semenovii,
Acer turkestanicum,Vitis
vinifera, Ampelopsis vi-
tifolia, Fraxinus potamo-
phila, Ostrowskia mag-
nifica (рис. 3), Malus
hissarica. Сюда же от-
носятся и наши виды
Roegneria, вероятно, и
Trachelanthus hissari-
cus, Lophanthus Lipsky-
anus, Lavatera cashemi-
riana. К типичнейшим
гималайским лесным ре-
ликтам, как выяснила
А. И. Пояркова (1950),
принадлежат и Ligularia
Thomsonii, L. persica,
L. altissima. К этой же
группе относится и
обнаруженный мною
(1950) в Кондаре Schoe-
noxiphium hissaricum
(рис. 4) (из сем. осоко-
вых) — прародитель ко-
брезий.
Аркто-третичное ре-
ликтовое ядро во флоре
Кондары, следователь-
но, играет довольно
большую роль. Его
удельный вес равен
3.5% от всего состава
флоры. Здесь уместно
отметить также, что во
флоре Кугитанга эта
группа состоит всего из
10—12 видов, т. е. ра-
вна 1.7% ее.
В составе аркто-тре-
тичной лесной реликто-
вой группы Средней
Азии А. И. Пояркова
(1950) рекомендует раз-
личать два разновозра-
стных элемента — более
молодой, гималайско-
гиссарский, и китай-
ский — более древний.
В предложенном выше
списке совершенно оче-
Флора ущелья Кондара
31
видно преобладание ги-
малайского элемента,
представленного к тому
же почти исключитель-
но травянистыми фор-
мами.
Это флористически
наиболее интересная
группа, и нет сомнений
в том, что дальнейший
систематико генетиче-
ский анализ флоры гор-
ной Средней Азии зна-
чительно увеличит ее
удельный вес.1
Все виды этой груп-
пы в своем распростра-
нении связаны с наи-
более влажными место-
обитаниями.
III. Средиземномор-
ские флористические
элементы в Кондаре
по числу видов зани-
мают господствующее
положение. В этом от-
ражается общая черта
среднеазиатской флоры,
в которой основное ядро
создают древнесреди-
земноморские ксеро-
фильные виды. Во всей
совокупности своей эта
группа представлена
почти 570 видами. При-
вожу выборочно часть
из них: Oryzopsis — 5
видов, Роа bulbosa, Lo-
lium cunеаturn и L. рег-
sicum, Taeniatherum —
2 вида, Hordeum — 5
видов, однолетние виды
Bromus, Vulpia, Aegi-
lops, Lophochloa phle-
oides, Carex, например,
1 Нельзя не сказать,
что в соседнем с Кондарой
ущелье—Т акобском — обна-
ружен и миндаль вязолист-
ный (Amygdalus ulmifolia) и
индо-гималаец Gymnocarpi-
um Fedtschenkoanum — па-
поротник, описанный А.И.
Поярковой (1950).
magnifica Rgl.
32
В. В. Письлукова
С. pachystilis, С. duriusculiformis, С. decaulescenes, С. diluta и др., Al-
lium, почти все Gagea, Ixiolirion, Ungernia, Gladiolus, Polygonum (кроме
перечисленных выше бореальных видов), Rheum, Eremurus, Celtis cau-
oasica, Atraphaxis pyrifolia, большая часть видов Silene, Dianthus,
Рис. 4. Схеноксифиум гиссарский — Schoenoxiphium hissaricum Pissjauk,
Delphinium, большая часть Ranunculus (напр. R. baldshuanicus, R. leptor-
rhynchus и др.), Clausia, Megacarpea gigantea, Roemeria, Papaver pavo-
ninum, виды Cotoneaster (кроме C. insignis), Crambe Kotschyana, Buchin-
gera axillaris, Astragalus, Onobrychis, Cicer, Haplophyllum, Euphorbia,
Tamarix, Eryngium, Prangos, Physocaulis nodosus, Scaligeria, Bunium,
Eremodaucus, Galagania fragrantissima, Ferula, Solenanthus, Linde-
Флора уъцелъя Кондара
33
lophia anchusoides, Trichodesma incanum, Echinospermum, Scutellaria,
Nepeta, Eremostachys, Salvia sclarea и S. virgata, Ziziphora, Origanum
tytthanthum, Phlomis fruticetorum, виды Asperula, Galium, Scabiosa
songorica, Phyteuma, Inula grandis, I. glauca, Cousinia, Scorzonera, Lac-
tuca, Barkhausia и многие другие виды. Общее число их составляет около
76% флоры Кондары, что несколько меньше, чем на Кугитанге (где их
около 80%). В большинстве своем виды, относящиеся к этой группе,
являются ксерофильными или ксерофилизированными в той или иной
степени. Они размещаются в Кондаре на открытых солнечных склонах
(преимущественно южных и юго-восточных), на каменистых осыпях
и карнизах скал во всех поясах.
При несколько более детальном анализе группы средиземноморских
элементов флоры Кондары можно отметить следующие ее особенности:
а) В Кондаре уже сравнительно бедно представлены средиземномор-
ские виды с родственными связями в пустынной гондванской флоре
(т. е. элементы флоры Вельвичии М. Г. Попова и — как части ее — «Флоры
Эфедры» С. А. Невского). Мне удалось выделить из списка всего около
40 видов (5.6%), в то время как на Кугитанге эта группа (I—III—IV,
по номенклатуре Невского) составляет шестую часть (16.7%) его флоры.
В Кондару заходят, например, следующие из таких видов:1 Psoralea dru-
расеа (только у ворот ущелья), Trichodesma incanum, Onosma, Tamarix
(только близ устья Кондары), Pistacia vera, Diarthron vesiculosum,
Avena Ludoviciana, виды Haplophyllum, Eremurus, Andropogon ischae-
mum (в нижней части ущелья), виды Thesium и несколько других. Сюда
не доходят (хотя некоторые еще встречаются в 5—6 км ниже по р. Вар-
зоб) такие южные пустынные формы, как Alhagi, Reamuria, Peganum,
Bibersteinia, Zygophyllum, Lycium, Capparis, Prosopis и многочисленные
Chenopodiaceae и Plumbaginaceae, составляющие основное ядро этой
группы на Кугитанге.
б) Что касается палеотропико-средиземноморского элемента Нев-
ского, то он в Кондаре оказался представленным 1G видами: Echinops
maracandicus, виды Eryngium, Cheilanthes persica, Celtis caucasica, Sage-
retia laetevirens, Asparagus, Arum и Eminium, а также оказалась у нас
и Agrostis verticillata. Любопытно отметить, что это растение встречается
только в 2—3 местах среднего течения Кондары, близ воды, вместе с Mueh-
lenbergia Hiigelii. По мнению Невского,1 2 3 это растение ничего общего с Ag-
rostis не имеет, а морфологически близко к тропическому роду Polypo-
gon, именно, к его многолетнему виду Р. demissus Steud. «и, вероятно,
произошло от него путем утраты остей».
в) Основная же масса средиземноморских видов Кондары — свыше
500 (как и вообще в области господства этой флоры) — связана в своем про-
исхождении с элементами голарктическими. Среди них Невский выделяет
два различных по возрасту генетических элемента — голарктико-среди-
земноморский и средиземноморско-голарктический. Первые — это само-
бытные средиземноморцы, со связями в голарктической флоре, вторые же
представлены видами — мигрантами из голарктики, но подвергшимися
ксерофилизации в новых для них условиях. Практическое разграниче-
ние этих двух категорий без тщательного флористического анализа видов
довольно затруднительно.
1 Большая часть из них встречается в пределах .до 1300—1400 м над ур. м.
и только некоторые виды Ferula, Oryzopsis, Crambe Kotschyana, Zozimia
.tragioides, Acantholimon alatavicum идут выше.
2 Acta Inst. Bot. Acad. Sci. URSS, ser. 1, 1936, вып. 3, стр. 143.
3 Ущелье Кондара
34
В. В. Нисъяукова
Однако даже и при очень относительной точности расчленения сохра-
няется довольно интересная особенность генетического строя средизем-
номорской группы во флоре Кондары.
Оказывается, что группа ксерофилизированных мигрантов голаркти-
ческой флоры в Кондаре больше чем на 100 видов превышает группу соб-
ственно средиземноморскую (первая состоит из 300 видов, а во второй
несколько больше 200). На Кугитанге, по данным Невского, эти группы
(V—VI) равноценны. Такое соотношение в обоих случаях легко понять,
если мы вспомним, что группа голарктико-средиземноморская, будучи
коренной средиземноморской, состоит из более ксероморфных видов,
чем группа вторая, состоящая из сравнительно поздних выходцев из
области мезофильной голарктической флоры.
К собственно средиземноморскому ксерофильному элементу в Кон-
даре относятся следующие роды (и виды): Taeniatherum, Hordeum, Aegi-
lops, Lolium, однолетние Bromus, Lophochloa, Phleum paniculatum,
Agropyrum trichophorum и A. intermedium, Eremopoa oxyglumis, Tulipa,
б. ч. Allium (кроме видов секции Rhiziridium и A. Oschanini), Ixiolirion,
Gladiolus, Ungernia, Juno, Atraphaxis, Vaccaria, Velezia, Roemeria,
Papaver pavoninum, Sysimbrium, Malcolmia, Pseudosedum, Rosularia,
Astragalus секции Xiphidium — A. baldshuanicus, A. maverranagri,
A. hissaricus (секции Proselius), Onobrychis, Gicer, Erodium, Linum,
Scandix, Torilis, Turgenia, Gaucalis, Eremodaucus, Prangos, Pimpinella,
Astomatopsis, Galagania, Korshynskia, Anchusa italica, Echium italicum,
Scutellaria, Nepeta, Eremostachys, Salvia, Origanum, Ziziphora, Linaria,
Leptorhabdos, Grucianella, Gallipeltis, Dipsacus, Scabiosa, Filago, Gou-
sinia, Onopordon, Koelpinia и некоторые другие. Всего — больше двухсот
видов. Остальные около 300 видов относятся к группе генетически более
молодой, названной G. А. Невским средиземноморско-голарктической.
В Кондаре эта группа составляет около 42% ее флоры (на Кугитанге —
30%).
В основном, как сказано выше, это менее ксерофильная группа и
большая часть ее видов в своем обитании связана с лесными формациями
различных типов, с розариями и травянистыми (особенно лигул яриевыми)
формациями Руидашта. Общий характер этой группы хорошо виден на
примере нескольких ее главных родов. Из Astragalus к этой группе в Кон-
даре относится 17 видов. Большая часть из них — члены таких мезо-
фильных секций, как: Myobroma (A. macronyx, A. glabrescens, A. var-
zobicus, A. andaulgensis), Euodmus (A. peduncularis), виды которой
распространены на Дальнем Востоке, в Сибири и Центральной Азии, сек-
ции Pendulina (A. macropodium, A. pauper, A. mirabilis, A. acormosus)
и др. Часть этих видов астрагала является эфемероидами, а боль-
шинство — мезофиты. Таковы же виды Hedysarum, Rosa Webbiana,
R. kokanica, Rhamnus dolichophylla, виды Gagea, Allium Barsczewskii,
A. tenuicaule, A. dolichomischum, Lepyrodiclis holosteoides, большая
часть наших видов Silene (все скальные формы Руидашта), Lonicera,
Adonis, почти все (кроме бореальных) виды Ranunculus, Delphinium
ternatum, D. biternatum, Solenanthus, виды Melandryum, Cotoneaster (осо-
бенно C. suavis, C. hissarica), виды Cicerbita, а также и более ксерофили-
зированные многочисленные виды, как Inula grandis, Роа (bactriana,
glabriflora, relaxa, bucharica и др.), многие виды Carex (diluta, Koshew-
nikovii, turkestanica, decaulescens), Clausia, Trigonella и др. Большая
часть этой группы относится к категории горных элементов флоры Гинкго^
в понимании М. Г. Попова (1941).
Флора ущелья Кондара
35
IV. Группа сорняков — мигрантов тропической флоры в Кондаре пред-
ставлена восемнадцатью видами. Это виды Sorghum, Eragrostis, Setaria,
Echinochloa, Verbena, Amaranthus, Portulaca, Chenopodium, Xanthium,
Cynodon.
Сравнивая флоры Малой Алмаатинки, Кондары и Кугитанга, можно
отметить следующее. Несмотря на заметное убывание бореальных эле-
ментов с северо-востока (от Заилийского Алатау) на юго-запад — к Гис-
сарскому хребту и Кугитангу, роль их в этих южных хребтах все же
остается довольно большой. Соотношение основных генетических элемен-
тов в трех сравниваемых участках горной Средней Азии в общем оказы-
вается следующим (табл. 1).
Таблица 1
Категории географо-генетических элементов флоры
Районы
бореальный
реликтовый
аркто-
третичный
I—VI группы средиземномор-
ской флоры Невского, обни-
мающие древнесредиземномор-
ский восточно- и центрально-
азиатский горный элемент
Попова (1941)
1. Бассейн р. Малой Алма-
атинки (Заилийский
Ала-тау)...............
2. Кондара (южные склоны
Гиссарского хребта)
3. Кугитанг
около 25%
» 18
» 13
около 1%
» 3.5
» 1.7
око;
76
80
И если, по замечанию М. Г. Попова, 25% бореальных элементов для
одного из участков Заилийского Алатау — это обильно, то 18% для
южных склонов Гиссарского хребта и 13% для Кугитанга — это, ко-
нечно, очень много.
Табл. 1 дает вообще довольно интересный материал для сопоставле-
ний. Даже если сделать все поправки на различное толкование каждым
автором границ и объемов географо-генетических элементов, то пред-
ставленные выше соотношения в основном, конечно, сохранятся.1 По
соотношению основных элементов флоры — средиземноморского и боре-
ального — Гиссарский хребет занимает промежуточное положение между
Заилийским Ала-тау и Кугитангом. Судя по удельному весу аркто-третич-
ных реликтовых форм во флоре Кондары, можно сказать, что южные
склоны Гиссарского хребта являются одной из основных областей их
сохранения. В составе флоры ущелья, как видим, сравнительно большое
место занимают все основные для Средней Азии географо-генетические
элементы. На эту контрастность и пестроту состава флоры горной Сред-
ней Азии уже давно обращали внимание все исследователи ее (особенно
М. Г. Попов во многих своих работах, Н. Ф. Гончаров и П. Н. Овчин-
ников и др.). Этим и объясняется необычайное богатство и разнообразие
флоры и растительного покрова Гиссарского и однотипных с ним хреб-
тов Западного Памиро-Алая.
1 О соотношении между собою различных групп средиземноморских элементов
сказано выше.
3
36
В. В. Писъяукова
Необычайно интересными оказываются сравнительные отношения
этих географо-генетических групп, взятых только в пределах наиболее
мезофильной лесной формации Кондары — в орехово-кленовых лесах
урочища Квак. Здесь зарегистрировано около 140 видов растений.1 По
группам элементов флоры они распределяются следующим образом
(табл. 2).
Таблица 2
Географо-генетические группы Количество видов % от общего числа видов формации
1. Бореальная . . 24 17.2
2. Реликтовая аркто-третичная .... 16 11.4
3. Ксерофильная средиземноморская (I—V группы Невского) . . . 21 15
4. Ксерофилизированные мигранты флоры Гинкго 79 56.4
Меньший, чем средний для Кондары, процент бореальных видов на
Кваке объясняется отсутствием в его списке прибрежных форм, соста-
вляющих большую часть бореального элемента в Средней Азии. Бореаль-
ные виды, обитающие в кленовых рощах, — лугово-лесные растения
Средней Европы.
Анализ лесных формаций Квака показывает, что десятая часть их
видового состава представляет собою третичные лесные реликты. Нако-
нец, совершенно очевидно, что основная масса видов этих формаций со-
стоит из сравнительно молодых (послеледниковых), лишь в той или
иной степени ксерофилизировавшихся элементов голарктической флоры.
Следует заметить, что такой же характер соотношений сохраняется
и в поясе выше лесного — на Руидаште. В составе флоры этого плато
первое и второе места принадлежат ксерофилизировавшимся мигрантам
голарктической флоры и элементам бореальным и лишь третье место за-
нимают представители ксерофильных средиземноморских групп.
В заключение остановимся на роли эндемичных видов во флоре Кон-
дары. Находясь в пределах горной Средней Азии, эта флора, конечно,
несет на себе печать высокого эндемизма, свойственного растительному
миру этой страны. Как сказано уже, основная часть видов флоры Кон-
дары принадлежит к большим полиморфным родовым циклам, находя-
щимся и сейчас в стадии интенсивного формообразования. В этом смысле
(что хорошо видно в списке флоры и в статистических сопоставлениях,
изложенных выше) свыше 30% видов Кондары оказываются эндемич-
ными, т. е. памиро-алайско-тяныпанскими. Тем не менее, в составе
флоры Кондары имеются виды с ареалом, в данное время очень ограни-
ченным. К ним, прежде всего, относятся новые, обнаруженные в Кон-
даре виды и известные пока только здесь или в 1—2 ближайших
ущельях: Ligularia altissima Pojark., известная только с Квака, Ra-
nunculus oligophyllus Pissjauk. — с Руидашта, Orobanche Solenanthi
1 Перечня их не даю, так как в списке флоры Кондары они все указаны для лесов
Квака или для «тенистых орехово-кленовых лесов».
Флора ущелья Кондара
37
Novopokr. et Pissjauk. — с Квака, Schoenoxiphium hissaricum Pissjauk. —
из Кондары, Такоба, Бигара, а также еще несколько намеченных к опи-
санию видов.
Кроме того, в Кондаре довольно велика группа видов, область рас-
пространения которых ограничивается только Гиссарским хребтом.
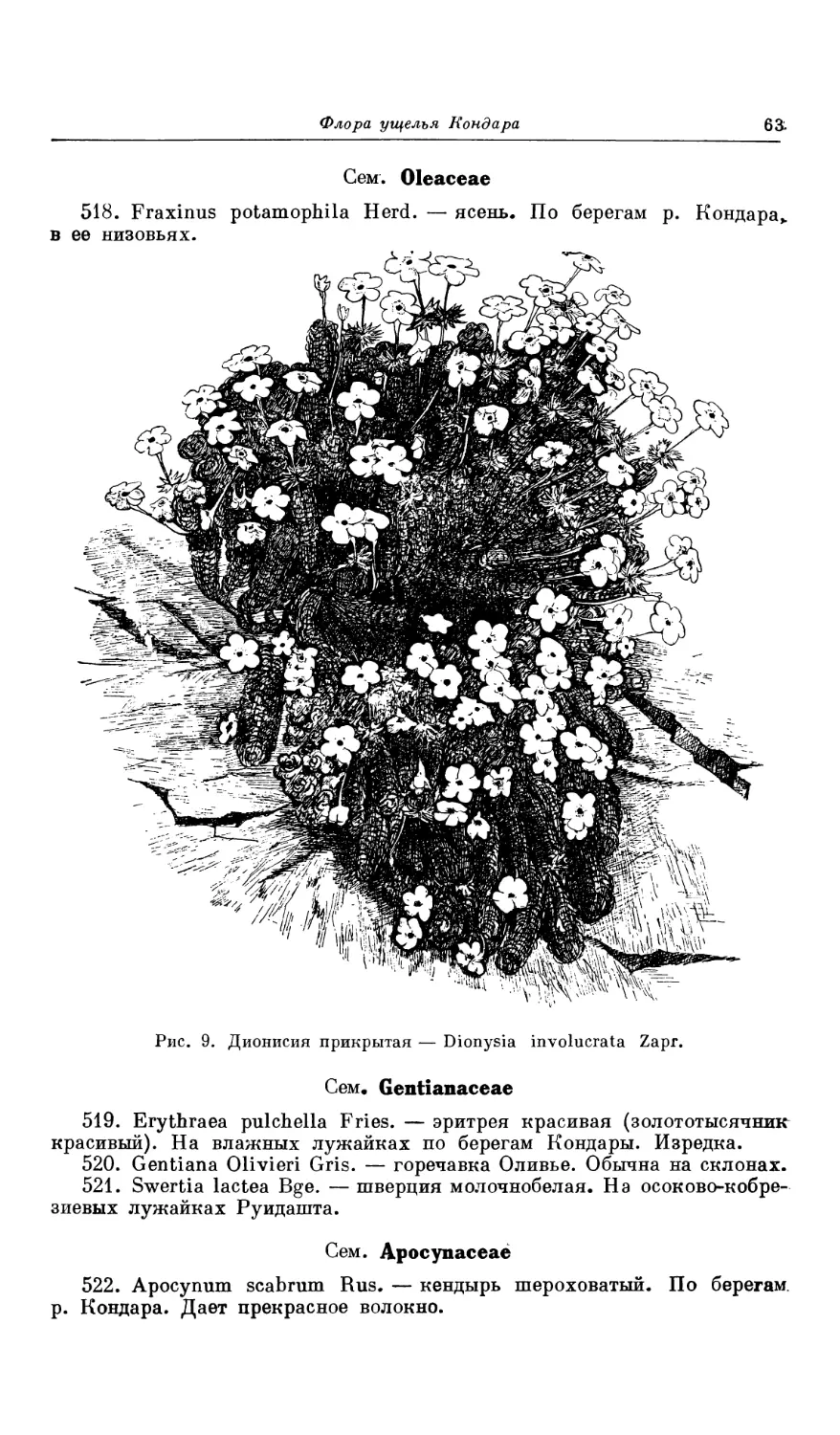
Например, Dionysia involucrata, Astragalus vegetior, A. Irinae, A. var-
zobicus, A. discessiflorus, Allium flavellum известны только из Вар-
зобского ущелья или бассейна Варзоба в целом. Eremostachys Popovii
известен только в бассейнах рек Варзоб и Сарда-и-Миона, Е. specta-
bilis—Варзоб и Вахш; Cousinia simulatrix—Варзоб и Каратаг. Ареал
некоторых видов, обитающих в Кондаре, локализирован только в запад-
ной части Гиссарского хребта, например у Trigonella Lipskyi, Oxytropis
leptophysa, Chesneya hissarica, Allium dolichomischum, Astomatopsis
galiocarpa. Пределами южных склонов Гиссарского хребта ограничи-
вается ареал Cousinia refracta и С. macilenta. Несколько больше группа
Гиссарских эндемиков в целом — Rosa hissarica, Astragalus mirabilis,
Allium hissaricum, Cousinia pulchella, C. acicularis, C. splendida, Astra-
galus acormosus, Trachelanthus hissaricus (последние 4 вида, кроме Гис-
сарского, встречаются еще только на Зеравшанском хребте). Необходимо
заметить, что среди перечисленных здесь видов с узким ареалом распро-
странения,1 в Кондаре имеются эндемики реликтовые. Это такие виды,
как Ligularia altissima и Schoenoxiphium hissaricum. Если к ним при-
бавить еще и описанный А. И. Поярковой (1950) Gymnocarpium Fed-
tschenkoanum — реликт той же третичной группы, живущий в соседнем
Такобском ущелье, то нужно сказать, что часть Варзобского ущелья от
Кондары и вверх до урочища Кобуты (включая ущелья Ходжа-оби-гарм
и Такобское) является редкой по своей флористической ценности.
Вследствие этого Кондара является базой, ценной для всякого рода
исследовательских ботанических работ и, особенно, работ флористи-
ческих.
СПИСОК РАСТЕНИИ УЩЕЛЬЯ КОНДАРА*
Отдел I. ARCHAEGONIATAE
FILICALES
Сем. Polypodiaceae
1. Cystopteris fragilis (L.) Bernh. — пузырник ломкий. В лесах,
среди кустарников, в тени скал. Обычен.
2. Asplenium ruta-muraria L. — костенец рута постенная. Близ
устья р. Кондары.
3. Ceterach officinarum Willd. — скребница аптечная. На скалах.
Обычен.
4. Gheilanthes persica Mett. — краекучник персидский. На скалах,
по южным и юго-восточным склонам.
5. Adianthum capillus veneris L. — адиантум венерин волос. У клю-
чей, водопадов, на влажных скалах. Обычен. 1 2
1 Большей частью эндемиков молодых.
2 Список составлен в результате личных сборов автора в летние сезоны 1945—
1947 гг. В небольшой степени список дополнен сборами Ю. С. Григорьева
Ф. Л. Запрягаева, А. С. Королевой и др., хранящимися в Гербарии Ботанического
института Таджикского филиала Академии Наук СССР в г. Сталинабаде.
38
В. В. Писъяукова
Сем. Equisetaceae
6. Equisetum ramosissimum Desf. — хвощ ветвистый. Обычен у рек
и ключей.
Отдел II. EMBRYOPHYTA SIPHONOGAMA
GYMNOSPERMAE
Сем. Cupressaceae
7. Juniperus seravschanica Кот. — можжевельник зеравшанский. На
склонах, от 1300—1400 м и выше.
Сем. Ephedraceae
8. Ephedra equisetina Bge. — хвойник хвощевый. На щебнистых и
скалистых склонах, от 1400 м и выше.
ANGIOSPERMAE
Сем. Gramineae
9. Imperata cylindrica (L.) P. В. — императа цилиндрическая. Близ
устья р. Кондара, на берегу р. Варзоб.
10. Andropogon ischaemum L. — бородач кровеостанавливающий. На
южных и юго-восточных склонах, в пределах усадьбы станции. Кор-
мовое.
И. Sorghum halepense (L.) Pers. — сорго алеппское. По берегам близ
устья р. Кондара. Злостный сорняк.
12. Echinochloa crus-galli (L.) Roem. — ежовник петушье просо.
Сорное.
13. Setaria glauca (L.) Р. В. — щетинник сизый. На южных склонах,
до 1000 м. Сорное.
14. S. viridis (L.) Р. В.—щ. зеленый. Вместе с предыдущим.
15. Digraphis arundinacea (L.) Trin. — двукисточник тростнико-
видный. По берегам р. Оби-сафед. Редко. Кормовое в молодом состоянии.
16. Lasiagrostis longearistata (Boiss.) Roshev. et Nevski —• чий
длинноостный. На каменистых северных склонах в древесно-кустарни-
ковом поясе, от 1300 м и выше.
17. Stipa caucasica Schmalh. — ковыль кавказский. Только перед
урочищем Квак, на каменистых осыпях.
18. Oryzopsis vicaria G. Grig. — рисовидка замещающая. На каме-
нистых склонах гор в древесно-кустарниковом поясе от 1300—1400 м
и выше.
19. О. latifolia Roshev. — р. широколистная. В кленовниках на се-
верных склонах, 1200—1600 м.
20. О. ferganensis Litw. — р. ферганская. В верхнем лесном поясе
и на Руидаште, 2800 м.
21. О. sogdiana G. Grig. — р. согдийская. На скалистых склонах
перед входом на плато Руидашт и на самом плато.
22. О. lateralis Stapf. — р. боковая. На скалах в долине Кондары,
от 1300 м, и на склонах холмов Руидашта.1
1 Все рисовидки — кормовые растения.
Флора ущелья Кондара
39
23. Milium vernale М. В. — бор весенний. В тени деревьев и кустар-
ников на склонах.
24. Muehlenbergia Hiigelii Trin. — мюленбергия Гюгеля. Только
в среднем течении р. Кондара, у ключа в ореховой роще.
25. Phleum paniculatum Huds. — тимофеевка метельчатая. На юж-
ных склонах.
26. Alopecurus seravschanicus Ovcz. — лисохвост зеравшанский. На
субальпийских лужайках Руидашта. Хорошее кормовое растение.
27. Agrostis canina L. — полевица собачья. На влажных осоковых
лужайках Руидашта.
28. A. hissarica Roshev- — п. гиссарская. Там же и в тени скал
в верховьях р. Кондара.
29. A. alba L. — п. белая. По берегам р. Кондара и ее притоков.
Кормовое растение. Обычное.
30. A. tianschanica N. Pavl.—п. тяныпанская. По берегам рек Кон-
дара и Оби-сафед, в их верховьях. Спорадично.
31. A. verticillata Vill. — п. мутовчатая. По берегам ключей, впа-
дающих в р. Кондара в ее среднем течении. Спорадично.
32. Calamagrostis epigeios (L.) Roth. — вейник наземный. Толь-
ко на Руидаште на каменистых склонах, в юго-восточной части
плато.
33. С. gigantea Roshev. — в. гигантский. На залежах по южному
склону левобережного хребта, от 1300 до 2000 м.
34. С. dubia Bge. — в. сомнительный. По берегам р. Кондара в ее
среднем и нижнем течении.
35. С. glauca (М. В.) Trin. — в. сизый. По берегу р. Кондара, в ее
верховьях и на опушках кленовников северного склона правобережного
хребта.
36. С. pseudophragmites (Hall, f.) Koeler — в. ложнотростниковый.
По берегам р. Кондара, в ее верховьях.
37. Avena Ludoviciana Dur. — овес Людовика. На щебнистых южных
склонах.
38. Cynodon dactylon (L.) Pers. — свинорой пальчатый. Там же,
в группировках эфемеров. Кормовое.
39. Phragmites communis Trin. — тростник обыкновенный. По бере-
гам р. Кондара и ее притоков.
40. Eragrostis minor Host. — полевичка малая. На мелкощебнистых
участках южного склона.
41. Е. Starosselskyi Grossh. — п. Старосельского. Урочище
Квак, в устье ключа и на песчаных отмелях близ устья р. Кон-
дара.
42. Lophochloa phleoides (Will.) Rchnb. — лофохлоа тимофеевковид-
ная. На щебнистых южных склонах левобережного хребта, близ урочища
Теги-камар.
43. Melica breviflora Boiss. — перловник короткоцветковый. На
скалистых обрывах, от 1300 м до Руидашта.
44. М. inaequiglumis Boiss. — п. неравночешуйный. На сухих щеб-
нистых склонах.
45. Dactylis glomerata L. — ежа сборная. Образует группировки,
а местами фон на северных склонах в древесно-кустарниковой полосе.
Очень хорошее кормовое растение.
46. Роа bulbosa L. — мятлик луковичный. Обычно всюду.
47. Р. bulbosa L. var. vivipara Koeler. — Там же.
40
В. В. Писъяукова
48. Р. bactriana Roshev. — м. бактрийский. Встречается в мезофит-
ных высокотравных кленовниках северных склонов урочища Квак и по
берегам р. Кондара, в ее верхнем течении.
49. Р. glabriflora Roshev. — м. гладкоцветковый. Обычно на склонах
холмов Руидашта.
50. Р. annua L. — м. однолетний. На склонах холмов Руидашта.
51. Р. bucharica Roshev. —м. бухарский. На склонах холмов Руидашта.
Обычен.
52. Р. trivialis L. — м. обыкновенный. По берегам р. Кондара, в ее
среднем течении. Кормовое.
53. Р. silvicola L. — м. лесной. Там же.
54. Р. pratensis L. — м. луговой. Часто на склонах холмов Руидашта
и в верхней части урочища Квак в древесно-кустарниковых зарослях.
Одно из лучших кормовых растений.
55. Р. angustifolia L. — м. узколистный. Там же.
56. Р. nemoralis L. — м. боровой. По берегам р. Кондара, в поясе
древесно-кустарниковой растительности. Обычен вплоть до Руидашта.
57. Р. nemoralis L. var vivipara Litw. — Только на Руидаште.
58. Р. relaxa Ovcz. — м. расползающийся. Обычен на склонах хол-
мов Руидашта и на скалистых его осыпях.
59. Eremopoa oxyglumis (Boiss.) Roshev. — эремопоа острочешуйная.
На каменистых склонах, главным образом, южной экспозиции, от низо-
вий р. Кондара до Руидашта.
60. Glyceria plicata Fries. — манник складчатый. По берегам р. Кон-
дара и ее притоков. Кормовое, особенно в молодом состоянии.
61. Festuca pratensis Huds. — овсяница луговая. По берегам р. Кон-
дара и ближайшим к ней склонам. Редко. Ценное кормовое растение.
62. F. gigantea (L.) Vill. — о. гигантская. В тенистых кленовниках
северного склона и по берегам р. Кондара.
63. Vulpia myuros (L.) Gmel. — вульпия мышехвостник. На южных
щебнистых склонах.
64. V. ciliata (Benth.) Link. — в. ресничатая. Там же.
65. Lolium persicum Boiss. et Hoh. — плевел персидский. На каме-
нистых южных склонах в пределах усадьбы Станции.
66. L. perenne L. — п. многолетний. На правом берегу р. Кондара,
вдоль тропы, перед первым водопадом. Кормовое.
67. L. cuneatum Nevski — п. клиновидный. Вдоль арыка на огороде.
68. Bromus inermis Leyss. — костер безостый. В зарослях кустар-
ников и на лесных опушках северных склонов правобережного хребта.
Ценнейший кормовой злак.
69. В. sterilis L. — к. бесплодный. В тени деревьев и кустарни-
ков и по берегам р. Кондара, в пределах усадьбы станции. Хорошее
кормовое растение.
70. В. tectorum L. — к. кровельный. Обычен в группировках эфемеров.
71. В. scoparius L. — к. метельчатый. На южных каменистых скло-
нах хребтов.
72. В. angrenicus Drob. — к. ангренский. На склонах холмов Руи-
дашта.
73. В. oxyodon Schrenk — к. острозубый. На южных каменистых
склонах. Обычен.
74. В. macrostachys Desf. — к. крупноколосый. Южный склон лево-
бережного хребта, в группировках эфемеров. Редко. Хорошее кормовое
растение.
Флора ущелья Кондара
41
75. В. Danthoniae Trin. — к. Дантонии. Там же. Хорошее кормовое
растение.
76. Brachypodium silvaticum (Huds.) Р. В. — коротконожка лесная.
В береговых зарослях р. Кондара и ее притоков. Очень распро-
странен.
77. Roegneria macrochaeta Nevski — регнерия крупнощетинистая.
В кленовниках на Руидаште на лужайках.
78. R. Drobovii Nevski — р. Дробова. На лесных опушках и по
берегам Кондары и ее притоков.
79. R. ugamica (Drob.) Nevski — р. угамская. На лесных лужайках
от урочища Квак и выше, а также и на Руидаште. Хорошее кормовое
растение.
80. Agropyrum intermedium (Host.) Р. В. — пырей средний. На
опушке кленовника у второго холма близ тропы на Квак.
81. A. trichophorum (Link.) Richt. — п. волосоносный. На сухих скло-
нах хребтов в долине Кондары и ее притоков. Кормовое.
82. A. repens (L.) Р. В. — п. ползучий. На залежах и опушках кленов-
ников.
83. Aegilops squarrosa L. — эгилопс оттопыренный. На каменистых
южных склонах.
84. A. triuncialis L. — э. трехдюймовый. Там же.
85. Taeniatherum crinitum (Schreb.) Nevski — лентоостник длинно-
волосый. Там же.
86. Т. asperum (Simk.) Nevski — л. шероховатый. На щебнистых
склонах.
87. Hordeum turkestanicum Nevski — ячмень туркестанский. На скло-
нах холмов Руидашта.
88. Н. bulbosum L. — я. луковичный. Образует ассоциации, местами
создает фон, на сухих открытых склонах в бассейне Кондары.
89. Н. leporinum Link. — я. заячий. На южном склоне левобережного
хребта в группировках эфемеров.
90. Н. hystrix Roth. — я. ощетиненный. На сухом глинистом склоне
в низовьях Кондары.
91. Н. geniculatum All. — я. коленчатый. В низовьях Кондары^
среди эфемеров.
Сем. Сурегасеае
92. Trichophorum pumilum (Vahl.) Schinz. — пухонос приземистый.
На осоково-кобрезиевых лужайках Руидашта.
93. Blysmus compressus (L.) Рапс. — блисмус сжатый. На осоковых
лужайк а х Р у и д ашта.
94. Heleocharis meridionalis Zinzerl. — болотница южная. Там же.
95. Heleocharis sp. — Там же.
96. Schoenoxiphium hissaricum Pissjauk. — схеноксифиум гиссар-
ский. В среднем течении Кондары, у водопадов.
97. Kobresia stenocarpa (К. et К.) Steud. — кобрезия узкоплодная.
На осоковых лужайках Руидашта.
98. К. pamiroalaica Iv. — к. памироалайская. Там же. Кормовое.
99. Carex duriusculiformis V. Krecz. — осока ложнотвердоватая. На
влажных лужайках Руидашта. Кормовое растение.
100. С. compacta Lam. — о. сжатая. На правом берегу Кондары
в ореховой роще у ключа.
42
В, В. Писъяукова
101. С. polyphylla Каг. et Kir. — о. многолистная. По берегам Кон-
дары и в кленовниках северных склонов.
102. С. pseudo-foetida Kuk. — о. ложновонючая. На влажных лу-
жайках Руидашта. Кормовое растение.
103. С. Slobodovii V. Krecz. — о. Слободова. Там же, вместе с пре-
дыдущим.
104. С. pachystylis Gay. — о. толстостолбиковая. На склонах по
ущелью, до 1300—1400 м, в группировках эфемеров. Хорошее кормовое
растение.
105. С. orbicularis Boott. — о. округлая. На сырых лужайках у клю-
чей на Руидаште.
106. С. decaulescens V. Krecz. — о. стеблеватая. По берегам ключей—
на Кваке и Руидаште.
107. С. Koshewnikowii Litw. — о. Кожевникова. Обычна на скалах
по ущелью Кондары и ее притокам.
108. С. turkestanica Rgl. — о. туркестанская. На мелкощебнистых
склонах хребтов, окружающих Кондару.
109. С. diluta М. В. — о. светлая. По берегам Кондары в нижнем ее
течении.
110. С. melanostachya М. В. — о. черноколосая. В кленовниках и ро-
зариях на юго-восточных склонах.
Сем. Агасеае
111. Arum Korolkowi Rgl. — аронник Королькова. Обычен в древес-
но-кустарниковом поясе.
112. Eminium Alberti (Rgl.) Engl. — эминиум Альберта. На сухих
склонах.
Сем. Juncaceae
ИЗ. Juncus bufonius L. — ситник лягушечий. На субальпийских осо-
ковых лужайках Руидашта.
114. J. minutulus V. Krecz. et Contsch. (J. bufonius v. p^rvulus
Hartm.) — с. мелкий. На влажных карнизах; по берегам ключей на
Руидаште.
115. J. lampocarpus Ehrh. — с. блестящеплодный. По берегам Кон-
дары и ее притоков.
116. J. glaucus Ehrh. — с. сизый. По берегам Кондары и ее притоков.
Сем. Liliaceae
117. Colchicum Kesselringii Rgl. — безвременник Кессельринга. На
склонах в урочище Квак.
118. Eremurus Regelii Vved. — эремурус Регеля. На каменистых скло-
нах правобережного хребта, от 1400 м и выше.
119. Е. brachystemon Vved. — э. короткотычинковый. На камени-
стых склонах выше уроч. Квак. 2400 м.
120. Е. comosus О. Fedtsch. — э. хохлатый. На южных и юго-восточ-
ных каменистых склонах в нижней части ущелья Кондары.
121. Е. Olgae Rgl. — э. Ольги. Там же. Поднимается до 1500 м.
Флора ущелья Кондара
43
122. Е. robustus Rgl. — э.
мощный (рис. 5). На горных
лугах и в лесах главным обра-
зом северных склонов, от 1400м
и выше. Корни съедобны. Де-
коративное, медонос.
123. Е. Aitchisonii Backer—
э. Ечисона. На северных и се-
веро восточных склонах в ниж-
ней части ущелья Кондары.
Выше сменяется предыдущим
видом.
124. Gagea pseudoerubescens
Pascher var. latifolia Grossh.—
гусиный лук ложнокраснова-
тый. На склонах холмов Руи-
дашта.
125. G. graminifolia Vved.—
г. л. злаколистный. Там же.
126. G. hiss rica Lipsky —
г. л. гиссарский. Там же.
127. G. vegeta Vved. — г.
л. крепкий. В разнотравном
кленовнике Квака. Редко.
128. Gagea sp.—На каме-
нистых осыпях в уроч. Квак.
129. G. gageoides (Zucc.)
Vved. — г. л. персидский. На
склонах всюду. Обычно.
130. G. minutiflora Rgl. var.
aphylla Grossh. — г. л. мелко-
цветный. На мелкоземистыми
щебнистых склонах холмов пла-
то Руидашт. Обычно.
131. G. capillifolia Vved. —
г. л. волосолистный. На карни-
зах и в трещинах скал. Обычное.
132. Allium Barsczewskii
Lipsky — лук Барщевского. На
щебнистых и каменистых скло-
нах от ни кнэй части ущелья
Кондары до Руидашта включи-
тельно. Обычен.
133. A. dolichomischum
Vved. — л. длинноцветоножко-
вый. На мелкощебнистых юж-
ных склонах левобережного
хребта, от 1200 до 1500 м, в
среднем течении Кондары.
134. A. tenuicaule Rgl. — л.
тонкостебельный. На щебнистых
и каменистых склонах холмов рис> 5. Эремурус мощный — Е rem u г us robustus
Руидашта, 2800 м. ‘ Rgl.
44
В. В. Письяукова
135. A. tytthanthum Vved. — л. мелкоцветный. На каменистых скло-
нах в восточной части плато Руидашт. Редко.
136. A. Oschanini О. Fedtsch. — л. Ошанина. На крупнокаменистых
осыпях перед входом на Руидашт, 2400 м. Редко.
137. A. verticillatum Rgl. — л. мутовчатый. На мелкощебнистых
южных склонах левобережного хребта близ урочища Теги-камар.
138. A. Alexejanum Rgl. — л. Алексея. На щебнистых склонах Руи-
дашта.
139. A. Suvorovi Rgl. — л. Суворова. На склонах.
140. A. stipitatum Rgl. — л. стебельчатый. На мягких северных
и северо-западных склонах в древесно-кустарниковом поясе ущелья Кон-
дары и ее притоков.
141. A. Rosenbachianum Rgl. — л. Розенбаха. В высокотравных
кленовниках на Кваке и в тени скал на Руидаште. Обычное.
142. A. flavellum Vved. — л. желтоватый. На щебнистых склонах
перед уроч. Квак.
143. A. seravschanicum Rgl. — л. зеравшанский. На каменистых
затененных склонах правобережного хребта и по берегам Кондары близ
ее устья.
144. A. darvasicum Rgl. — л. дарвазский. На каменистых склонах
уроч. Квак и на склонах холмов Руидашта. Обычное.
145. A. cupuliferum Rgl. — л. чашеносный. На каменистых и щеб-
нистых склонах правобережного хребта близ тропы на Квак.
146. A. hissaricum Vved. — л. гиссарский. На щебнистых склонах
перед уроч. Квак.
147. A. Pallasii Murr. — л. Палласа. Руидашт, на красных песча-
никах.
148. Rhinopetalum bucharicum (Rgl.) A. Los. — ринопеталум бухар-
ский. Обычно по склонам.
149. Fritillaria Eduardi Rgl. — рябчик Эдуарда. На горных склонах
от 1400 м и до Руидашта. Луковица используется местным населением для
приготовления крахмала, идущего в пищу.
150. Tulipa lanata Rgl. — тюльпан шерстистый. В древесно-кустарни-
ковом поясе, преимущественно в уроч. Квак.
151. Т. praestans Hoog. — т. превосходящий. На склонах в среднем
течении Кондары.
152. Т. hissarica М. Pop. — т. гиссарский. На склонах от 1300 м
и до Руидашта включительно.
153. Scilla puschkinioides Rgl. — пролеска пушкиниевидная. Всюду
на склонах.
154. S. bucharica Dessjat. — п. бухарская (рис. 6). На склонах всюду.
155. Asparagus bucharicus Iljin — спаржа бухарская. Всюду на
склонах хребтов.
156. Polygonatum Sewerzowi Rgl. — купена Северцова. Всюду в пре-
делах древесно-кустарникового пояса ущелья Кондары.
Сем. Amaryllidaceae
157. Ungernia tadshikorum Vved. — унгерния таджикская. На сухих
глинистых склонах от нижней части ущелья Кондары до уроч. Квак.
158. Ixiolirion tataricum (Pall.) Roem. et Schult. — иксиолириоп
татарский. На каменистых склонах.
Флора ущелья Кондара
45
Сем. Iridaceae
159. Crocus Korolkowi Rgl. — шафран Королькова. На склонах
всюду.
160. Iris Hoogiana Dykes. — касатик Гуга (рис. 7). На травянистых
склонах от 1400 м и выше. Очень красивое и душистое растение.
161. Juno bucharica (Foster) Vved. — юнона бухарская. Всюду на
склонах.
162. J. parvula Vved. — ю. мелкая. На склонах холмов Руидашта.
163. J. Rosenbachiana Rgl. — ю. Розенбаха. На склонах в кленов-
никах.
164. J. warleyensis (Foster) Vved. — ю. уорлийская. На склонах
холмов Руидашта.
Рис. 6. Пролеска бухарская — Scilla bucharica Dessjat.
165. Gladiolus segetum Ker-Gawl. — шпажник засоряющий. На юж-
ных склонах в долине Кондары, а также на залежах.
Сем. Orchidaceae
166. Epipactis Royleana Lindl. — дремлик Ройля. На влажных ска-
лах у сочащейся воды по Кондаре и ее притокам.
167. Orchis turkestanica Klinge — ятрышник туркестанский. На суб-
альпийских осоковых лужайках у ключей в южной части Руидашта.
Сем. Salicaceae
168. Salix babylonica L. — ива вавилонская. В нижней части ущелья.
Здесь культурное.
169. S. oxycarpa Andr. (=S. iranica Bornm.) — и. остроплодная.
В верховьях Кондары, в уроч. Квак и на Руидаште.
170. S. australior Anderss. — и. южная. По берегам Кондары.
171. S. tenuijulis Lab. — и. тонкосережчатая. По берегам Кондары
близ устья.
46
В. В. Письяукова
172. Populus pyramidalis Rozier. — тополь пирамидальный. По
берегам р. Кондара в двух-трех местах. Здесь культурное.
173. Р. tadshikistanica Кот. — т. таджикистанский. По берегам
в верхнем течении и близ устья Кондары.
Рис. 7. Касатик Гуга
Iris Hoogiana.
Сем. Juglandaceae
174. Juglans regia L. — орех грецкий. По
берегам р. Кондара и ее притоков на север-
ных склонах хребтов, до 1800 м.
Сем. Ulmaceae
175. Celtis caucasica Willd. — каркас кав-
казский. На сухих каменистых склонах в
долине Кондары.
Сем. Могасеае
175а. Morns alba L. — шелковица белая.
13 культуре.
Сем. Urticaceae
176. Urtica dioica L. — крапива двудомная.
По берегам в пределах усадьбы Станции.
177. Parietaria chersonensis (Lang, et Szov.)
Dorfler — постенница херсонская. В расщели-
нах камней, у ключей, в тени скал. Обыч-
ное.
Сем. Santalaceae
178. Thesium alatavicum Kar. et Kir. —
ленец алатавский. На травянистых склонах
холмов Руидашта. Часто.
179. Т. Gontscharovii Е. ВоЬг. — л. Гонча-
рова. На щебнистых склонах правобережного
хребта в среднем течении Кондары.
Сем. Polygonaceae
180. Rumex crispus L. — щавель курчавый. По берегам Кондары
в среднем ее течении, на галечнике.
181. R. Paulsenianus Recking. — щ. Паульсена. На травянистых
склонах в полосе древесно-кустарниковой. Обычен.
182. R. conglomeratus Мигг. — щ. скученноцветный. По берегам Кон-
дары и как сорное в пределах усадьбы Станции.
Флора ущелья Кондара
47
183. R. syriacus Meisn. — щ. сирийский. По тропам у берегов р. Кон-
дары и ее притоков.
184. Rheum Maximowiczi A. Los. — ревень Максимовича. На каме-
нистых склонах от 1300 м и выше.
185. R. Fedtschenkoi Maxim. — р. Федченко. На каменистых и щеб-
нистых склонах от 2000 м и выше (вместе с предыдущим).
186. Atraphaxis pyrifolia Bge. — курчавка грушелистная. На каме-
нистых южных склонах.
187. Polygonum paronychioides. С. А. М. — горец приноготковидный.
На сухих каменистых и щебнистых склонах. Обычен.
188. Р. biaristatum Aitch. et Hemsl. — г. двухостый. На щеб-
нистых склонах холмов Руидашта.
189. Р. fibrilliferum Кот. — г. волоконценосный. На сухих щеб-
нистых и каменистых склонах от 1400 м и выше. На плато Руидашт, обы-
чен на мелкоземистых склонах холмов.
190. Р. rupestre Kar. et Kir. — г. утесный. На щебнистых склонах
холмов Руидашта. Редко.
191. Р. heterophyllum Lindm. — г. разнолистный. На каменистых
склонах у троп левобережного хребта и на огороде станции.
192. Р. aviculare L. — г. птичий. Распространен от нижней части
ущелья до Руидашта включительно. Сорное.
193. Р. polycnemoides Jaub. et Spach. — г. подземковидный. На
щебнистых осыпях пред Кваком, по берегам Кондары на галечнике, на
залежи по южному склону левобережного хребта и близ устья квакского
ключа.
194. Р. persicaria L. — г. почечуйный. По берегам арыков в низовьях
Кондары. Краситель, лекарственное.
195. Р. hydropiper L. — г. водяной перец. По заболоченным берегам
р. Кондара и ее притоков. Обычное. Краситель.
196. Р. bucharicum G. Grig. — г. бухарский. На травянистых скло-
нах правобережного хребта, от 1400 м до Руидашта включительно. Дуби-
тель, краситель, медонос.
197. Р. bistorta L. — г. змеиный. На субальпийских осоковых лужай-
ках Руидашта. Медонос.
198. Р. convolvulus L. — г. вьющийся. В орехово-кленовом лесу
урочища Тангемур п на галечнике и огородах близ устья Кондары,
Медонос. Сорное.
Сем. Cbenopodiaceae
199. Chenopodium botrys L. — марь душистая. На галечниках по Кон-
даре.
200. Ch. foliosum (Moench.) Aschers — м. многолистная. На каме-
нистых склонах холмов Руидашта.
201. Ch. album L. — м. белая. Сорное.
Сем. Amarantaceae
202. Amaranthus retroflexus L. — ширица запрокинутая. Сорное.
203. A. blitum L. — щ. жминда. На террасах виноградника близ
метеорологических будок (правый берег Кондары). Сорное.
204. A. albus L. — щ. белая. Там же. Сорное.
-48
В, В. Письяукова
Сем. Portulacaceae
205. Portulaca oleracea L. — портулак огородный. Сорное, на огороде.
Съедобно.
Сем. Caryophyllaceae
206. Stellaria media (L.). Cyr. — звездчатка средняя. На огороде
близ устья Кондары. Медонос. Кормовое.
207. S. brachipetala Bge. — з. коротколепестная. На влажных лужай-
ках Руидашта.
208. S. fontana М. Pop. — з. приручейная. По берегам ключей на Руи-
даште.
209. S. alsinoides Boiss. — з. мокричная. Там же.
210. Cerastium cerastoides Britt. — ясколка трехстолбиковая. На
Руидаште у ключей.
211. С. inflatum Link. — я. вздутая. На каменистых и щебнистых
склонах в долине Кондары и ее притоков.
212. С. dichotomum L. — я. вильчатая. На каменистых склонах
в группировках эфемеров.
213. Lepyrodiclis holosteoides (С. А. М.) Fisch, et Меу. — пашенник
костенецовидный. В высокотравных кленовниках уроч. Квак, изредка
на Руидаште.
214. L. stellarioides Schrenk — п. звездочковидный. В поясе древес-
но-кустарниковой растительности на северных склонах хребтов Кон-
дары.
215. Minuartia Meyeri (Boiss.) Bornm. — минуарция Мейера. На
щебнистых склонах в нижней части ущелья.
216. Arenaria Griffithii Boiss. — песчанка Гриффица. На склонах
холмов Руидашта. Обычное.
217. A. turkestanica Schischk.—п. туркестанская. На склонах холмов
Руидашта.
218. A. serpyllifolia L. — п. чабрецолистная. По северным склонам
в кленовниках, по берегам Кондары на щебнистых склонах в тени.
219. A. leptoclados Guss. — п. тонкостебельная. В кленовниках на
северных склонах.
220. Herniaria glabra L. — грыжник гладкий. На скалистых склонах
восточного края Руидашта.
221. Н. hirsuta L. — г. волосистый. На тропинке у огорода на Кваке.
222. Agrostemma githago L.—куколь обыкновенный. Сорное, в посе-
вах. Редко.
223. Silene commutata Guss. — смолевка замещающая. В кленов-
никах правобережного хребта, 1200—2400 м.
224. S. samarkandensis Preobr. — с. самаркандская. На скалах
в западной части плато Руидашт.
225. S. conformifolia Preobr. — с. однороднолистная. На скалистых
склонах, от 1600 м и до Руидашта включительно.
226. S. baldshuanica В. Fedtsch. — с. бальджуанская. На каменистой
осыпи против дома Станции, около 1200—1300 м.
227. S. tachtensis Franch. — с. тахтинская.. На скалистых уступах
восточной и северо восточной окраин Руидашта.
228. S. guntensis В. Fedtsch. — с. гунтская. На скалах в северо-вос-
точной части Руидашта.
Флора ущелья Кондара
49
229. S. scabrifolia Кот. — с. шероховатолистная. На скалистых
склонах Руидашта.
230. S. Komarovii Schischk. — с. Комарова. В кленовниках уроч.
Квак, на опушках.
231. S. baldshuanica В. Fedtsch. var. Zaprjagaevii Schischk. — с. За-
прягаева. На каменистой осыпи против главного дома Станции.
232. S. bucharica М. Pop. — с. бухарская. На каменистой осыпи
против главного дома Станции и в кленовниках на северных склонах.
233. S. conica L. — с. коническая. На щебнистой вершине правобе-
режного хребта.
234. S. conoidea L. — с. конусовидная. В низовьях Кондары среди
эфемеров.
235. Coronaria coriacea (Moench.) Schischk. — горицвет кожистый.
На склонах хребтов и в долинах Кондары и ее притоков.
236. Melandrium ovalifolium Rgl. et Schmalh. — дрема овальнолист-
ная. В тенистых ущельях и по склонам на правом берегу р. Кондара
в среднем ее течении.
237. М. ferganicum (Preobr.) Schischk.—д. ферганская. В высокотрав-
ных кленовниках уроч. Квак.
238. М. erubescens Schischk. — д. краснеющая. На разнотравных
склонах уроч. Квак, до 2400 м.
239. М. longicarpophorum Кот. — д. длинноножковая. По склонам
холмов на Руидаште, 2700—2800 м.
240. Tunica stricta (Rgl.) Fisch, et Mey. — туника прямая. На щеб-
нистых сухих склонах, всюду.
241. Vaccaria segetalis (Neck.) Garcke — тысячеголов посевной. Сор-
ное. Семена ядовиты.
242. Dianthus tetralepis Nevski — гвоздика четырехчешуйная. Обычно
на скалах и каменисто-щебнистых склонах.
243. D. seravschanicus Schischk. — г. зеравшанская. На мелкощеб-
нистых осыпях уроч. Квак, 1500—2100 м.
244. Saponaria Griffithiana Boiss. — мыльнянка Гриффица. На мелко-
щебнистых осыпях уроч. Квак.
245. Velezia rigida L. — велеция жесткая. На щебнистых склонах.
Обычное.
Сем. Ranunculaceae
246. Paeonia anomala L. — пеон Марьин-корень. На склонах гор,
ют 1800 м и до Руидашта включительно.
247. Eranthis longistipitata Rgl. — весенник длинностебельчатый. На
склонах в уроч. Квак.
248. Nigella integrifolia Rgl. — чернушка цельнолистная. На скло-
нах у р. Кондара, по правому ее берегу, против главного дома Стан-
ции.
249. N. bucharica N. Schipcz.—ч. бухарская. На склонах в низовьях
Кондары.
250. Aquilegia lactiflora Kar. et Kir. — водосбор молочноцветный.
По ущельям в долине р. Кондара, у ключей, в тени.
251. Delphinium ternatum Huth. —живокость тройчатая. Верхняя
граница леса пред выходом на Руидашт, в розарии, 2400 м.
252. D. biternatum Huth. — ж. дваждытройчатая. В высокотравных
кленовниках северных склонов, от 1400 м до 2200 м.
Ущелье Кондара
50
В. В. Письяукова
253. D. semibarbatum Bienert — ж. полубородатая. На каменистых
южных склонах левобережного хребта. Красильное.
254. Delphinium sp. — вид, близкий к D. ternatum. Верхняя гра-
ница лесного пояса у входа на Руидашт, в зарослях кустарников.
255. Anemone bucharica Rgl. — ветреница бухарская. Обычное по
склонам.
256. A. Tschernjaewii Rgl. — в. Черняева. Обычное на травянистых
склонах.
257. Clematis orientalis L. — ломонос восточный. Среди кустарников
в долине р. Кондара и ее притоков.
258. Ranunculus baldshuanicus Rgl. — лютик бальджуанский. На
правом берегу Кондары, в среднем течении. Изредка.
259. R. laetus Wall. — л. красивый. По берегам Кондары и ее при-
токов, на влажных лужайках.
260. R. arvensis L. — л. полевой. На южных каменистых и щебнистых
склонах.
261. R. paucidentatus Schrenk — л. малозубчатый. На травянистых
склонах Руидашта и на щебнистых склонах уроч. Квак.
262. R. alajensis Ostenf. — л. алайский. На лужайках Руидашта,
у ключей.
263. R. afghanicus Aitch. et Hemsl. — л. афганский. На склонах
холмов Руидашта.
264. R. oligophyllus Pissjauk. — л. малолистный. На осоковых лу-
жайках Руидашта.
265. R. turkestanicus Franch. — л. туркестанский. На субальпийских
лужайках Руидашта.
266. R. mindshelkensis В. Fedtsch. — л. минджелькенский. На осо-
ковых лужайках Руидашта.
267. R. pinnatisectus М. Pop. — л. перистораздельный. В нижней
части ущелья Кондара.
268. R. tenuilobus Rgl. — л. тонколопастной. На склонах в лесной
полосе ущелья.
269. R. czimganicus Ovcz. — л. чимганский. В кленовниках Квака
на щебнистых осыпях.
270. R. Komarovii Freyn. — л. Комарова. В пределах древесно-
кустарникового пояса.
271. R. leptorrhynchus Aitch. et Hemsl. — л. тонконосиковый.
На склонах нижней части ущелья Кондара, до 1300 м.
272. Thalictrum sultanabadense Stapf. — василистник султанабад-
ский. На скалистых склонах, всюду.
273. Th. isopyroides G. А. М. — в. изопироидный. В расщелинах скал
нижней части ущелья.
274. Adonis turkestanicus (Korsh.) N. Adolf. — адонис туркестанский-
На северных склонах, от 2200 м. Лекарственное.
Сем. Berberidaceae
275. Leontice Albertii Rgl. — леонтица Альберта. На северных скло-
нах, в поясе древесно-кустарниковой растительности.
276. Bongardia chrysogonum (L.) Boiss.,— бонгардия золотистая. На
сухих склонах. Клубни съедобны.
277. Berberis integerrima Bge. — барбарис цельнокрайний. На
каменистых южных склонах долины р. Варзоб, близ ущелья Кондара.
Флора ущелья Кондара
51
Сем. Papaveraceae
278. Glaucium fimbrilligerum Boiss. — глауциум бахромчатый. На
щебнистых склонах в низовьях Кондары. Изредка.
279. Roemeria refracta (Stev.) DC. — рёмерия отогнутая. На склонах
всюду, до 1400 м. Обычное.
280. Papaver pavoninum Schrenk — мак павлиний. Обычен на скло-
нах в группировках эфемеров.
281. Р. Litwinowii Fedde — м. Литвинова. На щебнистой вершине
южного склона левобережного хребта.
282. Corydalis Ledebouriana Kar. et Kir. — хохлатка Ледебура. На
склонах. Обычное.
283. С. nudicaulis Rgl. — х. голостебельная. На склонах холмов
Руидашта.
284. Corydalis sp. — вид близкий к С. nudicaulis. Только в высоко-
травных кленовниках уроч. Квак.
Сем. Cruciferae
285. Alliaria officinalis Andrz. — чесночник лекарственный. Всюду
в древесно-кустарниковом поясе ущелья р. Кондара и ее притоков;
съедобно.
286. Sisymbrium Loeseli L. — гулявник Лёзелиев. На залежах.
287. Arabidopsis verna (С. Koch) N. Busch — резушка весенняя.
На скалистых склонах у выхода на Руидашт.
288. A. Thaliana (L.) Heynh. — р. Таля. На скалистых склонах
от устья Кондары до Руидашта.
289. Barbarea arcuata Rchb. — сурепка дуговидная. На галечниках
по берегам Кондары и ее притоков.
290. В. plantaginea DC. — с. подорожниковая. На северных скло-
нах. в тени леса в уроч. Квак и на Руидаште.
291. В. minor. С. Koch — с. малая. На субальпийских лужайках
Руидашта (по ключам).
292. Barbarea sp. — в орехово-кленовых рощах. Близкая к В. inte-
grifolia DC.
293. Cardamine densiflora N. Gontsch. — сердечник густоцветный. На
заболоченных лужайках у ключей в долине Кондары.
294. Nasturtium officinale (L.) R. Вт. — жеруха лекарственная. На
заболоченных лужайках у ключей. Изредка.
295. Arabis Montbretiana Boiss. — резуха Монбре. На скалистых и
щебнистых склонах правобережного хребта и уроч. Квак.
296. A. auriculata Lam. — р. ушастая. На скалах в устье Кон-
дары.
297. A. karategina Lipsky — р. каратегинская. На скалах правого
берега близ устья Кондары.
298. Clausia hispida (Rgl.) Lipsky — клаусия щетинистая. Обычное
на щебнистых склонах.
299. С. turkestanica Lipsky — к. туркестанская. На каменистых скло-
нах в долине Оби-сафед.
300. Malcolmia africana (L.) R. Br. —малькольмия африканская.
Каменистые южные склоны, до 1100 м.
301. Chorispora elegans Camb. — хориспора изящная. На лужайках
у ключей Руидашта.
4*
52
В. В. Письяукова
302. Ch. macropoda Trautv. — х. крупноногая. На склонах холмов
Руидашта.
303. Trichochiton inconspicuum Korn. — волосатик неприметный.
В розариях, на высоте 2300 м.
304. Alyssum campestre L. — бурачок полевой. На сухих склонах
в группировках эфемеров. Обычен.
305. A. dasycarpum Steph. — б. пушистоплодный. Там же.
306. Buchingera axillaris Boiss. — бухингера пазушная. В ореш-
никах. Образует с орехом ценозы, как Impatiens parviflora и Physocaulis
nodosus.
307. Erophila minima С. A. M. —веснянка карликовая. На щебнистых
склонах близ устья Кондары.
308. Е. verna (L.) Bess. — в. весенняя. На скалистых уступах в до-
лине Кондары.
309. Brassica sp. — Уроч. Квак, по ключам.
310. Eruca sativa Lam. — индау посевной. Сорное, на льняном поле
в долине р. Оби-сафед.
311. Crambe Kotschyana Boiss. —катран Кочи. На травянистых скло-
нах правобережного хребта против метеорологических будок. Корни
съедобны. Лучший медонос.
312. Stroganowia paniculata Rgl. — строгановия метельчатая. На
щебнистых сухих вершинах хребтов в долине Кондары.
313. Stubendorffia aptera Lipsky — штубендорфия бескрылая. На
каменистых склонах, от 1900 м и выше.
314. Megacarpaea gigantea Rgl. — крупноплодник гигантский. На
щебнистых склонах, от 1900 м. Корни съедобны.
315. Thlaspi Kotschyanum Boiss. et Hoh. — ярутка Кочи. На скали-
стых склонах у входа на Руидашт, около 2000—2400 м.
316. Th. perfoliatum L. — я. пронзенная. Обычное на склонах в груп-
пировках эфемеров. Сорняк.
317. Th. arvense L. — я. полевая. Сорное, на огороде Станции.
318. Neslia apiculata Fisch, et Mey. — неслия остроконечная. Только
на южном склоне левобережного хребта, у залежи.
319. Capsella bursa pastoris (L.) Med. — пастушья сумка. На склонах
в группировках эфемеров.
Сем. Crassulaceae
320. Pseudosedum longidentatum A. Bor. — ложноочиток длинно-
зубчатый. На каменистых и щебнистых склонах, от 1400 м и до Руи-
дашта.
321. Р. condensatum А. Вог. — л. плотный. Там же.
322. Rosularia hissarica А. Вог. — розеточница гиссарская. Обычное.
На скалах в долине Кондары, от 1100 м и до Руидашта.
323. R. paniculata (Rgl. et Schmalh.) Berger — p. метельчатая. Ha
мелкощебнистых осыпях правобережного хребта, от 1300 м и до Руидашта.
Сем. Platanceae
324. Platanus orientalior Dode — платан востока. В долине Варзоба
и его притоков. В Кондаре культурное.
Флора ущелья Кондара
53
Сем. Rosaceae
325. Spiraea baldshuanica В. Fedtsch. — таволга бальджуанская. На
скалах в долине р. Кондара.
326. Cotoneaster insignis Pojark. — кизильник замечательный. По
береговым зарослям р. Кондара, от устья до урочища Тангемур.
327. С. suavis Pojark. — к. привлекательный. В уроч. Квак, в кле-
новниках, на высоте 1800—1900 м. „
328. С. acutiuscula Pojark. — к. острый. Обычен в долине р. Кон-
дара и на скалистых склонах, ее ограничивающих (от уроч. Теги-камар
до уроч. Тангемур).
329. С. multiflora Bge. — к. многоцветковый. В средней части ущелья.
Изредка.
330. С. hissarica Pojark. — к. гиссарский. Там же и в ущельяхр. Оби-
сафед.
331. С. nummularioides Pojark. — к. монетовидный. Только на
скалистом уступе перед входом на Руидашт, 2400 м.
332. С. nummularia F. et М. — к. монетный. На каменистых и щеб-
нистых сухих склонах, от 1500 м и до скал Руидашта (уроч. Тангимур,
Квак, Тереакба).
333. Malus hissarica S. Kudr. — яблоня. Образует рощицы по скло-
нам у Кондары и ее притоков, а также растет по берегам. Приносит много
разнообразных по форме и довольно вкусных плодов.
334. Sorbus persica Hedl. — рябина персидская. На склонах (уроч.
Квак), от 1500 м до 2400. Изредка.
335. S. turkestanica Hedl. — р. туркестанская. В уроч. Квак.
336. Crataegus pontica С. Koch — боярышник понтийский. На сухих
склонах отдельными деревьями. Имеет крупные и вкусные плоды.
337. С. turkestanica Pojark. — б. туркестанский. Обычен на скло-
нах ущелья.
338. С. hissarica Pojark. — б. гиссарский. В долине Кондары, близ
ее устья и выше урочища Теги-камар (в среднем течении). Плоды
съедобны.
339. Rubus caesius L. var. turkestanicus Rgl. — ежевика. Обычное по
берегам Кондары и ее притоков, а также на северных склонах, на лесных
лужайках и опушках. Дает много вкусных плодов.
340. Potentilla kulabensis Th. Wolf. — лапчатка кулябская. Уроч.
Квак, в высокотравных, влажных кленовниках, по берегам Кондары
и ее притоков.
341. Р. transcaspia Th. Wolf. —л. закаспийская. На травянистых
склонах в долине Кондары.
342. Р. gelida С. А. М. — л. холодная. Обычное на склонах холмов
Руидашта.
343. Р. reptans L.—л. ползучая. По берегам Кондары.
344. Potentilla sp. — На Руидаште, по каменистым склонам холмов*
345. Geum urbanum L. — гравилат городской. На правом берегу
в среднем течении Кондары, у ключей, на влажных лужайках.
346. Orthurus kokanicus (Rgl. et Schmalh.) Juz. — г. коканский. Ha
каменистых склонах, от 2000 м.
347. Alchimilla sibirica Zam. — манжетка сибирская. Руидашт, на
лужайках у ключей.
348. Agrimonia asiatica Juz. — репейничек азиатский. Обычен в до-
лине Кондары, по берегам и на опушках кленовников северного склона
54
В. В. Письяукова
349. Poterium sanguisorba L. — черноголовник кровохлебковый. На
сухих щебнистых и каменистых, главным образом южных склонах,
в долине р. Кондара и ее притоков.
350. Rosa Webbiana Wall. — роза Уэбба. На скалистых обрывах по
краям Руидашта.
351. R. maracandica Bge. — р. самаркандская. На скалистых и щеб-
нистых сухих южных склонах левобережного хребта и близ устья Кон-
дары, на скалах. Витаминоносное.
352. R. hissarica Slobod. — р. гиссарская. На склонах холмов Руи-
дашта (северный край плато).
353. R. Есае Aitch. —р. Э. К. Эйчисон. Только на щебнистой вершине
правобережного хребта у тропы на Квак.
354. R. kokanica Rgl. — р. коканская. Обычна на склонах, образует
ассоциации. Витаминоносное.
355. R. canina L. — р. собачья. Обычна в полосе кленовников и
в ущельях. Дубитель.
356. R. corymbifera Borkh. — р. щитконосная. Там же, главным
образом у ключей, по берегам.
357. R. Beggeriana Schrenk — р. Беггера. У ручьев выше уроч. Квак
и в ущельях.
358. Amygdalus bucharica Korsh. — миндаль бухарский. На сухих
южных каменистых склонах, до 2300 м.
359. Prunus divaricate Ldb. — алыча. По берегам Кондары и ручьев
и в кленовниках на северных склонах, до 1800 м. Обычное. Плоды
съедобны и вкусны.
360. Gerasus mahaleb (L.) Mill. — вишня антипка. По берегам Кондары
и ручьев. Обычное. Имеет твердую хорошую древесину. Плоды дают
пурпурную окраску. Семена содержат много амигдалина.
361. С. verrucosa (French.) Nevski — в. бородавчатая. На каме-
нистых склонах хребтов.
Сем. Leguminosae
362. Thermopsis dolichocarpa V. Nikitin — термопсис длинноплод-
ный. На северных склонах по лесным опушкам. Ядовит.
363. Trigonella Lipskyi Sirjaew — пажитник Липского. На каме-
нистых склонах местами образует ассоциации.
364. Т. adscendens Aphan. et Gontsch. — п. восходящий. На красных
песчаниках Руидашта.
365. Т. Popovii Е. Ког. — п. Попова. На склонах холмов Руидашта.
Обычное.
366. Medicago lupulina L. — люцерна хмелевидная. На травянистом
склоне правого берега Кондары в среднем течении. Хорошее кормовое
растение.
367. М. sativa L. — л. посевная. На склонах. Сорное.
368. М. rigidula Desr. — л. жестковатая. На сухих щебнистых скло-
нах правобережного хребта, до 1400 м.
369. М. orbicularis (L.) АП. — л. округлая. На сухих щебнистых
склонах правобережного хребта, до 1400 м, в группировках эфемеров.
370. М. denticulate Willd. — л. зубчатая. Там же.
371. М. falcate L. — л. серповидная. Одичавшее, на северных скло-
нах, в пределах усадьбы Ботанической станции.
Флора ущелья Кондара
55
372. М. minima (L.) Bartal. — л. маленькая. На сухих юго-восточ-
ных щебнистых склонах, до 1200—1300 м.
373. Melilotus officinalis (L.) Desr. — донник лекарственный. На скло-
нах у залежей. Лекарственное, краситель, ароматическое.
374. Trifolium repens L. — клевер ползучий. По берегам Кондары.
Хорошее кормовое растение.
375. Т. campestre Schreb. — к. полевой. На травянистых склонах
правобережного хребта. Изредка. Кормовое растение.
376. Т. pratense L. — к. луговой. По берегам Кондары и ее притоков.
Кормовое.
377. Psoralea drupacea Bge. — псоралея костянковая. На щебнистых
склонах у входа в ущелье. Изредка.
378. Golutea canescens Shap. — пузырник седоватый. На склонах
правого берега Кондары, до 1200 м. Декоративное.
379. Caragana turkestanica Кот. — карагаиа туркестанская. На пра-
вом берегу Варзоба, близ кишлака Гажии, на склонах у вершины пра-
вобережного хребта Кондары.
380. Chesneya hissarica Boriss. — чезнейя гиссарская. На сухих щеб-
нистых склонах, до 1800—2000 м. Обычное.
381. Astragalus jagnobicus Lipsky — астрагал ягнобский. На скло-
нах холмов Руидашта.
382. A. macropodium Lipsky — а. крупноножковый. На щебнистых
склонах правобережного хребта, от 1400 до 2300 м.
383. A. pauper Bge. — а. бедный. На субальпийских лужайках Руи-
дашта. Обычен. Хорошее кормовое растение.
384. A. mirabilis Bge. — а. удивительный. На склонах, в пределах
кленовников и розариев, до 2000 м. Обычен.
385. A. acormosus N. Basil. — а. бесстебельный. Уроч. Квак на скло-
нах хребтов и плато Руидашт, где довольно обычен. Красивоцве-
тущее.
386. A. Sieversianus Pall. — а. Сиверса. На склонах хребтов в ущелье
Кондара, до 1400—1500 м. Обычен.
387. A. quisqualis Bge. — а. странный. В кленовниках в ущелье
р. Оби-сафед. Кормовое растение.
388. A. macronyx Bge. — а. самаркандский. На южном склоне лево-
бережного хребта выше урочища Теги-камар, в ячменно-разнотравных
группировках.
389. A. glabrescens N. Gontsch. — а. почти-голый. На склонах в до-
лине Кондары, в кленовниках и розариях.
390. A. varzobicus N. Gontsch. — а. варзобский. На скалистых скло-
нах Руидашта.
391. A. andaulgensis В. Fedtsch. — а. андаульгенский. В тенистых
кленовниках уроч. Квак и на склонах холмов Руидашта. Хорошее
пастбищное растение.
392. A. lasiosemius Boiss. — а. мохнатый. На Руидаште, в различ-
ных группировках кузиний.
393. A. peduncularis Boyle — а. цветоножечный. На склонах право-
бережного хребта, от 1400 м и до Руидашта, в злаково-разнотравных
кленовниках и розариях. Кормовое растение.
394. A. discessiflorus N. Gontsch. — а. рыхлоцветковый. На щебни-
стых склонах уроч. Квак. Редко.
395. A. Irinae В. Fedtsch. — а. Ирины. На красных песчаниках
Руидашта.
56
В. В. Пи'ъяукова
396. A. hissaricus Lipsky — а. гиссарский. Только на правом берегу
Кондары, против дома, в злаково-разнотравном кленовнике.
397. A. baldshuanicus М. Pop. — а. бальджуанский. Только на за-
лежи по южному склону левобережного хребта.
398. A. xanthomeloides М. Pop. et Е. Ког. — а. черножелтый. На щеб-
нистых склонах правобережного хребта Кондары.
399. A. maverranagri М. Pop. — а. маверанагрский. На Руидаште,
по склонам из мезозойских красных песчаников, на высоте 2350 м.
400. A. neurophyllus Franch. — а. жилковатолистный. На травя-
нистых склонах правобережного хребта в ущелье Кондара.
401. A. vegetior N. Gontsch.—а. рослый. На скалистых уступах под
Руидаштом, в верхней зоне древесно-кустарникового пояса, 2300 м.
402. Oxytropis Michelsoni В. Fedtsch. — остролодочник Михельсона.
На склонах холмов Руидашта.
403. О. leptophysa Bge. — о. тонкопузырчатый. На каменистых и щеб-
нистых склонах, от 1200 м до 2000 м.
404. О. Litwinowii В. Fedtsch. — о. Литвинова. Изредка на щебни-
стых и мелкоземистых склонах, от 1200 м до 2850 м (Руидашт).
405. Oxytropis sp. — Только на осыпи из красных песчаников в южной
части Руидашта.
406. Hedysarum flavescens Rgl. et Schmalh. — копеечник желтова-
тый. На склонах холмов Руидашта. Изредка. Кормовое.
407. Н. baldshuanicum В. Fedtsch. — к. бальджуанский. На север-
ных склонах, в кленовниках.
408. Н. denticulatum Rgl. — к. зубчатый. Руидашт, на склонах.
Красивоцветущее.
409. Onobrychis grandis Lipsky — эспарцет высокий. В светлых
кленовниках правобережного хребта Кондары, до 1800 м. Кормовое
и красивоцветущее растение.
410. О. schugnanica В. Fedtsch. — э. шугнанский. На залежах. Кор-
мовое растение.
411. Cicer songoricum Steph. — нут джунгарский. На склонах холмов
Руидашта.
412. Lens orientalis (Boiss.) М. Pop. — чечевица восточная. На скло-
нах в группировках эфемеров.
413. Vicia hyrcanica Fisch, et Mey. — вика гирканская. На склонах
в группировках эфемеров. Ценное кормовое растение.
414. V. sativa L. —в. посевная. На склонах в группировках эфемеров.
415. V. angustifolia L. — в. узколистная. Там же. Кормовое растение.
416. V. tenuifolia Roth. — в. тонколистная. Очень широко распро-
странен по берегам Кондары и в древесно-кустарниковых группировках,
часто фоновое. Одно из ценнейших кормовых растений.
417. V. kokanicica Rgl. et Schmalh. — в. коканская. На каменистых
склонах Руидашта. Хорошее кормовое растение.
418. V. ervilia (L.) Willd. — в. четкообразная. На залежах, всюду.
419. Lathyrus mulkak Lipsky — чина мулькак (рис. 8). Обычное
на склонах. Красивоцветущее растение.
420. L. inconspicuus L. — ч. незаметная. На сухих склонах в составе
эфемеровой растительности.
421. L. pratensis L. — ч. луговая. В зарослях кустарников по бере-
гам Кондары и ее притоков. Кормовое растение.
422. L. tuberosus L. — ч. клубненосная. На опушках кленовников
северного склона, против уроч. Теги-камар. Хорошее кормовое растение.
Рис. 8. Чина мулькак — Lathyrus mulkak Lipsky.
58
В. В. Письяукова
Сем. Geraniaceae
423. Geranium collinum Steph. — герань холмовая. На щебнистых
склонах, от 1900 м и до Руидашта включительно.
424. G. pusilium L.—г. маленькая. На галечнике близ устья р. Кон-
дара.
425. G. divaricatum Ehrh. — г. раскидистая. На каменистых скло-
нах в кленовниках и кустарниковых зарослях.
426. G. saxatile Kar. et Kir. — г. скальная. На северных склонах
холмов Руидашта. Красивоцветущее растение.
427. G. Robertianum L. — г. Роберта. В тенистых местах по берегам
рек Кондара и Оби-сафед.
428. G. Charlesii (Aitch. et Hemsl.) Vved. — г. Чарльза. Обычное
на склонах в долине Кондары.
429. Erodium cicutarium (L.) L’Herit. — журавельник. На сухих
склонах среди эфемеров.
Сем. Linaceae
430. Linum corymbulosum Rchb. — лен щиточковатый. На сухих
склонах.
431. L. macrorhizum Juz. — л. крупнокорневой. На южных скло-
нах левобережного хребта ущ. Кондара.
Сем. Rutaceae
432. Haplophyllum perforatum К. et К. — цельнолистник исколотый.
На сухих щебнистых склонах по Кондаре.
433. Н. Popovii Е. Ког. — ц. Попова. Среди разнотравья на опуш-
ках кленовников.
434. Dictamnus tadshikorum Vved. — ясенец таджикский. Среди раз-
нотравья на опушках кленовников.
Сем. Euphorbiaceae
435. Euphorbia seravschanica Rgl. — молочай зеравшанский. На скло-
нах холмов Руидашта. Обычное.
436. Е. transoxana Prokh. — м. заамударьинский. В кленово-орехо-
вых лесах ущелья р. Оби-сафед (от 1700—2100 м) и на Кваке.
437. Е. Franchetii В. Fedtsch. — м. Франше. На щебнистых скло-
нах в древесно-кустарниковом поясе.
438. Е. sogdiana М. Pop. — м. согдийский. На мелкоземистых южных
чжлонах.
439. Е. falcata L. — м. серповидный. Обычное на залежах.
440. Е. glomerulans Prokh. — м. скученный. На лужайках Руи-
дашта, по восточному его краю.
441. Е. poecilophylla Prokh. — м. пестролистный. По западному
краю плато Руидашт.
442. Е. Szowitsii Fisch, et Меу.—м. Совича. На щебнистой вершине
правобережного хребта.
443. Е. canescens L. — м. сереющий. На щебнистых склонах, на зале-
жах.
Флора уъцелъя Кондара
59
Сем. Anacardiaceae
444. Pistacea vera L. — фисташка. На сухих южных склонах лево-
бережного хребта Кондары. Плоды съедобны, цветы медоносны, древе-
сина идет на поделки.
445. Rhus coriaria L. — сумах. Образует ассоциации на юго-восточ-
ных склонах в соседнем ущелье ручья Гажни. Дубильное и красильное
растение.
Сем. Celastraceae
446. Euonymus Semenowii Rgl. et Herd. — бересклет Семенова.
Только в орехово-кленовом лесу уроч. Тереакба (по р. Оби-сафед).
Сем. Асегасеае
447. Acer turkestanicum Pax — клен туркестанский. Образует леса
на северных, северо-западных и северо-восточных склонах. Идет в ущелье
до 2400 м.
448. A. Regelii Pax — к. Регеля. На сухих южных и юго-восточ-
ных склонах, идет до 2100 м.
Сем. Balsaminaceae
449. Impatiens parviflora DC. — недотрога мелкоцветная. В тенистых
ореховых и кленовых рощах и в прибрежных зарослях. Обычное.
Сем. Rhamnaceae
450, Zizyphus jujuba Mill. — унаби, ююба. На сухих щебнистых
южных склонах, до 1400 м. Лекарственное, медоносное и с ценной дре-
весиной растение.
451. Rhamnus dolichophylla N. Gontsch. — жестер длиннолистный.
В кленовниках северного и северо-западного склонов. Лекарственное,
медоносное, дубильное, красильное растение и с крепкой древесиной.
452. Sageretia laetevirens (Korn.) Gontsch. — сажереция яркозеленая.
На скалах, до 1300 м.
Сем. Vitaceae
453. Vitis vinifera L. — виноград. В долине Кондары и ее притоков.
Обычное. Ягоды собираются жителями.
454. Ampelopsis vitifolia (Boiss.) Planch. — виноградовник виноградо-
листный. На каменистых склонах, до 1900—2000 м и по долинам.
Сем. Malvaceae
455. Lavatera caschemiriana Gambess. — хатьма кашмирская. В те-
нистых высокотравных кленовниках северных склонов (уроч. Квак и
60
В. В. Писъяукова
левобережный хребет, 2000 м); Красивоцветущее растение. Медонос и
волокнистое.
456. Althaea cannabina L. — алтей коноплевый. В среднем течении
Кондары, на лесных высокотравных полянках. Волокнистое растение
и медонос.
457. Alcea nudiflora (Lindl.) Boiss. — шток-роза голоцветковая.
Обычное на склонах, красивоцветущее растение. Медонос. Волокнистое.
458. Malva neglecta Wallr. — просвирник пренебреженный. Сорное,
от устья Кондары до Руидашта. Съедобно (салатный овощ).
Сем. Guttiferae
459. Hypericum scabrum L. — зверобой шероховатый. Обычное на
сухих каменистых склонах. Красильное растение.
460. Н. perforatum L. — з. продырявленный. Часто на опушках кле-
новников. Дает желтую краску. Лекарственное.
461. Н. elongatum Ldb. — з. вытянутый. На сухих щебнистых скло-
нах, до 2300 м. Обычен.
462. Elatine triandra Schkuhr. — повойничек трехтычинковый. В за-
стоявшихся водоемах Руидашта.
Сем. Tamaricaceae
463. Tamarix arceuthoides Bge. (=T. turkestanica Litw.) — гребен-
щик можжевеловый. В нижней части ущелья Кондары.
464. Myricaria alopecuroides Schrenk — мирикария лисохвостнико-
вая. По берегам Кондары. Лекарственное.
Сем. Violaceae
465. Viola Fedtschenkoana W. Bckr. — фиалка Федченко. На осыпях
близ устья Кондары.
466. V. turkestanica Rgl. et Schmalh. — ф. туркестанская. На юго-
западных склонах холмов Руидашта.
467. V. occulta Lehm. — ф. скрытая. На щебнистых склонах право-
бережного хребта, до 1200 м, в группировках эфемеров.
468. V. suavis М. В. — ф. приятная. В тенистых кленовых лесах,
у ручьев. Обычное.
Сем. Datiscaceae
469. Datisca cannabina L. — датиска коноплевая. Обычное по бере-
гам Кондары и ее притоков. Красильное, лекарственное и волокнистое
растение.
Сем. Thymeleace^ae
470. Diarthron vesiculosum (Fisch, et Mey.) С. A. M.—двучленник пу-
зырчатый. На щебнистых склонах.
Флора ущелья Кондара
61
Сем. Onagraceae
471. Epilobium hirsutum L. — кипрей мохнатый. Обычное по берегам.
472. Е. adnatum Gros. — к. сродный. На галечнике по берегам Кон-
дары и на карнизах скал у воды.
473. Е. nervosum Boiss. et Buse. — к. жилковатый. На склонах
у ключей.
474. Е. minutiflorum Hausskn. — к. мелкоцветковый. По берегам
Кондары, близ ее устья.
475. Е. palustre L. — к. болотный. В низовьях Кондары.
476. Ghamaenerium angustifolium (L.) Scop. — хаменериум узко-
листный. В высокотравных кленовниках уроч. Квак и на правом берегу
Кондары, от 1400 м.
Многие виды рода Epilobium — лекарственные и медоносные растения.
Сем. Umbelliferae
477. Eryngium Biebersteinianum Nevski — синеголовник Биберштейна.
В низовьях Кондары, на щебнистых склонах.
478. Е. macrocalyx Schrenk — с. крупночашечный. На лесных лу-
жайках северных склонов, в среднем течении Кондары.
479. Physocaulis nodosus (L.) Tausch. — вздутостебельник узло-
ватый. В ореховых рощах по берегам Кондары.
480. Torilis arvensis (Huds.) Link. — пузырник полевой. На южном
склоне среди кустарников.
481. Scandix pecten Veneris L. — скандикс венерин гребень. На скло-
нах в группировках эфемеров. Обычное.
482. S. stellata Soland. — с. звездчатый. На щебнистых солнечных
склонах и на залежах.
483. Caucalis leptophylla L. — прицепник тонколистный. На южном
склоне левобережного хребта в эфемерово-югановом миндальнике.
484. Turgenia latifolia Hoffm. — тургения широколистная. На щеб-
нистых склонах в группировках эфемеров.
485. Kozlovia paleacea (Rgl. et Schmalh.) Lipsky — козловия плен-
чатая. На вершине правобережного хребта, у залежей.
486. Scaligeria allioides (Rgl. et Schmalh.) Boiss. — скалигерия луко-
видная. Обычное на сухих щебнистых склонах в группировках эфемеров.
487. S. hirtula (Rgl. et Schmalh.) Lipsky — с. шершавая. На склонах
холмов Руидашта и в кленовниках северных склонов.
488. S. bucharica Е. Ког. — с. бухарская. На щебнистом склоне пра-
вобережного хребта, перед уроч. Квак, в кустах клена.
489. S. alaica (Lipsky) Е. Кот. — с. алайская. От устья Кондары
до Руидашта. Изредка.
490. Prangos pabularia Lindl. — прангос кормовой. Обычное на сухих
щебнистых и каменистых склонах. Образует ассоциации.
491. Р. bucharica В. Fedtsch. — п. бухарский. В нижней части
ущелья, до 1200—1300 м, по берегам Кондары и ближайшим к ним
склонам.
492. Bupleurum exalatum М. В. — володушка высокая. На скали-
стых склонах. Обычное. Медоносное и лекарственное.
493. Carum carvi L. — тмин обыкновенный. В нижней части ущелья,
у арыков.
62
В. В. Письяукова
494. Bunium Capusii (Franch.) Е. Kor. — буниум Кашо. На щебни-
стых склонах правобережного хребта, до 1300 м.
495. В. chaerophylloides (Rgl. et Schmalh.) Drude — б. бутеневидный.
В разнотравных кленовниках северных склонов. Обычное.
496. В. persicum (Boiss.) В. Fedtsch. — б. персидский. На травяни-
стых склонах холмов Руидашта.
Виды р. Bunium — лекарственные и медоносные растения.
497. Galagania fragrantissima Lipsky — галагания вонючая. На щеб-
нистых травянистых склонах. Обычное.
498. Korshinskya Olgae (Rgl. et Schmalh.) Lipsky — коржинския
Ольги. Обычное на склонах в кленовниках.
499. Pimpinella affinis Ldb. — бедренец родственный. На каменистых
склонах в долине Кондары в тени кустов.
500. Aegopodium tadshikorum Schischk. — сныть таджикская. В вы-
сокотравных кленовниках северных склонов от 1500 м (главным
образом на Кваке). Медонос, краситель, лекарственное.
501. Seseli macrophyllum Rgl. et Schmalh. — жабрица крупнолистная.
На склонах, от 1800 м и выше.
502. Ligusticum discolor Led. — лигустикум разноцветный. В высоко-
травных кленовниках уроч. Квак.
503. Ferula ovina Boiss. — ферула овечья. На каменистых осыпях
от Квака к Руидашту, от 1600 м.
504. F. Jaeschkeana Vatke — ф. Яшке. На склонах обычное. Лекар-
ственное.
505. Ferula sp. — На южном склоне, от 1900 до 2000 м.
506. Ferula sp. — На щебнистых склонах вместе с F. kokanica.
507. F. kokanica Rgl. et Schmalh. — ф. коканская. Обычное на скло-
нах.
508. Haracleum Lehmannianum Bge. — борщевик Лемана. По бере-
гам ключей у Кондары.
509. Zozimia tragioides Boiss. — зосимия. На скалистых уступах
(от 1400 м) Квака и восточного края Руидашта.
510. Daucus carota L. —морковь дикая. Сорное в низовьях Кондары.
Медоносное.
511. Astomatopsis galiocarpa Eug. Kor. — астоматопсис. На щеб-
нистых склонах Руидашта, от 2300 м.
512. Aulacospermum simplex Rupr. — бороздоплодник простой.
Там же.
513. Eremodaucus Lehmannii Bge. — эремодаукус Лемана. У огорода»
на усадьбе станции.
Сем. Primulaceae
514. Primula auricalata Lam. — примула ушастая. Руидашт. На суб-
альпийских осоковых лужайках. Медонос.
515. Dionysia involucrata Zapr. — дионисия прикрытая (рис. 9).
На скалах в долине Кондары.
516. Anagallis arvensis L. — очный цвет полевой. Часто, по склонам.
Медонос. Лекарственное.
Сем. Plumbaginaceae
517. Acantholimon alatavicum Bge. — акантолимон алатавский. На
склонах Руидашта.
Флора ущелья Кондара
6&
Сем. Oleaceae
518. Fraxinus potamophila Herd. — ясень. По берегам р. Кондара,.
в ее низовьях.
Рис. 9. Дионисия прикрытая — Dionysia involucrata Zapr.
Сем, Gentianaceae
519. Erythraea pulchella Fries. — эритрея красивая (золототысячник
красивый). На влажных лужайках по берегам Кондары. Изредка.
520. Gentiana Olivieri Gris. — горечавка Оливье. Обычна на склонах.
521. Swertia lactea Bge. — шверция молочнобелая. На осоково-кобре-
зиевых лужайках Руидашта.
Сем. Дросупасеаё
522. Apocynum scabrum Rus. — кендырь шероховатый. По берегам,
р. Кондара. Дает прекрасное волокно.
64
В. В. Писъяукова
Сем. Convolvulaceae
523. Cuscuta europaea L. — повилика европейская. Уроч. Квак,
в кленовниках.
524. С. Lehmanniana Bge. — п. Лемана. В огромном количестве
паразитирует всюду на травах, кустарниках и деревьях. Вреднейшее
растение.
525. С. indica (Englelm.) V. Petr. — п. индийская. В кленовниках,
выше уроч. Квак.
526. С. convallariaeflora N. Pavl. — п. ландышецветная. На травах,
по берегам Кондары.
527. С. cupulata Engelm. — п. бокальчатая. На залежи, по южному
склону левобережного хребта.
528. С. stenocalycina Palib. — п. узкочашечковая. На Руидаште.
529. Convolvulus subhirsutus Rgl. et Schmalh. — вьюнок полушерша-
вый. На сухих южных склонах в нижней части ущелья. Редко. Медонос.
530. С. arvensis L. — в. полевой. Сорное. Медонос.
531. С. pseudocantabrica Schrenk — в. ложнокантабрийский. На юж-
ных щебнистых склонах в нижней части ущелья. Обычное.
Сем. Boraginaceae
532. Trichodesma incanum (Bge.) DC. — триходесма серая. На сухих
щебнистых склонах.
533. Lindelophia anchusoides (Lindl.) Lehm. — линделофия анхузо-
видная. Так же, как и предыдущее. Обычное растение.
534. Solenanthus petiolaris DC. — трубкоцветник черешковый. Уроч.
Квак, в кленовниках.
535. S. coronatus Rgl. — т. увенчанный. Обычен в кленовниках север-
ных склонов.
536. S. Olgae Rgl. et Smirn. — т. Ольги. У тропы на Квак, в кустах
клена.
537. Trachelanthus hissaricus Lipsky — трахелантус гиссарский.
На травянистых лесных опушках правобережного хребта, у его вершины.
538. Cynoglossum denticulatum DC. — чернокорень зубчатый. На ка-
менистых южных склонах в нижней части ущелья.
539. Paracaryum himalayense (KL) Clarke — паракариум гималай-
ский. На сухих склонах у входа в ущелье.
540. Rindera cyclodonta Bge. — риндера. На скалистых уступах лево-
бережного хребта, от 1900 м.
541. ‘Echinospermum microcarpum Led. — липучка мелкоплодная.
На склонах в нижней части ущелья.
542. Е. Drobovii М. Pop. — л. Дробова. На склонах холмов Руи-
дашта. Обычна.
543. Asperugo procumbens L. — острица лежачая. Обычное в лесах.
Медоносное.
544' . Anchusa itsrlica Retz. — воловик итальянский. Обычное на сухих
склонах.
545. Myosotis caespitosa Schultz — незабудка дернистая. Обычна
на склонах. Медонос.
546. М. palu&tris (L.) With. — н. болотная. На заболоченных лужай-
ках, у ключей.
Флора ущелья Кондара
65
547. Onosma Barsczewskii Lipsky — оносма Барщевского. На скло-
нах, от 1400 м и выше. Красильное и медоносное.
548. Echium italicum L. — синяк итальянский. В нижней части
ущелья Кондара. Медоносное, красильное.
549. Rochelia disperma (L.) Wettst. — рохелия двусемянная. На мел-
коземистых склонах холмов Руидашта.
550. R. leiocarpa Led. — р. голоплодная. Сорное. На щебнистых
склонах.
551. R. peduncularis Boiss. — р. цветоножковая. Уроч. Квак, в кле-
новниках.
552. R. cardiosepala Bge. — р. сердцевидночашелистиковая. На юж-
ном склоне левобережного хребта Кондары.
Сем. Verbenaceae
553. Verbena officinalis L. — вербена лекарственная. В нижней части
ущелья. Медоносное.
Сем. Labiatae
554. Scutellaria orientalis L. — шлемник восточный. На сухих щеб-
нистых обнаженных склонах. Обычное.
555. S. hissarica В. Fedtsch. — ш. гиссарский. На склонах. Обычное.
556. S. phyllostachya Juz. — ш. листоколосый. На склонах холмов
Руидашта. Очень часто.
557. S. leptosiphon Nevski — ш. тонкотрубчатый. На скалах. Обычно.
558. Lophanthus Lipskyanus Jk.-Gal. et Nevski — лофантус Липского.
В розарии у окраины плато Руидашт, 2300—2400 м.
559. Nepeta podostachys Benth. — котовик ножкоколосковый. Обыч-
ное на склонах холмов Руидашта.
560. N. formosa Kudr. — к. прекрасный. В высокотравных кленов-
никах уроч. Квак.
561. N. micrantha Bge. — к. мелкоцветный. На каменистых северных
склонах в долине Кондары, в тени скал.
562. N. Severzowii Rgl. — к. Северцова. На склонах среди эфемеров.
Обычное.
563. Dracocephalum formosum N. Gontsch. — змееголовник прекрас-
ный. На склонах холмов Руидашта. Медоносное.
564. Siderites montana L. — железница горная. На щебнистых скло-
нах в нижней части ущелья.
565. Lallemantia Royleana (Walk.) Benth. — лалеманция Ройля.
На щебнистых склонах. Обычное в нижней половине ущелья. Медо-
носное.
566. Brunella vulgaris L. — Черноголовка обыкновенная. По берегам
р. Кондара, в нижней половине ущелья. Изредка. Медоносное, лекар-
ственное.
567. Eremostachys hissarica Rgl. — эремостахис гиссарский. На сухих
каменистых склонах. Часто.
568. Е. Lehmanniana Bge. — э. Лемана. На склонах. Обычное.
569. Е. seravschanica Rgl. — э. зеравшанский. На каменистых скло-
нах холмов Руидашта.
570. Е. Popovii N. Gontsch. — э. Попова. На северном склоне право-
бережного хребта.
Ущелье Кондара
6-
В. В. Письяукова
571. Е. labiosa М. Pop. — э. крупногубый. На глинистых склонах,
до 1400 м.
572. Е. spectabilis М. Pop. — э. великолепный. Вместе с предыдущим.
573. Е. speciosa Rupr. — э. прекрасный. На каменистых осыпях
у Руидашта, 2200 м.
574. Е. arctiifolia М. Pop. — э. лопухолистный. Уроч. Квак, на лес-
ных опушках.
575. Е. glanduligera М. Pop. — э. железистый. На каменистых юго-
восточных склонах холмов Руидашта.
576. Phlomis fruticetorum N. Gontsch. — зопник кустарниковый. На
каменистых склонах у Руидашта и на самом плато. Медоносное.
577. Ph. canescens Rgl. — з. седоватый. На западных склонах Руи-
дашта.
578. Lamium album L. — яснотка белая. По берегам Кондары и в кле-
новниках Квака. Медоносное.
579. Leonurus turkestanicus V. Krecz. et Kupr. — пустырник турке-
станский. В лесах уроч. Квак и Тереакба, 1500—1800 м. Медонос.
580. Stachyopsis oblongata (Schrenk) М. Pop. et Vved. — стахиопсис
продолговатый. Обычен на Руидаште.
581. Stachys hissarica Rgl. — чистец гиссарский. На каменистых
осыпях. Часто.
582. Salvia sclarea L. — шалфей мускатный. На сухих южных и
юго-восточных склонах в миндальниках. Обычнейшее растение.
583. S. virgata Ait. — ш. прутьевидный. На лесных сухих опушках.
Медоносное.
584. Ziziphora clinopodioides Lam. — зизифора. На щебнистых скло-
нах. Очень распространенное растение.
585. Melissa officinalis L. — мелисса лекарственная. На южных
склонах, обычное. Иногда образует фон (Яблоновый лес в уроч. Танге-
мур). Лекарственное и медоносное.
586. Satureja clinopodium (Garuel.) Spenner — чабер. В тенистых
орехово-кленовых лесах урочища Тереакба. Лекарственное и медоносное.
587. S. rotundifolia (Pers.) Briq. — ч. круглолистный. В группиров-
ках эфемеров в нижней части ущелья.
588. Hyssopus seravschanicus Pazij — гиссоп зеравшанский. На каме-
нистых и щебнистых склонах. Обычное. Лекарственное и медоносное.
589. Origanum tyttanthum N. Gontsch. — душица мелкоцветная.
Обычнейшее на мелкоземи. гшх склонах. Лекарственное и медо-
носное.
590. Lycopus europaeus L. — зюзник европейский. По берегам Кон-
дары в ее низовьях. Медоносное.
591. Mentha silvestris L. — мята лесная. Обычнейшее по берегам
Кондары и ее притоков.
Сем. Solanaceae
592. Hyoscyamus niger L. —белена черная. Уроч. Квак, близ огорода.
Сорное. Лекарственное.
Сем. Scrophulariaeeae
593. Verbascum songoricum Schrenk — коровяк джунгарский. На
сухих склонах.
Флора ущелья Кондара
67
594. V. bactrianum Bge. — к. бактрийский. На скалистых уступах
под Руидаштом.
595. V. turkestanicum Franch. — к. туркестанский. На щебнистых
склонах, при устье Кондары.
596. Gelsia heterophylla Desf. — цельсия разнолистная. По берегам
Кондары в ее низовьях.
597. Linaria Popovii Kupr. — льнянка Попова. На щебнистых скло-
нах. Изредка.
598. Scrophularia incisa Weinm. — норичник разрезной. На склонах
холмов Руидашта.
599. S. pinnata Каг. et Kir. — н. перистый. Вместе с предыдущим.
600. S. alata Gilib. — н. крылатый. В прибрежных зарослях по Кон-
даре. Медоносное.
601. Limosella aquatica L. — лужница водяная. На лужайках у клю-
чей в восточной части Руидашта.
602. Veronica verna L. — вероника весенняя. На Руидаште.
603. V. arvensis L. — в. полевая. По тропам у берегов р. Кондара.
604. V. nudicaulis Каг. et Kir. — в. голостебельная. На лужайках
у ключей Руидашта.
605. V. biloba L. — в. двулопастная. Обычна от Квака до Руидашта,
606. V. bucharica В. Fedtsch. — в. бухарская. На южных склонах
в группировках эфемеров.
607. V. cardiocarpa (Каг. et Kir.) Walp. — в. сердцевидноплодная.
На щебнистых склонах. Обычнейшее растение.
608. V. Tournefortii Gmel. — в. Турнефортова. На левом берегу
Кондары, близ метеорологической будки.
609. V. anagallis L. — в. водяная. На болотцах у ключей в долине
Кондары.
610. V. Beccabunga L. — в. поручейная. У ключей на Руидаште.
611. V. capillipes Nevski — в. волосовидная. На южных склонах
в группировках эфемеров. Обычное. Все представители рода Vero-
nica — медоносные растения.
612. Lagotis Korolkovii (Rgl. et Schmalh.) Maxim. — лаготис Король-
кова. На субальпийских лужайках Руидашта. Обычнейшее.
613. Leptorhabdos micrantha Schrenk — лепторабдос мелкоцветная.
На. травянистых склонах. Обычное.
614. Euphrasia tatarica Fisch. — очанка татарская. На лужайках
от Квака до Руидашта.
615. Pedicularis pycnantha Boiss. — мытник. На северных склонах
в древесно-кустарниковых зарослях.
Сем. Orobancbaceae
616. Orobanche pamiro-alaica N. Gontsch. et Novopokr. — заразиха
памиро-алайская. В кленовниках Квака.
617. О. lutea Baumg. — з. желтая. На люцерне в нижней части Кон-
дары.
618. О. coelestis Boiss. et Reut. — з. необыкновенная. На южных
щебнистых склонах в зарослях кровохлебки с редким миндалем.
619. О. gigantea (Beck.) N. Gontsch. — з. гигантская. Всюду на кор-
нях Ferula Jaeschkeana.
620. О. Kotschyi Reuter — з. Кочи. На Prangos pabularia и Ferula
Jaeschkeana. Обычная.
5*
68
В. В. Письяукова
621. О. solenanthi Novopokr. et Pissjauk. — з. трубкоцветниковая.
Уроч. Квак, на корнях трубкоцветника (Solenanthus).
Сем. Plantaginaceae
622. Plantago major L. — подорожник большой. По берегам Кон-
дары и Квакского ручья.
623. Р. lanceolate L. — п. ланцетный. Всюду на склонах гор.
Сем. Rubiaceae
624. Crucianella exasperate Fisch, et Mey. — крестовница. Обычное
на щебнистых склонах в группировках эфемеров.
625. С. gilanica Trin. — к. гиланская. На щебнистых восточных
склонах правобережного хребта, на высоте 1200—1400 м.
626. Asperula glomerate (М. В.) Gris. — ясменник скученный. На щеб-
нистых склонах. Обычное.
627. A. arvensis L. — я. полевой. На южных склонах в группиров-
ках эфемеров и на залежах. Обычное.
628. A. oppositifolia Rgl. et Schmalh. — я. супротивнолистный. На
щебнистых сухих склонах. Обычное.
629. A. aparine Schott. — я. цепкий. В кустарниковых зарослях,
в кленовниках, в тени скал у Кондары и ее притоков. Обычное.
Представители рода Asperula являются красильными, медоносными
растениями, а некоторые и лекарственными.
630. Galium verum L. — подмаренник настоящий. На лесных опуш-
ках. Весьма часто.
631. G. tricorne With. — п. трехрогий. На южном каменистом склоне
левобережного хребта, близ уроч. Теги-камар.
632. G. Vaillantii (DC.) Koch — п. цепкий Вайана. В кустах уроч.
Квак и на Руидаште.
633. G. aparine L. — п. цепкий. По кустам в долине Кондары и ее
притоков, в тени скал и на Кваке в кленовниках.
634. G. tenuissimum М. В. — п. тончайший. На щебнистых склонах
хребтов под кустами. Часто.
635. G. Decaisnei Boiss. — п. Декэна. На скалистых карнизах в до-
лине Кондары, в тени.
636. G. verticillatum Danth. — п. мутовчатый. На северо-восточном
каменистом склоне против дома Станции.
637. G. pedemontanum АН. — п. пиемонтский. На щебнистой вершине
правобережного хребта, до 1200 м, в кустах клена.
638. G. subreflexum М. Pop. — п. полуизогнутый. В тени скал
на южном склоне, выше урочища Теги-камар.
639. Gallipeltis cucullaria (L.) Stev. — каллипельтис колпачковая*
Весьма обычна на сухих щебнистых склонах среди эфемеров.
Сем. Caprifoliaceae
640. Lonicera Korolkovii Stapf. — жимолость Королькова. По бере-
гам Кондары и на склонах всюду. Медоносное.
Флора ущелья Кондара
69
641. L. seravschanica A. Pojark. — ж. зеравшанская. Скалистые
склоны у входа на Руидашт, 2400—2500 м. Медоносное.
642. L. persica Jaub. et Spach. — ж. персидская. В кленовниках и
розариях от 1800 м. Медоносное.
Сем. Valerianaceae
643. Valerianella dactylophylla Boiss. et Hohen. — валерьянница паль-
целистная. На каменистых склонах близ урочища Теги-камар.
644. V. truncata (Rchb.) Betcke — в. обрубленная. Там же.
645. V. coronata (L.) DC. — в. венечная. Весьма обычное по склонам
в группировках эфемеров.
646. Valeriana ficariaefolia Boiss. — валериана чистяковолистная.
Весьма обычное в нижней части ущелья в группировках эфемеров. Медо-
носное.
Сем. Dipsacaceae
647. Dipsacus azureus Schrenk — ворсянка лазоревая. На северном
склоне правобережного хребта против урочища Теги-камар. На лесной
лужайке. Медоносное.
648. Scabiosa songarica Schrenk — скабиоза джунгарская. Весьма
обычное на солнечных склонах.
Сем. Campanulaceae
649. Campanula incanescens Boiss. — колокольчик седеющий. Обыч-
ное на скалах.
650. Ostrowskia magnifica Rgl. — островския величественная. В кле-
новниках и кустарниках северных каменистых склонов, до 2500 м. Кра-
сивоцветущее.
651. Phyteuma argutum Rgl. — фитеума изящная. На склонах хол-
мов Руидашта.
652. Ph. baldshuanica В. Fedtsch. — ф. бальджуанская. Обычное
в поясе древесно-кустарниковой растительности. Медоносное.
653. Codonopsis clematidea С. В. Clarke — кодонопсис ломоносовая.
Обычна в ущельях по берегам Кондары и ручьев, в нее впадающих.
Сем. Compositae
654. Solidago virga aurea L. — золотарник обыкновенный. Часто
в лесном поясе. Медоносное и лекарственное.
655. Galatella punctata Lindl. — солонечник точечный. В кустах
на вершине правобережного хребта, у тропы на Квак.
656. Myriactis Gmelini DC. — мириактис Гмелина. В заболоченной
ореховой роще на правом берегу Кондары.
657. Erigeron canadensis L. — мелколепестник канадский. В нижнем
течении Кондары, по берегам.
658. Е. pseudoerigeron (Bge.) М. Pop. — м. ложный. На влажных
скалах у водопадов.
70
В. В. Писъяукова
659. Е. Lehmanni Boiss. — м. Лемана. На склонах холмов Руи-
дашта.
660. Е. alpinus L. — м. альпийский. В розариях, на высоте от 2300 м.
661. Е. Schmalhausenii М. Pop. — м. Шмальгаузена. На Руидаште.
662. Psychrogeton cabulicum Boiss. — психрогетон кабульский.
На склонах холмов Руидашта.
663. Р. leucophyllum (Bge.) — п. белолистный. Там же.
664. Gonyza khorassanica (Boiss.) Novopokr. — кониза хорассанская.
Обычное на Руидаште.
665. Filago arvensis L. — жабинец полевой. Обычнейшее на сухих
склонах.
666. Inula grandis Schrenk — девясил громадный. Обычное на скло-
нах северной и северо-западной экспозиций, до 1900 м. Медоносное.
Кормовое (в сухом виде).
667. I. rhizocephala Schrenk — д. корнеголовый. На влажных субаль-
пийских лужайках Руидашта.
668. I. glauca С. Winkl. — д. сизый. На скалах в ущелье реки Оби-
сафед (верховья р. Кондара, около 1800 м). Медонос.
669. Pulicaria salviaefolia Bge. — блошица иволистная. На берегу
р. Варзоб близ устья Кондары.
670. Р. dysenterica Gaertn. — б. дизентерийная. У арыков в нижней
части ущелья.
671. Vicoa Albertoregelia G. Winkl. — викоа Альберта-Регеля.
На скалах у реки Оби-сафед.
672. Achillea millefolium L. — тысячелистник обыкновенный. Квак
и Руидашт.
673. A. filipendulina Lam. — т. лабазниковый. Уроч. Квак, долина
Кондары и мелкоземистые склоны левобережного хребта.
674. A. trichophylla Schrenk — т. волосолистный. Обычное по скло-
нам.
675. Xanthium strumarium L. — дурнишник обыкновенный. Сорное.
676. Matricaria disciformis (G. А. М.) DG. — ромашка дисковидная.
На Руидаште и в уроч. Квак.
677. Pyrethrum parthenifolium Willd. — пиретрум партениелистный.
Обычнейшее в кленовниках, розариях и на лесных опушках.
678. Р. setaceum Rgl. at Schmalh. — п. щетинистый. На скалах в до-
лине Кондары.
679. Р. mucronatum Rgl. et Schmalh. — п. остроконечный. На скло-
нах холмов Руидашта. Обычнейшее, местами образует фон.
680. Р. hissaricum Krasch. — п. гиссарский. На скалах, от 1800 м.
681. Tanacetum Newesskianum G. Winkl. — пижма Невесского.
В уроч. Квак на склонах и по берегам ручьев. Обычное.
682. Т. pseudoachillea С. Winkl. — п. ложнотысячелистниковая.
В кленовниках уроч. Квак.
683. Т. tadshikorum S. Kudr. — п. таджикская. На каменистых скло-
нах в долине Кондары, до 1800 м.
684. Artemisia dracunculus L.—полынь — эстрагон. На Руидаште.
685. A. persica Boiss. — п. персидская. На гальке в долине Кондары
и на каменистых склонах холмов Руидашта.
686. A. absinthium L. — п. горькая. В высокотравных кленовниках
от 1500 м до уроч. Квак включительно.
687. A. vulgaris L. — п. обыкновенная, чернобыльник. Уроч. Квак,
на щебнистых гребнях у р. Оби-сафед.
Флора ущелья Кондара
71
688. Tussilago farfara L. — мать-и-мачеха. По берегам Кондары,
на гальке. Изредка.
689. Senecio Renardi G. Winkl. — крестовник Ренарда. На каме-
нистых склонах, от 2100 м, и на Руидаште. Медоносное.
690. Ligularia Thomsonii (Clarke) Pojark. — бузульник Томсона. В вы-
сокотравных кленовниках, от 1500 м и до Руидашта. Медоносное.
691. L. altissima Pojark. — б. высочайший. Только в высокотравных
кленовниках уроч. Квак.
692. L. persica Boiss. — б. персидский. Во влажных кленовниках
от 1600 м и на лужайках Руидашта. Местами фоновое. Медоносное.
693. Echinops maracandicus Bge. — мордовник самаркандский. На
щебнистых гривах у Кондары. Изредка. Медоносное.
694. Lappa major Gaertn. — лопух большой. Только в одном месте
на заболоченной лужайке перед урочищем Тангемур, в орешнике.
695. Cousinia umbrosa Bge. — кузиния теневая. Обычное по склонам.
696. С. pseudomollis С. Winkl. —к. лжемягкая. На щебнистой вер-
шине правобережного хребта (первый и второй холмы у тропы на
Квак).
697. С. fallax С. Winkl. — к. обманчивая. На мелкоземистых склонах
холмов Руидашта, от 2400 м.
698. С. refracta Juz. — к. преломленная. На южном склоне левобе-
режного хребта вокруг урочища Теги-камар. Фоновое.
699. С. macilenta С. Winkl. —к. тощая. На склонах холмов Руи-
дашта, где образует ассоциации. Появляется по склонам от 2200 м.
700. С. radians Bge. — к. лучевая. На склонах, до 1500 м. Обычна.
701. С. acicularis Franch. — к. игловидная. На Руидаште и у входа
на плато, от 2200 м. Часто.
702. С. integrifolia Franch. — к. цельнолистая. На склонах, часто.
703. С. pulchella Bge. — к. красивенькая. В разнотравных кленовни-
ках и на травянистых северных и северо-западных склонах Обычное.
704. С. simulatrix С. Winkl. — к. подражающая. На каменистых
осыпях выше Квака. Изредка.
705. С. commixta М. Kult. — к. смешанная. На каменистых осыпях
от Квака до Руидашта. Часто.
706. С. Fedtschenkoana Bornm. — к. Федченко. У восточного края
Руидашта.
707. С. splendida С. Winkl. — к. блестящая. На Руидаште. Образует
ассоциации.
708. С. candicans Juz. — к. белесоватая. На мелкощебнистых обна-
женных осыпях уроч. Квак и Оби-сафед, 1600—1800 м.
709. Cirsium turkestanicum Rgl. — бодяк туркестанский. На Руи-
даште.
710. С. glabrifolium (С. Winkl.) В. Fedtsch. — б. гололистный. По бе-
регам ключей на Руидаште.
711. С. lanceolatum (L.) Scop. — б. лапцетолистный. По берегам
Кондары.
712. Onopordon acanthium L. — татарник обыкновенный. На сухих
щебнистых склонах в нижней части ущелья. Редко.
713. Grupina vulgaris Cass. — крупина обыкновенная. На склонах.
714. Serratula chartacea С. Winkl. — серпуха бумажная. На Руи-
даште (скалы).
715. Acroptilon picris Fisch, et Mey. — василек-горчак. Обычное
на склонах. Сорное.
72
В. В. Писъяукова
716. Centaurea squarrosa Willd. — в. оттопыренный. На щебнистых
сухих склонах, весьма обычен.
717. Rhaponticum integrifolium G. Winkl. — рапонтикум цельно-
листный. Обычное в высокотравных кленовниках и на склонах.
718. Cnicus benedictus L. — бенедикт. В группировках эфемеров
в нижней части ущелья.
719. Gerbera kokanica Rgl. et Schmalh. — гербера коканская* На щеб-
нистых склонах. Обычное.
720. Gichorium inthybus L. — цикорий дикий. На склонах. Медо-
носное, съедобное.
721. Koelpinia linearis Pall. — кельпиния линейнолистная. Обычное
в группировках эфемеров.
722. Rhagadiolus hedypnois Fisch, et Mey. — рагадиолус. Обычное
на склонах в группировках эфемеров.
723. Acanthocephalus Benthamianus Rgl. — акантоцефалус Бентама.
На травянистых склонах.
724. Heteratia Szovitsii Fisch, et Mey. — гетерация Шовица. В груп-
пировках эфемеров. Обычное.
725. Pieris hieracioides L. — горчак ястребинковидный. На склонах
холмов Руидашта. Медоносное.
726. Р. japonica Thunb. — г. японский. На лесной высокотравной
опушке северного склона правобережного хребта перед урочищем Квак.
Медоносное.
727. Tragopogon paradoxum S. Nikit. — козлобородник парадоксаль-
ный. На высокотравной лужайке у вершины левобережного хребта,
выше урочища Теги-камар. Медоносное.
728. Т. seravschanicum S. Nikit. — к. зеравшанский. На щебнистых
осыпях близ устья Кондары.
729. Т. angustissimus S. Nikit. — к. узколистный. На каменистых
склонах близ устья Кондары, в кустах. Медоносное.
730. Scorzonera hissarica G. Winkl. — козелец гиссарский. На мелко-
щебнистых склонах хребта, у тропы в уроч. Квак.
731. S. bracteosa С. Winkl. — к. прицветниковый. Часто по скло-
нам.
732. S. tragopogonoides Rgl. et Schmalh. — к. козлобородниковид-
ный. На склонах холмов Руидашта.
733. S. acanthoclada Franch. — к. колючеветвистый, теке-сагыз. На
юго-западных склонах холмов Руидашта, образует ассоциации.
734. Ghondrilla juncea L. — хрящевница ситниковая. Обычное
на сухих щебнистых склонах.
735. Taraxacum montanum (С. А. М.) DC. — одуванчик горный. На
склонах холмов Руидашта, от 2500 м.
736. Т. officinale Wigg. — о. лекарственный. На лужайках Руидашта
между холмов.
737. Т. Fedschenkoi Hand. Mazz. — о. Федченко. На склонах холмов
Руидашта, обычное.
738. Т. Wallichii DG.? — о. Уоллича. На каменистых склонах холмов.
739. Т. syriacum Boiss. — о. сирийский. На Руидаште.
740. Gicerbita rosea (Pop. et Vved.) Н. Krasch. — цицербита розовая.
Обычное в древесно-кустарниковом поясе ir на лужайках Руидашта.
741. G. lutea Н. Krasch. et N. Gontsch. — ц. желтая. В кленовниках.
742. С. songorica (Rgl.) Krasch.—ц. джунгарская. Обычное по бере-
гам Кондары.
Флора ущелья Кондара
73:
743. С. crambifolia (Bge.) Krasch. — ц. капустнолпстная. На скали-
стой вершине левобережного хребта.
744. Sonchus palustris L. — осот болотный. На склонах с сочащейся
водой в долине Кондары. Медоносное.
745. S. oleraceus L. — о. огородный. Сорное в нижней части ущелья^
746. S. asper Vill. — о. колючий. Сорное в пределах усадьбы Станции.
747. Lactuca scariola L. — латук компасный. На сухих склонах^
обычное. Сорное.
748. L. auriculata DC. — л. ушкастый. Уроч. Квак и под Руидаш-
том, в розарии. Медоносное.
749. L. undulata Led. — л. поникающий. На южных склонах в тени
скал, до 1300 м.
750. L. orientalis Boiss.—л. восточный. На скалистых обрывах под,
Руидаштом. Медоносное.
751. Crepis pulchra L. — скерда красивая. Обычнейшее на склонах
в группировках эфемеров.
752. С. sibirica L. — с. сибирская. В лесах урочища Квак и на Руи-
даште.
753. Barkhausia trichocephala Krasch. — баркгаузия щетинистоголо-
вая. На щебнистом склоне в устье Кондары.
754. В. Bureniana (Boiss.) Krasch. — б. Буре. На щебнистых
склонах, изредка.
755. Hieracium procerum Fries. — ястребинка высокорослая. На скло-
нах в древесно-кустарниковых зарослях.
756. Н. virosum Pall. — я. ядовитая. На скалах с сочащейся водой^
на высоте 1200 м до 2000 м. Ядовитое растение.
ЛИТЕРАТУРА
Гончаров Н. Ф. и П. Н. Овчинников. 1935. Основные черты послетре-
тичной истории растительности зап. Памиро-Алая. Сов. ботан., № 6.
Невский С. А. 1937. Материалы к флоре Кугитанга и его предгорий. Тр. Инет,
ботаники АН СССР, сер. 1, вып. 4.
Павлов Н. В. и С. Ю. Липшиц. 1934. Эскиз флористических элементов Сыр-
дарьинского Каратау. Сов. ботан., К® 1.
Письяукова В. В. 1950. Schoenoxiphium hissaricum — представитель нового,
для флоры СССР рода. Бот. матер. Гербария Бот. инет. им. В. Л. Комарова
АН СССР, т. XII.
Попов М. Г. 1927. Основные черты истории развития флоры Средней Азии. Бюлл.
Ср.-Аз. Гос. унив., вып. 15.
Попов М. Г. 1938. Основные периоды формообразования и миграций во флоре
Средней Азии. Проблема реликтов во флоре СССР (Тезисы совещания).
Попов М. Г. 1940. Флора Алмаатинского Государственного заповедника. Тр.
Алмаат. заповеди., вып. III.
Попов М. Г. 1941. Географо-генетические элементы флоры Алмаатинского запо-
ведника. Растит. Казахстана, т. II.
Пояркова А. И. 1950. Новый вид папортника и вопрос о гималайских элемен-
тах во флоре Средней Азии. Сообщения Тадж. фил. АН СССР, вып. XXII.
Пояркова А. И. 1950. Материалы к систематике и истории рода Ligularia. Ботан.
матер. Бот. инет. им. В. Л. Комарова АН СССР, т. XII.
Рубцов Н.П. 1941. М. Г Попов. Флора Алмаатинского заповедника. Реферат,,
Сов. ботан., № 3.
А. С. КОРОЛЕВА
РАСТИТЕЛЬНЫЙ ПОКРОВ КОНДАРЫ
Растительность ущелья Кондара в общих чертах не представляет
исключения среди растительных ландшафтов южного склона Гиссар-
окого хребта. Однако широко распространенное в Гиссарском хребте
явление эндемизма определяет известную видовую обособленность расти-
тельности Кондары (что свойственно и другим участкам южного склона
Гиссарского хребта). Растительность древесно-кустарникового пояса
Кондары по своему общему характеру мало отличается от раститель-
ности аналогичного пояса смежных районов южного склона Гиссарского
хребта. Следует однако отметить, что в Кондаре среди древесных пород
отсутствует Exochorda Alberti Rgl., Fraxinus raibocarpa Rgl. и другие
виды, распространенные в районах, расположенных западнее и восточнее.
Что касается субальпийского пояса Кондары, то он является довольно
своеобразным участком южного склона хребта, характеризуясь присут-
ствием ряда кузиний, имеющих очень ограниченный ареал и не встре-
чающихся на большей части территории Гиссарского хребта (Gousinia
macilenta G. Winkl., G. splendida G. Winkl.).
Растительность Кондары развивается в климатических условиях,
характеризующихся наличием летней засухи. Общее количество осадков
в районе Кондары довольно значительно (достигает 1000 мм в год), но
выпадение их приурочено к зимне-весеннему периоду, летние же месяцы
и большая часть осени совершенно бездождны. Подобный характер рас-
пределения осадков обусловливает ряд своеобразных особенностей расти-
тельности. Большая часть травянистых растений отличается той или
иной степенью эфемеризации. При широком развитии эфемеров-однолет-
ников и типичных многолетних эфемероидов значительное число осталь-
ных многолетних видов носит эфемероидный характер и с наступлением
летней засухи более или менее быстро начинает отмирать. Во время
засушливого летнего периода продолжает вегетировать лишь очень
незначительная группа растений, теми или иными способами приспособ-
ленных к перенесению засухи. Благодаря засушливости лета, большая
часть древесных насаждений Кондары отличается разреженностью
древесного полога. Эта особенность свойственна на юге Средней Азии
почти всем ассоциациям сухих склонов, увлажняемых только атмосфер-
ной влагой (см. ст. В. И. Запрягаевой в этом сборнике). Сомкнутый дре-
весный полог наблюдается лишь в некоторых насаждениях долин, где
почва постоянно увлажняется грунтовыми «и речными водами.
Летняя засуха, сдвигающая вегетацию растительности на обильные
осадками весну—начало лета, наступает в различных частях ущелья
Кондары далеко не одновременно, что связано с довольно большими
Растительный покров Кондары
75
различиями абсолютных высот в отдельных частях склонов. В нижнем
течении р. Кондара (высота 1100 м1) разгар вегетации растительности
падает на апрель—май, в верхней части и на водоразделе между
р. Кондара и р. Лючоб (высота 2600 м) — на июнь—июль.
Склоны, на которых развивается растительность, характеризуются,
как и другие участки южного склона Гиссарского хребта, широким раз-
витием щебнистых и каменистых россыпей и скал, почти лишенных поч-
венного покрова. Мелкоземистые, более или менее глубокие почвы зани-
мают подчиненное положение.
Рис. 1. Северный склон ущелья Кондара
с широким распространением кленовиков
и южный склон с разреженной эфеме-
рово-югановой растительностью и единич-
ными деревьями миндаля.
(Фот. Варзобской ботанической станции).
Крутизна склонов, покрытых
растительностью, обычно колеб-
лется в пределах 30—35(40)°.
Нередко встречаются и более кру-
тые склоны и обнажения корен-
ных пород, где растительность
почти отсутствует.
Северный и южный склоны
ущелья Кондара довольно резко
отличаются друг от друга по ха-
рактеру мезоклиматических усло-
вий. Южные склоны нагреваются
сильнее, чем северные, и больше
испаряют, в результате чего к на-
чалу засухи запасы влаги на юж-
ных склонах оказываются значи-
тельно меньшими, чем на север-
ных. Поэтому южные склоны ха-
рактеризуются и более разрежен-
ным травяным покровом, что, в
свою очередь, не способствует
накоплению мелкозема. Исклю-
чительное развитие получают здесь
скелетные, щебнистые и камени-
стые почвы, растительный покров
которых представлен разрежен-
ными травянистыми ассоциациями
преимущественно эфемерного и эфе-
мероидного типа. Среди деревьев
и кустарников на южных скло-
нах преобладают ксерофитные породы, встречающиеся в виде единичных
или редких экземпляров и лишь иногда образующих светлые, несомкну-
тые насаждения. Мелкоземистые участки, покрытые мезофитными дре-
весно-кустарниковыми насаждениями, на южном склоне редки и приуро-
чены преимущественно к тальвегам и нижним частям склонов боковых
ущелий.
На северном склоне, при широком распространении щебнистых суб-
стратов, занятых травянистой эфемерной и эфемероидной раститель-
ностью, видное значение приобретают мелкоземистые почвы и связанные
с ними мезофитные древеснокустарниковые ассоциации (рис. 1).
Большая часть территории склонов ущелья Кондара находится
в поясе древесно-кустарниковой растительности, нижняя граница кото-
1 Здесь и в дальнейшем высота дана в метрах над ур. м.
76
А. С. Королева
рого лежит за пределами ущелья, на высоте 900—1000 м, а верхняя
поднимается до 2400 м. Лишь верхняя часть бассейна Кондары распо-
ложена в поясе субальпийской растительности, простирающейся от
высоты 2400 м до 3200 м.
ПОЯС ДРЕВЕСНО-КУСТАРНИКОВОЙ РАСТИТЕЛЬНОСТИ
Пояс древесно-кустарниковой растительности в районе Кондары,
так же как и по всему южному склону Гиссарского хребта, подразде-
ляется на две полосы — нижнюю и верхнюю. Нижняя полоса отличается
от верхней присутствием боярки дулоны, махалебской вишни, клена
Регеля, фисташки и др., не встречающихся в верхней полосе, измене-
ниями в составе травянистой растительности, а также более низкорослым
туркестанским кленом.
НИЖНЯЯ ПОЛОСА ПОЯСА ДРЕВЕСНО-КУСТАРНИКОВОЙ РАСТИТЕЛЬНОСТИ
Нижняя полоса пояса древесно-кустарниковой растительности под-
нимается до 1600м. На северных склонах среди древесных пород наиболее
широко распространен туркестанский клен (Acer turkestanicum Pax),
высота которого обычно не превышает здесь 13—14 м. К туркестан-
скому клену присоединяются алыча (Prunus divaricata Ldb.), боярка
туркестанская (Crataegus turkestanica Pojark.), махалебская вишня
[Cerasus Mahaleb (L.) Mill.], крушина длиннолистная (Rhamnus dolicho-
phylla N. Gontsch.), яблоня (Malus hissarica Kudr.), каркас (Celtis
caucasica Willd.), арча (Juniperus seravschanica Korn.).
На участках с разреженным древесным пологом нередко встречаются
миндаль бухарский (Amygdalus bucharica Korsh.), клен Регеля (Acer
Regelii Pax.).
На открытых местах изредка можно видеть боярку-дулону (Crataegus
pontica С. Koch), единично встречается фисташка (Pistacia vera L.).
Среди кустарников на северных склонах наиболее обычны: шиповник
кокандский (Rosa kokanica Rgl.), шиповник собачий (Rosa canina L.),
жимолость персидская (Lonicera persica Jaub. et Spach.), иргай (Coto-
neaster suavis Pojark.).
В нижней части ущелья изредка встречается пузырник (Colutea per-
sica Boiss.) и атрафаксис (Atraphaxis pyrifolia Bge.), приуроченный
преимущественно к долине Кондары. Грецкий орех (Juglans regia L.)
обитает в этой полосе почти исключительно в долине этой реки и в доли-
нах ее боковых притоков, лишь изредка заходя на нижние части склонов.
В долине Кондары и в долинах ее боковых притоков широко распро-
странен виноград (Vitis vinifera L.), часто встречаются различные виды
ив (Salix), ежевика (Rubus caesius L.), алыча, боярка, иргай, жимолость
Королькова (Lonicera Korolkowi Stapf.), шиповник собачий (Rosa cani-
na L.) и др.
Древесно-кустарниковые ассоциации северных склонов, состоящие
из клена с большей или меньшей примесью перечисленных пород, приуро-
чены преимущественно к склонам долин боковых ущелий и нижним частям
склонов к Кондаре.
Травяной покров этих ассоциаций варьирует в зависимости от сте-
пени сомкнутости древесного и кустарникового полога. Там, где почва
наиболее сильно затенена, развивается ассоциация ежово-разнотравно-
недотрогового кленовника, в травяном покрове которого на фоне пре-
Растительный покров Кондары
77
обладающей недотроги (Tmpatiens parviflora DC.) лишь в сравнительно
небольших количествах встречаются некоторые другие виды. Ежово-
разнотравно-недотроговые кленовники довольно часто встречаются на се-
верных склонах нижней полосы пояса древесно-кустарниковой расти-
тельности, где они занимают обычно нижние части склонов и тальвеги
сухих боковых долинок. Изредка на пониженных участках тальвегов
встречается ассоциация разнотравно-снытевого кленовника, характери-
зующегося преобладанием в травяном покрове сныти (Aegopodium tadshi-
corum Schischk.).
Наиболее широко распространенной древесно-кустарниковой ассоциа-
цией северных склонов являются ежово-девясилово-разнотравные кле-
новники. Эта ассоциация, отличающаяся сравнительно разреженным
древостоем, встречается в средних и верхних частях склонов боковых
ущелий и на склонах к Кондаре. В составе травяного покрова ежово-
девясилово-разнотравных кленовников недотрога отсутствует или встре-
чается в незначительных количествах у стволов деревьев, где больше
всего тени.
Наряду с видами, обычными и в ежово-разнотравно-недотроговых
кленовниках, здесь появляется ряд новых, из которых наиболее харак-
терен девясил (Inula grandis Schrenk), образующий эффектные, крупные
куртины. Нередко встречается и юган (Prangos pabularia Lindl.).
В верхних частях северного склона небольшое распространение имеет
юганово-разнотравная ассоциация, являющаяся переходной от ежово-
девясилово-разнотравных кленовников к разреженной эфемерово-югано-
вой растительности щебнистых участков.
В нижних частях склонов к Кондаре изредка встречается ассоциация
с преобладанием иргая (Gotoneaster suavis Pojark.), которая по составу
травяного покрова близка к ежово-девясилово-разнотравным кленовни-
кам. Иргайники такого типа развились, повидимому, вследствие уничто-
жения человеком клена, отдельные деревья и пни которого здесь можно
нередко видеть.
В нижних и средних частях склонов к Кондаре и ее боковым прито-
кам сравнительно небольшими пятнами встречается разнотравно-пажит-
никовая ассоциация с разреженным древостоем из туркестанского клена
и миндаля. На широко распространенных щебнистых и каменистых
участках северных склонов нижней полосы господствует разреженная
травянистая растительность с преобладанием эфемеров, эфемероидов
и многолетников ксерофитного характера. Изредка встречаются здесь
также отдельные деревья и кустарники, главным образом миндаль, кар-
кас, иргай, иногда арча. Часто можно видеть каменистые обнажения
и россыпи, почти лишенные растительного покрова, представленного
лишь единичными экземплярами ревеня (Rheum Maximoviczi A. Los.),
камоля (Ferula Jaeschkeana Vatke) и др.
На южных склонах нижней полосы пояса древесно-кустарниковой
растительности до высоты 1400—1500 м господствуют различные варианты
эфемерово-югановых ассоциаций, характеризующихся то обилием ширяша
(Eremurus comosus О. Fedtsch.) — эфемерово-ширяшево-югановая ассо-
циация, — то широким распространением кузинии преломленной (Сои-
sinia refracta Juz.). На более щебнистых участках развивается эфемерово-
чеснеево-югановая ассоциация, где наряду с чеснеей (Chesneya hissarica
А. Вог.) обычно присутствуют и другие элементы щебнистых субстратов,
как гвоздика (Dianthus tetralepis Nevski), шлемник (Scutellaria orientalis
L.) и др.
78
А. С. Королева
Эфемерово-чеснеево-югановая ассоциация поднимается и выше 1400 м,
до скал, завершающих на южных склонах нижнюю полосу пояса дре-
весно-кустарниковой растительности.
Деревья и кустарники, встречающиеся среди эфемерово-югановых
ассоциаций, представлены единичными экземплярами миндаля бухар-
ского, каркаса, клена Регеля (Acer Regeli Pax), шиповника самар-
кандского (Rosa maracandica Bge.)- В нижних частях долины часто
встречаются россыпи и скалы, в разреженном покрове которых часты
ширяш хохлатый (Eremurus comosus), камоль (Ferula Jaeschkeana Vatke),
ревень Максимовича, осока (Carex turkestanica Rgl.). Здесь же часто
встречаются единичные деревья миндаля. Подобного типа субстраты и
связанная с ними растительность спускаются и ниже указанной высоты
до нижнего предела пояса.
Мезофитные древесные и кустарниковые породы (клен туркестанский,
жимолость персидская и др.) распространены на южных склонах сравни-
тельно редко, преимущественно в нижних частях склонов к Кондаре
или по склонам боковых ущелий. Они образуют обычно небольшие,
сравнительно редкие насаждения, в составе которых постоянно присут-
ствуют и ксерофитные породы, как бухарский миндаль и каркас. Травя-
ной покров подобных ассоциаций (кленово-миндальная ассоциация
с ячменно-эфемерово-разнотравным травяным покровом) состоит из эфе-
меров и ряда многолетников, среди которых наиболее характерны луко-
вичный ячмень (Hordeum bulbosum L.), ежа сборная (Dactylis glome-
rata L.), девясил, иногда эремостахис гиссарский (Eremostachys hissarica
Rgl.). Лишь кое-где в долинах боковых ущелий и иногда на выходах
коренных пород и каменистых россыпях у гребня хребта ютятся неболь-
шие пятна густых кленовых и ореховых лесов, в травяном покрове кото-
рых преобладает недотрога, купена (Polygonatum Sewerzowi Rgl.), встре-
чаются островския (Ostrowskia magnifica Rgl.) и другие типично лес-
ные растения.
Растительность долины р. Кондара несколько отличается от расти-
тельности склонов. Долина Кондары, как и большей части горных рек,
очень не широка. Еще уже долины ее боковых притоков. Местами склоны
в виде отвесных обнажений сдвигаются, оставляя лишь узкое в 1—2 м
шириной ложе, где бежит река. Благодаря большому падению дна, ложе
реки часто образует уступы, по которым стремительно мчится поток
в виде живописных каскадов и водопадов. На нависших над водой ска-
лах, по окраинам водопадов, где нет потока, но куда постоянно дости-
гают водяные брызги, нередко встречаются куртины изящных папоротни-
ков, мхи и отдельные экземпляры покрытосемянных растений. У воды,
между скалами, куда почти не проникают лучи солнца, изредка образует
небольшие заросли ароматичная Geranium Robertianum L.
На большей части протяжения долины склоны раступаются, оставляя
неширокую пойму, 10—50 м. шириной. Здесь нередко встречаются
небольшие рощи грецкого ореха с травяным покровом из недотроги,
однолетних костров и некоторых других растений. Часты ивы, шипов-
ники, обильно плодоносящая алыча, яблони, густо перевитые дикой
виноградной лозой, образующие местами почти непроходимые заросли.
В среднем течении Кондары встречаются рощицы тополя (Populus tad-
shikistanica Кош.).
В полосе, прилегающей к руслу реки, в течение всего лета зеленеет
бордюр из коротконожки [Brachipodium silvaticum (Huds.) Roem. et
Schult.], ежевики, мяты (Mentha silvestris L.), высятся мощные заросли
Растительный покров Кондары
70
датиски (Datisca cannabina L.). На приподнятых, часто щебнистых тер-
расах в травяном покрове преобладают эфемеры и эфемероиды [однолет-
ние костры, бедренец (Pimpinella affinis Ld.), линделофия (Lindelophia
anchusoides (Lindl.) Lehm.J и др., выгорающие с наступлением летней
засухи. Здесь же часто встречаются кусты атрафаксиса. Местами зна-
чительные участки покрыты галечниками, почти лишенными раститель-
ности.
Переходим к описанию отдельных растительных ассоциаций.
Северный склон
1. Нсдотрогово-снытевые кленовники (Aceretum impatientoso-aego-
podiosum). Н едотрогово-снытевые кленовники являются наиболее мезо-
фитной древесно-кустарниковой ассоциацией северных склонов. В ниж-
ней полосе древесно-кустарниковой растительности они встречаются
крайне редко, занимая небольшие пониженные участки по тальвегам
сухих боковых долин. Почвы недотрогово-снытевых кленовников мелко-
земисты, нередко с крупными каменистыми глыбами. Господствующей
древесной породой является туркестанский клен, к которому обычна
примешиваются каркас, алыча, иргай и др. Густота древесного полога
достигает 60—100%. Травяной покров в разгар вегетаций в первой поло-
вине июня зелен и густ, покрывая 100% поверхности почвы. На фона
преобладающей сныти разбросаны немногочисленные другие виды. Среди
последних наиболее характерна недотрога.
2. Ежово-разнотравно-недотроговые кленовники (Aceretum dactyloso-
herboso-impatientosum). Ежово-разнотравно-недотроговые кленовники
довольно широко распространены в нижней полосе пояса древесно-
кустарниковой растительности. Эта ассоциация располагается на местах,,
повидимому, несколько менее увлажняемых весной, чем недотрогово-
снытевые кленовники, занимая ровные, более или менее мелкоземистыа
участки тальвегов сухих боковых ущелий и нижние части прилегающих
склонов. По сомкнутости древесного полога недотроговые кленовники
приближаются к недотрогово-снытевым кленовникам. Однако, как пра-
вило, сомкнутость полога здесь несколько меньше и не превышает
60—80%.
Среди древесных пород господствует туркестанский клен, к которому
иногда в значительных количествах присоединяется каркас, реже — алыча,,
крушина и др. Из кустарников наиболее часто встречаются шиповник
кокандский и иргай. Травянистая растительность образует почти сплош-
ной покров, одевающий почву на 80—100%. Фоновым растением здесь
является недотрога, среди которой в тех или иных количествах обычно-
разбросаны купена (Polygonatum Sewerzowi Rgl.), ежа сборная, ромаш-
ник (Pyrethrum parthenifolium Willd.), мятлик лесной (Роа nemoralis L.),
кузиния теневая (Cousinia umbrosa Rgl.), лук Розенбаха (Allium Rosen-
bachianum Rgl.), ясенец (Dictamnus tadshikorum Vved.), фитеума (Phy-
theuma baldshuanica B. Fedtsch.) и др.
Иногда встречается здесь и Ostrowskia magnifica Rgl., крупное, чрез-
вычайно эффектное колокольчиковое с большими белыми цветами. Сред-
няя высота травостоя 60—80 см. Выше поднимаются цветущие стебли ежи,
островскии, кузинии и некоторых других.
3. Ежово-девясилово-разнотравные кленовники (Aceretum dactyloso-
inuloso-herbosum) являются одной из наиболее широко распространенных
ассоциаций нижней полосы пояса древесно-кустарниковой раститель-
«О
А. С Королева
пости, заходя также в верхнюю полосу этого пояса. По сравнению с пре-
дыдущими ассоциациями ежово-девясилово-разнотравные кленовники
находятся в условиях увлажнения, повидимому, несколько менее благо-
приятных. Они располагаются на северных склонах Кондары и ее боко-
вых притоков, обычно выше мест обитаний, занятых недотрогово-сныте-
выми и ежово-разнотравно-недотроговыми кленовниками, нередко под-
нимаясь до водораздельного гребня. Почвы ежово-девясилово-разнотрав-
вых кленовников мелкоземисты, но иногда характеризуются большей
или меньшей примесью щебенки. Древесный полог описываемой ассо-
циации всегда сильно разрежен, сомкнутость его не превышает 20—
40%. Из древесных пород преобладает туркестанский клен, к которому
часто присоединяется махалебская вишня, клен Регеля, каркас и не-
которые другие. Из кустарников наиболее обыкновенны: иргаи, персид-
ская жимолость и кокандский шиповник. Травяной покров довольно
высок (90—100 см) и сравнительно густ, покрывая 80—90 (100%) поверх-
ности почвы.
В составе травостоя присутствуют многие растения, встречающиеся
и в ежово-недотрогово-разнотравных кленовниках, как недотрога, ежа,
фитеума, скалигерия [Scaligeria hirtula (Rgl. et Schmalh.) Lipsky], ясе-
нец, купена, островския и др. Однако в связи с большей разреженностью
древесного полога соотношения между растениями несколько изменяются.
Так, недотрога, фоновое для ежово-разнотравно-недотроговогокленовника
растение, здесь встречается лишь в небольших количествах и приурочена
к наиболее затененным местам у стволов деревьев, иногда и совсем от-
сутствует. Реже попадается и тенелюбивый ромашник. Напротив, значи-
тельно чаще можно встретить островскию, предпочитающую участки
светлых, разреженных ассоциаций. За счет уменьшения обилия или даже
исчезновения теневых форм появляется ряд светолюбивых видов, не встре-
чающихся под густым древесным пологом ежово-разнотравно-недотрого-
вых кленовников. Из их числа следует указать весьма обычные
для ежово-девясилово-разнотравных кленовников подмаренник (Galium
verum L.), скабиозу (Scabiosa songorica Schrenk), а также характерный
компонент этой ассоциации — девясил (Inula grandis Schrenk).
В тех случаях, когда ежово-девясилово-разнотравные кленовники
граничат с ассоциациями с обилием югана, в составе кленовников обычно
появляется и последний.
4. Недотрогово-разнотравные иргайники (Cotoneastretum impatien-
toso-herbosum) изредка встречаются в нижних частях северного склона
Кондары. Они — результат вырубки кленовников, на что указывают
единично сохранившиеся клены, а также старые пни. По составу травя-
ного покрова недотрогово-разнотравные иргайники приближаются
к ежово-девясилово-разнотравным кленовникам.
5. Разнотравно-югановая ассоциация (Prangetum herbosum) встре-
чается лишь в верхних частях северного склона нижнего течения реки,
на щебнисто-мелкоземистых почвах. Эта ассоциация является переходной
ют ежово-девясилово-разнотравных кленовников к эфемерово-гречихово-
югановой, занимающей водораздельный гребень и прилегающие части
склонов. В связи с почти полным отсутствием древесного полога в составе
травостоя становится еще меньше тенелюбивых видов, чем в ежово-девя-
силово-разнотравных кленовниках, за счет чего обильнее разрастаются
виды, не требующие или даже не выносящие затенения.
Среди деревьев встречаются единичные клены, миндали и, реже,
каркас. Часто встречаются отдельные кусты иргая.
Растительный покров Кондары
81
Травяной покров при средней высоте в 70—80 см занимает 80—90%
поверхности почвы. В составе травостоя по сравнению с ежово-девяси-
лово-разнотравными кленовниками происходят следующие изменения:
в значительных количествах появляется юган, который встречается
в ежово-девясилово-разнотравных кленовниках единично и не всегда,
увеличивается обилие эфемероидов (Hordeum bulbosum L.) и эфемеров
(Роа bulbosa L., Bromus oxyodon Schrenk и др.). Вместе с тем совершенно
исчезают такие типично лесные формы, как лесной мятлик (Роа nemo-
ralis L.), недотрога, почти не встречается фитеума. К растениям, харак-
терным как для ежово-девясилово-разнотравных кленовников, так и для
разнотравно-югановой ассоциации, относятся: ежа, тонколистная вика
(Vicia tenuifolia Roth.), скабиоза, купена. На некоторых участках этой
ассоциации пышно разрастается чина-мулькак (Latyrus mulkak Lipsky).
6. Разнотравно-пажитниковая ассоциация с редкими деревьями клена
и миндаля (Trigonelletum herbosum) встречается на северных склонах
нижнего течения р. Кондара, занимая преимущественно северо-восточ-
ные, реже северо-западные склоны боковых ущелий, а иногда и самые
нижние части склонов к Кондаре. Выше 1500 м эта ассоциация не под-
нимается. Почвы мелкоземисто-щебнистые и, повидимому, близки к поч-
вам разнотравно-югановой ассоциации и ежово-девясилово-разнотравных
кленовников. По составу растительности описываемая ассоциация также
имеет ряд близких черт с разнотравно-югановой. Немногочисленные
деревья представлены миндалем и кленами, среди кустарников часто
встречается шиповник кокандский и иргай. Так же как и в разно-
травно-югановой ассоциации, деревья настолько редки, что почти не
оказывают влияния на травостой, покрывающий поверхность почвы на
80—100%. Для травостоя характерны обычные и в разнотравно-югано-
вой ассоциации — ячмень луковичный, ежа, скабиоза, рапонтикум
(Rhaponticum integrifolium С. Winkl.), а также эфемеры — Роа
bulbosa L. и Bromus oxyodon Schrenk.
Основным отличием разнотравно-пажитниковой ассоциации от разно-
травно-югановой является почти полное отсутствие югана и обилие
пажитника Липского (Trigonella Lipskyi Sir.), создающего здесь фон
травостоя. Чаще, чем в разнотравно-югановой ассоциации, встречается
девясил.
7. Эфемерово-гречихово-югановая ассоциация (Prangetum ephemeroso-
polygonosum).
Встречается преимущественно в верхних частях северного склона,
у водораздельного гребня. Эта ассоциация очень близка к эфемерово-
ширяшево-югановой ассоциации южного склона, от которой отличается
меньшим обилием ширяша хохлатого (Eremurus comosus О. Fedtsch.),
присутствием единичных деревьев кленов, а также некоторых травяни-
стых растений (ежа, скабиоза и др.), заходящих из соседних,
расположенных ниже по склону кленовников и разнотравно-югановой
ассоциации. Среди деревьев и кустарников, помимо кленов, также
единично встречаются миндаль и иргай.
В относительно разреженном травяном покрове (степень покрытия
почвы 70%) преобладают юган, эфемеры [Bromus tectorum L. и В. oxyo-
don Schrenk, Роа bulbosa L., Vulpia ciliata (Danth.) Link., Alyssum cam-
pestreL.] и гречиха стелющаяся (Polygonum paronychioides C. A. Mey.).
8. Разреженная растительность щебнистых и каменистых осыпей.
На широко распространенных щебнистых и щебнисто-каменистых полу-
закрепившихся осыпях большей частью развивается не оформившаяся
6 Ущелье Кондара
82
А. С. Королева
в ценозы растительность с преобладанием травянистых эфемеров, эфеме-
роидов и ксерофитных многолетников. Из деревьев здесь встречаются
миндаль, каркас, махалебская вишня, арча, клены в единичных экзем-
плярах. Из кустарников, тоже единично, встречаются иргай (Gotoneaster
hissarica Pojark.), в нижних частях склонов — атрафаксис. Степень
покрытия почвы травянистой растительностью колеблется в очень
широких пределах — от 0 до 30—40%, редко — 50%. Наиболее обычными
представителями этой растительности являются юган, стелющаяся
гречиха, зверобой шероховатый (Hypericum scabrum L.), мятлик
луковичный, однолетние костры (Bromus tectorum L. и В. oxyodon
Schrenk), нередко встречаются крупные куртины камолей (Ferula koka-
nica Rgl. et Schmalh. и F. Jaeschkeana Vatke), ревеня, зизифоры (Ziziphora
clinopodiodes Lam.), невзрачная, но благоухающая необычайно тонким
ароматом гербера (Gerbera kokanica Rgl. et Schmalh.) и др.
Южный склон
9. Разнотравно-недотроговые кленовники (Aceretum herboso-impatien-
tosum). На южном склоне как нижней, так и верхней полосы пояса дре-
весно-кустарниковой растительности кленовники встречаются лишь
отдельными небольшими пятнами, изредка занимая более или менее
значительные территории в нижних частях склонов боковых ущелий.
Эти пятна, приуроченные к участкам мелкоземистых почв, представлены,
главным образом, разнотравно-недотроговыми кленовниками, являющи-
мися одной из наиболее мезофитных ассоциаций южного склона.
По общему характеру растительности эти кленовники довольно близки
к ежово-разнотравно-недотроговым кленовникам северных склонов.
Так же как и последние, разнотравно-недотроговые кленовники отли-
чаются довольно густым, хорошо развитым древостоем и богатым травя-
ным покровом мезофитного характера. Однако в составе травостоя обычно
отсутствуют столь типичные для кленовников северных склонов мятлик
лесной, ежа, ясенец, фитеума и др. В более густых группах
деревьев развит покров с преобладанием недотроги, на более разрежен-
ных местах встречаются девясил, кузиния теневая, купена, островския.
Почти всегда присутствует луковичный ячмень.
Мелкие участки разнотравно-недотроговой ассоциации часто встре-
чаются среди разреженной кленово-миндалевой ассоциации, описание
которой следует ниже.
10. Разреженная кленово-миндалевая ассоциация с ячменно-эфеме-
рово-разнотравным травяным покровом (Acereto-amygdaletum hordeoso-
ephemeroso-herbosum) является наиболее широко распространенной дре-
весно-кустарниковой ассоциацией южных склонов нижней полосы пояса
древесно-кустарниковой растительности. Она развивается на относительно
мелкоземистых почвах, почти всегда характеризующихся наличием круп-
ных, всюду разбросанных камней, а иногда и примесью щебенки.
Древостой кленово-миндалевой ассоциации, сложенный миндалем,
кленами и каркасом, чрезвычайно разрежен, сомкнутость крон не
превышает 10—30%.
В довольно густом травяном покрове (покрывающем 80—100% поверх-
ности почвы) характерны ячмень луковичный, однолетние эфемеры и в не-
больших количествах ежа. Из представителей многолетнего разнотравья
почти всегда присутствует девясил, нередко встречается алтея [Alcea
Растительный покров Кондары
83
nudiflora (Lindl.) Boiss.], тонколистная вика, кузиния теневая, на неко-
торых участках возвышаются стебли Eremostachys hissarica Rgl.
Ярусность травяного покрова выражена не ясно. Намечаются:
1-й ярус 150—200 см высоты, составленный генеративными побегами
девясила, ячменя, кузинии и некоторых других, и 2-й ярус 60—70 см
высоты, образованный злаковым подседом и основной массой вегетатив-
ных частей разнотравья.
11. Эфемерово-кузиниево-югановая ассоциация (Prangetum epheme-
roso-cousiniosum) является одной из наиболее широко распространенных
ассоциаций нижней полосы пояса древесно-кустарниковой раститель-
ности на южном склоне. Эта ассоциация, поднимающаяся до высоты
1500 м, развивается на относительно ровных щебнистых и мелкоземисто-
щебнистых участках. Деревья и кустарники встречаются здесь единично
и представлены миндалем и каркасом. Травяной покров разрежен, покры-
вая 50—70% поверхности почвы.
1-й ярус травостоя, 1.5—2 м высоты, образуют генеративные побеги
югана, кузинии и некоторых других, 2-й ярус, 60—80 см высоты, сло-
жен, главным образом, листьями югана, 3-й ярус, 15—30 см высоты, обра-
зует основная масса эфемеров.
Наряду с юганом, образующим фон травяного покрова, весьма харак-
терным растением является кузиния преломленная (Cousinia refracta
Juz.), а также эфемеры, среди которых наиболее обычны однолетние
костры (Bromus tectorum L. и В. oxyodon Schrenk), тимофеевка (Phleum
paniculatum Huds.), Taeniaterum crinitum (Schreb.) Nevski, Crepis pul-
chra L., Callipeltis cucullaria (L.) Stev. и др.
Постоянными компонентами эфемерово-кузиниево-югановой ассоциа-
ции являются ширяш хохлатый и камоль кокандский. Почти всегда при-
сутствует в небольших количествах и луковичный ячмень. Наличие
в большем количестве луковичного ячменя наблюдается на относи-
тельно мелкоземистых и нередко вогнутых участках.
12. Эфемерово-ширяшево-югановая ассоциация (Prangetum epheme-
roso-eremurosum) отличается от предыдущей только отсутствием или
наличием единичной кузинии преломленной. Встречается эта ассоциация
значительно реже, чем эфемерово-кузиниево-югановая.
13. Эфемерово-чеснеево-югановая ассоциация (Prangetum ephemeroso-
chesneyosum) связана с почвами более щебнистыми по сравнению с поч-
вами двух предыдущих ассоциаций. Она широко распространена в верх-
них частях южного склона ущелья под водораздельным гребнем, а также
на выпуклых участках в нижних и средних частях склонов.
Благодаря щебнистой почве травяной покров эфемерово-чеснеево-
югановой ассоциации значительно реже, чем травяной покров эфемерово-
кузиниево-югановой и эфемерово-ширяшево-югановой ассоциации, покры-
вает лишь 30—40, редко—60% поверхности почвы. По характеру
ярусности описываемая ассоциация близка к предыдущим: 1-й ярус,
150—200 см высоты, образуют генеративные побеги югана и иногда
кузинии преломленной, 2-й ярус, 60—70 см высоты, — листья югана
и генеративные побеги ширяша, 3-й ярус, 10—15 см высоты, —основ-
ная масса эфемеров, чеснея (Ghesneya hissarica А. Вог.), гвоздика
(Dianthus tetralepis Nevski). На фоне травянистой растительности в со-
ставе этой ассоциации почти всегда встречаются единичные деревья
миндаля, реже — каркаса, кусты шиповника самаркандского.
14. Растительность щебнистых и каменистых осыпей южных склонов.
Как уже упоминалось выше, на южных склонах исключительно широким
6*
84
А, С. Королева
развитием пользуются щебнистые и каменистые осыпи и почти отвесные
обнажения коренных пород в виде скал, огромных каменистых глыб
и т. д.
Разреженная скудная растительность подобных участков в общих
чертах мало отличается от растительности щебнистых и каменистых
северных склонов. Нередко можно здесь видеть единичные деревья мин-
даля, реже — каркаса, клена, арчи, кусты иргая (Cotoneaster acutiuscula
Pojark.), колючего самаркандского шиповника (Rosa maracandica Bge.).
Наиболее характерными представителями травянистой растительности
являются ревень Максимовича, ширяш хохлатый, камоли (Ferula
Jaschkeana Vatke и F. kokanica Rgl. et Schmalh.), Eremostachys
tadshikistanica B. Fedtsch., некоторые эфемеры (Bromus oxyodon
Schrenk, Crepis pulchra L., Crucianella exasperata F. A. M.) и иногда
юган.
На скалах лепятся причудливые куртины Dionysia involucrata Zapr.,
Scutellaria hissarica В. Fedtsch., S. leptosiphon Nevski, туркестанская
осока (Carex turkestanica Rgl.), кусты спиреи (Spiraea baldshuanica
B. Fedtsch.) и некоторые другие.
Долины
15. Недотроговые орешники (Juglandetum impatientosum) (рис. 2)
встречаются небольшими пятнами в долине Кондары и ее боковых при-
токов. Орех, достигающий здесь 20—25 м высоты, образует насаждения
с сомкнутыми или почти смыкающимися кронами. К ореху присоеди-
няется иногда туркестанский клен, каркас, в подлеске встречается ши-
повник собачий и иргай. В травяном покрове господствует недотрога,
среди которой лишь в небольших количествах кое-где встречаются группы
однолетних костров, лук Розенбаха, Ostrowskia magnifica Rgl., кузиния
теневая и некоторые другие виды. Чрезвычайно характерны для этой
ассоциации Physocaulis nodosus (L.) Tausch. и аллиария (Alliaria offici-
nalis Andrz.).
16. Недотроговые кленовники (Aceretum impatientosum), так же как
и недотроговые орешники, встречаются в долине р. Кондара и ее
боковых притоков, в виде отдельных небольших пятен. По характеру
местообитаний участки, занятые этой ассоциацией, близки к террито-
риям, занятым недотроговыми орешниками. По составу и характеру
травяного покрова они также чрезвычайно сходны с орешниками, основ-
ное отличие от которых заключается в замене ореха туркестанским кле-
ном, к которому нередко примешивается каркас. Единично встречается
и орех.
17. Недотроговые каркасники (Celtidetum impatientosum) очень
близки по характеру местообитаний и травяного покрова к недотрого-
вым кленовникам, от которых отличаются преобладанием в составе дре-
весного полога каркаса, а не клена.
18. Ежевиково-коротконожковые орешники (Juglandetum ruboso-bra-
chypodiosum) встречаются неширокими полосами вдоль речных русел. Орех
представлен здесь мощными деревьями, почти смыкающимися своими
кронами (сомкнутость 60—90%). К ореху обычно присоединяется ряд
других древесных пород (алыча, туркестанский клен, каркас, боярыш-
ник).
В подлеске господствует ежевика, образующая местами непроходи-
мые заросли, изредка встречаются шиповник собачий и иргай.
Растительный покров Кондары
85
В густом травяном покрове господствует коротконожка [Brachypo-
dium silvaticum (Huds.) P. В.], образующая вдоль берега потока сплош-
ной бордюр, зеленеющий в течение всего лета. К коротконожке приме-
шиваются немногие другие виды, количество которых увеличивается
по мере удаления от берега. У воды встречаются группы датиски.
Рис 2. Недотроговый орешник в долине р. Кондара.
(Фот. А. И. Иванова).
19. Ежевиково-коротконожковые тополевники (Populetum ruboso-bra-
chypodiosum) встречаются небольшими пятнами в среднем течении
долины р. Кондара. По составу подлеска и господствующих видов
травяного покрова близки к ежевиково-коротконожковым ореш-
никам. Сомкнутость древесного полога, образованного тополем (Populus
tadshikistanica Кот.), достигает 50—90%. К тополю присоединяются
единичные экземпляры ивы (Salix sp.). В подлеске преобладает ежевика
(Rubus caesius L.), нередко образующая заросли; встречается шиповник
собачий (Rosa canina L.). Основным элементом травяного покрова яв-
ляется коротконожка, в небольших количествах встречаются немногие
86
А. С. Королева
другие виды, как лесной мятлик, мелисса (Melissa officinalis L.), чина
(Lathyrus pratensis L.). У воды образует мощные заросли Datisca canna-
bina L. Здесь же часто растут мята (Mentha silvestris L.), Codonopsis
clematidea С. B. Clarke и некоторые другие.
20. Коротконожковые ежевичники (Rubetum brachypodiosum) неболь-
шими пятнами часто встречаются в долине Кондары и ее непересыхаю-
щих боковых притоков. Основным отличием этой ассоциации от двух
предыдущих является почти полное отсутствие древесных пород, пред-
ставленных лишь редкими ивами (Salix sp.), нигде не образующими со-
мкнутого полога. На местах, не занятых куртинами ежевики, в травяном
покрове господствует коротконожка, среди которой редко или единично
встречаются лесной мятлик, луговая чина; вдоль берегов высятся заросли
датиски, растет мята, кипрей (Epilobium hirsutum L.), лапчатка (Po-
tentilla reptans L.). Под деревьями можно видеть единичные группы
недотроги.
21. Болотистые орешники (Juglandetum paludosum) большею частью
встречаются в устьях боковых притоков, при наличии обильных выхо-
дов грунтовых вод. Среди деревьев, смыкающихся пологом на 50—80%,
преобладает орех, достигающий 20 м высоты. К ореху присоединяются
яблоня (Malus hissarica Kudr.), алыча и некоторые другие. Травяной
покров густ и зелен в течение всего лета.
22. Эфемерово-бедренцовая ассоциация (Pimpinelletum ephemerosum)
развивается на приподнятых мелкоземистых и мелкоземисто-каменистых
участках долины — на террасах, где уровень грунтовых вод настолько
опускается, что воды становятся недоступными для большей части травя-
нистых растений. Эта ассоциация нередко характеризуется присутствием
ряда деревьев и кустарников, которые обычно не образуют сомкнутого
полога. Здесь встречается каркас, алыча, тополь, грецкий орех, ивы,
иргай, шиповник собачий. На мелкоземистых участках травяной покров
покрывает почву на 90—100%, на каменистых — на 40—50%.
В составе травяного покрова преобладает бедренец (Pimpinella affi-
nis Led.), в значительных количествах встречается линделофия [Linde-
lophia anchusoides (Lindl.) Lehm.], алтея (Alcea nudiflora Lindl.), тыся-
челистник (Achillea filipendulina Lam. и A. trichophilla Schrenk), крово-
хлебка (Sanguisorba minor Scop.). Наряду с перечисленными многолет-
никами, богато представлены эфемеры, среди которых наиболее часты:
Crepis pulchra L., однолетние костры (Bromus oxyodon Schrenk, В. ste-
rilis L. и В. tectorum L.), Poa bulbosa L., Thaeniathaerum crinitum
(Schreb.) Nevski.
ВЕРХНЯЯ ПОЛОСА ПОЯСА ДРЕВЕСНО-КУСТАРНИКОВОЙ
РАСТИ ТЕЛ ЬНОСТ И
На северных склонах в этой полосе, как и в нижней, широко распро-
странен туркестанский клен, который отличается значительно лучшим
ростом, достигая 15—16 м высоты. Грецкий орех, в отличие от нижней
полосы, растет не только в долинах, но и на северных склонах; нередко
орех образует почти чистые насаждения, а также встречается в смешан-
ных ассоциациях. Как уже упоминалось выше, в верхней полосе отсут-
ствует махалебская вишня, клен Регеля, пузырник (Colutea persica
Boiss.), Atraphaxis pyrifolia Bge. Миндаль и каркас, широко распро-
страненные в нижней полосе, в верхней встречаются редко и только
Растительный покров Кондары
87
на южных склонах, причем каркас поднимается не выше 2000 м, а мин-
даль — до 2200 м.
В составе травянистой растительности верхней полосы также наблю-
даются существенные изменения. В травяном покрове древесно-кустар-
никовых ассоциаций появляется торон (Polygonum bucharicum Grig.),
Рис. 3. Клоповники верхней полосы пояса древесно-кустар-
никовой растительности на северном склоне уроч. Квак.
(Фот. А. И. Иванова).
бузульник Томсона [Ligularia Thomsoni (Clarke) Pojark.), Nepeta gran-
diflora M. B., Ligularia persica Boiss. и ряд других растений, не встре-
чающихся в нижней полосе. На лесных опушках и в светлых лесах вес-
ной цветут роскошные куртины ирисов (Iris Hoogiana Dikes.), изредка
можно увидеть эффектную Fritillaria Eduardi Rgl., которая раньше была
здесь довольно обычным растением, теперь же она истреблена из-за
съедобности клубней, содержащих крахмал. В верхней полосе отсутствуют
часто встречающиеся в нижней полосе: эспарцет (Onobrychis grandis
Lipsky), пажитник Липского (Trigonella Lipskyi Sir.) и некоторые эфе-
88
А. С. Королева
меры [Diarthron vesiculosum (Fisch, et Mey.) A. Mey.]; редко встречается
девясил. Древесно-кустарниковая растительность верхней полосы в ос-
новном приурочена к северным склонам, на южных склонах лишь изредка
встречаются небольшие лески.
На северных склонах древесно-кустарниковая растительность рас-
пространена довольно широко. Более или менее крупные массивы под-
нимаются только до 2000 м (рис. 3). Выше лишь кое-где можно видеть от-
дельные мелкие пятна розариев и разреженных кленовников, которые
с высоты 2400 м исчезают совершенно.
В пределах всей верхней полосы древесно-кустарниковая расти-
тельность распространена преимущественно на склонах боковых долин,
то более или менее крутых (до 35°), то сравнительно пологих и нередко
вогнутых. Почвы большей частью мелкоземисты, иногда каменисты.
Наиболее мезофитной древесно-кустарниковой ассоциацией северных
склонов верхней полосы являются высокотравно-снытевые кленовники,
приуроченные к депрессиям рельефа и встречающиеся преимущественно
в районе урочища Квак. Эти кленовники характеризуются хорошо раз-
витым древесным пологом и богатым травяным покровом с преобладанием
сныти (Aegopodium tadshikorum Schischk.), к которой присоединяется ряд
лесных видов.
На относительно ровных участках северных склонов к Кондаре,
ниже урочища Квак и на северных склонах к реке Оби-сафед, довольно
широко распространены бузульниково-недотроговые кленовники, отли-
чающиеся от ежово-разнотравно-недотроговых кленовников нижней
полосы присутствием бузулышка и некоторых других видов, свойствен-
ных верхней полосе. Древесный полог этой ассоциации, так же как и
в высокотравно-снытевых кленовниках, хорошо развит.
На крутых участках северных склонов изредка встречаются разно-
травно-мятликовые кленовые и кленово-ореховые ассоциации, в тра-
вяном покрове которых преобладает лесной мятлик и широко распростра-
нена душица (Origanum tyttanthum N. Gontsch.).
Изредка на северных склонах верхней полосы встречается ассоциация
ежово-девясилово-разнотравного кленовника (см. описание нижней
полосы).
Наиболее распространенной древесно-кустарниковой ассоциацией
верхней полосы являются разреженные высокотравные кленовники,
развивающиеся на ровных или слегка вогнутых участках преимущественно
северных склонов. В их густом травяном покрове чрезвычайно характерны
Ligularia Thomsoni (Clarke)Pojark., Rumex Paulsenianus Rech, f., Nepeta
grandiflora M. B., Tanacetum Newesskyanum C. Winkl. Под сенью дере-
вьев в заметных количествах встречается недотрога. В составе древес-
ной растительности, наряду с кленом, нередко видное участие принимает
грецкий орех. Среди разреженных высокотравных кленовников, осо-
бенно широко распространенных в районе урочища Квак, часто встре-
чаются открытые поляны с единичными деревьями и кустарниками. На
таких полянах травянистая растительность разрастается еще более
пышно, образуя ежово-тороново-разнотравнугр ассоциацию. В составе
ее, наряду с многими видами, обычными и для высокотравных кленов-
ников, весьма характерны и обильны некоторые растения, не встречаю-
щиеся или редко встречающиеся в кленовниках. К числу таких не
любящих затенения и процветающих здесь ’растений относятся торон
(Polygonum bucharicum Grig.), Ligusticum discolor Led. и некоторые
другие. Подобного типа ассоциации нередко встречаются и выше границы
Растительный покров Кондары 89
сплошного распространения древесно-кустарниковой растительности
(2000 м).
Высокотравные кленовники (ежово-тороново-разнотравная ассоци-
ация) благодаря сравнительно высокому обилию поедаемых трав
подвергаются наиболее сильному выпасу. Поэтому местами естественный
травяной покров сильно нарушен, представляя беспорядочные заросли
с пятнами преобладания то тех, то других видов. Такие участки особенно
часты вблизи прогонов скота из урочища Квак на пастбища Руидашта.
На небольших с пологими склонами выпуклых хребтиках, являю-
щихся водоразделами боковых притоков р. Уноу (верховья р. Кондара),
довольно часто прорезающих склоны, кленовые и ореховые насаждения
отсутствуют. Их заменяют розарии, в составе которых иногда встречаются
единичные деревья клена и арчи; орех сюда не поднимается. Почвы,,
по сравнению с почвами кленовых и ореховых насаждений, более сухи,
менее богаты перегноем и нередко щебнисты. В связи с относительной
сухостью почв и отсутствием древесного полога здесь встречаются лишь
немногие травянистые спутники лесов. В травяном покрове много эфе-
меров, эфемероидов и многолетников ксерофитного облика. Наиболее
обычной ассоциацией розариев являются югановые розарии, в травостое
которых господствует юган, широко распространены лапчатка (Poten-
tilla kulabensis Th. Wol.), луковичный мятлик, скабиоза, Scaligeria hir-
tula (Rgl. et Schmalh.) Lipsky, часто встречается гвоздика (Dianthus
seravschanicus Schischk.), неприятно пахнущая Galagania fragrantis-
sima Lipsky и др.
Значительно реже югановых розариев встречаются эфемеровые роза-
рии и разнотравные розарии.
Выше 2000 м кленовых и ореховых насаждений почти нет. Древесно-
кустарниковая растительность представлена отдельными небольшими
пятнами высокотравных розариев, среди которых иногда можно видеть
единичные клены. Розарии этой полосы занимают мелкоземистые, не-
редко вогнутые участки склонов. Они характеризуются красочным и
пышным травяным покровом, в составе которого преобладают лесные
формы.
Изредка на северных и восточных склонах верхней полосы древесно-
кустарникового пояса можно видеть небольшие пятна арчевников, сохра-
нившихся в настоящее время лишь на щебнистых крутых, удаленных от
троп участках. Арчевники характеризуются разреженным древесным
пологом и явно ксерофилизированным травяным покровом, в состав
которого часто входят Ziziphora clinopodioides Lam., Dianthus serav-
schanicus Schischk., Hypericum scabrum L. и другие растения, свой-
ственные открытым, щебнистым местообитаниям.
На южных склонах верхней полосы древесно-кустарниковые ассо-
циации редки. Кое-где встречаются небольшие участки разнотравно-
недотроговых кленовников (см. описание нижней полосы пояса древесно-
кустарниковой растительности) и сильно засоренных высокотравных
кленовников.
На щебнистых и щебнисто-мелкоземистых участках южных склонов
широко распространена эфемерово-тысячелистниково-югановая ассоци-
ация (часто с единичными деревьями миндаля, каркаса и некоторых дру-
гих), которая близка по своему составу и характеру к эфемерово-юга-
новым ассоциациям нижней полосы.
На щебнистых осыпях южных и восточных склонов нередко встре-
чаются камолево-югановые ассоциации, характеризующиеся разрежен-
so
А. С Королева
ным травяным покровом и присутствием ряда растений, специфических
для щебнистых и каменистых субстратов.
Большие территории на склонах всех экспозиций занимают щеб-
нистые и каменистые участки с редкой, большей частью не сформировав-
шейся травянистой растительностью, покрывающей не более 20—40%
поверхности почвы, или лишенные растительности. Участки подобного
типа особенно широко распространены на крутых, замыкающих долину
Кондары, преимущественно восточных склонах (в полосе от 2000 до
2400—2500 м высоты). Эти участки завершаются водораздельным гребнем
между реками Варзоб и Лючоб.
Здесь наиболее обычны: ревень Максимовича, хондрилла (Chondrilla
juncea L.), Polygonum paronychioides С. A. M. Нередко встречаются юган,
камоль. В отличие от щебнистых склонов нижней полосы часто наблю-
даются Astragalus macropodium Lipsky, иссоп (Hissopus officinalis L.),
Bupleurum exaltatum M. В. и некоторые другие.
Днища долин ручьев и речек узки и на большей части своего про-
тяжения покрыты галечниками. Мелкоземистые наносы встречаются редко,
и площади их очень невелики. На галечниках часто встречаются заросли
ив и богатая видами травянистая растительность, последняя по видовому
составу близка к растительности аналогичных местообитаний нижней
полосы.
23. Высокотравно-снытевые кленовники (Aceretum magno-herboso-
aegopodiosum) не имеют в верхней полосе пояса древесно-кустарниковой
растительности широкого распространения. Они встречаются сравни-
тельно небольшими пятнами на вогнутых участках северных склонов
в урочище Квак (рис. 4). От недотрогово-снытевых кленовников нижней
полосы они отличаются лучшим ростом клена, присутствием в высоком
и густом травяном покрове бузульника Томсона, торона, непеты
(Nepeta grandiflora М.В.), Ligusticum discolor Led., щавеля-шульхи и
некоторых других. Почвы мелоземистые глубокие. Сомкнутость крон
деревьев 50—90%. Помимо господствующего клена нередко встречается
орех, жимолость персидская, иногда арча и некоторые другие. Высота
травяного покрова колеблется в пределах 100—160 см.
24. Бузульниково-недотроговые кленовники (Aceretum ligularioso-
impatientosum) довольно широко распространены на северных склонах
среднего течения реки Кондара, в ее верховьях (реки Уноу и Оби-сафед)
и почти не встречаются в урочище Квак. Эта ассоциация развивается
на мелкоземистых коричневых почвах и характеризуется относительно
густым древесным пологом, смыкающимся на 50—80%. В составе древес-
ных пород преобладает туркестанский клен, к которому почти всегда
в различных количествах присоединяется грецкий орех. Иногда встре-
чаются единичные деревья крушины и арчи. В подлеске наиболее обычна
жимолость персидская, шиповник собачий, шиповник кокандский, иргай.
Травяной покров по своему составу близок к травяному покрову
разнотравно-недотроговых кленовников нижней полосы; он отличается,
главным образом, присутствием бузульника Томсона и иногда щавеля-
шульхи, которые не встречаются в нижней полосе.
Господствующее значение в травяном покрове принадлежит недотроге,
среди которой в тех или иных количествах разбросаны бузульник, ро-
машник, Scaligeria hirtula (Rgl. et Schmalh.) Lipsky, кузиния краси-
венькая (Cousinia pulchella Bge.), лук Розейбаха, душица, лесной мят-
лик. Высота основной массы травяного покрова (главным образом недо-
трога) 60—70 см. Степень покрытия почвы растительностью 80—100%.
Растительный покров Кондары
91
25. Б у зульниково-недотроговые орешники (Juglandetum ligularioso-
impatientosum) встречаются в условиях местообитания, аналогичных
с таковыми бузульниково-недотроговых кленовников, нередко чере-
дуясь с ними и переходя в смешанные кленово-ореховые насаждения. За
исключением замены клена орехом, описываемая ассоциация ничем не
отличается от бузульниково-недотроговых кленовников.
26. Разнотравно-мятликовые кленовники (Aceretum herboso-poaeosum).
Разнотравно-мятликовые кленовники встречаются изредка, неболь-
шими пятнами на крутых, преимущественно северных склонах ущелий
рек Оби-сафед и Кондары, с мелкоземистыми почвами. В составе древес-
Рис. 4. Высокотравно-снытевый кленовник на северном склоне уроч. Квак.
(Фот. А. И. Иванова).
ных пород преобладает клен, к которому часто, в значительных количе-
ствах, присоединяется орех. Изредка встречаются крушина, арча и не-
которые другие. Из кустарников наиболее обычны жимолость и иргай.
В травяном покрове преобладает лесной мятлик, довольно видную роль
играет душица, менее обильны, но довольно постоянны Scaligeria hirtula
(Rgl. et Schmalh.) Lipsky, вика тонколистная, ромашник.
27. Разреженные высокотравные кленовники (Aceretum magno-
herbosum) являются одной из наиболее распространенных древесно-
кустарниковых ассоциаций верхней полосы пояса древесно-кустарни-
ковой растительности. Они встречаются на северных, северо-западных
и северо-восточных (реже на южных) склонах с мелкоземистыми более
или менее глубокими почвами.
Древесный полог сильно изрежен, сомкнутость крон не превышает
30—50%, господствует туркестанский клен, к которому иногда присое-
диняется грецкий орех. Почти всегда присутствуют-единичные деревья
92
А. С. Королева
арчи, яблони, крушины. Из кустарников постоянно в небольшом коли-
честве встречаются персидская жимолость и иргай, несколько более
обилен кокандский шиповник (Rosa kokanica Rgl.)
Основную массу травяного покрова образуют высокие бузульник
Томсона, щавель-шульха (Rumex Paulsenianus Rech, f.), Scaligeria
hirtula (Rgl. et Schmalh.) Lipsky, Nepeta grandiflora M. В., пижма (Tana-
cetum Nevesskianum C. Winkl.), ромашник, вика тонколистная. Эти ра-
стения часто образуют неравномерные заросли, в которых преобладают
то одни, то другие виды. В небольших количествах постоянно встре-
чается ежа, Korshinskya Olgae (Rgl. et Schmalh.) Lipsky, Lavathera cas-
chemiriana Cambess., Conyza khorossanica (Boiss.) Novopokr. и др. Под
деревьями часты группы недотроги, Alliaria officinalis Andrz. Травяной
покров довольно густ, покрывает 90—100% поверхности почвы, в сред-
нем 100—150 см высоты.
28. Ежово-тороново-разнотравная ассоциация (Herbetum dactyloso-
polygonosum) довольно часто встречается на полянах северных склонов
среди кленовников урочища Квак и поднимается выше зоны сплошного
распространения кленовников (2000 м).
На фоне травянистой растительности постоянно встречаются единич-
ные деревья клена туркестанского и иногда рябины персидской (Sor-
bus persica Had!.), отдельные кусты кокандского шиповника, иргая
(Cotoneaster insignis Pojark.) и персидской жимолости.
В составе травяного покрова нет резко преобладающих видов.
Основную массу его образует группа двудольных растений, среди которых
наиболее характерны торон, Ligusticum discolor Led., Tanacetum Neves-
skyanum, Ligularia persica Boiss., смолевка (Silene commutata Guss.), Li-
gularia Tbomsoni, Scaligeria hirtula (Rgl. et Schmalh.) Lipsky, Phyt-
heuma baldshuanica B. Fedtsch. и др. В заметных количествах встречаются
злаки — ежа, мятлик лесной и пырей угамский. Все эти растения обра-
зуют красочный и густой покров, достигающий в среднем 100—150 см
высоты. Степень покрытия почвы травянистой растительностью 90—100%.
29. Разнотравно-югановые розарии (Rosarium herboso-prangosum)
широко распространены на мелкоземистых или слегка щебнистых поч-
вах в верхней полосе пояса древесно-кустарниковой растительности, где
занимают обычно выпуклые вершины водораздельных увалов, проре-
зающих склоны.
Эти розарии почти лишены деревьев; встречаются лишь единичные
экземпляры туркестанского клена, реже — крушины и некоторых дру-
гих. Среди кустарников преобладает кокандский шиповник, то рассеян-
ный в виде отдельных кустов, то образующий небольшие редкие заросли,,
высотой до 80—100 см. Травяной покров розариев довольно густ, покры-
вает 80—90% поверхности почвы. Высота его основной массы колеблется,
от 70—80 см, генеративные стебли югана и других растений поднимаются,
до 150—180 см.
Одним из наиболее характерных травянистых растений является юган,.
попутно в значительных количествах наблюдаются: лапчатка кулябская,
(Potentilla kulabensis Th. Wolf), луковичный мятлик, скабиоза, душица
и единичные — ежа и эремурус Регеля (Eremurus Regelii Vved.). Часто
к ним присоединяется скалигерия [Scaligeria hirtula (Rgl. et Schmalh.}
Lipsky], гвоздика (Dianthus seravschanicus Schischk.), Galagania frag-
rantissima Lipsky.
30. Разнотравные розарии (Rosarium herbosum) встречаются в место-
обитаниях, близких к местообитаниям юганово-разнотравных розариев,.
Растительный покров Кондары
93
но распространены значительно меньше, чем последние. По составу и
характеру травяного покрова разнотравные розарии также имеют ряд
общих черт с юганово-разнотравными розариями, основным отличием
от которых является отсутствие югана и лапчатки кулябской.
31. Ячменно-разнотравные розарии (Rosarium hordeoso-herbosum)
были отмечены лишь в нескольких местах на южных склонах ущелья
реки Уноу, где они встречаются на полянах среди кленовников. Травя-
ной покров этой ассоциации отличается резким преобладанием лукович-
ного ячменя, а также обилием сорных растений (Salvia sclarea L., Al-
cea nudiflora Lindl., Thermopsis dolichocarpa V. Nik. и др.), широко рас-
пространенных по всему склону, находящемуся на пути прогона скота
на пастбища Руидашта.
32. Ежово-разнотравные розарии (Rosarium dactyloso-herbosum)
встречаются небольшими пятнами в верхней части пояса древесно-кустар-
никовой растительности (выше 2000 м) на мелкоземистых участках север-
ных, восточных и западных склонов. Шиповник (Rosa kokanica Rgl.)
образует местами довольно густые заросли, среди которых развивается
пышный травяной покров. Иногда встречаются единичные клены и неко-
торые кустарники. В травяном покрове широко распространены лесной
мятлик, скалигерия и некоторые другие обычные спутники кленовников.
Встречаются также растения, не любящие затенения и отсутствующие
в кленовниках, как скабиоза джунгарская, торон, юган.
У границы субальпийского пояса в составе ежово-разнотравных
розариев начинают появляться и некоторые субальпийские растения,
как пион (Paeonia hybrida Pall.), горицвет [Adonis turkestanica (Korsh.)
N. Adolf], Cousinia splendida C. Winkl. и некоторые другие.
33. Зизифорово-разнотравные арчевники (Juniperetum ziziphoroso-
herbosum) встречаются редко, только на крутых, большей частью щеб-
нистых участках северных и восточных склонов. Древесный полог
арчевников обычно разрежен, сомкнутость крон не превышает 20—40%.
Среди господствующей арчи единично встречаются деревья клена, кусты
кокандского шиповника, иргая.
Травяной покров также не отличается густотой, покрывает 70—
80% поверхности почвы. В составе травостоя к господствующим видам
ксерофитного облика присоединяется ряд лесных растений и эфемеры.
Наиболее обильна зизифора, в меньших количествах встречаются
душица, зверобой шероховатый, гвоздика (Dianthus seravschanicus
Schischk.), скабиоза, вика тонколистная, лесной мятлик (Роа nemora-
lis. L.) и др. Под арчей всегда можно видеть группы ее постоянного спут-
ника — Leptorhabdos micrantha Schrenk.
34. Тысячелистниково-эфемерово-югановая ассоциация (Prangetum
achilleoso-ephemerosum) широко распространена на южных склонах
верхней полосы пояса древесно-кустарниковой растительности, где она
занимает мелкоземисто-щебнистые и щебнистые участки.
Эта ассоциация, весьма близкая к эфемерово-ширяшево-югановой
ассоциации нижней полосы, отличается от нее присутствием значитель-
ного количества тысячелистника (Achillea filipendulina Lam.), который
в нижней полосе встречается только в долинах рек, и отсутствием ряда
эфемеров — Gallipeltis cucullaria (L.) Stev., Diarthron vesiculosum
(F. etM.) С. A. M., Thaeniatherum crinitum (Schreb.) Nevski и некоторых
других.
Травяной покров данной ассоциации покрывает не более 70—90%
поверхности почвы. 1-й ярус, 150—200 см высоты, образован генератив-
94
А. С. Королева
ними побегами югана и других растений, 2-й ярус,. 60—70’ см высоты,—
листьями югана и некоторыми эфемерами.
35. Камолево-югановая ассоциация (Prangetum ferulosum) часто встре-
чается на южных и восточных склонах верхней полосы пояса древесно-
кустарниковой растительности и занимает более щебнистые участки,
чем эфемерово-югановая ассоциация, нередко представляющие полу-
закрепившиеся каменисто-щебнистые осыпи. Поэтому травостой здесь
разрежен и покрывает не более 40—60% поверхности почвы.
Так же как и в предыдущей ассоциации, широким развитием поль-
зуется юган, в меньших количествах распространены камоли (Ferula
Jaeschkeana Vatke, F. kokanica Rgl. et Schmalh.), нередко встречается
тысячелистник. В отличие от тысячелистниково-эфемерово-югановой ас-
социации эфемеры представлены только луковичным мятликом, но зато
в значительных количествах встречаются показатели щебнистых суб-
стратов, как ревень Максимовича, астрагал крупноножковый (Astra-
galus macropodium Lipsky), иссоп, гвоздика (Dianthus seravschanicus
Schischk.) и некоторые другие. У верхней границы пояса в составе камо-
лево-югановой ассоциации нередко появляются зопник (Phlomis fruti-
cetorum N. Gontsch.), горицвет [Adonis turkestanicus (Korsh.) N. Adolf],
торон, кузиния (Gousinia macilenta G. WinkL).
36. Растительность щебнистых и каменистых склонов верхней полосы
пояса древесно-кустарниковой растительности разрежена, покрывая
20—40% поверхности почвы, и часто имеет случайный характер.
Большая часть произрастающих здесь видов характерна и для
щебнистых участков нижней полосы, однако есть растения специ-
фичные для верхней полосы, некоторые же виды, обычные в нижней
полосе, здесь не встречаются. Деревья и кустарники на щебнистых скло-
нах редки и представлены большей частью единичными туркестанским
кленом, арчей и иргаем. Преобладают травянистые растения, среди ко-
торых наиболее обычны юган, камоли, ревень Максимовича, Chondrilla
juncea L., Polygonum paronychioides G. A. M., зверобой шероховатый,
Ziziphora clinopodioides Lam., гвоздика (Dianthus seravschanicus
Schischk.), иссоп, Orysopsis sogdiana Grig. На каменистых россыпях
у верхней границы пояса часто встречаются отдельные экземпляры и
группы торона, чина-мулькак (Lathyrus mulkak Lipsky), Fritillaria
Eduardi Rgl., Phlomis fruticetorum N. Gontsch. и некоторые другие.
37. Растительность речных долин верхней полосы пояса древесно-
кустарниковой растительности не имеет широкого распространения.
На галечниковых наносах часты заросли ив, среди которых встречаются
обычные и в долинах нижней полосы тысячелистник (Achillea filipen-
dulina Lam.), лапчатка (Potentilla reptans L.), кровохлебка (Sanguisorba
minor) и некоторые другие. В отличие от нижней полосы, здесь нередко
можно видеть персидскую полынь (Artemisia persica Boiss.).
У воды развиваются заросли датиски (Datisca cannabina L.), еже-
вики, встречается борщевик (Heracleum Lehmannianum Bge.), Codonop-
sis clamatidea Clarke, коротконожка [Brachyopodium silvaticum (Huds.)
P. В.]. Дикий виноград, столь обычный в речных долинах нижней полосы,
в верхнюю уже не заходит.
На сравнительно небольших мелкоземистых участках, в густом тра-
вяном покрове лугового типа преобладают': белая полевица (Agrostis
alba L.), клевер ползучий (Trifolim repens L.), лапчатка (Potentilla rep-
tans L.), подорожник (Plantago lanceolata L.), осоки, ситники и
другие.
Растительный покров Кондары
95
ПОЯС СУБАЛЬПИЙСКОЙ РАСТИТЕЛЬНОСТИ
Пояс субальпийской растительности в районе Кондары простирается
от 2400—2500 м до 2800 м, охватывая верхние части крутых и каменистых
склонов у верховьев Кондары и плато Руидашт.
Субальпийская растительность в районе Кондары относится только
к нижнему поясу этой растительности Гиссарского хребта. Верхняя часть
этого пояса в районе Кондары не выражена, так как абсолютные
высоты не превышают здесь 2800—2900 м.
Верхние части восточных склонов в Кондаре круты и каменисты.
Их растительный покров разрежен и представлен сочетаниями расте-
ний, несколько отличающимися от сочетаний на Руидаште.
На восточных склонах, в нижней полосе субальпийского пояса от
2400 до 2550 м, на щебнистых и щебнисто-каменистых участках господ-
ствует кузиниево (Cousinia macilenta С. Winkl.)-камолево-югановая ас-
социация. Выше ее сменяет разреженная ферулево-кузиниевая ассо-
циация, в которой преобладает та же кузиния тощая (Cousinia macilenta
С. Winkl.) в сочетании с ферулой (Ferula ovina Boiss.), ревенем Максимо-
вича, зверобоем (Hypericum scabrum L.) и некоторыми другими.
На относительно мелкоземистых участках восточных склонов, рас-
положенных в седловинах и впадинах между скал, изредка встречается
пажитниково-разнотравно-тороновая ассоциация, характеризующаяся
обилием торона (Polygonum bucharicum Grig.), сопровождаемого рядом
как субальпийских, так и лесных форм.
В пределах урочища Руидашт, на пологих склонах, щебнистые и ка-
менистые участки также играют весьма видную роль. Однако здесь до-
вольно широким распространением пользуются мелкоземистые и щеб-
нисто-мелкоземистые почвы, в значительной степени распаханные и
занятые под посевы ячменя или оставленные в виде залежей.
На щебнистых и щебнисто-каменистых склонах Руидашта господ-
ствует кузиниево-разнотравная ассоциация с преобладанием той же
кузинии тощей. Здесь отсутствует ревень, луковичный мятлик (Роа bul-
bosa L.) замещен мятликом голоцветным (Роа glabriflora Roshev.). Эта
ассоциация занимает около 30% всей площади Руидашта. Значительно
реже на каменисто-щебнистых участках южных и восточных склонов
встречается разреженная разнотравно-ромашниковая ассоциация с пре-
обладанием ромашника (Pyrethrum mucronatum Rgl. et Schmalh.) и
значительным количеством колючего подушкообразного астрагала (As-
tragalus lasiosemius Boiss.). В нижних крутых частях склонов нередки
каменистые и щебнистые осыпи, почти лишенные растительного покрова.
Последний отсутствует или крайне редок и на изредка встречающихся
обнажениях песчаников.
На мелкоземистых и щебнисто-мелкоземистых, преимущественно се-
верных и западных склонах широко распространена песчанково-лаго-
тисовая ассоциация с преобладанием песчанки (Arenaria Griffithii Boiss.)
илаготиса [Lagotis Korolkovi (Rai. et Schmalh.) Maxim.]. На красноцвет-
ных, нередко щебнистых почвах, песчанково-лаготисовая ассоциация
сменяется близкой кузиниево-песчанково-лаготисовой ассоциацией, ос-
новное отличие которой заключается в обилии Cousinia acicularis. В ниж-
них частях склонов, на пологих, часто вогнутых мелкоземистых участках,
где дольше залеживается снег, развивается кузиния блестящая (Gousi-
nia splendida С. Winkl.), среди которой в массе разрастается пажитник
96
А. С. Королева
Попова (Trigonella Popovii Е. Kor.), Polygonum fibrilliferum Korn, и
некоторые другие (пажитниково-кузиниевая ассоциация).
В верховьях саев, где образуются большие скопления снега или
близок уровень грунтовых вод, встречаются относительно влаголюби-
вые молочаево-ширяшево-бузульниковая и гречихово-бузульниковая ассо-
циации. Последняя часто встречается также узкими полосами вдоль
долин непересыхающих ручьев и речек.
Широкие долины пересыхающих летом речек, приподнятые террасы
рек с постоянным водотоком и небольшие впадины, т. е. места, где скоп-
ляются большие массы снега, обильно увлажняющие почву при таянии,
покрыты эфемероидной мятликово-осоковой ассоциацией с преобладанием
осоки (Carex duriusculiformis V. Krecz.) и большим количеством однолет-
него мятлика (Роа annua L.).
Вдоль русел непересыхающих летом небольших речек на торфянистых
почвах развивается зеленеющий в течение всего вегетационного пери-
ода густой ковер из осоки ложновонючей (Carex pseud of oetida Kiik.),
среди которой разбросаны многочисленные представители разнотравья
и злаков.
37. Кузиниево-камолево-югановая ассоциация (Prangetum feruloso-
cousiniosum) широко распространена на щебнисто-каменистых восточных
склонах в верховьях Кондары, в нижней части субальпийского пояса,
на высотах от 2400 до 2500 м. Разреженный травостой этой ассоциации
покрывает поверхность почвы на 60—70%. В травяном покрове гос-
подствует поедаемая форма югана (Prangos pabularia Lindl.), кузиния
тощая (Cousinia macilenta С. Winkl.) и камоль (Ferula Jaeschkeana
Vatke).
Остальные компоненты этой ассоциации представлены преиму-
щественно растениями, специфичными для субальпийского пояса, однако
встречается и ряд видов, распространенных в поясе древесно-кустарни-
ковой растительности и даже ниже, как Роа bulbosa L., Hypericum scab-
rum L., Scaligeria hirtula (Rgl. et Schmalh.) Lipsky и некоторые другие.
К числу таких растений относятся и юган с камолем, которые, так же
как и Роа bulbosa, на территорию Руидашта уже не заходят. 1-й, разре-
женный, ярус этой ассоциации, 150—200 см высоты, образуют генератив-
ные побеги камоля и югана, 2-й ярус, 60—170 см высоты,— листья югана
и стебли кузинии тощей и 3-й ярус, 20—25 см высоты,— листья кузинии
тощей.
38. Ферулево-кузиниевая ассоциация (Macilentae-cousinietum ferulosum)
господствует на крутых и щебнистых восточных склонах водораздела между
Кондарой и притоками Лючоба в полосе от 2500 до 2600 м. Травостой
этой ассоциации еще более редок, чем в кузиниево-камолево-югановой
ассоциации, покрывая всего лишь 40—50% поверхности почвы. В травя-
ном покрове резко преобладает кузиния тощая, среди которой весьма
характерно присутствие ферулы (Ferula ovina Bois.), ревеня (Rheum
Maximowiczi A. Los.), зверобоя (Hypericum scabrum L.). Намечаются:
1-й редкий ярус, 80—90 см высоты, образованный генеративными побе-
гами ферулы, 2-й ярус, 40—50 см, образованный генеративными побегами
кузинии тощей, и 3-й ярус, 20—25 см, составляющий основную массу
травостоя, сложенный листьями кузинии тощей.
39. Пажитниково-разнотравно-тороновая ассоциация (Polygonetum
trigonelloso-herbosum) изредка встречается на восточных склонах водо-
раздела между Кондарой и притоками Лючоба, где приурочена к неболь-
шим, зажатым между скалами ложбинкам с более или менее мелкозе-
Растительный покров Кондары
97
мистыми почвами. Растительность образует здесь высокий густой траво-
стой, покрывающий 90—100% поверхности почвы.
Господствующим растением является торон (Polygonum bucharicum
Grig.), среди которого, наряду с преобладающими элементами субаль-
пийского пояса (Trigonella Popowii Е. Kor., Psychrogeton cabulicum
Boiss., Роа bucharica Roshev., Euphorbia seravschanica Rgl. и др.),
встречаются некоторые растения, характерные для пояса древесно-кустар-
никовой растительности (Eremurus Regelii Vved., Geranium collinum
Steph., Astragalus peduncularis Royle и некоторые другие).
Ярусность выражена неясно, высота основной массы травостоя, обра-
зованного главным образом тороном,—90—100 см.
Рис. 5. Разнотравно-кузиниевая ассоциация в нижней части
субальпийского пояса.
(Фот. Варзобской ботанической станции).
40. Разнотравно-кузиниевая ассоциация (Macilentae-cousinietum
herbosum) широко распространена на щебнистых и щебнисто-каменистых
склонах урочища Руидашт, являясь господствующей в верхних частях
склонов, где преобладают щебнистые и щебнисто-каменистые почвы
(рис. 5).
Разнотравно-кузиниевая ассоциация развивается преимущественно
на склонах южной и восточной экспозиции, значительно реже встречаясь
также и на северных и западных склонах. Крутизна склонов колеблется
от 25 до 30°. Господствующим растением является кузиния тощая, серо-
ватые листья которой создают монотонный колорит травяного покрова.
На фоне кузинии — постоянно, но в сравнительно небольших количе-
ствах, встречаются мятлик голоцветный (Роа glabriflora Roshev.), зверо-
бой (Hypericum scabrum L.), кузиния иглистая (Cousinia acicularis
Franch.), Scaligeria hirtula (Rgl. et Schmalh.) Lipsky и др. Травяной по-
кров разрежен, покрывает 50—70% поверхности почвы: 1-й ярус траво-
стоя, 50—70 см высоты, образуют генеративные побеги кузинии тощей,
Ущелье Кондара
98
А. С. Королева
2-й ярус, 20—40 см высоты, сложен главным образом листьями той же
кузинии.
41. Разнотравно-ромашниковая ассоциация (Pyrethretum herbosum)
имеет сравнительно небольшое распространение в средней части Руи-
дашта, где занимает щебнистые и щебнисто-каменистые вершины и верх-
ние части южных и восточных склонов пологих увалов. По характеру
растительного покрова эта ассоциация близка к предыдущей, отличаясь,
главным образом, отсутствием или редкой встречаемостью кузинии то-
щей. В разнотравно-ромашниковой ассоциации нет резко преобладающих
видов — несколько чаще других растений встречается ромашник (Pyre-
thrum mucronatum Rgl. et Schmalh.), к которому присоединяются астра-
гал (Astragalus lasiosemius Boiss.), мятлик голоцветный (Роа glabriflora
Roshev.), Scaligeria hirtula (Rgl. et Schmalh.) Lipsky, Polygonum fibril-
liferum Кот. и другие растения, обычные и для разнотравно-кузиниевоп
ассоциации. Травяной покров разрежен, покрывает 50—60% поверх-
ности почвы. Средняя высота его 30—40 см.
42. Песчанково-лаготисовая ассоциация (Lagotidetum arenariosum)
весьма обычна в районе Руидашта, где приурочена к сравнительно поло-
гим (15—25°) мелкоземистым склонам северной, северо-западной и за-
падной экспозиции. Травяной покров сравнительно густ, покрывает
80—90% поверхности почвы. Господствующими растениями являются
лаготис [Lagotis Korolkovi (Rgl. et Schmalh.) Maxim.] и песчанка (Arenaria
Griffith!! Boiss.), несколько менее обильны теке-сагыз (Scorzonera acan-
thoclada Franch.) и Psychrogeton cabulicum Boiss. Почти всегда в тех или
иных количествах встречаются смолевка (Silene tachtensis Franch.),
мятлик голоцветный, пажитник Попова, астрагал андаульгенский
(Astragalus andaulgensis В. Fedtsch.), Phytheuma argutum Rgl. и неко-
торые другие. В травяном покрове намечается следующая ярусность:
1-й ярус, 30—40 см высоты, образуют генеративные побеги Silene tachten-
sis Franch., Phytheuma argutum Rgl. и некоторые другие, 2-й ярус, 10—
15 см высоты,— теке-сагыз, 3-й ярус, составляющий главную массу тра-
востоя, 2—5 см высоты, сложен песчанкой, лаготисом, Psychrogeton ca-
bulicum Boiss. и др.
43. Кузиниево-песчанково-лаготисовая ассоциация (Lagotidetum cou-
sinioso-arenariosum) довольно широко распространена в районе Руи-
дашта. Она встречается, главным образом, в северо-восточной части Руи-
дашта в окрестностях горы «Полосатая», характеризующейся преиму-
щественным развитием красноцветных почв. Склоны, занятые кузиниево-
песчанково-лаготисовой ассоциацией, мелкоземисты или мелкоземисто-
щебнисты и обращены на север, северо-запад или северо-восток. По
характеру растительности описываемая ассоциация близка к предыду-
щей, от,которой отличается, главным образом, обилием кузинии иглистой
(Cousinia acicularis Franch.) и значительно большей распространен-
ностью пажитника Попова. Как можно предполагать, обилие этих двух
растений (указывающих на места наиболее интенсивного выпаса) свя-
зано с тем, что выпас в окрестностях горы «Полосатая» интенсивнее, чем
в остальной части Руидашта, где распространена песчанково-лаготисовая
ассоциация. Кузиниево-песчанково-лаготисовая ассоциация приурочена
не только к красноцветным почвам; она, хотя и значительно реже, но
встречается и на других почвах.
44. Пажитниково-кузиниевая ассоциация (Splendidae-cousinietum tri-
gonellosum) встречается сравнительно редко на мелкоземистых, нередко
вогнутых участках в нижних частях склонов, где долго залеживается
Растительный покров Кондары
99
снег. Склоны, занятые пажитниково-кузиниевой ассоциацией, довольно
пологи и обращены преимущественно на север или запад, реже — на юг
и восток.
Фон ассоциации образует эффектная кузиния блестящая (Cousinia
splendida С. Winkl.), в нижнем ярусе всегда в больших количествах при-
сутствует мало заметный на первый взгляд, низкорослый пажитник
Попова (Trigonella Popovii Е. Kor.). Несколько менее обильны Polygo-
num fibrilliferum Кот., Роа glabriflora Roshev. и др.
1-й ярус травостоя, 70—80 см высоты, образуют генеративные побеги
кузинии блестящей, 2-й ярус, 40—50 см высоты. — генеративные побеги
Роа glabriflora Roshev., 3-й ярус, 3—18 см высоты, — пажитник Попова
и Polygonum fibrilliferum Кот.
45. Молочаево-ширяшево-бузульниковая ассоциация (Ligularietum
euphorbioso-eremurosum) встречается очень небольшими пятнами в вер-
ховьях саев юго-восточной части Руидашта. Условия для существования
относительно мезофитных представителей этой ассоциации (бузульник,
молочай, ширяш) создаются здесь, повидимому, благодаря более длитель-
ному залеживанию снега во впадинах, у верховий саев. Участки склонов,
занятых молочаево-ширяшево-бузульниковой ассоциацией, довольно по-
логи и характеризуются мелкоземистыми почвами.
Господствующими растениями являются бузульник (Ligularia рег-
sica Boiss.), молочай (Euphorbia glomerulans Prokh.) и ширяш (Ere-
murus Regelii Vved.). В больших количествах встречается также песчанка
(Arenaria Griffithii Boiss.).
Травостой покрывает 80—90% поверхности почвы. 1-й ярус, 120—
140 см высоты, образуют генеративные побеги ширяша, 2-й ярус, 50—
60 см высоты — молочай, листья бузульника, зопник (Phlomis frutice-
torum N. Gontsch.) и некоторые другие.
46. Гречихово-бузульниковая ассоциация (Ligularietum polygonosum)
встречается довольно часто узкими полосами вдоль речных долин, а также
в верховьях саев, где, видимо, неглубоко залегает уровень грунтовых
вод. Несколько ниже в долинах подобных саев можно видеть мочажины,
разделенные пересохшими участками русла; еще ниже струится уже
небольшой ручеек. Гречихово-бузульниковая ассоциация, резко выделя-
ющаяся светлой зеленью господствующего бузульника, образует довольно
густой (степень покрытия почвы 80—90%) и высокий травяной покров.
В травяном покрове заметное участие принимают щавель-шульха (Rumex
Paulsenianus Rech, f.), молочай зеравшанский (Euphorbia seravschanica
Rgl.), торон (Polygonum bucharicum Grig.), мятлик бухарский (Роа
bucharica Roshev.) и некоторые другие. В нижнем ярусе господствует
гречиха (Polygonum fibrilliferum Кот.).
Высота 1-го яруса, образованного генеративными стеблями бузуль-
ника и шульхи, 80—90 см, 2-й ярус, 50—60 см высоты, сложен в основном
листьями бузульника, 3-й ярус, 2—5 см высоты, составляет гречиха.
47. Мятликово-осоковая ассоциация (Duriusculiformi-caricetum рое-
osum) довольно часто встречается на территории урочища Руидашт
в виде небольших пятен или узких полос по сухим логам, на террасах
речек, в западинах, т. е. в местах, обильно увлажняемых весной тающим
снегом. Эта ассоциация, образующая сразу же после стаивания снега
густой зеленый ковер, носит эфемероидный характер, быстро засыхая
в первой половине лета. Господствующим растением является эфемероид-
ная осока (Carex duriusculiformis V. Krecz.). К осоке в значительных
количествах присоединяется однолетний мятлик (Роа annua L.), часто
7*
100
А. С. Королева
встречаются низкорослый подорожник (Plantago lanceolate L.), пажитник
Попова (Trigonella Popovii Е. Kor.), Scaligeria hirtula (Rgl. et Schmalh.)
Lipsky, лаготис и некоторые другие.
1-й ярус, 20 см высоты, образуют генеративные побеги мятлика и подо-
рожника, 2-й ярус, 6—7 см высоты, сложен осокой, пажитником и неко-
торыми другими.
48. Разнотравно-осоковая ассоциация (Pseudofoetidae-caricetum her-
bosum), зеленеющая в течение всего вегетационного периода, занимает
прирусловые полосы вдоль непересыхающих речек, а также окружает
выходы ключей. Эта ассоциация характеризуется невысоким (20—25 см
высоты) густым покровом (покрытие 100%), в котором преобладает осока
ложновонючая (Carex pseudofoetida Kiik.). Среди осоки в довольно зна-
чительных количествах разбросаны: клевер ползучий (Trifolium repens L.),
лапчатка (Potentilla gelida С. A. M.), лютик (Ranunculus alajensis
Ostenf.), ясколка [Cerastium cerastoides (L.) Brit.] и некоторые другие.
Иногда встречаются осока округлая (Garex orbicularis Boott.) и кобрезия
(Cobresia pamiroalaica). Два последние растения у воды местами обра-
зуют небольшие пятна зарослей, представляющих уже ассоциации
Orbiculari-caricetum и Cobresietum. Среди осоковых и кобрезиевых
ассоциаций вдоль берегов речек нередко встречаются узкие бордюры
из Ligularia persica Boiss., лисохвоста (Alopecurus seravschanicus
Ovcz.), щавеля-шульхи (Rumex Paulsenianus Rech, f.) и некоторых дру-
гих.
ЖИВОТНЫИ МИР
Б. С. ВИНОГРАДОВ
МЛЕКОПИТАЮЩИЕ
Млекопитающие Гиссарского хребта изучены очень мало. Имеющиеся
в литературе данные ограничиваются преимущественно немногими ука-
заниями о нахождении отдельных видов в некоторых пунктах хребта.
Приводимые здесь краткие сведения о млекопитающих южного склона
Гиссарского хребта — в долине р. Варзоб, ущелье Кондара, на плато
Руидашт н др. — основаны на исследованиях сотрудников Зоологического
института Академии Наук СССР.
Отр. Грызуны — Rodentia
1. Краснохвостый сурок — Marmota caudata Jackem. На территории
Горной ботанической станции и в смежных районах сурки не были най-
дены. В 1946 г., по словам А. И. Иванова, сурок был добыт в истоках
Кондары, в урочище Оби-сафед. Судя по сохранившимся норам на склоне
Руидашта, сурки здесь жили еще совсем недавно, что подтверждается
также указаниями местных жителей. Ближайший район, где сурок сей-
час еще не редок, — южные склоны главного хребта близ Анзобского
перевала.
2. Лесная соня — Dyromys nitedula Pall. Изредка встречается на
территории станции. Более обычен этот зверек в урочище Квак, в кле-
новом лесу, на высоте около 2000 м.
3. Дикобраз — Hystrix leucura Sykes (=hirsutirostris Br.). Широко рас-
пространен в долине Варзоба, от лёссовых предгорий (р. Гюльбиста) до
среднего пояса гор. В ущелье Кондара он также не редок и встречается
здесь всюду от устья Кондары до урочища Квак. Норы этого животного
приурочены преимущественно к крутым скалистым обрывам; отверстия
нор обычно располагаются в скалистых нишах или под защитой больших
обвалившихся обломков скал. Во многих местах на скалистых участках
в каменных нишах имеются также лёжки дикобраза, большие скопления
его помета и разбросанные иглы.
В противоположность большинству других грызунов, дикобраз яв-
ляется бродячим животным, уходящим на большие расстояния от нор.
Излюбленными путями его передвижения служат тропинки, протоптан-
ные человеком, вьючными животными или пасущимся скотом; на этих
тропинках можно каждое утро наблюдать свежие следы дикобраза,
находить его помет и отдельные иглы. В своих странствованиях дико-
браз нередко заходит в населенные места и даже подходит близко к жи-
лым постройкам; часто это животное проходит по трудно доступным и
недоступным для человека крутым скалам.
104
В. С. Виноградов
Дикобраз принадлежит к числу немногих грызунов, которые ну-
ждаются в регулярном питье воды. Это легко проследить по следам зверя,
ведущим к воде, и оставленным им у воды иглам. Весной и в начале лета
в районе Варзобской станции дикобраз питается преимущественно под-
земными частями разнообразных растений — корнями, корневищами и
луковицами. Это можно установить по остаткам поеденных им растений,
а также по составу помета; в частности, ранней весной можно находить
в большом количестве обгрызенные дикобразом корни камоля — Ferula
kokanica, тарана — Polygonum bucharicum, луковицы тюльпанов (осо-
бенно Tulipa hissarica) и др.
Летом и в начале осени дикобраз питается преимущественно плодами
диких и культурных растений. Тогда его помет состоит почти исключи-
тельно из косточек различных плодов, особенно дикого винограда, алычи,
дикой вишни (Prunus mahaleb) и др. В это же время года дикобраз при-
чиняет наиболее серьезные повреждения культурным растениям, осо-
бенно кукурузе, дыням, арбузам и тыквам.
В первую половину лета на участках Варзобской станции поврежде-
ния дикобраза были заметны, главным образом, на виноградниках:
дикобраз скусывал молодые побеги винограда и черешки листьев; ме-
стами на посадках богарного винограда были повреждены до 50% кустов.
Особенно значительный вред дикобраз причиняет во второй половине
лета после созревания огородных и бахчевых растений; в это время он
опустошает посевы кукурузы, скусывая за одну ночь до двух десятков и
более стеблей кукурузы, причем поедает лишь небольшую часть початков
и стеблей срезанных растений; в августе были отмечены повреждения
плодов тыкв, даже совершенно незрелых; большинство были надкусаны
и лишь немногие выедены до середины. По словам сотрудников Стан-
ции, дикобраз нанес также сильные повреждения клубням картофеля.
4. Туркестанская крыса — Rattus turkestanicus Sat. Места обитания
этого животного в ущелье Кондара чрезвычайно разнообразны. Наи-
более многочисленны крысы в ореховых лесах, которые здесь приурочены,
главным образом, ко дну ущелий; крысы встречаются здесь также в за-
рослях ив по берегам речек, на скалистых участках и в каменных рос-
сыпях; наконец, они живут также на приусадебных землях и в построй-
ках, где их поселение имеет ясно выраженный сезонный характер: в на-
чале лета крысы исчезают здесь из построек почти совсем, а в октябре
появляются вновь, возрастая в численности к началу зимы. В вертикаль-
ном направлении крысы распространены здесь по крайней мере до высоты
2000 м; во всяком случае, они еще многочисленны на урочище Квак
на высоте около 2000 м, а на южном склоне главного хребта прони-
кают по горным речкам значительно выше кишлака Зидды в субальпий-
скую зону.
Наиболее многочисленны поселения крыс в ореховых рощах; здесь
они роют норы под корнями орешника или селятся в дуплах. Нередко
в старых деревьях образуются большие полости, большая часть которых
бывает заполнена трухой, состоящей из помета крыс и скорлупы пое-
денных ими орехов.
Пища крыс в рассматриваемом районе чрезвычайно разнообразна.
Из растительных веществ они предпочитают семена и плоды различных
древесных пород, особенно грецкого ореха. Погрызенную крысами скор-
лупу ореха можно встретить в больших количествах не только в ореховых
рощах, но даже далеко от зарослей орешника, иногда до V2 км от ближай-
ших ореховых деревьев. Около нор крыс и в дуплах можно находить
Млекопитающие.
105
также погрызенную скорлупу косточек алычи, абрикоса, персика, дикой
вишни, каркаса (Celtis), фисташки и других плодов диких и культурных
деревьев. Крыса питается также животной пищей, о чем можно судить
по находимым среди остатков их пищи в дуплах костям мелких млекопи-
тающих и птиц со следами резцов крысы.
Туркестанская крыса принадлежит к числу наиболее вредных грызу-
нов в рассматриваемом районе; в наибольшей степени этот грызун вредит
в складочных помещениях, жилых постройках, приусадебных землях
(особенно огородах) и в ореховых рощах.
5. Домовая мышь — Mus musculus L. В ущелье Кондара домовая
мышь была добыта лишь однажды близ построек Горной ботанической
станции, среди больших камней, заросших диким виноградом. В лёс-
совых холмах предгорий Гиссарского хребта и на территории Сталин-
абада этот зверок очень обычен.
6. Лесная мышь — Apodemus sylvaticus L. Является наиболее много-
численным и наиболее широко распространенным грызуном ущелья Кон-
дара и прилежащих районов. Местообитания ее чрезвычайно разнооб-
разны. Наиболее высокая численность лесной мыши наблюдается на се-
верных склонах ущелья в зарослях клена, алычи, иргая, ореха, дикого
винограда и др. Лесная мышь многочисленна также в каменных осыпях
и скалах, часто лишенных древесной растительности. В меньшем коли-
честве она встречается на южных склонах ущелья с более разреженными
древесными зарослями и рано выгорающей травянистой растительно-
стью.
Основной пищей лесной мыши на южных склонах Гиссарского-
хребта являются плоды и семена различных диких п культурных дре-
весных пород, в том числе грецкого ореха, алычи, персика, абрикоса,
миндаля, фисташки, дикой вишни, каркаса (Celtis), клена и др.
Кроме вреда, причиняемого лесной мышью естественному возобнов-
лению леса, этот зверок вредит также богарным посевам пшеницы; здесь
лесная мышь селится под оставшимися среди пашни кустами и под кам-
нями и приносит значительный вред скусыванием колосьев и вылущива-
нием зерен.
7. Серый хомячок — Cricetulus migratorius Pall. В нижних частях
долины Кондары встречается редко, чаще попадается на более значи-
тельных высотах (например, на северных склонах ущелья, на высоте
1600—1700 м, и в урочище Квак, на высоте около 2000 м). В наибольшем
количестве был находим на южном склоне главного хребта, выше киш-
лака Зидды.
8. Арчевая полевка — Microtus carruthersi Thos. Очень характерна
для субальпийской зоны плато Руидашт, но встречается и на менее
значительных высотах, даже в урочище Квак, где этот зверек обитает
в необычной для него обстановке — среди кленового леса, и роет норы
под корнями деревьев.
9. Слепушонка — Ellobius talpinus Pall. В окрестностях Варзобской
станции встречается на высоте около 2000 м и выше, преимущественна
на мелкоземистых склонах, в частности, по гребню северного склона
ущелья Кондара; здесь норы этого зверька встречаются не только на
открытых безлесных местах, но даже среди кустарников и в кленовых
рощах. Более многочисленна слепушонка в субальпийской зоне Руи-
дашта, особенно на его пологих склонах, со стороны кишлака Ходжа-
оби-гарм. Весною первые следы роющей деятельности были замечены
7 марта.
106
Б. С. Виноградов
Отр. Насекомоядные — Insectivora
10. Землеройка-белозубка — Crocidura suaveolens Pall. В ущелье
р. Кондара редка. Встречается в разнообразных местообитаниях: среди
леса, в скалах, осыпях, прибрежных зарослях и пр.
Отр. Рукокрылые — Chiroptera
Летучие мыши в ущелье Кондара весьма немногочисленны, и в непо-
средственной близости от построек Станции можно было наблюдать по
вечерам не более 3—5 экземпляров. Видовой состав этих зверьков уста-
новить не удалось, так как ни один экземпляр здесь не был добыт.
Отр. Хищные — Carnivora
11. Лисица — Vulpes vulpes L. Была неоднократно наблюдаема на
территории Варзобской станции. Среди скал и осыпей здесь нередко
можно находить остатки растерзанных кекликов и других птиц, пови-
димому, добытых лисицами.
12. Волк — Canis lupus L. В Кондаринском ущелье не составляет
редкости. Летом держится в высокой зоне гор, в частности, на плато
Руидашт, где в это время пасутся стада овец. Зимой обычен в нижних
частях ущелья, посещая и район Горной ботанической станции.
13. Степная кошка — Felis ocreata Gmel. Встречающаяся изредка на
территории Варзобской станции дикая кошка, повидимому, принадлежит
к этому виду, свойственному преимущественно равнинам. Шкуры этой
кошки нередко поступают на Варзобский заготовительный пункт.
14. Медведь — Ursusarctos L. Принадлежит к довольно редким обита-
телям ущелья Кондара. Более стационарно он держится начиная с высоты
1500м, в частности, в кленовниках Квака и выше, в том числе и на открытом
плато Руидашт. Как только в нижних частях ущелья начинается созре-
вание плодовых деревьев, а позже — дикого винограда и диких яблок, мед-
веди начинают регулярные ночные посещения района станции, на день
•снова уходя в горы. Свои переходы животные совершают, пользуясь
пешеходными и вьючными тропами. День медведь проводит на лёжке,
по крайней мере, видеть его тогда не случается. Забравшись в культур-
ные сады, он нередко ломает деревья, например персиковые, поедает
виноград ит. д.; алычу или яблоки подбирает упавшими на землю, в за-
рослях дикого винограда производит целые опустошения, добираясь до
спелых гроздий. Так как дикий виноград по ущелью не поднимается выше
1300—1400 м, медведям, придерживающимся высот Руидашта, приходится
за ночь предпринимать спуск и подъем на 1500 м, что, видимо, не представ-
ляет для них затруднений. По окончании сезона фруктов жизнь животных
проходит в более высоких зонах, там же ложатся они и на зимнюю спячку.
15. Выдра — Lutra lutra L. В 1944 г. сотрудниками Варзобской стан-
ции экземпляр выдры был наблюдаем при впадении р. Кондара в р. Вар-
зоб, где выдра опустошала устроенные здесь ловушки для рыбы.
Из других хищников в верхних частях ущелья может быть найден
горностай (Mustela erminea L.), как об этом можно судить по заготовкам
из различных пунктов горного склона Гиссарского хребта шкурок гор-
ностая, поступающих на Варзобский заготовительный пункт.
Млекопитающие
107
Отр. Парнокопытные — Artiodactyla
16. Горный козел — Capra sibirica Meyer. Сотрудниками Варзобской
станции изредка был наблюдаем зимой на склонах ущелья Джеринод-
сой. Зимой 1944—1945 г. один экземпляр горного козла был добыт в Та-
ковском ущелье над поселком Такоб.
ЛИТЕРАТУРА
Виноградов Б. С. и А. И. Иванов. 1945. Грызуны Таджикистана. Сталин-
абад.
Соснина Е. Ф. 1949. Питание мышевидных грызунов южных склонов Гиссар-
ского хребта. Сообщ. Тадж. фил. АН СССР, вып. 12.
Соснина Е. Ф. 1949. О размножении некоторых мышевидных грызунов южных
склонов (Гиссарского хребта. Там же, вып. 18.
|А. я. тугаринов’
ПТИЦЫ
Состав орнитофауны южных склонов Гиссарского хребта выяснен
сравнительно полно. Если дальнейшие исследования, возможно, и допол-
нят список населяющих его видов новыми находками, то это существенно
не изменит имеющихся представлений об общем облике фауны и ее верти-
кальном распределении. Ущелье Кондара, изученное сейчас довольно
детально, может рассматриваться как населенное всеми характерными
видами птиц, по крайней мере, в пределах охватываемых им вертикаль-
ных зон. Иными словами, здесь отсутствует высокогорный элемент, хотя
частично птицы альпийской и нивальной зон появляются в различных
частях ущелья зимою.
Таким образом, состав и размещение птичьего населения в районе
Кондаринского ущелья дает картину, достаточно типичную для южных
склонов Гиссарского хребта.
Список птиц ущелья, по наблюдениям сотрудников Зоологического
института в 1944 г., пополненный данными А. И. Иванова, включает
135 видов и подвидов птиц. Имея ввиду такой сравнительно ограниченный
район, как само ущелье и его склоны, состав этот можно считать довольно
разнообразным. По характеру пребывания эти 135 форм распределяются
следующим образом: 29 принадлежат к оседлым обитателям района,
52 прилетают для гнездования, 29 появляются только на пролете и 25
проводят зиму. Последняя группа состоит из 16 видов, спускающихся из
более высоких частей Гиссарского хребта, и только 9 принадлежат к дальне-
прилетным с севера, проводящим здесь свое внегнездовое время. В этот
подсчет не входят некоторые виды, наблюдавшиеся хотя и в районе
ущелья, но лишь в соседних частях долины р. Варзоб. Так, во время
пролета на этой реке иногда появляются стайки уток: кряковых, шило-
хвостей, серух и чирков-свистунков, с трудом находящих спокойные за-
ливчики на этой бурной, стремительно несущейся реке.
Прежде чем перейти к характеристике размещения и образа жизни
гнездящегося, а также временно-оседлого населения, остановимся на явле-
ниях пролета. Поскольку ущелье Кондара лежит в окраинной зоне юж-
ных склонов Гиссарского хребта, пересекаемого здесь с севера на юг
долиной р. Варзоб, — в районе ущелья можно было бы ожидать обиль-
ного пролета как для видов, следующих к северу, так и для размещаю-
щихся во внутренних частях хребта. Наблюдения показали, что
сколько-нибудь обильного пролета по долине р. Варзоб не происходит.
Основная масса птиц следует рассыпным строем, появляясь то в большем,
то в меньшем числе особей. Так, некоторые, например варакушки, появ-
ляются весною (3 III), как и осенью (11 IX), буквально редкими оди-
Птицы
109
ночками, другие, например сибирские пеночки, пролетают очень обильно,
и их движение, начинаясь в середине марта, затягивается на два месяца.
То же и осенью: первые пеночки появляются в 20-х числах сентября,
лёт идет в течение октября и всего ноября, а одиночки задерживаются
до первой трети декабря. Поскольку эта пеночка в Средней Азии не гнез-
дится, можно установить, что в своем пролетном движении птичка пере-
секает Гиссарский хребет. Ограниченный пролет некоторых видов связан,
вероятно, с тем, что в районе ущелья появляются только те особи, кото-
рые затем будут гнездиться где-то в более внутренних частях хребта или
в более высоких его зонах. С 20-х чисел марта по склонам Кондаринского
ущелья появляются, например, отдельные особи чекана-плясуна или
рогатых жаворонков, держатся здесь десяток дней, а затем исчезают.
Вероятнее всего это те особи, которые поднимаются все выше в горы,
чтобы загнездиться затем на водораздельных субальпийских лугах.
Виды, гнездящиеся в лесной зоне, также некоторое время по прилете
вынуждены задерживаться в нижних частях ущелья, чтобы по мере тая-
ния снегов и развития растительности иметь возможность подняться на
свои горные стации. К ним относятся такие птицы, как обыкновенная
чечевица, лесной конек, пеночка Юма, серая славка и др.
Общий обзор птичьего населения ущелья Кондара и его распределе-
ния мы сделаем применительно к двум основным сезонам: зиме и лету.
В первом случае наша задача значительно облегчается тем, что зимою
птицы ограничиваются пребыванием в нижних частях ущелья — не выше
2000 м. Наибольшее разнообразие наблюдается в районе Горной ботани-
ческой станции, на высотах 1100—1400 м. Сосредоточенное здесь птичье
население состоит частью из оседлых, гнездящихся здесь видов, частью
из спустившихся из более высоких зон хребта, а также некоторого числа
зимующих, прилетных с севера. По самой р. Кондара, на скалистых обры-
вах, наблюдатель замечает чаще других синюю птицу, а на обтекаемых
водою камнях, среди стремнин, — изящную белоножку (рис. 1).
Неразлучные с водою оляпки по самой Кондаре не держатся, но по
Варзобу, на устье Кондары, оба вида оляпок зимою обычны. Там, где по
берегу речки есть мочажины, можно вспугнуть горного дупеля. Наиболее
оживлены древесные поросли по дну ущелья. Прежде всего здесь при-
влекают внимание кустарницы, скрывающиеся в густых зарослях,
и здесь же шмыгают в чаще многочисленные крапивники. Время от вре-
мени, обшаривая деревья, следуют смешанные стайки синиц — больших
серых, рыжешейных и желтогрудых. Обычны, хотя и не многочисленны,
черные дрозды.
Если, оставив дно ущелья, мы начнем подниматься по его южному
бесснежному склону, то прежде всего встретимся с кекликами. Не поки-
дая даже усадьбы Станции, в любой зимний день можно слышать голоса
этих красивых курочек. На скалистых обрывах, а то и на крыше дома,
случается заметить скалистого поползня (рис. 2), иногда здесь же мель-
кнет молчаливый стенолаз. Большее оживление замечается в боковых,
поросших деревьями ущельях. Здесь, прежде всего, привлекают вни-
мание кочующие шумливые стайки чернозобых дроздов. Едва они про-
следуют, как появляются общества дубоносов, задерживающиеся на
деревьях каркаса (Celtis), чтобы покормиться его ягодами. Также на
деревьях можно увидеть красноспинных горихвосток, юров, замешав-
шихся среди стаек щеглов, белошапочных овсянок. Если среди деревьев
есть достаточно старые, нередко можно заметить белокрылого дятла.
Особую колоритность придает древесным насаждениям присутствие
110
А. Я. Тугаринов
стаек розовых чечевиц. Всю зиму птички держатся строго оседло, изо
дня в день их можно встретить на одном месте, на тех же самых деревьях.
Если подниматься по склонам южных экспозиций еще выше, до гори-
зонталей порядка 1500—1700 м, мы достигнем границы, где снеговой по-
кров сохраняется всю зиму, а среди него выступают отдельные голые
скалы и россыпи крупных обломков. Здесь мы встретимся с небогатым
видами, но весьма характерным комплексом птиц. Дружными, много-
численными стайками, рассыпаясь по склону, торопливо перебегая
с места на место или перелетая с камня на камень, здесь кормятся алтай-
ские вьюрки. Так же держатся менее многочисленные памирские вьюрку
Рис 1. Белоножка — Microciclila scouleri (Vig.).
а изредка к ним присоединяются и альпийские вьюрки, образуя само-
стоятельные стайки. После снегопадов, когда граница сплошного снего-
вого покрова снижается временно до 1000 м и ниже, эти птички, гони-
мые бескормицей, спускаются до этих горизонталей. Зимний аспект
орнитофауны ущелья был бы не полон, если бы мы не упомянули обык-
новенную пустельгу, стационарно пребывающую на южных, пологих
склонах, где ее соседом оказывается, также строго оседлая, сорока.
В утренние часы над ущельем с удивительным постоянством можно ви-
деть пролетающие на юг, в предгорья, очевидно на пашни, стаи обыкно-
венных сизых голубей, в середине дня возвращающихся снова куда-то
в глубь хребта. И, наконец, с не меньшей регулярностью в первую поло-
вину дня над ущельем появляются стаи альпийских галок, очевидно,,
также совершающих свой дневной облёт в поисках корма.
На северных склонах ущелья птицы почти отсутствуют: сплошной
снеговой покров, облепленные снегом или ледяной корочкой деревья
Птицы
Ilf
не привлекают их сюда. Лишь изредка донесется позывка синичек или
постукивание дятла.
Картина распределения гораздо более разнообразного летнего насе-
ления значительно сложнее. Прежде всего прослеживается хорошо выра-
женная привязанность видов к определенным высотам и биотопам. Если
говорить о ближайших к Станции районах, лежащих в пределах 1000—
1200 м, то здесь наиболее характерный биотоп — сухие, незадернован-
ные каменистые осыпи и скалистые обрывы, с редкими деревьями мин-
даля. Только немногие птицы избирают эти места для гнездования. Чаще
Рис. 2. Скалистый поползень — Sitta tephronota Sharpe.
других здесь устраивается чекан-плешанка и овсянка Стюарта. Резкое
дневное нагревание склонов солнечными лучами трудно переносилось бы
этими гнездящимися на земле птицами, если бы свои гнезда они не
устраивали в тени, под защитой камней. Обитающая здесь же певчая
славка вьет свое гнездо на кустиках миндаля, так что, обвеваемое ветром,
оно оказывается гарантированным от перегрева.
Не часто обитает здесь синий дрозд, предпочитающий нагромождения
крупных обломков пород.
По сравнению с этой, бедной видами и числом особей, стацией дно
ущелья, а частью и его северные склоны полны оживления. На круп-
ных многолетних деревьях тута, ореха и других гнездятся иволги,
обычны щеглы, нередки дубоносы, многочисленны серые мухоловки,
славки-завирушки. По дну ущелья, в густых кустах, переплетенных
диким виноградом, находят надежное укрытие соловьи, черные дрозды,
кустарницы. Последних много и по всему северному облесенному склону.
112
А. Я. Тугаринов
Около воды, где есть скалистые выступы, по. теневой стороне привлекает
внимание синяя птица, здесь же, по соседству, устраивается горная тря-
согузка. Особенное своеобразие вносит присутствие райских мухоловок.
Как тени, мелькают они в густой листве громадных старых ореховых
деревьев. Странным кажется, при обилии мелких птичек, полное отсут-
ствие их преследователей из числа хищников, в виде, например, мелких
соколков или ястребов. Из первых изредка гнезддщийся выше, в лесной
зоне, чеглок появляется здесь исключительно редко, а ястреб-перепе-
лятник бывает только на пролете. Некоторое количество кекликов также
гнездится по северному склону. Для полноты перечня наиболее характер-
ных видов нельзя не упомянуть и домашнего воробья. Будучи пере-
летной птицей, поздно прилетающей, для гнездованья он устраивается
на деревьях, растущих по дну ущелья, а по выводе птенцов кочует
в лесной зоне.
Описываемые биотопы нижней части ущелья в основном остаются
теми же до высоты 1800 м. В связи с этим состав населения птиц как на
северных, так и на южных склонах изменяется мало. Выше, где начи-
нается господство кленовых насаждений, появляется ряд новых видов.
Прежде всего обращает внимание наличие кукушки, совершенно отсут-
ствующей в нижних частях ущелья (ниже 1800 м). Можно даже сказать,
что эта обычнейшая птица предпочитает здесь верхнюю границу леса
и ближайшие части субальпийской зоны. По ночам в кленовниках урочища
Квак дают о себе знать серая пясыть и сплюшка. Там, где много старых оре-
ховых деревьев, нередок белокрылый дятел. В верхней части лесного пояса
при наличии древовидного можжевельника (арчи) держится королько-
вый вьюрок, в этой же зоне и выше вместе с обыкновенной чечевицей
встречается краснокрылый чечевичник. Изредка гнездится гималайская
пищуха, обычны синицы — желтогрудая лазоревка и рыжешейная.
Подобно кукушке, почему-то только здесь, по окраинам кленовых рощ,
держится чернолобый сорокопут. По рекам Уноу и Оби-сафед А. И. Ива-
нову в этой же зоне удалось добыть рыжехвостую мухоловку и король-
ковую пеночку. В глухих участках леса в большом числе гнездятся
дрозды-дерябы, в скалах, у границы леса нередок крапивник. Из птиц,
гнездящихся в нижних частях ущелья, в зону кленовников поднима-
ются: иволга, щеглы, серые синицы, кустарницы, изредка райские мухо-
ловки, соловей, черный дрозд. Из этих перечней видно, что биотоп клено-
вых лесов имеет наиболее разнообразное птичье население. Некоторые
виды, например черный дрозд или соловей, очень многочисленны.
Выше лесной зоны начинается пояс субальпийского высокотравья,
«среди которого местами разбросаны пятна розариев. Здесь гнездятся
корольковый вьюрок, обыкновенная чечевица. Странно бывает видеть этих
лесных птичек качающимися на верхушках стеблей камоля (Ferula) или
других трав, достигающих громадного роста. К числу видов, связанных
только с этим поясом, относятся черноголовый чеканчик и серая славка.
Интересно также присутствие здесь удода, а при наличии скал —
горной овсянки (рис. 3) и горихвостки-чернушки. На подступах к Руи-
дашту, т. е. уже в зоне, где высокотравье сменяется горными лугами
(выше 2500 м) с ксерофитной флорой, держится степной конек, рогатый
жаворонок, кое-где каменный воробей; среди скалистых выступов очень
обыкновенна горная овсянка и горихвостка-чернушка. Здесь же среди
скал находит себе удобные для гнезд места скалистая ласточка.
Интересно, что некоторые виды оказываются совершенно не связанными
зональностью. От устья Кондары и до плато Руидашта — всюду можно
Птицы
113
обнаружить гнездящимся чекана-плешанку. От района Станции до роза-
риев под Руидаштом одинаково обыкновенны славка-завирушка, садовая
камышевка, синий каменный дрозд; даже желчная овсянка еще гнез-
дится в зоне высокотравья. Белогорлый соловей, гнездящийся в одном
из ущелий в долине Варзоба против устья Кондары, держится также до
верхней границы высокотравья под Руидаштом. Замечательно также,
что некоторые чисто лесные птицы, как, например, горлинка или дрозд-
деряба, проникают значительно выше предела леса, довольствуясь от-
дельными деревьями арчи
или приземистыми кустика-
ми жимолости.
Таковы, в основном, зо-
нальные аспекты гнездя-
щейся орнитофауны Конда-
ринского ущелья. Описанное
выше распределение птиц по
зонам и биотопам характер-
но для гнездового периода.
С прилета, а также по окон-
чании гнездования картина
размещения иная. Для ве-
сны хорошо выражен мо-
мент, когда, как уже упо-
миналось, прилетевшие пти-
цы, населяющие верхние зо-
ны хребта, вынуждены ожи-
дать возможности подняться
в гнездовые биотопы. Более
рельефно то же явление вер-
тикальных перемещений про-
слеживается осенью. Около
середины августа в нижней
части ущелья появляются
первые обыкновенные чече-
вицы — старые и молодые
птицы, видимо, семьи. Вслед
за ними спускаются серые
славки, чернолобые сороко-
путы, зеленые пеночки, при-
бывают желчные овсянки из
числа гнездившихся в более
глубоких и высоких частях
Рис. Горная овсянка — Kinberiza cia
par Hart.
хребта, тогда как местные отлетели еще ранее. В последней декаде коли-
чество перечисленных птичек заметно возрастает, появляются новые,
например желтоголовая трясогузка, черноголовый чеканчик и др. Из
субальпийской зоны в то же время скочевывает в лесную горихвостка-
чернушка. Короче говоря, идет неторопливый миграционный процесс,
при котором обитатели высоких зон прежде всего спускаются в низины
и, продержавшись на одном месте день-два, продвигаются далее.
В рамках настоящего очерка мы не имеем возможности останавли-
ваться на описании образа жизни отдельных видов. Поэтому ниже
дается общая характеристика условий существования, в которых про-
текает жизнь птиц в ущелье Кондара.
8 Ущелье Кондара
114
А. Я. Тугаринов
Как известно, период тепла наступает в Средней Азии рано. Расти-
тельность в лице эфемер начинает развиваться с конца февраля, в марте
предгорья Гиссарского хребта уже зеленеют, цветет миндаль, различ
ные косточковые и т. д. Если в один из теплых, ясных дней второй поло-
вины марта мы совершим экскурсию по ущелью, мы будем поражены
почти полным отсутствием птиц. Тогда как зимою они сосредоточивались
в нижних частях ущелья, теперь они разбрелись по предгорьям, высо-
когорные виды поднялись выше, к границе тающих снегов, а прилет ле-
тующих еще не начался. Это «бесптичье» продолжается весь март, и
только со второй декады апреля начинают появляться прилетные. От-
дельные виды прибывают далеко не одновременно, и весь процесс при-
лета растягивается по крайней мере на месяц, завершаясь тогда, когда,
по существу, уже наступило лето. Эфемеровая растительность окончила
вегетацию, деревья не только оделись листвой, но отцвели и покрыты
незрелыми плодами (тут, вишня и т. д.). Таким образом, совершенно
очевидно, что прилет птиц и их размещение по гнездовым биотопам не
стоит в прямой связи с фенологическими моментами весны, наступая
значительно позднее того времени, когда они могли бы найти себе и укры-
тие, и корм — как растительный, так и животный, в виде различных
многочисленных насекомых. Интересно, что и местные оседлые птицы
также не'спешат с гнездованьем. Кеклики, гнездящиеся на высотах
порядка 1500—1800 м, откладывают яйца не ранее конца апреля или
начала мая. Можно вообще считать, что гнездовой период для боль-
шинства птиц приходится на май и июнь месяцы, т. е. время наи-
более пышного развития растительности, ее плодоношения и обилия
насекомых. К июлю, т. е. моменту явной депрессии органической жизни
вследствие наступления засухи, птицы уже успевают вывести потомство.
На высотах более 2000 м те же сроки биологических моментов наступают
примерно на месяц позже. Состояние засушливости достигает и этих вы-
сот, — таким образом, период размножения также заканчивается ко вре-
мени выгорания растительности. В силу создавшейся обстановки пти-
цам становится невозможно оставаться на гнездовых биотопах. Даже
под пологом леса травяная растительность высыхает, поверхность почвы
распыляется. На высокотравных лугах побуревшие травы стоят покры-
тые слоем лёссовой пыли, приносимой горячими ветрами («афганец»).
Поверхность открытых склонов, высыхая, покрывается твердой, как
асфальт, коркой. Таким образом, единственными местами, где еще продол-
жают вегетировать растения и есть еще деятельные насекомые, остаются
ущелья с родниковыми водами илп речушками. Сюда-то и сваливает
птичье население. Ущелье Кондара, орошаемое потоком, заросшее густой
уремой с обилием плодоносящих деревьев, главным образом алычи, ди-
кого винограда и сочных трав, дает обильный и разнообразный корм
птицам. Интересно, что многие насекомоядные птицы в это время пере-
ходят на растительный корм. Можно увидеть серых и певчих славок,
серых мухоловок, поедающих ягоды тута, вишни и винограда. Послед-
ний особенно привлекает также домашних воробьев, которые выедают
мякоть, оставляя косточки и кожицу. Плоды каркаса дают обильную
пищу дубоносам; черный и синий дрозды, находившие ранее корм на по-
верхности земли — насекомых, моллюсков, — теперь также переходят
на питание ягодами. В августе и сентябре увлажняемые части Кондарин-
ского ущелья наиболее оживлены птицами.
Таковы основные черты жизненного уклада птичьего населения
ущелья. Само собой, в нем получают отражение общие условия, господ-
Птицы
115
ствующие в Средней Азии. Период размножения не связан, как в наших
широтах, с периодом тепла, что относится как к пролетному, так и осед-
лому населению (кроме высокогорного). Порядок жизни в значительно
большей степени складывается под воздействием периода летней засухи,
побуждая одни виды покидать гнездовую область, другие — сменять
биотопы. До некоторой степени этот уклад сходен с тем, который возни-
кает в субтропических широтах, например, Малой Азии или северной
Африки, хотя здесь он и приходится на другую часть года.
Нам осталось кратко охарактеризовать фаунистические связи
птичьего населения Гиссарского хребта и, следовательно, ущелья Кон-
дары. Наиболее яркую и своеобразную черту местной орнитофауны со-
ставляет присутствие видов китайско-гималайского происхождения.
Гималайская пищуха, рыжешейная синичка, райская мухоловка, кустар-
ница (тимелия), синяя птица, белоножка — все эти виды проникают
сюда из пригималайских стран, имея там немало родственных видов.
Другой характерной чертой является присутствие специфически горных
форм, из которых одни распространены в Ирано-Туркестанской провин-
ции, доходя на запад до Малой Азии, другие идут далеко на восток, вклю-
чая Центральную Азию. К первой группе относятся, например, такие,
как краснокрылый чечевичник, горная овсянка, ко второй — альпийский,
гималайский и туркестанский вьюрки, рыжехвостая мухоловка, горная
индейка (улар) и др. Далее, в рассматриваемой части Гиссарского хребта
мы имеем представителей группы видов, которых принято называть среди-
земноморскими, так как их ареал простирается далеко на запад
по странам Средиземноморья. Сюда принадлежат такие птицы, как
щурка, краснокрылый чечевичник, желчная овсянка, певчая славка
и др. Основную массу населения составляют виды широкого палеаркти-
ческого распространения. По отношению к ним интересно отметить, что
здесь многие из них становятся, так сказать, «горными». Если в северных
частях ареала, на Русской равнине, обыкновенная чечевица, степной
и лесной коньки, соловей, дрозд-деряба, крапивник и другие населяют
низменность, в Гиссарском хребте они поднимаются на вертикали 1500—
2000 м. Отчасти это может быть связано с тем, что только на этих высо-
тах названные виды находят нужную им жизненную обстановку. Но
в то же время это связано для них со сменой биотопов. Обыкновенная
чечевица и лесной конек поднимаются выше границы леса, в пояс высо-
котравья, крапивник живет в безлесных ущельях, среди нагроможде-
ния камней вдоль русел горных потоков, и т. д. Все эти «горные»,
широко населяющие Палеарктику птицы образуют местные подвиды.
СПИСОК ПТИЦ УЩЕЛЬЯ КОНДАРА
1. Ciconia nigra L. — черный аист. В мае встречен в районе уроч.
Квак.
2. Cerchneis t. tinnunculus (L.) — обыкновенная пустельга. Оседлая
птица ущелья.
3. Hypotriorchis subbuteo (L.) — чеглок. Гнездится в верхней лес-
ной зоне.
4. Milvus m. migrans В odd. — коршун. Гнездится до высоты 1500 м.
5. Accipiter n. nisus (L.) — ястреб-перепелятник. Изредка встре-
чается на весеннем и осеннем пролетах (IV—X).
6. Gyps f. fulvus (Gm.) — белоголовый сип. Эпизодически посещает
все зоны ущелья.
8*
116
А. Я. Тугаринов
7. Neophron percnopterus (L.) — стервятник. При облетах бывает
во всем ущелье, в районе Горной ботанической станции, видимо, гнез-
дится.
8. Aegvpius monachus (L.) — Черный гриф. Найден дважды: 25IV
1946 на Руидаште и 15 III 1947 на Кваке.
9. Gypaetos barbatus (L.) — бородач. Встречен в районе плато
Руидашт.
10. Aquila chrysaetus daphanea Sev. — беркут. Бывает только про-
летом.
И. Hieraetus pennatus (Gm.). — орел-карлик. Изредка в районе
Станции.
12. Hieraetus fasciatus (Vieill.) — ястребиный орел. То же.
13. Buteo rufinus Cretzschm. — канюк. Встречен на осеннем про-
лете.
14. Circus cyaneus (L.) — полевой лунь. То же.
15. Pandion haliaetus (L.) — скопа. Наблюдалась по р. Варзоб.
16. Coturnix coturnix (L.)—перепел. Отмечен 10 VI 1948 выше
Квака.
17. Alectoris k. kakelik (Falk.) — кеклик. Гнездится по всему ущелью.
18. Tetraogallus himalayensis subsp. — улар. Зимою бывает под Руи-
даштом и даже спускается в окрестности кишлака Гажни.
19. Grus grus (L.) — серый журавль. Пролетает, не останавливаясь,
весною.
20. Actitis hypoleucus (L.) — перевозчик. Гнездится по р. Варзоб
в районе устья р. Кондара.
21. Scolopax rusticola L. — вальдшнеп. Редок на осеннем про-
лете (X).
22. Capella solitaria (Hodgs.) — горный дупель. Встречается изредка
зимою по р. Кондара.
23. Columba livia neglecta Hume — сизый голубь. Оседлый в нижней
части ущелья.
24. Columba palumbus casiotus (Bp.) — вяхирь. Гнездится по верхней
границе леса.
25. Streptopelia orientalis meena (Sykes) — большая горлица. Много-
численна до зоны высокотравья, выводит птенцов дважды в лето.
26. Cuculus canorus telephonus Heine — кукушка. Гнездится только
в верхней лесной зоне.
27. Merops apiaster L. — щурка. В ущелье бывает только на пролете.
28. Upupa е. epops L. — удод. Прослежен гнездящимся до высоты
1800 м.
29. Coracias garrulus semenowi Loud, et Tschusi — сизоворонка.
Встречена на гнездовье только в районе уроч. Квак.
30. Apus apus subsp. — черный стриж. Отмечен на плато Руидашт.
31. Gaprimulgus europaeus subsp. — козодой. Редок от района Стан-
ции до уроч. Квак.
32. Asio otus L.—ушастая сова. Свежие перья ушастой совы най-
дены 14 X 1945 по р. Кондаре.
33. Strix aluco harmsi Zar. — серая неясыть. Гнездится в районе
уроч. Квак.
34. Otus scops pulchellus (Pall.) — совка-сплюшка. To же.
35. Bubo bubo subsp. — филин. Гнездится по всему ущелью.
36. Dryobates leucopterus leptorhynchus (Sev.) — белокрылый дятел.
Обычен в кленовниках уроч. Квак, р. Уноу и др.
Птицы 117
37. Jynx torquilla L. — вертишейка. Встречается только на пролете.
38. Corvus corax tibetanus Hodgs. — ворон. Летом придерживается
высокогорных районов.
39. Corvus corone orientails Ev. — черная ворона. Гнездится в районе
Станции, оседла.
40. Corvus cornix sharpei Oates — серая ворона. Единственный раз
(13 III) две пары встречены в районе уроч. Квак.
41. Corvus f. frugilegus L. — грач. Зимние кочующие стаи иногда
появляются на устье р. Кондара.
42. Coloeus m. monedula (L.) — галка. Пролетные стаи появляются
весной (II—III) в долине р. Варзоб на устье р. Кондара.
43. Pica pica bactriana Bp. — сорока. Оседла в районе Станции.
44. Pyrrhocorax pyrrhocorax centralis Str. — клушица. Стаями посе-
щает ущелье во внегнездовое время.
45. Pyrrhocorax graculus forsythi Stol. — альпийская галка. То же.
46. Sturnus vulgaris subsp. — скворец. Весною пролетает долиной
р. Варзоб.
47. Oriolus oriolus kundoo Sykes — иволга. Обыкновенна до высоты
1800 м.
48. Coccothraustes coccothraustes humei Sharpe — дубонос. Гнездится
во всей лесной части ущелья.
49. Mycerobas carnipes Hodgs. — арчевый дубонос. Отмечен зимою
1948/1949 г. в Кондаре.
Е0. Carduelis caniceps subcaniceps Zar. — щегол. Гнездится до зоны
высокотравья, оседлый.
51. Serinus pusillus (Pall.) — корольковый вьюрок. Гнездится в суб-
альпийской зоне.
52. Rhodopechys sanguinea (Gould) — краснокрылый чечевичник.
Свойствен верхней лесной и субальпийской зонам.
53. E’ythrina rubicilla (Giild.) — большая чечевица. Отмечена на ко-
чевке 5 V 1947 в ущ. Робот.
54. Erythrina rhodochlamys grandis (Blyth) — розовая чечевица.
Зимою стационарно держится в районе Станции.
55. Erythrina erythrina kubanensis (Laub.) — обыкновенная чечевица.
Гнездится в верхней лесной и субальпийской зонах.
56. Fringilla с. coelebs L. — зяблик. Зимующие (XII—II) стайки
посещают ущелье.
57. Fringilla montifringilla L. — юр. Зимою вместе с предыдущим.
58. Leucosticte nemoricola altaica (Ev.) — алтайский вьюрок. Зимою
многочислен в нижней части ущелья.
58. Leucosticte brandti pamirensis Sev.— памирский вьюрок. В общих
стаях с предыдущим, но реже.
60. Montifringilla nivalis alpicola (Pall.) — альпийский вьюрок.
Зимою у границы сплошных снегов.
61. Petronia petronia intermedia Hart. — каменный воробей. В мае
встречен на подступах к плато Руидашт.
62. Passer domesticus subsp. — домашний воробей. Гнездится
на деревьях около Станции, кочующим встречен в уроч. Квак.
63. Passer hispaniolensis transcaspicus Tschusi — испанский воробей.
По р. Кондара отсутствует, но гнездится по р. Варзоб в 1 км от
Станции.
64. Emberiza bruniceps Brandt — желчная овсянка. Гнездится
до высот плато Руидашт.
118
А. Я. Тугаринов
65. Emberiza leucocephala Gm. — белошапочная овсянка. В неболь-
шом числе зимует.
66. Emberiza stewarti Blyth — овсянка Стюарта. Обычна до суб-
альпийской зоны.
67. Emberiza cia par Hart. — горная овсянка. Обычна от границы
леса и выше.
68. Calandrella a. acutirostris Hume — тонкоклювый жаворонок. 22 V
встречена стайка на плато Руидашт.
69. Otocorys alpestris albigula Bonap. — рогатый жаворонок. Гнез-
дится на плато Руидашт.
70. Anthus campestris griseus Nicoll. — степной конек. Гнездится
на плато Руидашт.
71. Anthus trivialis sibirica Sushk. — лесной конек. Встречается
на пролете.
72. Anthus trivialis harringtoni Witherby — кашмирский лесной
конек. Субальпийская зона плато Руидашт.
73. Motacilla cinerea melanope Pall. — горная трясогузка. Спора-
дично гнездится по дну ущелья.
74. Motacilla alba personata Gould. — белая трясогузка. Гнездится
в районе Станции.
75. Motacilla alba dukhunensis Sykes — западно-сибирская белая
плиска. 5 V 1947 отмечена на пролете в Кондаре.
76. Budytes flava beema (Sykes) — желтая трясогузка. Встречены
6 IX пролетные стаи на плато Руидашт.
77. Budytes citreola calcarata Hodgs. — черноспинная трясогузка.
В конце августа и начале сентября пролетом в районе Станции и плато
Руидашт.
78. Certhia himalayana taeniura Sev. — пищуха. Гнездится в верх-
ней зоне леса.
79. Tichodroma muraria L. — стенолаз. Во время кочевок посещает
устье р. Кондара.
80. Sitta t. tephronota Sharpe — скалистый поползень. Обычен
по всему ущелью.
81. Parus major ferghanensis But. — большая синица. Характерна
для всей зоны леса.
82. Cyanistes cyanus flavipectus Sev. — желтогрудая лазоревка. Гнез-
дится в районе уроч. Квак.
83. Periparus г. rufonuchalis (Blyth) — рыжешейная синица. Гнездится
в кленовниках.
84. Remiz coronatus Sev. — ремез. Гнездится по дну ущелий, подни-
маясь до субальпийской зоны.
85. Regulus regulus (L.)—желтоголовый королек. Наблюдался 1 де-
кабря 1946 г. в Кондаре.
86. Lanius minor Gm. —чернолобый сорокопут. Только в кленовниках
уроч. Квак.
87. Lanius cristatus isabellinus Hempr. et Ehrenb.—рыжехвостый
сорокопут. Отмечен на пролете 18IV, при подъеме на Квак.
88. Bombicilla garrula L. — свиристель. Изредка прилетает на зи-
мовку.
89. Siphia р. parva (Bechst.) — малая мухоловка. Редка на про-
лете.
90. Muscicapa striata neumanni Poche — серая мухоловка. Много-
численна до верхней границы леса.
Птицы
119
91. Muscicapa ruficauda Swains. — рыжехвостая мухоловка. А. И. Ива-
новым найдена гнездящейся в кленовниках уроч. Квак.
92. Terpsiphone paradisi (L.)—длиннохвостая мухоловка. Обычна
на гнездовье.
93. Phylloscopus collybita tristis Blyth. — сибирская пеночка.
Обильна на пролете.
94. Phylloscopus nitidus viridanus Blyth. — зеленая пеночка. Гнез-
дится в верхней лесной зоне.
95. Phylloscopus occipitalis Blyth. — корольковая пеночка. Встре-
чена гнездящейся в районе уроч. Квак.
96. Phylloscopus inornatus humei (Brooks) — пеночка-зарничка.
Бывает только на пролете.
97. Phylloscopus griseolus Blyth. — индийская пеночка. Пролетной
добыта в районе Станции.
98. Phylloscopus neglectus Hume — закаспийская пеночка-кузнечик.
Гнездится в уроч. Оби-сафед и Квак.
99. Locustella naevia Bodd. — обыкновенный сверчок. Бывает на
весеннем пролете.
100. Cettia cetti albiventris Sev. — соловьиная камышевка. Редка
на пролете.
101. Acrocephalus dumetorum Blyth. — садовая камышевка. В леси-
стых частях ущелья нередка на кочевках и пролете.
102. Sylvia nisoria merzbacheri Schal. — ястребиная славка. Редка
на пролете.
103. Sylvia hortensis crassirostris Cretzsch. — певчая славка. Населяет
нижнюю часть ущелья.
104. Sylvia communis rubicola Stres. — серая славка. Пролетом
по всему ущелью.
105. Sylvia curruca affinis Blyth — славка-завирушка. Гнездится
по всему ущелью.
106. Garrulax lineatus bilkevitchi (Zar.). — кустарница. Оседла, на гне-
здовье поднимается до высоты 2000 м.
107. Turdus viscivorus bonapartei Cab. — дрозд-деряба. Верхняя лес-
ная и субальпийская зоны.
108. Turdus atrogularisTemm. — чернозобый дрозд. Обычен в качестве
зимующего.
109. Turdus merula intermedia (Richm.) черный дрозд. Оседлый, гнез-
дится до верхней границы леса.
110. Monticola saxatilis (L.) — каменный дрозд. Гнездится в субаль-
пийской зоне.
111. Monticola solitaria pandoo Sykes — синий дрозд. Изредка в рай-
оне Станции, гнездится.
112. Myophoneus coeruleus turcestanicus Zar. — синяя птица. Оседла,
на гнездовье не выходит за пределы лесной зоны.
113. Oenanthe oenanthe (L.) — обыкновенная каменка. Изредка встре-
чается на весеннем пролете.
114. Oenanthe р. pleschanka (Lep.) — чекан-плешанка. Всюду от устья
р. Кондара до плато Руидашт.
115. Oenanthe isabellina (Gretzchm.) — чекан-плясун. Бывает только
на пролете.
116. Saxicola torquata maura (Pall.) — черноголовый чеканчик. Свой-
ствен субальпийской зоне.
120
А. Я. Тугаринов
117. Phoenicurus ochruros phoenicuroides (Moore) — горихвостка-чер-
нушка. Характерна для субальпийской зоны.
118. Phoenicurus erythronota (Ev.) — красноспинная завирушка.
Обычна зимою в нижней части ущелья.
119. Phoenicurus erythrogastra grandis (Gould) — краснобрюхая гори-
хвостка. Редка зимою.
120. Phoenicurus coeruleocephala Vig. — синеголовая горихвостка.
Изредка гнездится.
121. Chaimarrornis leucocephalus pamirensis Zar. et Moltch. — водяная
горихвостка. Редка зимою по р. Варзоб, в районе Станции.
122. Luscinia megarhyncha golzii (Cab.) — соловей. Во всей лесной
зоне ущелья.
123. Irania gutturalis (Guer.) — белогорлый соловей. Изредка в рай-
оне Станции.
124. Cyanosylvia svecica subsp. — варакушка. Встречена в районе
Станции на весеннем (III) и осеннем (IX) пролетах, редка.
125. Microcichla scouleri (Vig.) — белоножка. Зимою по рекам
Кондара и Варзоб, нередко.
126. Prunella f. fulvescens Sev. — тяньшанская завирушка. Изредка
зимою в районе Станции.
127. Prunella atrogularis huttoni Moore — черногорлая завирушка.
Встречена на осеннем (X) пролете.
128. Piunella collaiis subsp. — альпийская завирушка. Изредка встре-
чается зимою на кочевках.
129. Troglodytes t. tianschanicus Sharpe — крапивник. Зимою в рай-
оне Станции, многочислен.
130. Cinclus с. leucogaster Вр. — белобрюхая оляпка. Зимою пор. Вар-
зоб, около устья р. Кондара.
131. Cinclus pallasi tenuirostris Bp. — бурая оляпка. Там же, значи-
тельно чаще предыдущей.
132. Hirundo г. rustica L. — деревенская ласточка. Бывает на про-
лете.
133. Hirundo daurica scullii Seeb. — рыжепоясничная ласточка. Гнез-
дится в скалах по р. Варзоб, в районе Станции.
134. Delichon urbica meridionalis Hart. — городская ласточка. Воз-
можно, гнездится на плато Руидашт, в районе Станции — пролетом.
135. Biblis rupestris Scop. — скалистая ласточка, Видимо, гнездится
в скалах на плато Руидашт.
ЛИТЕРАТУРА
Иванов А. И. 1940. Птицы Таджикистана. Тр. Тадж. базы АН СССР, т. X.
Иванов А. И. 1948. Новые данные о птицах Таджикистана. Сообщ. Тадж.
фил. АН СССР, вып. 6.
Иванов А. И. 1949. Зимняя орнитофауна окрестностей Сталинабада. Тр. Тадж.
фил. АН СССР, т. XIX.
Иванов А. И. 1950. Зимовки и пролет птиц на территории Памиро-Алая. Сборн.
пам. акад. П. И. Сушкина. Изд. АН СССР.
Козлова Е. В. 1949. Оседлые и кочующие птицы южных склонов Гиссарского
хребта. Тр. Зоол. инет. АН СССР, т. VIII.
С. А. ЧЕРНОВ
ПРЕСМЫКАЮЩИЕСЯ И ЗЕМНОВОДНЫЕ
Герпетофауна Гиссарского хребта и ее распределение, уже не говоря
об экологии, изучена очень слабо. Имевшиеся сведения ограничивались
случайными данными некоторых путешественников (Руссов, Грумм-
Гржимайло и др.), и в общем из пределов хребта было известно всего
5 видов пресмыкающихся. Сейчас, после работ сотрудников Зоологи-
ческого института АН СССР 1942—1944 гг., а частью Таджикского фили-
ала АН, главным образом в ущелье Кондара, список найденных видов
возрос до 11, из которых многие пока неизвестны из других районов
хребта. В тоже время виды — щитомордик и гималайская агама,— обна-
руженные в других частях Гиссара, в районе Кондары пока не встре-
чены. К известным 11 видам, повидимому, можно добавить голопалого
геккона, по некоторым сведениям также встречающегося в ущелье Кон-
дара. Из земноводных здесь может быть указана только зеленая жаба.
Найденные в районе Кондаринского ущелья представители класса
пресмыкающихся составляют, примерно, четверть всех известных для
Таджикистана видов. Все они встречаются в районе ущелья не часто,
а некоторые и исключительно редко. Более обыкновенными можно считать
туркестанскую агаму, живущую на скалистых склонах, и водяного ужа.
Связанный с речкой, он находит здесь для себя обильную пищу в виде молоди
маринок и туркестанских сомиков. Значительно реже встречаются широко
распространенные в Гиссарском хребте узорчатый и разноцветный полозы,
ядовитая гюрза (рис. 1), а местами в подходящих стациях азиатский
гологлаз (рис. 2). Невдалеке от ущелья по долине р. Варзоб была обна-
ружена слепозмейка, а по самому ущелью — степная агама, длинноно-
гий сцинк, восточный удавчик и краснополосый полоз. Все эти послед-
ние виды очень редки и известны по 1—2 экземплярам.
Редкость большинства видов пресмыкающихся лишает возможности
достаточно детально проследить их вертикальное распределение, и имею-
щиеся данные позволяют высказать лишь некоторые предварительные
соображения. Прежде всего, отметим, что в описываемом районе отсут-
ствуют пресмыкающиеся, характерные для высокогорья. Виды, извест-
ные с плато Руидашт (2300—2600 м), — разноцветный и узорчатый
полозы, уж, туркестанская агама и, возможно, азиатский гологлаз —
свойственны Кондаринскому ущелью и его устью (1100 м). Кроме агамы,
не отходящей далеко от Гиссарского хребта, все они широко распростра-
нены в Гиссарской долине, а оба полоза и уж также в Туркестанской
низменности, что указывает на их большую пластичность. Наибольшее
разнообразие герпетофауны мы имеем, таким образом, у места выхода
ущелья Кондара в долину Варзоба, где констатированы все 11 видов
122
С. А, Чернов
пресмыкающихся. Степная агама, характерная для пустынь и полупустынь
Средней Азии, длинноногий сцинк, принадлежащий к числу теплолю-
бивых видов, и слепозмейка — несомненно, находятся здесь у предела
Рис. 1. Гюрза среднеазиатская — Vipera lebetina turanica Cern.
своего вертикального распространения. Выше поднимаются восточный
удавчик, гюрза и краснополосый полоз. Первый обнаружен по ущелью
на высоте около 1800 м, кстати сказать, в совершенно не свойственной
Рис. 2. Азиатский гологлаз — Ablepharus brandti Str.
ому обстановке — среди древесной поросли. Гюрза на тех же высотах
по Кондаре не была встречена, но так как в Дарвазе и на Памире она
поднимается и выше, вероятно ее распространение в описываемом ущелье
по крайней мере до высоты 1800—2000 м. Здесь же найдется и красно-
полосый полоз, так как по южному склону Гиссарского хребта, несколько
западнее Кондары, он был встречен в арчевниках, на высоте около 2300 м.
П pec мыкающиеся и земноводные
123
СПИСОК ПРЕСМЫКАЮЩИХСЯ И ЗЕМНОВОДНЫХ УЩЕЛЬЯ КОНДАРА
Земноводные
1. Bufo viridis Laur. — зеленая жаба. Более или менее обычна;
встречается во всех высотных поясах. Полезна — уничтожает вредных
насекомых.
Пресмыкающиеся
1. Agama lehmanni (Nik.) — туркестанская агама. Самая распро-
страненная и обычная ящерица, встречается на скалах и осыпях во всех
высотных поясах.
2. Agama sanguinolenta sanguinolenta (Pall.) — степная агама. Очень
редка; обнаружена только на конусе выноса Кондары.
3. Ablepharus brandti Str. — азиатский гологлаз. Местами встре-
чается часто; распространен во всех высотных поясах.
4. Eumeces schneideri princeps (Eichw.) — длинноногий сцинк. Очень
редок; повидимому, выше 1200—1300 м в горы не поднимается.
5. Typhlops vermicularis Мегг. — слепозмейка. Относительно редка;
обнаружена лишь в долине р. Варзоб.
6. Егух tataricus subsp. nov. — восточный удавчик. Очень редок;
прослежен до высот в 1700—1800 м.
7. Natrix tessellata (Laur.) — водяной уж. Не представляет редкости
возле воды и в ней; встречается во всех высотных поясах. Вреден —
уничтожает молодь рыб.
8. Coluber rhodorhachis ladacensis Anders. — краснополосый полоз.
Редок; возможно, распространен во всех высотных поясах.
9. Coluber ravergieri ravergieri Menetr. — разноцветный полоз. Не
представляет редкости; распространен во всех высокогорных поясах.
Питаясь грызунами, уничтожает значительное число вредителей.
10. Elaphe dione (Pall.) — узорчатый полоз. Не представляет ред-
кости; распространен во всех высотных поясах. Полезен — уничтожает
мелких грызунов.
11. Vipera lebetina turanica Cern. — гюрза. Сравнительно редка. Ядо-
вита. Укус ее вызывает довольно длительную болезнь человека и домаш-
них животных, а иногда и их гибель.
ЛИТЕРАТУРА
Никольский А. М. 1899. Пресмыкающиеся и амфибии, собранные А. Н. Казна,-
ковым в путешествии в Шугнан и Рошан. Ежег. Зоол. музея Ак. Наук, IV :
172—178.
Никольский А. М. 1911. Материалы по герпетологии Восточной Бухары.
Ежег. Зоол. музея Ак. Наук, XVI 271—284.
Никольский А. М. 1915—1916. Фауна России и сопредельных стран. Пресмы-
кающиеся, т. I и II.
Царевский С. Ф. 1918. Пресмыкающиеся и земноводные, собранные В. Я.
Лаздиным в южной и восточной Бухаре и в с.-в. части Закаспийской области
летом 1915 года. Ежег. Зоол. музея Ак. Наук, XXII 79—90.
Чернов С. А. 1935. Ящерицы (Sauria) Таджикистана. Тр. Таджикской базы АН
СССР, 5 469—475.
Чернов С. А. 1944. Систематическое положение таджикистанской гюрзы. Изв.
Тадж. фил. АН СССР, 5 : 189—192.
Чернов С. А. 1945. Эколого-фаунистический очерк пресмыкающихся и земновод-
ных Гиссарской долины. Тр. Тадж. фил. АН СССР, XIV 53—68.
Г X. ШАПОШНИКОВА
РЫБЫ
Ихтиофауна реки Кондара представлена всего 3 видами рыб: форелью,
маринкой и туркестанским сомиком (см. рисунок). Их распростране-
ние в Кондаре ограничено водопадом, находящимся в 4 км от устья.
Все три вида рыб прекрасно приспособились к жизни в водоемах
с быстрым течением. Последнее, как известно, является основным абио-
тическим фактором, определяющим состав фауны горных рек. Форель
и маринка обладают торпедообразным телом, сильным хвостовым стеблем
Туркестанский сомик — Glyptosternum reticulatum McCli.
и являются прекрасными пловцами, способными преодолевать на своем
пути небольшие водопады. Мелкая чешуя не препятствует изгибам тела.
Сомик иначе приспособился к тем же экологическим условиям. Его тело
расплющено в дорзо-вентральном направлении, что дает ему возможность
как бы прилипать ко дну, передвигаясь между камнями. Зато, попав
в струю, он беспомощно сносится течением, пока ему снова не удастся
нырнуть на дно.
Для обитателей горных потоков характерна сравнительно небольшая
величина тела, замедленный рост и небольшая плодовитость. Это объяс-
няется тем, что большая часть энергии тратится на борьбу с быстрым
течением.
Питание форели и маринки довольно сходно. Пища добывается всеми
способами: с камней, со дна, в толще воды и с поверхности (падающие
в воду насекомые). Маринка с помощью заостренного рогового чехлика
на нижней челюсти соскабливает с камней водоросли и личинки ручей-
ников. Сомик питается, в основном, различными представителями лито-
реофильных биоценозов, не брезгуя, однако, и молодью рыб.
Рыбы
125
Форель принадлежит к видам стенотермным, икрометание происходит
осенью, в мелких горных потоках, при температуре воды ниже 10°.
Маринка относится к рыбам евритермным, мечет икру в летние месяцы
в сильно растянутые сроки. То же можно сказать и о туркестанском
сомике.
Являясь реликтом ледниковой эпохи, форель в бассейне Аму-дарьи
населяет горные притоки этой реки, не выходя ниже предгорий. Маринка
принадлежит к эндемикам нагорно-азиатской ихтиофауны. Туркестанский
сомик — индийского происхождения: населяя верховья Инда и Кабула,
он проник в верховья Аму-дарьи, где расселился в горных и предгор-
ных водоемах, не поднимаясь, однако, особенно высоко.
СПИСОК РЫБ РЕКИ КОНДАРА
1. Salmo trutta aralensis Berg m. fario L. — форель.
2. Schizothorax intermedins McCll — маринка.
3. Glyptosternum reticulatum McCll — туркестанский сомик.
ЛИТЕРАТУРА
Берг Л. С. 1935. Мальки рыб из бассейна р. Дюшамбинки (бассейн Аму-дарьи,
Таджикистан). Таджик, компл. эксп. 1932 г. Изд. Ак. Наук СССР, X.
Никольский Г. В. 1938. Рыбы Таджикистана. Тр. Таджик, базы Акад. Наук
СССР, III.
Турдаков Ф. А. 1941. Материалы по ихтиофауне рек Средней Азии. Сборн. тр.
Гос. Зоол. музея МГУ, VI.
Шапошникова Г X. 1951. Амударьинская форель. Тр. Карело-Финского
Отд. ВНИОРХ, т. III.
Е. П. ГОЛУБЦОВА
НАЗЕМНЫЕ МОЛЛЮСКИ
В ущелье Кондара и расположенных рядом с ним ущельях Гажни
и Заминдорос нами было собрано 26 видов моллюсков.
1. Cochlicopa lubrica (О. F. Miill.). — Найден только в Гажни и Замин-
дорос. Обитает на увлажняемых местах — вдоль арыков и ручейков,
при выходе родниковых вод, среди растительности, на поверхности почвы
и между камней. Широко распространенный вид.
2. Truncatellina strobeli (Gredl.). — Найден во всех трех ущельях;
в ущелье Кондара — как на северном, так и на южном склонах. Обитает
в растительной подстилке среди кустарников, встречаясь и на комках
совершенно сухой земли. Распространение средиземноморское — Кавказ,
Закаспий, южная Европа.
3. Vertigo antivertigo (Drap.). — В ущельях Заминдорос и Гажни,
в небольшом количестве при выходе родниковых вод, среди раститель-
ности и в воде. Широко распространенный вид.
4. Pupilla signata (Mouss.). — В ущелье Кондара, на каменистых рос-
сыпях, среди кустарников и полукустарников, на нижней поверхности
камней. Средняя Азия, Закавказье.
5. Orcula doliolum (Brug.).—В ущелье Кондара, чаще на южном
склоне. Среди лесной подстилки, в трещинах почвы, норах. Средняя
Азия, Кавказ, Крым, Европа.
6. Vallonia pulchella О. F. Mull. — В ущелье Кондара — на дне ущелья
и южном склоне, среди полукустарников, в кучах камней, трещинах
почвы, близ увлажненных мест. Широко распространенный в Палеарк-
тике вид.
7. Vallonia costata О. F. Mull. — В ущелье Кондара, часто вместе
с предыдущим видом. Среди растительности вдоль арыков, под кам-
нями каменистых россыпей. Широко распространенный в Палеарктике
вид.
8. Vallonia sp.? — ближе не определенный, характерный своей
структурой вид. Найден в ущелье Кондара, у подножья южного склона,
среди розария.
9. Achantinula aculeata О. F. Mull, (дефективная раковина). —
В ущелье Кондара, у родника. Распространен в средней полосе и на юге
Европейской части СССР, на Кавказе, в средней и южной Европе.
10. Pyramidula rupestris Drap. — По дну ущелья Кондара, среди
растительных остатков и под камнями каменистых россыпей. Немного-
числен. Северный Кавказ, Западная Грузия, Крым, Европа.
И. Subzebrinus sogdianus Mts.,
12. Subzebrinus sogdianus Mts. f. gracilior,
Наземные моллюски
127
13. Subzebrinus albiplicatus Mts.,
14. Subzebrinus kasnakovi West.,
15. Subzebrinus errans West. — Bee 5 видов в ущелье Кондара, пре-
имущественно на южном склоне (кроме S. kasnakovi, которая найдена
на северном склоне и по дну ущелья). Среди камней и на поверхности
земли. Большею частью пустые раковины, живые моллюски встре-
чаются редко. Средняя Азия.
16. Ena martensiana Ancey. — В ущелье Кондара, на северном
и южном склонах. Среди камней и растительных остатков. В небольшом
количестве. Средняя Азия.
17. Punctum pygmeum Drap. — В ущелье Заминдорос, на мокрых
бревнах изгороди. СССР (повсеместно), Европа.
18. Punctum sp.? (близка к Р. lederi Bttg.). — В ущелье Кондара,
на скале.
19. Macrochlamys retteri Rosen. — Пустые раковины в ущелье Кон-
дара, на южном склоне и по дну ущелья, среди крупного щебня. Средняя
Азия.
20. Zonitoides nitidus О. F. Mull.—В ущелье Кондара, на влажной
почве близ арыка. СССР, Европа, Азия, северная Африка, Северная
Америка.
21. Vitrina conoidea Mts. — В ущелье Кондара, на обоих склонах,
по дну ущелья, на уроч. Квак (1860 м над ур. м.). Среди растительности,
при выходе родниковых вод, среди камней, в лесной подстилке, на поверх-
ности почвы, под корой ореха. Средняя Азия.
22. Euconulus fulvus О. F. Mull.—В ущелье Кондара, до уроч. Квак.
На увлажненных местах — близ родников и ручьев, в трещинах
скал, среди растительности и под мхом. СССР, Европа, Азия, северная
Африка.
23. Cathaica indigena West. — В ущелье Кондара, на южном
склоне, единичными экземплярами между крупных камней. Таджи-
кистан.
24. Eulota rubens Mts. f. angulata West. — В ущелье Кондара, среди
камней каменистых осыпей, на глубине до 10—20 см от поверхности.
Средняя Азия.
25. Parmacella korshinskii Srth. — В ущелье Кондара и других
ущельях, на ур. Квак. Вредитель огородных культур и молодых древес-
ных насаждений. Весной очень многочислен. Средняя Азия.
26. Agriolimax agrestis (L.) turkestanus Srth. — В ущелье Кондара,
вдоль арыков и на поливных огородах. Вид широко распространен в СССР
и Европе. Подвид описан для Средней Азии.
Приведенный список наземных моллюсков, который, вероятно,
не исчерпывает всей фауны ущелья Кондара, показывает, что малако-
фауна данного участка Гиссарского хребта имеет неоднородный (в зоо-
географическом отношении) характер. Господствующим элементом фауны
здесь являются эндемики Средней Азии. Из моллюсков к ним принад-
лежат Subzebrinus sogdianus, S. albiplicatus, S. kasnakovi, S. errans, Ena
martensiana, Macrochlamys retteri, Vitrina conoidea, Cathaica indigena,
Eulota rubens f. angulata, Parmacella korshinskii. Кроме того, здесь
имеются виды средиземноморского распространения — Truncatellina
strobeli, Pupilia signata, .Orcula doliolum. Наконец, некоторая часть
фауны слагается видами, широко распространенными в Палеарктикеi
Cochlicopa lubrica, Vertigo antivertigo, Vallonia pulchella, V. costata,
Zonitoides nitidus., Euconulus fulvus, Punctum pygmeum.
128
Е. П. Голубцова
ЛИТЕРАТУРА
Жадин В. И. 1944. К изучению слизней Таджикистана. Изв. Тадж. фил. АН СССР.
№ 5.
Жадин В. И. 1945. Жизненный цикл одного среднеазиатского слизня. При-
рода, № 2.
Жадин В. И. 1946. К биологии Parmacella korshinskii Smrth. и о мерах борьбы
с ней в Таджикистане. Зоол. жури., XXV, 1.
Линдгольм В. 1928. Mollusca. Тр. Памирской экспедиции, VIII.
Мартенс Э. 1874. Слизняки. Путешествие в Туркестан А. П. Федченко, т. II,
ч. 1, вып. 1.
Simroth Н. 1901. Die Naktschneckenfauna des Russischen Reiches. St.-Petersb.
А. А. ШТАКЕЛЬБЕРГ
ДВУКРЫЛЫЕ
Кондаринское ущелье, благодаря многолетним исследованиям В. В.
Гуссаковского, является безусловно наилучше изученной в отношении
двукрылых точкой Средней Азии. Сборы Гуссаковского дают возмож-
ность весьма полно охарактеризовать фауну района; однако надо сказать,
что обработка этих материалов еще не закончена, и определение до вида
могло быть произведено далеко не всегда. Сборы по двукрылым Конда-
ринского ущелья таят в себе длинный ряд видов, исчисляющихся мно-
гими десятками, которые еще требуют выяснения и описания, а во многих
случаях — и монографической обработки соответствующей группы.
В силу этого настоящий очерк фауны двукрылых Кондаринского ущелья
имеет предварительный характер; следует иметь в виду, что к настоящему
моменту определено до вида никак не более 40—50% представленных в
коллекциях форм двукрылых кондаринской фауны. Полная обработка этой
фауны, несомненно, представит и большой систематический, и не менее
значительный зоогеографический интерес.
По своему генезису фауна Средней Азии представляет собою чрез-
вычайно сложный комплекс, составившийся из элементов весьма раз-
личного происхождения. Фауна двукрылых Кондары дает в миниатюре
ту же картину, что и фауна Средней Азии в целом. Разнохарактерность
стаций, сконцентрированных на небольшом пространстве ущелья, акку-
мулировала в Кондаре чрезвычайно богатую фауну двукрылых; здесь
есть и типичные панпалеаркты, группирующиеся по стациям более север-
ного облика, и настоящие средиземноморцы, избирающие для своего
обитания более «теплые» ксерофитные стации, и, наконец, среднеази-
атские эндемики, большинство которых связано со стациями, специфич-
ными именно для гор Средней Азии.
К настоящему моменту имеются коллекционные материалы примерно
по 600—650 видам двукрылых Кондаринского ущелья; из них определено
до вида всего 253. Из этого числа видов к панпалеарктам, охватывающим
в своем распространении большую часть палеарктической области, кроме
ее севера, относится примерно 120 видов; к средиземноморцам широ-
кого распространения — примерно 55 видов; к среднеазиатским энде-
микам — около 75 видов. Интересно отметить, что по сравнению с дру-
гими группами насекомых процент среднеазиатских эндемиков среди
двукрылых должен быть признан достаточно высоким; после обработки
всех имеющихся материалов по двукрылым едва ли он снизится, так
как среди необработанных групп остались такие богатые видами локаль-
ного распространения группы, как Empididae и Muscidae из мух, Ti pu-
li dae, Limoniidae и Tendipedidae — из комаров.
9 Ущелье Кондара
130
„1. .4. Штакельберг
Выше уже была дана краткая экологическая характеристика основ-
ных стаций, представленных в Кондаринском ущелье; для двукрылых
Кондары особенно типичными из них являются следующие.
Очень много своеобразного в отношении двукрылых дает сама речка
Кондара — бурный горный поток, с которым связано существование
в Кондаре особой богатой фауны (fauna hygropetrica). Набольших кам-
нях, лежащих в самом русло Кондаринки или в зоне водяных брызг,
обитают в очень большом количестве нежные длинноногие серые комары
из семейства Limoniidae, Antocha turkestanica; то сокращаясь, то уве-
личиваясь в своей численности, они встречаются в этих условиях в тече-
ние почти всего года, даже зимою. Вместе с Antocha на тех же камнях
сидят стройные хищные мушки Atalanta из семейства Empididae, охотя-
щиеся за мелкими насекомыми (Collembola); в Кондаре этот род пред-
ставлен несколькими еще не описанными видами. С речкой Кондаринкой
связаны широко известные гидробиологам комары семейства Dixidae,
которые иногда в массе попадаются на отвесных стенах камня, (‘пускаю-
щихся в речку, или на растениях, растущих по ее берегам; под вечер
поздней осенью иногда приходится наблюдать массовый лёт Dixidae над
речкой, где они образуют небольшие рои. Личинки Dixidae живут у уреза
воды. Интересно отметить, что наиболее обычный в Кондаре вид этого семей-
ства, Dixa christophersi Edw., является широко распространенным по на-
горьям Центральной Азии (Тибет) и по горам северной Индии (Кашмир).
Мезофильные стации собственно реки Кондара — затененные кустами
шиповника и ив берега речки, местами поросшие буйной травянистой
растительностью, имеют свою очень богатую по количеству видов, но мало
заметную фауну. В этих условиях встречаются многие Empididae, Doli-
chopodidae (Dolichopus, Hercostomus, Syntormon, у самой воды — Tachy-
trechus, Campsicnemus), Ephydridae, Drosophilidae. На цветах зонтич-
ных Prangos, растущих в тенистых местах, попадаются Sphegina sp. —
очень своеобразный, не описанный еще вид рода и некоторые тенелюби-
вые Syrphus. В траве по берегам Кондаринки обычен Chrysopilus clari-
cinctus — типичный мезофильный среднеазиатец, и целый ряд других
видов. Ксерофитные, бедные растительностью стации, в особенности
южного склона Кондаринского ущелья, привлекают особую фауну дву-
крылых, рекрутируемую по преимуществу за счет типичных средиземно-
морцев. К этой группе должны быть прежде всего отнесены большинство
Bombyliidae кондаринской фауны, а также некоторые ближе не опреде-
ленные виды ктырей (Asilidae).
Поднимаясь выше по склонам Кондаринского ущелья по пути на
уроч. Квак, мы вступаем в полосу более открытых ландшафтов, менее защи-
щенных от ветра; в связи с этим в составе фауны двукрылых этих мест
замечается преобладание форм, характеризующихся быстрым поле-
том и общей! значительной активностью; основу фауны двукрылых
этой полосы составляют многочисленные виды Syrphidae и Larvivoridae.
Расцвета эта фауна достигает в период цветения зонтичного Prangos
pabularia — в конце мая и в первой половине июня, когда на его ярко-
желтых шапках концентрируется огромное количество двукрылых самых
различных групп. Из наиболее характерных представителей этой фауны
можно назвать крупную эффектную львинку Stratiomyia nobilis и более
скромную — Stratiomyia bochariensis; встречающихся в массе, одетых
золотисто-бурыми густыми пушистыми волосками, похожих на пчел
журчалок из рода Lampetia, личинки которых развиваются в луковицах
лилейных (этот род представлен в Кондаре двумя новыми видами); чер-
Двукрылые
131
ных с яркожелтыми перевязями, крупных, напоминающих ос журчалок
из рода Chrysotoxum (Chr. parmense, Chr. kozhevnikovi); стройных,
с тонким брюшком, Conbpidae из рода Physocephala и других. Особенное
внимание привлекают на цветах Prangos крупные черные мухи с черными,
испещренными белыми точками крыльями и коренастым телом — Platy-
stoma elegans из сем. Platystomatidae, и также крупные, но стройные,
с узким брюшком, длинным яйцекладом и прозрачными со слабыми тем-
ными пятнами широкими крыльями Adapsilia из сем. Pyrgotidae. В огром-
ном количестве видов и особей на цветах Prangos попадаются Larvivori-
dae; из этой группы паразитических мух в условиях Кондары особенно
обычны крупные скоролетки из родов Linnaemyia, Winthemia и некото-
рых других; многие из этих видов несомненно являются новыми, не опи-
санными, а некоторые представляют собою формы новых, еще не извест-
ных родов.
Подъем на уроч. Квак представляет собою одно из самых увлекатель-
ных в дпптерологическом отношении предприятий, доступных энтомологу
в Таджикистане. После достаточно обедненной полосы, на протяжении
которой кроме массы саранчевых Gomphomastax gussakovskii не попа-
дается почти ничего, на высотах около 1800 м мы вступаем в зону настоя-
щего горного среднеазиатского леса, с его кленами, грецким орехом
и обильной травянистой растительностью. Многое в этом лесу напоми-
нает среднюю полосу Европы. Над тропинками здесь так же, как
и в Европе, перелетают металлически блестящие львинки Geosargus
cuprarius; в более влажных местах здесь также много плоских черных
львинок из рода Beris, хотя на уроч. Квак этот род представлен своим
видом. В этих же мезофильных стациях вместе со среднеазиатскими
видами Limoniidae, в большинстве, повидимому, новыми, встречаются
обычные повсюду в Европе Erioptera lutea. Габитуально похожа на евро-
пейскую, но так же своеобразна фауна двукрылых, свойственная соцве-
тиям крупных зонтичных Heracleum; в массе на них встречаются тем-
ные журчалки из рода Chilosia, представленные несколькими новыми
видами. В густой траве, в тенистых местах, попадаются средней вели-
чины мушки из рода Coelomyia (новый вид: из сем. Muscidae), с ногами
оригинального строения.
Подъем на плато Руидашт — сложная панорама стаций, быстро сме-
няющих одна другую. Помимо кленовых рощ на первых этапах подъема,
особенный интерес в дпптерологическом отношении представляют сильно
ннсолированные площадки с зонтичными и, особенно, сложноцветными, бо-
гатые всевозможными представителями семейства пестрокрылок (Trypeti-
dae), а также Phasiidae (Besseria) и Sarcophagidae (Sarcophila, Metopiinae).
Фауна двукрылых субальпики Руидашта едва затронута изучением,
но то, что известно, дает представление о большом ее своеобразии. Вместе
с панпалеарктамп, среди которых можно назвать хотя бы такие виды,
как Myopa testacea из Conopidae и, пожалуй, Blaesoxipha rossica —
паразита саранчевых, с Руидашта известны интереснейшие виды Ulidii-
dae (Clidia ruficeps и несколько новых видов того же рода) и некоторые
другие. Руидашт еще ждет своего исследователя.
СПИСОК ВИДОВ ДВУКРЫЛЫХ
Сем. Petauristidae
Petaurista maculipennis Mg. — Кондара, И Ш 1943 (1 Штакель-
берг). — Большая часть Палеарктики; Сев. Америка.
9*
132
А. А. Штакельберг
Сем. Limoniidae
Dicranomyia modesta Wd. — Кондара, 8 VI 1943 (1 6; Штакельберг)^.—
Большая часть Европы; Европ. часть СССР; Кавказ; Ср. Азия; Сибирь
до Приморского края.
Limonia inusta Mg. — Кондара, 8 VI 1943 (1 $; Штакельберг); Квак,
6, 7 VI 1943 (3 88; Штакельберг). — Большая часть Европы и Европ.
части СССР; Закавказье; Ср. Азия (Гиссарский хребет); Алтай.
Antocha turkestanica Meij. — Кондара, 8 10 III 1943, на камнях
в русле реки Кондара (6 88; Штакельберг). — Горные и предгорные
реки Ср. Азии и Семиречья; типичный представитель faunae hygropet-
ricae.
Erioptera lutea Mg. — Квак, 5 VI 1943 (4 88; Штакельберг). — Боль-
шая часть Европы; Европ. часть СССР; Закавказье; Ср. Азия (Гиссар-
ский хребет): Алтай. Мезофильные стации.
Erioptera (Symplecta) hybrida Mg. — Кондара, 21 VI, 28 VII 1938
(1 8, 1 2’» Гуссаковский). — Большая часть Европы; Сибирь; Ср. Азия.
Мезофильные стации.
Сем. Fungivoridae
Exechia confinis Winn. — Кондара, 10 III 1943 (1 8; Штакельберг). —
Большая часть Европы; Европ. часть СССР; горы Ср. Азии (Гиссарский
хребет).
Сем. Dixidae
Dixa christophersi Edw. — Кондара, 11 III 1943 (1 6, 1 2’> Штакель-
берг), 6 X1 1944 (36 SS, 22 22’ Штакельберг). Берег реки Кондара. —
Кашмир; Тибет; Ср. Азия (Гиссарский, Дарвазский хребты). На камнях
по берегам горных потоков.
Dixa lepnevae Stack. — Кондара, 11 III 1943 (1 8; Штакельберг). —
Таджикистан (Гиссарский хребет). По берегам горных потоков.
Dixa nebulosa Mg. — Кондара, 6 XI 1944 (3 88, 2 22» Штакельберг).
Сев. и ср. Европа; Ср. Азия (Гиссарский хребет). Берега водотоков.
Dixa platystyloides Stack. — Кондара, 11 III 1943 (3 88; Шта-
кельберг); Квак, 5 VI 1943 (1 <5; Штакельберг). — Гиссарский и Дар-
вазский хребты. По берегам горных потоков.
Dixa zernovi Stack. — Кондара, 6 XI 1944 (1 8; Штакельберг) ; Квак,
5 VI 1943 (4 88; Штакельберг). — Таджикистан (Гиссарский хребет). По
берегам горных потоков.
Сем. Blepharoceridae
Blepharocera fasciata turkestanica Brod. — Кондара, 7, 8 XI 1944
{3 88, 3 22» Штакельберг). — Горные потоки Ср. Азии. В Таджики-
стане обычен.
Сем. Cecidomyiidae
Perrisia medicaginis Rubs. (опр. Карпова). — Кондара (Карпова). —
Большая часть Европы, кроме севера; Ср. Азия. Вредит люцерне.
Двукрылые
133
Сем. Bibionidae
Bibio marci L. — Квак, 6 VI 1943 (1 Штакельберг). — Большая
часть Европы, кроме севера; горы Ср. Азии.
Сем. Stratiomyidae
Stratiomyia bochariensis Pl. (=S. turkestanica Pl. sec. typ., syn.
nov.). — Кондара, 5, 8 VI 1943 (2 $$, 2 99; Штакельберг); Квак,
6 VI 1943 (2 ; Штакельберг). — Горы Ср. Азии.
Stratiomyia nobilis Lw. — Кондара, 5, 8 VI 1943, на цветах Prangos
pabularia (2 3S, 5 22*, Штакельберг); Руидашт, 22 VII 1937 (1 25 Гус-
саковский). — Ср. Азия; Синьцзян.
Stratiomyia przewalskii Pl. — Кондара, 8 VI 1943 (1 9; Штакель-
берг). — Ср. Азия (распространение широкое).
Adoxomyia cinerascens Lw. — Кондара, 17, 24, 25, 27 VI, 3, 7 VII,
7, 14, 15, 16 IX 1937; 15 VI, 12, 13, 17, 20 VII, 13 IX 1939; 28 VII 1940
(13 3S, 9 22*’ Гуссаковский). — Туркмения; Таджикистан (Гиссарский,
Дарвазский хребты; зап. Памир); Иран. На камнях по берегам горных
потоков.
Adoxomyia obscuripennis Lw.—Кондара, 4, 5, 13, 15, 16, 24 VI 1937;
21 V, 4 VI 1939 (3 <5<5, 6 22’ Гуссаковский) ; 5 VI 1943, на цветах Pran-
gos pabularia (1 о, 1 9; Штакельберг); Квак, 6 VI 1943 (1 5, 1 Шта-
кельберг). — Ср. Азия: Таджикистан (Гиссарский хребет, долина
Ягноба); Узбекистан (Фергана).
Hermione aemula Lw. — Кондара, 13 VII 1939 (1 <5; Гуссаковский).—
Таджикистан (Гиссарский хребет); Узбекистан (Зеравшан).
Hermione meigeni fraterna Lw. — Кондара, 6, 17, 24, 28 VI, 9 IX
1937 (4 3S, 1 25 Гуссаковский). — Ср. Азия.
Hermione папа Lw. — Кондара, 18 VI 1939 (1 25 Гуссаковский). —
Таджикистан (Пенджикент, Гиссарский хребет).
Hermione nigriventris Lw. — Кондара, 24 VI 1937; 7 VI 1939 (2
Гуссаковский). — Таджикистан (Гиссарский хребет; Искандер-куль);
Алайская долина.
Hermione notata Lw. — Кондара, 4 VI 1939 (1 25 Гуссаковский).
— Таджикистан (Гиссарский хребет; Искандер-куль; Захматабад; Оббур-
ден).
Hermione tricolor Lw. — Кондара, 8 VIII 1937 (1 25 Гуссаковский).—
Таджикистан (Гиссарский хребет); Узбекистан.
Geosargus cuprarius L. — Кондара, 4, 24 VI, 7, 9, 15, 17 IX 1937
(1 3, 6 225 Гуссаковский). — Большая часть Европы и Европейской
части СССР; Закавказье; Ср. Азия (Гиссарский хребет); Монголия.
Beris heptapotamica Pl. — Квак, 10, 20 VI 1937 (2 S3, 1 9; Гусса-
ковский).— Алтай; Семиречье; Таджикистан (Гиссарский хребет). Мезо-
фильные стации.
Solva shnitnikovi Pl. — Кондара, 19 VI, 16 VII, 30 VIII 1938; 6 IX
1940 (2 6S, 4 225 Гуссаковский). — Семиречье; Таджикистан (Гиссар-
ский хребет; зап. Памир).
Сем. Rhagionidae
Chrysopilus auratus asiaticus Lind. — Квак, 7 VI 1943 (2 0(5; Штакель-
берг). — Ср. Азия. Мезофильные стации.
134
А. А. Штакельберг
Chrysopilus claricinctus Lind. — Кондара, 16, 24, 25, 26, 27, 28 VI,
2, 5, 7, 19 VII 1937; 17, 19, 21, 22 VI, 14, 16, 17 VII, 3, 7, 8, 20 VIII
1938; 7, 14, 15 VI, 13, 14, 15 VII, 13 1X 1939; 27, 28 VII, 7 VIII 1940
(32 $S, 18 22; Гуссаковский). — Ср. Азия. Мезофильные стации.
Symphoromyia crassicornis Pz. —Квак, 5, 6 VI 1943 (4 53, 1 9;
Штакельберг). — Большая часть Европы; Ср. Азия. Мезофильные стации.
Сем. Tabanidae
Chrysops (Heterochrysops) flavipes punctii’er Lw. — Кондара (Гусса-
ковский). — Южн. Европа; сев. Африка; М. Азия; Иран; Ср. Азия.
Silvius vitripennis Mg. — Кондара, 24 VI, 1 VII 1937 (2 33; Гуссаков-
ский). — Ср. и южн. Европа; Иран; Кавказ; Ср. Азия.
Tabanus (in sp.) leleani Aust. — Кондара (Гуссаковский). — Южн.
Европа; Иран; Кавказ; Ср. Азия.
Tabanus (in sp.) golovi golovi N. Ols. — Кондара (Гуссаковский).—
Семиречье; Кульджа; Таджикистан (Сталинабад, Кондара).
Tabanus (in sp.) bromius flavofemoratus Str. — Кондара (Гуссаков-
ский); Квак (Гуссаковский). — Южн. Европа; Передняя Азия; Кавказ;
Ср. Азия.
Tabanus (in sp.) semenovi N. Ols. —Кондара (Гуссаковский); Квак
(Гуссаковский); Руидашт (Гуссаковский). — Горы Ср. Азии (Ферганский,
Алайский п Гиссарский хребты).
Tabanus (in sp.) grandis Szil. — Кондара (Гуссаковский); Квак (Гус-
саковский). — М. Азия; горы Ср. Азии (Ферганский, Гиссарский, Дар-
вазский хребты).
Chrysozona bactriana N. Ols. —Квак, 12 VII—20 VIII 1937 (4 22»
Гуссаковский). — Гиссарский хребет; ? зап. Памир.
Сем. Nemestrinidae
Nemestrinus persicus Lichtw. — Руидашт, 20, 22 VII 1937 (3 33, 7 22;
Гуссаковский). — Мал. Азия; Египет; Ср. Азия.
Symmictoides simplex Lw. — Квак, 20, 21 VII 1937; 13 VII 1938;
4 VII 1940 (3 33, 13 22’ Гуссаковский); Руидашт, 20 VII 1938 (1 21
Гуссаковский). — Ср. Азия; Тибет.
Hirmoneura villosula Lw. — Кондара, 21, 23 VI, 10 VII 1938 (2 33,
1 2‘, Гуссаковский). — Таджикистан (Гиссарский хребет, Памир).
Сем. Therevidae
Psilocepliala nigrofemorata Krob. — Руидашт, 8 VII 1940 (2 33, 1 21
Гуссаковский). — Горы Ср. Азии.
Сем. Bombyliidae
Cyllenia giobiceps Lw. — Кондара, 27 VII 1940 (1 3; Гуссаковский).—
Ср. Азия (дол. Зеравшана, Гиссарский хребет, Дальвердинская степь).
Двукрылые
135
Amictus insignis Lw. —Кондара, 13 19 VI 1939; 4 VII 1940 (1 6,
2 2^; Гуссаковский). — Ср. Азия (дол. Зсравшана; Гиссарский хре-
бет).
Toxophora maculata ab. completa Par. — Кондара, 17 IX 1937 (1
Гуссаковский). —Южн. Европа; Закавказье; Туркмения (Копет-даг);
Узбекистан; Таджикистан (Гиссарский хребет).
Toxophora epargyra Herm. — Кондара, 23 VIII, 12 IX 1937 (1 о, 1 2;
Гуссаковский). —Сирия; Закавказье (Армения); Таджикистан (Гиссар-
ский хребет).
Phthiria vagans Lw. — Кондара, 7 VI 1939 (1 2; Гуссаковский). —
Южн. Европа; Иран; Ирак; юго-восток Европейской части СССР
(Сарепта); Ср. Азия.
Bombylius discoideus F. — Кондара, 9 VIII 1937; 29 VIII, 7 IX 1945
(1 <5, 2 2$; Гуссаковский). — Южн. Европа; Сев. Америка; юг Евро-
пейской части СССР; Ср. Азия.
Bombylius maculithorax Param. —Квак, 6 VI 1943 (1 (5; Штакель-
берг).—Таджикистан (Гиссарский хребет); Узбекистан (окр. Ташкента);
Китай (Гань-су).
Bombylius undatus Mikn. — Кондара, 8 VI 1943 (1 2< Штакельберг);
Квак, 6, 7 VI 1943 (2 65; 1 2» Штакельберг). — Ср. и южн. Европа;
средняя и южная полоса Европейской части СССР.
Antonia fedtschenkoi Lw. — Кондара, 31 VIII 1937 (1 6; Гуссаков-
ский). — Ср. Азия (окр. Мары; окр. Ташкента; Хива; Гиссарский
хребет).
Lomatia bella Lw. — Кондара, 25, 26, 27, 28 VI, 2, 3, 14, 15 VII
1937 (14 2$; Гуссаковский); Квак, 29 VI 1937 (4 22’ Гуссаковский). —
Таджикистан (дол. Ягноба, Гиссарский хребет).
Lomatia bella tibialis Lw. —Кондара, 19 VI 1939 (1 6; Гуссаков-
ский). — Таджикистан (Искандер-куль, Кондара).
Anthrax maculosa Sack. — Кондара, 22 VII 1940, 14 IX 1945 (3 22»
Гуссаковский). — Закавказье (Армения); Ср. Азия (Туркмения, Узбеки-
стан, Таджикистан).
Spongostylum (Satyramoeba) etruscum F. — Кондара, 15, 25, 28 VII,
3, 7 VIII, 9 IX 1937; 15, 22 VII 1939; 28 VII, 6 IX 1940; 27 VIII, 6, 10,
15 IX 1945 (И 66, 5 22; Гуссаковский). — Южн. Европа; сев. Африка.
Крым; Кавказ; Ср. Азия.
Exoprosopa grandis Wd. — Кондара, 23 VI, 15, 25 VII 1937; 12 VII
1939 (6 2$; Гуссаковский). — Южн. Европа; сев. Африка; Мал. Азия;
Закавказье; Ср. Азия.
Exoprosopa megerlei consanguinea Macq. — Кондара, 29 VIII 1945
(1 2J Гуссаковский). — Сев. Африка; Ср. Азия.
Exoprosopa melaena Lw. — Кондара, 13 VI, 9 IX 1937 (2 2$*, Гусса-
ковский); Квак, 10, 20 VI, 21, 22 VII 1937 (7 2$; Гуссаковский). —
Закавказье (Армения); сев. Иран; Ср. Азия (Кштут); Наманган; Гиссар-
ский и Дарвазский хребты; зап. Памир.
Thyridanthrax afer F. — Кондара, 27 VIII 1937 (1 2 J Гуссаковский).—
Ср. и южн. Европа; сев. Африка; Ср. Азия.
Hemipenthes morio L. — Кондара, 14 VI 1939 (1 2‘, Гуссаковский.) —
Ср. и южн. Европа; Аравия; горы Ср. Азии (Гиссарский хребет); Сев.
Америка.
Hemipenthes praecisus Lw. —Кондара, 10 1X 1945 (1 25 Гуссаков-
ский). — Ср. Азия, Семиречье, южн. Сибирь, Монголия.
136
А. А. Штакельберг
Сем. Asilidae
Eutolmus immaculatus Lw. — Кондара, 7 VI 1939 (1 (5; Гуссаков-
ский). — Ср. Азия (Сталинабад, Кондара, дол. Зеравшана, Пржевальск).
Сем. Dolichopodidae
Dolichopus signifer Hal. — Кондара, 20 IV 1944 (1 5; Штакельберг).—
Ср. и южн. Европа; Ср. Азия. Мезофильные стации.
Hercostomus blepharopus Lw. — Кондара, 24 VIII 1945 (1 5; Гусса-
ковский). — Туркмения (Копет-даг); Таджикистан (Гиссарский хребет);
Узбекистан (Зеравшан).
Hercostomus leptocercus Stack. — Кондара, 15 VI 1938 (1 <5; Гусса-
ковский). — Средняя зона Гиссарского и Дарвазского хребтов. На
камнях по берегам горных потоков.
Hercostomus longiventris Lw. — Кондара, 22 VI, 12, 14 VII, 18 IX
1938; 31 V 1939 (4 33, 2 jj; Гуссаковский). — Ср. и южн. Европа;
Ср. Азия (Гиссарский хребет).
Hercostomus ogloblini Stack. — Кондара, 16 VIII 1945 (1 3; Гусса-
ковский). — Гиссарский хребет (Кондара, Рахаты). По берегам гор-
ных потоков.
Hercostomus (Microhercostomus) dilatitarsis Stack. — Кондара, 16 IX
1938 (1 5; Гуссаковский). Вид известен пока только из Кондары.
Tachytrechus gussakovskii Stack. — Кондара, 29 VIII 1945 (1 3; Гус-
саковский). — Гиссарский и Дарвазский хребты. У воды.
Medetera lamprostoma Lw. — Квак—Руидашт, 2800 м, каменистые
россыпи, на камнях, 6 VI 1943 (2 33, 4 22» Штакельберг). — Ср. Азия.
Medetera (Oligochaetus) leucarista Stack. — Кондара, 19 VI, 9, 19,
20, 30 VIII, 13, 15, 19, 22 1X 1938 (6 33, 8 jj; Гуссаковский). —
Лесная зона Гиссарского хребта (Кондара, Рахаты).
Liancalus virens Scop. — Кондара, 24 VIII 1945 (6 33, 5 jj; Гус-
саковский). — Большая часть Палеарктики. У водопадов.
Syntormon cilitibia Stack. — Кондара, 17 VII, 18, 20 IX 1938; 13 IX
1939 (8 33; Гуссаковский). — Окр. Сталинабада; Кондара; зап. Памир.
У воды.
Diaphorus ussuriensis Stack. — Кондара, 17 21, 22 VI, 10, 12, 15, 17,
25 VII, 3, 9, 19, 20, 30 VIII, 6, 22 IX 1938; 15 VI, 13 IX 1939; 27 VIII
1940 (16 33, 9 22; Гуссаковский). — Лесная зона Гиссарского хребта;
Приморский край.
Campsicnemus barbitibia Stack. — Кондара, 24 VIII 1945(6 33; Гусса-
ковский). — Окр. Сталинабада; лесная зона Гиссарского и Дарвазского
хребтов; зап. Памир. На лужах.
Campsicnemus darvazicus Stack. — Кондара, 24 VIII 1945 (1 3; Гусса-
ковский). — Гиссарский и Дарвазский хребты. На лужах.
Campsicnemus umbripennis Lw. — Кондара, 11 VIII 1943 (1 3; Шта-
кельберг); 24 VIII 1945 (1 3, Гуссаковский). — Южн. Европа; Ср. Азия.
На лужах.
Сем. Dorylidae
Chalarus spurius Flln. — Кондара, 20 IX 1938 (1 3; Гуссаковский). —
Большая часть Палеарктики, кроме крайнего севера; в лесных стациях.
Двукрылые
13Г
Сем. Syrphidae
Liogaster splendidaMg.—Кондара, 2 VIII 1937 (1 5; Гуссаковский).—
Большая часть Европы, кроме севера; Ср. Азия.
Orthoneura nobilis FHn. — Квак, 31 VII 1939 (1 g; Гуссаковский).—
Большая часть Европы; Кавказ; горы Ср. Азии: Гиссарский хребет
(Квак, Гафильабад).
Paragus bicolor Flln. — Кондара, 23 VI 1938; 22, 24 VIII 1945 (1 5,
2 2$’, Гуссаковский), 2 V 1944 (1 g; Штакельберг); Квак, 17 VIII 1937
(1 g; Гуссаковский). — Большая часть Палеарктики, кроме севера.
Paragus tibialis Flln. — Кондара, 19 VI, 10 11, 25 VII, 15 1X 1938;
29 V, 15 VI 1939; 18, 21, 22, 24 VIII 1945 (15 55, 6 gg; Гуссаков-
ский). — Большая часть Палеарктики, кроме севера.
Melanostoma ambiguum Flln. — Кондара, 3 V 1944 (1 g; Штакель-
берг). — Большая часть Палеарктики, кроме севера.
Melanostoma mellinum L. — Кондара, 31 VIII 1937; 13, 16, 29 V 1939
(4 55, 1 g; Гуссаковский). — Большая часть Палеарктики.
Rohdendorfia dimorpha Smirn.—Кондара, 22 VII 1938 (3 (5(5; Гусса-
ковский). — Горы Ср. Азии.
Scaeva albomaculata Macq. —Кондара, 4 VI, 31 VIII 1937; 15 VII
1938 (1 5, 2 gg; Гуссаковский;) Квак, б VI 1943 (1 g; Штакельберг);
Руидашт, 22 VII 1937; 20 VII 1938 (1 (5, 1 g; Гуссаковский). —Среди-
земноморье; Ср. и Центр. Азия.
Scaeva pyrastri L. — Кондара, 20 IV 1944 (1 (5, 2 gg; Штакельберг);
Квак, 6 VI 1943 (1 g; Штакельберг). — Большая часть Палеарктики,
кроме севера.
Epistrophe balteata Deg. — Кондара, 20 IV 1944 (1 g; Штакель-
берг). — Большая часть Палеарктики, кроме севера; Ориентальная
область.
Syrphus corollae F. — Кондара, 2 VIII 1937 (1 (5; Гуссаковский);
Квак, 17 VIII 1937 (1 g; Гуссаковский); Руидашт, 22 VI 1937; 22 VII
1937 (1 5, 2 gg; Гуссаковский). — Большая часть Палеарктики, кроме
севера.
Syrphus vitripennis Mg. — Кондара, 3 V 1944 (2 55, 1 g; Штакель-
берг). — Большая часть Палеарктики.
Ischiodon scutellare F. —Кондара, 27 VI 1937 (1 g; Гуссаковский).
Sphaerophoria ruppeli Wd. —Кондара, 15 V, 2 VI 1939 (2 gg; Гус-
саковский). — Большая часть Палеарктики, кроме севера.
Sphaerophoria scripta L. — Кондара, 20 IV 1944 (1 5; Штакельберг).—
Большая часть Палеарктики.
Xanthogramma ornatum dives Rd. — Кондара — Квак, 2 V 1944
(1 5; Штакельберг); Квак, 6, 7 VI 1943 (3 55, 4 gg; Штакельберг). —
Ср. и южн. Европа; горы Ср. Азии.
Chrysotoxum intermedium Mg. — Кондара, 19, 21 IX 1938 (1 5,
2 gg; Гуссаковский). — Ср. Европа; Средиземноморье; Ср. Азия, где
широко распространен.
Chrysotoxum kozhevnikovi Smirn. — Кондара, 13 VII 1938 (1 5; Гус
саковский); 8 VI 1943 (1 5; Штакельберг); Квак, 17 VIII, 22 IX 1937,
16 VI 1938 (4 55, 3 gg; Гуссаковский), 7 VI 1943, на цветах Scaligeria
(4 55; Штакельберг); Руидашт, 22, 23 VII 1937 (3 gg; Гуссаковский).—
Горы Ср. Азии.
Chrysotoxum parmense Rd. — Кондара, 7, 17 VI, 9, 15 IX 1937 (1 5,
gg; Гуссаковский); 5 VI 1943, на цветах Prangos pabularia (1 5;
138
А. А. Штакельберг
Штакельберг); Квак, 5, 17 VIII, 22 IX 1937 (4 Гуссаковский). —
Средиземноморье, Закавказье, Иран, Ср. Азия.
Cerioides brunettii Shann. — Кондара, 5 VII, 9 IX 1937; 15 IX 1938;
4 VI, 12, 23 VII 1939 (2 55, 4 22’ Гуссаковский). — Белуджистан
(Кветта); Таджикистан (Гиссарский и Дарвазский хребты).
Cerioides sartorum Smirn. — Кондара, 7 VI, 15 VII 1937 (2 jj; Гус-
саковский). — Ср. и Центр. Азия.
Volucella zonaria Poda — Кондара, 19 VIII 1945 (1 j; Гуссаков-
ский). — Южн. Европа; юг Европейской части СССР; Кавказ: Ср.
Азия.
Eristalis arbustorum L. — Кондара, 2 V 1944 (1 5; Штакельберг). —
Вся Палерактика, кроме крайнего севера.
Eristalis tenax L. — Кондара, 27 VIII 1937; 18 VI, 19 IX 1938;
15 V 1939 (2 55, 2 22; Гуссаковский). — Вид распространен повсе-
местно по всему земному шару, кроме Арктики и Антарктики.
Myiatropa semenovi Smirn. — Кондара, 8 VI 1943; 20 IV 1944 (4 55;
Штакельберг). — Предгорья и горы Ср. Азии на северо-восток до Семи-
речья.
Tubifera continua Lw. — Кондара, 2 V 1944 (1 5; Штакельберг). —
Ср. и Центр. Азия; южн. Сибирь.
Tubifera trivittata F. —Руидашт, 18 VI 1938 (1 25 Гуссаковский). —
Большая часть Палеарктики, кроме севера.
Zelima ignava Pz. — Кондара, 2 V 1944 (2 22’ Штакельберг). —
Большая часть Палеарктики, кроме севера.
Eumerus amoenus Lw. — Кондара, 7, 14 IX 1937; 6, 16, 22 IX 1938;
17 VII 1939 (4 (5(5, 2 22’ Гуссаковский), 3 V 1944 (3 6S, 1 25 Штакель-
берг). — Средиземноморье; горные районы Таджикистана.
Eumerus coeruleus Beck. —Кондара, 24 VI 1937 (1 6; Гуссаков-
ский). — Окр. Ташкента; Гиссарский хребет (Кондара); Иран.
Eumerus gussakovskii Stack.—Квак, 18 IX 1945 (1 5, 2 qq; Гусса-
ковский).— Гиссарский хребет (Квак).
Eumerus latifrons Sack. — Кондара, 13 V 1939 (1 5; Гуссаковский). —
М. Азия; Гиссарский хребет (Кондара).
Eumerus pamirorum Stack. — Кондара, 16 IX 1937 (1 5; Гуссаков-
ский). — Гиссарский хребет (Кондара); зап. Памир.
Eumerus tadzhikorum Stack. — Кондара, 17 IX 1937 (1 5; Гусса-
ковский); Квак, 6 VI 1943 (1 25 Штакельберг). —Гиссарский хребет
(Кондара, Квак, Зидды).
Eumerus ursiculus Stack. — Руидашт, 23 VII 1936 (3 55; Гуссаков-
ский). — Гиссарский хребет (Руидашт).
Syritta pipiens L. — Кондара, 20 IX 1945 (1 5; Гуссаковский). —
Вся Палеарктика, кроме крайнего севера.
Сем. Conopidae
Brachyglossum capitatum Lw. —Кондара, 24 VI, 17 VII 1939 (2 55;
Гуссаковский). — Ср. Европа; юг Европейской части СССР; Ср. Азия:
Гиссарский хребет.
Physocephala rufipes F. — Кондара, 27 VI, 22 VII 1937; 20 VI 1938
(4 55; Гуссаковский); Руидашт, 20, 24 VIF1938 (3 55, 2 225 Гуссаков-
ский). — Большая часть Европы и Европ. части СССР, кроме севера;
Сибирь; Кавказ; Ср. Азия (Гиссарский хребет); Монголия.
Двукрылые
139
Physocephala vaginalis Rd. — Кондара, 7 VIII 1940 (1 $>; Гусса-
ковский). — Средиземноморье; Закавказье; Ср. Азия: Таджикистан
(Сталинабад, Кондара, Гиссар).
Physocephala vittata F. — Кондара, 13, 15, 16 V, 15, 27 VI, 2 VII,
2, 10, 15 VIII, 8 IX 1937 (17 55; Гуссаковский); 17, 20 VI, 10, 16 VII,
16 IX 1938 (6 55; Гуссаковский), 5 VI 1943, на цветках Prangos pabu-
laria (1 5; Штакельберг); Руидашт, 17 VI 1938 (1 5; Гуссаковский). —
Сф. и южн. Европа; юг Европ. части СССР; Ср. Азия; Сибирь (Иркутск).
Conops flavifrons Mg. (=Physocephala pugioniformis Beck. sec. typ.,
syn. nov.).— Кондара, 2, 14 VII, 3 VIII 1937 (3 55; Гуссаковский); 18 VII
1938, 13 VII 1939 (4 55; Гуссаковский). —Южн. Европа; сев. Африка;
М. Азия; юг Европ. части СССР; Ср. Азия: Таджикистан (Гиссарский
хр.; южн. Таджикистан).
Conops vesicularis L. — Кондара, 3, 6 VI 1939 (2 55; Гуссаковский);
8 VI 1943 (1 j; Штакельберг). —Большая часть Европы; Европ. часть
СССР, кроме севера; горы Ср. Азии (Гиссарский хребет); Сибирь.
Myopa buccata L. —Квак, 7 VI 1943 (1 5; Штакельберг); Руидашт,
18 VII 1938 (1 5; Гуссаковский). —Большая часть Европы; М. Азия;
Ср. Азия; Сибирь до Приморского края; Япония.
Myopa testacea L. — Руидашт, 17, 18 VI 1938 (9 55; Гуссаков-
ский). — Большая часть Европы; Сибирь до Байкала; горы <Ср. Азии;
сев. Иран.
Zodion cinereum F. — Кондара, 8, 9, 10, 15, 26 VI, 22 VII, 8 IX 1937;
14 VII 1938 (7 55, 2 2$; Гуссаковский); Руидашт, 22 VII 1937 (1 5;
Гуссаковский). — Большая часть Европы; Сибирь; Ср. Азия: Таджи-
кистан (Сталинабад, Гиссарский хребет).
Zodion erythrurum Rd. — Руидашт, 23 VII 1936 (1 5; Гуссаковский). —
Ср. и южн. Европа; юг Европ. части СССР; Ср. Азия: Таджикистан
(Гиссарский хребет).
Occemyia atra F. — Кондара, 5 VI, 9 1X 1937; 19 VI, 10, И VII,
13, 16, 19 IX 1938; 29 V 1939; 28 VII 1940 (9 55, 2 jj; Гуссаковский);
Руидашт, 23 VII 1936 (1 5; Гуссаковский). — Большая часть Европы;
М. Азия; сев. Африка; Ср. Азия: Таджикистан (Гиссарский хребет).
Occemyia distincta Mg. —Кондара, 7 VI 1937 (1 5; Гуссаковский);
Руидашт, 17 VI 1938 (1 5; Гуссаковский). — Большая часть Европы;
И. Азия; Ср. Азия: Таджикистан (Гиссарский хребет).
Сем. Platystomatidae
Platystoma canumPortsch. sec. typ. (= curvinerve Hendel, syn. nov .). —
Кондара, 10 IX 1945 (2 jj; Гуссаковский). — Закавказье; Туркмения;
Таджикистан.
Platystoma elegans Hendel. — Кондара, 8, 13 VI 1937 (3 55, 1 j;
Гуссаковский), 5 VI 1943, на цветах Prangos pabularia (2 55, 1 j; Шта-
кельберг); Квак, 20 VI 1937 (2 55, 2 jj; Гуссаковский). —Закавказье;
Туркмения (Копет-даг); Таджикистан (Гиссарский и Дарвазский хребты).
Сем. Ulidiidae
Physiphora demandata F. —Кондара, 15 VIII 1937 (1 5, 1 $>; Гус-
саковский); 15 VII, 13, 22 1X 1938 (2 55, 2 jj; Гуссаковский).—
140
А. А. Штакельберг
Европа и Азия, кроме крайнего севера; Африка; Сев. Америка, кроме
севера.
Ulidia erythrophthalma albidipennis Lw. — Кондара, 14 VII 1938 (1 3;
Гуссаковский); Квак, 4 VII 1940 (1 $>; Гуссаковский). —Юго-вост.
Европа; Закавказье; Иран; Таджикистан (Гиссарский хребет).
Ulidia melampodia Lw. — Кондара, 27 VI, 3 VII 1937; 15, 20 VII 1938
(2 5(5, 2 225 Гуссаковский). — Сев. Иран; Ср. Азия (дол. Зеравшана,
Гиссарский хребет).
Ulidia ruficeps Beck. — Руидашт, 23 VII 1936; 22 VII 1937; 16, 17 VI,
20 VII 1938 (6 33, 7 225 Гуссаковский). — Таджикистан (высоты Гиссар-
ского хребта); Персидский Белуджистан.
Ulidia salonikiensis Hennig. — Кондара, 5, 13 VI 1937; 13, 16 V 1939
(5 33, 3 gg; Гуссаковский); 5 VI 1943; 2, 3 V 1944 (4 33, 5 2$’» Шта-
кельберг). — Балканский п-ов; Ср. Азия: Таджикистан (Гиссарский
хребет).
Timia (Empyelocera) abstersa Lw. — Кондара, 15 VIII 1937 (1 25
Гуссаковский). — Юг Европ. части СССР; Туркмения; Таджикистан
(Сталинабад, Гиссарский хребет).
Timia (Empyelocera) nasuta Mik. — Кондара, 14 VIII 1937 (1 (5; Гус-
саковский). — Туркмения; Таджикистан (Кондара).
Timia (Empyelocera) nigrimana Lw. —Кондара, 13 VII 1938 (1 d;
Гуссаковский), 8 VI 1943 (1 3; Штакельберг). — Юго-восток Европ. части
СССР; Казахстан; Узбекистан; Таджикистан (Гиссарский хребет,
хр. Петра I).
Сем. Otitidae
Myennis octopunctata Coqueb. — Кондара (1 25 Гуссаковский). -
Ср. и южн. Европа; Украина; Ср. Азия (Таджикистан: Кондара);
Приморский край.
Сем. Lonchaeidae
Lonchaea (Dasyops) latifrons Mg. — Кондара, 20 IV, 3 V 1944
(3 2$5 Штакельберг). — Ср. и южн. Европа; Ср. Азия: Таджикистан
(Сталинабад, Кондара).
Сем. Trypetidae
Euribia stylata F. —Кондара, 15 VI, 15 VII 1939 (1 3, 1 25 Гусса-
ковский). — Европа.
Euribia tenuis Beck. — Кондара, 8 VI 1943 (1 3, 2 225 Штакельберг).
Ср. и Центр. Азия
Hypenidium roborovskii Beck. — Кондара, 16, 17, 20 IX 1938 (3 33:
Гуссаковский). — Кавказ; Средняя Азия: Таджикистан (Кондара; южн.
Таджикистан); Центр. Азия.
Aciura coryli Rossi — Кондара, 31 V, 15 VII 1939; 21 VIII, 10 IX 1945
(2 33, 3 225 Гуссаковский); 8 VI 1943 (1 25 Штакельберг). —Среди-
земноморье; сев. Африка; М. Азия; Ср. и Центр. Азия.
Oxyaciura tibialis R.-D. — Кондара, 12 VII 1938 (1 3; Гуссаков-
ский). — Южн. часть Ср. Европы; южн. Европа; сев. Африка; Иран;
Ср. Азия.
Двукрылые
141
Carpomyia schineri Lw. — Кондара, 14, 15 VII, 14, 15 1X 1938;
26 VIII 1945 (3 (55, 2 Гуссаковский). —Ср. Европа; Ср. Азия:
Таджикистан (Кондара, зап. Памир).
Carpomyia vesuviana Costa — Кондара, 29 VII 1940 (1 j; Гуссаков-
ский). — Южн. Европа; вост. Закавказье; Туркмения; Таджикистан
(Кондара).
Terellia longicauda Mg. —Кондара, 24 VII 1938 (1 j; Гуссаковский). —
Ср. и южн. Европа; сев. Африка; Ср. Азия: Таджикистан (Гиссарский
хребет).
Chaetorellia jaceae R.-D. — Кондара, 12 VIII 1937 (1 (5; Гуссаков-
ский); 8 VI 1943; 3 V 1944 (1 (5, 1 j; Штакельберг). — Большая часть
Европы; М. Азия; Иран; Ср. Азия: Таджикистан (Сталинабад, Кондара).
Chaetostomella onotrophes R.-D. — Квак, 7 VI 1943 (1 <5; Штакель-
берг).— Большая часть Европы; М. Азия: Ср. Азия—Таджикистан
(Сталинабад, Кондара).
Paroxyna tessellata Lw. — Кондара, 20 VIII 1938; 6 IX 1945 (1 (5,
1 о; Гуссаковский); Квак, 18 IX 1945 (1 (5; Гуссаковский). — Европа;
сев. Африка; М- Азия; Ср. Азия: Таджикистан (Гиссарский хребет);
Центр. Азия.
Tephritis cometa Lw. — Кондара, 20 VII 1940 (2 <5(5; Гуссаковский);
Квак—Руидашт, 2500 м, 6 VI 1943 (2 jj; Штакельберг); Руидашт,
17 VI 1938 (2 (5(5; Гуссаковский). —Ср. и южн. Европа; Малая Азия;
Ср. Азия: Таджикистан (Гиссарский хребет, южн. Таджикистан, зап.
Памир).
Tephritis dioscurea Lw. — Квак—Руидашт, 2500 м, 6 VI 1943 (1 (5;
Штакельберг). — Ср. и южн. Европа; Ср. Азия: Таджикистан.
Tephritis heiseri Lw. — Кондара, 10 VIII 1937 (1 <5; Гуссаковский). —
Ср. и южн. Европа; Ср. Азия: Таджикистан (Кондара); Куку-нор.
Tephritis poecilura Lw. —Квак—Руидашт, 2500 м, 6 VI 1943 (1 j;
Штакельберг). — Южн. Европа; сев. Африка; Ср. Азия: Таджикистан
(Гиссарский хребет).
Tephritis recurrens Lw. —Квак, 27 VIII 1937 (1 $>; Гуссаковский);
Руидашт, 20 VII 1938 (1 (5; Гуссаковский). — Средиземноморье; Ср. Азия:
Гиссарский хребет; Амурская обл.
Acanthiophilus helianthi Schrk. — Кондара, 21 VIII, 16 IX 1945 (2 jj;
Гуссаковский). — Большая часть Европы, кроме севера; сев. Африка;
М. Азия; Ср. и Центр. Азия.
Trypanaea amoena Frfld.—Кондара, 18, 26 VIII 1945 (2 jj; Гус-
саковский).— Ср. и южн. Европа; сев. Африка; Иран; Ср. и Центр.
Азия; Индия.
Trypanaea augur Frfld. — Кондара, 8 VI 1943 (2 jj; Штакельберг). —
Сев. и Ср. Африка; Иран; Ср. Азия: Таджикистан (Кондара, южн. Таджи-
кистан); Центр. Азия.
Trypanaea stellata Fuessly — Кондара, 8 VI 1943 (2 jj; Штакель-
берг); 15 IX 1945 (2 (5(5, 1 j; Гуссаковский). — Большая часть Европы,
кроме севера; сев. и вост. Африка; Иран; Ср. Азия: Таджикистан (Гис-
сарский и Дарвазский хребты, дол. Вахша).
Сем. Sciotoyzidae
Ditaenia cinerella Flln. — Руидашт, 17 VI 1938 (1 (5; Гуссаковский).
Большая часть Европы; сев. Африка.
142
А. А. Штакельберг
Сем. Lauxaniidae
Homoneura dentiventris Czerny— Кондара, 22 VI, 12, 14, 17 VII, 3,
19 VIII, 6, 17 IX 1938; 15 VII 1939 (4 53, 6 Гуссаковский). — Семи-
речье; Таджикистан (Гиссарский хребет).
Sapromyza biordinata Czerny — Кондара, 10, 15 VI, 9 VIII, 13, 14.
19, 20, 22 IX 1938; 29 V, 14 VI, 15 VII 1939; 27 VII, 7 VIII 1940 (10 33
10 22; Гуссаковский). — Ср. Азия (широко распространенный вид);
Пран.
Sapromyza simplex Lw. — Кондара, 14 IX 1938 (1 2» Гуссаковский).-
Большая часть Европы и Европ. части СССР: Ср. Азия.
Сем. Helomyzidae
Tephrochlamys rufiventris Mg. — Кондара, 11 III 1943 (1 3, 1 2;
Штакельберг). — Большая часть Европы; Ср. Азия: Таджикистан (Кон-
дара).
Сем. Trichoscelididae
Trichoscelis obscurella Flln. — Кондара, 16, 19 IX 1945 (2 33; Гус
саковский). — Большая часть Европы; Ср. Азия: Таджикистан (Кондара).
Сем. Chamaemyidae
Parochthiphila coronata Lw. — Кондара, 21 VIII 1945 (1 2: Гусса-
ковский). — Ср. и южн. Европа: Ср. и южн. Азия.
Сем. Opomyzidae
Geomyza tripunctata Mg. — Кондара, 17, 20 IX 1938 (1 3, 3 2$*, Гус-
саковский). — Большая часть Европы; Ср. Азия: Таджикистан (Кол-
дара).
Сем. Drosophilidae
Curtonotum anus Mg. — Кондара, 16 VII, 14 1X 1938 (3 2$; Гусса-
ковский). — Ср. и южн. Европа; Ср. Азия (Гиссарский хребет); Прпмо|
ский край.
Diastata unipunctata Zett. — Квак, 7 VI 1943 (1 25 Штакельберг).
Большая часть Европы; Ср. Азия (Гиссарский хребет).
Scaptomyza unipunctum Zett. —Квак, 5 VI 1943 (3 33, 1 д; Шта-
кельберг). — Сев. Европа; Сибирь (Иркутск); Камчатка; Семиречье;
Ср. Азия: Таджикистан (Гиссарский хребет). Мезофильные стации.
Drosophila littoralis Mg. — Кондара, 9 III 1943, на снегу (1 25 Шта-
кельберг). — Большая часть Европы; Ср. Азия (Гиссарский хребет).
Всюду редок.
Двукрылые
143
Drosophila obscura Flln. — Кондара, 11 III 1943 (1 Штакель-
берг). — Большая часть Европы: Ср. Азия (Гиссарский хребет); Сев.
Америка.
Сем. Sphaeroceridae
Sphaerocera curvipes Latr. — Кондара, 20 IV 1944 (2 <5d; Штакель-
берг). — Большая часть Европы; Ср. Азия; Сев. Америка. На навозе.
Crumomyia glabrifrons Mg. — Кондара, И III 1943 (1 j; Штакель-
берг); 20 IV 1944 (3 (55, 1 j; Штакельберг). — Большая часть Европы;
Ср. Азия (Гиссарский хребет). На навозе.
Paracollinella fontinalis Flln. — Кондара, 20 IV 1944 (1 (5, 1 Шта-
кельберг). — Большая часть Европы; Сев. Америка.
Сем. Ephydridae
Mosillus subsultans Flln. — Кондара, 20 VII, 7, 20 VIII, 20 IX 1938
(2 <5(5, 3 22; Гуссаковский). — Большая часть РЗвропы; Ср. и Центр.
Азия.
Psilopa nitidula compta Mg. — Кондара, 16, 17, 19 IX 1938; 3 VI 1939
(3 (5(5, 8 jj; Гуссаковский). — Большая часть Европы: сев. Африка;
Ср. Азия. Мезофильные стации.
Сем. Chloropidae
Elachiptera cornuta Flln. — Кондара, 6 IX 1944 (1 (j; Гуссаков-
ский). — Большая часть Европы; сев. Африка; Сибирь до Приморского
края; Ср. Азия (Гиссарский хребет).
Tricymba humeralis Lw. — Кондара, 15 IX 1945 (1 5, 1 Гуссаков-
ский). — Ср. и южн. Европа; сев. Африка; Ср. Азия (Гиссарский хребет).
Oscinella pusilia Mg. — Кондара, 14 IX 1938 (3 22’, Гуссаковский). —
Большая часть Палеарктики. Один из основных вредителей хлебных
злаков.
Meromyza pratorum Flln. — Кондара, 29, 31 VII 1940 (3 й(5, 3 22;
Гуссаковский). — Большая часть Европы; Сибирь до Приморского края;
Ср. Азия.
Meromyza saltatrix L. — Кондара, 15 IX 1945 (1 <5; Гуссаковский). —
Большая часть Европы; Сибирь; Ср. и Центр. Азия. Личинка вредит
хлебным злакам.
Thaumatomyia notata Mg. — Кондара, 27 VII 1940 (1 (5; Гуссаков-
ский). — Большая часть Европы; Сибирь; Ср. Азия.
Thaumatomyia sulcifrons Beck. — Кондара, 10, 14 V11, 14 IX 1938;
27, 29 VII 1940; 18, 19, 25 VIII 1945 (11 (5(5, 13 Гуссаковский).—
Юж и. Европа; сев. Африка; Ср. и Центр. Азия.
Сем. Odiniidae
Turanodinia coccidarum Stack. — Кондара, 18 VIII 1945 (1 2J Гус-
саковский). —Узбекистан (окр. Ташкента); Таджикистан (Кондара).
144
А. А. Штакельберг
Сем. Milicbiidae
Madiza glabra Flln. — Кондара, 20 VI, 16 IX 1938 (3 88, 4 Гус-
саковский). — Большая часть Европы; Сибирь; Ср. Азия; сев. Африка;
Сев. Америка.
Сем. Hippoboscidae
Hippobosca capensis Olf. — Кондара, 17 VII, 13, 14 IX 1938; 29 V,
22 VII 1939 (4 88, 4 22; Гуссаковский). — Южн. Европа: сев. Африка;
Ср. Азия.
Hippobosca equina L. — Кондара, 22 VI 1938 (1 25 Гуссаковский). —
Большая часть Европы; Ср. Азия.
Сем. Cordyluridae
Scopeuma stercorarium L. — Кондара, 15 V 1939 (2 88] Гуссаковский);
Руидашт, 17 VI 1938 (3 88; Гуссаковский). — Большая часть Европы;
Ср. Азия.
Сем. Muscidae
Schoenomyza littorella Flln. — Кондара, 20 IX 1938 (1 5; Гуссаков-
ский). — Большая часть Европы; Ср. Азия.
Coenosia atra Mg. — Кондара, 21 VIII 1945 (1 25 Гуссаковский). —
Большая часть Европы, кроме севера; Передняя Азия; горы Ср. Азии.
Calythea albicincta Flln. — Кондара; 10 VII, 30 VIII 1938; 13 IX 1939
(3 88; Гуссаковский); 29 VII, 7 VIII 1940 (8 55; Гуссаковский); Руи-
дашт, 17 VI 1938 (1 5; Гуссаковский). — Ср. и южн. Европа; Передн.
н Ср. Азия.
Lispa папа Macq. — Кондара, 28, 29 VII 1940 (3 55, 1 2’» Гуссаков-
ский). — Южн. Европа; сев. Африка; Ср. Азия.
Lispa tentaculata Deg. — Кондара, 14 IX 1938; 28, 29 VII 1940 (4 55,
2 2$; Гуссаковский). — Большая часть Европы; Ср. и Центр. Азия.
Fannia canicularis L. — Кондара, 19 IX 1945 (1 5; Гуссаковский). —
Европа; сев. Африка; Передн. и Ср. Азия. Синантроп.
Ophyra anthrax Mg. — Кондара, 19 VI, 10, 14, 18 VII, 15, 16, 17,
22 IX 1938; 13 VI 1939 (7 55, 3 22’ Гуссаковский). — Ср. и южн.
Европа; Ср. Азия (Туркмения, Таджикистан).
Muscina stabulans Flln. — Кондара 24 VIII 1945 (1 5; Гуссаков-
ский). — Большая часть Палеарктики, кроме крайнего севера.
Myiospila meditabunda Flln. — Квак, 7 VI 1943 (1 5; Штакельберг).—
Большая часть Европы; Передн. и Ср. Азия.
Stomoxys calcitrans L. (определил Зимин) — Кондара, 15 IX 1938
{1 2» Гуссаковский). — Европа; сев. Африка; Передн. и Ср. Азия.
Graphomyia maculata Scop, (определил Зимин) — Кондара, 30 VIII 1938;
24 VI, 12 VII 1939 (2 55, 1 25 Гуссаковский). — Большая часть Европы;
Сибирь; Ср. Азия.
Pyreilia cadaverina F. (определил Зимин) — Кондара, 15 VI, 16, 25 VII,
3, 19, 20, 30 VIII, 12, 13,17,18 1X 1938; 24 V, 13 VI, 31 VII 1939
(9 55, 9 225 Гуссаковский). —Большая часть Европы; Сибирь; Ср. Азия.
Двукрылые
145
Pyreilia secunda Zimin (определил Зимин) — Кондара, 17 VII 1938
(1 (5; Гуссаковский). — Ср. Азия.
Orthellia caesarion Mg. (определил Зимин) — Кондара, 15, 16 V 1939
(3 55, 1 Гуссаковский). — Большая часть Европы; Сибирь; Ср. Азия.
Dasyphora asiatica Zimin (определил Зимин) — Кондара, 17 VII 1938
(1 5; Гуссаковский). — Ср. и Центр. Азия.
Dasyphora gussakovskii Zimin (определил Зимин — Кондара, 13 IX 1937,
30 VIII, 6, 14, 15, 17, 18, 21, 22 IX 1938; 21 V 1939 (5 55, 10 Гус-
саковский). — Таджикистан (Гиссарский хребет, зап. Памир); зап. Каш-
гария.
Morellia simplex Lw. (определил Зимин) — Кондара, 15 VI, 16, 17,
26 VII, 3, 8, 20 VIII, 13, 18, 21 IX 1938; 15 VI, 14 VII 1939 (И 55, 7 2$5
Гуссаковский); 8 VI 1943 (1 5; Штакельберг). — Большая часть Европы,
кроме севера; Ср. Азия.
Musca autumnalis Deg. (определил Зимин) — Кондара, 9 IX 1937(2 55;
Гуссаковский). — Большая часть Европы, кроме крайнего севера; Кав-
каз; Ср. Азия.
Musca domestica vicina Macq. — Кондара, 21 VIII 1945 (3 55, 2 22*»
Гуссаковский). — Тропическая и субтропическая зона.
Musca larvipara Portsch. (определил Зимин) — Кондара, 7, 9 IX 1937
(2 55, 1 2$л Гуссаковский). — Ср. и южн. зоны Европейской части СССР;
Ср. Азия.
Musca osiris Wd. (определил Зимин) — Руидашт, 22 VI 1937 (1 25
Гуссаковский). — Ср. Азия.
Сем. Phasiidae
Besseria anthophila Lw. — Квак, 6 VI 1943 (1 25 Штакельберг). —
Ср. и южн. Европа; Ср. Азия.
Alophora pusilia Mg. — Кондара, 13 VII 1937 (1 25 Гуссаковский). —
Ср. и южн. Европа; Ср. Азия.
Phasia appendiculata Jacobs. — Кондара, 5 VI 1943, на цветах Pran-
gos pabularia (1 25 Штакельберг). — Гиссарский хребет; зап. Памир;
Джунгария.
Phasia crassipennis Mg. — Кондара, 2 V 1944 (1 5; Штакельберг). —
Ср. и южн. Европа; Ср. Азия.
Rhodogyne (Stylogymnomyia) nitens Mg. — Квак, 6 VI 1943 (1 5;
Штакельберг). — Ср. и южн. Европа; Ср. Азия.
Сем. Calliphoridae
Chrysomyia albiceps Wd. — Кондара, 11 IX 1937 (1 25 Гуссаков-
ский). — Сев. Африка; Ср. Азия.
Calliphora erythrocephala Mg. — Квак, 22 IX 1937 (1 5; Гуссаков-
ский). — Большая часть Европы; Ср. Азия.
Lucilia sericata Mg. — Кондара, 27 VI, 12 IX 1937 (2 55; Гуссаков-
ский). — Европа, кроме севера; Африка; Ср. Азия; Индия; Австралия.
Сем. Sarcophagidae
Blaesoxipha (Servaisia) rossica Vill. (определил Родендорф) — Квак—
Руидашт, 2800 м, каменистые россыпи, на камнях, 6 VI 1943 (5 55; Щта-
10 Ущелье Кондара
146
А. А. Штакельберг
кельберг). — Вост. Европа; Европейская часть СССР, кроме севера;
Сибирь на восток до Иркутска; Ср. Азия (центр. Тянь-шань, Гиссарский
хребет).
Blaesoxipha (in sp.) acuminata Vill. (определил Родендорф)— Кон-
дара, 22 VI, 10, 14 VII 1938, 8 IX 1945 (4 Гуссаковский). — Юго-
восток Европейской части СССР, Узбекистан, Таджикистан (Гиссарский
хребет).
Blaesoxipha (in sp.) cochlearis Pand. (определил Родендорф)—Кон-
дара, 11 VII, 7 VIII 1938 (2 55; Гуссаковский).—Юг Зап. Европы и
Европейской части СССР, Узбекистан, Таджикистан (Гиссарский хре-
бет) , Прибайкалье.
Blaesoxipha (in sp.) laticornis Mg. (определил Родендорф) — Квак,
29 VI 1937 (1 5; Гуссаковский). — Большая часть Европы, Ср. Азия,
Сибирь.
Blaesoxipha (in sp.) lineata Flln. (определил Родендорф) — Кон-
дара, 13 VI, 2 VII 1937, 7 VIII 1938 (3 9$; Гуссаковский); 3 V 1944 (1 5;
Штакельберг); Квак, 6 VI 1943 (1 $>; Штакельберг). — Средиземноморье;
лесостепи и степи Европейской части СССР, Крым, Кавказ; Ср. Азия;
южн. Сибирь.
Parasarcophaga portschinskyi Rohd. (определил Родендорф) — Кон-
дара, 23 VII 1938 (1 (5; Гуссаковский); экземпляр выведен из трупа
фаланги. — Юг Европейской части СССР, Ср. Азия, Монголия, Тибет.
Sarcophaga carnaria carnaria (L.) Rohd. (определил Родендорф) — Кон-
дара, 3 V 1944 (2 55; Штакельберг). — Сев. и ср. Европа; Европейская
часть СССР на юг до южн. Украины и южн. Урала; Сибирь; Ср. Азия
(Гиссарский хребет).
Coprosarcophaga haemorrhoidalis Flln. (определил Родендорф) — Кон-
дара, 26 VIII, 11, 12 IX 1937, 12, 17 VII, 7, 8 VIII, 15, 17 IX 1938, 29 V
1939 (4 55, 5 q$>; Гуссаковский); 20 IV 1944 (1 5; Шта ельберг).
Большая часть Европы, Ср. и Центр. Азия, Африка, Сев. Америка.
Pierretia plotnikovi Rohd. (определил Родендорф) —Кондара, 3 V 1944
(4 55; Штакельберг). — Таджикистан (Сталинабад, Гиссарский, Дар-
вазский хребты, зап. Памир); Узбекистан (окр. Ташкента).
Ravinia striata F. — Кондара, 5 IX 1945 (2 <5(5; Гуссаковский). —
Большая часть Палеарктики, кроме крайнего севера. На faeces.
Wohfahrtia magnifica Sehin, (определил Родендорф) — Кондара,
5 VI 1943 (1 j; Штакельберг). — Юг Европейской части СССР и Сибири,
Ср. Азия.
Miltogramma taeniatum Mg. — Кондара, 19 IX 1938 (2 55; Гуссаков-
ский). — Ср. и южн. Европа; Ср. Азия.
Miltogramma (Pseudomiltogramma) brevipilum Vill. (определил
Родендорф) — Кондара, 23 VI, 14, 15 VII 1938, 14, 17 VII 1939 (4 55,
6 Гуссаковский). — Южн. Европа, Ср. Азия.
Anacanthothecum testaceifrons Ros. (определил Родендорф).—Кон-
дара, 21 VI 1938 (1 9; Гуссаковский), 8 VI 1943 (2 55; Штакельберг). —
Ср. и южн. Европа, Ср. Азия.
Senotainia albifrons Rd. (определил Родендорф) — Кондара, 5, 26,
29 VIII, 9 1X 1937, 10, 17, 26 VII, 19 VIII 1938, 13 IX 1939, 22,
24 VIII 1945 (10 55, 12 оо; Гуссаковский).—Ср. и южн. Европа, Ср.
Азия.
Metopodia grisea Mg. (определил Родендорф). —Кондара, 24 VI 1937,
22 VIII 1945 (1 5, 1 j; Гуссаковский).—Ср. и южн. Европа, Ср.
Азия.
Двукрылые
147
Taxigramma (Eutaxigramma) multipunctatum Rd. — Кондара, 26 VIII
1945 (1 Гуссаковский). — Ср. и южн. Европа; Ср. Азия.
Metopia leucocephala Rossi — Кондара, 3 V 1944 (1 <8; Штакельберг);
24 VIII 1945 (1 8; Гуссаковский). — Большая часть Европы; Ср. Азия;
Сибирь до Приморского края.
Сем. Larvivoridae
Gonia bimaculata Wd. — Кондара, 9, 14 IX 1937 (2 88; Гуссаков-
ский). — Средиземноморье; Передн. и Средн. Азия.
Plagia ruricola Mg. — Кондара, 2 V 1944 (2 58; Штакельберг). —
Ср. и южн. Европа; Ср. Азия (в горах).
Voria ruralis Mg. — Кондара, 14 VIII 1937 (1 8; Гуссаковский). —
Большая часть Европы; горы Ср. Азии.
Stomatomyia filipalpis turanica Rohd. — Кондара, 3 V 1944 (1 8,
1 q; Штакельберг). — Ср. Азия. Типичная форма — Ср. и южн. Европа.
Viviania cinerea Flln. — Кондара, 8 VI 1943 (2 jj; Штакельберг). —
Большая часть Европы; Ср. Азия (Туркмения; Гиссарский хребет).
Meigenia mutabilis Flln. — Кондара, 14 IX 1937 (1 8; Гуссаков-
ский). — Большая часть Европы; Ср. Азия (Гиссарский хребет).
Exorista confinis Flln. — Кондара, 19 VII, 3 VIII 1937 (2 88; Гус-
саковский). — Большая часть Европы; Ср. Азия (Гиссарский хребет).
Linnaemyia compta Flln. (определил Зимин) — Кондара, 5 VI 1943, на
цветах Prangos pabularia (1 8, 3 22’ Штакельберг). — Большая часть
Европы, кроме севера; Сибирь; Ср. Азия.
Linnaemyia vulpina Flln. (определил Зимин) — Кондара, 20 IV 1944
(2 22; Штакельберг). Большая часть Европы, кроме севера; Крым ; Ср.
Азия (Сталинабад, Кондара).
Peletieria nigricornis Mg. (определил Зимин) — Кондара, 15 VI 1938,
24 VI 1939 (2 22’, Гуссаковский); Квак, 18 1X 1945 (1 25 Гуссаков-
ский). — Большая часть Палеарктики, кроме крайнего севера; Сев.
Америка.
Peletieria ruficeps Macq. (определил Зимин) — Кондара, 1 V 1939 (1 2‘,
Гуссаковский).—Сев. А{рика, южн. Европа, Крым, Закавказье, Ср.
Азия.
Peletieria sibirica Smirn. (определил Зимин) — Кондара, 19 VII 1937
(1 2’ Гуссаковский). — Гиссарский хр., хр. Петра I, зап. Памир,
Семиречье, Прибайкалье, Монголия.
Peletieria xenoprepes Lw. (определил Зимин)—Квак, 22 IX 1937
(3 22’ Гуссаковский). — Иран, Гиссарский и Дарвазский хребты, зап.
Памир, Алтай, вост. Сибирь (Иркутск).
Fabriciella ferox Pz. (определил Зимин) — Руидашт, 18 VI 1938 (2 88;
Гуссаковский). — Большая часть Европы, кроме севера; Закавказье;
Ср. Азия: Таджикистан (Гиссарский хребет, Памир); Сибирь; Монголия.
Cnephaotachina danilevskyi Portsch. (определил Зимин) — Кондара,
23 VI 1938 (1 2’, Гуссаковский). — Южн. Европа; юг Европейской части
СССР (Сарепта); Крым; Закавказье; Семиречье; Ср. Азия; Иран (Белуд-
жистан).
Tachina magnicornis Zett. (определил Зимин) — Кондара, 20 IV 1944
(1 2’, Штакельберг); Квак, 18 IX 1945 (1 2‘, Гуссаковский). — Большая
часть Палеарктики, кроме крайнего севера.
Tachina rohdendorfi Zim. (определил Зимин) — Кондара, 17 IX 1938;
19, 24 VI 1939 (3 88; Гуссаковский); Руидашт, 17, 18 VI, 20 VII 1938
10*
148
А. А. Штакельберг
(1 (5, 4 29; Гуссаковский). — Закавказье; Ср. Азия, Где широко распро-
странен.
Pareudora praeceps Mg. (определил Зимин) — Кондара, 24 VI 1939
(2 (5(5; Гуссаковский); 5 VI 1943; 2 V 1944 (1 (5, 3 2$; Штакельберг);
Квак, 7 VI 1943 (1 2‘, Штакельберг). —Ср. и южн. Европа; сев. Аф-
рика; юг Европейской части СССР; Кавказ и Закавказье; Ср. Азия;
Приморский край.
ЛИТЕРАТУРА
Ш т акельберг А. А. 1947. Новые виды Dolichopodidae из Таджикистана.
Энтом. обозр., XXIX, 1—2 93—102.
Штакельберг А. А. 1948. К познанию видов семейства Dixidae (Diptera, Nema-
tocera) Таджикистана. Памяти акад. С. А. Зернова [Сборник], Изд. АН СССР
236—241.
Штакельберг А. А. 1949. Виды рода Hercostomus Lw. (Diptera, Dolichopo-
didae) среднеазиатской фауны. Тр. Зоол. инет. АН СССР, VIII, 4 669—687.
Штакельберг А. А. 1949. Новые данные по роду Eumerus Mg. (Diptera, Syr phi-
dae) палеарктической фауны. Энтом. обозр., XXX, 3—4 : 426—439.
JJ. А. РУБЦОВ
МОШКИ
1. Eusimulium (Astega) persicum Rubz. — Иа выс. 1600—1800 м.
(Здесь и дальше высота в метрах над уровнем моря). Имаго в зоне арчи.
Редок.
2. Eusimulium (Astega) ovtshinnikovi Rubz. — p. Варзоб и ее истоки,
1200—3000 м. Массовый вид. Холоднолюбивый вид. В течение года —
две генерации. Кровосос.
3. Eusimulium (in sp.) montium Rubz. — мелкие притоки p. Кон-
дара.
4. Eusimulium (in sp.) brachyantherum Rubz. — Мелкие ручьи,
впадающие в р. Кондара на выс. до 1300 м.
5. Eusimulium nigrolusipes Rubz. — Мелкие прогреваемые ручейки
у Руидашта, на высоте около 2000 м.
6. Eusimulium (in sp.) inflatum Rubzov, sp. nov. — Личинки в хо-
лодном роднике, впадающем в р. Кондара.
7. Simulium (Odagmia) crassifilum Rubz. — Мелкие родники и ру-
чейки на высоте 1200—2500 м.
8. Simulium (Odagmia) ornatum var. caucasicum Rubz. — P. Кон-
дара (см. рис.).
9. Simulium (Odagmia) ferganicum Rubz. — Мелкие, относительно
теплые ручьи на выс. 2000 м (у Руидашта).
10. Simulium (Odagmia) alajense Rubz. — Широко распространенный
массовый вид, главный кровосос. Преобладает в р. Варзоб на выс. 1100—
3000 м.
11. Simulium (Odagmia) humerosum Rubz. — Изредка в Кондаре на
выс. около 1700 м. В основном распространен выше по Варзобскому
ущелью.
12. Wilhelmia veltistshevi Rubz. — Изредка в р. Варзоб против
Кондаринского ущелья. В основном вид распространен ниже, в р. Дю-
шамбинке против Сталинабада и ниже. Кровосос.
13. Simulium (Obuchovia) albellum Rubz. — Ущелье Кондара на
высоте около 1100 м. Кровосос. Основное распространение — выше,
район Ходжа-оби-гарма, в холодных ручьях.
14. Simulium (in sp.) angustifilum Rubz. — Ущелье p. Кондара на
выс. 1500 м. Редок в ущелье Кондара, но обычен выше (Ходжа-оби-
гарм, Зидды).
15. Simulium (in sp.) multistriatum Rubz. — Основной массовый
вид, населяющий р. Кондара и выплаживающийся на высоте от устья
р. Кондара до 2000 м. Злостный кровосос; нападает на человека и домаш-
них животных.
150
И, А. Рубцов
16. Simulium (in sp.) subornatoides Rubz. — Массовый вид в истоках
р. Кондара; встречается на выс. 1100—2500 м.
Перечисленными 16 видами, конечно, не исчерпывается перечень
видов мошек, которые могут встретиться в долине р. Кондара. Назван-
ные 16 видов — результат первого ориентировочного обследования. При
ближайшем исследовании здесь может быть обнаружено до десятка дру-
гих из числа 40 видов, обнаруженных в долине р. Варзоб и смежных
долинах Гиссарского хребта.
Экологически — это горная фауна, оригинальная в морфологическом
и биоценотическом отношениях. У большинства видов отчетливо выра-
в
Simulium ornatum Mg.
жены приспособления к условиям бы-
стротекущих горных ручьев. Все виды
встречаются в других долинах и бо-
лее или менее удаленных районах
Гиссарского хребта, по преимуще-
ству в вышележащих.
В зоогеографическом отношении
фауна носит сборный характер. Наи-
больший процент составляют сре-
диземноморские и туркменские эле-
менты. Целый ряд видов известен
из Кавказа: Simulium (Odagmia)
ornatum var. caucasicum Rubz.,
Eusimulium (Astega) montium Rubz.
и др. В смежных долинах имеются
виды общие с Ориентальной обла-
стью: Wilhelmia mediterraneum
Edw., Wilhelmia paraequina Puri,
Simulium (Eusimuluim) latipes Mg.
С другой стороны, имеются виды,
общие с Европой — Wilhelmia sa-
lopiensis (Edw.). Чрезвычайно ин-
тересным представляется нахожде-
ние видов, близких к европейским
[S. (Odagmia) ornatum Mg. и Eusi-
mulium aureum Fries], на высотах
около 2000 м. Несколько менее от-
А— взрослое насекомое, самка; Б— кукол- чвТЛИВЫ СВЯЗИ фауны Варзобской ДО-
ка; В—личинка. * J т~г *
лины и, в частности, Нондаринскои
с Индийской и Индокитайской подобластями. Таким образом, фауна
мошек Кондаринской долины, как и Таджикистана в целом, зоогеогра-
фически находясь на стыке четырех подобластей (Средиземноморской,
Туркменской, Индийской и Индокитайской), отражает в своем составе
их фаунистические элементы. Фауна в целом — молодая, эндемичные
виды маловероятны, и некоторые представляются таковыми вследствие
недостаточной изученности смежных областей.
Из биологических особенностей фауны мошек Кондаринского ущелья,
как и Гиссарского хребта вообще, следует прежде всего отметить следую-
щее. Несмотря на изобилие мошек в ручьях, как кровососы они мало
заметны. Основными кровососущими видами являются Simulium (Odag-
mia) alajense Rubz., Simulium (in sp.) multistriatum Rubz.
Для видов Кондаринского ущелья, далее, характерна фенология видов
северных широт, с резким перерывом развития зимой, что неясно выра_
Могик и
151
жено у видов, населяющих, например, Дюшамбинку и другие реки пред-
горий и выравненного рельефа Средней Азии.
Для наиболее массового вида Кондаринского ущелья — Simulium
(in sp.) multistriatum Rubz. характерно роение в утренние часы, над
речкой, что почти не свойственно видам северных широт.
Для видов, населяющих ручьи Кондаринского ущелья, как и вообще
видов Гиссарского и других хребтов, по сравнению с видами равнинных
областей Евразии, характерны некоторые общие морфо-физиологические
особенности.
Из них в первую очередь нужно отметить сильное развитие прикре-
пительных органов у личинки (заднее кольцо крючьев и «присоска»),
а у куколок — защитных приспособлений: укорочение нитей, усложне-
ние кокона, гладкая его поверхность. Эти морфо-физиологические при-
способления личинок и куколок находятся, очевидно, в связи с усло-
виями быстрого потока воды горных ручьев и носят по своему биологи-
ческому смыслу защитный характер. Сюда же можно отнести и преиму-
щественное расположение личинок некоторых видов не по ложу ручья,
а на стеблях прибрежных растений, свешивающихся в воду.
ЛИТЕРАТУРА
Рубцов И. А. 1947. Состав, связи, географическое распространение и вероятное
происхождение фауны мошек Средней Азии. Изв. АН СССР, сер. биолог.,
№ 1 87—116.
Рубцов И. А. 1951. К систематике мошек Средней Азии. Труды Зоологиче-
ского института АН СССР, том IX 439—556.
С. Г. ЛЕПНЕВА
РУЧЕЙНИКИ УЩЕЛЬЯ КОНДАРА
(Предварительный очерк)
Ручейникам Средней Азии А. В. Мартынов посвятил 4 работы (1914,
1927а, 19276, 1928).
Немногие дополнительные сведения, относящиеся к среднеазиатским
видам отряда Annulipalpia, вошли в определитель А. В. Мартынова
(1934).
Среди материалов, находившихся в распоряжении А. В. Мартынова,
сборы из Гиссарского хребта отсутствовали; фауна ручейников Горного
Туркестана освещалась этим автором главным образом по материалам,
относящимся к западной, северо-западной и северной его частям. Бли-
жайшими к Гиссарскому хребту местностями, фауны ручейников кото-
рых А. В. Мартынов коснулся в своих работах, являются окрестности
Самарканда, долина Зеравшана, долина Алая и Памир; сведения, со-
общенные названным автором о ручейниках этих мест, относятся к от-
дельным видам, из которых многие им описаны.
Автором настоящего очерка в 1943 и 1944 гг. исследовались ручей-
ники южного склона Гиссарского хребта в пределах бассейна р. Варзоб,
в интервале высот от 800 м1 (Дюшамбинка у Сталинабада) до 2800 м
(окрестности пос. Зидды).
То, что сделано, достаточно убедительно иллюстрирует тонкую диф-
ференцировку фауны ручейников в пределах исследованной местности,
обязанную неоднородности стаций, созданных высоко развитой гидро-
графической сетью в условиях резкого падения высот и сложно расчле-
ненного рельефа.
Ущелье Кондара, в частности, не может отразить всех особенностей,
присущих фауне ручейников южного склона Гиссарского хребта в целом,
а показывает лишь некоторые основные черты этой фауны. Ручейники
Кондаринского ущелья, как деталь общей фауны этой группы насекомых
южного склона Гиссарского хребта, обладают некоторыми, присущими
им особенностями, характерными для бокового ответвления Варзобской
долины, образованного малым притоком Варзоба в интервале умеренных
высот (1850—1100 м), лежащих ниже субальпийской зоны. Эта мест-
ная фауна, следовательно, представляет лишь часть общей фауны
ручейников южного склона Гиссарского хребта, сложившуюся в специ-
фических условиях малой поперечной долины.
Ручейники Кондары изучались от водопада на высоте около 1250 м
(«первого водопада») до устья реки (1100 м); ручейники родниковой зоны
1 Здесь и дальше высота в метрах над уровнем моря.
Ручейники ущелья Кондара
153
исследованы выше, от источника на уроч. Квак (1860 м) до родников
у нижнего течения реки. Сборы коснулись преимущественно водных
личиночных фаз этих насекомых, вследствие чего во многих случаях
оказалось невозможным определить видовую принадлежность личинки.
Нет основания думать, что выше первого водопада фауна ручейников
Кондары значительно меняется, так как на некотором расстоянии харак-
тер русла остается тем же. В верхнем течении, в районе водопадов и
у истока р. Кондара фауна ручейников, вероятно, значительно обеднена,
как это имеет место в примыкающем к водопаду участке одного из иссле-
дованных ближайших левых притоков Кондары и в родниках. Условий
для появления высокогорных видов, при отсутствии летующих снежников
и малой мощности Кондары у ее истока, нет.
СПИСОК РУЧЕЙНИКОВ УЩЕЛЬЯ КОНДАРА
Подотряд ANNULIPALPIA
Сем. Rhyacophilidae
Подсем. Rhyacophilinae
Rhyacophila obscura Mart. — Среднеазиатский горный холодновод-
ный вид. Известен (Мартынов, 1927) из Чимгана, в 75 км от Ташкента,,
из района 12 источников, личинки в речке Чимганка с температурой
воды 6°.
В Гпссарском хребте широко распространен в интервале высот 850—
1850 м; личинки этого вида населяют разнообразные потоки, начиная
от небольших родниковых ручьев, до р. Варзоб, в которой они наблю-
дались по всему течению до Сталинабада. В родниках и в Варзобе ли-
чинки R. obscura встречаются в небольшом количестве; наиболее обильно*
они представлены в потоках боковых ущелий Варзобской долины, подоб-
ных р. Кондара.
В этой речке личинки известны от пункта ниже первого водопада
(1250 м) до устья; обнаружены они и в родниковых ручьях Ореховой
рощи (1300 м, температура в июне 12°). Личинки живут на дне реки под
камнями, взрослые насекомые держатся в кустарниках и в траве на бе-
регах речки и родников. Обилием растительности, представляющей
удобные стации для жизни имаго, создаются условия оптимума вида для
взрослых его фаз в ущельях речек, подобных Кондаре; открытые берега
Варзоба такого рода убежищ для этих нежных насекомых не дают. Для
личинок Rhyacophila obscura в Кондаре и в подобных ей речках благо-
приятна их малая глубина, низкая температура воды (в августе 13.6°),
незначительная мощность, мелкокаменистое дно, довольно обильно на-
селенное, а, возможно, также и краткость периода половодья. Личинки
R. obscura — обитатели холодной и прозрачной воды.
Подсем. Glossosomatinae
Glossosoma dentatum McL. — Среднеазиатский горный вид. Изве-
стен из окрестностей Самарканда.
В р. Варзоб личинки Glossosoma широко распространены, но пока
еще нельзя уверенно сказать, относятся ли они к одному этому виду,
или же и к другим.
154
С. Г Лепнева
В ущелье Кондара видовая принадлежность личинок Glossosoma
установлена на основании определения полученного из них имаго (опре-
деление сделано О. М. Мартыновой).
Личинки G. dentatum встречаются в р. Кондара и в ее притоках вместе
с личинками Rh. obscura, но в большем числе. Они обитают под камнями,
нередко колониями. Имаго на растительности у берега в большем числе,
чем Rhyacophila. В р. Кондара личинки известны от первого водопада
до устья; в родниках, начиная от родников в Ореховой роще, — единично.
В Гиссарском хребте характерны для ущелий, подобных Кондарин-
скому.
Подсем. Agapeiinae
Agapetus sp. sp. Личинки видов этого рода обильно населяют Кон-
дару от первого водопада до устья. В меньшем числе, но широко, они
встречаются в связанных с нею родниках, поднимаясь в такого рода
водоемах выше всех прочих видов, до 1800 м (ур. Квак). Взрослые насе-
комые — на кустарниках у берегов в большом числе, во второй поло-
вине лета маленькими роями.
В р. Варзоб и в боковых ущельях его долины виды Agapetus широко
и обильно представлены. Эти мелкие насекомые (1.5—2 мм длины) в гор-
ных потоках Гиссарского хребта по своему количественному развитию
играют высокую роль, напоминая этим Hydroptilidae потоков предгорий
Алтая и Саян.
Сем. Philopotamidae
Dolophilodes sp. sp. Три вида этих характерных кренобионтов,
в нескольких экземплярах каждый, обнаружены по личинкам в род-
никах правого берега Кондары. Кроме этих водоемов в Гиссарском хребте
не констатированы.
Личинки отличаются исключительно крупными размерами головы.
Сем. Hydropsychidae
Hydropsyche sp. — Hydropsyche gracilis Mart, и H. exocellata McL.
известны из Варзоба; эти виды широко заселяют эту реку и, как харак-
терные потамобионты, представляют в ней господствующую часть фауны.
В ущелье р. Кондара, вследствие изменения характера текучеводных
стаций, фауна Hydropsyche резко снижается. Личинки Hydropsyche
в Кондаре и впадающих в нее родниках, начиная от родников Ореховой
рощи, встречаются единично.
Подотряд INTEGRIPALPIA
Сем. Sericostomatidae
Dinarthrum pugnax McL. Широко распространенный в горном Турке-
стане вид, известный из Киргизского хребта, восточного Каратау, из
окрестностей Алма-аты, из долины Зеравшана, из окрестностей Самар-
канда и Бухары; найден в окрестностях Искандер-куля.
Ручейники ущелья Кондара
155
В Гиссарском хребте D. pugnax широко распространен, встречаясь
в интервале высот от 900 до 1900 м. Личинки обитают преимущественно
в родниках. Кроме ущелья Кондара известны из реокренов:в 18 км выше
•Сталинабада (900 м), на берегу Сараджоу (1900 м) и у Ходжа-оби-гарм
(1900 м). В ущелье Кондара личинки этого вида населяют родники по пра-
вому берегу реки с постоянной температурой воды (9° — зимой, 12° —
летом), встречаясь в них в большом количестве в течение зимнего (I,
II, III) и летнего времен года.
Вид — чрезвычайно характерный для Кондаринского ущелья и ему
подобных. Имаго — среди кустарников; бросается в глаза своими мох-
натыми основаниями усиков, густо покрытыми волосками.
Сем. Limnophilidae
Подсем. Apatanlinae
Apatelia copiosa McL.(?). Определение сделано по личинке, бегло опи-
санной Мартыновым (1927). Широко распространенный вид горного Тур-
кестана. Известен из Алайского хребта, северного и восточного Каратау,
окрестностей Бухары и долины Зеравшана.
В Гиссарском хребте широко распространен, встречаясь в интервале
высот от 800 м (р. Варзоб у Сталинабада) до 1300 м (Гушары).
В ущелье Кондара постоянен в речке Кондара, населяя ее от первого
водопада до устья; личинки известны и из родников Ореховой рощи.
Подсем. Limnophilinae
Pseudostenophylax secretus Mart. Известен из района 12 источников
реки Чимганка, в 80 км к с.-в. от Ташкента. В Гиссарском хребте обна-
ружен в родниках на Кваке (1800 м).
Из относящихся к другим семействам видов в ущелье Кондара еди-
нично обнаружены личинки Psychomyidae и Leptoceridae, отмеченные
в родниках Ореховой рощи (5 VI 1944). Весьма вероятно присутствие
в родниках личинок Plectrocnemia sp. (сем. Polycentropidae), известных
из родников соседнего ущелья Гажни, а в речке Кондара — личинок
Hydroptilidae.
В ручье Джиринод-сой, ближайшем к Кондаре левом притоке Вар-
зоба с открытой южной экспозицией, обнаружен индийский вид Apsi-
lochorema indicum subsp. turanicum Mart. (Rhyacophilidae, подсем. Hydro-
biosinae). Из глубокого ущелья Кондара, с его закрытостью и северо-
восточной и восточной экспозицией, этот вид не известен.
Apsilochorema indicum Ulm. указана из Пенджаба (Индия); Ар. in.
subsp. turanicum Mart. — из окрестностей Бухары (Тенги-дувал).
Из приведенного списка видно, что систематический состав фауны ручей-
ников ущелья Кондара довольно пестр: указанные виды (в действительно-
сти их больше) относятся к 7 разным семействам. Подотряд Annulipalpia
господствует над Integripalpia и по числу семейств, и по количественному
развитию отдельных видов. Преобладание Annulipalpia над Integripalpia
представляет особенность, присущую всей фауне южного склона Гис-
сарского хребта; господство именно этого подотряда, состоящего из пер-
вично текучеводных семейств, совершенно естественно в фауне, почти
нацело относящейся к текучим водам.
156
С. Г. Лепнева
В ущелье Кондара из 8 семейств подотряда Annulipalpia отмечено 4;
вместе с предполагаемыми Hydroptilidae и Polycentropidae в нем должно
быть 6 семейств этого подотряда — то же число, что и во всем Гиссар-
ском хребте. Эти же 6 семейств Annulipalpia установлены А. В. Марты-
новым для всего горного Туркестана: семейства Arctopsychidae и Ste-
nopsychidae здесь отсутствуют. Первое из них в Советской Евразии
свойственно более северным широтам; второе распространено в пределах
узкой полосы предгорий горных цепей южной Сибири, на Дальнем Вос-
токе, в Японии, в Китае и в Индийской области. Отсутствие Stenopsy-
chidae на южном склоне Гиссарского хребта можно считать фактом,
твердо установленным, так как исключительно крупные личинки Ste-
nopsyche не могли бы остаться незамеченными рядом лиц, изучавших
фауну горных потоков этой местности.
Среди Annulipalpia Кондары преобладает семейство Rhyacophilidae^
виды которого (Rh. obscura, Gl. dentatum и Agapetus sp. sp.) наиболее
широко распространены в ущелье и наиболее богато количественно пред-
ставлены. В Кондаре это семейство преобладает над Hydropsychidae —
семейством, включающем в себя характерных потамобионтов, свойствен-
ных мощным потокам, в частности, широко населенному ими Варзобу,
особенно в его нижнем течении. Преобладание Rhyacophilidae над Hyd-
ropsychidae в Кондаре составляет характерную специфическую черту
фауны ручейников этого ущелья, как и других, подобных ему боковых
ущелий долины Варзоба, соответствующих небольшим притокам этой
реки. Эта черта существенно отличает фауну ручейников ущелья Кон-
дара от таковой долины Варзоба, особенно низовий ее, где Hydropsy-
chidae доминируют над Rhyacophilidae.
Второй специфической для ручейников района Кондары чертой
является отсутствие в нем среди Rhyacophilidae высокогорных форм:
Rhyacophila gigantea Mart, и близких к нему видов, личинки которых
обладают коническими жабрами (Лепнева, 1945); эта отрицательная черта
характеризует фауну ручейников ущелья Кондара, как свойственную
средним интервалам высот хребта (1100—1850), и отличает ее от фауны
ручейников горных долин более высоких горизонтов районов Ходжа-
оби-гарма (1500—2200 м) и Зидды (2200—2800 м).
Экологически фауна ручейников ущелья Кондара нацело слагается
из течениелюбивых форм; ни в редких прибрежных затишных стациях
речки Кондара, ни в разливах арычной сети, ни в родниках исследо-
ваниями 1943—1944 гг. видов, приуроченных к стоячим водам, не
обнаружено. В фауне ручейников всего бассейна Варзоба течениелюби-
вые виды ручейников также подавляюще господствуют, но и виды стоя-
чих вод в незначительном количестве присутствуют; местами их обита-
ния служат немногие стоячие водоемы поймы Варзоба и некоторых
других рек. Ущелье Кондара, следовательно, несколько утрирует основ-
ной реофильный характер фауны ручейников южных склонов Гиссарского
хребта, чрезмерно резко его представляя.
Зоогеографически ручейники ущелья Кондара в той их части, кото-
рая определена, всецело состоят из горных среднеазиатских видов; для
всех них Кондара является самой южной точкой местонахождений. В об-
щей фауне ручейников Гиссарского хребта гарный среднеазиатский эле-
мент также преобладает. Группу широко распространенных в палеарктике
видов представляют Cheumatopsyche lepida, Hydropsyche exocellata и
Hydropsyche gracilis; индийский элемент выражен присутствием вида
Apsilochorema indicum. Я полагаю, что при дальнейшем тщательном
Ручейники ущелья Кондара
157
исследовании палеарктические и в частности кавказские и закавказские
виды, подобные Hydropsyche gracilis, в ущелье Кондара обнаружены
будут, но не сомневаюсь, что эта часть фауны представлена в нем в зна-
чительно меньшей степени, чем в долине Варзоба, а следовательно, и на
южном склоне Гиссарского хребта в целом.
В биологическом сезонном цикле ручейников ущелья р. Кондара
наблюдаются некоторые уклонения по сравнению с хорошо изученными
европейскими видами. Для многих из этих последних, в частности для
обитателей мелких стоячих замерзающих водоемов, характерна зимовка
в одной из старших личиночных стадий. Весной, с освобождением водоема
от ледяной коры, наступает интенсивное питание, рост, последние линьки
и окукления личинок, а вслед за тем вылет взрослого насекомого
и откладка яиц. От времени откладки яиц зависит степень зрелости зи-
мующих личинок. В родниках, питающих Кондару, с их относительно
высокой зимней температурой (9—11° в январе—марте) и в незамерзаю-
щей Кондаре, с ее быстрым течением, такого рода цикличности не наблю-
дается. У Rhyacophila obscura, например, куколки обнаружены в тече-
ние круглого года. Начиная с мая, а может быть и апреля, в течение всего
теплого времени года по октябрь включительно происходит растянутый
вылет этого насекомого. У Glossosoma dentatum зимуют личинки всех
стадий и не успевшие осенью созреть до готовности к вылету куколки;
вылет перезимовавших куколок начинается в апреле; окукление, а вслед
за тем и вылет нового поколения происходят во второй половине лета
и осенью; запоздавшие куколки перезимовывают. У Agapetus sp. окук-
ление и вылет приурочены ко второй половине лета; в это время про-
исходит массовый лет этих насекомых, напоминающий это явление
у ручейников из сем. Hydroptilidae и у подёнок, когда эти мелкие насе-
комые, может быть, играют некоторую роль в пищевом бюджете насеко-
моядных птиц. В реках с ихтиофауной личинки всех видов ручейников
составляют существенный источник пищи рыб.
Настоящий очерк составлен автором на основании обработки материа-
лов, собранных в ущелье Кондара в 1943 и 1944 гг. А. В. Гриб, В. И. Жа-
диным, С. Г. Лепневой и К. Г. Ромадиной; часть взрослых насекомых по-
лучена в аквариумах и садках. Материал определен автором при уча-
стии О. М. Мартыновой; последней определены по имаго: Glossosoma
dentatum McL., Rhyacophila obscura Mart., Apsilochorema indicum Ulm.,
Hydropsyche gracilis Mart., Hydropsyche exocellata McL. Автор выражает
О. M. Мартыновой глубокую признательность.
ЛИТЕРАТУРА
Л е п н е в а С. Г. 1945. Замечательные личинки рода Rhyacophila из потоков Сред-
ней Азии. Энтомол. обозр., т. XXV 111, 3—4 : 64—74.
Мартынов А. В. 1914. К познанию Trichoptera Среднеазиатских владений Рос-
сии. Ежег. Зоол. муз. АН, XIX : 402—437.
Мартынов A. (Martynov А.) 1927а. Contributions to the aquatic entomofauna of
Turkestan. I. Trichoptera Annulipalpia. Ежег. Зоол. Муз. АН СССР, т. XXVIII:
162—193.
Мартынов А. 19276. Contributions to the aquatic fauna of Turkestan. II. Tri-
choptera Integripalpia. Ежег. Зоол. Муз. АН СССР, т. XXVIII: 457—497.
Мартынов А. В. 1928. Ручейники. Тр. Памирской экспедиции 1928 г., II. Зооло-
гия 79—80.
Мартынов А. В. 1934. Ручейники Trichoptera—Annulipalpia. Определители
по фауне СССР 1—343.
в. в. ПОПОВ
ПЧЕЛИНЫЕ
Пчелиные (Apoidea), одна из высших групп перепончатокрылых насе-
комых (Hymenoptera), богато представлены в Средней Азии (около 1200
видов); очень богато они представлены и в ущелье Кондара, урочище
Квак, на плато Руидашт, откуда известно уже свыше 200 видов (по сбо-
рам покойного В. В. Гуссаковского). Как и вся Средняя Азия, ущелье
Кондара еще далеко не достаточно изучено и к списку видов в будущем,
может быть, будет добавлено 70—80 видов. Во всяком случае и в настоящее
время Кондара в этом отношении является наиболее полно изученной
географической точкой в Средней Азии; в то же время здесь совершенно
не изучено распределение пчелиных по цветковой растительности, а как
раз с этой точки зрения Кондара—Руидашт представляет чрезвычайно-
удачное место.
Группа пчелиных была вызвана к жизни возникновением цветковой
(покрытосемянной) растительности, служа опылителями ее энтомофиль-
ных (мелитрофильных) видов; взаимосвязанная эволюция обеих групп
и их сопряженные взаимные приспособления общеизвестны. И морфо-
логические особенности пчелиных (например грызуще-лижущий рото-
вой аппарат, собирательный аппарат, богатый волосяной покров, пери-
стые волоски), и их образ жизни, гнездование, все сложные инстинкты,
связанные с гнездостроением, все поведение и, наконец, географическое
и стациальное распределение, — все в той или иной степени определяется
фактом питания личиночной фазы пчелиных пыльцой и нектаром опре-
деленных цветковых растений.
Флора Кондаринского ущелья очень богата; его разнообразные склоны
и лёссовые обрывы представляют обширные и удобные места для гнез-
дования тех видов, которые сооружают своп гнезда в земле или лепят их
в расселинах скал, а обильные деревья и кустарники — для тех пче-
линых, которые устраивают свои линейные гнезда в их стеблях или дре-
весине. Обильные запасы питания и разнообразные места гнездования,
расположенные на высоте 1100—2800 м,1 и определяют, в первую оче-
редь, богатство и состав фауны пчелиных Кондары—Руидашта.
Вслед за сезонной сменой растительных аспектов происходит значи-
тельная сезонная видовая и родовая смена фауны пчелиных. Весенняя
фауна, насчитывающая большее число моно- и олиготрофных форм,,
обильна видами с коротким лётным периодом; наоборот, летняя фауна
характерна преобладанием политрофных видов с растянутым пли даже
J Здесь и дальше высота над уровнем моря.
Пчелиные
159
сильно растянутым лётным периодом. Большинство видов дает одна
поколение в году и политрофно, лишь немногие имеют два поколения
(например Andrena flavipes Panz.).
Ранневесенняя фауна пчелиных ущелья Кондара не изучена, поздне-
весенняя немного известна и достаточно характерна. Со второй половины
мая или с начала его третьей декады в Кондаре отмечено свыше 30 видов.
В это время преобладают виды Andrena (рис. 1) и биологически свя-
занные с ним виды Nomada, немногие Halictus и виды Osmia (s. str).
Наоборот, раннелетняя фауна пчелиных характеризуется обилием видов
рода Osmia (в широком смысле), а летняя — обилием видов (и числен-
ностью особей) из родов Halictus, Anthophora и Megachile (и биологи-
чески связанные с ними паразитические Coelioxys и некоторые другие).
Рис. 1. Andrena lateralis F Mor. (j.
К началу сентября фауна пчелиных резко сокращается, хотя отдель-
ные виды активны и посещают цветы, — преимущественно для индиви-
дуального питания, — до последней декады октября.
В это время очень характерно обилие видов из рода Hylaeus (рис. 2),
а также ничтожное количество представителей Amegilla и Megachile,
и полное отсутствие таких обширных родов, как Anthophora, Halictus,
Andrena и Osmia (в широком смысле), закончивших активный период
своей жизни. Характерно присутствие Ceratina. Виды Ceratina (рис. 3 и 4)
отличаются чрезвычайно продолжительным лётным периодом и у них,
как почти полное исключение среди пчелиных, мать-основательница
гнезда доживает до вылета своих детей и встречается с ними; в сентябре
летает, в основном, молодое поколение, которое займется строительством
гнезд лишь с весны будущего года. Многие из сентябрьских пчелиных
представлены лишь подобным молодым поколением, что особенно пока-
зательно в тех случаях, когда летают лишь одни самцы (протерандрия,
как правило, преобладает у пчелиных) или оба пола совместно.
В тех же случаях, когда найдены лишь одни самки, всегда можно пред-
полагать, что исследователь имеет дело с запоздавшими представителями
160
В. В. Попов
летне-осенних видов; опасение это, однако, совершенно неосновательно
для тех видов, биология которых изучена (Xylocopa valga Gerst.).
Кормовые связи большинства пчелиных, и в частности среднеазиат-
ских, еще плохо изучены, и точное установление олиго- или монотрофизма
того или иного вида требует продолжительных и достаточно обильных
наблюдений. Кондаринские материалы собраны попутно и лишены ка-
Рис Hylaeus dolichocepha-
his F Mor.
Cera tin a laevifrons
F Mor. 6.
ких-либо экологических данных. Два вида из отмеченных в Кондаре—
Руидаште известны как монотрофы: Andrena fedtschenkoi F. Мог., по-
сещающие соцветия Tanacetum umbelliferum. и Tarsalia ancyliformis
Popov — преимущественно Centaurea solstitialis.
Обычными видами в Кондаре могут быть названы очень немногие:
♦Hylaeus dolichocephalus F. Mor., Megachile albidula Alfk., Osmia aenea L.,
♦Tetralonia fuscipes F. Mor. (рис. 5),
Amegilla albigena Lep.
Достаточно обильны также: Colletes
morawitzi Nosk., Hylaeus variegatus F.,
*H. pallidicornis F. Mor., Halictus subau-
ratus Rossi, H. smaragdulus Vach., H. hol-
tzi Schulz, H. albipes F. v. rubelloides
Bliithg., H. marginatus Brulle, *Andrena
laevifrons F. Mor., *A. mordax F. Mor.,
♦A. lateralis F’. Mor., *A. concava Lebed.,
Megachile argentata F., M. apicalis Spin.,
M. maritima Kby.,M. derasa Gerst., Chali-
codoma muraria nestorea Brulle, Hoplitis
simplicicornis F. Mor., Stelidomorpha na-
suta Latr., Ceratina laevifrons F. Mor.,
Рис. 4. Ceratina tibialis К Mor. q. tibialis F. Mor., Eucera tomentosa
F. Mor., Crocisa ramosa Lep.
За очень немногими исключениями, это все — виды, широко распро-
страненные и обычные в Средней Азии или даже в Средиземноморской
и Среднеазиатской подобластях одновременно. Лишь виды, отмеченные
знаком *, являются, повидимому, эндемиками горной системы хребтов
Туркестанского, Алайского, Зеравшанского и Гиссарского. Halictus
albipes v. rubelloides Bliithg. был известен только из Каратегина и Hop-
litis simplicicornis F. Mor. — только из Кызыл-кумов. Таким образом,
Пчелиные
161
эта группировка видов и их обыденность не являются оригинальными и
обычны для предгорной и значительной части нижней половины горной
зоны Средней Азии.
Фауну пчелиных Кондаринского ущелья следует сравнить с доста-
точно хорошо изученной фауной Зеравшана (Моравиц, 1875—1876,
1894). Верхний п средний Зеравшан (от Пенджикента до Кштута, Искан-
дер-куля, Ягноба и перевала Анзоб) вытянут приблизительно на 100 км
и образует горную страну с сильно пересеченным рельефом, расположен-
ную на высоте 1060 м (Пенджикент) — 2.800 м (Анзоб); район этот нахо-
дится в условиях, грубо говоря, достаточно сходных с физико-географи-
ческими, климатическими и иными условиями района Кондаринского
ущелья, который непосредственно к нему примыкает и с ним сопри-
касается.
Рис. 5. Tetralonia fuscipes F. Mor. g.
В пределах верхнего и среднего Зеравшана известно 223 вида пче-
линых. Из 203 видов, приводимых из Кондары—Руидашта, 104 являются
общими с указанной частью Зеравшана, 25 известны из других районов Зе-
равшана и так же, как 44 широко распространенных по всей Средней Азии
вида, лишь случайно еще не найдены в Зеравшане; 4 вида были известны
только из Ферганы, один из Ташкентского оазиса, один из Кызыл-кумов
идЗ из Туркмении, — невидимому, все они также достаточно широко рас-
пространены по всей Средней Азии. Только 8 нижеследующих видов или
являются эндемичными для Памирской горной системы, или в пределах
Средней Азии встречаются только здесь, повидимому, отсутствуя в Зе-
равшапе: Halictus mucidus Bliithg., Н. shestakovi Bliithg., H. albipes F. v.
rubelloides Bliithg., Icteranthidium tadzhicum Popov, Osmia pamirensis
Guss.. Habropoda tadzhica Popov, Anthophora vulpina Panz. (subsp. alti-
cola Hed.), A. borealis F. Mor.
’Изучение фауны пчелиных Кондаринского ущелья, помимо установ-
ления ряда новых форм, принесло несколько интересных видов, новых
для фауны Средней Азии: Melecta funeraria F. Sm. (Средиземноморская
подобласть), Nomada ruficornis L. (Европа), Megachile dacica Moes. (ю.-в.
-Европа), Dioxoides tridentata Nyl. (сев. и средн, часть Палеарктической
обл.) и др.
1 [ Ущелье Кондара
162
В. В. Попов
Результаты общего зоогеографического анализа фауны пчелиных
Кондаринского ущелья таковы:
Среднеазиатские эндемики . .
Средиземноморско-среднеазиатские .
Широко распространенные ....
Сибирские и вост.-туркестанские
45.8% (93 вида)
30.5% (62 вида)
22.7% (46 видов)
1.0% (2 вида)
Эти процентные отношения вероятно типичны для фауны пчелиных
большинства пунктов предгорной зоны Средней Азии; с этой точки зре-
ния фауна пчелиных Кондары мало оригинальна. Высокий видовой
эндемизм, значительные связи со Средиземноморьем и ничтожные —
с Восточным Туркестаном очень характерны для фауны среднеазиатских
пчелиных вообще; несомненно, что при дальнейшем изучении средне-
азиатских пчелиных восточные связи несколько возрастут количественно.
В. Гуссаковский (1940) отметил в фауне Sphecidae нижней зоны юж-
ного склона Гиссарского хребта присутствие элементов более южного
происхождения (Sceliphron deforme F. Sm., Crabro pugillata Kohl,
C. tuberculifer Kohl) и элементов северных (Crabro leucostoma L.,
C. quadrimaculata L. и др.); c другой стороны, им отмечены также эле-
менты явно реликтовые, — таков Hissarocephus stackelbergi Guss, из
сем. Cephidae (Гуссаковский, 1945).
Среди пчелиных, заселяющих южные склоны Гиссарского хребта,,
существуют подобные же элементы. К видам тожного происхождения сле-
дует прежде всего отнести Xylocopa (Nodula) bentoniCkll. (koltschi Hed.),
найденную в Ходжа-оби-гарме и известную ранее из Афганистана и Белу-
джистана; Nodula — довольно обширный эндемичный индийский подрод
Xylocopa, и Ходжа-оби-гарм — самое северное палеарктическое его
нахождение. Подобный же характер имеет Habropoda tadzhica Popov,,
представитель гималайской группы рода (Попов, 1948). Реликтовый ха-
рактер имеет и Dianthidium insulare F. Мог. (Попов, 1947). Все они при-
урочены к древесно-кустарниковому поясу и являются дериватами тре-
тичной фауны насекомых, корни которой, как и многих элементов флоры
Памиро-Алая, следует искать в юго-восточной Азии. К северным элемен-
там принадлежат: Colletes laevifrons F. Mor. (speculiferus Ckll.), известный
из СССР (Алтай) и сев. Японии, Stelis minuta Lep. (сев. и средн. Европа;
Чимбай) и Dioxoides tridentata Nyl. (сев. и средн, части Палеаркти-
ческой области) и др. Два последних названных вида — паразитические
и, как обычно бывает, характеризуются более узким распространением,
чем их хозяева. Dioxoides tridentata Nyl. паразитирует в гнездах Cha-
licodoma muraria Retz. — вида, нашедшего удобные места гнездования
на склонах Кондаринского ущелья, и Stelis minuta Lep. — в гнездах
мелких Osmiini и Eriades truncorum L., устраивающих свои гнезда
в стеблях Rubus, столь обильно растущих в ущелье Кондара.
Из 203 видов пчелиных, отмеченных в Кондаре—Руидаште, в Кон-
даре собрано 150, в уроч. Квак — 128 и на плато Руидашт — 78. Эти
цифры, однако, не отражают действительного содержания фауны пче-
линых при подъеме в горы. Тем не менее нет сомнения, что часть видов,
отмеченных для Квака и Руидашта или только для Руидашта, действи-
тельно отсутствует в Кондаре (например Colletes laevifrons F. Mor.,
некоторые Anthidiini, Anthophora bimaciilata Panz., A. vulpina Panz.,
A. borealis F. Mor.), а многие кондаринские виды отсутствуют на Руи-
даште.
Пчелиные
163
Наконец, следует упомянуть еще о видах, не найденных в Сталина-
баде, — даже в ближайшей к Кондаре его возвышенной части (Ботани-
ческий сад), где в течение полутора лет производились специальные интен-
сивные сборы пчелиных. Здесь следует отметить Andrena fedtschenkoi F.
Mor., Melitturga clavicornis Latr., Melitta leporina Panz., Xylocopa ver-
sicolor Alfk. Эти формы довольно обычны в степных и предгорных райо-
нах более северных частей Средней Азии.
Фауна пчелиных Кондаринского ущелья, численно богатая и разно-
образная по видовому составу, является в достаточной мере типичной
для Средней Азии по своим сезонным аспектам, видовому составу чи-
сленно преобладающих форм, сходству с фауной Зеравшана и процент-
ному отношению различных зоогеографических элементов, ее составляю-
щих; лишь незначительная примесь форм других зоогеографических груп-
пировок и ряд пока еще немногих эндемичных видов придают ей ориги-
нальные, местные черты.
СПИСОК ВИДОВ ПЧЕЛИНЫХ
Colletes morawitzi Nosk. (caspicus F. Mor.). Кондара (29 VIII—12 IX,
18 $). — Закавказье, Средняя Азия (Копет-даг, Фергана), сев. Иран.
Colletes laevifrons F. Mor. (speculiferus Ckll.). Руидашт (22 VII, 4 j),
Зидды (5 VIII, 3 j). — Алтай, Средняя Азия (Зеравшан), сев. Япония.
Colletes cunicularius L. var. Кондара (22 V—17 VII, 2 $>, 2 8), Квак
(8—21 VI, 6 1 8). — Типичная форма широко распространена по сев.
и средн. Палеарктике (до Кавказа и Балкан), приводимая форма встре-
чается только в Средней Азии (Зеравшан).
Colletes marginatus F. Sm. (floralis Eversm.). Кондара (2 VII—16 IX,
2 j), Квак (10 VI—5 IX, 7 j), Зидды (5 VIII, 2j). — Европейск. часть
СССР, Закавказье, Средняя Азия, Алтай, Зап. Европа, Малая Азия.
Colletes dorsalis F. Mor. (fasciata Rad., rossicus D.T.). Кондара
(26 VI, 1 J), Квак (16 VI—13 VII, 4 2 8), Анзоб (31 VII, 2 8). —
Кавказ, Средняя Азия (Токмак), Малая Азия.
Hylaeus dolichocephalus F. Мог. Кондара (20 VI—21 IX, 83 2» 2 8),
Квак (5 VIII—22 IX, 24 2), Руидашт (4 IX, 4 2), Зидды (5 VIII, 1 2>
1 8). — Среднеазиатский вид, известный из Ферганы и Зеравшана, в том
числе из Анзоба.
Hylaeus hungaricus Alfk. (affinis F. Mor.). Кондара (9 VI—15 IX,
37 8), Квак (20—23 VII, 5 8), Руидашт (22 VII, 4 8), Зидды (26 VII, 1 8).—
Юго-восток Европейск. части СССР, Средняя Азия (Зеравшан, Фергана,
Гиссарск. хр., сев. Памир, Копет-даг), Венгрия.
Hylaeus fedtschenkoi Ckll. (frontalis F. Mor.). Кондара (4 VII—
16 IX, 2 2> 6 8). — Средняя Азия (Аму-дарья, Зеравшан, Сталинабад,
Куляб, Кзыл-орда).
Hylaeus vulgaris F. Мог. Кондара (30 V—17 IX, 8 8), Руидашт (22 VII,
1 8), Анзоб (28 VII, 3 8). — Средняя Азия (Ташкентск. оазис, Зеравшан,
Фергана, Сталинабад, Куляб, Вахшск. долина).
Hylaeus distinguendus F. Мог. Кондара (27 VI, 1 2)- — Зеравшан.
Hylaeus atrocallosus F. Mor. Кондара (21 V—25 VII, 3 2 1 8), Квак
(21 VI—23 VII, 3 2), Руидашт (20 VII—2 IX, 3 2), Анзоб (31 VII, 1 2)-—
Зеравшан, зап. Памир.
Hylaeus angustifrons F. Мог. Кондара (29 V—16 IX, 10 8), Квак
(21 VII, 1 8), Гушары (14 VII, 1 8). — Зеравшан.
Hylaeus nigricollis F. Мог. Кондара (16 V, 1 2)- — Зеравшан.
11*
164
В. В. Попов
Hylaeus tibialis F. Мог. Анзоб (28 VII, 1 1 6). — Зеравшан (Анзоб),
Ашхабад.
Hylaeus bivittatus F. Мог. Квак (20 VII—17 VIII, 4 $>), Руидашт
(22 VII—4 IX, 4 2), Анзоб (31 VII, 1 д). — Фергана.
Hylaeus variolaris F. Мог. Кондара (5 VII—12 VIII, 8 j, 1 (5), Квак
(17 VII, 1 j). — Средняя Азия от Копет-дага до Памира.
Hylaeus pallidicornis F. Мог. Кондара (17 VI—7 IX, 31 (5), Квак
(20 VII—27 VIII, 4 6). — Фергана.
Hylaeus laticeps F. Mor. Кондара (9 VI—18 IX, 15 j), Квак
(17 VIII, 1 j). — Фергана.
Hylaeus (Navicularia) variegatus F. Кондара (12 VII—13 IX, 26 c,
10 8), Квак (13 VII—17 VIII, 13 ?, 3 8), Руидашт (22 VII, 3 8), Зидды
(26 VII—5 VIII, 3 2 8). — Средн, и южн. полосы Европейск. части
СССР, Средняя Азия, сев. Африка, Зап. Европа, Передн. Азия.
Andrena (Lepidandrena) fedtschenkoi F. Мог. Квак (20—29 VI, 5 j).—-
Средняя Азия (ю.-з. Казахстан, Узбекистан, Таджикистан). Монотрофный
вид, посещающий цветы Tanacetum umbelliferum.
Andrena (Andrenella) minutuloides Perk. v. sparsiciliata Alfk. Кон-
дара (30 V, 2 2), Квак (20 VII—4 VIII, 5 2), Р/вдашт (4 IX, 2 2)- “
Сев. и средн, полосы Европейск. части СССР, Средняя Азия, Зап.
Европа. Вероятно к этому виду относятся все указания о нахождении
в Средней Азии A. parvula Kby.
Andrena (Glyphandrena) carbonaria L. (pillpes F.). Кондара (22 V—
16 VII, 6 2> 4 8), Квак (16 VI—4 VIII, 5 2> 1 8), Руидашт (17 VI—4 IX,
2 2, 10 8), Зидды (26 VII, 1 2, 1 8), Гушары (14 VII, 1 8), Анзоб (28 VII,
2 о). — Широко распространенный обычный палеарктический вид, дохо-
дящий на юг до Кашмира включительно, где он также довольно обычен.
Andrena (Melanandrena) albopunctata Rossi. Кондара (5—25 VI, 2 2),
Квак (20 VII, 1 2), Зидды (26 VII, 1 2)- — Европейск. часть СССР, Сред-
няя Азия (Таджикистан), Зап. Европа, сев. Африка.
Andrena (Melanandrena) funebris Panz. (nigrobarbata F. Mor., macu-
laris Kriechb.). Кондара (28 V—17 VII, 6 2), Квак (21 VI, 1 2), Руидашт
(2—4 IX, 4 2)- — Юг Европейск. части СССР, Средняя Азия (Ташкентск.
оазис, Зеравшан, Фергана, Копет-даг), юг Зап. Европы, Малая
Азия.
Andrena (Chlorandrena) humilis Imh. (fulvescens F. Sm., cyanescens
Nyl., nasalis Thoms.). Квак (21 VI, 1 8). — Средняя, Азия (Ташкент),
Еврспейск. часть СССР, Зап. Европа.
Andrena (Gymnandrena) thoracica F. Кондара (1 VII, 1 2)’ Квак
(17 VII—5 IX, 2 2, 1 8), Руидашт (2 IX, 1 2)- — Широко распространен-
ный но Палеарктической обл. вид, в Средней Азии, известный из Таш-
кентск. оазиса, Зеравшана, Копет-дага.
Andrena (Gymnandrena) lateralis F. Mor. Кондара (4 VI—17 VII, 12 g,
2 8), Квак (8—29 VI, 6 2)- — Средняя Азия (сев. часть Средней Азии,
Зеравшан, Ашхабад).
Andrena (Zonandrena) flavipes Panz. (fulvicrus Kby.). Кондара
(19 VI—18 VII, 4 2, 3 8), Квак (8 VI—21 VIII, 5 5 8), Руидашт (18 VI —
4 IX, 4 2), Зидды (26 VII—5 VIII, 10 2), Гушары (14 VII, 1 8), Анзоб
(27 VII, 2 2)- — Европейск. часть СССР, Средняя Азия, Зап. Европа,
сев. Африка, Иран, Месопотамия, сев. Индия.
Andrena (Simandrena) combinata Christ. Кондара (5—19 V, 2 2)’
Руидашт (17 VI—4 IX, 6 2), Зидды (26 VII, 4 2)- — Средиземноморский
и понтийский вид, в Средней Азии известен из Зеравшана.
Пчелиные
165
Andrena (Simandrena) incisa Eversm. Кондара (16 V—19 VI, 4 j),
Квак (8 VI—20 VII, 12 $>), Руидашт (20 VII, 2 j). —Юго-восток Евpo-
пейск. части СССР, Средняя Азия (Зеравшан).
Andrena (Simandrena) proxima КЬу. Кондара (5 VI—4 VII, 5 $)>
Квак (8—21 VI, 7 g), Анзоб (27 VII, 1 $)). — Европейск. часть СССР,
Средняя Азия (Зеравшан), Зап. Европа.
Andrena (Simandrena?) aeneiventris F. Mor. Кондара (21 V—4 VII,
6 $>), Квак (8 VI, 2 $>). — Юг Европейск. части СССР, Средняя Азия
(Зап. Казахстан, Ашхабад), средн, и южн. полосы Зап. Европы.
Andrena (Cnemidandrena) jagnobensis F. Mor. Квак (21 VI, 1 j). —
Зеравшан (Ягноб), зап. Памир.
Andrena (Plastandrena) aulica F. Mor. Кондара (5 VI—2 VII, 9 $),
2 5). — Средняя Азия (юг Казахстана, Узбекистан, Туркмения, Таджики-
стан), Иран.
Andrena (Holandrena) labialis КЬу. Кондара (7 VI—3 VII, 6
5 Квак (10—29 VI, 4 <j). — Европейск. часть СССР, Средняя Азия
(Зеравшан), Зап. Европа.
Andrena fuscosa Erichs, (rutila atrocoerulea Gir.). Кондара (16 V—
11 Vll, 4 J, 2 8), Квак (20 VI—20 VII, 2 $, 1 <3), Руидашт (18 VI-
22 VII, 2 j). — Юг Европейск. части СССР, вся Средняя Азия, южн.
полоса Зап. Европы.
Andrena cineraria L. Руидашт (2 IX, 2 Руидашт—Кутарманба
(21 VI, 1 2). — Европейск. часть СССР, Средняя Азия (Фергана, Зерав-
шан, в том числе Анзоб), Зап. Европа, сев. Африка, Кашмир.
Andrena infirma F. Мог. Кондара (15 V, 1 2), Квак (21 VI, 2 2)- —
Средняя Азия (Зеравшан, Фергана).
Andrena mordax F. Мог. Кондара (14 V—5 VII, 10 2, 16 <5), Квак
(8—21 VI, 6 2, 2 3). — Средняя Азия (Зеравшан).
Andrena nigritula Ckll. (nigrita F. Mor.). Кондара (25 VII, 2 <5). —
Средняя Азия (Зеравшан).
Andrena pannosa F. Mor. Квак (20 VII—5 VIII, 4 2)- — Средняя Азия
(Фергана).
Andrena maculipes F. Mor. Кондара (19 V, 1 2)- — Средняя Азия
(Зеравшан).
Andrena convexiuscula Kby. Кондара (14 V—16 VII, 4 2), Руидашт
(18 VI, 1 2), Зидды (26 VII, 2 $), Анзоб (28 VII, 1 5)- — Средняя Азия
(Зеравшан, Фергана, Каракалпакия), Кашмир, Белуджистан, вся
Европа.
Andrena concava Lebed. Кондара (13 V—7 VI, 12 6), Квак (8—10 VI,
7 3). — Средняя Азия (Зеравшан, Фергана).
Andrena laeviventris F. Мог. Кондара (15 V—14 VII, 22 2» 5 <5), Квак
(8 VI—17 VIII, 7 2, 5 (5), Руидашт (22 VII—4 IX, 8 б). — Средняя Азия
(Зеравшан).
Andrena virescens F. Мог. Кондара (25 V, 1 2)- —Средняя Азия
(Ташкентск. оазис, Зеравшан, Марыйск. оазис).
Andrena aehnea F. Мог. Кондара (22 V, 1 2)- — Средняя Азия (Зерав-
шан).
Melitturga clavicornis Latr. Кондара (13 VI—4 VII, 2 2, 3 б), Квак
(30 VI—17 VII, 4 (5), Зидды (5 VIII, 4 5)- — Средн, и южн. полосы
Европейск. части СССР и Зап. Сибири, Средняя Азия (Зеравшан, Фер-
гана, ’зап. Памир, Тянь-шань), средн, и южн. полосы Зап. Европы.
Halictus annulipes F. Мог. Кондара (13 VI—4 VII, 2 2), Квак (10 VI,
1 $)• — Средняя Азия (Зеравшан, Дарваз, Алайск. хр., Ашхабад).
1В6
В. В. Попов
Halictus rhynchites F. Мог. Кондара (5 VI, 1 j), Руидашт (22 VII,
2 j). — Средняя Азия (Фергана, Алайск. хр., южн. Туркмения).
Halictus subauratus Rossi (meridionalis F. Mor.). Кондара (5 VI—
3 VIII, 22 j), Квак (10 VI—27 VIII, 7 2), Руидашт (22 VII—1 X, 2 о),
Зидды (5 VIII, 1 j), Гушары (14 VII, 1 j), Анзоб (28 VII, 1 j). — Средн,
и южн. полосы Европейск. части СССР, Средняя Азия (Ташкент, Фер-
гана, Зеравшан, Алайск. хр., Дарваз), Средн, и южн. полосы Зап. Европы,
Передняя Азия.
Halictus smaragdulus Vach. Кондара (5 VI—29 VIII, 20 q), Квак
(10 V—23 VII, 3 5), Руидашт (4 IX, 2 2), Гушары (14 VII, 2 2). —
Средняя Азия (Куляб), Средиземноморская подобласть.
Halictus croceipes F. Mor* (longiceps Bluthg.). Кондара (14 IX, 1 j). —
Средняя Азия (Узбекистан, Таджикистан, Туркмения).
Halictus pollinosus Sich. (cariniventris F. Mor.). Квак (5—17 VIII,
2 j), Анзоб (27 VII, 1 j). — Средняя Азия (Узбекистан, Таджикистан),
Белуджистан, Средиземноморье.
Halictus mucidus Bluthg. Кондара (3—15 VII, 3 j), Квак (20 VII,
2 j). — Средняя Азия (Дарваз, Каратегин, Копет-даг).
Halictus holtzi Schulz (sexcinctus F. Mor.). Кондара (5 VI—12 VIII,
17 ?, 10 (5), Квак (10 VI—27 VIII, 3 j, 3 3), Зидды (5 VIII, 1 3). —
Средняя Азия, вост, часть Средиземноморья, Белуджистан.
Halictus minor F. Mor. (fulvipes F. Mor.). Кондара (31 V—25 VII,
9 2), Квак (10 VI—20 VII, 4 j), Руидашт (17 VI—4 IX, 2 $), Зидды
(5 VIII, 1 2). — Средняя Азия (Ташкент, Зеравшан, Фергана, Туркмения).
Halictus subprasinus Bluthg. Кондара (15 IV, 1 j), Квак (20 VII, 1 $>).—
Средняя Азия.
Halictus quadricinctus F. (quadristrigatus Latr.). Квак (8 VI, 1 j);
v. aegyptiacus Friese, Кондара (7 VIII, 1 5), Квак (10 VI—6 VIII, 3 5),
Руидашт (17 VI, 2 о); v. brunescens Eversm. (duplocinctus Vach.), Кон-
дара (27 VII, 1 3), Квак (5 VIII, 1 3), Руидашт (2—4 IX, 6 3). — Ши-
роко распространен по всей средн, и южн. части Палеарктики.
Halictus palustris F. Мог. (pseudomaculatus Bliithg.). Квак (20 VI—
27 VIII, 4 5), Руидашт (22 VII, 6 5), Анзоб (28 VII, 2 5). — Средняя Азия
(Зеравшан, Фергана, Ашхабад).
Halictus funerarius F. Мог. Квак (8—16 VI, 3 $>). — Средняя Азия
(Ташкентск. оазис, Зеравшан).
Halictus longirostris F. Mor. (prognathus Per.). Кондара (14 VI—
15 VII, 7 $, 1 3), Квак (8 VI—23 VII, 2 5), Руидашт (22 VII 1 3). —
Средняя Азия (Зеравшан, Фергана).
Halictus leucozonius Schr. Кондара (21 V—7 VIII, 2 $>), Квак (23 VII,
1 j), Гушары (14 VII, 1 д). — Европейск. часть СССР, Средняя Азия,
вся Зап. Европа, сев. Африка.
Halictus morbillosus Kriechb. Кондара (14 V—29 VIII, 8 $>), Квак
(8—20 VI, 5 2)5 v- fertoni Vach. (r. glasunovi Bliithg.), Кондара (7 IX,
13). — Среднеазиатская и Средиземноморская подобл., Белуджистан.
Hafictus fallax F. Mor. (melanarius F. Mor.). Кондара (24 VI, 1 2)’
Квак (5 VIII, 1 о). — Средняя Азия (Фергана, Зеравшан, Джизак).
Halictus shestakovi Bluthg. (in litt.). Руидашт (17 VI, 1 $)). — Сред-
няя Азия (хр. Петра I).
Halictus xanthopus Kby. (fulvicrus Eversm.). Кондара (7 VI, 2 $>),
Квак (10 VI, 2 2), Руидашт (17 VI, 1 2)- —Средн, и южн. полосы
Европейск. части СССР, Средняя Азия, средн, и южн. полосы Зап.
Европы, сев. Африка, Передняя Азия.
Пчелиные
167
Halictus obscuratus F. Mor. (bentoni Ckll.). Кондара (15 VII, 1 j),
Квак (20 VII, 1 $>), Руидашт (22 VII, 1 $>). — Средняя Азия (Зеравшан,
Шуроабад, Копет-даг), сев. Индия.
Halictus albipes F. aff. v. rubelloides Bliithg. Руидашт (17—18 VI,
18 $>). — Типичная форма широко распространена по Европе, сев. и
Средней Азии, сев. Африке; v. rubelloides известна только из Средней
Азии (Каратегин).
Halictus marginatus Brulle (vulgaris F. Mor., riparius F. Mor.). Кон-
дара (И V—7 VI, 27 $>), Квак (10—20 VI, 3 9). —Южн. и средн, полосы
Европейск. части СССР, Средняя Азия (Каратау, Зеравшан, Ташкент),
южн. и средн, полосы Зап. Европы, Передняя Азия.
Halictus aschabadensis Bluthg. Кондара (21 V—24 VI, 3 j). — Сред-
дняя Азия (Зеравшан, Фергана, Куляб, Тахта-базар, Копет-даг).
Halictus lucidicollis F. Мог. Кондара (13 V—15 VII, 2 $>), Квак
(27 VIII, 1 5), Гушары (14 VII, 2 $), Анзоб (28 VII, 3 $).—Средняя
Азия.
Sphecodes fuscipennis Germ. Кондара (9 VI—17 IX, 3 } 2 d), Квак
(8—27 VIII, 2 2, 1 (3). — От сев. Африки до Монголии. Вероятный па-
разит Halictus quadricinctus F. или Colletes cunicularius L.
Sphecodes gibbus L. Кондара (15 VII, 1 d), Квак (17 VII, 1 d), Анзоб
(27 VII, 1 d). — От сев. Африки до Монголии. Паразит в гнездах Halic-
tus rubicundus Christ, и Н. quadricinctus F.
Sphecodes nigripennis F. Mor. Руидашт (2 IX, 4 6). — Средняя Азия
(Косарал, Зеравшан, южн. Туркмения).
Sphecodes divisus Kby. (similis Waesm.). Кондара (19 V—24 VI, 4 j),
Руидашт (18 VI, 1 2). — Средн, и южн. часть Палеарктической обл.
Sphecodes alternatus F. Sm. (gracilior F. Mor.). Кондара (12 VIII,
1 3). —Средняя Азия (Варзаминор, Куляб, Сары-чашма), Средиземно-
морье.
Nomioides minutissima Rossi. Кишл. Анзоб (28 VII, 1 j 2 d). —
Средиземноморская и Среднеазиатская подобл. (зап. Казахстан, Зерав-
шан, Туркмения).
Nomia diversipes Latr. Кондара (27 VI—2 VIII, 7 $>, 4 d), Квак
(23 VII, 1 d), Руидашт (22 VII, 1 d). — Средн, и южн. полосы Евро-
пейск. части СССР, Средняя Азия, средн, и южн. полосы Зап. Европы,
Передняя Азия.
Melitta leporina Panz. Квак (27 VIII, 1 d), Зидды (26 VII—5 VIII,
3 j, 1 d), Анзоб (27 VII, 2 d). — Европейск. часть СССР, Средняя
Азия (Ташкентск. оазис, Зеравшан, Фергана, Алайск. хр.), Зап. Европа,
сев. Иран.
Lithurgus fuscipennis Latr. Кондара (13—15 VII, 1 $, 1 d), Квак
(29 VI, 1 $>), Руидашт (22 VII, 1 d). — Юг. Европейск. части СССР,
Средняя Азия (зап. Казахстан, Ташкентск. оазис, Зеравшан, Копет-даг),
юг Зап. Европы.
Archianthidium forcipatum F. Мог. Руидашт (22 VII, 1 d). — Средняя
Азия (Зеравшан, в том числе Анзоб).
Megantidium (Asianthidium) glasunovi hissarense Mavrom. Квак
(10 VI, 1 9, 1 d). —Средняя Азия (Зеравшан, Ак-таш бл. Ташкента);
subsp. hissarense Mavrom. описана по приведенным экземплярам из Квака.
Dianthidium (Eoanthidium) insulare persicolum Mavrom. Кондара
(2 VI—25 VIII, 3 5, 1 d), Квак (4 VIII, 1 d), Руидашт (3 IX, 1 d). —
Средняя Азия; был известен ранее только из Ирана. Типичная форма—•
вост. Средиземноморье.
168
В. В. Попов
Anthidiellum strigatum luteum Friese. Квак (4—13 VII, 1 1 $). —
Типичная форма широко распространена по Палеарктической обл.;
subsp. luteum Friese известен из Средней Азии (Мургаб, Ташкентск.
оазис, южн. Туркмения), вост. Средиземноморья, Ирана, Месопотамии,
Вост. Туркестана.
Icteranthidium fedtschenkoi (F. Мог.). Квак (17—20 VII, 2 d). Средняя
Азия (Кизыл-кумы, Мары, Копет-даг).
Icteranthidium tadzhicum (Popov). Руидашт (3—4 IX, 2 $, 6 6). —
Средняя Азия (Дарваз).
Icteranthidium laterale auripes (Eversm.). Кондара (2 VI—29 Vr11,
j 2 c5). — Юго-восток Европейск. части СССР, сев.-зап. Казахстан,
Средняя Азия (Фергана), юг Зап. Европы.
Pseudoanthidium obscuratum (F. Мог.). Квак (4 VIII, 2 (5), Руидашт
(22 VII, 1 $>). — Средняя Азия (Зеравшан, Фергана).
Paranthidiellum cribratum (F. Мог.). Кондара (25 VIII—17 IX, 3
5 5). — Средняя Азия (Узбекистан, Таджикистан, Туркмения). Синь-
цзян.
Anthidium thomsoni F. Мог. Квак (21 VII, 1 $>). — Средняя Азия
(Ягноб, Памир).
Anthidium taschenbergi F. Мог. Кондара (3 VII, 1 j, 1 (5), Квак
(23 VII, 1 j). — Средняя Азия (Зеравшан).
Anthidium cingulatum Latr. Кондара (17 VI—15 IX, 2 1 (5), Квак
(29 VI—4 VIII, 4 5, 2 5), Руидашт (22 VII—4 IX, 1 $, 1 3). —
Юг Европейск. части СССР, сев. Казахстан, Сибирь, Средняя Азия
(Фергана, Алайск. хр., Тянь-шань, Таджикистан), юг Зап. Европы,
сев. Африка, зап. Азия.
Anthidium manicatum hissaricum Mavrom. Квак (20 VII, 1 $), 1 (5). —
Типичная форма, широко распространена в Средней Азии (Каракалпа-
кия), Европе, сев. Африке. Подвид описан из Квака по приведенным
экземплярам.
Antidium diadema Latr. Кондара (25 VI—17 VII, 6 д), Квак (29 VI—
17 VII, 1 $, 1 (5), Руидашт (4 IX, 1 $>). —Юг Европейск. части СССР,
Средняя Азия (Узбекистан, Таджикистан, Туркмения), юг Зап. Европы,
сев. Африка.
Anthidium florentinum F Кондара (10—21 IX, 2 j, 2 3), Квак
(27 VIII, 1 j). — Юг Европейск. части СССР, сев Казахстан, южн.
Сибирь, Средняя Азия (Ташкентск. оазис, Зеравшан, Фергана, Аму-дарья,
Сталинабад, Гиссарск. хр., Копет-даг), Передняя Азия, Белуджистан,
юг Зап. Европы.
Anthidium anguliventre F. Мог. Кондара (5 VII—3 VIII, 2 j).—Сред-
няя Азия (Таджикистан, Туркмения).
Anthidium gussakovskiji Mavrom. Кондара (9—15 VII, 2 j, 2 (5). —
Вид описан по приведенным экземплярам из Кондары.
Anthidium kvakicum Mavrom. Квак (4 VIII, 1 £, 1 (5). — Вид установ-
лен по приведенным экземплярам из Квака.
Anthidium punctatum oreophilum Popov. Руидашт (22 VII, 1 $>, 2 5). —
Основная форма широко распространена по сев. и средн. Европе и Си-
бири; subsp. oreophilum Popov приурочен к горам Средней Азии.
Anthidium trochantericum F. Мог. Кондара (4 VII, 1 5), Квак (22 IX,
1 (5). — Средняя Азия (Киргизск. хр., Зеравшан, Дарваз, зап. Па-
мир).
Stelidomorpha nasuta Latr. Кондара (20 VI—7 VII, 18 j). — Юг Евро-
пейск. части СССР, Кавказ, Закавказье, Средняя Азия (Зеравшан,
Пчелиные
169-
Фергана), средн, и южн. части Зап. Европы. Паразитирует в гнездах
Chalicodoma muraria Retz.
Stelis scutellaris F. Мог. Кондара (7 VII, 1 ^). — Средняя Азия
(широко распространен), Центр. Азия; subsp. inamoena Popov — юго-
восток Европейск. части СССР, Кавказ, Закавказье, сев. Иран.
Stelis minuta Lep. Кондара (9 VI, 1 (5). — Сев. и средн, часть Пале-
арктики; в Средней Азии был известен из Каракалпакии.
Dioxys cincta Jur. Кондара (4 VII, 1 $>). — Сев. и средн, часть Пале-
арктики; был известен с Памира.
Dioxoides tridentata (Nyl.). Руидашт (4 IX, 1 $>), Зидды (5 VIII, 1 2)-'—
Сев. и средн, часть Палеарктики. Новость для фауны Средней Азии..
Паразитирует в гнездах Chalicodoma muraria Retz., Megachile argentata F.
и некоторых видов Osmiini.
Heriades truncorum L. Квак (17 VII—5 VIII, 7 $), Зидды (5 VIII, 2 $>),
Гушары (12 VII, 4 j). — Европейск. часть СССР, Зап. Европа, сев.
Африка, Малая Азия. Новость для фауны Средней Азии.
Hoplitis simplicicornis (F. Мог.). Кондара (29 V—17 VII, 21 о, 9 3)b
Квак (20 VI—27 VIII, 10 j, 1 (5), Руидашт (22 VI, 1 j), Гушары (14 VII,
2 2 5). — Средняя Азия (Кызыл-кумы).
Hoplitis transcaspica (F. Мог.). Зидды (5 VIII, 1 $). — Средняя Азия
(Туркмения).
Hoplitis falcata (F. Мог.). Квак (21 VI, 1 5). — Средняя Азия (Зерав-
шан), сев. Иран.
Hoplitis (Liosmia) leucomelaena (Kby.) (claviventris F. Mor.). Кон-
дара (7—26 VI, 2 2), Анзоб (28 VII, 1 j). — Европейск. часть СССР,
Зап. Европа. Новость для фауны Средней Азии.
Anthocopa cyanescens (F. Мог.). Руидашт (18 VI, 3 3). — Средняя
Азия (Зеравшан).
Anthocopa glasunovi (F. Мог.). Кондара (20 VI—4 VII, 5 $, 1 3)^
Квак (29 VI—4 VIII, 3 $>). —Средняя Азия (Варзаут).
Anthocopa furcula (F. Мог.). Квак (23 VII, 1 3), Руидашт (22 VII,
1 $>), Анзоб (31 VII, 1 j). — Средняя Азия (Кызыл-кумы, Зеравшан).
Anthocopa sogdiana (F. Мог.). Кондара (1—19 VII, 7 $>), Квак (10—
30 VI, 5 $>), Руидашт (22 VII, 1 $>). —Средняя Азия (Зеравшан).
Anthocopa (Glossosmia) singularis (F. Mor.). Квак (20 VI, 1 j). Сред-
няя Азия (Фергана, Зеравшан).
Osmia fedtschenkoi F. Mor. Кондара (13—16 V, 6 $>).—Средняя Азия
(Зеравшан, Ашхабад).
Osmia melanocephala F. Mor. Квак (10—20 VI, 4 ^). — Средняя Азия
(Зеравшан, Алайск. хр., Ашхабад).
Osmia maxillaris F. Мог. Кондара (15 VI, 1 $), Квак (10—21 VI, 2 j,.
5 3). — Средняя Азия (Зеравшан, Туркмения).
Osmia difficilis F. Мог. Квак (20 VI, 2 j). — Средняя Азия (Зерав-
шан).
Osmia (Chalcosmia) aenea L. (caerulescens L.). Кондара (13 V—2 VIII,
32 5, 22 3), Квак (10 VI—13 VII, 4 2), Анзоб (28 VII, 5 $), Гушары
(14 VII, 1 j, 1 3). — Европейск. часть СССР, Средняя Азия (Ташкентск.
оазис, Зеравшан, Фергана, Тянь-шань), Зап. Европа, сев. Африка,
Передн. Азия.
Osmia (Chalcosmia) subcornuta F. Mor. Кондара (21 V, 1 j).—Средняя
Азия (Зеравшан).
Osmia (Melanosmia) pamirensis Guss, (rickmersi Alfk.). Руидашт (17—
18 VI, 2 2), Анзоб (27 VII, 1 5)- ~ Памир.
170
В. В. Попов
Chalicodoma muraria nestorea Brulle. Кондара (21 V—15 VII, 10
-3 5), Квак (20 VI, 1 $>), Руидашт (29 VII, 1 9). — Типичная форма, оби-
тает в средн, и южн. полосах зап. части Палеарктики; подвид nestorea
Brulle — в Средней Азии (Зеравшан, Фергана, Копет-даг), Белуджи-
стане, вост. Средиземноморье.
Chalicodoma desertorum F. Мог. Кондара (2 VI—3 VII, 5 — 9). Сред-
няя Азия (Кызыл-кумы, Туркмения), Белуджистан, Монголия;
в Египте — subsp. atrorufa Friese, в Китае — v. tsinanensis Ckll.
Chalicodoma difficilis F. Mor. Кондара (5 VI, 1 9), Квак (4—15 VI,
3 2 6). — Средняя Азия (Зеравшан).
Archimegachile derasa (Gerst.). Кондара (20 VI—17 VII, 6 j, 9 6),
Квак (13 VII, 2 ^). — Средняя Азия (Ташкентск. оазис, Зеравшан, Фер-
гана, Туркмения), вост. Средиземноморье.
Megachile (Eutricharaea) argentata F. Кондара ( 5 VI—19 IX, 12 5,
4 (5), Квак (29 VI—27 VIII, 6 $>). — Европейск. часть СССР, Средняя
Азия (Каракалпакия, Ташкентск. оазис, Кызыл-кумы, Зеравшан, Фер-
гана, Таджикистан, Туркмения), Зап. Европа, Сев. Африка, Передняя
Азия.
Megachile (Eutricharaea) albidula Alfk. Кондара (15 VI—21 IX, 15 9
54 <5), Квак (29 VI—5 VIII, 2 9, 4 (5), Руидашт (4 IX, 1 3), Зидды
(5 VIII, 1 9, 1 8), Анзоб (28 VII, 1 $>). — Средняя Азия (Таджикистан.
Памир).
Megachile (Eutricharaea) apicalis Spin. Кондара (26 VI—18 IX, 31 9),
Квак (4 VII—17 VIII, 7 9), Руидашт (2 IX, 1 9), Зидды (5 VIII, 1 9). -
Европейск. часть СССР, Средняя Азия (Ташкентск. оазис, Зеравшан.
Фергана, Таджикистан), средн, и южн. полосы Зап. Европы, сев.
Африка.
Megachile (Eutricharaea) terminata F. Mor. Кондара (8 VIII, 1 9),
Квак (23 VII, 1 9). — Средняя Азия (Зап. Казахстан, Зеравшан, Таджи-
кистан, Туркмения).
Megachile (Eutricharaea) dacica Moes. Кондара (25 VII, 1 9), Руи-
дашт (2—4 IX, 6 9). — Юг Европейск. части СССР, южн. Венгрия.
Новость для фауны Средней Азии.
Megachile (Eutricharaea) albisecta caucasica Lep. Кондара (15—16 VII,
1 9, 1 5), Квак (17 VII—17 VIII, 3 9), Зидды (26 VII—5 VIII, 3 9). —
Европейск. часть СССР, Средняя Азия (Ташкентск. оазис, Кызыл-кумы,
Таджикистан, вкл. Памир), Зап. Европа.
Megachile (Eutricharaea?) sanguinipes F. Mor. Кондара (20 VI—11 VII,
•8 5), Квак (17 VII, 1 6), Руидашт (22 VII, 1 (5). — Средняя Азия (Зерав-
шан, Дарваз).
Megachile (Anthemois) centuncularis L. Кондара (13—16 IX, 2 9). —
Европейск. часть СССР, Средняя Азия (Каракалпакия, Тянь-шань),
Зап. Европа, Передняя Азия.
Megachile (Anthemois) genalis F. Mor. Кондара (25 VIII, 1 9)- —
Европейск. часть СССР, Кавказ, Средняя Азия (Зеравшан), Синьцзян,
Зап. Европа.
Megachile (Xanthosaurus) lagopoda L. Кондара (7—15 VII, 2 9, 1 (5).
Квак (23 VII—27 VIII, 2 9), Руидашт (20 VII—4 IX, 4 9, 2 3). — Широко
распространенный палеарктический вид; Средняя Азия (Зеравшан, Фер-
гана, Тянь-шань, Туркмения).
Megachile (Xanthosaurus) maritima Kby. Кондара (21 V—21 IX,
22 9, 7 (5), Квак (4 VIII—22 IX, 2 9, 4 (5), Руидашт (22 VII—2 IX, 2 3),
Пчелиные
171
Анзоб (28 VII, 1 $). — Широко распространенный палеарктический вид;
-Средняя Азия (Фергана, Зеравшан, Памир, Тяныпань).
Coelioxys rufescens Lep. Кондара (20—27 VI, 1 $, 2 (5).—Широко рас-
пространенный палеарктический вид; Средняя Азия (Тянь-шань, Памир,
Каракалпакия). Паразит в гнезде Anthophora parietina F., и, вероятно,
Clisodon furcatus (Panz.).
Coelioxys conoidea Klug. Кондара (27 VI—17 IX, 1 j, 5 5), Квак
(13 VII, 1 j). Руидашт (2 IX, 1 6), Гушары (14 VII, 1 6). — Европейск.
часть СССР, Средняя Азия (северная часть, Туркмения), сев. и южн.
полосы Зап. Европы, Иран. Паразитирует в гнездах Megachile mari-
tima КЬу. и М. logopoda L.
Coelioxys afra Lep. Кондара (24 VI—6 VIII, 1 {, 5 (5). — Зап. часть
Палеарктики; Средняя Азия (Таджикистан, Туркмения). Паразитирует
у Megachile argentata F. и М. apicalis Spin.
Coelioxys emarginata Foerst. Кондара (8—25 VII, 1 j, 1 6), Квак
(23 VII, 1 ^). — Зап. часть Палеарктики; Средняя Азия (Фергана), Па-
разитирует в гнездах Megachile leucomalla Gerst.
Coelioxys brevis Eversm. Кондара (28 VI—13 VII, 4 $), 3 5). —
Широко распространенный палеарктический вид; Средняя Азия (Таш-
кентск. оазис, Фергана, Таджикистан). Паразитирует в гнездах Mega-
chile apicalis Spin, и, возможно, М. argentata F.
Coelioxys argentea Lep. Кондара (13—15 VII, 2 6), Зидды (5 VIII,
1 6). — Зап. часть Палеарктики; Средняя Азия (Ташкентск. оазис, Тад-
жикистан, Копет-даг).
Coelioxys sogdiana F. Мог. Кондара (27 VI—15 VII, 5 $>). Вост. За-
кавказье, Средн. Азия (Зеравшан, Ашхабад).
Ceratina nigra Handl, (nitidula F. Mor.). Кондара (10 VIII—17 IX,
4 j). — Средняя Азия (район Сыр-дарьи, Таджикистан, южн. Туркме-
ния).
Ceratina laevifrons F. Мог. Кондара (15 V—16 IX, 18 о, 9 (5), Квак
(8 VI-22 IX, 7 12 о), Руидашт (22 VII—4 IX, 3 5, 2 о), Руидашт—
Куторманба (21 VII, 1 5), Гушары (14 VII, 1 $>, 1 (5). — Вся Средняя Азия,
Месопотамия, Белуджистан.
Ceratina ferghanica F. Мог. Кондара (10—13 VII, 2 $>). — Средняя
Азия (Фергана, Таджикистан, Ашхабад).
Ceratina tibialis F. Мог. Кондара (14 V—29 IX, 25 j, 6 5). — Сред-
няя Азия, Месопотамия, Иран, Белуджистан.
Xylocopa valga Gerst. Кондара (4 VI—17 IX, 4 g), Квак (20 VI, 1 j),
Руидашт (22 VII, 5 $), 1 3). — Южная и средняя полосы Палеаркти-
ческой области, на восток до Центральной Азии включительно; вся
Средняя Азия.
Xylocopa turanica F. Мог. Варзоб (10 VI, 1 j). — Средняя Азия
(Зеравшан, Гиссар).
Xylocopa signata F. Мог. Кондара (17 VI, 1 5), Квак (29 VI, 1 (5). —
Средняя Азия (Чимган, Зеравшан, Фергана, горы Таджикистана, вклю-
чая зап. Памир, Копет-даг), Демавенд.
Xylocopa punctilabris F. Мог. Кондара (13—14 V, 1 3). — Средняя
Азия (Артух).
Xylocopa (Proxylocopa) versicolor Alfk. (lanata Per.). Кондара (4 VI—
19 VII, 2 j, 2 (5), Квак (29 VI, 2 $>). — Вост. Закавказье, средн, и южн.
полосы Средней Азии.
Nomada integricollis F. Мог. Квак (5 VIII, 2 (5), Гушары (14 VII,
1 (5), Анзоб (28 VII, 1 (5).—Зеравшан (Ягноб).
172
В. В. Попо»
Nomada flavopicta Kby. (jacobe^e Panz.). Зидды (26 VII, 3 3). Широко
распространенный палеарктический вид, в пределах Средней Азии из-
вестный из Ферганы и Зеравшана. Паразит в гнездах Melitta leporina
Panz. и некоторых видов Andrena.
Nomada pectoralis F. Мог. Кондара (9 VI—3 VII, 1 1 5), Квак (20—
29 VI, 1 £, 1 5).—Кавказ, Средняя Азия (Туркмения).
Nomada succincta Panz. Руидашт (17 VI—22 VII, 7 $>). — Широко рас-
пространенный палеарктический вид; Средняя Азия (Фергана, Копет-даг).
Nomada platyventris F. Мог. Кондара (15 V—17 VII, 5 j, 8 5), Квак
(16 VI—20 VII, 2 (5), Руидашт (18 VI, 1 $, 1 5), Зидды (26 VII, 1 $).
Анзоб (28 VII, 1 <3). — Зеравшан, в том числе Анзоб.
Nomada sarta F. Мог. Кондара (22—29 V, 2 <5), Анзоб (28 VII, 1 $). —
Средняя Азия (Фергана).
Nomada zonata Panz. Кондара (13—29 V, 4 5), Руидашт (18 VI,
1 (5). — Широко распространенный палеарктический вид; Средняя Азия
(Ташкентский оазис; Зеравшан, в том числе Анзоб; Копет-даг).
Nomada ruficornis L. Кондара (15 V—4 VII, 1 {, 1 <5), Квак (21 VI,
1 (3), Руидашт (17 VI—22 VII, 2 $>, 1 (5). — Западно-палеарктический
вид. Новость для фауны Средней Азии. Паразитирует в гнездах Andrena
varians Kby., A. helvola L., A. ovatula Kby. и др.; в Кваке, вероятно,
паразитирует в гнездах A. jagnobensis F. Мог.
Tarsalia ancyliformis Popov. Кондара (15 VII, 1 $, 1 (5), Квак (4—
6 VIII, 6 J, 1 (5). — Средняя Азия (Ташкенток, оазис, Таджикистан,
южн. Туркмения).
Pasites maculatus aschabadensis Rad. Кондара (16 VII, 1 $>). Средн,
и южн. части вост. Палеарктики, широко распространен по всей Сред-
ней Азии. Паразит Nomia diversipes Latr., и других видов этого рода.
Epeolus transitorius Eversm. Квак (13 VII—4 VIII, 1 $, 2 (5), Руидашт
'22 VII, 1 3), Зидды (5 VIII, 1 j). — Юго-восток Европейск. части СССР,
Средняя Азия (Зеравшан).
Tetralonia spectabilis F. Мог. Кондара (7 VI—27 VII, 7 $>), Квак
(10 VI, 1 $, 1 (3), Руидашт (18 VI, 1 (5). —Средняя Азия (Ташкенток,
оазис, Зеравшан, Фергана, Туркмения), Белуджистан.
Tetralonia fuscipes F. Мог. Кондара (28 VII—21 IX, 79 j, 16 (5),
Квак (17 VII, 1 j). — Средняя Азия (Зеравшан, Сталинабад, Куляб).
Tetralonia nigficeps F. Мог. Кондара (19 VI, 5 d), Квак (4—17 VII,
5 j, 1 <3). — Средняя Азия (Таджикистан, Туркмения).
Tetralonia malvae Rossi. Кондара (19 VI—15 VII, 3 j, 4(3).— Евро-
пейск. часть СССР, Средняя Азия (Зеравшан, Анзоб, Микоянабад),
средн, и южн. Европа, Передн. Азия.
Eucera tuberculata F. (longicornis Friese). Кондара (7—13 VI, 4 j),
Широко распространенный палеарктический вид, известный в Средней
Азии из «области Аму-дарьи».
Eucera tomentosa F. Мог. Кондара (13 V—15 VII, 10 j, 5 3), Квак
(10 VI—17 VII, 9 $, 4 (5), Руидашт (17 VI—22 VII, 5 $, 27 <3). — Южн. по-
лоса Европейск. части СССР, Средняя Азия, Иран, юго-восток Зап. Европы.
Eucera nigrifacies Lep. Квак (10—21 VI, 2 (5). Южн. полоса Евро-
пейск. части СССР, Кавказ, Средняя Азия (Каратау), средн, и южн.
полосы Зап. Европы, сев. Африка, Иран.
Eucera hispana.Lep. (melanocephala F. Mor.). Кондара (3 VII—3 VIII,
5 j), Квак (10 VI—4 VIII, 3 $), 3 <3). — Западно-палеарктический вид; сев.
Казахстан, Средняя Азия (Зеравшан, Туркмения), южн. Европа, сев.
Африка.
Пчелиные
173
Eucera chrysop^yga Per. Кондара (9 VI, 1 5), Квак (10 VI, 1 (5), Руи-
дашт (17 VI, 1 о). — Западно-палеарктический вид; Кавказ, юг Зап.
Европы, сев. Африка. Новость для фауны Средней Азии.
Habropoda tadzhica Popov. Квак (8 VI—20 VII, 8 6 (5). Вид опи-
сан по приведенным экземплярам из Квака, а также по 1 о с хр.
Петра I.
Amegilla quadrifasciata (Vill.). Кондара (1 VII—20 IX, 8 3 3),
Квак (4 VII—5 VIII, 4 $), 4 5), Руидашт (2 IX, 1 $). Средн, и южн. части
Палеарктической области, сев. Индия.
Amegilla albigena (Lep.). Кондара (18 VI—21 IX, 43 j, 24 5), Квак
(29 VI—22 IX, 13 j, 2 6), Руидашт (22 VII, 1 j), Зидды (5 VIII, 1 $).
Средиземноморская и Среднеазиатская подобласти, в Средней Азии
широко распространен.
Amegilla glycyrrhizae (Guss.). Кондара (28 VI—7 IX, 5 j). — Сред-
няя Азия (Ташкентск. оазис, Зеравшан, Таджикистан).
Anthophora deserticola F. Мог. Квак (29 VI, 2 j). — Средняя Азия
(Зеравшан, Сыр-дарья, Аму-дарья, Таджикистан).
Anthophora glasunovi F. Мог. Кондара (24 VI—19 VII, 6 j, 1 5), Квак
(17—21 VII, 3 (5), Руидашт (18 VI, 1 j). — Средняя Азия (Зерав-
шан, Гиссарск. хр.).
Anthophora kessleri Fedtsch. Кондара (7 VI—4 VII, 2 5), Квак (10—
29 VI, 3 2). — Средняя Азия (Ташкентск. оазис, Зеравшан, Гиссарск.
хр.), Месопотамия.
Anthophora uljanini Fedtsch. Руидашт (18 VI, 2 2)- — Средняя Азия
(Ташкентск. оазис, Зеравшан).
Anthophora fuliginosa F. Мог. Квак (10—21 VI, 1 2, 1 3), Руидашт
(22 VII, 2 2)- — Средняя Азия (Зеравшан: Ягноб; Памир — subsp.
pamiricola Hed.).
Anthophora turanica Fedtsch. (A. parietina Fedtsch., A. smirnovi
Kuzn.-Ug., syn.' nov). Кишл. Анзоб (28 VII, 1 2)- —Средняя Азия,
Закавказье.
Anthophora caucasica Rad. Кондара (13 VI, 1 5), Квак (29 VI, 1 2),
Руидашт (17 VI, 1 5). — Кавказ, Средняя Азия (Зеравшан), Средизем-
номорье.
Anthophora liturata Fedtsch. Кондара (27 VI, 1 2)» Квак (10 VI-
20 VII, 3 2, 1 3), Руидашт (22 VII, 1 2)- —Средняя Азия (Зеравшан,
Туркмения).
Anthophora radoszkowskyi Fedtsch. Кондара (7 VI—25 VII, 5 2)»
Квак (21 VII, 1 2), Руидашт (22 VII—2 IX, 6 2)- — Юго-восток Евро-
пейск. части СССР, Средняя Азия (Ташкентск. оазис; Зеравшан, в том
числе Анзоб, Сталинабад, Курган-тюбе), сев. Африка (?).
Anthophora semperi Fedtsch. Анзоб (27 VII, 5 2)- — Средняя Азия
(Зеравшан, Алайск. хр.), Иран (Шаху).
Anthophora vulpina Panz. Квак (27 VIII, 1 2), Руидашт (20 VII,
4 2)- — Европейск. часть СССР, Сибирь, Кавказ, Средняя Азия (Памир —
subsp. alticola Hed.), Зап. Европа.
Anthophora borealis F. Mor. Руидашт (2 IX, 1 6). Европейск. часть
СССР, сев. Казахстан, Кавказ, Тянь-шань, Монголия, Зап. Европа.
Новость для фауны Средней Азии.
Anthophora bimaculata Panz. Квак (29 VI—5 VIII, 5 2 2 (5) Руидашт
(2—4 IX, 4 2> 1 6), Зидды (5 VIII, 1 2)- — Европейск. часть СССР, Кав-
каз, Средняя Азия (Каракалпакия, Зеравшан, Фергана, Алайск. хр., Тад-
жикистан, Памир, южн. Туркмения), средн, и южн. полосы Зап. Европы.
474
В. В. Попов
Melecta plurinotata Brulle. Кондара (14 V, 1 q). Средиземноморская
и Среднеазиатская подобласти (в Средней Азии — Зеравшан, Туркмения).
Паразитирует в гнездах некоторых Anthophora (s. str.).
Melecta luctuosa Scop. Кондара (13 VI, 1 9), Квак (8 VI, 1 j), Руидашт
(22 VII, 1 9). — Европейск. часть СССР, Средняя Азия (Тянь-шань,
Заалайский хр., Памир, Зеравшан, южн. Туркмения), средн, и южн.
полосы Зап. Европы. Паразитирует в гнездах Anthopoia parietina F.
и других видов.
Melecta funeraria F. Sm. Кондара (26 VI, 1 9), Квак (10 VI, 1 9),
Руидашт (18 VI, 1 9). —Средиземноморская подобласть. Новость для
фауны Средней Азии. Паразит в гнездах Habropoda zonatula F. Sm.
(Европа). В Кваке, вероятно, паразитирует у Н. tadzhica Popov.
Crocisa scutellaris F. Кондара (7—17 VII, 2 9).—Средн, и южн. части
Палеарктической обл. (Туркмения). Паразит Anthophora vulpina Panz.,.
A. quadrimaculata F.
Crocisa truncata Per. Кондара (1 VII, 1 9). — Юг Европейск. части
СССР, Средняя Азия (Таджикистан), Средиземноморье.
Crosisa ramosa Lep. Кондара (26 VI—21 IX, 6 9, 16 5), Квак (5 IX,
1 9). — Юг Европейск. части СССР, Средняя Азия (Ташкентск. оазис,
Зеравшан, Фергана, Таджикистан, Туркмения), юг Зап. Европы, сев.
Африка, Нубия, Передняя Азия, срв. Индия.
Bombus (Bombus) lucorum L. Кондара (16 VII, 1 (5), Квак (4 VIIr
1 9). — Сев. и средн, части Палеарктики, горы Средней Азии.
Bombus (Pratobombus) leucurus Bisch. et Hed. (leucopygus F. Mor.). —
Кондара (13 VII, 1 6), Квак (17 VII, 1 6). — Горы Средней Азии.
Psithyrus (Ashtonipsithyrus) distinctus Per. Кондара (7 VI, 1 9). —
Сев. и средн, части Палеарктической обл., горы Средней Азии. Паразит
Bombus lucorum L.
Psithyrus (Metapsithyrus) morawitzianus 0. Vogt. Квак (18—20 VI,.
2 9). — Горы Средней Азии (Зеравшан, Каратегин, хр. Петра I).
Psithyrus (Metapsithyrus) ferghanicus Rad. Руидашт (22 VII, 1 9). —
Горы Средней Азии.
Apis mellifera L. Кондара (15 VI, 1 рабоч.), Квак (17 VI, 6 рабоч.). —
В Кондаре поставлена небольшая пасека и, несомненно, что здесь может
быть развернуто большое рентабельное пчеловодное хозяйство.
ЛИТЕРАТУРА
Гуссаковский В. В. 1940. К фауне перепончатокрылых Гиссарского хребта..
Природа, № 4 84.
Гуссаковский В. В. 1945. Новый род Cephidae из Средней Азии. Докл. Акад.
Наук СССР, XLVIII, № 7 : 558—560.
Моравиц Ф. Ф. 1875 и 1876. Пчелы в: Путешествие в Туркестан А. П. Федченко.
Изв. Общ. люб. естеств., антропол., этнограф., XIX, 2, 1875 II + 160:
3, 1876 161—304.
Моравиц Ф. Ф. (Morawitz F.). 1894. Supplement zur Bienenfauna Turkestans.
Horae Soc. Ent. Ross., XXVIII 1—87.
Попов В. В. 1947. О реликтовом характере Dianthidium insulare F. Mor. (Hyme-
noptera, Megachilidae) в фауне пчелиных Средней Азии. Докл. Акад. Наук
СССР, LVI1I, № 1 169—172.
Попов В. В. 1948. О реликтовом характере нового вида антофорид в фауне пчели-
ных Таджикистана. Сообщ. Тадж. фил. Акад. Наук СССР, VII : 31—34.
М. Н. НИКОЛЬСКАЯ
ОСЫ-БЛЕСТЯНКИ ЮЖНОГО СКЛОНА ГИССАРСКОГО ХРЕБТА
1. Ellampus (Dictenulus) hohlbecki Sem. — Кондара (14 VIII 1937x
1 2). — Ср. Азия (Таджикистан, Узбекистан, ю. Казахстан).
2. Ellampus (Dictenulus) dzhigit Sem. et Nik., in litt. — Кондара
(25 VIII 1938, 1 2).
3. Ellampus (Ellampus) bogdanovi Rad. — Кондара (16 VI 1929,
VIII 1937, 5 <5, 3 2)- — Ю.-в. Европейской части СССР, Ср. Азия
(Туркмения, Узбекистан, ю. Казахстан), в. Средиземноморье.
4. Ellampus (Ellampus) elongatus Sem. et Nik., in litt. — Кок-куль,,
истоки p. Лючоб (20 VIII 1940, 1 <5).
5. Ellampus (Ellampus) gussakovskii Sem. et Nik., in litt. — Кондара
(14 VII 1937, 2 2); Квак (20 VII 1937, 1 <5).
6. Ellampus (Ellampus) hirsutus Sem. — Кондара (31 VII 1937, 1 (5);
Руидашт (22 VII 1937, 1 5); Гиссарский хребет (12 VII 1912, 2 (5).
7. Ellampus (Ellampus) scrutator Sem. et Nik., in litt. — Кондара
(13—15 IX 1938, 2 2)-
8. Ellampus (Ellampus) subopacus Sem. et Nik., in litt. — Кондара
(2 VII—21 IX 1937; 14 VII—13 IX 1938, 3 S, 5 2)-
9. Hedychridium (Euchridium) trossulus Sem., in litt. — Перевал
Мура, Гиссарский хребет (15 VII 1912, 12, 1 (5).
10. Hedychridium (Hedychridium) alpestre Sem. et Nik., in litt. —
Кок-куль, истоки p. Лю1’об (20 VIII 1940, 6 5).
11. Hedychridium (Hedychridium) rhodinum Sem. et Nik., in litt. —
Руидашт (2 IX 1937, 1 $)-
12. Hedychridium (Hedychridium) turanicum Sem., in litt. — Кон-
дара (8 IX 1937, 1 2)- — Ср. Азия (Узбекистан).
13. Cyrteuchridium breviceps Sem., in litt. — Кондара (30 VIII 1938,
1 2)J — Закавказье, Ср. Азия (Таджикистан, Узбекистан).
14. Cyrteuchridium pusio Sem., in litt. — Кондара (12 VII 1938, 1 2). —
Ср. Азия (Туркмения, Узбекистан).
15. Euchrum turanum Sem., in litt. —Руидашт (22 VII 1937, 1 2);
Кондара (7 IX 1937, 1 2); Зидды (5 VIII 1938, 1 2)- — Ср. Азия (Таджи-
кистан, Узбекистан, Киргизия).
16. Zarudnidium laetum Sem. et Nik., in litt. — Кондара (2—
12 VIII 1937, 2 2)-
17. Zarudnidium sapphirinum Sem., in litt. — Кондара (4 VII 1937,
1 5). — Ср. Азия (Таджикистан, ю. Туркмения, Узбекистан).
18. Hedychrum frivaldskyi Moes. — Кондара (21 VI 1938, 1 (5). — Ср.
Азия (Таджикистан, ю. Туркмения, Узбекистан, ю. Казахстан).
19. Hedychrum sapphirinum Sem., in litt. — Кондара (9 VI 1937к
1 2)* — Ср. Азия (Узбекистан).
.'176
М. Н. Никольская
20. Holopyga amoenula Dahlb. — Кондара (3 VI—17 VII 1939, 2 (5,
1 $). — Юг ср. полосы Европейской части СССР и Сибири, Закавказье,
Ср. Азия (Таджикистан, Узбекистан, ю. Казахстан), 3. Европа,
с. Иран.
21. Holopyga chrysonota Foerst. — Руидашт (21 VII 1937, 1 3). —
Ю. ср. полосы Европейской части СССР и Сибири, Закавказье, Ср. Азия
(Таджикистан, Туркмения, Узбекистан, ю. Казахстан, Киргизия),
3. Европа, с. Иран.
22. Holopyga comosa Sem. et Nik., in litt. — перевал Анзоб
<31 VII 1937, 1 $).
23. Holopyga gloriosa Dahlb. — Кондара (22 VHI 1937, 1 j). —
Ю. ср. полосы Европейской части СССР и з. Сибири, Закавказье, Ср.
Азия (Таджикистан, Узбекистан ю. Казахстан), 3. Европа, с. Иран.
24. Holopygahirta Sem. et Nik., in litt. — Руидашт (20 VII 1938, 1 $)).
25. Holopyga punctatissima Dahlb. — Кондара (3 VIII—8 IX 1937,
21 VI 1938, 17—23 VII 1939, 4 3, 10 $); перевал Анзоб (28 VII 1938,
1 3). — Юг Европейской части СССР, Закавказье, Ср. Азия (Таджи-
кистан, Туркмения, Узбекистан, ю. Казахстан), Средиземноморье,
с. Иран.
26. Chrysis (Holochrysis) alticola Sem. — Гушары (14 VI 1938, 1 $)).—
Ср. Азия (горный Таджикистан).
27. Chrysis (Holochrysis) desidiosa Buyss. — Руидашт (22 VIII 1937.
4 2). — «Кавказ» (видимо, Закавказье).
28. Chrysis (Holochrysis) eos Sem., in litt. — Кондара (И VI 1937,
13 V 1939, 2 3). — Ср. Азия (Таджикистан, Узбекистан).
29. Chrysis (Holochrysis) fragaria Sem. et Nik., in litt. — Кондара
<1939, 2 ^).
30. Chrysis (Holochrysis) genalis Nees — Кондара (12 VI—8 VII 1937,
15 VI —12 VII 1938, 14 V—2 VII 1939, 17 j). — Ср. Азия (Таджикистан,
Узбекистан).
31. Chrysis (Holochrysis) korsakovi Sem. et Nik., in litt. — Руидашт
<16 VI 1939, 1 3).
32. Chrysis (Holochrysis) melpomena Sem. et Nik., in litt. — Кондара
<21 VI 1938, 4 о); Руидашт (16 V 1939, 1 3, 1 $>)
33. Chrysis (Holochrysis) persephone Sem. et Nik., in litt. — Кондара
(3—15 VII 1937, 30 V—14 VI 1939, 4 j). — Ср. Азия (Таджикистан,
Узбекистан).
34. Chrysis (Holochrysis) sogdiana Sem., in litt. — Кондара (13—
16 V 1939, 1 3, 3 2); перевал Анзоб (27 VII 1937, 1 5).—Ср. Азия (Тад-
жикистан, горный Узбекистан).
35. Chrysis (Holochrysis) umbra Sem. et Nik., in litt. — Квак
<26 VII 1939, 1 $).
36. Chrysis (Holochrysis) varicornis Spin. — Кондара (16 VI 1937,
1 3); Руидашт (23 VII 1937, 2 3). — Ср. Азия (Таджикистан, Узбекистан),
Средиземноморье.
37. Chrysis (Glossochrysis) matrona Sem., in litt. — Кондара
<15 IX 1937, 7 VIII 1938, 1 3, 1 $). — Ср. Азия (Таджикистан, Турк-
мения, У збеКистан).
38. Chrysis (Chrysidea) pumila KI. — Кондара (25 VIII—9 IX 1937,
-8 VIII 1938, 15 VI 1939, 4 j). — Юг Европейской части СССР, Закав-
казье, Ср. Азия (Таджикистан, Узбекистан, ю. Казахстан), 3. Европа.
39. Chrysis (Gonochrysis) gloriosa singula Rad. — Кондара (7 VIII 1937,
23 VI—26 VII 1938, 1 3, 3 j). — Ср. Азия (Туркмения, ю. Казахстан).
Осы-блестпянки, южного склона Гисса рского хребта
177
40. Chrysis (Gonochrysis) irenes Sem. et Nik., in litt. — Кондара
(17 1X 1937, 1 г); Квак (10 VI 1937, 1 $).
41. Chrysis (Gonochrysis) leachii Shuck. — Кондара (31 VII 1937,
12—26 VII 1938, 30 V—14 VI 1939, 2 3, 6 2). — Юг Европейской части
СССР, Закавказье, Ср. Азия (Таджикистан, Туркмения), 3. Европа.
42. Chrysis (Gonochrysis) succincta sparsepunctata Buyss. — Кондара
(15 VII 1937, 17 VII 1938, 2 $); Квак (17 VII 1937, 1 $). — Ср. Азия
(Таджикистан, Туркмения, Узбекистан).
43. Chrysis (Gonodontochrysis) dolens Sem. et Nik., in litt. — Квак
(6 VIII 1937, 1 3).
44. Chrysis (Chrysis) secernenda Moes. — Кондара (И VII—21 IX 1937,
5 j); Квак (20 VII 1937, 2 j). —Ср. Азия (Узбекистан, Киргизия,
ю. Казахстан), Синьцзян.
45. Chrysis (Tetrachrysis) analis turkestana Sem., in litt. — Квак
(20 VI—17 VIII 1937, 3 5); Кондара (7—22 VIII 1937, 2 $); Гушары
(28 VII 1930, 1 2). — Ср. Азия (Узбекистан).
46. Chrysis (Tetrachrysis) asiatica Rad. — Кондара (23 VII 1939;
1 2)- — Ср. Азия (Таджикистан, ю. Туркмения, Узбекистан).
47. Chrysis (Tetrachrysis) buechneri Sem. — Кондара (31 VIII 1937,
1 3). — Ср. Азия (Таджикистан, Узбекистан).
48. Chrysis (Tertachrysis) calypso Sem. et Nik., in litt. — Кондара
(30 VI 1939, 1 (5).
49. Chrysis (Tetrachrysis) csikiana Moc>_ — Кондара (19 VI 1936, 1 2)
Квак (29 VI 1937, 16 VI 1938, 2 2)- — Ср. Азия (Узбекистан).
50. Chrysis (Tetrachrysis) distincta Moes.— Кондара (4—26 VIII 1937,
И VII 1938, 4 2)‘> Руидашт (21 VII 1938, 1 2)- Юг Европ. части
СССР, в. Закавказье, Ср. Азия (Таджикистан, Туркмения, Узбекистан),
восток 3. Европы, с. Иран.
51. Chrysis (Tetrachrysis) eversmanni m. isiris Sem. in litt. — Кон-
дара (20 VI 1938, 1 2)- Ср. Азия (Таджикистан, Узбекистан).
52. Chrysis (Tetrachrysis) georgii Sem., in litt. — Кондара (9 VI 1937,
1 (5). Ср. Азия (Таджикистан, Туркмения, Узбекистан).
53. Chrysis (Tetrachrysis) gussakovskii Sem. et Nik., in litt. — Кон-
дара (21 VII 1938, 2 3); Руидашт (21 VII 1938, 1 3).
54. Chrysis (Tetrachrysis) fulgida L. — Кондара (14 VI 1937,
13 V—12 VII 1939, 1 3, 5 2)-—Палеарктика.
55. Chrysis (Tetrachrysis) inaequalis роёНса Sem. — Кондара
(24 VI—16 IX 1937, 20 VI—18 IX 1938, 8 <5 18 $); Руидашт (22 VII 1937,
1 3) — Закавказье, Ср. Азия (Таджикистан, Узбекистан).
56. Chrysis (Tetrachrysis) korbiana Moes.— Квак (20 VII 1937, 1 2)-—
Ср. Азия (Узбекистан).
57. Chrysis (Tetrachrysis) maculicornis KI. — Кондара (22 VII—
31 VIII 1937, 10—27 VII 1938, 6 3). — Ср. Азия (Таджикистан, Туркме-
ния, ю. Узбекистан), в. Средиземноморье, с. Иран, Монголия.
58. Chrysis (Tetrachrysis) mesembrina Sem. et Nik., in litt. — Кон-
дара (12 IX 1937, 25 VII —20 IX 1938, 9 $); Квак (17 VIII 1937, 16 VI-
1938, 1 3, 1 2)- — Ср. Азия (Таджикистан),
59. Chrysis (Tetrachrysis) neobulae Sem., in litt. — Кондара
(27 VIII 1937, 1 2)-—Ср. Азия (Таджикистан, ю.-з. Казахстан).
60. Chrysis (Tetrachrysis) remota Moks. — Кондара (19 VI 1939, 1 3).—
Закавказье, Ср. Азия (Таджикистан, Туркмения, Узбекистан, ю. Казах-
стан), с. Иран.
12 Ущелье Кондара
178
М. Н. Никольская
61. Ghrysis (Tetrachrysis) rutilans mesasiatica Sem. — Руидашт
(4 IX 1937, 1 2). — В. Закавказье, Ср. Азия (Таджикистан, Туркмения,
Узбекистан), с. Иран.
62. Chrysis (Tetrachrysis) scutellaris consobrina Moes. — Кондара
(12—26 VTII 1937, 21 VI — 26 VII 1938, 11 (5, 4 $); перевал Анзоб
(31 VII 1937, 1 (5). — Ср. Азия (Таджикистан, Туркмения, Узбекистан),
с. Иран, Синьцзян.
63. Chrysis (Tetrachrysis) uljanini Rad. — Кондара (14 VI-
28 VIII 1937, 20 VI—20 VIII 1938, 17 VII 1939, 4 5, 12 $); Квак
(17 VII 1937, 2 $). — В. Закавказье, Ср. Азия (Таджикистан, Узбеки-
стан), с. Иран.
64. Chrysis (Tetrachrysis) viridans Rad. — Кондара (25 VI—8 IX 1937,
10 VII—16 IX 1938, 19 V—17 VII 1939, 3 <5, 15 $).—Закавказье, Ср.
Азия (Таджикистан, ю. Туркмения, Узбекистан, ю. Казахстан), с.-в.
Иран, ю. Монголия.
65. Ghrysis (Tetrachrysis) viridula L. — Кондара (13 VII 1938,
1 2). — Европ. часть СССР, юг ср. Сибири, Закавказье, Ср. Азия
(Таджикистан, Туркмения, Киргизия, Казахстан), 3. Европа, с. Иран.
66. Chrysis (Hexachrysis) sexdentata Christ. — Кондара (17 VII 1936,
27 VI—26 VIII 1937, 15 VI—17 VIII 1938, 17 (3, 26 $); Квак (20 VII 1937,
1 (5). — Крым, Кавказ, Закавказье, Ср. Азия (Таджикистан, Узбеки-
стан, ю. Казахстан), 3. Европа.
67. Stilbum calens Spin. — Кондара (12 VII 1937, 3 VII 1937,
20 VII 1939 2 (5, 2 2)- — Юг Европ. части СССР и Сибири, Закавказье,
Ср. Азия (Таджикистан, Туркмения, Узбекистан, ю. Казахстан), 3. Ев-
ропа, Монголия.
68. Stilbum cyanurum Foerst. — Кондара (8 VII—11 VUI 1937,
15 VI—16 VII 1938, 13 VI—23 VII 1939, 13 5, И $); Квак (27 VIII 1937,
1 2)- — Юг Европ. части СССР и Сибири, Закавказье, Ср. Азия (Тад-
жикистан, Туркмения, Узбекистан, ю. Казахстан), 3. Европы, с. Иран.
М. Азия.
69. Spinolia (Pseudochrysis) humboldti Dahlb. — Кондара (15—VII 1937,
28 VI—7 VIII 1938, 15—20 VII 1939, 9 2)- — Юг Европейской части
СССР, Закавказье, Ср. Азия (Таджикистан, Туркмения, Узбекистан,
ю. Казахстан), 3. Европа, ю. Монголия.
70. Spinolia (Pseudochrysis) uniformis Dahlb. — Кондара (13 VII 1938,
1 2)- —Ср. Азия (Туркмения, Узбекистан), в. Средиземноморье, с.
Иран.
71. Parnopes glazunovi Sem. — Кондара (18 VII 1937, 1 3). Ср. Азия
(Таджикистан, Туркмения, Узбекистан, ю. Казахстан), в. Иран.
72. Parnopes grandior turkestanicus Sem., in litt. — Кондара (2—
24 VII 1937, 2 (5); Гушары (28 VII — 7 VIII 1930, 1 S, 1 $)« — Ср. Азия
(Таджикистан, Туркмения, Узбекистан, ю. Казахстан).
Вышеприведенный- список ос-блестянок, характеризующий фауну
южного склона Гиссарского хребта и ущелья Кондары, составлен по ру-
кописи «Фауна ос-блестянок. Таджикистана» А. П. Семенова-Тян-Шан-
скОго и М. Н. Никольской, предназначенной для сборника, издаваемого
Академией наук Тадж. ССР. В нее вошла обработка обширных сборов
Ctlrysididae, произведенных В. В. Гуссаковским в различных точках Тад-
жикистана, в частности -- из Кондары, Квака, Руидашта и других мест Гис-
сарского хребта, а также богатой коллекции А. П. Семенова-Тян-Шанского.
Осы-блестянки южного склона Гиссарского хребта
179
Из 125 видов, обнаруженных в пределах Таджикистана, 72 вошли
в настоящий список. Многие из этих видов известны пока только из Кон-
дары и других мест южного склона Гиссарского хребта, как то:
Ellampus dzhigit Sem. et Nik.
Ellampus elongatus Sem. et Nik.
Ellampus gussakovskii Sem. et Nik.
Ellampus scrutator Sem. et Nik.
Ellampus subopacus Sem. et Nik.
Hedychridium trossulus Sem.
Holopyga comosa Sem. et Nik.
Holopyga hirta Sem. et Nik.
Holochrysis fragaria Sem. et Nik.
Holochrysis korsakovi Sem. et Nik.
Holochrysis melpomene Sem. et Nik.
Holochrysis umbra Sem. et Nik.
Hedychridium alpestre Sem. et Nik.
Hedychridium rhodinum Sem. et Nik.
Zarudnidium laetum Sem. et Nik.
Gonochrysis irenes Sem. et Nik.
Gonodontochrysis dolens Sem. et Nik
Tetrachrysis gussakovskii Sem. et Nik
или только из пределов Таджикистана:
Holochrysis alticola Sem., Tetrachrysis mesembrena Sem. et ik.
Среди них есть типичные представители горной фауны, отличающиеся
длиной волосков, покрывающих тело:
Ellampus gussakovskii Sem.
Ellampus hirsutus Sem.
Hedychridium alpestre Sem.
Многие виды являются
в Средней Азии и иногда
Ellampus hohlbecki Sem.
Hedychridium turanicum Sem.
Gyrteuchridium pusio Sem.
Euchrum turanum Sem.
Zarudnidium sapphirinum Sem.
Hedychrum frivaldskii Moes.
Hedychrum sapphirinum Sem.
Holochrysis eos Sem.
Holochrysis genalis Nees
et Nik. Holopyga comosa Sem. et Nik.
Holopyga hirta Sem. et Nik.
et Nik. Holochrysis alticola Sem.
более или менее широко распространенными
представлены там особыми подвидами:
Holochrysis persephone Sem. et Nik.
Holochrysis sogdiana Sem.
Glossochrysis ma trona Sem.
Gonochrysis gloriosa singula Rad.
Gonochrysis succincta sparsepunctata
Buyss.
Tetrachrysis analis turkestana Sem.
Tetrachrysis eversmanni m. isiris Sem.
Tarnopes grandior turkestanicus Sem.
Таким образом, 45 из 72 видов эндемичны для Средней Азии, что
составляет 62.5%. Процент весьма высокий, который отчасти объясняется
относительно хорошей изученностью среднеазиатских Chrysididae. Несо-
мненно также, что Средняя Азия является центром видообразования
семейства.
Остальные 37.5% объединяют собой виды, общие у Ср. Азии с Закав-
казьем и с Ираном:
Gyrtenchridium breviceps Sem. Tetrachrysis rutilans mesasiatica Sem.
Holochrysis desidiosa Buyss. Tetrachrysis uljanini Rad.
Tetrachrysis inaequalis poetica Sem. Parnopes glazunovi Sem.
Tetrachrysis remota Moes.
Виды, распространяющиеся из Ср. Азии в ю.-в. направлении (Синь-
цзян, Монголия):
Tetrachrysis maculicornis KI.
Tetrachrysis scutellaris consobrina Moes.
Tetrachrysis viridans Rad.
Spinolia humboldti Dahlb.
Общими для Средней Азии и Средиземноморья являются: Holochrysis
varicornis Spin, и Spinolia uniformis Dahlb. (последняя встречается также
в с. Иране).
Западноевропейские виды, найденные на юге Европейской части
СССР и в Средней Азии, представлены следующими видами:
12*
180
М. Н. Никольская
Ellampus bogdanovi Rad.
Holopyga punctatissima Dahlb.
Chrysidea pumila KI.
Gonochrysis leachii Schuck.
Tetrachrysis distincta Moes.
Hexachrysis sexdentata Christ.
Еще более широкое в долготном направлении распространение имеют
виды, заселяющие как Западную Европу, так и юг Европейской части
СССР и Сибири; к ним принадлежат:
Holopyga amoenula Dahlb.
Holopyga chrysonota Foerst.
Holopyga gloriosa Dahlb.
Tetrachrysis viridula L.
Stilbum calens Spin.
Stilbum cyanurum Foerst.
Chrysis fulgida L. имеет своим ареалом почти всю Палеарктику и захо-
дит далеко на север, но была также обнаружена в Кондаре.
Сведения по биологии Chrysididae Кондары очень скудны.
A. H. КИРИЧЕНКО
НАСТОЯЩИЕ ПОЛУЖЕСТКОКРЫЛЫЕ И ЦИКАДОВЫЕ
Фауна настоящих полужесткокрылых, или клопов (Hemiptera), Кон-
даринского ущелья в настоящее время известна наиболее полно, чем
где бы то ни было в Гиссарском хребте. На небольшой территории от
впадения р. Кондара в р. Варзоб и до высокогорного плато Руидашт
уже констатировано нахождение 258 видов этого отряда, что составляет
почти половину числа видов всей гемиптерофауны Таджикской респу-
блики на современном уровне изучения с ее чрезвычайным разнообра-
зием географических ландшафтов. Тем не менее гемиптерофауну южного
склона Гиссарского хребта в районе р. Кондара нельзя считать богатой
пи в смысле видового многообразия, ни по численности популяций отдель-
ных видов (за некоторыми исключениями). Может быть, и вероятно вслед-
'твие отсутствия сведений по фауне других районов Гиссарского хребта,
кондаринская гемиптерофауна сейчас изобилует многими характерными
представителями отряда, но вряд ли они являются ее эндемиками, тем не
менее впервые открытыми только в самое последнее время в районе
Кондаринского ущелья.
Настоящие полужесткокрылые не принадлежат к тем представителям
животного мира, которые легко бросаются в глаза: они ютятся в детрите,
подчас в большом числе особей, главным образом у корней растений —
деревьев, кустарников и травянистых, где нередко превалируют над всем
макроскопическим животным населением, а отдельные представители их
занимают многие биотопы. Чтобы характеризовать гемиптерофауну
Кондаринского ущелья, сравнительно многочисленную по видовому
составу, но мало видимую взору туриста, остановимся на отдельных,
наиболее характерных биотопах от самого поворота из долины Варзоба
в Кондаринское ущелье до Руидашта.
На месте впадения р. Кондара в Варзоб, на крупных обкатанных ва-
лунах на берегу последнего живет очень изящный и красивый Leptopus
hispanus Ramb., мелкий хищник, как показывает его название, харак-
терный средиземноморский вид, в пределах СССР встречающийся в За-
кавказье и в Средней Азии. Это береговой обитатель из сем. Leptopo-
didae. Здесь уместно отметить чрезвычайную бедность описываемого рай-
она водными полужесткокрылыми, вплоть до Руидашта. Из них мы
пока знаем нахождение в Кондаре преимущественно родниковой и ти-
пично реофильной Velia rivulorum F. var. ventralis Put. и гладыша
Notonecta viridis mediterranea Hutsch., последнего в немногих особях.
Из водомерок здесь живет Gerris costai Ц. S. Эта бедность водными полу-
жесткокрылыми вполне объясняется водным режимом горной реки.
Как увидим ниже, высокогорное плато Руидашт, с его своеобразней-
шими типами водоемов, дает, наоборот, по крайней мере в конце весны,
182
A. H, Кириченко
необычайный расцвет видового разнообразия и многочисленности популя-
ций водных Hemiptera.
Перейдя через р. Варзоб и вступив в Кондаринское ущелье, мы
сразу встречаем характерную «кондаринскую» гемиптерофауну. Из от-
дельных биотопов нижнего течения реки мы можем остановиться на
главнейших, имеющих ландшафтный характер. Кондаринское ущелье —
в настоящее время, вероятно, наиболее сохранившийся облесенный
район Гиссарского хребта, в этом его интерес. И действительно, на
тополях, в небольшом числе растущих по краям дороги, идущей
Рис. 1. Dalpada pavlovskii Kir
Риг Xlu^lha biranovi Kir
через питомник Ботанической станции, мы находим характерную
фауну лесной зоны: крупных клопов из семейства щитников —
Pentatomidae: Apodiphus integriceps Horv., Dalpada pavlovskii Kir.
(рис. 1), Rhaphigaster brevispina Horv. Все они туранские эндемики,
но особого внимания заслуживает второй из них, открытый в самое по-
следнее время и являющийся характерным представителем третичной
фауны; сородичи его населяют южную п юго-восточную Азию, а отчасти
Меланезийскую подобласть Австралийской области. К трибе Haliini,
куда относятся приведенные выше роды Apodiphus и Dalpada, принадле-
жит и замечательнейший представитель кондаринской фауны Mustha
baranovi Kir. (рис. 2) — один из самых крупных по величине видов
палеарктической фауны, усаженный по краям тела шипами, но, к сожа-
лению, найденный до сих пор в немногих экземплярах: условия жизни его
пока остаются не известными. Специфическую гемиптерофауну находим
и на ивах, растущих в сильно увлажненных берегах нижнего течения
Настоящие полужесткокрылые и цикадовые
183
Рис 3. Aradus hissaricus Kir.
р. Кондара, это: Artheneis, Anthocoris, Lygus (Agnocoris) rubicundus Fall.,
Sthenarus perpulcher Kir., St. concinnus Kir., последние два—вновь от-
крытые эндемики туранской гемиптерофауны. На косточковых не обна-
ружены какие-либо специфические виды полужесткокрылых. Обитатель
и вредитель груш и яблонь — Stephanitis pyri F. живет ограниченными
популяциями. Из других древесных и кустарниковых обитателей упо-
мянем: Genocerus patellatus Kir. — главным образом на крушине, Нар-
loprocta pustulifera Stal — в большом числе во время плодоношения на
Atraphaxis вместе с Lygus (Orthops) foreli var. pilosulus Jak., Anthocoris
pilosus Jak. Первые два — характерные эндемики Средней Азии.
При подъеме из ущ. Кондара в уроч. Квак и выше на арче (Juniperus
seravschanica) живет ха-
рактерный для туранских
видов можжевельника —
Dichrooscytus pseud osabi-
naeReut., а на клене (Acer
turkestanicum) в Кваке об-
наружен в самое послед-
нее время один из самых,
пожалуй, эффектных ви-
дов отряда туранской фа-
уны — крупный и краси-
вый щитник Mesopriassus
vetustus Kir., которого
приходится считать харак-
терным элементом третич-
ной фауны, ранее извест-
ным только из ореховых
лесов Ферганского хребта
Тянь-шанской горной си-
стемы; в зоогеографичес-
ком отношении он являет-
ся аналогом вышеупомя-
нутой Dalpada pavlovskii
Kir.
Для кондаринской фа-
уны, конечно,
характерны
вают
наиболее
и заслужи-
особого внимания
в первую очередь представители дендробиоса и поэтому мы останав-
ливаемся на них более подробно. Здесь следует еще упомянуть о чисто
лесном элементе — мицетофаге Aradus hissaricus Kir. (рис. 3), найденном
под корой Celtis и ореха (Juglans) и также относящимся к реликтам тре-
тичного периода (кроме второго пункта на южном склоне Гиссарского
хребта — Ходжа-оби-гарм, вид этот до сих пор не был находим).
В отношении других биотопов мы не можем с такой подробностью
остановиться на их гемиптерофауне, как это сделано для лесных ее эле-
ментов. Фауна настоящих полужесткокрылых в вертикальном разрезе
от места впадения р. Кондара в Варзоб и до подножия Руидашта пре-
терпевает меньшие изменения по сравнению с ее видовым составом на
горных гребнях правого и левого берега р. Кондара. Дно самого ущелья
реки и склоны левого берега населяет в основном типичная средиземно-
морская фауна, характерная для предгорий и невысоких гор. (В. Ф. Оша-
184
Л. Н Ки,риченко
нин считал верхней границей этой зоны 1000 м, но в условиях южного
склона Гиссарского хребта эта фауна поднимается значительно выше). Если
наблюдать фауну отряда от низовий р. Кондара до уроч. Квак и выше,
то наиболее характерные представители ее: на широких листьях Rumex —
Coreus eversmanni Jak., вид сем. Coreidae с очень толстыми усиками;
в первую половину теплого времени, когда еще остаются зеленые листья
югана Prangos pabularia, — во множестве туранскпй эндемик Опсо-
tylus reuteri Rent, и в меньшем количестве Hissaritus dimorphus Kir.,
впервые описанный из Кондары (оба — сем. Miridae); на ксерофитных
склонах по правому берегу р. Кондара — Ellipsocoris tamerlani Kir»
Рис 4. Ellipsocoris tamerlani Kir.
Рис. Eurydema gussakovskii Kir
(Pentatomidae) (рис. 4) и из того же семейства — яркокрасочная Eury-
dema gussakovskii Kir. (рис. 5) — на крестоцветных, в особенности Barbarea
arcuata в уроч. Квак и верховьях р. Кондара — уроч. Оби-сафед. Здесь
были упомянуты только немногие, наиболее характерные и более или ме-
нее открыто живущие виды настоящих полужесткокрылых. Особый и
исключительно богатый мир настоящих полужесткокрылых представляют
герпетобии, ютящиеся в детрите у корней растений — травянистых, ку-
старников и деревьев, без большой специализации в отношении видового
состава растений, но в большом видовом разнообразии; из них отдельные
виды представлены многочисленными популяциями. Эта насыщен-
ность детрита растительности гемиптерофауной сильно возрастает в позд-
неосеннее, зимнее и ранневесеннее время, когда имеет место уход на зи-
мовку и зимовка многих видов настоящих полужесткокрылых, в другое
время года приуроченных к другим биотопам. Трудно дать какой-либо
список, характерных для этого биотопа видов — настолько он всеобщ.
Здесь впервые был констатирован для СССР монотипический представи-
тель подсемейства Bledionotinae — Bledionotus systellonotoides Reut.
Наспюягцие полужесткокрылые и цикадовые
.185
До сих пор мы описывали гемиптерологическое население долины
р. Кондара и горных склонов ее долины до высокогорного плато — Руи-
дашта. Руидашт, несмотря на кратковременность и эпизодичность иссле-
дования его энтомологической фауны (Гуссаковский, 22 VI, 22—23 VII,
2-4 1X 1937; 16, 17, 20 VII 1938; 16 VI 1939; 8 VII 1940; Кириченко,
2—4 VII 1943, 1 V 1944), представляется в гемиптерологическом отно-
шении особым зоологическим участком. Несмотря на высоту 2800 м над
ур. м., элементы высокогорья в фауне настоящих полужесткокрылых вы-
Рис. 6. Myrmecophyes nigripes Reut. $,
ражены очень слабо, это: Mimula maureri Jak. — крупный щитник,
известный ранее из высокогорий Памиро-Алая, найденный в одном эк-
земпляре и вторично на Руидаште X оджа-оби-гармском, Nysius ericae
obscuratus Horv., Lygus pachycnemis Reut., Myrmecophyes nigripes Reut.
(рис. 6), Scirtetellus, n. sp.
Если подниматься от истока р. Кондара на высокогорье Руидашта,
при самом подъеме на плато начинают встречаться отдельные кустики
Cousinia, которые выше, вместе с Adonis turkestanica, образуют специфи-
ческую фитогеографическую ассоциацию. Характернейшие ландшафтные
представители гемиптерофауны Руидашта: на Cousinia огромные попу-
ляции Myrmecophyes nigripes Reut. и Scirtetellus sp.1 Первый из них
1 Этот ландшафтный и в зоогеографическом отношении крайне важный вид был
мною наблюдаедг только в личиночной фазе, что не дает возможности его описания
и установления в системе
186
A. H, Кириченко
является характерным представителем ангарского зоогеографического
элемента в условиях Среднеазиатской горной области, давшего огром-
ный расцвет видообразования и видовой специализации. Горы Средней
Азии — хребты, отроги, северные и южные склоны их характеризуются
комплексом специфических форм: Parnassius (Lepidoptera), Carabus (Cole-
optera), Orgeriaria (Homoptera), Myrmecophyes (Heteroptera), Conophyma
(Orthoptera). В этом аспекте в фауне высокогорных плато Руидашта
крайне характерен упомянутый выше Myrmecophyes nigripes Rent., ко-
роткокрылый муравьиноподобный автохтонный элемент высокогорья
Памиро-Алая, он является характерным индикатором зоогеографических
и экологических подразделений Средней Азии и ее вертикальных гори-
зонтов. Вид рода Scirtetellus — также характерный обитатель высо-
когорий Средней Азии. К характеристике высокогорий склона Гиссар-
ского хребта нельзя не прибавить, что адонис-кузиниевая формация
Руидашта имеет своим компонентом клопа Spilostethus equestris — обыч-
ного транспалеаркта.
Как видно из таблицы распределения фауны полужесткокрылых
по высотным зонам (см. ниже), долина р. Варзоб ниже Кондары до Гис-
сарской долины в окрестностях Сталинабада в гемиптерологическом от-
ношении остается неизвестной. Поэтому не представляется возможным
осветить вопрос об инвазии и взаимном обмене отдельными компонен-
тами фаун Гиссарской долины и южного склона Гиссарского хребта.
Однако отдельные факты существуют и они достаточно ярки, чтобы о них
упомянуть. К ним я отношу нередкие и единичные нахождения в окрест-
ностях Сталинабада, т. е. в Гиссарской долине, горных видов — Alloeog-
lypta pretiosa Kir., Oxycarenus lacteus Kir., Hissaritus dimorphus Kir.,
характерных и, кроме первого, представленных в кондаринской фауне
многочисленнейшими популяциями. Обратно, в Кондаринском ущелье мы
встречаемся с крупным хищником — Ectomocoris ululans Rossi — харак-
терным галоксеном и полупустынным обитателем Гиссарской долины и
Южно-Таджикистанской депрессии.
Из особенностей биологии настоящих полужесткокрылых в Конда-
ринском ущелье следует отметить зимовку в имагинальной фазе и
в дневное время, во многих случаях в жизнедеятельном состоянии, очень
многих видов этого отряда, относящихся, однако, к определенным се-
мействам. Так, например, в середине февраля (7—15 II) во время кратко-
временного посещения ущелья Кондара можно было наблюдать до 70
видов полужесткокрылых принадлежащих к следующим семействам:
Thyraeocoridae, Pentatomidae, Coreidae, Aradidae, Neididae, Myodochidae,
Pyrrhocoridae, Tingitidae, Nabidae, Anthocoridae, Miridae, Hebridae,
Veliidae, Saldidae.
Перечень «зимней» фауны Кондаринского ущелья, заключающий
69 видов, констатированных в течение 9 дней, из которых в один из дней
{11 II) отмечен «сильный снегопад», свидетельствует о круглогодичной
жизнедеятельности многих видов настоящих полужесткокрылых и на
средних горизонтах Гиссарского хребта. К сожалению, подобных наблю-
дений не было произведено на Руидаште, где фенология его обитателей
должна претерпеть значительные изменения из-за краткости теплого
периода: при посещении Руидашта 1 V он представлял необозримое поле
снежного покрова с единичными площадками пригорков, освободившихся
от снега, населенными немногочисленными видами Heteroptera: Scioco-
ris consobrinus Kir., Coriomeris, Alia acuminata L., Berytus tipularius L.,
Ischnocoris punctulatus Fieb, Dichro61ytrus conjunctus Kir., Tingis valida
Настоящие полужесткокрылые и цикадовые
187
Put. var. dicessa Horv., Lygus pachycnemis Reut., Saldula orthochila
Fieb. и некоторых других.
Экономическое значение настоящих полужесткокрылых как вредите-
лей сельскохозяйственных культур и богарного садоводства невелико
в условиях Кондаринского ущелья и сводится, главным образом, к вре-
дителям богарной люцерны — Adelphocoris lineolatus Goeze, Lygus рга-
tensis L., Poeciloscytus, Carpocoris fuscispinus Boh., Dolycoris penicillatus
Horv., Codophila varia F., вызывающих сосанием свертывание листьев и за-
сыхание соцветий. Вредная черепашка (Eurygaster integriceps Put.), распро-
страненная почти до Руидашта, тем не менее и в годы массового размноже-
ния ее в Таджикистане (1941—1942) в Кондаринском ущелье значения
не имела. Вредитель яблонь и груш — Stephanitis pyri F. — наблю-
дался только два раза и в единичных особях. Грецкий орех (Juglans)
в Гиссарском хребте, как и в других островах своего произрастания
в Средней Азии, специфических вредителей из настоящих полужесткокры-
лых не имеет. Нет повода останавливаться на второстепенных и потен-
циально вредных видах, каковы, например: Graphosoma italicum Mull.,
Eurydema ornata L., Aelia melanota Fieb., Neottiglossa leporina H.—S.,
Eurydema ornata L., Acrosternum breviceps Jak., Mesocerus marginatus L.,
Liorhyssus hyalinus F., Nysius graminicola Koi., Stenodema turanicum
Reut., Trigonotylus ruficornis Geoffr. и мн. др., поскольку в Кондаринском
ущелье отсутствуют или занимают минимальную площадь культуры,
повреждаемые ими (огородные, технические) и поскольку мы не имеем
прямых указаний на приносимый вред (высокогорные посевы зерновых
на Руидаште). Вместе с тем здесь нужно подчеркнуть, что в условиях
богарного садоводства Кондары, связанного с интродукцией новых эле-
ментов флоры, всегда существует опасность перехода индиферентных
видов настоящих полужесткокрылых в разряд вредоносных, как это имело
место, например, в Азербайджане на южном побережье Каспийского
моря, где плантации тунгового дерева (Aleurites fordii, из сем. Euphorbi-
асеае) подверглись нападению большого щитника Apodiphus amygdali
Germ., обычно живущего на тополях, миндале и других древесных породах.
С этой точки зрения заслуживает особого внимания один вид кондарин-
ской фауны — Oxycarenus lacteus Kir., в изобилии живущий от уроч.
Квак до ущелья Кондара (единично встречен был и в Гиссарской долине)
на Cousinia, Centaurea и др. Это—ближайший в морфологическом отно-
шении вид к Oxycarenus hyalipennis Costa, являющегося, как известно,
одним из самых серьезнейших вредителей хлопчатника в Египте и эква-
ториальной Африке.
В Кондаринском ущелье до сих пор констатировано, как указано
выше, 258 видов настоящих полужесткокрылых (Hemiptera—Hetero-
ptera), относящихся к 164 родам. Цифра эта не очень значительна и ни
в коей мере не говорящая об истинной картине фауны. Поэтому зоогео-
графический анализ должен ограничиться пока немногими фактами.
Фауна настоящих полужесткокрылых Кондаринского ущелья харак-
теризуется немногими считающимися в настоящее время эндемичными
среднеазиатскими родами, это: Oplistochilus Jak., Alloeoglypta Kir., Mi-
mula Jak., Mesopriassus Kir., Hissaritus Kir.
Для зоогеографической характеристики гемиптерофауны Кондары
необходимо отметить род Dalpada Am. Serv., несомненно реликтового
характера, впервые отмечаемый для Средней Азии. Многочисленные дру-
гие виды этого рода в целом принадлежат индо-малайской фауне, а также
Меланезийской подобласти Австралийской области, встречаются и на
188
A. H. Кириченко
Мадагаскаре. В отношении истории происхождения гемиптерофауны
Памиро-Алая к роду Dalpada примыкают и вышеназванные эндемиче-
ские роды Alloeoglypta— очень близкий к Halyomorpha Мауг (эфиопская
и ориентальная области до Японии) и Mesopriassus Kir. — очень близ-
кий и Priassus Stal (ю.-в. Азия). Из предыдущего изложения видно, что
все они связаны с древесной растительностью: первые с чинаром и вто-
рично — с тополем, последние два — с кленом.
К древним палеотропическим эле-
ментам принадлежит и Stenolemus
bogdanovi Osh. (рис. 7), открытый
А. П. Федченко, но этот вид принад-
лежит к обитателям жилищ чело-
века.
Прежде чем перейти к численно
превалирующей в фауне ущелья Кон-
дара группе средиземноморских
родов, нужно отметить некоторые,
являющиеся общими для Средней и
Передней Азии, а в некоторых слу-
чаях доходящие до Закавказья (до-
лина Аракса) и Монголии, это роды:
Ellipsocoris Мауг (-{-Армения, Си-
рия), Tshingisella Kir. (-{-Нахиче-
ванская АССР), Dichroblytrus Kir.
[-{-СССР (Забайкалье), Иран, Монго-
лия].
Средиземноморских родов боль-
шинство:1 *()dontotarsus Lap., *Lep-
rosoma Baer., Ancyrosoma Am. Serv.,
Muis. R., *Tholagmas Stal., *Derula
Muis. R., *Apodiphus Spin., *Mus-
tha Am. Serv., Codophila Muis. R.,
*Haploprocta Stal., Maccevethus
Dall., Apterola Muis. R., Microplax
Fieb., Metopoplax Fieb., *Piezoscelis
Fieb., Bledionotus Reut., Icus Fieb.,
Lamprod ema Fieb., *Lasiocoris Fieb.,
Neurocladus Fieb., Ischnopeza Fieb.,
Рис. 7 Stenolemus bogdanovi Osh. *Monosteira Costa, Ploiaria Scop.,
Hyoidea Reut., Malthacosoma Reut.
Очень большой процент родового состава гемиптерофауны составляют
роды, имеющие представителей как в средиземноморской фауне, так и
других зоогеографических областях — Эфиопской, Индомалайской, но
отсутствующих в Европейско-сибирской; к ним относятся: Camptopus
Germ., Liorhyssus Stal, Agraphopus Stal, Paromius Fieb., Lethaeus Dall.,
Scantius Stal, Stenolemus Sign., Holotrichius Burm., Ectomocoris Mayr,
Sphedanolestes Stal.
Общее количество эндемичных туранских или турано-иранских ви-
дов — около 85, т. е. Vs всех зарегистрированных в Кондаринском
ущелье видов настоящих полужесткокрылых.
1 Многие из этих родов, отмеченные *. представлены в копдарипской фауне
эндемичными туранскпми видами.
Настоящие полужесткокрылые и цикадовые
189
СПИСОК НАСТОЯЩИХ ПОЛУЖЕСТКОКРЫЛЫХ И ЦИКАДОВЫХ
УЩЕЛЬЯ КОНДАРА
! 1 •11 П оДГ j Настоящие полужесткокрылые Hemiptera — Heteroptera 1 Долина p. Варзоб до 1100 м Кондара до 1800 м Квак до 2400 м Руидашт до 2800 м Другие нахождения
2 3 4 5 7
1 2 3 4 2 3 4 5а 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 20а 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 Сем. Thyreocoridae Ochetostethus nanus H.—S. Geotomus latiusculus Horv. Sehirus morio L. Canthophorus hissaricus m. Сем. Pentatomidae Odontoscelis fuliginosa L. Od. dorsalis F ... Irochrotus maculiventris Germ. Odontotarsus angustatus Jak. Od. impictus Jak. var. ranifer m. Od. armiger Kir. . . Ellipsocoris tamerlani Kir. Psacasta argillacea Horv Eurygaster integriceps Put. Tshingisella bella Kir. Leprosoma tuberculatum Jak. Oplistochilus pallidus Jak. Ancyrosoma leucogrammes Gmel. Tholagmus nigricornis Reut. Graphosoma consimile Horv. Gr. italicum Mull. Derula longipennis Osh. Mustha baranovi m. . . Apodiphus integriceps Horv Dalpada pavlovskii m. Sciocoris agnatus Jak. Sc. hospes m. Sc. consobrinus m. Sc. sulcatus Fieb. Sc. helferi Fieb. Sc. capitatus Jak. Aelia acuminata L. A. alticola Kir. A. melanota Fieb. A. furcula Fieb. . Neottiglossa leporina H.-S. Stagonomus bipunctatus L. St. amoenus Brulle Peribalus peltatus Jak. P. nitidus Kir. . . Alloeoglypta (n. gen.) pretiosa m. Carpocoris fuscispinus Boh. i 1 1 1 i l i - + Н—1—1—1—1—1—1——1—г+++ Н—Н‘++ ++'1"+-Н—Ь И 1 Ь 1 1 1 + 1 н и н 1 ++ -1 I-++ + +++ + ++++++ + + i 1 । 1 1 । | + + + пер. Анзоб пер. Анзоб Зидды Гушары, Зидды Гушары пер. Анзоб Гушары Гушары, Зидды Гушары
36 37 38 Mimula maureri Jak. Codophila varia F. Dolycoris penicillatus Horv. + 1 + +-Н- +++
190
A. H. Кириченцо
Продолжение
а и I Настоящие полужесткокрылые | Hemiptera — Heteroptera Долина 05 Р- Варзоб до 1100 м Кондара до 1800 м и- Квак до 2400 м Руидашт ДО 2800 м Другие нахождения 7
I 4 1 5 6
39 I Eurydema ornata L. + + г
40 E. festiva L. + 4- Зидды
41 E. gussakovskii m. + + + Зидды
42 E. oleracea L. + + Зидды
43 Acrosternum breviceps Jak. +
44 Rhaphigaster brevispina Horv + +
45 Mesopriassus vetustus m. + +
Сем. Coreidae 1 1
1 Gonocerus patellatus Kir. + + I 1
2 Mesocerus marginatus L. + Гушары,
Зидды
3 Syromastes rhombeus L. var. qua-
dratus F + + i
4 Haploprocta pustulifera Stal 1 + 1
5 Coreus eversmanni Jak. + + Гушары
6 . C. heinsi Osh. . 4- + + Гушары
7 Spathocera tenuicornis Jak.
8 Pseudophloeus waltli H.-S.
9 Bathysolen nubilus Fall. +
10 Ceraleptus obtusus Brulle +
11 Coriomeris sp. +
12 C. vitticollis Reut. + +
13 C. apricus m. . Кутарм-акба
14 C. pallidus Reut. +
15 Stenocephala ferghanensis Horv. + I +
16 Camptopus lateralis Germ. + + +
17 Corizus hyoscyami L. + + + , + Гушары,
I Зидды
18 C. limbatus Reut. var. subsimilis
Horv. . + + пер. Анзоб
19 C. altivolens m. + Кутарм-акба
20 Liorhyssus hyalinus F. ; . + + 4- Зидды
21 Rhopalus parumpunctatus Schill. + + +
22 Rhopalus var. lepidus Fieb. . 4- + +
23 Brachycarenus tigrinus Schill + +
24 Stictopleurus abutilon Rossi + + + Гушары
25 St. unicolor Jak. . . +
26 Maccevethus lineola F + + + j
27 M. persicus Jak. . . . + + + 1
28 Agraphopus lethierryi Stal +
29 Chorosoma schilling! Schill.
Сем. Aradidae 1 1 1
1 Aradus hissaricus m. т 1
Сем. Neididae ।
1 Berytus tipularius L. et var. favosus 1 1 -л
F ieb. । +- + Гушары,
i Зидды
Настоящие полужесткокрылые и цикадовые
191
П р одолжение
е с Настоящие полужесткокрылые llomiptera— Heteroptera дина Варзоб 1100 м ндара 1800 м ак 2400 м 1 адашт 2800 м Другие нахождения
® • о 5 о 5 © sc « §
1 2 3 4 5 6 7
2 Neiilеэ montivagus Mey + +
3 Gampsocoris punctipes Germ. + + +
Сем. Myodochidae
1 Spilostethus equestris I,. 4- + + иер. Анзоб
2 Sp. rubriceps Horv. + + + Кутарм-акба
3 Melanocoryphus albomaculatus Goeze + Гушары,
За M. tristrami Dgl. Sc. । l 4- Зидды
4 M. superbus Pollich +
5 M. curtipennis m. . + +
6 Apterola lowni Saund. . . . +
7 Lygaeosoma reticulatum H.-S. + 4-
8 N у si us ericae obscura tus Horv + ' + Зидды, Ку тарм-акба
9 N. graminicola Koi. +
10 N. ochraceus m. . . . 1 _L_ Г
11 Cymus claviculus Fall. -p-
12 C. melanocephalus Fieb. + i
13 Ischnodemus sabuleti Fall. + 1
14 Geocoris alaiensis Rent + +
15 G. fedtshenkoi Reut | +
16 G. apertus m. . 1 + + пер. Анзоб^
17 Holcocranum diminution Horv | “Г
18 Heterogaster cathariae Geoffr. 1 + +
19 H. parens Jak. . . 4- + +
20 H. artemisiae Schill. . . 1
21 Camptotelus montandoni Kir. + + +
22 Leptodemus minutus Jak. +
23 Cyproplax avenae Lindb. +
24 Microplax interrupta Fieb. 1 + 1 _l~
25 Metopoplax origani Koi. 1 + +
26 Oxycarenus lacteus Kir. + +
27 Ox. col laris M. R. + + 4-
28 Bledionotus systellonotoides Reut. ! । । 1 T
29 ! Paromius gracilis Ramb. + +
30 Rhyparochromus colon Put. + +
31 | Rh. praetextatus II.-S. Rh. sabulicola Thoms. 1 +
32 i +
33 Dichro61ytrus conjunctus Kir. +
34 Piezoscelis coarctata Kir. +
35 Icus angularis Fieb.. . . + 1 (
36 Jschnocoris punctulatus Fieb. ! I + “Г
37 Lamprodema maurum F ...
38 Plinthisus (Plinthisomus) dimidia-
tus m. +
39 Pl. (s. str.) mixtus n.
40 Pl. (s. str ) ferruginous m. _L_ l - —
41 Pl. (s. str.) brevipennis Latr +
42 Lasiocoris albomaculatus Jak.
43 L. sp. -1-
44 Peritrechu's gracilicornis Put. — । J
192
A. H. Кириченко
Про дол ж е н ие
• Настоящие полужесткокрылые 1 ф 2 сб ” о ж * си Ло © i 3 © Другие
с- 1 Hemiptera — Heteroptera И ж о ы сб сб © tjoo К г“| © сб сб © tf 00 а 01 нахождения
I До. Р- ДО 5 о ~и_ й « ° Рч W
1 j 2 1 3 ’ 4 1 5 1 6 7
45 Microtoma atrata Goeze 4 1 +
46 Trapezonotus arenarius L. 4 ' ' Зидды
4 7 Calyptonotus rolandri L. 4 : 4- : 4- Зидды
48 Aphanus (Plinthurgus) celeripes Kir. -f- ' i 1
49 Aphanus (Bleteogenus) currax Horv. -f- 1 ।
50 Aph. (s. str.) albomaculatus Goeze
var. flavatus Horv. + J- Гушары,
пер. Анзоб
51 Beosus 4-punctatus Mull. -j
5 2 Neuroclad us brachiidens Duf. 4-
53 Ischnopeza hirticornis II.-S. + + ,
54 Emblethis denticollis Horv — ' Гушары
5 г Embl. verbasci F. 4-
56 Embl. angustus Mont. 4-
57 Lethaeus picipes H.-S. + + |
58 Eremocoris procerus m. -I— 1
59 Drymus sylvaticus F 4- -4
60 Scolopostethus sp. + +
61 Sc. pilosus Reut. + +
62 Sc. lethierryi Jak. + 4
1 Сем. Pyrrhocoridae I 1 1
1 Scantius aegyptius L. 1 4
2 Pyrrhocoris apterus L. + + — Гушары,
Зидды,
j Кутарм-акба
Сем. Tingitidae
1 Dictyonota tricornis Schrck. 4-
1а Galeatus scrophicus Saund. 4
16 Stephanitis pyri F ... +
2 Tingis (s. str.) auriculata Costa 4-
2а г Г. (s. str.) adventoria m. +
3 T. (s. str.) zhadiana m. . . : 4"
4 T. (Tropidochila) valida Put. var. 1
biseriata Horv. . . ' + +
5 1 \ (Tr.) pauperata Put. +
6 Catoplatus citrinus Horv. -t- +
7 Monanthia nassata Put. —
•8 M. fulvescens m —
9 Monostira discoidalis Jak. -f-
Сем. Macrocephalidae
1 Phymata subinermis Horv. 4- +
Сем. Reduviidae
1 Stenolemus bogdanovi Osh. 1 +
1а Gardena insperata m. 1 4-
2 Ploiaria domestica Scop. +
/7астоящие полужесткокрылые и цикадовые
193
Продолжение
| № п.п. j 1 1 Настоящие полужесткокрылые Hemiptera — Heteroptera Долина p. Варзоб до 1100 м Кондара до 1800 м Квак до 2400 м Руидашт до 2800 м Другие нахождения
1 2 3 4 5 6 7
3 Oncocephalus plumicornis Germ. + 1
4 Holotrichius moestus Reut. +
5 Reduvius testaceus H.-S. 4-
6 R. disciger Horv. 4-
7 Ectomocoris ululans Rossi 4- +
8 Rhinocoris monticola Osh. 4- 4- -г Зидды
9 Rh. iracundus Poda + 4-
10 Rh. (Oncauchenius) abramovi Osh. | + +
11 Rh. (One.) nigrothorax m. + + 4- Гушары
12 Sphedanolestes lipskii Kir. + 4-
13 Coran us sp. i + 4-
Сем. Nabidae
1 Nabis guttula F . . +
2 N. sanguineus Rossi +
.3 Reduviolus (Aptus) maracandicus |
Reut. + 4- 4-
4 R. (s. str.) ferns L. + + 4- Гушары
5 R. (s. str.) rugosus L. ... пер. Анзоб
6 R. (Aspilaspis) viridis Brulle +
Сем. Cimicidae
1 Cimex lectularius L. । +
•2 Cimex sp. +
Сем. Anthocoridae
1 Xylocoris (s. str.) obliquus Costa i +
-2 Anthocoris pilosus Jak. + 4- + Гушары,
Зидды,
Кутарм-акбы
•3 An th. poissoni m. . +
4 Anth. angularis Reut. 4"
5 Orius niger Wolff +
6 Or. minutus L. + +
Сем. Miridae
1 Hissaritus (n. gen.) dimorphus m. + 4- +
2 Phytocoris hissariensis m. + + 4-
3 Ph. tener m. +
4 Adelphocoris lincolatus Goeze + 4- + Гушары,
Зидды,
пер. Анзоб
5 Вrachycoleus scriptus F. . +
6 Dichrooscytus pseudosabinae Reut. +
7 Lygus pachycnemis Reut. 4- + Гушары,
пер. Анзоб
Зидды
В L. pratensis L + 4-
9 L. (Orthops) sanguinolentus Reut. + + 4- пер. Анзоб
13 Ущелье Кондара
194 И. Н Кириченко
Продолжение
Е j Настоящие полужесткокрылые о s lg а© РчО сб о о о н ! S о сб О Другие
Hemiptera — Heteroptera а ! «00 х а <*• «00 а нахождения
% 1 5 2- X И О 0- « । —
“I 2 ! з 4 5 ' 6 7
10 L. (Orth.) foreli Fieb. var. pilosulus Jak "Г
11 L. (Orth.) kalmi L. г 4- пер. Анзоб
12 L. (Agnocoris) rubicundus Fall. +
13 Poeciloscytus brevicornis Reut 1 i
14 । P unifasciatus F
15 P vulneratus Panz. _i_
16 P. dissimilis Reut 4"
17 Charagochilus gyllenhali Fall. + -L
18 Camptobrochis pilipes Reut.
19 1 C. punctulatus Fall. . . -t- + Зидды
20 | Stenodema calcaratuni Fall. — 1— —J— —1—
21 (' St. turanicum Reut. . -- +
22 1 Notostira caucasica Koi. subsp. pop-
1 piusi Reut. . . “ +
23 । Trigonotylus ruficornis Geoffr + Газни
25 Miris dolobratus L.
26 27 M. ferrugatus Fall. Macrolophus nubilus H.-S. -L
28 Dicyphus testaceus Reut. - 4-
29 D. pallidior m. _L 4~
30 D. orientalis Reut. . . . —f—
31 Pilophorus sinuaticollis Reul
32 P. sp. . .
33 Myrmecophyes nigripes Reul —-
34 Hyoidea notaticeps Reut. _L_ 1 । I
35 Scirtetellus sp. . i 1
36 Orthocephalus niger Reut 1 4- ! Зидды
37 Halticus pusillus H.-.. + Зидды
38 Oncotylus setulosus H.-S.
39 One. reuteri Reut. “Г
40 Megalocoleus sp. . 1 1
41 Amblytylus concolor Jak: -i- -j-
42 Byrsoptera fuscicornis Reut + i
43 Psallus hiemalis m.
44 Ps. nebulosus Reut. i
45 Ps. ignitus m. —
46 . ? Criocoris J- i
47 ' Plagiognathus arbustorum F
48 Pl. chrysanthemi Wolff i +
49 ! Pl. elegaris m. +
50 1 Pl. bipunctatus Reut.
51 PL-sp. . . —
52 Eumecotarsus breviceps Reut
53 i Stheharus perpulcher m.
54 1 St. concinnus m. .• . —j—
ba ; ' Tuponia elegans Jak. . 4"
55 Atomoscelis onustus Fieb. 4-
Настоящие полужесткокрылые и цикадовые
195
Продолжение
Е П 1 Настоящие полужесткокрылые Hemiptera — Heteroptera Долина i р. Варзоб до 1100 м Кондара । до 1800 м | Квак ' °’ до 2400 м Руидашт до 2800 м j Другие нахождения 7
3 6
1 Сем. Hebridae llebrus montanus Koi. i 1 i 1 i Зидды
1 1 1 1 2 3 4 1 2 1 2 1 2 3 4 5 6 Сем. Veliidae Yelia rivulorum F var. ventralis Put Сем. Gerridae Gerris costai H.-S. Сем. Leptopodidae Leptopus hispanus Ramb. । Сем. Saldidae Saldula tadzhika Kir. S. orthochila Fieb. S. melanoscela Fieb. S. pallipes F . . Chartoscirta dilutipennis Rent. Сем. Notonectidae Notonecta viridis Dele. var. rnedi- | terannea Hutsch. N. obliqua Gall. Сем. Corixidae Corixa affinis Leach 1 Sigara lateralis Leach Цикадовые — Homoptera—Au ch e nor- ryncha 1 Сем. Cixiidae । Malemia bosnica Horv. Oliarius concolor Fieb. 01. 5-costatus L. Hyalestes obsoleus Sign. H. nigricollis V Kusn. Cixius bifasciatus Schr. 1 Слисок цикадовых составлен Г А. — 4- i 1 + -г + Сидорским, -4— 1 4- | Кутарм-акба I 1 _1_ i + 1 1 1 i 1 обработавшим сборы
В. В .. Гусе айовского.
13*
196
A. H. Кириченко
Н р о д о ляс е ние
й и % Настоящие полужесткокрылые Hemiptera — Heteroptera Долина р. Варзоб до 1100 м Кондара до 1800 м Квак до 2400 м g s ! Другие и О Л о 1 нахождения Он
1 2 3 4 1 5,61 7
Сем. Fulgoridae i 1
7 Chanithus avocetta Osh. . . + 1 4- ।
8 Nymphorgerius korolkovi Osh. . . 4- 4- 1 > ! •-<—
9 Phyllorgerius kusnezovi Sid. in litt. 4-
10 Orgerius ellipticus Osh. +
Сем. Issidae 1
11 Caliscelis gissarica V. Kusn. 4- + +
12 Aphelonema 4-virgata Fieb. . . +
13 Hystcropterum placophorum Horv. 4- i +
14 H. pictifrons Mel. + 4- 1 1 “Г
15 H. montanum Fieb. 4- i + 1
Сем. Tettigometridae 1 1
16 Tettigometra barani Sign. 4- + + 1
17 T. varia Fieb. . 4- F
18 T. obliqua Panz. v. platytaenia
Fieb + 4- 4-
T. v. bimaculata Fieb. 4- -4
T. v. tritaenia Fieb. + 1
T. v. thoracica Rey. . + 4- I
T. v. rosea Sid. in litt. +
T. v. nigrodorsalis Sid. in litt. + 1 4-
Сем. Delphacidae 1 1
19 Asiraca clavicornis F 4- [
20 Kelisia guttula Germ. 4- 4-
21 Delphax crassicornis F. 4-
22 Liburnia marginata F. . +
23 L. nigricornis Sid. in litt. +
24 L. albicollis Motsch. +
25 L. nigra Sid. in litt. +
26 L. aubei Perris 4-
27 L. brunneipennis Sid. in litt. +
28 Dicranotropis beckeri Fieb. + 1 I
Cicadidae 1 1 1
29 Paharia zevara V. Kusn. 4- I 4-
30 Cicadatra querula Pall. 4-
31 Melampsalta montana Scop. 4- + i +
Cercopidae
32 Lepyronia coleoptrata L. . 4- 1 4-
33 Neophilaenus notatus M. R. -1-
34 N. haupti Zachw. 4- + 1 +
Настоящие полужесткокрылые и цикадовые
197
Продолжение
•Ц п Настоящие полужесткокрылые Hemiptera — Heteroptera i 1 г долина p. Варзоб до 1100 м Кондара до 1800 м Квак до 2400 м Руидашт до 2800 м Другие нахождения
1 2 3 4 : 5 6 7
lassidae
35 Eupelix cuspidata F . . +
36 Euacanthus asiaticus Osh. — —1-
37 Batrachomorphus irroratus Lewis +
38 Agallia sinuata M. R. +
39 Idiocerus nobilis Fieb. +
40 Id. albicans Kirschb. . +
41 Macropsis fuscula Fieb. -1-
42 M. virescens F +
43 Platymetopius guttatus Fieb. +
44 Pl. breviceps Lindb. +
45 Balclutha chloris Horv. i 1 +
46 Phlepsius intricatus H.-S. ~г
47 Phi. pulcher Mel. . . 4- 1 -т-
48 Phi. kondarus Sid. in litt. + 1 +
49 Doratura salina Horv. +
50 Deltocephalus sp. (sect, striatus L.) + -f- 1 + 1 +
51 D. hypochlorus Fieb. . . +
52 Goniagnathus brevis H.-S. +
53 Euscelis interstitialis Germ. + 1 1
54 E. variegatus Kbm. + 1 +
55 E. procerus H.-S. + +
56 E. ignoscus Mel. + I +
57 Thamnotettix fenestratus H.-S. +
var. guttulatus Kbm. + +
var. hyalinus Sid. in litt. + +
58 Th. vittiventris Leth + +
59 Stenometopiellus sigillatus Haupt + + -1-
60 Opsius heydeni Leth. +
61 Ops. pallasi Leth. ; +
62 Ops. jucundus Leth. +
63 Chlorita flavescens F + 1
64 Chi. nervosa Fieb. . . + I
65 Empoasca smaragdula Fall. + 1
66 Eupteryx collina Flor +
67 Typhlocyba nitidula F. +
68 T. rosae L +
69 Zyginidia parvula Boh. i +
70 Z. jaxartensis Osh. . . . -j-
71 Erythroneura asiatica V. Kusn. + i
Psyllidae
72 Diaphorina propinqua F. Low + + 1
73 Psylla glycyrrhizae Beck. 1 1 -1- i 1
Л. Л. МИЩЕНКО
К ФАУНЕ УХОВЕРТОК, ТАРАКАНОВ, БОГОМОЛОВ, ПАЛОЧНИКОВ
И ПРЯМОКРЫЛЫХ ЮЖНОГО СКЛОНА ГИССАРСКОГО ХРЕБТА
Вследствие того, что изучение фауны южного склона Гиссарского
хребта в пределах Таджикистана началось всего лет 15 тому назад, ко-
нечно, еще рано говорить о ее полном видовом составе, но все же имею-
щиеся данные позволяют уже сейчас отметить ряд интересных ее осо-
бенностей.
В настоящее время из этого горного района Таджикистана известны:
2 вида уховерток, 6 тараканов, 5 богомолов, 1 палочник и 39 прыгающих
прямокрылых (из них 10 кузнечиков, 3 сверчка и 26 саранчевых).
Фауну вышеуказанных отрядов южного склона Гиссарского хребта
составляют преимущественно среднеазиатские виды, незначительная часть
которых только заходит пз Средней Азии в пограничные с ней страны;
общее число таких форм достигает 32, т. е. 60.3% всего видового состава
данного района, пз которых 13 видов являются пока его эндемиками.
15 видов, или 28.3% всего видового состава, — это виды, широко или
довольно широко распространенные в Средиземноморье; 2 пз них обитают
также в тропической Африке. 3 вида, или 5.7%, распространены, глав-
ным образом, в северных районах Палеарктики и, наконец, 3 вида, или
5.7%, заходят в этот район Таджикистана из Индомалайской области.
Если же проанализировать фауну южного склона Гиссарского хребта
с точки зрения происхождения родов, образующих ее состав, то на осно-
вании их географического распространения и, главным образом, на ос-
новании их родственных связей приходится сделать вывод, что эта фауна
состоит из следующих элементов.
а) древних, невидимому, домиоценовых среднеазиатских автохтонных
элементов, представленных родами: таракановые — Tartaroblatta B.-Bi-
enko; кузнечиковые — Glyphonotus Redt.; саранчевые Bienkoa Mistsh.,
Gonophyma Zub., Gomphomastax Br.-W. и Clinomastax B.-Bienko;
б) средиземноморских элементов: таракановые — Pbyllodromica Fieb.
и Polyphaga Brulle; богомоловые — Armene St., Iris Sauss и Rivetina
Berl. et Chop.: кузнечиковые — Phasgonura Steph., Ceraeocercus Uv.,
Paradrymadusa Herm., Tadzhikia Mistschenko (in litt.), Semenoviana Znr.,
Platycleis Fieb. и Eumetrioptera Mir.; саранчевые— Diciostaurns Fieb.,
Mizonocara Uv., Ramburiella I. Bol., Pyrgodera F.-W., Oedipoda Latr..
Sphingonotus Fieb. и Calliptamus Serv.;1
1 В число средиземноморских элементов я включаю эндемичные среднеазиатские
роды Tadzhikia Mistsh. (in litt.), Semenoviana Znr. и Eumetrioptera Mir., имеющие
ясное средиземноморское происхождение, вследствие своих тесных родственных
связей.
К фауне уховерток, тараканов, богомолов и прямокрылых
199
в) тропических элементов: таракановые роды — Ectobius Steph, и
Blatta L., богомоловые — Hierodula Burm. и Empusa Ill; приведенье-
вые — Gratidia St.; сверчковые — Gryllus L. и Oecanthus Serv.; саранче-
вые Acrida L., Locusta L., Oedaleus Fieb., Pyrgomorpha Serv., Oxya
Serv. и Paratettix I. BoL, из которых только Gryllus L., Oecanthus Serv.
и Paratettix I. Bol. широко распространены по всему тропическому поясу
земного шара, а остальные роды — эфиопского, индо-малайского или же
смешанного происхождения;
г) северных европейско-сибирских (ангарских) элементов; кожисто-
крылые— Oreasiobia Sem. и Anechura Scudd.; саранчевые — Chorthip-
pus Fieb.
Анализ же вертикального распространения видов, живущих на юж-
ном склоне Гиссарского хребта, также указывает на значительную неод-
нородность состава его фауны. Так, с одной стороны, в его состав входят
виды, широко распространенные в равнинах Средней Азии или даже
почти всей Палеарктики и только заходящие в горы, а с другой — спе-
цифически горные виды. Большинство равнинных видов заходит в горы
до 1100—2000 м над ур. м.: из таракановых представители рода Ectobius
Steph., черный таракан (Blatta orientalis L.) и черепашка Соссюра
[Polyphaga saussurei (Dohrn)J; из богомоловых — Hierodula tenuidentata
Sauss. и Empusa pennicornis (Pall.); из кузнечиковых — зеленый кузне-
чик [Phasgonura viridissima (L.)], Semenoviana tamerlana (Sauss.) и Gly-
phonotus thoracicus semenovi Mir.; из сверчковых — двупятнистый сверчок
(Gryllus bimaculatus De Geer), Tartarogryllus tartarus obscurior (Uv.)
и трубачик туранский (Oecanthus turanicus Uv.); из саранчевых — степ-
ная акрида [Acrida oxycephala (Pall.)], крестовичка пустынная [Docios-
tarus (s. str.) tartarus Uv.], туркменка [Ramburiella turcomana (F.-W.)],
гребневка [Pyrgodera armata (F.-W.)], полосатая кобылка [Oedaleus
decorus (Germ.)], полосатая пустынница (Sphingonotus rubescens fasciatus
Mistsh.), рисовая кобылка [Oxya fuscovittata (Marsch.)], полубогарный
прус (Calliptamus barbarus cephalotes F.-W.), обыкновенная остроголовка
[Pyrgomorpha conica (Ol.)] и южный прыгунчик [Paratettix meridionalis
(Ramb.)]; некоторые же виды, как, например, из кожистокрылых —
азиатская уховертка [Anechura (s. str.) asiatica asiatica Sem.], из при-
виденьевых — двубугорчатый палочник (Gratidia bituberculata Redt.),
из кузнечиковых — длиннохвостый кузнечик [Phasgonura caudata cau-
data (Charp.)] и туранский скачок (Platycleis intermedia turanica Znr.)
и из саранчевых — представители рода Chorthippus Fieb., широко рас-
пространены также и в горах, достигая 3000 и даже 3600 м (высота здесь
и дальше в метрах над уровнем моря). Главная масса горных форм приуро-
чена к зоне субальпийских лугов (2200—3600 м) и верхнего лесного пояса
(2000 м), и, как правило, ниже 1000 м не спускаются: из кожистокрылых
это уховертка Федченко [Oreasiobia fedtshenkoi fedtshenkoi (Sauss.)];
из таракановых — Phyllodromica sp. и Tartaroblatta tartara (Sauss.);
из богомоловых — Armene robusta Mistshenko (in litt.), Iris insolita Mis-
tshenko (in litt.) и Rivetina monticola Mistshenko (in litt.); из кузнечи-
ковых — Ceraeocercus fuscipennis hindukushanus Rme., Paradrymadusa
hissarica Mistshenko (in litt.), Tadzhikia pavlovskii Mistshenko (in litt.),
согдийский скачок [Platycleis sogdiana Mistshenko (in litt.)] и Eumet-
rioptera crassa Mistsh.; из саранчевых — Chorthippus curtus Mistshenko
(in litt.), Mizonocara kuznetzovae Um., кобылка Федченко (Oedipoda fed-
tshenkoi fedtshenkoi Sauss.), пустынницы (Sphingonotus pilosus Sauss.
и S. nebulosus discolor Uv.), украшенная кобылка [Bienkoa fedtshenkoi
200
Л. Л. Мищенко
ornata (Rme.).], представители рода Conophyma Zub., Gomphomastax
gussakovskii Mistsh., и только обыкновенный прус (Calliptamus italicus
reductus Rme.) и Clinomastax ninae (Mistsh.) заходят ниже этой границы,
выходя местами даже на возвышенную равнину (600 м).
По характеру своего обитания виды, живущие на южном склоне Гис-
сарского хребта, могут быть разбиты на несколько экологических групп.
Так, гигрофилами являются тараканы [Ectobius tadzhicus B.-Bienko
и Е. pavlovskii B.-Bienko (?)] и рисовая кобылка [Охуа fuscovittata
(Marsch.)], тесно связанные с растительностью по берегам различных
водоемов. К мезофилам принадлежат: из кожистокрылых — уховерткгг
Федченко [Oreasiobia fedshenkoi fedtshenkoi (Sauss.)], азиатская уховертка
[Anechura (s. str.) asiatica asiatica Sem.]; из таракановых — Phyllo-
dromica sp., Tartaroblatta tartara (Sauss.) и черный таракан (Blatta orien-
talis L.); из богомоловых — Hierodula tenuidentata Sauss., Iris insolita
Mistshenko (in litt.) и Empusa pennicornis (Pall.); из кузнечиковых —
представители рода Phasgonura Steph., Ceraeocercus fuscipennis hinduku-
shanus Rme., Paradrymadusa hissarica Mistshenko (in litt.), согдийский
скачок [Platycleis sogdiana Mistshenko (in litt.)] и Glyphonotus thoracicus
semenovi Mir.; из сверчковых — двупятнистый сверчок (Gryllus bima-
culatus De Geer) и туранский трубачик (Oecanthus turanicus Uv.); из са-
ранчевых — представители рода Chorthippus Fieb., украшенная кобылка
[Bienkoa fedtshenkoi ornata (Rme.)], конофимы зеленоватая и вооружен-
ная [Conophyma prasinum Mistshenko (in litt.) и C. armatum Mistsh.],
Gomphomastax gussakovskii Mistsh. и Clinomostax ninae (Mistsh.). Боль-
шинство этих видов приурочено к различного рода луговым стациям,
поросшим довольно густой и высокой растительностью, а также к зарос-
лям разнообразных кустарников и даже деревьев. Группу ксерофилов
образуют: из таракановых — черепашка Соссюра [Polyphaga saussurei
(Dohrn)J; из богомоловых — Armene robusta Mistshenko (in litt.) и Rive-
tina monticola Mistshenko (in litt.); из кузнечиковых — Tadzhikia pav-
lovskii Mistshenko (in litt.), Eumetrioptera crassa Mistsh.; из сверчковых—
Tartarogryllus tartarus obscurior (Uv.); из саранчевых — пустынная кре-
стовичка [Dociostaurus (s. str.) tartarus Uv.], Mizonocara kuznetzovae Um.,
гребневка [Pyrgodera armata (F.-W.)], полосатая кобылка [Oedaleus
decorus (Germ.)], кобылка Федченко (Oedipoda fedtshenkoi fedtshenkoi
Sauss.), представители рода Sphingonotus Fieb., конофимы Олсуфьева
и шероховатая (Conophyma olsufjevi Mistsh. и G. dumale Mistsh.),
полубогарный прус (Calliptamus barbarus cephalotes F.-W.). Боль-
шая часть ксерофилов обитает среди пустынных или полупустынных
открытых стаций, с редкой растительностью. Наконец, группу эвритоп-
ных видов, обладающих большой экологической пластичностью и обита-
ющих, вследствие этого, в самых разнообразных стациях, составляют:
из привиденьевых — двубугорчатый палочник (Gratidia bituberculata
Redt.); из кузнечиковых — Semenoviana tamerlana (Sauss.) и скачок
туранский (Platycleis intermedia turanica Znr.); из саранчевых — степ-
ная акрида [Acrida oxycephala (Pall.)], двупятнистый конек (Chorthip-
pus biguttulus meridionalis Mistsh.), туркменка [Ramburiella turcomana
(F.-W.)], обыкновенный прус (Calliptamus italicus reductus Rme.), обык-
новенная остроголовка [Pyrgomorpha conica (01.)] и южный прыгунчик
[Paratettix meridionalis (Ramb.)].
Почти все виды, составляющие фауну южного склона Гиссарскогс
хребта, зимуют в стадии яйца и имеют одну генерацию в году. В начале
или в середине весны происходит отрождение личинок, причем у саран-
К фауне уховерток, тараканов, богомолов и прямокрылых
201
чевых появление их происходит раньше, чем, например, у кузнечиков.
Примерно с апреля и до середины июня большинство видов проходит
постэмбриональное развитие, которое у саранчевых длится 40—45 дней,
у кузнечиков — около 80, у тараканов и богомолов — около 100 дней.
У некоторых видов, как, например, у представителей рода Chorthippus
Fieb., отрождение происходит в марте, первые взрослые особи по-
являются уже в мае, а иногда даже и в конце апреля. В зависимости от
высоты обитания сроки развития у одного и того же вида не одинаковы,
даты отрождения запаздывают и само постэмбриональное развитие, как
правило, идет несколько медленнее. Очень редкие виды, например, дву-
пятнистый сверчок (Gryllus bimaculatus De Geer) и, повидимому, дву-
бугорчатый палочник (Gratidia bituberculata Redt.), зимуют в личиночной
стадии, а у черепашки Соссюра [Polyphaga saussurei (Dohrn)] зимуют ли-
чинки и взрослые особи.
В настоящее время в качестве вредителей сельскохозяйственных ра-
стений на южном склоне Гиссарского хребта известны только 5 видов:
длиннохвостый кузнечик [Phasgonura caudata caudata (Charp.)], туран-
ский скачок (Platycleis intermedia turanica Znr.), украшенная кобылка
[Bienkoa fedtshenkoi ornata (Rme.)], обыкновенный прус (Galliptamus
italicus reductus Rme.), довольно сильно вредящие богарной люцерне,
и Gomphomastax gussakovskii Mistsh., значительно повреждающий бо-
гарную люцерну, многие плодовые деревья (урюк, персик, яблоню и др.),
культурный и дикий миндаль.
Повидимому, вышеуказанным перечнем не ограничивается число вред-
ных видов южного склона Гиссарского хребта, так как среди живущих
на нем видов многие сильно вредят полевым, огородным, бахчевым, тех-
ническим и лекарственным культурам, а также садоводству, виногра-
дарству, цветоводству, запасам, поделкам как на территории Средней
Азии, так и в других областях Палеарктики: из кожистокрылых это ухо-
вертка Федченко [Oreasiobia fedtshenkoi fedtshenkoi (Sauss.)] и азиатская
уховертка [Anechura (s. str.) asiatica asiatica Sem.],; из таракановых —
черный таракан (Blatta orientalis L.) и черепашка Соссюра [Polyphaga
saussurei (Dohrn)]; из кузнечиковых — зеленый кузнечик [Phasgonura
viridissima (L.)] и Semenoviana tamerlana (Sauss.); из сверчковых —
двупятнистый сверчок (Gryllus bimaculatus De Geer) и туранский труба-
чик (Oecanthus turanicus Uv.); из саранчевых — степная акрида [Acrida
oxycephala (Pall.)], коньки — обыкновенный и луговой [Chorthippus
biguttulus meridionalis Mistsh. и Ch. dorsatus dichrous (Ev.)], туран-
ская крестовичка [Dociostaurus (s. str.) tartarus Uv.], туркменка
[Ramburiella turcomana (F.-W.)], гребневка [Pyrgodera armata (F.-W.)],
полосатая кобылка [Oedaleus decorus (Germ.)], рисовая кобылка
[Oxya fuscovittata (Marsch.)], полубогарный прус (Galliptamus bar-
barus cephalotes F.-W.) и обыкновенная остроголовка [Pyrgomorpha
conica (01.)].
Черный таракан (Blatta orientalis L.) и черепашка Соссюра [Pylophaga
saussurei (Dohrn)], водясь в жилых домах и питаясь различными пищевыми
запасами, могут иметь эпидемиологическое значение для человека, раз-
нося на наружных частях тела и при помощи фекалий различные пато-
генные бактерии и, в частности, дизентерийные палочки, заражая ими
продукты питания.
Кроме того, черный таракан иногда нападает на спящих людей
и объедает эпидермис, щек, век, губ, шеи, пальцев, локтей и других
частей человеческого тела, вызывая заболевание.
202
Л. Л. Мищенко
СПИСОК ВИДОВ И ИХ РАСПРОСТРАНЕНИЕ ПО ЮЖНОМУ СКЛОНУ
ГИССАРСКОГО ХРЕБТА
DERMATOPTERA (КОЖИСТОКРЫЛЫЕ, ИЛИ УХОВЕРТКИ)
1. Oreasiobia fedtshenkoi fedtshenkoi (Sauss.) (уховертка Федченко) —
Кондара, Ходжа-обп-гарм, Квак, гора Мазор, Руидашт, Анзоб.
2. Anechura (s. str.) asiatica asiatica Sem. (уховертка азиатская) —
Руидашт.
BLATTODEA (ТАРАКАНОВЫЕ)
3. Ectobius tadzhicus B.-Bienko — Кондара, Квак, Руидашт.
4. Ectobius pavlovskii B.-Bienko — Кондара, Квак.
*5. Phyllodromica sp.1 — Кондара.
6. Tartaroblatta tartara (Sauss.) — Гушары, Квак. Руидашт.
7. Blatta orientals L. (черный таракан) — Кондара.
8. Polyphaga saussurei (Dohrn) (черепашка Соссюра) — Кондара.
MANTODEA (БОГОМОЛОВЫЕ)
*9. Armene robusta Mistshenko (in litt.) — Кондара, гора Мазор,
Руидашт.
10. Hierodula tenuidentata Sauss. — Кондара.
*11. Iris insolita Mistshenko (in litt.) — Кондара, Ходжа-оби-гарм,
Квак, гора Мазор.
*12. Rivetina monticola Mistshenko (in litt.) — Кондара.
13. Empusa pennicornis (Pall.) — Кондара.
PHASMATODEA (ПРИВИДЕНЬЕВЫЕ ИЛИ ПАЛОЧНИКИ)
14. Gratidia bituberculata Redt. (двубугорчатый палочник) — Кон-
дара, Квак, гора Мазор, Руидашт.?
SALTATORIA-ORTHOPTERA (ПРЫГАЮЩИЕ ПРЯМОКРЫЛЫЕ)
Tettigoniodena (кузнечиковые)
15. Phasgonura caudata caudata (Charp.) (длиннохвостый кузне-
чик) — Кондара, Квак, Руидашт.
16. Phasgonura viridissima (L.) (зеленый кузнечик) — Кондара.
17. Ceraeocercus fuscipennis hindukushanus Rme. — Кондара, гора
Мазор.
*18. Paradrymadusa hissarica Mistshenko (in litt.) — Ходжа-оби-гарм,
Квак, гора Мазор.
*19. Tadzhikia pavlovskii Mistshenko (in litt.) — Кондара.
20. Semenoviana tamerlana (Sauss.) — Кондара, Ходжа-оби-гарм.
21. Platycleis intermedia turanica Znr. (туранский скачок) — Кон-
дара.
*22. Platycleis sogdiana Mistshenko (in litt.) (согдийский скачок) —
Кондара, Гушары, Квак, Руидашт.
1 Звездочкой обозначены эндемики южного склона Гиссарского хребта.
К фауне уховерток, тараканов, богомолов и прямокрылых
203
*23. Eumetrioptera crassa Mistsh. — ущелье в 7 км на северо-запад
от Сталинабада.
24. Glyphonotus thoracicus semenovi Mir. — Кондара, Квак.
Gryllodea (сверчковые)
25. Gryllus bimaculatus De Geer (двупятнистый сверчок) — Кондара.
26. Tartarogryllus tartarus obscurior (Uv.) — Кондара, Квак.
27. Oecanthus turanicus Uv. (туранский трубачик) — Кондара, Квак,
гора Мазор.
Acridodea (саранчевые)
28. Acrida oxycephala (Pall.) (степная акрида) — Кондара, Руи-
дашт.
29. Chorthippus apricarius (L.) (бурый конек) — Кондара, Зидды,
Руидашт.
30. ChorthippusbiguttulusmeridionalisMistsh. (двупятнистыйконек)—
Кондара, Гушары, Ходжа-оби-гарм, Квак, гора Мазор, Руидашт, Анзоб.
31. Chorthippus curtus Mistshenko (in litt.) — Руидашт.
32. Chorthippus dorsatus dichrous (Ev.) (луговой конек) — Кондара,
Ходжа-оби-гарм, Зидды.
33. Dociostaurus (s. str.) tartarus Uv. (пустынная крестовичка) —
Кондара, Квак.
34. Mizonocara kuznetzovae Um. — Кондара, Квак.
35. Ramburiella turcomana (F.-W.) (туркменка) — Кондара.
36. Pyrgodera armata (F.-W.) (гребневка) — Кондара, Квак.
37. Oedaleus decorus (Germ.) (полосатая кобылка) — Кондара,
Ходжа-оби-гарм.
38. Oedipoda fedtshenkoi fedtshenkoi Sauss. (кобылка Федченко) —
Кондара, Ходжа-оби-гарм, Квак, гора Мазор, Руидашт.
39. Sphingonotus rubescens fasciatus Mistsh. (пустынница полоса-
тая) — Кондара, Квак.
40. Sphingonotus pilosus Sauss. (пустынница волосатая) — Кондара.
41. Sphingonotus nebulosus discolor Uv. (пустынница разноцвет-
ная) — Кондара, Квак.
42. Охуа fuscovittata (Marsch.) (рисовая кобылка) -- Кондара,
Ходжа-оби-гарм.
43. Bienkoa fedtshenkoi ornata (Rme.) (украшенная кобылка) —
Кондара, Ходжа-оби-гарм, Квак, Руидашт.
*44. Conophyma prasinum Mistshenko (in litt.) (конофима зеленова-
тая) — перевал Мира.
*45. Conophyma armatum Mistsh. (конофима вооруженная) —гора
Мазор.
46. Conophyma olsufjevi Mistsh. (конофима Олсуфьева) —ущелье
в 7 км на северо-запад от Сталинабада.
*47. Conophyma dumale Mistsh. (конофима шероховатая) — Руидашт.
48. Calliptamus italicus reductus Rme. (обыкновенный прус) —
Кондара, Ходжа-оби-гарм, Квак, гора Мазор, Зидды, Анзоб.
49. Calliptamus barbarus cephalotes F.-W. (полубогарный прус)—
Кондара.
204
Л. Л Мищенко
50. Pyrgomorpha conica (01.) (обыкновенная остроголовка)—Кон-
дара.
*51. Gomphomastax gussakovskii Mistsh. — Кондара, Гушары, Хо-
джа-оби-гарм, Квак, гора Мазор, Руидашт.
*52. Clinomastax ninae (Mistsh.) — ущелье в 7 км на северо-запад
от Сталинабада, Кондара.
53. Paratettix meridionalis (Ramb.) (южный прыгунчик) — Кон-
дара.
Видимо, вышеприведенным списком не исчерпывается фауна южного
склона Гиссарского хребта. С одной стороны, можно ожидать нахожде-
ния на нем новых, еще неизвестных науке форм, а с другой — наличие
в его фауне видов, живущих в пограничных с ним районах. Так, грани-
чащая с ним Гиссарская долина имеет значительно более богатую фауну.
В ней, кроме перечисленных выше равнинных видов, живущих как в до-
лине, так и на южном склоне Гиссарского хребта, обитают еще следую-
щие формы, часть которых, вполне возможно, живет также и на южном
склоне Гиссарского хребта и при дальнейших исследованиях может быть
обнаружена в его фауне: из кожистокрылых это прибрежная уховертка
[Labidura riparia (Pall.)] и малая уховертка [Labia minor (L.)]; из тара-
кановых— обыкновенный таракан [Blattella germanica (L.)]; из бого-
моловых — Armene pusilia (Ev.), обыкновенный богомол [Mantis reli-
giosa (L.)] и Rivetina crassa Mistsh.; из кузнечиковых — Anerota falcata
(Poda), конусоголовы [Conocephalus (Xiphidion) buxtoni Chop, и С. (X.)
fuscus (Fabr.)], Homorocorhyphus nitidulus (Scop.), Tessellana vittata
(Charp.), белолобый кузнечик [Tettigonia albifrons (Fabr.)] и Medecticus
assimilis (Fieb.); из сверчковых — однотипная медведка (Gryllotalpa
unispina Sauss.), Pteronemobius heydeni concolor (Walk.), сверчки — бор-
досский, степной и бухарский [Acheta burdigalensis (Latr.), A. deserta
(Pall.) и A. bucharica (B.-Bienko)] и Turanogryllus lateralis (Fieb.); из
триперстовых — триперст Савиньи [Tridactylus savignyi (Guerin)]; из
саранчевых — носатая кобылка [Truxalis nasuta (L.)], стреловидная ко-
былка [Gonista sagitta (Uv.)], кобылки согдийская и короткокрылая
(Duroniella sogdiana Mistsh. и D. brachyptera Um.), коньки туранский
и острокрылый (Chorthippus turanicus Tarb. и Ch. angulatus Tarb.), марокк-
ская саранча [Dociostaurus (s. str.) maroccanus (Thunb.)], крестовичка
Попова (Notostaurus popovi Mir.), Mizonocara robusta Mistsh. (обитает
только на каменистых склонах холмов в южной части Гиссарской до-
лины), Parapleurus alliaceus turanicus (Tarb.), азиатская саранча [Lo-
custa migratoria migratoria (L.)], Oedaleus senegalensis (Кг.), летуньи
обыкновенная и Савиньи [Aiolopus thalassinus thalassinus (Fabr.) и
A. savignyi (Kr.)], Hilethera turanica Uv., краснокрылая кобылка [Oedi-
poda miniata (Pall.)], Acrotylus insubricus insubricus (Scop.), пустынница
Савиньи (Sphingonotus savignyi Sauss.), египетская кобылка [Anacridium
aegyptium aegyptium (L.)], туранский прус (Galliptamus turanicus Tarb.),
ложный прусик [Metromerus coelesyriensis violaceipes Mistshenko (in litt.)],
бахчевая кобылка [Thisoicetrinus pterostichus (F.-W.)] и пустынный пры-
гунчик [Tetrix tartara (I. Bol.)].
ЛИТЕРАТУРА
(Б ей-Бпеико 'Г.) В e у - В i e n k о G. 1933. Records and descriptions of some
Orthoptera from U. S. S. R. Bol. Soc. Esp. Hist. Nat., XXXIII : 317—341.
(Б e й - Б it e н к о Г.) 1935. Descriptions of six new species of palaearctic Blattodea.
Konowia, XIV, 2: 117—134.
К фауне уховерток, тараканов, богомолов и прямокрылых 205
Б ей - Б и е нко Г. Я. 1936а. К фауне Blattodca Таджикистана. Тр. Тадж. базы
Акад. Наук СССР, 6 : 225—240.
Бей-Биенко Г. Я. 19366. Насекомые кожистокрылые. Фауна СССР, нов. сер.,
5, 240 стр.
Бей-Бпенко Г Я. 1948. Прямокрылые — Orthoptera и кожистокрылые —
Dermaptera. Животный мир СССР, II 270—291.
Бей-Биенко Г Я. 1949. Ревизия саранчевых подсем. Gomphomastacinae
(Orthoptera, Eumastacidae). Докл. Акад. Наук СССР, нов. сер., LXIV, 5
731—734.
Бей-Бпенко Г Я. 1950. Насекомые таракановые. Фауна СССР, нов. сер., 40,
343 стр.
Карпова А. И. 1945. Вредители богарной люцерны в условиях южного склона
Гиссарского хребта. Энтом. обозр., XXVIII, 1—2 1—7.
Мирам Э. Ф. 1935а. Blattodea (таракановые), Mantodea (богомоловые), Phasmodea
(привиденьевые) и Orthoptera (прямокрылые) Таджикистана. Тр. Тадж. базы
Акад. Наук СССР, 5 219—236.
(Мирам Э. Ф.) Miram Е. F. 19356. Beitrage zur naheren Kenntnis der Arten
der Gattung Glyphonotus Redt. (Tettigoniidae, Orthoptera). Konowia, XIV,
. 3 256—267.
Мирам Э. Ф. 1949. Богомоловые (Mantodea), привиденьевые (Phasmatodea) и пры-
гающие прямокрылые [Saltatoria (Orthoptera s. str.)] Таджикской ССР, по сбо-
рам Паразитологической экспедиции Академии Наук СССР 1934 г. Тр. Зоол.
инет. Акад. Наук СССР, МП : 714—719.
(Мищенко JI.) Mistshenko L. 1936—1937. Revision of palaearctic species
of the genus Sphingonotus Fieber (Orth., Acrid.), Eos, Xll : 65—282.
.(Мищенко Л.) 1937. Two new species of Acridodea (Orth.) from Tadjik S. S. R.
(form. E. Bokhara). Konowia, XVI, 2 130—136.
Мищенко Л. Л. 1947. Род Mizonocara Uv. [Saltatoria (Orthoptera sens, str.), Acri-
dodea]. Энтом. обозр. XXIX, 1—2 : 62—71.
Мищенко Л. Л. 1949а. Отряд Dermatoptera. Кожистокрылые, или уховертки.
Отряд Saltatoria (Orthoptera sens. str.). Прыгающие прямокрылые. Отряд
Blattodea. Таракановые. Вредные животные Средней Азии 147—169, 317,
345—347.
Мищенко Л. Л. 19496. Кожистокрылые (Dermatoptera), таракановые (Blattodea),
богомоловые (Mantodea), приведеньевые (Phasmatodea) и прыгающие прямо-
крылые [Saltatoria (Orthoptera s. str.)] Гпссарской долины (Таджикская ССР).
Тр. Зоол. Инет. Акад. Наук СССР, МП 735—749.
Мищенко Л. 1949в. Род Eumetrioptera Miram [Saltatoria (Orthoptera s. str.)
Tettigoniodea]. Энтом. обозр., XXX, 3—4 326—333.
Мищенко Л. Л. 1950а. Новые данные по среднеазиатской фауне вредных саран-
чевых (Saltatoria-Orthoptera, Acridodea). Докл. Акад. Наук СССР, нов. сер.,
LXXI, 4 : 789—792.
Мищенко Л. Л. 19506. Новые виды саранчевых (Saltatoria-Orthoptera, Acrididae)
из Средней Азии и Кашмира. Докл. Акад. Наук СССР, нов. сер., LXX11, 1 :
213—215.
(Тарбинский С. П.) Tarbinsky S. Р. 1927. New Asiatic grasshoppers of the genus
Chorthippus Fieb. Konowia, VI, 3 202—206.
Тарбинский, С. II. 1928. О некоторых новых и мало известных прямокрылых
насекомых из Палеарктической Азии. II. Изв. Инет. зоол. фитоп., 4 51—61.
Тарбинский, С. П. 1930. К познанию рода Calliptamus Serv. (Orthoptera, Acri-
didae). Изв. Акад. Наук, Отд. физ.-мат. наук 177—186.
(Умнов Н. Н.) U т.п о v N. N. 1931. Neue Heuschrecken-Arten aus Mittel-Asien.
(Orthopt. Acridiodea). Wien. Entom. Zeit., XLVIII, 3 : 132—137.
R a m m e W. 1930. Dermaptera und Orthoptera. Entomologische Ergebnisse der
Deutsch-Russischen Alai-Pamir Expedition 1928 (I). Mitt. Zool. Mus. Berlin,
XVI, 2 : 209—214.
R a m m e W 1931. Beitrage zur Kenntnis der palaearktischen Orthopterenfauna
(Tettig. et Acrid.). Mitt. Zool. Mus. Berlin, XVII, 1 166—200.
А. Н. ПОПОВА
СТРЕКОЗЫ
Из 40 видов стрекоз, известных для Таджикистана, в Кондаринском
ущелье фиксировано 9 видов, относящихся к 5 родам: Lestes (L. dryas
Kirby, L. Barbara Fabr.), Sympycna (S. fusca V. d. L.), Ischnura (I. pu-
milio Gharp., I. forcipata Mort.), Gordulegaster (G. coronatus Mort.) nSym-
petrum (S. fonscolombei Sei., S. striolatum pallidum Sei. и S. decoloratum
Sei.).
1. Lestes dryas Kirby — только Руидашт, 2700 м (здесь п дальше высота
в метрах над уровнем моря). Широко распространенный палеарктиче-
ский вид. По Бартеневу (1930), L. dryas, в общем вид северный, в южной
половине Палеарктики обитает только в горных районах, поднимаясь
до 2000 м. В Средней Азии вид очень редок, в Таджикистане также в,
вероятно, спорадичен.
2. Lestes bar-bara Farb. — южно-палеарктический вид, широко распро-
страненный в южной Европе; указан для Малой Азии, северной части
Индии, Алжира, заходит и в Центральную Азию. В Таджикистане вид
редкий, но распространенный более широко, чем L. dryas, и залетающий
в предгорья; известен с высоты 1200—2140 м.
Личинки этого и предыдущего вида — характерные обитатели неболь-
ших пересыхающих водоемов.
3. Sympycna fusca V.d.L.— обычная широко распространенная пале-
арктическая форма, по Бартеневу (1924), — выходец из средней Азии, а по
Шорыгину (1926) — выходец из Средиземноморья; вид заходит в предгорья
и поднимается приблизительно до 2000 м (Кавказ, Иссык-куль).
Для Таджикистана вид довольно обычный, предпочитающий пред-
горья, поднимающийся здесь до 2500 м (уроч. Хан-тахта, Хорог, Рушан-
ский район); в Кондаринском ущелье, повлдимому, редок.
Личинки относятся к лимнофилам, живут в мелких стоячих, рбычно
пересыхающих водоемах.
4. Ischnura pumilio Gharp. var. aurantiaca — южно-иалеарктический
вид; обычная форма в переднеазиатскпх странах, на Кавказе идет в горы
до высоты 2035 м.
В Таджикистане — один из обычных, часто встречаемых, повсюду
распространенных видов, летающих как в низменностях, так и в горах
до высоты 2500 м (западный Памир, Ванч).
5. Ischnura forcipata Mort.— характерный среднеазиатский вид, захо-
дящий в северную Индию.
В Таджикистане — самый обычный, самый распространенный и са-
мый обильный вид из всех пяти видов Ischnura, фиксированных для рес-
публики.
Стрекозы
207
Личинки этих двух видов Ischnura обитают в разнообразных биото-
пах, принадлежат к лимнофильным формам, но могут также обитать
в медленно текучих водах, являются фитофи ламп: заросли водных расте-
ний — непременное условие обитаемых ими водоемов.
6. Cordulegaster coronatus Mort. — один из интереснейших видов Таджи-
кистана. Относимые к нему литературные данные очень скудны. Впервые
этот вид был найден А. П. Федченко в 1875 г. при восхождении на гору
Аксай (830 м) в Шахризябском хребте (Зеравшан) и близ города Ургута
(1065 м) и неправильно Брауэром определен как insignis (Брауэр, 1877).
Имеются указания о нахождении его в Коканде [Мортон (Morton, 1915)].
В коллекциях Зоологического института Академии Наук СССР хра-
нятся 21 55 и 13 22 этого вида из следующих мест Средней Азии: Зерав-
шан (Урмитан, Агалык-сай), Ферганский хребет (река Падша-ата, 443 м),
Каратау (Кара-бас-тау), Киргизский хребет (ур. Кулан, Алмани, 1843 м),
Чаган (Акташ, р. Чимганка, около 777 м).
С. coronatus является, повидимому, представителем горной фауны,
именно нижнего пояса гор, пмеет вертикальное распространение при-
близительно от 500 м до 2000 м и может быть назван эндемиком Средней
Азии.
Помимо взрослых насекомых, в водоемах Кондаринского ущелья и
Ходжа-оби-гарма (1939, 1942—1944 гг.) были найдены неизвестные ранее
личинки этого вида.
Личинки живут в небольших, мелких, с очень слабым течением во-
доемах, расположенных у выходов родников, в мелких ручейках, выте-
кающих из родников, в небольших заводях горных речек, лужах, остав-
шихся от спада воды, и т. д.
Дно водоемов каменистое, но, как правило, сильно заиленное, покры-
тое толстым слоем отживших листьев, травой и корнями растений. Силь-
ное затенение водоемов является непременным условием обитания личи-
нок С. coronatus.
Личинки держатся, глубоко зарывшись в ил, пли среди подмытых
п покрытых илом корней трав и деревьев.
Вследствие этих особенностей поведения личинки слабо реагируют
на температурные изменения и были встречаемы в водоемах в различное
время года.
Личинки С. coronatus в большинстве случаев отмечались в условиях
температуры не выше 17—18° С; лишь в одном случае личинка была най-
дена в луже, имевшей температуру 29° С.
Личинки этого вида могут быть отнесены к слабо реофильным формам,
заходящим отчасти и в стоячие водоемы, о чем свидетельствует нахожде-
ние их в лужах, заводях со стоячей водой п т. д.
7. Sympetrum fonscolombei Sei. — эфиопский вид, распространен по
всей Эфиопской области, а также в южной и отчасти в средней Палеарк-
тике. На юге Средней Азии повсеместен и идет высоко в горы (Ак-басага,
2600 м); на Кавказе вид заходит высоко в горы, 2200 м — 2650 м (Акра-
мовский, 1948; Бартенев, 1915, 1930). В Таджикистане также заходит
высоко в горы (Руидашт, 2600 м).
Судя по литературным данным, в Средней Азии вид не очень част,,
в Кондаринском ущелье, по всей вероятности, редок.
Личинки этого вида относятся к лимнофилам.
8. Sympetrum striolatum pallidum Sei. — подвид приурочен к переход-
ной подобласти Палеарктики, известен из Средней и Центральной
Азии.
.208
A. H. Попова
В Таджикистане широко распространен самый многочисленный,
обыкновенный подвид из рода Sympetrum, летающий в низменностях
и предгорьях, от 1000 м до 2080 м (Хорог, 2080 м, Западный Памир, 1989 м,
Ванч, 1800 м).
В Кондаринском ущелье редок.
Личинки этого подвида живут в слабо проточных и стоячих водоемах
и могут быть отнесены к формам с широким экологическим диапазоном.
9. Sympetrum decoloratum Sei. — вид с достоверностью известен из вос-
точного Средиземноморья, Передней и Средней Азии.
В Средней Азии вид обычен на реке Сыр-дарье, а в районе станции
Чиназ (озеро Орумбай) был в массе найден в августе 1937 г. (Бартенев).
В Таджикистане вид довольно редок, в Кондаринском ущелье также.
9 видов стрекоз Кондаринского ущелья составляют 22.5% видов,
известных для Таджикистана.
Из них 3 вида принадлежат к эндемикам Средней Азии (Ischnura
forcipata Mort., Cordulegaster coronatus Mort, и Sympetrum striolatum
pallidum Sei.); остальные распространены в южной половине Палеарк-
тики, за исключением 1 вида (Lestes dryas Kirby), который широко рас-
пространен в северных частях Палеарктики, в южной половине ее оби-
тает только в горных районах.
ЛИТЕРАТУРА
А к р а м о в с к и й Н. Н. 1948. Фауна стрекоз Советской Армении. Зоол. сборы.
АН АрмССР, 5.
Артоболевский Г. В. 1930. К экологии стрекоз Теберды. Раб. Сев.-Кавк,
гидроб. ст., Щ, 1—3.
Бартенев А. Н. 1915. Насекомые ложносетчатокрылые. Фауна России и сопре-
дельных стран. I, 1.
Бартенев А. Н. 1924. К одонатофауне Кавказских гор. Bull. Mus. G£ogr., И.
Бартенев А. Н. 1930. Материалы к познанию западного Кавказа в одонатоло-
гическом отношении. Тр. Сев.-Кав. ассоц. научно-иссл. инет., I.
Бартенев А. Н. 1930. О палеарктических видах подрода Cordulegaster Leach
(Odonata). Раб. Сев.-Кавк, гидроб. ст., Ill, 1—3.
Б р а у е р Фр. 1877. Стрекозы (Odonata), в Федченко: Путешествие в Туркестан.
Изв. общ. люб. естеств., антр. и этногр., XXVI, 1.
Ill о р ы г и н А. 1926. К познанию рода Sympycna Charpentier 1840. Русск. энт.
обозр., XX.
Fraser F. С. 1929. A revision of the Tissilabioides (Cordulegasteridae, Patalliidae
and Petaluridae) (Order Odonata). Mem. Ind. Mus., 9, 3.
Morton K. J. 1915. Some Palaearctic species of Cordulegaster. Trans. Ent. Soc.
bond., P. Ill—IV.
Д. Е. ХАРИТОНОВ
ПАУКИ И СЕНОКОСЦЫ
Сборы пауков и сенокосцев из ущелья Кондара произведены
В. В. Гуссаковским в районе Ботанической станции в период с 29 июля
по 17 сентября 1945 г.
Всего в сборах оказалось 36 видов пауков и 1 вид сенокосцев и, кроме
того, некоторое количество молодых пауков и сенокосцев, не поддающихся
точному определению. Сенокосец относится к роду и виду, опи-
санному В. В. Редикорцевым1 с Гиссарского хребта. Из 36 форм пауков
8 (5 видов и 3 подвида) являются новыми, 16 относятся к средиземномор-
ским формам, половина из которых эндемична для Средней Азии, 4 —
более широко распространенные палеарктические формы, не заходящие
далеко на север, и 8 — транспалеарктические или голарктические формы.
Очень бедно представлены в сборах пауки из семейств Gnaphosidae и
Lycosidae, которые играют существенную роль в составе фауны всех
районов Средней Азии, и совсем отсутствуют представители некоторых
других характерных семейств, в частности, сем. Dipluridae. Недостаток
материала по этим семействам не позволяет подробно характеризовать
фауну, однако сравнительно большой процент новых форм в сборах
указывает на желательность дальнейшего изучения ее. Мы приводим ниже
список найденных видов с указанием дат сбора и описания новых форм.
Порядок OPILIONES
Сем. Phalangiidae
Dilophiocara bactriana Redik., 30 VII — 2 <5, 4 j.
Порядок ARANEI
Сем. Agelenidae
Coelotes major Cron., 30 VII — 2 j.
Сем. Lycosidae
Tarentula latifasciata Cron., 8 VIII — 1 j; 10 IX — 1 <5; 29 VII —
25 VIII — (5(5 et 22 inad.
Trochosa rustica Th.. 25 VIII — 1 j.
1 V Redikorzev (В. Редикорце в). Ein neuer Weberknecht aus
Buchara. z_ool. Anz., 97, H. 2.2 31—32.
14 Ущелье Кондара
210
Д. Е. Xаритонов
Сем. Oxyopidae
Oxyopes lineatus Latr., 25 VIII — 1 <5, 1 5.
Oxyopes maracandensis Charit., 25 VIII — 1 $•
Сем. Dictynidae
Dictyna pygmaea Th., 25 VIII — 1 i.
Сем. Theridiidae
Lithyphantes paykullianus (Walck.), 30 VII — 1 juv.
Dipoena tristis (Hahn) subsp. hissariensis Charitonov n. (рис. 1),
30 VII — 1 $, 1 juv.; 24 VIII — 1 juv.
$. Длина головогруди 1.0 мм, общая длина тела 2.8 мм. Относится
к группе tristis — braccata, но легко отличается от обоих этих видов
окраской ног. У взрослой самки только базальная часть IV бедра темно-
желтая и лапки всех ног светлее остальных члеников. У молодых экзем-
пляров базальная часть IV бедра и лапки всех ног светложелтые, резко
Рис. 1. Dipoena tristis (Hahn)
subsp. hissariensis n. 5: эпи-
гина.
отличающиеся от остальных темнокоричне-
вых члеников. Форма брюшка самки зани-
мает промежуточное положение между D. tri-
stis и D. braccata. Заднесредние глаза уда-
лены друг от друга на расстояние вдвое
меньшее, чем от боковых глаз того же
ряда. Передняя часть головогруди не вы-
ше заднесредних глаз. Вторая (задняя) пара
семеприемников видна снаружи в виде бе-
лых пятен или ямок, они чуть крупнее пе-
редних. Отсутствие самца не позволяет уточнить систематический ранг
данной формы, возможно, что она окажется самостоятельным видом.
Euryopis laeta (Westr.), 29 VII — 1 juv.
Theridium ovatum (CL), 30 VII — 1 j.
Сем. Argiopidae
Araneus redii (Scop.), 8 VIII — 1 juv.; 17 IX — 1 juv.
Araneus strandiellus Charitonov n. sp. (рис. 2), 30 VII — 1 j.
5. Длина головогруди 1.3, ширина 1.1, длина брюшка 2.0, ширина
1.7, высота 1.9 мм. Длина голени-|-колена I — 1.7, III — 0.85 мм. Голо-
вогрудь серо-желтая, в белых прилегающих волосках, задняя часть го-
ловного отдела желтая. Трапеция средних глаз шире сзади, чем спереди,
высота ее меньше основания. Диаметр передне-средних глаз в полтора
раза меньше, чем задне-средних. Боковые глаза обоих рядов сближены,
расстояние между ними меньше половины их диаметра. Они примерно
одинаковы по величине и вдвое меньше диаметром чем задне-средние
глаза. Верхний край желобка хелицер с 3 зубцами, из которых прокси-
мальный — вдвое меньше остальных; нижний край с 2 зубцами (дисталь-
ный немного крупнее). Стернум желтый с серой каймой по бокам и сзади.
Ротовые конечности и ноги желтые. Голени I и II имеют черно-серые,
прерванные сверху, базальное и медиальное кольца, которые по бокам
членика сливаются в общую продольную полосу. На голенях III и IV
Пауки и сенокосцы
211
этот рисунок недоразвит и имеются только пятна, соответствующие
частям колец. Предлапки I—IV с узким дистальным кольцом, кроме
того I затемнены по бокам в дистальной половине. Все лапки с узким
кольцом при основании и широким — в дистальной половине. Пальпы
имеют черно-серые пятна или неполные кольца: одно — на конце колена,
два — на голени и три — на лапке. Ноги вооружены белыми полупро-
зрачными иглами.
Брюшко в передней части высокое, с большими, толстыми коническими
буграми, расстояние между которыми меньше половины ширины бугра.
Основной цвет брюшка желтовато-белый. На переднем скате спинной
стороны, начинаясь от оснований бугров, идут кпереди 2 коричнево-
серых неровных полоски. Около основания бугров (между ними) 2 черно-
коричневых вдавленных точки, от которых отходят назад и в стороны
Рис. 2. Araneus strandiellus n. sp.
A— 9, общий вид паука; Б— 9, эпигина.
(заходя отчасти на бугры) 2 такого же цвета линии. На задней половине
брюшка виден слабо выраженный пирамидальный рисунок, в котором
лучше различаются 2 боковые зигзагообразные сходящиеся кзади линии
и посредине, одна за другой, несколько угловых линий, направленных
углом вперед. С каждой стороны от серо-желтых бородавок, находя-
щихся на сером участке хитина, — пара белых округлых пятнышек.
Срединное вентральное поле белое, оно ограничено по бокам серо-корич-
невыми линиями.
Mangora acalypha (Walck.), 20 VIII — 1 juv.; 24 VIII — 1 17 IX—
1 juv.
Hypsosinga albovittata (Westr.), 10 IX — 1 juv.
Zygiella kochi (Th.), 20 VIII — 1 j juv.; 24 VIII — 1 j juv.
Meta dentipalpis Cron., 30 VII — 2 juv.
Сем. Tetragnathidae
Tetragnatha extensa (L.) subsp. maracandica Charitonov, n., 8 VIII —
1 $ inad.; 17 IX — 1 $.
Отличается от основной формы следующими признаками: 1)стернум
без ясно выраженного светлого клиновидного пятна; 2) коготь хелицеры
самки с зубцом около основания, число зубчиков по краю желобка хели-
цер сверху и снизу 7—7 (иногда 8—7 или 7—6); 3) на хелицерах самца
14*
212
Д’. Е. Xаритонов
косой зубец сильно удален от главного, и расстояние его от концевого
бугорка основного членика хелицер меньше, чем от главного зубца, тогда
как у основной формы косой зубец расположен гораздо ближе к глав-
ному; 4) размеры 6: головогрудь 2.7—3.4 мм, брюшко 4.7—6.8 мм; раз-
меры головогрудь 3.4—3.5 мм, брюшко 7.5—9.0 мм.
Признаки этого подвида приведены по взрослым 66 и ££ из Узбе-
кистана (Ишкент Яккабагского* района, Кашка-дарьинской обл., 27—
28 VII 1942. Д. М. Федотов).
Сем. Gnaphosidae
Zelotes gussakovskyi Charitonov, n. sp. (рис. 3), 29 VII — 15.
Этот интересный вид довольно близок к Z. segrex Sim. Длина голово-
груди 2.5, ширина 1.9 мм; длина брюшка 3.0 мм. Головогрудь красновато-
коричневая с черно-серой краевой линией и мел-
Рис. 3. Zelotes gussa-
kovskyi n. sp., 6: ко-
пулятивный аппарат.
лапки и лапки I
кими пятнышками. От срединной бороздки расхо-
дятся тонкие линии. Поверхность матовая, очень
мелко морщинистая. Передне-боковые глаза вдвое
больше средних. Расстояние между передне-сред-
ними вдвое больше, чем между средним и боковым
глазом того же ряда. Задне-боковые глаза немного
больше задне-средних, расстояние между средни-
ми вдвое больше. Высота и ширина четыреуголь-
ника средних глаз примерно одинакова, он немного
уже спереди, чем сзади. Стернум красно-коричне-
вый, в мелких морщинках и точечных ямках, более
густых в средней части стернума. Ротовые органы
цвета головогруди, длина губы почти вдвое больше
ее ширины. Хелицеры сверху желобка с 3 зубцами,
снизу — с 1 более слабым, но хорошо выраженным
зубцом. Спереди, ближе к медиальной стороне,
хелицеры усажены длинными неровными торчащими
волосками. Бедра всех ног, а также колена и
голени I и IV темные, каштаново-бурые. Пред-
и II и лапки III и IV желтые. Остальные членики
красновато- или желто-коричневые; на передней стороне бедра I имеют
по продолговатому светлому пятну. Ноги I и II не вооружены, их лапки
и предлапки имеют scopulae, немного не доходящие до основания пред-
лапки. Размеры ног I II III IV =7.9: 6.9 5.9 : 8. 1. Длина колена-|-
4-голени I II III : IV = 2.4 2.1 1.6 2.5. Придаток тибиального
членика пальпуса в 2х/з раза длиннее своей ширины, длина придатка
примерно равна длине самого тибиального членика. Брюшко сверху
и снизу серо-желтое; с передней стороны, по бокам и сзади — черно-
серое. Паутинные бородавки черно-серые. В передней части брюшка ви-
ден небольшой красно-коричневый щиток из более твердого хитина.
Сем. Sparassidae
Olios sericeus (Cron.), 29 VII — 1 juv.; 8 VIII — 18; 24 VIII — In
inad.; 10 IX — 2 j juv.
Пауки и сенокосцы
213
Сем. Thomisidae
Philodromus pictus Cron., 8 VIII — 1 juv.; 10 IX — 1 juv.; 17 IX —
14 juv.
Philodromus aureolus (Cl.), 30 VIII — 2 juv.; 25 VIII — 3 juv.; 17 IX—
1 juv.
Diaea dorsata (F.), 8 VIII — 1 $ juv.; 24 VIII — 1 juv.; 17 IX—
1 juv.
Misumena vatia (Cl.)?, 30 VII — 1 juv.; 24 VIII — 1 9 inad.; 17 IX—
1 juv.
Thomisus albus (Gmel.), 29 VII — 1 o; 8 VIII — 1 9; 17 IX — 7 juv.
Xysticus obesus Th., 29 VII — 1 <5, 1 9; 10 IX — 1 j juv.
Сем. Clubionidae
Cheiracanthium pelagicum C. L. K., 29 VII — 1 9.
Cheiracanthium erraticum (Walck.), 30 VII — 1 9.
Micaria shadini Charitonov, n. sp. (рис. 4), 29 VII — lo.
9. Длина головогруди 1.7, ширина 1.2 мм. Индекс 75.88. Длина
брюшка 2.3, ширина 1.45 мм. Головогрудь желто-коричневая, очень
мелко морщинистая, шелковистая. Три пары
расходящихся линий (одна из них на границе
головного и грудного отделов) и четвертая,
неполная, начинающаяся несколько отступя
от середины головогруди, состоят из черно-
коричневого пунктира, сопровождающего тон-
Рис. 5. Micaria hissarica, n.
sp. $: копулятивный ап-
парат.
Рис. 4. Micaria shadini, n. sp. 9:
эпигина.
кие вдавления. Кроме того, вся головогрудь усеяна редкими черноватыми
точками и одета редким (может быть, отчасти стертым) покровом из бе-
лых чешуек. Стернум желтый, с сероватым налетом, мелко морщини-
стый, шелковистый. Ноги желтые, все членики ног III и IV и бедра I и
II имеют серо-коричневый налет. IV ноги очень длинные, конец их лапок
желтый. На ногах заметны отдельные белые чешуйки. Щупики желтые.
Брюшко золотисто-серое, с двумя широкими черными перевязями,
хорошо видимыми с боков и сверху. Золотисто-серая светлая поперечная
полоса между ними соответствует слабой перетяжке, опоясывающей
брюшко около его середины.
214
Д. Е. Харитонов
Micaria hissarica Charitonov, n. sp. (рис. 5), 29 VII — 1 3; 24 VIII —
1 § inad.
3. Длина головогруди 0.9, ширина 0.6 мм, длина брюшка 1.2 мм.
Головогрудь каштаново-бурая, шелковисто блестящая, в очень мелких
точках, с черным линейным рисунком, состоящим из продольной средин-
ной линии, тянущейся от середины глазного четыреугольника до заднего
конца головогруди, V-образного, ограниченного линиями, срединного
пятна (несколько более светлого по сравнению с остальной частью голо-
вогруди) и расходящихся линий, идущих к боковым краям головогруди.
Оба ряда глаз, при рассматривании сверху и несколько спереди, образуют
линии, загнутые концами вперед. Высота клипеуса равна диаметру
передне-боковых глаз. Боковые глаза крупнее средних. Расстояние между
передне-средними больше чем между средним и боковым. Глаза заднего
ряда расположены примерно на одинаковых расстояниях друг от друга.
Задне-средние глаза несколько удлинены (большие оси их направлены
назад и в стороны) и немного крупнее передне-средних. Четыреугольник
средних глаз длиннее своей ширины и спереди уже, чем сзади. Хели-
церы сверху и с боков каштаново-бурые, снизу желтоватые. Ноги жел-
тые, бедра всех ног, а также тазики и вертлуги I и II каштаново-бурые.
Бедра I и II более темные. Дистальные концы всех бедер желтые. Чле-
ники пальпуса, за исключением тарзального, темнокоричневые, тарзаль-
ный — светлее. Длина тарзального членика равна длине колена-(-голени.
Голень снабжена на конце шиповидным выростом, длина которого
меньше половины диаметра членика и меньше г/з его длины. Брюшко
коричнево-черное, покрыто довольно редкими переливчатыми чешуйча-
тыми волосками и имеет белый рисунок (из белых чешуек), состоящий
на спинной стороне из двух маленьких округлых пятнышек у переднего
края брюшка и короткой поперечной полоски в передней половине
брюшка около его середины. На вентральной стороне брюшка, непосред-
ственно позади эпигастральной складки, с каждой стороны имеется по
косой полоске из белых чешуек.
Сем. Salticidae
Cyrba algerina (Н. Luc.), 20 VIII — 1 3 inad.
Philaeus chrysops (Poda) v. haemorrhoicus (C. L. K.), 29 VIII—1 o;
10 IX — 1 juv.
Aelurillus variegatus Cron. ( = dorthesi And.?), 10 IX — 1 2 juv.
Euophrys thorelli Kulcz. subsp. asiatica Charitonov, n. (рис. 6),
29 VII — 1 2-
2- Длина головогруди 1.4, ширина 1.0 мм, брюшко 2.0 мм длиною.
Вооружение ног — как у типичной формы. Ноги темножелтые, все бедра
темнокоричневые, за исключением узкой более светлой продольной по-
лоски на спинной стороне, двух светлых (коричнево-желтых) пятнышек
на задней стороне бедер I и II и одного продольного пятнышка на задней
стороне бедер IV Все колена и голени с боков затемнены, предлапки
имеют слабый дымчатый налет по бокам. Эпигина отличается от
основной формы отсутствием хитиновой краевой пластинки на переднем
крае.
Salticus tricinctus (С. L. К.), 30 VII — 1 2; 20 VIII — 1 juv.
Hissarinus kondarensis Charitonov, n. g., n. sp. 29 VII — 1 о, 1
inad.; 30 VII — 1 2 juv; 24 VIII — 1 3, 2 juv.
Пауки и сенокосцы
215
Hissarinus Charitonov, n. g.
Относится к группе Chrysilleae и близок к роду Telamonia. Голень-}-
-4-колено III пары ног короче чем IV (у j — много короче). Глазной
четыреугольник в полтора раза шире своей длины и сзади сужен. Голов-
ной отдел вдвое короче грудного. Глаза
второго ряда расположены на оди-
наковом расстоянии от передне-боко-
вых и от глаз 3-го ряда, расстояние
их от 3-го ряда примерно равно
диаметру последних. Стернум впереди
сужен, но срезан прямо, ширина его
переднего края меньше диаметра тази-
ков. Срединная торакальная бороздка
довольно длинная.
Тип рода: Hissarinus kondarensis Рис 6. Euopbrys thorcdli Kulcz.
r subsp. asiatica, n., q: эпигина.
Charitonov, n. sp. 1 Y
Описание Hissarinus kondarensis Charitonov, n. sp. (рис. 7).
(5. Длина головогруди 3.6, ширина 2.7 мм, длина брюшка 4.0, ширина
2.5 мм. Наиболее высокая часть головогруди — немного позади задних
глаз, кпереди голова образует пологий скат, задняя половина имеет более
крутой скат. Основной цвет головогруди (клппеус, бока и часть дорзаль-
Рис. Hissarinus kondarensis. n. sp.:
A—d, общий вид паук
Б—<3, копулятивный аппарат; В— 5.
ной поверхности) — желтый, однако вся передняя половина спинной
стороны и 2 неправ иль но-четыреугольных участка в задней половине
имеют буроватый хитин, на фоне которого расположены 3 пары пятен,
образованных черными прилегающими густыми короткими волосками:
одна пара маленьких поперечных пятен в передней части позади глаз
и две пары крупных неправильно-четыреугольных пятен в середине
и в задней половине головогруди. Бока и желтые промежутки между
пятнами покрыты белыми прилегающими волосками. Базальная поло-
вина основного членика хелицер спереди и с внутренней стороны, а также
216
Д. Е. Харитонов
клипеус покрыты густой бородой из длинных желтовато-белых шел-
ковистых волосков. Реснички передне-средних глаз снизу и по бокам
белые или рыжевато-белые, сверху участок кольца ресничек имеет ярко-
оранжевый цвет. Реснички передне-боковых глаз беловатые, за исклю-
чением участка сверху, на котором реснички ярко оранжевые. Оранже-
вые реснички средних и боковых передних глаз соединены с каждой
стороны косым оранжевым пятном по бокам переднего конца голово-
груди. Глаза переднего ряда (при рассматривании сверху) образуют
линию, сильно загнутую назад. Верхние края передних глаз (при рас-
сматривании спереди) образуют линию, слабо загнутую вверх. Диаметр
средних глаз примерно вдвое больше диаметра боковых. Высота кли-
пеуса меньше половины диаметра средних глаз. Глаза 2-го ряда (очень
маленькие) расположены на одинаковом расстоянии от передних и зад-
них. Глаза 3-го ряда такого же размера, как передне-боковые. Расстоя-
ние между передне-средними глазами в 1.5 раза меньше, чем между сред-
ним и боковым. Расстояние между глазами 2-го и 3-го ряда примерно
равно диаметру глаза 3-го ряда. Стернум светложелтый в белых длинных
волосках. Тазики и вертлуги ног снизу желтые, с черными разреженными
волосками, более длинными на передних двух парах. Ноги желтые,
колена и голени I и II буровато-желтые. На бедрах всех ног спереди
и сзади черно-коричневые продольные пятна, занимающие не менее
половины ширины передней и задней стороны. Эти пятна у вершины
бедер соединены буроватыми поперечными перемычками. Бедра I и II
покрыты (кроме волосков) снизу и спереди белыми чешуйками. Колена
всех ног имеют по бокам в дистальной половине по треугольному черному
пятну. Голени II—IV снизу с узкой полоской из черных волосков, тяну-
щейся по всей длине членика, и концевым темным кольцом. Голени I
сплошь буро-коричневые, они густо покрыты черными длинными волос-
ками, особенно длинными снизу. Размеры ног I II : III : IV=
=9.9 : 8.8 9.65 10.45 мм. Колено+голень 111=2.9 мм, IV=3.0 мм.
Вооружение ног: бедра I—III сверху 1.1.1, спереди—сверху близ вер-
шины 2, сзади — сверху 1. 2. Вооружение бедер IV отличается только
отсутствием одиночной иглы сзади—сверху. Колена I—II спереди 1;
III—IV спереди и сзади по 1. Голени I—II снизу 2.2.2, спереди 1.0.1,
сзади 0.0.1 (или на II паре спереди 1.1.1 или сзади 1.0.1). Голени III—
IV сверху 1.2.0, снизу 1.0.2, спереди 0.1.0.1.0, сзади 0.1.0.1.0 (правая
IV голень снизу 2.0.2). Предлапки I снизу 2.2; II снизу 2.2, спереди 0.1;
III снизу 2.2, спереди и сзади по 0.1, сверху 2.2; IV снизу 2.2, спереди
0.0.1, сзади 0.0.1, сверху 2.2.2. Длина щеточек лапок превышает длину
коготков. Брюшко желтое. На спинной стороне 2 черных продольных
полосы, разделенных срединным светлым промежутком, образующим
пару небольших боковых выступов, заходящих в пределы черных полос.
Брюшная сторона желтая, с коричневым рисунком в виде V и продольной
тонкой коричневой полоской позади эпигастральной складки.
$). Длина головогруди 3.5, длина брюшка 4.6 мм. Рисунок голово-
груди менее дифференцирован, вся передняя половина ее имеет одно-
родную буро-коричневую окраску и покрыта коричневыми и редкими
беловатыми волосками. Границы задней пары пятен более расплывчаты.
Оранжевые пятна по бокам переднего конца головогруди хорошо выра-
жены. Окраска ног более бледная, густого покрова из черных длинных
волосков нет. Белые чешуйки на бедрах I и II имеются. Длина голени+
+колена III значительно меньше, чем IV. Продольные полосы на брюшке
более расплывчаты, чем у самца.
Б. В. ЛОТОЦКИЙ
ИКСОДОВЫЕ КЛЕЩИ
Изучение иксодофауны ущелья Кондара не ставилось в качестве само-
стоятельной задачи, но вопросы распространения этих клещей в Таджик-
ской республике изучались в различных высотных зонах; таким образом,
была исследована фауна иксодовых клещей и в Варзобском районе. По
природным особенностям и хозяйственному укладу этот район характе-
рен для районов, расположенных на южных склонах Гиссарского
хребта, и видовой состав иксодовых клещей здесь довольно полно отра-
жает общую картину распространения их в вертикально-зональном от-
ношении. Из И видов сем. Ixodidae и 2 видов сем. Argasidae, найденных
в Варзобском районе, к горным формам могут быть отнесены две:
Ixodes berlesei Bir. и Dermacentor marginatus Sulz., к равнинным также
две: Boophilus calcaratus Bir. и Hyalomma detritum P. Sch.; все про-
чие относятся к двузональным.
В зоогеографическом отношении (Померанцев, 1948) фауна иксодовых
клещей Варзобского района слагается из палеарктических видов и вклю-
чает наибольшее число представителей Средиземноморской подобласти,
куда входят такие виды, как Ixodes redikorzevi 01., Haemaphysalis
sulcata Can. et Fanz., Haem, numidiana Neum., Haem, caucasica 01.,
Boophilus calcaratus Bir., Rhipicephalus sanguineus Latr., Hyalomma
detritum P. Sch., H. anatolicum Koch и H. plumbeum Panzer: в мень-
шей доле сюда входят представители Европейской и Туранской подоб-
ластей.
В отношении специфичности видов к тем или иным классам своих,
хозяев, состав клещей Варзобского района может быть разделен на три
группы; к первой из них относятся такие виды, которые во всех актив-
ных фазах своего развития паразитируют на млекопитающих; сюда вхо-
дят Haemaphysalis caucasica 01., Dermacentor marginatus Sulz., Rhipi-
cephalus sanguineus Latr., Boophilus calcaratus Bir., Hyalomma detritum-
Р. Sch. и H. anatolicum Koch. Вторая группа включает паразитов птиц;
в Варзобском районе к ней относится лишь один вид — Ixodes berlesei
Bir. Третьей группой объединяются виды, паразитирующие на различ-
ных наземных позвоночных; в ее состав входят Ixodes redikorzevi 01.,
Haemaphysalis sulcata Can. et Fanz., Haem, numidiana Neum. и Hya-
lomma plumbeum Panzer.
В отношении сезонности паразитирования распределить клещей
по группам довольно трудно без допущения грубой схематизации. Боль-
шинство видов паразитирует в теплое время года; в зимний сезон на жи-
вотных равнинной части Варзобского района встречаются лишь нимфы
и личинки Hyalomma detritum. Возможно, что в этот сезон остаются*
218
В. В. Лотоцкий
активными некоторые норовые виды, как Ornithodorus papillipes Bir.,
Argas persicus Ok. и преимагинальные фазы Ixodes redikorzevi 01., однако
прямых наблюдений по этому вопросу не имеется.
Практический интерес представляет группирование видового состава
в отношении степени синантропности. К формам, связанным в своем жиз-
ненном цикле только с дикими животными, относятся 4 вида: Ixodes
berlesei Bir., I. redikorzevi 01., Haem, caucasica 01. и Haem, numidiana
Neum. К группе полусинантропных, паразитирующих преимущественно
на диких, но также и на домашних животных, можно отнести 5 видов:
Haem, sulcata Can. et Fanz., R. sanguineus Latr., H. plumbeum Panzer,
A. persicus Ok. и 0. papillipes Bir. К синантропным видам, паразити-
рующим по преимуществу на домашних животных и в меньшей степени —
на диких, относятся 4 вида: D. marginatus Sulz., В. calcaratus Bir., Н. de-
tritum P. Sch. и H. anatolicum Koch.
Наконец, прямой хозяйственный интерес представляет выделение
форм, вредящих здоровью человека и животных путем интоксикации или
переноса возбудителей болезней. Недостаточная изученность клещей
позволяет произвести подобное выделение лишь условно, так как виды,
считающиеся в настоящее время безопасными, могут при дальнейшем
исследовании оказаться вредными. Таким образом, к безусловно вредным
формам в Варзобском районе можно отнести 6 видов: D. marginatus
Sulz., R. sanguineus Latr., H. anatolicum Koch, H. plumbeum Panzer,
A. persicus Ok. и 0. papillipes Bir. Такие переносчики гемоспоридиозов
крупного рогатого скота, как Н. detri turn Koch и В. calcaratus Bir., почти
не имеют значения в животноводстве, так как распространены на очень
небольшой территории в южной равнинной части района.
Таковы общие черты иксодофауны Варзобского района. Более деталь-
ные сведения об отмеченных видах приводятся ниже.
Ixodes redikorzevi 01., 1927. В Варзобском районе найден в пределах
лесного пояса. В других районах республики встречается также и на равни-
нах. I. redikorzevi относится к средиземноморской фауне и известен из
Крыма, Закавказья и вне СССР из Палестины. Обнаружен на туркестанских
крысах, домовых мышах, на лисе, на горных куропатках (кекликах) и дру-
гих диких птицах. В равнинных районах республики был найден на зем-
ляных крысах, лесных мышах, слепушонках, зайцах и шакалах. Поло-
возрелые I. redikorzevi отмечались в январе, марте, июне, сентябре и
декабре; нимфы — в январе, марте, мае, октябре и декабре; личинки —
в мае и октябре. Сравнительно с другими представителями рода этот вид
наиболее многочислен и широко распространен. Биология и хозяйствен-
ное значение его не изучены.
В настоящее время установлен новый подвид этого вида — I. redi-
korzevi emberizae В. Pom., 1950, обнаруженный в сборах также из Гис-
сарского хребта, с горных куропаток (Померанцев, 1950).
Ixodes berlesei Bir., 1895. Принадлежит к видам, самки которых обла-
дают характерной сердцевидной формой; такая же форма тела свойственна
нимфам и личинкам. В Варзобском районе найден в верхнем лесном
и субальпийском поясах гор; ту же зональность ин обнаруживает и в дру-
гих районах республики. Находки I. berlesei известны из Восточной Си-
бири (Ангара) и из стран Западной Европы. Паразитирует на диких пти-
цах; в Варзобском районе половозрелые п нимфы встречены на стено-
лазе (Trichodroma muraria) и синем каменном дрозде (Monticola solitaria
pondoo), а нимфы и личинки — на горихвостке-чернушке (Phoenicurus
ochruros phoenicuroides). Взрослые клещи и нимфы отмечены в июле
Иксодовые клещи
219
и августе, а личинки — в июле. I. berlesei довольно малочисленный и
редкий вид. Биология и хозяйственное значение его неизвестны.
Haemaphysalis sulcata Can. et Fanz., 1877—1878. В Варзобском районе
обнаружен в пределах нижнего лесного пояса гор и в равнинной части;
в других районах встречается также в горах, но чаще — на равнине.
Н. sulcata сухолюбивый вид; он распространен в республиках Средней
Азии, на Кавказе; за пределами Советского Союза: в Китае (восточ-
ный Тибет), Индии, Иране и по всему Средиземноморью, населяя южные
леса и полупустыни. В половозрелой фазе паразитирует на крупных
млекопитающих, нападая обычно и на домашних животных; нимфы
и личинки обитают на мелких зверьках, птицах и очень часто — на
пресмыкающихся. Нимфы Н. sulcata встречались в Варзобском районе
на туркестанской крысе, а личинки — на лесной мыши и на полозе.
Взрослые клещи паразитируют в апреле, мае и июне, нимфы же
и личинки — в июне и июле. В естественных условиях развивается по
треххозяинному типу.
В Туркмении этот вид переносит пироплазмоз овец и может распро-
странять возбудителей бруцеллёза. Подозревается в способности вызы-
вать у животных параличи токсинами своей слюны. В Таджикистане
эпизоотологического значения не имеет.
Haemaphysalis numidiana Neum. 1897. В Варзобском районе обнару-
жен в нижнем лесном поясе гор, в других районах республики более
распространен на равнине, где обитает в оазисах, полупустынях и пу-
стынях. В СССР имеется в среднеазиатских республиках, в Казахстане,
в Крыму и на Кавказе. Этот сухолюбивый средиземноморский вид распро-
странен за пределами Советского Союза в Алжире, Тунисе и в северном
Иране, где обитает в пустынях, полупустынях, степях и горных лесах.
В Таджикистане распространен один из трех подвидов — Н. numidiana
turanica Pospelova-Strom, экологически связанный с норовыми живот-
ными.
Половозрелые клещи Н. numidiana обнаружены в Варзобском районе
на лисе, в других районах встречаются на ежах; нимфы и личинки пи-
таются на песчанках и фазанах. В литературе в качестве хозяев Н. numi-
diana отмечаются сельскохозяйственные животные, медведи и зайцы;
личинки и нимфы встречаются также на крысах и диких птицах. Сезон
паразитирования приходится на весенне-летний период — с марта по
июль.
Биология и хозяйственное значение этого вида не выяснены.
Haemaphysalis caucasica 01., 1928. Численно преобладает над про-
чими видами этого рода, встречаясь от предгорий до субальпийского
пояса гор. В других районах найден и в оазисах на равнине. В СССР
известен из Киргизии, с юга Украины, из Дагестана и восточного Закав-
казья. За пределами Советского Союза найден в северном Иране.
В половозрелой фазе Н. caucasica паразитирует, по преимуществу,
на зайцах, но встречается также на медведях, лисах и шакалах; нимфы
его нападают на ящериц. В Варзобском районе нимфы и личинки этого
клеща в большом количестве обнаруживались на туркестанских крысах
и лесных мышах (Соснина, 1950), а в других районах встречались на сено-
ставцах.
Половозрелые клещи паразитируют в мае, июне и августе, а нимфы
и личинки — с мая по октябрь.
Биология и хозяйственное значение Н. caucasica не изучены.
Dermacentor marginatus Sulz., 1776. Из двух видов рода Dermacentor,
220
Б. В. Лотоцкий
свойственных Таджикистану, в Варзобском районе, как и в других гор-
ных районах республики, распространен лишь D. marginatus, встре-
чающийся только в верхнем лесном и субальпийском поясах гор. Другой
вид — D. daghestanicus 01. обитает исключительно на равнинах южных
районов республики. Таким образом, ареалы обоих видов в Таджикистане
нигде не соприкасаются. В СССР D. marginatus известен из степей Укра-
ины и Дона, найден в Крыму, на Кавказе, в среднем и нижнем Поволжье,
в Казахстане, Западной Сибири и в горах среднеазиатских республик.
За пределами Советского Союза известен в странах южной Европы
и на островах Средиземного моря.
В половозрелой фазе D. marginatus питается на сельскохозяйственных
копытных, волках, собаках, зайцах и ежах; личинки и нимфы нападают
на различных мелких млекопитающих, главным образом, грызу-
нов.
Время паразитирования половозрелых клещей в Таджикистане —
с июля по октябрь; нимфы были обнаружены в сентябре; личинки появ-
ляются, видимо, раньше. Тип развития треххозяинный; в течение года
развивается одно поколение (Сердюкова, 1948).
D. marginatus известен как переносчик во втором поколении пиро-
плазмоза и нутталлиоза лошадей, что, по всей вероятности, имеет место
и в Таджикистане. Кроме того, он способен сохранять в своем организме
в течение длительного времени возбудителей и других заболеваний.
Эпидемиологическое и эпизоотологическое значение этого клеща в Таджи-
кистане тем не менее не велико вследствие ограниченности мест его оби-
тания лишь высокогорьем.
Rhipicephalus sanguineus Latr. 1806. Вопрос о видовом составе этого
рода в Таджикистане вообще требует выяснения. В Варзобском районе
найдены две формы, определяемые (в крайних вариантах) как R. san-
guineus Latr. и R. turanicus В. Pom.; обе обитают в одних и тех же ста-
циях, на одних и тех же хозяевах и паразитируют в одни и те же сезоны.
Вместе с тем наблюдается большое число экземпляров, обладающих
систематическими признаками обоих этих видов и не отличающихся от
них ни биологически, ни экологически. Некоторыми экспериментальными
данными подтверждается также и морфологическая идентичность соот-
ветствующих фаз развития этих форм, а поэтому дифференциация их
становится чрезвычайно трудной и искусственной. На основании этого
все подобные формы рассматриваются нами в дальнейшем как единый
полиморфный вид — R. sanguineus Latr.
В Варзобском районе этот вид встречается повсеместно до верхних
границ лесного пояса. В других районах республики он так же широко
распространен и в зоне равнин. В СССР он (R. sanguineus Latr.) обитает
на черноморском побережье в Закавказье и (R. turanicus В. Рот.) в рес-
публиках Средней Азии. За рубежом (R. sanguineus Latr.) известен по-
всеместно в странах с умеренным и жарким климатом.
Половозрелая фаза R. sanguineus паразитирует на различных млеко-
питающих (включая и всех сельскохозяйственных животных) особенно
на крупных хищных, главным образом на волках, собаках, лисах и ша-
калах. В Таджикистане встречен также на тиграх, барсах и на диких
свиньях (Лотоцкий, 1948). Личинки и нимфы питаются на мелких зверь-
ках, по преимуществу на грызунах. В Варзобском районе взрослые клещи
встречались на крупном и мелком рогатом скоте и непосредственно в при-
роде, а личинки и нимфы — на туркестанских крысах, домовых и лесных
мышах.
Иксодовые клегци
221
Половозрелая фаза R. sanguineus встречается в Таджикистане с марта
по октябрь, но в основном в весенний период, то же относится и к непо-
ловозрелым фазам, однако наибольшая численность их наблюдается
в июле.
Тип развития R. sanguineus треххозяинный; за год развивается одно
поколение.
Этот вид известен как переносчик очень многих заболеваний людей
и животных. В Таджикистане он имеет значение в распространении нут-
таллиоза лошадей.
Hyalomma plumbeum Panzer, 1795. Форма, распространенная в Та-
джикистане, относится к подвиду Н. plumbeum turanicum В. Pom.
(Померанцев, 1950). В Варзобском районе этот клещ встречается как
в предгорной южной части, так и в горной северной. В других районах
республики он обычен также и на равнине. В СССР Н. plumbeum tura-
nicum известен из сопредельных Таджикистану республик Средней Азии
и из южного Казахстана. Вне СССР найден в Иране, прочие подвиды рас-
пространены на юге Советского Союза и в Средиземноморье.
Хозяевами половозрелых Н. plumbeum служат крупные копытные
животные, главным образом домашние, а в Таджикистане также вер-
блюды, тугайный олень и заяц; нападает и на человека, но обычно не при-
сасывается. Личинки и нимфы в массе паразитируют на различных диких
птицах, ведущих или наземный образ жизни, или часто приземляющихся.
В Таджикистане список птиц-хозяев Н. plumbeum включает около 50
видов. Вместе с тем личинки и нимфы способны кормиться и на мелких
млекопитающих — зайцах и земляных крысах.
Развитие Н. plumbeum происходит по двуххозяинному типу; биоло-
гический цикл этого вида еще не вполне выяснен.
В горах все фазы развития встречаются с мая по сентябрь, а на рав-
нине — с марта по октябрь.
Н. plumbeum известен как переносчик некоторых заболеваний людей
и животных. В Таджикистане переносит нутталлиоз лошадей.
Hyalomma anatolicum Koch, 1844. В Таджикистане обитают два под-
вида: мелкий светлоокрашенный — Н. anatolicum anatolicum Koch и
более крупный темнопигментированный — Н. anatolicum excavatum
Koch; первый преобладает в горах, второй — на равнине. В Варзобском
районе, как и повсеместно в республике, встречаются обе формы. Поэтому
в дальнейшем мы не дифференцируем эти подвиды, относя их к еди-
ному виду Н. anatolicum Koch. В СССР Н. anatolicum известен из рес-
публик Средней Азии, южного Казахстана и Кавказа. За пределами
Советского Союза обнаружен в Иране, Ираке, Малой Азии и в
Африке.
Хозяевами взрослых клещей служат различные крупные млекопитаю-
щие, в том числе и домашние. В Таджикистане этот вид встречается
также на оленях, джайранах, диких свиньях и кошках (Felis ornata),
шакалах, хомячках и в норах сусликов. Личинки и нимфы паразитируют
на домашних животных и различных мелких диких зверьках. В Таджи-
кистане они были обнаружены на скоте, а также на домашних кошках
и кроликах, на зайцах, земляных крысах, песчанках, слепушонках,
ежах и на домовых мышах. В Варзобском районе личинки и нимфы
Н. anatolicum были найдены на туркестанских крысах и лесных мышах
(Соснина, 1950).
Взрослые клещи встречаются на своих хозяевах почти круглый год,
но, главным образом, с апреля по сентябрь, а личинки и нимфы — по
222
Б. В. Лотоцкий
преимуществу в июле—августе. Тип развития двух- и треххозяинный;
за год развивается одно поколение.
Н. anatolicum известен как переносчик тейлериоза крупного рогатого
скота и потому имеет значение в животноводстве республики.
Hyalomma detritum Р. Sch. 1919. Распространен только в южной рав-
нинной части Варзобского района. В других равнинных районах широко
распространен в пределах поливной площади, тугайной и оазисной рас-
тительности. Известен из соседних среднеазиатских республик, южного
Казахстана и Закавказья. За рубежом имеется в Иране, Китае и север-
ной Африке.
Хозяевами для всех активных фаз развития служат крупные млеко-
питающие, главным образом крупный рогатый скот; питается также на
овцах, козах, верблюдах и лошадях, а из числа диких животных в
Таджикистане встречается на тугайных оленях, диких свиньях и на
зайцах.
Половозрелая фаза паразитирует в массе с июля по сентябрь, единич-
ные экземпляры встречаются почти круглый год. Личинки появляются
в августе—сентябре и позднее; частью они нападают на своих хозяев
и в теплые дни зимою. Личинки линяют в нимф на том же хозяине; по-
следние остаются на нем до весны, отпадая в марте—апреле. Таким обра-
зом, развитие идет по двуххозяинному типу, образуя одно поколение
в год.
Н. detritum является основным переносчиком тейлериоза крупного
рогатого скота и потому в животноводстве Таджикистана вообще играет
значительную отрицательную роль, но в Варзобском районе значение
этого вида не велико, вследствие небольшого распространения — лишь
в самой южной его части, расположенной в предгорьях.
Boophilus calcaratus Bir., 1895. В фауне СССР существует как един-
ственный вид этого рода. В Варзобском районе чрезвычайно малочислен
и распространен только на небольшой площади равнины — в южной
части района. В других областях Таджикистана распространение этого
вида, подобно Н. detritum, связано с естественно или искусственно оро-
шаемыми долинами.
В СССР В. calcaratus известен из других среднеазиатских респуб-
лик, южного Казахстана, Северного Кавказа, Закавказья, Украины
и Крыма. За пределами СССР распространен в южной Европе, северной
Африке, в Малой Азии и в Иране.
Основным хозяином всех активных фаз развития В. calcaratus служит
крупный рогатый скот, но этот клещ может паразитировать и на диких
жвачных; в Таджикистане он обычен на тугайных оленях; в единичных
экземплярах встречается и на других млекопитающих. Развитие его
идет по однохозяинному типу. За год образуется до трех поколений;
в странах с более мягким и ровным климатом, как, например, в запад-
ном Закавказье, развитие В. calcaratus может итти, видимо, беспре-
рывно.
Сезон паразитирования во всех активных фазах охватывает в общем
все теплое время года, но максимум численности наблюдается весной
в апреле—мае, летом — в июле й осенью — в октябре.
В. calcaratus является основным переносчиком пироплазмоза, фран-
саиеллёза и анаплазмоза крупного рогатого скота, поэтому роль его
в животноводстве республики очень велика, однако в Варзобском районе
он не имеет такого значения по причине малочисленности и узости рас-
пространения.
Иксодовые клещи
223.
Argas persicus Ok., 1818. В Варзобском районе фактически обнаружен
пока только один вид этого рода, но допустимо существование здесь и двух
других, имеющихся в иксодофауне Таджикистана — A. reflexus Latr.
и A. vespertilionis Latr. Этот клещ не распространяется высоко в горы
и более характерен для фауны предгорной полупустыни, поэтому находки
его ограничиваются преимущественно более низкой южной частью
района. В аналогичных стациях, но также и в оазисах распространен
этот вид и в прочих районах Таджикистана. В СССР A. persicus известен
из других республик Средней Азии, южного Казахстана, Армении и Азер-
байджана. За рубежом распространен в Китае, Индии, Иране,Малой
Азии, Африке, Южн. и Сев. Америке и Австралии.
Хозяевами всех активных фаз развития A. persicus служат домашние
и дикие птицы. Он обитает в курятниках, на птичьих дворах или в гнез-
дах, прячась в различных щелях и трещинах или под корою деревьев.
В противоположность клещам семейства Ixodidae, присасывающимся
к своим хозяевам на срок в несколько дней, аргасовые клещи питаются
в течение очень короткого времени, измеряемого десятками минут.
В зависимости от климатических условий A. persicus может парази-
тировать либо в течение круглого года, либо в теплые сезоны. В Варзоб-
ском районе встречается в птичниках и в гнездах воробьев (Passer
domesticus).
Нападая на домашних птиц в больших количествах, вызывает инто-
ксикацию и гибель птиц, особенно молодняка. Кроме того, он является
переносчиком возбудителей спирохетоза птиц, вследствие чего относится
к вредителям птицеводства.
Ornithodorus papillipes Bir. 1895. Из трех видов этого рода, обитаю-
щих в Таджикистане, О. papillipes является, повидимому, наиболее*
распространенным. В Варзобском районе он отмечен в ряде населенных
пунктов (Соснина, 1945), начиная с предгорий и доходя до Анзобского
перевала. В других районах республики, например на Западном Памире,
он распространен и на больших высотах. В СССР известен из Узбекистана
и Туркмении, южного Казахстана и Закавказья, а за рубежом — из
Ирана и Индии.
Хозяевами всех активных фаз развития служат различные млеко-
питающие, а также и человек; способен питаться и на птицах. В процессе
постэмбрионального развития О. papillipes образует до пяти возрастов
нимф, но в половозрелых клещей могут линять уже третьи и четвертые
стадии нимф.
Обитает О. papillipes в жилых помещениях местного типа, прячась
в трещинах каменных стен или в лёссовой пыли полов, пещер и нор
различных животных, где и нападает на своих хозяев. Насасывается
крови и отпадает от своего хозяина О. papillipes столь же быстро, как
и A. persicus.
Сроки паразитирования этого вида также стоят в зависимости от
климатических условий. В естественной обстановке наибольшая актив-
ность наблюдается в теплое время года и почти совершенно замирает
в холодное.
О. papillipes имеет большое значение в эпидемиологии как перенос-
чик спирохет клещевого возвратного тифа, однако в Варзобском районе,
несмотря на обитание этого вида, заболевание не распространено, что
может объясняться разными причинами: иммунитетом местного насе-
ления, неинвазированностью переносчика, наконец, тем, что пришлое
население обычно не ночует в постройках местного типа.
-224 В. В. Лотоцкий
ЛИТЕРАТУРА
Лотоцкий Б. В. 1945. Материалы по фауне, биологии клещей надсемейства
Ixodoidea в Гиссарской долине Таджикистана, в связи с обоснованием мер
профилактики пироплазмозов крупного рогатого скота. Тр. ТФАН СССР,
т. XIV.
-Лотоцкий Б. В. 1948. Иксодовые клещи диких млекопитающих Таджикистана.
Сообщ. ТФАН СССР, вып. VIII.
П а вловский Е. Н. 1948. Руководство по паразитологии человека, т. 1.
Изд. АН СССР.
Померанцев Б. И. 1948. Географическое распространение клещей Ixodidae
и состав их фауны в палеарктической области. Тр. Зоол. инет. АН СССР,
т. VII.
Померанцев Б. И. 1950. Фауна СССР. Паукообразные, т. IV, вып. 2. Иксо-
довые клещи (Ixodidae). Изд. АН СССР.
‘Сердюкова Г. В. 1948. Метод определения продолжительности цикла развития
у клещей семейства Ixodidae. Паразитолог, сборн. Зоол. инет. АН СССР,
т. X.
Соснина Е. Ф. 1945. Испытание некоторых дезосредств для борьбы с клещами
Ornithodorus papillipes. Изв. ТФАН СССР, № 6.
Соснина Е. Ф. 1950. О сезоне паразитирования иксодовых клещей на мышевид-
ных грызунах. Сообщ ТФАН СССР, вып. XXIX.
Д. И. БЛАГОВЕЩЕНСКИЙ
ПУХОЕДЫ ПТИЦ
Приводимый список пухоедов (Mallophaga), собранных с птиц ущелья
Кондара, содержит 20 видов. Исследовано 57 птиц, относящихся к 31
виду 4 отрядов: 1 вид Gallinae, 2 вида Picariae, 1 вид Columbae и 27
видов Passeres. В эту коллекцию птиц входят как очень широко
распространенные виды (сокр. в списке — тп.), так и виды европей-
ской (ев.), средиземноморской (сзм.), ирано-туркестанской (ит.), индий-
ско-африканской (иа.), восточно-палеарктической (вп.), китайской (к.)
и туркестано-сибирской (тс.) групп фауны Палеарктики. Заражены
пухоедами 25 экземпляров 21 вида. Двойная инвазия, т. е. заражение
2 видами паразитов, отмечена у 5 форм: Upupa е. epops, Streptopelia
orientalis meena, Coccothraustes coccothraustes humei, Passer domesticus
bactrianus и Hirundo rufula scullii; тройная — у Alectoris kakelik.
Общая интенсивность заражения слабая — до 10 особей — у большин-
ства птиц, и умеренная — не превышающая 46 особей — у нескольких
экземпляров (Coracias garrulus semenowi, Coccothraustes coccothraustes
humei, Carduelis caniceps subcaniceps, Passer domesticus bactrianus,
Lanius m. minor, Oenanthe pleschanka, Hirundo rufula scullii).
Пухоеды
Подотряд AMBLYCERA KELLOGG.
Сем. Menoponidae Mjoberg
1. Menacanthus fertilis (Nitzsch):
1 $•
2. Menacanthus orioli, sp. n. (in
litt.): 1 j.
3. Myrsidea thoracica (Giebel):
1 $•
4. Myrsidearustica(Giebel): 1 5,
1 $, 12 L.
Птицы
Upupa e. epops L., тп., 5
3 VI 1944.
Oriolus oriolus kundoo Sykes,
тп., j, 27 V 1944.
Merula merula intermedia
Richm. ев., 3, 25 V 1944.
Hirundo rufula scullii (Seeb.),
тп., sex?, 24 V 1944.
Сем. Ricinidae Neumann
5. Ricinus tugarinovi, sp. n. (in
litt.): 2 2-
Terpsiphone paradisi turkesta-
nica (Zar. et Harms), иа., 6 VI
1944.
15 Ущелье Кондара
226
Д. И. Благовещенский
Подотряд ISCHNOCERA KELLOGG
Сем. Philopteridae Burmeister
6. Philopterus subflavescens
(Geoffroy): 3 <5, 11 j 12 L;
1 3, 3 2, 14 L;
3 L;
2 <5, 12;
1 $, 2 2, 6 L;
45, 13 L;
1 $•
7. Philopterus ruficeps(Nitzsch):
3 $, 7 2, 7 L;
1 S, 1 2, 1 L;
3 2> 2 L.
8. Philopterus cincli (Denny):
1 2-
9. Philopterus excisus Nitzsch:
1 <5.
10. Penenirmus albiventris
(Scop.): 1 5.
11. Penenirmus ? nirmoideus
Nitzsch: 7 3, 9 30 L.
12. Degeeriella subcuspidata
(Nitzsch): 12 3, 21 j
13. Degeeriella upupae (Denny):
1 3, 8 9.
14. Degeeriella juno (Giebel):
10 3, 16 J, 5 L.
15. Degeeriella cyclothorax
(Nitzsch): 1 3, 7 2
16. Cuclotogaster heterographus
(Nitzsch): 1 j
17. Columbicola columbae (Lin-
naeus): 1 3, 2 j.
18. Goniodes dispar Nitzsch:
3 L.
19. Goniodes piageti Johnston et
Harrison: 2
20. Goniocotes alatus Piaget: 2 $>.
2 Coccothraustes coccothraus-
tes humei Sharpe, тп., 5, 26 V
и 13 VI 1944;
Carduelis caniceps subcaniceps
Zar., ит., 5, 18 V 1944;
2 Serinus pusillus (Pall.), ит.,
3 и juv., 3 и 24 VI 1944;
Rhodopechys sanguinea (Gould),
сзм., 3, 24 VI 1944;
Erythrina e. erythrina (Pall.),
вп., 9, 15 V 1944;
Lanius m. minor Gm., ев.,
3, 3 VI 1944;
Acrocephalus d. dumetorum
Blyth, тс., 3, 15 V 1944.
3 Passer domesticus bactrianus
Zar. et Kudasch. тп., 3 и q, 15
и 30 V, 13 VI 1944;
Emberiza stewarti (Blyth), ит.,
3, 8 V 1944;
Emberiza bruniceps Brandt,
ит., 2, 6 VI 1944.
Cinclus cinclus L., тп., sex?,
2 XII 1940 (сбор E. Сосниной).
Hirundo rufula scullii (Seeb.).
(cm. 4).
Troglodytes troglodytes L., тп.,
sex.?, 2 XII 1940 (сбор E. Сосни-
ной).
Oenanthe pleschanka (Lep.),
сзм., 3, 17 V 1944.
Coracias garrulus semenowi
Loud, et Tsch., ев., 3, 21 V 1944.
Upupa e. epops L. (см. 1).
Coccothraustes coccothraustes
humei Sharpe (cm. 6, 26 V 1944).
Passer domesticus bactrianus
Zar. et Kudasch. (cm. 7, o, 13 VI
1944).
Alectoris kakelik (Falk), сзм.,
2, 22 V 1944.
Streptopelia orientalis meena
(Sykes), вп., 3, 25 V 1944.
Alectoris kakelik (Falk) (cm.
16).
Streptopelia orientalis meena
(Sykes) (cm. 17).
Alectoris kakelik (Falk.) (cm.
16).
Пухоеды птиц
227
Свободными от Mallophaga оказались птицы 10 видов Passeres:
Muscicapa striata neumanni Poche, ев., 2 5, 17 и 27 V 1944.
Phylloscopus nitidus viridanus Blyth, k., sex.?, 20 V 1944.
Phylloscopus inornatus humei (Brooks), k., 5 2, 24 V 1944.
Garrulax lineatus bilkewitchi (Zar.), k., 5 и j, 2 XII 1940; 5, 21 V 1944.
Lanius isabellinus phoenicuroides Schal., 5, 2 VI 1944.
Sylvia curruca blythi Tic. et Whist., ев., 5 и j, 18 V 1944.
Sylvia communis rubicola Stres., ев., j, 22 V 1944.
Sylvia nisoria merzbacheri Schal., ев., 5, 27 V 1944.
Sylvia hortensis crassirostris Cretzschm., сзм., 5, 2 VI 1944.
Irania gutturalis (Guer.), ит., S и juv., 23 VI 1944.
Я пользуюсь здесь случаем выразить признательность А. Я. Тугари-
нову и Е. В. Козловой-Пушкаревой за предоставление мне птиц для
осмотра во время работы на Кондаре в 1944 г.
15*
Е. Ф. СОСНИНА
ПАРАЗИТЫ МЫШЕВИДНЫХ ГРЫЗУНОВ
Фауна паразитов животных, обитающих в ущелье Кондара, еще не доста-
точно известна, так как лишь небольшая часть последних была подверг-
нута паразитологическому обследованию. Имеющийся материал позволяет
дать характеристику паразитофауны грызунов семейства Muridae: турке-
станской крысы Rattus turkestanicus Satun., лесной мыши Apod emus
sylvaticus L. и домовой мыши Mus musculus sewertzovi Kaschk.
Сбор материала производился автором в июле—сентябре 1945 г.
и с конца апреля до ноября 1946 г. Всего было обследовано 237 турке-
станских крыс, 199 лесных мышей и 39 домашних мышей. Грызуны отлав-
ливались, в основном, в ущелье Кондара, кроме того — в урочище Квак
и кишлаке Гажни, расположенном вблизи Горной ботанической станции,
в соседнем ущелье.
Черви определены автором, гамазовые клещи — Н. Г. Брегетовой,
иксодовые клещи — автором под руководством Б. В. Лотоцкого, красно-
телки — Е. Г. Шлугер, саркоптиды — А. А. Захваткиным, ногохвостки —
М. Н. Римским-Корсаковым, вши — автором под руководством Д. И.
Благовещенского, блохи — И. Г. Иоффом.
Помещенный ниже список включает не только паразитов грызунов,
но и некоторых непаразитических членистоногих, имеющих биологиче-
скую связь с грызунами. Всего здесь указано 48 видов червей и членисто-
ногих, чем, конечно, не исчерпывается паразитофауна обследованных
хозяев. Следует отметить, что в этом списке 17 видов оказались новыми,
из которых 15 относятся к клещам краснотелкам, по одному — к вшам
и блохам. Для круглого червя Rictularia amurensis найдены самцы и уста-
новлены хозяева. Ряд видов впервые обнаружен на территории Совет-
ского Союза. Многие виды указываются впервые для Таджикистана;
для некоторых паразитов установлены новые хозяева.
СОСАЛЬЩИКИ - TREMATODES
Сосальщики оказались редкими паразитами у обследованных грызу-
нов и представлены лишь двумя видами.
Сем. Dicrocoeliidae
Lyperosomum yitta (Dujardin). Найдены 4 экземпляра в желчных
протоках печени лесной мыши из ущелья Кондара. Цикл развития и про-
межуточные хозяева этого довольно редкого паразита не известны; веро-
ятно ими служат какие-то наземные моллюски.
Паразиты мышевидных грызунов
229
Сем. Bracbylaemidae
Brachylaemus recurvus (Dujardin). Обнаружен 21 экземпляр в кишеч-
нике двух туркестанских крыс, пойманных на берегу р. Варзоб. Про-
межуточные хозяева неизвестны, возможно, что ими являются слизни.
ЛЕНТОЧНЫЕ ЧЕРВИ - CESTODES
Ленточные черви, обнаруженные в ущелье в большом количестве,
представлены четырьмя видами. Для двух видов грызуны служат окон-
чательными хозяевами, для двух других — промежуточными.
Сем. Mesocestoididae
Mesocestoides lineatus (Goeze). В ущелье Кондара обнаружено 7998
личинок у 22 туркестанских крыс и 825— у трех лесных мышей. Турке-
станская крыса отмечена как новый промежуточный хозяин этого широко
распространенного паразита тонкого отдела кишечника хищных зверей.
Первым промежуточным хозяином служат панцырные клещи (Солда-
това, 1944); дальнейшее развитие личиночной стадии происходит в поло-
сти тела грызунов.
Сем. Hymenolepididae
Hymenolepis diminuta Rud. В ущелье Кондара найден у 5 туркестан-
ских крыс в количестве 78 экземпляров и у 9 лесных мышей 21 экзем-
пляр.
Обычный паразит кишечника крыс и мышей, имеющий всесветное
распространение, встречающийся и у других мелких грызунов. Проме-
жуточными хозяевами служат блохи, некоторые жуки, гусеницы бабо-
чек, тараканы, многоножки. Известны случаи паразитирования у чело-
века.
Сем. Taeniidae
Taenia taeniaeformis (Batsch). Цистицерки этого ленточного червя
найдены в печени у 10 туркестанских крыс в количестве 11 экземпляров,
у 7 лесных мышей 8 экземпляров и у 3 домовых мышей 4 экземпляра.
Половозрелые черви паразитируют у домашних и диких плотоядных
сем. Felidae.
Catenotaenia pusilia Goeze. Широко распространенный паразит кишеч-
ника домовой мыши, встречающийся и у других грызунов, представлен
21 экземпляром от 10 лесных мышей.
КРУГЛЫЕ ЧЕРВИ - NEMATODES
Круглые черви включают семь видов, большинство из которых обыч-
ные, повсюду обитающие паразиты грызунов, но некоторые являются
интересными и редкими находками.
230
Е. Ф. Соснина
Сем. Trichuridae
Trichuris muris (Schrank). Один самец этого широко распространен-
ного, но немногочисленного паразита грызунов найден в слепой кишке
лесной мыши из урочища Квак.
Сем. Tricbostrongylidae
Heligmosomoides dubius (Baylis). В ущелье Кондара у 112 лесных
мышей найдены 661 самец и 721 самка; у 14 туркестанских крыс 15 сам-
цов и 27 самок; в урочище Квак у 5 лесных мышей найдено 29 самцов
и 54 самки. Известен как паразит лесной мыши, обнаруженный на север-
ном Кавказе, в Закавказье, БССР, в Англии и Швейцарии. Отмечался
и у других грызунов.
Сем. Oxyuridae
Aspiculuris tetraptera (Nitzsch). Этот всесветно распространенный пара-
зит домовой мыши, а иногда и других грызунов, найден в ущелье Кон-
дара у 11 домовых мышей в количестве 37 самцов, 115 самок ; у двух дес-
ных мышей 19 самцов, 12 самок; у двух туркестанских крыс 4 самки.
Syphacia obvelata (Rudolphi). Обнаружен в ущелье Кондара: у 6 домо-
вых мышей в количестве 19 самок; у 26 лесных мышей 4 самца, 439 самок;
у туркестанской крысы 1 самка; в урочище Квак найдено 6 самок у лес-
ной мыши. Повсюду известный паразит слепой и толстой кишок мышей,
крыс, полевок и других грызунов, встречающийся иногда и у человека.
Сем. Heterakidae
Heterakis spurnosa Schneider. Всесветно распространенный паразит
крыс, мышей и других грызунов, найденный в ущелье Кондара: у 79
туркестанских крыс 123 самца, 232 самки; у домовой мыши 5 самцов,
2 самки; у трех лесных мышей 1 самец, 4 самки; в урочище Квак у крысы
8 самцов, 17 самок.
Сем. Spiruridae
Protospirura muris (Gmelin). Этот всесветно распространенный паразит
мышей, крыс и полевок встречен в ущелье Кондара у четырех лесных
мышей в количестве 20 экземпляров (4 самца и 16 самок).
Сем. Rictulariidae
Rictularia amurensis Schulz. В ущелье Кондара найден у 31 турке-
станской крысы 1 самец, 149 самок; у 39 лесных мышей 17 самцов, 126
самок. В урочище Квак обнаружен у 8 крыс 1 самец, 60 самок; у лесной
мыши 1 самец, 4 самки. Описан по самкам из коллекций Зоологического
института АН СССР от точно не указанного хозяина («мышь») из Пример-
Паразиты мышевидных грызунов
231
ской области. Таким образом, нами впервые найдены самцы и установ-
лены два хозяина для этого вида, цикл развития которого остается неиз-
вестным.
СКРЕБНИ - ACANTHOCEPHALA
Скребни обнаружены у грызунов ущелья лишь в редких случаях.
Всего отмечено два вида, для одного из которых грызуны являются
окончательным хозяином, для другого — промежуточным.
Сем. Oligacanthorbyncbidae
Macracanthorhynchus catulinus Kostylew. В брюшной полости турке-
станской крысы из ущелья Кондара обнаружена одна личинка этого
паразита кишечника хищников: собак, волков, лисиц и др. Как уста-
новлено В. Б. Дубининым (1948), личинки находятся в мышцах, тканях
внутренних органов и полости тела грызунов и мелких хищных. Пови-
димому, развитие происходит с двумя промежуточными хозяевами, при-
чем первым служат, вероятно, какие-то насекомые.
Сем. Monilif ormidae
Moniliformis moniliformis (Bremser). В ущелье Кондара встречен
у 3 лесных мышей в числе 3 самцов и 3 самок. Всесветно распространен-
ный паразит грызунов, в качестве промежуточных хозяев которого ука-
зываются тараканы и жуки чернотелки. Возможны случаи нахождения
скребня четковидного у человека.
КЛЕЩИ - ACARINA
Встреченные на мышевидных грызунах клещи относятся к трем под-
отрядам: Parasitiformes, Trombidiformes и Sarcoptiformes.
Подотряд PARASITIFORMES
Надсем. Parasiloidea
Гамазовые клещи, обнаруженные на грызунах ущелья, относятся
к 20 видам, часть которых в настоящее время не могла быть точно опре-
делена.
Сем. Parasitidae
Poecilochirus sp. Дейтонимфа этого рода найдена на туркестанской
крысе из ущелья Кондара 13 июня 1946 г.
Parasites consanguineus Oudms. et Vgts. Впервые отмечен в СССР
в числе двух самок и двух дейтонимф с туркестанской крысы, добытой
1 мая 1946 г. в ущелье Кондара.
232
Е. Ф, Соснина
Parasitidae, gen. sp. Одна дейтонимфа этого семейства снята с турке-
станской крысы, пойманной 3 мая 1946 г. в кишлаке Гажни.
Сем. Haemogamasidae
Haemogamasus sp. Две самки найдены в ущелье Кондара на турке-
станских крысах 19 августа и 30 октября 1946 г.
Euhaemogamasus oudemansi (Hirst). Две самки с домовой мыши, пой-
манной 28 октября 1946 г. на территории Горной ботанической станции,
оказались принадлежащими к этому виду, впервые указанному для Тад-
жикистана. Кроме того, вид был найден в Ленинграде. Известен из
Великобритании и южной Африки (Брегетова, 1949).
Сем. Laelaptidae
Laelaps turkestanicus Lange. Вид, описанный по сборам Н. Г. Бреге-
товой с туркестанской крысы из урочища Квак, оказался в наших сбо-
рах наиболее многочисленным из гамазовых клещей. Имеется 4102 самки,
227 самцов, 12 протонимф и 155 дейтонимф с двухсот туркестанских крыс
(в основном, из ущелья Кондара, а также из урочища Квак и кишлака
Гажни); 34 самки, 2 самца и 2 дейтонимфы с 22 лесных мышей (главным
образом из ущелья Кондара и немного из урочища Квак); 2 самки с двух
домовых мышей из ущелья Кондара. Отмечен также у грызунов в Стали-
набаде.
Laelaps agilis Koch. Имеется 248 экземпляров этого, впервые встре-
ченного в Таджикистане вида: 190 самок, 32 самца и 9 дейтонимф с 47
лесных мышей из ущелья Кондара; 2 самки и 4 дейтонимфы с двух лес-
ных мышей из урочища Квак; 8 самок и 2 самца с шести туркестанских
крыс из ущелья Кондара и 1 самка с крысы из кишлака Гажни.
Laelaps algericus Hirst. Из ущелья Кондара имеется 8 самок с пяти
домовых мышей, 2 самки с туркестанской крысы и 1 самка с лесной мыши.
Впервые найден в Таджикистане на домовой мыши в Сталинабаде.
Eulaelaps stabularis Koch. Имеется 39 самок с 24 туркестанских крыс
из ущелья Кондара, урочища Квак и кишлака Гажни и 2 самки с двух
лесных мышей из ущелья Кондара. Широко распространенный вид,
находимый свободно в природе, в хлевах, сеновалах, на грызунах, насе-
комоядных и в их гнездах, а также в гнездах береговых ласточек. Изве-
стен из различных областей Советского Союза (Брегетова и Высоцкая,
1949). В Таджикистане отмечен в Сталинабаде.
Mysolaelaps sp. Две самки сняты с лесной мыши из ущелья Кондара.
Gymnolaelaps sp. В ущелье Кондара найдены 2 самки: одна на лес-
ной мыши, другая — на домовой мыши.
Haemolaelaps pygmaeus (Muller). В Таджикистане обнаружен впер-
вые: по одной самке найдено на четырех туркестанских крысах из ущелья
Кондара и урочища Квак, 7 самок снято в ущелье с лесной мыши и 2
самки с домовой мыши. Обитает в гнездах грызунов и насекомоядных;
отмечался на крысах и полевках. В СССР найден в Ленинградской обла-
сти в гнездах серой полевки (Брегетова и Высоцкая, 1949).
Haemolaelaps megaventralis Str. Имеется 37 самок этого вида, впер-
вые обнаруженного в СССР: 16 самок с 14 туркестанских крыс из ущелья
Кондара; 1 самка с крысы из урочища Квак; 2 самки из кишлака Гажни;
Паразиты мышевидных грызунов
233
16 самок с домовой мыши, пойманной в сарае с саманом на Горной бота-
нической станции; 2 самки с двух лесных мышей из ущелья Кондара.
Androlaelaps sp. Имеются 3 самки с туркестанских крыс: 1 самка
из урочища Квак и 2 самки из ущелья Кондара.
Laelaptidae, gen. sp. I. Восемь самок клещей этого семейства найдены
на трех туркестанских крысах в ущелье Кондара в сентябре 1945 г.
Laelaptidae, gen. sp. II. Одна самка снята с туркестанской крысы
из ущелья Кондара в сентябре 1945 г.
Сем. Liponyssidae
Liponyssus bacoti Hirst. В Таджикистане обнаружен впервые в ущелье
Кондара: имеется 5 самок и 1 дейтонимфа с пяти туркестанских крыс
и 4 самки с четырех лесных мышей.
Тропический крысиный клещ, описанный из Египта; известен из мно-
гих далеко отстоящих друг от друга мест. Отмечены случаи нападения
на человека. Является переносчиком техасского тифа, резервуаром кото-
рого являются крысы.
Ichoronyssus sp. Из урочища Квак имеются 2 самки с двух турке-
станских крыс и 2 самки с двух лесных мышей; из ущелья Кондара —
2 самки с двух крыс и 3 самки с двух лесных мышей.
Сем. Dermanyssidae
Allodermanyssus sanguineus (Hirst). Имеется 35 самок, 3 самца, 39
протонимф и 50 дейтонимф с 44 туркестанских крыс из ущелья Кондара
и кишлака Гажни и 2 самки, 3 протонимфы и 1 дейтонимфа с пяти лес-
ных мышей из ущелья. Описан с грызунов Египта, известен также
из Вашингтона. В Таджикистане отмечен впервые в Сталинабаде на тур-
кестанских крысах и домовых мышах.
Надсем. Gamasoidea
В ущелье Кондара найдено 3 экземпляра (1 самка и 2 дейтонимфы)
точнее не определенных клещей на двух туркестанских крысах и 6 экзем-
пляров (2 самки, 3 самца, 1 дейтонимфа) на лесной мыши.
Надсем. Ixodoidea
Иксодовые клещи паразитируют на мышевидных грызунах почти
исключительно в неполовозрелых фазах развития. В ущелье Кондара
на грызунах обнаружены личинки и нимфы пяти видов иксодовых клещей.
Сем. Ixodidae
Ixodes redikorzevi Olen. В ущелье Кондара собраны личинки и нимфы
этого вида. Имеется 11 нимф и 114 личинок с 11 туркестанских крыс,
6 личинок с двух домовых мышей, 7 нимф и 28 личинок с 14 лесных
мьйпей.
234
Е. Ф. Соснина
Haemaphysalis sulcata Can. et Fanz. Имеется одна личинка этого вида
с лесной мыши, добытой 21 июня 1946 г. на территории Горной ботани-
ческой станции.
Haemaphysalis caucasica Olen. В сборах с мышевидных грызунов
найдены личинки и нимфы, относящиеся, повидимому, к этому виду:
из ущелья Кондара 59 нимф и 79 личинок с 40 туркестанских крыс, 40
нимф и 42 личинки с 36 лесных мышей; из урочища Квак 1 нимфа и 2
личинки с туркестанской крысы и 2 нимфы с лесной мыши.
Rhipicephalus turanicus В. Pom. Из ущелья Кондара имеется 46 нимф
и 59 личинок, принадлежащих, повидимому, к этому виду: 43 нимфы
и 50 личинок с 30 туркестанских крыс, 1 личинка с домовой мыши,
3 нимфы и 8 личинок с 6 лесных мышей.
Hyalomma anatolicum Koch. Из ущелья Кондара имеются 2 нимфы
и 5 личинок с 7 туркестанских крыс и 2 нимфы с 2 лесных мышей, отно-
сящиеся к этому виду.
Подотряд TROMBIDIFORMES
Собранный материал содержит хищных клещей, встречающихся
иногда на грызунах, некоторых постоянных паразитов грызунов, а также
многочисленных и богатых видами клещей краснотелок, паразитирую-
щих на грызунах в личиночной фазе развития.
Сем. Cheyletidae
Cheyletus sp. Хищные клещи найдены в ущелье Кондара на турке-
станских крысах: 22 августа 1946 г. — 1 экземпляр и 2 октября 1946 г. —
2 экземпляра.
Myobi a sp. В сборах имеется 5 клещей с пяти домовых мышей, 3 клеща
с лесной мыши и 2 клеща с двух крыс. Все развитие этих клещей
происходит на грызунах.
Сем. Trombidiidae
Клещи краснотелки, в личиночной фазе паразитирующие на различ-
ных животных (подсем. Trombiculinae — преимущественно на грызунах),
оказались представленными 18 видами, причем 15 видов являются новыми,
описанными Е. Г. Шлугер по настоящим сборам. Один из новых видов
обнаружен только на землеройке. Представители трех родов: Hannema-
nia, Leeuwenhockia и Walchia впервые найдены в СССР.
Trombicula manis Schluger. Из ущелья Кондара имеется 7 личинок
с четырех туркестанских крыс и 5 личинок с двух лесных мышей.
Trombicula pulchra Schluger. Обнаружено 3130 личинок с туркестан-
ских крыс, главным образом из ущелья Кондара, а также из урочища
Квак и кишлака Гажни, 248 личинок с лесных мышей, 3 личинки с домо-
вой мыши и 2 личинки с землеройки Crocidura sp. из ущелья.
Trombicula praemontanus Schluger. Из ущелья Кондара имеются 63
личинки с 17 туркестанских крыс, 76 с 23 лесных мышей, 3 с трех домо-
вых мышей, 3 с землеройки Crocidura sp.; из урочища Квак 3 личинки
с лесной мыши.
Паразиты мышевидных грызунов
235
Trombicula hirsutus Schluger. Из ущелья Кондара имеются 163 личинки
с туркестанских крыс, 9 личинок с лесных мышей; из урочища Квак
16 личинок с туркестанских крыс.
Trombicula raropinnis Schluger. Из ущелья Кондара имеется 583
личинки с туркестанских крыс, 110 личинок с лесных мышей и 52 с трех
землероек Crocidura sp.; из урочища Квак 183 личинки с крыс и 4 личинки
с лесной мыши; из кишлака Гажни 16 личинок с туркестанских крыс.
Trombicula talysini Schluger. В ущелье Кондара найдены 31 личинка
с 7 туркестанских крыс и 21 личинка с двух лесных мышей; в урочище
Квак 2 личинки с крысы и 2 личинки с лесной мыши; в кишлаке Гажни
1 личинка с туркестанской крысы.
Trombicula sosninae Schluger. Обнаружено 55 личинок в урочище
Квак в ушных раковинах двух туркестанских крыс.
Trombicula vulgaris Schluger. Из ущелья Кондара имеется 3690 личи-
нок с туркестанских крыс, 1177 с лесных мышей и 14 с домовых мышей;
из урочища Квак 12 личинок с туркестанских крыс.
Trombicula autumnalis Shaw. Из ущелья Кондара имеется 818 личи-
нок с туркестанских крыс, 1640 с лесных мышей, 7 с домовых мышей;
из урочища Квак 1 личинка с крысы и 8—с лесной мыши.
Neoschoengastia longior Schluger. В ущелье Кондара отмечены 2
личинки на туркестанских крысах и 9 личинок на лесных мышах.
Neoschoengastia intermedia Schluger. Найдены 204 личинки на турке-
станских крысах и 22 на лесных мышах в ущелье Кондара.
Neoschoengastia latyshevi Schluger. Имеется 185 личинок с турке-
станских крыс из ущелья Кондара; 22 личинки с крыс и 6 личинок с лес-
ной мыши из урочища Квак; 5 личинок с крысы из кишлака Гажни.
Впервые найден на бурундуке Eutamias asiaticus в Приморье (Шлугер,
1947).
Neoschoengastia rotundata Schluger. В ущелье Кондара обнаружено
3004 личинки с туркестанских крыс, 783 личинки с лесных мышей, 49
личинок с домовых мышей, 7 личинок с землеройки Crocidura sp.; в уро-
чище Квак 4 личинки с крыс, в кишлаке Гажни 2 личинки с крысы.
Личинки этого вида впервые найдены на грызунах в Приморском крае
(Шлугер, 1947).
Neoschoengastia sparsa Schluger. Представлен 177 личинками из ущелья
Кондара: 171 экземпляр с туркестанских крыс, 3 экземпляра с лесных
мышей и 3 экземпляра с домовой мыши.
Neoschoengastia olsufievi Schluger. Из ущелья Кондара имеется 875
личинок, из них 790 с туркестанских крыс и 85 с лесных мышей.
Hannemania oudemansi Schluger. В ущелье Кондара найдено236 экзем-
пляров: 22 личинки на туркестанских крысах, 213 на лесных мышах
и 1 личинка на домовой мыши.
Leeuwenhockia major Schluger. Имеется 90 личинок из ущелья Кон-
дара: 87 с туркестанских крыс и 3 личинки с лесных мышей.
Walchia parvula Schluger. Обнаружено шесть личинок на землеройке
Grocidura sp. из ущелья Кондара.
Подотряд SARCOPTIFORMES
Клещи этого подотряда представлены двумя семействами хлебных
и тремя семействами панцырных клещей. Хлебные клещи имеют древ-
нюю связь с грызунвми, обитая в их норах и пользуясь зверьками для
расселения (явление форезии). Панцырные клещи являются промежу-
236
Е. Ф. Соснина
точными хозяевами некоторых ленточных червей, паразитирующих у гры-
зунов, и нередко живут в норах последних.
Сем. Tyroglyphidae
Tyroglyphus farinae (L.). В ущелье Кондара обнаружено по одному
экземпляру на туркестанской крысе и лесной мыши. Этот всесветно рас-
пространенный мучной клещ встречается в природе в стогах сена, соломы,
в норах грызунов, а также на различных животных, являющихся распро-
странителями этого клеща.
Tyrophagus humerosus humerosus (Ouds.) A. Z. Один экземпляр найден
на туркестанской крысе на территории Горной ботанической станции.
Известен из различных районов СССР, Голландии, Германии; распро-
странен в полевой обстановке.
Сем. Glycyphagidae
Glycyphagus destructor Ouds. Из ущелья Кондара имеются два взрос-
лых клеща с туркестанской крысы и один гипопус с домовой мыши.
Один из наиболее распространенных и обычных видов амбарных кле-
щей; изредка находился в полевых условиях.
Glycyphagus domesticus (Deg.). В ущелье Кондара обнаружено
6 взрослых и 1 гипопус на туркестанских крысах, 2 экземпляра
на лесных мышах, 12 взрослых и 1 нимфа на домовых мышах.
Кроме того в тонких кишках двух лесных мышей найдено 3 экзем-
пляра хорошо сохранившихся клещей. Широко распространенный,
преимущественно домовый вид, встречающийся в поле в стогах и норах
грызунов.
Myacarus sp. Один экземпляр клеща, принадлежащего к этому роду,
обнаружен на туркестанской крысе из ущелья Кондара. Известны
только гипопусы двух видов этого рода, находимые на различных гры-
зунах и мелких насекомоядных.
Сем. Oribatulidae
Oribatula sp. обнаружен в ущелье Кондара на туркестанских крысах
в количестве 5 взрослых и 1 нимфы; на лесной мыши 1 нимфа.
Zygoribatula sp. отмечен на грызунах из ущелья: 7 клещей на крысах
и 2 на лесной мыши.
Zygoribates sp. найден 1 клещ на крысе из ущелья Кондара.
Сем. Ceratozetidae
Scheloribates sp. Имеется 1 клещ с крысы из ущелья Кондара.
Сем. Galumnidae
Galumna lanceatus kazakhstani Kriv. С трех крыс имеется 17 кле-
щей, с лесной мыши — 1 клещ.
Паразиты мышевидных грызунов
237
Allogalumna alifera Oudemans. Имеется 1 клещ с крысы из ущелья
Кондара.
НАСЕКОМЫЕ — INSECT А
НОГОХВОСТКИ — COLLEM BOLA
Ногохвостки, встреченные на грызунах ущелья Кондара в неболь-
шом количестве, представлены четырьмя видами. С. О. Высоцкой в окре-
стностях Ленинграда было отмечено довольно большое количество видов
ногохвосток в норах серой полевки и на самих зверьках. На основании
этого ею высказано предположение, что ногохвостки пользуются теплом
тела грызунов, а возможно и питаются омертвевшими частями кожи или
продуктами выделения последней (Высоцкая, 1949).
Сем. Poduridae
Hypogastrura armata Nic. Обнаружен на лесной мыши в количестве
6 экземпляров.
Pseudachorutes sp. Отмечен в одном экземпляре на лесной мыши.
Сем. Eutomobryidae
Eutomobrya sp. Найден в одном экземпляре на туркестанской крысе.
Orchesella sp. Обнаружен в одном экземпляре на лесной мыши.
ВШИ — ANOPLURA
Собранные в ущелье Кондара вши относятся к четырем видам одного
семейства, характерного для грызунов; вошь туркестанской крысы ока-
залась новым видом.
Сем. Haematopinidae
Hoplopleura affinis (Bunn.). Широко распространенный паразит лес-
ной мыши и других грызунов представлен 302 самками, 167 самцами
и 131 личинкой с 37 лесных мышей из ущелья Кондара и двух лесных
мышей из урочища Квак.
Hoplopleura hesperomydis (Osborn). Паразит домовой мыши, обнару-
женный на территории Горной ботанической станции на этом хозяине
в количестве 24 самок, 20 самцов и 1 личинки. Кроме того найдена 1 самка
на лесной мыши.
Hoplopleura pavlovskyi Sosnina. Этот вид впервые обнаружен на тур-
кестанских крысах в ущелье Кондара, урочище Квак и кишлаке Гажни.
Имеется 2001 самка, 1840 самцов и 2195 личинок.
Polyplax affinis Fahrenholz. Паразит лесной мыши, обнаруженный
в количестве 113 экземпляров (58 самок, 30 самцов и 25 личинок);
имеется также 1 самка с домовой мыши.
238
Е. Ф. Соснина
БЛОХИ — APHAXIPTERA
Блохи, собранные с грызунов ущелья, принадлежат к пяти видам,
один из которых оказался новым.
Сем. Ceratoph yllidae
Ceratophyllus (Nosopsyllus) simla J. et R. С туркестанских крыс
(главным образом из ущелья Кондара, а также из урочища Квак и киш-
лака Гажни) собрано 139 самок и 109 самцов; с лесных мышей 10 самок
и 8 самцов;-с домовой мыши 1 самка. Описана в Индии у крыс и домовой
мыши. В СССР найдена в Таджикистане и в Киргизии на туркестанской
крысе, может переходить и на других грызунов.
Neopsylla hissarica loff et Sosnina. Этот впервые обнаруженный вид
представлен 111 экземплярами, главным образом из ущелья Кондара,
а также из урочища Квак и кишлака Гажни. Имеется 67 самок и 38 самцов
с 67 туркестанских крыс и 4 самки и 2 самца с 6 лесных мышей.
Leptopsylla taschenbergi W. В ущелье Кондара собрано 111 самок
и 77 самцов с 80 лесных мышей и 6 самок и 6 самцов с 7 туркестанских
крыс. Паразит лесных мышей, встречающийся иногда и на других гры-
зунах.
Amphipsylla anceps W. В ущелье Кондара найден 1 самец на турке-
станской крысе. Является паразитом хомячков, но переходит и на дру-
гих грызунов; известен из Таджикистана, с Тянь-шаня и Алтая.
Сем. Pulicidae
Pulex irritans L. На туркестанской крысе, добытой на территории
Горной ботанической станции, найден 1 самец блохи человека и хищ-
ников.
ЛИТЕРАТУРА
Брегетова И. Г. 1949. Материалы по фауне клещей сем. Haemogamasidac в СССР.
Паразитолог, сборн. Зоолог, инет. АН СССР, т. XI.
Брегетова Н. Г. и С. О. Высоцкая. 1949. Гамазовые клещи (Gamasina,
Parasitiformes) — паразиты обыкновенной полевки (Microtus arvalis Pall.)
и обитатели ее гнезд в окрестностях Ленинграда. Паразитолог, сборн. Зоолог,
инет. АН СССР, т. XI.
Высоцкая С. О. 1949. Об одном из возможных путей происхождения эктопара-
зитизма. Докл. АН СССР, т. 64, вып. 3.
Дубинин В. Б. 1948. К вопросу о цикле развития скребня Machacanthorhynchu
catulinus Kostylew, 1927. Докл. АН СССР, т. 60, вып. 6.
Пофф И. Г 1949. Aphaniptera Киргизии. Эктопаразиты. Фауна, экология и эпидс
миологическое значение. Изд. Акад, медиц. наук СССР, вып. 1.
Пофф И. Г., О. И. Скалой, 3. М. Вовчинская, Н. Ф. Дарская
Н. Д. Емельянова, Э. В. Исаева-Гу рви ч, Б. А. Рости
гаев, Р Ф. Савенко, Е. Ф. Соснина, П. Т. Сы невский
1950. Новые виды блох (Aphaniptera). Сообщение II. Медицинская паразито-
логия и паразитарные болезни, т. XIX, вып. 3.
Солдатова А. П. 1944. К изучению цикла развития цестоды Mesocestoides
lineatus (Goeze, 1782), паразитирующей у хищных млекопитающих. Докл.
АН СССР, т. 45, вып. 7.
Соснина Е.Ф. 1949. К фауне сосальщиков мышевидных грызунов Таджикистана.
Сообщ. Тадж. фил. АН СССР, вып. XVIII.
Паразиты мышевидных грызунов
23<У
Соснина Е. Ф. 1950. К фауне ленточных червей мышевидных грызунов Таджи-
кистана. Там же, вып. XXII.
Соснина Е. Ф. 1950. К фауне скребней мышевидных грызунов Таджикистана.
Там же.
Соснина Е. Ф. 1950. К фауне круглых червей мышевидных грызунов Таджики-
стана. Там же, вып. XXIII.
Соснина Е. Ф. 1950. Исследование фауны клещей краснотелок в Таджикистане.
Там же, вып. XXV111.
Соснина Е. Ф. 1950. О сезоне паразитирования иксодовых клещей на мыше-
видных грызунах. Там же, вып. XXIX.
Ш л у г е р Е. Г 1947. Клещи подсемейства Troinbiculinae, зарегистрированные
в СССР. Новости медицины, вып. 5.
Е. С. КИРЬЯНОВА
ВОЛОСАТИКИ И КРУГЛЫЕ ЧЕРВИ
I
Плато Руидашт в районе Кондары изобилует волосатиками (Nema-
tomorpha), живущими во всех ручьях от самого истока и на всем протя-
жении их в районе Руидашта. Видовой состав волосатиков весьма разно-
образен и включает представителей 2 семейств: Chordodidae (роды: Рага-
chordodes, Gordionus, Paragordius) и Gordiidae (род Gordius). В ручьях
собственно ущ. Кондара волосатики встречаются довольно редко и ни разу
Рис. 1. Одновременный выход трех волосатиков в воду пз тела кузне-
чика Paradrymadusa hissaricola Mistshenko.
не были найдены в речке Кондара. Возможно, это объясняется чрезвы-
чайно крутым падением ручьев, впадающих в речку Кондара, и стреми-
тельным бурным течением самой речки, сносящей вниз попадающих
в нее волосатиков.
Личинки волосатиков паразитируют в различных водных и наземных
насекомых, в моллюсках и других беспозвоночных животных, покидая
их по достижении половозрелого состояния (рис. 1). Виды рода Gordius
размножаются в зимнее и ранневесеннее время (февраль—апрель), про-
изводя откладку яиц в виде длинных яйцевидных шнуров, значительно
превышающих длину их собственного тела (рис. 2). Самая ранняя яйце-
кладка волосатика Gordius sp. наблюдалась в соседнем ущелье Гажни
Волосатики и круглые черви
241
Рис. 2. Самка волосатика Gordius sp. в момент откладки ею яйцевого шнура.
(яйцевой шнур белого цвета).
10 февраля 1944 г., когда самка волоса-
тика вместе с отложенным ею яйцевым
шнуром была найдена под камнем у самого
истока ручья, где температура воды дости-
гала 8° С, несмотря на лежащий кругом
с нег.
II
Представители класса круглых червей
(Nematodes), обитающие в ущелье Кондара,
представлены 4 экологическими группами:
жители пресной воды, жители почвы, пара-
зиты растений и паразиты животных (бес-
позвоночных и позвоночных). Пресновод-
ная фауна нематод ущелья Кондара обсле-
довалась слабо. В данное время из числа
пресноводных нематод известно несколько
видов из родов Dorylaimus и Tripyla и 2—3
вида из сем. Mermithidae; последние в ли-
чиночном состоянии ведут паразитический
образ жизни.
Почвенная фауна нематод обследовалась
значительно более детально; она содержит
в своем составе несколько десятков червей,
большая часть которых представлена новы-
ми видами. В их число входят представи-
тели следующих родов: Mononchus (несколь-
ко видов), Rotylenchus (R. multicinctus),
Tylenchus (Т. filiformis и ряд других),
Рис. 3. Пшеница, сильно зараженная пшеничной
нематодой (а), и лист «василька» Rhaponticum
integrifolia С. Winkler (б), усыпанный галлами
нематоды Anguina sp.
16 Ущелье Кондара
242
Е. С. Кирьянова
Dorylaimus (D. intermedius, D. obtusicaudatus и много других), Alaimus
(A. primitivus и два новых вида), Longidorus, Nygolaimus, Monhystera,
Plectus, Rhabditis, Heterodera, Cephalobus, Eucephalobus (E. elonga-
tus) и некоторые другие. Многие виды почвенных нематод обладают
удивительной способностью переносить длительное высыхание, впадая
в состояние скрытой жизни. Так, например, в совершенно сухой поч-
ве, взятой на южном склоне горы в урочище Квак 30 июля 1942 г.,
нематоды пролежали 4.5 года без вреда для жизни и ожили 13—14 фев-
раля 1946 г. при увлажнении почвы, начав активно двигаться (Tylenchus
filiformis, Monhystera sp., Tylenchorhynchus sp.).
Паразиты растений обследовались также довольно детально, но мате-
риал в значительной степени не обработан. Широко распространенной
формой в Кондаре и близлежащих территориях является галлообразую-
щая нематода Anguina picridis Kirjanova, 1944 — паразит листьев и
стеблей горчака (Acroptilon picris С. А. М.) и Anguina sp., образующая
галлы на листьях различных видов кузиний, шалфея, скабиозы, василь-
ков и других растений (рис. 3). В пограничных с ущельем Кондара кол-
хозах встречается пшеничная нематода Anguina tritici — весьма серьез-
ный вредитель пшеницы в Таджикистане. В самом ущелье Кондара
на посевах пшеницы в 1942—1943 гг. эта нематода не была найдена.
Паразиты животных не изучались.
ЛИТЕРАТУРА
Кирьянова Е. С. 1944. Растениеядные нематоды Таджикистана. Пзв. Тадж.
филиала Ак. Наук СССР, 5 83—94.
Кирьянова Е. С. 1945. Растениеядные нематоды Таджикистана. Рефераты
работ учр. Отд. биолог, наук Ак. Наук СССР за 1941—1943 гг. 153.
Кирьянова Е. С. 1949. Новый вид волосатика из Средней Азии. Сообщ. Тадж.
филиала Ак. Наук СССР, 12 21—23.
Кирьянова Е. С. 1949. Первая находка новых видов волосатиков в Таджики-
стане. Там же, 14 : 19—21.
Кирьянова Е. С. 1950. Новый род волосатика (Progordius, gen. nov.) из Таджи-
кистана. Докл. Ак. Наук СССР, нов. сер. 71 (6) 1171—1173.
ntf о a
В. II. ЖАДИН, И. А. КИСЕЛЕВ, А. Г РОДИНА,
В. Я. ПАНКРАТОВА и |ЗС~В. ГРИБ ;
ЖИЗНЬ В ВОДАХ КОНДАРЫ
Воды Кондары, в том ее понимании, которое мы приняли для нашего
сборника, представляют собою три независимых друг от друга района.
Это, во-первых, воды субальпийского плато Руидашт, во-вторых, это
бассейн самой реки Кондара (в верхнем течении именуемой Уноу) и,
в-третьих, сюда же мы, присоединяем реку Варзоб, принимающую воды
Кондары у подножья ущелья.
1. ПЛАТО РУИДАШТ
О водах высокогорного плато Руидашт в настоящее время многого
сказать нельзя, потому что они изучены еще совершенно не достаточно.
Характерными особенностями водного режима Руидашта можно считать:
а) отсутствие вечных снегов, б) большое количество осадков, особенно
в виде снега, в) относительно малое количество постоянных, стабильных
водоемов и г) большое количество временных водоемов, питающихся
водами тающего снега и пересыхающих в середине лета.
Весной из-под тающего снега текут ручьи, образующие на своем
пути небольшие каскады, чередующиеся с участками полустоячей
воды — бочагами. В ледяной воде ручейков и согревшейся воде
бочагов в начале июня обитают личинки ортокладиин и ручейни-
ков и вместе с ними взрослые жуки — Helophorus brevipalpis и Hyd-
raena sp.
В обширных по площади и мелких по глубине пересыхающих лужах,
согревающихся до температуры, значительно превышающей температуру
воздуха, на дне часто можно встретить мох и синезеленые водоросли
Nostoc Linckia. В выжимке из мха и в планктоне луж найдены много-
численные водоросли, среди которых преобладают жгутиковые и зеле-
ные, в то время как диатомовые занимают второстепенное положение,
а синезеленые представлены всего 2 видами ностока. Руководящими
будут:
Flagellata — Euglena fusca, Euglena spirogyra, Euglena viridis, Phacus
acuminatus, Phacus orbicularis, Phacus pleuronectes, Trachelomonas his-
pida, Trachelomonas volvocina; Ghlorophyceae — Gonium pectorale, Pan-
dorina morum, Eudorina elegans, Volvox aureus, Phacotus lenticularis,
Oedogonium sp. sp., Bulbochaete sp., Glosterium pritchardianum, Gosma-
rium botrytis, Gosmarium calcareum, Gosmarium bireme, Gosmarium
asphaerophorum var. strigosum, Gosmarium phaseolus, Gosmarium subpro-
tumidum, Gosmarium Blyttii, Spirogyra sp.; Heterocontae — Tribortema
246 В, И. Жадин, И. А. Киселев, А. Г. Родина, В. Я. Панкратова и А, В. Гриб
sp.; Diatomeae — Synedra ulna, Neidium affine var. amphirhynchus,
Stauroneis anceps, Navicula cuspidata+var. Haribaudi, Pinnularia gibba,
Pinnularia pulchra, Pinnularia viridis, Hantzschia elongata; Hantzschia
amphioxys4-var. vivax; Cyanophyceae — Nostoc Linckia, Nostoc puncti-
forme.
Фауна очень богата и характерна для пересыхающих луж. В ней пре-
обладают различные группы насекомых, претерпевающих в теплой воде
луж быстрый метаморфоз, а также многочисленные ракообразные, обла-
дающие стойкими яйцами. В весенних лужах на Руидаште найдены
жуки и их личинки — Hydrophilus sartus, Deronectes airumulus, Coe-
lostoma transcaspicus, водяные клопы — Gerris costai, Notonecta viridis
subsp. mediterranea, личинки стрекоз Sympycna fusca, личинки тендипе-
дид — Orthocladiinae g.? larva adhaeria, larva dubia, Pelopia. Ракообраз-
ные представлены ветвистоусыми — Alonella excisa, Ceriodaphnia quad-
rangula, Rhynchotalona rostrata, Alona sp., Simocephalus vetulus, Chy-
dorus sphaericus, веслоногими — красным Diaptomus bacillifer и бесцвет-
ным Cyclops vernalis, и ракушковыми рачками — Eucypris lutaria, Cypris
pubera. Здесь также встречаются водяные клещи Eylais hamata, турбел-
ларии, нематоды и коловратки — Pedalia, Monostyla.
Летом, в бездождный период, водоемы Руидашта представлены пре-
имущественно маломощными ручейками и бочагами полустоячей воды.
Фауна такого рода водоемов состоит из насекомых и их личинок, немно-
гочисленных ракообразных и олигохет. Здесь найдены: личинки комара
Culex, личинки тендипедид Ablabesmyia ex gr. lentiginosa, Lauterbornia,
водяные жуки Gyrinus distinctus, Deronectes airumulus, Helophorus brevi-
palpis, Bidessus geminus, Gaurodytes sp., Hydroporus tesselatus, Ochthe-
bius depressus, Helodidae. Интересной чертой фауны ручьев Руидашта
является обилие волосатиков (см. статью Е. С. Кирьяновой в настоящем
сборнике).
В родниках Руидашта были обнаружены моллюски Pisidium caser-
tanum, веслоногие и ракушковые рачки — Cyclops fimbriatus, Candona
Candida, а также небольшое количество мелких турбелларий Seidlia,
жуков и личинок тендипедид.
2. БАССЕЙН РЕКИ КОНДАРА
а. Река Кондара
Начало свое река Кондара берет у подножья Руидашта в пределах
трех довольно обширных амфитеатров. Книзу долина суживается, пре-
граждается рядом неподдающихся размыву скал или скалистых нагро-
мождений, по которым река низвергается водопадами (рис. 1).
Истоки Кондары лежат на высоте 1860—1900 м над ур. м., а устье —
на высоте около 1120 м. Таким образом падение Кондары очень велико,
средний уклон ее равняется приблизительно 15 см на 1 м. Несмотря
на столь большое падение, скорость течения в Кондаре лишь при про-
хождении весенних паводочных вод достигает больших цифр. При межен-
ном состоянии реки большие скорости течения сохраняются лишь на во-
допадах, на обычных же местах, где вода, казалось бы, так же бурно
низвергается с камня на камень, измерения скоростей дают весьма невы-
сокие значения, порядка от 0.4 до 1.0 м в сек.
Жизнь в водах Кондары
247
Изредка, во время весенних обильных и длительных дождей, Кондара
вздувается и принимает характер потока, сметающего на своем пути
деревья и перемещающего камни.
Питается река Кондара преимущественно родниковыми водами. Чисто
родниковое питание продолжается все бездождное время летом и часть
зимы. Весной же, как говорилось, большая роль в питании Кондары
переходит к талым водам, осенью же некоторое дополнение к основному
родниковому питанию реки
дают дожди.
Термический режим Кон-
дары характеризуется значи-
тельной годовой амплитудой
температуры воды, достигаю-
щей 20.5°. Характерной чер-
той Кондары, как горной
реки, является большая су-
точная амплитуда темпера-
тур, особенно в летнее время.
Зимой суточная амплитуда
температуры воды колеблется
от 0 до 1.1°, весной она до-
ходит до 4.8, а летом до 6.5°.
Суточный ход температуры
воды здесь следует парал-
лельно ходу температуры
воздуха. Зимняя температу-
ра воды лишь редко дости-
гает 0°. В холодные зимние
дни кое-где у берегов появ-
ляется лед. При редких по-
холоданиях до —16.5° и
ниже, в Кондаре начинает
образовываться донный лед.
Подводные камни покрыва-
ются ледяными подушками,
по воде течет рыхлый лед
(шуга). В ряде мест он заби-
вает живое сечение реки и
образует плотины. Уровень
Рис. 1. Водопад на притоке Кондары.
(Фот. А. И. Иванова).
воды в местах таких подпруд
поднимается на 70—80 см;
река выходит из меженных
берегов, образуя бочаги, спо-
койная водная поверхность которых сейчас же сковывается льдом. Вода
реки, непрерывно поступающая из вышележащих участков, находит себе
дорогу поверх этих замерзших бочагов. Затем, чуть ослабнет мороз,
река, получающая согретую воду родников, просасывает себе путь
в ледяных плотинах и начинает течь как бы в ледяных каньонах под
куполом из тонкого льда.
Прозрачность воды в Кондаре на протяжении круглого года бывает
очень большой — дно реки видно отчетливо на всех ее участках. Лишь
после дождей вода реки становится слегка мутной, но и эта муть удер-
живается самое короткое время.
248 В. И. Жадин, И. А. Киселев, А. Г Родина, В. Я Панкратова и А. В. Гриб
Цвет воды чуть беловатый — в Кондаре не наблюдается той зеленовато-
голубой окраски, которая свойственна многим рекам Гиссарского хребта.
Химизм реки характеризуется умеренной жесткостью воды (см. Леп-
нева, 1944). Растворенный в воде кислород в течение всего года нахо-
дится в достаточном количестве, но колеблется на протяжении суток.
Суточная амплитуда содержания кислорода достигает 1.44 мг/л. Окис-
ляем ость воды незначительная.
Планктон р. Кондара состоит из бактерий, а кроме того, из дон-
ных животных и водорослей, отрывающихся течением воды и попадаю-
Рис. 2. Река Кондара.
(Фот. А. И. Иванова).
щих в речную струю. В один из осенних дней количество бактерий, вырос-
ших на мясо-пептонном агаре, колебалось от 28 до 56 на 1 см3 воды выше
Ботанической станции, достигая 217 ниже станции.
Донное население (бентос) Кондары группируется в 3 биоценоза:
1) биоценоз каменистого дна (литореофильный), 2) биоценоз песчаного
дна (псаммореофильный) и 3) биоценоз слегка заиленного дна (пелорео-
фильный). Господствующая роль принадлежит здесь биоценозу каме-
нистого дна, так как дно реки по меньшей мере на 95% состоит из кам-
ней и скал (рис. 2). Лишь в небольшом количестве мест позади камней
скапливается песок и совсем редко этот песок подергивается тонким
слоем наилка.
Растительная часть биоценоза каменистого дна Кондары состоит
из зеленых и синезеленых водорослей, прикрепляющихся к камням —
Spyrogyra, Phormidium uncinatum, Phormidium autumnale var. minor —
Жизнь в водах Кондары
249
и поселяющихся вместе с ними немногочисленных диатомей — Diatoma
elongatum var. tenuis, D. hiemale et var. mesodon, Synedra ulna typ. et
var. contracta, var. amphirhynchus, Synedra amphicephala, Synedra rum-
pens, Cocconeis placentula, Achnanthes linearis, Achnanthes minutissima
var. cryptocephala, Caloneis bacillum, Navicula cryptocephala et var.
veneta, Navicula radiosa, Navicula gracilis, Cymbella angustata var. minor,
Cymbella affinis, Cymbella naviculiformis, Cymbella Helvetica, Gompho-
nema parvulum var. subelliptica, var. exilis, Gomphonema intricatum
var. dichotoma, Nitzschia palea.
Животные, обитающие на камнях и скалах р. Кондара, принадлежат
преимущественно к классу насекомых (ручейники, веснянки, поденки,
двукрылые и жуки) и в небольшом количестве к водным клещам, червям
(турбелларии и олигохеты) и моллюскам.
Видовой состав фауны каменистого дна р. Кондара слагается сле-
дующими формами.
Oligochaeta
Lumbriculus variegatus
Nais pardalis
Nais bretscheri
Pristina rosea
Chaetogaster sp.
Tnrbellaria
Polycelidia receptaculosa
Seidlia sp.
Hydracarina
Sperchon plumifer
Sperchon glandulosus Cubanicus
Atrctides turkestan icus
Calonyx montanus
Dartia longispora
Megapus sp.
Lebertia sp.
Insecta
Dip ter a
Fam.? Gen?
Dixa
Blepharoceridae
Blepharocera fasciata asiatica
S i m u 1 i i d a e
Simulium (Odagmia) ornatum var. cauca-
sicum
Simulium (Odagmia) humerosum
Simulium (Obuchovia) albellum
Simulium (Simulium) multistriatum
Simulium (Simulium) subornatoides
Tendipedidae
Tendipedini G.? larva dulcis
Tanytarsus ex gr. exiguus
Microspectra
Lauterbornia
Ablabesmyia
Corynoneura
Orthocladiinae G.? larva condarensis
» principata
» sellata
» incurva
» oxiana
Eukiefferiella
Heptagia larva accomodata
Anatopynia
H e 1 e i d a e
Bezzia
Trichoptera (О пред. G. Г Лепневой)
Rhyacophila obscura
Glossosoma dentatum
Agapetus sp. sp.
Dolophilodes larva dolichocephala
Hydropsyche sp.
Dinarthrum pugnax
Apatelia copiosa
Leptoceridae G.? sp.?
Coleoptera
Staphylinidae (Lesteva fasciata?)
Ochthebius sp.
Plecoptera (indet.)
Ephemeroptera (Опред. О. А. Черновой,
рис. 3)
Rhithrogena
Ecdyonurus
Heptagenia
Ephemerella
Ordella
Baetis
Pseudocloeon?
Mollusca
Limnaea (Galba) truncatula
Рис. 3. Личинки поденок из р. Кондара:
А.— Ephemerella sp., Б—Rhithrogena sp., В—Iron sp.,r—Ecdyonurus sp. (Ориг.).
Жизнь в водах Кондары
251
Количество животных на камнях р. Кондара довольно велико.
На 0.1 м2 каменистого дна 1 здесь приходится весной в среднем 959 орга-
низмов, летом (в июле) 773, осенью (в октябре) 1069; среднее количество
за год составляет 934. Силевые потоки, иногда проходящие по Кондаре,
уменьшают количество животных — до 200—206 экз. на 0.1 м2.
Господствующее (по количеству) положение среди фауны биоценоза
каменистого дна Кондары занимают личинки ручейников, сидящие боль-
шею частью в своих чехликах, инкрустированных песчинками и камеш-
ками (рис. 4). Процент количества личинок ручейников к общему коли-
честву животных на камнях равнялся в среднем: в марте 61, в июле 86,
в октябре 47. В пробе, взятой после прохождения силя, ручейники состав-
ляли также 62%. Господствующее положение личинок ручейников
Рис. 4. Камень со дна р. Кондара с сидящими на нем
личинками и куколками ручейников.
в биомассе биоценоза каменистого дна будет еще яснее, если учесть,
что личинки ручейников представляют собою более крупные особи, чем
большинство остальных животных этого биоценоза. Следующее по оби-
лию место занимают личинки двукрылых (преимущественно тендипедид),
процент которых в пробах колеблется от 8 до 19 (рис. 5). Наконец, третье
(а иногда и второе) место принадлежит личинкам поденок, составляю-
щим от 1.2 до 29% всего количества животных биоценоза каменистого
дна. Прочие представители фауны (веснянки, клещи, олигохеты, турбел-
ларии, моллюски) играют здесь обычно незначительную роль.
По своим кормовым запросам большинство беспозвоночных, состав-
ляющих биоценоз каменистого дна Кондары, являются растениеядными.
Растительной пищей и бактериями (водорослями и приносимым течением
детритом) питаются все поденки, большая часть ручейников, почти все
двукрылые, олигохеты, моллюск. К хищникам можно, вероятно, отнести
лишь некоторых ручейников, турбелларию и клещей (?).
Из позвоночных к биоценозу каменистого дна в Кондаре следует при-
числить всех трех представителей ихтиофауны, найденных здесь, —
форель, маринку и горного сомика.. Все они используют в качестве пищи
L Описание метода определения количества животных на каменистом дне см.:
Жадин, 1940, стр. 529.
252 В. И. Жадин, И. А. Киселев, А. Г Родина, В. R. Панкратова и А. В. Гриб
составные элементы биоценоза каменистого дна: маринка с помощью
заостренного рогового чехлика на нижней челюсти соскабливает с кам-
ней водоросли и личинок ручейников, форель также питается живот-
ными, сидящими на камнях (но, помимо этого, и насекомыми, случайно
попадающими в воду с берега), сомик наряду с беспозвоночными загла-
тывает иногда и мальков рыб.
Подавляющее большинство организмов, населяющих биоценоз каме-
нистого дна, хорошо приспособлено к жизни на камнях в текучей воде.
Водоросли, найденные здесь, относятся к типичным литореофилам
(Киселев и Возженникова, 1950). Личинки ручейников или снабжены
домиками, построенными из песчинок и гравия (таковы Glossosoma,
Agapetia, Dinarthrum pugnax, Apatelia), или обладают крепкими крюч-
Рис. 5. Камень со дна р. Кондара с сидящими на нем личинками тендипедид,
ручейников и поденок и ползущей турбелларией.
ками, позволяющими цепляться за грунт (Rhyacophila, Hydropsyche)►
Куколки последней группы ручейников также заключены в прочный
песчано-гравелистый домик, плотно прикрепляющийся к камням.
Личинки поденок имеют сплющенную форму, плоскую нижнюю поверх-
ность, длинные хвостовые нити, увеличивающие контакт с грунтом;
жаберные листочки у некоторых форм, смыкаясь, образуют подобие
присоски. Ноги личинок расставлены в стороны, что дает им возможность,
двигаться, почти не отрывая тела от камня (см. рис. 3). Личинки мошек
снабжены сильно развитыми прикрепительными органами — задним
кольцом крючьев и «присоской». Куколки их имеют гладкую поверх-
ность кокона и укороченные нити. Личинки тендипедид или сидят в киле-
вых чехликах с нитями, прикрепляющихся к камешку (Tanytarsus ex
gr. exiguus, см. рис. 5), или обладают ничтожными размерами (2—4 мм)
и крепкими покровами тела, позволяющими им удерживаться за вся-
кими шероховатостями камней. Некоторые более крупные ортокладиины
(например, larva condarensis, имеющая длину 8 мм) снабжены на задних
ложных ножках крупными крючечками, сильно загнутыми на конце,
позволяющими личинке цепляться за камень. Укороченные задние лож-
ные ножки личинки Heptagia служат присосками, а пузыревидные вы-
ступы по краям всех сегментов тела служат, видимо, также для при-
крепления. Рыбы, обитающие в пределах литореофильного биоценоза.
Жизнь в водах Кондары
253
имеют также ясные приспособительные черты (см. статью Шапошниковой
в этом Сборнике).
Биоценоз песчаного дна занимает в Кондаре небольшое место и обла-
дает обычно кратковременным существованием. Субстратом для такого
биоценоза служит крупный песок, откладывающийся за камнями.
Среди песчинок живут мелкие олигохеты Nais variabilis, крупные
(длиною до 36 мм) личинки типулид, более мелкие, ближе не опре-
деленные личинки двукрылых (длиною до 9 мм), вздутые на конце
и обладающие способностью цепляться за песчинки, совсем мелкие
личинки тендипедид Tanytarsus ex gr. mancus. На поверхности песка
обитают очень мелкие личинки ручейников в песчаных чехликах и еди-
ничные мелкие личинки поденок.
Количество животных в биоценозе песчаного дна равнялось 535
на 0.1 м2; господствующее положение в нем (как по обилию, так и по
биомассе) занимали личинки двукрылых, составлявшие по количеству
31%, а по весу 99%.
Биоценоз заиленного дна на очень замедленном течении в Кондаре
можно встретить довольно редко. Наилок накапливается обычно поверх
песка. Господствующей группой фауны в нем являются личинки тенди-
педид Orthocladiinae G.?, Microspectra ex gr. trivalvis, Corynoneura, Lau-
terbornia. Здесь же обычны ближе не определенные личинки двукрылых,
а также мелкие личинки ручейников в песчаных домиках, единичные
экземпляры личинок поденок, единичные ракушковые рачки Eucypris sp.,
моллюск прудовик усеченный (Limnaea truncatula), довольно многочи-
сленные олигохеты Nais variabilis.
Обилие животных в биоценозе заиленного дна Кондары достигает
1600 экз. на 0.1 м2; 70% из них приходится на долю личинок тенди-
педид и 19% — на долю олигохет.
В местах затишья по спокойной водной поверхности стайками бегают
жуки-вертячки Gyrinus distinctus.
б. Источники в долине Кондары
Источники ущелья Кондара делятся на две высотные группы:
1) источники (родники) верхних частей ущелья (на выс. 1700—1900 м
над ур. м.) и 2) источники средних частей ущелья и его боковых вет-
влений.
Источники верхних частей ущелья в урочищах Квак и Оби-сафед
отличаются значительной мощностью и относятся к родникам, или лито-
реокренам (см.: Жадин. Жизнь в источниках. Жизнь пресных вод СССР,
т. 3, 1950). Выходя среди каменистого грунта из-под земли, они сразу же
получают большое падение и связанную с ним относительно большую
скорость течения. Родники при выходе на дневную поверхность сильно
затенены древесной и травяной растительностью, среди которой бросается
в глаза мощный борщовник и датиска.
Температура воды родников этой зоны держится в пределах 9.5—
9.7°. Вода довольно мягкая, слабо щелочная, со значительным насыще-
нием кислородом и небольшой окисляемостью.
Поскольку дно родников слагается преимущественно из камней,
жизнь в них, группируясь на камнях, составляет родниковый биоценоз
каменистого грунта. Растительные обрастания камней начинаются с са-
мого выхода родника на поверхность. Здесь появляются бурые скопле-
254 В. И. Жадин, И. А. Киселев, А. Г. Родина, В. Я. Панкратова и А. В. Гриб-
ния диатомей, а потом пленки синезеленой водоросли Phormidium unci-
natum. Всего здесь определено до 100 видов водорослей (Киселев и Воз-
женникова, 1950).
Фауна родников этой зоны не отличается видовым разнообразием.
Однако она представляет большой интерес как в экологическом, так
и зоогеографическом отношении. Среди нее мы видим амфибийного равно-
ногого рачка лигидиум, описанного Е. В. Боруцким (1948) как новый
вид — Ligidium shadini. Этот рачок живет во влажной среде между
камней в местах выхода родников, но заходит и в воду. Представители
рода Ligidium констатированы в Средней Азии впервые по нашим сбо-
рам. Другой вид этого рода Ligidium zernovi описан из Ферганы (Боруц-
кий, 1948). Типичный Ligidium shadini найден пока только в родниках
Гиссарского хребта, а разновидность его — subsp. gissarense — в род-
нике под старинной Гиссарской крепостью в Гиссарской долине.
В родниках Квака и Оби-сафеда собран также описанный Я. А. Бир-
штейном (1948) рачок-бокоплав Gammarus shadini. Этот вид относится
к подроду Sarothrogammarus, к которому, по Бирштейну, принадлежат
еще 2 вида, один из них обитает в ручьях Азорских островов, а дру-
гой — в горных ручьях северного склона Гиссарского хребта. Наш
бокоплав живет кроме верхней зоны ущелья Кондара также во многих
родниках Ходжа-оби-гарма. Он держится здесь при температуре от 6.5
до 13\
Найденная нами в роднике на Кваке личинка ручейника Pseudoste-
nophylax secretus (определенная С. Г. Лепневой) принадлежит к виду,
известному из района «12 источников» близ Ташкента.
Кроме перечисленных здесь видов в родниках верхней зоны были
найдены турбелларии, личинки мух, мошек, тендипедид, гелеид, блефа-
роцер, жуков, веснянок и поденок.
Помимо родников в верхней зоне ущелья Кондара местами встре-
чаются источники другого характера, которые можно обозначить как
топи. Они имеют заболачивающееся каменисто-гравелистое дно со слоем
ила, поросшее болотными растениями. Фауна топей бедная, среди нее
упоминания заслуживает личинка стрекозы Cordulegaster.
Источники следующей высотной зоны (1700—1300 м над ур. м.)
в ущелье Кондара обладают большею частью незначительным дебитом
и приурочены преимущественно к ореховым рощам. Часть их вытекает
на стыках коренных и наносных берегов реки Кондара. Источники эти,
выходя из-под земли, вначале тонкими пленками сочатся по крупному
гравию дна, на котором через некоторое расстояние появляются отдель-
ные камни. Несколько ниже уклон ложа источника делается еще меньше,
и на дне накапливается черный ореховый перегной, среди которого обо-
значается русло ручейка, текущего с небольшой скоростью. Прибли-
жаясь к реке, русло ручейка несколько углубляется, скорости течения
возрастают, признаки заболачивания утрачиваются. Такого рода «род-
никовые бассейны» занимают обычно небольшие площади, протяжением
в несколько десятков метров.
Родниковые бассейны зарастают пышной растительностью, целиком
закрывающей водную поверхность и образующей ассоциацию заболочен-
ного орешника, описанную в настоящем сборнике в главе о раститель-
ном покрове.
Температура воды источников этой зоны довольно постоянна — боль-
шую часть года она лежит около 12°. Годовая амплитуда температуры
при выходе источника из-под земли равняется всего 2°, а на участке за-
Жианъ в водах Кондары
255
пленного дна 6°. Вода источников мягкая, слабо щелочная, со значитель-
ной окисляемостью. Содержание растворенного кислорода в воде увели-
чивается по мере движения воды от выхода к реке.
Бактериологические исследования показали, что при выходе из-под
земли вода содержит всего от 6 до 22 бактерий в 1 см3 (методом выра-
щивания на агаре), или от 4000 до 6200 (методом прямого счета). При
продвижении воды от истока количество бактерий возрастает — до 53—
140 (посевом на агаре) и до 39 800—78 000 в 1 см3 (прямым счетом). По-
ставленные в воде предметные стекла после четырехсуточной экспозиции
показали, что бактериальная флора источника состоит из прикрепленных
нитчатых бактерий, значительного количества разнообразных палочек
и кокков, а также азотобактероподобных клеток. На стеклах, стоявших
вблизи выхода источника из-под земли, обрастание бактериями не носило
сплошного характера или характера колоний, зато на стеклах, стояв-
ших ниже по течению, образовались сплошные покрытия из многочис-
ленных мелких колоний разнообразных бактерий. В источниках были
обнаружены бактерии, обладающие протеолитическими ферментами, спо-
собными к глубокому расщеплению белковых веществ с выделением
сероводорода. В чашках на агаре часто развивался Bacillum mycoides,
указывающий на почвенное происхождение части бактерий. Особого
упоминания заслуживает факт постоянного наличия в источниках нитри-
фицирующих и азстфиксирующих бактерий (в том числе и азотобактера).
Изредка находились денитрифицирующие бактерии. Десульфурирующие
бактерии были находимы только в заиленной части источников.
Водная растительность имеет в источниках зональное распределение:
при выходе воды из-под земли растут лишайники, несколько ниже по те-
чению на камнях — мхи, а на илистом дне — многочисленные вероники.
Водоросли появляются уже среди мха на камнях близ выхода источ-
ника из-под земли. В самом начале это диатомеи, ниже к ним присоеди-
няются зеленые нитчатки. Всего из источников рассматриваемой высот-
ной зоны определено свыше 80 видов водорослей (Киселев и Возженни-
кова, 1950). При рассмотрении экологической природы водорослей, най-
денных в источниках, бросается в глаза значительное количество видов,
обитающих в почве. Характерны: Diatoma hiemale, Synedra ulna, Achnan-
thes lanceolata, Achnanthes minutissima, var. cryptocephala, Stauroneis
Smithii, Pinnularia appendiculata, Cymbella leptoceros, Gomphonema
angustatum, Gomphonema parvulum, Gomphonema gracile, Nitzschia micro-
cephala, Nitzschia amphibia, Hantzschia amphioxys и некоторые другие.
Фауна источников этой зоны состоит из многочисленных видов червей,
ракообразных, водяных клещей, насекомых и моллюсков, список кото-
рых дается ниже.
Oligochaeta
Lumbriculus variegatus
Tubifex tubifex
Limnodrilus sp.
Nais n. sp.
Enchytraeidae
Turbellaria
Seidlia sp.
Crustacea
Isopoda
Ligidium shadini
Ostracoda
Stenocypris sewelli (?)
Hydracarina
Tadjikothias connexa
Sperchon setiger tarnogradskii
Sperchon sp.
Panisus sp.
Insecta
Diptera
Dixa
Stratyomyidae
256 В. И. Жадин, И. А. Киселев, А. Г. Родина, В. Я. Панкратова и А. В. Гриб
Tipulidae
Psychodidae
Pericoma
S i m и 1 i i d а е
Eusimulium (Eusimulium) montium
Eusimulium (Eusimulium) brachyantherum
Simulium (Odagmia) crassifilum
Tendipedidae
Pelopiinae G.?
Orthocladiinae G.? larva sellata
» amphibia
» principata
Microspectra
Lauterbornia
Polypedilum
Procladius
Tanytarsus ex gr. exiguus
Diamesa larva spinosa
H e 1 e i d a e
Culicoides
Bezzia
Trichoptera
Glossosoma
Hydropsyche sp.
Dinarthrum pugnax
Agapetus sp.
Dolophilodes larva dolichocephala
» macrocephala
Rhyacophila obscura larva pallidiceps
Psychomyia sp.
Oecetis paula
Plecoptera (indet).
Ephemeroplera
Ordella
Odonata
Cordulegaster coronatus
Coleopter a
Helodidae
Staphylinidae — Lesteva fasciata, Stenus
cordatus
Molluscs
Amnicola raddei
Limnaea (Galba) truncatula
Pisidium casertanum
По участкам родникового бассейна фауна распределяется вполне
закономерно. Верхний отрезок источника с гравийным дном непосред-
ственно около выхода воды на дневную поверхность наиболее беден
видами, но зато имеет среди них весьма характерных представителей.
Здесь живет та же амфибийная мокрица Ligidium shadini, которая оби-
тает и в родниках более высокой зоны. Здесь же в большом количестве
встречается крошечный моллюск амникола — Amnicola raddei, для
которого высота 1500 м является, видимо, верхним пределом верти-
кального распространения.
Наиболее богат видами следующий (переходный) участок роднико-
вого бассейна, обладающий как гравийным и каменистым дном, так
и небольшими местными скоплениями илистого орехового перегноя.
Здесь встречаются турбелларии (вообще весьма редкие в источниках
данной высотной зоны), многочисленные и разнообразные двукрылые,
из которых особого упоминания заслуживает личинка Psychodidae,
относящаяся, видимо, к роду Peri сота, и ручейники. Только здесь обитает
личинка типичного родникового обитателя Dolophilodes, отличающаяся,
по данным С. Г. Лепневой, исключительно крупными размерами головы.
Наличие здесь биотопов илистого дна обусловливает нахождение наряду
с типичными родниковыми видами таких обитателей илистого дна, как
невзыскательный к условиям среды червь Tubifex tubifex.
На заиленном участке родникового бассейна иловые формы образуют
основной костяк фауны, типичные же родниковые обитатели, характер-
ные для вышележащих участков родникового бассейна, в значительной
мере выпадают. Общий список обитающих здесь видов становится очень
небольшим. Бросается в глаза частая встречаемость в илу моллюска
Pisidium casertanum, уже упоминавшейся олигохеты Lumbriculus varie-
Жизнь в водах Кондары
257
gatus, Т. tubifex и значительного количества ракушковых рачков. Заслу-
живает упоминания, что личинка ручейника Dolophilodes сохраняется
и в этом участке родникового бассейна, а личинка стрекозы Cordulegaster
так хорошо маскируется хлопьями ила, что представляется коренным
обитателем этих своеобразных условий.
Рис. 6. Камень из источника, поросший мхом, с сидящими на нем
личинками мошек, ручейников, мухи-львинки, турбеллариями,
дождевым червем и мокрицей Ligidium.
Количественное развитие фауны в источниках подвержено большим
колебаниям. Вычисляя обилие животных на 0.1 м2 дна, мы получили
для камня, обросшего мхом, 337 животных (рис. 6), для камня без мха —
155—1194 животных. Доминирующей группой на каменистом дне источ-
Рис. 7. Дождевой червь Dendrobaena sp., из источника.
(По Гриб, 1950).
ников этой высотной зоны оказались двукрылые, составлявшие 47—
55% обилия, за ними следовали личинки ручейников (9—42%). На одном
из обследованных камней значительную роль играли турбелларии,
составлявшие 39% обилия.
Интересной чертой фауны источников ущелья Кондара (равно как
и других мест Гиссарского хребта) является присутствие дождевых чер-
вей Eisenia foetida, Eiseniella tetraedra, Allolobophora yassyensis, Allo-
lobophora sp., Dendrobaena sp. (рис. 7, см. также: Гриб, 1950).
17 Ущелье Кондара
258 В. И. Ж ад ин, И. А. Киселев, А. Г. Родина, В. Я. Панкратова и А. В. Гриб
в. Поливные водоемы
Река Кондара, входя на территорию Ботанической станции, дает
начало целому ряду небольших водотоков, используемых для полива
расположенных здесь садов и огородов. На небольшом пространстве
низовьев ущелья в миниатюре воспроизводится картина зоны рассеива-
ния стока, столь характерная для гидрологии всей Средней Азии. Арыки,
орошающие территорию Ботанической станции, как правило, обладают
неустойчивым водным режимом. Вода пускается в них на несколько
часов в день, а затем перекрывается. Лишь в тех арыках, которые играют
роль головных, водный режим более постоянен. В периодически пересы-
хающих арыках развивается микроскопическая жизнь почвенно-водного
характера, в арыках же с устойчивым водным режимом развивается
биологическая картина, свойственная текучим водам. Сюда проникают
и рыбы, особенно горный сомик.
Там, где воды арыков сбрасываются вниз в виде каскадов, на обмы-
ваемых ими камнях разрастаются налеты водорослей Stratonostoc verru-
cosum, Tolypothrix distorta var. penicillata, Dichothrix Baueriana, Oedo-
gonium, Spirogyra, Mougeotia.
В некоторых местах арычные воды прорываются в пониженные котло-
винки и образуют лужи с довольно постоянным водным уровнем. В такого
рода лужах в большом количестве появляются моллюск Limnaea trun-
catula и личинки малярийного комара Anopheles superpictus.
3. РЕКА ВАРЗОБ
У подножья ущелья Кондара протекает бурная горная река Варзоб.
Дно реки сложено громадными глыбами гранита, а также камнями самой
различной величины (до гальки включительно). Местами, за каменными
глыбами или около берегов, наблюдаются скопления песка различной
крупности. Уровень воды в Варзобе, как типичной горной реке Средней
Азии, испытывает значительные колебания, приуроченные к определен-
ным сезонам. Скорости течения очень велики. Прозрачность воды нахо-
дится в зависимости от уровня воды — низкие меженные воды Варзоба
совершенно прозрачны, так что через красивую зеленовато-голубую
воду всюду видно дно. С повышением уровня, когда в реку поступает
большое количество мелкозема, вода становится мутной.
В термическом режиме Варзоба наблюдается ряд моментов, значи-
тельно отличающих его от термического режима р. Кондара. Прежде
всего, вода Варзоба значительно холоднее воды Кондары. В связи с этим
и годовая амплитуда температуры воды в Варзобе (=14°) на 6.5° меньше,
чем в Кондаре. Суточный ход температуры воды в Варзобе имеет также
свои особенности. Температура воды здесь как бы опаздывает за темпера-
турой воздуха: своего максимума она достигает в послеполуденные часы,
когда в воздухе наблюдается уже заметное похолодание. Это зависит
от того, что в р. Варзоб впадают притоки, обладающие (подобно Кондаре)
более высокой температурой, чем основная струя реки, и эти-то более
теплые воды доходят до района ущелья Кондара только к вечеру. Суточ-
ная амплитуда температуры воды Варзоба близ Кондары зимой равняется
0.5—1.5°, весной 2.3—4.6°, летом до 4.5°.
С химической стороны вода Варзоба характеризуется как умеренно
жесткая с нейтральной реакцией и малой окисляемостью (Глаголева,
Жизнь в водах Кондары
259
1945). Содержание растворенного кислорода находится примерно на том же
уровне, что и в р. Кондара. Суточный ход содержания кислорода показы-
вает максимум в утренние и дневные часы и значительное снижение ве-
чером.
Планктон р. Варзоб состоит из бактерий, закономерно развиваю-
щихся в воде, а также из организмов фито- и зообентоса, увлекаемых
в водную струю течением.
Бактерии весьма рельефно подчеркивают зависимость водных масс
Варзоба от вод его небольших притоков, на которых расположены насе-
ленные пункты. В меженное сухое время года эта зависимость прояв-
ляется в значительно меньшей степени, чем в дождливые периоды. Бакте-
риопланктон Варзоба реагирует на поступление вод из притоков, загряз-
няемых населением, возрастанием количества бактерий в воде как раз
у того берега, со стороны которого впадает приток. Приведем пример,
иллюстрирующий это положение. Ниже кишлака Гажни, расположен-
ного выше Кондары, у правого берега (где находится кишлак) в бездожд-
ное время количество бактерий, вырастающих на мясо-пептонном агаре,
было 319 в 1 см3, а у левого — 252, т. е. немногим меньше, чем у правого.
В дождливое же время количество бактерий у правого берега было больше
чем вдвое по сравнению с левым берегом: у правого — 7050, у левого —
3070 в 1 см3. Специфические показатели загрязнения воды — Escherichia
coli и Bacillus perfrigens также совершенно отчетливо отражают на себе
приток загрязнений с правого берега. Однако р. Варзоб довольно быстро
справляется с попадающими в нее загрязнениями. Этому способствует
мощное разбавление загрязненных вод основной струей реки, большое
количество перепадов, прекрасная аэрация воды и интенсивное солнеч-
ное освещение при малой глубине реки. На протяжении 1.5 км от Гажни
до Кондары Варзоб почти полностью освобождается от загрязнений.
В дождливое время количество бактерий от Гажни до Кондары падает
с 7050 до 1680 в 1 см3, титр Esch, coli изменяется в том же направлении,
а бактерии анаэробной кишечной группы (В. perfrigens) не обнаружи-
ваются в воде у ущелья Кондара совершенно.
Учет бактериопланктона методом прямого счета дал в р. Варзоб
в сухое время года величины от 26 000 до 128 000, а в дождливое —
до 161 000 бактерий в 1 см3 (Родина, 1944).
Настоящего фитопланктона в Варзобе нет, но в планктонную сетку
попадалось до 30 видов водорослей, принадлежащих к донному населе-
нию реки.
Дно р. Варзоб заселено бактериальными, растительными и живот-
ными организмами.
Среди бактерий, населяющих дно реки, особого упоминания заслу-
живает азотобактер, который был обнаружен в пробах как у кишлака
Гажни, так и у самого ущелья Кондара. Особого обилия азотобактер
достигал в другой реке — Лючоб, впадающей в Варзоб. Посевы грунта
из Лючоба на агаре Бейеринка в летние месяцы давали почти чистые
культуры азотобактера.
Растительные обрастания, покрывающие камни на дне и у берегов
реки, состоят из мхов, зеленых водорослей (Prasiola fluviatilis), хризо-
монады Hydrurus foetidus, нескольких видов синезеленых и коньюгат,
а также свыше 70 видов диатомовых (полный список микрофлоры Вар-
зоба см. у Киселева и Возженниковой, 1950).
Донная фауна р. Варзоб на участке от Такобского ущелья до выхода
из гор слагается видами, сведенными в нижеследующий список.
17*
260 В. И. /К ад ин, И. А. Киселев, А. Г Родина, В. Я. Панкратова и А. В. Гриб
Oligochaeta
Lumbriculus variegatus
Tur bell aria
Polycelidia receptaculosa
Hydracarina
Sperchon plumifer
Sperchon glandulosus cubanicus
Megapus sp.
Insecta
Diptera
Tipulidae
Atherix
S i m u 1 i i d a e
Simulium (Astega) ovtshinnikovi
Simulium (Odagmia) alajensis
Simulium (Wilhelmia) veltistschevi
Blepharoceridae
Blepharocera fasciata asiatica
Tendipedidae
Heptagia larva accomodater
Orthocladiinae G.? larva principata
» carbonaria
» sei lata
» fracta
Eukiefferiella larva oxiana
Lauterbornia
Corynoneura
Tanytarsus ex gr. exiguus
Tendipedini G.? larva dulcis
Trichoptera
Glossosoma sp.
Apatelia sp.? (copiosa?)
Hydropsyche sp.
Rhyacophila obscura larva pallidiceps
Plecoptera (indet.)
Ephemerop ter a
Rhitrogena
Iron
Cynigma?
Baetis
Ephemerella
Донная жизнь реки Варзоб группируется в биоценозы каменистого
дна и биоценоз песчаного дна, причем, как и в большинстве горных рек,
биоценозы каменистого дна занимают свыше 95% всего дна реки.
В Варзобе несомненно можно различать несколько биоценозов каме-
нистого дна, которые различаются между собою видовым составом как
растительного, так и животного населения. Здесь имеется, например,
биоценоз, растительная основа которого слагается мхом, а также биоце-
нозы в обрастаниях водорослей — празиолы и гидруруса. Крупным
глыбам скал, стоящих среди пенящейся воды на середине реки, свой-
ственны не те биоценозы, которые формируются на небольших камнях
берега в условиях замедленного течения. Однако до настоящего времени
мы не располагаем достаточными сведениями, чтобы сколько-нибудь полно
охарактеризовать все эти биоценозы каменистого дна.
Как и в р. Кондара, в Варзобе постоянным компонентом биоценозов
каменистого дна являются те же три вида рыб: форель, маринка и гор-
ный сомик.
В отношении количественного обилия фауны биоценоз каменистого
дна р. Варзоб (если судить по населению камней близ берега) почти
вдвое беднее соответствующего биоценоза р. Кондара. На 0.1 м2 дна
здесь приходится от 203 до 806 животных, а в среднем — 580. Как
и в Кондаре, господствующей группой животного населения здесь можно
считать личинок и куколок ручейников, которые в Варзобе составляют
в среднем 74%, доходя на отдельных камнях до 94%. За ручейниками
по обилию следуют личинки двукрылых и поденок, на долю которых
приходится в среднем по 11%. Остающиеся 6% делятся между другими
группами фауны — личинками веснянок, клещами и червями.
Жизнь в водах Кондары
261
Биоценоз песчаного дна в р. Варзоб, как говорилось, занимает неболь-
шие площади кое-где в русле реки за камнями или близ берегов —
в местах затишья и обратного течения. Песок, на котором развивается
биоценоз, принадлежит большею частью к категории мешаного, в кото-
ром различные фракции (от гравия до иловатой пыли) смешаны без замет-
ных закономерностей. В некоторых случаях поверхность песка покрыта
тонким слоем иловатого налета. Биоценоз песчаного дна в Варзобе
состоит всего из 3 форм — личинок ближе не определенных двукрылых
и тендипедид — Tendipedini larva dulcis, а также червя Lumbricuius varie-
gatus. Преобладали личинки тендипедид, найденные в одной пробе
в количестве 348 экз., в то время как две другие формы здесь же были
в единичных экземплярах.
Жизнь на поверхности воды в р. Варзоб не заметна без пристального
присматривания. Иногда где-нибудь у берега за камнем можно обнару-
жить жуков-вертячек. Зато на камнях, обмываемых бурными водами,
часто видишь резвую птичку-оляпку, временами в поисках пищи исче-
зающую под водой.
ЛИТЕРАТУРА
Акатова Н. А. 1950. К фауне Ostracoda Средней Азии. Тр. Зоолог, инет. АН
СССР, т. IX, вып. 1.
Бирштейн Я. А. 1948. Бокоплавы Таджикистана. Памяти акад. С. А. Зернова.
Изд. АН СССР
Боруцкий Е. В. 1948. К фауне родниковых Isopoda Средней Азии. Памяти
акад. С. А. Зернова. Изд. АН СССР.
Глаголева Е. М. 1945. Биотопы Anopheles superpictus Gr. и их химизм. Тр.
Инет, маляр, и медиц. паразитол., 1, Сталинабад.
Гриб А. В. 1950. Малощетинковые черви Средней Азии. Тр. Зоолог, инет. АН
СССР, т. II, вып. 1.
Жадин В. И. 1940. Фауна рек и водохранилищ. Тр. Зоолог, инет. АН СССР,
V, 3—4.
Жадин В. И. 1945. Результаты и следствия гидробиологических исследований
в Таджикистане. Тезисы докладов отчетной сессии Зоолог, инет. АН СССР.
Сталинабад.
Жадин В. И. 1950. Теоретическое и практическое значение гидробиологического
изучения бассейна Аму-дарьи. Тр. Зоолог, инет. АН СССР, т. IX, вып. 1.
Жадин В. И. 1950. Пресноводные моллюски бассейна Аму-дарьи. Там же.
Жадин В. И. 1950. Жизнь в источниках. Жизнь пресных вод СССР, т.Ш.
Лепнева С. Г. 1944. Биотопы Anopheles superpictus Grassi в водоемах окрестно-
стей Сталинабада и Варзобского района. Изв. Тадж. фил. АН СССР, № 5.
Киселев И. А. иТ. Ф. Возженникова. 1950. Материалы к изучению
флоры водорослей водоемов бассейна р. Аму-дарьи. Тр. Зоолог, инет. АН
СССР, т. IX, вып.1.
Панкратова В. Я. 1950. Фауна личинок Tendipedidae бассейна Аму-дарьи.
Там же.
Родина А. Г. 1944. Санитарное состояние воды р. Варзоб и основных арыков
г. Сталинабада. Изв. Тадж. фил. АН СССР. № 5.
Соколов И. И. 1948. Гидрахнеллы из Таджикистана. Памяти акад. С. А. Зер-
нова. Изд. АН СССР.
Шапошникова Г. X. 1950. Рыбы Аму-дарьи. Тр. Зоолог, инет. АН СССР
т. IX, вып. 1.
СЕЗОННЫЕ ЯВЛЕНИЯ
| А. Я. ТУГАРИНОВ
КАЛЕНДАРЬ ЖИЗНИ В УЩЕЛЬЕ КОНДАРА
(По материалам В. И. Запрягаевой и А. Я. Тугаринова)
Настоящая глава посвящена обзору годового хода явлений в орга-
нической природе в ущелье Кондара в их связи со сменами времен года.
Явления этого порядка, обычно называемые фенологическими, с одной
стороны, представляют безусловный интерес, как ближайшим образом
характеризующие среду обитания, а с другой, взятые комплексно, про-
ливают свет на многие стороны процессов, происходящих в органическом
мире. Словом, они подводят нас к вопросам ауто- и синэкологии. Для
наблюдателя не специалиста такие данные представляют тот интерес,,
что знакомят его с кругом явлений, происходящих в природе в тот или
иной момент, с их динамикой, последовательностью и т. д. Вот почему
включение в настоящий сборник соответствующей главы представлялось
коллективу авторов вполне целесообразным. Материалом для нее послу-
жили личные наблюдения сотрудников Зоологического института, рабо-
тавших в Кондаринском ущелье. Данные, касающиеся растительностиv
частью были предоставлены заведующей Горной ботанической станцией,
В. И. Запрягаевой, частью пополнены авторами настоящей главы. В по-
следнем случае растения были определены ботаником Таджикского^
филиала АН Ю. С. Григорьевым.
ЗИМА
Декабръ-февралъ. По метеорологическому календарю декабрь счи-
тается первым месяцем зимы. В нижней части Кондаринского ущелья
до высоты 1360—1400 м над ур. м. в начале декабря жизнь еще далеко*
не замирает. Среди голых осыпей и нагромождений обломков породы
зеленеют пятна свежей зелени. Деревья оголены, почва под ними
покрыта сухими опавшими листьями. Много летающих насекомых и среди
них бабочки-лимонницы, из-под ног прыгают кузнечики, ползают
муравьи (Cataglyphus bicolor). В лесных порослях кочуют стайки птиц.
Как насекомоядные — пеночки, горихвостки, так и зерноядные — дубо-
носы, юры, зяблики равно находят для себя обильный корм. Однако,
достаточно подняться несколько выше 1500 м — и картина резко меняется.
Здесь, как и выше, уже господствует настоящая зима.
В середине декабря органическая жизнь замирает, но вместе с тем
замечаются новые явления. Глубокие снега верхнего пояса гор вынудила
спуститься в низины некоторых высокогорных птиц: различных вьюрков
266
А. Я. Тугаринов
{Fringillauda nemoricola, Leucosticte brandti, Montifringilla nivalis),
краснокрылых чечевичников (Rhodopechys sanguinea), стенолазов; сюда
же перекочевали кеклики, появляющиеся теперь около построек Стан-
ции. Для зимней жизни горных вьюрков характерно их явное стремле-
ние при первой возможности снова подняться выше: едва по южным
склонам начнут появляться проталины, как эти птицы покидают низины
и остаются на грани сплошных снегов до нового снегопада.
В сменах ясных солнечных дней пасмурными, нередко с осадками,
иногда в виде дождя, проходит кондаринская зима. Тем не менее, в ниж-
ней части ущелья в это время на пригревах можно заметить проявление
деятельности животных. В любой солнечный день случается обнаружить
насекомых, например жуков, муравьев, бабочек-молей, мушек. Наиболь-
шее оживление вносят птицы. По дну ущелья у речки, строго придер-
живаясь раз избранного места, нередки синие птицы Myophonus caeru-
leus. Здесь же, в чаще кустов, скрытно держатся кустарницы Garrulax
lineatus. Их общество разделяют крапивники. На речке, там, где вода
тонким слоем стекает по камням, можно увидеть маленькую птичку,
бродящую среди струй воды; поминутно она склевывает что-то с мокрых
камней. Это — белоножка Microcichla scouleri, обитатель высоко-
горных потоков, сейчас скочевавший в низины. Обычные по Варзобу
оляпки на Кондару проникают редко, последняя для них, видимо, слиш-
ком мелка. Изредка у воды же можно вспугнуть горного дупеля, также
спустившегося в низменности со своих горных гнездовий. По обрывам
южного склона, среди скал, характерны поползни; подвижные и сует-
ливые, они старательно обшаривают трещины камней, время от времени
перекликаясь звонкой позывной. На территории Станции, среди посадок
фруктовых деревьев, кочуют щеглы. По вечерам, собравшись обществом
на дерево, они негромко поют. Здесь же время от времени появляются
стайки синичек: бухарских, желтогрудых князьков и рыжешейных;
обшаривая дерево за деревом, негромко перекликаясь, они так же неожи-
данно, как и появились, исчезают. Если к отмеченным, наиболее обыч-
ным зимним обитателям Кондаринского ущелья в его нижней части мы
прибавим прилетевших на зимовку чернозобых дроздов, юров, бело-
шапочных овсянок, красноспинных горихвосток и некоторых других,
то окажется, что зимнее птичье население ущелья достаточно разнооб-
разно. Среди него отсутствуют лишь птички, питающиеся деятельными
насекомыми. Очевидно, в те моменты, когда хотя бы всего на несколько
дней выпадают обильные снега, такие виды были бы обречены на голо-
данье. Все же названные выше виды являются или зерноядными, или
питающимися насекомыми, находящимися в состоянии зимнего покоя
или личиночных стадиях. Зимою почва не промерзает. На южных скло-
нах она нагревалась солнцем, на северных была укрыта снежным
покровом. А потому на местах, доступных солнечным лучам, уже в по-
следней декаде февраля начинается пробуждение растительности. Пер-
выми на пригорках появляются золотистые крокусы и почти одновре-
менно раннецветущие виды гусиного лука. В теплые зимы в конце месяца,
случается, зацветает дикий миндаль. Начинают вегетировать многолет-
ние травы, например юган Prangos pabularia, эремурусы, унгерния,
ферула и др. На крокусах можно заметить домашних пчел, берущих
первый взяток, летают лимонницы зимовавшей генерации. Выползают
из земли слизни. Наступает новый период деятельности, роста и раз-
множения для всех представителей животного и растительного
мира.
Календарь жизни в ущелье Кондара
267
ВЕСНА
Март. На устье р. Кондара, где ущелье расширено, среди наносов
гальки и крупных валунов попадаются площадки, покрытые мелкозе-
мом. С первых дней марта здесь появляются крохотные растеньица:
Erophila verna, Е. minima, приземистая осока (Carex pachystilis), гусиный
лук, изредка Colchicum Kesselringii. На южных склонах становится все
больше крокусов, местами образующих желтые пятна. Здесь еще с осени
начали свою вегетацию злаки: мятлик (Роа bulbosa), костер (Bromus),
ячмень (Hordeum bulbosum); теперь они быстро пошли в рост, покрывая
мелкоземистые склоны зелеными лужайками. К концу первой декады
начинается массовое цветение дикого миндаля. Осыпанные цветом, его
деревья как будто накрыты малиновым флером, а группы их по склонам
розовеют блеклыми пятнами. Под пологом деревьев, где больше перегноя,
показываются, первые ростки хохлатки (Corydalis Ledebouriana) и Leon-
tice Albertii. Через 3—5 дней они уже зацветут. В первой декаде месяца
лопаются почки на тополе, фруктовых деревьях, распускается листва
на ивах, шиповнике. Появляются бабочки-многоцветницы, много мух,
жуков Cicindella и других, молей, муравьев, около цветов летают шмели.
Пробудились от зимнего покоя слепушонки (Ellobius talpinus), всюду
на более мягких склонах заметны следы их роющей деятельности.
В птичьем мире перемен немного, господствует тот же зимний аспект.
Попрежнему кочуют стайки юров, горной и белошапочной овсянок,
обычны красноспинные горихвостки и другие из числа зимовавших.
Как и зимою, вскоре после восхода солнца можно увидеть пролетающих
над ущельем голубей (Columba livia); их стайки направляются на кор-
межку на пашни предгорий, возвращаясь обратно вскоре после полудня.
С такой же правильностью в утренние часы появляются над ущельем
стаи альпийских галок (Pyrrhocorax graculus). Они прилетают из более
глубоких частей Гиссарского хребта, совершая, очевидно, свой дневной
облет, связанный с посещением мест кормежек. Но уже невидно розовых
чечевичников (Erythrina rhodochlamys), укочевавших в горы. Изредка по
долине Варзоба проносятся стаи пролетных скворцов, галок. Появились
пролетом одиночные варакушки.
Во второй декаде в нижних частях ущелья массовое цветение кроку-
сов заканчивается. В затененных местах, среди кустов, зацветают фиалки
(Viola suavis), пролески (Scilla bucharica), Rhinopetalum bucharicurn, на-
ступает полный расцвет хохлатки, на пригревах появляются первые цвету-
щие экземпляры ветреницы (Anemone Tschernjaewii). Началось обильное
цветение миндаля по северным склонам. Буйно поднимается юган, раз-
росшиеся розетки унгернии, ферулы и эремурусов пестрят по южным
склонам. Травяная растительность развилась здесь уже настолько, что
служит пастбищем для скота. На северных склонах до высоты 1300—
1500 м начинается массовое цветение крокусов, а за ним и других ранне-
весенних растении. В составе птичьего населения попрежнему перемен
немного. Чаще попадаются на глаза белые трясогузки. Явно начался
их пролет: минуя устье ущелья, их маленькие общества в 5—7 особей
продвигаются вверх по Варзобу. Появились первые пеночки (Phylloscopus
collybita) и одиночки сорокопуты (Lanius cristatus isabellinus). У пено-
чек некоторые самцы поют. Голуби разбиваются на пары и приискивают
места для будущих гнездовий. В теплые солнечные дни, — а в это время
температура уже может достигать 20° в тени — можно заметить ящериц-
агам, вышедших из своих зимних убежищ, чтобы погреться на весеннем
268
А. Я, Тугаринов
солнышке. Однако выше по ущелью, особенно по северной стороне, весна
только что вступает в свои права.
Третья декада марта обогащает список цветущих растений целым
рядом новых видов. На южных склонах, на местах, богатых перегноем,
распускаются чудесные красные тюльпаны (Tulipa lanata), ирисы (Juno
Warleyensis), герань (Geranium Charlesii), ветреница (Anemone bucharica)
и др. Лопнули почки на большинстве деревьев, покрылись листочками
ивняки, зацветает алыча — ее деревья стоят как будто под белым поло-
гом. Также обильно покрывается цветом вишня-махалебка (Prunus Maha-
leb), набирает цвет дикая яблоня (Malus hissarica). В садах Станции зацве-
тает персик, смородина, появляются первые цветочки земляники-викто-
рии. Миндаль отцветает. К концу месяца почти все деревья, в том числе
и- грецкий орех, уже одеваются молодой листвой. На склонах южных
экспозиций растительность развилась настолько, что издали горы кажутся
зелеными. Заметных перемен в составе птичьего населения все еще нет.
Кеклики начинают образовывать пары, по-зимнему, стаями, держатся
щеглы, овсянки, альпийские галки, но уже не видно красноспинных
горихвосток, юров, белошапочных овсянок, покинувших свои зимние
ареалы. В самом конце месяца из прилетных появились чеканы-пле-
шанки и чеканы-плясуны, степные коньки и рогатые жаворонки. Три
последних вида зимовали на предгорных низменностях и теперь начали
движение вверх, поднимаясь по мере стаивания снега все выше, чтобы
на субальпийских лугах Руидашта занять свои гнездовые места. Пока-
зались редкие пролетные вальдшнепы, днем останавливающиеся на отдых
и кормежку в кленовниках по северным склонам. Летят журавли. Они
тянут прямо на север, на высоте, недоступной невооруженному глазу,
и только их крики доносятся с высоты. Их воздушные трассы проходят
так, что Гиссарский хребет с его высотами 3000—3500 м над ур. м.
на перевалах не является для них препятствием. Птицы летят «на про-
ход», лет их продолжается весь апрель до первых чисел мая. В эти же
дни конца марта нередко на границе тающего снега можно увидеть
маленькую ящерицу — азиатского гологлаза, начавшего свою деятельную
жизнь.
Апрель. В нижней части ущелья весна в разгаре. Летние солнечные
дни еще чередуются с пасмурными, дождливыми, хотя и теплыми. Однако
в верхних частях ущелья только наступает перелом к теплу.
Начавшийся в конце марта в нижней части ущелья интенсивный рост
трав продолжается нарастающими темпами. Зацветает тут (шелковица),
клены, несколько позднее их — каркас (Celtis), фисташка, орех. В пер-
вой же декаде наблюдается массовое цветение алычи, вишни, дикой
яблони, боярок. Сплошным розовым цветом покрылись деревья пер-
сика. Растения, развивавшиеся на южных склонах, расцветают теперь
на северных. Целыми полянками разрослись цветущие хохлатки,
ветреницы, пролески, красные тюльпаны, эспарцет (Onobrychis
grandis), герань. В середине месяца здесь же зацветает ревень (Rheum
Maximoviczi), выбирающий, кажется, самые недоступные места:
обрывы, трещины отвесных скал, осыпи. На южных склонах отдельные
растения достигают высоты 1 м. Цветут злаки — Hordeum bulbosum,
Dactylus glomerata, пышно разрослись кусты югана, ферулы, Eremo-
stachys и др. В зоне 1700—2000 м еще ранняя весна. Пятна тающих сне-
гов окаймлены желтым бордюром крокусов, частью их цветочные стрелки
пробиваются через тонкий слой снега. Позднее их сменяет пролеска.
В трещинах камней, давно прогреваемых солнцем, успела разрастись
Календарь жизни в ущелье Кондара
269
и цветет вшивица — (Pedicularis pycnantha). Деревья еще стоят голые,
только в последней декаде на них лопнут почки. В середине месяца в кле-
новниках Квака зацветает роскошная Fritillaria Eduardii. На Руидаште
еще господствует зимняя обстановка: и плато и склоны стоят сплошь
укрытые снегом.
В жизни птиц новые явления нарастают медленно. Из видов, приле-
тавших на зимовку, не осталось ни одного. Во второй половине месяца
из оседлых начинают токовать кеклики, строят гнезда и несутся вороны,
сороки, в конце месяца разбиваются на пары и приступают к устройству
гнезд синички. Прилет и пролет новых видов также идет замедленными
темпами. Как и везде в южных широтах, эти моменты не связаны с ранне-
весенними явлениями. Если у нас на севере прилет происходит в более
сжатые сроки и совпадает с другими фенологическими датами, то
в южных странах такие же явные совпадения трудно установимы. Неясно,
например, почему такая зерноядная птичка, как желчная овсянка,
в виде передовых особей появляется здесь лишь в самом конце апреля,
по существу — в летней обстановке, а массовый прилет приходится на пер-
вую декаду мая. Тогда же, в последней декаде апреля, появляются чече-
вичники (Erythrina erythrina). В отношении последнего известно, что
в горах он не гнездится ниже 2000 м. Таким образом, ущелье Кондара
является для него временной станцией, где прилетевшие птички ожи-
дают возможности подняться выше, в полосу розариев и субальпийского
высокогорья. С начала месяца появляются первые пролетные коньки
(Anthus trivialis). Больше становится пеночек (Phylloscopus collybita).
В общем прилет и пролет до середины апреля идет еще очень вяло, не внося
заметного оживления, вполне оправдывая таджикскую примету: «птица
придет, когда на туте лист будет», т. е. во второй половине апреля.1
Май. К концу второй декады устанавливается вполне летний тип
погоды: безоблачные, жаркие дни. Воздух потерял свою прозрачность,
не редки мглистые дни, случаются и ветры — «афганцы», приносящие
массы взвешенной мельчайшей пыли, скрывающей дали. По ущелью уста-
новилась правильная суточная смена ветров: с утра вниз по долине
тянет прохладный ветерок, стихающий к полудню. Он сменяется более
теплым ветром, дующим вверх по ущелью, — это поднимаются теплые,
нагретые массы воздуха с низин. С заходом солнца вновь начинает сте-
кать вниз более холодный воздух с верхнего пояса гор.
Ранневесенняя эфемеровая растительность в районе Станции к се
редине месяца почти выгорает. По склонам южных экспозиций в связи
с последними дождями почва еще достаточно напитана влагой, и расти-
тельность достигает наибольшего развития. Вместе с поднявшимися
до 1 м и более высоты злаками высятся стебли громадного камоля (Ferula
Jaeschkeana), Eremostachys hissarica, им немного уступают «василек»
(Rhaponticum integrifolia), пижма (Tanacetum tadshikorum), поповник
(Pyrethrum partenifolium), Mulgedium roseum. Здесь же дает желтеющие
пятна зверобой (Hypericum scabrum), обычна Onosma Barsczewskyi,
кровохлебка (Sanguisorba minor), изредка привлекают внимание ярко-
красные головки «мака» (Roemeria refracta). Пышно разросся, цветет
и плодоносит юган (Prangos pabularia). Все эти виды, как и ряд других,
не образуют на южных склонах, с грубой щебенистой почвой, сомкну-
1 Наблюдений над пролетом в течение апреля и первой половины мая у нас,
к сожалению, нет, так как в этот период орнитологи Зоологического института
на Кондаре не работали.
270
А. Я. Тугаринов
того травостоя, но растут отдельными кустами или дерновинами.
Из кустарников в начале мая цветет Atraphaxis pyrifolia, а в конце Zizi-
phus sativus. Незаметные цветочки этого колючего ксерофита издают
удивительно приятный, тонкий аромат. На северных склонах цветет
пузырник (Colutea canescens). Травянистая растительность дает здесь
более сомкнутый покров. Пажитник (Trigonella Lipskyi) придает жел-
теющий фон открытым лужайкам, обильны горечавки (Gentiana Olivieri),
выделяются крупные розовые соцветия эспарцета (Onobrychis grandis),
розово-фиолетовые стрелки шпажника (Gladiolus segetum). К концу
мая в теневых местах -реди этой пышной растительности поднимаются
полутораметровые стебли великолепного реликтового растения — Ostrow-
skia magnifica, несущие кисть громадных (10—12 см в диаметре) белых,
при отцветании розоватых или голубоватых колокольчиков. По дну
ущелья растительность также достигает теперь наиболее пышного раз-
вития. Ивняки, тополь, алыча, орех, кое-где яблоня, боярка стоят
в молодой, достигшей полного развития листве. Еще цветет кое-где жимо-
лость (Lonicera Korolkovi). Стволы и кроны деревьев густо обвиты
диким виноградом, образующим совершенно непролазные чащи. Здесь же
всюду раскинулись покрытые цветами крупные, колючие кусты ши-
повника (Rosa canina). Берега речки, как гигантская конопля, обступила
поросль Datisca cannabina. Там, где поток прижимается к отвесным
скалам, всегда затененным, среди подушек мхов свешиваются стебельки
венерина волоса (Adianthum capillus veneris).
В средней части ущелья, в кленовниках Квака, развитие раститель-
ности запаздывает. В первой половине в массе появляется красный тюль-
пан (Tulipa lanata), обильно цветут пролеска, хохлатки; целые заросли
шиповника (Rosa kokanica) стоят, усыпанные желтыми цветами. Цветет
иргай (Cotoneaster), боярка, еще есть отцветающие орехи, клены, в пол-
ном расцвете яблоня. Необычайно буйно идут в рост травы, еще не начав-
шие цветения. Во второй половине месяца появляются громадные лило-
вые ирисы (Iris Hoo^iana), зацветает таран (Polygonum bucharicum),
крупноцветный пунцовый «горошек» (Lathyrus mulkak), ясенец (Dictamnus
tadshikorum), Thermopsis dolichocarpa. Все эти растения образуют мощ-
ный сплошной покров на сильно увлажненной почве. Пробудилась жизнь
растений и на подступах к Руидашту, выше зоны кленовников. У ревеня,
из распластанных на скалах розеток гигантских листьев, поднимаются
цветущие стебли; под защитой скал, где есть скопления мелкозема, вытя-
нулись еще не зацветшие ирисы, Fritillaria Eduardii и ряд других
трав, ниже уже отцветших.
В первой половине мая прилет птиц еще продолжается. В 10-х числах
в районе Станции появляется замечательная райская мухоловка, скры-
ваясь в густой зелени ореховых и тутовых деревьев. Всюду заметны
зеленые пеночки, все больше становится желчных овсянок и чечевиц.
Все эти птички, кроме мухоловки, в сущности, уже закончили свой пере-
лет и держатся в лесной части ущелья в ожидании возможности под-
няться в горы — зону розариев и субальпийского высокотравья, где
и будут гнездиться. Носятся стаи домашних воробьев. Местные птицы
принадлежат к перелетной популяции — в районе Кондары они появ-
ляются в первой декаде мая. В середине месяца, разбившись на пары,
они занялись подыскиванием гнездовых мест и устройством гнезд, что
и продолжалось до конца месяца. В первой декаде началась кладка яиц
у кекликов, с середины мая — у щеглов; сидят на яйцах синички (Parus
flavipectus и Parus rufonuchalis). В 20-х числах последние уже выво-
Календарь жизни в ущелье Кондара
271
дят молодых. У желчных овсянок, разместившихся для гнездования
в нижней части ущелья, самцы усиленно поют, в токовом полете пере-
мещаясь с дерева на дерево. Загнездились серые мухоловки. Во второй
половине месяца летний аспект орнитофауны устанавливается вполне.
В связи с установившимся летним режимом с высокими температурами
дня, в полуденные часы деятельность птиц замирает.
У весьма обильного в ущелье слизня (Parmacella korshinskii) в мае
продолжается спариванье и происходит откладка яиц в еще влажную
почву, под камни и т. д.
ЛЕТО
Июнь. День за днем стоит жаркая сухая погода, случается, задует
«афганец», затянет небо серою мглою, сквозь которую солнце кажется
красным диском. Постепенно почва теряет запасы влаги. На южных
склонах до высоты 1500 м растительность кончает вегетацию и засыхает,
горы принимают бурый оттенок. На месте бывших зеленых лужаек под-
нимаются кусты грубых, жестких ксерофитов: Trichodesma incanum,
Salvia sclarea, Echium italicum, Lindelophia anchusoides, различные
Cousinia, аляповатая мальва (Alcea nudiflora). В начале месяца зацве-
тает нарядная Scabiosa songorica и невзрачный Eremurus comosus. Юган
плодоносит, достигает полного роста, и теперь население занято его сбо-
ром: прикорневые листья срезываются, здесь же, в снопах, высушиваются
и свозятся в кишлак. Это зимний корм для скота. Начинается созревание
плодовых деревьев. В середине месяца поспевают первые ягоды на туте,
вишне-махалебке, ежевике. С этого времени тут, частью и вишня, надолго
входят в пищевой рацион большого числа птиц. Сухость коснулась,
и северных склонов ущелья, и здесь травы блекнут, подсыхают. Даже
в таких затененных местах, как под пологом ореховых деревьев, нежная
недотрога — Impatiens parviflora, одно из немногих растений таких
мест, кончает вегетацию и увядает. Почва остается совершенно оголен-
ной. Зато выше, по склонам и в кленовниках Квака, в первой половине
месяца растительность достигает наиболее пышного развития. Травостой
в виде сплошного покрова поднимается до высоты IV2 м и более. Затеняя
друг друга, теснятся Ligularia peisic , щавель (Rumex Paulsenianus),
ясенец (Dictamnus tadshikorum), Thermopsis dolichocarpa, качаются
головки лука (Allium Rosenbachianum), местами фон дают белые зонтики
Aegopodium tadshikorum, а над всем этим мощным разнотравьем, как
свечи, поднимаются огромные стрелки беловато-розоватых Eremurus
robustus. Под Руидаштом еще цветет жимолость (Lonicera per. ica). Выше
лесной границы, на подъеме к Руидашту, растительность развилась
не менее пышно. Непролазные чащи желтого шиповника густо осыпаны
цветом, в их чаще краснеют громадные пионы (Paeonia hybrida), лило-
вые (Iris Hoogiana), по величине не уступающие крупным садовым сор-
там. Обилен и характерен для зоны горицвет (Adonis turkestanicus).
На каменистых россыпях кусты тарана (Polygonum bucharicum) благо-
ухают громадными белыми кистями цветов, рядом раскинулись плети
пунцового «горошка». Fritillaria Eduardii уже в плодах. На самом плата
Руидашт, освободившемся теперь от снега, по его пологим склонам зазе-
ленели дерновины мятликов (Роа annua, Р. glabriflora), цветут лютики
(Ranunculus afganicus, R. turkestanicus), пажитник (Trigonella Popovii)>
на грубых, травяных почвах распластались ветви Phlomis fruticetorumk
212
А. Я. Тугаринов
поднялись, хотя еще не цветут, щавель, лигулярия, ферула, юган и др.
.Для всей зоны от верхней границы леса и выше характерно заметное
ночное охлаждение. В середине месяца на плато пригоняют стада овец,
коз и табуны лошадей на летние пастбища.
В течение июня заканчивается период размножения у большинства
птиц. В начале месяца еще заняты гнездостроением такие поздно приле-
тающие виды, как желчные овсянки, домашние воробьи, райские мухо-
ловки, даурские ласточки и др. Хорошо прослеживается различие в сро-
ках гнездовых моментов, наступающих в районе Станции и выше
по ущелью. Так, в долине Варзоба у райской мухоловки 13 июня птенцы-
подлетки уже вышли из гнезда, а в уроч. Сангизард старые птицы имели
еще не полные кладки. У овсянки Стюарта на устье Кондары подлетки
появились около 20 июня, а на подступах к Руидашту 9 июня птицы
«еще только строили гнезда. Различие в сроках одних и тех же явлений
для птиц одного вида при разнице высот в 1000 м составляет 15—20 дней.
В районе Станции в последней трети июня появляются подлетки у домаш-
него воробья, сорокопута, серой мухоловки, садовой славки, Irania
gutturalis; у синего и каменного дроздов, краснокрылого чечевичника
(Rhodopechys sanguinea), чекана-плешанки, горного вьюрка (Serinus
pusillus) — в это время самостоятельные лётные молодые. Около 20-го
числа кончают петь соловьи.
Из отложенных в мае яиц слизня идет развитие молодых. Весь после-
дующий засушливый период они проводят под землей, скрывшись
в эмбриональной раковинке.
Июль, Весь месяц стоят томительно жаркие дни. В нижней части
ущелья даже ночь не приносит заметного охлаждения. Палящее солнце
иссушает почву, поверхность ее затвердевает. Накаленные за день камни
еще долго после заката солнца остаются горячими. Днем они трескаются,
разрушаются, скатываются вниз; частицы мелкозема и дресвы, нагре-
ваясь и расширяясь, на крутых склонах также приходят в движение,
медленно струясь по осыпи. Нередки мглистые дни: мельчайшие частицы
почвы восходящими токами поднимаются в воздух, засоряя атмосферу;
оседая, они покрывают пыльным слоем растения, листья деревьев, камни.
Случаются пыльные бури, приносимые «афганцем». Состояние засухи
коснулось теперь и средних частей ущелья — зоны кленовников. Травы
плодоносят, блекнут и вянут. В нижней части ущелья начался «летний
листопад»: часть листьев на деревьях сохнет и опадает. И только
по самому дну ущелья, около воды, деревья еще стоят в зеленой листве.
Здесь попрежнему мощно развиваются травы, громадного роста дости-
гает Datisca cannabina, цветут Epilobium hirsutum, Myricaria albopecu-
roides, начинает вызревать алыча.
На южных склонах травы совершенно выгорели, о прежней пышной
растительности напоминают лишь оголенные, полусухие стебли югана,
камоля, Eremostachys и других трав; прикорневые розетки эремурусов,
унгернии и других многолетников посохли. Кое-где еще цветут сухо-
любивые шалфей (Salvia sclarea), мальва (Alcea nudiflora), василек
(Centaurea squarrosa), Echium italicum, чаще других попадаются на глаза
различные кузинии (Cousinia refracta, С. radians). Колючие или шерсти-
стые, с серыми, либо сизыми листьями, все эти растения не оживляют
общего серо-бурого фона оголенной поверхности склонов. Деревца мин-
даля, почти лишившиеся листьев, стоят со зрелыми плодами и раскрыв-
шимися околоплодниками. На туте и вишне осыпаются поспевшие ягоды.
Созревают ранние сорта персиков.
Календарь жизни в ущелье Кондара
273
Для зоны кленовников отмечается постепенное угасание травяной
растительности. В первой половине месяца еще цветут запоздалые экзем-
пляры ясенца, Aegopodium tadshikorum, Ligulaiia Thomsonii, поближе
к августу от прежнего пышного травостоя остается лишь побуревшая,
наполовину полегшая растительная масса. И только на Руидаште еще
полный расцвет трав. Так, здесь цветут уже давно окончившие вегетацию
ниже: пажитник, зверобой (Hypericum scabrum), Gonyza khorassanica,
таран, иссоп (Hyssopus seravschanicus), а вместе с тем появились харак-
терные для субальпийской зоны различные кузинии, из которых особенно
обильна С. macilenta с ее седоватыми листьями, полынь (Artemisia
persica), астрагал (Astragalus lasiosemius), татарник (Cirsium turkesta-
nicum) и др. В понижениях и лощинах, где грунтовые воды близки к по-
верхности, мощное высокотравье (Ligularia, Rumex, Senecio и пр.) еще
в полном развитии. В начале месяца производится уборка посевов пше-
ницы и ячменя на пашнях, расположенных на высоте 1500 м и выше.
Созревание ягод алычи и фруктовых деревьев привлекает в район
Станции нежелательного посетителя — медведя. Его постоянным летним
обитанием служит зона кленовников и Руидашт как по склонам, так
и на плато. Сейчас он предпринимает ночные экскурсии до устья ущелья,
кормясь опавшими ягодами алычи; случается, ломает и персиковые
деревья, чтобы достать сочные плоды. Таким же ночным посетителем
становится теперь дикобраз, внимание которого привлекают поспеваю-
щие огородные овощи. Тутовые деревья дают обильный корм начавшим
кочевать семьям и стайкам птиц. В густой листве, выбирая спелые ягоды,
копошатся воробьи и другие зерноядные, время от времени появляются
вороны, иволги, поедают тут и некоторые насекомоядные, например
славки, серые мухоловки, не говоря о менее разборчивых дроздах —
синем и каменном, или синей птице. Мелкие птицы не съедают ягод цели-
ком, а выклевывают отдельные зернышки с частью мякоти. Все птицы
покинули гнездовые места и встречаются в чуждых им биотопах. Белые
трясогузки держатся семьями, родители еще кормят лётных молодых
(13 VII). В последних числах все гнездившиеся по ущелью желчные
овсянки начали продвижение на зимние квартиры; в верхней зоне,
на Руидаште, они — еще на местах, летные молодые следуют за родите-
лями, требуя пищи.
Август. До середины августа господствует тот же летний тип
погоды — знойные, безоблачные дни. В воздухе стоит пыльная мгла,
иногда она усиливается настолько, что солнце едва просвечивает.
На сухой побуревшей траве пыль осела в таком количестве, что вокруг
идущего человека она поднимается целым облачком. Со второй половины
месяца намечаются некоторые перемены. Днем над магистральной частью
хребта начинают появляться облачка, исчезающие к вечеру, даже в ниж-
ней части ущелья ночью становится более прохладно.
Растительная жизнь характеризуется продолжающимся угасанием
на всех высотных зонах. В нижней части ущелья еще кое-где доцветают
сухолюбы в виде шалфея, кузинии, но в основном все травы посохли.
Тем более интересно цветение унгернии, выпускающей цветочные стрелки,
тогда как прикорневые розетки листьев, вегетатировавшие ранней весной,
сейчас совершенно посохли. По дну ущелья, у реки, цветет ломонос
(Clematis orientalis), в более глубоких его частях идет дозревание алычи,
поспевают первые дикие яблоки, фисташка, поздние сорта персиков.
В середине месяца роняет последние ягоды тут, вызревает дикий вино-
град. В зоне кленовников созревание яблок начинается позднее, около
1S Ущелье Кондара
274
А. Я, Тугаринов
20-го числа. В это время и под лесным пологом все травы окончили пло-
доношение, засохли и полегли. Деревья потеряли значительную часть
листвы. На грани субальпийской зоны и выше все еще встречаются цве-
тущие растения: душица (Origanum tyttanthum), норичник (Scrophularia
pinnata), Senecio Renardi, Gerbera kokanica и др.. На Руидаште расти-
тельность в основной массе также закончила вегетацию, а вместе с тем
окончился и пастбищный сезон, стада перегнали на новые места.
Созревание яблок и винограда дало новый корм для медведя. Его
посещение виноградных зарослей приводит к полному их опустошению:
пытаясь достать высоко висящие кисти, зверь обрывает и топчет плети,
ломает деревья и т. д. Подбирая яблоки, медведь не довольствуется
упавшими, но стряхивает их с дерева. Виноград служит приманкой
и для многих птиц. Одни, как, например, воробьи, расклевывая ягоду,
выедают мякоть, не трогая кожуры и косточек, другие, как, например,
кустарницы, соловьи, едят с косточками. Продолжаются кочевки отгнез-
дившихся птиц. Исключение составляет горлинка. Еще можно услышать
воркующих самцов, а самки несутся вторичными кладками. Хорошо
выражено пролетное движение щурок. До нижних частей ущелья спу-
стились чечевичники, серые славки, на Кондаре появились Microcichla
sculerii, также прикочевавшие из верхней зоны хребта. Временами —
до середины месяца — попадаются молодые желчные овсянки: это отле-
тают последние из числа выведшихся в субальпийском поясе. В самом
конце месяца в районе Станции появляются желтоголовые трясогузки
(Motacilla citreola); летом их можно было встретить только на плате
Руидашт. В зоне кленовников к перечисленным бродячим стайкам птип
прибавляются семьи горихвосток (Phoenicurus ochruros), зеленых пено-
чек (Phyloscopus viridanus), малых сорокопутов (Lanius minor), дроздов-
деряб. В 10-х числах месяца здесь еще можно застать семьи серых мухо-
ловок, в которых родители кормят лётных молодых. Временами —
до середины месяца — случается услышать пение соловья, иволги, чече-
вичника и некоторых других, но это уже осеннее пение, не связанное
с давно окончившимся гнездовым периодом, частью это поют молодые
птицы. В первой декаде августа в р. Кондара можно заметить мальков
маринки, скатывающихся по течению в Варзоб.
ОСЕНЬ
Сентябрь, Первая половина месяца характеризуется еще летним
типом погоды. Вечера и ночи стали заметно прохладнее, воздух не засо-
рен пылью, открылись далекие горизонты. Над центральными частями
хребта все чаще собираются днем тучи, уже случаются проходящие шква-
листые дожди. С середины месяца короткие дожди начинают захватывать
и ущелье, по вершинам гор в таких случаях выпадает первый, скорс
стаивающий снежок, но в общем еще стоят ясные, нежаркие дни насту-
пающей осени.
На кленах, орехе и других деревьях появляется все более желтых
или побуревших листьев, частью они отпадают. Поспевают ягоды позд-
ники, шиповника, боярки (Crataegus pontica), вызревают яблоки на де-
ревьях у высотного предела распространения. С середины месяца,
по мере созревания, ранее — в нижних частях ущелья, начинается c6oj
орехов. Травяная растительность окончательно посохла. Только в верх-
ней части зоны кленовников в первую половину месяца попадаются за-
Календарь жизни в ущелье Кондара
275
поздавшие цветением экземпляры мальвы, да по дну ущелья еще есть
цветущие плети ломоноса. На Руидаште посохшая, вытравленная расти-
тельность также не радует глаза, и только в понижениях, где почва увлаж-
нена близкими грунтовыми водами, еще выделяются пятна свежей зелени.
Весь месяц идет постепенный отлет местных птиц и пролет некоторых,
не гнездившихся. В самых первых числах улетают иволги. Щеглы, дубо-
носы, синички кочуют, собравшись стайками. Чаще попадаются на глаза
семьи кустарниц; их стало значительно больше, очевидно, за счет при-
кочевавших из более глубоких частей ущелья. В первой декаде появ-
ляются пролетные общества коньков (Anthus trivialis), серых славок,
пеночек (Phylloscopushumei), горихвосток, чеканчиков (Pratincola maura).
К середине месяца исчезают райские мухоловки, чечевичники, показы-
ваются одиночки варакушек. На Руидаште бродят, сбившись в большие
стаи, рогатые жаворонки, желтые и желтоголовые трясогузки. Вслед
за перегоняемыми с летних пастбищ стадами овец появляются одиночки
и общества грифов и белоголовых сипов, рассчитывающих на поживу.
Местные черные вороны сбились в бродячие стаи. В конце второй декады
показываются передовые одиночки прилетающих на зимовку чернозобых
дроздов — начинается формирование зимнего комплекса орнитофауны.
Октябрь—ноябрь. Наметившийся в конце сентября переход к зим-
нему типу погоды в октябре определяется вполне. Ясные дни хотя и пре-
обладают, но их сменяют время от времени полосы ненастья. В ноябре
усиливается листопад, все деревья роняют уцелевшие от засухи листья
и к середине месяца совершенно оголяются. Вместе с тем увлажнение
почвы при преобладании положительных температур приводит к пробу-
ждению растительности. Под слоем посохшей и полегшей травы проби-
ваются листочки эфемеров, главным образом злаков — костра, мятлика
и др., появляются молодые листочки в розетках многолетников.
Те места, где злаки разрослись плотным ковром, кажутся зелеными
пятнами. На них можно встретить цветущие одуванчики. После каждого
дождя оживают лишайники, обильно покрывающие скалы и крупные
камни. Тогда их поверхность кажется испещренной зелено-бурыми пят-
нами. 1—2 дня бездождия снова приводят к высыханию лишайников.
Растения, начавшие осеннюю вегетацию, уходят под снег в зеленом со-
стоянии, не погибая. Низкие или даже отрицательные температуры лишь
задерживают их рост, и наступающее весною развитие является как бы
продолжением осенней деятельности.
В составе пернатого населения идет формирование зимнего комплекса.
Пришельцы с севера или из высоких поясов гор прибывают постепенно:
первые — по мере наступления зимних условий все далее к югу, гор-
ные — вслед за выпадением глубоких снегов. Из пролетных едва ли
не последними покидают ущелье пеночки (Phylloscopus collybita), когда
к декабрю исчезнут мелкие деятельные насекомые. В октябре, когда почва
увлажнится, не выходя на поверхность, начинают питаться и расти
молодые слизни.
18
хозяйственное освоение
БИОЛОГИЧЕСКИХ РЕСУРСОВ
В. И. ЗАПРЯГАЕВА
БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ДРЕВЕСНЫХ И КУСТАРНИКО-
ВЫХ ПОРОД В СВЯЗИ С МЕЛИОРАЦИЕЙ ЛЕСНОЙ РАСТИТЕЛЬНО-
СТИ ГИССАРСКОГО ХРЕБТА
I. ДРЕВЕСНО-КУСТАРНИКОВАЯ РАСТИТЕЛЬНОСТЬ УЩЕЛЬЯ
КОНДАРА
Древесно-кустарниковая растительность занимает почти все ущелье
Кондара, образуя резко выраженный пояс древесно-кустарниковой расти-
тельности. Этот пояс начинается, примерно, на высоте 900—950 м
над ур. м., т. е. несколько ниже ущелья Кондара; верхняя же граница
проходит на высоте около 2400 м.
Пояс древесно-кустарниковой растительности по своему флористиче-
скому составу является характерным для всего Гиссаро-Дарвазского
района, хотя некоторые древесные и кустарниковые растения здесь
и выпадают (Н. Ф. Гончаров, 1936).
В вертикальном направлении растительность распадается на 2 по-
лосы — нижнюю и верхнюю. Для нижней полосы характерно чрезвы-
чайное разнообразие древесных и кустарниковых пород, как Acer turke-
stanicum Pax, Acer Regelii Pax, Celtis caucasica Willd., Amygdalus
bucharica Korsh., Juglans regia L., Prunus sogdiana Vass., Cerasus
machaleb (L.) Mill., Crataegus pontica C. Koch, Crataegus turkestanica
A. Pojark., Malus hissarica Kudr., Fraxinus potamophila Herd., Juniperus
seravschanica Kom.
Из кустарников здесь встречаются несколько видов Cotoneaster,
несколько видов Rosa, Lonicera persica Jaub. et Spach., Lonicera Ko-
rolkowi Stapf. и др. Очень много дикого винограда Vitis vinifera L.
Верхняя полоса в ущелье Кондара начинается примерно с 1700 м
над ур. м., она обеднена древесными породами (рис. 1). Здесь
ярко выражен мезофильный травяной покров. Из древесных пород рас-
пространены Acer turkestanicum Pax, Juglans regia L., Malus hissarica
Kudr. и Juniperus seravschanica Kom., из кустарников — Rosa kokanica
Rgl., Lonicera persica Jaub. et Spach.
В настоящее время в ущелье Кондара древесно-кустарниковая расти-
тельность встречается фрагментарно, небольшими участками, разделен-
ными между собой скалами и осыпями. Возможно, что когда-то здесь
были более значительные по величине массивы древесной раститель-
ности. Подробное описание древесно-кустарниковой растительности
в ущелье Кондара дано авторами соответствующих разделов настоящего
сборника — В. В. Письяуковой и А. С. Королевой. Вследствие этого
я позволю себе остановиться здесь только на некоторых, не освещенных
280
В. И. Запрягаева
в указанных работах особенностях климата пояса древесно-кустарни-
ковой растительности, которые накладывают отпечаток на характер
растительного покрова.
Климат ущелья Кондара определяется его положением в Гиссаро-
Дарвазской горной стране, которая еще подчиняется распределению
сезонных явлений, характерных для всей субтропической зоны от Атлан-
тического океана на восток, но уже лежит близко к пределу, за которым
решающую силу приобретают континентальные особенности Азии как
таковой. Описываемый район доступен для юго-западных относительно
влажных токов воздуха, которые, задерживаясь на севере Гиссарским
хребтом и на востоке хребтами Петра I и Дарвазским, обусловливают
наличие здесь значительного количества зимне-весенних осадков.
В отличие от многих других районов Советского Союза выпадение
осадков здесь по временам года чрезвычайно неравномерно. Основная
Биологические особенности древесных пород
281
масса выпадает в течение зимне-весенних месяцев, в то время как летние»
месяцы остаются почти или совсем бездождными.
Из большого количества осадков, выпадающих в зимне-весенний
период, растениям, произрастающим по склонам гор, достается весьма
незначительная доля. Это объясняется тем, что во время зимне-весенних
месяцев осадки выпадают часто в виде ливней. Выпадающая влага, не-
успев проникнуть глубоко в почву, скатывается по крутым склонам
вниз, образуя часто силевые потоки, приносящие большой урон горному
сельскому хозяйству. Такие потоки воды, смешанной с мелкоземом
и камнями, с невероятной силой устремляются вниз, снося на своем пути
деревья, мелкие постройки и вырывая глубокие овраги. Древесная расти-
тельность по склонам гор является естественным закрепителем склонов
от размыва. В районе Кондары функцию закрепителя склона от размыва
и хранителя влаги древесная растительность, в силу своей разрежен-
ности, выполняет значительно хуже, чем в других горных районах.
Там, где участки расчищены из-под древесной растительности для пашен,
явления эрозии развиты чрезвычайно сильно. В горах можно наблю-
дать места бывших пашен, которые когда-то были разделаны на место
зарослей древесно-кустарниковой растительности, но в настоящее время
из-за смыва почвы вышли из употребления (Турский, 1932).
Небезинтересно проследить, как распределяется в течение года влага
в почве и насколько она доступна растениям. Влажность почвы на раз-
личных глубинах определялась на участках, расчищенных от расти-
тельности и ежегодно перепахиваемых, и на участках с естественными
насаждениями древесно-кустарниковой растительности.
Различие в количестве влаги в почве на различных высотах весьма
существенно, особенно в верхних горизонтах. Например, на глубине
5 см в марте на нижнем участке влажность почвы 29.13%, а на верхнем
в это же время — 55.20%. В июле на тех же глубинах: на нижнем участке
влажность почвы 8.14%, на верхнем — 16.50%.
Кривая накопления влаги в почве следует на обоих участках за коли-
чеством выпадающих осадков, иногда несколько отставая от него.
Дожди, как правило, начинают выпадать в значительном количестве
в октябре. С этого времени влажность почвы повышается, причем
на нижнем участке влажность повышается наиболее ощутимо тольке
в верхних горизонтах (до 30 см), в нижних же горизонтах (с 50 см) влаж-
ность остается низкой. На верхнем участке, где почва более рыхлая,,
влага проникает до 60 см. К декабрю почва успевает промокнуть на ниж-
нем участке до глубины 80 см, на верхнем — значительно глубже. В тече-
ние января и февраля на нижнем участке и в течение января—середины
апреля на верхнем почва остается под снегом; она не замерзает и,
оставаясь талой, в течение этих месяцев накапливает влагу. С апреля
количество влаги в почве начинает падать, а уже в мае кривая влажности
почвы до глубины 100 см быстро падает. Это объясняется тем, что в этом
месяце идут весенние ливни, влага которых, не успевая глубоко проник-
нуть в почву, скатывается вниз. Верхние же горизонты, насыщаемые
влагой во время таких ливней, под действием горячих лучей майского
солнца быстро пересыхает.
Начиная с мая, а на верхнем участке — с июня, количество влаги
в почве с глубиной, относительно верхних горизонтов, остается большим,
в то время как до мая запасы влаги в верхних горизонтах были значи-
тельно выше, чем в нижних. С июня начинается сухой, бездожд-
ный период, который длится до октября. Почва сильно пересыхает, в ее
282
В. И. Запрягаева
верхних горизонтах количество влаги приближается к мертвому запасу.
Этот период является критическим для растений.
Здесь было рассмотрено положение с влажностью почвы на участках,
расчищенных из-под зарослей древесно-кустарниковой растительности,
где иссушающая деятельность корневых систем очень ослаблена. В ассо-
циациях с нетронутой древесно-кустарниковой растительностью, в недо-
трогово-снытевом кленовнике уже в июне количество влаги в верхних
слоях почвы таково, что оно недоступно растениям; в разнотравно-
пажитниковом кленовнике период с недоступной растениям влагой насту-
пает уже в мае.
Несмотря на то, что почва чрезвычайно пересыхает в верхних гори-
зонтах, и почвенная влага остается недоступной растениям в течение
длительного периода, именно в этих горизонтах нами обнаружено боль-
шое скопление тонких корней, о чем будет сказано несколько ниже.
По био-экологическому характеру наиболее широко распространен-
ные в ущелье Кондара древесные породы можно разбить на 3 группы.
К первой группе следует отнести мезофильные широколиственные
породы: Acer turkestanicum Pax, Juglans regia L., Malus hissarica Kudr.,
сюда же отношу и Acer Regelli Pax, который занимает промежуточное
положение между первой и второй группами. Ко второй группе отно-
сятся ксерофильные лиственные породы, как Celtis caucasica Willd.,
Amygdalus bucharica Korsh., Crataegus pontica C. Koch, и, наконец,
к третьей группе — ксерофильные хвойные — Juniperus serawschanica
Кош.
1. МЕЗОФИЛЬНАЯ ШИРОКОЛИСТВЕННАЯ ДРЕВЕСНАЯ РАСТИТЕЛЬНОСТЬ
Эта растительность наиболее широко распространена в ущелье Кон-
дара, как и во многих других ущельях южного склона Гиссарского
хребта. Здесь она представлена 2 главнейшими формациями: кленов-
ников (Aceretum) и ореховых лесов (Juglandetum).
Особенно широко распространена формация клена Acer turkestanicum
Pax. Кленовики занимают склоны различных экспозиций, в то время
как ореховые леса приурочены к долине реки или местам выхода клю-
чей и родников и только как исключение поднимаются вверх по склонам.
Характерно, что древесно-кустарниковая растительность ущелья Кон-
дара не образует больших массивов сплошного леса, здесь можно наблю-
дать небольшие, по 2—4, реже — 6—8 гектаров, участки, перемежающие-
ся с пашнями или, чаще, скалистыми обнажениями материнских пород.
Кленовые и ореховые леса приурочены к более пониженным и, стало
'быть, более благоприятным в климатическом отношении частям рельефа,
с более или менее мощным почвенным покровом. Водораздельные гребни,
подверженные действию иссушающих ветров, почти всегда безлесны.
Как я уже отметила, можно полагать, что когда-то описываемое ущелье
было значительно больше облесено.
Кленовники
Формация кленовников, являясь основной зональной формацией,
наибольшего распространения достигает на мелкоземистых северных
склонах ущелья. Реже она распространена на склонах западных и восточ-
ных румбов и как исключение — на склоне южной экспозиции.
Кленовники представлены рядом группировок, приуроченных к раз-
личным экологическим условиям. Так как нижняя и верхняя полосы
Биологические особенности древесных пород
283
пояса древесно-кустарниковой растительности очень сильно отличаются
по климатическим и почвенным особенностям, а также по характеру
растительности, следует различать кленовники нижней и верхней полосы
этого пояса.
Необходимо отметить что Acer Regelli Pax встречается в ущелье
Кондара значительно реже Acer turkestanicum Pax и является второ-
степенным компонентом в ассоциациях, формируемых Acer turkestani-
cum.
В нижней части ущелья Кондара, на высоте 1200—1300 м, довольно
широко распространенной ассоциацией являются недотрогово-разно-
травные иргайники. Здесь туркестанский клен встречается либо единич-
ными деревьями, либо его нет совсем. Не вызывает никакого сомнения,
что участки, занятые в настоящее время недотрогово-разнотравными
иргайниками, были в свое время покрыты кленовниками.
Одной из основных причин отсутствия клена в иргайниках является
деятельность человека, который интенсивно вырубал клен.
Туркестанский клен и сопутствующие ему в ежово-разнотравно-
недотроговом кленовнике древесные породы, как Celtis caucasica Willd.,
Prunus sogdiana Vass., в иргайнике либо выпадают совсем (Prunus sog-
diana Vass.), либо встречаются отдельными деревьями (Acer turkesta-
nicum Pax, Celtis caucasica Willd.). Совсем исчезает в иргайнике Loni-
cera persica Jaub. et Spach., но зато иргай Cotoneaster hissaiica Pojark.
образует сплошные, трудно проходимые заросли. В травяном покрове
много общих видов, но встречаемость этих растений различна: клен,
вырубленный или уничтоженный каким-либо другим способом, на уча-
стках, занятых иргаем, не возобновляется. Мне не удалось встретить
ни одного всхода клена в иргайниках старше двух лет. Клен не возоб-
новляется здесь и вегетативно, как во многих других местах. Причину
этого явления следует искать в биологических и экологических особен-
ностях того и другого растения. Дело в том, что иргай имеет чрезвы-
чайно мощную корневую систему, которая располагается под поверх-
ностью почвы на небольшой глубине, от 0 до 120 см. Корневая система
у иргая не имеет стержневого корня, она очень разветвлена, и сосущие
корни ее подходят к самой поверхности (рис. 2, 3); в результате в почве,
284
В. И Запрягаева
на глубине 0—30 см, образуется густо переплетенная сетка из корней
иргая, которая перехватывает почти всю атмосферную влагу, попадаю-
щую на склоны.
В сплошных зарослях иргая верхние горизонты почвы (0—30 см)
относительно влажны и по запасам влаги приближаются к недотрогово-
снытевым кленовникам, т. е. самым влажным местообитаниям клена
в нижней полосе пояса древесно-кустарниковой растительности. Ниже
Рис. 3. Горизонтальная проекция корневой системы иргая Cotoneaslcr hissarica
Pojark.
30 см запасы влаги резко снижаются, и только на глубине 100 см начи-
нается некоторое повышение процента влажности.
Совершенно естественно, что корням клена, которые располагаются
тоже поверхностно, но несколько глубже иргая, не достается необходи-
мого для нормального развития количества влаги. Семена клена, кото-
рые заносятся в иргайники ветром, весной прорастают, но из-за недо-
статка влаги в почве к осени всходы отмирают, в то время как всходы
иргая продолжают нормально развиваться.
Одной из самых распространенных ассоциаций клена на северных
склонах нижней полосы пояса древесно-кустарниковой растительности
являются ежово-девясилово-разнотравные кленовники. Эта ассоциация
Биологические особенности древесных пород
285
располагается либо по северным склонам ущелья Кондара, либо по се-
веро-восточным или северо-западным склонам боковых ущелий, обычно
выше по склону местообитаний ежово-разнотравно-недотроговых кленов-
ников. Часто эта ассоциация поднимается до гребня водораздельных
гряд. Почва здесь мелкоземиста, но иногда с большой примесью щебенки.
Древесный полог всегда сильно разрежен, его сомкнутость не превышает
20—40%. Из древесных пород преобладает Acer turkestanicum Pax (sp.i),
часто примешивается Acer Regelii Pax (sol.), Cerasus machaleb (L.) Mill,
(sol.-sp.i), Celtis caucasica Willd. (sol.), Crataegus turkestanica Pojark.
(sol.), Juniperus seravschanica Kom. (sol.), Crataegus pontica C. Koch
(sol.), Malus hissarica Kudr. (sol.), Prunus sogdiana Vass, (sol.), Amyg-
dalus bucharica Korsh. (sol.). Обильно встречаются кустарники Coto-
neaster insignis Pojark. (sp.i), Lonicera persica Jaub. et Spach. (sol.-
sp.i), Rosa kokanica Rgl. Травяной покров чрезвычайно пестрый. Здесь
насчитывается около 60 видов различных растений.
У гребней водораздельных гряд, в местах, подверженных иссушаю-
щему действию солнца и ветра, располагается разнотравно-югановая
ассоциация, где туркестанский клен встречается единичными низкорос-
лыми деревьями; эта ассоциация не имеет широкого распространения
в ущелье Кондара, она является переходной от ежово-девясилово-разно-
травных кленовников к эфемерово-гречихово-югановой ассоциации, зани-
мающей гребень водораздельного хребта между Кондарой и Бигаром.
Из древесных пород в разнотравно-югановой ассоциации очень редко
встречаются Amygdalus bucharica Korsh. (sol.-sp.i), Acer Regelii Pax
(un.-sol.), Crataegus pontica C. Koch (sol.), Celtis caucasica Willd. (sol.-
sp.). Травяной покров не сплошной, он покрывает поверхность почвы
на 80—90%.
В аналогичных условиях у гребней водораздельных гряд или чаще
на самих грядах встречается эфемерово-гречихово-югановая ассоциация,
являющаяся крайней в экологическом ряду распространения клена.
Здесь Acer turkestanicum Pax либо присутствует в единичных, далеко
расставленных друг от друга экземплярах, либо часто его совсем нет.
Здесь же можно встретить экземпляры Acer Regelii Pax и Amygdalus
bucharica Korsh. Травяной покров очень редкий.
Довольно широко распространенной ассоциацией в нижней полосе
пояса древесно-кустарниковой растительности являются кленовники
разнотравно-пажитниковые, которые занимают небольшие по площади
участки на северных, северо-западных и северо-восточных склонах.
Почвы здесь мелкоземистые, но с очень большим количеством щебня,
напоминая в этом отношении почвы разнотравно-югановой ассоциации.
Эта ассоциация характеризуется очень разреженными зарослями Acer
turkestanicum Pax (sol.-sp.i), Amygdalus bucharica Korsh. (sol.-sp.i),
Acer Regelii Pax (sol.), среди которых довольно много кустарников
Rosa kokanica Rgl. (sol.-sp.i), Cotoneaster insignis Pojark. (sol.). В этой
ассоциации встречается около 70 видов травянистых растений.
По южным склонам, защищенным от неблагоприятного действия
ветров, на участках с относительно мощным почвенным покровом до-
вольно редко встречается разреженная миндально-кленовая ассоциация
с ячменно-эфемерово-разнотравным травяным покровом.
Древостой этой ассоциации чрезвычайно разрежен и состоит из
Amygdalus bucharica Korsh. (sp.i-sp.2), Acer turkestanicum Pax (sol.-
sp.i), Acer Regelii Pax, Celtis caucasica Willd. (sol.-sp.i), Crataegus ponti-
ca C. Koch (sol.), Malus hissarica Kudr. (sp.i), Pistacia vera L. (sol.).
286
В. И. Запрягаева
Травяной покров чрезвычайно разнообразен и включает около 70 видов
различных растений.
В верхней полосе пояса древесно-кустарниковой растительности тур-
кестанский клен образует целый ряд ассоциаций, которые в нижней по-
лосе не встречаются.
В наиболее благоприятных условиях по дну боковых ущелий с повы-
шенной влажностью почвы располагается самая мезофильная ассоциа-
ция высокотравных-снытевых кленовников. Почвы здесь лесные, сильно
гумусированные. Господствующей древесной породой является Acer tur-
kestanicum Pax (sp.2-сор.i). Часто к нему примешивается орех Juglans
regia L. (sol.-sp.i). Из кустарников распространена только Lonicera per-
sica Jaub. et Spach. (sol.). Травяной покров не разнообразен. Здесь тур-
кестанский клен достигает наиболее пышного развития, высота деревьев
15—16 м, он вместе с орехом часто образует покрытие до 50—90%.
Значительно более широкое распространение, чем предыдущая ассо-
циация, в пределах северных склонов верхней полосы древесно-кустар-
никового пояса имеет ассоциация высокотравных кленовников. Здесь
туркестанский клен развивается почти так же хорошо, как в предыдущей
ассоциации, хотя условия увлажнения почвы хуже. Господствующей
породой является, как и в предыдущей ассоциации, Acer turkestanicum
Pax, к нему примешивается орех Juglans regia L. (soL-sp.2), яблоня
Malus hissarica Kudr. (sol.-sp.i), Rhamnus dolichophylla N. Gontsch.
(sol.-sp.i), арча — Juniperus seravschanica Kom. (sol.). Из кустарников
встречаются Lonicera persica Jaub. et Spach., Cotoneaster insignisPojark»
(sol.), Rosa kokanica Rgl. (sp.). Травяной покров представлен высоко-
травьем и очень разнообразен по видовому составу, здесь встречается
около 70 видов различных травянистых растений.
Среди высокотравных и высок отрав но-снытевых кленовников можно
встретить на этих высотах, а иногда и несколько выше, ежово-тороново-
разнотравные кленовники, где клена очень мало (sol.-sp.i). К нему
иногда примешивается яблоня и рябина (Sorbus persica Held.). Травя-
ной покров по видовому разнообразию значительно беднее предыдущей
ассоциации.
Одной из наиболее распространенных ассоциаций в верхней полосе
пояса древесно-кустарниковой растительности являются бузульниково-
недотроговые кленовники. Эта ассоциация развивается на глубоких гуму-
совых буроземах и характеризуется более сомкнутым пологом (50—80%).
Преобладающей древесной породой является Acer turkestanicum Pax
(sp.2-сор.2), к которому почти всегда примешивается орех. Иногда встре-
чаются единичные экземпляры арчи и крушины. В подлеске Lonicera
persica Jaub. et Spach. (sol.), Rosa kokanica Rgl. (sp.i).
В верхней полосе пояса древесно-кустарниковой растительности
ущелья Кондара распространены очень широко розарии, чаще всего
разнотравно-югановые. Они занимают мелкоземистые и слегка щебни-
стые почвы по склонам северных румбов. Древесная растительность пред-
ставлена единичными, сильно угнетенными деревьями туркестанского
клена и еще реже арчей и яблоней, но зато кустарниковая растительность
богато представлена зарослями Rosa kokanica RgL, Cotoneaster sua-
vis Pojark., реже — Cotoneaster nummularia F. M., Lonicera persica Jaub.
et Spach. Желтоцветный шиповник Rosa kokanica Rgl. иногда образует
густые, непроходимые заросли.
Сравнивая флористический состав розариев и кленовников, легко
убедиться в том, что розарии являются вторичными ассоциациями,
Биологические особенности древесных пород
28?
появившимися на месте сведенных
кленовников. Шиповник создает та-
кие условия, при которых клен не
может развиваться нормально. Ши-
повник — Rosa kokanica Rgl.—об-
ладает способностью очень быстро
вегетативно размножаться. Размно-
жаясь корневыми отпрысками
(рис. 4), он очень быстро завоевы-
вает новые территории; из боль-
шого количества корней образует
густую сетку, которая располагается
под поверхностью почвы на глубине
О—15 см. Такая сетка, являясь не-
преодолимым заграждением для ат-
мосферной влаги, чрезвычайно силь-
но иссушает почву и не дает воз-
можности нормально развиваться
таким растениям, как туркестан-
ский клен, у которого корневая
система располагается несколько
глубже.
Естественное семенное возобнов-
ление туркестанского клена в роза-
риях почти прекратилось. Появив-
шиеся всходы клена ранней весной
развиваются только до прекращения
дождей, т. е. до июня, а затем засы-
хают. Нам удалось найти несколько
молодых растений клена в возрасте
10—12 лет; они имели высоту 12—
15 см.
Взаимоотношения между роза-
риями и кленовниками в верхней
полосе пояса древесно-кустарни-
ковой растительности совершенно
такие же, как между иргайниками
и кленовниками в нижней полосе
пояса. И в том и в другом случае
туркестанский клен заменяется при
активном содействии человека более
ксерофильными и более приспособ-
ленными к существующим условиям
кустарниками.
Если окинуть беглым взглядом
кленовые насаждения в ущелье Кон-
дара, то можно отметить, что, за ред-
ким исключением, когда клен растет
по дну боковых ущелий, они, так же
как и большинство других насажде-
ний в ущелье Кондара, очень^раз-
режены, не образуют сомкнутого
древесного полога. Отдельные
jPgc, 4. Вегетативное размножение шиповника Rosa kokani g
288
В. И. Запрягаева
деревья стоят на значительном расстоянии друг от друга, образуя на-
саждения паркового типа. Эта своеобразная особенность древесно-ку-
старниковых насаждений в горах Средней Азии была отмечена еще С.
Коржинским в 1896 г., затем В. И. Липским в 1896 и 1897 гг. и
после этого подтверждена многими другими исследователями.
Достаточное количество света в насаждении дает возможность форми-
роваться типичной для открытых мест широкой кроне деревьев. Стволы
у деревьев получаются сильно сбежистыми, кривыми, ветвление их
Рис. Профильная проекция бухарского миндаля Amygdalus bucharica Korsh,
на северном мелкоземистом склоне в ущелье Кондара.
начинается почти у основания. Деревья приобретают скорее вид мощных
кустов, высота которых достигает 6—12—14 м.
На основании многолетних наблюдений за древесно-кустарниковой
растительностью в естественных условиях, я и Ф. Л. Запрягаев пришли
к заключению, что основным фактором, обусловливающим своеобразную
структуру древесно-кустарниковых насаждений на южном склоне Гиссар-
ского хребта и, в частности, в ущелье Кондара, является недостаток
влаги в почве.
Растения, растущие по сухим склонам гор, чтобы обеспечить свое
нормальное развитие, протягивают корни на очень большие расстояния.
Наблюдения показали, что корни деревьев, отстоящих друг от друга
на 10—12 и даже 15 метров, смыкаются. У некоторых местных древес-
ных пород — яблони, клена, арчи, бухарского миндаля, иргая, шиповника
Биологические особенности древесных пород
289
и некоторых других — площадь проекции корневых систем превышает
площадь проекции крон в 10—20 раз (рис. 5). У большинства древесных
и кустарниковых пород основная масса корней располагается на глубине
до 100 см, а сосущие корни часто подходят к самой поверхности, распо-
лагаясь на глубине 0—2 см.
Недостаток влаги в почве в наших условиях вызывает развитие чрез-
вычайно мощных и поверхностных корневых систем. Это и является
основной причиной, обусловливающей своеобразную разреженность
насаждений. Здесь нет борьбы за свет, а есть борьба за влагу, происходя-
щая под поверхностью почвы. Кажущиеся «открытыми» древесно-кустар-
никовые ассоциации, так широко распространенные в описываемом рай-
оне, в действительности являются насыщенными.
Разреженность древесных насаждений, произрастающих по сухим
склонам гор, характерна не только для Гиссарского хребта, но и для
других районов южной части Средней Азии. В. И. Липский (1911) отме-
чал, что, благодаря разреженности и фрагментарности распределения,
среднеазиатские древесно-кустарниковые насаждения скорее похожи
на лесные рощи, чем на обычные леса.
Разреженность древесной растительности характерна для всего Гис-
саро-Дарваза (Н. Ф. Гончаров, 1937), для Копет-дага (Коржинский,
1896; Е. Г. Бобров, 1926; Е. А. Малюгин, 1931), для западного Тянь-
шаня (М. Г. Попов, 1940), для южных гор Средней Азии и для Турке-
станского хребта (Е. П. Коровин, 1934; В. Д. Дмитриев, 1938), для Зерав-
шанского хребта (В. Л. Комаров, 1896; Г. С. Сиворакша, 1938).
Сведения о разреженности древесных насаждений по сухим склонам
гор можно найти у М. П. Петрова (1944) при описании растительности
северного Ирана, у И. А. Линчевского и А. В. Прозоровского (1946)
для центрального массива Гиндукуша и предгорий Афганского Турке-
стана. Эти разреженные леса ксерофильного типа называют либо рощами,
либо редколесьем. И. А. Линчевский (1935) сравнивает насаждения
фисташки с континентальной саванной ю.-в. Африки, а П. Н. Овчин-
ников (1940) разреженные заросли фисташки с осоково-мятликовым
низкотравьем называет полусаваннами.
Разреженные насаждения широко распространены и в Закавказье,
особенно в восточной его половине, где кавказские ботаники называют
их то «светлыми лесами», то пустынным лесом, то древесной степью,
то аридным редколесьем или даже фриганой (Н. А. Буш, 1935;
А. К. Магакьян, 1941; А. В. Иванова, 1946; Д. И. Сосновский, 1943).
Можно отметить, что такие редколесья распространены еще дальше
на запад. В Малой Азии — в центральной части западной Анатолии
(П. М. Жуковский, 1933), в странах Средиземноморья (Rikli, 1912;
Maire, 1921; Andreanszky, 1939).
Разреженность древесно-кустарниковых насаждений некоторые
авторы пытаются объяснить биологической природой слагающих компо-
нентов, их неспособностью смыкаться кронами (С. Коржинский, 1896;
М. Г. Попов и Н. В. Андросов, 1937), большинство же исследователей —
воздействием человека, который хищнически уничтожал леса путем
порубок.
Наши наблюдения показывают, как уже было мною отмечено, что
смыканию крон препятствуют сильно развитые корневые системы
деревьев и кустарников, произрастающих в условиях недостаточного
почвенного увлажнения. Одни и те же деревья (туркестанский клен,
орех, яблоня, бухарский миндаль) в наших условиях могут образовывать
19 Ущелье Кондара
290
В. И. Запрягаева
и сомкнутые насаждения по дну балок и ущелий, и разреженные — на
сухих склонах.
Это же явление отмечает и Г. С. Сиворакша (1937) для фисташки,
которая иногда, в местах с повышенным увлажнением, образует рощи
с сомкнутым пологом.
О том, что в условиях недостаточного увлажнения растения разви-
вают мощные корневые системы, имеются довольно многочисленные ука-
зания в литературе (Н. А. Максимов, 1941; Г. Люндегорд, 1937;
Е. Я. Шефер-Сафонова, 1934; Т. К. Кварацхелия, 1927; А. В. Гурский,
1938; О. В. Заленский, 1940; Andreanszky, 1939).
Поверхностное распределение корневых систем по довольно много
численным исследованиям в этой области наблюдается не только у дре-
весных пород, но и у травянистых растений, произрастающих в условиях
недостаточного почвенного увлажнения, например в степях (Е. М. Лав-
ренко, 1940), полупустынях и пустынях (Е. П. Коровин, 1934; А. В. Тур-
ский, 1934; С. П. Кузьмин, 1929; В. М. Свешникова, 1944; Н. Walter,
1927; J. Е. Weaver, 1919, 1927; Jenny-Lips, 1930).
Описанная способность местных деревьев развивать корневые
системы, которые, соприкасаясь под поверхностью почвы, создают
разреженность насаждений, ввела в заблуждение сотрудников местных
лесхозов: они, желая получить в существующих условиях строевую
древесину, решили сгустить естественные насаждения путем посадки
и стали сажать различные деревья между кленами, орехом, каркасом
и другими с тем, чтобы занять эти, кажущиеся свободными, площадки. Но
посаженные молодые деревья со слабой корневой системой не могли
выдержать конкуренции с хорошо укоренившимися местными деревьями
или кустарниками, и. отмирание посаженных лесхозом саженцев про-
исходило либо в год посадки, либо на следующий год, и как исключение
отдельные экземпляры существовали несколько лет.
Специальные наблюдения за естественным возобновлением местных
древесных пород показали, что семенное возобновление многих из них
протекает с большим трудом. Основные причины этого явления, повиди-
мому, следующие: чрезвычайно высокий нагрев поверхности почвы днем
и охлаждение ночью, очень низкая влажность припочвенного слоя воз-
духа и сухость почвы. Летом разность между максимальной и минималь-
ной температурами почвы доходит до 40 и более градусов, что составляет
более годовой амплитуды средней суточной температуры воздуха. Ночью
температура в летние месяцы опускается до 10—18°, а днем повышается
до 66°, а на южных склонах до 74°.
Очень большое расхождение наблюдается летом между температу-
рами поверхности почвы и воздуха; на южном склоне оно доходит до
30°. С ноября по февраль и даже март максимальная температура поверх-
ности почвы северных склонов на высоте 1100 м и 1850 м над ур. м. ниже
соответствующей температуры воздуха, что объясняется их затенен-
ностью. Минимальные температуры поверхности почвы на высоте 1100 м
в течение круглого года ниже, чем в воздухе, в среднем на 3°.
Наряду с такими высокими температурами почвы, в припочвенном
слое наблюдается очень низкая относительная влажность воздуха.
Период летней засухи особенно губителен для молодых всходов древес-
ных и кустарниковых пород.
Неокрепшей и неразвитой корневой системе молодых растений неот-
куда черпать влагу в довольно продолжительный период летней засухи.
Если же ко всему этому прибавить иссушающее действие корневых
Биологические особенности древесных пород
291
систем укоренившихся растений, то станет ясным, почему мы находим
в кленовниках такое скудное возобновление клена. Немалую роль в от-
сутствии семенного возобновления играет выпас скота в лесу.
Из всех древесных пород ущелья Кондара наиболее неблагополучно
обстоит дело с естественным семенным возобновлением кленов Acer tur-
kestanicum Pax и Acer Regelii Pax (Ф. Л. Запрягаев, 1940).
Туркестанский клен довольно хорошо возобновляется только на ста-
рых заброшенных пашнях, причем возобновление клена мне удалось
видеть только на пашнях, расположенных в верхней полосе пояса дре-
весно-кустарниковой растительности.
Рис. 6. Вегетативное размножение Acer turkestanicum Pax. Кондара.
Из наблюдений над гибелью сеянцев клена может создаться впечатле-
ние, что в древесно-кустарниковом поясе описываемого района кленов-
ники, из-за того, что они не в состоянии произвести потомство, доживают
чуть ли не последние годы. В действительности это не так. Хотя по мно-
гим признакам мезофильная древесная растительность, которая является
остатком широколиственной древнетретичной мезофильной лесной расти-
тельности, деградирует (Е. М. Лавренко, 1938; Е. В. Вульф, 1944), но
в настоящее время она местами сохранилась еще в очень хорошем состоя-
нии. Такое явление в значительной степени объясняется тем, что клены,
орех, яблоня и др., как бы взамен почти утраченному семенному возоб-
новлению, приобрели способность возобновляться вегетативным путем
(рис. 6). Поскольку на вегетативном возобновлении древесных пород
на южном склоне Гиссарского хребта останавливается довольно по-
дробно в своих работах Ф. Л. Запрягаев, перейду к описанию дальнейшей
судьбы тех единичных растений, которые, несмотря на исключительно
неблагоприятные условия для своего развития, остаются живыми.
19*
292
В. И. Запрягаева
Наблюдения за характером роста путем измерений приростов на де-
реве в течение ряда лет и подсчет годичных колец прироста по спилам
показали, что клены на сухих склонах растут чрезвычайно медленно,
особенно в первые годы (рис. 7),
Из графика видно, что молодое дерево, выросшее из семени, растет
чрезвычайно медленно. Только на 33-й год своей жизни оно достигает
высоты 1м. Высоты 2 м клен достигает к 58, а 5м — только к 100 го-
Рис. 7. Скорость роста древесных пород на южном склоне Гиссарского хребта.
1— Acer turkestanicum Pax; 2— Acer Regelii Pax; 3— Juglans regia L.; 4— Populus pyra-
mldalis Rozier.
дам. Дальше его рост несколько ускоряется, оставаясь в общем весьма
замедленным.
Годичные приросты взрослого туркестанского клена чрезвычайно
невелики и не превышают 8—11 см в год. Период роста клена связан
с периодом выпадания осадков и наличия влаги в почве.
Рост туркестанского клена, как и целого ряда других пород, происхо-
дит, примерно, до конца июня — начала июля, т. е. до того времени,
когда влага, накопленная за зимне-весенний период, еще не успела
израсходоваться из нижних горизонтов почвы. В начале июля прирост
почти у всех деревьев прекращается. Вскоре после прекращения роста
туркестанский клен, так же как и ряд других древесных растений, начи-
нает сбрасывать листву.
Биологические особенности древесных пород
293
Сопоставив данные о возобновлении естественным семенным путем
клена со скоростью его роста в естественных условиях, можно притти
к определенному выводу о возможном характере эксплоатации кленовых
лесов. Эксплоатация кленовых насаждений даже и на топливо была бы
затруднена, если бы не пришла на помощь в этом отношении уже указан-
ная раньше способность туркестанского клена возобновляться вегетатив-
ным путем.
В насаждениях редко можно встретить деревья клена, состоящие
из одного ствола. Обычно клен растет своеобразным «гнездом», состоя-
щим из 5—20 и более стволов различного возраста. Все эти побеги, как
правило, кривые, ветвление их начинается почти у самого основания,
что делает невозможным использование их для строительных целей.
Рост клена Регеля (Acer Regelii Pax) несколько отличается от
роста туркестанского клена. Предельная высота деревьев этого вида
всегда несколько ниже и достигает 7—10 м. Высоты одного метра
этот вид клена достигает за 4—6 лет. Рост его замедляется к 30 годам
жизни. Рост побегов связан, так же как и у туркестанского клена,
с количеством выпадающих осадков и влажностью почвы. У взрослого
дерева этого клена годичные приросты значительно меньше, чем у тур-
кестанского клена. Клен Регеля, так же как и туркестанский, имеет
склонность возобновляться вегетативным путем.
Ореховые леса
Ореховые леса не имеют в ущелье Кондара такого широкого распро-
странения, как кленовники. Они, как правило, сосредоточены в местах
с близким стоянием воды, по дну ущелий, у ручьев и родников. В средней
части ущелья, где климатические условия наиболее благоприятны, орех
поднимается высоко по северным склонам с мощными мелкоземистыми
почвами. Если в нижней части древесно-кустарникового пояса он встре-
чается отдельными деревьями или маленькими группами, то выше он
образует настоящие ореховые рощи.
Если проследить распределение ореха в вертикальном направлении
по ущелью Кондара, то можно сказать, что хотя он произрастает и до
2000 м над ур. м., т. е. переходит границу нижней полосы пояса древесно-
кустарниковой растительности, но наилучшего своего развития дости-
гает у верхнего предела нижней полосы пояса, т. е. примерно на высоте
1400—1700 м над ур. м. Здесь, как я уже отмечала, он образует настоя-
щие ореховые рощи.
В отличие от туркестанского клена, орех не образует такого разно-
образия растительных ассоциаций. В нижней полосе пояса древесно-
кустарниковой растительности орех ютится около воды, образуя по
долине реки Кондара ореховые леса с недотрогой, где орех достигает
высоты 20—25 м и образует сомкнутое или почти сомкнутое насажде-
ние. Участки орехового леса с недотрогой часто перемежаются с насажде-
ниями ореха, в которых присутствует ежевика, образующая часто непро-
ходимые заросли.
Иногда орех образует густые насаждения на маленьких болотцах
в устьях боковых ущелий. В атом случае орех также достигает огромных
размеров 20—25 м и часто образует сомкнутый полог. Помимо этих ассо-
циаций, орех часто встречается единичными деревьями по дну ущелья
Кондара.
294
В. И. Запрягаева
В верхней полосе пояса древесно-кустарниковой растительности
орех можно встретить только по склонам, на дно ущелий он не спускается.
Здесь орех никогда не образует чистых насаждений, он растет всегда
в смеси с туркестанским кленом, яблоней, реже арчей. Из наиболее
распространенных ассоциаций на этих высотах следует отметить орехо-
вые леса с бузульниково-недотроговым травяным покровом, разрежен-
ные высокотравные и высокотравно-снытиевые кленовники, где орех
играет подчиненную роль.
Ореховые леса с недотрогой, как я уже отметила, встречаются неболь-
шими пятнами в долине Кондары и ее боковых притоков. Орех здесь
образует сомкнутый полог. К ореху иногда присоединяется туркестан-
ский клен, каркас. В подлеске шиповник Rosa canina L. В травяном по-
крове господствует недотрога.
Вдоль речных русел, перемежаясь с предыдущей ассоциацией, до-
вольно часто встречаются ореховые леса с ежевикой и коротконожкой.
Здесь высокие деревья ореха часто образуют сомкнутый полог. К ореху
присоединяются алыча, туркестанский клен, каркас, боярка. В под-
леске очень густые заросли ежевики Rubus caesius L.
Значительно реже описанных ассоциаций встречаются ореховые
лесочки на болотах. Орех здесь иногда образует сомкнутые насаждения.
К ореху присоединяются — яблоня (sp.i), алыча (sp.i). В подлеске
довольно много ежевики Rubus caesius (sp.i).
Очень часто орех совместно с яблоней, алычей, каркасом встречается
отдельно стоящими деревьями на приподнятых участках долины р. Кон-
дара. Здесь же, между деревьями, широко распространен шиповник Rosa
canina L. и иргай.
В верхней полосе пояса древесно-кустарниковой растительности
орех, как я уже отметила выше, не образует чистых насаждений, он встре-
чается как подчиненная или равнозначущая порода в разреженных высо-
котравных и высокотравно-снытиевых кленовниках, располагающихся
по северным склонам.
Помимо этих ассоциаций можно назвать еще ореховые леса с бузуль-
никово-недотроговым травяным покровом, которые всеречаются в сред-
нем течении р. Кондара по северным склонам на богатых гумусом бурозе-
мах. Здесь наряду с орехом формирует первый ярус Acer turkestanicum
Pax (sp.i), реже встречается арча (sol.), крушина (sol.), яблоня (sol.),
туркестанская боярка (sol.); из кустарников распространены Rosa koka-
nica Rgl. (sol.), Gotoneaster insignis Pojark. (sol.) и G. suavisPojark. (sol.).
Орех, в отличие от клена, который почти всегда образует разрежен-
ные насаждения паркового типа, встречается в виде густых с сомкнутым
пологом насаждений, но такие сомкнутые насаждения наблюдаются
исключительно в местах с постоянным увлажнением. По сухим склонам
орех, так же как и клен, образует разреженные насаждения. Орех,
так же как и клен, является в настоящее время реликтовым растением,
ареал которого все больше сокращается.
Одной из причин постепенного уменьшения площадей распростране-
ния ореха является чрезвычайно ослабленное семенное возобновление
его. О том, что орех в Средней Азии имеет слабое семенное возобновление,
много писалось различными исследователями. Основными причинами
такого явления большинство авторов считает деятельность человека
и животных, направленную на сбор плодов, и пастьбу скота в насажде-
ниях ореха. В. П. Учайкин (1934) и Д. И. Прутенский (1940) в своих
работах, помимо вышеуказанных причин слабого семенного возобновле-
Биологические особенности древесных пород
295
ния ореха, приводят еще один фактор, ограничивающий распространение
грецкого ореха по склонам южной, восточной и западной экспозиций, —
это недостаток влаги в почве.
Наши наблюдения дают основание полагать, что основными причи-
нами отсутствия семенного возобновления ореха, так же как и туркестан-
ского клена, на склонах гор являются: недостаток влаги в почве, высокие
температуры поверхности почвы и низкая относительная влажность
в припочвенном слое воздуха.
Несмотря на то, что плоды ореха довольно тщательно собираются,
часть из них остается несобранной, попадает в почву, и весной из само-
сева начинается развитие молодых растений. До июля развитие всхо-
дов протекает нормально, а затем начинается их увядание и отмира-
ние.
Условия для нормального существования молодых, еще не окреп-
ших сеянцев мало благоприятны. Имеющиеся запасы влаги в верхних
горизонтах в течение июля—сентября таковы, что недоступны растениям.
Помимо крайнего недостатка влаги, отрицательным явлением для раз-
вития молодых растений является сильный нагрев почвы. Максималь-
ные температуры поверхности почвы в высокотравном кленовнике до-
стигают 58.1°.
Конечно, при всем этом нельзя не считаться с губительным дей-
ствием выпаса скота, который вытаптывает молодые всходы. В результате
действия всех этих факторов у ореха, растущего на сухих склонах,
наблюдается значительный отпад сеянцев, доходящий до 70—80%
в первый же год развития.
В последующие годы в силу тех или других обстоятельств происхо-
дит еще некоторый отпад, и в результате выживают только единичные
экземпляры.
Небезинтересно проследить, как развиваются те экземпляры, которые
сохраняются от самосева. Скорость роста ореха зависит, так же как и его
семенное возобновление, от условий местообитания дерева. Орех, про-
израстающий на сухих склонах, растет чрезвычайно медленно (см.
рис. 7), при выходе родников — быстрее.
В начале, т. е. до 10—12 лет, рост их идет, примерно, одинаково,
но в 12 лет дерево, растущее около ручья, благодаря постоянному увлаж-
нению, начинает быстро расти, в 1—3 года достигая прироста 1 м. Де-
рево же ореха, растущее на сухом склоне, очень сильно замедляет свой
рост; для того чтобы вырасти до 1 м, ему требуется 6—8, а иногда и 29
лет. Примерно к 35 годам дерево, растущее в условиях достаточного
увлажнения, имеет высоту 12 м, а дерево, растущее в условиях недоста-
точного увлажнения, имеет такую же высоту только к 136 годам.
Орех, так же как и туркестанский клен, обладает способностью веге-
тативно размножаться: этим обеспечивается существование деревьев
на чрезвычайно долгое время.
По сравнению с деревьями семенного возобновления деревья от порос-
левых побегов развиваются значительно быстрее. Так, например, дерево
семенного возобновления, достигшее 10 м высоты, имеет возраст 124 года,
в то время как дерево порослевого происхождения в однородных усло-
виях достигает такой же высоты к 29 годам. Разница в характере роста
между отдельными деревьями порослевого происхождения, произрастаю-
щими в различных условиях, хотя и имеется, но она не столь велика,
как это было отмечено для деревьев семенного возобновления. Молодые
порослевые побеги ореха, выросшего на сухом склоне, имеют, в отличие
296
В. И. Запрягаева
от побегов кроны, более удлиненный период роста. Свой рост они закан-
чивают на 10—15 дней позднее конца роста побегов кроны. В этом слу-
чае наши наблюдения можно сравнивать с наблюдениями Н. К. Вехова
(1934), проведенными на лесостепной опытной станции над искусствен-
ными посадками ореха, где разница в окончании роста между побегами
порослевого характера и побегами кроны достигала в некоторых слу-
чаях 30 дней.
К мезофильной древесной растительности ущелья Кондара можно
отнести дикую яблоню Malus hissarica Kudr. Она здесь не образует
чистых насаждений, а встречается всегда в тех или других ассоциациях,
формируемых Acer turkestanicum Pax или Juglans regia L., которые
описаны выше.
Мною не производилось полной раскопки корневой системы яблони,
но отдельные наблюдения говорят о том, что яблоня в наших условиях,
так же как и на западных склонах Ферганского хребта, по описанию
Ал. А. Федорова, П. К. Красильникова и А. А. Никитина, имеет поверх-
ностную корневую систему, далеко уходящую за пределы проекции
кроны. Семенное возобновление яблони в условиях сухих склонов почти
отсутствует. Можно встретить очень редкие всходы только по берегам
ручьев. Довольно часто яблоня в наших условиях возобновляется веге-
тативным путем. Яблоня, так же как клен и орех, в условиях сухих
склонов растет чрезвычайно медленно, что связано, повидимому, тоже
с недостатком влаги в почве.
В начале своего роста она растет по сравнению с кленом довольно
быстро и достигает высоты 1 м за 3 года. Но затем примерно с 5 лет
рост ее замедляется, и для того чтобы вырасти на 1 м, ей необходимо
затратить 6 лет, а позднее еще больше — до 17 лет.
2. КСЕРОФИЛЬНАЯ ЛИСТВЕННАЯ ДРЕВЕСНАЯ РАСТИТЕЛЬНОСТЬ
Эта растительность в ущелье Кондара очень широко представлена.
Здесь обычны бухарский миндаль Amygdalus bucharica Korsh., каркас
Celtis caucasica Willd. и боярышник Crataegus pontica C. Koch.
Миндаль и каркас очень широко распространены в поясе древесно-
кустарниковой растительности, занимая склоны всех экспозиций в ниж-
ней полосе пояса до высоты 1700 м над ур. м.
Боярышник имеет в этом ущелье значительно меньшее распростране-
ние, хотя поднимается до высоты 1700 м (см. рис. 1), отмечая своим верх-
ним пределом распространения границу между двумя полосами пояса
древесно-кустарниковой растительности. Для всех трех пород деревьев
характерно, что они никогда не образуют чистых насаждений и никогда
не встречаются порознь; последнее относится к миндалю и каркасу,
которые во всех ассоциациях встречаются вместе.
Миндальники
Одной из самых распространенных ассоциаций, формируемых Amygda-
lus и Celtis, являются эфемерово-югановые миндальники. Они занимают
южные склоны до высоты 1500 м, со щебнистыми или мелкоземисто-щеб-
нистыми почвами. Здесь Amygdalus bucharica Korsh. совместно с карка-
сом, который встречается несколько реже, образуют чрезвычайно разре-
женные насаждения, с расстояниями между деревьями около 20—25 м.
Биологические особенности древесных пород
297
Несколько выше по склонам, заходя часто на водораздельную гряду,
располагаются миндальники эфемерово-чеснеево-югановые. Почвы под
этой ассоциацией сильно щебнисты.
Amygdalus bucharica Korsh., так же как и в предыдущей ассоциации,
встречаются в виде отдельно стоящих деревьев, расстояние между кото-
рыми доходит до 20—25—40 м. Миндаль перемежается с Celtis cauca-
sica Willd. и иногда арчей Juniperus seravschanica Кот. Иногда в этих
миндальниках можно встретить единичные кустарники Atraphaxis pyri-
folia Bge, Rosa maracandica Bge. Травяной покров здесь очень редкий*
В средней части ущелья, примерно на высотах 1450—1600 м, по юж-
ному склону ущелья Кондара можно встретить небольшие по площади
участки, занятые разреженными зарослями миндально-кленовой ассоциа-
ции с ячменно-эфемерово-разнотравным травяным покровом.
Очень широко миндаль распространен по скалистым участкам, где
он поселяется вместе с каркасом в расщелинах скал. В значительно
меньшем количестве бухарский миндаль встречается по северным или
близким к ним склонам. Здесь он, повидимому, вытесняется более мезо-
фильными породами. Наиболее распространенными ассоциациями север-
ных склонов, где произрастает миндаль, будут описанные выше кленов-
ники ежово-девясилово-разнотравные, кленовники разнотравно-пажит-
никовые и разнотравно-югановая и эфемерово-гречихово-югановая ассо-
циации.
Для всех насаждений, образованных бухарским миндалем, каркасом
и боярышником, характерна разреженность; травяной покров не изме-
няет своего состава в зависимости от наличия или отсутствия древесных
пород.
Как показали наши исследования, бухарский миндаль, каркас
и боярышник имеют чрезвычайно мощные корневые системы, обеспечи-
вающие им возможность существования на таких участках, где развитие
других деревьев немыслимо.
Интересно, что корневые системы этих деревьев не всегда имеют по-
верхностное распределение: в некоторых случаях, например в скалах,
корни деревьев опускаются на значительную глубину. На каменистых
субстратах бухарский миндаль имеет корневую систему, часть которой
располагается непосредственно под поверхностью почвы на глубине
до 50 см (рис. 8), другая же часть опускается по трещинам скал до глу-
бины 200 и более сантиметров. Такое распределение корней дает воз-
можность миндалю использовать влагу из верхних горизонтов почвы
в ранневесенний период, и из более глубоких горизонтов в летнюю
засуху, когда влага остается в расщелинах скал на значительной глу-
бине. На мелкоземистых почвах вся корневая система бухарского мин-
даля располагается в основном до глубины 100 см (рис. 9).
В этих условиях, как видно на прилагаемой схеме, площадь проекции
корневой системы во много раз превышает площадь проекции кроны.
Основные корни тянутся от ствола миндаля на большие расстояния, рас-
полагаясь до глубины 50—60 см, в то время как сосущие корни подходят
к самой поверхности — глубина их залегания до 5 см. Несколько отлична
корневая система бухарского миндаля, растущего по долине реки,
с близко подходящим постоянным увлажнением. В этом случае одна часть
корней, главным образом сосущих, располагается на глубине 0—50 см,
часто подходя к самой поверхности почвы, другая часть опускается,
в зону повышенного увлажнения, достигая самого сильного разветвления
сосущих корней у уровня грунтовых вод, на глубине около 200 см..
298
В. И. Запрягаева
Просматривая корневые системы бухарского миндаля, можно убедиться
в том, что значительная часть их распределяется на небольшой глубине
под поверхностью почвы. Очень часто сосущие корни подходят к самой
поверхности, располагаясь на глубине всего 0—1 см. Схема (рис. 10)
дает представление, как распределяются сосущие корни бухарского
миндаля в поверхностном слое почвы. Можно думать, что эти корни, под-
ходящие к самой поверхности почвы, имеют специальное назначение —
использовать конденсационную влагу, или служат для улавливания
атмосферной влаги только в осенне-весенний период.
Корневые системы боярышника и каркаса аналогичны только что
описанной корневой системе бухарского миндаля. У каркаса (Celtis
caucasica Willd.), по описанию Ф. Л. Запрягаева (1937), корни распо-
Рис. 8. Профильная проекция бухарского миндаля Amygdalus bucha-
rica Korsh. на южном каменистом склоне ущелья Кондара.
лагаются на глубине 20—80 см, а мочки подходят к самой поверхности.
Иногда, когда каркас растет в скалах, его корни опускаются в поисках
воды на очень большую глубину, доходящую в некоторых случаях, как
это приходилось мне наблюдать, до 2.5—3 м. О способности каркаса
опускать свои корни глубоко по расщелинам скал говорит также в своей
работе по Закавказью П. Виноградов-Никитин (1928).
Если посмотреть, как возобновляются естественным семенным путем
растения, принадлежащие к этой группе, станет совершенно очевидным,
что, в противоположность растениям предыдущей группы, им не угро-
жает опасность уменьшения ареала.
В противоположность развитию сеянцев туркестанского клена,
сеянцы бухарского миндаля на склонах гор развиваются нормально,
хотя и очень медленно. Средняя высота их к концу первого года дости-
гает 15.4 см. Как исключение, отдельные сеянцы достигают 39.1 см.
Количество отмерших растений в первый год всего только 18%, в то время
как у туркестанского клена оно часто доходит до 100о/о.
Биологические особенности древесных пород
299
Примерно так же благополучно обстоит дело и с естественным семен-
ным возобновлением каркаса (Ф. Л. Запрягаев, 1940). «Примерно с сере-
Рис. 9. Профильная проекция бухарского миндаля Amygdalus bucharica
Korsh. по дну ущелья Кондара.
дины апреля появляются мелкие всходы каркаса 1.5—2 см. к началу
июня достигают 7 см высоты, дальше рост, а вместе с ним и развитие
постепенно замедляются и к середине июля прекращаются вовсе.
Рис. 10. Расположение сосущих корней Amygdalus bucharica
Korsh. у поверхности почвы.
Массовое отмирание сеянцев за этот период (первый год жизни)
наблюдается ранней весной, и связано оно, главным образом, с механи-
ческими повреждениями, которые вызываются объеданием улитками
300
В. И, Запрягаева
нежных, еще не одеревеневших частей молодого растения. В продолже-
ние всего остального периода отмирания сеянцев не наблюдается».
Каркас по сравнению с кленом размножается хорошо семенным путем,
даже в условиях самых сухих горных склонов.
Дальнейшее развитие молодых растений — представителей этого типа
растительности — протекает совершенно своеобразно, особенно у бухар-
ского миндаля.
Чтобы вырасти бухарскому миндалю из семени до высоты одного метра,
требуется 10—15 лет, но затем, когда молодое деревцо развило доста-
точно мощную корневую систему, оно начинает расти более быстро. По
сравнению с другими древесными породами он растет быстрее, но, так же
как и остальные растения, для своего роста не использует всего теплого
периода года. Его рост продолжается до середины июля, т. е. до того
периода, когда в почве сохраняется еще достаточное количество влаги.
Первым предвестником наступающей почвенной засухи является пожел-
тение листьев. Это обычно бывает во вторую половину июня. В июне же,
как только прекращается рост, наблюдается массовое сбрасывание
листьев, причем у бухарского миндаля это явление выражено зна-
чительно более резко, чем у других пород. Поведение его в этом слу-
чае можно сравнивать с миндалем в условиях Туркмении, где, по данным
В. К. Василевской (1941), миндали, культивируемые в Копет-даге, сбра-
сывают листву уже в мае.
Рост бухарского миндаля происходит значительно интенсивнее на
склонах южной экспозиции, чем на всех остальных. Например, средний
годовой прирост побегов в кроне бухарского миндаля на южном склоне
составляет 10.1 см, на восточном склоне всего только 3.5 см. На южном
склоне миндаль растет значительно быстрее, что, невидимому, влечет
за собой и более быстрое наступление старости деревьев.
Бухарский миндаль почти на протяжении всей своей жизни образует
у корневой шейки дерева или на главном стволе порослевые побеги,
которые появляются почти ежегодно. За год они достигают 103—129,
а иногда и 150 см. Такие побеги в плодоводстве называют «жировыми»
и для того, чтобы они не мешали нормальному развитию кроны, их выре-
зают, не давая им развиться до естественного предела. Образование
«жировых» побегов у бухарского миндаля в условиях культуры (на
Копет-даге) отмечает В. К. Василевская (1941), но, повидимому, в ее
опытах «жировым» побегам также не давали развиться до конца. Нам же,
совместно с Л. Ф. Запрягаевым, удалось проследить, какое значение для
жизни дерева в естественных условиях имеют эти «жировые побеги».
На схеме (см. рис. 8 и 9) показано образование порослевых побегов
у бухарского миндаля. Обычно появление таких побегов происходит
в то время, когда дерево имеет еще вполне нормальное развитие, когда
его крона распределяется, примерно, в плоскости, перпендикулярной
главному стволу. Но вот появились порослевые побеги; быстро разви-
ваясь, они достигают высоты кроны. Своей новой разветвленной кроной
они начинают угнетать крону материнского дерева, которая постепенно
отклоняется в сторону. Затем можно наблюдать, как основной ствол,
имевший когда-то вертикальное положение, изгибаясь, отклоняется
и, под влиянием развивающегося нового побега, отмирает. На смену
старой развивается новая крона, у основания ствола которой вскоре
после завоевания ею господствующего положения появляются новые
порослевые побеги, которые через несколько лет готовят новую смену.
И так продолжается в течение всей жизни дерева. Нам удалось наблю-
Биологические особенности древесных пород
301
дать у бухарского миндаля до 7—8 циклов в развитии кроны (под циклом
здесь понимается время от начала появления побега до его отмирания).
Возраст всего дерева из-за того, что бухарский миндаль периодически
сменяет крону, чрезвычайно трудно установить. В отличие от многих
других древесных пород, обладающих способностью вегетативного возоб-
новления, бухарский миндаль меняет всю крону вместе с главным стволом.
Крона бухарского миндаля существует, как правило, не более 15—
20 лет. Говорить же о чрезмерной недолговечности всей надземной части
бухарского миндаля нельзя, ибо нами наблюдались случаи, когда ниж-
няя часть ствола дерева, примерно до высоты 100—120 см, сохранялась
на протяжении 30—50 лет. Вообще же бухарский миндаль, повидимому,
не очень долговечен. Мне не удалось встретить деревьев старше 75 лет.
Каркас
Каркас (Celtis caucasica Willd.), в отличие от бухарского миндаля,
не имеет в процессе своего роста таких периодически сменяющихся цик-
лов в развитии кроны. У него, как у большинства древесных пород,
с самого раннего возраста определяется главный ствол. Характер роста
каркаса на сухих склонах отличается во многом от описываемых выше
представителей мезофильной растительности. Семенное возобновление
каркаса протекает более или менее нормально. Тяжелые условия поч-
венной засухи не являются для него такими катастрофическими, как,
например, для кленов. После некоторого отпада сеянцев в первый
и второй год жизни каркас продолжает развиваться вполне удовле-
творительно.
Высоты 1 м каркас достигает за 6 лет, высоты 3 м — за 12 лет,
высоты 7 м — за 25 лет и, наконец, в 65 лет он имеет высоту 13 м,
в то время как туркестанский клен в таком же возрасте достигает только
2.5 м. В возрасте около 30 лет каркас замедляет свой рост, его годичные
приросты становятся очень небольшими и достигают всего 6—8—15 см.
Динамика роста побегов в течение сезона находится в тесной зависи-
мости от количества выпадающих осадков и влажности почвы.
Фисташка
К описываемой группе ксерофильных древесных пород по своим био-
логическим особенностям близко подходит фисташка (Pistacia vera L.).
Основные массивы фисташковых зарослей сосредоточены в низкогор-
ном поясе. Нижняя граница ее распространения проходит около 600—
500 м над ур. м. В районе ущелья Кондара фисташка находится у верх-
ней границы своего распространения. Здесь она встречается небольшой
рощицей на высоте 1300 м по южному склону. Выше 1300 м нам не уда-
лось встретить фисташки. Небольшая рощица по ущелью Кондара со-
стоит исключительно из деревьев порослевого происхождения.
Порослевый побег, отходящий от корневой шейки, быстро развивается
только в первые годы своей жизни, но затем, примерно с 5 лет, рост его
чрезвычайно замедляется. За 28—34—43 года фисташка вырастает всего
на 1 м в высоту. В этом отношении она даже превосходит своим замед-
ленным ростом туркестанский клен. Проанализированный нами 155-лет-
ний побег фисташки ничего еще не говорит о возрасте всего дерева,
302
В. И. Запрягаева
который трудно установить в наших условиях, но, повидимому, он очень
велик.
3. КСЕРОФИЛЬНАЯ ХВОЙНАЯ ДРЕВЕСНАЯ РАСТИТЕЛЬНОСТЬ
В ущелье Кондара представителем этой растительности является
только арча — Juniperus seravschanica Korn.
Арча
Арча, как видно на рис. 1, распространена в ущелье Кондара по всему
поясу древесно-кустарниковой растительности — от 1100 до 2400 м над
ур. м. Высота 2400 м не является пределом распространения арчи: в дру-
гих местах Гиссаро-Дарвазского района арча поднимается вплоть до
нижней границы альпийского пояса.
Арча по ущелью Кондара большей частью встречается по сухим ка-
менистым склонам различных экспозиций, включая сюда и южные;
здесь она, не встречая себе конкурентов, развивается в мощные деревья
высотой до 18 м.
В нижней полосе пояса древесно-кустарниковой растительности арча
никогда не образует чистых насаждений, встречаясь единичными
деревьями в кленовниках и миндальниках. Из наиболее распространенных
ассоциаций, в которых произрастает арча, можно отметить кленовники
с ежово-разнотравно-недотроговым и с ежово-девясило-разнотравным
травяным покровом и миндальники с эфемерово-чеснеево-югановым по-
кровом. Эти ассоциации описаны выше. На этих высотах чаще всего
арчу можно встретить на каменистых участках и на скалах, мало доступ-
ных человеку.
В верхней полосе пояса арча распространена значительно шире.
Здесь она встречается в разреженных высокотравных и высокотравно-
снытиевых кленовниках и розариях в виде незначительной примеси
и даже образует настоящие арчевники с зизифорой и разнотравьем;
правда, такие арчевники встречаются довольно редко и приурочены
к щебнистым восточным и северным склонам. Среди арчи Juniperus serav-
schanica Кот. (эрг) встречаются только единичные деревья Acer turkesta-
nicum Pax. Деревья, как и везде по склонам, не образуют сомкнутого
полога. Травяной покров довольно густой и представлен растениями
ксерофильного типа.
Значительно чаще, чем в нижней полосе пояса, арча встречается
здесь на южных скалистых склонах, где она вырастает в мощные деревья
свыше 20 м в высоту.
По всей вероятности, арча в прежние времена в ущелье Кондара
занимала большие площади, чем сейчас, и, повидимому, была значительно
шире распространена на мелкоземистых почвах в нижней части древесно-
кустарникового пояса. Но, поскольку эти места являются более доступ-
ными, человек хищнически уничтожил ее здесь и непроизвольно сохра-
нил в местах, недоступных ему, — на скалах. Возможно, что ограни-
чивающим распространение арчи на мелкоземистых склонах в нижней
части древесно-кустарникового пояса фактором является конкуренция
других произрастающих здесь деревьев и кустарников (П. Н. Овчин-
ников, 1940). О более широком распространении арчи в наших условиях
Биологические особенности древесных пород
303
можно судить по сохранившимся пням. Так как древесина арчи пре-
красно противостоит гниению, то ее пни остаются не сгнившими в тече-
ние долгого времени.
В ущелье Кондара, так же как и в других районах Средней Азии, ар-
чевые насаждения так же сильно разрежены, в них почти отсутствует
связь между травяным покровом и пологом деревьев. На скалистых
Рис. 11. Профильная проекция
Juniperus seravschanlca Korn.
склонах арча разрежена в силу того, что она селится только в местах,,
где имеются хотя бы минимальные запасы питательного субстрата,
а таких мест чрезвычайно мало. На мелкоземистых же склонах разрежен-
ность арчевых насаждений объясняется тем, что здесь, так же как и в опи-
санных раньше древесных насаждениях, основным фактором, опреде-
ляющим структуру насаждений, является не свет, а недостаток влаги
в почве, в связи с которым арча уже в самом раннем возрасте развивает
мощную корневую систему, ее сосущие корни подходят к самой поверх-
ности, располагаясь часто на глубине 0—5 см. Мне приходилось наблю-
дать у старых деревьев арчи тонкие сосущие корешки, сплошь покрываю-
304
В. И. Запрягаева
щие корни арчи, диаметром в 7—8 см, на глубине до 12 см под поверх-
ностью почвы.
На схемах (рис. 11, 12) представлена корневая система молодого
дерева арчи, выросшего на мелкоземистом склоне. У этого дерева корне-
Рис. 12. Горизонтальная проекция корневой системы Juniperus seravschanica Kom.
вая система двояка: часть корней протягивается непосредственно под
поверхностью почвы, а часть уходит вглубь до 2 м. У взрослых экземпля-
ров мне удавалось наблюдать только поверхностные корни, но, воз-
можно, что у арчи, растущей в скалах, часть корней проникает, в поисках
Биологические особенности древесных пород
305
запасов влаги, глубоко по расщелинам, напоминая в этом отношении
корневую систему миндаля и каркаса.
Естественное семенное возобновление зеравшанской арчи в условиях
ущелья Кондара почти отсутствует. Можно пройти километры, не
встретив молодого подроста. Очень редкие всходы, которые удавалось
мне наблюдать, погибают в первый же год.
Специальные исследования, которые проводились мной в 1946 г.
над естественным семенным возобновлением, показали, что арча дает
здоровых семян не больше 3%, все же остальные поражены.
То, что арча очень плохо воз-
обновляется семенами в естест-
венных условиях, отмечает Е. А.
Малюгин (1931) для Копет-дага.
Возобновление зеравшанской арчи
в Туркестанском хребте происхо-
дит значительно хуже чем других
видов арчи (А. Ф. Радзиевский,
1934; М. Г. Попов и Н. В. Анд-
росов, 1937; В. Д. Дмитриев, 1930,
1938; Г С. Сиворакша, 1938), ее
подрост там встречается редко и
только под пологом материнских
деревьев. Отсутствие возобновле-
ния арчи семенным путем указан-
ные авторы объясняют пастьбой
скота. В наших же условиях этот
фактор почти не имеет значения,
и отсутствие возобновления можно
объяснить, с одной стороны, боль-
шой повреждаемостью семян и, с
другой стороны, неблагоприят-
ными климатическими условиями,
при которых развиваются молодые
всходы.
Рис. 13. Вегетативное возобновление арчи
Juniperus seravschanica Кот. Ущелье
Кондара.
(Фот. Ф. Л. Запрягаева).
землей, и в том случае, если они
Арча, так же как и ряд других
местных древесных пород, очень
хорошо размножается вегетатив-
ным путем. У арчи нижние ветки,
как это видно на фотографии
(рис. 13), часто соприкасаются с
бывают засыпаны землей, старыми листьями или прижаты осыпаю-
щимися камнями, то у них на месте соприкосновения с поверхностью
почвы появляются корни, которые дают начало новому растению. При бег-
лом осмотре древесных насаждений в ущелье Кондара может создаться
ложное впечатление, что здесь встречается молодой подрост арчи. В дей-
ствительности же это большей частью отводки. Часто бывает трудно вос-
становить, откуда появляется тот или другой отводок. На северном склоне
в ущелье Кондара мною был отмечен старый пень арчи (судя по пню, это
было очень большое дерево арчи, выросшее из семени); от него на расстоя-
нии 2 м по склону находится второй пень, который с предыдущим пнем не
соединяется. Возможно, что это тоже было семенное дерево, выросшее под
пологом материнского. На расстоянии 1.5 м от второго пня обнаружен
мною третий пень, который на глубине 25 см соединялся со вторым
20 Ущелье Кондара
306
В, И. Запрягаева
пнем веткой, погребенной когда-то под поверхностным слоем почвы. Вся
эта ветка сплошь покрыта мелкими корнями, подходящими к самой по-
верхности почвы. От третьего пня еще ниже по склону растет очень моло-
дое на вид дерево арчи, в действительности же — это естественно обра-
зовавшийся отводок дерева, от которого в настоящее время остался
только пень (третий). И так как ветка, давшая жизнь новому деревцу,
погребена, а старых пней вокруг почти не заметно, то получается впечат-
ление, что это вполне самостоятельное дерево. Нижние ветки дерева,
пригибаясь к земле, образуют новые отводки, которые пока еще не совсем
укоренились. Если бы арча не уничтожалась человеком, то можно
было бы наблюдать на месте ныне существующего маленького деревца
целую рощу из арчи. Способность арчи возобновляться вегетативным
путем дает ей возможность сохраняться по склонам гор.
Изученные выше особенности древесно-кустарниковой раститель-
ности не являются присущими только одному ущелью Кондара, они свой-
ственны растительности всего Гиссаро-Дарвазского района, т. е. значи-
тельной части древесно-кустарниковой растительности Таджикистана,
что дает право сделать несколько общих выводов.
Древесная растительность Гиссаро-Дарвазского района, как видно
на примере ущелья Кондара, в своем естественном виде не может обеспе-
чить республику ни строительной, ни поделочной древесиной. В наших
лесах не формируются гладкие, малосбежистые стволы деревьев, так как
леса очень разрежены. Слабое семенное возобновление наиболее техни-
чески ценных древесных пород, их крайне замедленный рост не дают
возможности организовать правильную эксплоатацию. Этим собственно
и объясняется то, что республика ежегодно ввозит из других областей
Советского Союза на много миллионов рублей строевой и поделочной
древесины.
Древесно-кустарниковая растительность требует улучшения и при-
способления ее для более рационального использования в хозяйстве
республики.
Улучшение существующих древесно-кустарниковых насаждений
можно организовать в двух направлениях.
А. Прежде всего нужно привести в порядок уже существующие наса-
ждения. Нужно убрать старые, перестоявшие деревья, причем рубку
производить очень осторожно, помня, что многие удаленные из этих
мест деревья никогда не смогут появиться здесь вновь без активного
вмешательства человека. Для некоторых старых деревьев, может быть,
нужно создать такие условия, при которых они могли бы, нормально
развиваясь, давать обильное вегетативное потомство, и т. д. Особо нужно
выделить деревья, приносящие хорошие плоды — орех, яблоню, алычу,
фисташку, виноград, которых в ущелье Кондара не мало. Здесь имеются
формы яблони, алычи, винограда, приносящие высококачественные
плоды, мало отличимые от культурных сортов. В ущелье Кондара можно
встретить весьма ценные формы ореха; кондаринские орехи отличаются
значительным весом, уступая в этом отношении только более крупным
кавказским формам; по абсолютному весу ядра они очень мало отли-
чаются от культурных сортов и, наконец, по содержанию жира стоят
значительно выше киргизских и французских. Даже худшие образцы
кондаринских орехов имеют процент жира в ядре (63—65%), рав-
ный проценту жира лучших французских сортов (64%), лучшие же
образцы превышают средний процент жира французских орехов на 18%
(С. С. Печникова, 1940).
Биологические особенности древесных пород
307
Выделенные плодовые деревья должны быть охраняемы и служить
маточными деревьями для последующего размножения. Те же плодовые
деревья, которые приносят низкосортные плоды (яблоня, алыча, боя-
рышник, дикие вишни), могут с успехом использоваться как подвои
для лучших сортов плодовых.
Б. Помимо мероприятий по благоустройству существующих необхо-
димо создавать новые насаждения, способные давать строевую и поде-
лочную древесину. Но для развития новых насаждений в описываемых
нами условиях нужна своя, специально приспособленная к существую-
щим условиям методика. Одно из основных условий для создания про-
дуктивного леса на сухих склонах — обеспечение почвы влагой. Без
него любые мероприятия не смогут принести эффекта.
II. БИОЛОГИЧЕСКИЕ ОСНОВЫ ГОРНОГО БОГАРНОГО
ЛЕСОРАЗВЕДЕНИЯ И ПЛОДОВОДСТВА
Изучение лесной растительности ущелья Кондара — биологии важ
нейших древесных и кустарниковых пород — дало возможность вскрыть
ряд закономерностей в развитии представителей этой растительности
и на основе их построить систему мероприятий, направленных на ее видо-
изменение и улучшение, следуя принципам мичуринской биологии.
Ниже излагаются результаты экспериментальных работ за 10 лет
по богарному лесоразведению, богарному плодоводству и виноградар-
ству, а также по использованию дикорастущих плодовых деревьев
в лесосадах.
1. ОПЫТ БОГАРНОГО ЛЕСОРАЗВЕДЕНИЯ
Одним из путей, по которым должна быть направлена мелиорация
древесных насаждений, является искусственное древоразведение. Созда-
ваемые насаждения должны, в отличие от естественных, формировать
деревья с малосбежистыми, гладкими и высокими стволами. Это может
быть достигнуто тем, что на единице площади будет размещено значи-
тельно больше деревьев, чем это наблюдается в естественных условиях.
Но, учитывая климатические и почвенные условия ущелья, всякое сгу-
щение растений ио склонам гор может быть достигнуто только в том
случае, если почва получит дополнительные запасы влаги.
Создание древесных насаждений, отвечающих требованиям лесного
хозяйства, в условиях орошаемых склонов или по долинам узких горных
ущелий не представляет особых трудностей. Здесь же я остановлюсь
только на методах разведения искусственных насаждений на неорошае-
мых склонах гор, которые в Средней Азии принято называть богарными.
Разведение древесных насаждений в ущелье Кондара и в аналогич-
ных по условиям других горных районах Таджикистана мыслится
не в виде создания сплошных лесных массивов по склонам гор, а в виде
отдельных небольших (0.5—1 га) участков. В хозяйственном отношении
это неудобно, но отсутствие в горах больших площадей с достаточно
мощно развитым почвенным покровом заставляет приспосабливаться
к имеющимся условиям.
В нашем районе древесные насаждения можно располагать прежде
всего по участкам, расчищенным от естественных насаждений, но в силу
тех или иных условий не используемым под полевые культуры. Можно
20*
308
В. И. Запрягаева
также расчищать территории, занятые в настоящее время зарослями
кустарников, абсолютно не используемых в хозяйстве.
Попытки разведения леса в горных условиях делались и раньше
и были связаны с работами по закреплению склонов от эрозии. «Лес в дан-
ном случае не самоцель, а средство» (А. С. Карасев, 1934). Так же, при-
мерно, пишет и В. Ф. Морозов (1944). Мне кажется, что в Таджикистане
создание высокопродуктивного леса на склонах гор должно быть также
и целью во всех тех местах, где осуществляется чрезвычайно важное
для народного хозяйства республики мероприятие по сохранению почвы
от смыва.
В практике лесоразведения на горных склонах применялись различ-
ные методы посадки и посева и, естественно, получились различные ре-
зультаты; деревья высаживались по склонам гор или без всякой под-
готовки последних, или иногда на перекопанных площадках различных
размеров, расчищенных от всяких посторонних растений.
Лучшим способом мелиорации горных склонов, довольно широко
применяемым в некоторых районах Советского Союза и за границей,
является террасирование. Оно сводится к проведению на склонах гор
горизонтальных канав или террас, которые задерживают выпадающую
на данную территорию и стекающую по склонам влагу и аккумулируют
ее в почве.
В России вопросами горной агромелиорации впервые начали зани-
маться в 1878 г. в Крыму, на склонах хребта Тете-оба, прилегающих
к Феодосии. Здесь эту работу произвел А. Ф. Вещицкий (Ходоровский,
1904). Параллельно подошве горы по склону устраивались канавы на
расстоянии 2 м друг от друга, шириной в 1 м и глубиной 2 м. Из канав
почва вынималась, но затем разрыхленная вновь укладывалась в канаву,
но не до верха, а чуть ниже, так, чтобы в ней оставалось место для сбора
воды. Деревья высаживались прямо в канаву. Недостатком этих работ
было отсутствие всяких расчетов, канавы были часто не горизонталь-
ными, и вода скапливалась только в наиболее низкой части их. Из-за
постоянного заиливания самой канавы сползающей со склона почвой
и отсутствием возможности расчистить террасу (мешали посаженные
деревья) растения не могли нормально развиваться, отсюда—некоторые
неудачи в опытах.
Примерно к тому же времени (1880 г.) относятся опыты, заложенные
Н. И. Корольковым в Аман-кутане на северном склоне хребта Кыр-
тау, в 40 км от Самарканда, в пределах 1200—1800 м над ур. м. Работы
Н. И. Королькова имели целью ослабить силевые потоки, разру-
шающие оросительную систему Зеравшанского оазиса. Н. И. Корольков
положил в основу горизонтальность террас, но террасы раздвинул на
расстояние 12—15 м. Террасы Королькова проводились на склонах гор
следующим способом: по горизонтали склона почва вынималась и склады-
валась с нижней стороны канавы так, чтобы получился вал разрыхлен-
ной почвы. В эту разрыхленную почву сажались растения. Сама же
канава выкапывалась треугольной формы, с уклоном в сторону горы.
Несмотря на ряд преимуществ, система террасирования Н. И. Король-
кова имеет и недостатки: большие расстояния между террасами не дают
возможности формировать сомкнутые насаждения, и треугольное сече-
ние канавы затрудняет обработку почвы.
В 1898 году горномелиоративные мероприятия были организованы
в урочище Ак-таш, 60 км. к северо-востоку от Ташкента, в зоне от 1000
до 1800 м над ур. м. С. Ю. Раунером. Его работы также были предпри-
Биологические особенности древесных пород
309
няты с целью защитить оросительные каналы от разрушения силями.
В работах Раунера использовался опыт Н. И. Королькова, но они про-
водились на основе расчетов, и террасирование проводилось по строго
горизонтальным линиям, определяемым инструментально. Расстояние
между террасами и величина выемки террасы определялись по принципу:
чем круче склон, тем больше выемка. Сами террасы делились глухими
перемычками через каждые 20 м с тем, чтобы в случае размыва террасы
вода могла бы вылиться только из одного отделения, а не из всей канавы.
Расстояние между террасами Раунер допускал 6 м на крутых скло-
нах и 16—18 м — на пологих, что так же, как и у Королькова, не давало
возможности формировать густые насаждения.
В 90-х годах прошлого столетия, в целях ослабления разрушитель-
ной деятельности горных потоков на склонах Телетского хребта в Гру-
зии, агролесомелиоративные работы проводил Лисневский. Принципы
его террасирования были те же, что и у Раунера, но он сблизил террасы
до расстояния 2 м, создав тем самым условия для формирования сомкну-
тых насаждений.
В Советском Союзе агромелиоративные работы получили значительно
большее распространение. Так, начиная с 1928 г., горномелиоративные
работы развертываются в Узбекистане, в бассейне реки Шахимардан,
в 1927—1928 гг. — в горах Копет-дага (Малюгин, 1931); позднее,
с 1930 г., — в бассейне реки Аман-кутан в Гиссарском хребте и с 1932 г.,—
в урочищах Миндон-сельхане, Кассай-сай в Чаткальском хребте. Задачей
этих работ также являлась выработка метода борьбы с силевыми пото-
ками (А. С. Карасев, 1934).
В Таджикистане агромелиоративные исследования начались в 1932—
1933 г. в Сильбурсае (отроги Гиссарского хребта) на высоте 1400 м
над ур. м. Здесь применялись для посадки лесных пород террасы типа
Раунера. Следующими по времени работами в этой области были наши
работы* начатые в 1935 г. Несколько позднее опытные посадки древесных
пород были заложены Лесной опытной станцией на высоте 850—900 м
над ур. м. в предгорьях Гиссарского хребта, прилегающих к Сталина-
баду. Наконец, были организованы работы по созданию искусственных
древесных насаждений на склоне хребта Арук-тау, на высоте 700 м над
ур. м. (В. Ф. Морозов, 1944).
Свои опытные работы по созданию древесных насаждений на неоро-
шаемых склонах гор Варзобская станция проводила в поясе древесно-
кустарниковой растительности ущелья Кондара на двух основных
участках.
Первый участок располагается на высоте 1100 м над ур. м. Здесь
опытные посадки производились в трех пунктах: 1) на склоне северной
экспозиции с мощными мелкоземистыми почвами, занятом в свое время
насаждениями туркестанского клена, бухарского миндаля и другими,
но затем расчищенном под пашню; 2) на склоне южной экспозиции
с мелкоземистыми почвами и, наконец, 3) тоже на склоне южной экспози-
ции, но на каменисто-песчаной осыпи, занятой разреженными зарослями
бухарского миндаля.
Второй участок располагается в урочище Квак, на высоте 1850 м
над ур. м. Здесь опытные посадки производились на северном склоне,
расчищенном из-под разреженного высокотравного кленовника.
Опытные посадки производились на склонах, мелиорированных тер-
расами. Как исключение производилась посадка древесных пород на
площадках. Расчет террас производил, в основном, инженер Народного
310
В. И. Запрягаева
Комиссариата водного хозяйства Таджикской ССР П. Ф. Иванов
(1938).
Террасы, применявшиеся в наших работах, были горизонтальными,
непрерывными, с перемычками и вмещали весь сток воды с межтеррас-
ного пространства. С целью сохранить накопленную во время дождей
влагу, терраса имела профиль, обладающий наименьшим коэффициен-
том испарения. Профиль террасы был сконструирован так, чтобы, пере-
ходя на склон большой крутизны, терраса уменьшала свою емкость
пропорционально уменьшающейся поверхности стока и его коэффи-
циента. Изменение емкости достигалось изменением глубины террасы.
Нижеприводимая схема (рис. 14) характеризует профиль террас, приме-
няемых для посадки древесных и плодовых пород.
Для посадок деревьев наиболее целесообразно применять террасы
с расстоянием между ними в 6—8 м. На крутых склонах, обычных для
горного Таджикистана, сближать террасы больше, чем на 6 м, как пока-
Рис. 14. Поверочный профиль террасы.
зал опыт, нельзя. Так как при указанном расстоянии между рядами
деревьев нельзя будет получить густых насаждений, то межтеррасные
пространства используются также для посадки деревьев. В этом случае
деревья располагаются горизонтальными по склону рядами.
Определение расстояний между террасами производится на основа-
нии расчета по формулам, в основе которого лежала задача предотвра-
тить смыв почвы со склонов (П. Ф. Иванов, 1938).
Отсутствие собственных наблюдений по критической скорости в наших
условиях вызвало необходимость использовать данные Ф. К. Кочерга
(1933), полученные им для Узбекистана.
Сооружавшиеся нами террасы по всей своей длине через каждые
20 м делились глухими перемычками, расположенными в шахматном
порядке. Во избежание утечки в виде подпочвенных токов аккумули-
рованной в почве воды, для террасирования выбирались участки с мелко-
земистым слоем почвы, не менее 2—3 м мощности.
На нижнем участке описанные террасы выполнили свою роль по
задержанию атмосферной влаги на склонах с большим успехом. После
сильных дождей приемная часть террасы примерно на 3/4 своего объема
заполняется водой, которая постепенно просачивается в глубину
Биологические особенности древесных пород
311
На втором участке, расположенном на высоте 1850 м (в верхней полосе
пояса древесно-кустарниковой растительности), метод террасирования
сплошными, абсолютно горизонтальными террасами оказался не совсем
пригодным; основной дефект заключался в недостаточной емкости их
для задержания снеговой воды. Дело в том, что на этих высотах в тече-
ние зимы накапливается снег мощностью до 2—2.5 м. В конце апреля
он начинает очень быстро таять и дает такую массу воды, что описанные
выше террасы не в состоянии вместить ее. Вода, переливаясь через край
террасы, производит разрушения. Устраивать же террасы более емкими
нельзя, так как вся масса снеговой воды, получающаяся от таяния в тече-
ние 15—20 дней, может произвести перенасыщение почвы и вызвать
оползни. Для того чтобы избежать этого, Ф. Л. Запрягаев (1940) пред-
ложил применять на высоте 1850 м «самоочищающиеся» террасы, придав
им уклон 2 см на 1 м длины. На таких, очень слабо наклонных террасах
устраивались перемычки до половины террасы с тем, чтобы с террасы
уходил только излишек воды. Как только таяние снега оканчивалось,
все террасы для того, чтобы удержать выпавшие весной атмосферные
осадки, закрывались через каждые 10 м высокими, до краев террасы,
перемычками.
Нижеприведенная таблица (табл. 1) демонстрирует запасы влаги
в почве на нижнем участке в течение критических для растений месяцев.
Цифры в таблице показывают, насколько велика разница во влажности
почвы на склонах, мелиорированных водозадерживающими террасами
и без них. Если еще в июне максимальное расхождение в процентах
влаги на одной и той же глубине достигает 3%, то в июле, когда расте-
ния наиболее сильно ощущают недостаток влаги в почве, разница во
влажности почвы на участках с террасами и без них достигает 12.4%.
Так же велика разница во влажности почв в сентябре, когда она доходит
на определенных глубинах до 11.5%. Характерно, что чем глубже, тем
сильнее вырисовывается эта разница.
Таблица 1
Сравнительная динамика влажности почвы на участках с террасами и без террас
(Кондара. Выс. 1100 м над ур. м., склон северной экспозиции, 1937 г.)
Влажность почвы (в %)
Ен 15 VI 1 16 VII 20 IX
Глубина (в Коэффицие] увядания без террас с террасами разница во влаж- ности без террас. 1 с террасами| разница во влаж- ности без террас с террасами разница во влаж- ности
0— 5 । 9.9 1 14.6 16.8 2.2 6.3 7.7 1.4 3.0 5.9 2.9
10— 15 9.9 15.8 17.4 1.6 8.0 15.7 /. / 4.8 9.0 4.2
20— 25 10.3 16.0 19.1 3.1 8.3 17.4 1 9.1 6.5 14.5 8.0
40— 45 10.7 18.1 20.4 2.3 9.9 19.2 9.3 8.0 17.3 9.3
50— 55 10.8 20.0 22.2 2.2 . 10.0 19.3 9.3 8.1 19.4 ] 1.3
95—100 9.2 21.2 22.5 1.3 10.2 I 21.6 11.4 8.3 19.4 11.1
120—125 7.8 21.6 23.2 1.6 I 10.3 1 1 22.7 12.4 8.7 20.2 11.5
312
В. И. Запрягаева
На склоне без террас влажность почвы в течение летне-осенних меся-
цев катастрофически быстро падает даже в более глубоких горизонтах.
Так, разница между влажностью почвы на глубине 120—125 см между
июнем и сентябрем достигает 12.9%, в то время как на террасированном
участке на этой же глубине и за такой же период разница во влажности
почвы доходит всего до 3%.
Полученные путем террасирования склонов дополнительные запасы
влаги в почве дали возможность нормально развиваться на склонах гор
целому ряду местных и иноземных древесных пород. Для того, чтобы
подчеркнуть, насколько террасирование улучшает развитие растении,
позволю себе привести несколько фактов, наблюдавшихся мною в ущелье
Кондара.
Выше уже отмечалось, что местные лесхозы, желая привести лесное
хозяйство горных склонов в нормальное состояние, решили сгустить
естественные насаждения путем посева и посадки различных древесных
пород. Они делали это без всяких мелиоративных мероприятий. На
склонах выкапывались площадки, около 0.5 кв. м площадью, и туда либо
опускалось несколько семян, либо сажалось молодое деревцо. Иногда
посев производился между деревьями, но большей частью на участках,
расчищенных от древесных насаждений. Много труда вкладывали лес-
хозы в свои агролесомелиоративные мероприятия, но без всякого успеха.
Как правило, растения погибали в первый же год, как исключение, —
оставались, но их состояние было весьма плачевным. Фисташка, напри-
мер, высеянная по склонам северной экспозиции в 1934 г., достигла
к 1945 г., т. е. за 11 лет, высоты 10—18 см. Миндаль бухарский, высеян-
ный там же, достиг за тот же период высоты 38—41—60 см. Все эти
«деревья» в возрасте 10—11 лет имели вид кустиков, с большим количе-
ством побегов, среди которых было много отмерших, поэтому высота
растений, которая мною здесь приводится, относится только к некоторым
сохранившимся побегам 3—5-летнего возраста.
На одном из участков, где лесхоз проводил свои работы с посевом
семян древесных растений, впоследствии (через 1 год после посева) были
проведены террасы. Поскольку всходы фисташки и миндаля, скрытые
травою, были очень малы, их не заметили и оставили между террасами.
Таким образом им была предоставлена возможность развиваться само-
стоятельно, без каких-либо элементарных мер ухода, и с ними про-
изошли весьма замечательные перемены. Поскольку террасы на данном
участке были сближены до 6 метров, почва межтеррасного пространства,
где росли эти растения, получила дополнительные запасы влаги. И это
дало возможность растениям развиваться более нормально. Высота
фисташки на И год после посева достигла в межтеррасном пространстве
175—190—330 см, т. е. была больше, чем в 10 раз, высоты кустиков
фисташки этого же возраста, оставшихся расти без террас. Характерно,
что фисташка начала расти быстро не сразу после проведения террас,
а только через 3 года, т. е. с 1938—1939 гг. Аналогичная картина полу-
чилась и с миндалем: высота растений на 11-й год после посева на участке
без террас достигала 38—41—60 см, в то время как высота растений,
попавших между террасами, равняется 230—350—400 см.
В наших опытах древесные породы высаживались однолетними сажен-
цами или высевались семенами на внутренний откос вала, как это
указано в схеме (см. рис. 14). Посадочный материал брался из питомни-
ков, расположенных в долине. Уход за растениями сводился, в основном,
к оправке террас от осыпавшейся почвы, к рыхлению почвы и очень незна-
Биологические особенности древесных пород
313
чительной обрезке деревьев. Посадка производилась на расстоянии
в ряду 1—2 м с последующим прореживанием. Как правило, каждого
вида древесных высаживалось от. 20 до 50 экземпляров.
Сравнивая результаты, полученные от посадки и посева различных
древесных пород на террасах, можно отметить, что наилучший резуль-
тат дают посадки растений саженцами. Культура большинства видов
древесных пород, высеянных на богаре, не удается без особого притене-
ния. Молодые всходы гибнут либо от ожогов корневой шейки, которые
получаются в результате сильного нагрева почвы, либо от интенсивного
испарения в результате низкой влажности воздуха при высоких темпера-
турах, часто доходящих до критических пределов. Сеянцы, имеющие
по 3—5 листиков, засыхают так быстро, что не успевают даже потерять
зеленой окраски. Из многих испытанных растений положительные
результаты получены только с посевом бухарского миндаля и гледичии,
но рост их в первые 2 года замедлен. Наш опыт показал, что вполне
рационально пользоваться поливными питомниками для выращивания
посадочного материала. Создавать же богарные питомники не следует,
так как молодые растения развивают на богаре очень мощную корневую
систему, затрудняющую ее выкапывание при пересадке.
Подбор растений для создания продуктивных древесных насаждений
производился нами путем изучения опыта предшествующих исследовате-
лей. Подбирались древесные породы быстрорастущие, дающие хорошую
древесину. Для опытов использовались местные и иноземные древес-
ные породы.
Перейдем теперь к изложению результатов опытов по богарному
лесоразведению.
Каркас (Celtis caucasica Willd.)
Каркас в естественных условиях достигает высоты 1 м к 6 годам;
из местных пород — это наиболее быстро растущее дерево. На террасах
он достигает такой же высоты на второй год (включая сюда время, про-
веденное в питомнике). Десятилетнее дерево каркаса в естественных
условиях имеет высоту около 2.5 м, а на террасированном склоне его
высота в 10 лет доходит до 3—4 м. Диаметр ствола каркаса в десятилет-
нем возрасте на террасах значительно превышает таковой у дерева,
выросшего в естественных условиях. Годичные приросты каркаса, расту-
щего в естественных условиях, достигают 6.1 см, а у каркаса, растущего
на террасах, могут достигать 152 см.
Клен туркестанский (Acer turkestanicum Pax)
В естественных условиях туркестанский клен — медленно растущая
порода, достигающая предельной высоты 10—13—14 м. Стволы клена,
как правило, кривы, сильно сбежисты и мало пригодны для строитель-
ных целей.
Наш опыт с туркестанским кленом был направлен к тому, чтобы
создать условия, при которых он мог бы развиваться в стройное, быстро
растущее дерево. Для этого сеянцы из поливного питомника были пере-
несены на склон северной экспозиции, т. е. в обычные для жизни клена
условия, но с измененным режимом почвенной влаги, созданным
водозадерживающими террасами.
314
В. И. Запрягаева
Здесь высота семилетних деревьев доходит до 3.3 м. Годичные при-
росты достигают на 7-й год, в среднем, 22 см, а иногда доходят и до
ИЗ см. Туркестанский клен на террасах начинает формировать гладкий
ствол только на 4—5-й год, а некоторые растения даже на 7-й год после
посадки все еще имеют форму куста. В естественных условиях для того,
чтобы молодому клену вырасти из семени до высоты 1 м, требуется около
33 лет, на террасированном же склоне — всего только 3 года. В естест-
венных условиях, до 3 м клен вырастает за 74 года, а на террасирован-
ном склоне только за 7 лет. Таким образом туркестанский клен на тер-
расах отнюдь не является медленно растущей породой и с успехом может
быть использован для создания высокопродуктивных лесов на богар-
ных, мелиорированных соответствующим образом склонах. Разводить
клен туркестанский можно и чистыми насаждениями, и в смеси с дру-
гими породами.
Клен Регеля (Acer Regelii Pax)
Развивается на богарных склонах совершенно так же, как туркестан-
ский клен.
Шелковица (Morns alba L.)
Шелковица является одной из самых распространенных пород в Сред-
ней Азии, занимая, как правило, искусственно орошаемые территории
близ селений. В горах Таджикистана шелковица поднимается иногда
довольно высоко. Здесь можно наблюдать, как она хорошо развивается
не только на поливных землях, но и на удаленных от мест орошения,
что позволяет считать шелковицу породой засухоустойчивой. Засухо-
устойчивость шелковицы способствовала тому, что опыты по ее разведе-
нию на богаре многократно производились в различных районах Средней
Азии (Федоров, 1932; Киселев, 1936).
В Таджикистане первые опыты по разведению шелковицы на богаре
были заложены на Варзобской Горной ботанической станции в 1935 г.,
затем в 1936 г. Лесная опытная станция НКЗ Таджикской ССР поставила
опыты по испытанию шелковицы на сухих склонах в предгорьях южного
склона Гиссарского хребта. Позднее, с 1938 по 1941 г., вопросу разве-
дения шелковицы на богаре уделяется еще более серьезное внимание,
л эта работа включается в тематический план Таджикского филиала Ака-
демии Наук СССР Лесная опытная станция ТФАН (Морозов, 1944)
организует работы на высотах 600—800 м над ур. м. Эти работы приводят
к заключению о том, что в полупустынных предгорьях на высоте 600 м
над ур. м. недостаток почвенной влаги ограничивает возможность разве-
дения шелковицы, в то же время на высоте 800 м шелковица развивается
хорошо, при сравнительно высокой приживаемости растений после
посадки.
Результаты наших работ по разведению шелковицы на богаре в усло-
виях пояса древесно-кустарниковой растительности сводятся к следую-
щему. Развитие шелковицы протекает здесь весьма удовлетворительно,
хотя отпад растений после посадки был довольно значительным (послед-
нее можно объяснить, главным образом, плохим качеством посадочного
материала). В своем росте в высоту шелковица не делает резких скачков,
кривая ее роста плавно поднимается. Годичные приросты даже на 10-й
год только несколько уменьшаются, оставаясь все же, по сравнению
Биологические особенности древесных пород
315
•с другими растениями, довольно большими. В 1945 г. был впервые по-
ставлен опыт эксплоатации шелковицы, высаженной на богарном склоне.
Богарная шелковица показала прекрасную способность к восстановле-
нию обрезанных частей. Несколько хуже, чем на северном склоне, шелко-
вица развивается на южном, мелкоземистом, тоже террасированном
«склоне; здесь ее годичные приросты значительно меньше. Высота деревьев
на 3-й год после посадки достигает всего 85—120 см, в то время как на
•северном склоне — 163—232 см. Здесь процент погибших растений дохо-
дит до 55—70. Опыт с посадкой шелковицы на южном каменистом склоне
•совсем не удался. Растения погибли в год посадки, за исключением
4 экземпляров,, которые отмерли на следующий год. Таким образом
вопрос культуры шелковицы на богаре, в пределах 800—1200 м над ур. м.,
в условиях, аналогичных описываемым, является разрешенным. Расши-
рение площади под культуру шелковицы на богаре даст возможность
•освободить поливные земли под другие культуры.
Орех (Juglans regia L.)
Грецкий орех можно рассматривать, с одной стороны, как садовую
культуру, дающую высококачественные, богатые витаминами плоды,
с другой стороны, — как лесную породу с очень хорошей поделочной
и строевой древесиной.
В отличие от описанных выше растений, по ореху имеется весьма
обширная литература, где значительное место уделяется вопросу его
культуры вообще и на богаре — в частности. О распространении ореха
в Средней Азии, о некоторых его биологических особенностях имеются
многочисленные данные в работах В. Л. Некрасовой (1927), М. Г. По-
пова (1939), X. Г. Дьяченко (1934), А. В. Гурского (1932), Л. А. Смолья-
ниновой (1936) и, наконец, уже непосредственно по Таджикистану,
в работах Г П. Викторовского (1935), В. И. Запрягаевой (1940).
В естественных условиях орех обычно селится по дну ущелий
и у родников, т. е. там, где имеется достаточное количество влаги.
Но орех довольно часто поднимается и вверх по склонам, произрастая на
мощных мелкоземистых почвах.
Впервые в Средней Азии опыт с посадкой ореха на неполивных землях
был заложен в Аман-кутане в конце прошлого века (А. И. Федоров,
1929). Позднее эти опыты повторялись в других местах Средней Азии
и, в частности, в Таджикистане, на участке Лесного управления
НКЗ в Сильбурсае. По описанию В. М. Киселевпча (1936), опыт Амап-
кутана оказался не совсем удачным. Такого же мнения придерживается
и П. И. Троицкий (1936), объясняя это тем, что орех в Аман-кутане
высаживался на мало удобных землях, часто по южным щебнистым скло-
нам, открытым для действия юго-западных ветров, в результате чего
деревья ореха принимали карликовый вид. К тому же за посадками
ореха не было никакого ухода. В опытах Раунера в Ак-ташо получились
значительно лучшие результаты (А. И. Федоров, 1929); здесь орех вполне
нормально рос и плодоносил даже в неблагоприятных условиях. Непло-
хие результаты получились и на участке в Сильбурсае, где состояние
ореховых насаждений, по моим наблюдениям в 1938 г., было вполне удо-
влетворительным. Растения, высаженные в канаву по способу Раунера,
дали довольно высокий процент приживаемости. Мною же наблюдались
посадки ореха, сделанные Лесхозом Варзобского района по склонам
316
В. И. Запрягаева
ущелья Кондара. Так как орех высаживался здесь без предварительной
подготовки склона, то молодые растения погибали на 2—3-й год после
посадки.
Наши опытные посадки ореха были сделаны на высоте 1100 м над
ур. м., на склоне северной экспозиции. Орех высаживался на полотне
водозадерживающей террасы.
Для ореха характерно начало интенсивного роста не в первые годы
после посадки, а через несколько лет (примерно, через 4—5 лет). Если
в первые 2 года орех давал годичные приросты всего по 8—25 см, то на
5-й год прирост доходил до 40—91 см. К концу 5-го года орех имел вы-
соту 78—108 см, а к концу 10-го года его высота достигала 424—550 см.
Диаметр ствола ореха значительно превышает диаметр многих других
описанных выше древесных культур в том же возрасте. Орех, посажен-
ный на богаре, на 10-й год вступил в пору плодоношения.
Небезинтересно сравнить скорость роста ореха на террасированном
склоне со скоростью его роста в обычной для него естественной обстановке.
До 5 лет орех, высаженный на террасированном склоне, развивается
примерно так же, как орех, растущий в самых неблагоприятных усло-
виях, т. е. за 5 лет достигает высоты всего 1 м. Но затем, в отличие от
деревьев, растущих на сухом, немелиорированном склоне, на террасах
в его росте наблюдается скачок и за последующие 5 лет он вырастает
на 3.5—4.5 м в высоту, достигая к концу 10-го года 4.2—5.5 м. Таким
образом он оставляет позади себя медленно растущие деревья ореха
на сухих склонах, приближаясь по интенсивности роста к орехам, расту-
щим в наиболее благоприятных условиях, по берегам ключей. Из
этого можно сделать заключение, что те запасы влаги, которые аккуму-
лируются в почве с помощью террас, являются вполне достаточными
для нормального развития ореха и поэтому вполне целесообразно вклю-
чить его в список древесных пород для агролесомелиорации в горных
условиях, аналогичных южному склону Гиссарского хребта. Разводить
орех в горах можно в пределах 1000—1900 м (зона его естественного рас-
пространения), на склонах северных экспозиций с мощными мелкозе-
мистыми почвами. Если орех предназначается для создания лесных
насаждений, с целью получить ценную строительную и поделочную
древесину, то посадки его следует сгустить. В случаях же, когда орех
предназначается для получения плодов, расстояние между террасами
следует довести до пределов, описанных в следующем разделе настоя-
щего очерка, соответственно увеличивая расстояние между деревьями
в ряду; при этом следует высаживать только привитые культурными сор-
тами саженцы.
Помимо местных древесных пород, в опытах по богарному лесоразведе-
нию участвовали иноземные древесные породы. Привлечение последних
для создания полноценных лесных насаждений необходимо, ибо ассорти-
мент местных древесных пород, пригодных для практических целей,
чрезвычайно беден.
Айлант (Ailanthus altissima L.)
Очень быстрорастущая и засухоустойчивая порода, достигающая
в обычных условиях своего существования высоты 20 м. Древесина его
невысокого качества. Особое значение айлант приобретает на горных
склонах, где его быстрорастущие деревья могут использоваться как ты-
Биологические особенности древесных пород
317
•чины для богарного винограда. До сих пор Таджикистан ввозит много
этих кольев из других мест Советского Союза.
Айлант как порода, пригодная для облесения горных склонов, из-
вестен уже давно. Его высаживали в Крыму, близ Феодосии, в 80-х
годах прошлого столетия (Карасев, 1934). Очень хорошо сохранилась
роща айланта в Аман-кутане, на северных склонах западной оконечности
Зеравшанского хребта. Здесь айлант высаживался в конце прошлого
•столетия по южным щебнистым склонам на террасах типа Раунера (Троиц-
кий, 1936). В окрестностях кишлака Багрин у подножья Зеравшанского
хребта диаметр богарного айланта достигал 30 см. Айлант применялся
Рис. 15. Айлант (Ailanthus altissima), на террасах в ущелье Кондара.
Слева — возраст 1 год (фот. Ф. Л. Запрягаева); справа — возраст 10 лет
(фот. А. И. Иванова).
и в работах Раунера в Ак-таше, но Раунер, в отличие от многих
исследователей, не считает его породой засухоустойчивой, пригодной
для облесения горных склонов. Работы Варзобской станции с айлантом
ни в какой мере не подтверждают мнения, высказанного Раунером.
В наших условиях айлант показал себя как одна из лучших пород, при-
годных для облесения горных склонов (рис. 15).
Склоны северной экспозиции с карбонатными серо-бурыми, более
или менее глубокими почвами — лучшее для его развития местообита-
ние. После посадки айлант, без всякого отпада, начинает быстро разви-
ваться. Уже на 2-й год он дает годичные приросты до 96—188 см, его
высота достигает 2 м. К концу 4-го года отдельные деревья айланта до-
стигают 5 м, годичные приросты все так же велики; но затем начинается
некоторое замедление роста, и на 10-м году годичные приросты не пре-
вышают 54 см. На 10-й год после посадки, при высоте 6—7—9 м, толщина
ствола у основания равна 14.7—24 см, на высоте 1 м—9.3—14.2 см. Это
весьма характерно для айланта, у которого формируется гладкий, мало-
сбежистый ствол.
318
В. И. Запрягаева
Одновременно айлант был высажен на склоне южной экспозиции
с каменистыми почвами. Эти условия оказались для него совсем непри-
годными. К концу второго года высаженный айлант погиб на 93.4%.
Айлант очень хорошо развивается в предгорьях южного склона Гис*
сарского хребта, где лесная опытная станция Таджикского филиала Ака-
демии Наук СССР организовала опытные посадки на высоте 900 м над
ур. м. В 1945 г. 7-летние деревья айланта имели среднюю высоту 5.5 м,
а максимальная высота деревьев доходила до 8 м.
Необходимо отметить способность айланта к порослевому возобновле-
нию. Об этом уже говорили многие исследователи (Троицкий, 1936).
В наших условиях айлант стал давать первые порослевые побеги на
4-й год после посадки. Поросль развивается чрезвычайно быстро, за
3 года порослевые побеги достигают 3 м высоты, годичные приросты таких
побегов доходят до 100—135 см. На южном склоне у айланта поросле-
вых побегов не образуется. Посев айланта семенами не дал хороших
результатов. Обильные всходы с наступлением жарких дней быстро засы-
хали и к концу первого же года погибали. Отмирание всходов связано-
с почернением и омертвлением молодого стебелька у корневой шейки,
что, невидимому, вызывается сильным нагревом поверхности почвы. Все
изложенное выше говорит, что в наших условиях айлант пригоден для
культуры только по склонам северной или близкой к ней экспозиции.
Белая акация (Robinia pseudoacacia L.)
Быстрорастущая порода. Высота дерева может достигать 30 м. Вет-
роустойчива, к почве очень нетребовательна. Древесина ее ценна для
строительства, используется для телеграфных столбов, а также для
железнодорожных шпал, так как чрезвычайно стойка против гниения,
что дает основание сравнивать ее в этом отношении с дубом (Киселе-
вич, 1936). Имеет применение в изготовлении виноградных тычин.
В Средней Азии белая акация распространена по орошаемым оазисам.
Опыт перенесения белой акации в условия горной богары был произ-
веден еще в 80-х годах прошлого столетия в Крыму, близ Феодосии
(Карасев, 1934), и в Аман-кутане, а несколько позднее повторен ферган-
ской и кашка-дарьинской агролесомелиоративными партиями. Во всех
случаях посадки белой акации на богарные склоны результат получился
очень хороший. Совершенно отличное от всех исследователей мнение
по вопросу культивирования белой акации на горных склонах высказы-
вает Раунер (1901), который считает, что белую акацию, так же как
и айлант, даже нельзя назвать лесной породой, так как она, якобы,
страдает от засухи и мороза. Судя по нашим опытам в ущелье Кондара,
белая акация лучше всего развивается на высоте 1100 м над ур. м., на
северном склоне (рис. 16).
Рост белой акации на северном склоне мало чем отличается от роста
айланта. И общая высота и годичные приросты так же велики, как
у айланта. Так же, как айлант, белая акация бурно развивается впервые
5 лет, затем ее годичные приросты уменьшаются. Значительно хуже раз-
вивается белая акация в других экологических условиях.
Как показал наш опыт, разводить белую акацию в верхней части
древесно-кустарникового пояса (высота 1850 м над ур. м.) малоцеле-
сообразно, там она сильно страдает от поломки снегом, особенно в первые-
годы. Почти каждую зиму сползающим снегом уничтожаются все побеги
Биологические особенности древесных пород
319
предшествующего года. Когда же белая акация вырастает выше уровня
снегового покрова, ее ствол под тяжестью того же сползающего снега
сильно изгибается и становится непригодным для использования в стро-
ительном деле.
Опыт с белой акацией в предгорьях Сталинабада, проводившийся
Лесной опытной станцией Таджикского филиала Академии Наук, пока-
зал, что на высоте 900 м над ур. м. белая акация развивается очень
хорошо. В 1945 г. 7-летние деревья имели здесь среднюю высоту 4.5 м,
а отдельные экземпляры достигали 7.4 м.
Таким образом можно считать, что лучшими для разведения белой
акации па горных склонах являются склоны северной экспозиции или
Рис. 16. Белая акация Robinia pseudoacacia L. на террасах в ущелье Кондара.
Слева — возраст 1 год; справа—возраст 4 года (фот. Ф. Л. Запрягаева).
близкие к ней, с развитыми мелкоземистыми карбонатными серо-бурыми
почвами в пределах 900—1400 м над ур. м. Белую акацию можно здесь
разводить либо чистыми насаждениями, либо в смеси с другими, более мед-
ленно растущими породами, например с дубом или кленом, с тем, чтобы
через 10—15 лет акацию можно было вырубить и тем дать возможность
свободнее расти посаженным вместе с ней породам. Однако ее ни в коем
случае не следует сажать вместе с кустарниками, так как последние
будут только конкурентами в потреблении почвенной влаги.
Гледичия (Gleditschia triacanthos L.)
Быстрорастущая, засухоустойчивая порода, достигающая высоты
40 м. Дает прочную,' пригодную для поделок древесину.
В Средней Азии распространена в орошаемых оазисах. Впервые
в условиях богары испытывалась на Кавказе на склонах Телецкого
хребта Лисневским.
•320
В. И. Запрягаева
В опытах Варзобской ботанической станции гледичия дала очень
хорошие результаты. Ее посадка производилась в различных экологи-
ческих условиях нижней части пояса древесно-кустарниковой расти-
тельности.
Рост ее, хотя и отстает от роста айланта и белой акаций, но все же
происходит довольно быстро. На 10-й год она достигает высоты 4—5 м.
На склоне северной экспозиции гледичия, в отличие от многих пород,
прекрасно развивается из семян. Примерно на 3-й год после посева моло-
дые растения достигают в высоту такого же размера, как растения, пере-
саженные однолетними саженцами из питомника. Несколько хуже гле-
дичия развивается на южном склоне с мелкоземистыми почвами, ее высота
здесь к концу 5-го года после посадки достигает только 92—128 см,
в то время как на северном склоне — 310—430 см. На южном склоне
с каменистыми почвами гледичия продержалась только в течение 3 йет,
а затем погибла полностью. Сделанные нами попытки разведения гле-
дичии на высоте 1850 м были неудачны, так как здесь она очень сильно
страдала от поломок снегом.
На основании опыта Варзобской станции можно считать, что произ-
водственного эффекта от посадки гледичии можно ожидать только при
культуре ее в нижней части пояса древесно-кустарниковой раститель-
ности, на северных склонах с мелкоземистыми почвами.
Карагач (Ulmus foliacea Gilib. v. campestris L.)
Быстрорастущая, засухоустойчивая порода, достигающая 40 м
высоты, не морозоустойчива. В Таджикистане карагач встречается как
в орошаемых оазисах, так и на сухих горных склонах. Первые искус-
ственные посадки карагача на богаре были сделаны в Аман-кутанской
даче (Киселевич, 1936). Затем опыт повторен кашкадарьинской агро-
лесомелиоративной партией. И в том и в другом случае результат опытов
получился очень хороший. ]
Наши первые опытные посадки карагача были произведены в 1935 г.
в ущелье Кондара на высоте 1100 м над ур. м., на склонах различных
экспозиций.
Рост карагача на террасированном склоне очень хороший. Он усту-
пает в росте только айланту и белой акации. У карагача, так же как
и у предыдущих древесных пород, наблюдается чрезвычайно бурное
развитие в первые годы после посадки, когда годичные приросты дости-
гают 122 см. Примерно на 4-м году после посадки рост карагача начи-
нает замедляться, что выражается в сравнительно небольших годичных
приростах, которые на 10-й год достигают всего 14—28 см. Вполне нор-
мально развивается и толщина ствола, на 10-й год она достигает у кор-
невой шейки 10—16 см, а на высоте 1м — 6.4—11 см. Отмерших расте-
ний после посадки в этих условиях совсем не наблюдалось. Много хуже
чувствует себя карагач на склоне южной экспозиции с мелкоземистыми
почвами. Посаженный в 1935 г. карагач вырос к 1938 г., т. е. к концу
4-го года, всего только на 60—79 см, в то время как его высота на север
ном склоне достигала к этому же времени 188.4—272.0 см. В последую-
щие годы карагач очень плохо развивался; его годичные приросты дости-
гали всего 10—12 см. Процент погибших растений на этом склоне очень
велик; к 5-му году после посадки он достигал 63.1 см. Еще хуже чув-
ствует себя карагач на каменистых почвах южного склона, здесь он
Биологические особенности древесных пород
321
не выдерживает суровых условий и погибает на 4-й год. Таким образом,
лучшими условиями для культуры карагача на богаре являются склоны
северной или близкой к ней экспозиции с мелкоземистыми почвами.
Лесная опытная станция ТФАН организовала посадку карагача
на мелиорированном водозадерживающими террасами склоне в пред-
горьях южного склона Гиссарского хребта, на высоте 900 м над ур. м.
Здесь семилетние деревья к 1945 г. достигли высоты 3.5—5.5 м.
Вяз (Ulmus effusa Willd.)
Это дерево может достигать высоты 20—25 м. Очень долговечно,
доживает до 400 лет. По данным М. Б. Колпикова (1944), засухоустойчив,
но к почве требователен. Древесина вяза очень ценится из-за того, что
может гнуться, и широко применяется в кустарном производстве и маши-
ностроении.
Первый опыт использования вяза для горной агролесомелиорации
был сделан Зибольдом в 1897 г. близ Феодосии, но был не совсем удачен
(Ходоровский, 1904).
В наших опытах вяз высаживался на северных склонах на высотах
1100 и 1850 м над ур. м., т. е. в нижней и верхней полосах пояса дре-
весно-кустарниковой растительности. На нижнем участке вяз разви-
вался значительно лучше, чем на верхнем, что, однако, не является пре-
пятствием для его разведения в более высоких областях.
Вяз развивается и растет в условиях горной богары значительно
быстрее карагача. Высоты 411—615 см он достигает за 4 года, в то время
как примерно такой же высоты (447—645 см) карагач достигает за 10 лет.
То же самое можно сказать и о толщине ствола. Отмирание посаженных
на горной богаре растений не наблюдалось совсем. Таким образом, хотя
вяз и является новой культурой в горной агролесомелиорации, но весьма
перспективен.
Ясень европейский (Fraxinus excelsior L.)
Может достигать 40 м высоты. Растет быстро, особенно в молодом
возрасте. По данным М. В. Колпикова (1944), растет довольно хорошо
на сухих степных карбонатных почвах. Древесина крепка, упруга и очень
ценится в мебельном деле, идет также на изготовление ободьев для колес.
В последнее время имеет широкое применение в авиационной промыш-
ленности. О возможности использования ясеня в агролесомелиоратив-
ных работах высказываются противоречивые мнения. В. Н. Киселевич
(1936) считает ясень весьма требовательным к почве, особенно к ее влаж-
ности, и, помимо того, подверженным нападениям растительных и живот-
ных вредителей. Напротив, Н. Н. Степанов (1944) считает ясень глав-
ной породой в степном лесоразведении. Десятилетний опыт Варзобской
станции с культурой ясеня на богаре показал, что Fraxinus excelsior L.
прекрасно развивается на склоне северной экспозиции в ущелье р. Кон-
дара, на высоте 1100 м над ур. м.
Ясень растет очень быстро и рост его продолжается в течение про-
должительного срока. Замедление в росте, выражающееся в уменьшении
годичных приростов, замечается только на 6—7-й год. Его высота на 10-й
год достигает 521—670 см, а диаметр ствола у корневой шейки 10.7—
21 Ущелье Кондара
322
В. К Запрягаева
16.5 см. До сих пор никаких признаков угнетения деревьев и их повре-
ждений обнаружено не было. Процент погибших деревьев доходит до 25,
но отмирание растений наблюдалось только в первый год после посадки
и было, повидимому, связано с плохим качеством посадочного материала.
Так же, как и все прочие древесные породы, ясень плохо развивается
на южном склоне даже на местах с мелкоземистыми почвами. Рост его
здесь чрезвычайно замедлен; выживают после посадки только единич-
ные экземпляры. Главный отпад растений наблюдается на следующий
год после посадки. Очень значительный процент отпада наблюдался нами
при посадке этого вида ясеня на богарном склоне в пределах верхней
полосы пояса древесно-кустарниковой растительности, где ясень поги-
бал от поломок снегом. Рост в этих условиях был несколько замедлен;
так, деревья 3-летнего возраста, посаженные на северном склоне, на
высоте 1850 м над ур. м., имели высоту 76.9—156 см, в то время как
на высоте 1100 м на северном склоне ясень имел в этом же возрасте
высоту 179—278 см.
Посадка ясеня без соответствующей подготовки склонов не дает
эффекта. Нами проанализирован рост ясеня на склоне северной экспо-
зиции соседнего с Кондарой ущелья — Джиринодсой, где ясень был
высажен лесхозом в 1933 г.
На протяжении всех лет замечался большой процент отпада; 1939 г.
на всем участке, где высаживался ясень, нельзя было найти ни одного
живого растения.
Вопреки сложившемуся мнению, автор, руководствуясь собственным
опытом, считает вполне целесообразным использовать ясень в горных
агролесомелиоративных работах, на склонах северной экспозиции с мощ-
ными мелкоземистыми почвами.
Опыт лесной опытной станции Таджикского филиала Академии Наук
СССР с посадкой ясеня на террасированном склоне в предгорьях южного
склона Гиссарского хребта, на высоте 900 м над ур. м., показал, что
культура ясеня может быть распространена в агромелиоративных рабо-
тах в пределы 900—1500 м над ур. м. На участке этой станции 7-летние
деревья ясеня достигали в 1945 г. средней высоты в 286 см, а отдель-
ные деревья имели высоту до 4 м.
Ясень американский (Fraxinus americana L.)
В Средней Азии и в Таджикистане встречается редко и только в оро-
шаемых оазисах. В. Н. Киселевич (1936) высказывается в очень катего-
рической форме против культуры американского ясеня на горной богаре,
ссылаясь на то, что он к 10 годам жизни замедляет свой рост, чахнет
и нередко погибает. Совершенно противоположное мнение высказывает
Н. И. Степанов (1922). Изучив 80-летний опыт степного лесоразведения,
он пришел к убеждению, что американский ясень зарекомендовал себя
как порода быстрорастущая и мало требовательная к влаге. Американ-
ский ясень образует с самого раннего возраста очень стройные деревца.
Опыта разведения его на горной богаре до работ Варзобской станции,
повидимому, не было.
В наших опытах американский ясень высаживался на склоне север-
ной экспозиции, на высоте 1100 м над ур. м. В 8-летнем возрасте амери-
канский ясень здесь не несет никаких признаков угнетения, хотя прирост
его в последние 2 года замедляется. На всем протяжении жизни ясеня
нд богаре отпада не наблюдалось.
Биологические особенности древесных пород
323
Ясень американский, высаженный на южный склон ущелья Кондара
на высоте 1100 м, развивается очень плохо, здесь срок его жизни огра-
ничивался всего 3 годами. Неплохо развивается американский ясень
в верхней части пояса древесно-кустарниковой растительности, на высоте
1850 м над ур. м. Высота 4-летних деревьев достигает 243 см, при тол-
щине ствола, доходящей у корневой шейки до 3.7 см. Годичные при-
росты ясеня в этих условиях достигают 73—100 см.
На основании опыта Варзобской станции американский ясень можно
рекомендовать разводить во всем поясе древесно-кустарниковой расти-
тельности, отводя для него только склоны северной экспозиции.
Ясень ланцетный (Fraxinus lanceolata Borkh.)
Впервые в СССР опыт использования этого ясеня в горной агролесоме-
лиорации был поставлен, повидимому, на Варзобской горной ботанической
станции, где он высаживался на высоте 1100 и 1850 м над ур. м. При
незначительном отпаде в первые годы после посадки ясень развивался
вполне нормально. Высота деревьев ясеня на верхнем участке, на высоте
1850 м, достигает на 3-й год после посадки 85—155 см. Нет никаких
оснований полагать, что этот вид ясеня не может быть использован более
широко в горной аромелиорации.
Ясень остроплодный (Fraxinus oxycarpa Willd.)
Так же как и предыдущий, этот вид ясеня, повидимому, нами испы-
тывался на горной богаре впервые в СССР Опыты с ним были заложены
в нижней полосе пояса древесно-кустарниковой растительности на высоте
1100 м, на северном мелкоземистом склоне ущелья Кондара. Резуль-
таты получены вполне удовлетворительные. 8-летние деревья достигают
высоты 170—230 см, при диаметре ствола у корневой шейки 1.7—4.7 см.
Ясень приречный (Fraxinus potamophila Herd)
Широко распространен по горным долинам Средней Азии. В Таджики-
стане встречается в Гиссарском хребте и Придарвазье в пределах 1000—
1800 м над ур. м. В речных поймах достигает 20 м высоты и 80 см в диа-
метре.
Был высажен нами на террасированный склон северной экспозиции
с мелкоземистыми почвами на высоте 1100 м над ур. м. В первые 2 года
развивался очень медленно. Его годичные приросты достигали всего
8—35 см, но на 3-й год жизни он стал быстро расти, достигнув к концу
6-го года высоты 92—121 см. На 10-й год после посадки этот ясень имел
вид стройных деревьев высотой до 280—405 см, с диаметром ствола у кор-
невой шейки до 13.1 см.
Таким образом этот вид ясеня развивается вполне удовлетворительно
на богарных склонах северной экспозиции, мелиорированных водозадер-
живающимп террасами.
Дуб летний (Quercus pedunculata Ehrh.)
Теплолюбивая порода, требует хорошо развитых почв. К влаге
менее требователен. Дерево первой величины, очень долговечно, имеет
21*
324
В. И. Запрягаева
прекрасную, прочную и гибкую древесину, находящую широкое при-
менение в различных производствах.
Первые опыты по использованию летнего дуба при облесении горных
склонов были поставлены в Крыму, близ Феодосии, в 80-х годах про-
шлого столетия. Примерно тогда же дуб был высажен на горные склоны
в Аман-кутане. Позднее опыт с дубом был повторен ферганской партией.
По описанию В. М. Киселевича (1936), опыт с культурой дуба в Аман-
кутане оказался не совсем удачным. В одном случае дуб высаживался
на высоте 1200—1300 м на западных и юго-западных щебнистых склонах
и здесь почти весь погиб. В другом случае дуб высаживался на высоте
1700 м вблизи гребня водораздельного хребта и тоже почти вся посадка
погибла; оставшиеся единичные экземпляры имели вид слабых, болез-
ненных растений. Дубки страдали от систематического обмерзания вер-
хушечных почек.
В опытах Варзобской станции дуб высаживался на 2 участках на
высоте 1100 и 1850 м над ур. м; в обоих случаях по северным мелко-
земистым склонам.
С первого же года посадки дуб принял стелющуюся форму, он начал
ветвиться почти у основания ствола, поэтому большинство дубков имело
форму распростертого куста, но некоторые из них в настоящее время
довольно ясно образовали главный ствол с кроной, находящейся в верх-
ней части ствола, нижние же ветки, появившиеся в первые годы, так
и остаются стелющимися по земле. Высота- 10-летних деревьев дуба
достигает 347—550 см. В этих условиях 10-летние дубки вступили в пору
плодоношения. Дуб при весенних посадках дает весьма значительный
процент отпада, доходящий в некоторых случаях до 70%, поэтому его
лучше сажать поздней осенью; тогда его отпад не превышает 20—25%.
Одновременно с посадкой на высоте 1100 м над ур. м. дуб высаживался
на высоте 1850 м, на северном склоне, в верхней части древесно-кустар-
никового пояса. Здесь весенние посадки 1936 г., так же как и на нижнем
участке, не дали никаких результатов, 75% саженцев погибло в пер-
вый же год после посадки, остальные деревца погибли на 3-й год,
влача жалкое существование в течение 2 лет. Значительно лучше раз-
вивались деревца при осенних посадках. Высота их на 3-й год после
посадки достигала 40—88 см. Годичные приросты доходили до 14—56 см.
Посадка дуба на южных склонах как на высоте 1100 м, так и на высоте
1850 м, не дала никаких положительных результатов; растения погибали
в первый же год после посадки.
Исходя из опыта Варзобской станции, можно рекомендовать дуб для
использования в горной агролесомелиорации, но отводить для его посадки
следует только склоны северной экспозиции с хорошими глубокими
почвами в пределах 1000—2000 м над ур. м. Свойство дуба стелиться
и давать искривленный ствол в первые годы жизни не является препят-
ствием для его разведения, так как в последующие годы он выпрямляется.
Конский каштан (Aesculus hippocastanum L«)
В Средней Азии разводится, так же как и в других местах, как пар-
ковое декоративное дерево в городах. Довольно морозоустойчив, что
позволяет его разводить даже в Ленинграде. Древесина конского каш-
тана, по данным В. Д. Городецкого (1934), мягкая, пригодная для резьбы,
для подделки под красное дерево. Это — быстрорастущая порода, но, по
Биологические особенности древесных пород
325
указанию того же автора, редко достигает высоты 15 м. Указаний
на использование конского каштана в горной агролесомелиорации
в пределах Советского Союза не имеется. Возможно, что опыт Варзоб-
ской станции в этом отношении был первым.
Конский каштан растет в условиях горной богары значительно
медленнее, чем многие другие древесные породы в аналогичных условиях,
но кривая его роста в течение указанных лет непрерывно поднимается.
9-летние деревья достигают высоты 450 см, при толщине ствола у осно-
вания в 5—11 см. Конский каштан при посадке в нашем опыте дал про-
цент отпада, доходящий до 42. Возможно, что эта неудача относится
за счет поздней весенней посадки.
Таким образом конский каштан может с успехом занять место в списке
лесных пород для горной агролесомелиорации в пределах 1000—1200 м
над ур. м.
Софора японская (Sophora japonica L.)
В наших опытах софора высаживалась на высоте 1100 м над ур. м.
на северном и южном склонах. В связи с большим процентом отмерших
в первый же год растений можно считать, что склоны южной экспозиции
не пригодны для разведения софоры. Склоны северной экспозиции на ука-
занной высоте более пригодны; здесь высота софоры на 4-й год после
посадки достигала 182—333 см. Несмотря на то, что в первые годы софора
имела тенденцию к кущению, путем своевременной обрезки лишних
ветвей удалось сформировать гладкоствольное дерево, высота которого
на 10-й год достигает 431—560 см, прп толщине ствола у основания
в 11—16 см.
Липа (Tilia parvifolia Ehrh., Т. grandifolia Ehrh.)
Оба вида, достигающие 25 м в высоту, имеют очень ценную, мягкую
древесину, широко используемую на резьбу, токарные и столярные
изделия. Помимо того, липа является прекрасным медоносом. Оба вида
лип высаживались нами на высоте 1850 м над ур. м., на мощных мелко-
земистых почвах северного склона в урочище Квак. Высота 3-летних
деревьев доходит до 130—136 см. На 7-й год после посадки высота
деревьев достигает 500—550 см. Годичные приросты на 3-й год довольно
значительны и достигают 83—84 см, в то время как в первые годы они
не превышали 10—42 см.
Осина (Populus tremula L.)
В опытах Варзобской станции осина высаживалась на высоте 1850 м
на северном мелкоземистом склоне. Высота деревьев 3-летнего возраста
достигает 108—143 см. К концу 7-го года после посадки высота деревьев
достигала 630—750 см.
Лещина (Corylus avellana L.)
Лещина высаживалась нами на высоте 1100 и 1850 м над ур. м.
на северных мелкоземистых склонах. В обоих случаях результат полу-
чился вполне удовлетворительный. На высоте 1850 м лещина, при срав-
326
В. И. Запрягаева
нительно невысоком проценте отпада, который наблюдался только в пер-
вый год после посадки, развивается в кусты, достигающие на 3-й год
высоты 58—111 см. На нижнем участке лещина развивается медленнее
и высоты 110—150 см достигла только на 8-й год.
Помимо посадки лиственных деревьев мы делали опыты с посадкой
хвойных — видов родов Pinus, Picea, Larix, Pseudotsuga, — но резуль-
таты получили весьма скромные, и то только с сосной Pinus Pithyusa
Strangw. В наших опытах этот вид сосны высаживался на высоте 1100
и 1850 м. На обоих участках процент отпада достигал 80, т. е., по суще-
ству, оставались живыми только единичные экземпляры, которые и
в настоящее время развиваются еще очень медленно. Сосенки на высоте
как 1100 м, так и 1850 м на З-й год после посадки достигли 44—57 см
высоты, на 9-й год высота их возросла до 137—150 см. Опыты с хвой-
ными были заложены, помимо указанных участков, и на высоте 2400 м
над ур. м., на высокогорном плато Руидашт, в поясе субальпийской
растительности. Это плато лишено древесной растительности, но есть
основания полагать, что она в эту зону когда-то заходила. У посеянных
здесь растений, как правило, весной появлялись обильные всходы, кото-
рые погибали летом в первый же год, повидимому, от сильного нагрева
поверхности почвы. Опыты с посевом хвойных повторялись безуспешно
в течение ряда лет. Причина неудачи не ясна; повидимому, мы чего-то
не учли. Судя по опыту в Аман-кутане, сосна должна была бы занять
определенное место в облесении горных склонов Таджикистана. В Аман-
кутане хорошо развиваются деревья крымской сосны на сухих склонах
гор, достигая к 39 годам высоты 10 м.
Подводя итоги нашим опытным работам в области богарного лесо-
разведения, можно сделать следующие практические выводы:
А. Наиболее целесообразным методом разведения древесных пород
на горной богаре, в условиях, аналогичных описываемым, для создания
сомкнутых насаждений надо считать посадки на террасированных скло-
нах с расстоянием между террасами до 6—8 м.
Б. При разведении древесных пород на богаре следует признать пре-
имущество посадки перед посевом. Высаживать лучше всего саженцы
одно-двухлетнего возраста, выращенные в поливных питомниках.
В. Межтеррасные пространства в создаваемых насаждениях можно
занимать только рядами древесных пород, это дает возможность сгустить
насаждения, но ни в коем случае не следует занимать их кустарниками,
так как всякое постороннее растение будет являться конкурентом в потре-
блении столь дефицитной почвенной влаги.
Г За всеми посаженными растениями, в целях создания высоко-
продуктивных древесных насаждений, необходим регулярный уход,
особенно в первые годы. Уход должен состоять в оправке террас, рыхле-
нии почвы, подрезке растений.
2. ОПЫТ БОГАРНОГО САДОВОДСТВА1
Варзобская ботаническая станция разрабатывает методы разведения
искусственных насаждений не только ради получения строевой и поде-
лочной древесины, но также и для получения плодов. На сухих склонах
1 Работы по настоящему разделу проводились коллективом сотрудников: науч-
ными сотрудниками В. И. Запрягаевой и Ф. Л. Запрягаевым, садовником И. Я. Зен-
тарским, техниками-плодоводами А. Г Тошаевой и Е. А. Базовой.
Биологические особенности древесных пород
327
гор выращиваются богарные сады из персика, абрикоса, яблони, груши,
сладкого миндаля, инжира, граната и некоторых других плодовых и ягод-
ных культур.
На южном склоне Гиссарского хребта и в других, аналогичных ему
горных районах Таджикистана сады хотя и разводятся колхозами,
но в весьма ограниченных размерах. Чаще встречаются единичные пло-
довые деревья около домов или небольшие садики в нижней части гор-
ного пояса, с 10—15 деревьями. Все эти сады и отдельные деревья встре-
чаются только на искусственно-орошаемых территориях, которых в горах
очень мало. С высоты же 1800—2000 м над ур. м. никаких фруктовых
деревьев вообще не разводится; поэтому расширение площадей под сады
за счет использования огромных богарных территорий по склонам гор
имеет чрезвычайно важное значение, особенно для высокогорных рай-
онов.
Разведение плодовых пород, так же как п лесных, на горных склонах
основано на принципе обогащения почвы влагой путем террасирования
склонов водозадерживающими террасами. Опыт местного лесхоза, пытав-
шегося в ущелье Кондара разводить персик без террас, потерпел неудачу.
Нами разработаны две системы богарных садов на горных склонах —
сплошных и комбинированных (Ф. Л. Запрягаев и В. И. Запрягаева,
1941). Сплошные сады занимают весь отведенный для них участок, так
что для посторонних культур не остается места. В сплошном саду рас-
стояния между террасами и между деревьями в ряду определяются осо-
бенностями крон и корневых систем высаживаемых плодовых деревьев.
Как правило, расстояние между террасами равняется 8—10 м; межтер-
расные пространства не засеиваются никакими другими культурами.
Под такие сады в горах могут быть использованы территории, либо заня-
тые в настоящее время кустарниками, не имеющими никакого хозяй-
ственного значения, либо участки, уже расчищенные от древесно-кустар-
никовой растительности, но почему-либо не использованные под зерно-
вые культуры и находящиеся в состоянии залежей на протяжении мно-
гих лет. Крутизна склонов, отводимых для сплошных садов, не имеет
значения; осваиваться могут склоны с наклоном до 40° Территории,
которые могут осваиваться под сады, располагаются обычно по склонам
разрозненными участками, прерываемыми осыпями и обнажениями скал.
Это не дает возможности говорить о разведении в горах больших мас-
сивов плодовых насаждений.
В комбинированных садах плодовые деревья высаживаются по тер-
расам, располагающимся на большом расстоянии друг от друга, а меж-
террасные пространства засеваются зерновыми и полевыми культурами.
Такой сад исполняет сразу два назначения: дает урожай плодов и спо-
собствует прекращению эрозии на склонах, что имеет громадное значе-
ние для повышения урожайности зерновых культур и сохранения пахот-
ных угодий. В таких комбинированных садах расстояния между тер-
расами лимитируются скоростью поверхностного стока воды и могут
быть доведены, в зависимости от крутизны склона, до 20—30 м. Под
комбинированные сады целесообразно отводить склоны не круче 25°.
Расчеты террас, применявшихся нами в опытах по богарным садам, осно-
ваны на тех же принципах, что п для лесных пород. Межтеррасные посевы
ни в какой степени не будут мешать развитию плодовых деревьев, так
как должна засеваться только часть межтеррасного пространства, уда-
ленная по внешнему откосу террасы на 2 метра от деревьев, т. е. зона,
в пределы которой корневые системы деревьев, высаженных на террасах,
328
В. И. Запрягаева
не заходят (рис. 17). Раскопка корневой системы персика в возрасте
7 лет, посаженного на террасе в богарном саду, показала, что вся масса
его корней уходит вглубь почвы под полотном террасы. Никаких при-
Ненарушенный с/игом
Рис. 17. Корневая система 7-летнего персика, на террасе в ущелье Кондара.
А — вертикальная проекция; Б — горизонтальная проекция.
знаков поверхностного распределения корней, свойственного деревьям
и кустарникам, растущим в естественных условиях на таких же склонах,
здесь нет; повидимому, такое распределение корневых систем у деревьев
на террасах связано с наличием достаточного количества влаги в почве.
Биологические особенности древесных пород
329
Из плодовых пород в нижней полосе пояса древесно-кустарниковой
растительности, в пределах 1000—1600 м над ур. м., как показали наши
опыты, на богарных склонах очень хорошо развиваются: орех, о кото-
ром уже говорилось выше, персик, абрикос, яблоня, груша, сладкий
миндаль, гранат, инжир, виноград, малина. Совсем не удались опыты
с вишней и черешней, а из ягодных — с земляникой. В верхней полосе
пояса древесно-кустарниковой растительности, начиная примерно с 1600—
1700 м над ур. м. и до высоты 1900 м, из плодовых может культивиро-
ваться яблоня и груша, да и то при особом формировании кроны. Все же
остальные перечисленные выше плодовые не выносят суровых условий
высокогорной зимы с ее мощным снеговым покровом.
Для разведения плодовых в горах имеет большое значение экспози-
ция склона и высота по склону от дна ущелья. Более подробно об этом
будет сказано несколько ниже, при описании опытов по культуре отдель-
ных плодовых пород.
Особо важным является вопрос о выращивании посадочного матери-
ала для описываемых условий. Как показал наш опыт, наиболее целе-
сообразно переносить в богарный сад саженцы, выращенные и заокули-
рованные в орошаемых питомниках. Этим, однако, не исключается воз-
можность посева некоторых плодовых, в частности миндаля и персика,
семенами, но сеять нужно тогда сразу на постоянное место, без пере-
садки. Выращиванию посадочного материала в питомниках должно быть
уделено очень большое внимание. Здесь имеет значение не только общее
состояние саженца, который должен быть вполне здоровым, с хорошо
развитой корневой системой, но и то, на чем он привит. Вопрос о под-
боре подвоев для культивируемых на богаре плодовых требует к себе
особого внимания. Подвой должен быть вынослив к почвенной засухе
и к морозам. Необходимо, чтобы между подвоем и привоем была физио-
логическая совместимость; это обеспечит прочное срастание подвоя
с привоем, нормальный рост и плодоношение. Для того, чтобы плодовое
дерево нормально развивалось, подвой должен обладать быстрым ростом
и долговечностью. Во всех случаях надо исходить прежде всего из воз-
можностей использования для подвоев местных дикорастущих плодовых;
это, с одной стороны, уменьшит расходы, а с другой — увеличит
эффективность культур.
Наши опытные работы по богарному садоводству были развернуты
на участках, описанных в предыдущей главе и использовавшихся для
опытных работ с лесными культурами, в связи с чем непосредственно
перехожу к описанию опытов с богарными садами.
Персик (Persica vulgaris Mill.)
В Средней Азии персик чрезвычайно широко распространен на оро-
шаемых территориях. Из-за своего быстрого роста, прекрасных плодов
заслуженно пользуется большой популярностью. Это — теплолюбивая
порода, к почвам крайне нетребовательная. Многие авторы отмечают
засухоустойчивость персика. В. Н. Киселевич (1936), наоборот, харак-
теризует персик как культуру влаголюбивую и предостерегает от исполь-
зования его на богаре, считая, что он может быть использован здесь,
только будучи привит на миндале. Опыт Варзобской ботанической стан-
ции ни в какой степени не подтвердил этого мнения и показал, что персик
очень засухоустойчив и прекрасно развивается на горной богаре на своем
330
В. И. Запрягаева
собственном подвое. О росте и развитии персика на богаре дает пред-
ставление рис. 18. Для решения вопросов не только о возможности куль-
тивирования персика на горной богаре, но также и о подборе сортов
Рис. 18. Персик на террасах в ущелье Кондара.
Вверху— возраст 1 год; внизу— возраст 3 года.
для определенных высот горного пояса, нами было испытано довольно
большое количество сортов персика. На склоны высаживались не только
привитые в питомнике сеянцы, но и дички, которые на 2—3-й год после
посадки, когда они уже были достаточно укоренившимися, подвергались
Биологические особенности древесных пород
331
облагораживанию на месте. Как правило, на богару высаживались
саженцы персика, привитые на собственном подвое. В литературе есть
многочисленные указания, что для персика хорошими подвоями являются
миндаль, абрикос, алыча, терн и другие (Белохонов, Курындин и др.,
1939), но это в большинстве случаев все же не засухоустойчивые расте-
ния; исключение представляет миндаль, который и нами был исполь-
зован как подвой для персиков. Однако персик, привитый на бухарском
миндале, получается маленьким и слабым.
Посадка персика в богарных садах производилась как комбинирован-
ного, так и сплошного типа. Разницы в развитии персика в обоих типах
садов не было никакой. Поэтому в дальнейшем я буду говорить о раз-
витии персика вообще на богаре, не упоминая о том, в каком типе сада
он был посажен. То же относится и ко всем другим испытывавшимся
плодовым. Персик высаживался и чистыми насаждениями, и между
деревьями яблони (как промежуточная культура), с тем чтобы быть уда-
ленным при наступлении старости, которая в поливных условиях, по мне-
нию многих исследователей, наблюдается на 10-м году. Расстояние в ряду
между деревьями допускалось до 4 м, а междурядья доходили до 8—
15—25 м. Опытные посадки персика производились на различных участ-
ках, располагающихся на высотах от 850 до 1850 м над ур. м., но пол-
ные наблюдения за его развитием имеются только по участку, располо-
женному в ущелье Кондара на высоте 1100 м. Опыт культуры персика
в более высоких районах (1850 м) не удался. Здесь персик ежегодно
страдал от снега. Опыт с персиком в нижней части горного пояса (850 м)
уже вне пределов ущелья Кондара был удачным, деревья развивались
в первые годы очень хорошо. Однако через некоторое время они были
уничтожены пастьбой скота. О возможности культивирования персика
на этих высотах можно судить по данным Лесной опытной станции, кото-
рая производила посадку персика на участке Заргар. Здесь персик пре-
красно развивается, дает в год до 50—90 см прироста и приносит еже-
годно урожай плодов. Ниже я излагаю свои наблюдения над развитием
персика, посаженного на высоте 1100 м над ур. м. в ущелье Кондара
(табл. 2).
Таблица 2
Скорость роста персика, посаженного на террасированном склоне в ущелье
Кондара
(Высота — 1100 м над ур. м.; посадка — декабрь 1935 г.; средние данные по сортам: Инжирный
белый, Александр ранний, Роугани-Гоу и Гринсборо)
Г О Д ы
1937 1938 1939 । 1940 I 1942 1 1943 1 1944 1945
Саженцы
Высота средн, (в см) заокули- 102 156 218 341 430 494 494
То же максим. рованы на месте 153 235 300 388 1 500 520 520
Год. приросты средн . 94 64 87 47 i 35 30 26
То же максим. % приживаемости сеян- 139 92 134 116 105 82 77
цев 100 100 100 100 100 100 100
332
В. И. Запрягаева
Так как разницы в росте отдельных сортов персика не наблюдалось,,
то наши данные относятся к сортам: Инжирный белый, Александр ран-
ний, Роугани-гоу и Гринсборо. Совершенно так же развиваются деревья
сортов: Чиляги красный, Мупр, Майский цветок, Ак-Шафтолю, Амсден,
Риверс, Победитель, Эльберта, № 1411 и др. Возраст персика к моменту
окулировки (1937) достигал 3 лет, общий же возраст деревьев — 10 лет.
Деревья к концу 10-го года не несут признаков старости, пх годичные
приросты хотя и уменьшились, но все еще остаются довольно значитель-
ными, достигая 30—82 см. Значительные по величине приросты у персика,
посаженного на террасированном склоне, объясняются не только тем.
что он быстро растет, но еще и тем, что он имеет значительно более удли-
ненный период роста, который, по сравнению, например, с кленом, мин-
далем и каркасом, растущим в естественных условиях, увеличен на 15—
20 дней. В табл. 3 приведены данные по скорости роста персика в тече-
ние вегетационного периода.
Таблица 3
Скорость роста персика Чиляги красный на террасированном склоне в ущелье
Кондара
Возраст деревьев — 3 года. 1941 г.
Даты
наблюдений
Длина
прироста
(в см)
I
0.1
I
5.4 10.2 20.7
36.7 I 51.6 : 70.2
!
81.0 89.0 | 90.8 | 94.8 I 94.8 | 94.8
В данном случае персик окончил свой рост в первых числах августа,
в то время как в естественных условиях древесная растительность закан-
чивает свой рост обычно в начале июля, а начало приостановки роста
падает на 1—2-ю декаду июня.
Как правило, персик-однолетка, посаженный на террасированном
склоне, начинает цвести через год, т. е. не уступает в этом отношении
персику в обычных поливных условиях. Наблюдения за цветением
и плодоношением персиков, высаженных на богаре, показали, что цве-
тение персика происходит дружнее других плодовых пород, напри-
мер сладкого миндаля. Разница в сроках цветения по отдельным сортам
не превышает 2—3 дней. Цветет персик довольно поздно: 7—16 IV —
19 IV—2 V Это дает возможность персику избегать весенних замороз-
ков и, в отличие от сладкого миндаля и абрикоса, плодоносить ежегодно -
Первые плоды персика появляются либо в год начала цветения, либо,
чаще всего, на следующий год. Урожай плодов с наших богарных пер-
сиков был, как правило, не велик. Это, в основном, объясняется отсут-
ствием достаточного количества удобрений, которые требуются для пер-
сика, но которые мы не могли вносить из-за недостатка рабочей силы.
У персика наблюдалось массовое опадайте завязей в раннелетнпй период.
Причина этого явления не выяснена, но^можне-пблагать, что оно свя-
зано не с недостатком влаги в почве, а скорее с недостатком какого-либо
элемента минерального питания.
Биологические особенности древесных пород
333
Урожай с дерева бо-
гарного персика полу-
чается несколько меньший
по сравнению с урожаем
поливного персика, вы-
ращенного в том же
ущелье Кондара. Плоды
тоже несколько мельче
(рис. 19 и 20), но зато
почти нет расхождений в
проценте выхода мякоти.
Как видно из таблицы,
плоды богарных персиков
являются мясистыми и с
небольшой косточкой. Раз-
меры плодов колеблются
в зависимости от погод-
ных условий года, но ко-
лебание это не велико.
Только для некоторых
сортов оно доходит до 32 г
(сорт Роугани-Гоу), ма-
ксимальный вес в 1940 г.—
93 г, а в 1942 г. — 125 г.
Выводы о перспекти-
вах использования этой
плодовой породы при ос-
воении горных богарных
склонов сводятся к сле-
дующему: а) культивиро-
вать персик следует
только на водозадержи-
вающих террасах; б) луч-
шим способом разведения
персика является посадка
однолетними саженцами,
которые подготовляются
в питомнике; в) лучшим
подвоем для персика,
предназначенного для бо-
гары, является персик;
г) подбирать сорта надо
таким образом, чтобы в
ассортименте были пер-
сики раннего, среднего и
позднего сроков созрева-
ния, но основной упор дол-
жен быть сделан на ран-
ние сорта, которые со-
Рис. 19. Персик Инжирный красный. Ущелье Кон-
дара. Богарный сад.
зревают в сроки, когда
в почве еще имеется достаточный запас влаги; персики дают при этих
условиях более сочные плоды; д) за высаженными растениями должен
быть систематический, тщательный уход, сводящийся к внесению
334
В. И. Запрягаева
удобрений (только в осенние сроки, под зимние дожди), рыхлению почвы,
тщательной формировке кроны, борьбе с вредителями; е) персик должен
О
Рис. 20. Персик Майский цветок. Ущелье Кондара. Богарный сад.
высаживаться в богарных садах как промежуточная культура среди
яблонь, груш, ореха с тем, чтобы его можно было впоследствии, при
наступлении старости, удалить.
Биологические особенности древесных пород
335
Сладкий миндаль (Amygdalus communis L.)
Основной областью культуры миндаля является Средиземноморье:
Турция, Иран и пр., в Америке — Калифорния. В СССР площадь
под посадками сладкого миндаля весьма ограничена и сосредоточена
в Средней Азии, Крыму и Закавказье, но ни в одном из районов СССР
культура сладкого миндаля не имеет промышленного значения. В зоне
своего распространения сладкий миндаль культивируют, в основном,
в орошаемых долинах, что отнюдь не является необходимостью для него,
так как, будучи растением весьма засухоустойчивым, он может расти
даже в полупустыне. Очень влажных земель избегает и, как правило,
вытесняется на них другими, более влаголюбивыми породами. Все это
говорит о том, что сладкий миндаль не должен занимать места на полив-
ных землях.
Наши опыты показали, что сладкий миндаль может быть разводим
в горной части Таджикистана. Посаженные на богару однолетние
саженцы сладкого миндаля прекрасно развиваются. Сладкий миндаль
в своем росте не делает таких резких скачков, как персик. Кривая его
роста медленно, но непрерывно поднимается вверх. Годичные приросты
значительно меньше, чем у персика. Разница в росте по отдельным сор-
там настолько незначительна, что ею можно пренебречь.
Сладкий миндаль, высаженный на террасе, прекращает свой рост
значительно раньше персика. У него начало замедления приростов падает
на конец мая, а полное прекращение роста совпадает с концом июня,
но иногда у Алты-арыкского сорта рост продолжается и дальше. Резкий
скачок в росте у миндаля во вторую половину июня объясняется про-
буждением почки, заложенной в этом же месяце.
У миндаля возобновление роста летом, как правило, появляется после
рыхления почвы.
Хотя годичные приросты миндаля по сравнению с персиком и очень
уменьшены, но зато по сравнению с бухарским миндалем (Amygdalus
bucharica Korsh.), произрастающим в естественных условиях, значи-
тельно больше и превышают последние на 10—19 см.
Сладкий миндаль, высаженный на богарный склон, впервые зацве-
тает на 2—3 год после посадки, иногда в этом же году дает первые плоды.
Нам не удалось собирать с миндаля, выращенного на богаре, полного
урожая из-за массового опадания завязей в раннелетний период, что,
повидимому, связано не с недостатком влаги, а с недостатком питатель-
ных веществ в почве (удобрения почвы в миндальном саду не произво-
дились). В дальнейшем, при разработке агротехники для богарного мин-
даля, должен быть разработан вопрос о сохранении урожая.
Спорным является вопрос о выборе подвоя для сладкого миндаля.
Подвой должен быть безусловно очень засухоустойчивым, особенно если
он предназначается для богары. Некоторые исследователи (А. В. Тур-
ский) считают из миндалей более засухоустойчивым Amygdalus communis
L., другие — Amygdalus bucharica Korsh. Нами одновременно высажи-
вались на богаре саженцы сладкого миндаля, привитые на Amygdalus
bucharica и A. communis L. Существенной разницы в росте деревьев
не было, но, как правило, средняя высота дерева была несколько выше
у миндаля, привитого на Amygdalus bucharica Korsh. Например, сорт
Лангедок имел среднюю высоту дерева, будучи привит на Amygdalus
bucharica Korsh., на пятый год — 252 см, а на Amygdalus communis
336
В. И. Запрягаева
L. — 195 см. У сорта Алты-арыкский средняя высота дерева, привитого
на Amygdalus bucharica, на пятый год после посадки равнялась
304 см, а у привитого на Amygdalus communis — 222 см. С точки зре-
ния целесообразности использования того или другого миндаля в каче-
стве подвоя, в данных условиях надо отдать предпочтение Amygdalus
bucharica Korsh., который растет в диком виде в горах в чрезвычайно
большом количестве. Принято считать, что в Таджикистане насчиты-
вается около 400 000 га зарослей бухарского миндаля. Нет смысла
ввозить дорогостоющие семена Amygdalus communis L., в то время как
в горах можно собрать семена Amygdalus bucharica Korsh. в неограни-
ченном количестве.
Спорным является также вопрос о том, как разводить миндаль: выса-
живать ли саженцами, либо высевать сразу на постоянное место. Мне
кажется, что не следует в такой категорической форме, как это делает
В. Н. Киселевич (1936), отрицать посадку как метод разведения миндаля
на склонах гор. Наш опыт показал, что миндаль вполне удовлетвори-
тельно переносит пересадку на богару, не давая, как правило, отпада.
Основным условием, обеспечивающим успех посадки, является хорошо
подготовленная корневая система саженца. Способ разведения миндаля
на богаре посевом очень трудоемок. Семена, высеянные весной, быстро
прорастают. Молодые деревца довольно быстро развиваются, но гото-
выми к окулировке они становятся не раньше чем через 3 года. Как пока-
зал опыт, окулировку посеянного миндаля гораздо лучше производить
не на сеянце, стволик которого бывает очень тонким и ветвистым,
а на поросли, которую можно вызвать, срезая на второй год после посева
надземную часть молодого растения. Образовавшиеся таким путем побеги
более удобны для окулировки и развитие на них молодых культур-
ных побегов начинается через несколько дней после окулировки (при
условии производства окулировки в июне) и протекает очень быстро,
достигая к концу этого же года 20—25 см.
Значительно более серьезным обстоятельством, которое решает благо-
получный исход дела, является умелый подбор сорта сладкого миндаля.
Здесь надо учитывать не только то, что миндаль требует перекрестного
опыления соответствующим сортом, но и то, что не все имеющиеся сорта
могут быть разводимы на горных склонах.
Несколько ниже, в том разделе моей работы, где описывается исполь-
зование бухарского миндаля как подвоя для сладкого мпндаля в лесо-
садах, указан перечень сортов сладкого миндаля, пригодных для раз-
множения в средней части горного пояса.
Размещать посадки сладкого миндаля следует в местах, удаленных
от токов холодного воздуха, особенно от ложа горных ущелий, оставляя
эти места для более поздно цветущих плодовых — персика, яблони, груши
и винограда. В весеннее время пониженные части рельефа наиболее
опасны, так как сюда стекают массы холодного воздуха, и температура
часто опускается значительно ниже нуля, в то время как на склонах
она значительно выше нуля.
Яблоня (Malos pumila Mill.) и груша (Pirns communis L.)
Эти ценные плодовые породы наиболее широко распространены
в Советском Союзе. До 70% площади, занятой под плодовые культуры,
приходится на долю яблони. В Средней Азии яблоня и груша имеют
Биологические особенности древесных пород
337
ограниченное распространение. Особенно редко встречаются культур-
ные сады из яблони и груши в горах Средней Азии, в частности, в Таджи-
кистане. Местное население предпочитает собирать яблоки с дикорасту-
щих яблонь, среди которых имеются деревья, дающие прекрасные плоды.
Однако большинство яблонь дает мелкие, малосъедобные плоды, при-
годные, может быть, только для сушки.
Одной из задач научно-исследовательской работы в области плодо-
водства в Таджикистане является разработка методов культивирования
плодовых в более высоких горных районах. Из них первыми кандидатами
на продвижение вверх являются яблоня и груша как одни из самых моро-
зостойких плодовых культур. В горах Таджикистана дикорастущие
яблоня и груша занимают не только места с повышенным увлажнением
по дну ущелий, но и поднимаются высоко по сухим склонам (Викторов-
ский, 1935; Богушевский, 1935); это дало нам основание использовать
грушу и яблоню в горных богарных садах.
Яблоня и груша высаживались на двух участках по северным мелко-
земистым склонам. Посадка производилась 2-летними саженцами, выра-
щенными и заокулированными в питомниках. Посадка произведена
в 1939 г., но к 1945 г. еще ни один сорт яблони и груши не вступил в пору
плодоношения, поэтому пока несколько преждевременно делать оконча-
тельные выводы; но результаты шестилетних наблюдений говорят, что
яблоня и груша могут быть с успехом разводимы на богарных склонах.
Яблоня и груша близки по характеру роста на богаре.
Характер кривой роста яблони и груши в течение года аналогичен
кривой роста сладкого миндаля, посаженного на террасах, и значительно
отличается от персика, у которого рост продолжается до августа.
Опыт с посадкой яблони и груши на высоте 1850 м обычным способом,
т. е. вертикально стоящими деревьями, не удался; молодые деревца под-
вергались ежегодным поломкам от снега. Тогда был применен опыт
с посадкой растений наклонно, под углом в 45° Этот способ оказался
наиболее применимым, но пользоваться им для выращивания садов нужно
с большой осторожностью, так как поверхность почвы нагревается
до 64°.
При такой высокой температуре у поверхности почвы наблюдается
очень низкая влажность, падающая в некоторых случаях до 9%. Расте-
ние, попадая в такую обстановку, погибает. Посаженное под углом дерево
нельзя летом держать прижатым к земле; его надо приподнимать на 20 см
от поверхности почвы, где температура воздуха значительно ниже, а отно-
сительная влажность заметно выше. Примененный нами метод посадки
садов в этой части горного пояса заимствован у А. Д. Кизюрина (1938),
который применял стелющуюся форму плодовых деревьев в Омской
области, спасая их от зимних ветров и морозов.
Значение выбора подвоя для яблони и груши, предназначенных для
посадки на богаре, очень велико. Подвой должен быть засухоустойчи-
вым. Нет смысла искать новых подвоев и ввозить семена яблони и груши
из дальних мест, когда можно использовать семена местной дикорасту-
щей, хорошо приспособленной к данной обстановке яблони [Malus his-
sarica Kudr.]. Несколько сложнее с подвоями из местных груш.
В Таджикистане встречается, по описанию Г. П. Викторовского (1935)
и М. Г. Попова (1934), несколько видов: Pirus heterophylla Rgl. рас-
пространена на верхнем Зеравшане, в пределах 1000—2000 м над ур. м;
груша Коржинского (Р. Korshinskyi Litw.) распространена в пределах
1200—1700 м, реже — Р. Vavilovii М. Pop. и Р. bucharica Litw. Распро-
22 Ущелье Кондара
338
В. И. Запрягаева
стрднение этих видов, так же как и количество деревьев по отдельным
видам, незначительно. Хотя все эти виды применяются как подвои для
р/азводимых в этих местах груш (Викторовский, 1935; Линчевский, 1940),
они не получили еще достаточного признания в плодоводстве республики.
В условиях горной богары груша, как показала наша работа, очень
хорошо развивается и на подвое айвы.
Из наших опытов можно сделать выводы о том, что яблоню и грушу
разводить в богарных садах описываемого нами района можно в пре-
делах 1000—2000 м над ур. м., с условием формирования в более высо-
ких районах стелющейся формы дерева. При посадке яблони и груши
в нижней части горного пояса можно использовать в качестве проме-
жуточной быстро растущей культуры, персик, который можно убрать
к тому времени, когда наступит плодоношение яблони. Для выращивания
подвоя яблони надо использовать, прежде всего, семена местной яблони.
Инжир (Ficus carica L.)
Инжир — растение сухих субтропиков. Зоной распространения куль-
турного инжира в СССР являются Средняя Азия, Закавказье и Крым.
В Средней Азии инжир встречается, помимо того, и в диком виде.
По Таджикистану инжир в диком виде распространен в южной части
республики — в Кулябской области (Линчевский, 1937) и по Пянджу,
где он, вырастая до 5—6 м в высоту, поднимается до 1400 м (близ селе-
ния Хирманджоу Даштиджумского района) или даже до 1750 м над
ур. м. В центральном Таджикистане инжир в диком виде не встречается.
В горах Таджикистана, как правило, инжир встречается единичными
экземплярами или, реже, маленькими группами деревьев. Инжир засу-
хоустойчив и крайне нетребователен к почве (В. Н. Киселевич, 1936:
Б. С. Розанов, 1940), может произрастать почти на голых скалах. Это
дало основание к испытанию инжира на богарных склонах ущелья Кон-
дара. Помимо того, назрел вопрос о подборе для инжира более благо-
приятных в климатическом отношении мест культуры. До сих пор инжир
культивируется в долинах. Ферганская долина является крупнейшим
центром его культуры, а между тем климатические ее особенности далеко
не отвечают требованиям для инжира. Здесь минимальная температура
опускается до 25°, в то время как мороз —18° вызывает уже обмерзание
инжира, а при температуре 20—22° инжир обмерзает до корневой шейки.
Горные же районы центрального Таджикистана, в частности описывае-
мый район, не подвержены резким колебаниям температур, здесь нет
таких низких минимумов. Необходимо считать, что наиболее подходя-
щим районом для культуры инжира являются горные районы. Самая
низкая температура в ущелье Кондара (за период с 1936 по 1941 г.), отме-
ченная в 1937 г., —15.6°; хотя для инжира это почти критическая тем-
пература, связанная с обмерзанием надземной части, но даже слабое
укрытие может предотвратить гибель. Необходимо, помимо того, отме-
тить, что умелым размещением инжира по рельефу можно предохранить
его от обмерзания. Для инжира должны быть отведены по склонам гор
места, защищенные от холодных токов воздуха. В горах ни в коем слу-
чае нельзя сажать инжир по дну ущелий, куда стекает весь холодный
воздух.
Разница в температуре воздуха (термометры установлены в будке)
между дном ущелья и склоном его на высоте до 100 м от дна ущелья
достигает в некоторых случаях 6°. Особое значение эта разница в тем-
Биологические особенности древесных пород
339
пературах приобретает в период заморозков, когда температура воздуха
по пониженным частям рельефа опускается намного ниже 0° и вызывает
обмерзание растений.
Опыты с посадкой инжира на богарных склонах в соседнем с Кон-
дарой ущелье Джиринодсой показали, что инжир может развиваться
без всякого полива на богарных склонах с водозадерживающими тер-
расами. В отличие от многих других плодовых пород, инжир хорошо
развивается не только на склонах северной экспозиции, но даже на южных
склонах с мелкоземистым почвенным слоем. Инжир, посаженный одно-
летними саженцами в 1941 г., дал уже в 1942 г. первые плоды, которые
по размерам не велики (вес 7—13 г), но высокого вкусового качества.
Небольшие опыты Варзобской станции с инжиром не решают, конечно,
задачи перенесения инжира в горы. Нужна дальнейшая работа по раз-
работке агротехнических мероприятий, среди которых важнейшими
являются вопросы предохранения инжира от вымерзания и повышения
плодородия почвы путем внесения соответствующих удобрений.
Гранат (Punica granatum L.)
Гранат, так же как и инжир, является растением субтропическим.
В СССР гранат в диком виде довольно широко распространен в Закав-
казье и республиках Средней Азии, особенно в Узбекистане. В Таджик-
ской ССР гранатовые насаждения сосредоточены в Кулябском, Шуро-
абадском и Даштиджумском районах (Крейдик, 1940), где он, по данным
Линчевского (1937), поднимается до высоты 2000 м над ур. м., но пло-
щади, занятые гранатом в пределах Таджикистана, крайне незначительны.
Гранат, как и инжир, весьма нетребователен к почве, растет на ска-
листых сухих склонах, лишенных почвы, иногда даже селится на осыпях
щебня (М. Г. Попов, 1939). Гранат дает не только вкусные, питательные
плоды, но и ряд других технических продуктов: в коре ствола, корней
и веток содержится много таннидов, идущих на дубление кож. Плодо-
вые корки используются для окрашивания материй и в медицине.
Состояние граната на богарных склонах вполне удовлетворительно.
Первые цветы граната, посаженного в 1941 г. на богарном склоне одно-
летними саженцами, появились в 1943 г., а первые плоды — в 1944 г.
Наши опыты не дают еще права окончательно решить вопрос о куль
туре граната в описываемом или аналогичных ему горных районах. Необ-
ходима дальнейшая работа по предохранению граната от обмерзания.
Подводя итоги настоящему разделу статьи, можно сказать, что в пре-
делах района Варзобской ботанической станции и аналогичных ему
по физико-географическим условиям садоводство может развиваться
не только на поливных землях долин, но и на богарных склонах гор,
мелиорированных водозадерживающими террасами. Для перенесения
нашего опыта по богарному плодоводству в широкую практику горных
колхозов в доработке нуждаются вопросы агротехники богарного садо
водства.
3. ОПЫТ БОГАРНОГО ВИНОГРАДАРСТВА 1
Вопрос о перенесении культуры винограда с орошаемых земель
на богарные давно назрел. Еще в прошлом веке в Ташкентском и Самар-
1 Работы по настоящему разделу вела техник-плодовод Е. А. Базова, под руко-
водством Ф. Л. Запрягаева.
340
В. И. Запрягаева,
кандском оазисах русские поселенцы делали попытки разведения вино-
града на богаре, но эти попытки оказались неудачными (Киселевич, 1936).
Более удачным был опыт М. А. Тупикова в 1926 г. на Аккавакской стан-
ции; посаженный им богарный виноград развивался вполне удовлетво-
рительно, годовые приросты его достигали 150 см. Успех опыта Тупи-
кова объясняется хорошей подготовкой почвы перед посадкой винограда.
Еще позднее, в 1933 г., виноград высаживался на богаре в Аман-кутане.
Опыт здесь хотя и не совсем удался, но результат получился явно поло-
жительный, свидетельствующий о том, что виноград на горной богаре
развиться может (М. Г. Попов, 1934).
В горах Средней Азии, и в Таджикистане в частности, дикий виноград
Vitis vinifera L. распространен довольно широко. Он поднимается в горы
до высоты 2000 м, занимая, как правило, места по долинам рек или вблизи
водоемов, но иногда он поднимается довольно высоко и по сухим скло-
нам гор. То же самое можно сказать и о культурном винограде. Иногда
в горных кишлаках виноградники не поливаются долгое время, но, не-
смотря на это, дают хороший урожай. В холмистых предгорьях южного
склона Гиссарского хребта существуют виноградники, которые поли-
ваются только раз в год, но их урожай поражает своим обилием.
Варзобская Горная ботаническая станция свои опыты с богарным
виноградом начала в 1941 г. Для этого были выделены три уча-
стка:
1-й участок — южный склон в ущелье Джиринодсой (это соседнее
с Кондарой ущелье, аналогичное ему по характеру растительности).
Участок располагается на старой заброшенной пашне. Крутизна склона
колеблется от 30 до 40° Над участком обнажаются гранитные скалы
с разреженными зарослями Amygdalus bucharica Korsh. С других сторон
к участку прилегают насаждения Amygdalus bucharica Korsh. и Celtis
caucasica Willd. Почва на участке по составу не однородная; в основ-
ном мелкозем с примесью кварца и полевого шпата. При высыхании
сильно цементируется. Подстилающей почву породой является сильно
выветренный гранит.
2-й участок располагается на северном склоне в ущелье Кондара,
на месте старой пашни, которая, по всей вероятности, была заложена
в свое время на месте сведенных кленово-миндальных насаждений, кото-
рые теперь окружают участок. Все климатические, почвенные и расти-
тельные особенности этого участка совершенно такие же, как там, где
проводились опыты с посадками лесных и плодовых пород.
3-й участок располагается по дну ущелья Кондара и, в отличие
от предыдущих, искусственно орошается. Он является контрольным для
богарных. Почвенные условия его очень неблагоприятны (наносный песок
с незначительной примесью мелкозема). Участок несколько удобрен перед
посадкой винограда.
На всех трех участках высаживались однолетние саженцы винограда
различных сортов. На первых двух посадка производилась по террасам,
с расстоянием между кустами 4 м, на последнем участке, в долине
реки, посадка произведена обычным способом — рядами, с тем же самым
расстоянием между кустами.
Каждый сорт винограда высаживался одновременно на трех участках.
Всего в опыте участвовало 12 сортов. На основании пятилетних наблю-
дений можно отметить следующее.
На всех участках приживаемость винограда велика. На богарных
склонах только у некоторых сортов отмерло по 1 экземпляру, все осталь-
Биологические особенности древесных пород
341
ные живы. Годичные побеги очень велики, что указывает на обеспечен-
ность растений влагой.
Виноград гораздо лучше развивается на южном склоне, чем на север-
ном, несмотря на то, что почвенные условия здесь менее благоприятны.
Если взять данные средних приростов, то разница между приростами
на северном и южном склонах по одному и тому же сорту достигает 121 см.
Еще более велика разница по максимальным приростам. Максимальные
приросты на южном склоне больше приростов на северном склоне
на 275 см (сорт Султани), но для некоторых сортов приросты на обоих
склонах примерно одинаковы. Значительно лучше, как и надо было
ожидать, развивается виноград на 3-м участке с частыми поливами. Здесь
длина плетей винограда за один год достигает 250—620 см (Мускат вен-
герский), 212—590 см (Кишмишный белый) и 310—730 см (Султани).
Многие сорта зацвели на,3-й год после посадки, и некоторые дали в этот же
год первые кисти винограда. Наблюдение за плодоношением винограда
так и не удалось полностью провести потому, что каждый год уро-
жай винограда уничтожался медведями, которые спускаются из
ущелья к самым участкам станции и наносят им большой вред, ломая
плодовые деревья и уничтожая урожай не только винограда, но и
персика.
Формирование винограда на участках станции производится по чаше-
видной форме: на кусте оставляется, в зависимости от его силы, от 2 до 4
побегов, которые режутся на 3—4 глазка. Плети винограда подвязы-
ваются на тычины, расположенные вокруг куста.
Небольшой опыт Варзобской станции с богарными посадками вино-
града еще раз подтверждает возможность перенесения культуры вино-
града на богарные горные склоны.
4. ОПЫТ ИСПОЛЬЗОВАНИЯ ДИКОРАСТУЩИХ ДЕРЕВЬЕВ И КУСТАРНИКОВ
В ЛЕСОСАДАХ
Из дикорастущих плодовых или близких к ним, в ущелье Кондара
можно встретить довольно много ореха (Juglans regia L.), горького бу-
харского миндаля (Amygdalus bucharica Korsh.), фисташки (Pistacia
vera L.), яблони (Malus hissarica Kudr.), которая образует большое
разнообразие форм, боярышника (Crataegus pontica С. Koch, С. turke-
stanica A. Pojark.), алычи (Prunus sogdiana Vass.), дикорастущей вишни
[Cerasus machaleb L., Cerasus verrucosa (Franch.) Nevski] и винограда
(Vitis vinifera L.).
За исключением миндаля и ореха, плодовые деревья не образуют
сплошных насаждений, а встречаются единичными экземплярами в наса-
ждениях других древесных пород или образуют, как, например, яблоня,
небольшие рощицы.
Коллективом сотрудников Варзобской горной ботанической станции
под руководством автора были поставлены опыты по использованию
дикорастущих плодовых как подвоев для прививки на них культурных
сортов плодовых, таким образом на месте диких лесов путем облаго-
раживания деревьев создавались весьма своеобразные сады, которые мы
называем лесосадами.
В литературе понятие «лесосад» применяется различно. Примерно
Другие же — В. П. Дробов (1936), Н. Я. Череватенко (1936) и А. Ф. Рад-
342
В. И. Запрягаева
зиевский (1934) — называют лесосадами дикорастущие заросли плодовых.
М. Г. Попов (1934) называет лесосадами посаженные плодовые деревья
на неорошаемых территориях, за которыми проводятся упрощенные
меры ухода. М. П. Сеславин (1936) определяет лесосад, как «такой хозяй-
ственный комплекс, в котором имеются все характерные признаки леса,
но в то же время основной продукцией является не древесина, а плоды».
Совершенно особое понятие в термин «лесосад» вкладывает В. Н. Киселе-
вич (1936). Он говорит: «под именем лесосада понимаем такое насаждение
естественного, искусственного или смешанного происхождения, кото-
рое включает в свой состав систематически или периодически эксплоати-
руемые плодовые и технические древесно-кустарниковые породы с при-
месью, в качестве вспомогательных, пород лесного характера».
Идея облагораживания дикорастущих плодовых не нова. П. П. Гусев
(1938) и Н. Я. Череватенко описывают опыт облагораживания дикорасту-
щих плодовых в лесах Кавказа, где в настоящее время сохранились
столетние деревья яблони, когда-то привитые на дикорастущих яблонях
черкесами. Эти деревья приносят до сих пор богатый урожай, который
достигает 7.5—14 центнеров с дерева. Следуя примеру черкесов, рус-
ские переселенцы в окрестностях своих селений также проводили облаго-
раживание дикорастущих. Необходимо отметить большого энтузиаста
лесосадов на Черноморском побережье (Геленджикский район) Н. Я.
Череватенко, который, начав свои опыты еще в 1904 г., проделал очень
большую работу и опубликовал несколько ценных по своим практиче-
ским указаниям статей. С 1931 г. вопросами облагораживания дикорасту-
щих начинает заниматься Майкопская опытная станция в Краснодарском
крае.
Возможно, что в Средней Азии история облагораживания дикорасту-
щих плодовых насчитывает не меньшее количество лет, но, к сожалению,
исследований в этой области не производилось. Интересные наблюдения
были сделаны И. А. Линчевским (1940) в 1935 г. Он обнаружил на южной
оконечности Дарвазского хребта в Даштиджумском и Шуроабадском райо-
нах Таджикистана грушевые лесосады, которые, по рассказам дехкан,
были созданы на месте сведенной древесно-кустарниковой растительности.
Произрастающая здесь в диком виде груша (Pirus Korshinskyi Litw.) была
затем дехканами облагорожена. С 1934 г. вопросами организации лесо-
садов начинают интересоваться в различных пунктах Средней Азин.
Известно, что были сделаны попытки облагораживания дикорастущих
в Аман-кутане. В 1936 г. начаты работы по этой же линии Всесоюзным
Институтом сухих субтропиков на Арсламбобском опорном пункте и,
наконец, к этому же периоду относятся наши опыты. В районах, при-
легающих к нашим опытным участкам, среди дехкан облагораживание
дикорастущих не практиковалось.
Наши работы по использованию дикорастущих плодовых для созда-
ния лесосадов явились воплощением в жизнь идеи покойного Владимира
Леонтьевича Комарова, появившейся у него во время путешествия
по Зеравшанскому хребту.
Объектами работ были все указанные выше дикорастущие плодовые,
за исключением ореха и фисташки. Кроме того, использовался как под-
вой для культурных сортов плодовых кустарник иргай (Cotoneaster his-
sarica Pojark.), который не относится к плодовым, но в систематическом
отношении близок к некоторым из них. Ниже излагаются результаты
работ с облагораживанием отдельных видов дикорастущих плодовых
ущелья Кондара.
Биологические особенности древесных пород
343
Бухарский миндаль (Amygdalus bucharica Korsh.)
В естественном виде бухарский миндаль находит лишь небольшое
применение в хозяйстве. Это, прежде всего, источник хорошей, плотной
древесины, идущей у дехкан исключительно на дрова. Орех бухарского
миндаля находит очень ограниченный спрос в кондитерской промышлен-
ности и в медицине. В естественном виде он почти не съедобен и даже
ядовит из-за наличия в ядре синильной кислоты. Но, несмотря на это, дех-
кане собирают орехи миндаля и используют их, правда, в весьма незна-
чительных количествах, для приготовления съедобных блюд. В горах
Каратага А. И. Толмачевым был найден бухарский миндаль со сладкими
плодами, который из-за своего ограниченного распространения мало
известен даже местному населению.
Наиболее рационально бухарский миндаль можно использовать как
подвой для культурных сортов сладкого миндаля, персика, сливы и абри-
коса. Нам удалось разработать простую методику облагораживания
бухарского миндаля в местах его естественного произрастания. В общих
чертах она сводится к следующему: выбрав участок под будущий лесо-
сад, поздней осенью приступают к подготовке деревьев. Для этого сре-
зается вся надземная часть дерева, на высоте 10—15 см от корневой
шейки. Весной следующего года на пне миндаля образуется целая щетка
порослевых побегов. Периодическим уничтожением части из них и оста-
влением наиболее развитых мы достигаем того, что они к июню стано-
вятся готовыми для окулировки. Из испытанных нами способов облаго-
раживания лучшими оказалась окулировка глазком в Т-образный над-
рез коры молодого порослевого побега. Этот способ наиболее прост
по выполнению и не требует особых материалов, которые нужны для
прививки. Помимо того, при прививке инкрустацией, которую мы также
применяли для облагораживания бухарского миндаля, наблюдается боль-
шой процент отмирающих растений за счет своеобразного повреждения,
которое получается как бы от ожога коры. Совершенно такое же явление
наблюдалось при прививке дикорастущих в Арсламбобском опорном
пункте Всесоюзного Института сухих субтропиков, где проводил свою
работу С. М. Момот (1940). Явления такого ожога С. М. Момот объясняет
отмиранием ткани на обрезанном стволе, которое происходит вследствие
прекращения нормального тока воды и других питательных веществ
в данном месте.
В литературе встречается довольно много указаний по методике обла-
гораживания дикорастущих, но все авторы, как правило, рекомендуют
производить окулировку или прививку либо в высокий штамб, либо
в крону облагораживаемого дерева. Для бухарского миндаля оба метода
абсолютно неприемлемы, потому что он периодически меняет свою крону,
и если не учитывать этого обстоятельства, то можно встретиться с таким
фактом, что привитое в высокий штамб Amygdalus bucharica Korsh.
дерево персика или сладкого миндаля может оказаться недолговечным.
Это подтверждалось и нашей практикой. Один и тот же сорт персика
прививался на двух деревьях бухарского миндаля. В одном случае окули-
ровка производилась в порослевые побеги, отходящие от самой корне-
вой шейки, а в другом — на побеги, отходящие на высоте 100 см от осно-
вания ствола миндаля. Деревья персика развивались по-разному. В пер-
вом случае в течение 7 лет дерево сохранило вид нормального куста, без
признаков старости, в то время как у дерева персика, развивающегося
344
В. И. Запрягаева
на высоком штамбе, признаки старости появились уже на 3-й год. На 4-й
год плодоношение прекратилось, а на 8-й год дерево имело жалкий вид,
с несколькими листочками на концах прошлогодних побегов.
В том случае, когда окулировка сделана в мае—июне, развитие куль-
турных побегов начинается в этом же году, когда же срок окулировки
более поздний, например в июле—августе, пробуждение глазка начи-
нается на следующий год.
Рис. 21. Сладкий миндаль Большой сул-
тан европейский, привит на Amygdalus
bucharica Korsh. Ущелье Кондара.
(Фот. 3. С. Калединой).
Уход за развивающимися побе-
гами довольно прост и совсем не
отличается от обычных мероприя-
тий по уходу за окулированными
растениями. Здесь только особое
внимание уделяется быстрорасту-
щим культурным побегам, которые
легко обломаются, если не будут
своевременно подвязаны. В тече-
ние первых двух лет деревья пер-
сика, сладкого миндаля, сливы и
абрикоса очень интенсивно рас-
тут, но затем годичные приросты
резко сокращаются, дерево оста-
навливает свой рост в высоту,
оставаясь во все последующие
годы своеобразным карликом,
высотой до 1.5—2 метров. Эта
своеобразная карликовая форма
дерева характерна для всех пло-
довых, привитых на бухарском
миндале (рис. 21) в условиях су-
хих склонов.
Нижеприводимые данные яв-
ляются результатами большого
количества опытов с облагоражи-
ванием бухарского миндаля. Все-
го на Amygdalus bucharica Korsh.
нами было привито 49 сортов
сладкого миндаля, 39 сортов пер-
сика, 26 сортов сливы, 27 сортов абрикоса, 8 сортов черешни и 5 сортов
вишни. Заокулировано различными сортами плодовых 10 563 побега
бухарского миндаля.
Сладкий миндаль
Лучшим методом облагораживания бухарского миндаля сладким мин-
далем является окулировка либо растущим глазком, окулированным
как можно раньше в мае—июне, либо спящим глазком, окулированным
в июле—августе. При окулировке растущим глазком уже в первый год
молодые культурные побеги вырастают до 90 см. При окулировке спя-
щим глазком побеги достигают таких же размеров или несколько боль-
ших на второй год. Уменьшение приростов наблюдается на третий год,
когда растение начинает формировать большое количество укороченных
побегов с цветочными почками.
Биологические особенности древесных пород
345
Предельной высоты сладкий миндаль, привитый на Amygdalus bucha-
rica Korsh., достигает, в условиях сухих склонов, на 2—3 год после оку-
лировки, оставаясь не-
большим кустиком вы-
сотой в 1.5—2.5 м
(рис. 22). Если бухар-
ский миндаль произра-
стает в условиях доста-
точного увлажнения, то
сладкий миндаль, при-
витый на нем, выраста-
ет в нормальное дерево
высотой до 5—7 м. Эти
явления дают основа-
ние карликовый рост
плодовых, привитых на
Amygdalus bucharica
Korsh., отнести исклю-
чительно за счет недо-
статка влаги в почве.
Первое цветение у
сладкого миндаля, при-
витого на бухарском
миндале, начи нается,
как правило, на 3-й год
после окулировки; пло-
доношение — либо в год
начала цветения, либо
на следующий.
Сладкий миндаль —
одно из плодовых де-
ревьев, которое подвер-
жено губительному дей-
ствию весенних замо-
розков. Запоздалые ве-
сенние заморозки — ча-
стое явление в горах,
особенно по дну уще-
лий, куда стекают хо-
лодные массы воздуха.
Поэтому при организа-
ции лесосадов подбор
сортов сладкого минда-
ля имеет исключитель-
ное значение. Из испы-
танных нами сортов
сладкого миндаля, у
которых период цвете-
ния проходит позднее
Рис. 22. Сладкий миндаль Перлес, привит на Ainyg-
dalus bucharica Korsh. (Ориг.).
весенних заморозков, можно рекомендовать совсем незначительное ко-
личество.
За исключением одного сорта сладкого миндаля (Иранский 1375),
у цсех сортов период цветения наступает значительно позднее, чем
346
В. И. Запрягаева
у бухарского миндаля, на корнях которого воспитываются эти сорта.
Различные сорта сладкого миндаля имеют различные сроки цветения
иногда интервал во времени между началом цветения у отдельных
сортов достигает 30 дней, что дает возможность подбирать сорта
для того или другого микрорайона. Характерно, что, чем холоднее
весна, чем больше колебаний в температурах воздуха (например в
1940 г.), тем более растянут период цветения сладкого миндаля и, наобо-
рот, чем теплее и дружнее весна, тем меньше расхождения в сро-
ках начала цветения у различных сортов сладкого миндаля (1941).
В наших условиях все испытанные нами сорта сладкого минда-
ля можно разбить на 2 группы. К первой группе относятся такие,
у которых период цветения попадает в полосу весенних заморозков.
Сюда, как правило, попадают сорта местного происхождения и сорта,
вывезенные из Ирана. Ко второй группе относятся сорта с поздним
сроком цветения, уходящим целиком или частично из полосы весенних
заморозков; это большей частью сорта западноевропейские и амери-
канские.
Принято считать, что мороз в —4° уничтожает цветы сладкого миндаля.
Если посмотреть на рис.23,на котором изображен период цветения различ-
ных сортов сладкого миндаля, привитых на Amygdalus bucharica Korsh.
в ущелье Кондара, то станет очевидным, что все те сорта сладкого миндаля,
которые зацвели до 15 марта 1940 г., в этом году не плодоносили; сюда
относятся: Иранский 1375, Алты-арыкский, Мягкоскорлупный № 138,
Иранский 1365; в то же время сорта Нек-плюс-ультра, Перлес, Нон-
парейль и другие, зацветшие после последнего заморозка, сохранили
плодоношение.
Из изложенного видно, что для наших условий приходится, вопреки
агроуказаниям, составленным Всесоюзным Научно-исследовательским
институтом сухих субтропиков (1939), рекомендовать для размноже-
ния такие сорта, как Большой султан европейский, Дрейк-никитский,
Лангедок, Нонпарейль, Перлес, Нек-плюс-ультра, Сары-бодоми. Все-
союзный Институт сухих субтропиков признает их как временно допу-
стимые, основными же считает местные сорта.
В таблице (см. рис. 23) помещены результаты наблюдений за цвете-
нием сладкого миндаля на склоне южной экспозиции. На северных
склонах цветение у различных сортов миндаля запаздывает на 3—6,
а иногда и 10 дней.
Плодоношение сладкого миндаля, привитого на Amygdalus bucharica
Korsh., начинается на 3—4-й год. О том, как велик может быть урожай
плодов с дерева, сейчас трудно сказать. Безусловно, это будет зависеть
от ухода за деревом и, главным образом, от количества вносимых удобре-
ний. Возможно, что недостаток питательных веществ вызывает массовое
опадание завязей у сладкого миндаля, привитого на Amygdalus bucharica
Korsh. Иногда опадание завязей достигает катастрофических размеров.
Например на дереве сорта Большой султан европейский 13 IV 1944
было 829 завязей, 3 V их осталось всего 64 шт. На другом дереве этого
же сорта 19 IV было 2623 завязи, 3 V—498, а 20 V — только 389 шт.
Дерево Нонпарейль 13 IV 1944 имело 1818 завязей, 3 V — 283. Дерево
сорта Нек-плюс-ультра 13 IV 1944 имело 6831 завязь, a3V — только
431 шт. Таких примеров можно привести много; эти факты заставляют
обратить на них серьезное внимание.
На рис. 24 изображены орехи бухарского миндаля и различных сор-
тов сладкого миндаля, привитых на Amygdalus bucharica Korsh. Размер
Биологические особенности древесных пород
347
орехов в наших опытах мало отличается от принятых стандартов. Напри-
мер сорт Нонпарейль, привитый на бухарском миндале, дает средний
вес ореха 1.3—1.4 г, иногда вес доходит до 2.5 г, с выходом ядра
до 55%.
Рис. 23. Период цветения привитых в ущелье Кондара различных сортов сладкого миндаля
По данным Киселе-
вича (1936), орех минда-
ля этого же сорта, вы-
ращенного в нормаль-
ных условиях, т. е. на
поливе и на собственном
подвое, весит в среднем
1.93 г, с выходом ядра
в 66.6%. В орехах мин-
даля этого сорта, при-
витого на бухарском
миндале, жира содер-
жится 52.25%; по дан-
ным А. А. Рихтера (1938),
в орехах сорта Нонпа-
рейль, выращенного в
обычных условиях, —
58%. Сорт Нек-пл юс-
ультра дал в наших опы-
тах средний вес ореха
1.9—2.0 г, с выходом
ядра 45.0—60.7%. По
данным Киселевича, у
этого сорта, выращенно-
го в нормальных усло-
виях, средний вес ореха
2.6 г, с выходом ядра
56%. В орехах сорта
Нек-плюс-ультра, со-
бранных с бухарского
миндаля, содержится
жира 54.03%, а в оре-
хах этого же сорта, со-
бранных в нормальных
условиях, по данным
А. А. Рихтера, — 57%.
Следует отметить,что
установить какую-либо
закономерность в раз-
мере орехов сладкого
миндаля и количестве
жира в ядре, в зависи-
мости от экологических
условий района Кон-
дары, не удалось. У од-
ного и того же сорта сладкого миндаля, привитого на бухарском миндале,
вес орехов примерно один и тот же, независимо оттого, где рос бухарский
миндаль: на богарных ли склонах южной или северной экспозиции, или
в условиях орошения по долине реки. Процент жира в ядре у одних сор-
348
В. И. Запрягаева
тов значительно выше в условиях полива (Нек-плюс-ультра) растений,
у других (Сары-Бодоми) выше при произрастании на сухих южных скло-
нах. В этом отношении сладкий миндаль, привитый на бухарском мин-
дале, отличается от персика, у которого разница в размере плодов
и в их вкусовых качествах довольно значительна и зависит от места про-
израстания деревьев. Поэтому при организации лесосада из сладкого
миндаля можно рекомендовать использовать дикорастущий бухарский
Рис. 24. Плоды
1 — орех Amygdalus bucharica Korsch.; 2— орехи сладкого миндаля Нек-плюс-
ультра, привитого на Amygdalus bucharica Korsh.; з—Большой султан европей-
ский; 4—Лангедок.
минадаль по склонам южной или близкой к ней экспозиции, где сладки)!
миндаль может развиваться вполне нормально, а для более требователь-
ного персика отводить места с более благоприятными почвенными усло-
виями.
Персик
Развитие персика, привитого на бухарском миндале, протекает при-
мерно так же, как и развитие сладкого миндаля. Если окулировка сде-
лана в конце мая или июне, развитие культурного побега начинается
в этом же году, если в более цоздний срок (июль—август), развитие побе-
Биологические особенности древесных пород
349
тов начинается ранней весной следующего года. В первом случае высота
побегов уже в первый год достигает 24—112 см, во втором случае такой
же или несколько большей высоты побег достигает к концу следующего
лета. Буйное развитие молодых побегов прекращается на 3—4 год, что
ювязано, как у сладкого миндаля, с формированием плодовых почек.
Несмотря на то, что персик в последующие годы продолжает образо-
вывать довольно значительные по размерам побеги, его рост в высоту
прекращается. Персик достигает своей предельной высоты в 2—2.5 м
на 2—3-й год после окулировки.
Цветение персика, привитого на бухарском миндале, начинается,
как правило, на 3-й год после окулировки, но иногда и на 2-й год. Плодо-
Рис 25. Персик Александр ранний, привит на Amygdalus bucharica Korsh.
ношение наступает на 3—4-й год. Урожай плодов с дерева небольшой
п достигает в среднем 2.5—3.5 кг, но иногда доходит до 12 кг (рис. 25).
Надо отметить, что почва под облагороженными деревьями бухарского
миндаля в наших опытах не удобрялась на всем протяжении опыта.
Можно полагать, что систематически удобряемые участки безусловно
дадут значительно больший урожай. Недостатком питательных веществ
в почве, повидимому, можно объяснить и массовое опадение завязей
у персика, привитого на бухарском миндале. Например на дереве сорта
Александр ранний имелось завязей 25 IV — 124 шт., а к 3 V — только
34 шт. Дерево сорта Роугани-гоу 25 IV имело 812 завязей, к 3 V их
осталось 286 шт., а к 20 V — только 161 шт. Опадающие завязи не имеют
каких-либо видимых повреждений. Это явление, отмеченное также
у сладкого миндаля, привитого на Amygdalus bucharica Korsh., безу-
словно должно быть устранено в дальнейшем.
В том случае, если Amygdalus bucharica Korsh. произрастает в более
нормальных в отношении полива условиях, размер плодов несколько
повышается, причем средний вес плода часто остается таким же, как
350
В. И. Запрягаева
у дерева, растущего на сухом склоне, но отдельные плоды по своему
весу значительно превосходят максимальный вес плодов с сухого склона;
0 1 2 3 4 5 6 см
1____1 1 к 1----1 Л
Рис. 26. Контур плода персика Роу-
гани-Гоу, привитого на Amygdalus
bucharica Korsh.
иногда это расхождение достигает
20—80 г. Если сравнить получаемые
нами плоды персика, привитого на
бухарском миндале, произрастающе-
го на сухом каменистом склоне, с
плодами персика, выращенного в
обычных условиях, т. е. привитого
на собственном подвое, в условиях
нормального водоснабжения, то раз-
ница более ощутима. Например
сорт Роугани-Гоу в нормальных ус-
ловиях имеет средний вес плода
193 г, в то время как на южном
склоне на подвое бухарского мин-
даля он весит только 75—126 г
(рис. 26). У сорта Амсден в нормаль-
ных условиях средний вес плода
53 г, а у привитого на бухарском
миндале 33—40 г. Сорт Александр
ранний в нормальных условиях
имеет средний вес плода 70 г, мак-
симальный вес 101 г, а на бухарском миндале на южном склоне средний
вес плода привитого персика 43—68 г, максимальный — 88 г.
Слива
Облагораживанпе бухарского миндаля сливой, по сравнению с пре-
дыдущими привоями, сложнее, так как процент приживающихся после
окулировки растений значительно ниже, и сам период окулировки значи-
Рис. 27. Слива Итальянская венгерка, привита на Amygdalus bucharica Korsh.
тельно короче (май—середина июля). В отличие от миндаля и персика,,
у сливы, привитой на бухарском миндале, развитие побегов, даже при
ранних сроках окулировки, начинается, как правило, на следующий год»
Биологические особенности древесных пород
351
Годичные приросты сливы, хотя и резко сокращаются на 3-й—4-й год
после окулировки, но все же остаются довольно большими. Высота
дерева сливы, привитой на бухарском миндале, остается всегда около
2—2.5 м независимо от того, в каких условиях увлажнения растет бухар-
ский миндаль.
Цветение сливы наступает значительно позднее,чем у других плодовых,
привитых на бухарском миндале. Плодоношение иногда наступает на 4-й
год окулировки, а иногда даже на 8-й год. Урожай сливы не велик
и достигает с одного дерева на южном склоне 2—7 кг (рис. 27). Возможно,
что при внесении удобрений в почву урожай будет значительно выше.
Качество плодов вполне
удовлетворительное, пло-
ды получаются мясистыми
и сочными (рис. 28).
Абрикос
Нами были сделаны
многочисленные попытки
использовать бухарский
миндаль как подвой для аб-
рикоса, но результаты в
общем получены плохие.
Абрикос дает очень пло-
хое срастание с бухар-
ским миндалем. Окулиро-
вка, сделанная даже в са-
Рис. 28. Контуры плодов сливы, привитой на
Amygdalus bucharica Korsh.
МЫе ранние сроки, часто j—яичная желтая; 2—итальянская венгерка,
не удается, число непри-
нявшихся растений доходит до 100; иногда же абрикос приживается,
и в этих случаях происходит очень быстрый рост культурного побега,
который на 2-й год после окулировки достигает 74—119 см. На
3-й год годичные приросты все еще велики, но затем наступает их
уменьшение. Предельной высоты абрикос, привитый на бухарском мин-
дале, достигает на 3-й год после окулировки. Абрикос на бухарском мин-
дале недолговечен. Несмотря на то, что развитие культурных побегов
происходит как будто нормально, очень часто на 3—4—5-й год происхо-
дит обламывание на месте прививки. Цветение наступает на 3-й год после
окулировки. Плодоношение абрикоса на Amygdalus bucharica Korsh. —
редкое явление, потому что ежегодно наблюдается массовое опадание
цветов и завязей.
Нами были поставлены опыты по использованию персика как проме-
жуточной культуры между бухарским миндалем и абрикосом, но абри-
кос, привитый на персике, который в свою очередь был привит на Amygda-
lus bucharica Korsh., развивался чрезвычайно слабо и на протяжении ряда
лет не вступил в пору плодоношения.
Вишня и черешня
Испытывались как привои на Amygdalus bucharica Korsh., но почти
во всех случаях опыты дали резко отрицательный результат. Вишня
и черешня не срастаются с Amygdalus bucharica Korsh. Только однажды
352
В. И. Запрягаева
черешня, привитая на нем, прижилась, образовала на следующий год
побег высотой до 60 см, но в дальнейшем не развивалась. В результате
плохого срастания с миндалем побег обломился на 3-й год.
В итоге наших исследований можно считать бухарский миндаль
вполне пригодным для облагораживания сладким миндалем, персиком
и сливой.
Иргай (Cotoneaster hissarica Pojark.)
Иргай относительно близок в систематическом отношении к яблоне,
груше и айве, что дало основание использовать его как подвой для этих
плодовых (Запрягаева, 1944).
Опыты с облагораживанием иргая заложены в нижней полосе пояса
древесно-кустарниковой растительности на высоте 1100 м. На иргае
было привито различными способами очень большое количество сортов
яблони, груши и айвы. Прививка производилась либо в многолетние
побеги у корневой шейки, либо в однолетние порослевые побеги, полу-
ченные после срезания всей надземной части куста. В отличие от других
испытываемых нами подвоев, иргай очень плохо срастается с яблоней
и грушей, иногда процент отпада после окулировки достигает 60—80.
Значительно лучше развивается на иргае айва.
Для яблони и груши, привитых на иргае, характерно то, что они до-
стигают предельной высоты в течение первых 3 лет. На 4-й год после
окулировки прирост в высоту почти приостановился. Растение остается
в последующее время небольшим деревцом высотой около 2 м. Стимули-
ровать рост в высоту яблони или груши, привитых на иргае, не удалось.
Яблоня, привитая на иргае, затем переведенная на свои собственные
корни, стала очень интенсивно расти в высоту, достигая за один год
увеличения на 80 см. Этот опыт показал, что карликовый рост плодовых,
привитых на иргае, объясняется исключительно влиянием подвоя.
Отрицательным моментом в использовании иргая как подвоя для
яблони и груши является плохое срастание подвоя с привоем. Одним
из возможных путей устранения этого является использование айвы
как промежуточной культуры между иргаем и грушей. Айва, как пока-
зал наш опыт, очень хорошо срастается с иргаем.
По данным И. В. Мичурина (1936, 1938), некоторые культурные сорта
груши, будучи привитыми на айве, прекрасно развиваются и обладают
значительно более слабым ростом, чем обычно, что в данном случае может
считаться одной из положительных черт этого привоя для иргая.
Цветение яблони и груши, привитых на иргае, начинается на 3—
4-й год. У многих сортов в этот же период начинается и плодоношение.
Общий урожай плодов у яблони и груши, привитых на иргае, колеблется
от 2—3 до 16 кг с дерева (рис. 29). Урожайность в основном зависит от
места произрастания иргая; на мелкоземистой почве урожай с дерева
выше, на скалах и осыпях урожай мал, и плоды получаются меньше.
Контуры плодов яблони Ранет шампанский (рис. 30), полученных
с деревьев, произрастающих в различных условиях, дают представление
о колебании в размере плодов.
Размер плодов яблони колеблется по годам. Максимальный вес
полученного плода яблони Ранет шампанский доходил до 186.9 г. То же
самое можно отметить и в отношении груши, у которой наблюдались
большие колебания в размере плодов не только в связи с различными
условиями произрастания иргая, на котором привита груша, но и в за-
Биологические особенности древесных пород
353
висимости от климатических условий того или другого года. У груши
Лесная красавица, привитой на иргае, средний вес плодов 179.7 г, макси-
Рис. 29. Плодоносящая ветка яблони Ранет шампанский, привитой на
иргае. Кондара.
(Фот. Ф. Л. Запрягаева).
7
2
О 1 2 3 « 5 6 7 8 см
----1-----1----1-----1-----1-----।_____1____i
Рис. 30. Яблоня Ранет шампанский, привита на иргае:
1— с дерева, растущего на осыпи у воды; 2— с дерева, растущего в скалах.
мальный вес — 185.2 г (рис. 31).У груши Фердинанда, привитой на иргае,
плоды достигали значительно больших размеров*. средний вес плода —
23 Ущелье Кондара
354
В. И. Запрягаева
283 г, а максимальный доходил до 345 г, но общий вес плодов груши
с дерева в наших опытах был не велик и колебался в пределах 2—3 кг.
Исходя из нашего первого опыта, сейчас трудно сделать какие-либо
окончательные выводы о возможности разведения в Таджикистане лесо-
садов из яблони и груши на иргае, но совершенно бесспорным является
то, что иргай заслуживает весьма серьезного внимания. Нужно дорабо-
тать методику его облагораживания. Применение иргая в садоводстве
Таджикистана имеет большие перспективы. Помимо того, что его можно
использовать в естественных условиях как базу для организации ябло-
невых и грушевых лесосадов, он может быть с успехом использован
как источник получения огромного количества семян для нового карли-
кового плодоводства семечковых. Наш опыт с окулировкой однолетних
О 12 3 4 5 6/ 8 см
растений иргая в питомнике яблоней
и грушей дал более положительные
результаты приживаемости после
окулировки, чем в естественных
условиях. В настоящее время про-
должаются наблюдения за ябло-
нями, привитыми на иргае в питом-
нике. В 1946 г. они, будучи 6-лет-
ними деревцами, высотой в 100—
150 см, цвели, но плодов еще не
дали.
Боярышник (Crataegus turkestanica
A. Pojark.)
Указания на использование этого
вида боярышника как подвоя для
яблони и груши имеются в работе
С. М. Момот (1940). В опыте Мо-
мот не получалось хорошего сраста-
Рис. 31. Груша Лесная красавица, ния подвоя с привоем, отчего рост
привита на иргае. привоев получался очень слабым.
Но, несмотря на это, как отмечает
автор, яблоня, привитая на боярышнике, начала плодоносить уже
на 2-й год.
Примерно так же вела себя яблоня, привитая на Crataegus turkesta-
nica A. Pojark., и в наших опытах. При всех испытанных нами способах
облагораживания яблоня приживается на боярышнике только в единич-
ных случаях. Несколько лучше срастается с боярышником груша. При
прививке в штамб дерева боярышника яблоня и груша ведут себя при-
мерно одинаково. Они в первый же год дают прирост культурного по-
бега до 1 м. При окулировке в порослевые побеги таких больших годич-
ных приростов не наблюдается. Рост в высоту яблони и груши, привитых
на боярышнике, в нашем опыте прекратился на 3-й год. Деревья, всту-
пившие в пору плодоношения, имеют вид небольшого деревца, высотой
в 2—2.5 м. Во всех наших опытах прочного срастания яблони и груши
с боярышником не было. Обламывание на месте прививки происходит
на 2—3-й год. Как исключение в опыте сохранились 7—8-летние деревья.
Яблоня и груша, привитые на боярышнике, начинают цвести на 3—
4-й год. Плодов у яблони не удалось собрать ни разу, так как завязи
Биологические особенности древесных пород
355
осыпались вскоре после цветения, у груши же плодоношение начинается
поздно, на 7—8-й год. Урожай с дерева груши высотой в 2 м достигает
2.5 кг. Плоды груши получаются вполне нормальными, например, сред-
ний вес плода груши Оливье-де-Серр — 163, а максимальный — до 235 г.
Опыт Варзобской станции с облагораживанием боярышника подтверждает
данные Момота и говорит о возможности использования боярки при
создании лесосадов. Для Таджикистана, где боярышник довольно ши-
роко распространен, это имеет немалое значение; очередной задачей
является создание способов облагораживания боярышника, при кото-
рых было бы достигнуто прочное срастание подвоя с привоем.
Боярышник-дулона (Crataegus pontica С. Koch)
Плоды этого вида боярышника используются местным населением
в пищу. Нас интересовал вопрос об использовании этого вида боярыш-
ника в лесосадах как подвоя для яблони и, особенно, для груши.
Облагораживание производилось окулировкой в порослевые побеги
и прививкой в высокий штамб дерева. Одновременно с опытом в естествен-
ных условиях производился посев семян понтийского боярышника
в питомнике с последующим облагораживанием молодых сеянцев. В есте-
ственных условиях опыт облагораживания боярышника не удался;
ни яблоня, ни груша, привитые на боярышнике, нормально не разви-
ваются. Помимо того, Crataegus pontica С. Koch совершенно не перено-
сит удаления надземной части дерева, засыхая на 2-й год после обреза-
ния кроны.
Иная картина получилась в питомнике. Здесь яблоня и груша довольно
хорошо приживаются после окулировки; процент прижившихся расте-
ний достигает 90—95. На 2-й год после окулировки высота заокули-
рованных в питомнике растений достигает 75—79 см. На 3-й год рост
в высоту замедляется, высота растений увеличивается всего на 25—30 см.
В последующие годы яблоня и груша остаются очень маленькими, сла-
быми деревцами, высотой в 1.5—2 м. Цветения и плодоношения у при-
витых шестилетних деревьев еще не наблюдалось.
Яблоня (Malus hissarica Kudr.)
Об использовании дикорастущих яблонь в лесосадах имеется обшир-
ная литература. Методика облагораживания яблони довольно проста,
прививка производится с одинаково хорошим успехом и в порослевые
побеги и в штамб дерева.
Нами были заложены опыты по облагораживанию яблони, произрас-
тающей в различных экологических условиях. Получился вполне удо-
влетворительный результат, который говорит о том, что дикорастущая
яблоня ущелья Кондара может быть с успехом использована как компо-
нент в лесосадах. Достаточна привести хотя бы один из наших примеров,
чтобы убедиться, насколько хороши плоды яблони, выращенной на дико-
растущей яблоне; яблоня сорта Кальвиль красный, например, дает плоды,
которые весят в среднем 225 г, а максимальный вес плода достигает 257—
264 г.
23*
356
В. И. Запрягаева
Алыча (Prunus sogdiana Vass.)
Очень широко распространена в ущелье Кондара, но плоды ее мест-
ным населением почти не используются. В наших опытах алыча исполь-
зовалась как подвой для абрикоса, сливы, вишни и черешни. Облагора-
живание алычи производилось прививкой в высокий штамб и окулиров-
кой в порослевые побеги. Лучшие результаты были получены с прививкой
сливы. Опыты с вишней и черешней не удались совсем. Абрикос, при-
витый на алыче, не дает прочного срастания и обламывается на месте
окулировки. Слива очень хорошо срастается с алычей [что совпадает
с данными Момот (1940)], хотя наблюдается большой процент непри-
нявшихся растений после окулировки или прививки.
Слива, привитая на алыче, в первый же год своего развития образует
приросты до 83—192 см. В последующие годы хотя размеры годичных
приростов и снижаются, но все же остаются довольно значительными
(в пределах 60—100 см). Цветение сливы, привитой на алыче, начинается
на 6—7-й год после окулировки, плодоношение на 7-й год. Плоды полу-
чаются совершенно нормальными и достигают, например у сорта Итальян-
ская венгерка, 37—42—50 г.
При организации лесосадов в ущелье Кондара или в другом месте,
аналогичном нашему ущелью, встает вопрос и об облагораживании,
помимо растений, с которыми мы работали, ореха и иногда фисташки,
постоянно сопутствующих яблоне, боярышнику, алыче и бухарскому
миндалю. На Станции не ставилось специальных работ по облагора-
живанию ореха и фисташки, так как эти вопросы разрешены: разра-
ботанные методы облагораживания этих плодовых могут быть с успе-
хом применены и в наших условиях.
Суммируя все изложенное в настоящей главе, можно сказать, что
использование дикорастущих плодовых или близких к ним растений
для создания лесосадов имеет очень большие перспективы не только
в ущелье Кондара, но и по всему горному Таджикистану; достаточно
взглянуть на карту распространения бухарского миндаля по Таджики-
стану, чтобы убедиться в этом. Лесосады — это наиболее простая форма
создания плодоводства в горах. Несмотря на разбросанность плодовых
деревьев по мало удобным для работы склонам, лесосады имеют ряд
преимуществ перед садами. Используя местные хорошо укоренившиеся
растения как подвои, мы достигаем того, что привитые культурные пло-
довые вступают в пору плодоношения значительно раньше, чем в обыч-
ных условиях. Кроме того, под лесосады используются так называемые
«бросовые земли» (крутые склоны, осыпи, скалы), где культура других
сельскохозяйственных растений немыслима.
ЛИТЕРАТУРА
Агролесомелиорация, под редакцией Горшенина Н. М. 1941 г.
Бейдеман И. Н. 1938. К программе изучения корневых систем. Тр. Ботан. инет.
Азербайдж. филиала АН СССР, т. 3.
Берг Л. С. 1938. Климат СССР. Растительность СССР, I.
Б е л о х о н о в И. В., И. И. Курындин и др. 1939. Плодоводство.
Бобров Е. Г. 1926. К вопросу о поясах растительности среднего Копет-дага.
Сб. Географ.-экономич. исследов. инет.
Богушевский П. Н. 1935. Культурная яблоня Зеравшана. Плодовые Сред-
него Таджикистана.
Биологические особенности древесных пород
357
Буш Н. А. 1935. Ботанико-географический очерк Кавказа.
Вальтер Г. и В. В. Алехин. 1946. Основы ботанической географии.
Василевская В. К. 1941. Анатомическое строение плодовых Копет-дага в раз-
личных условиях культуры. Сов. ботан., № 1.
В е х о в Н. К. 1934. К биологии ореха рода Juglans. Орехи, биология, культура
хозяйства, в. III.
Викторовский Г. П. 1935. Грецкий орех Juglans regia L. ssp. fallax (Dode)
M. Pop. Плодовые среднего Таджикистана.
Викторовский Г. П. 1935. Груша среднего Таджикистана. Плодовые Сред-
него Таджикистана.
Виноградов-Никитин II. 1928. Плодовые и пищевые деревья лесов Закав-
казья. Тр. по прикл. ботан., генет. и селекции, т. XXII, вып. 5.
Вульф Е. В. 1944. Историческая география растений.
Гончаров Н. Ф. 1936. Очерк растительности Центрального Таджикистана.
Гончаров Н.Ф. 1937. Районы флоры Таджикистана и их растительность. Флора
Таджикистана, т. 5.
Городецкий В. Д. 1934. Пособие по дендрологии для Средней Азии.
Григорьев Ю. С. 1944. Очерк растительности бассейна Среднего Зеравшана.
Изв. Таджик, фил. Акад. Наук СССР, № 7.
Гурский А. В. 1932. Лесоводственные исследования. Таджикская комплексная
экспедиция.
Гурский А. В. 1932. Орехи западного Копет-дага. Тр. по прикл. ботан., генет.
и селекции, т. VIII, I.
Гурский А. В. 1934. Экологические черты лессовых полупустынь западного
Копет-дага. Сов. субтропики, № 3.
Гурский А. В. 1938. Корневые системы древесных растений в связи с их эколо-
гией и культурой на пустынных почвах.
Гурский А. В. 1939. Опыт неорошаемого древоводства в пустыне. Докл.
ВАСХНИЛ, вып. 2—3.
Гусев П. П. 1938. Лесосады.
Дмитриев В. Д. 1930. Кипарисовые Средней Азии (арча).
Дмитриев В. Д. 1938. Состав, биология и современное состояние арчевников
Узбекистана.
Дробов В. П. 1936. Лесосады Средней Азии. Тр. Узбекск. лесокультурной и агро-
лесомелиоративной ст., вып. 1.
Дьяченко А. Я. 1934. Грецкий орех в Южной Киргизии. ВНИЛАМИ, вып. 3.
Жуковский П. М. 1933. Земледельческая Турция.
Заленский О. В. 1940. Распространение и экологические особенности фисташки
(Pistacia vera L.) и миндаля (Amygdalus communis L.) в западном Копет-даге
(Туркменская ССР). Ботан. журн., № 9.
Запрягаев Ф. Л. 1937. Древесно-кустарниковая растительность Таджикистана.
Сов. ботан., № 6.
Запрягаев Ф. Л. 1940. Естественное семенное возобновление некоторых дре-
весных пород на южных склонах Гиссарского хребта. Тр. Таджикск. базы АН
СССР, т. VIII, Ботаника.
Запрягаев Ф. Л. иВ. И. Запрягаева. 1940. Облагораживание дико-
растущих древесных и кустарниковых пород Таджикистана. Тр. Таджикск.
базы АН СССР, т. VIII, Ботаника.
Запрягаев Ф. Л. и В. И. Запрягаева 1941. Перспективы горного богар-
ного плодоводства в Таджикистане. Изв. Таджикск. фил. Акад. Наук СССР,
№ 1.
Запрягаева В. И. 1940. Орех Juglans regia L. в бассейне реки Кондара. Тр.
Таджикск. базы АН СССР, т. VIII, Ботаника.
Запрягаева В. И. 1941. Лесосады на базе диких зарослей бухарского миндаля.
Запрягаева В. И. 1944. Опыт использования иргая как подвоя для культур-
ных плодовых. Изв. Таджикск. фил. Акад. Наук СССР, № 3.
Иванов Г. И. 1931. Опытные лесокультурные работы в районе Чулийской дачи.
Тр. Туркменской лесокультурной опытной станции.
Иванов П. Ф. 1938. Пояснительная записка к проектированию террас под богар-
ные сады.
Иванова А. В. 1946. Можжевеловые редколесья южной Армении. Тр. Ботан.
инет. Акад, наук Армянской ССР, т. IV.
Карасев А. С. 1934. Лесные мелиорации.
Карасев А. С. 1934. Освоение гор и пустынь Узбекистана. Сб. Узбекистан.
Кварацхелия Т. К. 1927. Материалы к биологии корневой системы плодовых
деревьев.
358
В. И. Запрягаева
Керн Э. Э. 1925. Деревья и кустарники.
Кизюрин А. Д. 1938. Стелющийся сад и кустовая система.
Киселев и ч В. Н. 1936. О выборе пород для горной агромелиорации.
К о л п и к о в М. В. 1944. Общее лесоводство.
Комаров В. Л. 1896. Материалы по флоре Туркестанского нагорья. Бассейн
Зеравшана. Тр. СПб. общ. естествоиспыт.
Комаров В. Л. 1896. Краткий очерк растительности горного Зеравшана. Тр.
СПб. общ. естествоиспыт., т. XXVI.
Коржинский С. 1896. Очерки растительности Туркестана. Зап. Акад. Наук,
серия IV, VIII, № 4.
Коровин Е. П. 1934. Растительность Средней Азии.
Коровин Е. П. и А. Н. Розанов. 1938. Почвы и растительность Средней Азии
как естественная производительная сила. Тр. Средне-Азиатского Гос. унив.,
серия ХПа, География, вып. 17.
Королева А. С. 1940. Очерк растительности центральной части южного склона
Гиссарского хребта и ее естественные кормовые ресурсы. Тр. Таджикск. базы
АН СССР, т. VI.
Королева А. С. 1951. Растительный покров Кондары. Ущелье Кондара.
Кочерга Ф. К. 1933. Борьба с силевыми потоками в Узбекистане.
Красильников П. К. 1941. О корневых системах Parrotia persica, Carpinus
betulus, Quercus castanefolia в низменности и предгорьях Талыша. Сов. бота-
ника, № 4.
К р е й д и к Б. М. 1940. Гранат. Культура инжира и граната.
Кузьмин С. П. 1929, 1930. Водный баланс и засухоустойчивость растений Апше-
рона в связи с особенностями строения их корневой системы. Тр. по прикл.
ботан., генет. и селекции, т. XXIII.
Лавренко Е.М. 1938. История флоры и растительности СССР но данным совре-
менного распространения растений. Растительность СССР, I.
Лавренко Е. М. 1940. Степи СССР Растительность СССР, II.
Линчевский И. А. 1935. Растительность Бадхыза. Сб. «Раст, ресурсы Туркмен-
ской СССР».
Линчевский И. А. 1937. Микрорайоны сухих субтропиков. Сов. субтропики,
№ 4.
Линчевский И. А. 1940. Лесосады и перспективы их развития в Таджикистане.
Тр. Таджикск. базы АН СССР, т. V111, Ботаника.
Линчевский И. А. и А. В. Прозоровский. 1946. Основные законо-
мерности распределения растительности Афганистана. Сб. научи, тр. Ботан.
инет. им. В. Л. Комарова АН СССР.
Липский В. И. 1911. Лесная растительность в Туркестане.
Липский В. И. 1897. Гиссарская экспедиция 1896 года.
Липский В. И. 1902. Горная Бухара, ч. I—III.
Липский В. И. 1915. Ботанические исследования в Закаспийской области
1912 года.
Люндегорд Г. 1937. Влияние климата и почвы на жизнь растений.
Магакьян А. К. 1941. Растительность Армянской ССР.
Максимов Н. А. 1941. Краткий курс физиологии растений.
Малюгин Е. А. 1931. Кара-калинский район. Тр. Туркм. лесокультурно-опыт-
ной ст.
Малюгин Е. А. 1931. Работы по акклиматизации и натурализации. Тр. Туркм.
лесокультурно-опытной ст.
Мичурин И. В. 1936. Итоги шестидесятилетних работ.
Мичурин И. В. 1939. Принципы и методы работы.
М о м о т С. М. 1940. Использование плодовых, сопутствующих грецкому ореху.
Сб. «Грецкий орех южной Киргизии».
Морозов В. Ф. 1944. Культура шелковицы на богаре. Изв. Тадж. фил. Акад.
Наук СССР, № 3.
Некрасова В. Л. 1927. Род Juglans L. в Туркестане. Тр. по прикл. ботан., генет.
и селекции, т. XV111.
Овчинников П. Н. 1940. К истории растительности юга Средней Азии. Совет-
ская ботаника, № 3.
Петров М. П. 1939. О диких плодовых горной Туркмении. Природа, № 5.
Петров М. П. 1944. Очерк растительности северного Ирана. Сов. ботан.,
4—5.
Печникова С. С. 19407 Внутривидовая изменчивость Таджикистанских орехов
Juglans regia L. в ущелье Кондара. Тр. Таджикск. базы АН СССР, т. VIII.
Ботаника.
Биологические особенности древесных пород
359
Попов М. Г. 1922. О растительности гор Сары-тау и урочища Сель-рохо в Коканд-
ском уезде Ферганской области. Тр. Туркест. Гос. унив., вып. 4.
Попов М. Г 1934. Дикие плодовые леса Узбекистана. Сб. «Узбекистан».
Попов М. Г 1939. Дикие плодовые деревья и кустарники Средней Азии. Тр.
по прикл. ботан., генет. и селекции, т. 22, вып. 3.
Попов М. Г. 1940. Растительность Казахстана.
Попов М. Г. и Н. В. Андросов. 1937. Растительность заповедника Гуралаш
и Зааминской лесной дачи.
Прутенский Д. И. 1940. Естественное семенное возобновление грецкого ореха.
Сб. «Грецкий орех Южной Киргизии».
Радзиевский А. Ф. 1934. Лесное хозяйство Узбекистана. Сб. «Узбекистан».
Р а у н е р С. Ю. 1901. Горные леса Туркестана и их значение для водного хозяйства
края.
Рихтер А. А. 1938. Культура миндаля в республиках Средней Азии.
Розанов Б. С. 1940. Инжир. Культура инжира и граната.
Свешникова В. М. 1944. Водный режим почв и корневая система растений
Восточного Памира. Изв. Тадж. фил. Акад. Наук СССР, № 7.
Сеславин П. М. 1936. Лесосадовые насаждения Узбекистана.
Сиворакша Г. С. 1938. Арча и ее использование. Арча.
Сиворакша Г. С. 1937 Фисташковые рощи Баба-тага. Соц. наука и техника,
№ 12.
Смольянинова Л. А. 1936. Орех. Культурная флора СССР. XVII. Орехо-
плодные.
Соснове кий Д. И. 1943. Опыт хронологизации основных фитоценозов Восточ-
ной Грузии. Сообщ. Академии наук Грузинской ССР, т. IV, № 2.
Степанов Н.П. 1944. Степное лесоразведение.
Сукачев В. Н. 1934. Дендрология с основами лесной геоботаники.
Тольский А. 1911. К вопросу о влиянии типа почв на строение корней сосны.
Троицкий П. И. 1936. Опыты лесоразведения в Аман-кутанской даче. Тр. Узбекск.
лесокультурной и агролесомелиоративной ст., вып. 1.
Учайкин В. П. 1934. Культура грецкого ореха и освоение гор Средней Азии.
Федоров А. И. 1929. Освоение культуры ореха в Средней Азии. Тр. Средне-
азиатск. Гос. унив., сер. 18, Ботаника, вып. 8.
Федоров А. И. 1932. Шелковица и ее культура.
Федоров А. А., П. А. Красильников, А. А. Никитин. 1945.
К биологии яблони Сиверса (Malus Sieversii (Ldb.) М. Roem. Сов. ботан.,
т.ХШ, № 6.
Федченко Б. А. 1925. Горный Туркестан. Ботан.-географ, сб.
Ходоровский К. К. 1904. Горно-культурные работы в Азиатской и Европей-
ской России.
Череватенко Н. Я. 1936. Облагораживание диких фруктовых зарослей. Сов.
субтропики, № 3.
Шефер-Сафонова Е. А. 1934. Влияние влажности почвы на развитие неко-
торых древесных пород. ВНИЛАМИ, вып. II.
Andreanszky. 1939. Der Baumwuchs und seine klimatischen Grenzen in Nordaf-
rica. Bot. Jahrb., Bd. 70, H. 2.
J enny-Lips. 1930. Vegetationsbedingungen und Pflanzengesellschaften auf Fels-
schutt. Bot. Zentralbl., 16.
Maire Rene. 1921. Etudes sur la Vegetation et la Flore du Grand Atlas et du Moyen
Atlas Marocains.
Rikli M. 1912. Lebensbedingungen und Vegetationsverhaltnisse der Mittelmeergeb.
und der Atlantischen Inseln.
Walter H. 1927. Einfiihrung in die allgemeine Pflanzengeographie Deutschlands.
Weaver J.E. 1919. The ecological relations of roots. Carnegie Institution Publ. 286.
Weaver J.E. and P. Clements. 1929. Plant Ecology.
А. С. КОРОЛЕВА
КУЛЬТУРА КОРМОВЫХ ТРАВ НА БОГАРЕ
Работа с кормовыми травами, производившаяся на Варзобской гор-
ной ботанической станции в 1938—1941 гг., ставила своей целью первич-
ное интродукционное испытание ряда культурных и диких кормовых
растений в различных условиях высоты и экспозиции склонов горной
богары пояса древесно-кустарниковой растительности.
В нижней части субальпийского пояса на территории урочища Руи-
дашт, примыкающего к верховьям бассейна р. Кондары, в 1936 г.
Ф. Л. Запрягаевым были проведены опытные посевы люцерны. Испыта-
ние ряда кормовых трав в верхней части субальпийского пояса в вер-
ховьях бассейна р. Лючоб (за пределами бассейна Кондары) ведется
с 1939 г. А. П. Саверкиным. Результаты испытаний в субальпийском
поясе затрагиваются в настоящем очерке поверхностно.
До работ Варзобской станции испытания кормовых трав на богаре
среднегорной полосы Таджикистана не проводилось. Культурой кормо-
вых трав (главным образом, люцерны) на богаре в течение ряда лет зани-
малась в Таджикистане Богарная селекционная станция Министерства
земледелия, расположенная в окрестностях Сталинабада в поясе эфеме-
ровой растительности. Природные условия богарного пояса, занимаю-
щего область предгорий, значительно отличаются от условий в районе
Кондары, лежащего в среднегорной области. Эти различия природных
условий создали необходимость постановки работ на Варзобской ботани-
ческой станции.
В поясе древесно-кустарниковой растительности, где проводит работу
Варзобская станция, вопрос разведения кормовых трав стоит очень
остро. Под выпас весной и летом здесь используются участки, занятые
древесно-кустарниковой растительностью, и старые залежи. Запасы
кормов здесь очень невелики и их хватает для выпаса лишь небольшой
части скота. Основное поголовье угоняется на субальпийские пастбища
высокогорной области. Площади поливных люцерников в поясе древесно-
кустарниковой растительности ничтожны. Хорошие естественные сено-
косы представлены небольшими полянами среди древесно-кустарниковых
зарослей и залежами. Основную массу сена, заготовляемого в поясе
древесно-кустарниковой растительности, составляет крупное ароматиче-
ское зонтичное — юган (Prangos pabularia Lindl.), не поедаемый в све-
жем состоянии и дающий сено, пригодное лишь для рогатого скота.
Таким образом количество сена, заготовляемого в поясе древесно-кустар-
никовой растительности, далеко не удовлетворяет растущих требований
животноводства, вследствие чего создание на богаре этого пояса искус-
ственных пастбищ и сенокосов является насущной необходимостью.
Культура кормовых трав на богаре
361
При этом нужно иметь в виду, что люцерновые и некоторые другие траво-
стои на богаре могут быть использованы в зеленом состоянии до конца
августа, в то время как окружающая естественная травянистая расти-
тельность засыхает большей частью уже в июне.
Кормовые травы испытывались в культуре на двух участках Варзоб-
ской станции, в нижней и верхней полосе пояса древесно-кустарниковой
растительности. Посевы кормовых трав производились на более или менее
мелкоземистых склонах как северной, так и южной экспозиции.
В нижней полосе (Кондара) мелкоземистый участок с посевами на
северном склоне лежал на высоте 1230—1370 м над ур. м., крутизна его
была 25—40°. Участок с посевами на южном склоне Кондары характе-
ризовался мелкоземисто-щебнистыми почвами. Высота его была 1310—
1330 м, крутизна склонов 25—30°.
В верхней полосе (уроч. Квак) участок с посевами на северном склоне
лежал на высоте 1700—1750 м, крутизна его была 25—30°. Высота
участка на южном склоне достигала 1800 м, крутизна склонов была
здесь 15—20°. Почвы как на северном, так и на южном склонах участка
Квак были мелкоземистые.
На обоих участках посевы кормовых трав проводились на террито-
риях комбинированных садов (см. ст. В. И. Запрягаевой в настоящем
сборнике). Под культуры кормовых трав использовались вытянутые
поперек склонов полосы 6—20 м шириной, протягивающиеся между водо-
задерживающими террассами, вдоль которых были высажены плодовые
деревья. Посевы производились преимущественно весной. Испытывались
и осенние сроки посевов, показавшие в большинстве случаев отрицатель-
ные результаты.
Почти во всех случаях приходилось довольствоваться лишь предпо-
севной вспашкой. Удобрения, за немногими исключениями, не вноси-
лись. Однако и при таких сравнительно примитивных приемах агротех-
ники культура ряда кормовых трав дала хорошие результаты и показала
полную рентабельность их возделывания на богаре пояса древесно-
кустарниковой растительности.
Ниже дается краткое изложение основных опытов.
На Варзобской горной ботанической станции в поясе древесно-кустар-
никовой растительности испытывалось около 20 сортов синих культур-
ных люцерн, 2 сорта желтых культурных люцерн, дикая синяя люцерна
из трех пунктов Таджикистана и из западного Тянь-шаня, культурные
донники — белый и люцерновидный донник Альфа, дикий желтый дон-
ник, культурные эспарцеты, культурные красный и розовый клевера,
дикая тонколистная вика, дикие пажитники (пажитник Липского,
пажитник Попова и пажитник Гриффица), культурные однолетние вики,
чина, горохи, культурный костер безостый, овсяница луговая, жит-
няки, тимофеевка, дикие ежа сборная и ячмень луковичный, 3 культур-
ные сорта суданки и некоторые другие.
Среди бобовых культур наилучшие результаты показали люцерны,
на которых следует остановиться несколько подробнее, чем на других
культурах.
Люцерны
Люцерны высевались на северном и южном склонах верхней и ниж-
ней полосы пояса древесно-кустарниковой растительности. Для люцерн
испытывались как весенние, так и осенние сроки посевов, причем поло-
жительные результаты дали только весенние сроки.
362
Л. С. Королева
А. НИЖНЯЯ ПОЛОСА ПОЯСА ДРЕВЕСНО-КУСТАРНИКОВОЙ
РАСТИТЕЛЬНОСТИ (КОНДАРА)
Северный склон
На северном склоне нижней полосы испытание началось с 1938 г.
4—5 мая на делянках в 20 кв. м в двух повторностях были высеяны 12
сортов синих культурных люцерн и одна дикая местная люцерна. Посев
был рядовой с междурядьями шириной в 25 см и при норме высева 15 кг.
Ряды проводились поперек склонов (табл. l).j
Таблица 1
Урожайность люцерн посева 1938 г. на северном склоне участка Кондара
1938 1939 1 1940 1941 Сумма за 4 года
Сорт люцерны 1-й укос 15 VII 1-й укос 4 V 2-й укос 7 VII 3-й укос 4 VIII 1-й УКОС 3 VI 2-й укос 2 VII Сумма 1 1-й укос 27 V 2-й укос 2 VII л Б
Люцерна ферганская 3.3 18.7 10.3 3.6 32.6 28.6 17.9 46.5 20.9 9.5 30.4 112.8
Л. самаркандская 4.3 18.8 10.7 3.7 33.2 22.4 , 15.5 37.9 14.9 8.6 23.5 98.9
Л. семиреченская 2.9 19.8 12.0 5.4 37.2 26.1 ! 18.5 44.6 18.7 11.7 30.4 115.1
Л. алмаатинская 4.1 15.2 11.4 4.4 31.0 24.6 11.7 36.3 15.5 9.5 25.0 96.4
Л. хивинская 3.9 18.0 10.6 3.3 31.9 , 20.7 13.1 33.8 10.5 7.7 18.2 87.8
Л. афганская 3.5 11.1 9.0 4.2 24.3 21.1 12.9 34.0 13.2 8.8 22.0 83.8
Л. сирийская 5.1 17.8 16.0 5.1 38.9 1 28.3 19.9 43.2 17.3 3.7 13.0 113.2
Л. малоазиатская 2.5 16.7 8.3 4.0 29.0 21.2 । 12.5 33.7| 21.7 10.9 32.6 97.9
Л. арабская 3.6 21.6 12.3 6.3 40.2 24.8 14.9 39.7, 19.0 11.6 30.6 114.1
Л. венгерская 3.4 21.9 12.2 5.9 40.0 25.2 19.3 44.5 18.6 13.4 32.0 119.9
Л. французская 3.5 14.0 11.0 3.7 28.7 22.7 14.8 37.51 18.8 9.0 27.8 97.5
Л. Гримм-Зайкевича 3.4 15.5 7.6 2.8 25.9 21.0 12.8 33.8: 16.5 I 8.3 24.8 87.9
Как видно из таблицы, в первый год жизни люцерны дали только
один укос 1 с очень небольшой урожайностью. Высота травостоя была
при этом 20—30 (50) см. Различия в росте и урожайности по отдельным
сортам в этом году не выявились. На втором году жизни люцерны дали
3 укоса, общей урожайностью 25.6—40.2 ц на га. Высота травостоя во
нремя 1-го укоса была 50—70 (100) см, во время 2-го укоса—30—
40 (60) см, 3-го укоса — 20—40 (50) см (рис. 1). Различия между сортами
в этом году выявились достаточно определенно: на первое место по
урожайности выдвинулись люцерны ферганская, самаркандская, семи-
реченская, сирийская, арабская и венгерская. Дикая местная люцерна
характеризовалась низким (25—30 см выс.) травостоем и распростертыми
мелколистными побегами, вследствие чего была непригодна для укоса.
На третьем году жизни люцерны дали за 2 укоса максимальный урожай:
от 33.7 до 46.5 ц на га. При этом высота люцерн во время 1-го укоса
1 Укосы люцерны брались в начале массового цветения, за некоторыми исклю-
чениями, которые оговорены особо.
Культура кормовых трав на богаре
363
была 60—90 см, 2-го укоса — 40—60 см. Первые места по урожайности
заняли люцерны, бывшие лучшими и на втором году жизни. На четвер-
том году жизни урожайность всех люцерн, давших 2 укоса, снизилась.
На пятом году урожайность не учитывалась, но общее состояние лю-
церны значительно ухудшилось и при учете урожайность их, несомненно,
была бы снижена по сравнению с четвертым годом жизни.
В 1939 г. на северном склоне нижней полосы пояса древесно-кустар-
никовой растительности испытывался дополнительно ряд культурных
и диких синих люцерн и желтая люцерна. Испытание велось при увели-
ченной до 50 см ширине междурядий и сокращенной до 10 кг норме вы-
Рис. 1. Люцерна второго года жизни на северном склоне участка Кондара.
(Фот. Варзобской ботанической станции).
сева. Увеличение ширины междурядий было обусловлено тем, что на всех
делянках люцерн посева 1938 г. срединная часть делянки была заметно
угнетена, по сравнению с краями. Это обстоятельство вызвало предполо-
жение, что при увеличении ширины междурядий угнетенность может
исчезнуть или уменьшиться. Кроме того, увеличение ширины междуря-
дий сильно облегчило бы уход за люцерной на крутых склонах и дало
бы возможность этот уход механизировать (табл. 2).
Результаты широкорядных посевов не показали здесь преимуществ
по сравнению с посевами с междурядьями в 25 см. Правда, угнетенность
срединной части делянки при широкорядном посеве была выражена
менее резко, но урожайность люцерн почти во всех случаях уступала
таковой в посевах с междурядьями в 25 см.
Принимая во внимание, что сравнивались посевы разных годов (узко-
рядный посев 1938 г. и широкорядный посев 1939 г.) при малых делян-
ках и отсутствии повторностей, данные о преимуществах узкорядного
364
А. С. Королева
посева в нижней полосе пояса древесно-кустарниковой растительности
требуют проверки. Испытание желтой люцерны дало значительно худ-
шие результаты, чем испытание синих культурных люцерн. Желтая
люцерна все время отставала в росте от синих и давала меньшие урожаи.
Таблица 2
Урожайность люцерн посева 1939 г. на северном склоне участка Кондара
Сорт люцерны 1940 1941 Сумма за 3 года
1-й укос 3 VI 2-й укос 2 VII 1 Сумма 1-й укос 28 V 2-й укос 3 VII Сумма
Люцерна Перу 7.7 3.6 11.3 11.7 5.1 16.8 28.1
Л. канзасская 6.8 4.9 11.7 8.6 5.5 14.1 25.8
Л. арабская 721 8.0 4.8 12.8 13.0 5.5 18.5 31.3
Л. ферганская 700 7.0 5.2 12.2 14.0 5.4 19.4 31.6
Л. армянская 4.5 — — 30.5 9.3 39.8 —
Л. мархаматская 13.0 9.8 22.8 19.9 13.3 33.2 56.0
Л. Триполи 12.9 10.2 23.1 19.1 13.3 32.4 55.5
Л. желтая кубанская 14.1 5.1 19.2 17.1 9.6 26.7 40.8
Л. дикая из зап. Тянь-шаня 18.8 8.5 27.3 21.9 18.9 40.8 68.1
Л. дикая из Хаита 12.5 — — — — — —
Л. дикая из Кондары 9.6 — — — 1 — — —
Л. дикая из Квака 6.2 i — — — — — —
Из диких люцерн показала хорошую урожайность лишь форма из запад-
ного Тянь-шаня. Остальные дикие формы были мелколистны, низко-
рослы и почти непригодны для укоса.
Южный склон
На южном склоне нижней полосы пояса древесно-кустарниковой
растительности люцерны испытывались с 1939 г.
Нужно заметить, что участок на южном склоне был не вполне благо-
приятным для испытаний по причине значительной щебнистости его
почв. Вследствие этого можно предполагать, что состояние и урожай-
ность люцерн на более мелкоземистых почвах южного склона могло бы
быть лучшим.
Люцерны высевались на южном склоне на делянках в 20 кв. м, без
повторностей, при междурядьях 50 см шириной. Посев был произведен
3 мая. В год посева люцерны укоса не дали, так же как и люцерны
посева того же года на северном склоне. В 1940 г. первый укос был страв-
лен скотом, урожайность 2-го укоса синих культурных люцерн колеба-
лась от 7.5 до 13.0 ц на га, т. е. несколько превышала урожайность
2-го укоса люцерн посева того же года на северном склоне. В 1941 г.,
т. е. на третий год жизни, синие культурные люцерны дали на южном
склоне за 2 укоса 20—24 ц на га, урожайность тех же сортов на северном
склоне была 25.8—28 ц на га.
Культура кормовых трав на богаре
365
Б. ВЕРХНЯЯ ПОЛОСА ПОЯСА ДРЕВЕСНО-КУСТАРНИКОВОЙ
РАСТИТЕЛЬНОСТИ
Северный склон
На северном склоне верхней полосы пояса древесно-кустарниковой
растительности люцерны испытывались с 1938 г. 11 мая на делянках
в 25 кв. м, без повторностей, были высеяны 12 сортов синих культурных
люцерн. Посев был рядовой, с междурядьями в 25 см, при норме высева
15 кг на га. Несмотря на внешнюю однородность участка с посевами,
почвенные условия в разных его частях оказались неодинаковыми, что
обусловило резкое различие в состоянии одних и тех же сортов люцерн
в разных его частях. Благодаря небольшим размерам делянок и отсут-
ствию повторностей, некоторые сорта полностью оказались в обедненной
части участка, другие, наоборот, в лучшей его части. Поэтому говорить
о преимуществах того или иного сорта представилось здесь совершенно
невозможным, и приводимые цифры урожайности являются лишь при-
мерами, иллюстрирующими возможные колебания урожаев люцерн
при данных условиях.
В первый год жизни в начале августа люцерны на северном склоне
участка Квак были скошены и показали урожайность 4—9 ц на га. В тече-
ние первой зимы все люцерны пострадали от каких-то роющих млеко-
питающих, которые местами на протяжении целых рядов объели корне-
вые шейки, вследствие чего значительная часть растений погибла.
В 1939 г. дружное отрастание люцерн началось в мае, сразу же после
стаивания снега. В этом году люцерны дали 2 укоса, общей урожайностью
37—51 ц на га. Первый укос был произведен 11 VII, второй — 15 VIII.
Средняя высота люцерн во время 1-го укоса достигала 60—90 см, во время
2-го укоса —40—70 см. В следующем 1940 г., на третьем году жизни,
урожай люцерн был почти равен урожаю 1939 г., т. е. колебался в преде-
лах 35—60 ц на га. Нужно иметь в виду, что срок взятия 1-го укоса был
по техническим причинам в 1940 г. несколько запоздалым (он был взят
в начале отцветания люцерн), вследствие чего, возможно, была снижена
и урожайность 2-го укоса. В 1941 г. урожайность люцерн значительно
возросла. Колебания урожаев за 2 укоса были 58—90 ц на га. В 1942 г.
урожайность не учитывалась, но внешнее состояние люцерн было пре-
красное и никаких признаков вырождения они не показывали.
Таким образом урожаи люцерн на северном склоне участка Квак
во все годы превышали урожаи люцерн на северном склоне участка Кон-
дара. В отличие от последнего, где на четвертый год жизни урожаи всех
люцерн, по сравнению с третьим годом жизни, упали, на участке Квак
на четвертом году наблюдалось возрастание урожаев. Состояние люцерн
продолжало быть хорошим и в 1945 г. Большая по сравнению с нижней
полосой пояса древесно-кустарниковой растительности долговечность
люцерны в верхней полосе объясняется, видимо, большим количеством
осадков здесь и, как показывают соответствующие данные, большей
влажностью почв. Как известно, долговечность люцерны на поливе зна-
чительно превышает таковую на богаре. Продолжительность жизни
травостоя богарной люцерны считается в среднем 3—4 года (Голодков-
ский В. Л. и Михайловская М. А., 1936) (здесь имеются в виду нижние
зоны богары), между тем как травостой люцерны на поливе остается
хозяйственно годным в течение 6—7 лет. В то время как срок стояния
люцерны в нижней полосе пояса древесно-кустарниковой растительности
366
А. С. Королева
не должен превышать обычного срока стояния богарной люцерны, т. е.
3—4 лет (что и подтверждают данные Кондары), в верхней полосе этого
пояса, где богара очень богата осадками, продолжительность жизни
люцерновых полей может быть значительно большей.
В 1939 г. люцерны испытывались на северном и южном скло-
нах участка Квак. Посев был рядовой при междурядьях 50 см шири-
ной и норме высева 10 кг на га. На северном склоне урожайность синих
культурных люцерн на втором году жизни была за 2 укоса 40—50 ц
на га, т. е. приблизительно равнялась урожайности люцерн посева
1938 г., при междурядьях 25 см. Таким образом в два раза большее
увеличение междурядий в верхней полосе пояса древесно-кустарниковой
растительности не оказало на урожайность отрицательного влияния.
Учет урожайности семян показал, что семенная продукция на Кваке
в 2х/г раза превышала таковую в Кондаре, равняясь 0.5 ц на га (в Кон-
даре урожайность семян была 0.2 ц на га). При сравнении веса семян
одних и тех же сортов в Кондаре и на Кваке вес кваковских семян
оказался несколько большим.
Южный склон
На южном склоне участка Квак с 1939 г. испытывались синяя и жел-
тая культурные люцерны. В 1939 и 1940 гг. люцерны на южном склоне
участка Квак очень сильно страдали от потравы скотом, вследствие чего
учесть их урожайность не представлялось возможным. В 1941 г., на
третьем году жизни, урожай 1-го укоса синих люцерн равнялся 24—
37 ц с га, урожай желтой люцерны — 38 ц на га (второй укос был страв-
лен скотом). Как видно, урожайность первого укоса на южном склоне
почти не уступала таковому на северном склоне (30—40 ц на га). Инте-
ресно отметить, что желтая люцерна на южном склоне Квака, в отличие
от участка Кондары, не отставала в росте и урожайности от синих,
а в некоторых случаях превышала их.
Все изложенные опыты с люцерной относятся к весенним срокам
посевов. Что касается осенних посевов люцерны, то они испытывались
на обоих участках в 1939—1940 гг., но дали отрицательные результаты.
Семена, посеянные в конце лета или начале осени в сухую землю, всхо-
дили только после первых осенних дождей в октябре—ноябре, незадолго
до начала ночных заморозков, которые убивали всходы и не успевшие
окрепнуть молодые растения. К концу ноября, еще до выпадения более
или менее постоянного снегового покрова, уже не оставалось ни одного
живого растения. Все это показывает, что люцерна, всходы и молодые
растения которой очень нежны, может высеваться осенью только в тех
районах, где после осенних дождей наступает теплый период, достаточ-
ный для того, чтобы растения могли окрепнуть до наступления замороз-
ков. Наши наблюдения над гибелью осенних посевов люцерны полностью
подтверждаются данными богарной селекционной станции Министерства
земледелия Таджикистана и данными опытной станции Узбекистана
(Димитриев, 1935).
Посевы люцерны как в нижней, так и в верхней полосе пояса древесно-
кустарниковой растительности во все годы поражались мучнистой росой
Культура кормовых трав на богаре
367
(Leveillula taurica Arn.). Однако в виду того, что мучнистая роса обильно
распространялась лишь после 2-го укоса, заметного влияния на урожай
она не оказывала. В некоторые годы осенью люцерну довольно сильно
поражала люцерновая златка, которая, разрушая корни в верхних их
частях, послужила причиной гибели многих растений. В зиму 1938/1939 г.
большой ущерб посевам люцерн на Кваке нанесли какие-то роющие
млекопитающие. Однако в последующие годы они не вредили. В 1941 г.
все люцерны в Кондаре в начале цветения были поражены бурой пят-
нистостью листьев [Pseudopeziza medicaginis (Lib.) Saccard.], которая
Рис. 2. Участок с посевами люцерны в нижней части субальпийского пояса.
(Фот. Варзобской ботанической станции).
несколько снизила урожай как первого, так и второго укосов. Семена
и бобы люцерны во все годы очень сильно страдали от повреждения раз-
личными насекомыми. На борьбу с перечисленными болезнями и вреди-
телями нужно в первую очередь обратить внимание при дальнейшей
работе с люцерной.
В заключение следует сказать несколько слов о культуре люцерны
на богаре пояса субальпийской растительности, лежащего выше пояса
древесно-кустарниковой растительности и простирающегося от 2400
до 3000—3200 м высоты над ур. м. (рис. 2).
Нижняя часть субальпийского пояса является зоной богарного
земледелия с широким распространением культуры ячменя, реже — пше -
ницы и ряда зерновых бобовых. Однако при существующей системе дли-
368
А. С. Королева
тельных залежей большая часть территории субальпийского пояса
используется под выпас. Верхняя часть субальпийского пояса лежит
выше современной границы земледелия и используется только как лет-
ние пастбища. Естественная растительность субальпийского пояса пред-
ставлена исключительно травянистыми группировками. Бедность в кор-
мовом отношении этих группировок, а также сильная засоренность их
грубыми непоедаемыми растениями обусловливают их низкую продук-
тивность и ставят с особенной остротой вопрос о создании в субальпий-
ском поясе искусственных высокопродуктивных сенокосов и пастбищ.
В нижней части субальпийского пояса опытные посевы люцерны
были проведены в 1936 г. в урочище Руидашт Ф. Л. Запрягаевым. Эти
посевы, несмотря на крайне примитивные условия применявшейся агро-
техники и полное отсутствие ухода, дали интересные результаты. Хотя
урожайность люцерн здесь не учитывалась, так как травостой подвер-
гался систематическому стравливанию, все же в 1938 г., на третьем году
жизни, отдельные экземпляры люцерн достигали 70—90 см высоты,
образовывая мощные, густо олиственные кусты, и пышно цвели. В 1939 г.
на участке с посевами все время пасся скот, что не дало возможности
люцернам достичь цветения. В 1940 г., на пятом году жизни, люцерны,
не показывая признаков вырождения, достигали 70—80 см высоты и нор-
мально цвели. Несмотря на обильное цветение и образование завязей,
семена, за исключением единичных бобиков, не успевали созревать
и убивались ранними заморозками в сентябре—октябре.
Испытание А. П. Саверкиным культуры люцерны в верхней части
пояса субальпийской растительности также дало, в общем, положитель-
ные результаты. Люцерны, хотя и отличающиеся здесь значительно худ-
шим ростом, чем в нижней полосе субальпийского пояса, существовали
здесь в удовлетворительном состоянии с 1940 по 1946 г., почти лишенные
ухода. _______
Основным выводом в результате первичных испытаний люцерны на
богаре пояса древесно-кустарниковой растительности и субальпийского
пояса является несомненная рентабельность возделывания здесь этой
культуры.
В колхозной практике люцерна уже в течение ряда лет культивируется
на богаре пояса древесно-кустарниковой растительности. Однако она
еще не завоевала себе должного признания. Основным бичом культуры
люцерны на богаре является сильнейшая засоренность ее посевов, благо-
даря чему в сене люцерников большой, а часто и преобладающий про-
цент составляют сорняки. Между тем никаких мер для уничтожения
сорняков не принимается, необходимая для люцерны зяблевая вспашка
в большинстве случаев отсутствует, полка и другие способы уничтоже-
ния сорняков не имеют места.
Поднятие уровня культуры богарной люцерны в производстве на
должную высоту является важной сельскохозяйственной задачей бли-
жайших лет. Необходима еще большая опытная работа. Основные агро-
технические правила для богары зоны предгорий разработаны богарной
селекционной станцией Министерства сельского хозяйства Таджик-
ской ССР. Однако в среднегорной области эти правила могут быть приме-
нимы лишь частично. Принимая во внимание специфику горных районов,
целый ряд агротехнических вопросов следует решать здесь иначе, чем в зоне
предгорий. Наиболее существенными пунктами агротехники горных райо-
нов должно явиться изучение сроков весеннего посева, способов посева,
Культура кормовых трав на богаре
369
норм высева, норм и сроков внесения удобрений. Кроме того, необходимо
в зональном разрезе провести сортоиспытание люцерн.
Не менее важной задачей богарного травосеяния является подбор
злаковых компонентов для травосмесей с люцерной. Дальнейшая опыт-
ная работа по травосеянию должна также включить в свой план подроб-
ное изучение ряда других культурных и диких бобовых, совершенно
не известных на богаре Таджикистана, если не считать культуры ряда
зерновых бобовых (горох, чечевица, нут), которые в некоторых
районах идут на корм скоту. При развертывании опытной работы со зла-
ковыми и бобовыми культурами некоторой наметкой могут служить
результаты посевов кормовых бобовых и злаков на Варзобской станции,
излагаемые ниже.
Эспарцеты культурные
Испытывались в нижней и верхней полосах пояса древесно-кустарни-
ковой растительности. Посевы эспарцетов производились весной.
Эспарцет песчаный, испытывавшийся на северных и южных склонах
Кондары, показал результаты значительно худшие, чем люцерны. Уро-
жайность его на северном склоне Кондары за 2 укоса на втором году
жизни была 16 ц на га, на третьем году — 23 ц на га. На южном склоне
Кондары состояние эспарцета было совершенно неудовлетворительным,
и урожайность его даже не учитывалась.
На северном склоне Квака испытывались песчаный эспарцет и эспар-
цет неизвестного сорта. Урожаи их на втором году жизни за 2 укоса
достигали 15—20 ц на га, на третьем году 20—30 ц на га, т. е. значительно
уступали урожаям люцерны.
На южном склоне Квака состояние эспарцета так же, как и в Кондаре,
было неважным, и укоса он не давал.
Приведенные данные Варзобской станции говорят о том, что культура
эспарцета в поясе древесно-кустарниковой растительности является
менее перспективной, чем культура люцерны. Работа с эспарцетом Бо-
гарной селекционной станции Министерства земледелия Таджикистана
также показала, что эспарцет по урожайности значительно уступает
люцерне.
Культура эспарцета, более засухоустойчивого чем люцерна, явится,
повидимому, рентабельной лишь для более сухих зон богарного земледе-
лия, где его урожаи превосходят урожаи люцерны (Голодковский
и Михайловская, 1936).
Донники
В верхней и нижней полосах пояса древесно-кустарниковой расти-
тельности на северных и южных склонах испытывался культурный белый
донник, на южных склонах обоих участков — дикий донник лекарствен-
ный, и на северном склоне Кондары — культурный донник люцерновидный
Альфа. Все донники высевались весной.
Донник белый на северном склоне Кондары в первый год жизни дал
урожай 12 ц на га за один укос, на второй год жизни — за 2 укоса 29 ц
на га (1-й укос 23 ц, 2-й укос 6 ц). На южном склоне Кондары
донник в год посева урожая не дал, на втором году жизни он был страв-
лен скотом, но по состоянию сохранившихся растений было видно, что
урожай его не уступал бы урожаю на северном склоне.
24 Ущелье Кондара
370
А. С. Королева
На северном склоне участка Квак донник белый в год посева урожая
не дал, на втором году жизни урожай его в первом укосе достигал 40 ц
на га. Отрастание после первого укоса было незначительное и второй укос
не снимался.
На южном склоне участка Квак донник так же, как и на северном
склоне, в год посева урожая не дал, на втором году жизни урожай его
в первом укосе был 37 ц на га, т. е. почти равнялся урожаю на северном
склоне.
Донник люцерновидный отличается от белого донника хорошо олист-
венными, более нежными стеблями и меньшим содержанием кумарина.
При испытании на северном склоне участка Кондара донник люцерно-
видный, благодаря плохой всхожести семян, характеризовался крайне
разреженным травостоем, что не дало возможности учесть его урожай-
ность. По росту он все время отставал от белого донника.
Донник лекарственный дал на втором году жизни на южном склоне
Кондары урожай 20 ц на га, на южном склоне участка Квак — 30 ц на га.
Сравнивая урожаи донников на всех участках с урожаями люцерн,
мы видим, что в большинстве случаев урожаи донников не превышали
урожаев люцерн. Вследствие грубости стеблей донника и присутствия
в нем значительного количества кумарина, сено его, по питательности
почти не уступающее люцерновому, поедается животными гораздо менее
охотно. Поэтому в тех зонах богары, где люцерна удается не хуже дон-
ника, культура его вряд ли имеет широкие перспективы.
Вики
Из многолетних вик на Варзобской станции на северных склонах
обоих участков испытывалась дикая тонколистная вика (Vicia tenui-
folia Roth.). Эта вика, образующая большую надземную массу, ценится
местным населением как корм высокого качества и заготовляется всюду,
где встречается в более или менее значительных количествах. На участке
Кондара посевы тонколистной вики не дали хороших результатов. При
весеннем посеве нормально развивавшиеся сначала молодые растения по-
чти полностью отмерли в середине лета, с наступлением засухи. При осен-
нем посеве вика, в противоположность люцерне, хорошо перенесла осень
и зиму, но прирост ее с весны был крайне-замедленным, ив середине лета
она начала подсыхать и частично отмерла. Повидимому вика, развиваю-
щаяся в нижней полосе, обычно под пологом деревьев и кустарников,
плохо переносит в молодом состоянии лето без прикрытия. Это — вопрос,
требующий дальнейшего исследования.
На участке Квак вика тонколистная широко распространена в диком
состоянии, нередко образуя заросли, даже без полога деревьев. Культи-
вируемая на северном склоне Квака тонколистная вика при весеннем
посеве прекрасно развивалась, не уступая в этом отношении вике в есте-
ственной обстановке. На второй год жизни она дала укос урожайностью
25—32 ц на га. После укоса отрастание было незначительным, от чего
общая урожайность вики значительно уступала урожайности люцерны.
Из однолетних вик на северном склоне Кондары и Квака испытыва-
лась культурная вика посевная (Vicia sativa L.). В Кондаре культура
однолетней вики не дала положительных результатов. Урожай вики,
испытывавшейся 2 года, не превышал 3—4 ц на га. На участке Квак
урожай вики был значительно выше, достигая 10—17 ц на га.^
Культура кормовых трав на богаре
371
Однолетние вики, так же как и другие однолетние бобовые (см. ниже),
повидимому,. мало перспективны для культуры в нижней полосе пояса
древесно-кустарниковой растительности, в верхней же полосе, в сево-
оборотах, однолетние бобовые могут представить значительный интерес.
Пажитники
Из многолетних диких пажитников на северном склоне Кондары
испытывался пажитник Липского, на северном склоне Квака — пажит-
ник Попова и пажитник Гриффица.
Пажитник Липского, являющийся хорошим кормовым растением,
образует в Кондаре довольно крупные естественные заросли по северным
и восточным склонам. Он рано вегетирует и достигает укосной
зрелости раньше большей части других кормовых трав. При весеннем
посеве пажитник вел себя так же, как и тонколистная вика, т. е. погиб
почти полностью в середине лета. При посеве осенью пажитник Липского
после дождей дал всходы, хорошо перенесшие осень и зиму и развивав-
шиеся, правда, довольно замедленными темпами, в первой половине
лета. Со второй половины лета растения начали желтеть, тем не менее
до наступления зимы были живы. К сожалению, весной 1942 г. участок
с посевами был перепахан, и дальнейшие наблюдения не проводились.
Во всяком случае из вышеизложенного ясно, что культура пажитника
Липского (так же как и вики тонколистной), если она и окажется рен-
табельной, возможна только при осенних посевах.
Пажитник Гриффица и пажитник Попова являются прекрасными
кормовыми травами на пастбищах субальпийского пояса, характери-
зуясь быстрым отрастанием после стравливания. При культивировании
этих пажитников на северном склоне участка Квак развиваемая ими
масса была слишком ничтожна для укоса, вследствие чего урожайность
их не учитывалась.
Таким образом культура этих пажитников, могущая представить
лишь некоторый интерес при селекционной работе, непосредственного
практического значения в поясе древесно-кустарниковой растительности
иметь не будет.
Однолетний культурный пажитник (Trigonella foenum graecum L.)
культивировался на северном склоне участка Кондара. Однако разви-
ваемая им травяная масса была так ничтожна, что учет ее урожайности
не производился.
Клевера
Клевер красный испытывался на северных склонах обоих участков
Варзобской станции. Вследствие разреженности его травостоя на всех
делянках (по причине плохой всхожести семян) урожайность его не учи-
тывалась. Интересно отметить, что состояние клевера на обоих участках
было очень хорошим, высота его достигала 60—70 см, причем он обильно
цвел и плодоносил.
Клевер розовый на обоих участках чувствовал себя также неплохо;
вследствие разреженного травостоя. урожайность его не учитывалась.
На основании полученных данных культура клевера в поясе древесно-
кустарниковой растительности несомненно заслуживает некоторого
внимания.
24*
372
А. С. Королева
Однолетние зерновые бобовые
Из однолетних зерновых бобовых на Варзобской станции испыты-
вались на северном склоне участка Квак культурные горох и чина. Уро-
жайность чины была 11 ц на га, гороха 30 ц на га. В Кондаре эти куль-
туры не испытывались. Интересно, что в поясе эфемеровой растительности
культура местного сорта гороха-пелюшки, испытанная богарной селек-
ционной станцией Министерства земледелия в Таджикистане, дала поло-
жительные результаты. Возможно, что и в нижней полосе пояса древесно-
кустарниковой растительности горох окажется перспективным кормовым
растением.
Многолетние злаки
На Варзобской станции, в нижней и в верхней полосах пояса дре-
весно-кустарниковой растительности, испытывался ряд многолетних зла-
ков как возможных компонентов для травосмесей с люцерной и другими
бобовыми.
В нижней полосе результаты культуры многолетних злаков оказа-
лись пока мало удовлетворительными, но, несомненно, требуют дополни-
тельной проверки.
Здесь на северном склоне испытывались при весеннем посеве культур-
ные костер безостый и житняки, дикие ежа сборная и ячмень лукович-
ный. Все эти злаки давали весной хорошие всходы, которые при очень
слабом приросте развивались до половины лета, а затем погибали.
В 1940 г. были произведены осенние посевы злаков. Все злаки, взошед-
шие после дождей, хорошо перенесли осень и зиму. За вегетационный
период они развивались очень слабо, давая незначительный прирост,
и не достигли далее генерации. Тем не менее к зиме все растения оста-
лись живыми. Дальнейшие наблюдения не проводились. Тот факт,
что некоторые из испытывавшихся злаков (ежа, ячмень луковичный)
прекрасно развиваются в диком виде на окружающих склонах, говорит
о том, что успех их культуры зависит лишь от разрешения вопросов
агротехники.
В верхней полосе пояса древесно-кустарниковой растительности
культура многолетних злаков удается значительно лучше. В этой полосе
на северном склоне участка Квак испытывались культурные костер
безостый, тимофеевка, овсяница луговая, дикие ежа.сборная и пырей
Дробова. Среди всех злаков наибольшую урожайность пока-
зал костер безостый, урожайность которого на втором году жизни дости-
гала 12 ц на га, на третьем 65 ц на га, на четвертом 63 ц на га. Такие
высокие цифры урожайности костра показывают, что его культура будет
здесь весьма успешной. По указанию ряда авторов, костер безостый
может быть использован лишь для долгосрочных травосмесей и совер-
шенно непригоден для кратковременных, так как корневища его начи-
нают в верхнем слое почвы отмирать лишь в 6—7-летнем возрасте и при
более ранней распашке сильно засоряют почву (Е. К. Алексеев
и Н. А. Мочалов 1939; П. Н. Константинов, 1936; Д. Н. Прянишников
и И. В. Якушкин, 1938). Это обстоятельство необходимо учитывать при
введении костра в культуру.
Ежа сборная развивалась также очень хорошо, но урожаи ее (34—
39 ц на га) значительно уступали урожаям костра.
Культура кормовых трав на богаре
373
Тимофеевка луговая в течение трех лет развивала очень небольшую
массу травостоя и лишь на четвертом году дала хороший урожай в 44 ц
на га.
Овсяница луговая образовала травостой, слишком редкий для укоса.
Однолетние злаки
Из однолетних злаков на богаре нижней полосы пояса древесно-
кустарниковой растительности на северном склоне участка Кондара
испытывалась суданка. В 1938 г. суданка за 3 укоса дала урожай 30—
33 ц на га. При повторных испытаниях в последующие годы урожаи
суданки были значительно ниже, достигая лишь 10—13 ц на га. Тем
не менее хорошие урожаи 1938 г. заставляют обратить на нее внимание
в дальнейшей опытной работе.
ЛИТЕРАТУРА
Алексеев Е. Н. и Н. А. Мочалов. 1939. Люцерна в Воронежской области.
Г о л о д к о в с к и й В. Л. и М. А. Михайловская. 1936. Кормовые куль-
туры Узбекистана.
Димитри е в. 1935. Результаты агротехнических опытов с люцерной на богаре.
Бюлл. Всесоюзн. Инет, по хлопководству, № 2.
Константинов И. Н. 1936. Житняк.
Прянишников Д. Н. и И. В. Я к у ш к и н. 1938. Растения полевой куль-
туры.
ВРЕДИТЕЛИ И БОЛЕЗНИ
РАСТЕНИЙ
A. E. СЕМЕНОВ
ВРЕДНАЯ ЭНТОМОФАУНА БОГАРНОГО САДОВОДСТВА В КОНДАРЕ
Работа по изучению вредителей богарного садоводства — насеко-
мых, переходящих с дикой растительности на культурную в связи с ос-
воением горных лесов под подвои, была организована в 1939 г. на Вар-
зобской ботанической станции Таджикского филиала Академии Наук
СССР.
Проблема богарного земледелия в Таджикской ССР является одной
из актуальнейших проблем народного хозяйства республики.
При организации богарного садоводства нужно учитывать следующее.
1. Вредная энтомофауна богарного садоводства горной лесной
зоны довольно специализована и заметно отличается от видового
состава вредных насекомых тех же культур в поливных районах Гиссар-
ской долины. То же можно сказать в отношении численности и хозяйст-
венного значения отдельных видов. Видовой состав вредных насекомых
в условиях Гиссарской долины оказался значительно беднее, чем это
имеет место на садовых насаждениях в условиях горной лесной зоны
южного склона Гиссарского хребта.
2. Переделка природы в горах Таджикистана путем освоения местной
древесной растительности под подвои изменяет экологические условия
этих лесов, что, в свою очередь, сказывается на мире насекомых. Изме-
няя экологические условия местных лесов, мы тем самым лишаем многих
насекомых их типичные кормовых растений; при этом мы прежде всего
сталкиваемся с переходом насекомых с дикой растительности на культур-
ную п значительным вредом, приносимым ими богарному садоводству.,
В качестве примера можно привести большую белокрылую цикаду (Tibi-
cina zevara V Kuzn.), которая в новых условиях, созданных богарным
садоводством, стала серьезным вредителем плодовых культур (Семе-
нов, 1940).
Известно, что многие вредители в массе размножаются периодически^
и состав вредных насекомых сильно изменяется. Поэтому приведенные
ниже данные, основанные на наблюдениях 1939—1941 гг., следует рас-
сматривать как предварительные.
3. Интересную и достаточно характерную картину дает и анализ-
списка вредителей с точки зрения географического распространения
и экологии. Бросается в глаза несомненное численное преобладание
типичных среднеазиатских видов над видами, распространенными
в Европе и в Средней Азии.
Пожалуй, еще более характерную картину дает разбивка нашего
списка на экологические группировки: в группе среднеазиатских видов
наблюдается резкое количественное преобладание специфически лесных
378
A. E. Семенов
(следовательно, в современных условиях, и специфичных горных) видов
над видами, безразлично относящимися к условиям рельефа и связанным
•с ним характером растительности; таких специфически горно-лесных
видов 21.1
4. При защите урожая богарного садоводства от вредных насекомых
нужно учитывать следующее: а) своеобразие рельефа не позволяет при-
менять мощную аппаратуру; б) метод опрыскивания инсектисидамп,
по возможности, заменяется опыливанием; в) гусеничный клей ней-
тральных марок, не вызывающий ожогов в обычных условиях, в горной
местности на южных склонах дает ожоги коры молодых деревьев; г) за-
щитные мероприятия в горных условиях являются более дорогими, по-
скольку требуют значительно большей затраты рабочей силы; д) необхо-
димость считаться с постоянным переходом насекомых с дикой рас-
тительности на культурную, так как все перечисленные в списке на-
секомые жили на местных растениях.
СПИСОК ВРЕДНЫХ НАСЕКОМЫХ
1. Большая персиковая тля (Pterochloroides persicae Sulz.). Как от-
мечает Мордвилко (1927), широко распространена на персике в Закав-
казье, Средней Азии, Палестине и Пенджабе. Кроме персика, как
указывает Невский (1929), этот вид наблюдается реже на абрикосе и
яблоне.
Обычно сосет большими колониями на стволе и нижней стороне тол-
стых ветвей. Зимует в стадии неоплодотворенного яйца; вылупление
из яиц основательниц происходит в начале марта; эта тля одна из самых
первых отрождающихся весною тлей. Основательницы, по Архангель-
скому (1917), дают до 11 поколений бескрылых тлей в год. Среди колоний
с начала мая и до сентября можно встретить крылатых расселительниц.
В конце октября появляются бескрылые яйцекладущие самки, которые
откладывают неоплодотворенные яйца на кору ветвей и стволов. Самцов
не бывает.
Персиковая тля, как подчеркивает Невский (1929), для Узбекистана
не может быть отнесена к разряду серьезных вредителей, так как она
нападает на второстепенные части растений. То же самое отмечает Бо-
денгеймер (Bodenheimer, 1930) и для Палестины.
В Таджикистане большая персиковая тля наблюдалась в огромном
количестве на стволах миндаля и персика. В результате значительной
потери сока нередки были случаи, когда развитое дерево персика усыхало.
Это происходит потому, что богарное садоводство и особенно лесосады
в большой степени лимитированы водой. В данных условиях, в силу
специфики богарного садоводства и особенно лесосадов, вопрос о вред-
ности тли на персике необходимо пересмотреть.
2. Персиковая тля (Myzodes persicae Sulz.) является здесь одним из
-самых вредных видов и принадлежит к числу многоядных тлей (Невский,
1929). Основным растением для этого вида тли является персик, на кото-
ром самки откладывают яйца. Основательницы вылупляются в апреле
и сосут развертывающиеся почки, а потом цветы и листья с нижней
стороны. В результате листья скручиваются.
1 В списке они отмечены звездочкой (*).
Вредная энтомофауна богарного садоводства в Кондаре
379
В мае, затем в течение всего лета появляются крылатые девственницы,
которые перелетают на другие растения (деревья и травы). В октябре
крылатые полоноски и самцы с промежуточных растений возвращаются
на персик. Полоноски на персике дают самок, которые, после спаривания
в ноябре, откладывают яйца.
Распространен этот вид тли во всех среднеазиатских республиках,
в Европе и других частях света.
3. Миндальная тля (Myzaphis amigdalina Nevsk.)* Как вредителя
миндаля ее отмечает Невский (1929) для Узбекистана. В Таджикистане
тля наблюдалась и в Гиссарской до-
лине и в Варзобском районе повсе-
местно там, где произрастал миндаль.
Тля большими колониями на-
блюдалась на нижней стороне ли-
стьев миндаля. Под влиянием со-
сания листья свертываются. Было
повреждено около 10—15% деревьев
миндаля. Тля появилась в апреле
и в значительных количествах
встречалась до конца августа.
Распространена в Средней Азии:
Узбекистан и Таджикистан.
*4. Большая белокрылая цикада
(Tibicina zevara V. Kuzn.). Как
нами выяснено (Семенов, 1940),
широко распространена в лесной
зоне горного Таджикистана. Перво-
начально этот вид был найден в
1928 г. Неуструевым в горах южного
Таджикистана (Кизил-мазар), близ
г. Куляба, и Кузнецовой — на южном
склоне Гиссарского хребта близ
селения Зевар, в честь которого и
было дано видовое название. Нами
Рис. 1. Большая белокрылая цикада
(По А. Е. Семенову).
эта цикада наблюдалась в массовых
количествах в Кондаре и прилежа-
щих соседних кишлаках в долине
р. Варзоб.
Большая белокрылая цикада (рис. 1) оказалась серьезным вредите-
лем богарного садоводства. Эта цикада повреждала следующие древес-
ные породы: яблоню, абрикос, сливу, персик, алычу, вишню, миндаль
культурный и дикий, иргай, боярышник. с
Взрослое насекомое, хотя и питается соком перечисленных растений
путем прокалывания коры и высасывания сока из растения, однако суще-
ственного вреда этим не причиняет. То же самое и личинка цикады,
живущая в почве, питаясь соком корневой системы, заметного вреда
не приносит. Вредит при яйцекладке, пробуравливая своим огромным
яйцекладом древесину веток. Полости имеют значительные размеры;
проходят глубоко в древесину, достигая сердцевины веток; цикада
в полость откладывает 10—20 яиц, затем заливает отверстие особым
клейким веществом. Снаружи такое поражение веток хорошо заметно
по вывернутой наружу в виде щепы древесине. Раны цикада наносит
обычно строчкой, 15—20 штук в строгом порядке одна за другой, с про-
380
A. E. Семенов
межутками около 0.5 см. Яйцекладку цикада совершает с нижней части
ветки и постепенно подвигается вверх. Образованные цикадой полости
в древесине сильно понижают прочность веток. Такие ветки под влиянием
постоянного горного ветра в большинстве случаев ломаются (рис. 2).
Искривление ствола молодых плодовых деревьев является также резуль-
татом деятельности цикады. В условиях Кондаринского ущелья на север-
ном склоне и внизу нами были проведены учеты повреждений, причинен-
ных большой цикадой 3-летним деревьям персика и абрикоса. Процент
повреждения цикадой деревьев персика и абрикоса внизу ущелья значи-
тельно ниже, чем на северном склоне, где она обитает в значительно
больших количествах.
Начало лёта наблюдалось 1 VI. Нимфы перед последней линькой
выходили из земли, взбирались на стебли травянистых растений или
кустарников, где претерпевали дальнейшее превращение. Появление
Рис. 2. Ветка миндаля, поврежденная цикадой.
Сверху— внешний вид повреждения, снизу— разрез поврежденной части. (По А. Е. Семенову).
взрослых цикад стало заметно по громкому и резкому пению самцов.
В десятых числах июня самки приступили к яйцекладке, массовая
яйцекладка отмечена с 19 VI. В самом конце июня лёт цикад быстро
начал падать, последняя самка была зарегистрирована 3 VII. Как видно,
взрослая цикада живет только около одного месяца.
Развитие яиц белокрылой цикады совершается очень медленно. Уда-
лось выяснить, что из яиц, отложенных цикадой 17 VI, начали вылуп-
ляться личинки 31 VII. Таким образом развитие яиц происходило
44 дня. Вышедшая из яйца личинка уже в первом возрасте имеет харак-
терные большие копательные передние ноги (рис. 3). Личинка вылезает
из камеры, где лежали яйца, падает и быстро закапывается в землю.
Дальнейшее, чрезвычайно длительное развитие личинок происходит
в земле на корнях деревьев, соком которых они питаются.
Цикл развития большой белокрылой цикады, повидимому, очень дли-
телен и, вероятно, продолжается несколько лет. Вопрос длительности
развития певчих цикад еще очень плохо изучен. Для наглядного представ-
ления считаю уместным привести как пример американскую цикаду
(Madicicada septendicim L.), развитие которой длится 17 лет (Marlatt,
1907 и Felt, 1913).
Необходимо подчеркнуть, что склоны гор, покрытые лесом, в Таджи-
кистане являются типичными местами обитания белокрылой певчей
Вредная энтом офауна богарного садоводства в Кондаре
381
цикады. Можно предполагать, что в связи с расширением в горах богар-
ного садоводства и поливного плодоводства большая певчая цикада
будет приспособляться к плодовой древесной растительности, и вред
от нее станет более значительным
5. Длиннохвостый кузнечик (Tettigonia caudata Charp.). Длинно-
хвостый кузнечик приносит ежегодно на Северном Кавказе и в Гру-
зии заметный вред разнообразным сельскохозяйственным культурам.
В Узбекистане кузнечик повреждает вишню, яблоню и другие плодо-
вые деревья.
В Таджикистане этот вид питается, в основном, травянистой расти-
тельностью, а древесным широколиственным породам вредит случайно.
В Варзобском районе кузнечик объедал листья плодовых деревьев.
Рис. 3. Личинка белокрылой цикады.
Вид с верхней (2) и с боковой (2) стороны; передняя копательная лапа (3). (По А. Е. Семенову).
*6. Зеленая бескрылая кобылка (Gomphomastax gussakovskii Mistch.)
является типичным полифагом. Вредит разнобразной травянистой расти-
тельности, а также и древесной, объедая листья.
В противоположность кузнечику, вред от бескрылой кобылки на
деревьях не случаен. Благодаря длинным лапкам с хорошо развитыми
на них присосками (рис. 4 и 5), бескрылая кобылка легко взбирается
на самые высокие деревья, являясь постоянным обитателем крон широко-
лиственных деревьев.
В богарном садоводстве это насекомое серьезно повреждает персик,
абрикос, сливу и другие. Особенно сильно объедает листья дикого и слад-
кого миндаля.
Биология этой своеобразной и интересной группы — бескрылых кобы-
лок— еще совершенно не изучена. Бескрылая зеленая кобылка обитает
во всей лесной горной зоне и в огромной численности наблюдается у верх-
382
A. E. Семенов
него пояса произрастания древесной растительности 1900—2500 м над
УР- м-
*7. Бурая бескрылая кобылка (Gomphomastax ninae Mistch.). Как
вредитель этот вид отмечается нами впервые; повреждает он миндаль,
яблоню, абрикос, персик.
Рис. 4. Зеленая бескрылая кобылка, самец. (Ориг.).
Среди насекомых, повреждающих богарные сады в Таджикистане,
большое экономическое значение имеют жуки-златки рода Capnodis,
из них наибольший вред фруктовым деревьям наносят два вида: шести-
пятнистая — С. sexmaculata Ball, и дымчатая — С. tenebricosa 01.
Два указанных вида златок широко распространены в лесной зоне гор
и предгорий. Биология их очень сходна и только в экологическом отно-
Рис. 5. Зеленая бескрылая кобылка, самка. (Ориг.).
шении они сильно разнятся между собой. Так, шестипятнистая златка
чрезвычайно термофильная, явно предпочитает солнечные места и оби-
тает исключительно на южных голых каменистых склонах. Дымчатая же
златка встречается только на северных склонах гор.
Жизненный цикл шестипятнистой и дымчатой златок в наших
условиях, повидимому, завершается в два года. Жуки наблюдаются
в течение всего вегетационного периода — с начала апреля и до конца
октября. Яйцекладка происходит с начала июля по сентябрь. Жуки
Вредная энтомофауна богарного садоводства в Ванд а ре
3831
откладывают яйца группами по 20—40 штук, склеенные места наблю-
даются у корневой шейки растения в трещинах коры. Из отложенных
яиц через 12—15 дней вылупляются личинки, которые проникают под
кору и, повидимому, питаются камбием растения. Как правило, личинки
вредят корням и корневой шейке растений. При питании они окольцовы-
вают корни и тем самым прекращают питание растения. Поэтому личинки
златок очень опасны, особенно в питомниках и школках. Продолжитель-
ность развития личинок еще не ясна; в условиях лесной зоны гор и пред-
горий личинки развиваются, повидимому, 1.5 года.
8. Дымчатая златка (Gapnodis tenebricosa 01.) (рис. 6) как вреди-
тель впервые была отмечена в 1884 г. Мокржецким в Крыму на абрикосе.
Затем как вредителя груши ее указывает в 1926 г. Плотников дляТуркме-
Рнс. 6. Дымчатая златка.
(По А. Е. Семенову).
Рис. 7 Шестипятнистая златка.
(По А. Е. Семенову).
нии. В 1930 г. Боденгеймер (Bodenheimer, 1930) отмечает этот вид
как наиболее вредный для Палестины; вредит он там преимущественно
в горной области, в плодовых школках и питомниках. В Таджикистане-
этот вид повреждает миндаль дикий, миндаль культурный, иргай,.
боярышник, яблоню, персик и абрикос.
*9. Шестипятнистая златка (Capnodis sexmaculata Ball.) (рис. 7) яв-
ляется также многоядным насекомым. Как вредитель плодовых культур
этот вид впервые был отмечен нами. Известен он из Копет-дага в Турк-
мении и из гор Таджикистана (Гиссарский хребет, Дарваз, западный
Памир). Вероятно, эти два ареала связаны друг с другом через горы
Афганистана. В условиях Кондары эта златка зарегистрирована на.
диком и культурном миндале, яблоне, алыче, вишне, персике. Оба вида
златок явно предпочитают дикий и культурный миндаль.
Принято считать, что вред приносят только личинки златок,
а жуки, как правило, значительного вреда не приносят. Согласно
нашим наблюдениям, в силу специфических условий богарного садовод-
ства в Таджикистане основной вред растениям приносят именно жуки.
Продолжительность вредной деятельности жуков-златок в условиях
Таджикистана чрезвычайно велика и охватывает весь вегетационный.
384
A. E. Семенов
период. В зависимости от времени года характер вреда изменяется.
В весенний период (с начала и до конца мая) жуки наблюдаются на крове
миндальных деревьев, где они питаются главным образом черешками
листьев. В это время земля под деревом бывает усыпана пластинками
листьев без черешков. По этому признаку очень легко обнаружить вреди-
теля.
В начале июня, с наступлением сухой и жаркой погоды, жуки поки-
дают крону, переходя на сочную молодую поросль, которая в это время
обильно отрастает от корней миндаля. Жуки питаются молодыми листьями
поросли и охотно объедают сочную молодую кору до древесины, тем
самым вызывают гибель молодых растений, уничтожают результаты
прививки («зеленый глазок»), причиняя таким образом очень существен-
ный вред.
10. Фисташковая златка (Gapnodis parumstriata Pall.) отмечена как
вредитель фисташки Мориц-Романовой (1927). Как вредитель богар-
ных садов нами приводится впервые. Вредят жуки в мае—июле, также
как два предыдущих вида златок.
11. Миндальная златка (Gapnodis cariosa Pall.). Личинки вредят
древесине фисташки, миндаля и персика в Передней Азии. В СССР этот
вид мы отмечаем как вредителя фисташки и миндаля впервые.
Жуки наблюдаются в течение всего вегетационного периода, преимуще-
ственно на фисташке, вредят так же, как предыдущие виды златок.
*12. Малая миндальная златка (Sphenopteia kaznakovi В. Jak.). Био-
логия златок рода Sphenoptera изучена крайне слабо. Только для видов,
обитающих на травянистой растительности, известно, что их личинки
развиваются в корнях растений. Цикл жизни малой миндальной златки
представляет интерес, поскольку ее личинки повреждали дикий миндаль,
сладкий миндаль, персик, абрикос (Семенов, 1944).
Нам удалось наблюдать в конце мая вылупление и выход жука этого
вида златки из пенька бухарского миндаля. Жуки (рис. 8) в течение
июня—июля откладывали яйца в трещины коры растений. Личинки
этого вида златки питаются под корой стволов деревьев. Поврежденное
дерево заметно по многочисленным истечениям камеди, обычно в нижней
части ствола.
Жуки малой миндальной златки обычно не летают, и их можно видеть
бегающими по стволу миндаля, персика, абрикоса, в который они откла-
дывают яйца.
*13. Волосистая шпанка (Teratolytta pilosella Sols.). Ранней весной
на распускающиеся цветы различных плодовых деревьев нападают
в больших количествах жуки волосистой шпанки (рис. 9). Жуки в
Кондаре в апреле—мае повреждают все цветущие плодовые деревья,
выгрызая половые органы цветка, и тем самым в какой-то степени влияют
на урожай.
Биология волосистой шпанки не прослежена. Как указывает Плот-
ников (1926), личинки ее являются паразитами в гнездах диких пчел,
подобно обыкновенной шпанке.
14. Большой усач (Aeolestes sarta Sols.) (рис. 10). Большой усач
в Средней Азии нападает главным образом на тополя, карагачи и ивы,
но нередко повреждает и фруктовые деревья — яблоню, грушу, сливу
и алычу (Плотников, 1926). Вредит личинка. Первоначально молодая
личинка образует червоточину в коре, затем, при дальнейшем развитии,
вгрызается в древесину, образует многочисленные широкие ходы. Взро-
слая личинка, питаясь древесиной, достигает 6 см в длину. Бывает
Вредная энтомофауна богарного садоводства в Кондаре
385
достаточно только одной личинки, чтобы вызвать полную гибель плодо-
вого дерева. Окукление личинок происходит в древесине; там же из
куколки в конце лета появляется жук. Только весной жук прогрызает
лётное отверстие и выходит наружу. Весь цикл развития от яйца
до выхода жука большой усач проходит в 2 года.
Распространен большой усач в Туркмении и Таджикистане. В усло-
виях Кондары лёт жуков отмечается нами в июне. Дикие яблони
и алыча повреждались личинками, а деревья тутовника и грецкого ореха
подвергались нападению жуков.
*15. Миндальный опушенный листоед (Lefevrella amygdali D. Ogl.).
Этот вредитель широко распространен в нижней лесной зоне гор Таджи-
кистана; в условиях Кондары он наблюдался в значительных количе-
ствах только на южном склоне ущелья.
Опушенный миндальный листоед в течение года имеет одно поколе-
ние, зимует в фазе жука (рис. 11). В условиях Кондары жуки появля-
лись с мест зимовки рано весной; в конце апреля—начале мая жуки
в значительных количествах нападали на сладкий миндаль (культур-
ный). Вредили только жуки, объедая листья миндаля. Жуки хорошо
летают, на деревьях быстро передвигаются по веткам и, если к ним при-
близиться, они притворяются мертвыми и падают на землю, где их почти
невозможно обнаружить. Кроме листьев сладкого миндаля, жуки также
питались листьями сливы. В садках жуки охотно питались листьями дикого
25 Ущелье Кондара
386
A. E. Семенов
миндаля, но в природных условиях на этом виде деревьев их удавалось
встречать очень редко.
Жуки наблюдались в заметных количествах в течение всего мая
и только в середине июня совершенно исчезли. Интенсивное спаривание
происходило в начале—середине мая. В условиях садка с 6 VI наблюда-
лась яйцекладка. Развитие яиц протекало очень долго (24 дня), 30 VI
вылупились личинки.
Где живет и чем питается личинка миндального опушенного листо-
еда, еще не ясно.
Рис. 10. Большой усач. (Ориг.). Гис. И. Миндальный опушенный листо-
грыз. (По А. Е. Семенову).
* 16. Черный миндальный листоед (Luperus flavilabris Ogl.). Черный
миндальный листоед, как и миндальный опушенный листоед, широко
распространен в нижней лесной зоне гор Таджикистана. По условиям
обитания эти два вида листоедов различны. Черный миндальный листоед
наблюдался только на северном склоне ущелья, обильно покрытом тра-
вянистой и древесной растительностью.
Жуки черного миндального листоеда (рис. 12) появились с середины
мая и держались до конца июня на листьях дикого бухарского миндаля
и сладкого культурного миндаля. Жуки выгрызали листья небольшими
участками. Яйцекладка происходила в первых числах июня, но личинок
вывести не удалось.
* 17. Листоед-красношейка (Gynandrophthalma discolor Sols.). Жуки
листоеда-красношейки (рис. 13) на территории Варзобской станции и сосед-
В редкая энтомофаун-а богарного садоводства в Кондаре
387
них селений наблюдались в незначительных количествах в течение-
всего июня; они питались листьями абрикоса и персика. В середине июня
происходила яйцекладка. Яйца развивались 13—15 дней.
* 18. Миндальный плодовый долгоносик (Rhynchites zeravshanicus
Voss.) (рис. 14) является специфичным вредителем миндаля, широко
распространенным в лесной зоне гор Таджикистана. Миндальный пло-
довый долгоносик — типичный монофаг, специфичный вредитель плодов
миндаля. Личинки этого жука питаются только ядром миндаля (рис. 15)
и тем самым
Биология
наносят непосредственный вред урожаю.
миндального долгоносика очень тесно связана с феноло-
гией развития дикого миндаля —
Amygdalus bucharica.
Слоник, повидимому, имеет одно-
и двухгодичную генерацию.
Зимовка происходит в земле под
кроной деревьев или там, куда
Рис. 12. Миндальный черный листо-
грыз. (Ориг.).
Рис 13. Лнстогрыз-краспотпейка.
(Ориг.).
могли случайно закатиться упавшие плоды, поврежденные личинками
этого насекомого. В пахотном слое почвы зимуют на глубине 8—10 см.
Зимуют личинки, куколки и жуки.
В отличие от других долгоносиков рода Rhynchites, вредящих плодо-
вому садоводству в СССР, миндальный долгоносик появляется с мест
зимовки очень поздно, в то время когда плоды дикого миндаля достигают
почти предельной величины; в условиях Варзобского района это первая
декада мая. После зимовки жуки сразу же поселяются на миндальных
деревьях, плодами которых питаются; яйца откладывают в плоды.
Жуки прекрасно летают, заселяют крону миндаля в верхней ее части.
Продолжительность периода яйцекладки миндального долгоносика
немного превышает месячный срок. Развитие яйца в плоде миндаля
происходит 10—12 дней. Яйцекладка совершается в период, когда мин-
дальный плод достигает зелено-молочной спелости.
25*
388
A. E„ Семенов
Личиночная фаза вредителя проходит в период от восковой до полной
зрелости миндального ядра. Развившаяся личинка долгоносика, как пра-
вило, покидает ядро миндаля и окукляется в почве. Период развития
личиночной фазы определяется, примерно, пятью декадами. Отродив-
шиеся молодые жуки до весны из почвы не выходят. Часть личинок обычно
диапаузирует. Из диапаузирующих личинок жуки, повидимому, появ-
ляются лишь на вторую весну.
* 19. Большой персиковый долгоносик (Leptomias tadzhikistanicus L.
Агп.). До нашего исследования биология этого долгоносика была совер-
шенно не изучена. Нами раньше он ошибочно назывался L. bimaculatus
Fst. Как вредитель плодового сада этот вид приводится впервые.
Рис. 15. Повреждение миндаля
личинкой плодового долгоносика.
(Ориг.).
7 — вскрытый поврежденный плод в
кожуре; 2—без кожуры.
Жуки (рис. 16) в больших количествах появились на северных скло-
нах гор с середины июня и повреждали листья персика и абрикоса.
В двадцатых числах июня жуки в массе спаривались; в конце июня—
начале июля самки, в природных условиях, откладывали большое коли-
чество белых продолговатых яиц. Развитие яиц в садках происходило
16—18 дней, после чего появлялись личинки. Чем питаются личинки,
еще не выяснено. В середине августа жуки встречались в незначительных
количествах.
* 20. Малый абрикосовый слоник [Myllocerus sp. (рг. rufitarsis Reih.)].
Биология малого абрикосового слоника до нашего исследования также
не была изучена.
В условиях Кондары жуки (рис. 17) появились в середине июня
и повреждали листья абрикоса, персика и иргая. Спаривание жуков
наблюдалось в двадцатых числах июня. Отложенные яйца (в условиях
садка) развивались 20—25 дней. Яйцекладка растянутая, начало отме-
чено в конце июня, конец в середине августа. Чем питается личинка,
не установлено.
* 21. Полосатый листовой слоник (Polydrosus obliquatus Fst.). Согласно
литературным данным, — вредитель листьев яблони, груши, вишни,
Вредная энтомофауна богарного садоводства в Кондаре
389
черешни, сливы, абрикоса, персика и фисташки (Плотников, 1926,
и Яхонтов, 1929).
В Кондаре наблюдался на различных плодовых деревьях.
* 22. Щетинистый листовой слоник (Polydrosus pilifer Hohn.). Как
вредителя богарного садоводства в лесной зоне Таджикистана мы его
отмечаем впервые.
Вредят жуки различным плодовым деревьям.
* 23. Персиковый цветоед (Anthonomus sibiricus Desbr.) (рис. 18).
Никаких данных по биологии этого вида опубликовано до сих пор
Рис. 16. Большой персиковый долго- Рис. 17. Малый абрикосовый слоник,
носик. (Ориг.). (Ориг.).
не было, как вредитель он отмечается здесь впервые. Появление жуков
в условиях Кондары происходит в начале апреля. Самки откладывают
яйца по одному в нераспустившиеся зеленые бутоны персика. Отродив-
шиеся личинки поедают тычинки и пестики, склеивая экскрементами
изнутри лепестки поврежденного бутона. Поврежденные бутоны не рас-
пускаются и засыхают. Личинки в поврежденных бутонах превращаются
в куколок. Выход жуков наблюдается в период сбрасывания избыточ-
ных завязей.
В третьей декаде апреля наблюдалось массовое повреждение цветов
персика личинками этого вредителя. С 4 мая личинки начали превра-
щаться в куколок. Первые жуки появились из бутонов 13 V. В 1941 г.
это насекомое вредило цветам персика в массовом количестве.
* 24. Скелетирующий долгоносик (Magdalis myochroa Rchdt). В лите-
ратуре имеется единственное краткое указание, сделанное Рейхардтом
390
A. E. Семенов
(1927), на то, что скелетирующий долгоносик повреждал листья алычи
в Гузаре (Узбекистан). В условиях Кондары, по нашим наблюдениям,
эти мелкие долгоносики в большом количестве выгрызали круглыми
дырочками паренхиму листьев бухарского миндаля. Поврежденные
листья казались сетчатыми и прозрачными. Жуки (рис. 19) появились
на северном склоне ущелья в массовом количестве в начале мая и наблю-
дались до середины июля. Алыча ими не повреждалась.
25. Туранская оленка (Epicometis turanica Rtt.). Отмечена Плотнике-
вым (1926) как вредитель плодовых для Средней Азии.
Рис 18. Персиковый цветоед. (Ориг.).
Рис 19. Скелетирующий до;
носик. (Oj)lir.).
Вредят только жуки (рис. 20). Они повреждают цветы различных
плодовых растений. До цветения и после цветения фруктовых деревьев
они питаются цветами травянистых растений.
В наших условиях с начала апреля и до начала мая жуки в массе
нападали на цветущий миндаль, персик, абрикос, сливу и другие плодо-
вые деревья. Жуки питаются различными частями цветов и преимущест-
венно половыми органами цветка (тычинками и пестиками), в результате
чего поврежденные цветы не плодоносят. Жуки не летают высоко, а по-
тому от них страдают особенно сильно карликовые деревья. Для богар-
ного садоводства туранская оленка является постоянным серьезным
вредителем.
Туранская оленка имеет в году одно поколение. Личинки пре-
вращаются в куколки в земле, а затем в жуков, которые там и зимуют.
Весною жуки появляются на поверхности земли и начинают вредить
цветущим плодовым деревьям.
26. Рябая бронзовка (Oxythyrea cintella Schaum.) (рис. 21) очень
сходна по образу жизни с туранской оленкой. Вредят только жуки,
питаясь также цветами различных плодовых деревьев и травянистых
Вредная энтомофауна богарного садоводства в Кондаре
391
растений. Появление этого насекомого происходит несколько позднее
оленки. Первые жуки начали появляться с начала мая и наблюдались
на цветах различных растений до июля—августа.
Вред богарному садоводству от этого насекомого меньший, чем от
туранской оленки, вследствие того, что основные фруктовые деревья
успевают отцвести до массового появления жуков. В 1939 г. рябая брон-
зовка очень сильно вредила розам в Ботаническом саду в Сталинабаде.
Распространен этот вредитель на юге Европейской части СССР, на
Кавказе и в Средней Азии.
27. Зеленая бронзовка (Potosia marginicollus Ball.). В отличие от
рябой бронзовки, зеленая бронзовка в фазе жука повреждает цветы,
побеги и плоды яблони, груши и других фруктовых деревьев, как это
отмечает Плотников (1926) для Средней Азии.
В условиях Гиссарского хребта зеленая бронзовка вредила цветущим
деревьям богарного садоводства.
Рис. 20. Тура некая о/юпка. (Ориг.). Рис 21. Рябая бронзовка. (Ориг.).
28. Вредный хрущ (Polyphylla adspersa Mots.) (рис. 22). Личинки
вредного хруща считаются наиболее опасными из всех известных хрущей,
обитающих в Средней Азии. Эти личинки обгрызают корневую шейку
и корни молодых плодовых деревьев, вызывая этим нередко, как сооб-
щает Плотников (1926), гибель плодоносящих деревьев. Наибольший
вред личинки приносят плодоводству в школках и питомниках. Личинки
этого вида в Кондаре нами были обнаружены в двадцатых числах мая
и повреждали персик, абрикос и миндаль. Лёт жуков в больших количе-
ствах происходил около месяца — с начала и по* конец июля.
29. Трехзубчатый хрущ (Polyphylla tridentata Rtt.) отмечен Плотни-
ковым (1926) как вредитель корней плодовых деревьев. В Кондаре лёт
жуков наблюдался с первого июля. Распространен в Средней Азии.
30. Июньский хрущ (Amphimalon solstitiale L.) (рис. 23) широко
распространен в СССР и хорошо известен как вредитель полевых
культур в степных местностях.
В условиях района Кондары интенсивный лёт жуков наблюдался
с 2 по 15 VI. Личинки вредили в школках молодым посадкам культурного
миндаля.
392
A. E. Семенов
*31. Таджикский майский хрущ—(Melolontha clypeata Rtt.). Таджик-
ский майский хрущ (рис. 24) как вредитель нами отмечался впервые
в списке вредителей богарного садоводства (Семенов, 1944). Личинки
этого вида хруща повреждали различные плодовые деревья. Массовый
лёт жуков в условиях верхней границы лесной зоны (кленовник) Квака
на высоте2000мнад ур. м. происходите 10по 27 V. Этот вредитель в усло-
виях района Кондары кроме большого вреда плодовым насаждениям
приносит вред широколиственным древесным породам.
*32. Горный майский хрущ (Melolontha gussakovskii Med.). Нахожде-
ние в горах нового вида хруща (рис. 25) представляет большой интерес,
Рис. 23. Июньский хрущ. (Ориг.).
тем более что этот вид насекомого зарегистрирован нами как вредитель
плодовых деревьев в лесной горной зоне южных отрогов Гиссарского
хребта (Семенов, 1944).
Биология этого насекомого совершенно еще не изучена. Его ареал,
возможно, узко локализован. (Медведев, 1945).
В Кондаре этот вид наблюдался в гораздо меньших количествах, чем
Melolontha clypeata Rtt. Лёт жуков горного майского хруща происходил
в условиях Кондары с первых чисел мая по 23 мая.
33. Морщинистый заболонник (Eccoptogaster rugulosus Ratz.). Вредит,
живет под корой тонких стволов и ветвей всех видов плодовых деревьев.
Повреждает грушу, яблоню, сливу, вишню, персик, абрикос, черешню,
боярышник, тутовое дерево и др.
*34. Пилильщик-ткач—(Pamphiliidae). В середине мая наблюда-
лась яйцекладка пилильщика-ткача с нижней стороны листьев мин-
даля. Зеленые личинки пилильщика скручивали паутинкой листья
миндаля и объедали их. Вред был незначительный. Окукление про-
исходило в десятых числах июня в земле. Зимуют куколки.
Вероятно это — Neurotoma similis К. Ug. — единственный вид этого
семейства, найденный в Кондаре.
Вредная энтомофауна богарного садоводства в Кондаре
393
35. Боярышница (Aporia crataegi L.) широко распространена по всей
Палеарктике, кроме крайнего севера. Везде она является обычным вре-
дителем и в годы массового размножения приносит большие опустоше-
ния плодовым садам.
Рис. 24. Таджикский майский хрущ.
(Ориг.).
Рис. 25. Горный майский
хрущ. (Ориг.).
В Узбекистане, как отмечает Плотников (1926), заметного вреда
боярышница не приносит.
В Кондаре с весны наблюдаются гусеницы на шиповнике и боярыш-
нике. В первой половине июня происходит лёт бабочек. Яйцекладка
Рис. 26. Туркестанская желтогузка. (Ориг.).
наблюдается в середине и в конце июня, в начале июля происходит
отрождение гусениц, которые впоследствии зимуют в гнездах из листьев,,
склеенных паутиной, на разнообразных деревьях.
36. Непарный шелкопряд (Porthetria dispar L.). Непарный шелко-
пряд — известный, весьма серьезный вредитель плодовых деревьев
394
A. E. Семенов
и лесных лиственных пород. В Средней Азии этот вредитель имеет боль-
шое значение в предгорных районах и прилегающих к ним долинах.
По нашим наблюдениям в Кондаре, гусеницы непарного шелкопряда в
заметных количествах начали выползать из яиц в конце апреля и вначале
Рис. 2/ Горный кольчатый шелкопряд, самец. (Ориг.).
мая. Гусеницы повреждали абрикос, персик, сливу, яблоню, иву, тополь,
фисташку. Окукление гусениц началось с 17 VI, первые бабочки выле-
тели 8 VII. Бабочки откладывали яйца преимущественно на кору
деревьев, яйцекладки имеют вид желтоватой овальной лепешечки.
Рис 28. Горный кольчатый шелкопряд, самка. (Ориг.).
в каждой из них до 500 яиц. Сверху такая лепешечка покрыта слоем волос-
ков с брюшка самки, образующих светлобурый войлочек. Развитие яиц
заканчивается к концу осени, и зимуют уже совершенно сформировав-
шиеся гусенички внутри яйцевой оболочки.
*37. Туркестанская желтогузка (Euproctis kargalica Moore). Турке-
станская желтогузка, как пишет Плотников (1926), в Средней Азии раз-
Вредная энтомофауна богарного садоводства в Кондаре
395
вивается в горах на диких плодовых деревьях; как вредитель большого
значения не имеет.
В Кондаре в течение всего мая и до середины июня гусеницы желто-
гузки питались диким миндалем, сладким миндалем, яблоней, перси-
ком, абрикосом и др. Лёт бабочек (рис. 26) начался 30 VI и закончился
Рис 29. Кольцо яиц горного кольчатого шелкопряда па бухарском
миндале. (Ориг.).
20 VII. Яйцекладка происходила 17 VII Начало отрождения из яиц
гусениц летнего поколения произошло 2 VIII.
Зимует желтогузка, повидимому, в фазе гусеницы в развилках веток,
в зимних гнездах, образованных из нескольких листьев, склеенных пау-
тинками.
*38. Горный кольчатый шелкопряд (Malacosoma parallela Stgr.) обитает
в горах Средней Азии. Биологически он очень близок к европейскому
виду кольчатого шелкопряда (Malacosoma neustria L.), причем последний
396
A. E. Семенов
известен как серьезный вредитель фруктовых деревьев. Горный кольча-
тый шелкопряд отличается от европейского по окраске бабочек и гусе-
ниц (рис. 27 и 28) и тем, что его яйца, отложенные на ветки, всегда по-
крыты белой массой (рис. 29).
По Плотникову (1926) горный кольчатый шелкопряд в Средней Азии
большого значения не имеет. По нашим наблюдениям в Таджикистане,
этот вид шелкопряда в горной лесной зоне южного склона Гиссарского
хребта — бич плодового садоводства и некоторых диких плодовых, как то:
дикого миндаля, яблони, иргая, боярышника, алычи и других. В долинах
шелкопряд не обитает, ниже Сталинабада в Гиссарской долине на плодо-
вых насаждениях отсутствует совершенно. В горной зоне распространен
до высоты 2000 м над ур. м.
рис. 30. Бабочка Павлиноглазка Шенка.
(По А. Е. Семенову).
Зимует горный кольчатый шелкопряд сформировавшейся гусеничкой
внутри яйцевой оболочки, аналогично непарному шелкопряду. Вылу-
пление гусениц весною из яиц совпадает с моментом распускания листьев
бухарского миндаля. В условиях Кондары гусеницы шелкопряда с конца
апреля до конца июня приносят чрезвычайный вред вышеперечислен-
ным растениям. Окукление гусениц происходит в середине июня. В конце
июня наблюдается лёт бабочек и яйцекладка, яйца откладывает на ветки
различных деревьев. Из отложенных яиц гусеницы вылупляются только
весной. В одной кладке бывает от 100 до 400 штук яиц. Для полного
уничтожения листвы одного дерева достаточно 4—5 кладок. В годы раз-
множения шелкопряда в условиях Кондары на одном дереве наблюдается
до 16 кладок при среднем числе — 5. Уничтожив листья одного дерева,
гусеницы шелкопряда, как правило, переползают на соседние деревья.
39. Павлиноглазка Шейка (Neoris huttoni schenki Stgr.) (рис. 30)
является крупной и очень красивой бабочкой, относящейся к семейству
Saturniidae. Этот вид распространен только в горных районах Средней
Азии.
Павлиноглазка широко распространена в лесной зоне гор Таджики-
стана и является постоянным вредителем древесных широколиственных
Вредная энтомофауна богарного садоводства в Кондаре
397
пород (Семенов и Герасимов, 1941). В Кондаре павлиноглазка имеет
в году одну генерацию. Лёт этой ночной бабочки наблюдается в конце
сентября и в начале октября. Яйцекладку бабочки совершают
осенью; зимуют яйца. С наступлением апреля из яиц появляются моло-
дые гусенички, которые быстро растут, и достигая огромных размеров
(рис. 31), становятся очень прожорливыми. Гусеницы этого насекомого
в районе Варзобской станции значительно повреждали яблоню, алычу,
вишню, миндаль дикий и культурный, персик, абрикос, боярышник,
иргай, ясень, клен и другие.
Период вредной деятельности гусениц наблюдается обычно с начала
.апреля и продолжается до середины июня.
Окукление наблюдается с середины июля. Перед окуклением гусе-
ница плетет большой шелковистый сетчатый кокон из шелка с редкими
Рис. 31. Гусеница-Павлиноглазка Шенка.
(По А. Е. Семенову).
неправильными отверстиями. Окукление происходит открыто у земли,
на различных растениях.
Фаза куколки продолжается около четырех месяцев. Первые куколки
наблюдались 3 VI, массовое окукление имело место 10 VI. Вылет бабо-
чек из куколок в лабораторных условиях начался с 27 IX и продолжался
до 15 X. Вылупление бабочек начиналось с 16 час. и к 18—19 часам,
когда становилось темно, бабочки успевали расправить крылья и начи-
нали совершать полет. В природных условиях лёт бабочек наблюдался
с 20 до 23 часов. Бабочки хорошо летят на свет. Лёт бабочек наблюдался
немного больше двух недель (27 IX—14 X). Температурные условия
в октябре позволяют бабочкам спариваться и откладывать яйца.
40. Яблонная плодожорка (Garpocapsa pomonella L.) по своему эко-
номическому значению считается наиболее крупным вредителем плодо-
водства. Этот вредитель распространен повсюду, где только встре-
чается яблоня в диком или культурном виде. Плодожорка нападает
главным образом на яблоню, грушу и айву, а другие плоды, как отме-
чает для Средней Азии Плотников (1926), она повреждает как исключе-
ние. Вредная деятельность плодожорки заключается в том, что гусе-
ницы этого насекомого выгрызают ход внутри плода, питаясь семенами,
и вызывают опадание плодов. В условиях района Кондары плодожорка
в течение вегетационного периода, по всей вероятности, имеет 3 полных
генерации.
41. Сливовая плодожорка (Grapholitha funebrana tr.) широко распро-
странена в Европейской части СССР, в Крыму, на Кавказе, в Средней
398
A. E. Семенов
Азии, на Дальнем Востоке, повреждает плоды сливы, алычи, абрикосаг
черешни. Значение ее как вредителя сливы очень велико.
В Таджикистане сливовая плодожорка имеет в году несколько гене-
раций, но сколько — еще не установлено. Повидимому, сливовая плодо-
жорка, подобно яблонной плодожорке, развивается здесь в трех гене-
рациях.
Зимует сливовая плодожорка в фазе гусеницы, в коконах, в тре-
щинах коры и в дуплах деревьев. Во время образования и роста завя-
зей сливы гусеницы окукляются. Бабочки откладывают яйца на зеленые
плоды сливы и абрикоса. Гусеницы вгрызаются в плод и внутри плода
питаются мякотью, устраивая там длинные ходы вокруг косточки, при
Рис. 32. Повреждения различных сортов плодожоркою.
Слева направо: алы мелкоплодная слива; ренглод зеленый.
этом гусеницы подъедают основные сосудистые стволы, которые питают
ядро (рис. 32). Плоды сливы, поврежденные гусеницами первой генера-
ции, падают на землю.
Перед окуклением взрослые гусеницы покидают поврежденный плод
или вместе с поврежденным плодом падают на землю. Гусеницы всегда
покидают плоды и окукляются в земле. Вылет бабочек второй гене-
рации совершается в период, когда слива достигает значительной
величины.
Развитие второго поколения сливовой плодожорки происходит, при-
мерно, так же, как и первого. Вылетевшие бабочки второго поколения
(рис. 33) откладывают на плоды сливы яйца, которые и дают гусениц
и бабочек третьего поколения. Эти гусеницы не окукляются в земле,,
а коконируются в трещинах коры деревьев и остаются на зимовку.
Где совершается яйцекладка и где питаются гусеницы третьей генера-
ции, пока еще не ясно.
В Таджикистане, в условиях лесной зоны Гиссарского хребта, пло-
дожорка нами была отмечена как серьезный вредитель красноплодной
Вредная энтомофауна богарного садоводства в Кондаре
399
алычи (85—100% поражения), обильно произрастающей в диком состоя-
нии по ущельям.
Этот вредитель распространен и в долинах.
В Кондаре ежегодно очень сильно повреждает сливу; этому способ-
ствует красноплодная алыча, являющаяся резерватором данного вре-
дителя .
42. Чехликовая моль (Eupista hemerabiola Fil.) в Таджикистане рас-
пространена в горах и в долинах. В долинах чехликовая моль повре-
ждает только яблоню, в горных районах она встречается на персике,
абрикосе, миндале, сливе, груше и на других.
Чехликовая моль повреждает почки и листья яблони. В марте—
апреле гусеницы чехликовой моли питаются набухшими почками. Повре-
жденные почки снаружи имеют кругловатые дырочки. Гусенички вгры-
заются внутрь почек, при сильном повреждении почки совсем отмирают,
при слабом — дают уродливые листья.
Наибольший вред дереву, как считает Плотников (1926), чехликовая
моль наносит ранним, еще не тронувшимся в рост, почкам. При обиль-
Рис. 34. фруктовая полосатая моль.
(Ориг.).
Рис. 33. Сливовая плодожорка.
(Ориг.).
ном размножении чехликовой моли, что случается иногда на яблонях,
все листовые и цветовые почки повреждаются и отмирают, а дерево долго
остается без листьев.
После распускания почек гусеницы повреждают листья; они вгры-
заются в мякоть листа и выедают небольшими участками паренхиму
между верхней и нижней кожицей. Сильно испещренный такими повре-
ждениями лист желтеет, скручивается, частично листья осыпаются или
остаются висеть на дереве.
Затем гусеницы окукливаются. В конце мая, в начале июня начи-
нают летать бабочки. В условиях Кондары лёт бабочек наблюдается
в начале июля. Бабочки откладывают яйца на нижнюю сторону
листьев яблони вдоль жилок. Вылупившиеся из яиц гусенички сильно
повреждают листья яблони — это происходит в начале июля и продол-
жается до конца лета, а затем гусеницы перестают питаться и уходят
на зимовку.
43. Плодовая моль (Hyponomeutha padellus L.) является серьезным
вредителем косточковых плодовых деревьев,, широко распространена
в Крыму, на Кавказе и в Средней Азии.
Плодовая моль по внешнему виду мало чем отличается от всем хорошо
известной яблонной моли Hyponomeutha malinellus L. Отличие заклю-
чается в том, что плодовая моль многоядна и развивается на многих
диких древесных растениях и на яблоне, в то время как яблонная моль
живет только на яблоне (Витковский, 1914). Гусеницы плодовой моли
400
A. E. Семенов
в Кондаре и в Зиддах повреждали листья боярышника, алычи и яблони.
В Кондаре гусеницы повреждали перечисленные растения в течение мая,
•окукление гусениц началось с 1 V Начало лёта бабочек наблюдалось
с И VI.
44. Фруктовая полосатая моль (Anarsia lineatella L.) (рис. 34) широко
распространена на Кавказе и в Средней Азии. В Средней Азии ее отметил
Плотников (1926), который сообщил, что гусеницы этой бабочки питаются
в плодах и почках абрикоса и персика. Под Ташкентом Яхонтову (1929)
удалось вывести один экземпляр бабочки этого вида из гусеницы, оби-
тавшей в побегах миндаля.
В условиях Кондары нам удалось наблюдать в июле гусениц фрук-
товой моли, в больших количествах повреждавших плоды дикого
миндаля. Поврежденные плоды миндаля, кроме гусениц A. lineatella
L., содержали в большом количестве гусениц другого вида — плодо-
жорки (Laspeyresia sp.). Из 500 проанализированных плодов миндаля
466 плодов (92.4%) было повреждено одновременно двумя видами.
По всей вероятности, эти же виды плодожорок в условиях Кондары
повреждают плоды персика и абрикоса.
Окукление гусениц фруктовой полосатой моли началось с 12 VII.
Лёт бабочек наблюдался с 22 VII по 10 VIII.
ЛИТЕРАТУРА
Архангельский П. П. 1917. К биологии персиковой тли — Pterochloroides
persicae Col. Ташкент.
Витковский И. В. 1914. Яблонная и плодовая моль. Кишинев.
Медведев С. И. 1945. Новый вид хруща (Coleoptera, Scarabaeidae) из Таджи-
кистана. Энтомол. обозр., XXVIII, 1—2.
Мордвилко А. К. 1927. Кормовые растения тлей СССР и сопредельных стран.
Мориц-Романова. 1927. Обзор вредителей сельского хозяйства Туркмении.
Невский В. П. 1929. Тли Средней Азии. Ташкент.
Плотников В. И. 1926. Насекомые, вредящие хозяйственным растениям в Сред-
ней Азии. Ташкент.
Рейхардт А. Н. 1927. Описание нового вредного вида слоника из Бухары. Защита
растений, IV
Семенов А. Е. 1940. Большая белокрылая цикада — Tibieina zevara V Kuzn.,
как вредитель богарного садоводства. Докл. ВАСХНИЛ, № 10.
Семенов А. Е.иД. А. Оглоблин. 1941. Два новых вида жуков-листоедов.
Докл. ВАСХНИЛ, № 1.
Семенов А. Е. 1944. Значение подвоев богарного садоводства как резерваций
вредных насекомых в Таджикистане. Изв. Таджикск. филиала Академии Наук
СССР, № 5.
Семенов А. Е. 1945. Жуки-златки рода Capnodis, вредящие богарному садовод-
ству в Таджикистане, и меры борьбы с ними. Энтомолог, обозр., № 1—2.
Семенов А. Е. иА. М. Герасимов. 1941. Новый вредитель богарного садо-
водства павлиноглазка Шенка. Докл. ВАСХНИЛ, № 6.
Яхонтов В. В. 1929. Список вредителей хозяйственных растений и зарегистри-
рованных на них хищников и паразитов. Тр. Ширабудск. оп. станции. № 2.
Bodenheimer F. S. 1930. Die Schadlingsfauna Paliistinas. 217—221.
Felt E. P. 1913. Periodical Cicada (Tibieina septendecim Linn.). Journ. Ent. Soc.,
XXI. New York.
Marlatt G. Z. 1907. A successful seventeennyear breeding record for the periodical
cicada. Proc. Ent. Soc. Washington. IX.
Marlatt G. Z. 1907. The Periodical cicada. U. S. A. Dept. Agr. Bur. Ent. Bull.,
LXXI.
В. Ф. КУПРЕВИЧ
БОЛЕЗНИ РАСТЕНИЙ УЩЕЛЬЯ КОНДАРА
В ущелье Кондара насчитывается свыше 750 видов явнобрачных
растений. Подавляющее большинство их поражается грибными, бактери-
альными или вирусными болезнями. Сведения о болезнях растений Гис-
сарского хребта, как, впрочем, для всей горной части Таджикистана,
весьма скудны. В 1892 —1893 гг. в Самаркандской области путешествовал
В. JI. Комаров; в эти годы им собрана значительная коллекция паразитных
грибов, преимущественно ржавчинных, с Зеравшанского и Гиссарского
хребтов. В списке, опубликованном Комаровым (1894), приводится 106
видов грибов, из них 76 ржавчинных и 19 мучнисторосяных; описано
13 новых видов. Наиболее полный перечень грибов Средней Азии опу-
бликован Н. Г. Запрометовым (1926, 1928). Автор использовал обшир-
ный гербарный материал, собранный разными исследователями, и опу-
бликованные ранее списки (Сорокин, 1884; Комаров, 1894; Спешнев,
1901, и др.). В списке упоминается несколько видов из горной части
Таджикистана. В последние годы наиболее обстоятельные исследования
микрофлоры Средней Азии, в частности Памира, производились П. Н. Го-
ловиным (1941, 1944).
Первые наблюдения над болезнями растений и сборы гербарного
материала в ущелье датированы 1939—1941 гг.; производились они
фитопатологами Н. Олтаржевским и Т. Роткевич. В последующие годы,
1943 —1946, болезни растений изучались более планомерно. Накопленный
за истекшее время материал сравнительно невелик — около 300 гербар-
ных листов. Сопоставляя эту цифру с общим количеством явнобрачных —
свыше 750 видов, можно видеть, насколько недостаточны проведенные
наблюдения и сборы. Многие паразиты, особенно растений, имеющих
сравнительно небольшой ареал и недавнюю известность, изучены очень
слабо. Наблюдения над болезнями и сборы пораженных растений произ-
водились большею частью эпизодически, вследствие чего паразитные
грибы на рано отцветающих эфемерах могли остаться незамеченными.
Основное содержание настоящего краткого обзора составляют болезни
сельскохозяйственных культур и древесных пород, наиболее обстоя-
тельно изученные. Обзор болезней дикорастущих травянистых растений
ограничен небольшим числом видов. Сравнительно хорошо изучены
ржавчинные грибы, занимающие по количеству видов и частоте встре-
чаемости в ущелье одно из первых мест. Перечень видов паразитных
грибов составлен по нашим наблюдениям и сборам 1944—1946 гг.
Состав болезней сельскохозяйственных культур в ущелье и в сосед-
ней Гиссарской долине примерно одинаков. Различия незначительны;
состоят они в том, что некоторые культуры, в частности помидоры, пора-
26 Ущелье Кондара
402
В. Ф. Купревич
жаются в долине гораздо сильнее. Сильно поражаются в ущелье неко-
торые плодовые деревья. Так, основное насаждение абрикоса на тер-
ритории Станции настолько страдало вследствие камедетечения, что
в 1943 г. его пришлось вырубить. В 1944 г. наблюдалось сплошное пора-
жение мучнистой росой и «курчавостью листьев» многих сортов персика.
В ущелье повсеместно распространен ржавчинник Tranzschelia pruni-
spinosae, поражающий бухарский миндаль; на сладком миндале гриб
не наблюдался. Эцидии этого ржавчинника ежегодно в большом коли-
честве наблюдаются на Anemone Tschernjaewii Rgl. На абрикосе и мин-
дале сладком сильно распространен кластероспориоз; в середине лета
листья этих деревьев становятся настолько изрешеченными грибом, что
остаются лишь центральная и одна-две боковые жилки с полузасохшими
обрывками ткани между ними.
Большой стойкостью отличаются яблоня и груша; в течение послед-
них четырех лет на них наблюдалась только обыкновенная парша; между
тем в долине, в частности в окрестностях Сталинабада, яблоня и груша
сильно поражаются паршой, плодовой гнилью и другими грибами.
Успешно культивируется виноград, представленный на опытных участках
Станции многими превосходными сортами, местными и европейскими.
На листьях отдельных лоз изредка наблюдалась пятнистость (Cercospora,
Gloeosporium), никогда не достигавшая сильного развития. В двух слу-
чаях замечена мучнистая роса в виде очень слабо развитого налета
на молодых листьях. Вовсе не обнаружен бактериальный рак, являю-
щийся бичом ряда виноградников в окрестностях Сталинабада.
Поразительно стоек здесь помидор. Между тем в долине эта куль-
тура жестоко поражается вирусными болезнями, чаще всего скручива-
нием листьев и болезнью, патографически сходною со столбуром. Так,
в 1944 г. крупные плантации помидора во многих колхозах (в долине)
были запаханы вследствие сплошного поражения и гибели растений.
В том же году на большой плантации, расположенной возле Станции,
ни одного больного помидорного растения не наблюдалось. В последую-
щие годы на отдельных листьях и плодах, получивших механические
повреждения, замечена бактериальная гниль, и в одном случае Clado-
sporium sp.
Огурцы и другие тыквенные из года в год поражаются в слабой сте-
пени бактериозами. Свекла и капуста проявляют признаки сильного
угнетения, растут плохо, вероятно вследствие неблагоприятных почвен-
ных условий. Не поддавался культуре картофель. В 1943 г. он оказался
сплошь зараженным вирусными болезнями и в последующие годы не
высаживался. Впрочем, в соседних кишлаках, Гажни и Бигар, кар-
тофель растет неплохо.
Из дикорастущих ценных пород особенно сильно поражается орех.
Ежегодно в середине лета на его листьях наблюдается обильное разви-
тие несовершенного гриба Marssonia juglandis Magn., вызывающего пят-
нистость. Нередко гриб переходит на молодые плоды, вызывая их прежде-
временное засыхание и опадание. Особенно сильно поражаются деревья,
расположенные в тенистых и сырых местах или непосредственно на берегу
р. Кондара. В этих условиях орех довольно часто поражается также
рядом других паразитов, в частности — многими видами трутовиков. Чаще
других встречается Inonotus hispidus, причиняющий значительный
вред.
На листьях фисташки наблюдалась пятнистость, вызванная грибом
Septoria pistaciae. Особенно сильно поражались деревья, расположен-
Болезни растений ущелья Кондара
403
ные на склоне южной экспозиции. Пятна, вначале небольшие и слабо
заметные, постепенно разрастаются, распространяясь на всю пластинку.
Отмершие и засохшие листья обычно не опадают. Они сохраняются
на ветвях до осени. Трутовый гриб Phellinus rimosus, дважды собранный
на старых деревьях, вероятно большого значения не имеет.
Широко распространены в ущелье виды Gymnosporangium, поражаю-
щие иргай и боярышник. Нередко листья и особенно плоды этих пород
сплошь покрываются густой «щетиной» спороношений ржавчинника (см.
список). Телейтоспоры гриба образуются на Juniperus seravschanica;
их можно наблюдать глубокой осенью, в период дождей, когда они при-
обретают вид небольших скоплений оранжево-желтой студенистой массы
на ветвях и молодых побегах.
Высокой устойчивостью к болезням характеризуются в изобилии
произрастающие дикие алыча и виноград. Дикий виноград представляет
основное и нередко господствующее растение во многих местах, в част-
ности, боковых ущельях южной экспозиции. Одно из этих ущелий, полу-
чившее название «Виноградное», отличается изумительным разнообра-
зием произрастающего в нем дикого винограда. Наиболее распространен-
ная разновидность имеет сравнительно мелкие черные ягоды в неболь-
ших довольно рыхлых гроздьях. Однако здесь же встречаются лозы, несу-
щие грозди до 1 кг весом. Цвет и величина ягод исключительно разно-
образны. Так, на отдельных лозах или группах их наблюдались светло-
зеленые, зеленовато-желтые, золотисто-желтые, желтые с розовым или
красноватым основанием, наконец, красные и красновато-бурые ягоды.
Величина их варьирует от 0.5 до 2.5 см в диаметре; форма также под-
вержена сильной изменчивости, — чаще всего ягоды широко эллипсо-
идальные, реже — яйцевидные и шаровидные, у некоторых лоз — с высту-
пающим в виде сосочка носиком, в одном случае — асимметричные,
с боков как бы сдавленные, напоминающие плоды фисташки. Вкус вино-
града в зрелом виде сладкий (черные и желтые разновидности), кисло-
вато-сладкий (зеленовато-желтые), медово-сладкий (желтые с розовым
основанием); кислые разновидности отсутствуют. Некоторые дикорасту-
щие лозы по своим качествам не уступают многим культурным сортам
и с упехом могли бы войти в культуру. Все эти многочисленные разно-
видности отличаются, как указывалось выше, высокой устойчивостью
к заболеваниям. Единственное заболевание, наблюдавшееся преимуще-
ственно на лозах, растущих у р. Кондара, имеет, вероятно, функцио-
нальную природу. Проявляется оно изменением окраски и последующим
отмиранием участков ткани листа (см. Vitis vinifera, 64).
Некоторые болезни, как, например, мучнистая роса персика и кла-
стероспориоз абрикоса, причиняют значительный ущерб. Борьба с ними
на опытных участках Станции велась в соответствии с правилами, пери-
одически издаваемыми Министерством сельского хозяйства СССР. Резуль-
таты ее далеко не всегда оправдывали возлагаемые надежды. Несо-
мненный успех имело опрыскивание персика бордоской жидкостью против
«курчавости листьев» и опыливание серными препаратами против муч-
нистой росы. Малой эффективностью отличалась борьба с кластероспори-
озом. Почти все мероприятия по борьбе с болезнями проводились на участ-
ках, расположенных на территории, примыкающей к р. Варзоб, и отчасти
на участках, смежных со зданием Станции, на дне ущелья. В меньшей
мере страдали от болезней опытные посадки на склонах южной и север-
ной экспозиций. Это счастливое обстоятельство освобождало сотруд-
ников от необходимости обрабатывать фунгисидами труднодоступные
26*
404
В. Ф. Купревич
террасы и, особенно, мелкие каменные площадки, лепящиеся, напо-
добие ласточкиных гнезд, по крутым склонам.
Желательна разработка мероприятий против болезней ореха, в частно-
сти против пятнистости листьев и плодов. Возможно проведение элемен-
тарных мероприятий, уборка захламленных валежником участков (район
Танге-мура) и удаление отмерших или отмирающих частей кроны старых
деревьев. Эти мероприятия частично осуществляются в процессе заготовки
топлива. Некоторые очень старые, полузасохшие деревья, являющиеся
источником инфекции, целесообразно удалить. Весьма желательна орга-
низация элементарного ухода за молодым подростом ореха и охрана его.
Обзор паразитных грибов (и болезней) изложен по питающим расте-
ниям, в порядке их системы. По числу видов первое место занимают
ржавчинники — 56 видов из общего количества 106. Несовершенные
(12 видов) и сумчатые (10 видов) изучены недостаточно; дальнейшие
исследования несомненно увеличат список представителей этих грибов.
Из ущелья, включая плоскогорие Руидашт, описано 5 новых видов ржав-
чинных грибов, 2 новые формы мучнисторосяных и один вид из группы
несовершенных. Новый вид ржавчинника на реликтовой Ostrowskia
magnifica, широко распространенный в ущелье Кондара, назван по имени
его — Aecidium kondariense Кпрг. Помимо болезней, вызываемых гри-
бами, обнаружены бактериозы ореха и некоторых других растений.
О функциональной болезни виноградной лозы упоминалось выше. Вирус-
ные болезни, обнаруженные на малине и других растениях, в список
не включены ввиду слабой изученности их и трудности определения;
необходимы систематические наблюдения в течение хотя бы одного веге-
тационного сезона.
Гербарный материал из ущелья и плато Руидашт обработан в Отделе
споровых растений Ботанического института Академии Наук СССР, где
и оставлен на хранение; дублеты переданы в гербарий Ботанического
института Таджикского филиала АН СССР. В определении ряда дико-
растущих питающих растений существенную помощь оказали мне бота-
ники А. И. Пояркова и В. В. Письяукова. Коллекция трутовых грибов
определена А. С. Бондарцевым.
СПИСОК ПАРАЗИТНЫХ ГРИБОВ (И БОЛЕЗНЕЙ) УЩЕЛЬЯ КОНДАРА
(По семействам питающих растений)
Сем. Cupressaceae
1. Juniperus seravschanica Korn. 1. Gymnosporangium fusispo-
rum Ed. Fisch. (Траншель, 75).Cno-
роношение (телейтоспоры) на вет-
вях, в виде оранжево-желтой сту-
денистой массы (в намокшем состоя-
нии). — Вблизи Станции, северный
склон, 4 XI 1944.
Сем. Gramineae
2. Imperata cylindrica (L.) 2. Sphacelotheca Schweinfurthi-
P. B. ana (Thum.) Sacc. (Гутнер, 136).
Гриб разрушает соцветия. — У р.
Варзоб, 2 IX 1945. Часто.
Болезни растений ущелья Кондара
405
3. Andropogon ischaemum L.
4. Calamagrostis pseudophrag-
mites (Hall.) Koeler.
5. Gynodon dactylon (L.) Pers.
6. Phragmites communis Trin.
7. Poa nemoralis L.
8. Triticum cult.
9. Hordeum cult.
10. Hordeum bulbosum L.
3. Sphacelotheca andropogonis
(Opiz) Bubak (Гутнер, 137). Гриб
полностью разрушает соцветия. —
У р. Варзоб, 2 IX 1945.
4. Puccinia coronata Corda.
(Траншель, 112). Уредо- и телейто-
споры на листьях и листовых вла-
галищах. — Квак, 30 VII 1945.
5. Puccinia cynodontis Desm.
(Траншель, 98). На листьях и вла-
галищах. — У р. Варзоб, 2 1X1945.
6. Ustilago cynodontis Р Henn.
(Гутнер, 66). Гриб разрушает соцве-
тия. Часто. — У р. Варзоб, 2 IX
1945.
7. Puccinia phragmitis (Schum.)
Korn. (Траншель, 94).—Ущелье,
14 IX 1944. Сравнительно редко.
8. Uromyces роае Rabh. (Тран-
шель, 108). — Ущелье, в трещине
скалы, 7 IX 1945.
9. Puccinia triticina Erikss.
(Траншель, 108). — Ущелье, юж-
ный склон, в посеве, VI 1943.
10. Ustilago hordei (Pers.) Kel-
lerm. et Sw. (Гутнер, 95). — Руи-
дашт, 16 IX 1945. Довольно часто.
11. Erysiphe graminis DC.
(Ячевский, 141). — Ущелье, 27 V
1944.
Сем. Cyperaceae
11. Carex decaulescens V.Krecz.
12. Carex melanostachya M. B.
13. Carex Koshewnikowii Litw.
12. Puccinia caricis A. Kleb.
(Траншель, 123). — Руидашт, 16
IX 1945.
13. Cintractia caricis (Pers.)
Magn. (s. 1.; Гутнер, 157). Квак,
9 VIII 1944.
14. Puccinia sp.--Ущелье, 27
IV 1944.
Сем. Liliaceae
14. Eremurus comosus O.
Fedtsch.
15. Allium sp.
16. Rhinopetalum bucharicum
(Rgl.) A. Los. (Fritillaria bucha-
rica Rgl.).
15. Puccinia eremuri Kom.
(Траншель, 144). — Ущелье, 29 V
1941, собр. H. Олтаржевский.
16. Caeoma alliorum Link. (Тран-
шель, 139). — Ущелье, 10 IV 1944.
17. Uromyces mogianensis Bu-
bak (Траншель, 143). — Ущелье,
3—28 IV 1944. Часто.
406
В. Ф. Купревич
17. Tulipa praestans Hoog. 18. Puccinia tulipae Schrdt.
(Траншель, 143). — Квак, 9 V 1944.
19. Ustilago Heufleri Fuck. (Гут-
нер, 101). — Квак, 24 V 1944.
Сем. Salicaceae
18. Salix sp. 20. Melampsora salicina Lev. (s.
1.; Траншель, 156). — Ущелье, VIII
1946. Часто.
19. Populus tadshikistanica 21. Melampsora populina (Pers.)
Kom. Lev. (Траншель, 154). — Ущелье,
28 IX 1944. Часто.
20. Populus pyramidalis Rosier. 22. Fomes fomentarius (Fr.)
Gill. На стволе живого дерева. —
Усадьба, 8 IV 1944.
Сем. Juglandaceae
21. Juglans regia L. 23. Inonotus hispidus (Bull.)
Karst. На стволе живого ореха. —
У р. Кондара, 11 IV 1944. Гриб
встречается довольно часто; наибо-
лее вредоносный из встречающихся
в ущелье трутовиков.
Fomesfomentarius (Fr.) Gill. На
стволе живого ореха. — Ущелье,
9 IX 1944.
24. Ganoderma applanatum
(Wollr.) Pot. На отмерших ство-
лах. — Ущелье, 23 X 1944.
25. Cerrena unicolor (Bull.)
Murr. На мертвом стволе. —
Ущелье, 10 IX 1945; Квак, 12 IX
1945.
26. Marssonia juglandis Magn.
(Gnomonia leptostyla Ces. et De
Not.). (Oudemans, II, 289). Гриб
вызывает пятнистость листьев; пят-
на сначала мелкие, разбросанные,
желтовато-бурые; затем распростра-
няются на всю пластинку; пора-
женные листья отмирают и опа-
дают. Гриб переходит на молодые
плоды, вызывая их преждевремен-
ное отмирание и опадание. —
Ущелье, 27 V 1944. Но всему
ущелью, часто.
27. Microstroma juglandis Sacc
(Oudemans, II, 289). Гриб вызы-
вает беловатые, неправильные или
Болезни растений ущелья Кондара
407
угловатые пятна на листьях. —
Ущелье, уроч. «Ферма», 27 V 1944.
Сравнительно редко.
28. Pseudomonas (Phytomonas)
Juglandis Pierce (Ячевский, Бак-
териозы, 338). Бактериоз. На
листьях молодого (4—5 лет) ореха
болезнь проявляется в виде неболь-
ших округлых или угловатых чер-
но-бурых пятен (с беловатой точ-
кой в центре), окруженных по пери-
ферии светложелтым полупрозрач-
ным (в проходящем свете) колеч-
ком. В пораженной ткани в зна-
чительном количестве встречаются
амилопласты, 8—30а диам. Скопле-
ние бактерий обычно наблюдается
в клетках, расположенных не-
посредственно возле отмершей тка-
ни, на границе пятна. На листьях
старых деревьев бактериоз чаще
всего проявляется побурением жи-
лок, создающим картину штрихо-
ватости, и образованием неболь-
ших некротических пятен. Болезнь
переходит на плоды. — Ущелье,
9 V 1945, 7 IX 1945; особенно силь-
ное развитие бактериоза наблю-
далось в 1946 г., в августе и сен-
тябре.
Сем. Ulmaceae
22. Celtis caucasica Willd. Cerrena unicolor (Bull.) Murr. Ha
коре живого дерева. — уроч. Танге-
мур, 10 IX 1945. (См. Juglans).
Сем. Santalaceae
23. Thesium Gontscharowii 29. Puccinia thesii-decurrentis
Bobr. (P. Henn.) Diet. (Траншель, 164). Эцидии и телейтоспоры. — Квак, 2 VIII 1945, собр. В. Письяукова.
Сем. Polygonaceae
24. Atraphaxis pyrifolia Bge.
25. Polygonum paronychioides
C. A. Mey.
30. Leptothyrium sp. — Ущелье,
9 VII 1941, собр. T. Роткевич.
31. Uromyces polygoni (Pers.)
Fuck. (Траншель, 169).—Ущелье,
южные склоны, 31 VIII 1945.
408
В. Ф. Купревич
26. Polygonum bucharicum 32. Puccinia monticola Кот.
G. Grig. (Траншель, 171). — Квак, 20 VIII
1944; по тропе на Руидашт, 16 IX
1945.
Сем. Caryophyllaceae
27. Arenaria Grilfithii Boiss. 33. Puccinia arenariae-Griffithii
Kuprewicz (sp. n.). На живых
листьях. Уредоспоры шаровидные
или широко эллипсоидальные,
светлобурые, гладкие, с 2 порами,
18—24 ел диам., телейтоспоры в рых-
лых кучках, светлобурые, глад-
кие, 18—24 х 26—35 и. — Руи-
дашт, 16 IX 1945.
Сем. Ranunculaceae
28. Anemone Tschernjaewii Rgl. 34. Tranzschelia pruni-spinosae
(Pers.) Diet. (Траншель, 237). —
Северный склон, 10 IV 1944. Весь-
29. Ranunculus ма часто. Эцидиальная стадия; гриб переходит на виды Amygdalus. tenuilobus Rgl. 35. Aecidium ranunculacearum
30. Thalictrum Stapf. DC. (s. 1. Траншель, 195). — Ущелье, 28 IV 1944. sultanabadense 36. Aecidium thalictri-flavi Wint. (Траншель, 195). — Ущелье, 4 IV 1944.
31. Cotoneaster jark. Сем. Rosaceae insignis A. Po- Gymnosporangium fusisporum Ed. Fisch, (cm. Juniperus seravscha- nica Kom.). — Ущелье, 1 VIII1944. Часто.
32. Cotoneaster jark. hissarica A. Po- Gymnosporangium fusisporum Ed. Fisch. — Ущелье, 1 VIII 1944. Часто.
33. Cotoneaster A. Pojark. 34. Cotoneaster et M. nummularioides Gymnosporangium fusisporum Ed. Fisch. 17 X 1944. Часто. nummularia F. Gymnosporangium fusisporum Ed. Fisch. 16 IX 1945. Ржавчин- ник на видах Cotoneaster весьма
распространен в ущелье; особенно
поражаются виды, растущие на дне
ущелья. Исключением является Co-
toneaster с малиново-красными пло-
дами, по определению А. И. Пояр-
4
Болезни растений
ущелья Бонда pa 409
35. Pyrus communis L. Груша.
36. Malus hissarica Kudr. Дикая
яблоня.
37. Malus domestica Borkh.
Яблоня.
38. Crataegus pontica C. Koch.
Боярышник.
39. Crataegus turkestanica A. Po-
jark.
40. Crataegus hissarica A. Po-
jark.
41. Rubus caesius L. var. turke-
stanicus Rgl.
42. Fragaria ananassa Duch.
Земляника.
43. Potentilla transcaspica Th.
Wolf.
ковой, — C. multiflora Bge., вовсе
не поражающийся грибом. Этот вид
занимает небольшую площадь на
северном склоне ущелья, непосред-
ственно перед первым поворотом
тропы на северный склон (перед
«Фермой»). Ржавчинник несомненно
связан с Juniperus serawschanica
Кот., принадлежность его к Gym-
nosporangium fusisporum сомни-
тельна. Необходимы опыты с искус-
ственными культурами гриба на
обоих питающих растениях.
37. Fusicladium pirinum Fuck.
(Venturia pirina Aderh.). Преиму-
щественно на плодах. — Усадьба,
17 VIII 1944.
38. Fusicladium dendriticum
Fuck. (Venturia inaequalis Wint.).
Преимущественно на плодах. —
Ущелье, 17 VIII 1944; 21 VIII1944.
Часто. Inonotus hispidus (Bull.)
Karst. На живом стволе. — Квак,
25 IX 1944.
Fusicladium dendriticum Fuck.—
—Усадьба, 21 VIII 1944. Преиму-
щественно на плодах.
39. Gymnosporangium confusum
Plowr. (Траншель, 76). Эцидии на
листьях и плодах. — По тропе на
Квак, 4 VII 1941, соор. Т. Ротке-
вич; там же, 24 V 1944; 17 IX 1944.
Очень часто сильно поражаются
листья и плоды. Телейтоспоры,
повидимому, на Juniperus serav-
schanica Kom.
Gymnosporangium confusum
Plowr. — Ущелье, 17 IX 1944.
Gymnosporangium confusum
Plowr. — Ущелье, 24 V 1944.
40. Phragmidium rubi (Pers.)
Wint. (Траншель, 233). —Ущелье,
24 VIII 1945. Сравнительно часто.
41. Ramularia Tulasnei (Mycos-
phaerella fragariae Lind.). Пятнис-
тость листьев; пятна округлые»
красноватые, затем беловатые с тем-
нокрасным ободком. — Усадьба,
5 VI 1944, VIII 1945.
42. Phragmidium potentillae
(Pers.) Karst. (Траншель, 231). На
листьях. Часто. — Ущелье, 27 V
1944.
410
В. Ф. Купревич
44. Geum kokanicum Rgl. et Schmalh. 43. Phragmidium circumvalla- tum Magn. (Траншель, 232). — Руи- дашт, 18 VI 1945, собр. В. В. Письяукова.
45. Poterium sanguisorba L. 44. Phragmidium sanguisorbae (DC.) Schrot. (Траншель, 230). — Ущелье, 15 IX 1945.
46. Rosa dumetorum Thuill. 45. Phragmidium tuberculatum J. Miill. (Траншель, 235). — Ущелье, 31 VIII 1944, 24 VIII 1945.
47. Rosa kokanica Rgl. Phragmidium tuberculatum J. Miill. На листьях. — Северный склон, 27 V 1944. 46. Phragmidium devastatrix Sorok. (Траншель, 233). Гриб пора- жает целые побеги, вызывая их де- формацию. Спороношение обиль- ное, иногда сплошь по всему побе- гу. — Ущелье, 24 VIII 1945.
48. Prunus domestica L. Слива. 47. Polystigmina rubra Sacc.; сумчат. стадия Polystigma rubrum DC. (Oudemans, III, 725). Красно- ватые подушковидные пятна на листьях. — Усадьба, X 1945. 48. Clasterosporium carpophi- lum (Lev.) Aderh. (Oudemans, III, 698). На листьях; вследствие выпа- дения отмерших участков ткани листья становятся продырявлен- ными. — Усадьба, IX 1945.
49. Prunus divaricata Ldb. Алыча. 49. Coriolus versicolor (L.) Quel. На мертвом стволе. — Ущелье, 24 VIII 1945.
50. Amygdalus bucharica Korsh. Миндаль горький. Tranzscheliapruni-spinosae (Pers.) Diet. На листьях черные раз- бросанные подушечки телейтоспор. Эцидии на Anemone Tschernjae- wii Rgl. — Ущелье, 10 IX 1945. Очень часто. На сладкий миндаль гриб в обычных условиях не пере- ходит. Polystigmina rubra Sacc. — Ущелье, 10 IX 1945. Часто.
51. Amygdalus communis L. Миндаль сладкий. Polystigmina rubra Sacc.— Усадьба, 10 IX 1945. Сравнительно часто. Clasterosporium carpophilum (Lev.) Aderh. —Усадьба, 19 IX 1945. Часто, поражение сильное.
52. Cerasus mahaleb (L.) Mill. 50. Phellinus pomaceus (Pers.) Quel. На живом стволе. — Вино- градное ущелье, 19 IX 1946.
Болезни растений ущелья Кондара
411
53. Armeniaca vulgaris Lam. Аб- рикос. Clasterosporium carpophilum (Lev.) Aderh. — Усадьба, 19 IX 1945. Сильное поражение; еже- годно. 51. Камедетечение. На пора- женных деревьях наблюдается обильное выделение камеди, обра- зующей наплывы разной величины. В местах усиленного образования и выделения камеди, под наплыва- ми, всегда наблюдались механиче- ские повреждения (трещины, ссадины, повреждения насекомыми, некроз ткани, вызванный гриба- ми, и т. п.). Болезнь распростра- нилась по всему насаждению, вы- звав массовое усыхание абрикоса. Усадьба, 1942—1946 гг.
54. Persica vulgaris Mill. Персик. 52. Exoascus deformans (Berk.) Fuck. На листьях и цветах. — Усадьба, 5 IV 1944. Сильное пора- жение, вызвавшее массовое опада- ние цветов. 53. Sphaerotheca pannosa Lev. var. persicae Woronichin. Мучни- стая роса. — Усадьба, 23 VII 1940, собр. Н. Олтаржевский; там же, 10 V 1944. Сильное поражение листьев.
Сем. Leguminoseae
55. Medicago sativa L. Люцер- на. 54. Phyllosticta medicaginis Rab. et Desm. — Квак, 9 IX 1941, собр. T. Роткевич.
56. Trifolium repens L. Клевер ползучий. 55. Uromyces nerviphilus (Gro- gnot) Hotson (Траншель, 235). — Ущелье, 6 VII 1943, собр. T. Рот- кевич; Квак, 18 IX 1944. 56. Uromyces trifolii-repentis (Cast.) Liro. (Траншель, 251). Руи- дашт, 16 IX 1945.
57. Astragalus lasiosemius Bo- iss. 57. Uromyces astragali-lasiose- mi Kuprewicz (sp. n.). На живых листьях; телейтокучки темнобу- рые, эллипсоидальные, 1—2 мм длиной. Телейтоспоры светлобу- рые, широко эллипсоидальные, ша- ровидные, реже продолговатые, 18 —21 X 19—26 а, оболочка около 2 (л толщины, у вершины не утол- щенная или слабо утолщенная,
гладкая. Уредоспоры светлобурые,
шаровидные или широко эллипсо-
идальные, 15—18 х 17—21 а. -
Руидашт, 16 IX 1945. Часто.
Сем. Anacardiaceae
58. Pistacia vera L. Фисташка. 58. Phellinus rimosus (Berk.)
Bond, et Sing. На живом стволе. —
Южный склон ущелья, 6V 1944;
там же, 31 VIII 1945.
59. Septoria pistaciae Desm.
(Oudemans, III, 1113). Пятнистость
листьев. — Южный склон, 31 VIII
1945. Болезнь широко распростра-
нена.
Сем. Асегасеае
59. Acer turkestanicum Pax. 60. Rhitisma acerinum Fries
(Melasmia acerina Lev.) (Oudemans,
III, 1155). На листьях. — По тропе
на Квак, 7 IX 1945. Сильное пора-
жение, повсеместно.
60. Acer Regelii Pax. Rhitisma acerinum Fries. — Се-
верный склон, 14 IX 1945. Пора-
жается слабее предыдущего.
Сем. Balsaminaceae
61. Impatiens parviflora DC. 61. Puccinia Komarovii Tranz.
(Траншель, 247). — Квак, 15 IX
1944. Обильно, повсеместно.
Сем. Rhamnaceae
62. Rhamnus dolichophylla 62. Puccinia rhamni Kuprewicz
Gontsch. (sp. n.). На живых листьях. Эци-
дии на нижней стороне листьев.
Телейтокучки преимущественно на
нижней стороне листьев, выступаю-
щие из-под разорванного эпидер-
миса, округлые, реже неправиль-
ные, продолговатые, расположен-
ные вдоль жилок, черные. Телей-
тоспоры с 1—4 пальцевидными при-
датками у вершины, верхняя
клетка более или менее округлая,
нищняя продолговатая, светло
окрашенная; 36—66 х 15—21 jx.—
Квак, «Заххана», 6 IX 1944; там же
15 IX 1944. — Усадьба, 8 IX 1944.
Болезни растений ущелья Кондара
413
Сем. Vitaceae
63. Vitis vinifera культурный. L. Виноград 63. Gloeosporium ampelopha- gum Sacc. (Oudemans, III, 1275). На листьях и молодых побегах. — Усадьба, 12 IX 1940, собр. Н. Ол- таржевский. 64. Cercospora Roesleri Sacc. (Ou- demans, III, 1280). На листьях.— У p. Варзоб, VI—VIII 1944. 65. Oidium Tuckeri Berk. (Unci- nula necator Burr; Ячевский, 370). На листьях, очень слабое пораже- ние. — У р. Варзоб, V 1944.
64. Vitis vinifera дикий. L. Виноград 66. Функциональная болезнь. Отдельные листья, реже — почти все листья побега приобретают светлофиолетовую, иногда желто- ватую или красноватую окраску; пятна постепенно распространяют- ся от периферии к центру, захва- тывая все большую площадь. От- мершие участки бледнеют, мацери- руются и опадают. Болезнь встре- чается преимущественно на лозах, растущих непосредственно у р. Кон- дара. На культурных лозах сход- ное заболевание также наблюда- лось, но гораздо реже. 22—24 VIII 1945; IX 1946. Сем. Malvaceae
65. Alcea nudiflora Lindl. 67. Leveillula taurica Arnaud.
forma altheae Jacz. (Ячевский, 411). Клейстокарпии 160—200 [х диам.; сумки 75 х 30 и, на короткой ножке, в числе 24 в клейстокарпии (в 8 рядах, по 3 сумки в каждом). — Усадьба, 16 VIII 1944. Сем. Datiscaceae
66. Datisca cannabina L. 68. Leveillula taurica Arnaud,
forma datiscae Jacz. Клейстокарпии 160—180 jx диам.; сумки в числе 8—10, на короткой ножке. На листьях, повсеместно и обильно. — Ущелье, 14 IX 1944. 69. Coleosporium datiscae Tranz. (Траншель, 283). Оби-сафед, 5 IX 1945. Часто.
414
В. Ф. Купревич
Сем. Onagraceae
67. Epilobium hirsutum L. 70. Puccinia vagans (DC.) Artb.
(Траншель, 286). Уредо- и телейто-
споры; эцидии отсутствуют. —
Усадьба, 5 IX 1945.
Сем. Umbelliferae
68. Prangos pabularia Lindl. 71. Puccinia libani Magn. (Тран-
Юган. шель, 301). Различают ядовитую
и неядовитую разновидности юга-
на; первая произрастает преиму-
щественно внизу, на высоте при-
мерно до 1600 м, тогда как вторая,
неядовитая, чаще встречается на
высоте 1600 м и выше. Обе разно-
видности поражаются ржавчинни-
ком, причем форма на ядовитом
югане отличается от типа более
правильными, вверху закруглен-
ными телейтоспорами. — По тропе
на Квак, VI 1944.
69. Ferula Jaeschkeana Vatke. 72. Aecidium sarcinatum Lindr.
(Траншель, 296). — Усадьба, 27 IV
1944.
73. Puccinia sogdiana Kom.
(Траншель, 398). Телейтокучки
многочисленные, рассеянные по
всему листу. — По тропе на Квак,
19 V 1941.
74. Puccinia Litvinovii Tranz.
et Erem. (Траншель, 306). Нижняя
поверхность листьев сплошь покры-
та черно-бурым слоем телейто-
спор. — Квак, на вершине хребта,
по тропе, 9 V 1944.
Сем. Oleaceae
70. Fraxinus potamophila Herd. 75. Septoria fraxini-potamophili
Kuprewicz (sp. n.). Пикнидии на
верхней стороне листьев на угло-
ватых, сначала желтых, затем кра-
сновато-бурых пятнах, рассеянные,
черные, в проходящем свете фио-
летовые, погруженные, 160—180 и.
диам. Стилоспоры бесцветные, с 1—
2 невнятными перегородками, ци-
линдрические, согнутые или пр я-
Болезни растений ущелья Кондара
415.
мне, на концах закругленные, 30—
53 х 2—3 [л. Растущие у здания
Станции экземпляры ясеня еже-
годно очень сильно поражаются
грибом. Сначала на листьях обра-
зуются многочисленные светложел-
тые угловатые пятна; впоследствии
пятна становятся красноватыми или
светлобурыми. Ободок вокруг пя-
тен отсутствует. Пораженные
листья преждевременно засыхают
и опадают. — Усадьба. 21 VIII
1945.
76. Cylindrosporium sp. На
листьях. — Усадьба, 24 VII 1945.
Сем. Аросупасеае
71. Apocynum scabrum Russ. 77. Melampsora apocyni Tranz..
(Траншель, 320). На листьях. —
Усадьба, 22 IX 1946. Обильно.
Сем. Boraginaceae
72. Trichodesma incanum (Bge.) 78. Aecidium trichodesmae Kup-
DC. rewicz (sp. n.). Эцидии преимуще-
ственно на нижней стороне листьев.
Эцидиоспоры шаровидные или ши-
роко эллипсоидальные, бесцвет-
ные, 18—27 [л диам., тонко штри~
ховатые. Клетки перидия бесцвет-
ные, угловатые, 33—38 [л шири-
ной; со стороны, обращенной
внутрь, покрыты многочисленными
плотно стоящими шипиками. — На
южном склоне, за «Фермой», 27 V
1944.
Сем. Labiatae
73. Eremostachys seravschanica 79. Leveillula taurica Arnaud.
RgL forma phlomidis Jacz. (Ячевский,
410). Клейстокарпии 160—200 [л
диам.; сумки многочисленные >
60 х 36 и; сумкоспоры 33x21 и. —
Руидашт, 5 IX 1944.
74. Stachys hissarica Rgl. 80. Puccinia Hariotii Lagerh.
(Траншель, 331). — Руидашт, 6 IX
1944.
75. Mentha silvestris L. 81.Puccinia menthae Pers. (Тран-
шель, 331). — Усадьба, 5 IX 1945.,
Повсеместно.
416
В. Ф. Купревич
Сем. Solanaceae
76. Solanum Томаты. lycopersicum L. 82. Cladosporium sp. На повре- жденных листьях и плодах. Усадь- ба, VIII 1945. 83. Bacillus carotovorus Jones (?). Гниль недозрелых плодов, пре- имущественно поврежденных. — Усадьба, VIII—IX 1945. Сем. Scrophulariaceae
77. Veronica biloba L. 84. Ramularia veronicae Fuck.
(Oudemans, IV, 690). — Руидашт,
16 IX 1945.
Сем. Rubiaceae
78. Galium verum L. 85. Puccinia valantiae Pers.
(Траншель, 343). Телейтоспоры,
у вершины утолщенные до 12 а,
светлобурые, 48—54 х 16—23 а.—
Танге-мур, 23 VIII 1945.
79. Galium tricorne With. 86. Puccinia punctata Link.
(Траншель, 344). — Ущелье, 17
VIII 1945.
80. Lonicera Spach. Сем. Caprifoliaceae persica Laub, et 87. Lasiobotrys lonicerae Kunze et Schm. (Oudemans, IV, 829). — Северный склон, 9 VII 1941, собр. T. К. Роткевич.
81. Lonicera Pojark. 88. Phellinus lonicerinus (А. Bond.) Bond, et Sing. На живом стволе. — Квак, 9 VIII 1944. seravschanica A. Phellinus lonicerinus (A. Bond.) Bond, et Sing. — По тропе на Квак, у подъема на Руидашт, 16 IX 1945.
82. Scabiosa Сем. Dipsacaceae songarica Schrenk. 89. Sphaerotheca fuliginea Pal- lacci forma scabiosae Jacz. (Ячев-
ский, 104). Клейстокарпии 75—
90 и диам., сумки 72—80 х 57—
63
болезни растений ущелья Кондара
41>
Сем. Cucurbitaceae
83. Cucumis sativus L. 90. Bacterium lachrymans E. F.
SM. et Br. На листьях беловатые
или темносерые угловатые пятна.—
Усадьба, VI—VII 1945.
Сем. Campanulaceae
84. Ostrowskia magnifica Rgl. 91. Aecidium kondariense Kup-
rewicz. (sp. n.). На живых листьях
и стеблях. Эцидии на нижней сто-
роне листьев, на желтоватых пят-
нах, тесно скученными группами
или концентрическими кругами;
также на стеблях. Клетки перидия
угловатые, 18—30 [л, с внутрен-
ней стороны грубо бородавчатые.
Эцидиоспоры шаровидные, широко
эллипсоидальные или яйцевидные,
бесцветные, тонко бородавчатые,
почти гладкие, 16—25 [л диам. От
видов Aecidium на Campanula и
Adenophora отличается бесцветны-
ми спорами. — Северный склон, 27
V 1944.
Сем. Compositae
85. Psychrogeton cabulicus Во- 92. Erysiphe cichoracearum DC.
iss. forma psychrogetonis Kuprewicz (f.
n.). Клейстокарпии 100—120 u
диам., с рассеянными по всей по-
верхности бесцветными у основа-
ния коричневатыми простыми при-
датками. Сумки мешковидные, без
ножки, 51 х 33 в числе 8; сум-
коспоры яйцевидные, 21 х 15 а.—
Руидашт, 6 IX 1944.
86. Tanacetum Nevesskyanum 93. Puccinia tanaceti DC. (Тран-
C. Winkl. шель, 397). — Квак, 24 VIII 1941,
собр. H. Олтаржевский.
87. Artemisia persica Boiss. 94. Puccinia cinae Tranz. et
Kuprew. (Траншель, 397). Телейто-
кучки рыхлые; телейтоспоры тем-
нобурые, у вершины утолщенные,
ясно бородавчатые, 39—60 х 24—
33 [л. — Руидашт, 6 IX 1944.
88. Echinops maracandicus Bge. 95. Puccinia echinopis DC.
(Траншель, 398). — У p. Варзоб,
22 VIII 1945.
27 Ущелье Кондара
418
В. Ф. Купревич
89. Ligularia Thomsoni (Clarke) 96. Puccinia expansa Link.
A. Pojark. (Траншель, 387). — Руидашт, VII 1945
90. Cousinia radians Bge. Д. . 97. Puccinia cousiniae Syd. (Траншель, 398). — Ущелье, 10 VIII 1941; по тропе на Квак, 24 V 1944.
91. Cousinia pulchella Bge. Puccinia cousiniae Syd.— По тропе на Квак, 24 V 1944.
92. Cousinia umbrosa Bge. Puccinia cousiniae Syd. —Квак
9 V 1944.
93. Cousinia splendida C. Winkl. 94. Cirsium arvense (L.) Scop. Puccinia cousiniae Syd. Руи- дашт, 16 IX 1945. 98. Puccinia suaveolens (Pers.)
Rostr. (Траншель, 389). — У p. Варзоб, IX 1945.
95. Serratula chartacea C. 99. Puccinia heterophyllae Co-
Winkl. oke (Траншель, 389). — У p. Вар- зоб, 10 IV 1944, собр. А. И. Пояр-
кова.
96. Rhaponticum integrifolium 100. Puccinia centaureae DC.
Winkl. (Траншель, 401). — По тропе на Квак, 24 V 1944.
97. Cichorium intybus L. 101. Puccinia cichorii (DC.)
Bell. (Траншель, 402). — Усадьба, 27 VIII 1945.
98. Scorzonera acanthoclada 102. Leveillula taurica Arnaud.
Franchet. forma scorzonerae Kuprewicz (f. n.). Клейстокарпии 130—180 [л диам., темнобурые, с многочисленными бесцветными ветвящимися придат- ками, выходящими из нижней части; в засохшем виде вогнутые наподо- бие чашечки. Сумки 80—90 X 33 it, по 8 в клейстокарпии; сумкоспоры яйцевиднйе или широко эллипсо- идальные, бесцветные, 30 X 25 и.— Руидашт, 16 IX 1945. 103. Ustillago scorzonerae (Alb. et Schw.) Schr. (Гутнер, 135).— Руидашт, 16 IX 1945.
99. Chondrilla juncea L. 104. Puccinia chondrillina Bubak
et Syd. (Траншель, 404).5VII 1943, собр. T. К. Роткевич. По тропе на Квак, 15 VIII 1945.
100. Taraxacum sp. 105. Puccinia taraxaci (Rebent) Plowr. (Траншель, 403), —Руи- дашт, 6 IX 1944.
101. Mulgedium roseum Pep. et 106. Puccinia mulgedii Syd.
Vved. ssp. flavum Krasch. (Траншель, 393).— Северный склон, 28 IV 1944.
В ущелье широко распространены виды Cuscuta, особенно С. Lehman- niana Bge. Поселяясь на молодой поросли древесных пород, в частности
Болезни растений ущелья Кондара 419^
клена, этот вид причиняет значительный вред. Действие С. Lehmanniana
на хозяина можно видеть на следующем примере. В июне 1944 года побе-
гам повилики, в изобилии растущей на склоне, удалось зацепиться за ветви
молодых персиков на одной из террас. Спустя 2 недели пораженные ветви
отмерли и засохли. Молодые деревца клена и небольшие кусты, под-
вергшиеся нападению, отмирают и засыхают спустя 1—2 месяца.
В ущелье, помимо указанного вида, встречаются С. europaea L., С. аррго-
ximata Engelm. и С. convallariaeflora N. Pavl. Эти виды менее распро-
странены. Из них довольно часто встречается С. convallariaeflora, пара-
зитирующая на травянистых растениях на дне ущелья. Изредка этот
вид наблюдается на лозах дикого винограда. Борьба с заразихой (С. Leh-
manniana) велась преимущественно в районе усадьбы. Пораженные
участки тщательно скашивались, а высохшие растения использовались
в качестве топлива.
ЛИТЕРАТУРА
Г о л о в и н П. Н. 1941. Грибы песчаных пустынь Средней Азии. Тр. Узб. фил.
Акад. Наук СССР, вып. 1.
Головин П. Н. 1944. Закономерности распределения микологической флоры
на Памире. Изв. Тадж. фил. Акад. Наук СССР, № 8.
Гутнер Л. С. 1941. Головневые грибы. Ленинград.
Запрометов Н.Г Материалы по микофлоре Средней Азии, вып. 1, 1926; вып. 2,
1928, Ташкент.
Комаров В. Л. 1894. Паразитные грибы горного Зеравшана. Изд. СПб. унив.
Сорокин Н. В. 1884. Материалы для флоры Средней Азии. Bull. Soc. Nat.
de Moscou.
Спешнев H. H. 1901. Грибные паразиты (новые и малоизвестные) Закаспийской
области и Туркестанского края. Тр. Тифлисск. бот. сада, V, Тифлис.
Траншель В. Г. 1939. Обзор ржавчинных грибов СССР. М.—Л.
Я ч е в с к и й А, А. 1927. Карманный определитель грибов, вып. 2. Мучнисто-рося-
ные грибы. Ленинград.
Я чсвский А. А. 1935. Бактериозы растений. Ленинград.
Oudemans С. А. 1919—1924. Enumeratio systematica Fungorum. I—V.
27*
СОДЕРЖАНИЕ
Стр.
Акад. Е. Н. Павловский и В. И. Жадин. Введение . . . 5
Акад. Е. Н. Павловский. Возникновение и развитие деятельности Варзобской
горной ботанической станции Таджикского филиала Академии Наук
СССР 9
Растительный мир
В. В. Письяукова. Флора ущелья Кондара 23
А. С. Королева. Растительный покров Кондары 74
Животный мир
Б. С. Виноградов. Млекопитающие 103
|л. Я. Тугаринов.\ Птицы 108
С. А. Чернов. Пресмыкающиеся и земноводные 121
Г. X. Шапошникова. Рыбы 124
Е. П. Голубцова. Наземные моллюски 126
А. А. Штакельберг. Двукрылые 129
И. А. Рубцов. Мошки 149
С. Г. Лепнева. Ручейники ущелья Кондара 152
В. В. Попов. Пчелиные . 158
М. Н. Никольская. Осы-блестянки южного склона Гиссарского хребта 175
А. Н. Кириченко. Настоящие полужесткокрылые и цикадовые 181
Л. Л. Мищенко. К фауне уховерток, тараканов, богомолов, палочников и пря-
мокрылых южного склона Гиссарского хребта 198
А. Н. Попова. Стрекозы 206
Д. Е. Харитонов. Пауки и сенокосцы 209
Б. В. Лотоцкий. Иксодовые клещи 217
Д. И. Благовещенский. Пухоеды птиц 225
Е. Ф. Соснина. Паразиты мышевидных грызунов. 228
Е. С. Кирьянова. Волосатики и круглые черви 240
Воды
В. И. Жадин, И. А. Киселев, А. Г Родина, В. Я. Панкратова и |Л. В. Гриб |.
Жизнь в водах Кондары 245
Сезонные явления
\А. Я. Тугаринов.\ Календарь жизни в ущелье Кондара 265
Хозяйственное освоение биологических ресурсов
В. И. Запрягаева. Биологические особенности древесных и кустарнико-
вых пород в связи с мелиорацией лесной растительности Гиссарского
хребта . 279
А. С. Королева. Культура кормовых трав на богаре 360
Вредители и болезни растений
А. Е. Семенов. Вредная энтомофауна богарного садоводства в Кондаре 3/7
В. Ф. Купревич. Болезни растений ущелья Кондара 401
Печатается по постановлению
Редакционно-издательского совета
Академии Наук СССР
*
Редактор издательства Н. С, Ворхсениус
Художник М. П. Цыбасов
Технический редактор Д. М. Крол.
Корректоры О. Г Крючевская и
И. И. Удимое
*
РИСО АН СССР № 4534 М-00892
Подписано к печати 9 VI 1951 г.
Бумага 70ХЮ8/16. Бум. л. 13V4
Печ. л. 36,3. Уч.-изд. л. 31.5
Тираж 1500. Заказ. № 1830.
Цена в переплете 28 руб.
1-я тип. Издат. Академии Наук СССР.
Ленинград, В. О., 9 линия, 12