



































































































Текст
Издатель ст во “Kiman ”
выпустило следующие книги по энтомологии
(на казахском и руском языках):
ЭНТОМОЛОГИЧЕСКИЙ КАЛЕЙДОСКОП
МУРАВЬИ
ПЧЭОИг ITVM ИИ\ПЧкИ 1ПМП1' 'I UP
логический кллейдоскоп
эНТ0М°
йГ /
11
нкугап-Д9^^
Изда^^Л-2008
ББК
М
Рецензенты: доктор биологических наук, профессор В. Л. Казенас,
кандидат биологических наук М. К. Чильдебаев
Тимоханов С.А. Муравьи, богомолы, стрекозы. Алматы: Изд-во "К1тап-Дэу1р",
2008.96 с. Илл.: 136.
В книге рассматриваются основные особенности биологии, строения и
систематики муравьев, богомолов и стрекоз на примере фауны Казахстана. Текст
иллюстрирован большим количеством цветных рисунков. Книга рассчитана на
широкий круг читателей интересующихся природой, а также для учащихся и
учителей в качестве дополнительного материала по биологии.
Timokhanov S. A. The ants, mantises, dragonflies. Almaty: "К1тап-
Д yip", 2008.96 pp. ill.: 136.
This book gives a concise and brief illustration of basic biology, structures and
systematics of ants, mantises, and dragonflies of the Kazakhstanian fauna. The book is
directed towards wide range of readers who are interested in the nature, it can also be
used as resource material for both students and teachers of biology.
Тимоханов С. А. К;умырск;алар, дэуггтер, инелжтер. Алматы: "KiTan-fleyip"
баспасы, 2008.96 б. Суреттер: 136.
Ютапта Казахстан фаунасындагы кумьрска, деу!т, инелжтердщ непзп
биологиялык; ерекшел!ктер1, курылысы жене жуйелену! бер!лген. Мэт1н кептеген
турл! туст! суреттермен керкемделген. К!тап табигатты тамашалайтын жалпы
к,ауымга, сонымен к,атар окушылар мен мугал!мдерге косымша к,урал релнде
усынылады.
Выражаем искреннюю благодарность за помощь в работе и консультации
Казенасу В. Л., Кащееву В. А., Кадырбекову Р. Х.,Чильдебаеву М. К., Громову А.В.,
Пачикину К. М., Ященко Р. В. и Мариковскому П. И.
М
ББК
ISBN
© ТОО Издательство “KiTan-flayip”
© Тимоханов С. А.
© Иллюстрации: Тимоханов В. А.
© Фото: Тимоханов С. А.
ПРЕДИСЛОВИЕ
Перед вами книга, посвященная трем группам насекомых: муравьям, богомолам и
стрекозам. Несмотря на то, что насекомые этих групп резко отличаются друг от друга
морфологически и по образу жизни и на эволюционной лестнице стоят далеко друг от
друга, выбор их не случаен и определился задачей автора показать размах
разнообразия насекомых в морфологическом, экологическом и биологическом
аспектах и вызвать интерес к этой группе животных у молодых читателей,
начинающих биологов.
Муравьи - экологически очень важная группа насекомых, имеющая большое
практическое значение. Многие виды какхищники являются активными регуляторами
численности других насекомых, среди которых немало вредителей сельского и
лесного хозяйства. Некоторые виды являются потребителями семян растений.
Устраивая подземные гнезда, муравьи способствуют аэрации почвы и участвуют в
почвообразовательных процессах. Собирая мертвых беспозвоночных, они
выполняют роль санитаров природы. Иногда являются в природе промежуточными
хозяевами паразитов человека и животных. Являясь общественными насекомыми,
муравьи представляют собой удобный модельный объект для различных научных
экспериментов и решения сложных теоретических проблем, связанных с
происхождением и эволюцией социальности в животном мире. Знание фауны,
экологии, биологии и распространения муравьев имеет большое значение для
разработки мер по регулированию процессов, происходящих в природных
экосистемах. Между тем, муравьи остаются еще недостаточно изученной группой
насекомых в Казахстане.
Богомолы немногочисленны в нашей стране, но своей своеобразной формой и
охотничьими повадками неизменно вызывают большой интерес у натуралистов,
специалистов и любителей. Несмотря на древность своего происхождения, они
обладают важными специализированными приспособлениями, позволяющими им
сосуществовать с более прогрессивными формами насекомых.
То же можно сказать и о третьей группе. Стрекозы в личиночной фазе ведут
водный образ жизни, а во взрослой - живут в воздушной среде. Высокая численность,
активное хищное поведение и широкое распространение определили важное место
этих насекомых в водных и прилегающих к водоемам наземных биоценозах.
Материал книги основан на рассмотрении фауны Республики Казахстан и
содержит обширную научную информацию обо всех трех группах, в ней по
возможности подробно рассмотренны вопросы наружной морфологии, анатомии,
физиологии, систематики, фаунистики и биологии этих насекомых. Неся
просветительский, учебный характер, отличается обилием иллюстративного
материала с простыми краткими пояснениями, что делает ее привлекательной как
для самых молодых читателей, так и биологов широкого профиля. Наличие
определительных таблиц и систематически точных, отражающих особенности
строения рисунков делают ее "универсальной". Книга может использоваться в
качестве источника информации для учителей-биологов и студентов высших и
средних учебных заведений.
3
«да«
ВВЕДЕНИЕ В СИСТЕМАТИКУ
При рассмотрении как комплексов видов животных, обитающих на определенной
территории, так и особей животных, входящих в какое либо сообщество животного
населения, невозможно без системы научных названий для организмов и их групп,
связанных той или иной степенью родства и достаточно обособленных, чтобы им можно
было присвоить определенную таксономическую категорию того или иного ранга (вид, род,
семейство и тд.).
Задачей систематики является описание всех существующих и вымерших организмов,
присвоение им научных названий, а также их упорядочение (классификация) и
распределение всего множества живых организмов по определенной системе иерархически
соподчиненных групп (таксонов), для создания возможности ориентирования во множестве
существующих видов. Таксономические категории подразумевают не реальные организмы,
а определенный ранг или уровень классификации, ступени определенной иерархии.
Принципы систематики являются примером логической категоризации понятий, логической
системой, позволяющей охватить все разнообразие огромного класса насекомых.
Современные биологические номенклатуры ведут начало от классических работ К. Линнея
(сер. 18 в.), впервые применившего бинарные (биноминальные) названия для всех
известных ему видов. Эти названия состоят из названий рода и собственно вида. К. Линней
впервые установил твердую и стройную классификационную схему из основных категорий:
класс, отряд, род, вид. Название в которой всем таксонам дают на латинском языке, при этом
для каждого таксона устанавливается только одно название. Латинская номенклатура всюду
должна быть краткой, удобопроизносимой и рациональной. Выбор единственно
правомочного названия определяется правилами приоритета, согласно которому
действительным (валидным) считается старейшее из названий, обнародованное в
соответствии с правилами Международного кодекса биологической номенклатуры. Отряды
крылатых насекомых обычно имеют окончание -ptera (от греч. pteron - «крыло»). Окончание -
oidea применяется при обозначении надсемейств или надотрядов, -idae, -inae - соответ-
ственно семейств и подсемейств, -ini - триб.
Муравьи* наряду с другими группами насекомых - термитами (Isoptera), настоящими
осами (Vespidae), некоторыми пчелиными (Apoidea) - являются общественными, или
социальными, насекомыми, образующими постоянные (сезонные или многолетние)
объединения в природе - семьи, состоящие из репродуктивных (размножающихся) и
рабочих особей. У многих из них наблюдается временная (возрастная) или постоянная
специализация рабочих особей для выполнения определенных функций. Регуляция состава
и жизнедеятельности семьи осуществляется путем постоянного перераспределения между
всеми особями пищи, гормонов и феромонов, обуславливая координацию поведения
особей насекомых образующих семьи.
На основе современных представлений о систематике муравьев надсемейство
Formicoidea (муравьи) представлено современным единственным семейством Formicidae,
включающим в себя шестнадцать подсемейств, три из которых - Nothomyrmeciinae,
Myrmeciinae, Ponerinae - составляют группу “низших” в филогенетическом (историческом)
отношении и близких к исходной линии муравьев, которые характеризуются целым рядом
архаичных (древних) для них признаков. Все остальные подсемейства - Dorylinae,
Myrmicinae, Pseudomyrmecinae, Dolichoderinae, Formicinae - относятся к “высшим",
филогенетически более продвинутым группам муравьев. Мирмикофауну Казахстана
составляют представители пяти подсемейств: понерин (Ponerinae), дорулин (Dorylinae),
мирмицин (Myrmicinae), долиходерин (Dolichoderinae) и формицин (Formicinae),
включающих двадцать пять родов и около двухсот видов.
5
Совокупность всех фаз развития, пройдя которые организм достигает
зрелости и становится способным дать начало следующему поколению,
составляет цикл развития. Его длительность зависит от числа поколений (генера-
ций), развивающихся в течение года, она также определяется продолжительностью
претерпеваемого организмом обязательного периода покоя, или диапаузы. При раз-
витии со сложным жизненным циклом - метаморфозом (метаболией), при котором
происходит глубокое преобразование организма, в течение развития одной особи
прослеживается следующий цикл: яйцо-личинка-куколка-имаго (взрослое насеко-
мое). А в онтогенезе (индивидуальном развитии особи) происходит резкое разделе-
ние основных функций (питание в стадии личинки, расселение и размножение во
взрослой стадии). Данный тип метаморфоза относитсится к полному превращению,
или голометаболии.
Личинка (larva).
В своем развитии претерпевает пять
личиночных возрастов разделенных линьками. У
нее различаются слабо хитинизированная
головная капсула, несущая ротовые части,
представленные различимыми жвалами, нижними
челюстями и губой, а также - три грудных и *
десять брюшных сегментов.
Яйцо (ovum).
У большинства муравьев, как и у всех
жалящих перепончатокрылых, из
оплодотворенных яиц развиваются самки и
рабочие, а из неоплодотворенных - самцы.
Однако у нескольких родов муравьев,
являющихся исключением, из
неоплодотворенных яиц могут развиваться как
самцы, так и самки.
6
ЖИЗНЕННЫЙ ЦИКЛ
У насекомых с полным превращением червеобразная личинка не похожа на
взрослое насекомое и переход во взрослую форму осуществляется через стадию
куколки, которая не питается и (обычно) неподвижна. В ней происходит интенсивная
внутренняя перестройка, в процессе которой насекомое превращается из личинки во
взрослую особь, при этом большинство личиночных органов разрушается, а органы
имаго формируются заново из особых зачатков - имагинальных дисков. У свободных
(открытых) куколок крылья, усики и ноги плотно прижаты к телу, но не сцеплены с ним.
В целом особенности метаморфоза и жизненного цикла являются важнейшими
диагностическими признаками насекомых. При этом различия по метаморфозу
присущи преимущественно высшим систематическим категориям - семействам,
отрядам и др., а пожизненному циклу-отдельным видам.
Предкуколка (praepuppa).
Взрослое насекомое
(imago)
Куколка (puppa, сосоп).
Куколка муравьев относится к свободному
типу и обычно покрыта коконом. Но
имеется много исключений, и один и тот же
вид при одних условиях может делать кокон,
при других - нет. Стадии куколки
предшествует стадия предкуколки, а
куколки разных каст хорошо различаются.
7
нога
усик
скапус
глазки
просома,
включающая
голову
мезосома,
включающая
три грудных
и первый
абдоминальный
сегмент
метасома,
включающая
сегменты
брюшка
бедро
вертлуг
голень
гребенчатая
шпора
базитарзус
Передняя нога рабочего
Camponotus lameerei Emery, 1898.
пятичлениковая
лапка
тазик
Общий вид самки муравья - жнеца Messor
clivorum (Ruzsky),1905.
Строение узелка у подсемейств Myrmicinae,
Dolichoderinae, Formicinae.
8
ВНЕШНЕЕ СТРОЕНИЕ (МОРФОЛОГИЯ)
стрекотательный
напильничек
головная капсула
сложный глаз
окулярный гребень
лобный валик
лобная бороздка
лобная площадка
усиковые ямки
передняя ямка тенториума
клипеус (личной щиток, наличник)
верхняя губная пластинка
мандибула (верхняя челюсть)
нижняя губа
максилла (нижняя челюсть)
максиллярный щупик
лабиальный щупик
Голова солдата
стрекотательный
и ротовые органы (гнатиты) у муравья Camponotus
herculeanus Linnaeus, 1758.
слияние заднегруди с
морфологически первым,
истинным сегментом брюшка
образует эпинотум.
Строение стридуляционного аппарата
представителей подсемейства
Myrmicinae и Ponerinae.
В некоторых группах муравьев в нижней
части задней вертикальной поверхности
эпинотума имеется острый край эпинотального
склерита стрекотательный кантик, служащий
для издания звуков фрикционным (трением)
способом. Звуковое излучение производится
при трении стрекотательного кантика по
верхней гофрированной поверхности петиоля,
образованной рядом параллельно идущих
ребрышек, называемых стрекотательным
напильничком, при движении его вверх - вниз.
При этом кантик образующий связку между
эпинотумом и петиолем, устроен таким
образом, что позволяет петиолю двигаться или
со звуком, или без него (Myrmicinae). У других
муравьев (Ponerinae) звук извлекается путем
сдвижения - раздвижения следующего за
петиолем сегмента брюшка, отграниченного от
идущего далее сегмента глубокой бороздой.
9
Муравьи относятся к жалящим перепончатокрылым. И у самок и у
рабочих на конце брюшка имеется жало, которое развито слабо имея
различную степень изменения строения и редукции отдельных частей вплоть до
полного исчезновения (рудиментации).
органы
секреции
органы
пищеварения
нервная
система
железа Дюфура
10
ВНУТРЕНЕЕ СТРОЕНИЕ МУРАВЬЯ (АНАТОМИЯ)
Уникальна избирательная способность муравьев по отношению к пище,
связанная с образованием фильтрационного аппарата на стенках глотки в виде
множества мелких волосков, направленных наружу, определяющих возможность
поглощать взрослыми муравьями только жидкую пищу, а не проглатывать твердые
кусочки.
слюнная железа
грудные
нервные узлы
надглоточный ганглий
брюшная
нервная
цепочка
подглоточный ганглий
глоточная железа
нижнечелюстные
железы
глотка
предротовая камера
нижняя губа
максиллярная железа
Внутреннее строение муравья рода Formica.
11
12
СОСТАВ СЕМЬИ, ПОЛИМОРФИЗМ И РАЗДЕЛЕНИЕ ФУНКЦИЙ
Самка (слева), самец, рабочая особь и солдаты
(диморфизм рабочих особей)
Pheidole pallidula (Nylander),1848.
У муравьев имеются определенные виды работ выполняемые различными
группами рабочих. В общем случае одни обеспечивают уход за тлями, и сбор пади,
другие - сбор строительного материала и охоту на беспозвоночных, перенос рабочих
и обмен молодью между гнездами. Для обозначения фиксированных различий в
выполнении разными рабочими определенного круга функций введено понятие
полиэтизма.
Полиэтизм особей носит возрастной и кастовый характер. В первом случае
наблюдается смена выполняемых функций, зависящая от возраста рабочих особей.
Рабочие сначала выполняют работы в гнезде и занимаются уходом за молодью. В это
время у них развиты хорошо функционирующие максиллярные железы и яичники.
После затухания их функционирования рабочие особи становятся «фуражирами» и
строителями. При этом скорость прохождения разных периодов неодинакова и
зависит от потребности семьи. Во втором периоде наблюдается слабая размерная
дифференциация (различие) рабочих особей, определяющая различие в
выполнении разных функций вне гнезда. В пределах общей размерной изменчивости
резко выделяется группа рабочих, собирающих падь тлей, и обычно средние размеры
этих особей меньше, чем у особей всего гнезда. Вторую группу рабочих образуют
особи, переносящие мертвых муравьев и гнездовой материал. Группа рабочих,
защищающих гнездо, не отличается размерами от особей всей семьи, поскольку эту
функцию может выполнять все население гнезда.
13
*- _r*
Спаривание муравья Formica
cunicularia glauca Ruzsky,1895.
Как уже отмечалось, муравьи
живут семьями, насчитывающими
от нескольких десятков до сотен
тысяч особей, строя сложные гнез-
да. В состав семьи входят рабочие
особи, бесплодные, с недоразвиты-
ми половыми органами, модифици-
рованные самки и одна самка -
основательница (т.н. моногинные
семьи). Самок - основательниц в
гнезде у отдельных видов иногда
может быть несколько (в случае
полигинии семьи).
Каждая особь в семье муравьев выполняет строго определенные функции. Самцы
выполняют только функции, связанные с размножением, а после оплодотворения
самок (копуляции) они погибают. Функция самок, меняясь в зависимости от времени,
после оплодотворения состоит в расселении, основании новой семьи и воспитании
первого потомства, а в дальнейшем заключается только в откладке яиц. Заботу о
потомстве несут рабочие особи, которые обеспечивают уход за яйцами, личинками и
куколками внутри гнезда.
Переход к размножению, или созревание семьи, возникает при высокой числен-
ности рабочих особей, когда царица (репродуктивная самка) не в силах эффективно
сдерживать выращивание плодовитых самок и подавлять плодовитость всех рабо-
чих, откладывающих яйца (из которых появляются самцы, что характерно далеко не
для всех видов). А максимальный размер семей зависит от продуктивности царицы,
выживаемости расплода и выживаемости взрослых особей различных каст. Эволюци-
онирование в направлении увеличения репродуктивности ведет к появлению у
муравьев физогастрии (увеличения объема брюшка царицы) и полигинии, когда в
одном гнезде живет свыше одной репродуктивной самки. Половые особи выходят из
коконов, как правило, осенью, зимуют, а весной покидают гнездо, тогда происходит их
лет. После копуляции (спаривания) самки теряют крылья и основывают самосто-
ятельные гнезда, или их принимают в старые гнезда, где нужны самки.
14
РАЗМНОЖЕНИЕ, ОБРАЗОВАНИЕ СЕМЬИ
У муравьев наиболее распространенным типом основания новых гнезд является
самостоятельное образование новых семей. При этом молодая оплодотворенная
самка, сбросив крылья, выкапывает в земле ход и делает одну камеру, в которой
производит первую откладку яиц. Вылупившихся личинок она кормит выделениями
ротовых желез и частью откладываемых яиц. В этот период она не получает
дополнительного питания и расходует собственное жировое тело и крыловые мышцы,
которые рассасываются. Первые рабочие резко отличаются от последующих партий
потомства мелкими размерами; они начинают кормить самку и добывать пищу, а
самки с этого времени только откладывают яйца.
Молодая оплодотворенная самка Messor aralocaspius (Ruzsky),1902,
в подземном гнезде ухаживает за кладкой яиц.
В процессе эволюции у ряда видов муравьев выработались такие способы
основания гнезд, при которых уязвимый период одиночного существования самки
сокращен до минимума. Один из таких способов - проникновение самки муравья в
гнездо другого вида, или временный социальный паразитизм.
У многих видов, основывающих гнезда по типу социального паразитизма, имеется
еще один способ увеличения колоний, который называют «делением», или
«почкованием», при этом группа особей вместе с самкой и молодью уходит из старого
гнезда и основывает новое гнездо, которое может в дальнейшем отделиться,
постепенно утрачивая связи со старым гнездом.
15
Гнезда большинства видов в той или иной степени расположены в
земле. Хотя строение гнезд даже в пределах одного вида муравьев
сильно изменяется в зависимости от местности, все же возможно
свести все их разнообразие к определенной системе.
Земляное гнездо и схема (в
разрезе) гнезда муравья Messor
denticulatus (Kusnezov -
Ugamskij),1927.
Гнездо
Formica fusca Linnaeus, 1758.
Гнездо, устраиваемое
муравьями Camponotus lameeri
Emery, 1898 под корой дерева.
16
Основным, наиболее примитивным типом являют-
ся земляные гнезда, состоящие из входного отверстия
и поверхностной горизонтальной системы ходов, где
весной находится основная масса расплода. Иногда
вокруг входа заметны выбросы почвы, а у некоторых
видов он окружен земляным валиком, который бывает
очень высоким и напоминает кратер. Многие, особен-
но обитающие в пустыне виды на время своего отсу-
тствия закрывают вход частичками почвы, мелкими
камешками или сухими растительными остатками. В
этих гнездах температура не отличается от темпера-
туры окружающей почвы, и муравьи вынуждены пере-
носить личинок и куколок в камеры, расположенные
на различных горизонтах гнезда, с оптимальной тем-
пературой и влажностью для развития потомства.
Часто гнезда муравьев располагаются под камнями,
кусками древесины и другими естественными укрыти-
ями, хорошо аккумулирующими тепло.
Гнезда с куполообразной земляной постройкой не
испытывают резких колебаний температуры, как это
отмечается на поверхности почвы, а повышенный
нагрев поверхности гнезда во многом обусловлен
ориентацией гнезда относительно инсоляции (потока
солнечного тепла). Форма купола гнезда меняется в
зависимости от условий и может быть на открытых
участках невысокой и плоской, а на затененных -
конусообразной. Во влажных стациях постройка при-
обретает цилиндрическую форму, а с высоким тра-
востоем - конусообразный вид. При преобладании
вынесенной наружу в виде комочков почвы с мелкими
растительными остатками в качестве основного мате-
риала одни гнезда имеют плотное строение и обильно
зарастают травой и мхами, другие имеют рыхлую
структуру и лишены растительности.
Для ряда других видов муравьев характерны гнез-
да, расположенные в пнях, сучьях, под корой или в
других полостях, связанных с отмершей древесиной.
Затрагивая лишь верхние слои заболони между корой
и древесиной или проникая по поврежденным учас-
ткам в более глубокие слои древесины, ходы и каме-
ры занимают готовые подходящие полости.
СТРОЕНИЕ ЖИЛИЩА (ГНЕЗДО)
гнезда земляной холмик
Гнездо и схема строения гнезда
муравьев подрода Coptoformica.
Гнезда, построенные с использованием расти-
тельных остатков, куполообразной формы, являются
наиболее сложными по конструкции постройками.
Основой подземной части является кратер из плот-
ной почвы, выброшенной из ходов. Снаружи края
кратера имеют форму вала, обычно поросшего тра-
вой. Над кратером возвышается густо пронизанный
травинками внутренний холмик из рыхлой земли, на
вершине которого находится своеобразная «под-
ушка» из семян злаков, мелких травинок и стебель-
ков мха. Сверху все, кроме наружных краев кратера,
покрыто плотным куполом из стеблей и листьев
травы и мха, реже из тонких веточек и игл. Между
«подушкой» и верхней частью купола помещается
главная камера, в которой обычно находится при-
плод, а в периоды интенсивной откладки яиц - и
репродуктивные самки.
В другом случае внутренний холмик образо-
ван крупными палочками, принимает форму
конуса. В нем, как правило, происходит развитие
молоди. Сверху гнездо прикрыто толстым
наружным куполом из разнообразнейшего мате-
риала, состоящего из мелких веточек, игл,
черешков листьев, толстых стеблей травы.
Материал гнездового холмика муравейника
постоянно перемешивается, и муравьи, посто-
янно поднимая мелкий гнездовой материал из
внутренних частей наружного купола, спосо-
бствуют формированию внутреннего конуса
вследствие опускания крупных фрагментов
гнездового материала, слишком громоздких для
своего поднятия наверх. Изменяя форму вер-
хней части купола, муравьи регулируют темпе-
ратурный режим гнезда. В основе строения
других муравейников служит пень, разрушенный
ходами ксилофагов (животных, питающихся
древесиной), при этом наружная часть гнезда
почти целиком помещается в пне, к которому
принесено небольшое количество растительных
остатков.
Кроме основных гнезд, муравьи делают
небольшие временные гнезда без сложной,
развитой структуры, служащие как проме-
жуточные постройки при переселении, и кормо-
вые гнезда.
Гнездо и схема строения гнезда
муравьев подрода Formica s. str.
14°С
16°С
18°С
20°С
22°С
23°С
24°С
25°С
Температура окружающего
воздуха и в гнезде.
17
18
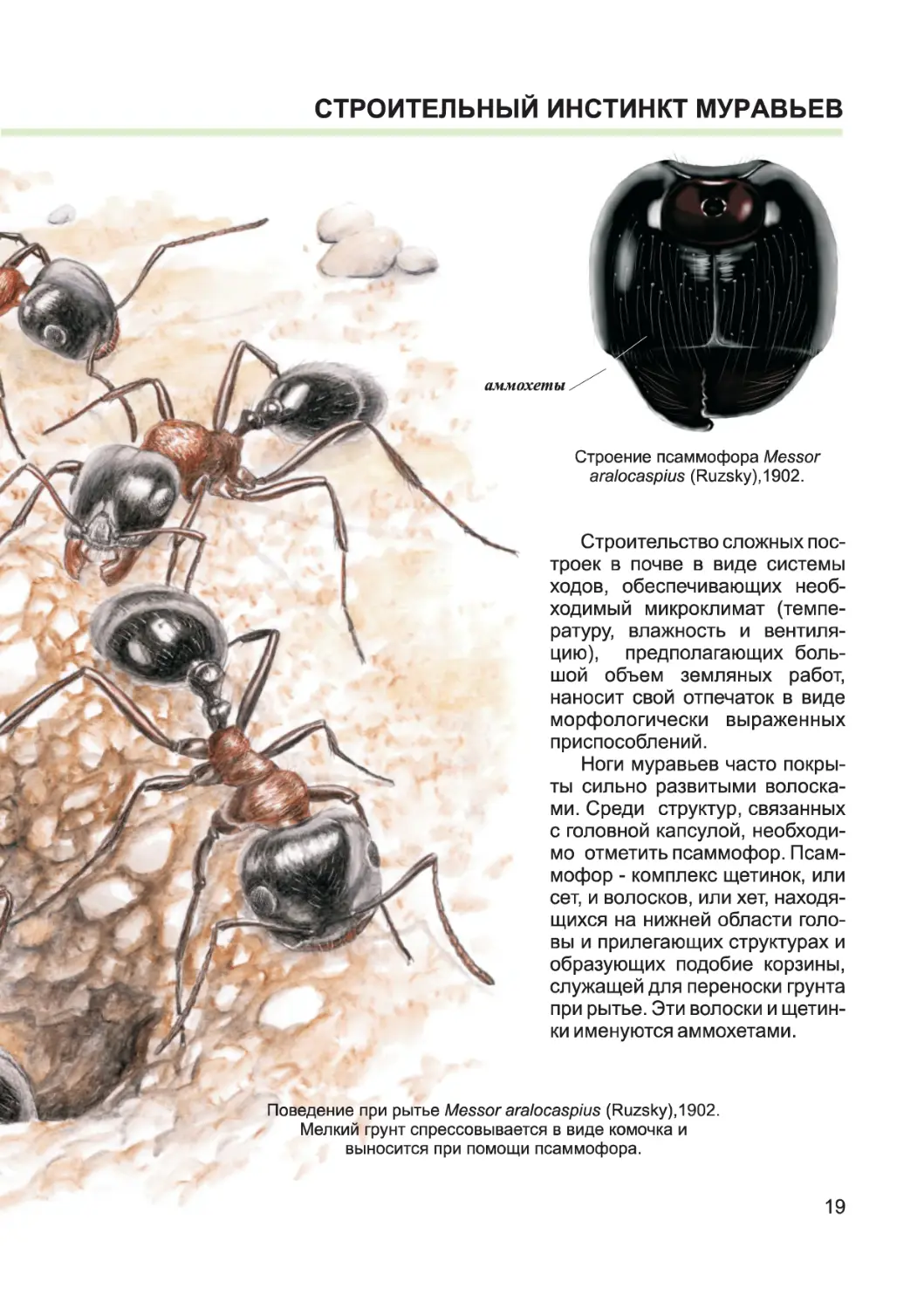
СТРОИТЕЛЬНЫМ ИНСТИНКТ МУРАВЬЕВ
аммохеты
Строение псаммофора Messor
aralocaspius (Ruzsky),1902.
Поведение при рытье Messor aralocaspius (Ruzsky),1902.
Мелкий грунт спрессовывается в виде комочка и
выносится при помощи псаммофора.
Строительство сложных пос-
троек в почве в виде системы
ходов, обеспечивающих необ-
ходимый микроклимат (темпе-
ратуру, влажность и вентиля-
цию), предполагающих боль-
шой объем земляных работ,
наносит свой отпечаток в виде
морфологически выраженных
приспособлений.
Ноги муравьев часто покры-
ты сильно развитыми волоска-
ми. Среди структур, связанных
с головной капсулой, необходи-
мо отметить псаммофор. Псам-
мофор - комплекс щетинок, или
сет, и волосков, или хет, находя-
щихся на нижней области голо-
вы и прилегающих структурах и
образующих подобие корзины,
служащей для переноски грунта
при рытье. Эти волоски и щетин-
ки именуются аммохетами.
19
Охотясь на улиток, муравьи Formica pratensis
Retzius, 1783 растворяют раковину,
выбрызгивая из ядовитой железы кислоту.
Большинство муравьев имеет сме-
шанную диету, состоящую главным обра-
зом из насекомых и других беспозвоноч-
ных, пади и выделений растений, а обмен
кормом происходит путем трофоллаксиса
(взаимного обмена жидкой пищей между
особями). В общем случае пищу в рационе
муравьев можно разделить на углеводную
и белковую, а муравьев характеризовать
как высокоспециализированных и неспе-
циализированных хищников с высокой
общественной специализацией, тяготею-
щих выбирать более мелкую жертву, кото-
рую можно одолеть без помощи жала, но
зато в большем числе.
Каждый муравей, принесший жидкую
пищу, не использует ее для себя, а распре-
деляет между другими особями внутри гнезда. Большую роль при этом имеет строе-
ние провентрикулюса, который не позволяет пище проходить из зоба в желудок, и
лишь незначительное количество добытой пищи рабочие поедают сами. При обмене
пищеи происходит не только распределение питательных веществ, но распределяют-
ся и феромоны, регулирующие поведение особей в гнезде. Активация трофоллакси-
са происходит тактильными, при прикосновении усиков и лапок стимулами. Направ-
ление обмена пищей зависит от интенсивности движений какого - либо из партнеров.
Прикосновения наблюдаются чаще у акцептора, чем у донора. После насыщения
акцептор в свою очередь становится донором. Личинки и царицы либо совсем ничего
не получают при трофоллаксисе, либо получают лишь ничтожное количество пищи, в
виде секрета, выделяемого лабиальны-
ми железами рабочих.
Выделения тлей (Aphididae) и других
равнокрылых хоботных (кокцид, листо-
блошек и цикад) занимают важное место
в питании муравьев. Они носят название
«падь», или «медвяная роса», и характе-
ризуются большим содержанием углево-
дов. Тли, потребляя большое количество
сока растений, получают необходимое
количество белка и компенсируют повы-
шенный расход влаги из - за тонких покро-
вов и отсутствия защитных волосков и
лакового слоя, выделяя углеводы в виде
экскрементов. Выделение экскрементов,
богатых углеводами, является результа-
том длительной эволюции, направленной
на приспособление тлей к симбиозу с
муравьями.
Муравей Formica fusca Linnaeus, 1758
собирающий падь тли Metopeurum
fuscoviride Stroyan.
20
ПИТАНИЕ
Помимо равнокрылых хоботных (Homoptera), муравьи связаны с гусеницами
голубянок (Lepidoptera, Lycaenidae). Гусеницы голубянок имеют железистые
специальные органы, выделяющие сладкий экскрет.
Транспортировка семян
в гнездо рабочими муравья-жнеца
Messor aralocaspius (Ruzsky),1902.
Плеэргат (вверху справа),
крупный и мелкий рабочий Proformica
epinotalis Kusnezov-Ugamskij,1927.
У некоторых муравьев есть
особая каста - плеэргаты (рабочие,
способные сохранять большие
запасы жидкой пищи в зобу), в
отличии от солдат не являющейся
морфологической кастой. Их ана-
томическое строение не отличает-
ся от строения нормальных рабо-
чих такого же размера, и потенци-
ально плеэргатом может стать
любой крупный молодой рабочий.
Многие виды переходят к пита-
нию собираемыми семенами (кар-
пофагия) и созданию запасов. Эти
муравьи еще питаются беспозво-
ночными, когда их можно добыть,
но семена представляют такой
неистощимый и питательный вид пищи, что привычки к собиранию и запасанию семян
в гнездах становятся высоко развитыми. Питание семенами зарегистрировано у мно-
гих видов муравьев, но они чаще поедают не сами семена, а съедобные различные
придатки (эласмосомы). Возникновение таких придатков - результат приспособления
растений к расселению муравьями (мирмекохория).
21
Обмен пищей Formica truncorum Fabricius, 1804. Пища распределяется между рабочими
путем своеобразного обмена - трофоллаксиса.
Главными отличительными особенностями общественного (социального) образа
жизни является наличие обязательного разделения труда. На первой стадии происхо-
дит разделение на половых особей и бесплодных рабочих, в дальнейшем происходит
появление полиэтизма, приводящего в ходе эволюционирования у муравьев к поли-
морфизму. Между всеми членами происходит постоянный обмен информацией, и ни
один из индивидуумов семьи (кроме самцов и молодых самок-основательниц, служа-
щих для размножения и расселения вида) не может существовать длительное время
отдельно.
Проявляя эти особенности, социальные насекомые приобретают ряд преиму-
ществ в виде возможности создания условий, являющихся оптимальными для разви-
тия молоди: возможности создания запасов пищи либо в специальных хранилищах,
либо в зобиках определенной группы рабочих, а также способности к быстрой регене-
рации семьи при гибели части особей. Семьи наиболее высокоорганизованных
общественных насекомых,
обладая наличием большого
количества плодовитых самок и
возможностью их замены, су-
ществуют до тех пор, пока им
позволяют внешние условия.
Как и другим социальным
насекомым, муравьям присуща
развитая система коммуника-
ции, позволяющая поддержи-
вать стабильность структуры
семьи и обмен информацией.
(слева) и угрожающая поза муравья
atogaster subdentata Мауг, 1877.
ВЗАИМООТНОШЕНИЯ В СЕМЬЕ
На основе иерархии и
доминирования у муравьев
возникла функциональная
иерархия, включающая в
себя разделение функций
рабочих особей (группы
муравьев-фуражиров, гнез-
довых рабочих, муравьев-
нянек, царицы или цариц).
Поддержание стабильности
структуры семьи (гомеоста-
за) и регуляция происходит
как при помощи развитой
системы желез и рецепторов,
расположенных на теле
муравьев экзогенно (внеш-
не), так и с помощью внутрен-
них (эндогенных) лабиаль-
ных (нижнегубных), постфа-
рингеальных (заглоточных) кормовых желез, ядовитой железы и мн. др. факторов,
поддерживающих химическую коммуникацию, а также осуществляется с помощью
зрительных сигналов, звуковых, тактильных сигналов, сигнальных поз, движений,
характерных (видоспецифичных) многим группам муравьев.
Механизмы коммуникации поддерживают стабильность основных свойств, увя-
зывая части в целое, при этом система обладает выраженной индивидуальностью и
своеобразием строения, поддерживая неизменное численное соотношение между
особями различных каст, рабочими и личинками.
Транспортировка рабочего Formica cunicularia
glauca Ruzsky, 1895.
Обычная (вверху) и угрожающая поза
муравья Formica truncorum Fabricius, 1804.
При наличии благоприят-
ных условий семья муравьев
разрастается настолько, что
одно гнездо уже не вмещает
всего населения. Это явление
приводит к появлению коло-
ний, когда семьи муравьев,
обитающих одновременно в
нескольких муравейниках,
производят постоянный обмен
между особями разными гнез-
дами. Он может осущест-
вляться как постоянно, так и
периодически, по сети подзем-
ных ходов в первом случае
путем транспортировки осо-
бей, выполняющих работы
внутри гнезда, по наружным
тропам во втором.
23
Обитающий в подстилке
стафилинид синехтр
Xantholinus longiventris Heer.
Синойка Clytra atraphaxidis
Pallas, личинка, которой
обитает в муравейниках,
питаясь растительными
остатками.
Как правило, обитающие в гнездах муравьев и боль-
ше нигде не встречающиеся беспозвоночные, называют-
ся мирмекофилами. По общепринятой классификации
различаются пять биологических категорий мирмекофи-
лии: синехтрия, синойкия, симфилия, трофобиоз и пара-
зитизм.
Отношения между муравьями и мирмекофилами,
когда муравьи враждебно относятся к своим сожителям и
пытаются изгнать их из своего гнезда, называются синех-
трией. Такие сожители обычно питаются мертвыми
муравьями и их личинками и всевозможными пищевыми
отбросами. Эту небольшую по числу видов и количеству
особей категорию представляют в основном стафилини-
ды (Staphylinidae), обитающие в подстилке.
Безразличное отношение муравьев к своим сожите-
лям называется синойкией (метохия) и является наибо-
лее распространенным видом отношений между муравь-
ями и мирмекофилами. Нейтральные синойки безраз-
лично относятся к муравьям и их молоди и питаются
гнездовым разлагающимся
пищевыми отходами или
материалом. Этими сожи-
телями являются встреча-
ющиеся в каждом гнезде
ногохвостки и клещи
орибатиды, тироглифиды,
плоские своеобразные
личинки сирфид из рода
Microdon, личинки жуков
бронзовок (Cetonia), жуки
перистокрылки (Ptiliidae) и
многие другие. Часто в
гнездах можно увидеть
личинок жуков листоедов
рода Clytra, питающихся
преимущественно расти-
тельными остатками.
Миметичные и симфилоидные синойки имеют
подражательное сходство формы и окраски с муравья-
ми, у которых живут, либо имеют волосковидные желе-
зы, так называемые трихомы, выделяющие вещества,
слизываемые муравьями.
Миметичный паук мирмекофил
Micaria pulicaria (Sundevall).
24
ВЗАИМООТНОШЕНИЯ С ДРУГИМИ ОРГАНИЗМАМИ В ПРИРОДЕ
Мирмекоклепты - это мирмекофилы-воры. К ним
муравьи относятся отрицательно, но благодаря
чрезвычайной подвижности им всегда удается скрыться.
Мирмекофильные щетинохвостки рода Atelura питаются,
подбираясь к передающим друг другу каплю пищи
муравьям, и кормится вместе с ними.
Муравей Myrmica bergi Ruzsky, 1902
и настоящий симфил Homoeusa acuminata Maerklin.
Муравей мирмекоклепт
Formicoxenus nitidulus
(Nylander),1846.
Настоящие сожители муравьев - это симфилы, за которыми муравьи ухаживают
Эти мирмекофилы имеют железы, которые выделяют привлекающие муравьев
вещества, они состоят из собранных в пучки волосков (трихом), расположенных на
брюшке (Staphylinidae), элитрах (надкрылья жуков) (Pselaphidae), антеннах
(Paussidae).
25
sanguinea Latreille, 1798.
У муравьев в результа-
те закрепления разделения
функций в семьях, состоя-
щих из нескольких видов,
развилась своеобразная
форма паразитизма - «ра-
бовладение», или «куколоч-
ный паразитизм». Парази-
тизм определяется как
форма взаимоотношений
двух различных организ-
мов, когда один из них (па-
разит) использует другого
(хозяина) в качестве среды
обитания или источника
пищи, возлагая на него
регуляцию своих отноше-
ний с внешней средой.
При куколочном парази-
тизме, развившемся незави-
симо у различных муравьев,
рабочие вида-паразита пери-
одически совершают набеги
колонной рабочих на гнезда
других видов муравьев и
похищают куколок. Вышед-
шие из этих куколок рабочие
выполняют различные функ-
ции в гнезде, как это происхо-
дит обычно в родном гнезде,
выполняя в основном работы
внутри гнезда.
proformicarum К. Arnoldi, 1928.
26
РАБОВЛАДЕНИЕ
Проявление куколочно-
го паразитизма имеет раз-
личный уровень организа-
ции. При наиболее прими-
тивной организации отсу-
тствуют морфологические
изменения и присутствует
упрощенность поведения
(Raptiformica). Муравьи не
совершают далеких «экспе-
диций» за куколками и
нападают лишь на гнезда,
находящиеся на кормовом
участке их гнезда. Часто
встречаются семьи, живу-
щие без «рабов».
christophi Emery, 1889.
Высокоспециализиро-
ванных «рабовладельцев»
отличают, прежде всего,
морфологические различия.
Жвалы этих муравьев име-
ют лишь крошечные зубчики
(у Rossomyrmex) либо и
вовсе лишены развитых
зубцов, они имеют серпо-
видную форму (у Polyergus,
Strongylognathus) и не при-
способлены к размельчению
пищи и гнездовым работам.
27
28
ОРГАНИЗАЦИЯ ПОХОДОВ
Грабительский набег Polyergus rufescens (Latreille),1798
на~гнездо Formica cuniculaha glauca Ruzsky, 1895.
Сдавливая саблевидными челюстями голову
противников, солдаты муравьев-амазонок временно
их обездвиживают и уносят куколок в свое гнездо.
29
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 .Стебелек брюшка одночлениковый и состоит из петиоля, несущего чешуйку..........3
- Стебелек брюшка двучлениковый. Состоит из петиоля и постпетиоля.................2
2 .Клоакальное отверстие в виде поперечной щели. Анальные железы отсутствуют.
Жало имеется. Куколки без кокона......................................2.Myrmicinae.
3 .Покровы тела жесткие. Первый сегмент брюшка отграничен от второго глубокой
перетяжкой. Жало имеется..........................................................4
- Покровы тела мягкие. Брюшко без перетяжки. Жало отсутствует.......................5
4.Отверстие клоаки в виде поперечной щели. Анальные железы отсутствуют. Куколки
всегдав коконах.........................................................1 .Ponerinae.
5.Отверстие клоаки круглое, выдающееся, окруженное венчиком волосков. Лобная
площадка хорошо отделена от клипеуса. Клипеус не вдается треугольником между
лобными валиками. Анальных желез нет. Куколки чаще с коконом..........4.Formicinae.
-Отверстие клоаки имеет вид поперечной щели. Клипеус вдается треугольником
между лобными валиками. Лобная площадка необозначенная. Анальные железы
имеются. Куколки без коконов.......................................3.Dolichoderinae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ MYRMICINAE
1 .Постпетиоль причленяется в верхней части первого сегмента брюшка. Брюшко
сердцевидное, сплюснутое, заостренное, снизу выпуклое....................................5.Crematogaster.
-Постпетиоль причленяется по центру первого сегмента брюшка. Брюшко иной
формы...................................................................................................2
2 .Постпетиоль широкий, в 2 раза шире петиоля...........................................4.Cardiocondyla.
- Постпетиоль не шире петиоля (см.сверху)...............................................................3
3. Петиоль снизу с большим широким зубцом, постпетиоль с длинным
шипом...................................................................................................4
- Постпетиоль без шипа снизу (см. в профиль)...........................................................5
4 .Лобные валики короткие, почти прямые, параллельные. Усики вместе со скапусом
насчитывают 11 члеников у самок и рабочих, 12 члеников усамцов 11 .Formicoxenus.
- Лобные валики длинные, немного изогнутые, расходящиеся. Усики вместе со
скапусом насчитывают 12 члеников у самок и 13 члеников у самцов. Рабочие
отсутствуют..........................................................................10.Leonomyrma.
5 . Усики с двучлениковой булавой..........................................................7.Solenopsis.
- Булава усиков неявственная или имеет более двух члеников................................6
6. Жвалы узкие, серповидные, без зубцов на жевательном Kpae...3.Strongylognathus.
- Жвалы широкие, треугольные или трапециевидные, с зубчиками.........................7
7. Боковые части клипеуса килевидно приподняты, резко ограничивая усиковую
впадину...................................................................................12.Tetramorium.
- Боковые части клипеуса не приподняты килевидно...................................................8
8. Шипы эпинотума в виде неявственных бугорков..........................................................9
- Эпинотум с явно выраженными шипами....................................................................10
9. Брюшко овальное, приплюснутое сверху, спереди притупленное. Мандибулы
узкие...................................................................................6.Monomorium.
- Брюшко шаровидное. Мандибулы широкие.......................................................2.Messor.
30
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ
10. Вершина средней и задней голени с крупной вершинной гребенчатой
шпорой.....................................................................................f.Myrmica.
- Вершины голеней без гребенчатых шпор.................................................................. 11
11. Мандибуллярные и максиллярные щупики двучлениковые.................................3.Pheidole.
- Мандибуллярные щупики пятичлениковые, максиллярные щупики
трехчлениковые.....................................................................................12
12. Усики у самок и рабочих вместе со скапусом 12 члениковые, у самцов 13 члени-
ковые................................................................................8. Leptothorax.
-Усики у самок и рабочих 11 члениковые, а у самцов 12 члениковые 9.Temnothorax.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ DOLICHODERINAE
1. Эпинотум с сильно вогнутой покатой поверхностью (см. в профиль). Скульптура
тела грубая. Клипеус плоской формы, без киля................................1 .Dolichoderus.
- Эпинотум с ровной или покатой поверхностью. Скульптура тела тонкая и
нежная. Клипеус выпуклый..................................................................2
2. Чешуйка рудиментарная, еле заметная. Клипеус прикрывает основания жвал и
выступает между лобными валиками. Лобные валики параллельные...................2.Tapinoma.
- Чешуйка маленькая, но всегда заметная. Клипеус с ровным передним краем, мало
высту-пающий между лобными валиками......................................3.Bothriomyrmex.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ FORMICINAE
1. Мандибулы саблевидные, с очень мелкими зубчиками............................8 Polyergus.
- Мандибулы не саблевидные, с крупными зубцами............................................2
2. Эпинотум с резко вогнутой покатой поверхностью, у рабочих несет
зубцы.......................................................................2.Acantholepis.
- Эпинотум с выпуклой покатой поверхностью, без зубцов....................................3
3. Скапус расположен на значительном расстоянии от клипеуса. Усиковая ямка
отделена от клипеальной.......................................................3.Camponotus.
- Скапус причленяется близко от края клипеуса. Усиковая и клипеальная ямки
слиты.....................................................................................4
4. Лобная площадка резко ограничена......................................................5
- Лобная площадка не ясная................................................................6
5. Лобные валики прямые, параллельные, лишь немного расходящиеся
5.Cataglyphis.
-Лобные валики расходящиеся........................................................................ 6.Formica.
6. Голова с выемкой на заднем крае. Скапус короче заднего края
головы.......................................................................9.Rossomyrmex.
- Голова без выемки. Скапус длиннее заднего края головы...................................7
7. Глазки рабочих всегда развиты. Рабочие полиморфны...........................7.Proformica.
- Гпазки у рабочих рудиментарные или отсутствуют. Рабочие мономорфны......................8
8. Чешуйка маленькая, узкая, закругленная, наклоненная вперед, без
вырезки........................................................................1 .Plagiolepis.
- Чешуйка крупная, довольно широкая, прямая, нередко с вырезкой на верхнем
крае........................................................................................................................4.Lasius
31
Род Myrmica Latreille, 1804
Род Leptothorax Mayr, 1855
Род Crematogaster Lunder, 1831
Род Tetramorium Mayr, 1855
Род Messor Forel, 1890
Род Monomorium Mayr, 1855
Род Strongulognathus Mayr, 1853
Род Pheidole Westwood, 1840
Род Solenopsis Westwood, 1841
Род Cardiocondyla Emery, 1869.
Род Dolichoderus Lunder, 1831.
32
ПРИЛОЖЕНИЕ
Род Tapinoma Foerster, 1850
Род Formica Linnaeus, 1758
Род Camponotus Mayr, 1861
Род Bothriomyrmex Emery, 1869
Род Proformica Ruzsky, 1903
Род Lasius Fabricius, 1804
Род Plagiolepis Mayr, 1861
Род Rossomyrmex K.Arnoldi,1928
Род Cataglyphis Foerster, 1850
Род Acantholepis Mayr, 1861.
Род Polyergus Latreille, 1802.
33
воптяы
I орпйл
Самка богомола Mantis religiosa Linnaeus, 1758.
ХАРАКТЕРИСТИКА ГРУППЫ
Это преимущественно крупные дневные теплолюбивые насекомые с удлиненным
телом и особенно сильно развитым передним грудным сегментом, несущим
характерные для богомолов хватательные конечности. Характерная структура тела
богомола наглядно свидетельствует о приспособлениях к хищному образу жизни. А их
поведение представляет собой классический пример «подстерегающих хищников»,
иначе называемых засадниками.
У богомолов очень подвижная голова, имеющая треугольную форму и несущая с
боков большие выпуклые глаза, а у основания усиков три простых глазка. Ротовой
аппарат грызущего, гипогнатического типа (голова направлена ротовыми органами
вниз и немного назад). Усики обычно имеют нитевидную форму, но у некоторых видов
перистые, или гребенчатые. Бедро и голень лапки, складываясь вместе, образуют
мощный хватательный аппарат. Бедро по нижнему краю усажено тремя рядами
сильно развитых шипов. Голень короче бедра и по нижнему краю тоже усажена
острыми шипами, а лапка богомолов тонкая, пятичлениковая, не отличается от лапок
на остальных ногах. Конец брюшка с короткими членистыми придатками (церками), у
самок яйцеклад отсутствует. Крылья часто укороченные, или с различной степенью
рудиментации. Передние сильнее склеротизированные, а задние перепончатые
веерообразно складывающейся вдоль тела. Богомолы бывают полнокрылыми,
короткокрылыми или полностью лишенными крыльев. Многие окрашены в зеленые,
бурые или пегие криптические (сливающиеся с окружающим фоном) тона. Некоторые
виды расцвечены ярче, иногда с выраженным рисунком, в виде перевязей и пятен,
"переливающейся" окраской, зависящей от преломления световых лучей.
Личинка богомолов имея внешнее сходство со взрослыми насекомыми, также
являктся активным хищником, охотящимся на более мелкую добычу.
Богомолы наряду с таракановыми (Blattoptera), приведеньевыми (Phasmoptera) (к
которым относятся палочники и листовидки), прямокрылыми (Orthoptera, Saltatoria) и
рядом других образуют группу ортоптероидного комплекса. Этот комплекс - один из
самых древних по происхождению и характеризуется целым рядом архаичных
признаков, таких как неполное превращение, имагообразное строение личинок,
гипогнатическая голова (направленная ротовыми органами вниз) относящаяся к
грызущему типу. В отряд богомоловые, или богомолы (Mantoptera, Mantodea),
включают семейство настоящих богомолов (Mantidae, Manteidae) и семейство эмпуз,
или эмпузид (Empusidae).
Богомолы - сравнительно небольшая группа насекомых по количеству видов
представленных в умеренных широтах, имеющая древнее происхождение и
обладающая интересным поведением. На южных широтах группе присуще большее
разнообразие форм, особенно в тропических регионах. Фауна богомолов территории
Казахстана состоит из около десятка представителей семи родов, а мировая фауна
насчитывает свыше двух тысяч видов. Два вида этих насекомых: Bolivaria brachyptera
Pallas, 1773 (боливария короткокрылая) и Hierodula tenuidentata Saussure, 1869
(древесный богомол) внесены в Красную книгу Республики Казахстан и подлежат
охране.
35
У богомолов, в онтогенезе (индивидуальном развитии особи) строение
и образ жизни ранних стадий и имаго сходен, личинка похожа на взрослое
насекомое, а изменения организации сопровождаются в основном постепенным
Богомолы яйца
откладывают на
различные предметы
группами, покрывая
пенистым секретом
анальных желез,
который, застывая
образует своеобразную
капсулу - оотеку.
В своем развитии претерпевает семь
личиночных возрастов (разделенных линьками).
Она напоминает миниатюрную копию взрослых
богомолов.
36
ЖИЗНЕННЫЙ ЦИКЛ
ростом, с каждой линькой личинки, зачатков крыльев. Данный тип метаморфоза
(процесс превращения личинки во взрослую особь) называют неполным
превращением, или гемиметаболией.
Схема метаморфоза богомола рода Hierodula transcaucasica Brunner von Wattenwyl, 1878.
37
Для богомолов характерно не только специфическое строение
передних ног, на заднегруди расположены ультразвуковые органы слуха.
Имея очень подвижную голову, с крупными сложными глазами, они обладают
надкрылье
крыло
переднегрудь
тергиты
анальная
пластинка
передняя
хватательная
нога
средняя
бегательная,
или ходильная
нога
стерниты
церк
задняя
бегательная, или
ходильная нога
Строение тела богомола Iris
oratoria Linnaeus,1758.
38
ВНЕШНЕЕ СТРОЕНИЕ (МОРФОЛОГИЯ)
очень большым углом обзора, позволяющим видеть происходящее сзади насекомого.
Пронотум, включающий переднегрудь, не покрывает большую часть головы, в чем
богомолы сходны с тараканами по общим признакам и строению.
Строение передней (хватательной) ноги Iris oratoria Linnaeus, 1758.
усик
сложный глаз
Строение головы
Iris oratoria Linnaeus, 1758.
верхняя губа
челюстные щупики
Строение головы
Empusa pennicornis Pallas, 1773.
39
органы
секреции
органы
пищеварения
нервная
система
слепые придатки
средней кишки
прямая кишка
тонкая кишка
яйцевод
анальная
железа "
жевательный
желудок
зоб
парные яичники
40
ВНУТРЕННЕЕ СТРОЕНИЕ (АНАТОМИЯ)
железа
грудные
Схема внутреннего строения богомола.
41
42
РАЗМНОЖЕНИЕ
Спаривание богомолов Mantis rehgiosa Linnaeus, 1758.
Самки богомолов значительно крупнее самцов.
У самцов помимо хрупкого телосложения, в отли-
чие от самок, на кончике брюшка располагаются
кроме церок, парные придатки - грифельки. Про-
цесс спаривания у богомолов происходит доста-
точно долго и нередко заканчивается для самца
гибелью - самка отгрызает ему голову и перед-
нюю часть туловища. При этом нервные центры,
управляющие функциями гениталий самца, рас-
положены на задних участках его тела, что
позволяет не прерывать процесса копуляции.
43
Откладка яиц богомолом Bolivaria brachyptera Pallas, 1773.
Богомолы откладывают яйца, располагая рядами и склеивая их вместе пенистым,
застывающим секретом крупными массами определенной конфигурации. Эти массы,
или оотеки с яйцами, приклеиваются к ветке, камню или другому предмету. Окраска
оотеки варьирует от светло - желтой до коричневой или серой. Число яиц в кладке
колеблется в зависимости от вида и составляет от 50 до 300. В северных областях
ареала у богомолов зимовка происходит на стадии яйца и развивается одно поколе-
44
РАЗМНОЖЕНИЕ
Выход личинок богомола
из оотеки.
ние в год. Весной из яиц вылупляются
личинки. Личинка одолевает выходное
отверстие и выбирается наружу, а упру-
гие края отверстия оотеки, сокращаясь,
зажимают хвостовые нити. В таком по-
ложении личинка приступает к линьке.
Перелиняв, она становится похожей на
взрослого богомола с зачаточными
крыльями.
Богомол Empusa pennicornis Pallas, 1773 -
подстерегающий добычу.
Все виды богомолов ведут хищный образ жизни, питаясь насекомыми и другими
членистоногими, которых они ловят, подстерегая на растениях, или подкрадываясь
среди растительности, при помощи хватательных передних ног. Размер добычи уве-
личивается с ростом насекомого. Если в начале жизни у вновь отродившейся личинки,
основу рациона составляют тли и другие мелкие животные, то позднее добычей
46
ПИТАНИЕ
более взрослой особи богомола становятся крупные, порой небезобидные
членистоногие - различные прямокрылые, двукрылые, пчелы, осы, пауки. Излишняя
активность богомола как хищника иногда приводит к его гибели. Нападая на добычу, с
которой богомол не может справиться (крупные кузнечики, или другие богомолы),
порой он сам становится их добычей и бывает съеден.
Paravespula.
47
48
ПОВЕДЕНИЕ В ПРИРОДЕ
Полет у богомолов неуклюжий и неу-
стойчивый, поэтому многие виды пред-
почитают передвигаться среди расти-
тельности при помощи ног, лишь изредка
перелетая, или планируя на крыльях.
Обследуя территорию в поисках пищи
степные виды способны преодолевать
значительные расстояния. Часто, осо-
бенно в период размножения богомолы
приходят, или прилетают ночью к
источникам искусственного света.
Обитатель степей - богомол Rivetina папа Mistshenko, 1967.
1
49
Самка богомола Mantis religiosa Linnaeus,
1758, поедает окончившего копуляцию
самца.
50
КАННИБАЛИЗМ
Каннибализм животных - это поедание
особей своего вида, внутривидовое
хищничество. Чаще наблюдается при
неблагоприятных условиях, возника-
ющих при переуплотнении популяции,
или недостатке корма. При этом самки
более склонны к каннибализму, чем
самцы.
Среди богомолов широко распро-
страненное и совершенно “нормальное”
явление - поедание самкой самца после
окончания спаривания. Каннибализм у
богомолов носит постоянный (обли-
гатный) характер и возник в процессе
эволюции как полезное для вида при-
способление, которое, выступает в ка-
честве природного регулятора числен-
ности популяции вида, или фактора,
стабилизирующего популяцию, уста-
навливающего число особей в соответ-
ствии с кормовыми ресурсами и способ-
ствующего выживанию.
51
Покровительственная окраска и форма богомолов в ходе эволюции
достигла совершенства и для них характерно проявление всех ее типов -
маскировки, демонстрации и мимикрии (подражательного сходства). Окраска и
форма этих животных делает их менее заметными на фоне окружающей среды,
маскируя с окружающим фоном. Мимикрия у богомолов выражается внешним
сходством с предметами окружающей среды и растениями - мимезией. Для многих
Богомол - крошка Armene pussila Eversmann, 1859, прячется среди камней имитируя сухую
опавшую веточку.
52
МИМИКРИЯ
характерно проявление дизруптивной, или расчленяющей, окраски с наличием
контрастных пятен и полос, нарушающих зрительное впечатление о контурах тела,
часто в сочетании с криптической, когда пятна в окраске гармонируют с фоном.
Личинка Empusa pennicornis Pallas, 1773, при передвижении покачивается, имитируя движение
окружающей растительности от ветра.
53
Богомола, сидящего в засаде во время охоты, незаметным делает не
только его неподвижная поза, но также сама форма его тела и окраска.
Различно окрашенные цветовые формы, или морфы, встречаются в популяциях вида
совместно и обозначаются словом аберрации.
54
ЗАЩИТНАЯ ОКРАСКА
Окраска тела обычно носит защитный характер. Преобладают зеленый, желтый и
бурый тона. В пределах одного и того же вида могут встречаться особи,
отличающиеся по цвету. В течении сезона тон и насыщенность окраски меняются в
зависимости от фона окружающей растительности.
55
Отпугивая птиц и других крупных хищников, потревоженные богомолы
принимают угрожающие позы. Размахивая передними ногами, распа-
хивают веер крыльев, демонстрируя скрывающийся при сложенных в покое и
неброско окрашенных надкрыльях резко бросающиеся в глаза яркие крупные рисунки
в виде пятен, образующих переливающиеся на свету глазки.
Отпугивая потенциальных хищников богомол Iris
polystictica Fischer de Waldheim, 1846,
разворачивает крылья, неожиданно демонстрируя
переливающийся рисунок, образованный
перевязями, напоминающий глаза.
56
ВЗАИМООТНОШЕНИЯ В ПРИРОДЕ
Угрожающая поза может сопровождаться устрашающими звуками, возникающи-
ми при раздражении насекомого от шуршания крыльев и щелчков голени по бедру.
Одновременно богомол начинает раскачиваться из стороны в сторону и размахивает
передними ногами, часто несущими на внутренней стороне бедер рисунок.
Потревоженный самец богомола Mantis
religiosa Linnaeus, 1758, издает
движением крыльев о тергиты
волнообразно изгибающегося брюшка
шуршащие звуки, напоминающие
шипение.
57
У некоторых богомолов развились в процессе эволюции удивительные
приспособления для охоты. Вогнутое зеркальце у рода Empusa,
расположенное на голове, при попадании на него солнечного света создает иллюзию
Kai
58
УЛОВКИ БОГОМОЛОВ
фоне выгоревшей на солнце бурой растительности, росинкой попадают в “лапы”
коварного хищника. Похожие зеркальца и тактика имеются у богомолов рода
Hierodula. Они находятся у них на бедре между рядами острых щипов.
Богомол Hierodula transcaucasica Brunner von Wattenwyl, 1878.
59
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ И ПОДСЕМЕЙСТВ
Строение головы характерное для богомолов, типичной треугольной формы,
несущей, у самок и самцов нитевидные усики.................1. Manteidae.
- Голова имеет выступающий конический отросток, иногда раздвоенный на конце,
усики у самцов перистые....................................2. Empusidae.
1. Переднеспинка длиннее тазиков передних ног. Расширенная часть переднеспинки
находится заметно ближе к переднему концу. Преимущественно крупные формы.
Крылья иногда с рисунком. Мелкие формы безкрылы............1. Manteinae.
- Переднеспинка короче тазиков передних ног. Расширенная часть переднеспинки
находится примерно посередине. Усики самцов нитевидные. Крылья, если есть,
всегда без рисунка, хотя могут быть окрашены. Преимущественно мелкие
формы.......................................................3. Amelinae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ MANTEINAE
Внутренняя поверхность средних и задних бедер и голеней с рядами упорядоченных
шипов. Крылья, если есть, с рисунком, первый членик задних лапок длиннее всех
остальных взятых вместе.............................................1
- Внутренняя поверхность средних и задних бедер и голеней без рядов шипов, но
иногда покрыта неупорядоченными очень мелкими шипиками. Крылья, если есть, с
рисунком или без него...............................................2
1. Крылья дымчатые, с сероватым костальным полем, у внешнего края непрозрачные,
у самцов и самок укороченные...................................Bolivaria.
- Крылья дымчатые с круглым темным (белым в центре) глазком в апикальной
вершинной) области, у самцов нормально развиты..............Rivetina.
2. Передние бедра с четырьмя внешними шипами, лобовой склерит без бугорков.
Анальная пластинка короткая, закругленная, крылья прозрачные без темного
рисунка.............................................................3
- Передние бедра с пятью внешними шипами, лобовой склерит с двумя бугорками.
Анальная пластинка продолговатая, крылья с темным рисунком в виде пятен, первый
членик задних лапок короче всех остальных взятых вместе..........Iris
3. Ширина лобового склерита заметно больше высоты, бедра средних и задних ног без
шипа на вершине, глазок на надкрыльях неявственный............Mantis.
- Ширина лобового склерита примерно равна его высоте, бедра средних и задних ног с
шипом на вершине, глазок на надкрыльях резко выраженный, белый.Hierodula.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ AMELIDAE
Глаза заострены заметно в стороны........................Oxyothespis.
- Глаза с заострением несколько вперед, либо округлые................1
1. Корневой членик задней лапки не короче остальных члеников вместе взятых.
Сложные глаза округлой формы, крылья у самок нормальной длины..Armene.
- Корневой членик задней лапки немного короче, равен или немного длиннее
следующего членика. Сложные глаза конические, крылья у самок укороченные...
..............................................................Ameles.
60
ПРИЛОЖЕНИЕ
Ameles decolor
Charpentier, 1825.
Строение надкрылии
Hierodula tenuidentata
Saussure, 1869.
Голова Empusa
pennicornis Pallas,
1773.
Строение надкрылии
Mantis religiosa Linnaeus ,
1758.
Строение члеников
задней ноги Rivetina
папа Mistshenko,
1967.
Строение
переднегрудного отдела
Armene pussila
Eversmann, 1859.
Строение члеников
задней ноги Iris polystictica
Fischer de Waldheim, 1846.
Строение
переднегрудного отдела
Bolivaria brachyptera
Pallas, 1773.
анальная пластинка
Строение анальной
пластинки у Iris
polystictica Fischer
de Waldheim, 1846.
Строение анальной
пластинки у Mantis
religiosa Linnaeus,
1758.
Голова Ins polystictica
Fischer de Waldheim.,
1846.
61
СТРЕКОЗЫ
ODONhTUPTERA
Стрекоза Sympetrum sanguineum sanguineum Muller, 1764.
ХАРАКТЕРИСТИКА ГРУППЫ
Отряд стрекоз, занимая особое место среди насекомых, отличается одной из важ-
ных особенностей - высокой морфологической специализацией.
Современный отряд стрекоз принято считать древним по происхождению, учиты-
вая палеозойский возраст ископаемых стрекозоподобных. Тем не менее, стрекозам
присущи многие прогрессивные черты, выражающиеся в широком распространении,
большом видовом многообразии (около 6 тысяч рецентных (известных и описанных
видов)), обилии в водных и о кол о водных формациях.
Даже из одной яйцекладки стрекозы имеют разную скорость развития, что позво-
ляет сохраниться хотя бы части яиц в случае гибели остальных вследствие измене-
ния внешних условий, поэтому в водоемах одновременно присутствует большое
количество личинок разных возрастов одного вида.
Личинки стрекоз ведут водный образ жизни и в общей своей массе обитают среди
водных растений, где они медленно ползают по стеблям или по дну. У семейства
Epallagidae они живут под камнями в быстрых потоках. А у настоящих стрекоз (семей-
ство Libellulidae) держатся у поверхности дна (рода Leucorrhinia Brittinger, Sympetrum
Newman) или сидят, зарывшись у самой поверхности (рода Libellula Linnaeus, Cordulia
Leach). Личинки булавобрюхов (семейство Cordulegasteridae) и дедок, или речников
(семейство Gomphidae) - также обитатели дна водоемов, закапывающиеся довольно
глубоко в грунт. Форма и строение ног находится в тесной связи со специальным обра-
зом жизни личинок. Передняя пара ног, обыкновенно употребляется для удерживания
пищи во время еды. Стрекозы дышат не только через любой тонкий участок покровов,
но в основном через поверхность специальных дыхательных органов. У личинок круп-
ных стрекоз дыхание осуществляется только путем «вдохов» и «выдохов» воды через
анальное отверстие в заднюю (прямую) кишку. С силой выталкивая воду, личинка
делает резкие движения в водной среде по принципу действия реактивного двигателя.
У личинок более мелких равнокрылых стрекоз дыхательными органами служат листо-
видные трахейные жабры, находящиеся на заднем конце брюшка, служащие и орга-
ном передвижения, как хвостовой плавник рыбы. Даже у этих стрекоз, имеющих тра-
хейные жабры, большая часть потребляемого кислорода поступает в организм через
стенки задней кишки. Число личиночных стадий у различных видов непостоянно,
например, известно, что виды рода Lestes Leach линяют от девяти до одиннадцати
раз, а виды рода Sympetrum Newman - семь, иногда восемь раз.
Сложное поведение, наличие высокоадаптивных жизненных форм и активное
хищничество определяют важную роль стрекоз в трофических сетях биоценозов. При
чередовании водной и наземной фаз развития и большой биомассе стрекозы вносят
существенный вклад в процессы круговорота и трансформации вещества в природ-
ных экосистемах.
В фауне насекомых Республики Казахстан к настоящему времени зарегистрирова-
но около ста видов, из которых семь отмечены как нуждающиеся в охране и внесены в
Красную книгу Казахстана.
63
Стрекозы, за немногими исключениями, относятся к дневным насе-
комым, летающим преимущественно в ясную погоду. Разнокрылые стрекозы (подо-
тряд Anisoptera Selys, 1840) характеризуются быстрым резким полетом и способнос-
Подотряд Zygoptera Selys, 1840.
Подотряд Anisoptera Selys, 1840.
64
ОБЩЕЕ СТРОЕНИЕ
тью отлетать на далекие расстояния от водоемов. Равнокрылые стрекозы (подотряд
Zygoptera Selys, 1840) - медленно порхающие стрекозы, не отлетающие далеко от
водоемов.
узелок
крыловой треугольник
птеростигма
перепоночка
анальное поле
предузелковые
поперечные жилки
крыловой
четырехугольник
добавочная
(интеркалярная)
продольная жилка 1R2
Строение (жилкование) крыльев у
стрекоз различных групп.
65
Жизненный цикл у стрекоз характеризуется неполным прев-
ращением. Личинки стрекоз имеют внешнее морфологическое сходство с имаго, имея
укороченное, грубое тело, крылья в виде зачаточного органа и более длинные ноги.
Яйца стрекоз с про-
долговатым телом -
удлиненной формы, а у
стрекоз с более коротким
и широким телом -
округлой.
Из предличинок выходят маленькие, всего около одного
миллиметра длиной плоские, светло - зеленые или
кремовые личинки, из которых с ростом происходят при
каждой линьке изменения всех органов. Обычно число
личиночных стадий у различных видов непостоянно.
66
ЖИЗНЕННЫМ ЦИКЛ
Размещение в биоценозе у стрекоз разделяется не только по территориальному
принципу, но и по вертикальному: они занимают травяной, кустарниковый и
древесный ярусы растительности.
У
Взрослые стрекозы в умеренных широтах летают на протяжении всего теплого периода,
при этом их видовой состав все время меняется, а осенью они исчезают. В фазе имаго
перезимовывают лишь немногие виды.
Схема метаморфоза стрекозы Calopteryx spledens Harris, 1782.
67
Стрекозы способны не только на простые формы зрительного ана-
лиза, такие как стабилизация полета относительно объекта, обнаружение и преследо-
вание добычи, и защитное бегство. Им присущи и более сложные, такие как опозна-
ние брачного партнера, запоминание и охрана
территории, возврат к охраняемой территории с
расстояния нескольких километров, а также
дальние миграции.
верхний анальный
придаток
крыло
Внешнее строение стрекозы Anax imperator Leach, 1815.
68
ВНЕШНЕЕ СТРОЕНИЕ (МОРФОЛОГИЯ)
У стрекоз - преследователей (разнокрылые), захватывающих добычу в полете,
поля зрения обоих глаз перекрываются всего несколькими рядами фасеток, поэтому
они не могут эффективно оценивать глубину пространства. Стрекозы - собиратели
(равно-крылые) летают медленно и схватывают добычу среди растительности,
обладают большей бинокулярной зоной, позволяющей им выделять характерные
объекты на сложном фоне, например мелких двукрылых насекомых, летающих среди
береговой растительности, окружающей водоем.
Семейство Aeschnidae Bianchi, 1905
(коромысла).
первый боковой
сложный
глаз
Семейство Gomphidae Banks, 1892
(дедки, или речники).
Семейство Lestidae Bianchi, 1905 (лютки).
69
органы
секреции
органы
пищеварения
нервная
система
пищевод
гло
грудные ганглии
слепые придатки
средней кишки
жевательный
желудок
слюнная
железа
подглоточный
ганглий
надглоточный
ганглий
70
ВНУТРЕНЕЕ СТРОЕНИЕ (АНАТОМИЯ)
непарный яйцевод
гонопофизы
(яичники)'
железа семяприемника
тонкая кишка
средняя кишка
мальпигиевы сосуды
прямая кишка
Внутреннее строение равнокрылой стрекозы (подотряд Zygoptera Selys, 1840).
71
Большую часть жизни стрекозы проводят в стадии личинки, ведущей
водный образ жизни, предпочитая преимущественно стоячие или медленно текущие
водоемы. Только некоторые из них живут в реках с более или менее сильным
Общий вид личинки стрекозы
Erythromma najas Hansemann, 1823.
Общий вид личинки стрекозы
Sympetrum danae Sulzer, 1776.
Строение хватательной нижней губы-
плоской маски у личинки стрекозы рода
Erythromma Charpentier, 1840.
Строение хватательной нижней губы-
шлемовидной маски у личинки рода
Sympetrum Newman, 1833.
Строение ротовых органов у личинки
стрекозы poflaAeschna Fabricius, 1775.
72
СТРОЕНИЕ ЛИЧИНКИ
течением выбирая для своей жизни слабопроточные участки. Личинки встречаются не
только в пресных, но и в солоноватых водах, обитая в прибрежных лагунах, заливах и
бухтах. Личинки стрекоз - хищники-полифаги и питаются теми объектами, которые
являются для них более доступными в соответствующих условиях, а во время охоты
стрекозы отдают предпочтение более крупным насекомым. При захвате добычи
личинка выбрасывает маску - видоизмененную нижнюю губу, а пойманная добыча
крепко зажимается крючками лопастей и подносится ко рту, при складывании маски в
прежнее положение.
Охота личинки стрекозы Anaciaeschna isosceles isosceles Muller, 1767 на малька рыбы.
73
Спаривание Апах imperator Leach, 1815.
74
РАЗМНОЖЕНИЕ (СПАРИВАНИЕ)
Когда у стрекоз завершается период
созревания, они возвращаются к водое-
мам для спаривания и откладки яиц. Спа-
ривание стрекоз происходит в воздухе на
лету, самец откладывает сперматофор в
ямку на особом выступе третьего членика
своего брюшка. Затем он хватает самку
клешнеобразными придатками заднего
конца брюшка и летает с ней до тех пор,
пока она не поднимет к сперматофору
задний конец брюшка, на котором нахо-
дится половое отверстие, и не произойдет
копуляция. Такие попарно летающие
стрекозы принимают в это время вид
кольца.
Самка
75
Оплодотворенная самка откладывает яйца поодиночке прямо в воду,
во влажную почву, в ткани водяных растений, или же кучками или шнурами,
окруженными слизью. Иногда откладывает их в подводные части растений, опускаясь
по стеблям до самого дна, причем по мере движения самка все время откладывает
яйца.
Спаривание у стрекоз Coenagrion puella Linnaeus, 1758.
76
РАЗМНОЖЕНИЕ (ОТКЛАДКА ЯИЦ)
Некоторые виды откладывают яйца в надводные части растений (стебли, ветки) и
из яиц выходят предличинки, еще не сформировавшиеся личинки, которые
Откладка яиц стрекоз Calopteryx spledens Harris, 1782.
77
Ко времени превращения во взрослое насекомое личинки начинают
дышать атмосферным воздухом, для чего они по какой - либо опоре (стебли растений,
сваи, камни или любой грунт дна) вылезают наполовину из воды и остаются в таком
положении от нескольких часов до нескольких суток. Для окончательного
превращения во взрослое насекомое они выбираются из воды совсем, поднимаясь по
опоре или выходя прямо на берег. Выходя из воды, они сидят совершенно открыто и
свободно, крепко держась при помощи ног и коготков за свою опору, некоторое время
отдыхают, не двигаясь, а после отдыха начинают постепенно освобождаться от
Выход на сушу личинки стрекозы Aeschna cyanea Muller, 1764.
78
МЕТАМОРФОЗЫ
личиночной шкурки. После освобождения от шкурки стрекоза, не достигшая своих
окончательных размеров, сидит неподвижно, затем начинает делать колебательные
движения брюшком, с каждым движением увеличивая длину крыльев и брюшка,
расправляя их от основания.
Выход из личинки взрослой стрекозы Sympetrum danae Sulzer, 1776.
79
Взрослые крупные стрекозы хищники - преследователи, охотятся
только на подвижные жертвы, их основная добыча крупные хорошо летающие
насекомые (комары, мошки, мухи, поденки и ручейники).
80
ПИТАНИЕ (ИМАГО)
Большинство более мелких стрекоз относятся к группе подстерегающих
хищников, питающихся как у берегов водоемов, так и вдали от них, мелкими
малоподвижными насекомыми, придерживающимися прибрежной растительности.
Питание и патрулирование территории у стрекозы
Aeschna cyanea Muller, 1764.
81
Стрекозы сидят на выступающих частях растений и различных
предметов и взлетают за пролетающими насекомыми. Схватив добычу, они
возвращаются к месту старта. Питаются они на определенных участках, которые
активно защищают от других стрекоз. Выбор места для посадки или места
Platycnemis pennipes Pallas, 1771
Erythromma najas Hansemann, 1823
(семейство Coenagrionidae Tillyard,
1926[Agrionidae], или стрелки).
Sympecma fusca Vanderlinden, 1823
(семейство Lestidae Bianchi, 1905
(Coenargionidae), или лютки).
Enallagma cyathigerum Charpentier, 1840
(семейство Coenagrionidae Tillyard,
1926[Agrionidae], или стрелки).^^
(семейство Platycnemidae Tillyard et Fraser,
1938, или стрекозы - плосконожки).
82
ХАРАКТЕРНЫЕ ПРЕДСТАВИТЕЛИ
патрулирования над охраняемым участком в первую очередь зависит от видовой
принадлежности. Почти у всех групп стрекоз размер охраняемого участка
определяется размерами насекомого и в значительной мере зависит от плотности
особей на территории.
Ischnura pumilio Charpentier, 1825
(семейство Coenagrionidae Tillyard
1926[Agrionidae], или стрелки).
Nehalennia speciosa Charpentier, 1840
(семейство Coenagrionidae Tillyard,
1926[Agrionidae], или стрелки).
Lestes dryas Kirby, 1890 (семейство Lestidae Bianchi, 1905
(Coenargionidae), или лютки).
83
Среди взрослых особей, как и среди их личинок, нередко наблюда-
ются случаи каннибализма. Часто, мелкие и небольшие являются жертвами других,
более крупных хищных стрекоз.
Gomphus vulgatissimus Linnaeus, 1758
(семейство Gomphidae Banks, 1892,
или дедки, речники).
Calopteryx virgo virgo Linnaeus, 1758
(семейство Calopterygidae Buckecker, 1876,
или красотки).
84
ХАРАКТЕРНЫЕ ПРЕДСТАВИТЕЛИ
Somatochiora flavomaculata flavomaculata
Vanderlinden, 1825 (семейство Corduliidae
Karach, 1894, или бабки, патрульщики).
Hemianax ephippiger Sjostedt, 1925
(семейство Aeschnidae Bianchi,
1905, или коромысла).
85
Orthetrum cancellatum
cancellatum Linnaeus, 1758
(семейство Libellulidae
Stephens, 1836, или настоящие
стрекозы).
Sympetrum flaveolum Linnaeus, 1758
(семейство Libellulidae Stephens,
1836, или настоящие стрекозы).
86
ХАРАКТЕРНЫЕ ПРЕДСТАВИТЕЛИ
Sympetrum danae Sulzer, 1776
(семейство Libellulidae Stephens,
1836, или настоящие стрекозы).
Libellula depressa Linnaeus, 1758
(семейство Libellulidae Stephens,
1836, или настоящие стрекозы).
87
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДОТРЯДОВ
Жилкование и форма передних и задних крыльев одинаковое, крылья в покое
складываются (за исключением рода Lestes) над брюшком. Анальное поле развито
слабо, либо редуцированное. Имеется крыловой четырехугольник, а перепоночка
отсутствует. Глаза полушаровидные, выступающие, разделенные промежутком,
который шире глаза. У самцов по одной паре верхних и нижних анальных придатков. У
самок всегда есть яйцеклад...............(Подотряд Zygoptera - Равнокрылые
стрекозы).
- Передние и задние крылья не одинаковы по форме и жилкованию, в покое держатся
распростертыми. Анальное поле, особенно задних крыльев, хорошо развито, у осно-
вания они шире передних крыльев. Имеется крыловой треугольник и обычно перепо-
ночка. Глаза соприкасаются; если разделены, то промежуток между ними уже отдель-
ного глаза. У самцов одна пара верхних анальных придатков и единственный нижний
анальный придаток. Яйцеклад иногда отсутствует................................
..................................(ПодотрядАп1зор1ега - Разнокрылые стрекозы).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ ПОДОТРЯДА ZYGOPTERA
Крылья окрашены. Предузелковых поперечных жилок много. Четырехугольник вытя-
нут, подразделен поперечными жилками на многие ячейки. Анальное поле развито
слабо. Птеростигмы у самцов нет, у самок на ее месте светлое пятно («ложная пте-
ростигма»). Первый боковой шов вполне развит ..............................1.
Calopterygidae.
- Крылья не окрашенные, совершенно прозрачные. Предузелковых поперечных жи-
лок только две. Четырехугольник не подразделен поперечными жилками, состоит
из одной ячейки. Анальное поле отсутствует. Птеростигма всегда имеется.
Первый боковой шов неполный.................................................1
1. Птеростигма сравнительно длинная, равна по длине, по крайней мере, двум
ячейкам. Добавочная продольная жилка 1R2 искривленная. На темени за усиками
имеется борозда. Тело с металлическим блеском. Верхние анальные придатки самца
заметно длиннее десятого сегмента брюшка, клещеобразные .................. 2.
Lestidae.
- Птеростигма сравнительно короткая, длиной в одну ячейку. Добавочная
продольная жилка 1R2 прямая. Борозды за усиками нет. Тело редко и то тогда
лишь местами с металлическим блеском. Верхние анальные придатки обычно
короче десятого сегмента брюшка и не являются клещеобразными........................2
2. Четырехугольник правильный, длина его передней стороны примерно равна длине
задней стороны. Предузелковых дискоидальных ячеек две. Голени двух задних пар
ногу самцов расширены........................................3. Platycnemidae.
- Четырехугольник неправильный, передняя его сторона значительно короче, чем
задняя. Предузелковых дискоидальных ячеек, по крайней мере, три. Полени не рас-
ширены ......................................................4.Coenagrionidae.
88
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ ПОДОТРЯДА ANISOPTERA
Треугольники на всех крыльях расположены одинаково - вытянуты по длине крыла.
Первичные предузелковые поперечные жилки имеются (в виде двух утолщенных жи-
лок). Все лопасти нижней губы примерно одинаковой величины, боковые лопасти с
подвижными концевыми крючками..........................................1
- Треугольники на крыльях расположены неодинаково: на передних крыльях они вы-
тянуты поперек крыла, на задних - вдоль крыла. Первичных предузелковых попе-
речных жилок нет. Средняя лопасть нижней губы гораздо меньше боковой,
последняя без подвижных концевых крючков................................3
1. Глаза не соприкасаются, раздвинуты. Яйцеклад отсутствует..................................5. Gomphidae.
- Гпаза соприкасаются. Яйцеклад имеется....................................................................2
2. Глаза соприкасаются в одной точке. Яйцеклад большой, значительно длиннее
анальных придатков. Черные с желтыми пятнами стрекозы................................7. Cordulegasteridae.
- Глаза соприкасаются на некотором протяжении. Яйцеклад не длиннее конца
брюшка. Окраска другая........................................................................6. Aeschnidae.
3. Задний боковой край глаз посредине сильно изогнут. Основание задних крыльев
самцов вырезанное. Голени самцов на нижней внешней стороне с удлиненным, плас-
тинообразным килем. На втором сегменте брюшка самцов имеются ушки. Тело
обычно с металлическим блеском................................................................3. Corduliidae.
- Задний боковой край глаз прямой. Основание задних крыльев самцов закругленное.
Голени самцов без киля. Ушек у самцов нет. Тело без металлического блеска........................................
........................................................................................................................9.Libellulidae.
средняя лопасть нижней
Строение нижней губы у различных стрекоз.
род Sympetrum Newman, 1833
(Libellulidae Stephens, 1836).
род Cordulegaster Leach, 1815
(Cordulegasteridae Banks, 1892).
poflAeschna Fabricius, 1775
(Aeschnidae Bianchi, 1905).
Семейство Libellulidae
Stephens, 1836
(стрекозы настоящие).
89
Азональные - территории луговых формаций.
Аллометрия-пропорциональное изменение размеров тела муравьев.
Анатомия-внутреннее строение тела.
Аридные-пустынные (засушливые).
Биоценоз - совокупность животных, совместно населяющих участок суши или водоема.
Временный социальный паразитизм - проникновение самки муравья в гнездо
другого вида.
Гомеостаз - стабильность структуры семьи.
Гормоны-биологически активные вещества, выделяемые экзогенными железами.
Гумидные- лесные достаточно увлажненные биоценозы.
Железы - органы, вырабатывающие и выделяющие специфические вещества,
участвующие в физиологических отправлениях организма.
Инсоляция - поток солнечного тепла.
Карпофагия-питание семенами.
Кастовый диморфизм - наличие рабочих и особей-солдат.
Коадаптивный комплекс - совокупность родственных групп видов, связанных
конкурентными и мутуалистическими взаимоотношениями и заселяющих один
биоценоз.
Конкуренция - соревнование групп животных за одни и те же ресурсы внешней среды
при их недостатке.
Копуляция-спаривание половых особей.
Куколочный паразитизм - поведение социальных насекомых, при котором рабочие
вида-паразита периодически совершают набеги колонной рабочих на гнезда других
видов муравьев и похищают куколок.
Ландшафты - исторически сложившиеся, устоявшиеся формации из характерных
абиотических и биотических элементов.
Местообитание вида - совокупность отвечающих экологическим требованиям вида
участков.
Метаморфоз (метаболия) - развитие организма в онтогенезе со сложным жизненным
циклом.
Мобилизация - передача тем или иным способом информации о местонахождении
источника пищи и организация фуражировки.
Моновольтиннаягенерация- метаморфоз с годичным циклом развития.
Морфология - внешнее строение тела.
Мутуализм - симбиотические взаимоотношения группы организмов, возлагающих
друг на друга функцию регуляции своих отношений с внешней средой.
90
КРАТКИЙ СЛОВАРЬ ТЕРМИНОВ
Нефиксированный мономорфизм -тип изменчивости, при котором размеры рабочих
в одной семье могут сильно варьировать.
Общественные, или социальные, насекомые - насекомые, образующие
постоянные (сезонные или многолетние) объединения.
Онтогенез - индивидуальное развитие особи.
Падь - экскременты тлей.
Плеэргаты- каста рабочих, способных сохранять большие запасы жидкой пищи в зобу
Поливольтинная генерация - генерация, при которой метаморфоз занимает
несколько лет.
Полигиния семьи - обитание в одном гнезде насекомых свыше одной репродуктивной
самки.
Полиэтизм - фиксированные различия в выполнении разными рабочими
определенного круга функций.
Половой диморфизм - наличие репродуктивных и бесплодных самок и самцов
Провентрикулюс - жевательный желудок.
Семиаридные - полузасушливые степные области.
Трофоллаксис- взаимный обмен жидкой пищей между особями.
Феромоны -биологически активные вещества, выделяемые эндогенными железами.
Физогастрия-увеличения объема брюшка царицы.
Фиксированная вариабельность - тип изменчивости с унимодальным (равно-
встречающимся) частотным распределением особей по размерным классам и
монофазной аллометрией.
Фиксированный мономорфизм - тип изменчивости, при котором все рабочие особи
отдельной семьи имеют одинаковые размеры, но в зависимости от обеспеченности
пищей размер рабочих в разных семьях будет различаться.
Цикл развития - совокупность всех фаз развития, пройдя которые организм достигает
зрелости и становится способным дать начало следующему поколению.
Экзогенные - внутренние.
Экспозиция-направление ориентации склонов относительно частей света.
Эласмосомы- съедобные различные придатки семян.
Эндогенные - внешние.
Эргатоформные половые особи - рабочеобразные самки и самцы.
Прим. *Слово «муравей» образовано от древнерусского слова “мурава"(луговая трава).
Прим.** Богомолы - название группы насекомых происходит от строения передних лапок
насекомого и характерного поведения хищника (охотника) - засадника, словно застывшего в
молитвенной позе.
91
АЛФАВИТНЫЙ УКАЗАТЕЛЬ НАЗВАНИЙ НАСЕКОМЫХ
Aeschna cyanea Muller, стр. 78, 80, 81.
Ameles decolor Charpentier, 61.
Anaciaeschna isosceles isosceles Muller, стр. 73.
Anax imperator Leach, стр. 68, 69, 74, 75
Armene pusilia Eversmann, стр. 52, 61.
Bolivaria brachyptera Pallas, стр. 44, 61.
Calopteryx spledens Harris, стр. 66, 67,77.
Calopteryx virgo virgo Linnaeus, стр. 84.
Camponotus herculeanus Linnaeus, стр. 9,12.
Camponotus lameerei Emery, стр. 8,16.
Cataglyphis setipes (Forel), стр. 4.
Clytra atraphaxidis Pallas, стр. 24.
Coenagrion puella Linnaeus, стр. 76
Crematogaster subdentata Mayr, стр. 22.
Empusa pennicornis Pallas, стр. 39, 46, 53, 58.
Enallagma cyathigerum Charpentier, стр. 82.
Erythromma najas Hansemann, стр. 72, 82.
Formica cunicularia glauca Ruzsky, стр. 14, 23,
28, 29.
Formica fusca Linnaeus, стр. 16, 20.
Formica pratensis Retzius, стр. 20.
Formica sanguinea Latreille, стр. 26
Formica truncorum Fabricius, стр. 22, 23.
Formicoxenus nitidulus (Nylander), стр. 25.
Gomphus vulgatissimus Linnaeus, стр. 84.
Hemianax ephippiger Sjostedt, стр. 85.
Hierodula tenuidentata Saussure, стр. 36, 37,
59.
Hierodula transcaucasica Brunner von Wattenwyl,
61.
Homoeusa acuminata Maerklin, стр. 25.
Iris oratoria Linnaeus, стр. 38,
Iris polystictica Fischer de Waldheim, стр. 56, 60,
61.
Ischnura pumilio Charpentier, стр. 83.
Lasius niger (Linnaeus), стр. 12.
Lestes dryas Kirby, стр. 83.
Libellula depressa Linnaeus, стр. 87.
Mantis religiosa Linnaeus, стр. 34, 42, 43, 47, 50,
51, 54, 55, 57, 61.
Meropeurum fuscoviride Stroyan, стр.20.
Messor aralocaspius (Ruzsky), стр. 15, 18, 19, 21.
Messor clivorum (Ruzsky),1905. стр. 8.
Messor denticulatus (Kusnezov-Ugamsky), стр.
16.
Micaria pulicaria (Sundevall). стр. 24.
Myrmica bergi Ruzsky, стр. 25.
Nehalennia speciosa Charpentier, стр. 83.
Orthetrum cancellatum cancellatum Linnaeus,
стр. 86.
Pheidole pallidula (Nylander), стр. 13.
Platycnemis pennipes Pallas, стр. 82.
Polyergus rufescens (Latreille), стр. 27, 28, 29.
Proformica epinotalis Kyznetzow-Ugamsky, стр.
21.
Rivetina nana Mistshenko стр.48, 49.
Rossomyrmex proformicarum K. Arnoldi, стр. 26.
Somatochiora flavomaculata flavomaculata
Vanderlinden, стр. 85.
Strongylognathus christophi Emery, стр. 27.
Sympecma fusca Vanderlinden, стр. 82.
Sympetrum danae Sulzer, стр. 72, 79, 87.
Sympetrum flaveolum Linnaeus, стр. 86.
Sympetrum sanguineum sanguineum Muller,
стр. 62
Xantholinus longiventris Heer, стр. 24.
Бартенев A. H. Libellulidae. Фауна России и сопредельных стран. Насекомые ложносетчатокрылые.
Вып. 1 -2. Пгр., 1915-1919.
Бей-Биенко Г.Я. Общая энтомология. М.,1980.416 с.
Белышев Б. Ф., и др. Фауна и экология стрекоз. Новосибирск, 1989, 207 с.
Белышев Б.Ф,, Харитонов А.Ю. Одределитель стрекоз по крыльям (роды Бореального фаунистичес-
кого царства и сопредельных земель, виды фауны СССР) Новосибирск: Наука. Сиб. отд-ние,1970,
397 с.
Белышев В.Ф. Стрекозы - Odonata. 1/ Руководство к определению. Алма-Ата. Изд - во Казах, ин -
та, 1968. 12 с.
Белышев Б.Ф, Шенченко В.В. Фауна стрекоз (Odonata распределение ее компонентов в Казахста-
не). 1/ Биол. науки. 1971. Вып. 2. С. 73 - 77.
Белышев Б.Ф. Стрекозы Сибири. Новосиб.: Наука. Сиб. отд-ние, 1973.Т. 1. 620 с.
92
и список рекомендуемой литературы
Белышев Б.Ф. Стрекозы Сибири. Новосиб.: Наука. Сиб. отд-ние, 1974,Т. 2. 351 с.
Богданов-Катьков Н. Н. Руководство к практическим занятиям по общей энтомологии. М - Л., 1947.
356 с.
Бондаренко Н. В., Глущенко А. Ф. Практикум по общей энтомологии. Л.,1972. 344 с.
Гребенщиков В. С. В стране насекомых: записки и зарисовки энтомолога и художника. М., 1979, 168с.
Длусский Г. М. Муравьи рода формика. М.-Л., «Наука»: 1967, 236 с.
Длусский Г. М., и др., Муравьи Туркменистана, Ашхабад, 1990 г., 273 с.
Дмитриенко В. К., Петренко Е. С. Муравьи таежных биоценозов Сибири. Новосиб., 1976, 220 с.
Жизнь животных. Т. 2. М.,1968, 563 с.
Жизнь животных. Т. 3. М.,1969, 576 с.
Захаров А. А. Внутривидовые отношения муравьев. М., «Наука»: 1972, 216 с.
Захаров А. А. Муравей, семья, колония. М.,1978, 144 с.
Захаров, А. А. Экология муравьев. Итоги науки и техники. Зоол. беспозвоночных, М., 1980, т 7, с.
132-205.
Захваткин Ю. А., Исаичев В.В. Словарь - справочник энтомолога. М.,1992. 334 с.
Караваев В. А. Фауна родины Formicidae (мурашки) Укран!. Ки!в; I (1934): 1-62; II (1936): 1-316.
Кипятков В. Е. Мир общественных насекомых. Л., 1991,408 с.
Кипятков В. Е. Происхождение общественных насекомых. М., 1985, 64 с.
Кнауер Фридрих. Муравьи.(Под ред. В. А. Фаусека), СПб., 1905,188 с.
Козлов М., Нинбург Е. Ваша коллекция. Сбор и изготовление зоологических коллекций: Пособие
для учащихся. М., 1971.159 с.
Красная книга Казахской ССР. Редкие и находящиеся под угрозой исчезновения виды животных и
растений. Т. 1. Животные. Алма-Ата, 1981.560 с.
Купянская А. Н. Муравьи (Hymenoptera, Formicidae) Дальнего Востока СССР ДВО АН СССР, 1990,
257 с.
Леббок Дж. Муравьи, осы, пчелы. СПб., 1884, вып. 16, 491 с.
Мамаев Б.М. Определитель насекомых по личинкам. М.,1972. 400 с.
Мамаев Б.М., Медведев Л.Н., Правдин Ф.Н. Определитель насекомых европейской части СССР.
М.,1976. 304 с.
Мариковский П.И. Муравьи пустынь Семиречья. Алма-Ата, 1979. 264 с.
Негробов О. П., Черненко Ю. И. Определитель семейств насекомых. Воронеж, 1990. 181 с.
Определитель насекомых европейской части СССР. М., Л.,1948.1128 с.
Плавильщиков Н.Н. Определитель насекомых. М.,1957. 548 с.
Попона А.Н. Личинки стрекоз фауны СССР (Odonata) М.; Л.: Изд-во АН СССР, 1953а. 235 с.
Правдин Ф.Н. Экологическая география насекомых Средней Азии. М.: Наука, 1978. 271 с.
Резникова Ж. И. Межвидовые отношения муравьев. Новосибирск, 1983, 206 с.
Рузский М. Д. Муравьи России. Казань: 1905-1907, 1-799.
Руководство по энтомологической практике. Л.,1983, 230 с.
Спурис 3. Д. Стрекозы Латвийской ССР. Рига, 1956.
Тарбинский Ю. С. Муравьи Киргизии. Фрунзе, 1976, с 1 - 217.
Фабр Ж. А. Инстинкт и нравы насекомых. СПБ.д 1,1914, 590 с.
Фабр П. Насекомые. М., 1976.192 с.
Фасулати К.К. Полевое изучение наземных беспозвоночных. М.,1971.424 с.
Халифман И. А. Муравьи. М., 1964, 303 с.
Харитонов А.Ю. Отряд Odonatoptera - стрекозы. Определитель насекомых дальнего Востока
СССР. Л.: Наука. Ленингр. отд-ние, 1986. Т. 1. С. 142 -162.
Шванвич Б.Н. Курс общей энтомологии. М.; Л.: Сов, наука, 1949. 900 с.
Якобсон Г. Г. и В. Л. Бианки. Прямокрылые и ложносетчатокрылые Российской империи и сопре-
дельных стран. СПб.,1905. 592 с.
93
СПИСОК ЛАТИНСКИХ НАЗВАНИЙ ЧАСТЕЙ ТЕЛА
антеннальная борозда (sulcus antenalis)
антеннальные впадины (cavitas antennalis)
антеннальный склерит (sclerites antennalis)
антенны (antennae)
базитарзус (basitarsis)
бедро (femora)
брюшко (abdomen, gaster)
вертлуг (trochanter)
внутренние орбиты (orbita interior)
волселла (volsella)
гениталии (genitalia)
гипопигий (hypopigium)
глазки (ocellus)
глоссы (glossa)
глотка (pharynx)
голень (tibia)
голова (caput)
головная капсула (epicranium)
горизонтальная (дорсальная) поверхность
эпинотума (facies horizontalis epinotei)
грудь (thorax)
губа (labrum)
дигитус (digitus)
дистальный (пятый концевой) членик
лапки (distapsus)
длинный первый членик (metatarsis)
дыхальце (stigmata)
жало (aculeus)
железы (glandulae)
заднегрудь (metathorax)
задняя поверхность эпинотума (facies verticalis
posterior epinotei)
зоб (ingluvies)
кантик (feniculus)
кишечник (intenstium)
клипеальная борозда (sulcus clypealis)
клипеус (clypeus)
коготки (unguis),
крыловые крышечки (tegulae)
куспис (cuspis)
лабиальные щупики (palpes labiales)
лапки (tarsus).
латеральная(боковая) поверхность эпинотума
(facies laterales epinotei)
латеральные глазки (ocellus lateralis)
лациния (lacinia)
лицо (facies)
лоб (fronts)
лобная бороздка (sulcus frontalis)
лобная площадка (areae frontalis)
лобные валики (laminae frontalis)
максиллы (maxilla)
максиллярный комплекс (complex maxillaris)
малярные промежутки (areae malaris)
мандибулы (mandibulae)
медиальный глазок (ocellus medialis)
мезосома (mesosoma)
метасома (metasoma)
нижняя губа (labium)
ноги(pedes)
окулярные гребни (crista ocularis)
параглосса (paraglossa)
парамеры (paramera)
параокулярные участки (areae paraoculares)
петиоль (petiol)
пигидий (pygidium)
пищевод (oesophagus)
подушечки (pulvilli)
половые щупики (palpus genitalis, cerki)
постпетиоль (postpetiol)
прементум (prementum)
придатки головы (appendiculae epicraniales)
провентрикулюс (proventriculus)
просома (prosoma)
птеростигма (pterostigma)
радиальная ячейка (radialis cellae)
саггита (saggita)
сеты (setae)
сквамула (scvamula)
сложные глаза (oculi)
срединный сегмент (propodeum)
среднегрудь (mesothorax)
стернум (sternum)
стипес (stipes)
стрекотательный кантик (plectrum, scrapper)
стрекотательный напильничек (pars strideris,
file)
тазик(coxa)
тазиковые впадины (cavitas coxalis)
тело (soma)
темя (vertex)
тенториум (tentorium)
хеты (hetae)
шипы (spinae)
шпоры (calcaria)
эдиагус (aedeagus)
94
СОДЕРЖАНИЕ
OOs|-lf>CDOOOC\lsrCDOOOCXIsrCDCOOC\l^rinCDCOOC\ICDOOOC\lsrCDOOOT-CXlCOsrCDCOOCXlsrCDOOOC\|COOC\l^riD
: Т- Т- Т- Т- Т- C\l CXI CXI CXJ CXJ СО СО СО 00 СО 00 sr sr LO ю Ю Ш Ю CD CD CD CD CD CD CD I4- N- h- Is*- Is- СО 00 СО О) О СП О)
Предисловие...................................................................
МУРАВЬИ (FORMICOIDEA)......................................................
Введение в систематику...................................................
Жизненный цикл...........................................................
Внешнее строение (морфология)............................................
Внутреннее строение (анатомия)...........................................
Состав семьи, полиморфизм и разделение функции...........................
Размножение, образование семьи...........................................
Строение жилища (гнездо).................................................
Строительный инстинкт муравьев...........................................
Питание..................................................................
Взаимоотношение в семье..................................................
Взаимоотношения с другими организмами в природе..........................
Рабовладение.............................................................
Организация походов......................................................
Определительные таблицы..................................................
Приложение...............................................................
БОГОМОЛЫ (MANTOPTERA (MANTODEA, MANTOIDEA))................................
Характеристика группы....................................................
Жизненный цикл...........................................................
Внешнее строение (морфология)............................................
Внутреннее строение (анатомия)...........................................
Размножение..............................................................
Питание..................................................................
Поведение в природе......................................................
Каннибализм..............................................................
Мимикрия.................................................................
Защитная окраска.........................................................
Взаимоотношения в природе................................................
Уловки богомолов.........................................................
Определительные таблицы..................................................
Приложение...............................................................
СТРЕКОЗЫ (ODONATOPTERA (ODONATA))..........................................
Характеристика группы....................................................
Общее строение...........................................................
Жизненный цикл...........................................................
Внешнее строение (морфология)............................................
Внутреннее строение (анатомия)...........................................
Строение личинки.........................................................
Размножение (спаривание).................................................
Размножение (откладка яиц)...............................................
Метаморфозы..............................................................
Питание (имаго)..........................................................
Характерные представители................................................
Определительные таблицы..................................................
Краткий словарь терминов......................................................
Алфавитный указатель названий насекомых и список рекомендуемой литературы.....
Список латинских названии частей тела.........................................
Содержание....................................................................
95
ЭНТОМОЛОГИЧЕСКИЙ КАЛЕЙДОСКОП
МУРАВЬИ, БОГОМОЛЫ, СТРЕКОЗЫ
Автор:
Сергей Анатольевич Тимоханов
Художник:
Владимир Анатольевич Тимоханов
Редактор:
Сергей Анатольевич Тимоханов
Рецензенты:
доктор биологических наук, профессор Владимир Лонгинович Казенас,
кандидат биологических наук М. К. Чильдебаев.
Дизайн и верстка: В. А. Тимоханов,
О.С. Любовицкая
Формат 6Ох9О7в. Гарнитура "Arial".
Подписано в печать 12.11.07.
Печать офсетная. Бумага офсетная.
Усл. печ. л. 12. Тираж .
Заказ l\i .
ТОО Издательства "Кйтап-Д yip"
г.Алматы, пр.Гагарина, 93
Тел.: 269-93-89, 269-94-39