






































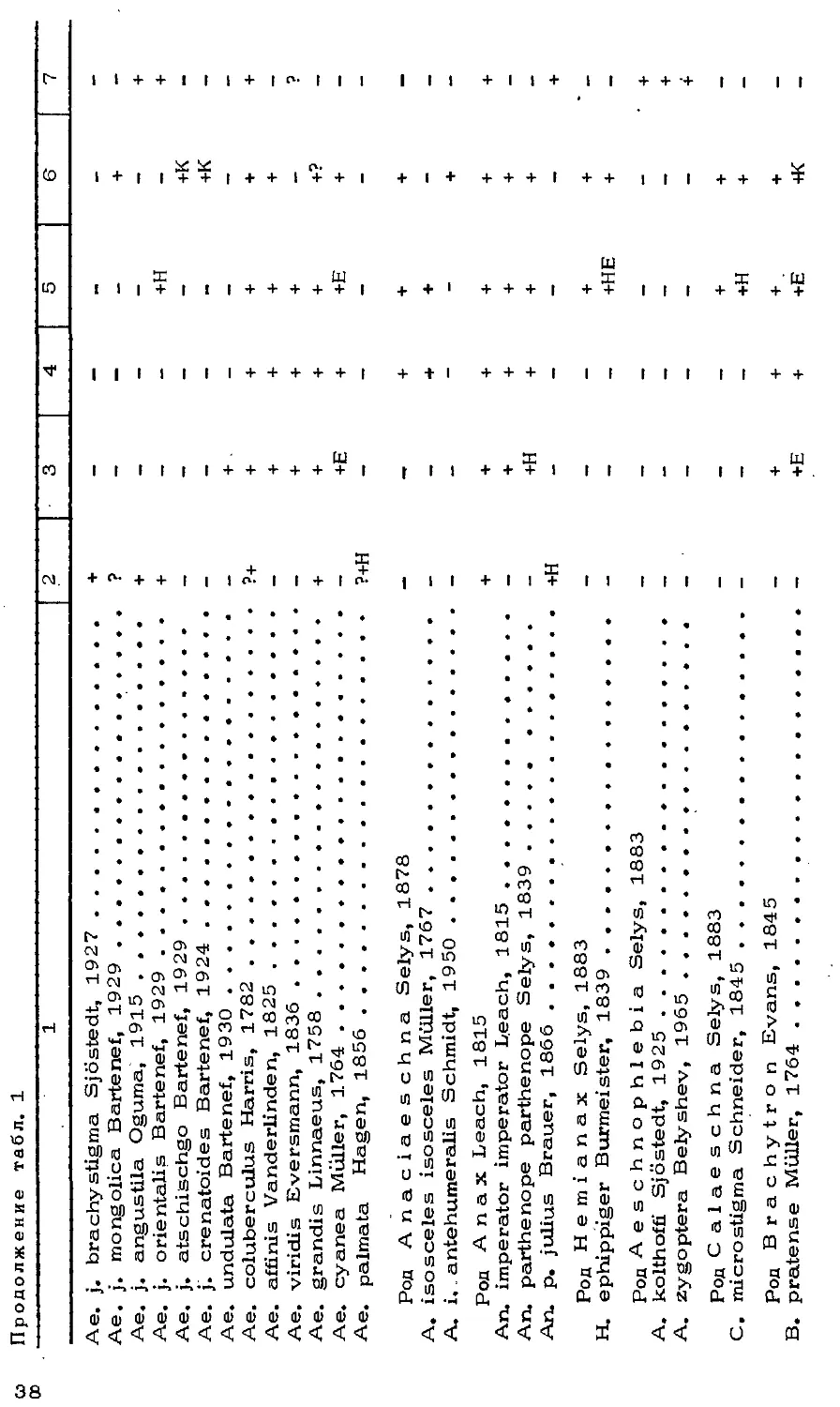


































































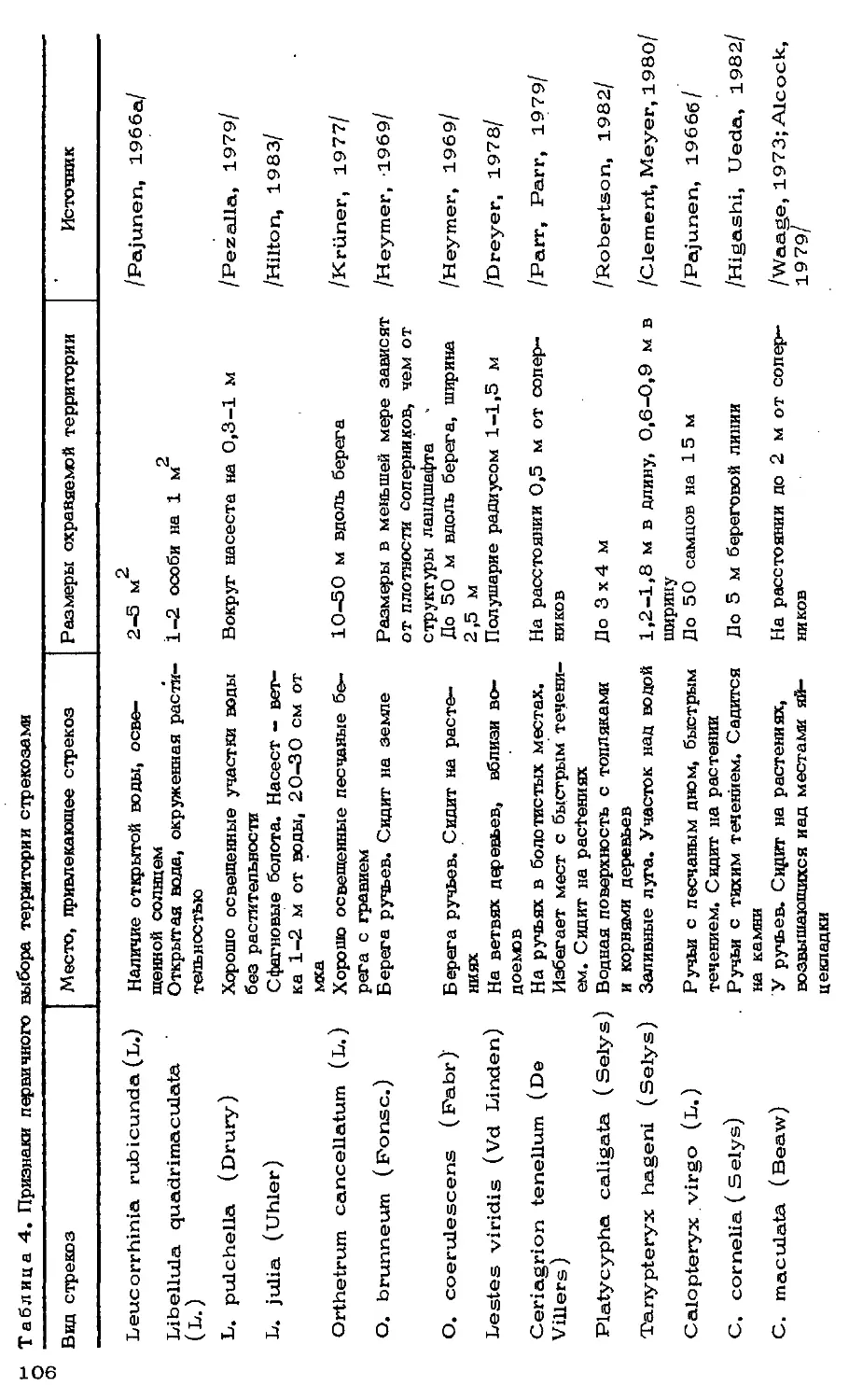

























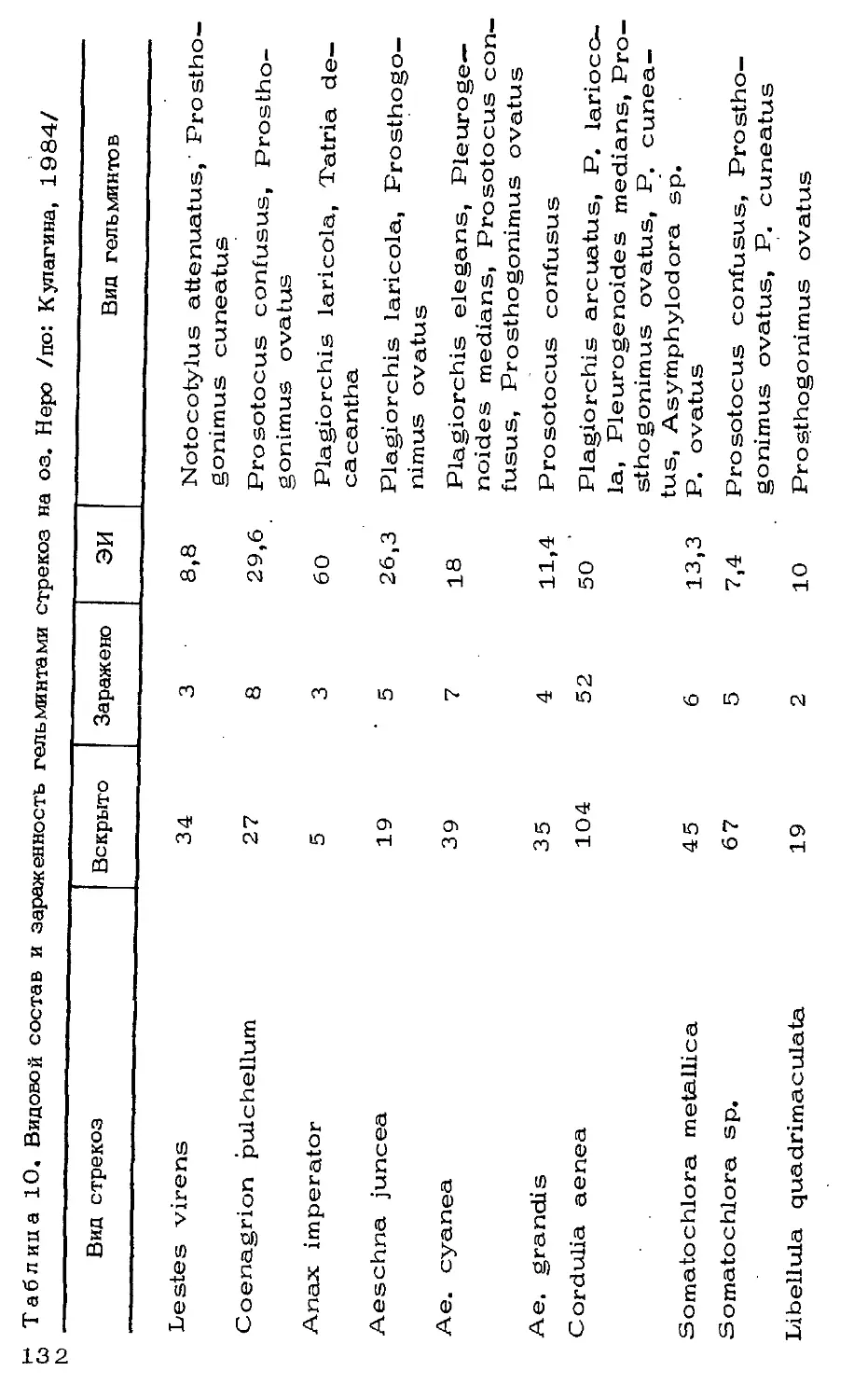


















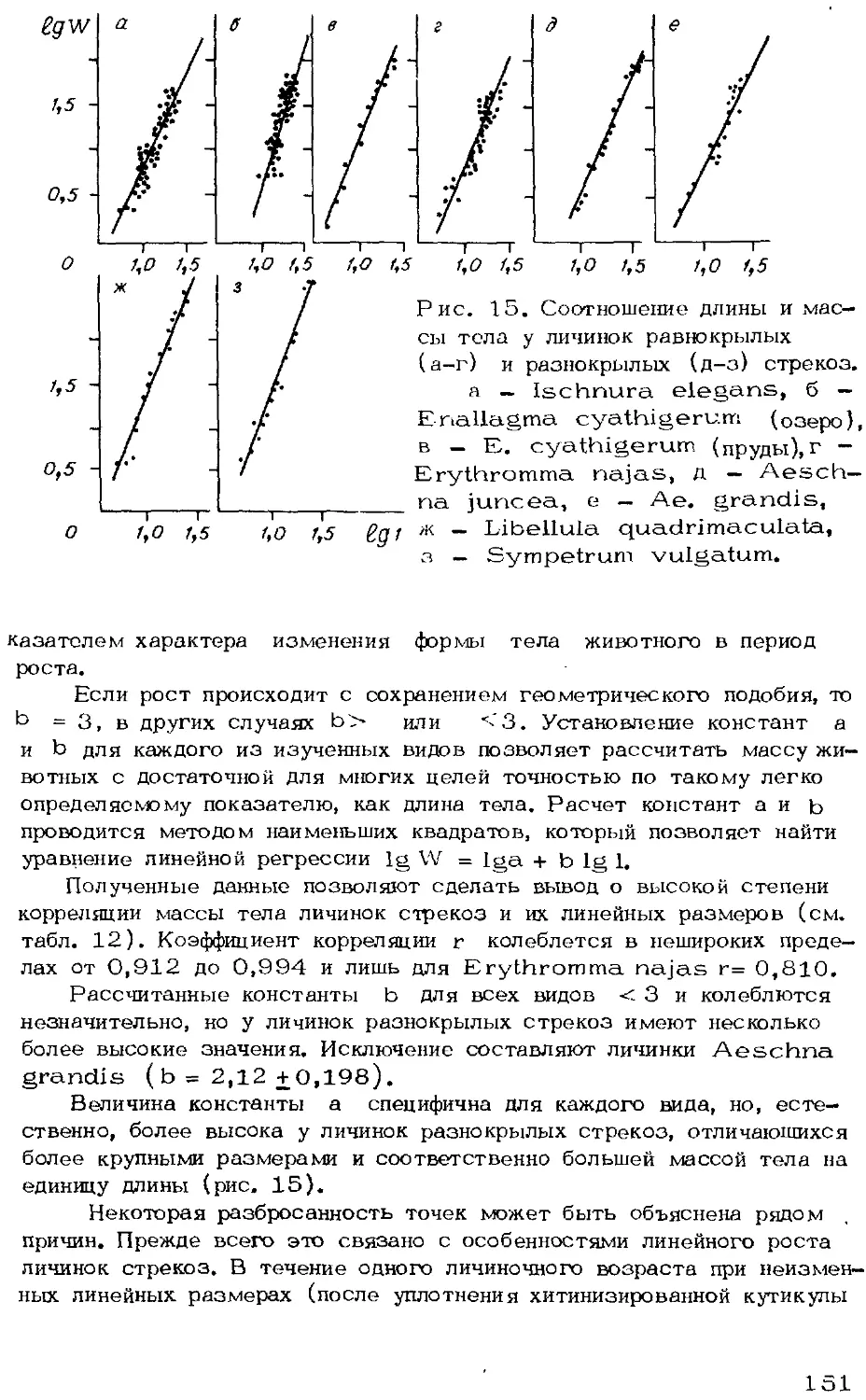
































































Автор: Мордкович В.Г.
Теги: articulata (членистые) биология экология насекомые монография фауна стрекозы исследования стрекоз биоценоз
ISBN: 5—02—028931—0
Год: 1989




АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
БИОЛОГИЧЕСКИЙ ИНСТИТУТ
ФАУНА
И
ЭКОЛОГИЯ
СТРЕКОЗ
Ответственный редактор
доктор биологических наук В.Г. Мордкович
в
НОВОСИБИРСК
«НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
1989
УДК 595. 733
Авторы
Б. Ф. Белышев, А.Ю, Харитонов, С.Н. Борисов, З.Д, Спурис, Г.А. Мазохин-
Поршняков, П.А. Мокрушов, Р.С. Павлюк, Л.Н. Притыкина, Г.И. Рязанова,
Е.С. Шалопенок, А.Д. Писаненко, Г.А. Сухачева. И.Н, Харитонова. В.В. Заика.
Л. И, Францевич
Фауна и экология стрекоз / Белышев Б.Ф., Харитонов А.Ю., Борисов С.Н.
и др. - Новосибирск: Наука. Сиб. отд-ние, 1989. - 207 с.
ISBN 5—02—028931—0.
Монография обобщает результаты исследований стрекоз и характеризует
состояние изученности этого отряда насекомых в нашей стране. Основу книги
составили материалы докладов участников Первого Всесоюзного симпозиума,
одонатологов, состоявшегося 1-3 июля 1986 г. в Новосибирске. Дан анализ изу-
ченности фауны стрекоз различных регионов СССР, освещены аспекты их эко-
логии, показано значение этих насекомых как'модельных объектов для разно-
образных биологических исследований. Особое внимание уделено оценке роли
стрекоз в биоценозах, в частности, их паразитарным связям.
Книга рассчитана на энтомологов, экологов, этологов, паразитологов, гид-
робиологов, специалистов в области охраны природы, студентов-биологов.
Табл. 15. Ил. 19. Фототабл. 7. Библиогр.: 887 назв.
Рецензенты
кандидаты биологических наук А.В. Баркалов. Т.В. Лобанова
Утверждено к печати
Биологическим институтом СО АН СССР
ф LggTQPPOQQ-.eig 59-_8„ кн 2
Ф 055(02)-89 595 891 кн- 2
ISBN 5-02-028931-0
© Коллектив авторов, 1989
ПРЕДИСЛОВИЕ
Отряд стрекоз занимает особое место в классе насекомых. Одна
из важных его особенностей - высокая морфологическая специализация,
благодаря которой многие авторы /Шванвич, 1949; Родендорф, 1,980;
и др./ относят стрекоз к особому отделу или инфраклассу, противопо-
ставляя их остальным крылатым насекомым. Учитывая палеозойский
возраст ископаемых стрекозоподобных, современный отряд стрекоз
принято считать древним по происхождению. Тем не менее стрекозы
отличаются многими прогрессивными чертами, такими, как широкое
распространение, видовое многообразие (около 6 тыс. репентных ви-
дов), обилием в водных и околоводных биоценозах умеренных и юж-
ных широт. Сложное поведение, наличие высокоадаптивных жизнен-
ных форм, активное хищничество определяют их важную роль в тро-
фических сетях биоценозов. При чередовании водной и наземной фаз
развития и большой биомассе стрекозы вносят существенный вклад в
круговорот веществ в биогеопенозах.
Широко известна положительная роль стрекоз в массовом истре-
блении кровососущих насекомых, а иногда и вредителей сельского и
лесного хозяйств. Делаются обнадеживающие попытки использования
их для биоиндикации качества природных вод. В отдельных случаях
стрекозы могут приносить и вред, в частности, составляя пишевую
конкуренцию молоди рыб в рыбоводных прудах, распространяя гельмин-
тозы птип, истребляя пчел в районах пасек. Но этот вред локален и
несопоставим с большим полезным значением стрекоз в природе. Из-
вестна и роль стрекоз как носителей уникального генофонда, что по-
будило взять под охрану редкие виды этих насекомых и включить их
в Красные книги МСОП, СССР и союзных республик. Стрекозы широ-
ко используются в качестве модельных объектов в самых различных
биологических исследованиях.
Все эти причины привлекают к стрекозам внимание многих специ-
алистов. С одонатологией в настояшее время связано более 600 ис-
следователей почти из 60 стран мира. Для улучшения координации одо-
натологических исследований и повышения их эффективности в октябре
1971 г. в Бельгии (г. Гент) был созван Первый европейский симпо-
зиум одонатологов, учредивший Международное общество одонатопогов -
The Societas Internationalis Odonatologica (S.I.O.) co штаб-
квартирой при Утрехтском университете в Нидерландах.
С марта 1972 г. S.I.O. начало издавать свой ежеквартальный
3
журнал Odonatologica, а с июня 1978 г. в дополнение к нему -
бюллетень Notulae Odonatologicae. Создание S.I.O. и его пе-
чатного органа резко повысило интерес к одонатологии. Во многих
странах были организованы национальные отделения S.I.O., начавшие
издавать свои одонатологические бюллетени. Концентрация информации
в специализированных изданиях, регулярные симпозиумы S.I.O. и его
национальных отделений, публикация S.I.O. каталогов, обзоров и дру-
гой справочной литературы, создание службы определения и библиогра-
фического обслуживания способствовали заметному прогрессу одонато-
погии в последние годы.
Значительный вклад в развитие одонатологии внесли русские и со-
ветские ученые. Работавший в первой половине текущего столетия
А.Н. Бартенев по оценке экспертов S.I.O. отнесен к первой десятке
крупнейших специалистов в истории мировой одонатологии. Мировое
признание получили работы доктора биологических наук Б.Ф. Белыше-
ва и его учеников по географическому распределению и гене'зису фа-
уны стрекоз. Б.Ф. Белышев принимал деятельное участие в создании
S.I.O. и сейчас является его почетным членом.
Вместе с тем в работе отечественных специалистов, связанных в
своих исследованиях со стрекозами, не было организационно четких
контактов, в связи с чем по инициативе Сибирского и Латвийского
отделений Всесоюзного энтомологического общества было решено про-
вести Первый Всесоюзный одонатологический симпозиум, который сос-
тоялся в Новосибирске 1-3 июля 1986 г. В резолюции, принятой уча-
стниками симпозиума, было, в частности, отмечено, что в области одо-
натологии в СССР за послевоенный период достигнуты существенные
результаты - издана капитальная двухтомная монография Б.Ф. Белы-
шева по стрекозам Сибири, региональные определители по европейской
части СССР, Сибири, Дальнему Востоку, определитель личинок фауны
СССР, атлас-определитель крыльев родов мировой фауны и видов фауны
СССР, двухтомная География стрекоз /Белышев, Харитонов, 1981, 1983/,
монография и каталог по стрекозам Латвийской ССР, раздец по стре-
козам Красной книги СССР. По стрекозам защищены докторская и де-
сять кандидатских диссертаций. Получены существенные результаты по
изучению ископаемых стрекоз. Расширились исследования по экологии,
поведению, паразитарным связям, оценке роли стрекоз в водных и око-
ловодных биоценозах.
Однако многие интересные в научном и практическом плане вопро-
сы жизнедеятельности стрекоз пока не-затронуты исследованиями,
Крайне неравномерно изучена одонатофауна различных регионов СССР,
многие сведения по распространению и экологии стрекоз очень непол-
ны и фрагментарны, а подчас и противоречивы. Пель настоящей книги -
подытожить результаты и основные достижения отечественной одона-
тологии, охарактеризовать современное состояние исследований и на-
метить их перспективы. Список использованной в монографии литера-
туры представляет собой довольно полную отечественную одонатологи-
ческую библиографию, поэтому книга будет особенно полезна начина-
ющим исследователям.
А.Ю. Харитонов
4
Г лава I
ИСТОРИЯ ИЗУЧЕНИЯ, СОСТАВ И ГЕНЕЗИС ФАУНЫ
Первые сведения об отдельных видах стрекоз с территории нашей
страны появились в научной литературе в последней четверти XVIII в.
На протяжении последующих 100 лет одонатологические материалы
пополнялись незначительно и их публикация представляет в основном
исторический интерес. Первой крупной и содержательной работой по
стрекозам России явилась статья П. Иванова /1876/ с довольно по-
дробными сведениями по морфологии и биологии 36 видов. В начале
XX в. вышла в свет работа Г.Г. Якобсона и В.Л, Бианки /1905/, со-
держащая очень полные для своего времени одонатологическую библи-
ографию и определительные таблицы стрекоз, не утратившие своего
значения до настоящего времени.
В 1907 г. была опубликована первая статья А.Н. Бартенева
(1882-1946), которой начался новый период истории отечественной
оДонатологии. .За попрека исключительно плодотворной деятельности
А.Н. Бартенев опубликовал около 100 работ преимущественно по фа-
уне и систематике стрекоз. Его фаунистические работы охватывают
почти всю нашу страну, материалы по некоторым группам стрекоз
обработаны в объеме палеарктической и даже мировой фауны.
В 1951 г. выход в свет статьи Б.Ф. Белышева ознаменовал но-
вый этап развития одонатологии. Автор наряду с фаунистическими ра-
ботами проводит исследования по морфологии, систематике и особен-
но географическому распространению стрекоз, опубликовав по этим во-
просам около 200 работ. Начав с изучения одонатофауны Сибири, он
вскоре приступает к анализу распространения стрекоз мировой фауны,
результатом которого явилась обширная серия зоогеографических пу-
бликаций /1956-1986/, завершившаяся изданием двух написанных
совместно с А.Ю. Харитоновым монографий, составляющих в сумме ру-
ководство по географии стрекоз мира.
Работы А.Н. Бартенева и Б.Ф. Белышева составили золотой фонд
не только отечественной, но и мировой одонатологии и получили заслу-
женное международное признание.
Более подробно история одонатологических исследований и изучен-
ность фауны стрекоз СССР будет охарактеризована далее по регио-
нальному принципу.
5
ЕВРОПЕЙСКАЯ ЧАСТЬ СССР
Сведения о составе фауны стрекоз европейской части СССР впер-
вые были опубликованы в XVIII в. учеником К. .Линнея-рижанином
Я. Фишером /Fischer, 1778/. В своей книге о природе Лифляндии
он привел пять видов, обнаруженных предположительно в окрестностях
Риги. Судя по кратким описаниям, два из них были определены невер-
но. Через несколько лет он /Fischer, 1784/ дополнил этот список
пятью видами и все эти сведения повторил во втором издании упомя-
нутой книги /Fischer, 1791/. Следовательно, можно считать, что
в конце XVIII в. в Латвии было выявлено восемь видов стрекоз (Са—
lopteryx splendens, G-omphus vulgatissimus, Cordulia
aenea, Orthetrum cancellatum, Libellula depressa, L. qua-
drimactilata, Sympetrum flaveolum, Leucorrhinia rubicun-
da). Эти данные являлись первыми и для всей территории СССР. В
самом конце века И. Цедерхьелм опубликовал некоторые данные по
стрекозам Ленинградской области /G-ederhjelm, 1798/.
В XIX в. опубликовано сравнительно много работ, особенно на
Украине. Так, стрекозам Харьковской губернии посвящены четыре
крупные статьи /Иванов, 1876; Ярошевский, 1881; Родзянко, 1889,
1895/. Были опубликованы данные по Полтавской /Родзянко, 1887,
1889, 1895/ и Киевской /Belke, 18бб/ губерниям, по Подолии
/Belke, 1859; Lomnicki, 1877/. В начале XX в. к ним добавля-
ются статьи А. Браунера /1902/ по стрекозам Херсонской губернии
и Крыма, Н.М. Арнольда- /1902/ по Могилевской губернии, Дзедзе-
левича /Dziedzielewicz, 1902/ по Галипии. Последняя работа вы-
полнена весьма основательно и содержит список 60 видов.
Особого повторного упоминания заслуживает работа П. Иванова,
резко и положительно выделяющаяся среди работ своего времени. В
ней даны весьма подробные описания всех обнаруженных в окрестно-
стях г. Купянска 36 видов, приведены определительные таблицы, ри-
сунки и русские названия родов и видов. Следует отметить, что все
родовые названия, данные П. Ивановым, сохранились до наших дней.
Источники родовых названий не указаны, за исключением слова "дед-
ка", которое заимствовано у рыбаков, однако этим словом они назы-
вали как виды рода Gomphus, так и рода Aeschna. Наконец, рус-
ские названия родов совершенно правильно стоят в множественном чис-
ле.
По Прибалтике упоминания заслуживают статьи Ратке /Rathke,
1832/, по Курляндии - Кавалла /Kawa.ll, 1864/, по Лифляндии и
Эстонии - Бруттана /Bruttan, 1878, 1881а, б/. Статья Ратке ин-
тересна тем, что в ней впервые на материале из России рассматри-
вается анатомия стрекоз: сборы стрекоз в окрестностях Тарту, по су-
ществу, не.представляли фаунистических целей, так как они (7 видов)
были использованы для изучения гениталий. В статьях для Латвии ука-
зано 26 видов, т.е. около половины фактического видового состава, а
для Прибалтики в целом - 33 вида.
По Петербургской губернии опубликована лишь одна специальная
фаунистическая работа - статья О. Полетаевой /1880/ со списком
47 видов.
V
6
По Московской губернии некоторые сведения даны уже в 1802 г.
Двигубским /Dwigubsky, 1802/. Хорошие для того времени рабо-
ты по сетчатокрылым и прямокрылым Московского учебного округа
опубликовал В. Ульянин /1867, 1869/, указав в них 40 видов стре-
коз (один определен недостаточно точно). Судя по описанию, автор,
возможно, имел также Calopteryx splendens var. ancllla.
Для всех видов указано количество собранных экземпляров, притом
раздельно по Полам, что совсем необычно для работ XIX в. В 1869 г.
он дал дополнительную таблицу с фенопогичебкими данными по 34 ви-
дам. Кроме того, с учетом литературных данных им дан список видов
Европейской России, включающий 51 вид. Для сравнения укажем, что
для Азиатской России В. Ульянин перечислил 26 видов, для Кавказа -
44. Небольшие дополнения по Московской губернии в начале XX в.
дал Г.А. Кожевников /1902/.
По Приволжским территориям некоторые данные сообщались в
классических работах Эверсманна /Eversmann, 1836, 1837/.
Следует вспомнить, что он описал новые для науки виды Lestes
macrostigma и Aeschna viridis. Другие данные имеются в публи-
кациях В.Е. Яковлева /lakowleff, 1870/ и А.А. Силантьева /1894/.
В целом можно сказать, что работы XIX в. содержат много цен-
ных сведений, однако большинство из них сегодня имеют лишь истори-
ческий интерес. Депо в том, что фаунистические работы того времени,
в особенности первой половины столетия, обычно представляют собой
только списки видов, притом неполные, без каких-либо дополнительных
данных, или с отрывочными указаниями мест сбора и т.п. Экологичес-
кая характеристика мест сбора отсутствует. Это естественно, так как
работы прошлого столетия написаны в соответствии требованиям и тра-
дициям своего времени. Их авторами во многих случаях были энтузи-
асты-любители, нередко достаточно квалифицированные. Например, на-
иболее ценные работы по стрекозам Прибалтики в прошлом веке опу-
бликовали упомянутые выше И. Кавалп и А. Бруттан. И. Кавалл был
священником в сельской местности Курляндской губернии. Он натура-
лист-любитель широкого профиля, член ряда научных обществ, в том
числе Московского общества испытателей природы, стрекозами зани-
мался наряду с изучением многих других насекомых. А. Бруттан ра-
ботал в Эстонии (Тарту) учителем и увлекался главным образом бо-
таникой (хорошо знал мхи и лишайники).
В начале нашего века в одо'натологию вступает А.Н. Бартенев -
яркая фигура не только отечественной науки. Однако по европейской
части СССР од. опубликовал мало. Первая его статья касается стрекоз
Полесья и окрестностей Вильнюса /Бартенев, 1907/. В некоторых
последующих статьях упоминаются стрекозы с периферии европейской
части СССР, например Крыма /1912/, окрестностей Ростова-на-До-
ну /1916в/.
С начала века до второй мировой войны опубликовано немало ра-
бот других авторов-, заслуживающих упоминания. Прежде всего следует
назвать несколько статей Г.В. Артоболевского по украинской фауне -
стрекозам Полтавской области /1917, 1929 а/, окрестностей Киева
/1927а/, Подолии /19276/, Крыма /19296/. Кроме того, он опу-
7
бликовал данные по Калужской области /1923/. По фауне и экологии
стрекоз Московской области две содержательных статьи есть у В.Г. Ко-
лесова /1930а, б/.
А.М. Дьяконов более известен как специалист по иглокожим и пя-
деницам, однако он написал статью о стрекозах карельского оз. Сан-
дала и его окрестностей /1922а/, составил определитель стрекоз Ле-
нинградской области /1926/. Кроме того, он описал четыре новых ви-
да из СССР, правда, лишь один из европейской части - Aeschna
elisabethae /19226/.
Фауну стрекоз крупнейшего эстонского о. Сааремаа выявил Л. Мер-
жеевский /Ь. Mierzejewski, 1913/. В. Мержеевский описал с это-
го острова новый вид Aeschna osiliensis /w. Mierzejewski,
1.913/, считающийся ныне подвидом Ае. serrata. В 30-х годах
стрекоз Эстонии изучали работники Тартусского университета: Юрйс
/juris, 1938/, опубликовавший обобщающую статью, и Каури /Ka-
uri, 1949/, защитивший диссертацию по фауне стрекоз Эстонии. Со-
ответствующие данные были опубликованы лишь после войны.
Обстоятельную статью по стрекозам Латвии, в которой приведены
также разные их промеры, в 1942 г. опубликовал латышский гидроби-
олог, позднее крупный специалист по коловраткам Берзиньш /Ber-
zin^, 1942/. Он привел 47 видов, среди них южные виды Ischnu—
га pumilio, Aeschna mixta, Ае. isosceles, Anax impera—
tor.
З.Д. Спурис в течение 3-петних наблюдений на площади в несколь-
ко квадратных километров в южной части Латвии около р. Свете вы-
явил 35 видов /Spuris, 1943/. Среди них Lestes virens, Sym-
pecma paedisca, Aeschna mixta. Позднее на этом участке най-
дены также Coenagrion armatum, Ischnura pumilio и Aeschna
isosceles. Приведенные данные показывают значение длитель-
ных стационарных наблюдений для выявления полного состава фауны.
В 20-х годах супругами Прюфферами впервые опубликованы срав-
нительно полные данные о стрекозах Литвы /Priiffer, 1923; Zna—
mierowska, 1923; Znamierowska— Prtifferowa, 1927/. Рас-
смотрена восточная часть Литвы. Всего указано 39 видов. Стреко-
зам нынешней Калининградской области посвящена работа Роя /Roi,
1911/.
О фауне стрекоз Белоруссии обобщающую работу опубликовал Вну-
ковский /Wnukowsky, 1937/, указав 48 видов.
Следует назвать еще некоторые работы этого периода: Ю. Зографа
/1909/ по Украине, Б.К. Григорьева /1906/ по Валдайской возвышен-
ности, Ю.М. Колосова /1925, 1929а, 1930, 1932/ по Ярославской
губернии и Белоруссии, Л. Круликовского /1907/ по Вятской губер-
нии, Б.А. Редько /1913/ по окрестностям Саратова, А.И. Радкевича
/1928/ по Белоруссии, В. Меландера /1927/ по Смоленской губер-
нии. Если еще упомянуть работу С.В. Солодовникова /1929/ по ли-
чинкам стрекоз р. Донца и отметить, что А.Л. Бенинг /1924/ в сво-
ей известной монографии по Волге привел данные также по стрекозам,
то обзор одонатопогиче<рких публикаций периода до второй мировой вой-
ны можно считать завершенным.
8
Фаунистические исследования стрекоз европейской части СССР в
послевоенные годы удобнее рассмотреть по союзным республикам.
В Эстонии одонатофаунистических исследований проведено немно-
го. Можно назвать лишь одну небольшую статью, специально посвящен-
ную стрекозам /Спурис, 1968/. В ней рассматривается в основном
юго-восточная часть Эстонии и приведен 31 вид. Интересно отметить,
что в послевоенном периоде эстонскими энтомологами впервые вы-
явлены южные виды Sympecma annulata ( = S. paedisca),
Ischnura pumilio, Aeschna mixta /Н. Remm, 1966/. Всего в
республике выявлено 53 вида. На эстонском языке опубликовано не-
сколько определителей стрекоз республики как по имаго /Н. Remm,
1957, 1966/, так и по личинкам /Е. Remm, 1963/.
В Латвии одонатофаунистические исследования проведены почти
исключительно Э.Д. Спурисом. В 1956 г. опубликована обзорная ра-
бота по стрекозам региона с определительными таблицами имаго. Все-
го в ней указано 53 вида. Впоследствии найден лишь один дополни-
тельный вид - Coenagrion concinnum /Спурис, 19646/. Всего
в республике выявлено 54 вида /Спурис, 1980/ - 53 местных вида
и один очень редкий иммигрант (Sympetrum fonscolombei). Сре-
ди них установлен ряд южных видов /Спурис, 1951/.
Следует отметить, что изучение стрекоз в Латвии проводилось
почти в 2 00 пунктах, и по густоте сети мест сборов и наблюдений
она занимает одно из первых мест в СССР. Пункты исследований по
территории республики распределены сравнительно равномерно, однако
совершенно недостаточно до сих пор изучена фауна верховых болот. В
1956 г. для облегчения описания распространения стрекоз в пределах
республики территория ее по ландшафтному принципу была разделена
на 15 фаунистических экологических округов. Для каждого из них ука-
зано количество выявленных видов, что неплохо охарактеризовало фа-
унистическую изученность округов. Впоследствии было опубликовано
много дополнительных данных по встречаемости стрекоз, а количество
округов увеличено до 16 /Спурис, 1965/. Перечисление видов стре-
коз по округам можно рассматривать как начальный этап картирова-
ния их распространения. Определитель взрослых стрекоз на латышском
языке издан в 1957 г. /Spuris, 1957/.
В Литве фаунистикой стрекоз занимались А.П. Станёните и
З.Д. Спурис. А.П. Станёните /1962а, б, 1963а-в, 1964, 1968, 1985/
изучала стрекоз главным образом восточной части Литвы, притом она
исследовала личинок и их паразитарные связи. З.Д. Спурис опублико-
вал две статьи /1959, 1971/ по распределению стрекоз в Литве.
Всего в республике достоверно выявлено 54 вида, не считая Coe-
nagrion concinnum, присутствие которого окончательно не дока-
зано. Небезынтересно отметить, что в Литве обнаружены как север-
ные виды Coenagrion armatum и Somatophlora arctica, так
и южные Anax parthenope, Sympetrum pedemontanum, S.
depressiusculum.
В Белоруссии стрекоз изучал А.И. Радкевич /1957/. Для северо-
восточной части республики он указал 43 вида. Однако его данные о
Coenagrion ornatum и С. concinnum требуют подтверждения.
9
С 1977 г. фауну стрекоз региона изучает А.Д. Писаненко. Результа-
ты первых лет его исследований опубликованы в виде крайне сжатой
обзорной статьи /Писаненко, 1985/, в которой по литературным и
собственным данным для республики указано 53 вида, в том числе
названные выше два сомнительных вида. Однако территория республики
обследована неравномерно, некоторые крупные районы, в особенности
в восточной части, остались неизученными. Существенные сведения по
личинкам стрекоз оз. Нарочь содержатся в работах гидробиологичес-
кого характера /Шалопенок, 1963; Камлюк, 1973; Шалопенок, Па-
вельева, 1973; Гигиняк, Павельева, 1974; Шалопенок, Камлюк, 1975/.
На Украине еще в начале века весьма подробно была изучена ее
западная часть. Так, например, польский энтомолог Дзедзелевич
/Dziedzielewicz, 1902/ в крупной работе по стрекозам Галиции
привел 60 видов. В послевоенное время большой вклад в изучение
стрекоз западной части Украины внес Р.С. Павлюк /1975а/. Основ-
ной его задачей было паразитологическое исследование стрекоз, од-
нако при сборе паразитов добыт богатый фаунистический материал,
относящийся к 58 видам. Всего в семи западных областях (Волынской,
Ровенской, Львовской, Тернопольской, Закарпатской, Иваново-Франков-
ской и Черновипкой) обнаружено 65 видов. Среди них числится ука-
занная до 1941 г. редкая в СССР стрекоза Cordulegaster biden—
tatus.
Особый интерес представляют сведения о распределении стрекоз
по физико-географическим районам западной части Украины. По дан-
ным Р.С. Павлюка, в фауне стрекоз Украинских Карпат выявлено 38 ви-
дов, но следует признать, что эта фауна изучена пока недостаточно. В
равнинных частях Закарпатья зарегистрировано 46 видов, а в Пред-
карпатье - 57.
Фауну стрекоз юго-восточной части Украины в 1971-1974 гг.
изучал А.И. Олигер в основном в Донецкой области, частично в Воро-
шиловградской. Обнаружено 47 видов /Олигер, 1980, 1985/, среди
них Aeschna serrata, что требует подтверждения.
Всего на Украине выявлено около 70 видов, но их территориаль-
ное распределение внутри республики определено недостаточно: ряд
областей вовсе не затронут исследованиями, например нет современ-
ных данных по Крыму. Нелишне отметить, что в гидробиологических
работах для Украины по личинкам указаны Coenagrion mercu-
rials и С. scitulum, но эти данные крайне сомнительны, поэтому
источники здесь не указываются.
Стрекозы Молдавии изучены очень плохо. По данным книги 'Жи-
вотный мир Молдавии" /1983/, в республике зарегистрировано толь-
ко 20 видов.
Фауна стрекоз огромной территории европейской части РСФСР в
послевоенный период практически не исследовалась. Опубликованы
лишь пять статей, заслуживающих упоминания. В одной из них дается
беглый обзор фауны Коми АССР /Седых, 1974/, в которой приведено
35 видов. В статье по фауне Кировской области указано 47 видов
/Леви, 1968/. Имеются работы пО стрекозам Чувашии, в которой ука-
зано 29 видов, Астраханского заповедника /А. Олигер, 1970/ и Куй-
10
бышевской области /Крыжановский, 1982/. Все они сравнительно не-
большие, новых крупных работ нет.
По стрекозам европейской части СССР опубликовано два опреде-
лителя /Дьяконов, 1948; Спурис, 1964а/, причем второй из них вклю-'
чает не только обнаруженные, но и ожидаемые виды. Всего в этом
регионе выявлено 76 видов стрекоз. В целом хорошо изучена запад-
ная его часть, включая Прибалтику, Белоруссию, западные районы
Украины.
АЗИАТСКАЯ ЧАСТЬ СССР
Этот регион по территории огромен и обследован неравномерно,
поэтому его следует рассмотреть по отдельным районам, относу к
ним и такие пограничные территории, как Урал и Кавказ.
УРАЛ И ПРИУРАЛЬЕ
Первое указание на нахождение отдельных видов стрекоз на Ура-
ле появилось немногим более 100 лет назад в статье Бергрота
/Bergroth, 1881/. Последующие публикации по фауне стрекоз Ура-
ла и Приуралья относятся уже к началу нашего века. Фрей-Гесснер
/1907/ упомянул о находке одного вида на Среднем Урале, П. Во-
ронцовский /1909, 1912, 1913/ опубликовал небольшие списки ви-
дов стрекоз окрестностей г. Оренбурга.
Большой вклад в изучение уральской одонатофауны внес А.Н. Бар-
тенев. В 1908 г. он опубликовал статью "Коллекция стрекоз из окре-
стностей оз. Увильды", в которой приводится список 28 видов, от-
дельные биологические наблюдения, делается попытка зоогеографичес-
кого анализа фауны. Годом позже появилась его "Заметка о стреко-
зах Зоологического отдела музея Уральского общества любителей есте-
ствознания" /Бартенев, 1909а/, в которой приводится список 14 ви-
дов стрекоз Среднего Урала с примечаниями по морфологии некоторых
из них. Затем он /Бартенев, 1911г, 1915г/ опубликовал критические
обзоры первых статей по одонатофауне Урала и Приуралья П. Ворон-
цовского и Ю.М. Колосова, в которых показал ошибочность описания
некоторых новых видов и форм стрекоз. В 1930 г. в Германии выш-
ла большая статья А.Н. Бартенева /Bartenef, 1.930е/ со списком
стрекоз Ильменского заповедника, включающим 2 6 видов. В этой ра-
боте, в частности, дано описание нового вида Aeschna undulata,
найденного на оз. Ильменское.
Значительная заслуга в изучении фауны стрекоз Урала и Приу-
ралья принадлежит Ю.М. Колосову. Им опубликована серия "Матери-
алов к познанию энтомофауны Урала" /Колосов, 1915а, 1916а,б,
1922, 1929а/, которые содержат указания на находки отдельных, не
отмечавшихся прежде в этом регионе, видов стрекоз. Среди них осо-
11
бого внимания заслуживают находки Anax parthenope и Aesch-
па суапеа. В "Заметке о стрекозах Челябинского округа" /Коло-
сов, 1927/ содержится список 23 видов, часть из которых была от-
мечена личными наблюдениями автора летом 1925 г. в окрестностях
Челябинска и часть обнаружена в коллекциях Челябинского краевед-
ческого музея.
Первые сведения о фауне стрекоз северной половины Восточного
Приуралья приведены в работе К. Валле /Valle, 1932/, указавшим
несколько видов для р. Конды. Пять видов, по определению А.М. Дья-
конова, отмечены В.Ю. Фридолиным /1936/ для полярных районов
восточного склона Урала. Среди них особого интереса заслуживает на-
ходка Somatochlora sahlbergi.
Отдельные сведения о стрекозах главным образом Южного Урала
и лесостепи Восточного Приуралья содержатся в гельминтологических
публикациях в связи с выяснением роли стрекоз в поддержании цик-
лов развития некоторых видов гельминтов /Ткачев, 1970, 19716,в,
1973; Окороков, Ткачев, 1973, 1974; Баянов, 1974, 1986/, а
также в работах гидробиологов по изучению кормовой базы рыб /Гран-
дилевская-Дексбах, 1961, 1966а,б; Грандилевская-Дексбах, Шилкова,
1971; Ковалькова, 1971/.
С 1968 г. планомерные исследования уральской одонатофауны про-
водятся А.Ю. Харитоновым. На протяжении полных пяти полевых сезо-
нов он ведет фаунистическое обследование Южного Урала и Приуралья,
в 1973 г. на Полярном Урале и в 1974 г. на Среднем Урале. В по-
следующие годы проводятся эпизодические маршрутные обследования
многих районов южной части региона. Результаты исследований публи-
куются в серии статей как специально посвященных Уралу, так и в
более широких зоогеографических и экологических работах с привле-
чением материалов по уральской одонатофауне /Окороков, Харитонов,
1971; Харитонов, 1972, 1974а,б, 1975а-ж, 1976а-г- 1980а,б;
Белышев, Харитонов, 1973-1975а, 1977а-в-1985а; Belyshev,
Haritonov, 1973, 1978, 1980/.
В итоге исследований на Урале и в Приуралье выявлена богатая
и разнообразная фауна, насчитывающая 60 видов стрекоз. Наибольшим
видовым разнообразием отличаются предгорные леса Южного Урала и
лесостепь Восточного Приуралья, где найдено 52 вида стрекоз. В та-
ежной зоне региона отмечен 31 вид, а в лесотундровой - 11 видов.
Значительному видовому богатству уральской одонатофауны, веро-
ятно, способствует ландшафтное разнообразие и обилие водоемов, осо-
бенно в южной части региона. Наиболее богата видами фауна стрекоз
мелководных заливов больших олиго- и мезотрофных предгорных озер
(Увильды, Ильменское, Миассово и т.п.), насчитывающая 44 вида.
Это объясняется, видимо, благоприятными и относительно стабильны-
ми условиями кислородного режима и химизма воды в озерах такого
типа в сочетании с высокой кормностью, обилием водной растительно-
сти и достаточной прогреваемостью мелководных заливов. Следует от-
метить, что десятки южно-уральских озер необследованы и здесь воз-
можны новые интересные фаунистические находки. Например, на неко-
торых озерах ильменской группы А.Ю. Харитонов обнаружил один из
Ч
12
редчайших видов стрекоз Ischnura aralensis, первоначально счи-
тавшийся среднеазиатским эндемиком. Буквально на соседних озерах
обнаружен восточно-сибирский вид Somatochlora graeseri. Тре-
буют подтверждения указания для южно-уральских озер таких видов,
как Aeschna cyanea, Ае. undulata, Anax imp erat or.
Мало обследованы водоемы степной зоны, в частности Оренбург-
ской области. Почти полностью отсутствуют материалы с Северного
Урала. Необходимы стационарные наблюдения за составом фауны, под-
верженным большим межгодовым колебаниям, особенно в родах
Anax, Orthetrum, Leucorrhinia, некоторые виды которых в одни
годы почти исчезают, в другие становятся обильными. Такие наблюде-
ния целесообразно организовать прежде всего на охраняемых террито-
риях, из которых для этих целей наиболее удобен Ильменский запо-
ведник.
СИБИРЬ
Это громадная территория от Уральских гор до берегов Тихого
океана. История одонатологических исследований в регионе дана в мо-
нографии Б.Ф. Белышева/1973а/. Поэтому данный вопрос здесь да-
ется в кратком изложении.
Первые сведения о фауне стрекоз Сибири появились в литературе
в середине прошлого века. Так, Hagen /1856/ по материалам раз-
ных коллекторов опубликовал данные о находках ряда видов стрекоз
в различных местах Сибири: Томске (1 вид), Иркутске (17 видов),
Забайкалье (2 вида), Охотском побережье (4 вида), на Камчатке
(8 видов) и некоторых других. В другой работе /Hagen, 1858/ со-
держатся материалы для долины р. Вилюй по сборам участника изве-
стной Амурской экспедиции Р.К. Маака. В ней приводится всего де-
вять видрв, но ее значение несомненно, так как эти данные явились
первыми для северной части Сибири. Мочульский /Motschulsky,
1859/ в работе по Приамурью упоминает три вида стрекоз, но пра-
вильность его определения вызывает сомнение. Два крупнейших
одонатолога Сели Лоншан и Мак Лахлан /Selys Longchamps, Mac
Lachlan, 1872/ опубликовали работу по одонатофауне северной ча-
сти Азии, преимущественно востоку Сибири. В ней приведено 44 вида,
среди которых много новых описаний. Часть упоминаемых видов, по
всей вероятности, определена ошибочно, так как повторно они никем
не обнаружены. Особенно это относится к видам с побережья Охотско-
го моря. Указывались, например, некоторые американские виды, кото-
рые, вероятно, попали в коллекции из русских колоний в Америке. В
1887 г. вышла в свет работа Сели Лоншана по восточной части Сиби-
ри.
Шведский ученый Ф. Трибом изучал энтомофауну на р. Енисей в
1876 г. Его исследования охватили большой отрезок долины реки от
г. Енисейска до пос. Дудинка. Материалы исследования были опубли-
кованы в 1889-г. В работе приведено 20 видов, в том числе несколь-
ко новых для науки, из которых Coenagrion hylas и Somato-
13
chlora sahib ergi оказались реально существующими, а остальные
были сведены в синонимы. Эта работа, написанная 100 лет назад, до
сих пор остается единственной публикацией по одонатофауне такого
крупного.района, как бассейн низовий Енисея.
Таким образом, начали исследовать одонатофауну Сибири не рус-
ские энтомологи, а немцы, бельгийцы, англичане и шведы, но из них
только один Ф. Трибом сам работал в Сибири, а все остальные лишь
обрабатывали и публиковали данные, собранные' главным образом рус-
скими коллекторами, имена которых для нас не известны.
В первой четверти XX в. приоритет в изучении стрекоз Сибири
переходит к отечественным исследователям. Немногочисленные рабо-
ты иностранных авторов /Klapalek, 1901; Ris, 1909-1916;
Oguma, 1915; Matsumura, 1.917/ в этот период связаны в ос-
новном с дальневосточными островами. В 1906 г. Б. Григорьев опу-
бликовал заметку по югу Алтая по сборам А.Г.. Якобсона. В ней опи-
саны некоторые интересные находки, в том числе Libellula dep—
ressa. Ю.М. Колосов в 1915 г. опубликовал статью о распростра-
нении и биологии Aesahna gigas Bart; (=Ае. crenata Hag.),
а также список видов окрестностей городов Кургана и Ялуторовска.
Основное место в изучении одонатофауны Сибири в этот период при-
надлежит А.Н. Бартеневу, опубликовавшему около 20 работ по фауне
и систематике стрекоз с широким привлечением сибирских материалов.
К наиболее интересным исследованиям в 20-40-е годы относятся
работы по фауне Камчатки шведа Шостеда /Sjdstedt, 1927/, япон-
цев Огума /Oguma, 1926/, Коно" и Тамануки/Kono, Tamanuki,
19 28/, Окумара /Okumura, 1942/, Асахина /Asahina, 1949/
по фауне Сахалина, финна Валле /Valle, 1932/по материалам немно-
гих мест Северной Азии по старым сборам, хранящимся в финляндских
коллекциях. Из отечественных авторов прежде всего необходимо упо-
мянуть А.М. Дьяконова ‘/Djakonov, 1926/, описавшего три новых
вида, В.В. Внуковского /1928/, опубликовавшего материалы по За-
падной Сибири, А.Н. Попову /1933/, обработавшую сборы с Алтая, а
главное - А.Н. Бартенева /1921а, 1929а, 1930ж, 19316/, включив-
шего в свои обобщающие работы много сведений по сибирской одона-
тофауне. В 1956 г. посмертно вышла в свет последняя работа А.Н. Бар-
тенева по стрекозам Дальнего Востока.
В 50-е годы, как уже отмечалось, впервые началось планомерное
изучение одонатофауны Сибири. Б.Ф. Белышев /1951-1955а-в и др./
обследует Алтай, Нарымский край, Кулундинскую степь, Прибайкалье,
Приморье, низовье р. Лены. Помимо собственных материалов он обра-
батывает многочисленные сборы из многих мест Сибири энтомологов
СО АН СССР и др. В этот же период публикуются статьи К.Б. Город-
кова /1956/ о находках стрекоз на крайнем севере Сибири, Асахина
/Asahina, 1958/ по одонатофауне Курильских островов и небольшое
число менее значимых для познания сибирской фауны работ других ав-
торов.
В 70-80-е годы изучение сибирской фауны продолжает главным
образом А.Ю. Харитонов /19746, 19756, 1976а-в, 197 86, 1981,
1984а,б, 1986; Белышев, Харитонов, 19756, 1978в; Харитонов, Ха-
14
ритонова, 1982/. Он проводит экспедиционное обследование Зауралья,
юга Западной Сибири, Саян, юга Приморского края, Камчатки. Обра-
батывает многочисленные коллекционные материалы, поступившие в
Зоологический музей Биологического института СО АН СССР.
В этот же период несколько работ по югу Западной Сибири и о. Ку-
нашир. публикует В.В. Заика /1974, 1980, 1982/. Изучение стрекоз
лесостепной зоны Западной Сибири, в том числе стапионарные эколо-
гические работы, продолжает Г.А. Смирнова /Смирнова, Харитонов,
1987/, а Верхнего Приобья в целом - О.Э. Костерин /1987; Заика,
Костерин, 1987/.
В итоге исследований в Сибири (включая Дальний Восток) выяв-
лено 114 видов стрекоз. Число это довольно условно, поскольку так-
сономическое положение отдельных видов нуждается в уточнении, а
многие крупные территории Сибири слабо изучены или совсем нё за-
тронуты исследованиями. Наиболее богата фауна стрекоз Приморского
края и Приамурья, насчитывающая более 80 видов. Однако .этот реги-
он, особенно Среднее Приамурье, изучен еще крайне недостаточно.
Неполно выяснен состав фауны дальневосточных островов, почти пол-
ностью отсутствуют материалы по северо-востоку Сибири, включая
Охотское побережье, и огромному пространству Средне-Сибирского
плоскогорья. Мало обследованы горы юга Сибири, низовье Оби, весь
бассейн Енисея, север Сибири, Даже в наиболее изученном регионе
Сибири - в Верхнем Приобье и на Алтае - до настоящего времени
случаются неожиданные фаунистические находки. Так, на оз. Манжерок
и в приобском Сузунском бору обнаружены локальные популяции вида
Sympetrum croceolum, считавшегося эндемическим для Дальнего
Востока.
За составом фауны и динамикой численности отдельных видов, осо-
бенно в районах интенсивного хозяйственного освоения и на охраняе-
мых территориях, необходимы стационарные наблюдения. Нет сведений
по видовому составу стрекоз подавляющего большинства сибирских за-
поведников, за исключением Кедровой пади и Алтайского, для которых
составлены относительно полные списки одонатофауны.
КАВКАЗ
Благодаря пограничному положению на стыке Голарктической и
Субголарктической областей Бореального фаунистического царства,
сложно расчлененному рельефу и теплому климату, Кавказ представ-
ляет значительный фаунистический и зоогеографический интерес.
Первые упоминания о стрекозах Кавказа появились в работах пре-
имущественно европейских авторов в середине XIX в. Так, Коленати
/Kolenati, 1846/ приводит 4 вида, Хаген /Hagen, 1856/ - 13
видов, в том числе довольно редкий для региона - Libellula fulva.
Одна из значительных работ прошлого века принадлежит В. Ульянину
/1869/, опубликовавшему список 24 видов стрекоз из разных мест
Кавказа. Заслуживают упоминания статьи Сели Лоншана /Selys
Longchamps, 1.887/ и Г.И. Радде/1899/, включающие'в список 18
15
видов. Но большинство работ прошлого века содержат лишь отрывоч-
ные сведения и не представляют существенного интереса.
В начале текущего столетия была опубликована статья А. Брау-
нера /1903/ с небольшими списками стрекоз, собранных между Но-
вороссийском и Адлером. В этот же период начинает исследование
одонатофауны Кавказа А.Н. Бартенев /19096/, внесший выдающийся
вклад в изучение стрекоз региона. Первые обстоятельные сборы одо-
натологических материалов (29 видов) проведены им летом 1907 г.
на Карском плато. Затем А.Н. Бартенев /1910г, 19126/ обрабаты-
вает частную коллекцию барона О.В. Розена, собранную в юго-запад-
ной части Кубанской области, и коллекции Кавказского музея в Тиф-
лисе. В последующие годы А.Н. Бартенев обследует сам и обрабаты-
вает сборы других исследователей из Закавказья /1912д, 1919в,
1929в/, Кавказских Минеральных Вод /1915д/, окрестностей-Сиг-
нахи /19166/, Ростова—на—Дону /1916в/, Ставропольской губернии
/19216/, окрестностей Нальчика /1918/, Тбилиси /1928в/ и дру-
гих мест. В обшей сложности около 30 его работ полностью посвя-
щены или содержат материалы по одонатофауне собственно Кавказа,
Предкавказья и Закавказья. Многие труды ученого содержат не толь-
ко фаунистические списки с замечаниями по систематике, но и ценные
наблюдения по биологии стрекоз. Так, в 1921 г. в окрестностях Ста-
рой Мацесты он впервые в мировой Одонатологии применил протоко--
пирование репродуктивного поведения стрекоз /Бартенев, 1924а/. В
Грузии им изучена фенология и проанализированы периоды лёта имаго
/Бартенев, 1930в/:
Большие заслуги в изучении одонатофауны Закавказья принадле-
жат Н.Н. Акрамовскому /1939, 1948/. В статье о стрекозах Нахи-
чеванской республики приводится 23 вида и подвида. В обстоятельной
работе по Армении опубликован аннотированный список 47 видов, дан
зоогеографический анализ фауны, описаны новые для науки подвиды
Agrion ( = Calopteryx) splendens erevanense и Sympet—
rum flaveolum austrinum. В серии "Животный мир СССР" /1958а/
Н.Н. Акрамовский - автор раздела по стрекозам Кавказа. В нём упо-
минается 47 видов, дан анализ их распространения, краткие сведения
об экологии с учетом вертикальной поясности горной страны. В этом
же году Н.Н. Акрамовский /19586/ опубликовал описание личинки
стрекозы Onychogomphus flexuosus, а в 1964 г. - дополнение
к фауне стрекоз Восточного Закавказья, увеличив список одонатофауны
Армении еще на восемь видов.
Н.Н. Акрамовский первым в нашей стране поставил вопрос о не-
обходимости охраны стрекоз, выделил особо интересные виды Кавказа
и Закавказья и попытался проанализировать их распространение по ох-
раняемым территориям /1975а-в/. Материалы по фауне стрекоз вы-
сокогорья Кавказа были заметно пополнены работами Е.С. Шенгелия
/1953, 1964, 1966; Akramowski, Schengelia, 1967/.
Помимо упомянутых авторов небольшие работы по одонатофауне
региона принадлежат Ю.М. Колосову /19156/ по Кубанской области,
П.В. Виноградову /1926, 1928/ по Ставропольскому краю, Г.В. Ар-
тоболевскому /1929в, 1930/ по Дагестану и экологии стрекоз Те-
берды, Ш, Акопяну /1939/ по личинкам стрекоз окрестностей Ерева-
16
на, А.В. Богачеву /1937, 1951/ по Азербайджану, А.Н. Поповой
/1928, 1947/ По сборам Северо-Кавказской гидробиологической
станции и описанию личинки Epallage fatime, А.Л. Бенингу и
А.Н. Поповой /1947/ по р. Занге. Зоогеографический анализ одонато-
. фауны Кавказа на основе всех известных литературных данных дан
Б.Ф. Белышевым и А.Ю. Харитоновым /1983в/.
В фауне стрекоз региона зарегистрировано 80 видов; 57 в Пред-
кавказье и 75 в Закавказье. Несмотря на относительно полное выяв-
ление видового состава, степень изученности стрекоз Кавказа пока
недостаточна: нуждается в уточнении распространение ряда видов, не-
достает сведений по биологии, неясен статус некоторых таксонов. Осо-
бенно большой интерес представляет изучение представителей родов
Calopteryx и Cordulegaster, исследование формообразователь-
ных процессов в условиях горной изоляции.
СРЕДНЯЯ АЗИЯ И ЮЖНЫЙ КАЗАХСТАН
Мы характеризуем территорию, ограниченную на юге и востоке
Государственной границей СССР, на западе - восточным побережьем
Каспийского моря, на севере граница условно проходит по линии: се-
верное побережье Аральского моря — оз. Балхаш - оз. Алакуль.
Первые сведения по стрекозам этого региона появились в науч-
ной литературе в конце XIX в. Ф. Брауер /1877/ по сборам экспе-
диции А.П. Федченко по Туркестану в 1868-1871 гт. опубликовал
небольшой список стрекоз. В начале XX в. указания на находки от-
дельных видов стрекоз в Средней Азии приведены в работах Б. Гри-
горьева /1905/ и Г.Г. Якобсона и ВЛ. Бианки /1905/. Значительный
интерес представляют работы А.Н. Бартенева /1911в - 1 913,
1915а, 1916а, 19196, 1929в, 19306, г, ж, 1933/, в которых со-
держатся данные по среднеазиатским стрекозам.
Сведения о единичных находках стрекоз рода Sympecma в
Средней Азии приводятся в работе А. Шорыгина /1926/. И.И. Соко-
лов /1933/ на основании определений А. Дьяконова описал фауну
стрекоз водоемов Старой Бухары и ее окрестностей.
Заметный вклад в изучение стрекоз Средней Азии внесла А.Н. По-
пова /1936, 1.953/, опубликовавшая, в частности, большую статью о
стрекозах Таджикистана /1951/, в которую вошли ее собственные
материалы и данные из неопубликованной рукописи А.Н. Бартенева о
стрекозах Палеарктической Азии. ИЛ. Старостин /1951/ в статье
по сернистым источникам Копетдага указал девять видов стрекоз,
В.Р. Садыкова /1959/ охарактеризовала одонатофауну рисовых полей
Ташкентского оазиса. Б.Ф. Белышев /1958а, 1 9б1е/ привел список
стрекоз окрестностей Ташкента к Прибалхашья, а также Западного
Тянь-Шаня /Белышев, Шевченко, 1958/ и Казахстана в целом /Бе-
лышев, Шевченко, 1971/.
С конца 60-х гг. появилось несколько работ о стрекозах Кирги-
зии и Казахстана /Крылова, 1968а, 6, 1969, 1972; Чибиченко,
Крылова, 1.968; Крылова, Чиров, 1971; Кумачев, 1973; Харитонов,
2 Здк. 611
17
1979, 19806; Ионычев, Ибадулаев, 1981/. О находках единичных
видов стрекоз на Памире упоминается А.И. Янковской /1950, 1965/
и К£. Городковым /1961/. Ископаемым стрекозам Средней Азии по-
священы работы Л.Н. Притыкиной /1968, 1970/. В последнее время
проведены целенаправленные исследования и опубликован ряд статей
по одонатофауне Таджикистана, Туркмении и сопредельных районов
/Харитонов, Борисов, 1981, 1985; Борисов, 1983-1987; Павлюк,
Курбанова, 1984; Борисов, Харитонов, 1986/.
Значительный интерес представляют публикации по пограничным
со Средней Азией районам Афганистана /Schmidt, 1961; Asahina,
1963, 1966; Dumont, 1975/, Ирана /Schmidt, 1954/, Ирака
/Asahina, 1973/ и Кашмира /Valle, 1932; Ander, 1944;
Asahina, 19 78/.
Оценивая степень изученности одонатофауны, нужно отметить, что
она крайне неравномерна в различных районах Средней Азии. К на-
стоящему времени относительно полно выявлен видовой состав стре-
коз в Киргизии и Таджикистане. Для всей остальной территории име-
ются лишь фрагментарные сведения из отдельных районов.
Непосредственно нашими исследованиями были охвачены большая
часть территории Таджикистана; окрестности городов Нукус, Хива,
Ташкент, Бекабад, Самарканд в Узбекистане; Суусамырская долина,
Сары-Челекский заповедник, окрестности городов Мин-Куш и Достук в
Киргизии; оз. Ай-Бугур на севере Туркмении и окрестности г. Кызыл-
Орды в Южном Казахстане.
С учетом литературных данных и наших материалов в фауне Сред-
ней Азии и Южного Казахстана в настоящее время известен 91 вид
стрекоз. В этот список мы включаем также таксоны, указанные
А.Н. Бартеневым /1916а/ для Астрабада (ныне Горган) в Северном
Иране, находящегося в непосредственной близости от Государственной
границы СССР. Учитывая отсутствие географических и экологических
барьеров, можно предполагать, что эти виды обитают в Юго-Западной
Т уркмении.
В целом одонатофауна Средней Азии изучена хуже, чем любого
другого крупного региона СССР. Для этой территории отсутствует оп-
ределитель стрекоз, а имеющиеся определители по европейской части
СССР, Сибири и Дальнему Востоку не пригодны для идентификации
среднеазиатской фауны. Кроме того, в систематике ряда среднеазиат-
ских видов много неясного и разные авторы неоднозначно трактуют
таксономический статус отдельных форм, например, видов рода Ca-
lo pteryx.
С учетом этих обстоятельств фауна стрекоз Средней Азии и Южного
Казахстана характеризуется нами более подробно. Ниже дается обзор
всех таксонов, отмеченных для региона и пограничных с ним терри-
торий.
Семейство Gomphidae в фауне Средней Азии включает виды пя-
ти родов. Представитель еще одного рода Paragomphus — Р. linia—
tus Selys — отмечен в Северо-Западном Афганистане /Schmidt,
1961/, поблизости от Государственной границы СССР. Возможно его
нахождение на юге Т уркмении и Узбекистана.
18
Род Gomphus включает три вида. Из Северо-Западного Аф-
ганистана (г. Герат) описан G. amseli /Schmidt, 1961/, находка
которого также возможна в пределах Средней Азии.
Наиболее обычен для рода G. flavipes liniatus, распростра-
ненный локально в низменных районах. Шмидт /Schmidt, 1961/ экземп-
ляры из Термеза, Сирии и Афганистана относит к иному роду и виду-
Stylurus liniatus. Некоторые авторы /Carfi et al., 1982/
также считают G. flavipes Charp. и S. liniatus Bart, раз-
ными таксонами, что требует специального исследования.
G. schneideri приводится А.Н. Бартеневым /1916а/ для
Горгана в Северном Иране. Одни авторы /Schmidt, 1954/ считают
эту форму самостоятельным видом, другие - подвидом /Бартенев,
1929в/.
G. davidi приводится А.Н. Поповой /1951/ для Туркмении
без конкретного указания мест находок. Вероятно, данные заимство-
ваны ею из неопубликованной рукописи А.Н. Бартенева. Один экземп-
ляр самки G. davidi из окрестностей г. Ашхабада хранится в кол-
лекции Зоологического института АН СССР.
Род Onychogomphus представлен четырьмя видами. О. fon-
cipatus unguilatus наряду с G. v. schneideri отмечен в Се-
верном Иране (Горган) А.Н. Бартеневым /1916а/ и найден нами в
Западном Копетдаге. А.Н. Поповой /1951/ без ссылок на источники
приводится для Южного Казахстана О. forcipatus. Если последнее
верно, то на севере Средней Азии вид, возможно, представлен номи-
нативной формой. Э. Шмидт / Schmidt, 1954/ описал из Ирана под- .
вид О. f. lucidostriatus, к которому относит и экземпляры из Аст—
рабада (Горган), что требует уточнения.
О. flexuosus обычен в долинах и предгорьях Таджикистана и
Узбекистана до высоты 800-900 м. Отмечен для Туркмении, Южного
Казахстана /Попова, 1951/, Юго-Восточного Казахстана /Белышев,
19б1е/ и Киргизии /Крылова, 1969/.
О. lefebvrei известен по единичным находкам в Юго-Восточ-
ной Туркмении из района хр. Кугитанг /Борисов, Харитонов, 1986/,
в Северном Иране /Бартенев, 1916а/, окрестностях Ташкента (в кол-
лекции Биологического института СО АН СССР находятся 2 сГ и 1
собранные Р.Н. Мёкленбурцевым в 1954 г.), в массе найден в Запад-
ном Копетдаге. Преимагинальные фазы О. lefebvrei не известны.
О. assimilis assimilis, по нашим материалам, обычен в'
Западном Копетдаге.
О. assimilis fulvipennis отмечен А.Н. Поповой /1951/
для Туркмении (вероятно, по рукописи А.Н. Бартенева). Систематика
вида дискуссионна. Преимагинальные фазы не известны.
Род Ophiogomphus включает один обычный на всей терри-
тории вид - О. reductus. При наличии подходящих по термическим ус-
ловиям проточных водоемов он поднимается довольно высоко в горы -
до 2000 м над ур. м., например в Сары-Челекском заповеднике.
Возможно, на северо-востоке региона будет найден О. cecilia, кото-
рый отмечен в Прииртышье /Белышев, 19б1е/.
Род Anormogomphus также включает единственный
19
вид - A. kiritschenkoi. С момента описания вида А.Н. Барте-
невым /1913/ из Термеза оставался малоизвестным по единичным
находкам в Юго-Западном Таджикистане и Узбекистане. Нами в
1988 г. в массе обнаружен на Каракумском канале в окрестностях
Ашхабада.
Род Lindenia, вероятно, монотипичен, так как вид Ь.in-
kiti вызывает сомнение в реальности существования. Вид L. tetra—
phylla обычен в долинах Таджикистана и Узбекистана. Отмечен в
Южной Туркмении /Павлюк, Курбанова, 1984/, Киргизии /Крылова,
1969/ и Южном Казахстане /Попова, 1951/. В Западном Тянь-Ша-
не найден на высоте 1300 м над ур. м, /Белышев, Шевченко, 1958/.
Семейство Cordulegasteridae представлено одним родом
Cordulegaster, систематика которого весьма дискуссионна. Коли-
чество видов в пределах Средней Азии точно не установлено.
В.Н. Крылова /1969/ приводит для Киргизии три вида: С. insignis,
С. bidentatus, С. coronatus. Возможно, в Копетдаге
будет найден С. brevistigma volia Fraser, отмеченный в Се-
верном Афганистане в верховье р. Теджен /Carfi et al., 1982/.
А.Н. Бартенев /цит. по: Попова, 1951/ считал, что в Средней Азии
обитает только один вид рода - С. coronatus, а Г.В. Артоболевский
определял все среднеазиатские экземпляры (коллекции Зоологического
института АН СССР) как С. insignis.
Также требует подтверждения обитание в Киргизии С. bidenta—
tus.
С учетом последних исследований по систематике рода и данных
систематического списка видов стрекоз мира /Davies, Tobin,
1984/ мы относим все известные нам среднеазиатские экземпляры
представителей рода к подвиду С. insignis coronatus. Этот
подвид известен в предгорьях и горах Тянь-Шаня и Памиро-Апая на
абсолютных высотах от 720 до 2400 м. В Афганистане отмечен до
высоты 2700 м над ур. M./Schmidt, 1961/. Указание АЛ. Поповой
/1951/ о находке стрекоз этого вида на высоте 443 м над ур. м.
на Ферганском хребте представляется нам ошибочным, так как северо-
восточная часть Ферганской долины и тем более прилегающие горы
расположены на значительно больших высотах.
Семейство Aeschnidae в фауне Средней Азии представлено
шестью родами. Возможно, в Бадахшане будет найден представитель
еще одного рода - Cephalaeschna — С. klapperichi, описан-
ный из Северо-Восточного Афганистана /Schmidt, 1961/.
Род Brachytron монотипический, распространен в
Европе, Малой и Передней Азии, представители его проникают в За-
падный Копетдаг. Вид В. pratense отмечен А.Н. Бартеневым
/1916а/ для Горгана в Северном Иране, в непосредственной близости
от территории Юго-Западной Туркмении, и поэтому включается нами в
список стрекоз Средней Азии.
Род Calaeschna также монотипичен, распростра-
нен в восточной части Средиземноморья, С. microstigma обычен
в Западном Копетдаге. Количество, видов рода Aeschna в Сред-
ней Азии точно не установлено. Пока, следуя, данным А.Н. Поповой
/1951/, мы оставляем в списке региона шесть видов.
20
Ae. serrata приводится А.Н. Поповой /1951/ без ссылки на
источники в списке стрекоз Средней Азии для Туркмении. По сборам
Е. Гавриловой, единственная самка известна из окрестностей оз. Ала-
Куль.
Ae. viridis и Ae. grandis указываются А.Н. Поповой
/1951/ для Южного Казахстана, что требует подтверждения.
Ae. juncea mongolica в Средней Азии обитает только в
горах, на высотах примерно от 800 до 2500 м над ур. м. Возмож-
но, личинки способны развиваться в водоемах и на большей высоте.
В Афганистане залеты имаго отмечены до абсолютной высоты 5000 м
над ур. м. Вид многочислен в среднегорье Тянь-Шаня, значительно
реже встречается в Памиро-Алае. Здесь нами найден в окрестностях
г. Хорог, а А.Н. Поповой /1951/ упоминается об одной личинке из
окрестностей г. Душанбе, "'по систематическим признакам ближе все-
го подходящей к этому виду". В горах Копетдага не найден. Вероятно,
по мере продвижения на юг места обитания смешаются во все более
высокогорные. Так, в Афганистане вид отмечен не ниже 2000 м над
ур. м. /Schmidt, 1961/.
Ae. coluberculus на всей территории Средней Азии, кроме
Туркмении, обычен как в долинах, так и в горах до высоты 2300 м
над ур. м. Возможно, .преимагинальные фазы способны развиваться и
на больших высотах. ВЛ. Крылова /1969/ упоминает, что в Кирги-
зии вид достигает "верхнего предела" распространения стрекоз (до
3000 м над ур. м.).
Ae. affinis отмечен в Прибалхашье /Белышев, 1961е/, Юж-
ном Казахстане, Северном Иране /Бартенев, 1916а/. В Киргизии
указывается В.Н. Крыловой /1969/ на высоте более 2500 м над
ур. м. В Таджикистане единично отмечен в заповеднике Тигровая бал-
ка. Вероятно, на юге Средней Азии локален.
Род А п а х включает в Средней Азии два вида. В Северо-Восточ-
ном Афганистане отмечен индийский A. immaculifrons Ramb. /Scmidt,
19 61/. Учитывая великолепные лётные качества представителей
этого рода, можно ожидать его находки в Бадахшане.
A. imperator обычен на большей части территории Средней
Азии. Отсутствует в долинах Юго-Западного Таджикистана. По-види-
мому, у южных границ своего распространения обитает только в го-
рах, где известен до высоты 2300 м над ур. м.
A. parthenope parthenope обычен на всей территории. В
горы поднимается до 2000 м над ур. м.
Род Hemianax включает единственный вид -Н. ephip-
piger. Отмечен для Киргизии /Крылова, 1969/, Южного и Юго-Во-
сточного Казахстана /Белышев, Шевченко, 1971/, локально распро-
странен в Узбекистане и Таджикистане. Редок.
Род Anaciaeschna также включает единственный вид -
A. isosceles anthehumeralis. Известен из Юго-Восточного
Казахстана /Белышев, 1961е/, отмечен как редкий в Киргизии /Кры-
лова, 1969/, в Узбекистане и Таджикистане локален в долинах, более
обычен в предгорьях на высоте 700-1000 м над ур. м. АЛ. Поповой
/1951/ в списке стрекоз Средней Азии указывается для всех средне-
азиатских республик и Южного Казахстана.
21
Семейство Corduliidae представлено двумя родами.
PoflSomatochlora — бореальный, незначительно
проникающий в северные районы Средней Азии, где известен один вид-
S. rr.etallica, указанный А.Н. Поповой /1951/ для Южного Казах-
стана' без конкретного указания места.
Род С о г dulia включает один вид - С. аепеа. От-
мечен ВЛ. Крыловой /1969/ для Тянь-Шаня как редкий. Вероятно,
реликт ледникового периода, локально распространенный в горах.
Семейство Libellulidae в Средней Азии представлено семью
родами. В Северо-Восточном Афганистане отмечен вид эфиопско-
ориентального рода Palpopleura — Р. sexmaculata, широко рас-
пространенный в Ориентальной области /Schmidt, 19614 Возможно
его нахождение в Бадахшане.
Род Orthetrum в таксономическом отношении разра-
ботан недостаточно. В список стрекоз Средней Азии нами включаются
шесть видов. Упомянутый в работе А.Н. Поповой /1951/ "Orthet—
rum fragmentatum Bart., in litt", название которого заимст-
вовано из неопубликованной рукописи А.Н. Бартенева, нами во внима-
ние пока не принимается, так как какие-либо сведения об описании,
сохранности и местонахождении типовых экземпляров этого вида от-
сутствуют. В Северо-Восточном Афганистане отмечены О. taeniola—
turn, О. pruinosum, О. triangular© /Schmidt, 1961 /,
нахождение которых возможно на юге Средней Азии. О. martensi
описан с термальных источников Кашмира /Asahina, 1978/ и мо-
жет быть найден'в аналогичных условиях на Памире.
О. brunneum Широко распространен по всей территории. В го-
рах достигает максимальных высот, на которых способны развивать-
ся преимагинальные фазы стрекоз: термальный источник Шаймак на
Памире - 3900 м над ур. м. /Янковская, 1950, 1965; Борисов,
19856; Борисов, Харитонов, 1986/.
О. cancellatum cancellatum обычен в низменных север-
ных районах Средней Азии примерно до широты Ташкента. Южнее еди-
нично найден в Ферганской долине /Ленинабад). Из работы В.Н. Кры-
ловой /1969/ не ясно, встречается ли в Киргизии в горах. Э. Шмидт
/Schmidt, 1954/ экземпляры из Северного Ирана относит к подвиду
О. с. kraepelini Ris.
О. albistylum albistylum широко распространен в долинах
Средней Азии. В южных районах обитает также в предгорьях, где от-
мечен до абсолютной высоты 1350 м.
О. sabina распространен в низменных районах Средней Азии.
В предгорьях Таджикистана отмечен на высоте 1350 м над ур. м.
Вопрос о подвидовой принадлежности - номинативный это подвид или
О. s. nigrescens - пока не ясен.
О. ransonneti для территории СССР до настоящего времени не
указан, но отмечен А.Н. Бартеневым /1916а/ для Горгана в Север-
ном Иране. Ожидая находок стрекоз этого вида на юго-западе Турк-
мении, мы включаем его в список стрекоз Средней Азии.
О. anceps обычен на всей территории в долинах и предгорьях
до высоты 1500 м над ур. м. В горах на юге отмечен до абсолют-
22
ной высоты 2400 м. В систематике вида не все ясно. А.Н. Барте-
нев /1929в/ описывает по двум самкам из пос. Мигры в Южной Ар-
мении О. translatum, близкий к О. anceps, и как один из основ-
ных признаков указывает сильное развитие желтой окраски на крыльях.
В.Н. Крылова /1969/ приводит О. translatum для Киргизии. Про-
смотр более 100 экз. из различных мест Средней Азии показал, что
сильное пожелтение крыльев характерно для самок О. anceps, при-
чем этот признак хорошо проявляется у молодых, только что выпло-
дившихся особей. С возрастом желтая окраска у большей части самок
исчезает и хорошо заметной остается лишь у единичных экземпляров.
Для окончательного решения вопроса требуется сравнение материала
из Средней Азии и Закавказья. Вероятнее всего, что О. translatum
окажется синонимом О. anceps.
PoflLibellula включает два вида.
L, depressa отмечен в Южном и Юго-Восточном Казахстане
/Белышев, Шевченко, 1971/, как редкий указан для Киргизии /Крыло-
ва, 1969/. А.Н. Попова /1951/ приводит его для Таджикистана по
находке единственной личинки возле пос. Куйбышев на юго-западе рес-
публики. А.Н. Бартенев /1916а/ отмечает в Северном Иране. Подви-
довая принадлежность среднеазиатских экземпляров - номинативный
это подвид или L. d. taurica Beutler, 1984 — не установлена.
L. quadrimaculata quadrimaculata широко распросв-
ранен по всей территории. Мнение /Белышев, 1958а/ о том, что в
южной половине Средней Азии четырехпятнистая стрекоза обитает
только в горах, оказалось ошибочным. Вид обычен как в долинах, так
и в горах, где отмечен до высоты 2800 м над ур. м. Э. Шмидт
/Schmidt, 1961 / по экземплярам из Северо-Восточного Афгани-
стана описывает подвид L. q. grigorievi, отождествляя афганских
стрекоз с самцом, описанным из Семиречья /Григорьев, 1905/. Ос-
новным признаком для выделения подвида послужило изменение рисун-
ка брюшка: общее посветление, наличие полулунных пятен на IX сег-
менте. А.Н. Бартенев /19126/ упоминает об экземплярах из Казалин-
ска и с р. Тарим с сильно развитыми желтыми пятнами по бокам IX
сегмента брюшка. Такие же отличия экземпляров с Гиссарского хреб-
та находит Б.Ф. Белышев /1958а/, но здесь же оговаривается, что
среди типичных встречаются почти аналогичные экземпляры. Просмотр
большой серии стрекоз этого вида из различных мест Таджикистана,
Киргизии и Узбекистана показал, что наряду с типичными обычны и
более светлые экземпляры, идентичные описанию L. q. grigorievi,
и все переходы между ними. Несмотря на то что усматривается обшая
тенденция к посветлению экземпляров по направлению с севера на юг
и в горах по вертикали сверху вниз, мы считаем нецелесообразным
выделение среднеазиатских стрекоз в особый подвид, тем более,
что ареал вида, как показали наши исследования, оказался
сплошным. Этот циркумбореальный вид приспособился к обитанию в до-
линах даже на юге Средней Азии, сместив репродуктивную часть жиз-
ненного цикла на короткий весенний интервал своего температурного
оптимума. Например, в долинах Юго-Западного Таджикистана четырех-
пятнистая стрекоза летает с середины марта до середины мая.
23
Как в долинах, так и в горах обычна аберрация "praenubila",
свойственная в основном самкам.
Род Crocothemis насчитывает два вида.
С. erythraea chaldaeorum распространен на большей ча-
сти территории. На юге обычен в предгорьях и редок в горах до аб-
солютной высоты 2000 м. В Киргизии /Крылова, 1969/ в горах не
отмечен.
По систематическим признакам экземпляры с юга Средней Азии
соответствуют описанию С. е. chaldaeorum в работе Шнайдера
/Schneider, 1985/, несколько отличаясь деталями, например, хотя
слабым, но заметным базальным пятном на передних крыльях у неко-
торых самцов и иногда наличием у обоих полов трех рядов ячеек в
дискоидальном поле передних крыльев.
С. servilia (ssp. ?) обычен в Таджикистане, Узбекистане и
Южном Казахстане. Отмечен в Северном Иране /Бартенев, 1916е/.
Мнение о его редкости в Средней Азии /Попова, 1951/, на наш
взгляд, ошибочно. Пересмотр коллекционного материала Института зо-
ологии и паразитологии АН Таджикской ССР показал, что А.Н. Попо-
вой экземпляры С. servilia ошибочно определены как С. erythraea.
В горах Таджикистана отмечен до высоты 2500 м над ур. м.
В систематике вида много неясного. Цитотаксономический анализ
/цит. по: Lohmann, 1981/ показал значительные различия между
японским (номинативная форма) и остальным азиатским материалом.'
Г.Г. Якобсон и В.П. Бианки /1905/ в определительной таблице для
видов рода Crocothemis как один из основных признаков для самок
С. servilia указал более короткую и менее закругленную гениталь-
ную пластинку, что совершенно не подходит к среднеазиатским экземп-
лярам. У самок из Средней Азии генитальная пластинка значительно
больше, чем у С. erythraea, оттянута вниз и более заострена. По
этим признакам среднеазиатские экземпляры близки к индонезийским
и новогвинейским, т.е. к С. s. novogvinensis, и резко отличны
от номинативной формы.
Ломанн /Lohmann, 1981/ серию экземпляров из Индонезии от-
носит к особому подвиду, оставляя его без названия и со знаком во-
проса. Для выяснения систематики среднеазиатской С. servilia тре-
буется сравнительный материал из разных мест ареала, в первую оче-
редь из Японии, Китая, Новой Гвинеи, Индонезии. Очень желательно
пере исследование забытой С. soror Rambur, 1842, описанной из Ги-
малаев и упоминаемой В.П. Бианки /Якобсон, Бианки, 1905/ и
А.Н. Бартеневым /19196/, и С. indica Sahni, 1965, описанной так-
же из предгорий северо-западных отрогов Гималаев /Sahni, 196 5/.
Род Diplacodes включает единственный эфиопско-
мадагаскарский вид D. lefebvrei, проникающий на севере в Среди-
земноморскую подобласть Субголарктической области. Обычен в доли-
нах Юго-Западного Таджикистана и Юго-Западном Копетдаге. Найден
в Гарме (Таджикская ССР) на высоте 1450 м над ур. м. Возможны
находки и в других районах Средней Азии.
Род Trithemis также включает единственный вид
Т. festiva, широко распространенный в Ориентальной области и прони-
24
кающий в пределы Средиземноморской подобласти Субголарктики, от
Афганистана до Малой Азии /Schmidt, 1961/. В Средней Азии из-
вестен по единичным находкам на юго-востоке Т уркмении из района
хр. Кугитанг /Харитонов, Борисов, 1985/.
Род Sympetrum самый обширный, представлен 13
видами.
S. flaveolum flaveolum обычен в горах Тянь-Шаня и
Памиро-Алая на абсолютных высотах от 1300 до 2800 м. У ниж-
ней границы своего распространения редок. Б.Ф. Белышевым /1961е/
отмечен для предгорий на юге и юго-востоке Казахстана, А.Н. Бар-
теневым /1929в/ - для Семиречья. Вероятно, на северо-востоке
Средней Азии ареал вида сплошной, без разрыва в долинах, с выхо-
дом в Прииртышье. Для среднеазиатских экземпляров обычны абер-
рации typica, flaveolata, typica heterochroma. А.Н. Бар-
теневым /1929в/ из Семиречья описана аберрация pishpe-
hi. Нами для оз. Искандер-куль в Таджикистане (по сборам Е. Луп-
повой в 1951 г.) отмечена самка с признаками аберрации innomi-
nate.
S. fonscolombei широко распространен по всей территории.
В горах обычен до 2300 м над ур. м., выше редок. Находки на Па-
мире на абсолютной высоте 4100 м (р. Маркансу), вероятно, связа-
ны с залетом этих стрекоз /Борисов, 19856/.
S. pedemontanum спорадичен в долинах и в горах до высоты
2000 м над ур. мо Местами многочислен, особенно на рисовых чеках,
Не отмечен для относительно хорошо обследованных долин Юго-Запад-
ного Таджикистана. Б.Ф. Белышев /1958а/ экземпляры из Ташкента
относит к подвиду S. р. intermedium, а экземпляры из Прибалха-
шья /Белышев, 1961е/ затрудняется идентифицировать с каким-либо
подвидом. Мы также не смогли отнести стрекоз из Таджикистана и
Киргизии к какому-либо подвиду, так как экземпляры, собранные да-
же в одном месте, сочетают в себе признаки и номинативного подви-
да, и S. р. intermedium.
S. depressiusculum распространен аналогично предыдущему
виду. Не указан для малоизученной Туркмении. Вероятно, как и пре-
дыдущий вид, на юге Средней Азии уже редок и находится у южной
границы своего распространения, тем более, что оба вида отсутству-
ют в Афганистане / Schmidt, 1961; Carfi и др., 1982/.
S. tibiale, по нашим наблюдениям, очень обычен в южных
районах Казахстана. Указания на находки вида на Памире /Борисов,
19856; Борисов, Харитонов, 1986/ оказались ошибочными. Вероятно,
S. tibiale не обитает и в Афганистане. На наш взгляд, и Шмидт
/Schmidt, 1961/, и Дюмонт /Dumont, 19 75/ ошибочно приняли за
этот вид тогда еще не описанный S. haritonovi /Борисов, 1983/.
S. vulgatum flavum — циркумбореальный вид. В Средней
Азии известен из Тянь-Шаня с высот 1500-2500 м над ур. м. Ука-
зан для Южного /Попова, 1951/ и Юго-Восточного Казахстана /Бе-
лышев, Шевченко, 1971/.
S. vulgatum flavum обычен в предгорной и среднегорной
частях Тянь-Шаня и Памиро-Алая. В Киргизии /Крылова, 1969/ и
25
Юго-Восточном Казахстане /Белышев, 1961е/ приводится и для до-
лин. На юге Средней Азии, южнее Ташкента, приурочен только к го-
рам. Верхний предел распространения, по-видимому, около 3000 м.
Б.Ф. Белышев /1961е/ упоминает об экземпляре S. v. flavum
из Заилийского Алатау с "яркими переходными признаками" к номи-
нативному подвиду. Все просмотренные нами экземпляры (80 шт.) из
Таджикистана и Киргизии уклонений от описываемого подвида не
имеют.
S. haritonovi обычен в Гиссаро-Дарвазе выше 1300 м над
ур. м., многочислен на термальных источниках Памира /Джеланды -
3550 м, Джаушангоз - 3306 м над ур. м.), отмечен как обычный
на южных склонах Чаткальского хребта (1400-1600 м) в Киргизии.
Личинка не описана.
S. striolatum pallidum широко распространен по всей тер-
ритории Средней Азии. В горах известен до высоты 2700 м над ур. м.
S. striolatum striolatum обычен в Копетдаге. Отмечен
в Афганистане /Schmidt, 1961/. А.Н. Бартенев /цит. по: Попова,
1951/ указывает, что в Средней Азии встречаются все переходы от
S. s. striolatum до S. s. pallidum.
S. decoloratum обычен в долинах и предгорьях. Верхний пре-
дел распространения в горах - 1500 м над ур. м. Личинка не описана.
S. sanguineum обычен в горах Памиро-Алая и Тянь-Шаня на
высотах от 800 до 2400 м над ур. м. В Киргизии приводится и для
долин /Крылова, 1972/. А.Н. Бартеневым /цит. по: Попова, 1951/
указывается для Ташкента и Намангана, т.е. с высот менее 800 м
над ур. м. В низменностях Прибалхашья /Белышев, 1961е/ вид уже
обычен. Вероятно, южнее широты Ташкента S. sanguineum обитает
только в горах. Подвидовая принадлежность среднеазиатских стрекоз
точно не установлена. А.Н. Попова /1951/ отмечает, что экземпля-
ры из Таджикистана ближе всего к подвиду S. s. armeniacum.
Б.Ф. Белышев /1961е/ стрекоз из Прибалхашья относит к подвиду
S. s. obsoletum, оговариваясь, что их систематическое положение
не ясно и название сохраняется только ввиду того, что место сбора -
terra typica для подвида.
S. meridionale обычен на большей части территории. В до-
линах на юге спорадичен. В горах отмечен до абсолютной высоты
2500 м.
Род Leucorrhinia бореальный, незначительно прони-
кающий в Среднюю Азию, где отмечено 2 вида: L. pectoralis и
L. caudalis, указаны А.Н. Поповой /1951/ для Южного Казахста-
на, что требует подтверждения. 1'. pectoralis обычен на оз. Ала-
куль.
Род Pantala представлен единственным циркумтропиче-
ским видом Р. flavescens, склонным к широким миграциям. Обы-
чен на большей части территории Средней Азии. Не отмечен для ма-
лоизученных Туркмении и Юго-Восточного Казахстана. В горах за-
летные особи встречаются до высоты 5000 м над ур. мр. /Борисов,
19856/.
Семейство Macrodiplactidae включает монотипический
26
род Selysiothemis - эндемик Средиземноморской подобласти
Бореального фаунистического царства /Белышев, Харитонов, 1981/.
S. nigra обычен в низменных районах Средней Азии до абсо-
лютной высоты 700 м, изредка встречается и на высотах до 1000 м
над ур. м. Обладает значительной сезонной изменчивостью популяций
в размерах /Борисов,' 1984/.
Семейство Platycnemidae в Средней Азии также известно
по одному роду. В Северо-Восточном Афганистане /Schmidt, 1961/
отмечен Calicnemis eximia, проникающий из Индостана. Возмож-
на его находка в Бадахшане.
Род Platycnemis представлен тремя видами.
P.pennipes отмечен для Киргизии /Крылова, 1969/ и Узбе-
кистана /Попова, 1951/. Вероятно, вид лишь незначительно проника-
ет в северные районы Средней Азии.
Р. dealbata обычен в долинах Юго-Западного Таджикистана и
на юге Туркмении. Отмечен для юго-западных предгорий Зеравшан-
ского хребта под названием Р. aedipus /Белышев, 1958а/. При-
урочен к проточным водоемам искусственных оросительных систем
/Борисов, 1985а/. Статус среднеазиатских представителей таксона
разными авторами трактуется по-разному. Шмидт /Schmidt, 1961/
выделяет их в особый вид Р. dealbata, чего придерживаемся и мы.
Преимагинальные фазы не описаны.
Р. insularis известен по находке двух самок в Южной Турк-
мении /Павлюк, Курбанова, 1984/, что требует подтверждения.
Семейство Coenagrionidae объединяет четыре рода. На севе-
ро-востоке Афганистана отмечен род Agriocnemis (a. pygmaea),
который, возможно, будет найден в Бадахшане.
Род Coenagrion насчитывает семь видов.
С. armatum отмечен В.Н. Крыловой /1969/ для Киргизии. Ве-
роятно, это изолированное от основного, более северного ареала ме-
стонахождение в горах. Подвидовая принадлежность не известна.
C.hastulatum приводится А.Н. Поповой /1951/ в списке стре-
коз Средней Азии для Южного Казахстана, где находится у южных
пределов Своего распространения.
С. pule helium asiaticum распространен спорадично. Нами
найден в Таджикистане (окрестности Душанбе и Ховалинг), Узбеки-
стане (окрестности Самарканда, Красногвардейска, район "Ворота
Тимурлена"), Киргизии (Сары-Челекский заповедник). Отмечен для
западной оконечности Гиссарского хребта /Белышев, 1958а/, Кирги-
зии /Крылова, 1969/, Южного Казахстана /Попова, 1951/. На юге
явно приурочен к предгорной полосе и горам. Вероятно, более обы-
чен в северной половине Средней Азии. На оз. Зайсан, сравнительно
недалеко от среднеазиатского региона, этот вид один из самых мно-
гочисленных среди стрекоз /Белышев, 1961е/.
С. puella отмечен для Южного Казахстана /Попова, 1951/,
Киргизии /Крылова, 1969/, юго-востока Казахстана /Белышев,
1961е/. Подвидовая принадлежность не установлена. Б.Ф. Белышев
/19б1е/ отметил отличие самца из окрестностей г. Алма-Аты от
типичных.
27
C.syriacum отмечен А.Н. Бартеневым /1916а/ для Горгана в
Северном Иране под названием Agrion puella. Ожидая находок ви-
да на юго-западе Туркмении, мы включаем его в фауну региона.
С. ornatum отмечен нами для Юго-Западного КопеТдага.
С. vernale указан А.Н. Поповой /1951/ в списке стрекоз
Средней Азии для Южного Казахстана.
С. scitulum отмечен в Киргизии для долин и предгорий /Кры-
лова, 1972/, в Северном Иране /Бартенев, 1916а/. В Таджикиста-
не спорадичен в горах на высотах от 1300 до 2300 м над ур. м.
По—видикюму, на юге Средней Азии обитает только в горах.
Род Erythromma включает два вида.
Е. najas отмечен нами как обычный в Южном Казахстане.
А.Н. Попова /1951/ приводит его для Узбекистана и Киргизии без
указания мест находок. Подвидовая принадлежность среднеазиатских
экземпляров не установлена.
Е. viridulum распространен в низменных районах региона, вы-
ше 800 м над ур. м. не найден. В северных районах редок. По-види-
мому, в Средней Азии обитает подвид Е. v. orientate, что требу-
ет специального исследования.
Род I schnura в Средней Азии представлен семью
видами.
I. senegalensis не включается нами в фауну региона, так
как указания на его находки здесь оказались ошибочными /Белышев,
Харитонов, 1981/. Не принимаются во внимание также упомянутые
А.Н. Поповой /1951/ в работе по стрекозам Таджикистана L ке—
lesi, L bidentata, L calicis, L sinuata, названия которых
заимствованы ею из неопубликованной рукописи А.Н. Бартенева, а
описание и сведения о сохранности и местонахождении типовых эк-
земпляров отсутствуют. В Кашмире (Северо-Западная Индия) в высо-
когорье на термальных источниках обитает L inarmata /Asahina,
1978/. Вполне вероятно нахождение этого вида в аналогичных
условиях на Памире и в Бадахшане.
L pumilio широко распространен по всей территории как в до-
линах, так и в горах. Многочислен на термальных источниках Памира
до абсолютной высоты 3550 м /Борисов, 19856/. Обычны две фор-
мы самок - типичная и aurantiaca.
I. elegans - один из самых обычных видов стрекоз на всей
территории. В горах отмечен до высоты 2500 м над ур. м. Можно
ожидать находок и на больших высотах. Характерна сильная индиви-
дуальная изменчивость, что не позволяет нам отнести среднеазиат>-
ских стрекоз к какому-либо из подвидов, выделенных Шмидтом
/Schmidt, 19 54 /. Требуется специальное исследование изменчи-
вости. Возможно, выделение подвидов окажется нецелесообразным. От-
мечена сезонная изменчивость популяций в окраске и размерах /Бо-
рисов, 1984/. Самки трех форм: типичной, "самцовой" окраски и
aurantiaca. Последняя, редкая на большей части территории, най-
дена нами в массе в Сары-Челекском заповеднике.
L fountainei распространен р низменных районах на всей
территории, в горах как редкий найден до абсолютной высоты 1300 м.
28
К северу численность убывает. Отмечена сезонная изменчивость по-
пуляций по окраске и размерам. Самки трех форм: aurantiaca,
зеленая и "самцовой" окраски. Последняя относительно редка. Из бас-
сейна р. Сырдарьи описана гинандроморфная самка /Харитонова, 1987/.
Преимагинальные фазы не описаны.
L forcipata обычен в южной половине Средней Азии. Север-
ная граница распространения проходит примерно по 42° с.ш. Вид на-
ми не найден в Каракалпакии и Южном Казахстане, хотя обычен под
Ташкентом. К сожалению, ВЛ. Крылова /1969/ не указывает кон-
кретных мест нахождения в Киргизии. В горах на юге отмечен до
абсолютной высоты 2800 м. Таксономическое положение не совсем
ясно, необходим материал для сравнения из других мест ареала, в
первую очередь из Ориентальной области. Обычны две формы самок -
типичная и aurantiaca.
L intermedia описан по трем самцам и самке из Турции/Du-
mont, 1974/, после чего до недавнего времени нигде не был отме-
чен. Нами обнаружен на юге Туркмении, где особенно обычен в Запад-
ном Копетдаге. Несколько экземпляров .этого вида из разных мест
юга Средней Азии идентифицировано нами в коллекциях Зоологическо-
го института АН СССР среди серий I. forcipata. Преимагинальные
фазы не известны.
I. evansi обычен в низменных районах на север до широты Таш-
кента. В.Н. Крыловой /1969/ отмечен для Киргизии. В горах найден
до высоты 1300 м над ур. м. Отмечена сезонная изменчивость попу-
ляций по окраске и размерам. Обычны самки трех форм: светлая, ко-
ричневая и "самцовой" окраски. Очень редка вариация aurantiaca.
Преимагинальные фазы не описаны.
I. aralensis описан из бассейна р. Сырдарьи /Харитонов,
1979/. Позднее найден нами на Южном Урале. Ареал в целом и в
пределах Средней Азии не ясен. Преимагинальные фазы не известны.
Род Enallagma представлен единственным видом Е. суа-
thigerum. Распространен в низменностях на севере региона и только
в горах на юге. Обычен в Памире-Алае на высотах от 1300 до
2300 м над ур. м. Самая высокогорная из известных находка на вы-
соте 3550 м над ур. м. - термальный источник Джеланды на Памире.
Систематическое положение среднеазиатской формы не ясно. Шмидт
/Schmidt, 1961/ описал из Афганистана Е. risi, к которому очень
близки экземпляры из Таджикистана и Киргизии. Необходимы допол-
нительные материалы и ревизия рода. Самки двух форм: типичные и
светлые.
Семейство Lestidae включает два рода.
Род Lestes насчитывает в Средней Азии 5 видов.
L. rracrostigma распространен спорадично, более обычен в се-
верных районах региона.
L. dryas спорадичен в горах Памиро-Алая на высотах от 1300
до 2800 м над ур. м. В Тянь-Шане отмечен не ниже 800 м над ур.
м. /Крылова, 1972/. Редок в долинах на юго-востоке Казахстана
/Бартенев, 1929в; Белышев, 1961е/. В Средней Азии явно приуро-
чен к горам, например в среднегорье Киргизии это один из самых
29
обычных видов стрекоз, и только в северных районах региона спосо-
бен обитать в долинах.
L. sponsa изредка встречается в горах на юге на высотах от
800 до 2800 м над ур. м. В Киргизии обитает и в долинах /Крыло-
ва, 1972/, обычен в Южном Казахстане /Харитонов, 19806/.
L. Barbara обитает в горах и низменностях северной половины
региона примерно до 42° с.ш. Южнее обычен в горах до высоты
2300 м над ур. м. и очень редок в долинах. АЛ. Бартенев /1916а/
приводит его для Горгана в Северном Иране, Р.С. Павлюк, Т. Курба-
нова /1984/ - для окрестностей Ашхабада и Иолотани в Туркмении.
Не найден нами в относительно хорошо изученных долинах Юго-Запад-
ного Таджикистана, хотя в горах вокруг обычен.
L. virens vestalis распространен в низменностях на севере
региона, южнее широты Ташкента найден только в горах. Обычен в
диапазоне абсолютных высот от 1300 до 2300 м, но ниже редок.
Б.Ф. Белышев /1961е/ по экземплярам из Юго-Восточного Казах-
стана описал подвид L. v. marikovskii, отличающийся от типа мел-
кими размерами, что вряд ли целесообразно, так как у L. v. vesta—
размеры тела, по данным промеров экземпляров из Таджикистана,
заметно варьируют и среди них мелкие особи обычны.
Род Sympecma включает три вида.
S. paedisca paedisca обычен на всей территории в долинах,
редок в горах, где отмечен до абсолютной высоты 1900 м /Белышев,
Шевченко, 1958/. Вид подвержен сильной клинальной изменчивости,
выражающейся в потемнении окраски по направлению с юга на север.
На севере региона отмечены переходные экземпляры к подвиду S. р.
braueri. Вид S. annulata Selys, 1827 сводится нами в сино-
нимы описываемого подвида. Экземпляров, переходных к S. gobica
и упомянутых А.Н. Бартеневым /1912в/ и А. Шорыгиным /1926/,
нами не найдено. Все они могут быть отнесены либо к одному, либо
к другому виду.
S. gobica распространен в регионе аналогично предыдущему
виду. Более обычен в горах до высоты 2200 м над ур. м. Преима-
гинальные фазы не известны.
S. fusca обычен на всей территории в долинах и горах, отме-
чен на высотах до 2800 м над ур. м.
Семейство Е pallagidae монотипично.
Род E pallage с недостаточно разработанной системати-
кой. Количество видов разными авторами понимается по-разному. Од-
ни /Белышев, Харитонов, 1981/ считают род монотипическим, другие
/Якобсон, Бианки, 1905; Бартенев, 1916а/ - состоящим из несколь-
ких видов.
Е. fatime известен на юге Туркмении. А.Н. Бартенев /1916а/
приводит его для Северного Ирана (Шахруд).
Семейство Calopterygidae также включает один род - Са-
lopteryx, систематика которого очень запутана. Разные авторы
по-разному понимают таксономический статус форм группы splen-
dens. До окончательного решения вопроса мы пока оставляем
большинство среднеазиатских форм в ранге видов. Видовая принадлеж-
30
ность имеющихся у нас экземпляров из окрестностей Пархара на юго-
западе Таджикистана пока неясна. Возможно, они принадлежат к осо-
бому виду.
С. virgo отмечен ВЛ. Крыловой /1969/ для Киргизии. АЛ. По-
пова /1951/ в списке стрекоз Средней Азии приводит его для Южно-
го Казахстана, Киргизии и Узбекистана. Вероятно, вид незначительно
проникает лишь в северные районы региона. АЛ. Бартенев /1929в/
самку из Семиречья относит к номинативному подвиду.
С. maracandica спорадичен в Таджикистане и южных районах
Узбекистана и Киргизии. При'' наличии подходящих по термическим ус-
ловиям проточных водоемов проникает довольно высоко в горы. В Са-
ры-Челекском заповеднике отмечен на высотах 1300-1500 м над
ур. м. Шмидт /Schmidt, 1961/ для Афганистана приводит его с высо-
ты до 2500 м над ур. м. Преимагинальные фазы не описаны.
С. splendens спорадичен на всей территории. В долинах при-
урочен к проточным водоемам искусственной оросительной системы. В
горах в массе найден в Сары-Челекском заповеднике на теплой одно-
именной реке. Как отмечалось выше, таксономический статус группы
splendens во многом неясен.
С. intermedia найден в окрестностях г. Бекабада в Узбеки-
стане, где летает совместно с С. splendens. Личинка не известна.
С. orientalis распространен по южному побережью Каспийско-
го моря от Ленкорани до Красноводска /Бартенев, 1912а/. Личинка
не известна.
С. transcaspica описан из Геок-Тепе на юге Туркмении. При-
водится для окрестностей Ашхабада /Бартенев, 1912а; Павлюк, Кур-
банова, 1985/. Личинка не известна.
С. unicolor описан из Нового Маргелана в Туркестане. По
сборам РЛ. Мекленбурцева (коллекции Биологического института СО
АН СССР), отмечен для Ташкента. Личинка не известна.
СОСТАВ ФАУНЫ СТРЕКОЗ СССР
В составе одонатофауны СССР в настоящее время зарегистриро-
вано 172 вида и 69 подвидов стрекоз, объединяемых в 52 рода, 10
семейств и 3 подотряда. Число видов довольно условно И по мере на-
копления и анализа материалов по таксономическим причинам может
быть изменено на 10-15. Учитывая недостаточную изученность мно-
гих районов, особенно юга Дальнего Востока и Средней Азии,, можно
предполагать обнаружение на территории страны еще одного-двух де-
сятков видов стрекоз.
Внутривидовая изменчивость стрекоз изучена пока совершенно не-
достаточно, поэтому трудно дать оценку реального числа подвидов в
составе фауны. Работу по разработке внутривидовой систематики стре-
коз можно считать находящейся на самом начальном этапе развития.
Необходим поиск и обоснование критериев для выделения подвидовых
таксонов.
Более 10 видов и подвидов, сведение которых в синонимы мы счи-
31
Рис. 1. Зоогеографическое районирование СССР на основании распро-
странения стрекоз (Odonata) .
Голарктическая область: I - Сибирская, II - Европейско-Сибир-
ская, III - Европейская, IV - Монголо—Казахская подобласти; Суб—
голарктическая область: V - Средиземноморская, VI - Восточно-
Азиатская (Амурская) подобласти.
таем доказанным, не включены нами в предлагаемый ниже список. По
сравнению с опубликованным ранее перечнем стрекоз СССР /Белышев,
19766/ в данный список внесено около 40 изменений и уточнений си-
стематического, номенклатурного и географического характера. В ча-
стности, в последнее время в составе фауны страны обнаружены два
новых рода и шесть видов стрекоз, в том числе два вида новых для
науки. Для отдельных таксонов уточнено распространение и таксоно-
мический статус.
Для облегчения представления о пространственном размещении ви-
дов список дан в виде таблицы (табл. 1) с указанием принадлежности
таксонов к зоогеографическим областям и подобластям, схематические
границы которых приведены на карте (рис. 1). Ареалы большинства
видов стрекоз нашей страны выяснены пока весьма приблизительно.
Отсутствие достаточно подробных материалов по распространению ви-
дов и их комплексов не позволяет провести четкое зоогеографическое
районирование территории и наметить реальные, а не схематические
границы подразделений.
Территория страны входит в состав двух областей Бореального
фаунистического царства: Голарктической и Субголарктической. Первая
включает четыре подобласти: Сибирскую, Европейско-Сибирскую, Евро-
32
пейскую и Монголо-Казахскую, вторая - две: Средиземноморскую и
Восточно-Азиатскую, незначительно входящие в пределы СССР, но
фаунистически наиболее богатые. Для Сибирской, Европейской, Среди-
земноморской и Восточно-Азиатской подобластей характерны автох-
тонные фаунистические комплексы. Подобласти Европейско-Сибирская
и Монголо-Казахская практически не имеют автохтонных элементов и
их гетерогенные по составу фаунистические комплексы сформированы
выходцами из соседних подобластей.
ПАЛЕОНТОЛОГИЯ И ЭВОЛЮЦИЯ СТРЕКОЗ
Изучение геологической истории стрекоз необходимо для эволю-
ционных построений и обоснования классификационной схемы, совре-
менное состояние которой требует пересмотра и уточнения. В свою
очередь, такая система и распространенность стрекоз в осадочных
отложениях делают возможным использование сведений о них для па-
леоэкологических и палеогеографических реконструкций, а также для
биостратиграфии.
В предлагаемом разделе, естественно, невозможно решить все
сложные и многообразные проблемы палеонтологии и эволюции стрекоз.
В основном здесь в самых общих чертах будет дана только макроси-
стема и макроэволюция инфракласса. Отдельные аспекты темы неиз-
бежно освещаются неравномерно.
Основные сведения о системе, филогении и палеонтологии стрекоз
содержатся в работах Гандлирша /Handlirsch, 1906—1908/, Тильяр-
да /Tillyard, 1929/, Фрэзера /Fraser, 1957/, А.В. Мартынова
/1938/, Б.Б. Родендорфа /1962/ и Л.Н. Притыкиной /1980/. Кро-
ме литературных источников использованы некоторые неопубликован-
ные результаты изучения богатой коллекции ископаемых стрекоз, хра-
нящейся в Палеонтологическом институте АН СССР. Она содержит
большую часть мировых сборов: несколько тысяч образцов, собранных
в 140 местонахождениях Евразии и Северной Африки. Основное до-
стоинство нашей коллекции - присутствие в ней не только отпечатков
крыльев, но и значительного количества остатков личинок и имаго. С
их помощью возможно изучение структур и органов, которых обычно
недостает палеонтологу. Крылья сохраняются чаще всего, они дают
много сведений для эволюционных и классификационных построений, но
в отдельных случаях, когда основное ветвление родословного древа
связано не с модификацией летательного аппарата, а с экологической
дивергенцией личинок, как у эсхноидов, система, основанная на при-
знаках крыла, в значительной мере формальна.
В геологической истории стрекоз немало географических и стра-
тиграфических пробелов. Так, мезозойская фауна известна почти'ис-
ключительно по находкам в Евразии. Для полноценного описания исто-
рии стрекоз очень недостает триасового, особенно раннетриасового
материала. Без него картина начала жизни современных стрекоз, "ис-
тинных" Odonata, остается все еще туманной. Сведения о фауне ран-
немелового возраста в основном ограничены Восточной Азией. Почти
3 Зак. 611
33
w Таблица 1. Стрекозы фауны СССР
Л -----------------------------
Состав фауны
1
ПОДОТРЯД ANISOPTERA SELYS, 1840
Сем. LibeUulidae Stephens, 1836
Род Pantala Hagen, 1860
Р. flavescens Fabricius, 1798 ...................
Род Li b ellula Linnaeus, 1758
L. depressa Linnaeus, 1758.......................
L. fulva fulva, Milller, 1764 ...................
L. f. pontica Albarda, 1887 .....................
L. relicta Belyshev et Kiauta, 1968 .............
L. quadrimaculata quadrimaculata Linnaeus, 1758 . .
L. q. orientalis Belyshev, 1956 .................
L. q. asahini Schmidt, 1957 .....................
Род Lyriothemis Brauer, 1868
Ly. pachigastra Selys, 1878 .....................
Род Leucorrhinia Brittinger, 1850
L. albifrons albifrons Burmeister, 1839 .........
L. a. obensis Belyshev, 1973 ....................
L. orientalis Selys, 1887 .......................
L. dubia dubia Vanderlinden, 1825
L. d. circassica Bartenef, 1929 .................
L. pectoralis Charpentier, 1825 ....................
L. rubicund^ Linnaeus, 1758 .....................
L. intermedia Bartenef, 1911 ....................
L. caudalis Charpentier, 1840 ...................
Голарктическая область
Субгол арктиче
ская область
Подобласти
Сиби]>- ская Европей- ско-Си- бирская Европей- ская Монголо- Казах- ская Среди- земно- морская Восточ но-Ази- атская
2 3 4 5 6 7
+н - — — + + +
+ + + + + +
— +н 4- + + —
— +НЕ + +Е — —
Род Sympetrum Newman, 1833
S. eroticum eroticum Selys, 1863 ..............
S. kunckeli Selys, 1884 ........................
S. baccha matutinum Ris, 1911...................
S. flaveolum flaveolum Linnaeus, 1758 ............
S. f. luteola Selys, 1850 ......................
S. f. austrinum Akramovski, 1948 ...............
S. fonscolombei Selys, 1840 .................
S. pedemontanum pedemontanum Allioni, 1766 . . .
S. p. intermedia Belyshev, 1955 ................
S. p. kurenzovi Belyshev, 1956 .................
S. p. elatum Selys, 1883 .......................
S. cordulegaster Selys, 1883 ...................
S. infuscatum Selys, 1883 ...........
S. parvulum Bartenef, 1912...............
S. risi risi Bartenef, 1914 ........ ...........
S. frequens Selys, 1833 ........................
S. depressiusculum Selys, 1841..................
S. tibiale Ris, 1897 ...........................
S. scoticum scoticum Donovan, 1811..............
S. s. matrix Bartenef, 1915.....................
S. vulgatum vulgatum Linnaeus, 1758 ............
S. v. imitans Selys, 1886 ......................
S. v. flavum Bartenef, 1915........................
S. v. fuscopterum Belyshev, 1971 ..........
S. uniforme Selys, 1883 ........................
S. croceolum fuscoatrum Belyshev, 1964 .........
S. striolatum striolatum Charpentier, 1840 ........
S. s. pallidum Selys, 1887 .....................
S. s. imitoides Bartenef, 1915..................
S. decoloratum Selys, 1884 .............
S. sanguineum sanguineum Muller, 1764 ......
S. s. armeniacum Selys, 1884 ...................
S. s. obsqletum Bartenef, 1915...............
S. s. sykinia Belyshev, 1955....................
S. meridionale meridionale Selys, 1841 .......
S., haritonovi Borisov, 1983 ................
+ + + + + +
— -
•• - +
••
а—
— +
+ + + + -»-Ср. А +
— — +Ср. А —
— -i м> +К
•* — +? + 4- 1
— + + +Е + м
+ + м> +А >_— _
+н — — — +
— — —
аа- . а
••
•• ” +
—
— ~ +
ж— а
***
+
+-Н - -к + 4.
— +А, НЕ + —
+ + — + +
+н —
+Н + +
— — -> +
— — — + Н +
-
““ — *** 4*
+н — м> — +
+л л>-
— + НЕ +• + + _
— — — +н + м
4-Н — — — ц.
•* — — +А +
— + + + Е • м»
•* — — — +к
— — +А + Ср. А -
* ? + — -(НА -
— +НЛ + м
— — — + —
Продолжение табл. 1
1
Род Neurothemis Brauer, 1867
N. palliata palliata Rambur, 1342 ..........
Род Crocothemis Brauer, 1868
C. erythraea erythraea Brulle, 1832 ........
C. e. chaldaeorum Morton, 1920 .............
C. servilia Drury, 1773 .....'..............
Род D e i e 1 i a Kirby, 1889
D. phaon Selys, 1883 .................
Род В rachithemis Brauer, 1868
B. leucostica Burmeister, 1839 .............
Род Orthetrum Newman, 1833
Or. coerulescens coerulescens Fabricius, 1798 ...
Or. brunneum brunneum Fonscolombe, 1837 .....
Or. cancellatum cancellatum Linnaeus, 1758 ..
Or. c. kraepelini Ris, 1897 ...............
Or. c. orientalis Belyshev, 1958 .........
Or. albistylum albistylum Selys, 1842 ......
Or. a. speciosum Uhler, 1858 ..............
Or. sabina sabina Drury, 1773 ..............
Or. s. nigrescens Bartenef, 1929 ...........
Or. anceps Schneider, 1845 ...............
Or. triangulare melania Selys, 1883 ..........
Род Diplacodes Kirby, 1889
D. lefebvrei Rambur, 1842 ................
Род Tri t hemi s Brauer, 1868
T*. festiva Rambur, 1842 .................
Сем. Macrodiplactidae Fraser, 1957
Род Selysiothe mis Ris, 1897
S. nigra Vanderlinden, 1825 ...........
2 3 4 5 . 6 7
— — м. 4-
* • • • - - - - - +
- - + + + -
• • • • mb — + — — —
• • • • мв м 4- +н + —
4 • ♦ • - - - - -Юр. А -
- - — - - +
• Я • • - - - - - +
мв м. м. + —
• • • • - - - - +к -
4- + ’ 4- + + +
« • • • — 4- — +? —
* * ♦ ♦ ? — — + + —
МВ 4- 4- + 4- —
• • • • • м. м. м. !А 1Ср. А —
4- +НА — !Н — —
4* — + + + —
• • • * — — — — — +
• • • • — — — — + —
• • • • — м> МВ — + —•
♦ • • • - - - +А, НЕ + -
• « • • • • — — — — — +
— — — — + —
• • • • - - — — + , -
— — — — + —
« • « • — - — — + —
— м. +НА 4- м.
• • • • м. — — +НА 4- —
Сем. Corduliidae Karsch, 1894
Род Epitheca Burmeister, 1893
Ep. bimaculata bimaculata Charpentier, 1825 .
Ep. b. sibirica Selys, 1887 . .......
Ep. b. altaica Belyshev, 1951 .............
Род S omatochlora Selys, 1871
S. metallica metallica Vanderlinden, 1825
S. m. exuberata Bartenef, 1911 ..........
S. m. abacanica Belyshev, 1955 ............
S. m. japonica Matsumura, 1911 ..............
S. flavomaculata flavomaculata Vanderlinden, 1825 .
S. f. minor Belyshev, 1955 ................
S. viridiaenea (?) viridiaenea Uhler, 1858 ......
S. arctica Zettersted, 1840 ...............
S. graeseri graeseri Selys, 1887 ...........
S. uchidai Foerster, 1909 ..............
S. sahlbergi sahlbergi Trybom, 1889 .........
S, s. relicta Belyshev, 1971 ..............
S. alpestris Selys, 1840 ....................
Род C ordulia Leach, 1815
C. aenea aenea Linnaeus, 1758 ....... .......
C. a. amurensis Selys, 1887 ........
Род Macromia Rambur, 1842
M. sibirica Djakonov, 1926 ............
M. bartenevi Belyshev, 1973..............
Род E po phthal mi a Burmeister, 1839
Ep, elegans Brauer, 1865 ....................
Сем. Aeschnidae Bianchi, 1905
Род Aeschna Fabricius, 1775
Ae. squamata squamata Muller, 1764 ..........
Ae. crenata crenata Hagen, 1856 ..............
Ae. c. wnukovskii Belyshev, 1967 ...........
Ae. serrata serrata Hagen, 1856 .............
Ae. s. osiliensis Mirzejewski, 1913 ..........
Ae. subarctica elisabethae Djakonov, 1922 ......
Ae. juncea juncea Linnaeus, 1758 ...........
+НК
Сем. Gordulegasteridae Banks, 1892
Род A n о t о g a s t e r Selys, 1854
An. sieboldi Selys, 1854 .............
Род C ordulegaster Leach, 1815
C. boltonii boltonii Donovan, 1807 ........
C. princeps Morton, 1915 .............
C. insignis charpentieri Kolenati, 1846 .
C. i. coronatus Morton, 1916 ............
C. i. nobilis Morton, 1915..........
C. mzymtae Bartenef, 1929 ...............
C. bidentatus Selys, 1843 .............
Сем. Gomphidae Banks, 189 2
Род Nihonogomphus Oguma, 1926
N. ruptus Selys, 1857 ...................
Род Onychogomphus Selys, 1854
On. forcipatus forcipatus, Linnaeus, 1758 . . . .
On. f. unguiculatus Vanderlinden, 1820 .....
On. flexuosus Schneider, 1845 ..........
On. assimilis assimilis Schneider, 1845 . . . . .
On. a. fulvipennis Bartenef, 1912. . . .
On. lefebvrei Rambur, 1842 ...............
Род Ophiogomphus Selys, 1854
Oph. serpentinus Charpentier, 1825 .......
Oph. obscurus Bartenef, 1930 . .. .
Oph. reductus Calvert, 1898 .............
Oph. spinicornis Selys, 1878..............
Род Ani sogomphus Selys, 1857
An. maakii Selys, 1872 .........
An. amurensis Bartenef, 1930 ...........
Род Anormogomphus Selys, 1854
A. kiritschenkoi Bartenef, 1913 ..........
Род Trigomphus Bartenef, 1911
Co Tr. nigripes Selys, 1887..................
+ м> — +
+н - - - - +
— + + + + —
+ + !Е т. —
— — — ?+к —
• •••••• — — -ч — +к —
- - +АН +Ср.А -
—• —• — +
т. — — +к —
- + ? +к -
+н — — — +
+н - — - +
- + + + + —
+ + — + —
м> — 4-
- - - +А, НЕ + -
— — — + —
— — — +к —
— — — + -
+ + + + + +
+л + + 4- — т.
»•••••• + +НА — — — +
— - +НА + —
- - -. - +
+ т. — т. — +
+? - - - — -
- — - - +
- - - - + -
- — - + -
1 - - - - +
. . ! •W м> •W м> +
w
№
Продолжение табл. 1
-
Ae. j. brachystigma Sjostedt, 1927 .......
Ae. j. mongolica Bartenef, 1929 ..........
Ae. j. angustila Oguma, 1915............... .
Ae. j. orientalis Bartenef, 1929 ..........
Ae. j. atschischgo Bartenef, 19 29 ........ .
Ae. j. crenatoides Bartenef, 1924 ..........
Ae. undulata Bartenef, 1930 ................
Ae. coluberculus Harris, 1782 ...........
Ae. affinis Vanderlinden, 1825 .............
Ae. viridis Eversmann, 1836 ................
Ae. grandis Linnaeus, 1758..................
Ae. cyanea Muller, 1764 ..............
Ae. palmata Hagen, 1856 ....................
Род Anaciaeschna Selys, 1878
A. isosceles isosceles Muller, 1767 . .
A. i.. antehumeralis Schmidt, 1950 .........
Род A пах Leach, 1815
An. imperator imperator Leach, 1815 .......
An. parthenope parthenope Selys, 1839 .....
An. p. julius Brauer, 1866 ..........
Род Hemianax Selys, 1883
H. ephippiger Burmeister, 1839 ..........
Род Aeschnophlebia Selys, 1883
A. kolthoffi Sjostedt, 1925 ...........
A. Zygoptera Belyshev, 1965 ............
Род Calaeschna Selys, 1883
C. microstigma Schneider, 1845 ..........
Род Brachytron Evans, 1845
B. pratense Muller, 1764 ..............
12 3 1 4 5 6 7
+ м. —
• •••••« ? — — — + —
• •••••• 4- — 4-
— — +н — 4-
+К »>
— — - +к —
+ . — —
?+ 4- 4- 4- + 4-
+ + 4- 4-
4- 4- 4- —
4- 4- 4- +? —
+Е + +Е 4- —
?+н — — - - -
— - + + + —
— -к + - —
- - + -
+ 4- 4- 4- 4- 4-
4- 4- 4- 4-
+н + 4- 4- —
+н - - - - +
— — _ 4- 4- —
— — +НЕ + -
— — - ' 4.
— —» 4-
— — - - +
— — — 4- 4-
— — +н + -
- 4- 4- 4- 4- —
+Е + +Е +к
C. v. altaica Belyshev, 1955 .................
C. v. japonica Selys, 1869 ....................
C. maracandica Bartenev, 1913..................
C. splendens splendens Harris, 1782 ...........
C. s. erevanense Akramovski, 1948 .............
C. s. tshaldirica Bartenef, 1912...............
C. s. cartvelica Bartenef, 1930 . •............
C. s. taurica Selys, 1853 .....................
C. s. mingrelica Selys, 1868 ..................
C. s. ciscaucasica Bartenef, 1912..............
C. transcaspica Bartenef, 1911.................
C. orientalis Selys, 1887 .................
C. intermedia Selys, 1887 .....................
C. iohanseni Belyshev, 1955 ...................
C. unicolor Bartenef, 1912.....................
ПОДОТРЯД ZYGOPTERA SELYS, 1840
Сем. Lestidae Bianchi, 1905
Род L e s t e s Leach, 1815...............
L. macrostigma Eversmann, 1836 .............
L. dryas Kirby, 1890 .......................
L. sponsa Hansemann, 1823 ..................
L. temporalis Selys, 1883 ..................
L. barbara Rabricius, 1798 .................
L. virens vestalis Rambur, 1842 ............
L. v. marikovskii Belyshev, 1961............
L. viridis parvidens Artobolevskij, 1929 ....
Род Sympecma Burmeister, 1839
S. paedisca paedisca Brauer, 1877 ......... ,
S. p. braueri Bianchi, 1905 ................
S. gobica Rorster, 1900 ....................
S. fusca Vanderlinden, 1823 ................
Сем. Coenagrionidae Tiltyard, 1926
Род Coenagrion Kirby, 1890
C. hylas hylas Trybom, 1889 ................
+ +н —
+н
— “ — — +Ср. А
+ + + + —
— — — — +К
— « — — +к
*Е +к
““ — _ _ +к
— “ — — ?+к
“ — — — +Ср.А
!НЕ + - -Ж +Ср. А +
+ + + + +
— +н — + +
. . + + + + +
....... + + + + +
— — — —
• — + + + +
> . •••••• — + + +ЕА +
- — — +А +
— - + +Е ?+к
+ + + + +
— — — +
+ + + + +
— — — + +Ср. А
+Е + + +
+НА
д
о
Продолжение табл. 1
1
Род G о m р hu s Leach, 1815
G. flavipes flavipes Charpentier, 1825 ........
G. f. sibirica Bartenef, 1909 .................
G. f. liniatus Bartenef, 1929..................
G. vulgatissimus vulgatissimus Linnaeus, 1758 . .
G. schneideri Selys, 1850 .....................
G. epophthalmus Selys, 1872 ...................
G. chankae Bartenef, 1956 .....................
G. davidi Selys, 1887 .........................
род Dav idius Selys, 1878
D. annulatus Djakonov, 1926 ...................
Род Hologomphus Bartenef, 1914
H. lunatus Bartenef, 1914......................
Род Hagenius Selys, 1854
H. albardae Selys, 1886 .......................
Род Ictinus Rambur, 1842
Ic. clavatus Fabricius, 1775 ..................
Род Lindenia Selys, 1840
L. tetraphylla Vanderlinden, 1825 .............
L. inciti Bartenef, 1929 ......................
ПОДОТРЯД CALOPTERA ZALESSKU, 1932
Сем. Epallagidae Tillyard et Fraser, 1938
Род E pallage Charpentier, 1840
Ep. fatime Charpentier, 1840 ......................
Сем. Calopterygidae Buckecker, 1876
Род Calopteryx Leach, 1815
C. atrata Selys, 1853 .........................
C. virgo virgo Linnaeus, 1758 .................
C. v. festiva Briille, 1832 ...................
2 3 4 5 6 7
4- 4- 4- 4- 4- 4-
— 4- 4- 4- +К —
+? 4- — — — +?
— — — 4- —
• ••••• — + 4- 4- - -
— — — 4- —
4- +н - - - 4-
• ••••• — — — — — 4-
- - - + -
— - - - — 4-
- — т- - +
- - - - - 4-
- - - - +
- - - - — 4-
— - - - +
— - - — — 4-
- - - - - 4-
- - 4- 4- —
— — 4- 4- —
...... — — — +к —
4-
..... - - +Ср. А
4- 4- 4- 4- 4- 4-
— — — — — 4-
««•••• — 4- 4- 4- 4- —
!Н
En. belyshevi Haritonov, 1975 . • • • — — — — ’ — +
En. risi Schmidt, 1961 ... ? — — ! 1 —
En. circulatum Selys, 1883 . . . . . . . • — — — — — +
En. nigrolineatum Belyshev et Haritonov, 19?5 • • • + - - - -
Род Ischnura Charpentier, 1840 + + + + + 4-
I. aralensis Haritonov, 1979 . - - — + — 4- 4- —
I. pumilio Charpentier, 1825 +H + + + —
I. elegans elegans Vanderlinden, 1823 • « • + + + + + +
L evansi Morton, 1919 . • л . — — — +A, HE + —
L fountainei Morton, 190 5 . . . - — — +A, HE 4- —
I. forcipata Morton, 1907 . . . — — — I A +Cp. A —
I. intermedia Dumont, 1974 жж. — — — + —
I. lobata Needham, 1930 . • . ~ - - - - +
Род Nehalennia Selys, 1850 + + + + 4-
N. speciosa Charpentier, 1840 • ♦ • 4 + + + + ?+H +
Род Mortonagrion Eraser, 1920 — — — -f.
M. selenion Ris, 1911 • 4 4 4 - - - - — +
Сем. Platycnemidae Tillyard et Eraser, 1938 Род Denticnemis Bartenef, 1956 — 4-
D. bicolor Bartenef, 1956 . « . 4 — - - - - +
Род Platy cnemis Charpentier, 1840 + 4- + + 4- 4-
Pl. pennipes Pallas, 1771... .... +H + + 4- +K, Cp.A? —
Pl. phyllopoda Djakonov, 1926 — — — — +
Pl. dealbata Selys, 1836 . • ж ж — — — +H 4- —
Pl. insularis Selys, 1863 » 4 . • — — - + -
Примечания. + присутствие вида; - отсутствие вида; ? - сомнительные данные; ! - вид не указан, но должен быть; Н -
незначительный заход нз соседних областей; Л - локальные очаги; А - азиатская, Е - европейская части области; К - Кавказ,
Ср. А. - Средняя Азия.
Л Окончание табл. 1
№ . .
1
С. h. ussuriensis Belyshev, 1956 ................
С. concinnum concinnum lohanssen, 1859 ............
C. c. bartenevi Belyshev, 1955 ....................
C.' armatum armatum Charpentier, 1840..............
C. glaciale glaciate Selys, 1872 ..................
C. g. orientalis Belyshev et Haritonov, 1973 . . . .
C. hastulatum Charpentier, 1825 ...................
C. lanceolatum Selys, 1872 ..........................
C. pulchellum interruption Charpentier, 1825 ......
C. p. asiaticum Belyshev, 1964 ......................
C. p. sibiricum Belyshev, 1964 ....................
C. p. saisanicum Belyshev, 1964 .....................
C. puella puella Linnaeus, 1758 ...................
C. syriacum Morton, 1924 .........................
C. vernale Hagen, 1839 ............................
C. ecprnutum Selys, 1872 ..........................
C. mercuriale mercuriale Charpentier, 1840.........
C. ornatum Selys, 1850 ........... .......
C. brevicauda Bartenev, 1956 . . ...........
C. v—nigrum Needham, 1930 .........................
C. scitulum Rambur, 1842 ..........................
C. lindeni Selys, 184b.............................
Род Erythromma Charpentier, 1840
Er. najas najas Hansemann, 1823 ...................
Er. n. humerale Selys, 1887 .......................
Er. n. baicalensis Belyshev, 1964 .................
Er. viridulum viridulum Charpentier, 1840 .........
Род Pyrrhosoma Charpentier, 1840
P. nymphula Sulzer, 17 76..........................
Род Enallagma Charpentier, 1840
En. cyathigerum cyathigerum Charpentier, 1840 . . ,
En. c. rotundatum Bartenev, 1929 ................
2 3 4 5 6 7
• + — — +
— + + — — —
+ — — — — +
— + + +н + -
• ••••• + — — — — —
* • • • • +? — — — — +
+ + + + — —
+ — — — — +
• ••••• — + + +Е +к —
• • • • • - - - - +Ср. X -
• « • • • — + — — — —
— +н — А + —
— + + + + —
— — — — +к —
+ + + +н +к —
+ +н — — — +
— — — — +к —
• • • • • — — + +н + —
• « • • • — — — — — +
— — — — — +
- - +? +Е + —
• • • • • - - - - +к -
+ + + + + +
+ + + + + —
+ — — — — +
> • • > • + — — — — —
• • • • • - - + +НЕ + -
— +Е + +Е + —
- +НЕ + +Е +к -
+ + + + + +
+ + + + + —
+к
Рис. 2. Схема филогенетических отно-
шений стрекоз.
нет фаун позднего мела и т.д. Палеон-
тологическая летопись о многом умал-
чивает, и это постоянно приходится
иметь в виду, делая широкие обобще-
ния. Например, смена таксонов в раз-
резе одного региона может отражать
последовательность миграций (это ти-
пичный случай), а не ветвлений родо-
словного древа.
В комплексах остатков ископаемых
животных (ориктоценозах) закономерно
искажается реальный экологический
спектр древних фаун. Они непропорцио-
нально обогащены эврибионтами, круп-
ными, хорошо летающими формами, ми-
грантами, а в мезозое - специфичес-
кими озерными группами с плавающи-
ми личинками. В основном стрекозы сохраняются в осадках озер, ре-
же лагун и в прибрежно-морских фациях. В составе ориктоценозов
представлены стрекозы с самой разной экологией, но перечисленные
группы имеют наибольший удельный вес. Обитатели рек, ручьев, гор-
ных потоков, болот и излюбленных стрекозами небольших и недолго-
вечных водоемов имеют мало шансов на захоронение и по тафономи-
ческим причинам не оставляют следов в палеонтологической летописи.
Например, древнее и вместе с тем обширное семейство Gomphidae
на протяжении всей своей истории, с поздней юры до настоящего вре-
мени, крайне бедно представлено в ориктоценозах.
Необходимо отметить, что стрекозы по своим физическим и биоло-
гическим особенностям имеют тафономические преимущества перед
другими насекомыми, и поэтому их история несравненно полнее отра-
жена палеонтологической летописью. Для того чтобы в этом убедить-
ся, достаточны несложные подсчеты, основанные на актуалистических
данных по питанию и продолжительности жизни имаго и сведениях об
участии стрекоз в ориктоценозах. Оказывается, фактическое участие
стрекоз в озерных ориктоценозах на несколько порядков выше ожидае-
мого, особенно в морских и лагунных фациях.
При всей неполноте летописи ископаемый материал не теряет свое-
го уникального значения, оставаясь документальным свидетельством
развития таксонов и фаунистических комплексов во времени. Вряд ли
можно согласиться с радикальным скепсисом Хеннига / Hennig,
1969/ и его последователей, признающих из всего палеонтологиче-
ского материала достойными изучения только инклюзы в ископаемых
смолах. В нашей коллекции есть немало отпечатков стрекоз прекрас-
ной сохранности, с доступными для изучения разнообразными и весь-
ма тонкими структурами, в том числе и во внутренних органах -
бранхиальной камере и провентрикулусе.
44
Ниже приводится в виде списка принятая система старших таксо-
нов и схема их филогенетических отношений (рис. 2). Названия унифи-
цированы в соответствии с рекомендациями Родендорфа /1977/ и
частично опубликованы ранее /Притыкина, 1980, 1981/. В скобках
даны общепринятые названия старших таксонов.
ИНФРАКЛАСС LIBELLULONES
Отряд Meganeurida (= Fbotodanata + Odonata р. р.)
Подотряд Meganeurina (= Protodonata)
Подотряд Ditaxineurina (= Protanisoptera + Permanisoptera)
Подотряд Kennedyina (= Protozygoptera + Archizygoptera)
Подотряд Triadophlebiina
Отряд Libellulida (= Odonata p. p.)
Подотряд Libellulina (= Anisoptera)
Подотряд Epiophlebiina (= Anisozygoptera p. p.)
Подотряд Heterophlebiina (= Anisozygoptera p. p.)
Подотряд Calopterygina Selys, 1850 /nom. transl. Prity—
kina, nic (ex Calopterygines Selys, 1850 )/(= Zygoptera)
В настоящее время нет единого мнения о рангах высших таксо-
нов, относимых к стрекозам, их объемах и родственных связях. Не
оспаривается только монофилия, происхождение всех вымерших и сов-
ременных стрекоз от гипотетического общего предка. Однако обосно-
вание монофилии ни одним автором не приводится.
Наиболее популярны представления о том, что гигантские стреко-
зы палеозоя из-за отсутствия у них узелка и птеростигмы образуют
самостоятельный отряд Protodonata (фототабл. I, фиг. 1, 2), противо-
стоящий всем прочим стрекозам отряду Odonata /Handlirsch,
1906-1908; Tillyard, 1917, 1929, 1938; Carpenter, 1931,
1977; Мартынов, 1938; Carle, 1982а/. Ф. Фрэзер /Fraser,
1957/ счел доводы в пользу отрядового ранга неубедительными и
включил Protodonata в состав Odonata как подотряд Megani—
soptera; ему последовали Б.Б. Родендорф /1962/ и Л.Н. Притыкина
/1980/.
Основные противоречия связаны с различной оценкой классифика-
ционного веса признаков и полярности некоторых морфоклин. Так, по
мнению одних авторов, исходным было стебельчатое крыло с редуци-
рованным типом жилкования. Другие считали более примитивным кры-
ло с широким основанием и полным набором жилок. Р. Тильярд пола-
гал, что пермские стебельчатокрылые Protozygoptera, примитив-
нейшие из стрекоз, дали начало подотряду Zygoptera, предку всех
других подотрядов стрекоз с более широким крылом и полным жилко-
ванием /Tillyard, 19 29/. В рамках концепции Карпентера /Carpen-
ter, 1931/ от Protozygoptera происходят только Zygoptera,
тогда как другой палеозойский подотряд, Protanisoptera, дал на-
чало подотряду Anisozygoptera —предку Anisoptera. Позже эту
версию поддержал Тильярд /Tillyard, 1938-1940/. Ф. Фрезер - ав-
тор последней ревизии отряда - принял первую, "монистическую", кон-
45
цепцию Р. Тильярда, признав всех мезо-кайнозойских стрекоз потом-
ками кеннедиид /Fraser, 1957/.
Фактически все авторы в палеозойских группах видят предков ме-
зо-кайнозойских "настоящих" Odonata; их построения отличаются
лишь в деталях. Только А.А. Мартынов считал палеозойских стрекоз
предшественниками, но не предками современных, предлагая разделить
отряд Odonata на две группы - Permodonata и Neodonata,
дивергировавших в карбоне от неизвестного предка. С его схемой со-
гласился Карле / Carle, 1982а/, а предположение о первичности
крыла с широким основанием и наибольшей примитивности Anisopte-
га среди Neodonata нашло поддержку у автора настоящих строк
/Притыкина, 1980/.
Новые данные об ископаемых и современных стрекозах позволя-
ют рассматривать макросистему и филогению инфракласса существен-
но иначе, чем предшественники. Кроме того, автору приходится отка-
заться от некоторых собственных прежних /Притыкина, 1980/ пред-
ставлений.
РОДСТВЕННЫЕ ВЗАИМООТНОШЕНИЯ ОТРЯДОВ
MEGANEURIDA И LIBELLULIDA
И РАННИЕ ЭТАПЫ ЭВОЛЮЦИИ СТРЕКОЗ
Монофилия отряда Meganeurida поддерживается боль-
шим количеством общих признаков. Из них наиболее важными
представляются следующие: ушек нет, задний край задних крыль-
ев самцов и самок одинаково округленный; de нет; дистальное, на
уровне конца Sc, положение развилка RS (при нестебельчатом крыле
вблизи его середины); наличие свободного основания Си до слияния
с 1А; вогнутая жилка 2А (если она есть); 1А отходит от Си +1А
проксимальнее дужки; лапка 4- или 5-члениковая с длинным базаль-
ным члеником; копулятивный аппарат без семенной капсулы, оплодот-
ворение, вероятно, сперматофорное; развитие наземное.
Свободное основание Си не сохранилось только у протомирме—
леонтид в связи с крайней специализацией жилкования. Соматические
признаки триадофлебииновых не известны из-за отсутствия остатков
тел. Строение копулятивного аппарат^ установлено на примере дитак-
синевриновых (неопубликованные данные aBTopaj см. фототабл. I,
фиг. 3). О наземном развитии меганевридовых можно судить по почти
полному отсутствию в захоронениях личинок при обилии имаго. Един-
ственная меганевридная личинка, известная по фрагментарному остат-
ку из среднего карбона Северной Америки /Kukalova—Peck, 1983/,
не имеет каких-либо морфологических особенностей, указывающих на
водный образ жизни. Напротив, несомненно водные личинки отряда
Libellulida появляются в захоронениях одновременно с имаго в
триасе /Rozefelds, 1985/ и обычны в более молодых отложениях.
Единство происхождения подотрядов, объединяемых в отряд Libel—
lulida, и вместе с тем обособленность от Meganeurida пред-
ставляются несомненными. ОтряД характеризуют следующие признаки:
ушки есть, задние крылья самцов с вырезкой, угловатые, у самок -
округленные; de есть; проксимальное положение развилка RS, на-
много проксимальнее вершины Sc, в базальной половине крыла; от-
46
сутствие свободного основания Си; все анальные жилки выпуклые;
1А отходит от Си дистальнее дужки, от постеродистального угла de;
лапка 3-члениковая с коротким базальным члеником; копулятивный
аппарат с семенной капсулой — хранилищем семенной жидкости; разви-
тие водное.
Некоторые из перечисленных признаков в отдельных таксонах мо-
гут исчезать. Например, у некоторых мегаподагрионид развилок RS
находится на уровне конца Sc, но при этом жилкование в целом ока-
зывается весьма специализированным. Вообще, в любых вариантах фи-
логении Calopterygina архетипическое крыло имеет развилок RS
проксимальнее узелка. Ушки и коррелирующая с ними вырезка на кры-
ле калоптеригиновых, по-видимэму, исчезли вследствие ранней специа-
лизации копулятивного аппарата (подробнее об этом признаке несколь-
ко позже).
Распространение признаков в отрядах не дает оснований для опре-
деленного вывода о большей примитивности Meganeurida.
В некоторых отношениях более молодые Libellulida даже при-
митивнее. Например, у них нет вогнутых ветвей анальной жилки, а на-
бор жилок более полный, если справедливо предположение А.В. Марты-
нова /1938/ о наличии у Anisoptera рудиментов СиА в составе
de. К числу несомненных плезиоморфий Meganeurida относятся
свободное основание Си на крыле имаго и наземное развитие. В ос-
новном же полярность характерных признаков отрядов остается неясной.
Но дело даже не в том, что Meganeurida по каким-то призна-
кам примитивнее, чем Libellulida. Важно отсутствие между отря-
дами переходных групп, где бы эти признаки встречались совместно.
Палеонтологам известно, что именно сложная мозаика признаков в пе-
реходных группах затрудняет проведение границы между таксонами
предком и потомком. Появления сходства с Libellulida можно бы-
ло бы ожидать у более поздних Meganeurida, но ничего подобного
не наблюдается. У более поздних, позднепермских, Meganeurida
формообразование заторможено, сколько-нибудь заметный морфологиче-
ский прогресс отсутствует. Крайняя специализация немногих раннеме-
зозойских представителей отряда - Triadophlebiina и Protomyr—
meleontidae - исключает их из числа возможных предков Libel—
lulida.
Теоретически единство инфракласса можно было бы обосновать
иначе, допустив существование неизвестной примитивной группы стре-
коз, от которой в раннем карбоне ответвились Meganeurida, a Libel—
lulida одновременно с ними /Притыкина, 1980/ или в неизвестное
время до триаса. Но этот путь сейчас кажется ненадежным. Реконст-
рукция гипотетического общего предка явно требует слишком многих
допущений. Трудно себе представить, по каким причинам таксон, от-
носимый к стрекозам, будь то общий предок инфракласса или только
отряда, никак не обнаруживал себя два геологических периода (около
90 млн лет), не оставив следов в летописи, а тафономические досто-
инства стрекоз относительно велики.
Приведенные факты и рассуждения наводят на мысль о возможно-
сти дифилетического происхождения стрекоз. В этом случае момент
47
первой дивергенции смещается вниз, в пределы таксона, не относяще-
гося к стрекозам. Тогда инфракласс Libellulones выделен,
скорее, по горизонтальному принципу, хотя объединяемые им отряды
очень близки. Прямых доказательств независимого происхождения
двух отрядов стрекоз у нас нет. Поэтому целесообразно сохранить
инфракласс Libellulones, его мэнофилия под сомнением и нужда-
ется в более углубленном анализе.
Попытаемся представить себе начало эволюции стрекоз с нетра-
диционных позиций, в самых общих чертах, ибо для детализации пока
нет достаточного материала.
Прежде всего нужно отказаться от привычных представлений о
какой-то необыкновенной примитивности, архаичности стрекоз. Прими-
тивны ли постоянно распростертые в стироны крылья или закономер-
ное чередование выпуклых и вогнутых жилок? Сейчас ни один серьез-
ный исследователь не поддержит идеи о подобном архетипическом кры-
ле Pterygota. Можно ли считать признаками особой архаичности
сомнительную "простоту" (скорее, упрощенность) аксиллярного аппа-
рата или редукцию продольных мышц птероторакса? Совершенный по-
лет стрекоз - вершина эволюции четырехкрылого полета. И едва ли
кому-нибудь покажутся примитивными ноги, утратившие локомоторную
функцию, нижняя губа личинки, преобразованная в ловчую маску, или
сложнейший вторичный копулятивный,аппарат. Вероятно, высказывания
о примитивности связаны с действительно большой древностью находок
стрекоз. Как предполагают Рик и Кукалова-Пек /Riek, Kukalova—
Peck, 1984/, описанные ими стрекозы из намюра Аргентины, воз-
можно, древнейшие среди крылатых насекомых вообще. Древность же,
естественно, ассоциируется с примитивностью.
Можно предположить, что неожиданно раннее появление стрекоз
скорее следствие их тафономических преимуществ перед другими на-
секомыми. Поэтому их сравнительный эволюционный возраст оказал-
ся завышенным. Обратимся к ценотическому аспекту возникновения
стрекоз. Ясно, что специализированный воздушный хищник, прожорли-
вый и долгоживущий, мог появиться не прежде, чем сформировалось и
стало высокопродуктивным сообщество летающих насекомых. Кроме
того, возникновение стрекоз - сложный процесс освоения новой адап-
тивной эоны, и естественно предположить, что не все их многочис-
ленные специализации возникли сразу, а отчасти унаследованы от
предшественника в эволюционном ряду.
Предки стрекоз, по всей вероятности, были тоже крупными хищ-
никами, но подстерегающими жертву на субстрате. От них могли быть
унаследованы такие, например, признаки, как ноги, вооруженные хищ-
ными шипами, подвижная голова, хорошо развитое зрение, короткие
церки у самцов, приспособленные для фиксации на теле самки. По-
следний признак весьма важен: по-видимому, церки-щипцы имели ре-
шающее значение при возникновении уникального способа копуляции
стрекоз /Carle, 1982b/. Предстерегаюших хищников немало среди
карбоновых и болеё поздних насекомых, но подходящих на роль пред-
ка стрекоз среди них пока не найдено.
Отряды (прежде подотряды) и почти все объединяемые ими под-
отряды (прежде инфраотряды) кратко охарактеризованы в прежних
48
публикациях /Притыкина, 1980, 1981/. Ниже приводятся диагнозы
Heterophlebiina и Calopterygina, первоначально отнесенных
к инфраотряду Lestomorpha.
Подотряд Heterophlebiina ( = Anisozygoptera р. р.)
Табл. IV-VI
Типовое семейство - Heterophlebiidae Handl., 1906; юра
Евразии.
Диагноз. Имаго. Голова шаровидная, глаза без тенденции к ла-
теральному смещению; верхняя часть интерплеврального шва редуци-
рована. Задний край заднего крыла самца угловатый, с вырезкой: de
переднего и заднего крыла почти всегда существенно различны. Если
на переднем крыле de не замкнута, то на заднем имеет вид просто-
го неразделенного четырехугольника; если на переднем крыле de
замкнута, то на заднем разделена на t и st. Только в родах Iso—
phlebia и Stenophlebia de обеих пар крыльев одинаковы: со-
ответственно простые четырехугольники и разделенные на t и st; у
Turanophlebia при незамкнутой de переднего крыла de задне-
го разделена на t и st Ушки есть. Копулятивный аппарат самца с
развитой лигулой и задними крючочками; пенис образован членистым
оральным отделом семенной капсулы (неопубликованные данные авто-
ра). Нижний каудальный отросток самца непарный, эпипрокт.
Личинка. Бранхиальная камера есть. Анальная пирамида не раз-
вита; пластинчатые каудальные отростки покрыты плотной кутикулой,
лишены трахей, функционально образуют хвостовой плавник.
Состав. Десять вымерших мезозойских семейств: Heterophle—
biidae Handl., 1906; Archithemistidae Till., 1917; Tarso—
phlebiidae Handl., 1906; Liassophlebiidae Till., 1925;
Oreopteridae Pritykina, 1968; Triassolestidae Till., 1918
( sensu/Притыкина, 1981/); Campterophlebiidae Handl., 1925;
Karatawiidae Mart., 1925; Isophlebiidae Handl., 1906 и
Stenophlebiidae Handl., 1906.
Сравнение. Heterophlebiina близки одновременно к
подотрядам Libellulina и Epiophlebiina. Их объединяет сходст-
во в форме головы и глаз, тип редукции интерплеврального шва, на-
личие вырезки на заднем крыле, развитых задних крючочков копуля-
тивного аппарата и нижнего каудального отростка у самцов. Личинки
в этой группе подотрядов с внутренним органом водного дыхания -
бранхиальной камерой. Признаки, обшие только с Libellulina, —
наличие ушек (неопубликованные данные автора) и идентичный тип ко-
пулятивного аппарата. От Libellulina (фототабл. VII ) отличаются бо-
лее узким основанием крыльев, некоторой редукцией жилкования и
строением de. Тип копулятивного аппарата совершенно иной, чем у
Epiophlebia (подробно о копулятивном аппарате эпиофлебий см.
/Asahina, 1954; Pfau, 1971/),
Уникальная особенность гетерофлебиин - развитый у личинок хво-
стовой плавник. Отдельные особенности жилкования характерны также
исключительно для этого подотряда.
4 Зак. 611
49
Замечание. Строение копулятивного аппарата изучено только у
Isophlebiidae и tTarsophlebiidae (неопубликованные данные
автора).
Подотряд Calopterygina Selys, 1850
(nom. transl. Pritykina, hie
(ex Calopterygines Selys, 1850))
( = Zygoptera auct)
Типовое семейство - Calopterygidae Selys, 1850; нео-
генные.
Диагноз. Имаго. Голова поперечная, с широким промежутком
между глазами; интерплевральный шов полный или редуцированный в
средней и нижней частях. Крылья стебельчатые, задние не угловатые,
без вырезки; de обеих лар крыльев почти всегда простые - замк-
нутые четырехугольники. Незамкнутые de передних крыльев только
у представителей родов Hemiphlebia и Chorismagrion. Ушек
нет, пенис - модифицированная лигула, семенная капсула простая, зад-
ние крючочки копулятивного аппарата развиты слабо. Нижние каудаль-
ные отростки самца, если они есть, парные - парапрокты.
Личинки. Бранхиальной камеры нет, хотя частично газообмен
осуществляется дыхательным эпителием ректума (кровяные жабры);
анальная пирамида отсутствует; пластинчатые каудальные отростки
пронизаны трахеями и лишены эпикутикулы, функционально являются
жабрами. У реобионтов пластинчатые каудальные жабры могут быть
различным образом модифицированы, утрачивая дыхательную функцию.
Взамен развиваются боковые брюшные жабры или пучки нитевидных
жабр на околоанальных пластинках, увеличивается участие в газооб-
мене переднего отдела ректума, но без образования трахейных жабр.
Состав. Три современник надсемейства - Coenagrionoidea,
Lestoidea и Calopterygoidea. Два первых представлены в иско-
паемом состоянии, начиная с поздней юры, третье - с палеоцена.
С ра внен и е. Calopterygina отличаются от остальных трех
подотрядов гораздо резче, чем последние между собой. Основные от-
личия касаются формы головы и положения глаз, типа редукции ин-
терплеврального шва, отсутствия ушек и вырезки на задних крыльях,
а также природы нижних каудальных отростков у самцов. Только у
Calopterygina пенис является производным лигулы, а задние крю-
чочки копулятивного аппарата сильно редуцированы. Уникальная осо-
бенность подотряда - отсутствие бранхиальной камеры у личинок и
наружнье жабры разных типов.
По жилкованию и форме крыла наиболее далеки от Libellulina,
обнаруживая некоторое поверхностное сходство с более узкокрылыми
гетерофлебииновыми и эпиофлебиями. По-видимому, это сходство воз-
никло параллельно и независимо при сужении оснований крыльев.
50
ИСТОРИЧЕСКОЕ РАЗВИТИЕ ОТРЯДА MEGANEURIDA
В отряде примитивными считаются гигантские стрекозы Mega—
neurina из-за отсутствия таких важных структур стрекозиного кры-
ла, как узелок и птеростигма. Они раньше других стрекоз появляются
в истории развития насекомых в конце раннего карбона. Основное на-
правление эволюции меганевриновых - увеличение размеров, хотя са-
мые древние и примитивные Их представители из семейства Eugero—
pteridae были невелики - длина крыльев 4-5 см /Riek, Kuka—
lova—Peck, 1984/.
В ранней перми среди стрекоз отряда появились настоящие гиган-
ты: у Meganeuropsis размах крыльев превышал 70 см. Это са-
мые крупные из всех известных насекомых. Отсутствие узелка и пте-
ростигмы указывает на меньшее совершенство их полета в сравнении
с другими стрекозами. Узкое и длинное крыло с острой вершиной счи-
тается характерным признаком планера. Такой важный фактор, в стиму-
лировании развития полета, как бегство от врага /Расницын, 1969/,
для гигантских стрекоз не существовал - они были самыми крупными
и сильными воздушными хищниками палеозоя. Для полета таким круп-
ным насекомым были необходимы большие пространства, верхнего яру-
са растительности.
В позднем карбоне - ранней перми меганеврины достигли наивыс-
шего расцвета и широко расселились в тропических болотистых лесах
Еврамерии. К концу перми их разнообразие снижается и в триас пере-
ходит единственное семейство Triadotypidae.
Меганевриновые стрекозы дали начало двум группам потомков,
преимущественно пермской - Ditaxineurina + Kennedyina и
эндемичной для триаса - Triadophlebiina. Наиболее близки к ги-
гантским стрекозам дитаксиневриновые. Их ноги образуют "ловчую
корзину", основания крыльев широкие, базальный развилок RS при-
близительно в середине крыла, жилка А^^., начинается из корня кры-
ла, ветвиста, с вогнутой 2А. Несомненные апоморфии этого подотря-
да - узелок и птеростигма. Его обособление, по-видимому, связано с
совершенствованием лётных качеств крыла. Вершина его у дитакси-
невриновых становится закругленной, что можно связывать с увеличе-
нием маневренности, уменьшением элементов планирования, активиза-
цией полета. Фактор бегства от врага для дитаксиневриновых так же,
как и для более поздних стрекоз, приобрел очень важное значение.
Первые вынуждены были спасаться от гигантских стрекоз, второе - от
летающих рептилий и птиц. По совершенству крыла дитаксиневриновые
сравнимы с ширококрылыми Libellulida и, видимо, занимали сходные
экологические ниши в биоценозах палеозоя и мезо-кайнозоя.
Полного расцвета дитаксиневриновые достигли в ранней перми, ши-
роко распространились в Еврамерии, но в основном тяготели к запад-
ной окраине Ангариды - Приуралью и Кузнецкому бассейну. К концу
периода разнообразие дитаксиневриновых снизилось, ио одновременно
шло расселение подотряда на континенты Южного полушария в соот-
ветствии с общим оживлением связей между биотами северных и юж-
ных континентов. В послепермских отложениях дитаксиневриновые не
обнаружены.
51
Одновременно с дитаксиневриновыми в ранней перми появились
их стебепьчатокрылые потомки - подотряд Kennedyina (фототабл. Ц}
фиг. 1, 2). Преемственность между ними очевидна. Некоторые осо-
бенности в строении узелка, ветвлении главных продольных жилок яв-
но унаследованы кеннедииновыми от предка. Другие, например прокси-
мальное смешение N и развилка RS, узкое дискоидальное поле, зна-
чительная редукция анальных жилок, коррелируют с сужением основа-
ний крыльев до стебельчатых.
В экологическом аспекте начало эволюции кеннедииновых можно
связывать с освоением более укрытых, чем у предка, стаций и пере-
ходом к питанию сидящими на растениях насекомыми. Необходимость
перемещения в тесных пространствах вызывала некоторое мельчание
стрекоз и предъявляла новые требования к свойствам полета. Крыло
стебельчатого типа, как известно, позволяет выполнять маневр оп-
тимальным способом, без поворота тела. Еще одно достоинство сте-
бельчатых крыльев - компактное складывание. Оно позволяет исполь-
зовать для отдыха и ночевки более защищенные укрытия. Адаптивная
ценность стебельчатых крыльев подтверждается независимым появле-
нием стебельчатокрылых в неродственных группах стрекоз.
Другая особенность кеннедииновых уникальна и не имеет анало-
гий среди стрекоз: их ноги не образуют "ловчей корзины" и не при-
годны для охоты й воздухе. Вместе с переходом к жизни в зарослях
изменился и объект питания. По-видимому, кеннедииновые питались
малоподвижными сосущими филлофагами, характерными для пермских
о риктоценозо в.
Раннепермские кеннедииновые были несравненно многочисленнее
и разнообразнее меганевридовых с широким основанием крыла. Основ-
ной расцвет подотряда приходится на вторую половину перми. Кенне—
дииновые из Meganeurida исчезли последними: небольшое мезозой-
ское семейство Protomyrmelepntidae просуществовало до ранне-
го мела включительно.
Во второй половине перми, с появлением семейства Triadoty—
pidae, среди Meganeurida наметилась вторая эволюционная линия,
развитие которой привело к появлению еще одного подотряда стрекоз
со стебельчатыми крыльями - Triadophlebiina(фототабл.II,фиг. 3,
фототабл. 1П, фиг. 1-3). Экологически они были близки к Kennedyina,
но в отличие от последних питались малоподвижными, но не мелкими
насекомыми, на что указывают крупные размеры большинства пред-
ставителей подотряда, не намного уступающих в этом отношении сво-
им предкам - триадотипидам. По разнообразию триадофлебииновые
сравнимы с кеннедииновыми, но отличались от них локальностью рас-
пространения и недолговечностью, будучи эндемиками среднеазиатской
провинции во второй половине триаса.
Отраженная палеонтологической летописью последовательность по-
явлений и расцветов основных ветвей отряда Meganeurida совпа-
дает с эволюционной. Возможно,' это происходит потому, что конти-
ненты Северного полушария, Еврамерия и Ангарида, лучше всего оха-
рактеризованные палеонтологически, были областью первичного формо-
образования и расселения этого отряда.
52
ИСТОРИЧЕСКОЕ РАЗВИТИЕ ОТРЯДА LIBELLULIDA
Принятая здесь схема филогенетических отношений подотрядов
Libellulida во многом соответствует схеме, предложенной
А.В. Мартыновым /1938/, но при этом его группировки Permodo—
nata (= Ditaxineurina + Kennedyina) и Neodonata (=Li—
bellulida) не принимаются за сестринские. Кроме того, в свете по-
следних палеонтологических данных выделяемый обычно подотряд Ani—
sozygoptera - таксон полифилетический.
Обособленность по жилкованию крыла мезозойских Anisozygo—
ptera и Epiphlebia ранее отмечалась Карпентером / Carpen-
ter, 1931/. Однако отдельных названий для них не предлагалось, и
оба таксона до сих пор фигурируют в специальной литературе как
Ani so zygo ptera.
Приступая к обсуждению родственных отношений подотрядов Li—
bellulida,попытаемся оценить полярность характерных признаков. Пле-
зиоморфиями представляются следующие особенности: крыло с широким
основанием и полным набором главных продольных жилок, включающим
рудименты МР и Сиа; шарообразная голова без латерального смеще-
ния сложных глаз; полный интерплевральный шов; наличие ушек и кор-
релирующей с ними вырезки на задних крыльях самцов; пенис, обра-
зованный тремя элементами второго брюшного сегмента - лигулой и
задними крючочками; водное дыхание личинки, осуществляемое только
дыхательным эпителием ректума, без образования трахейных жабр;
каудальные отростки личинки - простые трехгранные выросты. С не-
которым сомнением к числу плезиоморфий можно отнести непарный
нижний каудальный отросток самцов, морфологически эпипрокт.
Представление о плезиоморфности комплекса взаимосвязанных
признаков ушки + угловатое крыло основано на общей картине его
распространения среди членов отряда. Он кажется архаичным, так как
всегда встречается совместно с другими архаичными особенностями.
Напротив, в различных концевых ветвях, в группах так или иначе
сильно отклоняющихся от типичного состояния, ушки и угловатость
задних крыльев исчезают. Таковы, например, все Calopterygina.
Среди гетерофлебииновых, судя по имеющимся материалам, углова-
тость задних крыльев исчезает только в аберрантном семействе 'Гаг—
sophlebiidae и только у более молодых позднеюрских и раннеме-
ловых представителей, тогда как у раннеюрского рода Myopophle-
bia заднее крыло с вырезкой. Нет ушек у эпиофлебий, но слабая
угловатость крыльев сохранилась. Среди Libellulina ушек и угло-
ватых крыльев нет: у Libellulidae, некоторых высших Aesch—
nidae, а также у вымерших мезозойских Aeschnidiidae, харак-
теризующихся резким вторичным расширением крыльев и крайней спе-
циализацией жилкования.
Большинство плезиоморфий, по-видимому, одновременно встреча-
ются у либеллюлиновых. К числу специализаций имаго относится не-
полный интерплевральный шов и членистый пенис - производное
третьего стернита, а у личинок - высокоэффективный внутренний ор-
ган водного дыхания - бранхиальная камера.
53
Естественную монофилетическую группировку образуют подотряды
Libellulina, Heterophlebiina и Epiophlebiina. Их объе-
диняют следующие синапоморфии: наличие бранхиальной камеры у ли-
чинок, редукция интерплеврального шва выше метастигмы и специали-
зированный эпипрокт у самца. Правда, выдержанность последних двух
признаков у гетерофлебииновых не бесспорна. У представителей раз-
личных семейств гетерофлебииновых, изученных нами, верхняя часть
интерплеврального шва всегда редуцирована. В то же время у двух
экземпляров Anisozygoptera, неясного систематического положения
из лейаса Англии, обнаружен полный интерплевральный шов /Asahi—
па, 1957/.
Непарный нижний каудальный отросток имели самцы Liasso-
phlebiidae /Tillyard, 19 25/, а также, по наблюдениям автора,
Isophlebiidae и Tarsophlebiidae. Хаген / Hagen, 1866/,
напротив, изобразил изофлебиид и тарэофлебиид из Зольнгофена с пар-
ными нижними каудальными отростками. Подобные морфологические
различия представителей одного и того же семейства кажутся мало-
вероятными, и точность рисунков эольнгофенских стрекоз вызывает
сомнение.
На фоне этих специализаций обсуждаемая группа подотрядов со-
храняет ряд примитивных черт: шаровидную голову без латерального
смещения глаз, относительно широкое крыло и полное жилкование,
ушки и вырезку на заднем крыле. Присутствие вырезки у Epiophle—
bia, не имеющего ушек, можно считать унаследованным от предка,
обладавшего последней структурой. Здесь мы исходим из предположе-
ния, что вырезка возникла для предохранения крыла от травм при
контакте с ушками.
Наиболее обособленное положение в отряде занимают Caloptery—
gina. Они существенно отклоняются от исходного типа по форме го-
ловы, по форме и жилкованию крыла, отсутствию ушек и вырезки на
крыльях. Кроме того, к числу апоморфий относятся наружные трахей-
ные жабры личинок и, по всей вероятности, редукция задний крючоч-
ков копулятивного аппарата. Единственная несомненная плезиоморфия
подотряда - полный интерплевральный шов, развитый у калоптеригои-
дов. Не исключено, что отсутствие у некоторых калоптеригиновых ниж-
них каудальных отростков тоже примитивная особенность.
В фундаментальном исследовании, посвященном сравнительной и
функциональной морфологии копулятивного аппарата /Pfau, 1971/,
состояние этого органа у Zygoptera оценивается как примитив-
нейшее, исходное для более продвинутых Anisozygoptera и Aniso—
ptera. Считается, что первичным пенисом у стрекоз была лигула,
сериальный гомолог каудальных отростков типичных брюшных стерни-
тов /Schmidt, 1915; Pfau, 1971/. Но зигоДтерный пенис хотя и
является производным лигулы, все же слишком сложен морфологиче-
ски и функционально для начального звена морфологического ряда. При
этом оказываются необъяснимыми наличие примитивной длинной, про-
никающей в вагину, лигулы yAnisoptera и полная ее редукция,
впервые отмеченная X. Пфау у Epiophlebia. Кроме того, представ-
ление о примитивности Zygoptera плохо согласуется с отсутствием
54
именно у них каудальных отростков типичных стернитов, тогда как у
Epioplebia, Anisopteran мезозойских Anisozygoptera (неопуб-
ликованные данные автора) они развиты.
Можно предполагать, что никакие из известных стрекоз не сохра-
нили вполне примитивного копулятивного аппарата. Скорее всего, в
начале эволюции копулятивного процесса пенис состоял из лигулы и
длинных задних крючочков, вместе составлявших желобок для семен-
ной жидкости. Из этого исходного типа гораздо легче выводятся все
основные типы копулятивного аппарата, в том числе и зигоптерный.
Картина распределения плезио- и апоморфий позволяет восстано-
вить последовательность основных ветвлений родословного древа от-
ряда. Неизвестный сейчас общий предок Libellulida, вероятно,
морфологически и экологически был подобен Libellulina, отлича-
ясь от них большей примитивностью копулятивного аппарата и отсут-
ствием трахейных жабр у личинок, которые тогда еще не были посто-
янными жителями водоемов.
При первой дивергенции от исходной группы ответвились общий
предок Libellulina + Heterophlebiina + Epiophlebiina и об-
щий предок Calopterygina. Первая группа меньше отклонилась
от исходного состояния, подобно предку продолжая осваивать откры-
тые пространства. Жизненная форма имаго не изменилась, лишь совер-
шенствовались лётные качества крыла с широким основанием и бога-
тым набором жилок. Возможно, тогда же началась редукция интерпле-
врального шва - отражение возрастающего функционального единства
птероторакса. Копулятивный аппарат на этой стадии эволюции сохра-
нял примитивное состояние, но, вероятно, усовершенствовался способ
фиксации самца на теле самки, и к церкам, охватывающим передне-
спинку, добавился помещаемый на голове самки специализированный
эпипрокт.
Личинки прогрессировали сильнее, чем имаго. В триасе, когда
континентальные водоемы заселились целым комплексом водных насе-
комых, постоянными жителями их стали и личинки стрекоз. При этом
первостепенное значение приобрела проблема водного дыхания. В об-
суждаемой группе стрекоз на основе преадаптации кишечного эпитё—
лия к газообмену в переднем отделе ректума сформировался высоко-
эффективный орган водного дыхания - бранхиальная камера. Первич-
ная функция анальной пирамиды, по-видимому, - фильтрация воды, по-
ступающей в бранхиальную камеру.
Предки калоптеригиновых гораздо дальше отклонились от исход-
ного состояния. Крылья их сузились и стали стебельчатыми, исчезли
различия в форме обеих пар крыльев, а жилкование в большинстве так-
сонов частично редуцировалось. В экологическом аспекте начальную
историю калоптеригиновых, как и стебельчатокрылых меганевридовых,
можно связывать с заселением зарослей растений. Несомненные пре-
имущества стебельчатых крыльев при полете в тесных пространствах
и использовании защищенных укрытий уже отмечались при обсуждении
кеннедииновых.
В отличие от своих пермских аналогов стебельчатокрылые Libel—
lulida унаследовали от предка "ловчую корзину", а с нею и способ-
55
ность схватывать насекомых не только на субстрате, но и в воздухе.
Возможно, латеральное смещение глаз, из-за которого голова стала
поперечной, как-то связано со спецификой новых местообитаний. Ко-
пулятивный аппарат усовершенствовался. Лигула путем дополнительной
дифференциации превратилась в пенис. Ее апикальный отдел приобрел
способность заякориваться в вагине, а на поверхности образовался
канал для спермы, вследствие чего задние крючочки утратили свое зна-
чение и уменьшились, сохранив только сенсорную функцию. Не исклю-
чено, что ранняя специализация копулятивного аппарата - одна из при-
чин редукции ушек у калоптеригиновых. Возможно, эта структура име-
ет какое-то, не вполне ясное отношение к копуляции /Carle, 1982b/.
Водное дыхание личинок совершенствовалось иначе, чем в преды-
дущей группе. Частично газообмен осуществлялся, как у предка, ды-
хательным эпителием ректума, но теперь его дополняли наружные
пластинчатые жабры - производные эпи- и парапроктов. Позже, при
колонизации текучих вод, первичные жабры видоизменялись, утрачивая
способность к газообмену, и их заменяли наружные жабры иной
природы.
Первые калоптеригиновые сохраняли полный интерплевральный шов.
В более поздней истории он частью редуцируется, но в отличие от
предыдущих подотрядов исчезает его нижняя и средняя часть.
Следующая дивергенция поделила ствол Libellulina + Hetero—
phlebiina на общего предка первых двух подотрядов и предка Epio-
phlebiina. Первая ветвь сохранила широкие основания крыльев и
полный набор жилок. Личинки по-прежнему заселяли стоячие воды кон-
тинентов.
Наиболее важная синапоморфия, возникшая в этом стволе, касает-
ся копулятивного аппарата. Из орального отдела семенной капсулы
развился членистый пенис, а лигула и задние крючочки сохранились,
изменив функцию (подробнее см. / Pfau, 1971/).
Обособление эпиофлебииновых, несомненно, было обусловлено пе-
реходом к развитию в текучих водах. Такое допущение хорошо объяс-
няет и крайнюю реобионтность единственного современного рода, и от-
сутствие ископаемых представителей этого явно древнего подотряда.
Крылья предков эпиофлебий сузились до образования короткого
стебелька, число продольных жилок сократилось. Копулятивный аппа-
рат приобрел уникальные черты: лигула исчезла, а гипертрофирован-
ные задние крючочки образовали пенис. Таким образом, хотя копуля-
тивный процесс у эпиофлебий кажется несовершенным' /Pfau, 1971/,
структуры второго стернита у них дальше от примитивного типа, чем
в других подотрядах.
Самая молодая ветвь отряда - подотряд Heterophlebiina.
Эти стрекозы явно превосходят либеллюлиновых по общему уровню
организации, оказываются во всех отношениях эволюционно более про-
двинутыми и потому могут считаться их прямыми потомками. Личин-
ки гетерофлебииновых в большей степени водные животные, чем ли-
чинки либеллюлиновых, потому что имеют больше адаптаций к водно-
му образу жизни, обладая одновременно органом водного дыхания -
бранхиальной камерой и органом водной локомоции - хвостовым плав-
56
ником. Вероятно, и обособление подотряда, и его ранний расцвет за-
висели от успеха в заселении континентальных водоемов, которые в
мезозое имели свою специфику.
Другая специализация гетерофлебииновых состояла в сужении ос-
нований крыльев, которое повлекло за собой некоторую редукцию жил-
кования, вначале несущественную. Примитивнейшие из них, раннеюр-
ские гетерофлебииды, отличались от либеллюлиновых главным обра-
зом неразделенной de передних крыльев. Позже в некоторых семей-
ствах редукция жилкования независимо усиливается. Еще одна прогрес-
сивная черта гетерофлебииновых - способность складывать крылья, да-
же при относительно широких их основаниях. Адаптивная ценность это-
го свойства вполне очевидна. Судя по имеющимся ископаемым мате-
риалам, положение крыльев у гетерофлебииновых, даже у особей одно-
го вида, в покое могло быть произвольным: распростертым, полурас-
крытым или компактно сложенным.
Обратимся теперь к геологической истории. Первые Libellulida
известны по немногим находкам в триасовых отложениях Южного по-
лушария: в Австралии, Южной Африке и Америке, где Meganeurida
практически уже вымерли. В фаунах Евразии этого времени преобла-
дали Meganeurida, сильно изменившиеся, но еще многочисленные и
разнообразные. Libellulida здесь исключительно редки и на фоне
коренного населения этих территорий кажутся пришельцами.
Картина резкой географической дифференциации Триасовых фаун
отражает, по всей вероятности, положение древних генетических оча-
гов отрядов. Однако мезо-кайнозойская история стрекоз так же, как
и палеозойская, известна почти исключительно по находкам в Север-
ном полушарии. Поэтому можно предполагать, что палеонтологическая
летопись отражает не столько последовательность ветвлений родослов-
ного древа Libellulida, сколько последовательность заселения ими Евра-
зии. Кроме того, известно, что величина ареала связана со скоростью
расселения. Расселительные потенции зависят от таких свойств, кото-
рые- не всегда доступны для изучения у ископаемых. В частности, для
мигрантных современных стрекоз отмечаются сложные поведенческие
и физиологические адаптации /Corbet, 196 2/.
Стратиграфическое описание последовательности развития подот-
рядов с реконструированным по данным сравнительной морфологии не
совпадает, что объясняется искажением летописи реального хода со-
бытий. Первыми в позднем триасе появились Heterophlebiina,
за ними в ранней юре - Libellulina и только в поздней юре - пер-
вые несомненные Calopterygina. Ключ к пониманию неожиданно
раннего появления и расцвета гетерофлебииновых, историческая судьба
которых отчасти напоминает динозавров, дает своеобразная экология
их личинок.
Основу комплекса водных насекомых мезофита составляли подвиж-
ные, плавающие формы /Калугина, 1980/. Есть основания предпола-
гать, что в мезозое, как и сейчас, важную роль в питании стрекоз
играли водные двукрылые. Поэтому особый интерес представляют ука-
зания на подвижный, плавающий образ жизни личинок юрских и мело-
вых водных двукрылых (см. /Калугина, 1974, 1986; Калугина, Ко-
57
вапев, 1985/). Учитывая это обстоятельство, можно предполагать,
что подвижные, активно плавающие личинки гетерофлебииновых с са-
мого начала получили огромное адаптивное преимущество перед бен-
тосными личинками либеллюлиновых и калоптеригиновых и-успешно
колонизировали пресные воды континентов на обширных территориях.
Уместно будет вспомнить также, что основной тип насекомоносных от-
ложений - осадочные толщи больших озер. Причем в мезозое озерная
литораль не имела зарослевой зоны погруженных цветковых растений,
аналогичной современной. В таких условиях способность к плаванию
должна была быть особенно полезной, помогая активно разыскивать
благоприятные стации, хорошо прогреваемые и аэрируемые, защищен-
ные от волн и богатые кормом. Действительно, личинки гетерофлеби-
иновых обычны в озерных ориктоценозах мезозоя, особенно юрских.
Быстрое развитие подотряда началось не позднее второй полови-
ны триаса, но распространение его в то время ограничивалось конти-
нентами Южного полушария. С наступлением раннеюрской эпохи, ког-
да начавшаяся в триасе гумидизация достигла максимума и на обшир-
ных территориях установился влажный и жаркий климат, гетерофле-
бииновые достигли вершины своей эволюции. Они заняли господствую-
щее положение среди стрекоз Евразии, определяя весь облик фауны
раннего мезофита. В то время существовали 8 из 10 семейств под-
отряда. Судя по разнообразию морфологических форм имаго, больше-
му, чем у остальных Libellulida, вместе взятых, важным направ- •
пением эволюции гетерофлебииновых было увеличение экологического
спектра взрослых стрекоз,
С началом позднеюрской аридизации гетерофлебииновые приходят
в упадок, уступая господствующее положение либеллюлиновым. В Во-
сточной Азии они просуществовали до раннего мела включительно,
тогда как на западе Евразии, по всей вероятности, вымерли раньше,
до наступления мелового периода.
Libellulina ( Anisoptera ) появились в ранней юре на
западе Евразии и до начала поздней оставались малозаметными в фау-
нах стрекоз. В поздней юре и неокоме произошло расселение подотря-
да на восток, а участие в фаунах заметно возросло. Ветвление родо-
словного древа либеллюлиновых на уровне семейств завершилось, по-
видимому, уже в поздней юре. Самое молодое семейство подотряда -
Libellulidae - найдено в берриасовых отложениях Испании /Whal-
ley, Jarzembowsky, 1985/. Тогда же подотряд достиг своего на-
ивысшего расцвета, включая, кроме современных, еще пять вымерших
семейств. Позже его разнообразие идет на убыль и во второй полови-
не мела состав подотряда на семейственном уровне почти соответству-
ет современному. Только одно из мезозойских семейств - Aeschni-
diidae — продержалось на тихоокеанском побережье Азии до начала
позднего мела (сеноман Приохотья).
Позднее появление в разрезе калоптеригиновых, не соответствую-
щее представлениям об их эволюционной древности, вероятно, обуслов-
лено тем, что область их первичного формообразования и расселения
находилась за пределами Евразии и долго оставалась ограниченной. В
Евразию они проникли только в конце юры, но уже на довольно позд-
58
ней стадии развития, когда не только существовали, но и далеко ди-
вергировали их основные ветви - надсемейства Coenagrionoidea
и Lestoidea. К этому времени должны были бы появиться и калоп-
теригоиды, но их история, как и других реофилов, вообще плохо отра-
жена в летописи. Древнейшая находка калоптеригоидов датируется па-
леоценом, но, судя по высокому уровню специализации первых пред-
ставителей надсемейства, нет никаких оснований связывать момент их
появления с возрастом находки.
Суммарный объем коллекции раннемеловых стрекоз Евразии очень
велик, но среди них нет калоптеригиновых, а находки позднемеловых
единичны. Только с палеогена удельный вес подотряда в фаунах стре-
коз становится значительным. Не только появление калоптеригиновых
в Евразии отделено значительным временным промежутком от предпо-
лагаемого момента возникновения, но и расцвет их начался с запозда-
нием более чем на период относительно первой находки. Может быть
причины этого явления, кроме предполагаемой удаленности первичного
фауногенетического очага от палеонтологически изученных территорий,
заключались в низкой скорости расселения древних калоптеригиновых.
ПУТИ ФОРМИРОВАНИЯ БОРЕАЛЬНОЙ ФАУНЫ
В отечественной одонатологической литературе значительное вни-
мание уделено проблемам географического распространения и генези-
са фауны стрекоз. Накопление фаунистического материала позволило
осуществить зоогеографический анализ региональных и мировой фаун
стрекоз. По этим вопросам имеется большое количество публикаций,
в том числе обобщающих сводок по географии и истории одонатофауны
/Белышев, Харитонов, 1981, 1983а/. Поэтому ниже будут даны лишь
некоторые основные положения представлений о формировании бореаль-
ной фауны и критические замечания в их адрес.
В распространении стрекоз имеются существенные отличия от мно-
гих привычных хрестоматийных положений зоогеографии. Географиче-
ское районирование на основании распространения стрекоз в отдельных
деталях больше соответствует схемам геоботаников, чем многих зоо-
географов. Такое положение складывается в результате сочетания осо-
бенностей, свойственных этому отряду насекомых - древности проис-
хождения, амфибионтному и хищному образу жизни, развитию в интра-
зональных водных биотопах, большим возможностям при расселении.
Своеобразие распространения и наличие палеонтологических материалов
способствуют реконструкции истории фауны стрекоз.
Данные географии стрекоз позволяют разделить мировую сушу на
два фаунистических царства - Бореальное и Меридиональное, террито-
риально близко совпадающие с бывшими суперматериками Лавразией'
и Гондваной, которые были разделены древним морем Тетис (рис. 3).
Бореальное царство по занимаемой площади сходно с Голарктической
областью в понимании большинства зоогеографов, но не подменяет его,
так как имеет иную структуру. В частности, оно не может быть разде-
лено на Пале— и Неоарктическую подобласти, поскольку, например, Ка-
59
05
О
Рис. 3. Зоогеографическое районирование мировой суши на основании распространения стрекоз.
нада по составу одонатофауны резче отличается от юга Северной Аме-
рики, чем от Сибири. Неприемлема в традиционном объеме и Палеарк-
тическая подобласть, объединяющая слишком разнородные фауны Сиби-
ри, Средиземноморья, Приамурья. Весь юг этой совершенно искусст-
венной с одонатогеографической точки зрения подобласти целесообраз-
но изменить в ранге и выделить в качестве Субголарктической обла-
сти, неоднотипной по составу на востоке и западе, но структурно од-
нородной. Вся остальная территория Северного полушария объединяет-
ся в Голарктическую фаунистическую область, которая включает более
или менее близкие и одновозрастные фауны.
Фауна Бореального царства сложилась на территории Лавразии в
относительной изоляции от Гондваны, в пределах которой сформирова-
лась фауна Меридионального царства. При этом южная полоса Боре-
ального царства - Сонорская область на американском континенте и
Субголарктическая на евразиатском - представлена богатой и более
древней одонатофауной, исходной для Северного полушария в целом.
Более обширная северная полоса Бореального царства - Голарктиче-
ская область - отличается значительно более бедной и молодой по
времени формирования фауной.
Особое значение в формировании бореальной фауны имеет выясне-
ние возможных миграционных путей, зависящих, в частности, от па-
леогеографической обстановки соответствующих исторических периодов.
Однотипность голарктической фауны свидетельствует об очень тес-
ных контактах и исходном единстве американской и евразиатской одо—
натофаун. Сходство северной биоты в целом и наличие у некоторых
групп животных и растений так называемых берингийских ареалов
привели к признанию многими биогеографами Берингии как моста для
обмена фаунами между Северной Америкой и Азией и даже центра
формообразования голарктической фауны и флоры. Вероятно, для ряда
групп, прежде всего субарктических животных и растений, эти пред-
ставления вполне обоснованны, но далеко не для всех. Палеогеогра-
фия Берингии во многом дискуссионна, хотя она в определенные пе-
риоды представляла собой обширную сушу, климат в этом районе по
большинству реконструкций с конца палеогена был крайне суровым.
Это явление обусловлено глобальной дифференциацией климата в кай-
нозое и смешением климатических эон, связанном с перемешением Се-
верного полюса по приберенгийской территории к его современному
местонахождению.
Очевидно, Берингия послужила мостом-фильтром, пропускавшим од-
них животных и задерживающим других, более теплолюбивых, в том
числе стрекоз. Действительно, в современном распространении стрекоз
нет ни одного факта, подтверждающего их берингийские трансконтинен-
тальные миграции, но есть такие, которые свидетельствуют в пользу
североатлантических фаунистических связей двух континентов по былой
климатически благоприятной суше, называемой иногда Арктидой и сое-
динявшей Северную Америку, Гренландию, Шпицберген, Северные Евро-
пу и Азию в единое целое. Множество палеогеографических и биогео-
графических данных подтверждают важную роль этого канала транскон-
тинентальной связи фаун и флор.
61
Изоляция Тетисом и объединение Арктидой обусловливают основной
и наиболее древний тип формирования и принципиальное единство бо-
реальной одонатофауны. Дальнейшее ее формирование произошло на
фоне дифференциации и колебаний климата, антропогеновых оледенений
и морских трансгрессий, перекрывавших крупные регионы евразиатской
суши и приводивших к дизъюнкциям ареалов.
В пределах Евразии наиболее древние дизъюнкции ареалов, просле-
живающиеся в современной фауне, возникли еще в палеогене. К ним, в
частности, относятся значительные долготные разобщения ареалов ро-
дов Pyrrhosoma и Boyeria. Виды первого из них встречаются в
Западной Европе (Р. nymphula) и Южном Китае (Р. tinctipennis),
второго - в Западной Европе (В. irene) и Японии (В. maclach—
lani ). Представители рода Boyeria (В. grafiana и В. vinosa)
обитают и в Северной Америке, где они распространены широко. Ве-
роятно, разрыв ареалов этих родов начался еще в палеогене под воз-
действием обширных морских трансгрессий и развития аридности и
повышения континентальности климата в результате поднятия Цент-
ральной Азии.
Очень своеобразен дизъюнктивный ареал рода Epiophlebia,
который относится к процветавшему в мезозое реликтовому подотря-
ду Anisozygoptera, представленному в настоящее время единст-
венным родом с двумя видами: Е. superstes, обитающим в Японии,
и Е. laidlawi, известным из Гималаев. Интересно, что оба вида
этих живых ископаемых сохранились в горах и отличаются исключи-
тельной реобионтностью, обитая в холодных горных потоках с относи-
тельно стабильными гидрологическими и биотическими условиями.
Наиболее большой разрыв видового ареала наблюдается у Сое-
nagrion hylas - сибирского вида, имеющего изолированный локаль-
ный ареал в Альпах. Очевидно, это ледниковое разобщение и остаточ-
ный европейский участок ареала имеют реликтовый характер. Вообще
ледниковое время наложило глубокий отпечаток на формирование ре-
цептной евразиатской одонатофауны. Резкие похолодания и затопления
в течение этого времени уничтожили аборигенную фауну Западной Си-
бири. По мнению большинства палеогеографов, на протяжении большей
части плейстоцена на юго-западе Западно-Сибирской равнины существо-
вал обширный слабопроточный водный бассейн, возникший вследствие
ледниковой запруды рек бассейна Оби. Уровень бассейна поднимался
до 100 м и более, в результате чего происходил сток вод в бассейны
Тургая и Арала в виде широкого пролива, полностью изолировавшего
европейско—уральские и сибирские части ареалов стрекоз.
Восстановлению фауны в периоды регрессий водного бассейна и
после его исчезновения препятствовали климатические колебания. В
частности, в позднее постгляциальное время этот процесс был невоз-
можен вследствие развития ксеротермической климатической фазы, во
время которой произошло остепнение территории, резкое сокращение
количества пресноводных водоемов, т.е. мест обитания стрекоз, и
сильная минерализация оставшихся. Так, по мере удаления от горной
части Урала к Васюганью, где в это время развились лесостепные
пространства, резко возрастала засоленность озер. Даже в отложени-
62
ях глубоких, тектонического происхождения предгорных озер появились
диатомовые водоросли, характерные для солоноватых вод /Благове-
щенский, 1943/. Еще сильнее процесс осолонения развивался в бо-
лее южных районах, что создавало неблагоприятные экологические ус-
ловия для существования стрекоз.
Восстановление одонатофауны на этой территории стало возмож-
ным сравнительно недавно и происходило преимущественно за счет
миграций с запада, о чем свидетельствует современная структура
ареалов и принадлежность экземпляров западно-сибирских стрекоз к
европейским подвидам. При этом у ряда видов так и не произошло
полного восстановления ареалов, и на пространстве между Приураль-
ем и Приобьем они не обнаруживаются совсем или прерывисто распро-
странены в основном по долинам рек Тобол, Ишим и Иртыш.
Одонатофауна Восточной Европы испытала, возможно, не столь
катастрофическое, как в Западной Сибири, но все-таки сильное влия-
ние похолоданий и затоплений ледникового времени. Если равнины не-
однократно перекрывались ледниковыми водами, то горные территории
оказывались в более благоприятных условиях, и населявшая их фауна
могла хотя бы частично сохраниться с доледникового времени. Об
этом свидетельствует большее по сравнению с прилегающими равнина-
ми видовое разнообразие и наличие реликтовых видов стрекоз на Ал-
тае, Урале и в Карпатах. Например, в горах Алтая и возвышенных
приалтайских землях сохранились реликтовые изолированные местона-
хождения видов Libellula depressa, Anax parthenope, Sympet—
rum croceolum. В примыкающих к Алтаю ленточных борах бассейна
Оби (также, кстати, сохранившихся с неогена) сосредоточены популя-
ции видов Leucorrhinia pectoralis, L. albifrons, L. cauda-
lis, Somatochlora flavomaculata и некоторых других, основ-
ные ареалы которых расположены в европейской части континента.
Сходная ситуация наблюдается на Урале. Так, в предгорьях и го-
рах Южного Урала насчитывается 53 вида стрекоз в отличие от 35
на сопредельных равнинных территориях, причем уральские популяции
ряда из этих видов изолированы от основных ареалов.. К ним отно-
сятся, например, Brachytron pratense, Anax parthenope, So-
matochlora graeseri, Ischnura aralensis и некоторые другие.
Еще более заметные следы ледниковых разобщений и последую-
щей аридиэации климата обнаруживаются в широтных дизъюнкциях
ареалов. Значительное распространение среди евразиатских стрекоз
имеют ареалы бореалъно- и даже арктоальпийского типа, когда изо-
лированные участки ареалов локализуются в южных горах, где сохра-
няются популяции видов, широко распространенных на более северных
широтах. Один из наиболее ярких примеров такого разобщения - рас-
пространение стрекозы Somatochlora sahlbergi. Основной
ареал этого вида лежит в субарктической полосе Евразии и Америки,
а изолированные места нахождений известны в горах Южной Сибири
от Алтая до Прибайкалья.
В горах Таджикистана отмечено не менее 10 видов стрекоз,
среднеазиатские участки ареалов которых изолированы от основных,
более северных областей распространения. Примерно столько же юж-
63
ных горных иэолятов известно для Кавказа. Большая часть этих ви-
дов в изолированных горных очагах морфологически не отличается от
популяций из основного ареала, но в отдельных случаях, как напри-
мер у Leucorrhinia dubia, они образовали особые подвиды.
Изредка наблюдается и обратное явление - изолированность на
севере фрагментов ареалов южных видов, представляющих собой, по-
видимому, реликты ксеротермических периодов. Иногда они связаны
с горячими источниками, например Orthetrum albistylum, ос-
новной ареал которого лежит в пределах Субголарктической обла-
сти, а северные изолированные очага известны на выходах термаль-
ных вод на северо-востоке Байкала и в долине р. Чары.
Относительно мало были затронуты ледниковыми пертурбациями
в Бореальном фаунистическом царстве территории Субголарктической
области, что повлияло на сохранение здесь богатой, разнообразной и
достаточно древней одонатофауны. Так, из 19 эндемичных для царст-
ва монотипических родов 18 оказались сосредоточеными в Субгол—
арктической области и лишь 1 - в Голарктической, крупнейшей по
площади, но наиболее молодой и бедной по видовому составу фауни-
стической области Земли.
Следует отметить, что все построенные на основании распростра-
нения стрекоз принципиальные зоогеографические схемы и масштаб-
ные по времени историко-фаунистические реконструкции хорошо со-
гласуются с теорией горизонтальных перемещений материков. Учиты-
вая геологическую древность отряда стрекоз, следует признать, что
отражение в их распространении долговременных геофизических про-
цессов вполне естественно. Вместе с тем апелляции к идеям моби—
лизма в зоогеографических работах вызывают иногда критические за-
мечания. Особенно энергично высказаны они в рецензии К.Ю. Есько-
ва /1984/ на монографии по географии стрекоз /Белышев, Харито-
нов, 1981, 1983а/. К сожалению, рецензия мало касается сути мо-
нографий, явствующей уже из их названий, а именно географии стре-
коз, и сводится главным образом к критике палеогеографических ос-
нов исторических реконструкций, занимающих в этом труде подчинен-
ное положение и составляющих незначительную часть его объема. Ре-
цензент упрекает авторов в приверженности к теории Вегенера, поче-
му-то беря ее в кавычки, хотя здесь же сообщает, 'что ныне среди
биогеографов предметом обсуждения является не факт перемещения
материков (признаваемый почти всеми), а масштабы и характер его
влияния на современное распространение живых организмов". От вни-
мания рецензента, видимо, ускользнуло то обстоятельство, что авто-
ры подходят к данной теории не как к непогрешимой догме и отдают
себе отчет в том, что не все в ней бесспорно, делая, например, та-
кие замечания: "Конечно, на теорию А. Вегенера и теперь приходит-
ся смотреть как на принципиальную схему, где необходима доработка
многих деталей, уточнение частностей и во времени, и в пространст-
ве..." /Белышев, Харитонов, 1981, с. 125/. "...Следует подчеркнуть
и то обстоятельство, что классическая схема А. Вегенера не отлича-
ется точностью. Она в принципе верна, удостоверена данными геоло-
гии, палеомагнитологии, палеонтологии и биогеографии, но, конечно,
требует дальнейших доработок" /Там же, с. 128/.
64
Вместе с тем в литературе, в том числе и наиболее современ-
ной, существует столь большой и общеизвестный разнобой в датиров-
ках и оценке масштабов движения земной коры, что "все это застав-
ляет нас отдавать пока предпочтение линиям дрейфа Вегенера - Кеп-
пена, построенным в основном на палеонтологическом материале" /Гам
же, с. 125/. Казалось бы, авторская позиция оговорена достаточно
четко, тем не менее рецензент отмечает, что 'авторы используют в
конкретных построениях картину перемещения материков и полюсов,
воссозданную самим Вегенером, между тем последние десятилетия
внесли столь существенные коррективы в первоначальные представле-
ния о континентальном дрейфе (см., например, /Красилов, 1977;
Уэда, 1980/), что пользоваться реконструкциями 1924 г. по меньшей
мере странно'. Кстати, В.А. Красилов, к работе которого апеллирует
рецензент, отмечает, что 'теория Вегенера открыла новую страницу
исторической биогеографии. Гипотеза дрифта континентов органически
вошла в биогеографические построения многих исследователей... По
современным представлениям, дрифт континентов - это частный аспект
тектоники плит' /Красилов, 1977, с. 63-64/.
Принципиальную правильность построений А. Вегенера даже на фо-
не самых современных данных подчеркивают многие авторы. Так, Д.
и М. Тарлинг /1973, с. 15/ отмечают, что 'схема реконструкции
Вегенера основывалась на фактах из многих областей науки; она уди-
вительно сходна с нашими современными представлениями об эволю-
ции южных материков'. Известные советские геологи С.А. Ушаков и
Н.А. Ясаманов в капитальной сводке "Дрейф материков и климаты зем-
ли' /1984, с. 5/ подчеркивают, что «Теория тектоники литосферных
плит представляет собой современное развитие того направления науки
о Земле, которое родилось еще в начале XX в. в виде гипотезы дрей-
фа континентов. Факты, приводившиеся мобилистами 50-60 лет назад
в защиту 'сумасбродной' гипотезы, выдержали проверку временем; ны-
не эти факты получили полное подтверждение и дальнейшее развитие».
Академик А., Л. Яншин в одном из своих выступлений в прессе отме-
тил, что 'сейчас гипотеза Вегенера, получившая в ее современном ви-
де название теории движения тектонических плит, или новой глобаль-
ной тектоники, принята, с незначительными поправками, геологами все-
го мира, лишь немногие ученые упорствуют в своем с ней несогласии"
/Известия, 08. 08. 84 г./.
Критикуя выводы авторов по генезису фауны, рецензент не обра-
тил внимания на то, что они даны как сугубо предположительные по-
строения и во введении к книгам подчеркнуто, что 'в основу историко-
фаунистического анализа положено современное распространение стре-
коз, морфологическая близость таксонов и палеогеографические дан-
ные, которые опираются на теорию континентальных перемещений, ос-
новы которой были заложены А. Вегенером... В вопросах истории и
времени расселения много условностей, поскольку палеонтологический
материал почти отсутствует и мы должны опираться на косвенные дан-
ные и логические построения. Следовательно, все эти вопросы пока
находятся на уровне рабочих гипотез. Но высказать эти гипотезы мы
должны, так как это продвинет изучение вопроса независимо от того,
5 Зак. 611
65
будут они приняты в свете новых данных или отвергнуты. То и дру-
гое равноценно, так как подтолкнет к подобным исследованиям, даст
для них опорные точки' /Белышев, Харитонов, 1981, с. 4/.
В начале рецензии сообщено, что "мы посвяшаем рецензию наи-
более крупным зоогеографическим обобщениям, выдвигаемым автора-
ми', в ней в основном дан разбор частных палеогеографических во-
просов, причем далеко не во всем аргументация рецензента бесспор-
на и точна. Например, рецензент пишет, что "авторы говорят о су-
ществовании Пангеи еще в середине третичного периода... тогда как
по современным представлениям раскрытие Тетиса, отделившего Лав-
разию от Гондваны, и одновременный раскол последней на восточную
и западную части произошли уже в конце триаса - начале юры". Та-
кой упрек основан на явном недоразумении. В разделе рецензируемой
книги, посвяшенном общим вопросам формирования голарктической фау-
ны, написано: 'История Северного полушария в свете теории А. Веге-
нера рисуется следующим образом. До юры существовала общая кон-
тинентальная глыба. С этого времени она начала располагаться, ее
части раздвигались, таким образом наметился Атлантический океан"
/Там же, с. 156/. Ясно сказано, что целостность Пангеи сохранялась
до юры, но никак до середины третичного периода, применительно к
которому речь идет уже лишь о фрагментах бывшей Пангеи.
Упрекая авторов в излишней приверженности к теории Вегенера в
том, что они 'всецело полагаются" на его реконструкции, рецензент
сокрушается по поводу того, что в необходимых случаях они обраща-
ются к современной литературе, уточняющей схемы Вегенера. Так,
К.Ю. Еськов пишет, что "для обоснования сильной обособленности
одонатофауны Индии и Юго-Западной Азии они пользуются современ-
ными представлениями о северном дрифте Индостана как фрагмента
Гондваны и о его совсем недавнем контакте с Азией, что не согласу-
ется с построениями Вегенера, у которого Индия движется как часть
Евразии".
Не вдаваясь более в дискуссию по достаточно частным и далеко
выходящим за рамки биологического труда проблемам, в заключение
еще раз отметим, что все наши зоогеографические и историко-фау-
нистические выводы базируются прежде всего не на отвлеченных рас-
суждениях и не на спорных палеогеографических реконструкциях, а на
неоспоримых фактах распространения стрекоз, при этом они ни в коей
мере не претендуют на всеобщее зоогеографическое значение.
Глава II
ЭКОЛОГИЯ И ПОВЕДЕНИЕ
До 30-х годов текущего столетия вопросы экологии и поведения
стрекоз не составляли предмета специального изучения. Сведения по
аутэкологии отдельных видов’ на уровне натуралистических наблюдений
собирались попутно с фаунистическими исследованиями. Первые спе-
66
циальные работы и обобщения по экологии стрекоз, прежде всего фе-
нологии, были опубликованы А.Н. Бартеневым /19306, д, е; 1932а/
и В.Г. Колесовым /19306/. В 50 -60-х годах появилась серия пре-
красных работ по экологии стрекоз Корбета /Corbet, 1956, 1957,
1962/, в том числе результаты экспериментального изучения фотопе-
риодической регуляции жизненных циклов стрекоз, впервые полученные
для беспозвоночных животных вообще. В этот же период Б.Ф. Белышев
/1959а, 1960а-г, 1961в, 1963а, д, 1964в, 19656, 1966г, д,
19676/ выполняет исследования по фенологии, суточной активности,
биотопическому распределению стрекоз в разных природных зонах Си-
бири.
В последующие годы количество работ по экологии и этологии
стрекоз возрастает, по отдельным проблемам появляются солидные
обзоры /Рязанова, Мазохин-Поршняков, 1984/. Многообразие эколо-
гической и этологической проблематики не позволяет дать ее подроб-
ный анализ, поэтому в данную главу включены лишь отдельные вопро-
сы, приоритетные в отечественной одонатологии.
ФЕНОЛОГИЯ СТРЕКОЗ
Сезонные ритмы жизнедеятельности стрекоз входят в число важ-
нейших составляющих их экологических характеристик. Сведения о фе-
нологии стрекоз очень неполны в связи со слабой изученностью жиз-
ненных циклов этих нссекомых. Для территории СССР относительно
точные данные по жизненным циклам имеются всего лишь по 13 ви-
дам, изученным в Восточном Приуралье /Харитонов, 1977а/, Прибал-
тике /Станёните, 1966/ и в окрестностях Рыбинского водохранилища
/Таранова, 1980/. Сведения по подавляющему большинству остальных
видов фрагментарные и нередко сводятся из разных регионов воедино
/Попова, 1953а/, вследствие чего действительная картина искажается.
Существенный экологический интерес представляет общая продол-
жительность активности имаго, специфичная для каждого вида, но спо-
собная значительно изменяться в разных частях ареала. В настоящее
время периоды лёта стрекоз относительно хорошо изучены в ряде рай-
онов европейской части СССР /Дьяконов, 1922а; Колосов, 1925; Бар-
тенев, 1930д; Акрамовский, 1948; Спурис, 1956; Станёните, 1963а,
1968; Олигер, 1980; Павлюк, 19806; Таранова, 1980/, в Зауралье
и Казахстане /Харитонов, 1975ж, 19806/, в некоторых районах Си-
бири /Белышев, 1960а, б, 1963а, 19656, 1966г, д/, в Киргизии
/Чибиченко, Крылова, 1968; Крылова, 1969, 1972/ и Таджикистане
/Попова, 1951; Борисов, 1986, 1987/. Ряд обобщающих работ по-
священ анализу географической изменчивости фенологии /Бартенев,
1930ц; Белышев, 1974а; Харитонов, 19806/.
Сезонная ритмика лёта — один из механизмов адаптации стрекоз
к сезонным изменениям среды. Период активного имаго может варьи-
ровать в зависимости от погодных особенностей года. На начало лёта
в сильной мере влияет температура воздуха. Выход имаго из личинок
возможен при достижении среднесуточными температурами определен-
67
Рис. 4. Изменения среднесуточных температур воздуха в весенние пе-
риоды 1978, 1980-1982 гг. и даты начала выплода стрекоз в запо-
веднике "Тигровая балка".
Виды стрекоз: 1 — 1. evansi; 2 — 1. fountainei; 3 — A. parthe—
nope; 4 — L. quadrimaculata; 5 — P. dealbata; 6 — L. tet—
raphylla; 7 - S. nigra; 8 - C. erythraea.
ных видоспецифичных пороговых значений. Корреляция начала выплода
первых весенних видов с температурами установлена в различных при-
родных зонах умеренных широт /Белышев, 1974а; Харитонов, 19806;
Corbet, 1956, 19627. Например, на юго-западе Таджикистана вы-
ход из личинок L evansi, L fountainei и A. parthenope в
1978 г. начался на две недели позже, чем в 1981 г., но в том и
другом случае при одинаковой среднесуточной температуре около
12 °C (рис. 4). Имаго L. tetraphylla в 1978 г. появились на три
недели позже, чем в 1981 г., но в обоих случаях при одинаковой тем-
пературе - около 25 °C. L. quadrimaculata начал летать в 1981 г.
13 марта, в 1982 г. 19 марта, а в 1980 г. только с 1 апреля,
но во всех случаях при одинаковой среднесуточной температуре — око-
ло 14 °C (см. рис. 4). В Латвии отмечено, что разница между на-
чальными датами вылета первых весенних видов в разные годы может
быть больше месяца /Спурис, 195 6/. Необычайно раннее появление
имаго A. junius (4 апреля — на 1,5 мес раньше обычного), свя-
занное с теплой погодой, отмечено в Канаде /Butler et al., 1975/.
68
Фотопериодическая реакция имеет сигнальное значение для нача-
ла выплода, вероятно, в большей степени для тех видов, имаго кото-
рых начинают летать во второй половине лета при высоких устойчи-
вых температурах. Важность фотопериода в регуляции развития стре-
коз в областях с умеренным климатом предполагается и для некото-
рых видов стрекоз, летающих осенью /Procter, 1973/. Строгая
ритмичность в развитии каждого вида в умеренных широтах обуслов-
лена эндогенными ритмами, сложившимися под длительным влиянием
конкретных климатических условий. Фотопериод выступает здесь как
счетчик времени и сигнал предстоящих изменений климатических фак-
торов.
С продвижением на юг сезонная ритмика у некоторых видов на-
рушается. Так, на юго-западе Таджикистана выплод С. servilia,
I. elegans (и, вероятно, других видов) возможен зимой, в январе
/Борисов, 19866, 1987/. В данном случае главную роль играет тем-
пературный фактор воздушной среды и значение фотопериодической
реакции отодвигается на задний план.
Несомненно, что и условия развития преимагинальных фаз играют
определенную роль в регуляции выплода. А.Н. Бартенев /1910а/ и
Б.Ф. Белышев /1959в/ отмечают разные сроки лёта на степных и
таежных озерах в верховьях Енисея и Оби. В аридной зоне Таджики-
стана имаго С. servilia на проточных водоемах с более суровым
термическим режимом появляются значительно позже, чем на непро-
точных /Борисов, 11187/. Аналогичное явление отмечено в Индии
/Kumar, 1979/. В Бельгии описан зимний вылет Ае. суапеа из
теплого из-за промышленных сточных вод водоема /Hinnekint,
1972/. В последнем случае, видимо, выплоду благоприятствовала и
температура воздуха.
Время окончания лёта также определяется эндогенным ритмом,
корректируемым теми же факторами, что и его начало, - температу-
рой воздуха и фотопериодом. У весенних видов с особенно четким эн-
догенным ритмом в умеренных широтах окончание лёта происходит в
довольно стабильные по годам сроки. В пустынной зоне Таджикистана
северный вид Ъ. quadrimaculata прекращает лёт обычно в середине
мая, когда среднесуточные температуры превысят 24 °C. У видов с
продолжительным лётом репродуктивный период растянут, постоянно
происходит возобновление имаго за счет выплода новых особей, и пре-
кращение активности взрослых стрекоз связано с ухудшением погод-
ных условий. В зависимости от конкретных условий среды сроки окон-
чания их лёта могут значительно варьировать: на юге Таджикистана
у ряда видов с начала ноября до"конца декабря, т.е. с разницей поч-
ти в 2 мес.
В зависимости от календарных сроков лёта в конкретном регионе
в работах по фенологии стрекоз принято выделять сезонные группы.
Время лёта каждого вида и отдельной группы ограничивается крайни-
ми датами находок имаго. Периоды лёта многих видов, особенно на
юге, сильно растянуты, внутри них существуют большие и закономер-
ные изменения численности. Например, на юге Средней Азии многим
видам стрекоз свойственно резкое снижение численности в жаркую
69
середину лета. У ряда эвритопных бивольтинных видов с увеличением
проточности водоемов возрастает численность стрекоз второго поко-
ления и соответственно уменьшается численность первого. Виды, уни-
вольтинные в более холодных проточных водоемах, переходят на би-
вольтинный тип развития в более теплых непроточных водах. В этом
случае главными факторами, влияющими на развитие стрекоз, служат
термический режим и аэрация, сочетание которых различно в разных
водоемах и видоспецифично воздействует на личинок стрекоз. Подоб-
ное явление отмечено также в Индии /Kumar, 1972, 1979/ и Ка-
наде /Leggott, Pritchard, 1985/.
Фенология стрекоз подвержена закономерным географическим из-
менениям. Такие изменения прослеживаются в широтном, долготном
и высотном (в горах) направлениях. Наши материалы /Харитонов,
19806/ дают возможность проследить широтные изменения фенологии
стрекоз от Заполярного Урала до южных пустынь Средней Азии при-
мерно на одной долготе между 60 и 70° в.д. Сравнительно полные
данные по фенологии на этой более чем 3-тысячекилометровой тран-
секте получены в пяти точках: лесотундра - стационар Института эко-
логии растений и животных УРО АН СССР, левобережье Оби немного
севернее Полярного круга (66° с.ш.); тайга - окрестности г. Верхо-
турье, верховье р. Туры (59° с.ш.); лесостепь - окрестности горо-
дов Челябинск и Курган, реки Миасс и Тобол (55° с.ш.); полупусты-
ня - окрестности г. Кызыл-Орда, нижнее течение р. Сырдарьи (45°
с.ш.); пустыня - заповедник "Тигровая балка" на юго-западе Таджи-
кистана, верховье р. Амударьи (37° с.ш.). Все перечисленные рай-
оны находятся на абсолютной высоте не более 350 м. Общий пери-
од лёта стрекоз на этом расстоянии изменяется от круглогодичного
на юге Средней Азии до 50-55 дней в Заполярье.
В лесотундре лёт стрекоз короткий и почти одновременный у всех
видов, продолжающийся около двух месяцев: с начала июля до конца
августа. В тайге общая продолжительность активности имаго немно-
го превышает 4 мес: с конца мая до начала октября. По времени по-
явления и длительности лёта здесь выделяются пять сезонных групп.
Начало лёта летне-осенних видов накладывается на период исчезнове-
ния первых весенних, поэтому количество одновременно летающих ви-
дов в середине лета максимальное (рис. 5). В лесостепи общий пе-
риод лёта стрекоз составляет около 6 мес: с конца апреля до на-
чала октября. Здесь выделяются восемь сезонных групп /Окороков,
Харитонов, 1971/ и два основных периода лёта, как и в тайге, — ве-
сенний и летний (см. рис. 5).
В полупустыне общий период активности имаго около 7 мес: с
начала апреля по октябрь. У большинства видов лёт длительный, свы-
ше 4 мес. Сезонные изменения в фауне менее заметны. Из пяти вы-
деленных здесь сезонных групп четко обособлена только зимующая
(два вида рода Sympecma). Количество одновременно летающих
видов нарастает постепенно и не дает резкого пика в середине лета
(см. рис. 5).
На самом юге Средней Азии в пустынной зоне при благоприятных
условиях имаго некоторых видов активны круглый год /Борисов, 19866,
70
Рис. 5. Сезонные изменения количества видов стрекоз в имагиналь—
ной фазе в разных широтных зонах.
1 - пустыня; 2 - полупустыня; 3 — лесостепь; 4 — тайга; 5 —
лесотундра.
1987/. Лёт подавляющего большинства видов длительный, свыше
6 мес. Сезонная ди. реренциация одонатофауны очень слабая. Большая
часть выделенных здесь девяти сезонных групп весьма условна, за
исключением весенней и осенней, представленных каждая одним ви-
дом -L. quadrimaculata и Ае. coluber cuius соответст-
венно.
Таким образом, продолжительность лёта и характер сезонного раз-
вития стрекоз изменяются на разных широтах. Сроки появления первых
весенних видов в природе с продвижением на юг смешаются к весне
с начала июля в Заполярье до начала марта (по усредненным много-
летним данным) на юге Средней Азии. Но лёт начинается на всех ши-
ротах примерно при одинаковых среднесуточных температурах в 10—
12 °C, которые можно считать пороговыми. При этом на севере они
немного ниже (10 °C), чем на юге (11-12 °C). Сроки начала лёта
видов летней фауны также изменяются и заметно сдвигаются с про-
движением на юг к весне. Последнее противоречит выводам Б.Ф. Бе—
лышева /1974а, с. 332/, согласно которым с изменением широты
местности сроки развития весенних видов меняются, а у летне-осен-
них остаются постоянными на одном меридиане. Видимо, такое явле-
ние не закономерность, а скорее исключение из правил, обусловленное
своеобразием климатических условий северо-восточного побережья Бай-
кала и Тункинской долины, где Б.Ф. Белышев проводил исследования.
Четкая сезонная ритмика в развитии стрекоз характерна для уме-
ренных широт и нарушается в экстремальных условиях севера, а так-
же на юге из-за очень продолжительного теплого периода.
71
.Град'С.ш. у Рис. 6. Периоды лёта стрекозы L. qu—
S9 । adrimaculata в разных широтных
55 5 ---------- зонах’
1 - пустыня; 2 - полупустыня; 3 -
45- 2 ----------1 лесостепь; 4 - тайга.
57- 1'------
—,—,—,—,—। । . ।—r-r-i—г—->— Особый интерес представляет ана-
/ III V VII IX XI А,
Месяц лиз изменчивости фенологии отдельных
широко распространенных видов, напри-
мер циркуМэореального L. quadrimaculata, доходящего от лесо-
тундры до пустынных областей юга Средней .Азии. Существование ви-
да в таком большом диапазоне климатических условий возможно бла-
годаря высокой экологической пластичности, один из механизмов ко-
торой - смена сезонов активности имаго по известному правилу /Го-
родков, 1986/.
Лёт этого вида в пустынной зоне Таджикистана начинается в кон-
це I декады марта. С продвижением на север сроки появления имаго
постепенно сдвигаются в сторону лета и в таежной зоне Зауралья че-
тырехпятнистая стрекоза начинает лёт только с 1П декады мая
(рис. 6). Оптимальные условия для развития этого вида, по—видимо-
му, находятся примерно па 45° с.ш., где период его лёта наиболее
продолжительный — более 4 мес. С продвижением на север время ак-
тивности имаго сокращается до 2 мес (в таежной зоне). Интересная
картина наблюдается на юго-западе Таджикистана, где дет прекраща-
ется уже в середине мая, задолго до наступления жаркого периода,
т.е. в крайних для него термических условиях L. quadrimaculata
почти вдвое, по сравнению с оптимальным вариантом, сокращает пери-
од активности в имагинальной фазе. Таким путем этот вид приспосаб-
ливается к аридному климату, совмещая репродуктивную часть жизнен-
ного цикла с коротким весенним интервалом своего температурного
оптимума. Если в таежной зоне Зауралья в середине мая первые осо-
би только появляются в воздушной среде, то в заповеднике "Тигровая
балка" на юге Средней Азии развитие L. quadrimaculata уже за-
канчивается. В связи с этим интересно упоминание Д. Проктера /Pro-
cter, 1973/ о том, что в Канаде четырехпятнистая стрекоза отно-
сится к группе осенних видов, в то время как в умеренных широтах
Восточного полушария это один из фоновых видов весенней фауны.
Приведенный пример широтной смены сезонов активности - один
из наиболее ярких, но аналогичное явление свойственно большинству
распространенных видов. При этом в наиболее южных частях ареала
периоды их лёта могут изменяться по-разному в зависимости от эко-
логического, в первую очередь температурного, стандарта вида. Так,
у летних реофильных видов Gomphus flavipes и Calopteryxsplen-
dens периоды лёта на 37° с.ш. удлиняются на 1-1,5'мес по срав-
нению с 45° с.ш., а максимум активности так же, как и северный,
приходится на середину сезона. У1. elegans, в этом же диапазоне
широт, на юге лёт удлиняется на 2 мес и более, но с резким спадом
численности в жаркую середину лета, что достигается переходом на
72
бивольтинное развитие. Период лёта Anax partnenope в пустынной
эоне (37° с.ш.) на месяц больше, чем в полупустынной (45° с.ш.),
но максимальной интенсивности на юге он достигает только в конце
апреля - начале мая. Aeschna coluberculus в Верхнем Приобье
летает с середины июля до I декада! октября, а в заповеднике "Тиг-
ровая балка' на юге Таджикистана - с середины сентября до сере-
дины декабря. Как и у L. quadrimaculata лёт этого вида сильно
смещен в связи с 'избеганием' жаркого периода, но только в сторону
осени.
Изменение фенологии стрекоз в долготном направлении выражено
менее ярко, чем по широте, но все же прослеживается. В направлении
с запада на восток уменьшается обший период лёта стрекоз и смеша-
ются сроки появления первых весенних видов в сторону лета. В ГДР
имаго стрекоз активны 7 мес: с 20 апреля до 10 ноября / Schie—
menz, 1953/, в Латвии (расположенной немного севернее) -
6 мес; с начала мая по октябрь /Спурис, 1956/, на юге Западной
Сибири (примерно на одной широте с ГДР) - 5 мес; с 20 мая до
20 октября. В Южном Прибайкалье лёт стрекоз начинается только в
начале июня /Белышев, 1974а/. Закономерно изменяются и сроки лё-
та отдельных видов. Так, L. quadrimaculata в ГДР начинает ле-
тать в первых числах мая, на юге Западной Сибири с 20 мая, а в
Южном Прибайкалье лишь с 5 июня. Прекращается лёт во всех этих
регионах примерно в одно время - в середине августа, т.е. с продви-
жением на восток сезон активности четырехпятнистой стрекозы отод-
вигается в сторону осени в результате смещения сроков появления
имаго и уменьшается с 3,5 в европейской части до 2,5 мес в При-
байкалье.
Период лёта летнего вида Ophiogomphus serpentinus за-
метно уменьшается с продвижением на восток вследствие более позд-
него появления и более раннего исчезновения имаго: в ГДР этот вид
летает с середины июня до начала октября, в Латвии - с 20 июня до
начала сентября, на юге Западной Сибири - только с июля до начала
сентября. Имаго осеннего вида Ае. coluberculus в ГДР активны
с конца июля до начала ноября - более 3 мес, в Приобье лёт этого
вида начинается и заканчивается раньше - с середины июля до октяб-
ря - 2,5 мес.
Значительный интерес представляет выяснение особенностей фено-
логии стрекоз в горах, где даже у незначительно удаленных Друг от
друга популяций на разных абсолютных высотах наблюдаются сущест-
венные различия в сезонном развитии. До последнего времени этот
вопрос был мало изучен. Отрывочные данные имеются для Кавказа
/Бартенев, 1928в, 1930в/. В Тянь-Шане установлено, что общая про-
должительность и сезонная ритмика лёта стрекоз значительно отлича-
ются в долинах и горах /Крылова, 1969/. Квентин / Quentin,
1959/ для Западной Европы проанализировал изменение сроков лёта в
Разных частях ареалов у двух видов с бореоальпийским распространени-
ем. Наши исследования фенологии стрекоз в Таджикистане позволяют
проследить изменчивость этого явления от низменных юго-западных
73
районов (350 м над ур. м.) до высокогорья Памира (33 60-3 900 м
над ур. м.).
Особенности сезонного развития фауны стрекоз в пустынной зоне
отмечены выше: круглогодичная активность имаго при благоприятных
погодных условиях и слабая сезонная дифференциация одонатофау-
ны, за исключением двух резко отличных по срокам лёта видов - ве-
сеннего L. quadrimaculata и осеннего Ae. coluberculus.
В предгорье Гис capo-Дарваза (до 1500 м над ур. м.) общий пе-
риод лёта стрекоз при благоприятных погодных условиях составляет
около 6,5 мес: с середины апреля до конца октября. По характеру лё-
та выделяются семь групп стрекоз. Из них довольно четко обособле-
на только осенняя, представленная одним видом S. haritonovi,
имаго которого появляются в начале августа и летают до холодов -
около 3 мес. Период, в который сохраняется максимальное количест-
во одновременно летающих видов, в 2 раза короче, чем в долинах на
юго-западе Таджикистана,- около 3 мес и аналогичен лёту стрекоз
в полупустыне Южного Казахстана (рис. 7). В среднегорье Гиссаро-
Дарваза (до 2500 м над ур. м.) лёт стрекоз начинается позднее, чем
в предгорье, - в середине I декады, мая и продолжается до конца сен-
тября.
В высокогорье Памира (до 3 900 м над ур. м.) стрекозы обита-
ют только на термальных источниках, где их лёт сдвинут на вторую
половину лета - с июля до середины сентября, охватывая наиболее
теплый период. Все четыре отмеченных здесь вида летают одновремен-
но и очень короткий срок: 2—2,5 мес, что аналогично фенологии стре-
коз в Заполярье (см. рис. 7).
Таким образом, сезонная активность имаго стрекоз значительно
отличается на разных абсолютных высотах. Общая продолжительность
лёта постепенно уменьшается от круглогодичной в долинах до 2-
2,5 мес в высокогорье. Сроки появления первых весенних видов с
увеличением абсолютной высоты смешаются в сторону лета, но почти
при одинаковых среднесуточных температурах — 10—12 °C (см. рис. 5).
Исчезновение взрослых стрекоз происходит также примерно при одина-
ковых температурах 7-9 °C (рис. 8).
Главенствующую роль в регуляции лёта стрекоз в горах играют
среднесуточные температуры воздуха, так как здесь фактически при
одинаковом фотопериоде сезоны активности имаго резко отличаются на
разных абсолютных высотах. Развитие личинок стрекоз в благоприят-
ных по термическому режиму термальных источниках на Памире не
оказывает заметного влияния на время появления и исчезновения има-
го. Начало и- окончание лёта зависят здесь от благоприятных усло-
вий в воздушной среде.
Значительным высотным изменениям подвержена фенология отдель-
ных видов стрекоз, широко распространенных по высотным поясам. Вид
L. quadrimaculata в пустынной зоне (350 м над ур. м.) летает
только весной — с середины марта, когда среднесуточные температуры
достигают 13-14 °C, до середины мая, когда температуры превысят
24 °C. В предгорье (1300-1500 м над ур. м.) лёт этого вида при-
урочен к летнему периоду. Появляясь в начале мая, при такой же по-
74
Рис. 7. Сезонные изменения количества видов стрекоз в имагиналь-
ной фазе в разных высотных поясах.
1 - Вахшская долина (350 м над ур. м.); 2 - предгорье Гис-
саро-Дарваза (1300-1500 м над ур. м.); 3 - высокогорье Памира
(3360—3550 м над ур. м.).
роговой температуре, имаго летают почти все лето, так как макси-
мальные среднесуточные температуры здесь не превышают 24 °C. Ви-
ды I. pumilio и О. brunneum в долинах (350 м) имеют очень
продолжительный период лёта (март - ноябрь); в предгорье (1300-
1500 м над ур. м.), сохраняя температурные пороги появления и от-
мирания, они летают только с конца апреля до середины сентября, а
в высокогорье Памира (33 60-3550 м) - с июля до середины сентяб-
ря. Таким образом, продолжи-
тельность лёта этих видов
уменьшается с 8,5 мес в доли-
нах до 2,5 мес в высокогорье
и четко прослеживается верти-
кальная смена сезонов актив-
ности имаго.
Рис. 8. Изменения среднесу-
точных температур воздуха и
начало выплода первых весен-
них (a-в) и отмирание имаго
осенних видов (г-е).
1, а, е - Вахшская доли-
на (350 м над ур. м.); 2, б,
д - предгорье Гиссаро-Дарваза
(13 50 м над ур. м.); 3, в, г-
предгорье Памира (3350 м
над ур. м.).
75
Интересными особенностями отличается фенология типичного гор-
ного вида S. haritonovi. В предгорье он начинает летать с середи-
ны I декады августа, а в среднегорье - с середины июля, становясь
многочисленным в начале августа. В высокогорье Памира лёт начина-
ется в начале июля, максимума достигает в середине августа и пре-
кращается в середине сентября, в то время как в предгорье макси-
мум лёта приходится на конец сентября, а лёт прекращается в конце
октября, т.е. в более теплом вертикальном поясе время лёта смеща-
ется в сторону осени.
Из изложенного можно сделать следующие выводы:
1. Ритм сезонной активности стрекоз относительно постоянен в
одном биотипе, но подвержен значительным географическим и высот-
ным изменениям.
2. Широтная изменчивость фенологии аналогична высотной: с про-
движением на север и с увеличением абсолютной высоты в горах по-
степенно сокращается общий период лёта от круглогодичного в низмен-
ных южных районах Средней Азии до 2—2,5 мес в Заполярье и у верх-
ней границы распространения в южных горах. Менее четко прослежива-
ются долготные изменения сезонов активности в умеренных широтах:
с продвижением на восток общий период лёта сокращается от 7 мес в
Западной Европе до 5 мес в Средней Сибири.
3. Главным экзогенным фактором, определяющим начало и прекра-
щение лёта, а также его продолжительность, является среднесуточная
температура воздуха. Сроки появления первых весенних видов с про-
движением на север и с увеличением абсолютной высоты в горах сме-
щаются в сторону лета, но при одинаковых температурных порогах в
10-12 °C. Другим важным фактором, регулирующим фенологию стре-
коз, служит фотопериод.
4. Для широко распространенных видов стрекоз в разных частях
ареалов характерны изменения сезонов активности имаго, адекватные
градиенту оптимальных климатических условий; по мере удаления . от
географического и высотного оптимума в сторону потепления периоды
лёта сокращаются и сдвигаются на весну у весенних видов или осень
у летне-осенних видов или, наоборот, удлиняются с резким сокраще-
нием численности в жаркий период; с продвижением от оптимума в
сторону похолодания сезоны активности имаго изменяются однозначно:
сокращаются и приходятся на наиболее теплый период.
Благодаря лабильной фенологии, позволяющей выбирать оптималь-
ные условия в воздушной среде, многие виды стрекоз в умеренных
широтах Северного полушария имеют обширные ареалы, охватывающие
разные природные зоны и вертикальные пояса. Фенологические сдвиги
не требуют сложных морфологических и физиологических перестроек
/Городков, 1986/ и представляют для этих насекомых наиболее быст-
рый и прогрессивный путь адаптации к макроклимату.
76
СУТОЧНЫЕ РИТМЫ АКТИВНОСТИ СТРЕКОЗ
Изучение суточных ритмов активности стрекоз - одно из необхо-
димых условий для оценки их роли в трофических сетях биоценозов,
например, в регуляции численности гнуса или вредителей сельского и
лесного хозяйств, а также представляет интерес с точки зрения вы-
яснения адаптивных возможностей этих насекомых.
Крупные размеры и особенности поведения стрекоз делают их
удобным модельным объектом в полевых исследованиях суточных рит-
мов. Тем не менее на общем фоне публикаций по суточной активности
насекомых стрекозам уделяется очень мало внимания. Отрывочные
сведения об изменении активности в течение дня получены для несколь-
ких европейских видов /Corbet, 1957; Parr, 1973/. В СССР иссле-
дования по суточной активности стрекоз и влиянию на нее погодных
условий проведены на о. Кунашир /Заика, 1980/, в Южной Сибири
/Белышев, 19676, 1974а; Заика, Воронова, 1977/, Восточном При-
уралье /Харитонов, 1975а, б/, на юге Украины /Опигер, 1980,
1985/ и юго-западе Таджикистана,/Борисов, 1985в, 1987/. Накоп-
ленный материал фрагментарен и невелик по объему, тем не менее по-
зволяет обобщить имеющиеся сведения и опыт в изучении суточной
ритмики стрекоз, что может послужить предпосылкой дальнейшего раз-
вития исследований в этом направлении.
В своих исследованиях мы придерживались терминологии, пред-
ложенной в монографии BJB. Чернышева /1984/ "Суточные ритмы
активности насекомых", - первой сводке в мировой литературе. Преж-
де чем перейти к обсуждению методов изучения суточной активности,
поясним, что она означает по отношению к стрекозам (рассматрива-
ется только для имаго). По определению В£. Чернышева, активность-
это любая деятельность насекомого. Средством для ее осуществления
служит подвижность. Б.Ф. Белышев /19676/ признаком активности
стрекоз считает полёт, так как с ним целиком связано питание и раз-
множение - важнейшие жизненные функции. Представления о лёте и
активности отождествляют и другие авторы /Заика, Воронова, 1977;
Заика, 1980; Олигер, 1980, 1985; Борисов, 1985в, 1987/, что в
целом неоправданно сужает диапазон понятия активности стрекоз, фак-
тически ограниченный периодом с момента начала движения после ноч-
ного покоя до полного прекращения двигательной активности перед но-
чевкой. Иначе, между началом или прекращением активности и непо-
средственно лётом существуют переходные периоды, когда насекомые
проявляют двигательную активность (например, выползают из укры-
тий), но из-за действия каких-либо факторов еще или уже не способ-
ны к полету. Эти периоды соответствуют времени потенциальной го-
товности - ВПГ /Чернышев, 1973, 1981, 1984/. Знание ВПГ имеет
непосредственно практическое значение при профилактике гельминтоз-
ных заболеваний, переносчиками которых являются стрекозы. Во вре-
мя потенциальной готовности стрекозы наиболее уязвимы и могут в
массе поедаться птицами.
Суточный ритм складывается из разных видов частной активности.
Например, летом возле водоемов обычно наблюдается репродуктивная
77
активность: охрана участков и поиск или поджидание самок самцами,
спаривание, откладывание яиц, выход имаго из личинок. При этом у
многих разнокрылых стрекоз, например видов рода Crocothemis,
Orthetrum, более активны и учитываются в основном самцы (до
98%), самки же благодаря скрытному поведению и биотопической ра-
зобщенности с сакщами в отдельные часы суток отмечаются лишь
единично. В удалении от водоемов учитываемое соотношение полов
близко к естественному и общее время активности преимущественно
трофической здесь больше, чем возле водоемов.
Некоторые виды частной активности наблюдать сложно, например,
ночной выход имаго из личинок, ночной охотничий лёт у Anax
parthenope Selys. Неясны пока причины лёта на искусственный
свет некоторых видов стрекоз. Таким образом, суточная ритмика стре-
коз - явление сложное, по всей вероятности, каждому виду активности
свойствен свой ритм и общая ритмическая картина складывается из
их совокупности. Несомненно при этом, что лёт стрекоз и связанные с
ним виды активности имеют основное значение в формировании суточ-
ных ритмов^ Эндогенный суточный ритм подвержен корректировке по-
годными факторами. Впервые количественная оценка воздействия по-
годных факторов на активность стрекоз произведена Б.Ф. Белышевым
/19676/. А.Ю. Харитонов /1975а, б/ в дополнение к этому разрабо-
тал методы количественной оценки уровня активности.
В экспериментальных условиях с заранее заданными параметрами
среды исследование ритмики стрекоз сложно из-за их большой подвиж-
ности и проявлении эффекта 'замкнутого пространства' (повышенное
возбуждение, стремление вырваться из ограниченного пространства,
затем потеря ориентации и, как следствие этого, невозможность полё-
та). В полевых условиях необходимо параллельно учитывать изменение
уровня активности и изменение факторов среды.
Начало и прекращение активности можно фиксировать визуальными
наблюдениями, перемещаясь в местах ночевок или укрытий стрекоз
при неблагоприятных погодных условиях. Преимущество этого метода
заключается в возможности наблюдать сразу многие виды и на значи-
тельном пространстве. Основной недостаток - необходимость отличать
типичный лёт от спровоцированного наблюдателем во время потенциаль-
ной готовности стрекоз. Более трудоемкий, но и более объективный
метод - наблюдение за поведением стрекоз на точно оконтуренных пло-
щадках или маршрутах.
На учетных площадках подсчитываются пролетающие стрекозы или
количество поведенческих актов в единицу времени (10-15 мин). Же-
лательно отдельно учитывать самцов, самок, копулирующие пары, са-
мок, откладывающих яйца, и т.д. Наблюдения за активностью мелких
равнокрылых стрекоз целесообразно проводить на небольших по разме-
рам площадках (4-6 м^), а учеты разнокрылых стрекоз - на площад-
ках 6-10 м^ и более. Учеты на площадках более результативны в
местах концентрации стрекоз, как правило, возле водоемов. Показа-
телем активности в этом случае служит количество поведенческих ак-
тов (взлеты, патрульные облеты и др.), но последнее в сильной мере
зависит от количества активных особей, причем не только конспеци-
78
фичных. Известно, что с ростом плотности популяций до определенно-
го предела у стрекоз прогрессирует территориальное поведение /Ма-
зохин-Поршняков, Рязанова, 1984; Борисов, 1985в/.
На учетных маршрутах, протяженность которых определяется в
зависимости от обилия исследуемых видов (50-500 м), показателем
уровня активности служит количество активных особей в момент про-
хождения маршрута. Удобны кольцевые или параллельные маршруты,
проложенныее в местах, различающихся условиями, например, на солн-
це и в тени, в заветрии и на открытом месте.
Изучение суточных ритмов невозможно без постоянного контроля
за факторами среды. Необходимо через определенные промежутки вре-
мени непосредственно в местах работы учитывать время наблюдений
и основные показатели: температуру и относительную влажность воз-
духа, прямую солнечную радиацию, освещенность, давление, скорость
ветра, наличие осадков. Интервалы времени, через которые проводят
наблюдения, выбирают в зависимости от вариабельности показателей.
Например, в моменты начала и прекращения активности стрекоз и при
резких изменениях погодных условий наблюдения проводят с периодич-
ностью 10-15 мин, в других случаях промежутки можно увеличивать
до 1 ч.
При обработке результатов наглядную картину дает графическое
изображение изменения показателя активности в сочетании с графи-
ками изменения внешних факторов. По оси абсцисс откладывается
время суток, по оси ординат - показатели уровня активности и факто-
ров среды. При помощи таких графиков удобно проводить анализ влия-
ния факторов на ритм и уровень активности. Составление термогигро-
грамм, совместных значений температуры и относительной влажности
воздуха, при которых зарегистрирована активность стрекоз, позволяет
выделить пороговые значения этих факторов и характеризует отноше-
ние к ним в общем исследуемого объекта. Последнее возможно при
достаточном количестве наблюдений при условии большой вариабельно-
сти факторов. Например, при 600 наблюдениях на юго-западе Таджи-
кистана удалось построить такие термогигрограммы для восьми ви-
дов (рис. 9).
В настоящее время суточные ритмы стрекоз наиболее полно изу-
чены в аридных условиях на юго-западе Таджикистана. В зависимости
от распределения максимумов здесь выделяются три основных типа
активности: преимущественно дневная - для термоксерофильных видов
на протяжении всего сезона и гигрофильных в холодное время года
(ранней весной, поздней осенью и зимой), преимущественно утренне-
вечерняя - для гигрофильных видов в теплое время года и преимуще-
ственно вечерняя (сумеречная) - для ряда видов сем. Aeschnidae
в весенне-летний период.
По отношению к гигротермическим факторам все виды стрекоз
можно разделить здесь на две группы: ксерофильную и гигрофильную.
В группу ксерофильных видов входят наиболее теплолюбивые виды
стрекоз S. nigra, О. sabina, О. albistylwn, О. brwneum,
С. erythraea, Р. dealbata и эвритермный С. servilia.
К этой группе, вероятно, относится большинство раз-
79
Рис. 9. Температура и относительная влажность воздуха, зафиксиро-
ванные на юго-западе Таджикистана и их значения (заштрихованы),
при которых возможна активность стрекоз.
а - S. nigra, б— О. sabina, в — С. serythraea, г —
С. servilia, д — A. parthenope, е — I. fountainei, ж —
I. evansi, з — I. elegans.
30 -
22 -
40
20 -
t’C
Зв -
14 J
Экз.
60 -
-во
- во
140
20
8 10 12 14 16 18 20
20Л
Экз.
Лк 60-
-700 п
-56040'
-420
-210 го'
-440
t’C
II
28-
24-
-80
-60
16 18 20 22
8 10 42 44
-20
лк
-700
-560
-420
\-2в0
\'-140
40
Г'в
8ремя,ч
Рис. 10. Суточная динамика активности стрекоз (экз.) и погодные
условия.
I - 19. V 1982 г., II — 18. V 1980 г. Вид: а - ксёрофиль-
ный С. erythraea; б - гигрофильный I. evansi; в- A. parthe-
nope; г - О. sabina. 1- температура; 2 - относительная влажность, %;
3 - наличие солнца; 4 - освещенность, лк.
80
нокрылых стрекоз (исключая сем. Aeschnidae). Макси-
мум активности этих видов приходится на середину дня при наиболь-
ших значениях температуры и низких влажностях воздуха (рис. .10, 1а).
К гигрофильным видам относятся представители рода Ischnura
(L elegans, I. evansi, L fountainei ) и, видимо, еще не-
которые равнокрылые стрекозы. Для видов этой группы в теплый пе-
риод года характерен бимодальный ритм с максимумами активности в
утренние и вечерние часы. Таким образом, они избегают воздействия
высоких температур при низкой влажности воздуха (рис. 10, 16). К
этой же группе относится своеобразный по типу ритм Anax parthe—
поре и, вероятно, некоторые другие виды сем. Aeschnidae
(Ае. coluberculus, Н. ephippiger) . у A. parthenope
наблюдаются два типа суточного лёта: репродуктивный и охотничий.
Мощные вечерние пики охотничьей активности приурочены к сумеркам
и совпадают с максимумами активности гнуса (рис. 10, II ). Отмечена
также активность утром, в сумерках и ночью, чего не наблюдается у
стрекоз в умеренных широтах. Аналогичное поведение у близкого вида
A. imperator описывает Корбет/Corbet, 1957/ На юге Украины ве-
черний охотничий лёт наблюдается у Brachytron. hafniense ( = pra—
tense), Ae. affinis, Ae. coluberculus, Cordulia aenea
/Олигер, 1985/. Для видов рода Aesc-hna активность с одним ве-
черним максимумом отмечена в лесостепи Восточного Приуралья и За-
падной Сибири /Харитонов, 1975а, б/.
Строгого разграничения между различными по типу активности ви-
дами стрекоз нет. В зависимости от конкретных погодных и микрокли-
матических условий ритмика у одного и того же вида может законо-
мерно изменяться. В холодное время года максимумы активности всех
видов стрекоз приходятся на самое теплое время суток.
В лесостепи Восточного Приуралья отмечено также три типа ак-
тивности стрекоз /Харитонов, 1975а, б/: с одним максимумом в се-
редине дня (для большинства видов при температуре не выше 28-
30 °C и влажности не ниже 30 %), с двумя пиками активности в на-
чале и конце дня (для большинства видов при температуре выше
30 °C и влажности воздуха менее 30 %) и с одним вечерним макси-
мумом (для видов рода Aeschna, охотящихся на скоплениях двукры-
лых насекомых). Следует отметить, что принципиальных различий в
суточных ритмах в пустынной эоне и лесостепи почти нет, но на юге
в жаркую середину дня активность некоторых видов падает почти до
нуля, а в лесостепи происходит ее незначительный спад.
В более северной таежной зоне стрекозам присущ лишь один тип
активности с максимумом в самое теплое время дня /Харитонов,
19756/. В лесотундре в условиях непрерывной освещенности полярно-
го дня и большой изменчивости других факторов среды суточные ритмы
стрекоз выражены слабо и описывающие их кривые отличаются нетипич—
ностью /Харитонов, 19756/.
На юге Украины А.И. Олигер /1980/ в зависимости от числа от>-
ловленных экземпляров на протяжении дня все виды стрекоз по типу
активности распределяет на пять групп: отловленные в основном в
первой половине дня, более активные в утренние и вечерние часы, с
6 Зак. 611
81
равномерной активностью на протяжении дня, с максимумом активно-
сти в середине дня, с максимумом активности во второй половине дня.
Этот же автор /Олигер, 1985/ для ряда видов стрекоз описывает ве-
черние пики активности. Метод отлова при изучении суточных ритмов
по многим причинам малопригоден применительно к стрекозам. Тем
не менее по этим данным также вырисовываются три типа активности,
аналогичные таковым в лесостепи Восточного Приуралья и пустынной
зоне Таджикистана.
По наблюдениям В.В. Заики /1980/, на о. Кунашир выделяются
виды с одним максимумом активности в дневное время и с ее спадом
в середине дня.
Таким образом, в южных районах СССР, где летние дневные тем-
пературы могут превышать 28-30 °C, для стрекоз характерны три
основных типа суточной активности: с максимумом в середине дня,
с двумя максимумами утром и вечером и с одним вечерним.
Время активности стрекоз закономерно изменяется в зависимости
от конкретных условий среды, времени года и географического поло-
жения. На юго-западе Таджикистана общая продолжительность актив-
ности варьирует от практически круглосуточной у A. parthenope
в мае - июне до 5-7 ч у С. servilia и S. decoloratum в декаб-
ре. У большинства термэксерофильных видов в теплый период года
максимальное время активности 15-16 ч в удалении от водоемов и
12-13 (репродуктивная активность) - возле водоемов, у гигрофиль-
ных 14-15 ч возле водоемов. Период суточной активности у полисе-
зонных видов (С. servilia, I. elegans, Sympecma sp.)
зимой примерно в 3 раза короче, чем летом.
В лесостепи Восточного Приуралья лёт большинства видов стре-
коз длится около 15 ч, а у видов рода Aeschna превышает 17 ч
/Харитонов, 19756/. В Южном Прибайкалье при благоприятных по-
годных условиях лёт обычно продолжается не более 12 ч, а макси-
мальное время лёта не более 15 ч /Белышев, 19676/. По данным
А.И. Олигер /1980/, на юге Украины общая продолжительность лёта
не превышает 14 ч, что кажется несколько заниженным. В условиях
островного климата на Дальнем Востоке длительность лёта стрекоз
значительно меньше, чем на материке, и составляет на о. Кунашир
5-6 ч на побережье и 8-9 ч в кальдере вулкана, что обусловлива-
ется высокой влажностью и сильными туманами в утренние и вечер-
ние часы /Заика, 1980/.
Стрекозы в умеренных широтах - дневные насекомые. Исключе-
ние составляют некоторые виды сем. Aeschnidae, активные также
в сумерках и ночью. Из всех видов двигательной активности полет
больше всего нуждается в зрительной ориентации и связан с опреде-
ленными уровнями освещенности.
Освещенность - главный фактор, определяющий начало и прекра-
щение активности, имеет значение только при благоприятнь!х гигро-
термических условиях. Активность большинства ксерофильных видов
зарегистрирована при освещенности не менее 150-200 лк. Гигрофиль-
ные виды при прочих благоприятных условиях летают в сумерках и
активность их прекращается при минимальных значениях освещенности.
82
Сумеречная и даже ночная активность A. parthenope связана с
активностью гнуса, время роения которого четко связано с освещен-
ностью /Чернышев, 1984/. Выход имаго из личинок у многих видов
происходит ночью, но первый полет - только утром, при высоких ее
уровнях.
В лесостепи Восточного Приуралья освещенность является сигна-
лом к началу и прекращению активности. Лёт стрекоз отмечен здесь
при освещенности не менее 100-200 лк, а у видов рода Aes ch-
па - в глубоких сумерках при минимальных ее уровнях /Харитонов,
1975а/.
В Южном Прибайкалье из-за неблагоприятных сочетаний других
факторов в моменты наблюдений лёт стрекоз отмечен только при вы-
соких значениях освещенности /Белышев, 19676/. Примерно также
обстоит депо, судя по данным BJ3. Заики /1980/, на о. Кунашир.
Температура и влажность воздуха - важнейшие факторы в регуля-
ции суточных ритмов стрекоз. Активность каждого вида ограничена
определенными сочетаниями этих показателей (см. рис. 9). Особен-
но сильно проявляется их воздействие на ритм в пустынной Зоне, где
суточный размах температуры в местах обитания стрекоз превышает
25 °C и значительно изменяется влажность воздуха (от 10 до 98 %).
Температурные пороги активности специфичны для каждого вида и
значительно отличаются. Так, например, активность С. servilia
возможна в пределах от 7,2 до 39,6 °C (вероятно выше), A. par—
thenope- от 12,2 и только до 34 °C, S. nigra - лишь от 23 до
40 °C и более, L evansi - от 13 до 35,6 °C.
Температурные пороги лёта выше, чем пороги обшей подвижности,
например у видов рода Ischnura на 2-3 °C. Начало и прекра-
щение лёта у гигрофильных видов определяется преимущественно тем-
пературой, а у ксерофильных в значительной мере и влажностью. Воз-
ле водоемов время лётной активности ксерофилов меньше на 2-4 ч,
чем у гигрофилов, и ограничено высокой влажностью в утренние и ве-
черние часы. Изменение уровня активности на протяжении дня также
находится в прямой зависимости от гигротермических факторов. С
увеличением температуры и падением влажности (до известных преде-
лов) снижается активность гигрофильных видов и повышается у ксеро-
фильных и наоборот. При чрезмерно высоких значениях температуры
(37-40 °C) и низкой влажности воздуха лёт гигрофилов прекращает-
ся, а у самых теплолюбивых ксерофильных видов (например, S. ni-
gra) уровень активности максимален.
Температурные пороги активности у стрекоз на более северных
широтах ниже, что и естественно. Так, в Южном Прибайкалье лёт
многих видов возможен при температуре воздуха выше 10 °C /Бе-
лышев, 19676/, в лесостепи Восточного Приуралья - 13,5 °C /Ха-
ритонов, 1975а/. Определяющее значение температуры (наряду с ос-
вещенностью) в регуляции лётной активности отмечено в лесостепи и
тайге, но особенно ярко проявляется оно в лесотундре /Харитотов,
19756/. Высокая влажность воздуха и тесно связанные с ней роса и
туман, по данным Б.Ф. Белышева /19676/, оказывают сильное сдер-
живающее влияние на активность стрекоз в Южном Прибайкалье. Та-
83
кое же явление отмечено для о. Кунашир /Заика, 1980/. В лесостепи
Восточного Приуралья при низкой влажности (<.30 %) и высокой тем-
пературе ( > 28-30 °C) происходит снижение уровня активности /Ха-
ритонов, 1975а/, аналогичное-таковому у гигрофильных видов в пу-
стынной эоне.
Прямая солнечная радиация. Особенно сильное влияние этого фак-
тора на активность стрекоз отмечено в пустынной зоне и в горах
Средней Азии. В холодное время года и при незначительном превы-
шении температурой воздуха пороговых величин лёт стрекоз возможен
только при воздействии на них лучистой энергии солнца. Так, зимой
и ранней весной в пустынной зоне лёт С. servilia и S. decolora-
tum возможен при температуре 7-1 О °C только в ясную погоду при-
мерно с 11 ч при достаточно высоком положении солнца. В пасмурные
дни эти стрекозы не летают даже при температуре воздуха 16-18 °C.
В весенний период резко снижается уровень активности ксерофильных
видов в моменты, когда солнце скрывается за тучами (см. рис. 10).
В высокогорье Памира полностью прекращается лёт S. haritonovi,
О. brunneum и I. pumilio даже при незначительной облачности,
не сопровождающейся заметным падением температуры воздуха.
Летом в пустынной зоне при высокой температуре и низкой влаж-
ности воздуха прямая солнечная радиация оказывает сдерживающее
влияние на активность стрекоз. При этом гигрофильные виды или пол-
ностью прекращают летать, или перемешаются в затененные места.
На более северных широтах отмечено, что облучение стрекоз солнцем
всегда содействует их активности и часто играет роль решающего
фактора начала лёта /Харитонов, 1975а/.
Воздействие ветра на активность стрекоз тесно связано с темпе-
ратурой воздуха. Чем больше скорость ветра, тем большие значения
температуры (и освещенности) требуются для начала лёта. В лесо-
тундре скорость ветра - один из главнейших факторов в регуляции
лётной активности /Харитонов, 19756/.
При высоких температурах воздуха сдерживающее активность стре-
коз влияние ветра значительно ослабляется. Так, в лесостепи стреко-
зы способны летать при скорости ветра до 4,5 м/с /Харитонов,
1975а/, а в аридных областях Таджикистана отдельные виды ( S. ni-
gra) - до 8 м/с.
Отрицательное действие на активность стрекоз изменения атмо-
сферного давления отмечено в Южном Прибайкалье /Белышев, 19676/
и в лесостепи Зауралья /Харитонов, 1975а/. Даже в разгар лёта рез-
кое падение давления (например, в течение 45 мин с 759 до 756 мм
рт. ст) способно вызвать полное его прекращение.
В дождь обычно лёт стрекоз прекращается, но при слабых и крат-
ковременных осадках многие виды сохраняют активность.
В целом сведения о суточной активности стрекоз очень фрагмен-
тарны. Необходимо изучение ритмики отдельных видов с учетом всех
видов частной активности и факторов, влияющих на них. Важно выяс-
нить особенности ритмов стрекоз в экстремальных условиях среды,
например, в высоких широтах, в высокогорье, зимой на юге, а у рас-
пространенньк видов - в разных ландшафтно-географических районах.
84
В последнем случае, в частности, интересно выявить различия в форми-
ровании суточных ритмов в разных частях ареалов у бореоальпийских
видов*
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ ПОПУЛЯЦИЙ ЛИЧИНОК
(на примере рода Calopteryx)
Поведение стрекоз привлекает внимание многих исследователей.
Сравнительно многочисленные данные по периоду репродукции демонст-
рируют его многообразие и сложность /Рязанова, Мазохин-Поршняков,
1984/ и дают возможность рассматривать действие самцов многих
видов как модель территориального поведения у насекомых вообще.
Большой интерес представляет очень слабо затронутое исследованиями
поведение личинок, тем более, что их лабораторное содержание срав-
нительно просто и приближено к естественным условиям. Кроме того,
образ жизни личинок стрекоз - это хороший пример распространенно-
го среди насекомых изолированного от имаго существования преимаги-
нальных фаз, ’ обладающих оригинальным поведением, не осложненным
репродукцией. Наконец, изучение личинок стрекоз перспективно в пла-
не хозяйственного использования пресных вод, где они нередко явля-
ются одним из наиболее многочисленных и функционально важных ком-
понентов сообщества беспозвоночных.
Немногочисленные работы по поведению личинок стрекоз затрагива-
ют такие вопросы, как охотничье поведение, его модификации в зависи-
мости от поведения жертв, развитие в онтогенезе и сенсорное обеспе-
чение /Мокрушов, 1975; Францевич, 1980; Хорхордин, 1985; Cail—
ier, 1974; Sherk, 1977; Etienne, 1978; Weber, Caillere,
1978; Williams, 1979; Folsom, Collins, 1984/. Есть работы,
посвященные пространственным взаимоотношениям кон— и гетероспеци-
фических особей, агонистическому поведению / Baker, 1980, 1981,
1983; Rowe, 1980, 1985/ и взаимоотношениям стрекоз с рыба-
ми /Pierce, Crowley, 1985/. Описано существование у личинок ряда
видов стрекоз долговременной привязанности к месту. Так, личинки
Xanthocnemis zealandica в опыте при содержании по 2 и 8
особей оставались на месте более 50 сут безотлучно /Rowe, 1980/.
Обнаружено, что взаимоотношения конспецифических особей отличают-
ся от взаимоотношений стрекоз с их жертвами /Baker, 1983/. Для
вида X. zealandica описано территориальное поведение с защитой
индивидуального участка путем угрожающих демонстраций и прямых
столкновений / Rowe, 1980, 1985/. Однако роль поведения во внут-
ривидовых взаимоотношениях и, в, частности, в формировании структуры
популяции еще не ясна, хотя и существуют предположения о вытесне-
нии младших особей с лучших кормовых участков как о механизме, ре-
гулирующем сроки развития особей и плотность популяции /Масап,
1964, 1977; Corbet, 1980; Baker, 1980/.
В данном разделе рассмотрена роль взаимоотношений у личинок
Calopteryx splendens в построении пространственной структуры
популяций: закономерности размещения личинок в местах обитания, их
передвижения и взаимоотношения с конспецифическими особями. Ли-
85
чинки С. splendens избраны как малоподвижные исключительно
тактильные охотники /СаШег, 1974; Sherk, 1977/, чье прост-
ранственное поведение предположительно должно быть проще, чем у
видов подвижных и обладающих хорошо развитыми сенсорными систе-
мами. В то же время образ жизни имаго С. splendens хорошо
изучен и служит образцом совершенного территориального поведения
/ Heymer, 1973/. Рассмотрение пространственного поведения личи-
нок этого вида, возможно, позволит оценить взаимосвязь преимагиналь-
ного поведения с имагинальным.
Опыты проводили в Москве и Закарпатской области в 1983-
1985 гг. Использовали индивидуально меченых личинок, собранных в
Р = 5— F возрастах. Содержали их в аквариумах с отстоенной водо-
проводной водой, песком и растениями, при постоянной принудительной
аэрации и естественном освещении. Объем воды 6 и 3 л, толщина слоя
воды 7-15 см. Летом аквариумы находились в боксе с крышей и сет-
чатыми стенками, затененными вьющимися растениями. Температура
колебалась от 15 до 28 °C. Кормом служили Daphnia, олигохеты
Tubificidae, личинки Ephemeroptera, Chironomidae и
Culicidae. Корм предлагали в избытке во всех опытах. Личинок со-
держали группами в 3-8 особей одного или разных возрастов. На про-
зрачную крышку аквариума наносили сетку со стороной ячеи 5 см (дли-
на тела F личинки 3,7-4,5 см). Местонахождение каждой особи от-
мечали на плане аквариума, руководствуясь положением проекции пе-
редней половины ее тепа на сетке. Местоположения личинок в зависи-
мости от задач опыта регистрировали 1-4 раза в сутки.
Эксперименты показали, что при наличии в аквариуме 7-33 см
побегов элодеи на одну особь личинки основное время проводили на
растениях (около 90 % регистраций). Регистрация каждой личинки в
ячейках аквариума за все время наблюдений свидетельствует о до-
стоверно неслучайных перемещениях ( У2 = 66,9, Р < 0,01 для 28
особей за 40 наблюдений в аквариумах с 15 условными ячейками).
Оказалось, что личинки С. splendens возрастов F = 5 — F при-
вязаны к однажды избранному месту. В разных условиях эксперимен-
тов каждую особь отмечали в одной и той же ячейке в 34 + 1,3 - 47 +
+3,5 % регистраций. При этом, как правило, личинка находилась не
просто в одной ячейке, но на одном и том же листе или на одном и
том же участке ствола растения. Достоверных различий в степени
локализации личинок младших и старших возрастов ( Е ~ 5 - F =
= 4 и F - 1- F ) не обнаружено.
Привязанность личинок С. splendens к месту высока, одна-
ко они не остаются в одном пункте значительное время. Более того,
опыты показали, что большинство их передвигается ежесуточно. Из
78 особей, наблюдаемых в июле - сентябре 1985 г., лишь 15 % ос-
тавались на месте 3-6 сут подряд (при четырех регистрациях в сут-
ки). При этом обнаружено достоверное численное превосходство ноч-
ных прогулок над дневными (для восьми аквариумов критерий Вилко—
ксона для сопряженных рядов Z = О, Р < 0,01). Сравнение часто-
ты перемещений личинок разных полов дает основание говорить о
большей активности самок в ночных передвижениях /Мазохин-Поршня-
86
ков, Рязанова, 1987/. Количественная оценка перемещений восьми ли-
чинок в разные периоды линочного цикла показала, что 5-6 сут перед
линькой и 5-6 сут в середине линочного цикла стрекозы гораздо менее
подвижны, чем 5-6 сут сразу после линьки: передвижения составили
38+18, 30+16 и 84+18 % ночных регистраций на особь соответ-
ственно. Различие высоко достоверно: U = 4 (критерий Вилко—
ксона - Манна - Уитни), Р = 0,001 и U = О, Р <.0,001. Часто-
та перемещений личинок от плотности их в аквариуме обнаруживает
прямую корреляцию между относительной длиной элодеи (основного
места пребывания личинок) и числом перемещений особей: чем мень-
ше плотность личинок в траве, тем больше они ходят ( = 0,772,
Р < 0,05). В среднем разницы в числе передвижений старших ( В4 =
= 1- F ) и младших ( Б'=5 - F = 4) личинок не выявлено.
Сочетание привязанности личинок С. splendens к месту и
ежесуточных прогулок большинства из них свидетельствует о том, что,
постоянно передвигаясь, личинки регулярно возвращаются на излюб-
ленные места. Установлено, что такие возвраты после прогулок ха-
рактерны для всех особей с частотой, в среднем значительно превы-
шающей случайную ( .X2 = 216,9, Р <0,001). Самцы и самки не
различаются по этой характеристике. Наиболее часто личинки возвра-
щаются утром в место, оставленное вечером (в сравнении с возвра-
тами из дневных прогулок Z = О, число пар 8, Р < 0,01). Мы за-
фиксировали такие возвраты и после вынужденных отлучек, вызван-
ных изъятием личинок из аквариума для нанесения новой метки в
связи с очередной линькой. В этих случаях возврат личинок тем бо-
лее удивителен, что насекомое претерпевает столь серьезные воздей-
ствия, как ночйая линька, поимка пинцетом, пребывание на воздухе
на сухом фильтре до 3 мин, нанесение метки лаком и затем помеще-
ние в середину аквариума. Тем не менее из 14 личинок, изъятых ут-
ром, три при вечерней регистрации оказались на месте, откуда их
брали для мечения, что достоверно отличается от случайного попада-
ния в ту же ячейку ( X2 = 8,5, Р <0,01). Еще больше возвратов
произошло при мечении вечером - из восьми личинок четыре верну-
лись на старое место: одна через несколько минут, две ко времени
ночной регистрации, одна - к утренней.
Нами сделана попытка раскрыть механизм возвратов личинок в
"убежище", т.е. место, где они без движения проводят большую часть
времени. Оценено влияние на поведение личинок освещения, локализо-
ванного звука, течения, а также возможности заполнения маршрута
по оставленным химическим меткам, тактильным признакам или с ис-
пользованием моторной памяти /Мазохин-Поршняков, Рязанова, 1987/.
К сожалению, слишком мало сведений о сенсорных системах личинок
стрекоз, чтобы a priori нацелить поиск на выявление определен-
ных ориентационных стимулов, кроме визуальных.
В опыте, чтобы лишить стрекоз зрительных стимулов, аквариумы
затемняли черной бумагой с вечерней до утренней регистрации (за
исключением нескольких минут ночной регистрации в красном свете).
Устраняя стимулы, связанные с течением, изолировали в аквариуме
распылитель воздуха, помешая его в пробирку с водой. Выключая
87
микрокомпрессор, убирали одновременно течение и звук. При лишении
личинок связи с пройденным маршрутом их извлекали после вечерней
регистрации на 3-5 мин в чашку Петри с водой, затем выпускали
всех одновременно в середину уже затемненного или лишенного тече-
ния (в зависимости от условий опыта) аквариума, Контроль и опыт
чередовали ежесуточно.
Результаты опытов свидетельствуют о несомненном влиянии ос-
вещения на передвижение личинок. В полной темноте достоверно мень-
ше личинок оказываются утром в месте вечерней регистрации, чем в
незатемненных аквариумах, имеющих сумеречное и слабое ночное ос-
вещение ( Z = 45, число пар 24, Р <0,01). При этом значение ос-
вещения в ориентации повышается с возрастом личинок. Опыты также
показали, что достоверно большее число особей успешно ориентир уют-
с я при возможности использовать стимулы, даваемые течением ( 2 =
= 35, число пар 12, Р <0,01). Сравнение между собой числа воз-
вратов спонтанно и принудительно передвигающихся личинок в отсутст-
вии света, течения и звука свидетельствует о достоверной зависимо-
сти успешной ориентации от факторов, которые запоминает личинка,
однажды пройдя маршрут ( Z =21, число пар 16, Р = 0,01). Уди-
вительно, что часть личинок не случайно возвращалась в убежище и в
отсутствие перечисленных выше ориентационных факторов: в полной
темноте, без течения и без связи с пройденным маршрутом, хотя воз-
вращение в таких случаях затруднено. Может быть, у них хранится в
памяти представление о топографии аквариума, сложившееся при его
обследовании в предыдущие дни?
Эксперименты с 33 личинками F“ = .1 - F возрастов выявили
ограниченность периодических передвижений каждой из этих особей
определенным участком. При этом пребывание личинок в эонах участ-
ка, по разному удаленных от его центра, неодинаково по времени и
достоверно зависит от плотности особей. Рассмотрено распределение
личинок по аквариуму при различной длине веток элодеи - от 8,5 до
33,5 см на особь. Эксперимент выявил существование прямой корре-
ляции между плотностью популяции и временем, проводимым личинкой
в центре своего участка (одна ячейка): коэффициент корреляции ран-
гов 0,661, Р < 0,0005. Оценка корреляции между плотностью особей
и временем пребывания стрекоз в эонах, разноудаленных от центра,
позволяет утверждать, что при высокой плотности личинки основное
время проводят в зоне с R = 7,5 см, при низкой - они меньше бы-
вают в центре, чаше и дальше от него расходятся /Рязанова, Мазо—
хин-Поршняков, 1985/. Для взаимного расположения центров пребы-
вания личинок F* = 1 - F возрастов характерно их разобщение ие
менее чем на 5-6 см или при высокой плотности поочередное исполь-
зование одного места для длительного пребывания. Изучение законо-
мерностей контактов личинок в аквариуме показало, что частота крат-
ковременных посещений чужих центров в присутствии хозяина зависит
от плотности особей: 3,5+0,9 случаев на насекомое при одной и бо-
лее особей на 15 см элодеи (за 40 регистраций) и 0,4+0,3 - при
одной и менее особей на 25 см элодеи ( = 3,1, Р < 0,01). По-
сещение центров в отсутствие хозяина соответственно 3,9 и 3,1 слу-
88
чаев. Предполагается, что при малой плотности присутствие хозяина
в центре сдерживает посещение его другими особями. Были выполне-
ны эксперименты с изъятием хозяина центра, какие обычно использу-
ют при изучении территориального поведения животных для выявле-
ния охраны территории. В двух из семи таких опытов (только в ус-
ловиях высокой плотности особей) освобожденный центр был немед-
ленно занят для длительного пребывания другой особью, чего не про-
исходило за 30 предыдущих дней в присутствии хозяина. По-видимо-
му, у личинок двух последних возрастов С. splendens существуют ин-
дивидуальные территории, недоступные для занятия какой-либо иной
особью в присутствии хозяина. Несмотря на отсутствие взаимных аг-
рессивных действий, такое поведение, на наш взгляд, может быть на-
звано территориальным в соответствии с наиболее приемлемым, по
нашему мнению, определением территории, данным Уилсоном /Wil-
son, 1975/: 'Территория - область, занятая более или менее ис-
ключительно животными или группами животных посредством явной
агрессии или демонстрации".
Изучение пространственного распределения личинок разных воз-
растов с учетом пола насекомого позволило выявить возрастные и по-
ловые особенности этого поведения. Всего в эксперименте использова-
ли 68 особей F = 5 - Е“ возрастов, регистрируя их положение в ак-
вариуме один раз в сутки - около захода солнца. Обнаружено, что во
всех восьми аквариумах личинки разделяются на две когорты. Пред-
ставители одной склонны к группированию и совместно занимают од-
ну и ту же ячейку, несмотря на обилие (около 60 см элодеи на ак-
вариум) растительности. В случаях, когда в такую группу входят
3-5 личинок, они нередко подолгу находятся в тактильном контакте,
не проявляя признаков агрессии. Личинки другой когорты ведут оди-
ночный образ жизни, не сближаясь с конспецифическими особями. В
группах отмечены особи обоего пола и всех изучаемых возрастов. Од-
нако среди группирующихся личинок резко преобладают младшие ( F* =
= 5 — F = 2 с длиной тела менее 30 мм). Они составляют 70 % об-
щего числа особей этой когорты. Среди одиночных особей младших
личинок всего 36 % при общем преобладании их в опытах (40 из 68).
Различие в возрастном составе когорт высоко достоверно ( X'2 =
= 6,8, Р <0,01) и свидетельствует о снижении у личинок старших
возрастов ( F = 1 - F* с длиной тела более 30 мм) стремления к
агрегации.
Среди одиночных личинок значительно преобладают самцы старшего
возраста (11 из 22 особей). Всего из наблюдаемых старших самцов
(19 экз.) 58 % были одиночными, тогда как среди младших самцов
(25 экз.) одиночными были лишь 20 %. Это различие в поведении
самцов старшего и младшего возрастов высоко ( ,Х2 = 6,7, Р <0,01).
Для самок такого различия в поведении особей разных возрастов не
обнаружено ( У!2 = 0,5, Р < 0,05, п = 24). Интересно, что оди-
ночные старшие самцы превосходят группирующихся старших самцов по
размерам (длине) тела: U = 18, Р < 0,05, п =11, п^ = 8. Од-
нако увеличение процента одиночных особей среди личинок старших воз-
89
растов, очевидно, связано не просто с их физическим ростом. С од-
ной стороны, часто еще более крупные самки остаются в группах, с
другой - самки с увеличением возраста сохраняют в эксперименте
индивидуальные особенности пространственного поведения, тогда как
у самцов с возрастом достоверно растет число одиночных особей.
Можно думать, что пол личинки влияет на характер ее пространствен-
ного поведения. Наблюдаемое избегание скоплений преимущественно
старшими особями свидетельствует о вероятном антагонизме их по
отношению друг к другу и хорошо согласуется с существованием опи-
санных выше элементов территориального поведения у F1 = 1 — F
личинок.
Таким образом, рассмотрены некоторые характеристики индиви-
дуального пространственного поведения и взаимоотношений с конспе-
цифическими особями у личинок С. splendens. Мы подтвердили пред
полагавшуюся другими авторами высокую степень их привязанности к
однажды избранному месту. Однако в отличие от существующих пред-
ставлений о личинках С.splendens, как о сидячих подстерегающих
хищниках /Sherk, 1977/, нами показано, что пребывание их на из-
бранном месте перемежается с передвижениями. В наших опытах это
происходило при избытке корма. Поэтому можно усомниться в абсо-
лютности высказанного Кроулеем / Crowley, 1979/ для Ischnura
и Маканом / Масап, 1977/ для Pyrrhosoma предположения о
том, что передвижения личинок стрекоз - это рассеивание при боль-
шой плотности особей, вызванное недостатком пищи. Тем более что
увеличение частоты и дальности передвижений находится в обратной
связи с увеличением плотности особей. Возможно, у С. splendens
такие передвижения связаны с поиском нового вида корма, с иссле-
довательской активностью или, наконец, с изменением взаимоотноше-
ний особей после линьки. Обнаруженное нами возвращение личинок
после прогулок на облюбованное место вызывает сомнение в том, что
у С. splendens передвижения — рассеивание, ведущее к снижению
плотности популяции. Отсутствие различий в частоте передвижений у
личинок разных возрастов и размеров позволяет утверждать, что у
С.splendens, по крайней мере при обилии корма, мелкие особи
не вытесняются в худшие условия, как это описано Маканом /1977/
для Pyrrhosoma. Обнаруженная корреляция плотности особей с
частотой и дальностью их перемещений и со степенью привязанности
к месту свидетельствует о несомненном взаимном влиянии особей на
взаимное поведение.
Обнаружено различие во взаимоотношениях с конспецифическими
особями у личинок разных возрастов и полов. Основная масса младших
личинок и старших самок склонны к агрегации, большинство стар-
ших самцов - к одиночному существованию. Рассматривая эту осо-
бенность поведения личинок С. splendens,а также описанные для
личинок двух старших возрастов элементы территориального поведе-
ния, кажется заманчивым предположить, что у личинок изучаемого вш
да в последних перед метаморфозом возрастах начинают появляться
черты сходного с имагинальным пространственного поведения. Вид
С. splendens хорошо известен /Неутег,! 973/ характерным' для
90
части имаго самцов территориальным поведением, которое наблюда-
ется одновременно с существованием стай разновозрастных самцов
и самок. В настоящее время мы не можем определить природу и зна-
чение обнаруженных у старших личинок С. splendens элементов тер-
риториального поведения. Возможно, они выработались у личинок и
являются лишь аналогом поведения имаго. Они могут определять успех
развития личинки в условиях дефицита ресурсов, например корма или
укрытий при обилии хищников. Для выявления такой роли поведения
необходимы дополнительные исследования. Можно лишь отметить, что
условия существования личинок С. splendens способствуют созданию
такого рода дефицитов.
Так, изучаемая нами закарпатская популяция состоит из групп
личинок, заселяющих небольшие, часто не более 0,5 м^, куртины вод-
ных растений у берега реки при глубине около 50 см. Куртины эти
разделены между собой голыми участками глинистого дна по 10-30 м
вдоль берега, которые, возможно, препятствуют переходам личинок.
Численность личинок на растениях может быть очень велика, -судя по
количеству яйцекладущих имаго. Специальных учетов мы не проводили,
но на двух-трех таких участках в июле вылавливали по 100-620 экз.
F = 1- F возрастов. Проявление элементов территориального пове-
дения личинками С. splendens может представлять собой лишь
отражение начального этапа развития физиологических механизмов со-
ответствующего поведения имаго и не иметь собственного значения в
жизни личинок. Такую трактовку территориального поведения личинок
подтверждает одиночное существование преимущественно старших сам-
цов, что хорошо согласуется с территориальным поведением имаго
самцов и кажется неоправданным в случае повышения в результате
этого поведения успеха развития личинок.
Итак, наши исследования показали, что индивидуальное простран-
ственное поведение и территориальные взаимоотношения с конспецифи-
ческими особями у личинок С. splendens сложнее, чем можно бы-
ло ожидать от сидячего подстерегающего хищника, использующего лишь
тактильную сенсорную систему в процессе охоты. Поведение названных
личинок наряду с другими экологическими факторами имеет значение
в формировании структуры популяции этого хищника и в его взаимооту-
ношениях с жертвами. В частности, поведение в какой-то степени влия-
ет на распределение в пространстве мест долговременного пребывания
особей и в связи с этим определяет плотность и границы обитания по-
пуляции. Передвижения особей, коррелирующие по частоте и дальности
с плотностью популяции, обеспечивают контакт с жертвами во всем
местообитании хищников, даже при невысокой их плотности. Данные о
способности личинок С. splendens к ориентированию заставляют ду-
мать о значительно больших, чем предполагалось, возможностях их
сенсорных систем, по-видимому, используемых в передвижениях и
охоте.
91
РОЛЬ ЗРЕНИЯ В ПОВЕДЕНИИ СТРЕКОЗ
Многочисленные поведенческие и физиологические исследования
показывают, что зрение насекомых достигло высокой степени совер-
шенства. В то же время зрительный анализатор насекомых имеет
более простое по сравнению с позвоночными устройство. Миниатюр-
ность зрительного анализатора насекомых сочетается с разнообрази-
ем решаемых им задач. Несмотря на относительно низкую разрешаю-
щую способность сложного глаза, членистоногие чрезвычайно чувстви-
тельны к видимому движению. Для насекомых с хорошо развитым зре-
нием восприятие движения лежит в основе формирования поведенческих
актов. В естественных условиях движение объекта, несомненно, слу-
жит сигналом о приближении добычи, опасного объекта или брачного
партнера. Кроме того, движение оптического изображения способству-
ет выделению признаков формы объекта и повышает остроту зрения.
Стрекозы, ведущие дневной подвижный образ жизни, - удобны для
исследования зрительного анализатора у насекомых. Они способны не
только к таким простым формам зрительного анализа, как стабилиза-
ция движения относительно источника света, обнаружение и преследо-
вание добычи, бегство от угрожающих объектов, но и таким, которые
не уступают по сложности актам позвоночных животных - опознание
брачного партнера, запоминание и охрана территории, возврат к охра-
няемой территории с расстояния нескольких километров. Электрофи-
зиологическими исследованиями установлено, что в зрительном анали-
заторе взрослых стрекоз и их личинок имеются интернейроны, специ-
фически чувствительные к некоторым признакам оптического окруже-
ния. Предполагается, что некоторые из них могут тем или иным об-
разом участвовать в запуске определенных поведенческих реакций у
стрекоз и их личинок. Несомненно, что запуск и поддержание таких
поведенческих актов - бегство, обнаружение добычи, выбор места для
посадки — могут осуществляться нейронами, которые способны выде-
лять одиночные крупные и мелкие объекты.
Многолетние работы со стрекозами дали возможность сопоставить
результаты электрофизиологических и поведенческих опытов и объяс-
нить физиологические механизмы поведения стрекоз, управляемые зре-
нием.
ПРОСТЫЕ ФОРМЫ ЗРИТЕЛЬНОГО АНАЛИЗА
Специфическое поведение стрекоз позволяет использовать количе-
ственные методы регистрации в поведенческих опытах и подробно опи-
сать способности зрительного анализатора стрекоз и их личинок. На
этих же объектах выполнены классические работы гистологов по Строе-
нию оптических и нервных структур у насекомых. Электрофизиологи
зарегистрировали в нервных центрах стрекоз клетки, отвечающие эле-
ктрическим разрядом на изменения оптического окружения. Некоторые
92
из систем нейронов, возможно, участвующие в запуске поведенческих
реакций, приведены в табп. 2. Далее в разделах эти явления будут
описаны более подробно.
Органы зрения. Сложный глаз. Зрительный анализатор среди орга-
нов чувств стрекоз занимает особое место. Ему принадлежит главная
роль в управлении основными жизненными функциями насекомого: пи-
тания, размножения и т.п. Органы зрения стрекоз представлены про-
стыми и сложными глазами. Основой зрительного анализатора насеко-
мых и их личинок являются сложные глаза - очень крупные парные
образования, занимающие почти всю поверхность головы. У многих
групп стрекоз Anisoptera они сливаются в области лба.
Сложный глаз стрекозы состоит из множества (1000-28000)
длинных цилиндрических зрительных единиц - омматидиев. В оммати-
дии выделяются три типа элементов: оптический (бветопреломляющий),
сенсорный (светочувствительный), пигментный (с вето изолирующий).
Подробнее описание устройства и принцип работы глаза насекомого
приведены в работах /Мазохин-Поршняков, 1965; Францевич, 1980;
Грибакин, 1981/.
Разрешающая способность. Острота зрения, или разрешающая спо-
собность, - это свойство органа зрения устанавливать наименьший
угол между двумя точками. Такая способность в первую очередь опре-
деляется строением сетчатки: чем больше чувствительных клеток на
единицу телесного угла сетчатки, тем выше острота зрения. Для насе-
комых, у которых воспринимающей единицей является целый оммати-
дий, разрешающая способность глаза будет тем выше, чем меньше
угол между оптическими осями соседних омматидиев (угол расхожде-
ния). Для взрослых стрекоз Anax, Aeschna, Hemicordulia, Li—
bellula угол расхождения вычислен в 0,4—1,2° /Pritchard, 1966;
Horridge, 1969; Laughlin, 1973/. У стрекоз Libellula вент-
ральная часть глаза обладает более высокой разрешающей способ-
ностью.
Спектральная чувствительность. Способностью различать излуче-
ния по спектральному составу, кроме человека, обладают многие пред-
ставители животного мира, в том числе и большинство насекомых.
Вопрос о восприятии насекомыми цвета освещен в литературе /Мазо-
хин—Поршняков, 1965, 1973/. Возможности цветового зрения стрекоз
показаны разными методами /Мазохин-Поршняков, 1973; Мокрушов,
Францевич, 1976а/. Глаз стрекоз имеет двойную функцию — верхняя
его часть чувствительна к ультрафиолету или синим лучам, а нижняя
обычно обладает полным набором светочувствительных приемников.
Причем у некоторых стрекоз, кроме обычных для насекомых чувстви-
тельных в синезеленой области спектра приемников, имеются и красно-
чувствительные /Meinertzhagen et al., 1983/ Спектральные харак-
теристики зрительного анализатора стрекоз представлены в табл. 3.
Бинокулярное зрение. Стрекозы и их личинки обладают способно-
стью воспринимать пространственное положение потенциальной жертвы
и определять расстояние до нее. Достигается это в результате пере-
крещивания зрительных осей глаз, что позволяет видеть некоторую
часть поля зрения обоими глазами (поля зрения обоих глаз перекры—
93
CD д Таблица 2. Типы центральных зрительных нейронов у стрекоз
Тип нейрона Вид стрекоз Локализация ней- рона Адекватная поведен- ческая реакция Источник
Нейроны, чувствитель- ные к направлению освещения S утр strum vulga- tum Брюшная нервная цепочка Фототропическая ре- акция /Золотов, 1982/
Нейроны, чувствитель- ные к размеру объ- ектов и направлению движения (ДР-ней- роны) S. vulgatum, Aeschna viridis, S. flaveolum Коннектива Обнаружение добычи /Зенкин, Пигарев, 1969, 1971; Мок- рушов, Францевич, 1976а/
Высокочастотные де- текторы движения ' (ВДД) Aeschna cyanea, род Sympetrum Третий оптиче- ский ганглий Обнаружение опасных объектов, реакция бегства /Мокрупюв, Фран- цевич, 1973; Фран- цевич, Мокрушов, 1974/
Дирекционно-чувстви- тельные (ДЧ) ней- роны Ae. viridis S. flaveolum Брюшная нервная цепочка Управление оптомо- торной реакцией /Зенкин, Пигарев, 1969, 1971/
ДЧ-нейроны, чувстви- тельные к движению расчлененных узоров S. vulgatum Коннектива Управление оптомотор- ной реакцией в скоро- стном полете /Францевич и др., 1978/
ДЧ-нейроны (мульти- сенсорные) Anax j unius Между про— и ме- заторакальными ганглиями Управление полетом /Olberg, 1981/
Таблица 3. Спектральные характеристики светоприемников некото-
рых видов стрекоз /по: Мазохин—Поршняков, 1973; Lavoie— Dornik
et al., 19 81; Meinertzhagen et aL, 19 83/
Вид M аксимальная чувстви- тельность светоприем— ника, нм Часть глаза Источник
Libellula quadri— maculata 420 515; 610 Верхняя Нижняя /Мазохин- Поршняков, 1965/
L. pulchella 420 380; 518; 550 Верхняя Нижняя /Ruck, 1965/
L. needhami 415; 430 450; 550 Верхняя Нижняя /Horridge, 1969/
Sympetrum rubi- cundulum 340; 410 4 90; 540; 620 /Meinertz— hagen et al., 1983/
Hemicordulia tau 350; 420; 450; 500; 520; 540 Нижняя /Laughlin, 1975,1976/
Anax j unius 370; 3 90; 420-440 420-520 Верхняя Нижняя /Horridge, 1969/
Aeschna cyanea Ae. cyanea 356; 445-458 519 412-432; 519; 356; 615; 336 Верхняя Нижняя /Eguchi, 1971/ /Autzum, Kolb, 1968/ Там же
ваются). Бинокулярную эону зрения у некоторых стрекоз методами на-
блюдения ложного зрачка или свечения омматидиев при освещении гла-
за изнутри измеряли Л.И. Францевич и В.Е. Пичка /1976/. Одновре-
менно авторы подсчитывали количество омматидиев, захватываемых
бинокулярной зоной. Они показали, что размер и организация биноку-
лярного поля зрения у стрекоз соответствуют особенностям их пове-
дения. Так, у стрекоз-преследователей (Aeschna), захватывающих
добычу в полете, поля зрения обоих глаз перекрываются всего не-
сколькими рядами фасеток. Поэтому стрекозы этой группы не могут
эффективно оценивать глубину пространства и при быстром полете ата-
ка на цель и облет препятствий у них осуществляется по другим при-
знакам. Стрекозы-собиратели (равнокрылые), которые летают медлен-
но и схватывают добычу среди растительности, обладают большей би-
нокулярной зоной, насчитывающей несколько тысяч омматидиев. Такое
большое бинокулярное перекрытие позволяет стрекозам этой группы
выделять характерные объекты на сложном фоне.
95
Морфологические основы зрения. Впервые описание тонкого строе-
ния зрительных центров у насекомых осуществлено А.А. Заварзиным
/1914/ на примере личинки стрекозы. Светочувствительной единицей
омматидия является ретинальная (зрительная) клетка, преобразующая
оптическое изображение в нервный сигнал. Своими аксонами ретиналь-
ные клетки связаны с парными, очень крупными у стрекоз, оптически-
ми долями, которые являются частью протоцеребрума. В каждой зри-
тельной доле имеются обособленные зоны - нейропили, образованные
переплетением отростков нервных клеток. Каждая такая эона носит
название оптического ганглия. У стрекоз их три, они осуществляют
последовательную переработку зрительной информации. Структурной
единицей оптических ганглиев является нейро мматидий — дискретный
пучок нервных волокон. Растр нейромматидиев строго соответствует
растру омматидиев. Нейромматидии образуют проекцию сетчатки с
сохранением разрешающей способности оптики глаза.
Строение и функциональное назначение отдельных элементов нерв-
ной системы стрекоз и их личинок дано в работах /Цвилинева, 1950,
1951; Плотникова, 1979; Satja, 1958; Mill, 1965; Mathur,
1972; и др./. Развитие сложных глаз и перестройка нервных связей
в зрительных центрах личинок стрекоз при имагинальной линьке ис-
следованы /Mouze, 1972; Sherk, 1977, 1978/.
Фототропизм. Жизнедеятельность насекомых, особенно с разви-
тым зрением, определяется влитием света. Посредством света у них
запускаются ориентационные механизмы и суточная активность, через
органы зрения поступает информация об окружающей обстановке. Для
громадного количества насекомых свойственно явление положительно-
го фототропического поведения — перемещение в область более высо-
кой освещенности. Положительный фототропизм - причина лёта ночных
насекомых на искусственные источники света /Мазохин-Поршняков,
1965; Горностаев, 1984/.
Ориентирующее воздействие света известно и для стрекоз. Для
Anax imperator экспериментально показано /Mittelsfaedt,
1950/ влияние реакции "свет сверху" на стабилизацию полета. Так,
стрекозы, закрепленные в специальном устройстве, позволяющем им
взмахивать крыльями, в темноте характеризуются нестабильным, вра-
щающимся вдоль продольной оси тела полетом. Направленное освеще-
ние подопытного насекомого вызывает вначале поворот головы к све-
ту, а затем и всего туловища. Необходимо отметить, что управление
моторными центрами осуществляется не только оптическими сигнала-
ми, но и от других сенсорных входов, в частности от волосковых ме—
ханорецепторов. Так, после перерезки чувствительных нервов, контро-
лирующих положение головы относительно тела или удаления волоско-
вых механорецепторов у Anax imperator, стрекоза реагирует на
источник света поворотом головы, но неуправляемое вращение туло-
вища сохраняется. Одноглазые насекомые в закрепленном полете так-
же вращаются вдоль продольной оси тела до тех пор, пока зрячий
глаз не будет стимулирован светом до определенного уровня. Таким
образом, реакция "свет сверху" действенна только в том случае, ес-
ли возбуждению подверглось заданное количество омматидиев. Суще-
96
ствуют ли специализированные участки глаза, возбуждение которых
управляет реакцией 'свет сверху*, точно не установлено. Глаз стре-
козы в дорсальной своей части имеет более крупные фасетки, чем в
вентральной. Причем у видов, летающих на открытых пространствах
(Anax, Aeschna, Libellula ), граница между крупными и мел-
кими фасетками смещена сильнее в сторону вентральной части глаза,
в сравнении со стрекозами, обитающими в закрытых пространствах
(zYg°ptera)* Но такое увеличение дорсальной эоны глаза у крупных
стрекоз не влияет на реакцию 'свет сверху*. Скорее всего подобное
устройство глаза связано с пишедобыванием. У закрепленных стрекоз
при диффузном освещении в полете наблюдается вращательное движе-
ние в том случае, если закрашивали часть мелких фасеток в дорсола-
теральной эоне одного из глаз.
Электрофизиологическими исследованиями в брюшной нервной це-
почке стрекоз Sympetrum vulgatum обнаружено несколько типов
нейронов, чувствительных к направлению освещения /Золотов, 1982/.
Описанные нейроны обладают широкими, перекрывающимися рецептив-
ными полями, с разными диаграммами направленности. Предполагает-
ся, что некоторые из них могут управлять ориентационными реакция-
ми стрекоз. Так, нейроны с равномерными (моно-’и бинокулярными)
диаграммами направленности, обладающие высокой светочувствитель-
ностью, могут управлять фототропическими реакциями насекомых при
низких интенсивностях освещенности. Группа нейронов, характерной
особенностью которых являются низкая светочувствительность и не-
равномерные диаграммы направленности (типа синусоидальных), спо-
собны, вероятно, управлять такими поведенческими реакциями, как
поляротропизм, астроориентация и т.п.
Охотничье поведение. В опытах по изучению охотничьего поведе-
ния хищных насекомых часто исследуются реакции броска на добычу.
Неоднократно опыты ставились на личинках стрекоз рода Aeschna,
обладающих хорошим зрением и ведущих открытый образ жизни. При
захвате добычи личинка выбрасывает маску, что легко фиксируется
наблюдателем. Очень важно и то, что личинка может пытаться захва-
тить не только мелких водных животных, но в условиях опыта и моде-
ли жертвы - небольшие движущиеся объекты. Использование моделей
позволяет предъявлять личинкам объекты с заданными параметрами:
размер, цвет и т.п.
• Специальные опыты с личинками стрекоз показали, что реакция
на добычу у последних возникает и сохраняется до тех пор, пока объ-
екты подходящего размера воспринимаются как подвижные. Фогт
/Vogt, 1964/ показывал личинкам стрекоз два подсвечиваемых лам-
почками пятна (диаметр 1,6 мм),, которые могли вспыхивать одно-
временно или поочередно. Скачок пятна на небольшие расстояния вос-
принимается насекомыми как движение. В других опытах /Hoppen—
heit, 1964/ личинкам предъявлялись объемные (шарики) и плос-
кие (диски) объекты. Было установлено, что с увеличением моделей
от 3 до 15 мм активность реакции нападения у личинок убывает.
При захвате добычи личинка предпочитает плоские предметы объем-
ным и компактные расчлененным. Оптимальная скорость движения
7 Зак. 611
97
приманки в опытах Хоппенхайта приближалась к скорости ползания
личинок (немногим более 1 см/с). Очень эффективным раздражите-
лем для личинок являются зигзагообразные движения светлой точки
со скоростью по вертикали 2,5 см/с (амплитуда вертикальных коле-
баний 0,2—4 см), а по горизонтали 0,3 см/с /Etienne, 1969/. Па-
раметры модели могли меняться в зависимости от физиологического
состояния личинки.
Взрослые стрекозы родов Aeschna, Libellula, Sympetrum
преследуют только подвижные жертвы. Стрекозы Libellula quadri—
maculata относятся к группе подстерегающих хишьиксв /Заи-
ка и др., 1977/. Они могут питаться как у берегов водоемов, так и,
вдали от них. Стрекозы сидят на выступающих частях субстрата (вер-
хушки трав, кустарников и т.п.) и взлетают за пролетающими насеко-
мыми. Схватив добычу, они возвращаются к месту старта. Такое по-
ведение стрекоз этой группы позволяет исследователю приблизиться к
ним и с помощью удилища и тонкой лески показывать разные модели.
В наших опытах /Мокрушов, 1972/ при одновременном предъявлении
белого и черного шариков, расположенных в 5 см один под другим,
стрекозы L. quadrimaculata предпочитали захватывать нижний ша-
рик, независимо от его цвета. Стрекозы чаще всего бросаются на мо-
дель при ее движении снизу вверх, особенно поднимающуюся над силу-
этами деревьев. Охотящееся насекомое обнаруживает цели, движущие- .
ся с угловой скоростью до 400 град/с и с угловыми размерами око-
ло 5 мин. Ниже представлена зависимость реакции преследования от
размеров модели:
Размер цели, Продолжение
мм учета, мин
1,5 40
3 50
6 50
12 45
Число брос- Максимальная
ков в мину- дальность об-
ту наружения, м
6,8 1
5,2 1.5
3,3 3,0
0,44 —
При отведении электрической активности в грудных ганглиях стре-
коз Sympetrum flaveolum регистрируются нейроны, избиратель-
но чувствительные к размеру объектов и направлению их движения
/Зенкин, Пигарев, 1969/. Нами /Мокрушов, Францевич, 1976а/ бы-
ли подробно исследованы такие характерные свойства этих нейронов,
как дирекциональная чувствительность, чувствительность к размеру
движущегося объекта и локальное привыкание к повторной стимуляции.
Нейроны этого класса не отвечают на изменение общей освещенности
и движение узора из черно-белых полос. Мощный разряд следует в
момент движения в поле зрения одиночных небольших объектов снизу
вверх. Оптимальные размеры стимула (плоский диск или светлая по-
лоса на экране электронно-лучевой трубки) составляют 5-7 градусов.
Знак контраста движущегося стимула не имеет существенного значе-
ния: белые и черные диски на сером фоне вызывают в среднем одина-
98
ковую реакцию (12,0+1,1 и 10,8+1,3 имп на предъявление). Кро-
ме того, нейроны этого класса особо чувствительны к быстрым дви-
жениям. Исследованные нейроны обладают симметричными монокуляр-
ными рецептивными полями, вытянутыми узкой лентой по горизонтали
от медиального края глаза на 120-150 град. Чувствительность к
движению в рецептивном поле убывает к латеральной части глаза.
Латеральное торможение. Изучая электрические реакции оммати-
дия в латеральном глазу мечехвоста /Hartleine et al., 1956/,
установили, что если рядом с исследуемым омматидием освещать дру-
гой, то это оказывает существенное влияние на первый. Причем это
влияние чисто тормозное и суть его заключается в том, что возбуж-
дение соседнего омматидия повышает порог чувствительности и сни-
жает спонтанную частоту импульсов исследуемого омматидия. Тормоз-
ные влияния взаимны, т.е. каждый омматидий тормозит соседние ом-
матидии и, в свою очередь, сам тормозится ими. Тормозные эффекты
нескольких омматидиев определяются суммой тормозных влияний, каж-
дого из них. Описанное взаимодействие в сложном глазу мечехвоста
получило название латерального торможения. Оно происходит между
особыми модифицированными фоторецепторами - эксцентрическими
клетками. В. Рейхард /1968/ провел теоретический анализ нервного
торможения в глазу мечехвоста и на основании данных Хартлайна и
Ратлиффа показал, что процесс латерального торможения компенсиру-
ет эффекты перекрывания зрительных попей смежных омматидиев и
тем самым повышает разрешающую способность сложного глаза. Функ-
циональная роль латерального торможения состоит в том, что оно уси-
ливает контраст "изображения", передаваемого нервными структурами,
что, в свою очередь, способствует выделению контуров или границы
объекта. Латерально-тормозные взаимодействия лежат в основе выде-
ления точечных объектов.
В глазу насекомых латеральное торможение между омматидиями
отсутствует. Объясняется это тем, что в отличие от системы эксцент-
рических клеток мечехвоста в зрительном анализаторе насекомых меж-
ду фоторецепторными аксонами нет непосредственных морфологических
связей. Электрофизиологи ческие исследования на мухах Calliphora
erythocephala /Zettler, Jarvilehto, 1972/, Boettcherisca pe—
rygrina /Mimura, 1972/, Phaenicia sericata /Arnett, 1971/
свидетельствуют о том, что латеральное торможение у этих насеко-
мых происходит на уровне первого оптического ганглия. Данные по-
веденческих опытов с насекомыми, несомненно, указывают на роль
латерального торможения в выделении объектов минимального разме-
ра. Из наших опытов по охотничьему поведению видно, что стрекозы
L, quadrimaculata яростно преследуют модели жертв маленьких
размеров. Модели большого диаметра вызывают более слабую реакцию.
Сравнительно крупные шарики, диаметром более 1 см, не привлекают
насекомых. Резкое предпочтение стрекозами мелких целей можно объ-
яснить, допуская, что сигналы от соседних омматидиев в рецептивных
полях нейронов, выделяющие цель, тормозят друг друга.
К такому же выводу приводят и результаты опытов с предъявле-
нием двойных и тройных целей. Если три шарика подвесить один под
99
другим на расстоянии 2-5 см, то количество нападений на них со
стороны стрекоз распределяется следующим образом: 71 % нападений
(83 из 117) стрекозы совершают на нижний шарик, 21 % - на верх-
ний и только 8 % - на средний. Возможно, что такое выделение край-
них элементов можно объяснить процессами латерального торможения.
В одной из серий опытов мы постепенно увеличивали расстояние
от О до 2 см по горизонтали между двумя шариками диаметром
1,5 мм. Одновременно в 30 см от двойной цели показывали конт-
роль - два вместе склеенные шарики тех же размеров. Увеличение
расстояния между шариками до 2 см подавляло реакцию преследова-
ния. В опытах / Meyer,1971/ на водомерке Velia caprai пока-
зано, что если расстояние между двумя небольшими темными диска-
ми, имитирующими добычу, увеличивать до такого, что каждый из
них начинает попадать в поле зрения одного из двух соседних омма-
тидиев, то число выборов данной модели в сравнении с одиночным
диском уменьшалось: оно становилось минимальным при расстоянии,
равном углу расхождения омматидиев.
Наблюдая за поведением стрекоз родов Libellula, Sympetrum
в природе, можно отметить одну особенность: очень часто, подстере-
гая добычу, они выбирают для посадки узкие, вертикально протяжен-
ные или горизонтально расположенные предметы. Привлечение к вер-
тикально протяженным предметам свойственно и насекомым-фитофа-
гам. Стрекоз особенно привлекают светлые объекты и их вершины.
Так, в наших опытах при одновременном предъявлении черных и бе-
лых палочек, имитирующих насесты, стрекозы L. quadrimaculata
совершили 77 % посадок на белые палочки. Полосатые черно-белые
папочки со светлой верхушкой выбирались вдвое чаше, чем с черной.
Реакцию посадки тормозят большие диски или поперечные элементы,
перекрывающие верхушку. Если выставить три палочки рядом, то до-
ля посадок на среднюю из них возрастает с увеличением расстояния
между палочками. Как уже отмечалось, нервной основой выделения
краев и линий являются процессы латерального торможения в рецеп-
тивном поле. Можно предполагать, что у стрекоз Libellula, Sym—
petrum имеются нейроны со специфической геометрией тормозных
связей в рецептивном поле, которые позволяют выделять линии опре-
деленной ориентации. В частности, выделение вертикальной пинии мо-
жет осуществляться в рецептивном поле из нейронов, описанных у мух
/Arnett, 1971/. Эти нейроны возбуждаются при освещении фоторецеп-
торов своего нейромматидия и тормозятся соседними, расположенны-
ми справа и слева в том же горизонтальном ряду.
Оборонительная реакция. В поведенческих опытах с личинками
стрекоз /Hoppenheit, 1964/ было показано, что диски диаметром
свыше 8 мм могут вызывать реакцию бегства и тем сильнее, чем
больше размер модели. В наших опытах /Мокрушов, Золотов, 1973/
мы показывали личинкам стрекозы Ае. суапеа модели, имитирую-
щие жертву. Моделями служили белые и черные диски диаметром от
2,5 до 60 мм, которые предъявляли на контрастном фоне: белые на
черном, черные на белом. В одной из серий опытов измеряли расстоя-
ние, с которого черные диски разного размера вызывали реакцию бег-
100
ства, и вычисляли критический угловой размер модели у нормальных
личинок и у личинок, один глаз которых был закрашен черным ла-
ком. Критический размер черного диска на белом фоне для половины
подопытных насекомых превышает 53 град. Но реакция личинок на
диске большого и малого абсолютного размера существенно различа-
ется. Так, средний критический угловой размер 1,5-сантиметрового
диска равен 58,1+2,1 град, а 6-сантиметрового - 38,8+0,83. Та-
кое поведение личинок свидетельствует о том, что они могут оцени-
вать не только угловые размеры объектов, но и их действительные
размеры.
Одноглазые личинки стрекоз пытаются схватить объекты неболь-
ших угловых размеров на расстоянии значительно большем, чем нор-
мальное расстояние захвата /Baldus, 1926/. В наших опытах бег-
ство одноглазых личинок вызывали критические угловые размеры 6-
сантиметрового диска 88+3,2 и 1,5-сантиметрового - 71,1 +
+ 2,4 град.
В электрофизиологических опытах в проксимальной части треть-
его оптического ганглия личинок Ае. cyanea регистрируются
нейроны, отвечающие залпом разрядов на движение контрастных объ-
ектов в рецептивном попе, без проявления чувствительности к на-
правленному движению. Эти нейроны - высокочастотные детекторы
движения (ВДД) разряжаются со значительно большей частотой
(150-200 имп./с) в ответ на движение черных дисков на сером фо-
не, чем на движение белых дисков такого же размера. Приближение
черного диска к голове пичинки вызывает сильный разряд, удаление
(сразу после приближения или через 15—20 с) не вызывает или поч-
ти не вызывает реакции нейронов. Частота разряда таких нейронов
при движении одиночного черного объекта прямо зависит от разме-
ров этого объекта. Так, для стандартного стимула (проход на рас-
стоянии 40 град со скоростью 50-60 град/с) чиспо импульсов в
реакции на черный диск угловыми размерами 15 град было в 2-3
раза больше, чем на диск диаметром 2 град.
Признаки простейших моделей, вызывающих оборонительную реак-
цию личинок стрекоз в поведенческих опытах, весьма похожи на при-
знаки стимулов, которые возбуждают реакцию ВДД. Поведенческие
реакции испуга (бегства) и упомянутые нейроны сильнее всего воз-
буждаются движением темного диска на контрастном фоне. Очень
сильно действует приближение темного объекта. Темные диски мало-
го углового размера почти не возбуждают эти нейроны и соответст-
венно вызывают не реакцию бегства, а реакцию атаки. Слабо дейст-
вуют на нейрон удаление черных объектов и движение светлых. Круп-
ные белые диски не вызывают у личинок стрекоз реакции бегства, ли-
чинки даже могут нападать на них.
Роль детектора в формировании поведения животного можно про-
верить в поведенческих экспериментах. Так, в электрофизиологиче-
ских опытах /Зенкин, Пигарев, 1969/ 'детектор пятна' в сетчатке
лягушки сильно реагировал на вхождение в репептивное поле тонкой
темной палочки, ширина которой была меньше диаметра поля. В по-
веденческих экспериментах животное хватало кончик вдвигаемой в
101
попе зрения попоски так же хорошо, как и темное пятно, что, несом-
ненно, свидетельствует о том, что указанный детектор может запу-
скать реакцию захвата добычи.
Из наблюдений над высокочастотными детекторами движения из-
вестно, что этот нейрон слабо реагирует на движения оптически бо-
гатого фона, быстро привыкает к нему и на его фоне не отвечает на
движение одиночного объекта. Можно было ожидать, что приближение
черного диска на фоне непрерывно движущегося узора будет с боль-
шим трудом запускать оборонительную реакцию, чем движение этого
же диска на неподвижном фоне. В поведенческих опытах личинкам
демонстрировали приближение 3-сантиметрового черного диска на
фоне вертикальных узоров двух типов: один с шириной полос 5 см,
второй - 1 см. Диск приближался к одной и той же пичинке на фо-
не неподвижного и возвратно-поступательного движения узора. В этих
экспериментах мы не получили однозначного ответа. Критические уг-
ловые размеры черного диска на белом фоне и неподвижном полоса-
том примерно одинаковы. Не отличалась реакция от контроля и при
приближении диска на фоне движущегося растра с шириной попос 1 см.
И только движение узора с шириной полос 5 см подавляло реакцию
бегства. Поэтому еще нельзя утверждать, что именно ВДД запускают
реакцию бегства у личинок стрекоз.
Опыты Хоппенхайт /Hoppenheit,1964/ и наши /Мокрушов, Зо-
лотов, 1973/ показывают, что модели жертв, привлекающие личинок
с увеличением их размеров, начинают вызывать оборонительную реак-
цию у насекомых. Более подробно взаимодействие этих двух реакций-
атаки и бегства - было исследовано в поведенческих и электрофизио-
логических опытах Л.И. Францевичем /1982/. Автор исследовал ре-
акцию ДР-нейронов в ответ на стимуляцию их обоими типами специ-
фических раздражителей. В электрофизиологических опытах стрекозам
рода Sympetrum показывали движение белого круга с угловым диа-
метром 4-5°, который двигался снизу вверх со скоростью 300 град/с.
Одновременно с началом движения белого диска или с заданной за-
держкой начиналось движение черного круг» угловыми размерами 30-
35°. Черный диск двигался со скоростью 135 град/с. В поведенче-
ских опытах стрекозам показывали примерно такие же по характери-
стикам стимулы. Моделью жертвы служил светлый шарик диаметром
1—2 мм, а опасность имитировали темным флагом размером 30 х
х40 см. Результаты электрофизиологических опытов свидетельствуют
о том, что взаимодействие детекторных нейронов, управляющих реак-
циями атаки и бегства, проявляется в кратковременном торможении
реакции нападения, если в этот момент возбужден нейрон, управляю-
щий бегством. Предполагается, что взаимодействием этих двух типов
детекторных систем определяется в естественных условиях конкурен-
ция между поведенческими реакциями атаки и бегства. Так, во время
бегства, вызванного приближением темного флага, стрекозы не атаку-
ют модель жертвы (светлый шарик). В свою очередь, в момент пере-
хвата добычи чувствительность насекомого к движению опасного объ-
екта понижается. В 18 случаях из 22 (82 %) стрекозы преследова-
ли цель, если им показывали темный флаг с некоторой задержкой пос-
102
ле старта, стрекозы на движущийся шарик. Взаимодействие между ней-
ронами, управляющими реакциями атаки и бегства у стрекоз, по всей
вероятности, происходит на уровне нейронов с большими рецептивны-
ми полями.
Оптомоторная реакция. Многие животные обладают способностью
реагировать на перемещение в их поле зрения различных объектов
движением глаз, головы, приближением тела или поворотом всего жи—.
вотного в сторону видимого движения. Такое компенсирующее движе-
ние в ответ на зрительные раздражители получило название опто мю-
торной реакции (ОМР).
ОМР наблюдались у ракообразных, головоногих мюллюсков, рыб,
пресмыкающихся, птиц и млекопитающих (в том числе и у человека).
Зрительно-двигательная реакция свойственна и насекомым. Впервые
ОМР у насекомых описана Радлем / Radi, 1902/. Личинки стрекоз
Aeschna в опытах /Salzle, 1932/ в ответ на движение опти-
ческого узора сначала поворачивали голову, а затем и сами начина-
ли двигаться в сторону движения. Было установлено / Gaffron,
1934/, что личинки фиксируют контрастные линии в неподвижном оп-
томоторном барабане. Если оптомоторный барабан начинает медленно
двигаться, то свободноползущие личинки следят за его движением,
перемещаясь на арене. Закрепленные в стеклянной трубке личинки
/Топпег, 1938/ отвечают на движение изображения характерными
асимметричными положениями тела. В этих опытах показано, что эф-
фект ОМР наблюдается только при движении всего оптического окру-
жения. Поведенчески' работы по ОМР у взрослых стрекоз нам не из-
вестны. Функциональ ое значение ОМР для стрекоз, как и для других
насекомых с хорошим полетом, состоит в корректировке курса при
непроизвольных отклонениях в прямом полете.
Нервной основой поддержания ОМР, по всей вероятности, служат
нейроны, чувствительные к направленному движению (дирекционно-
чувствительные, ДЧ-нейроны). Для этих нейронов характерна реакция
возбуждения на движение объекта в определенном направлении (пред-
почитаемом). При движении в противоположном (нулевом) направле-
нии активность нейрона уменьшается или полностью отсутствует. Де-
текторы направления описаны на разных уровнях зрительного анали-
затора позвоночных, некоторых ракообразных и у насекомых: саранчи,
мух, пластинчатоусых жуков и у стрекоз.
Обстоятельное изучение ДЧ-нейронов у насекомых показало, что
они обладают некоторым набором общих свойств для многих изученных
видов. Главным свойством ДЧ-нейронов, как уже отмечалось, являет-
ся способность возбуждаться на движение стимула в предпочитаемом
направлении. Реакция ДЧ-нейронов у насекомых не зависит от знака
контраста между объектом и фоном. На предпочитаемое направление
не оказывают влияния ни форма и площадь, ни яркость движущегося
объекта. Так, сложные предметы, перемещающиеся в предпочитаемом
направлении, вызывают реакцию такого же типа, как и простое тем-
ное пятно. Движение в направлении, перпендикулярном к предпочитае-
мому, реакции не вызывает.
У всех изученных объектов выделено четыре основных направле-
103
ния дирекционной чувствительности, которые, как и у позвоночных,
примерно совпадают с горизонтальной и вертикальной осями головы
насекомого. Так, у 15 ДЧ-нейронов, которые были обнаружены в
Lamiganglionaris стрекоз Aeschna viridis, Sympetrum fla—
veolum /Зенкин, Пигарев, 1969/, рецептивные поля занимали об-
ласть диаметром 40-60 град, а их ориентация распределялась сле-
дующим образом: 6 нейронов были чувствительны к движению снизу
вверх, 5 - сверху вниз и 4 - от головы к хвосту (каудально). Кро-
ме того, в коллективах стрекоз Libellula, Sympetrum, Leucor-
rhinia отводятся ДЧ-нейроны, которые в 1,5-2 раза и более
чувствительны к движению расчлененного узора (шахматным, случай-
ным), чем к узору из вертикальных полос /Францевич и др., 1978/.
Выделение направленного движения происходит в результате тор-
мозящего взаимодействия между соседними нейро мматидиями, причем
процесс торможения распространяется односторонне - в нулевом на-
правлении от возбужденного участка /Францевич, 1969/.
Рецептивные поля ДЧ-нейронов у насекомых могут быть органи-
зованы по-разному. Как показано в исследованиях на мухах /Collet,
King, 1975; Dvorak et al., 1975; Eckert, 1978/, их основу
составляют монокулярные нейроны с небольшими рецептивными поля-
ми. Монокулярные нейроны взаимодействуют между собой, а их реак-
ции суммируются на более крупных нейронах, имеющих бинокулярные
рецептивные поля, ответы которых легко регистрируются на выходе
в брюшную цепочку или в противоположную сторону мозга. Наиболее
часто встречаются случаи, когда рецептивное поле одного волокна
охватывает сетчатки обоих глаз, но предпочитаемые направления их
ориентированы противоположным образом. ДЧ-нейроны с такой орга-
низацией рецептивного поля выделяют специфический признак: враще-
ние оптического окружения, который вызывает ОМР. Также показа-
но /Bischop, Keehn, 1966 /t что в зависимости от угловой
скорости движения и периода узора частота импульсации нейронов
мух изменяется по зависимости такого же вида, как и двигательный
ответ в ОМР. Поведенческое значение дирекциональной чувствитель-
ности стрекоз к движению сложных узоров может состоять в реше-
нии задач, связанных с контролем направления и скорости полета,
обнаружения препятствий.
СЛОЖНЫЕ ФОРМЫ ПОВЕДЕНИЯ СТРЕКОЗ, УПРАВЛЯЕМОГО ЗРЕНИЕМ
Сложные формы поведения стрекоз (выбор и охрана территории,
распознавание брачных партнеров и т.п.) на нейронном уровне в прин-
ципе не исследованы, хотя в последние годы значительная часть по-
веденческих работ посвяшена этим вопросам. Существуют определен-
ные трудности при исследовании сложных форм поведения и в поле-
вых условиях, поскольку метод, условных рефлексов к стрекозам пока
применить не удалось. Работы по территориальному поведению стре-
коз носят описательный характер. В исследованиях репродуктивного
поведения стрекоз и, в частности, по распознаванию брачных партне-
104
ров применяются такие приемы, как перекраска живых насекомых
или их трупов, показ синтетических моделей и т.п.
Территориальное поведение и ориентация на местности. После ок-
рыления, и до начала репродуктивного периода стрекозы интенсивно
питаются, ведя хищнический образ жизни. Питаются они на определен-
ных участках, которые можно защитить от других стрекоз. Охрана ин-
дивидуальных участков питания от особей этого же вида или особей
другого вида рассматривается как охотничий территориализм. Терри-
ториальное поведение при добывании пищи проявляется у стрекоз, об-
ладающих мощным крыловым аппаратом, - Anax, Aeschna, Cor—
dulia /Заика, 1977; Sakagami et al.,1974/.
Особенно заметно территориальное поведение проявляется у сам-
цов некоторых видов стрекоз в репродуктивный период. Участки мест-
ности, где ожидается появление самок, самцы могут охранять как в
полете (Anax, Aeschna), так и сидя на выступающих частях субстра-
та (Calopteryx, Libellula, Sympetrum). Выбор места для по-
садки или места патрулирования над охраняемым участком в первую
очередь зависит от видовой принадлежности. Почти у всех групп стре-
коз размер охраняемого участка определяется размерами насекомого
/Corbet, 1962; Parr, 1980/ и в значительной мере зависит от
количества самцов, присутствующих в местах спаривания, т.е. от их
плотности /Мокрушов, 1982; Pajunen, 1966а; Heymer, 1969;
Ubukata, 1975; Hassan, 1978; Parr, 1980 /. Как прави-
ло, с увеличением количества особей на единицу площади уменьшает-
ся размер охраняемой территории. Для некоторых видов стрекоз (Ca-
lopteryx maculata /Waage, 1973/, Leucorrhinia rubicunda
I Pajunen, 1966a/) при высокой плотности насекомых тер-
риториальное поведение подавляется. Плотность активных самцов на
определенном участке зависит не только от их физиологического со-
стояния и погодных условий, но и от таких факторов, как топография
местности и растительность на ней, степень освещенности солнцем и
т.п. Некоторые данные об оптических признаках территории, выбирае-
мой самцами, ее размерах приведены в табл. 4.
Роль зрения при территориальном поведении у стрекоз сводится
не только к выбору по оптическим признакам участка, пригодного для
размножения. Есть сведения о том, что самцы Plathemis lydia
могут неоднократно возвращаться к своим участкам с мест охоты или
ночлега за несколько километров / Jacobs, 1955/. Самцы L. qu—
adrimaculata, пролетев несколько метров над водой в погоне за со-
перником или самкой, безошибочно возвращаются к месту старта
/Мокрушов, 1982/. Стрекозы Lestes barbara после разлета с
мест окрыления способны снова находить родные водоемы /Utzeri
et al., 1976/. Строгих доказательств о том, каким образом стреко-
зы решают эти довольно сложные ориентационные задачи, пока не су-
ществует. Можно только предполагать, что самцы многих видов стре-
коз, обладающих территориальным поведением, при возврате к водое-
мам, выборе на этих водоемах участка, пригодного для яйцекладки и
насеста на охраняемой территории, используют местные ориентиры.
Неоднократные возвраты из разных точек территории к месту старта
105
106
Таблица 4. Признаки первичного выбора территории стрекозами
Вид стрекоз Место, привлекающее стрекоз Размеры охраняемой территории Источник
Leucorrhinia rubicunda (L.) Libellula quadrimaculata (L.) Наличие открытой воды, осве- щенной солнцем Открытая вода, окруженная расти- тельностью 2-5 м2 2 1-2 особи на 1 м /Pajunen, 1966а/
L. pulchella (Drury) L. julia (Uhler) Хорошо освещенные участки воды без растительности Сфагновые болота. Насест - вет- ка 1-2 м от воды, 20-30 см от мха Вокруг насеста на 0,3-1 м /Pezalla, 1979/ /Hilton, 1983/
Orthetrum cancellatum (L.) Хорошо освещенные песчаные бе- рега с гравием 10-50 м вдоль берега /Krilner, 1977/
O. brunneum (Fonsc.) Берега ручьев. Сидит на земле Размеры в меньшей мере зависят от плотности соперников, чем от структуры ландшафта /Heymer, 1969/
O. coerulescens ( Fabr) Берега ручьев. Сидит на расте- ниях До 50 м вдоль берега, ширина 2,5 м /Неутег, 1969/
Lestes viridis (Vd Linden) На ветвях деревьев, вблизи во- доемов Полушарие радиусом 1-1,5 м /Dreyer, 1978/
Ceriagrion tenellum (De Villers ) На ручьях в болотистых местах. Избегает мест с быстрым течени- ем. Сидит на растениях На расстоянии 0,5 м от сопер- ников /Рагг, Parr, 1979/
Platycypha caligata ( Selys) Водная поверхность с топляками и корнями деревьев До 3 х 4 м /Robertson, 1982/
Tanypteryx hageni (Selys) Заливные луга. Участок над водой 1,2-1,8 м в длину, 0,6-0,9 м в ширину /Clement, Meyer, 1980/
Calopteryx virgo (L.) Ручьи с песчаным дном, быстрым течением. Сидит на растении До 50 самцов на 15 м /Pajunen, 19666 /
C. Cornelia ( Selys) Ручьи с тихим течением. Садится на камни До 5 м береговой линии /Higashi, Ueda, 1982/
C. maculata (Beaw) У ручьев. Сидит на растениях, возвышающихся иад местами яй- цекладки На расстоянии до 2 м от сопер- ников /Waage, 1973;Alcock, 1979/
стрекозы могут осуществлять по астроориентирам. Подробно теорети-
ческие вопросы ориентации животных рассматриваются в книге
Л.И. Францевича /1986/.
Репродуктивное поведение. У стрекоз существует некоторый набор
механизмов (оптических и неоптических), обеспечивающих репродуктив-
ную изоляцию между видами. К неоптическим механизмам можно от-
нести:
1) сезонную изоляцию, которая может быть между видами, похо-
жими по поведению и местам обитания, но размножающимися в разное
время сезона. Например, стрекозы родов Sympetrum, Leucorrhi—
nia;
2) временная изоляция - у многих видов стрекоз активность при-
урочена к разному времени светового периода;
3) изоляция на местности - близкие виды стрекоз предпочитают
разные места встречи брачных партнеров, хотя яйцекладку эти виды
могут осуществлять в одних и тех же местах. Наблюдая за поведе-
нием стрекоз S. flaveolum и S. sanguineum на небольшом сыром
лугу, мы практически не обнаружили у этих видов приуроченности к
каким-либо участкам луга. В то же время в выборе места посадки
виды несколько отличались. Самцы S. sanguineum предпочитали верх-
ний ярус;
4) механическая изоляция - несоответствие размеров самих жи-
вотных. У некоторых групп стрекоз обнаруживается морфологическое
несоответствие в строении анальных придатков самцов структуре пе-
реднегруди и головы самки.
Оптическая изоляция. Органам зрения принадлежит ведущее место
в выборе и распознавании брачного партнера у стрекоз. Возможно,
что для отправления таких функций, управляемых зрением, как охот-
ничье поведение, ориентация на местности и т.п., может осуществлять-
ся по яркости, и наличие цветового зрения не обязательно.
Половой диморфизм у стрекоз играет определенную роль в репро-
дуктивном поведении этих насекомых. Необходимо отметить, что цвет
стрекоз меняется с возрастом и окончательно устанавливается к мо-
менту половой зрелости. Примерно 10 % видов стрекоз имеют крыло-
вой узор, что также, вероятно, способствует видовому распознаванию.
Рассмотрены лишь некоторые вопросы, касающиеся поведения
стрекоз, управляемого зрением. Данные многочисленных опытов сви-
детельствуют о том, что в результате специализированного взаимо-
действия (возбуждения, латерального или унилатерального торможения)
между нервными элементами зрительной системы стрекоз происходит
выделение специфических признаков в оптическом окружении насекомо-
го, что, в свою очередь, запускает и поддерживает многие простые
поведенческие реакции. Несмотря на то что поведение стрекоз посто-
янно привлекает внимание исследователей разных специальностей, на
наш взгляд, еще недостаточно освещена роль зрительного анализа в
решении этими насекомыми таких задач, как ориентация на местности
(в том числе и миграции), способность самок некоторых видов стре-
коз находить субстрат для откладывания яиц. Так, яйцекладущие тан-'
демы Eryfhromma najas очень привлекаются к плавающим на по-
107
верхности воды плодам кубышки желтой ( Nuphar lutea Sibth. et Sm.)
Особенность такого поведения отмечена для стрекоз Lestes
barbara, откладывающих в условиях Сибири яйца только в сусак зон-
тичный Butoneus umbellatus L. /Заика, 1977/; охотничье пове-
дение, в частности, такое часто встречающееся явление, как "роевое
питание". Стрекозы Aeschna, Libellula собираются в многочис-
ленные стаи в местах скопления мелких насекомых. Не ясно, чем ру-
ководствуются хищники в этом случае. Привлекают ли их топографи-
ческие особенности местности, наличие жертвы или элементы подра-
жательного поведения, уже отмеченные у таких насекомых, как пчелы
/Мазохин-Поршняков и др., 1984/. Заслуживают внимания и некото-
рые другие акты поведения стрекоз. Так, только что окрылившиеся
самки Aeschna grandis, еще не окрепшие, отлетев от воды, как
правило, садятся для отдыха на растения, имеющие бурую окраску. По
всей вероятности, выбор стрекозами субстрата для посадки растения
такого цвета не был случайным.
ЭТОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ СТРЕКОЗ РОДА SYMPETRUM
Оптические признаки занимают определенное место среди различ-
ных факторов межвидовой изоляции у стрекоз. Роль их в узнавании
самцами самок своего вида у стрекоз исследовалась неоднократно
(табл. 5). Ме1Гьше опытов и наблюдений посвящено зрительному рас-
познаванию территориальными самцами самцов-соперников своего ви-
да (табл. 6). Для территориальных самцов важна способность диетант^
но узнавать особь своего вида, что позволяет им избегать как беспо-
лезных стычек с самцами, так и экономить силы, не преследуя самок
других видов. Полевые наблюдения многих исследователей свидетельст-
вуют, что очень часто самцы некоторых видов плохо различали видо-
вые признаки и проявляли агрессию к стрекозам других видов, родов
и семейств - Lestes viridis /Dreyer, 1978/, T'anypteryx ha-
geni /Clement, Meyer, 1980/, Cordulia aenea /Ubukata,
1975/, Leucorrhinia caudalis /Pajunen, 1964/, Libellula
quadrimaculata /Заика, 1977/, Orthetrum cancellatum /Kru—
ner, 1977/ и др. Несмотря на высокую способность самцов разли-
чать самок своего вида, в природе неоднократно наблюдались случаи
образования гетероспецифических пар, особенно у Libellulidae
и мелких Zygoptera.
Удобным объектом для исследования роли зрительных стимулов в
распознавании самцов самок своего вида у стрекоз являются стре-
козы рода SympetrunxСамцы S. sanguineum, S. flaveolum,
S. vulgatum, S. pedemontanum, S. danae обладают террито-
риальным поведением, а их распределение по сезону и на местности
широко перекрывается. В некоторых местах могут одновременно при-
сутствовать самцы сразу 4-5 видов. По нашим наблюдениям, в местах
скопления самцов можно было встретить одиночных самок и тандемы.
Так, 18. VIII 1982 г. на небольшом сыром лугу в период с
12 ч 20 мин до 13 ч 50 мин зарегистрировано 52 самца S. fla-
1 08
Таблица 5. Оптические признаки самок своего вида для стрекоз
Вид стрекоз Способ исследо— вания Существенные признаки Источник
Тело Цвет крыла X арак- тер полета Д емонстраци- онные позы
раз- мер цвет
брюш- ка крыль- ев
109
Calopteryx virgo
С. b.aemorrhoida-
Полевые наблюдения
Перекраска стрекоз
lis
С. maculata Полевые наблюдения
Neurobasis chinensis " *
Enallagma cyathige—
rum
Platycnemis penni- Перекраска стрекоз
pes, P. dealbata
Ischnura elegans,
I. pumilio
Nesciothemis nige— ’
riensis
Leucorrhinia dubia, Показ стрекоз на
L. rubicunda удочке
Belonia croceipen— Полевые наблюдения
nis
Platycypha caligata Показ моделей
Plathemis lydia
Perithemis tene'ra
Cordulia aenea
Полевые наблюдения
Перекраска стрекоз
Показ моделей
/Pajunen, 1966b/
/Неутег, 1973/
/Waage, 1973/
/Kumar, Prasad, 1977/
/Doerksen, 1980/
/.Buchholta, 195б/
/Krieger, Krieger—
Loibl, 1958/
/Parr, Parr, 1974/
/Pajunen, 1964/
/Williams, .1977/
/Robertson, 1982/
/Jacobs, 1955/
tf
/Ubukata, 1983/
Таблица 6. Оптические признаки самцов своего вида для стрекоз
Вид стрекоз 1 Способ иссле- дования С ущественные признаки Источник
Тело Цвет крыла Движе- ние кры- ла Демонстрацион- ные позы
раз- мер цвет
брюшка крыльев
Hetaerina americana,
Н. titia
Calopteryx haemor-
rhoidalis
Libellula pulchella
L. luctuosa, Erythe-
mis simplicicollis
Leucorrhinia dubia,
L. rubicunda,
L. caudalis
Orthetrum albistylum
Perithemis tenera
Plathemis lydia
Полевые наблюдения
Перекраска стрекоз
Полевые наблюдения +
Перекраска стрекоз
Полевые наблюдения
ж г
Перекраска стрекоз
/Johnson, 1961/
/Heymer, 1973/
/Pezalla, 1979/
/Andrew, 1966/ '
/Pajunen, 1962, 1964/
Nesciothemis nigeri— ' * + +
ensis
Pachydiplax longipen—Полевые наблюдения, +
nis спектрофотометрия
Cordulia aenea
Показ моделей
Наврав- Форма
ление брюш-
полета ка
/Ио, 1966/
/Jacobs, 1955/
/Jacobs, 1955; Campa-
nella, Wolf, 1974/
/Рагг, Parr, 1974/
/Johnson, 1962; Robey,
3 975/
/Ubukata, 19B3I
veolum и 34 тандема этого же вида. Здесь же присутствовало 117
самцов S. sanguineum и 24 тандема.
Полевые опыты проводились в окрестностях Киева в июле - сен-
тябре с 12 до 16 ч местного времени, в солнечную погоду по сле-
дующей методике. Модели стрекоз демонстрировали, применяя техни-
ку удочки /St Quentin, 1934/. Учтено 6144 реакции более чем
на 80 различных моделей. Модели 'самцов* изготовляли из синтети-
ческих материалов: резисторов, цветной изоляции проводов, крылья
имитировали прозрачной или окрашенной триацетатной пленкой. В опы-
тах по распознаванию самок использовали пять серий окрашенных мо-
делей (высушенные тела самок S. vulgatum с натуральными крыль-
ями) по четыре образца осветленности в каждой серии. Тело окра-
шивали масляными красками, близкими по окраске к монохроматиче-
скому освещению с длиной волны 540 (А), 560 (Б), 580 (В), 600
(Г) или 620 (Д) нм (рис. 11). Градации осветленности создавали
путем добавления черной краски. Коэффициент отражения самой тем-
ной (четвертой) серии составлял 14-15 %. В этих же опытах самцам
предъявляли синтетические модели разной формы, окрашенные масля-
ной краской, которая при спектрофотометрическом измерении наиболее
соответствовала натуральному цвету. Моделям приклеивали крылья са-
мок S. vulgatum. Наблюдатель показывал модель, идя от сам-
ца к самцу, до 10 раз, затем заменял ее другой. Чтобы выровнять
сезонные и погодные изменения активности самцов, в каждом опыте
показывали одну модель, эталонную для данного вида. Контролем в
опытах с моделями 'самок* служили тепа самок S. vulgatum.
Модели 'самца* проводили на расстоянии 2-0,5 м от сидящего
территориального самца исследуемого
вида. Модель 'самки* старались опус- S
тить ниже и впереди сидящего самца /
S. flaveolum с таким расчетом,
чтобы он мог видеть ее сверху. Сам-
цы S. flaveolum были выбраны для 0$
опытов потому, что в предыдущих эк-
спериментах они были самыми актив-
ными. г /
Реакции самцов классифицировались
Рис. 11. Спектральные характерис-
тики зрения и оптического окружения
стрекоз.
а - спектральная чувствительность
( S ) фоторецепторов стрекозы S. ru—
bicundulum /по: Meinertzhagen
et aL, 1983/; б - усредненные
спектры отражения (г) тепа самца
и самки четырех видов стрекоз рода
Sympetrum; в - спектры отражения
красок для моделей А-Д: А- длина
волны, нм.
111
Таблица 7. Видимые различия стрекоз рода Sympetrum
Вид стрекоз Размер самца, мм Цвет тела
длина тела размах крыльев самец
sanguineum 33,6+1,2 55,0+2,6 Кроваво-красный vulgatum 39,3+2,0 58,8 + 1,4 Вишнево-красный flaveolum 33,6+0,7 57,3+2,4 Оранжево-красный danae 31,7+0,4 49,4+1,3 Черный pedemonta- 31,8 + 0,6 53,7 £0,7 Кораллово-красный num
:на типы: 1) атака - подлет к модели на расстояние менее 5 см, до
физического контакта, повторение маневров модели в воздухе; 2) ос-
мотр - прямой полет к модели на расстояние до 10-15 см н более с
последующим возвратом; 3) бегство - полет без возврата на терри-
торию; 4) безразличие - стрекоза не взлетает; 5) тандем - захват
модели в тандем.
- Спектры пропускания крыльев записывали на спектрофотометре
SPECORD. Спектры отражения образцов применявшихся красок для
моделей и тел стрекоз (самцов и самок) (см. рис. 11, б, в) изме-
ряли с помощью спектрофотометра СФ-4 в отражающем режиме и фо-
тоумножителя ФЭУ-31. Белым стандартом служила баритовая бумага.
Так как тела стрекоз и фон (зеленые растения) слабо отражают ульт-
рафиолетовое излучение, мы ограничились анализом окрасок в видимой
части спектра. Цветовой треугольник для стрекоз Sympetrum рас-
считан по опубликованным характеристикам фоторецепторов /Meiner—
tzhagqn et al., 1983 /. За белый цвет принят равнокванто-
вый поток в видимай и ближней ультрафиолетовой части спектра. Для
расчета их характеристики спектр чувствительности отдельных типов
рецепторов нормировали к одинаковой площади под графиком (см.
рис. 11, а). Различия между распределениями реакций на разные мо-
дели устанавливали критерием .X2.
В итоге получены следующие результаты.
Распределение видов в пространстве и во времени. В местах на-
блюдения стрекоз Sympetrum первыми окрыляются и созревают
самцы S. flaveolum, затем следуют S. sanguineum, S.pedemon—
tanum, S. danae, S. vulgatum Период размножения всех пяти
видов перекрывается в августе - сентябре. Самцы S. flaveolum,
S. sanguineum занимают участки на берегах небольших водоемов,
на лугах и пересыхающих болотах. Обычно территориальные самцы
S. sanguineum занимают более высокий ярус и сидят на травинках,
кустиках, в то время как более агрессивные S. flaveolum пред-
почитают дороги, широкие листья. На растениях вдоль каналов с чис-
той водой встречаются совместно S. pedemontaniimn S. sangui—
112
Цвет тела Цвет крыловой, пластинки Рисунок крыловой пластинки
самка
Желтоватый Слегка дымчатый Отсутствует
Серо-бурый Буроватый №
Б уро-желтый Прозрачный Желтое пятно в основании крыльев
Желто-бурый № Отсутствует
Желтый tt Предвершинная бурая перевязь
neum. На дорогах пойменных лугов наиболее часто можно встре-
тить самцов S. vulgatum. Другие виды Sympetrum в этих
местах бывают редко. Территориальных самцов S. danae в боль-
шом количестве нам не приходилось встречать, хотя массовый выплод,
охоту и яйцекладку у этого вида наблюдать легко.
Видовые различия. Признаки самцов и самок пяти видов Sym—
petrum указаны в табл. 7. Наблюдение крыльев стрекоз в желтом и
ультрафиолетовом свете показало, что узор крыльев S. flaveolum
и S. pedemontanum выглядит для стрекоз гораздо контрастнее,
чем для человека-наблюдателя; крылья S. vulgatum и S. sangui—
neum заметно темнее прозрачных крыльев S. danae.
Размер и форма тела. Самцам S. sanguineum и S. vulga—
turn, имеющим прозрачные крылья, но несколько отличающимся по
размеру, показывали серии моделей с пропорциональным изменением
их длины от 27 до 50 мм. Связи между размером модели и типом
реакции не установлено.
Наиболее важный признак, привлекающий самцов, - темный центр
определенного размера (6x12 мм) и крылья вокруг него. Самцы
реагируют и на крыло без тела. В опытах с моделями "самца" мате-
риал крыла для атакующих самцов был безразличен. Пленочное крыло
можно было с успехом заменить несколькими витками тонкой прово-
локи. Подвеска модели вниз или вверх головой не имеет значения.
Стрекозы одинаково реагируют на модели без "груди". Кольцевая мо-
дель и модели с множественными элементами (тело или хвост) не
запускают атаки или вызывают бегство. Наиболее привлекательной
моделью "самца", поведенческая эффективность которой превышает
90-95 %, - пленочный диск с темным компактным центром
(рис. 12, I ).
Результаты опытов с моделями "самок" разной формы представ-
лены на рис. 12, П. Значимого развития между распределениями по-
ложительных реакций (Т и О) на контроль и последующими пятью мо-
делями при помощи критерия X-квадрат не установлено. Снижают
привлекательность моделей такие их искажения, как утолщение брюш-
- 8 Зак. 611 113
Рис. 12. Реакция самцов S. flaveolum и S. sanguineum на
модели "самцов" ( I ) и самцов S. flaveolum на модели "самок"
(II ) разной формы. Образцы моделей показаны сверху.
А - атака; О - осмотр; Б - безразличие; б - бегство; К - конт-
II
□ 6
роль - мертвый самец; - контроль - мертвая самка
turn; Т - тандем.
S. vulga—
ка или добавочная голова. Перемещение таких моделей самцы сопро-
вождают движениями головы, но не взлетают.
Узор крыла. Для самцов S. flaveolum необычайно привлека-
тельны модели с оранжевым прикорневым пятном. Крыло без тела,
но с оранжевым пятном в центре вызывает агрессию со стороны сам-
цов S. flaveolum. Сверхоптимальными были модели с желтым пят—
ном, которое занимало половину крыла (рис. 13). Самцы S. flaveo—
lum надежно отличали модели с прикорневым оранжевым пятном от
моделей без пятна, имитирующих S. sanguineum.
Для самцов S. pedemontanum 100 % атак вызывали модели с
телом и полосами по краям крыльев. Модели с прикорневым пятном и
вообще без узора не вызывали реакции преследования со стороны стре-
коз этого вида. Сверхоптимальной была черная модель с крыловым
диском, краевым секторным узором и компактным темным центром
(рис. 14). В местах обитания самцы S.pedemontanum четко отлича-
ли модели с полосами на крыльях от моделей, имитирующих стрекоз
S. sanguineum.
114
Рис. 13. Различение видоспецифичной модели по крыловому узору у
S. flaveolum (а) и S. sanguineum (б).
Образны окраски крыльев показаны сверху. Ниже основной линии
показано бегство. Усл. обозн. см. рис. 12.
б
Рис. 14. Различение видоспецифичной модели по крыловому узору у
S. pedemontanum (а) и S. sanguineum (б).
Образцы окраски крыльев показаны сверху. Ниже основной линии
показано бегство. Усл. обозн. см. рис. 12.
Цвет. Разноцветные модели 'самца* имитировали форму тела
S. sanguineum. Длина равнялась 33 мм, размер 'груди* 6x12 мм,
крыловая пластинка 40x10 мм. В этих опытах самцы плохо разли-
чали цвета, наблюдалось предпочтение желтых и красных моделей
только в сравнении с белыми и синими. В дальнейшем предъявлялись
модели с 'телом' красного цвета.
Для самцов S. pedemontanum цвет полосы на крыльях - оран-
жевый, коричневый, угольно-черный, пикриново-желтый ("ультрачер-
ный', окраска пикриновой кислотой, поглощающей УФ-пучи), пурпур-
ный (окраска сафранином, пропускающим УФ-пучи) не имел значения.
Самцы S. flaveolum отличали модели с желтым прикорневым пяг-
113
Таблица 8. Реакция самцов стрекоз Sympetrum на модели раз-
ного цвета
Уровень осветлен- ности Процент тандемов для серий окрасок
А Б В Г Д
1 9,1 17,8 51,8 13,6 О
2 12,5 37,5 37,0 27,2 0
3 9,1 31,8 30,6 66,6 9,1
4 15,0 26,1 22,0 34,6 15,0
ном, сделанным анилиновой краской, от пятна, окрашенного пикрино-
вой кислотой.
Количественные учеты реакций самцов S. flaveolum на цвет-
ные модели "самок" выявили (табл. 8), что положительная реакция
(высокий процент тандемов) на модели В. и Г приближается к та-
1 о
ковой на контрольную самку. Высокую привлекательность этих моде-
лей можно объяснить оптимально подобранной краской; желто-зеленые
и оранжевые окраски высокой осветленности резко тормозят положи-
тельную реакцию, более сильнее действуют зеленые и красные тона.
Из поведенческих опытов известно, что окраска тела и крыльев
у стрекоз может служить для распознавания особей своего вида /Ви—
chholtz, 1951, 1956; Krieger, Krieger—Loibl, 1958; He—
ymer, 1973/. У стрекоз родов Leucorrhinia и Cordulia
нет полового диморфизма по окраске. Очевидно, по этой причине сам-
цы этих родов распознают самок от самцов по характеру полета и по
форме тела /Pajunen, 1966а; Ubukata, 1983/.
В наших опытах с моделями 'самца' у стрекоз Sympetrum
ключевой стимул, запускающий атаку с высокой эффективностью, сво-
дится к простейшим концентрическим конфигурациям из компактного
тела, диска крыла и узоров на нем. Казалось бы, что для стрекоз
не менее важным ключевым стимулом должно служить удлиненное
брюшко. Но, как показали опыты с моделями, для атакующего сам-
ца Sympetrum положение брюшка и даже его отсутствие не влияют
на его действия. Именно в таком ракурсе, без выдающегося брюшка,
атакующий самец наблюдает своего соперника во время погони или
спирального полета головой к голове.
Поведение самки на территории резидентного самца отличается
от поведения самца-нарушителя. Она не вступает в борьбу, а пытает-
ся спастись бегством. Преследуя самку, самец занимает верхний эше-
лон и ориентирует свой полет вдоль тела самки. В этом случае суще-
ственную роль играют элементы формы тела (например, голова - ме-
сто захвата в тандем) и его цвет.
В том и другом случае важнейшим положительным признаком мо-
дели является наличие крыльев. С точки зрения оптики, крылья - это
материал, задерживающий или пропускающий около 10 % световых лу-
116
чей. Самцы, подлетая к сопернику снизу, рассматривают его на фоне
неба и поэтому плохо различают цвета, а крылья видят как тень.
Самку самцы наблюдают сверху, в отраженном свете. Здесь может
играть определенную роль отражение ультрафиолетовых и синих лучей
от крыльев. Возможно, из-за последнего эффекта нам и не удалось
смоделировать крылья для самки. При распознавании конспецифичной
особи сверху становится важным наличие у стрекоз цветового зрения.
Сравнение возможностей цветового зрения человека и стрекоз на гра-
фических моделях (треугольниках смешения цветов) показывает, что
стрекоза видит чистые излучения в отличие от человека менее интен-
сивно окрашенными; реальные объекты — самец, самка, фон — окраше-
ны для стрекоз и для человека одинаково ненасыщенно; самцы и сам-
ки довольно близки по окраске, различие их по цвету требует у че-
ловека и стрекоз примерно равной тонкости анализа.
Наличие у стрекоз Sympetrum красночувствительного при-
емника само по себе не обеспечивает узнавания самца на фоне расти-
тельности, но в результате определенного взаимодействия между крас-
но— и зеленочувствительными приемниками такое выделение может
осуществляться. Так, признаком цвета самца может служить возбуж-
дение красночувствительного приемника, вдвое и более сильное, чем
возбуждение зеленочувствительного рецептора. Одинаковое возбужде-
ние красного и зеленого приемников служит сигналом об однородном
фоне. Самка может быть опознана в том случае, если возбуждение
обоих рецепторов будет находиться на промежуточном уровне. Такое
взаимодействие красно- и зеленочувствительных приемников могло бы
быть основано на работе цветооппонентных нейронов, например, воз-
буждающихся красным и тормозящихся зеленым цветом. Такие нейро-
ны хорошо известны у млекопитающих, описаны у бабочек / Swi—
hart, 1970/, а механизм цветооппонентного анализа предполагался
нами для объяснения поведения Erythromma najas /Мокрушов,
Францевич, 19766/.
Виды рода Sympetrum, которые имеют заметный узор крыль-
ев, а именно S. flaveolum и S. pedemontanum, надежно
отличают по этому признаку особей своего вида от других стрекоз.
Эта способность особенно важна для редкого в исследуемом районе
S. pedemontanum, который вынужден выискивать партнеров свое-
го вида в превосходящем по численности населении других стрекоз.
S. sanguineum, лишенный узора на крыле, обладает гораздо мень-
шей избирательностью реакций. Наличие чуждых виду признаков умень-
шает привлекательность, но не уничтожает ее.
Расчеты показывают, что глаз S. pedemontanum может раз-
решить три элемента модели, а именно центр тела, прозрачную и ок-
рашенную часть крыла, только в том случае, если изображение проек-
тируется в диаметре не менее чем на пять фасеток. Критическое рас-
стояние, на котором тело и краевая полоса S. pedemontanum вид-
ны раздельно, составляет всего полметра. Поэтому для дальнего рас-
познавания особей своего вида, имеющих узор на крыле, выгодно рас-
полагать их элементы как можно ближе или как можно дальше от тела.
Было интересно узнать, существуют ли в природе стрекозы с не-
117
выгодными (срединными) элементами узора. Анализ 190 видов и
подвидов мировой фауны стрекоз (фонд ЗИН АН СССР) с заметным
рисунком на крыле показал, что подавляющее большинство стрекоз
имеет четко выраженные прикорневые или краевые элементы узора. У
восьми видов, или 4 %, темная или матово-белая полоса приближена
к середине крыла, из них только у двух проходит ровно по середине;
20 видов (10 %) имеют пестрый узор.
Роль элементов узора крыльев во взаимном распознавании стре-
коз с пестрым или срединным узором не известна. Необходимо ли им
для этого различать детали или достаточно видеть общее уменьшение
светопропускания? Также неизвестно, с какими другими видами стре-
коз им приходится сосуществовать на занимаемых территориях. Во
всяком случае дальнее узнавание особи своего вида по рисунку на
крыльях у них может быть затруднено. Таких видов немного -12-
15 % от всех стрекоз с рисунком на крыльях.
Глава III
РОЛЬ В БИОЦЕНОЗАХ
ПАРАЗИТЫ СТРЕКОЗ
Стрекозы находятся в тесных биоценотических отношениях с мно-
гочисленными организмами из разных систематических групп. Одной
из форм этих взаимосвязей является паразитизм — использование
стрекоз различными организмами в качестве хозяев в цикле своего
развития.
Стрекозы привлекли внимание паразитологов сравнительно недав-
но, но к настоящему времени они уже стали одной из наиболее изу-
ченных в паразитологическом отношении групп насекомых. Одонатоло—
гов прежде всего интересует комплекс паразитов, связанных со стре-
козами, но таких работ сравнительно немного. Эти насекомые обыч-
но служат предметом изучения более узких специалистов—паразитоло-
гов: протозоологов, трематодологов, цестодологов и т.д. Поэтому в
главе будет рассмотрено состояние изученности зараженности стре-
коз отдельными группами паразитов в разных районах европейской и
азиатской частей СССР, т.е. по систематически-территориалыюму
принципу.
ПАРАЗИТИЧЕСКИЕ ПЕРЕПОНЧАТОКРЫЛЫЕ
Некоторые виды водных перепончатокрылых из семейств Мута—
ridae Eulophidae и Trichogrammatidae откладывают
яйца в яйца стрекоз, в которых развиваются их личинки и куколки.
Впервые это явление в нашей стране описал И.И. Мечников /Metsch—
nikoff, 1866/. М.Н. Римский-Корсаков /1916, 1940/ указывал,
118
что Anagrus incarnatus Forster (сем. Mymaridae ) является
паразитом яиц стрекоз, откладывающих яйца в ткани растений: Ca-
lo pteryx, Coenagrion и Lestes. Зараженные яйца автор нахо-
дил в Бологовском озере Калининской области и в Княжем Дворе на
р. Углянка. Свои исследования он проводил также на р. Донец в
Харьковской области.
По данным А.Н. Поповой /1953а/, для СССР отмечены следую-
щие виды перепончатокрылых, паразитирующих в яйцах стрекоз: Pre—
stwichia solitaria Ruschka (сем. Trichogrammatidae),
Anagrus incarnatus Hal. ( = Polynema natans Lubb.),веро-
ятно, Polynema bergi Ashm. и P. wagneri R.-Kors. (сем.
Mymaridae).
B.H. Фурсов /1984/ впервые для фауны СССР привел четыре ви-
да хальпид из рода Tetrastichus без указания видовой принадлеж-
ности, паразитирующих в яйцах стрекоз. Позже В.Н. Фурсов и
В.В. Костюков /1987/ дали описание и привели некоторые сведения
по биологии трех новых для науки видов хальцид из этого рода: Tet—
rastichus rimskykorsakovi из яиц Zygoptera (Воронежский
заповедник), Т. zerovae из яиц Zygoptera и Aeschnidae (Там-
бовская обл.) и Т. natans из яиц стрекоз (Воронежская обл.).
В целом видовой состав перепончатокрылых, развивающихся в яй-
цах стрекоз, их роль в снижении численности хозяев и видовая при-
надлежность заражаемых яиц стрекоз еше очень мало изучены.
ЛИЧИНКИ ВОДЯНЫХ КЛЕЩЕЙ HYDRACARINA
М.Е. Крендовский /1878, 1879/ исследовал зараженность стре-
коз (и других насекомых) личинками водяных клещей в окрестностях
Харькова, в Кременчугском и Кобеляцком районах Полтавской области.
На нижней стороне груди Sympetrum meridionale, S. flaveo—
lum и некоторых видов Coenagrion и Lestes обнаружены ли-
чинки Arrenurus papillator Mull. Автор сообщает, что труд-
но было встретить стрекозу S. meridionale, на крыльях которой
было бы менее полсотни этих клещей. В течение лета одна стрекоза
S. meridionale и S. flaveolum способна вынашивать по край-
ней мере 100-200 личинок, причем передние крылья обычно несут
больше паразитов, чем задние. У разных видов стрекоз из родов
Coenagrion и Lestes автор находил личинок A. emarginator
Mull. и Limnochares aquaticus Dug. Личинки A. emar-
ginator прикрепляются кучками на нижней стороне груди между но-
гами стрекоз Libellula depressa и Anax.Переход личинок кле-
щей на имаго стрекоз осуществляется на водных растениях.
А. Ицкал /1928/ исследовал стадии развития клешей рода Ar-
re nurus в окрестностях Ленинграда. Он отметил, что зараженность
стрекоз зависит от водоема и колеблется в очень больших пределах.
• У Coenagrion pulchellum и С. hastulatum паразитировали
личинки клещей A. kjerrmani Neuman, A. maculator O.F. Mill.
(= A. cuspidator O.F. Mill.), A. glcbator O.F. Mill., A. tet—
119
racyphus Piesing; у Libellula quadrimaculata ( на одной бы-
ло до 300 экз.) - A. pustulator O.F. Mill.; у Erythromma na-
jas — A. bicuspidator Berlese и A. affinis Koenike. Изу-
чая зараженность стрекоз Литовской ССР (окрестности Вильнюса и
оз. Жувинтас) паразитами, А.П. Станёните /1962а, б, 1964 и др./
у личинок и взрослых стрекоз находила водяных клещей и их личинок
из рода Arrenurus. Они были зарегистрированы у 23 видов из
48 исследованных, т.е. у 44,2 %. Больше всех были заражены Сог—
dulia aenea — ЭИ 80,3 %, Erythromma najas — 68,0, Enal—
lagma cyathigerum - 61,2, Ischnura elegans - 45,0. Ре-
же личинки клещей встречались у Calopteryx splendens —
ЭИ 2,8; Lestes dryas — 35,0; L. sponsa — 1,0; Sympycna
annulata — 5,4; Platycnemis pennipes — 12,1; Nehalen—
nia speciosa — 60,0; Coenagrion hastulatum — 25,2;
C. pulchellum - 40,5; C. puella - 21,6; C. vernale -
33,3; Aeschna viridis — 25,0; Libellula quadrimaculata —
23,2; L. fulva — 50; Sympetrum flaveolum — 4,5; S. vul—
gatum — 18,1; Leucorrhinia albifrons — 37,5; L. pectora—
lis — 5,8; L. rubicunda — 31,6< L. dubia - 25,0. На одной
особи стрекозы насчитывалось клещей от единичных экземпляров до
нескольких сотен (максимально 930). Их личинок находили с начала
мая до сентября. У личинок стрекоз они встречались всегда непо-
средственно перед линькой на имаго.
В западных областях Украины личинки водяных клещей рода Аг—
renurus найдены на покровах тела 25 видов стрекоз (17 видов
Zygoptera и 8 - Anisoptera), личинки клешей A. papilla-
tor - у 10 видов /Павлюк, 1968, 1972в, 1975а/. ЭИ стрекоз ли-
чинками клещей составляла 11,2 %. Наиболее высокая зараженность
личинками клещей в дельте Дуная отмечена у Ischnura elegans —
ЭИ 68,1 %. I. elegans и С. pulchellum впервые зарегистриро-
ваны в качестве хозяев для клещей A. papillator /Павлюк,
19806/.
В Донбассе обследования стрекоз (5836 экз., 47 видов) на за-
раженность личинками клещей сем. Arrenuridae проводил
А.И. Олигер /1975/. Клещи были найдены на 15 видах стрекоз, что
составляет 32 % от всех обследованных. Описаны экстенсивность за-
раженности отдельных видов стрекоз, а также численное соотношение
клещей, локализующихся на крыльях, груди и брюшке у разных видов.
Б.Ф. Белышев и В.В. Шевченко /1958/ сообщают, что при обработ-
ке коллекционных материалов из Западного Тянь-Шаня много красных
личинок водяных клещей из рода Arrenurus обнаружено на крыльях
стрекоз Sympetrum vulgatum Bart. В Сибири, по многочисленным
данным Б.Ф. Белышева и А.Ю. Харитонова, личинки водяных клещей
встречаются повсеместно почти на всех видах стрекоз, кроме обитаю-
щих в личиночной фазе в проточной воде. Они обнаружены от Запо-
лярья до крайнего юга и от Урала до Приморья.
Часто личинки клещей остаются прикрепленными к покровам тела
стрекоз, хранящихся в коллекциях. Так, личинки водяных клещей об-
наружены на сухих стрекозах, много лет хранившихся на ватных мат-
120
расиках: Sympetrum tibiale, Ischnura fountainei и Sym—
pycna gobica из Кзыл-Ордынской области, Coenagrion concin-
num из Приамурья, оз. Ханка и низовьев р. Лены, С. hylas из
Алтая и п-ова Ямал, С. lanceolatum и Leucorrihinia inter-
media из Южного Прибайкалья и др. /Павлюк, 1981/.
Видовой состав паразитирующих на стрекозах личинок клешей, при-
уроченность их к хозяевам, экологические условия инвазии и ее по-
следствия, географические закономерности связей ареалов стрекоз и
клещей и многие другие стороны этого интересного явления почти не
исследованы.
ГРЕГАРИНЫ
Грегарины - наиболее распространенные и массовые, но малоизу-
ченные паразиты кишечника личинок и взрослых стрекоз. Немецкий
протоэоолог Вельмер /Wellmer, 1911/ на территории нынешней Ка-
лининградской области в кишечнике стрекоз Calopteryx virgo и
С. splendens находил грегарин Hoplorhynchus oligacan—
thus (Sieb.), а у стрекоз Coenagrion puella- грегарин Meno—
spora polyacantha L^ger.
B.H. Цветков /1929/ исследовал зараженность насекомых в ок-
рестностях Петергофа. В кишечнике личинок Aeschna sp. он на-
ходил большое число грегарин G eneiorhynchus aeschnae Grawley,
у личинок Coenagrion sp. и имаго С. hastulatum - грега-
рин Hoplorhynchus oligacanthus, у личинок и имаго С. ри—
ella — грегарин Menospora polyacantha. Для указанных ви-
дов грегарин автор дал морфологическое описание.
Г.А. Штейн /1960/ исследовала зараженность водных членистоно-
гих, в том числе личинок стрекоз, грегаринами на озерах южной части
Карелии (Пертозеро, Кончозеро, Сямозеро и Святозеро). У личинок
стрекоз рода Aeschna были найдены грегарины Geneiorhynchus
aeschnae. Автор отмечает, что меньше всего заражены самые
крупные личинки, готовые к метаморфозу. К этому времени грегарины
обычно инцистируются и выводятся во внешнюю среду. Личинки С. ha—
stulatum и С. pulchellum были заражены грегаринами Me—
nospora polyacantha на 22 % и 50 % соответственно. На этих
же озерах Карелии Е.Н. Фролова /19657 у личинок Coenagrion sp.,
С. hastulatum и С. pulchellum отметила грегарин Me—
nospora polyacantha, у личинок Aeschna sp., Anax sp. и
Sympetrum sp-. — Geneiorhynchus aeschnae; у Cordulia
aenea - Gregarina sp. 1 и у Platycnemis pennipes —
Gregarina sp. 2.
А.П. Станёните /1964/ в Литовской ССР обнаружила грегарин у
10 видов стрекоз: Enallagma cyathigerum, Coenagrion hastu—
latum, C. armatum, Erythromma najas, Cordulia aenea, Li—
bellula quadrimaculata, Somatochlora metallica, Aeschna
grandis, Leucorrhinia rubicunda и L. albifrons. Более
интенсивно были заражены равнокрылые стрекозы: С. hastulatum —
121
32,7 %, E. najas - 25,0, Е. cyathigerum - 12,5 %. Личинки
заражены более интенсивно, чем имаго. Грегарины Hoplorhynchus
oligacanthus чаще всего встречались у стрекоз подотряда Zygo—
ptera, a Menospora polycantha и Geneiorhynchus ae—
schnae — Anisoptera.
В западных областях Украины с 1965 по 1985 г. нами вскрыто
17 620 стрекоз, в основном имаго, отловленных у 170 водоемов. Из
58 видов стрекоз у 35 обнаружены грегарины Hoplorhynchus
oligacanthus, Menospora polyacantha и шесть новых для
науки видов: Hoplorhynchus tenuis, Bothryopsid.es mirabilis,
Acanthospora (?) calopterygis, Anthorhynchus (?) pul—
cherrimus, Geneiorhynchys (?) shuravnicus и Acantho—
spora (?) гага /Павлюк, 1971, 1972b, 1975b, 1980в/. Ввиду
того что спороцист от последних четырех видов со знаком (?) рас-
смотреть не удалось (форма их является важным диагностическим
признаком рода), к указанным родам они пока зачислены условно. На-
иболее высокая ЭИ отмечена у Calopteryx virgo, в среднем
55,68 + 5,3 %, но у стрекоз из разных водоемов она колеблется от
О до 88,88+6,0 %; у С. pulchellum - 42,55 + 1,4 (от 12,50 +
+ 5,8 до 90,36+3,2), у С. puella - 41,17_+1,3 и у С. hastu—
latum - 38,52+3,1 %. Средняя экстенсивность инвазии стрекоз из
подотрядов Zygoptera и Caloptera составляла 26,51 -
28,25 %, а из подотряда Anisoptera — лишь 1,78-2,72 %, т.е.
у последних она в 10 раз ниже.
В результате вскрытия 566 стрекоз 22 видов из Килийской дель-
ты Дуная у шести видов обнаружены грегарины Н. oligacanthus
и Н. tenuis. Для Anisoptera в целом экстенсивность инвазии
составляла менее 1 %, а для Zygoptera - более 9, т.е. соотно-
шение близко к указанному выше /Павлюк, 19806/.
Из Киевского водохранилища Д.П. Курандина /1981, 1982/ вскры-
ла 244 личинки стрекоз, 16,1 % из них были заражены грегаринами
Н. oligacanthus и G. aeschnae. Автор указала также вид Acti—
nocephalus sieboldi, но он является синонимом Н. oligacanthus.
Грегаринами были заражены личинки Lester sponsa, Erythromma
najas, Enallagma cyathigerum, Aeschna grandis, Sympet-
rum sanguineum.
При обследовании 34 стрекоз 10 видов из пос. Висим Свердлов-
ской области у Lestes sponsa и Enallagma cyathigerum
нами найдены грегарины Н. oligacanthus.
На юге Туркмении были вскрыты 411 стрекоз 16 видов /Павлюк,
Курбанова, 1986/. Грегаринами были заражены три вида: Calopte—
ryx splendens transcaspica(28,6 %) - главным образом гре-
гариной Acanthospora (?) calopterygis (максимальная интен-
сивность до 500 экз.), реже грегариной Bothriopsides sp. и
др.; Ischnura elegans - грегаринами Н. oligacanthus и
Н. tenuis (34,4 %, максимальная интенсивность до 50 экз.);
Epallage fatime - не определенными до вида грегаринами (4,7%).
Итак, в СССР в качестве хозяев грегарин зарегистрированы мно-
гие виды стрекоз и описано шесть новых для науки видов. Однако гре-
122
гаринофауна стрекоз изучалась пока на сравнительно небольшой тер-
ритории. Дальнейшее изучение ее, безусловно, обещает много инте-
ресных находок, в том числе новых видов паразитов.
ГЕЛЬМИНТЫ
Трематоды. Первым отечественным исследователем личинок тре-
матод - метацеркарий, развивающихся в организме стрекоз, был
Д.Ф. Синицын /1905/. Опыты по выяснению жизненных циклов тре-
матод у рыб и лягушек он проводил в окрестностях Варшавы. Скарм-
ливая церкарии нескольких видов трематод личинкам водных насеко-
мых, главным образом стрекоз, он выяснил, что дополнительными
(или вторыми промежуточными) хозяевами трематод рода Gorgode—
га являются личинки стрекоз рода Epitheca, хозяевами трема-
тод Sicrjabinoeces similis (автор ошибочно привел ее под на-
званием Haematoloechus variegatus ) и Halipegus ovo-
caudatus — личинки и имаго стрекоз Calopteryx virgo; мета—
перкарии трематоды Prosotocus confusus Д.Ф. Синицын находил
в личинках и взрослых Aeschna sp.; Pleurogenoides medians —
в жировом теле и полости тела Coenagrion sp. Для всех обнару-
женных метацеркарий трематод автор дал морфологическое описание
и рисунки.
После исследований Д.Ф. Синицына долгие годы никто тремато-
дами не занимался, а его данные с некоторыми неточностями вошли
в ряд солидных отечественных и зарубежных работ. Лишь почти пол-
века спустя MJO. Паскальская /1954/ начала изучать цикл развития
трематоды Plagiorchis arcuatus Strom, паразитирующей в
яйцеводах и фабрициевой сумке кур. По словам автора, в одной из
центральных областей СССР, в очаге заболевания и гибели кур, ли-
чинки и имаго стрекоз Coenagrion hastulatum и Lestes
sponsa были инвазированы метацеркариями Р. arcuatus, ЭИ была
0,98 %, максимальная ИИ 21 экз. Автор выяснила, что проникнове-
ние церкарий Р. arcuatus в личинки стрекоз в эксперименте
идет активно через наружные покровы тела с помощью стилета в те-
чение 30-50 мин. Возможно и пассивное внедрение через рот и во
время кишечного дыхания через прямую кишку.
Церкарии в данном случае с помощью стилета проникают сквозь
стенки кишечника в полость тела личинки стрекозы. Спустя 12 ч с
момента внедрения церкарии инцистируются в брюшных мышцах насе-
комого. К 60-му дню циста значительно увеличивается в диаметре и
метацеркария становится инвазионной. В 1953-1954 гг. в окрестно-
стях Москвы в очагах плягиорхоза кур М.Ю. Паскальской /195 9/
вскрыты следующие виды стрекоз: С. hastulatum — 4047 экз.,
L. sponsa — 1740, С. pulchellum — 9 2, Platycnemis pen—
nipes — 55, Libellula quadrimaculata — 416, Aeschna vi—
ridis - 268, Sympetrum vulgatum - 338 экз. Метацерка-
рии P. arcuatus были выявлены лишь у равнокрылых стрекоз.
Очень жаль, что в результате вскрытия только из окрестностей Моск—
123
вы 6878 стрекоз кроме метацеркарий Р. arcuatus автор не упо-
минает другие виды, которые при вскрытии должны были встретиться.
На основании обработки собранного в 1952-1955 гг. материала
на некоторых озерах Карелии Г.А. Штейн /1957/ сообщила, что у
13,3 % личинок Cordulia aenea из Пертозера (исследовано
45 экз.) выявлены метацеркарии Prosthogonimus pellucidus
(= Р. cuneatus), а в одной из шести личинок Sympetrum sp.
из Кончозера найдена метацеркария Pleurogenoides medians Ols-
son. В личинках стрекоз на юге Карелии автор находила и грегарин.
Некоторые из приведенных здесь данных повторяет в своей работе
Е.Н. Фролова /1965/. Почти одновременно появились два небольших
сообщения В.И. Здуна /1957, 1959/ о роли насекомых, в том числе
стрекоз как дополнительных хозяев трематод в условиях западных об-
ластей Украины. Эти работы показали, что круг дополнительных хозя-
ев трематод не ограничивается насекомыми, указанными Д.Ф. Синицы-
ным. Они вызвали интерес к паразитологическим исследованиям вод-
ных и амфибионтных насекомых, среди которых стрекозы занимают
видное место.
В Ковровском районе Владимирской области Т.А. Краснолобова
/1958, 1962/ проводила исследования биологии трематоды Prost—
hogonimus pellucidus (= Р. cuneatus), возбудителя заболева-
ния яйцеводов домашних птиц - простогонимоза. Из 11 исследованных
видов стрекоз метацеркариями этой трематоды были заражены: Сог—
dulia aenea, Libellula quadrimaculata, Leucorrhinia rubi—
cunda, Somatochlora metalliaca. Выявленные у стрекоз
метацеркарии были представлены тремя типами: в двухслойной, одно-
слойной цисте и свободные (неинцистированные). Автор дала описание
всех трех типов метацеркарий. В дальнейшем Т.А. Краснолобовой
/1968/ установлено, что на территории СССР существуют два вида
из рода Prosthogonimus — Р. cuneatus и Р. ovatus, а все
другие названия являются лишь их синонимами.
В Горьковской области, по данным Т.А. Краснолобовой /1961/,
метацеркариями трематоды Plagiorchis arcuatus были зара-
жены Coenagrion hastulatum, С. pulchellum и Lestes
sponsa, а метацеркариями Prosthogonimus cuneatus — Cor—
dulia aenea, Libellula quadrimaculata и Leucorrhinia
dubia. Видовая принадлежность метацеркарий из стрекоз Aesch-
na grandis и Sympetrum flaveolum не установлена.
В Литовской ССР в 1959-1962 гг. паразитологические иссле-
дования стрекоз проводила А.П. Станёните /1963а, б, 1964/. Из
379 вскрытых личинок стрекоз 125 (33,3 %) были заражены просто-
гонимами. Метацеркарии простогонимов обнаружены у 9 из 41 вида
личинок и имаго стрекоз, найденных в республике: Coenagrion
hastulatum — ЭИ 6,8 %; С. armatum - 16,6; Aeschna
grandis - 58,3; Anax parthenope—28,5; Cordulia aenea—
37,8; Somatochlora metallica — 40,0; Libellula ’quadrima—
culata — 44,1; Leucorrhinia rubicunda - 22,0 и L. albif—
rons - 12,5; максимальная ИИ 50 экз. Метацеркарии в теле личи-
нок локализовались в разных органах и частях тела, чаще всего в
124
мускулатуре брюшка, иногда встречались в голове и даже в маске и
крыловых чехлах. Метацеркарий обнаружены трех типов, как и по дан-
ным Т.А. Красно лобовой. Экспериментальное заражение подопытных
цыплят метацеркариями из стрекоз показало, что они относились к
Prosthogonimus cuneatus и реже к Р. ovatus. В. Киселене
/1967/ отмечает Aeschna grandis в качестве нового дополни-
тельного хозяина трематоды Porosthogonirrus cuneatus.
В 1977-1978 гг. на озерах Игналинской системы Литовской
ССР Т«П. Илюшина /1980/ вскрыла 1011 ?ез. стрекоз семи видов.
Из них только у двух найдены метацеркарий трематод: у Aeschna
cyanea (4 эка. из 28 вскрытых) - Prosthogonimus ova—
tus, ЭИ 14,28 %; у Sympetrum flaveolum (190 из 347 вскры-
тых) - Prosotocus confusus, ЭИ 62 %. Массовые виды
Ischnura elegans (вскрыт 151 экз.), Enallagma cyathigerum.
( 266 экз. ), Erythromma najas (203 экз.) в гельминто-
логическом отношении оказались стерильными.
В целом в паразитофауне стрекоз Литовской ССР зарегистриро-
ваны три вида трематод и общий объем паразитологических исследо-
ваний в республике пока невелик.
Из Калининградской области имеется лишь сообщение МЛ. Голи-
ковой /1960а/, которая в 1954-1957 гг. на территории птицефер-
мы совхоза "Львовский" вскрыла 211 личинок стрекоз пяти видов.
Среди них 4,3 % (5 экз. из 117 вскрытых) Lestes viridis
были заражены метацеркариями невыясненной систематической при-
надлежности, для которых дается краткое описание и рисунок. Однако
правильность определения данного вида стрекоз вызывает сомнение,
так как этот средиземноморский вид в северных областях не встре-
чается.
В Латвии проводились паразитологические исследования стрекоз
в трех пунктах центральной и северной частей республики. В 195 6-
1963 гг. из поймы р. Лиелупе близ г. Елгавы и оз. Буртниеку были
вскрыты 118 стрекоз, относящихся к шести родам (до вида они не
определялись). В них Э.Р. Ланге /1969/ нашла четыре вида мета-
церкарий. В личинках Aeschna sp. обнаружены метацеркарий
Pleurogenoides medians и, точнее, не определенные плягиорхид-
ные метацеркарий. В личинках Coenagrion sp., Libellula sp.
nCordulia sp. обнаружены метацеркарий рода Prosthogoni-
mus.
В 1970 г. появилась работа Т.А. Краснолобовой "Зараженность
стрекоз Латвийской ССР метацеркариями трематод" на основании ис-
следований 700 личинок и 7680 имаго стрекоз 18 видов, отловлен-
ных в 1963-1968 гг. на оз. Энгурес (табл. 9). Однако данные по
одному озеру не могут характеризовать паразитологическую ситуацию
во всей республике.
Здесь интересно, что у стрекоз L. sponsa найдены метацер-
карии одного лишь вида трематоды - Skrjabinoeces similis,
которые ни у какого другого вида из 8280 исследованных стрекоз не
обнаружены. Также лишь один вид трематоды - Plagiorchis lari-
cola ( = Р. elegans) —обнаружен у 400 экз. из 2 тыс. вскрытых
125
126
Таблица 9. Видовой состав и зараженность метацеркариями стрекоз на оз. Энгурес /по: Краснолобова, 1970/
Вид стрекоз Вскрыто Заражено Вид трематод
Calopteryx virgo 5 —
Lestes sponsa 100 12 Skrjabinoeces similis (Looss)
Coenagrion pulchellum 20 — —
Enallagma cyathigerum 2000 400 Plagiorchis laricola Skrjab.
Erythromma najas 15 — —
Aeschna grandis: имаго 300 90 P. laricola, Opisthoglyphe sp.?
личинки 200 28 Prosthogonimus cuneatus Rud., Prosotocus confusus
Ae. viridis 15 1 Looss, Pleurogenoides medians Olsson P. confusus
Anax imperator 25 1 Prosthogonimus sp.
Cordulia aenea: имаго 3000 1200 P. ovatus
личинки 100 24 P. cuneatus
Libellula quadrimaculata: имаго 1500 350 P. ovatus
личинки 400 28 P. cuneatus
L. depressa 50 1 Prosthogonimus sp.
Orthetrum cancellatum 100 7 Prosthogonimus sp., Plagiorchis laricola
Sympetrum flaveolum 250 82 Skrjabinoeces sp., Prosotocus confusus, Pleurogenes
S. danae 50 2 claviger Rud. P. confusus, Pleurogenoides medians
Leucorrhinia caudalis 100 12 P. cuneatus, Prosotocus sp.
L. pectoralis 100 3 P. cuneatus, Prosotocus sp.
L. rubicunda 25 8 P. cuneatus
L. dibia 25 3 P. cuneatus
Примечание. ТЛ. Краснолобова /19716/ и Л.А. Шардило, В.П. Шарпило /1972/ Р, laricola свели в синонимы
Р. elegans, в дальнейшем изложении они упоминаются как синонимы. В данной таблице в случаях смешанной инвазии да-
ется общее количество зараженных стрекоз метацеркариями всех видов трематод.
E. cyathigerum. Довольно странно, что у 1200 имаго С. ae-
nea и 350 имаго L. quadrimaculata найдены метацерка-
рии одного вида трематоды - Prosthogonimus ovatus, а у ли-
чинок тех же видов стрекоз - только Р. cuneatus. Если личинки
заражены только трематодой Р. cuneatus, а взрослые стреко-
зы - только Р. ovatus. то неясно, куда деваются метацеркарии
Р. cuneatus после метаморфоза личинок стрекоз, если у имаго их
уже нет и, наоборот, откуда у имаго стрекоз берутся метацеркарии
Р. ovatus, если их не было у личинок.
Т.А. Краснолобова дает краткое описание морфологии всех видов
выявленных метацеркарий, отмечая при этом, что метацеркарии Pro—
sthogonimus cuneatus и Р. ovatus в инцистированном состоя-
нии отличить друг от друга невозможно, а наиболее надежным спосо-
бом определения их видовой принадлежности является эксперимент.
Что касается трематод Opisthoglyphe sp. и Pleurogenes clavi—
ger, то автор не находила их в организме стрекоз, но делает вывод
о возможности включения стрекоз в качестве дополнительных хозяев
этих трематод на том основании, что они были найдены при вскрытии
лягушек, которым за несколько дней до этого скармливались целые,
не подвергавшиеся исследованию на наличие паразитов стрекозы. Све-
дений о паразитировании в стрекозах метацеркарий этих двух видов
трематод в мировой литературе к настоящему времени не было.
На материале оз. Энгурес Т.А. Краснолобова /1971а/ проводила
экспериментальное изучение жизненного цикла трематоды Plagior—
chis laricola ( = Р. elegans). Оказалось, что цикл развития
этой трематоды может проходить двумя путями: 1) с участием одно-
го хозяина - моллюска, в котором проходит одновременное развитие
спороцист, содержащих церкарий и инцистированные метацеркарии,
2) с участием моллюска как промежуточного хозяина и разных бес-
позвоночных: насекомых (в том числе стрекоз) и ракообразных в ка-
честве дополнительных хозяев. У церкарий этой трематоды отсутству-
ет избирательная способность в выборе дополнительного хозяина, но
из 18 видов стрекоз зараженных ими было лишь три.
Т.Г. Никулин /1967, 1969, 1971/ сообщает, что при обследова-
нии 15 озер и одной реки в северной часта Белоруссии у 31 % личи-
нок стрекоз обнаружены личинки гельминтов. При экспериментальном
скармливании подопытным утятам личинок стрекоз L. sponsa и
S. vilgatum утята заразились эхиносто матицами (Echinostoma
revolutum и Hypodaereum conoideum), от А. суапеа - тре-
матодой Catatropis verrucosa из сем. Notocotylidae.
Особый интерес, представляет впервые зарегистрированный автором
факт заражения стерильных в гельминтологическом отношении утят
трематодами Е. revolutum, Н. conoideum и с. verrucosa по-
средством личинок стрекоз. По-видимому, личинки стрекоз в данном
случае играют роль резервуарных хозяев. Автор считает, что метацер-
карии нотокотилид могли втянуться с илом в "хвостовую часть", т.е.
в бронхиальную камеру прямой кишки личинок стрекоз и задержаться
вплоть до поедания их птицами. Из 1310 стрекоз (775 имаго и 535
личинок), отловленных в окрестностях птицеферм Витебской и Грод-
127
ненской областей, метацеркариями простогонимид (в основном Pro—
sthogonimus cuneatus) были инвазированы 12 видов стрекоз:
Calopteryx splendens, Lestes dryas, L. sponsa, Gomph-
us vulgatissimus, Aeschna cyanea, Ae. grandis, Epitheca
bimaculata, Cordulia aenea, Libellula depressa, L. quadri—
maculata, L. fulva и Sympetrum vulgatum. Наибольший про-
цент инвазии наблюдался в мае - 3 6,3 %, к ноябрю он снижался до
15 %. Метацеркарий простогонимид локализуются чаще всего в брюш-
ном (до 25,5 %) и в грудном отделах (до 21,1 %), значительно ре-
же - в головном (до 2,8 %).
Е.А. Литвиенковой /1970/ из Глубокского, Городокского и Ви-
тебского районов Витебской области вскрыто 180 имаго девяти ви-
дов и 72 личинки трех видов стрекоз. Метацеркариями простогониму—
сов были заражены 38,3 % имаго (Ае. grandis - ЭИ 77,7 %;
L. quadrimaculata — 75,0; L. depressa — 72,2; L. fulva —
5,0; G. vulgatissimus — 66,6; E. bimaculata — 57,1;
C. splendens — 27,5) и 25 % личинок: S. scoticum —
27,6 и Gomphus sp. — 50,0 %. Максимальная ИИ у разных
видов составляла 12-19 экз.
Таким образом, по гельминтофауне стрекоз Белоруссии имеются
лишь фрагментарные сведения, причем большая часть ее территории
не обследована.
С севера Волынской области Украинской ССР В.И. Здуном /1957,
1959/ вскрыты 150 личинок и имаго Calopteryx virgo, Aesch—
па sp., Cordulia aenea и Epitheca bimaculata. Най-
дены метацеркарий четырех видов трематод. Из района среднего тече-
ния р. Северского Донца и пойменных водоемов Харьковской области
вскрыты 1425 личинок стрекоз 10 видов /Шевченко, Барабашова,
1960; Шевченко, Вертун, 1961; Шевченко, 1962, 1976/. Наиболее
высокая инвазия метацеркариями трематод отмечена у S. flaveo—
lum — ЭИ 40,4 %; Ае. isosceles - 28,5; Ае. grandis —
19,5 и С. aenea - 16,1 %. Установлено, что личинки стрекоз на-
ряду с ручейниками являются второстепенными дополнительными хо-
зяевами трематоды Pleurogenoides medians, главные допол-
нительные хозяева ее - жуки-плавунцы. Единичные метацеркарий этой
трематоды найдены у личинок S. flaveolum, Ае. isosceles, Ае.
grandis, Е. najas и L. sponsa. Личинки S. flaveolum,
Ае. isosceles, Ае. grandis, Ае. viridis, С. aenea и
С. puella — главные дополнительные хозяева трематоды Prosoto—
cus confusus, а личинки жуков - второстепенные. У личинок
С. aenea, Е. cyathigerum, С. puella и Е. najas Н.Н. Шев-
ченко находила неопределенные метацеркарий со слабо выраженными
морфологическими признаками, с оболочкой цист, покрытой волосками.
У одного вида личинок стрекоз (вид не указан) автор находила мета—
церкарии Pleurogenes claviger.
У личинок Aeschna sp. из дельты Дуная Т.И. Комарова
/1968/ выявила метацеркарий трематоды Pleurogenoides me-
dians.
Из 566 вскрытых нами /Павлюк, 19806/ взрослых стрекоз 22
128
видов из Килийской части дельты Дуная метацеркариями трематод
были инвазированы 22,08 % стрекоз 15 видов. Наиболее высокая ЭИ
отмечена у S. vulgatum - 83,33 %, сильная зараженность наблю-
далась метацеркариями Skrjabinoeces similis — ЭИ 11,66 %
(10 видов стрекоз) и Prosotocus confusus - 6,89 (12 видов).
Реже они поражались метацеркариями Plagiorchis elegans —
ЭИ 2,65 % (5 видов), Prosthogonimus sp. - 1,41 (6 видов) и
Pleurogenoid.es medians—1,41 (2 вида). Довольно редкими па-
разитами были метацеркарии Halipegus ovocaudatus — ЭИ 0,54 %
(2 вида) и Corgodera sp.—0,54 (1 вид). В качестве дополнитель-
ных хозяев впервые зарегистрированы: S. meridionale — для
трематод Pleurogenoides medians, Prosotocus confusus
и Prosthogonimus sp.; C. erythraea — для P. confusus,
Skrjabinoeces similis, Plagiorchis elegans и Prostho—
gonimus sp.; E. viridulum — для P. confusus; C. ornatum-
для P. confusus и Prosthogonimus sp.; I. elegans - для
Prosthogonimus sp.
На территории семи западных областей Украины (Волынская, Ро-
венская, Львовская, Тернопольская, Ивано-Франковская, Закарпатская
и Черновицкая) с 1965 по 1985 г. из более 170 водоемов отловле-
ны и вскрыты 17 620 стрекоз 58 видов в основном в имагинапьной
фазе /Павлюк, 1972-1976, 1978, 1980а и др.; Павлюк, Стражник,
1985/. Предварительные вскрытия показали, что зараженность обоих
полов стрекоз одинакова. Принимая меры по охране стрекоз, вскры-
вали преимущественно самцов (около 80 %). В случаях вскрытия са-
мок более редких видов, яйца, извлеченные из них, помещали в со-
суды с водой и выводили из них личинок, так как у многих самок
часть яиц была уже оплодотворена и готова к развитию. Личинки на
первых порах подкармливались культурой инфузорий и планктонных ра-
кообразных, а затем выпускались в водоем. Редких, заслуживающих
охраны видов стрекоз вовсе не вскрывали. Личинок для вскрытия бра-
ли лишь последнего возраста, содержащих более полный набор парази-
тов. Данные вскрытия личинок стрекоз без учета их возраста не мо-
гут дать достоверной картины паразитологической ситуации, так как
многие личинки младших возрастов еще не ’ успели заразиться. К тому
же дефинитивные хозяева заражаются личинками гельминтов в основ-
ном при поедании взрослых стрекоз, содержащих и больший процент
инвазионных метацеркарий, чем личинки.
Метацеркарии рода Prosthogonimus обнаружены у 17 видов
стрекоз, 9 из которых в качестве дополнительных хозяев зарегистри-
рованы впервые. Найденные у 15 видов стрекоз метацеркарии на ос-
новании неточного рисунка и описания MJO. Паскальской вначале были
определены нами как Plagiorchis arcuatus, в дальнейшем выясни-
лось, что они относятся к виду Prosthogonimus ovatus, а ос-
тальные метацеркарии рода Prosthogonimus — к виду Р. cu-
neatus. У 35 видов стрекоз обнаружены метацеркарии Plagiorchis
elegans, причем 31 вид стрекоз в качестве дополнительных
хозяев зарегистрирован впервые. Максимальная ИИ 34 экз. отмечена
у Anax imperator. У 30 видов стрекоз (из них у 25 видов впер-
9 Зак. 611
129
вые) обнаружены метацеркарий Skrjabinoeces similis.
Максимальная ИИ (117 экз.) отмечена у S.sanguineum. У 28 ви-
дов (из них у 19 впервые) - метацеркарий Prosotocus confu—
sus. Как правило, у равнокрылых стрекоз метацеркарий Р. confu—
sus имеют лишь одну прозрачную цисту, а у разнокрылых в боль-
шинстве случаев они снаружи покрыты еще плотной коричневой обо-
лочкой и в 2-3 раза мельче, чем у первых. По-видимому, наружная
оболочка является соединительно—тканным выделением стрекозы—хо-
зяина как защитная реакция организма, изолирующая метацеркарий и
сдерживающая поступление к ним питательных веществ, вследствие
чего такие метацеркарий не достигают больших размеров.
Максимальная ИИ до 60 экз. отмечена у Sympetrum flaveo—
Lum и S. sanguineum. У 18 видов стрекоз (из них у 13 впервые)
зарегистрированы метацеркарий Pleurogenoides medians; мак-
симум ИИ (28 экз.) - у S. flaveolum. У 25 видов стрекоз (из
них у 23 впервые) найдены метацеркарий рода Gorgodera poss;
максимальная ИИ (91 экз.) - у S. vulgatum. В кишечнике 17 ви-
дов стрекоз впервые отмечены свободные, очень подвижные метацер-
карии Halipegus ovocaudatus. ЭИ и ИИ довольно низкие, мак-
симальная интенсивность была у С. pulchellum. У 22 видов стре-
коз найдены также метещеркарии трех видов трематод, видовую при-
надлежность которых выяснить не удалось.
Очень часто наблюдалось сопаразитирование в одном хозяине раз-
ных видов, составляющих паразитоценоэы всех возможных комбинаций.
Исследования показали, что множественные инвазии стрекоз паразита-
ми, в частности, метацеркариями разных видов трематод - явление
довольно обычное /Павлюк, 1983а/. При этом соотношения видов при
множественных инвазиях колеблются в определенных пределах, что
можно проиллюстрировать на двух наиболее сильно поражающихся ме-
тацеркариями видах: Sympetrum sanguineum и S. vulgatum.
Из числа инвазированных особей метацеркариямй одного вида трема-
тод S. sanguineum составляли 68,55, a S. vulgatum- 70,02 %,
двумя видами - соответственно 23, 38 и 23,34 %, тремя - '8,46 и
6,36 %, четырьмя - 1,61 и 1,06 % и пятью - 0,40 и 0,00 %, т.е.
эти соотношения для обоих видов стрекоз очень близки. Если принять
всех инвазированных стрекоз за 100 %, то инвазированные одним ви-
дом трематод составят около 70 %, двумя - примерно в 3 раза мень-
ше, тремя - также примерно в 3 раза меньше, чем двумя, и четырь-
мя - в 5-6 раз меньше, чем тремя. Эти соотношения позволили выя-
снить некоторые формы взаимоотношений паразитов со стрекозами-хо-
зяевами /Павлюк, 19836/.
Выяснилось, что метацеркариями трематод поражаются почти все
(а возможно, и все) виды стрекоз, и гостальная специфичность их по
отношению к своим хозяевам колеблется в широких пределах /Павлюк,
1986/. Степень инвазированности стрекоз метацеркариями различна,
что обусловлено в основном экологическими факторами. Численное со-
отношение паразитов у стрекоз различно в разных биотопах и в раз-
ные годы /Павлюк, Стражник, 1985/, но в целом стрекозы западных
областей УССР в наибольшей степени поражаются метацеркариями
130
Skrjabinoeces similis, затем Prosotocus confusus,
Plagiorchis elegans, Gorgodera sp., Prosthogonimus sp.,
Pleurogenoides medians, Halipegus ovocaudatus и мета-
церкарии невыясненной видовой принадлежности.
Для. выяснения паразитологической ситуации в окрестностях Кие-
ва вскрыто 148 личинок последнего возраста и имаго в основном
двух наиболее сильно поражающихся трематодами видов стрекоз -
Sympetrum sanguineum и S. vulgatum /Стражник, Павлюк,
1986/. У них зарегистрированы Prosotocus confusus —
20 %, Skrjabinoeces similis — 12, Prosthogonimus sp. —
7,7, Pleurogenoides medians — 6,7, Gorgodera sp. — 4,8
и единично Plagiorchis elegans,
В Ярославской области из оз. Неро О.П. Кулагиной /1984/
вскрыто 384 личинки и 157 имаго стрекоз 10 видов, которые на
17,5 % были заражены метацеркариями семи видов трематод
(табл. 10). Видовая принадлежность личинок гельминтов была уста-
новлена экспериментально путем скармливания их позвоночным живот-
ным и последующим их вскрытием.
Среди личинок гельминтов на оз. Неро первое место занимают
виды рода Prosthogonimus, обнаруженные у восьми видов стре-
коз. Наибольшее число видов гельминтов (б) обнаружено у Cordu—
lia аепеа. Заслуживают внимания случаи обнаружения метацер-
карий Asymphylodora sp. у С. аепеа и адолескарий Notoco—
tylus attenuatus у Lestes virens.
A.E. Жохов /1984/ в Ярославской области установил ЭИ у бес-
позвоночных животных двух водоемов. В числе беспозвоночных были
вскрыты 32 особи трех видов стрекоз из городского загрязненного
водоема (они оказались незараженными) и 130 стрекоз из удален-
ного на 30 км от города водоема, где стрекозы были заражены ме—
тацеркариями Prosotocus confusus: Aeschna grandis и Ae.
cyanea — no 40 %, Coenagrion armatum - 2,3 и Ischnu—
ra pumilio — 9,5 %.
Из окрестностей Казани, Волжско-Камского заповедника Татар-
ской АССР с 1973 по 1978 г. исследовано 786 личинок и 162
имаго стрекоз 28 видов /Любарская, Галеева, 1980а, б; Любарская,
Соколина, 1983/. Общая зараженность стрекоз составила 46,8 %.
Имаго были заражены на 35,1 (+3,7), а личинки - на 49,2
(+1,8) %. У разнокрылых стрекоз ЭИ (56,3+1,4 %) была почти в
3 раза выше, чем у равнокрылых (18,8+2,4 %). Наиболее заражен-
ными оказались Cordulia аепеа: 74,8 % - личинки и 96,8 % -
имаго; Libellula quadrimaculata — 52,9 и 85,7 % соответст-
венно; Aeschna grandis - 75,0 % (личинки),Coenagrion ha-
stulatum - 33,3 (личинки), Lestes sponsa - 3 7,5 % (имаго).
Выявлены метацеркарии восьми видов трематод: Prosotocus con-
fusus, Pleurogenoides medians, Skrjabinoeces similis,
Plagiorchis elegans, Prosthogonimus sp., Halipegus sp.,
Gorgodera sp., Laterotrema (?) sp. Последний в стрекозах за-
регистрирован впервые. При скармливании стерильной в отношении
трематод.лабораторной мышы двух экземпляров Libellula quadrima—
131
132
Таблица 10. Видовой состав и зараженность гельминтами стрекоз на оз. Неро /по: Кулагина, 1984/
Вид стрекоз Вскрыто Заражено ЭИ Вид гельминтов
Lestes virens 34 3 8,8 Notocotylus attenuatus,' Prostho— gonimus cuneatus
Coenagrion pulchellum 27 8 29,6 Prosotocus confusus, Prostho- gonimus ovatus
Anax imperator 5 3 60 Plagiorchis laricola, Tatria de- ca cantha
Aeschna juncea 19 ' 5 26,3 Plagiorchis laricola, Prosthogo— nimus ovatus
Ae. cyanea 39 7 18 Plagiorchis elegans, Pleuroge— noides medians, Prosotocus con- fusus, Prosthogonimus ovatus
Ae. grandis 35 4 11,4 Prosotocus confusus
Cordulia aenea 104 52 50 Plagiorchis arcuatus, P. lariocc- la, Pleurogenoides medians, Pro— sthogonimus ovatus, P. cunea- tus, Asymphylodora sp.
Somatochlora metallica 45 б 13,3 P. ovatus
Somatochlora sp. 67 5 7,4 Prosotocus confusus, Prostho- gonimus ovatus, P. cuneatus
Libellula quadrimaculata 19 2 10 Prosthogonimus ovatus
culata получена половозрелая трематода Prosthodendrium
chilostomum (Mehl., 1931) из сем. Lecithodendriidae.
Степень зараженности стрекоз метацеркариями варьирует в зави-
симости от сезона. Наиболее высокая она в мае, к середине лета
уменьшается и вновь возрастает к осени. Часто отмечалась множест-
венная инвазированность стрекоз. Особенно это характерно для ряда
видов с высокой ЭИ -С. аепеа, Ae. grandis, L. quadrimaculata.
Восемь видов из подотряда Anisoptera были заражены на 55,8 %
метацеркариями рода Laterotrema (?) sp. Основным дополнитель-
ным хозяином у них оказалась стрекоза Cordulia аепеа - 28,8 %
от общего числа зараженных стрекоз при максимальной интенсивности
до 116 экз., второстепенными хозяевами были стрекозы родов Ae-
schna и Libellula. Десять видов из подотряда Anisoptera и
один из отряда Zygoptera (43,6 % стрекоз) были заражены мета-
церкариями Prosotocus confusus, основным хозяином для которо-
го служат стрекозы рода Aeschna (22,7 %), второстепенными -
виды родов Libellula и Sympetrum. Метацеркариями Skrja—
binoeces similis были заражены 21,4 % стрекоз (6 видов из под-
отряда Anisoptera и 6 - Zygoptera), основные его хозяева
Cordulia аепеа (9,8 %) и Coenagrion hastulatum (4,9 %).
Метацеркариями Plagiorchis elegans были заражены 12,6 %
разнокрылых стрекоз, основной хозяин - Cordulia аепеа (10,1 %),
второстепенный - Aeschna grandis. По одному виду равнокрылых
стрекоз были заражены метацеркариями Gorgodera sp. (0,9 %)
и Halipegus sp. (0,3 %).
В окрестностях Воронежа ПА. Положенцевым и В.П. Негробовым
/1958, 1967/ в 1956 г. отловлены и вскрыты 1207 личинок и
имаго стрекоз. Личинки Aeschna isosceles — 122, Ischnura
elegans - 58, Cordulia sp. - 513, Libellula quadrimacu-
lata — 62, Lestes sp. — 280; имаго Calopteryx virgo —
115, Libellula depressa - 57. Метацеркариями единственного
вида трематод Gorgodera sp. были заражены лишь Les—
tes sp. — ЭИ 60 %, C. virgo — 54,8 и L. depressa — 15,
с максимальной - ИИ 180 экз. Метацеркарии локализовались в груд-
ной полости.
В.Е. Судариков и Е.М. Карманова /1964/ сообщают, что в дель-
те Волги на территории Астраханского заповедника в числе разных
видов беспозвоночных были вскрыты стрекозы (92 личинки) четырех
видов, из которых у Enallagma cyathigerum и Gomphus sp. най-
дено по одной метацеркарии без указания видовой принадлежности.
Т.Л. Илюшиной /1979/ на этой же территории отловлены и вскрыты
2274 стрекозы десяти видов. Метацеркариями трематод были зара-
жены: Erythromma najas - ЭИ 12,7 %; Enallagma cyathige-
rum - 1,7; Ischnura elegans - 20,7; Platycnemis sp. -
64,8; Cordulia aenea — 62,7; Sympetrum vulgatum — 63 и
Aeschna cyanea - 60. Метацеркариями Prosthogonimus ova-
tus - Ae. cyanea и S. vulgatum, ЭИ 3.5%, ИИ 1-3 экз.;
метацеркариями Prosotocus confusus — Ae. cyanea ( ЭИ
30 %), S. vulgatum (17,8), C. aenea (62,7); Gorgodera
sp. I — Platycnemis sp. ( ЭИ 7,5 %) и I. elegans (42,3),
а метацеркариями Gorgodera sp. II - Platycnemis sp. (ЭИ
2,8 %).
Впервые в мускулатуре тела Е. najas найдена метацер—
кария Asymphylodora sp. — паразита рыб (ЭИ 0,17 %), а
в мускулатуре брюшка личинки S. vulgatum — две инцистирован-
ные метацеркарий Halipegus occidualis, причем данный вид
трематоды зарегистрирован впервые на территории СССР. У
S. vulgatum, I. elegans, Е. najas, Е. cyathigerum, Platy—
cnemis sp. найдены метацеркарий Plagiorchis elegans -
ЭИ 6,9 %, ИИ 1—7 экз. Метацеркарий Skrjabinoeces si—
milis найдены в мускулатуре тела L elegans и Platycnemis
sp. _ ЭИ 3,5 %, ИИ 1-2 экз.
На территории Астраханского заповедника Т.А. Краснолобова
/1979/, проводившая исследования цикла развития трематоды Р1а—
giorchis elegans, выяснила, что найденные у Е. cyathigerum
метацеркарий полностью идентичны описанным В.Е. Судариковым и
Е.М. Кармановой от слепня под названием Р. laricola.
В 1970—1974 гг. П.П. Дремковой /1976/ исследовано 956 ли-
чинок стрекоз семи видов, отловленных в пойменных озерах, криках и
маленьких временных водоемах Волго-Ахтубинской поймы. У 12,9 %
личинок обнаружены метацеркарий трематод, цистицеркоиды цестод и
личинки нематод. Каждый вид личинок стрекоз оказался дополнитель-
ным хозяином только одного вида гельминта. По два вида плагиорхид-
ных и горгодеридных метацеркарий найдены у фитофильных личинок
равнокрылых стрекоз из мелководных пойменных озер и временных во-
доемов. ЭИ плагиорхидными метацеркариями составляла 0,5 %, ИИ
1-3 экз. Горгодеридные метацеркарий встречались у 10,6 % личинок
стрекоз, ИИ 1—5 экз. У личинок равнокрылых и разнокрылых стрекоз
находили метацеркарий Prosthogonimus ovatus и Р. cunea—
tus. Видовая принадлежность личинок стрекоз и найденных у них па-
разитов, за исключением простогонимусов, автором не указаны.
На Южном Урале (Учалинский р-н Башкирской АССР, озера Бол.
Учалы, Карагайлы и Ургун) М.Г. Баяновым /1974, 1986/ вскрыты
902 стрекозы - 3 93 личинки и 509 имаго 13 видов. Найдены мета-*
церкарии Plagiorchis elegans: у Coenagrion armatum,
С. pulchellum, Enallagma cyathigerum-ЭИ 31,25 %, ИИ
1-3 экз., у Anax imperator, Cordulia aenea -ЭИ 26,8, ИИ
до 22 и у Libellula quadrimaculata- ЭИ 2,5 %, ИИ 1-3 экз. В
подопытном цыпленке, которому были скормлены имаго С. pulchel—
lum, найдены по одной трематоде Schistogonimus rarus и Echi—
noparyphium aconiatum. У двух видов стрекоз (виды не указа-
ны) найдена Echinostoma revolutum и Plagiorchis папа.
Автор отмечает, что паразитирование у стрекоз эхиносто матид носит,
скорее всего, случайный характер. Метацеркарий Prosthogonimus
cuneatus найдены у Cordulia aenea - ЭИ 43,5 %, ИИ до
70 экз.; Libellula quadrimaculata — ЭИ 23,8, ИИ до 79 и
Aeschnidae gen. sp.; Р. ovatus - у Anax imperator—ЭИ
15,8 %, ИИ 25 экз., L. quadrimaculata и Aeschnidae gen.
134
sp. ; а метацеркарии Skrjabinoeces similis — у Cordulia
aenea.
М.Г. Баянов /1974/ наблюдал в оэ. Карагайлы массовое зара-
жение стрекоз сем. Aeschnidae: Ae. grandis, Ae. juncea, Ае.
cyanea, Anax imperator, прогенетическими метацеркариями
Prosotocus confusus при отсутствии там окончательных хозяев
этой трематоды - бесхвостых амфибий. ЭИ достигала 63,5 %, ИИ
2-73 экз. Большинство стрекоз (58,3 %) было заражено метацер-
кариями только этого вида трематоды. Искусственное заражение про-
генетическими метацеркариями лягушек показало, что такие метацер-
карии не способны к длительному паразитированию в организме амфи-
бий. Причину перехода метацеркарий Р. confusus к прогенетиче-
скому размножению автор объясняет исчезновением на озере оконча-
тельного хозяина. В искусственно зараженной личинке Ae. juncea
обнаружены в голове 24, груди 104, брюшке 165, конечностях 38
(по 6—7 в каждой) метацеркарий. Опыты показали, что заражение
моллюска происходит при проглатывании яиц.
На Среднем Урале в Свердловской области НДЗ. Чуриной /1964/
вскрыто 364. личинки и 145 имаго стрекоз и установлено, что
L. quadrimaculata и Е. najas являются дополнительными хо-
зяевами трематоды Prosthogonimus cuneatus, причем для
Е. najas это отмечается впервые. Автором установлен факт развития
трематоды Notocotylus attenuatus двумя путями: с образованием
адолескарий и с образованием метацеркарий, т.е. с включением допол-
нительных хозяев, играющих роль резервуара инвазии в водоемах. В
условиях Среднего Урала взрослые стрекозы были заражены метацер-
кариями трематод на 28,2 %, а личинки - на 10,1 %. Личинки боль-
ше заражены в естественных прудах (17,9 %), значительно слабее в
пойменных (1,3 %), а в озере оказались стерильными. Высокая ин-
вазированность (43,1 %) личинками трематод отмечена и у взрослых
стрекоз, отловленных вблизи естественных прудов. Стрекозы, пойман-
ные у озера и пойменных прудов, были свободны от инвазии.
В.И. Окороков и В.А. Ткачев /1974/ исследовали зараженность
стрекоз гельминтами на оз. Песчаное в Челябинской области. Вскры-
то 2700 личинок семи видов. Зараженность гельминтами составляла:
весной 12,2, летом 4,2 и осенью 1 %. В личинках Coenagrion
vernale найдены метацеркарии Plagiorchis elegans (у ав-
торов под названием Р. laricola), ЭИ 0,2 %; у Libellula quad—
rimaculata — Prosthogonomus cuneatus, 1,6 %; у Cordulia
aenea — P. elegans, P. cuneatus и Echinostoma sp.;
у Aeschna viridis — P. cuneatus, P. ovatus, Echinosto—
ma revolutum и P. elegans; у Ae. grandis — P. cunea-
tus, P. elegans и Plagiorchis sp.
На озерах Карасукской системы - Титове, Кусган и Кротова Ля-
га в Новосибирской области - Т.Л. Илюшиной /1973, 1975/ иссле-
довано 8767 стрекоз семи видов. Метацеркариями трематод были за-
ражены имаго Aeschna juncea - ЭИ 27,7 %, Sympetrum
vulgatum — 52,0, S. flaveolum - 47,6, Lestes sponsa —
5,88 и Enallagma cyathigerum — 2,2 %. Метацеркариями Pla—
135
giorchis laricola (= P. elegans) — Aeschna juncea, Sym-
petrum flaveolum, S. vulgatum, Lestes sponsa и
Enallagma cyathigerumНаиболее высокая ЭИ отмечена у S. vul—
gatum (46,7 %) и S. flaveolum (44 %). Характерно, что в роде
Sympetrum зараженными были только самки. Последнюю особенность
приходится считать либо удивительной случайностью (из 4197 особей
рода Sympetrum около половины были заражены), либо ошибкой в
определении пола стрекоз, так как наш обширный фактический матери-
ал свидетельствует, что степень инвазированности любого вида стре-
коз для обоих полов примерно одинакова.
Метацеркарии Opisthoglyphe sp. зарегистрированы у S. vul—
gatum (ЭИ 0,32 %, ИИ 1-3 экз.) и S. flaveolum (0,1 и 1);
Pneumonoeces sp.- в цистах у одной стрекозы S. flaveolum
(0,05 и 2); Prosthogonimus ovatus - у S. flaveolum (ЭИ
5 %), S. vulgatum (o,9), Ae. juncea (1,8); Prosotocus
confusus - у S. flaveolum ( ЭИ 1,41 %), S. vulgatum
(o,19) и у Ae. juncea ( ЭИ 0,6 %); Pleurogenes clavi—
ger - у личинок S. vulgatum (ЭИ o,3 %, ИИ 4-7 экз.);
Pleurogenoides medians - у S. vulgatum (ЭИ 2 %). В
1970 и 1971гг. в единичных случаях обнаружены метацеркарии
Asymphylodora sp. у S. flaveolum - ЭИ 0,14, которые впер-
вые зарегистрированы вообще у насекомых. В среднем ЭИ стрекоз
метацеркариями трематод составляла 30,54 %, максимальная ИИ
38 экз. По сравнению с другими группами насекомых зараженность
стрекоз была наиболее высокой.
С.В. Карпенко и В.В. Заика /1979/ в Северной Кулунде изуча-
ли экологию трематоды Plagiorchis elegans. Наибольшая экс-
тенсивность инвазии метацеркариями отмечена у стрекоз S. vulga—
turn - 46,7 и S. flaveolum - 44 %, т.е. точно такая же, как и
по данным Т.Л. Илюшиной /1973/ для Карасукских озер.
В 1979 г. нами вскрыто 114 стрекоз шести видов из с. Майма,
расположенного на высоте 830-850 м над ур. м. в Горно-Алтайской
АО (материал не опубликован). У Sympetrum vulgatum найдены
метацеркарии Prosthogonimus sp., ЭИ 21,7 %, у S. flaveolum —
метацеркарии Skrjabinoeces similis — 1,7; Plagiorchis slo-
gans — 1,7; Prosotocus confusus — 8,6 и Gorgodera sp. —
1,7 %.
Из Кургальджинских озер Целиноградской области Казахской ССР
вскрыто 216 стрекоз восьми видов /Климишин, Павлюк, 1972/. Сре-
ди них у пяти видов найдены метацеркарии трематод (ЭИ 22,18):
Prosthogonimus sp. - у Aeschna coluberculus, ЭИ 18,18,
Sympetrum vulgatum- 10 и S. flaveolum - 0,86; Plagiorchis
elegans - у Aeschna juncea (у четырех из шести), Ae.
coluberculus - 18,18, S. flaveolum - 15,5, S. scoticum-
24,4, S. vulgatum - 22,66; Prosotocus sp. - у S. flaveolum -
5,17. Этот район Д. Кукашев /1979/ отмечает как очаг простогони-
моза, где в качестве дополнительных или вторых промежуточных хо-
зяев отмечены стрекозы Libellula quadrimaculata — ЭИ
65 %, Anax parthenope — 63, Aeschna coluberculus (=Ae.
136
mixta ) - 62 %, реже Sympetrum flaveolum и Coenagrion
sp ИИ у отдельных насекомых превышала 100 экз. Зараженность
метацеркариями личинок стрекоз была очень низкой.
В.Я. Панин /1957, 1958/, изучая цикл развития трематод рода
Prosthogonimus в районе оз. Зайсан в Казахстане, вскрыл 11 ви-
дов: Libellula quadrimaculata — ЭИ 39,46 % (имаго) и
5,95 (личинки), Sympetrum depressiusculum - соответственно
23,7 и 2,75 и Anax parthenope - 17,35 и 3,33 %. Как допол-
нительные хозяева трематод отмечены Prosthogonimus ovatus
и Р. cuneatus, причем первый из них служит основным переносчи-
ком простогонимоза в районе оз. Зайсан. В этом природном очаге и
в радиусе 40-50 км от него (г. Зайсан, с. Карабулак и Актер) до-
машние куры заражались просто гони мозом (ЭИ до 20).
Из четырех типов водоемов Иссык-К улье кой котловины Киргизской
ССР исследовано 1943 стрекозы восьми видов /Чибиченко, 1966,
1968; Чибиченко, Крылова, 1968; Токобаев, Чибиченко, 1976/. В
пресных водоемах и болотах зараженность стрекоз личинками трема-
тод достигала 43,7-80 %, в других водоемах она была относительно
низка - 2,8-17 %. Метацеркариями плягиорхид (их видовая принад-
лежность, к сожалению, не указана) были заражены стрекозы из мел-
ководных заливов оз. Иссык-Куль: Lestes dryas - ЭИ 6,6 %,
Enallagma cyathigerum- 4,0, Orthetrum cancellatum -
5,0; из озера подсобного хозяйства Ко'г-Сары: L. dryas — 27,5
(плягиорхиды - 15,8 и нотокотилиды - 11,7) и Е. cyathigerum —
10,9; из пресноводных озерков: L. dryas - 70,6, Sympetrum
flaveolum — 48,0 и S. scoticum - 44,3; из болот: L. dry—
as — 71,0 и s. flaveolum — 50,0 %.
Исследователи сообщают, что метацеркарии плягиорхид и нотоко-
тилид были обнаружены в кишечнике стрекоз. Данное сообщение очень
интересно, так как такая необычная локализация метацеркарий пляги-
орхид больше никем не зарегистрирована. Перкарин Notocotylus
attenuatus, проглоченные личинками стрекоз, попадали в пищева-
рительный тракт и там инцистировались. Таким образом, возникло но-
вое звено в цикле развития N. attenuatus с участием стрекоз, ко-
торые могут аккумулировать в себе нотокотилидную инвазию и быть
резервуарными хозяевами, служащими источником заражения птиц.
В.Н. Крылова /1969/ для этой же территории в качестве допол-
нительных хозяев трематод рода Prosthogonimus отмечает Les—
tes dryas, Sympetrum flaveolum, S. scoticum и E. cy—
athigerum, т.е. те же виды, в которых вышеуказанные авторы нахо-
дили плягиорхид. ЭИ личинок стрекоз в разных водоемах составляла
от 10 до 70 %. Автор сообщает, что максимумы интенсивности и
экстенсивности инвазии приходятся на июнь и июль, т.е. на время ли-
чиночного развития стрекоз.
В Восточной Туркмении в среднем течении р. Амударьи Н.М. Ми-
найловой и М.Г. Мушкамбаровой /1982/ в числе других насекомых
были вскрыты 29 стрекоз Aeschna sp., в гельминтологическом
отношении они оказались стерильными. В южных районах Туркмении,
окрестности Ашхабада, были вскрыты 411 стрекоз, относящихся к
137
16 видам /Павлюк, Курбанова, 1986/. Метацеркарий рода Prostho—
gonimus найдены у Calopteryx splendens transcaspica —
ЭИ 4,5 %; Skrjabinoeces similis — у Sympetrum striolatum
и Epallage fatime, 9-10; Plagiorchis elegans - у Isch—
nura elegans, 1,6 %. Впервые в стрекозах СССР выявлены мета-
церкарии рода Eumegacetes, которыми были инвазированы Epal—
lage fatime - 13,5, С. splendens transcaspica — 0,9,
Sympetrum meridionale - 2,2 и Orthetrum anceps - 30,4,
максимальная ИИ 34 экз.
Для паразитологических исследований нами /Павлюк, 1981/ были
использованы также сухие стрекозы, многие годы хранившиеся на ват-
ных матрасиках, а перед вскрытием размоченные в теплой воде. Об-
наруженные личинки некоторых гельминтов оказались пригодными для
определения. В результате исследования длительно хранившихся стре-
коз впервые выявлены метацеркарий рода Prosthogonimus в таких
видах, как Cordulegaster coronatus из заповедника “Рамит" в
Таджикистане, Aeschna crenata из г. Иркутска, Ае. serrata
из р. Тобол Курганской области, Somatochlora graeseri из
оз. Лобозино Амурской области, Grocothemis erythraea из
окрестностей Самарканда, Leucorrhinia orientalis из г. Абаза
(Западный Саян), L. intermedia из Южного Прибайкалья, Coenag—
rion concirmurn из Приамурья и С. vernale из Кургальджин-
ских озер Целиноградской области.
Цестоды. В полости тела личинок Coenagrion hastulatum
из двух водоемов Озерского района Калининградской области М.Н. Го-
ликова /19606, 1961/ впервые нашла цистицеркоиды (личинки) це-
стоды Tatria decacantha (ЭИ 12,8 %) - паразита кишечника
поганок. В Ярославской области цистицеркоиды этого же вида цесто-
ды впервые найдены О.П. Кулагиной /1984/ у Anax imperator.
П.П. Дремкова /1976/ сообщает, что в полости тела двух личи-
нок разнокрылых стрекоз из Волго-Ахтубинской поймы Астраханской
области обнаружены по 1-2 цистицеркоида, похожих на Т. decacan-
tha. Цистицеркоиды этого вида найдены в полости тела 12 видов
стрекоз из 40 пунктов западных областей Украины /Павлюк, 19736,
1980г; Павлюк, Стражник, 1985/. Стрекозы Sympetrum depres-
siusculum, S. vulgatum, S. sanguineum, Coenagrion pul—
chellum, C. pueUa, C. vernale, Ischnura pumilio, I. ele—
gans, Lestes dryas, L. sponsa, L. virens в ка-
честве хозяев зарегистрированы впервые, ЭИ 0,15-7,02 %, средняя-
0,42, максимальная ИИ 7 экз. В полости тела L. sponsa из ок-
рестностей Львова обнаружен цистицеркоид Tatria acanthorhyn—
cha Wedl.
H.B. Чурина /1964/ отмечает, что в условиях Среднего Урала
для цестоды Micro soma canthus paracompressa резервуарны-
ми хозяевами наряду с моллюсками являются и личинки Coenagri—
on sp. Благодаря большой плотности птиц на единицу, площади во-
доема и высокой численности различных водных беспозвоночных, со-
здается частый контакт паразитов всех стадий развития с разными
138
видами хозяев и в цикл развития некоторых гельминтов вовлекаются
неспецифические для них хозяева.
Из водоемов Южного Урала В.А. Ткачевым /1969а, б -
1971а, б/ вскрыто 4830 личинок стрекоз: Lestes sponsa — 340,
Coenagrion pulchellum — 290, Calopteryx splendens —
320, Sympetrum scoticum - 230, Aeschna grandis - 150
и Ae. cyanea — 3500. В результате исследований только в ли-
чинках Ae. cyanea обнаружены цистицеркоиды Т. decacantha.
ИИ спонтанно зараженных личинок 2—45 экз., а в лабораторных усло-
виях - до 80. Учитывая практическое отсутствие достоверных находок
Ae. cyanea на водоемах восточного склона Южного Урала, приходит-
ся предположить, что автором допущена ошибка в определении вида
стрекозы и он имел дело с личинками Ae. serrata, описание кото-
рой было впервые опубликовано только в 1973 г. /Белышев, Харито-
нов, 1973/. В.А. Ткачевым в природных и лабораторных условиях
проводились наблюдения над развитием этой цестоды.
По поводу работ В.А. Ткачева К.М. Рыжиков и Л.М. Толкачева
/1981/ отмечают: "... наиболее подробно биологический цикл цестоды
Tatria decacantha изучал Ткачев. Следует, однако, отметить, что
достоверность некоторых данных Ткачева вызывает сомнение. В част-
ности, приведенное им описание личинки цестоды существенно отлича-
ется от описаний, даваемых другими авторами. Видимо, Ткачев допус-
тил ошибку. Мы думаем, что найденная и описанная им личинка при-
надлежит какой-то другой цестоде". Также в личинках Coenagrion
pulchellum на озерах Челябинской области В.И. Окороков и
В.А. Ткачев /1973/ нашли цистицеркоидов Schistotaenia mathe—
vossianae, отнесенных ими к оказавшимся синонимом Tatria ju-
bilaea.
В Учалинском районе Башкирии М.Г. Баянов /1974/ нашел цис—
тицеркоиды Т. decacanthay двух из 27 вскрытых личинок Anax
imperator из оз. Карагайлы. В кишечнике подопытного цыпленка, ко-
торому были скормлены имаго Enallagma cyathigerum из этого
же озера, была обнаружена цестода Dilepidoidea gen. sp.
В Казахстане Д.Ш. Кукашев /1985, 1986/ из стрекоз впервые
выявил и описал личинок трех видов цестод. В брюшной полости на по-
верхности средней кишки личинки Ischnura elegans из солоновато-
го озера близ с. Борки Уральской области (бассейн р. Кушум) им об-
наружен цистицеркоид Schistotaenia ( Para schistotaenia ) со—
lymba, ЭИ 0,71 % (у 1 экз. из 140 вскрытых I. elegans ).
В брюшной полости личинки Coenagrion hastulatum из оз. Кургальд-
жин в Центральном Казахстане обнаружен цистицеркоид Tatria du—
bininae, ЭИ 0,93 % (у 1 экз. из вскрытых С. hastulatum ).
Цистицеркоиды Schistotaenia (Paraschistotaenia) srivastavi
обнаружены в личинках 8 видов стрекоз: Sympycna sp., Coenag-
rion hastulatum, C. armatum, Coenagrion sp., Erythromma na-
jas, Enallagma cyathigerum, Ischnura pumilio, Ischnura sp.,
собранных на оз. Бол. Шошкалы (бассейн р. Убаган в Северном Ка-
захстане) и оз. Кургальджин, ИИ 1-6 экз. Локализация — наружная
поверхность средней кишки в области расположения мальпигиевых со-
судов.
139
В одной особи Sympetrum meridionale из пруда Ашхабада
найдено /Павлюк, Курбанова, 1976/ около 250 цистицеркоидов Tat—
ria decacantha, все тело личинки буквально было набито цистицер-
коидами - явление, не наблюдавшееся даже в эксперименте.
Нематоды. Литературных сведений об инвазии стрекоз нематода-
ми очень мало, что связано с относительной редкостью самих парази-
тов. А.Н. Попова /1953а/ сообщает, что неоднократно находила личи-
нок стрекоз сем. Libellulidae, полость тела которых была заполне-
на нематодами, но систематическое положение их автором не дается.
К.И. Скрябин /1923/ указывает, что в полости тела личинок Со-
enagrionsp.napaaHTHpyioT личинки нематоды Camallanus lacust—
ris. Место их обнаружения не указано. А.М. Супряга /1971, 1972/
сообщает, что в южных районах СССР личинки стрекоз, наряду с маль-
ками рыб и головастиками амфибий, могут принимать участие в разви-
тии нематоды Avioserpens mosgovoji (Camallanata, Dracun—
culidae) некоторых водоплавающих птиц в качестве резервуарных
хозяев.
У личинок Coenagrion hastulatum из озер южной части Ка-
релии (Пертозеро и Кончозеро) Е.Н. Фролова /1965/ находила нема-
тод Contracaecum sp. (Ascaridata, Anisakidae).
A.A. Мозговой, В.И. Шахматова и М.К. Семенова /19686/ экспе-
риментально установили, что личинки стрекоз родов Coenagrion и
Calopteryx, наряду с рыбами, являются дополнительными хозяева-
ми нематоды Contracaecum spiculigerum домашних и промыс-
ловых птиц. Эти же авторы /Мозговой и др., 1968а/ в числе допол-
нительных хозяев Contracaecum microcephalum— паразита мно-
гих рыбоядных птиц (наряду с мальками рыб и хирономидами) называ-
ют также личинок стрекоз. В южной части Аграханского залива (Да-
гестан) М.К. Семенова /1972-1974/ проводила заражение экспери-
ментальных животных, в том числе 233 личинок стрекоз, личинками
III стадии Contracaecum micropapillatum- паразита желудка пе-
ликанов. При этом установлено, что в число факультативных резерву-
арных хозяев этой нематоды входят личинки стрекоз - Calopteryx
sp., Coenagrion sp., Ischnura elegans, Lestes sp., Anax
imperator, Aeschna grandis, Cordulia sp., а в роли
резервуарного хозяина С. microcephalum выступают личинки
A. imperator, Ae. grandis и L elegans. Автор отмечает, что
среди беспозвоночных наиболее восприимчивыми к контрацекозной ин-
вазии оказались молодые личинки равнокрылых стрекоз. Несколько
труднее заражаются в опытах личинки разнокрылых стрекоз.
Личинки Contacoecum spiculigerum находил М.Г. Баянов
/1974/ в личинках стрекоз Erythromma najas, Aeschna jun—
cea и Anax imperator в Учалинском районе Башкирской АССР. На
оз. Песчаное Челябинской области В.И. Окороков и В.А. Ткачев
/1974/ у личинок Coenagrion puella, Sympetrum scoticum
и Cordulia aenea обнаружили личинок Mermis sp.
В стрекозах западных областей Украины, главным образом Во-
лынской и Львовской, найдены и описаны личинки новых видов мерми-
тид /Рубцов, Павлюк, 1972/: Hexamermis (?) pachydermis
140
из полости брюшка Lestes virens; Agamomermis coenagrio—
nis из Coenagrion pulchellum, Ischnura elegans и Sym-
petrum vulgatum; Agamomermis lestis из Lestes sponsa
и L elegans; Paramermis zygopterorum из разнокрылых стре-
коз (этикетка вида утеряна), а также ближе не определенная личинка
из Orthetrum aibistylum из окрестностей Ивано-Франковска. За-
тем найденную нами в имаго Lestes sponsa личинку мермити-
ды у г. Нововолынска И.А. Рубцов /1976/ описал как новый вид
Paramermis (?) foveata.. Личинку мермитиды не установленной ви-
довой принадлежности мы нашли также у Calopteryx splendens
из окрестностей Львова и у С. s. transcaspica из Туркмении. Во
всех случаях зараженность стрекоз личинками мермитид не превыша-
ла 1 %. В стрекозах Западной Украины изредка встречались также очень
мелкие личинки нематод невыясненного систематического положения.
П.П. Дремкова /1976/ в нимфах трех видов стрекоз из водоемов
Волго-Ахтубинской поймы нашла 11 мермитид. И.А. Рубцов и А.И.Ки-
селев /1975/ описали новый вид мермитиды Odonatomermis аге—
naria по личинке из стрекозы (вид не указывается), отловленной
на оз. Песчаное Челябинской области. В данной работе И.А. Рубцов
отмечает, что все известные до сих пор личинки мермитид из стре-
коз, определявшиеся как Hexamermis и Agamomermis, из-за недо-
статочности материала относятся к одному роду Odonatomermis.
Н.Н. Шевченко/1962/ отмечает редкий случай паразитирования
личинки волосатика Gordius aquaticus в личинке стрекозы Enal-
lagma cyathigerum в Харьковской области.
Итак, паразитологические исследования стрекоз в. нашей стране
начаты сравнительно недавно, но по масштабам исследований (в це-
лом вскрыто свыше 80 тыс. особей) и достигнутым результатам они
далеко превосходят таковые любой другой страны. Среди паразитиче-
ских организмов, найденных в стрекозах, описан ряд новых для науки
видов. Для многих видов паразитов значительно уточнены сведения о
круге их хозяев, о их гостальной специфичности. В то же время па-
разитологическими исследованиями стрекоз охвачена лишь незначи-
тельная часть территории нашей огромной страны. Основная работа
еще впереди. Есть основание сказать, что в стрекозах еще будут най-
дены и описаны многие новые для науки виды паразитов из разных
систематических групп. А вопрос о видовом составе, сезонной дина-
мике численности, значении поселяющихся на покровах тела личинок
стрекоз сидячих инфузорий, мшанок, коловраток, низших водорослей и
других эпибионтов еще почти не затронут исследованиями.
Непреодоленным пока недостатком в паразитологических исследо-
ваниях стрекоз остается ориентация специалистов на изучение отдель-
ных узких групп паразитов, но не всего их комплекса в целом. Бога-
тый фактический материал по паразитам стрекоз требует не только
дальнейшего пополнения, но и теоретических обобщений, имеющих и
практическое значение, так как они касаются важных звеньев в слож-
ных биоценотических отношениях.
141
ОБЗОР ИЗУЧЕННОСТИ ТРОФИЧЕСКИХ СВЯЗЕЙ СТРЕКОЗ
Стрекозы - массовый компонент многих водных и наземных биоце-
нозов. По типу питания это активные хищники как в личиночной, так
и имагинальной фазах, вносящие существенный вклад в процессы круго-
ворота и трансформации вещества в экосистемах. Однако в литературе
почти отсутствуют сведения об оценке хищничества стрекоз и их био-
ценотическом значении.
Первые исследования, посвященные изучению питания личинок
стрекоз, появились в 20-х гг. нашего столетия /Попова, 1925/. В
40-е гг. опубликовано еще несколько статей /Березина, 1940, 1946;
Wright, 1946/. Начиная с 60-х гг. изучение трофических связей личи-
нок стрекоз с другими представителями водных биоценозов заметно
оживилось. Личинки стрекоз сравнительно легко содержатся в лабора-
торных условиях и являются удобным объектом Для построения мате-
матических моделей системы "хищник - жертва", поэтому вопросам их
хищничества к настоящему моменту посвящено примерно 70 % всех
работ, касающихся питания стрекоз.
Такая неравноценность связана еще и с тем, что для изучения
трофических связей личинок существует ряд простых и не очень трудо-
емких методик, позволяющих выявить рацион, суточную и сезонную ак-
тивность питания и некоторые другие вопросы. Наиболее часто исполь-
зуемый метод - анализ содержимого фекалий личинок, который приме-
няли В.И. Попченко /1971/, Pandian, Mathavan /1974/, Thomp-
son /1975, 1978/, Cloarec /1977/, Blois/1985/ и некото-
рые другие авторы. С помощью этого метода Притчард /Pritchard,
1964/ выяснил рацион личинок стрекоз различных видов. Основную
долю в нем составляли личинки хирономид, равнокрылых стрекоз, мел-
кие ракообразные, моллюски. Всего же он идентифицировал 17 групп
беспозвоночных, остатки которых встречались в фекалиях личинок.
При изучении состава жертв личинок Anax imperator в есте-
ственных условиях установлено, что они питаются главным образом
личинками поденок, равнокрылых стрекоз, хирономид и водных клопов
рода Plea /Cloarec, 1977/. Другие же виды беспозвоночных, такие
как водные клещи, ракообразные, личинки и взрослые жуки, некоторые
водные клопы, широко представленные в исследованном автором био-
топе, в рационе личинок отсутствовали. Это свидетельствует о том,
что использование хищником-полифагом в пищу жертв определенного
вида зависит от многочисленных факторов: сезонной и-суточной актив-
ности жертв, их биотопического распределения, различных поведенче-
ских реакций на одни и те же изменения биотических и абиотических
факторов. Н.А. Березина /1946/ дает более узкую трактовку анало-
гичного вывода: личинки стрекоз - полифаги и питаются теми объек-
тами, которые являются для них более доступными в соответствую-
щих условиях. Полифагию личинок, г зволяющую им менять вид жерт-
вы, Cloarec /1977/ рассматривает в качестве возможного меха-
низма стабилизации численности популяций как хищника, так и жерт-
вы. На наш взгляд, это свидетельствует о широкой адаптации стрекоз
142
к нестабильным биотическим факторам среды - существенному коле-
банию численности всех видов жертв по сезонам и годам.
Однако если полифагия личинок стрекоз, их роль в регуляции чис-
ленности некоторых членистоногих, например комаров, мошек, дафний
и других, изучалась целым рядом исследователей /Jonson, 1973;
Ezenwa, 1974; El Amin el Rayah, 1975; El Amin el Rayah,
Abu Shama,1977; и др./, то для имаго стрекоз этот вопрос практиче-
ски не изучен. Имеются лишь отдельные разрозненные данные о пи-
тании имаго на уровне случайных наблюдений. Например, Ларсен
/Larsen, 1981/ описывает поедание стрекозами Orthetrum
austenti бабочек разных видов, но все наблюдения были проведе-
ны в .течение одного дня и лишь над четырьмя стрекозами. Ларид
/ Larid, 1977/ в своей работе дает ссылки на некоторых авторов,
наблюдавших случаи поимки мухи це-це стрекозами родов Cacerga—
tes и Orthetrum. Киеллберг / Kjellberg, 1972/ рассматривает
стрекоз как основных хищников взрослых поденок. Известны случаи
массового поедания термитов стрекозами Anax junius и неко-
торыми либеллюлидами / G-illaspy, 1971/, истребления мух Ino—
pus rubriceps сем. Stratiomyidae стрекозами Heminax pupu—
ensis и Notoaeschna sagittata /Osborn, Forteath, 1974/.
Среди активных хищников, поедающих крылатых самок муравьев в
момент их массового выхода, отмечены стрекозы Pachydiplax
longipennis, Tramea Carolina, Anax junius и Somatochlo-
ra provacans /Whitcomb et al., 1973/.
Лишь несколько работ отличаются системным подходом к изуче-
нию трофических связей имаго стрекоз, хотя они и не лишены неко-
торых методических недостатков. Так, в статье по стрекозам агро-
биоценозов сои во Флориде / Neal, Whitcomb, 19721 проанализи-
рован видовой состав стрекоз с одновременной регистрацией времени
лета, характерных особенностей питания и рациона. Всего в агробио-
ценозах сои отмечено 35 видов стрекоз, 9 из которых были массо-
выми и потенциально могли участвовать в регуляции численности от-
дельных групп вредителей. Однако полевые наблюдения показали, что
только три из них (Erythemis simplicicollis, Pachydiplax lon-
gipennis и Perithemis tenera seniinole ) оказыва-
ли влияние на численность некоторых экономически важных вредите-
лей. Но авторы отмечают, что все эти виды активно питались и в
других биоценозах. Лишь одна равнокрылая стрекоза Anomalagri—
on hastatum была постоянно связана с культурой сои. Какие-
либо количественные данные по оценке экономической значимости
стрекоз в биоценозах сои в этой работе отсутствуют, осталось неяс-
ным их место в системе "хищник — жертва", взаимоотношения и со-
существование с другими хищниками.
Следует также отметить работу японского одонатолога Хигаши
/Higashi, 1973/. Автором проведены наблюдения за суточной ак-
тивностью двух видов: Calopteryx Cornelia и Sympetrum fre—
quens, основное внимание уделялось пищевому и репродуктивному
поведению. Изучалась частота фуражировочных полетов, эффективность
поимок, частота действительного питания, ежедневное потребление пи-
143
щи и соотношение между пищевым потреблением и массой стрекозы.
Однако в работе отсутствуют сведения о том, какую роль играют два
вышеуказанных вида в исследованном биотопе, почему именно на них
был остановлен выбор. К тому же необходимо отметить, что в дан-
ном случае все наблюдения были проведены в естественных условиях,
в то время как большинство авторов использует лабораторные экспе-
рименты, особенно по элективности питания.
В 1973 г. опубликована первая отечественная работа специально
по питанию имаго 'Роль стрекоз и ос в снижении численности гнуса
в бассейне реки Или' /Кумачев, 1973/. Полевые исследования про-
водились автором в местах массового выплода кровососущих комаров
и слепней. Плотность мелких видов стрекоз учитывалась на участках
1x10 м, а разнокрылых и ос - на площадках 5x20 м. Однако метод
учета вызывает сомнение: количество стрекоз учитывалось при вспу-
гивании их веткой во время прохождения по учетным площадкам. Пи-
тание исследовалось несколькими методами. Так, для равнокрылых
регистрировались их жертвы в течение 30 мин на учетных площадках,
а в трех точках площадки брались пробы насекомых сачком для коше-
ния. Сопоставление жертв стрекоз с видовым составом отловленных
насекомых позволяло судить о предпочитаемости тех или иных жертв.
Аналогичным образом изучался выбор жертв у крупных видов стрекоз.
Среди объектов питания автор указывает некровососущих комаров,
мокрецов, цикадок и их личинок, комаров-долгоножек, мух-пестрокры-
пок, чешуекрылых, слепней, тахин, поденок, мелких и средних жестко-
крылых, а для стрекоз родов Anax и Aeschna - еще и кровосо-
сущих комаров. К сожалению, автором не были сделаны определения
жертв хотя бы до рода, а также не приведены данные по численности
стрекоз и их основных жертв, т.е. в работе практически отсутствуют
конкретные количественные данные, что придает ей описательный ха-
рактер. Несмотря на это, автор делает вывод о том, что существен-
ное влияние на численность гнуса в бассейне р. Или оказывает огра-
ниченный круг видов стрекоз. Практическая значимость их, в свою
очередь, в значительной мере зависит от их численности.
Нам представляется более существенным в этой работе замеча-
ние о том, что во время охоты стрекозы отдают предпочтение бокее
крупным насекомым. И это становится вполне, естественным, если
рассмотреть энергетический баланс хищника при ловле мелких и круп-
ных насекомых.
И еще одна работа, на наш взгляд, заслуживает внимания, по-
скольку авторами сделана попытка оценить влияние стрекоз на чис-
ленность кровососущих насекомых. Она была выполнена во Флориде
/ Edman, Haeger,1974/, где в естественных условиях был постав-
лен интересный эксперимент. На слегка залесенный остров площадью
100 акров выпустили почти одновременно 1,2 млн 2-3-дневных ко-
маров Aedes taeniorhynchus. После этого наблюдалась быстрая
аккумуляция меченых комаров в желудках Anax junius. Полевые
наблюдения показали селективное выедание этой искусственно создан-
ной повышенной концентрации жертвы стрекозами. Однако авторы не
привели данных о том, какой процент в пишевом рационе Anax ju—
144
nius составляют комары этого вида при их фоновой плотности в
изучаемом биотопе, не пытались провести сравнение пищевой избира-
тельности стрекоз при одинаковой плотности двух потенциальных жертв,
например, кровососущих и некровососущих комаров.
Обобщая сказанное, можно констатировать, что до сих пор не про-
ведено ни одного системного комплексного исследования трофических
связей имаго стрекоз. Отдельные элементы изучения питания некото-
рых видов и натуралистические наблюдения не позволяют оценить по-
тенциальную роль стрекоз в различных типах экосистем. Одна из при-
чин такого положения - отсутствие подходящих методов для изучения
рациона стрекоз и количественных оценок потребления пиши. Исполь-
зуемые ранее методы - наблюдение, протоколирование, анализ содер-
жимого желудков и фекалий - трудоемки и не всегда дают статисти-
чески достоверные результаты. Вне поля зрения исследователя оста-
ются мелкие и спабохитинизированные объекты питания, а ошибки в.
определении их, как правило, обусловлены опытом и квалификацией ис-
следователя. Очевидно, необходимо привлекать в исследования подоб-
ного рода новые современные методы.
В 1986 г. нами предпринята попытка использовать серологиче-
ский метод для анализа пищевого спектра имаго стрекоз. Основа это-
го метода заключается в постановке реакции преципитации, которая
служит для выявления белков жертвы в пищеварительном тракте хищ-
ника. Применение серологического метода в энтомологии началось при-
мерно с 50-х гг. Т.К. Сергеева /1970а/ приводит краткий обзор ра-
бот в этой области и в последующих, публикациях /Сергеева, 19706,
19$2а, б, 1983 и др./ подробно описывает методику. Несколько мо-
дифицировав методики, используемые для почвенных насекомых, мы на-
чали работу со стрекозами.
Для получения антисывороток были выбраны четыре тестовые груп-
пы: Tabanidae, Culicidae, Chironomidae, стрекозы рода Sym—
petrum. Выбор тестовых групп на данном этапе работы преследо-
вал цель выяснить, как часто в рационе стрекоз встречаются кровосо-
сущие насекомые и могут ли стрекозы при известной плотности в
околоводных биоценозах оказывать регулирующее воздействие на их
численность. Полевые наблюдения 1985-1987 гг. показали, что для
массовых видов равнокрылых стрекоз (Coenagrion vernale,
С. pulchellum, Lestes sponsa, L. dryas) наиболее часто
добычей служат представители сем. Chironomidae, а такие виды,
как Aeschna serrata и Libellula quadrimaculata, пре-
имущественно питаются стрекозами родов Sympetrum и Coenag—
rion соответственно.
Для иммунизации кроликов в 1986 г. в Новосибирской области
(окрестности оз. Чаны, Панове кая опорная база Биологического инсти-
тута СО АН СССР) собран антигенный материал. В качестве антигена
использовали водорастворимые белки потенциальных жертв. Собранных
насекомых выдерживали в стеклянных садках объемом 500-1000 см^
на увлажненной фильтровальной бумаге 2-3 дня для полной очистки
желудочно-кишечного тракта. Затем предварительно приморенных эфи-
ром и взвешенных насекомых одного вида растирали чистым скальпе-
10 Зак. 611
145
Таблица 11. Сравнительная характеристика процентного содержа-
ния водорастворимых белков в некоторых группах насекомых
Группа, вид Дата сбора Сырая биомас- са, мг Кол-во экстра- гента, мл Выде- ленный белок, %
Tipula scripta* Pogo nognathell us 18. VI 1979 3170 10 0(,13
flavescens* 16. VIII 1979 900 8 0,35
1. VIII 19 79 850 6 9,22
Allolobophora cali- ginosa* 18. VI 1979 3000 5 1,80
Aedes excruciens 2. VI 1986 1326 7 4,90
Chironomidae 20. VII 1986 1111 10 6,10
25. VIII 1986 681 10 4,20
Sympetrum flaveolum 15. VII 1986 975 10 5,50
Tabanidae 29. VI 1987 452 5 8,50
Lestes sponsa 26. VII 1987 467 5 5,30
Coenagrion vernale 5. VI 1987 201 4 7,20
* /по: Сергеева, 19826/.
Лем на фильтре, высушивали при комнатной температуре и хранили в
стеклянных бюксах над слоем обезвоженного CaCl^. Далее, в лабора-
торных условиях экстрагировали водорастворимые белки с фильтров
по Конышеву /Лабораторные методы..., 1967/ и определяли их кон-
центрацию по Лоури. Концентрация водорастворимых белков не варьи-
ровала у насекомых различных систематических групп в таких широ-
ких пределах, как об этом сообщает Т.К. Сергеева /19826/. Срав-
нительные данные приведены в табл. 11.
Из данных табл, if видно, что пропент выделенного белка для
наших тестовых групп варьировал в пределах 5 %.
Исходя из определенных концентраций белка, готовили антиген с
адьювантом Фрейнда для иммунизации кроликов. Иммунизацию прово-
дили по следующей схеме: первая, на 14-й день - вторая, на 28-й -
третья;"на 35-й - взятие 40 мл крови для определения титра, на
42-й день - четвертая, на 56-й - пятая, на 70-й - шестая, на 77-й
день можно брать кровь для определения окончательного титра. Анти-
ген вводили подкожно в шести точках в области лопаток по 0,1 мг
белка (в 0,5 мл физиологического раствора и 0,5 мл адьюванта) за
одну иммунизацию. На каждый антиген иммунизировались два живот-
ных—самца массой 3 кг. Данная схема иммунизации по длительности
несущественно превышает схему, предлагаемую Т.К. Сергеевой
/1970а/, однако позволяет существенно сократить количество вводи-
мого белка, а следовательно, уменьшить количество насекомых, по-
146
тенциапьных жертв, необходимых для приготовления антигена.
В 1986 г. нам удалось получить две антисыворотки с рабочим
титром: анти-Culicidae ’ (1:32) и анти-Chironomidae (1:32).
С их помощью в 1987 г. было поставлено несколько тестов в поле-
вых условиях. Для тестирования содержимого желудков стрекоз исполь-
зовали реакцию двойной иммунодиффузии в гель по Оухтерлони и усо-
вершенствованную Л.А. Зильбером и Г.П. Абелевым /1962/. Предвари-
тельная проверка видоспецифичности полученных сывороток показала,
что сыворотка, полученная против одного вида хирономид, дала пере-
крестные реакции со всеми другими видами данного семейства, кото-
рые были представлены в биотопах во время сбора антигенного мате-
риала (июнь - июль 1986 г.). Аналогичная ситуация наблюдалась для
сыворотки против кровососущих комаров. Это позволило назвать сыво-
ротки анти-Chironomidae и анти-Culicidae и идентифицировать
объекты питания стрекоз на данно'м этапе исследований до семейства.
На наш взгляд, это уже является существенным преимуществом
перед методиками, используемыми в работах подобного рода ранее. Со-
поставление хитиновых остатков в желудках стрекоз с видами, обитаю-
щими в исследуемых биотопах, их идентификация требует значительно-
го времени. Полевые наблюдения и отнимание добычи в момент поим-
ки не позволяют за 1-2 полевых сезона набрать материал, достаточ-
ный для статистической обработки и обобщений. Идентификация насеко-
мых-жертв, как правило, проводится до семейства, в лучшем случае
до рода и лишь в единичных случаях до видов.
26 июля 1987 г. в естественных местообитаниях собрано около
40 экз. Lestes sponsa и 20 экз. Sympetrum scoticum. У
стрекоз отпрепарировали желудочно-кишечные тракты. Их содержимое
поместили в лунки на специальном плато, залили физиологическим
раствором из расчета.0,2 мл на один желудок и поместили в холо-
дильник на 12 ч. Стеклянные пластины залили 1,5 %-м агаром, после
остывания вырезали лунки. Через 12 ч в центральные лунки разлили
экстракт содержимого желудков, а в периферические - антисыворотки.
Пластины поместили во влажную камеру. Дальнейшая суть метода за-
ключается в том, что в агаре белки, являющиеся антигенами, из со-
держимого желудков и антитела к ним, находящиеся в сыворотках, на-
чинают диффундировать навстречу друг другу. В зоне оптимальной кон-
центрации реагентов образуется преципитат, т.е. комплекс "антиген —
антитело". При боковом освещении он хорошо виден в виде мутных
полос на прозрачном агаре. По образованию линий преципитации мож-
но судить о том, какая жертва была съедена стрекозой. Анализ содер-
жимого желудков стрекоз, собранных 26/VII 1987 г., показал, что дву-
крылыми сем.Chironomidae питалось 80 % L. sponsa и 20 %
S. scoticum, а кровососущих комаров удалось обнаружить лишь в
желудке одного S. scoticum. Количественные учеты насекомых-
хортобионтов, являющихся потенциальными жертвами стрекоз, подтвер-
дили доминирование представителей сем. Chironomidae во всех ис-
следуемых биотопах. Полученные к настоящему моменту данные не по-
зволяют пока сделать существенных обобщений и выводов, но они уже
раскрывают преимущества серологического метода диагностики питания
147
стрекоз и возможность использования его в качестве экспресс-мето-
да при наличии достаточного количества сывороток с рабочими титра-
ми. Их видоспецифичность будет определяться цепью исследований.
Для оценки регулирующего воздействия стрекоз на численность крово-
сосущих насекомых, видимо, будет достаточно 1-2 сывороток против
доминантных видов.
В целом трофические связи стрекоз и их роль в трансформации
вещества в экосистемах изучены слабо. Известны лишь объекты до-
бычи по отдельным видам, и есть некоторые теоретические расчеты,
построенные на единичных полевых наблюдениях. Однако существуют
методы, используемые в смежных областях биологии, а с недавних
пор и в энтомологии, позволяющие успешно решать этот вопрос.
ОПЫТ ОЦЕНКИ РОЛИ СТРЕКОЗ В ВОДОЕМАХ БЕЛОРУССИИ
Опенка роли стрекоз в природных биоценозах проводилась многи-
ми исследователями. Однако до последнего времени в большей части
публикаций даются лишь общие указания на хищный образ жизни стре-
коз и потребление ими различных наземных и водных животных. Вме-
сте с тем в литературе, особенно гидробиологической, накоплен зна-
чительный материал, позволяющий глубже оценить роль стрекоз в при-
родных сообществах.
В фауне зарослей различных пресных водоемов большое значение
приобретают личинки>стрекоз, из которых многие относятся к типи-
чным хищникам, питающимся крупными подвижными водными животны-
ми. Крупные личинки разнокрылых стрекоз представляют интерес как
прямые потребители личинок и мальков рыб и как конкуренты рыб в
питании ракообразными (в первую очередь дафниями) и личинками ко-
маров /Березина, 1946/. Значительный вред рыбному хозяйству при-
носят крупные личинки Aeschna grandis и Anax imperator,
которые в массе потребляют в прудах мальков карпа и линя. По дан-
ным Н.А. Березиной /1947/, личинка A. imperator массой
971,3 мг съедает до 1226,8 мг мальков карпа. По мнению различ-
ных авторов, в нерестово-выростных прудах рыбхозов наибольший вред
приносят личинки стрекоз из семейства Aeschnidae и Libellu—
lidae.
Для выяснения роли личинок стрекоз в водных экосистемах в те-
чение ряда лет на одном из крупнейших водоемов Белоруссии оз. На-
речь проводилось изучение количественных закономерностей роста,
питания, интенсивности обмена.
Сообщество личинок стрекоз, обитающих на литорали оз. Нарочь,
включает, по данным Е.С. Шалопенок /1963/, 10 видов. Позднее при
обследовании литорали озера и основных нерестилищ отмечено 11 ви-
дов личинок стрекоз. Основными фоновыми видами на различных уча-
стках литорали оказались Enallagma cyathigerum, Ischnura ele—
gans, Lestes sponsa, Sympetrum vulgatum, Cordulia
aenea. В некоторых биотопах среди доминанте в отмечены личинки
Aeschna grandis.
148
Удельный вес личинок стрекоз в общей биомассе бентоса в лет-
нее время составляет около 30 % (3 г сырой массы на 1 м^ лито-
рали при общей биомассе бентоса 9,8 г/м£) /Арабина, 1968/. Сум-
марная биомасса комплекса хищных насекомых, включающего помимо
личинок стрекоз хищных клопов, жуков, личинок поденок, ручейников
и другие группы, достигает на некоторых участках литорали
4,6 г/м^.
Наблюдения за изменением численности и биомассы личинок стре-
коз на различных участках литорали озера показали, что предпочитае-
мыми и наиболее благоприятными для них являются биотопы с заилен-
ным грунтом и обильными зарослями макрофитов.
Результаты многолетних наблюдений позволили установить, что
максимальная численность личинок стрекоз наблюдается в июне. Ак-
тивный лёт происходит в июле (стрекозы летней группы). В этот пе-
риод в популяциях преобладают крупные особи с большой индивидуаль-
ной массой тела. Аналогичные результаты дает и анализ весенней по-
пуляции, основу которой составляют перезимовавшие особи с высоки-
ми весовыми и линейными показателями. С середины лета появляются
личинки новой генерации, численность которых возрастает к осени.
Для установления возрастного состава популяций личинок стрекоз,
а также при постановке длительных физиологических экспериментов
возникает необходимость получить данные о массе тела насекомых.
Многократное взвешивание и связанное с этим предварительное обсу-
шивание личинок затруднительны, часто сопровождаются травмирова-
нием насекомых, поэтому большое значение приобретает возможность
получения констант для вычисления массы организмов по легко уста-
навливаемым линейным размерам тела.
Нами проведено определение соотношения между длиной тела и
сырой массой личинок восьми массовых видов, обитающих на литорали
оз. Нарочь и в прудах рыбхоза "Изобелино' Минской области
(табл. 12). У личинок разнокрылых стрекоз за длину тела принимали
расстояние от переднего края головной капсулы до конца анальной
пирамиды, у личинок равнокрылых стрекоз длину тела измеряли без
листовидных хвостовых жабр, которые нередко обламываются. Сырую
массу личинок определяли на торзионных весах взвешиванием после
предварительного обсушивания на фильтровальной бумаге до исчезнове-
ния влажных пятен.
Исследованы линейно—весовые характеристики четырех видов раз-
нокрылых стрекоз: Ae. grandis, Ae. juncea, L. quadrimacula—
ta, S. vulgatum и трех видов раЪнокрылых: Е. najas, L elegans
и Е. cyathigerum.
Для анализа последнего вида исследованы особи из озерной и пру-
довой популяций.
Вопрос о соотношении массы тела и линейных размеров у различ-
ных групп животных достаточно широко освещен в литературе. Извест-
но, что между указанными параметрами существует степенная зависи-
мость, выражающаяся формулой W = а 1^, где W - масса тела, 1 -
длина тела, а - специфическая для каждого вида константа, равная
массе животного при единице длины; Ь - константа, являющаяся по-
149
Таблица 12. Параметры степенных уравнений, связывающих весовые и линейные показатели у личинок
стрекоз
Вид Пределы колебаний массы, мг Число изме- рений <^х Г й>а а bi&b
Aeschna juncea* 23-750 24 0,453 0,184 0,990 0,045 0,105 2,45+0,050
Ае. grandis 2-55 60 0,438 0,189 0,912 0,178 0,046 2,12 +0,198
Libellula quadrimaculata 5-620 28 0,580 0,215 0,993 0,069 0,105 2,67+0,060
Sympetrum vulgatum* 10-000 18 0,542 0,207 0,991 0,072 0,123 2,60+0,082
Enallagma cyathigerum* (пруды ) 1-105 20 0,534 0,216 0,994 0,058 0,03 9 2,46-^0,060
E. cyathigerum* (озеро) 4-44 120 0,385 0,140 0,993 0,045 0,025 2,2 9+0,213
Erythromma najas 2-55 101 0,480 0,211 0,810 0,279 0,033 2,44+0,2 97
Ischnura elegans 1-45 99 0,515 0,1 99 0,924 0,197 0,020 2,39+0,168
Примечания. Среднее квадратичное отклонение - по фактору веса, & - по фактору линейных раз-
у X
меров. Звездочкой отмечены виды, для которых расчеты проводили по усредненным данным массы и длины
тела.
1,0 1,5 1,0 1,5 1,0 1,5 1,0 1,5 1,0 1,5
3 7
/ Рис. 15. Соотношение длины и мае—
£ сы тела у личинок равнокрылых
г (а-г) и разнокрылых (д-з) стрекоз.
/• а — Ischnura elegans, б —
.г Enallagma cyathigerum (озеро),
/’ в — Е. cyathigerum (пруды),г -
' Erythromma najas, д — Aesch—
____________па juncea, е — Ae. grandis,
1,0 1,5 (?д 1 ж — Libellula quadrimaculata,
з — Sympetrum vulgatum.
казателем характера изменения формы тела животного в период
роста.
Если рост происходит с сохранением геометрического подобия, то
b = 3, в других случаях Ь> или <3. Установление констант а
и Ь для каждого из изученных видов позволяет рассчитать массу жи-
вотных с достаточной для многих целей точностью по такому легко
определяемому показателю, как длина тела. Расчет констант а и ь
проводится методом наименьших квадратов, который позволяет найти
уравнение линейной регрессии 1g W = Iga + b 1g 1.
Полученные данные позволяют сделать вывод о высокой степени
корреляции массы тела личинок стрекоз и их линейных размеров (см.
табл. 12). Коэффициент корреляции г колеблется в нешироких преде-
лах от 0,912 до 0,994 и лишь для Erythromma najas r= 0,810.
Рассчитанные константы b для всех видов < 3 и колеблются
незначительно, но у личинок разнокрылых стрекоз имеют несколько
более высокие значения. Исключение составляют личинки Aeschna
grandis ( b = 2,12 +0,198).
Величина константы а специфична для каждого вида, но, есте-
ственно, более высока у личинок разнокрылых стрекоз, отличающихся
более крупными размерами и соответственно большей массой тела на
единицу длины (рис. 15).
Некоторая разбросанность точек может быть объяснена рядом
причин. Прежде всего это связано с особенностями линейного роста
личинок стрекоз. В течение одного личиночного возраста при неизмен-
ных линейных размерах (после уплотнения хитинизированной кутикулы
151
покровов) продолжается возрастание индивидуальной массы тела. Так,
для личинок Ischnura elegans (см. рис. 15, а) при длине тела
10 мм (1g 1 = 1,0000) отмечено колебание массы тела от 3 до
7 мг (lg W = 0,4771-0,9031). Сходные результаты получены для
личинок других равнокрылых стрекоз (см. рис. 15, д—з).
С другой стороны, масса тела личинок стрекоз одного возраста
с одинаковыми линейными размерами иногда оказывается различной в
связи с отсутствием или наличием воды в ректальном отделе кишеч-
ника. Это возможно прежде всего для личинок разнокрылых стрекоз,
хотя исключение не составляют личинки и равнокрылых стрекоз.
Для выяснения значения личинок стрекоз в водных биоценозах
важно знать некоторые характеристики их питания. Это касается как
механизма захвата пищи, что изучено достаточно подробно различны-
ми авторами, так и состава и величины рациона, усвояемости корма,
элективности питания.
Для определения величины суточного рациона личинок стрекоз бы-
ла поставлена серия опытов с массовыми видами: Cordulia aenea,
Lestes sponsa, Ischnura elegans и Enallagma cy—
athigerum. Отбирались животные приблизительно одинаковой мас-
сы. Личинки обсушивались на фильтровальной бумаге до исчезновения
влажных пятен на фильтре, после чего взвешивались на торзионных ве-
сах. Количество животных в одной кювете колебалось от 4 до 22 осо-
бей в зависимости от их размеров. Экспозиция опыта 5-8 дней. Опыт-
ные животные активно двигались, линяли. Смертности личинок за вре-
мя опытов не наблюдалось. В качестве кормовых объектов личинкам
стрекоз предлагались мальки рыб, олигохеты, хирономиды и водяные
ослики. Такой выбор определен был тем, что указанные группы гид-
робионтов доминируют в водоеме и являются основными кормовыми
объектами хищников. В опытах использовались в основном мальки плот-
вы. Из олигохет в качестве корма отбирались Limnodrilus udeke—
mianus и Lumbriculus variegatus: хирономиды были
представлены тремя видами: Cricotopus sp., Chironomus plumo—
sus, Ch. thummi.Последний вид отбирался в очистных прудах, где
численность его была очень высокой. В зарослях хары отлавливались
водяные ослики - Asellus aquaticus.
До начала опытов личинок стрекоз и их кормовые объекты в те-
чение 2-3 ч содержали в сосудах с озерной водой для адаптации, за-
тем помещали в опытные кюветы с отфильтрованной озерной водой.
Кормление стрекоз проводилось один раз в сутки. Навеска корма
обычно была не менее 30 % от массы личинок, в связи с характер-
ным для водных хищников избыточным питанием, отмеченным у гид-
робионтов не только в опытах, но и в естественных условиях.
Количество потребляемой пищи определяли как разность между за-
данным и оставшимся кормом. Величина рациона выражена в весовых
и энергетических (кал) показателях. Для этого методом бихроматного
окисления определяли калорийность личинок стрекоз, корма и фекалий^-.
1В опытах принимали участие А.П. Остапеня, А.П. Павлютин,
Ю.Г. Гининяк, Е£. Павельева.
152
Таблица 13. Среднесуточный рацион для личинок стрекоз
W R R/W-lOO %
мг кал мг кал
сырая сухая сырая сухая
Подотряд Zygoptera
10,9 1,97 7,84 2,67 0,322 1,53 19,52
13,4 2,53 10,12 3,00 0,360 1.71 16,90
13,4 2,53 10,12 . 2,17 0,260 1,23 12,16
14,6 2,665 10,64 4,75 0,570 2,70 25,38
14,8 2,74 10,94 4,50 0,540 2,56 23,32
24,5 4,65 18,16 5,08 0,610 2,89 15,54
25,8 4,83 19,42 4,33 0,520 2,47 12,79
25,7 4,82 19,42 3,06 0,3 67 1,71 8,65
44,0 8,31 33,24 6,00 0,720 3,42 10,20
61,9 11,26 45,04 11,50 1,380 6,55 14,54
70,5 ' 13,13 52,52 7,54 0,905 4,29 6,25
71,5 13,5 54,00 10,00 1,200 5,70 10,56
72,3 13,67 54,68 7,23 0,868 4,12 7,54
Род Aeschna
55,65 9,07 42,63 12,84 1,54 7,32 14,50
75,95 12,38 58,18 27,00 3,25 15,44 21,30
92,09 15,01 70,55 33,00 3,96 18,81 26,26
148,34 24,18 113,64 34,25 4,11 19,52 13,57
172,15 28,06 131,84 30,20 3,62 17,14 13,00
449,69 73,30 344,50 68,40 8,20 98,95 9,05
707,06 115,25 541,67 110,25 13,23 62,84 9,08
Род Cordulia
48,5 6,305 27,11 16,92 2,030 9,643 35,57
48,5 6,305 27,11 10,03 1,203 5,714 21,08
102,0 13,26 57,02 27,25 3,270 15,53 27,24
102,0 13,26 57,02 22,28 2,675 12,71 22,29
152,0 19,76 84,97 50,00 6,000 28,50 33,54
152,0 19,76 84,97 45,42 5,450 25,89 30,47
Как видно из приведенных данных (табл. 13), абсолютные вели-
чины рационов у личинок стрекоз разных родов и подотрядов обнару-
живают отчетливую тенденцию к повышению по мере возрастания ин-
дивидуальной массы животного (рис. 16).
Рацион, выраженный в процентах от массы тела (кал), законо-
мерно падает (рис. 17). Таким образом, количество пиши, потребляе-
мое личинками стрекоз в расчете на 1 экз., возрастает с увеличени-
153
---! ! !---- ! ! !— ! ! (------
/,2 2,0 2,6--1,4 1,6 2,2 1,0 1,4 1,6 SgW
Рис. 16. Зависимость суточного рациона ( R, кал) от массы тела
(W, кал) у личинок стрекоз.
а - род Aeschna, б - род Cordulia, в - Zygoptera.
ем массы тела, а в расчете на единицу веса - снижается. Это хоро-
шо прослеживается по результатам оценки рационов личинок стрекоз
рода Aeschna. При массе тела личинок 55,65-92,09 мг (r/w х
х 100) составляет 14,5-26,26 %.
У личинок, масса тела которых равна 148,34-707,06 мг, ука-
занный показатель снижается до 13,57-9,05 %.
Установлена зависимость величины среднесуточного рациона (R )
от массы тела ( W) для разных групп личинок стрекоз и для всего
отряда в целом (рис. 18).
Величина рациона находится в параболической зависимости от мас-
сы тела. Эта зависимость может быть выражена уравнением R =
=р • Wm , где R - суточный рацион (в единицах массы или энергии);
W - масса животного; р - константа, определяющая уровень потреб-
ления пищи в данных условиях за единицу времени при W = 1; т-
коэффициент, определяющий скорость изменения рациона при увеличении
массы организма.
Рис. 17. Изменение величины суточного рациона ( R) от массы те-
ла ( W) в процентах у личинок стрекоз подотрядов Zygoptera (а)
и Anisoptera (б).
154
Рис. 18. Зависимость рациона от мас-
сы тела.
1 - О donate. в целом, 2 -
Anisoptera, 3 - Zygoptera. Шка-
лы логарифмические.
На основании экспериментальных
данных методом наименьших квадра-
тов рассчитаны уравнения зависимости
и равнокрылых стрекоз.
рациона от массы тела личинок равнокрылых
Для Zygoptera уравнение имеет вид
R = 0,449 • АлР’7х3,
где R - суточный рацион 1 экз. в калориях, W - эквивалентная
массе энергетическая ценность одной особи.
Для Anisoptera уравнение имеет вид
R = 1,062 • АлР'657.
Суммарно для Odonata уравнение имеет вид
R = 0,298 • W0,88.
Коэффициенты последнего уравнения близки к величинам, получен-
ным для Zygoptera. Если учесть, что в сообществе личинок стре-
коз литорали оз. Нарочь основная роль принадлежит равнокрылым
стрекозам, то с достаточным приближением для определения величины
рационов по массе тела можно использовать формулу зависимости R
от W с общими для равнокрылых и разнокрылых стрекоз коэффици-
ентами.
Таким образом, зная линейные размеры личинок стрекоз, мы мо-
жем рассчитать массу их тела, а по показателям массы тела опреде-
лить величину рациона.
Изучая питание хищных личинок стрекоз с точки зрения оценки
их роли в водоеме, мы исследовали элективность питания в отношении
таких кормовых объектов, как личинки рыб, хирономиды, олигохеты и
водяные ослики.
Степень избирательности в выборе кормового объекта оценивается
как отношение концентрации его в рационе к концентрации в среде
/Ивлев, 1955/ и рассчитывается по следующей формуле:
г. - р.
Е = г. + р. ,
1 1
где Г£ — концентрация пищевого компонента в рационе, р — то же, в
среде. 1
Результаты экспериментов показали, что личинки Cordulia
aenea при наличии в пищевом комплексе личинок хирономид, олиго-
хет, водяных осликов и мальков рыб потребляют все пищевые объекты,
но явное предпочтение отдают личинкам хирономид (Е = 0,13) и оли-
155
156
Таблица 14. Элективность питания личинок некоторых видов стрекоз
Вид r. , % 1 Pi О/ , ЛЭ Е
Личин- ки хи- роно- мид Олиго- хеты Водя- ные осли- ки Личин- ки рыб Личин- ки хи- роно- мид Олиго- хеты Водя- ные осли- ки Личин- ки рыб Личин- ки хи- роно- мид Олиго- хеты Водя- ные осли- ки Личин- ки рыб
Cordulia 45,1 8,6 25,0 21,3 35,6 9,2 28,6 31,9 0,11 -0,04 -0,06 -0,18
аепеа 52,4 13,2 17,4 17,2 36,4 12,0 27,1 24,2 0,20 0,05 -0,21 -0,16
40,7 13,6 23,5 12,2 34,4 6,9 28,0 30,7 0,09 0,33 -0,09 -0,43
Среднее . . . 46,0 11,8 21,6 16,9 35,4 9,4 27,9 28,9 0,13 0,11 -0,12 -0,19
Lestes 37,6 15,6 10,6 36,2 33,5 11,0 28,4 27,0 0,05 0,18 -0,46 0,18
sponsa 33,8 23,0 24,5 28,7 26,9 9,7 2 9,0 34,4 0,11 0,58 -0,08 -0,09
31,6 34,1 20,0 24,3 26,5 8,8 36,3 28,0 0,09 0,61 -0,27 -0,07
Среднее . . . 34,3 24,2 18,4 29,7 28,9 9,8 31,2 29,8 0,08 0,45 -0,25 -0,001
Ischnura 52,9 14,4 21,6 11,1 42,4 8,4 22,9 28,3 0,11 0,26 -0,03 -0,43
elegans 40,6 10,8 27,5 21,1 40,0 7,0 27,8 . 25,2 0,01 0,21 -0,01 -0,09
Среднее . . . 46,7 12,6 24,3 16,1 41,2 7,7 25,3 26,7 0,06 0,24 -0,02 -0,24
Enallagma 54,6 15,7 25,7 4,0 38,8 6,0 34,5 20,7 0,16 0,45 -0,12 -0,68
cvathigerum 31,4 20,6 25,9 22,1 29,3 7,7 33,7 29,3 0,03 0,45 -0,13 -0,14
Среднее . . . 43,0 18,1 25,8 13,0 34,0 6,8 34,1 25,0 0,11 0,45 -0,13 -0,31
Таблица 15. Вероятность выедания молоди рыб личинками стрекоз
Вид Количество серий опытов Продолжительность опыта, сут Среднее кол-во уничтоженных ли- чинок (в сутки) Размер личинок, мм Среднее кол-во хищ- ников на 1 м^ Среднее кол-во ли- чинок рыб, уничто- женных за сутки, экз.
„ 2 на 1 м на 1 га
Aeschna
grandis Cordulia 1 10 12 10-20 2,2 24,4 244 000
aenea Sympetrum 3 24 5 10-20 4,3 21,5 215 000
vulgatum Enallagma 2 12 4 10-18 3,1 12,4 124 000
cyath. Ischnura 6 19 2 7-14 4,5 9,0 90 000
elegans Lestes 5 24 6 9-15 2,1 12,6 126 000
sponsa 5 12 3 8-15 11,2 33,6 336 000
Итого... — — — — — 113,5 1 135 000
гохетам (Е = 0,11), доля которых в рационе составляет соответст-
венно 46,0 и 11,8 %. Водяные ослики, мальки и личинки рыб, доля
которых в рационе составляет 21,6 и 16,9 %, в эксперименте избе-
гаются, индекс элективности (или выборочного питания, когда в ра-
ционе пищевые объекты представлены. в иной пропорции, чем в окру-
жающей среде) отрицателен (-0,12 и -0,19 соответственно)
(табл. 14). Сходные результаты получены для Ischnura elegans
и Enallagma cyathigerum. Lestes sponsa, один из самых
массовых видов личинок в водоеме, имеет индекс элективности при
питании личинками рыб, близкий к О. В отдельных опытах Е= 0,18.
В экспериментах, где личинкам стрекоз для питания предлагались
лишь личинки рыб, что близко к условиям на нерестилищах, получены
следующие результаты (табл. 15). В пересчете на единицу площади
(1 и 1 га) с учетом средней численности личинок стрекоз мы по-
лучили величину потребления хищниками стрекоз мальков рыб, что
позволяет судить о возможной степени выедания молоди рыб на нере-
стилищах.
Личинка стрекозы Aeschna grandis за сутки способна уничто-
жить до 12 личинок плотвы размером 7-10 мм, 3-5 личинок разме-
ром 15-25 мм. Личинка Cordulia аереа уничтожала в опытах
ежесуточно до 5 личинок рыб размером 10-20 мм, более крупные
особи для нее недоступны. Личинка Sympetrum vulgatum потреб-
157'
ляла в сутки 8-10 личинок плотвы размером 7-12 мм или 3—4 раз-
мером 14-15 мм. Для личинок Enallagma cyathigerum, Ischnu-
ra elegans, Lestes sponsa суточный рацион в условиях
эксперимента включал 9-10 личинок рыб размером 7-8 мм или
3 экз. размером 14-15 мм. Личинки рыб более 20 мм недоступны
для этих хищников.
Анализируя полученные опытным путем данные по потреблению
личинками стрекоз молоди рыб, можно определить приблизительный
ущерб, наносимый рыбному хозяйству в период размножения рыб на
нерестилищах оз. Нарочь.
Следует, однако, принять во внимание, что в естественных усло-
виях хищные личинки стрекоз уничтожают меньшее количество рыб,
чем в эксперименте.
В опытах нет водной растительности, служащей личинкам рыб ук-
рытием, а на белом дне опытной кюветы жертва более заметна, по-
этому для расчетов мы приняли допущение, что в природных место-
обитаниях личинки потребляют в 10 раз меньше молоди рыб, чем в
опытных условиях. По расчетным данным, средняя величина потребле-
ния молоди рыб личинками стрекоз 12 экз./м^, что в пересчете на
гектар составляет 100 тыс. личинок и мальков рыб за сутки. В те-
чение месяца на той же площади уничтожается 346 экз./См^ • месяц),
что составляет 3 460 000 экз./(га • месяц).
Сходные данные приведены В.В. Шерстюком /1967/ для Кремен-
чугского' водохранилища.
ОПЫТ ИССЛЕДОВАНИЯ НАСЕЛЕНИЯ СТРЕКОЗ
В БИОЦЕНОЗАХ ЗАПАДНОЙ СИБИРИ
Для интегральной оценки роли стрекоз в биогеоценозе необходимо
учитывать всю совокупность видов, появляющихся в течение сезона на
данной территории. Такую совокупность особей всех видов определен-
ной систематической группы, занимающих определенную территорию,
принято называть населением /Дажо, 1975/.
Материалов по изучению населения стрекоз пока очень немного.
В последние годы они получены для некоторых районов азиатской ча-
сти СССР, в частности Карасукских озер в Кулундинской степи, и
выполнялись В.В. Заикой в 1973-1975 гг.
Учеты стрекоз проводили в разные периоды их жизни и в разных
биотопах. Взрослые стрекозы учитывали в местах выплода возле во-
доемов на трансектах, проходящих вдоль берега и на маршрутах протя-
женностью 4-10 км.
Формирование населения стрекоз будет рассмотрено нами для сле-
дующих территорий: возле водоемов в озерных котловинах, в колках и
на открытых пространствах полей.
Население озерных котловин. Котловины озер в районе исследова-
ний занимают понижения между гривами и другие депрессии с малым
водосборным бассейном. На дне котловин располагаются озера и тро-
стниковые займища, а на склонах - березово-осиновые колки, в цент-
ральной части которых за счет таяния снега и дождей обычно застаи-
вается вода и создается заболоченность.
158
Вблизи озер проводят всю свою жизнь только равнокрылые стре-
козы, разнокрылые же прилетают к водоемам лишь для спаривания и
откладки яиц. Естественно, что доля последних в населении стрекоз
приозерной территории невелика и не превышает 10 % (исключение
представляет лишь период выплода стрекоз). Поэтому население озер-
ных котловин состоит в основном из видов подотряда Zygoptera,
которые относятся к жизненной форме "собирателей", охотящихся на
концентрирующихся в прибрежной растительности мелких двукрылых.
Стрекозы других жизненных форм не могут охотиться в толше тра-
востоя.
Население озерных котловин формируется в начале весеннего пе-
риода. Во второй половине мая появляются С. armatum и С. ver—
nale, которые не встречаются в других биотопах, а держатся толь-
ко у водоемов. Одновременно с ними появляются L. quadrimacu—
lata, которые по численности в момент выплода составляют пример-
но пятую часть населения, но вскоре их доля уменьшается и разно-
крылые остаются в незначительном количестве. В июне - июле проис-
ходит выплод видов родов Enallagma и Lestes, которые в се-
редине лета достигают максимальной плотности у водоемов.
В целом население озерных котловин на протяжении всего сезона
представлено последовательно сменяющими друг друга массовыми
видами равнокрылых стрекоз.
Население колков. Колки располагаются в межгривных понижени-
ях. Нередко в них имеются поляны с богатой энтомофауной, многие
представители которой служат пишей для стрекоз. На полянах и опуш-
ках колков происходит концентрация стрекоз, которые относятся к
жизненной форме "подстерегателей", выплаживающихся в водоемах,
расположенных от колков на значительном расстоянии, иногда до не-
скольких километров. Население формируется в основном за счет ви-
дов родов Sympetrum, Leucorrhinia и Libellula. Небольшое
участие особей "преследователей" и "собирателей" (С,2-5 %) проис-
ходит в моменты их массового выплода, а также за счет рассеива-
ния этих видов в конце лета, когда их доля возрастает до 20 % у
"преследователей" и 12 % у "собирателей". В состав населения вхо-
дят сначала весенне—летние виды: L. pectoralis, L. rubicunda
и L. quadrimaculata, связанные трофически с разлетающимися от
водоемов двукрылыми насекомыми и концентрирующиеся вслед за ни-
ми в колках и влажных западинах. Затем эти виды сменяются летне-
осенними и осенними рода Sympetrum:S.flaveolum, S. scoticum,
S. sanguineum, S. vulgatum. Средний (кустарниковый) ярус биогеоце-
ноза, так же как и нижний (травяной), в озерных котловинах занят
массовыми видами стрекоз в течение всего сезона.
Население открытых пространств. Водораздельные территории, сво-
бодные от колков, заняты полями. На этих пространствах преобладают
стрекозы "преследователи", основную массу которых составляют виды,
появляющиеся с середины лета, - Ae. serrata и Ae. grandis.
Они способны успешно охотиться на крупную добычу, такую, например,
как слепни, которые достигают максимума численности в конце июня -
150
середине июля. Таким образом, в .отличие от травяного и кустарни-
кового ярусов верхний (воздушный) занимается стрекозами только с
середины лета.
Итак, особенности формирования населения стрекоз южной части
Западно-Сибирской равнины позволяют этим насекомым охотиться в
течение всего сезона во всех ярусах биогеоценозов, создавая тем
самым сложную структуру пресса хищников, который влияет на чис-
ленность мелких, в том числе кровососущих насекомых.
Стрекозы уже в момент выплода начинают играть немаловажную
биогеоценотическую роль, вынося из водоема накопленные в теле ли-
чинок минеральные и органические вещества, которые тут же начина-
ют свой путь в наземной части биогеоценоза в качестве пищи для
птиц и хищных беспозвоночных. После выплода стрекозы разлета-
ются по территории и начинают охотиться в местах локализации мел-
ких двукрылых возле водоемов, в колочных западинах и на полях,
обеспечивая пресс хищников на межводных пространствах. Сфера де-
ятельности в биоценозе у разных жизненных форм стрекоз разделя-
ется не только по территориальному принципу, но и по вертикальному:
они занимают травяной, кустарниковый и древесный ярусы. Благодаря
этому, весь приз емный слой воздуха, заселенный летающими насеко-
мыми, контролируется разными видами стрекоз в течение всего теп-
лого сезона.
Когда у стрекоз завершается период созревания, они воз-
вращаются к водоемам для спаривания и откладки яиц. В
этот период они вновь становятся жертвами птиц. Откладывая
яйца в больших количествах (до 2 тыс. шт. у четырехпят-
нистой стрекозы), стрекозы тем самым компенсируют свою
высокую смертность и обеспечивают существование вида. Яйца даже
из одной яйцекладки имеют разную скорость развития, это позволяет
сохраниться хотя бы части яиц в случае гибрли остальных вследст-
вие изменения внешних условий. Из-за неравномерного созревания
яиц выплод личинок растянут во времени, и в водоемах одновремен-
но присутствует большое количество личинок разных возрастов одно-
го вида. Личинки разных жизненных форм практически все время при-
сутствуют в водоеме и поедают многих доступных им гидробионтов
вплоть до мальков рыб. В водоеме же происходит заражение личинок
церкариями трематод, нематодами, цестодами и личинками клешей.
Личинки стрекоз, сами будучи хищниками, служат пищей для других
хищных беспозвоночных, а также рыб и птиц.
Итак, стрекозы являются массовым компонентом всех ярусов вод-
ных и наземных биогеоценозов, имеют трофические связи с птицами,
рыбами и многими группами беспозвоночных животных, существенно
влияя на численность некоторых насекомых, в первую очередь двукры-
лых. Будучи связующим звеном водных и наземных частей биогеоце-
нозов, они участвуют в перераспределении веществ между водоемами
и сушей.
160
СПИСОК ЛИТЕРАТУРЫ
Акимов М.П. Биоморфологический метод изучения биоценозов // Бюл. МОИП.
Отд. биол. - 1954. - №3. - С. 27-36.
Акрамовский Н.Н. Стрекозы Нахичеванской республики // Нахичеванские тр.
Ереван, гос. ун-та. - 1939. - № 9. - С. 47-53.
Акрамовский НЛ. Фауна стрекоз Советской Армении // Зоол. сб. АН .
АрМОСР, - Ереван, 1948. - Вып. 5. - С. 117-188.
Акрамовский Н.Н. Систематический обзор животных Кавказского перешейка.
9. Стрекозы - Odonata // Животный мир СССР. Горные области евро-
пейской части СССР. - М.; Л.: Изд-во АН СССР, 1958а. - Т. 5. -
С. 457-463.
Акрамовский Н.Н. Личинка стрекозы Onychogomphus flexu'osus
(Schneider) (ins., Odonata, Gomphidae)// Изв. АН АрмССР.- 19586.-
T. 11, № 5. - С. 87-90.
Акрамовский Н.Н. Дополнение к фауне стрекоз Восточного Закавказья //
Там же. - 1964. - Т. 17, № 10. - С. 99-101.
Акрамовский Н.Н. Новый общебиологический и энтомологический журнал
"Odonatologica", журнал международного общества одонатологов //
Общая биология. - 1973. - Т. 34, № 2. - С. 319.
Акрамовский НЛ. К вопросу о цепях охраны малочисленных и исчезающих
видов насекомых и беспозвоночных животных вообще 11 Тез. докл. II совещ.
об охране насекомых. - Ереван, 1975а. - С. 18-20.
Акрамовский Н.Н. Некоторые особо интересные виды стрекоз Кавказа и За-
кавказья и предложения по их охране II Там же. - 19756. - С. 14-17.
Акрамовский Н.Н. О типах фауны и биогеографическом районировании (на
примере современной фауны моллюсков и стрекоз АркССР) // Тез. докл.
VI Всесоюэ. зоогеогр. конф. - 1975в. - С. 6-7.
Арабина И.П. Сезонная, годовая динамика и продукция зообентоса озер На-
рочь, Мястро н Баторин: Автореф. дис. ... канд. биол. наук. - Минск,
1968.
Арнольд Н.М. Каталог насекомых Могилевской губернии. - Спб., 1902. -
С. 73-74.
Артоболевский Г.В. Заметки о стрекозах Полтавской губернии Н Материа-
лы к познанию фауны Юго-Западной России. - 1917. - Т. 2. - С. 17-29.
Артоболевский Г.В. Материалы к фауне стрекоз окрестностей г. Калуги И
Фауна насекомых Калужской губернии. - Калуга, 1923. - Вып. 1. -
С. 18-20.
Артоболевский Г.В. Стрекозы окрестностей Киева // 36ipH£K прац Зоологи-
ческого музея Украинской АН. - 1927а. - Т. 3. - С. 65-118.
Артоболевский ГЛ. До вивчения бабок Подиппя II Там же. - 19276 . -
С. 159-162.
Артоболевский Г.В. Нотатки про бабок Поптавшини // Ф1з.-мат. siflfliny
Всеукраинской АН. - 1929а. - Т. 13. - С. 3-20.
.11 Зак. 611 161
Артоболевский Г.В. Стрекозы Крыма // Бюл. о-ва натуралистов и друзей
природы. - 19296. - Т. 11. - С. 139-150.
Артоболевский Г.В. Стрекозы Дагестана // Рус. энтомол. обозрение. -
1929в. - Т. 23, № 3/4. - С. 225-240.
Артоболевский Г.В. К экологии стрекоз Теберды // Работы Сев.-Кавк,
гидробиол. станции. - 1930. » Т. 3, вып. 1-3. - С. 33-43.'
Бартенев А.Н. Odonata Полесской и Виленской экспедиций // Тр. /
Студ. кружок дли исследовании русской природы при Моск, ун-те. - 1907.-
Т. 3. - С. 133-146.
Бартенев А.Н. Коллекции стрекоз из окрестностей оз. Увильды Екатеринбург-
ского уезда Пермской губернии // Тр. о-ва / О-во естествоиспыт. при Ка-
зан. ун-те. - 1908. - Т. 41, вып. 1. - С. 1-40.
Бартенев А.Н. Заметка о стрекозах Зоологического музеи Уральского общест-
ва любителей естествознании // Зап. Урал, о-ва любителей естествознания.-
1909а. - Т. 29. - С. 142-144.
Бартенев А.Н. Odonata Карской экскурсии // Тр. /Студ. кружок для
исследовании русской природы при Моск, ун-те. - 19096. - Т. 4. -
С. 63-75.
Бартеиев АЛ. Материалы по фауне стрекоз Сибири, № 1-5 // Изв. Том.
ун-та. - 1909в. - Т. 37. - С. 17-56.
Бартенев АЛ. .Материалы по фауне стрекоз Сибири, № 6-14 // Работы лаб.
эоол. кабинета Варшав. ун-та. - 1910а. - С. 1-24.
Бартенев АЛ. Коллекция стрекоз из Мацуими (Японии), пожертвованнаи
братьями М.А., С.А. Сапожниковыми (Москва) // Изв. Том. ун-та. -
19106. - С. 1-16.
Бартенев АЛ. Материалы по фауне стрекоз Сибири, № 15 // Работы лаб.
эоол. кабинета Варшав. ун-та. - 1910в. - С. 1-78.
Бартенев АЛ. К фауне Odonata Кубанской области // Рус. энтомол.
обозрение. - 1910г. - № 10, вып. 1/2. - С. 27-38.
Бартенев А.Н. К фауне стрекоз Польши // Там же. - 1910д. - С. 124-125.
Бартенев АЛ. К фауне стрекоз Закавказья И Работы лаб. эоол. кабинета
Варшав. ун-та. - 1911а. - Т. 1. - С. 1-20.
Бартенев А.Н. К составу фауны Кавказа // Протокол о-ва естествоиспыт.
при Варшав. ун-те. - 19116. - Т. 23, № 1/2. - С. 211-239.
Бартенев А.Н. Материалы по стрекозам палеарктической Азии нэ коллекции
Зоологического музеи Имперской АН, 1 // Ежегод. Зоол. музеи АН. -
1911в. - Т. 16. - С; 409-448.
Бартенев А.Н. /Рецензия/ // Рус. энтомол. обозрение. - 1911г. - Т. 11,
вып. 1. - С. 23-24. - Рец. на ст.: Воронцовский П. Материалы к фауне
стрекоз окрестностей г. Оренбурга. - Изв. Оренбург, отд. Рус. геогр. о-ва.-
Оренбург, 1909. - Вып. 21. - С. 113-120.
Бартенев АЛ. Палеарктические и восточно-азиатские виды и подвиды рода
Calopteryx Leach (Odonata, Calopterygidae) ll Работы лаб. эо-
ол. кабинета Варшав. ун-та. - 1912а. - Т. 1. - С. 163-257.
Бартенев А.Н. О коллекции стрекоз Кавказского музеи // Изв. Кавк, му-
зеи. - 19126. - Т. 7, вып. 1. - С. 107-116.
Бартенев АЛ. Материалы по стрекозам палеарктической Азии из коллекций
Зоологического музеи АН, 2 // Ежегод. Зоол. музея. - 1912в. - Т. 17.-
С. 289-310.
Бартенев А.Н. К фауне стрекоз Крыма // Там же. - 1912г. -С. 281-288.
Бартенев А.Н. Сборы стрекоз в Закавказье летом 1911 г. // Работы лаб.
эоол. кабинета Варшав. ун-та. - 1912д. - С. 132-161.
Бартенев А.Н. Заметки о стрекозах Черногории II Рус. энтомол. обозрение.-
1912е. - № 12, вып. 1. - С. 76-80.
Бартенев АЛ. О коллекциях стрекоз из Южной Бухары (Туркестан) // Там
же. - 1913. - № 13. - С. 176-189.
162
Бартенев А.Н. Материалы по фауне стрекоз Сибири, № 16-17 // Тр. о-ва /
Рус. энтомол. о-во. - 1914а. - Т. 41, № 2. - С. 1-32.
Бартенев А.Н. К вопросу о принципах зоогеографических делений // Варшав.
университет, изв. - 19146. - № 6. - С. 1-35.
Бартенев А.Н. К вопросу о виде и его составе // Протокол о-ва естество-
испыт. Варшав. ун-та. - 1914в. - Т. 25, № 3/4. - С. 31-32.
Бартенев А.Н. Насекомые ложносетчатокрылые. Libellulidae // Фауна
России и сопредельных стран. - Пг.: Изд-во АН, 1915а. - Т. 1, вып. 1,-
352 с.
Бартенев А.Н. Американские представители рода Sympetrum // Варшав.
университет, изв. - 19156. -№ 5. - С. 1-26.
Бартенев А.Н. Sympycna paedisca в Европейской России // Рус. энто-
мол. обозрение. - 1915в. - Т. 15, вып. 2. - С. 293-^94.
Бартенев АЛ. /Рецензия/ // Там же. - 1915 г. - Т. 15, вып. 3. -
С. 446-448. - Рец. на ст.: Колосов Ю.М. Заметки о насекомых Тобольской
губернии. - Зап. Урал, о-ва любителей естествозн. - Екатеринбург, 1914. -
Т. 34, вып. 1/2. - С. 13-36.
Бартенев А.Н. Стрекозы Кавказских Минеральных Вод // Рус. энтомол. обоз-
рение. - 1915д. - Т. 15, вып. 4. - С. 604-612.
Бартенев А.Н. К фауне стрекоз Северной Персии // Там же. - 1916а. -
Т. 16, № 1/2. - С. 38-45.
Бартенев А.Н. Заметки о стрекозах окрестностей Сигнаха Тифлисской губер-
нии // Там же. - 19166. - Т. 16, № 3/4. - С. 343-345.
Бартенев А.Н. Одонатологические экскурсии 1916 г. в окр. Ростова-на-До-
ну // Там же. - 1916в. - Т. 16, № 3/4. - С. 319-327.
Бартенев А.Н. О классификации и конституции рода Sympetrum Newm.
(Odonata, Libellulidae ) // Варшав. университет, изв. - 1916г. -
№ 5. - С. 1-19.
Бартенев А.Н. Стрекозы Нальчика Терской области // Изв. Кавк, музея. -
1918. - Т. 11, вып. 3/4. - С. 228-244.
Бартенев АЛ. Лёт стрекоз 1914 г. и о лётах стрекоз вообще // Протокол
о-ва естествоиспыт. при Дон. ун-те, 1916-1918. - 1919а. - Вып. 1,-
С. 49-66.
Бартенев А.Н. Насекомые ложносетчатокрыпые. Libellulidae // Фауна
России и сопредельных стран. - Пг.: Изд-во АН, 19196. - Т. 1, вып. 2. -
С. 353-577.
Бартенев АЛ. Заметки о стрекозах Закавказья И Изв. Кавк, музея. -
1919в. - Т. 12. - С. 196-199.
Бартенев А.Н, Материалы по фауне стрекоз Сибири, № 16 // Рус. энтомол.
обозрение. - 1921а. - Т. 17. - С. 129-134.
Бартенев А.Н. Список стрекоз Ставропольской губернии // Там же. - 19216.-
С. 144-151.
Бартенев А.Н. Одонатологические наблюдения зимой 1921 г. в Сочи, Куба-
но-Черно морс кой области // Там же. - 1924а. - Т. 18, № 4. - С. 223-^28.
Бартенев А.Н. К одонатофауне Кавказских гор И Бюл. Гос. музея Грузин. -
19246. - Т. 2. - С. 28-86.
Бартенев А.Н. Заметки о стрекозах Кавказа // Работы Сев.-Кавк. гидробиол.
ст. при Горском с.-х. ин-те. - 1928а. - Т. 2, вып. 2/3. - С. 124-125.
Бартенев АЛ. Определительная таблица родов всего света подсемейства
Libellulidae (Odonata) // Изв. Сев.-Кавк. ун-та. - 19 2 86. - Т. 3,
вып. 15. - С. 152-198.
Бартенев А.Н. Odonata окрестностей гор. Тифлиса и других мест Гру-
зии // Бюл. Гос. музея Грузии. - 1928в. - Т. 5. - С. 65-86.
Бартенев А.Н. С видовых группах Aeschna juncea и Aeschna clep-
sydra (Odonata, Aeschnidae) в Палеарктике //Тр. Сев.-Кавк. ассо-
циации н.—и. ин-тов. — Ростов—н/Д., 1929а. — № 54. — С. 64—79. — (Тр.
163
Ин-та математики и естествознания при Сев.-Кавк. гос. ун-те; Вып. 8).
Бартенев А.Н. Материалы к познанию Западного Кавказа в одоиатологическом
отношении // Там же. - 19296. - № 72. - С. 1-138. - (Тр. Ин-та мате-
матики н естествознания цри Сев.-Кавк. гос. ун-те; Вып. 14).
Бартенев А.Н. Новые данные о стрекозах Закавказья, Персии и Туркестана И
Рус. энтомол. обозрение. - 1929в. - Т. 23, № 1/2. - С. 124-131,
Бартенев А.Н. Заметка о сборах стрекоз Северо-Кавказской гидробиологиче-
ской станции в 1928 г. в Кабарде и Балкарии // Работы Сев.-Кавк. гидро-
бнол. ст. при Горском с.-х. ин-те. - 1930а. - Т. 3, вып. 1-3. - С. 85-86.
Бартеиев А.Н. К вопросу о зимовках imago стрекоз // Рус. эоол. ж урн. -
19306. - № 10, вып. 1. - С. 65-92.
Бартенев А.Н. О весенней и осенней одонатофауне Грузии й других мест За-
кавказья И Бюл. музея Грузии. - 1930в. - Т. 6. - С. 85-96.
Бартенев АЛ. О палеарктических видах подрода Cordulegaster Leach
(Odonata) // Работы Сев.-Кавк. гидробиол. ст. при Горском с.-х. ин-те. -
1930г. - Т. 3, вып. 1-3. - С. 1-32.
Бартенев А.Н. Опыт биологической группировки стрекоз европейской части
СССР. Ч. 1 // Рус. эоол. журн. - 1930а. - Т. 10, вып. 4. - С. 57-131.
Бартенев А.Н. О случаях определенной ориентировки стрекоз по отношению к
странам света // Там же. - 1930е. - № 10, вып. 1. - С. 53-64.
Бартенев АЛ. По поводу коллекции стрекоз из Восточной Сибири и Туркеста-
на о роде Ophiogomphus Sei. в Палеарктике // Рус. энтомол. обозрение. -
1930ж. - Т. 24, № 1/2. - С. 115-127.
Бартенев АЛ. Изменение сроков лёта стрекоз в пределах Палеарктики // Тр.
IV Всесоюз. съезда зоологов, анатомов, гистологов. Киев, 6-12 мая
1930 г. - Киев; Харьков, 1931а. - С. 46-47.
Бартенев А.Н. Опыт биологической группировки видов стрекоз Европейской
России // Там же. - 19316. - С. 48.
Бартенев А.Н. Стрекозы Западного Кавказа // Там же. - 1931в. - С. 47-48.
Бартенев АЛ. Опыт биологической группировки стрекоз европейской части
СССР. Ч. 2 // Зоол. журн. - 1932а. - Т. 11, вып. 1. - С. 3-60.
Бартенев АЛ. О некоторых основных положениях зоогеографии // Там же.-
19326. - Т. 11, вып. 2. - С. 23-38.
Бартенев А.Н. Заметка о представителях подсемейства Lestinae и Ag—
rioninae (Odonata) Кавказа и Туркестана // Бюл. Гос. музея Грузии. -
1933а. - Т. 8. - С. 42-44.
Бартенев АЛ. О некоторых положениях истории и сложения палеарктической
фауны // Зоол. журн. - 19336. - Т. 12, вып. 1. - С. 101-129.
Бартенев АЛ. О некоторых положениях истории и сложения палеарктической
фауны // Там же. - 1933в. - Т. 12, вып. 3. - С. 121-149.
Бартенев А.Н. К характеристике сложения фауны Западного Предкавказья //
Там же. - 1934а. - Т. 13, вып. 1. - С. 139-147.
Бартенев А.Н. К вопросу об основах зоогеографических характеристик . живот-
ных групп Палеарктики // Там же. - 19346. - Т. 13, вып. 4. -
С. 762-772.
Бартенев А.Н. О понтийской фауне // Там же. - 1935. - Т. 14, вып. 4. -
С. 762-784.
Бартенев АЛ. Стрекозы - Odonata // Животный мир СССР. - М.; Л.:
Изд-во АН СССР, 1936. - Т. 1. - С. 503-507.
Бартенев А.Н. О зоогеографическом анализе перелетов палеарктических птиц //
Учен. зап. Казах. ун-та. Биология. - 1939а. - Т. 1. - С. 9-27.
Бартенев А.Н. К теории ареалов и границ в зоогеографии // Там же. -
193 96. - Т. 1. - С. 29-43.
Бартенев А.Н. К теории зоогеографических делений // Там же. - 1939в,-
Т. 1. - С. 45-65.
164
Бартенев А.Н. К вопросу о зоогеографии севера Карагандинской области //
Там же. - 1939г. - Т. 1. - С. 67-80.
Бартенев А.Н. Фенология как наука И Сов. наука. - 1940. - № 3/4. -
С. 126-144;
Бартенев А.Н. Материалы по одонатологической фауне ( Odonata, Insects)
Дальнего Востока // Тр. / ДВ фил. им, Комарова АН СССР. Сер. эоол. -
1956. - Т. 3. - С. 202-238.
Бартенев А.Н., Попова А.Н. Материалы по фауне стрекоз Палеарктики // Рус.
энтомол. обозрение. - 1928. - Т. 22, № 3/4. - С. 235-239.
Баянов М.Г. Стрекозы Башкирии как промежуточные хозяева гельминтов //
Гельминты животных, человека и растений на Южном Урале. - Уфа, 1974. -
Вып. 1. - С. 77-87.
Баянов М.Г. Водные беспозвоночные животные Южного Урала как носители
метацеркарий // Паразиты и болезни водных беспозвоночных. Тез. докп. IV
Всесоюз. снмпоз. - М., 1986. - С. 13.
Бей-Биенко Г.Я., Мищенко Л.Л. Саранчовые фауны СССР и сопредельных
стран. Определители по фауне СССР. - М.; Л.: Изд-во АН СССР, 1951. -
Т. 1-2. - 667 с.
Белышев Б.Ф. К биологии и систематике Epitheca bimaculata Charp.
(Odonata) в Купундинской степи // Энтомол. обозрение. - 1951. - Т. 31,
№ 3/4. - С. 504-505.
Белышев Б.Ф. Материалы по фауне стрекоз северной части Томской области //
Учен. зап. Том. ун-та. - 1952. - № 18. - С. 89-94.
Белышев Б.Ф. Стрекозы на мысе Челюскина // Природа. - 1953а. - № 4.-
С. 40.
Белыйев Б.Ф. К фауне стрекоз Юго-Восточного Алтая // Заметки по фауне и
флоре Сибири. - Томск, 19536. - Вып. 17. - С. 49-52.
Белышев Б.Ф. Таинственная стрекоза на Алтае // Природа. - 1954. - № 11. -
С. 119.
Белышев Б.Ф. К изучению стрекоз Верхнего Приобья // Тр. ун-та / Том.
ун-т. - 1955а. - Т. 131. - С. 387-398.
Белышев Б.Ф. Неизвестные формы и стадии стрекоз из Сибири // Томск,
19556. - № 18. - С. 27-29.
Белышев Б.Ф. /Рецензия/ // Зоол. журн. - 1955в. - Т. 34, вып. 3. -
С, 697-698. - Рец. на кн.: Попова А.Н. Личинки стрекоз фауны СССР (Odo-
nata). - М.; Л.: Изд-во АН СССР, 1953. - 235 с.
Белышев Б.Ф. Южный внд стрекоз (Odonata, Insects) на горячих ис-
точниках Северного Забайкалья // Там же. - 1956а. - Т. 35, вып. 11. -
С. 1735-1736.
Белышев Б.Ф. Материалы к познанию дальневосточной фауны // Тр. ДВ фил.
им. Комарова АН СССР. - Владивосток, 19566. - Т. 3(6). - С. 181-199.
Белышев Б.Ф. Основные принципы географического распространения стрекоз
в Палеарктике // Тр. ун-та / Том. ун-т. - 1956в. - Т. 142. -
С. 185-194.
Белышев Б.Ф. Карликовая форма Lestes uncatus Kirby (Odonata, In-
sects) с горячих ключей Северного Забайкалья // Энтомол. обозрение. -
1957а. - Т. 36, № 1. - С. 161-162.
Белышев Б.Ф. /Рецензия/ // Зоол. журн. - 19576. - Т. 36, вып. 6. -
С. 957. - Рец. на кн.: Спурис З.Д. Стрекозы Латвийской ССР. - Рига:
Изд-во АН ЛатвССР, 1956. - 96 с.
Белышев Б.Ф. О пределах Палеарктической области в Восточной Азии на ос-
новании распространения стрекоз ( Odonata ) // Тез. докп. III совета.
ВЭО. - М.; Л., 1957в. - С. 4-6.
Белышев Б.Ф. О некоторых закономерностях в распределении окраски у
стрекоз // Докл. VII науч. конф. - Томск: Изд-во Том, ун-та, 1957г. -
С. 65-66.
165
Белышев Б.Ф. К систематике Ophiogomphus и Somatochlora (Odonata)
из Северной Азии Н Тез. докл. совеш. зоологов Сибири (ноябрь, 1957). -
Новосибирск, РИС СО АН СССР, 1957д. - С. 14-15.
Белышев Б.Ф. К фауне ( Odonata) Средней Азии //Fragm. faunist -
Warszawa, 1958а. - Т. 8, № 5. - С. 34-40.
Белышев Б.Ф. Стрекозы Верхнего Приобья: Автореф. дис. ... канд. биол. наук.-
Томск, 19586. - 11 с.
Белышев Б.Ф. Новые материалы по фауне стрекоз Алтайского края И Учен,
зап. Бийского пед. ин-та. - 1958в. - Вып. 3. - С. 85-111.
Белышев Б.Ф. Некоторые особенности вертикального распространения стрекоз
в горах Алтая // Природа и природные ресурсы Алтайского края. - Бийск,
1959а. - С. 114-115.
Белышев Б.Ф. Одонатологическая фауна Монголии в связи с фауной Южного
Алтая // Там же. - 19596. - С. 115-117.
Белышев Б.Ф. Одонатологическая фауна боровых озер южной оконечности
Верхне-Обского борового массива // Fragm. faunist — Warszawa,
1959в. - Т. 8, № 11. - С. 191-206.
Белышев Б.Ф. Палеарктическая область и ее подобласти на основании распро-
странения стрекоз (Odonata) // Тез. докл. IV съезда ВЭО. - М.; Л.,
1959г. - Т. 1. - С. 18-20.
Белышев Б.Ф. Фенология лёта стрекоз (Odonata)в Приалтайских степях н
некоторые общие закономерности этого явления // Энтомол. обозрение. -
1960а. - Т. 3 9, № 2. - С. 3 95-403.
Белышев Б.Ф. Некоторые особенности фенологии одонатологнческой фауны в
Приалтайских степях // Вопр. регион, фенол, и биогеограф. Бюл. Вост.-Сиб.
фен. Комисе. - Иркутск, 19606. - № 1. - С. 46-47.
Белышев Б.Ф. Горячий источник как среда обитания личинок стрекоз // Тр.
зап. I Баргузин, заповедник. - 19б0в. - Вып. 2. - С. 131-133.
Белышев Б.Ф. Условия существования личинок реликтовой стрекозы Orthet-
rum albistylum Selys в горячем ключе Северо-Восточного Прибай-
калья И Зоол. журн. - 1960 г. - Т. 39, вып. 9. - С. 1432-1433.
Белышев Б.Ф. Основные подразделения Палеарктической области на основании
распространения стрекоз (Odonata, Insecta) // Изв. СО АН СССР. -
1960д. - № 10. - С. 94-102.
Белышев Б.Ф. К познанию одонатологической фауны северо-востока Сибири //
Биологический сб. - Иркутск, 1960е. - С. 222-226.
Белышев Б.Ф. Сельскохозяйственное значение стрекоз II Сел. хоз-во Сиби-
ри. - 1961а. - № 7. - С. 65-66.
Белышев Б.Ф. О некоторых закономерностях в распределении окраски у севе-
ро-палеарктических стрекоз // Тр. Вост.-Сиб. фил. АН СССР. Сор. биол. -
19616. - Т. 36. - С. 102-105.
Белышев Б.Ф. Вертикальные пределы распространения Стрекоз в горах Алтая//
Зоол. журн. - 1961b. - Т. 40, вып. 7. - С. 1103-1104.
Белышев Б.Ф. К познанию одонатологической фауны Баргузинского заповедни-
ка и некоторых прилежащих земель // Тр. зап. / Баргузин, заповедник. -
1961 г. - Вып. 3. - С. 181-185.
Белышев Б.Ф. Зоогеография Верхнего Приобья на основании распространения
стрекоз (Odonata, Insecta) // Изв. СО АН СССР. - 1961д. - № S.-
С. 93-104.
Белышев Б.Ф. К фауне стрекоз (Odonata) Восточного Казахстана //
Fragm. faunist. — Warszawa, 1961е. - Т. 9, № 4. - С. 27-43.
Белышев Б.Ф. Граница-палеарктической Азии на основании распространения
Odonata // Ann. zool. - Warszawa. 1961ж. - T. 19, № 12.-
С. 437-453.
Белышев Б.Ф. О некоторых элементах третичной одонатологической фауны в
166
боровых массивах южной части Западно-Сибирской равнины (Odonata, In—
secta) И Изв. СО АН СССР. - 1962а. - № 4. - С. 115-120.
Белышев Б.Ф. О вероятных, миграциях одонатопогической фауны на островах
н полуостровах Восточной Азии в четвертичное время П Материалы научно-
го совещания географов Сибири и Дальнего Востока. - Владивосток, 19626,-
С. 3-5. '
Белышев Б.Ф. К познанию одонатопогической фауны Западно-Сибирской равни-
ны // Изв. Вост.-Снб. отд. геогр. о-ва СССР. - 1962в. - Т. 60. -
С. 56-59.
Белышев Б.Ф. Одонатопогическая фауна северо-восточной части Кулундннской
степи И Там же. - 1962г. - Т. 60. - С. 60-66.
Белышев Б.Ф. Пределы Палеарктической области в Новом Свете на основании
распространения стрекоз ( Odonata ) // Зоол. журн. - 1962д. - Т. 41,
вып. 11. - С. 1743-1745.
Белышев Б.Ф. Некоторые вопросы условий обитаний личиночных фаз Sympet—
rum flaveolum L. (Odonata, Insecta ) в пересохших временных
водоемах // Учен. зап. Бурят, пед. нн—га. - 1962е. - Вып. 24. - С. 57-61.
Белышев Б.Ф. К фенологии стрекоз (Odonata) в Южном Прибайкалье и не-
которые общие вопросы развития фауны И Сезонная и вековая динамика при-
роды. - Иркутск, 1963а. - С. 57-66. - (Бюл. Вост.-Снб. фенолог, комис-
сии; № 2/3).
Белышев Б.Ф. История формирования одонатопогической фауны Верхнего Прн-
обья И Там же. - 19636. - С. 67-73.
Белышев Б.Ф. Определитель стрекоз Сибири по нмагннапьным и личиночным
фазам. - М.; Л.: Изд-во АН СССР, 1963в. - 114 с.
Белышев Б.Ф. Структура и история формирования островных и полуостровных
одонатопогическнх фаун Северо-Восточной Азин // Зоол. журн. - 1963г. -
Т. 42, вып. 11. - С. 1638-1645.
Белышев Б.Ф. Одонатопогическая фауна Сибири: Автореф. дне. ... д-ра бноп.
наук. - Иркутск, 1963д. - 34 с.
Белышев Б.Ф. Основные вопросы взаимоотношений фаун стрекоз (Odonata,
Insecta ) Сибири и Америки в пределах Палеарктической области // Изв.
СО АН СССР. - 1963е. - Вып. 3, № 12. - С. 66-75.
Белышев Б.Ф. Материалы к познанию одонатофауны высокогорных частей Цент-
рального и Западного Алтая // Изв. Алтай, отд. геогр. о-ва СССР. - 1963ж.
Вып. 3. - С. 72-76.
Белышев Б.Ф. Особенности в структуре одонатопогической фауны Прнаптайской
степи в 1953 н 1954 годах // Там же. - 1963з. - Вып. 3. - С. 77-78.
Белышев Б.Ф. К познанию одонатопогической фауны Сибири //Fra gm. fau—
nist. —Warszawa, 1964a. - T. 11, № 5. - C. 53-74.
Белышев Б.Ф. Одонатопогическая фауна Верхнего Прнобья // Стрекозы Сиби-
ри. - Л., 19646. - С. 4-70. - (Тр. Вост.-Снб. фил. АН СССР. Сер. бноп.;
Вып. 40).
Белышев Б.Ф. Распределение стрекоз по различным типам водоемов Верхнего
Приобья н некоторые вопросы профилактики простогоннмоза птиц // Изв. СО
АН СССР. Сер. бноп.-мед. наук. - 1964в. - № 8, вып. 2. - С. 131-13 9.
Белышев Б.Ф. Личинка Leucorrhinia orientalis Selys и некоторые
вопросы систематики и генезиса вида // Изв. Вост.-Сиб. отд. геогр. о-ва
СССР. - 1964. - Т. 62. - С. 3-7.
Белышев Б.Ф. Новый вид и новый подвид стрекоз (Odonata, Insecta)
из Южного Приморья // Там же. - 1964д. - Т. 62. - С. 7-10.
Белышев Б.Ф. О происхождении некоторых биологических особенностей сибир-
ских стрекоз // Там же. - 1964е. - Т. 62. - С. 11-17.
Белышев Б.Ф. О случаях заселения Сибири восточными по происхождению ви-
дами стрекоз с запада // Там же. - 1964ж. - Т. 62. - С. 18-20.
167
Белышев Б.Ф. Новые для фауны СССР роды и виды стрекоз (Odonata,
Insecta ) с Дальнего Востока // Зоол. журн. - 1965а. - Т. 44, вып. 4.-
С. 611-613.
Белышев Б.Ф. Фенология лёта стрекоз (Odonata, Insecta) в Заполяр-
ной Сибири и некоторые общие закономерности этого явления на севере Па-
леарктики // Там же. 19656. - Т. 44, вып. 7. - С. 1014-1017.
Белышев Б.Ф. О вероятном центре проискождения и пути расселения основного
ядра голарктической одонатофауны (Odonata, Insecta)// Изв. Вост.-Сиб.
отд. геогр. о-ва СССР. — 1965в. — Т. 64. — С. 3-14.
Белышев Б.Ф. Голарктическая фауна стрекоз (Odonata, Insecta),ее струк-
тура, пределы распространения и вероятные пути формирования // Изв. СО
АН СССР. Сер. биоп.-мед. наук. - 1965г. - № 12, вып. 3. - С. 76-83.
Белышев Б.Ф. Новый вид стрекоз (Odonata, Insecta) из Южного При-
морья И Зоол. жури. - 1965д. - Т. 44, вып. 12. - С. 1864-1866.
Белышев Б.Ф. О формировании ареала и о постоянстве морфологических особен-
ностей у Pantala flavescens Fabr. (Insecta, Odonata) // Там
же. - 1965e. - T. 47, вып. 6. - С. 945-947.
Белышев Б.Ф. Формообразующие территории в Северной Палеарктике // Изв.
Алт. отд. геогр. о-ва СССР. - 1965ж. - Вып. 5. - С. 15-18.
Белышев Б.Ф. К познанию одонатопогической фауны Сибири ( II. Материалы
к фауне и экологии стрекоз Тунинской долины в Южном Прибайкалье) //
Fra gm. faunist - Warszawa, 1966а. - Т. 12, № 23. - С. 385-411.
Белышев Б.Ф. Неизвестные и малоизвестные фазы стрекоз (Odonata, In—
secta) ио Юго-Восточной Сибири // Гидробиол. журн. - 19666,-
Т. 11, вып. 15. - С. 10-23.
Белышев Б.Ф. Об истории возникновения китайского и камчатского дизъюнк-
тивных ареалов американского дозорщика Anax junius Drury (Odo—
nata, Insecta) // Зоол. журн. - 1966в. - T. 45, вып. 8. - С. 1,159-1163.
Белышев Б.Ф. Особенности в сезонном развитии одонатофауны на водоемах се-
веро-восточного побережья оз. Байкал // Фауна и экология членистоногих
Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1966г. - № 1. - С. 203-205.
Белышев Б.Ф. Сезонное развитие одонатопогической фауны в условиях муссон-
ного климата Южного Приморья // Изв. СО АН СССР. Сер. биоп.-мед. наук.-
1966д. - № 4, вып. 1. - С. 120-128.
Белышев Б.Ф. Стрекозы как компоненты воднык и сухопутных биоценозов //
Охрана и рациональное использование дикой живой природы. - Алма-Ата,
1966е. - С. 95-97.
Белышев Б.Ф. Интересная закономерность в возникновении цветных аберраций
у некоторых видов стрекоз (Odonata, Insecta) // Зоол. журн. - 1967а.-
Т. 46, вып. 8. - С. 1258-1259.
Белышев Б.Ф. Условия погоды и суточная активность стрекоз (Odonata) //
Энтомэл. обозрение. - 19676. - Т. 46, № 4. - С. 778-783.
Белышев Б.Ф. К познанию одонатопогической фауны Сибири ( III. К фауне и
биологии стрекоз (Odonata) долины реки Монхухай в Южном Приморье)//
Fra gm. faunist - Warszawa, 1968а. - Т. 12, № 26. - С. 457-492.
Белышев Б.Ф. Стрекозы - Odonata // Руководство к определению. -
Алма-Ата: Изд-во Казах, ун-та, 19686. - 12 с.
Белышев Б.Ф. К познанию одонатопогической фауны Сибири (География стре-
коз Сибири) /1 Fragm. faunist. - Warszawa, 1968в, - Т. 14, № 13.-
С. 407-536.
Белышев Б.Ф. К познанию одонатофауны Забайкалья // Изв. Забайк. фил. геогр.
о-ва СССР. - 1968г. - Т. 4, вып. 1. - С. 112-114.
Белышев Б.Ф. Некоторые закономерности в современном географическом рас-
пространении репиктовык подотрядов и семейств стрекоз (Odonata, Insec—
ta ) по мировой суше // Изв. СО АН СССР. Сер. биоп.-мед. наук. -
1968д. - № 15, вып. 3. - С. 32-39.
168
Белышев Б.Ф. К истории рода Lestes Leach. (Odonata, Insecta)
в пределах Голарктики и на сопредельных землях // Там же. - 1968е. -
№ 10, вып. 2. - С. 104-107.
Белышев Б.Ф. Фрагменты к зоогеографии стрекоз (Odonata) мира II Ann.
zool. —Warszawa, 1969а. — Т. 27, № 1. — С. 1—27.
Белышев Б.Ф. Моллюски или стрекозы? II Вопросы малакологии Сибири. -
Томск, 19696. - С. 165-167.
Белышев Б.Ф. Некоторые спорные вопросы классификации и эволюции стрекоз
(Odonata, Insecta) П Изв. СО АН СССР. Сер. биол. наук. - 1969в. -
№ 15, вып. 3. - С. 109-115.
Белышев Б.Ф. Фаунистические связи Чилийской зоогеографической провинции
Неотропической области с другими фаунистическими областями мировой су-
ши Н Изв. Всесоюз. геогр. о-ва. - 1969г. - Т. 101, вып. 6. -
С. 555-557.
Белышев Б.Ф. Геофизическая теория Вегенера как основа к пониманию совре-
менного распространения одонатофауны и ее история II Изв. Забайк. фил.
геогр. о-ва СССР. — 1969ц. - Т. 5, № 2. — С. 28—34.
Белышев Б.Ф. Чем вызван перелет стрекоз? // Природа. - 1970а. - № 4. -
С 32.
Белышев Б.Ф. Особенности распределения стрекоз (Odonata) по типам
водоемов в Южном Приморье // Энтомол. обозрение. - 19706. - Т. 4 9,
№ 2. - С. 304-306.
Белышев Б.Ф. Структурные особенности в одонатофауне приалтайских районов
в 1951 г. // Изв. Алтайск. отд. геогр. о-ва. - 1970в. - Вып. 2. -
С. 131-132.
Белышев Б.Ф. О зоогеографических соотношениях в районе Карибского моря
на основании распространения стрекоз (Odonata, Insecta) // Изв. СО
АН СССР. Сер. биол. наук, 1970г. - № 10, вып. 2. - С. 109-113.
Белышев Б.Ф. О зональном распределения стрекоз (Odonata, Insecta)//
Фауна Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1970д. - С. 48-52.
Белышев Б.Ф. Одонатофауна окрестностей г. Бийска // Изв. Алтайск. отд.
геогр. о-ва. - 1970е. - С. 128-130.
Белышев Б.Ф. Mortonagrion selenion Ris (Odonata, Insecta) —
новый вид в фауне СССР И Новые и малоизвестные виды фауны Сибири. -
Новосибирск: Наука. Сиб. отд-ние, 1970ж. - Вып. 3. - С. 17-19.
Белышев Б..Ф. Закономерность географической изменчивости размеров голарк-
тических одонат (Odonata) в пределах Евразии II Изв. Алт. отд. геогр.
о-ва. - 1970з. - Вып. 2. - С. 125-127.
Белышев Б.Ф. Проблема Тихого океана с точки зрения одонатологии // Изв.
Забайк. фил. геогр. о-ва СССР. - 1971а. - Т. 5, вып. 6. - С. 107-115.
Белышев Б.Ф. Одонатофауна заповедников Средней Сибири и некоторых со-
предельных с ними земель II Тр. зап. / Гос. заповедник "Столбы'. -
19716. - Вып. 8. - С. 80-84.
Белышев Б.Ф. Новая географическая форма стрекозы (Odonata) из Южной
Сибири II Новые и малоизвестные виды фауны Сибири. - Новосибирск:
Наука. Сиб. отд-ние, 1971в. - Вып. 4. - С. 5-6.
Белышев Б.Ф. Некоторые своеобразные черты в структуре одонатофауны (Odo—
nata, Insecta) Цейлона и вероятное их происхождение // Изв. СО АН
СССР. Сер. бнол. наук. - 1971г. - № 10, вып. 2. - С. 110-115.
Белышев Б.Ф. Зоогеографическое районирование Австралийской фаунистической
области на основании распространения стрекоз (Odonata, Insecta) //
Там же. - 1971д. - № 5, вып. 1. - С. 66-72.
Белышев Б.Ф. О линии разграничении Ориентальной и Австралийской фаунисти-
169
ческих областей Меридионального царства на основании распространения
стрекоз // Там же. - 1972а. - № 10, вып. 2. - С. 94-97.
Белышев Б.Ф. О зависимости распространения стрекоз (Odonata, Insecta)
в условиях Северной Евразии от экологического фактора - влажности II Там
же. - 19726. - № 15, вып. 3. - С. 131-134.
Белышев Б.Ф. Заметки о географической изменчивости стрекоз и морфология
личинки Somatochlora alpestris Selys II Тр. ин-та / Биол. ин-т
СО АН СССР. Фауна и экология членистоногих Сибири. - 1972в. - Вып. 2. -
С. 43-45.
Белышев Б.Ф. Стрекозы Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1973а,-
Т. 1. - 620 с.
Белышев Б.Ф. Стрекозы как компоненты водных биоценозов и некоторые об-
щие вопросы хищничества в природе // Проблемы экологии. - Томск, 19736.-
Т. 3. - С. 131-134.
Белышев Б.Ф. О зоогеографических соотношениях в экваториальной части при-
атлантических стран на основании распространения стрекоз (Odonata, In—
secta) II Изв. СО АН СССР, Сер. биол. наук. - 1973в. - № 10, вып. 2.-
С. 109-114,
Белышев Б.Ф. Одонатофауне долины реки Норды в Заполярной Сибири II Тр.
ин-та / Биол. ин-т СО АН СССР. Фауна Сибири. - 1973г. - Ч. 2,
вып. 16. - С. 24-31.
Белышев Б.Ф. Об атлантическом разрыве в распространении стрекоз (Odona—
ta) Бореального фаунистического царства II Зоол. журн. - 1973д. - Т. 52,
выд. 7. - С. 1013-1016.
Белышев Б.Ф. Зоогеографическое районирование Сибири иа основании распро-
странения стрекоз (Odonata, Insecta) // Тр. ин-та / Биол. ин-т СО АН
СССР. Фауна Сибири. - 1973е. - Ч. 2, выц. 16. - С. 32-51.
Белышев Б.Ф. Вымершие и вымирающие стрекозы (Odonata) и вероятные
причины, обусловливающие вымирание таксонов // Проблемы эволюции. - Но-
восибирск: Наука. Сиб. отд-ние, 1973ж. - Т. 3. - С. 119-131.
Белышев Б.Ф. Стрекозы Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1974а.-
Т. 2.-351 с.
Белышев Б.Ф. Распространение реликтовых стрекоз, тихоокеанский рудный по-
яс и теория дрейфа континентов // Изв. СО АН СССР. Сер. биол. паук. -
19746. - № 10, вып. 2. - С. 144-145.
Белышев Б.Ф. Новые и малоизвестные Odonata Сибири И Биология и
морфология новых и малоизвестных видов фауны Сибири. - Новосибирск: Нау-
ка. Сиб. отд-ние, 1974в. - Вып. 8. - С. 29-32.
Белышев Б.Ф. Зоогеографические соотношения в одонатофауне ( Odonata, In—
secta) тропической Африки II Изв. СО АН СССР. Сор. биол. наук. - 1974г.-
№ 10, вып. 2. - С. 76-81.
Белышев Б.Ф. Некоторые моменты в современном направлении эволюции ок-
раски стрекоз Сибири и сопредельных земель И Вопросы биологии. - Томск:
Изд-во Том. ун-та. - 1975. - С. 123-128.
Белышев Б.Ф. Трофические связи личинок стрекоз рода Aeschna (Odonata)
с моллюсками // Охрана и преобразование природы лесостепей Западной Си-
бири. - Новосибирск: Наука. Сиб. отд-ние, 1976а. - С. 177-179.
Белышев Б.Ф. Одонатофауна (insecta, Odonata) СССР и территориаль-
ное распределение ее компонентов И Фауна гельминтов и членистоногих Си-
бири. - Новосибирск: Наука, Сиб. отд-ние, 19766. - С. 134-150. - (Тр.
ин-та / Биол. ин-т СО АН СССР; Вып. 18).
Белышев Б.Ф. О количественных несоответствиях в одонатофаунах приатлан-
тических стран Голарктической области Бореального фаунистического царства И
Изв. СО АН СССР. Сер. биол. наук. - 1977а. - К» 10, вып. 2. - С. 87-89.
Белышев Б.Ф. Экологические факторы и географическое распространение стре-
170
коз (insects, Odonata) // Проблемы зоогеографии и истории фауны. -
Новосибирск: Наука. Сиб. отд-ние, 1980а. - С. 84-94. - (Тр. ин-та / Биол.
ин-т СО АН СССР; Вып. 40) .
Белышев Б.Ф. Одонатофауне (insecta, Odonata) Бореального царства и
территориальное распределение ее компонентов II Фауна и экология расти-
тельноядных и хищных насекомых Сибири. - Новосибирск: Наука. Сиб. отд-
ние, 19806. - С. 7-24. - (Тр. ин-та / Биол. ин-т СО АН СССР; Вып. 43).
Белышев Б.Ф. Особенности распространения реликтового рода Mortonagrion
Fras. (Odonata, Insecta) // Фауна Сибири. - Новосибирск: Наука.
Сиб. отд-ние, 1985а.
Белышев Б.Ф. Об основных биогеографических подразделениях мировой суши
на основании распространения растений, позвоночных животных и стрекоз //
Изв. Всесоюз. геогр. о-ва СССР. - 19856. - Т. 117, вып. 1. - С. 31-34.
Белышев Б.Ф., Белышев НЪ. Anax parthenope Selys - реликтовый
вид в одонатофауне Алтая II Вопросы энтомологии Сибири. - Новосибирск:
Наука. Сиб. отд-ние, 1974. - С. 35-37.
Белышев Б.Ф., Белышев Н.Б. Anax parthenope Selys - реликтовый
вид в одонатофауне Горного Алтая // Вопросы географии Сибири. - Томск,
1976а. - Вып. 9. - С. 151.
Белышев Б.Ф., Белышев HJ5. Своеобразный перелет стрекоз (Odonata)
через г. Иркутск // Проблемы экологии. - Томск, 19766. - Т. 4. -
С. 187-188.
Белышев Б.Ф., Бессолицина Е.П., Костина Н.С., Полякова П.Е. Новые
данные по одонатофауне (insecta, Odonata) Северо-Восточной Азии II
Членистоногие Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1978. -
Вып. 34. - С. 3 9-46.
Белышев Б.Ф., Вержутский БЛ. Сравнительная оценка берингийских и ат-
лантических связей фауны Голарктики // Изв. Забайк. фил. геогр. о-ва
СССР. - 1969. - Т. 3, вып. 2. - С. 82-93.
Белышев Б.Ф., Гагина Т.Н. К фауне стрекоз (Odonata) Прибайкалья II
Fragm. faunist.—Warszawa, 1959. - Т. 8, № 7. - С. 115-117.
Белышев Б.Ф., Дошидорджи А. К фауне стрекоз (Odonata ) Монголии И
Зоол. журн. - 1958. - Т. 37, вып. 1. - С. 34-40.
Белышев Б.Ф., Дошидорджи А. Новые материалы к познанию одонатологи-
ческой фауны Монголии II Биол. сб. - Иркутск, 1960. - С. 217-221.
Белышев Б.Ф., Дулькейт ГЛ. Одонатологическая фауна Восточного Алтая//
Стрекозы Сибири. - Л., 1964. - С. 81-97. - (Тр. Вост.-Сиб. фил. АН
СССР. Сер. биол.; Вып. 40).
Белышев Б.Ф., Золотаренко Г.С., Велижанин А.Г. Одонатофауне Южных
Курильских островов и некоторые вопросы ее структуры и формирования //
Фауна гельминтов и членистоногих Сибири. - Новосибирск: Наука. Сиб. отд-
ние, 1976. - С. 165-173. - (Тр. ин-та / Биол. ин-т СО АН СССР;
Вып. 18).
Белышев Б.Ф., К оршунов Ю.П. Новые материалы к познанию одонатофауны
(Insecta,Odonata) Заполярной Сибири // Там же, 1976в. -С. 151-156.
Белышев Б.Ф., Куренцов А.И. К познанию стрекоз Приамурья // Стрекозы
Сибири. - Л., 1964. - С. 71—80, - (Тр. Вост.-Сиб. фил. АН СССР. Сер.
биол.; Вып. 40).
Белышев Б.Ф., Наси мо вич АЛ., Ракилин В.К. Одонатофауне островов Ми-
рового океана // Островная фауна Мирового океана. - М„ 1972. -
С. 57-61.
Белышев Б.Ф., Оводов Н.Д. Somatochlora sahlbergi Trybom (Odona—
ta, Insecta ) в Южной Сибири И Зоол. журн. - 1961. - Т. 40,
вып. 12. - С. 1892-1893.
Белышев Б.Ф., Ремм X. Личиночная фаза Agrion lanceolatum Selys
171
II Изв. Вост.-Сиб. отд. геогр. о-ва СССР. - 1969. - Т. 66. - С. 155-156.
Белышев Б.Ф., Ремы Х„ Панкратьев А.Г. К познанию одонатологической
фауны Уссурийского края II Живая природа Дальнего Востока. - Таллин,
1971. - С. 162-170.
Белышев Б.Ф., Степанчук Е.С. Суточные миграции у четырехпятнистых стре-
коз // Изв. Вост.-Сиб. отд. геогр. о-ва СССР. - 1964. - Т. 62. -
С. 20-22.
Белышев Б.Ф., Степанчук Е.С. Интересные в зоогеографическом отношении
находки стрекоз (Odonata) в Южном Приморье // Там же. - 1965. -
Т. 64. - С. 15-18.
Белышев Б.Ф., Томилов А.А. К фауне стрекоз (Odonata ) Восточной
Сибири I! Тр. фил. / Вост.-Сиб. фил. АН СССР. Сер. биол. - 1961. -
Т. 36. - С. 69-92.
Белышев Б.Ф., Харитонов А.Ю. Личинка Aeschna serrata Hag. (Odo—
nata, Insecta) // Новые и малоизвестные виды фауны Сибири. - Новосибирск:
Наука. Сиб. отд-ние, 1973. - Вып. 7. - С. 27-30.
Белышев Б.Ф., Харитонов А.Ю. Agrion hylas Trybom (Odonata, In—
secta ), его подвиды и близкие к нему виды в Сибири // Биология и мор-
фология новых и малоизвестных видов фауны Сибири. - Новосибирск: Наука.
Сиб. отд-ние, 1974. - Вып. 8. - С. 21-29.
Белышев Б.Ф., Харитонов А.Ю. О систематическом и номенклатурном поло-
жении северного евразиатского вида Sympycna braueri Bianch (Odo—
nata, Lestidae) Ц Вопросы экологии. - Красноярск, 1975a. - С. 193-
198. - (Тр. гос. заповедника "Столбы"; Вып. 10).
Белышев Б.Ф., Харитонов А.Ю. О японо-гималайских ареалах стрекоз Юго-
Восточной Азии Ц Зоол. журн. - 19756. - Т. 54, вып. 3. - С. 471-472.
Белышев Б.Ф., Харитонов А.Ю. Зоогеографические и морфологические груп-
пировки вида Sympetrum Newmann (1833) и история нх расселения И
Изв. СО АН СССР. Сер. биол. наук. - 1977а. - № 5, вып. 1. - С. 51—56.
Белышев Б.Ф., Харитонов А.Ю. К истории бореального вида стрекозы Ne—
halennia speciosa Charp. (1840) и о центре происхождения рода
Nehalennia Selys (i860) (Odonata, Coenagrionidae) // Энто-
мол. обозрение. - 19776. - Т. 56, № 4. - С. 776-780.
Белышев Б.Ф., Харитонов А.Ю. Определитель стрекоз по крыльям (роды
Бореального фаунистического царства и сопредельных земель, виды фауны
СССР) Ц Новосибирск: Наука. Сиб. отд-ние, 1977в. - 3 97 с.
Белышев Б.Ф., Харитонов А.Ю. О двух ископаемых стрекозах (insecta,
Odonata) с верховий р. Васюган II Изв. СО АН СССР. Сер. биол. наук.-
1978а. - № 10, вып. 2. - С. 138.
Белышев Б.Ф., Харитонов А.Ю. О причинах резкой изоляции фаун стрекоз
полуострова Индостан и Юго-Западной Азии И Зоол. журн. - 19786. -
Т. 47, вып. 1. - С. 140-141.
Белышев Б.Ф., Харитонов А.Ю. С широтных н долготных дизъюнкциях ареа-
лов евраэиатских стрекоз (Odonata, Insecta) и их происхождении II Изв.
СО АН СССР. Сер. биол. наук. - 1978в. - № 5, вып. 1. - С. 114-118.
Белышев Б.Ф., Харитонов А.Ю. Об атлантическом разрыве ареалов стрекоз
(Odonata) южных частей Африки и Америки // Зоол. журн. - 1979а. -
Т. 58, вып. 4. - С. 610-611.
Белышев Б.Ф., Харитонов А.Ю. С распространении рода Orthetrum Newm.
(Odonata, Libellulidae ) // Вести, зоологии. - 19796. - № 3. -
С. 81-84.
Белышев Б.Ф., Харитонов А.Ю. Заполярная одонатофауна (insecta, Odo—
nata) Северного полушария и вероятные пути ее формирования // Изв. СО
АН СССР. Сер. биол. наук. - 1980а. - N° 15, вып. 3. - С. 35-38.
Белышев Б.Ф., Харитонов А.Ю. С поздних фаунистических связях Эфиопской
172
и Австралийской областей на примере стрекоз (Odonata) // Энтомол.
обозрение. - 19806. - Т. 59, № 1. - С. 89-91.
Белышев Б.Ф., Харитонов А.Ю. Особенности географического распростране-
ния рода Enallagma Charp., 1840 (insecta, Odonata)// Проблемы зоогеогра-
фии и истории фауны. - Новосибирск: Наука. Сиб. отд-ние, 1980в. -
С. 80-84. - (Тр. ин-та / Б иол.- ин-т СО АН СССР; Вып. 40).’
Белышев Б.Ф., Харитонов А.Ю. Распространение по мировой суше и центры
развития семейства Cordulegastridae Calv. (insecta, Odonata)
// Там же. - 1980г. - Вып. 40. - С. 76-80.
Белышев Б.Ф., Харитонов А.Ю. География стрекоз (Odonata) Бореаль-
ного фаунистического царства. - Новосибирск: Наука. Сиб. отд-ние, 1981. -
279 с.
Белышев Б.Ф., Харитонов А.Ю. Enallagma cyathigerum Charp. (in-
secta, Odonata) и проблема беренгийских фаунистических связей // Изв.
СО АН СССР. Сер. биол. наук., 1982. - Вып. 3. - С. 84-86.
Белышев Б.Ф., Харитонов А.Ю. География стрекоз (Odonata) Меридио-
нального фаунистического царства. - Новосибирск: Наука. Сиб. отд-ние,
1983а. - 153 с.
Белышев Б.Ф., Харитонов А.Ю. Зоогеографическая структура фауны стрекоз
Средиземноморской подобласти Субголарктики // Вести, зоол. - 19836. -
№ 5. - С. 9-15.
Белышев Б.Ф., Харитонов А.Ю. О зоогеографических соотношениях в районе
Кавказского хребта // Изв. СО АН СССР. Сер. биол. наук. - 1983в. -
Вып. 1. - С. 85-89.
Белышев Б.Ф., Харитонов А.Ю. Монотипические таксоны стрекоз (insec-
ta, Odonata ) Бореального фаунистического царства // Фауна Сибири. -
Новосибирск: Наука. Сиб. отд-ние, 1985а. - С. 44-47.
Белышев Б.Ф., Харитонов А.Ю. Новый реликтовый подотряд стрекоз (in-
sects, Odonata) // Изв. СО АН СССР. Сер. биол, наук^ - 19856. -
Вып. 3. - С. 51-53.
Белышев Б.Ф., Хусаинова Н.З. Памяти Александра Николаевича Бартенева
(1882-1946) И Биол. науки. - 1971, - Вып. 3. - С. 213-225.
Белышев Б.Ф., Шевченко В.В. К фауне стрекоз (Odonata) Таласского
Алатау и Каратау II Тр. ин-та / Зоол. нн-т АН КазССР. - 1958. - Т. '8. -
С. 222-223.
Белышев Б.Ф., Шевченко В.В. Фауна стрекоз (Odonata, Insecta) и распре-
деление ее компонентов в Казахстане И Биол. науки. - 1971. - Вып. 2. -
С. 73-77.
Бенинг А.Л. К изучению придонной жизни реки Волги // Монографии Волжской
биологической станции Саратовского общества естествоиспытателей. - 1924,-
Т. 1. - 398 с.
Бенинг А.Л., Попова АЛ. Материалы по гидробиологии р. Занги, от истока
до г. Еревана // Тр. / Севенская гидробиол. станция. - 1947. - Т. 8. -
С. 5-75.
Березина Н.А. Питание личинок стрекоз подотряда Zygoptera // Зоол. журн,-
1946. - Т. 25, вып. 6. - С. 509-516.
Березина НА. Питание личинок стрекоз из подотряда Anisoptera // Бюл.
МОИП. Отд. биол. - 1947. - Т. 52, № 6. - С. 21-32.
Березина НА. Роль некоторых представителей Odonata, Hemiptera и
Coleopters в трофике пресных водоемов //, Трофология водных жи-
вотных. Итоги и задачи. - М.: Наука, 1973. - С. 206-211.
Благовещенский ГА. История лесов восточного склона Среднего Урала //
Сов. ботаника. - 1943. - № 6. - С. 3-16.
Богачев А.В. Стрекозы, собранные в Кахе ком районе АзССР // Тр. Азерб.
фил. АН СССР. Сер. эоол. - 1937. - Т. 20. - С. 33-44.
173
Богачев А.В. Животный мир Азербайджана. - Баку, 1951. - С. 275-278.
Борисов С.Н. Новый вид стрекозы рода Sympetrum Newman (Odonata,
Libellulidae) из Таджикистана // Изв. АН ТаджССР. Сер. биол. - 1983.-
№ 2(91). - С. ,68-70.
Борисов С.Н. Изменчивость поколений у стрекоз (insecta^ Odonata)//
Там же. - 1984. - № 4 (97). - С. 68-70.
Борисов С.Н. Искусственные оросительные системы как стации личинок стре-
коз (insecta, Odonata) в долинах Таджикистана // Докл. АН ТаджССР.-
1985а. - Т. 28, № 9. - С. 541-543.
Борисов С.Н. Стрекозы (insecta, Odonata) Памира// Материалы рес-
публиканской научно-теоретической конференции молодых ученых и специали-
стов Таджикской ССР. - Душанбе: Дониш, 19856. - С. 87-88.
Борисов С.Н. Суточный ритм активности Anax parthenope Selys (Odo—
nata, Libellulidae) в условиях аридной зоны // Докл. АН ТаджССР. -
1985в. - Т. 28, № 10. - С. 603-606.
Борисов С.Н. Численность некоторых видов стрекоз в долинах Юго-Западного
Таджикистана // Веста, зоологии. - 1986а. - № 2. - С. 38—42.
Борисов С.Н. Фауна и экология стрекоз (Insecta,Odonata) заповедника
'Тигровая балка' // Докл. АН ТаджССР. - 19866. - Т. 29, № 9, -
С. 560-564.
Борисов С.Н. Об экологии двух близких видов стрекоз в Таджикистане // Эко-
логия. - 1987. - № 1. - С. 85-87.
Борисов С.Н., Харитонов А.Ю. Фауна стрекоз (insecta, Odonata) Тад-
жикистана // Изв. АН ТаджССР. Сер. биол. - 1986. - № 1(102). —
С. 46-49.
Брауер Фр. Стрекозы (Odonata). Путешествие в Туркестан А.П. Федчен-
ко // Изв. о-ва любителей естествознания. - Спб.; М., 1877. - Вып. 14,
т. 2. Зоогеографические исследования, ч. 5. - С. 2-11.
Браунер А. Заметки о стрекозах Херсонской губернии и Крыма // Зап. Ново-
рос. о-ва натуралистов. - 1902. - Т. 24. - С. 73-102.
Браунер А. Заметки о стрекозах (Odonata) // Рус. энтомол. обозрение. -
1903. -№2. - С. 89-91.
Виноградов П. Дополнение к списку ставропольских стрекоз // Изв. Ставро-
пол. энтомол. о-ва. - 1926. - Т. 2.
Виноградов П. Заметки о стрекозах // Там же. - 1928. - Т. 4, вып. 2. -
С. 50-52.
Внуковский В.В. К фауне стрекоз (Odonata) Западной Сибири // Тр. Си-
бирского ин-та сельского хоз-ва и лесоводства. - Омск, 1928. - Т. 10. -
С. 3-9.
Воронов А.Г. Биогеографии. - М.: Изд-во МГУ, 1963. - 340 с.
Воронцовский П. Материалы к фауне стрекоз (Odonata) окрестностей
г. Оренбурга // Изв. Оренбург, отд. Рус. геогр. о-ва. - 1909. - Вып. 21.-
С. 113-120,
Воронповский П. Материалы к изучению фауны стрекоз (Odonata) окрест-
ностей г. Оренбурга // Там же. - 1912. - Вып. 23. - С. 100-115.
Воронцовский П, Материалы к изучению фауны стрекоз (Odonata) ок-
рестностей г. Оренбурга (сборы 1912 г.) // Там же. - 1913. - Т. 24. -
С. 111-113.
Вульф Е.В, Введение в историческую географию растений. - Л., 1932. -
355 с.
Гентнер В.Г. Общая зоогеография. - М.; Л.: Биомедгиэ. 1936. - 548 с.
Гигиняк Ю.Г., Павельева Е.Б. Весовая и энергетическая характеристика
личинок стрекоз озера Нарочь // Вести. Белорус, ун-та. Сер. 11. - 1974. -
Вып. 1. - С. 37-40.
Голикова М.Н. К биологии некоторых видов ленточных червей водоплавающих
174
птиц // Докл. АН СССР. - 1960а. - Т. 131, № 5. - С. 1222-1224.
Голикова МЛ. Эколого-паразитологическое изучение биоценоза некоторых
озер Калининградской области // Фауна трематод беспозвоночных животных.-
Л., 19606. - С. 80-94. - (Вестн. Ленингр. ун-та. Сер. биол.; № 21,
вып. 4).
Голикова МЛ. Эколого-паразитологическое изучение биоценоза некоторых озер
Калининградской области: Автореф. дис. ... канд. биол. наук. - Л., 1961. -
23 с.
Горностаев Г.Н. Введение в этологию насекомых>тфотоксенов // Этология на-
секомых. - Л.: Наука. Ленингр. отд-ние, 1984. - С. 101-167. - (Тр. ВЭО;
Т. 66).
Городков KJ5. Некоторые данные о распространении стрекоз на севере // Эн-
томол. обозрение. - 1956. - Т. 35, № 1. - С. 120-123.
Городков К.Б. Pantala flavescens Fabr. (Odonata, Libellulidae)
в высокогорье Восточного Памира // Зоол. журн. - 1961. — Т. 40, вып. 4.-
С. 610-611.
Городков К.Б. Трехмерная климатическая модель потенциального ареала и не-
которые ее свойства // Энтомол. обозрение. - 1986. - Т. 65, вып. 1. -
С. 81-95.
Граидилевская-Дексбах МЛ. Основные черты донной фауны и питания рыб
Камского водохранилища (1955-1959) // Тр. отд-ния // Урал отд-ние
НИОРХ. - 1961. - Т. 5. - С. 131-175. •
Грандилевская-Дексбах М.Л. Дониая фауна и питание рыб Аргазинского во-
дохранилища // Там же. - 1966а. - Т. 7. - С. 25-38.
Грандилевская-Дексбах МЛ. Донная фауна и питание рыб озер Исетское,
Шарташ, Балтым и Половинное // Там же. - 19666. - Т. 7. - С. 133-165.
Грандилевская-Дексбах МЛ., Шилкова Е.В. Материалы по бентосу
р. Пышмы в пределах Свердловской области // Там же. - 1971. - Т. 8. -
С. 195-199.
Грибакин Ф.Г. Механизмы фоторецепции насекомых. - Л.: Наука. - Ленингр. отд-
ние, 1981. - 216 с.
Григорьев Б. Список стрекоз Семиречья // Рус. энтомол. обозрение. - 1905. -
№ 5/6. - С. 216-220.
Григорьев Б. Список Ortoptera, Odonata Болога (Новгородская губерния)
и его окрестностей // Тр. / Пресноводная биол. ст. С.-Петербург о-ва есте-
ствоиспытателей. - 1906а. - Т. 2. - С, 65-69.
Григорьев Б. Одонатологические заметки // Рус. энтомол. обозрение. -
19066. - Т. 6, Ns 3/4. - С. 205-207.
Дажо- Р. Основы экологии. - М.: Прогресс, 1975. - 415 с.
Дремкова П.П. О гельминтофауне личинок стрекоз в водоемах Волго-Ахтубин-
ской поймы // Современные проблемы зоологии и совершенствование методи-
ки ее преподавания в вузе и школе. - Пермь, 1976. - С. 64^65,
Дьяконов А.М. К фауне Odonata озера Сандала и его окрестностей //
Тр. / Олонецкая науч, экспедиция. - 1922а. - Т. 4, № 1. - С. 1-37.
Дьяконов А.М. Новый вид стрекозы рода Aeschna из Северной и Севе-
ро-Западной России (Aeschna elisabethae, sp. п.) // Изв. Сев. обл.
ст. защиты растений от вредителей. - 19226. - Т. 3. - С. 123-130.
Дьяконов А.М. Наши стрекозы. Определитель стрекоз и их личинок. - М.; Л.:
Гос. изд-во, 1926. - 72 с.
Дьяконов А.М. Отряд Odonata - стрекозы // Определитель насекомых
европейской часта СССР. - М,; Л.: Сельхозгиз, 1948. - С, 42-55.
Еськов К.Ю. /Рецензия/ // Зоол. журн. - 1984. - Т. 63, вып. 4. - С. 637-
640. - Рец. на кн.: Белышев Б.Ф., Харитонов А.Ю. География стрекоз (Odo-
nata) Бореального фаунистического царства. - Новосибирск: Наука. Сиб.
отд-ние, 1981. - 280 с. Белышев Б.Ф., Харитонов А.Ю. География стрекоз
175
(Odonata) Меридионального фаунистического царства. - Новосибирск: Нау-
ка. Сиб. отд-ние, 1983. - 152 с.
Животный мир Молдавии. Насекомые . - Кишинев, 1983.
Жохов А.Е. Гель минто фаун а животных двух водоемов, различающихся по степе-
ни антропогенного воздействия И Вопросы экологической гельминтологии. -
Ярославль, 1984. - С. 31-35.
Заварзин А.А. Избранные труды. - М.; Л.: Изд-во АН СССР, 1952. - Т. 1.
Заика В.В. Фауна стрекоз (Odonata) северной части Купундинской степи tf
Вопросы энтомологии Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1974. -
С. 43-44.
Заика В.В. Поведение стрекоз Северной Кулунды в репродуктивный период //
Этологические проблемы экологии насекомых Сибири. - Новосибирск: Новосиб.
ун-т, 1977. - С. 106-125.
Занка В.В. Население стрекоз наземной части биогеоценозов Северной Кулун,-
ды // Вопросы экологии. - Новосибирск: Новосиб. ун-т, 1979. - С. 87-111.
Заика В.В. Экология и поведение стрекоз (Odonata) острова Кунашир //
Там же. - 1980. - С. 73-88.
Заика В.В. Стрекозы южной части Западно-Сибирской равнины: Автореф. дис....
канд. биол. наук. - Новосибирск, 1982. - 18 с.
Заика В.В., Воронова ИА. Поведение стрекоз в озерной степи Западной Си-
бири // Этологические проблемы экологии насекомых Сибири. - Новосибирск:
Новосиб. ун-т, 1977. - С. 82-105.
Заика В.В., Костерин О.Э. Структура населения и этологические аспекты
'стрекоз Манжерокского озера (Алтай) // Экология И география членистоно-
гих Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1987. - С. 73-74.
Заика В.В., Стебаев И.В., Резникова Ж.И. Опыт изучения поведения насе-
комых в связи с ярусной структурой биогеоценоза ( Odonata Acridoidea,
Formicidae) // Этология насекомых и клещей. - Томск: Изд-во Том.
ун-та, 1977. - С. 7-39.
Здун В.И. Наоекомые - промежуточные (добавочные) хозяева дигеиетических
трематод // Ц1 совещание всесоюз. энтомол. о-ва. - Л., 1957. -
С. 194-196.
Здун В.И. Матери 1али до вивчення комах як додаткових хазя1в трематод в умо-
вах зайдних областей УРСР 11 Проблемы ентомологи на yxpaiHi АН УРСР.-
Киев, 1959. - С. 120-121.
Зенкин Г.М., Пигарев И.Н. Специализированные элементы в зрительной си-
стеме стрекоз // Зоол. журн. - 1969. - Т. 48, № 11. - С. 1629—1633.
Зенкин Г.М., Пигарев И.Н. Зрительно обусловленная активность в шейной
нервной цепочке стрекозы // Биофизика. - 1971. - Т. 16, № 2, -
С. 299-306.
Зильбер ЛА., Абелев Г.И. Вирусология и иммунология рака. - М.: Меди-
цина, 1962. - С. 1—458.
3 о граф Ю. К фауне Odonata Донской области II Тр. студенческого круж-
ка для исследования русской природы. - 1909. - Т. 4. - С. 94-96.
Золотов В.В. Пространственная организация рецептивных попей светочувстви-
тельных нейронов брюшной цепочки насекомых // Эволюционная биохимия и
физиология. - 1982. - Т. 18, № 4. - С. 366-372.
Иванов П. Материалы для энтомологии Харьковской губернии. Описание стре-
коз окрестностей г. Купянска // Тр. / О-во испытателей природы Харьков,
ун-та. - 1876. - Т. 10. - С. 4-113.
Ивлев В.С. Экспериментальная экология питания рыб. - М.: Пищепромиэдат,
1955.
Илюшина ТЛ. Водные насекомые Карасукской системы озер как дополнитель-
ные хозяева трематод И Экология и таксономия гельминтов. - М.: Наука,
1973. - С. 55-64. - (Тр. / Гепьмиитол. паб. АН СССР; Т. 23),
176
Илюшина ТЛ> Роль водных насекомых в жизненных циклах трематод // Па-
разиты в природных комплексах Северной Кулунды. - Новосибирск: Наука.
Сиб. отд-ние, 1975. — С. 53—94. — (Тр. ин-та / Биол. ин-т СО АН СССР;
Вып. 17).
Илюшина ТЛ. Фауна метацеркарий трематод водных насекомых Астраханского
заповедника // Тр. Гельминтол. лаб. АН СССР. - 1979. - Т. 29. -
С. 59-66.
Илюшина ТЛ. Зараженность водных насекомых метацеркариями трематод на
озерах Игналинской системы в Литовской ССР // Гельминты насекомых. -
М.: Наука, 1980. - С. 46-49.
Ионычев С.С., Ибад улаев И .А. Фауна и биотопическое распределение стре-
коз низовий р. Сырдарьи // Фауна и екопогия животных Казахстана. - Алма-
Ата: Наука, 1981. - С. 6-10.
Ицкал А. К биологии рода Arrhenurus Dug. (Hydracarina) ll Тр. I
Ленингр. о-во естествоиспытателей. - 1928. - Т. 58, вып. 2. - С. 3-26.
Калугина Н.С. Изменение подсемейственного состава хирономид (Diptera,
Chironomidae) как показатель возможного евтрофирования водоемов в
конце мезозоя // Бюп. МОИП. Отд. биол., 1974, - Т. 79, № 6. - С, 45-55.
Калугина Н.С. Насекомые в водных экосистемах прошлого // Историческое
развитие класса насекомых. - М.; Л„ 1980. - С. 224-240. - (Тр. нн-та /
Палеонтол. ин—т АН СССР; Т. 175).
Калугина Н.С. Двукрылые. Muscidae («• Diptera). Инфраотряды Tipuld—
morpha и Culicomorpha // Насекомые в раннемеловых экосистемах
Западной Монголии. - М.: Наука, 1986. - С. 112-125.
Калугина Н.С., Ковалев В.Г. Двукрылые насекомые юры Сибири. - М.: Неуз-
ка, 1985. - 198 с.
Камлюк Л.В. Энергетический баланс сообщества личинок стрекоз в литорали
озера Нарочь // Вопросы рыбного хозяйства Белоруссии. - Минск, 1973. -
Т. 9. - С. 124-135.
Карпенко С.В., Заика В.В. Материалы по екологин трематоды Plagiorchis
elegans (Rudolphi, 1802) в Северной Кулунде // Экология и морфоло-
гия гельминтов Западной Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1979.-
С. 131-138.- (Тр. ин-та/Биол, ин-т СО АН СССР; Вып. 38).
К ис ел ей е В. Aeschna grandis L. - новый дополнительный хозяин Pro—
sthogonimus cuneatus (Rud., 1809) 11 Acta Parasitol. Lithua—
nica. - 1967. - Vol. 7. - C. 67-69.
Климишнн A.C., Павлюк Р.С. Биотопы зарослей прибрежной растительности
водоемов К ургадь джинс кого озерно-равнинного района как основные очаги ин-
вазии некоторых животных паразитами // Сб. тр. геоботанической експеди-
пии. - Львов: Изд-во Львов, ун-та, 1972. - С. 116-121.
Ковалькова МЛ. Донная фауна заморного озера Вашты // Тр. / Урал,
отд-ние СибНИИРХ. - 1971. - Т. 8. - С. 189-193.
Кожевников Г.А. Дополнения к спискам животных Московской Губернии.
№ 4 // Дневник зоол. отд-ния Имперского о-ва любителей естествознания. -
1902. - Т. 3, вып. 4. - С. 16-17.
Колесов ВЛ. Основные черты фауны стрекоз Московской губернии И Тр. /
О-во изучения Моск. обл. - 1930а. - Т. 4. - С. 1-20.
Колесов ВЛ. Экология Odonata Московской губернии // Зап. биол. ст.
в Большеве. - 19306. - Т. 4. - С. 59-129.
Колосов Ю.М. Заметки о насекомых Тобольской губ. // Зап. Урал, о-ва лю-
бителей естествознания. - Екатеринбург, 1914, - Т. 34, вып. 1/2. -
С. 13-36.
Колосов Ю.М. Энтомологические заметки. III. О нахождении в Вятской губер-
нии G-omphus flavipes Charp. // Там же. - 1915а. - Т. 35,
вып. 8/10. - С. 145-146.
12 Зак. 611
177
Колосов Ю^1, Дополнение к фауне Odonata Кубанской области // Рус.
энтомол. обозрение. - 19156. - Т. 15, № 3. - С. 468.
Колосов Ю.М. О географическом распространении и биологии Aeschna gi—
gas Bart // Там же. - 1915в. - Т. 15, № 2. - С. 292-293.
Колосов Ю.М. Материалы к познанию энтомофауны Среднего Урала. V. So-
matochlora arctica Zett, в Пермской губернии // Зап. Урал, о-ва лю-
бителей естествознания. - 1916а. - Т. 36, вып. 5-8. - С. 71-73.
Колосов Ю.М. Материалы к познанию энтомофауны Урала. VI. Aeschna суа-
nea Mull, в Пермской губернии // Там же. - 19166. - С. 73-78.
Колосов Ю.М. Материалы к познанию энтомофауны Урала. VII. Anax parthe-
nope Selys в Пермской губернии // Там же. - 1922. - Т. 38, вып. 1,-
С. 21-23.
Колосов Ю.М. Стрекозы Ярославской губернии // Тр. / Ярослав, естеств.-
ист. и краевед, о-во. - 1925. - Т. 4, вып. 1. - С. 39-44.
Колосов Ю.М. Заметки о стрекозах Челябинского округа // Сборник материа-
лов по изучению Челябинского округа. - Челябинск, 1927. - Вып. 1. -
С. 7-13.
Колосов Ю.М. Материалы к познанию энтомофауны Урала. XIV. Наиболее ин-
тересные стрекозы, собранные в Вятской и Пермской губерниях Камской эк-
спедицией В.Б.С. в 1925 г. // Работы Волжской биол. ст. - Саратов,
1929а. - Т. 10, вып. 5. - С. 86-92.
Колосов Ю.М. Определительная таблица встречающихся в СССР видов р. Leu-
corrhinia // Зап. Бел. дзярж. акад. сел. лясн. гаспад. - 19296. - Т. 15.
Колосов Ю.М. Материалы да паказаньня энтамафауны Беларуси // Там же. -
1930. - Т. 11. - С. 75-78.
Колосов Ю.М. Показыпк л!таратуры па пытаньнях тэорытычнай i прикладной
энтомолёги Беларус! // Матар'ялы да вывучэньня фпёры i фауны Беларусь -
1932. - Т. 7. - С. 61-98.
Комарова Т.Н. Метацеркарии дигенетических трематод бентических членисто-
ногих водоемов дельты Дуная // Вести, зоологии. - 1968. - № 6. -
С. 7-14.
Костерин Ой. Находка восточно-азиатской стрекозы (Odonata, Libellu-
lidae ) на Манжерокском озере (Алтай) // Насекомые, клеши и гель-
минты. - Новосибирск: Наука. Сиб. отд-ние, 1987. - С. 57-63.
Красилов ВЛ. Эволюция и биостратиграфия. - М.: Наука, 1977. - 256 с.
Краснолобова ТЛ. К биологии Prosthogonimus pellucidus (Lunst., 1873)-
возбудителя простогонимоза домашних птиц // Работы по гельминтологии к
80-летию акад. К.И. Скрябина. - М.: Изд-во АН СССР, 1958. - С. 173-175.
Краснолобова ТЛ. Жизненный цикл возбудителя заболевания домашних птиц -
Prosthogonimus cuneatus (Rudolphi, 1809 ) // Helminthologia. -
M.: Изд-во АН СССР, 1961. - Т. 3, № 1-4. - С. 183-192.
Краснолобова ТЛ. Биологический цикл трематоды Prosthogonimus pel—
lucidus (Lunstow, 1873), возбудителя заболевания птиц // Helmintholo—
gia. - М.: Изд-во АН СССР. - 1962. - Т. 4. - С. 217-229.
Краснолобова ТЛ. Влияние некоторых биотических факторов иа морфологию-
трематод рода Prosthogonimus (Ltihe, 1839). Гельминты человека,
животных и растений и борьба с ними. - М.: Наука, 1968. - С. 115-125.
Краснолобова ТЛ. Экспериментальные доказательства идентичности родов
Prosthogonimus Liihe, 1899 и Schisthogonimus (ДЗуаип, 1901),
Liihe, 1909 (Trematoda, Prosthogonimidae) // Тр. / Гепьминтол,
лаб. АН СССР. - 1969. - Т. 20. - С. 79-87.
Краснолобова ТЛ. Зараженность стрекоз Латвийской ССР метацеркариями
трематод // Зоол. жури. - 1970. - Т. 49, вып. 9. - С. 12 90-1297.
Краснолобова ТЛ. Биологические особенности рода Plagiorchis (Liihe,
1899) Plagiorchidae. Экспериментальное изучение жизненного цикла
178
трематоды Plagiorchis laricola (Skrjabin, 1924). 4. 1. // Вопро-
сы биологии, физиологии и биохимии гельминтов животных и растений. - М.:
Наука, 1971а. - Т. 21. - С. 43-57.
Краснолобова Т.А. Биологические особенности трематод рода Plagiorchis.
Развитие трематоды Plaricola в окончательных хозяевах. Ч. 2 //
Тр. / Гельминтол. лаб. АН СССР. - 19716. - Т. 22. - С. 92-118.
Краснолобова Т.А. О развитии Plagiorchis elegans Lilhe, 1899
в условиях дельты Волги // Гельминты животных и растений. - М.: Наука,
1979. - С. 75-80.
Крендовский М.Е. О явлениях превращения у водяных акарид // Тр. I О-во
испытателей природы при Импер. Харьк. ун-те. - Харьков, 1878. - Т. 12.-
С. 221-286.
Крендовский М.Е. О паразитизме акарид на насекомых Н Там же. - 1879.-
Т. 13. - С. 169-212.
Криволуцкий Д.А. Понятие "жизненная форма* в экологии животных Н Журн.
общ, биологии. - 1967. - Т. 28, № 2. - С. 153-162.
Крупиковский Л. Стрекозы Малмыжского и Уржумского уездов Вятской губер-
нии Ц Зап. Урал, о-ва любителей естествознания. - 1907. - Т. 26, N» 52,-
С. 179-187.
Крыжановский В.В. Редкие виды стрекоз Куйбышевской области // Экология
и охрана животных. - Куйбышев, 1982. - С. 137-140.
Крылова В.Н. Некоторые особенности фенологии и способов сбора стрекоз в
условиях Средней Азии // Энтомологические исследования в Киргизии. —
Фрунзе: Илим, 1968а. - С. 57-59.
Крылова В.Н. Распределение стрекоз фауны Киргизии по типам водоемов //
Тез. докл. конф, по вопросам рыбного хозяйства республик Средней Азии и
Казахстана. - 19686. - С. 90-91.
Крылова В.Н. Стрекозы Киргизии: Автореф. дис. ... канд. биол. наук. -
Фрунзе, 1969. - 18 с.
Крылова В.Н. Вертикальные пределы распространения стрекоз в Тянь-Шане //
Энтомологические исследования в Киргизии. - Фрунзе: Илим, 1972. -
С. 20-25.
Крылова В.Н., Чиров П.А. О роли стрекоз в снижении численности некоторых
кровососущих насекомых Тянь-Шаня // Материалы по членистоногим энтомо-
фагам Киргизии. - Фрунзе: Илим, 1971. - С. 92-94.
Кукашев Д. К природной очаговости простогонимоза в Центральном Казахста-
не // X Всесоюз. конф, по природной очаговости болезней. - Алма-Ата: Нау-
ка, 1979. - С. 194-195.
Кукашев Д. Ш. К фауне и биологии стрекоз (Odonata, Insecta) бассейна
озера Кургальджин // Изв. АН КаэССР. Сер. биол. - 1982, № 6. -
С. 110-116.
Кукашев Д.Ш. Стрекозы (Odonata) - промежуточные хозяева цестод се-
мейства Amabiliidae в Казахстане // Паразитология. - 1985. - Т. 9,
вып. 6. - С. 479-482.
Кукашев Д.Ш. Зараженность стрекоз и водных клопов личинками цестод сем.
Amabiliidae в некоторых водоемах Казахстана 7/ Паразиты и болез-
ни водных беспозвоночных. (Тез. докл. IV Всесоюз. симпоз). - М., 1986.-
С. 76-77.
Кулагина О.П. Зараженность стрекоз озера Неро личинками гельминтов // Во-
просы экологической гельминтологии. - Ярославль, 1984. - С. 49-52.
Кумач ев И.С. Роль стрекоз и ос в снижении численности гнуса в бассейне
реки Или // Регуляторы численности гнуса на юго-востоке Казахстана. - Ал-
ма-Ата: Изд-во АН КаэССР, 1973. - С. 78-87.
Курандина Д.П. К изучению паразитов и симбионтов членистоногих Киевского
водохранилища // IV съезд Всесоюз. гидробиологического о-ва. (Тез. докл.
симпоз. гидропаразитологов). - Киев: Наук, думка. - 1981. - С. 27-28.
179 .
Курандииа Д.П. Грегарины водных членистоногих Киевского водохранилища И
Современные проблемы протозоологии. - Вильнюс, 1982. - С. 193.
Кухарчук Л.П. Кровососущие комары (Culicidae)//Биологические основы
борьбы с гнусом в бассейне Оби. - Новосибирск, 1966. - С. 9-53.
Лабораторные методы исследования в неинфекционной иммунологии. - М.:
.Медицина. - 1967. - С. 288-289.
Ланге Э.Р. Фауна метацеркарий в водных личинках насекомых И Вопросы фи-
зиологии и зоологии. - Рига, 1969. - Т. 100. - С. 95-113.
Леви Э.К. К фауне стрекоз Кировской области //Latvijas Ent — 1968. -
Вып. 12. - С. 77-84.
Литвенкова Е.А. Зараженность стрекоз метацеркариями простогоиимусов //
Животный мир Белорусского Поозерья. - Минск, 1970. - Вып. 1. -
С. 174-177.
Любарская О.Д., Галеева Л.Х. К вопросу о специфичности у метацеркарий
трематод // IX конф. Украинского республиканского паразитологического об-
щества. - Киев: Наук, думка, 1980а. - Ч. 3. - С. 39-41.
Любарская О.Д., Галеева Л.Х. О зараженности стрекоз метацеркариями тре-
матод в Татарской АССР И Вопросы паразитологии водных беспозвоночных
животных. - Вильнюс, 19806. - С. 68-70.
Любарская О.Д., Соколина Ф.М. Роль беспозвоночных в распространении
трематод и предпосылки формирования очагов трематодозов И Биологические
основы борьбы с гельминтами животных и растений. (Тез. докл. Всесоюз.
о-ва гепьминтол.). - М.: Наука, 1983. - С. 40-51.
Мазохин-Поршняков Г.А. Основные приспособительные типы чешуекрылых
Lepidoptera .// Зоол. журн. - 1954. - Т. 33, вып. 4. - С. 822-840.
Мазохин-Поршняков Г.А. Зрение насекомых. - М.: Наука, 1965. - 264 с.
Мазохин-Поршняков Г.А. Механизмы цветораэличения у насекомых //Био-
ника. Т. 1. (Итоги науки и техники ВИНИТИ АН СССР). - М.: 1973. -
С. 126-162.
Мазохин-Поршняков Г.А., Рязанова Г.И. Поиск убежища личинками стре-
козы Calopteryx splendens (Harris) (Zygoptera ) // Журн. общ.
биологии. - 1987. - Т. 48, Xs 2. - С. 248-253.
Мазохин-Поршняков ГЛ., Семенова С.А., Любарский Г.Ю. Анализ груп-
пового визуального поведения медоносных пчел при фуражировке // Там же. -
1984. - Т. 45, № 1. - С. 79-88.
Мамаев Б.М. Эволкция галлообразующих насекомых - галлин. - Л.: Наука.
Ленингр. отд-ние, 1968. - 237 с.
Мартынов А.В. Основные черты географического распространения ручейников
(Trichoptera ) // Докл. Российской АН. - Пг., 1922. - С. 48-51.
Мартынов А.В. Очерки геологической истории и филогении отрядов насекомых.
Ч. 1 // Тр. ин-та / Палеонтол. ин-т АН СССР. - 1938. - Т. 7, вып. 4,-
149 с.
Меландер В. Очерк феуны стрекоз (Odonata)Смоленской губернии // Тр.
Смоленское о-во естествоиспытателей и врачей. - 1927. - Т. 2. -
С. 157-188.
Минайлова Н.М., Мушкамбарова М.Г. Насекомые равнинной части Восточ-
ной Туркмении как промежуточные хозяева гельминтов // Изв. АН ТССР.
Сер. биол. - 1982. - № 1. - С, 29-35.
Мозговой А.А., Семенова М.К., Шахматова В.И. Жизненный цикл Cont-
racaepum microcephalum (Ascaridata, Anisakidae) - парази-
та рыбоядных птиц // Гельминты человека, животных и растений и борьба с
ними. - М.: Наука, 1968а. - С. 262-272.
Мозговой А.А., Шахматова В.И., Семенова М.К. Жизненный цикл Cont—
racaecum spiculigerum (Ascaridata, Anisakidae) - паразита
домашних и промысловых птиц // Там же. - 19686. - С. 129-136.
180
Мокрушов ПЛ. Зрительные стимулы в поведении стрекоз. I. Охота и посадка
у стрекозы четырехпятнистой (Libellula quadrimaculata L.) // Вести,
зоологии. - 1972. - № 4. - С. 46-51.
Мокрушов ПЛ. Зрительные стимулы в поведении личинок и взрослых стрекоз //
Поведение водных беспозвоночных. - Борок, 1975. - С. 52-53. '
Мокрушов П.А. Территориальное поведение четырехпятнистой стрекозы (Libel—
lula quadrimaculata) (Odonata, Anisoptera) // Вести, зоологии. -
1982. - № 2. - С. 58-62.
Мокрушов ПЛ., Золотов В.В. Зрительные стимулы в поведении стрекоз. П.
Охота и бегство у личинок коромысла синего Aeschna cyanea Mull. //
Там же. - 1973. - № 6. - С. 75-77.
Мокрушов П.А., Франпевич Л.И. Нейроны, чувствительные к движению конт-
растных объектов, у личинок стрекозы Aeschna cyanea // Журн. эволкц.
биохимии и физиологии. - 1973. - Т. 9, № 2. - С. 189-194.
Мокрушов П.А., Францевич Л.И. Зрительные интернейроны, чувствительные
к размеру объектов и направлению движения, у стрекоз рода Sympetrum И
Там же. - 1976а. - Т. 12, № 4. - С. 341-346.
Мокрушов П.А., Франпевич Л.И. Зрительные стимулы в поведении стрекоз.
III. Выбор места для посадки у стрелки-наяды Erythromma najas Han-
sem. // Вести, зоологии. - 19766. - № 4. - С. 20-24.
Нарэикулов М.Н. О жизненных формах насекомых, их становление и эволю-
ция // Фауна и экология насекомых Средней Азии. - Душанбе: Ифрон,
1970. - С. 3-25.
Никулин Т.Г. К вопросу гельминтологической оценки водоемов и выяснению
промежуточных хозяев гельминтов северной зоны Белоруссии // Проблемы
паразитологии. - Киев: Наук, думка, 1967. - С. 180-181.
Никулин Т.Г.. Моллюски. и личинки стрекоз водоемов БССР как промежуточные
и резервуарные хозяева трематод и цестод птиц. - Витебск, 1969. -
С. 51-52. - (Тез. докп. итогов науч, коиф, Витеб. вет. ин-та, 1968 г.).
Никулин Т.Г. Стрекозы поозерья Белоруссии и их роль в распространении тре-
матодозов птиц // Сб. работ по гельминтологии. - М.: Колос, 1971. -
С. 284-289.
Окороков В.И., Ткачев В.А. К обнаружению цистицеркоида рода Tatria
из личинок стрекоз иа Южном Урале // Вопр. зоологии. - Челябинск, 1973.-
Вып. 2. - С. 79-80.
Окороков В.И., Ткачев ВЛ. Заражение личинок стрекоз гельминтами на озе-
ре Песчаное в Челябинской области // Гельминты животных, человека и ра-
стений на Южном Урале. - Уфа, 1974. - Вып. 1. - С. 142-144.
Окороков В.И., Харитонов А.Ю. Фауна и биология стрекоз на Южном Урале
и их роль как промежуточных хозяев гельминтов // Вопр. зоологии. - Челя-
бинск, 1971. - Вып. 2. - С. 32-40.
Олигер А.И. О стрекозах Астраханского заповедника // Учен. зап. Чуваш; гос.
пед. ин-та. - 1970. - Вып. 31. - С. 100-107.
Олигер А.И. К вопросу о зараженности стрекоз (Odonatoptera) клещами се-
мейства Arrhenuridae в Донбассе И Проблемы паразитологии. - Киев:
Наук, думка, 1975. - Ч. 2. - С. 78-80.
Олигер А.И. Фенология и суточная активность стрекоз (Odonata) в юго-
восточной части Украины // Зоол. журн. - 1980. - Т. 59, вып. 9. -
С. 1425-1427.
Oh иге р А.И. О биотопнческом распределении стрекоз Донецкой области // Бюл.
МОИП. Отд. биол. - 1985. - Т. 90, № 5. - С. 25-33.
Павлюк Р.С. Об эктопаразитах стрекоз (Odonata) - личинках водяных
клещей рода I rhenurus // V Всесоюз. совещ. по болезням и парази-
там рыб и водн. х беспозвоночных. - Л.: Наука. Ленингр. отд-ние, 1968. -
С. 91-92.
181
Павлюк Р.С. Грегарины, поражающие стрекоз (insecta, Odonata) западных
областей Украинской ССР // Материалы I съезда всесоюз. о-ва протозооло-
гов. - Баку: Элм, 1971. - С. 294-295.
Павлюк Р.С. К биологии метацеркариев трематоды Prosotocus confusus
(Looss, 1894) Ц Проблемы паразитологии. - Киев: Наук, думка, 1972а, -
Ч. 2. - С. 100-101.
Павлюк Р.С. Итоги изучения параэитофауны стрекоз (insecta, Odonata)
западных областей Украинской ССР Ц Там же, 19726. - Ч, 2. -
С. 102-103.
Павлюк Р.С. К вопросу о видовой принадлежности не инцистиро ванных метацер-
карий плягиорхидного типа из полости тела личинок и имаго стрекоз // Па-
разиты водных беспозвоночных животных. - Львов, 1972в. - С. 68-69.
Павлюк Р.С. Новые виды дополнительных хозяев трематод рода Gorgodera
Loose, 1901 Ц Гидробиол. журн. - 1972г. - Т. 8. № 3. - С. 105-107.
Павлюк Р.С. Новые данные о метацеркариях Halipegus ovocaudatus
Vulp., 1958 (Trematoda, Halipegidae)// Вести, зоологин. - 1973a. -
№ 2. - С. 33-37.
Павлюк Р.С. О цистицеркоидах'Icitriadecacantha Fuhrmann, 1913 (Cestoda:
Amabiliidae) из стрекоз западных областей Украины // Паразитология. -
19736. - Т. 7, вып. 4. - С. 353-356.
Павлюк Р.С. Стрекозы (Odonata) - дополнительные хозяева трематод рода
Prosthogonimus Liihe // Материалы VII съезда Всесоюз. энтомол.
о-ва. - Л., 1974. - Ч. 1. - С. 100-101.
Павлюк Р.С. Стрекозы (insecta, Odonata) западных областей УССР, их
паразиты и враги: Автореф. дис. ... каид. биол. наук. - Львов, 1975а. - 29 с.
Павлюк Р.С. Новые дополнительные хозяева трематоды Plagiorchis ele-
gans Rud., 1802 И Проблемы паразитологии. - Киев: Наук, думка,
19756. - Ч. 2. - С. 88-89.
Павлюк Р.С. Новые виды грегарин из стрекоз (insecta, Odonata) запад-
ных областей Украины // Паразиты и паразитоэы животных и человека. -
Киев: Наук, думка, 1975в. - С. 88-99.
Павлюк Р.С, О зараженности личинок и имаго стрекоз (insecta, Odonata)
западных областей УССР метацеркариями трематод Pleurogenoides me-
dians (Olsson, 1876) //II Воесоюз. симпоз. по болезням и парази-
там водных беспозвоночных: Тез. докл. - Л., 1976. - С. 56-57.
Павлюк Р.С. Структура параэитоценоэов комплексов стрекоз (insecta, Odo—
nata), приуроченных к различным типам водоемов западных областей
УССР // I Всесоюз. съезд паразитоценологов. - Киев; Наук, думка, 1978.-
Ч. 2. - С. 38-40.
Павлюк Р.С. Грегаринофауна бабок (insecta, Odonata) водоемов западных
областей Украинской ССР // Mani водойми У крайни та питания ix охорони. -
Киев: Наук, думка. - 1980а. - С. 38-42.
Павлюк Р.С. К фенологии стрекоз Западной лесостепи // Исследования по эн-
томологии и акарологии на Украине. - Киев, 19806. - С. 51-52. - (Тез.
докл. 2-го съезда УЭО, Ужгород, 1980).
Павлюк Р.С. Материалы по паразитам стрекоз (insecta, Odonata) дельты
Дуная И IX конференция Украинского республиканского параэитол. о-ва. -
Киев: Наук, думка, 1980в. - Ч. 3. - С. 133-134.
Павлюк Р.С, О поражаемости личинок стрекоз (insecta, Odonata) личинка-
ми цестод в западных областях Украины // Вопросы паразитологии водных
беспозвоночных животных. - Вильнюс, 1980г. - С. 81-83.
Павлюк Р.С. О пораженности стрекоз в западных областях УССР метацерка-
риями Skrjabinoeces similis (Trematoda, Plagiorchiidae) 11
Вести, зоологии. - 1980д. - № 1. - С. 81-84.
Павлюк Р.С, Паразитологические исследования стрекоз (insecta, Odonata),
182
хранящихся в энтомологических коллекциях II Там же. - 1981. - № 2. -
С. 90-92.
Павлюк Р.С. Материалы о грегарииах, поражающих стрекоз (insecta, Odo—
nata) дельты Дуная II Современные проблемы протозоологии. - Вильнюс,
1982. - С. 275.
Павлюк Р.С. Соотношения множественных инвазий у стрекоз II Й Всесоюз.
съезд паразитоценологов. - Киев:' Наук, думка, 1983а. - С. 254-256.
Павлюк Р.С. Взаимоотношения паразитов стрекоз с хозяевами и факторы ок-
ружающей среды // Там же. - Киев: Наук, думка, 19836. - С. 256-257.
Павлюк Р.С. О пределах гостальной специфичности личиночных стадий гельмин-
тов, развивающихся в организме стрекоз (insecta, Odonata) И X коиф.
Украинского республиканского параэитол. о-ва. - Киев: Наук, думка, 1986. -
Ч. 2. - С. 88.
Павлюк Р.С., Курбанова Т.М. К изучению фауны стрекоз (insecta, Odo-
nata) Туркмении II Изв. АН ТССР. Сер. биол. - 1984. - № 4. -
С. 72-74.
Павлюк Р.С., Курбанова Т.М. Параэитофауна стрекоз южных районов Турк-
мении П X коиф. Украинского параэитол. о-ва. - Киев: Наук, думка, 1986.-
Ч. 2. - С. 89.
Павлюк Р.С., Стражник Л.В. Гельминтоценоэы стрекоз (insecta, Odona-
ta) водоемов Западных областей Украины // Вести, зоологии. - 1985. -
24 с. - Деп. в ВИНИТИ № 1921-85.
Павлюк Р.С., Харитонов А.Ю. Номенклатура стрекоз (insecta, Odonata)
СССР Ц Полезные и вредные насекомые Сибири. - Новосибирск: Наука. Сиб.
отд-ние, 1982. - С. 12—42.
Пании В.Я. Распространение трематод рода Prosthogonimus среди диких
птиц и природная очаговость простогонимоэа // Тр. ин-та / Ии-т зоологии
АН КазССР. - 1957. - Т. 7. - С. 216-226.
Панин В.Я. К фауне стрекоз - переносчиков простогонимоэа в Зайсанской кот-
ловине II Там же. - 1958. - Т. 9. - С. 164-166.
Паскальская М.Ю. Изучение цикла развития трематоды Plagiorchis arcua-
tus Strom, 1924, паразитирующей в яйцеводе и фабрициевой сумке кур Ц
Докл. АН СССР. - 1954. - Т. 97, № 3. - С. 561.
Паскальская М.Ю. Цикл развития Plagiorchis arcuatus Strom,
1924 // Тр. ин-та / Всесоюз. ин-т гельминтологии им. Скрябина. - 1959.-
Т. 6. - С. 24-30.
Писаненко А.Д. Фаунистический очерк стрекоз (insecta, Odonata) Бело-
руссии // Вести. Белорус, ун-та. Сер. 2 (химия, биология, география). -
1985. - № 3. - С. 37-41.
Плотникова С.И. Структурная организация центральной нервной системы иа-
секомых. - М.: Наука, 1979. - 118 с.
Полетаева О. Петербургские Odonata И Тр. / Рус. энтомол. о-во. -
1880. - Т. 11. - С. 97-119.
Положеицев П.А., Негробов В.П. К исследованию насекомых как промежу-
точных хозяев биогельмиитов в Воронежской области // Работы по гельмин-
тологии к 80-петию акад. К.И. Скрябина. - М.: Изд-во АН СССР, 1958, -
С. 274-278.
Положенцев П.А., Негробов В.П. О насекомых-хозяевах паразитических
червей человека и животных // Вредные и полезные насекомые. - Воронеж,
1967. - С. 3-159.
Попова А.Н, К вопросу питания личинок стрекоз // Работы Волжской биологи-
ческой станции. - 1925. - Т. 8, № 1-3.
Попова А.Н. Odonata, собранные Сев.-Кавказской гидробиологической стан-
цией в 1921-1923 и 1925 гг. // Работы Сев.-Кавк. гидробиол. ст. -
1928. - Т. 2, вып. 2/3. - С. 120-123.
183
Попова А.Н. К фауне стрекоз бассейна Телецкого озера // Исследование озер
СССР. - Л., 1933. - Вып. 3. - С. 131-134.
Попова А.Н. Материалы к фауне и биологии личинок стрекоз реки Чу Ц Тр. /
Киргизская комплексная экспедиция, - 1936. - Т. 3, вып. 1. - С. 215-
219.
Попо ва А.Н. Личинки стрекозы Epallage fatime (Charp.) среди дон-
ных обитателей реки Занги // Тр. / Севанская гидробиол. ст. - 1947. -
Т. 8. - С. 77-86.
Попова А.Н. Стрекозы - Odonata // Животный мир СССР. - М.; Л.: Изд-
во АН СССР, 1950. - Т. 3. - С. 288-294.
Попова АЛ, Стрекозы (Odonata) Таджикистана // Тр. ин-та / Зоол. ин-т
АН СССР. - 1951. - Т. 9. - С. 861-894.
Попова А.Н. Личинки стрекоз фауны СССР (Odonata). - М.; Л.: Изд-во АН
СССР, 1953а. - 235 с.
Попова А.Н. Стрекозы - Odonata // Животный мир СССР. - М.; Л.:
Изд-во АН СССР, 19536. - Т. 4. - С. 552-555.
Попченко В.И. Потребление малощетинковых червей рыбами и беспозвоночны-
ми // Вопр. ихтиологии. - 1971. - Т. 2, № 1. - С, 96-102.
Правдин ФЛ. Экологическая география насекомых Средней Азии. - М.: Наука,
1978. - 271 с.
Притыкина Л Л. Материалы к морфоэкологической классификации личинок стре-
коз (Odonata) 7/ Энтомол. обозрение. - 1965. - Т. 44, № 3. - С. 503-
519.
Притыкина ЛЛ. Стрекозы Каратау (Odonata) // Юрские насекомые Ка-
ратау. — М.: Наука, 1968. — С. 26—54.
Притыкина ЛЛ. Стрекозы семейства Liassophlebiidae из триаса и юры
Средней Азии 7/ Палеонтол. журн. - 1970. - Вып. 1. - С, 104-117.
Притыкина Л.Н. Отряд Libellulidae. Стрекозы // Историческое развитие
класса насекомых. - М.: Наука, 1980. - С. 128-134. - (Тр. ин-та / Палеон-
тол. ин-т АН СССР; Т. 178).
Притыкина Л.Н. Новые триасовые стрекозы Средней Азии II Новые ископае-
мые насекомые с территории СССР. - М.: Наука, 1981. - С. 5-42. - (Тр.
ии-та / Палеонтол. ин-'t АН СССР; Т. 183).
Пузанов И.И. Зоогеография. - М., ,1938. - 357 с.
Радде Г.И. Коллекции Кавказского музея. I. Тифлис, 1899. - 540 с.
Радкевич А.И. Стрелк! В'щебшчыны (Odonata). - Вщебшчына, 1928. -
Т. 2, - С. 85-91.
Радкевич А.И. Материалы к изучению эитомофауны БССР. Стрекозы северо-
восточной части Белорусской ССР // Учеи. зап. Витеб. гос. пед. ин-та. -
1957. - Вып. 6. - С. 71-86.
Расиипыи А.П. Происхождение и эволюция низших перепончатокрылых. - М.:
Наука, 1969. - 196 с. - (Тр. ин-та 7 Палеонтол. ин-т АН СССР; Т. 123).
Редько Б.А. Материалы к фауне стрекоз окрестностей города Саратова //
Работы Волжской биол. станции. - 1913. - Т. 4. - С, 19-33.
Рейхардт В. Обработка сенсорной информации нервной системы И Теоретиче-
ская и математическая биология. - М„ 1968. - С. 373-400.
Римский-Корсаков МЛ. Биологические наблюдения над водными перепон-
чатокрылыми 7/ Рус. эитомол. обозрение. - 1916. - Т. 16, № 3/4. -
С. 208-225.
Римский-Корсаков М.Н. Перепончатокрылые (Hymenoptera) 7/Жизнь
пресных вод СССР. - М., 1940. - Т. 1. - С. 227-232,
Родендорф Б.Б. Надотряд Odonatoidea // Основы палеонтологии. Тра-
хейные и хелицеровые. - М.: Наука, 1962. - С. 73-85.
Родендорф Б.Б. О рационализации названий таксонов высокого ранга в зооло-
гии // Палеонтол, журн., 1977. - № 2. - С. 14-22.
Родендорф БЛ. Инфракласс Libellulones. Стрекоэообразные // Ио-
184
торическое развитие класса насекомых. - М.: Наука, 1980, - 256 с.
Родзянко В. Список стрекоз, водящихся в Полтавской губернии // Тр. о-ва /
О-во испытателей природы Харьков, ун-та. - 1887. - Т. 20. - С. 97-104.
Родзянко В. К сведению об одонатологической фауне Харьковской и Полтав-
ской губерний // Там же. — 1889, - Т. 22. - С. 209-224.
Родзянко В. Новые сообщения о фауне стрекоз Полтавской и Харьковской гу-
берний // Бюл. МОИП. - 1895. - № 1. - С. 119-127.
Рубцов И.А. Новые виды мермитид // Зоол. журн. - 1976. - Т. 55. -
С. 1292-1298.
Рубцов И.А., Киселев А.И. Новые виды мермитид из водных жуков и стре-
коз // Гельминты животных и растений Южного Урала. - Челябинск, 1975. -
С. 28-33.
Рубцов И.А., Павлюк Р.С. Мермитиды (Nematoda, Mermithidae) —
паразиты стрекоз западных областей УССР // Вести, зоол. - 1972. - № S.-
С. 34-42.
Рыжиков К.М., Толкачева Л.М. Аколеаты - ленточные гельминты птиц //
Основы цестодологии. - М.: Наука, 1981. - Т. 1 О. - 214 с.
Рязанова Г.И., М аз о х ин-П о рш н яко в Г.А. Поведение как механизм оптими-
зации репродукции у насекомых (на примере стрекоз) И Усаетм соврем, био-
логии. - 1984. - Т. 98, вып. 3. - С. 451—463,
Рязанова Г .И., М азохин-Поршняко в Г.А. Пространственное взаимодействие
личинок Calopteryx splendens (Odonata, Calopterygidae )
в лабораторном эксперименте И Зоол. журн. - 1985. - Т. 64, вып. 9. -
С. 1360-1367.
Садыкова В.Р. К одонатофауне рисовых полей Ташкентского оазиса. - Таш-
кент: Изд-во АН УзССР, 1959. - Вып. 3. - С. 59-66.
Свешников В.Г. Рефлекторные механизмы, запускающие и поддерживающие по-
лет стрекозы Aeschna grandis // Журн. эволкц. биохимии и физиологии. -
1970. - Т. 6, № 4. - С. 472-473.
Седых К.Ф. Отряд стрекозы - Odonata // Животный мир Коми АССР. Бес-
позвоночные. - Сыктывкар: Коми кн. изд-во, 1974. - С. 68-72.
Семенова М.К. К изучению биологии нематоды Contracoecum micropapil-
latum (Stossich, 1890) Baylis, 1920 (Ascaridata; Anisa-
kidae) // Пробл. паразитологии. - Киев: Наук, думка, 1972. - Ч. 2. -
С. 238-240.
Семенова М.К. Нематоды Contracoecum micropapillatum (Stossich,
1890) Baylis, 1920 (Ascaridata: Anisakidae) // Экология и так-
сономия гельминтов. - М.: Наука, 1973. - С. 55-64.
Семенова М.К. Изучение биологии Contracoecum micropapillatum и
Contracoecum microcephalum (Ascaridata: Anisakidae) -
нематод рыбоядных птиц: Автореф. дис, ... канд. биол. наук. - М., 1974.
Сергеева Т.К. Методика получения антисыворотки к белкам вредной черепаш-
ки Eurygaster integriceps для изучения ее хищников // Зоол. журн.-
1970а. - Т. 49, вып. 3. - С. 440-444.
Сергеева Т.К. Фиксация и обнаружение белков вредной черепашки (Eurygaster
integriceps ) в желудках хищных членистоногих // Там же. - 19706. -
Т. 49, вып. 11. - С. 1715-1719.
Сергеева Т.К. Метод серологической диагностики питания хищных беспозвоноч-
ных для изучения трофической структуры сообщества // Биоиндикация состоя-
ния окружающей среды Москвы и Подмосковья. - М.: Наука, 1982а. -
С. 105-112.
Сергеева Т.К. Методы и современное состояние изучения трофических связей
хищных почвенных беспозвоночных: серологический анализ питания // Зоол.
журн. - 19826. - Т. 61, вып. 1. - С. 109-119.
Сергеева Т.К. Изучение хищничества почвообитающих проволочников серологи-
185
ческим методом // Там же. - 1983. - Т. 62, вып. 10. - С. 1510-
1513.
Силантьев А.А. Фауна Падовь, имения В.Л. Нарышкина Балашовского уезда
Саратовской губернии. - Спб., 1894. - Т, 19. - С. 141-142,
Синицын Д.Ф. Материалы по естественной истории трематод: Дистомы рыб и
лягушек окрестностей Варшавы // Изв. Варшав. ун-та, 1905. - Вып. 7-9.—
С. 1-96.
Скрябин К.И. Паразитические Nematodes пресноводной фауны Европейской
и отчасти Азиатской России // Пресноводная фауна Европейской России. -
М„ 1923. - С. 1-98.
Смирнова Г.А., Харитонов А.Ю. Стрекозы бассейна озера Чаны // Экология
и география членистоногих Сибири. - Новосибирск, 1987. - С. 96-97.
Соколов И.И. К познанию фауны водоемов Старой Бухары и ее окрестностей //
Тр. ин-та / Узбекистан, ин-т тропической медицины, - 1933. - Т. 1, вып. 4,-
С. 4-16.
Солодовников С.В. Материалы к фауне и биологии личинок стрекоз р. Донца и
некоторых притоков // Тр. Харьковского тов. доел, природы. - 1929. -
Т. 52. - С. 249-268.
Спурис ЗЛ. Южные элементы в фауне стрекоз (Odonata) Латвийской
ССР // Энтомол. обозрение. - 1951. - Т. 31, вып. 3/4. - С. 500-503.
Спурис З.Д. Стрекозы Латвийской ССР. - Рига: Изд-во АН ЛатвССР, 1956. -
96 с.
Спурис З.Д. О фауне стрекоз Литовской ССР // Тр. ин-та / Ин-т биологин
АН ЛатвССР. - 1959. - Т. 12. - С. 87-93.
Спурис З.Д. Отряд Odonatoptera (Odonata) - стрекозы // Определитель
насекомых европейской части СССР. - М.; Л.: Наука, 1964а. - Т. 1. -
С. 137-161.
Спурис З.Д. Стрекоза Coenagrion concinnum Joh. в Латвийской ССР //
Latvijas Ent. — 19646. - Вып. 9. - С. 77-86.
Спурис З.Д. Новые данные о распространении стрекоз в Латвии И Там же. -
1965. - Вып. 7. - С. 21-40.
Спурис З.Д. Заметки о распространении стрекоз в Эстонии // Там же. -
1968. - Вып. 12.-С. 72-76.
Спурис З.Д. Заметки о распространении стрекоз в Южной Литве И Там же. -
1971. - Вып. 13. - С. 27-34.
Спурис З.Д. Каталог насекомых Латвии. I. Стрекозы (Odonata) И Лат-
вийский энтомолог. - 1980, - № 23. - С. 5-19.
Станёните А.П. К материалам по экологии стрекоз юго-восточной части Лит-
вы // Вторая зоологическая конференция Белорусской ССР: Тез. докл. -
Минск: Изд-во АН БССР. - 1962а. - С; 185,
Станёните А.П. Фауна личинок стрекоз в водных бассейнах окрестностей
г. Вильнюса // Тр. АН ЛитССР. Сер. Б. - 19626. - Т. 1(27). -
С. 153-160.
Станёните А.П. Биология и паразиты стрекоз (Odonata) Лиговской ССР //
Автореф. дис. ... канд. биол. наук. - Вильнюс, 1963а. - 22 с.
Станёните А.П. Зараженность стрекоз (Odonata) паразитами в Литов-
ской ССР // Материалы науч. конф. Всесоюз. о-ва гельминтологов. - М.,
19636. - Ч. 2.
Станёните А.Н. Некоторые данные о стрекозах (Odonata) Литовской ССР //
Тр. АН Литовской ССР. Сер. Б. - 1963в, - Вып. 1(30). - С. 51-63.
Станёните А.П. Паразиты стрекоз Литовской ССР // Там же. Сер. В. -
1964. - Т. 3. - С. 83-91.
Станёните А.П. Биологическое исследование личинок некоторых видов стрекоз //
Там же. - 1966. - № 3(41). - С. 113-120.
Станёните А.П. Фауна стрекоз (Odonata) озера Жувинтас и его окрестно-
стей И Заповедник Жувинтас. - Вильнюс, 1968. - С. 239-242,
186
Станёните А.П. Изученность стрекоз Литовской ССР // Новые и редкие для
ЛитССР виды насекомых: сообщение и описание 1985 г. - Вильнюс, 1985.-
С. 99-103.
Старостин И.Б. Сернистые источники Копетдаге (фауна и флора) // Тр. /
Мургабская гидробиол. ст. - Ашхабад, 1951. - Вып. 1. - С. 127-138.
С т е б а е в И.В. Жизненные формы и половой диморфизм саранчовых Т увы и Юго-
Восточного Алтая // Зоол. журн. - 1970. - Т. 49, вып. 3, - С. 325-338.
Стебаев И.В., Никитина С.И. Особенности поведения саранчовых разных
жизненных форм из степей и полупустынь Тувы. I // Там же. - 1975. -
Т. 54, № 5. - С. 688-700.
Стражник Л.В., Павлюк Р.С. Материалы к паразитофауне личинок стрекоз
(insecta, Odonata) в водоемах окрестностей г. Киева // Паразиты и
болезни водных беспозвоночных. - М., 1986. - С. 137-139.
Судариков В.Е., Карманова Е.М. Работа Астраханского отряда 320-й со-
юзной гельминтологической экспедиции 1962 г. // Тр. / Астраханский эапо—
вед. - 1964. - Вып. 9. - С. 32-39.
Супряга А.М. Биологический цикл Avioserpens mosgovoyi (Camallana—
ta, Dracunculidae ) - нематоды водоплавающих птиц // Сборник ра-
бот по гельминтологии. - М.: Колос, 1971. - С. 374-383.
Супряга А.М. К изучению очага авиосерпенсоэа водоплавающих птиц // Про-
блемы паразитологии. - Киев: Наук, думка, 1972. - Ч. 2. - С. 304-306.
Таранова В.М. Сезонное развитие фауны стрекоз (Odonata) прибрежья
Рыбинского водохранилища и прилегающих к нему водоемов // Тр. ин-та /
Ин-т биол. внутр, вод АН СССР. - 1980. - № 41/44. - С. 145-154.
Тарлинг Д., Тарлинг М. Движущиеся материки. - М.: Мир, 1973. - 104 с.
Ткачев В.А. Развитие цестоды Tatria decacantha Fuhrmann, 1913
в организме дефинитивного хозяина // Вопр. экологии. - 1969а. -
С. 49-55.
Ткачев В.А. Экспериментальное исследование развития ленточного паразита
Tatria decacantha Fuhrmann, 1913 в организме промежуточного хо-
зяина // Там же. - 19696. - С. 56-63.
Ткачев В.А. К обнаружению цистицеркоида Tatria decacantha Fuhrmann,
1913 в личинках стрекоз // Материалы IV науч. конф, зоологов
пед. ин-та. - Горький, 1970. - С. 136-137.
Ткачев В.А. Влияние личинок цестод на промежуточного хозяина и развитие их
в условиях переуплотненной популяции // Вопр. зоологии. - 1971а. -
С. 81-82.
Ткачев В.А. Вопросы выяснения заражения водных организмов личинками це-
стоды Т. decacantha Fuhrmann, 1913 на озерах Южного Урала // Там
же. - 19716. - Вып. 2. - С. 63-64.
Ткачев В.А. Сезонная динамика заражения птиц отряда Podicipitiformes
цестодой Tatria decacantha Fuh., 1913 // Там же. - 1971в. -
Вып. 2. - С. 51-53.
Ткачев В.А. Некоторые данные питания большой поганки на озере Дергуново,
Горетое, Калинкино в Челябинской области // Там же. - 1973. - Вып. S.-
С. 45-47.
Токобаев М.М., Чибиченко Н.Т. Фаунистические комплексы личинок гель-
минтов у воднь!х беспозвоночных Киргизии // II Всесоюз. симпозиум по бо-
лезням и паразитам водных беспозвоночных: Тез. докл. - Л.: Наука. Ленингр.
отд-ние, 1976. - С. 69-70.
Ульянин В, Список московских сетчатокрылых и прямокрылых. - М., 1867. -
111 с.
Ульянин В. Материалы для этомэлогии губерний Московского учебного округа.
Вып. 2. Список сетчатокрылых и прямокрылых насекомых // Изв. Импер.
о-ва любителей естествознания, антропологии и этнографии. - 1869. - Т. 6,
вып. 2. - 120 с.
187
Ушаков С.А., Ясаманов Н.А. Дрейф материков и климаты Земли // М.:
Мысль, 1984. - 206 с.
Уэда С. Новый взгляд на Землю. - М.: Прогресс, 1980. - 213 с.
Францевич Л.И. Нейроны зрительных долей, чувствительные к видимому дви-
жению, у пластинчатоусых жуков И Журн. эволюц. биохимии и физиологии. -
1969. - Т. 5, № 6. - С. 577-582.
Францевич Л.И. Зрительный анализ пространства у насекомых. - Киев: Наук,
думка, 1980. - 288 с.
Францевич Л.И. Взаимодействие оптических ключевых стимулов, вызывающих
у стрекоз атаку и бегство // Журн, эволки. биохимии и физиологии. - 1982,-
Т. 18, № 2. - С. 150-154.
Ф ра нц е ви ч Л.И. Пространственная ориентация животных. - Киев: Наук, дум-
ка, 1986. — 200 с.
Францевич Л.И., Мокрушов П.А. Ответы нейронов - высокочастотных де-
текторов движения у личинок стрекозы на движение одивочных объектов //
Нейрофизиология. - 1974. - Т. 6, № 1. - С. 68-74.
Францевич Л.И., Мокрушов П.А., Дуб В.А. Реакция дирекционно-чувстви-
тельных оптических интернейронов насекомых на движение двумерных узоров /1
Журн. эволюц. биохимии и физиологии. - 1978. - Т. 14, № 3. - С. 251-258.
Францевич Л.И., Пичка В.Е. Размеры бинокулярной зоны поля зрения у насе-
комых // Там же. - 1976. - Т. 12, № 5. - С. 461-465.
Ф рн е й-Гесс не р. Материалы к энтомологии Урала. Насекомые, собранные в
Екатеринбургском уезде Вл. и Мод. Клер в 1896-1897 гг, // Зап. Урал,
о-ва любителей естествознания. - 1907. - Т. 26. - С. 75-80.
Фридолин В.Ю. Фауна Северного Урала как зоогеографическая единица и как
биоценотическое целое // Тр. / Ледниковые экспедиции. Вып. 4. Урал. При-
полярные районы. - Л., 1936. - С. 245-270.
Фролова Е.Н. Паразиты членистоногих и моллюсков в озерах Карелии // Фауна
озер Карелии. - М.; Л.: Изд-во АН СССР, 1965. - С. 311-324.
Фурсов В.Н. К изучению фауны и биологии хальцид (Hymenoptera, Chalci-
doidae) - паразитов яиц водных насекомых // IX съезд Всесоюз. энтомол.
о-ва. - Киев: Наук, думка, 1984. - Ч. 2. - С. 214.
Фурсов В.Н., Костюков В.В. Новые виды рода Tetrastichus (Hymeno-
ptera, Eulophidae ) - паразиты яиц стрекоз и жуков-плавунцов // Зоол.
журн. - 1987. - Т. 66, вып. 2. - С. 217-228.
Харитонов А.Ю. Распространение стрекоз на территории Челябинской области И
Тез. докл. науч.-краевед. конф. Челяб. отд. геогр. о-ва СССР. - Челябинск,
1972. - С. 51-52.
Харитонов А.Ю. Заенисейские стрекозы на Полярвом Урале И Вопросы энто-
мологии Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1974а. - С. 68-69.
Харитонов А.Ю. О некоторых закономерностях в изменении биологических осо-
бенностей сибирских стрекоз (Odonata, Insecta) в зависимости от ге-
ографического положения изучаемых фаун // Изв. СО АН СССР. Сер. биол.
наук. - 19746. - № 10, вып. 2. - С. 146-147.
Харитонов А.Ю. К биологии малоизвестного вида стрекоз Somatochlora
sahlbergi Trybom ( Odonata, Corduliidae) // Таксономия и экология
животных Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1975а. - С. 21-23.-
(Новые и малоизвестные виды фауны Сибири; Вып. 9).
Харитонов А.Ю. Североевразиатские виды рода Enallagma Charp. (Odo-
nata, Insecta)// Там же. - С. 11-20.
Харитонов А.Ю, Стрекозы как массовые компоненты водных биоценозов //
Биологические ресурсы Западной Сибири и их охрана (материалы науч. конф,
молодых ученых). - Новосибирск: Наука. Сиб. отд-ние, 1975в. - С. 63-64.
Харитонов А.Ю. Стрекозы Ильменского заповедника // Вопр. зоологии, -
1975г. - Вып. 4. - С. 63-65.
188
Харитонов А.Ю. Суточная активность стрекоз и влияние на нее погодных ус-
ловий // Там же. - 1975д. - Вып. 4. - С. 66-75.
Харитонов А.Ю. Условия обитания личиночных фаз стрекоз в Заполярье //
Экология. - 1975е. - № 3. - С. 96-99. <
Харитонов А.Ю. Стрекозы Урала и Зауралья (фауна, экология, зоогеография):
Автореф. дис. ... канд. биол. наук. - Новосибирск, 1975ж. - 25 с.
Харитонов А.Ю. Основные направления эволкиии репродуктивного поведения у
равнокрылых стрекоз и его таксономическое значение // Групповое поведение
животных: Докл. участников II Всесоюз. конф, по поведению животных. - М.:
Наука, 1976а. - С. 409-412.
Харитонов AJO. К вопросу о разрыве ареалов у некоторых видов стрекоз (in—
secta, Odonata) в районе Тургайской ложбины // Фауна гельминтов и
членистоногих Сибири. - Новосибирск: Наука. Сиб. отд-ние, 19766. -
С. 162-164. - (Тр. ин-та / Биол. ин-т СО АН СССР; Вып. 18).
Харитонов А.Ю. Таксономическое положение малоизвестных видов группы
Agrion concinnum Joh. (Odonata, Insecta) // Новости фауны Си-
бири. - Новосибирск: Наука. Сиб. отд-ние, 1976в. - Вып. 10. - С. 52-56. -
(Новые и малоизвестные виды фауны Сибири; Вып. 10).
Харитонов А.Ю. Фауна стрекоз (insecta, Odonata) Урала и Восточного
Приуралья // Фауна гельминтов и членистоногих Сибири. - Новосибирск: Нау-
ка. Сиб. отд-иие. - 1976г. - С. 157-161. - (Тр. ин-та / Бнол. ии-т СО
АН СССР; Вып. 18).
Харитонов AJO. Жизненные пиклы некоторых видов стрекоз в Восточном При-
уралье // Изв. СО АН СССР. Сер. биол. наук. - 1977а. - № 5. - С. 55-59.
Харитонов А.Ю. Описание реликтовых видов стрекоз ( Insecta, Odonata )
из южных районов Западной Сибири // Таксоны фауны Сибири. - Новосибирск:
Наука. Сиб. отд-ние, 19776, - С. 117-124. - (Новые и малоизвестные виды
фауны Сибири).
Харитонов А.Ю. Зоогеографическое районирование Восточного Приуралья на
основании распространения стрекоз (Insecta, Odonata ) И Членистоногие
Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1978а. - С. 47-54. - (Тр. ии-
та / Биол. ии-т СО АН СССР; Вып. 34),
Харитонов А.Ю. Новые сведения о морфологии, экологии и поведении двух ма-
лоизвестных видов стрекоз (insecta, Odonata) // Таксономия и экология
членистоногих Сибири. - Новосибирск: Наука. Сиб. отд-ние, 19786. -
С. 70-78.
Харитонов А.Ю. Новый вид стрекозы рода Ischnura Chard. (Odonata,
Coenagrionidae) из Южного Казахстана // Новые виды насекомых. - Но-
восибирск: Наука. Сиб. отд-ние, 1979. - С. 5-7. - (Тр. ВЭО; Т. 61).
Харитонов А.Ю. Первый опыт установления контактов одонатологии и археоло-
гии // Новые методы в археологии. - Томск: Изд-во Том. ун-та, 1980а. -
С. 46.
Харитонов А.Ю. Широтные изменения фенологии стрекоз (insecta, Odonata)
Зауралья и Казахстана // Экология. - 19806. - С. 93-96.
Харитонов А.Ю. Морфология и биология малоизвестного вида стрекоз Gom-
phus epophthalmus Selys (Odonata, Gomphidae) // Насеко- 1
мые и клещи Сибири. - Новосибирск: Наука. Сиб. отд-ние, 1981. -
С. 111-113. - (Новые и малоизвестные виды фауны Сибири).
Харитонов А.Ю. Совершенствование аэрозольной технологии применения инсек-
тицидов и проблемы охраны природы // XXVI съезд КПСС и управление
научно-техническим и социальным прогрессом (Сб. тез. докл. обл. иауч,-
практ. коиф. - Новосибирск. - 1982. - С. 141-144.
Харитонов А.Ю. Опыт опенки экологической роли стрекоз в пойменных водое-
мах и о коло водных биоценозах // Тез. докл. IX съезда Всесоюз. энтомол.
о-ва. - Киев: Наук, думка, 1984а. - Ч. 2. - С. 296.
189
Харито нов А.Ю. Отряд Odonata — стрекозы. Красная книга СССР //
Редкие и находящиеся под угрозой исчезновения виды животных и растений.-
М.: Лесн. пром-сть, 19846. - Т. 1. - С. 217-224.
Харитонов А.Ю. Отряд Odonata - стрекозы. Определитель насекомых
Дальнего Востока СССР. - Л.: Наука. Ленингр. отд-ние, 1986. - Т. 1. -
С. 142-162.
Харитонов А.Ю., Белышев Б.Ф. Особенности географического распростране-
ния рода Enallagma Charp., 1840 (insecta, Odonata) // Пробле-
мы зоогеографии и истории фауны. - Новосибирск: Наука. Сиб. отд-
ние, 1980. - С. 80-84. - (Тр. ин-та /Биол. ин-т СО АН СССР; Вып. 40).
Харитонов А.Ю., Борисов С.Н. Diplacodes Kirby - новый для фауны
СССР род стрекоз (Odonata, Libellulidae ) // Энтомол. обозрение,-
1981. - Т. 60, № 3. - С. 604-606.
Харитонов AJO., Борисов СЛ. Trithemis Brauer - новый для фауны
СССР род стрекоз (Odonata, Libellulidae ) // Докл. АН ТаджССР. -
1985. - Т. 28, № 8. - С. 482.
Харитонов А.Ю., Харитонова И.Н. Личинка G-omphus epophthalmus
Selys (Odonata, Insecta) // Гельминты, клещи и насекомые. - Ново-
сибирск: Наука. Сиб. отд-ние, 1982. - С. 20-22. - (Новые и малоизвест-
ные виды фауны Сибири).
Харитонов А.Ю., Харитонова И.Н. Опыт использования садков для оценки
смертности насекомых под воздействием инсектицидных аэрозолей Н Опти-
мизация технологии применения инсектицидных аэрозолей. - Новосибирск,
1983. - С. 63-73.
Харитонова ИЛ. Первый случай обнаружения гинандроморфной равнокрылой
стрекозы // Экология и география членистоногих Сибири. - Новосибирск:
Наука. Сиб. отд-ние, 1987. - С. 119-120.
Хорхордин Е.Г. Интенсивность хищничества личинок Aeschna affinis (©do-
nate, Aeschnidae) разных возрастов в зависимости от стадии развития
жертв и величины экспериментального пространства И Зоол. журн. - 1985. -
Т. 64, вып. 9. - С. 1426-1428.
Цветков ВЛ. К фауне грегарин насекомых Петергофа и окрестностей Н Тр.
ин-та / Петергофский естеств.-науч. ин-т. - 1929. - № 6. - С. 155-198.
Цвилинева ВЛ. Брюшной мозг Aeschna (по материалам акад, АЛ. За-
варзина). (Сообщение I. Строение ганглиев грудного отдела) // Изв. АН СССР.
Сер. биол. - 1950. - № 2. - С. 91-128.
Цвилинева В.А. Брюшной мозг Aeschna (по материалам акад. А.А. За-
варзина). (Сообщение П. Строение ганглиев брюшного отдела) // Там же. -
1951. - № 2. - С. 66-116.
Чернышев ВЛ. Поведение животных и циркадные ритмы // Журн. общ. биоло-
гии. - 1973. - Т. 34, № 2. - С. 284-293.
Чернышев ВЛ. Взаимосвязь суточных ритмов активностей насекомых // Тр. /
Всесоюэ. энтомол. о-во. - 1981. - Т. 63. - С. 159-162.
Чернышев В.Б. Суточные ритмы активности насекомых. - М.: Изд-во МГУ,
1984. - 218 с.
Чибиченко Н.Т. Водные беспозвоночные животные Северной Киргизии как
промежуточные хозяева гельминтов птиц И- Гельминты животных Киргизии
и сопредельных территорий. - Фрунзе: Илим, 1966. - С. 66-75.
Чибиченко Н.Т. Гельминтологическая характеристика обследованных водоемов
в связи с перспективами использования их для разведения птиц // Гельминты
животных и растений Киргизии. - Фрунзе: Илим, 1968. - С. 99-100.
Чибиченко Н.Т., Крылова В.Н. Стрекозы водоемов Иссык-Кульской котло-
вины и их зараженность паразитами // Там же. - С. 111-118.
Чурина Н.В. Гельминты водоплавающих птиц Среднего Урала и гельминтологи-
ческое обследование водоемов: Автореф. дис. ... канд. биол. наук. - М.,
1964. - 17 с.
190
Шалопенок Е.С. Фауна водных насекомых литорали озера Нарочь // Биологи-
ческие основы рыбного хозяйства на внутренних водоемах Прибалтики. -
Минск: Наука и техника, 1963. - С. 196-201.
Шалопенок Е.С., Камлюк Л.В. Размерно-весовая характеристика личинок не-
которых видов стрекоз // Биол. науки. - 1975. - № 9. - С. 128-131.
Шалопенок Е.С., Павельева Е£. Роль личинок стрекоз в трансформации
энергии в литоральной эоне оз.' Нарочь // Вести. Белорус, ун-та. Сер. П. —
1973. - Вып. 1. - С. 41-45.
Шарова ИЛ. Жизненные формы и значение конвергенций и параллелизмов в их
классификации // Журн. общ. биологии. - 1973. - Т. 34, № 4. -
С. 563-570.
Шарова ИЛ. Жизненные формы личинок жужелиц ( Coleopters, Carabidae)
и их эволюционные взаимоотношения П Эволюционная морфология личинок на-
секомых. - М.: Наука, 1976. - С. 56-80.
Шарова И.Х. Жизненные формы жужелиц. - М.: Наука, 1981. - 340 с.
Шарпило Л.Д., Шарпило В.П, Отношение некоторых форм рода Plagior-»
chis Luhe, 1909, паразитирующих у рептилий и грызунов, к виду
Plagiorchis elegans (Rud., 1802); Trematoda, Flagiorchidae//
Пробл. паразитологии. - Киев: Наук, думка, 1972. - С. 425-42 7.
Шванвич Б.Н. Курс общей энтомологии. - М.; Л.: Сов. наука, 1949. - 900 с.
Шевченко Н.Н. О личинках гельминтов у водных насекомых реки и пойменных
водоемов Северского Донца // Докл. АН СССР. - 1962. - Т. 142, № 4. -
С. 972-975. .
Шевченко Н.Н. Динамика инвазии водных беспозвоночных Северского Донца
гельминтами в разные годы // И Всесоюз. симпозиум по болезням и парази-
там водных беспозвоночных (Тез. докл.). - Л.: Наука. Ленингр. отд-ние,
1976. - С. 73.
Шевченко Н.Н., Барабашова В.М. Водщ членистоног! як прокйжн! хазя!
гельм1нт1в // Допов1д! АН УРСР. - 1960. - № 11. - С. 1555-155 9.
Шевченко Н.Н., Вергун Г.И. О жизненном цикле трематоды амфибий Pro—
sotocus confusus (Looss, 1894) Looss, 1899 // Helmintholo-
gies. - 1961. - T. 3, вып. 1-4. - C. 294-298.
Шенгелия Е.С, Стрекозы (Odonata) Грузии ll Тр. ин-та / Ин-т зоологии
АН ГЮСР. - 1953. - Т. 11. - С. 147-165.
Шенгелия Е.С. Стрекозы (Odonata) в фауне высокогорья Большого Кав-
каза в Грузии // Фауна высокогорья Большого Кавказа в пределах Гру-
зии. - Тбилиси, 1964. - С. 15-19.
Шенгелия Е.С. Стрекозы - Odonata // Фауна беспозвоночных Триапетско-
го хребта. - Тбилиси, 1966.
Шерстюк В.В. Роль хищников в поедании икры и мзлодн рыб на нерестилищах
Кременчугского водохранилища: Автореф. дис. ... каид. биол. наук. - Киев,
1967. - 20 с.
Шорыгнн А, К познанию рода Sympycna Charpentier, 1840И Рус. эн-
томзл. обозрение. - 1926. - Т. 20, № 1/2. - С. 57-63.
Штейн Г.А. Материалы по паразитологии водных членистоногих некоторых озер
Карелии // Учен. зап. Петрозавод. ун-та. Сер. биол. - 1957. - Т. 8,
вып. 3. - С. 120-139.
Штейн ГЛ. Грегарины водных членистоногих Карельских озер // Зоол. журн. -
1960. - Т. 39, вып. 8. - С. 1135-1144.
Якобсон Г.Г., Бианки В.П. Прямокрылые и ложносетчатокрылые Российской
империи и сопредельных стран. - Спб., 1905. - 836 с.
Янковская А.И. По водоемам Памира // Природа. - 1950. - № 2. - С. 46-49
Янковская А.И. Фауна теплых родников Восточного Памира // Фаунистика и
экология животных. - М.; Л.: Наука, 1965. - Т. 35. - С. 43-56.
191
Ярошевский А. Перечень сетчатокрылых насекомых (Neuroptera ),
встречающихся в Харьковской губернии // Тр. / О-во испытателей природы
Харьков, ун-та. - 1881. - Т. 15. - С. 95-99.
Akramowski N.N., S'che ngelia E.S. Neue Angaben ilber
Cordulegaster mzymtae Barteneff, 1930 (Odonata, Cordulegaste-
ridae) // Dtsch. entomoL Z. (N.F.). — 1967. — Bd 14, N 3/4. —
S. 313-321.
Akramowski N.N., Zhiltzova L.A. Obituary. Ariadna Niko-
laevna Popova /I Odonatologica. - 1973. - Vol. 2, N 1. — P. 45—47.
Alcock J. Multiple mating Calopteryx maculata (Odonata; Calop-
terigidae) and the adoantage of non-contact guarding by ma-
les // J. Natur,' Hist. - 1979. - Vol. 13, N 4. - P. 439-446.
A n d e r K. Einige Odonata aus Kasmir und Ostturkestan Ц
Kungl. Fysiogr. Sbllsk. F6rh. - 1944. - N 14.. — P. 168-178.
Andrew C.Gr. Sexual recognition in adult Erythemia simplicicol—
lis (Odonata) // Ohio J. Sci. — 1966. — Vol.' 66. — P. 613—617.
Arnett D.W. Receptive Field Organization of Units in the First
optic ganglion of Diptera // Science. - 1971. — Vol. 173,
N 4000. - P. 929-931.
Asahina S. Odonata of Sachalin. Transactions Kansai // En—
tom. Soc. — Osaka, 1949. — Vol. 14, pt 2. — P. 23—32.
Asahina S. A morphological study of a relic dragonfly Epio-
phlebia superstes Selys (Odonata, Anisoptera) // The Japan Soc.
for promotion of Sci. — Tokyo, 1954. — 153 P.
Asahina S. On the pterotoracic interpleural suture of Mesozoic
and recent Odonata // J. Fac. Sci. Hokkaido Univ., 1957. —
Ser. VI, ZooL 13. - P. 1-7. :
Asahina S. A revision of the Odonata of the Kurile islands //
Insecta Matsumur, — Tokyo, 1958. — Vol. 22, N 3/4. — P. 63—70.
Asahina S. Odonata Taken by Japanese Expeditions to Kara-
korum, Afghanistan, Iran and Pakistan // PubL Kyoto Univ. —
Kyoto, 1963. - P. 45-50.
Asahina S. The Afghan Odonata taken by Dr. Josii in 1960 //
Ibid. - 1966. - P. 159-163.
Asahina S. The Odonata of Iraq Ц Japan J. Zool., 1973. —
Vol. 17, N 1. - P. 17-36.
Asahina S. A new and some known species of Odonata from
Kashmir // Senckenberg. biol. - 1978. - Vol. 59 (1/2) . - P. 115-120.
Baker R.L. Use of space in relation to feeding areas by Zy—
gopteran nymphs in captivity // Canad. J. Zool. — 1980. —
Vol. 58, N 6. - P. 1060-106 5.
Baker R. L. Behavioural interactions and use of feeding areas
by nymphs of Coenagrion resolutum (Coenagrionidae; Odo—
nata) // Oecologia. - 1981. - Vol. 49. -P. 353-358.
Baker R.L. Spacing behaviour by larval Ischnura cervila Se-
lys: effects of hunger, previous interactions and familarity with
an area (Zygoptera: Coenagrionidae) Ц Odonatologica. — 1983,—
Vol. 12, N 3. - P. 201-207.
В a 1 d u s K. Experimented Untersuchung ilber die Entfernungs—
lokalisation der Libellen (Aeschna cyanea) // Z. vergleichen—
de Physiol - 1926. - Bd 3, N 4. - P. 475-505.
Bartenef A.N. Date relating to Siberian Dragonflies // Zool.
Anz. - 1910. - Bd 35, N 9/10. - P. 270-278.
Bartenef A.N. Contributions to the knowledge of the Species
of the genus Sympycna Charpentier 1840 and of their Subdivi-
sions j/ Ежегод. Зоол, музея АН. - 1912а. - № 17. - С. 144-164.
Bartenef A.N. Materialien zur Odonatenfauna Sibiriens // ZooL
Jahrb. System, geogr. und baologic. Tiere. - 19126. — Bd 32,
H. 3. - S. 221-284.
Bartenef A.N, Die Bestimmungs—tabell der Gattungen der Un—
terfamilie Libellulidae (Odonata, Fam. Libellulidae) // Zool. Jahrb.
Abt. System. - 1929a. - Bd 56, H. 5/6. - S. 357-524.
Bartenef A.N. Testudo ibera Pall, in Pitzunda, Kaukasus, Ufer
des Schwarzen Meeres Ц Zool. Anz. — 19296. — Bd 81, N 1—4.—
S. 94-95.
Bartenef A.N. New Arten und Varetaten der Odonata des
West—Kaukasus // Ibid. — 1929b. — Bd 85, N 3/4. — S. 54—68.
Bartenef A.N. Uber Calopteryx splendens (Odonata) und ihre
Biotypen, besonders die westasiatischen // Zool. Jahrb. Abt.
System. — 1930a. — Bd 58, H. 5/6. — S. 521—540.
Bartenef A.N. Uber die Aberrationen von Libellula quadrimacu—
lata L. (Odonata) // Zool. Anz. — 19306. — Bd 87, N 7/8. —
S. 191-198.
Bartenef A.N. Uber eine kleine Odonatensammlung aus Japan
and Nordchina // Ibid. - 1930b. - Bd 88, N 11/12. - S. 326-329.
Bartenef A.N. Zur Systematic der palaarktischenen Aeschna—
Arten (Odonata, Aeschninae) // Ibid. — 1930г. — Bd 89, N 1/2,—
S. 39-56.
Bartenef A.N. Noch einmal uber die Artehgruppe Aeschna jun-
cea (Odonata, Aeschninae) in der Palaearktik // Ibid. — 1930д. -
Bd 89, N 7-10. - S. 229-245.
Bartenef A.N. Altaigomphus heterostylus g. n., sp. n. (Odona-
ta, Gomphidae) aus dem Slid—Altaigebirge // Ibid. — 1930e. —
Bd 89, N 11/12. - S. 327-331.
Bartenef A.N. Sind Orthetrum anceps Schn. und Orthetrum
coerulescens Fabr. (Odonata, Libellulinae) selbstandige Arten//
Ibid. - 1930 ж. — Bd 91, N 1-4. - S. 6 7-71.
Bartenef A.-N. Uber das Artenpaar Orthetrum albistylum Selys
und Orthetrum cancellatum L. (Odonata, Libellulinae) // Ibid. —
1930a. - Bd 92, N 3/4. - C. 79-89.
Bartenef A.N. Uber die geographische Verbreitung von Fla—
vescens Fabr. (Odonata) // Zool. Jahrb. Abt. System. — 1931a.».
Bd 60, H. 5/6. - S. 471-488.
Bartenef A.N. Die geographisch—biologische Charakteristik
und die Artenpaarilngen der Gattung Sympetrum Newn, 1883
(Odonata) // Ibid. - 19316. - Bd 61. - S. 347-360.
Bartenef A.N. Uber einige Tupen von Arten—und Rassenkrei—
sen in der Palaarktik // Zool. Anz. — 1932a. — Bd 97, N 7, —
S. 236-238.
Bartenef A.N. Uber die Tageskurven des Fluges von Aesch-
na juncea L. (Odonata) an Hochgebirgsgewassern // Ibid. —
19326,- Bd 98, N 3/4. - S. 91-94.
Bartenef A.N. Tibet die Veranderung der Libellenfauna Von
Krasnodar ( Nor.dkaukasus) im Lanf von 25 Jahren // Ibid. —
1932b. - Bd 98, N 5/6. - S. 128-131.
Bartenef A.N. Uber die Verschiedenheit der Verbreitungagren—
zen der Odonatenarten in der Palaarktik nach Norden // Ibid.—
1932г. - Bd 98, N 9/10. - S. 267-271.
Bartenef A.N. Ubersicht der Leucorrhinia (Britt., 1850). Arten
und Varietaten unter geographischen, biologischen und morpho—
13 Зак. 611
193
logischen Gesichtspunkten Ц Arch. Naturgesech. — 1933a. —
Bd 2, N 1. - S. 1-53.
Bartenef A.N. Zur Frage der Migrationstypen von Tieren in
Vorderasien // Zool. Anz. — 19336. — Bd 103, N 9/10. —
S. 235-244.
Bartenev A.N. Uber einige Grundlagen der Geschichten und
des Bestanddes der palaarktischen Fauna // Arch. Naturgesch.
(N.F.). - 1934. - Bd 3. - S. 158-181.
Bartenef A.N. Uber die Gattung Libellula und Besonders uber
ihre palaarktischen Arten // Ibid. — 1935. — Bd 4. - S. 2 74— 290.
Bartenef A.N. Uber Verschwinden der Odonaten in Herbst //
Entomol. tidskr. — 1937. — Bd 58. — S. 66—74.
В e 1 к e G. Esquisse de 1’histoir naturelle de Kamenetz—Podolski,
precedee d'un cop—d’ oeil sur les travaux der naturalistes de
povinces occidentales de la Russie et du Rayame de Polonge
an XIX circle // Bui. Soc, Natur. Moscou. — 1859. — P. 64—66.
В e 1 к e G. Notice sur I'hirtoire naturelle du district de Radomysl
(Gouv. de Kief). 2. Catalogue des animaux qui habitent le di-
strict de Radomysl // Ibid. — 1866. — P. 510-511.
Belyshev B.F. Altaigomphus heterostylus Bart. (Odonata, In-
sects) // Ann. zool. — Warszawa, 1957. — T. 16, N 25. —
S. 475-486.
Belyshev B.F. Die Larve von Agrion hylas Trybom (Odona-
ta, Agrionidae) // Dtsch, entomol. Z., 1957. - Bd 4, N 3/4. -
S. 191-192.
Belyshev B.F. Die Verbreitung Odonata in Nordasien // Ibid.-
1958. - Bd 5, N 1/2. - S. 79-85.
Belyshev B.F. Die Larve von Somatochlora graeseri Selys
(Odonata, Corduliinae) // Entomol. Z,, 1959. — Bd 69, N 18.—
S. 200-202.
В elyshev B.F. Aeschna septentrionalis Burm. — eine fur Eura-
sien neue Libellenart (Odonata, Insecta) // Mitt. Zool. Muse-
um Univ. - Berlin, 1966. - Bd 42, N 1. - S. 153-154.
Belyshev B.F, Biologische Gesetzmassigkaiten der aurasiati—
schen Libellen fauna in borealen Bereich (Odonata) // Dtsch.
entomol. Z. - 1968. - Bd 15, N 4/5. - S. 317-320.
Belyshev B.F. Ozeanische Libellen // Ibid. — 1968. - Bd 15,
N 1/3. - S. 9-13.
В elyshev B.F. To the Participants in the Meeting to Europe-
an Odonatologists. Abstracts of papers read at the First Euro-
pean Symposium on Odonatology // Genetica. - 1971. —P. 10—11.
Belyshev B.F. Individual and geographic variation in Coenag-
rion glaciale (Selys, 1872), with description of a new subspe-
cies (Zygoptera: Coenagrionidae) // Odonatologica. - 1973. -
Vol. 11, N 2. - P. 99-103.
Belyshev B.F., К i a u t a B. To Nicolai Nicolaevich Akramow—
ski on his 70th birthday // Ibid. — 1976. — Vol. 4. —P. 309—312.
Belyshev B.F., Haritonov A. Yu. The distribution of the
Cosmopolitan Genus Anax Leach and its probable origin (Ani—
soptera: Aeschnidae) // Ibid. — 1978. — Vol. 7, N 2. - P. 115-
121.
Belyshev B.F., Haritonov A. Yu. On the reasons for a
sharp Curve in the Western boundary of ranges of some
eastern dragonfly species in the north of Western Sibirica //
Ibid. - 1980. - Vol. 9, N 4. - P. 317-319.
194
Bergroth E. Zur geographischen Verbreitund einiger Odona-
ten // Entomol. Nachr. — 1881. — Bd 7. _ S. 85-88.
Berzins B. Beitrag zur Kenntnis der Odonatenfauna Lettlands.//
Folia zool. hydrobioL - 1942, Bd 11, N 2. - S. 329-350.
Bishop L.G., К e e h n D.G. Two types of neurones sensitive
to motion in the optic lobe of the fly // Nature. - 1966. —
Vol. 212, N 5068. - P. 1374-1376.
Blois C. The larval diet of three anisopteran (Odonata) Spe-
cies // Freshwater BioL — 1985. — Vol. 15, N 4. — P. 505—514.
Bruttan A. Die Odonaten Liv— und Ehstlands. // Sitz.—Ber. Na—
turf. Ges. Dorpat — 1878. - Bd 4, N 3. — S. 420—426.
Bruttan A. Nachtrag ilber Odonaten Liv— und Ehstlands //Ibid.—
1881a. - Bd 5, N 1. - S. 114-115.
Bruttan A. Zwei fiir das Gebiet neue Odonaten // Ibid. —
18816. - Bd 5, N 3. - S. 379.
Buchholtz C. Untersuchungen an der Libellengattung Calop—
teryx Leach, unter besonderer Berilcksichtigung ethologischer
Fragen // Z. TierpsychoL — 1951. — Bd 8, N 2. — S. 273—293.
Buchholtz C. Eine Analyse des Paarungsverhaltens und der
dabei wirkenden Ausldser bei den Libellen Platycnemis penni—
pes Pall, und Pl. dealbata Klug // Ibid. — 1956. — Bd 13,
N 1. - S. 13-25.
В u 11 e r T., Peterson J.E., Corbet Ph. S. An exceptional-
ly early and informive arrival of adult Anax junius in Ontario
(Odonata: Aeshnidae) // Canad. entomologist. — 1975. — Vol.
107, N 12. - P. 1253-1254.
C a i 11 e r L. Ontogenese du corr.portement de capture chez la
larve d'Agrion (Calopteryx Auct) splendens Harris (Odonato—
pteres) // Behaviour. — 1974, — Vol. 51, N 3/4. - P. 166—194,
Campanella P.J., Wolf L.L. Temporal Lakes as a mating
system in a temperate zone dragonfly (Odonata; Anisoptera)
I. Plathemis lydia (Drury) // Ibid. — 1974. — Vol. 51, N 1/2,-
P. 49-87.
C a r f i S., C e n t i n a P., T e r z a n i F« Contributo alia conos—
cenza della odonatofauna dell'Afghanistan // Redia. — 1982. -
Vol. 65. - P. 207-214.
Carle F.L. The wing vein homologies and phylogeny of the
Odonata: a continuing debate // S.LO. Rapid Communications. —
1982a. - N 4. - P. 1-66.
Carle F.L. Evolution of the Odohate copulatory process //
Odonatologica. — 19826. — Vol. 11, N 4. — P. 271—286.
Carpenter F.M. The Lower Permian insects of Kansas.
Pt 2: The orders Palaeodictyoptera, Protodonata and Odona-
ta // Amer. J. Sci. - 1931. - Vol. 21, N 5., - P. 97-139.
Carpenter F.M. Geological history and evolution of the in-
sects // Proc. 15 Int. Congr. Entomol. — 1977. — P, 63—70.
Cederhjelm J. Faunae Ingricae Prodromus. — Lipsiae, 1798. -
348 p.
Clement S.L., Meyer R.P. Adult biology and behaviour of
the dragonfly Tanypteryx hageni (Odonata: Petaluridae) //
J. Kansas Entomol. Soc. — 1980. — Vol. 53, N 4. — P. 711—719.
Cloarec A. Alimatation de larves d'Anax imperator Leach dan-
sum milien (Anisoptera: Aeschnidae) // Odonatologica. — 1977. —
Vol. 6, N 4. - P. 227-243.
C ollett T.S., King A.J. Visual during fliegt // Compaund Eya
and Vision Insects. — Oxford. — 1975. — P. 434—466.
195
Corbet Ph.S. Environmental factors influencing the induction
and termination of diapause in the emperor dragonfly, Anax im—
perator Leach (Odonata: Aeschnidae) // J. Exptl Biol. — 1956,—
Vol. 33, N 1. — P. 1-14.
Corbet Ph.S. The Life—History of the Emperor Dragonfly Anax
imperator Leach (Odonata, Aeschnidae) // J. Animal EcoL —
1957. - Vol. 26, N 1. - P. 1-68.
Corbet Ph.S. A Biology of Dragonflies. — L., 1962. — 247 p.
Corbet Ph.S, Biology of Odonata // Ann. Rev. EntomoL —
1980. - Vol. 25. - P. 189-217.
Cothran M.L., Thorp J.H. Test of prey preference and swi-
tching behaviour of the dragonfly Celithemis fasciata // Oikos. —
1985. - Vol. 44. - P. 350-355.
Crowley P.H. Behaviour of zygopteran nymphs in a simulated
weed bed // Odonatologica. —1979. — VoL 8, N 1. — P. 91—101.
Davies D., Tobin P. The dragonflies of the world; A sy-
stematic list of the extant species of Odonata. — Utrecht, 1984,—
Vol; 1. Zygoptera, Anisozygoptera. — 127 p. — (Soc. Inter. Odo—
natologica Rapid Communications (Supplements); No. 3).
Djakonov A.M. Drei neue Odonaten — Arten aus dem Paia—
arctischen Faunengebiet // Рус, энтомол. обозрение. - 1926. - Bd 20,
N 3/4. - S. 2 28-234.
Doerksen G.P. Notes on the reproductive behaviour of Enal—
lagma cyathigerum (Charpentier) (Zygoptera: Coenagrionidae) //
Ibid. - 1980. - Vol. 9, N 4. - P. 293-296.
Dreyer W. Etho — bkologische Untersuchungen an Lestes viri—
dis (v. d. Lenden) (Zygoptera: Lestidae) // Ibid. - 1978. -
Vol. 7, N 4. - P. 309-322.
Dumont H.J. Jschnura intermedia spec. nov. from Turkey, and
its relations to J. forcipata Morton, 1907 and J. pumilio ( Char-
pentier, 1825) (Anisoptera: Coenagrionidae) // Odonatologica.—
1974. - Vol. 3, N 3. - P. 153-165.
Dumont H.J. A note on some dragonflies from Afghanistan //
Ibid. - 1975. - Vol. 4, N 4. - P. 234-248.
Dwigubsky L Primitiae faunae Mosquensis. — M., 1802. —
215 p.
Dvorak D.R., Bishop L.G., E с к e r t H.E. On the identifica-
tion of movement detectors in the fly optic lobe // J. Compar.
Physiol. - 1975. - VoL 100, N 1. - P. 5-23.
Dziedzielewicz T. Wazki Galicyi i prryleglych krajiw pol—
skich (Odonata Haliciae religuarumque provinciarum Poloniae)//
Muzeum Dzieduszyckich. — 1902. — N 5. — S. 175.
Eckert H.E. Response properties of dipteran giant visual inter-
neurones involved in control of optomotor behaviour // Nature.—
1978. - Vol. 271, N 5643. - P. 358-360.
Edman J.D., H a e g e r J.S. Dragonflies attracted to and selec—
tivly feeding on mosquitoes // Fla Ent — 1974. — VoL 57, N 4.—
P. 408.
El Amin el Rayah. Dragonfly nymphs as activ predators of
mosquito larvae // Mosquito News. — 1975. — Vol. 35, N 2. —
P. 229-230.
El Amin el Rayah, Abu Shama F.T. The predatory ca—
pasity of nymphs of the dragonflies: Trithemis annulata scorte—
cii Nielsen (Odonata: Anisoptera) and Ischnura senegalensis
Rambur (Odonata: Zygoptera) // Rev. zooL et bob afric, —
1977. - Vol. 91, N 2. - P. 381-384.
196
Etienne A.S. Analyse der schlagauslosenden Bewegunspara-
meter einer punktformigen Beuteattrappe bei der Aeschnalarve //
Z. vergleichende Physiol. — 1969. — Vol. 64, N 1. — S. 71—110.
Etienne A.S. Energy versus time dependent parameters in the
determination of a behavioural sequens in the Aeschna larva //
J. compar. and physiol. — 1978. — Vol. 127, N 1. — P. 89-96,
Eve r s m a n n E. Libellulae Wolgam fluvium inter et montes Ura—
lenses observatae et desciptae spec. nov. // Bub Soc. Natur.
Mosqou. — 1836. — P. 233—248.
Eversmann E. Insecta Wolgam fluvium inter et montes Uralen—
ses observata // Ibid. — Mosqou. — 1837. — P. 33—39.
E z e n w a A.O. Ecology of Simuliidae, Mermithidae and Microspo—
rida in Newfoundland freshwater // Canad. J. Zool. — 1974. —
Vol, 52, N 5. - P. 557-565.
Fischer J.B. Versuch einer Naturgeschichte von Livland. —
Leipzig, 1778. - 390 S. (Odonata: S. 155-156).
Fischer J.B. Zusatze zu "Versuch einer Naturgeschichte von
Livland" // J.J. Ferbers Anmerkungen zur physischen Erdbesch—
reibung von Kurland, nebst J.B. Fischers Zusatzen zu seinem
Versuch einer Naturgeschichte von Livland. — Riga, 1784. —
305 P. (Odonata; S. 84—85).
Fischer J.B. Versuch einer Naturgeschichte von Livland. —
Zweite Aufl. — Ktinigsberg, 1791. — 826 S. (Odonata:
S. 335-337).
Folsom T.C., Collins N.C. The diet and foraging behaviour
of the larval dragonfly Anax junius (Aeschnidae), with an as-
sessment of the role of refuges and prey activity // Oikos. —
1984. - Vol. 42. - P. 105-113.
Fraser F.S. A reclassification of the order Odonata // Roy. Zo-
ol. Soc. N.S.W. — Sydney, 1957. — 133 p.
G a f f г о n M. Untersuchungen ilber das Bewegungssehen bei Li—
bellenlarven, Eleigen und Fischen // Z. vergleichende PhysioL —
1934. - Vol. 20, N 3. - S. 299-337.
G i 11 a s p у J.E. A spectacula case of termite predation by dra-
gonflies // Taius. - 1971. - Vol. 4, N 1. - P. 87-88.
Hagen H. Odonates de la Russie. Etud Entom. Red. Motschul—
sky, V. Helsingfors, 1856. — P. 5 2—5 9.
Hagen H. Beitrag zur Odonaten — Fauna des Russischen
Reichs. - Stettind, 1858.- Bd 19. - P. 96-101.
Hagen H.A. Die Neuroptera des Lithographischen Schiefers in
Bayern // Cassel. — 1866. — VoL 15. — S. 1—40.
Handlirsch A. Die fossilen Insecten und die Phylogenie der
rezenten Formen. — Leipzig, 1906-1908. — Bd 1—3.
Hartleine H.K., Wagner H.G., Ratliff F. Enhibition in the
eye of Limulus Ц J. Gen. Physiol. - 1956. - N 39. - P. 651.
Hassan A.T. Reproductive behaviour of Acisoma panoproides
inflatum Selys (Anisoptera: Libellulidae) // Odonatologica. —
1978. - Vol. 7, N 3. - P. 237-245.
Heinrich B., Casey T.M. Heat transfer in dragonflies: "fli-
ers" and "perchers" // J. Exptl. Biol. - 1978. - VoL 74. -
P. 17-36.
Hennig W. Die Stammesgeschichte der Insekten. — Frankfurt:
Shenckenberg—Buch, 1969.
H e у m e r A. Fortpflanzungsverhalten und Territorialitat bei Ort—
hetrum coerulescens (Fabr., 1798) und O. brunneum (Fonsc.,
197
183?) (Odonata, Anizoptera) // Rev. Comp. Animal. — 1969. —
Vol. 3, N 2. - S. 1-24.
Heymer A. Etude du comportement reproductur et analyse des
mec^nismes dedencheurs innes (MDl) optiques chez les Ca—
lopterygidae (Odonata, Zygoptera) // Ann. Soc. entomol. — 1973.—
Vol. 9, N 1. - P. 219-255.
Higashi K. Estimation of the food consumption for some speci-
es of dragonflies. 1. Estimation by observation for the frequen—
sy of feeding flights of dragonflies // Reports Ebino BioL Lab.
Kyushu Univ. - 1973. - Vol. 1. - P. 119-129.
Higashi K., Ueda T. Territoriality and movement pattern in a
population of Calopteryx Cornelia (Selys) (Zygoptera: Calopte—
rygidae) // Odonatologica. — 1982. — Vol. 11, N 2. — P. 129-
137.
Hilton D.F.J. Territoriality in Libellula julia Uhler (Anisoptera:
Libellulidae) // Ibid. - 1983. - Vol. 12, N 2. - P. 115-124.
Hinnenkint B.O. Thermal pollution as a probable cause of
a winter ecdysis of Aeschna cyanea (Muller) (Anisoptera:
Aeshnidae) // Ibid. - 1972. - Vol. 1, N 3. - P. 163-164.
Hoppenheit M. Beobachtungen zum Beutefangverhalten der
Larve von Aeschna cyanea Mull. (Odonata) // Zool. Anz. —
1964. - Vol. 172, N 3. - S. 216-232.
Horridge G.A. Unit studies of the retina of dragonflies // Z.
vergleichende Physiol. — 1969. — Vol. 62, N 1. — P. 1—37.
lakowleff V. Materialen zur entomologischen Fauna der Wol—
ga—Gegend. 2. Verzeichnis der Neuropteren der Wolga—Gegend//
Тр. / Рус. энтомол. о—во. - 1870. - T. 6. - C. 121-122.
Ito V. Territorial!sm and residentalyty in a dragonfly Orthetrum
albistylum speciosum Uhler // Ann. Entom. Soc. America. —
1960. - Vol. 53, N 6. - P. 851-853.
Jacobs M.E. Studies on territorialism and sexual selection in
dragonflies // Ecology. — 1955. — V. 36, N 1—4. — P. 566—586.
Johnson C. Breeding behaviour and oviposition in Hetaerina
americana (Fabricius) and H. titia (Drury) (Odonata: Agrii-
dae) // Canad. entomologist. — 1961. — Vol. 93, N 4. —
P. 260-266.
Johnson C. A study of territoriality and breeding behaviour
in Pachydiplax longipennis Burm. (Odonata: Libellulidae) //
Southwest. Naturalist. — 1962. — N 7. — P. 191—197.
Jonson D.M. Predation by damselfly naiads on cladoceran po-
pulation fluctuating intensity // Ecology. — 1973. — Vol. 54,
N 2. - P. 251-268.
Juris A. Erganzende Bemerkungen zur estlandischen Fauna
der Odonata // Tartu Ulik. juures oleva Loodusuurijate seltsi
aruannetest - 1938. — Vol. 43, fasc. 3/4. — P. 297—306.
К a r a m a n B.S. Contribution a la connaissance de 1'ecologie
des larves d'Odonates dans I'ecosysteme du lac de Doiran //
Ann. Fac. BioL Univ. Scopje. — 1979. — Vol. 32. — P. 197—205.
К a w a 1 1 J.H. Die Orthopteren und Neuropteren Kurlands //Korr.—
Blatt Naturf.—Ver. — Riga, 1864. — Bd 14, H. 11. — S. 155—168.
Kauri H. Die Odonaten—Fauna von Estland // A pophoreta Tar—
tuensia. — Stockholm, 1949.
Kjellberg G. Autecological studies of Lephophlebia vesperti-
ne (Ephemeroptera) in a small forest pond from 1966—1968 //
Entomol. tidskr. - 1972. - Vol. 93, N 1-3. - P. 130-133.
198
К la pal ek FY. Neuropteren in: Graf E. Zichy, Dritte Asiatische
Fordchungereise Ц Zool. Ergeben. — Budapest, 1901. — Bd 2. —
P. 203-231. '
К olenati F.A. Insecta Caucasi 11 Meletemata Entomologica. —
Petropoli, 1846. - Vol. 5. - P. 113-115.
Kono H., Tamanuki K. Insecten — Ausbeute aus Nord—Sa—
chalin I) Insecta Matsumur, — Tokyo, 19 28. — Vol. 2. —
P. 128-129.
Krichevskii R.E., R у a b k о Ya., Haritonov A.Yu. Opti-
mal key for taxons ordered in accordance with their frequenci-
es // Appl. Mathematics. — 1981. — Vol. 3. — P. 67—72.
Krieger F., К r i e g e r-L о i b 1 E. Beitrage zum Verhalten
von Ischnura elegans und Ischnura pumilio (Odonata) // Z. Ti—
erpsychol. - 195.8. - VoL 15, N 1. - S. 82-93.
К run er U. Revier— und Fortpflanzungsverhalten von Orthetrum
cancellatum (Linnaeus) (Anisoptera: Libellulidae) // Odonatilo—
gica. - 1977. - Vol. 6, N 4. - P. 263-270.
К u k a 1 о v a-P e c k J. Origin of the insect wing and wing arti-
culation from the arthropodan leg // Canad. J. Zool. — 1983,—
Vol. 61, N 7. - P. . 1618-1669.
Kumar A. The phenology of dragonflies in the Dahra Dun Val-
ley, India // Odonatologica. — 1972. — VoL 1, N 4. — P. 199-
20 7.
Kumar A. On the occurrence of multivoltine generations in so-
ma Indian dragonflies // Sci. and Culture. — 1979. — VoL 45,
N 3. - P. 126-127.
Kumar A., Prasad M. Reproductive behaviour in Neuroba-
sis chinensis chinensis (Linnaeus). (Zygoptera: Calopterygi—
dae) // Odonatologica. — 1977. — VoL 6, N 3. — P. 163—177.
L a r i d M. Ed. Tsetse. The future for biological methods in integ-
rated controL — Ottawa: InL Develop. Res. Cent, 1977. — 220 p.
Larsen T.E. Butterflies as prey for Orthetrum austenti (Kirby)
(Anisoptera: Libellulidae) // Notulae Odonatologicae. — 1981. —
VoL 8, N 1. - P. 130-133.
Lavovie — Dornik J., Pilon J.—G., A 1 i М,—A., M о u z e M.
Revue critique la vision chez les ©donates: ^lectrophysiologie//
Rev. Canad. Biol. — 1981. — VoL 40, N 3. —P. 287—304.
Laughlin S.E. Neural integration in the first optic neuropile of
dragonflies. 1. Bignala amplification in dark—adapted second or-
der neurons // J. Compar. PhysioL — 19 73. — VoL 84, N 4. —
P. 335-356.
Leggott M., Pritchard G. The effect of temperature on ra-
te of egg and larval development in populations of Argia vivi—
da Hagen (Odonata: Coenagrionidae) from habitats with diffe-
rent thermal regimes // Canad. J. Zool. — 1985. — VoL 63,
N 11. - P. 2578-2582.
Lomnicki M. Sprawozdanie z wycieczki zoologiczej podbytej
na Podolu w r. 1876 pomieddzy Seretem, Zbruczem a Dniest—
rem // Sprawozd. Komisyi fizyjogr. — 1877. — VoL 11. —
P. 128-151.
Lutz P. Effect of temperature and photoperiod on larval deve-
lopment in Tetragoneuria cynosure (Odonata: Libellulidae) //
Ecology. - 1974. - VoL 55, N 2. - P. 370-377.
M a с a n T.T, The Odonata of a moorland fishpond Ц Internet
rev. ges. HydrobioL - 1964. - VoL 49. - P. 325-360.
199
M a c a n T.T. The influence of predation on the composition of
freshwater animal communities // Biol. Rev. — 1977. — Vol. 52;—
P. 45-70.
M a t h u r B.N. The central nervous system of Crocothemis servi—
lia erythraea (Bridle) (Odonata: Libellulidae) // Entomol. Mon—
they Mag. - 1972. - Vol. 108, N 1. - P. 40-45.
Matsumura S. Erster Beitrag zur Insecten — Fauna von Sa—
chalin II J. Coled. of Agricult. Tohoku Univ. — Sapporo, 1911,—
Vol. 4, N 1. - P. 1-144.
M a 11 e w s LV. Tertiary land briges and their climat; backdrop
for development of the present Canadian insect fauna // Canad.
Entomol. - 1980. Vol. 112, N 11. - P. 1089-1103.
McCrae A.W.R., Corbet Ph.S. Oviposition behaviour of Tet—
rathemis polleni (Selys) : a possible adaptation to life in tur-
bid pools (Anisoptera: Libellulidae) // Odonatologica. — 1982.—
VoL 11, N 1. - P. 23-31.
Meinertzhagen LA., Menzel R., Kahle G-. The identi-
fication of spectral Receptor Types in the Retina and Lamina
of the Dragonfly Sympetrum rubicundulum // J. Compal. PhysioL—
1983. - A151, N 3. - P. 295-310.
Metschnikoff E, Embryologische Studien an Insecten. — Le-
ipzig, 1866.
Meyer H.W. Schliisselreise ftir die Auslosung der Beutefang—
handlung beim Bachwasserlaufer Velia caprai (Hemiptera, Hete—
roptera). Untersuchung der Wirkung zeitlicher Reizmuster mit
Flimmerlicht // Z. vergl. Physiol. — 1971. — VoL 72, N 3. —
P. 298.
Mierzejewski W. Aeschna osiliensis nov. spec. Ц BuL Acad.
Sc. Cracovi, Cl: Sc. Math. Nat., Ser. В (Sc. Nat). — 1913. —
P. 79-87.
Mierzejewski L. Die Libellen (Odonata) der Insel О esel (Liv-
land, Rusland) // VerhandL zool.-boL Ges. Wien. - 1913. -
Bd. 53, H. 7-8. - S. 300-307.
Mill P.J. An anatomical study of the abdominal nervous and mus-
cular system of dragonfly (Aeschnidae) nymphs 11 Proc. Zool.
Soc. - L., 1965. - VoL 145. - P. 57-73.
M i m u r a K. Neural mechanisms, subserving directional selecti-
vity of movement in the optic lobe of the fly // J. Compar. Phy—
sioL - 1972. - Vol. 80, N 4. - P. 409-437.
Mittelstaedt H. Physiologie des gleichgewichtsinnes bei flie—
genden Libellen // Z. Vergl. PhysioL — 1950. — VoL 32, N S.-
Р. 422-463.
Morin P.J. Odonate guild composition: experiments with coloniza-
tion history and fish predation // Ecology. — 1984. — VoL 65,
N 6. - P. 1866-1872.
Motschulsky V. Catalogue des insecter repportes des envi-
rons du fleuve Amur, depuis la Schilka jusqu e Nikolaevsk,
examines et enumeres // Bull. Soc. NaL Mose. — Moscou,
1859. - P. 487-507.
M о u z e M. Croissame et metamorphose de Lapparell visuel des
Aeschnidae (Odonata) // InL J. Insect MorphoL and EmbryoL—
1972. - VoL 1, N 2. - P. 181-200.
Neal T.M., Whitcomb W.H. Odonata in the Florida soybean
agroecosystem // Fla Ent. — 1972. — Vol. 52, N 2. — P. 107-
114.
200
Oguma К. Japanese dragonflies of the subfamily Aeschninae //
Ent. Mag, Kyoto. — Kyoto, 1915a. — N 1 (з). — P. 120—131.
Oguma K. List of Libellulidae of Japan // Ibid. — Kyoto, 19156. —
N 1 (1). - P. 5-14.
Oguma K. Japanese Corduliinae // Zool. Mag. Tokyo. — Tokyo,
1915b. _ N 25. - P. 439-454.
Oguma K. The Japanese Aeschnidae // Insecta Matsumur. - Tokyo,
19 26. - Vol. 1, N 2. - P. 78-100.
Okumura T. Odonata from the Kutile Islands (ll) // Kontyu. —
Tokyo, 1942. - VoL 1б(1). - P. 12-19.
О 1 b e r g R.M. Parallel eucoding of direction of wind, head abdo-
men, and visual pattern movement by single interneurons in the
dragonfly // J. Compar. Physiol. — 1981. — A142, N 1. —
P. 27-41.
Osborn A.W., Eorteath Gr.N.R. Biology, dispersion and na-
tural enemis of Inopus rubriceps // Environ. Ent. — 19 74, —
VoL 3, N 1. - P. 29-32.
Pajunen V.L A description of agressive behaviour between
males of Leucorrhinia dubia v.d. Lind. (Odonata, Libellulidae)//
Ann. Ent Penn. - 1962. - VoL 28. - P. 108-118.
Pajunen V.L Agressive behaviour in Leucorrhinia caudalis
Charp. (Odonata: Libellulidae) // Ann. ZooL Penn. — 1964« —
VoL 1. - P. 357-369.
Pajunen V.L The influence of population density on the terri-
torial behaviour of Leucorrhinia rubicunda L. (Odonata: Libellu—
lidae) // Ibid. — 1966a. — VoL 3, N 1. — P. 40—52.
P a j u n e n V.L Agressive behaviour and territoriality in a popula-
tion of Calopteryx! virgo L. (Odonata, Calopterygidae) // Ibid. —
19666. - VoL 3, N 3. - P. 201-214.
P a n d i a n T.J., Mathavan S. Patterns of energy utilization
in the tropical dragonfly, Diplacodes trivialis (Rambur) and so-
me other aquatic insects (Anisoptera: 1-ibellulidae) // Odonatolo—
gica. - 1974. - VoL 3, N 4. - P. 241-248.
Parr M.J. Ecological studies of Ischnura elegans (van der Lin-
den) (Zygoptera: Coenagrionidae). 1. Age groups, emergence
patterns and numbers // Ibid. — 1973. — VoL 2, N 3. —
P. 139—157^
Parr M.J. Territorial behaviour of the African libellulid Orthet—
rum julia Kirby (Anisoptera) // Ibid. — 1980. — VoL 9, N 1, —
P. 75-9 9.
Parr M. J., Parr M. Studies on the behaviour and ecology of
Nesciothemis nigeriensis Gambles (Anisoptera: Lebellulidae) //
Ibid. - 1974. - VoL 3, N 1. - P. 21-47.
Parr M.J., Parr M. Some observations on Coenagrion tenel—
lum (de Villers) in southern England (Zygoptera; Coenagrioni—
dae) // Ibid. - 1979. - VoL 8, N 3. - P. 171-194.
P e z a 1 1 a V.M. Behaviour ecology of the dragonfly Libellula pu—
Ichella Drury (Odonata: Anisoptera) // Amer. Midland Natura-
list. - 1979. - VoL 102, N 1. - P. 1-22.
Pfau H.K. Struktur und Funktion des sekundaren Kopulationsap—
parates der Odonata (insecta: Palaeoptera), ihre Wandlung in
der Stammesgeschichte und Bedeutung fur die adaptive Entfal—
tung der Ordnung // Z. Morphol. Tiere. — 1971. — VoL 70,
N 4. - S. 281-371.
Pierce C.Z., Crowley P.H. Behaviour and ecological inte—
201
Factions of larval Odonata // Ecology. — 1985. - Vol. 66, N 5.—
P. 1504-1512.
Pritchard G. On the morphology of the compound eyes of
dragonflies (Odonata: Anisoptera), with special reference to
their in prey capture // Proc. Roy. Ent. Soc. London A. — 1966.—
Vol. 41, N 1-3. - P. 1-8.
Pritchard G. The prey of dragonfly larvae (Odonata: Aniso—
ptera) in ponds in northern Alberta // Canad. J. Zool. - 1964.—
Vol. 42, P. 785-800.
Procter D. The effect of temperature and photoperiod on lar-
val development in Odonata // Ibid. — 1973. — Vol. 51, N 11. —
P. 1165-1170.
Priiffer J. Untersuchungen iiber die bei Wilna vorkommenden
Formen der Gattung Leucorrhinia // Trav. Soc. Sc. Lettr. Vilno,
Cl. Sc. Math. Nat - 1923. - Vol. 1. - S. 1-16.
Quentin D.St Beobachtuhgen und Versuchen an Libellen in ih—
ren Jagdrevieren // Konovia. — 1934. — VoL 13, S. 275—282.
Quentin D.St Der Eangapparat der Odonaten Ц Osterr. zool.
Z. - 1953. - N 3. - S. 375-390.
Quentin D.St Zum Problem der borealpinen Verbreitung // En-
tomol. Nachr. Osterr. Schwez, Ent — 1959. — Bd 11, N 3. —
S. 82—84.
Radi E. Uber die Lichtreaktionen der Artropoden auf der Dreh—
scheibe // Biol. ZbL - 1902. - Vol. 22. - S. 728-732.
Rathke M. Miscelanea anatomico—physiologica. Ease. 1: De li—
bellarum partibus genitalibus. — Regimonti, 1832.
R e m m E. Eesti Kiilivastsete Maaraja (= Определитель личинок стре-
коз Эстонии). - Tartu, 1963. — 41 Р.
R е m m H. Eesti NSV kiilid (= Определитель стрекоз Эстонской ССР). -
Tartu, 1957. - 50 Р.
R е m m H, Putukate valimaraja (= Полевой определитель насекомых).—
Tartu, 1966. — 184 р. (Odonata: Р. 20—28).
R i е k E.P., Kukalova — Peck J. A new interpretation of dra-
gonfly wing venation based upon Early Upper Carboniferous
fossils from Argentina (insecta: Odonata) and basic character
states in pterygote wings // Canad. J. Zool. — 1984. - VoL 62.-
P. 1150-1166.
Ris Fr. Coll. Selys. Libellulinae, fas. IX—XVI. — Bruxelles. —
1909-1916.
Robertson H.M. Mating behaviour and its relationship to ter-
ritoriality in Platycypha caligata Selys (Odonata: Calopte -
Chloro cyphidae) // Behaviour. — 1982. - VoL 79, N 1. —
P. 11-27.
Robey C.W. Observation on breeding behaviour of Pachydiplax
longipennis (Odonata: Libellulidae) // Psyche. — 1975. —
Vol. 82, N 1. - P. 79-96.
R о i O. Die Odonaten von Ostpreussen // Schr. phys.—dkon. Ges.
Kd’nigsberg. — 1911. — Bd 52. — S. 13—30.
Rowe R.J. ’Territorial behaviour of a larval dragonfly Xantho—
cnemis zealandica (McLachlan) (Zygoptera: Coenagrionidae) //
Odonatologica. — 1980. — Vol. 9, N 4. — P. 285—292.
Rowe R.J. Intraspecific interactions of New Zealand damselfly
larvae L Xanthocnemis zealandica, Ischnura aurora and Aust—
. rolestes colenonis (Zygoptera: Coenagrionidae: Lestidae) //
N.Z.J. ZooL - 1985. - Vol. 12. - P. 1-15.
202
Rozefelds A.S. A fossil Zygopteran Nymph (insecta: Odona-
ta) from the Late Triassic Aberdare Conglomerate, Southeast
Queensland // Proc. Roy. Soc. Od. — 1985. — Vol. 96. —
P. 25-32.
Sakagami S.E., Ubukata H., Ida M., Toda M.J. Observa-
tions on the behaviour of some Odonata in the Bonin Islands,
with considerations on the evolution of reproductive behaviour
in Libellulidae // J. Fac, Sci. Hokkaido Univ. Ser. 6. — 1974. —
Vol. 19, N 3. - P. 722-757.
S a 1 z 1 e. Untersuchungen an Libelienlarven Uber das sehen be—
wegter Objekte // Z. vergL Physiol. — 1932. — Vol. 18. —
S. 347-368.
S a t i j a R.C. A histological study of the brain and thoracic ner-
ve cord of Aeschna nympha with special reference to the de-
scending nervous pathways // Res. Bui. Panjab, Univ. — 1958.—
Vol. 138. - P. 33-47.
Schiemenz H. Die Libellen unserer Heimat — Jena, 1953. —
154 S.
Schmidt Er. Vergliichende Morphologie des zweiten und drit—
ten Abdominalsegments bei m&nnlichen Libellen // Zool. Jahrb.
Abt. Anat. - 1915. - Vol. 39. - S. 87-200.
Schmidt Er. Die Libellen Irans. Ergeb. der Osterreih. Iran—Ex—
pedit. 1949/50 // Osterr. Akad. Wissensch. — Wien, 1954. —
Bd 16 3, N 4/5. - S. .223-260.
Schmidt Er. Ergebnisse der Deutsvhen Afghanistan—Expedition
1956 der Landessamlungen fur Naturkunde Karlsruhe sowie
der Expeditionen J. Klapperich, Bonn 1952—53 und Dr. K.
Lindberg, Lund (Schweden), 1957—60 // Beitr. natur. K. Forsch,
SW—Deutschl. — Karlsruhe, 1961. — Bd 19, N 3. — S, 399—435.
Selys Longchamps Ed. Cdonates de L'Asia mineure et re-
vision de ceux des auters parties de la fauna dite europiene//
Ann. Soc. Entom. Belgique. — Bruxelles, 1887. — Bd 31. —
P. 1-85.
Selys Longchamps Ed., Mac Lachlan R. Materiaux
pour une faune Nevropterologique de L'Asie septentrionale //
Ann. Soc. Entom. Belgique. - Bruxelles, 1872. — Bd 15. —
P. 25-45.
Sherk T.E. Development of the compound eyes of dragonflies
(Odonata). Larval compound eyes // J. Expti. Zool. — 1977. —
Vol. 201, N 3. - P. 391-416.
Sherk T.E. Development of the compound eyes of dragonflies
(Odonata). IV. Development of the adult compound eyes //
Ibid. - 1978. - Vol. 203, N 2. - P. 183-199.
Sjostedt L Entomologischen Ergebnisse der Schvedischen..
Kamtschatka — Expedition 19 20—19 22. B. Odonata // Arkif for
Zoologi. — Lund, 19 27. — Bd 19, ab. 2, N 16.
Spuris Z. Quelques donnees nouvelles sur la faune odonato—
logique de la Lettonie // Eolia zool. hydrobiol. — Riga, 1943. —
Vol. 12, fasc. 1. - P. 87-91.
Spuris Z. Odonata — spares // Latvijas PSP dzivnieku notei—
сё}з. — Riga, 1957. — Vol. 1. Bezmugurkaulnieki. — P. 2 8 9— 299.
S w i h a r t S.L, The neural basis of colour vision in the butter-
fly Papilio troilus // J. Inst. Phys. — 1970. — Vol. 16, N 8. —
P. 1623-1636.
Thompson D.J. Towards a predator—prey model incorporating
203
age structure: The effects of predator and prey sise on the
predation of Daphnia magna by Ischnura elegans // J. Anim.
Ecol. - 1975. - VoL 44, N 3. - P. 907-916.
Thompson D.J. The natural prey of larvae of damselfly, Isch—
nura elegans (Odonata: Zygoptera) // Freshwater Biol. — 1978,—
VoL 8. - P. 377-384.
Tillyard R.J. The biology of dragonflies. — Cambridge Univ.
Press, 1917.
Tillyard R.J. The British Liassic Dragonflies (Odonata) //
Fossil Insects. — Brit Mus. Nat Hist, 1925. — N 1. — P. 3—40.
Tillyard R.J. The evolution of the order Odonata // Proc. 4th
Intern. Congr. EntomoL (Ithaca, 1928). — Hamburg, 1929. —
P. 543-545.
Tillyard R.J., Fraser F.C. A reclassification of the order
Odonata // Austral. Zool. - 1938-1940. - Vol. 9. - P. 125-169,
195-221, 359-390.
T о n n e r F. Habsreflexe und Bewequngssehen bei Arthropoden //
Z. vergL Physiol. — 1938. — VoL 25, N 4. — S, 427—454.
Ubukata H. Life and behaviour of a Corduliid dragonfly, Cor-
dulia aenea amurensis Selys. IL Reproductive period with spe-
cial reference to territory // J. Fac. Sci. Hokkaido Univ. Ser.
VI Zool. - 1975. - Vol. 19, N 4. - P. 812-833.
Ubukata H. An experimental study Of sex recognition in Cor-
dulia aenea amurensis Selys (Anisoptera: Corduliidae) // Odo—
natologica. — 1983. — VoL 12, N 1. — P. 71—81.
Utz e r i C., Falchetti E., С a r c h i n i G. Acuni acpetti eto—
logici della ovideposizione di Lestes barbarus (Fabricius)
presso pozze tern poranee (Zygoptera: Lestidae) // Ibid. —
1976. - VoL 5, N 2. - P. 175-179.
Valle K.L. Nordasiatische Odonaten // Ann. Zool. Soc.—Botan.
Fennicae. — 19 32. — VoL 1, N 2. — P. 27—48.
Vogt P. Uber die optischen Schlusselreize beim Beutewerb der
Larven der Libelle Aeschna cyanea Mull // Zool. Jahrb. Phy-
siol. - 1964. - VoL 71, N 1. - S. 171-180.
W a a g e J.K. Reproductive behaviour and its relation to territo-
riality in Calopteryx maculata (Beanvois) (Odonata: Calopte—
rygidae) // Behaviour. — 1973. — VoL 47, N 3/4. — P. 240—256.
Weber T., Caillere L. Thermistor telemetry of ventilation
during prey capture by dragonfly larvae (Cordulegaster bolto—
ni. Odonata) // J. Compar, Physiol. — 1978. — VoL 128, N 4. —
P. 341-345.
W e 1 1 m e r L. Sporozoen ostpreussischer Arthropoden // Schrif—
ten der Phys.—okoL Ges. zu Konigsberg in Pr. — Leipzig; Ber-
lin, 1911. - S. 103-164.
Whalley P.E.S., Jarzembowsky E.A. Fossil insects from
the lithographic limestone of Montsech (Late Jurassic—Early
Cretaceous), Lerida province, Spain // BuL Brit Mus. Nat
Hist GeoL - 1985. - VoL 38, N 5. - P. 381-412.
Whitcomb W.H., В h a t c a r A., Nickerson J.C. Preda-
tors of Solenopsis invicta queens prior to successful colony
establishment // Environ. Ent — 1973. — VoL 2, N 6. —
P. 1101-1103.
Williams C.E. Courtship display in Belonia croceipennis (Se-
lys), with notes on copulation and oviposition (Anisoptera: Li-
204
bellulidae) 11 Odonatologica. — 1977, — Vol. 6, N 4. — P. 281-
287.
W illiams C.E. Notes on the behaviour of the late instar nym-
phs of four Macromia species under natural and laboratory
conditions (Anisoptera: Macromiidae) // Notulae Odonatol. —
1979. - Vol. 1, N 2. - P. 27-28.
Wilson E.O. Sociobiology: the new synthesis. — Cambridge,
1975. - 696 P. ~
Wnukowsky W. Beitr&ge zur Kenntnis der Odonaten — Fauna
Weiss—Rus stands (Weiss — Rutheniens) // Festschrift 60. Ge—
burtstage Prof. Dr. Embrik Strand. — Riga, 1937. — Bd 3. —
S. 568-581.
Wright M. The economic importance of dragonflies (Odonata)//
J. Tennessee Acad. Sci. — 1946, - Vol. 21. - P. 60—71.
Zamberti G.A., Resh V.H. Seasonal patterns of invertebrate
predators and prey in coyote Hills Marsh // Proc, and Pap.
52nd Ann. Conf. Calif. — Mosquito and Vector Contr. Assoc.,
Sacramento, Calif., 1985. — P. 126-128.
Z e tt 1 e r F., Jarvilehto M. Lateral inhibition in an insect
eye // Z. vergL Physiol. — 1972. — Vol. 76, N 3. — P. 233—244.
Znamierowska M. Die Odonaten fauna der Umgebung von
Wilna II Trav. Soc. Sc. Lettr. Vilno, Cl. Sc. math, nat., 1923.—
Bd 1. - S. 29-39.
Z namierows ka-Pruffe rowa M. Materialien zur Kennt-
nis der Odonatenfauna des nordbstlichen Polen // Ibid. — 1927,—
Bd 3. - S. 181-189.
ФОТОТАБЛИЦЫ И ОБЪЯСНЕНИЯ К НИМ
Таблица I
Подотряды Meganeurina и Ditaxineurina
Фиг. 1. Arctotypus sinuatus Martynov (Meganeuridae), экз< № 53/117,ПИН, перед
нее крыло (х0,8); Архангельская область, верхняя пермь.
Фиг. 2. Permaeschna dolloi Martynov (P.ermaeschnidae), ©кз. № 3353/75, ПИН,
переднее крыло (х1,8); Архангельская область, верхняя пермь.
Фиг. 3. Ditaxineurina fam. gen. sp., экз. № 1700/325, ПИН, неполный остаток тела и
крыльев (х1,1); Урал, нижняя пермь.
Таблица II
Подотряды Kennedyina и Triadophlebiina
Фиг. 1. Permolestes sp. (Permolestidae ), экз. № 3353/63, ПИН, крылья (х2); Ар-
хангельская область, верхняя пермь.
Фиг. 2. Permolestidae gen. sp., экз. № 1700/477, ПИН, неполный остаток тела и
крыльев (х1,5); Урал, нижняя пермь.
Фиг. 3. Neritophlebia elegans Pritykina (Triadophlebiidae), экз. № 2785/3, ПИН,
фрагменты крыльев (х2,6); Средняя Азия, триас /Притыкина, 1981/.
Таблица Ш
Подотряд Triad о р hie b i i па
Фиг. 1. Paurophlebia lepida Pritykina (Triadophlebiidae), голотип № 2785/7,
ПИН, крыло (х4,2); Средняя Азия, триас /Притыкина, 1981/.
Фиг. 2. Cyrtophlebia sinuosa Pritykina (Zygophlebiidae), голотип № 2785/25,
ПИН, фрагменты двух крыльев (х4,3); Средняя Азия, триас /Притыкина, 1981/.
Фиг. 3. Xamenophlebia ornata Pritykina (Xamenophlebiidae), экз. № 2555/2074,
ПИН, фрагмент крыла (х2,5); Средняя Азия, триас /Притыкина, 1981/.
Таблица IV
Подотряд Heterophlebiina
Isophlebiidae gen. sp., колл. ПИН, верхняя юра Сибири и МНР: 1 - экз. № 2815/41,
личинка (х!,9); 2 - экз. Ns 3795/426, хвостовой плавник личинки (х4); 3 - экз. № 3688/230,
птероторакс (х2,2); 4 - экз. 3063/914, фрагменты второго и третьего брюшных сегментов с
копулятивным аппаратом (хб,4); 5 - экз. № 3795/427, фрагменты тела и крыльев (х1,1).
Таблица V
Подотряд Heterophlebiina
Фиг. 1. Karatawia shurabica Pritykina (Karatawiidae ), голотип № 2068/55,
ПИН, фрагмент переднего крыла (х4); Средняя Азия, нижняя юра /Притыкина, 1980/.
Фиг. 2. Triassolestodes asiaticus Pritykina (Triassolestidae) ,-голотип
№ 2240/1783, ПИН, заднее крыто (х4,б); Средняя Азия, триас/Притыкина, 1981/.
Фиг. 3. Sibirioneura angarica Pritykina ( Campterophlebiidae), голотип
№ 2375/16, ПИН, фрагмент крыла (х2,8); Сибирь, нижняя юра /Притыкина, 1985/.
Фиг. 4. Sogdopteron leve Pritykina (Oreopteridae), голотип № 2389/15, ПИН,
заднее крыто (-х5,1); Средняя Азия, нижняя юра /Притыкина, 1980/.
Таблица VI
Подотряд Heterophlebiina
Фиг. 1. Cyclothemis sagulica Pritykina (Archithemistidae ), голотип Ns 2389/40,
ПИН, фрагмент заднего крыла (хб,9); Средняя Азия, нижняя юра /Притыкина, 1980/.
Фиг. 2,3. Молодые личинки Samarura gigantea Br., Rdtb., Gglb. (Campterophle—
biidae), нижняя - средняя юра Сибири: 2 - экз. № 1604/7, ПИН, (х4); 3 - экз. № 1670/143,
ПИН, (х5,1).
Фиг. 4. Shurabiola папа Pritykina (Archithemistidae), экз. № 508/11, ПИН, ли-
чинка (х4,4); Сибирь, нижияя - средняя юра /Притыкина, 1965/.
Фиг. 5. Turanophlebia sp. ( Tarsophle biidae ), экз. № 3790/1, ПИН, личинка
(хЗ,6); МНР, нижний мел.
Таблица VII
Подотряд Libellulina (= Anisoptera)
Фнг. 1, 2. Hemeroscopus baissicus Pritylina (Hemeroscopidae), нижний мел
Забайкалья: 1 - голотип Ns 3064/141, ПИН, заднее крыло (х2,3)/Притыкина, 1977/; 2 - экз.
№ 1989/1941, ПИН, взрослая личинка (х!,б).
Фиг. 3. Sinaeschnidia sp. ( Aeschnidiidae ), Ns 2385/878, ПИН, фрагмент заднего
крыла (х2,2); Забайкалье, нижний мел.
Таблица I
2
Таблица П
Таблица Ш
1
Таблица IV
Таблицах/
Таблица VI
£
г
ПЛ в Пи Lt g в J,
ОГЛАВЛЕНИЕ
Предисловие (А.Ю. Харитонов)...............•............................. 3
Глава I.' История изучения, состав и генезис фауны....................... 5
Европейская часть СССР (З.Д. Спурис)................................. 6
Азиатская часть СССР (Б.Ф. Белышев, А.Ю. Харитонов, СЛ. Бори-
сов) ............................................................... 11
Состав фауны стрекоз СССР (Б.Ф» Белышев, AJO. Харитонов, И.Н. Ха-
ритонова, С.Н. Борисов)............................................. 31
Палеонтология и эволюция стрекоз (Л.Н. Притыкина)................... 43
Пути формирования бореальной фауны (Б.Ф. Белышев, А.Ю. Харито-
нов) ............................................................ 59
Глава Ц. Экология и поведение............................................ 66
Фенология стрекоз (А.Ю. Харитонов, СЛ. Борисов)..................г 67
Суточные ритмы активности стрекоз (А.Ю. Харитонов, СЛ. Борисов) 77
Пространственная организация популяций личинок (на примере рода Ca-
lopteryx) (Г.И. Рязанова, Г.А. Маэохин-Поршняков)................... 85
Роль зрения в поведении стрекоз (П.А. Мокрушов)..................... 92
Этологические исследования стрекоз рода Sympetrum (П.А. Мок-
рушов, Л.И. Францевич)...............................................Ю8
Глава и. Роль в биоценозах..............................................118
Паразиты стрекоз (Р.С. Павлюк).................................... -
Обзор изученности трофических связей стрекоз (Г.А. Сухачеве) .... 142
Опыт опенки роли стрекоз в водоемах Белоруссии (Е.С. Шалопенок,
А.Д. Писаиеико).................................................. 148
Опыт исследования населения стрекоз в биоценозах Западной Сибири
,(В.В. Заика) . ................................................. 158
Список литературы................................................... 161
Научное издание
Белышев Борис Федорович
Харитонов Анатолий Юрьевич
Борисов Сергей Николаевич и др.
ФАУНА И ЭКОЛОГИЯ СТРЕКОЗ
Редакторы издательства В.В. Волковинцер,
Л.И. Шпаковская
Художник В.В. С ед уно в
Технический редактор Н.М. Остроумова
Корректоры Н.М. Горбачева, С.В. Блинова
ИБ № 34371
Сдано в набор 25.04.89. Подписано в печать 29.08.89.
МН-01460, Формат 60x90 1/16. Бумага кн.-журн.
Усл. печ. л. 13 + 0,5 п.л. на мел. бум. Усл. кр.-отт. 13,5.
Уч.-изд. л. 16,5. Тираж 900 экз. Заказ № 611,
Нена 3 р. 7 О к.
Ордена Трудового Красного Знамени издательство 'Наука',
Сибирское отделение. 630099 Новосибирск, ул. Советская, 18.
4-я типография издательства 'Наука*. 630077 Новосибирск,
ул. Станиславского, 25.