















































































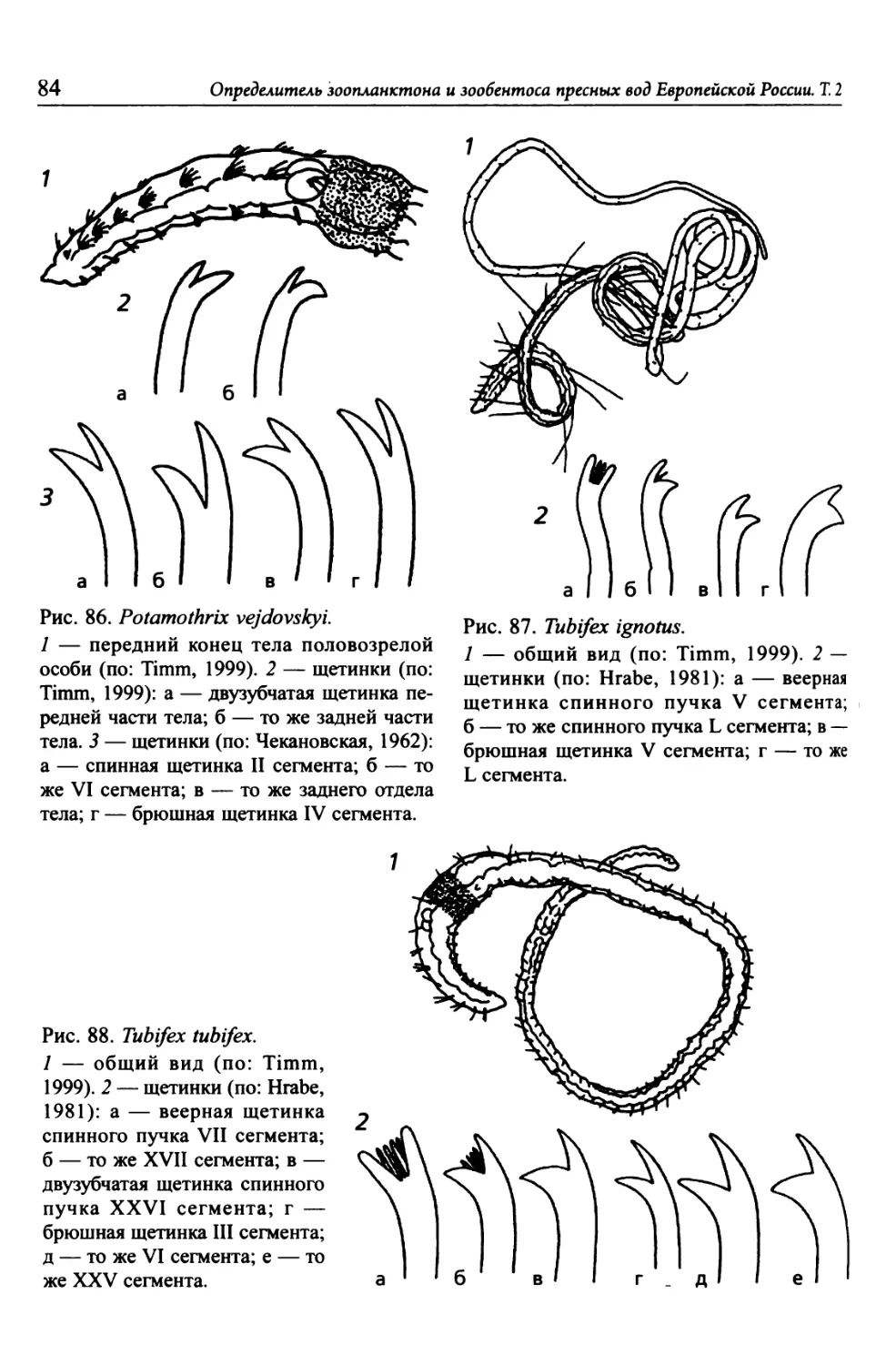


























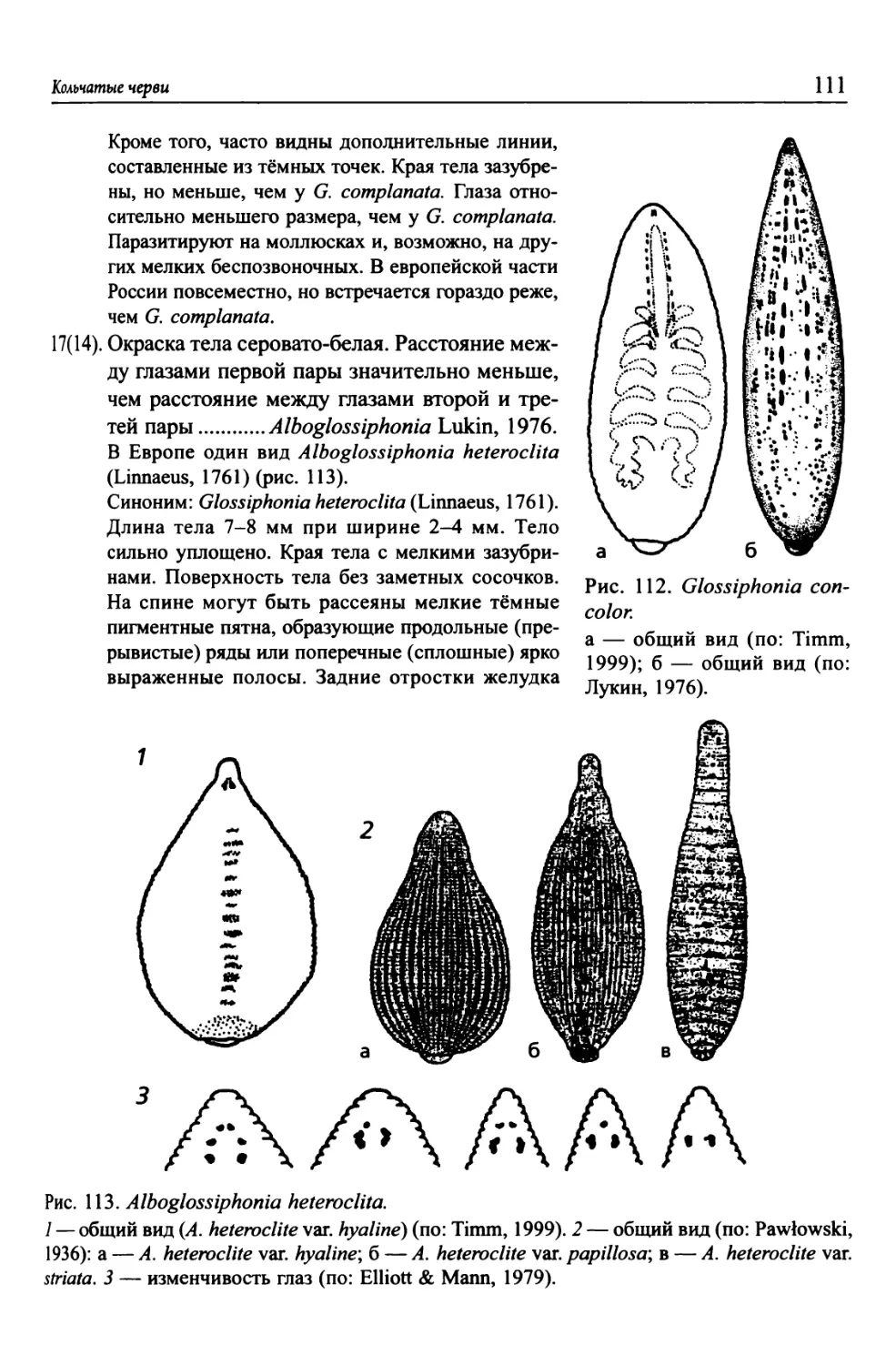

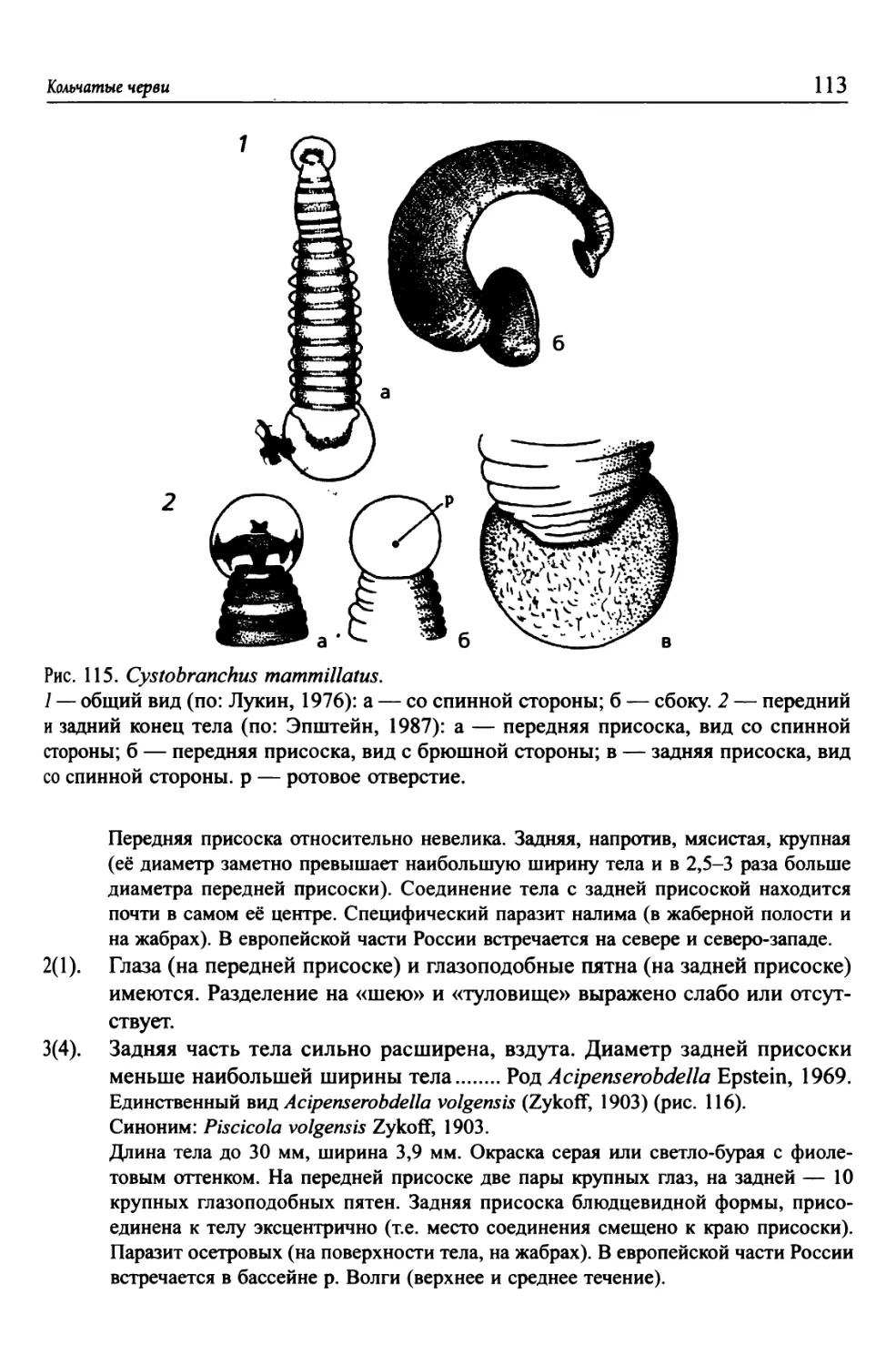


















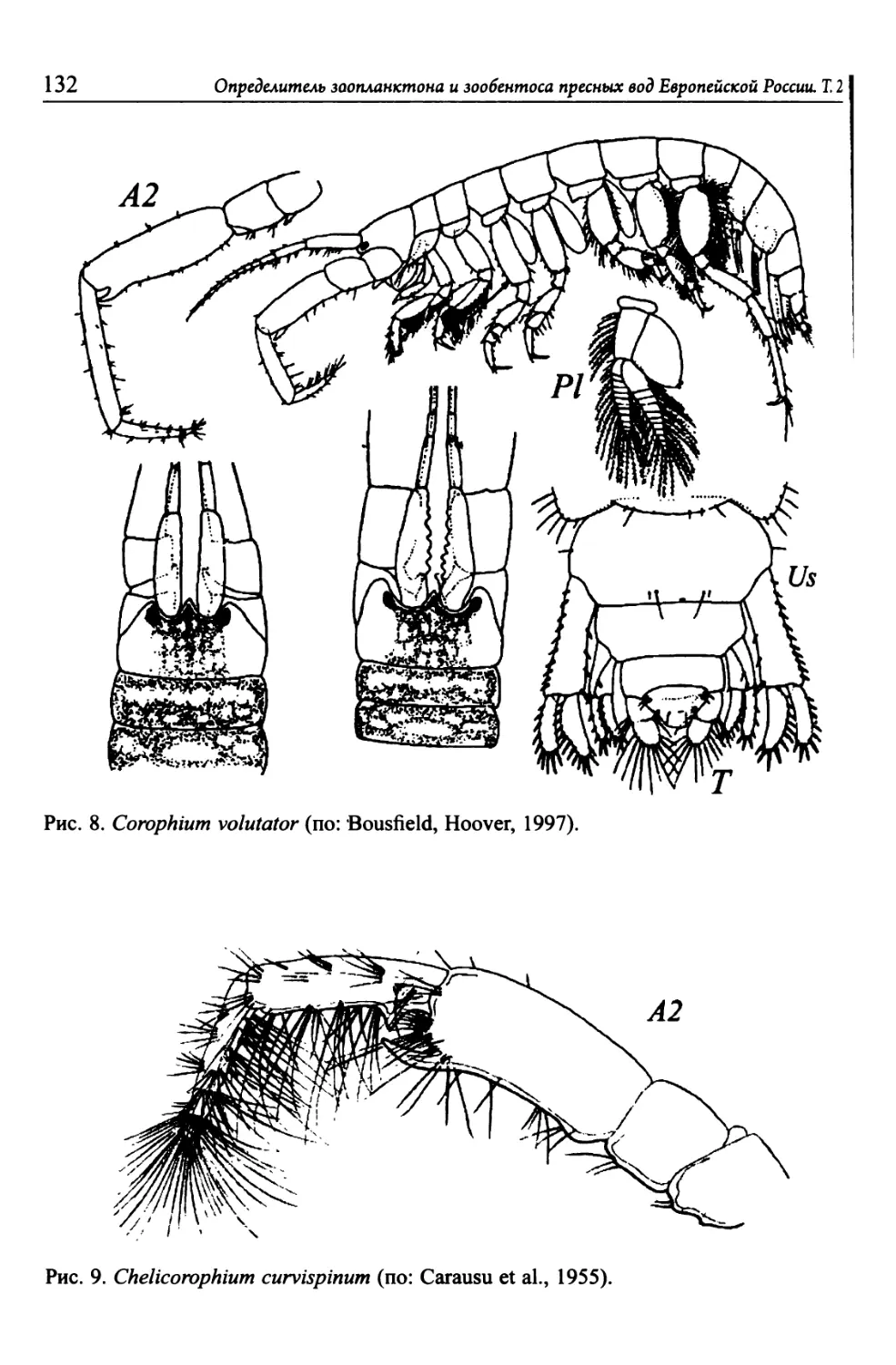





























































































































































































































































































































































Автор: Цалолихин С.Я. Алексеев В.Р.
Теги: общая экология биоценология гидробиология биогеография систематика животных специальные зоологические науки биология экология беспозвоночные
ISBN: 978-5-9907572-4-0
Год: 2016




ОПРЕДЕЛИТЕЛЬ
зоопланктона и зообентоса
пресных вод
Европейской России
Том 2
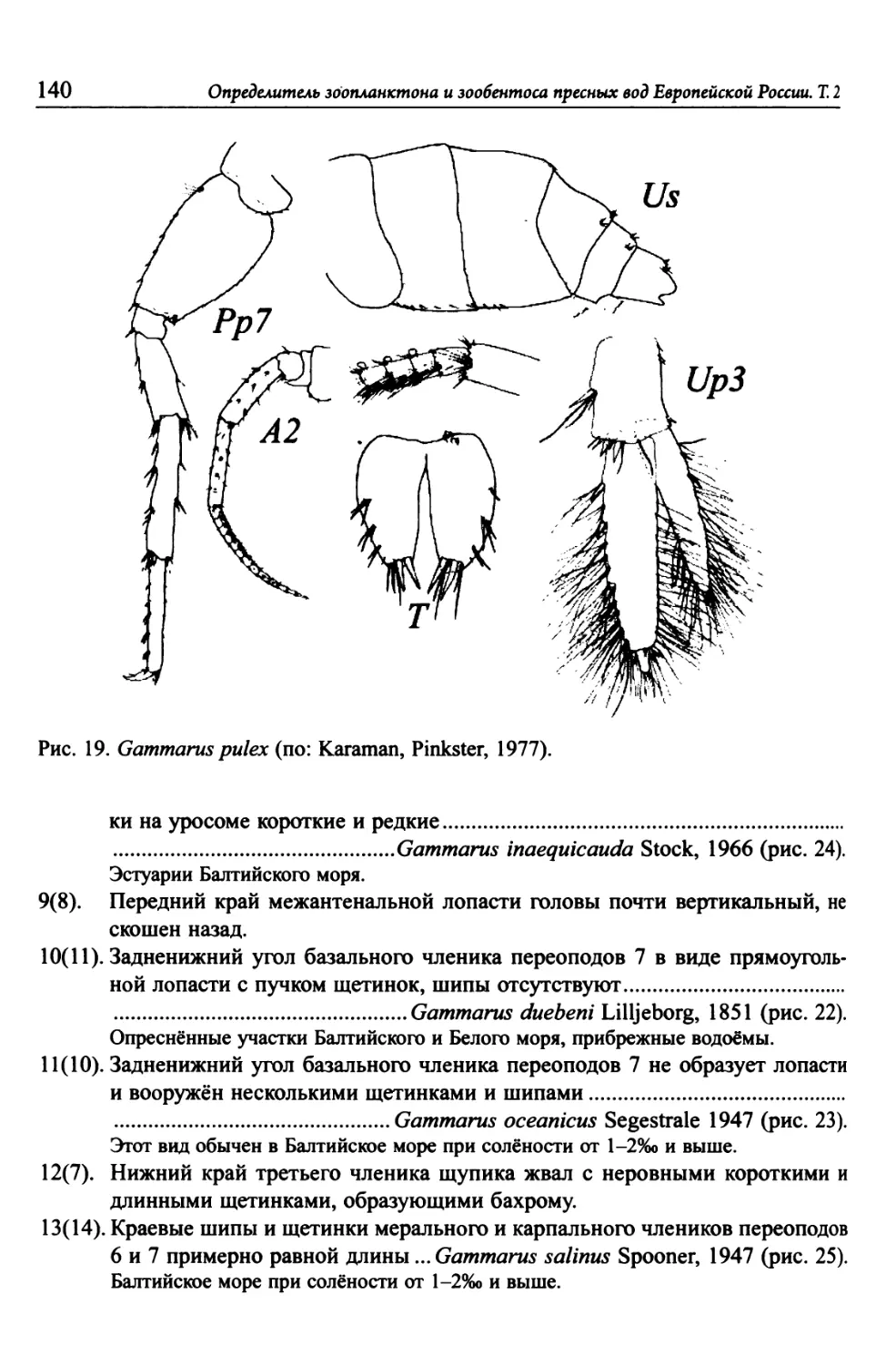
Зообентос
российская академия наук
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
ОПРЕДЕЛИТЕЛЬ
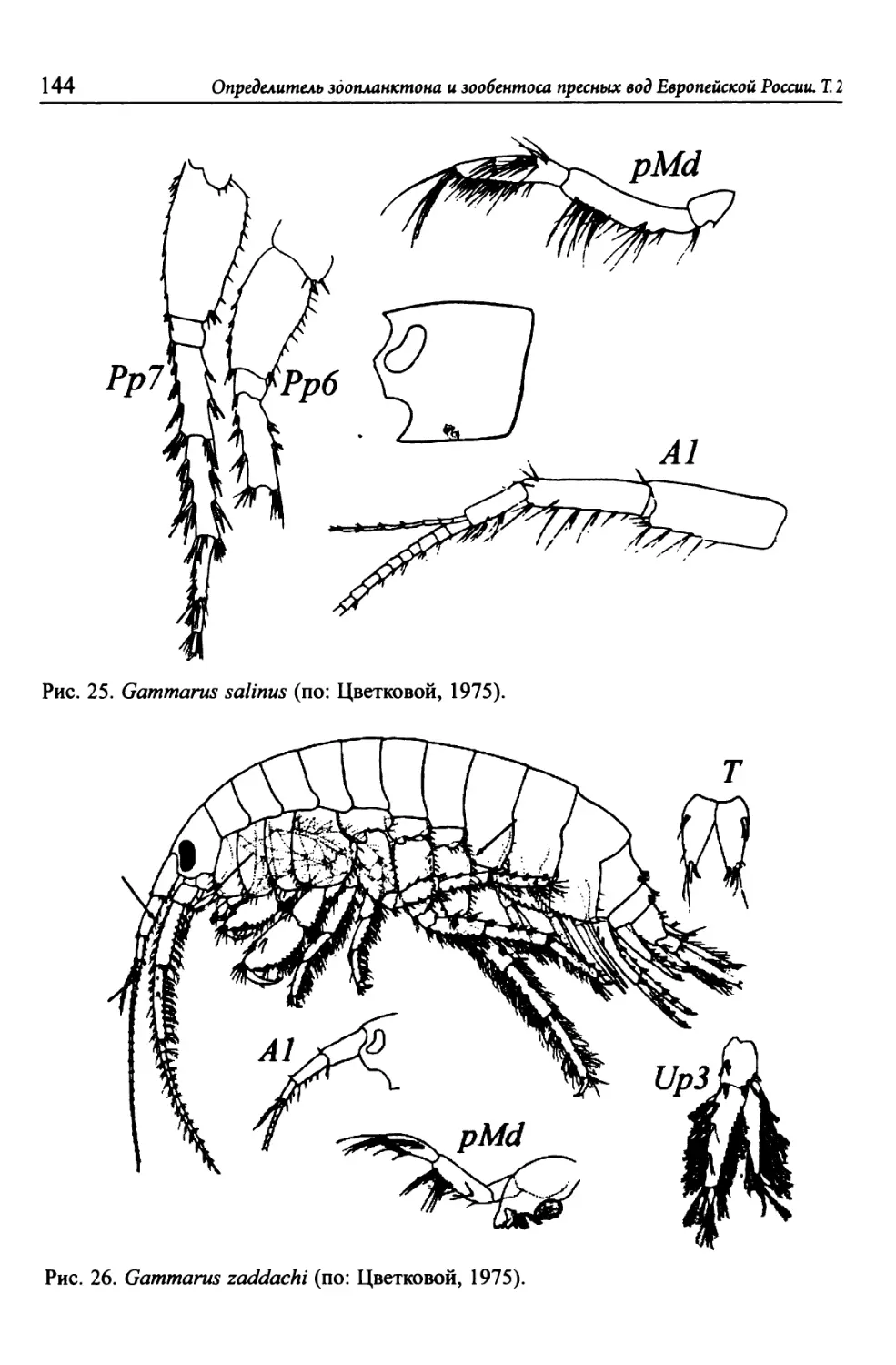
зоопланктона и зообентоса
пресных вод
Европейской России
Под редакцией
В.Р. Алексеева, С.Я. иалолихина
*
Том 2
Зообентос
Редактор тома
С.Я. иалолихин
Товарищество научных изданий КМК
Москва - Санкт-Петербург ❖ 2016
УДК 574.583(470X083.71)
ББК 28.082(235)я28+28.691я28
0-62
Главный редактор
член-корреспондент РАН О.Н. Пугачев
Редакционная коллегия
Н.Б. Ананьева, А.Ф. Алимов, Н.В. Аладин, А.О. Аверьянов, ТА. Асанович, А.В. Балушкин,
В.Я. Бергер, С.Д. Гребельный, М.В. Крылов, Е.Л. Мархасёва, С.Ю. Синев, С.Я. Резник,
М.К. Станюкович, А.Н. Тихонов, В.В. Хлебович, С.Я. Цалолихин
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2.
О_62 Зообентос / Под редакцией В.Р. Алексеева и С.Я. Цалолихина. — М.-СПб.: Товари-
щество научных изданий КМК, 2016. — 457 с., ил., 6 с., 18 цв. вкл.
Определитель предназначен для исследователей водных экосистем, а также для студентов
биологических специальностей высших учебных заведений. В Определителе, в разной
степени подробности, представлены все основные группы донных беспозвоночных жи-
вотных, встречающихся в пресных водоёмах и водотоках Европейской России с акцентом
на фауну северо-западных регионов.
Редактор тома
С.Я. Цалолихин
Рецензенты тома
С.М. Голубков, А.В. Шошин
Издание осуществлено при финансовой поддержке
Программы фундаментальных исследований Отделения общей биологии РАН
"Биологическиересурсы России"
и, частично, при поддержке РФФИ (грант Xs 14-04-00932)
ISBN 978-5-9907572-4-0
© Зоологический институт РАН, 2016
© Коллектив авторов, текст, иллюстрации,
подбор иллюстраций, 2016
© Товарищество научных изданий КМК, издание, 2016
ПРЕДИСЛОВИЕ
Предлагаемый 2-й том «Определителя зоопланктона и зообентоса пресных
эд Европейской России» включает основные группы гидробионтов, населяющих
но водоёмов от крупных озёр до небольших прудов. Главной целью при состав-
ении этого тома было научить начинающих исследователей, в первую очередь
гудентов-биологов, пользоваться определительными ключами и дать возмож-
ость ознакомиться с типичной пресноводной фауной России. Такая задача сто-
ла и при составлении ранее опубликованного 6-ти томного «Определителя пре-
новодных беспозвоночных России и сопредельных территорий» (1994-2004),
котором бентосным животным было посвящено более 70% общего объёма из-
дания. Однако надежда себя не оправдала в том смысле, что «Определитель» ока-
ался сложен для первокурсников, и от руководителей соответствующих кафедр
юступило предложение выпустить упрощённое издание, ограниченное одним
омом, который можно было бы использовать на летней практике студентов. К со-
калению, включить в одну книгу весь объём шеститомника, где только одни дву-
срылые насекомые занимают 1000 страниц, конечно невозможно.
При составлении тома «Бентос» в основу был положен принцип: «лучше до-
стоверно определить род, чем сомнительно вид», тем более, что система многих
рупп водных беспозвоночных продолжает совершенствоваться на видовом уров-
4е, многие названия переходят в категорию синонимов, отдельные виды разделя-
ются на ряд новых, уточняются биогеографические границы видов.
Внутренняя структура тома «Бентос» отличается от традиционной «филогене-
тической». Последовательность разделов в этом томе следует от наиболее часто
встречающихся и легко попадающихся на глаза обитателей дна водоёмов к встре-
чающимся не часто и требующих специальных способов «охоты».
Во всех тех случаях, когда возникает интерес или необходимость в более точ-
ном определении материала исследователю следует обращаться к специальным
определителям, в том числе и к приложению «Пресноводные моллюски севера и
северо-запада Европейской России» (ред. П.В. Кияшко), данному в этой книге и к
нашему «Определителю пресноводных беспозвоночных России и сопредельных
территорий». Особенно можно рекомендовать 3, 4 и 5 тома, где приводятся под-
робные многоступенчатые ключи для определения имаго и личинок насекомых.
4 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
«Определитель пресноводных беспозвоночных России и сопредельных
территорий» (под редакцией С.Я. Цалолихина)
Том 1. 1994. Низшие беспозвоночные (ред. С.Я. Цалолихин): Губки, Книда-
рии, Турбеллярии, Коловратки, Гастротрихи, Нематоды, Волосатики, Олигохеты,
Пиявки, Мшанки, Тихоходки. 396 с.
Том 2. 1995. Ракообразные (ред. В.Р. Алексеев): Листоногие, Ветвистоусые,
Веслоногие, Остракоды, Кумойые, Мизиды, Изоподы, Декаподы, Амфиподы.
628 с.
Том 3. 1997. Паукообразные и низшие насекомые (ред. Э.П. Нарчук, Д.В. Ту-
манов, С.Я. Цалолихин): Акариды, Орибатиды, Галакариды, Гидрахниды, Пауки,
Ногохвостки, Подёнки, Веснянки, Стрекозы, Клопы. 448 с.
Том 4. 1999. Высшие насекомые (ред. Э.П. Нарчук): Двукрылые насекомые.
1000 с.
Том 5. 2001. Высшие насекомые (ред. Э.П. Нарчук): Ручейники, Бабочки,
Жуки, Сетчатокрылые. 836 с.
Том 6. 2004. Моллюски, Полихеты, Немертины (ред. В.В. Богатов, С.Я. Цало-
лихин): Двустворчатые, Брюхоногие, Полихеты, Немертины. 527 с.
Издание предлагаемого Определителя осуществлено благодаря поддержке и
помощи председателя Научного совета по изучению, охране и рациональному ис-
пользованию животного мира, президента Гидробиологического общества, ака-
демика А.Ф. Алимова и куратора программы фундаментальных исследований
Отделения общей биологии РАН «Биологические ресурсы России», члена-кор-
респондента РАН Б.Р. Стригановой, за что выражаю им глубокую благодарность.
Кроме этого, при подготовке к печати данного тома частично были использованы
средства гранта РФФИ 14-04-00932.
С.Я. Цалолихин
ДОННЫЕ БЕСПОЗВОНОЧНЫЕ ПРЕСНЫХ ВОД
Таблица для определения групп пресноводных беспозвоночных,
наиболее часто встречающихся в сборах бентоса
(длина тела не менее 3 мм)
ЕСТЬ УДЛИНЁН Н Ы ЕЧЛ ЕН ИСТЫЕ КОНЕЧНОСТИ
___________________ТРИ ПАРЫ ЧЛЕНИСТЫХ НОГ____________________
Голова с парой крупных (фасеточных) глаз по бокам. Грудные сегменты
с двумя парами крыловых зачатков (в виде лопастей, направленных назад)
или же задняя половина тела покрыта плотными надкрыльями.
EPHEMEROPTERA — ПОДЕНКИ (личинки).
Тело удлинённое. Задний конец тела с 3 «нитями».
Сегменты брюшка (многие или некоторые) с листо-
видными или нитевидными жабрами по бокам. Есть
2 пары крыловых зачатков. Могут активно плавать.
PLECOPTERA — ВЕСНЯНКИ (личинки).
Тело удлинённое. Задний конец тела с 2 «нитями».
Есть 2 пары крыловых зачатков. Как правило, не пла-
вают.
ODONATA — СТРЕКОЗЫ (личинки).
Тело удлинённое или компактное. Задний конец тела
с 3 удлинённо-листовидными пластинками или без
придатков; в последнем случае заострён, с шипами на
вершине. Голова снизу прикрыта складной «маской»,
которая может выдвигаться вперёд. Есть 2 пары крыло-
вых зачатков. Могут активно плавать.
HETEROPTERA — ПОЛУЖЕСТКОКРЫЛЫЕ
(личинки и имаго).
Тело удлинённое или компактное. Ротовой аппарат в
виде непарного хоботка, удлинённого и заострённо-
го (обычно направлен назад между основаниями ног);
если хоботок не виден, то голова снизу треугольная.
Надкрылья, если имеются, накладываются друг на
друга вершинами (разделены диагональным «швом»).
Если надкрыльев нет, то обычно есть 2 пары крыловых
зачатков. Бегают по поверхности воды или плавают.
COLEOPTERA — ЖЕСТКОКРЫЛЫЕ (имаго).
Тело компактное, с плотными покровами. Жёсткие
парные надкрылья плоско покрывают сверху заднюю
половину тела, смыкаясь по средней линии (без диаго-
нального «шва»). Ротовой аппарат с развитыми парны-
ми челюстями. Многие активно плавают.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Голова без крупных глаз; по бокам головы обычно по несколько глазков
в виде группы мелких тёмных пятен. Грудные сегменты без следов крыловых
зачатков, с более-менее прямыми задними краями; надкрылий нет.
MEGALOPTERA — ВИСЛОКРЫЛКИ (личинки).
Тело удлинённое. Задний конец тела с длинным за-
острённым непарным выростом. Сегменты брюшка по
бокам с членистыми нитевидными жабрами. Вырост и
жабры с бахромой волосков. Могут активно плавать.
TRICHOPTERA — РУЧЕЙНИКИ (личинки).
Тело удлинённое, облик обычно слегка напоминает гу-
сениц. Головная капсула и грудные сегменты (один-два
передних или все три) плотные; остальная часть тела
мягкая, более-менее цилиндрическая. Вершина тела с
парой коротких выростов, несущих по крючку. Чаще
всего в переносных домиках. Как правило, не плавают.
COLEOPTERA — ЖЕСТКОКРЫЛЫЕ (личинки).
Тело удлинённое, при этом может быть уплощено. Не-
редко задний конец тела вытянут и заострён, может
нести парные «нити» (они могут быть в длинных воло-
сках). Сегменты брюшка могут нести по бокам жабры в
виде нечленистых лопастей или «нитей», которые могут
иметь бахрому из волосков. Многие активно плавают.
ЧЕТЫРЕ ПАРЫ ЧЛЕНИСТЫХ НОГ
HYDRACHNIDIA — ВОДЯНЫЕ КЛЕЩИ.
Тело компактное, округлое или овальное, без перетяж-
ки. Длина до 8 мм (редко более 4 мм). Многие активно
плавают, попеременно двигая ногами.
ARANEI — ПАУКИ.
Тело из двух компактных отделов, разделённых пере-
тяжкой. Неплавающие; могут бегать по воде или под
водой.
БОЛЕЕ ЧЕТЫРЁХ ПАР ЧЛЕНИСТЫХ НОГ
AMPHIPODA — БОКОПЛАВЫ.
Тело сжатое с боков, сегментированное. Активно пла-
вают.
ISOPODA — РАВНОНОГИЕ РАКИ.
Тело уплощённое, удлинённо-овальное, сегментирован-
ное. Не плавают.
NOTOSTRACA — ЩИТНИ.
Передняя часть тела покрыта овальным несегментиро-
ванным спинным щитом, из-под которого торчит удли-
нённый сегментированный брюшной отдел с 2 «нитя-
ми» на конце. Могут активно плавать.
Донные беспозвоночные пресных вод
7
ЧЛЕНИСТЫХ КОНЕЧНОСТЕЙ НЕТ
ТЕЛО ЗАКЛЮЧЕНО В ТВЁРДУЮ РАКОВИНУ
GASTROPODA — БРЮХОНОГИЕ МОЛЛЮСКИ.
Раковина спиральная или колпачковидная. Не плавают.
BIVALVIA — ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ.
Раковина двустворчатая, створки примерно равного раз-
мера и одинаковой формы. Не плавают.
ТЕЛО БЕЗ РАКОВИНЫ
HIRUDINEA — ПИЯВКИ
Тело с большой присоской на заднем конце (обычно
также видна присоска на переднем конце), уплощённое
(удлинённо-лентовидное или листовидное), кольчатое
(более 20 коротких сегментов). Ползают, используя при-
соски и активно плавают, волнообразно изгибая тело в
вертикальной плоскости.
TURBELLARIA — РЕСНИЧНЫЕ ЧЕРВИ.
Тело уплощённое (лентовидное или удлинённо-ли-
стовидное), не кольчатое, без головной капсулы и
придатков. Передний край прямой, угловатый или за-
круглённый, часто с тёмными глазными пятнами. Не-
плавающие; двигаются по субстрату, плавно скользя.
OLIGOCHAETA — МАЛОЩЕТИНКОВЫЕ ЧЕРВИ.
Тело тонкое, нитевидное, не уплощённое, кольчатое
(более 20 коротких сегментов), без головной капсулы и
придатков. Как правило, способны ненаправленно пла-
вать, быстро изгибаясь.
DIPTERA — ДВУКРЫЛЫЕ (личинки).
Тело более-менее цилиндрическое или веретеновидное,
кольчатое (не более 13 сегментов). Передний конец с
уплотненной головной капсулой (бывает частично втя-
нута) или с просвечивающими тёмными продольными
структурами. Некоторые сегменты могут нести бугор-
ковидные нечленистые ложноножки; задний конец тела
может нести длинные щетинки, парные или непарные
придатки. Многие могут плавать, резко изгибаясь.
DIPTERA — ДВУКРЫЛЫЕ (куколки).
Тело с вздутым цельным передним отделом (с нижней
стороны могут просвечивать неподвижные чехлы ног)
и с удлинённым кольчатым брюшком (не более чем из
9 сегментов). Передний конец тела может нести пару
придатков различной формы (веретеновидные, трубко-
видные или в виде ветвящихся нитей). Многие могут
плавать, резко изгибая брюшко.
моллюски
Пресноводные моллюски — одна из наиболее крупных, таксономически раз-
нообразных и широко распространённых групп гидробионтов. Эти животные,
являющиеся специальными объектами изучения для специалистов-малакологов,
нередко попадают в сферу научных интересов биологов других специальностей
при проведении комплексных фаунистических, биогеографических, гидробиоло-
гических, паразитологических, ихтиологических и палеонтологических исследо-
ваний, как теоретической, так и прикладной направленности.
Система пресноводных моллюсков базируется на сложном комплексе разно-
родных морфологических признаков и во многом ещё далека от завершённости.
Совершенствование системы, связанное с привлечением к анализу новых призна-
ков и таксономическими перестановками, закономерно приводит к устареванию
фаунистической информации и усложнению процедуры идентификации видов.
Поэтому, учитывая интерес к пресноводным моллюскам специалистов разных об-
ластей знаний, представляется актуальным периодическое переиздание на основе
современных данных пособий для определения брюхоногих (Gastropoda) и дву-
створчатых (Bivalvia), обитающих в пресных водах.
В представленном томе такое пособие для определения моллюсков состоит из
двух блоков, позволяющих идентифицировать животных с разной степенью под-
робности. В основном тексте тома даны ключи, позволяющие определять пресно-
водных моллюсков до рода. Они предваряются кратким морфологическим очер-
ком, дающим общее представление о внешнем облике, диагностически значимых
признаках и об основных методах сбора и камеральной обработки двустворчатых
и брюхоногих пресноводных моллюсков. Ключи построены на основе данных,
публикуемых в приложении к данному тому ’’Определителя пресноводных бес-
позвоночных...” и снабжены изображениями типичных для каждого рода раковин.
Более подробный определитель, позволяющий идентифицировать виды пресно-
водных моллюсков, обитающих на севере и северо-западе Европейской России,
дан в виде приложения в конце тома.
Приложение. Пресноводные моллюски севера и северо-запада Европейской
России.
Класс Двустворчатые моллюски — Bivalvia Linnaeus, 1758 (В.В. Богатов,
П.В. Кияшко).
Класс Брюхоногие моллюски — Gastropoda Cuvier, 1797 (П.В. Кияшко,
Е.В. Солдатенко, М.В. Винарский).
Краткая морфологическая характеристика
пресноводных моллюсков
Беспозвоночные с мягким несегментированным телом, у представителей
большинства классов подразделяющимся на три отдела: голову (отсутствует
Моллюски
9
у двустворчатых), туловище и ногу. Нога находится на брюшной стороне тела
и в некоторых случаях способна претерпевать значительные изменения соот-
ветственно образу жизни и характеру движений животного. Чаще всего нога
представлена плоской ползательной подошвой (брюхоногие моллюски). У дву-
створчатых моллюсков нога образует плоский мускулистый вырост, иногда ре-
дуцирована. У большинства брюхоногих моллюсков, дышащих с помощью жабр,
а также у некоторых «лёгочных» моллюсков на задней части ноги имеется кон-
хиолиново-известковая крышечка, закрывающая устье раковины при втягивании
внутрь мягкого тела животного.
Туловище моллюсков билатерально (двусторонне) симметрично или спираль-
но закручено. На большем или меньшем его протяжении имеется свешивающа-
яся книзу складка — мантия, наружная поверхность которой образует раковину.
У двустворчатых моллюсков края лепестков мантии могут срастаться, образуя
три отверстия: одно, расположенное примерно посередине брюшного края, слу-
жит для высовывания ноги и два (вводное и выводное) в задней части животно-
го — для обеспечения тока воды в мантийной полости. Иногда участки мантии
окружающие вводное и выводное отверстия разрастаются, образуя сифоны. Меж-
ду внутренней поверхностью мантии и телом животного находится мантийная
полость, сообщающаяся с внешней средой. Мантийная полость содержит жабры,
анальное отверстие, отверстия выделительных органов и иногда половых прото-
ков. У некоторых брюхоногих моллюсков жабры редуцируются, а участок стенки
мантии преобразуется в «лёгкое» (Pulmonata sensu lato).
Тело пресноводных моллюсков заключено в раковину, образованную монолит-
ной спирально завитой трубкой или двумя створками.
У двустворчатых моллюсков на каждой створке выделяют спинной, брюш-
ной, передний и задний края, плавно переходящие друг в друга или образующие
на границе смежных краёв закруглённый уголок. Створки на спинной стороне
связаны эластичной перемычкой — лигаментом. Основная, наиболее заметная
часть лигамента всегда лежит позади макушек, что определяет положение перед-
него и заднего концов створки. Над спинным краем в той или иной степени воз-
вышается макушка — начальная часть каждой створки. Она может быть сдвину-
та вперёд, назад или располагаться более или менее посередине спинного края.
Форма поверхности раковины около макушек (степень её приподнятости над кра-
ем створки) имеет таксономическое значение.
У некоторых групп двустворчатых спинной край за макушками может разрас-
таться, закрывая лигамент и образуя наклонённое к переднему краю крыло. На
наружной поверхности створки видны линии нарастания (роста), а реже — кон-
центрические валики — морщины. Наиболее выделяющиеся своей глубиной и
чёткостью линии роста обычно рассматриваются как годовые кольца. У некоторых
раковин макушечная скульптура может быть не развита или не видна из-за корро-
зии. Часто за макушками имеется линия в виде резкого перегиба, проходящего от
макушки до нижней части заднего края створки — задний килевой перегиб.
10 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
У многих Bivalvia внутри раковины имеется замок, состоящий из расширен-
ного участка спинного края — замочной площадки и расположенных на нём не-
скольких выступов различной формы — зубов. Замок предназначен для точного
совмещения створок при их смыкании, поскольку каждому зубу одной створки
соответствует углубление в другой. Замки бывают различных типов и имеют
большое значение при выделении таксонов высокого ранга. У представителей
Unionidae и Margaritiferidae замок схизодонтный, относящийся к группе прегете-
родонтных, у Sphaeriidae — гетеродонтный.
На внутренней поверхности раковины в её передних и задних частях имеются
по две пары мускульных отпечатков — мест прикрепления мускулов-замыкате-
лей (аддукторов) и мышц, втягивающих ногу (ретракторов) к створкам. Поверх-
ность отпечатков обычно гладкая, с заметными линиями роста. У представителей
Margaritiferidae передние отпечатки, хотя бы частично, имеют морщинистый ре-
льеф. Рядом с отпечатками аддукторов (или сливаясь с ними) помещаются отпе-
чатки ретракторов ноги. В некоторых случаях аддукторы крепятся к особым вну-
тренним примакушечным выростам — септам (например, у представителей рода
Dreissena). Вдоль краёв створок тянется след от прикрепления мышц мантии —
мантийная линия. В тех случаях, когда имеются развитые сифоны, мантийная
линия в задней половине створки образует более или менее глубокий изгиб —
мантийный синус. Участок между мантийной линией и брюшным краем створ-
ки имеет вид слабовыпуклого валика, называемого прикраевым валиком. У пред-
ставителей Margaritiferidae мантийные мускулы прикрепляются кроме мантийной
линии почти по всей внутренней поверхности створки.
У брюхоногих моллюсков раковина представляет собой замкнутую с одного
конца коническую трубку, в разной степени скрученную в спираль, либо преобра-
зованную в невысокий конус (колпачок), сформированный увеличенным послед-
ним оборотом.
Узкий замкнутый конец раковинной трубки именуется вершиной. На проти-
воположном конце раковина открывается устьем. Совокупность оборотов, рас-
положенных выше устья, называется завитком. Внутренние стенки оборотов
обычно сливаются, образуя столбик (колумеллу), который часто снабжён полым
внутренним каналом, открывающимся у основания завитка раковины отверстием
(пупком). Пупок может быть открытым, либо в различной степени или целиком
прикрытым отворотом колумеллярного края устья. Первые 1,5-2,0 оборота рако-
вины (редко 3,0), образующиеся ещё на зародышевой стадии, называются эмбри-
ональными или протоконхом. Нередко протоконх отличается цветом или микро-
скульптурой от остальной (дефинитивной) части раковины. Воображаемая линия,
вокруг которой происходит навивание оборотов называется осью раковины. Ли-
ния, разделяющая обороты, именуется швом. Шов может быть мелким (у раковин
с уплощёнными оборотами) или в той или иной степени углублённым (у раковин
с выпуклыми оборотами), прямым (ориентированным более или менее перпенди-
кулярно к оси раковины) или скошенным. Иногда по краю спирально закручен-
Моллюски
11
ных раковин развита резкая угловатость — киль. Поверхность оборотов может
быть гладкой или с различными скульптурными элементами: рёбрами, морщина-
ми, бороздками, вмятинами, шипами и волосками. Рёбра, морщины и бороздки
могут быть спиральными (ориентированными вдоль оборотов) или радиальными/
осевыми (ориентированными поперёк оборотов).
По направлению навивания раковины брюхоногих моллюсков могут быть
лево- или правозавитыми. Чтобы установить это направление необходимо распо-
ложить раковину замкнутым концом спиральной трубки (верхушкой) вверх так,
чтобы плоскость устья была перпендикулярна оси зрения наблюдателя. При этом
расположение устьевого контура относительно оси раковины оси покажет сторо-
ну навивания оборотов.
В пределах устьевого контура различают: париетальный край устья, образо-
ванный стенкой предпоследнего оборота; колумеллярный край, образованный
столбиком; свободный палатальный край устья, причём его нижняя часть иногда
выделается как самостоятельный базальный край. В верхней части устья обычно
имеется явственно различимый парието-палатальный угол.
Важное значение для диагностики многих семейств и родов брюхоногих мол-
люсков имеет форма раковины. Формы раковин брюхоногих моллюсков, исполь-
зуемые в ключах данного определителя, приведены на рис. 1.
Методы сбора и камеральной обработки пресноводных моллюсков
Двустворчатые моллюски
Крупные двустворчатые моллюски собираются вручную или также как и мел-
кие — с использованием специальных орудий лова (зубчатый сачок, драги раз-
личных конструкций, черпаки, небольшие бредни с привязанной к нижней подбо-
ре цепью, невода, широкие грабли с решёткой на тыльной стороне, выполненной
из металлических стержней и проволоки).
Первую фиксацию двустворчатых моллюсков следует проводить 96% этило-
вым спиртом, который через сутки следует заменить на 70% спирт. Пустые рако-
вины лучше всего сохранять в сухом виде. Чтобы раковины не растрескивались
при высыхании, их смазывают глицерином.
Для изучения строения мягкого тела моллюсков их раковины вскрывают.
У крупных экземпляров скальпелем или тонким ножом как можно аккуратнее пе-
ререзают аддукторы на участке между мантией и створкой, после чего раковина
раскрывается. Мелких моллюсков при таком способе легко повредить, поэтому
их перед вскрытием выдерживают в течение 1-2 суток в 40-50% растворе эти-
лового спирта. При такой процедуре створки раскрываются за счёт слабой маце-
рации в зоне прикрепления аддукторов. После этого материал фиксируют в 70%
спирте.
Обычно раковина двустворчатых моллюсков измеряется по трём стандартным
промерам: длина — расстояние по продольной оси раковины между наиболее
12
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 1. Формы раковин пресноводных Gastropoda.
А — колпачковидная; Б — неритоидная (полушаровидная, полуяйцевидная); В — дис-
ковидная турбоспиральная; Г — дисковидная плоскоспиральная; Д — коническая;
Е — низкоконическая (1 — прижатая, 2 — не прижатая); Ж — высококоническая; 3 —
овально-коническая; И — башневидно-коническая (1 — для представителей большинства
групп пресноводных гастропод, 2 — для представителей сем. Lymnaeidae по Хохуткин и
др., 2009); К — башневидная (1 — для представителей большинства групп пресноводных
гастропод, 2 — для представителей сем. Lymnaeidae по Хохуткин и др., 2009); Л — оваль-
ная; М — яйцевидная (овально-башневидная); Н — уховидная; О — шаровидная; П —
плоскоспиральная линзовидная (в виде двояковыпуклой линзы); Р — плоскоспиральная
линзовидная [в виде односторонне выпуклой (плосковыпуклой) линзы]; С — плоскоспи-
ральная шайбовидная; Т — кубаревидная.
Моллюски
13
Рис. 2. Основные морфологические особенности и промеры раковин крупных дву-
створчатых моллюсков.
I. — раковина перловицы Unio pictorum Linnaeus, 1758 (по: Жадин, 1952); II. — раковина
сем. Unionidae [по: Старобогатов и др., 2004 (с изменениями); III. — раковина сем. Dreis-
senidae [по: Богуцкая и др., 2013 (с изменениями)].
1 — задние мускульные отпечатки; 2 — латеральные зубы; 3 — кардинальные зубы; 4 —
передние мускульные отпечатки; 5 — мантийная линия; 6 — макушка; 7 — лигамент; 8 —
киль (килевой перегиб); 9 — септа; A-Aj — длина раковины; Б-Bj — высота раковины;
Г-Fj — выпуклость (толщина) раковины; АВ — передняя часть раковины; BAj — задняя
часть раковины; вк — высота раковины у крыла; лмвк — линия максимального выпуклого
контура на боковой поверхности створки; мт — расстояние между макушкой и наиболее
выступающей точкой боковой поверхности раковины; т — наиболее выступающая точка
боковой поверхности раковины.
удалёнными точками переднего и заднего краёв; высота — расстояние по перпен-
дикуляру к продольной оси раковины между макушкой (высота у макушек), либо
лигаментом (высота у лигамента), либо вершиной крыла (высота у крыла) и наи-
более удалённой точкой брюшного края; выпуклость — расстояние по перпенди-
куляру к комиссуральной плоскости между наиболее удалёнными от неё точками
створок (рис. 2). Высота у лигамента обычно измеряется в случаях сильной изъ-
еденности (коррозии) макушек, сломанном крыле или когда макушка значительно
смещена к переднему краю и расположена заметно ниже лигамента. При измере-
нии высоты раковины необходимо указывать, в каком месте было проведено из-
мерение (у макушки, лигамента или крыла). Степень выпуклости раковины (ин-
декс выпуклости) обычно оценивается соотношением выпуклости раковины (В) к
14 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
её высоте (Н). Если значение индекса В/Н меньше 0,3-0,4 — раковина считается
плоской или слабовыпуклой, при значении индекса от 0,3-0,4 до 0,5-0,6 — уме-
ренно выпуклой, от 0,5-0,6 до 0,7 — выпуклой, и свыше 0,7 — сильновыпуклой.
Через соотношение высоты раковины (Н) к её длине (L) определяется общая
форма раковины. Например, если у моллюсков с неразвитым крылом соотноше-
ние H/L не превышает 0,45, то раковина считается удлинённо-овальной, от 0,5
до 0,6 — овальной, и около 0,7-0,8 — яйцевидной. Наличие островершинного
крыла в совокупности с узким передним и задним краями придаёт раковине тре-
угольную форму, а у моллюсков без крыла, но с заметными резкими переходами
или уголками между, как минимум, тремя краями раковины (например: передним
и спинным, спинным и задним, задним и нижним) раковина оценивается как тра-
пециевидная или овально-четырёхугольная. Суженная за макушками задняя часть
раковины придаёт ей коническую форму, а суженная передняя часть раковины —
каплевидную или клювовидную.
Брюхоногие моллюски
Простейшим способом отлова брюхоногих моллюсков является сбор моллю-
сков руками на мелководье и в осушённой зоне по берегам водоёма. Вручную или
пинцетом производится сбор моллюсков или их кладок с поверхности водных
растений и погружённых в воду предметов.
Основными орудиями качественного сбора бентосных моллюсков являются
скребок и драга. Если скребки применяются на мелководьях, то драгами, вле-
комыми по дну на верёвке, можно облавливать как мелководье, так и глубокие
участки водоёмов. Драги бывают треугольной или четырёхугольной формы, они
могут быть снабжены зубчатыми или режущими краями, захватывающими грунт.
Количественные сборы моллюсков проводятся с применением дночерпателей
различных типов и модификаций.
Первичная фиксация собранных животных проводится в полевых условиях с
помощью 96% этанола. При этом следует учитывать, что в мантийной полости
моллюсков может содержаться некоторое количество воды, выделяющейся после
гибели, что ведёт к снижению исходной концентрации фиксатора. Спустя сутки,
животные переводятся в 70% этанол для длительного хранения. Для облегче-
ния анатомирования перед фиксацией моллюсков может применяться анастезия
с помощью различных химических веществ: хлоралгидрата, нембутала, пенто-
барбитала, ментола, хлороформа, эфира и т.п. Также умерщвление собранных
животных можно производить крутым кипятком, после чего тело моллюска срав-
нительно легко извлекается (выкручивается) из раковины и переносится в спирт.
Раковина при этом высушивается и хранится отдельно.
Промеры раковины обычно осуществляются с точностью до 0,1 мм при помо-
щи штангенциркуля или окуляр-микрометра микроскопа. Для каждого из морфо-
логических типов раковины разработана особая схема измерений (рис. 3), строгое
Моллюски
15
Рис. 3. Схемы промеров раковин брюхоногих моллюсков, используемые в определи-
тельных таблицах.
А, Б — по: Андреева и др., 2010; В — по: Старобогатов и др., 2004. А — турбоспиральная
раковина, Б — плоскоспиральная раковина, В — колпачковидная раковина. Обозначения
промеров см. в тексте.
следование которой необходимо для сопоставимости полученных данных со зна-
чениями, приводимыми в определительных ключах.
Измерение турбоспиральных раковин (семейств Bithyniidae, Lymnaeidae, Phy-
sidae, Valvatidae и др.; рис. 2А) предполагает шесть основных промеров: высота
раковины (ВР), измеряемая строго вдоль оси раковины от вершины до нижнего
(базального) края устья; ширина раковины (ШР) — расстояние между наиболее
удалёнными от оси точками палатального края устья и поверхности последнего
оборота на противоположной устью (в проекции на линию, перпендикулярную
оси раковины); высота завитка (ВЗ) — расстояние от вершины до верхней точки
устья (в проекции на ось раковины); высота последнего оборота (ВПО) — рассто-
16
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
яние от базального края устья до расположенного над устьем участка шва меж-
ду последним и предпоследним оборотами; высота устья (ВУ) — расстояние от
верхней точки устья до наиболее удалённой от него точки базального края (изме-
ряется строго в плоскости устья); ширина устья (ШУ) — расстояние между наи-
более удалёнными точками устья, измеренное перпендикулярно ВУ У раковин с
развитым колумеллярным отворотом оно может измеряться двояко: включая ко-
лумеллярный отворот (ШУСКО) или без него. Большая часть указанных промеров
применима и к неритоидным раковинам (род Theodoxus семейства Neritidae).
У плоскоспиральных раковин (семейство Planorbidae, некоторые Valvatidae;
рис. 2Б) промеры ВЗ и ВПО выполнить невозможно, зато вводятся несколько до-
полнительных измерений. Это — диаметр внутренних оборотов (ДВО) и ширина
трубки последнего оборота (ШПО), соотношение которых даёт индекс внутрен-
них оборотов (ИВО). ИВО может определяться как с апикальной (вершинной),
так и с базальной поверхности раковины и отражает скорость нарастания обо-
ротов раковины. Иногда необходимо знать ширину предпоследнего оборота
(ШППО), измеряемую аналогично ШПО.
При измерении колпачковидных раковин (семейства Acroloxidae и Ancylidae;
рис. 2В) высотой (ВР) является расстояние от самой верхней точки раковины до
плоскости устья. Длина раковины измеряется между двумя наиболее удалённы-
ми точками, расположенными на переднем и заднем крае устья соответственно.
Ширина раковины измеряется перпендикулярно длине. Эти промеры у колпачко-
видных раковин соответствуют длине и ширине устья у спиральных форм. Затем,
измеряют удаление вершины от переднего края (УПК) в проекции на линию про-
мера длины; удаление вершины от левого края (УПК) в проекции на линию про-
мера ширины и длину хорды переднего склона (ДХ) — расстояние от вершины
до самой передней точки устья.
В качестве дополнительного показателя у турбоспиральных раковин исполь-
зуется величина апикального угла измеряемого между двумя касательными, про-
ходящими через наиболее выступающие точки всех или нескольких оборотов
завитка. Измерение апикального угла проводится по рисунку или фотографии
раковины. В последнее время для этой же цели стали применяться различные
устройства для оцифровки изображений.
КЛАСС ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ —
BIVALVIA LINNAEUS, 1758
Ключ для определения семейств и некоторых родов
пресноводных двустворчатых моллюсков
1(4). Внутренняя поверхность створки перламутровая. У европейских пресно-
водных макушки сдвинуты вперёд, но не на самый передний конец рако-
вины.
Моллюски
17
2(3).
3(2).
4(1).
5(6).
6(5).
1(4).
2(3).
Передние зубы хорошо развиты на обеих
створках, а задние полностью отсутству-
ют или представлены едва заметными ва-
ликами. Отпечаток переднего аддуктора,
хотя бы частично, неправильно морщини-
стый...Сем. Margaritiferidae Haas, 1940.
В европейской части России семейство пред-
ставлено одним родом Margaritifera Schum-
acher, 1816 (рис. 4).
Хорошо развиты и передние и задние
зубы, или зубы замка полностью отсут-
ствуют. Отпечаток переднего аддуктора
гладкий или с тонкими линиями.........
..........Сем. Unionidae Fleming, 1828.
Внутренняя поверхность створки не пер-
ламутровая, матовая. Форма раковины
другая.
Макушки сдвинуты на самый передний
Рис. 4. По: Старобогатов и др.,
2004.
Рис. 5.
конец раковины, что придаёт ей треуголь-
ную, каплевидную или клювовидную форму; часто
имеется более или менее выраженный килевой перегиб................
........................................Сем. Dreissenidae Gray, 1840.
В европейской части России семейство представлено одним родом Dreissena Van
Beneden, 1835 (рис. 5).
Раковина округлая, округло-овальная или округло-треугольная. Макушки
располагаются посередине спинного края створок или могут быть слегка
смещены вперёд или назад. Килевой перегиб отсутствует.................
................................Сем. Sphaeriidae Deshayes, 1855 (1820).
Ключ для определения родов семейства Unionidae
Передние и задние зубы имеются.
Раковина удлинённо овальная, овальная
или овально-коническая. Макушечная
скульптура представлена двумя расходя-
щимися рядами бугорков, либо бугорками
и валиками в виде букв V или W, либо
валиками в виде буквы W. Передние зубы
пластинчатые........................
Рис. 6. По: Старобогатов и др.,
2004.
... Unio Philipsson in Retzius, 1788 (рис. 6).
3(2). Раковина коротко-овальная, овальная или трапециевидная. Спинной край
часто выгнут в виде широкого горба. Макушечная скульптура представлена
вложенными друг в друга тонкими валиками в виде греческой буквы й
18
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 7. По: Старобогатов и др.,
2004.
Рис. 8. По: Старобогатов и др.,
2004.
Рис. 9. По: Старобогатов и др.,
2004.
Рис. 10. По: Старобогатов и
др., 2004.
(омега), либо двумя рядами спрямлённых
валиков, сдвинутых по отношению друг к
другу. Передние зубы треугольные или тра-
пециевидные ...........................
..........Crassiana Servain, 1882 (рис. 7).
4(1). Передние и задние зубы отсутствуют.
5(8). Раковина разной степени выпуклости. Вы-
сота отпечатков задних мускулов-замыкате-
лей обычно меньше трети высоты раковины
за лигаментом. Мантийная линия у заднего
края узкая, слабозаметная. Макушечная
скульптура в виде тонких спрямлённых ва-
ликов, либо тонких концентрических расхо-
дящихся валиков. Передний край раковины
широкий, если узкий, то макушечная скуль-
птура в виде тонких спрямлённых валиков.
6(7). Раковины очень крупные, длиной до 20 см
и более. Наружная поверхность створок
чаще всего морщинистая. Макушечная
скульптура в виде концентрических расхо-
дящихся тонких валиков......................
..........Anodonta Lamarck, 1799 (рис. 8).
7(6). Раковины средних размеров, длиной до
10-14 см. Боковая поверхность чаще всего
гладкая. Макушечная скульптура в виде
ряда спрямлённых тонких валиков.............
...Colletopterum Bourguignat, 1876 (рис. 9).
8(5). Раковина плоская или умеренно выпуклая.
Высота отпечатков задних мускулов за-
мыкателей около или больше трети высоты
раковины за лигаментом. Мантийная линия
у заднего края раковины широкая, обычно
хорошо заметная. Макушечная скульптура в
виде крупных коротких валиков или отсут-
ствует. Передний край раковины узкий........
Pseudanodonta Bourguignat, 1876 (рис. 10).
Ключ для определения подсемейств и родов
семейства Sphaeriidae
1(4). Длина раковины взрослых особей не менее
7-8 мм. Макушка занимает срединное поло-
Моллюски
19
жение, иногда наклонена вйерёд.......
..............Подсем. Sphaeriinae s. str.
2(3). Раковина тонкостенная, хрупкая. Макуш-
ки очень узкие, выступающие («сосцевид-
ной» формы). Эмбриональная раковина
очень маленькая, в виде «шапочки»; ино-
гда она обособлена от остальной части
створки заметной бороздой. Замочная
площадка резко сужена под макушкой.........
.....Род Musculium Link, 1807 (рис. 11).
3(2). Раковина толстостенная, относительно
прочная. Макушки широкие. Эмбрио-
нальная раковина крупнее, не имеет вид
шапочки резко обособленной бороздой.
Как правило, она отграничена от осталь-
ной части створки линией нарастания или
цветом. Замочная площадка под макушкой
не сужена..................................
....Род Sphaerium Scopoli, 1777 (рис. 12).
4(1). Макушка отчётливо смещена к заднему
краю, если это смещение не выражено,
то раковина маленькая (длина раковины
меньше 5 мм).
5(20). Выводковая сумка расположена ближе к
брюшному краю створок. Если раковина
больше 5,5 мм в длину и лигамент виден
со спинной стороны закрытой раковины,
Рис. 11. По: Корнюшин, 1996.
Рис. 13. По: Корнюшин, 1996.
то внешняя поверхность створок гладкая, с тонкими линиями нарастания,
тонко исчерченная или с тонкими рёбрышками..........................
..............................Подсем. Euglesinae Pirogov et Starobogatov, 1974.
6(7). Раковина относительно крупная (длина створок не менее 7 мм). Лигамент
хорошо виден, если смотреть на закрытую раковину сверху...................
.........................................Род Lacustrina Sterki, 1916 (рис. 13).
7(6). Раковина меньше 7 мм в длину.
8(19). Раковина небольшая (длина створок от 2,5 до 5,5 мм). Лигамент закрыт
тонкой известковой стенкой и снаружи не виден (или заметен, просвечи-
вая через стенку).
9(16). Участок сросшихся листков мантии между нижним сифоном и не срос-
шимися листками мантии в передней части тела, образующими педальное
отверстие для высовывания ноги, не удлинён и составляет не более 0,4
длины свободного края каждого из мантийных листков.
20
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 14. По: Корнюшин, 1996.
Рис. 15. По: Корнюшин,
1996.
Рис. 16. По: Корнюшин,
10(11). Лигаментная ямка узкая и длинная. Ма-
кушки узкие, отчётливо смещены к задне-
му краю створок, благодаря чему раковина
приобретает округло-треугольную форму.
На макушках, как правило, формируются
косо расположенные складки. Если склад-
ки отсутствуют, то макушки располага-
ются ближе к центру спинной стороны
створок и не придают раковине округло-
треугольную форму.............................
....Род Henslowiana Fagot, 1892 (рис. 14).
11(10). Лигаментная ямка короткая и широкая.
Макушки широкие или узкие, но шире,
чем у предыдущего рода, всегда без скла-
док, сильно или слабо сдвинутые к за-
днему краю створок (в последнем случае
спинной край створок выгнутый). Ракови-
на округло-овальная или неправильно-че-
тырёхугольная, если округло-треугольная,
то макушки широкие.
12(15). Раковина округло-овальная с выраженной
концентрической исчерченностью или
ребристостью. Если гладкая, то непра-
вильно-четырёхугольной, округло-треу-
гольной или трапециевидной формы. От-
печатки внутренних радиальных мантий-
1996. ных мускулов не отделены от мантийной
линии.
13(14). Раковина округло-овальная с узкими макушками и выраженной концен-
трической исчерченностью наружной поверхности створок (особенно в
области макушки). Внутренние радиальные мантийные мускулы развиты
слабо и малозаметны...........................Род Hiberneuglesa Starobogatov
in Dolgin, 1983 (рис. 15).
14(13). Раковина с широкими макушками. Если округло-овальная, то с концентри-
ческой ребристостью наружной поверхности створок, если гладкая, то не-
правильно-четырёхугольной формы. Внутренние радиальные мантийные
мускулы хорошо развиты....................................................
...............Род Cingulipisidium Pirogov et Starobogatov, 1974 (рис. 16).
15(12). Раковина гладкая, округло-овальной, реже трапециевидной или округло-
треугольной формы. Отпечатки внутренних радиальных мантийных му-
скулов (все или частично) отделены от мантийной линии.....................
............................Род Euglesa Leach in Jenyns, 1832 (рис. 17).
Моллюски
21
16(9). Участок сросшихся листков мантии между
нижним сифоном и не сросшимися листка-
ми мантии в передней части тела заметно
удлинён и составляет около половины (или
больше) длины свободного края каждого
из мантийных листков.
17(18). Раковина удлинённо-четырёхугольная
(трапециевидная), с ясно выраженными
углами и спрямлённым брюшным краем.
Макушки приближены к середине спин-
ного края. Наружная поверхность створок
гладкая, блестящая. Передний аддуктор
очень большой, бобовидной формы. Почти
все отпечатки мантийных мускулов (кроме
двух в срединной части створок) слиты с
мантийной линией............................
...........Род Tetragonocyclas Pirogov et
Starobogatov, 1974 (рис. 18).
18(17). Раковина округло-треугольная, клиновид-
ная, гладкая, тонко ребристая или исчер-
чена. Брюшной край выгнутый. Макушки
сильно смещены к заднему краю створок
и наклонены назад. Передний аддуктор
не увеличен, овальной формы. Отпечатки
мантийных мускулов отделены на значи-
тельное расстояние от мантийной линии...
.................Род Pseudeupera German, 1913
(рис. 19).
19(8). Раковина очень маленькая (длина створок
не более 2,0 мм). Сифональное отверстие
одно, наружная полужабра отсутствует........
....Род Conventus Pirogov et Starobogatov,
1974 (рис. 20).
20(5). Выводковая сумка расположена ближе к
спинному краю створок. Если раковина
больше 5,5 мм в длину и лигамент виден
со спинной стороны закрытой раковины,
то внешняя поверхность створок с концен-
трическими резкими рёбрами или грубо
концентрически исчерчена....................
...........Подсем. Pisidiinae Gray, 1857.
Рис. 17. По: Корнюшин, 1996.
Рис. 19. По: Корнюшин, 1996.
Рис. 20. По: Корнюшин, 1996.
22
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 21. По: Корнюшин, 1996.
Рис. 22. По: Корнюшин, 1996.
Рис. 23. По: Корнюшин, 1996.
21(22). Длина створки достигает 11 мм. Сифональ-
ных отверстий два..............................
Род Pisidium Pfeiffer, 1821 (рис. 21).
22(21). Длина створки, как правило, не превышает
5 мм. Сифональное отверстие одно.
23(24). Раковина очень маленькая (длина створки
1,5-2,3 мм). Лигамент находится в верхней
части замочной площадки (закрытый тип
лигамента). Лигаментная ямка удлинена,
ланцетовидной формы...........................
...Род Neopisidium Odhner, 1921 (рис. 22).
24(23). Раковина более крупная. Лигамент глубоко
погружён в толщу раковины и достигает
нижнего края замочной площадки (инвер-
тированный тип лигамента). Лигаментная
ямка треугольной формы........................
.........Род Odhneripisidium Kuiper, 1962
(рис. 23).
КЛАСС БРЮХОНОГИЕ МОЛЛЮСКИ —
GASTROPODA CUVIER, 1797
Ключ для определения семейств и некоторых родов
пресноводных брюхоногих моллюсков
1(4). Раковина колпачковидная, не завита в спираль.
2(3). Контур устья (или раковины при взгляде сверху) округло- или яйцевидно-
овальный. Вершина в виде небольшого загнутого рожка располагается на
продольной оси раковины или немного смещена вправо...............
Рис. 24.
.................Сем. Ancylidae Rafinesque, 1815.
В европейской части России семейство представлено
одним родом Ancylus O.F. Muller, 1773 (рис. 24).
3(2). Контур устья (или раковины при взгляде
сверху) продолговто-овальный. Вершина в
виде небольшого загнутого рожка явствен-
но смещена влево от продольной оси рако-
вины.......Сем. Acroloxidae Thiele, 1931.
Моллюски
23
В европейской части России семейство представлено
одним родом Acroloxus Beck, 1837 (рис. 25).
4(1). Раковина в виде замкнутой с одного конца трубки,
закрученной в спираль различной формы.
5(30). Устье раковины закрывается крышечкой, как прави-
ло, не врезано стенкой последнего оборота. Устье-
вые края внутри не имеют наплывов и складок.
6(7). Раковина неритоидная (полушаровидная или полу-
яйцевидная), “уховидная” если смотреть со сто-
роны устья; с немногочисленными (до 3), быстро
нарастающими оборотами. Наружная поверхность
раковины с рисунком из светлых пятен на тёмном
фоне или зигзагообразных линий, реже полностью
чёрная. Завиток почти не выступающий или сла-
бо выступающий. Устье полукруглое, с широкой
хорошо выраженной колумеллярной (устьевой)
площадкой. Крышечка с двумя внутренними от-
ростками (апофизами)................................
.................Сем. Neritidae Rafinesque, 1815.
В европейской части России семейство представлено
одним родом Theodoxus Montfort, 1810 (рис. 26).
7(6). Раковина иная.
Рис. 25.
Рис. 26.
8(13). Раковина при 3-8 оборотах очень маленькая. Высо-
та раковины до 6,5 мм, ширина до 2,7 мм.
9(10). Раковина овальная, яйцевидная (овально-башневидная),
башневидная с высотой не меньше 2,2 мм и шириной не
меньше 1,5 мм или овально-коническая с высотой не более
3,0 мм и шириной не более 1,7 мм............................
...........................Сем. Amnicolidae Tryon, 1863.
В европейской части России семейство представлено одним ро-
дом Marstoniopsis Regteren Altena, 1936 (рис. 27).
10(13). Раковина кубаревидная, башневидно-коническая, высоко-
коническая или яйцевидная. Если раковина овальная или
башневидная, то её высота не больше 2,1 мм, ширина не
больше 1,4 мм. Если раковина овально-коническая, то её
высота не менее 3,1 мм, ширина не менее 1,8 мм..............
.........................Сем. Hydrobiidae Troschel 1857.
11(12). Раковина коническая или высококоническая, обороты
сильно выпуклые, разделённые глубоким швом. Вершина
парието-палатального угла (угловатости в верхней части
устья) резко выражена.......................................
...................Род Ventrosia Radoman, 1977 (рис. 28).
Рис. 28.
24
Определитель, зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 30.
Рис. 29.
Рис. 31.
Рис. 32.
12(11). Раковина высоко коническая, обороты умеренно или сла-
бо выпуклые, шов не глубокий. Вершина парието-пала-
тального угла (угловатости в верхней части устья) плавно
скруглена............Род Potamopyrgus Stimpson 1865 (рис. 29).
13(8). Раковина при 3-8 оборотах, как правило, более крупная.
В тех случаях, когда высота раковины меньше 6,6 мм, её
ширина всегда не менее 2,8 мм.
14(17). Раковина не менее 25 мм высотой, кубаревидная, редко
коническая. Поверхность оборотов, как правило, с 2-3
спиральными бурыми полосами на светлом фоне. Иногда
эти полосы настолько широкие, что могут сливаться, об-
разуя однотонную бурую окраску раковины....................
...........................Сем. Viviparidae Gray, 1847.
15(16). Раковина толстостенная, кубаревидная или овально-кони-
ческая (отношение ширины раковины к её высоте — не
более 0,78), пупок полностью закрыт отворотом устьевого
края, либо в виде очень узкой щели. Обороты умеренно
выпуклые, на периферии могут быть уплощённые. Устье
округло-каплевидное, как правило, с отчётливым углом
между париетальным и палатальным краями. Вершина за-
витка куполообразно сглаженная, притуплённая...............
...................Род Viviparus Monfort, 1810 (рис. 30).
16(15). Раковина относительно тонкостенная, кубаревидная или
коническая (отношение ширины раковины к её высоте
превышает 0,80), пупок широко-щелевидный. Обороты
сильновыпуклые, на периферии равномерно закруглённые,
расположенные ступенчато. Устье овальное или округло-
каплевидное со сглаженной угловатостью между парие-
тальным и палатальным краями. Вершина завитка — за-
острённая...........Род Contectiana Bourguignat, 1880 (рис. 31).
17(14). Раковины не превышает 25 мм в высоту, Различной
формы: от плоскоспиральной, дисковидной и низкокони-
ческой до высококонической и кубаревидной. Бурые спи-
ральные полосы на поверхности оборотов отсутствуют.
18(21). Раковина кубаревидная, с гладкой, фарфоровидной поверхностью или с
очень тонкой радиальной (осевой) исчерченностью оборотов. Последний
оборот сильно вздут.
19(20). Раковина очень маленькая, при 4 оборотах её высота 2,5-4,5 мм, ширина
3,5-5,5 мм. Колумеллярная (устьевая) площадка отсутствует; пупок округ-
лый, проколовидный, менее чем на половину прикрыт отворотом колумел-
лярного края устья....................................................
....Сем. Valvatidae Gray, 1840, род Borysthenia Lindholm, 1914 (рис. 32).
Моллюски
25
20(19). Раковина крупнее, при 4 оборотах её высота 7,0-9,0 мм,
ширина 6,5-8,0 мм. Утолщённые и отвёрнутые колу-
меллярный и париетальный края устья образуют узкую
колумеллярную (устьевую) площадку. Пупок полностью
закрыт отворотом колумеллярного края устья................
....................Сем. Lithoglyphidae Troschel, 1857.
В европейской части России семейство представлено одним
родом Lithoglyphus Megerle von Miihfeld in Hartmann, 1821
(рис. 33).
21(18). Раковина не кубаревидная: дисковидная (плоско- или тур-
боспиральная), низко-, овально- или башневидно-кониче-
ская, коническая. Последний оборот не вздут, расширяет-
ся более или менее равномерно.
22(27). Раковина коническая или овально-коническая, с заострён-
ной вершиной, поверхность оборотов в общем гладкая
(блестящая или матовая), иногда со спиральной исчерчен-
ностью и/или тонкой радиальной (осевой) морщинисто-
стью. Пупок щелевидный, в разной степени открытый,
иногда полностью закрытый отворотом колумеллярного
края устья. Устье овальное или каплевидно-овальное, за-
крывается обызвествлённой концентрической крышечкой
.................................Сем. Bithyniidae Gray, 1857.
23(24). Раковина маленькая, при 4,0-5,5 оборотах её высота не
превышает 6 мм. Ширина предпоследнего оборота не пре-
вышает 2,6 мм.............................................
......Род Codiella Monterosato in Locard, 1894 (рис. 34).
24(23). Раковина крупнее, при 4,0-5,5 оборотах её высота не ме-
нее 6,1 мм. Ширина предпоследнего оборота более 2,8 мм.
25(26). Обороты слабовыпуклые, иногда на периферии уплощён-
ные, разделённые мелким швом. Пупок закрытый или
в виде очень узкой щели...................................
..............Род Bithynia Leach in Abel, 1818 (рис. 35).
26(25). Обороты сильновыпуклые, иногда ступенчатые, разделён-
ные глубоким швом. Пупок каплевидный в разной степени
открытый (иногда довольно узкий, но не узкощелевид-
ный)......................................................
Рис. 33.
Рис. 34.
Рис. 35.
Рис. 36.
.........Род Opisthorchophorus Beriozkina, Levina, Starobogatov, 1995 (рис. 36).
27(22). Раковина дисковидная (плоско- или турбоспиральная), низко- или баш-
невидно-коническая с притуплённой вершиной; поверхность оборотов
гладкая или со спиральными, либо радиальными (осевыми) рёбрышками.
Всегда имеется пупок округлой формы, широко развёрнутый, перспек-
тивный или узкий проколовидный, открытый или в различной степени
26
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
прикрытый отворотом колумеллярного края устья. Устье округлое, иногда
с лёгкой угловатостью в париетально-палатальной области, закрывается
необызвествлённой спиральной многооборотной крышечкой.............
.............................................Сем. Valvatidae Gray, 1840.
Рис. 37.
Рис. 38.
28(29). Обороты раковины в сечении круглые, разделённые
глубоким швом. Устье круглое или слегка овальное,
но всегда без резкого уголка или более или менее
плавной угловатости в верхней части. Ширина ра-
ковины у большинства видов при 3-4 оборотах не
превышает 4 мм. Если раковина плоскоспиральная,
то имеющиеся радиальные рёбрышки не несут по
кромке пластинчатых гребней, сформированных за
счёт периостракума.......................................
...................Род Valvata Muller, 1774 (рис. 37).
29(28). Обороты раковины в сечении овальные, разделённые
умеренно глубоким швом. Устье округлое, непра-
вильно-округлое, реже овальное, но всегда с резким
уголком или более или менее плавной угловатостью
в верхней части. Ширина раковины у большинства
видов при 3—4 оборотах превышает 4 мм. Если рако-
вина плоскоспиральная, то её поверхность покрыта
радиальными рёбрышками, несущими по кромке
пластинчатые гребни, сформированные за счёт пери-
остракума.............Род Cincinna Hubner, 1810 (рис. 38).
30(5). Крышечка, закрывающая устье, отсутствует. Колу-
меллярный и париетальный края устья внутри могут
иметь утолщения, складки или складкообразные на-
Рис. 39.
плывы. Устье нередко врезано стенкой последнего
оборота.
31(40). Раковина турбоспиральная, уховидная, шаровидная, яйцевид-
ная, башевидная, овально-, башневино- или высококоническая.
32(35). Раковина правозавитая.....Сем. Lymnaeidae Lamarck 1812.
33(34). Раковина очень стройная, башневидной формы, с большим
числом (6-7) медленно нарастающих плоских оборотов. Мешок
пениса состоит из двух частей — проксимальной (с тонкими
стенками) и дистальной (с железистыми стенками)................
.Род Aenigmomphiscola Kruglov et Starobogatov, 1981 1 (рис. 39).
1 У рода Aenigmomphiscola отсутствует конхологический диагноз. По признакам раковины
виды этого рода невозможно отличить от представителей подрода Omphiscola рода Lymnaea,
поэтому изучение анатомического строения моллюсков необходимо для надёжной диагностики
их родовой принадлежности (см. Приложение к тому: «Пресноводные моллюски севера и севе-
ро-запада Европейской России»: 279 с.).
Моллюски
27
34(33). Раковина иной формы; если,,башневидной, то
мешок пениса простого строения, не подразде-
лённый на проксимальную и дистальную части
...............Род Lymnaea Lamarck, 1799 (рис. 40).
35(32). Раковина левозавитая, овально- или высоко-
коническая............Сем. Physidae Fitzinger, 1837.
36(39). Раковина овально-коническая. Завиток невысо-
кий, его высота меньше высоты устья. Обороты
выпуклые.
37(38). Вершина раковины заострённая. Стенка ра- рИс. 40.
ковины прочная, не просвечивающая. Высота
раковины достигает 15-17 мм....................................
..........................Род Costatella Dall, 1870 (рис. 41).
38(37). Вершина раковины притуплённая. Стенки раковины тонкие,
полупрозрачные. Размеры меньше (высота раковины может до-
стигать 12 мм, но обычно — не более 10 мм).....................
.......................Род Physa Drapamaud, 1801 (рис. 42).
39(36). Раковина высококоническая. Завиток высокий и его высота пре-
вышает высоту устья. Обороты уплощённые........................
.........................Род Aplexa Fleming, 1820 (рис. 43).
40(31). Раковина плоскоспиральная. Если турбоспиральная, то диско-
видная (с едва возвышающимся над последним оборотом завит-
ком) или в различной степени прижатая кубаревидная (высота
завитка примерно равна половине высоты устья или меньше).
41(42). Раковина левозавитая (верхняя сторона раковины определяется
по меньшей погружённости/вдавленности начальных оборотов
в области верхушки по сравнению с более глубоко погружённы-
ми начальными оборотами в области пупка), плоскоспиральная,
крупная (при 4-5 оборотах ширина раковины больше 20 мм)....
..............................Сем. Helisomatidae ЕС. Baker, 1928.
В европейской части России семейство представлено одним родом
Planorbarius Dumeril, 1806 (рис. 44).
42(41). Раковина правозавитая, плоскоспиральная или турбо-
Рис. 41.
Рис. 42.
Рис. 43.
спиральная (дисковидная, линзовидная или кубаревид-
ная), относительно или очень маленькая (при 4-5 обо-
ротах ширина раковины меньше 20 мм).
43(46). Раковина плоскоспиральная. При 4-5 оборотах её ши-
рина не больше 8,6 мм. Контур раковины в поперечном
сечении может быть: симметрично выпуклым с верхней
и нижней сторон, так, что раковина имеет вид двояковы-
пуклой линзы, выпуклым сверху и уплощённым снизу
так, что раковина имеет форму от плоско-выпуклой
Рис. 44.
28
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 46.
Рис. 48.
линзы до полушаровидной................................
.....................Сем. Segmentinidae ЕС. Baker, 1945.
44(45). Последний оборот внутри содержит 2-3 группы радиально
ориентированных пластинок.....................................
..................Род Segmentina Fleming, 1817 (рис. 45).
45(44). Поперечные пластинки внутри последнего оборота отсутст-
вуют .........Род Hippeutis Agassiz in Charpentier, 1837 (рис. 46).
46(43).Раковина турбоспиральная (дисковидная или кубаревидная).
Если плоскоспиральная, то с уплощёнными сверху и снизу
оборотами, не придающими раковине облик двояко-, пло-
ско-выпуклой линзы или полушаровидную форму. При 4-5
оборотах ширина раковины больше 8,7 мм. В центральных
и северных областях европейской России виды с кубаревид-
ной раковиной неизвестны......................................
.......................Сем. Planorbidae Rafinesque, 1815.
47(48). Раковина крупная, твёрдостенная, при 4 оборотах её шири-
на не менее 9,0 мм, высота около 3,0 мм.......................
..................Род Planorbis O.F. Mtiller, 1773 (рис. 47).
48(47). Раковина средних размеров или маленькая, при 4 оборотах
её ширина не более 8,0 мм, высота около 1,8 мм.
Рис. 47.
49(50). При взгляде со стороны устья контур раковины в виде
высокой шайбы. При взгляде сверху тесно навитые
медленно нарастающие обороты напоминают скручен-
ный ремень.......................Род Bathyomphalus Agassiz
in Charpentier, 1837 (рис. 48).
50(49). Раковина плоскоспиральная в виде уплощённого дис-
ка, плоского диска или турбоспиральная.
51(52). Верхняя и нижняя стороны раковины при визуальном
осмотре (без увеличения) выглядят практически оди-
наковыми. Высота оборотов раковины увеличивается
медленно и равномерно относительно оси навивания, поэто-
му завиток и пупок раковины почти не погружённые (верх-
ние обороты и околопупочное поле сильно не вдавленные)
и имеют равные диаметры...........
.............Род Anisus Studer, 1820 (рис. 49).
52(51). Верхняя и нижняя поверхности ракови-
ны, за редким исключением, легко раз-
личимы (без увеличительных приборов).
Высота оборотов раковины увеличивает-
ся быстро и часто неравномерно относи-
тельно оси навивания, поэтому завиток
и пупок раковины погружены (верхние
Рис. 49.
Моллюски
29
Рис. 50. Рис. 51. Рис. 52.
обороты и околопупочное поле вдавленные) и имеют различные диа-
метры.
53(54). Устье не врезано стенкой предпоследнего оборота. Часто на поверхности
раковины развиты периостракальные рёбра...............................
...............................Род Armiger Hartmann, 1840 (рис. 50).
54(53). Устье врезано стенкой предпоследнего оборота, периостракальные рёбра
отсутствуют.
55(56). Число дефинитивных оборотов раковины больше 4.................
....................Род Gyraulus Agassiz in Charpentier, 1837 (рис. 51).
56(55). Число дефинитивных оборотов раковины не превышает 4...........
........................Род Choanomphalus Gerstfeldt, 1859 (рис. 52).
КОЛЬЧАТЫЕ ЧЕРВИ
Ключ для определения подклассов и отрядов кольчатых червей
1(4). Присоски и челюсти отсутствуют. Анус расположен терминально на за-
днем конце тела. Щетинки имеются у подавляющего большинства видов.
Передний конец тела несёт головную лопасть (за исключением видов рода
Сhae togas ter).
2(3). Волосные щетинки имеются только в спинных пучках или волосные ще-
тинки отсутствуют совсем.........................................
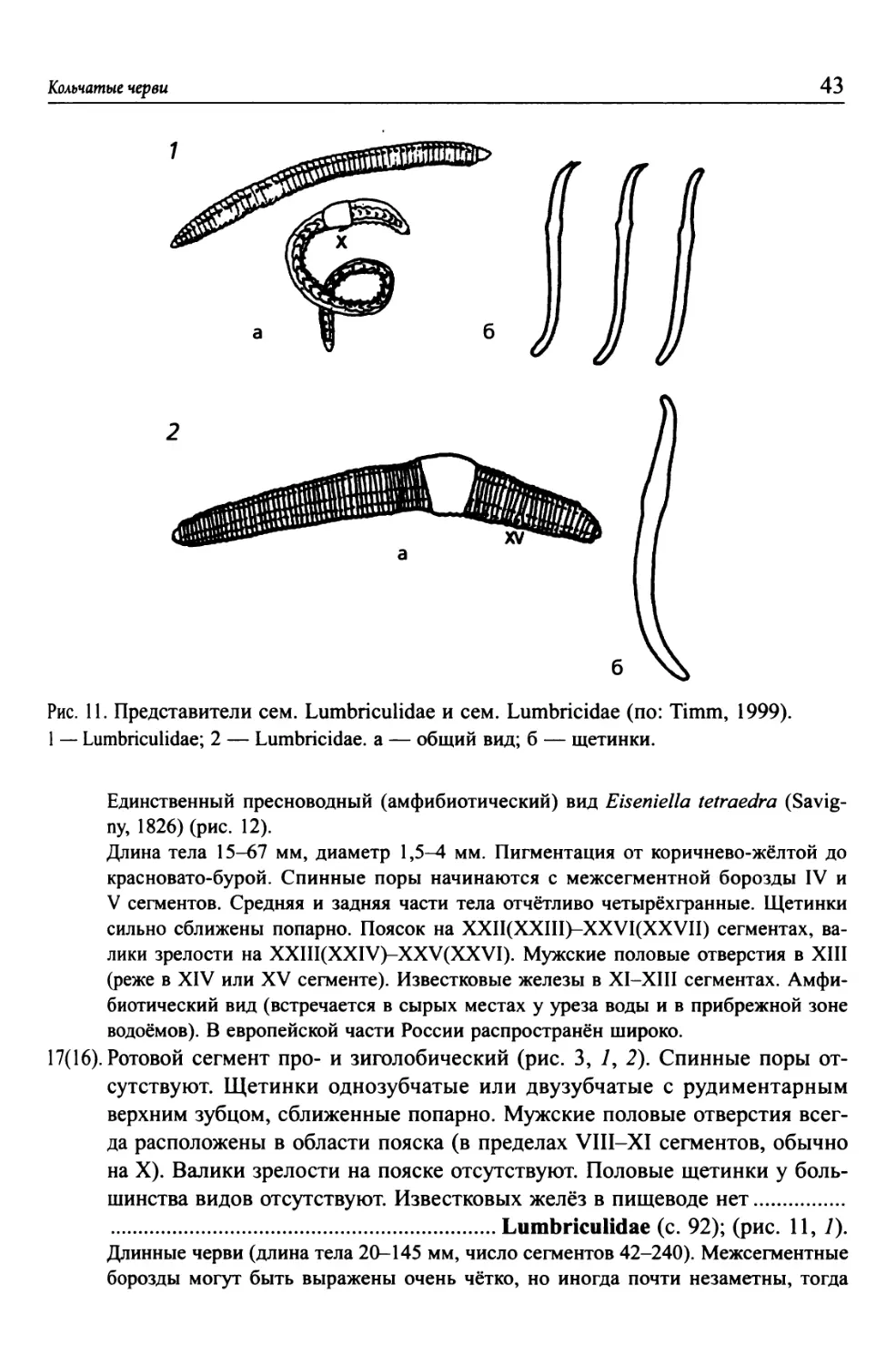
............Подкласс Oligochaeta (класс Clitellata) (с. 32) (рис. 3-94).
3(2). Волосные щетинки имеются и в спинных, и в брюшных пучках..
.............Отряд Aphanoneura (класс Polychaeta) (с. 30) (рис. 1; 2).
4(1). Присоски имеются хотя бы на заднем конце тела. Анус расположен на
спинной стороне у основания задней присоски. Головная лопасть отсут-
ствует. Щетинки отсутствуют (за исключением подкл. Acanthobdellida).
5(6). Присоски имеются на обоих концах тела (за исключением вторичной ре-
дукции у Archaeobdella esmonti)..................................
.................Подкласс Hirudinea (класс Clitellata) (с. 98) (рис. 99-125).
6(5). Присоска имеется только на заднем конце тела.
7(8). Первые 4 сегмента тела слиты в единый головной отдел. Щетинки отсут-
ствуют. Мелкие, прозрачные черви (несколько мм длиной)...........
.......Подкласс Branchiobdellida (класс Clitellata) (с. 94) (рис. 95-97).
8(7). Первые сегменты не слиты в единый отдел. Щетинки имеются на брюш-
ной стороне переднего конца тела (на I-V сегментах). Покровы тела пиг-
ментированы. Черви среднего размера (несколько см длиной)........
..........Подкласс Acanthobdellida (класс Clitellata) (с. 96) (рис. 98).
Класс МНОГОЩЕТИНКОВЫЕ ЧЕРВИ — POLYCHAETA
Отряд АФАНОНЕУРА — APHANONEURA
Семейство AEOLOSOMATIDAE (рис. 1; 2)
(ранее сем. Aeolosomatidae включали в состав
подкласса Малощетинковых червей (Oligochaeta))
Эолосаматиды — очень мелкие черви (длина тела от 0,1 до 4-6 мм, редко
10 мм). Тело короткое (состоит из 5-14, редко 19-22 сегментов), обычно сплю-
щено в спино-брюшной плоскости. Кутикула очень тонкая, благодаря чему фор-
ма тела весьма изменчива. Характерная особенность покровов — наличие сфе-
рических эпидермальных телец, цвет которых определяет окраску червя. Само
тело прозрачное. Глаза отсутствуют. Сегментация почти не выражена: межсег-
ментные борозды отсутствуют, диссепименты (межсегментные перегородки) раз-
Кольчатые черви
31
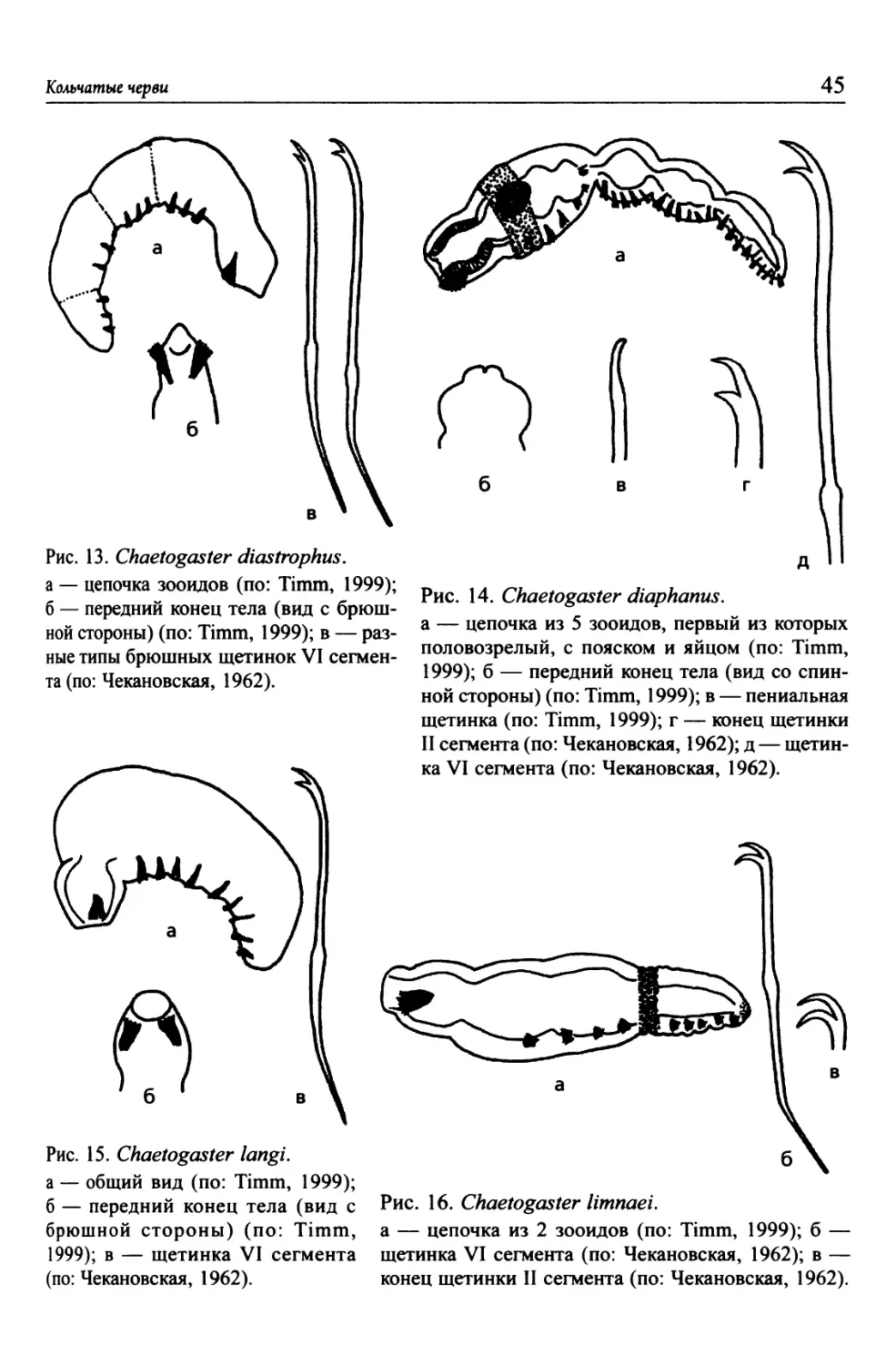
Рис. 1. Aeolosoma hemprichi (по: Чеканов-
ская, 1962).
А — цепочка зооидов; Б — пучок щетинок.
Рис. 2. Aeolosoma tenebrarum (по: Чека-
новская, 1962).
А — цепочка зооидов; Б — пучки щетинок.
виты слабо или отсутствуют. Головная лопасть часто шире передних сегментов.
На её брюшной стороне и вокруг ротового отверстия располагаются реснички.
Щетинки начинаются со II сегмента, чаще волосные, иногда встречаются дву-
зубчатые или пильчатые. В некоторых случаях щетинки отсутствуют. Половых
щетинок нет. Спинной кровеносный сосуд развит только в передней части тела.
Кровь бесцветная. Гермафродиты. Обособленные гонады у большинства видов
отсутствуют. Ни мужских, ни женских половых протоков нет (сперма выводит-
ся через нефридии). Семенные и яйцевые мешки отсутствуют. Семяприёмники
имеются только у некоторых видов. Особи с развитой половой системой встреча-
ются крайне редко. Большинство видов размножается почти исключительно бес-
полым путём (паратомией). В водоёмах встречаются в основном в виде цепочек
из 2-8 особей. Обитают только в пресных водоёмах (в поверхностных горизонтах
грунтов и среди водной растительности). Способны перемещаться по субстрату
характерным скользящим движением, используя реснички, расположенные на го-
ловной лопасти и вокруг рта. Некоторые виды плавают.
Мировая фауна включает около 30 видов эолосоматид, 25 из них обитает в
Палеарктике. В пресных водоёмах европейской части России отмечено 10 видов
этих организмов.
Для адекватного определения представителей сем. Aeolosomatidae необходи-
мо работать с живым материалом. В данном пособии мы не приводим ключей
для идентификации родов и видов.
32
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Класс ПОЯСКОВЫЕ — CLITELLATA
Характерной особенностью поясковых является образование на теле пояска
(clitellum) — железистого разрастания эпидермиса — клетки которого участвуют
в формировании коконов при откладке яиц. Поясковым свойственен гермафроди-
тизм. Гонады располагаются лишь в отдельных строго определённых сегментах
тела. Развитие прямое. Щетинки немногочисленны или отсутствуют. Головная
лопасть невелика и, как правило, лишена придатков.
Подкласс Малощетинковые черви — Oligochaeta
Строение тела
Число сегментов в теле малощетинковых червей никогда не бывает постоян-
ным даже в пределах одного вида, обычно оно колеблется от 7-14 до нескольких
сотен у представителей разных видов. (Номера сегментов принято обозначать
римскими цифрами). Глубина и выраженность наружных межсегментных борозд
различна у разных видов. Встречается также вторичная кольчатость (обычно в пе-
редних сегментах тела). Первый сегмент на брюшной стороне несёт ротовое от-
верстие. Спереди к нему присоединена головная лопасть. Выделяют 4 различных
типа сочленения головной лопасти с первым сегментом (рис. 3). Форма головной
лопасти может быть различной, у некоторых видов она вытянута в хоботок.
Прижизненная окраска малощетинковых червей очень разнообразна (как
правило, сильнее пигментирована спина и передний конец тела). При фиксации
окраска часто обесцвечивается, и черви приобретают сероватый оттенок.
'ДДДДД
Рис. 3. Типы сочленения головной лопасти малощетинковых червей с ротовым сег-
ментом (по: Попченко, 1988).
А — вид сверху; Б — вид сбоку. 1 — зиголобический; 2 — пролобический; 3 — эпилоби-
ческий открытый; 4 — эпилобический закрытый; 5 — танилобический
Кольчатые черви
33
В покровном эпителии у всех малощетинковых червей в большом количестве
имеются железистые клетки. У некоторых видов они расположены группами,
имеющими вид эпидермальных сосочков.
Все сегменты, кроме первого, несут по 2 спинных и по 2 брюшных пучка ще-
тинок. Иногда спинные щетинки отсутствуют во всех или в нескольких передних
сегментах. У отдельных видов щетинки отсутствуют совсем. Форма щетинок
очень разнообразна (рис. 4). Двузубчатые, веерные и однозубчатые щетинки, как
правило, имеют утолщение, называемое узелком. Однако узелок может и отсут-
ствовать. Особой модификацией брюшных щетинок являются половые щетинки
(рис. 4, 10-11). Они располагаются на брюшной стороне (вместо обычных локо-
моторных) и бывают двух разновидностей: пениальные (рядом с мужскими поло-
выми отверстиями) и сперматекальные (рядом с отверстиями семяприёмников).
Очень редко у представителей сем. Tubificidae половые щетинки располагаются
в сегментах, соседних с несущими половые отверстия. У дождевых червей по-
ловые щетинки не связаны топографически с половыми отверстиями — распо-
ложены в некоторых передних сегментах на железистых сосочках. Щетинки —
образования непрочные, легко ломаются, быстро снашиваются, выпадают и
заменяются новыми. Поэтому в пучках помимо сформированных щетинок можно
видеть мелкие растущие щетинки. (Их не надо учитывать при подсчёте щети-
нок в пучке!)
Целом (вторичная полость тела) у малощетинковых червей хорошо развит.
Он выстлан перитонеальным эпителием и разделён посегментно перегородками
(диссепиментами). Хлорагогенная ткань, одевающая со всех сторон большую
часть кишечника, а часто и спинной кровеносный сосуд с прилегающими коль-
цевыми сосудами, является видоизменением перитонеального эпителия. Полость
тела может сообщаться с наружной средой посредством специальных спинных и
головных пор. В полости тела в целомической жидкости содержатся полостные
тельца (целомоциты). У отдельных групп олигохет (наидиды, некоторые тубифи-
циды) они крупные хорошо различимые в световой микроскоп, у других — мел-
кие.
Пищеварительный тракт разделён (морфологически и функционально) на ро-
товую полость, мускулистую глотку, пищевод, пищеварительный и концевой от-
делы. Пищевод при переходе в пищеварительный отдел (желудок) расширяется
резко или постепенно. Все отделы кишечника, за исключением концевого, снаб-
жены секреторными железами разных типов. В области глотки и пищевода рас-
положены многоклеточные септальные железы. У дождевых червей в пищеводе
развиты известковые железы. Стенки пищевода в этих местах имеют пластин-
чатую многослойную структуру, сам пищевод расширен. При сильном развитии
пластин известковых желёз образуются карманообразные выпячивания пищево-
да — дивертикулы.
Отдельные виды семейств Naididae и Tubificidae обладают специализирован-
ными органами дыхания — наружными жабрами, расположенными либо на рас-
34
Кольчатые черви
35
ширении заднего конца тела — жаберном ложе, либо попарно в каждом сегмен-
те.
У многих наидид по бокам головной лопасти располагаются примитивные
глаза, состоящие из 5-6 увеличенных клеток кожного эпителия, подостланных
несколькими пигментными клетками.
Положение половой системы различно у представителей разных семейств.
Половой аппарат состоит из гонад (семенников и яичников), мужских и женских
половых протоков (гонодуктов) с их железами и совокупительным органом, се-
менных и яйцевых мешков и семяприёмников. Самым сложным и разнообразным
строением характеризуются мужские гонодукты. В наиболее полном варианте
мужской гонодукт состоит из семенной воронки, семяпровода, атрия с проста-
тической железой (железами), семяизвергательного канала и совокупительного
органа (рис. 71). Обычно мужские гонодукты парные на всём протяжении. Од-
нако у отдельных видов происходит сближение концевых отделов с образовани-
ем непарного мужского полового отверстия. Простатические железы могут быть
2-х типов: 1) диффузная простатическая железа (поверхность атрия равно-
мерно покрыта простатическими клетками); 2) обособленная простатическая
железа, свободно лежащая в целоме и впадающая в определённую часть атрия.
Совокупительные органы {пениальные булъбусы, пенисы, псевдопенисы) име-
ются у многих малощетинковых червей. У некоторых видов тубифицид пенисы
заключены в толстые хитиноидные трубки (жёлтого или бурого цвета), хорошо
различимые под микроскопом даже при небольшом увеличении. У большинства
олигохет половые отверстия открываются наружу в области пояска, у дождевых
червей — впереди него. Семяприёмники состоят из ампулы разнообразной фор-
мы и выводного протока различной длины. Обычно они парные. У немногих
видов (например, виды рода Bothrioneurum, отдельные виды дождевых червей)
семяприёмники отсутствуют. В этом случае передача спермы осуществляется
посредством сперматофоров, прикрепляемых к поверхности тела партнёра при
спаривании. У дождевых червей на брюшной стороне пояска развиты валики, или
бугорки зрелости, являющиеся утолщениями железистого эпителия.
Оплодотворение и развитие яиц происходит внутри коконов, образуемых же-
лезами пояска. Форма коконов характерна для каждого вида (рис. 5). Число яиц,
заключённых в коконе, различно у представителей разных семейств (у наидид —
1, у представителей других семейств — до нескольких десятков). Коконы откла-
Рис. 4. Типы щетинок малощетинковых червей (по: Чекановская, 1962).
1-2 — оперённые волосные; 3 — пильчатая волосная; 4 — зубчатая; 5 — весловидная;
6 — двузубчатая с редуцированными зубцами; 7 — однозубчатая палочковидная; 8 —
однозубчатая игловидная; 9 — однозубчатая крючковидная; 10-11 — половые щетинки;
12—13 — двузубчатые щетинки с равными зубцами; 14-15 — веерные; 16 — двузубчатая
щетинка с редуцированным нижним зубцом; 17 — двузубчатая щетинка с редуцирован-
ным верхним зубцом.
36
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 5. Коконы малощетинковых червей.
1 — Chaetogaster diaphanus (по: Чекановская, 1962): а — стенка кокона; б — загустевшая
слизь, приклеивающая кокон к субстрату; в — задняя трубка; г — яйцо на стадии двух
бластомеров; д — передняя трубка. 2 — Rhynchelmis limosella (по: Чекановская, 1962).
3 — Tubifex tubifex (ювенильный червь выползает из яйцевого кокона) (по: Чекановская,
1962). 4 — Limnodrilus hoffineisteri (по: Архипова, 1983): а — кокон, покрытый грунтом;
б — кокон, частично освобождённый от грунта. 5 — Potamothrix moldaviensis (по: Архи-
пова, 1983): а — кокон, покрытый грунтом; б — кокон, освобождённый от грунта.
Кольчатые черви
31
дываются в грунт или прикрепляются к подводным растениям. Развитие прямое,
коконы покидают уже сформировавшиеся черви, отличающиеся от взрослых
только размерами. Многим малощетинковым червям свойственно бесполое раз-
множение в форме архитомии (отдельные виды тубифицид, энхитреид и люмбри-
кулид) или паратомии (все наидиды и пристиниды). При архитомии происходит
деление червя на две (иногда более) части с последующей регенирацией недоста-
ющего переднего и заднего конца. При паратомии вначале происходит образова-
ние морфологически отдельных дочерних особей, которые до завершения форми-
рования остаются связанными с материнским организмом и друг с другом, что
приводит к образованию цепочки особей.
Малощетинковые черви широко распространены в пресных, морских и соло-
новатоводных (эстуарных) водоёмах, а также во влажных почвах всех материков.
Отсутствуют только в гиперсолёных континентальных водоёмах, горячих источ-
никах и водоёмах полностью лишённых кислорода. Среди донных беспозвоноч-
ных пресных водоёмов они занимают одно из ведущих мест по видовому разноо-
бразию и функциональной роли в сообществах.
Большинство малощетинковых червей по способу питания являются мирны-
ми грунтоедами (тубифициды, энхитреиды, люмбрикулиды, дождевые черви) или
собирателями (большинство наидид, пристиниды). Ripistes parasita может быть
отнесена к фильтраторам, т.к. использует свои гигантские щетинки для сбора
взвеси из толщи воды. Хищники очень немногочисленны — это некоторые виды
рода Chaetogaster.
Мировая фауна насчитывает почти 5000 видов малощетинковых червей, из ко-
торых около 1700 видов являются водными (пресные водоёмы населяют 1100 ви-
дов). В пределах Палеарктики обитает более 600 пресноводных видов. В пресных
водоёмах европейской части России отмечено более 300 видов малощетинковых
червей.
Методы сбора и обработки
Для сбора малощетинковых червей, обитающих в толще грунта и на его по-
верхности, подходят традиционные гидробиологические орудия лова (дночерпа-
тели, драги, сачки, скребки и т.п.). Образцы грунта промывают по стандартной
методике (через серию сит с разным размером ячеи). Для учёта мелких видов
следует использовать сита и промывалки из мельничного газа №№ 38—49. При
промывке проб необходимо соблюдать осторожность, т.к. малощетинковые чер-
ви имеют тонкие покровы и легко повреждаются, кроме того при грубой отмывке
могут быть выломаны щетинки, что осложнит дальнейшее определение матери-
ала. Обитатели зарослей и перифитона добываются путём смывов со срезанных
водных растений и камней. Полученные смывы фильтруют через мельничный
газ с ячеёй как можно более мелкого размера, т.к. в данном биотопе присутствует
много субмикроскопических представителей олигохет.
38
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
После промывки пробы можно сразу зафиксировать 4% формалином и после
этого выбирать из них животных. Однако, если позволяют условия, предпочти-
тельнее выбирать малощетинковых червей живыми и затем уже фиксировать их
(4% формалином или 96° этиловым спиртом). Во-первых, исследование червей с
целью определения вида целесообразно начинать с прижизненных наблюдений.
Во-вторых, свежезафиксированным червям можно придать положение удобное
для дальнейшего исследования. В то время как червей, фиксированных вместе с
грунтом и пролежавших в таком состоянии какое-то время, уже невозможно рас-
править, и это значительно осложняет изучение их признаков (при воздействии
фиксирующей жидкости черви, особенно длинные, изгибаются, скручиваются,
завязываются в узлы). Просмотр проб следует производить под бинокуляром,
чтобы обнаружить всех мелких червей. Хранить материал лучше в 70° спирте.
Формалин, являясь хорошим первичным фиксатором, не подходит для длительно-
го хранения, т.к. черви становятся хрупкими. После первичной фиксации форма-
лином червей необходимо, сполоснув водой, перенести в спирт. При фиксации и
дальнейшем хранении нужно следить, чтобы объём фиксирующей жидкости был
достаточно большим для фиксируемого (хранимого) материала.
Определение видов малощетинковых червей осуществляют под микроскопом.
При изучении живого материала мелких червей помещают в каплю воды на пред-
метном стекле, прикрыв покровным стеклом, крупных — в чашку Петри или за-
жав между двумя предметными стёклами. Тотальные препараты фиксированных
червей изготавливают, поместив объект на предметное стекло в каплю глицери-
на и накрыв его покровным стеклом с легким надавливанием. Детали строения
щетинок и пениальных трубок удобнее рассматривать, когда червь лежит на боку
(т.е. сагитально). Пениальные трубки обычно удаётся рассмотреть при неболь-
шом надавливании на покровное стекло. Экземпляры с очень плотными непро-
зрачными покровами можно для просветления выдержать в глицерине (в течение
суток). Если же это не помогает, необходимо вскрыть червя в области пояска и
отпрепарировать гонодукт. Для этого фиксированного червя (его передний ко-
нец с пояском) прикалывают тонкими энтомологическими булавками к специ-
альной пробковой подставке спиной вверх. Подставка представляет собой проб-
ковую пластину небольшой толщины, покрытую сверху слоем чёрной туши для
контрастности. Для очень крупных экземпляров можно использовать ванночку с
дном из парафино-восковой смеси. Вскрытие лучше всего производить тонкими
препаровальными иглами, заточенными на конце; также можно воспользовать-
ся лезвием безопасной бритвы. Разрезанную стенку тела отводят в обе стороны
и закрепляют тонкими энтомологическими булавками. После этого очень тонки-
ми препаровальными иглами осторожно распутывают и извлекают мужской го-
нодукт, который помещают в каплю глицерина на предметное стекло и, прикрыв
покровным, изучают под микроскопом. Наиболее пригоден для вскрытия свеже-
зафиксированный материал.
Кольчатые черви
39
Основные пособия для определения малощетинковых червей
Чекановская О.В. 1962. Водные малощетинковые черви фауны СССР. М.; Л.: Изд-во АН
СССР. 411 с.
Попченко В.И. 1988. Водные малощетинковые черви (Oligochaeta limicola) Севера Евро-
пы. Л.: Наука. 287 с.
Timm Т. 1999. A Guide to the Estonian Annelida. Tartu - Tallinn: Estonian Academy Publishers.
208 p.
Ключ для определения семейств и видов малощетинковых червей
1(18). Щетинки имеются.
2(11). В каждом сегменте, хотя бы в передней части тела, более 2 щетинок в
пучке.
3(4). Спинные щетинки начинаются с III-VI сегмента, иногда они отсутствуют.
У многих видов имеются глаза. Поясок у половозрелых особей располага-
ется на V-VII сегментах. Многие виды способны плавать.....................
...........................................Naididae (с. 44); (рис. 6, /).
Рис. 6. Представители сем. Naididae и сем. Pristinidae (по: Timm, 1999).
1 — Naididae (общий вид). 2 — Pristinidae (общий вид). 3 — щетинки: а — брюшные;
б — спинные.
40
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Размножение преимущественно бесполое путём паратомии с образованием це-
почек особей. Мелкие черви (длина цепочки особей редко превышает 20 мм;
число сегментов одиночной особи редко больше 50, обычно гораздо меньше).
Прозрачные, бесцветные, жёлтые, коричневатые, часто с пигментными пятнами и
полосами. Половые щетинки имеются. Половозрелые особи чаще встречаются в
конце лета и осенью.
4(3). Спинные щетинки начинаются со II сегмента. Глаза отсутствуют. Поясок
располагается кзади от VI сегмента тела. К плаванию не способны.
5(6). Размножение преимущественно бесполое путём паратомии с образовани-
ем цепочек особей. У половозрелых особей поясок всегда расположен в
области VII-IX сегментов.......................Pristinidae (с. 65); (рис. 6, 2).
Мелкие и очень мелие черви (длина цепочки особей 1,5-8,5 мм). Прозрачные,
молочно-белые, светло-жёлтые. Головная лопасть некоторых видов снабжена хо-
ботком.
6(5). Размножение преимущественно половое. Бесполое в виде исключения,
путём архитомии. Паратомия не встречается. Цепочки особей никогда
не образуются. Поясок обычно располагается кзади от IX сегмента тела
(в редких случаях на VII—VIII или VIII—IX).
7(8). Щетинки разных типов — двузубчатые, веерные, волосные. Половая си-
стема и поясок, как правило, располагаются в X-XI сегментах тела (реже
на VII—VIII или VIII—IX). Мужские половые отверстия в XI сегменте (реже
в VII или IX). Семяприёмники находятся в сегментах, соседних с несущи-
ми мужские половые отверстиями. Половые щетинки имеются у многих
видов............................................Tubificidae (с. 69); (рис. 7).
Черви нитевидные. Длина тела обычно 10-65 мм, иногда более 120-150 мм.
Число сегментов 30-120. Окраска от светло-розовой до красной. Некоторые виды
способны к бесполому размножению путём архитомии.
8(7). Щетинки однозубчатые (игловидные или палочковидные) или двузубчатые
с рудиментарным верхним зубцом. Поясок расположен на XI-XIII(XIV)
сегментах. Мужские половые отверстия в XII сегменте. Семяприёмники
Рис. 7. Представители сем. Tubificidae (по: Timm, 1999).
а — общий вид; б — концы спинных щетинок.
Кольчатые черви
41
Рис. 8. Представители сем. Enchytraeidae (по: Timm, 1999).
а — общий вид; б — щетинки.
отделены от остальной половой системы несколькими сегментами. По-
ловые щетинки отсутствуют.
9(10). Щетинки только однозубчатые: прямые или S-образно изогнутые, с узел-
ком или без, одинаковой или разной длины в пучке. Отверстия семяприём-
ников в межсегментной борозде IV и V сегментов. У большинства видов
в передней части тела (в IV-IX сегментах) по бокам от глотки хорошо
различимы крупные септальные железы......................................
.....................................Enchytraeidae (частично) (рис. 8).
Черви относительно короткие (число сегментов редко превышает 50, обычно
25-40; длина тела 2—40 мм). Преимущественно белого цвета с гладкой кутику-
лой. Некоторые виды способны к бесполому размножению путём архитомии.
Большинство видов ведёт почвенный образ жизни, предпочитая увлажнённые
богатые органикой субстраты. Встречаются также в пресных и морских водоёмах.
Немало среди них амфибиотических видов. В пресных водоёмах наиболее бога-
то представлены в прибрежье и зоне уреза воды. Определение представителей
этого семейства достаточно затруднительно и требует специальных методов.
В данном пособии мы не приводим ключей для идентификации родов и видов.
10(9). Щетинки двузубчатые с рудиментарным верхним зубцом. Отверстия
семяприёмников в межсегментной борозде III и IV сегментов. Крупные
септальные железы в передней части тела отсутствуют.........Propappidae.
Единственный род Propappus Michaelsen, 1905.
В европейской части России единственный вид Propappus volki Michaelsen, 1916
(рис. 9).
Длина тела 6-12 мм, диаметр 0,2-0,3 мм. Молочно-белого цвета. Головная ло-
пасть удлинённая с небольшим хоботком. В пучке 3—4 щетинки. В европейской
части России распространён широко.
11(2). В брюшных и спинных пучках не более, чем по 2 щетинки.
12(13). Мелкие полупрозрачные черви, преимущественно белого цвета.......
....................................Enchytraeidae (частично) (рис. 8).
13(12). Черви средних и крупных размеров. Розового, красного и тёмных цветов.
42
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 9. Propappus volki (по: Timm, 1999).
а — общий вид; б — щетинка.
Рис. 10. Haplotaxis gordioides.
а — общий вид (по: Чекановская, 1962); б —
щетинки (по: Попченко, 1988).
14(15). Тело длинное, тонкое, аскаридообразное. Щетинок в средней и задней
частях тела по 2 или по 1 в пучке. Спинные пучки щетинок могут отсут-
ствовать....................................................Haplotaxidae.
Единственный род Haplotaxis Hoffmeister, 1843.
В европейской части России единственный вид Haplotaxis gordioides (Hartmann,
1821) (рис. 10).
Очень длинные, тонкие черви, по форме напоминающие волосатиков. Длина тела
275-310 мм, диаметр 1-1,2 мм. Щетинки однозубчатые, крючковидные. Брюшные
щетинки крупнее спинных. Поясок кольцевидный на Х-ХШ сегментах. Половые
отверстия находятся в области пояска. Стенотермный холодлюбивый вид. В евро-
пейской части России в Карелии и Республике Коми. Очень редкий.
15(14). Тело не выделяется своей длиной по отношению к ширине. В брюшных и
спинных пучках всех сегментов по 2 щетинки.
16(17). Ротовой сегмент эпи- и танилобический (рис. 3, 3-5). На спинной стороне
в межсегментных бороздах имеются поры. Щетинки всегда однозубчатые,
S-образно изогнутые, сближенные попарно или расставленные пооди-
ночке. Мужские половые отверстия всегда расположены впереди пояска
(чаще в XV сегменте), имеют вид поперечных щелей на высоких желези-
стых подушках. Поясок на XXII-XXXVII сегментах. На брюшной стороне
пояска располагаются валики зрелости. Половые щетинки имеются. Они
более длинные, чем локомоторные, острые, иногда с желобком. Пищевод
в X-XIV сегментах снабжён известковыми железами...........................
............................Lumbricidae (Дождевые черви) (рис. 11, 2).
Кольчатые черви
43
Рис. 11. Представители сем. Lumbriculidae и сем. Lumbricidae (по: Timm, 1999).
1 — Lumbriculidae; 2 — Lumbricidae. а — общий вид; б — щетинки.
Единственный пресноводный (амфибиотический) вид Eiseniella tetraedra (Savig-
ny, 1826) (рис. 12).
Длина тела 15-67 мм, диаметр 1,5-4 мм. Пигментация от коричнево-жёлтой до
красновато-бурой. Спинные поры начинаются с межсегментной борозды IV и
V сегментов. Средняя и задняя части тела отчётливо четырёхгранные. Щетинки
сильно сближены попарно. Поясок на XXII(XXin)-XXVI(XXVII) сегментах, ва-
лики зрелости на XXIII(XXIV)-XXV(XXVI). Мужские половые отверстия в ХШ
(реже в XIV или XV сегменте). Известковые железы в XI—XIII сегментах. Амфи-
биотический вид (встречается в сырых местах у уреза воды и в прибрежной зоне
водоёмов). В европейской части России распространён широко.
17(16). Ротовой сегмент про- и зиголобический (рис. 3, /, 2). Спинные поры от-
сутствуют. Щетинки однозубчатые или двузубчатые с рудиментарным
верхним зубцом, сближенные попарно. Мужские половые отверстия всег-
да расположены в области пояска (в пределах VIII—XI сегментов, обычно
на X). Валики зрелости на пояске отсутствуют. Половые щетинки у боль-
шинства видов отсутствуют. Известковых желёз в пищеводе нет.............
....................................Lumbriculidae (с. 92); (рис. 11, /).
Длинные черви (длина тела 20-145 мм, число сегментов 42-240). Межсегментные
борозды могут быть выражены очень чётко, но иногда почти незаметны, тогда
44
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 12. Eiseniella tetraedra (по: Попченко, 1988).
1 — передний и 2 — задний конец тела. <3 — мужское половое отверстие; $- женское
половое отверстие; по — поясок; вз — валик зрелости.
червь приобретает «аскаридную» внешность. Нередко хорошо выражена вторич-
ная кольчатость. Щетинки на концах тупые или тонко заострённые. У отдельных
видов отмечено бесполое размножение путём архитомии.
18(1). Щетинки отсутствуют полностью во всех сегментах тела................
.....................Lamprodrilus achaetus palearcticus Popchenko, 1988
(род Lamprodrilus Michaelsen, 1901; сем. Lumbriculidae).
Длина тела 9-12 мм, диаметр 0,4-0,5 мм. Бесцветные черви. Поверхность тела
гладкая, межсегментные борозды совсем не выражены. Вторичной кольчатости
нет. Мужских половых отверстий 2 пары на выступающих бугорках (папиллах)
в X и XI сегментах. Карелия, Архангельская обл. Очень редкий.
Ключ для определения родов и видов семейства Naididae
1(2). Спинные щетинки отсутствуют. Головная лопасть у большинства видов
отсутствует или рудиментарна. II и (или) III сегменты тела удлинены по
сравнению с остальными сегментами..........................................
...........................Chaetogaster Baer, 1827 (с. 58); (рис. 13-17).
Мелкие прозрачные черви с небольшим числом сегментов (не более 20 у одиноч-
ной особи). Глаза отсутствуют. Брюшные щетинки во II сегменте длиннее, чем в
остальных. В III—V сегментах брюшные щетинки отсутствуют.
2(1). Спинные щетинки имеются. Головная лопасть хорошо развита. II и III
сегменты тела не удлинены.
3(10). Волосные щетинки в спинных пучках отсутствуют.
4(5). Спинные щетинки начинаются с III сегмента... Amphichaeta Tauber, 1879.
В пресных водоёмах европейской части России единственный вид Amphichaeta
leydigi Tauber, 1879 (рис. 18).
Длина тела одиночной особи до 1,7 мм, цепочки особей — до 4 мм. Головная ло-
пасть треугольная заострённая. Глаза отсутствуют. Брюшные и спинные щетинки
Кольчатые черви
45
а — цепочка зооидов (по: Timm, 1999);
б — передний конец тела (вид с брюш-
ной стороны) (по: Timm, 1999); в — раз-
ные типы брюшных щетинок VI сегмен-
та (по: Чекановская, 1962).
Рис. 14. Chaetogaster diaphanus.
а — цепочка из 5 зооидов, первый из которых
половозрелый, с пояском и яйцом (по: Timm,
1999); б — передний конец тела (вид со спин-
ной стороны) (по: Timm, 1999); в — пениальная
Рис. 15. Chaetogaster langi.
а — общий вид (по: Timm, 1999);
б — передний конец тела (вид с
щетинка (по: Timm, 1999); г — конец щетинки
II сегмента (по: Чекановская, 1962); д — щетин-
ка VI сегмента (по: Чекановская, 1962).
Рис. 16. Chaetogaster limnaei.
брюшной стороны) (по: Timm,
1999); в — щетинка VI сегмента
(по: Чекановская, 1962).
а — цепочка из 2 зооидов (по: Timm, 1999); б —
щетинка VI сегмента (по: Чекановская, 1962); в —
конец щетинки II сегмента (по: Чекановская, 1962).
46
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 18. Amphichaeta leydigi (по: Timm, 1999).
а — цепочка из 2 зооидов; б — щетинка.
Рис. 17. Щетинки Chaetogaster setosus (по: Чекановская, 1962).
а — щетинки II сегмента; б — то же середины тела.
Ja U б
Рис. 19. Paranais frici (по: Timm, 1999).
а — общий вид; б — щетинка.
одинаковые по форме: двузубчатые с
зубцами равной длины, расходящи-
мися в разные стороны. В брюшных
пучках II и III сегментов по 4 щетин-
ки, IV и V по 2, в последующих по 4
(иногда по 1-3) щетинке. В спинных
пучках III сегмента по 5 щетинок, в
последующих — подобно брюшным
тех же сегментов. Пищевод расширя-
ется в V сегменте. Половые щетинки
отсутствуют. В европейской части
России на севере, северо-западе и в
центре. Очень редкий.
5(4). Спинные пучки щетинок начина-
ются далее III сегмента.
6(7). Спинные пучки с V сегмента. Глаза отсутствуют........................
............................................Paranais Cemiavsky, 1880.
В пресных водоёмах европейской части России единственный вид Paranais frici
НгаЬё, 1941 (рис. 19).
Длина фиксированных червей 2,7-6 мм, живых — до 9 мм. Брюшные и спинные
щетинки однотипные: тонкие двузубчатые с более длинным верхним зубцом (во
II сегменте длина верхнего зубца превосходит длину нижнего в 2 и более раз).
В брюшных пучках II сегмента по 2-4 щетинки, в спинных пучках V сегмента
по 3 щетинки, во всех остальных — по 1-2. Желудок явственный, в VII сегменте.
Пениальные щетинки по форме не отличаются от обычных локомоторных щети-
Кольчатые черви
41
Рис. 21. Ophidonais serpentina.
а — общий вид (по: Timm, 1999); б — спинные
щетинки (по: Чекановская, 1962); в — брюшная
щетинка (по: Timm, 1999).
Рис. 20. Uncinais uncinata.
а — цепочка из 2 зооидов (по: Timm,
1999); б — щетинка (по: Чеканов-
ская, 1962).
нок, но более глубоко погружены в стенку тела, по 2-3 в пучке. В европейской
части России в пресных и слабосолоноватых участках эстуариев и низовьях рек
бассейна Понто-Каспия и Балтийского моря. Находится в состоянии быстрого рас-
селения при помощи человека.
7(6). Спинные пучки с VI сегмента. Глаза имеются.
8(9). Спинные и брюшные щетинки одинаковой формы — двузубчатые................
.................................................Uncinais Levinsen, 1884.
В европейской части России единственный вид Uncinais uncinata (Oersted, 1842)
(рис. 20).
Длина цепочки особей 5-18 мм. Головная лопасть толстая, тупая, округлая. Ще-
тинки с верхним зубцом длиннее нижнего. В брюшных пучках по 2-4 щетинки,
в спинных — по 2-7. Желудочное расширение начинается в VIII сегменте. Пени-
альные щетинки на конце крючковидно загнуты, по 2-3 в пучке. В европейской
части России повсеместно.
9(8). Спинные щетинки толстые, прямые, палочковидные, брюшные — двузуб-
чатые................................................Ophidonais Gervais, 1838.
Единственный вид Ophidonais serpentina (Muller, 1774) (рис. 21).
Длина цепочки особей 5-36 мм. В переднем отделе тела несколько поперечных
пигментных полос коричневого цвета. Поверхность тела усеяна чувствительными
сосочками, расположенными поперечными рядами. Кожный эпителий покрыт тон-
ким панцирем из частиц грунта. Головная лопасть короткая закруглённая. В брюш-
ных пучках по 2-6 щетинок с более длинным верхним зубцом, в спинных — по
1 щетинке. У некоторых экземпляров спинные щетинки отсутствуют. Желудок в
VIII-X, иногда IX-XI сегментах. Половые щетинки по 2-3 в пучке с крючковид-
ным или расширенным концом. В европейской части России повсеместно.
10(3). Волосные щетинки в спинных пучках имеются.
11(12). Спинные щетинки начинаются с III сегмента......Bratislavia Kosel, 1976.
Единственный вид Bratislavia palmeni (Munsterhjelm, 1905) (рис. 22).
Синонимы: Pristina elegans Finogenova, 1966; Pristina napocensis Pop, 1973.
48
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 22. Bratislavia palmeni (по:
Timm, 1999).
а — передний конец тела; б —
брюшная щетинка II сегмента; в —
то же задней части тела; г — игло-
видная щетинка спинного пучка.
Длина тела 2-11 мм. Покровы имеют эпи-
дермальные сосочки. Головная лопасть
короткая треугольной формы. Глаза от-
сутствуют. Брюшные щетинки по 2-7 в
пучке с зубцами равной длины. Спинные
пучки содержат по 1-3 гладких волосных
щетинки и по 1-3 игловидных щетинки с
двумя очень длинными зубцами, верхний
из которых немного длиннее нижнего.
Игловидные щетинки в передних сегмен-
тах тела тоньше и короче, чем в последую-
щих. Желудочное расширение начинается
в VIII сегменте. Пениальные щетинки 4-6
в пучке с двумя короткими зубцами или
без них. В европейской части России на
севере. Очень редкий.
12(11). Спинные щетинки начинаются далее
III сегмента.
13(32). Спинные щетинки начинаются с IV-
VI сегмента.
14(19). Головная лопасть спереди вытянута в
хоботок.
15(16). Хоботок длинный. Волосных щетинок по 1-3 в каждом сегменте начиная
с VI...............................Stylaria Lamarck, 1816 (с. 59); (рис. 23, 24).
Передний конец тела пигментирован. Глаза обычно имеются. Брюшные щетинки
двузубчатые с длинным изогнутым верхним зубцом и укороченным нижними.
Пениальные щетинки имеются.
Рис. 23. Stylaria lacustris (по: Чеканов-
ская, 1962).
а — передний конец тела; б — брюшная
щетинка.
Рис. 24. Stylaria fossularis (по: Чека-
новская, 1962).
а — передний конец тела (вид с брюш-
ной стороны); б — конец брюшной
щетинки.
Кольчатые черви
49
16(15). Хоботок короткий. Волосных ще-
тинок больше 3-х в пучке, по край-
ней мере в VI—VIII сегментах.
17(18). Волосные щетинки VI—VIII сег-
ментов примерно одинаковой дли-
ны с таковыми последующих сег-
ментов. Во всех сегментах, начи-
ная с VI, большое число волосных
щетинок (от 8 до 18).................
..........Arcteonais Piguet, 1928.
Единственный вид Arcteonais lomondi
(Martin, 1907) (рис. 25).
Длина цепочки особей 8-12 мм. Глаза
имеются, но могут отсутствовать у
отдельных экземпляров. В брюшных
пучках по 3-8 щетинок с верхним зуб-
цом длиннее нижнего. В спинных пуч-
ках помимо длинных волосных щети-
нок, расположенных веерообразно,
имеется 9-12 коротких игловидных
щетинок. Желудочное расширение на-
чинается в VIII сегменте. Пениальные
щетинки на конце крючковидные, по 4
в пучке. В европейской части России
повсеместно.
Рис. 25. Arcteonais lomondi (по: Чека-
новская, 1962).
а — передний конец тела половозрелой
особи (вид сбоку); б — брюшная щетин-
ка II сегмента; в — то же XXVIII сегмен-
та; г — спинной пучок щетинок.
18(17). Волосные щетинки VI-
VIII сегментов во много раз
длиннее, чем в последу-
ющих. Большое число во-
лосных щетинок (до 16) на-
блюдается только VI—VIII
сегментах, в остальных со-
держится по 1-3 волосные
щетинки.........................
....Ripistes Dujardin, 1842.
Единственный вид Ripistesраг-
asita (Schmidt, 1847) (рис. 26).
Длина цепочки особей 2-8 мм.
Глаза имеются. Брюшные ще-
тинки в IV и V сегментах от-
сутствуют, во И-Ш сегментах
по 2-7 щетинок с более длин-
ным верхним зубцом в пучке,
в последующих (начиная с VI
Рис. 26. Ripistes parasita.
а — передний конец тела (по: Чекановская,
1962); б — брюшная щетинка II сегмента (по:
Sperber, 1948); в — то же III сегмента (по: Sper-
ber, 1948); г — то же задней части тела (по:
Sperber, 1948).
50 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
берные придатки с ресничками.
Рис. 27. Aulophorus furcatus (по: Timm, 1999).
а — общий вид; б — различный вид жаберного
ложа; в — брюшная щетинка II сегмента; г — то
же задней части тела; д — игловидная щетинка
спинного пучка.
сегмента) по 3-8 щетинок с более коротким верхним зубцом в пучке. Пищевод
расширяется в желудок резко в VI или VIII сегментах. Пениальные щетинки с
крючковидными концами по 2 в пучке. Строит мягкие трубки. В европейской
части России повсеместно.
19(14). Головная лопасть треугольная или округлая без хоботка.
20(23). На заднем конце тела, расширенном в виде жаберного ложа, имеются жа-
12). Кроме жаберных придатков
с ресничками жаберное ложе
несёт 2 длинных придатка, ли-
шённых ресничного покрова.
Спинные щетинки начинаются
с V сегмента......Aulophorus
Schmarda, 1861.
В европейской части России
единственный вид Aulophorus fur-
catus (Oken, 1815) (рис. 27).
Длина цепочки особей 6-20 мм.
Головная лопасть закруглённая.
Глаза отсутствуют. Жаберное
ложе в виде бочонка с 3-4 парами
пальцевидных жабр. Брюшные
пучки содержат по 2-6 двузубча-
тых щетинок. Во II—IV сегментах
зубцы их длинные, почти равного
размера, с V сегмента зубцы ста-
новятся короче, нижний из них
длиннее верхнего. В спинных
пучках по 1 гладкой волосной и
по 1 игловидной двузубчатой ще-
тинке с более коротким верхним
зубцом. Желудок начинается с VII
или VIII сегмента. Пениальные щетинки отсутствуют. Строит мягкие трубки.
В европейской части России распространён широко. Редкий.
22(21). Длинных нересничных придатков на жаберном ложе нет. Спинные щетин-
ки начинаются с IV или VI сегмента........................................
...................................Dero Oken, 1815 (с. 59); (рис. 28-30).
Головная лопасть округлая. Глаза отсутствуют. Половые щетинки отсутствуют.
Строят мягкие трубки.
23(20). Жаберные придатки отсутствуют.
24(25). Волосные щетинки в VI сегменте в 3—4 раза длиннее, чем в последующих.
Кожный эпителий с клейкими сосочками и рядами чувствительных бугор-
ков, снаружи обычно покрыт частицами грунта. Окрашен в оранжевый
цвет................................................Slavina Vejdovsky, 1884.
Кольчатые черви
51
Рис. 28. Dero dorsalis (по: Timm, 1999).
а — общий вид; б — передний конец тела; в — различный вид жаберного ложа; г —
брюшная щетинка II сегмента; д — то же задней части тела; е — игловидная щетинка
спинного пучка.
Рис. 29. Dero digitata (по: Timm, 1999).
а — передний конец тела; б — различный вид жаберного ложа; в — брюшная ще-
тинка II сегмента; г — то же задней части тела; д — игловидная щетинка спинного
пучка.
Рис. 30. Dero obtusa (по: Timm, 1999).
а — передний конец тела; б — различный вид жаберного ложа; в — брюшная ще-
тинка II сегмента; г — то же задней части тела; д — игловидная щетинка спинного
пучка.
52
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 31. Slavina appendiculata (по: Timm, 1999).
а — общий вид; б — брюшная щетинка; в —
игловидная щетинка спинного пучка.
Рис. 32. Щетинки Vej-
dovskyella macrochae-
ta (по: Чекановская,
1962).
а — брюшная щетинка
X сегмента; б — то же
VII сегмента; в — ко-
нец брюшной щетин-
ки VII сегмента; г —
пениальная щетинка.
В европейской части России единственный вид
Slavina appendiculata (Udekem, 1855) (рис. 31).
Длина цепочки особей 2-22 мм. Головная
лопасть короткая округлая. Глаза имеются.
Брюшные щетинки двузубчатые по 2-5 в пучке
с верхним зубцом немного тоньше и длиннее
нижнего. В спинных пучках, начинающихся с
VI сегмента, по 1-2 волосных и 1-2 игловидных щетинки. Игловидные щетинки
очень тонкие с едва заметным вздутием на концах. Желудочное расширение в
VII-VIII сегменте. Пениальные щетинки с острыми изогнутыми концами. В евро-
пейской части России повсеместно.
25(24). Волосные щетинки VI сегмента не удлинены. Кожный эпителий гладкий
(без клейких сосочков и чувствительных бугорков); покров из частиц
грунта отсутствует.
26(27). Волосные щетинки толстые, пильчатые.................................
........Vejdovskyella Michaelsen, 1903 (с. 60); (рис. 32-34).
Головная лопасть короткая, округлая. Благодаря мощным
волосным щетинкам спинных пучков имеют своеобраз-
ный «лохматый» вид. Пениальные щетинки имеются.
27(26). Волосные щетинки тонкие, гладкие.
28(29). Игловидные щетинки спинных пучков очень тон-
кие, одно- и двузубчатые, по форме резко отличаю-
щиеся от брюшных щетинок (если же игловидные
щетинки толстые, то они однозубчатые заострён-
ные)..............Nais Muller, 1774 (с. 61); (рис. 35—45).
Головная лопасть округлая или коническая. Глаза обыч-
но имеются (могут отсутствовать у отдельных экземпля-
ров в порядке индивидуальной изменчивости). Пениаль-
ные щетинки имеются.
29(28). Игловидные щетинки спинных пучков толстые
двузубчатые, по форме и характеру изгиба напоми-
нают брюшные щетинки.
30(31). Глаза есть. Зубцы игловидных щетинок спинных
пучков одинаковой длины с таковыми брюшных.
Волосные щетинки очень тонкие, короткие (не
более чем в 1,5 раза длиннее игловидных), в коли-
честве 0-3 в пучке..............Piguetiella Sperber, 1939.
В европейской части России единственный вид Pigueti-
ella bland (Piguet, 1906) (рис. 46).
Кольчатые черви
53
Рис. 33. Vejdovskyella intermedia.
а — передний конец тела (по: Timm, 1999); б — брюшная щетинка II сегмента (по: Timm,
1999); в — брюшная щетинка VI сегмента (по: Чекановская, 1962); г — конец брюшной
щетинки VI сегмента (по: Timm, 1999); д — спинной пучок щетинок (по: Чекановская,
1962); е — игловидная щетинка спинного пучка (по: Timm, 1999).
Рис. 34. Vejdovskyella comata.
ъ — одиночная особь целиком (по: Чекановская, 1962); б — брюшная щетинка II сегмента
(по: Чекановская, 1962); в — то же из середины тела (по: Чекановская, 1962); г — волос-
ная щетинка спинного пучка (по: Чекановская, 1962); д — фрагмент волосной щетинки
(по: Timm, 1999); е — игловидная щетинка спинного пучка (по: Чекановская, 1962).
54
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
а б и в
Рис. 36. Щетинки Nais alpina (по:
Timm, 1999).
а — брюшная щетинка II сегмен-
та; б — то же задней части тела;
в — игловидная щетинка спинного
Рис. 35. Щетинки Nais behningi (по: Timm, 1999).
a-в — различные варианты брюшных щетинок
II сегмента; г — брюшная щетинка задней части
тела; д — игловидная щетинка спинного пучка.
а — брюшная щетинка II сегмен-
та; б — то же задней части тела;
в — игловидная щетинка спинно-
го пучка.
пучка.
Рис. 38. Nais barbata (по: Timm, 1999).
а — половозрелая особь; б — брюшная щетинка
II сегмента; в — то же задней части тела; г — игло-
видная щетинка спинного пучка.
Кольчатые черви
55
Рис. 39. Nais pseudobtusa (по: Timm, 1999).
а — передний конец тела; б — брюшная щетинка II сегмента; в — то же задней части
тела; г — игловидная щетинка спинного пучка.
Рис. 40. Щетинки Nais pardalis (по: Timm, 1999).
а — брюшная щетинка II сегмента; б — увеличенная брюшная
щетинка; в — брюшные щетинки задней части тела; г — игло-
видная щетинка спинного пучка.
Рис. 41. Щетинки Nais bretscheri (по: Timm, 1999).
а — брюшная щетинка II сегмента; б — то же VI сегмента;
в — гигантские брюшные щетинки; г — брюшная щетинка
задней части тела; д — игловидная щетинка спинного пучка.
56
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 42. Nais elinguis.
а — передний конец тела половозрелой особи с пояском и яйцом (по: Timm, 1999); б —
брюшная щетинка II сегмента (по: Чекановская, 1962); в — то же из середины тела (по:
Чекановская, 1962); г — игловидная щетинка спинного пучка (по: Чекановская, 1962).
Рис. 43. Щетинки Nais christinae (по:
Timm, 1999).
а — брюшная щетинка II сегмента; б —
то же задней части тела; в — игловид-
Рис. 44. Щетинки Nais communis (по:
Timm, 1999).
а — брюшная щетинка II сегмента; б —
то же задней части тела; в — игловид-
ная щетинка спинного пучка.
ная щетинка спинного пучка.
Рис. 45. Щетинки Nais variabilis (по: Timm, 1999).
а — брюшная щетинка II сегмента; б — то же зад-
ней части тела; в — игловидная щетинка спинного
пучка.
Кольчатые черви
57
тинка; в — спинная щетинка; г — волосная
щетинка; в — спинная щетинка; г — во-
щетинка.
лосная щетинка.
Длина цепочки особей 3-7 мм. Го-
ловная лопасть округлая. Брюшные
пучки содержат по 3-9 двузубчатых
щетинок с зубцами равной длины.
Во II сегменте брюшные щетинки
немного длиннее, чем в последую-
щих. Желудок в VIII—IX сегментах.
Пениальные щетинки отсутствуют.
Сперматекальные имеются в V сег-
менте, по 2 в пучке, просто заост-
рённые или с небольшим нижним
зубцом. В европейской части России
повсеместно.
31(30). Глаз нет. Зубцы игловидных ще-
тинок спинных пучков короче,
чем таковые брюшных. Волосные
щетинки относительно толстые,
длинные (превышают длину игло-
видных в 2 раза), в количестве 2-6
в пучке........Specaria Sperber, 1939.
Единственный вид Specaria josinae
(Vejdovsky, 1884) (рис. 47).
Длина цепочки особей 3-10 мм. Го-
ловная лопасть слегка удлинённая.
Брюшные щетинки с более длинным
верхним зубцом, по 5-10 в пучке.
Желудок в VII-VIII сегментах. Пе-
Рис. 48. Haemonais waldvogeli (по: Timm,
1999).
а — передний конец тела; б — брюшная
щетинка II сегмента; в — то же задней
части тела; г — спинной пучок щетинок.
58
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
ниальные щетинки однозубчатые с крючковидными концами, по 1-3 в пучке.
В европейской части России распространён широко.
32(13). Спинные щетинки начинаются с XVIII-XX сегментов....................
...........................................Haemonais Bretscher, 1900.
Единственный вид Haemonais waldvogeli Bretscher, 1900 (рис. 48)
Длина цепочки особей 4-20 мм. Головная лопасть заострённая. Глаза отсутствуют.
В брюшных пучках по 2-4 двузубчатые щетинки. Во П-XVI сегментах они тонь-
ше, длиннее и прямее, чем в последующих, верхний зубец их длиннее нижнего.
Брюшные щетинки остальных сегментов с более коротким и тонким верхним
зубцом. В спинных пучках по 1 короткой волосной щетинке и 1 двузубчатой с
удлинённым верхним зубцом. Хорошо выраженного желудка нет. Пениальные ще-
тинки двузубчатые сильно изогнутые, по 1-3 в пучке. В европейской части России
распространён широко. Очень редкий.
Ключ для определения видов рода Chaetogaster Baer, 1827
1(2). Головная лопасть хорошо выражена. У фиксированных экземпляров она
треугольной формы, заострённая, у живых — овальной формы с длинны-
ми чувствительными волосками по краю.......................................
............................Ch. diastrophus (Gruithuisen, 1828) (рис. 13).
Длина цепочки особей 1-5 мм. В брюшных пучках II сегмента по 4-8 щетинок, в
остальных — по 3-7. Пениальные щетинки по 2-3 в пучке. В европейской части
России распространён широко.
2(1). Головная лопасть рудиментарная или отсутствует.
3(8). Щетинки двузубчатые с хорошо развитыми верхним и нижним зубцами.
4(7). Верхний зубец щетинок длиннее нижнего. Свободноживущие черви.
5(6). Относительно “крупные” черви: длина цепочки особей 2,5-25 мм; длина
щетинок II сегмента 145-350 мкм............................................
............................Ch. diaphanus (Gruithuisen, 1828) (рис. 14).
Головная лопасть отсутствует; у некоторых экземпляров в центре верхней губы
имеется вырезка. В брюшных пучках II сегмента по 6-13 щетинок, в осталь-
ных — по 4-10 менее длинных и более тонких. Пищевод очень короткий. Пени-
альные щетинки тупоконечные или с рудиментарным верхним зубцом, по 3-5 в
пучке. Хищники. В европейской части России повсеместно.
6(5). Очень мелкие черви: длина цепочки особей 0,8-2 мм; длина щетинок II
сегмента 63-100 мкм........................Ch. langi Bretscher, 1896 (рис. 15).
Головная лопасть рудиментарная. В брюшных пучках II сегмента по 3-9 щетинок,
в остальных — по 3-6. Пищевод хорошо развит, одинаковой длины с глоткой. Пе-
ниальные щетинки по 2 в пучке. В европейской части России в бассейне Белого
моря, Карелии, Ленинградской обл., бассейне р. Волги.
7(4). Зубцы щетинок равной длины или верхний зубец немного короче нижне-
го. Оба зубца сильно изогнуты, имеют вид крючков. Комменсалы брюхо-
ногих моллюсков (обитают на поверхности тела или внутри мантийной
полости)......................................Ch. limnaei Baer, 1827 (рис. 16).
Кольчатые черви 59
Длина цепочки особей 0,7-5 мм. Менее прозрачны, чем другие виды этого рода.
Головная лопасть слабо развита, едва заметна. В брюшных пучках II сегмента по
5-20 щетинок, в остальных — по 4-20. Пищевод очень короткий. Пениальные ще-
тинки двузубчатые или однозубчатые заострённые, по 2-4 в пучке. В европейской
части России распространён широко.
8(3). Щетинки однозубчатые или с рудиментарным верхним зубцом.
9(10). Щетинки однозубчатые...............Ch. setosus Svetlov, 1925 (рис. 17).
Длина цепочки особей 0,7-2 мм. Головная лопасть редуцирована. В брюшных
пучках II сегмента по 3-10 щетинок, в остальных — по 4-10. Пищевод хорошо
развит. Карелия, Архангельская область, Республика Коми, Невская губа Финско-
го залива, бассейн р. Камы. Редкий.
10(9). Щетинки с рудиментарным верхним зубцом............................
....................................Ch. krasnopolskiae Lastockin, 1937.
Длина цепочки особей 1,2-1,4 мм. Головная лопасть выражена слабо. Пищевод
хорошо развит. Карелия, Архангельская область, Республика Коми, реки: Москва,
Скалба, Днепр, Хопёр, Волга. Очень редкий.
Ключ для определения видов рода Stylaria Lamarck, 1816
1(2). Головная лопасть угловатая широкая с ямкой, в глубине которой прикреп-
лён хоботок............................S. lacustris (Linnaeus, 1767) (рис. 23).
Длина цепочки особей 5-18 мм. Глаза всегда имеются. В брюшных пучках по 4—7
щетинок с двумя угловатыми изгибами в проксимальной части. В спинных пучках
по 1-3 тонких длинных волосных щетинки и по 3—4 коротких игловидных одно-
зубчатых. Желудок начинается в VII или VIII сегменте. Пениальные щетинки с
крючковидными концами, по 2 в пучке. В европейской части России повсеместно.
Один из самых распространённых видов.
2(1). Головная лопасть округлая, хоботок прикреплён к её вершине.........
....................................S. fossularis Leidy, 1852 (рис. 24).
Длина тела одиночной особи 6-10 мм, цепочки особей — 14-18 мм. Глаза обыч-
но имеются. В брюшных пучках по 5-15 двузубчатых щетинок, во II сегменте
они длиннее, чем в последующих. В спинных пучках по 1-2 длинных (в 2 раза
превышающих диаметр тела) волосных щетинки и по 1-3 коротких игловидных
однозубчатых. Желудок начинается в VIII сегменте. Пениальные щетинки с крюч-
ковидными концами, по 2-3 в пучке. В европейской части России обнаружен пока
только в Республике Коми (более характерен для азиатской части страны — Сиби-
ри, Алтая, Дальнего Востока).
Ключ для определения видов рода Dero Oken, 1815
1(2). Спинные пучки щетинок начинаются с IV сегмента. Жабр 5 пар.........
..................................D. dorsalis Ferroniere, 1899 (рис. 28).
Длина цепочки особей 10-32 мм. Жаберное ложе с округлыми или угловатыми от-
ростками на заднебоковых краях. Форма жабр разных пар различна. В брюшных
пучках II—V сегментов по 5-7 двузубчатых щетинок с верхним зубцом в 1,5-2 раза
длиннее нижнего. В последующих сегментах количество щетинок уменьшается,
60
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
а зубцы становятся одинаковыми или верхний короче нижнего. Длина брюшных
щетинок убывает по мере удаления кзади. Спинные пучки содержат по 1-2 во-
лосных щетинки и по 1-2 игловидных двузубчатых с верхним зубцом немного
длиннее нижнего. Желудок начинается в IX-XI сегментах. В европейской части
России распространён широко. Редкий.
2(1). Спинные пучки начинаются с VI сегмента. Жабр 3-4 пары.
3(4). Жабр в норме 4 пары. Верхний зубец спинных игловидных щетинок длин-
нее нижнего почти в 2 раза.................D. digitata (Muller, 1774) (рис. 29).
Длина цепочки особей 5-36 мм. Отдельные пары жабр иногда редуцируются и
имеют вид небольших бугорков. Спинной край жаберного ложа может быть с вы-
емкой. В брюшных пучках II—V сегментов по 3-6 двузубчатых щетинок с верхним
зубцом вдвое длиннее нижнего. В последующих сегментах по 2-5 щетинок в пуч-
ке, более толстых и сильно изогнутых, с верхним зубцом лишь немного длиннее
нижнего. Спинные пучки содержат по 1 волосной и по 1 игловидной двузубчатой
щетинке. Желудок в IX-X или X-XI сегментах. В европейской части России по-
всеместно.
4(3). Жабр не более 3 пар. Зубцы спинных игловидных щетинок равной длины,
короткие....................................D. obtusa Udekem, 1855 (рис. 30).
Длина цепочки особей 5-19 мм. Боковая пара жабр имеет более широкое осно-
вание, чем прочие. Пара жабр, примыкающая к спинному краю жаберного ложа,
может быть редуцирована и представлена небольшими вздутиями. В брюшных
пучках II-V сегментов по 2-5 двузубчатых щетинок, более длинных и тонких,
чем последующие, с верхним зубцом вдвое длиннее нижнего. В последующих
сегментах по 3-6 щетинок в пучке, более коротких толстых и сильно изогнутых.
По направлению кзади длина верхнего зубца убывает, а толщина нижнего возрас-
тает. Спинные пучки содержат по 1 волосной и по 1-2 игловидных двузубчатых
щетинки. В европейской части России распространён широко.
Ключ для определения видов рода Vejdovskyella Michaelsen, 1903
1(4). Брюшные щетинки VI и нескольких последующих сегментов гигантских
размеров (значительно утолщены и удлинены) по сравнению с остальными.
2(3). Глаза имеются. Гигантские брюшные щетинки VI-VII (редко и VIII-IX)
сегментов двузубчатые, по форме сходные с остальными.......................
..............................V. macrochaeta (Lastodkin, 1921) (рис. 32).
Длина тела одиночной особи 1-3 мм. В IV-V сегментах брюшные щетинки отсут-
ствуют. Гигантские щетинки в количестве 1-2 в пучке, вместо щетинок обычного
размера (иногда вместе с ними). В спинных пучках содержатся волосные пильча-
тые и игловидные однозубчатые щетинки. Пениальные щетинки однозубчатые с
серповидно загнутым концом. Карелия, Республика Коми, Ленинградская и Ива-
новская обл. Редкий.
3(2). Глаза отсутствуют. Гигантские брюшные щетинки VI (часто и VII-IX
сегментов) трех-, четырёхзубчатые с очень толстым нижним зубцом и 2-3
тонкими верхними зубцами. Гигантские щетинки сильно изогнуты в от-
личие от остальных....................V. intermedia (Bretscher, 1896) (рис. 33).
Кольчатые черви
61
Длина цепочки особей 1,3-3 мм. В брюшных пучках II-V сегментов по 2-4 дву-
зубчатые щетинки с более длинным верхним зубцом. Щетинки II сегмента длин-
нее прочих. Начиная с VI сегмента в брюшных пучках по 1 щетинке. В спинных
пучках по 4-9 пильчатых волосных и по 3-9 тонких игловидных однозубчатых
щетинок. В европейской части России распространён широко.
4(1). Брюшные щетинки всех сегментов примерно одинакового размера. Ги-
гантские щетинки отсутствуют...........V. comata (Vejdovsky, 1884) (рис. 34).
Длина цепочки особей 2-8 мм. Глаза имеются. В брюшных пучках по 3-6 двузуб-
чатых щетинок с верхним зубцом чуть длиннее нижнего. Во II сегменте щетинки
длиннее и толще, чем в остальных, в V отсутствуют. В спинных пучках по 4-8
саблевидно изогнутых волосных щетинок, выпуклый край которых по всей длине
усажен 2 рядами мелких зубчиков, и по 1-8 тонких игловидных однозубчатых
щетинок. Желудок в VIII сегменте. Пениальные щетинки крючковидные по 1 в
пучке. В европейской части России повсеместно.
Ключ для определения видов рода Nais Muller, 1774
1(10). Игловидные щетинки спинных пучков однозубчатые.
2(3). Нижний зубец брюшных щетинок II-V сегментов рудиментарный, плот-
но прижат к верхнему или вовсе отсутствует, верхний — серповидной
формы, сильно вытянут в длину, с острым концом, загнутым под прямым
углом к оси щетинки....................N. behningi Michaelsen, 1923 (рис. 35).
Длина цепочки особей 3-5 мм. Головная лопасть длинная. В брюшных пучках II-V
сегментов по 4-10 двузубчатых щетинок, почти прямых, значительно более длинных
и вдвое более тонких, чем в последующих сегментах. Начиная с VI сегмента в пучках
по 2-7 S-образно изогнутых щетинок, их нижний зубец нормально развит, но короче
верхнего в 2 раза. В спинных пучках по 1 волосной и по 1-2 игловидных щетинки.
Пищевод при переходе в желудок расширяется резко. Пениальные щетинки крючко-
видные, по 2-4 в пучке. В европейской части России распространён широко. Редкий.
3(2). Нижний зубец брюшных щетинок II—V сегментов хорошо развит, хотя и
короче верхнего.
4(7). Игловидные щетинки спинных пучков с тупыми концами, по форме резко
отличаются от волосных.
5(6). Зубцы брюшных щетинок II—V сегментов тонкие, почти параллельные;
нижний зубец образует с осью щетинки острый угол. Во всех сегментах
тела верхний зубец брюшных щетинок вдвое длиннее нижнего. Брюшные
щетинки II—V сегментов заметно длиннее и тоньше, чем в последующих.
Узелок спинных игловидных щетинок располагается на расстоянии Уд-’Л
от верхнего конца щетинки...................N. alpina Speber, 1948 (рис. 36).
Длина цепочки особей 3,6-4,8 мм. Длина головной лопасти немного превышает
её ширину у основания. В брюшных пучках II—V сегментов по 2-8 двузубчатых
щетинок, начиная с VI сегмента — по 3-7. В спинных пучках по 1-2 волосных и
по 1-2 игловидных щетинки. Пищевод при переходе в желудок расширяется рез-
ко. Пениальные щетинки просто заострённые или крючковидные, по 2-3 в пучке.
В европейской части России на севере и северо-западе. Редкий.
62
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
6(5). Зубцы брюшных щетинок II—V сегментов образуют между собой почти
прямой угол. Верхний зубец брюшных щетинок заметно (в 1,6-2 раза)
длиннее нижнего только в передних сегментах (II—V), начиная с VI сег-
мента зубцы брюшных щетинок почти равной длины (верхний зубец лишь
слегка длиннее нижнего). Брюшные щетинки только II сегмента заметно
длиннее и тоньше, чем в последующих. Узелок спинных игловидных ще-
тинок располагается на расстоянии V3 от верхнего конца щетинки...........
.....................................N. simplex Piguet, 1906 (рис. 37).
Длина цепочки особей 4-9 мм. Головная лопасть удлинённая. В брюшных пучках
по 2-6 двузубчатых щетинок. В VI и последующих сегментах брюшные щетинки
более толстые и изогнутые, чем в передних. Их нижний зубец в 2-3 раза толще
верхнего. В спинных пучках по 1-2 волосных и по 1-2 игловидных щетинки.
Пищевод при переходе в желудок расширяется резко. Пениальные щетинки крюч-
ковидные, по 2-3 в пучке. В европейской части России распространён широко.
7(4). Игловидные щетинки спинных пучков с острыми тонко оттянутыми кон-
цами, по форме сходны с волосными.
8(9). Волосные щетинки спинных пучков толстые, длина их превосходит дли-
ну игловидных менее чем в 3 раза. Верхний зубец брюшных щетинок
длиннее нижнего только во II—V сегментах; начиная с VI сегмента зубцы
брюшных щетинок равной длины................N. barbata Muller, 1774 (рис. 38).
Длина цепочки особей 3,5-6,5 мм. Головная лопасть чуть удлинённая. В брюшных
пучках по 2-5 двузубчатых щетинок, во II-V сегментах они длиннее, тоньше и
прямее, чем в последующих. Начиная с VI сегмента щетинки значительно толще и
сильнее изогнуты, их нижний зубец в 2-3 раза толще верхнего. В спинных пучках
по 1-5 волосных и по 2-5 игловидных щетинок. Пищевод при переходе в желудок
(в VII сегменте) расширяется резко. Пениальные щетинки крючковидные, по 2-3
в пучке. В европейской части России распространён широко.
9(8). Волосные щетинки спинных пучков тонкие, длинные (их длина превосходит
длину игловидных в 3-3,5 раза). Верхний зубец брюшных щетинок длиннее
нижнего во всех сегментах тела...........N. pseudobtusa Piguet, 1906 (рис. 39).
Длина цепочки особей 1.5-6 мм. Головная лопасть удлинённая. В брюшных пучках
по 2-5 двузубчатых щетинок. Во II—V сегментах они длиннее и тоньше, чем в по-
следующих, с зубцами одинаковой толщины. Начиная с VI сегмента щетинки ста-
новятся более крупными, нижний зубец толще верхнего. В спинных пучках по 1-3
волосных и по 1-3 игловидных щетинки. Пищевод при переходе в желудок (в VII
сегменте) расширяется резко. Пениальные щетинки с рудиментарным верхним зуб-
цом, по 2-3 в пучке. В европейской части России распространён широко. Редкий.
10(1). Игловидные щетинки спинных пучков двузубчатые.
11(14). Среди брюшных щетинок VI и последующих сегментов имеются значи-
тельно увеличенные с удлинённым верхним зубцом.
12(13). Увеличенные брюшные щетинки начинаются с VI сегмента, гигантских
среди них нет (максимальная толщина щетинок 4 мкм). Их нижний зубец
имеет обычную форму. Игловидные щетинки спинных пучков с коротки-
ми тонкими зубцами, расходящимися под некоторым углом. Пищевод при
Кольчатые черви
63
переходе в желудок (в VII сегменте) расширяется резко..............
.......................•.............N. pardalis Piguet, 1906 (рис. 40).
Длина цепочки особей 2,5-9 мм. В брюшных пучках II-V сегментов по 2-5
двузубчатых щетинок, более тонких и длинных, чем в последующих, с верхним
зубцом в 1,5-2 раза длиннее нижнего. Начиная с VI сегмента, брюшные пучки
содержат по 1-5 щетинок. Помимо увеличенных щетинок с верхним зубцом в
2-3 раза длиннее нижнего имеются щетинки обычного размера с верхним зуб-
цом чуть длиннее, равным или чуть короче нижнего. В спинных пучках по 1-2
коротких волосных и по 1-2 игловидных щетинки. В месте перехода пищевода
в желудок имеется скопление вытянутых коричнево-красных клеток, вдающихся
своими концами в полость кишечника (хорошо различимо у живых экземпляров).
Пениальные щетинки прямые, тупоконечные, по 2-3 в пучке. В европейской части
России распространён широко.
13(12). Увеличенные брюшные щетинки начинаются с VII сегмента, обычно име-
ются гигантские щетинки толщиной до 7,5 мм. Их нижний зубец может
отсутствовать или быть рудиментарным. Игловидные щетинки спинных
пучков с относительно длинными параллельными зубцами. Пищевод при
переходе в желудок расширяется постепенно...............................
...............................N. bretscheri Michaelsen, 1899 (рис. 41).
Длина цепочки особей 3-7 мм. Головная лопасть конусообразная, удлинённая.
В брюшных пучках II-V сегментов по 2-7 двузубчатых щетинок, более тонких
и прямых, чем в последующих сегментах, с сильно изогнутым верхним зубцом
вдвое длиннее нижнего. Начиная с VI сегмента, брюшные пучки содержат по 2-5
щетинок разных типов. Большинство из них обычного размера с верхним зубцом
более тонким и длинным, чем нижний. Наряду с ними имеются утолщённые
щетинки с верхним зубцом в 2-3 раза длиннее нижнего и гигантские щетинки с
рудиментарным нижним зубцом (по 1-2 в пучке). В спинных пучках по 1 (реже
2) короткой волосной щетинке (превышает длину игловидных щетинок в 2 раза) и
по 1-2 игловидных щетинки. Пениальные щетинки крючковидные, по 2 в пучке.
В европейской части России распространён широко.
14(11). Все щетинки VI и последующих сегментов приблизительно одинакового
размера.
15(16). Зубцы спинных игловидных щетинок длинные, почти параллельные.
Верхний зубец брюшных щетинок вдвое длиннее нижнего во всех сегмен-
тах тела. Зубцы брюшных щетинок образуют между собой прямой угол во
всех сегментах тела.......................N. elinguis Muller, 1774 (рис. 42).
Длина цепочки особей 2,2-12 мм. Головная лопасть короткая конусовидная.
В брюшных пучках по 2-5 двузубчатых щетинок, мало отличающихся по длине
и толщине в разных отделах тела. В спинных пучках по 1-4 волосных и по 1-4
игловидных щетинок. Пищевод при переходе в желудок расширяется постепенно.
Пениальные щетинки крючковидные по 2-4 в пучке. В европейской части России
повсеместно.
16(15). Зубцы спинных игловидных щетинок короткие, расходящиеся под неко-
торым углом. Верхний зубец брюшных щетинок длиннее нижнего только
64 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т.
во II—V сегментах, начиная с VI сегмента зубцы брюшных щетинок при
близительно равной длины.
17(20). Брюшные щетинки II—V сегментов с тонкими параллельными у основани
зубцами; верхний зубец длиннее нижнего в 1,5-2 раза; нижний зубец об
разует острый угол с осью щетинки.
18(19). В спинных пучках волосные щетинки превосходят по длине игловидные
3 и более раз. Зубцы игловидных щетинок очень мелкие, едва различимы!
(как правило, хорошо видны лишь при использовании иммерсии). Брюш
ные щетинки начиная с VI сегмента короткие, очень толстые (до 4-5 мкм)
изогнутые. Их нижний зубец в несколько раз толще верхнего.................
..................................N. christinae Kasprzak, 1973 (рис. 43)
Длина цепочки особей 5-8 мм. Головная лопасть широкая, округлая. В брюшньп
пучках по 2-4 двузубчатых щетинки. В спинных по 1 (реже 2) волосной щетинке
и по 1-2 игловидных. Пениальные щетинки крючковидные, по 2-3 в пучке. От
мечен в Ленинградской области. Редкий.
Возможно, имеет более широкое географическое распространение. Его отсутствие
в видовых списках объясняется тем, что он относительно недавно описан и от-
сутствует в определителях по фауне малощетинковых червей России. При опре-
делении экземпляры N. christinae можно перепутать с другими похожими на неп)|
видами рода Nais (например, N. variabilis или N. pardalis).
19(18). В спинных пучках волосные щетинки превосходят по длине игловидные
менее чем в 3 раза (обычно не более, чем в 2,75 раза). Зубцы игловидных
щетинок хорошо различимы. Брюшные щетинки никогда не бывают тол
ще 3 мкм.............................................N. pardalis Piguet, 1906
Экземпляры без увеличенных брюшных щетинок.
20(17). Зубцы брюшных щетинок II—V сегментов расходятся от основания по;
острым углом; верхний зубец лишь немного длиннее нижнего; нижнш
зубец образует прямой угол с осью щетинки.
21(22). Пищевод расширяется в желудок постепенно (в VII сегменте). Спинньи
игловидные щетинки изогнуты на конце, их зубцы хорошо различимы
Брюшные щетинки всех сегментов практически не различаются по длине
толщине и степени изгиба...................N. communis Piguet, 1906 (рис. 44
Длина цепочки особей 1,5-12 мм. Головная лопасть удлинённая. В брюшных пуч-
ках по 2-7 двузубчатых щетинок. В спинных по 1-2 волосных и по 1-2 игловид-
ных. Пениальные щетинки крючковидные или с рудиментарным верхним зубцом,
по 2-3 в пучке. В европейской части России повсеместно.
22(21). Пищевод расширяется в желудок резко (в VI или VII сегменте). Зуб-
цы спинных игловидных щетинок очень мелкие, едва различимые (как
правило, хорошо видны лишь при использовании иммерсии). Брюшные
щетинки, начиная с VI сегмента более короткие, толстые и сильнее изо-
гнуты, чем в передних сегментах. Их зубцы приблизительно равной дли-
ны, но верхний всегда тоньше нижнего......................................
.....................................N. variabilis Piguet, 1906 (рис. 45).
Кольчатые черви
65
Длина цепочки особей 4-10 мм. Головная лопасть удлинённая. В брюшных пучках
по 2-7 двузубчатых щетинок. В спинных по 1-2 волосных и по 1-2 игловидных
щетинки. Пениальные щетинки крючковидные, по 3-4 в пучке. В европейской
части России повсеместно.
Ключ для определения видов семейства Pristinidae
Единственный род Pristina Ehrenberg, 1828. Ранее виды этого рода включали
в сем. Naididae.
Хоботок имеется.
1(4).
2(3).
3(2).
Волосные щетинки III сегмента в несколько раз длиннее, чем во всех
остальных. Игловидные щетинки спинных пучков однозубчатые...........
..................................Р longiseta Ehrenberg, 1828 (рис. 49).
Длина цепочки особей 3,5-6 мм. Брюшные пучки содержат по 3-9 двузубчатых
щетинок, верхний зубец которых длиннее нижнего. Во II сегменте щетинки длин-
нее и тоньше, чем в последующих, с верхним зубцом в 2 раза длиннее нижнего.
В спинных пучках по 1-4 волосных пильчатых щетинки и по 2-5 игловидных
просто заострённых. Желудок начинается в VII-VIII сегментах. Сперматекальные
щетинки в VI сегменте, изогнутые одно- и двузубчатые, по 1-2 в пучке. В евро-
пейской части России распространён широко.
Волосные щетинки III сегмента не удлинены по сравнению с остальными.
Игловидные щетинки спинных пучков двузубчатые
....Р aequiseta Bourne,
1891 (рис. 50).
Синоним: Pristina foreli Pi-
guet, 1906.
Длина цепочки особей 1,8—
8,5 мм. В брюшных пучках
по 2-8 двузубчатых ще-
тинок. Во II сегменте они
длиннее и тоньше осталь-
ных, с верхним зубцом
вдвое длиннее нижнего; в
III—VII сегментах верхний
зубец лишь немного длин-
нее нижнего. У некоторых
особей брюшные щетинки
IV-V сегментов крупнее,
чем в остальных сегментах;
их верхний зубец длин-
Рис. 49. Pristina longiseta (по: Чекановская, 1962).
а — передний конец тела сбоку; б — брюшная ще-
тинка II сегмента; в — то же III сегмента; г — то
же IV сегмента; д — игловидная щетинка спинного
пучка; е — волосная щетинка спинного пучка.
нее нижнего более чем в
2 раза. После VII сегмента
щетинки становятся толще
и короче, более изогнуты,
с зубцами равной длины.
66
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 51. Щетинки Pristina lasto-
tshkini (по: Попченко, 1988).
а — брюшная щетинка II сегмента;
б — то же V сегмента; в — игло-
видная щетинка спинного пучка.
Рис. 50. Pristina aequiseta (по: Чекановская, 1962).
а — передний конец тела сбоку; б — брюшная ще-
тинка II сегмента; в — то же VIII сегмента; г — то
же IV сегмента; д — игловидная щетинка спинного
пучка.
В спинных пучках по 1-4 тонких волосных оперённых щетинки и по 1-4 тонких
игловидных двузубчатых. Желудок начинается в VIII сегменте. В европейской
части России распространён широко.
Вид представлен двумя формами: 1) форма, имеющая крупные брюшные щетинки
в IV-V сегментах; 2) форма, у которой они отсутствуют. Последняя форма ранее
считалась самостоятельным видом Pristina foreli Piguet, 1906.
4(1). Хоботок отсутствует.
5(6). Поверхность тела покрыта палочковидными эпидермальными сосочками
в 2 ряда на каждом сегменте..........Р lastotshkini Popchenko, 1971 (рис. 51).
Длина тела одиночной особи 1,5-2,8 мм. Головная лопасть короткая шлемовид-
ная, длина её значительно меньше ширины у основания. В брюшных пучках по
2-4 двузубчатых щетинки с более длинным и тонким верхним зубцом. В спинных
пучках по 1-3 волосных щетинки и по 2-4 игловидных. Волосные щетинки глад-
кие и толстые, длина их равна поперечнику тела или превосходит его. Игловидные
щетинки с двумя расходящимися под углом 45° очень короткими зубчиками оди-
наковой длины. Желудок начинается в VII сегменте. Река Печора. Очень редкий.
6(5). Поверхность тела гладкая.
7(12). У игловидных щетинок спинных пучков верхний зубец короче нижнего;
волосные щетинки гладкие.
8(9). Желудочное расширение начинается в VI сегменте.......................
...............................Р amphibiotica Lastockin, 1927 (рис. 52).
Синоним: Pristinella amphibiotica (Lastockin, 1927).
Длина цепочки особей 3-6 мм. Головная лопасть удлинённая, на конце овальная.
В брюшных пучках по 3-6 двузубчатых щетинок. В передних сегментах верх-
ний зубец длиннее или равен нижнему, в последующих он значительно короче.
Кольчатые черви
67
Рис. 52. Щетинки Pristina amphibiotica (по: Попченко, 1988).
а — брюшная щетинка II сегмента; б — то же задней части тела; в —
игловидная щетинка спинного пучка.
В спинных пучках по 1-2 волосных щетинки и 1-2 игловидных
двузубчатых с верхним зубцом значительно короче нижнего.
В IV или V сегменте игловидные щетинки длиннее и толще, чем
в остальных. Пениальные щетинки с сильно загнутым зубцом
по 1 в пучке в IX сегменте. В европейской части России на севе-
ре, северо-западе и в центре. Очень редкий.
9(8). Желудочное расширение начинается в VII или VIII сег-
менте.
10(11). Верхний зубец брюшных щетинок в передней и средней
частях тела длиннее нижнего, в задней — зубцы щетинок
равной длины. Игловидные щетинки спинных пучков
всегда двузубчатые, слабоизогнутые............................
.........................Р rosea (Piguet, 1906) (рис. 53).
Синоним: Pristinella rosea (Piguet, 1906).
Длина цепочки особей 4—6 мм. Головная лопасть слегка удлинённая. В брюшных
пучках по 2-5 двузубчатых щетинок. В спинных пучках по 1-2 волосных и по 1-2
игловидных двузубчатых с тонкими зубцами. Желудок начинается в VII или VIII
сегменте. В европейской части России распространён широко.
11(10). Верхний зубец брюшных щетинок длиннее нижнего только в передней ча-
сти тела, в задней он короче нижнего. Игловидные щетинки спинных пучков
а V б
Рис. 53. Pristina rosea.
1 — общий вид (по: Timm, 1999). 2 — щетинки (по: Timm, 1999): а — брюшная II сегмен-
та; б — то же задней части тела; в — спинная игловидная. 3 — щетинки (по: Чекановская,
1962): а — брюшная; б — игловидная. 4 — щетинки (по: Попченко, 1988): а — брюшная;
б — игловидная.
68
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 54. Щетинки Pristina menoni (по: Sperber, 1948).
а — конец брюшной щетинки II сегмента; б — то же задней части тела;
в — игловидная щетинка спинного пучка.
просто заострённые или двузубчатые с рудиментарным верх-
ним зубцом, сильно изогнутые на конце.....................
...........................Р menoni (Aiyer, 1930) (рис. 54).
Синоним: Pristinella menoni (Aiyer, 1930).
Длина цепочки особей 3,7-7 мм. В брюшных пучках по 2-5 двузуб-
чатых щетинок. В спинных пучках по 1-2 волосных щетинки и по
1-2 игловидных. Желудок начинается в VII сегменте. В Карелии,
Республике Коми, Ленинградской и Московской обл. Очень редкий.
12(7). Игловидные щетинки спинных пучков с зубцами равной
длины или с верхним зубцом немного длиннее нижнего; во-
лосные щетинки оперённые или пильчатые.
13(14).Игловидные спинные щетинки с очень короткими зубцами,
верхний длиннее нижнего; на конце щетинки имеется боль-
шой излом. Волосные щетинки оперённые (покрыты воло-
сками с 2-х сторон). Желудок начинается в VI сегменте............
..........................Р komi Popchenko, 1988 (рис. 55).
Длина тела одиночной особи 1,8 мм. Го-
В
Рис. 55. Щетинки Pristina komi (по:
Попченко, 1988)
а — брюшная щетинка II сегмента;
б — то же IV сегмента; в — то же
VII сегмента; г — игловидная ще-
тинка спинного пучка; д — волосная
щетинка спинного пучка.
ловная лопасть вытянута (длина её больше
ширины у основания). Передние сегменты
(II—V) значительно длиннее последующих.
В брюшных пучках по 3-4 двузубчатых
S-образно изогнутых щетинки. В передних
сегментах верхний зубец более тонкий
и чуть короче или равен нижнему, в за-
дних — длиннее нижнего. В спинных
пучках по 1-3 волосных щетинки, по дли-
не превосходящих поперечник тела, и по
1-2 игловидных двузубчатых. Республика
Коми (р. Кайгородка — приток р. Печоры).
Очень редкий.
14(13).Игловидные спинные щетинки с зуб-
цами равной длины; конец щетинки
без излома. Волосные щетинки пиль-
чатые (покрыты волосками только с
одного края). Желудок начинается в
VIII сегменте........................
..Р bilobata (Bretscher, 1903) (рис. 56).
Синоним: Pristinella bilobata (Bretscher,
1903).
Кольчатые черви
69
Длина цепочки особей 4-7 мм. В брюш-
ных пучках по 3-8 двузубчатых щетинок.
В спинных пучках по 1-2 волосных ще-
тинки и по 1-2 игловидных двузубчатых
с параллельно направленными зубцами.
В европейской части России на севере,
северо-западе и в центре.
Рис. 56. Pristina bilobata.
а — передний конец тела сбоку (по:
Чекановская, 1962); б — конец игло-
видной щетинки спинного пучка
(по: Попченко, 1988).
Ключ для определения родов и видов семейства Tubificidae
1(34). Жаберных нитей на поверхности тела нет.
2(7). Тело покрыто тонким слоем вязкого секрета с прилипшими к нему ча-
стицами грунта, которые образуют плотно прилегающую к телу оболочку
(панцирь). Поверхность тела имеет зернистую структуру благодаря на-
личию эпидермальных сосочков и прилипших частиц грунта. Головная
лопасть короткая, обычно втянута внутрь.
3(4). Брюшные щетинки в сегментах за пояском значительно крупнее, чем в
передних, сильно изогнуты, с очень мощным загнутым нижним зубцом
и коротким верхним. Сперматекальные щетинки у половозрелых особей
отсутствуют (в X сегменте в брюшных пучках обычные локомоторные
щетинки). Веерные (двузубчатые) щетинки спинных пучков хотя и тонкие,
но хорошо различимы..................Spirosperma ferox Eisen, 1879 (рис. 57).
Единственный представитель рода Spirosperma Eisen, 1879 в европейской части
России.
Синоним: Peloscolex ferox (Eisen, 1879).
Длина тела 15-58 мм, диаметр 0,4-1,2 мм. В эпителии расположены чувствитель-
ные папиллы в 2-3 ряда на каждом сегменте. В брюшных пучках по 2-5 двузубча-
тых щетинок, в передних сегментах с более длинным и тонким верхним зубцом.
В спинных пучках по 2-8 толстых, саблевидно изогнутых, относительно коротких
волосных и по 2-5 тонких веерных (в задних сегментах — двузубчатых) щетинок.
После откладки коконов область пояска временно освобождается от оболочки из
секрета желёз и частиц грунта. Кроме того, в неблагоприятных условиях (напри-
мер, при токсическом загрязнении водоёма) эпидермальные железы слабо развиты
и на поверхности тела (местами или на всём протяжении) не образуется панциря
из секрета и частиц грунта, так что черви выглядят «голыми». В европейской ча-
сти России повсеместно.
4(3). Брюшные щетинки в сегментах за пояском не выделяются своими раз-
мерами и наличием мощного нижнего зубца по сравнению с таковыми
70
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
2
Рис. 57. Spirosperma ferox.
1 — передний конец тела (по: Чекановская, 1962). 2 — щетинки: а — брюшная щетинка
II сегмента (по: Timm, 1999); б — то же V сегмента (по: Timm, 1999); в — то же задней
части тела (по: Timm, 1999); г — веерная щетинка спинного пучка (по: Timm, 1999); д —
конец веерной щетинки (по: Грабье, 1962); е — волосная щетинка спинного пучка (по:
Timm, 1999). 3 — передний конец половозрелой особи (по: Timm, 1999).
передних сегментов. У половозрелых экземпляров в X сегменте имеются
сперматекальные щетинки характерной формы. Веерные (двузубчатые)
щетинки спинных пучков очень тонкие и короткие (различимые лишь при
сильном увеличении).
Рис. 58. Embolocephalus velu-
tinus (по: Чекановская, 1962).
а — веерная щетинка спинно-
го пучка IV сегмента; б-г —
брюшные щетинки разных
типов.
-5(6).
В брюшных пучках щетинки трёх типов:
однозубчатые заострённые, двузубчатые с
укороченным (рудиментарным) верхним
зубцом, двузубчатые с зубцами равной
длины.....Embolocephalus Randolph, 1892.
В европейской части России единственный вид
Embolocephalus velutinus (Grube, 1879) (рис. 58).
Синоним: Peloscolex velutinus (Grube, 1879).
Длина тела 18-62 мм, диаметр 0,8-2,0 мм.
В брюшных пучках по 2-4 щетинки. В спинных
пучках по 1—4 волосных и до 4 почти не вы-
дающихся над поверхностью тела двузубчатых
щетинок с зубцами, расходящимися под острым
углом. Сперматекальные щетинки по 1-2 в пуч-
ке, гладкие, тонкие, просто заострённые или
немного расширенные с углублением на конце.
В европейской части России на севере, северо-
западе и юге. Редкий.
Кольчатые черви
71
6(5). В брюшных пучках только двузубчатые щетинки. Их верхний зубец в
передних сегментах длиннее и тоньше нижнего, в задних сегментах зубцы
примерно равны по длине и толщине....................Alexandrovia НгаЬё, 1962.
Единственный вид Alexandrovia ringulata (Sokolskaja, 1961) (рис. 59).
Синоним: Alexandrovia onegensis НгаЬё, 1962.
Длина тела 7-24 мм, диаметр в области пояска 0,6-0,9 мм. Передние сегменты
двукольчатые. В эпителии имеются крупные чувствительные папиллы, располо-
женные в 2-3 ряда на каждом сегменте. В брюшных пучках передних сегментов
(до пояска) по 3-5 стройных, S-образно изогнутых двузубчатых щетинок, за по-
яском — по 1-2 щетинки. В передних сегментах верхний зубец брюшных щетинок
длиннее и тоньше нижнего (во II
сегменте — почти вдвое). За пояском
зубцы постепенно выравниваются по
длине и толщине. У некоторых осо-
бей брюшные щетинки IV сегмента
имеют тонкий промежуточный зуб-
чик. У половозрелых экземпляров
отсутствуют брюшные щетинки в
XI сегменте. В спинных пучках по
3-7 (до 10) длинных, относительно
толстых (однако более тонких, чем у
Spirosperma ferox), гладких волосных
и по 1-6 веерных щетинок с неболь-
шим изгибом на конце. Кишечник
покрыт хлорагогенной тканью начи-
ная с V сегмента. Сперматекальные
щетинки по 1 в пучке, однозубча-
тые, узкие с продольным желобком,
слегка загнутые на конце. На севере
европейской части России. Редкий.
7(2). Оболочка из секрета и инородных
частиц отсутствует. Поверхность
тела гладкая, без эпидермальных
сосочков. Головная лопасть хоро-
шо развита, не втянута внутрь.
8(11). В полости тела имеются крупные
зернистые полостные тельца.
Обособленные простатические
железы отсутствуют. Пенисов нет.
9(10). Очень тонкие черви. Голов-
ная лопасть с чувствительной
ресничной ямкой на спинной
стороне. Мужское половое от-
верстие непарное, на брюшной
Рис. 59. Alexandrovia ringulata.
1 — передний конец тела (по: Грабье,
1962). sp — чувствительные папиллы.
2 — щетинки (по: Попченко, 1988): а —
брюшная щетинка II сегмента; б — то же
IV сегмента; в — то же V сегмента; г — то
же из середины тела; д — веерная щетинка
спинного пучка; е — конец веерной ще-
тинки; ж — волосная щетинка спинного
пучка; з — сперматекальная щетинка.
72
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
стороне XI или IX сегмента, по центру. Семяприёмники отсутствуют.
У червей, прошедших копуляцию, на поверхности тела имеются сперма-
тофоры. Характерно бесполое размножение путём архитомии..........
..........................................Bothrioneurum Stoic, 1886.
В европейской части России единственный вид Bothrioneurum vejdovskyanum
Stoic, 1886 (рис. 60).
Длина тела 35 мм, диаметр 0,3-0,4 мм. В брюшных и спинных пучках передних
сегментов по 3-5 щетинок, верхний зубец которых тоньше и почти вдвое длиннее
нижнего. В пучках задних сегментов по 2 щетинки. Зубцы щетинок средних и за-
дних сегментов равной длины. Кишечник покрыт хлорагогенной тканью начиная
с VI сегмента. Пениальные щетинки рядом с мужским половым отверстием, по
2-6 в пучке, залегают в железистом щетинковом мешке. Их концы имеют вид
сильно загнутых крючков, под которыми располагаются два ряда мелких зубчи-
ков. В европейской части России на юге, а также в Чудском оз. и на Валдайской
Рис. 60. Bothrioneurum vejdovskyanum.
а — общий вид (по: Timm, 1999); б — го-
ловная лопасть с чувствительной ямкой
(по: Timm, 1999); в — несколько сегмен-
тов передней части тела со сперматофо-
рами, приклеенными к кутикуле (по: Че-
кановская, 1962); г — щетинка передних
сегментов (по: Timm, 1999); д — то же
задних сегментов (по: Timm, 1999); е —
конец пениальной щетинки (по: Чеканов-
ская, 1962).
возвышенности.
Возможно, имеет более широкое географиче-
ское распространение, но редко распознаётся
исследователями, т.к. размножается преиму-
щественно бесполым путём.
Рис. 61. Rhyacodrilus falciformis (по:
Timm, 1999).
а — общий вид половозрелой особи;
б — конец локомоторной щетинки; в —
конец пениальной щетинки.
Кольчатые черви
73
10(9). Черви обычного облика. Чув-
ствительная ресничная ямка на
головной лопасти отсутствует.
Мужские половые отверстия
парные. Семяприёмники име-
ются. Сперматофоры отсут-
ствуют. Размножение только
половое.........................
.......Rhyacodrilus Bretscher,
1901 (с. 85); (рис. 61,62).
Сперматекальные щетинки в X
сегменте отсутствуют, имеются
пениальные в XI сегменте.
11(8). Крупные полостные тельца
отсутствуют. На атрии одна
или несколько обособленных
простатических желёз. Пенисы
имеются.
12(13). Зона образования новых сег-
ментов расположена на значи-
тельном расстоянии от заднего
конца тела. Позади неё на-
ходится несегментированный
сократимый отдел, функциони-
рующий как орган дыхания........
.....Aulodrilus Bretscher, 1899
(с. 86); (рис. 63-65).
Черви небольших размеров с про-
зрачным телом. Головная лопасть
короткая. Верхний зубец двузуб-
чатых и верных щетинок заметно
короче нижнего, сами щетинки в
дистальной части (непосредствен-
но под зубцами) могут быть фрон-
тально расширены в виде вздутия
или латеральных «крыльев». Во-
лосные щетинки короткие (их
длина превышает длину двузуб-
чатых примерно в 2 раза). Поясок
у большинства видов расположен
на VI—VIII сегментах. Размноже-
ние преимущественно бесполое
(путём архитомии), половозрелые
особи встречаются редко.
Рис. 62. Rhyacodrilus coccineus.
а — передний конец тела половозрелой
особи (по: Timm, 1999); б — полостные
тельца в головной лопасти (по: Timm, 1999);
в — пучок пениальных щетинок (по: Timm,
1999); г — конец брюшной щетинки VIII
сегмента (по: Чекановская, 1962); д — типы
пениальных щетинок (по: Чекановская,
1962); е — конец веерной щетинки спинного
пучка (по: Чекановская, 1962).
Рис. 63. Aulodrilus limnobius (по: Timm,
1999).
а — общий вид; б — щетинка.
74
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Рис. 65. Aulodrilus pluriseta.
а — общий вид (по: Timm, 1999);
б — латеральный и фронтальный вид
щетинки (по: Timm, 1999); в — дву-
зубчатая щетинка спинного пучка II
сегмента (по: Чекановская, 1962); г —
брюшная щетинка IV сегмента (по:
Чекановская, 1962).
Рис. 64. Aulodrilus pigueti.
1 — брюшная щетинка XXI сегмента (по: Че-
кановская, 1962). 2 — задний конец тела (по:
Чекановская, 1962). 3 — спинные щетинки раз-
ной формы из одного пучка XV сегмента (по:
Чекановская, 1962). 4 — весловидные щетинки
XXV сегмента (по: Чекановская, 1962). 5 —
латеральный и фронтальный вид весловидных
щетинок (по: Timm, 1999).
13(12). Зона образования новых сегментов расположена на заднем конце тела
(терминально). Несегментированного отдела позади неё нет.
14(23). Волосные щетинки в спинных пучках отсутствуют. Щетинки в спинных и
брюшных пучках однотипные — двузубчатые.
15(20). Верхний зубец щетинок длиннее или равен (изредка немного короче) ниж-
нему.
16(19). Пенисы одеты твёрдыми хитиновыми трубками. Пениальные щетинки в
XI сегменте отсутствуют.
17(18). Пениальные трубки (в XI сегменте) грибовидной формы, сильно расши-
ренные на обоих концах, с толстыми складчатыми стенками. Их длина
почти равна максимальной ширине. В X сегменте имеются сперматекаль-
ные щетинки, тонкие однозубчатые с узким жёлобом в дистальной части.
Черви мелкие........................Peipsidrilus pusillus Timm, 1977 (рис. 66).
Кольчатые черви
75
Длина тела 6-8 мм, диаметр в об-
ласти пояска 0,2-0,4 мм. Голов-
ная лопасть удлинённая, на конце
коническая. Передние сегменты
двукольчатые. Пучки содержат по
1-6 щетинок, зубцы которых равной
длины или верхний немного длин-
нее нижнего. Кишечник покрыт
хлорагогенной тканью начиная с
V или VI сегмента. В европейской
части России в Псковско-Чудском
и Онежском озёрах, Невской губе
Финского залива. Редкий.
18(17). Пениальные трубки цилиндриче-
ские с небольшим терминальным
расширением. Длина трубки
превосходит её максимальную
ширину в 4 и более раз. Сперма-
текальные щетинки отсутствуют
(в X сегменте обычные локомо-
торные щетинки). Черви относи-
тельно крупные.....................
....Limnodrilus Claparede, 1862
(с. 87); (рис. 67-72).
19(16). Пенисы мягкие, без хитиновых
трубок. В XI сегменте рядом с
мужскими половыми отверсти-
ями располагаются пениальные
щетинки (по 1 в пучке). Их верх-
ний зубец прямой и значительно
длиннее и тоньше нижнего...........
...Potamothrix moldaviensis Vej-
dovsky et Mrazek, 1903 (рис. 73).
Рис. 66. Peipsidrilus pusillus (по: Timm,
1999).
а — передний конец половозрелой особи;
б — локомоторная щетинка; в — сперма-
текальная щетинка; г — латеральный вид
пениальной трубки; д — апикальный вид
пениальной трубки
Рис. 67. Limnodrilus udekemianus (по:
Timm, 1999).
а — передний конец половозрелой особи;
б — участок хвостового отдела тела; в —
щетинка II сегмента; г — то же V сегмен-
та; д — то же заднего отдела.
Рис. 68. Щетинки Limnodrilus claparedea-
nus (no: Timm, 1999).
a — щетинка переднего отдела тела; б — то
же заднего отдела тела.
76
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 69. Limnodrilus hoffineisteri.
а — передний конец половозрелой
особи (по: Timm, 1999); б — брюшная
щетинка переднего отдела тела (по:
Чекановская, 1962); в — то же XV сег-
мента (по: Чекановская, 1962).
Рис. 70. Limnodrilus profundicola (по: Timm,
1999).
а — передний конец половозрелой особи; б —
щетинка.
Рис. 72. Пениальные трубки представителей
рода Limnodrilus (по: Чекановская, 1962).
а — Limnodrilus profundicola (= Limnodrilus
helveticus)\ б — Limnodrilus udekemianus', в —
Limnodrilus hoffineisteri', г — Limnodrilus clapa-
redeanus.
Рис. 71. Мужской гонодукт Limnodrilus hoffineisteri (по: Чекановская, 1962).
a — семенная воронка; б — семяпровод; в — простатическая железа; г — атрий; д — пе-
нис; е — пениальная сумка; ж — мужское половое отверстие.
Кольчатые черви
77
Синоним: Ilyodrilus moldavien-
sis (Vejdovsky et Mrazek, 1903).
Длина тела 10-40 мм. Головная
лопасть короткая закруглён-
ная. Межсегментные борозды
глубокие, хорошо различи-
мые, особенно в заднем отделе
тела. Верхний зубец щетинок
длиннее и тоньше нижнего.
В переднем отделе тела по 5-9
щетинок в пучке, в заднем от-
деле число щетинок уменьша-
ется. Кишечник покрыт хлора-
гогенной тканью начиная с VI
сегмента. В X сегменте имеют-
ся сперматекальные щетинки,
однозубчатые с продольным
жёлобом в дистальной части и
слабо загнутым кончиком, по
1 в пучке (по форме сходны со
сперматекальными щетинкам
других представителей рода
Potamothrix). В европейской
части России распространён
широко.
20(15). Верхний зубец щетинок
значительно короче ниж-
него (по крайней мере, в
переднем отделе тела).
21(22). У половозрелых экземпля-
ров в брюшных пучках IX,
X и XII сегментов по 1 по-
ловой щетинке с верхним
зубцом в 2 раза длиннее
нижнего, изогнутым под
прямым углом к оси щетин-
ки. Зубцы локомоторных
щетинок образуют между
собой острый угол. Хлора-
гогенная ткань на кишечни-
ке начинается с V сегмента.
Пенис мягкий без хитино-
идного чехлика................
...Isochaetides НгаЬё, 1965.
Рис. 73. Potamothrix moldaviensis.
а — общий вид половозрелой особи (по: Timm,
1999); б — конец спинной щетинки II сегмента
(по: Чекановская, 1962); в — концы пениальных
щетинок (по: Чекановская, 1962); г — спермате-
кальная щетинка (по: Чекановская, 1962).
Рис. 74. Isochaetides michaelseni.
1 — передний конец тела с кровеносной систе-
мой (по: Чекановская, 1962). 2 — локомоторные
щетинки (по: Чекановская, 1962): а — брюшная
щетинка V сегмента; б — то же VIII сегмента;
в — то же XIII сегмента; г — то же задней части
тела; д — спинная щетинка III сегмента; е — то
же VIII сегмента. 3 — половая щетинка — вид с
разных сторон (по: Timm, 1999).
78
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 75. Tubifex newaensis.
а — передний конец половозрелой особи (по:
Timm, 1999); б — брюшные щетинкл переднего
отдела тела (по: Чекановская, 1962).
В европейской части России единственный
вид Isochaetides michaelseni (Lastodkin, 1936)
(рис.74).
Синоним: Limnodrilus michaelseni Lastodkin,
1936.
Длина тела 25-80 мм. Головная лопасть короткая округлая. Передние сегменты
(II—IV) двукольчатые. В переднем отделе тела по 4-6 щетинок в пучке, за по-
яском — 3-4. В сегментах за пояском разница в величине зубцов щетинок менее
ощутима, чем в передних. В XI сегменте брюшные щетинки часто отсутствуют.
Половые щетинки примерно в 1,5 раза длиннее локомоторных. В европейской
части России распространён широко.
22(21). Половых щетинок нет. Все щетинки двузубчатые с укороченным верхним
зубцом. Угол между зубцами щетинок близок к прямому. Хлорагогенная
ткань на кишечнике начинается с VI сегмента. Пенис одет толстым хити-
ноидным чехликом..................Tubifex newaensis (Michaelsen, 1903) (рис. 75).
Синонимы: Limnodrilus newaensis Michaelsen, 1903, Isochaetides newaensis (Mi-
chaelsen, 1903).
Длина тела 32-93 мм, диаметр 0,6-3,0 мм. Гигант среди тубифицид. Головная
лопасть закруглённая. Щетинки S-образно изогнуты. Иногда их зубцы стёрты и
имеют округлые концы. В переднем отделе тела брюшных щетинок по 3-4 в пуч-
ке, спинных — по 4-8. Позади пояска во всех пучках по 2 щетинки. В XI сегменте
у половозрелых червей брюшные щетинки отсутствуют. Межсегментные пере-
городки (диссепименты) очень толстые, хорошо различимы сквозь покровы тела.
В европейской части России распространён широко.
23(14). Волосные щетинки в спинных пучках имеются.
24(25). За пояском двузубчатые и веерные щетинки заметно толще и грубее, чем в
передних сегментах тела.............................................
.......................Psammoryctides НгаЬё, 1964 (с. 88); (рис. 76-78).
25(24). Двузубчатые и веерные щетинки примерно одинаковые во всех сегментах
тела: щетинки за пояском не выделяются своей толщиной по сравнению с
таковыми переднего отдела тела.
26(31). Пенисы заключены в пениальные трубки.
27(28). Пениальные трубки воронкообразные с широким основанием и сужен-
ным, немного изогнутым и слепо замкнутым концом. Отверстие трубки
расположено на боковой поверхности её конца...............................
.......................................Tasserkidrilus Holmquist, 1985.
В пресных водоёмах европейской части России единственный вид Tasserkidrilus
kessleri (НгаЬё, 1962) (рис. 79).
Кольчатые черви
79
Рис. 76. Psammoryctides barbatus.
1 — передний конец тела половозрелой осо-
би (по: Timm, 1999). 2 — участок заднего
отдела тела (по: Timm, 1999). 3 — щетинки
(по: Чекановская, 1962): а — брюшная ще-
тинка III сегмента; б — то же средней части
тела; в — веерная щетинка передней части
тела; г — то же задней части тела; д — спер-
матекальная щетинка.
Рис. 77. Psammoryctides albicola.
1 — передний конец тела половозрелой
особи (по: Timm, 1999). 2 — участок за-
днего отдела тела (по: Timm, 1999). 3 —
щетинки: а — спинная веерная щетинка
III сегмента (по: Чекановская, 1962); б —
брюшная щетинка V сегмента (по: Чека-
новская, 1962); в — фрагмент волосной
щетинки (по: Timm, 1999) ; г — спермате-
кальная щетинка (по: Timm, 1999)
Рис. 78. Щетинки Psammoryctides moravicus (по:
Чекановская, 1962).
а — спинная веерная щетинка VI сегмента; б — спин-
ная двузубчатая щетинка области за пояском; в, г —
концы брюшных щетинок III сегмента; д — конец
брюшной щетинки IV сегмента.
80
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 79. Tasserkidrilus kessleri (по: Грабье,
1962).
а — конец брюшной щетинки IX сегмента;
б — брюшная щетинка и щетиночная же-
леза; в — пениальная трубка; г — продоль-
ный разрез пениального влагалища.
Синоним: Tubifex kessleri НгаЬё, 1962.
Длина тела 18-34 мм, диаметр 0,4-0,9 мм.
Головная лопасть небольшая закруглённая.
В брюшных пучках передних сегментов
тела по 3-5 двузубчатых щетинок с верхним
зубцом немного длиннее нижнего; в не-
которых щетинках имеется 1 дополнитель-
ный промежуточный зубчик. У основания
брюшных щетинок имеется грушевидная
железа с эозофильным секретом. В спин-
ных пучках передних сегментов тела по
1-2 коротких волосных и по 3-5 веерных
щетинок с 1-2 промежуточными зубчиками.
Волосные щетинки имеются только в V, VI, VIII и X сегментах. Половые щетинки
отсутствуют. Онежское оз. (Петрозаводская губа). Очень редкий.
28(27). Пениальные трубки конусообразные, более широкие у основания и сужи-
вающиеся к концу. Отверстие расположено терминально на конце трубки.
29(30). Межсегментные борозды неглубокие, слабо выражены. Мужское половое
отверстие в XI или в IX сегменте, поясок на X-XI или на VIII-IX сегмен-
тах. Длина пениальных трубок превышает их максимальную ширину ме-
нее, чем в 2 раза. Кишечник покрыт хлорагогенной тканью начиная с VIII
сегмента. Черви мелкие............Peipsidrilus saamicus (Timm, 1978) (рис. 80).
Синоним: Umbadrilus saamicus Timm, 1978.
Длина тела до 6 мм, диаметр 0,2 мм. Головная лопасть коническая закруглённая
на конце. В брюшных пучках по 1-5 двузубчатых щетинок, верхний зубец кото-
рых немного длиннее и тоньше или равен нижнему. Многие брюшные щетинки
содержат очень мелкий промежуточный зубчик, видимый лишь при очень боль-
шом увеличении (под иммерсией). В сегментах с половыми отверстиями (X и
XI или VIII и IX) брюшные щетинки отсутствуют. В спинных пучках до пояска
по 1-6 гладких волосных и по 1-4 веерных щетинки, за пояском по 1-3 гладких
волосных и по 1-2 веерных щетинки. Сдвиг половой системы вперёд косвенно
указывает на возможное бесполое размножение путём архитомии. В европейской
части России в Карелии и на Кольском п-ове. Редкий.
Возможно, имеет более широкое географическое распространение, но остаёт-
ся неопределённым из-за редкой встречаемости половозрелых экземпляров.
30(29). Межсегментные борозды глубокие, хорошо выражены, особенно в заднем
конце тела. Мужское половое отверстие всегда в XI сегменте, поясок на
Кольчатые черви
81
Рис. 80. Peipsidrilus s a amicus.
1 — передний конец тела (по: Timm, 1999). 2 — щетинки (по: Попченко, 1988): а — брюш-
ная щетинка II сегмента; б — то же VIII сегмента; в — то же IX сегмента; г — то же задней
части тела; д — веерная щетинка спинного пучка VII сегмента; е — то же VIII сегмента;
ж — то же задней части тела. 3 — пенис с пениальной трубкой (по: Тимм, Попченко, 1978).
п — пенис; п.т. — пениальная трубка.
IX-XIL Длина пениальных трубок превосходит их максимальную ширину
в 2 раза. Кишечник покрыт хлорагогенной тканью начиная с VI сегмента.
Черви среднего размера..........................Ilyodrilus Eisen, 1879.
В европейской части России единственный вид Ilyodrilus templetoni (Southern,
1909) (рис. 81).
Синоним: Tubifex templetoni Southern, 1909.
Длина тела 10-30 мм. Передние сегменты двукольчатые, заднее кольцо шире
переднего. В брюшных пучках передней части тела по 3-4 двузубчатых щетинки с
более длинным и тонким верхним зубцом. У половозрелых экземпляров брюшные
щетинки в XI сегменте отсутствуют. В спинных пучках передней части тела по 1-4
волосных, тонких и длинных, и по 3-4 веерных щетинки. Половые щетинки отсут-
ствуют. В европейской части России распространён широко. Относительно редкий.
31(26). Пенисы мягкие или покрыты тонкой коричневатой кутикулой, не образую-
щей трубок.
32(33). В брюшных пучках X сегмента имеются сперматекальные щетинки.......
..............Potamothrix Vejdovsky et Mrazek, 1903 (с. 89); (рис. 82-86).
33(32). Сперматекальные щетинки отсутствуют — в брюшных пучках X сегмента
обычные локомоторные щетинки...............................................
.............................Tubifex Lamarck, 1816 (с. 91); (рис. 87-89).
82
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 81. Ilyodrilus templetoni.
а — общий вид половозрелой особи (по:
Timm, 1999); б — брюшная щетинка
передней части тела (по: Timm, 1999);
в — то же задней части тела (по: Timm,
1999); г — веерная щетинка спинного
пучка (по: Timm, 1999); д — пенис с
пениальной трубкой (по: НгаЬе, 1981).
Рис. 82. Potamothrix bedoti.
1 — общий вид (по: Timm, 1999). 2 — особь
в состоянии архитомического деления (пе-
редний конец тела в стадии регенирации)
(по: Timm, 1999). 3 — щетинки: а — веерная
щетинка спинного пучка (по: Timm, 1999);
б — брюшная щетинка передней части тела
(по: Timm, 1999); в — то же задней части
тела (по: Timm, 1999); г — сперматекальная
щетинка (по: Чекановская, 1962).
34(1). На заднем конце тела в каждом сегменте имеются спинные и брюшные
жаберные нити......................................Branchiura Beddard, 1892.
Единственный вид Branchiura sowerbyi Beddard, 1892 (рис. 90).
Длина тела 40-185 мм, диаметр 1-2,5 мм. Головная лопасть заострённая. Сегмен-
ты переднего отдела тела двукольчатые. Отдел, несущий жабры, занимает ’/3-2/5
всей длины тела. В брюшных пучках переднего отдела тела до 16 двузубчатых
щетинок с более коротким (иногда рудиментарным) верхним зубцом, в среднем
отделе — до 4, в заднем — не более 2. В спинных пучках переднего отдела тела
по 1-8 коротких волосных и по 4-12 двузубчатых (иногда однозубчатых) щетинок,
сходных с брюшными. Длина волосных щетинок превосходит длину двузубчатых
в 3 раза. В жаберном отделе волосных щетинок нет. Последние сегменты тела
вообще лишены щетинок. В полости тела содержатся крупные, зернистые по-
лостные тельца круглой формы. В европейской части России на юге, а также в
теплицах ботанических садов центральных и северных районов.
Кольчатые черви
83
Рис. 83. Potamothrix heuscheri (по:
Timm, 1999).
а — передний конец тела половозрелой
особи; б — брюшная щетинка передней
части тела; в — то же задней части тела;
г — веерная щетинка спинного пучка;
д — сперматекальная щетинка.
Рис. 84. Potamothrix bavaricus.
а — передний конец тела половозрелой осо-
би (по: Timm, 1999); б — сперматекальная
щетинка (по: Чекановская, 1962); в — ве-
ерная щетинка спинного пучка (по: Timm,
1999); г — брюшная щетинка передней ча-
сти тела (по: Timm, 1999); д — то же задней
части тела (по: Timm, 1999).
Рис. 85. Potamothrix hammoniensis (по: Timm, 1999).
а — общий вид половозрелой особи; б — брюшная щетинка передней части тела; в — то
же задней части тела; г — веерная щетинка спинного пучка; д — сперматекальные ще-
тинки.
84
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 86. Potamothrix vejdovskyi.
1 — передний конец тела половозрелой
особи (по: Timm, 1999). 2 — щетинки (по:
Timm, 1999): а — двузубчатая щетинка пе-
редней части тела; б — то же задней части
тела. 3 — щетинки (по: Чекановская, 1962):
а — спинная щетинка II сегмента; б — то
же VI сегмента; в — то же заднего отдела
тела; г — брюшная щетинка IV сегмента.
Рис. 87. Tubifex ignotus.
1 — общий вид (по: Timm, 1999). 2 —
щетинки (по: НгаЬе, 1981): а — веерная
щетинка спинного пучка V сегмента;
б — то же спинного пучка L сегмента; в —
брюшная щетинка V сегмента; г — то же
L сегмента.
Рис. 88. Tubifex tubifex.
1 — общий вид (по: Timm,
1999). 2 — щетинки (по: НгаЬе,
1981): а — веерная щетинка
спинного пучка VII сегмента;
б — то же XVII сегмента; в —
двузубчатая щетинка спинного
пучка XXVI сегмента; г —
брюшная щетинка III сегмента;
д — то же VI сегмента; е — то
же XXV сегмента.
Кольчатые черви
85
Рис. 89. Tubifex smimowi (по: Timm,
1999).
а — передний конец тела половозре-
лой особи; б — брюшная щетинка;
в — веерная щетинка спинного пучка.
Рис. 90. Branchiura sowerbyi (по: Чекановская, 1962).
а — общий вид; б — двузубчатая щетинка передней ча-
сти тела; в — то же задней части тела.
Ключ для определения видов рода Rhyacodrilus Bretscher, 1901
1(2). Волосные щетинки в спинных пучках отсутствуют. Пениальные щетинки
крупные (в 1,5-2 раза длиннее локомоторных брюшных щетинок), сильно
изогнутые, серповидной или крючковидной формы, по 1-2 в пучке.............
...............................Rh. falciformis Bretscher, 1901 (рис. 61).
Длина тела 8-10 мм, диаметр 0,3 мм. Головная лопасть закруглённая. Щетинки в
брюшных и спинных пучках во всех отделах тела однотипные. Их верхний зубец
почти вдвое длиннее и немного тоньше нижнего. В брюшных пучках переднего
отдела по 3-6 щетинок, заднего — по 2. В спинных пучках по 3-4 щетинки. В Ле-
нинградской области. Очень редкий.
2(1). Волосные щетинки в спинных пучках имеются. Пениальные щетинки
одинакового размера с локомоторными брюшными щетинками, S-образно
изогнуты на концах, однозубчатые или с редуцированным верхним зуб-
цом, по 2-5 в пучке. Дистальные концы пениальных щетинок сближены,
так что пучок образует «веер»........Rh. coccineus (Vejdovsky, 1875) (рис. 62).
86
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Длина тела 15-35 мм, диаметр 0,4-0,6 мм. В брюшных пучках 2-8 двузубчатых
щетинок. Зубцы щетинок в переднем отделе тела равной длины, в среднем и за-
днем — нижний зубец чуть толще и длиннее верхнего. В спинных пучках до XX
сегмента по 3-8 волосных и по 3-7 веерных щетинок, в последующих сегментах
веерные щетинки заменяются двузубчатыми (по 3-4 в пучке) с более толстым и
длинным нижним зубцом. В европейской части России распространён широко.
Ключ для определения видов рода Aulodrilus Bretscher, 1899
1(2). Волосные щетинки в спинных пучках отсутствуют......................
..................................A. limnobius Bretscher, 1899 (рис. 63).
Длина тела 8-16 мм, диаметр 0,2-0,3 мм. Головная лопасть короткая закруглённая.
Спинные и брюшные щетинки однотипные — двузубчатые, S-образно изогнутые,
в количестве до 10 в пучке. Кишечник расширяется в VIII сегменте, хлорагоген-
ные клетки на кишечнике начинаются с VII сегмента. Поясок на VII—VIII (IX) сег-
ментах. Половые щетинки отсутствуют. В европейской части России повсеместно.
2(1). Волосные щетинки в спинных пучках имеются.
3(4). В спинных пучках, начиная с VI-IX сегментов двузубчатые щетинки
заменены на весловидные уплощённые щетинки, конец которых может
быть закруглённым или двузубчатым.........................................
.................................A. pigueti Kowalewski, 1914 (рис. 64).
Длина тела 5-14 мм, диаметр 0,2-0,3 мм. Головная лопасть слабо выражена, ко-
ническая. В брюшных пучках по 3-6 двузубчатых щетинок. В спинных пучках
передних сегментов по 1-4 волосных и по 2-6 двузубчатых щетинок. Кишечник
заметно расширяется в VII сегменте, хлорагогенные клетки начинаются с VII
сегмента. Поясок на VII—VIII сегментах. Пениальные щетинки в VII сегменте,
изогнутые, заострённые на концах, с продольной бороздой, по 1-2 в пучке. В ев-
ропейской части России повсеместно.
4(3). Весловидных щетинок в спинных пучках нет.
5(6). Поясок у половозрелых экземпляров расположен на VII—VIII сегментах.
Мужское половое отверстие в VII сегменте. Имеются только двузубчатые
щетинки, веерные отсутствуют. Кишечник расширяется в VIII сегменте...
..........................................A. pluriseta (Piguet, 1906) (рис. 65).
Длина тела 12-40 мм, диаметр 0,3-0,7 мм. Головная лопасть короткая конусо-
видная. В брюшных пучках передних сегментов по 13-17 двузубчатых щетинок,
сильно S-образно изогнутых. В спинных пучках передних сегментов до 8 волос-
ных и до 10 почти прямых двузубчатых щетинок. В задних сегментах число ще-
тинок уменьшается. Половые щетинки отсутствуют. В европейской части России
распространён широко.
6(5). Поясок у половозрелых экземпляров расположен на IX-XI сегментах.
Мужское половое отверстие в X сегменте. В спинных и брюшных пучках
помимо двузубчатых имеются веерные щетинки с очень мелкими допол-
нительными боковыми зубчиками (от 1 до нескольких по обеим сторонам
щетинки). Кишечник расширяется в X-XI сегментах...........................
........................................A. japonicus Yamaguchi, 1953.
Кольчатые черви
87
Длина тела 20-26 мм, диаметр 0,3-т0,4 мм. Головная лопасть короткая коническая.
В брюшных пучках по 5-14 двузубчатых (веерных) щетинок. В спинных пучках
по 2-8 волосных и по 4-10 двузубчатых (веерных) щетинок. Во II сегменте и в
нескольких последних сегментах число щетинок минимально. Кишечник плотно
покрыт хлорагогенными клетками начиная с X сегмента, но небольшое количе-
ство этих клеток можно обнаружить в VI-IX сегментах. Половые щетинки от-
сутствуют. Отмечен пока только в Невской губе Финского залива и Рыбинском
водохранилище.
Вероятно, данный вид имеет более широкое географическое распространение
(сравнимое с распространением других представителей рода). Его отсутствие в
видовых списках можно объяснить тем, что неполовозрелые экземпляры этого
вида легко принять за A. pluriseta (дополнительные латеральные зубчики на дву-
зубчатых щетинках видны только при очень большом увеличении), а половозре-
лые особи редки вследствие преимущественно бесполого размножения.
Ключ для определения видов рода Limnodrilus Claparide9 1862
1(2). Верхний зубец щетинок в передних сегментах значительно длиннее ниж-
него и крючковидно загнут. Длина пениальной трубки превосходит её
ширину у основания не более чем в 4 раза..................................
............................L. udekemianus Claparede, 1862 (рис. 67; 726).
Длина тела 20-90 мм, диаметр 0,5-1,3 мм. Относительно длинные черви (до 160
сегментов в составе тела). В заднем отделе тела имеются поперечные пигментные
полосы (жёлтого цвета). Головная лопасть короткая, слегка заострённая. Сегмен-
ты в переднем отделе тела двукольчатые (переднее кольцо шире заднего). Сегмен-
ты заднего отдела удлинены. В передних сегментах по 5-8 щетинок в пучке, в
средних — до 4, в задних — по 2. Зубцы щетинок заднего отдела равны по длине.
Тёмная хлорагогенная ткань на кишечнике начинается с VI сегмента. В европей-
ской части России повсеместно.
2(1). Верхний зубец щетинок немного длиннее, равен или чуть короче нижнего.
Длина пениальных трубок превосходит их ширину у основания в большее
число раз.
3(4). Пениальная трубка очень длинная — отношение её длины к ширине у
основания составляет 23-33 : 1 ...........................................
............................L. claparedeanus Ratzel, 1868 (рис. 68; 72г).
Длина тела 20-112 мм (отдельные экземпляры до 135 мм), диаметр 0,6-1,3 мм.
Головная лопасть удлинённая. В пучках переднего отдела по 5-10 щетинок, верх-
ний зубец которых равен или слегка длиннее нижнего. В заднем отделе тела число
щетинок уменьшается. Тёмная хлорагогенная ткань на кишечнике начинается с V
сегмента. В европейской части России повсеместно.
4(3). Пениальная трубка короче.
5(6). Длина пениальной трубки превосходит ширину у основания в 8-14 раз....
.............................L. hoffmeisteri Claparede, 1862 (рис. 69; 71; 72в).
Длина тела 12-66 мм. Головная лопасть короткая тупая. В переднем и среднем
отделах тела по 4-8 щетинок в пучке, в заднем — по 2-3. Зубцы щетинок рав-
88
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
ной длины или верхний слегка короче нйжнего. Тёмная хлорагогенная ткань на
кишечнике начинается с V сегмента. В европейской части России повсеместно.
Один из самых распространённых видов.
6(5). Длина пениальной трубки превосходит ширину у основания в 5-6 раз......
.............................L. profundicola (Verrill, 1871) (рис. 70; 72а).
Синоним: L. helveticus Piguet, 1913.
Длина тела 20-50 мм, диаметр 0,5-1,2 мм. Головная лопасть короткая, чуть за-
острённая. В передних сегментах в пучках по 5-9 щетинок, в задних — по 1-2.
Зубцы щетинок равной длины или верхний немного длиннее нижнего. Тёмная
хлорагогенная ткань на кишечнике начинается с V-VI сегмента. В европейской
части России распространён широко.
Ключ для определения видов рода Psammoryctides НгаЬё, 1964
1(2). Веерные щетинки спинных пучков в передних сегментах тела по форме
сильно отличаются от двузубчатых брюшных (напоминают садовые веер-
ные грабли). Их краевые зубцы одинаковы по форме и размеру, широко
расставлены и расположены симметрично по отношению к продольной
оси щетинки (верхний и нижний зубцы неразличимы). Между краевыми
зубцами имеется большое число промежуточных зубчиков, по размерам
практически не отличающихся от краевых. Волосные щетинки гладкие,
без оперения...............................Р barbatus (Grube, 1861) (рис. 76).
Синоним: Psammoryctes barbatus (Grube, 1861).
Длина тела 19-70 мм, диаметр 0,7-1,3 мм. Головная лопасть треугольная за-
острённая. Первые 3 сегмента тела двукольчатые. В брюшных пучках передних
сегментов по 3-4 двузубчатые щетинки с более длинным верхним зубцом. В сег-
ментах среднего и заднего отделов по 2 щетинки, верхний зубец которых тоньше
и короче нижнего, а сами щетинки значительно более толстые и грубые, чем
в переднем отделе тела. В спинных пучках по 1-2 волосных и по 3-8 веерных
щетинки. Веерные щетинки VIII и последующих сегментов по форме близки к
двузубчатым брюшным с тонкими промежуточными зубчиками. Сперматекальные
щетинки в X сегменте, прямые заострённые, в дистальной части полые, немно-
го длиннее локомоторных. В европейской части России распространён широко,
встречаемость постепенно уменьшается в направлении на север и восток.
2(1). Веерные щетинки спинных пучков сходны с брюшными двузубчатыми во
всех сегментах тела. Их промежуточные зубчики значительно короче и
тоньше краевых зубцов. Волосные щетинки оперённые.
3(4). В спинных пучках всех сегментов тела имеются волосные и веерные ще-
тинки. Веерные щетинки однотипные во всех отделах тела, их нижний
зубец заметно толще верхнего. Оперение волосных щетинок хорошо раз-
личимо.................................Р albicola (Michaelsen, 1901) (рис. 77).
Синоним: Psammoryctes albicola (Michaelsen, 1901).
Длина тела 24-41 мм, диаметр 0,6-1,1 мм. Передние сегменты двукольчатые.
В брюшных пучках по 1-3 двузубчатых щетинки. В спинных по 1-3 волосных
и по 2-3 веерных щетинки. Сперматекальные щетинки в X сегменте, прямые
Кольчатые черви
89
заострённые, в дистальной части полые и косо срезанные, по длине равны локо-
моторным. В европейской части России распространён широко, на севере встре-
чается редко.
4(3). Волосные щетинки в спинных пучках имеются только до XXI сегмента
тела, веерные — только до XIV(XV). Начиная с XIV(XV) сегмента веер-
ные щетинки заменены двузубчатыми. Веерные и двузубчатые щетинки
спинных пучков различны по форме: краевые зубцы веерных щетинок
прямые, равные по длине и толщине, у двузубчатых щетинок нижний
зубец в несколько раз толще и длиннее верхнего. Оперение волосных ще-
тинок заметно только при сильном увеличении.................................
....................................Р moravicus (НгаЬё, 1934) (рис. 78).
Синоним: Psammoryctes moravicus НгаЬё, 1934.
Длина тела 20-30 мм. В брюшных пучках впереди пояска по 2-4 двузубчатые
щетинки, позади пояска — по 2, в задней части тела — по 1. Верхний зубец
брюшных щетинок длиннее нижнего (во II и III сегментах в 3-4 раза). Брюшные
щетинки II и III сегментов меньшего размера, чем в последующих. В спинных
пучках впереди пояска по 1-5 волосных и 2-3 веерных щетинок, позади пояска по
1 волосной и по 1 веерной щетинке, после XX сегмента только по 1 двузубчатой
щетинке. Сперматекальные щетинки в X сегменте прямые с дистальным концом
в виде полукруглого долота. В европейской части России в р. Оке и нижнем тече-
нии р. Нарвы. Очень редкий.
Ключ для определения видов
рода Potamothrix Vejdovsky et MrAzek, 1903
(кроме Potamothrix moldaviensis Vejdovsky et Mrazek, 1903; cm. c. 000)
1(2). Сперматекальные щетинки в VII-VIII сегментах (в обоих или в одном),
асимметричной формы, напоминающие птичий клюв. В IX сегменте рас-
положены пениальные щетинки по форме сходные со сперматекальными.
Поясок на VIII — IX сегментах. Способны к бесполому размножению.............
........................................Р bedoti (Piguet, 1913) (рис. 82).
Синоним: Ilyodrilus bedoti Piguet, 1913.
Длина тела 4-20 мм, диаметр 0,3-0,7 мм. Головная лопасть удлинённая, слегка
заострённая. В брюшных пучках по 2-6 двузубчатых щетинок с более длинным и
тонким верхним зубцом, иногда зубцы равной длины. В спинных пучках по 3-5
волосных и по 3-5 веерных щетинки с более длинным и тонким верхним зубцом.
В середине тела веерные щетинки заменяются двузубчатыми. Сперматекальные
и пениальные щетинки крупные, по 1-2 в пучке. В европейской части России на
севере, северо-западе и юге.
Вероятно, имеет более широкое географическое распространение, но редко рас-
познаётся исследователями, т.к. размножается преимущественно бесполым путём.
Неполовозрелые экземпляры Р bedoti неотличимы от Р hammoniensis.
2(1). Сперматыкальные щетинки в X сегменте, края их более или менее парал-
лельны. Пениальные щетинки (в XI сегменте) отсутствуют. Поясок на X-
XI сегментах. Размножение только половым путём.
90
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
3(8). В спинных пучках имеются веерныё щетинки. Зубцы веерных и двузуб-
чатых щетинок равной длины или верхний немного длиннее нижнего.
Волосные щетинки относительно длинные (их длина превышает длину
веерных в 2,5 и более раз).
4(5). Сперматыкальные щетинки узкие, тонкие, без расширения в дистальной
части, загнутые на концах..............Р. heuscheri (Bretscher, 1900) (рис. 83).
Синоним: Ilyodrilus heuscheri (Bretscher, 1900).
Длина тела 6-15 мм. В брюшных пучках по 3-5 двузубчатых щетинок. В передних
сегментах верхний зубец щетинок длиннее и тоньше нижнего, за пояском зубцы
равной длины. В спинных пучках по 2-5 волосных и по 2-5 веерных щетинок.
В европейской части России в реках и эстуариях бассейнов Понто-Каспия и Бал-
тийского моря.
5(4). Сперматыкальные щетинки расширены в дистальной части, их концы не
загнуты.
6(7). Сперматыкальные щетинки копьеобразной формы, с коротким жёлобом,
сильно расширенным у основания и заканчивающимся остриём..................
................................Р bavaricus (Oschmann, 1913) (рис. 84).
Синоним: Ilyodrilus bavaricus Oschmann, 1913.
Длина тела 12-35 мм, диаметр 0,3-0,8 мм. Головная лопасть закруглённая.
В брюшных пучках по 2-5 двузубчатых щетинок. В спинных пучка по 1-5 волос-
ных и по 2-5 веерных щетинок. В задних сегментах (после ХХХ-ХХХП сегмен-
тов) веерные щетинки заменяются двузубчатыми с более толстым нижним зубцом.
В европейской части России в реках, эстуариях и водохранилищах бассейнов
Понто-Каспия и Балтийского моря.
7(6). Сперматыкальные щетинки более узкие, с удлинённым и слабо расширен-
ным в дистальной части жёлобом............................................
...........................Р hammoniensis (Michaelsen, 1901) (рис. 85).
Синоним: Ilyodrilus hammoniensis Michaelsen, 1901.
Длина тела 10-49 мм, диаметр 0,4-1,0 мм. Головная лопасть округлая. В брюш-
ных пучках по 3-6 двузубчатых щетинок. В спинных пучках по 1-5 волосных и по
2-6 веерных щетинок; в середине тела веерные щетинки заменяются двузубчаты-
ми. В европейской части России повсеместно. Один из самых распространённых
видов.
8(3). Веерные щетинки в спинных пучках отсутствуют, вместо них имеются
двузубчатые, верхний зубец которых немного короче нижнего. Волосные
щетинки относительно короткие (их длина превышает длину двузубчатых
не более чем в 2 раза)....................Р vejdovskyi (НгаЬё, 1941) (рис. 86).
Синоним: Ilyodrilus vejdovskyi НгаЬё, 1941.
Длина тела 10-21 мм, диаметр (в области пояска) 0,7 мм. Головная лопасть окру-
глая. В брюшных пучках 3-10 двузубчатых щетинок с верхним зубцом немного
короче или равным нижнему. В спинных пучках по 2-5 волосных и по 2-6 двузуб-
чатых щетинки, сходных с брюшными. Сперматекальные щетинки по форме напо-
минают таковые Р hammoniensis. В европейской части России в реках, эстуариях
и водохранилищах бассейнов Понто-Каспия и Балтийского моря.
Кольчатые черви
91
Ключ для определения видов рода Tubifex Lamarck, 1816
(кроме Tubifex newaensis (Michaelsen, 1903); см. с. 78)
1(4). Волосные щетинки содержатся во всех сегментах тела. Зубцы брюшных
щетинок равной длины или верхний длиннее нижнего, по крайней мере, в
переднем отделе тела. Задние сегменты тела не шире передних.
2(3). Черви очень длинные и тонкие, нитевидные. Волосные щетинки за по-
яском в 4-5 раз длиннее поперечника тела....................................
.......................................Т. ignotus (Stoic, 1886) (рис. 87).
Длина тела 39-80 мм, диаметр (в области пояска) 0,4-0,9 мм. Головная лопасть
закруглённая. Передние сегменты (II-VI) двукольчатые, переднее кольцо уже
заднего. В брюшных пучках по 1-4 двузубчатых щетинок с более длинным и
тонким верхним зубцом в передних сегментах; в задних сегментах зубцы равной
длины, нижний значительно толще верхнего. В спинных пучках передних сегмен-
тов по 2-5 волосных и по 2-3 веерных щетинки; в сегментах за пояском волос-
ных щетинок по 1-3 в пучке; в задних сегментах веерные щетинки заменяются
двузубчатыми. Пениальные щетинки отсутствуют. В европейской части России
распространён широко.
3(2). Пропорции тела обычные для тубифицид, тело не удлинённое, не ните-
видное. Длина волосных щетинок не более, чем в 2 раза превосходит по-
перечник тела...............................Т. tubifex (Muller, 1774) (рис. 88).
Длина тела 10-100 мм, диаметр 0,5-1,2 мм. Головная лопасть слегка удлинённая.
В брюшных пучках переднего отдела тела по 3-6 двузубчатых щетинок (иногда
с 1-2 промежуточными зубчиками), верхний зубец которых немного длиннее и
тоньше нижнего. В спинных пучках переднего отдела тела по 1-6 длинных, тон-
ких, оперённых волосных и по 2-5 веерных щетинок с зубцами равной длины.
За пояском веерные щетинки заменяются двузубчатыми, число щетинок сокраща-
ется (1-2 волосных и 1-3 двузубчатых). В заднем отделе тела спинные и брюшные
двузубчатые щетинки с более тонким и коротким верхним зубцом, нижний зубец
толстый и изогнутый. Пениальные щетинки отсутствуют, в брюшных пучках
XI сегмента содержится по 2-6 (чаще по 4—6) двузубчатых щетинок. В европей-
ской части России повсеместно. Один из самых распространённых видов.
4(1). Волосные щетинки имеются только в передних сегментах тела. Верхний
зубец брюшных щетинок короче нижнего во всех отделах тела. Сегменты
заднего отдела тела шире передних...........................................
....................................Т. smirnowi Lastockin, 1927 (рис. 89).
Длина тела 7-12 мм, диаметр 0,3-0,4 мм. Головная лопасть вытянутая и заострён-
ная на конце. В брюшных пучках по 2-6(7) двузубчатых щетинок. В спинных
пучках по 1-3 оперённых волосных и по 1-4 веерных щетинки. Зубцы веерных
щетинок почти равной длины (верхний чуть короче), между ними 1-3 проме-
жуточными зубчиками. Пениальные щетинки отсутствуют. В европейской части
России на севере, северо-западе и в центре. Очень редкий.
92 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Ключ для определения родов и видов семейства Lumbriculidae
Определение большинства родов и видов этого семейства основано на иссле-
довании строения внутренних органов и требует специальных методов. Ниже при-
ведены виды, легко идентифицируемые по особенностям внешней морфологии.
1(6). Головная лопасть с длинным хоботком.......Rhynchelmis Hoffineister, 1843.
2(3). Щетинки во всех пучках только однозубчатые, слегка S-образно изогну-
тые, тупые на концах...................Rh. limosella Hoffineister, 1843 (рис. 91).
Длина тела 32-140 мм, диаметр 1-3 мм. Поверхность тела с радужным блеском.
Сегменты двукольчатые с более широким передним кольцом. В поперечном сече-
нии передние сегменты круглые, остальные — четырёхгранные трапециевидные;
задний конец тела уплощён. Одна пара мужских половых отверстий в X сегменте
и одно непарное мужское половое отверстие в IX сегменте. В европейской части
России распространён широко. Редкий.
3(2). Щетинки двузубчатые или разных типов (однозубчатые и двузубчатые).
4(5). Щетинки в брюшных и спинных пучках только двузубчатые с рудиментар-
ным верхним зубцом, сильно S-образно изогнутые, заострённые на конце
.......................................Rh. tetratheca Michaelsen, 1920 (рис. 92).
Длина тела 15-40 мм, диаметр 0,8-1,5 мм. Двукольчатость сегментов хорошо вы-
ражена. Одна пара мужских половых отверстий в X сегменте и одно непарное
мужское половое отверстие в IX сегменте. В европейской части России на севере
(повсеместно) и северо-западе.
5(4). Щетинки брюшных пучков однозубчатые, лишь некоторые из них имеют
едва заметный верхний зубец. Щетинки спинных пучков двузубчатые с
рудиментарным верхним зубцом..............Rh. granuensis onegensis НгаЬё, 1962.
Длина тела 32-50 мм, диаметр 0,8-1,3 мм. Сегменты двукольчатые. Мужских по-
ловых отверстий одна пара в XI сегменте. Карелия (Онежское оз.; р. Уница; ключ
у с. Кончозеро). Очень редкий.
6(1). Головная лопасть без хоботка.
7(8). У половозрелых экземпляров в X сегменте тела имеются невтягивающие-
ся длинные пенисы. Щетинки с очень коротким и тонким верхним зубцом
(у некоторых щетинок верхний зубец редуцирован полностью). Головная
лопасть небольшая, конической формы, закруглённая на конце. Межсег-
ментные борозды глубокие, хорошо выраженные. Сегменты, начиная с IV,
Рис. 91. Rhynchelmis limosella (по: Timm,
1999).
а — передний конец тела; б — щетинка.
Рис. 92. Rhynchelmis tetratheca (по: Timm,
1999).
а — передний конец тела; б — щетинка.
Кольчатые черви
93
Рис. 93. Stylodrilus heringianus.
1 — передний конец тела половозрелой особи (по: Timm, 1999). 2 — разные типы щети-
нок (по: Timm, 1999). 3 — щетинки (по: Попченко, 1988): а — спинная щетинка VI сег-
мента; б — спинные щетинки II сегмента; в — брюшные щетинки VIII сегмента.
Рис. 94. Lumbriculus variegatus.
1 — передний конец тела (по: Timm, 1999). 2 — особь с регенерирующими передним и
задним концами тела (по: Timm, 1999). 3 — щетинка: а — по: Чекановская, 1962; б — по:
Timm, 1999; в — по: Попченко, 1988.
94 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
двукольчатые с широким передним и узким задним кольцом. Спинной со-
суд не имеет слепых ветвящихся придатков. Размножение только половое
......................Stylodrilus heringianus Claparede, 1862 (рис. 93).
Длина тела 20-60 мм, диаметр в области пояска 0,7-1,4 мм. Щетинки в пучке не-
равной длины: одна короче другой. В европейской части России на севере, северо-
западе и в центре. Предпочитает водоёмы с холодной и чистой водой.
8(7). Невтягивающиеся пенисы у половозрелых экземпляров отсутствуют.
Верхний зубец щетинок короче нижнего, но хорошо различим. Головная
лопасть большая, удлинённая (её длина больше ширины у основания).
Межсегментные борозды выражены слабо. Сегменты однокольчатые.
Спинной сосуд, начиная с XI сегмента, снабжён слепыми сократимыми
придатками (по 1 паре в каждом сегменте). В передней части тела они
простые, мешковидные, в задней — гроздевидно ветвятся, образуя харак-
терный рисунок в виде пары клубочков по бокам от кишечника. Придатки,
как и спинной сосуд, покрыты хлорагогенной тканью и хорошо видны
сквозь стенку тела. Размножение в основном бесполое (путём архитомии).
Живые экземпляры при прикосновении легко распадаются на отдельные
фрагменты, каждый из которых способен полностью регенерировать....
......................Lumbriculus variegatus (Muller, 1774) (рис. 94).
Длина тела 13-90 мм, диаметр 1-1,5 мм. Брюшные и спинные щетинки однотип-
ные, S-образно изогнутые. Мужских половых отверстий одна пара в VIII (иногда
IX) сегменте. В европейской части России повсеместно.
Подкласс Бранхиобделлида — Branchiobdellida
Отряд БРАНХИОБДЕЛЛИДА — BRANCHIOBDELLIDA
Семейство BRANCHIOBDELLIDAE («рачьи пиявки»)
Ранее сем. Branchiobdellidae включали в подкласс Малощетинковых червей
(Oligochaeta).
Данное семейство включает в себя паразитов и комменсалов речных ра-
ков (живут на поверхности тела и жабр). Некрупные черви (длиной 4-13 мм) с
уплощённым коротким телом. Окраска белая, розовая, красная или светло-корич-
невая. Тело состоит из трёх отделов: нерасчленённого снаружи головного (из 4
сегментов), туловищного (из 11 двукольчатых сегментов) и мускулистой задней
присоски (расположена на XV сегменте). Головная лопасть и щетинки отсутству-
ют. Глаз нет. Около ротового отверстия имеется мышечная складка в виде губ с
сосочками. Ротовая полость вооружена хитиноидным челюстным аппаратом, со-
стоящим из пары челюстей с зубцами или без. Полость тела развита только в ту-
ловищном отделе. Анальное отверстие находится на спинной стороне у основа-
ния задней присоски. Размножение только половое. Поясок на IX-XI сегментах.
Кольчатые черви
95
Парные семяпроводы объединяются в один непарный семяпровод, впадающий в
непарный атрий. Атрий на конце переходит в пенис, способный выворачиваться
наружу. Мужское половое отверстие непарное (на брюшной стороне X сегмента).
Семяприёмник также непарный (в IX сегменте).
Всего известно 150 видов бранхиобделлид. В пресных водоёмах европейской
части России обитают представители только одного рода — Branchiobdella Odier,
1823.
Ключ для определения видов рода Branchiobdella Odier, 1823
1(4). Челюсти треугольной формы.
2(3). Обе челюсти приблизительно одинаковой величины. По бокам от боль-
шого среднего зуба имеется по 2—4 маленьких боковых зубчика. Пенис
на конце покрыт многочисленными крючьями, направленными своими
остриями назад. Обитает преимущественно на поверхности тела (очень
редко на жабрах) узкопалого и широкопалого раков. Окраска от серовато-
белой до жёлтой с красным оттенком.... В. parasita (Braun, 1805) (рис. 95).
Длина тела 4-6 - 10-12 мм. Головной отдел такой же ширины как первые сегмен-
ты туловищного отдела. Сегменты тела двукольчатые. Переднее кольцо сегментов
туловищного отдела шире заднего. Присоска несколько шире сегментов, лежащих
перед ней. В европейской части России на северо-западе.
3(2). Челюсти разного размера: одна из них значительно больше другой. Воору-
жены челюсти единственным крупным зубом. Боковые зубчики отсутству-
Рис. 95. Branchiobdella parasita.
а — общий вид (по: Timm, 1999); б — пара
челюстей (по: Timm, 1999); в — форма че-
люсти (по: Чекановская, 1962).
Рис. 96. Branchiobdella astaci.
1 — пара челюстей (по: Timm, 1999):
а — верхняя челюсть; б — нижняя че-
люсть. 2 — форма челюсти (по: Чеканов-
ская, 1962).
96
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 97. Branchiobdella pentodonta.
а — общий вид половозрелой особи (по: Timm, 1999); б — две разные пары челюстей (по
Timm, 1999); в — форма челюсти (по: Чекановская, 1962).
ют или рудиментарны. Пенис без крючьев. Обитает на жабрах узкопалого
и широкопалого раков. Окраска красновато-коричневая....................
...........................................В. astaci Odier, 1823 (рис. 96).
Длина тела 3,5-5 - 8-12 мм. Головной отдел почти не обособлен от туловищного
Около ротового отверстия можно различить спинную и брюшную губы. Присоска
развита слабо. Наиболее патогенный из видов, обитающих в европейской части
России. В европейской части России распространён широко.
4(1). Челюсти четырёхугольной формы (прямоугольные или трапециевидные),
удлинённые в поперечном направлении...........................................
...................................В. pentodonta Whitman, 1882 (рис. 97).
Длина тела 2-6 мм. Тело прозрачное, бесцветное или беловатое. Головной коней
значительно уже туловищного. Челюсти снабжены 5 (иногда 4) крупными зубца-
ми на свободном крае. Форма и число зубчиков варьирует. Средний зубец всегда
длиннее краевых. Пенис совершенно прямой, гладкий (без крючков) или вздутый
на конце. Обитает на поверхности тела широкопалого рака. В европейской части
России на северо-западе и в центре.
Подкласс Щетинконосные пиявки — Acanthobdellida
Отряд ЩЕТИНКОНОСНЫЕ ПИЯВКИ — ACANTHOBDELLIDA
Ранее семейства щетинконосных пиявок включали в подкласс Пиявок (Him-
dinea).
Число сегментов (сомитов) в теле постоянно — 30. Вторичная кольчатость хо-
рошо выражена, полный сомит состоит из 4-х колец.
Кольчатые черви
97
Рис. 98. Щетинконосная пиявка Acanthobdella peledina.
а — общий вид фиксированного экземпляра (по: Лукин, 1977); б — передний конец тела
с брюшной стороны (по: Лукин, 1977); в — передний конец тела (по: Эпштейн, 1987).
I-VI — сомиты; 1 — ротовое отверстие; 2 — щетинки, г — щетинки из последнего и
первого рядов (по: Эпштейн, 1987).
Отличительной особенностью является наличие щетинок на брюшной сторо-
не переднего конца тела (на первых пяти сегментах) (рис. 98). Щетинки образуют
поперечные ряды и сближены попарно.
Передняя присоска отсутствует или слабо развита. Задняя присоска мала, рас-
положена перпендикулярно продольной оси тела.
Желудок и кишка не имеют слепых отростков, но отчётливо разделены на ка-
меры.
Размножение только половое. Мужское и женское половые отверстия располо-
жены в области пояска и отделены друг от друга тремя кольцами. Позади женско-
го полового отверстия имеется углубление — сперматека, к которой прикрепляет-
ся сперматофор.
Эктопаразиты рыб.
98
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Всего известно два вида щетинконосных пиявок, относящихся к двум моно-
типическим семействам. На территории европейской части России обитает един-
ственный вид — Acanthobdella peledina Grube, 1851 (семейство Acanthobdelli-
dae) (рис. 98).
Длина тела 30-40 мм, ширина 2-3,5 мм. Головная лопасть и передняя присо-
ска отсутствуют. Передний конец тела закруглён. Тело цилиндрическое, у фик-
сированных экземпляров часто задняя его часть вздута. Окраска изменчива (от
буровато-серой до тёмно-зелёной с жёлтыми пятнами или серой с красноватым
оттенком). На спинной стороне можно различить более тёмные поперечные сег-
ментальные полосы (числом до 24). Фиксированные экземпляры имеют белова-
тую, желтоватую, иногда ярко-жёлтую окраску. Щетинки крючковидной формы
с закруглёнными концами. Три пары мелких глаз. Паразиты лососевых, сиговых
и хариусов (на поверхности тела и плавниках). Постоянные эктопаразиты, поки-
дают хозяина только на период размножения. В европейской части России встре-
чается в бассейне р. Уса (Республика Коми), водоёмах Кольского п-ова, озёрах
Онежском и Имандра.
Подкласс Пиявки — Hirudinea
Отряд ПИЯВКИ — HIRUDINIDA
Строение тела
Число сегментов (сомитов) в теле постоянно — 34 (рис. 99).
Отличительной чертой внешнего облика пиявок является наличие двух присо-
сок (у некоторых видов могут подвергаться редукции). Другая характерная осо-
бенность — вторичная кольчатость тела: каждый сегмент неглубокими перетяж-
ками делится на ряд дополнительных колечек. Число колец в сомите различно у
представителей разных видов.
Форма тела большинства пиявок более или менее уплощённая, цилиндриче-
ская встречается гораздо реже. У одних видов она постоянна, у других подвер-
жена значительной изменчивости. Края тела могут быть ровными или иметь за-
зубрины.
Пиявки, как правило, интенсивно окрашены (исключение составляют посто-
янные эктопаразиты и обитатели пещерных водоёмов). Поверхность кожи у од-
них видов пиявок гладкая, у других на спинной стороне располагаются сосочки
(папиллы), часто образующие правильные ряды.
На спинной стороне переднего конца тела у большинства пиявок расположены
глаза (рис. 100, 103). У некоторых видов рыбьих пиявок по окружности задней
присоски располагаются глазоподобные пятна.
Передний отдел пищеварительного тракта (глотка и часть пищевода) у пи-
явок семейств Glossiphoniidae и Piscicolidae превращён в выдвижной хоботок
(рис. 101, 117). У челюстных пиявок в ротовой полости развиты валики, несущие
Кольчатые черви
99
Рис. 99. Расчленение тела пиявки (по: Лукин, 1976).
а — вид со спинной стороны; б — вид с брюшной стороны. I-XXXIV — сомиты; —
мужское половое отверстие; $ — женское половое отверстие.
зубы. У глоточных валики и зубы редуцированы. Желудок и кишка кровососущих
видов имеют боковые выросты (рис. 101). Анальное отверстие расположено на
спинной стороне впереди задней присоски.
Пиявки размножаются только половым путём. Половые отверстия открывают-
ся на пояске: сначала мужское, за ним женское (рис. 99, 101). Поясок образован
у пиявок тремя сомитами (XI, XII, XIII). У челюстных пиявок конечный отдел
атриума превращен в копулятивный орган.
100
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 100. Расположение глаз у пиявок (по: Лукин, 1977).
1 — Glossiphonia complanatcr, 2-4 — G. concolor, 5, 6 — G. heteroclite\ 7 — Helobdella
stagnalis', 8 — Boreobdella verrucata; 9 — Batracobdella paludosa\ 10 — Theromyzon\ 11 —
Hemiclepsis marginatcr, 12 — Cestobranchus и Piscicola\ 13 — Erpobdellcr, 14 — Hirudo и
Haemopis.
Оплодотворение внутренние при помощи копулятивных органов (челюстные
пиявки) или сперматофоров (все остальные). Оплодотворённые яйца помещают-
ся в коконы, которые прикрепляются к различным подводным субстратам или за-
рываются во влажном грунте выше уреза воды. Для плоских пиявок характерна
забота о потомстве: яйца и (или) молодь, вышедшая из коконов, прикрепляются к
брюшку матери. Строение коконов различно у представителей разных семейств.
Развитие прямое (плоские пиявки) или с образованием скрытой личинки (прочие
пиявки). В последнем случае личинка развивается внутри кокона, питаясь белко-
вой жидкостью.
Пиявки широко распространены во всех зоогеографических зонах и в океа-
не от высоких широт обоих полушарий до тропических морей. В основном, это
Кольчатые черви
101
Рис. 101. Внутреннее строение пиявки (на примере Glossiphonia complanata) (по:
Лукин, 1977).
— мужское половое отверстие; $ — женское половое отверстие. 1 — ротовое от-
верстие; 2 — хоботок во втянутом состоянии; 3 — семенник (первый); 4 — семяпровод;
5 — кишечник; 6 — семенник (десятый); 7 — задняя кишка; 8 — анальное отверстие (на
спинной стороне тела); 9 — боковой вырост желудка (первый); 10 — яичник; 11 — бо-
ковой вырост кишки (первый); 12 — боковой вырост желудка (шестой); 13 — боковой
вырост кишки (четвёртый).
водные организмы, однако, некоторые виды приспособлены к наземному образу
жизни в условиях повышенной влажности тропических лесов.
Пиявки — исключительно плотоядные животные. По способу питания они
или кровососы, паразитирующие на различных видах беспозвоночных и позво-
ночных животных, или хищники. Постоянных эктопаразитов, покидающих своих
хозяев лишь на короткие периоды размножения, немного (большинство рыбьих
пиявок). Остальные виды, в т.ч. и кровососущие, ведут активный образ жизни.
102
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Всего на земном шаре известно приблизительно 680 видов пиявок, 480 из них
ведут пресноводный образ жизни. В континентальных (пресноводных) водоёмах
Палеарктики обитает 185 видов пиявок. В пресных водоёмах европейской части
России распространено около 30.
Методы сбора и обработки
При сборе пиявок следует учитывать их поведенческие особенности. В при-
брежных зарослях, где фауна пиявок наиболее богата и разнообразна, для мало-
подвижных форм наиболее эффективен сбор руками с поверхности различных
подводных предметов. Для подвижных форм и активных пловцов неплохие ре-
зультаты даёт сачок. Виды, обитающие в грунте, хорошо улавливаются скребком.
При сборе пиявок, обитающих вдали от берега на больших глубинах, подходят
традиционные гидробиологические орудия (дночерпатель, драга). При лове меди-
цинских пиявок широко пользуются методом приманок (свежий вскрытый труп
кролика или лягушки; похлопывание палкой по поверхности воды, имитирующее
движение жертвы). Данный метод может применяться и при сборе других кро-
вососущих видов. Стационарных паразитов, покидающих своих хозяев лишь на
короткие периоды размножения, лучше собирать, осматривая их потенциальных
жертв вскоре после вылова (т.к. пиявки быстро оставляют погибших хозяев).
Фиксируют пиявок 4% формалином или 70° спиртом. В формалине лучше со-
храняется окраска и глаза. Однако, черви, фиксированные формалином, сильно
сжимаются, половые отверстия в этом случае плохо различимы. При фиксации
и хранении в спирте пиявки сохраняют большую гибкость тела, на таком матери-
але легче изучать кольчатость и расположение половых отверстий, но он быстро
обесцвечивается. Непосредственное погружение в крепкие растворы фиксаторов
может вызвать сильное сжатие тела, что затруднит дальнейшее исследование,
поэтому целесообразно прибегать к анестезии живых червей слабым раство-
ром спирта или хлоралгидратом, и только после того, как пиявки окончательно
перестали реагировать на раздражители, переносить их в формалин или крепкий
спирт. Фиксированный материал следует хранить в темноте, т.к. свет губителен
для пигментов.
Изучение внешних деталей строения пиявок проводят при помощи лупы, би-
нокуляра и микроскопа как в падающем, так и в проходящем свете. При этом по-
лезно зажимать пиявок между двумя предметными стёклами. Такая манипуляция
содействует выпрямлению червя и уменьшению его толщины. Для выявления не-
которых признаков (например, чтобы лучше рассмотреть глаза) целесообразно
просветлять пиявок в глицерине.
Подсчёт числа глаз и изучение особенностей их расположения весьма полезен
при определении пиявок. Глаза хорошо сохраняются у фиксированных экземпля-
ров. Однако следует помнить, что глаза у пиявок подвержены значительной из-
менчивости (редукции, слиянию). Чисто формальное отношение к этому призна-
Кольчатые черви
103
ку может привести к ошибкам. Поэтому при вынесении окончательного решения
о видовой принадлежности исследуемого экземпляра необходимо обратить вни-
мание и на остальные характерные признаки.
Подсчёт колец в полном сомите облегчается тем, что окраска тела пиявок ча-
сто носит сегментарный характер или кольца сомита имеют разную длину и чере-
дуются в разном порядке.
Пособия для определения пиявок
Лукин Е.И. 1976. Пиявки пресных и солоноватых вододоемов. Серия: Фауна СССР. Пияв-
ки. Т. 1. Л.: Наука. 484 с.
Лукин Е.И. 1977. Класс Пиявки Hirudinea // Определитель пресноводных беспозвоночных
европейской части СССР (планктон и бентос) / Ред. Л.А. Кутикова, Я.И. Страробога-
тов. Л.: Гидрометеоиздат. С. 201-212.
Эпштейн В.М. Тип Кольчатые черви — Annelida. Класс Пиявки — Hirudinea // Определи-
тель паразитов пресноводных рыб фауны СССР / Ред. О.Н. Бауер. Т. 3. Паразитиче-
ские многоклеточные (вторая часть). Л.: Наука. С. 340-372.
Timm Т. 1999. A Guide to the Estonian Annelida. Tartu - Tallinn: Estonian Academy Publishers.
208 p.
Elliott J.M., Mann K.H. 1979. A key to the British freshwater Leeches. Freshwater Biological
Association, Scientific Publication. No. 40. 72 p.
Ключ для определения семейств и видов
1(8). Задняя и передняя присоски хорошо развиты. Глаза имеются.
2(3). Присоски дисковидной формы, резко отделены от прилегающих частей
тела......................Сем. Piscicolidae (Рыбьи пиявки) (рис. 1026) (с. 112).
Синоним: Ichthyobdellidae.
Тело или цилиндрическое, или уплощённое и разделённое на два отдела: перед-
ний более узкий и короткий («шею») и задний более длинный и широкий («туло-
вище»). По краям тела расположены боковые пузыри (обычно 11 пар), величина
которых варьирует от очень большой до совсем незначительной. Коконы мелкие с
прочными хитиноидными стенками; содержат по одному мелкому яйцу. Прикреп-
ляются к подводным предметам.
3(2). По крайней мере, передняя присоска не имеет дисковидной формы и не
отделена или мало отделена от остальной части тела.
4(5). Тело уплощённое, листовидное. Глаза (1—4 пары) расположены посере-
дине переднего конца тела. Ротовое отверстие маленькое. Передняя часть
пищеварительной трубки превращена в хобот (который, как правило,
трудно увидеть). Пиявки малой и средней величины (длина до 50 мм)............
.............Сем. Glossiphoniidae (Плоские пиявки) (рис. 102а) (с. 105).
Коконы студенистые тонкостенные бесформенные; содержат крупные богатые
желтком яйца. Прикрепляются к брюшной стороне материнского организма или к
подводной растительности.
104
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 102. Форма тела пиявок разных семейств (по: Лукин, 1976).
а — сем. Glossiphoniidae; б — сем. Piscicolidae; в — сем. Erpobdellidae; г — сем. Hirudinidae.
5(4). Тело червеобразное. Глаза (4-5 пар) расположены по краям переднего
конца тела. Ротовое отверстие большое. Хобота нет. Пиявки средней и
крупной величины.
6(7). Глаз 5 пар, расположены в виде дуги. В передней части глотки находят-
ся челюсти, вооруженные зубчиками. Имеется совокупительный орган.
Крупные черви (длина до 100 мм и более)....................................
.......Сем. Hirudinidae (Челюстные пиявки) (рис. 102 г, 103а, б) (с. 117).
Коконы имеют губчатые стенки и откладываются в сыром грунте прибрежной
полосы. Размеры коконов относительно крупные, содержат большое количество
мелких яиц.
7(6). Глаз 4 пары (у некоторых видов подвергаются частичной или полной ре-
дукции). Челюсти отсутствуют (сохранились лишь рудименты в виде трёх
складок). Совокупительных органов нет (оплодотворение при помощи
сперматофоров). Черви среднего размера (длина до 30-50 мм).................
.......Сем. Erpobdellidae (Глоточные пиявки) (рис. 102в, 103в) (с. 119).
Синоним: Herpobdellidae.
Кольчатые черви
105
Рис. 103. Расположение глаз у представителей семейств Hirudinidae и Erpobdellidae
(по: Elliott & Mann, 1979).
а — Haemopis sanguisuga: вид переднего конца тела сбоку; б — Haemopis sanguisuga: вид
переднего конца тела со спины; в — Erpobdella octoculata: вид переднего конца тела со
спины.
8(1)-
Коконы имеют хитиноидные стенки и при-
крепляются к подводным субстратам. Коконы
среднего размера, число яиц в них относи-
тельно невелико.
Задняя присоска редуцирована и пре-
вращена в пластинку, прижатую к за-
днему концу брюха. Передний конец тела
сильно сужен в виде небольшого хоботка.
Глаза отсутствуют......................
......Archaeobdella esmonti Grimm, 1876
(рис. 104).
Род Archaeobdella Grimm, 1876; сем. Erpobdel-
lidae.
Длина тела 15-30 мм, ширина 0,5-3,5 мм.
Рис. 104. Archaeobdella esmonti.
Общий вид (по: Лукин, 1976).
Покровы не содержат пигмента. Тело довольно сильно уплощено. Обитатели ила.
Хищники. Каспийское и Азовское моря и устья впадающих в них рек.
Ключ для определения родов и видов семейства Glossiphoniidae
1(2). Передний конец сильно расширен и явно отделён от остальной части тела,
напоминая «голову».............................Род Hemiclepsis Vejdovsky, 1884.
Единственный вид Hemiclepsis marginata (Muller, 1774) (рис. 105).
Длина тела обычно 10-15 мм, ширина 2-3 мм (встречаются экземпляры до 30 мм
при ширине 3-8 мм). Окраска зеленовато-коричневая, у многих экземпляров на
спинной стороне расположены 7 продольных рядов бледно-жёлтых пятнышек.
Поверхность тела гладкая. Края тела прозрачные со слабовыраженной зазубрен-
ностью. Глаз 2 пары, из которых передние значительно меньше. Задняя присоска
относительно большая. Желудок имеет 7 ветвящихся отростков. Сосут кровь рыб
106
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 105 Hemiclepsis marginal а.
а — общий вид (по: Timm, 1999); б —
общий вид (по: Лукин, 1977); в — перед-
ний конец тела, вид со спины (по: Elliott
& Mann, 1979).
Рис. 106. Helobdella stagnalis.
а — общий вид (по: Timm, 1999); б —
общий вид (по: Лукин, 1976); в — перед-
ний конец тела, вид со спины (по: Elliott
& Mann, 1979). c.s. — хитиноидная пла-
стинка (callous scute).
и земноводных (тритонов и головастиков бесхвостых амфибий). В европейской
части России повсеместно, однако на крайнем севере и северо-востоке редок. Ни-
где не достигает массового развития.
2(1). Передний конец не расширен.
3(4). На спине, позади глаз, имеется желтоватая или коричневая хитиноидная
пластинка......................................Род Helobdella Blanchard, 1896.
В европейской части России единственный вид Helobdella stagnalis (Linnaeus,
1758) (рис. 106).
Длина тела редко достигает 8-10 мм, обычно не более 5-6 мм, ширина 2-4 мм.
У фиксированных экземпляров часто передний конец загнут кверху. Окраска се-
ровато-белая, иногда со слабым зеленоватым оттенком. Поверхность тела гладкая.
Края тела с хорошо выраженными зазубринами. Глаза большие, одна пара. Из 6
пар отростков желудка хорошо развита только последняя. (В то же время все от-
ростки кишки выражены очень хорошо.) Сосут кровь, соки тела и разжиженные
ткани личинок насекомых и других мелких беспозвоночных. Своих жертв выса-
сывают целиком, способны заглатывать отдельные куски добычи, поэтому могут
Кольчатые черви
107
Рис. 107. Theromyzon tessulatum.
а — общий вид со спинной стороны тела (по: Timm, 1999); б — общий вид с брюшной сто-
роны тела (по: Timm, 1999); в — положение половых отверстий (по: Timm, 1999); г — общий
вид (по: Лукин, 1977); д — передний конец тела, вид со спины (по: Elliott & Mann, 1979).
быть отнесены скорее к хищникам. Один из самых распространённых видов пия-
вок в пресных водоёмах России.
4(3). Хитиноидной пластинки на спинной стороне нет.
5(10). Тело очень мягкое, желеобразное. При жизни способны к значительным
изменениям формы тела. Края тела заворачиваются на брюшную сторону.
6(9). Глаз 4 пары. Спина почти гладкая..........Род Theromyzon Philippi, 1867.
Синоним: Protoclepsis Livanov, 1902.
Форма тела живых пиявок в состоянии покоя имеет миндалевидную, овальную
форму. При движении весьма изменчива. Грубая фиксация может исказить форму
тела до почти шарообразной. Желудок снабжён 7 парами лопастных отростков.
Паразиты водоплавающих птиц (сосут кровь из слизистых оболочек ротовой по-
лости и верхних дыхательных путей).
7(8). На спине имеются продольные ряды мелких жёлтых пятнышек. Мужское
и женское половые отверстия разделены четырьмя кольцами. Задняя при-
соска относительно крупная (её диаметр равен или несколько превышает
половину наибольшей ширины тела пиявки, находящейся в состоянии по-
коя) ..........................Theromyzon tessulatum (Muller, 1774) (рис. 107).
Синоним: Protoclepsis tessulata (Muller, 1774).
Длина тела 10-50 мм, ширина 4—15 мм. Окраска тела очень изменчива: от светлой
до тёмной (зелёных, оливковых и коричневых тонов). В европейской части России
чаще встречается в умеренных и северных широтах, чем на юге.
108
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Рис. 108. Theromyzon maculosum.
1 — общий вид (по: Timm, 1999): а — со спинной стороны тела; б — с брюшной стороны
тела. 2 — расположение половых отверстий (по: Timm, 1999). 3 — изменение окраски и
формы тела (по: Лукин, 1976): а, г — вытянутое тело; б — пиявка в спокойном состоянии
(просвечивает кишечник); в — сильно сокращённое тело.
8(7). На спине кроме рядов мелких пятнышек имеются крупные светлые лин-
зообразные пятна. Половые отверстия разделены двумя кольцами. Задняя
присоска невелика (её диаметр значительно меньше половины наиболь-
шей ширины тела пиявки, находящейся в состоянии покоя)..........................
.........................Theromyzon maculosum (Rathke, 1862) (рис. 108).
Синоним: Protoclepsis maculosa (Rathke, 1862).
Длина тела 20-50 мм, ширина 5-10 мм. Основной фон тёмный (от тёмно-коричне-
вого до тёмно-оливкового). Количество крупных светлых пятен различно у разных
экземпляров (от небольшого числа до практически сплошного светлого окраса). В
европейской части России в умеренных и северных широтах.
9(6). Глаз 3 пары (из которых первая может редуцироваться). На спине 4 пары
продольных рядов очень крупных сосочков.........................................
..........................................Род Boreobdella Johansson, 1929.
Единственный вид Boreobdella verrucata (Muller, 1844) (рис. 109).
Длина тела 20-30 мм, ширина 6-12 мм. Окраска спины тёмно-зелёная. Вдоль сре-
динной части спины (парамедиально) проходят две пунктирные тёмные полоски
(прерываются в местах расположения сосочков). Брюшная сторона светлая (бело-
ватая). Глаза довольно крупные. Глаза первой пары проявляют очень сильную
тенденцию к редукции. Могут редуцироваться и обе передние пары глаз. Желудок
имеет 7 пар укороченных отростков. Паразиты брюхоногих моллюсков. Питаются
соками тела, иногда высасывая своих жертв целиком (т.е. по способу питания мо-
Кольчатые черви
109
Рис. 109. Boreobdella verrucata.
а — общий вид (по: Лукин, 1976); б — три
кольца из средней части тела, демонстри-
рующие папиллы (сосочки) (по: Elliott &
Mann, 1979); в — передний конец тела (по:
Elliott & Mann, 1979); г — желудок (по:
Лукин, 1976).
Рис. ПО. Haementeria costata.
1 — общий вид (по: Timm, 1999). 2 — об-
щий вид (по: Лукин, 1976). 3 — общий вид
(по: Лукин, 1977). 4 — передний конец тела
(по: Elliott & Mann, 1979): а — вид со спин-
ной стороны (положение сосочков и глаз);
б — вид с брюшной стороны (положение
рта и половых отверстий), е — глаза (eyes);
m — рот (mouth); — мужское половое
отверстие; $ — женское половое отверстие.
Рис. 111. Glossiphonia сот-
planata.
1 — общий вид (по: Timm,
1999): а — вид со спинной
стороны; б — вид сбоку. 2 —
общий вид (по: Лукин, 1976).
3 — желудок (по: Лукин,
1976). 4 — изменчивость глаз
(по: Elliott & Mann, 1979).
по
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
гут быть отнесены скорее к хищникам). В европейской части России отмечен на
территории Республики Коми.
10(5). Тело более или менее плотное.
11(12). На спинной стороне многочисленные мелкие сосочки, образующие про-
дольные ряды, один из которых идет вдоль средней линии спины. Глаз
1 пара........................................Род Haementeria de Filippi, 1849.
В Европе единственный вид Haementeria costata (Muller, 1846) (рис. ПО).
Длина тела 25-40 мм, редко до 72 мм; ширина 5-13 мм, редко до 26 мм. Окраска
тела очень изменчива (зелёная, светло- или тёмно-коричневая и совсем тёмная).
Посередине спины проходит тёмная широкая пунктирная полоса. По краям тела
имеются светлые пятна. Края тела зазубрены. Желудок снабжён 7 парами хорошо
развитых и ветвистых отростков. Паразиты болотной черепахи. Факультативно
могут сосать кровь птиц и человека, а также, возможно, рыб и амфибий. Обычен
в бассейнах рек Волги и Дона и прилегающих районах Кавказа. Известны отдель-
ные находки из Московской и Рязанской обл., Ладожского озера.
12(11). Сосочки на спинной стороне, если имеются, никогда не располагаются
вдоль средней линии. Глаз в норме более одной пары.
13(18). Консистенция тела очень плотная. Края тела зазубрены. Желудок с ше-
стью парами отростков. Глаз 3 пары, иногда частично редуцированных
или слитых между собой.
14(17). Окраска тела коричневато-зелёная или зеленовато-коричневая. Расстояние
между глазами первой пары практически равно расстоянию между глаза-
ми второй и третей пары.......................Род Glossiphonia Johnson, 1816.
15(16). Тело грушевидной формы. На спине три (иногда четыре) пары продоль-
ных рядов сосочков. В наружном слое кожи имеются скопления коричне-
вого пигмента, образующие продольные и поперечные ряды. Глаза первой
пары обычно хорошо выражены................................................
.....................Glossiphonia complanata (Linnaeus, 1758) (рис. 111).
Длина тела 10-30 мм, ширина 4-10 мм. Окраска от светло-зеленоватой до интен-
сивно коричневой. Вдоль срединной части спины (парамедиально) проходят две
пунктирные тёмные линии (признак очень характерный для данного вида). Края
тела с крупными зазубринами, поверхность которых также имеет зазубрины. Гла-
за неправильной почти треугольной формы. Отростки желудка не разветвлены.
Сосут кровь моллюсков, личинок насекомых и некоторых других мелких беспоз-
воночных. Кроме того, способны высасывать икру рыб и коконы пиявок, не про-
являющих заботу о потомстве. Один из самых распространённых видов пиявок в
пресных водоёмах России.
16(15). Тело узкое, миндалевидное. Поверхность тела гладкая (сосочки на спине
незаметны). В наружном слое кожи скоплений коричневого пигмента нет.
Глаза первой пары обычно сильно, а иногда совсем редуцированы..............
.........................Glossiphonia concolor (Apathy, 1888) (рис. 112).
Длина тела 10—15 мм. Окраска кофейного цвета. Вследствие отсутствия поверх-
ностного пигмента тело прозрачное даже у крупных экземпляров. Продольные
пунктирные линии на спинной стороне выражены также как у G. complanata.
Кольчатые черви
111
Кроме того, часто видны дополнительные линии,
составленные из тёмных точек. Края тела зазубре-
ны, но меньше, чем у G. complanata. Глаза отно-
сительно меньшего размера, чем у G. complanata.
Паразитируют на моллюсках и, возможно, на дру-
гих мелких беспозвоночных. В европейской части
России повсеместно, но встречается гораздо реже,
чем G. complanata.
17(14). Окраска тела серовато-белая. Расстояние меж-
ду глазами первой пары значительно меньше,
чем расстояние между глазами второй и тре-
тей пары..............Alboglossiphonia Lukin, 1976.
В Европе один вид Alboglossiphonia heteroclita
(Linnaeus, 1761) (рис. 113).
Синоним: Glossiphonia heteroclita (Linnaeus, 1761).
Длина тела 7-8 мм при ширине 2-4 мм. Тело
сильно уплощено. Края тела с мелкими зазубри-
нами. Поверхность тела без заметных сосочков.
На спине могут быть рассеяны мелкие тёмные
пигментные пятна, образующие продольные (пре-
рывистые) ряды или поперечные (сплошные) ярко
выраженные полосы. Задние отростки желудка
Рис. 112. Glossiphonia con-
color.
а — общий вид (по: Timm,
1999); б — общий вид (по:
Лукин, 1976).
А А А
Рис. 113. Alboglossiphonia heteroclita.
1 — общий вид (A. heteroclite var. hyaline) (по: Timm, 1999). 2 — общий вид (по: Pawlowski,
1936): а — A. heteroclite var. hyaline’, б — A. heteroclite var. papillosa’, в — A. heteroclite var.
striata. 3 — изменчивость глаз (no: Elliott & Mann, 1979).
112
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
’А А
Рис. 114. Batracobdella paludosa.
1 — общий вид: а — по: Timm, 1999; б —
по: Лукин, 1976. 2 — желудок (по: Elliott
& Mann, 1979). 3 — изменчивость глаз
(по: Elliott & Mann, 1979).
имеют добавочные ответвления. Со-
сут кровь мелких беспозвоночных
(малощетинковых червей, моллю-
сков, личинок насекомых). Мелких
моллюсков высасывают полностью.
Способны заглатывать куски тканей
добычи, самых мелких жертв могут
глотать целиком (т.е. по способу пи-
тания могут быть отнесены скорее
к хищникам). В европейской части
России повсеместно.
18(13). Консистенция тела менее плот-
ная. Края тела гладкие, без за-
зубрин. Желудок с семью парами
отростков. 2 пары больших глаз
(иногда 3 пары, при этом глаза
третей пары очень мелкие), кото-
рые могут сливаться в различных
сочетаниях вплоть до образова-
ния единственного крестообраз-
ного пятна..........................
..Род Batracobdella Viguier, 1879.
В европейской части России един-
ственный вид Batracobdella palu-
dosa (Сагепа, 1824) (рис. 114).
Длина тела 7-20 мм, ширина 3-5 мм.
Окраска взрослых экземпляров зеленовато-коричневая, однотонная без тёмных пя-
тен или полос. Тело узкое, края почти параллельны друг другу. Поверхность тела
без сосочков. Покровы взрослых особей довольно прозрачны, поэтому заполнен-
ный кровью кишечник хорошо заметен. Молодые пиявки совершенно прозрачны,
голубоватого цвета. Боковые отростки передней пары желудка направлены вперед
и раздвоены на концах. Паразиты земноводных и моллюсков. В европейской части
России встречается, главным образом, в южных районах. Известны отдельные на-
ходки (сомнительные, по мнению Е.И. Лукина) из Московской области, Карелии,
устья р. Невы.
Ключ для определения родов и видов семейства Piscicolidae
1(2). Глаза и глазоподобные пятна отсутствуют. Разделение на «шею» и «туло-
вище» чётко выражено............................Род Cystobranchus Diesing, 1859.
В европейской части России единственный вид Cystobranchus mammillatus (Malm,
1863) (рис. 115).
Длина до 11-35 мм, ширина 2-7 мм. Окраска от серой до грязно-розовой. На брюш-
ной и спинной поверхностях тела имеются сегментальные поперечные тёмно-се-
рые полосы с нечёткими краями. Боковые пузыри невелики, но хорошо заметны.
Кольчатые черви
113
Рис. 115. Cystobranchus mammillatus.
1 — общий вид (по: Лукин, 1976): а — со спинной стороны; б — сбоку. 2 — передний
и задний конец тела (по: Эпштейн, 1987): а — передняя присоска, вид со спинной
стороны; б — передняя присоска, вид с брюшной стороны; в — задняя присоска, вид
со спинной стороны, р — ротовое отверстие.
Передняя присоска относительно невелика. Задняя, напротив, мясистая, крупная
(её диаметр заметно превышает наибольшую ширину тела и в 2,5-3 раза больше
диаметра передней присоски). Соединение тела с задней присоской находится
почти в самом её центре. Специфический паразит налима (в жаберной полости и
на жабрах). В европейской части России встречается на севере и северо-западе.
2(1). Глаза (на передней присоске) и глазоподобные пятна (на задней присоске)
имеются. Разделение на «шею» и «туловище» выражено слабо или отсут-
ствует.
3(4). Задняя часть тела сильно расширена, вздута. Диаметр задней присоски
меньше наибольшей ширины тела..................Род Acipenserobdella Epstein, 1969.
Единственный вид Acipenserobdella volgensis (Zykoff, 1903) (рис. 116).
Синоним: Piscicola volgensis Zykoff, 1903.
Длина тела до 30 мм, ширина 3,9 мм. Окраска серая или светло-бурая с фиоле-
товым оттенком. На передней присоске две пары крупных глаз, на задней — 10
крупных глазоподобных пятен. Задняя присоска блюдцевидной формы, присо-
единена к телу эксцентрично (т.е. место соединения смещено к краю присоски).
Паразит осетровых (на поверхности тела, на жабрах). В европейской части России
встречается в бассейне р. Волги (верхнее и среднее течение).
114
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
4(3). Задняя часть тела не вздута.
5(6). Диаметр задней присоски приблизительно равен наибольшей ширине
тела. Пиявки малой величины.......................Род Caspiobdella Epstein, 1966.
В пресных водоёмах европейской части России один вид Caspiobdella fadejewi
(Epstein, 1961) (рис. 117).
Синоним: Piscicola fadejewi Epstein, 1961.
Рис. 116. Acipenserobdella volgensis.
1 — общий вид (по: Лукин, 1976). 2 — передний и задний конец тела (по: Эпштейн, 1987):
а — передняя присоска, вид со спинной стороны; б — передняя присоска, вид с брюшной
стороны; в — задняя присоска, вид со спинной стороны.
Рис. 117. Caspiobdella fadejewi.
1 — общий вид (по: Лукин, 1976): а — со спинной стороны; б — с брюшной. 2 — перед-
ний и задний конец тела (по: Эпштейн, 1987): а — передняя присоска, вид со спинной
стороны; б — передняя присоска, вид с брюшной стороны; в — задняя присоска, вид
сбоку, х — хобот.
Кольчатые черви
115
Рис. 118. Piscicola fasciata.
1 — общий вид (по: Лукин, 1976): а — со спинной стороны; б — с брюшной. 2 — окра-
ска спины (по: Лукин, 1976). 3 — передний и задний конец тела (по: Эпштейн, 1987):
а — передняя присоска, вид со спинной стороны; б — передняя присоска, вид с брюшной
стороны; в — задняя присоска, вид со спинной стороны, р — ротовое отверстие.
Длина тела до 13 мм, обычно менее 10 мм, наибольшая ширина 2,5 мм. Спинная
сторона светлая, брюшная тёмная. По бокам тела узкие, поперечные тёмные по-
лоски. На задней присоске радиальные пигментные полосы и глазоподобные
пятна. Соединение тела с задней присоской находится почти в самом её центре.
Боковые пузыри невелики, но хорошо заметны. Паразитируют на многих видах
рыб (обычно в ротовой полости, реже на жабрах и в жаберной полости), особенно
часто поражает леща и щуку. В европейской части России в реках и водохранили-
щах Азово-Черноморского бассейна. Сейчас обнаруживается в Волге и волжских
водохранилищах в большом количестве. В различных частях ареала вытесняет
Piscicola geometra.
116
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 119. Piscicola geometra.
1 — общий вид (по: Timm, 1999). 2 — общий вид (по: Лукин, 1976). 3 — передний (а)
и задний (б) конец тела, вид со спинной стороны (по: Лукин, 1976). 4 — передний и за-
дний конец тела (по: Эпштейн, 1987): а — передняя присоска, вид со спинной стороны;
б — передняя присоска, вид с брюшной стороны; в — задняя присоска, вид со спинной
стороны, х — хобот.
6(5). Диаметр задней присоски значительно превосходит наибольшую ширину
тела. Пиявки среднего и крупного размера....Род Piscicola Blainville, 1818.
7(8). Диаметр задней присоски в 2 раза превосходит наибольшую ширину тела.
Тело несколько уплощённое и удлинеённое. Длина тела превышает шири-
ну обычно в 5-6 раз......................Piscicola fasciata Kollar, 1842 (рис. 118).
Синоним: Cystobranchus fasciatus (Kollar, 1842).
Кольчатые черви
117
Одна из самых крупных рыбьйх пиявок Палеарктики. Длина тела может дости-
гать 75 мм при ширине 8 мм (обычно длина тела 25-50 мм, ширина 3-4,5 мм).
Окраска живых червей серо-бурая, иногда с фиолетовым оттенком. На теле че-
редуются широкие тёмные полосы с узкими светлыми. На передней присоске
имеется крестообразное пятно. На задней 14 радиальных пигментных полос и 10
глазоподобных пятен. Характерная особенность задней присоски — чрезвычайно
эксцентричное соединение с “туловищем” (место соединения сильно смещено к
переднему краю присоски). Боковые пузыри невелики, но хорошо заметны. Пер-
вая пара глаз заметно крупнее второй. Специфический паразит сома (чаще всего
на голове); иногда на синце, рыбце и подусте. В европейской части России в реках
и водохранилищах бассейнов Чёрного, Азовского и Каспийского морей; известны
находки из притоков Западной Двины.
8(7). Диаметр задней присоской 1,3-1,6 раза превосходит наибольшую ширину
тела. Тело длинное, округлое в поперечном сечении (у фиксированных
пиявок форма тела может сильно изменяться). Длина тела превышает ши-
рину обычно в 10-12 раз........Piscicola geometra (Linnaeus, 1761) (рис. 119).
Длина тела до 50 мм, обычно 15-30 мм; ширина 2-5 мм. Вдоль средней линии
спины проходит светлая полоса, пересекаемая поперечными светлыми полосками.
Брюшная сторона светлая. На передней присоске пигментные пятна располагают-
ся в виде креста. На задней присоске 10-14 радиальных пигментных полос и 6-12
глазоподобных точек. Передняя присоска почти круглая, задняя — эллиптическая.
Паразитируют на самых разнообразных рыбах (в особенности на карповых) и на
круглоротых. Обычно на поверхности тела, плавниках, а также в ротовой и жабер-
ной полостях. Данный вид не является постоянным эктопаразитом и вскоре после
насыщения кровью покидает хозяина. В европейской части России повсеместно, в
том числе в опреснённых участках Балтийского, Азовского и Каспийского морей.
Ключ для определения родов и видов семейства Hirudinidae
1(2). На спинной стороне имеется узор из двух продольных полос оранжевого
цвета. Каждая челюсть несёт один ряд острых зубчиков (в количестве от
69 до 91). Задняя присоска большая (диаметр её значительно больше по-
ловины ширины тела). Анус маленький, плохо различимый.....................
............................................Род Hirudo Linnaeus, 1758.
В европейской части России единственный вид Hirudo medicinalis Linnaeus, 1758
(рис. 120).
Длина тела взрослой пиявки 100-150 мм, ширина 10 мм. Окраска тёмно-корич-
невая, варьирует в широких пределах, однако сегментарный узор виден всегда.
Поверхность тела покрыта мелкими сосочками. Желудок с 10 парами хорошо
развитых отростков. Сосут кровь позвоночных животных (главным образом, мле-
копитающих и лягушек). В европейской части России в южных районах повсе-
местно (численность в природе во многих частях ареала сокращается в результате
варварского вылова и загрязнения водоёмов).
2(1). Окраска спины чёрная иногда с рассеянными ещё более тёмными пят-
нами, сегментарный узор из полос отсутствует. На каждой челюсти два
118
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 120. Hirudo medicinalis.
1 — общий вид (по: Лукин, 1976): а — вид со спинной стороны; б — вид с брюшной сто-
роны. 2 — передний конец тела (вид со спины) (по: Timm, 1999). 3 — задний конец тела
(вид со спины) (по: Лукин, 1976).
Рис. 121. Haemopis sanguisuga.
1 — общий вид (по: Лукин, 1976). 2 — передний конец тела (по: Timm, 1999): а — вид со
спинной стороны; б — вид с брюшной стороны. 3 — задний конец тела (вид со спины)
(по: Лукин, 1976). 4 — положение половых отверстий (по: Timm, 1999).
Кольчатые черви
119
ряда тупых зубчиков (в количестве от 7 до 18). Задняя присоска маленькая
(диаметр её меньше половины ширины тела). Анус большой, явно разли-
чимый......................................VonHaemopis Savigny, 1822.
В Европе единственный вид Haemopis sanguisuga (Linnaeus, 1758) (рис. 121).
Длина тела обычно 60-100 мм, иногда до 160 мм; ширина 10-15 мм. Окраска
тела очень изменчива. Спина чёрная, чёрно-коричневая, чёрно-зелёная. Брюшная
сторона светло-серая, зеленовато-серая, иногда с небольшими чёрными пятныш-
ками. Поверхность тела гладкая. Желудок имеет только заднюю пару отростков.
Хищники, поедающие различных мелких животных (малощетинковых червей,
пиявок, личинок насекомых, мелких рыб, головастиков и др.). Заглатывают до-
бычу целиком или отрывая крупные куски. Могут вести амфибиотический образ
жизни — выползают в прибрежную полосу для охоты, переносят пересыхание во-
доёмов в случае сохранеи^! влажного грунта на дне. В европейской части России
повсеместно, кроме крайнего Севера. Особенно многочислен в южных районах.
На севере и северо-востоке европейской части России редок.
Ключ для определения видов рода Erpobdella Blainville, 1818
семейства Erpobdellidae
1(2). Заднее (пятое) кольцо сомита заметно длиннее остальных. На спине две
сплошные узкие чёрные полоски...............Е. lineata (Muller, 1774) (рис. 122).
Синонимы: Herpobdella lineata (Muller, 1774); Dina lineata (Muller, 1774).
Длина тела 16-45 мм, ширина 3-8 мм. Окраска спины тёмная, коричнево-серая.
Поверхность тела гладкая. Глаза подвержены большей изменчивости, чем у дру-
гих представителей этого рода. Средние глаза переднего ряда обычно крупнее
остальных. Иногда могут появляться добавочные, но более мелкие глаза. Половые
Рис. 122. Erpobdella lineata (по: Лукин, 1976).
а — общий вид (со спинной стороны); б — положение половых отверстий;
в — изменчивость глаз.
120
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 21
Рис. 123. Erpobdella octoculata.
а — передний конец тела со спинной стороны (по: Timm, 1999); б — общий вид тела
сбоку (по: Timm, 1999); в — общий вид со спинной стороны (по: Лукин, 1976); г — по-
ложение половых отверстий (по: Timm, 1999).
отверстия разделены обычно 2, редко 2!4-3 кольцами. Хищники. Ареал вида про-
стирается до южной Карелии, однако наиболее многочислен на юге европейской
части России. На северо-востоке европейской части России отсутствует.
2(1). Сомит состоит из пяти равных колец. Чёрные сплошные полосы на спине
либо отсутствуют, либо имеется одна полоса с неровным краем.
3(4). Половые отверстия разделены 2/4-3 кольцами..............................
.................................Е. octoculata (Linnaeus, 1758) (рис. 123).
Синоним: Herpobdella octoculata (Linnaeus, 1758).
Длина тела 15-80 мм (обычно 30-40 мм), ширина 3-10 мм. На поверхности тела
имеются поперечные ряды сосочков, но они очень мелкие. Окраска тела очень
сильно варьирует. У типичной формы на спинной стороне каждого кольца имеет-
ся поперечный ряд жёлтых пятнышек на тёмном фоне. У других происходит ре-
дукция тёмного пигмента до почти полного исчезновения, и спинная поверхность
выглядит одноцветной (светлой) с редкими тёмными пятнами. Хищники с широ-
ким спектром питания — потребляют тот вид пищи, который наиболее доступен
в данный момент. Питаются мелкими беспозвоночными (малощетинковыми чер-
вями, личинками насекомых, ракообразными), икрой брюхоногих моллюсков и
рыб, также иногда занимаются каннибализмом, могут поедать падаль. Самый рас-
пространённый и многочисленный вид пиявок в пресноводных водоёмах России.
4(3). Половые отверстия разделены 4, иногда 5 кольцами.
5(6). Покровы тела в значительной степени прозрачны, вследствие чего видны
тёмные головные нервные узлы, узлы брюшной нервной цепочки и неко-
торые части лакунарной системы целома.........................................
.................................Е. nigricollis (Brandes, 1900) (рис. 124).
Синоним: Herpobdella nigricollis (Brandes, 1900).
Кольчатые черви
121
Рис. 124. Erpobdella nigricollis.
а — передний конец тела со спинной стороны (по: Лукин,
1976); б — положение половых отверстий (по: Лукин,
1977).
Самая мелкий палеарктический представитель рода
Erpobdella (длина тела обычно составляет 15-25 мм,
редко 35 мм, максимальная ширина 3-5 мм). Спина
однотонная (коричневая или зеленоватая). Тело тон-
кое с заострёйным, почти цилиндрическим передним
концом. Поверхность тела гладкая. Хищники, поеда-
ют мелких животных (к|к и прочие представители
этого рода). В ЕвропейсмРчасти России повсемест-
но, особенно многочислен на северо-востоке.
6(5). Покровы непрозрачны. На спинной стороне у
многих экземпляров проходит тёмная полоса
с неровными краями; могут быть разбросаны
тёмные пятна.....................................
..........Е. testacea (Savigny, 1822) (рис. 125).
Синоним: Herpobdella testacea (Savigny, 1822).
Длина тела 19-45 мм (обычно 20-30 мм), ширина 2-5 мм. Окраска спины одно-
тонная (коричневато-серая, зеленовато-серая) или с узкой тёмной полосой вдоль
средней линии спины, иногда заметны тёмные пятнышки. Брюшная сторона не-
много светлее. Хищники, поедают мелких животных. Ареал во многом совпадает
с ареалом предыдущего вида (Е. nigricollis), однако на северо-востоке европей-
ской части Е. testacea очень редок.
Рис. 125. Erpobdella testacea.
а — передний конец тела со спинной стороны (по: Timm, 1999); б — общий вид тела
сбоку (по: Timm, 1999); в — общий вид со спинной стороны (по: Pawlowski, 1936); г —
положение половых отверстий (по: Timm, 1999).
АМФИПОДЫ
Отрад Amphipoda Latreille, 1816, или бокоплавы (класс Malacostraca Latreille,
1802) — подвижные ракообразные, способные к свободному перемещению, как
в придонных слоях, так и в толще воды. Внешняя морфология тела амфипод при-
ведена на рис. 1. При измерении амфипод за длину тела принимается расстояние
вдоль линии спины (при выпрямленном теле рачка) от переднего края головы до
основания тельсона, т.е. до конца последнего сегмента тела.
Тело состоит из 18 сегментов и делится на три отдела: голову, включающую
четыре головных и слившийся с ними один грудной сегмент; грудь (мезосома,
Л&) составленную семью сегментами З^эрюшко, состоящее из шести свобод-
ных сегментов (рис. 1). Брюшной отдел подразделяется на два отдела — перед-
ний (метасома, Mis, или плеосома) и задний отдел (хвостовой, или уросома, Us).
Спинная сторона последних грудных и (или) первых трех брюшных сегментов
нередко несёт срединный продольный (медиальный) киль. На спинной стороне
уросомальных сегментов у большинства видов имеются бугорки, как правило,
усаженные группами шипов и щетинок.
Передний край головы имеет лобный вырост — рострум. Боковые выросты
головы оттянуты в межантеннальную лопасть. Нижняя часть головы образует
глубокий межантенальный синус, в котором помещаются основания нижних ан-
тенн. Голова несёт пять парных и три непарных придатка (рис. 2): верхние ан-
тенны (антенна 1, Л7), нижние антенны (антенна 2, Л2), жвалы (мандибулы, Md),
Ms
Рис. 1. Внешняя морфология представителя отряда Амфипода (по: Barnard, Barnard,
1983).
Амфиподы
123
Рис. 2. Голова и придатки головы бокоплава (по: Bousfield, 1973).
верхняя (максилла 1, Мх1) и нижняя челюсти (максилла 2, Мх2), верхняя губа (/),
нижняя губа (Z) и ногочелюсти (максиллипеды, Мхр). Антенна 1 и 2 состоят из
стебелька (рА1,рА2), и основного жгутика (fAl,fA2). Антенна 1 также несёт до-
бавочный жгутик (af). Второй сегмент стебелька антенны 2 с конусовидной же-
лезой. Все членики стебелька и жгутика обеих антенн несут пучки длинных или
коротких щетинок. Расположение, количество, форма и длина щетинок имеет
важное систематическое значение. На сегментах жгутика антенны 1 также рас-
124
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
рт
Рис. 3. Строение грудных и брюшных конечностей бокоплава (по: Bousfield, 1973).
Амфиподы
125
положены булавовидные хеморецепторы — эстетаски. У самцов на члениках
жгутика и дистальных сегментах стебелька антенны 2 имеются видоизмененные
щетинки — кальцеолы (cal), выполняющие функцию органов чувств. Ротовой
аппарат включает верхнюю и нижнюю губы (выросты кутикулы вокруг ротово-
го отверстия), жвалы, челюсти и ногочелюсти. Жвалы состоят из тела и щупика
(pMd), вооруженного щетинками. Расположение и длина щетинок третьего чле-
ника щупика жвал — важный в таксономическом отношении морфологический
признак у представителей рода Gammarus.
Два первых грудных сегмента несут специализированные хватательные конеч-
ности — гнатоподы (Gpl и Gp2), грудные сегменты 3-7 несут пять пар ходиль-
ных ножек, или переоподов (РрЗ-7, рис. 3). Грудные конечности представляют
собой придатки, состоящие из (фш члеников: коксального членика в виде расши-
ренной пластины (Со), базального членика (Ь) и пяти хорошо развитых линейной
формы члеников эндоподита — ишиального (i), мерального (т), карпального (с),
проподального (р) и коготка (d). Проподус гнатопод (шестой членик) имеет вид
ладони, которая вместе с коготком (седьмым члеником) образует ложную клеш-
ню, или субхелю. Передний край проподуса обычно слабо вооружен щетинками,
его задний край усажен густыми пучками щетинок, а пальмарный край (рт) не-
сёт и шипы, и щетинки. Гнатоподы самцов обычно более массивны, чем у самок,
так как они выполняют функцию прекопуляторных хватательных придатков. Ба-
зальные членики трех последних пар переоподов (Рр5-7) расширены в разной
степени. Жаберные мешки помещаются на грудных сегментах в основании ко-
нечностей, представляют собой простые выросты коксальных члеников грудных
ног, покрытые тонким слоем кутикулы и снабжённые кровеносными сосудами
(рис. 3).
Выросты трех метасомальных сегментов (передних брюшных сегментов) об-
разуют эпимеральные пластины (Epl-З). Три сегмента метасомы несут парные
двуветвистые плавательные конечности, или плеоподы (РП-З). Движение плеопо-
дов обеспечивает постоянный ток обогащенной кислородом воды к расположен-
ным впереди жабрам и выводковой (марзупиальной) камере самок.
Хвостовой отдел включает три задних брюшных (абдоминальных) сегмента
и хвостовую пластинку — тельсон (Т). Сегменты уросомы несут парные пры-
гательные ножки, или уроподы (Upl-З). Уроподы состоят из стебелька (pUp) и
двух ветвей — экзоподита, или наружной (го) и эндоподита, или внутренней (ri)
ветвей.
Методы сбора и обработки
В мелководье озёр, рек и эстуариев амфипод собирают ручным сачком с мел-
коячеистым планктонным газом в толще воды или вручную с камней или расту-
щих макрофитов, а также вручную у уреза воды в водорослевых выбросах или
сыром песке. Субстрат переносят в небольшое пластиковое ведро или таз и затем
126
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
сливают воду вместе с освободившимися от растений и других субстратов рач-
ками в сито. Из сита амфипод осторожно выбирают, используя для этого пинцет,
или вручную, и переносят в пластиковые баночки, содержащей фиксирующую
жидкость (70% этилового спирта и несколько капель уксусной кислоты). Пробы
снабжают этикеткой со сведениями о месте и времени сбора материала.
Для изучения популяционных характеристик амфипод, требующих учета чис-
ленности, животных собирают с определённой площади дна водоёма (Методи-
ческие рекомендации, 2005). Участок дна ограничивают квадратной рамкой или
цилиндрическим пробоотборником с известной площадью покрытия (сечения).
Твёрдые субстраты вместе с прикреплёнными к ним водорослями (растениями)
собирают с этого ограниченного участка и переносят в таз с чистой водой. Затем
при помощи сачка отлавливают всплыви^ животных и собирают верхний слой
мягкого грунта в течение нескольких минут. Все части пробы помещают в сита
с размером ячеи (0,25 мм), промывают чистой водой и фиксируют 70% спиртом,
или 4% формалином.
Нектобентосных амфипод собирают тралом. В некоторых случаях могут быть
использованы специальные ловушки, представляющие собой садок с мелкоячеи-
стой сетью. Ловушки подвешивают в водоёме на определённой глубине на ночь,
а утром вынимают. Морских блох семейства Talitridae, которые обитают в вы-
бросах водорослей и песке в супралиторали морей и эстуариев рек, обычно со-
бирают при помощи небольшого железного ящика с острыми краями. Ящиком
покрывают соответствующую площадку песка и загоняют его на 5-10 см в глу-
бину. Под ящик подсовывают немного больших размеров крышку и быстро пере-
ворачивают. Собранный грунт вместе с рачками пересыпают в емкости и транс-
портируют в лабораторию. Амфипод, обитающих в подземных водах, собирают в
местах естественного (источники, родники, ключи) или искусственного (колодцы,
колонки, буровые скважины) выхода вод на поверхность. В источниках и родни-
ках рачки скапливаются у самого выхода вод, поэтому, прежде всего, необходимо
провести ловлю сачком у самого выхода родника и обследовать нижнюю поверх-
ность крупных камней, дерновин мха и травянистого покрова, нависающих над
берегами источника. Для исследования фауны интерстициальных вод использует-
ся весьма простой, но эффективный прием Шаппуи (Chappuis, 1942). В 1-2 м от
берега реки (с песчаным или мелко галечным грунтом) вырывают яму, достигаю-
щую глубины уреза речной воды. Затем фильтруют заполнившую яму грунтовую
воду через сачок. Также для сбора амфипод в грунтовых водах используют руч-
ные колонки для получения питьевой воды.
Основные пособия по определению
Бирштейн Я.И. 1940. Высшие раки (Malacostraca). Жизнь пресных вод СССР. Т. 1. М.; Л.
Мордухай-Болтовской Ф.Д., Грезе И.И., Василенко С.В. 1969. Отряд амфиподы или раз-
ноногие — Amphipoda Latreille, 1816-1817. Определитель фауны Чёрного и Азовского
морей. Т. 2. Киев.
Амфиподы
127
Цветкова Н.Л. 1975. Прибрежные гаммариды северных и дальневосточных морей СССР и
сопредельных вод. Л.
Barnard J.L., Barnard С.М. 1983. Freshwater Amphipoda of the World. II. Handbook and Bib-
liography. Virginia.
Список обозначений к рисункам
Ms — грудной отдел, мезосома
Mts — передний хвостовой отдел, метасома
Us — задний хвостовой отдел, уросома
А1 — верхние антенны, или антенны 1
А2 — нижние антенны, или антенны 2
рА1,рА2 — стебелёк (стержень) ^^енны 1
и антенны 2
af—добавочный жгутик антенны 1
cal — кальцеолы
Gp 1, Gp 2 — хватательные конечности,
или гнатоподы 1 и 2
Md— жвалы, или мандибулы
pMd— щупик жвал
Мх 1 — верхняя челюсть, или максилла 1
Мх 2 — нижняя челюсть, или максилла 2
Мхр — ногочелюсти, или максиллипеды
рМхр — щупик максиллипед
/ — верхняя губа
L — нижняя губа
Pl I-III — плавательные конечности, или
плеоподы
рР1 — стебелёк (стержень) плеопода
Рр III-VII— ходильные ножки,
или переоподы
Up I-III — прыгательные ножки,
или уроподы
pUp — стебелёк (стержень) уропода
го — наружная ветвь уропода
ri — внутренняя ветвь уропода
Т — хвостовая пластинка, или тельсон
Ер I-III — эпимеральные пластины
Со — коксальный членик,
В — базального членик
I — ишиальный членик
m — моральный членик
с — карпальный членик
р — проподальный членик
d— коготок
pm — пальмарный край проподуса
гнатопод
Ключ для определения семейств
1(2). Тело сжато в спинно-брюшном направлении. Стебелёк антенны 2 массив-
ный, с коротким трёхчлениковым жгутиком и крючковидными апикальны-
ми шипами............................Сем. Corophiidae Dana, 1849 (с. 131).
2(1). Тело сжато с боков. Стебелёк антенны 2 немассивный.
3(6). Уроподы 3 обычно с одной короткой ветвью. Тельсон сросшийся или
только слегка расщеплён.
4(5). Антенны 1 длиннее антенн 2. Сегменты уросомы слиты, ветвь уроподов 3
короткая конической формы............Сем. Crangonyctidae Bousfield, 1973.
В подземных водах, ручьях и временных водоёмах) распространён один вид
Synurella ambulans Muller 1846 (син. Stygobromus ambulans) (рис. 4).
5(4). Антенны 1 много короче антенн 2, без добавочного жгутика. Уросомаль-
ные сегменты раздельные. Ветвь уропода 3 почти такой же длины, что и
стебелёк...........................Сем. Talitridae Rafinesque, 1815 (с. 134).
6(3). Уроподы двуветвистые. Добавочный жгутик антенны 1 развит в разной
степени. Тельсон глубоко расщеплён, хотя бы на 14 длины.
128
Определитель зоопланктона и зообентоса пресных вод Европейской России, Т. 2
Рис. 4. Synurella ambulans (по: Barnard, Barnard, 1983).
7(8). Добавочный жгутик антенны 1 рудиментарный, одночленниковый. Второй
членик переопода VII в основании широкий и покрыт множеством щети-
нок....................................Сем. Micruropidae Kamaltynov 1999.
В озёрах, водохранилищах, реках Северо-Запада России, эстуариях Балтийского
моря встречается один вид Gmelinoides fasciatus (Stebbing 1899) (рис. 5; вкл. рис. 1).
8(7). Добавочный жгутик антенны 1 состоит из двух и более члеников.
9(10). Антенны 1 и 2 почти равной длины. Базиоподит переопода 7 расширен
в крупную закруглённую по нижнему краю пластину, так что его шири-
на равна или немного меньше длины. Его длина превышает длину всех
остальных члеников переоподов. Переоподы (особенно, шестой и седьмой
Амфиподы
129
Рис. 5. Gmelinoides fasciatus (по: Гурьяновой, 1951).
пары) снабжены шипами, иглами и перистыми щетинками. Задний край
первого сегмента уросомы гладкий без шипиков.....................
....................................Сем. Pontoporeiidae Dana, 1853.
В озёрах Карелии, опреснённых районах эстуариев рек Балтийского и Белого
моря встречается один вид Monoporeia affinis (Lindstrom, 1855) (рис. 6).
10(9). Антенна 1 длиннее антенны 2. Базиоподит 7 переоподов иной формы и
заметно короче всех остальных члеников вместе взятых.
11(14). Тело с хорошо развитыми спинным и боковым рядами вооружений в виде
зубцов или вздутий, которые распространяются до сегментов метазомы
12(13). Голова с длинным заострённым рострумом. Вдоль всего тела развит спин-
ной киль, зубцы которого увеличиваются в задней части тела. Переоподы
6 заметно длиннее переоподов 7. Уроподы 3 мощные, с ветвями оваль-
130
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1
Рис. 6. Monoporeia affinis (по: Гурьяновой, 1951).
Рис. 7. Pallasea quadrispinosa (по: Barnard, Barnard, 1983).
но-листовидной формы равной длины, густо окаймленными перистыми
щетинками.............Сем. Gammaracanthidae Bousfield, 1989 (с. 134).
13(12). Голова с небольшим рострумом. Срединный ряд возвышений отсутству-
ет или слабо развит. Боковой ряд возвышений на седьмом сегменте тела
отсутствует, либо в виде вздутия. Переоподы 6 не длиннее или немного
Амфиподы
131
длиннее переоподов 7. Экзоцодит уроподов 3 длиннее эндоподита в два
раза..................................Сем. Pallaseidae Tachteew, 1995.
В озёрах бореальной зоны, эстуариях рек Балтийского моря встречается один вид
Pallasea quadrispinosa Sars, 1967 (син. Pallasiopsis quadrispinosa, Pallasiola quad-
rispinosa) (рис. 7).
14(11). Тело гладкое без кутикулярного вооружения.
15(16). Эндоподит уроподов 3 составляет около !6 длины экзоподита. Базальный
членик переопода 7 с лопастью на задненижнем углу, часто выступающей
за границу с ишиальным члеником...........................................
........................Сем. Pontogammaridae Bousfield, 1977 (с. 136).
16(15). Внутренняя ветвь уроподов III составляет не менее половины наружной
ветви или редуцирована*go чешуйковидного придатка. Базальный членик
переоподов VII без лопасти на задненижнем углу и с суживающимся
книзу крыловидным краем. Эпимеральные пластинки хорошо развиты с
прямым или заострённым задненижним углом..................................
................................Сем. Gammaridae Leach, 1813 (с. 138).
Ключ для определения видов сем. Corophiidae Dana, 1849
1(2). Первый членик стебелька антенны 1 с мелкими зубчиками вдоль нижнего
внутреннего края (самец и самка сходны). Отросток второго членика сте-
белька антенны 2 хорошо развит, направлен вперёд. Третий членик этой
антенны несёт один дистальный зубец, который выступает немного за его
конец. Четвёртый членик без шипов и зубцов................................
..Род Corophium Latreille, 1806: Corophium volutator (Pallas, 1766) (рис. 8).
Североатлантический вид, встречается в опреснённых заливах и эстуариях рек
Балтийского моря (в Ленинградской и Калининградской областях). Морфоло-
гически близкий вид С. orientate Schellenberg, 1928, изначально описанный как
С. volutator orientate (Stock, 1960), обитает в Средиземноморском и Азово-Черно-
морском бассейнах. Отличается от С. volutator наличием диморфизма у самцов и
самок в строение антенны 2 (у самцов имеются мелкие зубчики вдоль нижнего
внутреннего края первого сегмента). Также, у С. orientate внешний край стебелька
уропода 1 опушён на всём протяжении, а ветвь уропода 3 опушена бедно.
2(1). Внутренний край первого членика стебелька антенны 1 незубчатый, с
проксимальными щетинками. На третьем членике стебелька антенны 2
кроме главного зубца имеются еще два зубчика. Четвёртый членик антен-
ны 2 с одним проксимальным зубцом в виде воротникообразного выроста
у самцов, либо с двумя зубцами (проксимальным и дистальным) у самок,
причём дистальный зубец меньше проксимального зубца и направлен впе-
рёд .............................Род Chelicorophium (Bousfield, Hoover, 1997),
Chelicorophium curvispinum (Sars, 1895) (рис. 9; вкл. рис. 2).
Вид Понто-Каспийского происхождения, встречается в опреснённых заливах и
эстуариях рек Балтийского моря (в Ленинградской и Калининградской областях).
132
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 8. Corophium volutator (по: Bousfield, Hoover, 1997).
Рис. 9. Chelicorophium curvispinum (no: Carausu et al., 1955).
Амфиподы
133
Рис. 10. Gammaracanthus loricatus (по: Гурьяновой, 1951).
Рис. 11. Gammaracanthus lacustris (по: Гурьяновой, 1951).
134 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Ключ для определения видов сем. Gammaracanthidae Bousfield, 1989
1(2). Длина пальмарного края проподуса гнатоподов 2 существенно больше,
чем у проподуса гнатоподов 1. Нижний край первой и второй эпимераль-
ных пластин с крепкими короткими шипами................................
.....Gammaracanthus loricatus (Sabine, 1821) (= G. loricatus aestuariorum
Lomakina, 1952) (рис. 10; вкл. рис. 3).
Бассейн Белого и Баренцева морей, в устья рек и прибрежных озёр
2(1). Длина пальмарного края проподуса гнатоподов 2 не больше, чем у пропо-
дуса гнатоподов 1. Нижний край второй и третьей эпимеральных пластин
с длинными изогнутыми иголочками.......................................
.......................... ft*...Gammaracanthus lacustris Sars, 1867
(син. Gammaracanthus relictus Sars 1895) (рис. 11).
В ледниковых озёрах Карелии, Скандинавии.
Ключ для определения видов сем. Talitridae Rafinesque, 1815
8(9). Проподус гнатоподов 1 без ладони. Гнатоподы 2 с маленьким проподусом
и пальцем. Ветвь уропода 3 с одним большим шипом сходным по длине с
ветвью..................Talitrus saltator (Montagu, 1808) (рис. 12; вкл. рис. 4).
Песчаные пляжи у уреза воды: побережье Балтийского моря (Польша, Прибал-
тика).
Рис. 12. Talitrus saltator.
А — внешний вид по: Carausu et al., 1955; Б — уропод 3 по: Spicer, Janas, 2006.
Амфиподы
135
9(8). Проподус гнатоподов 1 расширен в маленькую ладонь. Гнатоподы 2 сам-
цов с сильно расширенным проподусом и с крупным коготком. Мерус
(четвёртый членик) гнатоподов 1 и 2 с лопастью в нижней части. Ветвь
уропода 3 без шипа...............Orchestia cavimana (Heller, 1865) (рис. 13).
Встречается у уреза воды, в выбросах растений на побережье Балтийского моря.
Рис. 14. Dikerogammarus villosus (по: Carausu et al., 1955).
136 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 21
Ключ для определения видов сем. Pontogammaridae Bousfield, 1977
1(4) Длина антенны I не меньше половины тела.
2(3). На спинной стороне первого и второго сегментов уросомы чётко выражен-
ные бугорки не менее чем с тремя шипиками, антенны 2 и гнатоподы 1 и
2 с густыми пучками длинных волосков.....................................
...........Dikerogammarus villosus (Sowinsky 1894) (рис 14; вкл. рис. 5).
Понто-Каспийский вселенец в реках и эстуариях Балтийского моря.
3(2). На спинной стороне первого и второго сегментов уросомы есть округлые
выступы........................Dikerogammarus haemobaphes (Eichwald, 1941)
(син. D. haemobaphes fluviatilis, D. fluviatilis Martynov, 1919) (рис. 15).
Понто-Каспийский вселенец. Обитает в реках центральной России, опреснённых
участках эстуариев и реках БалтийЛъго моря.
Рис. 15. Dikerogammarus haemobaphes.
А — внешний вид по: Konopacka, 1998; Б — детали по: Carausu et al., 1955.
Амфиподы
137
Рис. 16. Obesogammarus crassus (по: Carausu et al., 1955).
4(1). Длина антенны 1 меньше половины тела. Антенны 1 и 2 и уроподы 3 уко-
рочены и расширены. Жгутик антенны 1 короче, чем её стебелёк.
5(6). Задний край меруса переопода 6 прямой с пучками коротких щетинок,
пятый членик этого же переопода прямоугольный. Базальный членик
переопода 7 с сильно развитой нижнезадней лопастью, заходящей за
конец ишиального членика. Коксальные пластины 1-4 с небольшим коли-
чеством коротких волосков вдоль заднего края. На спиной стороне сегмен-
тов уросомы обычно два-три шипа.........Obesogammarus crassus (Sars, 1894)
(син. Pontogammarus crassus) (рис. 16).
Понто-каспийский вселенец в прибрежных водах Балтийского моря, реках, эстуа-
риях рек Балтийского бассейна, Калининградской области.
6(5). Задний край меруса переопода 6 выгнутый с веером длинных щетинок, а
пятый членик того же переопода яйцевидный или овальный. Задненижняя
лопасть базального членик переопода 7 не достигает конца ишиального
членика. Коксальные пластины 1-4 с многочисленными длинными во-
лосками вдоль заднего края. Второй сегмент уросомы с 4—7 шипами или
волосками, расположенными в ряд на спинной стороне.....................
............Pontogammarus robustoides (Sars, 1894) (рис. 17; вкл. рис. 6).
Понто-Каспийский вселенец. Этот вид широко распространён в прибрежных во-
дах Балтийского моря, реках, эстуариях рек Балтийского бассейна, Ленинградской
и Калининградской областях.
138
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 17. Pontogammarus robustoides (по: Carausu et al., 1955).
Ключ для определения видов сем. Gammaridae Leach, 1813
1(2). Эндоподит уроподов составляет менее чем !4 длины экзоподита. Базаль-
ный членик переопода 7 расширен в верхней части. Добавочный жгутик
антенны 1 имеет не более двух члеников. Все членики антенны 2 с груп-
пой длинных и закрученных щетинок вдоль вентрального края..............
............................Chaetogammarus warpachowskyi (Sars, 1894)
(син. Echinogammarus warpachowskyi) (рис. 18).
Устья рек Балтийского моря, озёра и водохранилища его бассейна, Прибалтика.
2(1). Длина эндоподита уроподов 3 составляет не менее половины длины эк-
зоподита, или около % первого членика экзоподита. Добавочный жгутик
антенны 1 имеет более двух члеников.
3(4). Антенна 1 составляет половину длины тела. Эпимеральные пластинки
хорошо развиты с прямым или слегка заострённым задненижнем углом...
..................................Gammarus pulex (Linnaeus, 1758) (рис. 19).
Амфиподы
139
Рис. 18. Chaetogammarus warpachowskyi (по: Carausu et al., 1955).
Ручьи, реки, озёра в Ленинградской, Псковской области, Европейская часть Рос-
сии.
4(3). Антенна 1 составляет менее Уз длины тела. Нижний край первой эпиме-
ральной пластины закруглённый, а вторая и третья эпимеральные пласти-
ны с вытянутым острым задненижнем углом.
5(6). Конический отросток на вентральной стороне второго членика стебелька
антенны 2 прямой. Переоподы 3-7 с длинным коготком. Вдоль нижнего
края двух последних эпимеральных пластин содержится несколько шипи-
ков и коротких щетинок...................................................
......................Gammarus lacustris Sars, 1863 (рис. 20; вкл. рис. 7).
Озёра Карелии, Ленинградской, Псковской области, центральная Россия.
6(5). Конический отросток второго членика стебелька антенны 2 закручен.
Переоподы 3-7 с коротким коготком. Вторая и третья эпимеральные пла-
стины несут вдоль нижнего края только шипики.............................
......................Gammarus varsoviensis Jazdzewskyi, 1975 (рис. 21).
Обитает в реках и ручьях.
7(12). Нижний край третьего членика щупика жвал с рядом коротких одинако-
вой длины щетинок, часто в виде щеточки.
8(9). Межантенальная лопасть головы треугольной формы. Задний край тре-
тьей эпимеральной пластины без щетинок или с одной щетинкой. Щетин-
140
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 19. Gammarus pulex (по: Karaman, Pinkster, 1977).
ки на уросоме короткие и редкие.....................................
............................Gammarus inaequicauda Stock, 1966 (рис. 24).
Эстуарии Балтийского моря.
9(8). Передний край межантенальной лопасти головы почти вертикальный, не
скошен назад.
10(11). Задненижний угол базального членика переоподов 7 в виде прямоуголь-
ной лопасти с пучком щетинок, шипы отсутствуют............................
...........................Gammarus duebeni Lilljeborg, 1851 (рис. 22).
Опреснённые участки Балтийского и Белого моря, прибрежные водоемы.
11(10). Задненижний угол базального членика переоподов 7 не образует лопасти
и вооружён несколькими щетинками и шипами.................................
............................Gammarus oceanicus Segestrale 1947 (рис. 23).
Этот вид обычен в Балтийское море при солёности от 1-2%о и выше.
12(7). Нижний край третьего членика щупика жвал с неровными короткими и
длинными щетинками, образующими бахрому.
13(14). Краевые шипы и щетинки мерального и карпального члеников переоподов
6 и 7 примерно равной длины ...Gammarus salinus Spooner, 1947 (рис. 25).
Балтийское море при солёности от 1-2%о и выше.
Амфиподы
141
Рис. 20. Gammarus lacustris (по: Karaman, Pinkster, 1977).
Рис. 21. Gammarus varsoviensis (no: Karaman, Pinkster, 1977).
142
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 22. Gammarus inaequicauda (по: Цветковой, 1975).
Амфиподы
143
Рис. 23. Gammarus duebeni (по: Цветковой, 1975).
Рис. 24. Gammarus oceanicus (по: Цветковой, 1975).
144
Определитель зоопланктона и зообентоса пресных вод Европейской России, Т. 2
Рис. 25. Gammarus salinus (по: Цветковой, 1975).
Рис. 26. Gammarus zaddachi (по: Цветковой, 1975).
Амфиподы
145
14(13). Краевые щетинки мерального и карпального члеников переоподов 6 и 7
у самцов намного длиннее шипов. Нижний край стебелька антенны 1 и
задний край базиоподита переопода 7 самцов с густыми закрученными
щетинками.
15(16). Стебелёк антенны 1 с 14-24 группами щетинок по нижнему краю........
..............................Gammarus zaddachi Sexton, 1912 (рис. 26).
Эстуарии Балтийского моря, выдерживает распреснение.
16(15). Стебелёк антенны 1 с менее чем 10 группами щетинок по нижнему краю
..........................Gammarus tigrinus Sexton, 1939 (рис. 27; вкл. рис. 8).
Вселенец из Северо-Атлантического бассейна. Обитает в эстуариях Балтийского
моря, Финском заливе, озёрах и реках Европы.
Рис. 27. Gammarus tigrinus (по: Bousfield, 1973).
изоподы
Изоподы (отряд Isopoda) имеют уплощённое дорзо-вентрально тело, которо
достигает размеров от 0,6 мм до 46 см. Выделяются три отдела тела: голова, пере
он и плеон. Голова включает сегменты собственно головы и слитый с ней первы
сегмент груди. Обе пары антенн у разных видов имеют неодинаковую длину. Ан
тенуллы короче антенн и состоят из 5 членикового стебелька и многочлениковоп
жгутика.
Грудной отдел (переон) состоит из 7 одинаковых по форме подвижно сочле
ненных сегментов. Каждый сегмент снабжён парой грудных ног — переоподов
представляющих собой одноветвисты^конечности. У половозрелых самок на пе-
редних пяти или четырёх парах переоподов с внутренней стороны имеются пла
стинки — оостегиты, образующие выводковую камеру.
Плеон полностью соответствует абдомену (брюшку) и включает шесть сег-
ментов и терминальную пластину — тельсон. У изопод, обитающих в пресны)
водах, тельсон слит с последним сегментом абдомена и образует плеотельсон
Каждый сегмент абдомена несёт по паре абдоминальных ног, за исключение*
Asellota, у которых передняя пара трансформирована или отсутствует. Плеопод!
служат для дыхания и плавания.
Последняя пара абдоминальных ног — уроподы состоит из проподита (основ
ного членика) и двух ветвей — экзоподита и эндоподита.
Равноногие ракообразные в пресных водах северо-запада России представле
ны несколькими видами семейств Asellidae, Janiridae и Chaetiliidae.
Основные пособия по определению
Бирштейн Я.А. 1940. Высшие раки (Malacostraca). Жизнь пресных вод СССР. Т. 1, M.;J
С. 405—430.
Кусакин О.Г. 1988. Морские и солоноватоводные равноногие ракообразные (Isopoda) х(
лодных и умеренных вод Северного полушария. III. Подотряд Asellota, ч. 1.
Ключ для определения семейств и видов
1(2). Тело овальной формы с относительно короткими ногами и уроподами.1
Последний абдоминальный сегмент с маленькой выемкой, внутри которой
Рис. 1. Внешний вид Jaera sarsi.
Рис. 2. Jaera sarsi. Конец абдомена
с уроподами.
Изоподы
147
Рис. 3. Внешний вид Asellus aquaticus. Рис. 4. Asellus aquaticus. Конец абдомена
с уроподами.
находятся очень короткие уроподы, лишь немного выступающие за край
абдомена............................................Сем. Janiridae.
Во внутренних водах Европы и опреснённых участках рек Балтийского, Баренце-
ва и Белого морей встречаются морфологически трудноразличимые виды из рода
Jaera Leach 1814 (J. sarsi Valkanov 1936 (рис. 1, 2), J. praehirsuta Forsman 1949,
J. ischiosetosa Forsmann 1949; J. albifrons Leach 1814).
2(1). Тело продольно вытянуто. Последний сегмент абдомена закруглён с длин-
ными ясно выделяющимися разветвлёнными уроподами, заметно высту-
пающими за край абдомена..................................Сем. Asellidae.
В пресных водоёмах европейской части России широко распространён 1 вид
Asellus aquaticus Linnaeus 1758 (рис. 3, 4).
В Ладожском озере и солоноватых водах Фин-
ского залива встречается крупная бентическая
изопода из семейства Chaetiliidae Dana, 1849, мор-
ской таракан Saduria entomon (L., 1758). Ранее
этот вид был также известен как Nfesidotea ento-
mon. Эта изопода имеет плоское, овальное, серо-
го или серо-коричневого цвета тело длиной свы-
ше 20 мм у половозрелых особей (рис. 5). Самые
крупные экземпляры достигают в длину 9 см. По
бокам головы вблизи глаз имеются отчетливые
V-образные, глубокие вырезки. Щупик ногочелю-
стей у S. entomon пятичлениковый. На дорсальной
поверхности третьего и четвёртого грудных сег-
Рис. 5. Saduria entomon. Самец. Внешний вид.
По: Kaestner, 1959.
148
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
ментов имеются продольные кили,
дорсальная поверхность четвёртых
коксальных пластин на четвёртых
грудных сегментах плоская без киля.
Заднебоковые углы первого грудно-
го сегмента тупые и закруглённые
(рис. 6).
Рис. 6. Saduria entomon. Самец, голова и
I грудной сегмент.
По: Gruner, 1965.
ГУБКИ
Губки (Porifera) — чрезвычайно своеобразная группа низших многоклеточ-
ных, они не имеют нервной системы, и основную часть их тела составляет ме-
зохил (паренхима), где располагаются различно дифференцированные клетки,
которые можно свести к 2 морфологическим типам: амебоидным и жгутиковым
(хоаноцитам). Снаружи губка покрыта дермальной мембраной, состоящей из
2 слоев клеток — наружного (экзопинакодермы) и внутреннего (эндопинакодер-
мы), между которыми тонкая прослойка мезоглеи, содержащая коллагеновые во-
локна, немногочисленные амебоидные клетки и специальные дермальные ске-
летные элементы. Эндопинакодерма формирует внутренний слой дермальной
мембраны, а также выстил^г субдермальных полостей и каналов водоносной си-
стемы. Снизу, со стороны поверхности субстрата, губка одета базопинакодермой,
состоящей из клеток продуцирующих коллагеновые волокна и комплекс веществ,
прикрепляющих губку к субстрату. Наличие в базопинакоцитах сократимых эле-
ментов позволяет молодой губке перемещаться по субстрату на небольшие рас-
стояния. Экзопинакодерма пронизана многичисленными порами, пропускаю-
щими воду в субдермальные полости и далее в каналы водоносной системы с
воротничковыми камерами. Каждая такая камера состоит из хоаноцитов, жгутики
и воротнички которых направлены в просвет, в сторону единственного выводного
отверстия — оскулюма.
Скелет пресноводных губок построен из кремниевых спикул и белкового ве-
щества спонгина, который соединяет спикулы в точках их пересечения или по-
крывает спикулы так, что они оказываются заключенными в спонгиновый фут-
ляр. Определённые комбинации спикул, их расположение в теле, а также размер и
форма макро- и микросклер (крупных и мелких спикул), содержание спонгина и
строение геммосклер специфичны для крупных таксонов и используются в видо-
вой диагностике губок.
Методы сбора и обработки
Собирать губок следует в ясную погоду около полудня, когда лучше освещается
дно водоёма. Со свай губку можно срезать вместе со слоем дерева. Срезая губку с
камня, следует обратить внимание на слой, непосредственно прилегающий к суб-
страту. Мелкие экземпляры лучше брать вместе с субстратом, у крупных необхо-
димо брать наиболее типичную часть, содержащую как дермальную мембрану, так
и базальный слой, прилегающий к субстрату. С глубины губок можно добывать с
помощью «кошки» или сачка. На больших глубинах применяются драги и тралы.
Собранный материал фиксируют 80-90° спиртом или 10% формалином.
В крайнем случае допускается сушка нефиксированных губок. Определение ве-
дётся под микроскопом, для чего приготавливается микропрепарат спикул следу-
ющим образом.
150
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. Спикулы губок.
1-5 — амфиоксы, 6—13 — амфистронгили, 14-19 — амфидиски.
1. Небольшой, расщеплённый препаровальными иглами кусочек губки
(2-4 мм), включающий участок дермальной мембраны с подлежащим мезохилом
и базальный участок, содержащий геммулы, помещают на предметное стекло с
лункой или в мелкую пробирку и заливают любым хозяйственным отбеливате-
лем, содержащим хлорку, например «Белизна».
2. После растворения тканей жидкость сливают или отсасывают пипеткой;
осадок несколько раз тщательно промывают водой, после чего лишнюю воду от-
сасывают и осадок заливают 90° спиртом.
3. На предметное стекло пипеткой переносят несколько капель осадка, содер-
жащего спикулы, спирт отсасывают фильтровальной бумагой и препарат подсу-
шивают на воздухе или над пламенем спиртовки и заключают в глицерин-жела-
тин или бальзам, накрывая покровным стеклом. Спикулы на препарате должны
лежать не слишком густо, чтобы не затруднять просмотр.
Губки
151
Иконографический ключ для определения видов губок
Вид Макросклеры (Ma) Микросклеры (Mi) Геммосклеры (Gs)
Spongilla lacustris
Eunapius fragilis
e,
Ephidatia muelleri
Ephidatia fluviabHs jmn. IU„ , , _ |_ |J I 1
Trochospongilla horrida
r i г •,:
Важно следить за чистотой предметных стекол и пробирок, в которых про-
водится разрушение тканей губки, чтобы не занести спикулы из одной пробы
в другую. Для изучения строения геммулы или скелета губки приготавливают
срезы толщиной от 0,5 до 1 мм, положив губку или геммулу на пробку. Срезы
можно окрасить насыщенным раствором основного фуксина с сафронином в 90°
спирте.
Ключ для определения родов семейства Spongillidae, Gray, 1867
1(2). Дермальные и мезенхимные микросклеры присутствуют. Геммосклеры
представлены амфистронгилями или амфиоксами...............................
...........................................Spongilla Lamarck, 1816.
2(1). Дермальные и мезенхимные микросклеры отсутствуют.
3(4). Геммосклеры представлены амфиоксами..............Eunapius Gray, 1867.
4(3). Геммосклеры представлены амфидисками.
5(6). Края диска рассечены более или менее глубоко на лучи, диск имеет вид
звезды или зубчатого колёсика.....................Ephydatia Lamoroux, 1816.
152 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
6 (5). Диски у амфидисков цельнокрайные в виде неправильных кружков, глад-
кие или с тонкой радиальной исчерченностью, амфидиски имеют вид
катушек....................................Trochospongilla Vejdovsky, 1883.
Ключ для определения видов рода Ephydatia Lamoroux, 1816
1(2). Стержень амфидиска короче или равен диаметру диска. Диск глубоко рас-
сечён на небольшое число лучей (4-15). Преобладают шиповатые макро-
склеры, гладкие встречаются реже. В редких случаях присутствуют только
один тип макросклер...................Ephydatia muelleri (Lieberkuhn, 1856).
2(1). Стержень амфидиска длиннее радиуса диска, он тонкий, часто сужен в
средней части. Иногда стержень ^меет один-два шипа, более коротких,
чем лучи диска. Диск неглубоко рассечён на многочисленные лучи; обыч-
но несколько более глубоких вырезок делят диск на секторы с зубчатыми
краями. Преобладают гладкие макросклеры. Шиповатые макросклеры не-
сут мелкие многочисленные шипики.............Ephydatia fluviatilis (L., 1758).
Ключ для определения видов рода Eunapius Gray, 1867
1(2). Геммосклеры по форме и строению похожи на макросклеры. Те и другие
представлены гладкими амфиоксами...Eunapius carteri (Bowerbank, 1863).
2(1). Геммосклеры по строению отличаются от макросклер и представлены
редко шиповатыми амфистронгилями.............Eunapius fragilis (Leidy, 1851).
Ключ для определения видов рода Spongilla Lamarck, 1816
1(2). Шипы в средней части микросклер (мезенхимных и геммульных) на своих
вершинах рассечены на 2-^ крючка, загнутых в разные стороны. У концов
спикулы шипы простые, прямые или изогнутые.............................
...................................Spongilla biseriata Weltner, 1895.
2(1). Шипы у микросклер простые, не рассечённые. Мезенхимные микроскле-
ры покрыты частыми мелкими шипиками. Геммульные микросклеры с
более редкими и крупными шипами.................Spongilla lacustris L., 1758.
МШАНКИ
(Bryozoa, Polyzoa, Ectoprocta)
Мшанки — колониальные животные, веду-
щие, как правило, прикреплённый образ жизни.
В континентальных водах России и сопредель-
ных государств встречаются представители 2
надклассов мшанок — Покрыторотые (Phylac-
tolaemata) и Голоротые (Gymnolaemata) (А.В.
Виноградов повысил ранг Phylactolaemata jyo
типа). Покрыторотые обитает в пресных во-
доёмах, хотя отдельные виды могут перено-
сить временное осолонение. Среди голоротых
мшанок большинство составляют морские и
солоноватоводные виды, но имеются семей-
ства, обитающие в пресных водах. Настоящи-
ми пресноводными мшанками можно считать
только покрыторотых и два семейства ктено-
стомных мшанок (Paludicellidae и Hislopiidae).
Phylactolaemata обнаружены всюду в пресных
водах во всём мире, за исключением полярных
областей, в широкой амплитуде от уровня моря
до высокогорных озёр, и на глубинах варьирую-
щих от мелководья мелких потоков и луж до дна
глубоких озёр. Gymnolaemata распространены в
морских, солоноватоводных условиях, и иногда
в пресноводных бассейнах. Bryozoa. являются
важной частью обрастаний и могут создавать
мшанково-губковые биоценозы. Колонии мша-
нок или зоарий могут быть разной величины,
формы и веса. Величина колоний меняется от
нескольких миллиметров до десятков сантимет-
ров. Живые колонии могут весить от несколь-
ких миллиграммов до нескольких килограммов.
Зоарии могут проявлять полимофизм или, что
то же самое, обладать различной формой у од-
ного и того же вида, и это вызвано различиями
в условиях существования. Форма колонии мо-
жет зависеть также от ее возраста. Зооиды или
индивидуумы, из которых образована колония,
состоят из 2 отделов — нежного и прозрачно-
Строение мшанки.
1 — щупальца, 2 — лофофор,
3 — ротовое отверстие, 4 — эпи-
стом, 5 — пищевод, 6 — аналь-
ное отверстие, 7 — щупальцевое
влагалище, 8 — кутикула, 9 —
слепой мешок, 10 — спермин,
11 — статобласты, 12 — фуни-
кулюс.
154
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
го полипида, и хитинизированного, желатинизированного или обызвествленного
цистида (домика), куда втягивается полипид. Полипид несёт венчик щупалец на
специальном щупальценосце или основании — лофофоре. У мшанок из надклас-
са Gymnolaemata лофофор кольцевой. Большинство же филактолемат обладает
подковообразным лофофором с двойным рядом щупалец. У некоторых филакто-
лемат лофофор круглый или овальный; среди них примитивный вид Fredericella
sultana. Щупальца у филактолемат соединены внизу межщупальцевой перепон-
кой, они полые, снабжены наружным мерцательным эпителием; внутри венчика
щупалец находится ротовое отверстие, которое только у филактолемат прикрыто
языкообразным выростом — эпистомом. Пищеварительный тракт петлеобраз-
ный, порошица находится вне венчика^щупалец рядом с ротовым отверстием.
За ротовым отверстием следует глотка, затем пищевод, расширенный мешкоо-
бразный желудок и тонкая кишка, заканчивающаяся порошицей. От дна желуд-
ка отходит тонкий канатик — фуникулюс, на котором образуются статобласты
и семенники. Яичники и семенники диффузного типа, представлены небольшим
скоплением оогониев и сперматогониев и не имеют выводных половых прото-
ков. Яичники формируются на брюшной стороне, а семенники, кроме фуникулю-
са, — на стенке полипида. Полипид заключен в секретированный стенкой тела
экзоскелет, называемый цистидом. Стенка цистида утолщена за счёт мягкого эла-
стичного хитина, который у высших филактолемат способен ослизняться (жела-
тинизироваться). Филактолематы — гермафродиты. При половом размножении
появляются свободноплавающие личинки, которые не являются личинками в
полном смысле слова, так как уже на ранних стадиях развития представляют со-
бой малозооидные колонии (за исключением рода Fredericella). Личинка у Phy-
lactolaemata, на самом деле, является свободно плавающей анцеструлой (первый
зооид колонии) или маленькой колонией, состоящей из нескольких аутозооидов,
которая позднее прикрепляется к субстрату, что присуще нескольким видам, или
дает свободно движущуюся колонию, как у Cristatella. Колонии Phylactolaemata
могут использовать «бегущую стратегию», а также пресноводные Gymnolaema-
ta, таких как Paludicella и Victorella. Phylactolaemata, в основном, не способны
менять место после того как колония прикрепилась к субстрату, но Cristatella
ползает на протяжении всей своей жизни, а молодые колонии Lophopus, Lopho-
podella, и Pectinatella также могут двигаться. Движение в этих родах часто свя-
зано с разделением колонии на части и последующим отделением друг от друга
дочерних колоний. Филактолематам свойственен оригинальный способ размно-
жения с помощью статобластов, представляющих собой продукт внутреннего
почкования. Различают 2 основных типа статобластов: прикреплённые (сессо-
бласты) и свободные (фрибласты). Фрибласты в свою очередь делятся на 2 ос-
новные группы: флотобласты и спинобласты. Сессобласты обычно имеют только
развитую капсулу; флотобласты — капсулу и развитое плавательное кольцо; спи-
нобласты — капсулу, плавательное кольцо и прикрепительный аппарат (шипы,
крючки). Мелкие ячейки плавательного кольца заполнены газом. Наружная по-
Мшанки
155
верхность статобластов имеет тонкую скульптуру, хорошо видную в элекгрон-
но-сканирующий микроскоп и специфичную для разных видов филактолемат.
Состав стенки капсулы статобластов — хитин, но более прочной модификации,
чем содержащийся в кутикуле трубок зоариев. Размеры статобластов в настоящее
время являются необходимым систематическим признаком как для современ-
ных, так и ископаемых филактолемат. Особенно важно отношения абсолютных
величин, например, отношение длины статобласта к его ширине, отношение по-
лярной ширины плавательного кольца к латеральной ширине кольца (боковой) —
L/В и pol/lat. Статобласты некоторых видов филактолемат имеют различия в
строении дорзальной и вентральной стороны. Для Phylactolaemata свойственна
высокая степень колониальной изменчивости, поэтому желательно использовать
для определения вместе с формой колонии и статобласты, особенно свободные.
В составе надкласса Gymnolaemata имеется 2 класса: Stenolaemata (един-
ственный представитель из ныне живущих — исключительно морские, отряд
Cyclostomata) и Eurystomata. Эвристомные мшанки так же, как и филактолем-
ные, как правило, сидячие колониальные животные, представленные в континен-
тальных водоёмах двумя отрядами — Ctenostomata и Cheilostomata (некоторые
представители последнего отряда могут факультативно переносить опреснение
или жить в солоноватоводных условиях). Венчик щупалец у аутозооидов круго-
вой, однорядный. Внутри него находится ротовое отверстие, а вне его — поро-
шица. Целом включает два отдела — туловищный и щупальцевый. При втягива-
нии переднего конца тела щупальца помещаются в полости тела в щупальцевом
влагалище. Стенка тела у Ctenostomata по сравнению с филакголематами упро-
щена, состоит из хитиновой кутикулы, полигональных эпидермальных клеток и
слоя перитонеума. У кгеностомных мшанок пищеварительная система усложне-
на также наличием жевательного желудка — гизарда, форма и размеры которого
имеют таксономическое значение. Яичники, как правило, возникают на спинной
стороне и перитонеуме полипида, семенники — на фуникулюсе и стенке ци-
стида, и обычно развиваются одновременно, хотя у некоторых видов выражена
протерандрия, т.е. созревание мужских гонад происходит ранее женских. Вну-
треннее оплодотворение осуществляется перекрёстно. Оплодотворенные яйца
могут выбрасываться в воду, но у большинства хейлостомных, кгеностомных и
циклостомных мшанок их развитие до личиночной стадии происходит либо в
целоме аутозооидов, либо в специальных выводковых камерах — овицеллах (у
Cheilostomata). Яйца выбрасываются через отверстие в стенке тела, помещаю-
щееся у основания щупалец, — супраневральную пору, или целомопору, либо
через терминальные поры, расположенные на кончиках щупалец, или же через
межщупальцевый орган. Для перенесения неблагоприятных условий, в первую
очередь холодной зимы, некоторые виды пресноводных кгеностомных мшанок,
в их числе Paludicella и Victorella, образуют стадии покоя — гибернакулы, пред-
ставляющие собой автономные, замкнутые фрагменты трубок зоария, что пред-
ставляет некоторую аналогию со статобластами филактолемат. Paludicella мо-
156
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
жет также продуцировать так называемые bagged (инкапсулированные) почки
(Вгает, 1914, 1951). Цистид у кгеностомных мшанок состоит из хитина, у хей-
лостомных — из кальцита. Зооиды могут быть обособленными, но чаще сопри-
касаются стенками с соседними зооидами, или даже имеют обобщенные стенки
цистидов. Связь между отдельными зооидами осуществляется или через поры,
или у хейлостомных мшанок через более сложные структуры — поровые пла-
стинки, или септулы, поровые камеры или диеталии, расположенные в стенках
цистидов. У Bryozoa, Gymnolaemata отверстие цистида или зоеция называется
орифиций, и у Cheilostomata часто снабжено закрывающей его крышечкой —
оперкулюмом. Пространство между вершинами щупалец, втянутыми во влага-
лище, и орифицием называется атриум, или вестибулюм.Часто в колониях хей-
лостомных, и иногда ктеностоМных мшаЬок, зооиды полиморфные, т.е. наряду
с зооидами типичного строения — аутозооидами — имеются видоизменённые
морфологически, и физиологически специализированные — гетерозооиды, в их
числе овицеллы, где происходит созревание половых продуктов, кенозооиды,
несущие часто опорную функцию, авикулярии (защитная функция), вибракуля-
рии (по-видимому, обеспечивающие ток воды над колонией) и некоторые другие
типы.
Субстратом для мшанок служат камни, водные растения, затонувшая древеси-
на, раковины моллюсков, панцири раков, а также металл, пластмасса, бумага, бе-
тон и другие объекты, подводные конструкции. Bryozoa пресноводных водоёмов
часто могут иметь вид выростов на погружённых растениях. Мшанки из Frederi-
cellidae селятся на любом субстрате, в том числе и на илах, а высокоорганизо-
ванные плюмателлиды и кристателлиды обитают преимущественно в зарослях
макрофитов, на древесных остатках, раковинах моллюсков и камнях. Когда по-
липид высовывается из цистида (домик, в котором находится полипид), его ло-
фофор поворачивается и изгибается во всех направлениях, создавая своими рес-
ничками, расположенными на щупальцах лофофора, ток воды ко рту, который
может быть обнаружен в 2 мм от него (Brooks, 1929). Как и у Gymnolaemata, этот
поток является механизмом ловли пищи у Phylactolaemata, которые таким обра-
зом питаются с помощью ресничек, или как часто называют, являются фильтра-
торами. Поток проходит между щупальцами, расположенными на лофофоре и
частички пищи удерживаются и направляются в рот. Пища состоит из крошеч-
ных организмов, таких как бактерии, диатомовые водоросли, рогатории, десми-
ды, и другие одноклеточные водоросли, простейшие, мелкие ракообразные, но,
несомненно, без детрита. Riische (1938), изучивший питание Plumatella fungosa,
отметил почти 40 крошечных планктонных организмов в месте обитания этого
вида, главным образом десмиды, зелёные флагелляты, рогатории и мелкие рако-
образные. Он обнаружил, что большинство из них были проглочены без разбора,
за исключением очень больших и слишком громоздких по форме. Щупальца мо-
гут быть согнуты друг другу, чтобы предотвратить выход пищевых организмов
или широко расставлены, для того чтобы слишком большие или нежелательные
Мшанки
157
частицы были удалены. Удаление также достигается внезапным сокращением.
Коловратки и ракообразные могут сбегать при помощи энергичных движений
и часто в любом случае не могут быть проглочены из-за их размера. Щупальца
могут двигаться независимо друг от друга. В связи с прозрачностью стенок по-
липида у аутозооидов цвет пищи может определять их окраску. Кроме того, окра-
ска наблюдается в случаях симбиоза с бактериями и синезелёными водорослями.
Большинство Phylactolaemata предпочитают более или менее затенённые водные
условия для жизни. Fredericella sultana может быть обнаружена в водопроводных
трубах в полной темноте. Состав воды, по всей вероятности, важен. Phylactolae-
mata, в основном, в том числе и их личинка, имеют положительный стереотаксис,
хотя есть тенденция у н^соторых видов и варьететов давать прямостоячие ветви,
что указывает на обратный ответ по отношению к стереотаксису. Щупальца обла-
дают отрицательным стереотаксисом и имеют тенденцию избегать касания друг
друга и щупалец соседних лофофоров. Plumatellidae с толстой кутикулой могут,
вероятно, противостоять в определённой степени обсушиванию на короткий пе-
риод. Kraepelin (1887) часто находил Phylactolaemata, растущих в мелких ручьях
на плоских камнях и банках, которые неминуемо в той или иной степени обсыха-
ли летом. Температура воды, без сомнения, главный фактор среды, контролирую-
щий рост и выживание колоний Phylactolaemata. В основном, молодые колонии
и молодые полипиды более чувствительны ко всем факторам среды, чем старые.
Обычно в умеренных широтах колонии умирают зимой из-за падения темпера-
туры, и выживают только за счёт статобластов. Колонии развиваются весной из
переживших зиму статобластов, когда температура достаточно повысится, и до-
стигают наибольшего развития летом и ранней осенью, затем умирают, когда
температура снижается поздней осенью. Следовательно, обычный ежегодный
цикл зависит от температур в этих широтах. Статобласты, созданные в начале
лета, будут развиваться в колонии без изменения летних условий обитания в то
же самое лето, и продуцировать другое поколение статобластов. Статобласты, об-
разовавшиеся поздним летом, обычно остаются перезимовывать, и развиваются
следующей весной. Следовательно, большинство видов умеренных широт имеют
ежегодно два поколения колоний, возникающих из статобластов. Все статобла-
сты должны оставаться в покоящемся состоянии определённый период, различ-
ный у разных видов, прежде чем они начнут развиваться. Влияние температуры
на время развития статобластов хорошо изучено. Свет не влияет на процент раз-
вивающихся статобластов. Содержание кислорода наиболее важно, поскольку
статобласты, например, Pectinatella не развиваются в воде с низким содержани-
ем кислорода или при отсутствии кислорода, хотя сохраняют жизнеспособность.
Летние статобласты, удалённые из живых колоний, не выдерживают более одной
недели осушки. Процент развивающихся статобластов снижается после увели-
чения времени осушки, но практически нет изменений во времени развития. До-
ступная информация указывает, что статобласты адаптированы для переживания
неблагоприятных условий, но недостаточно осушки и замораживания, чтобы их
158
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
способность выдерживать эти условия изменялась у вида, и, в основном, осушка
более губительна, чем заморозка. Число продуцируемых статобластов огромно.
Brown (1933) подсчитал, что колонии Plumatella repens на растениях на 1 кв. м в
Douglas Lake, Мичиган, США могут производить 800 000 статобластов в осенний
период. Вдоль прибрежья этого озера он нашёл дрейфующий пояс, состоящий из
статобластов, от 1 до 4 футов шириной, простирающийся на пол мили у берега.
Этот пояс статобластов, главным образом, состоял из мёртвых статобластов или
створок, оставшихся после развившихся статобластов. В обычном годовом цикле
колонии развиваются из статобластов, переживших зиму, и быстро приступают к
половому размножению, которое происходит только один раз в году и длится в
природе 3 или 4 недели. Половое размножение часто выпадает в северном кли-
мате или в альпийских озёрах, возможно,4 из-за короткого сезона роста. Бесполое
почкование продолжается весь сезон роста во всех колониях, которые возникли
или из личинок, или из статобластов, до его остановки при падении температуры.
Два типа размножения, соответственно, характерны для Gymnolaemata и Phylac-
tolaemata (Ectoprocta) — половое и бесполое. Дополнительный тип встречается,
например, у Cristatellz, когда её колония разделяется на две новые дочерние. Ко-
лонии, как правило, в наших широтах переживают только одно лето, хотя иног-
да в мягком климате существуют в течение зимы или зим. Подобно другим при-
креплённым животным, колонии Phylactolaemata неизбежно служат местом для
прикрепления и обитания многочисленных мелких животных, таких как простей-
шие, гидры, планарии, коловратки, гастротрихи, нематоды, копеподы, аннелиды,
тардиграды, и личинки насекомых. Мшанки могут срастаться с пресноводными
губками. Личинки хирономид, по-видимому, одни из наиболее постоянных парт-
нёров колоний филактолемат. Многие из квартирантов Phylactolaemata также их
постоянные враги, питающиеся полипидами и статобластами. Полипиды едят
плоские черви, аннелиды, улитки, орибатиды и личинки насекомых. Личинки хи-
рономид самые худшие враги, собирающиеся на колониях в орды и питающиеся
не только полипидами, но и статобластами, которые они разрушают своим грызу-
щим ротовым аппаратом.
В прошлом Phylactolaemata часто вступали в прямые экономические отноше-
ния с человеком как нарушители в городской водной системе. Не только трубы
становились забитыми растущими эктопроктами («трубный мох») и ордами со-
провождающих питающихся детритом и фильтрующих животных, но и фрагмен-
ты разрушенных колоний блокировали водные решётки и счётчики. Предположи-
тельно с установкой в наше время более эффективной системы фильтров таких
трудностей не возникает.
Методы сбора и сохранения в коллекциях
Сбор зоариев, плавающих и прикреплённых статобластов в континентальных
водоёмах проводится по общепринятой методике гидробиологических исследова-
Мшанки
159
ний перифитона, бентоса и планктона. Просмотр воды под бинокуляром, взятой
любой посудой в прибойной зоне, особенно в зарослях макрофитов, даёт возмож-
ность обнаружить статобласты и их фрагменты, а просмотр разного рода субстра-
тов, извлечённых из воды — зоарии.
Мшанок следует фиксировать 70° спиртом или 4%-ным формалином, причём
второе предпочтительней, если зоарии хитиновые, так как в спирте эти колонии
и статобласты становятся хрупкими. Обызвествленные мшанки, наоборот, лучше
хранятся в спирте. Крупные зоарии филактолемат или Eurystomata высушивают-
ся на субстратах.
Основные пособия по определению
Клюге Г.А. 1962. Мшанки северных морей СССР. М.; Л. 584 с.
Lacourt A. W. 1968. A Monograph of the Freshwater Bryozoa Phylactolaemata Leiden. 159 p.
(Zoologische Verhandlungen. No. 93).
Wood, T.S. and Beth Okamura. 2005. A new kew to the freshwater bbryozoans of Britain, Ire-
land and continental Europe, with notes on their ecology. Titus, Wilson and Son, Kendal,
Cumbria. 113 p.
Woss, E. 2005. Moostiere (Bryozoa). Linz, Austria. 369 p.
Определительная таблица мшанок континентальных водоёмов
европейской части России
1(2). Эпистом отсутствует. Статобласты отсутствуют. Могут иметься гиберна-
кулы. Зоарии однорядные, линейные.........................................
...................Надкласс Gymnolaemata (Голоротые) Allman, 1856.
Зоарий мелкий, колония од-
норядная, в виде цепочки,
отдельные цистиды аутозоо-
идов удлинённой конусовид- ' Т r
ной или веретенообразной г
формы, разделены внутрен-
ними перегородками, ответ- рис i Paludicella articulata (колония),
вляются друг от друга под
косым или почти прямым углом. Цистид не имеет шипов по краю отверстия,
орифиций цистида четырёхугольный. Венчик щупалец на лофофоре круглый.
Щупальцев от 10 до 20.
Род Paludicella Gervais, 1836 — Болотница; Р articulata Ehrenberg, 1810 — Б. чле-
нистая (рис. 1).
Встречается преимущественно в реках, реже в озёрах.
2(1). Ротовое отверстие прикрывается эпистомом. Статобласты имеются. Зре-
лые зоарии преимущественно многорядные, либо с явно выраженной
хитиновой кутикулой, тёмно-коричневые, либо просвечивающие, стекло-
видные, либо желатинизированные...........................................
................Надкласс Phylactolaemata (Покрыторотые) Allman, 1856.
160
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Рис. 2. Lophopus crystallinus.
1 — колония, 2 — статобласт, 3 — колония на ряске.
Рис. 3. Lophopodella
carteri (статобласт).
3(10). Статобласты (спинобласты) с прикрепительными образованиями (шипа-
ми). Зоарий неветвящийся, желатинизированный.
4(7). Шипы у спинобластов располагаются преимущественно на полюсах пла-
вательного кольца.
5(6). Спинобласт овальной формы (похож на лимон), но сбоку кажется верете-
новидным, так как каждый полюс спинобласта вытянут в один заострён-
ный шип. Крючки на шипах отсутствуют (L = 1 мм, В = 0,5 мм, отношение
L/B = 1,5)........................Род Lophopus Dumotier, 1835 — Полковник;
L. crystallinus (Pallas, 1768) — П. кристальный (рис. 2).
Колонии небольшие. Колония простирается по субстрату, веерообразная. Лофо-
фор подковообразный с двумя рядами щупалец. Каждый аутозооид образует толь-
ко один статобласт. Встречается в реках и проточных водоёмах.
6(5). Спинобласт широкоовальной формы со слегка усеченными или широко
округлыми полюсами, имеет сбоку вид седла. Каждый полюс спинобла-
ста с несколькими шипами (от 7 до 14), несущими обычно крючки (М
пары). Срединные шипы длиннее боковых. Длина спинобластов (L) в
пределах 1070-1150 мкм, ширина (В) 705-784 мкм, соотношение L/B
1,48-1,52. Длина срединных шипов 65-78 мкм, боковых 44-52 мкм.............
.....................Род Lophopodella Rousselet, 1904 — Подкововидка;
L. carteri (Hyatt, 1866) — П. Картера (рис. 3).
Колония мешковидная, иногда шарообразная или дольчатая. Лофофор подковоо-
бразный с двумя рядами щупалец. Встречается в реках и озёрах.
7(4). Шипы у спинобласта расположены вне его полюсов.
8(9). Спинобласты круглые, примерно 1 мм в диаметре (отношение L/B = 1). На
периферии центральной капсулы плавательного кольца спинобласта рас-
Мшанки
161
положены дорзальное и вентральное кольцо
шипов с крючками............................
.....Род Cristatella Cuvier, 1798 — Хохлатка;
С. mucedo Cuvier, 1798 — X. слизистая (рис. 4).
Зоарий свободно ползающий, гусеницевидный (це-
нозоеций), билатерально симметричный, обычно по
краям своим из двух-трёх рядов полипидов, окру-
жающих прозрачное пространство внутри ценозо-
еция. Лофофор подковообразный с двумя рядами
щупалец. Внутри ценозоеция могут просвечивать
формирующиеся спинобласты. Встречается в про-
точных водоёмах, например, в Ропшинских прудах
и водотоках. .
9(8). Спинобласт с якоревидным шипом в центре
капсулы спинобласта. Шип расположен пер-
пендикулярно к его поверхности....................
.....Род Gelatinella Toriumi, 1955; G. toanensis
(Hozawa et Toriumi, 1940) — Ж. тоанская
(рис. 5).
Колония Gelatinella toanensis отличается сложными
ветвями, каждая из которых состоит из осевой ветви,
составленной рядом первых зооидов и низких боко-
вых мелких веточек. Лофофор подковообразный с
двумя рядами щупалец. Редко встречающийся вид.
Встречается в лентических и логических водоёмах.
Рис. 4. Cristatella mucedo
(статобласт).
Рис. 5. Gelatinella toanen-
sis (статобласт).
10(3). Статобласты без прикрепительных образований (шипов, крючков). Зоарий
прикреплён к субстрату, или ветвящийся, или грибовидный.
11(12). Лофофор округлый........................Род Fredericella Gervais, 1836.
Зоарий ветвящийся. Лофофор с одним рядом щупалец и эпистомом. Сессобла-
сты вытянутые, овальные или почковидные, или очень удлиненные, со светлой,
тонкой, гибкой кутикулой, в сухом виде
выглядят гладкими и блестящими. Все
сессобласты прикреплены к стенкам
цистида. Их форма и размер зависит от
ширины цистида зооида.
Fredericella sultana (Blumenbach, 1779)—
Фредерицелла венценосная (рис. 6).
Встречается в реках, реже в озёрах и
протоках.
12(11). Лофофор подковообразный.
13(20). Имеются сессобласты и флотобла-
сты.
Рис. 6. Fredericella sultana.
1 — колония, 2 — статобласты.
14(15). Кутикула трубок аутозооидов жела-
тинизирована. Колонии линейные,
162
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 7. Hyalinella punctata.
I — колония, 2 — статобласты.
Рис. 8. Plumatella fungosa (колония).
радиальные или компактные пузыревидные, желатинизированные, бесц-
ветные, изредка коричневатые................Род Hyalinella Jullien, 1885.
Сессобласты имеются. Флотобласты округло-овальные, дисковидные, почти оди-
наковые с дорзальной и вентральной стороны. Размер флотобластов значительно
варьирует (L от 0,23 до 0,50 мм).
Род Hyalinella Jullien, 1885 — Стекловидка; Н. punctata Jullien, 1885 — X. точеч-
ная (рис. 7).
У Hyalinella punctata множественное почкование не обязательно приводит к об-
разованию ветвей, а аутозооиды из различных почкующихся рядов могут сосуще-
ствовать в одной ветви. Кутикула трубок желатинизирована. Колонии линейные,
радиальные или компактные пузыревидные, желатинизированные, бесцветные,
изредка коричневатые. Лофофор подковообразный с двумя рядами щупалец. Ча-
сто встречается в водоёмах и водотоках.
15(14). Кутикула трубок аутозооидов не желатинизирована. Колонии ветвящиеся
или грибовидные, или ползучие..................Род Plumatella Lamarck, 1816.
16(19). Отношение длины к ширине у флотобластов L/В менее 1,5.
17(18). L/В флотобласта = 1,45. У флотобластов дорзальная сторона равномерно
покрыта мелкими пузырьковидными образованиями. Имеются сессобла-
сты, покрытые мелкими выростами (tubercules). L/B у сессобластов 1,42..
..........Plumatella fungosa (Pallas, 1768) — Плюмателла грибовидная (рис. 8).
Зоарий (взрослый) крупный, компактный, грибовидный, губкообразный, вначале
прозрачный, затем становится янтарного цвета, позднее коричневеет. Молодые ко-
лонии могут либо «ползти» по субстрату (running strategy), либо ветвиться близко
к субстрату. Цистиды у зооидов могут сливаться. Наиболее распространённый вид
в водоёмах Европейской части России.
18(17). L/В флотобласта = 1,3-1,4, чаще 1,3. Сбоку дорзальная и вентральная
стороны флотобласта почти одинаково выпуклые. Имеются сессобласты с
Мшанки
163
сетчатой структурой узкого плавательного кольца...................
..................Plumatella repens (L., 1758) — Плюмателла ползучая.
Цистиды зооидов прикреплены к субстрату на большей части своей длины.
Стенки колонии прозрачные с легкой инкрустацией. Встречается в лентических и
логических водоёмах.
19(16). Отношение длины к ширине у флотобластов L/В более 1,5. Имеются сес-
собласты и флотобласты, последние с тонкой плёнчатой кутикулой (леп-
тобласты). Средние размеры флотобластов составляют: длина 370 мкм,
ширина 245 мкм, отношение L/B — 1,51....................................
...............Plumatella casmiana Oka, 1907 — Плюмателла касмиана.
Колония компактная, с короткими, густо ветвящимися трубочками, может быть
полупрозрачной или ^товой. Аутозооиды по всей своей длине прикреплены к
субстрату, но в особенно густых местах могут быть вертикальными. Лофофор от-
носительно небольшой. Редко встречающийся вид.
20(13). Имеются только флотобласты.
21(24). Отношение длины к ширине (L/В) у фло-
тобласта 1,65 или более.
22(23). L/В флотобласта = 1,65. Дорзальная сторо-
на флотобласта почти плоская, а вентраль-
ная сильно выпуклая и немного больше
дорзальной. На дорзальной стороне плава-
тельное кольцо с мелкой пузырьковидны-
ми структурой сильно заходит на капсулу,
кольцо серебристое.........................
...........Plumatella emarginata Allman,
1844 — Плюмателла окаймлённая (рис. 9).
Колонии могут быть компактными, или вет-
вистыми со свободными ветвями. Встречается
в проточных водоёмах, преимущественно в
малых реках.
23(22). Флотобласты широкоовальные или поч-
ти круглые. L/В флотобласта = 2,0-2,8.
Дорзальная сторона флотобластов мало
отличается от вентральной. Сбоку флотобласт выглядит как невысокое
куполовидное образование................................................
............Plumatella fruticosa Allman, 1844 — Плюмателла кустистая.
Колония с длинными, тонкими, свободными ветвями. Аутозооиды удалены друг
от друга. Встречается в лентических и логических водоёмах.
24(21). Средние размеры флотобластов менее или равны 1,4. Они составляют око-
ло: длина 340 мкм, ширина 245 мкм, отношение L/B — 1,39-1,4.............
.......Plumatella coralloides Allman, 1850 — Плюмателла кораллоидная.
Зоарий обычно имеет кустовидные выросты. Редко встречающийся вид.
КЛЕЩИ
Клещами называется несистематическая группа мелких хелицеровых, крайне
разнообразных как морфологически, так и экологически. В пресных водах могут
быть обнаружены лишь представители отряда Acariformes (Акариформные кле-
щи). Тело водных клещей компактное, не подразделённое на сегменты, лишь у
представителей подотрядов Acaridida и Oribatida оно может быть разделено по-
перечной бороздой на два отдела — протеросому и гистеросому (табл. 1, 7; 16).
Покровы часто несут склеротизированные щитки, число и расположение которых
может быть различным. У большинства представителей подотряда Oribatida по-
кровы всего тела сильно склеротизиров^ны. На переднем конце тела находятся
ротовые придатки: хелицеры и педипальпы (пальпы), объединённые в специфи-
ческое образование — гнатосому (гнатему). У водных форм всегда 4 пары ног,
обычно с парными терминальными коготками, между которыми часто распола-
гается непарная щетинковидная или коготковидная структура — эмподий; реже
парные коготки отсутствуют и имеется только эмподиальный коготок. С половым
отверстием обычно связано различное число половых присосок. В целом внеш-
нее строение клещей исключительно разнообразно, поэтому описание морфоло-
гических особенностей необходимых для определения отдельных групп этих жи-
вотных будет дано в соответствующих разделах.
Методы сбора и обработки
Для сбора водных клещей используются стандартные гидробиологические
инструменты: сачки, скребки, различные дночерпатели. Пробы предпочтитель-
но фиксировать спиртом (до 70%), однако лучше исследовать живых клещей.
Дальнейшая обработка материала может несколько отличаться для различных
групп клещей. Для сбора крупных форм (преимущественно клещей группы
Hydrachnidia) содержимое пробы следует сразу же промыть и поместить в ем-
кость с водой, а затем просматривать в белой кювете или тарелке небольшими
частями. Для отделения от субстрата мелких форм (Halacaridae, Oribatida и др.)
можно промывать субстрат через два сита с размером отверстий 2 и 0,1 мм. Со-
держимое тонкого сита смывается в чашку Петри и просматривается под бино-
куляром. Для извлечения из субстрата представителей групп Oribatida и Acaridida
используется термоэклекторная установка Берлезе-Тулльгрена, состоящая из
воронки, сетки (на которую кладется субстрат) и верхнего обогревателя (обыч-
но используются лампы накаливания 15—40 Вт) (табл. 1, 9). Клещей собирают в
80%-ный спирт или в воду. Учитывая повышенную влажность субстрата, требу-
ется выдерживание его в установке не менее 5 дней. Для фиксации всех групп
водных клещей можно использовать 70%-ный спирт, однако для Hydrachnidia
наилучшим средством для фиксации и хранения является жидкость Удеманса
(87 частей 75%-ного спирта, 8 частей уксусной кислоты и 5 частей глицерина).
Клещи 165
Для приготовления необходимых- для определения микроскопических препа-
ратов обычно используется жидкость Фора-Берлезе, состоящая из следующих
весовых частей: глицерин — 20, хлоралгидрат — 200, гуммиарабик или вишне-
вый клей — 30, дистиллированная вода — 50. Бесцветные и слабоокрашенные
формы (часть Halacaridae, некоторые Acaridida) можно использовать для приго-
товления препаратов без предварительной фиксации. Сильно пигментированные
формы (Oribatida, некоторые Halacaridae) перед заключением в препарат необхо-
димо просветлить. Для просветления может использоваться слегка подогретая
молочная кислота (с последующей отмывкой в 70%-ном спирте), 15%-ный КОН
(отмывка дистиллированной водой) или жидкость Несбитта (хлоралгидрат 40 г,
дистиллированная вода 25 мл, НС1 (конц.) 2 капли; не более часа). Для просвет-
ления клещей семейстда Anoelijae, накапливающих в теле большое количество
экскретов (кристаллы гуанина) так же желательно использовать щелочь. Опреде-
ление обычно ведется на тотальных препаратах клещей. Для большинства групп
необходимо дорсовентральное расположение животного в препарате, однако для
Oribatida часто необходимо изучение клеща в латеральном положении (можно ис-
пользовать временный препарат в капле молочной кислоты на стекле с лункой).
При приготовлении препаратов из Hydrachnidia и некоторых Halacaridae необхо-
димо отделить гнатему и расположить её латерально. Для Oribatida можно реко-
мендовать отделение нотогастрального щита по циркумгастральному шву. Каче-
ство препарата улучшается, если после изготовления выдержать его в термостате
при температуре 45-50° в течение нескольких суток.
Ключ для определения крупных таксонов клещей
1(8). Хелицеры колющие или колюще-режущие, с редуцированным неподвиж-
ным пальцем. Подвижный палец стилетовидный или ножевидный, глад-
кий, реже с мелкими зубчиками (табл. 1,6)................................
.............................Подотряд Actinedida (= Trombidiformes).
2(3). Эмподий тонкий, щетинковидный (табл. 1, 2). Тело полусферической фор-
мы, сильно склеротизировано. Под покровами имеются воздухоносные
полости (мешковидные или трубковидные)...................................
..............Когорта Eleutherengona, сем. Homocaligidae Wood, 1969.
1 род Homocaligus Berlese, 1910. Я. scapularis (Koch, 1838) широко распространён
в Европе; Н. amphibius Wainstein, 1975 известен из европейской части России
(табл. 1, /). Амфибионтные формы, встречаются по берегам рек и во временных
водоёмах.
3(2). Эмподий утолщённый, коготковидный или отсутствует. Тело либо уплоще-
но дорсовентрально, либо мешковидной или сферической формы; в раз-
личной степени склеротизировано. Воздухоносные полости отсутствуют.
4(5). Тело уплощено дорсовентрально, обычно ромбовидной формы. В составе
пальп не более четырёх члеников..........................................
..............Когорта Eupodina, сем. Halacaridae Murray, 1876 (с. 190).
166
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
5(4). Тело обычно мешковидное или сферическое, реже уплощено дорсовен-
трально. В составе пальп не менее пяти члеников (исключая Calyptostomi-
dae и некоторых Limnocharidae).........................Когорта Parasitengona.
Таблица 1
7 — Homocaligus amphibius Wainstein, 1975, общий вид, латерально; 2 — Homocaligus
scapularis (C.L. Koch, 1838), предлапка с коготками и эмподием; 3 — Calyptostoma sp.,
общий вид, дорсально; 4 — Calyptostoma brevirostris Wainstein, 1977, гнатема; 5 —
грызущая клешневидная хелицера Oribatida; 6 — Litarachna communis Walter, 1925,
хелицера: общий вид самки, дорсально; 7, 8 — Schwiebea danielopoli Fain, 1982, об-
щий вид самки, вентрально; 9 — схема аппарата для извлечения клещей из субстрата.
1 — по: Вайнштейн, 1975; 2 — по: Wood, 1969; 3 — по: Kranz, 1970; 4 — по: Вайнштейн,
1977; 5 — по: Гиляров, 1975; 6 — по: Тузовский, 1997; 7, 8 — по: Fain, 1982; 9 — по: За-
хваткин, 1953.
Клещи
167
6(7). Тело несёт неопределённо большое число щетинок. 2 пары половых при-
сосок. Вся гнатема (включая пальпы) способна втягиваться в тело......
............Фаланга Trombidia, сем. Calyptostomidae Oudemans, 1923.
1 род Calyptostoma Cambridge, 1875. В России 2 вида: С. brevirostris Wainstein,
1977 и С. longirostris Wainstein, 1977 (табл. 1, 3, 4), различающихся относительной
длиной гнатемы. Амфибионтные формы, встречающиеся на болотах, по берегам
рек, во временных водоёмах.
7(6). Тело несёт небольшое строго определённое число щетинок. У взрослых
особей 3 или более пар половых присосок, реже они отсутствуют........
.....................................Фаланга Hydrachnidia (с. 167).
8(1). Хелицеры обычно грызущие, клешневидные (табл. 1, 5). Реже (семейство
Anoetidae) подвиэедый палец редуцирован, а неподвижный палец обычно
несёт ряд крупных зубцов.
9(10). В составе пальп не менее 3 члеников, обычно они пятичлениковые. Ноги
с 1 или 3 (редко 2) коготками. Покровы взрослых особей сильно склероти-
зированы.....................Подотряд Oribatida (= Cryptostigmata) (с. 196).
10(9). Пальпы двучлениковые. Ноги всегда с одним эмподиальным коготком.
Покровы мягкие, не склеротизированные...........................
..................................Подотряд Acaridida (= Astigmata).
В пресных водах могут быть обнаружены представители семейств Anoetidae Oude-
mans, 1904 и Acaridae Leach, 1816, род Schwiebea Oudemans, 1916 (табл. 1, 7, 8).
HYDRACHNIDIA
Клещи, относящиеся к данной группе (часто их называют гидракаринами, ги-
драхнеллами, гидрахнидиями или пресноводными клещами), распространены по-
всеместно и заселяют все типы пресных водоёмов и морскую среду.
Клещи проходят сложный цикл развития, включающий три активных фазы
(стадии): личинку, дейтонимфу и имаго. Личинки гидрахнидий паразитируют
преимущественно на имаго водных насекомых, а последующие активные фазы
хищничают, питаясь различными мелкими беспозвоночными. Водяные кле-
щи раздельнополы. После копуляции самка откладывает яйца — от нескольких
штук до многих сотен у разных групп и видов. Вылупившаяся из яйца личинка
(табл. 2, 7) имеет три пары ног, у неё отсутствуют какие-либо генитальные при-
датки. Все личинки гидрахнидий довольно сходны между собой, но по особен-
ностям морфологии их подразделяют на две группы: низшие и высшие. Покровы
личинок первой группы не смачиваются и после выхода из яйца они всплывают
на поверхность воды, бегают по ней и выступающим частям водных растений,
где и отыскивают хозяина. Личинки второй группы не могут сами преодолеть
силы поверхностного натяжения воды и отыскивают хозяев в водной среде. Насо-
савшись гемолимфы хозяина, личинка переходит в покоящееся состояние и затем
линяет в нимфу. Наружное строение нимфы (табл. 2, 7, 8) существенно отличает-
168
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
169
^лещи
до от личинки. Во-первых, у нимфы 4 лары ног; во-вторых, появляются наруж-
ные генитальные придатки (2 или более пар присосок, створки или пластинки),
хотя половое отверстие отсутствует; в-третьих, форма, пропорции и размеры всех
органов совершенно непохожи на личиночные. После активного периода, напи-
тавшись, нимфа также переходит в покоящееся состояние, после чего превраща-
ется во взрослого клеща. Внешний облик большинства низших клещей, многих
мягкотелых высших (особенно самок), настолько сходен с нимфальной стадией,
что зачастую только исследование генитального поля позволяет установить фа-
зовую принадлежность особи. Половое отверстие у взрослых клещей продоль-
ное, по бокам его располагаются три или более пар присосок, которые полностью
или частично прикрыты створками, покоятся на пластинках или размещаются
непосредственно на покровах. Спереди и сзади половое отверстие часто ограни-
чено опорными телами. Половые присоски отсутствуют лишь у Pontarachnidae
и Hydrovolziidae. Тазики ног обычно хорошо развиты, слиты с вентральной по-
верхностью тела и могут группироваться самым различным образом, вплоть до
слияния их в один блок. Передние тазики, как правило, имеют отростки, погру-
жённые под кутикулу (передние аподемы), а тазики IV нередко — задние аподе-
мы. Ноги, помимо тазиков, состоят из 6 свободных члеников: вертлуга, бедра I,
бедра II, колена, голени и лапки. Последние членики ног на конце имеют выемку,
в которую при плавании втягиваются парные коготки. Ротовой аппарат состоит из
гипостомальной пластинки (сросшиеся тазики педипальп), хелицер и педипальп.
Передняя часть гипостомальной пластинки (хоботок, рострум) нередко удлине-
на и в той или иной мере развернута на вентральную сторону. Хелицеры колю-
ще-режущего типа и состоят из 2 члеников: крупного базального (основного) и
подвижного маленького, заострённого на конце (хела). Педипальпа представле-
на 5 свободными члениками: вертлугом, бедром, коленом, голенью, лапкой. Тело
обычно округлой или яйцевидной формы, часто сплющено в дорсовентральном
Таблица 2
1 — Arrenurus sp., общий вид личинки, дорсально; 2 — Mideopis orbicularis (Muller,
1776), общий вид самки дорсально; 3 — Eylais meridionalis Thon, 1899, общий вид
дорсально;^ — Arrenurus globator (Muller, 1776), общий вид самца дорсально; 5 —
Hydrachna geographica Muller, 1776, общий вид дорсально; 6 — Unionicola crassipes
(Muller, 1776), общий вид вентрально; 7, 8 — общий вид дейтонимфы Hydrachnidia
(схематично): 7 — вентрально, 8 — дорсально.
АН — анальная пластинка, ГП — гипостомальная пластинка, ГЕН — генитальные при-
соски, ГС — генитальные створки, ЗА — задняя аподема, ПА — передняя аподема, ПЕ —
педипальпы, Н1-Н4 — ноги 1-4, В — вертлуг, Б1 — бедро 1, Б2 — бедро 2, К — колено,
Г — голень, Л — лапка, КГ — коготки, Т1-ТЗ — тазики 1-3, ХЕЯ — хелицеры.
/ — по: Kranz, 1970; 2 — по: Soar & Williamson, 1929; 3, 6 — по: Павловский, Лепнева,
1948; 4 — по: Wesenberg-Lund & Storch, 1939; 5 — по: Ланге, 1969; 7, 8 — по: Тузовский,
1997.
170
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
направлении, иногда сжато с боков и несёт определённый набор щетинок. Покро-
вы мягкие или со щитками, которые могут разрастаться, сливаясь между собой
вплоть до образования панциря (табл. 2, 2-6).
Гидрахнидии ведут придонный образ жизни, обитают в озёрах, водохранили-
щах, прудах, реках, ручьях, родниках. Заселяют они также подземные воды, го-
рячие источники, а клещи семейства Pontarachnidae — морскую среду. Наиболее
многочисленны они в прибрежной зоне водоёмов, в зарослях высшей раститель-
ности, исключая жёсткую (камыш, тростник), на илистых грунтах. Бедностью ка-
чественного и количественного состава клещей отличаются открытые прибойные
участки крупных водоёмов с песчаными грунтами.
К настоящему времени в мире известно около 4000 видов и подвидов гидрах-
нидий. В России известно свмше 500 видов и 60 родов, относящихся к 8 надсе-
мействам. Лишь одно надсемейство Hydrovolzioidea пока не обнаружено на тер-
ритории России. Находки представителей этой группы вполне вероятны, так как
они найдены почти всюду, от Скандинавии до Японии и Северной Америки.
Основные пособия по определению
Вайнштейн Б.А. 1980. Определитель личинок водяных клещей. Л. 237 с.
Соколов И.И. 1940. Hydracarina — водяные клещи. Фауна СССР. М.; Л. 510 с.
Тузовский П.В. 1990. Определитель дейтонимф водяных клещей. М. 236 с.
Szalay L. 1964. Viziatkak Hydracarina. Fauna Hungary, 72. Budapest. 380 p.
Ключ для определения надсемейств
1(4). Генитальные присоски отсутствуют.
2(3). Тело уплощено. На туловище имеются хорошо развитые дорсальные и
вентральные щитки. Латеральные части тазиков III—IV выдаются за бо-
ковую линию тела, задний край тазиков IV без отростков. Пресноводные
формы.......................................................Hydrovolzioidea.
Одно семейство Hydrovolziidae Thor, 1905. В России представители этой группы
клещей пока не обнаружены. Вполне вероятно нахождение видов рода Hydrovolzia
Thor, 1905 (табл. 3, 1, 2), обычных для западной, южной Европы и Скандинавии.
Обитают в ручьях, родниках и подземных водах.
3(2). Тело не уплощено. Настоящие туловищные щитки или пластинки отсут-
ствуют. Тазики III—IV не выдаются за боковую линию тела, задний край
тазиков IV с 2 длинными узкими отростками. Морские формы.....................
.....................................................Pontarachnoidea.
Включает одно семейство Pontarachnidae Koenike, 1910, насчитывающее около
20 видов. В России известно 2 рода: Pontarachna Philippi, 1840 и Litarachna Walter,
1925. Обитают исключительно в морской среде.
4(1). Генитальные присоски имеются.
5(6). Тело сферической формы. Хелицеры одночленистые стилетовидные. Пе-
редняя часть гипостомальной пластинки сильно вытянута в длину, образуя
Клещи
171
желобовидный хоботок нацодобие клюва, сопоставимый по величине с
педипальпами.........................................Hydrachnoidea.
Включает одно семейство Hydrachnidae Leach, 1815, богатое видами. В России
1 род Hydrachna Muller (табл. 3, 3, 4), 1776 и около 30 видов. Обитатели различно-
го рода временных водоёмов, озёр, прудов и слабопроточных заросших водотоков.
6(5). Тело иной формы. Хелицеры состоят из 2 раздельных члеников. Передняя
часть гипостомальной пластинки не образует хоботка или он значительно
короче педипальп.
Таблица 3
1,2 — Hydrovolzia sp.: 1 — дорсально, 2 — вентрально; 3, 4 — дорсальные щитки и
пластинки Hydrachna: 3 — Hydrachna cruenta Muller, 1776, 4 — Hydrachna conjecta
Koenike, 1895; 5, 6 — Piersigia palustris Tuzovskij, 1986: 5 — дорсально, 6 — вен-
трально; 7-9 — Limnochares aquatica (Linne, 1758): 7 — передняя часть вентральной
поверхности, 8 — генитальные присоски, 9 — дорсальный щиток; 10 — Eylais mutila
Koenike, 1897, глазные пластинки (очки).
1—10— по: Тузовский, 1997.
172
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
7(12). Глаза расположены на переднем дорсальном щитке или соединены узкой
поперечной пластинкой. Передняя часть ротового аппарата образует круп-
ный присасывательный диск.
8(9). Глазные капсулы широко расставлены и расположены в латеральных ча-
стях довольно крупного переднего дорсального щита, имеющего задний
медиальный отросток......................................Piersigioidea.
Представлено одним семейством Piersigiidae Oudemans, 1902. В России 1 род Pier-
sigia Protz, 1896 и 2 вида: Р palustris Tuzovskij, 1986 (табл. 3, 5, 6) и Р limnophila
Protz, 1896. Первый вид обитает в осоково-сфагновых болотах, второй — в озёрах
и слабопроточных водотоках.
9(8). Глазные капсулы сближены между собой в срединной части переднего от-
дела туловища и соединены продольной или поперечной пластинками.
10(11). Глаза находятся по бокам продольной пластинки, образуя крестообразную
фигуру. Ноги без плавательных волосков. Ротовой диск не развернут на
вентральную сторону.....................................Limnocharoidea.
Включает одно семейство Limnocharidae Grube, 1859. В России 1 род и вид Limno-
chares aquatica (Linne, 1758) (табл. 3, 7-9). Обитает на илистых грунтах в непро-
точных и слабопроточных водоёмах.
11(10). Глазные капсулы соединены более или менее развитой узкой поперечной
пластинкой, образуя так называемые «очки» (табл. 3, 10). Ноги I—III с
многочисленными плавательными волосками. Ротовой диск обычно раз-
вернут на вентральную сторону...............................Eylaioidea.
Включает одно семейство Eylaidae Leach, 1815. В России 1 род — Eylais Latreille,
1776 и свыше 40 видов. Обитают преимущественно в заросших непроточных и
слабопроточных водоёмах.
12(7). Глаза находятся по бокам переднего отдела тела и не соединены между со-
бой, хотя между ними могут располагаться щиток или пластинки. Перед-
няя часть ротового аппарата не образует крупного диска.
13(14). Красные клещи, часто с фронтальным органом. Туловище, ходильные
конечности и педипальпы без половых отличий. Голень педипальпы с дор-
содистальным отростком, если отросток не развит, то на голени имеется
дорсодистальная шиловидная щетинка...............Hydryphantoidea (с. 172).
14(13). Окраска клещей самая разнообразная. Фронтальный орган, как правило,
отсутствует. Туловище, ходильные конечности и педипальпы часто различ-
ные у самцов и самок. Голень педипальп без дорсодистального отростка..
...................................................Hygrobatoidea (с. 176).
Надсемейство HYDRYPHANTOIDEA
Цвет красный или жёлтый с самыми разнообразными оттенками. Покровы с
сосочками, с тенденцией к образованию пластинок или щитков, которые могут
разрастаться и сливаться, образуя иногда панцирь. Реже, настоящие пластинки
не развиты, но сохраняются их рудименты. Боковые глаза обычно имеются, ча-
Клещи
173
сто они заключены в капсулы, занимают латеральное положение и не соединены
медиально пластинками. Медиальный глаз (фронтальный орган) обычно имеется,
хотя у отдельных видов он может отсутствовать. Тазики ног чаще всего объеди-
нены в 4 группы. Ноги с плавательными волосками или без них. Коготки про-
стые, часто с надкоготковой пластинкой.
Ключ для определения семейств
1(4). Передние и задние группы тазиков на небольшом удалении друг от друга.
Наружные гениталии расположены между задними группами тазиков или
несколько сзади от них.
2(3). Боковые глаза в капсулах. Большей частью имеются более или менее раз-
витее туловЙЪцные щитки или пластинки. Голень педипальпы с коротким
дорсодистальным отростком, если отросток не развит, то имеется шипо-
видная дорсодистальная щетинка.... Hydryphantidae Piersig, 1896 (с. 173).
3(2). Боковые глаза не в капсулах, находятся на удалении друг от друга. Покро-
вы без щитков и пластинок. Голень педипальпы с длинным, величиной с
лапку, дорсодистальным отростком..............Hydrodromidae Viets, 1936.
В России 1 род и вид Hydrodroma despiciens (Muller, 1776) (табл. 4, 1, 2). Один из
самых обычных видов. Распространён повсеместно.
4(1). Передние и задние группы тазиков разделены большим промежутком, в
котором расположены наружные гениталии..........Protziidae Koenike, 1909.
В России известно 2 рода: Protzia Piersig, 1896 (табл. 4, 3, 4) и Calonyx Walter,
1907. Последний отличается от первого тем, что наружные гениталии имеют
длинные лентовидные пластинки, тогда как у видов рода Protzia они отсутствуют.
Семейство HYDRYPHANTIDAE
На дорсальной поверхности тела обычно хорошо развит 1 или несколько щит-
ков. Глаза обычно попарно объединены в капсулы. Половой орган с подвижными
створками.
Ключ для определения родов
1(2). Сосочки на покровах тела оканчиваются на вершине пучком тонких отро-
стков.................................................................
.....Подсем. Pseudohydryphantinae, род Pseudohydryphantes Viets, 1907.
В России 2 вида. Обитают в озёрах и проточных водоёмах. Наиболее распростра-
нён Р parvulus Viets, 1907 (табл. 4, 5, 6).
2(1). Сосочки на покровах тела без отростков на вершине.
3(6). На туловище хорошо выражен только передний дорсальный щиток. Ноги
с плавательными волосками.......................Подсем. Hydryphantinae.
4(5). Наружный генитальный аппарат обычно с 3 (иногда с 4-7) парами присо-
сок. На дорсальном щитке хорошо выражены только задние отростки.......
...........................Род Hydryphantes Koch, 1841 (табл. 4, 7, 8).
174
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Таблица 4
7,2 — Hydrodroma despiciens (Muller, 1776): 1 — вентральная поверхность, 2 — пе-
дипальпа; 3, 4 — Protzia eximia (Protz, 1896): 3 — передняя часть вентральной по-
верхности, 4 — коготки; 5, 6 — Pseudohydryphantes parvulus Viets, 1907: 5 — тазики
ног и генитальный орган, 6 — кутикулярные сосочки; 7, 8 — Hydryphantes placationis
Thon, 1899: 7 — дорсальный щиток, 8 — генитальные пластинки; 9, 10 — Georgella
koenikei (Maglio, 1906): 9 — тазики ног и генитальный орган, 10 — дорсальный щи-
ток; 77 —Thyopsis cancellata (Protz, 1896), дорсальная поверхность.
7-77 — по: Тузовский, 1997.
Клещи
175
5(4). Наружный генитальный аппарат с многочисленными присосками. Дор-
сальный щиток имеет хорошо выраженные задние и боковые отростки и
передний выступ................................Род Georgella Koenike, 1907.
В России известно 3 вида. Наиболее распространён G. koenikei (Maglio, 1906)
(табл. 4, 9, 10).
6(3). Туловище со многими дорсальными и вентральными щитками, которые
могут объединяться самым различным образом, вплоть до образования
монолитного щита. Иногда щитки слабо выражены. Ноги без плаватель-
ных волосков...........................................Подсем. Thyasinae.
7(8). Все дорсальные пластинки объединены в 1 крупный щит..............
...............................Род Thyopsis Piersig, 1898 (табл. 4, 11).
8(7). Дорсальные плаегинки не объединены в 1 щит.
9(18). Передний дорсальный щит (пропельтидий) монолитный, не расчленен на
фрагменты.
10(15). Пропельтидий не несёт на себе щетинок и не слит с другими пластинками.
11(12). Пропельтидий ромбовидной формы, его длина больше, чем ширина. Вто-
рая пара присосок находится близ середины генитального органа..........
.................................Род Euthyas Piersig, 1898 (табл. 5, 1).
12(11). Пропельтидий округло-угловатой формы, его ширина примерно равна
длине. Вторая пара присосок находится на заднем конце генитального
органа.
13(14). Латеральные пластинки и задняя пара дорсоцентральных щитков очень
крупные. Медиальный глаз не пигментирован...............................
.............................Род Parathyas Lundblad, 1926 (табл. 5, 2).
14(13). Латеральные и задняя пара дорсоцентральных пластинок умеренной вели-
чины. Медиальный глаз пигментирован.....................................
.............................Род Thyasides Lundblad, 1926 (табл. 5, 3).
15(10). Пропельтидий несёт на себе 1 пару щетинок и слит с передней парой дор-
соцентральных пластинок.
16(17). Задние дорсоцентральные пластинки срослись между собой в 1 щит. Зад-
няя часть переднего дорсального щита лишь немного уже передней части
.......................................Род Panisus Koenike, 1896 (табл. 5, 4).
17(16). Задние дорсоцентральные пластинки не срослись между собой. Задний
отросток переднего дорсального щита вдвое уже передней части............
.................................Род Panisopsis Viets, 1926 (табл. 5, 5).
18(9). Пропельтидий расчленен на 2 или 4 маленьких фрагмента. Иногда они
очень плохо заметны.
19(20). Дорсоцентральные и дорсолатеральные пластинки более или менее выра-
жены. Генитальные присоски не срастаются со створками...................
....................................Род Thyas Koch, 1835 (табл. 5, 6).
20(19). Настоящие дорсальные щитки или пластинки отсутствуют. Присоски
срослись со створками...............Род Zschokkea Koenike, 1892 (табл. 5, 7).
176
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Таблица 5
1 — Euthyas truncata (Neuman, 1875), передняя часть дорсальной поверхности; 2 —
Parathyas thoracata (Piersig, 1896), дорсальная поверхность; 3 — Thyasides dentata
(Thor, 1897), передняя часть дорсальной поверхности; 4 — Panisus major Tuzovskij,
1994, дорсальная поверхность; 5 — Panisopsis sp., дорсальная поверхность; 6 —
Thyas fastus Tuzovskij, 1980, дорсальная поверхность; 7 — Zschokkea oblonga (Koe-
nike, 1892), дорсальная поверхность.
ДЦ1-5 — дорсоцентральные пластинки 1-5, ДЛ1-4 — дорсолатеральные пластинки 1-4,
ПДЩ — передний дорсальный щит (пропельтидий), ФР — фронтальный орган.
1-7 — по: Тузовский, 1997.
Надсемейство HYGROBATOIDEA
Покровы обычно мягкие, нередко с пластинками или щитками, а порой при-
обретают характер панциря. Боковые глаза занимают латеральное положение.
Наружные гениталии с присосками, располагающимися прямо на покровах, при-
крытых створками или на 2(4) пластинках. Хелицеры двучленистые. Педипаль-
Клещи
177
пы обычно состоят из 5 члеников, но иногда отдельные членики могут срастаться
между собой. Голень педипальпы без дорсодистального отростка. Для многих ви-
дов характерны половые различия в строении различных органов и систем. Са-
мая богатая родами и видами группа гидрахнидий.
Ключ для определения семейств
1(10). Половой орган с более или менее выпуклыми и подвижными створками.
2(5). Тазики ног обычно объединены в 4 группы. Передние группы тазиков мо-
гут соприкасаться или даже срастаться медиальными краями.
3(4). Ноги с плавательными волосками. Лапки IV без коготков. На голени педи-
пальпы 1 шип....................................Teutoniidae Koenike, 1910.
Включает 1 род Teutonia Koenike, 1899. В России известно 3 вида, наиболее рас-
пространён Т cometes (Koch, 1837) (табл. 6, 1, 2). Обитает как в проточных, так и
в непроточных водоёмах.
4(3). Ноги без плавательных волосков, лапки IV с коготками. На голени педи-
пальпы 2 шипа или бугор (во втором случае без шиловидных щетинок)....
...........................................Sperchonidae Thor, 1900 (с. 180).
5(2). Тазики ног объединены в одну группу.
6(9). Покровы на дорсальной поверхности мягкие, без щитков или пластинок.
Половой орган с 3 парами присосок.
7(8). Лапки всех ног с коготками....................Lebertiidae Thor, 1900.
Обитатели преимущественно проточных водоёмов. Изредка в озёрах, водохрани-
лищах и других непроточных водоёмах. В России 1 род Lebertia Neuman, 1880 и
свыше 50 видов (табл. 6, 3).
8(7). Лапки ног IV без коготков..................Oxidae Viets, 1926 (с. 180).
9(6). Покровы сильно склеротизованы и приобрели характер панциря, лишь
узкая интерскутальная мембрана отделяет дорсальный щит от остальной
склеротизованной части тела. Половой орган с 6 парами присосок..........
.......................................Torrenticolidae Piersig, 1902.
В России 1 род Torrenticola Piersig, 1896 (табл. 6, 4, 5) и более 10 видов.
10(1). Половой орган без подвижных створок.
11(24). Голень педипальпы тоньше, чем бедро и колено, без вентродистального
выступа.
12(13). Боковые глаза отделены друг от друга широким промежутком. Лапки ног
IV без коготков....................................Limnesiidae Thor, 1900.
В России 1 род Limnesia Koch, 1836 (табл. 6, 6, 7) и около 15 видов, которые оби-
тают в разного рода непроточных и слабопроточных водоёмах и являются одними
из самых многочисленных групп гидракарин в озёрах, прудах и водохранилищах.
13(12). Глаза попарно с каждой стороны туловища, сближены между собой. Лап-
ки ног IV с коготками.
14(17). Задние группы тазиков более или менее треугольной формы, с узкими
медиальными и широкими латеральными сторонами.
178
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Таблица 6
1,2 — Teutonia cometes (Koch, 1837): 1 — тазики ног и генитальный орган самки, 2 —
тазики ног III—IV и генитальный орган самца; 3 — Lebertia dubia Thor, 1899, тазики
ног и генитальный орган; 4,5 — Torrenticola amplexa (Koenike, 1908): 4 — дорсальная
поверхность, 5 — вентральная поверхность; 6, 7 — Limnesia connata Koenike, 1895:
6 — вентральная поверхность, 7 — педипальпа; 8 — Feltria minuta Koenike, 1892, дор-
сальная поверхность самки; 9, 10 — Acalyptonotus violaceus Walter, 1911: 9 — педи-
пальпа, 10 — вентральная поверхность самца; 11, 12 — Krendowskia latissima Piersig,
1895: 11 — педипальпа нимфы, 12 — вентральная поверхность самки.
1—12 — по: Тузовский, 1997.
Клещи
179
15(16). Тазики ног большей частью расположены в 3 группы: тазики I—II объ-
единены в один блок, а тазики III-IV — в две группы. Тазики IV без
переднелатерального отростка. Задние группы тазиков покоятся прямо на
покровах..................................Hygrobatidae Koch, 1842 (с. 182).
16(15). Тазики ног объединены в 4 группы. Тазики IV с крупным переднелате-
ральным отростком, охватывающим полностью или частично тазики III.
Задние группы тазиков покоятся на вентральных щитках, иногда все про-
межутки между тазиками склеротизованы..............Feltriidae Viets, 1926.
Богатая видами группа клещей. В России пока известен 1 род Feltria Koenike,
1892 (табл. 6, 5) и около 20 видов, большинство из которых найдено в восточных
районах страны. Самые мелкие из гидракарин (0,3-0,5 мм). Реофилы, обитают на
каменистых участках и моэдвых обрастаниях водотоков.
17(14). Задние тазики иной формы, латеральные края тазиков III—IV такой же дли-
ны, как и медиальные или немногим длиннее их.
18(19). Передняя пара ног обычно утолщена, с крупными, сидящими на бугорках,
мечевидными зазубренными щетинками. Передние тазики с хорошо раз-
витыми аподемами. Медиальные края задних тазиков такой же ширины
как и латеральные. Амбулакры (коготки) без коготковых пластинок..........
...............................Unionicolidae Oudemans, 1909 (с. 182).
19(18). Передние ноги обычно не утолщены, без бугров и зазубренных щетинок.
Тазики I—II с мелкими аподемами или они не развиты вовсе. Медиальные
края задних тазиков несколько уже латеральных. Амбулакры с коготковой
пластинкой.
20(23). Заднемедиальные части тазиков IV вогнуты и образуют генитальную вы-
емку, в которой или близ которой расположены наружные гениталии. Зад-
ние края тазиков IV с угловатым выступом или выпуклые.
21(22). Задний край тазиков IV с угловатым выступом и маленькой аподемой. По-
кровы большей частью мягкие, с небольшими дорсальными пластинками.
У самцов иногда развит крупный дорсальный щит............................
.........................................Pionidae Thor, 1900 (с. 184).
22(21). Задний край тазиков IV без угловатого выступа и аподемы. Покровы
большей частью сильно склеротизованы, имеется лишь узкая бороздка,
отделяющая дорсальный щиток от вентрального щита.........................
..................................Mideopsidae Koenike, 1910 (с. 187).
23(20). Тазики IV не образуют генитальную выемку. Задние края тазиков IV пря-
мые или с небольшими выемками.................Aturidae Thor, 1900 (с. 188).
24(11). Голень педипальпы обычно такой же толщины и высоты как бедро и коле-
но, с вентродистальным выступом, несущим крупную щетинку.
25(26). Тазики ног и наружные гениталии покоятся на общем вентральном щите,
дорсальный щиток отсутствует..................Acalyptonotidae Walter, 1911.
1 род Acalyptonotus Walter, 1911. В России 1 вид A. violaceus Walter, 1911 (табл. 6,
9, 10). Озёрная форма, характерен для умеренных широт и больших глубин (до
125 м). Распространён в Скандинавии, европейской и азиатской частях России.
180
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
26(25). Покровы склеротизованы полностью. Лишь дорсальная бороздка разгра-
ничивает дорсальный щиток от вентрального щита.
27(28). Медиальные края тазиков IV вогнутые и образуют генитальную выемку.
Половых присосок 3-4 пары, створки узкие продолговатые...............
.......................................Krendowskiidae Viets, 1926.
В Европе и России известен 1 род Krendowskia Piersig, 1895 и 1 вид К. latissima
Piersig, 1895 (табл. 6, 11, 12). Проточные водоёмы.
28(27). Медиальные края тазиков IV не вогнуты и не образуют генитальную
выемку. Половые присоски многочисленны и расположены на узких по-
перечных пластинках..............................Arrenuridae Thor, 1900.
Клещи с хорошо выраженными половыми различиями. Самки обычно яйцевид-
ной формы, половое отверстие у нихФрупное, прикрыто подвижными створками,
генитальные пластинки довольно широкие и не переходят на боковые и дорсаль-
ную стороны тела (табл. 7, 1). Самцы же могут иметь самую разнообразную фор-
му тела, которое часто снабжено различными выступами, буграми и придатками;
генитальное отверстие маленькое без створок, пластинки узкие и часто переходят
на латеродорсальную поверхность (табл. 2, 7, 2, 3). Одна из самых богатых
видами групп гидрахнидий. В России 1 род Arrenurus Duges, 1834 и свыше 70
видов. Обитатели преимущественно заросших высшей водной растительностью
непроточных и слабопроточных водоёмов.
Семейство SPERCHONIDAE
Обитатели проточных водоёмов.
Ключ для определения родов
1(2). Голень педипальпы на вентральной стороне с бугром, но без шиловидных
щетинок (штифтов). Выходные отверстия кутикулярных желёз размеща-
ются на высоких конусовидных буграх..................................
............................Sperchonopsis Piersig, 1896 (табл. 7, 4, 5).
2(1). Голень педипальпы с 2 маленькими, обычно широко расставленными
шипиками, но бугор отсутствует. Отверстия кутикулярных желёз распола-
гаются на плоских пластинках..........Sperchon Kramer, 1877 (табл. 7, 6, 7).
Богатый видами род. В России известно около 30 видов.
Семейство OXIDAE
Обитатели непроточных и слабопроточных водоёмов.
Ключ для определения родов
1(4). Тело сильно сжато с боков, его высота гораздо больше ширины. Тазики
ног чрезвычайно увеличены в размерах и занимают большую часть по-
верхности тела.
Клещи
181
Таблица 7
1 —Arrenurus globator (Muller, 1776), вентральная поверхность самки; 2 — Arrenurus
absurdus Wainstein et Tuzovskij, 1971, дорсальная поверхность самца; 3 — Arrenurus
sinuator (Muller, 1776), дорсальная поверхность самца; 4, 5 — Sperchonopsis verrucosa
(Protz, 1896): 4 — вентральная поверхность, 5 — педипальпа; 6 — Sperchon glan-
dulosus Koenike, 1885, вентральная поверхность; 7 — Sperchon tridentatus Sokolow,
1940, педипальпа; 8 — Frontipoda musculis (Muller, 1776), туловище в профиль; 9 —
Gnaphiscus ekmani Thor, 1913, туловище в профиль; 10 — Oxus ovalis (Muller, 1776),
вентральная поверхность.
ЛО — лентовидный отросток. 1—10 — по: Тузовский, 1997.
182
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
2(3). Анальное отверстие открывается на продолговатой пластинке. Передний
конец тазиков I с 2 короткими щетинками, но без лентовидного отростка.
...............................................Frontipoda Koenike, 1891.
В России 1 вид F musculis (Muller, 1776) (табл. 7, 8).
3(2). Анальное отверстие без особой склеротизации. На переднем конце тази-
ков I имеется, помимо 2 щетинок, длинный лентовидный отросток........
.........................................Gnaphiscus Koenike, 1898.
В России известно 4 вида, наиболее распространён G. ekmani Thor, 1913 (табл. 7,9).
4(1). Тело сигарообразной формы, его ширина примерно равна высоте. Тазики
ног развиты умеренно и занимают только вентральную или вентролате-
ральную стороны тела......................Oxus Kramer, 1877 (табл. 7,10).
В России насчитывается около 10 вийов.
Семейство HYGROBATIDAE
Богатая родами и видами группа клещей, предпочитающая проточные водоёмы.
Ключ для определения родов
1(4). Передние ноги не длиннее ног второй пары, лапки I не изогнуты, голень I
без длинных мечевидных щетинок. Гипостомальная пластинка срослась с
тазиками I.
2(3). Тазики IV с подкожным заднемедиальным зубовидным выростом. Колено
педипальпы без вентральных зубцов..................Mixobates Thor, 1905.
В России 3 вида. Наиболее распространён М. uncatus (Sokolow, 1930) (табл. 8,1-4).
3(2). Тазики IV без подкожного заднемедиального зубовидного выроста. Коле-
но педипальпы по вентральному краю с мелкими зубцами.................
...............................Hygrobates Koch, 1837 (табл. 8, 5-7).
В России найдено 15 видов.
4(1). Передние ноги длиннее и толще ног второй пары, лапка I изогнута, голень
I с 2 длинными вентродистальными мечевидными щетинками. Гипосто-
мальная пластинка не срослась с тазиками I...........................
...............................Atractides Koch, 1837 (табл. 8, 8-11).
В России известно свыше 20 видов.
Семейство UNIONICOLIDAE
Обитатели непроточных и слабопроточных водоёмов. Богатая родами и ви-
дами группа клещей. В России широко распространены представители 2 родов,
каждый из которых насчитывает по 10 видов.
Ключ для определения родов
1(2). Передние тазики с узкими короткими аподемами. Шов между тазиками
III—IV неполный и косо направлен к переднемедиальному углу тазиков
Клещи
183
Таблица 8
1-4 — Mixobates uncatus (Sokolow, 1930): 1 — вентральная поверхность самки, 2 —
генитальная пластинка самца, 3 — педипальпа самки, 4 — голень и лапка ноги I;
5 — Hygrobates fluviatilis (Strom, 1768), педипальпа; 6, 7 — Hygrobates trigonicus Koe-
nike, 1895: 6 — генитальный орган самки, 7 — генитальная пластинка самца; 8-11 —
Atractides ovalis Koenike, 1883: 8 — тазики ног самца, 9 — генитальный орган самки,
10 — генитальная пластинка самца, 11 — голень и лапка ноги I; 12—14 — Unionicola
figuralis (Koch, 1836): 12 — тазики ног, 13 — педипальпа, 14 — генитальные пластин-
ки; 75, 16 — Neumania papillosa (Soar, 1902): 75 — вентральная поверхность самца,
16 — генитальный орган самки.
1-16 — по: Тузовский, 1997.
184
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
III. Вентральные щетинки и штифт на голени педипальп обычно сидят на
крупных буграх или отростках......................................
...........................Unionicola Haldeman, 1842 (табл. 8, 12-14).
2(1). Передние тазики с широкими длинными аподемами. Шов между тазиками
III—IV обычно полный и перпендикулярен к их медиальному краю. Вен-
тральные щетинки и штифт покоятся на крошечных буграх или прямо на
поверхности голени педипальп..........Neumania Lebert, 1879 (табл. 8, 15-16).
Семейство PIONIDAE
Очень богатая родами и видами группа клещей, обитающих в разного рода не-
проточных и слабопроточных водоёмах^
Ключ для определения родов
1(2). Покровы с сосочками. Гипостомальная пластинка с хорошо выраженным
рострумом. Ротовой аппарат прикреплён к эластичной трубке, которая мо-
жет выпячивать его вперёд........................Nautarachna Moniez, 1888.
В России 1 вид N. crassa (Koenike, 1908) (табл. 9, 1, 2).
2(1). Покровы без сосочков. Гипостомальная пластинка без рострума. Ротовой
аппарат без эластичной трубки и не может выпячиваться вперёд.
3(10). Наружные гениталии с 3 парами присосок.
4(5). Педипальпы и ноги очень длинные. Самцы на заднем конце тела имеют
палочковидный петиолус, состоящий из 3 члеников.........................
.............................Hydrochoreutes Koch, 1837 (табл. 9, 3-5).
5(4). Педипальпы и ноги обычных размеров. Самцы без петиолуса.
6(7). Передняя пара ног короткая, с сильно вздутым концевым члеником, снаб-
жённым крупными коготками. Швы между тазиками III—IV и задний край
тазиков IV направлены под острым углом к продольной оси тела. Задние
ноги самцов без вторичных отличий.....................Wettina Piersig, 1892.
В России 1 вид W. podagrica (Koch, 1837) (табл. 9, 6-8), обитает преимущественно
в проточных водоёмах.
7(6). Передняя пара ног обычных размеров, без увеличенных концевых члени-
ков и коготков. Швы между тазиками III—IV и задний край тазиков IV на-
правлены под прямым углом к продольной оси тела. Задние ноги самцов
со вторичными отличиями.
8(9). Задний выступ тазиков IV у самок хорошо выражен, с острой или прямо-
угольной вершиной и довольно длинной аподемой. Колено ноги IV сам-
цов большей частью расширено или искривлено, лапка IV прямая............
.....................................Fiphys Koch, 1836 (табл. 9, 9-11).
В России 15 видов. Обычны для временных водоёмов и слабопроточных зарос-
ших водотоков.
9(8). Задний выступ тазиков IV у самок сглажен, с тупоугольной или овальной
вершиной и короткой аподемой. Колено и голень ноги IV самцов несколь-
Клещи
185
Таблица 9
1, 2 — Nautarachna crassa (Koenike, 1908): 1 — вентральная поверхность самки,
2 — вентральная поверхность самца; 3-5 — Hydrochoreutes krameri Piersig, 1895:
J — вентральная поверхность самки, 4 — вентральная поверхность самца, 5 — коле-
но и голень ноги IV самца; 6-8 — Wettina podagrica (Koch, 1837): 6 — вентральная
поверхность самца, 7 — голень и лапка ноги I, 8 — генитальный орган самки; 9-11 —
Tiphys ensifer (Koenike, 1895): 9 — вентральная поверхность самки, 10— вентральная
поверхность самца, 11 — колено голень и лапка ноги IV самца.
1-11 — по: Тузовский, 1997.
186
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Таблица 10
7-3 — Pionacercus leuckartii Piersig, 1894: 1 — вентральная поверхность самки, 2 —
вентральная поверхность самца, 3 — лапка ноги IV самца; 4, 5 — Forelia variegator
(Koch, 1837): 4 — вентральная поверхность самки, 5 — вентральная поверхность сам-
ца; 6-8 — Huitfeldtia rectipes Thor, 1898: 6 — вентральная поверхность самца, 7 — гени-
тальный орган самки, 8 — педипальпа; 9 — Piona conglobata (Koch, 1836), педипальпа;
10—13 — Piona coccinea Koch, 1836:10 — генитальный орган самки, 77 — тазики III-IV
и генитальный орган самца, 72 — коготки III самца, 73 — коготок I самца.
7-73 — по: Тузовский, 1997.
Клещи 187
ко утолщены к дистальному концу, лапка IV серповидно изогнута.....
...........................................Pionacercus Piersig, 1894.
В России известно 5 видов. Наиболее распространён Р leuckartii Piersig, 1894
(табл. 10, 7-3). Обитатели непроточных водоёмов.
10(3). Наружные гениталии с многочисленными присосками.
11(12). Медиальный край тазиков IV не выражен. Задние тазики имеют более или
менее треугольную форму. Шов между тазиками III—IV и задний край та-
зиков IV направлены под острым углом к продольной оси тела.............
..................................Forelia Haller, 1882 (табл. 10, 4, 5).
12(11). Медиальный край тазиков IV хорошо развит. Задние тазики имеют форму
многоугольника. Шов между тазиками III—IV и задний край тазиков IV на-
правлены более или мейЬе перпендикулярно к продольной оси тела.
13(14). Колено педипальпы с длинной, примерно вдвое длиннее самого членика,
наружной щетинкой. Тазики и ноги у самцов без вторичных отличий........
............................................HuitfeldtiaVbQx, 1898.
1 вид Н. rectipes Thor, 1898 (табл. 10, 6-8), обитает преимущественно в непро-
точных водоёмах.
14(13). Колено педипальпы с короткой (короче или немногим длиннее самого
членика) наружной щетинкой. Задние тазики и ноги III—IV самцов со вто-
ричными отличиями........................Piona Koch, 1842 (табл. 10, 9-13).
Род очень богат видами, которые характеризуются эвритопностью, эвритермно-
стью и заселяют как непроточные (озёра, болота, пруды, водохранилища), так и
слабопроточные водоёмы. Многие виды достигают высокой численности. В Рос-
сии известно свыше 20 видов.
Семейство MIDEOPSIDAE
Довольно богатая родами и видами группа клещей. В России пока найдено
4 рода. Обитатели непроточных и слабопроточных водоёмов.
Ключ для определения родов
1(2). Тазики ног образуют 4 группы (тазики I не слиты между собой, хотя у
самцов они нередко тесно сближены). Общего вентрального щита нет.
Голень педипальпы с 2 крупными шиловидными щетинками...................
.............................................Momonia Halbert, 1906.
В России 2 вида, наиболее распространён М. falcipalpis Halbert, 1906 (табл. 11,
1, 2).
2(1). Тазики I слиты между собой и расположены на общем вентральном щите.
Вентральные щетинки на голени педипальпы тонкие волосовидные.
3(4). Голень педипальпы короткая толстая, короче двух предыдущих члеников,
вместе взятых, с вентральным бугром, несущим волоски. Генитальных
присосок 3 пары...................................Mideopsis Neuman, 1880.
188
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Таблица 11
1,2 — Momonia falcipalpis Halbert, 1906: 1 — вентральная поверхность самки, 2 —
педипальпа; 3,4 — Mideopsis orbicularis (Muller, 1776): 3 — вентральная поверхность
самца, 4 — педипальпа; 5-7 — Midea orbiculata (Muller, 1776): 5 — вентральная по-
верхность самца, 6 — педипальпа, 7 — генитальный орган самки.
1-7 — по: Тузовский, 1997.
В России 2 вида, наиболее распространён М. orbicularis (Muller, 1776) (табл. 11,
3, 4).
4(3). Голень педипальпы длинная тонкая, не короче двух предыдущих члени-
ков, без вентрального бугра. Генитальные присоски довольно многочис-
ленны — 8-14 пар.........................................Midea Bruzelius, 1854.
В России 1 вид М. orbiculata (Muller, 1776) (табл. 11, 5-7).
Семейство ATURIDAE
Богатая родами и видами группа клещей. В России зарегистрировано 10 ро-
дов, наиболее распространены из них Brachypoda, Neobrachypoda, Ljania, Axo-
nopsis, Aturus, Kongsbergia. Остальные встречаются редко. Обитают преиму-
щественно в разного рода водотоках, но нередко встречаются в непроточных
водоёмах и подземных водах.
Клещи
189
Таблица 12
1,2 — Brachypoda versicolor (Muller, 1776): 1 — вентральная поверхность самца, 2 —
вентральная поверхность самки; 3-5 — Neobrachypoda ekmani (Walter, 1911): 3 — вент-
ральная поверхность самца, 4 — вентральная поверхность самки, 5 — педипальпа;
6—Ljania macilenta Koenike, 1908, вентральная поверхность самки; 7 — Axonopsis sp.,
вентральная поверхность самки; 8, 9 — Aturus natangensis Protz, 1900: 8 — вентраль-
ная поверхность самки, 9 — вентральная поверхность самца; 10-11 — Kongsbergia
vshivkovae Tuzovskij, 1990: 10 — вентральная поверхность самки, 11 — педипальпа.
1-11 — по: Тузовский, 1997.
190 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Ключ для определения родов
1(8). Наружные гениталии с 3^4 парами присосок.
2(5). Бедро педипальпы с вентральным бугром.
3(4). Наружные гениталии с 3 парами присосок. Четвёртый и пятый членики
ног IV самцов видоизменены.......................Brachypoda Lebert, 1879.
В России 2 вида. Наиболее распространён В. versicolor (Muller, 1776) (табл. 12,
1, 2).
4(3). Наружные гениталии с 4 парами присосок. Задняя пара ног самцов без
вторичных отличий............................Neobrachypoda Koenike, 1913.
1 вид N. ekmani (Walter, 1911) (табл. 12, 3-5). Озёрная форма.
5(2). Бедро педипальпы без вентрального бугра.
6(7). Границы всех танков, расположенных на общем вентральном щите, чётко
обозначены. Тазики IV с заднемедиальной угловатой выемкой.............
......................................Ljania Thor, 1898 (табл. 12, 6).
7(6). На общем вентральном щите намечены контуры лишь тазиков I—III, грани-
цы тазиков IV не обозначены............Axonopsis Piersig, 1893 (табл. 12, 7).
8(1). Наружные гениталии с многочисленными присосками.
9(10). Генитальные присоски довольно крупные, расположены обычно узкой по-
перечной лентой в 1 ряд. Бедро педипальпы с 1 бугром..................
.................................Aturus Kramer, 1875 (табл. 12, 8, 9).
10(9). Генитальные присоски очень мелкие и многочисленные, образуют не-
сколько поперечных рядов в виде широкой ленты. Бедро педипальпы с
несколькими буграми................Kongsbergia Thor, 1899 (табл. 12, 10, 11).
В России известно 3 вида. Реофилы.
HALACARIDAE
Галакариды — небольшая группа клещей, обитающих исключительно в во-
дной среде. Основная масса видов галакарид — морские организмы, однако су-
ществует несколько таксонов, в ранге от подсемейства до вида, представители ко-
торых обитают исключительно в пресных водах.
Тело галакарид, длиной 0,15-1,7 мм, округлой, эллиптической или ромбо-
видной формы, обычно сильно уплощено в дорсо-вентральном направлении
(табл. 13, 1, 2). На спинной стороне обычно имеются 4 склеротизированных
пластинки: передняя дорсальная, пара окулярных и задняя дорсальная. В слу-
чае, если имеются глаза, окулярные пластинки, как правило, имеют связанные с
ними линзовидные структуры (роговицы). Число спинных пластинок может быть
увеличено за счёт появления мелких дополнительных склеритов или разделения
основных, либо уменьшено в результате их слияния или редукции. На брюшной
стороне имеются: передние эпимеральные пластинки, связанные с основани-
ями двух первых пар ног, обычно слитые в единый щит; задние эпимеральные
пластинки, связанные с основаниями задних пар ног, и, обычно единая, генито-
Клещи
191
анальная пластинка. Все склеротизированные участки разделены зонами тонкой
мембранозной кутикулы, строение поверхности которой может использоваться
для различения некоторых видов. Как на пластинках, так и на участках тонкой
кутикулы располагаются туловищные щетинки, число и расположение которых
используется в систематике.
С расположенным на генито-анальной пластинке половым отверстием обычно
связана пара генитальных склеритов (створок) и половые присоски. Анальное от-
верстие обычно расположено субтерминально или терминально на заднем конце
тела, иногда с ним связана пара анальных склеритов.
На переднем конце тела галакарид находятся ротовые органы, объединённые
в гнатосому (гнатему). Гнатосома состоит из основания, с расположенным вен-
трально гипостомом (Рострумом), парные лопасти которого образуют желоб, в
котором движутся двучлениковые колющие хелицеры. Также к основанию гнато-
сомы прикрепляется пара пальп (педипальпы). Пальпы обычно состоят из 4 чле-
ников (Р1-Р4), форма, относительная длина и особенности состава и расположе-
ния щетинок которых играют важную роль в систематике галакарид. В некоторых
группах число члеников пальп может быть уменьшено за счёт слияния или недо-
развития.
Ноги взрослых галакарид (I-IV) состоят из 6 члеников. Все членики ног несут
щетинки, которые могут быть шиловидными, оперенными или ветвистыми. При-
знаки, связанные с составом, строением и расположением щетинок конечностей
играют исключительно важную роль в систематике группы. К вершине лапки
каждой ноги прикреплены 2 латеральных коготка. Для некоторых видов характер-
но наличие также непарного медиального коготка, выраженного в различной сте-
пени. Латеральные коготки могут быть гладкими, или могут нести дополнитель-
ные образования: гребенчатость, состоящую из шипиков, расположенных обычно
на внутреннем крае коготка, и придаточный зубец, расположенный у вершины ко-
готка на наружном крае.
Постэмбриональное развитие галакарид включает несколько стадий. Из яйца
выходит личинка, имеющая лишь 3 пары пятичлениковых ног. За личинкой сле-
дует протонимфальная стадия, которая, как и все последующие, имеет 4 пары ног,
но отличается от них тем, что ноги IV пары состоят лишь из 5 члеников. За про-
тонимфой могут следовать одна или две нимфальные стадии (дейтонимфа и три-
тонимфа соответственно), обычно отличающиеся от половозрелой стадии мень-
шим числом половых присосок и щетинок конечностей. Особенности строения
туловищных склеритов у преимагинальных стадий обычно близки к наблюдае-
мым у взрослых форм, однако могут наблюдаться и различия (у видов, для имаги-
нальных стадий которых характерны слияние или редукция склеритов).
Гал акариды обитают исключительно в бентосе различных водоёмов (случай-
но могут быть встречены в планктонных пробах). Большинство видов являются
хищниками, поедающими различных мелких беспозвоночных. Европейские виды,
встречаемые в жаберной полости речных раков, видимо являются комменсалами.
Клещи
193
Основные пособия по определению
Соколов И.И. 1952. Водяные клещи — Halacarae. Фауна СССР. М.; Л. 201 с.
Green J., Macquitty М. 1987. Halacarid Mites. Synopses of the British Fauna, 36. London.
178 p.
Ключ для определения подсемейств и родов
1(2). Передняя и задняя дорсальные пластинки слиты, окулярные пластинки
отсутствуют. Вентральные пластинки объединены в общий щит. Пальпы
короткие........................................Ropohalacarinae Bartsch, 1989.
1 род Ropohalacarus с единственным видом R. uniscutatus (Bartsch, 1982) (табл. 13,
3-5). Известен из пресных вод Северной Америки, Германии. В России R. cfr.
uniscutatus обнаружен в водо^ах Ленинградской области.
2(1). Передняя и задняя дорсальные пластинки не слиты. Окулярные пластинки
обычно имеются. Пальпы удлинённые.
3(10). Пальпы прикреплены к гнатосоме дорсально, расстояние между их осно-
ваниями не превышает ширины Р1.
4(5). Длина рострума почти равна длине пальп, вместе они образуют хвата-
тельный аппарат. Коготки ног I со слабо развитой гребенкой из мелких
зубчиков..........................................Lohmannellinae Viets, 1927.
В пресных водах встречаются представители рода Porolohmannella Viets, 1933.
В России 1 вид — Р. violacea (Kramer, 1879) (табл. 13 6-8). Реки, озёра, сфагновые
болота; встречен на жабрах речных раков.
5(4). Рострум короче пальп, они не образуют хватательного органа. Коготки ног
I с развитой гребенчатостью.....................Limnohalacarinae Viets, 1927.
6(7). Коготки ног I дистально не расширены.....Limnohalacarus Walter, 1917.
L. wackeri (Walter, 1914) (табл. 14 1-4) широко распространён в Европе; болоти-
стые лужи, пруды, озёра; L. и/. astacicolus Viets, 1927 описан из жаберной полости
речного рака.
Таблица 13
1,2 — внешний вид Halacaridae: 1 — вентрально, 2 — дорсально; 3 — Ropohalacarus
uniscutatus (Bartsch, 1982), гнатема вентрально; 4, 5 — Ropohalacarus cf. uniscutatus'.
4 — вентральная поверхность самки, 5 — дорсальная поверхность самки; 6-8 — Po-
rolohmannella violacea (Kramer, 1879): 6 — гнатема латерально, 7 — дорсальная по-
верхность самки, 8 — вентральная поверхность самки.
АН — анальное отверстие, ГА — генито-анальная пластинка, ГО — половое отверстие,
ЗД — задняя дорсальная пластинка, ОК — окулярные пластики, П — педипалыты, ПД —
передняя дорсальная пластинка, ПЭ — передняя эпимеральная пластинка. 1-6 — членики
ног.
1-3 — по: Bartsch, 1989; 4, 5 — по: Туманов, 1997; 6--по: Teschner, 1969; 7, 8 — по:
Abe, 1990.
194
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Таблица 14
1-4 — Limnohalacarus wackeri (Walter, 1914): 1 — дорсальная поверхность самца,
2 — вентральная поверхность самца, 3 — вершина ноги I, 4 — педипальпа; 5-8 —
Parasoldanellonyx parviscutatus (Walter, 1917): 5 — дорсальная поверхность, 6 —
вентральная поверхность, 7 — педипальпа, 8 — нога I; 9-12 — Soldanellonyx chap-
puisi Walter, 1917: 9 — дорсальная поверхность самки, 10 — вентральная поверхность
самки, 11 — гнатема латерально, 12 — нога I.
1—3 — по: Green & Macquitty, 1987; 4, 7, 8-12 — по: Соколов, 1952; 5, 6 — по: Viets, 1959.
Клещи
195
Таблица 15
/, 2 — Lobohalacarus weberi (Romijn et Viets, 1924): 1 — дорсальная поверхность,
2 — вентральная поверхность; 3, 4 — Porohalacarus alpinus (Thor, 1910): 3 — дор-
сальная поверхность самки, 4 — вентральная поверхность самки.
/, 2 — по: Green & Macquitty, 1987; 3, 4 — по: Morselli & Mari, 1979.
7(6). Коготки ног I дистально с дисковидным расширением, по периметру ко-
торого сидят зубцы, направленные проксимально, параллельно стержню
коготка.
8(9). Третий и пятый членики ног I утолщены, значительно шире четвёртого
членика. В области сочленения Р1 и Р2 пальпы изогнуты под прямым
углом...........................................Parasoldanellonyx 1929.
Р parviscutatus (Walter, 1917) (табл. 14, 5-5) в России известен из водоёмов Коль-
ского полуострова и из Байкала.
9(8). Третий и пятый членики ног I не утолщены, почти такой же толщины, что
и четвёртый членик. В области сочленения Р1 и Р2 пальпы не изогнуты
под прямым углом....................................Soldanellonyx Walter, 1917.
Из пресных вод России известны 5. monardi Walter, 1919 и 5. chappuisi Walter,
1917 (табл. 14, 9-12).
10(3). Пальпы прикреплены к гнатосоме латерально, расстояние между ними
превышает ширину Р1.
11(14). РЗ с волосовидной или шиловидной щетинкой.....Halacarinae Viets, 1927.
12(13). Все вентральные пластинки слиты. Передний край спинной поверхности
вытянут в шиловидный отросток.......................Lobohalacarus Viets, 1939.
L. weberi (Romijn et Viets, 1924) (табл. 15, 1, 2) и L. w. quadriporus (Walter, 1947)
широко распространены в Европе; В России L. w. quadriporus известен из озера
на о. Средний (Белое море).
13(12). Вентральные пластинки не слиты.............Porohalacarus Thor, 1923.
196
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
R alpinus (Thor, 1910) (табл. 15, 3, 4) широко распространён в Европе; Россия —
р. Тверда близ Твери, Днепр, мелкие водоёмы Северо-Запада, сильно распреснён-
ный участок финского залива в районе Лужской губы. Эвритопный вид, встреча-
ется в жаберной полости речных раков.
14(11). РЗ без щетинки...............................................
.....Copidognathinae Bartsch, 1983, род Copidognathus Trouessart, 1888.
Преимущественно морской род, из пресных вод Европы известны С. tectiporus
(Viets, 1935) и С. dactyloporus Benfatti, Mari et Morselli, 1989. В России не обна-
ружены.
ПАНЦИРЩДЕ КЛЕЩИ
Панцирные клещи (Orobatida) представляют собой, по-видимому, сборную
группу клещеобразных хелицеровых, обладающих во взрослом состоянии скле-
ротизированным панцирем. Из более чем 7000 описанных видов лишь около 100
можно рассматривать как собственно гидробионтных, хотя известно, что многие
виды орибатид способны переносить длительное затопление и даже откладывать
под водой яйца (Rostrozetes ovulurri). Представители нескольких близких семейств
(Fortuyniidae, Podacaridae, Ameronothridae) населяют морскую супра- и эули-
тораль. Панцирные клещи чрезвычайно широко распространены в водоёмах со
стоячей и проточной водой, где входят в состав мейоэпифитона, будучи связаны
с макрофитами и скоплениями нитчатых водорослей. Также они встречаются в
болотах и некоторых других специфических местообитаниях (заполненные водой
дупла деревьев и т.д.). Известны ассоциации с мшанками.
Размеры пресноводных орибатид не превышают 1 мм. Тело отчётливо разде-
лено сеюгальной бороздой на передний (протеросома) и задний (гистеросома) от-
делы. Дорсальная часть протеросомы покрыта щитом (продорсумом), передняя
часть которого образует рострум. На продорсуме могут быть продольные греб-
ни (ламеллы), часто со свободными дистальными концами — кусписами и пере-
мычкой (трансламеллой), другие структуры, например, туберкулы (возвышения).
На продорсуме располагается пара трихоботрий — органов, служащих для вос-
приятия колебаний. Основания трихоботрий погружены в чашевидные ботридии.
У гидробионтных орибатид трихоботрии могут быть щетинковидными, колбовид-
ными, либо отсутствовать. Кроме трихоботрий, на продорсуме имеется еще 4-5
пар щетинок. Спинной щит, или нотогастр, более или менее явственно отделён
с боков и сзади от остальной части тела циркумгастральной бороздой. Спереди
по бокам нотогастра у взрослых клещей нескольких семейств имеются крыловид-
ные разрастания (птероморфы), которые могут загибаться на вентральную сторо-
ну. Они обычно неподвижные, но у некоторых видов способны прижиматься к
боковым сторонам тела. По бокам нотогастра у большинства орибатид имеется
пара опистосомальных желёз, открывающихся наружу порами, а также несколь-
ко пар щелевидных органов чувств. В нескольких родах гидробионтных орибатид
Клещи
197
(Hydrozetes, Aquanothrus) спереди на нотогастре у взрослых клещей вторично раз-
вивается глаз (лентикулюс). На вентральной стороне выделяют три отдела: гнато-
сому, эпимеральную область (ограниченную коксостернитами (эпимерами) ног),
аногенитальную область (позади кокс четвёртой пары ног). Генитальные и аналь-
ные крышки несут генитальные и анальные щетинки, соответственно.
Комплекс ротовых частей помещается в камеростоме и включает хелицеры,
оканчивающиеся клешней, состоящей из дорсального неподвижного и вентраль-
ного подвижного пальцев; 3-5-члениковые пальпы и гипостомальную пластинку,
имеющую одну пару рутеллумов, представляющих собой видоизмененные кок-
сальные щетинки.
Ноги состоят из 5 свободных члеников: вертлуга, бедра, колена, голени, лап-
ки. У многих клещей Вертлуги I и II очень малы. Предлапка с одним или тремя
(очень редко двумя) коготками. Количество, расположение и строение щетинок
(хетотаксия) чрезвычайно важны для видовой диагностики.
Активно плавать ни один из известных видов панцирных клещей не спо-
собен. Большинство видов ползает по водной растительности, некоторые спо-
собны передвигаться и по поверхностной плёнке воды (Heterozetes palustris),
другие — под ней (род Hydrozetes). Расселение этих клещей по суше маловеро-
ятно. У большинства видов орибатид имеется церотегумент — гидрофобная
оболочка, удерживающая припокровный слой воздуха, которая формируется це-
ментным и восковым слоями эпикутикулы и имеется не только у взрослых, но и
у ювенильных фаз, что может рассматриваться как адаптация к условиям пере-
менной влажности. Пластрон — слой воздуха, удерживаемый с помощью спе-
циальных кутикулярных микроструктур и обеспечивающий газообмен, у гидро-
бионтных орибатид развит очень хорошо и во многих случаях (роды Hydrozetes,
Aquanothrus) его локализация видима при малых увеличениях стереомикроскопа.
Видимо, пластрон также служит источником для формирования внутрикишечно-
го воздушного пузыря (пузырей) у клещей рода Hydrozetes, благодаря чему про-
исходит их «парение» (левитация) в воде. Это обеспечивает эффективное рассе-
ление клещей.
Почти все гидробионтные орибатиды размножаются партеногенетически (те-
литокия). Самцы либо неизвестны, либо составляют крайне незначительный про-
цент в популяции. Обычно у панцирных клещей половой диморфизм выражен
слабо. Онтогенез включает фазы яйца, предличинки, личинки, прото-, дейто- и
тритонимфы, имаго. Яйца обычно откладываются в аэренхиму водных растений,
в пустоты стеблей, в пазухи листьев и другие укромные места. Известны случаи
живорождения (вивипария) с выходом личинок (Trimalaconothrus). Ювенильные
фазы ряда видов (р. Hydrozetes) ведут полускрытый образ жизни, вбуравливаясь
в листья и стебли водных растений или используя естественные укрытия. У нимф
заднекрайние щетинки превращены в длинные нити, имеющие, видимо, псевдо-
церкальную функцию.
198
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Основные пособия по определению
Гиляров М.С. (ред.). 1975. Определитель обитающих в почве клещей. Sarcoptiformes. М.
491 с.
Balogh J., Mahunka S. 1983. Primitive oribatids of the Palaearctic region. Budapest. 372 p.
Ключ для определения семейств и родов
1(6). Генитальное и анальное отверстия не разъединены участком вентрального
щита; если разъединены, то незначительно: менее чем на треть длины ге-
нитального отверстия.
2(3). Предлапка с 1 коготком...................Malaconothridae Berlese, 1916,
Malaconothrus Berlese, 19Q$ (табл. 16, 7).
В пресных водах европейской части России 3 вида. Болота, заливные луга, при-
брежные участки водоёмов.
3(2). Предлапка с 3 коготками.
4(5). Окраска светло-жёлтая, сероватая или беловатая.......................
............Malaconothridae Berlese, 1916, Trimalaconothrus Berlese, 1916
(табл. 16, 2).
В пресных водах европейской части России 4 вида. Болота, прибрежные участки
водоёмов.
5(6). Окраска светло-коричневая............................................
..Trhypochthoniidae Willmann, 1931, Trhypochthoniellus Willmann, 1928.
В пресных водах европейской части России 1 вид: Т. setosus Willmann, 1928
(табл. 16, 3).
6(1). Генитальное и анальное отверстия широко разъединены участком вент-
рального щита.
7(8). Циркумгастральная борозда не отделяет нотогастр и вентральный щит в
задней части цилиндрического туловища. Продорсум в задней части несёт
пару туберкул в форме зубца или ряда зубцов с общим основанием............
........................................Nanhermanniidae Sellnick, 1928.
Клещи этого семейства населяют очень влажные субстраты, обычны в болотах; в
открытых водоёмах не встречаются.
8(7). Нотогастр чётко отделён от вентрального щита циркумгастральной бо-
роздой. Тело не цилиндрическое. Туберкулы в задней части продорсума
отсутствуют.
9(10). Плечевые выступы нотогастра (птероморфы) не развиты.................
........................................Hydrozetidae Grandjean, 1954.
Наиболее хорошо изученное семейство пресноводных орибатид. Содержит один
род: Hydrozetes Berlese, 1902 (табл. 16, 4). Взрослые клещи размером 450-700 мкм.
Распространены всесветно. Обычные обитатели пресных вод: прибрежной зоны
озёр, рек, а также болот. В аквариумах. С территории России известно 7 видов.
10(9). Птероморфы имеются.
11(16). Птероморфы неподвижные.
Клещи
199
12(13). Птероморфы загибаются лцтерально. Трихоботрии колбовидные (могут
обламываться), либо редуцированные до щетинки. Бедро и вертлуг IV без
вентральных листовидных расширений. Предлапки всех ног с 3 когот-
ками.............................................Limnozetidae Grandjean, 1954.
В семействе 1 род: Limnozetes Hull, 1916 (табл. 16,5, 6). Обычны в сфагновых боло-
тах, прибрежье озёр. Достоверно зарегистрированы на территории России 4 вида.
Таблица 16
1 — Malaconothrus gracilis van der Hammen, 1952, общий вид дорсально; 2 — Trimala-
conothrus foveolatus Willmann, 1931, общий вид дорсально; 3 — Trhypochthoniellus
setosus Willmann, 1928, общий вид дорсально; 4 — Hydrozetes parisiensis Grandjean,
1948, общий вид дорсально; 5, 6 — Limnozetes onondaga Behan-Pelletier, 1989: 5 —
общий вид дорсально, 6 — трихоботрия; 7 — Heterozetes palustris (Willmann, 1917),
общий вид дорсально; 8 — Zetomimus furcatus (Pearce et Warburton, 1906), общий вид
дорсально.
1,2 — no: Balogh & Mahunka, 1983; 3 — no: Kuriki & Aoki, 1989; 4 — no: Grandjean, 1948;
5 — no: Behan-Pelletier, 1989; 6 — по: Толстиков, 1997; 7, 8 — по: Шалдыбина, 1975.
200 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
13(12). Птероморфы не загибаются латерально. Трихоботрии веретеновидные.
Бедро и вертлуг IV с вентральными листовидными расширениями. Пред-
лапки I с 1 коготком..............................Zetomimidae Shaldybina, 1966.
14(15). Предлапки I однокоготковые, II-IV — трехкоготковые...................
............................................Heterozetes Willmann, 1917.
В фауне России 1 вид: Н. palustris (Willmann, 1917) (табл. 16, 7). Европейская
часть России, Западная Сибирь.
15(14). Предлапки I, II — однокоготковые, III, IV — трехкоготковые...........
...................................................Zetomimus Hull, 1916.
В фауне России 1 вид: Z. furcatus (Pearce et Warburton, 1906) (табл. 16, 8). Европей-
ская часть России, Западная Сибирь.
16(11). Птероморфы подвижные.......................Galumnidae Grandjean, 1936.
Некоторые виды семейства (например Orthogalumna terebrantis) могут рассматри-
ваться как пресноводные. На территории России не зарегистрированы.
ОСТРАКОДЫ
Ракушковые рачки (класс Ostracoda) широко распространены во всех типах
пресноводных водоёмов. Мировая фауна пресноводных остракод насчитывает
около 2000 видов из более чем 200 родов. Наибольшее число видов описано для
Палеарктического региона.
Остракоды — типичные представители мейобентоса водных экосистем. Мно-
гочисленны они также среди зарослей водной растительности. Остракоды обита-
ют как в постоянных, так и во временных водоёмах, в поверхностных и подзем-
ных водах. Размеры большинства видов пресноводных остракод не превышают
1-1,5 мм, однако имеются формы больших размеров (до 3,5 и более мм). Несмот-
ря на повсеместное распространение, эта группа водных беспозвоночных про-
должает оставаться одной из наименее изученных среди пресноводных низших
ракообразных, хотя их численность и биомасса часто достигает весьма высоких
величин. Они играют существенную роль в процессах трансформации вещества
и энергии в водоёмах, являясь, в частности, кормовым ресурсом для многих ви-
дов рыб, особенно для молоди. Остракоды являются уникальными биоиндикато-
рами состояния не только современных водных экологических систем, но и усло-
вий далеких эпох, будучи надежными элементами стратиграфии.
Несегментированное тело остракод, подразделяемое на головной отдел и ту-
ловище, заключено в двустворчатую раковинку с латерально расположенными
створками (рис. 1). Створки раковинки образованы двойной складкой кожи, сво-
бодно покрывающей тело рачка, кроме места прикрепления на границе головно-
го и туловищного отделов. Наружный листок этого кожного образования с двух
сторон покрыт обызвествленной кутикулой (рис. 1). Центральные части вну-
треннего листка не обызвествлены. Зона соединения наружного и внутреннего
листка называется зоной сращения. Между уплотненной и неуплотненной ча-
стями внутреннего листка находится, как правило, хорошо заметная (особенно с
внутренней стороны створки) граница, называемая внутренним краем створки.
Проксимальная граница зоны сращения (линия сращения) может совпадать с вну-
тренним краем створки, однако на переднем и заднем концах створок внутренний
край расположен намного ближе к центру створки.
Зону сращения пронизывают простые или разветвлённые поровые канальца,
которые могут заканчиваться щетинками или бородавчатыми возвышеньицами.
Свободные края створок часто снабжены особой каемкой кожи, покрытой тонким
слоем непропитанного известью хитина — кутикулярной каемкой. Она может
быть расположена как на наружной границе зоны сращения, так и проксимально
от неё (рис. 1).
Места прикрепления к створкам замыкающего мускула хорошо заметны в
виде отпечатков замыкающего мускула. Впереди отпечатков замыкающих мышц
расположены 2 отпечатка мандибулярных мышц. Сверху створки соединены эла-
202
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
V»
ктракоды 203
Яичной связкой. Как правило, на створках в этом месте развиты различного рода
шступы, зубчики и углубления, формирующие замок раковинки. Створки редко
ывают совершенно симметричными. Чаще наблюдается асимметрия различных
Встей створок, которая является важным систематическим признаком. Поверх-
ость створок может быть гладкой или скульптурированной различного рода
дублениями, шипами, зубцами и бугорками.
В пределах головного отдела остракод различают парные: первые антенны
штеннулы), вторые антенны, мандибулы и максиллы (рис. 2). Одноветвистые
ервые антенны состоят из 4-7 члеников. Три терминальных членика вооружены
ибо длинными оперенными щетинками (плавающие формы), либо эти щетинки
олее-менее укорочены и неоперены (ползающие и роющие формы). У предста-
ителей подотрядов Cytherocopina и Darwinulocopina эти щетинки, как правило,
оготковидны.
Двухветвистые вторые антенны состоят из 2-членистого протоподита, 4-чле-
истого эндоподита и экзоподита, который достигает значительной длины только
Cytherocopina, а у Cypridocopina и Darwinulocopina редуцирован до малозамет-
ен чешуйки с несколькими (2-3) щетинками (рис. 2).
Мандибулы и максиллы (табл. 18) остракод состоят из жевательной части
протоподита), щупальца (эндоподита) и жаберного придатка (экзоподита).
Строение туловищной ножки первой пары связано с её различным функ-
(иональным использованием (движение, питание, размножение) и различает-
to. 1.1-3 — общий план строения и расположения конечностей у остракод из подо-
рядов Cypridocopina (7 — Herpetocypris reptans (Baird, 1835)), Darwinulocopina (2 —
)arwinula stevensoni (Brady & Robertson, 1870)) и Cytherocopina (3 — Limnocythere
anctipatricii Brady & Robertson, 1869); 4 — Darwinula stevensoni (Brady & Robert-
юп, 1870), раковинка сверху; 5-7 — фурки у Cypridocopina (5 — Candona caudata
(aufinann, 1900, 6 — рудиментарная фурка у рода Potamocypris} и Cytherocopina (7 —
Imnocythere inopinata (Baird, 1850)); 8 — эякуляционный орган; 9 — поперечный
>азрез через раковинку (схематично); 10 — Eucypris lilljeborgi (G.W. Muller, 1900),
левая створка самки сбоку.
[—первые антенны; II — вторые антенны; III — мандибулы; IV — максиллы; V — туло-
вищные конечности первой пары; VI — туловищные конечности второй пары; VII — ту-
ловищные конечности третьей пары; VIII — фурка; IX — жаберный придаток; X — кау-
дальный придаток; XI — копулятивный орган; XII — щупальце мандибуллы. СТ — ствол,
ЗЩ — задняя щетинка, ЗК (Щ) — задний коготок (щетинка), ПК (ак) —передний (апи-
кальный) коготок, ПЩ — передняя щетинка, ГП — генитальный придаток. 3 — замок,
ОНЛ — обызвествленный наружный листок, ЗМ — замыкающие мускулы, ММ — ман-
дибулярные мышцы, ОММ — отпечаток мандибулярных мышц, М — мандибула, НВЛ —
необызвествленный внутренний листок, ОВЛ — обызвествленный внутренний листок,
ВКС — внутренний край створки, ЛС — линия сращения, КК — кутикулярная каемка,
ЗС — зона сращения, Л С — линия сращения.
1-4,9,10 — по: Henderson, 1990; 5-7 — по: Бронштейн, 1947; 8 — по: Tressler, 1959.
21
23
24
16
Оопракоды
205
а у Cytherocopina, Cypridocopina и Darwinulocopina. У Cytherocopina все три
юры туловищных ножек ходильные (рис. 1, 2) сходны по строению и состоят из
2-3-членистого эндоподита, одной или нескольких оперенных щетинок, являю-
щихся рудиментом жаберного придатка (экзоподита) и протоподита. У самцов
Cytherocopina правая ножкка первой пары может трансформироваться в хвата-
тельную, а правая ножка второй пары становится рудиментарной, при этом нож-
и левой стороны не отличаются от таковых у самок.
У Cypridocopina ножка первой пары, являясь частью приротового аппарата,
достоит из жевательной лопасти (протоподита), жаберного придатка с одним или
^сколькими лучами (экзоподита) и щупальца (эндоподита). У самцов щупальце
трансформировано в орган для захватывания самок при копуляции (хватательное
упальце) (рис. 1, 2). Асимметрия и (^рма левого и правого хватательного щу-
Вльца самцов — важный диагностический признак.
[ У Darwinulocopina ножка первой пары отличается от таковой у Cypridocopina
жсутствием коготка на эндоподите и более развитым жаберным придатком
не. 1,2).
Ножки второй и третьей пары Darwinulocopina являются ходильными, сходны
) строению и похожи на конечности второй пары у Cypridocopina. Они состо-
: из протоподита и четырёхчленистого эндоподита. Число члеников эндоподита
йпример, род Cypris) может быть сокращено до трех.
ас. 2. 1—3 — вторые антенны Ostracoda: 1 — Cypridocopina, 2 — Darwinulocopina
larwinula stevensoni (Brady & Robertson, 1870)), 3 — Cytherocopina (Cytherissa lacus-
is (G.O. Sars, 1882)); 4-6 — мандибулы Ostracoda: 4 — Cypridocopina (Cypris pubera
F. Muller, 1776), 5 — Darwinulocopina (Darwinula stevensoni (Brady & Robertson,
870)), 6—Cytherocopina (Cytherissa lacustris (G.O. Sars, 1882)); 7 — максилла; 8-13 —
отовищные конечности Cypridocopina: 8 — туловищная ножка первой пары самки (че-
остная ножка), 9 — челюстная ножка самца с хватательными щупальцами, 10, И —
и4-членистые ходильные ножки второй пары, 12 — чистильная ножка с разделен-
ии 3-м члеником, 13 — апикальная часть чистильной ножки с щипцевидным при-
пособлением; 14-16 — туловищные конечности Darwinulocopina: 14 — конечность
ервой пары (челюстная ножка), 15, 16 — туловищные ходильные ножки второй и
ретьей пары; 17-24 — туловищные конечности Cytherocopina: 17-19 — ходильные
южки первой, второй и третьей пары у самки Cytherissa lacustris (G.O. Sars, 1882),
О— ножка третьей пары самца Cytherissa lacustris (G.O. Sars, 1882), 21, 22 — левая
правая (хватательная) ножки первой пары самца, 23, 24 — левая и правая (рудимен-
арная) ножки второй пары самца.
IP — протоподит, ЭК — экзоподит, ЭН — эндоподит, ПЛЩ — плавательные щетинки,
ЧЩ — чувствительные щетинки, ЖП — жаберный придаток (экзоподит), Щ — щупальце
(эндоподит), СТ (ПР) — ствол (протоподит), ЖЛ — жевательная лопасть, ХЩ — хвата-
тельные щуп аль цы.
1-9,13-16, 17-24 — по: Бронштейн, 1947; 10—12 — по: Henderson, 1990.
206 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
4-5-членистая ножка третьей пары у Cypridocopina (рис. 1, 2) трансформиро-1
вана в так называемую чистильную ножку. Она обращена назад и вверх и служит
для очистки внутренней поверхности раковинки и тела. Её строение — важный
систематический признак.
Важными диагностическими признаками являются также строение фурки, ге-
нитального отдела тела самок (генитальные придатки) (рис. 1) и копулятивного
органа самцов (рис. 38), в строении которого выделяют наружный, внутренний и
серединный придатки.
Методы сбора и обработки
Для качественных сборов остракод пригодны любые гидробиологические
орудия лова (сачки, драги, сети, дноче^Ьатели). В ходе оценки количественного
развития остракод в донных биоценозах для взятия проб рекомендуется исполь-
зовать трубчатые и коробчатые пробоотборники и дночерпатели (первые предпоч-
тительнее, т.к. они меньшего диаметра и лучше улавливают бентическую фауну).
Для количественных сборов среди зарослей применяют пробоотборники и зарос-
лечерпатели специальных конструкций.
Для избавления от лишнего грунта и детрита отобранные пробы лучше всего
промывать через газ с диаметром ячеи не более 100-125 мкм, что позволяет поч-
ти на 100% избежать потери остракод.
Промытые пробы в «живом» или фиксированном состоянии просматривают
или под бинокуляром, или в кювете с использованием (или без использования)
лупы.
Фиксироваать остракод рекомендуется в 70-80-градусном спирте. Фиксация
формалином весьма нежелательна, т.к. происходит декальцинация раковинки
и искажение её формы, которая является важным диагностическим признаком.
Фиксация в спирте позволяет получить рачков с приоткрытыми створками, что
облегчает доступ к телу рачка для дальнейшего его препарирования. Для получе-
ния сомкнутых раковинок (это важно для точной зарисовки их формы) фиксиро-
вать рачков следует в 5-10% растворе глицирина в 80-90-градусном спирте.
Препаровку остракод лучше всего производить в глицерине или слегка разбав-
ленном водой глицерин-желатине при помощи тонких игл. Препаровка в подогре-
том разбавленном глицерин-желатине удобна, т.к. дает возможность после препа-
ровки сразу получить постоянный препарат. Как вариант может быть применена
препаровка в глицерине, с перенесением в её процессе створки и отделённые от
тела конечности в подогреваемый на предметном стекле глицерин-желатин, где
они могут быть расположены удобным образом и накрыты покровным стеклом
После охлаждения границы покровного стекла рекомендуется замазать лаком
Такой препарат может храниться очень долго. Для получения постоянных препа
ратов конечностей и раковинок можно использовать также канадский бальзам г
жидкость Фора-Берлезе. Для изготовления последней 30 частей сухого гуммиа
Остракоды
207
рабика растворяют в 50 частях (по массе) дистиллированной воды, добавляют 20
частей глицерина и 20 частей хлоралгидрата. Смесь подогревают в бюксе (баноч-
ка с притертой пробкой) на водяной бане до полного растворения, фильтруют и
охлаждают до комнатной температуры.
Основные пособия по определению
Бронштейн З.С. 1947. Ostracoda пресных вод. Фауна СССР. Ракообразные. М.; Л. Т. 2.
Вып. 1. 371 с.
Курашов Е.А. 2012. Определительные ключи остракод внутренних водоёмов Европейской
части России. М.: Т-во науч, изданий КМК. 152 с.
Meisch С. 2000. Freshwater Ostracoda of Western and Central Europe. Suesswasserdauna von
Mitteleuropa 8/3, J. Schwoerbel a^l P. Zwick, eds.: Spektrum Akademischer Verlag, Hei-
delberg, Germany. 522 p.
Sywula T. 1974. Malzoraczki (Ostracoda). Fauna Slodkowodna Polski. 24. Panstwowe Wydaw-
nictwo Naukowe. Warszawa-Poznan. 316 p.
Классификация и список родов Ostracoda
внутренних водоёмов европейской части России
Тип Arthropoda
Надкласс Crustacea Pennant, 1777
Класс Ostracoda Latreille, 1802
Подкласс Podocopa Muller, 1894
Отряд Podocopida Muller, 1894
Подотряд Cypridocopina Jones, 1901
Сем. Ilyocyprididae Kaufmann, 1900
Род Ilyocypris Brady & Norman, 1889
Сем. Cypridopsidae Kaufmann, 1900
Подсемейство Cypridopsinae Bronstein, 1947
Род Sarscypridopsis McKenzie, 1977
Род Plesiocypridopsis Rome, 1965
Род Cypridopsis Brady, 1868
Род Cavemocypris Hartmann, 1964
Род Potamocypris Brady, 1870
Сем. Cyprididae Baird, 1845
Подсемейство Cyprettinae Hartmann, 1963
Род Cypretta Vavra, 1898
Подсемейство Notodromatinae Kaufmann, 1900
Род Notodromas Lilljeborg, 1853
Подсемейство Cyproidinae Hartmann, 1963
Род Cyprois Zenker, 1854
Подсемейство Scottinae Bronstein, 1947
Род Scottia Brady & Norman, 1889
208
Определитель, зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Посемейство Hungarocypridinae Bronstein, 1947
Род Hungarocypris Vavra, 1906
Подсемейство Herpetocypridinae Kaufmann, 1900
Род Ilyodromus Sars, 1894
Род Stenocypria G.W. Muller, 1901
Род Herpetocypris Brady & Norman, 1889
Подсемейство Stenocyprinae Ferguson, 1964
Род Stenocypris Sars, 1889
Подсемейство Cyprinotinae Bronstein, 1947
Род Cyprinotus Brady, 1886
Род Heterocypris Claus, 1892
Подсемейство Cypriditfae Baird, 1845
Род Cypris Muller, 1776
Подсемейство Eucypridinae Bronstein, 1947
Род Cypricercus Sars, 1895
Род Eucypris Vavra, 1891
Род Strandesia Stuhlmann, 1888
Подсемейство Dolerocypridinae Triebel, 1961
Род Dolerocypris Kaufmann, 1900
Подсемейство Isocypridinae Rome, 1965
Род Isocypris G.W. Muller, 1908
Семейство Candonidae Kaufmann, 1900
Подсемейство Cyclocypridinae Kaufmann, 1900
Род Cypria Zenkeri, 1854
Род Cyclocypris Brady & Norman, 1889
Род Physocypria Vavra, 1897
Подсемейство Candoninae Kaufmann, 1900
Род Candona Baird, 1845
Род Fabaeformiscandona Krstic, 1972
Род Cryptocandona Kaufmann, 1900
Род Paracandona Hartwig, 1899
Род Candonopsis Vavra, 1891
Род Pseudocandona Kaufmann, 1900
Подотряд Cytherocopina Baird, 1850
Надсемейство Cytheroidea Baird, 1850
Сем. Limnocytheridae Klie, 1938
Подсемейство Metacypridinae Danielopol, 1965
Род Metacypris Brady & Robertson, 1870
Подсемейство Limnocytherinae Klie, 1938
Род Limnocythere Brady, 1868
Род Leucocythere Kaufmann, 1892
Сем. Cytherideidae Sars, 1925
Остракоды
209
Род Cytherissa G.O. Sars, 1925
Род Cyprideis Jones, 1857
Сем. Hemicytheridae Puri, 1953
Род Tyrrhenocythere Ruggieri, 1955
Сем. Leptocytheridae Hanai, 1957
Род Leptocythere Sars, 1925
Подотряд Darwinulocopina Sohn, 1988
Сем. Darwinulidae Brady & Norman, 1889
Род Darwinula Brady & Robertson in Jones 1885
КЛЮЧИ ДЛЯ ОПРЕДЕЛЕНИЯ ОСТРАКОД КОНТИНЕНТАЛЬНЫХ
ВОДОЁМОВ ЕВРОПЕЙСКОЙ ЧАСТИ РОССИИ
Ключ для определения подотрядов пресноводных
Ostracoda Latreille, 1802
1(2, 3). Все три пары туловищных конечностей у самок сходного строения —
ходильные ножки. У самцов ножки 1-й, а также 2-й пары, могут быть
превращены в хватательные, или они такие же как у самок. Экзоподит II
антенны в виде членистой щетинки.................Cytherocopina Baird, 1850.
В пресных водах европейской части России представители надсемейства Cythe-
roidea Baird, 1850.
2(1, 3). Первая пара туловищных конечностей — ногочелюсти, вторая и третья —
ходильные ножки. Фурка отсутствует............Darwinulocopina Sohn, 1988.
Одно семейство Darwinulidae Brady & Norman, 1889; в Европейской части России
один род — Darwinula Brady & Robertson in Jones 1885; один вид — Darwinula
stevensoni (Brady & Robertson, 1870) (рис. 1, 2).
Длина самки: 0,68-0,79 мм.
Обитает в постоянных водоёмах на заиленном песке, илистом грунте, среди рас-
тительности. Эвритермичен. Широко распространён.
3(1, 2). Все три пары туловищных конечностей имеют разное строение: I пара —
ногочелюсти, II пара — ходильные ножки, III пара — чистильные ножки.
Фурка имеется....................................Cypridocopina Jones, 1901.
В пресных водах европейской части России представители надсемейства Cypri-
doidea Baird, 1845.
Ключ для определения семейств и родов Cytheroidea Baird, 18S0
1(2). На апикальном членике первых антенн 4 щетинки: 3 простых и 1 чувстви-
тельная...................................Сем. Limnocytheridae Klie, 1938.
2(1). На апикальном членике первых антенн 3 щетинки: 2 простых и 1 чувстви-
тельная.
3(4). Жаберный придаток мандибул сильно редуцирован, с 2 лучами. Глаза
слившиеся.................................Сем. Leptocytheridae Hanai, 1957.
210
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 3. Leptocythere pediformis Schomikov, 1966.
1,2 — раковинки самца и самки сверху; 3 — правая створка самца сбоку; 4 — левая
створка самки сбоку; 5 — правая створка самки изнутри сбоку; 6 — вторая антенна;
7-9 — ходильные ножки; 10 — фурки; 11 — копулятивный орган.
1-6, 11 — по: Шорников, 1969; 7-10 — по: Коваленко, 1976..
Во внутренних водах европейской части России один род Leptocythere G.O. Sars,
1925 (рис. 3).
4(3). Жаберный придаток мандибул хорошо развит.
5(6). Жаберный придаток мандибул с 4-5 щетинками...........................
........................................Сем. Cytherideidae Sars, 1925.
6(5). Жаберный придаток мандибул только с одной хорошо развитой оперенной
щетинкой и 1 рудиментарной....................Сем. Hemicytheridae Puri, 1953.
Во внутренних водах европейской части России один род Tyrrhenocythere Ruggieri,
1955; 1 вид — Tyrrhenocythere donetziensis Dubowsky, 1926 (синоним Hemicythere
sicula (Brady, 1902)) (рис. 4).
Рис. 4. Tyrrhenocythere donetziensis Dubowsky, 1926.
1,2 — левая и правая створки раковинки самца; 3,4 — левая и правая створки раковинки
самки; 5 — структура поверхности створок; 6 — фурка.
М — по: Коваленко, 1976; 5, 6 — по: Daday, 1910.
Длина самки: до 0,85 мм, длина самца: до 1,00 мм.
Устья рек, пресные и осолонённые озёра и другие водоёмы.
Ключ для определения родов семейства Limnocytheridae Klie, 1938
1(2). Раковинка при взгляде сверху очень широкая и короткая. Соотношение
длины к ширине для самок около 1,25, для самцов около 1,5. Первые ан-
тенны 6-членистые.........................Metacypris Brady & Robertson, 1870.
1 вид — Metacypris cordata Brady & Robertson, 1870) (рис. 5).
Длина самки: 0,56-0,63 мм, длина самца: 0,50-0,54 мм.
Обитатель озёр. Часто в прибрежной зоне среди растительности.
2(1). При взгляде сверху раковинка более сжата латерально. Соотношение дли-
ны к ширине около 2 или более. Первые антенны 5-членистые.
212
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 5. Metacypris cordata Brady & Robertson, 1870.
7,2 — раковинки самки и самца сбоку; 3,4 — раковинки самки и самца сверху; 5 — пер-
вая антенна.
7, 2 — по: Muller, 1912; 3,4 — по: Sywula, 1974; 5 — по: Henderson, 1990.
3(4). Апикальная щетинка второго членика первых антенн значительно не дохо-
дит до дистального края предпоследнего членика и превышает длину тре-
тьего членика примерно в 2 раза..............Limnocythere Brady, 1868 (рис. 1, 6).
4(5). Апикальная щетинка второго членика первых антенн длинная и достигает
у самок основания последнего членика, а у самцов апикальной его части.
.................................................Leucocythere Kaufmmann, 1892.
Во внутренних водах европейской части России 1 вид — Leucocythere mirabilis
Kaufmmann, 1892) (рис. 7).
Длина самки: 0,78-0,81 мм, длина самца: 0,89-0,92 мм.
Дно озёр.
Остракоды
213
*ис. 6. Limnocythere inopinata (Baird, 1843).
2 — раковинки самки и самца сбоку; 3 — раковинка сверху; 4 — первая антенна; 5 —
торая антенна; 6 — ходильные ножки первой, второй, третьей пары и фурка.
-3 —по: Henderson, 1990; 4-6—по: Sywula, 1974.
Рис. 7. Leucocythere mirabilis Kaufrnmann, 1892.
/ — правая створка раковинки самки сбоку; 2 — левая створка самца сбоку; 3 — раковин
ка сверху; 4 — фурка. 1,2,4 — по: Kaufmann, 1900; 3 — по: Ekman, 1914.
214
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Рис. 8. Cytherissa lacustris (G.O. Sars, 1863).
1,2 — раковинка сбоку и сверху; 3 — мандибула; 4 — первая антенна.
По: Henderson, 1990
Рис. 9. Cyprideis torosa (Jones, 1850).
1,2 — правая и левая створки раковинки самки; 3 — левая створка раковинки самца; 4,
5 — раковинки самки и самца сверху; 6 — вторая антенна; 7 — копулятивный орган.
По: Sywula, 1974.
Остракоды
215
Ключ для определения родов семейства Cytherideidae Sars, 192S
1(2). Апикальный членик II антенн несёт 3 коготка...Cytherissa G.O. Sars, 1925.
Во внутренних водах европейской части России один вид — Cytherissa lacustris
(G.O. Sars, 1863) (табл. 2, 8).
Длина самки: 0,89-0,95 мм, длина самца: до 1,1 мм.
Обитает на дне довольно крупных озёр. Индикатор ненарушенных антропоген-
ным воздействием условий. Оксифилен.
2(1). Апикальный членик II антенн несёт 2 коготка............................
..........................................Cyprideis Jones, 1856 (рис. 9).
Ключ для определения семейств и родов Cypridocopina Jones, 1901
1(2). На поверхности* створок^имеются хорошо очерченные ячеистые углубле-
ния. Спиной край прямой. Эндоподит первой пары ног (ногочелюстей)
у самок членистый...............................Ilyocyprididae Kaufmann, 1900.
Один род — Ilyocypris Brady & Norman, 1889) (рис. 10).
Рис. 10. Ilyocypris gibba (Ramdohr, 1808).
1,2 — раковинка самки сбоку и сверху; 3 — вторая антенна; 4 — челюстная ножка самки;
5 — ходильная ножка; 6 — чистильная ножка; 7 — фурка самки; 8,9 — левое и правое
хватательные щупальца челюстных ножек самца; 10 — копулятивный орган.
1, 3 —по: Henderson, 1990; 2, 4-10 — по: Sywula, 1974.
216
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
2(1). На поверхности створок отсутствует ячеистая структура. Спиной край
дугообразно изогнут. Эндоподит первой пары ног у самок не членистый.
3(4). Фурка редуцирована. Состоит из короткого ствола и бича. На переднем, а
иногда и на заднем краях створок могут присутствовать особые колонко-
видные, радиально расположенные образования...............................
........................................Cypridopsidae Kaufmann, 1900.
4(3). Фурка хорошо развита, вооружена коготками. Колонковидные, радиально
расположенные образования на краях створок отсутствуют.
5(6). Как правило 4-й членик чистильной ножки очень маленький, и эта ко-
нечность с щипцевидной структурой. При отсутствии щипцевидной
структуры длина последнего членика вторых антенн более, чем в 3 раза
превосходит его ширину. Семевввдящие органы самцов не менее, чем с 9
розетками, либо они с многочисленными лучами, которые не формируют
правильных розеток...................................Cyprididae Baird, 1845.
6(5). Четвёртый членик чистильной ножки больших размеров, цилиндриче-
ский, вооружён 3 щетинками. Щипцевидная структура всегда отсутствует.
Семевводящие органы самцов с 7-8 хитиновыми розетками.....................
...........................................Candonidae Kaufmann, 1900.
Ключ для определения родов Cypridopsidae Kaufmann, 1900
1(4). При закрытых створках с брюшной стороны левая створка перекрывает
правую.
2(3). Плавательные щетинки вторых антенн редуцированы, едва достигают или
слегка превосходят основание предпоследнего членика вторых антенн.
При взгляде со спиной стороны раковинка удлинённая: длина отчётливо
более, чем в два раза превышает ширину ....Cavernocypris Hartmann, 1964.
Один вид — С. subterranea (Wolf, 1920) (рис. 11).
Длина самки: 0,58-0,72 мм.
Реки, ручьи, подземные воды, колодцы. Предпочитает холодные чистые воды. Из-
вестен из Европы. Нахождение в пределах России весьма вероятно.
3(2). Плавательные щетинки вторых антенн хорошо развиты. При взгляде со
спиной стороны раковинка вздутая, её ширина составляет около половины
длины.......................................Cypridopsis Brady, 1868 (рис. 12).
4(1). При закрытых створках с брюшной стороны правая створка перекрывает
левую.
5(6). Конечный членик щупальца максилл трапециевидный. Створки раковинки
сильно асимметричны: правая превосходит левую со спиной стороны...........
....................................Potamocypris Brady, 1870 (рис. 13).
6(5). Конечный членик щупальца максилл цилиндрический, не трапециевид-
ный. Створки раковинки слабо асимметричны.
7(8). Ствол фурки цилиндрический, дистально резко сужен...................
........................................Plesiocypridopsis Rome, 1965.
Остракоды
217
Один вид — Р. newtoni (Brady & Robertson, 1870). Правая створка превышает ле-
вую спереди и менее отчётливо сзади. Цвет створок коричнево-зелёный, (рис. 14).
Длина самки: 0,62-0,85 мм, длина самца: 0,62-0,70 мм.
В разнообразных стоячих и текучих пресных и солоноватых водоёмах).
8(7). Ствол фурки постепенно сужается от основания к дистальному концу..........
.........................................Sarscypridopsis McKenzie, 1977.
Один вид S. aculeata (Costa, 1847). Створки покрыты кроме обыкновенных ще-
тинок шиповидными щетинками. Раковинка тёмного голубовато-зелёного цвета
(рис. 15).
Длина самки: 0,65-0,75 мм.
Солоноватоводная форма. Встречается также и в пресных водоёмах.
Рис. 11. Cavemocypris subterranea (Wolf, 1920).
1,2 — правая и левая створки сбоку; 3 — раковинка сверху; 4 — ножка первой пары;
5 — вторая антенна; б — максилла и шипы первого жевательного придатка; 7 — хо-
дильная ножка второй пары; 8 — чистильная ножка; 9 — фурки.
1, 2, 4, 6-9 — по: Marmonier et al., 1989; 3, 5 — по: Henderson, 1990.
218
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 12. Cypridopsis vidua (O.F. Muller, 1776).
1, 2 — левая и правая створки самки сбоку; 3 — раковинка сверху; 4 — вторая антенна;
5 — челюстная ножка с жаберным придатком; 6 — ходильная ножка; 7 — чистильная
ножка; 8 — фурка.
7, 2,5,8 — по: Sywula, 1974; 3, 7 — по: Henderson, 1990; 4, 6 —по: Marmonier et al., 1989.
Остракоды
219
Рис. 13. Potamocypris pallida Alm, 1914.
1,2 — левая и правая створки самки; 3 — раковинка сверху; 4 — вторая антенна; 5 — че-
люстная ножка; 6 — терминальный членик щупальца максилл; 7 — фурка.
По: Henderson, 1990.
Рис. 14. Plesiocypridopsis newtoni (Brady & Robertson, 1870).
1,2 — раковинка самки сбоку и сверху; 3 — фурка; 4, 5 — левое и правое хватательные
щупальца самца; 6 — копулятивный орган.
1, 3 — по: Muller, 1900; 2 — по: Henderson, 1990; 4-6 —по: Sywula, 1974.
220
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 15. Sarscypridopsis aculeata (Costa, 1847).
1 — левая створка сбоку; 2 — общий вид рачка; 3 — раковинка сверху; 4 — максилла;
5 — челюстная ножка; 6 — фурка.
1-4 — по: Henderson, 1990; 5,6 — по: Muller, 1900.
Ключ для определения подсемейств и родов Cyprididae Baird, 1845
1(2). Фурка с тонким и слабым стволом дегенеративного вида, лишенная перед-
ней щетинки....................................Cyprettinae Hartmann, 1963.
Один род Cypretta Vavra, 1898; один вид С. dubiosa (Daday, 1901) (рис. 16).
2(1). Фурка хорошо развита.
3(4). Фуркальные ветви с выраженной асимметрией: правая, как правило, более
широкая и сильнее изогнута, чем левая, которая почти прямая...............
.........................................Stenocyprinae Ferguson, 1964.
Один род Stenocypris Sars, 1889 (рис. 17).
Остракоды
221
Рис. 16. Cypretta dubiosa (Daday, 1901).
1 — раковинка самки сверху; 2, 3 — правая и левая створки самки с наружной стороны;
4 — фурка; 5 — терминальная часть чистильной ножки.
По: Бронштейн, 1947.
4(3). Фуркальные ветви без выраженной асимметрии.
5(8). Первая жевательная лопасть ствола максилл с 6 шиповидными щетинками.
6(7). Щипцевидная структура на чистильной ножке имеется. Длина последнего
членика вторых антенн менее чем в 3 раза превосходит его ширину.........
........................................Cyproidinae Hartmann, 1963.
Один род Cyprois Zenker, 1854 (рис. 18).
7(6). Щипцевидная структура на чистильной ножке отсутствует. Длина послед-
него членика вторых антенн более чем в 3 раза превосходит его ширину..
...........................................Notodromatinae Kaufmann, 1900.
Один род Notodromas Lilljeborg, 1853 (рис. 19).
8(5). Первая жевательная лопасть ствола максилл с 2 шиповидными щетинками.
9(10). Туловищные конечности 2-й пары (ходильные ножки) с 2 коготками на
конце...........................................Scottinae Bronstein, 1947.
Один род — Scottia Brady & Norman, 1889); один вид — S. pseudobrowmaria
Kempf, 1971 (синоним — S. browniana (Jones, 1856) (рис. 20).
222
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 17. Stenocypris bronsteini Semenova, 2006.
1, 2 — правая и левая створки раковинки самки; 3 — вторая антенна; 4 — шипы максилл;
5 — ходильная ножка; 6 — чистильная ножка; 7, 8 — левая и правая фурки.
По: Семенова, 2006.
Длина самки: 0,65-0,85 мм, длина самца: 0,65-0,85 мм.
Ключи, родники.
10(9). Туловищные конечности 2-й пары (ходильные ножки) с 1 коготком на конце.
11(12). Спинной (задний) край фурки с 2 щетинками.........................
...................................Hungarocypridinae Bronstein, 1947.
Один род — Hungarocypris Vavra, 1906; один вид — Н. madaraszi (Orley, 1886)
(рис. 21).
Длина самки: 4,5-5,0 мм, длина самца: 4,5-5,0 мм.
Реки, ильмени, старицы, болота, прибрежные заросли озёр.
12(11). Спинной (задний) край фурки с 1 щетинкой, или щетинка отсутствует.
13(14). На поверхности створок бугорки 2-х типов: многочисленные мелкие без
волосков и более крупные с волосками.......................................
...................................Herpetocypridinae Kaufmann, 1900.
)апракоды
223
Рис. 18. Cyprois marginata (Strauss, 1821).
1,2 — левая створка самки и самца сбоку; 3 — раковинка сверху; 4 — фурка; 5 — шипо-
видные щетинки первой жевательной лопасти ствола максилл; 6 — чистильная ножка; 7,
5 — правое и левое хватательные щупальца челюстных ножек самца.
И — по: Henderson, 1990; 5-8 — по: Sywula, 1974.
224
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1
Рис. 19. Notodromas monacha (O.F. Muller, 1776).
7, 2 — левая створка самки и самца сбоку; 3, 4 — раковинка самки сверху и снизу; 5 —
вторая антенна; 6 — первая жевательная лопасть ствола максилл; 7 — чистильная ножка;
£ — фурка; 9, 10 — левое и правое хватательные щупальца самца; 11 — копулятивный
орган.
1-4, 8 — по: Henderson, 1990; 5-7, 9-11 — по: Sywula, 1974.
14(13). На поверхности створок бугорки только одного типа (окончания поровых
канальцев) с волосками или без них. Если присутствуют более мелкие
бугорки, то единично.
15(16). Передний и задний край более короткой створки с многочисленными
округлыми или зубчатыми бугорками.............Cyprinotinae Bronstein, 1947.
16(15). Округлые или зубчатые бугорки на переднем и заднем крае более корот-
кой створки отсутствуют.
17(18). Туловищные конечности второй пары 4-членистые (2 предпоследних чле-
ника сращены)......................................Cypridinae Baird, 1845.
Один род — Cypris Muller, 1776 (рис. 2, 22).
Остракоды
225
Рис. 20. Scottia pseudobrowniana Kempf, 1971.
1 — левая створка самки сбоку; 2 — правая створка раковинки самца сбоку; 3 — раковин-
ка самки сверху; 4 — ходильная ножка; 5 — фурка; б, 7 — левое и правое хватательные
щупальца самца; 8 — копулятивный орган.
1-3 — по: Henderson, 1990; 4-8 — по: Sywula, 1974.
Рис. 21. Hungarocypris madaraszi (Orley, 1886).
1 — правая створка самки сбоку; 2 — фурка.
По: Daday, 1900.
226
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 22. Cypris pubera O.F. Muller, 1776.
7, 2 — раковинка сбоку и сверху; 3 — шипы максилл; 4 — ходильная ножка; 5 — чистиль-
ная ножка; 6 — фурка.
По: Henderson, 1990.
18(17). Туловищные конечности второй пары 5-членистые (2 предпоследних чле-
ника не сращены).
19(20). На задних краях створок внутренний край слит с зоной сращения. Створ-
ки покрыты длинными волосками, на заднем краю каждой створки имеет-
ся по 2 особенно длинных волоска..................Isocypridinae Rome, 1965.
Один род — Isocypris G.W. Muller, 1908 (рис. 23).
20(19). Внутренний край створок на задних краях створок расположен прокси-
мально от линии сращения.
Остракоды
227
Рис. 23. Isocypris beauchampi (Paris, 1919).
1 — левая створка самки сбоку; 2 — раковинка сверху; 3 — вторая антенна; 4 — ходиль-
ная ножка; 5 — чистильная ножка; 6 — фурка.
1,2 — по: Henderson, 1990; 3-5 — по: Meisch, 2000; 6 — по: Sywula, 1974.
228
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 24. Dolerocypris fasciata (O.F. Muller, 1776).
7,2 — створки самки сбоку; 3 — раковинка сверху; 4 — вторая антенна; 5 — шипы мак-
силл; 6 — челюстная ножка; 7 — ходильная ножка; 8 — чистильная ножка; 9 — фурка.
1—3 — по: Sars, 1925; 4, 9 — по: Henderson, 1990; 5 — по: Sywula, 1974; 6-8 — по: Victor,
Fernando, 1979.
Остракоды
229
21(22). Передняя щетинка фурки явственно меньше Уг длины её переднего ко-
готка........................................Eucypridinae Bronstein, 1947.
22(21). Передняя щетинка фурки не менее Уг длины её переднего коготка. Коготки
фурки глубоко зазубрены......................Dolerocypridinae Triebel, 1961.
Один род — Dolerocypris Kaufmann, 1900 (рис. 24).
Ключ для определения подсемейств Candonidae Kaufmann, 1900
1(2). Плавательные щетинки на вторых антеннах имеются.....................
......................................Cyclocypridinae Kaufmann, 1900.
2(1). Плавательные щетинки на вторых антеннах отсутствуют.................
...........................................Candoninae Kaufmann, 1900.
Ключ для определения родов Herpetocypridinae Kaufmann, 1900
1(2). Задняя (спинная) щетинка фурки толстая, когтевидная.................
........................................Ilyodromus Sars, 1894 (рис. 25).
2(1). Задняя (спинная) щетинка фурки обычная, не когтевидная.
3(4). Апикальный членик щупальца максилл цилиндрический, дистально не
расширен.....................................Stenocypria G.W. Muller, 1901.
Один вид — S. fischeri (Lilljeborg, 1883) (рис. 26).
Длина самки: 1,90-2,15 мм.
Озёра и мелкие водоёмы.
4(3). Апикальный членик щупальца расширен в дистальной части..............
........................Herpetocypris Brady & Norman, 1889 (рис. 1, 27).
Ключ для определения родов Cyprinotinae Bronstein, 1947
1(2). Левая створка всегда превосходит правую спереди и со спиной стороны...
.............................................Cyprinotus Brady, 1886 (рис. 28).
2(1). Левая створка превосходит правую спереди и с брюшной стороны........
.....................................Heterocypris Claus, 1892 (рис. 29).
Ключ для определения родов Eucypridinae Bronstein, 1947
1(4). Передняя щетинка фурки явственно меньше !4 длины её переднего коготка.
2(3). На скелетном хитиновом тяже, поддерживающем фурку, в верхней прок-
симальной части хитиновые ответвления образуют характерную овальную
петлю. Фурка длинная, очень тонкая...........Cypricercus Sars, 1895 (рис. 30).
3(2). Овальная петля на скелетном хитиновом тяже, поддерживающем фурку,
отсутствует. Фурка обычного типа, довольно широкая, средней длины.........
........................................Eucypris Vavra, 1891 (рис. 1, 31).
4(1). Передняя щетинка фурки не менее !6 длины её переднего коготка. Коготки
фурки не зазубрены: голые или покрыты тонкими волосками....................
...................................Strandesia Stuhlmann, 1888 (рис. 32).
230
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1
Рис. 25. Ilyodromus olivaceus (Brady & Norman, 1889).
1,2 — левая и правая створки самки сбоку изнутри; 3 — раковинка сверху; 4 — шипы
максилл; 5 — апикальный членик щупальца максилл; 6 — чистильная ножка; 7 — конеч-
ные членики чистильной ножки с щипцевидным приспособлением; 8 — фурка; 9, 10 —
левое и правое хватательные щупальца челюстных ножек самца; 11 — копулятивный
орган.
1—3, 6-8 — по: Henderson, 1990; 4, 5 — по: Sywula, 1974; 9-11 — по: Ярвекюльг, 1959.
Остракоды
231
Рис. 26. Stenocypria fischeri (Lilljeborg, 1883).
1,2 — левая и правая створки раковинки самки сбоку; 3 — раковинка сверху; 4 — апи-
кальный членик щупальца максилл; 5 — фурка; 6 — один из шипов максилл
1-5 — по: Sywula, 1974; 6 — по: Vavra, 1891.
Рис. 27. Herpetocypris reptans (Baird, 1835).
1 — левая створка самки сбоку; 2 — раковинка сверху; 3 — вторая антенна; 4 — фурка;
5 — расположение шипиков на заднем краю фурки.
/, 2,4, 5 — по: Henderson, 1990; 3 — по: Sywula, 1974.
232
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1
Рис. 28. Cyprinotus salinus (Brady, 1868).
1,2 — раковинка сбоку и сверху; 3 — чистильная ножка; 4 — фурка.
1,2,4 — по: Sywula, 1974; 3 — по: Коваленко, 1976.
Рис. 29. Heterocypris incongruens (Ramdohr, 1808).
1 — раковинка самки сбоку; 6 — раковинка самки сверху; 3 — часть мандибулы с шипами
первой лопасти; 4 — фурка; 5, 6 — левое и правое хватательные щупальца самца; 7 —
копулятивный орган.
1, 2 — по: Henderson, 1990; 3, 4 — по: Sywula, 1974; 5-7 — по: Muller, 1900.
Остракоды
233
Рис. 30. Cypricercus affinis (Fischer, 1851).
1 — раковинка сбоку; 2 — раковинка сверху; 3 — раковинка при взгляде сзади; 4 — фур-
ка; 5 — шипы первого жевательного придатка максилл; 6,7 — левое и правое хвататель-
ные щупальца челюстных ножек самца; 8 — копулятивный орган.
1 — по: Henderson, 1990; 2-5 — по: Sywula, 1974; 6,7, 8 — по: Alm, 1915.
Ключ для определения родов Cyclocypridinae Kaufmann, 1900
1(2). Более короткая створка (чаще правая) на заднем и переднем краю (иногда
только на одном) с небольшими зубчиками с закруглёнными вершинами..
.....................................................Physocypria Vavra, 1897.
Один вид — Р fadeewi Dubowsky, 1927) (рис. 33).
Длина самки: 0,65-0,68 мм, длина самца: 0,53-0,58 мм.
Реки, старицы, озёра.
2(1). Края створок без закруглённых шипиков.
3(4). Длина последнего членика чистильной ножки равна или немного превы-
шает его ширину.................................Cypria Zenkeri, 1854 (рис. 34).
4(3). Длина последнего членика чистильной ножки не менее, чем в 2 раза пре-
вышает его ширину..................Cyclocypris Brady & Norman, 1889 (рис. 35).
234
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 31. Eucypris virens (Jurine, 1820).
1,2 — раковинка самки сбоку и сверху; 3 — вторая антенна; 4 — ходильная ножка;
5 — чистильная ножка; 6 — конечный членик щупальца максилл; 7 — шипы максилл;
5 — фурка; 9,10 — левое и правое хватательные щупальца челюстных ножек самца; 11 —
копулятивный орган.
1-5 — по: Henderson, 1990; 6-8 — по: Sywula, 1974; 9-11 — по: Gauthier, 1928.
Остракоды
235
Рис. 32. Strandesia obliqua (Brady, 1868).
1,2 — левая и правая створки раковинки самки сбоку изнутри; 3 — раковинка сверху;
4 — вторая антенна; 5 — один из шипов максилл; 6 — ходильная ножка; 7, 8 — фурка.
7, 2, 4-7 — по: Karanovic, 2005; 3 — по: Sars, 1925; 8 — по: Muller, 1900.
Рис. 33. Physocypria fadeewi Dubowsky, 1927.
7, 2 — левая и правая створки раковинки самки; 3 — раковинка сверху; 4 — фрагмент
заднего края правой створки; 5 — фурка; 6,7 — левое и правое хватательные щупальца
самца; 8 — копулятивный орган. По: Sywula, 1974.
236
Определитель зоопланктона и зообентоса пресных вод Европейской России, Т 2
Рис. 34. Cypria ophtalmica (Jurine, 1820).
1,2 — раковинка самки и самца сбоку; 3 — раковинка сверху; 4 — вторая антенна; 5 —
фурка и генитальный отдел тела самки; 6, 7 — левое и правое хватательные щупальца
челюстных ножек самца; 8 — копулятивный орган.
1,2,4 — по: Henderson, 1990; 3, 5-8 — по: Sywula, 1974.
Рис. 35. Cyclocypris laevis (Muller, 1785).
1, 2 — раковинка сверху и сбоку; 3 — чистильная ножка; 4 — фурка; 5, 6 — левое и
правое хватательные щупальца челюстных ножек самца; 7 — копулятивный орган.
1-6 — по: Henderson, 1990; 7 — по: Sywula, 1974.
Остракоды
237
Рис. 36. Candonopsis kingsleii (Brady & Robertson, 1870).
/, 2 — левая и правая створки раковинки самки; 3 — левая створка раковинки самца;
4 — раковинка сверху; 5 — чистильная ножка; 6 — фурка самки; 7, 8 — левое и правое
хватательные щупальца челюстных ножек самца; 9 — копулятивный орган.
По: Sywula, 1974.
Рис. 37. Paracandona euplectella (Brady & Norman, 1889).
1 — раковинка сбоку; 2 — раковинка сверху; 3 — чистильная ножка; 4 — фурка; 5, 6 —
левое и правое хватательные щупальца челюстных ножек самца; 7 — копулятивный орган.
1,2 — по: Henderson, 1990; 3-7 — по: Sywula, 1974.
238
Определитель зоопланктона и зообентоса пресных вод Европейской России, Т. 2
Ключ для определения родов Candoninae Kaufmann, 1900
1(2). Фурка без задней щетинки...............Candonopsis Vavra, 1891 (рис. 36).
2(1). Задняя щетинка на фурке имеется.
3(6). Сетчатая или ямчатая скульптура на поверхности створок имеется.
4(5). По всей поверхности створок сетчатая скульптура и бородавчатые возвы-
шения с волосками. Длина самок до 0,7 мм..........Paracandona Hartwig, 1899.
Один вид — Р euplectella (Brady & Norman, 1889) (рис. 37).
Длина самки: до 0,7 мм, длина самца: 0,75-0,77 мм.
В болотах, старицах, реках и других постоянных водоёмах.
5(4). Створки густоволосисты и скульптурированы ямочками только в своей
центральной части. Длина самок до 0,95 мм..................................
...................v..................Pseudocandona Kaufmann, 1900.
Один вид — Р insculpta (G.W. Miiller, 1900) (рис. 38).
Длина самки: 0,82-0,94 мм Длина самца: 0,82-0,94 мм.
В довольно крупных озёрах на илистых грунтах.
6(3). Поверхность створок гладкая.
Рис. 38. Pseudocandona insculpta (G.W. Miiller, 1900).
1, 2 — левая и правая створки самки сбоку; 3 — раковинка самки сверху; 4 — щупальце
мандибулы; 5 — чистильная ножка; 6 — фурка; 7 — копулятивный орган; 8, 9 — левое и
правое хватательные щупальца челюстных ножек самца.
1-5 — по: Henderson, 1990; 6-9 — по: Sywula, 1974.
Остракоды
239
Рис. 39. Cryptocandona vavrai Kaufmann, 1900.
1,2 — левая и правая створки самки сбоку; 3 — раковинка самки сверху; 4 — чистильная
ножка; 5 — фурка и генитальный отдел тела самки; 6 — копулятивный орган.
1-5 — по: Sywula, 1974; 6 — по: Klie, 1938.
Рис. 40. Candona Candida (Muller, 1785).
1,2 — левая и правая створки самки сбоку; 3 — раковинка самки сверху; 4 — мандибу-
лярное щупальце; 5 — чистильная ножка; 6 — фурка и генитальный отдел самки; 7 — ко-
пулятивный орган; 8, 9 — левое и правое хватательные щупальца самца.
1, 2, 4-6 — по: Sywula, 1974; 3 — по: Henderson, 1990; 7-9 — по: Muller, 1900.
240
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 41. Fabaeformiscandona fabaeformis (Fischer, 1851).
1,2 — правая створка раковинки самки и самца сбоку; 3 — раковинка сверху; 4 — фурка
и генитальный придаток самки; 5 — мандибулярное щупальце; 6 — чистильная ножка; 7,
8 — левое и правое хватательные щупальца челюстных ножек самца; 9 — копулятивный
орган.
1—3 — по: Henderson, 1990; 4-9 — по: Sywula, 1974.
Остракоды 241
7(8). На серединё разделённого или неразделённого предпоследнего членика
чистильной ножки имеется щетинка.......................................
..............................Cryptocandona Kaufmann, 1900 (рис. 39).
8(7). На середине разделённого или неразделённого предпоследнего членика
чистильной ножки щетинка отсутствует.
9(10). Со спиной стороны карапакс субовальной формы. Отношение его ширины к
длине более 0,4. Медиальная щетинка на предпоследнем членике щупальца
мандибул оперенная. На втором членике щупальца мандибул на внутреннем
крае группа из 4-5 щетинок.................Candona Baird, 1845 (рис. 1, 40).
10(11). Карапакс со спинной стороны довольно сильно сжат. Отношение его ши-
рины к длине менее 0,4 (за исключением Fabaeformiscandona levanderi
Hirschmihn, 1912, соотношение ширины и длины около 0,42). Медиаль-
ная щетинка на предпоследнем членике щупальца мандибул гладкая. На
втором членике щупальца мандибул на внутреннем крае группа из 3-5
щетинок............................Fabaeformiscandona Krstic, 1972 (рис. 41).
тихоходки
Тихоходки (Tardigrada) — небольшая группа микроскопических сегментиро-
ванных целомических животных, сочетающих в своем строении признаки анне-
лид и членистоногих. Широко распространены в природе. Наибольшего разно-
образия достигают в наземных биотопах, содержащих капельно-жидкую воду:
куртинах мхов и лишайников, опаде, почве. В постоянных и временных водоёмах
тихоходки встречаются практически в любых типах субстратов, однако их излю-
бленным местообитанием является крупный заиленный песок.
Тело тихоходок обычно вальковатое, выпуклое со спинной стороны и упло-
щённое с брюшной (табл. 1, 1, 2; 2, 2). Слабыми перетяжками оно разделено на
головную лопасть, на которой расположены ротовое отверстие и органы чувств
и 4 следующих за ней сегмента, несущих по паре ножек. Размеры большинства
тихоходок находятся в интервале 90-800 мкм, однако некоторые представители
семейства Macrobiotidae, а также отдельные экземпляры Milnesium tardigradum
могут достигать длины 1000-1200 мкм, в то время как вышедшие из яиц особи
некоторых видов не превышают в длину 50 мкм. В большинстве своем тихоходки
бесцветны, но встречаются и окрашенные виды с тремя вариантами окраски: за
счёт просвечивающего содержимого кишечника, за счёт пигментированных эле-
ментов полостной жидкости и за счёт окрашенной гиподермы и кутикулы.
По особенностям строения покровов всех тихоходок можно разделить на две
не систематических группы: «панцирных» и «голых». «Панцирными» называют-
ся тихоходки, кутикула которых образует утолщённые участки — спинные и (ред-
ко) брюшные пластинки (табл. 1, 2). К группе «голых» тихоходок относятся виды,
кутикула которых может нести различную скульптуру, но не образует правильно
расположенных щитков (табл. 1, 7; 2, 7, 2).
Тело тихоходок несёт 4 пары бугорчатых ножек. Ножки 4-й пары часто сраста-
ются своими основаниями, направлены назад и в движении не участвуют, а ис-
пользуются для прикрепления к субстрату. Каждая ножка на своей вершине не-
сёт коготки, форма и расположение которых играют важную роль в систематике
группы.
Ножки представителей класса Eutardigrada несут по 2, так называемых, «двой-
ных коготка» (табл. 1, 4-14). Каждый из них состоит из общего основания и двух
ветвей: первичной (главной) и вторичной. Форма, расположение и характер диф-
ференцировки ветвей каждого «двойного коготка» и самих коготков в пределах
каждой лапки играют важную роль в систематике тихоходок. К числу дополни-
тельных кутикулярных структур, которые могут присутствовать на ножках, отно-
сятся лунулы — кутикулярные образования, связанные с основаниями коготков
(табл. 1, 7, 12), а также кутикулярные полоски, иногда развитые вблизи основа-
ний коготков (табл. 1, 9).
Уникальной особенностью тихоходок, отсутствующей у других групп живот-
ных, является их рото-глоточный аппарат (табл. 1,15).
Тихоходки
243
Ротовое отверстие у большинства тихоходок находится на вершине терми-
нального или субтерминального ротового конуса. У представителей некоторых
родов Eutardigrada оно окружено кутикулярными складками — околоротовыми
ламеллами. За ротовым отверстием следует ротовая трубка, обычно жёсткая и
прямая. Однако в нескольких группах тардиград независимо возникает разделе-
ние её на 2 части: жёсткую собственно ротовую трубку и гибкую глоточную труб-
ку. Стенки глоточной трубки всегда несут спиральные утолщения (рото-глоточ-
ный аппарат типа Diphascon) (табл. 1, 16). Для Eutardigrada характерно наличие
различных дополнительных структур в передней части ротовой трубки, а именно:
вентрального мостика — продольного кутикулярного гребня (табл. 1, 75; 2, 3),
либо дорсальной^ вентрального гребней для прикрепления мышц — протракто-
ров стилетов (табл. 2, 3-7). Форма этих гребней играет важную роль в системати-
ке некоторых семейств Eutardigrada.
По сторонам от ротовой трубки лежат 2 стилета — органы перфорации. Пе-
редние концы стилетов способны выдвигаться через ротовое отверстие и прока-
лывать пищевые объекты тихоходок. Задние их концы обычно связаны с ротовой
трубкой при помощи, так называемых, опор стилетов.
Проксимальный конец ротовой трубки переходит в мускулистую сосательную
глотку. В месте перехода иногда присутствуют клапаноподобные структуры (апо-
физы). У большинства тихоходок просвет глотки снабжён различными кутику-
лярными образованиями. Существует 2 основных варианта их организации: тон-
кие непрерывные кутикулярные полоски (большинство Heterotardigrada) и ряды
обособленных пластинок — плакоидов (большинство Eutardigrada). Наиболее
полный набор плакоидов состоит из 3 макроплакоидов и 1 микроплакоида (ука-
зывается число плакоидов в одном ряду); иногда позади ряда плакоидов имеется
особая кутикулярная структура, называемая септулой, вероятно, играющая роль
клапана.
В систематике некоторых родов тихоходок (Macrobiotus, Dactylobiotus,
Ramazzottius) используется такой признак, как характер скульптуры хориона от-
ложенных яиц. Зачастую их изучение — это единственный способ, дающий воз-
можность различать виды, взрослые особи которых практически идентичны.
Используются такие признаки, как форма выростов хориона (полусферическая,
коническая, филаментозная, грибовидная) и особенности строения поверхности
яиц между ними (грануляция, ретикуляция, соотношение размеров ячеек и т.д.).
Методы сбора и определения
Для взятия проб в глубоких участках водоёмов используются различные типы
бентометров, на мелководье можно использовать сачок из мельничного газа
№ 78. В случае, если невозможно разобрать пробы на месте в течение 1-2 дней,
их можно зафиксировать: вначале ледяной уксусной кислотой (1-2 мл на пробу),
затем, через 2-3 минуты, 4-5%-ным формалином.
244
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Для извлечения тихоходок из пробы материал процеживают через 2 сита: с га-
зом № 9 (для удаления крупных частиц) и с газом № 78. Просматривая под би-
нокуляром в чашке Петри содержимое сита, обнаруженных тихоходок и их яйца
извлекают пипеткой и переносят в стекла с лунками.
Определение ведется только на постоянных препаратах, для приготовления ко-
торых может быть рекомендована следующая методика.
1. Зафиксировать уксусной кислотой. В стекло с лункой или солонку добавить
2-3 капли ледяной уксусной кислоты и подождать несколько минут, до расправ-
ления зафиксированных животных.
2. Перенести растравившихся животных пипеткой на предметное стекло и
полностью высушить. Большинство экземпляров следует располагать дорсовен-
трально, но для некоторых родов тйхЬходок важно иметь возможность исследо-
вать строение рото-глоточного аппарата в латеральном положении.
3. Заключить в жидкость Фора. Для изучения препараты готовы через 1-2 дня.
При определении необходимо использовать фазово-контрастную оптику.
Основные пособия по определению
Ramazzotti G., Maucci W. 1983. Il Philum Tardigrada. 1012 p.
Dastych H. 1988. The Tardigrada of Poland. 288 p.
Maucci W. 1986. Tardigrada d’Italia. 388 p.
Ключ для определения тихоходок
1(4). Головной конец несёт щетинковидные придатки — латеральные циррусы
(табл. 1).
2(3). В глотке отсутствуют оформленные плакоиды, могут быть лишь тонкие
непрерывные кутикулярные полоски................... Класс Heterotardigrada.
Из представителей этого класса пресноводная Carphania fluviatilis Binda, 1978
(«голая» форма; длина тела 186-290 мкм) известна из Сицилии. Отдельные виды
семейства Echiniscidae (панцирные формы) были встречены в пресноводных
местообитаниях, по-видимому, оказываясь там в результате случайного заноса
(табл. 1, 2, 3).
3(2). В глотке имеются макроплакоиды..................Класс Mesotardigrada.
Единственный представитель Thermozodium esakii Rahm, 1937 (360-490 мкм) об-
наружен в сернистом термальном источнике. Япония, остров Кюсю.
4(1). Головные латеральные циррусы отсутствуют, могут присутствовать сен-
сорные структуры в виде коротких папилл.................Класс Eutardigrada.
5(6). Головные папиллы имеются (6 околоротовых и 2 головных папиллы),
глотка без кутикулярных структур, главная и боковая ветви коготков
полностью разделены, боковая ветвь с двумя разнонаправленными базаль-
ными ветвями (табл. 1, 4)...................................Отряд Apochela.
Из представителей этого отряда в России встречается лишь Milnesium tardigradum
Doyere, 1840 (500-1000 мкм). Часто встречается в наземных биотопах, в воде редок.
Тихоходки
245
6(5). Головные папиллы отсутствуют (у представителей некоторых родов может
быть пара слабо выраженных латеральных головных бугорков). В глотке
обычно имеются плакоиды. Главная и боковая ветви образуют единый
коготок (табл. 1, 4-14).....................................Отряд Parachela.
7(12). Коготки каждой ножки одинаковой формы и размера, расположены сим-
метрично (последовательность ветвей: боковая-главная-главная-боковая)
(табл. 1, 5-7).
8(9). Коготки типа тасгопух (общее основание короткое, первичные ветви силь-
но удлинены, вторичные — слабо развиты. Лунулы отсутствуют, основа-
ния коготков соединены кутикулярными полосками) (табл. 1, 5)................
..................................Род Dactylobiotus Nelson et al., 1980.
Пресноводный родРВ России встречаются виды: D. haplonyx Maucci, 1980, D. sele-
nicus Bertolani, 1981, D. ambiguus (Murray, 1907), D. dispar (Murray, 1907).
9(8). Коготки другие (табл. 1, 6-7).
10(11). Коготки типа hufelandi (оба коготка небольших размеров, общее основание,
первичная и вторичная ветви хорошо развиты. Ветви расходятся примерно
на середине высоты коготка (Y-форма). Основание коготка соединено с
лунулой филаментозной структурой) (табл. 1, 7), макроплакоидов 2 или 3
.........Роды Macrobiotus Schultze, 1834 и Paramacrobiotus Guidetti et al, 2008.
Более 150 видов. Большинство видов приурочены к наземным местообитаниям.
Многие виды эвритопны и часто встречаются в прибрежной зоне водоёмов. Род
Paramacrobiotus недавно выделен из рода Macrobiotus на основании данных
молекулярно-гентичесгого анализа. К нему отнесены виды, ранее входившие в
состав групп видов richtersi и areolatus. В пресных водах России наиболее широ-
ко распространены виды, относящиеся к группам видов Macrobiotus hufelandi и
М. harmsworthi и Paramacrobiotus richtersi. Точное определение видов этих родов
возможно только при наличии отложенных яиц.
11(10). Коготки типа pullari (общее основание слабо выражено, ветви хорошо раз-
виты, расходятся от основания коготка (V-форма). Основание коготка со-
единено с лунулой пластинчатой структурой (табл. 1, 6), макроплакоидов
2, без микроплакоидов....................Род Murrayon Bertolani et Pilato, 1988.
В пресных водах России встречаются виды: М. hastatus (Murray, 1907), М. pullari
(Murray, 1907), М. ovoglabellus (Biserov, 1988).
12 (7). Коготки каждой ножки, за редкими исключениями, имеют разную форму
и размеры, всегда расположены асимметрично относительно продольной
оси ножек; последовательность ветвей боковая-главная-боковая-главная
(табл. 1, 8-14).
13(14). Коготки типа Eohypsibius (коготки каждой ножки близки по форме и раз-
мерам, состоят из трёх сегментов каждый: базальной части, вторичной и
первичной ветвей, соединённых последовательно и отчётливо разделён-
ных перегородками) (табл. 1, 10)............................................
....................Сем. Eohypsibiidae, род Bertolanius Ozdikmen, 2008
(ранее Amphibolus Bertolani, 1981).
246
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Тихоходки
247
Из представителей семейства один вид В. markevichi Biserov, 1992 известен из
Байкала. В Западной Европе и Скандинавии из пресных вод известны также
В. smreczinskii (Weglarska, 1970) и В. weglarskae (Dastych, 1972).
14(13). Коготки иного типа.
15(16). Коготки типа Microhypsibius (коготки каждой ножки близки по форме,
общее основание коготка образовано только базальной частью первичной
ветви, вторичная ветвь коготков жёстко причленяется к основанию, в ме-
сте соединения имеется поперечная перегородка) (табл. 1, 14), скульптура
кутикулы в виде поперечных рядов бугорков (табл. 2, 1).........................
.....................Сем. Microhypsibiidae, род Fractonotus Pilato, 1998.
Из пресных вод России известен Е caelatus (Marcus, 1928).
16(17). Кбготки иного типа. Кутикула гладкая или скульптурирована.
17(20). Коготки типа Calohypsibius (коготки каждой ножки близки по форме, не-
большого размера, с ветвями, базальные части которых вместе образуют
общее основание коготка, сохраняя разделяющую их продольную перего-
родку) (табл. 1, 13). Вторичная ветвь коготков часто сильно редуцирована
...........................................................Сем. Calohypsibiidae.
18(19). Кутикула дорсально несёт скульптуру в виде поперечных рядов шипиков
(табл. 2, 2). Каждая ножка несёт пару двойных коготков.........................
..........................................Род Calohypsibius Thulin, 1928.
В пресных водах России отмечен С. omatus (Richters, 1900).
19(18). Кутикула гладкая. Ножки 4-й пары несут лишь по одному двойному когот-
ку каждая.......................................Род Parhexapodibius Pilato, 1969.
В пресных водах России отмечен Р pilatoi (Bernard, 1977).
20(17). Коготки типа Hypsibius, Isohypsibius или Ramazzottius (см. ниже) (табл. 1,
8, 9, 11, 12).
21(26). По крайней мере наружные коготки каждой ножки типа Isohypsibius
(коготки каждой ножки близки по форме и размерам, первичная ветвь
связана с основанием гибкой связкой. Основание коготка образует со вто-
ричной ветвью прямой угол) (табл. 1, 8).
Таблица 1
1 — Macrobiоtus sp. (Eutardigrada, Macrobiotidae), «голая» форма, общий вид, вен-
трально; 2 — Pseudechiniscus suillus (Heterotardigrada, Echiniscudae), «панцырная»
форма, общий вид, дорсально; 2 — Echiniscus sp., ножка с коготками; 4 — Milnesium
tardigradum, коготки; 5 — Dactylobiotus sp., коготки; 6 — Murrayon sp., коготки; 7 —
Macrobiotus sp., коготки; 8 — Isohypsibius sp., коготки; 9 — Hypsibius sp., коготки;
10 — Bertolanius sp., коготки; 11 — Mixibius sp., коготки; 12 — Ramazzottius sp., ко-
готки; 13 — Calohypsibius ornatus, коготки; 14 — Fractonotus caelatus, коготки; 15 —
рото-глоточный аппарат Eutardigrada, схема строения; 16 — рото-глоточный аппарат
типа Diphascon.
1, 4-12, 16 — ориг.; 2, 3, 15, 13 — по: Marcus, 1936; 14 — по: Pilato, 1998.
248
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т2'
22(23). Наружные коготки каждой ножки типа Isohypsibius, внутренние — про-
межуточного между типами Isohypsibius и Hypsibius (табл. 1, 11). Два
макроплакоида.......................................Род Mixibius (Pilato, 1992).
В пресных водах России отмечены М. saracenus (Pilato, 1973) и M ninguidus Bi-
serov 1999.
23(22). Все коготки типа Isohypsibius.
24(25). Ротовая трубка с вентральным мостиком (табл. 2, 3). Макроплакоидов 2,
без микроплакоидов.............................Род Doryphoribius Pilato, 1969.
В пресных водах России отмечены D. macrodon Binda, Pilato et Dastych, 1980 и
D. polynettae Biserov, 1988.
25(24). Ротовая трубка без вентрального мостика. На переднем конце ротовой
трубки имеются лишь выросты, ^Служащие для прикрепления мышц сти-
летов в форме гребней (при виде сбоку) (табл. 2, 4)........................
.................Роды Isohypsibius Thulin, 1928, Thulinius (Bertolani, 1981),
Pseudobiotus Nelson, 1980.
Достоверно различаются только деталями строени ротового аппарата: у предста-
вителей рода Isohypsibius околоротовые ламеллы отсутствуют, виды рода Thulinius
имеют 12 околоротовых ламелл, у видов рода Pseudobiotus — 30 околоротовых ла-
мелл. При исследовании в световой микроскоп, даже при использовании фазового
контраста, этот признак крайне труден для наблюдения.
26(21). Коготки типа Hypsibius или Ramazzottius (см. ниже) (табл. 1, 9, 12).
27(32). Ротовая трубка жёсткая на всём протяжении.
28(29). Коготки типа Ramazzottius (наружный и внутренний коготки каждой нож-
ки очень сильно отличаются друг от друга, первичная ветвь наружного
коготка очень Длинная и тонкая, соединена гибкой связкой с массивным
комплексом «основание + вторичная ветвь») (табл. 1, 12). Дорсальный и
вентральный гребни для прикрепления протракторов стилетов различной
формы (табл. 2, 5). Яйца орнаментированы, откладываются свободно...........
...................................Род Ramazzottius Binda & Pilato, 1986.
На территории России виды этого рода были обнаружены во временных водоёмах
и в прибрежной зоне постоянных водоёмов.
29(28). Коготки типа Hypsibius (наружный и внутренний коготки каждой ножки
значительно отличаются друг от друга, первичная ветвь связана с ос-
нованием гибкой связкой. Основание коготка не образует со вторичной
ветвью отчётливого угла, а формирует единую плавно изогнутую кривую)
(табл. 1, 9). Яйца (за редкими исключениями) гладкие, откладываются при
линьке в пустой экзувий.
30(31). Дорсальный и вентральный гребни для прикрепления протракторов
стилетов со вздутыми каплевидными дистальными концами (табл. 2, 6).
В глотке 2 толстых макроплакоида, оба с перетяжками........................
.......................................Род Borealibius Pilato et al., 2006.
Монотипический род. В. zetlandicus (Murray, 1907) известен из пресных вол
России.
Тихоходки
249
Таблица 2
1 — Fractonotus caelatus, общий вид, дорсально; 2 — Calohypsibius ornatus, об-
щий вид, латерально; 3 — Murrayon sp., рото-глоточный аппарат типа Macrobiotus
(с вентральным мостиком), латерально; 4 — Isohypsibius sp., рото-глоточный аппарат,
латерально; 5 — Ramazzottius sp., передний конец ротовой трубки, латерально; 6 —
Borealibius zetlandicus, передний конец ротовой трубки, дорсально; 7 — Hypsibius
dujardini, передний конец ротовой трубки, латерально.
1,2 — по: Marcus, 1936; 3-7 — ориг.
31(30). Дорсальный и вентральный гребни для прикрепления протракторов сти-
летов одинаковые, в форме тонких крючков (табл. 2, 7). Если глотка с дву-
мя толстыми макроплакоидами, второй не имеет отчётливой перетяжки ...
..............................................Род, Hypsibius Ehrenberg, 1848
Род включает около 40 видов, многие из которых часто втречаются в пресных во-
дах. На территории России наиболее широко распространён Н. dujardini (Doyere,
1840).
32(27). Между жёсткой ротовой трубкой и глоткой имеется гибкая глоточная
трубка, со спиральным утолщением стенок (рото-глоточный аппарат типа
Diphascon) (табл. 1, 16).
33(36). Ротовая и глоточная трубки широкие, свыше 3 мкм. В глотке два тонких
длинных макроплакоида.
34(35). Микроплакоиды отсутствуют. Опоры стилетов крепятся на ротовой трубке
в месте перехода её в гибкую глоточную трубку............................
...........................................Род Platicrista Pilato, 1987.
Во влажных мхах часто встречается вид Р. angustata (Murray, 1905). В воде редок.
250 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
35(34). Микроплакоиды имеются. Опоры стилетов крепятся на ротовой трубке
кпереди от перехода её в гибкую глоточную трубку............................
.............................................Род Mesocrista Pilato, 1987.
Во влажных мхах севера России широко распространён вид М. spitzbergensis
(Richters, 1903). В воде редок.
36(33). Ротовая и глоточная трубки узкие: 1-3 мкм...Род Diphascon Plate, 1889.
Род включает более 70 видов тихоходок приуроченных преимущественно к назем-
ным местообитаниям. В пресных водах России обнаружены D. belgicae Richters,
1911, D. arduifrons (Thulin, 1928), D. scoticum Murray, 1905.
КНИДАРИИ
Тип Cnidaria подразделяется на 2 подтипа Anthozoa и Medusozoa и включает
преимущественно морские организмы. Однако известно небольшое количество
книдарий подтипа Medusozoa, обитающих либо исключительно в пресных водо-
ёмах (представители класса Hydrozoa — отряды Hydrida, Limnomedusae), либо в
сильно опреснённых участках моря, в том числе в интерстициали (Hydrozoa —
отряды Anthoathecata, Leptothecata).
Жизненный цикл большинства книдарий подтипа Medusozoa в классическом
варианте предполагает наличие двух стадий — стадию полипа, и стадию медузы,
каждая из которых может быть развита в большей или меньшей степени. У неко-
торых представителей стадия Йедузы может быть редуцирована.
Самая характерная черта книдарий — наличие в их эпителиях особых образо-
ваний — стрекательных клеток (нематоциты или книдоциты), в которых форми-
руются стрекательные капсулы (нематоцисты или книдоцисты — книды). Это —
уникальная, самая сложноустроенная клеточная структура Metazoa, которая и
дала название типу животных — Cnidaria или Стрекающие. Набор типов капсул,
характерный для той или иной группы книдарий принято называть книдомом.
Наиболее широко известные пресноводные представители книдарий — гидры
(отряд Hydrida, сем. Hydridae). В России и в Европе насчитывается 6 видов, при-
надлежащих к 3 родам гидр, это — бесскелетные одиночные полипы, которые
могут образовывать временные колонии. Форма тела полипа зависит от окружаю-
щих условий и от характера фиксации: тело может быть удлинённым, округлым,
боченковидным, со стебельком и без него. Конусовидная оральная часть полипа
называется гипостомом, вокруг которого располагается 4-8 полых нитевидных
щупелец разной длины — от очень длинных, превышающих длину тела полипа,
до очень коротких. Наличие или отсутствие стебелька, длина щупалец, соотноше-
ние длины тела полипа и длины щупалец видоспецифичны для гидр. Размножа-
ются гидры почкованием, половым и бесполым путем. Стадия медузы у гидр от-
сутствует. Оплодотворенные яйца гидры приступают к дроблению и гаструляции,
находясь, как правило, на теле материнского организма. Здесь же, на зародыше
развивается наружная оболочка — эмбриотека, покрытая шипами. Вскоре после
образования эмбриотеки зародыш либо отваливается от материнского полипа и
падает на дно водоёма, либо остаётся прикреплённым к телу материнского орга-
низма, вплоть до развития из него молодого полипа. На дне водоёма зародыш на-
ходится всю зиму, до весны, когда из него возникает новый организм.
Книдом всех родов и видов гидр одинаков и представлен 4 типами капсул:
стенотёлами, десмонёмами, изоризами голотрихами и изоризами атрихами.
Видоспецифичными могут считаться лишь размеры капсул, их форма и харак-
тер расположения нити внутри невыстреленной капсулы. Но пользоваться строе-
нием капсул нематоцист как диагностическим признаком непросто — для этого
нужна специальная оптика и навык распознавания структур капсулы.
252
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Кроме гидр, типичными для пресных водоёмов оказывается следующие пред-
ставители класса Hydrozoa. Два вида рода Cordylophora — С. lacustris (Европа и
Азия) и С. caspia (понто-каспийский регион) (отряд Anthoathecata, подотряд Fili-
fera, семейство Clavidae). Представители этой группы отличаются колониальны-
ми полипами, тело которых покрыто перисаркальным скелетом. Головки полипов
этих организмов имеют веретеновидную форму и снабжены многочисленными
нитевидными щупальцами, беспорядочно распределенными по всей поверхности
головки. Скелетной чашечки, в которую может прятаться сокращенная головка
полипа, нет. Жизненный цикл кордилофор характеризуется наличием медузоид-
ной стадии, редуцированной до состояния споросака, заключенного в гонофор,
внутри которого развиваются планулы. Свободные медузы отсутствуют. В кни-
доме известен единственный тип нематоцист — эврителы микробазические. Род
Craspedacusta (Hydrozoa, Trachylinae, Limnomedusae, Olindiasidae) представлен в
относительно холодных водах пресных водоёмов северного и южного полушарий
одним видом С. sowerbyi Lankester, 1880 (рис. 1). В жизненном цикле предста-
вителей этой группы хорошо известны свободные медузы. Они имеют полусфе-
Рис. 1. Craspedacusta sowerbyi Lankester, 1880.
Половозрелая медуза из водоёма близ оз. Байкал (по Степаньянц и др., 2009).
Книдарии
253
рический или уплощённый колокол, 4 радиальных канала и множество краевых
щупалец, расположенных в несколько рядов чуть выше края колокола, со сторо-
ны эксумбреллы. Щупальца разной длины, но особенно хорошо развиты 4 пер-
радиальных щупальца. Гонады — на радиальных каналах. Рот — на достаточно
длинном манубриуме, окружен 4 лопастями. Статоцисты (конструкции, играю-
щие роль органов чувств) погружены в мезоглею. Полипы (там, где известны)
представлены маленькими червеобразными организмами, без щупалец, но име-
ют оральное скопление нематоцист. Могут образовывать временные колонии или
дают фрустулы.
Методы сбора и обработки
Гидр собирают «вслепую». Это значит, что из водоёма вылавливают водные
растения (элодею, кувшинки и др.) или ветки деревьев, куски древесины, камеш-
ки и т.п. и помещают их в емкости с водой. Спустя час-другой, если гидры есть,
они расправляют щупальца и становятся видны с освещённой стороны банки.
Изучать гидр лучше «живьём», так как в фиксированном состоянии они сильно
сокращаются, меняют форму тела, их щупальца укорачиваются и полипы труд-
но идентифицируются. Обычный способ фиксации гидр — 4% формалин или 76°
спирт. Колонии Cordylophora можно собирать сачком или вручную, соскребая их
с твёрдого субстрата, например, с погружённых в воду деревянных свай причала,
или собирая вместе с водными растениями.
Медузы ловят сачком (со шлюпки) или сетью Джеди (с борта судна). Чаще
всего используется сеть Джеди с диаметром входного отверстия 37 мм; конус
сети изготовляется из газа N 38. Для анестезии пойманных медуз помещают в
банку с водой, в которую добавляют несколько кристаллов MgCl или MgSO4. По-
сле того как медуза «расслабленно замирает», в жидкость добавляют 40% форма-
лин, доведя раствор до 4%.
Ключ для определения родов и видов гидр
1(2). Прижизненный цвет полипов ярко-зелёный. Капсула голотрих изориз за-
метно заужена базально..................................................
................Род Chlorohydra, Ch. viridissima (Schulce, 1914) (рис. 2).
2(1). Полипы имеют иную прижизненную окраску. Капсулы голотрих изориз не
заужены базально.
3(6). Тело полипа со стебельком. Нить в невыстреленной капсуле голотрих изо-
риз уложена вертикальными витками по всей своей длине...................
..................................................Род Pelmatohydra.
4(5). Длина капсул голотрих изориз превышает в 2-3 раза их ширину.......
.....................................Р oligactis (Pallas, 1766) (рис. 3).
5(4). Длина капсул голотрих изориз превышает их ширину меньше, чем в
2 раза.............................Р baikalensis (Swarczewsky, 1923) (рис. 4).
254
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Рис. 2. Chlorohydra viridissima
(Schulze, 1914).
А — полип гидры; Б — голотриха
изориза, невыстреленная капсу-
ла, зауженная базально. Рисунок
Б.А. Анохина.
Рис. 3. Pelmatohydra oligactis (Pallas, 1766).
А — полип со стебельком; Б — невыстреленная
капсула голотрихи изоризы с нитью, уложенной
внутри капсулы, по всей её длине вертикальны-
ми витками. Рисунок Б.А. Анохина.
Рис. 4. Pelmatohydra baikalensis (Swar-
czewsky, 1923).
A — тело полипа co стебельком; Б — не-
выстреленная капсула голотрихи изоризы с
нитью, уложенной внутри капсулы, по всей
её длине вертикальными витками. Рисунок
Б.А. Анохина.
Книдарии
255
Рис. 5. Hydra circumcincta Schulze, 1914.
A — полипа; Б — невыстреленная кап-
сула голотрихи изоризы с нитью, уло-
женной внутри капсулы, в апикалной её
части горизонтальными витками. Рисунок
Б.А. Анохина.
Рис. 7. Hydra vulgaris Pallas, 1766.
А — полип, длина щупалец которого больше
длины тела полипа; Б — невыстреленная капсула
голотрихи изоризы. Рисунок Б.А. Анохина.
Рис. 6. Hydra oxycnida Schulze, 1914.
А — полип, длина щупалец которого
не превышает половину длины тела;
Б — невыстреленная капсула голо-
трихи изоризы с нитью, уложенной
внутри капсулы, в апикальной её
части горизонтальными витками. Ри-
сунок Б.А. Анохина.
256
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Рис. 8. Cordylophora caspia (Pallas, 1766).
А — фрагмент колонии из Каспия; Б — схема ветвления колонии; В — молодая колония.
Изменено по рукописи Тарасова.
Книдарии
257
Рис. 9. Cordylophora lacustris Allman, 1844.
Полипы с гонофорами. Рисунок Б.А. Анохина.
258 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
6(3). Тело полипа без стебелька. Верхняя часть нити голотрих изориз в невы-
стреленной капсуле уложена чаще всего горизонтально, или косыми гори-
зонтальными витками...............................................Род Hydra.
7(10). Щупальца в расслабленном остоянии не превышают длину полипа.
8(9). Полипы маленькие (редко достигают 15 мм). Ширина капсул их голотрих
изориз превышает длину таковых....//, circumcincta Schulze, 1914 (рис. 5).
9(8). Полипы крупные (достигают 28 мм в длину). Ширина капсул голотрих
изориз не превышают половину их длины....................Н. oxycnida (рис. 6).
10(7). Щупальца в расслабленном состоянии превышают длину полипа...........
...................................................Н. vulgaris (рис. 7).
Ключ для определения видов рода Cordylophora
1(2). Колонии полипов крупные и достигают 40 см в высоту; ветвление — до
2^4 порядков. Перисарк колонии членистый по всей длине ствола и вет-
вей ..........................................С. caspia (Pallas, 1771) (рис. 8).
2(1). Колонии маленькие, достигают в высоту несколько десятков миллиме-
тров; ветвление, как правило, не более, чем второго порядка. Перисарк
имеет членистость только в основании ствола и ветвей.......................
....................................С. lacustris (Allman, 1844) (рис. 9).
Представители отрядов Anthoathecata и Leptothecata,
встречающиеся в водоёмах с солёностью воды до 8%о
Anthoathecata: Filifera: Pandeidae
Halitholus cirratus Hartlaub, 1914 (рис. 10-12).
Колокол молодой медузы (около 2 мм в диаметре) овальный, с короткой апи-
кальной проекцией, с коротким и широким манубриумом и 4 короткими лопа-
стями вокруг рта; 4 радиальных канала и 4 краевых щупальца. Сформированная
медуза (около 20 мм в диаметре) имеет очень высокую апикальную проекцию, го-
нада — на манубриуме, четырёхлопастная, складчатая; р зооида отовые лопасти
бахромчатые.
Морфология полипов у представителей этого и близких семейств не имеет
четких диагностически признаков — все они представлены мелкими зооидами,
отходящими от сетки гидроризы, с перисарком, окружающим ножку полипа и до-
ходящим до основания его головки, с небольшим числом щупалец вокруг гипо-
стома. Гонофоры располагаются тоже на гидроизе.
Описан из Балтийского моря, в районе бухты Данцигер, где были встречены и
медузы, и полипы.
Число краевых щупалец обычно соответствует числу радиальных каналов.
Каждое щупальце ветвится, часто до 5 раз. Каждое щупальце и все его ветви по-
крыты нематоцистами. В основании каждого щупальца имеется удлинённая кра-
евая бульба, в гастродермисе которой имеется множество конкреций, а с её внеш-
Книдарии
259
ней стороны — по одному глаз^су. На бульбе имеются также по 2-6 булавовидных
щупальца — т.н. «ходильные ноги», на которых медуза не ходит, а стоит на грунте.
Тепловодный вид, обитающий при температурах до 25 °C и довольно высокой
солёности — до 35%о. Но часто встречается и в распреснённой воде (как в приро-
де, так и в аквариумах). Например, известен в Балтийском море.
Рис. 10. Halitholus cirratus Hartlaub, 1914.
Колония полипов из Балтийского моря. По:
Hartlaub, 1914.
Рис. 11. Halitholus cirratus Hartlaub, 1914.
Молодая медуза из Балтийского моря. По:
Hartlaub, 1914.
Рис. 14. Halitholus cirratus Hartlaub, 1914.
Половозрелая медуза из Балтийского моря. По: Hartlaub, 1914
НЕМАТОДЫ
Нематоды (Nematoda) — одна из самых распространённых групп беспозво-
ночных. В пресных водах отмечено более 100 родов нематод с 600 видами. Длина
большинства пресноводных видов не превышает 2 мм. Тело покрыто плотной ку-
тикулой, либо гладкой, либо так или иначе орнаментированной — это могут быть
кольца, склероции, продольные рёбра. Довольно обычны и короткие соматиче-
ские щетинки. Ротовое отверстие расположено терминально и может быть окру-
жено головными щетинками. По бокам головы на уровне стомы располагаются
органы хеморецепции — амфиды. Форма амфидов у представителей разных от-
рядов различна. Круглые амфиды характерны для отряда Monhysterida, спираль-
ные — для отряда Chromaddfeida, карманообразные — для Enoplida, Mononchida
и Dorylaimida. Пищеварительная система состоит из ротовой полости (стомы),
пищевода, средней кишки и ректума, которому иногда предшествует преректум.
Стома — трёхстенное кутикулярное образование. Зубовидные выросты стенок
стомы называются онхами; наибольшего развития обычно достигает дорсальный
онх. Некоторые таксономические группы нематод в качестве вооружения стомы
имеют подвижные кутикулярные зубы, примером которых может служить копьё
дорилаймид или крюковидные зубы иронид. Строение пищевода в разных от-
рядах различно. Так, для многих представителей отрядов Chromadorida, Агаео-
laimida и Rhabditida характерно наличие бульбуса — расширенного основания
пищевода с особо развитой кутикулярной выстилкой, образующей дробильный
аппарат. Некоторые виды имеют 2 бульбуса — передний (метакарпальный) и за-
дний (кардиальный). Пищевод соединяется со средней кишкой кардием — клапа-
ном, который открывается только в передне-заднем направлении. У многих видов
из отрядов Enoplida и Monhysterida в области кардия развиты 3 пищеварительные
железы, которые не следует путать с бульбусом.
Нематоды — раздельнополые животные. Женская половая система (чаще пар-
ная) состоит из гонад, маток с протоками, мышечной вагины и вульвы, которая
расположена приблизительно в середине тела. Если система непарная, то вульва
может быть сдвинута в сторону ануса. Первичные половые органы самцов плохо
различимы, зато вторичные — спикулы, супплементы, бурса — видны хорошо и
играют существенную роль в диагностике видов (табл. 1).
Методы сбора и обработки
Наибольшей численности нематоды достигают на литорали озёр, предпочитая
слабозаилённые пески и ризосферу растений. Основная масса нематод сосредото-
чена в верхнем слое грунта до 1 см. Для качественных исследований достаточно
любым способом собрать верхний слой грунта с площади до 0,1 м2 и зафикси-
ровать его 4%-ным формалином. Перед лабораторной обработкой фиксирован-
ную пробу необходимо промыть. Промывка осуществляется в кристаллизаторе
Нематоды
261
(простоквашнице, миске) диаметром 15-20 см. Проба заливается чистой водой
и взбалтывается кругообразными движениями. После такого «центрифугирова-
ния» лёгкая фракция, куда входят и нематоды, сливается в чистый сосуд, затем
процедура повторяется. Промытую пробу проливают через газ с диаметром ячеи
100-200 мкм, нематод смывают с ситечка и разбирают под бинокуляром. Опре-
деление следует вести на постоянных глицерин-желатиновых или временных
глицериновых препаратах под микроскопом. Для изготовления препаратов прово-
дится предварительная обработка материала: в часовое стекло с водой, куда поме-
щены нематоды, добавляют несколько капель чистого глицерина, через сутки ещё
несколько капель и так до полного испарения воды и замещения её глицерином.
Определение до вида трудоёмко т.к. требует расчёта ряда индексов и, в большин-
стве случаев, •использования иммерсионной оптики, однако в общих гидробиоло-
гических исследованиях бывает достаточно довести определение до рода, что не
составляет особого труда.
Для таксономической ориентировки ниже приводится поотрядный список (по
алфавиту) всех включённых в «Определитель» родов.
Enoplida
Brevitobrilus, Epitobrilus, Eutobrilus, Neotobrilus, Paratrilobus, Prismatolaimus,
Semitobrilus, Tobrilus, Trischistoma, Tripyla, Tripylella, Tripilina.
Dorylaimida
Alaimus, Aporcelaimellus, Aporcelaimus, Aquatides, Chrysonema, Crocodorylai-
mus, Dorylaimoides, Dorylaimus, Ecumenicus, Epidorylaimus, Eudorylaimus, Ironus,
Labronema, Laevides, Laimydorus, Mesodorylaimus, Neoactinolaimus, Nygolaimus,
Paractinolaimus, Paramphidelus, Paravulvus, Prodorylaimus, Tylencholaimus.
Mononchida
Anatonchus, Clarkus, lotonchus, Miconchus, Mononchus, Mylonchulus, Para-
mononchus, Prionchuloides, Prionchulus.
Monhysterida
Eumonhystera, Monhystera.
Chromadorida
Achromadora, Chromadorina, Chromadorita, Ethmolaimus, Рагасуatholaimus,
Punctodora.
Desmodorida
Prodesmodora
Araeolaimida
Aphanolaimus, Bastiania, Chronogaster, Cylindrolaimus, Paraphanolaimus, Rhab-
dolaimus.
Plectida
Plectus, Paraplectonema.
Rhabditida
Diplogaster, Fictor, Koemeria, Mononchoides, Pelodera.
262
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Основные пособия по определению
Гагарин В.Г. 1992-1993. Свободноживущие нематоды пресных вод СССР. СПб.
Andrassy I. 2005-2009. Free-living Nematodes of Hungary. Budapest.
Ключ для определения родов нематод
1(38). Стома вооружена копьём (табл. 1, 5)..............Отряд Dorylaimida.
2(31). Копьё расположено в середине стомы по ходу главной оси тела (аксиаль-
ное копьё).
3(6). Вокруг копья имеется обширный вестибулюм с крупными онхами (табл. 2,
I, 3).
4(5). Кроме макроонхов вестибулюме имеются мелкие онхи, напоминающие
рашпиль................................Paractinolaimus Meyl, 1957 (табл. 2,7).
5(4). Рашпильных онхов не имеется.......................................
.............................Neoactinolaimus Thome, 1967 (табл. 2, 3).
6(3). Вестибулюм не выражен; других типов вооружения стомы, кроме копья,
нет.
Таблица 1
Элементы строения нематод.
1,2 — схема строения самца (/) и самки (2) (1 — стома, 2 — пищевод, 3 — нервное
кольцо, 4 — прикардиальные железы, 5 — кардий, 6 — средняя кишка, 7 — ректум, 8 —
вульва, 9 — женские гонады с яйцами, 10 — супплементы, 11 — спикулы, 12 — рулёк,
13 — хвост).
3-7 — строение головы, стомы и пищевода: 3, 7 — рабдитоидный тип, 4 — тобрилоидный
тип, 5 — дорилаймоидный тип, 6 — мононхоидный тип (14 — губы с папиллами, 15 —
головные щетинки, 16 — амфиды, 17 — буккальная полость, 18 — карманы стомы, 19 —
онхи, 20 — ротовое отверстие, 21 — хейлостома, 22 — простома, мезостома . метастома,
23 — телостома, 24 — зубы, 25 — просвет пищевода, 26 — копьё, 27 — отверстие копья,
28 — приставка копья, 29 — направляющее кольцо, 30 — дорсальный онх, 31 — ребро,
32 — гребень с пиловидными микроонхами, 33 — рашпильные микроонхи, 34 — суб-
вентральные онхи, 35 — кутикулярные поры, 36 — кольчатость кутикулы, 37 — корпус
пищевода, 38 — истмус пищевода, 39 — метакарпальный бульбус, 40 — кардиальный
бульбус).
8-15 — типы амфидов (верхний ряд — латерально, нижний ряд — дорсально): 8, 9 —
круглый (монхистеридный), 10, 11 — спиральный (хромадоридный), 12, 13 — плетевид-
ный (плектидный), 14-15 — бокаловидный-карманообразный (эноплидный).
16 — кутикулярные рёбра, характерные для некоторых дорилаймид (поперечный срез
тела).
17, 18 — строение хвоста (41 — хвостовые железы, 42 — спиннерета, 43 — субтерми-
нальная щетинка, 44 — бурса).
19-22 — типы супплементов: 19 — трубчатые (дорилаймоидные), 20 — альвеолярные
(этмоляймоидные), 21 — тобрилоидные-погружённые, 22 — тобрилоидные-ежевидные.
По: Цалолихин, 1994.
Нематоды
263
264
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
7(10). Копьё массивное, короткое, отверстие копья достигает или превышает ’Л
его длины.
8(9). Длина тела 4,5-10 мм; кутикула с продольной и поперечной (крест-на-
крест) исчерченностью.....................................................
....................Aporcelaimus Thome et Swanger, 1936 (табл. 2, 5, 7).
9(8). Длина тела 1,5-3,5 мм; кутикула с тонкой поперечной исчерченностью....
...........................Aporcelaimellus Heyns, 1965 (табл. 2, 8-10).
10(7). Отверстие копья не превышает, или незначительно превышает Уз его дли-
ны.
11(12). Копьё очень короткое, несколько искривлённое, вентральная сторона его
короче дорсальной.........................................................
................^Dorylaimoides Thome et Swanger, 1936 (табл. 2, 11-13).
12(11). Копьё правильной формы.
13(14). Передняя часть пищевода очень тонкая, мускулатура пищевода выражена
слабо; мелкие нематоды, обычно около 1 мм.................................
...........................Tylencholaimus de Man, 1880 (табл. 2, 14-16).
14(13). Мускулатура пищевода хорошо выражена; в подавляющем большинстве
крупные нематоды длиной более 1,5 мм.
15(22). Хвост у самца и самки различной формы; у самца, как правило, короткий,
часто закруглённый, у самки удлинённый, тонкий, плетевидный.
16(17). Кутикула с хорошо выраженными продольными рёбрами (табл. 1, 16)...
.............................Dorylaimus Dujardin, 1845 (табл. 2, 17-19).
17(16). Кутикула без продольных рёбер, гладкая или слобокольчатая.
18(19). В основании копья имеется 4 веретеновидных желтоватых образования....
.................................Crocodorylaimus Andrassy, 1988 (табл. 2, 20).
19(18). В основании копья желтоватых образований нет.
20(21). Длина тела, как правило, менее 2 мм; преректум самца не выходит или
незначительно выходит за пределы супплементарной области..................
...........................Mesodorylaimus Andrassy, 1959 (табл. 3, 1-3).
Таблица 2
1,2 — Paractinolaimus macrolaimus (de Man, 1880): 1 — голова, 2 — задняя часть
тела самца; 3, 4 — Neoactinolaimus duplicidentatus (Andrassy, 1968): 3 — голова, 4 —
хвост самки; 5-7 — Aporcelaimus eurydorys (Ditlevsen, 1911): 5 — голова, 6 — хвост
самца, 7 — хвост самки; 8-10 — Aporcelaimellus obscurus (Thome et Swanger, 1936):
8 — голова, 9 — задняя часть тела самца, 10 — хвост самки; 11-13 — Dorylaimoides
limnophilus (de Man, 1880): 11 — голова, 12 — вульварный отдел, 13 — хвост самки;
14-16 — Tylencholaimus sp.: 14 — пищевод, 15 — голова, 16 — хвост самки; 17-19 —
Dorylaimus stagnalis Dujardin, 1845: 17 — голова, 18 — задняя част тела самца, 19 —
хвост самки; 20 — Crocodorylaimus flavomaculatus (Linstow, 1876) — голова.
По: Цалолихин, 1994.
Нематоды
265
19
266
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
21(20). Длина тела более 2 мм; преректум самца значительно выходит за пределы
супплементарной области.................Laimydorus Siddiqi, 1969 (табл. 3, 4-6).
22(15). Хвост у самца и самки одинаковой формы.
23(24). Хвост у самца и самки истончённый до нитевидного..................
...........................Prodorylaimus Andrassy, 1959 (табл. 3, 7-9).
24(23). Хвост у самца и самки короткий, конический, никогда не истончённый до
плетевидного.
25(26). Тело очень тонкое, стройное, длина его не менее, чем в 60 раз превосходит
толщину..............................Chrysonema Thome, 1929 (табл. 3, 10, 11).
26(25). Длина тела не более, чем в 50 раз (обычно в 20-40) превосходит толщину.
27(28). У самок развита только одна задняя гонада (опистодельфные самки); сам-
цы неизвестны...................*....ESnmenicus Thome, 1974 (табл. 3, 12-14).
28(27). Самки дидельфные.
29(30). Хвост относительно длинный, его длина соответствует 4-8 диаметрам
тела в области ануса..............Epidorylaimus Andrassy, 1986 (табл. 3, 15-16).
30(29). Хвост короткий, его длина соответствует 1-3 диаметрам тела в области
ануса.............................Eudorylaimus Andrassy, 1959 (табл. 3, 17-19).
31(2). Копьё смещено от главной оси тела к брюшной стороне, обычно прижато
к левой субвентральной стенке стомы.
32(33). Вульва продольная, в области вульвы имеются маленькие папиллы или
поры.................................Paravulvus Heyns, 1968 (табл. 3, 20-22).
33(32). Вульва поперечная, в области вульвы папиллы или поры отсутствуют.
34(35). Длина копья заметно меньше ширины головы..........................
.................................Laevides Heyns, 1968 (табл. 3, 23-25).
35(34). Длина копья равна или почти равна ширине головы.
Таблица 3
1—3 — Mesodorylaimus meyli (Andrassy, 1958): 1 — голова, 2 — задняя часть тела сам-
ца, 3 — хвост самки; 4-6 — Laimidorus pseudostagnalis (Micoletzky, 1927): 4 — голова,
5 — задняя часть тела самца, 6 — хвост самки; 7-9 — Prodorylaimus longicaudatoides
Altherr, 1978: 7 — голова, 8 — задняя часть тела самца, 9 — хвост самки; 10, 11 —
Chrysonema holsaticum (Schneider, 1925): 10 — голова, 11 — задняя часть тела самца;
12-14 — Ecumenicus monohystera (de Man, 1880): 12 — голова, 13 — вульварный
отдел, 14 — хвост самки; 15, 16 — Epidorylaimus consobrinus (de Man, 1918): 75 —
голова, 16 — хвост самки; 17-19 — Eudorylaimus carteri (Bastian, 1865): 17 — голова,
18 — задняя часть тела самца, 19 — хвост самки; 20-22 — Paravulvus hartingii (de
Man, 1880): 20 — голова, 27 — хвост самца, 22 — хвост самки; 23-25 — Laevides
laevis (Tome, 1939): 23 — голова, 24 — задняя часть тела самца, 25 — хвост самки;
26-28 — Nygolaimus brachyuris (de Man, 1880): 26 — голова, 27 — хвост самки, 28 —
хвост самца.
По: Цалолихин, 1994; Andrassy, 2009.
Нематоды
267
268 Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
36(37). Копьё заметно расширятся к основанию, дельтовидное..............
...............................Nygolaimus Cobb, 1913 (табл. 3, 26-28).
37(36). Копьё тонкое, игловидное......Aquatides Heyns, 1968 (табл. 4, 11-13).
38(1). Стома либо лишена вооружения, либо вооружена крупными или мелкими
онхами или зубами; форма стомы различна, или стома отсутствует (визу-
ально отсутствует).
39(58). Стома обширная, бочкообразная, иногда несколько сужающаяся к основа-
нию, вооружена в разной степени развитыми онхами, пищевод всегда без
бульбусов.
40(57). Головные щетинки отсутствуют.
41(44). Стома вооружена 1 дорсальным макроонхом и микроонхами, напоминаю-
щими рашпиль.
42(43). Дорсальный макроонх расположен в передней части стомы, рашпильные
микроонхи многочисленны................Mylonchulus Cobb, 1916 (табл. 4, 10).
43(42). Дорсальный макроонх расположен в средней части стомы, микроонхи
малочисленны.......................Prionchuloides Mulvey, 1963 (табл. 4, 8).
44(41). Стома вооружена 1 или 3 макроонхами, микроонхи, напоминающие раш-
пиль отсутствуют.
45(46). Напротив дорсального макроонха расположены микроонхи в виде пилы...
........................................Prionchulus Cobb, 1916 (табл. 4, J).
46(45). Микроонхи в виде пилы отсутствуют.
47(50). Стома вооружена 3 макроонхами.
48(49). Все 3 макроонха расположены в основании стомы и направлены вперёд...
.................................Miconchus Andrassy, 1958 (табл. 4,1).
49(48). Все 3 макроонха расположены в передней части стомы и (у взрослых жи-
вотных) направлены назад.................Anatonchus Cobb, 1916 (табл. 4, 9).
50(47). Стома вооружена только 1 дорсальным онхом.
51(52). Макроонх расположен ниже середины стомы.........................
....................................lotonchus Cobb, 1916 (табл. 4, 2).
52(51). Макроонх расположен выше середины стомы.
Таблица 4
1-10 — строение стомы мононхид: 1 — Miconchus, 2 — lotonchus, 3 — Prionchulus,
4 — Paramononchus, 5 — Clarkus, 6 — Mononchus, 7 — Coomansus, 8 — Prionchuloides,
9 — Anatonchus, 10 — Mylonchulus^ 11-13 — Aquatides aquaticus (Tome, 1930): 11 —
голова, 12 — хвост самца, 13 — хвост самки; 14, 15 — Paratrilobus grandipapilloides
Micoletzky, 1922: 14 — голова, 75 — общий вид самца; 16-19 — Ironus tenuicaudatus
de Man, 1876: 16 — передний отдел тела, 17 — яйцо в половых путях самки, 18 —
хвост самки, 19 — хвост самца; 20 — Rhabdolqimus sp.: 20 — пищевод.
По: Цалолихин, 1994.
Нематоды
269
16 17
270
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1
53(54). Напротив макроонха имеется продольный зазубренный гребень; самцы
неизвестны...........................Paramononchus Mulvey, 1978 (табл. 4, 4\.
54(53). Напротив макроонха продольного зазубренного гребня нет; самцы имеются.
55(56). Спикулы самцов широкие и короткие, супплементы папилловидные......
.......................................Clarkus, Jairajpuri (табл. 4, 5i.
56(55). Спикулы самцов тонкие и длинные, супплементы трубчатые............
..................................Mononchus Bastian, 1865 (табл. 4, 6}.
57(40). Головные щетинки имеются...Paratrilobus Micoletzky, 1922 (табл. 4, 14}.
58(39). Стома воронковидная, бокаловидная, чашевидная, трубковидная или
стома отсутствует (визуально отсутствует); пищевод без бульбусов или
с бульбусами (1 или 2).
59(62). Стома в виде узкой трубки с 3 хорошо развитыми зубами в передней ча-
сти.
60(61). Длина тела не менее 1 мм; пищевод без бульбусов...................
.................................Ironus Bastian, 1865 (табл. 4, 16-19}.
61(60). Длина тела не более 0,5 мм; пищевод с 1 бульбусом.................
...............................Rhabdolaimus de Man, 1880 (табл. 4, 20}.
62(59). Стома устроена иначе, с иным вооружением или не вооружена.
63(72). Стома цилиндрическая.
64(65). Головные щетинки отсутствуют; у самцов на хвосте имеется бурса; пище-
вод с 1 бульбусом.......................Отряд Rhabditida, сем. Rhabditidae.
В загрязнённых органикой водоёмах в большинстве случаев могут быть обнару-
жены представители рода Pelodera Schneider, 1866 (табл. 5: 1-4).
65(64). Головные щетинки имеются; хвост у самцов не имеет бурсы.
66(67). Кутикула гладкая; глубина стомы примерно в 20 раз превышает её шири-
ну; в основании стомы имеются маленькие онхи.............................
................................Cryptonchus Cobb, 1913 (табл. 5, 5-6).
67(66). Кутикула кольчатая; глубина стомы либо равна её ширине, либо превы-
шает её ширину незначительно; или превышает её ширину примерно в
15 раз.
Таблица 5
1-4 — Pelodera sp.: 1 — голова, 2 — хвост самки, 3,4 — хвост самца с бурсой; 5, 6 —
Cryptonchus tristis (Ditlevsen, 1911): 5 — голова, 6 — хвост самки; 7 — Cylindrolaimus
communis (голова); 8-9 — Paraphanolaimus behningi Micoletzky, 1923: 8 — общий
вид самки с эмбрионами, 9 — голова; 10 — Aphanolaimus aquaticus (голова); 11-
13 — Paramphidelus dolichurus'. 11 — передний отдел тела, 12 — вульварный отдел.
13 — хвост самки; 14-16 — Alaimus primitivus de Man, 1880: 14 — передний отдел
тела, 75 — хвост самца, 16 — задний отдел пищевода; 77, 18 — Tripylella intermedia
(Butschli, 1873): 77 — голова, 18 — хвост самки; 19-21 — Tripylina arenicola (de Man.
1880): 19 — голова, 20 — вульварный отдел, 27 — хвост самки.
По: Цалолихин, 1994; Andrassy, 2009.
Нематоды
271
272
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
68(69). Глубина стомы примерно в 15 раз превышает её ширину; стома лишена
вооружения............................Cylindrolaimus de Man, 1889 (табл. 5, 7).
69(68). Глубина стомы приблизительно соответствует её ширине или превышает
её не более, чем в 4 раза.
70(71). Глубина стомы приблизительно равна её ширине; кутикула тонкокольча-
тая............................Paraphanolaimus Micoletzky, 1923 (табл. 5, 8, 9).
71(70). Глубина стомы превышает её ширину не более, чем в 4-5 раз; кутикула
грубокольчатая........................Aphanolaimus de Man, 1880 (табл. 5, 10).
72(63). Стома иной формы или визуально отсутствует.
73(86). Стома визуально отсутствует.
74(77). Головные щетинки отсутствуют.
75(76). Амфиды огромные, хорошо различимые.................................
..........................Paramphidelus Andrassy, 1977 (табл. 5, 11-13).
76(75). Амфиды едва заметные, поровидные...................................
.................................Alaimus de Man, 1880 (табл. 5, 14-16).
77(74). Головные щетинки имеются.
78(81). Головные щетинки расположены в 1 круг.
79(80). Женская половая система парная.....................................
.........Tripylella Brzeski et Winiszewska-Slipinska, 1993 (табл. 5, 17, 18).
80(79). Женская половая система непарная, продельфная (развита только перед-
няя гонада)...........................Tripylina Brzeski, 1963 (табл. 5, 19-21).
81(82). Головные щетинки расположены в 2 круга.
82(83). Кутикула гладкая.................Trischistoma Cobb, 1913 (табл. 6, 5).
83(82). Кутикула явственно кольчатая.
84(85). В сжатой до просвета пищевода стоме имеется маленький онх треуголь-
ной формы..................................Tripyla Bastian, 1865 (табл. 6, 1-4).
85(84). Стома не вооружена..............Bastiania de Man, 1876 (табл. 6, 6, 7).
86(73). Стома воронковидная, бокаловидная, чашевидная или сложной конфигу-
рации.
Таблица 6
1,2 — Tripyla glomerans Bastian, 1865: 1 — голова, 2 — хвост самца; 3, 4 — Tripyla
setifera Butschli, 1873: 3 — голова, 4 — хвост самца; 5 — Trischistoma monohystera (de
Man, 1880) — голова; 6, 7 — Bastiania gracilis de Man, 1886: 6 — голова, 7 — хвост
самки; 8, 9 — Diplogaster rivalis (Leydig, 1854): 8 — голова, 9 — спикула с рульком;
10—12 — Korneria lepida (Andrassy, 1958): 10 — голова, 11 — хвост самца, 12 — хвост
самки; 13-15 — Fictor fictor (Bastian, 1865): 13 — голова, 14 — хвост самца, 75 —
хвост самки; 16-18 — Mononchoides sp.: 16 — голова, 17 — хвост самца, 18 — хвост
самки; 19-21 — Eumonhystera sp.: 19 — голова, 20 — спикула, 21 — хвост самки;
22-24 — Monhystera paludicola de Man, 1881: 22 — голова, 23 — хвост самки, 24 —
спикула.
По: Цалолихин, 1994; Andrassy, 2005.
Нематоды
273
274
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. L
87(94). Стома сложной конфигурации; пищевод с 2 бульбусами.
88(91). Хорошо развит только 1 дорсальный зуб (другие зубы, если имеются, зна-
чительно меньше дорсального).
89(90). Дорсальный зуб расположен в основании стомы......................
..............................Diplogaster Schultze, 1857 (табл. 6, 8, 9
90(89). Дорсальный зуб расположен в передней части стомы.................
................................Koerneria Meyl, 1961 (табл. 6, 10—12
91(88). Хорошо развиты 2 зуба — дорсальный и субвентральный
92(93). Общая глубина стомы превышает её ширину; зубы расположены в сред-
ней части стомы.....................Mononchoides Rahm, 1928 (табл. 6, 16-18
93(92). Общая глубина стомы равна или несколько меньше её ширины; зубы рас-
положены ближе к основанию стомы.........................................
..............................Fictor Paramonov, 1952 (табл. 6, 13-15
94(87). Стома воронковидная, бокаловидная, чашевидная; пищевод с 1 бульбусом
или без бульбусов.
95(98). Стома в виде мелкой воронки; амфиды хорошо выраженные, круглые.
96(97). Расстояние от переднего конца тела до центра амфида превышает ширин;»
головы; спикулы самцов короткие..........................................
..........................Eumonhystera Andrassy, 1981 (табл. 6, 19—21
97(96). Расстояние от переднего конца тела до центра амфида меньше или равнс
ширине головы; спикулы самцов длинные....................................
..............................Monhystera Bastian, 1865 (табл. 6, 22-24 •
98(95). Стома хорошо выражена, бокаловидная, чашевидная или иной формы.
99(118). Пищевод с 1 бульбусом.
100(111). Амфиды хорошо выражены, в виде спирали, незамкнутые круглые или
петлеобразные.
101(102). Амфиды незамкнутые круглые, расположены на уровне основания
стомы..............................Prodesmodora Micoletzky, 1923 (табл. 7. 1 <
102(101). Амфиды спиральные или петлеобразные.
Таблица 7
1 — Prodesmodora circulata (Micoletzky, 1913) — передний отдел тела; 2, 3 — Ethmo-
laimus pratensis de Man, 1880: 2 — голова, 3 — задняя часть тела самца; 4, 5 — Рага-
cyatholaimus intermedius (de Man, 1880): 4 — голова, 5 — хвост самки; 6, 7 — Achro-
madora terricola (de Man, 1880); 8, 9 — Punctodora ratzeburgensis (Linstow, 1876'.
10 — Chromadorina viridis (Linstow, 1876) — голова; 11-13 — Chromadorita leuckarr.
(de Man, 1876); 14, 15 — Paraplectonema pedunculatum (Hofmaenner, 1913); 16 — Plec-
tus cirratus Bastian, 1865; 17 — Plectus aquatilis Andrassy, 1985; 18, 19 — Prismato-
laimus verrucosus Hirschmann, 1952: 19 — терминус хвоста; 20-22 — Chronogaster
typica\ 20 — передний отдел тела, 21 — задний отдел пищевода, 22 — хвост самки.
По: Алексеев, 1983; Цалолихин, 1994; Micoletzky, 1925; Zullini, 1982.
Нематоды
275
276
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
103(108). Амфиды спиральные.
104(105). Стома с параллельными стенками, окружена капсулой; в верхне части
стомы расположены 3 небольших зуба примерно одинаковых размеров...........
..............................Ethmolaimus de Man, 1880 (табл. 7, 2, 3).
105(104). Стома сложной конфигурации, в задней части воронкообразная, дор-
сальный зуб крупный, субвентральные очень маленькие.
106(107). Дорсальный зуб крупный, прямой, направлен вперёд; ректум не превы-
шает диаметра тела на уровне ануса........................................
......................Paracyatholaimus (Micoletzky, 1922) (табл. 7, 4, 5).
107(106). Дорсальный зуб крупный, когтевидный; ректум заметно превышает
диаметр тела на уровне ануса...........Achromadora Cobb, 1913 (табл. 7, 6, 7).
108(103). Амфиды петлеобразные.
109(110). Пищевод с бульбусообразным расширением в передней части; кардий
цилиндрический, «раздвигающий» пищевод и среднюю кишку....................
..........................Paraplectonema Strand, 1934 (табл. 7, 14-15).
110(109). Пищевод без бульбусообразного расширения в передней части; кардий
расположен в полости средней кишки........................................
.................................Plectus Bastian, 1865 (табл. 7, 16, 17).
111(100). Амфиды неразличимы или различимы плохо.
112(113). Бульбус отделён от средней кишки утончённой частью пищевода; тер-
минус хвоста с 3^4 шипиками.... Chronogaster Cobb, 1913 (табл. 7, 20-22).
113(112). Бульбус прилегает к средней кишке; терминус хвоста без шипиков.
114(117). Сильно развит только дорсальный зуб, субвентральные либо очень ма-
ленькие, либо отсутствуют.
115(116). Дорсальный сектор пищевода в области стомы асимметрично вздут..
............................Chromadorita Filipjev, 1922 (табл. 7, 11-13).
116(115). Вздутия в области стомы нет.... Punctodora Filipjev, 1930 (табл. 7, 8, 9).
117(114). Дорсальный зуб ненамного крупнее субвентральных................
...............................Chromadorina Filipjev, 1918 (табл. 7, 10).
118 (99). Пищевод без бульбуса.
Таблица 8
1-3 — Epitobrilus medius (G. Schneider, 1916): 1 — голова вентрально, 2 — голова
латерально, 3 — задний конец тела самца; 4-6 — Neotobrilus longus (Leidy, 1852):
4 — общий вид самца, 5 — голова, 6 — вагина; 7-9 — Semytobrilus pellucidus (Bastian,
1865): 7 — голова, 8 — задний конец тела самца, 9 — средняя часть тела самки; 10,
11 — Brevitobrilus stefanskii (Micolotzky, 1925): 10 — голова, 11 — задняя часть тела
самца; 12, 13 — Eutobrilus Andrassy (Altherr, 1963): 12 — голова, 13 — задняя часть
тела самца; 14-16 — Tobrilus gracilis (Bastian, 1865): 14 — голова, 75 — задняя часть
тела самца, 16 — хвост самки.
По: Цалолихин, 1995; 2000; 2001; 2005; 2009.
Нематоды
277
12
11
278
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
119(120). Кутикула кольчатая; стома узкая, с параллельными стенками; в основа-
нии стомы имеется маленький онх...........................................
..........................Prismatolaimus de Man, 1880 (табл. 7. 18, 19).
120(119). Кутикула гладкая или едва заметно кольчатая; стома бокаловидная с 2
карманообразными расширениями в основании, где располагаются 2 онха.
121(128). Стома с 2 карманами, разделёнными протоком и расположенными вдоль
оси тела.
122(123). Передний карман слит с буккальной полостью......................
..........................:.Epitobrilus Tsalolikhin, 1981 (табл. 8, 1-3).
123 (122). Передний карман отчётливо отделён от буккальной полости.
124(125). Вагина в виде луковицы; самцы с 3 очень крупными и 3 маленькими
супплементами; спикулы длинные и тонкие...................................
............................Neotobrilus Tsalolikhin, 1981 (табл. 8, 4-6).
125(124). Вагина узкая (не в виде луковицы); супплементы самцов одинаковых
размеров.
126(127). Вагина расположена перпендикулярно к поверхности тела; все 6 суп-
плементов самцов с узкой шейкой, одинаковых размеров; спикулы корот-
кие и широкие...................Brevitobrilus Tsalolikhin, 1981 (табл. 8, 10, 11).
127(126). Вагина расположена под углом к поверхности тела; супплементы сам-
цов редуцированы до едва заметных; спикулы длинные и тонкие...............
............................Semitobrilus Tsalolikhin, 1981 (табл. 8, 7-9).
128(121). Оба кармана расположены почти на одном уровне, налегая друг на дру-
га; онхи также расположены почти на одном уровне.
129(130). Супплементы самцов ежевидные, число их, как правило, не превыша-
ет 6...............................Eutobrilus Tsalolikhin, 1981 (табл. 8, 12, 13).
130(129). Супплементы самцов без шипиков, с уплощённой ампулой, число их,
как правило, более 6..................Tobrilus Andrassy, 1959 (табл. 8, 14-16).
Самки Eutobrilus и Tobrilus практически неразличимы.
ПРИЛОЖЕНИЕ
ПРЕСНОВОДНЫЕ МОЛЛЮСКИ СЕВЕРА
И СЕВЕРО-ЗАПАДА ЕВРОПЕЙСКОЙ РОССИИ
Отдельные части раздела подготовлены следующими авторами:
П.В. Кияшко (ЗИН РАН): Общая характеристика типа Mollusca; Ключ для
определения семейств брюхоногих моллюсков; Семейство Neritidae; Семейство
Viviparidae Gray, 1847; Семейство Bithyniidae Gray, 1857; Семейство Hydrobi-
idae Troschel 1857; Семейство Amnicolidae Tryon, 1863; Семейство Lithoglyphidae
Troschel, 1857; Семейство Valvatidae Gray, 1840; Семейство Dreissenidae Gray,
1840; совместно с M.B. Винарским — Класс брюхоногие моллюски Gastropoda
Cuvier, 1797 (общая характеристика); совместно с В.В. Богатовым — Класс дву-
створчатые моллюски Bivalvia Linnaeus, 1758 (общая характеристика); Ключ для
определения семейств пресноводных двустворчатых моллюсков.
М.В. Винарский (Омский гос. пед. университет): Методы сбора и камераль-
ной обработки пресноводных брюхоногих моллюсков; Семейство Acroloxidae
Thiele, 1931; Семейство Physidae Fitzinger, 1833; Семейство Lymnaeidae Rafin-
esque, 1815; совместно с П.В. Кияшко — Класс брюхоногие моллюски Gastropo-
da Cuvier, 1797 (общая характеристика).
Е.В. Солдатенко (Смоленский гос. пед. университет): Семейство Ап-
cylidae Rafinesque, 1815; Семейство Helisomatidae F.C. Baker, 1945; Семейство
Planorbidae Rafinesque 1815; Семейство Segmentinidae F.C. Baker, 1945.
В.В. Богатов (Биолого-почвенный институт ДВО РАН): Методы сбо-
ра и камеральной обработки пресноводных двустворчатых моллюсков; Семей-
ство Margaritiferidae Haas, 1940; Семейство Unionidae Fleming, 1828; совместно
с П.В. Кияшко — Класс двустворчатые моллюски Bivalvia Linnaeus, 1758 (общая
характеристика); Ключ для определения семейств пресноводных двустворчатых
моллюсков.
Общая редакция приложения — П.В. Кияшко (ЗИН РАН).
ПРЕДИСЛОВИЕ
Из всего многообразия пресноводных животных, моллюски: брюхоногие (Gas-
tropoda) и двустворчатые (Bivalvia), являются одной из наиболее крупных, широ-
ко распространённых и сложных для изучения групп гидробионтов. Они играют
важную роль на разных уровнях функционирования пресноводных экосистем: от
средообразующего действия крупных скоплений двустворчатых моллюсков до
280
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
элементарного значения особей гастропод и бивальвий в трофических сетях, транс-
формации веществ и потоках энергии, а также в качестве промежуточных хозяев
паразитических червей. Трудности в изучении пресноводных моллюсков отчасти
обусловлены небольшими размерами многих видов, затрудняющими их поиск в
водоёмах, последующее анатомирование и исследование морфологических струк-
тур. Кроме того относительная простота строения раковин и важных для диагно-
стики органов мягкого тела, как правило, лишённых у пресноводных моллюсков
ярких видовых особенностей, в сочетании со значительным размахом изменчи-
вости большинства признаков создают существенные проблемы для идентифика-
ции видов. Это вынуждает специалистов-малакологов привлекать к диагностике
таксонов новые методики, анализировать новые признаки или новые комбинации
признаков. Получаемые в результате данные обуславливают быстрые изменения в
системе пресноводных моллюсков, набирающие темп с конца XX века.
В России такие изменения были произведены Я.И. Старобогатовым и его уче-
никами. Не вдаваясь в обсуждение использованных методик и привлечённых при-
знаков, следует отметить, что созданная этими специалистами система пресно-
водных моллюсков, характеризуется значительно возросшим количеством родов
и видов, представленных на территории России — около 140 родов и более 1000
видов (Старобогатов и др., 2004). Большое количество установленных видов, от-
личающихся только небольшими изменениями пропорций раковины и/или фраг-
ментов половой системы, а также «бурлящий котёл» из, вовлекаемых в система-
тику группы всё новых признаков, и быстро сменяющих друг друга изменений в
таксономии и номенклатуре, обуславливают закономерное усложнение процедуры
определения таксонов и, как следствие, замедляют получение новых фаунисти-
ческих данных. Нередко всё это просто отпугивает специалистов из прикладных
областей биологии и студентов биологического профиля от детального изучения
пресноводных моллюсков при проведении фаунистических, гидробиологических,
паразитологических, экологических и биомониторинговых исследований.
При подготовке данного определителя авторами предпринята попытка хотя бы
частично изменить сложившуюся ситуацию. При этом основной задачей явилось
создание подробного пособия для идентификации пресноводных моллюсков, по-
зволяющего биологам различного профиля и квалификации определять по при-
знакам раковины все рода и хорошо очерченные виды. Такой подход, по замыслу
авторов, с одной стороны будет способствовать возрождению интереса к группе,
с другой позволит исследователям при обработке массового материала быстро
отделить легко определяемые виды (включённые в настоящий определитель) от
сложных и спорных таксонов, требующих углублённого изучения (с привлечени-
ем разных диагностических методик и специальной литературы) или обращения
за консультацией к малакологам — специалистам по конкретной группе.
Определитель включает: краткую характеристику типа Mollusca и две части,
посвящённые двум классам пресноводных моллюсков — Bivalvia и Gastropoda.
Каждая часть содержит краткую морфологическую характеристику класса, опи-
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России 281
сание методов сбора и обработки животных и собственно определительные
ключи. Для семейств и родов указаны номенклатурные типы, в виде отдельных
аннотаций даны краткая морфологическая характеристика и распространение
представителей. Для видов в тексте ключей указаны: ссылка на первоописание,
синонимы, некоторые морфологические характеристики, не вошедшие в ключи
(но необходимые для точного определения), размерные диапазоны и особенности
распространения. Там где необходимо основной текст дополнен краткими ком-
ментариями. Для некоторых видов двустворчатых моллюсков указаны экземпля-
ры, хранящиеся в коллекции ЗИН РАН. При этом после указания места хранения
приведены номера, под которыми моллюски хранятся в систематической коллек-
ции. Если экземпляры в коллекции ЗИН РАН указаны под другими названиями,
то эти названия приведены перед номером хранения.
При подготовке определителя нам не удалось найти специалиста по двуствор-
чатым моллюскам семейства Sphaeriidae, который бы согласился принять участие
в написании книги. Поэтому, чтобы не оставлять группу без внимания, в опреде-
лителе приводятся переработанные с учётом результатов более поздних таксоно-
мических исследований ключи для определения подсемейств, родов и подродов
шаровок из работы А.В. Корнюшина1 «Двустворчатые моллюски надсемейства
Pisidioidea Палеарктики (фауна, систематика, филогения)», изданной в Киеве в
1996 году тиражом в 300 экземпляров. Эта работа, ставшая на сегодняшний день
библиографической редкостью, является последней наиболее всесторонней обоб-
щающей сводкой по морфологии, анатомии, систематике и распространению
мелких пресноводных двустворчатых моллюсков.
При пользовании определителем необходимо помнить, что приведённые в
ключах для определения моллюсков семейств Margaritiferidae и Unionidae морфо-
метрические индексы не универсальны и применимы лишь к раковинам со стан-
дартными формами без видимых дефектов. В ключах не учтены промеры раковин
неполовозрелых и старых особей (если их параметры специально не оговарива-
ются), раковин с нарушениями роста в результате внешних воздействий, раковин
гибридов, а также уродливых, деформированных и сильно корродированных ра-
ковин. В то же время, для представителей этих семейств в иллюстрированные та-
блицы определителя помещены фотографии раковин с наиболее часто встречаю-
щимися отклонениями от типовых (стандартных) форм.
1 А.В. Корнюшин (1996) относил палеарктических шаровок к трём семействам: Sphaeriidae,
Euglesidae и Pisidiidae надсемейства Pisidioidea. По современным представлениям эти моллю-
ски относятся к надсемейству Sphaerioidea Deshayes, 1855 (1820), включающему наряду с ис-
копаемыми одно рецентное семейство Sphaeriidae с четырьмя подсемействами: Sphaeriinae,
Euglesinae; Euperinae и Pisidiinae (Bouchet, Rocroi, 2010). Внесённые в репринт ключей допол-
нения и изменения взяты из более поздних работ «Taxonomic revision of the genus Sphaerium
sensu lato in the Palaearctic Region, with some notes on the North American species» (Komiushin,
2001) и из главы «Моллюски» в «Определителе пресноводных беспозвоночных России и со-
предельных территорий» (Старобогатов и др., 2004).
282
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Каждый вид моллюсков проиллюстрирован в определителе фотографиями ра-
ковин, выполненными П.В. Кияшко, Е.В. Солдатенко, М.В. Винарским, В.В. Бога-
товым с экземпляров, хранящихся в малакологической коллекции Зоологического
института РАН (ЗИН РАН, Санкт-Петербург), Биолого-почвенного института ДВО
РАН (БПИ ДВО РАН, Владивосток), Омского государственного педагогического
университета (ОмГПУ, Омск). Ряд фотографий любезно предоставлены коллега-
ми: Т.Я. Ситниковой из Лимнологического института СО РАН (ЛИН СО РАН, Ир-
кутск), И.О. Нехаевым из Мурманского морского биологического института КНЦ
РАН (ММБИ КНЦ РАН, Мурманск) и Р. Gloer (Hetlingen, Германия).
Авторы выражают признательность Л.Л. Ярохнович (ЗИН РАН) за помощь в
работе с коллекциями моллюсков Зоологического института РАН, И.О. Нехаеву
(Мурманский морской биологический институт) за предоставленные фотографии
Lymnaea fulva, L. mucronata, L. subangulata, P. Gloer за предоставленные фотогра-
фии Acroloxus lacustris, Physa taslei), Т.Я. Ситниковой (ЛИН CO РАН) за предо-
ставленные фотографии раковин ряда видов родов Valvata и Cincinna, а также
И.Е. Аракчееву (дер. Конищево, Тверская обл.) за помощь в организации экспеди-
ции в бассейн р. Медведица.
П.В. Кияшко
ОБЩАЯ ХАРАКТЕРИСТИКА ТИПА
МОЛЛЮСКИ — MOLLUSCA
П.В. Кияшко
Беспозвоночные с мягким несегментированным телом, у представителей
большинства классов подразделяющимся на три отдела: голову (отсутствует у
двустворчатых), туловище и ногу (отсутствует у каудофовеат). На голове распо-
ложены рот, щупальца и глаза. Нога находится на брюшной стороне тела и в не-
которых случаях способна претерпевать значительные изменения соответственно
образу жизни и характеру движений животного. Чаще всего нога представле-
на плоской ползательной подошвой (брюхоногие моллюски). У двустворчатых
моллюсков нога образует плоский мускулистый вырост, иногда редуцирована.
Туловище моллюсков билатерально (двусторонне) симметрично или спираль-
но закручено. На большем или меньшем его протяжении имеется свешивающа-
яся книзу складка — мантия, наружная поверхность которой образует раковину.
У двустворчатых моллюсков края лепестков мантии могут срастаться, образуя
три отверстия: одно, расположенное примерно посередине брюшного края, слу-
жит для высовывания ноги и два (вводное и выводное) в задней части животно-
го — для обеспечения тока воды в мантийной полости. Иногда участки мантии,
окружающие вводное и выводное отверстия, разрастаются, образуя сифоны.
Между внутренней поверхностью мантии и телом животного находится мантий-
ная полость, сообщающаяся с внешней средой. Мантийная полость содержит жа-
бры, анальное отверстие, отверстия выделительных органов и иногда половых
протоков. У некоторых брюхоногих моллюсков жабры редуцируются, а участок
стенки мантии преобразуется в «лёгкое» (Pulmonata sensu lato).
У большинства моллюсков имеется раковина, образованная монолитной спи-
рально завитой трубкой, рядом пластинок или двумя створками. Стенка ракови-
ны состоит из органического конхиолинового наружного слоя (периостракум) и
одного-двух слоёв углекислого кальция (остракум и гипостракум).
Пищеварительная система представлена передней, средней и задней кишками.
Передняя кишка образует расширение («глотку») внутри которого помещается
особый орган захвата и измельчения пищи — радула (отсутствует у двустворча-
тых моллюсков). В глотку или переднюю кишку впадают протоки слюнных же-
лёз. Средняя кишка может формировать сложно структурированное вздутие —
желудок, кроме того в неё впадают протоки пищеварительной железы («печени»).
Задняя кишка имеет форму простой трубки или снабжена слепым карманом или
отростком («слепой кишкой»). В разных группах моллюсков кишечник проходит
либо почти прямо через тело, либо петлеобразно скручен перед терминально рас-
положенным анальным отверстием, либо, перекручиваясь, образует крутой изгиб
с перемещением ануса ближе к головному отделу. Органами выделения служат
284
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
видоизменённые метанефридии. Кровеносная система хорошо развитая, незам-
кнутая, однако с выраженной тенденцией к образованию обособленных крове-
носных сосудов. Имеется заключённое в перикард (околосердечную сумку) серд-
це, состоящее из желудочка и одного, двух или четырёх предсердий, количество
которых исходно связано с числом жабр. От желудочка отходят две аорты — к
голове и к висцеральному мешку. Отходящие от аорт артерии переходят в лаку-
ны. Нервная система — или с выраженными нервными стволами, или разбросан-
но-узлового типа — представлена нервными узлами (ганглиями), соединёнными
нервными волокнами. Большинство моллюсков раздельнополые, остальные —
гермафродиты. Представленные одной парой гонады обладают собственными по-
ловыми протоками и могут становиться непарной структурой путём слияния или
редукции. Оплодотворение наружное или внутреннее. В последнем случае фор-
мируется сложный копулятивный аппарат. Развитие прямое или с метаморфозом,
при котором имеются планктонные личиночные стадии.
В современной фауне насчитывается около 150 000 видов моллюсков, при-
надлежащих восьми классам (Кантор, Шилейко, 1994). Моллюски представляют
важный (часто доминирующий) компонент бентосной фауны морских и конти-
нентальных водоёмов, а их личинки играют заметную роль в планктонных со-
обществах. Многие группы этих животных являются ценными кормовыми объ-
ектами для ряда беспозвоночных, рыб (включая многих промысловых), птиц и
млекопитающих. Отдельные виды употребляются человеком в пищу; использу-
ются в качестве удобрений и в составе комбикормов в сельском хозяйстве и при
разведении рыб. Велика роль моллюсков в качестве промежуточных хозяев пара-
зитических червей — трематод. В пресноводных водоёмах обитают моллюски,
относящиеся к двум классам: Bivalvia Linnaeus, 1758 и Gastropoda Cuvier, 1797.
КЛАСС ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ —
BIVALVIA LINNAEUS, 1758
В.В. Богатов, П.В. Кияшко
Двустворчатые моллюски (класс Bivalvia) — одна из самых древних групп
донных беспозвоночных (наиболее ранние находки известны из верхнекембрий-
ских слоёв). Современные представители класса насчитывают около 20000 видов,
обитающих в Мировом океане, краевых морях, а также в солоноватых и пресных
водоёмах.
Тело пресноводных двустворчатых моллюсков одето раковиной, состоящей из
двух, как правило, симметричных створок (рис. 1). Плоскость их смыкания на-
зывается комиссуральной плоскостью. На каждой створке выделяют спинной,
брюшной, передний и задний края, плавно переходящие друг в друга или обра-
зующие на границе смежных краёв закруглённый уголок. Створки на спинной
стороне связаны эластичной перемычкой — лигаментом. Основная, наиболее
заметная часть лигамента всегда лежит позади макушек, что определяет поло-
жение переднего и заднего концов створки (рис. 1). Часто лигамент расположен
на известковых пластинках (нимфах), которые начинаются под макушкой и резко
Рис. 1. Раковина перловицы Unio pictorum L. По: Жадин, 1938.
1 — задние мускульные отпечатки; 2 — латеральные зубы; 3 — кардинальные зубы; 4 —
передние мускульные отпечатки; 5 — мантийная линия; 6 — щит; 7 — щиток; 8 — сину-
лус; 9 — макушка (верхушка); 10 — лигамент; 11 — синус. Схема промеров: АА1 — длина
раковины; ББ} — высота раковины; ГГ{ — выпуклость (толщина, ширина) раковины;
АВ — передняя часть раковины; BAt — задняя часть раковины.
286
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
обрываются на заднем крае, образуя уголок у основания нимфы. Над спинным
краем в той или иной степени возвышается макушка — начальная часть каждой
створки. Она может располагаться посредине спинного края или может быть в
той или иной степени сдвинутой вперёд или назад. Форма поверхности раковины
около макушек (степень её приподнятости над краем створки) имеет таксономи-
ческое значение.
У некоторых групп моллюсков спинной край за макушками может разрастать-
ся, закрывая лигамент и образуя наклонённое к переднему краю крыло. Впереди
макушек у ряда видов имеется уплощённый участок ланцетовидной формы —
щиток (рис. 1). Аналогичный, но более вытянутый участок, называемый щитом,
формируется и позади макушек. Позади лигамента расположено углубление, за-
крытое плёнкой, которое носит название лигаментного синуса. Впереди лигамен-
та и макушек между створками находится ланцетовидное отверстие, прикрытое
плёнкой — синулус. На наружной поверхности створки видны линии нарастания
(роста), а реже — концентрические валики — морщины. Наиболее выделяющиеся
своей глубиной и чёткостью линии роста обычно рассматриваются как годовые
кольца. Для представителей Unionidae характерна макушечная скульптура в виде
валиков или бугорков разной толщины и конфигурации. Макушечная скульптура
важна при определении родовой принадлежности этих моллюсков. У некоторых
раковин макушечная скульптура может быть не развита или не видна из-за корро-
зии. Часто за макушками имеется линия в виде резкого перегиба, проходящего от
макушки до нижней части заднего края створки — задний килевой перегиб.
У многих Bivalvia внутри раковины имеется замок, состоящий из расширен-
ного спинного края — замочной площадки и, расположенных на ней, нескольких
выступов различной формы — зубов. Замок предназначен для точного совмеще-
ния створок при их смыкании, поскольку каждому зубу одной створки соответ-
ствует углубление в другой. Замки бывают различных типов и имеют большое
значение при выделении таксонов высокого ранга. У представителей Unionidae и
Margaritiferidae замок схизодонтный, относящийся к группе прегетеродонтных, у
Sphaeriidae — гетеродонтный.
На внутренней поверхности раковины в её передних и задних частях имеют-
ся по две пары мускульных отпечатков — мест прикрепления мускулов-замыка-
телей (аддукторов) и мышц, втягивающих ногу (ретракторов), к створкам. По-
верхность отпечатков у Unionidae обычно гладкая, с заметными линиями роста.
У представителей Margaritiferidae передние отпечатки, хотя бы частично, имеют
морщинистый рельеф. Задний аддуктор прикрепляется близ спинного края створ-
ки. Рядом с отпечатками аддукторов (или сливаясь с ними) помещаются отпечат-
ки ретракторов ноги. В некоторых случаях аддукторы крепятся к особым вну-
тренним примакушечным выростам — септам (например, у представителей рода
Dreissend). Вдоль краёв створок тянется след от прикрепления мышц мантии —
мантийная линия. В тех случаях, когда имеются развитые сифоны, мантийная
линия в задней половине створки образует более или менее глубокий изгиб —
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
287
Рис. 2. Поперечный шлиф ра-
ковины беззубки Anodonta. По:
Жадин, 1933.
1 — периостракум; 2 — остракум
(призмы); 3 — гипостракум (перла-
мутр); 4 — зёрнышки конхиолина.
мантийный синус. Участок между мантийной
линией и брюшным краем створки имеет вид
слабовыпуклого валика, называемого прикра-
евым валиком. У представителей Margaritiferi-
dae мантийные мускулы прикрепляются кроме
мантийной линии почти по всей внутренней
поверхности створки.
Стенка раковины образована тремя слоя-
ми (рис. 2). Внешний слой — периостракум,
представленный органическим веществом —
конхиолином, может достигать значительной
толщины. Он обладает определённой устойчи-
востью к химическим воздействиям и служит
для защиты подлежащих слоёв углекислой
извести. В некоторых случаях конхиолин на-
столько тонок, что может стираться (главным
образом в области макушек). Остракум обра-
зует основную стенку раковины и представлен
призматическими кристаллами углекислой из-
вести. Гипостракум (внутренний слой) — это пластинчатые кристаллы углекис-
лой извести, расположенные параллельно поверхности створки и способные ир-
радировать, образуя характерный перламутровый блеск. У некоторых моллюсков
перламутр отсутствует, а гипостракум представляет собой матовый фарфоровид-
ный слой с перекрёстно-пластинчатой структурой известковых кристаллов.
Листки мантии, покрывающие мягкое тело моллюска, срастаются в несколь-
ких местах, оставляя, как правило, три отверстия: большее для ноги и два мень-
ших сифональных отверстия — нижнее (жаберный сифон) для входа воды и верх-
нее (анальный сифон) для её выброса (рис. 3). Край и внутренняя поверхность
нижнего сифонального отверстия, а иногда и край верхнего (например, у некото-
рых Nodulariinae) снабжены папиллами. Их количество, форма и порядок распо-
ложения могут иметь таксономическое значение. Иногда дорсально от сифональ-
ных отверстий раположено супрасифональное отверстие. В мантийной полости
имеются полужабры (жабры) по две пары с каждой стороны. Часто внутренние
листки полужабр срастаются, образуя горизонтальную перегородку, разделяю-
щую мантийную полость на верхнюю (супрабранхиальную) и нижнюю части.
Нижняя часть является дыхательной полостью, а через верхнюю происходит уда-
ление отработанной воды. Нога в большинстве случаев топоровидная или кли-
новидная, хорошо развита и направлена концом вперёд. На верхней поверхности
заднего края ноги иногда формируется плоский листок, расположенный между
жабрами — педальный киль. У моллюсков рода Dreissena нога сильно укорочена
в связи с прикреплённым образом жизни. У её основания имеется железа, форми-
рующая нити биссуса, которыми моллюск прикрепляется к твёрдому субстрату.
288
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 3. Внутреннее строение беззубки Anodonta (мантия и жабры правой стороны
удалены). По: Жадин, 1933.
I — головной, II — ножной, III — висцеральный нервные узлы; 1 — передний мускул-за-
мыкатель раковины; 2 — задний мускул-замыкатель раковины; 3 — анальный сифон; 4 —
жаберный сифон; 5 — задний мускул-ретрактор; 6 — верхний отдел боянусова органа;
7 — нижний отдел боянусова органа; 8 — предсердие; 9 — желудочек сердца; 10 — киш-
ка; 11 — сообщение почки с околосердечной сумкой; 12 — прикрепление двух пластинок
правого внутреннего жаберного листа; 13 — ротовые щупальца; 14 — отверстие протока
печени; 15 — желудок; 16 — передний мускул-ретрактор; 17 — нога; 18 — половая желе-
за; 19 — внутренняя жабра; 20 — печень.
С двух сторон от ротового отверстия расположены своеобразные мускулистые
ресничные лопасти — ротовые щупальца. Пищеварительная система состоит из
короткого пищевода, сложно устроенного мешкообразного желудка, средней и
задней кишок. В желудок открываются пищеварительные дивертикулы (протоки
«печени») и отверстие мешка, содержащего «кристаллический стебелёк» (сту-
денистую массу, состоящую из белкового вещества типа глобулина). Слюнные
железы отсутствуют. Кровеносная система незамкнутая, сердце расположено
на спинной стороне, оно состоит из охватывающего заднюю кишку желудочка
и двух боковых предсердий, заключённых в околосердечную сумку — перикар-
дий. От желудочка отходят передняя и задняя аорты. Выделительная система
представлена парными почечными мешками или боянусовыми органами, сооб-
щающимися посредством реноперикардиальных каналов с околосердечной сум-
кой и посредством нефропоров с мантийной полостью. Кроме того имеются экс-
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
289
креторные перикардиальные железы. Нервная система в виде трёх пар ганглиев
(головных, ножных и висцеральных), соединённых между собой длинными кон-
нективами. Половая система образована парными гонадами, открывающимися в
супрабранхиальное пространство мантийной полости, или впадающими в прото-
ки выделительной системы. Пресноводные двустворчатые моллюски — раздель-
нополые, за исключением мелких двустворок из семейства Sphaeriidae, которые
являются гермафродитами. Иногда и у крупных двустворчатых (беззубок и перло-
виц) в условиях стоячих замкнутых водоёмов могут появляться гермафродитные
особи. Кроме этого у крупных двустворчатых моллюсков часто наблюдаются слу-
чаи межвидового скрещивания. Гибридам обычно свойственны признаки обоих
родителей, что затрудняет определение моллюсков.
Индивидуальное развитие у представителей семейств Unionidae и Margaritifer-
dae протекает с образованием своеобразной личинки (глохидия) определённое
время паразитирующей на рыбе. Формирующиеся из оплодотворённых яиц гло-
хидии, сначала сохраняются в особым образом изменённых участках жабр —
марсупиях. У Margaritiferidae марсупии образуются как в наружных, так и вну-
тренних полужабрах, у Unionidae — только в наружных полужабрах. Выброс
зрелых глохидиев происходит через выводной сифон моллюска. У представите-
тей рода Dreissena из яйцевых оболочек выходит личинка, которая через очень
кроткое время преобразуется в планктонную двустворчатую стадию — ротигер
велигер). В толще воды такая личинка перемещается с помощью особого двига-
тельного аппарата — велюма (паруса). По мере роста личинка приобретает ма-
<ушки, но сохраняет велюм. Такая личинка называется великонхом. Достигнув
зпределённого возраста, личинка опускается на дно, велюм редуцируется, и мо-
тэдой моллюск переходит к образу жизни, характерному для взрослых особей.
Двустворчатые моллюски ведут малоподвижный образ жизни, а особи рода
Z^eissena прикрепляются к твёрдому субстрату нитями биссуса, при этом они
зтособны образовывать обширные многоярусные скопления — друзы (рис. 4).
Малоподвижный образ жизни, отсутствие головы и специализированных органов
гснаружения пищи определяют способ питания двустворчатых за счёт взвешен-
-ых в воде пищевых частиц, подгоняемых к ротовому отверстию мерцательным
тзижением ресничек ротовых лопастей или жабр. Основными объектами питания
сляются взмученный детрит и микропланктон. Ток воды, обусловленный филь-
-тационной активностью двустворчатых моллюсков, улучшает кислородный ре-
и санитарное состояние водоёмов.
Двустворчатые моллюски разных возрастных стадий активно поедаются ры-
гами и птицами (утиные, гусеобразные). Агглютинированные фекалии двуствор-
чатых моллюсков являются ценным кормом для многих детритофагов. Некоторые
•лты Bivalvia используются человеком в гастрономических и промышленных це-
Мясо потребляемых моллюсков обладает высокой калорийностью, содержит
♦:-огие витамины и микроэлементы (такие как йод, медь, железо, цинк). Толчё-
ные раковины и тела входят в состав удобрений, а также кормовых смесей, ис-
290
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 4. Друза из дрейссен Dreissena polymorpha (Pallas) различного возраста на ра-
ковине двустворчатого моллюска. Оз. Дуглас (Douglas Lake), Мичиган, США. Фото
В. Богатова.
пользуемых при разведении рыб и домашней птицы. В настоящее время больше
половины добываемых в мире двустворчатых моллюсков получают в результате
промышленного разведения.
Методы сбора и камеральной обработки пресноводных
двустворчатых моллюсков
Методы изучения пресноводных двустворчатых моллюсков описаны в ряде
специальных руководств и монографий (Жадин, 1938; Алимов, Богатов, 1975;
Старобогатов, 1977; Шкорбатов, Старобогатов.
1990; Богатов и др., 2005).
Крупные двустворчатые моллюски собира-
ются вручную или также как и мелкие — с ис-
пользованием специальных орудий лова, среди
которых наиболее эффективен зубчатый сачок
(рис. 5). Кроме того, для сбора моллюсков при-
меняют драги различных конструкций, черпа-
Рис. 5. Зубчатый сачок для ловли Unionidae.
По: Жадин, 1938.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 17S8
291
Рис. 6. Сбор двустворчатых моллюсков с помощью граблей. Фото И. Аракчеева.
ки, небольшие бредни с привязанной к нижней подборе цепью, невода. Для разо-
вых сборов можно использовать широкие грабли с решёткой на тыльной стороне,
выполненной из металлических стержней и проволоки (рис. 6). Мелкие моллю-
ски имеют довольно большую плотность поселения и поэтому обильны в дночер-
пательных пробах.
Первую фиксацию двустворчатых моллюсков следует проводить 96% этило-
вым спиртом, который через сутки следует заменить на 70% спирт. Пустые рако-
вины лучше всего сохранять в сухом виде. Чтобы раковины не растрескивались
при высыхании, их смазывают глицерином.
Для изучения строения мягкого тела моллюсков их раковины вскрывают.
У крупных экземпляров скальпелем или тонким ножом как можно аккуратнее пе-
ререзают аддукторы на участке между мантией и створкой, после чего раковина
раскрывается. Мелких моллюсков при таком способе легко повредить, поэтому
их перед вскрытием выдерживают в течение 1-2 суток в 40-50% растворе эти-
лового спирта. При такой процедуре створки раскрываются за счёт слабой маце-
пации в зоне прикрепления аддукторов. После этого материал фиксируют в 70%
;лирте.
Обычно раковина двустворчатых моллюсков измеряется по трём стандартным
промерам: длина — расстояние по продольной оси раковины между наиболее
.палёнными точками переднего и заднего краёв; высота — расстояние по перпен-
дикуляру к продольной оси раковины между макушкой (высота у макушек), либо
пигаментом (высота у лигамента), либо вершиной крыла (высота у крыла) и наи-
:елее удалённой точкой брюшного края; выпуклость — расстояние по перпен-
292
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 7. Основные промеры раковин крупных двустворчатых моллюсков (А — вид
сбоку, Б — вид спереди в позиции максимально выпуклого сечения): м — макушка,
т — наиболее выступающая точка боковой поверхности раковины, вк — высота
раковины у крыла, вм — высота у макушки, д — длина раковины, вп — выпуклость
двух створок, мт — расстояние между макушкой и наиболее выступающей точкой
боковой поверхности раковины, лмвк — линия максимально выпуклого контура на
боковой поверхности створки, мвк — высота максимально выпуклого контура. По:
Старобогатов и др., 2004 (частью).
дикуляру к комиссуральной плоскости между наиболее удалёнными от неё точ-
ками створок (рис. 1; рис. 7). Высота у лигамента обычно измеряется в случаях
сильной изъеденности (коррозии) макушек, сломанном крыле или когда макушка
значительно смещена к переднему краю и расположена заметно ниже лигамента.
При измерении высоты раковины необходимо указывать, в каком месте было про-
ведено измерение (у макушки, лигамента или крыла). Степень выпуклости рако-
вины (индекс выпуклости) обычно оценивается соотношением выпуклости рако-
вины (В) к её высоте (Н). Если значение индекса В/Н меньше 0,3-0,4 — раковина
считается плоской или слабовыпуклой; при значении индекса от 0,3-0,4 до 0,5-
0,6 — умеренно выпуклой; от 0,5-0,6 до 0,7 — выпуклой и свыше 0,7 — сильно-
выпуклой. Через соотношение высоты раковины (Н) к её длине (L) определяется
общая форма раковины. Например, если у моллюсков с неразвитым крылом соот-
ношение Н/L не превышает 0,45, то раковина считается удлинённо-овальной; от
0,5 до 0,6 — овальной, и около 0,7-0,8 — яйцевидной. Наличие островершинного
крыла в совокупности с узким передним и задним краями придаёт раковине тре-
угольную форму, а у моллюсков без крыла, но с заметными резкими переходами
или уголками между, как минимум, тремя краями раковины (например: передним
и спинным, спинным и задним, задним и нижним) раковина оценивается как тра-
пециевидная или овально-четырёхугольная. Суженная за макушками задняя часть
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
293
Рис. 8. Раковины перловицы (левые створки) Unio protractus Lindholm.
а — раковина, близкая к стандартной форме (задний край створки частично недоразвит),
б — раковина с неразвитой задней частью (р. Ивица, Тверская обл.; коллекция БПИ ДВО
РАН), в — раковина с неразвитой задней частью спинного края (Дельта Волги, Белинский
канал; коллекция ЗИН РАН). Масштаб 3 см.
раковины придаёт ей коническую форму, а суженная передняя часть раковины —
каплевидную или клювовидную.
Важно помнить, что у видов, обладающих аллометрическим типом роста, с
увеличением размеров моллюсков соотношения основных промеров раковины
могут значительно изменяться. В таких случаях в определительных ключах ука-
зываются значения индексов, рассчитанные только для средневозрастных моллю-
сков, при этом результаты промеров молодых и старых раковин не учитываются,
либо их значения оговариваются специально. Кроме того, в процессе развития
двустворчатых моллюсков может наблюдаться резкое замедление роста (рис. 8а,
б) или, наоборот, разрастание (рис. 9а, б) задней части спинного и/или заднего
краёв раковины, т.е. тех её частей, которые не погружены в грунт. В результате
общая форма створок может заметно искажаться. При определении таких моллю-
сков, необходимо обращать внимание на то, что поверхность раковины на участ-
ках с угнетённым ростом становится морщинисто-шероховатой, а её призматиче-
ский и перламутровый слои — тонкими. У экземпляров с разросшимся задним
краем линии роста наиболее раннего времени образования, как правило, очерчи-
вают контуры близкие к стандартной форме раковины (рис. 9в). Подобные иска-
жения формы моллюсков не имеют систематического значения и, по-видимому,
связаны с воздействием на край мантии, выделяющей раковину, метаболитов со-
:юшества перифитона, развивающегося на омываемых водой участках створок, и
:остоящего, в основном, из бактерий, водорослей, грибов, мелких беспозвоноч-
ных и др. организмов.
294
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 9. Раковины перловицы (левые створки) Crassiana crassa (Philipsson in Retzius).
a — стандартная форма (p. Ока у г. Калуга, коллекция ЗИН РАН); б — раковина с раз-
росшимся задним краем створки (из коллекции Вестерлюнда, коллекция ЗИН РАН);
в — часть той же раковины: вид створки близ макушки с линиями роста раннего времени
образования. Масштаб 1 см.
При видовой идентификации двустворчатых моллюсков иногда применяют
компараторный метод, позволяющий сравнивать рисунки внешних контуров по-
перечных максимально выпуклых сечений створок (МВС). Это сечения, которые
проходят через макушку и точки, максимально удалённые в разные моменты об-
разования раковины от комиссуральных плоскостей (обычно МВС проходит
под углом к продольной оси раковины) (рис. 7)1. Существует точка зрения, что
1 Применявшиеся в 1970-1990 годах прошлого столетия для таксономических целей конту-
ры фронтальных сечений створок (Старобогатов, 1977; Шкорбатов, Старобогатов, 1990), т.е.
сечений, проведённых через макушку, перпендикулярно продольной оси раковины, или пер-
пендикулярно линиям, соединяющим нижние края передних и задних зубов (у перловиц), или
перпендикулярно середине отпечатков мускулов замыкателей (у беззубок) — не подходят для
видовой идентификации двустворчатых моллюсков. У большинства видов Bivalvia необходи-
мое для сравнение фронтальное сечение не совпадает с внешним фронтальным контуром створ-
ки. При установке раковины передним краем вверх в микроскоп видны контуры максимально
выпуклого сечения, но не перпендикулярного продольной оси раковины, а наклонённого до 40
градусов. У моллюсков с развитым крылом при такой позиции видно закилевое поле (закиле-
вую поверхность). Следует также иметь в виду, что у большинства крупных Bivalvia в процессе
роста задняя часть раковины с большей скоростью отодвигается от комиссуральной плоскости
по сравнению с передней частью. Если в этом случае рассматривать контуры фронтальных се-
чений у разных по размеру раковин, то чем крупнее моллюск, тем видимый в микроскоп внеш-
ний контур створки будет дальше смещаться к заднему краю. Фактически в данной ситуации
мы будем сопоставлять разные участки створок, т.е. иметь дело с разными кривыми (изменение
формы тела в процессе роста наблюдается и по другим характеристикам, например, по изме-
нению траектории нарастания створок и др.). Ситуация становится ещё более сложной, если
<ласс двустворчатые моллюски — Bivalvia Linnaeus, 17S8
295
> многих таксономических групп
моллюсков форма (степень кри-
визны) контуров МВС видоспе-
пифична. Здесь следует иметь в
виду, что раковины двустворчатых
моллюсков растут от макушки, и
ождая из траекторий нарастания
<меет собственную форму кривой/
:лирали. Направления траекторий
взрастания можно проследить, на-
пример, на некоторых створках по
расходящимся от макушки тёмным
полосам (рис. 10). Контуры МВС
Рис. 10. Левая створка перловицы Crassiana
папа (Lamarck) с расходящимися от макушки
полосами (р. Жаксыкар в окр. г. Актюбинска,
коллекция ЗИН РАН). Масштаб 1 см.
представляют собой максимально выпуклую траекторию, по которой происходит
нарастание раковин. Контуры таких траекторий наиболее удобны при сопостав-
лении разных по размеру моллюсков.
Для прорисовки контуров МВС применяют микроскоп (увеличение 6 * 0,6) и
рисовальный аппарат. Створку под микроскопом устанавливают передним концом
ззерх в положение, при котором полярные оптические оси микроскопа лежали бы
з комиссуральных плоскостях разного времени образования, в том числе и наибо-
лее раннего (подобные плоскости очерчены линиями роста). За начальную точку
развертывания кривой МВС (полярный центр) берут точку схождения у макушки
хорошо видимых в микроскоп линий роста (если макушка изъедена, то линии ро-
рта прорисовывают до точки их схождения). Считается, что в некоторых случаях
полученные контуры могут быть близки к логарифмической или равноугольной
опирали. Отличительное свойство логарифмической спирали заключается в том,
^по угол (его обозначают греческой буквой а) между касательной к любой точке
спирали и полярным радиусом (линией, проведённой из полярного центра к точке
осания) остаётся для данной спирали постоянным (рис. Па). Характер кривиз-
ны контуров логарифмических спиралей определяется величиной угла а, а также
ттношением ширины спирали к её высоте (рис. 116). Если контур максимально
выпуклого сечения отличается от логарифмической спирали, то угол а в процессе
тоста может увеличиваться (тогда с возрастом раковина становится более выпу-
клой) или уменьшаться (тогда с возрастом раковина становятся более плоской).
В некоторых случаях траектории нарастания створок представлены иными кривы-
ми или ломаными линиями, не имеющими отношение к спиралям.
галивину устанавливают по линии, проходящей через нижние края зубов, либо через середины
скальных отпечатков. В результате можно получить разные контуры даже у близких по раз-
<е?у моллюсков одной и той же генерации, т.к. зубы/отпечатки по отношению к продольной
тег? раковины могут находиться под разными углами (разные контуры у одноразмерных особей
<сгут получаться при фронтальном положении моллюска и при условии, если максимально вы-
~ клые сечения створок находятся под разными углами к продольной оси раковины).
296
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 11. Логарифмические спирали.
А — постоянный угол логарифмической спирали, а. (О — полярный центр; ОА, ОВ и ОС —
полярные радиусы); Б — соотношение между выпуклостью логарифмической спирали S
к её высоте Н в двух произвольно взятых позициях — Sp S, и Нр Н2, соответственно.
В определительных ключах для приблизительной характеристики степени
кривизны максимально выпуклых контуров, близких к логарифмическим спира-
лям, используют индекс выпуклости МВС, представляющий собой соотношение
выпуклости раковины к высоте/длине МВС, т.е. линии между макушкой и брюш-
ным краем раковины, проведённой через точку максимально удалённую от ко-
миссуральной плоскости (рис. 7, лмвк). Иногда используют отношение выпукло-
сти раковины к расстоянию между макушкой и точкой максимально удалённой
от комиссуральной плоскости. Чем ближе кривая соответствует логарифмической
спирали, тем более точно проводится определение моллюсков с помощью индек-
сов выпуклости МВС. Если максимально выпуклое сечение у разных по разме-
ру моллюсков наклонено к продольной оси раковины примерно под одинаковым
углом — в определительных ключах используют обычный индекс выпуклости
(отношение выпуклости раковины к её высоте). Такой подход особенно актуален
для моллюсков с удлинённо-овальной или эллиптической формой раковины, для
которых точное определение высоты/длины МВС проблематично. Важно пом-
нить, что при заметном отклонении кривой контура МВС от логарифмической
спирали, обычный индекс выпуклости и индекс выпуклости МВС оцениваются
для средневозрастных моллюсков, а при значительном изменении угла а индексы
выпуклости оцениваются для разных произвольно выбранных размерных групп,
либо приводятся уравнения зависимости индексов выпуклости от каких-либо ли-
нейных параметров раковины.
\.асс двустворчатые моллюски — Bivalvia Linnaeus, 1758
297
Список литературы
Алимов А.Ф., Богатов В.В. 1975. Рост беззубки Anodonta piscinalis в водохранилищах Ка-
лининской области // Зоол. ж. Т. 54. Вып. 1. С. 27-31.
Андрусов Н.И. 1897. Ископаемые и живущие Dreissensidae Евразии // Тр. СПб. общ-ва
естествоисп. Отд. геол, и мин. Т. 25. С. i-iv, 1-683. (Докт. дис. Н.И. Андрусова.)
Еюгатов В.В., Старобогатов Я.И., Прозорова Л.А. 2005. Моллюски рода Colletopterum (Апо-
dontinae, Bivalvia) России и сопредельных территорий // Зоол. ж. Т. 84. № 9. С. 1050-1063.
эсгуцкая Н.Г., Кияшко П.В., Насека А.М., Орлова М.И. 2013. Определитель рыб и бес-
позвоночных Каспийского моря. Т. 1. Рыбы и моллюски. СПб.; М.: Т-во науч, изданий
КМК. 543 с.
аддин В.И. 1933. Пресноводные моллюски СССР. Л.: Ленснабтехиздат. 232 с.
Кддин В.И. 1938. Семейство Unionidae. Фауна СССР. Моллюски. Т. 4. Вып. 1. М.; Л.:
Изд-во АН СССР. 170 с.
р’-иин В.И. 1952. Моллюски пресных и солоноватых вод СССР. Определители по фауне
СССР, издаваемые Зоологическим институтом академии наук СССР. М.; Л.: Изд-во
.АН СССР. 346 с.
Зевина Г.Б. 1958. Моллюски Mytilaster lineatus (Gmelin) и Dreissena polymorpha Pall, в об-
растаниях на Каспийском море // Изв. АН Туркм. ССР. № 4. С. 57-62.
<±нтор Ю.И., Шилейко. А.А. 1994. О разнообразии моллюсков // В.Е. Соколов, Ю.С. Ре-
шетников (ред.). Биоразнообразие. Степень таксономической изученности. М.: Наука.
С. 86-97.
«Стрнюшин А.В. 1996. Двустворчатые моллюски надсемейства Pisidioidea Палеарктики
(фауна, систематика, филогения). Киев. 175 с.
Тиндгольм В.А. 1932. Пресноводные моллюски из плиоценовых отложений по р. Ирты-
шу // Тр. Всесоюз. геол.-развед. объединения НКТП СССР. Вып. 239. С. 3-27 + 7 табл.
’гхнович В.П., Каратаев А.Ю., Ляхов С.М., Андреев Н.И., Андреева С.И., Афана-
сьев С.А., Дыга А.К., Закутский В.П., Золотарева В.И., Львова А.А., Некрасова М.Я.,
Осадчих В.Ф., Плигин Ю.В., Протасов А.А., Тищиков Г.М. 1994. Условия обита-
ния // Я.И. Старобогатов (ред.). Дрейссена, Dreissena polymorpha (Pall.) (Bivalvia,
Dreissenidae). Систематика, экология и практическое значение. М.: Наука. С. 109-119.
Старобогатов Я.И. 1970. Фауна моллюсков и зоогеографическое районирование водоёмов
земного шара. Л.: Наука. 372 с.
Старобогатов Я.И. 1977. Класс двустворчатые моллюски Bivalvia // Определитель пресно-
водных беспозвоночных европейской части СССР (планктон и бентос). Л.: Гидроме-
теоиздат. С. 123-151.
Старобогатов Я.И. 1994. Систематика и палеонтология // Я.И. Старобогатов (ред.). Виды
фауны России и сопредельных стран. Дрейссена Dreissena polymorpha (Pall.) (Bivalvia,
Dreissenidae). M.: Наука. C. 18-46.
Старобогатов Я.И., Андреева С.И. 1994. Ареал И Я.И. Старобогатов (ред.). Виды фауны
России и сопредельных стран. Дрейссена Dreissena polymorpha (Pall.) (Bivalvia, Dreis-
senidae). M.: Наука. C. 47-53.
Старобогатов Я.И., Корнюшин А.В. 1986. Особенности яйцеживорождения и систематика
сфериид (Bivalvia Pisidioidea Sphaeriidae) // Тр. Зоол. ин-та АН СССР. Т. 152. С. 30-41.
Старобогатов Я.И., Прозорова Л.А., Богатов В.В., Саенко Е.М. 2004. Моллюски // С.Я. Ца-
лолихин (ред.). Определитель пресноводных беспозвоночных России и сопредельных
территорий. Т. 6. Моллюски, полихеты, немертины. СПб.: Наука. С. 9-^491.
298
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Шевченко Т.К. 1949. О движении дрейссены // Природа. № 9. С. 82-83.
Шкорбатов Г.Л., Старобогатов Я.И. (ред.). 1990. Методы изучения двустворчатых моллю-
сков И Тр. Зоол. ин-та АН СССР. Т. 219. Л. 208 с.
Bogatov V.V., Prozorova L.A., Starobogatov Y.I. 2003. The family Margaritiferidae (Mollusca:
Bivalvia) in Russia // Ruthenica. V. 13. No 1. P. 41-52.
Bouchet P., Rocroi J.-P., Fryda J., Hausdorf B., Ponder W., Valdes A., Waren A. 2005. Classifica-
tion and nomenclator of gastropod families // Malacologia: International Journal of Mala-
cology. 47 (1-2). Hackenheim. Germany: ConchBooks. 397 p.
Gmelin J.F. 1791. Caroli a Linne Systema Naturae per regna tria naturae, secundum classes, or-
dines, genera, species; cum characteribus, differentiis, synonymis, locis. Editio decimo ter-
tia, reformata. Tomus 1, pars 6. Lipsia (Leipzig): G.E. Beer. P. 3021-3910.
Holandre J. 1836. Faune de departement de la Moselle. Mollusques ou coquilles terrestres et flu-
viatiles des environs de Metz. Metz, Thiel, 58 p.
Kantor Yu.I., Vinarski M.V., Schileyko A.A. 2010. Catalogue of the continental mollusks of
Russia and Adjacent territories. Version 2.3. Электронная публикация: http://www.
ruthenica.com/documents/Continental_Russian_molluscs_ver 2-3-1.pdf
Komiushin A.V. 2001. Taxonomic revision of the genus Sphaerium sensu lato in the Palaearctic
Region, with some notes on the North American species (Bivalvia: Sphaeriidae) // Archiv
fur Molluskenkunde. Band, 129 (1/2). P. 77-122.
Kiister H.C. 1838. Die Gattung Anodonta nebst den iibrigen Najaden mit unvollkommenem Schloss
Systematisches Conchylien-Cabinet von Martini und Chemnitz. Bd. 9. Abth. 1. P. 41-64.
Lamarck J.P.B.A. de Monet de. 1819. Histoire naturelie des animaux sans vertebres, present-
ant les caracteres generaux et particuliers de ces animaux, leur distribution, leurs classes,
leurs families, leurs genres, et la citation des principales especes qui s’y rapportent; precede
d’une introduction offrant la determination des caracteres essentiels de Г animal, sa distinc-
tion du vegetal et des autres corps naturals, enfi n, 1’exposition des principes fondamentaux
de la zoologie. Tome sixieme. Premiere partie. Paris: Verdiere. P. i-vi + 1-343.
Linnaeus C.1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera,
species, cum characteribus, differentiis, synonymis, locis. Tomus 1. Editio decima, refor-
mata. Holmia (Stockholm): Laurentii Salvii. iv + 824 p.
Nilsson S. 1822. Historia Molluscorum Sveciae terrestrium et fluviatilim breviter delineata.
Lund, 124 p.
Pallas P.S. 1771. Reise durch verschiedene Provinzen des Russischen Reichs. Erster Theil.
Physicalische Reise durch verschiedene Provinzen des Russischen Reichs im 1768 und
1769sten Jahre. St. Petersburg: Kaiserlichen Akademie der Wissenschaften. [12] + 504 p.
Tabs. 1-11 + Tabs. A-L.
Pfeiffer C. 1825. Naturgeschichte Deutcher Land- und Susswasser-Mollusken, 2. Weimar, Verla-
ge des GrossherzogL Sachs, 40 p.
Retzius A. J. 1788. Dissertatio historico-naturalis sistens nova Testaceorum genera. Quam venia
ampliss. facult. philosophicae preside D. M. Andr. J. Retzio ... defert L. M. Philipsson ...
in-4°, Lundae, 23 p.
Rossmassler E.A. 1835. Iconographie der Land- und Susswasser-Mollusken Europa’s, mit
vorzuglicher Beriicksichtigung der Europaischen noch nicht abgebildeten Arten. Dresden,
Leipzig, 1(1). P. 1-132.
Rossmassler E.A. 1836a. Iconographie der Land- und Susswasser-Mollusken Europa’s, mit
vorzuglicher Beriicksichtigung der Europaischen noch nicht abgebildeten Arten. Dresden.
Leipzig, 1(3). P. 1-33.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758 299
Rossmassler Е.А. 1836b. Iconographie der Land- und Siisswasser-Mollusken Europa’s, mit
vorziiglicher Beriicksichtigung der Europaischen noch nicht abgebildeten Arten. Dresden,
Leipzig, 1(4). P. 1-27.
Spengler L. 1793. Beskrivelse over et nyt Slaegt af de toskallede Konkylier, forhen af mig kaldet
Chaena, saa og over det Linneiske Slaegt Mya, hvilket noiere bestemmes, og inddeles I
tvende Slaegter. Skrivter af Naturhistorie-Selskabet, Kiobenhavn, 3(1). P. 16-69.
Westerlund C.A. 1871. Expose critique des mollusques de terre et d‘eau douce de la Suede et de
laNorvege. Nova Acta Societatis Regiae Scientiarum Upsaliensis, ser. 3, 8(1). P. 1-200.
Ключ для определения семейств пресноводных двустворчатых моллюсков
1(4). Внутренняя поверхность створки перламутровая. У европейских пресно-
водных представителей макушки сдвинуты вперёд, но не на самый перед-
ний конец раковины.
2(3). Передние зубы хорошо развиты на обеих створках, а задние полностью
отсутствуют или представлены едва заметными валиками. Отпечаток пе-
реднего аддуктора, хотя бы частично, неправильно морщинистый. Вына-
шивание глохидий осуществляется как в наружных, так и во внутренних
полужабрах....................................Margaritiferidae Haas, 1940 (с. 299).
В европейской части России семейство представлено одним родом Margaritifera
Schumacher, 1816.
3(2). Хорошо развиты и передние и задние зубы, или зубы замка полностью
отсутствуют. Отпечаток переднего аддуктора гладкий или с тонкими ли-
ниями. Вынашивание глохидий осуществляется в наружных полужабрах.
..................................................Unionidae Fleming, 1828 (с. 301).
4(1). Внутренняя поверхность створки не перламутровая, матовая. Форма рако-
вины другая.
5(6). Макушки сдвинуты на самый передний конец раковины, что придаёт ей
треугольную, каплевидную или клювовидную форму. Угол, образованный
макушкой — острый; часто имеется более или менее выраженный киле-
вой перегиб.........................Dreissenidae Gray, 1840 (с. 313).
6(5). Раковина округлая, округло-овальная или округло-треугольная. Макушки
округлые, располагаются посередине спинного края створок или могут
быть слегка смещены вперёд или назад. Килевой перегиб отсутствует.....
...........................Sphaeriidae Deshayes, 1855 (1820) (с. 315).
Семейство MARGARITIFERIDAE Haas, 1940
Типовой род: Margaritifera Schumacher, 1815.
Замок состоит только из кардинальных зубов (один кардинальный зуб в пра-
вой створке и два — в левой), или из кардинальных и слабо выраженных лате-
ральных зубов. Задние края мантии раздельные, супраанальное отверстие не вы-
ражено. Жабры в задней части не сращены с мантией. Вынашивание глохидий
осуществляется как в наружных, так и во внутренних полужабрах.
300
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
В европейской части России только один род Margaritifera Schumacher, 1816,
представители которого обитают в речках, характеризующихся быстрым течени-
ем и чистотой воды, протекающих среди кристаллических пород.
Род Margaritifera Schumacher, 1816
Типовой вид: Муа margaritifera Linnaeus, 1758.
Раковина прочная, овальная, овально-вытянутая или почковидная. В послед-
нем случае брюшной край вогнутый. В зависимости от внешних условий задний
край раковины может быть укороченным или вытянутым. Длина жемчужниц в
южной части ареала достигает 15 см и более. Макушечная скульптура представ-
лена слабыми концентрическими морщинками или отсутствует. Замок состоит
только из кардинальных зубов (один кардинальный зуб в правой створке и два —
в левой створке). На замочном крае под лигаментом имеются следы редуциро-
ванных латеральных зубов в виде узких валиков. Внутри мантии может образо-
вываться жемчуг. Моллюски малоподвижны. Обитая на быстром течении среди
камней, у жемчужниц в процессе роста часто формируется искривлённая ракови-
на. В редких случаях может замедляться рост заднего края раковины, в результате
чего моллюски приобретают яйцевидную форму.
Комментарии. В пределах Севера европейской части России обнаружено 3 вида
жемчужниц — Margaritifera margaritifera (Linnaeus, 1758), М. elongata (Lamarck, 1819)
и M. borealis (Westerlund, 1871), хорошо различимых по выпуклости максимально-выпу-
клого сечения (МВС) створки. В определительном ключе для этих видов приводятся соот-
ношения выпуклости раковины (В) к её максимальной высоте (Н). Поскольку показатели
В/Н жемчужниц увеличиваются с увеличением линейных размеров раковин, приведённые
в тексте значения В/Н относятся только к среднеразмерным моллюскам с длиной створок
7-11 см.
В настоящее время у части специалистов, занимающихся биологией европейских жем-
чужниц, нет уверенности в самостоятельности М. elongata и М. borealis. Действительно,
компараторный метод не даёт ответа на вопрос, являются ли выделенные с его помощью
формы МВС доказательством самостоятельности видов или это пример внутривидовой
изменчивости. По-видимому, таксономические проблемы, связанные с составом рода Mar-
garitifera, будут решаться в рамках будущих исследований с применением иных современ-
ных методов (например, молекулярно-генетического).
Ключ для определения видов рода Margaritifera
1(2). Раковина выпуклая. Отношение выпуклости раковины к её максимальной
высоте не меньше 0,65.........................................................
.........Margaritifera borealis (Westerlund, 1871) (вкл. табл. I, рис. 1 а-з).
— borealis Westerlund, 1871: 168 (Margaritana margaritifera f.).
Редкий вид, обитающий на севере Европы. Моллюски встречаются в реках на
быстром течении.
2(1). Раковина уплощённая или умеренно выпуклая. Отношение выпуклости
раковины к её максимальной высоте меньше 0,62.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
301
3(4). Раковина уплощённая: отношение выпуклости раковины к её максималь-
ной высоте не превышает 0,56................................................
......Margaritifera margaritifera (Linnaeus, 1758) (вкл. табл. I, рис. 2а-д).
— margaritifera Linnaeus, 1758: 651 (Муа).
Массовый вид, распространённый на севере Европы. Моллюски обитают в реках
на участках с быстрым течением.
4(3). Раковина умеренно выпуклая. Отношение выпуклости раковины к её мак-
симальной высоте 0,58-0,62..................................................
..........Margaritifera elongata (Lamarck, 1819) (вкл. табл. I, рис. За-в).
— elongata Lamarck, 1819: 70 (Unio).
Массовый вид, распространённый на севере Европы. Моллюски обитают в реках
на участках с быстрым течением.
Семейство UNIONIDAE Fleming, 1828
Типовой род: Unio Philipsson in Retzius, 1788.
Замок выражен или отсутствует. Вводной и выводной сифоны разделены диа-
фрагмой. Наружный жаберный листок сращён с мантией. Вынашивание глохидий
осуществляется в наружных полужабрах.
Ключ для определения подсемейств семейства Unionidae
1(4). Передние и задние зубы имеются.
2(3). Раковина удлинённо овальная, овальная или овально-коническая. Маку-
шечная скульптура представлена двумя расходящимися рядами бугорков,
либо бугорками и заметными валиками в виде букв V или W, либо валика-
ми в виде буквы W. Передние зубы пластинчатые. Яйца окрашены в ярко-
жёлтые или бледно-жёлтые цвета.......................Unioninae s. str. (с. 302).
В европейской части России подсемейство представлено одним родом Unio Phil-
ipsson in Retzius, 1788.
3(2). Раковина коротко-овальная, овальная или трапециевидная. Спинной край
часто выгнут в виде широкого горба. Макушечная скульптура представле-
на вложенными друг в друга тонкими валиками в виде греческой буквы
омега, либо двумя рядами спрямлённых валиков, сдвинутых по отноше-
нию друг к другу. Передние зубы треугольные или трапециевидные. Яйца
окрашены в красный цвет...............Psilunioninae Starobogatov, 1970 (с. 304).
В европейской части России подсемейство представлено родом Crassiana Servain,
1882.
4(1). Передние и задние зубы отсутствуют.
5(6). Раковина разной степени выпуклости. Высота отпечатков задних муску-
лов-замыкателей обычно меньше трети высоты раковины за лигаментом.
Мантийная линия у заднего края узкая, слабозаметная. Макушечная
скульптура в виде тонких спрямлённых валиков, либо тонких концентри-
ческих расходящихся валиков. Передний край раковины широкий, если
302
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
узкий, то макушечная скульптура в виде тонких спрямлённых валиков.....
..................................Anodontinae Rafinesque, 1820 (с. 306).
В европейской части России подсемейство представлено двумя родами: Anodonta
Lamarck, 1799 и Colletopterum Bourguignat, 1876.
6(5). Раковина плоская или умеренно выпуклая. Высота отпечатков задних му-
скулов замыкателей около или больше трети высоты раковины за лигамен-
том. Мантийная линия у заднего края раковины широкая, обычно хорошо
заметная. Макушечная скульптура в виде крупных коротких валиков или
отсутствует. Передний край раковины узкий....................................
............................Pseudanodontinae Stadnichenko, 1984 (с. 311).
В европейской части России подсемейство представлено родом Pseudanodonta
Bourguignat, 1876
Подсемейство Unioninae s. str.
Типовой род: Unio Philipsson in Retzius, 1788.
Раковина удлинённо овальная, овальная или овально-коническая, средних раз-
меров. Длина раковины 5-8 см, отдельные раковины могут достигать 10-12 см.
Задний край на конце образует суженный клюв или конус. Окраска периостра-
кума от жёлтой и жёлто-зелёной, до коричневой и почти чёрной. Макушечная
скульптура представлена двумя расходящимися рядами бугорков, либо бугорками
и заметными валиками в виде букв V или W, либо валиками в виде буквы W, при-
чём вершины этих валиков выступающие (вкл. табл. I, рис. 4, 5, 11). Кардиналь-
ные и латеральные зубы пластинчатые, расположены на одной слабо выгнутой
линии, или под малозаметным углом друг к другу. Папиллы вводного сифона тон-
кие, упругие, сформированы в 4-5 рядов (вкл. табл. I, рис. 6, 10). Яйца окрашены
в ярко жёлтые или бледно жёлтые цвета.
Комментарии. В разделе, посвящённом двустворчатым моллюскам, определителя
пресноводных беспозвоночных России (Старобогатов и др., 2004) род Unio был представ-
лен четырьмя видами: Unio limosus Nilsson, 1822, U rostratus Lamarck, 1799, U. protractus
Lindholm, 1922 и U. pictorum (Linnaeus, 1758). Кроме того, там же был выделен род Tumi-
diana Servain, 1882, в состав которого были включены три вида, ранее относимые к роду
Unio\ Tumidiana conus (Spengler, 1793), T. tumida (Philipsson in Retzius, 1788) и T. mulleri
(Rossmassler, 1838). В результате таксономической ревизии было установлено, что между
представителями родов Unio и Tumidiana (sensu Старобогатов и др., 2004) нет различий
достаточных для отнесения их к разным родам. Целесообразнее в пределах рода Unio вы-
делить два подрода: Unio s. str. и Tumidiana, представители которых отличаются формой
раковины и взаиморасположением передних и задних зубов.
Также установлено, что название Unio rostratus является синонимом U. pictorum, а на-
звание Tumidiana mulleri — синонимом названия U. protractus (часть особей, хранящихся
в коллекции ЗИН РАН и определённых ранее как U. mulleri, по кривизне контура МВС
оказались неотличимы от U. tumida). Кроме того, подтверждена самостоятельность вида
U. longirostris в составе подрода Tumidiana. Ранее название U. longirostris считалось сино-
нимом названия U. pictorum.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
303
Род Unio Philipsson in Retzius, 1788
Типовой вид: Mya pictorum Linnaeus, 1758
Ключ для определения подродов рода Unio
1(2). Раковина удлинённо-овальная. Отношение высоты раковины к её длине
обычно меньше 0,47 (чаще всего находится в пределах 0,42-0,45). Передние
и задние зубы расположены на одной слабо выгнутой линии. Макушечная
скульптура представлена двумя расходящимися рядами бугорков, либо ком-
бинацией бугорков и валиков в виде букв V или W, либо почти незаметна.
Периостракум обычно окрашен в жёлтый цвет....................Unio s. str. (с. 304).
2(1). Раковина овальная или овально-коническая. Отношение высоты раковины
к её длине обычно выше 0,5 (чаще всего находится в пределах 0,52-0,55).
Передние зубы заметно опущены. Макушечная скульптура представлена
валиками в виде буквы W, либо комбинацией бугорков и валиков в виде
букв V или W, либо почти незаметна. В окраске периостракума преобла-
дают зелёные или коричневые тона.........................Tumidiana Servain, 1882.
Подрод Tumidiana Servain, 1882
Типовой вид: Unio tumidus Philipsson in Retzius, 1788.
Ключ для определения видов подрода Tumidiana
1(2). Раковина овально-коническая, умеренно выпуклая. Отношение В/Н не
превышает 0,65.................................................................
......Unio tumidus (Philipsson in Retzius, 1788) (вкл. табл. I, рис. 12 a-в).
— tumidus Philipsson in Retzius, 1788: 17.
[= Unio ovalis (Montagy, 1803) partim, Unio mulleri Rossmassler, 1836 partim].
Вид распространён в Европе (кроме севера и северо-востока) и на юге Восточной
Сибири (река Иртыш, бассейн Оби).
2(1). Раковина овальная или овально-коническая, выпуклая или сильновыпук-
лая. Отношение В/Н превышает 0,65.
3(4). Раковина овальная, выпуклая. Лигамент крупный, заметно возвышается
над спинным краем. Отношение В/Н составляет 0,65-0,71..........................
.............Unio longirostris Rossmassler, 1836 (вкл. табл. I, рис. 13 а-д).
— longirostris Rossmassler, 1836а: 26.
Вид распространён в Европе (кроме севера и северо-востока) и на юге Восточной
Сибири.
Форма контура максимально-выпуклого сечения сходна с таковой у U. pictorum.
4(3). Раковина овально-коническая, сильновыпуклая. Лигамент обычно не воз-
вышается над спинным краем, а если возвышается, то только своей перед-
ней частью. Отношение В/Н превышает 0,68.......................................
...................Unio conus (Spengler, 1793) (вкл. табл. I, рис. 14 а-д).
— conus Spengler, 1793: 60-61.
304
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
[= Unio ovalis (Montagy, 1803) partim].
Вид распространён в Европе (кроме севера и северо-востока).
Подрод Unio s. str.
Типовой вид: Муа pictorum Linnaeus, 1758.
Ключ для определения видов подрода Unio
1(4). Раковина выпуклая или умеренно выпуклая. В левой створке выемка
между внутренним и внешним передними зубами хорошо выражена. От-
ношение В/Н меньше 0,70.
2(3). Раковина умеренно выпуклая. Отношение В/Н 0,58-0,64 ......................
.................Unio protractus Lindholm, 1932 (вкл. табл. I, рис. 7 а-в).
— protractus Lindholm, 1932: 15-16, pl. 5, fig. 1, 2, 4-7.
[= Unio mulleri Rossmassler, 1836 partim].
Вид распространён в Европе (кроме севера и северо-востока) и на юге Восточной
Сибири (возможно до Алтая).
Комментарии. В разделе, посвящённом крупным двустворчатым моллюскам,
определителя пресноводных беспозвоночных России (Старобогатов и др., 2004)
на таблице 18, рис. 5-6 под названием Tumidiana muelleri приведён рисунок взрос-
лого экземпляра U. protractus.
3(2). Раковина выпуклая. Отношение В/Н 0,65-0,70................................
.................Unio pictorum (Linnaeus, 1758) (вкл. табл. I, рис. 8 а-в).
— pictorum Linnaeus, 1758: 671 (Муа) [= Unio rostratus Lamarck, 1799].
Ареал вида охватывает Европу (кроме севера и северо-востока), Западное Закавка-
зье и юго-восточную часть Сибири. Интродуцирован в окрестностях города Чита.
Комментарии. Форма контура максимально выпуклого сечения (МВС) сходна с
таковой у U. longirostris.
4(1). Раковина сильновыпуклая, почти цилиндрическая. В левой створке вы-
емка между внутренним и внешним передними зубами малозаметна. От-
ношение В/Н больше 0,74 (чаще всего 0,74-0,82)..................................
......................Unio limosus Nilsson, 1822 (вкл. табл. I, рис. 9 а-в).
— limosus Nilsson, 1822: 110-111.
Вид распространён в Европе (кроме севера и северо-востока), интродуцирован в
окрестностях города Чита.
Подсемейство Psilunioninae Starobogatov, 1970
Типовой род: Psilunio Stefanescu, 1896.
Род Crassiana Servain, 1882
Типовой вид: Unio crassus Philipsson in Retzius, 1788.
Раковина коротко-овальная, овальная, овально-четырёхугольная или эллип-
тическая, прочная, средних размеров (5-8 см, некоторые раковины достигают
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
305
10-12 см). Спинной край часто заметно выгнут, задний край опущен к низу. Ма-
кушечная скульптура представлена вложенными друг в друга тонкими округлыми
или слегка изогнутыми валиками в виде греческой буквы омега, либо двумя ря-
дами спрямлённых валиков, сдвинутых по отношению друг к другу (вкл. табл. II,
рис. 15 а, б). Кардинальные и латеральные зубы расположены под заметным
углом друг к другу. Кардинальный зуб правой створки массивный, округло-тре-
угольный, конический или трапециевидный, зазубренный. Латеральный зуб не-
сколько изогнутый, зазубренный по гребню. Кардинальные зубы левой створки
массивные, зазубренные, лежащие один за другим. Латеральные зубы прямые, по
периферии с насечками. Папиллы вводного сифона тонкие, упругие (вкл. табл. II,
рис. 16). Яйца окрашены в красный цвет.
Комментарии. В разделе по моллюскам определителя пресноводных беспозвоночных
России (Старобогатов и др., 2004) род Crassiana был представлен шестью видами, отне-
сёнными к двум подродам: Crassiana s. str. и Bataviana Servan, 1882. Последующие ис-
следования, проведённые на основе коллекции ЗИН РАН, а также новые многочисленные
сборы из бассейна реки Медведица (Тверская обл.) и р. Великой (окр. Пскова), хранящие-
ся в коллекции БПИ ДВО РАН, показали несостоятельность подразделения рода Crassiana
на два подрода.
Также установлено, что названия Crassiana fuscula (Rossmassler, 1836), С. cyprinorum
(Bourguignat in Locard, 1882) и C. irgizlaica (Lindholm, 1904) являются синонимами на-
звания C. musiva (Spengler, 1793). Выявлено, что представители рода Crassiana могут об-
разовывать гибриды с особями рода Unio подрода Tumidiana. При этом форма раковины
оказывается почти неотличимой от представителей подрода Tumidiana, однако кардиналь-
ные зубы гибридных раковин обычно сохраняют пирамидальную форму, характерную для
рода Crassiana.
Ключ для определения видов рода Crassiana
1(2). Раковина яйцевидная или овальная, умеренно выпуклая. Примакушечная
область плоская, почти не выступает над контуром створки. Отношение
В/Н меньше 0,6..............................................................
..............Crassiana папа (Lamarck, 1819) (вкл. табл. II, рис. 17 а-г).
— папа Lamarck, 1819: 76 (Unio).
Вид распространён в Европе (кроме северных районов). Моллюски обитают в ре-
ках на участках, характеризующихся выраженным течением. Иногда обитают в
речных затонах.
2(1). Раковина овальная, удлинённо-овальная или четырёхугольная, выпуклая
или сильновыпуклая. Примакушечная область раковины выгнута, заметно
выступает над контуром створки. Отношение В/Н превышает 0,6.
3 (4) Раковина овальная или удлинённо-овальная, у крупных особей массивная,
выпуклая. Отношение В/Н 0,60-0,66...........................................
............Crassiana musiva (Spengler, 1793) (вкл. табл. II, рис. 18 а-ж).
— musivus Spengler, 1793: 57 (Unio).
[= Crassiana fuscula (Rossmassler, 1836), C. cyprinorum (Bourguignat in Locard,
1882), C. irgizlaica (Lindholm, 1904)].
306
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Вид распространён в Европе (кроме северных районов). Моллюски обитают в
реках на участках, характеризующихся выраженным течением.
Комментарии. Полиморфный массовый вид. Для С. musiva характерна высокая
изменчивость соотношения высоты и длины раковины в пределах одного и того
же водоёма/биотопа (вкл. табл. II, рис. 18 а-г). Известны гибриды Crassiana mu-
siva и Unio longirostris, сходные по общей форме раковины с U. longirostris, а по
строению кардинальных зубов — с С. musiva.
4(3). Раковина овальная или овально-четырёхугольная, у крупных особей мас-
сивная, сильновыпуклая. Отношение В/Н у молодых особей более 0,67, а
у взрослых особей — более 0,70...............................................
....Crassiana crassa (Philipsson in Retzius, 1788) (вкл. табл. II, рис. 19 а-д).
— crassus Philipsson in Retzius, 1788: 17 (Unio).
[= Unio pseudolittoralis Clessin, 1875 partim].
Редкий вид, распространённый в Европе (кроме северных районов). Моллюски
обитают в реках на участках, характеризующихся выраженным течением.
Комментарии. Некоторые специалисты считают, что сильная выпуклость рако-
вины, не является видоспецифичным признаком С. crassa, однако, в отличие от
остальных видов рода сильная выпуклость створок не только у взрослых (вкл.
табл. II, рис. 19 в-д), но и у молодых особей С. crassa (вкл. табл. II, рис. 19 а, б)
может служить подтверждением самостоятельности данного вида.
Подсемейство Anodontinae Rafinesque, 1820
Типовой род: Anodonta Lamarck, 1799.
Зубы отсутствуют. Макушка смещена к переднему краю и расположена на
одном уровне с лигаментом или ниже. Спинной край у молодых особей спрям-
лённый. Некоторые представители подсемейства имеют развитое крыло. Самая
задняя точка раковины расположена близ середины высоты раковины, или немно-
го ниже. Макушечная скульптура в виде концентрических расходящихся тонких
валиков или широкого ряда спрямлённых тонких валиков. Высота отпечатков за-
дних мускулов-замыкателей обычно меньше трети высоты раковины. Мантийная
линия у заднего края узкая, слабозаметная.
В пределах европейской части России подсемейство представлено 2 родами:
Anodonta Lamarck, 1799 и Colletopterum Bourguignat, 1876.
Ключ для определения родов подсемейства Anodontinae
1(2). Раковины очень крупные, длиной до 20 см и более. Наружная поверхность
створок чаще всего морщинистая. Макушечная скульптура в виде кон-
центрических расходящихся тонких валиков.....................................
........................................Anodonta Lamarck, 1799 (с. 207).
2(1). Раковины средних размеров, длиной до 10-14 см. Боковая поверхность
чаще всего гладкая. Макушечная скульптура в виде ряда спрямлённых
тонких валиков.........................Colletopterum Bourguignat, 1876 (с. 308).
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
307
Род Anodonta Lamarck, 1799
Типовой вид: Mytilus cygneus Linnaeus, 1758.
Раковины очень крупные, разной степени выпуклости, длиной до 20 см и бо-
лее. Боковая поверхность чаще всего морщинистая. Высота отпечатков задних
мускулов-замыкателей обычно меньше трети высоты раковины за лигаментом.
Мантийная линия у заднего края узкая, слабозаметная. Макушечная скульптура
в виде тонких концентрических расходящихся валиков (вкл. табл. II, рис. 20 а,
б). Передний край раковины широкий. Вводной сифон эллиптический. Папиллы
вводного сифона канатообразные, упругие, при заборе воды не вывернуты на-
ружу.
В малакофауне России установлены три вида данного рода.
Ключ для определения видов рода Anodonta
1(2). Спинной край у молодых и взрослых раковин чаще всего параллелен
брюшному краю. Если смотреть на створку сверху под углом около 20
градусов к вертикальной оси, то примакушечная поверхность значительно
приподнята. Раковина сильновыпуклая. Отношение В/Н превышает 0,7....
...................Anodonta zellensis (Gmelin, 1791) (вкл. табл. II, рис. 21 а-в).
— zellensis Gmelin, 1791: 3362 (Mytilus).
Вид распространён в Европе (кроме крайнего севера и северо-востока). Моллюски
обитают в реках, озёрах и прудах на илисто-песчаном или илистом грунте.
2(1). Спинной край у молодых раковин чаще всего наклонен вперёд, у взрослых
раковин он обычно выгнутый или параллелен спинному краю. Отношение
В/Н меньше 0,6, а если больше, то примакушечная область плоская.
3(4). Если смотреть на створку сверху под углом около 20 градусов к верти-
кальной оси, то примакушечная поверхность очень плоская. У раковин
длиной до 4-5 см, отношение В/Н не превышает 0,3. С возрастом вы-
пуклость створок заметно увеличивается, в результате чего особи длиной
больше 10 см становятся очень выпуклыми. Отношение В/Н при длине
раковины 10-11 см составляет около 0,5, а при длине 17-20 см превышает
0,7.......................A. stagnalis (Gmelin, 1791) (вкл. табл. II, рис. 22 а-д).
— stagnalis Gmelin, 1791: 3362 (Mytilus).
Вид распространён в Европе (кроме крайнего севера и северо-востока). Моллюски
обитают в озёрах и прудах на заиленном грунте.
4(3). Спинной край у молодых раковин чаще всего наклонен вперёд, у взрос-
лых раковин спинной край обычно выгнутый. Если смотреть на створку
сверху под углом до 20 градусов к вертикальной оси, то примакушечная
поверхность умеренно приподнята. Раковина выпуклая. Отношение В/Н
0,5-0,6............Anodonta cygnea (Linnaeus, 1758) (вкл. табл. II, рис. 23 а-в).
— cygnea Linnaeus, 1758: 706 (Mytilus).
Вид распространён в Европе (кроме крайнего севера и северо-востока). Моллюски
обитают в реках, озёрах и прудах на илисто-песчаном или илистом грунте.
308 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Род Colletopterum
Типовой вид: Anodonta letourneuxi Bourguignat, 1880.
Раковина овально-треугольная, овальная или удлинённо-овальная, средних
размеров, длиной до 10-14 см. Крыло хорошо выражено или отсутствует. Боковая
поверхность обычно гладкая. Макушечная скульптура в виде ряда спрямлённых
тонких валиков (вкл. табл. II, рис. 24 а, б). Вводной сифон эллиптический. Папил-
лы вводного сифона тонкоконические, упругие, при заборе воды не вывернуты
наружу (вкл. табл. II, рис. 25).
В составе рода насчитывается 11 видов, сгруппированных в два подрода: Col-
letopterum s. str. и Piscinaliana Bourguignat 1881. На севере и северо-западе ев-
ропейской части России обитают виды подрода Piscinaliana, распространённого
также в Сибири и Средней Азии.
Комментарии. В разделе, посвящённом двустворчатым моллюскам, определителя
пресноводных беспозвоночных России (Старобогатов и др., 2004) указано, что на тер-
ритории европейской части России было зафиксировано семь видов рода Colletopterum'.
С. subcirculare (Clessin, 1873), С. apollonicum (Bourguignat, 1880), С. nilssoni (Kiister, 1842),
C. ponderosum (C. Pfeiffer, 1825), C. anatinum (Linnaeus, 1758), C. piscinale (Nilsson, 1822)
и C. rostratum (Rossmassler, 1836). Вскоре в южных областях европейской части России
было обнаружено еще 4 вида рода: С. baeri Bogatov, Starobogatov et Prozorova, 2005, C. os-
tiarium (Drouet 1881), C. milaschevichi Bogatov, Starobogatov et Prozorova, 2005 и C. con-
vexum (Drouet 1888) (Богатов, Старобогатов, Прозорова, 2005). Все вновь обнаруженные
виды имели высокое или хорошо выраженное крыло и по этому признаку были объеди-
нены в подрод Colletopterum s. str., куда были также отнесены С. subcirculare и С. apol-
lonicum. Остальные пять видов рода — С. nilssoni, С. ponderosum, С. anatinum, С. piscinale
и С. rostratum — вошли в состав подрода Piscinaliana Bourguignat 1881 [типовой вид С.
piscinale (Nilsson, 1822)].
Подрод Colletopterum s. str.
Типовой вид: Anodonta letourneuxi Bourguignat, 1880.
Раковина овально-треугольная с высоким крылом. У взрослых или сильно вы-
пуклых раковин крыло может быть низкое, однако всегда хорошо выраженное.
Ключ для определения видов подрода Colletopterum
1(6). Раковина умеренно выпуклая или выпуклая, так что отношение выпукло-
сти раковины к высоте максимально выпуклого сечения створок (индекс
выпуклости МВС) не менее 0,47. Если смотреть на створку сверху под
углом около 20 градусов к вертикальной оси, то примакушечная поверх-
ность раковин заметно приподнята. Наиболее выступающая точка боковой
поверхности раковины лежит выше середины её высоты.
2(3). Раковина умеренно выпуклая, так что индекс выпуклости МВС у средне-
размерных особей не превышает 0,50. У взрослых особей в срединной
части контура МВС имеется заметный спрямленный участок........................
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
309
......Colletopterum milaschevichi Bogatov, Starobogatov et Prozorova, 2005
(вкл. табл. II, рис. 26 a-e).
— milaschevichi Богатов, Старобогатов, Прозорова, 2005: 1056, рис. 3 н-р, рис. 4в.
Вид известен в Европе из Нижней Волги.
3(2). Раковина в большей степени выпуклая, так что индекс выпуклости МВС у
среднеразмерных особей не менее 0,51. В срединной части контура МВС
спрямленный участок отсутствует.
4(5). Индекс выпуклости МВС не превышает 0,55.................................
........Colletopterum convexum (Drouet, 1888) (вкл. табл. II, рис. 27 а-г).
— convexa Drouet, 1888: ПО (Anodonta).
Вид распространён в южных районах Европы.
5(4). Индекс выпуклости МВС не менее 0,60.....................................
.. Colletopterum apollonicum (Bourguignat, 1880) (вкл. табл. II, рис. 28 а-г).
— apollonica Bourguignat, 1880: 91, Taf. 15, Fig. 1-3 (Anodonta).
[= Anodonta piscinalis var. volgensis Жадин, 1938 (partim); Anodonta ponderosa vol-
gensis Старобогатов, Пирогов, 1970 (partim)].
Вид распространён в южной Европе и Малой Азии. Обитает в реках и озёрах.
6(1). Раковина в различной степени уплощённая, так что индекс выпуклости
МВС не превышает 0,46. Если смотреть на створку сверху под углом
около 20 градусов к вертикальной оси, то примакушечная поверхность
раковины плоская и даже у крупных раковин (длиной больше 7 см) не
выступает над контуром створки. Наиболее выступающая точка боковой
поверхности раковины лежит против середины её высоты.
7(8). Раковина плоская или очень плоская, так что индекс выпуклости МВС не
превышает 0,38................................................................
......Colletopterum subcirculare (Clessin, 1873) (вкл. табл. Ill, рис. 29 а-в).
— subcircularis Clessin, 1873: 8, Taf. 22, Fig. 3—4 (Anodonta).
[= Anodonta piscinalis var. volgensis Жадин, 1938 (partim); Anodonta subcircularis
astrachanica Старобогатов, Пирогов, 1970; Anodonta ponderosa volgensis Старо-
богатов, Пирогов, 1970 (partim)].
Южная часть центральной и восточной Европы, Причерноморье, нижнее течение
Волги и Дона. Вид обитает в озерах и реках на медленном течении.
8(7). Раковина слабо выпуклая, так что индекс выпуклости МВС не менее 0,40.
9(10). Крыло очень высокое.........................Colletopterum baeri Bogatov,
Starobogatov et Prozorova, 2005 (вкл. табл. Ill, рис. 29 a-в).
— baeri Богатов, Старобогатов, Прозорова, 2005: 1055, рис. 3 л, м, рис. 4 д.
Вид распространён в низовьях Волги. Редок.
10(9). Крыло относительно низкое, но хорошо выраженное........................
........Colletopterum ostiarium (Drouet, 1881) (вкл. табл. Ill, рис. 31 а-в).
— ostiaria Drouet, 1881: 26 (Anodonta).
[= Anodonta piscinalis var. ostiaria Жадин, 1938; Anodonta piscinalis var. volgensis
Жадин, 1938 (partim); Anodonta piscinalis shadini Старобогатов, Пирогов, 1970].
Вид распространён в Причерноморье, нижних и центральных частях бассейна
Волги.
310 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Подрод Piscinaliana Bourguignat 1881
Типовой вид: Colletopterum piscinale (Nilsson, 1822).
Раковина разной формы. Крыло даже у молодых особей низкое или почти не
выражено.
Ключ для определения видов подрода Piscinaliana
1(8). Раковина яйцевидная, овальная, овально-четырёхугольная или клювовид-
ная, разной степени выпуклости. У молодых особей спинной край пря-
мой, наклонён вперёд. Если смотреть на створку сверху под углом около
20 градусов к вертикальной оси, то примакушечная поверхность в виде
высокого или низкого поднятия, занимающего только часть спинного края
створки. Наиболее выступающая точка боковой поверхности створки рас-
положена примерно на уровне середины высоты раковины или выше.
2(3). Раковина яйцевидная, уплощённая, так что индекс выпуклости МВС не
превышает 0,46...............................................................
.........Colletopterum nilssoni (Kiister, 1838) (вкл. табл. Ill, рис. 32 а-в).
— nilssoni Kiister, 1842: 61 (Anodonta).
Редкий вид, распространённый в Европе и Сибири. Моллюски обитают в озёрах и
реках на участках с медленным течением.
3(2). Раковина обычно овальная, овально-четырёхугольная (у особей, обитаю-
щих в местах с выраженным течением, может быть удлинённо-овальная)
или клювовидная, более выпуклая по сравнению с предыдущим видом.
Индекс выпуклости (МВС) более 0,50.
4(5). Раковина овально-четырёхугольная, массивная. Индекс выпуклости (МВС)
не менее 0,62. Макушки у взрослых раковин заметно выступают над кон-
туром створки. В окраске периостракума преобладают тёмные тона...............
...Colletopterum ponderosum (С. Pfeiffer 1825) (вкл. табл. III, рис. 33 а-г).
— ponderosa С. Pfeiffer, 1825: 31-32, pl. 4, figs. 1-6 (Anodonta).
Редкий вид, распространённый в Европе и Сибири. Моллюски обитают в озёрах и
реках на участках с медленным течением.
5(4). Раковина клювовидная или овальная, не массивная, умеренно выпуклая,
так что индекс выпуклости (МВС) не превышает 0,60. Макушки у взрос-
лых раковин слабо выступают над контуром створки. В окраске раковины,
особенно её задней части, преобладают зелёные тона.
6(7). Раковина средних размеров (обычно не превышает 8-9 см в длину), клю-
вовидная, так как передний край узкий, а задняя часть раковины (сразу за
лигаментом) при переходе в задний край заметно выгнута.......................
......Colletopterum anatinum (Linnaeus 1758) (вкл. табл. Ill, рис. 34 а-д).
— anatinus Linnaeus, 1758: 706 (Mytilus).
Вид распространён в реках и озёрах Европы и Сибири.
Комментарии. За С. anatinum часто принимают тугорослые небольшие экземпля-
ры С. ponderosum и С. piscinale. Не исключено также, что за С. anatinum могут
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
311
быть приняты гибриды от видов из подрода Piscinaliana и рода Pseudanodonta
(подсемейство Pseudanodontinae). Молодые раковины С. anatinum из-за их клю-
вовидной формы можно спутать с представителями рода Pseudanodonta. В мала-
кологическом разделе определителя пресноводных беспозвоночных Европейской
части СССР (Старобогатов, 1977) С. anatinum был ошибочно отнесён к роду Pseu-
danodonta, от видов которого он отличается заметно более выпуклой раковиной
(особенно у взрослых особей) и макушечной скульптурой, характерной для рода
Colletopterum.
7(6). Раковина может достигать больших размеров (до 15 см), овальная, так
как её передний край обычно широкий. На течении или в прибойной зоне
больших озёр форма раковины может быть удлинённо-овальной. У моло-
дых раковин имеется небольшое крыло. Участок при переходе спинного
края в задний вогнут, за исключением форм с неразвитым задним краем..
..............Colletopterum piscinale (Nilsson, 1822) (вкл. табл. Ill, рис. 35 а-г).
— piscinalis Nilsson, 1822: 116-117 (Anodonta).
Вид широко распространён в озёрах и реках Европы, Сибири и Средней Азии.
Комментарии. В стоячих водоёмах центральной России могут отмечаться формы
С. piscinale с неразвитым задним краем. Установлено, что на глубине число таких
форм увеличивается и достигает максимума в зоне, куда свет практически не про-
никает (Алимов, Богатов, 1975).
8(1). Раковина удлинённая и сильновыпуклая, поэтому индекс выпуклости
(МВС) не менее 0,65. Спинной край, как у молодых, так и у взрослых осо-
бей, почти параллелен брюшному краю. Если смотреть на створку сверху
под углом около 20 градусов к вертикальной оси, то примакушечная по-
верхность в виде невысокого очень широкого поднятия, проходящего по
всему спинному краю створки. Наиболее выступающая точка боковой
поверхности раковины расположена в районе середины её высоты...............
.... Colletopterum rostratum (Rossmassler, 1836) (вкл. табл. Ill, рис. 36 а-ж).
— rostrata Rossmassler, 18366: 25, pl. 20, fig. 284 (Anodonta).
Редкий вид, распространённый в реках и озёрах Европы и Сибири.
Подсемейство Pseudanodontinae Stadnichenko, 1984
Типовой род: Pseudanodonta Bourguignat, 1877.
Зубы отсутствуют. Макушка значительно смещена к переднему краю и рас-
положена заметно ниже лигамента. Спинной край выгнутый. Самая задняя точка
раковины расположена близ середины высоты заднего края или ниже. Высота от-
печатков задних мускулов-замыкателей около или больше трети высоты раковины.
Мантийная линия у заднего края раковины широкая, обычно хорошо заметная.
В пределах европейской части России подсемейство представлено родом Pseu-
danodonta Bourguignat, 1876.
Род Pseudanodonta Bourguignat, 1876
Типовой вид: Anodonta complanata Ziegler in Rossmassler, 1835.
312
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Раковина клювовидная, часто с заострённым передним краем, реже эллип-
тическая, средних размеров (длина 5-7 см, высота 3-4 см, наиболее старые ра-
ковины достигают в длину 9 см и в высоту 5 см). Моллюски, обитающие на
участках с выраженным течением или в прибойной зоне, могут иметь удлинён-
но-овальную или эллиптическую раковину. Периостракум молодых и среднераз-
мерных раковин обычно блестящий, в его окраске преобладают зелёные тона,
крупные раковины могут быть окрашены в тёмно-коричневые тона. Под воздей-
ствием внешних факторов задний край раковины часто не развивается, в резуль-
тате чего раковина становится треугольной. В более редких случаях задний край
сильно разрастается, придавая раковине удлинённо-овальную форму. Макушеч-
ная скульптура в виде крупных коротких валиков или отсутствует (вкл. табл. III,
рис. 37 а-г). Вводной сифон овальный, папиллы вводного сифона уплощённые,
широкие у основания, не упругие, при открытом сифоне вывернуты наружу (вкл.
табл. III, рис. 38).
Комментарии. В разделе по моллюскам определителя пресноводных беспозвоночных
России (Старобогатов и др., 2004) род Pseudanodonta был представлен четырьмя видами:
Pseudanodonta elongata (Hollandre, 1836), Р complanata (Rossmassler, 1835), P nordenski-
oldi Bourguignat, 1881 и P kletti (Rossmassler, 1835). Наши последующие исследования
показали, что название Р kletti является синонимом Р complanata. Кроме того, уточнён
диагноз вида Р. elongata, поскольку в работе Старобогатова и др. (2004) на таблице 21
(рис. 1-2) под названием Р. elongata ошибочно приведён рисунок молодого экземпляра
Colletopterum rostratum (Rossmassler, 1836). Пока остаётся под вопросом нахождение на
территории России Р. nordenskioldi Bourguignat, 1881, имеющего более вздутую, по срав-
нению с Р. elongate, околомакушечную поверность раковины.
Иногда к подсемейству Pseudanodontinae ошибочно относят некоторые изменённые
формы видов Colletopterum anatinum и С. rostratum. Особенно в тех случаях, когда задняя
часть их спинного края и/или верхняя часть заднего края отстают в росте, в результате
чего самая задняя точка раковины опускается. В таких случаях для точного определе-
ния моллюсков необходимо внимательно осматривать линии нарастания на проблемных
участках раковины (на участках замедления роста, на внешней стороне раковины обычно
формируются шероховатые борозды и складки, а на внутренней стороне истощаются при-
зматический и перламутровый слои), рисунок макушечной скульптуры, а также отпечатки
задних мускулов-замыкателей и мантийной линии.
Ключ для определения видов рода Pseudanodonta
1(2). Примакушечная область раковины, как и сама раковина плоские. Отноше-
ние В/Н составляет 0,35-0,45 .............................................
Pseudanodonta complanata (Rossmassler, 1835) (вкл. табл. IV, рис. 39 а-е).
— complanata Ziegler in Rossmassler, 1835a: 112, pl. 3, fig. 68 (Anodonta).
[= Pseudanodonta kletti (Rossmassler, 1835)].
Моллюски широко распространены в Европе. В России встречаются в бассей-
нах Волги, Чёрного и Балтийского морей, Северной Двины. Обитают в реках на
участках с выраженным течением и в затонах на песчано-илистом или заиленном
грунте, проникают в пресные и солоноватые воды внутренних морей.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
313
2(1). Примакушечная область в виде заметного широкого поднятия. Раковина
умеренно выпуклая. Отношение В/Н более 0,50..............................
.....Pseudanodonta elongata (Hollandre, 1836) (вкл. табл. IV, рис. 40 а-к).
— elongata Holandre, 1836: 54 (Anodonta).
Как и предыдущий вид, эти моллюски широко распространены в Европе. В Рос-
сии они обитают в бассейнах Волги, Северной Двины, Чёрного и Балтийского
морей. Населяют песчано-илистые или заиленные грунты на участках с выражен-
ным течением и в затонах. Проникают в пресные и солоноватые воды внутренних
морей.
Семейство DREISSENIDAE Gray, 1840
Типовой род: Dreissena Van Beneden, 1835.
Раковина равно- или неравностворчатая, ромбическая, каплевидная (иногда
клювовидная) или округло-треугольная, лишённая перламутрового слоя. Килевой
перегиб выражен в разной степени, расположен ближе к брюшному или к спин-
ному краю. Макушки очень сильно сдвинуты на переднюю сторону. Замочный
край без зубов; внутри в передней части раковины имеется перегородка (септа), к
которой прикрепляется передний мускул-замыкатель. Иногда имеется связанный
с септой пластинчатый вырост внутренней поверхности спинного края — апо-
физа (у видов родов Congeria Partsch, 1836 и Mytilopsis Conrad, 1858). К апофизе
крепится передний педальный ретрактор. Два мускульных отпечатка. Лигамент
наружный. Мантийная линия цельная или с синусом. Мантийный край сраста-
ется, образуя на брюшной стороне отверстие для ноги, и сзади два коротких си-
фона, из которых нижний (вводной) более длинный, изнутри по краю с тонкими
папиллами. Нога маленькая, служит преимущественно для прикрепления; биссус
имеется. Развитие с пелагической личинкой.
Ранние представители семейства известны с эоцена из тропических широт по
обеим сторонам Атлантического океана. Наиболее разнообразно они были пред-
ставлены в Паратетисе в миоцен-плиоценовый период (Старобогатов, 1994).
Современные виды широко распространились, нередко за счёт интродукций,
в континентальных водах Европы и Передней Азии, в водоёмах, относящихся к
бассейну Атлантического океана в Северной и, частично, Южной Америке, на
островах Карибского архипелага и в Западной Африке (Старобогатов, 1970; Ста-
робогатов, Андреева, 1994). Семейство включает пресноводные и солоноватовод-
ные виды. Наличие биссуса определяет прикреплённый образ жизни дрейссенид.
В настоящее время известно около 30 рецентных видов семейства Dreissenidae,
сгруппированных в 3 рода. В европейской части России обитают представители
одного рода — Dreissena Van Beneden, 1835
Род Dreissena Van Beneden, 1835
Типовой вид: Mytilus polymorphic Pallas, 1771.
314
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Раковина от клювовидной и каплевидной до округло-треугольной, реже оваль-
ная, зеленовато-жёлтая, беловатая или коричневато-роговая, часто с радиальными
или зигзагообразными тёмными полосами. Макушки настолько сильно смещены
вперёд, что передний край створок полностью отсутствует. На левой створке под
макушкой имеется выраженный в различной степени зубовидный вырост, входя-
щий в соответствующую выемку на правой створке. Зияния между створок нет.
септа хорошо развита, мантийная линия без синуса.
Моллюски рода Dreissena — представители эпибиоса, прикрепляются нитями
биссуса к твёрдому субстрату (раковинам, скалам и камням, растениям, гидро-
техническим конструкциям и т.п., или к уплотнённым илистым, илисто-камени-
стым, ракушечным, илисто-ракушечным, плотным илисто-песчаным и песчаным
грунтам) в диапазоне глубин от 0 до нескольких сотен метров. Молодые и, реже,
взрослые особи способны к небольшим перемещениям (до 7 см в сутки). При
этом они отбрасывают биссус и движутся при помощи ноги (главным образом в
ночное время) (Шевченко, 1949; Зевина, 1958). Дрейссены характеризуются эври-
галинностью, их солевой диапазон колеблется в пределах от 0 до 14%о (Ляхнович
и др., 1994). По способу питания — фильтраторы, сестонофаги; основные пище-
вые объекты — фитопланктон и взвешенный в воде детрит. Особи раздельнопо-
лые, размножаются в тёплое время года. Количество яиц, продуцируемых одной
самкой, исчисляется сотнями тысяч. Развитие с пелагической личинкой.
Область распространения рода: Европа, черноморско-каспийский бассейн, Пе-
редняя Азия; отдельные непреднамеренно интродуцированные инвазионные по-
пуляции известны в Северной Америке (Великие озёра) (Богуцкая и др., 2013).
На севере и северо-западе в европейской части России распространены два вида.
Ключ для определения видов рода Dreissena
1(2). Раковина более или менее равносторонняя, угловато-каплевидной или
клювовидной формы; с отчётливым килем, смещённым ближе к уплощён-
ной (иногда вогнутой) брюшной поверхности.................................
..........Dreissena polymorpha (Pallas, 1771) (вкл. табл. IV, рис. 41 а, б).
— polymorphic Pallas, 1771: 368, 435 (Mytilus).
[= Mytilus volgae Chemnitz, 1795; Mytilus volgensis Gray, 1825; Mytilus hagenii
Baer, 1825; Mytilus area Kickx, 1834; Dythalmia danubii Jay, 1835 (nomen nudum);
Tichogonia chemnitzi Rossmassler, 1835; Dreissena fluviatilis Bourgignat, 1856; Dreis-
sensia anatolica Locard, 1893; Dreissensia arnouldi Locard, 1893; Dreissensia bedoti
Locard, 1893; Dreissensia belgrandi Locard, 1893; Dreissensia bourguignati Locard.
1883; Dreissensia chantrei Locard, 1883; Dreissensia complanata Locard, 1893; Dreis-
sensia curta Locard, 1893; Dreissensia elongata Locard, 1893; Dreissensia eximia
Locard, 1893; Dreissensia gallandi Locard, 1893; Dreissensia hellenica Locard, 1893:
Dreissensia hermosa Locard, 1893; Dreissensia lacunosa Locard, 1893; Dreissensia
letourneuxi Locard, 1893; Dreissensia locardi Locard, 1893; Dreissensia lutetiana
Locard, 1893; Dreissensia magnifica Locard, 1893; Dreissensia occidentalis Locard,
1893; Dreissensia paradoxa Locard, 1893; Dreissensia recta Locard, 1893; Dreissensia
Класс двустворчатые моллюски — Bivalvia Linnaeus, 17S8
315
servaini Locard, 1893; Dreissensia siouffi Locard, 1893; Dreissensia sulcata Locard,
1893; Dreissensia thiesae Locard, 1893 (как thiesa)', Dreissensia tumida Locard, 1893;
Dreissensia ventrosa Locard, 1893; Dreissensia westerlundi Locard, 1893; Dreissena
polymorpha var. angustiformis Колесников, 1950; Dreissena cornuliformis Али-Заде,
1961; Dreissena gubkini Али-Заде, 1961; Dreissena kambai Али-Заде, 1961; Dreis-
sena kopetdagica Али-Заде, 1961; Dreissena lunata Али-Заде, 1961; Dreissena na-
livkini Али-Заде, 1961].
Раковина зеленовато- или желтовато-бурая с тёмными зигзагообразными полоса-
ми. Линии нарастания резкие. Спинной край сильно выгнутый, часто угловатый.
Форма, выпуклость и скульптура раковин подвержены изменчивости. Встречают-
ся как сильно вытянутые, узкие, так и короткие, широкие раковины. Биссусная
выемка всегда хорошо выражена.
Размеры: ДР до 50 мм.
Вид обитает в пресноводных водоёмах Европы (кроме крайних северных райо-
нов) и севера Малой Азии, в Балтийском, Чёрном, Каспийском, Аральском морях
и их бассейнах. Во второй половине XX века интродуцирован в Северную Амери-
ку (Великие озёра). Моллюски прикрепляются биссусом к твёрдому субстрату и
является важным компонентом обрастаний.
2(1). Раковина не равностворчатая, округло-треугольная. Киль отсутствует или
выражен в виде плавного перегиба только в области макушек. Брюшной
край выгнутый................................................................
.........Dreissena bugensis (Andrusov, 1897) (вкл. табл. IV, рис. 42 а, б).
— bugensis Андрусов, 1897: 285-286, табл. 15, рис. 31-37 (Dreissensia).
Раковина не равностворчатая, окраска коричневато-бурая, иногда с рисунком из
тёмных слабоизогнутых концентрических линий. Створки неравномерно выпук-
лые; от центра к брюшному краю скошены более или менее отвесно, к спинно-
му — полого. Спинной край выгнутый, образует плавно закруглённую, реже угло-
ватую, дугу. Линии нарастания тонкие. Макушки сглаженные, невыступающие,
тесно сближенные. Биссусная выемка имеется или отсутствует.
Размеры: ДР до 50 мм.
Исходный ареал вида охватывает опреснённые участки Днепровско-Бугского ли-
мана Чёрного моря, Южный Буг и нижнее течение Днепра. За последние несколь-
ко десятилетий вид широко расселился в крупные реки черноморско-каспийского
бассейна (дельта Дуная, Днестр, Днепр с верхними притоками, Дон, Волга), про-
ник в Таганрогский залив Азовского моря, Северный Каспий авандельта Волги)
и в Финский залив Балтийского моря. Изолированные популяции, связанные с
антропохорным заносом особей, известны в северной части Западной Европы
(бассейн Рейна) и в Северной Америке (Великие озёра). В озёрах, реже в реках,
на медленном течении. Прикрепляется биссусом к твёрдому субстрату, но менее
прочно, чем предыдущий вид.
Семейство SPHAERIIDAE Deshayes, 1855 (1820)
Типовой род: Sphaerium Scopoli, 1777.
Длина раковины наиболее крупных видов не превышает 25 мм, большинство
шаровок имеют размеры не более 10 мм, а длина самых мелких раковин состав-
316
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
ляет около 2-5 мм. Раковина характеризуется срединным (иногда наклонённым
вперёд) или задним положением макушки, пронизана порами. Замок гетеродонт-
ный, кардинальные и латеральные зубы хорошо развиты; в правой створке име-
ется один кардинальный и по два латеральных зуба, в левой, соответственно, два
кардинальных и по одному латеральному зубу.
Мантийные листки в задней части тела срастаются, образуя в типичном слу-
чае два сифона: анальный (верхний) и бранхиальный (нижний). Первый снабжён
одной парой ретракторов, второй — одной или двумя. Нижний сифон обнаружи-
вает склонность к редукции. Наружная полужабра всегда значительно меньше
внутренней, иногда полностью редуцирована. Выделительные органы (нефридии)
хорошо развиты, имеют трубчатое строение и характеризуются сложной конфи-
гурацией. Развитие прямое (без планктонной личинки), протекает во внутренних
полужабрах материнской особи, в особых органах — выводковых сумках.
Ареал семейства охватывает все континенты кроме Антарктиды. В Палеаркти-
ке представлено тремя подсемействами: Sphaeriinae s. str., Euglesinae Pirogov et
Starobogatov, 1974 и Pisidiinae Gray, 1857.
Комментарии. В это семейство входят маленькие двустворчатые моллюски, палеар-
ктических представителей которых А.В. Корнюшин (1996) относил к трём семействам:
Sphaeriidae, Euglesidae и Pisidiidae надсемейства Pisidioidea
Ключ для определения подсемейств семейства Sphaeriidae
1(2). Длина раковины взрослых особей не менее 7-8 мм. Макушка занимает
срединное положение, иногда наклонена вперёд. Оба сифона трубчатой
формы. Наружная полужабра составляет не менее Уз высоты внутренней.
Во внутренней полужабре несколько выводковых сумок.........................
............................................Sphaeriinae s. str. (с. 317).
2(1). Макушка отчётливо смещена к заднему краю, если это смещение не вы-
ражено, то раковина маленькая (длина раковины меньше 5 мм). Анальный
сифон имеет вид короткой трубки или воронки, бранхиальный редуциро-
ван до простого отверстия мантии, иногда отсутствует. Наружная полужа-
бра составляет менее Уз высоты внутренней или отсутствует. Во внутрен-
ней полужабре одна выводковая сумка.
3(4). Выводковая сумка смещена вентрально. Если раковина больше 5,5 мм в
длину и лигамент виден со спинной стороны закрытой раковины, то внеш-
няя поверхность створок гладкая, с тонкими линиями нарастания, тонко
исчерченная или с тонкими рёбрышками.......................................
.......................Euglesinae Pirogov et Starobogatov, 1974 (с. 321).
4(3). Выводковая сумка расположена во внутренней полужабре дорсально.
Если раковина больше 5,5 мм в длину и лигамент виден со спинной сто-
роны закрытой раковины, то внешняя поверхность створок с концентри-
ческими резкими рёбрами или грубо концентрически исчерчена.................
........................................Pisidiinae Gray, 1857 (с. 332).
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
317
Подсемейство Sphaeriinae s. str.
Типовой род: Sphaerium Scopoli, 1777.
Раковина относительно крупная: длина створки от 7-8 до 20-25 мм. Макуш-
ки занимают срединное положение, иногда наклонены вперёд. Сифонов два, оба
трубчатой формы, верхний сифон снабжён одной парой ретракторов, нижний
двумя парами. Наружная полужабра относительно хорошо развита (составля-
ет не менее Уз высоты внутренней) и имеет две нормально развитые ламеллы.
Во внутренней полужабре одновременно формируется несколько выводковых
сумок.
Подсемейство характеризуется всесветным распространением. В Палеарктике
представлено семью родами, шесть из которых представлены в европейской ча-
сти России.
Ключ для определения родов подсемейства Sphaeriinae
1(2). Раковина тонкостенная, хрупкая. Макушки очень узкие, выступающие
(«сосцевидной» формы). Эмбриональная раковина очень маленькая, в
виде «шапочки»; иногда она обособлена от остальной части створки за-
метной бороздой. Замочная площадка резко сужена под макушкой............
..............................................Musculium Link, 1807.
2(1). Раковина толстостенная, относительно прочная. Макушки широкие. Эм-
бриональная раковина крупнее, не имеет вид шапочки резко обособленной
бороздой. Как правило, она отграничена от остальной части створки линией
нарастания или цветом. Замочная площадка под макушкой не сужена.........
...................................Sphaerium Scopoli, 1777 (с. 318).
Род Musculium Link, 1807
Типовой вид: Tellina lacustris O.F. Muller, 1774.
Раковина, как правило, небольшая, лишь у немногих видов длина створок до-
стигает 10 мм. Лигамент никогда не выступает над спинным краем створки. Кар-
динальные зубы небольшие, почти прямые, иногда редуцированы. Отпечаток
верхнего сифонального мускула имеет вид выступа на передне-спинном секторе
отпечатка заднего аддуктора. Наружная полужабра составляет не более /з высоты
внутренней.
Ареал рода охватывает Европу, Северную, Центральную и Восточную Азию,
Северную Америку и Юго-Восточную Азию. Моллюски населяют различные
типы водоёмов — от небольших луж и каналов до крупных рек и озёр.
В составе рода насчитывается до 19 видов. На севере и северо-западе европей-
ской части России обитают 5 видов: Musculium creplini (Dunker, 1845), М. lacus-
tre (O.F. Muller, 1774), M. ryckolti (Normand, 1844), M. terverianum (Dupui, 1834) и
M. mucronulatum (Moquin-Tandon, 1855).
318
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Род Sphaerium Scopoli, 1777
Типовой вид: Tellina cornea Linnaeus, 1758.
Раковина по сравнению с моллюсками предшествующего рода более толсто-
стенная, с более выраженной скульптурой. Эмбриональная часть створки отно-
сительно велика (что связано с крупными размерами молоди, длительное время
развивающейся под защитой материнского организма). Настоящей «шапочки», как
у Musculium, не образуется, поскольку рост раковины молодого животного начина-
ется сразу после отрождения. Однако замедление или остановка роста на поздних
стадиях онтогенеза, вызванная неблагоприятными условиями, может привести к
появлению в верхней части створки резкого перегиба или глубокой борозды. Замок
хорошо развит. Наружная полужабра достигает Vi высоты внутренней.
Ареал рода — Голарктика. В составе рода различают от 4 до 6 подродов (Ког-
niushin, 2001).
Ключ для определения подродов рода Sphaerium
1(2). Раковина относительно крупная (длина створки достигает 20 мм и более).
Лигамент заметно выступает над спинным краем раковины. Верхние ре-
тракторы нижнего сифона очень длинные, их отпечатки далеко отстоят от
отпечатков задних аддукторов...............Rivicoliana Servain, 1888 (с. 320).
2(1). Раковина меньших размеров. Лигамент не выступает над спинным краем
створки, хотя может быть хорошо виден со спинной стороны. Сифональ-
ные ретракторы более короткие, их отпечатки соединяются с отпечатками
аддукторов, если обособлены, то не удалены от последних.
3(8). Верхние ретракторы нижнего сифона длинные, их отпечатки удалены от
отпечатков аддукторов.
4(7). Раковина удлинённо-овальная. Наружная поверхность створок покрыта
концентрическими рёбрами или с исчерченностью. Поры раковины не-
большие, редко расположенные, малозаметные.
5(6). Макушки выступающие. Наружная поверхность створок равномерно по-
крыта крупными отчётливыми рёбрами.........................................
................................Cyrenastrum Bourguignat, 1854 (с. 319).
6(5). Макушки мало выступающие. Наружная поверхность створок исчерчена.
....................................Amesoda Rafinesque, 1820 (с. 319).
7(4). Раковина округлая или коротко-овальная. Наружная поверхность створок
тонко исчерченная или гладкая, со специфическим шелковистым блеском.
Створки пронизаны многочисленными крупными порами..........................
....................Nucleocyclas Alimov et Starobogatov, 1968’ (с. 319).
1 А.В. Корнюшин (Komiushin, 2001) рассматривает Nucleocyclas Alimov et Starobogatov, 1968
и Parasphaerium Alimov et Starobogatov, 1968 в качестве видовых групп неопределённого ранга.
За неимением лучшего решения, в определителе эти таксоны пока оставлены в качестве под-
родов рода Sphaerium.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
319
8(3). Верхние ретракторы нижних сифонов короткие, их отпечатки слиты с от-
печатками аддукторов.
9(10). Раковина небольшая (длина не более 7-8 мм). Эмбриональная часть
створки несёт правильную концентрическую исчерченность....................
....................Parasphaerium Alimov et Starobogatov, 1968 (с. 320).
10(9). Раковина относительно крупная (длина более 10 мм). Эмбриональная
часть створки гладкая........................Sphaerium Scopoli, 1777 (с. 321).
Подрод Amesoda Rafinesque, 1820
Типовой вид: Cyclas similis Say, 1816.
Раковина средних размеров или более или менее крупная (длина створок наи-
более крупных экземпляров, как правило, от 8 мм до 13 мм), более или менее тол-
стостенная, прочная. Форма раковины удлинённо-овальная или неправильно че-
тырёхугольная. Макушки мало выступающие. Скульптура в виде исчерченности,
иногда довольно резкой. Зубы замка хорошо развиты. Кардинальный зуб правой
створки изогнут под тупым углом, его задняя ветвь утолщена или расщеплена.
Внутренний кардинальный зуб левой створки угловатый, Наружный плавно вы-
гнут. Его передняя часть может далеко заходить вперёд, полностью перекрывая
внутренний зуб.
Ареал подрода охватывает восточную Палеарктику. На севере европейской ча-
сти России встречается один вид Sphaerium asiaticum Martens, 1884.
Подрод Cyrenastrum Bourguignat, 1854
Типовой вид: Cyclas solida Normand, 1854.
Раковина средних размеров (длина створок, как правило, не превышает
10 мм), овальная, толстостенная прочная. Поверхность створок покрыта хорошо
выраженными концентрическими рёбрами. Макушки выступающие. Кардиналь-
ные зубы хорошо развиты, угловатые. Кардинальный зуб правой створки на за-
днем конце расщеплён. Наружный кардинальный зуб левой створки может быть
заметно короче заднего.
Монотипический подрод. Типовой вид распространён в западной Палеарктике
вплоть до Урала. Sphaerium solida является типичным реофилом, характерна для
крупных рек с сильным течением, встречается в водохранилищах. Предпочитает
песчаные грунты.
Подрод Nucleocyclas Alimov et Starobogatov, 1968
Типовой вид: Cyclas nucleus Studer, 1820.
Раковина крупная (длина створки до 12 мм), округлая или широкоовальная,
иногда удлинённая, относительно толстостенная, гладкая, со специфическим
шелковистым блеском. Макушки более или менее широкие, мало выступающие.
320
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Раковина пронизана многочисленными равномерно расположенными порами,
расстояние между ними составляет 30-45 мкм. Внутренний кардинальный зуб ле-
вой створки резко изогнут, наружный зуб располагается косо, его передний конец
не заходит за точку перегиба внутреннего зуба. Кардинальный зуб правой створ-
ки тоже изогнут, его задний конец расщеплён. Лигамент длинный.
Ареал подрода скорее всего голарктический (Старобогатов, Корнюшин, 1986).
В Палеарктике обитают три вида, два из которых: Sphaerium nucleus (Studer,
1820) и S. radiatum (Clessin in Westerlund, 1877) встречаются в европейской части
России. Моллюски приурочены к заболоченным водоёмам (небольшие озерца,
мелиоративные канавы, залитые водой луга), а также к медленно текущим рекам,
старицам и крупным озёрам (S. radiatum).
Подрод Parasphaerium Alimov et Starobogatov, 1968
Типовой вид: Sphaerium rectidens Starobogatov et Streletzkaja, 1967.
Раковина широкоовальная или сферическая, небольшая (длина створки, как
правило, не превышает 7,5 мм). Характерен прямой спинной край створки с выра-
женными передними и задними углами. Поверхность гладкая, с чёткими линиями
нарастания. Эмбриональная часть створки несёт ярко выраженную концентри-
ческую скульптуру. Макушки относительно узкие, выступающие. Кардинальные
зубы обоих створок тонкие, почти прямые. Кардинальный зуб правой створки не
расщеплён, наружный кардинальный зуб левой створки смещён назад по отноше-
нию к внутреннему. Лигамент короткий, не выходит за контур макушки.
В составе подрода различают два вида, по крайней мере в Палеарктике (Кор-
нюшин, 1996; Komiushin, 2001). Информация об обитании этих моллюсков в Се-
верной Америке нуждается в подтверждении. В европейской части России встре-
чается один вид Sphaerium nitidum (Clessin in Westerlund, 1876), ареал которого
охватывает арктические и субарктические широты от Скандинавии до Чукотки.
Моллюски обитают в небольших тундровых озёрах.
Подрод Rivicoliana Servain, 1888
Типовой вид: Cyclas rivicola Leach in Lamarck, 1818.
Наиболее крупные представители надсемейства (длина створок до 23 мм, вы-
сота до 18 мм, выпуклость до 14 мм). Раковина толстостенная, поверхность ство-
рок резко, но неравномерно исчерчена. Макушки широкие и уплощённые, мало
выступающие, отчётливо наклонены вперёд. Кардинальные зубы резко изогнуты,
задняя часть кардинального зуба правой створки расщеплена.
Монотипический подрод. Единственный представитель Sphaerium rivicola
(Leach in Lamarck, 1818) распространён в Европе: к северу до Балтики и к югу до
Альп, на Южном Урале и на юге Западной Сибири. Реофильные моллюски, оби-
тают в реках и недавно отделившихся от русла пойменных водоёмах. Встречают-
ся как на песчаных, так и на илистых грунтах, на глубинах до 2,5-3,0 м.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
321
Подрод Sphaerium s. str.
Типовой вид: Tellina cornea Linnaeus, 1758.
Раковина крупная (длина створки до 12 мм), от широкоовальной до почти сфе-
рической, вздутая, тонкостенная, гладкая или неравномерно исчерченная, обычно
с чёткими линиями нарастания. Макушки широкие, выступающие. Поры скон-
центрированы в макушечной части, по направлению к наружному краю плот-
ность пор быстро убывает. Кардинальные зубы левой створки почти одинаковой
длины, располагаются параллельно друг другу и краю замочной площадки, плав-
но выгнуты. В правой створке кардинальный зуб тонкий, плавно изогнут, на за-
днем конце слегка расщеплён.
Представители подрода распространены в Голарктике. В Палеарктике выделя-
ют 2 вида, один из которых Sphaerium baicalense известен исключительно из Бай-
кала (Komiushin, 2001). Второй — S. согпеит — широко распространён в Европе,
Сибири и на Алтае. Типичными местообитаниями для них являются медленно те-
кущие реки, пойменные водоёмы и крупные озёра.
Подсемейство Euglesinae Pirogov et Starobogatov, 1974
Типовой вид: Euglesa Jenyns, 1832.
Раковина небольшая или средних размеров. Макушки смещены к заднему
краю створок. Сифоны редуцированы: верхний (анальный) образует короткую
широкую трубку или воронку, нижний (бранхиальный), если имеется, представ-
ляет собой простое отверстие; каждый сифон снабжён парой ретракторов. Иногда
бранхиальное отверстие отсутствует, в этом случае его ретракторы тоже редуци-
руются. Наружная полужабра в различной степени редуцирована, как правило,
состоит из одной ламеллы (наружной или восходящей), у некоторых представи-
телей сохраняется рудиментарная нисходящая ламелла, иногда наружная полужа-
бра отсутствует. Выводковая сумка одна, развивается из утолщений филаментов в
нижней трети внутренней полужабры.
Подсемейство распространено по всем континентам.
Ключ для определения родов подсемейства Euglesinae
1(2). Раковина относительно крупная (длина створок не менее 7,0 мм). Лига-
мент хорошо виден, если смотреть на закрытую раковину сверху. Наруж-
ная полужабра имеет небольшую нисходящую ламеллу, которая налегает
на внутреннюю полужабру.....................Lacustrina Sterki, 1916 (с. 330).
2(1). Раковина небольшая (длина створок не более 5,5 мм). Лигамент закрыт
тонкой известковой стенкой и снаружи не виден (или заметен, просвечи-
вая через стенку). Наружная полужабра (если имеется) состоит из одной
ламеллы и не налегает на внутреннюю полужабру.
3(14). Раковина маленькая (длина створок 2,5-5,0 мм). Сифональных отверстий
два, наружная полужабра имеется.
322
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
4(11). Пресифональная спайка (участок сросшихся листков мантии между ниж-
ним сифоном и не сросшимися листками мантии в передней части тела,
образующими педальное отверстие для высовывания ноги) не удлинена и
составляет не более 0,40 длины свободного края каждого из мантийных
листков.
5(6). Лигаментная ямка узкая и длинная. Макушки узкие, отчётливо смещены
к заднему краю створок, благодаря чему раковина приобретает округло-
треугольную форму. На макушках, как правило, формируются косо распо-
ложенные складки. Если складки отсутствуют, то макушки располагаются
ближе к центру спинной стороны створок и не предают раковине округло-
треугольную форму. В средней части внутренней полужабры утолщённые
филаменты, образующие сумку, чередуются с неизменёнными...................
.....................................Henslowiana Fagot, 1892 (с. 327).
6(5). Лигаментная ямка короткая и широкая. Макушки широкие или узкие,
но шире, чем у предыдущего рода, всегда без складок, сильно или слабо
сдвинутые к заднему краю створок (в последнем случае спинной край
створок выгнутый). Раковина округло-овальная или неправильно-четырёх-
угольная, если округло-треугольная, то макушки широкие. Утолщённые
филаменты, образующие сумку, расположены подряд.
7(10). Раковина округло-овальная с выраженной концентрической исчерченно-
стью или ребристостью. Если гладкая, то неправильно-четырёхугольной,
округло-треугольной или трапециевидной формы. Отпечатки внутренних
радиальных мантийных мускулов не отделены от мантийной линии.
8(9). Раковина округло-овальная с узкими макушками и выраженной концен-
трической исчерченностью наружной поверхности створок (особенно в
области макушки). Внутренние радиальные мантийные мускулы развиты
слабо и малозаметны.......................................................
.....................Hiberneuglesa Starobogatov in Dolgin, 1983 (с. 329).
9(8). Раковина с широкими макушками. Если округло-овальная, то с концентри-
ческой ребристостью наружной поверхности створок, если гладкая, то не-
правильно-четырёхугольной формы. Внутренние радиальные мантийные
мускулы хорошо развиты....................................................
...................Cingulipisidium Pirogov et Starobogatov, 1974 (с. 323).
10(7). Раковина гладкая, округло-овальной, реже трапециевидной или округло-
треугольной формы. Отпечатки внутренних радиальных мантийных му-
скулов (все или частично) отделены от мантийной линии.....................
.................................Euglesa Leach in Jenyns, 1832 (с. 325).
11(4). Пресифональная спайка заметно удлинена и составляет около половины
(или больше) длины свободного края каждого из мантийных листков.
12(13). Раковина удлинённо-четырёхугольная (трапециевидная), с ясно выражен-
ными углами и спрямлённым брюшным краем. Макушки приближены к
середине спинного края. Наружная поверхность створок гладкая, блестя-
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
323
щая. Передний аддуктор очень большой, бобовидной формы. Внутренние
радиальные мантийные мускулы каждой стороны собраны в один мощный
пучок. Почти все их отпечатки (кроме двух в срединной части) слиты с
мантийной линией... Tetragonocyclas Pirogov et Starobogatov, 1974 (с. 331).
13(12). Раковина округло-треугольная, клиновидная, гладкая, тонко ребристая
или исчерчена. Брюшной край выгнутый. Макушки сильно смещены к за-
днему краю створок и наклонены назад. Передний аддуктор не увеличен,
овальной формы. Внутренние радиальные мантийные мускулы каждой
стороны собраны в несколько пучков (не менее трёх). Их отпечатки от-
делены на значительное расстояние от мантийной линии.......................
..........................................Pseudeupera German, 1913 (с. 330).
14(3). Раковина очень маленькая (длина створок не более 2,0 мм). Сифональное
отверстие одно, наружная полужабра отсутствует.............................
........................Conventus Pirogov et Starobogatov, 1974 (с. 324).
Род Cingulipisidium Pirogov et Starobogatov, 1974
Типовой вид: Pisidium nitidum Jenyns, 1832.
Раковина 2,5-4,5 мм длиной, овальная или неправильно-четырёхугольная. Ма-
кушка, как правило, узкая, немного смещена назад. Поверхность ребристая или
грубо исчерченная. Поры редкие. Замочная площадка развита в разной степени,
кардинальные зубы прямые или слабо изогнуты. Отпечатки мантийных мускулов
заметно выступают над мантийной линией. Бранхиальное отверстие жабры име-
ется.
Ареал рода охватывает Европу, Северную Азию и Северную Америку.
В составе рода выделяют три подрода, два из которых распространены в евро-
пейской части России.
Ключ для определения подродов рода Cingulipisidium
1(2). Раковина овальная, брюшной край выгнут. Замочная площадка нормаль-
но развита, иногда расширена. Скульптура представлена несколькими
рёбрышками, окаймляющими макушку или равномерно расположенными
концентрическими рёбрами...............................Cingulipisidium s. str.
2(1). Раковина неправильно-четырёхугольная, гладкая, брюшной край спрям-
лённый. Замочная площадка очень узкая......................................
.............................Pseudosphaerium Komiushin, 1990 (с. 324).
Подрод Cingulipisidium s. str.
Типовой вид: Pisidium nitidum Jenyns, 1832.
Раковина небольшая (длина створки, как правило, не превышает 3,5 мм, но
иногда достигает 4,3 мм), овальная. Наружная поверхность створок вся покрыта
концентрическими рёбрышками, либо могут быть развиты только 2-3 рёбрышка,
324
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
окаймляющих макушку. Макушки умеренно выступающие. Поры редкие или от-
сутствуют. Замочная площадка относительно широкая. Кардинальные зубы пря-
мые или дугообразно изогнутые.
Ареал подрода в целом соответствует ареалу рода. Моллюски населяют не-
большие медленно текущие реки, пойменные водоёмы и прибрежные зоны озёр.
Встречаются на заиленных и песчаных грунтах.
В Палеарктике подрод представлен 11 видами, 7 из которых могут быть встре-
чены в европейской части России: Cingulipisidium cingulatum (Anistratenko et Sta-
robogatov, 1990 [1991]), C. crassum (Stelfox, 1918), C. depressinitidum (Anistraten-
ko et Starobogatov, 1990[1991]), C. feroense Komiushin, 1991, C. nitidum (Jenyns,
1832), C. splendens (Bandon, 1857), C. tanugaTmm, 1975.
Подрод Pseudosphaerium Komiushin, 1990
Типовой вид: Pisidium pseudosphaerium Favre, 1927.
Раковина небольшая (длина створки составляет до 3 мм), неправильно-четы-
рёхугольной формы, гладкая. Макушки мало выступающие, слегка смещены к за-
днему краю створок. Замочная площадка очень узкая, зубы замка несколько недо-
развиты, кардинальные зубы в виде тонких пластинок.
Подрод характеризуется европейским ареалом и включает два вида: Pseudo-
sphaerium favrei (Kuiper, 1947) и Р pseudosphaerium (Favre, 1927). Оба вида при-
урочены к болотным водоёмам и водотокам: небольшим озёрам и медленно те-
кущим рекам с заболоченными берегами, черноольховым топям, мелиоративным
канавам. Встречаются в зарослях среди корней растений, в залитом водой расти-
тельном опаде или в сплавине.
Род Conventus Pirogov et Starobogatov, 1974
Типовой вид: Pisidium conventus Clessin, 1877.
Раковина небольшая (длина створки до 2 мм), округло-овальная или непра-
вильно-четырёхугольная, гладкая. Макушки широкие, уплощённые, мало высту-
пающие, незначительно смещены к заднему краю створок. Поры редкие. Ширина
замочной площадки варьирует, кардинальные зубы прямые или слабоизогнутые.
Латеральные зубы тонкие. Имеется только верхнее сифональное отверстие. Ниж-
нее сифональное отверстие и его мускулатура редуцированы. Наружная полужа-
бра отсутствует.
Ареал рода охватывает Северную и Центральную Европу, север Азии и Север-
ную Америку. Моллюски приурочены к озёрам и являются характерными пред-
ставителями европейской глубоководной фауны. В озёрах Центральной Европы
отмечены на глубине 350 м. В Карелии (Умб озеро) — до 100 м.
В Палеарктике род представлен по меньшей мере 6 видами, три из которых
являются эндемиками Байкала. На севере европейской части России обитают:
Conventus conventus (Clessin, 1877) и С. urinator (Clessin, 1877).
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
325
Род Euglesa Leach in Jenyns, 1832
Типовой вид: Tellina henslowiana Leach in Sheppard, 1823.
Длина створок раковины варьирует от 2,5-3,5 мм до 5,0-6,0 мм. Форма рако-
вины округло-овальная, реже округло-треугольная или трапециевидная. Гладкая.
Макушки широкие, заметно смещены к заднему краю створок. Ширина замочной
площадки, конфигурация и взаимное расположение зубов варьируют в широких
пределах. Отпечатки внутренних мантийных мускулов, как правило, отделены от
мантийной линии. Бранхиальное (нижнее) сифональное отверстие мантии име-
ется. Размеры и положение наружной жабры крайне изменчивы даже в пределах
одного вида.
Род распространён по всем континентам. В Палеартике представлен тремя
подродами.
Ключ для определения подродов рода Euglesa
1(2). Раковина небольшая (длина створки не превышает 3,5 мм). Макушки вы-
ступающие, замочная площадка в подмакушечной части резко сужена.
Латеральные зубы укорочены. Передние пучки внутренних радиальных
мышц мантии, прилегающие к аддуктору, ослаблены и трудно различимы
.............................................Cyclocalix Dall, 1903 (с. 326).
2(1). Раковина 3,0-6,0 мм длиной. Макушки мало выступающие. Замочная пло-
щадка не сужена или сужена равномерно. Латеральные зубы относительно
длинные, если укорочены, то раковина более 4,0 мм длиной. Передние
пучки внутренних радиальных мышц мантии хорошо выражены.
3(6). Раковина толстостенная, прочная. Замочная площадка не сужена. Карди-
нальные зубы резко изогнуты. Внутренние радиальные мантийные муску-
лы мощные, число их пучков больше 7.
4(5). Латеральные зубы укорочены. Поры раковины многочисленные. Наруж-
ная полужабра крупная, у взрослых экземпляров её наружный край нахо-
дится на уровне пятого или шестого филаментов внутренней.................
..........................................Roseana Fagot, 1892 (с. 327).
5(4). Латеральные зубы не укорочены. Поры раковины малочисленные, редкие.
Наружная полужабра небольших размеров, её передний край расположен
не ближе, чем уровень десятого филамента внутренней полужабры............
...........Euglesa s. str. partim (? секция Casertiana Fagot, 1892) (c. 326).
6(3). Раковина тонкостенная, хрупкая. Замочная площадка узкая, кардинальные
зубы слабо изогнуты или почти прямые. Внутренняя радиальная мантий-
ная мускулатура несколько редуцирована (особенно в задней области) за
счёт исчезновения и/или ослабления отдельных пучков. Число мышечных
пучков, как правило, меньше 7 (обычно 5-6). Если больше 7, то мозоле-
видный наплыв правой створке отсутствует.
326 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
7(8). В правой створке имеется зубообразный мозолевидный наплыв — каллюс.
Внутренние радиальные мышцы мантии образуют 5-6 мощных пучков....
.........................Euglesa s. str. partim (? секция Euglesa s. str.) (c. 326).
8(7). Каллюс отсутствует. Пучков внутренних радиальных мантийных мышц 7,
но они ослаблены. Задние пучки, прилегающие к пресифональной спайке,
трудноразличимы..........Euglesa s. str. partim
(? секция Potamopisidium Komiushin, 1991).
Подрод Cyclocalix Dall, 1903
Типовой вид: Pisidium scholtzi Clessin, 1871.
Раковина небольшая (2-3 мм длиной), широкоовальная или округлая, выпук-
лая, иногда вздутая. Макушки заметно выступающие. Замок очень короткий,
особенно укорочены латеральные зубы. Мантийная мускулатура несколько ос-
лаблена, особенно пучки, прилегающие к пресифональной спайке и переднему
аддуктору. Число пучков 5-6. Средние пучки наиболее длинные и обнаруживают
тенденцию к слиянию.
Подрод распространён в Европе, Северной, Центральной и Восточной Азии,
Северной Америке. Моллюски характеризуются значительной экологической
пластичностью. Наиболее характерны для заболоченных водоёмов, часто встре-
чаются на заливных лугах и в родниковых топях, особенно под пологом леса. Не-
редки и в крупных озёрах, где приурочены к участкам берега, характеризующим-
ся некоторой заболоченностью, или к зарослям. Встречаются в медленнотекущих
ручьях и канавах.
В европейской части России обитают 7 видов: Euglesa dubrueilii (Baudon,
1872), E.fragilis (Clessin in Westerlund, 1873), E. hinzi (Kuiper, 1975), E. lapponicus
(Clessin in Westerlund, 1873), E. magnificus (Clessin in Westerlund, 1873), E. obtusa-
lis (Lamarck, 1818), E.scholtzi (Clessin, 1871).
Подрод Euglesa s. str.
Типовой вид: Tellina henslowiana Leach in Sheppard, 1823.
Раковина 3,0-5,5 мм, овальная, реже округлая, треугольная или трапециевид-
ная, относительно плоская, с широкими мало выступающими макушками. Замок
развит в разной степени, но всегда замочная площадка относительно длинная.
Пучки внутренних радиальных мышц мантии развиты в разной степени, несколь-
ко усиливаются в направлении сзади наперёд. Их число варьирует от 5-6 до 8-10.
Подрод распространён по всем континентам. Точное количество видов подро-
да установить затруднительно из-за запутанности систематики группы и неясного
статуса многих описанных форм. Более или менее обоснованно можно говорить
об обитании на севере и северо-западе европейской части России около 14 видов.
Большинство из них приурочены к небольшим речкам и ручьям, реже к озёрам.
Это: Euglesa buchtarmensis Krivosheina, 1978, Е. curta (Clessin, 1874), E. hoyeri
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
327
(Clessin in Esmark et Hoyer, 1886), E. intermedia (Gassies, 1855), E. jaudouiniana
(Gassies, 1855), E. obliquata (Clessin in Martens, 1874), E. ponderosa (Stelfox, 1918),
E. rivularis (Clessin, 1874), E. steenbuchi (Moller, 1842), E. subponderosa Krivo-
sheina, 1976. Некоторые виды: Euglesa eichwaldi Stadnichenko, 1983 и E. personata
(Malm, 1855) — обитают главным образом в родниках и родниковых топях. Ино-
гда встречаются в колодцах. Euglesa likharevi (Komiushin, 1991) приурочен к про-
фундали крупных озёр.
Подрод Roseana Fagot, 1892
Типовой вид: Pisidium roseum Scholtz, 1843.
Раковина до 5 мм длиной, округлая или широкоовальная, умеренно или силь-
новыпуклая, относительно тонкостенная. Макушки широкие, выступающие, рас-
положенные недалеко от середины спинного края створок. Раковина пронизана
многочисленными относительно крупными порами, которые расположены осо-
бенно тесно в подмакушечной части (здесь расстояние между ними не превыша-
ет 40 мкм). Замочная площадка сужена, особенно под макушками. Кардинальные
зубы отчётливо изогнутые, латеральные — сильно укороченные. Вдоль края ман-
тии насчитывается не менее семи пучков внутренних радиальных мышечных во-
локон.
Ареал подрода охватывает Евразию, кроме, вероятно, самых южных районов.
Моллюски приурочены к болотным водоёмам. Встречаются в небольших прудах
и заболоченных лесных озёрцах, на заливных лугах, в мелиоративных канавах
и ручьях с медленным течением. В европейской части России обитают не ме-
нее трёх видов: Roseana globularis (Clessin in Westerlund, 1873), R. rosea (Scholtz,
1843) и R. silesiaca (Starobogatov et Komiushin in Komiushin, 1991).
Род Henslowiana Fagot, 1892
Типовой вид: Pisidium henslowianum Fagot, 1892.
Раковина небольшая, длина створки как правило не превышает 5 мм. Форма
раковины овальная, округло-треугольная иногда неправильно-четырёхугольная.
Скульптура выражена в разной степени. Макушки узкие, отчётливо смещены
назад, выступающие. Ширина замочной площадки варьирует, характерны от-
чётливый изгиб кардинального зуба правой створки (кроме популяций, харак-
теризующихся общей редукцией замка), а также относительно узкая и длинная
лигаментная ямка. Пресифональная спайка мантии короткая. Пучки внутренних
радиальных мантийных мускулов всегда несколько ослаблены, но передние 3-4
могут быть хорошо выражены.
Виды рода известны из Европы, Северной и Восточной Азии, а также в Север-
ной Америке. Палеарктические представители рода группируются в четыре под-
рода, три из которых представлены в европейской части России.
328
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Ключ для определения подродов рода Henslowiana
1(2). Раковина четырёхугольная, коротко- или округло-овальная. Макушечные
складки отсутствуют. Пучки внутренних радиальных мантийных муску-
лов хорошо выражены..........................................................
.......................Arcteuglesa Pirogov et Starobogatov, 1974 (с. 328).
2(1). Раковина округло-треугольная. Макушечные складки имеются, в случае
их отсутствия раковина имеет отчётливо удлинённую форму. Пучки вну-
тренних радиальных мантийных мускулов ослаблены, малозаметны и
могут распадаться на отдельные волокна.
3(4). Раковина удлинённо-овальная. Замочная площадка не расширена. Перед-
ние пучки внутренних радиальных мантийных мускулов не распадаются
на отдельные волокна. Мантийная линия расположена близко к краю ство-
рок ...............................................Henslowiana s. str. (с. 328).
4(3). Раковина округло-треугольная. Замочная площадка отчётливо расширена.
Все пучки внутренних радиальных мантийных мускулов распадаются на
отдельные волокна. Мантийная линия далеко отстоит от краёв створок...........
....................................Supiniana Komiushin, 1996 (с. 329).
Подрод Arcteuglesa Pirogov et Starobogatov, 1974
Типовой вид: Pisidium lilljeborgi Clessin, 1886.
Раковина четырёхугольная, коротко- или округло-овальная, относительно тон-
костенная, гладкая или исчерченная, иногда очень тонко ребристая. Макушки без
складок. Поры раковины многочисленные, обычно хорошо заметны. Замочная
площадка от умеренно широкой до очень узкой. В последнем случае кардиналь-
ные и наружные латеральные зубы обнаруживают склонность к редукции. Отпе-
чатки внутренних радиальных мантийных мускулов выступают над мантийной
линией, передние могут быть обособлены.
Ареал подрода бореально-альпийский. Моллюски приурочены к озёрам.
Встречаются как на литорали, так и в профундали, на песчаных и заиленных
грунтах. На севере и северо-западе европейской части России обитают три вида:
Henslowiana lilljeborgii (Clessin in Esmark et Hoyer, 1886), H ruut (Timm, 1975),
H waldeni (Kuiper, 1975).
Подрод Henslowiana s. str.
Типовой вид: Pisidium henslowianum Fagot, 1892.
Раковина удлинённо-овальная, умеренно выпуклая, относительно тонкостен-
ная, гладкая или слабо исчерченная. Макушки обычно несут косо расположенные
складки. Поры раковины редкие. Замочная площадка не расширена. Отпечатки
передних внутренних радиальных мантийных мускулов выступают над мантий-
ной линией, но не обособляются от неё.
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
329
Ареал подрода соответствует ареалу рода. Моллюски являются лимнофилами.
Населяют пойменные водоёмы средних и крупных рек, встречаются также в не-
больших реках и озёрах. В европейской части России представлены пять видов:
Henslowiana dupuyana (Normand, 1854), Н. henslowana (Leach in Sheppard, 1823),
H. ostroumovi (Pirogov et Starobogatov, 1974), H. polonica (Anistratenko et Starobo-
gatov, 1990 [1991]), H. suecica (Clessin in Westerlund, 1873).
Подрод Supiniana Komiushin, 1996
Типовой вид: Pisidium supinumk. Schmidt, 1850.
Раковина округло-треугольная, умеренно выпуклая, толстостенная, отчётливо
регулярно исчерченная или ребристая. Макушки со складками, поры раковины
имеются, но обычно они малозаметные. Отпечатки внутренних радиальных ман-
тийных мускулов не выступают над мантийной линией, мантийная линия ровная
или слегка волнистая. Замочная площадка расширена. Латеральные зубы очень
мощные.
Подрод известен только в Палеарктике. Его ареал охватывает Европу и при-
легающие части Западной Сибири (бассейн реки Иртыш). Моллюски относятся
к реофилам. Обитают в русловой части крупных рек, реже встречаются в озёрах.
Предпочитают песчаные или слабо заиленные грунты. На севере и северо-запа-
де европейской части России обитают три вида: Henslowiana baudonii (Clessin,
1873), Н. conica (Baudon, 1857) и H. supina (A. Schmidt, 1850).
Род Hiberneuglesa Starobogatov in Dolgin, 1983
Типовой вид: Pisidium hibernicum Westerlund, 1894.
Раковина небольшая (длина створки до 3 мм), округло-овальной формы. При
переходе спинного края в передний и задний края, как правило, имеются уголки
(«плечики»), что иногда придаёт контурам створок неправильно-четырёхугольный
облик. Макушки узкие, заметно выступающие, приближены к середине спинного
края. Довольно хорошо выражена концентрическая исчерченность, особенно вбли-
зи макушки. Замочная площадка узкая. Кардинальные зубы прямые или дугообраз-
но изогнутые, латеральные зубы короткие, с чётко обособленными выступающими
верхушками. Лигаментная ямка короткая. Отпечатки внутренних радиальных ман-
тийных мускулов выступают над мантийной линией. Бранхиальное сифональное
отверстие имеется, пресифональная спайка мантии не удлинена. Число пучков вну-
тренних радиальных мантийных мускулов достигает 7-8, при этом пучки, приле-
гающие к передним аддукторам, выделяются крупными размерами и хорошо раз-
личимы на окрашенных препаратах. Остальные пучки слабые, трудно различимые.
Ареал рода охватывает Северную и Среднюю Европу, а также Западную Си-
бирь. Находки представителей рода в Восточной Сибири и Северной Америке
нуждаются в подтверждении. Моллюски являются лимнофилами, встречают-
ся в крупных ручьях, небольших реках и пойменных водоёмах более крупных
330
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
рек. Обычны также в небольших озёрах. В крупных водоёмах они приурочены к
участкам берега, защищённым от действия прибоя и заросшим растительностью.
В европейской части России, исключая южные районы, обитают пять видов: Hi-
berneuglesa bodamica (Starobogatov et Komiushin, 1989), H. normalis (Stelfox,
1929), H. parvula (Clessin in Westerlund, 1873), H. portentosa (Stelfox in Ellis, 1940),
H. subhibernica (Starobogatov et Komiushin, 1989).
Род Lacustrina Sterki, 1916
Типовой вид: Pisidium idahoense Roper, 1890.
Раковина овально-треугольная, гладкая, с широкими мало выступающими
макушками, заметно сдвинутыми назад. Замочная площадка широкая, карди-
нальные зубы изогнуты. Бранхиальное сифональное отверстие имеется. Преси-
фональная спайка мантии короткая. Пучки внутренних радиальных мантийных
мускулов мощные, равномерно расположенные вдоль края педального отверстия
мантии. Отпечатки этих пучков обособлены от мантийной линии.
Род в целом имеет голарктическое распространение. На большей части Пале-
арктики (исключая Чукотский, Камчатский полуострова и острова Курильской
гряды) представлен одним видом — Lacustrina dilatata (Westerlund, 1897). Этот
вид приурочен к холодноводным озёрам.
Род Pseudeupera German, 1913
Типовой вид: Pisidium landeroini Germain, 1909.
Раковина 3-4 мм длиной, овальная или округло-треугольная, гладкая или ре-
бристая. Макушка более или менее сужена, значительно смещена к заднему краю
створок. Поры раковины редкие. Ширина замочной площадки варьирует. Карди-
нальные зубы замка спрямлены. Лигаментная ямка сравнительно короткая. От-
печатки внутренних радиальных мантийных мускулов обособлены от мантийной
линии, передние удалены в большей степени, чем задние. Бранхиальное сифо-
нальное отверстие имеется. Внутренние радиальные мантийные мускулы хорошо
развиты, сконцентрированы в области педального отверстия мантии. Мощность
пучков увеличивается сзади наперёд, передние пучки проявляют тенденцию к
слиянию. Хорошо развита концентрическая мускулатура мантии.
Род имеет голарктический ареал и представлен двумя подродами.
Ключ для определения подродов рода Pseudeupera
1(2). Раковина гладкая или тонко исчерченная. Пресифональная спайка состав-
ляет более половины длины педального отверстия мантии. Пучки вну-
тренних радиальных мантийных мускулов сконцентрированы в передней
части мантийного края.............................Pseudeupera s. str. (с. 331).
2(1). Раковина ребристая. Пресифональная спайка составляет менее полови-
ны длины педального отверстия мантии. Пучки внутренних радиальных
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
331
мантийных мускулов располагаются вдоль краёв мантии оконтуривающих
педальное отверстие и равномерно усиливаются сзади наперёд...........
...................Pulchelleuglesa Starobogatov in Dolgin, 1983 (с. 331).
Подрод Pseudeupera s. str.
Типовой вид: Pisidium landeroini Germain, 1909.
Раковина округло-треугольная, гладкая или тонко исчерченная. Макушки уз-
кие, смещены к заднему краю створки и наклонены назад. Число отпечатков вну-
тренних радиальных мантийных мускулов невелико, все они, особенно передние,
далеко отставлены от мантийной линии. Внутренние радиальные мантийные му-
скулы объединены в 4-5 мощных пучка, иногда позади них можно наблюдать ещё
1-2 слабых пучка.
Ареал подрода палеарктический. В конце XX века отечественные специали-
сты различали в составе подрода 13 видов. По мнению западноевропейских авто-
ров подрод представлен в Палеарктике одним видом — Pseudeupera subtruncata
(Malm, 1855). Моллюски характеризуются значительной экологической пластич-
ностью, обитают в ручьях, малых и средних реках, пойменных водоёмах, мелио-
ративных каналах, в озёрах разной величины.
Учитывая сложность в определении видов (как данного подрода, так и вообще
шаровок в целом), а также отсутствие в русскоязычной литературе ключей для
определения видов семейства Euglesidae, при работе с определителем относи-
тельно количества видов подрода рекомендуется предварительно принимать точ-
ку зрения западноевропейских авторов.
Подрод Pulchelleuglesa Starobogatov in Dolgin, 1983
Типовой вид: Pisidium pulchellum Jenyns, 1832.
Раковина овальная. Макушки относительно широкие, маловыступающие, за-
метно смещены к заднему краю створок. Поверхность раковины, за исключени-
ем макушечной части, равномерно покрыта резкими концентрическими рёбрами.
Отпечатки внутренних радиальных мантийных мускулов обособлены, но не уда-
лены от мантийной линии. Вдоль краёв мантии, оконтуривающих педальное от-
верстие расположено 6-7 мощных мускульных пучков, постепенно увеличиваю-
щихся сзади наперёд.
Два вида подрода распространены в Европе и Сибири. Это: Pulchelleuglesa
acuticostata (Starobogatov et Komiushin, 1989) и P pulchella (Jenyns, 1832). Мол-
люски являются реофилами, обитают в ручьях, небольших реках и пойменных
водоёмах, реже в озёрах. Чувствительны к загрязнению воды и могут быть ис-
пользованы для биоиндикации чистоты водоёмов.
Род Tetragonocyclas Pirogov et Starobogatov, 1974
Типовой вид: Pisidium milium Held, 1836.
332
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Раковина небольшая (длина створок 2,5-3,5 мм), удлинённо-четырёхугольная
(трапециевидная), с ясно выраженными углами и спрямлённым брюшным кра-
ем. Макушки приближены к середине спинного края створок, узкие, мало вы-
ступающие. Наружная поверхность створок гладкая, обычно блестящая. Замоч-
ная площадка узкая. Кардинальные зубы почти прямые, параллельны друг другу.
Латеральные зубы вытянуты. Лигаментная ямка короткая. Сифональных отвер-
стий два. Пресифональная спайка мантии практически равна по длине педаль-
ному отверстию. Внутренние радиальные мантийные мускулы каждой стороны
образуют 1-3 мощных сближенных пучка. Кроме того, имеются более слабо раз-
витые пучки (1-2) и отдельные мышечные волокна в области пресифональной
спайки. Очень характерны крупные размеры и бобовидная форма переднего ад-
дуктора.
Виды подрода, по-видимому, широко распространены в Палеарктике, однако
находки из бассейна реки Амур и из Северной Африки нуждаются в подтвержде-
нии. В европейской части России представлен один вид — Tetragonocyclas baudo-
niana (de Cessac, 1855). Моллюски обитают в озёрах, небольших реках, ручьях и
пойменных водоёмах, где могут служить индикаторами некоторой заболоченно-
сти. Также населяют и настоящие болота, отмечены в сплавинах.
Подсемейство Pisidiinae Gray, 1857
Типовой род: Pisidium С. Pfeiffer, 1821.
Размеры раковины варьируют в широких пределах (длина створки от 1,5—
2,0 мм до 7,0-11,0 мм). Макушки смещены к заднему краю створки. Сифоны
устроены так же, как в подсемействе Euglesidae: верхний сифон в виде короткой
и широкой трубки, снабжён парой ретракторов; нижний представляет собой про-
стое отверстие и также имеет пару ретракторов. У представителей многих так-
сонов нижнее сифональное отверстие мантии сливается с педальным вследствие
редукции пресифональной спайки. Сифональные ретракторы при этом сохраня-
ются. Наружная полужабра если имеется, то относительно небольшая, сильно
смещена назад и имеет только одну (восходящую) ламеллу. Исчезновение преси-
фональной спайки сопровождается полной редукцией наружной полужабры. Вы-
водковая сумка одна; развивается в задней части внутренней полужабры и вклю-
чает большое число филаментов. Образованию сумки предшествует разрастание
филаментов по всей длине, либо развитие утолщений в их верхней трети.
Виды подсемейства распространены на всех континентах и сгруппированы в
три рода.
Ключ для определения родов подсемейства Pisidiinae
1(2). Раковина средних размеров (длина створки достигает 11 мм). Сифональ-
ных отверстий два. Наружная полужабра имеется...............................
......................................Pisidium С. Pfeiffer, 1821 (с. 334).
Класс двустворчатые моллюски — Bivalvia Linnaeus, 1758
333
2(1). Раковина небольшая (длина створки, как правило, не превышает 5 мм).
Сифональное отверстие одно. Наружная полужабра отсутствует.
3(4). Раковина очень маленькая (длина створки 1,5-2,3 мм). Лигамент находит-
ся в верхней части замочной площадки (закрытый тип лигамента). Лига-
ментная ямка удлинена, ланцетовидной формы................................
...........................................Neopisidium Odhner, 1921.
4(3). Раковина более крупная. Лигамент глубоко погружён в толщу раковины и
достигает нижнего края замочной площадки (инвертированный тип лига-
мента). Лигаментная ямка треугольной формы................................
.................................Odhneripisidium Kuiper, 1962 (с. 333).
Род Neopisidium Odhner, 1921
Типовой вид: Pisidium torquatum Stelfox, 1918.
Раковина очень маленькая, длина створок не превышает 2,3 мм, треуголь-
ной или трапециевидной формы, исчерченная, иногда неравномерно ребристая.
Макушки широкие, мало выступающие, заметно смещены назад, как правило,
окаймлённые резкими полулунными складками. Замочная площадка относитель-
но широкая, дугообразно выгнута. Кардинальные зубы слабоизогнутые или поч-
ти прямые. Кардинальный зуб правой створки расщеплён на заднем конце. Лига-
мент закрытого типа. Внутренние радиальные мантийные мускулы образуют 6-7
слабых трудноразличимых пучков. Их отпечатки не выступают над мантийной
линией.
Виды рода распространены в Европе и Западной Сибири. По разным данным
в составе рода насчитывается три или четыре вида (Корнюшин, 1996; Kantor et
al., 2010). Реофильные моллюски, населяют реки, где держатся в русловой части,
а также озёра, как в прибрежных биотопах, так и на значительных глубинах. Оби-
тают на песчаных или слегка заиленных грунтах.
На севере и северо-западе европейской части России представлены три вида:
Neopisidium moitessierianum (Paladilhe, 1866), N. torquatum (Stelfox, 1918), N. tri-
gonum (Bourguignat in Locard, 1893).
Род Odhneripisidium Kuiper, 1962
Типовой вид: Pisidium steward Preston, 1909.
Раковина небольшая или средних размеров, овальная или округло-треуголь-
ная. Макушки умеренно широкие, не выступающие. Поверхность гладкая или
слабо исчерченная, иногда тонко ребристая. Признаки замка варьируют. Лига-
мент погружён под замочную площадку, лигаментная ямка треугольной формы.
Пучки внутренних радиальных мантийных мускулов развиты в разной степени.
Род насчитывает 4 подрода и объединяет виды распространённые большей ча-
стью в Азии и Южной Европе (Средиземноморье). На севере Западной Европы
обитают 3 вида, объединяемые Корнюшиным (1996) в подрод Europisidium Staro-
334
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
bogatov in Stadnichenko, 1984. Учитывая теоретическую возможность обнаруже-
ния этих видов на севере и северо-западе европейской части России, в определи-
теле приводится описание подрода Europisidium.
Подрод Europisidium Starobogatov in Stadnichenko, 1984
Типовой вид: Pisidium tenuilineatum Stelfox, 1918.
Раковина небольшая (длина створки до 2,5 мм), округло-треугольная или кли-
новидная, с ясно выраженными уголками («плечиками») при переходе спинного
края створок в передний и задний края. Скульптура представлена отчётливыми
тонкими концентрическими рёбрышками. Замок хорошо развит, замочная пло-
щадка относительно широкая, кардинальные зубы отчётливо изогнутые, лате-
ральные — крупные, мощные. Встречаются, однако, экземпляры с ослабленным
замком. Обычно выражен инвертированный тип лигамента, лигаментная ямка
треугольная. Отпечатки внутренних радиальных мантийных мускулов, как прави-
ло, нечёткие, но у некоторых экземпляров удаётся различить несколько отпечат-
ков, заметно выступающих над мантийной линией. Внутренние радиальные ман-
тийные мускулы образуют 6-7 слабых, но оформленных пучков.
В подрод сгруппированы виды достоверно известные из Западной Европы
(Британские острова, Скандинавия, Альпы, территории Германии и Польшы).
Это три вида: Europisidium alpinum (Odhner, 1938), Е. stelfoxi (Pirogov et Starobo-
gatov in Timm, 1976), E. tenuilineatum (Stelfox, 1918). Моллюски известны из род-
ников и озёр, прежде всего горных. Встречаются также в небольших реках, про-
текающих по более или менее возвышенным территориям.
Род Pisidium С. Pfeiffer, 1821
Типовой вид: Cyclas obliqua Lamarck, 1818.
Раковина от удлинённо-овальной до округло-треугольной. Макушки узкие,
мало выступающие, отчётливо смещены к заднему краю створок. Поверхностная
скульптура хорошо выражена: от резкой неравномерной исчерченности до пра-
вильной ребристости. Поры раковины хорошо заметны, но сравнительно немно-
гочисленны. Замочная площадка широкая. Кардинальный зуб правой створки и
внутренний кардинальный зуб левой очень резко дуговидно изогнуты (у молодых
особей с отчётливым изломом). Пресифональная спайка мантии короткая. Пуч-
ки внутренних радиальных мантийных мускулов (8-10) хорошо развиты и распо-
ложены равномерно вдоль края педального отверстия. Их отпечатки удалены от
мантийной линии.
Ареал рода в общем охватывает Голарктику, но находки в Северной Аф-
рике нуждаются в проверке. В Европе обитает один вид — Pisidium amnicum
(O.F. Muller, 1774). Эти реофильные моллюски в основном приурочены к рекам и
прирусловым пойменным водоёмам. В озёрах встречаются на песчаных грунтах
открытых участков, часто в зоне прибоя.
КЛАСС БРЮХОНОГИЕ МОЛЛЮСКИ —
GASTROPODA CUVIER, 1797
П.В, Кияшко, Е.В. Солдатенко, М.В. Винарский
Брюхоногие моллюски (Gastropoda) составляют крупнейший из всех классов
моллюсков. Видовое богатство ныне живущих гастропод оценивается в 110-120
тысяч видов (Старобогатов, 1994; Шилейко, Кантор, 1994). Будучи изначально
морскими животными, брюхоногие в ходе широкой адаптивной радиации осво-
или также континентальные водоёмы (пресные и солоноватоводные), а также
наземно-воздушную среду обитания. Видовое разнообразие современных пре-
сноводных моллюсков составляет 7000-8000 видов, из которых более половины
приходится на долю гастропод (Старобогатов, 1994; Strong et al., 2008). Согласно
современным филогенетическим построениям на уровне высших таксонов Gas-
tropoda брюхоногие моллюски, включённые в данный определитель, рассматри-
ваются в составе «клад» Neritimorpha, Caenogastropoda и Heterobranchia, которым
ближе всего соответствует ранг подклассов (Aktipis et al., 2008).
Тело большинства гастропод состоит из головы, ноги и внутренностного (вис-
церального) мешка, покрытого мантией и заключённого в спирально закрученную
или колпачковидную раковину. Голова несёт ротовое отверстие, а также пару не-
втягиваемых щупалец, у основания которых расположены глаза (рис. 12). Нога
Рис. 12. Строение мягкого тела Anisus albus (O.F. Muller, 1774) [Planorbidae] (no:
Meier-Brook, 1983).
a — глаз; 6 — щупальце; в — мужское половое отверстие; г — мантия; д, и — кишечник;
е — лёгкое; ж — почка; з — пищеварительная железа; к — нога.
336
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
располагается вентрально, имеет форму мускулистой ползательной подошвы и
выполняет локомоторную функцию. Внутренностный мешок в основном содер-
жит органы пищеварительной и половой систем. Его основание покрыто особой
кожной складкой —мантией, свободный край которой прилегает к нижнему краю
раковины и участвует в её формировании (Иванов, 1940). Между складкой мантии и
внутренностным мешком имеется мантийная полость, в которой располагаются жа-
бры, анальное отверстие, отверстия выделительных органов и иногда половых про-
токов. У некоторых брюхоногих моллюсков жабры редуцируются, а участок стенки
мантии преобразуется в лёгкие [лёгочные моллюски — Pulmonata sensu lato]. У на-
земных брюхоногих большая часть свободного края мантии срастается со стенка-
ми внутренностного мешка. Для сообщения мантийной полости с внешней средой
остаётся небольшое отверстие — пневмостом, снабжённое сфинктером.
Раковина брюхоногих моллюсков формируется в результате секреционной де-
ятельности эпителия внешней поверхности мантии и представляет собой замкну-
тую с одного конца коническую трубку, в разной степени скрученную в спираль,
либо преобразованную в невысокий конус (колпачок), сформированный увели-
ченным последним оборотом. Она состоит преимущественно из карбоната каль-
ция и органического матрикса, доля которого в разных группах лёгочных моллю-
сков варьирует от 0,01% до 21,4% (Круглов, 2005). По направлению навивания
раковины брюхоногих моллюсков
могут быть лево- или правозави-
тыми. Чтобы установить это на-
правление необходимо располо-
жить раковину замкнутым концом
f А спиральной трубки (верхушкой)
\ I ввеРх так> чтобы плоскость устья
была перпендикулярна оси зрения
А Б наблюдателя. При этом располо-
Рис. 13. Лево-(А) и правозавитая (Б) раковины жение палатального края устья
(по: Burch, 1898). покажет сторону навивания обо-
ротов (рис. 13).
У пресноводных гастропод стенка раковины образована тремя слоями (пе-
риостракум, остракум и гипостракум), однако последний развит слабо. В раз-
ных таксонах класса Gastropoda можно наблюдать различные степени редукции
раковины вплоть до её полного исчезновения (Лихарев, Миничев, 1983). Узкий
замкнутый конец раковинной трубки именуется вершиной. На противоположном
конце раковина открывается устьем. Совокупность оборотов, расположенных
выше устья, называется завитком. Внутренние стенки оборотов обычно сливают-
ся, образуя столбик (колумеллу), который часто снабжён полым внутренним кана-
лом, открывающимся у основания завитка раковины отверстием (пупком). Пупок
может быть открытым, либо в различной степени или целиком прикрытым отво-
ротом колумеллярного края устья. Первые 1,5-2,0 оборота раковины (редко 3,0),
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
337
образующиеся ещё на зародышевой стадии, называются эмбриональными или
протоконхом. Нередко протоконх отличается цветом или микроскульптурой от
остальной {дефинитивной) части раковины. Воображаемая линия, вокруг которой
происходит навивание оборотов называется осью раковины. Линия, разделяющая
обороты, именуется швом. Шов может быть мелким (у раковин с уплощёнными
оборотами) или в той или иной степени углублённым (у раковин с выпуклыми
оборотами), прямым (ориентированным более или менее перпендикулярно к оси
раковины) или скошенным. При сильновыпуклых оборотах их верхние части мо-
гут образовывать пришовную площадку — плечо. Иногда по краю спирально за-
крученных раковин развита резкая угловатость — киль. Поверхность оборотов
может быть гладкой или с различными скульптурными элементами: рёбрами,
бороздками, вмятинами, шипами, волосками и т.п. Рёбра и бороздки могут быть
спиральными (ориентированными вдоль оборотов) или радиальными/осевыми
(ориентированными поперёк оборотов). У многих видов можно обнаружить мал-
леатную скульптуру (от латинского malleus — молот) в виде хаотично разбросан-
ных или в той или иной мере упорядоченных вдавлений, напоминающих следы
от ударов молотка. Касательная, проведённая к наиболее выступающим точкам
оборотов, называется тангент-линией. Если по мере формирования раковины
ширина оборотов возрастает равномерно — тангент-линия прямая. Если обороты
нарастают неравномерно — тангент-линия становится выгнутой или вогнутой.
Устье раковины бывает округлое, овальное или неправильной формы. У боль-
шинства брюхоногих моллюсков, дышащих с помощью жабр, а также у некото-
рых лёгочных моллюсков на задней части ноги имеется конхиолиново-известко-
вая крышечка, закрывающая устье раковины при втягивании внутрь мягкого тела
животного.
Край устья, образованный стенкой предпоследнего оборота называют пари-
етальным', край, образованный столбиком — колумеллярным; свободный край
устья называется палатальным, причём его нижняя часть иногда выделается как
самостоятельный базальный край. В верхней части устья обычно имеется яв-
ственно различимый парието-палатальный угол.
Все пресноводные моллюски имеют хорошо развитую раковину, разнообра-
зие форм которой может быть сведено к четырём основным типам: колпачковид-
ной, неритоидной (полушаровидной или полуяйцевидной), турбоспиральной и
плоскоспиральной (рис. 14). В свою очередь турбоспиральные и плоскоспираль-
ные раковины разнообразны по внешнему облику и могут быть разделены на не-
сколько форм, номенклатура которых в отечественной литературе ещё не вполне
устоялась. Например, для моллюсков семейства Lymnaeidae чаще всего выделяют
башневидные, конические, яйцевидно-конические, яйцевидные и уховидные ра-
ковины (Андреева и др., 2010). Для наземных моллюсков используют более под-
робную классификацию форм раковины (Лихарев, Раммельмейер, 1952). Формы
раковин брюхоногих моллюсков, используемые в ключах данного определителя,
приведены на рис. 15.
338
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 14. Морфологические типы форм раковин пресноводных Gastropoda (по: Burch,
1989).
А — плоскоспиральная; Б — колпачковидная; В — неритоидная; Г — разновидности тур-
боспиральных раковин, различающихся вершиной апикального угла.
Мускулатура брюхоногих моллюсков включает несколько слоёв подкожных
мышечных волокон, сложную систему мышц ноги и специфический колумел-
лярный мускул с его дериватами. Последний — наиболее крупный мускул в теле
большинства раковинных моллюсков. При сокращении этого мускула тело жи-
вотного втягивается в раковину, а у видов, имеющих крышечку, с его помощью
крышечка прижимается к устью.
Внутренние органы брюхоногих моллюсков представлены пищеварительной,
дыхательной, кровеносной, выделительной, нервной и репродуктивной системами.
Пищеварительная система включает в себя ротовое отверстие, открывающееся
в эктодермальную глотку (буккальную полость), снабжённую собственной муску-
латурой и содержащую конхиолиновую пластину — челюсть, одонтофор — сво-
его рода внутренний опорный скелет радулярного аппарата, собственно радулу
(тёрку), представленную отдельными зубами, располагающимися на одонтофоре.
Сюда же впадают протоки парных слюнных желёз. Далее следует пищевод —
также эктодермального происхождения. Он впадает в энтодермальную среднюю
кишку, часто образующую мешковидное мускулистое расширение — желудок.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
339
Рис. 15. Формы раковин пресноводных Gastropoda.
А — колпачковидная; Б — неритоидная (полушаровидная, полуяйцевидная); В — дис-
ковидная турбоспиральная; Г — дисковидная плоскоспиральная; Д — коническая;
Е — низкоконическая (1 — прижатая, 2 — не прижатая); Ж — высококоническая; 3 —
овально-коническая; И — башневидно-коническая (1 — для представителей большинства
групп пресноводных гастропод, 2 — для представителей сем. Lymnaeidae по: Хохуткин и
др., 2009); К — башневидная (1 — для представителей большинства групп пресноводных
гастропод, 2 — для представителей сем. Lymnaeidae по Хохуткин и др., 2009); Л — оваль-
ная; М — яйцевидная (овально-башневидная); Н — уховидная; О — шаровидная; П —
плоскоспиральная линзовидная (в виде двояковыпуклой линзы); Р — плоскоспиральная
линзовидная [в виде односторонне выпуклой (плосковыпуклой) линзы]; С — плоскоспи-
ральная шайбовидная; Т — кубаревидная.
340
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
В желудок открываются проток или парные протоки пищеварительной железы
(гепатопанкреаса, «печени»), занимающей большую часть внутренностного меш-
ка. Позади желудочного расширения могут располагаться несколько петлеобраз-
ных извивов остальной средней кишки, впадающей в заднюю кишку, открываю-
щуюся анальным отверстием в мантийную полость.
Дыхательная система представлена парными или единственной жаброй, либо
«лёгким» — видоизменённым участком крыши мантийной полости, пронизан-
ным многочисленными кровеносными сосудами. Также возможен газообмен че-
рез поверхность тела.
Кровеносная система незамкнутая, сердце состоит из одного желудочка или
одного или двух предсердий. От сердца отходят передняя аорта, снабжающая
кровью голову, передние отделы пищеварительного тракта, мантию и ногу; за-
дняя аорта, снабжающая кровью внутренностный мешок и заключённые в нём
органы.
У большинства гастропод выделительная система состоит из единственной
левой почки, представляющей собой видоизменённый метанефридий. Он сооб-
щается с перикардом посредством рено-перикардиального протока, и с внешней
средой посредством простого отверстия, открывающегося в мантийную полость
или специализированного мочеточника, расположенного в мантийной полости и
развитого в разной степени.
Нервная система разбросанно-узлового типа. Её основу составляют ганглии,
сконцентрированные, главным образом, в области глотки и пищевода. Ганглии
соединены продольными и поперечными перемычками (коннективами и комиссу-
рами соответственно). Органы чувств представлены парой глаз, а также чувстви-
тельными клетками, разбросанными по поверхности тела и особенно многочис-
ленными на щупальцах, по краям рта, мантии и ноги. Имеются парные органы
равновесия — статоцисты, расположенные в передней части ноги и органы хемо-
рецепции — осфрадии (исходно парные), лежащие в мантийной полости у осно-
вания жабр.
Брюхоногие моллюски могут быть раздельнополыми (большинство жабер-
ных) и гермафродитными (все лёгочные). Репродуктивная система представлена
половой железой, снабжённой в разной степени развитыми протоками, открыва-
ющимися либо в почку, либо в мантийную полость или в копулятивные органы,
расположенные в передней части цефалоподиума. У гермафродитных животных
половые протоки приобретают сложное строение за счёт формирования специ-
ализированных желёз и внутренних структур.
Методы сбора и камеральной обработки пресноводных
брюхоногих моллюсков
Методы изучения пресноводных брюхоногих моллюсков многообразны и опи-
саны в ряде специальных руководств и монографий (Жадин, 1960; Березкина,
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
341
Старобогатов, 1988; Круглов, 2005; Стадниченко, 2006; Мордухай-Болтовской,
1975; Шаповалова, Вологдин, 1979; Андреева и др., 2010). Здесь мы остановимся
только на способах сбора животных в водоёмах и на тех методиках их изучения,
которыми необходимо владеть для пользования определительными таблицами.
Методы сбора пресноводных моллюсков. Простейшим способом является
сбор моллюсков руками на мелководье и в осушенной зоне по берегам водоёма.
Вручную или пинцетом производится сбор моллюсков или их кладок с поверхно-
сти водных растений и погружённых в воду предметов.
При необходимости получить крупные выборки раковин следует искать резко
мелеющие или даже пересыхающие в сухой сезон водоёмы. На обсохших участ-
ках дна таких водоёмов часто образуются настоящие «кладбища» моллюсков,
когда поверхность покрыта сотнями и тысячами пустых раковин различных ви-
дов, а в небольших углублениях или под остатками водной растительности, где
ещё сохраняется вода, концентрируются живые моллюски.
Нередко скопления раковин обнаруживаются в береговых выбросах рек и
крупных озёр. Однако экологическая и фаунистическая информация, полученная
при изучении таких скоплений, не всегда достоверна, так как зачастую сложно
определить истинное место обитания моллюсков. Особенно это касается речных
выбросов, в которых встречаются раковины животных, обитавших в самых раз-
ных придаточных и пойменных водоёмах, возможно, даже весьма удалённых от
места обнаружения.
Основными орудиями качественного сбора бентосных моллюсков являются
скребок и драга. Если скребки применяются на мелководьях, то драгами, вле-
комыми по дну на верёвке, можно облавливать как мелководье, так и глубокие
участки водоёмов. Драги бывают треугольной или четырёхугольной формы, они
могут быть снабжены зубчатыми или режущими краями, захватывающими грунт.
Количественные сборы моллюсков проводятся с применением дночерпателей
различных типов и модификаций (рис. 16).
Первичная фиксация собранных животных проводится в полевых условиях с
помощью 96% этанола. При этом следует учитывать, что в мантийной полости
моллюсков может содержаться некоторое количество воды, выделяющейся после
гибели, что ведёт к снижению исходной концентрации фиксатора. Спустя сут-
ки, животные переводятся в 70% этанол для длительного хранения. Допускается
применение в качестве фиксатора формалина (нейтрализованного добавлением
мела), но при долгом нахождении моллюсков в формалине их мягкие ткани ста-
новятся жёсткими и ломкими, что затрудняет анатомирование животных. Кроме
того, формалин разрушает раковину при длительном хранении. Для облегчения
анатомирования моллюсков перед фиксацией рекомендуется умерщвлять, поме-
щая в небольшой, плотно закрытый сосуд с кипячёной водой, содержащей мало
растворенного кислорода, в результате чего животное погибает от асфиксии.
С этой же целью может применяться анестезия моллюсков с помощью различных
химических веществ: хлоралгидрата, нембутала, пентобарбитала, ментола, хло-
342
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 16. Орудия количественного и качественного сбора пресноводных моллюсков (из:
Андреева и др., 2010 по: Иоганзен, 1959; Жадин, 1960; Мордухай-Болтовской, 1975).
А — скребок; Б — ковшёвый дночерпатель; В — трубчатый дночерпатель; Г — коробча-
тый дночерпатель; Д — драга.
роформа, эфира и т.п. (Meier-Brook, 1976а, 1983; Barker, 1981; Kunigelis, Saleud-
din, 1984; Стадниченко, 2006). Также умерщвление собранных животных можно
производить крутым кипятком, после чего тело моллюска сравнительно легко из-
влекается (выкручивается) из раковины и переносится в спирт. Раковина при этом
высушивается и хранится отдельно.
Промеры раковины брюхоногих моллюсков. В большинстве случаев табли-
цы для определения таксономической принадлежности пресноводных гастропод
основаны на признаках раковины. Это могут быть как качественные признаки
(форма раковины, наличие определённого типа скульптуры и т.п.), так и коли-
чественные. Среди последних наиболее часто используются число оборотов ра-
ковины, а также специальные линейные промеры (высота и ширина раковины,
высота и ширина устья и т.п.), на основе которых могут рассчитываться морфо-
метрические индексы (значения отношений линейных промеров), характеризу-
ющие пропорции раковины. Несмотря на то, что использование этих индексов
имеет ряд ограничений (Терентьев, 1970), они чаще применяются в определи-
тельных таблицах, нежели линейные промеры раковины и её частей. Причина в
том, что размер раковины — это довольно пластичный признак, который может
изменяться в зависимости от условий обитания, например, от трофики водоёма.
При использовании морфометрических индексов также нужно учитывать высо-
кую степень их онтогенетической изменчивости, возникающей вследствие алло-
метрического роста (Амитров, 1971; Круглов, 2007).
Промеры раковины обычно осуществляются с точностью до 0,1 мм при по-
мощи штангенциркуля или окуляр-микрометра микроскопа. Для каждого из мор-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
343
Рис. 17. Схемы промеров раковин брюхоногих моллюсков, используемые в определи-
тельных таблицах (А, Б — по: Андреева и др., 2010; В — по: Старобогатов и др., 2004).
А — турбоспиральная раковина, Б — плоскоспиральная раковина, В — колпачковидная
раковина. Обозначения промеров см. в тексте.
фологических типов раковины разработана особая схема измерений (рис. 17),
строгое следование которой необходимо для сопоставимости полученных данных
со значениями, приводимыми в определительных ключах. В настоящем издании
представлены брюхоногие моллюски, принадлежащие четырём морфологиче-
ским типам раковины: турбоспиральному, неритоидному (полушаровидная рако-
вина с широкой колумеллярной/устьевой площадкой), плоскоспиральному и кол-
пачковидному.
Измерение турбоспиральных раковин (семейств Bithyniidae, Lymnaeidae, Phy-
sidae, Valvatidae и др.; рис. 17А) предполагает шесть основных промеров: высота
раковины (ВР), измеряемая строго вдоль оси раковины от вершины до нижнего
(базального) края устья; ширина раковины (ШР) — расстояние между наиболее
удалёнными от оси точками палатального края устья и поверхности последне-
го оборота на противоположной устью стороне (в проекции на линию, перпен-
дикулярную оси раковины); высота завитка (ВЗ) — расстояние от вершины до
344
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
верхней точки устья (в проекции на ось раковины); высота последнего оборота
(ВПО) — расстояние от базального края устья до участка шва между последним
и предпоследним оборотами, расположенного над устьем; высота устья (ВУ) —
расстояние от верхней точки устья до наиболее удалённой от него точки базаль-
ного края (измеряется строго в плоскости устья); ширина устья (ШУ) — рассто-
яние между наиболее удалёнными точками устья, измеренное перпендикулярно
линии, по которой измерялась ВУ У раковин с развитым колумеллярным отворо-
том оно может измеряться двояко: включая колумеллярный отворот (ШУСК0) или
без него. Большая часть указанных промеров применима и к неритоидным рако-
винам (род Theodoxus семейства Neritidae).
У плоскоспиральных раковин (семейство Planorbidae, некоторые Valvatidae;
рис. 17Б) промеры ВЗ и ВПО выполнить невозможно, зато вводятся несколько до-
полнительных измерений. Это — диаметр внутренних оборотов (ДВО) и ширина
трубки последнего оборота (ШПО), соотношение которых даёт индекс внутрен-
них оборотов (ИВО), предложенный Л.А. Прозоровой и Я.И. Старобогатовым
(Prozorova, Starobogatov, 1996). ИВО может определяться как с апикальной (вер-
шинной), так и с базальной поверхности раковины и отражает скорость нараста-
ния оборотов раковины. Этот индекс приложим и к турбоспиральным раковинам,
но измеряется у них только с базальной стороны и называется индексом пупка
(Старобогатов и др., 2004). Иногда необходимо знать ширину предпоследнего
оборота (ШППО), измеряемую аналогично ШПО.
При измерении колпачковидных раковин (семейства Acrloxidae и Ancylidae;
рис. 17В) высотой (ВР) является расстояние от самой верхней точки раковины до
плоскости устья. Длина раковины измеряется между двумя наиболее удалённы-
ми точками, расположенными на переднем и заднем крае устья соответственно.
Ширина раковины измеряется перпендикулярно длине. Эти промеры у колпачко-
видных раковин соответствуют длине и ширине устья у спиральных форм. Затем,
измеряют удаление вершины от переднего края (УПК) в проекции на линию про-
мера длины; удаление вершины от левого края (УЛК) в проекции на линию про-
мера ширины и длину хорды переднего склона (ДХ) — расстояние от вершины
до самой передней точки устья (Старобогатов и др., 2004).
В качестве дополнительного показателя у турбоспиральных раковин использу-
ется величина апикального угла измеряемого между двумя касательными, прохо-
дящими через наиболее выступающие точки всех или нескольких оборотов завит-
ка. Для измерения апикального угла контур раковины зарисовывается с помощью
рисовального аппарата, и необходимые измерения проводятся по рисунку. В по-
следнее время для этой же цели стали применяться различные устройства для
оцифровки изображений.
У некоторых жаберных брюхоногих таксономическую значимость имеют так-
же признаки крышечки. В частности, у битиниид используются такие признаки,
как форма крышечки, размер и положение её центрального ядра, выраженность
линий нарастания (Анистратенко, Стадниченко, 1995). Сопоставление размеров
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
345
Рис. 18. Схема подсчёта числа
оборотов раковины (по: Burch,
1989).
таксономического положения
конхиолиновой и известковой частей крышечки
может использоваться для видовой диагностики
представителей рода Theodoxus из водоёмов юга
Восточной Европы (Анистратенко, Анистратенко,
2001).
Число оборотов раковины определяется по спе-
циальной схеме (рис. 18) с точностью до ХА или !4.
Использование морфометрических индексов наи-
более эффективно при сравнении раковин с при-
мерно одинаковым числом оборотов. Это связано
с изменением пропорций раковины и отражающих
их морфометрических индексов в онтогенезе мол-
люсков (Терентьев, 1970).
Определение видовой принадлежности пре-
сноводных брюхоногих моллюсков. В пода-
вляющем большинстве случаев для определения
пресноводных гастропод используются признаки раковины. Признаки внутрен-
него строения используются в ключах реже, преимущественно для определения
лёгочных моллюсков. При этом анатомические признаки зачастую рассматрива-
ются как более значимые для систематики и диагностики, поскольку они значи-
тельно меньше, чем признаки раковины, подвержены модифицирующему воздей-
ствию среды обитания. В некоторых группах пресноводных лёгочных, например
в подроде Stagnicola рода Lymnaea надёжная идентификация ряда видов возмож-
на только путём изучения строения половой системы (Jackiewicz, 1993; Гарбар,
2001; Андреева и др., 2010). Поэтому при работе с пресноводными пульмонатами
рекомендуется для подтверждения определений, полученных по раковине, по воз-
можности использовать результаты вскрытий мягкого тела.
Для видовой диагностики пресноводных лёгочных моллюсков чаще всего ис-
пользуются признаки строения копулятивного аппарата. В некоторых случаях по-
лезны сведения о строении простаты, семеприемника и его протока1. Для изуче-
ния этих признаков необходимо хорошо представлять топографию внутренних
органов моллюсков, в частности, расположение половой системы относительно
других систем органов (рис. 19).
Широкое применение в видовой диагностике пресноводных Pulmonata по-
лучил индекс копулятивного аппарата (ИКА): соотношение длин препуциума и
мешка пениса (Jackiewicz, 1998; Круглов, 2005) (рис. 20). Перед измерением ме-
шок пениса и препуциум должны быть расправлены, но при этом необходимо
избегать чрезмерного растягивания этих органов, что может исказить результа-
ты. Следует также учитывать возможное изменение пропорций копулятивно-
1 Методы вскрытия брюхоногих моллюсков подробно описаны в многочисленных пособиях
и специальных работах (Шилейко, 1984; Иванов и др., 1985; Стадниченко, 1990, 2006; Gloer,
2002; Круглов, 2005; Солдатенко, Стыгар, 2006).
346
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 19. Расположение органов половой системы Lymnaea balthica (L., 1758) относи-
тельно частей тела вскрытого животного (из: Хохуткин и др., 2009 по: Jackiewicz, 1954).
бж — белковая железа, в — вагина, гж — гермафродитная железа, гт — грушевидное
тело, ич — извитая часть яйцевода, мп — мешок пениса, нж — нидаментальная железа,
пр — простата, пре — препуциум, сп — семяпровод, ст — семяприёмник.
Рис. 20. Строение копулятивного
аппарата (на примере Lymnaea
fragilis).
ПП — препуциум, МП — мешок
пениса.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
347
го аппарата в зависимости от способа фиксации (Hubendick, 1955; Meier-Brook,
1983; Круглов, 2005), а также изменчивость гениталий, связанную с возрастом
животного. Информацию об этом можно найти в монографии ГВ. Берёзкиной
и Я.И. Старобогатова (1988). Для диагностики на уровне родов и подродов ис-
пользуются такие признаки как длина протока семеприемника, число внутренних
складок в простате (у прудовиков), строение пениса, в частности, наличие или от-
сутствие стилета (у Planorbidae) и другие.
Список литературы
Акрамовский Н.Н. 1976. Фауна Армянской ССР: Моллюски (Mollusca). Ереван: Изд-во
АН Арм. ССР. 272 с.
Амитров О.В., 1971. Применение количественных методов в изучении раковин гастропод.
Современные проблемы палеонтологии. М.: Наука. С. 231-245.
Андреев Н.И., Андреева С. И. 2014. Моллюски семейства Bithyniidae (Gastropoda, Pectini-
branchia) водоемов Коргалжынекого заповедника // Успехи современного естествозна-
ния. №5(1). С. 227-227.
Андреева С.И., Андреев Н.И., Винарский М.В. 2010. Определитель пресноводных брюхо-
ногих моллюсков Западной Сибири. Ч. 1. Gastropoda: Pulmonata. Вып. 1. Семейства
Acroloxidae и Lymnaeidae. Омск, 200 с.
Анистратенко В.В. 1998. Определитель гребнежаберных моллюсков (Gastropoda, Pectini-
branchia) фауны Украины. Ч. 2. Пресноводные и наземные // Вести, зоологии. Отд.
вып. Прил. 8. С. 67-117.
Анистратенко В.В., Анистратенко О.Ю. 2001. Класс Панцирные или Хитоны. Класс Брю-
хоногие — Cyclobranchia, Scutibranchia и Pectinibranchia (часть) // Фауна Украины.
Моллюски. 29 (1, кн. 1). Киев: Велес. 240 с.
Анистратенко В.В., Стадниченко А.П. 1994 (1995). Литторинообразные, риссоиобраз-
ные // Фауна Украины. Моллюски. 29 (1, кн. 2). Киев: Паукова думка. 175 с.
Анистратенко О.Ю., Старобогатов Я.И., Анистратенко В.В. 1999. Моллюски рода Theo-
doxus (Gastropoda, Pectinibranchia, Neritidae) Азово-Черноморского бассейна // Вести,
зоологии. Т. 33. № 3. С. 11-19.
Анистратенко В.В., Черногоренко Е.В. 1989. Фауна и экология брюхоногих моллюсков
бассейна Среднего Днепра // Вести, зоологии. Т. 23. № 2. С. 3-6.
Берёзкина Г.В., Старобогатов Я.И. 1988. Экология размножения и кладки яиц пресновод-
ных легочных моллюсков. // Тр. Зоол. ин-та АН СССР. 174. 306 с.
Бекман М.Ю., Старобогатов Я.И. 1975. Байкальские глубоководные моллюски и род-
ственные им формы // М. Ю. Бекман, Я. И. Старобогатов (ред.). Новое о фауне Бай-
кала. Тр. Лимнологического ин-та СО АН СССР Т. 18(38), ч. 1. Новосибирск: Наука.
С. 92-111.
Гарбар А.В. 2001. Комплексный анализ морфологических признаков представителей под-
родов Lymnaea (Corvusiana) и L. (Stagnicola) (Gastropoda, Pulmonata, Lymnaeidae) и
оценка их диагностической значимости // Вести, зоологии. Т. 35. № 2. С. 15-22.
Градовский В.М. 2000. Распространение и некоторые особенности экологии моллюсков
семейства Bithyniidae (Gastropoda, Pectinibranchia) в водоемах Правобережной Украи-
ны // Вести, зоологии. № 14. С. 11-22.
348
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Жадин В.И. 1952. Моллюски пресных и солоноватых вод СССР // Определители по фауне
СССР, издаваемые Зоологическим институтом АН СССР. 46. М.; Л.: Изд-во АН СССР.
376 с.
Жадин В.И. 1960. Методы гидробиологического исследования. М.: Высшая школа.
191 с.
Иванов А.В. 1940. Класс Брюхоногих моллюсков (Gastropoda). Руководство по зоологии.
Т. 2. М.; Л.: Изд-во АН СССР. С. 323-455.
Иванов А.В., Полянский Ю.И., Стрелков А.А. 1985. Большой практикум по зоологии бес-
позвоночных. Т. 3. М.: Высшая школа. 390 с.
Иззатуллаев 3. 1972. К фауне пресноводных моллюсков Гиссарского хребта и сопре-
дельных районов Таджикистана // Изв. АН Тадж. ССР, отд-ние биол. наук, № 3(48).
С. 44-Л9.
Иззатулаев З.И. 1980. Двустворчатые моллюски семейства Corbiculidae Средней Азии //
Зоол. ж. Т. 59. Вып. 8. С. 1130-1136.
Кантор Ю.И., Сысоев А.В. 2005. Каталог моллюсков России и сопредельных стран. М.:
Т-во науч, изданий КМК. 627 с.
Кантор Ю.И., Сысоев А.В. 2006. Морские и солоноватоводные брюхоногие моллюски
России и сопредельных стран: иллюстрированный каталог. М.: Т-во науч, изданий
КМК. 371 с., 140 табл.
Коробков И.А. 1954. Справочник и методическое руководство по третичным моллюскам.
Брюхоногие. Л.: Гостоптехиздат. 786 с.
Круглов Н.Д. 2005. Моллюски семейства прудовиков Европы и Северной Азии. Смоленск:
Изд-во СГПУ. 508 с.
Круглов Н.Д. 2007. Особенности стабилизации основных таксономических параметров
у легочных моллюсков (Mollusca: Gastropoda: Pulmonata) и ошибки систематиков //
Моллюски: Морфология, таксономия, филогения, биогеография и экология. 7-е (XVI)
совещ. по изучению моллюсков. Сб. науч, работ. СПб. С. 143-145.
Круглов Н.Д. 2008. Две системы моллюсков семейства прудовиков (Gastropoda, Pulmonata,
Lymnaeidae): европейская и российская. Где истина? Ч. 1. Анализ системы на родовом
и подродовом уровнях // Изв. Смоленск, гос. ун-та. № 2. С. 33-51.
Круглов Н.Д. 2009. Две системы моллюсков семейства прудовиков (Gastropoda, Pulmonata,
Lymnaeidae): европейская и российская. Где истина? Ч. 2. Жизненные формы прудо-
виков и проблема вида в малакологии (Gastropoda Pulmonata Lymnaeidae) // Изв. Смо-
ленск. гос. ун-та. № 6. С. 5-24.
Круглов Н.Д., Солдатенко Е.В. 1997. Ревизия рода Segmentina (Planorbidae) // Ruhtenica:
7(2). С. 111-132.
Круглов Н.Д., Солдатенко Е.В. 2000. Новые данные о видах подрода Gyraulus рода Ani-
sus европейской части России и сопредельных территорий (Gastropoda Planorbidae) //
Ruhtenica: 10(2). С. 113-120.
Круглов Н.Д., Старобогатов Я.И. 1981. Новый род лимнеид и система подрода Omphiscola
рода Lymnaea (Gastropoda, Pulmonata) // Зоол. ж. Т. 60. Вып. 7. С. 965-972.
Круглов Н.Д., Старобогатов Я.И. 1984. Особенности морфологии и систематика моллю-
сков подрода Corvusiana рода Lymnaea (Gastropoda Pulmonata Lymnaeidae) // Бюл.
Моск, общ-ва испыт. прир. Отд. биол. Т. 89. Вып. 2. С. 58-70.
Круглов Н.Д., Старобогатов Я.И. 1985. Объем подрода Galba и сходных с ним подродов
рода Lymnaea (Gastropoda, Pulmonata) // Зоол. ж. Т. 64, вып. 1. С. 24-35.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
349
Круглов Н.Д., Старобогатов Я.И. 1986. Моллюски подрода Stagnicola рода Lymnaea фауны
СССР (Gastropoda, Pulmonata) // Бюл. Моск, общ-ва испыт. прир. Отд. биол. Т. 91(2).
С. 59-72.
Круглов Н.Д., Старобогатов Я.И. 1987. Жизненные формы лимнеид и некоторые пробле-
мы построения системы // Моллюски. Результаты и перспективы их исследований.
Всесоюз. совещ. по изучению моллюсков. Автореф. докл. 8. Л.: Наука. С. 68-70.
Круглов Н.Д., Старобогатов Я.И. 1991. Родовой состав семейства Acroloxidae (Gastropoda,
Pulmonata) и виды рода Acroloxus, обитающие в СССР // Зоол. ж. Т. 70. Вып. 2. С. 66-
79.
Лазуткина Е.А., Андреева С.И., Андреев Н.И. 2011. Находки Digyrcidum bourguignati
(Paladilhe, 1869) в водоёмах Западной Сибири и Урала // Омский научный вестник.
№ 1(104). ОмГПУ. С. 217-220.
Лазуткина Е.А., Андреева С.И., Андреев Н.И. 2014. Моллюски рода Digyrcidum Locard,
1882 (Gastropoda, Pectinibranchia) в водоемах Западной Сибири и Урала // Ruthenica.
Vol. 24, № l.C. 15-23.
Лихарев И.М., Миничев Ю.С. 1983. Слизни как один из главных типов организации брюхо-
ногих моллюсков // Моллюски: систематика, экология и приспособления к размноже-
нию. Всесоюз. совещ. по изучению моллюсков. Автореф. докл. 7. Л.: Наука. С. 32-36.
Лихарев И.М., Раммельмейер Е.С. 1952. Наземные моллюски фауны СССР // Определи-
тели по фауне СССР, издаваемые Зоологическим институтом АН СССР. 43. М.; Л.:
Изд-во АН СССР. С. 511 с.
Марковский Ю.М. 1955. Фауна беспозвоночных низовьев рек Украины. Ч. 3. Водоемы Ки-
лийской дельты Дуная. Киев: Изд-во АН УССР. 280 с.
Мордухай-Болтовской Ф.Д. (ред). 1975. Методика изучения биогеоценозов внутренних во-
доёмов. М.: Наука. 240 с.
Першко I.O. 2006. Систематична структура родин Bithyniidae, Lithoglyphidae, Melanop-
sidae (Mollusca: Gastropoda: Pectinibranchia) за результатами конхюлопчних, анатом!ч-
них та кардюлопчних дослщжень. Автореф. дис. ... канд. бюл. наук. Ки!*в: Б.в. 20 с.
Першко I.O. 2010. Особливосп карюлогп представниюв родини Bithyniidae (Mollusca:
Gastropoda: Pectinibranchia) фауни УкраГни // Вюник Харювського нацюнального yni-
верситету 1меш В.Н. Каразша. Сер. бюлопя. Вип. 12 (№ 920). С. 98-103.
Прозорова Л.А. 2003. Таксономическая структура и видовой состав рода Anisus Studer
(Gastropoda, Planorbidae) в фауне России // Бюл. Дальневосточного малакологического
общ-ва. Вып. 7. С. 99-115.
Ситникова Т.Я. 1983. Система байкальских эндемичных видов рода Megalovalvata и неко-
торые вопросы систематики семейства Valvatidae (Gastropoda, Pectinibranchia) // Зоол.
ж. Т. 62. Вып. 1. С. 32-44.
Ситникова Т.Я., Старобогатов Я.И. 1982. Объем и систематический статус группы Archi-
taenioglossa (Gastropoda, Pectinibranchia) // Зоол. ж. Т. 61. Вып. 6. С. 831-842.
Ситникова Т.Я., Старобогатов Я.И., Черногоренко Е.В. 1986. Род Borysthenia (Gastropoda,
Valvatidae), его систематическое положение и видовой состав // Вести, зоологии. Т. 20.
№ 1.С. 9-14.
Ситникова Т.Я., Старобогатов Я.И., Широкая А.А., Шибанова И.В., Коробкова Н.В., Адов
Ф.В. 2004. Брюхоногие моллюски (Gastropoda) // О.А. Тимошкин (отв. ред.). Анноти-
рованный список фауны озера Байкал и его водосборного бассейна. Т. 1. Озеро Бай-
кал. Кн. 2. Новосибирск: Наука. С. 935-1002.
350
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Солдатенко Е.В. 2004. Особенности морфологии вида Anisus (Vorticulus) charteus (Gastro-
poda Pulmonata) // Н.Д. Круглов (отв. ред.). 4-е научные чтения памяти В.В. Станчин-
ского. Вып. 4. Смоленск: Изд-во Смоленск, гос. пед. ун-та. С. 480-482.
Солдатенко Е.В. 2008. Кутикулярные образования полового аппарата моллюсков семей-
ства Planorbidae (Gastropoda: Pulmonata), их разнообразие и систематическое значе-
ние // Отчётная научная сессия по итогам работы 2008 г. Зоол. ин-та РАН, 7-9 апреля
2009 г. Тез. докл. СПб.: ЗИН РАН. С. 36-37.
Солдатенко Е.В. 2010. Особенности морфологии некоторых балканских видов рода Апсу-
lus (Gastropoda, Pulmonata) // Зоол. ж. Т. 89. Вып. 9. С. 1037-1051.
Солдатенко Е.В. 2011. Развитие копулятивного аппарата у моллюсков семейства Planorbi-
dae (Gastropoda: Pulmonata) и его значение в систематике группы // О.В. Зайцева (отв.
ред.). Сб. докл. 2-й Всерос. конф, с международным участием «Современные пробле-
мы эволюционной морфологии животных», 17-19 октября 2011 г. / СПб.: ЗИН РАН.
С. 318-321.
Солдатенко Е.В., Старобогатов Я.И. 2004. Род Ancylus Muller, 1774 (Gastropoda Planorbi-
dae) // Ruthenica: 14(1). C. 37-56.
Солдатенко E.B., Стыгар Л.В. 2006. Методика анатомирования моллюсков семейств Pla-
norbidae и Succineidae // Естественные науки и экология: Ежегодник ОмГПУ. Вып. 10.
Омск: Изд-во ОмГПУ. С. 76-80.
Сон М.О. 2007. Моллюски-вселенцы в пресных и солоноватых водах Северного Причер-
номорья. Одесса: Друк. 132 с.
Стадниченко А.П. 1968. Galhz occulta Jackiewicz, 1959 (Gastropoda, Pulmonata) — новый
вид фауны СССР // Зоол. ж. Т. 47. Вып. 6. С. 942-944.
Стадниченко А.П. 1990. Прудовикообразные (пузырчиковые, витушковые, катушковые) //
В.И. Монченко (ред.). Фауна Украины. Т. 29. Моллюски. Вып. 4. Киев: Наукова думка.
292 с.
Стадниченко А.П. 2004. Прудовиковые и чашечковые (Lymnaeidae, Acroloxidae) Украины.
Киев: Центр учебной литературы. 327 с.
Стадниченко А.П. 2006. Lymnaeidae и Acroloxidae Украины: Методы сбора и изучения,
биология, экология, полезное и вредное значение. Житомир: Рута. 168 с.
Старобогатов Я.И. 1967. К построению системы пресноводных легочных моллюсков //
Я.И. Старобогатов (ред.). Моллюски и их роль в биоценозах и формировании фаун.
Тр. Зоол. ин-та АН СССР. Т. 42. Л.: Наука. С. 280-304.
Старобогатов Я.И. 1970. Фауна моллюсков и зоогеографическое районирование континен-
тальных водоёмов земного шара. Л.: Наука. 372 с.
Старобогатов Я.И. 1976. Система и филогения Lymnaeidae // Проблемы зоологии. Л. С. 79-81.
Старобогатов Я.И. 1977. Класс брюхоногие моллюски Gastropoda // Л.А. Кутикова,
Я.И. Старобогатов (ред.). Определитель пресноводных беспозвоночных европейской
части СССР: (планктон и бентос). Л.: Гидрометеоиздат. С. 152-174.
Старобогатов Я.И. 1994. Биологическое разнообразие моллюсков континентальных водоё-
мов и состояние его изученности в Российской Федерации и соседних независимых
государствах. Биоразнообразие: Степень таксономической изученности. М.: Наука.
С. 60-65.
Старобогатов Я.И., Будникова Л.Л. 1976. О фауне пресноводных брюхоногих моллюсков
крайнего Северо-Востока СССР // Тр. биол.-почв, ин-та ДВНЦ АН СССР. Т. 36(139).
Владивосток. С. 72-88.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
351
Старобогатов Я.И., Затравкин М.Н. 1987. Bithynioidea (Gastropoda, Pectinibranchia) фауны
СССР // Я.И. Старобогатова, А.Н. Голикова, И.М. Лихарева (ред.). Моллюски, резуль-
таты и перспективы их исследований. 8-е всесоюз. совещ. по изучению моллюсков.
Автореф. докл. Л : Наука. С. 150-153.
Старобогатов Я.И., Прозорова Л.А., Богатов В.В., Саенко Е.М. 2004. Моллюски // С.Я. Ца-
лолихин (ред.). Определитель пресноводных беспозвоночных России и сопредельных
территорий. Т. 6. СПб.: Наука. 492 с.
Старобогатов Я.И., Прозорова Л.А., Затравкин М.Н. 1989. Состав семейства Physidae
(Gastropoda, Pulmonata, Lymnaeiformes) Сибири и Дальнего Востока СССР (с замеча-
ниями о европейских физидах) // Бюл. Моск, общ-ва испыт. прир. Отд. биол. Т. 94(1).
С. 62-76.
Сушкина А.П., 1949. Питание и рост некоторых брюхоногих моллюсков // Тр. Всесоюз.
гидробиол. общ-ва. Т. 1. С. 118-131.
Тейлор Д.В., Ситникова Т.Я. 2004. Изучение брюхоногих моллюсков семейства Physidae
(Gastropoda: Hygrophila) Сибири, Украины и Монголии // Еколого-функцюнальш та
фаунютичш аспекти дослщження молюсюв, ix роль у биошдикацн стану навколиш-
нього середовища: ЗбАрник наукових праць. Житомир: Волынь. С. 217-219.
Терентьев П.В. 1970. Географическая изменчивость раковины большого прудовика //
Вести. ЛГУ. Биология. № 14. С. 146-154.
Хохуткин И.М., Винарский М.В., Гребенников М.Е. 2009. Моллюски Урала и прилегаю-
щих территорий. Семейство Прудовиковые Lymnaeidae (Gastropoda, Pulmonata, Lym-
naeiformes). Ч. 1. Екатеринбург: Гощицкий. 156 с.
Хохуткин И.М., Винарский М.В. 2013. Моллюски Урала и прилегающих территорий.
Семейства Acroloxidae, Physidae, Planorbidae (Gastropoda, Pulmonata, Lymnaeiformes).
4. 2. Екатеринбург: Гощицкий. 184 с.
Цихон-Луканина E.A. 1961. К вопросу о фильтрационном способе питания у Bithynia ten-
taculata (L.), Valvata piscinalis (Muller) (Gastropoda, Prosobranchia) // Бюл. Ин-та био-
логии водохранилищ АН СССР. № 10. С. 28-30.
Цихон-Луканина Е.А. 1987. Трофология водных моллюсков. М.: Наука. 176 с.
Черногоренко Е.В. 1988 О видовом составе вивипарид (Gastropoda, Viviparidae) Европы и
Западной Азии // Зоол. ж. Т. 67. Вып 5. С. 645-655.
Черногоренко Е.В., Старобогатов Я.И. 1987. Обзор современных Viviparidae Европы //
Я.И. Старобогатова, А.Н. Голикова, И.М. Лихарева (ред.). Моллюски, результаты и
перспективы их исследований. 8-е всесоюз. совещ. по изучению моллюсков. Автореф.
докл. Л : Наука. С. 145-147.
Шаповалова И.М., Вологдин М.П. 1979. К методике количественного учёта погружённой
растительности и фитофильной фауны // Гидробиол. ж. Т. 15. № 4. С. 111-113.
Шилейко А.А. 1967. К вопросу об изменчивости лимнеид (Gastropoda, Mollusca) Кольско-
го полуострова // Гидробиол. ж. Т. 3. С. 77-80.
Шилейко А.А. 1984. Наземные моллюски подотряда Pupillina фауны СССР (Gastropoda,
Pulmonata, Geophila) // Фауна СССР. Моллюски. Т. 3. Вып. 3. 399 с.
Шилейко А.А., Кантор Ю.И. 1994. О разнообразии моллюсков. Биоразнообразие: Степень
таксономической изученности. М.: Наука. С. 60-65.
Широкая А.А., 2005. Байкальские эндемичные моллюски семейства Acroloxidae (Gastro-
poda: Pulmonata): Морфология, таксономия, распределение и формирование фауны.
Автореф. дис. ... канд. биол. наук. СПб. 28 с.
352
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Шихова Т.Г. 2004. Фауна моллюсков бассейна реки Вятки и Вятско-Двинской водораз-
дельной области. Автореф. дис. ... канд. биол. наук. СПб. 26 с.
Aktipis S.W., Giribet G., Lindberg D.R., Ponder W.F. 2008. Gastropod phylogeny: an overview
and analysis. Phylogeny and Evolution of the Mollusca. Berkeley etc.: California University
press. P. 201-237.
Albrecht C., Kuhn K., Streit B. 2007. A molecular phylogeny of Planorboidea (Gastropoda, Pul-
monata): insights from enhanced taxon sampling // Zoologica Scripta. 36. P. 27-39.
Alder J. 1838. Supplement to a catalogue of the land and freshwater testaceous Mollusca, found
in the vicinity of Newcastle // Transactions of the Natural History Society of Northumber-
land. 2. P. 337-342.
Baker F.C. 1945. Molluscan family Planorbidae. Urbana: University of Illinois Press. 530 p.
Baker H.B. 1956. Family names in Pulmonata // Nautilus. Vol. 69. P. 128-139.
Bargues M.D., Vigo M., Horak P., Dvorak J., Patzner R.A., Pointier J.P., Jackiewicz M., Meier-
Brook C., Mas-Coma S., 2001. European Lymnaeidae (Mollusca: Gastropoda), intermediate
hosts of trematodiases, based on nuclear ribosomal DNA ITS-2 sequences // Infection, Ge-
netics and Evolution. Vol. 1. P. 87-107.
Bargues M.D., Horak P., Patzner R.A., Pointier J.P., Jackiewicz M., Meier-Brook C., Mas-Co-
ma S. 2003. Insights into relationships of Palearctic and Nearctic lymnaeids (Mollusca:
Gastropoda) by rDNA ITS-2 sequencing and phylogeny of stagnicoline intermediate host
species of Fasciola hepatica // Parasite. Vol. 10. P. 243-255.
Bargues M.D., Artigas P., Jackiewicz M., Pointier J.P., Mas-Coma S. 2006. Ribosomal DNA
ITS-1 sequence analysis of European stagnicoline Lymnaeidae (Gastropoda) // Heldia.
Vol. 6(1-2). P. 57-68.
Bargues M.D., Mas-Coma S. 2005. Reviewing lymnaeid vectors of fascioliasis by ribosomal
DNA sequence analyses // Journal of Helminthology. Vol. 79. P. 257-267.
Barker G.M. 1981. Nembutal for narcotisation of mollusks // Veliger. Vol. 24(1). P. 76.
Beran L., Gloer P. 2006. Gyraulus chinensis (Dunker, 1848) — a new greenhouse species for
the Czech Republic (Gastropoda: Planorbidae) // Malacologica Bohemoslovaca. Vol. 5.
P. 25-28.
Beriozkina G.V, Levina O.V, Starobogatov Ya.I. 1995. Revision of Bithyniidae from European
Russia and Ukraine // Ruthenica: 5(1-2). P. 27-39.
Boeters H. D., Falkner G. 1998. Valvatapulchella S. Studer und Valvata studeri n. sp. (Gastrop-
oda, Ectobranchia: Valvatidae) // Heldia. Vol. 2(5-6). P. 113-122.
Bouchet P., Rocroi J.-P., Fryda J., Hausdorf B., Ponder W., Valdes A., Waren A. 2005. Classifica-
tion and nomenclator of gastropod families // Malacologia. Vol. 47(1-2). P. 1-397.
Bourguignat J.R. 1860. Malacologie terrestre et fluviatile de la Bretagne. Paris. J.B. Bailliere.
178 p.
Bourguignat J.R. 1862. Notice monographique sur les Limnees d’Europe du groupe de la Lim-
naea stagnalis // Revue et Magasin de Zoologie. Ser. 2. 14. P. 54-63.
Bourguignat J.R. 1880. Resensement des Vivipara du systeme europeen. Paris. Imp. Bouchard-
Huzard. 52 p.
Burch J.B. 1989. North American freshwater snails. Malacological Publications, Hamburg,
(Michigan): Malacological publications, 366 p.
Cristofori J., Jan, G. 1832. Catalogue in IV. Sectiones divisus rerum naturalium in museo exstan-
tium Josephi De Cristofori et Georgii Jan plurium Acad. Scient. et Societ. Nat. Cur. Soda-
lium complectens adumbrationem oryctognosiae et geognosiae atque prodrumum faunae et
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
353
floriae Italiae Superioris. Sectio II. — Pars I. — pp. [1-7], [Conchylia] 1-8, [1], [Mantissa]
1—4, [Excerptum] 1-4, [1], [Conchylia fossilia] 1-16, [1-2]. Parmae.
Clessin S. 1882. Die Familie der Ancylinen // Systematisches Conchylien-Cabinet von Martini
und Chemnitz. Numberg. Bauer und Raspe. 60 p.
Clessin S. 1884. Die Familie der Limnaeiden enthaltend die Genera Planorb is, Limnaeus, Physa
und Amphepeplea // Systematisches Conchylien-Cabinet von Martini und Chemnitz. Num-
berg. Bauer und Raspe. P. 111-222.
Climo F.M., Pullan N.B. 1972. A taxonomic review of the family Lymnaeidae (Mollusca: Gas-
tropoda) in New Zealand // Journal of the Royal Society of New Zealand. Vol. 2(1). P. 5-13.
Da Costa E.M. 1778. Historia naturalis testaceorum Britanniae, or, the British Conchology; con-
taining the descriptions and other particulars of natural history of the shells of Great Britain
and Ireland: illustrated with figures. London. Millan. White. Elmsley & Robson, printed for
the author, xii +254+vii p.
Drapamaud J.P.R. 1805. Histoire naturelle des mollusques terrestres et fluviatiles de la France.
Paris. L. Colas, vii+164 p.
Kiister H.C., Dunker W. 1850. Die Familie der Limnaeiden enthaltend die Genera Planorbis,
Limnaeus, Physa und Amphepeplea И Systematisches Conchylien-Cabinet von Martini und
Chemnitz. Numberg, Bauer und Raspe. P. 35-62.
Dupuy D. 1850. Histoire naturelle des mollusques terrestres et d’eau douce qui vivent en France.
Paris. Fascicule 4. 331^458.
Durand P., Pointier J.P., Escoubeyrou K., Arenas J.A., Yong M., Amarista M., Bargues M.D.,
Mas-Coma S., Renaud F. 2002. Occurrence of a sibling species complex within neotropical
lymnaeids, snail intermediate hosts of fascioliasis //Acta Tropica: 83. P. 233-240.
Ehrmann P. 1933. Mollusca // Die Tierwelt Mitteleuropas Vol. 2(1). 264 p.
Falkner G., Bank R.A., Proschwitz T. von. 2001. Check-list of the non-marine molluscan spe-
cies-group taxa of the states of Northern, Atlantic and Central Europe (CLECOM I) // Hel-
dia. Vol. 4. P. 1-76.
Falniowski A., 1990. Anatomical characters and SEM structure of radula and shell in the species
level taxonomy of freshwater prosobranchs (Mollusca: Gastropoda: Prosobranchia): a com-
parative usefulness study // Folia Malacologica. V 4. P. 53-142.
Ferussac J.B.L. 1807. Essai d’une metode conchyliologique appliquee aux Mollusques terrestres
et fluviatiles. Paris, xvi+142 p.
Fretter V, Graham A. 1962. British Prosobranch Molluscs. Their Functional Anatomy and Eco-
logy. Ray Society. London 755 p.
Fromming E. 1956. Biologie der mitteleuropaischen Siisswasserschnecken. Duncker & Hum-
bolt. Berlin. 313 p.
Geyer D. 1927. Unsere Land- und Susswasser-mollusken. Stuttgart: Lutz, xi+224 s.
Gittenberger E., Janssen A.W., Kuijper W.J., Kuiper J.G.J., Meijer T, Velde G. van der, Vries
J.N. de. 1998. De Nederlandse zoetwatermollusken. Recente en fossiele weekdieren uit zoet
en brak water // Nederlandse Fauna. Vol. 2. Leiden: Nationaal Natuurhistorisch Museum
Naturalis, KNNV Uitgeverij & EIS-Nederland. 288 p.
Gloer P. 2002. Die Stisswassergastropoden Nord- und Mitteleuropas: Bestimmungschlussel,
Lebenweise, Verbreitung. Hackenheim: Conchbooks. 327 s.
Gloer P., Bossneck U. 2007. Zur Identitat von Gyraulus piscinarum (Bourguignat, 1852) mit
der Beschreibung von G. bekaensis n. sp. (Gastropoda: Planorbidae) // Mollusca: 25(2).
P. 139-146.
354
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Gloer Р., Bouzid S. 2008. Redescription of Planorbis agraulus Bourguignat, 1864 (Gastropoda:
Planorbidae) // Journal of Conchology. Vol. 39 (5). P. 487^491.
Gloer P., Meier-Brook C. 2003. Susswassermollusken. Ein Bestimmungschliissel fur die Bun-
desrepublik Deutschland. 13 Auflage. Hamburg: Deutscher Jugendbund fiir Naturbeobach-
tung. 134 s.
Gloer P., Pesic V. 2012. The freshwater snails (Gastropoda) of Iran, with descriptions of two new
genera and eight new species // ZooKeys. Iss. 219. P. 11-61.
Gloer P., Vinarski M.V. 2009. Taxonomical notes on Euro-Siberian freshwater molluscs: 2. Re-
description of Planorbis (Gyraulus) stroemi Westerlund, 1881 (Mollusca: Gastropoda: Pla-
norbidae) // Journal of Conchology. Vol. 39 (6). P. 717-725.
Gloer P., Zettler M.L. 2009. Redescription of Planorbis moquini Requien, 1848 (Gastropoda:
Planorbidae) // Journal of Conchology. Vol. 39 (6). P. 727-732.
Gmelin J. F. 1791. Caroli a Linne, systema naturae. Tom. I. Pars VI. P. 3021-3910. Lipsiae.
(Beer).
Gray J. E. 1843. Notes on the materials at present existing towards a fauna of New Zealand.
P. 177-181. In: Dieffenbach, E.: Travels in New Zealand; with contributions to the geog-
raphy, geology, botany, and natural history of that country. In two volumes: Vol. II. pp. i-iv
[= 1-4], 396, pl. [1]. London. (Murray).
Gredler VM. 1859. TiroPs Land- und Siisswasser-Conchylien. II. Die Siisswasser-Conchylien //
Verhandlungen der Kaiserlich-koniglichen zoologisch-botanischen Gesellschaft in Wein. 5.
S. 213-308.
Hartmann J.D.W. 1821. System der Erd und FluBschnecken der Schweiz // Neue Alpina. 1.
S. 194-268.
Hartmann J. D. W. 1840-1844. Erd- und Siisswasser-Gasteropoden der Schweiz. Mit Zugabe ei-
niger merkwiirdigen exotischen Arten. I. Band. St. Gallen. (Scheitlin & Zollikofer). pp. i-xx
[= 1-20], 1-227, Tab. I-XII [= 1-12], I-XII [sic, = 13-24], 25-84.
Held F. 1836. Aufzahlung der in Bayern lebenden Mollusken // Isis (Von Oken). Vol. 29(4).
S. 271-282.
Hodgson A.N., Healy J.M. 1998. Comparative sperm morphology of the pulmonate limpets Tri-
musculus costatus, T. reticulatus (Trimusculidae) and Bumupia stenochorias and Ancylus
Xuviatilis (Ancylidae) // Journal of Molluscan Studies. Vol. 64. Iss. 4. P. 447-460.
Hubendick B. 1951. Recent Lymnaeidae. Their variation, morphology, taxonomy, nomenclature
and distribution // Kunglike Svenska Vetenskapsakademiens Handlingar. Series 4. Vol. 3(1).
P. 1-223.
Hubendick B. 1954. On the anatomy of Choanomphalus (Moll. Pulm.) // Arkiv for Zoologi:
(2)6. P. 503-509.
Hubendick B. 1955. Phylogeny in the Planorbidae // Transactions of the Zoological Society of
London. Vol. 28. P. 453-542.
Hubendick B. 1957. On the male copulatory organ in Anisus // Proceedings of the Malacological
Society of London. Vol. 32(5). P. 208-212.
Hubendick B. 1960. Faunistic review of the Ancylidae of Lake Ohrid //Arhiv Biol. Nauka. Vol.
13. № 3-4. P. 89-97.
Hubendick B. 1962. Studies on Acroloxus (Mollusca, Basommatophora) // Goteborgs Kungliga
Vetenskaps-och Vitterhets-Samhalles Handlingar. Vol. 9. № 2. P. 1-68.
Hubendick B. 1964. Studies on Ancylidae, the subgroups // Goteborgs Kunglige Vetenskaps-och
vitterhets-sam-halles handlingar. Vol. 9. № 6. Goteborg. P. 1-75.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
355
Hubendick В. 1978. Systematics and comparative morphology of the Basommatophora. In: Pul-
monates. Vol. 2A / V. Fretter, J.F. Peake eds. Academic Press: London. P. 1-47.
Hudec V. 1967. Bemerkungen zur Anatomie von Arten aus der Gattung Anisus Studer, 1820
aus slowakischen Populationen (Mollusca, Pulmonata) // Biologia (Bratislava). Vol. 22.
P. 345-364.
Inaba A. 1969. Cytotaxonomic studies of lymnaeid snails // Malacologia. Vol. 7(2-3). P. 143-168.
Jackiewicz M. 1993. Phylogeny and relationships within european species of the family Lym-
naeidae (Gastropoda: Pulmonata: Basommatophora) // Folia Malacologica. Vol. 5. P. 61-96.
Jackiewicz M. 1998. European species of the family Lymnaeidae (Gastropoda Pulmonata
Basommatophora) // Genus. Vol. 9(1). P. 1-93.
Jackiewicz M. 2000. Blotniarki Europy, Gastropoda, Pulmonata, Lymnaeidae. Wydawnictwo
Kontekst, Poznan. 116 p.
Jaeckel, S. G. A. (1962). Erganzungen und Berichtigungen zum rezenten und quartaren Vorkom-
men der mitteleuropaischen Mollusken // Brohmer, Ehrmann & Ulmer (Hrsg.), Die Tierwelt
Mitteleuropas. 2 (Lief.l, Erg.). Leipzig [Quelle & Meyer]. P. 25-294.
Jordaens K., Dillen L., Backeljau Th. 2007. Effects of mating, breeding system and parasites
on reproduction in hermaphrodites: pulmonate gastropods // Animal Biology. Vol. 57(2).
P. 137-195.
Jorgensen A., Kristensen T.K., Stothard J.R. 2004. An investigation of the ,,Ancyloplanorbidae“
(Gastropoda, Pulmonata, Hydrophila): preliminary evidence from DNA sequence data //
Molecular Phylogenetics and Evolution. Vol. 32. P. 778-787.
Komiushin A.V 1999. New records of Lymnaea (Stagnicola) species in the West Ukraine
(Gastropoda: Basommatophora: Lymnaeidae) // Malakologische Abhandlungen. Vol. 19.
P. 281-286.
Kruglov N.D., Starobogatov Ya.I. 1993a. Annotated and illustrated catalogue of species of the
family Lymnaeidae (Gastropoda Pulmonata Lymnaeiformes) of Palaearctic and adjacent
river drainage areas. Pt 1 // Ruthenica: 3(1). P. 65-92.
Kruglov N.D. Starobogatov Ya.I. 1993b. Annotated and illustrated catalogue of species of the
family Lymnaeidae (Gastropoda Pulmonata Lymnaeiformes) of Palaearctic and adjacent
river drainage areas. Pt 2 // Ruthenica: 3(2). P. 161-180.
Kunigelis S.C., Saleuddin A.S.M. 1984. Anesthetization of Planorbidae (Mollusca): methodol-
ogy and pharmacology // Experientia. Vol. 40. P. 584-585.
Kiister H.C. 1862. Die Gattungen Limnaeus, Amphepeplea, Chilina, Isidora und Physiopsis 11
Systematisches Conchylien-Cabinet von Martini und Chemnitz. Numberg, Bauer und Ras-
pe. 1-48.
Lamarck J.B.P.A. de M. de. 1822. Histoire naturelle des animaux sans vertebres. 6(2). Paris,
Verdiere. 252 p.
Lilly M.M. 1953. The mode of life and the structure and functioning of the reproductive ducts
of Bithynia tentaculata (L.) // Proceedings of the Malacological Society of London. Vol. 30.
P. 87-110.
Lindholm W.A. 1911. Uber mollusken aus dem Ladogasee und der Nevabucht // Ежегодник
Зоологического музея Императорской Академии наук. (Annuaire du Musee Zoologique
de I’Academie Imperiale des Sciences de St.-Petersbourg). T. 16. C. 285-310.
Linnaeus C. 1758. Sy sterna naturae per regna tria naturae, secundum classes, ordines, genera,
species, cum characteribus, differentiis, synonymis, locis. T. 1. Editio decima, reformata.
Holmia (Stockholm): Laurentii Salvii. iv+824 p.
356
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Mattice J. S. 1972. Production of a natural population of Bithynia tentaculata L. (Gastropoda:
Mollusca) // Ekologia Polska. Ekologia Polska Vol. 20. P. 525-539.
Meier-Brook C. 1964. Gyraulus acronicus und G. rossmaessleri, ein anatomischen Vergleich
(Planorbidae) //Archiv fur Molluskenkunde. Bd. 93 (5/6). S. 233-242.
Meier-Brook C. 1976a. The influence of varied relaxing and fixing conditions on anatomical
characters in a Planorbis species // Basteria. Vol. 40(4/6). P. 101-106.
Meier-Brook C. 1976b. The generic position of Planorbis (Gyraulus) intermixtus Mousson,
1874, and Planorbis presbensis Sturany, 1894 (Gastropoda, Basommatophora) // Basteria.
Vol. 40(4/6). P. 107-118.
Meier-Brook C. 1983. Taxonomic studies on Gyraulus (Gastropoda:Planorbidae) // Malacologia.
Vol. 24(1/2). P. 1-113.
Menke K.T. 1846. Kritische Ubersicht der lebenden Hz/vata-Arten // Zeitschrift fur Malakozoo-
logie. Voll. 2 (1845). P. 115-130.
Menzel H. 1904. Zwei neue Arten von Valvata Muller (Gruppe Cincinna Hiibner) // Nachrichts-
blatt der Deutschen Malakozoologischen Gesellschaft. Vol. 36(2). S. 77-79.
Millet P.A. 1813. Mollusques terrestres et fluviatiles, observees dans le departement de Maine et
Loire. Angers. Imp. L. Pavie. 82 p.
Morch O.A. 1864. Synopsis Molluscorum terrestrium et fluviatilium Daniae. Kjobenhavn.
105 s.
Morgan J.A.T., DeJong R.J., Jung Y., Khallaayoune K., Kock S., Mkoji G.M., Loker E.S. 2002.
Schistosoma parasites of humans and domestic animals did not co-speciate with their pla-
norbid snail hosts; a story of host switching and recent diversification // Molecular Phyloge-
netics and Evolution. Vol. 25. P. 477-488.
Morris J., Sowerby G.B. 1838. I. On the Deposits containing Carnivora and other Mammalia
in the Valley of the Thames (Morris). II. Comparison of Cyrena, Valvata, and Unio found
fossil at Grays, with recent species (Sowerby) // The Magazine of Natural History (N. S.).
Vol. 2. P. 539-546 (I), 546-548 (II).
Mozley A. 1934. New fresh-water mollusks from northern Asia // Smithsonian Miscellaneous
Collections. Vol. 92(2). P. 1-7.
Muller O.E 1774. Vermium terrestrium et fluviatilium seu animalium infusoriorum, helminthi-
corum et testaceorum, non marinorum, succincta historia. 1(2). Hafhiae et Lipsia. Heineck
et Faber, xxxvi + 214 s.
Nordsieck H. 1990 (1993). Pylogeny and system of the Pulmonata (Gastropoda) // Archiv fur
Molluskenkunde. Bd. 121. S. 31-52.
Odhner N. 1956. Distinctions between Anisus (Armiger) crista (L.) and Anisus (Gyraulus) ri-
parius (West.) // Proceedings of the Malacological Society. Vol. 32. P. 127-131.
Paasch A. 1842. Beschreibungen einer neuer bei Berlin gefundenen Paludina // Archiv fur Na-
turgeschichte. Vol. 8. S. 300-301.
Paladilhe A. 1867. Nouvelles miscellanees malacologiques // Revue et Magasin de Zoologie.
Ser. 2. Vol. 19. P. 38-53, 88-95.
Paladilhe A. 1869. Descriptions du quelques Paludinidees, Assiminidees et Melanidees nou-
velles // Revue et Magasin de Zoologie. Vol. 2 (21). P. 225-237.
Paraense W.L. 2003. A bird’s eye survey of central American planorbid molluscs // Memorias do
Institute Oswaldo Cruz, Rio de Janeiro: 98. P. 51-67.
Paraense W.L. 2004. Planorbidae, Lymnaeidae, and Physidae of Ecuador (Mollusca: Basomma-
tophora) // Mem. Inst. Oswaldo. Cruz.: 99. P. 357-362.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
357
Paraense W.L., Pointier J.-P. 2003. Physa acuta Drapamaud, 1805 (Gastropoda: Physidae): a
study of topotypic specimens // Memorias do Institute Oswaldo Cruz. Vol. 98(4). P. 513-517.
Pesic V, Gloer P. 2013. A new freshwater snail genus (Hydrobiidae, Gastropoda) from Montene-
gro, with a discussion on gastropod diversity and endemism in Skadar Lake // ZooKeys. Iss.
281. P. 69-90.
Pfeiffer C. 1821. Systematische Anordnung und Beschreibung deutscher Land- und Siisswasser-
Schnecken, mit besonderer Riicksicht auf die bisher in Hessen gefimdenen Arten. Cassel.
Verlag des Verfassers. 134 p.
Pfeiffer C. 1828. Naturgeschichte Deutscher Land- und Siisswasser Mollusken. 3. Weimar, 84 p.
Piaget J. 1929. L’adaptation de la Limnaea stagnalis aux milieux lacustres de la Suisse ro-
mande // Revue Suisse de Zoologie : 36(17). P. 263-531.
Piechocki A. 1979. Mi^czaki (Mollusca). Slimaki (Gastropoda). // Fauna slodkowodna Polski;
Vol. 7. Warszawa. Poznan: Panstwowe Wydawn. Naukowe. 188 s.
Ponder W.F., Waterhouse J. 1997. A new genus and species of Lymnaeidae from the lower
Franklin River, south western Tasmania // Journal of Molluscan Studies. Vol. 63. P. 441—
468.
Prosorova L.A., Starobogatov Ya.I. 1996. Guide to Recent molluscs of northen Eurasia. 6. Ge-
nus Armiger Hartmann, 1840 (family Planorbidae) // Ruthenica: 5(2). P. 167-175.
Prozorova L.A., Starobogatov Ya.I. 1997. New species of the subgenus Gyraulus of the genus
Anisus and Gyraulus-simiiar forms from Asiatic Russia (with notes on the classification of
the genera Anisus and Choanomphalus) (Gastropoda Planorbidae) // Ruthenica: 7(1). P. 39-
50.
Roffiaen F. 1868. Mollusques terrestres et fluviatiles recuiellis en Suisse //Annales de la Societe
malacologique de Belgique. Vol. 3. P. 65-84.
Rossmassler E.A. 1835a. Iconographie der Land- und Susswasser-Mollusken Europa’s, mit
vorzuglicher Beriicksichtigung der Europaischen noch nicht abgebildeten Arten. Dresden.
Leipzig. 1(1). P. 1-132.
Schafer H. 1952. Ein Beitrag zur Emahrungsbiologie von Bithynia tentaculata II Archiv fur
Molluskenkunde. Bd. 82. S. 67-70.
Schmidt A. 1851. Malakologische Mittheilungen // Zeitschrift fur Malakozoologie. 8. S. 179—
192.
Schmidt A. 1856. Verzeichniss der Binnenmolluscken Norddeutchlands mit kritischen Bemer-
kungun // Zeitschrift fur die Gesammter Naturwissenchaften. 8. P. 120-169.
Schrank F. von Paula 1803. Fauna Boica. Durchgedachte Geschichte der in Baiem einheimi-
schen und zahmen Thiere, 3(2). Lanshut. Philipp Krull. 372 S.
Servain G. 1881. Histoire malacologique du lac Balaton en Hongrie. Poissy. Imp. S. Lejay et Co.
125 p.
Sheppard, 1823 Sheppard R. 1823. Description of seven new British land and fresh-water shells,
with observations upon many other species, including a list of such as have been found in
the country of Suffolk // Transactions of the Linnean Society of London. Vol. 14(1). P. 148—
170.
Sitnikova T.Ya., Ropstorf P. 1999. A new species of planorbid gastropod of the genus Choanom-
phalus fron Lake Baikal // Ruthenica: 9(2) P. 123-128.
Soldatenko E.V 2009. New diagnostic characters of Kolhymorbis angarensis (B. Dybowski
et Grochmalicki, 1925) (Gastropoda: Pulmonata: Planorbidae) // Zoosystematica Rossica.
Vol. 18. Iss. 2. P. 191-195.
358
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Soldatenko E.V., Petrov А.А. 2009. Cuticular structures in the copulatory apparatus of Planorbis
planorbis (Linnaeus, 1758) (Gastropoda: Pulmonata: Planorbidae) // Zoosystematica Ros-
sica. Vol. 18. Iss. P. 11-16.
Soldatenko E.V Petrov A.A. 2012. Mating behaviour and copulatory mechanics in six species
of Planorbidae (Gastropoda: Pulmonata) // Journal of Molluscan Studies. Vol. 78. Iss. 1.
P. 185-196.
Soldatenko E.V, Petrov A.A. 2013. Mating and morphology of the copulatory apparatus in Pla-
norbarius corneus (Linnaeus, 1558) (Gastropoda: Pulmonata) // Zoosystematica Rossica.
Vol. 22. Iss. 2. P. 153-164.
Soldatenko E.V, Shatrov A.B. 2013. Comparative ultrastructure of the terminal portions of the
male copulatory apparatus in Planorbidae (Gastropoda: Pulmonata) // Invertebrate Repro-
duction & Development. Vol. 57. Iss. 3. P. 224-236.
Soldatenko E.V, Starobogatov Ya. I. 2000. Genus Planorbis (Gastropoda Pulmonata Planorbi-
dae) // Ruthenica: 10(1). P. 23-35.
Soos A. 1935. Morphologie and Histologie des Geschlechtsapparates der ungarischen Planorbis-
Arten //Allattani Kozlemenyek (Budapest). Bd. 32. S. 21^46.
Strong E.E., Gargominy O., Ponder W.F., Bouchet Ph. 2008. Global diversity of gastropods
(Gastropoda; Mollusca) in freshwater// Hydrobiologia: 595. P. 149-166.
Studer, 1820 Studer, S. 1820. Kurzes Verzeichniss der bis jetzt in unserm Vaterlande entdeckten
Conchylien // Naturwissenschaftlicher Anzeiger der Allgemeinen Schweizerischen Gesell-
schaft fur die Gesammten Naturwissenschaften. Bern. Bd. 3(11). S. 83-90, 91-94.
Taylor D. W., Sohl N.F. 1962. An outline of gastropod classification // Malacologia. Vol. 1.
P. 7-32.
Taylor D.W. 2003. Introduction to Physidae (Gastropoda: Hygrophyla): Biogeography, classifi-
cation, morphology // Revista de biologia tropica. Vol. 51 (suppl. 1). P. 1-300.
Те G.A. 1978. Systematic study of the family Physidae (Basommatophora: Pulmonata). PhD
Thesis. Michigan University. 324 p.
Thiele J. 1931. Handbuch der Systematischen Weichtierkunde. Bd. 1. Jena: 1-77,8.
Troschel F.H. 1834. De Limnaeaceis seu de gasteropodis pulmonatis quae ndstris in aquis viv-
unt. Berolini. Typis Academiae Regiae Scientiarum. 65 p. /
Vinarski M.V 2013. One, two, or several? How many lymnaeid genera are there? // Ruthenica:
23(1). P. 41-58.
Vinarski M.V, Schniebs K., Gloer P., Son M.O., Hundsdoerfer A.K., 2Ш2. The steppe relics:
taxonomic study on two lymnaeid species endemic to the former USSR (Gastropoda: Pul-
monata: Lymnaeidae) //Archiv fur Molluskenkunde. Bd. 141(1). P. 67-85.
Wesenberg-Lund C. 1939. Biologie der SiiBwassertiere — Wirbellose TiereAWien (Springer).
xi + 818s.
Westerlund C.A. 1865. Sveriges land- och sotvatten-mollusker. Lund W.K. GleerupsX(42 p.
Westerlund C.A. 1871. Expose critique des mollusques de terre et d‘eau douce de la Suede et de la
Norvege // Nova Acta Societatis Regiae Scientiarum Upsaliensis. Ser. 3. Vol. 8(1). P. 1-200.
Westerlund C.A. 1873. Fauna molluscorum terrestrium et fluviatilium Sueciae, Norvegiae et Da-
niae. 2. Sotvatten Mollusker. Stockholm. O.W. Backmann. i-v, 297-651 p.
Westerlund C.A. 1875. Malakologische Studien, Kritiken und Notizen // Malakozoologische
Blatter Vol. 22. S. 98-117.
Westerlund C.A. 1881. Malakologiska bidrag. II. For Vetenskapen nya Land- och Sotvatten
Mollusker. Ofversigt af Kongliga Vetenskaps-Akademiens Forhandlingar. Vol. 4. P. 50-70.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
359
Westerlund С.А. 1885а. Fauna der in der Palaarktischen Region (Europa, Kaukasien, Sibirien,
Turan, Persien, Kurdistan, Armenien, Mesopotamien, Kleinasien, Syrien, Arabien, Egypten,
Tripolis, Tunisien, Algerien und Marocco) lebenden Binnenconchylien. V. Fam. Succineidae,
Auriculidae, Limnaeidae, Cyclostomidae und Hydrocenidae. Lund. H.Ohlsson. 135+14 p.
Westerlund C.A. 1886. Fauna der in der Palaarktischen Region (Europa, Kaukasien, Sibirien,
Turan, Persien, Kurdistan, Armenien, Mesopotamien, Kleinasien, Syrien, Arabien, Egypten,
Tripolis, Tunisien, Algerien und Marocco) lebenden Binnenconchylien. VI. Fam. Ampullari-
idae, Paludinidae, Hydrobiidae, Melapiidae, Valvatidae & Neritidae. Lund, 156+13 p.
Westerlund C.A. 1897. Synopsis molluscorum extramarinorum Scandinaviae (Sueciae, Norvegi-
ae, Daniae & Fenniae) //Acta Societatis pro Fauna et Flora Fennica. Vol. 13(7). 238 p.
Wilke T, Davis G. M., Falniowski A., Giusti F., Bodon M., Szarowska M. 2001. Molecular sys-
tematics of Hydrobiidae (Mollusca: Gastropoda: Rissooidea): testing monophyly and phy-
logenetic relationships // Proceedings of the Academy of Natural Sciences of Philadelphia.
151. P. 1-21.
Zilch A. 1959-1960. Teil 2: Euthyneura // O.H. Schindewolf, ed. Handbuch der Palaozoologie.
Band 6. Gastropoda. Bomtraeger. Berlin, xii+835 p.
Ключ для определения семейств пресноводных брюхоногих моллюсков
1(4).
2(3).
3(2).
4(1).
Раковина колпачковидная, не завита в спираль.
Контур устья (или раковины при взгля-
де сверху) округло- или яйцевидно-
овальный. Вершина в виде небольшо-
го загнутого рожка в задней части ра-
ковины располагается на продольной
оси раковины или немного смещена
вправо. Тело левозавитое. Половые,
выделительное и дыхательное отвер- рис. 21.
стия расположены с левой стороны...
...........Ancylidae Rafinesque, 1815
(с. 413; рис. 21).
Контур устья (или раковины при
взгляде сверху) продолговто-овальный.
Вершина в виде небольшого загнутого
рожка в задней части раковины яв-
ственно смещена влево от продольной
оси раковины. Тело правозавитое. По-
ловые, выделительное и дыхательное
отверстия расположены с правой сто-
роны.........Acroloxidae Thiele, 1931
(с. 391; рис. 22).
Раковина в виде замкнутой с одного
конца трубки, закрученной в спираль
Рис. 22.
различной формы.
360
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
5(22). Устье раковины закрывается крышечкой, как правило, не вырезано стенкой
последнего оборота. Устьевые края внутри не имеют наплывов и складок.
6(7). Раковина неритоидная (полушаровидная или полуяйцевидная), “ухо-
видная” если смотреть со стороны устья; с немногочисленными (до 3),
быстро нарастающими оборотами. Наружная поверхность раковины с
рисунком из светлых пятен на тёмном фоне или зигзагообразных линий,
реже полностью чёрная. Завиток почти не вы-
ступающий или слабо выступающий. Устье
полукруглое, с широкой хорошо выраженной
колумеллярной (устьевой) площадкой. Кры-
шечка с двумя внутренними отростками (апо-
физами) ..................................
... Neritidae Rafinesque, 1815 (с. 364; рис. 23).
Рис. 23.
7(6). Раковина иная.
8(13). Раковина при 3-8 оборотах очень маленькая. Высота раковины до 6,5 мм,
ширина до 2,7 мм.
9(10). Высота завитка составляет чуть больше половины высоты устья. Раковина
низкоконическая, сильно прижатая. Пупок округлый, перспективный, в
виде широкой воронки.......................Hydrobiidae Troschel 1857 (с. 378).
Роды Hauffenia Pollonera, 1899 и Islamia Radoman, 1973. В евро-
х—х пейской части России не известны.
Ю(9). Высота завитка больше высоты устья или равна ей. Ракови-
f на другой формы. Пупок не перспективный, каплевидный,
j щелевидный, иногда в виде очень узкой щели или полно-
( j стью закрытый отворотом колумеллярного края устья.
I И(12). Раковина овальная, яйцевидная (овально-башневидная),
\. "ЯИГ | башневидная с высотой не меньше 2,2 мм и шириной не
J меньше 1,5 мм или овально-коническая с высотой не бо-
лее 3,0 мм и шириной не более 1,7 мм...........................
Рис- 24. ..............Amnicolidae Tryon, 1863 (с. 380; рис. 24).
12(11).Раковина кубаревидная, баш-
невидно-коническая, высококо-
ническая или яйцевидная. Если
раковина овальная или башне-
вцДная, то её высота не боль-
шие 2,1 мм, ширина не больше
1,4 мм. Если раковина оваль-
но-коническая, то её высота не
менее 3,1 мм, ширина не менее
к8 мм........................
Hydrobiidae Troschel 1857
(с. 378; рис. 25).
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
361
Рис. 26.
13(8). Раковина при 3-8 оборотах, как прави-
ло, более крупная. В тех случаях, когда
высота раковины меньше 6,6 мм, её ши-
рина всегда не менее 2,8 мм.
14(15). Раковина не менее 25 мм высотой,
кубаревидная, редко коническая. По-
верхность оборотов, как правило, с 2-3
спиральными бурыми полосами на свет-
лом фоне. Иногда эти полосы настолько
широкие, что могут сливаться, образуя однотонную бурую окраску рако-
вины ...............................Viviparidae Gray, 1847 (с. 365; рис. 26).
15(14). Раковины не превышает 25 мм в высоту, Различной формы: от плоскоспи-
ральной, дисковидной и низкоконической до высококонической и кубаре-
видной. Бурые спиральные полосы на поверхности оборотов отсутствуют.
16(19). Раковина кубаревидная, с гладкой, фарфоровидной поверхностью или с
очень тонкой радиальной (осевой) исчерченностью оборотов. Последний
оборот сильно вздут.
17(18). Раковина очень маленькая, при 4 оборотах её высота
2,5-4,5 мм, ширина 3,5-5,5 мм. Колумеллярная (устье-
вая) площадка отсутствует; пупок округлый, проколо-
видный, менее чем на половину прикрыт отворотом
колумеллярного края устья.............................
....Valvatidae Gray, 1840; род Borysthenia Lindholm, рис 27.
1914 (с. 382; рис. 27).
18(17). Раковина крупнее, при 4 оборотах её высота 7,0-9,0 мм, ширина 6,5-
8,0 мм. Утолщённые и отвёрнутые колумеллярный и париетальный края
устья образуют узкую колумеллярную (устьевую) площадку. Пупок пол-
ностью закрыт отворотом колумеллярного края устья.......................
........................Lithoglyphidae Troschel, 1857 (с. 381; рис. 28).
19(16). Раковина не кубаревидная: дисковидная
(плоско- или турбоспиральная), низко-,
овально- или башневидно-коническая,
коническая. Последний оборот не вздут,
расширяется более или менее равно-
мерно.
20(21). Раковина коническая или овально-ко-
ническая, с заострённой вершиной,
поверхность оборотов в общем гладкая
(блестящая или матовая), иногда со спи-
ральной исчерченностью и/или тонкой
радиальной (осевой) морщинистостью.
Пупок щелевидный, в разной степени Рис. 28.
362
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 30.
Рис. 29.
открытый, иногда полностью закрытый
отворотом колумеллярного края устья.
Устье овальное или каплевидно-овальное,
закрывается обызвествлённой концентри-
ческой крышечкой......................
....Bithyniidae Gray, 1857 (с. 370; рис. 29).
21(20). Раковина дисковидная (плоско- или тур-
боспиральная), низко- или башневидно-
коническая с притупленной вершиной;
поверхность оборотов гладкая или со спи-
ральными, либо радиальными (осевыми)
рёбрышками. Всегда имеется пупок окру-
глой формы, широко развёрнутый, пер-
спективный или узкий проколовидный,
открытый или в различной степени при-
крытый отворотом колумеллярного края
устья. Устье округлое, иногда с лёгкой
угловатостью в париетально-палатальной
области, закрывается необызвествлённой
спиральной многооборотной крышечкой..
.........................Valvatidae Gray, 1840
(с. 382; рис. 30).
22(5). Крышечка, закрывающая устье, отсутствует. Колумеллярный и париеталь-
ный края устья внутри могут иметь утолщения, складки или складкоо-
бразные наплывы. Устье нередко вырезано стенкой последнего оборота.
23(26). Раковина турбоспиральная, уховидная, шаровидная, яйцевидная, баше-
видная, овально-, башневино или высококоническая.
24(25). Раковина правозавитая.....Lymnaeidae Lamarck 1812 (с. 398; рис. 31).
Рис 31.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
363
Рис. 33.
25(24). Раковина левозавитая, овально- или высококони-
ческая.........................Physidae Fitzinger, 1833
(с. 392; рис. 32).
26(23). Раковина плоскоспиральная. Если турбоспираль-
ная, то дисковидная (с едва возвышающимся над
последним оборотом завитком) или в различной
степени прижатая кубаревидная (высота завит-
ка примерно равна половине высоты устья или
ч Рис. 32.
меньше).
27(28). Раковина левозавитая (верхняя сторона
раковины определяется по меньшей по-
гружённости/вдавленности начальных
оборотов в области верхушки по срав-
нению с более глубоко погружёнными
начальными оборотами в области пуп-
ка), плоскоспиральная, крупная (при
4-5 оборотах ширина раковины больше
20 мм)......................................
........Helisomatidae ЕС. Baker, 1928
(с. 415; рис. 33).
28(27). Раковина правозавитая, плоскоспиральная или турбоспиральная (диско-
видная, линзовидная или кубаревидная), относительно или очень малень-
кая (при 4-5 оборотах ширина раковины меньше 20 мм).
29(30). Раковина плоскоспиральная. При 4-5
оборотах её ширина не больше 8,6 мм.
Контур раковины в поперечном сечении
может быть: симметрично выпуклым с
верхней и нижней сторон, так, что рако-
вина имеет вид двояковыпуклой линзы,
выпуклым сверху и уплощённым снизу
так, что раковина имеет форму от плоско-
выпуклой линзы до полушаровидной....
...............Segmentinidae ЕС. Baker, 1945
(с. 433; рис. 34).
30(29). Раковина турбоспиральная (дисковидная
или кубаревидная). Если плоскоспи-
ральная, то с уплощёнными сверху и
снизу оборотами, не придающими рако-
вине облик двояко-, плоско-выпуклой линзы или полушаровидную форму.
При 4-5 оборотах ширина раковины больше 8,7 мм. В центральных и
северных областях Европейской России виды с кубаревидной раковиной
неизвестны......................Planorbidae Rafinesque, 1815 (с. 417; рис. 35).
Рис. 34.
364
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Семейство NERITIDAE Rafinesque, 1815
Типовой род: Nerita O.F. Muller, 1774.
Раковина толстостенная (в большинстве случаев), неритоидная (полуяйце-
видная, “уховидная” если смотреть со стороны устья), с немногочисленными
быстро нарастающими гладкими, либо морщинистыми или ребристыми оборо-
тами. Завиток малозаметный, почти не выступающий или слабо выступающий.
Пупок отсутствует. Устье относительно небольшое, полукруглое, закрывающее-
ся обызвествленной малооборотной крышечкой с двумя внутренними отростка-
ми (апофизами). Колумеллярный край сильно расширен, образуя так называемую
колумеллярную (устьевую) площадку. Раздельнополые животные. Своеобразная
женская половая система представлена двумя связанными протоками, идущими
от половой железы и открывающимися самостоятельными отверстиями. У сам-
цов пенис расположен на внутренней стороне правого щупальца. Поперечный
ряд радулы включает 1 рахидальный зуб, 2 субцентральных (по одному с каждой
стороны от рахидального), 4 парацентральных (по 2 с каждой стороны), 2 иници-
альных (по 1 с каждой стороны) и п маргинальных. Формула поперечного полу-
ряда радулы: R-S-2P-I-nM1 (Анистратенко, Анистратенко, 2001).
Древнейшие неритиды известны из отложений триаса. В континентальных во-
доёмах разных материков (искл. Антарктиду) они начинают появляться с палео-
гена, образуя современные пресноводные и солоноватоводные роды. Современ-
ные представители неритид это обитатели прибрежных участков морей, пресных
и солоноватоводных водоёмов, а также супралиторальные амфибиотические фор-
мы. В составе семейства известно более 10 родов. В Евразии обитают представи-
тели одного рода Theodoxus Montfort, 1810.
1 Здесь и далее по тексту даны следующие обозначения зубов радулы: R — рахидальный зуб,
S — субцентральные, Р — парацентральные, I — инициальные, М — маргинальные.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
365
Род Theodoxus Montfort, 1810
Типовой вид: Neritina fluviatilis Linnaeus, 1758.
Раковина, полушаровидная, оборотов 2-3, с почти не выступающим завитком
(высота устья практически равна высоте раковины) и широкой колумеллярной
площадкой. Обороты гладкие или с радиальными линиями нарастания, обычно
ярко окрашены, часто с рисунком из многочисленных тёмных ломанных или изо-
гнутых линий на светлом фоне, либо светлых округлых пятен на тёмном фоне.
Крышечка с отростком на внутренней стороне.
Представители рода известны с олигоцена. Ныне распространены в Европе,
Передней Азии и Северной Африке. Около 40 видов. Обитают в пресноводных
и солоноватоводных водоёмах. В водоёмах северной части Европейской России
обитает только 1 вид — Theodoxus fluviatilis (Linnaeus, 1758).
Комментарии. Раковина теодоксусов бедна признаками, поэтому многие исследовате-
ли, описывая новые таксоны и формы, опирались преимущественно на особенности из-
менчивой окраски и характер рисунка поверхности оборотов. Вопрос о таксономическом
значении окраски у теодоксусов до сих пор остаётся предметом дискуссий. Однако, как
показали данные О.Ю. Анистратенко и В.В. Анистратенко (основанные на оригинальных
наблюдениях и анализе литературы), определённый таксономический вес имеет только
характер рисунка (наличие полос или пятен, однотонный фон), тогда как варианты цвета
окраски диагностического значения не имеют и могут варьировать даже в пределах одной
популяции (Анистратенко др., 1999; Анистратенко, Анистратенко, 2001).
Theodoxus fluviatilis (Linnaeus, 1758) (вкл. табл. IV, рис. 43).
—fluviatilis Linnaeus, 1758: 777 (Nerita).
При взгляде со стороны устья контур раковины прижато-яйцевидный, раковина
толстостенная, относительно крупная (высота до 7 мм, ширина до 12 мм) со слабо
выступающим или практически не выступающим завитком. Обороты разделены
мелким швом, линии нарастания выражены в разной степени, иногда присут-
ствует неясная спиральная исчерченность. Окраска от чёрной до оливково-бурой.
Рисунок образован обильными светлыми округлыми пятнами на тёмном фоне.
Нередко сгущение пятен преобразует рисунок таким образом, что можно разли-
чить тёмные спиральные волнистые линии на относительно светлом фоне. Устье
заметно скошенное, с отогнутым верхним краем.
Размеры: ВР до 7 мм, ШР до 12 мм.
Вид обитает в водоёмах Западной и Центральной Европы, в бассейнах рек Бал-
тийского и Чёрного морей. Нередко встречается в морских лиманах при солёности
до 5-7%о, питается губками и зелёными водорослями.
Семейство VIVIPARIDAE Gray, 1847
Типовой род: Viviparus Montfort, 1810.
Раковина крупная (высота до 60 мм), от кубаревидной и овально-конической
до конической (в разной степени вытянутой), поверхность оборотов неравномер-
но радиально морщинистая, реже почти гладкая. Окраска от зеленовато- до крас-
новато-бурой. Часто имеются 3 коричневых спиральных полосы, наиболее выра-
366
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
женные на последнем обороте раковины. На раковинах молодых особей имеются
периостракальные щетинки, расположенные в 3 спиральных ряда. С возрастом
они утрачиваются. На поверхности дефинитивных оборотов (кроме последнего)
может присутствовать своеобразная маллеатная (“удары молотка”) скульптура,
имеющая упорядоченный характер. Пупок щелевидный или отсутствует. Устье
овальное с неотвернутыми краями, закрывающееся тонкой роговой концентри-
ческой крышечкой. Щупальца длинные, у самцов правое — функционирует как
копулятивный орган. Глаза расположены на небольших бугорковидных утолще-
ниях у оснований щупалец. Позади щупалец из-под мантийного края выступают
наружу две лопасти, правая из которых формирует жёлоб для подачи воды к жа-
бре. Жабра гребневидная. Самки несколько крупнее самцов и отличаются более
вздутой раковиной. У самцов от двулопастного семенника (слабо обособленно-
го от пищеварительной железы) отходит семепровод, расширяющийся в длин-
ную трубку с мышечно-железистыми стенками — пенис (эякуляторная камера
в работах Анистратенко, 1998 и Анситратенко, Анистратенко, 2001). У самок от
яичника отходит короткий яйцевод, впадающий в объёмную матку с коротким
влагалищем, открывающимся в мантийную полость. В яйцевод впадает проток
белковой железы. Животные яйцеживородящие. Радула короткая, зубов в попе-
речном ряду — 7. Формула поперечного полуряда радулы R-I-2M (Анистратен-
ко, Анистратенко, 2001).
В ископаемом состоянии представители семейства известны с миоцена Евро-
пы. Ныне живущие моллюски Viviparidae относятся к 4 родам, распространены
как в заболоченных водоёмах, так и в проточной воде в Евразии и Северной Аме-
рике (Ситникова, Старобогатов, 1982; Черногоренко, 1988). Обитают на песча-
ных, илисто-песчаных грунтах и в зарослях водных растений на мелководьях (до
2 м) затопленных речных пойм, а также на илистых грунтах в крупных проточ-
ных водоёмах на глубинах до 7 м. Зимуют в неактивном состоянии, зарывшись
в ил. Из-за периодических зимних диапауз на поверхности оборотов раковины
формируются резкие радиальные морщины (линии нарастания/роста). Однако ис-
пользовать их для определения возраста моллюсков не следует. Подобные линии
формируются также в экстремальные периоды, не зависящие от сезона, а также
при недостаточном питании (Fromming, 1956; Gloer, 2002).
В европейской части России представлены 2 рода.
Ключ для определения родов семейства Viviparidae
1(2). Раковина толстостенная, кубаревидная или овально-коническая (отноше-
ние ширины раковины к её высоте — не более 0,78), пупок полностью
закрыт отворотом устьевого края, либо в виде очень узкой щели. Обороты
умеренно выпуклые, на периферии могут быть уплощённые. Устье округ-
ло-каплевидное, как правило, с отчётливым углом между париетальным и
палатальным краями. Вершина завитка куполообразно сглаженная, при-
тупленная...................................Viviparus Monfort, 1810 (с. 367).
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
367
2(1). Раковина относительно тонкостенная, кубаревидная или коническая (от-
ношение ширины раковины к её высоте превышает 0,80), пупок широ-
ко-щелевидный. Обороты сильновыпуклые, на периферии равномерно
закруглённые, расположенные ступенчато. Если ступенчатость оборотов
выражена слабо, все равно они имеют отчётливую пологую область вбли-
зи шва (подшовное плечо). Устье овальное или округло-каплевидное со
сглаженной угловатостью между париетальным и палатальным краями.
Вершина завитка — заострённая .... Contectiana Bourguignat, 1880 (с. 368).
Род Viviparus Montfort, 1810
Типовой вид: Helix vivipara Linnaeus, 1758.
Раковина кубаревидная или овально-коническая, толстостенная, с 5,5-7,0 сла-
бовыпуклыми оборотами часто уплощёнными на периферии. Окраска изменчи-
вая: желтовато-роговая, зеленовато-коричневая или чёрная, иногда однотонная
(без полос). Пупок практически полностью закрыт отворотом устьевого края,
либо в виде очень узкой щели. Щетинки на раковинах молодых особей короткие
(не длиннее 0,2 мм). Формула полуряда радулы R-I-1M-2M. Большинство зубов
радулы (рахидальные R, инициальные I и первые маргинальные 1М) имеют круп-
ный трёхвершинный срединный зубец. Крайние зубы (вторые маргинальные 2М)
с 5 примерно одинаковыми небольшими зубчиками по режущему краю. Яйцевые
капсулы крупные, располагаются в матке в 1 ряд.
Представители рода известны с миоцена Европы. Ныне они широко распро-
странены в водоёмах Европы, Западной Азии и востока Северной Америки. Оби-
тают в реках и озёрах на мягких грунтах и в зарослях водных растений.
Количество видов рода колеблется, по мнению разных специалистов от 4-5,
до 9. В малакофауне севера европейской части России насчитывается не менее
3 видов.
Комментарии. Отечественные малакологи в работах конца XX начала XXI в. (Черно-
горенко, Старобогатов, 1987; Анистратенко, 1998; Анистратенко, Анистратенко, 2001;
Старобогатов и др., 2004) различали в составе рода 3 подрода (Viviparus s. str., Hazayipal-
udina Chemogorenko et Starobogatov, 1987 и Balcanipaludina Starobogatov, 1986). В опре-
делителе не поддерживается это точка зрения ввиду нечёткости подродовых диагнозов,
предложенных вышеназванными авторами на основании крайне изменчивых признаков
раковины.
Ключ для определения видов рода Viviparus
1(4). Раковина крупная (при 4,5 оборотах высота раковины ЗСМ-5 мм; шири-
на 24-30 мм; высота последнего оборота 26-29 мм, ширина последнего
оборота без устья не меньше 19 мм). На поверхности последнего и пред-
последнего оборотов, кроме радиальной морщинистости, присутствует
отчётливая маллеатная (“удары молотка”) скульптура из вмятин в сово-
купности образующих правильный решетчатый узор.
368
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
2(3). Обороты вздутые, в профиле плавно закруглённые. Наиболее выпуклая
область последнего оборота практически не смещена к околопупочной об-
ласти............Viviparus sphaeridius (Bourguignat, 1880) (вкл. табл. IV, рис. 46).
— sphaeridia Bourguignat, 1880: 47 {Vivipara).
Размеры: ВР до 45,0 мм, ШР до 30 мм.
Вид распространён в Южной и Центральной Европе. Населяет водоёмы бассей-
нов Дуная, Днестра, Западной Двины. Часто встречается совместно с Viviparus
viviparus и Viviparus at er, крупных скоплений не образует (Черногоренко, Старо-
богатов, 1987; Анистратенко, Черногоренко, 1989; Анистратенко, 1998; Анистра-
тенко, Анистратенко, 2001; Старобогатов и др. 2004; Кантор, Сысоев, 2005).
3(2). Обороты не вздутые, на периферии уплощённые. Последний оборот в
профиле округло-тупоугольный. Его наиболее выпуклая область сильно
смещена к околопупочной области (иногда настолько сильно, что ракови-
на приобретает почти правильные конические очертания).........................
..........Viviparus ater (Christophori et Jan, 1832) (вкл. табл. IV, рис. 47).
— atra Christophori et Jan, 1832: 3 {Paludina).
Размеры: BP до 45,0 мм, ШР до 35,0 мм.
Вид распространён в Южной и Центральной Европе, а также в Северном При-
черноморье. Моллюски населяют водоёмы бассейнов Дуная, Днестра, Западной
Двины, Нижнего Дона. Относительно недавно вид обнаружен в водоёмах южного
берега Крыма, окрестностей Тольятти и Пятигорска. Часто встречается совместно
с Viviparus viviparus, образуя крупные скопления.
4(1). Раковина относительно небольшая (при 4,5 оборотах высота раковины
26-30 мм; ширина 21,0-22,5 мм; высота последнего оборота 21,6-23,0 мм,
ширина последнего оборота без устья меньше 19 мм). Маллеатная скуль-
птура отсутствует, если имеется, то представлена слабо (выше предпо-
следнего оборота) и не образует правильного решетчатого узора.................
...............Viviparus viviparus (Linnaeus, 1758) (вкл. табл. IV, рис. 48).
— vivipara Linnaeus, 1758: 772 {Helix).
[= Viviparus vistulae (Kobelt, 1900)].
Размеры: BP до 35 мм, ШР до 25 мм.
Вид повсеместно распространён в Европе за исключением крайнего севера и юга,
а также в восточном, южном Причерноморье и в западном Закавказье. В 1990-е гг.
XX века интродуцирован в Новосибирское водохранилище (Западная Сибирь).
Моллюски обитают в реках, озёрах и водохранилищах; образуют плотные по-
селения.
Род Contectiana Bourguignat, 1880
Типовой вид: Cyclostoma contectum Millet, 1813.
Раковина кубаревидная или коническая, от умеренно толстостенной до тон-
костенной и просвечивающей, с острой вершиной завитка (макушкой). Обороты
(5,5-7,0) сильно вздутые, закруглённые на периферии, ступенчатые, разделён-
ные глубоким швом. Пупок широко-щелевидный. Устье слабо вырезано стенкой
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
369
предпоследнего оборота, в сечении овальное или округло-каплевидное со сгла-
женной угловатостью между париетальным и палатальным краями, слабо ско-
шенное. Щетинки на раковинах молодых экземпляров относительно длинные
(0,9-1,2 мм). Формула полуряда радулы R-I-1M-2M Большинство зубов радулы
(рахидальные R, инициальные I и первые маргинальные 1М) имеют крупный
прямоугольный срединный зубец по сторонам от которого располагаются по 4
мелких зубчика. Крайние зубы (вторые маргинальные 2М) с 8 примерно одина-
ковыми небольшими зубчиками по режущему краю. Яйцевые капсулы располага-
ются в матке в несколько рядов.
Представители рода известны в Европе с плиоцена. Ныне распространены по
всей Европе и на юге Западной Сибири (до русла Оби на восток). Количество ви-
дов рода по представлениям разных специалистов колеблется от 3 до 9. В цент-
ральных и северных областях европейской части России обитают 2 вида.
Ключ для определения видов рода Contectiana
1(2). Раковина кубаревидная, всегда умеренно толстостенная, не просвечива-
ющая. Последний оборот сильно вздут и по периферии правильно за-
круглён. Отношение ширины раковины к её высоте 0,9 и более. Высота
возвышающейся над последним оборотом части завитка не превышает
0,20 высоты последнего оборота................................................
...............Contectiana ladogensis Chemogorenko et Starobogatov, 1987
(вкл. табл. IV, рис. 45).
— ladogensis Черногоренко, Старобогатов, 1987: 147; 1988: 653, рис. 4з
(Contectiana).
Размеры: ВР 29,2 мм, ШР 25,5 мм (голотип).
Вид пока известен только из типового местонахождения в северной части Ладож-
ского озера.
2(1). Раковина тонкостенная, просвечивающая, иногда хрупкая; реже умеренно
толстостенная. Обороты более стройные (контур раковины коническиий).
Последний оборот часто неравномерно закруглённый (наиболее выпуклая
часть оборота расположена выше периферии, ближе к шву). Отношение
ширины раковины к её высоте всегда меньше 0,9. Высота возвышающейся
над последним оборотом части завитка превышает 0,21 высоты последне-
го оборота............Contectiana contecta (Millet, 1813) (вкл. табл. IV, рис. 44).
— contectum Millet, 1813: 5 (Cyclostoma).
Размеры: ВР до 45 мм, ШР до 35 мм.
Вид распространён в водоёмах Западной, Центральной и Восточной Европы,
исключая крайние южные районы, а также на юге Западной Сибири (на восток
до русла Оби). Моллюски обитают в сильно заросших мелководных заводях при
практически полном отсутствии течения, а также в озёрах. Держатся преимуще-
ственно на грунте и в зарослях растений. Плотных скоплений не образуют.
Комментарии. Полиморфный вид. Размеры, толщина стенок, окраска раковины, а
также степень вздутости последнего оборота подвержены изменчивости.
370
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Семейство BITHYNIIDAE Gray, 1857
Типовой род: Bithynia Leach in Abele, 1818.
Раковина относительно небольшая (высота не превышает 20 мм); от вытяну-
той в разной степени овально-конической до конической и кубаревидной форм.
Иногда в русскоязычной литературе (Анистратенко, Стадниченко, 1994[1995])
встречаются такие определения формы раковины, как яйцевидная, башневидная
и бочонковидная, но они (в приложении к представителям данного семейства)
представляются менее удачными. Окраска однотонная, от тёмно-коричневой
до светло-роговой, с зеленоватым, желтоватым или сероватым оттенками. По-
верхность оборотов гладкая (блестящая или матовая) иногда со спиральной ис-
черченностью и/или тонкой частой морщинистостью (радиальными линиями
нарастания). Пупок щелевидный, в разной степени открытый, у некоторых пред-
ставителей полностью закрыт отворотом колумеллярного края устья. Устье оваль-
ное, иногда с выраженным париетально-палатальным углом (широко-каплевид-
ное). Закрыто обызвествлённой крышечкой. У ряда представителей семейства
выражен половой диморфизм, проявляющийся в том, что самцы имеют более
стройную раковину, чем самки. Передняя часть головы выдаётся подобно ко-
роткому хоботу. Щупальца (в расправленном состоянии) длинные, тонкие и за-
острённые. У основания щупалец расположены глаза. Подошва ноги короткая,
уплощённая спереди и вытянуто-суженная сзади. Раздельнополые животные.
У самцов с правой стороны из-под мантии выступает пенис с развитым боко-
вым выростом (пениальным аппендиксом). Остальная часть мужских гениталий
представлена половой железой, от которой отходит семепровод, пронизывающий
перед впадением в пенис простату. В пениальный вырост впадает в слепо зам-
кнутый извитой придаток — флагеллюм. У самок от яичника отходит длинный
яйцевод, расширяющийся в объёмную матку. Белковая железа отчётливо не вы-
ражена. Матка, сужаясь, переходит во влагалище. На границе матки и влагалища
расположен семеприемник в виде небольшого карманообразного выпячивания.
В поперечном ряду радулы 7 зубов (формула поперечного полуряда: R-I-2M). Ра-
хидальный зуб трапециевидный с зазубренным режущим краем и крупным сре-
динным зубцом.
Естественная область распространения битиниид охватывает все континен-
ты за исключением Северной и Южной Америк. Во второй половине XX века в
Северную Америку была занесена В. tentaculata (Linnaeus, 1758) (Jaeckel, 1962).
Моллюски населяют заиленные медленно текущие реки, озёра, пруды, боло-
та. Питаются растительным детритом, нитчатыми и диатомовыми водорослями
(Wesenberg-Lund, 1939; Lilly, 1953; Fretter, Graham, 1962; Цихон-Луканина, 1987);
в отдельных случаях отмечено фильтрационное питание (Schafer, 1952; Цихон-
Луканина, 1961; Mattice, 1972).
Семейство включает около 130 видов, сгруппированных в два десятка родов
(Strong et al., 2008). В европейской части России обитают 3 вида, относящиеся к
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
Ъ1\
3 родам: Bithynia Leach in Abel, 1818; Opisthorchophorus Beriozkina, Levina, Staro-
bogatov, 1995 и Codiella Monterosato in Locard, 1894 [или подродам одного рода
Bithynia (sensu Gloer, 2002)].
Комментарии. Европейские моллюски семейства по морфологии раковины делятся
на 2 чётко очерченные группы. Одни обладают раковиной с уплощёнными на перифе-
рии оборотами, разделёнными мелким швом и закрытым либо узкощелевидным пупком.
Другие имеют раковину с сильновыпуклыми (иногда ступенчатыми) оборотами, раз-
делёнными глубоким швом и с широко-щелевидным (иногда каплевидным) пупком. По-
следние разделяются на маленьких моллюсков с раковиной, не превышающей 6,0 мм в
высоту (при 4,0-5,5 оборотах), и более крупных животных с раковиной, превышающей
6,0 мм в высоту (при таком же количестве оборотов). Крупные (более 6,0 мм.) раковины,
в свою очередь, бывают с высоким (значительно превышающим высоту устья) и относи-
тельно низким (практически равным высоте устья) завитком. До последнего десятилетия
XX века эти различия позволяли относить европейских моллюсков семейства к 4 видам
рода Bithynia'. В. tentaculata (Linnaeus, 1758), В. leachi (Sheppard, 183), В. troschelii Paasch,
1842, В. inflata Hansen, 1845 (Старобогатов, 1977).
В ревизии битиний европейской России и Украины Г.В. Березкиной, О.В. Левиной,
Я.И. Старобогатова (Beriozkina, Levina, Starobogatov, 1995) и в книге В.В. Анистратенко,
А.П. Стадниченко (1994[ 1995]), частично предваряющей эту ревизию, система Bithyni-
idae, предложенная Я.И. Сторобогатовым (1977), была значительно изменена за счёт вали-
дизации ряда старых (синонимичных) названий видов битиний и описания новых таксо-
нов (родов, подродов и видов). Эти изменения были основаны на результатах применения
компараторного метода (в данном случае метода сравнения контуров завитка раковины),
сравнительного изучения линейных параметров копулятивного аппарата (представителей
рода Bithynia sensu Beriozkina, Levina, Starobogatov, 1995) и размеров ядра крышечек. При
этом анализировался фрагментарный материал без учёта индивидуальной и географиче-
ской (межпопуляционной) изменчивости признаков раковины. В результате европейские
битинии были отнесены к 5 родам [Codiella Monterosato in Locard, 1884; Bithynia Leach in
Abele, 1818; Digyrcidum Locard, 1882; Opisthorchophorus Beriozkina, Levina et Starobogatov,
1995; Paraelona Beriozkina et Starobogatov in Anistratenko et Stadnichenko, 1994(1995)] и 15
видам [Codiella leachi, C. kickxii (Westendorp, 1835), Bithynia tentaculata, B. curta (Gamier
in Picard, 1840), B. decipiens (Millet, 1843), B. producta Moquin-Tandon, 1855, Digyrcidum
bourguignati (Paladilhe, 1869), Opisthorchophorus troschelii, O. baudonianus (Gassies, 1859),
O. hispanicus (Servain, 1880), O. valvatoides Beriozkina et Starobogatov in Beriozkina, Levi-
na et Starobogatov, 1995, Paraelona majewskyi (Frauenfeld, 1862), P boissieri (Charpentier
in Kiister, 1852), P milachevitchi Beriozkina et Starobogatov in Anistratenko et Stadnichenko,
1994[ 1995] и P socialis (Westerlund, 1886)].
Позднее И.А. Першко (2006, 2010), синонимизировала род Digyrcidum с родом Bithyn-
ia, а вид Bithynia decipiens с В. curta. В определителе принимается её точка зрения каса-
тельно рода Digyrcidum, а из моллюсков рода Bithynia признается обитание в пределах
европейской России только одного вида — В. tentaculata (см. комментарии к роду Bithy-
nia). Род Paraelona считается синонимом рода Opisthorchophorus, представленного в евро-
пейской России одним видом — О. troscheli (см. комментарии к роду Opisthorchophorus).
В составе рода Codiella также признается один вид — С. leachi (см. комментарии к роду
372
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Ключ для определения родов семейства Bithyniidae1
1(2). Раковина маленькая, при 4,0-5,5 оборотах её высота не превышает 6 мм.
Ширина предпоследнего оборота не превышает 2,6 мм.........................
...........................Codiella Monterosato in Locard, 1894 (с. 372).
Обороты всегда выпуклые, разделённые глубоким швом. Пупок всегда в виде
широкой щели.
2(1). Раковина крупнее, при 4,0-5,5 оборотах её высота не менее 6,1 мм. Шири-
на предпоследнего оборота более 2,8 мм.
3(4). Обороты слабовыпуклые, иногда на периферии уплощённые, разделённые
мелким швом. Пупок закрытый или в виде очень узкой щели...................
.................................Bithynia Leach in Abel, 1818 (с. 374).
4(3). Обороты сильновыпуклые, иногда ступенчатые, разделённые глубоким
швом. Пупок каплевидный в разной степени открытый (иногда довольно
узкий, но не узкощелевидный)..............................................
........Opisthorchophorus Beriozkina, Levina, Starobogatov, 1995 (с. 377).
Раковина вздутостью оборотов, глубоким швом и формой пупка схожа с предста-
вителями рода Codiella, однако разница в размерах позволяет хорошо различать
моллюсков этих родов. Кроме того, представители этих родов чётко различаются
морфологией копулятивного аппарата (Falniowski, 1990).
Род Codiella Monterosato in Locard, 1894
Типовой вид: Turbo leachi Sheppard, 1823.
Раковина самая маленькая в семействе (при 4,0-5,5 оборотах её высота не пре-
вышает 6,0 мм), овально-коническая или коническая с вытянутым разной степе-
ни завитком. Тонкостенная, просвечивающая, желтовато- или зеленовато-рого-
вая, реже коричневая (часто поверхность раковины покрыта плотным тёмным
налётом). Обороты (4,0-5,5) сильно выпуклые, ступенчатые, разделённые глубо-
ким швом. Высота завитка (у взрослых моллюсков), как правило, больше высо-
ты устья, реже эти значения равны. Поверхность оборотов тонко неравномерно
радиально исчерченная. Устье овальное, его края образуют цельный (замкнутый)
контур. Париетально-палатальный угол, если имеется, — тупой, очень слабо вы-
раженный. Крышечка тонкая концентрическая, с острым краем. Пупок широко
щелевидный, наполовину прикрыт отворотом устьевого края. Радула типичная
для семейства. Пениальный аппендикс ответвляется от дистальной трети пениса.
На конце аппендикса имеется небольшая папилла (Falniowski, 1990).
Представители рода распространены в Западной и Центральной Европе север-
нее Альп, в Восточной Европе в бассейне Балтийского моря и верховьях Волги
1 При определении родов битиний необходимо учитывать степень сформированности рако-
вин. С высокой степенью достоверности можно различать только раковины половозрелых мол-
люсков. Значения линейных параметров раковин у ювенильных моллюсков, как правило, пере-
крываются. Раковины ювенильных моллюсков отличаются небольшим числом оборотов (менее
4), тонким ломким устьевым краем и высотой устья, превышающей высоту завитка.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797 373
(Старобогатов, Затравкин, 1987; Анистратенко, Стадниченко, 1994[1995]; Berioz-
kina et al., 1995; Старобогатов и др., 2004; Gloer, 2002; Кантор, Сысоев, 2005).
В европейской части России обитает 1 вид — Codiella leachii (Sheppard, 1823).
Комментарии. В конце XX века российские и украинские малакологи отнесли к роду
Codiella два вида: Codiella leachii (Sheppard, 1823) и Codiella kickxii (Westendorp, 1835),
указав, что эти виды встречаются совместно, различаются по размерам раковины и не
имеют переходных форм (Анистратенко, Стадниченко, 1994[1995]; Beriozkina, Levina,
Starobogatov, 1995; Старобогатов, Прозорова, Богатов и др., 2004). Однако четыре экземп-
ляра битиниид, определённые Я.И. Старобогатовым как С. kickxii и послужившие осно-
вой для валидизации этого названия, были выделены (Я.И. Старобогатовым) из пробы,
содержащей ещё 6 экземпляров, оставшихся под названием — leachii {“Bithynia leachii
№ 1, Suecia, Kristianstad”, сбор и определение — Westerlund, коллекция ЗИН РАН). Все 10
вероятно разновозрастных экземпляров этой пробы демонстрируют плавный спектр пере-
ходных по размеру форм в диапазонах: высота раковины 3,8-5,0 мм, ширина 2,7-3,4 мм,
отношение высоты завитка к высоте устья 0,9-1,2 при количестве оборотов 3,5-4,5. Эк-
земпляры, характеризующиеся крайними размерными значениям, в общем, соответству-
ют описаниям С. leachii и С. kickxii [= С. celtica (Paladilhe, 1870)] в работе В.В. Анистра-
тенко, А.П. Стадниченко (1994 [1995])', но также они соответствуют описанию Bithynia
leachi Sheppard, 1823, приведённому в работе К. Вестерлюнда (Westerlund, 1886). В этой
же работе К. Вестерлюнд указал Paludina kickxii Westendorp, 1835 как синоним В. leachi.
С другой стороны, он (Westerlund, 1886) даёт описание В. leachi var. kickxii (Nyst in Ktister,
1852) (non Westendorp, 1835) из Голландии, отличающейся маленькими размерами ракови-
ны (высота раковины 4,5 мм, ширина 3,0 мм, без указания количества оборотов). Однако,
кроме совпадения размеров, раковины из коллекции ЗИН РАН не соответствуют другим
специфическим особенностям этого вариетета: устье с выраженным тупым париетально-
палатальным углом, наличие радиальных жёлтых полос на поверхности оборотов, распро-
странение — Голландия. До окончательно прояснения ситуации с валидностью С. kickxii
мы считаем это название синонимом С. leachii.
Codiella leachii (Sheppard, 1823) (вкл. табл. IV, рис. 50).
— leachii Sheppard, 1823: 152 {Turbo).
[= Paludina kickxii Westendorp, 1835; Bithynia ventricosa Gray in Turton, 1840; Bithyn-
ia celtica Paladilhe, 1870; Bithynia leachi var. kickxii Nyst in Ktister, 1852; Bithynia
leachi Westerlund, 1886; Paraelona hellenica Kobelt, 1892; Paraelona socialis Wester-
lund, 1886; Codiella kickxii Анистратенко, Стадниченко, 1994(1995); Beriozkina,
Levina, Starobogatov, 1995; Старобогатов, Прозорова, Богатов и др., 2004; Paraelona
milachevitchi Beriozkina, Starobogatov in Anistratenko et Stadnichenko, 1995].
Признаки раковины такие же, как у рода. Высота раковины и завитка у молодых
экземпляров могут варьировать.
Размеры: ВР до 4,5-6,0 мм, ШР до 3,0-4,5 мм.
Вид распространён в Западной и Центральной Европе севернее Альп, в Восточ-
ной Европе в бассейне Балтийского моря и верховьях Волги. Моллюски населяют
заросли макрофитов по берегам медленно текущих рек, каналов, озёр и прудов.
В работах Г.В. Березкиной, О.В. Левиной, Я.И. Старобогатова (Beriozkina, Levina, Starobo-
gatov, 1995) и Я.И. Старобогатова, Л.А. Прозоровой, В.В. Богатова и др. (2004) эти виды приво-
дятся без морфологических описаний.
374
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Род Bithynia Leach in Abel, 1818
Типовой вид: Helix tentaculata Linnaeus, 1758.
Раковина овально-коническая или кубаревидная. Твердостенная, зеленова-
то-роговая или желтовато-роговая, просвечивающая (поверхность раковины мо-
жет быть покрыта плотным тёмным илистым налётом). Обороты (4,0-6,0) сла-
бовыпуклые, часто на периферии уплощённые, разделены неглубоким швом.
Поверхность оборотов тонко радиально исчерченная, со слабыми спиральными
струйчатыми линиями, реже гладкая или с резкими широко расставленными ли-
ниями нарастания. Последний оборот может быть сильно вздутым. Высота за-
витка у взрослых особей, как правило, больше, редко равна высоте устья (иногда
может быть слегка меньше — у представителей формы — curta sensu Beriozkina
et al., 1995). Устье грушевидно-овальное, слабо, но отчётливо вырезанное стен-
кой последнего оборота. Его края образуют цельный (замкнутый) контур иногда
усилены светлым губообразным наплывом. Парието-палатальный угол хорошо
выражен, прямой или немного более 90°. Крышечка по центру вдавленная, кон-
центрическая. Пупок закрытый или в виде очень узкой щели. Радула типичная
для семейства. Пениальный аппендикс ответвляется от середины либо от дис-
тальной трети пениса.
Род известен с юрского времени. Ныне живущие представители рода рас-
пространены в Европе и Западной Сибири (Старобогатов, 1970; Старобога-
тов, Затравкин, 1987; Анистратенко, Стадниченко, 1995; Beriozkina et al., 1995;
Старобогатов и др., 2004; Gloer, 2002; Кантор, Сысоев, 2005). Во второй половине
XX века в Северную Америку была занесена Bithynia tentaculata (Linnaeus, 1758)
(Jaeckel, 1962). Моллюски населяют проточные и непересыхающие стоячие водо-
ёмы, лиманы (Анистратенко, Стадниченко, 1994[ 1995]). Обитают на разных грун-
тах от илистых осадков до каменистых и щебнистых перекатов; нередко встре-
чаются на водной растительности. Живут около 2 лет, выдерживают значительно
осолонение — до 17%о. Начало размножения приурочено к весеннему периоду
(при температуре воды около 10°), вымётывание кладок прекращается к концу
сентября — началу октября (Анистратенко, Стадниченко, 1994 [1995]). Кладки
прикреплены к нижней поверхности водных растений и к камням. Выход молоди
приурочен к началу мая.
В составе рода выделяют от 1 до 4 видов (Beriozkina et al., 1995, Першко,
2006, 2010; Gloer, 2002). Анатомические и конхологические признаки этих видов
перекрываются. Поэтому до проведения ревизии рода Bithynia в определителе
принимается точка зрения, близкая к взглядам малакологов из Западной Евро-
пы, согласно которой в пределах центральных и северных областей Европей-
ской России обитает 1 вид — Bithynia tentaculata (Linnaeus, 1758).
Комментарии. В ревизии битиниид европейской России и Украины Г.В. Березки-
ной, О.В. Левиной, Я.И. Старобогатова (Beriozkina, Levina, Starobogatov, 1995) и в кни-
ге В.В. Анистратенко, А.П. Стадниченко (1994[ 1995]) в составе рода Bithynia приводятся
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
375
4 вида, отнесенные к двум подродам: Bithynia s. str. (В. tentaculata, В. curta, В. producta)
и Milletelona Beriozkina et Starobogatov, 1995 (B. decipiens). В качестве диагностических
признаков подродов указаны: форма тангент-линии раковины (прямая или изогнутая),
выпуклость оборотов (уплощенные или слегка выпуклые) и выраженность парието-па-
латального угла (резкий или округлый). Анализируя коллекционные материалы ЗИН РАН
(послужившие основой для вышеназванных работ), ни по одному из диагностических
признаков не удалось однозначно разделить виды битиний на два подрода. Кроме того,
не обнаружены какие-либо существенные отличия типового вида подрода Milletelona —
В. decipiens от В. tentaculata (ни в строении раковины и гениталий; ни по ключевым при-
знакам, предложенным в работах: Анистратенко, Стадниченко 1994[1995]; Beriozkina,
Levina, Starobogatov, 1995; Старобогатов, Прозорова, Богатов и др., 2004). Учитывая от-
сутствие типовых экземпляров и крайне неконкретное первоописание В. decipiens, валид-
ный статус этого вида и соответственно подрода Milletelona представляется сомнитель-
ным. К такому же заключению на основе изучения морфологических и кариологических
признаков украинских битиний приходит И.А. Першко (2006; 2010). В своих работах она
синонимизирует В. decipiens с В. curta и относит к роду Bithynia вид Digyrcidum bour-
guignati (Paladilhe, 1869), не считая Digyrcidum самостоятельным родом. Точка зрения
И.А. Першко в отношении рода Digyrcidum справедлива. Г.В. Березкина с соавторами
(Beriozkina, Levina, Starobogatov, 1995) восстановили род Digyrcidum Locard, 18821 (ранее
считавшийся синонимом Bithynia), отнеся «странных битиний» из Каневского водохрани-
лища (окр. Триполья, Украина, 1 экз.), дельты Волги (Россия, 4 экз.) и Голландии (1 экз.)
к типовому виду этого рода — Bithynia bourguignati Paladilhe, 1869, описанному по 1 су-
хому экземпляру из окрестностей Перпиньяна (Франция). Основаниями для этого послу-
жили: наличие крышечки со спиральным ядром и совпадение (при сравнении компаратор-
ным методом) контура завитка «странных битиний» с таковым на рисунке В. bourguignati,
данном А. Паладиле (A. Paladilhe). К сожалению, Г.В. Берёзкина с соавторами (Beriozkina,
Levina, Starobogatov, 1995) не учли, что раковина голотипа В. bourguignati (согласно пер-
воописанию) при пяти оборотах имеет высоту не более 5 мм, а спиральное ядро её кры-
шечки крупное (максимальный диаметр более 1 мм) и образовано двумя оборотами. Тогда
как все экземпляры, изученные Г.В. Берёзкиной при подготовке ревизии, имеют при том
же количестве оборотов высоту раковины не менее 9 мм, а спиральные ядра их крышечек
существенно меньше (максимальный диаметр менее 0,9 мм) и образованны всего одним
оборотом (рис. 36: 3, 6 б, в). Кроме того, контуры завитка Digyrcidum bourguignati sensu
Beriozkina et al. (1995) не соответствуют таковым на рисунке A. Paladilhe (1869) (рис. 36).
Всё это не позволяет отнести раковины, изученные Г.В. Берёзкиной и др. (Beriozkina,
Levina, Starobogatov, 1995) к В. bourguignati и таким образом, следует признать произве-
дённое ими восстановление валидного статуса рода Digyrcidum неправомерным. Весьма
вероятно, что А. Паладиле (A. Paladilhe) описал таксон -bourguignati, посчитав видоспе-
цифичными признаки ювенильной раковины какой-то уже известной битинии (вероятно
В. tentaculata). Об этом косвенно свидетельствует тот факт, что за 125 лет прошедших с
момента описания -bourguignati и до выхода работы Г.В. Березкиной и др. (Beriozkina,
Levina, Starobogatov, 1995) других находок моллюсков этого вида известно не было. Све-
дения об обнаружении D. bourguignati на территории России и Украины, опубликован-
1 Несколько ранее В.В. Анистратенко и А.П. Стадниченко (1994[1995]) указали род Digirci-
dum как валидный со сноской: «Диагноз рода составлен Г.В. Берёзкиной и Я.И. Старобогато-
вым».
376
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Рис. 36. Различные представления о виде Bithynia bourguignati Paladilhe, 1869.
1 — В. bourguignati по: Paladilhe, 1869; 2 — В. bourguignati рисунок A. Paladilhe по:
Beriozkina, Levina, Starobogatov, 1995; 3 — Digyrcidum bourguignati no: Beriozkina, Levina,
Starobogatov, 1995; 4 — D. bourguignati по: Анистратенко, Стадниченко, 1994[1995]; 5 —
D. bourguignati по: Лазуткина, Андреева, Андреев, 2014; 6 — сравнение спиральных ядер
крышечек: а — по: Paladilhe, 1869; б — по: Beriozkina, Levina, Starobogatov, 1995; в — по:
Лазуткина, Андреева, Андреев, 2014. Масштаб: 1, 2, 3 — 3 мм; 4, 5, 6 — 1 мм.
ные в конце XX - начале XXI века (Beriozkina, Levina, Starobogatov, 1995; Анистратенко,
А.П. Стадниченко, 1994[1995]; Анистратенко, 1998; Градовский, 2000; Лазуткина, Андре-
ева, Андреев, 2011; 2014; Андреев, Андреева, 2014 и нек. др.), скорее всего, относятся к
какому-то другому виду рода Bithynia.
Согласно данным И.А. Першко (2006, 2010), корректирующим взгляды Г.В. Березки-
ной др. (Beriozkina, Levina, Starobogatov, 1995) и В.В. Анистратенко, А.П. Стадниченко
(1994[1995]), на территории европейской России и на Украине обитают 4 вида битиний:
В. tentaculata, В. curta (= В. decipiens), В. producta и В. bourguignati. Однако сравнительное
изучение раковин этих видов показало, что значения их линейных параметров (высота ра-
ковины ВР, ширина раковины ШР, высота завитка ВЗ, высота устья ВУ, высота последнего
оборота ВПОб, ширина последнего оборота без устья ШПОбУ) и морфометрических ин-
дексов (ШР/ВР, ВЗ/ВУ, ВПОб/ВР, ШПОбУ/ВПОб, ВЗ/ВПОб), а также признаки, предло-
женные в определительных ключах (Анистратенко, Стадниченко, 1994[1995]; Beriozkina,
Levina, Starobogatov, 1995; Старобогатов, Прозорова, Богатов и др., 2004), — перекрыва-
ются. Результаты сравнительного анализа пропорций копулятивного аппарата (линейные
промеры: длина пениса, ширина пениса, длина пениального аппендикса, ширина пени-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
377
ального аппендикса)1 и кариологического анализа (Першко, 2006; 2010) демонстрируют
похожую картину. Хотя обнаруженные ею на фоне общего кариотипического сходства
различия морфологии отдельных хромосом у разных видов украинских битиний небезын-
тересны, сами по себе они недостаточны для обоснования самостоятельности видов.
Учитывая изложенное выше, до проведения более подробной ревизии рода Bithynia в
определителе принимается точка зрения, близкая к взглядам малакологов западноевропей-
ской школы. Согласно ей в пределах центральных и северных областей европейской Рос-
сии обитает 1 вид — В. tentaculata.
Bithynia tentaculata (Linnaeus, 1758) (вкл. табл. IV, рис. 49)
— tentaculata Linnaeus, 1758: 774 {Helix).
[= Bithynia curta (Gamier in Picard, 1840); Bithiynia codia Bourgignat, 1862; Bithynia
decipiens (Millet, 1843); Bithynia producta Moquin-Tandon, 1855; Paludina impura
var. producta Menke, 1830, nom. nud.; Bithynia bourguignati Paladilhe, 1869; Bithynia
potamica Locard, 1893; Bithynia tentaculata f. producta Menke, 1828].
Признаки раковины такие же, как у рода, однако линейные размеры могут сильно
варьировать.
Размеры: ВР 6,0-14,8 мм, ШР 5,0-8,5 мм.
Распространение и экология такое же, как у рода.
Род Opisthorchophorus Beriozkina, Levina et Starobogatov, 1995
Типовой вид: Paludina troscheli Paasch, 1842.
Раковина коническая с вытянутым в разной степени завитком, серовато-, жел-
товато- или зеленовато-роговая, умеренно твёрдостенная, просвечивающая (ча-
сто покрытая плотным слоем илистого налёта). Обороты (до 6) сильно выпуклые
I иногда ступенчатые), разделённые глубоким швом. Поверхность оборотов тонко
морщинистая, как правило, с разрозненными резкими линиями нарастания. Ча-
сто присутствуют тонкие спиральные струйчатые линии, лучше выраженные на
поверхности последнего оборота и хорошо заметные при увеличении х25 и бо-
лее. Высота завитка раковин взрослых особей всегда больше высоты устья. Устье
овальное (яйцевидное, суженное в парието-палатальной области). Иногда наблю-
дается плавный тупой парието-палатальный угол. Устьевой контур замкнутый,
иногда усиленный размытым, светлым наплывом наподобие губы. Крышечка тон-
кая, концентрическая, ровная, если в её центре имеется вдавление, то оно слабое,
едва заметное. Пупок широко-щелевидный, иногда каплевидный. Радула типич-
ная для семейства. Пениальный аппендикс ответвляется от проксимальной трети
пениса. На конце аппендикса папилла отсутствует (Falniowski, 1990).
Как показывает практика анатомирования моллюсков, пропорции частей копулятивных
аппаратов подвержены значительной индивидуальной изменчивости и малоприменимы в си-
стематике. При жизни моллюска они сильно варьируют в зависимости от возраста и физио-
логического состояния животного (стадии репродуктивного цикла), от наличия патологий. При
сиксации они непредсказуемо изменяются в результате неравномерного сокращения (сжатия)
типичных участков половых протоков. Кроме того, следует учитывать уменьшение объема и
деформацию мягких тканей моллюсков при их длительном (несколько десятков лет и более)
оанении в спирте.
378
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
В ископаемом состоянии представители рода в Европе известны, по меньшей
мере, с голоцена. Современные моллюски рода распространены в Восточной
Европе и Западной Сибири; населяют сильно заросшие озёра, слабопроточные
речные и временные (иногда пересыхающие и промерзающие) водоёмы. Предпо-
читают песчано-илистые грунты. Неблагоприятные периоды пережидают зарыв-
шись в грунт.
Комментарии. ГВ. Берёзкиной и Я.И. Старобогатовым в 1995 году на основе исклю-
чительно конхологических признаков были описаны два новых для науки рода битини-
ид: Opisthorchophorus и Paraelona, включавшие по пять видов каждый (Beriozkina, Levi-
na, Starobogatov, 1995). Изучение экземпляров, отнесённых этими авторами (Beriozkina,
Levina, Starobogatov, 1995), а также В.В. Анистраенко (Анистратенко, Стадниченко, 1994
[1995]) к данным родам показало, что значения диагностических признаков перекрыва-
ются у всех 10 видов, несомненно, относящихся к одному роду, отличающемуся от родов
Codiella и Bithynia. В качестве приоритетного для этого рода мы оставляем имя, начина-
ющееся на букву, более близкую к началу алфавита — Opisthorchophorus. Проведённый
анализ раковин также показал, что в пределах европейской части России к роду Opisthor-
chophorus следует отнести всего один вид — О. troschelii, обладающий весьма вариабель-
ной по размерам раковиной. Виды, отнесённые к роду Paraelona, представляют собой
причудливую смесь моллюсков О. troschelii и С. leachii.
Opisthorchophorus troschelii (Paasch, 1842) (вкл. табл. IV, рис. 51).
— troscheli Paasch, 1842: 300-301, textfigs а-d (Paludind).
[= Bithynia similis Stein, 1850; Bithynia leachii (Sheppard, 1823) Lehmann, 1873 partim;
Bithynia goldfussi Kobelt, 1891; Bithynia limosus troscheli (Paasch, 1842) S.H. Jaeckel,
1961; Bithynia transsilvanica Reischiitz, 1998; Bithynia leachi var. inflata Hansen,
1845; Bithynia leachi var. majewskyi Frauenfeld, 1862; Opisthorchophorus baudonianus
(Gassier, 1859) Beriozkina, Levina, Starobogatov, 1995; Opisthorchophorus hisphanicus
(Servain, 1880) Beriozkina, Levina, Starobogatov, 1995; Opisthorchophorus valvatoides
Beriozkina et Starobogatov, 1995; Paraelona majewsky (Frauenfeld, 1862) Beriozkina,
Levina, Starobogatov, 1995; ?Paraelona fausseki Beriozkina et Starobogatov, 1995;
Paraelona boissieri (Kiister, 1852) Beriozkina, Levina, Starobogatov, 1995; Paraelona
socialis (Westerlund, 1886) Beriozkina, Levina, Starobogatov, 1995 partim; Paraelona
milachevitchi Beriozkina et Starobogatov, 1995 partim]
Признаки раковины такие же, как у рода, однако линейные размеры могут сильно
варьировать.
Размеры: ВР 6,2-13,6 мм, ШР 4,5-8,8 мм.
Распространение и экология такое же, как у рода.
Семейство HYDROBIIDAE Stimpson, 1865
Типовой род: Hydrobia Hartmann, 1821.
Раковина в большинстве случаев гладкая, реже ребристая или исчерченная,
маленькая (при 3-8 оборотах её высота не более 6,5 мм, ширина до 2,7 мм), раз-
личной формы от низкоконической, овальной и овально-конической до башне-
видной и кубаревидной. Устье овальное, иногда заострённое в верхней части,
устьевой край сомкнутый, иногда утолщённый. Крышечка роговая, спиральная.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
379
Пупок выражен в разной степени. Передняя часть головы вытянута наподобие
хобота или рыла. Щупальца нитевидные. Глаза располагаются у основания щу-
палец. Нога широкая, сзади может быть округлая или заострённая. Рахидальный
зуб радулы с крупными зубцами по режущему краю и 1^4 дополнительными зуб-
чиками по бокам основания. Животные раздельнополые, яйцекладущие или яйце-
живородящие.
Семейство известно с олигоцена Европы (Старобогатов, 1970). Современные
представители населяют моря и эстуарии у побережий Европы, а также европей-
ские пресные и солоноватые быстротекущие водоёмы. Систематика и объём се-
мейства являются предметом дискуссий. Согласно более или менее современным
данным, в составе семейства выделяют не менее пяти подсемейств и около полу-
сотни родов (Wilke et al., 2001; Bouchet et al., 2005).
В пресных водоёмах европейской части России встречаются представители
двух родов: Potamopyrgus Stimpson 1865 и Ventrosia Radoman, 1977.
Ключ для определения родов и видов семейства Hydrobiidae
1(2). Раковина коническая или высококоническая, обороты сильно выпуклые,
разделённые глубоким швом. Вершина парието-палатального угла (угло-
ватости в верхней части устья) резко выражена.................................
...................................Род Ventrosia Radoman, 1977 (рис. 37);
Ventrosia stagnorum (Gmelin, 1791) (вкл. табл. IV, рис. 53).
— stagnorum Gmelin, 1791: 3653 (Helix).
[= Hydrobia staglnalis Van Benthem-Jutting, 1933].
Размеры: BP до 6,3 мм, ШР до 2,9 мм. JAk
Вид распространён на атлантическом побережье Европы, в Среди-
земноморье и опреснённых участках Балтийского моря. Моллюски Л
обитают в солоноватых водах (2-8%о) среди водной растительности. /
2(1). Раковина высоко коническая, обороты умеренно или слабо eL
выпуклые, шов не глубокий. Вершина парието-палатального Ш
угла (угловатости в верхней части устья) плавно скруглена ....
.....Род Potamopyrgus Stimpson 1865 (рис. 38); Potamopyrgus рис 37
antipodarum (Gray, 1843) (вкл. табл. IV, рис. 52).
— antipodarum Gray J.E., 1843: 241 (Amnicold). А
[= Hydrobia ventrosa var. carinatus Marshall, 1889; Hydrobiajenkinsi
Smith, 1889].
Размеры: BP до 6 мм, ШР до 3 мм. J
Новозеландский вид, непреднамеренно завезённый в Европу в пер- [ ]
вой половине XIX века. В настоящее время широко распространён V ~~ j
в пределах европейской части материка. На севере и северо-западе t
европейской части России известен в солоноватых участках Балтий- \ 1
ского моря. Моллюски обитают в пресной, солоноватой и солёной /
воде (в диапазоне солёности 0-17 %о), как правило, держатся на по-
верхности и в толще грунта, реже среди растительности. Рис. 38.
380
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Семейство AMNICOLIDAE Tryon, 1863
Типовой род: Amnicola Gould et Haldeman, 1840.
Раковина маленькая (высота до 4 мм, ширина до 2,5 мм) овально-коническая,
яйцевидная (овально-башневидная) или овальная. Если яйцевидная, то очень
маленькая (высота до 3 мм, ширина до 1,7 мм), умеренно твёрдостенная или
хрупкая, тонкостенная, почти прозрачная; беловатая, сероватая, желтовато- или
зеленовато-роговая. Обороты (около 5) выпуклые, разделены глубоким швом. По-
верхность оборотов очень тонко радиально морщинистая. Пупок щелевидный,
иногда в виде очень узкой щели или практически закрытый. Устье яйцевидно-
овальное, практически не вырезанное стенкой последнего оборота, иногда с лёг-
кой угловатостью в суженной парието-палатальной области. Крышечка роговая,
спиральная. Нога спереди угловато заострённая, сзади широко закруглённая. Щу-
пальца тонкие, нитевидные. Рахидальный зуб радулы широкий, с одним или не-
сколькими боковыми и двумя базальными зубцами. Его режущий край с крупным
зубцом посередине и мелкими зубчиками по бокам. Остальные зубы неправиль-
но-ромбические с многочисленными зубчиками. Раздельнополые животные. Ко-
пулятивный орган самцов с крупным железистым придатком.
Семейство известно с карбона или юры. В его составе насчитывается око-
ло 200 видов, относящихся к 17 рецентным и ископаемым родам, сгруппиро-
ванным в 3 подсемейства (Amnicolinae Tryon, 1863; Baicaliinae Р. Fischer, 1885;
Emmericiinae Brusina, 1870). Большинство видов распространены в Северной
Америке. В Европе обитают представители трёх родов. На севере и северо-запа-
де европейской части России представлен один род — Marstoniopsis Van Regteren
Altena, 1936, относящийся к подсемейству Amnicolinae.
Род Marstoniopsis Regteren Altena, 1936
Типовой вид: Hydrobia steinii Martens, 1858.
Раковина овально-коническая или башневидная, тонкостенная, слабо блестя-
щая или матовая, с притуплённой вершиной. Обороты сильно выпуклые, разделе-
ны глубоким швом. Устье овальное или яйцевидное, с сомкнутым краем. Пупок в
виде широкой щели. У представителей рода выражен половой диморфизм. Сам-
цы имеют более стройную раковину по сравнению с самками. Рахидальный зуб
радулы имеет пару базальных зубцов с каждой стороны.
Представители рода распространены в Европе. По данным разных специали-
стов в составе рода насчитывается от одного до трёх видов (Анистратенко, 1998;
Gloer, 2002; Старобогатов и др., 2004). На севере и северо-западе европейской ча-
сти России обитает один вид.
Marstoniopsis scholtzi (A. Schmidt, 1856) (вкл. табл. V, рис. 54).
— scholtzi A. Schmidt, 1856: 158 (Hydrobia).
[= Hydrobia steinii Martens, 1858].
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
381
Раковина овально-коническая, тонко и неравномерно исчерченная, тонкостенная,
просвечивающая, с притуплённой верхушкой. Обороты (до 4,5) сильно выпуклые,
разделённые глубоким швом. Часто поверхность раковины может быть покрыта
илистым налётом, маскирующим толщину стенок и скульптуру.
Размеры: ВР до 3 мм, ШР до 2 мм.
Вид распространён в бассейне Балтийского моря и в верхней части волжского бас-
сейна. Моллюски обитают в реках (на участках, характеризующихся медленным
течением) и в озёрах.
Комментарии. В конце XX - начале XXI века в литературных источниках на
русском языке для севера и северо-запада европейской части России приводилось
два вида: М. scholtzi и Marstoniopsis steinii (Martens, 1858). При этом в качестве
основного межвидового различия утверждалась разная высота завитка относи-
тельно высоты устья. Изучение коллекционных материалов, собранных в раз-
личных районах Европы и хранящихся в Зоологическом институте РАН, показало
непригодность этого признака для разграничения видов. Поэтому в определителе
принимается точка зрения близкая к взглядам западноевропейских малакологов,
согласно которой М. steinii является синонимом М. scholtzi.
Семейство LITHOGLYPHIDAE Troschel, 1857
Типовой вид: Lithoglyphus Menke, 1830.
Раковина кубаревидная, толстостенная, с гладкой, фарфоровидной поверхно-
стью или с очень тонкой радиальной исчерченностью оборотов. Последний обо-
рот сильно вздут и составляет большую часть корпуса раковины. Устье капле-
видное или округло-треугольное. Утолщённые и отвёрнутые колумеллярный и
париетальный края устья образуют колумеллярную (устьевую) площадку (более
узкую по сравнению моллюсками семейства Neritidae). Пупок полностью закрыт
отворотом колумеллярного края устья или в виде очень узкой щели. Крышечка
роговая, спиральная. Рахидальный зуб радулы с крупным базальным зубцом. Жи-
вотные раздельнополые, развитие прямое, протекает в яйцевых капсулах.
Семейство объединяет около 20 рецентных и два ископаемых рода (Анистра-
тенко, 1998). На севере и северо-западе европейской части России обитают мол-
люски одного рода — Lithoglyphus Menke, 1830.
Род Lithoglyphus Megerle von Muhfeld in Hartmann, 1821
Типовой вид: Lithoglyphus eburneus Megerle von Muhfeld in Hartmann, 1821
(nom. nud. = Paludina naticoides Ferussac in C. Pfeiffer, 1828).
Раковина кубаревидная, твёрдостенная, поверхность оборотов гладкая матовая
или очень тонко неравномерно радиально исчерченная. Обороты мало выпуклые,
разделённые слабо углублённым швом. Крышечка с двумя или тремя оборотами
спирали. Рахидальный зуб радулы с крупным и острым центральным зубцом и
несколькими мелкими боковыми. Кроме того по бокам основания имеются два
ряда мелких зубчиков. Латеральные и маргинальные зубы длинные, дуговидно
изогнутые с мелкими зубчиками (числом от 6 до 12).
382
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Род насчитывает до 6 рецентных видов (Анистратенко, 1998). На севере и се-
веро-западе европейской части России обитает один вид.
Lithoglyphus naticoides (С. Pfeiffer, 1828) (вкл. табл. V, рис. 55 а, б).
— naticoides С. Pfeiffer, 1828: 45-46, pl. 8. fig. 1, 2, 4 (Paludina).
Размеры: BP до 9 мм, ШР до 7 мм.
Вид распространён в бассейне Балтийского моря и в верхней части волжского бас-
сейна. Моллюски обитают в реках (на участках, характеризующихся медленным
течением) и в озёрах.
Семейство VALVATIDAE Gray, 1840
Типовой род: Valvata O.F. Muller, 1774.
Раковина от кубаревидной или низкоконической (в различной степени прижа-
той) до дисковидной (плоско- или турбоспиральной); с выраженным округлым (в
разной степени перспективным) пупком, иногда частично прикрытым приустье-
вой частью последнего оборота. Обороты (3-5) гладкие, либо со спиральными
линиями, рёбрами или радиальными рёбрышками. Устье округлое, с цельным
краем; крышечка тонкая, роговая, с центральным ядром и относительно узкими
оборотами спирали. Передний край ноги широкий с загнутыми углами, щупальца
длинные, относительно сближенные, глаза расположены позади щупалец. Ман-
тия справа несёт нитевидный придаток, а слева вытянута в перистую жабру. Име-
ется парная челюсть, радула короткая. В поперечном ряду радулы 7 зубов. Фор-
мула поперечного полуряда радулы R-I-2M (Анистратенко, Анистратенко, 2001).
Рахидальный зуб (R) спереди суженный, сзади расширенный с треугольным зазу-
бренным режущим краем. Остальные зубы могут быть зазубренные или гладкие.
Гермафродиты. Женский половой проток расширяется в мешкообразную камеру
оплодотворения (бурсу) в которую впадают железы (белковая, оотекальная), да-
лее следует относительно короткая вагина, открывающаяся наружу женским по-
ловым отверстием. Мужской половой проток направляется к пенису, удалённому
от основания правого щупальца и расположенному на спинной стороне. Простата
прилегает к проксимальному участку белковой железы.
Вальватиды населяют пресные водоёмы Северного полушария. Семейство
подразделяется на 2 подсемейства и 6 родов (Ситникова, 1983; Ситникова и др.,
1986). В европейской части России обитают представители 3 родов из 2 подсе-
мейств.
Ключ для определения подсемейств и родов
1(4). Поверхность раковины с выраженной скульптурой в виде радиальных
морщинок или рёбрышек иногда с размытыми спиральными рёбрами или
исчерченностью. Обороты сильновыпуклые, разделены глубоким швом.
Последний оборот около устья менее чем в 2 раза шире предпоследнего.
Устье умеренно скошенное, округлое или в верхней части со слабым па-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
383
рието-палатальным углом. Пупок округлый, перспективный, открытый
или менее чем на ’/з прикрыт последним оборотом....................
......................................Подсем. Valvatinae Gray, 1840 (с. 383).
2(3). Обороты раковины в сечении круглые, разделённые глубоким швом. Устье
круглое или слегка овальное, но всегда без резкого уголка или более или
менее плавной угловатости в верхней части. Ширина раковины у боль-
шинства видов (искл. V pusilia и V. andreaei) при 3^4- оборотах не превы-
шает 4 мм. Если раковина плоскоспиральная, то имеющиеся радиальные
рёбрышки не несут по кромке пластинчатых гребней, сформированных за
счёт периостракума..........................Род Valvata O.F. Miiller, 1774 (с. 384).
3(2). Обороты раковины в сечении овальные, разделённые умеренно глубоким
швом. Устье округлое, неправильно-округлое, реже овальное, но всегда с
резким уголком или более или менее плавной угловатостью в верхней ча-
сти. Ширина раковины у большинства видов (искл. С. studeri и С. discors)
при 3-4 оборотах превышает 4 мм. Если раковина плоскоспиральная, то
её поверхность покрыта радиальными рёбрышками, несущими по кромке
пластинчатые гребни, сформированные за счёт периостракума................
..........................................Род Cincinna Hubner, 1810 (с. 386).
4(1). Поверхность раковины блестящая, очень тонко нитевидно радиально ис-
черчена, иногда скульптура выражена настолько слабо, что поверхность
выглядит матовой, фарфоровидной. Спиральная скульптура отсутствует.
Обороты умеренно выпуклые, шов умеренно глубокий. Последний оборот
сильно вздут (в приустьевой области более чем в 2 раза шире предпо-
следнего). Устье сильно скошено, у раковин с возвышающимся завитком
с отчётливым (парието-палатальным) углом в верхней части. Пупок на Уз
прикрыт приустьевой частью последнего оборота............................
................Подсем. Borystheniinae Starobogatov in Sitnikova, 1983;
род Borysthenia Lindholm, 1914 (c. 390).
Подсемейство Valvatinae Gray, 1840
Типовой род: Valvata O.F. Muller, 1774.
Форма раковины от кубаревидной и в различной степени прижатой низкоко-
нической до дисковидной (плоско- или турбоспиральной). Обороты сильновы-
пуклые с радиальными морщинами или рёбрышками, иногда с размытыми спи-
ральными рёбрами или исчерченостью; в сечении правильно круглые, близ устья
может присутствовать лёгкая угловатость. Половая система типичного для семей-
ства строения. Яйцеживородящие животные, продуцируют кладки, из которых
выходят вполне сформировавшиеся особи.
Моллюски подсемейства известны с юрского периода, ныне распространены
в Евразии, Северной Африке и Северной Америке. В европейской части России
обитают представители 2 родов.
384
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Род Valvata O.F. Muller, 1774
Типовой вид: Valvata cristata O.F. Muller, 1774.
Проток, соединяющий яйцевод с камерой оплодотворения (бурсой) длинный,
полость камеры оплодотворения обширная, оотекальная железа прилегает к бур-
се с дорзальной стороны, белковая железа в виде короткой массивной петли; про-
стата относительно небольшая, прилегает к белковой железе, незначительно вы-
ступая за её дистальную часть.
В европейской части России, вероятно, обитают 6 видов 2 подродов.
Комментарии. Представления об объёме рода сильно варьируют в отечественных и
зарубежных малакологических сводках. В настоящий определитель включены виды, само-
стоятельность которых и их обитание в пределах европейской части России не вызывает
сомнений.
Ключ для определения подродов и видов рода Valvata
1(4). Раковина дисковидная, завитая практически в одной плоскости, как пра-
вило, с плоским не выступающим завитком или иногда с очень слабо воз-
вышающимся предпоследним оборотом. Пупок в виде широкой воронки,
перспективный...........................................Подрод Valvata s. str.
2(3). Раковина относительно тонкостенная, просвечивающая, блестящая; обо-
роты (3-4) покрыты густыми радиальными морщинками (отчётливо замет-
ными при увеличении *25 и более). При трёх оборотах ширина раковины
2,5-3,0 мм. Отношение ширины трубки последнего оборота в области устья
к суммарной ширине предшествующих оборотов (с верхней стороны) 0,7
и более. Пупок в виде широкой углублённой (за счёт вздутости последнего
оборота) воронки... Valvata cristata O.F. Muller, 1774 (вкл. табл. V, рис. 56).
— cristata O.F. Muller, 1774: 198.
Размеры: BP до 1,2 мм, ШР до 3,0 мм.
Вид распространён в Европе и Западной Сибири. Обитает на песчаных грунтах
мелководий рек и водохранилищ, а также в мелких заросших водоёмах.
3(2). Раковина меньших размеров (её ширина при трёх оборотах 2,2-2,3 мм),
относительно твёрдостенная, чаще всего непросвечивающая. Обороты
(3,0-3,5) тонко радиально исчерченные. При трёх оборотах отношение
ширины трубки последнего оборота в области устья к суммарной ширине
предшествующих оборотов (с верхней стороны), как правило, не превы-
шает 0,7. Пупок в виде расширенной неуглублённой воронки....................
..............Valvata planorbulina Paladilhe, 1867 (вкл. табл. V, рис. 57).
— planorbulina Paladilhe, 1867: 50, pl. 21, fig. 23-26.
Размеры: BP 1,0-1,2 мм, ШР до 2,4 мм.
Вид распространён в Европе; в России в бассейне Балтийского моря. Обитает на
погружённой растительности рек и водохранилищ.
4(1). Раковина с явственно возвышающимся завитком, в различной степени
прижатая низкоконическая или кубаревидная. Пупок округлый, проколо-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
385
видный, в различной степени перспективный, слегка прикрыт стенкой
последнего оборота......................................................
.............................Подрод Microcincinna Starobogatov in Sitnikova, 1983
5(6). Раковина низкоконическая, при 3,5 оборотах высота завитка меньше или
равна высоте устья.....Valvata elatior Menzel, 1904 (вкл. табл. V, рис. 58).
— latior Menzel, 1904: 77 (Valvata andreana var.).
Раковина тонкостенная, роговая, просвечивающая, сильно прижатая, обороты
(3,0-4,0) с радиальными рёбрышками. Возвышение верхних оборотов над по-
следним оборотом может быть небольшим (около 0,2 мм). Пупок перспективный,
слегка прикрыт стенкой последнего оборота.
Размеры: ВР до 3,0 мм, ШР до 4,0 мм.
Вид распространён в бассейне Балтийского моря, обитает в озёрах.
6(5). При 3,5 и более оборотах высота завитка больше высоты устья, если равна
ей, то раковина кубаревидная.
"(8). Ширина третьего оборота от устья составляет больше половины ширины
предпоследнего оборота, вершина раковины уплощённая............................
.......................Valvata geyeri Menzel, 1904 (вкл. табл. V, рис. 59).
— geyeri Menzel, 1904: 78.
Эмбриональные обороты практически невыступающие, поэтому контур раковины
при взгляде со стороны устья имеет форму усечённого конуса или высокой трапе-
ции. Обороты с сильно сглаженными радиальными морщинками, иногда практи-
чески гладкие; пупок уже, чем у предыдущего вида (диаметр ~ 0.4 мм).
Размеры: ВР до 3,1 мм, ШР до 2,7 мм.
Вид распространён в бассейне Балтийского моря и Среднем Днепре. Обитает сре-
ди водорослей в озёрах с прозрачной водой.
N 7). Ширина третьего оборота от устья меньше или равна половине ширины
предпоследнего оборота, вершина раковины более или менее заострённая.
10). Раковина кубаревидная. Обороты с тонкой радиальной ребристостью. При
3,5-4,0 оборотах высота завитка не превышает 1,5 мм..........................
.................Valvata pusilia (O.F. Muller, 1774) (вкл. табл. V, рис. 60).
— pusilia O.F. Muller, 1774: 171 (Nerita).
Раковина умеренно толстостенная. Высота раковины почти равна, либо равна её
ширине (отношение ВР к ШР 0,9-1,0).
Размеры: ВР до 4,8 мм, ШР до 5,0 мм.
Вид распространён в бассейнах Балтийского и Северного морей, в Западном Буге.
Обитает в озёрах с чистой водой.
.0(9). Раковина низкоконическая, обороты с тонкой радиальной исчерченно-
стью, при 3,0-4,0 оборотах высота завитка 1,6 мм и более.......................
.....................Valvata andreaei Menzel, 1904 (вкл. табл. V, рис. 61).
— andreaei Menzel, 1904: 77 (Valvata).
Обороты круглые в сечении, но устье иногда с очень лёгкой угловатостью в верх-
ней части. Высота раковины чуть больше её ширины (отношение ВР к ШР, как
правило 1,07-1,12).
Размеры: ВР до 3,0-4,5 мм, ШР до 4,0 мм.
386
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Вид распространён в бассейне Балтийского моря. Обитает среди водорослей в
озёрах с прозрачной водой.
Род Cincinna Hubner, 1810
Типовой вид: Nerita piscinalis O.F. Muller, 1774.
Проток, соединяющий ренальный яйцевод с камерой оплодотворения (бурсой)
короткий, полость камеры обширная, но заметно меньше, чем у представителей
рода Valvata. Оотекальная железа прилегает к бурсе с дорзальной стороны, бел-
ковая железа в виде очень длинной узкой петли; простата клиновидная, в боль-
шинстве случаев вытянутая и значительно выступает за петлю белковой железы,
иногда относительно короткая (Ситникова, 1983).
В европейской части России, вероятно, обитают 11 видов.
Комментарии. Представления об объёме рода сильно варьируют в отечественных и
зарубежных малакологических сводках. В настоящий определитель включены виды, само-
стоятельность которых и их обитание в пределах европейской части России не вызывает
сомнений.
Ключ для определения видов рода Cincinna
1(4). Раковина в общем дисковидная (но не плоскоспиральная), над последним
оборотом слегка возвышается один оборот, второй, если и возвышается,
то едва заметно. Высота завитка меньше 1 мм. Пупок широкий, перспек-
тивный (ширина пупка не меньше 0,3 ширины раковины), сквозь него
просматриваются все обороты.
2(3). Отношение высоты раковины (ВР) к её ширине (ШР) не превышает 0,6;
отношение высоты завитка (ВЗ) к высоте устья (ВУ) не более 0,36.............
............Cincinna macrostoma Morch, 1864 (вкл. табл. V, рис. 62 а, б).
— macrostoma Morch, 1864: 59 (Valvata).
[= Valvata umbilicata Fitzinger, 1833].
Отношение ширины внутренних оборотов к ширине трубки последнего оборота
возле устья (индекс внутренних оборотов) с нижней стороны раковины не менее
0,63. Обороты (3,0-4,0) густо покрыты тонкими рёбрышками, окраска раковины
зеленовато- или коричневато-роговая.
Размеры: ВР 2,2-3,0 мм, ШР 3,5-5,0 мм.
Вид распространён в Европе. Предпочитает мелководные пересыхающие водоёмы
с илистым дном.
3(2). Отношение высоты раковины (ВР) к её ширине (ШР) не менее 0,61; от-
ношение высоты завитка (ВЗ) к высоте устья (ВУ) не менее 0,37...............
...............Cincinna frigida (Westerlund, 1873) (вкл. табл. V, рис. 63).
—frigida Westerlund, 1873: 436 (Valvata).
Отношение ширины внутренних оборотов к ширине трубки последнего оборота
возле устья (индекс внутренних оборотов) с нижней стороны раковины не ме-
нее 0,5. Обороты раковины с радиальными рёбрышками, несущими по кромке
пластинчатые гребни, сформированные за счёт периостракума. При 3 оборотах
ширина раковины не больше 2,7 мм, а при 3,5 — всегда меньше 4 мм.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
387
Вид распространён в Северной Европе и Сибири (до бассейна Охотского моря).
Обитает в озёрах лесо-таёжной зоны.
Комментарии. По общему облику и размерам вид сходен с предыдущим, однако
хорошо отличается пропорциями раковины и наличием периостракальных греб-
ней по кромке рёбрышек на поверхности оборотов.
4(1). Раковина кубаревидная или низкоконическая в различной степени при-
жатая. В любом случае над последним оборотом явственно возвышаются
два предшествующих. Высота завитка больше 1 мм. Пупок уже, чем у
предыдущих видов (ширина пупка не превышает 0,25 ширины раковины).
В редких случаях, когда над последним оборотом возвышается только
один оборот — высота и ширины раковины не менее 5 мм.
5(8). Высота завитка при 3,5-4,0 оборотах не превышает 1,29 мм. Пупок окру-
глый, полностью открытый.
6(7). При 3,5—4,0 оборотах с верхней стороны раковины отношение ширины
внутренних оборотов к ширине трубки последнего оборота возле устья
(индекс внутренних оборотов) не меньше 1,9. Отношение высоты ракови-
ны к её ширине не меньше 0,8...................................................
...........Cincinna studeri Boeters et Falkner, 1998 (вкл. табл. V, рис. 64).
— studeri Boeters et Falkner, 1998: 113, Taf. 14-16.
[= Valvata pulchella Studer, 1820].
Раковина низкоконическая, коричневатая, блестящая; обороты (3-4) разделены
глубоким швом.
Размеры: ВР до 2,3 мм, ШР до 3.0 мм.
Вид распространён в Европе и Западной Сибири. Обитает в мелководных пере-
сыхающих водоёмах с илистым дном.
7(6). При 3,5^4,0 оборотах с верхней стороны раковины отношение ширины
внутренних оборотов к ширине трубки последнего оборота возле устья
(индекс внутренних оборотов) не более 1,88. Отношение высоты ракови-
ны к её ширине не превышает 0, 79..............................................
...............Cincinna depressa (С. Pfeiffer, 1821) (вкл. табл. V, рис. 65).
— depressa С. Pfeiffer, 1821: 100, Taf. 4, fig. 33.
Раковина низко коническая, желтовато-роговая; обороты (3,5-4,0) покрыты тонкой
радиальной исчерченностью или несут густо расположенные тонкие рёбрышки.
Размеры: ВР 2,8-4,5 мм, ШР до 4,0-5,5 мм.
Вид распространён в Европе и Западной Сибири (преимущественно в северной
части). Обитает в реках, озёрах, крупных прудах и водохранилищах.
8(5). Высота завитка при 3,5-4,0 оборотах составляет 1,30 мм и более. Пупок
округлый но, как правило, в той или иной степени прикрытый участком
стенки последнего оборота. Если открытый, то высота завитка меньше 2 мм.
9(14). Высота завитка не превышает 2 мм и она заметно меньше высоты устья
(отношение высоты завитка (ВЗ) к высоте устья (ВУ) 0,44-0,80). Отноше-
ние ширины внутренних оборотов к ширине трубки последнего оборота
возле устья (индекс внутренних оборотов) составляет 2,3 или меньше.
388
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
10(11). Раковина с относительно низким, прижатым завитком. Отношение высо-
ты завитка (ВЗ) к высоте устья (ВУ) не превышает 0,6. Высота верхних
оборотов, возвышающихся над последним оборотом не более 0,4 мм. От-
ношение ширины внутренних оборотов к ширине трубки последнего обо-
рота возле устья (индекс внутренних оборотов) 1,6-2,0.........................
..............Cincinna ambigua (Westerlund, 1873) (вкл. табл. V, рис. 66).
— ambigua Westerlund, 1873: 439 (Valvata).
Раковина зеленовато-роговая, тонкостенная, оборотов 3,5-4,5. Вершина настолько
прижата, что часто над последним возвышается практически только предпо-
следний оборот. Последний оборот сильно вздут, покрыт тонкими радиальными
рёбрышками. Устье яйцевидное с несколько суженной верхней частью.
Размеры: ВР до 4,2 мм, ШР до 4,7 мм.
Вид распространён в Европе и Западной Сибири. Обитает в реках и водохранили-
щах на плотных и слегка заиленных грунтах.
11(10). Раковина с более высоким, не сильно прижатым завитком. Отношение вы-
соты завитка (ВЗ) к высоте устья (ВУ) составляет 0,70-0,85. Высота верх-
них оборотов, возвышающихся над последним 0,45-0,67 мм. Отношение
ширины внутренних оборотов к ширине трубки последнего оборота возле
устья (индекс внутренних оборотов) 2,1-2,3.
12(13). Раковина тонкостенная, с выпуклыми оборотами, разделёнными глубоким
швом и покрытыми тонкой радиальной ребристостью. Отношение высоты
завитка к высоте устья 0,76-0,85. Отношение ширины внутренних оборо-
тов к ширине трубки последнего оборота возле устья (индекс внутренних
оборотов) не менее 2,25. Отношение ширины пупка к ширине раковины
0,16-0,17. Через пупок просматривается весь предпоследний оборот. Вы-
сота верхних оборотов, возвышающихся над последним не менее 0,6 мм
.......................Cincinna skorikovi (Lindholm, 1911) (вкл. табл. V, рис. 67).
— skorikovi Lindholm, 1911: 299, n. 22 (Valvata).
Раковина коричневато-роговая, обороты (3,5-4,5) выпуклые, расположены не-
сколько ступенчато. Устье яйцевидное с несколько угловатой верхней частью.
Размеры: ВР до 4,0 мм, ШР до 4,5 мм.
Вид распространён по всей Европе, в Причерноморье и в дельте Волги. Обитает в
реках и озёрах на илистых грунтах.
13(12). Раковина толстостенная, очень тонко радиально ребристая. Верхние обо-
роты (начиная с предпоследнего) мало выпуклые, разделённые мелким
швом. Завиток ниже по сравнению с предыдущим видом. Отношение вы-
соты завитка к высоте устья не более 0,75. Отношение ширины внутрен-
них оборотов к ширине трубки последнего оборота возле устья (индекс
внутренних оборотов) не более 2,2. Пупок, перспективный у молодых
особей, при достижении раковиной 3 оборотов (и более) он сужается. От-
ношение ширины пупка к ширине раковины у взрослых особей 0,12-0,15.
Через пупок просматривается меньше половины предпоследнего оборота
................Cincinna piscinalis (O.F. Muller, 1774) (вкл. табл. V, рис. 68 а, б).
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
389
— piscinalis O.F. Muller, 1774: 172 (Nerita).
Раковина зеленовато-роговая, образованная 4,0-4,5 оборотами, покрытыми очень
тонкой и густой ребристостью. Устье округлое или овальное с небольшим окру-
глым уголком в верхней части.
Размеры: ВР до 4,5 мм, ШР до 5,0 мм.
Вид распространён в Европе и Западной Сибири. Обитает в реках и озёрах.
14(9). Высота завитка больше, чем 2,1 мм, она примерно равна или заметно пре-
вышает высоту устья (отношение высоты завитка (ВЗ) к высоте устья (ВУ)
0,87-1,23). Отношение ширины внутренних оборотов к ширине трубки
последнего оборота возле устья (индекс внутренних оборотов) больше,
чем 2,36.
15(16). Раковина одна из наиболее крупных среди моллюсков рода. У взрослых
животных её высота может достигать 9 мм, ширина — 8 мм. Ширина ра-
ковины при трёх оборотах не меньше 4,6 мм..........................................
..........Cincinna lilljeborgi (Westerlund, 1897) (вкл. табл. V, рис. 69 а, б).
— lilljeborgi Westerlund, 1897а: 137 (Valvata).
Раковина башневидно-коническая, с притуплённой вершиной. Желтовато- или
зеленовато-роговая. Обороты (4,0-4,5) выпуклые, разделённые глубоким швом,
с тонкой густой радиальной ребристостью. Пупок относительно узкий (ширина
пупка 0,11-0,23 ширины раковины), проколовидный. Устье округлое, с выражен-
ным уголком в верхней части.
Размеры: ВР до 9,8 мм, ШР до 8,0 мм.
Вид распространён в северных районах Западной Европы, в России — в бассейне
Балтийского моря. Обитает в реках, проточных прудах, водохранилищах и озёрах;
предпочитает плотные или слабо заиленные грунты.
16(15). Раковина меньших размеров. Её ширина при трёх оборотах меньше 4,5 мм.
17(18). Последний оборот сильно вздут. Его высота вместе с устьем (измеренная
со стороны устья) составляет 0,86 и более высоты раковины..........................
Cincinna antiqua (Morris in Morris et Sowerby, 1838) (вкл. табл. V, рис. 70).
— antiqua Morris — Morris, Sowerby, 1838: 547, fig. 26 (Valvata).
Раковина от кубаревидной до прижатой низкоконической, умеренно толстостен-
ная, светлая, практически гладкая (имеющиеся сглаженные радиальные морщин-
ки становятся заметны при увеличении х20). Обороты (4,0—4,5) уплощённые,
разделены неглубоким швом. Пупок узкий, проколовидный, приблизительно на
Уз прикрыт отворотом колумеллярного края. Устье яйцевидное, с выраженным
резким уголком в верхней части.
Размеры: ВР и ШР до 4,5 мм
Вид распространён в Европе (исключая Средиземноморье) и Западной Сибири.
Обитает в реках и водохранилищах, на слегка заиленных грунтах.
18(17). Последний оборот менее вздутый. Его высота вместе с устьем (измерен-
ная со стороны устья) составляет 0,85 и менее высоты раковины.
19(20). Возвышение верхних оборотов над последним оборотом не превышает
0,9 мм...........................Cincinna falsifluviatilis Starobogatov in Anistratenko
et Anistratenko, 2001 (вкл. табл. V, рис. 71).
390
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
—falsifluviatilis Starobogatov in В. Anistratenko et О. Anistratenko, 2001: 139-140,
fig. 110 (Cincinna) [nom. nov. pro Valvata fluviatilis sensu Westerlund, 1886; non
Colbeau, 1859].
Раковина кубаревидная, твёрдостенная, светлая, тонко и густо радиально ребри-
стая. Обороты (4,0—4,5) выпуклые, разделены глубоким швом. Пупок проколовид-
ный, но не узкий (сквозь него видно около половины предпоследнего оборота).
Устье яйцевидное, с угловатостью в верхней части.
Размеры: ВР до 4,7 мм, ШР до 5,3 мм.
Вид распространён в Европе; в России — в бассейне Балтийского моря. Обитает
в реках.
20(19). Возвышение верхних оборотов над последним оборотом составляет
0,95 мм и более.................................................................
...............Cincinna discors (Westerlund, 1886) (вкл. табл. VI, рис. 72).
— discors Westerlund, 1886: 133 (Valvata).
Раковина низкоконическая, от прижатой до весьма высокой, иногда почти оваль-
ной (выраженный овальный облик раковины достигается за счёт вздутости
последнего оборота) серовато-, желтовато- или коричневато-роговая. Обороты
(3,5-4,5) покрыты радиальными рёбрами. Высота раковины примерно равна её
ширине.
Размеры: ВР и ШР до 3-3,5 мм.
Вид обитает в озёрах и реках бассейна Балтийского моря, центральной части Дне-
провского бассейна, и в Западной Сибири (бассейн реки Иртыш).
Подсемейство Borystheniinae Starobogatov in Sitnikova, 1983
Типовой род: Borysthenia Lindholm, 1914.
Раковина кубаревидная с прижатым (в различной степени) завитком; умеренно
толстостенная. Обороты (3-4) слабо выпуклые, уплощённые на периферии, очень
тонко нитевидно радиально исчерчены, иногда скульптура выражена настолько
слабо, что раковина выглядит гладкой, блестящей или матовой, фарфоровидной.
Последний оборот сильно вздут (в приустьевой области более чем в 2 раза шире
предпоследнего). Устье округло-каплевидное, сильно скошенное. Колумеллярный
и париетальный края устья могут быть утолщены мозолью. Животные живоро-
дящие, их женские половые протоки с массивной выводковой камерой. Внешние
маргинальные зубы радулы узкие, серповидно изогнутые, не зазубренные. В со-
ставе подсемейства один род.
Род Borysthenia Lindholm, 1914
Типовой вид: Valvata naticina Menke, 1845.
Признаки рода совпадают с диагнозом подсемейства. Ареал рода охватывает
Восточную Европу, Средний Дунай до Причерноморья. На севере и северо-запа-
де европейской части России распространён один вид.
Borysthenia naticina (Menke, 1846) (вкл. табл. VI, рис. 73).
— naticina Menke, 1846: 129 (Valvata).
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
391
Размеры: ВР 2,5-4,5 мм; ШР 3,5-5,5 мм.
Область распространения совпадает с ареалом рода. В России находки живых
особей вида известны из небольших рек в Калининградской области. Ископаемые
раковины вида обнаружены в четвертичных слоях в Ленинградской области.
Семейство ACROLOXIDAE Thiele, 1931
Типовой род: Acroloxus Beck, 1837.
Раковина небольших размеров (до 8-9 мм в длину), колпачковидная, тонко-
стенная, с продолговато-овальным или округлым устьем. Вершина в виде неболь-
шого загнутого рожка явственно смещена влево от продольной оси раковины,
покрыта радиальной микроскульптурой. Поверхность раковины с микроскопи-
ческими конхиолиновыми волосками (Hubendick, 1962). Тело правозавитое. Щу-
пальца цилиндрической формы. Внутренностный мешок блюдцеобразной или
пирамидальной формы, спиральная скрученность не выражена. Мантийная по-
лость сильно редуцирована. Дыхание растворённым в воде кислородом осущест-
вляется с помощью адаптивной жабры.
Семейство известно с мелового периода, включает 6 современных родов и 53
вида, большинство из которых являются эндемиками древних гигантских озёр —
Байкал, Охрид и др. (Широкая, 2005). Ареал семейства охватывает Палеарктику,
северную часть Сино-Индийской области, а также Неарктику [единственный се-
вероамериканский вид Acroloxus coloradensis (Henderson, 1930)]. В водоёмах ев-
ропейской части России отмечены представители одного рода Acroloxus.
Комментарии. Число видов в составе рода Acroloxus является предметом дискуссии.
Зарубежные исследователи относят всех европейских представителей рода к одному виду
Acroloxus lacustris (Linnaeus, 1758), широко распространённому в западной части Палеар-
ктики (Hubendick, 1962; Gittenberger et al., 1998; Gloer, 2002). В ходе ревизии, проведён-
ной с помощью компараторного метода, Н.Д. Круглов и Я.И. Старобогатов (1991) пришли
к выводу о том, что в водоёмах Европы (кроме южной её части) обитают как минимум
шесть видов: A. lacustris s. str., A. oblongus (Lightfoot, 1786), A. lindholmi Kruglov et Staro-
bogatov, 1991; A. okaensis Kruglov et Starobogatov, 1991; A. rossicus Kruglov et Starobogatov,
1991 и A. shadini Kruglov et Starobogatov, 1991. Определительные ключи, предложенные
для разграничения этих видов, основаны исключительно на конхологических признаках, а
межвидовые различия сводятся к очень небольшим отличиям пропорций раковины (Кру-
глов, Старобогатов, 1991). При этом по крайней мере для трёх видов (A. lacustris s. str.,
A. oblongus, A. shadini) было показано отсутствие статистически достоверных различий по
диагностически значимым признакам раковины (Хохуткин, Винарский, 2013). Анатомиче-
ское строение некоторых видов до сих пор не изучено.
Учитывая это, мы полагаем, что вопрос о числе видов Acroloxus в фауне Европы до
сих пор открыт и может быть окончательно решён с применением большего числа мето-
дов изучения, включая молекулярно-генетические. До тех пор, пока это не сделано, всех
представителей рода из водоёмов севера европейской части России рекомендуется обозна-
чать как Acroloxus lacustris s. lato.
Acroloxus lacustris (Linnaeus, 1758) (вкл. табл. VI, рис. 74).
— lacustris Linnaeus, 1758: 783 (Patella).
392
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Размеры: ДР до 9 мм, ВР до 2 мм, ШР до 3,5 мм.
Вид распространён в Европе и на юге Западной Сибири. Моллюски обитают в
стоячих и слабопроточных водоёмах; среди растительности и на камнях.
Семейство PHYSIDAE Fitzinger, 1833
Типовой род: Physa Drapamaud, 1801.
Раковина турбоспиральная, левозавитая, маленькая или средних размеров (до
20 мм высотой); окраска раковины в пределах семейства может варьировать от
светлой (грязно-белой) до тёмно-коричневой. Форма раковины от овально-кони-
ческой до башневидной; поверхность её матовая или гладкая, блестящая; скуль-
птура развита слабо и обычно представлена линиями нарастания. Стенка ракови-
ны непрозрачная или полупрозрачная. Пупок отсутствует.
Рис. 39. Physa sp. с отвернутыми на
поверхность раковины краями ман-
тии (по: Стадниченко, 1990).
ные. Челюсть многопластинчатая
Тело левозавитое, тёмно-окрашенное (от
желтовато-серого до практически чёрного
цвета). У представителей большинства ро-
дов (кроме Aplexa Fleming, 1820 и Sibirenau-
ta Starobogatov et Streletzkaja, 1967) бахром-
чатые края мантии в той или иной степени
отвернуты на поверхность мантии (рис. 39),
выполняя функцию адаптивной жабры (ис-
тинная жабра у представителей семейства
отсутствует). Щупальца длинные, нитевид-
с редуцированными латеральными частями
(Старобогатов, 1967). Зубы радулы многозубчатые, скошенные, расположены в
виде Х-образных поперечных рядов (Стадниченко, 1990).
Половая система в целом характерная для отряда Lymnaeiformes (Берёзкина,
Старобогатов, 1988; Jordaens et al., 2007), имеет ряд примитивных черт. Гонада
состоит из неправильных дивертикул. Простата состоит из пальцеобразных ди-
вертикул, впадающих непосредственно в семепровод. Копулятивный аппарат
разделён на препуциум и мешок пениса, придатки копулятивного аппарата от-
сутствуют. У многих видов на совокупительном органе имеется фиксаторное
утолщение.
Представители семейства населяют преимущественно небольшие постоянные
или временные водоёмы, богатые растительностью, со стоячей или полу проточ-
ной водой. Некоторые виды обитают в ручьях (Strong et al., 2008). Питаются во-
дорослями и тканями высших водных растений (Цихон-Луканина, 1987). Могут
выступать промежуточными хозяевами некоторых видов трематод.
Представители семейства известны со всех континентов, кроме Антарктиды
(Taylor, 2003). Большинство видов и родов отмечено в водоёмах Северной и Цен-
тральной Америки, Европы и Северной Азии (Старобогатов, 1970). Многие виды
в историческое время расширили свой ареал благодаря расселению человеком; в
частности, в водоёмах Африки, Австралии, некоторых островов Океании обита-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
393
ют только интродуцированные с других континентов виды (Taylor, 2003). Мак-
симальное таксономическое разнообразие физид наблюдается ныне в водоёмах
тихоокеанского побережья Америки, что, вероятно, указывает на американское
происхождение семейства (Taylor, 2003). В геологической летописи семейство из-
вестно с юрского периода (Старобогатов, 1970).
В состав семейства входят 23 рода и около 80 видов, включая несколько ви-
дов неясного таксономического положения (Taylor, 2003; Strong et al., 2008). В со-
ставе аборигенной малакофауны Европы насчитывается 3 рода — Physa, Aplexa и
Costatella (= Phy sella partim).
Комментарии. Имеется несколько различных вариантов системы семейства Physidae
водоёмов Голарктики (Те, 1978; Старобогатов и др., 1989; Taylor, 2003). Здесь мы прини-
маем разработанную в отечественной литературе систему (Старобогатов, 1967; Старобо-
гатов и др., 1989, 2004), в соответствии с которой семейство разделяется на два подсемей-
ства: Physinae Fitzinger, 1833 и Aplexinae Starobogatov, 1967.
Ключ для определения подсемейств и некоторых родов семейства Physidae
1(4). Раковина овально-коническая. Завиток невысокий, его высота меньше вы-
соты устья. Обороты выпуклые.................................................
..............................Подсем. Physinae Fitzinger, 1833 (с. 394).
2(3). Вершина раковины заострённая. Стенка раковины прочная, не просвечи-
вающая. Высота раковины достигает 15-17 мм...................................
.....Род Costatella Dall, 1870 (с. 000); Costatella acuta (Drapamaud, 1805)
(вкл. табл. VI, рис. 77).
— acuta Drapamaud, 1805: 55 (Physa).
Размеры: BP до 14 мм, ШР до 8 мм.
Вид описан из водоёмов Франции, имеет очень широкий ареал. Обитает преиму-
щественно в стоячих водоёмах, в Европе (кроме самой северной части), Передней,
Южной и Юго-Восточной Азии, Африке, Северной Америке и Австралии, на
Дальнем Востоке и Гавайских островах (Paraense, Pointier, 2003). Столь широкий
ареал обусловлен, по-видимому, высоким инвазионным потенциалом вида и уча-
стием человека в его расселении. В местностях с менее благоприятным климатом
(например, в Западной Сибири), С. acuta может образовывать популяции в водоё-
мах-охладителях, куда сбрасываются подогретые воды теплоэлектростанций.
Комментарии. В отечественной литературе (Старобогатов и др., 1989, 2004;
Стадниченко, 1990) вид помещается в состав рода Costatella. Однако зарубежные
авторы относят его либо к роду Physella Haldeman, 1842 (Gloer, 2002), либо к роду
Haitia Clench et Aguayo, 1932 (Taylor, 2003).
3(2). Вершина раковины притупленная. Стенки раковины тонкие, полупро-
зрачные. Размеры меньше (высота раковины может достигать 12 мм, но
обычно — не более 10 мм)..................Род Physa Drapamaud, 1801 (с. 394).
4(1). Раковина высококоническая. Завиток высокий и его высота превышает
высоту устья. Обороты уплощённые.............................................
.. Подсем. Aplexinae Starobogatov, 1967; род Aplexa Fleming, 1820 (с. 394).
394
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Подсемейство Physinae Fitzinger, 1833
Типовой род: Physa Drapamaud, 1801.
Раковина овально-коническая, обычно тонкостенная, с относительно неболь-
шим завитком и крупным, иногда сильно вздутым, последним оборотом. Обо-
роты раковины как правило более или менее выпуклые. Мантия с пальцевидны-
ми отростками, в расправленном состоянии охватывающими раковину снаружи.
В стенке препуциума имеется особая препуциальная железа.
Род Physa Drapamaud, 1801
Типовой вид: Bulla fontinalis Linnaeus, 1758.
Раковина некрупная (высота её не превышает 12 мм, обычно не более 10 мм),
тонкостенная, овально-коническая, последний оборот заметно вздут. Поверх-
ность раковины совершенно гладкая или со слабо выраженной спиральной скуль-
птурой, матово блестящая. Вершина раковины закруглена, устье относительно
крупное и широкое. Мешок пениса целиком или почти целиком железистый. Пе-
нис тонкий, с ланцетовидным стилетом на терминальном конце.
Общее распространение — северная часть Евразии, Северная Америка. По
данным разных исследователей, в составе рода насчитывается от 9 (Taylor, 2003)
до 14 видов (Старобогатов и др., 1989).
Ключ для определения видов рода Physa
1(2). Раковина широко овально-коническая, с очень низким тупым завитком.
Последний оборот сильно вздут.................................................
..................Physa fontinalis (Linnaeus, 1758) (вкл. табл. VI, рис. 78).
—fontinalis Linnaeus, 1758: 727 (Bulla).
Размеры: ВР до 12 мм, ШР до 5 мм.
Общее распространение вида — Европа и Сибирь. Моллюски обитают преимуще-
ственно в небольших стоячих водоёмах.
2(1). Раковина овально-яйцевидной формы, с относительно высоким заострён-
ным завитком. Последний оборот умеренно вздут.................................
....................Physa taslei Bourguignat, 1860 (вкл. табл. VI, рис. 79).
— taslei Bourguignat, 1860: 70 (Physa).
[= Physa bulla sensu Starobogatov et al., 1989].
Размеры: BP до 12 мм, ШР до 4,5 мм.
Вид распространён в Европе (кроме крайнего севера), Западной Сибири. Преиму-
щественно в стоячих и полупроточных водоёмах. Вероятно, именно к этому виду
относятся указания североамериканского видаР/?. skinneri Taylor, 1954, с недавних
пор отмечаемого в водоёмах Украины и Европейской России (Тэйлор, Ситникова,
2004; Сон, 2007).
Подсемейство Aplexinae Starobogatov, 1967
Типовой род: Aplexa Fleming, 1820.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
395
Раковина высококоническая, довольно тонкостенная, завиток сравнительно
высокий, его высота заметно превышает высоту устья. Высота раковины дости-
гает 20 мм. Поверхность раковины гладкая, с мягким «шелковистым» блеском,
окраска золотисто-коричневая или тёмно-коричневая. Обороты раковины упло-
щённые, шов мелкий.
Половая система характеризуется отсутствием препуциальной железы (имеет-
ся у представителей подсемейства Physinae). Мешок пениса может быть цельным
(железистым), а может состоять из двух или даже трёх частей (железистой и му-
скульных).
Подсемейство включает 12 родов, 4 трибы и не менее 34 видов, распростра-
нённых в Северной и Южной Америке и на севере Евразии (Taylor, 2003). Мол-
люски обитают преимущественно во временных водоёмах.
Род Aplexa Fleming, 1820
Типовой вид: Bulla hypnorum Linnaeus, 1758.
Раковина некрупная (высота до 15 мм), высококоническая, коричневая или зо-
лотисто-коричневая, стройная, матово-блестящая, гладкая. Завиток заострённый,
обороты завитка уплощённые. Устье составляет менее половины высоты ракови-
ны. Мешок пениса целиком железистый, заметно короче препуциума.
Общее распространение: Европа (кроме юга), Западная Сибирь, Дальний Вос-
ток (бассейн Амура)1. Число видов рода, по разным данным, составляет от одного
(Gloer, 2002; Taylor, 2003) до шести (Старобогатов и др., 1989).
Ключ для определения видов рода Aplexa
1(2). Раковина относительно стройная, так что ширина её последнего оборота,
измеренная без учёта устья, составляет менее половины высоты послед-
него оборота. Препуциум длинный, так что ИКА составляет около 1,2............
................Aplexa turrita (O.F. Muller, 1774) (вкл. табл. VI, рис. 76).
— turrita O.F. Muller, 1774: 169 (Planorbis).
Размеры: BP до 15 мм, ШР до 5 мм.
Общее распространение вида — Европа и Западная Сибирь. Моллюски обитают
преимущественно во временных водоёмах.
Комментарии. В отечественной литературе (Старобогатов и др., 1989, 2004) для
водоёмов Европейской России указывается ещё один вид со сходными пропорци-
ями раковины, A. ataxiaca Fagot, 1844, выделяемый в особый подрод Paraplexa
Starobogatov in Starobogatov, Prozorova et Zatravkin, 1989. Тэйлор (Taylor, 2003) по-
высил ранг этого таксона до родового, однако по данным этого автора в пределах
Российской Федерации представители вида A. ataxiaca не обитают, так что ареал
рода Paraplexa ограничен юго-востоком Европы. Моллюски, описанные Я.И. Ста-
1 Дальневосточные представители выделены в подрод Amuraplexa Starobogatov, Prozorova et
Zatravkin, 1989, который Тэйлор (Taylor, 2003) считает самостоятельным родом.
396
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
робогатовым и др. (1989) как A. ataxiaca, по мнению Тэйлора (2003) должны быть
отнесены к A. hypnorum.
2(1). Раковина менее стройная, так что ширина её последнего оборота, изме-
ренная без учёта устья, составляет более половины высоты последнего
оборота. Препуциум короткий, так что ИКА составляет около 0,50..............
.............Aplexa hypnorum (Linnaeus, 1758) (вкл. табл. VI, рис. 75).
— hypnorum Linnaeus, 1758: 727 (Bulla).
Размеры: ВР до 14 мм, ШР до 6 мм.
Вид распространён в Европе и на юге Западной Сибири. Моллюски обитают пре-
имущественно во временных водоёмах.
Комментарии. Виды рода Aplexa довольно близки по пропорциям раковины, по-
этому для определения видовой принадлежности моллюсков рекомендуется по
возможности использовать значения ИКА.
Семейство LYMNAEIDAE Rafinesque, 1815
Типовой род: Lymnaea Lamarck, 1799.
Животные обычно малых и средних размеров (у представителей подрода Lym-
naea s. str. высота раковины достигает 70 мм). Раковина турбоспиральная, у всех
европейских представителей семейства правозавитая (иногда могут быть встре-
чены особи прудовиков с левозавитой раковиной, возникающей в результате до-
статочно редкой мутации). Форма и пропорции раковины значительно варьируют
в пределах семейства. Известны виды с уховидной, шаровидной, овально-кониче-
ской, и башневидной раковиной.
Тело правозавитое. Дыхание осуществляется атмосферным воздухом, или рас-
творённым в воде кислородом. В последнем случае лёгкое заполняется водой, а
край мантийной полости функционирует как адаптивная жабра (Иванов, 1940).
Большое значение имеет также кожное дыхание. Все прудовики — гермафроди-
ты, однако способны к самооплодотворению. Половая система представлена не-
парной гонадой (гермафродитной железой), мужскими и женскими протоками.
Мужские протоки заканчиваются копулятивным аппаратом, который у большин-
ства видов состоит из двух частей: препуциума и мешка пениса, внутри которо-
го находится пенис (рис. 20). У представителей рода Aenigmomphiscola Kruglov
et Starobogatov, 1981 копулятивный аппарат более сложного строения, в котором
мешок пениса подразделён на две части (тонкостенную и железистую), а внутри
препуциума, близ его границы с мешком пениса, развит так называемый препу-
циальный орган (рис. 40).
Семейство распространено практически по всему земному шару, однако мак-
симального видового разнообразия достигает в континентальных водах умерен-
ной зоны Северного полушария. Наименьшее число видов представлено в Афри-
ке, Южной Америке, Австралии и на островах Тихого океана (Strong et al., 2008).
Прудовики населяют пресные, реже солоноватые воды, термальные источники,
разного рода увлажнённые поверхности. Фитофаги, крупные формы всеядны, мо-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
397
гут питаться трупами головастиков, тритонов и
рыб (Сушкина, 1949). Известны случаи канниба-
лизма (Цихон-Луканина, 1987).
Общее число современных видов не ме-
нее 200-250, в фауне Палеарктики насчиты-
вается около 140 видов (Kruglov, Starobogatov,
1993а, b). Семейство возникло, предположитель-
но, в самом конце палеозоя (Inaba, 1969; Старо-
богатов, 1976), в геологической летописи извест-
но с юры (Коробков, 1954).
Наибольшую ценность для таксономии лим-
неид имеют признаки половой системы, в пер-
вую очередь — строение копулятивного аппара-
та, простаты, длина протока семеприемника и др.
(Jackiewicz, 1998; Круглов, 2005). В частности,
для видовой диагностики широко используется
индекс копулятивного аппарата, ИКА. Ракови-
на прудовиков весьма изменчива, поэтому кон-
хологические признаки имеют относительную
меньшую систематическую ценность, к тому же
в пределах семейства отмечены многочисленные
случаи конвергенции, когда виды, не находящи-
еся в близком родстве, имеют сходную раковину
(Круглов, Старобогатов, 1987). Поэтому опреде-
ление видовой принадлежности лимнеид реко-
Рис. 40. Копулятивный аппа-
рат Aenigmomphiscola uvalievae
(по: Круглов, Старобогатов,
1981).
а — нежелезистая проксималь-
ная часть мешка пениса; б — же-
лезистая дистальная часть мешка
мендуется по возможности проводить с привле-
чением признаков строения половой системы.
Комментарии. Общепринятой системы семейства
Lymnaeidae не существует. По мнению Н.Д. Кругло-
пениса; в — пенис; г — семяиз-
вергательный канал; д — препу-
циум; е — препуциальный орган.
ва и Я.И. Старобогатова (1981), семейство включает два рода: Lymnaea Lamarck, 1799 и
Aenigmomphiscola Kruglov et Starobogatov, 1981. Другие авторы принимают значительно
большее число родов в составе семейства (Ponder, Watehouse, 1997; Vinarski, 2013). Чис-
ло признаваемых видов также неодинаково в разных версиях системы. В странах Запад-
ной Европы придерживаются традиционной системы, в рамках которой признается от-
носительно небольшое число полиморфных видов (Hubendick, 1951; Jackiewicz, 1993,
1998; Falkner et al., 2001; Gloer, 2002). В странах бывшего СССР большее распростране-
ние получила система семейства, разработанная Н.Д. Кругловым и Я.И. Старобогатовым
(Kruglov, Starobogatov, 1993a,b; Круглов, 2005, 2008, 2009), в которой принят более дета-
лизированный подход к выделению видов, в результате чего их число значительно боль-
ше, чем в современной западноевропейской системе. Близкородственные виды в этой
системе разграничиваются по тонким различиям в геометрии раковины, а также по коли-
чественным различиям пропорций частей копулятивного аппарата. В связи с этим тради-
ционное определение видов по морфометрическим индексам раковины бывает затрудне-
398 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
но. Некоторые виды, относящиеся к роду Aenigmomphiscola и подроду Stagnicola Leach in
Jeffreys, 1831 рода Lymnaea можно достоверно определить только по анатомическим при-
знакам. Также представителей подродов Corvusiana и Stagnicola рода Lymnaea надёжнее
всего отличать друг от друга именно по строению мужской половой системы; при работе
с пустыми раковинами возможны ошибки. Таксономическая самостоятельность ряда ви-
дов, указываемых для водоёмов Европы, также нуждается в подтверждении. В настоящий
определитель включены виды, самостоятельность которых и их обитание в пределах ев-
ропейской части России не вызывают сомнений. Ключи для определения родов, подро-
дов и видов Lymnaeidae составлены на основе конхологических признаков, однако в ряде
случаев даны и анатомические признаки, позволяющие более точно установить таксоно-
мическую принадлежность животных1. Подготовка части определителя, посвящённой се-
мейству Limnaeidae, была завершена в 2011 году. Поэтому в ней не учтены некоторые пу-
бликации, вышедшие позднее. В том числе работа автора (Vinarski, 2013), отражающая его
новый взгляд на систему семейства.
Ключ для определения родов семейства Lymnaeidae
1(2). Раковина очень стройная, башневидной формы, с большим числом (6-7)
медленно нарастающих плоских оборотов. Мешок пениса состоит из двух
частей — проксимальной (с тонкими стенками) и дистальной (с желези-
стыми стенками).............Aenigmomphiscola Kruglov et Starobogatov, 19812.
2(1). Раковина иной формы; если башневидной, то мешок пениса простого
строения, не подразделённый на проксимальную и дистальную части.........
......................................Lymnaea Lamarck, 1799 (с. 399).
Род Aenigmomphiscola Kruglov et Starobogatov, 1981
Типовой вид: Aenigmomphiscola europaea Kruglov et Starobogatov, 1981.
Раковина средних размеров (высота раковины до 17 мм), башневидная, очень
стройная. Основным отличием этого рода от рода Lymnaea является уникаль-
ное строение копулятивного аппарата: наличие особого препуциального органа,
располагающегося в проксимальной части препуциума и представляющего со-
бой сильно разросшийся велюм (Круглов, Старобогатов, 1981). Мешок пениса у
Aenigmomphiscola подразделён на две части (рис. 40), чего нет (как нет и препу-
циального органа) у видов рода Lymnaea. Простата с одной внутренней складкой.
1 Подробные определительные ключи европейских Lymnaeidae, составленные в соответствии
с системой Н.Д. Круглова и Я.И. Старобогатова можно найти в целом ряде монографий (Стад-
ниченко, 2004; Круглов, 2005; Хохуткин и др., 2009; Андреева и др., 2010). Система семейства,
принятая в странах Западной Европы, наиболее полно изложена в публикациях М. Яцкевич
(Jackiewicz, 1998, 2000) и П. Глоэра (Gloer, 2002; Gloer, Meier-Brook, 2003), где также имеются
ключи для определения видов прудовиков.
2 У рода Aenigmomphiscola отсутствует конхологический диагноз (Круглов, 2008). По при-
знакам раковины виды этого рода невозможно отличить от представителей подрода Lymnaea
(Omphiscold), поэтому изучение анатомического строения моллюсков необходимо для надёж-
ной диагностики их родовой принадлежности.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
399
По строению раковины виды Aenigmomphiscola очень близки к представите-
лям подрода Omphiscola рода Lymnaea, но ареалы этих таксонов, по-видимому, не
перекрываются. Общее распространение рода Aenigmomphiscola — Европейская
часть России, Южный Урал, степные районы на юге Западной Сибири, Горный
Алтай. Род представлен в современной фауне тремя видами, эндемичными для
территории бывшего СССР, два из которых представлены в фауне Европейской
России. Моллюски обитают преимущественно во временных водоёмах.
Ключ для определения видов рода Aenigmomphiscola
1(2). Линия, проведённая через верхний угол устья параллельно оси раковины,
делит просвет устья на две слегка неравные части (не более 3:2 или 3,2:2).
Тонкостенная и толстостенная части мешка пениса почти равны по длине
........................Aenigmomphiscola europaea Kruglov et Starobogatov, 1981
(вкл. табл. VI, рис. 80).
— europaea Круглов, Старобогатов, 1981: 969, рис. 1,7, 7, 72; За; 4а.
Индекс копулятивного аппарата составляет 0,5-0,7.
Размеры: ВР до 17 мм, ШР до 5 мм.
Вид известен из водоёмов Подмосковья, Нижегородской и Ярославской областей,
республики Башкирия, а также, вероятно, обитает в бассейнах р. Дон и Печора.
Возможно, вид широко распространён по Восточно-Европейской равнине. Обита-
ет во временных водоёмах.
2(1). Линия, проведённая через верхний угол устья параллельно оси раковины,
делит просвет устья на две резко неравные части (не менее 4:1). Тонко-
стенная часть мешка пениса почти втрое короче толстостенной.................
.................Aenigmomphiscola uvalievae Kruglov et Starobogatov, 1981
(вкл. табл. VI, рис. 81).
— uvalievae Круглов, Старобогатов, 1981: 971-973, fig. 1, 2, 8, 13; 2; ЗВ; 4В; 5.
Индекс копулятивного аппарата составляет 0,4-0,5.
Размеры: ВР до 12,0 мм, ШР до 4,5 мм.
Вид описан из водоёмов Северного Казахстана. Известен также из одного место-
обитания в Республике Башкирия, расположенного в 230 км к востоку от г. Уфы
(Хохуткин и др., 2009). Вероятно, ареал вида охватывает восточную часть Евро-
пейской России и юг Западной Сибири.
Род Lymnaea Lamarck, 1799
Типовой вид: Helix stagnalis Linnaeus, 1758.
Раковина разнообразная по размерам, форме и числу оборотов. В составе рода
различают более 200 рецентных видов, сгруппированных в 26 подродов (Kruglov,
Starobogatov, 1993а, b). Моллюски населяют пресные воды всего земного шара,
кроме Антарктики. От рода Aenigmomphiscola отличается просто устроенным ко-
пулятивным аппаратом, без препуциального органа и железистых образований в
стенках мешка пениса.
400 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Ключ для определения подродов и некоторых видов рода Lymnaea
1(2). Раковина коническая или уховидная, крупная (при 5,5-6,0 оборотах её
высота составляет как правило, не менее 30 мм) со слабовыпуклыми или
совершенно плоскими оборотами. Простата с несколькими (5-10) развет-
влёнными внутренними складками................Подрод Lymnaea s. str. (с. 403).
2(1). Раковина иной формы, если коническая или уховидная, то число оборотов
не превышает 5,0. Высота раковины, как правило, меньше 30 мм, если
крупнее, то раковина башневидная или овально-коническая с относитель-
но некрупным устьем, либо уховидная с заметно выпуклыми оборотами.
Простата без внутренних складок, либо с одной или несколькими склад-
ками (в последнем случае они неразветвлённые).
3(12). Высота завитка больше высоты устья или приблизительно равна ей.
4(7). Раковина маленькая (при 4,5-5,0 оборотах её высота не превышает 10-
12 мм; если крупнее, то раковина башневидной формы с плоскими или
слабовыпуклыми оборотами и очень слабо вздутым последним оборотом).
Общее число оборотов не менее 6,0.
5(6). Форма раковины от башневидной и башневидно-конической до овально-
конической. Раковина умеренно стройная, последний оборот умеренно
или сильно вздут. Обороты сильновыпуклые или почти ступенчатые............
.................................Подрод Galba Schrank, 1803 (с. 404).
6(5). Форма раковины башневидная. Раковина очень стройная, последний обо-
рот очень слабо вздут. Обороты плоские или слабовыпуклые..................
..................................Подрод Omphiscola Rafinesque, 1819;
Lymnaea clavata (Westerlund, 1885) (вкл. табл. VI, рис. 82).
— clavata Westerlund, 1885: 49 (Limnaea glabra var.).
Размеры: BP до 14 мм, ШР до 5 мм.
В Европейской России подрод Omphiscola представлен единственным видом
L. {О.) clavata, обитающим в водоёмах бассейна Балтийского моря. Этот вид ис-
ключительно редок: его единственная достоверная находка в России относится
к 1895 году и с того времени в водоёмах нашей страны вид не найден (Круглов,
Старобогатов, 1981). Вид также известен из водоёмов бассейна Западного Буга на
Украине. Обитает в озёрах, прудах и старицах.
Комментарии. Европейские систематики рассматривают L. clavata как синоним
вида L. (О.) glabra (O.F. Muller, 1774).
7 4). Раковина средних размеров (при 4,5-5,0 оборотах её высота составляет не
менее 12 мм; если меньше, то обороты плоские или слабовыпуклые).
8(9). Раковина сравнительно крупная (высотой до 32-35 мм). Мешок пениса
очень короткий, составляет около !4 длины препуциума. Простата с 5-10
внутренними неразветвлёнными складками....................................
.............................Подрод Corvusiana Servain, 1881 (с. 406).
9(8). Раковина меньших размеров, её высота, как правило, не превышает 25 мм,
если крупнее, то высота устья заметно меньше половины высоты ракови-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
401
ны, а обороты плоские или слабовыпуклые. Мешок пениса, как правило,
равен по длине препуциуму или длиннее его. Если мешок пениса короче
препуциума, то не более чем вдвое. Простата с одной внутренней складкой.
10(11). Раковина башневидная, с плоскими или слабовыпуклыми оборотами, по-
следний оборот очень слабо расширен в сравнении с предшествующими.
Отворот колумеллярного края устья молочно-белого цвета, довольно ши-
рокий, толстый..........................Подрод Ladislavella В. Dybowski, 1913
Lymnaea terebra (Westerlund, 1885) (вкл. табл. VI, рис. 83).
— terebra Westerlund, 1885: 46 (Limnaea palustris var.).
[= L. occulta Jackiewicz, 1959; L. vulnerata sensu Kruglov, Starobogatov, 1986 non
Kiister, 1862].
Ширина мешка пениса лишь незначительно меньше, чем ширина препуциума.
У сходных по раковине видов из подродов Omphiscola и Stagnicola препуциум
светлоокрашенный и заметно шире, чем мешок пениса.
Размеры: ВР до 25 мм. (обычно менее 22 мм), ШР до 12 мм.
Вид известен из водоёмов Восточной и Южной Европы, Скандинавии, Сибири.
Указывался из западных областей Украины под названием L. occulta (Стадничен-
ко, 1968; Komiushin, 1999). В коллекции ЗИН РАН имеются раковины вида из
окрестностей Архангельска и бассейна Печоры (сборы начала XX в). Возможно
нахождение в других областях севера Европейской России. Важным анатомиче-
ским признаком для диагностики L. terebra служит тёмно-окрашенный препуциум
и примерно равный ему по длине мешок пениса. L. terebra обитает преимуще-
ственно во временных водоёмах, также может быть встречен в ручьях и малых
реках.
11(10). Раковина башневидно-коническая или овально-коническая, если башне-
видная, то отворот колумеллярного края устья неширокий и тонкий. По-
следний оборот заметно расширен по сравнению с предшествующими..............
........................Подрод Stagnicola Leach in Jeffreys, 1831 (с. 407).
12(3). Высота завитка заметно меньше высоты устья. Последний оборот относи-
тельно крупный и расширенный по сравнению с оборотами завитка.
13(14). Раковина округлая, очень хрупкая, просвечивающая, гладкая, её завиток
настолько мал, что создаётся впечатление, будто вся раковина состоит из
одного последнего оборота. У живого моллюска раковина окружена на-
ружными выростами мантии....................................................
......Подрод Myxas Sowerby, 1822; Lymnaea glutinosa (O.F. Muller, 1774)
(вкл. табл. VI, рис. 84).
— glutinosum O.F. Muller, 1774: 129 (Buccinum).
Размеры: BP до 20 мм (обычно до 15 мм), ШР до 18 мм.
Вид широко распространён в водоёмах Европейской России. Общее распростра-
нение L. glutinosa — Европа, Урал, Западная Сибирь, единичные находки в бас-
сейне Енисея.
1 В западноевропейской систематике эта группа рассматривается как род Catascopia Meier-
Brook et Bargues, 2002 (Gloer, Meier-Brook, 2003; Bargues et al., 2003, 2006).
402 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Комментарии. Отечественные систематики (Kruglov, Starobogatov, 1993b; Кру-
глов, 2005) помимо L. glutinosa приводят для водоёмов Европейской России ещё
два вида, L. (М.) dupuyi Locard, 1893 и L. (М) mabillei Locard, 1893, Отличия
между ними и от L. glutinosa сводятся к тонким различиям в относительной высо-
те завитка, пропорциях копулятивного аппарата и длине протока семеприемника1.
Экология всех этих видов сходна. Они обитают в постоянных полноводных водоё-
мах, обычно на листьях водных растений.
14(13). Раковина иной формы, если округлая — то стенки её непрозрачные, а за-
виток даже при небольших размерах заметно возвышается над последним
оборотом. Раковина живого моллюска не окружена наружными складками
мантии.
15(16). Раковина уховидная. Завиток с неравномерно выпуклыми (грушевидны-
ми) оборотами. На колумеллярном крае устья имеется сильное вдавление,
прикрывающее пупок. Проток семеприемника длинный, его длина значи-
тельно превышает длину резервуара.............................................
.......Подрод Radix Montfort, 1810; Lymnaea auricularia (Linnaeus, 1758)
(вкл. табл. VI, рис. 85).
— auricularia Linnaeus, 1758: 774 (Helix).
Размеры: BP до 34 мм, ШР до 30 мм.
Вид повсеместно распространён в водоёмах Европейской России. Общее распро-
странение — Европа, Северная Африка, Ближний Восток, Средняя и Восточная
Азия, Сибирь на восток до Берингии; интродуцирован в Северную Америку.
Обитает в озёрах, прудах, старицах, реже ручьях и протоках; на растительности
или у дна.
Комментарии. Может также быть обнаружен родственный вид L. (R.) parapsilia
(Vinarski et Gloer, 2009) [= L. (R.) psilia sensu Kruglov et Starobogatov, 1993a], име-
ющий восточно-палеарктическое распространение и встречающийся более редко.
От L. auricularia он отличается меньшими размерами раковины (высота раковины
до 28 мм), более высоким завитком, придающим раковине уховидно-коническую
форму2. Обитает в тех же биотопах, что и L. auricularia.
16(15). Раковина уховидной, яйцевидной или иной формы. Обороты почти пло-
ские, либо равномерно выпуклые; если неравномерно выпуклые, то рако-
вина не уховидная. Вдавление на колумеллярном крае развито слабо, так
что париетальная площадка плавно переходит в отворот колумеллярного
края. Пупок открытый. Проток семеприемника короткий...........................
...............................Подрод Peregriana Servain, 1881 (с. 409).
Подрод Lymnaea s. str.
Типовой вид: Helix stagnalis Linnaeus, 1758.
1 В случае сомнений в точности определения видовой принадлежности моллюсков подрода
Myxas рекомендуем обозначать их как L. (М.) glutinosa s. lato.
2 В случае сомнений в точности определения видовой принадлежности моллюсков подрода
Radix рекомендуем обозначать их как L. (R.) auricularia s. lato.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
403
Раковина самая крупная в пределах семейства (высотой до 70 мм), с высоким
и широким устьем, стенка раковины обычно умеренной толщины. Форма рако-
вины варьирует у разных видов подрода от уховидной до высококонической, с
очень высоким и стройным завитком. Окраска раковины обычно светло-роговая,
иногда тёмно-бурая либо почти белая с розоватым оттенком. Проток сперматеки
длинный. Представители подрода распространены в северной Голарктике, интро-
дуцированы в Тасманию и Новую Зеландию (Geyer, 1927; Climo, Pullan, 1972).
Ключ для определения видов подрода Lymnaea s. str.
1(4). Раковина высококоническая или коническая, дефинитивная раковина име-
ет более 6,0 оборотов (до 8,25). Завиток относительно высокий, составля-
ет (при 6,0 и более оборотах) не менее 0,45 высоты раковины.
2(3). Завиток шиловидной или высококонической формы, очень стройный, вы-
сокий, обороты нарастают в высоту очень медленно. Высота раковины,
как правило, в два и более раза превышает её ширину1.........................
................Lymnaea fragilis (Linnaeus, 1758) (вкл. табл. VI, рис. 86).
—fragilis Linnaeus, 1758: 774 (Helix).
[= L. stagnalis var. producta Colbeau, 1859; L. raphidia Bourguignat, 1860; L. stagna-
te var. subulata Westerlund, 1873].
Индекс копулятивного аппарата составляет не менее 3,75 (обычно не менее 3,95).
Размеры устья и последнего оборота раковины подвержены сильной изменчиво-
сти. Особи с сильно вздутым последним оборотом иногда рассматриваются как
особый «подвид» L. f producta (Colbeau, 1859).
Размеры: ВР до 65 мм, ШР до 45 мм.
Вид распространён по всей Палеарктике, от атлантического побережья на западе,
до бассейна Колымы на востоке; в водоёмах Европейской России встречается по-
всеместно. Населяет водоёмы различного типа, от мелких эфемерных водоёмов до
проток и затонов крупных рек. Наибольшей численности и биомассы достигает в
небольших хорошо прогреваемых и богатых органикой озёрах и прудах. Взрослые
особи держатся преимущественно у поверхности воды или на растениях.
3(2). Завиток конической формы, относительно невысокий, обороты нарастают
в высоту довольно быстро. Высота раковины, как правило, менее чем в
два раза превышает ширину раковины...........................................
..............Lymnaea stagnalis (Linnaeus, 1758) (вкл. табл. VI, рис. 87).
— stagnate Linnaeus, 1758: 774 (Helix).
[= L. turgida Menke in Hartmann, 1840].
Индекс копулятивного аппарата составляет не более 3,75 (обычно не более 3,50).
Размеры: ВР до 60 мм, ШР до 45 мм.
Вид распространён по всей Европе и Северной Азии на восток до Берингии,
в Северной Африке, Северной Америке, интродуцирован в Новую Зеландию.
1 При использовании индексов раковины следует учитывать, что в ряде случаев виды L. stag-
nalis и L. fragilis неотличимы по этим параметрам и для определения следует привлекать анато-
мические данные.
404
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
В водоёмах Европейской России встречается повсеместно; на севере региона
сравнительно редок. Наиболее обычен в непроточных или слабо проточных водо-
ёмах, богатых водной растительностью, как пересыхающих, так и постоянных.
В поймах рек и крупных глубоких озёрах редок. Обычно держится в прибрежье
среди гидрофитов.
Комментарии. Наиболее изменчивой является высота завитка раковины. Экзем-
пляры с особенно низким завитком иногда рассматриваются как особый «подвид»
L. s. turgida (Menke in Hartmann, 1840).
4(1). Раковина яйцевидная или почти уховидная, дефинитивная раковина имеет
до 5,5 оборотов (как исключение до 6,0). Завиток невысокий, составляет
не более 0,4 высоты раковины....................................................
.............Lymnaea doriana (Bourguignat, 1862) (вкл. табл. VI, рис. 88).
— doriana Bourguignat, 1862: 60 {Limnaea).
[= L. lacustris (Studer, 1820) non Leach, 1819].
Индекс копулятивного аппарата составляет около 4,0.
Размеры: ВР до 40 мм, ШР до 30 мм.
Вид распространён в Европе (кроме крайнего севера), на Кавказе, в Горном Алтае
и на юге Восточной Сибири (верховья Енисея). В водоёмах Европейской России
вид известен из ряда местообитаний, расположенных в Ленинградской области и
республике Карелия (по материалам коллекции ЗИН РАН). Моллюски обитают в
прибойной зоне крупных озёр среди растительности.
Комментарии. В крупных озёрах Восточной Европы (Карелия) и Альп (Боден-
ское озеро) встречаются представители подрода Lymnaea s. str. с очень низким
завитком и расширенным устьем, раковина которых имеет уховидную форму и
несколько напоминает по форме раковину уховидного прудовика L. {Radix) auricu-
laria. Европейские авторы (Geyer, 1927; Piaget, 1929; Ehrmann, 1933) традиционно
рассматривали таких моллюсков, как «озёрную форму» вида L. stagnalis, возник-
шую в результате адаптации к обитанию в прибойной зоне крупных озёр. Этого
же мнения придерживались и некоторые отечественные авторы (Жадин, 1952;
Шилейко, 1967). В системе Н.Д. Круглова и Я.И. Старобогатова (Kruglov, Staro-
bogatov, 1993а) «озёрные формы» рассматриваются как самостоятельные виды
L. (£.) bodamica (Miller, 1873) и L. (£.) media (Kobelt, 1877), характеризующиеся
относительной низкой высотой раковины (отношение высоты раковины к её ши-
рине составляет 1,4 и менее).
Подрод Galba Schrank, 1803
Типовой вид: Galbapusilia Schrank, 1803.
Раковина маленькая (до 10-12 мм высотой), форма её варьирует у разных ви-
дов от башневидной до овально-конической. Обороты, как правило, сильно выпу-
клые, иногда почти ступенчатые, тангент-линия выгнутая. Простата имеет одну
неразветвлённую внутреннюю складку, проток семеприемника длинный.
Представители подрода характеризуются как обитатели разного рода увлаж-
нённых поверхностей, например, мокрых берегов небольших рек и ручьев, мо-
чажин; ряд видов адаптирован к жизни в родниках. Ареал подрода охватывает
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
405
Палеарктику, исключая северо-восточную часть Азии; вид L. truncatula интроду-
цирован в Южную Америку и Новую Зеландию (Hubendick, 1951; Climo, Pullan,
1972; Durand et al., 2002; Круглов, 2005).
Ключ для определения видов подрода Galba
1(2). Раковина высококоническая или башневидная. Завиток высокий и строй-
ный, его высота равна или превышает высоту устья..............................
.............Lymnaea truncatula (O.F. Muller, 1774) (вкл. табл. VI, рис. 89).
— truncatulum O.F. Muller, 1774: 130 (Buccinum).
[=£. minuta Drapamaud, 1801].
Обороты сильновыпуклые медленно нарастающие, иногда почти ступенчатые.
Индекс копулятивного аппарата составляет около 3,0.
Размеры: ВР до 12 мм (обычно не превышает 10 мм), ШР до 5,5 мм.
Вид распространён в Европе, на Кавказе, в Сибири (на восток до бассейна Лены),
Средней Азии, Северной Африке, на Аравийском полуострове; интродуцирован
в Боливию, Перу и Новую Зеландию. В водоёмах европейской части России,
по-видимому, распространён повсеместно. Амфибионтный вид. Встречается в
прибрежной зоне озёр, в реках (в зоне заплеска), а также в различных мелких не-
постоянных водоёмах (лужах, мочажинах, на сырых лугах в заполненных водой
отпечатках копыт скота).
Комментарии. Отечественные малакологи (Круглов, Старобогатов, 1985; Кру-
глов, 2005) выделяли ещё два близких по морфологии и экологическим особен-
ностям к L. truncatula вида, обладающих раковинами башневидной или высоко-
конической формы с относительно высоким и стройным завитком. Это L. goupili
(Moquin-Tandon, 1856) и L. oblonga (Puton, 1847). Они отличаются от L. truncatula
геометрией роста раковины, её пропорциями, а также различными значениями
индекса копулятивного аппарата. Европейские специалисты рассматривают эти
виды как синонимы L. truncatula'.
2(1). Раковина овально-коническая, завиток относительно низкий и широкий,
высота его составляет не более 0,9 высоты устья...............................
............Lymnaea subangulata (Roffiaen, 1868) (вкл. табл. VI, рис. 90).
— subangulata Roffiaen, 1868: 78, pl. 1, fig. 9 {Limnaea truncatula var.).
[= L. ventricosa Moquin-Tandon, 1856 non Hartmann, 1841].
Индекс копулятивного аппарата составляет 2,83. От L. truncatula вид отличается
также большим значением апикального угла.
Размеры: ВР до 8,0 мм, ШР до 4,5 мм.
Вид распространён в Европе (кроме её северной части), Центральной Азии, Се-
верной Африке. Обитает в пересыхающих водоёмах и мелких родниковых моча-
жинах.
1 В случае сомнений в точности определения видовой принадлежности моллюсков группы
L. truncatula рекомендуем обозначать их как L. (G.) truncatula s. lato.
406
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Подрод Corvusiana Servain, 1881
Типовой вид: Helix corvus Gmelin in Linnaeus, 1791.
Раковина твёрдостенная, относительно крупная (до 35 мм высотой), форма её
варьирует у разных видов от овально-конической до башневидной. Устье крупное
и сравнительно широкое. Проток семеприемника длинный. Препуциум крупный,
мешковидный, плотный, мешок пениса значительно короче препуциума, заметно
вздут в области проксимальных камер.
По мнению Я.И. Старобогатова (1976), подрод Corvusiana является древней-
шим из подродов рода Lymnaea, возникшим, предположительно, в конце палео-
зоя. В современной фауне насчитывается 5 видов подрода, распространённых в
Европе и Казахстане.
Ключ для определения видов подрода Corvusiana
1(2). Раковина башневидно-коническая или овально-коническая. Обороты пло-
ские или слабовыпуклые ......................................................
.......Lymnaea corvus (Gmelin in Linnaeus, 1791) (вкл. табл. VI, рис. 91).
— corvus Gmelin in Linnaeus, 1791: 3665 {Helix).
Мешок пениса узкий. Индекс копулятивного аппарата составляет около 3,5.
Размеры: ВР до 35 мм, ШР до 15 мм.
Область распространения вида — Европа, кроме крайнего севера и юга. На восток
вид распространён до бассейна р. Урал (в пределах Прикаспийской провинции).
Моллюски обитают во временных стоячих водоёмах, а также в малых реках на
участках с замедленным течением.
Комментарии. Ряд отечественных малакологов (Круглов, Старобогатов, 1984;
Круглов, 2005) признавали существование ещё двух близких по морфологии и
экологическим особенностям к L. corvus видов, отличающихся более низким
завитком и более крупным последним оборотом, а также пропорциями копуля-
тивного аппарата. Это L. gueretiniana (Servain, 1881) и L. curtacorvus Kruglov et
Starobogatov, 1984. Первый из них широко распространён в водоёмах Европей-
ской России и Украины, второй достоверно известен из некоторых озёр Карелии
(Пилджское, Святозеро). Европейские специалисты рассматривают эти виды в
составе вида L. corvus ’.
2(1). Раковина башневидная. Обороты сильно выпуклые, иногда почти ступен-
чатые...........Lymnaea taurica kazakensis Mozley, 1934 (вкл. табл. VI, рис. 92).
— palustris kazakensis Mozley, 1934: 3, pl. 1, 7 {Lymnaea).
Мешок пениса широкий. Индекс копулятивного аппарата составляет около 2,5-2,7.
Размеры: ВР до 34 мм, ШР до 14 мм.
Вид распространён на юге Восточной Европы, юге Западной Сибири, в Северном
и Южном Казахстане. В Европейской России известен из водоёмов Оренбургской
области. Возможно обитает в Астраханской области. Номинативный подвид,
L. taurica taurica (Clessin, 1880) описан из Крыма; недавно установлено его оби-
1 В случае сомнений в точности определения видовой принадлежности моллюсков группы
L. corvus рекомендуем обозначать их как L. (С.) corvus s. lato.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
407
тание на юге Украины, в Одесской области (Vinarski et al., 2012). L. t. kazakensis
обитает в степных и полупустынных районах во временных водоёмах1.
Подрод Stagnicola Leach in Jeffreys, 1831
Типовой вид: Buccinum palustre O.F. Muller, 1774.
Раковина стройная, твёрдостенная, башневидная, высококоническая либо
овально-коническая, малых и средних размеров (до 25-28 мм высотой). Пропор-
ции раковины, форма и степень выпуклости оборотов, скорость их нарастания
значительно варьируют в пределах подрода. Простата с одной внутренней склад-
кой. Проток семеприемника длинный. Для видов подрода характерна высокая
степень изменчивости раковины, поэтому в ряде случаев надёжное определение
видовой принадлежности моллюсков возможно только путём изучения копуля-
тивного аппарата.
Общее распространение подрода — голарктическое. Большинство видов Stag-
nicola обитают во временных или полупостоянных водоёмах, некоторые ведут
амфибионтный образ жизни.
Ключ для определения видов подрода Stagnicola
1(6). Обороты раковины плоские или слабо и равномерно выпуклые. Завиток
конической или яйцевидно-конической формы. Колумеллярная складка
выражена слабо.
2(3). Раковина очень стройная, башневидная, тангент-линия выгнутая. Высота
раковины не более 23 мм.....................................................
......................Lymnaea archangelica Kruglov et Starobogatov, 1986
(вкл. табл. VI, рис. 93).
— archangelica Круглов, Старобогатов, 1986: 60, рис. 1, 7; 2, 7 (Lymnaea).
Индекс копулятивного аппарата составляет чуть более 1,0.
Размеры: ВР до 23 мм, ШР до 8 мм.
Вид распространён на севере Европы, в Подмосковье, на юге Урала, в Западной
Сибири. Населяет временные водоёмы.
3(2). Раковина менее стройная, башневидно-коническая или высококоническая,
тангент-линия прямая или очень слабо выгнутая. Раковина крупнее, чем у
предыдущего вида, её высота достигает 28-30 мм.
4(5). Раковина башневидно-коническая с завитком правильной конической фор-
мы. Завиток относительно узкий, так что апикальный угол составляет не
более 40°..........Lymnaea palustris (O.F. Muller, 1774) (вкл. табл. VI, рис. 94).
— palustre О.F. Muller, 1774: 131 (Buccinum).
Индекс копулятивного аппарата составляет 0,7-0,8.
Размеры: ВР до 28 мм, ШР до 14 мм.
1 Данные молекулярной систематики указывают на принадлежность вида L. taurica к подроду
Lymnaea s. str. (Vinarski et al., 2012).
408
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Вид распространён в Европе, Передней и Малой Азии, в Западной Сибири. Мол-
люски обитают преимущественно в небольших временных водоёмах, часто в той
или иной степени заболоченных.
5(4). Раковина высококоническая, с относительно низким и широким завитком.
Апикальный угол составляет 45° и более.........................................
.....................Lymnaea atra (Schrank, 1803) (вкл. табл. VI, рис. 95).
— atrum Schrank, 1803: 288 (Buccinum).
[= L. palustris var. corviformis (Bourguignat in Servain, 1881)].
Индекс копулятивного аппарата составляет около 0,91.
Размеры: ВР до 32 мм, ШР до 16 мм.
Вид рапространён в Европе (на восток до Башкирии). Моллюски обитают в непо-
стоянных водоёмах.
Комментарии. В водоёмах крайнего северо-востока Европы (бассейн р. Печоры)
обитает близкий вид L. zebrella (В. Dybowski, 1913), который может быть встречен
и западнее, в бассейнах рек Белого и Баренцева морей. От L. atra он отличается
значительно более мелкими размерами (высота раковины не превышает 15 мм), а
также большими значениями Индекса копулятивного аппарата (0,97-1,15). L. ze-
brella, описанный из оз. Байкал, распространён также на севере Западной Сибири
и в Восточной Сибири.
6(1). Обороты раковины явственно выпуклые. Завиток высококонический, ко-
нический или башневидный. Колумеллярная складка обычно хорошо вы-
ражена.
7(8). Раковина очень стройная, башневидная. Завиток узкий и высокий. По-
следний оборот некрупный, его высота составляет не более 0,65 от высо-
ты раковины.................................Lymnaea callomphala (Servain, 1881)
(вкл. табл. VI, рис. 96).
— callomphala Servain, 1881: 78 (Limnaea).
[= L. palustris var. transsylvanica Kimakowicz, 1884].
Индекс копулятивного аппарата составляет около 0,75-0,90.
Размеры: ВР до 24 мм, ШР до 10 мм.
Вид распространён в Восточной Европе. По данным Н.Д. Круглова (2005) в ев-
ропейской части России вид распространён во временных пойменных водоёмах к
югу от линии Санкт-Петербург-Москва-Нижний Новгород. На крайнем юго-вос-
токе Европы эти моллюски отсутствует.
8(9). Раковина менее стройная, яйцевидно-коническая. Завиток широкий и
сравнительно низкий. Последний оборот высокий, его высота составляет
не менее 0,7 от высоты раковины.
9(10). Завиток коротко-башневидный. Апикальный угол не более 45°...............
.....................Lymnaea turricula (Held, 1836) (вкл. табл. VI, рис. 97).
— turricula Held, 1836: 278 (Limnaed).
Индекс копулятивного аппарата составляет около 1,0.
Размеры: ВР до 16 мм, ШР до 7 мм.
Вид распространён в Центральной и Восточной Европе. Моллюски обитают во
временных водоёмах, включая небольшие лужи.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
409
Комментарии. При использовании современной западноевропейской малаколо-
гической литературы (Jackiewicz, 1998; Gloer, 2002) следует иметь в виду, что в
ней под названием Lymnaea turricula или Stagnicola turricula фигурирует совсем
другой вид, отличающийся длинным мешком пениса и соответственно меньшими
значениями индекса копулятивного аппарата (0,20-0,33). Пока неясно, обитает ли
этот вид в водоёмах европейской части России.
10(9). Завиток правильной конической формы. Апикальный угол не менее 50°...
........................Lymnaea fusca (С. Pfeiffer, 1821) (вкл. табл. VI, рис. 98).
—fuscus С. Pfeiffer, 1821: 92, pl. 4, fig. 25 (Limnaeus).
Индекс копулятивного аппарата составляет около 0,61-0,66.
Размеры: ВР до 22 мм, ШР до 14 мм.
Вид распространён в постоянных водоёмах бассейна Балтийского моря, включая
опреснённые участки моря, откуда представители вида известны под названием
L. fusca maritima (Clessin, 1878).
Комментарии. При использовании современной западноевропейской малаколо-
гической литературы (Gloer, 2002; Gloer, Meier-Brook, 2003) следует иметь в виду,
что в ней под названием Stagnicola fusca фигурирует совсем другой вид, имеющий
не одну, а две внутренних складки в простате и специфическое строение копуля-
тивного аппарата, близкое к таковому у L. corvus. В работах М. Яцкевич (Jackie-
wicz, 1993, 1998) этот же вид именуется L. vulnerata (Kiister, 1862) non L. vulnerata
sensu Kruglov et Starobogatov, 1986. Пока неясно, обитает ли этот вид в водоёмах
европейской части России.
Подрод Peregriana Servain, 1881
Типовой вид: Buccinum peregrum O.F. Muller, 1774.
Раковина средних размеров (до 30-35 мм высотой), различной формы, от ухо-
видной до яйцевидной, относительно тонкостенная, со сравнительно некрупным
коническим завитком и более или менее вздутым последним оборотом. Колумел-
лярное вдавление развито слабо или отсутствует. Столбик с внутренним каналом.
Проток семеприемника короткий. Простата с одной внутренней складкой. Пре-
пуциум плотный, мешковидный, мешок пениса значительно более узкий; индекс
копулятивного аппарата заметно варьирует в пределах подрода.
Комментарии. Подрод Peregriana был выделен из состава подрода Radix Я.И. Старо-
богатовым и Л.Л. Будниковой (1976) на основе конхологических (слабое развитие колу-
меллярного вдавления, столбик с внутренним каналом) и анатомических (короткий проток
семеприемника) признаков. Зарубежными систематиками все виды подрода включаются в
состав (под)рода Radix s. lato (Piechocki, 1979; Jackiewicz, 1993; Ponder, Waterhouse, 1997;
Falkner et al., 2001; Gloer, 2002). Эта точка зрения по результатам некоторых исследований
подтверждается методами молекулярной систематики (Bargues et al., 2001; Bargues, Mas-
Coma, 2005).
Общее распространение подрода — палеарктическое, большинство видов обитают в
Европе и Северной Азии.
410
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Ключ для определения видов подрода Peregriana
1(6). Раковина уховидной формы. Устье очень крупное, широкое. Завиток чрез-
вычайно мал в сравнении с размерами последнего оборота.
2(3). Верхняя точка устья расположена выше верхней точки завитка или при-
близительно на одном уровне с ней.............................................
..............Lymnaea monnardi (Hartmann, 1841) (вкл. табл. VI, рис. 99).
— monnardi Hartmann, 1841: 71, tab. VI, fig. 18, 1-3 {Gulnaria}.
Обороты завитка сильно и равномерно выпуклые, нарастают быстро. Индекс ко-
пулятивного аппарата составляет 1,35-1,50.
Размеры: ВР до 25 мм, ШР до 22 мм.
Вид распространён в Центральной и Восточной Европе, на восток проникает до
бассейна Печоры и гор Южного Урала. Моллюски населяют постоянные водоёмы.
Комментарии. В водоёмах европейской части России может быть встречен ещё
один конхологически близкий вид, L. hartmanni (Studer, 1820), который отличается
от L. monnardi чуть меньшими значениями индекса копулятивного аппарата (око-
ло 1,2) и умеренно выпуклыми оборотами. Обороты завитка L. hartmanni нарас-
тают чуть быстрее, чем у L. monnardi.
3(2). Верхняя точка устья расположена ниже верхней точки завитка.
4(5). Завиток очень небольшой, едва возвышается над последним оборотом.
Высота его составляет, как правило, менее 0,2 высоты раковины.................
...............Lymnaea ampla (Hartmann, 1821) (вкл. табл. VI, рис. 100).
— ampla Hartmann, 1821: 250, tab. II, f. 29 (Limneus auricularius var.).
[= L. patula sensu Kruglov, Starobogatov, 1993b non Da Costa, 1778; L. tobolica Laza-
reva, 1967].
Индекс копулятивного аппарата составляет 0,7-1,0.
Размеры: ВР до 30 мм, ШР до 28 мм.
Вид распространён в Центральной, Южной и Восточной Европе, на Урале, в За-
падной Сибири, на юге Восточной Сибири (на восток до Тывы и, возможно, оби-
тает в верховьях рек Лена и Нижняя Тунгуска). Реликтовая часть ареала находится
в низовьях Енисея. Моллюски обитают в постоянных водоёмах со стоячей или
слабо проточной водой.
5(4). Завиток более крупный, заметно возвышается над последним оборотом.
Высота его составляет, как правило, 0,2-0,3 высоты раковины...................
...................Lymnaea tumida (Held, 1836) (вкл. табл. VII, рис. 101).
— tumida Held, 1836: 278 (Limnaea).
Индекс копулятивного аппарата составляет 1,5-2,0.
Размеры: ВР до 25 мм, ШР до 18 мм.
Вид распространён в Европе, горной части Центральной Азии, на юге Сибири (на
восток до Байкала). Реликтовая часть ареала находится в низовьях Енисея. Встре-
чается преимущественно в постоянных водоёмах.
6(1). Раковина иной формы, от яйцевидной до овально-конической. Устье уме-
ренных размеров, если крупное и широкое, то его палатальный край от
точки соединения с париетальным краем всегда идёт несколько вниз, что
придаёт раковине яйцевидную форму. Завиток сравнительно хорошо развит.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
411
7(10). Обороты завитка сильно и равномерно выпуклые, иногда даже ступенча-
тые, разделены глубоким швом.
8(9). Устье очень крупное, его высота в 2,5-3,0 раза превышает высоту завитка.
Форма раковины широкояйцевидная или близка к шаровидной......................
................Lymnaea mucronata (Held, 1836) (вкл. табл. VII, рис. 102).
— mucronata Held, 1836: 278 (Limnaea).
Индекс копулятивного аппарата составляет 0,85-0,86.
Размеры: ВР до 20 мм, ШР до 14 мм.
Вид распространён в Европе (бассейн Балтийского и Северного морей, атлантиче-
ское побережье). Моллюски обитают в крупных водоёмах.
9(8). Устье сравнительно некрупное, его высота менее чем в 2 раза превышает
высоту завитка. Форма раковины яйцевидная....................................
...........Lymnaea fulva (Ziegler in Ktister, 1862) (вкл. табл. VII, рис. 103).
—fulva Ktister, 1862: 16, taf. 4, fig. 19-21 (Limnaeus).
Индекс копулятивного аппарата составляет около 1,13.
Размеры: ВР до 20 мм, ШР до 14 мм.
Вид распространён в юго-восточной части Западной Европы, на юге и западе
Восточной Европы, включая южную часть бассейна Балтийского моря. Населяет
постоянные и временные водоёмы.
Комментарии. В водоёмах севера европейской части России может быть встречен
ещё один конхологически близкий вид, L. carelica Kruglov et Starobogatov, 1983,
который отличается несколько большей высотой устья и меньшей высотой завит-
ка. Изменчивость, экология и географическое распространение L. carelica плохо
изучены. До сих пор этот вид известен только из двух местонахождений: из Каре-
лии (Круглов, 2005) и на севере Кировской области (Шихова, 2004).
10(7). Обороты завитка уплощённые, слабовыпуклые или сильно, но неравно-
мерно выпуклые (грушевидные).
11(12). Раковина широкояйцевидная (иногда почти уховидная), последний оборот
очень сильно вздутый и высокий (его высота составляет около 0,9 высоты
раковины). Обороты завитка прижатые: предпоследний оборот очень ши-
рокий, значительно шире предшествующих; его ширина примерно в 4-5
раз превышает его высоту.....................................................
.......Lymnaea ampullacea (Rossmassler, 1835) (вкл. табл. VII, рис. 104).
— ampullaceus Rossmassler, 1835: 19, taf. 7, fig. 124 (Limnaeus).
Индекс копулятивного аппарата составляет 1,05-1,15.
Размеры: ВР до 25 мм, ШР до 20 мм.
Вид распространён в Европе, на юге Сибири (на восток до озера Байкал). Моллю-
ски населяют преимущественно постоянные непроточные водоёмы.
Комментарии. Особи этого вида с наиболее широкими раковинами иногда трудно
отличить от раковин L. ampla. В этом случае следует обращать внимание на фор-
му оборотов завитка (у L. ampullacea они прижатые, у L. ampla — не прижатые).
12(11). Раковина иной формы, если широкояйцевидная, то обороты завитка не
прижатые, так что ширина предпоследнего оборота в 3 и менее раза пре-
вышает ширину предшествующих. Последний оборот умеренно вздут, его
высота составляет обычно менее 0,9 высоты раковины.
412 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
13(16). Раковина яйцевидная с относительно низким завитком ширококонической
формы, мало возвышающимся над последним оборотом.
14(15). Оборот завитка, предшествующий последнему, заметно вздут и расширен
в сравнении с начальными оборотами. Апикальный угол не превышает
80°..................Lymnaea ovata (Drapamaud, 1805) (вкл. табл. VII, рис. 105).
— ovatus Drapamaud, 1805: 50, pl. 2, fig. 30-31 (Limneus).
Индекс копулятивного аппарата составляет около 0,95.
Размеры: ВР до 25 мм, ШР до 20 мм.
Вид распространён в Европе, Сибири (на восток до озера Байкал). Моллюски на-
селяют постоянные непроточные водоёмы и реки.
15(14). Оборот завитка, предшествующий последнему, почти не вздут и слабо
расширен в сравнении с вышележащими начальными оборотами. Апи-
кальный угол не менее 90°.....................................................
..............Lymnaea balthica (Linnaeus, 1758) (вкл. табл. VII, рис. 106).
— balthica Linnaeus, 1758: 775 (Helix).
[= L. ovata var. inflata Kobelt, 1870].
Индекс копулятивного аппарата составляет около 1,2.
Размеры: ВР до 28 мм, ШР до 22 мм.
Вид распространён в Европе и Южной Сибири (на восток до Прибайкалья и
оз. Байкал); реликтовая часть ареала — низовья Енисея. Моллюски населяют во-
доёмы различного типа. Наиболее часто встречаются в постоянных непроточных
водоёмах.
16(13). Раковина овально-коническая с относительно высоким завитком, заметно
возвышающимся над последним оборотом.
17(20). Завиток очень высокий, его высота составляет 0,7 и более от высоты рако-
вины.
18(19). Обороты завитка слабо и равномерно выпуклые, тангент-линия прямая
или очень слабо выгнутая. Мешок пениса заметно длиннее препуциума...
....................Lymnaea peregra (O.F. Muller, 1774) (вкл. табл. VII, рис. 107).
— peregrum O.F. Muller, 1774: 130 (Buccinum).
[=£. labiata (Rossmassler, 1835)].
Индекс копулятивного аппарата составляет 0,65-0,85.
Размеры: ВР до 22 мм, ШР до 14 мм.
Вид распространён в Европе и на юге Западной Сибири. Моллюски населяют пре-
имущественно временные водоёмы.
19(18). Обороты завитка неравномерно выпуклые (грушевидные), тангент-линия
вогнутая. Мешок пениса заметно короче препуциума..............................
................Lymnaea lagotis (Schrank, 1803) (вкл. табл. VII, рис. 108).
— lagotis Schrank, 1803: 290 (Buccinum).
Индекс копулятивного аппарата составляет 1,3-1,5.
Размеры: ВР до 25 мм, ШР до 15 мм.
Вид распространён в Европе, Западной Сибири, оз. Байкал, горах Центральной
Азии. Моллюски населяют преимущественно временные водоёмы, могут жить в
ручьях и малых реках.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
413
20(17). Завиток умеренно высокий, его высота составляет менее 0,6 от высоты
раковины.
21(22). Обороты завитка равномерно выпуклые, последний оборот умеренно
вздут. Высота завитка составляет 0,5-0,6 высоты устья. Мешок пениса
короткий (индекс копулятивного аппарата составляет 1,75-2,00)..................
.............Lymnaea intermedia Lamarck, 1822 (вкл. табл. VII, рис. 109).
— intermedia Lamarck, 1822: 162 {Lymnaea).
Размеры: ВР до 20 мм, ШР до 12 мм.
Вид распространён в Европе и на севере Азии (на восток до бассейна Колымы).
Моллюски населяют преимущественно временные водоёмы.
22(21). Обороты завитка неравномерно выпуклые, последний оборот сильно
вздут. Высота завитка составляет менее 0,48 высоты устья. Мешок пениса
равен по длине препуциуму или немного короче (индекс копулятивного
аппарата составляет 1,0-1,2)...................................................
...............Lymnaea fontinalis (Studer, 1820) (вкл. табл. VII, рис. ПО).
—fontinalis Studer, 1820: 27 {Limneus).
[= L. acutalis Morelet, 1845].
Размеры: BP до 22 мм, ШР до 15 мм.
Вид распространён в Европе, Сибири, оз. Байкал. Моллюски населяют водоёмы
различного типа, часто встречаются в ручьях и малых реках, а также некрупных
временных водоёмах.
Семейство ANCYLIDAE Rafinesque, 1815
Типовой род: Ancylus O.F. Muller, 1774.
Раковина маленькая (высота до 4 мм), колпачковидная. Поверхность может
быть ребристая или почти гладкая с тонкой исчерченностью. Окраска от роговой
до тёмно-коричневой. Эмбриональная раковина (протоконх) имеет форму кепки
или корнета, всегда ребристая (Hubendick, 1960; Солдатенко, 2010). Вершина (за-
виток) располагается на продольной оси раковины или немного смещена вправо,
в пределах контура устья, или вне его. Устье округлое, овальное или яйцевидное.
Тело левозавитое. Половые, выделительное и дыхательное отверстия рас-
положены последовательно, с левой стороны тела. Щупальца короткие, не пиг-
ментированные. Имеется маленькая адаптивная жабра. Гермафродитная железа
кистевидная, простата имеет небольшое число (до 7) неразветвлённых диверти-
кулов. Простатический проток отсутствует. Копулятивный аппарат, состоит из 2
отделов. Проксимальный отдел — мешок пениса с пенисом внутри и мышечно-
железистым придатком, который открывается в дистальной части мешка пениса,
образуя единую структуру. Дистальный отдел — препуциум (мышечный мешок с
мощными стенками), проксимальная часть которого у взрослых особей погруже-
на в дистальную. Препуциальный орган, препуциальная железа и внешний про-
ток отсутствуют. Пенис без стилета. Копулятивный аппарат очень компактный за
счёт частичного погружения его проксимального отдела в дистальный.
414
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Челюсть многопластинчатая. Радула насчитывает от 80 до 200 поперечных ря-
дов зубов. Зубов в поперечном ряду от 19 до 95. Формула поперечного полуряда
радулы С-пМ.
В ископаемом состоянии представители семейства известны из олигоцена
(эоцена) Европы, Северной Африки, Передней Азии, а также из неогена Север-
ной Америки (Старобогатов, 1970). Современные виды семейства распростране-
ны в проточных водоёмах и крупных озёрах (Охрид, Преспа, Селигер, Долосец)
Евразии, Северной Африки, Передней Азии и Северной Америки (Старобога-
тов, 1970). Моллюски обитают на крупных камнях и гальке, на илисто-песчаных
и илистых грунтах. Встречаются в зоне заплеска на камнях, на мелководьях (до
0,5 м), в озёрах на глубине до 40 м. Зимуют в неактивном состоянии, зарывшись
в грунт.
В пределах семейства Ancylidae выделяют 3 рода. В европейской части России
представлен 1 родЯису/ws' О.Е МйПег, 1774.
Комментарии. В отечественной и западной литературе не существует общепринято-
го мнения относительно таксономического статуса группы. Анцилиды неоднократно при-
нимались малакологами в ранге семейства (Thiele, 1931; Baker, 1956; Zilch, 1959-1960;
Taylor & Sohl, 1962; Hubendick, 1964; Акрамовский, 1976; Piechocki, 1979; Burch, 1989),
однако его границы были очерчены по-разному. Я.И. Старобогатов (1967) поместил род
Ancylus в семейство Planorbidae и эта точка зрения в конце XX века стала доминирующей
в отечественной (и некоторой зарубежной) малакологической литературе. Её разделяли
Стадниченко (1990) Солдатенко (Солдатенко, Старобогатов, 2004) и Глёер (Gloer, 2002).
Однако, в настоящий момент, опираясь на систему Бёрча (Burch, 1989) и анализируя весь
комплекс данных известных о пресноводных анцилюсах [с учётом морфологии сперми-
ев (Hodgson & Healy, 1998) и молекулярных данных (Jorgensen et al., 2004; Albrecht et al.,
2007)] выделение самостоятельного семейства Ancylidae представляется более обосно-
ванным.
Род Ancylus О.Е Muller, 1773
Типовой вид: Ancylus fluviatilis О.Е Muller, 1774.
Раковина маленькая, колпачковидная, часто полупрозрачная, рогового, жёлто-
го или коричневого цвета. Поверхность телеоконха (дефинитивной части рако-
вины) может быть ребристой или почти гладкой с продольной (спиральной) или
кольцевой (коллаборальной) исчерченностью. Вершина (завиток) располагается
на продольной оси раковины.
Для копулятивного аппарата характерен мешок пениса с пенисом внутри и
длинным мышечно-железистым придатком (7-8 мм) который открывается в дис-
тальной части мешка пениса, в месте его впадения образуются мышечные склад-
ки аналоги саркобеллума у Planorbidae.
Число поперечных рядов, количество зубов и их величина сильно варьируют в
пределах рода.
Род Ancylus обитает только в проточных водоёмах в переделах Палеарктики:
Европа до Урала, Северная Африка, Эфиопия, Передняя Азия. В палеонтологиче-
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
415
ской летописи известен с олигоцена и неогена на выше перечисленных террито-
риях (Старобогатов, 1970).
В настоящее время признаны в пресных водах европейской части России и на
прилегающих территориях 8 видов. Для севера европейской части России харак-
терны два вида.
Ключ для определения видов рода Ancylus
1(2). Раковина низкая (высота составляет менее 0,38 длины)..................
.........Ancylus subcircularis Clessin, 1882 (вкл. табл. VII, рис. 111 а-в).
— subcircularis Clessin, 1882: 33 {Ancylus).
Размеры: ВР до 2,4 мм, ШР (перпендикулярно продольной оси тела) до 5,6 мм, ДР
(вдоль продольной оси тела) до 6,8 мм.
Вид распространён в Европе. Моллюски обитают в проточных водоёмах в бассей-
нах Балтийского и Северного морей.
2(1). Раковина высокая (высота её составляет не менее 0,40 длины)...........
.........Ancylus fluviatilis O.F. Muller, 1774 (вкл. табл. VII, рис. 112 а-в).
—fluviatilis O.F. Muller, 1774: 201-202 {Ancylus).
Размеры: BP до2,9 мм, ШР до 5,2 мм, ДР до 6,1 мм.
Вид распространён в Европе (на восток до Урала). Не встречается в Средиземно-
морье и крайних северных областях. Моллюски обитают в проточных водоёмах.
Семейство HELISOMATIDAE ЕС. Baker, 1928
Типовой род: Helisoma Swainson, 1840.
Раковина крупная (ширина до 40 мм), плоскоспиральная или турбоспираль-
ная, гиперстрофная1 [как правозавитая {Planorbella pilsbryi, Helisoma anceps),
так и левозавитая {Planorbella duryi)]. Поверхность оборотов может быть ребри-
стой {Planorbella pilsbryi), или гладкой {Planorbella duryi). Окраска от роговой и
желтоватой до тёмно-коричневой. Оборотов 5-6. Периферия последнего оборота
может быть округлой {Planorbarius corneus), иметь острый уголок {Menetus орег-
cularis) или киль {Menetus brogniartiana). Пупок разного диаметра и глубины.
Форма устья от округлой {Planorbarius corneus) до овальной {Planorbella magnifi-
сит) или трапециевидной {Carinifex newberryi). Внутри последнего оборота по-
перечных пластин нет. Эмбриональная раковина асимметричная (имеет смещение
начальных оборотов).
Тело левозавитое. Половые, выделительное и дыхательное отверстия рас-
положены компактно в районе головы, с левой стороны. Щупальца нитевидные,
пигментированные. Адаптивная жабра хорошо развита. Гермафродитная железа
многорядная. У большинства видов простата с многочисленными веерообразны-
Гиперстрофия — явление, при котором у плоскоспиральных раковин нарастающие обороты
в верхней части расширяются сильнее, чем в нижней так, что каждый последующий оборот
оказывается как бы чуть выше предыдущего. Такая раковина, если она правозавитая, выглядит
ложно левозавитой и наоборот. Пример гиперстрофной раковины см. рис. 17Б.
416
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
ми дивертикулами, впадающими в семяпровод (без простатического протока). Ко-
пулятивный аппарат состоит из двух (мешок пениса и препуциум) или из трёх
(эякуляторный проток, мешок пениса и препуциум) отделов. Мешок пениса не
имеет железистых придатков. Пенис различной длины, семяизвергательный ка-
нал открывается субтерминально. Кончик пениса мягкий или вооружён стилетом
различной формы и размера. Внутри препуциума находятся мышечно-желези-
стые структуры (атриальный и препуциальный органы). Многие представители
семейства имеют дополнительный наружный проток, соединяющий мешок пени-
са и препуциум.
Челюсть подковообразная, преимущественно трех-раздельная или многопла-
стинчатая (подрод Carinifex). Формула поперечного полуряда радулы С-nM. Зу-
бов в поперечном ряду от 17 до 45. Зубы мелкие (их длина до 18 мкм).
В ископаемом состоянии моллюски семейства известны с юры (Старобогатов,
1970). Современные виды распространены в постоянных и временных пресно-
водных водоёмах Европы, в Северной Африке, Северной и Центральной Амери-
ке, Азии. В Европе моллюски населяют мелководные (глубиной до 1 м) участки
в различных водоёмах (озёра, реки, пруды, лиманы) с обильным листовым опа-
дом и зарослями водных растений. Зимуют в неактивном состоянии, зарывшись в
грунт (до 2 м).
Комментарии. Состав семейства окончательно не определён. В европейской части
России представлен только один род семейства — Planorbarius. Интродуцированные
виды, периодически появляющиеся в европейских водоёмах, не рассматриваются.
Род Planorbarius Dumeril, 1806
Типовой вид: Helix cornea Linnaeus, 1758.
Раковина большая (ширина до 40 мм), плоскоспиральная, левозавитая, твёр-
достенная, красноватого, коричневого или чёрного цвета. Высота раковины со-
ставляет 0,37-0,57 ширины. Верхняя (апикальная) поверхность раковины незна-
чительно погружена относительно последнего оборота. Обороты (5-6) выпуклые,
быстро нарастающие, хорошо просматриваются с обеих сторон раковины. Пери-
ферия последнего оборота округлая. Устье округлое или овальное, вырезанное
стенкой предпоследнего оборота, внутри без образований. Поверхность раковины
иногда со спиральной исчерченностью, особенно хорошо заметной на последнем
обороте, или с маллеатной скульптурой.
Половые протоки массивные, не тонкие. Копулятивный аппарат представите-
лей рода массивный (до 25 мм в длину). Он состоит из толстостенного эякулятор-
ного протока (выполняющего функцию изгнания спермы во время копуляции),
мешка пениса и препуциума, соотношение длин последних (индекс копулятивно-
го аппарата) — 7,0-9,0. Пенис очень маленький, семяизвергательный канал от-
крывается субтерминально. На конце пениса имеется маленькая папилла, стилет
отсутствует. Внутри препуциума находятся атриальный и препуциальный органы
(мышечно-железистая выросты стенки препуциума) (Soldatenko & Petrov, 2013)
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
417
Длина копулятивного аппарата равна длине глотки половозрелых особей и при-
мерно в 4 раза меньше длины тела фиксированного моллюска.
Радула широкая, число поперечных рядов зубов 160-173. Формула поперечно-
го пол уряда радулы до 045 М.
Род Planorbarius известен с эоцена (Старобогатов, 1970). Современные пред-
ставители рода распространены в Европе, Северной Африке, Азии (на восток до
бассейна Енисея). Моллюски обитают в зарослях водной растительности времен-
ных и постоянных водоёмов равнинных районов Палеарктики (Старобогатов,
1977; Стадниченко, 1990; Старобогатов и др., 2004).
Разные специалисты различают в составе рода от 1 до 8 видов (Старобогатов
и др., 2004; Gloer, 2002). Для севера европейской части России достоверно уста-
новлено обитание двух видов.
Ключ для определения видов рода Planorbarius
1(2). При взгляде со стороны устья контур раковины трапециевидный (области
периферии последнего оборота и устья скошены). При 5 оборотах высота
устья составляет Vi ширины раковины. Обороты слабовыпуклые, разде-
лённые умеренно углублённым швом...........................................
......Planorbarius corneus (Linnaeus, 1758) (вкл. табл. VII, рис. 113 а-в).
— corneus Linnaeus, 1758: 770 (Helix).
Размеры: ВР до 14 мм, ШР до 35 мм. Тело моллюсков чёрное, у молодых особей
коричневато-чёрное.
Вид распространён в Европе, Западной Сибири (до бассейна Оби). Моллюски
обитают в крупных стоячих и проточных водоёмах на растительности.
2(1). При взгляде со стороны устья контур раковины овально-прямоугольный
(области периферии последнего оборота и устья более или менее равно-
мерно округлые). При 5 оборотах высота устья составляет менее Yi шири-
ны раковины. Обороты сильновыпуклые (особенно последний), разделён-
ные глубоким швом..........................................................
..................Planorbarius grandis (Dunker in Ktister et Dunker, 1850)
(вкл. табл. VII, рис. 114 a-в).
— grandis Dunker — Ktister et Dunker, 1850: 35, pl. 7, figs. 1-3 (Planorbis).
Размеры: BP до 20 мм, ШР до 40 мм. Тело моллюсков серое.
Вид распространён в северо-западной части Причерноморья. Возможно обнару-
жение вида в бассейне Балтийского моря. Моллюски живут в заросших участках
озёр и рек.
Семейство PLANORBIDAE Rafinesque 1815
Типовой род: Planorbis О.Е Muller, 1773.
Раковина маленькая (ширина до 20 мм), плоскоспиральная или турбоспи-
ральная, правозавитая (Bathyomphalus contortus), гиперстрофная (Planorbis pla-
norbis), реже левозавитая, условно правозавитая (не имеет смещения эмбрио-
418
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
нальных оборотов как у Planorbis planorbis). Поверхность оборотов может быть
ребристой, гладкой, иногда с периостракальными рёбрышками, щетинками и
даже шипами (род Armiger). Окраска от роговой и желтоватой до тёмно-корич-
невой. Оборотов 3-9. Периферия последнего оборота может быть округлой (Ani-
sus spirorbis), иметь острый уголок (Gyraulus stelmachoetius) или киль (Planorbis
planorbis). Диаметр внутренних оборотов (ДВО) не менее !4 ширины раковины.
Пупок разного диаметра и глубины. Форма устья от серповидной (Bathyomphalus
contortus) до округлой (Choanomphalus rossmaessleri) или ромбовидной (Anisus
vortex). Эмбриональная раковина симметричная (не имеет смещения начальных
оборотов).
Тело левозавитое. Половые, выделительное и дыхательное отверстия располо-
жены компактно в районе головы, с левой стороны тела. Щупальца нитевидные,
могут быть пигментированными. Адаптивной жабры нет. Гермафродитная желе-
за может состоять из одного ацинуса (Choanomphalus annuliformis), а также быть
одно- (Choanomphalus grachevi) или двухрядной (Planorbis planorbis). У боль-
шинства видов простата с многочисленными (до 50) неразветвлёнными, располо-
женными в один ряд (Planorbis planorbis) или в несколько рядов (Carinogyraulus
crenophilus) дивертикулами, впадающими в простатический проток. У наиболее
примитивных групп семейства простатический проток отсутствует. Копулятив-
ный аппарат состоит из двух отделов (мешок пениса с пенисом и препуциум).
Мешок пениса может иметь один-два свободных железистых придатка. Кончик
пениса мягкий или вооружён стилетом различной формы и размера (Baker, 1945;
Soldatenko, 2009). Челюсть подковообразная многопластинчатая (род Planorbis)
или трёхраздельная (род Biomphalaria). Радула длинная. Формула поперечного
полуряда радулы С-nM. Зубов в поперечном ряду от 17 до 113. Зубы мелкие (их
длина не более 14 мкм).
В ископаемом состоянии моллюски семейства известны с юры (Старобогатов,
1970). Современные виды распространены в постоянных и временных пресно-
водных водоёмах всех континентов. В Европе они населяют участки с обильным
листовым опадом и заросли водных растений обычно на мелководьях (до 0,5 м).
Зимуют в неактивном состоянии, зарывшись в грунт (до 2 м).
В составе семейства выделяют два подсемейства: Biomphalariinae Watson,
1954 и Planorbinae Rafinesque, 1815. В европейской части России представлено
только одно подсемейство Planorbinae.
Комментарии. Состав семейства значительно менялся на протяжении последних 150
лет (Thiele, 1931; Soos, 1935; Baker, 1945; Zilch, 1959; Taylor & Sohl, 1962; Hudec, 1967;
Старобогатов, 1967; Hubendick, 1978; Piechocki, 1979; Burch, 1989; Nordsieck, 1992). Рабо-
ты последних лет (Morgan et al., 2002; Jorgensen et al., 2004; Albrecht et aL, 2007; Солдатен-
ko, 2011; Soldatenko & Petrov, 2012; Soldatenko & Shatrov, неопубликованные данные 2016)
показали, что в объёме, принимаемом до конца XX века, семейство являлось сборным.
В результате бывшие подсемейства Ancylinae Rafinesque, 1815, Helisomatinae Baker. 1928
и Segmentinae Baker, 1945 были выделены из состава Planorbidae в ранге самостоятельных
семейств.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
419
Подсемейство Planorbinae Rafinesque 1815
Типовой род: Planorbis O.F. Muller, 1773.
Раковина маленькая (ширина до 20 мм), плоскоспиральная или турбоспираль-
ная, правозавитая, реже левозавитая, гиперстрофная. Плоскоспиральные ракови-
ны могут иметь облик плоского диска (Anisus vortex), утолщённого диска (Anisus
spirorbis) или шайбы (Bathyomphalus contortus). Турбоспиральные раковины мо-
гут быть кубаревидными (Choanomphalus amauronius) или низкоконическими
(всегда сильно прижатыми как у Choanomphalus lindholmi). Эмбриональные обо-
роты, как правило, покрыты тонкими спиральными линиями. Дефинитивные обо-
роты с брюшной стороны могут быть не вдавленными или вдавленными (погру-
жёнными). В последнем случае можно говорить о наличии пупка в виде широкой
воронки (не менее Уг ширины раковины). Иногда пупок узкий (до проколовидно-
го). Устье различной формы.
Копулятивный аппарат состоит из двух отделов (мешок пениса с пенисом и
препуциум), не имеющих дополнительных мышечных или железистых структур.
Кончик пениса со стилетом.
Радула выступает за пределы глотки, но составляет не больше !4 её длины.
В поперечном ряду радулы от 17 до 113 мелких зубов (длина зуба 3-14 мкм).
В ископаемом состоянии моллюски подсемейства известны с юры (Старобо-
гатов, 1970). Ныне живущие представители подсемейства относятся к девяти ро-
дам, шесть из которых представлены в европейской части России.
Ключ для определения родов подсемейства Planorbinae
1(2). Раковина крупная, твёрдостенная, при 4 оборотах её ширина не менее
9,0 мм, высота около 3,0 мм.........Род Planorbis O.F. Muller, 1773 (с. 431).
2(1). Раковина средних размеров или маленькая, при 4 оборотах её ширина не
более 8,0 мм, высота около 1,8 мм.
3(4). При взгляде со стороны устья контур раковины в виде высокой шайбы.
При взгляде сверху тесно навитые медленно нарастающие обороты напо-
минают скрученный ремень................................................
................Род Bathyomphalus Agassiz in Charpentier, 1837 (с. 424).
4(3). Раковина плоскоспиральная в виде уплощённого диска, плоского диска
или турбоспиральная.
5(6). Верхняя и нижняя стороны раковины при визуальном осмотре (без уве-
личения) выглядят практически одинаковыми. Высота оборотов раковины
увеличивается медленно и равномерно относительно оси навивания, по-
этому завиток и пупок раковины почти не погружённые (верхние обороты
и околопупочное поле сильно не вдавленные) и имеют равные диаметры.
...........................................Род Anisus Studer, 1820 (с. 420).
6(5). Верхняя и нижняя поверхности раковины, за редким исключением, легко
различимы (без увеличительных приборов). Высота оборотов раковины
420
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
увеличивается быстро и часто неравномерно относительно оси навивания,
поэтому завиток и пупок раковины погружены (верхние обороты и около-
пупочное поле вдавленные) и имеют различные диаметры.
7(8). Устье не врезано стенкой предпоследнего оборота. Часто на поверхности
раковины развиты периостракальные рёбра....................................
...................................Род Armiger Hartmann, 1840 (с. 423).
8(7). Устье врезано стенкой предпоследнего оборота, периостракальные рёбра
отсутствуют.
9(10). Число дефинитивных оборотов раковины больше 4......................
.......................Род Gyraulus Agassiz in Charpentier, 1837 (с. 428).
10(9). Число дефинитивных оборотов раковины не превышает 4................
............................Род Choanomphalus Gerstfeldt, 1859 (с. 426).
Род Anisus Studer, 1820
Типовой вид: Helix spirorbis Linnaeus, 1758.
Раковина маленькая (ширина не более 11,0 мм), плоскоспиральная (в виде
уплощённого диска или блюдца), условно правозавитая, гиперстрофная, тонко-
стенная, невысокая (высота раковины 0,11-0,28 ширины). Поверхность её глян-
цевая, гладкая, часто прозрачная, или с осевой, либо спиральной исчерченностью
(Anisus spirorbis). у Anisus strauchianus с радиальными рёбрышками. Цвет варьи-
рует (от светло-рогового и желтоватого до коричневого). Верхняя и нижняя сто-
роны раковины выглядят почти одинаково, но поверхность оборотов на нижней
стороне более плоская. Оборотов 5-9, медленно нарастающих. Все они хорошо
просматриваются с обеих сторон раковины. Периферия последнего оборота мо-
жет быть гладкой, угловатой или с хорошо выраженным килем. Устье ромбовид-
ное, овальное или округлое, вырезанное стенкой предпоследнего оборота.
Половые протоки тонкие и длинные. Копулятивный аппарат состоит из мешка
пениса и препуциума примерно равных по длине и ширине. Семяизвергательный
канал открывается субтерминально, кончик пениса представлен мышечно-желе-
зистой папиллой и всегда заканчивается трубчатым стилетом. Длина копулятив-
ного аппарата в два раза превышает длину глотки. Радула длинная, число по-
перечных рядов зубов до 140-195. Формула поперечного полуряда радулы до
С-20М (С-16М, по Baker, 1945).
В ископаемом состоянии моллюски рода известны из миоцена и плиоцена Ев-
ропы и Северной Африки, плиоцена запада США (Старобогатов, 1970). Совре-
менные представители распространены в Палеарктике. В европейской части Рос-
сии обитают 6 видов из 4 подродов.
Комментарии. Разные специалисты указывали разное число видов в составе рода.
В работах на русском языке приводится от 7 (Жадин, 1952) до 56 (Старобогатов и др.,
2004). В европейских научных источниках признаны валидными не более 7 видов (Gloer,
2002; Bouchet et al., 2005). Такая ситуация объясняется, отсутствием значительных раз-
личий в строении копулятивного аппарата у представителей родов Anisus Studer, 1820,
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
421
Bathyomphalus Agassiz in Charpentier, 1837 и Gyraulus Agassiz in Charpentier, 1837, на ос-
новании чего, Старобогатов и др. (2004) объединили их в один род Anisus в ранге подро-
дов. Авторы определителя не разделяют точку зрения Старобогатова и др. (2004), насчи-
тывая в составе рода 10 видов, сгруппированных в 4 подрода.
Ключ для определения подродов и некоторых видов рода Anisus
1(4). Последний оборот раковины с килем или хорошо выраженным углом, что
отражается на форме палатального края устья.
2(3). Периферия последнего оборота раковины посередине с углом. Последний
оборот с верхней стороны раковины практически не выступает над пред-
шествующими...................Подрод Vorticulus Prozorova et Starobogatov, 1997;
Anisus vorticulus (Troschel, 1834) (вкл. табл. VII, рис. 115 а-в).
— vorticulus Troschel, 1834: 51 (Planorbis).
Периферийный угол стенки последнего оборота может быть смещён ниже середи-
ны высоты раковины, если обороты снизу погружённые.
Размеры: ВР до 0,8 мм, ШР до 5,5 мм.
Вид распространён в Европе и на юге Западной Сибири. Редкий вид. Моллюски
пока известны только из озёр.
Комментарии. В начале XXI столетия на основе положения и выраженности
периферийного угла последнего оборота раковины было предложено различать в
составе подрода 3 вида: A. vorticulus, A. charteus (Held, 1837) и Anisus bavaricus
(Westerlund, 1885) (Прозорова, 2003; Старобогатов и др., 2004). При подготовке
определителя у предложенных видов достоверных различий по этим признакам
не выявлено. Положение и выраженность периферийного угла сильно изменчивы.
Вид A. charteus имеет анатомические особенности (Солдатенко, 2004). Копулятив-
ный аппарат и сперматека у половозрелых особей очень маленькие (около 1 мм).
3(2). Периферия последнего оборота с углом или килем, расположенным на
границе с нижней стороной раковины. Последний оборот с верхней сто-
роны раковины заметно выступает над предшествующими, придавая рако-
вине облик блюдца...........................Подрод Disculifer С. Boettger, 1944;
Anisus vortex (Linnaeus, 1758) (вкл. табл. VII, рис. 116 а-в).
— vortex Linnaeus, 1758: 770 (Helix).
Если нижняя сторона раковины выпуклая, то киль может смещаться ближе к сере-
дине периферии последнего оборота.
Размеры: ВР до 1,3 мм, ШР до 10,2 мм.
Вид распространён в Европе (кроме крайнего севера) и Западной Сибири. Мол-
люски обитают в постоянных водоёмах (озёрах, реках и их старицах) и некоторых
временных водоёмах, имеющих связь с постоянными (небольшие болота или при-
дорожные канавы).
Комментарии. В составе подрода различают от двух (Gloer, 2002) до шести ви-
дов (Прозорова 2003; Старобогатов и др., 2004). Результаты наших исследований
раковины и анатомии моллюсков подрода не подтвердили видовую самостоятель-
ность Anisus carinea (Westerlund, 1897); A. discoides (Reinhardt, 1870); A. johanseni
(Mozley, 1935) и A. nummulus (Held, 1837).
422
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
4(1). Последний оборот раковины без киля или сильно выраженного угла, пала-
тальный край устья закруглённый или с лёгкой угловатостью на границе с
базальным краем.
5(6). Поверхность раковины покрыта радиальными рёбрышками.....................
......Подрод Costorbis Lindholm, 1926; Anisus strauchianus (Clessin, 1884)
(вкл. табл. VII, рис. 117 а-г).
— strauchianus Clessin, 1884: 204-205, pl. 31, fig. 5 {Planorbis).
Размеры: BP до 0,9 мм, ШР до 5,0 мм.
Вид распространён в Европе, кроме крайнего севера. Вид встречается редко. Мол-
люски живут во временных водоёмах.
Комментарии. Статус подрода спорный, так как характерных анатомических осо-
бенностей, отличающих его представителей от моллюсков подрода Anisus s. str. не
установлено.
6(5). Поверхность раковины лишена ребристости, только с тонкими линиями
нарастания.................................................Подрод Anisus s. str.
Поверхность раковины с тонкими линиями нарастания. Устье, округлое, овальное
или округло-квадратное (ширина примерно равна высоте), не вырезанное стенкой
предпоследнего оборота. Копулятивный аппарат взрослых моллюсков длинный
(2-3 мм).
Комментарии. В пресных водах европейской части России и на прилегающих
территориях в настоящее время известно 5 видов подрода. В данном определителе
приведены только 3 хорошо различимых вида.
Ключ для определения видов подрода Anisus s. str.
1(2). Обороты нарастают неравномерно, ширина последнего оборота в два раза
больше ширины предпоследнего. Последний оборот с верхней стороны
раковины заметно возвышается над предшествующими, раковина имеет
облик блюдца..................................................................
............Anisus spirorbis (Linnaeus, 1758) (вкл. табл. VII, рис. 118 а-в).
— spirorbis Linnaeus, 1758: 770 {Helix).
Размеры: ВР до 1,5 мм, ШР до 7,2 мм.
Вид распространён в Европе и Западной Сибири. Моллюски обитают во времен-
ных водоёмах.
2(1). Обороты нарастают равномерно, ширина последнего оборота больше
ширины предпоследнего примерно в 1,5 раза. Каждый оборот с верхней
и нижней сторон раковины незначительно возвышается над предыдущим
(исключая эмбриональные обороты), поэтому раковина имеет облик вы-
пуклого (в различной степени) диска.
3(4). Обороты на верхней и нижней сторонах раковины практически одинаково
выпуклые...........Anisus leucostoma (Millet, 1813) (вкл. табл. VII, рис. 119 а-в).
— leucostoma Millet, 1813: 1617 {Planorbis).
[= Planorbis rotundatus W. Dybowski, 1913; Planorbis {Anisus) leucostoma var. niko-
lensis Kozhov, 1936].
Размеры: BP до 1,5 мм, ШР до 9,0 мм.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
423
Вид распространён в Европе и Сибири. Моллюски обитают во временных водо-
ёмах.
Комментарии. Крупные особи этого вида часто относят к виду Anisus septem-
gyratus (Ziegler in Rossmassler, 1835), обитающему только в южной Европе и не
проникающему севернее территорий Чехии и Украины. A. leucostoma достоверно
отличается от A.septemgyratus скоростью нарастания оборотов.
4(3). Обороты на верхней стороне раковины выпуклые, а на нижней — пло-
ские ...Anisusperezii (Graells in Dupuy, 1850) (вкл. табл. VII, рис. 120 a-в).
— perezii Graells — Dupuy, 1850: 441 (Planorbis).
Размеры: BP до 1,5 мм, ШР до 8,0 мм.
Вид распространён на северо-западе и юге Европы. В России встречается в бас-
сейнах Балтийского и Белого морей. Моллюски живут в мелководных временных
водоёмах.
Комментарии. В определительных ключах, изданных на русском языке во второй
половине XX - начале XXI веков, в качестве характерного признака для иденти-
фикации A. perezii предлагалось использовать наличие белого утолщения/губы
внутри устья или беловатых полос на последнем обороте (Старобогатов, 1977;
Прозорова, 2003; Старобогатов и др., 2004). Этот признак не является диагно-
стическим, так как такие утолщения вблизи устьевого края и внутри последнего
оборота, выглядящие как белые радиальные полосы, формируются у всех видов
данного подрода в период приостановки роста раковины в неблагоприятных ус-
ловиях.
Род Armiger Hartmann, 1840
Типовой вид: Nautilus crista Linnaeus, 1758.
Раковина маленькая (ширина до 3 мм), плоскоспиральная, правозавитая, тон-
костенная, рогового, жёлтого или коричневого цвета. Верхняя сторона раковины
плоская, или с несколько выступающим завитком. Оборотов 3^4, быстро нараста-
ющих. Последний оборот к устью опущен. Периферия последнего оборота угло-
ватая или округлая. Устье овальное, не вырезанное стенкой предпоследнего обо-
рота. Поверхность оборотов может нести периостракальные рёбрышки, иногда
переходящие в шипы.
Копулятивный аппарат состоит из примерно равных по длине отделов, но ме-
шок пениса уступает по ширине препуциуму. Пенис длинный, семяизвергатель-
ный канал открывается субтерминально, кончик пениса заканчивается мышеч-
но-железистой папиллой и всегда имеет колпачковидный стилет (Odhner, 1956;
Hubendick, 1957). Копулятивный аппарат взрослой особи втрое длиннее глотки.
Радула имеет до 150 поперечных рядов зубов. Формула поперечного полуряда
С-12М (С-16М по Baker, 1945).
Моллюски рода Armiger в ископаемом состоянии известны из миоцена и плио-
цена Европы и Алтая. Современные представители рода широко распространены
в водоёмах Европы, северных областей Азии и Северной Америки (Старобогатов,
1970). На севере и северо-западе европейской части России обитают 2 вида.
424
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Комментарии. Западноевропейские систематики (Gloer, 2002) признают валидным на
территории Европы только 1 вид — Armiger crista (Linnaeus, 1758). Ряд отечественных ма-
лакологов различают от четырёх (Старобогатов и др., 2004) до семи (Иззатуллаев, 1972;
Prosorova et Starobogatov, 1996) видов.
Ключ для определения видов рода Armiger
1(2). Раковина светлая, серовато-жёлтая, прозрачная, пупок широкий (диаметр
пупка составляет 0,70-0,95 ширины последнего оборота)...........................
.............Armiger crista (Linnaeus, 1758) (вкл. табл. VII, рис. 121 а-в).
— crista Linnaeus, 1758: 709 (Nautilus).
[= Turbo nautileus Lnnaeus, 1767; Planorbis imbricatus O.F. Muller, 1774: 165; Pla-
norbis cristatus Drapamaud, 1805; Planorbis crista var. spinulosus Clessin, 1873; Ar-
miger jagellonensis B. Dybowski et Grochmalicki, 1922; Armiger sobieskianus B. Dy-
bowski et Grochmalicki, 1922; Armiger gedyminensis B. Dybowski et Grochmalicki,
1922; Planorbis crista var. inermis Lindholm, 1926, partim].
Угол на периферии последнего оборота (если имеется) может быть смещён к
верхней стороне раковины, в результате чего контур устья приобретает овально-
четырёхугольную форму. Последний оборот может нести мощные, редко рас-
ставленные рёбра (числом не больше 18), переходящие на периферии раковины в
шипы. Встречаются особи с раковиной, лишённой рёбер, с плавно закруглённой
периферией последнего оборота и овальным контуром устья.
Размеры: ВР до 1 мм, ШР до 3 мм.
Вид распространён в Европе и Западной Сибири. Моллюски обитают в постоян-
ных стоячих и слабопроточных водоёмах. Обычно держатся в зарослях водной
растительности стариц и затонов рек.
2(1). Раковина роговая, серовато-коричневая или красноватая, полупрозрачная,
пупок уже, чем у предыдущего вида (диаметр пупка 0,55-0,67 ширины
последнего оборота).............................................................
..........Armiger bielzi (Kimakowicz, 1884) (вкл. табл. VII, рис. 122 а-в).
— crista var. bielzi Kimakowicz, 1884: 79 (Planorbis).
[= Armiger vitoldianus B. Dybowski et Grochmalicki, 1922; Armiger paravitoldianus
B. Dybowski et Grochmalicki, 1922; Planorbis crista var. inermis Lindholm, 1926,
partim].
Угол на периферии последнего оборота расположен на середине его высоты, устье
овально-ромбическое. Рёбра (если имеются), более тонкие и частые, чем у пре-
дыдущего вида (их число не менее 20), шипов не образуют и не увеличиваются в
высоту к периферии последнего оборота.
Размеры: ВР до 0,9 мм, ШР до 2,7 мм.
Вид распространён в Европе и Западной Сибири. Моллюски живут в постоянных
стоячих и слабопроточных водоёмах. Вид обычен в зарослях водной раститель-
ности стариц и затонов рек.
Род Bathyomphalus Agassiz in Charpentier, 1837
Типовой вид: Helix contorta Linnaeus, 1758.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
425
Раковина маленькая (ширина до 6 мм), плоскоспиральная, условно правозави-
тая или правозавитая, относительно высокая (напоминает шайбу или свернутый
в спираль ремень, её высота составляет 0,3-0,4 ширины), окраска варьирует от
светло-роговой и жёлтой, до коричневатой. Иногда раковина прозрачная. Верхняя
сторона раковины уплощённая, нижняя — с воронковидным пупком. Оборотов
7-9, медленно нарастающих. Все они хорошо просматриваются с обеих сторон
раковины. Периферия последнего оборота гладкая. Скульптура эмбриональных и
дефинитивных оборотов представлена тонкими линиями нарастания. Устье сер-
повидное или овальное, вырезанное стенкой предпоследнего оборота.
Половые протоки тонкие и длинные. Копулятивный аппарат состоит из двух
отделов, мешка пениса (более длинного) и препуциума. Индекс копулятивного
аппарата половозрелых особей 0,50-0,89. Пенис длинный, семяизвергательный
канал открывается субтерминально, кончик пениса заканчивается мышечно-же-
лезистой папиллой и трубчатым стилетом. Копулятивный аппарат взрослой особи
в три раза длиннее глотки. Радула насчитывает до 240 поперечных рядов зубов.
Формула поперечного полуряда С-18М.
В палеонтологической летописи моллюски рода Bathyomphalus появляются с
миоцена и плиоцена Европы и Северной Африки (Старобогатов, 1970). Современ-
ные представители распространены в водоёмах Европы и севера Азии (Старобо-
гатов и др., 2004). На севере европейской части России распространены 3 вида.
Комментарии. Европейские малакологи (Hudec, 1964; Gloer, 2002) признают на тер-
ритории Европы один вид — Bathyomphalus contortus (Linnaeus, 1758), объединяя внутри
него несколько форм, существенно различающихся особенностями раковины (например,
скоростью нарастания оборотов). Отечественные малакологии выделяют до пяти видов
Bathyomphalus (Старобогатов и др., 2004).
Ключ для определения видов рода Bathyomphalus
1(2). При 5 оборотах высота раковины не превышает 1,4 мм, устье относитель-
но широкое (ширина просвета устья больше половины высоты последнего
оборота)...................................................................
...Bathyomphalus dispar (Westerlund, 1871) (вкл. табл. VII, рис. 123 а-в).
— dispar Westerlund, 1871: 131 (Planorbis).
Обороты с верхней стороны раковины — выпуклые, шов между ними глубокий.
Размеры: ВР до 1,4 мм, ШР до 4,5 мм.
Вид распространён в Европе. Моллюски обитают в постоянных водоёмах, как
правило, в озёрах.
2(1). При 5 оборотах высота раковины не меньше 1,45 мм, устье узкое, серпо-
видное (ширина просвета устья не более половины высоты последнего
оборота).
3(4) Обороты нарастают быстро, на верхней стороне раковины они сильно-
выпуклые, разделены глубоким швом, начальные обороты (около двух)
заметно погружены..........................................................
...Bathyomphalus crassus (Da Costa, 1778) (вкл. табл. VIII, рис. 124 а-в).
426
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
— crassa Da Costa, 1778: 66 (Helix).
[= Planorbis (Bathyomphalus) contortus sensu Lindholm, 1909; B. Dybowski, 1912;
Kozhov, 1936, partim, non Linnaeus, 1758].
Размеры: BP до 2 мм, ШР до 6 мм.
Вид распространён в Европе и Сибири (до реки Колымы). Моллюски обитают в
постоянных водоёмах.
4(3). Обороты нарастают медленно, на верхней стороне раковины они слабо-
выпуклые, шов между ними мелкий, начальные обороты не погружённые
(верхняя сторона раковины плоская)............................................
....Bathyomphalus contortus (Linnaeus, 1758) (вкл. табл. VIII, рис. 125 а-в).
— contorta Linnaeus, 1758: 770 (Helix).
Размеры: ВР до 2 мм, ШР до 6 мм.
Вид распространён в Европе и Сибири. Моллюски обитают в постоянных про-
точных и стоячих водоёмах.
Род Choanomphalus Gerstfeldt, 1859
Типовой вид: Choanomphalus maacki Gerstfeldt, 1859.
Раковина маленькая (ширина до 7 мм), турбоспиральная (кубаревидная или
сильно прижатая низкоконическая), реже плоскоспиральная (дисковидная); пра-
возавитая, обычно твёрдостенная, непрозрачная [исключение Choanomphalus ri-
parius (Westerlund, 1865)], коричневого, желтоватого или рогового цвета. Завиток
сильно изменчив (от вдавленного до значительно возвышающегося), пупок от
широко открытого и воронковидного до проколовидного и закрытого. Оборотов
3-5. Периферия последнего оборота угловатая или округлая. Устье различной
формы, слабо вырезанное стенкой предпоследнего оборота.
Копулятивный аппарат состоит из двух отделов: узкого, но хорошо отличимо-
го от семяпровода мешка пениса (различной длины) и массивного препуциума.
Индекс копулятивного аппарата варьирует от 0,33 до 3,00. Пенис длинный, семя-
извергательный канал открывается субтерминально или терминально. Имеется
конусовидный стилет. Копулятивный аппарат половозрелых особей в 1,5-2,0 раза
больше глотки. Радула включает 132-195 поперечных рядов зубов. Формулы по-
перечного полуряда радулы у разных видов от С-8М до С-31М.
В ископаемом состоянии моллюски рода Choanomphalus известны из верхнего
мела (?), и миоцена севера Азии (Старобогатов, 1970). Современные представи-
тели рода распространены на территории Европы и на севере Азии. Моллюски
обитают в зарослях водной растительности во временных и постоянных водоёмах
на глубинах от 0 до 580 м. По современным данным в составе рода насчитывает-
ся 11 подродов, включающих 34 вида. Большинство видов обитают в озёрах Бай-
кал и Хубсугул, а также в реке Ангаре (Бекман, Старобогатов, 1975; Prozorova,
Starobogatov, 1997; Sitnikova, Ropstorf, 1999; Старобогатов и др., 2004; Ситникова
и др., 2004). Для севера европейской части России характерны два вида из подро-
дов Lamorbis Starobogatov, 1967 и Pseudogyraulus Prozorova et Starobogatov, 1997.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
427
Комментарии. В малакологической литературе стран Западной Европы в настоящее
время предлагается относить Choanomphalus (Lamorbis) riparius и Choanomphalus (Pseu-
dogyraulus) rossmaessleri к роду Gyraulus Agassiz in Charpentier, 1837 на основании сход-
ства признаков раковины и отсутствия надёжных различий в строении выделительной и
половой системы (Gloer, 2002; Bouchet et al., 2005). Однако для моллюсков из родов Choa-
nomphalus и Gyraulus характерны принципиально разные стилеты (конусовидный и труб-
чатый соответственно). В последнее время показано, что такие типы закладки и, опреде-
ляемые ими формы стилета, позволяют хорошо различать обсуждаемые рода (Солдатенко,
2008b).
Ключ для определения подродов и некоторых видов рода Choanomphalus
1(2). Раковина светлая, прозрачная, тонкостенная, плоскоспиральная с углова-
то-суженным на периферии последним оборотом. Устье овальное, сильно
оттянутое вбок, таким образом, что его ширина заметно превосходит вы-
соту.....................................Подрод Lamorbis Starobogatov, 1967.
Обороты раковины нарастают равномерно, Эмбриональная раковина со
скульптурой из тонких спиральных линий. Линии нарастания дефинитив-
ных оборотов едва заметны. Индекс копулятивного аппарата 0,89-1,31.
Длина стилета — 25 мкм. Монотипический подрод......................
..Choanomphalus riparius (Westerlund, 1865) (вкл. табл. VIII, рис. 126 а-в).
— riparius Westerlund, 1865: 106 (Planorbis).
Размеры: ВР до 0,7 мм, ШР до 3,5 мм.
Вид распространён в Европе (кроме крайнего севера) и Западной Сибири. Моллю-
ски обитают в постоянных и временных водоёмах. Редкий вид, но его популяции
всегда многочисленны. Обитает не глубже 0,5 м, преимущественно в хорошо
прогреваемых частях озёр, стоячих водоёмах и подтопленных территориях, в за-
рослях водной растительности.
2(1). Раковина коричневая, непрозрачная, твёрдостенная, плоскоспиральная с
плавно закруглённым на периферии последним оборотом. Устье округло-
овальное, его высота практически равна ширине............................
................Подрод Pseudogyraulus Prozorova et Starobogatov, 1997.
Верхняя сторона раковины, как правило, почти плоская. Только у некото-
рых видов, встречающихся за пределами европейской части России, по-
следний оборот смещается вниз, делая заметным предпоследний. Пупок
в виде широкой воронки. Поверхность оборотов практически гладкая.
Слабые линии нарастания хорошо заметны на нижней стороне раковины.
Индекс копулятивного аппарата 2,4-3,0. Подрод имеет в своём составе
не менее трёх видов распространённых в Палеарктике. В пресных водах
европейской части России встречается 1 вид.........................
...........Choanomphalus rossmaessleri (Auerswald in A. Schmidt, 1851)
(вкл. табл. VIII, рис. 127 а-в).
— rossmaessleri Auerswald — A. Schmidt, 1851: 179 (Planorbis).
Размеры: ВР до 1,8 мм, ШР до 6,2 мм.
428 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Вид распространён в Европе (кроме крайнего севера) и Сибири. Моллюски оби-
тают в стоячих и временных водоёмах. Редкий вид, как и Ch. riparius обитает не
глубже 0,5 м. Обычен в лесных болотах и мочажинах с большим количеством
растительного детрита.
Род Gyraulus Agassiz in Charpentier, 1837
Типовой вид: Planorbis albus Muller, 1774.
Раковина маленькая (ширина до 7 мм), плоскоспиральная, правозавитая, ино-
гда условно правозавитая, часто полупрозрачная, коричневого, желтоватого или
рогового цвета. Верхняя и нижняя стороны раковины имеют вдавления (сверху
могут быть погружены только начальные обороты). Оборотов 4,5-6,0, быстро на-
растающих, хорошо просматривающихся с обеих сторон раковины. Поверхность
оборотов выпуклая с обеих сторон. Периферия последнего оборота может быть
гладкой, угловатой или с хорошо выраженным килем и периостракальным кан-
том. Устье угловато-овальное (сердцевидное), овальное или округлое, вырезанное
стенкой предпоследнего оборота.
Половые протоки компактные, не тонкие. Копулятивный аппарат состоит из
мешка пениса и препуциума. Индекс копулятивного аппарата не превышает 2,0.
Пенис длинный, семяизвергательный канал открывается субтерминально. На кон-
це пениса имеется трубчатый стилет. Соотношение длин копулятивного аппарата
и глотки сильно варьирует (от 1,2 до 3,5). Радула включает 122-176 поперечных
рядов. Формула поперечного полуряда радулы варьирует от С-14М до С-56М
(Hubendick, 1954; Meier-Brook, 1983; Soldatenko, 2009).
Моллюски рода Gyraulus в ископаемом состоянии известны с юры (Старобо-
гатов, 1970). Современные представители распространены на всех континентах,
кроме Южной Америки. В составе рода насчитывают около 60 видов из 4 подро-
дов (Meier-Brook, 1983; Prozorova, Starobogatov, 1997; Круглов, Солдатенко, 2000;
Прозорова, 2003; Paraense, 2003; Paraense, 2004; Старобогатов и др., 2004; Beran,
Gloer, 2006; Gloer & Bossneck, 2007; Gloer, Vinarski, 2009; Gloer, Pesic, 2012; Pesic,
Gloer, 2013) На севере европейской части России встречаются шесть видов из двух
подродов: Gyraulus Agassiz in Charpentier, 1837 и Torquis Dall, 1905. Эти моллюски
обитают в зарослях водной растительности во временных и постоянных водоёмах.
Ключ для определения подродов и некоторых видов рода Gyraulus
1(2). Наивысшая точка устья расположена почти посередине его верхнего края
................................................Подрод Gyraulus s. str. (с. 429).
2(1). Наивысшая точка устья значительно приближена к шву....................
..................Подрод Torquis Dall, 1905; Gyraulus laevis (Alder, 1838)
(вкл. табл. VIII, рис. 128 а-в).
— laevis Alder, 1838: 337 {Planorbis).
Размеры: BP до 1,5 мм, ШР до 5,6 мм.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
429
Вид распространён в Европе (кроме юга и юго-востока). Моллюски обитают в не-
больших водоёмах с родниковым питанием. Редкий вид.
Подрод Gyraulus s. str.
Типовой вид: Planorbis albus Muller, 1774.
Раковина плоскоспиральная, дисковидная, средних размеров (ширина до
9 мм), светлая (конхиолиновый слой, покрывающий раковину, может быть свет-
лым и тёмным), часто прозрачная. Периферия последнего оборота закруглённая,
закруглённо-угловатая или с килем. Угол или киль (если имеются) всегда окайм-
ляют последний оборот по центру (не смещаются к околошовной области), у не-
которых видов (G. draparnaudi, G. centrifugus, G. stroemi) угловатость или киль
несут периостракальный кант. Обороты раковины хорошо просматриваются с
обеих сторон. На эмбриональной раковине всегда имеется тонкая скульптура, с
возрастом у одних видов (G. albus, G. draparnaudi) она становится более отчётли-
вой, а у других (G. borealis, G. stroemi) исчезает совсем (Meier-Brook, 1983).
Устье разнообразной формы, при наличии угла, киля или канта на периферии
последнего оборота, оно приобретает более удлинённую форму (овальную или
сердцевидную). Значения индекса копулятивного аппарата сильно варьируют от
0,75 до 1,80. Размер стилета от ПО до 368 мкм.
Общее распространение подрода — все континенты, кроме Южной Америки.
Ключ для определения видов подрода Gyraulus s. str.
1(6). Поверхность раковины с чёткой хорошо заметной спиральной скульпту-
рой.
2(5). Обороты с нижней стороны раковины более выпуклые, по сравнению с
верхней стороной. Верхняя сторона раковины практически плоская, ино-
гда начальные обороты могут быть очень слабо погружёнными. Область
пупка представляет собой широкую воронку (вдавление оборотов здесь
выражено сильнее, чем на верхней стороне раковины).
3(4). Обороты нарастают быстро и равномерно. Каждый последующий оборот
вдвое больше предыдущего. Диаметр внутренних оборотов (ДВО) равен
или немного превышает ширину последнего оборота. При 3,5 оборотах
ширина раковины не менее 3,5 мм.............................................
.............................Gyraulus stelmachoetius (Bourguignat, 1860)
(вкл. табл. VIII, рис. 129 а-в).
— stelmachoetius Bourguignat, 1860b: 139, pl. 2, figs. 10-13.
[= Planorbis stelmachoetius var. notatus Westerlund, 1885].
Скульптура сильнее выражена на нижней стороне раковины. Высота раковины
составляет 0,3-0,4 ширины.
Размеры: ВР до 1,8 мм, ШР до 6,2 мм.
Вид распространён в Европе и Западной Сибири. В постоянных водоёмах (как
стоячих, так и проточных).
430
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
4(3). Обороты нарастают медленно и неравномерно. Диаметр внутренних обо-
ротов (ДВО) в 1,5-1,7 раз больше ширины последнего оборота. При 3,5
оборотах ширина раковины не превышает 3,0 мм....................................
.........Gyraulus albus (O.F. Muller, 1774) (вкл. табл. VIII, рис. 130 а-г).
— albus (O.F. Muller, 1774: 164 (Planorbis).
[= Planorbis albus var. cinctutus Westerlund, 1871].
Спиральная скульптура одинаково хорошо выражена на всей поверхности ракови-
ны, часто имеются ряды длинных периостракальных щетинок. Высота раковины
меньше, чем у G. stelmachoetius (высота раковины составляет 0,26-0,30 ширины).
Размеры: ВР до 1,4 мм, ШР до 5,0 мм.
Вид распространён в Европе. В постоянных водоёмах.
5(2). Обороты одинаково выпуклые с обеих сторон, нарастают равномерно. Вы-
сота оборотов раковины увеличивается медленно, поэтому обороты как на
верхней, так и на нижней стороне раковины погружены одинаково слабо.
................Gyraulus acronicus (Ferussac, 1807) (вкл. табл. VIII, рис. 131 а-в).
— acronicus Fdrussac, 1807: 105 (Planorbis).
[= Planorbis devianus var. euphratica Mousson, 1874; Planorbis rossmassleri Hart-
wich, 1914].
Встречаются особи с матовой раковиной рогового цвета и с блестящей, у первых
скульптура отчётливо выражена с обеих сторон, у вторых преимущественно на
нижней (базальной) стороне. У представителей ряда европейских популяций
имеется широкий периостракальный кант или киль, что придает им сходство с
G. stroemi.
Размеры: ВР до 2,2 мм, ШР до 7,0 мм.
Вид распространён в Северной и Центральной Европе, а также в Сибири. Моллю-
ски обитают в постоянных водоёмах.
6(1). Поверхность раковины лишена спиральной скульптуры, даже линии на-
растания выражены слабо.
7(8). Раковина обычно имеет киль и/или большой периостракальный кант, ко-
торые придают устью сердцевидную форму..........................................
.........Gyraulus stroemi (Westerlund, 1881) (вкл. табл. VIII, рис. 132 а-в).
— stromi Westerlund, 1881: 63 (Planorbis).
[= Planorbis (Gyraulus) angasolensis B. Dybowski, 1912].
Раковина тонкостенная, даже после очистки раковины от конхиолинового слоя
по периферии последнего оборота чётко просматривается тонкий киль или угол.
Размеры: ВР до 1,5 мм, ШР до 9,0 мм.
Вид распространён в Северной Европе, Сибири (на восток до Охотского моря),
Приморье, Северном Казахстане и Монголии. Моллюски обитают в постоянных
водоёмах.
8(7). Раковина без киля и периостракального канта. Устье округлое или округ-
ло-овальное.........................Gyraulus borealis (Loven in Westerlund, 1875)
(вкл. табл. VIII, рис. 133 а-в).
— borealis Lovdn - Westerlund, 1875: 112-113, pl. 2, figs. 23-25 (Planorbis).
[= Anisus (Gyraulus) acronicus, sensu Starobogatov, 1970, partim, non Fdrussac, 1807;
Gyraulus gredleri var. borealis Lovdn, 1845].
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
431
Раковина твёрдостенная, обычно тёмно-жёлтого или красновато-бурого цвета.
Размеры: ВР до 2,0 мм, ШР до 7,0 мм.
Вид распространён в Северной Европе, Приуралье, на севере Западной Сибири.
Моллюски обитают в постоянных водоёмах. Часто в небольших озёрах и речных
разливах.
Род Planorbis O.F. Muller, 1773
Типовой вид: Helixplanorbis Linnaeus, 1758.
Раковина крупная (ширина до 20 мм), плоскоспиральная, условно правозави-
тая гиперстрофная или правозавитая; в большинстве случаев дисковидная. Вы-
сота раковины составляет 0,17-0,42 ширины. Стенки раковины непрозрачные,
прочные, желтоватого или коричневого цвета. Верхняя сторона раковины пло-
ская или слегка погружённая, нижняя — от плоской до воронковидно-вогнутой.
Оборотов 4-6, медленно нарастающих. Все они хорошо просматриваются с обе-
их сторон раковины. Периферия последнего оборота может быть с килем или
без него. Устье округлое, овальное или ромбическое, слегка вырезанное стенкой
предпоследнего оборота.
Копулятивный аппарат состоит из двух отделов: узкого, почти не отличимо-
го от семяпровода мешка пениса (различной длины) и массивного препуциума.
Индекс копулятивного аппарата варьирует от 0,44 до 5,39. Пенис длинный, семя-
извергательный канал открывается субтерминально, кончик пениса лишен папил-
лы. Имеется маленький внутренний стилет (длина 12-18 мкм), который создан
покрывающими эпителиальными клетками пениса (Soldatenko, Petrov, 2009; Sol-
datenko, Shatrov, 2013). Копулятивный аппарат половозрелых особей массивный
(1,0-6,7 мм), примерно в 1,5-2 раза больше глотки. Радула насчитывает 127-209
поперечных рядов зубов. Формула полуряда радулы С-18М (Baker, 1945), или
С-26М (у крупных особей).
В ископаемом состоянии моллюски рода известны с олигоцена на территории
Европы и с неогена в Северной Африке, Передней и Центральной Азии (Старо-
богатов, 1970). Современные представители рода Planorbis распространены в Ев-
ропе, Северной Африке, Передней, Средней, Центральной и Северной Азии (на
восток до бассейна Енисея включительно). Моллюски населяют заросли рипали
рек, водоёмы их придаточной системы, литоральную зону озёр, пруды, мелиора-
тивные рвы, лужи, встречаются на рисовых чеках, лиманах и в дельтах рек (Мар-
ковский, 1955; Стадниченко, 1990; Soldatenko, Starobogatov, 2000).
Комментарии. В настоящее время малакологи (Meier-Brook, 1976b; Soldatenko, Staro-
bogatov, 2000; Gloer, 2002; Bouchet et al., 2005; Gloer, Bouzid, 2008; Gloer, Zettler, 2009)
выделяют в пределах рода 2 подрода {Crassiplanorbis и Planorbis s. str.), которые насчи-
тывают 16 видов. Подрод Crassiplanorbis выделен Мейер-Бруком (Meier-Brook, 1976b)
на конхологической основе и включает только один эндемичный вид Planorbis presbensis
Sturany, 1894 из озера Преспа (Македония).
432 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Подрод Planorbis s. str.
Типовой вид: Helix planorbis Linnaeus, 1758.
Раковина плоскоспиральная, дисковидная, её нижняя сторона плоская или по-
гружённая. Выпуклость оборотов с обеих сторон раковины может быть одинако-
вой или различной, но скорость их нарастания одинаковая. Диаметр внутренних
оборотов на нижней стороне раковины (ширина пупка) равен или значительно
больше ширины последнего оборота измеренной около устья. Периферия послед-
него оборота гладкая, угловатая или с ярко выраженным килем. Устье горизон-
тальное, округлой, овальной или ромбической формы.
Подрод включает 15 видов. В пресных водах севера европейской части России
обитают не менее трёх видов.
Ключ для определения видов подрода Planorbis s. str.
1(2). Верхняя и нижняя стороны раковины одинаково выпуклые, в результате
просвет устья имеет ромбовидную форму. Киль проходит посередине по-
следнего оборота..............................................................
.......Planorbis carinatus O.F. Muller, 1774 (вкл. табл. VIII, рис. 134 а-в).
— carinatus O.F. Muller, 1774: 157-158 (Planorbis).
Киль может быть смещен к нижней стороне раковины, особенно у ювенильных
особей.
Размеры: ВР до 3 мм, ШР до 17 мм.
Вид распространён в Европе (на восток до Урала). Моллюски обитают в посто-
янных водоёмах. Обычно встречаются в прибрежных участках рек, протоках с
медленным течением и на примыкающих к ним заливных лугах.
Комментарии. Вид исключительно редкий. Тело практически не имеет пигмента-
ции (имеются только серые участки в виде мантийного кольца и двух, отходящих
от него коротких продольных полос). Индекс копулятивного аппарата составляет
1,05-2,11.
2(1). Верхняя сторона раковины выпуклая, нижняя сторона практически пло-
ская. В результате просвет устья имеет скошено-квадратную или округлую
форму.
3(4) Киль проходит по периферии последнего оборота ближе к нижней стороне
раковины. При взгляде сверху он хорошо заметен................................
.......Planorbis planorbis (Linnaeus, 1758) (вкл. табл. VIII, рис. 135 а-в).
— planorbis Linnaeus, 1758: 769 (Helix).
[= Planorbis marginatus Drapamaud, 1805; Planorbis (Tropidiscus) marginatus
latistoma Puzanov, 1925].
Тело серого цвета. Длина почки равна сумме длин копулятивного аппарата и се-
мяпровода.
Размеры: ВР до 4 мм, ШР до 20 мм.
Один из самых распространённых видов в Европе и Западной Сибири. Обитает
преимущественно в проточных постоянных и временных водоёмах.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
433
4(3). Киль (если имеется) проходит по периферии последнего оборота ближе к
нижней стороне раковины, но при этом он не заметен при взгляде сверху
............Planorbis umbilicatus O.F. Miiller, 1774 (вкл. табл. VIII, рис. 136 а-в).
— umbilicatus СР. Muller, 1774: 160-161 (Planorbis).
[^Planorbis marginatus Drapamaud, 1805].
Размеры: BP до 4 мм, ШР до 20 мм.
Один из самых распространённых видов в Европе и Западной Сибири. Обитает
преимущественно в стоячих постоянных и временных водоёмах.
Комментарии. По конхологическим признакам практически не отличим от Р pla-
norbis. Тело чёрного цвета. Длина почки в 1,5-2,0 раза меньше суммы длин копу-
лятивного аппарата и семяпровода.
Семейство SEGMENTINIDAE ЕС. Baker, 1945
Типовой род: Segmentina Fleming, 1817.
Раковина маленькая (ширина до 8,6 мм), плоскоспиральная, правозавитая,
при взгляде со стороны устья контур раковины имеет линзовидный облик. Рако-
вина мало вариабельна в пределах семейства. Выделяют виды с двояковыпуклой
(обороты выпуклые как на верхней, так и на нижней сторонах раковины), пло-
ско-выпуклой (обороты выпуклые на верхней стороне раковины и уплощённые
на нижней), полушаровидной (обороты сильно выпуклые на верхней стороне ра-
ковины и плоские на нижней) раковиной. Поверхность оборотов гладкая, блестя-
щая. Окраска от беловато-прозрачной до красновато-коричневой. Эмбриональная
раковина имеет тонкую скульптуру из частых спиральных линий. Оборотов 4-5.
Последний оборот составляет большую часть ширины раковины, его периферия
всегда округло-заострённая, киль отсутствует. Пупок воронковидный, умеренно
широкий (не более Vi ширины раковины) или узкий (до проколовидного). Устье
поперечно вытянутое, сердцевидное, глубоко вырезано стенкой предпоследнего
оборота. Внутри последнего оборота вблизи устья обычно имеются поперечные
пластины, число и размеры которых сильно варьируют у разных видов семейства.
Эмбриональная раковина правозавитая с сердцевидным устьем.
Тело левозавитое. Половые, выделительное и дыхательное отверстия рас-
положены компактно в районе головы слева. Щупальца нитевидные, не пигмен-
тированные. Адаптивная жабра отсутствует. Гермафродитная железа состоит из
ацинусов, расположенных в два ряда. Простата имеет до 9 двулопастных или не-
разветвлённых, дивертикул, расположенных в один ряд и впадающих в простати-
ческий проток. Копулятивный аппарат имеет 2 отдела (мешок пениса с пенисом
и препуциум с препуциальным органом). В проксимальной части мешок пениса
имеет один-два свободных железистых придатка или вздутие. На конце пени-
са у представителей некоторых родов (Segnetillia, Kolhymorbis) имеется стилет.
У представителей рода Helicorbis копулятивный аппарат с внешним протоком.
Радула длинная, насчитывает от 120 до 275 поперечных рядов зубов. Формула по-
луряда радулы от С-15М до С-28М.
434
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
В ископаемом состоянии представители семейства известны с палеоцена
(Старобогатов, 1970). Современные виды семейства Segmentinidae относятся
к 11 родам. Эти моллюски встречаются в заболоченных временных и постоян-
ных (участки затопленных речных пойм, озёра) водоёмах в Евразии, Африке и
Австралии (Старобогатов, 1970). Моллюски обитают преимущественно на или-
сто-песчаных, илистых и глинисто-илистых грунтах, среди листового опада и в
зарослях водных растений на мелководьях (до 0,5 м). Зимуют в неактивном со-
стоянии, зарывшись в грунт (на глубину до 2 м). В европейской части России
представлены 2 рода.
Комментарии. Зарубежные и отечественные малакологи неоднократно отмечали спец-
ифичные морфологические черты подсемейства Segmentinae (по Baker, 1945) или трибы
Segmentinini (согласно: Zilch А. 1959-1960; Старобогатов, 1967; Hubendick, 1978) в рамках
семейства Planorbidae. Применение современных методик электронной (трансмиссионной
и сканирующей) и конфокальной микроскопии при проведении ультраструктурных иссле-
дований спермиев и копулятивных органов позволило выявить новые признаки, на основе
которых группу следует рассматривать в ранге самостоятельного семейства Segmentinidae
(Soldatenko & Shatrov, неопубликованные данные 2016).
Ключ для определения родов семейства Segmentinidae
1(2). Последний оборот внутри содержит 2-3 группы радиально ориентирован-
ных пластинок...............................Segmentina Fleming, 1817 (с. 435).
2(1). Поперечные пластинки внутри последнего оборота отсутствуют............
.........................Hippeutis Agassiz in Charpentier, 1837 (с. 437).
Род Segmentina Fleming, 1818
Типовой вид Nautilus lacustris Lightfoot, 1786.
Раковина маленькая (ширина до 8,6 мм) в виде двояковыпуклой или плоско-
выпуклой линзы, глянцевая, часто прозрачная, тонкостенная, окраска от свет-
ло-роговой и желтоватой до красно-коричневой. Периферия последнего оборота
угловатая. Пупок узкий (0,16-0,24 ширины раковины) Устье поперечно-вытяну-
тое, сердцевидное, сквозь стенку последнего оборота хорошо видны внутренние
пластинки.
Копулятивный аппарат характеризуется длинным и тонким мешком пениса с
двумя железистыми придатками разной длины (больший около 1,0-1,5 мм). Пе-
нис без стилета, семяизвергательный канал открывается субтерминально, кон-
чик пениса представлен мышечной заострённой папиллой. Препуциум обычно
грушевидной или овальной формы. Препуциальный орган у большинства видов
связан с боковой стенкой структуры на значительном расстоянии, и лишь у S. dis-
tinguenda удерживается на двух проксимальных мышечных тяжах. Препуциаль-
ный орган не имеет внешнего протока. Копулятивный аппарат половозрелых осо-
бей примерно в 3 раза больше глотки. Радула насчитывает 210-275 поперечных
рядов зубов. Формула полуряда радулы С-21М.
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
435
Род известен с палеоцена на территории Европы и Северной Африки (Старо-
богатов, 1970). Современные виды рода Segmentina распространены в Европе,
Передней Азии и Западной Сибири. Соглашаясь с мнением Бекера (Baker, 1945),
считаем, что в Северной Африке моллюски этого рода отсутствуют.
Комментарии. В пределах рода выделено два подрода: Segmentina s. str. и Paraseg-
mentina Kruglov et Soldatenko, 1997 (Круглов, Солдатенко, 1997). Виды, ранее относимые
к подроду Subsegmentina Kruglov et Soldatenko, 1997 являются представителями рода Hip-
peutis. Западноевропейские систематики признают в составе рода только один вид — S. ni-
tida (Baker, 1945; Gloer, 2002; Bouchet, Rocroi, 2005). Между тем, существуют чёткие кон-
хологические и анатомические особенности, позволяющие идентифицировать большее
количество видов этого рода. В отечественных малакологических работах приводятся до
14 видов, однако статус азиатских и некоторых европейских представителей Segmentina,
описанных только по конхологическим признакам, требует уточнения (Иззатуллаев, 1980;
Круглов, Солдатенко, 1997).
Ключ для определения подродов и некоторых видов рода Segmentina
1(2). Раковина в виде двояковыпуклой линзы. Периферический угол располо-
жен на уровне середины высоты раковины. Внутренние обороты нарас-
тают относительно медленно; диаметр внутренних оборотов (ДВО) со-
ставляет 0,62-0,68 ширины последнего оборота. Ширина предпоследнего
оборота меньше ширины последнего в 5-7 раз.................................
.....................Подрод Parasegmentina Kruglov et Soldatenko, 1997;
Segmentina distinguenda (Gredler, 1859) (вкл. табл. VIII, рис. 137 а-в).
— distinguenda Gredler, 1859: 217 (Planorbis nitidus var.).
Подрод представлен одним видом. У некоторых особей раковина может не иметь
форму идеальной двояковыпуклой линзы. Цвет раковины может варьировать от
рогового до бледно-оранжевого, последний вариант наиболее характерен. Пре-
пуциальный орган копулятивного аппарата связан двумя мышечными тяжами с
проксимальным участком стенки препуциума.
Размеры: ВР до 1,50 мм, ШР до 4,75 мм.
Вид распространён в Европе (кроме крайнего севера), на юге Западной Сибири,
возможно, в северном Казахстане и на Алтае. В пределах европейской части Рос-
сии эти моллюски встречаются очень редко в постоянных водоёмах.
2(1). Раковина в виде плосковыпуклой линзы. Периферический угол располо-
жен заметно ниже середины высоты раковины..................................
..............................................Подрод Segmentina s. str.
Подрод Segmentina s. str.
Типовой вид: Nautilus lacustris Lightfoot, 1786
Раковина маленькая (ширина до 8,6 мм), в виде плоско-выпуклой линзы. Пе-
риферический угол расположен заметно ниже середины высоты раковины. У не-
которых видов подрода (Segmentina nitida, S. montgazoniana) наблюдается вы-
сокая степень изменчивости раковины, поэтому встречаются особи, имеющие
436
Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
форму почти двояковыпуклой линзы. В ряде случаев надежное определение ви-
довой принадлежности моллюсков возможно только по признакам половой систе-
мы. Копулятивный аппарат половозрелых особей длинный (до 3 мм), примерно в
3 раза больше глотки. Длина копулятивного аппарата примерно, в 3 раза меньше,
длины тела фиксированного моллюска. Препуциальный орган всегда связан с бо-
ковой стенкой препуциума на значительном участке. Индекс копулятивного аппа-
рата варьирует от 0,67 до 1,44.
Общее распространение подрода — палеарктическое, большинство видов оби-
тают в Европе и Северной Азии. В малакофауне севера европейской части России
насчитывается не менее четырёх видов. Моллюски этого подрода населяют вре-
менные и постоянные водоёмы.
Ключ для определения видов подрода Segmentina s. str.
1(4). Внутренние обороты раковины нарастают быстро (расширяются почти в
геометрической прогрессии), при измерении на одном уровне каждый по-
следующий оборот в 2 и более раза шире предшествующего.
2(3). Ширина последнего оборота с верхней стороны превышает таковую пред-
последнего оборота не менее чем в 5 раз. Отношение радиуса раковины
(учитывающего ширину устья) к ширине предпоследнего оборота с верх-
ней стороны не меньше 7,0...................................................
.....Segmentina nitida (O.F. Muller, 1774) (вкл. табл. VIII, рис. 138 а-в).
— nitidus O.F. Muller, 1774: 163-164 {Planorbis).
Размеры: BP до 2,1 мм, ШР до 5,1 мм.
Вид распространён в Европе, кроме крайнего северо-востока и Средиземноморья.
Моллюски обитают в постоянных водоёмах (затоны рек, старицы, запруды).
Редкий вид.
3(2). Ширина последнего оборота с верхней стороны превышает ширину пред-
последнего оборота не более чем в 4 раза. Отношение радиуса раковины
(учитывающего ширину устья) к ширине предпоследнего оборота с верх-
ней стороны не больше 6,0...................................................
....................Segmentina starobogatovi Kruglov et Soldatenko, 1997
(вкл. табл. VIII, рис. 139 а-в).
— starobogatovi Круглов, Солдатенко, 1997: 122-124, fig. 4В, 7В, ЮС {Segmentina).
Раковина по сравнению с S. nitida более высокая и с округлённым контуром, если
смотреть со стороны устья.
Размеры: ВР до 2,1 мм, ШР до 5,3 мм.
Вид распространён в реках бассейнов Балтийского и Северного морей. Моллюски
обычно держатся на прогреваемых мелководьях (глубиной до 1 м). Изредка встре-
чаются во временных водоёмах.
4(1). Внутренние обороты раковины нарастают медленно, первые два дефини-
тивных оборота равны по ширине.
5(6). Внутренние обороты нарастают неравномерно, ширина предпоследнего
оборота, больше ширины предыдущего не менее чем в 1,5 раза. Раковина
Класс брюхоногие моллюски — Gastropoda Cuvier, 1797
437
светло-жёлтого или серого цвета........................................
...Segmentina oelandica (Westerlund, 1885) (вкл. табл. VIII, рис. 140 а-в).
— oelandicus Westerlund, 1885: 86 (Planorbis nitidus var.).
Размеры: BP до 2,2 мм, ШР до 5,3 мм.
Вид распространён в Европе, кроме крайнего севера-востока, а также на юге
Западной Сибири. Моллюски обитают в мелких постоянных (реки, запруды) и
временных водоёмах с большим количеством детрита. Наиболее обычный вид на
подтопленных и заболоченных территориях.
6(5). Внутренние обороты нарастают равномерно, ширина предпоследнего
оборота с верхней стороны почти равна ширине предыдущего оборота.
Раковина жёлтого или красновато-жёлтого цвета, хрупкая........................
...................Segmentina montgazoniana Bourguignat in Servain, 1881
(вкл. табл. VIII, рис. 141 а-в).
— montgazoniana Servain, 1881: 89-90 (Segmentina).
Раковина наиболее уплощённая, по сравнению со всеми приведёнными выше
видами рода.
Размеры: ВР до 1,3 мм, ШР до 5,0 мм.
Вид распространён в Европе, кроме крайнего севера и Средиземноморья. Мол-
люски обитают преимущественно в озёрах, затонах рек и протоках. Наибольшей
численности и биомассы достигают в небольших хорошо прогреваемых и богатых
органикой участках.
Род Hippeutis Agassiz in Charpentier, 1837
Типовой вид: Helix complanatus Linnaeus, 1758.
Раковина маленькая (ширина до 5 мм) в виде двояковыпуклой или плоско-
выпуклой линзы, глянцевая, прозрачная, тонкостенная, окраска желтоватого или
рогового цвета. Периферия последнего оборота угловатая, без киля. Пупок во-
ронковидный, узкий (0,16-0,25 ширины раковины). Устье поперечно-вытянутое,
сердцевидное, на периферии суженное, сильно вырезанное стенкой предпослед-
него оборота, без пластинок внутри.
Копулятивный аппарат короткий (до 1,5 мм) с коротким тонким мешком пени-
са, имеющим два коротких одинаковых (до 0,3 мм) железистых придатка. Пенис
без стилета, отверстие семяизвергательного канала расположено субтерминально,
папилла маленькая (40 мкм). Препуциум грушевидной формы. Препуциальный
орган связан с боковой стенкой на значительном расстоянии и не имеет внешнего
протока. Копулятивный аппарат и глотка у половозрелых особей примерно рав-
ной длинны. Радула насчитывает 120-150 поперечных рядов зубов. Формула по-
луряда радулы С-16М.
Род предположительно известен с эоценового времени на территории Европы
(Baker, 1945). Современные представители распространены в Европе и на юге За-
падной Сибири. Моллюски обитают во временных и постоянных водоёмах. На
севере и северо-западе европейской части России обитает один вид — Hippeutis
complanatus (Linnaeus, 1758).
438 Приложение. Пресноводные моллюски Севера и Северо-Запада Европейской России
Комментарии. Отечественные малакологи различают в составе рода три вида {Н. dia-
phanella, Н. euphaea и Н. f on tana), обитающих на территории России. Все они выделены
на основе конхологических признаков без ознакомления с типовым материалом осталь-
ных видов рода (Старобогатов, 1977; Старобогатов и др., 2004). Мы не отрицаем, что со-
став рода на территории России может насчитывать больше одного вида, но анализ ра-
ковин, хранящихся в коллекции ЗИН РАН и собственных сборов, показали отсутствие
хиатуса по признакам, использованным для разграничения Н. diaphanella, Н. euphaea и
Н. fontana. Анатомические различия также не обнаружены.
Hippeutis complanatus (Linnaeus, 1758) (вкл. табл. VIII, рис. 142 а-в).
— complanatus Linnaeus, 1758: 769 {Helix).
[= Planorbis {Hippeutis) colchicus Lindholm, 1913].
Размеры: BP до 1,5 мм, ШР до 5,0 мм.
Вид распространён практически по всей Европе (кроме крайнего севера) и на юге
Западной Сибири. Моллюски обитают в постоянных водоёмах; преимущественно
в озёрах, затонах и старицах рек.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
А
absurdus, Arrenurus 181
Acalyptonotidae 179
Acalyptonotus 179
Acanthobdella 97
Acanthobdellida 30, 96
Acanthobdellidae 98
Acaridida 164, 167
Acarididae 167
Acariformes 164
achaetus, Lamprodrilus 44
Achromadora 261
Acipenserobdella 113
Acroloxidae 16,22,359,391
Acroloxus 23, 391
acronicus, Anisus 430
acronicus, Gyraulus 430
acronicus, Planorbis 430
Actinedida 165
aculeate, Sarscypridopsis 217, 220
acuta, Costatella 393
acuta, Physa 393
acutalis, Lymnaea 413
acuticostata, Pulchelleuglesa 331
Aenigmomphiscola 26, 396
Aeolosoma 31
Aeolosomatidae 30
aequiseta, Pristina 65, 66
aestuariorum, Gammaracanthus 134
affinis, Cypricercus 233
affinis, Monoporeia 129,130
Alaimus 261
albicola, Psammoryctes 88
albicola, Psammoryctides 79, 88
albifrons, Jaera 147
Alboglossiphonia 111
albus, Anisus 335
albus, Gyraulus 429, 430
albus, Planorbis 428, 429, 430
Alexandrovia 71
alpina, Nais 54,61
alpinum, Europisidium 334
alpinus, Porohalacarus 195,196
amauronius, Choanomphalus 419
ambigua, Cincinna 388
ambigua, Valvata 388
ambiguus, Dactylobiotus 245
ambulans, Stygobromus 127
ambulans, Synurella 127,128
Ameronothridae 196
Amesoda 318,319
Amnicola 380
Amnicolidae 23, 360, 380
amnicum, Pisidium 334
amphibiotica, Pristina 66, 67
amphibius, Homocaligus 165,166
Amphibolus 245
Amphichaeta 44
Amphipoda 6, 122
ampla, Limneus auricularius var. 410
ampla, Lymnaea 410
amplexa, Torrenticola 178
ampullacea, Lymnaea 411
ampullaceus, Limnaeus 411
Amuraplexa 395
anatinum, Colletopterum 308, 312
anatolica, Dreissensia 314
Anatonchus 261
anceps, Helisoma 415
Ancylidae 16,22,359,413
Ancylus 22,413,414,415
andreana, Valvata 385
angasolensis, Planorbis 430
angustata, Platicrista 249
angustiformis, Dreissena polymorpha var.
315
Anisus 28,419,420,421,422
annuliformis, Choanomphalus 418
Anodonta 18, 287, 288, 302, 306, 307
Anodontinae 302, 306
Anoetidae 165, 167
Anthoathecata 252, 258
Anthozoa 251
antipodarum, Amnicola 379
antipodarum, Potamopyrgus 379
antiqua, Cincinna 389
440
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
antiqua, Valvata 389
Aphanolaimus 261
Aphanoneura 30
Aplexa 27, 392, 393, 394, 395
Aplexinae 393, 394
Apochela 244
apollonica, Anodonta 309
apollonicum, Colletopterum 308
Aporcelaimellus 261,262
Aporcelaimus 261
appendiculata, Slavina 52
Aquanothrus 197
aquatica, Limnochares 171,172
Aquatides 261
aquaticus, Asellus 147
Araeolaimida 260,261
Aranei 6
area, Mytilus 314
Archaeobdella 30, 105
archangelica, Lymnaea 407
Arcteonais 49
Arcteuglesa 328
arduifrons, Diphascon 250
Armiger 29, 420, 423,424
amouldi, Dreissensia 314
Arrenuridae 180
Arrenurus 169, 180
articulata, Paludicella 159
Asellidae 146, 147
Asellus 147
asiaticum, Sphaerium 319
astaci, Branchiobdella 95, 96
Astigmata 167
astrachanica, Anodonta subcircularis
ataxiaca, Aplexa 395
ater, Viviparus 368
atra, Lymnaea 408
atra, Paludina 368
Atractides 182
atrum, Buccinum 408
Aturidae 179, 188
Aturus 188, 190
Aulodrilus 73, 86
Aulophorus 50
auricularia, Helix 402
auricularia, Lymnaea 402
Axonopsis 188,189, 190
В
baeri, Colletopterum 308
baicalense, Sphaerium 321
baicalensis, Pelmatohydra 253, 254
Balcanipaludina 367
balthica, Helix 412
balthica, Lymnaea 346, 412
barbata, Nais 54, 62
barbatus, Psammoryctes 88
barbatus, Psammoryctides 79, 88
Bastiania 261
Bataviana 305
Bathyomphalus 28,419,424,425
Batracobdella 112
baudoniana, Tetragonocyclas 332
baudonianus, Opisthorchophorus 371
baudonii, Henslowiana 329
bavaricus, Anisus 421
bavaricus, Ilyodrilus 90
bavaricus, Potamothrix 83, 90
beauchampi, Isocypris 227
bedoti, Dreissensia 314
bedoti, Ilyodrilus 89
bedoti, Potamothrix 82, 89
behningi, Nais 54,61
belgicae, Diphascon 250
belgrandi, Dreissensia 314
Bertolanius 245, 247
bielzi, Armiger 424
bielzi, Planorbis crista var. 424
bilobata, Pristina 68, 69
bilobata, Pristinella 68
Biomphalaria 418
biseriata, Spongilla 152
Bithynia 25, 370, 371, 372, 374
Bithyniidae 15,25,370,371,372
Bivalvia 7, 10, 16,279,285
blanci, Piguetiella 52, 57
bodamica, Hibemeuglesa 330
bodamica, Lymnaea 404
boissieri, Paraelona 371,378
Borealibius 248
borealis, Gyraulus 430
borealis, Gyraulus gredleri var. 430
Указатель латинских названий
borealis, Margaritana margaritifera f. 300
borealis, Margaritifera 300
borealis, Planorbis 430
Boreobdella 108
Borysthenia 24,361,383,390
Borystheniinae 383, 390
Bothrioneurum 72
bourguignati, Bithynia 375, 376
bourguignati, Digyrcidum 371, 375, 376
bourguignati, Dreissensia 314
Brachipoda 188, 190
Branchiobdella 95
Branchiobdellida 30, 94
Branchiobdellidae 94
Branchiura 82
Bratislavia 47
bretscheri, Nais 55, 63
brevirostris, Calyptostoma 166,167
Brevitobrilus 261
brogniartiana, Menetus 415
bronsteini, Stenocypris 222
browniana, Scottia 221
Bryozoa 153
Buccinum 407
buchtarmensis, Euglesa 326
bugensis, Dreissena 315
bugensis, Dreissensia 315
Bulla 394
bulla, Physa 394
C
caelatus, Fractonotus 247, 249
callomphala, Lymnaea 408
Calohypsibiidae 247
Calohypsibius 247
Calonyx 173
Calyptostoma 166, 167
Calyptostomidae 166, 167
cancellata, Thyopsis 174
Candida, Candona 239
Candona 208,241
Candonidae 208,216,229
Candoninae 208, 229, 238
Candonopsis 238
carelica, Lymnaea 411
carinatus, Hydrobia ventrosa var. 379
carinatus, Planorbis 432
carinea, Anisus 421
Carinogyraulus 418
Carphania 244
carteri, Eunapius 152
carteri, Lophopodella 160
Casertiana 325
casmiana, Plumatella 163
caspia, Cordylophora 252, 256, 258
Caspiobdella 114
Catascopia 401
caudata, Candona 203
Cavemocypris 216
cavimata, Orchestia 135
celtica, Bithynia 373
celtica, Codiella 373
centrifugus, Gyraulus 429
Cestobranchus 100
Chaetiliidae 146
Chaetogammarus 138
Chaetogaster 30, 37, 44, 58
chantrei, Dreissensia 314
chappuisi, Soldanellonyx 194, 195
charteus, Anisus 421
Cheilostomata 155
Chelicorophium 131
chemnitzi, Tichogonia 314
Chlorohydra 253
Choanomphalus 29, 420, 426, 427
Chrisonema 261
christinae, Nais 56, 64
Chromadorida 260,261
Chromadorina 261
Chromadorita 261
Chronogaster 261
Cincinna 26, 386, 386
cinctutus, Planorbis albus var. 430
cingulatum, Cingulipisidium 324
Cingulipisidium 20, 322, 323
circumcincta, Hydra 255, 258
cirratus, Halitholus 258, 259
claparedeanus, Limnodrilus 75, 87
Clarkus 261
clavata, Limnaea glabra var. 400
Clavidae 252
Clitellata 30, 32
442
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Cnidaria 251
coccinea, Piona 186
coccineus, Rhyacodrilus 73, 85
Codiella 25,371,372
colchicus, Planorbis 438
Coleoptera 5, 6
Colletopterum 18, 302, 306, 308
Colohypsibius 247
coloradensis, Acroloxus 391
comata, Vejdovskyella 53, 61
cometes, Teutonia 177,178
communis, Litarachnia 166
communis, Nais 56, 64
complanata, Anodonta 311
complanata, Dreissensia 314
complanata, Glossiphonia 100, 101,109, 110,
111
complanata, Pseudanodonta 312
complanatus, Helix 437
complanatus, Hippeutis 437, 438
concolor, Glossiphonia 100, 110, 111
Congeria 313
conglobata, Piona 186
conica, Henslowiana 329
conjecta, Hydrachna 171
connata, Limnesia 178
contecta, Contectiana 369
Contectiana 24, 367, 368, 369
contectum, Cyclostoma 368,
contorta, Helix 424
contortus, Bathyomphalus 417, 419, 425
conus, Tumidiana 302
conus, Unio 303
Conventus 21, 323, 324
conventus, Conventus 324
conventus, Pisidium 324
convexa, Anodonta 309
convexum, Colletopterum 308
Copidognathinae 196
Copidognathus 196
coralloides, Plumatella 163
cordata, Metacypris 211,212
Cordylophora 252
cornea, Helix 416
cornea, Tellina 321
comeum, Sphaerium 321
comeus, Helix 417
comeus, Planorbarius 415, 417
comuliformis, Dreissena 315
Corophiidae 127, 131
Corophium 131
corviformis, Lymnaea palustris var. 408
corvus, Helix 406
corvus, Lymnaea 406, 409
Corvusiana 400, 406
costata, Haementeria 109,110
Costatella 27, 393
Costorbis 422
Crangonyctidae 127
Craspedacusta 252
crassa, Crassiana 294, 306
crassa, Helix 426
crassa, Nautarachna 184,185
Crassiana 18,301,305
crassipes, Unionicola 169
Crassiplanorbis 431
crassus, Bathyomphalus 425
crassus, Unio 304, 306
crassus, Obesogammarus 137
crassus, Pontogammarus 137
crenophilus, Carinogyraulus 418
creplini, Musculium 317
crissum, Cingulipisidium 324
crista, Anisus 424
crista, Armiger 424
crista, Nautilus 423, 424
cristata, Valvata 384
Cristatella 154, 158, 161
cristatellus, Lophopus 160
cristatus, Planorbis 424
Crocodorylaimus 261,262
cruenta, Hydrachna 171
Cryptocandona 241
Cryptostigmata 167
crystallinus, Lophopus 160
Ctenostomata 155
curta, Bithynia 371, 375
curta, Dreissensia 314
curta, Euglesa 326
curtacorvus, Lymnaea 406
curvispinum, Chelicorophium 131,132
Cyclas 319
Указатель латинских названий
443
Cyclocalix 325, 326
Cyclocypridinae 208, 229
Cyclocypris 208, 233
Cyclostoma 368
Cyclostomata 155
cygnea, Anodonta 307
cygnea, Mytilus 307
Cylindrolaimus 261
Cypretta 207, 220
Cyprettinae 207, 220
Cypria 208, 233
Cypricercus 208, 229
Cyprideis 209,215
Cyprididae 207,216,220
Cypridinae 208, 224
Cypridocopina 203, 205, 206, 207, 209, 215
Cypridopsidae 207,216
Cypridopsinae 207
Cypridopsis 207,216
cyprinorum, Crassiana 305
Cyprinotinae 208, 224, 229
Cyprinotus 208, 229
Cypris 208, 224
Cyproidinae 207,221
Cyprois 207,221
Cyrenastrum 318
Cystobranchulus 112
Cytherideidae 208, 210, 215
Cytherissa 209
Cytherocopina 203, 205, 208, 209
Cytheroidea 208, 209
D
Dactylobiotus 243, 245, 247
dactyloporus, Copidognathus 196
danielopoli, Schwiebia 166
danubii, Dythalmia 314
Darwinula 209
Darwinulidae 209
Darwinulocopina 203, 205, 209
decipiens, Bithynia 371,375
dentata, Thyasides 176
depressa, Cincinna 387
depressinitidum, Cingulipisidium 324
Dero 50, 59
Desmodorida 261
despiciens, Hydrodroma 173, 174
devianus, Planorbis 430
diaphanella, Hippeutis 438
diaphanus, Chaetogaster 36, 45, 58
diastropus, Chaetogaster 45, 58
digitata, Dero 51, 60
Digyrcidum 371
Dikerogammarus 135
dilatata, Lacustrina 330
Diphascon 243, 250
Diplogaster 261
Diptera 7
discoides, Anisus 421
discors, Cincinna 383, 390
discors, Valvata 390
Disculifer 421
dispar, Bathyomphalus 425
dispar, Planorbis 425
dispar, Dactylobiotus 245
distinguenda, Planorbis nitidus var. 435
distinguenda, Segmentina 435
Dolerocyprinae 208, 209, 229
Dolerocypris 208, 209, 229
donetziensis, Tyrrhenocythere 210, 211
doriana, Lymnaea 404
dorsalis, Dero 51, 59
Dorylaimida 260,261,262
Dorylaimoides 261,262
Dorylaimus 261,262
Doryphoribius 248
draparnaudi, Gyraulus 429
Dreissena 10, 17,286,287,289,313,314
Dreissenidae 17, 299, 313
Dreissensia 314
dubia, Lebertia 178
dubiosa, Cypretta 220, 221
dubrueilii Euglesa 326
duebeni, Gammarus 140,143
dujardini, Hypsibius 249
duplicidentatus, Neoactinolaimus 264
dupuyana, Henslowiana 329
dupuyi, Lymnaea 402
duryi, Planorbella 415
E
ebumeus, Lithoglyphus 381
444
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Echiniscudae 244
Echiniscus 247
Ectoprocta 153
Ecumenicus 261
eichwaldi, Euglesa 327
Eiseniella 43
ekmani, Gnaphiscus 181
ekmani, Neobrachypoda 189,190
elatior, Valvata 385
elegans, Pristina 47
Eleuterengona 165
elinguis, Nais 56, 63
elongata, Anodonta 313
elongata, Dreissensia 314
elongata, Margaritifera 300
elongata, Pseudanodonta 312
emarginata, Plumatella 163
Embolocephalus 70
Enchytraeidae 41
Enoplida 260, 261
ensifer, Tiphys 185
entomon, Mesidotea 147
entomon, Sadura 147, 148
Eohypsibiidae 245
Ephemeroptera 5
Ephydatia 151, 152
Epidorylaimus 261
Epitobrilus 261
Erpobdellidae 104, 105
Erpopbdella 100, 119, 121
esakii, Thermozodium 244
esmonti, Archeobdella 30, 105
Ethmolaimus 261
Eucypridinae 208, 229
Eucypris 208,229
Eudorylaimus 261
Euglesa 20,321,325
Euglesinae 19,316,321
Eumonhystera 261
Eunapius 151, 152
euphaea, Hippeutis 438
euphratica, Planorbis devianus var. 430
euplectella, Paracandona 237, 238
Eupodina 165
eurydoris, Aporcelaimellus 264
Euristomata 155
europaea, Aenigmomphiscola 398
Europisidium 334
Eutardigrada 242, 243, 244
Euthyas 175
Eutobrilus 261
eximia, Dreissensia 314
eximia, Protzia 174
Eylaidae 172
Eylaioidea 172
Eylais 172
F
fabaeformis, Fabaeformiscandona 240
Fabaeformiscandona 208,241
fadeewi, Physcypria 233, 235
fadejewi, Caspiobdella 114
fadejewi, Piscicola 114
falciformis, Rhyacodrilus 72, 85
falcipalpis, Momonia 187,188
falsifluviatilis, Cincinna 389, 390
fascheri, Stenocypria 229
fasciata, Dolerocypris 228
fasciata, Piscicola 115,116
fasciatus, Cystobranchus 116
fasciatus, Gmelinoides 128, 129
fastus, Thyas 176
favrei, Pseudosphaerium 324
Feltra 179
Feltriidae 179
feroense, Cingulipisidium 324
ferox, Peloscolex 69
ferox, Spirosperma 69, 70, 71
Fictor 261
figuralis, Unionicola 183
Filifera 252, 258
fischeri, Stenocypria 229, 230
flavomaculatus, Crocodorylaimus 264
fluviatilis, Ancylus 414, 415
fluviatilis, Carphania 244
fluviatilis, Dreissena 314
fluviatilis, Ephidatia 151,152
fluviatilis, Hygrobates 183
fluviatilis, Nerita 365
fluviatilis, Neritina 365
fluviatilis, Theodoxus 365
fluviatilis, Valvata 390
Указатель латинских названий
445
fluviatilis, Dikerogammarus 136
fontana, Hippeutis 438
fontinalis, Bulla 394
fontinalis, Limneus 413
fontinalis, Lymnaea 413
fontinalis, Physa 394
foreli, Pristina 65
Forelia 187
Fortuyniidae 196
fossularis, Stylaria 48, 59
foveolatus, Trimalaconothrus 199
Fractonotus 247
fragilis, Euglesa 326
fragilis, Helix 403
fragilis, Lymnaea 403
fragilis, Eunapis 151,152
Fredericella 154, 161
Fredericellidae 156
frici, Paranais 46
frigida, Cincinna 386
frigida, Valvata 386
Frontipoda 182
fruticosa, Plumatella 163
fulva, Limnaeus 411
fulva, Lymnaea 411
fungosa, Plumatella 156,162
furcatus, Aulophorus 50
furcatus, Zetomimus 199
fusca, Lymnaea 409
fusca, Stagnicola 409
fuscula, Crassiana 305
fuscus, Limnaeus 409
G
Galba 400, 404, 405
gallandi, Dreissensia 314
Galumnidae 200
Gammaracanthidae 130, 134
Gammaracanthus 134
Gammaridae 131, 138
Gammarus 125, 138
Gastropoda 7,12, 22, 279, 335, 338, 339, 343
gedyminensis, Armiger 424
Gelatinella 161
geographica, Hydrachna 169
geometra, Piscicola 115,116, 117
Georgella 175
geyeri, Valvata 385
gibba, Ilyocypris 215
glabra, Lymnaea 400
glandulosus, Sperchon 181
globator, Arrenurus 169,181
globularis, Roseana 327
Glossiphonia 110
Glossiphoniidae 98, 103,104, 105
glutinosa, Lymnaea 401,402
glutinosum, Buccinum 401
Gmelinoides 128
Gnaphiscus 182
gordioides, Haplotaxis 42
goupili, Lymnaea 405
grachevi, Choanomphalus 418
gracilis, Malaconothrus 199
grandis, Planorbarius 417
grandis, Planorbis 417
granuensis, Rhynchelmis 92
gubkini, Dreissena 315
gueretiniana, Lymnaea 406
Gymnolemata 153, 154, 159
Gyraulus 29, 420, 428, 429
H
Haementeria 110
haemobaphes, Dikerogammarus 136
Haemonais 58
Haemopis 100, 119
hagenii, Mytilus 314
Halacaridae 165, 190
Halitholus 258
hammoniensis, Ilyodrilus 90
hammoniensis, Potamothrix 83, 89, 90
haplonyx, Dactylobiotus 245
Haplotaxidae 42
Haplotaxis 42
harmsworthi, Macrobiotus 245
hartmanni, Lymnaea 410
hastatus, Murrayon 245
Hazayipaludina 367
Helicorbis 433
Helisoma 415
Helisomatidae 27,363,415
hellenica, Dreissensia 314
446
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
hellenica, Paraelona 373
Helobdella 106
helveticus, Limnodrilus 88
Hementeria 109, 110
Hemiclepsis 105
Hemicythere 210
Hemicytheridae 209,210
hemprichi, Aeolosoma 31
henslowana, Henslowiana 329
Henslowiana 20, 327, 328
henslowiana, Tellina 326
henslowianum, Pisidium 327, 328
heringianus, Stylodrilus 93, 94
hermosa, Dreissensia 314
Herpetocypridinae 208, 222, 229
Herpetocypris 208, 229
Heterobranchia 335
heteroclita, Alboglossiphonia 100, 111
Heterocypris 208, 209
Heteroptera 5
Heterotardigrada 244, 247
Heterozetes 197,200
heuscheri, Ilyodrilus 90
heuscheri, Potamothrix 83, 90
Hibemeuglesa 20, 322, 329
hibemicum, Pisidium 329
hinzi, Euglesa 326
Hippeutis 28, 434, 437
Hirudinea 7, 30, 96, 98
Hirudinida 98
Hirudinidae 104,105, 117
Hirudo 100, 117
Hislopiidae 153
hisphanicus, Opisthorchophorus 371
hoffineisteri, Limnodrilus 36, 76, 87
Homocaligidae 165
Homocaligus 165
horrida, Trochospongilla 151
hoyeri, Euglesa 326
hufelandi, Macrobiotus 245
Huitfeldtia 187
Hungarocypridinae 208, 209
Hungarocypris 208, 222
Hyalinella 162
Hydra 258
Hydrachna 171
Hydrachnidae 171
Hydrachnidia 6, 164, 165, 167
Hydrachnoidea 171
Hydrida 251
Hydridae 251
Hydrobia 378
Hydrobiidae 23, 360, 378, 379
Hydrochoreutes 184
Hydrodroma 173
Hydrodromidae 173
Hydro volzia 170,171
Hydrovolziidae 170
Hydro volzioidea 170
Hydrozetes 197, 198
Hydrozetidae 198
Hydrozoa 251,252
Hydryphantes 173
Hydryphantidae 173
Hydryphantinae 173
Hydryphantoidea 172
Hygrobates 182
Hygrobatidae 179
Hygrobatoidea 172, 176
hypnorum, Aplexa 396
hypnorum, Bulla 395, 396
Hypsibius 247, 249
I
Ichtiobdellidae 103
idahoense, Pisidium 330
ignotus, Tubifex 84, 91
Ilyocyprididae 207,215
Ilyocypris 207,215
Ilyodrilus 81
Ilyodromus 208, 229
imbricatus, Planorbis 424
imnophila, Piersigia 172
inaequicauda, Gammarus 140, 142
incongruens, Heterocypris 232
inermis, Planorbis crista var. 424
inflata, Bithynia 371
inflata, Lymnaea ovata var. 412
inopinata, Limnocythere 203, 213
insculpta, Pseudocandona 238
intermedia, Euglesa 327
intermedia, Lymnaea 413
Указатель латинских названий
447
intermedia, Vejdovskyella 53, 60
lotonchus 261
irgizlaica, Crassiana 305
Ironus 261
ischiosetosa, Jaea 147
Isochaetides 77
Isocypridinae 208, 226
Isocypris 226
Isohypsibius 247, 248, 249
Isopoda 6, 146
J
Jaera 146, 147
jagellonensis, Armiger 424
Janiridae 146, 147
japonicas, Aulodrilus 86
jaudouiniana, Euglesa 327
jenkinsi, Hydrobia 379
johanseni, Anisus 421
josinae, Specaria 57
К
kambai, Dreissena 315
kazakensis, Lymnaea palustris 406
kazakensis, Lymnaea taurica 406
kessleri, Tasserkidrilus 78, 80
kessleri, Tubifex 80
kickxii, Bithynia leachi var. 373
kickxii, Codiella 371,373
kickxii, Paludina 373
kingsleii, Candonopsis 237
kletti, Pseudanodonta 312
koenikei, Georgella 174,175
Koemeria 261
Kolhymorbis 433
komi, Pristina 68
Kongsbergia 188, 190
kopetdagica, Dreissena 315
krameri, Hydrochoreutes 185
krasnopolskiae, Chaetogaster 59
Krendowskia 180
Krendowskiidae 180
labiata, Lymnaea 412
Labronema 261
lacunosa, Dreissensia 314
lacustre, Musculium 317
Lacustrina 19,321,330
lacustris, Acroloxus 391
lacustris, Cordylophora 252, 257, 258
lacustris, Cytherissa 205, 214, 215
lacustris, Gammaracanthus 133
lacustris, Gammarus 139,141
lacustris, Lymnaea 404
lacustris, Nautilus 434, 435
lacustris, Patella 391
lacustris, Spongilla 151, 152
lacustris, Stylaria 48, 59
lacustris, Tellina 317
Ladislavella 401
ladogensis, Contectiana 369
Laevides 261
laevis, Cyclocypris 236
laevis, Gyraulus 428
laevis, Planorbis 428
lagotis, Buccinum 412
lagotis, Lymnaea 412
Lamorbis 426, 427
Lamprodrilus 44
landeroini, Pisidium 330,331
langi, Chaetogaster 45, 58
longirostris, Calyptostoma 167
lapponicus, Euglesa 326
lastotschkini, Pristina 66
latior, Valvata andreana var.
latissima, Krendowskia 178,179
latistoma, Planorbis marginatus 432
leachi, Bithynia 371
leachii, Codiella 372, 373
leachi, Turbo 372
Lebertia 177
Lebertiidae 177
Leptocythere 210
Leptocytheridae 209
Leptothecata 251,258
letoumeuxi, Anodonta 308
letoumeuxi, Dreissensia 314
leuckartii, Pionacercus 186,187
Leucocythere 212
leucostoma, Planorbis 422
leucostoma, Anisus 422
448
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
levanderi, Fabaeformiscandona 241
leydigi, Amphichaeta 44, 46
likharevi, Euglesa 327
lilljeborgi, Cincinna 389
lilljeborgi, Pisidium 328
lilljeborgi, Valvata 389
lilljeborgi, Eucypris 203
lilljeborgii, Henslowiana 328
limnaei, Chaetogaster 45, 58
Limnesia 177
Limnesiidae 177
limnobius, Aulodrilus 73, 86
Limnochares 172
Limnocharidae 166, 172
Limnocharoidea 172
Limnocythere 208,212
Limnocytheridae 208,209,211
Limnocytherinae 208
Limnodrilus 75, 87
Limnohalacarinae 193
Limnohalacarus 193
Limnomedusae 252
limnophylus, Dorylaimoides 264
Limnozetes 199
Limnozetidae 199
limosella, Rhynchelmis 36, 92
limosus, Unio 302, 304
lindholmi, Acroloxus 391
lindholmi, Choanomphalus 419
lineata, Erpobdella 119
lineata, Herpobdella 119
Litarachna 170
Lithoglyphidae 25,361,381
Lithoglyphus 25,381
Ljania 188, 190
Lobohalacarus 195
locardi, Dreissensia 314
Lohmannellinae 193
lomondi, Arcteonais 49
longirostris, Calyptostoma
longirostris, Unio 302, 303
longiseta, Pristina 65
Lophopodella 154, 160
Lophopus 154, 160
loricatus, Gammaracanthus 133
Lumbricidae 42, 43, 92
Lumbriculidae 43, 44
lunata, Dreissena 315
lutetiana, Dreissensia 314
Lymnaea 26, 27, 345, 396, 399, 400
Lymnaeidae 12, 15, 26, 362, 396
M
maacki, Choanomphalus 426
mabillei, Lymnaea 402
macilenta, Ljania 189
Macrobiotus 243, 245, 247
macrochaeta, Vejdovskyella 52, 60
macrodon, Doryphoribius 248
macrolaimus, Paractinolaimus 264
macrostoma, Cincinna 386
macrostoma, Valvata 386
maculosa, Protoclepsis 108
maculosum, Theromyson 108
madaraszi, Hungarocypris 222, 225
magnifica, Dreissensia 314
magnificum, Planorbella 415
magnificus, Euglesa 326
majewskyi, Paraelona 371
major, Panisus 176
Malaconothridae 198
Malaconothrus 198
Malacostraca 122
mammilatus, Cystobranchulus 112,113
Margaritana 300
Margaritifera 17, 299, 300
margaritifera, Margaritifera 300, 301
margaritifera, Mya 300
Margaritiferidae 10, 17, 286, 289, 299
marginata, Cyprois 223
marginata, Hemiclepsis 100,105,106
marginatus, Planorbis 432, 433
maritima, Lymnaea fusca 409
markevichi, Bertolaimus 247
Marstoniopsis 23, 380
media, Lymnaea 404
medicinalis, Hirudo 117,118
Medusozoa 251
Megaloptera 6
menoni, Pristina 68
menoni, Pristinella 68
meridionalis, Eylais 169
Указатель латинских названий
449
Mesidotea 147
Mesocrista 250
Mesodorylaimus 261,262
Mesotardigrada 244
Metacypridinae 208
Metacypris 208,211
michaelseni, Isochaetides 77
michaelseni, Limnodrilus 77
Miconchus 261
microcephalus, Planorbis
Microcincinna 385
Microhypsibiidae 247
Micruropidae 128
Mideopis 187
Mideopsidae 179, 187
milachevitchi, Paraelona 371, 373, 378
milaschevichi, Colletopterum 308
milium, Pisidium 331
Milletelona 375
Milnesium 244
minuta, Lymnaea 405
minuta, Feltra 178
mirabilis, Leucocythere 212,213
Mixibius 247, 248
Mixobates 182
moitessierianum, Neopisidium 333
moldaviensis, Ilyodrilus 77
moldaviensis, Potamothrix 36, 75, 77
Momonia 187
monacha, Notodromas 224
monardi, Soldanellonyx 195
Monhystera 261
Monhysterida 260,261
monnardi, Gulnaria 410
monnardi, Lymnaea 410
Mononchida 260,261
Mononchoides 261
Mononchus 261
montgazoniana, Segmentina 435, 437
moravicus, Psammoryctes 89
moravicus, Psammoryctides 79, 89
mucedo, Cristatella 161
mucronata, Lymnaea 411
mucronulatum, Musculium 317
muelleri, Ephydatia 151,152
mulleri, Tumidiana 302, 304
mulleri, Unio 302, 303, 304
Murrayon 245, 247, 249
musculis, Frontipoda 181
Musculium 19,317
musiva, Crassiana 305
musivus, Unio 305
mutila, Eylais 171
Mya 301
Mylonchulus 261
Mytilopsis 313
Mytilus 314
Myxas 401
N
Naididae 33, 39, 44, 65
Nais 52,61
nalivkini, Dreissena 315
nana, Crassiana 295, 305
nana, Unio 305
Nanhermanniidae 198
napocensis, Pristina 47
natangensis, Aturus 189
naticina, Borysthenia 390
naticina, Valvata 390
naticoides, Lithoglyphus 382
naticoides, Paludina 381
Nautarachna 184
nautileus, Turbo 424
Nautilus 424
Nematoda 260
Neoactinolaimus 261,262
Neobrachipoda 188, 190
Neopisidium 22, 333
Neotobrilus 277, 278
Nerita 364, 385
Neritidae 16,23,344,360,364
Neumania 184
newaensis, Isochaetides 78
newaensis, Limnodrilus 78
newaensis, Tubifex 78,91
newberryi, Carinifex 415
newtoni, Plesiocypridopsis 217,219
nigricollis, Erpobdella 120,121
nigricollis, Herpobdella 120
nikolensis, Planorbis leucostoma var. 422
nilssoni, Anodonta 310
450
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
nilssoni, Colletopterum 308
ninguidus, Mixibius 248
nitida, Segmentina 435, 436
nitidum, Cingulipisidium 324
nitidum, Pisidium 323
nitidum, Sphaerium 320
nitidus, Planorbis 436
nordenskioldi, Pseudanodonta 312
normalis, Hibemeuglesa 330
notatus, Planorbis stelmachoetius var. 429
Notodromas 207, 221
Notodromatinae 207, 221
Notostraca 6
Nucleocyclas 318
nucleus, Cyclas 319
nucleus, Sphaerium 320
nummulus, Anisus 421
Nygolaimus 261
О
Obesogammarus 137
obliqua, Cyclas 334
obliqua, Strandesia 235
obliquata, Euglesa 327
oblonga, Lymnaea 405
oblonga, Zschokkea 176
oblongus, Acroloxus 391
obscurus, Aporcelaimellus 264
obtusa, Dero 51, 60
obtusalis, Euglesa 326
occidentals, Dreissensia 314
occulta, Lymnaea 401
oceanicus, Gammarus 140,143
octocullata, Erpobdella 105,120
octocullata, Herpobdella 120
Odhneripisidium 22, 333
Odonata 5
oelandica, Segmentina 437
oelandicus, Planorbis nitidus var. 437
okaensis, Acroloxus 391
oligactis, Pelmatohydra 253, 254
Oligochaeta 7, 30, 32, 94
Olindiasidae 252
olivaceus, Ilyodromus 230
Omphiscola 26, 399, 400, 401
onegensis, Alexandrovia 71
onondaga, Limnozetes 199
opercularis, Menetus 415
Ophidonais 47
ophtalmica, Cypria 236
Opisthorchophorus 25,371,372,377
orbicularis, Mideopis 169, 188
orbiculata, Midea 188
Orchestia 135
Oribatida 164, 165, 167
orientale, Corophium 131
omatus, Calohypsibius 247, 249
Orthogalumna 200
ostiaria, Anodonta 309
ostiaria, Anodonta piscinalis var. 309
ostiarium, Colletopterum 308
Ostracoda 201,207
ostroumovi, Henslowiana 329
ovalis, Atractides 183
ovalis, Oxus 181
ovalis, Unio 303, 304
ovata, Lymnaea 412
ovatus, Limneus 412
ovoglabellus, Murrayon 245
ovulum, Rostozetes 196
Oxidae 177, 180
Oxus 182
oxycnida, Hydra 255, 258
P
Pallasea 130
Pallaseidae 131
Pallasiola 131
Pallasiopsis 131
pallida, Potamocypris 219
palmeni, Bratislavia 47, 48
Paludicella 154, 159
Paludicellidae 153
Paludina 368
paludosa, Batracobdella 100,112
palustre, Buccinum 407
palustris, Lymnaea 401
palustris, Heterozetes 197,199, 200
palustris, Piersigia 171,172
Pandeidae 258
Panisopsis 175, 176
Panisus 175
Указатель латинских названий
451
papillosa, Neumania 183
Paracandona 208, 238
Parachela 245
Paractinolaimus 261,262
Paracyatholaimus 261
paradoxa, Dreissensia 314
Paraelona 371,378
Paramacrobiotus 245
Paramononchus 261
Paramphidelus 261
Paranais 46
Paraphanolaimus 261
Paraplectonema 261
Paraplexa 395
parapsilia, Lymnaea 402
Parasegmentina 435
parasita, Branchiobdella 95
parasita, Ripistes 37, 49
Parasitengona 166
Parasoldanellonyx 195
Parasphaerium 319,320
Parathyas 175
Paratrilobus 261
paravitoldianus, Armiger 424
Paravulvus 261
pardalis, Nais 55, 63, 64
Parhexapodibius 247
parisiensis, Hydrozetes 199
parviscutatus, Parasoldanellonyx 194, 195
parvula, Hibemeuglesa 330
parvulus, Pseudohydryphantantes 173,174
patula, Lymnaea 410
Pectinatella 154, 157
pediformes, Leptocythere 210
Peipsidrilus 80
peledina, Acanthobdella 97, 98
Pelmatohydra 253
Pelodera 261
Peloscolex 69
pentodonta, Branchiobdella 97, 98
peregra, Lymnaea 412
Peregriana 402,409,410
peregrum, Buccinum 409, 412
perezii, Anisus 423
perezii, Planorbis 423
personata, Euglesa 327
Phylactolaemata 153, 159
Physa 27, 392, 394
Physella 393
Physidae 15,27,363,392,393
Physinae 394
Physocypria 208, 233
pictorum, Mya 302
pictorum, Unio 13, 285, 302
Piersigia 172
Piersigioidea 172
pigueti, Aulodrilus 74, 86
Piguetiella 52
pilatoi, Parhexapodibius 247
pilsbryi, Planorbella 415
Piona 187
Pionacercus 187
Pionidae 179, 184
Piscicola 100, 116
Piscicolidae 98, 103,104, 112
piscinale, Colletopterum 308
Piscinaliana 308
piscinalis, Anodonta 308, 309
piscinalis, Cincinna 386, 388
piscinalis, Nerita 389
Pisidiinae 21,316,332
Pisidium 22, 332, 334
placationes, Hydryphantes 174
Planorbarius 27,416,417
Planorbidae 16, 28, 344, 363, 364, 417
Planorbinae 419
Planorbis 28, 417, 419, 431, 432
planorbis, Helix 431,432
planorbis, Planorbis 417,418,432
planorbulina, Valvata 384
Platicrista 249
Plecoptera 5
Plectida 261
Plectus 261
Plesiocypridopsis 207,216
Plumatella 162
Plumatellidae 157
pluriseta, Aulodrilus 74, 86, 87
Podacaridae 196
podagrica, Wettina 184,185
Podocopa 207
Podocopida 207
452
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
polonica, Henslowiana 329
Poly chaeta 30
polymorpha, Dreissena 290,314
polymorphus, Mytilus 313
polynette, Doryphoribius 248
Polyzoa 153
ponderosa, Anodonta 308
ponderosa, Euglesa 327
ponderosum, Colletopterum 308
Pontarachna 170
Pontarachnidae 170
Pontarachnoidea 170
Pontogammaridae 131, 136
Pontoporeiidae 129
Porifera 149
Porohalacarus 195
Porolohmannella 193
portentosa, Hibemeuglesa 330
potamica, Bithynia 376
Potamocypris 203,207,216
Potamopisidium 326
Potamopyrgus 24 379
Potamothrix 77,81,89
praehirsuta, Jaera 147
presbensis, Planorbis 431
Prionchuloides 261
Prionchulus 261
Prismatolaimus 261
Pristina 65
Pristinella 66
Pristinidae 39,40, 65
Prodesmodora 261
Prodorylaimus 261
producta, Bithynia 371,375
producta, Bithynia tentaculata f. 377
producta, Lymnaea f. 403
producta, Lymnaea stagnalis var. 403
profundicola, Limnodrilus 76, 88
Propappidae 41
Propappus 41
Protoclepsis 107
protractus, Unio 293, 302, 304
Protzia 173
Protziidae 173
Psammoryctes 78
Psammoryctides 88
Pseudanodonta 18, 311, 312
Pseudanodontinae 302, 311
Pseudechiniscus 247
Pseudeupera 21, 323, 330
Pseudobiotus 248
pseudobrowniana, Scottia 221, 225
pseudobtusa, Nais 55, 62
Pseudocandona 238
Pseudogyraulus 426, 427
Pseudohydryphantantes 173
Pseudohydryphantinae 173
pseudolittoralis, Unio 306
Pseudosphaerium 324
pseudosphaerium, Pisidium 324
pseudosphaerium, Pseudosphaerium 324
psilia, Lymnaea 402
Psilunio 304
Psilunioninae 301,304
pubera, Cypris 205, 226
pulchella, Pulchelleuglesa 331
pulchella, Valvata 387
Pulchelleuglesa 331
pulchellum, Pisidium 331
pulex, Gammarus 138,140
pullari, Murrayon 245
Pulmonata 9
punctata, Hyalinella 162
Punctodora 261
pusilia, Galba 404
pusilia, Nerita 385
pusilia, Valvata 385
pusillus, Pepsidrilus 74, 75
Q
quadrispinosa, Pallasea 130
quadrispinosa, Pallasiola 131
quadrispinosa, Pallasiopsis 131
R
radiatum, Sphaerium 320
Radix 402, 409
Ramazzottius 243, 247, 248, 249
raphidia, Lymnaea 403
recta, Dreissensia 314
rectidens, Sphaerium 320
rectipes, Huitfeldtia 186
Указатель латинских названий
453
relictus, Gammaracantus 134
repens, Plumatella 158, 163
reptans, Herpetocypris 203, 231
Rhabditida 260, 261
Rhabdolaimus 261
Rhyacodrilus 73, 85
Rhynchelmis 92
richtersi, Paramacrobiotus 243
ringulata, Alexsandrovia 71
riparius, Choanomphalus 426,427, 428
riparius, Planorbis 427
Ripistes 49
rivicola, Cyclas 320
rivicola, Sphaerium 320
Rivicoliana 318,320
rivularis, Euglesa 327
robustoides, Pontogammarus 137,138
Ropohalacarinae 193
Ropohalacarus 193
rosea, Pristina 67
rosea, Pristinella 67
rosea, Roseana 327
Roseana 325, 327
roseum, Pisidium 327
rossicus, Acroloxus 391
rossmaessleri, Choanomphalus 427
rossmaessleri, Gyraulus
rossmaessleri, Planorbis 418, 427, 430
rostrata, Anodonta 311
rostratum, Colletopterum 308,312
rostratus, Unio 302
Rostrozetes 196
rotundatus, Planorbis
ruut, Henslowiana 328
ryckolti, Musculium 317
S
saamicus, Peipsidrilus 80, 81
saamicus, Umbadrilus 80
Sadura 147
salinus, Cyprinotus 232
salinus, Gammarus 140,144
saltator, Talitrus 134
sanctipatricii, Limnocythere 203
sanguisuga, Haemopis 105,118, 119
saracenus, Mixibius 248
Sarscypridopsis 207, 217
sarsi, Jaera 146
sawerbyi, Branchiura
scapularis, Homocaligus 165, 166
scholtzi, Euglesa 326
scholtzi, Hydrobia 380
scholtzi, Marstoniopsis 380,381
scholtzi, Pisidium 326
Schwiebia 167
scoticum, Diphascon 250
Scottia 207,221
Scottinae 207,221
Segmentina 433, 434, 435, 436
Segmentinae 434
Segmentinidae 28, 363, 433, 434
Segnetillia 28, 433
selenicus, Dactylobiotus 245
Semitobrilus 261
septemgyratus, Anisus 423
serpentina, Ophidonais 47
servaini, Dreissensia 315
setosus, Chaetogaster 46, 59
setosus, Trhypochthoniellus 199
shadini, Acroloxus 391
shadini, Anodonta piscinalis
Sibirenauta 392
sicula, Hemicythere 210
similis, Cyclas 319
simplex, Nais 54, 62
sinuator, Arrenurus 181
siouffi, Dreissensia 315
skorikovi, Cincinna 388
skorikovi, Valvata 388
Slavina 50
smerczinskii, Bertolaimus 247
smimowi, Tubifex 85, 91
sobieskianus, Armiger 424
socialis, Paraelona 371,373,378
Soldanellonyx 195
solida, Cyclas 319
solida, Sphaerium 319
sowerbyi, Branchiura 82, 85
sowerbyi, Craspedacusta 252
Specaria 57
Sperchon 180
Sperchonidae 177, 180
454
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2
Sperchonopsis 180
sphaeridius, Viviparus 368
Sphaeriidae 10, 17, 18, 315, 316
Sphaeriinae 19, 316, 317
Sphaerium 19,315,317,318,319,321
spinulosus, Planorbis crista var. 421
spirorbis, Anisus 417, 419, 420, 422
spirorbis, Helix 420, 422
Spirosperma 69
spitzbergensis, Mesocrista 250
splendens, Cingulipisidium 324
Spongilla 151,152
Spongillidae 151
stagnalis, Anodonta 307
stagnalis, Dorylaimus 264
stagnalis, Helix 402, 403
stagnalis, Hydrobia 379
stagnalis, Lymnaea 399, 403
stagnalis, Mytilus 307
stagnalis, Helobdella 100,106
Stagnicola 345, 398, 401,407
stagnorum, Helix 379
stagnorum, Ventrosia 379
Standesia 235
starobogatovi, Segmentina 436
steenbuchi, Euglesa 327
steinii, Hydrobia 380
steinii, Marstoniopsis 381
stelfoxi, Europisidium 334
stelmachoetius, Gyraulus 429, 430
stelmachoetius, Planorbis 417
stenochorias, Bumupia
Stenocypria 208, 229
Stenocyprinae 208, 220
Stenocypris 220
Stenolaemata 155
stevensoni, Darwinula 203, 205, 209
steward, Pisidium 333
Strandesia 208, 229
strauchianus, Anisus 420, 422
strauchianus, Planorbis 422
stroemi, Gyraulus 430
stroemi, Planorbis 430
studeri, Cincinna 383, 387
studeri, Valvata 387
Stygobromus 127
Stylaria 48, 59
Stylodrilus 93, 94
subangulata, Limnaea truncatula var. 405
subangulata, Lymnaea 405
subcirculare, Colletopterum 308
subcircularis, Ancylus 415
subcircularis, Anodonta
subhibemica, Hibemeuglesa 330
subponderosa, Euglesa 327
Subsegmentina 435
subterranean, Cavemocypris 216,217
subtruncata, Pseudeupera
subulata, Lymnaea stagnalis var. 403
suecica, Henslowiana 329
suillus, Pseudechiniscus 247
sulcata, Dreissensia 315
sultana, Fredericella 154, 157,161
supina, Henslowiana 329
Supiniana 329
supinum, Pisidium 329
Synurella 128
Talitridae 127, 134
Talitrus 134
tanuga, Cingulipisidium 324
Tardigrada 242
tardigradum, Milnesium 244, 247
taslei, Physa 394
Tasserkidrilus 78
taurica, Lymnaea 406
tectiporus, Copidognatus 196
Tellina 317,321
templetoni, lyiodrilus 81,82
templetoni, Tubifex 81
tenebrarum, Aeolosoma 31
tentaculata, Bithynia 370, 374, 375, 377
tentaculata, Helix 374
tenuilineatum, Europisidium 334
tenuilineatum, Pisidium 334
terebra, Limnaea palustris var. 401
terebra, Lymnaea 401
terebrandis, Orthogalumna 200
terverianum, Musculium 317
tessulata, Protoclepsis 107
tessulatum, Theromyson 107
Указатель латинских названий
455
testacea, Erpobdella 121 testacea, Herpobdella 121 tetraedra, Eiseniella 43, 44 Tetragonocyclas 21, 323, 331 tetratheca, Rhynchelmis 92 Teutonia 177 Teutoniidae 177 Theodoxus 16,23,344,364,365 Thermozodium 244 Theromyzon 100, 107 thiesa, Dreissensia thiesae, Dreissensia 315 thoracata, Parathyas 176 Thulinius 248 Thyas 175 Thyasides 175 Thyasinae 175 Thyopsis 175 Tichogonia 314 tigrinus, Gammarus 145 Tiphys 184 toanensis, Gelatinella 161 tobolica, Lymnaea 410 Tobrilus 261 torosa, Cyprides 214 torquatum, Neopisidium 333 torquatum, Pisidium 333 Torquis 428 Torrenticola 177 Torrenticolidae 177 Trombidiformes 165 Trombidida 166 Tropidiscus 432 troscheli, Bithynia limosus 371 troscheli, Paludina 371 troschelii, Bithynia 371 troschelii, Opisthorchophorus 371 truncata, Euthyas 176 truncatula, Lymnaea 405 truncatulum, Buccinum 405 Tubifex 81,91 tubifex, Tubifex 36, 84, 91 Tubificidae 33, 40, 69 tumida, Dreissensia 315 tumida, Lymnaea 410 tumida, Tumidiana 302 tumida, Unio 302 Tumidiana 302, 303 tumidus, Unio 303 Turbellaria 7 Turbo 373 turgida, Lymnaea 403 turgida, Lymnaea stagnalis 403, 404 turricula, Lymnaea 408 turricula, Stagnicola turrita, Aplexa 395 turrita, Planorbis 395 Tylencholaimus 261,262,264 Tyrrhenocythere 209,210
transsilvanica, Bithynia transsylvanica, Lymnaea palustris var. 408 Trhypochthoniellus 198 Trhypochthoniidae 198 Trichoptera 6 tridentatus, Sperchon 181 trigonicus, Hygrobates 183 trigonum, Neopisidium 333 Trimalaconothrus 197, 198 Tripyla 261 troschelii, Opisthorophorus 371, 378 troschelii, Paludina 377 Tripylella 261 Tripylina 261 Trischistoma 261 Trochospongilla 152 U udekemianus, Limnodrilus 75, 87 umbilicata, Valvata 386 umbilicatus, Planorbis 433 uncatus, Mixobates 182,183 Uncinais 47 uncinata, Uncinais 47 Unio 17,301,302,303 Unionicola 184 Unionicolidae 179 Unionidae 10, 17, 286, 289, 299 Unioninae 301,302 uniscutatus, Ropohalacarus 193 urinator, Conventus 324 uvalievae, Aenigmomphiscola 397, 399
456
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т 2
Valvata 26, 382, 383, 384
Valvatidae 15, 16, 24, 26, 344, 361, 362,
382
Valvatinae 383
valvatoides, Opisthorchophorus 371
variabilis, Nais 56, 64
variegator, Forelia 186
variegatus, Lubriculus 93, 94
varsoviensis, Gammarus 139,141
vavrai, Cryptocandona 239
vejdovskyanum, Bothrioneurum 72
Vejdovskyella 52, 60
vejdovskyi, Ilyodrilus 90
vejdovskyi, Potamothrix 84, 90
velutinus, Embolocephalus 70
velutinus, Peloscolex 70
ventricosa, Bithynia 374
ventricosa, Lymnaea 374
ventrosa, Dreissensia 315
ventrosa, Hydrobia 379
Ventrosia 23, 379
verrucata, Boreobdella 100, 108,109
verrucosa, Sperchonopsis 181
versicolor, Brachypoda 189, 190
Victorella 154
vidua, Cypridopsis 218
villosus, Dikerogammarus 135,136
violaceus, Acalyptonotus 178, 179
violacia, Porolohmannella 193
virens, Eucypris 234
viridissima, Chlorohydra 253, 254
vistulae, Viviparus 368
vitoldianus, Armiger 424
Vivipara 368
vivipara, Helix 367, 368
Viviparidae 24, 361, 365, 366
Viviparus 24, 365, 366, 367
viviparus, Viviparus 368
volgae, Mytilus 314
volgensis, Anodonta piscinalis var. 309
volgensis, Anodonta ponderosa 309
volgensis, Mytilus 314
volgensis, Acipenserobdella 113,114
volgensis, Piscicola 113
volki, Propappus 41, 42
volutator, Corophium 131,132
vortex, Anisus 418
vortex, Helix 418
Vorticulus 421
vorticulus, Anisus 421
vorticulus, Planorbis 421
vshivkovae, Kongsbergia 189
vulgaris, Hydra 255, 258
vulnerata, Lymnaea 401,409
W
wackeri, Limnohalacarus 193,194
waldeni, Henslowiana 328
waldvogeli, Haemonais 57, 58
warpachowskyi, Chaetogammarus 138,139
warpachowskyi, Echinogammarus 138
weberi, Lobohalacarus 195
weglarskae, Bertolaimus 247
westerlundi, Dreissensia 315
Wettina 184
Z
zaddachi, Gammarus 144, 145
zebrella, Lymnaea 408
zellensis, Anodonta 307
zellensis, Mytilus 307
zetlandicus, Borealibius 248, 249
Zetomimidae 200
Zetomimus 200
Zschokkea 175
Рис. 1. Gmelinoides fasciatus (фото H.A. Березиной).
Рис. 2. Chelicorophium curvispinum (фото H.A. Березиной).
Рис. 3. Gammaracanthus loricatus (фото H.A. Березиной).
Рис. 4. Talitrus saltator (no: Spicer, Janas, 2006).
Рис. 5. Dikerogammariis villosus (фото H.A. Березиной).
Рис. 6. Pontogammarus robustoides (фото H.A. Березиной).
Рис. 7. Gammarus lacustris (фото НА. Березиной).
Рис. 8. Gammarus tigrinus (фото H.A. Березиной).
Spongilla lacustris (p. Оредеж).
Ephydatia muelleri (p. Оредеж).
Cristatella mucedo. Статобласт.
Cristatella mucedo. Колония co статобластами.
Cristatella mucedo. Статобласт.
Cristatella mucedo. Молодая колония.
\
Fredericella sultana. Часть колонии.
Hyalinella punctata. Колония.
Lophopus. Колонии.
Lophopus. Спинобласт.
Paludicella. Колония.
Paludicella. Часть колонии.
Plumatella emarginata. Колонии.
Plumatella emarginata. Колонии. Показа-
ны margin.
Plumatella fruticosa. Колонии.
Plumatella repens. Колония.
Plumatella fungosa. Колония.
Plumatella fungosa. Колония.
Damvinula stevensoni (Brady & Robertson,
1870). По: E. Day, 2004.
Limnocythere inopinata (Baird, 1843).
По: E. Day, 2004.
Sarscypridopsis aculeata (Costa, 1847).
По: E. Day, 2004.
Potamocypris pallida Alm, 1914.
По: E. Day, 2004.
Cypridopsis vidua (O.F. Muller, 1776).
По: E. Day, 2004.
Cyprois marginata (Strauss, 1821).
По: E. Day, 2004.
Cyclocypris ovum (Jurine, 1820).
По: E. Day, 2004.
Candona rostrata (Brady & Norman, 1889).
По: E. Day; 2004.
Cyprois occidentalis Sars, 1926.
По: E. Day, 2004.
Candona rectangulata Alm, 1914.
По: E. Day, 2004.
Candona Candida (Muller, 1785).
По: E. Day, 2004.
Таблица I
Рис. 1 а-з. Margaritifera borealis (Westerlund, 1871): а, б — из-под г. Архангельск
(ЗИН, №1); в — искривленный экз. без указания места сбора (ЗИН, № 5); г, д,
е — экз. с неразвитым задним краем, р. Лямукея, Архангельская обл. (ЗИН, № 4):
ж, з — р. Муна, Мурманская обл. (ЗИН, № 6). а, г, д — вид сбоку и изнутри: з —
вид сбоку; б, в, е, ж — вид спереди. Масштабная линейка 3 см.
Рис. 2 а-д. Margaritifera margaritifera (Linnaeus, 1758): а, б — экз. с неразвитым
задним краем, р. Варзуга, Кольский п-ов (ЗИН, № 12); в, г, д — экз. из коллекции
Михалкова без указания места сбора (ЗИН, № 48): в, г — вид сбоку и изнутри;
б — вид сбоку; а, д — вид спереди. Масштабная линейка 3 см.
Рис. 3 а-в. Margaritifera elongata (Lamark, 1819): озерцо у р. Плюссы, дер. Сер-
бино (ЗИН, №1): а, б — вид сбоку и изнутри; в — вид спереди. Масштабная
линейка 3 см.
Рис. 4. Макушечная скульптура Unio pictorum (Linnaeus, 1758).
Рис. 5. Макушечная скульптура Unio limosus Nilsson, 1822.
Рис. 6. Сифоны Unio pictorum (Linnaeus, 1758): оз. Сапшо, Смоленская обл.
Рис. 7 а-в. Unio protractus Lindholm, 1922: старина р. Ока, Московской обл.
(ЗИН, = U. muelleri, № 3). а — вид спереди; б, в — вид сбоку и изнутри. Мас-
штабная линейка 3 см.
Рис. 8 а-в. Unio pictorum (Linnaeus, 1758): Швеция (ЗИН, = U. rostratus, №1). а,
б — вид сбоку и изнутри; в — вид спереди. Масштабная линейка 3 см.
Рис. 9 а-в. Unio limosus Nilsson, 1822: Низовья Волги у г. Астрахань, пос. Ки-
ровский (ЗИН, = U. limosus bemingi, № 1): а, б — вид сбоку и изнутри; в — вид
спереди. Масштабная линейка 3 см.
Рис. 10. Макушечная скульптура Unio longirostris Rossmaessler, 1836.
Рис. 11. Сифоны Unio longirostris Rossmaessler, 1836: оз. Сапшо, Смоленская обл.
Рис. 12 а-в. Unio tumidus (Philipsson in Retzius, 1788): p. Ока, Прилуки (ЗИН,
№ 78): а, б — вид сбоку и изнутри; в — вид спереди. Масштабная линейка 3 см.
Рис. 13 а-д. Unio longirostris Rossmaessler, 1836: а — оз. Сапшо, Смоленская обл.
(БПИ); б, в — басе. р. Медведица, Саратовская обл. (ЗИН, № 13); г, д — р. Иви-
ца, Тверская обл. (БПИ): а — вид сбоку; б, г — вид сбоку и изнутри; в, д — вид
спереди. Масштабная линейка 3 см.
Рис. 14 а-д. Unio conus (Spengler, 1793): а, б, в — р. Днепр (ЗИН, хранилась под
названием U. tumidus, № 21); г, д — оз. Селигер, Тверская обл. (ЗИН, = U. ovalis,
№ 3): а, б, г — вид сбоку и изнутри; в, д — вид спереди. Масштабная линейка 3 см.
Таблица II
Рис. 15 а, б. Примеры макушечной скульптуры видов рода Crassiana.
Рис. 16 Сифоны Crassiana musiva (Spengler, 1793): р. Ивица, Тверская обл.
Рис. 17 а-г. Crassiana папа (Lamarck, 1819): р. Жаксыкар, окр. г. Актюбинск
(ЗИН).
Рис. 18 a-ж. Crassiana musiva (Spengler, 1793): а, б, в, г — экземпляры разной
степени вытянутости, собранные в одном биотопе р. Ивица, Тверская обл. (БПИ);
д, е, ж — р. Ока у г. Калуга (ЗИН).
Рис. 19 а-д. Crassiana crassa (Philipsson in Retzius, 1788): a, 6 — p. Великая у
г. Пскова (БПИ); в, г, д — р. Ока у г. Калуга (ЗИН).
Рис 20 а, б. Примеры макушечной скульптуры видов рода Anodona.
Рис. 21 а-в. Anodonta zellensis (Gmelin, 1791): оз. Тайное, Брестская обл. (ЗИН,
№51).
Рис. 22 а-д. Anodonta stagnalis (Gmelin, 1791): а, б, в — оз. Башкирское у г. Стер-
литамак (ЗИН); г, д — оз. Жушэр, Республика Мэрий Эл (ЗИН).
Рис. 23 а-в. Anodonta cygnea (Linnaeus, 1758): дельта Волги (ЗИН).
Рис. 24 а, б. Примеры макушечной скульптуры видов рода Colletopterum.
Рис. 25. Сифоны Colletopterum sp.
Рис. 26 а-е. Colletopterum milaschevichi Bogatov, Starobogatov et Prozorova, 2005:
a, 6 — молодой экз., паратип, дельта Волги у д. Оли (ЗИН); г, д, е — голотип,
дельта Волги, Кабаний Ильмень (ЗИН, № 1).
Рис. 27 а-г. Colletopterum convexum (Drouet 1888): р. Дон (ЗИН).
Рис. 28 а-г. Colletopterum apollonicum (Bourguignat 1880): оз. Апполония, Малая
Азия (ЗИН, № 1).
Рис. 21 в, 22в, 23в, 26в, 27в, 28в — вид на створку сверху под углом около 20° к вертикаль-
ной оси. Ориентация остальных раковин такая же, как на таблице I. Масштабная линейка
3 см.
Таблица III
Рис. 29 а-в. Colletopterum subcirculare (Clessin, 1873): р. Дон у г. Павловск (ЗИН,
№ 10).
Рис. 30 а-в. Colletopterum baeri Bogatov, Starobogatov et Prozorova, 2005: голотип,
p. Волга у Астрахани (ЗИН, № 1).
Рис. 31 а-в. Colletopterum ostiarium (Drouet 1881): р. Днепр, ниже г. Кременчук
(ЗИН, № 7).
Рис. 32 а-в. С. nilssoni (Kiister 1842): р. М. Истра у г. Истра, Московская обл.
(ЗИН, № 1).
Рис. 33 а-г. Colletopterum ponderosum (С. Pfeiffer 1825): оз. Узьминское, Псков-
ская обл. (ЗИН, № 36).
Рис. 34 а-д. Colletopterum anatinum (Linnaeus 1758): р. Ивица, Тверская обл. (БПИ).
Рис. 35 а-г. Colletopterum piscinale (Nilsson, 1822): а — сифоны (р. Ивица, Твер-
ская обл., фото В. Богатова); б, в, г — дельта Волги у п. Барановка (ЗИН).
Рис. 36 a-ж. Colletopterum rostratum (Rossmassler, 1836): а, б, в, г — Германия
(ЗИН, № 1); д, е, ж — р. Ивица, Тверская обл. (ЗИН).
Рис. 37 а-г. Примеры макушечной скульптуры видов рода Pseudanodonta.
Рис. 38 Сифоны Pseudanodonta elongata (Hollandre, 1836): р. Ивица, Тверская
обл.
Рис. 33г, 34г, 36г — вид на створку сверху под углом около 20° к вертикальной оси. Ори-
ентация остальных раковин такая же, как на таблице 1. Масштабная линейка 3 см.
Таблица IV
Рис. 39 а-е. Pseudanodonta complanata (Rossmaessler, 1835): а, б, в, г, д — Кали-
нинградская обл., Куршская коса (ЗИН); е — экз. с неразвитым задним краем,
устье Днестра (ЗИН, № 35).
Рис. 40 a-к. Pseudanodonta elongata (Hollandre, 1836): а, б, в, г, д — Калининград-
ская обл., Куршская коса (ЗИН, № 4); е, ж, з — р. Ивица, Тверская обл. (БПИ);
и — р. Сев. Двина (ЗИН, № 2); к — экз. с неразвитым задним краем, басе. р. Мед-
ведица, Саратовская обл. (ЗИН, № 22); л — экз. с разросшимся задним краем,
р. Днепр, низовья (ЗИН).
Рис. 41 а, б. Dreissena bugensis (Andrusov, 1897): Днепровское водохр., у г. Дне-
пропетровск (ЗИН, № 2).
Рис. 42 а, б Dreissena polymorpha (Pallas, 1771): без указания места сбора (ЗИН).
Рис. 43. Theodoxus fluviatilis (Linnaeus, 1758) (по: Кантор, Сысоев, 2006). Мас-
штабная линейка — 5 мм.
Рис. 44. Contectiana contecta (Millet, 1813).
Рис. 45. Contectiana ladogensis Chemogorenko et Starobogatov, 1987.
Рис. 46. Viviparus ater (Christophori et Jan, 1832).
Рис. 47. Viviparus sphaeridius (Bourguignat, 1880).
Рис. 48. Viviparus viviparus (Linnaeus, 1758).
Рис. 49. Bithynia tentaculata Linnaeus, 1758).
Рис. 50. Codiella leachii (Sheppard, 1823).
Рис. 51. Opisthorchophorus troschelii (Paasch, 1842).
Рис. 52. Potamopyrgus antipodarum (Gray, 1843).
Рис. 53. Ventrosia stagnorum (Gmelin, 1791).
Рис. 39д, 40д — вид на створку сверху под углом около 20° к вертикальной оси. Ориента-
ция остальных раковин такая же, как на вклейке 1. Масштабная линейка 3 см. Рис. 41, 42:
а — вид сбоку и изнутри; б — вид спереди. Масштабная линейка 1 см. Рис. 44—48 — мас-
штабная линейка 10 мм; рис. 49 — масштабная линейка 3 мм; рис. 50-53 — масштабная
линейка 2 мм.
Таблица V
Рис. 54. Marstoniopsis scholtzi (A. Schmidt, 1856).
Рис. 55 а, б. Lithoglyphus naticoides (С. Pfeiffer, 1828).
Рис. 56. Valvata cristata O.F. Muller, 1774.
Рис. 57. Valvata planorbulina Paladilhe, 1867.
Рис. 58. Valvata elatior Menzel, 1904.
Рис. 59. Valvata geyeri Menzel, 1904.
Рис. 60. Valvata pusilia (O.F. Muller, 1774).
Рис. 61. Valvata andreaei Menzel, 1904.
Рис. 62 а, б. Cincinna macrostoma Morch, 1864.
Рис. 63. Cincinna frigida (Westerlund, 1873).
Рис. 64. Cincinna studeri Boeters et Falkner, 1998 (no: Gloer, 2002: Abb. 224 — Val-
vata (Cincinna) studeri Boeters et Falkner, 1998).
Рис. 65. Cincinna depressa (C. Pfeiffer, 1821).
Рис. 66. Cincinna ambigua (Westerlund, 1873).
Рис. 67. Cincinna skorikovi (Lindholm, 1911).
Рис. 68 a, 6. Cincinna piscinalis (O.F. Miiller, 1774).
Рис. 69 a, 6. Cincinna lilljeborgi (Westerlund, 1897).
Рис. 70. Cincinna antiqua (Morris in Morris et Sowerby, 1838).
Рис. 71. Cincinna falsifluviatUis Starobogatov in Anistratcnko et Anistratenko, 2001.
Рис. 54-63, 65-71 — масштабная линейка I мм.
Таблица VI
Рис. 72. Cincinna discors (Westerlund, 1886).
Рис. 73. Borysthenia naticina (Menke, 1846).
Рис. 74. Acroloxus lacustris (Linnaeus, 1758).
Рис. 75. Aplexa hypnorum (Linnaeus, 1758).
Рис. 76. Aplexa turrita (O.F. Muller, 1774).
Рис. 77. Costatelia acuta (Drapamaud, 1805).
Рис. 78. Physa fontinalis (Linnaeus, 1758).
Рис. 79. Physa taslei Bourguignat, 1860.
Рис. 80. Aenigmomphiscola europaea Kruglov et Starobogatov, 1981.
Рис. 81. Aenigmomphiscola uvalievae Kruglov et Starobogatov, 1981.
Рис. 82. Lymnaea clavata (Westerlund, 1885).
Рис. 83. Lymnaea terebra (Westerlund, 1885).
Рис. 84. Lymnaea glutinosa (O.F. Muller, 1774).
Рис. 85. Lymnaea auricularia (Linnaeus, 1758).
Рис. 86. Lymnaea fragilis (Linnaeus, 1758).
Рис. 87. Lymnaea stagnalis (Linnaeus, 1758).
Рис. 88. Lymnaea doriana (Bourguignat, 1862).
Рис. 89. Lymnaea truncatula (O.F. Muller, 1774).
Рис. 90. Lymnaea subangulata (Roffiaen, 1868).
Рис. 91. Lymnaea corvus (Gmelin in Linnaeus, 1791).
Рис. 92. Lymnaea taurica kazakensis Mozley, 1934.
Рис. 93. Lymnaea archangelica Kruglov et Starobogatov, 1986.
Рис. 94. Lymnaea palustris (O.F. Miiller, 1774).
Рис. 95. Lymnaea atra (Schrank, 1803).
Рис. 96. Lymnaea callomphala (Servain, 1881).
Рис. 97. Lymnaea turricula (Held, 1836).
Рис. 98. Lymnaea fusca (C. Pfeiffer, 1821).
Рис. 99. Lymnaea monnardi (Hanmann, 1841).
Рис. 100. Lymnaea ampla (Hartmann, 1821).
Рис. 72-74, 76, 79—81, 89, 90 — масштабная линейка 1 мм: рис. 75, 77, 78. 82. 93-98 —
масштабная линейка 2 мм; рис. 83, 84 — масштабная линейка 4 мм: рис. 85-88, 91, 92, 99,
100 — масштабная линейка 5 мм.
Таблица VII
Рис. 101. Lymnaea tumida (Held, 1836).
Рис. 102. Lymnaea mucronata (Held, 1836).
Рис. 103. Lymnaea fulva (Ziegler in Kiister, 1862).
Рис. 104. Lymnaea ampullacea (Rossmassler, 1835).
Рис. 105. Lymnaea ovata (Drapamaud, 1805).
Рис. 106. Lymnaea balthica (Linnaeus, 1758).
Рис. 107. Lymnaeaperegra (O.F. Muller, 1774).
Рис. 108. Lymnaea lagotis (Schrank, 1803).
Рис. 109. Lymnaea intermedia Lamarck, 1822.
Рис. 110. Lymnaea fontinalis (Studer, 1820).
Рис. Ill а-в. Ancylus subcircularis Clessin, 1882.
Рис. 112 а-в. Ancylus fluviatilis O.F. Muller, 1774.
Рис. 113 а-в. Planorbarius corneas (Linnaeus, 1758 '.
Рис. 114 а-в. Planorbarius grandis (Dunker in Kiister et Dunker, 1850).
Рис. 115 а-в. Anisus vorticulus (Troschel, 1834).
Рис. 116 а-в. Anisus vortex (Linnaeus. 1758).
\ J J
Рис. 117 а-г. Anisus strauchianus (Clessin, 1884).
Рис. 118 а-в. Anisus spirorbis (Linnaeus, 1758).
Рис. 119 а-в. Anisus leucostoma Millet, 1813).
Рис. 120 а-в. Anisus perezii (Graells in Dupuy, 1850).
Рис. 121 а-в. Armiger crista (Linnaeus, 1758).
Рис. 122 а-в. Armiger bielzi (Kimakowicz, 1884).
Рис. 123 а-в. Bathyomphalus dispar (Westerlund, 1871).
Рис. 101, 109, 110, 113 и 114 — масштабная линейка 5 мм; рис. 102-108 — масштабная
линейка 2 мм; рис. 111, 112, 115-123 — масштабная линейка 1 мм. Рис. 111, 112: а — вид
сверху, б— вид справа, в — вид сзади: рис. 113-123: а — вид спереди, б — вид сверху,
в — вид снизу; рис. 117 г — скульптура последнего оборота.
Таблица VIII
Рис. 124 а-в. Bathyomphalus crassus (Da Costa, 1778).
Рис. 125 а-в. Bathyomphalus contortus (Linnaeus, 1758).
Рис. 126 а-в. Choanomphalus riparius (Westerlund, 1865).
Рис. 127 а-в. Choanomphalus rossmaessleri (Auerswald in A. Schmidt, 1851).
Рис. 128 а-в. Gyraulus laevis (Alder, 1838).
Рис. 129 а-в. Gyraulus stelmachoetius (Bourguignat, 1860).
Рис. 130 а-г. Gyraulus albus (O.F. Muller, 1774).
Рис. 131 а-в. Gyraulus acronicus (Ferussac, 1807).
Рис. 132 а-в. Gyraulus stroemi (Westerlund, 1881).
Рис. 133 а-в. Gyraulus borealis (Loven in Westerlund, 1875).
Рис. 134 а-в. Planorbis carinatus O.F. Muller, 1774.
Рис. 135 а-в. Planorbis planorbis (Linnaeus, 1758).
Рис. 136 а-в. Planorbis umbilicatus O.F. Muller, 1774.
Рис. 137 а-в. Segmentina distinguenda (Gredler, 1859).
Рис. 138 а-в. Segmentina nitida (O.F. Muller, 1774).
Рис. 139 а-в. Segmentina starobogatovi Kruglov et Soldatenko, 1997.
Рис. 140 а-в. Segmentina oelandica (Westerlund, 1885).
Рис. 141 а-в. Segmentina montgazoniana Bourguignat in Servain, 1881.
Рис. 142 а-в. Hippeutis complanatus (Linnaeus, 1758).
Рис. 124, 125, 127-133, 137-142 — масштабная линейка 1 мм; рис. 126 — масштабная
линейка 2 мм; рис. 134-136 — масштабная линейка 3 мм. Рис. 124-142: а — вид спереди,
б — вид сверху, в — вид снизу; рис. 130 г — скульптура последнего оборота.
Таблица I
Ml
Таблица II
Таблица III
Таблица IV
Таблица V
Таблица VI
Таблица VII
Таблица VIII
СОДЕРЖАНИЕ
Предисловие (С.Я. Цало.наин).....................................................................3
Донные беспозвоночные пресных вод (А.А. Пржиборо)................................................5
Моллюски (77. В. Кияшко).........................................................................8
Кольчатые черви (ИГ. Циплёнкина)...........................................................................30
Многощетинковые черви.......................................................................30
Малощетинковые черви........................................................................32
Бранхиобделлиды.............................................................................94
Щетинконосные пиявки........................................................................96
Пиявки......................................................................................98
Амфиподы (Н.А. Березина).......................................................................122
Изоподы (Н.А. Березина)........................................................................................... 146
Губки (Л. В. Иванова)..........................................................................149
Мшанки (В.И. Гонтарь)..........................................................................153
Клещи (Д.В. Туманов)...........................................................................164
Остракоды (Е.А. Курашев).......................................................................201
Тихоходки (Д.В. Туманов).......................................................................242
Книдарии (С.Д. Степаньянц) .................................................................................251
Нематоды (С.Я. Цалолихин)......................................................................260
Приложение. Пресноводные моллюски Севера и Северо-Запада
Европейской России.........................................................................279
Предисловие (П.В. Кияшко)..................................................................279
Общая характеристика типа Моллюски (П.В. Кияшко)...........................................283
Класс Двустворчатые моллюски (В.В. Богатов, П.В. Кияшко)...................................285
Класс Брюхоногие моллюски (П.В. Кияшко, Е.В. Солдатенко,
М. В. Винарский)..................................................................................................... 335
Указатель латинских названий...................................................................439
Научное издание
ОПРЕДЕЛИТЕЛЬ
зоопланктона и зообентоса пресных вод
Европейской России
Под редакцией
В.Р. Алексеева. С.Я. Цалолихина
Том 2
Зообентос
Редактор тома С.Я. Цалолихин
СПб. - М.: Товарищество научных изданий КМК. 2016.
457 с.. 6 с., 18 цв. вкл.
при участии ИП Михайлова К.Г.
Редактор издательства К.Г. Михайлов
Оригинал-макет: ТА. Горлина
Для заявок:
123100, Москва, а/я 16, Издательство КМК
электронный адрес mikhailov2000@gmail.com
http://avtor-kmk. ш
Подписано в печать 21.12.2015. Заказ № 2088
Формат 70x100/16. Объём 28,5 + 1,5 п.л. Усл. печ.л. 39.
Бумага офсетн. и мелов. Тираж 800 экз.
Отпечатано в ППП «Типография «Наука»,
121099, Москва, Шубинский пер., 6
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 2. Зообентос /
Под ред. В.Р. Алексеева, С.Я. Цалолихина. — М.: Товарищество научных изданий КМК, 2016. —
457 с., ил., 6 с., 18 цв. вкл.
Иллюстрированный определитель организмов зоопланктона и зообентоса внутренних вод Евро-
пейской России предназначен для исследователей водных экосистем. Он также может быть ис-
пользован для обучения и самообучения студентов биологических специальностей вузов. В соот-
ветствии с делением биологических специальностей в гидробиологии он состоит из двух томов,
посвященных анализу видового состава зоопланктона и зообентоса. Во втором томе представле-
ны, в разной степени подробности, все основные группы донных беспозвоночных животных, с
акцентом на фауну северо-запада региона.
Книга предназначена для зоологов, лимнологов, экологов, водных токсикологов, биологов.