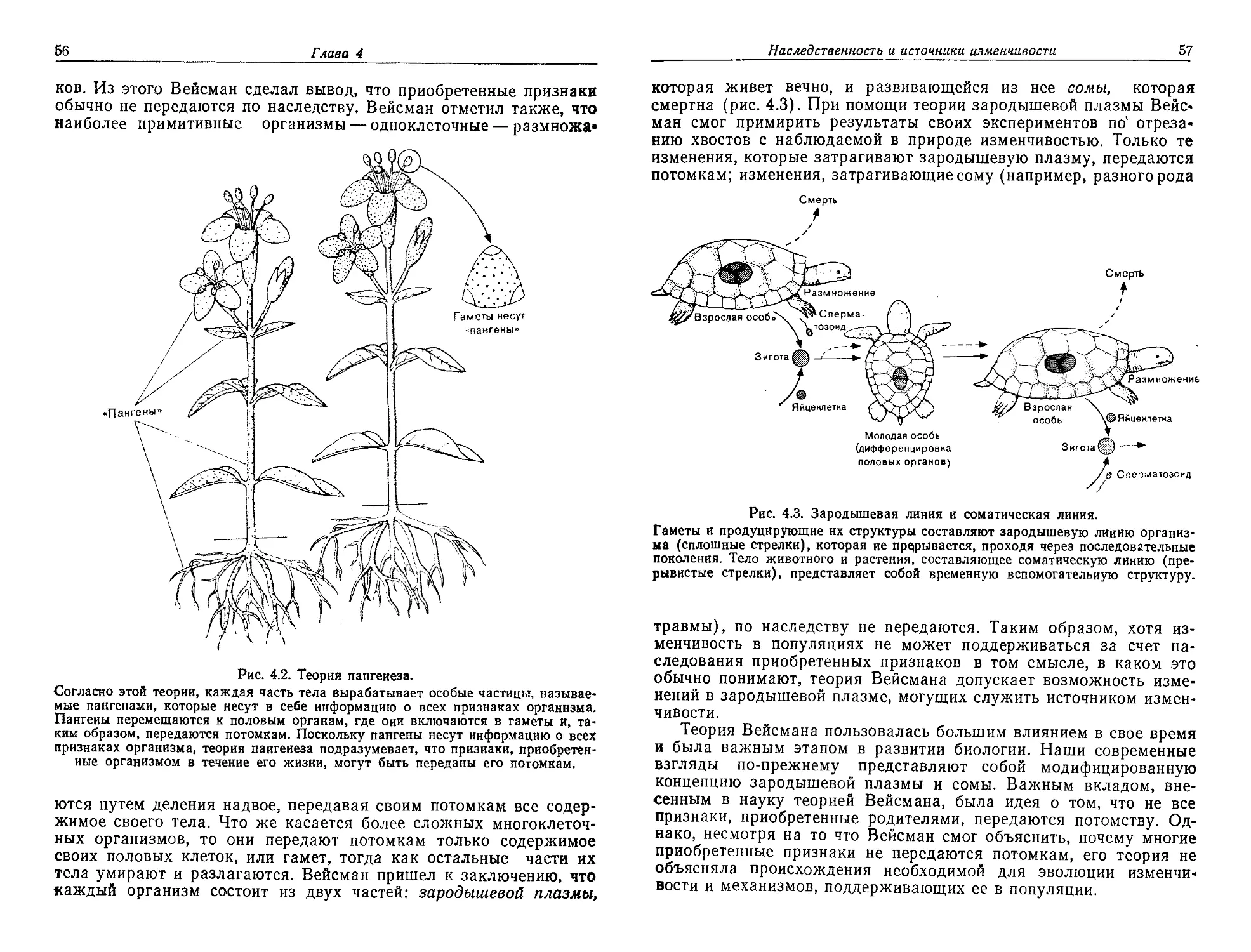
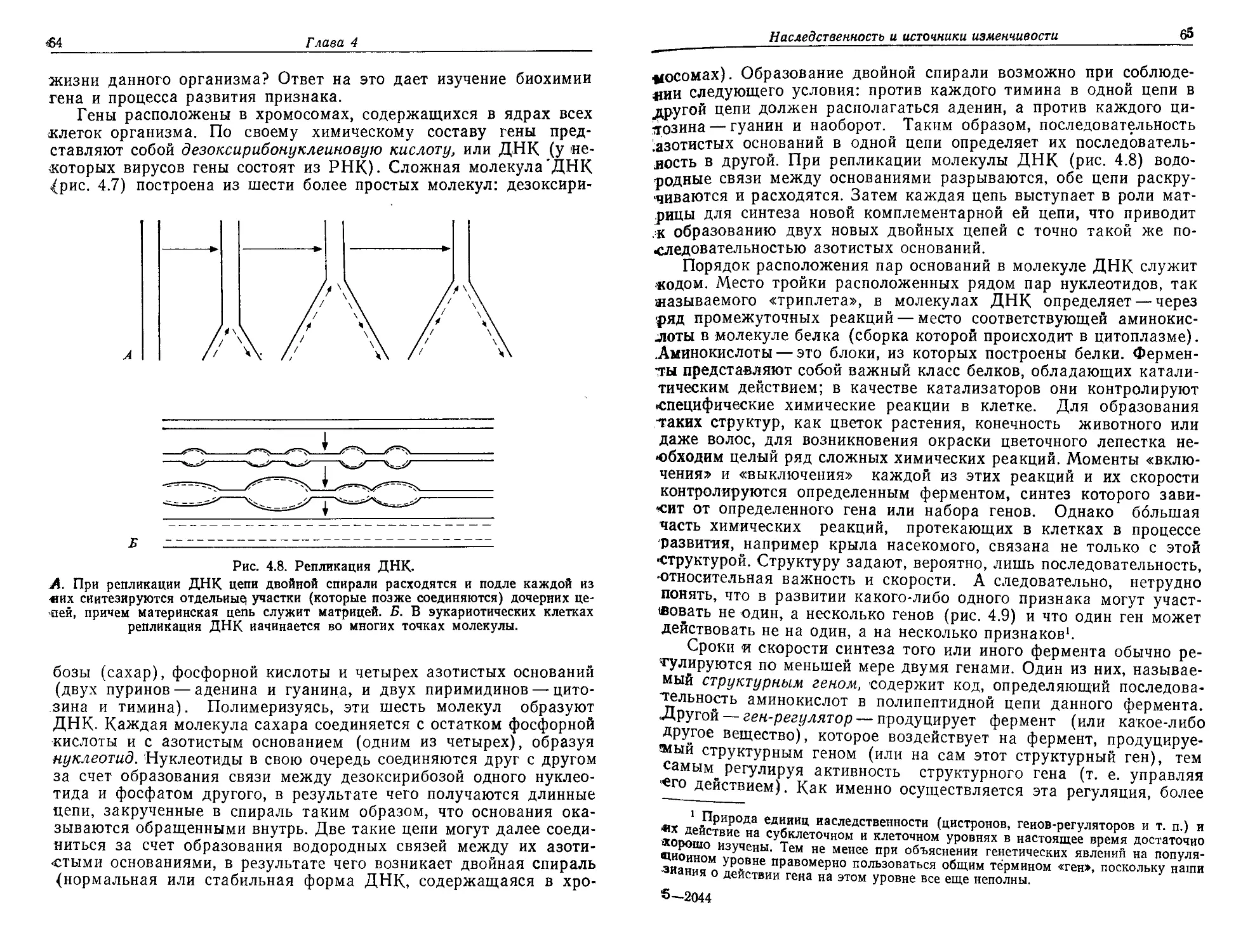
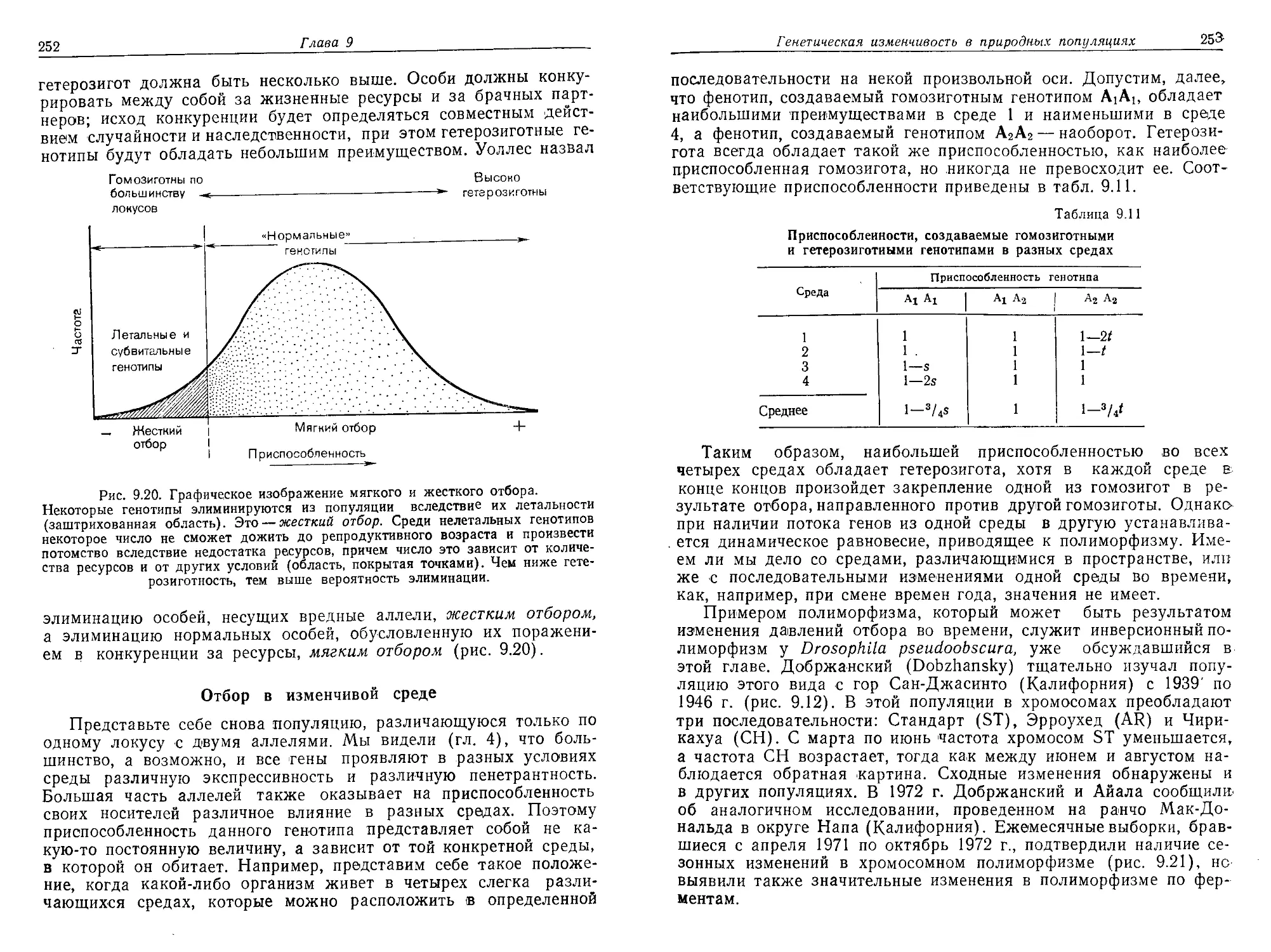
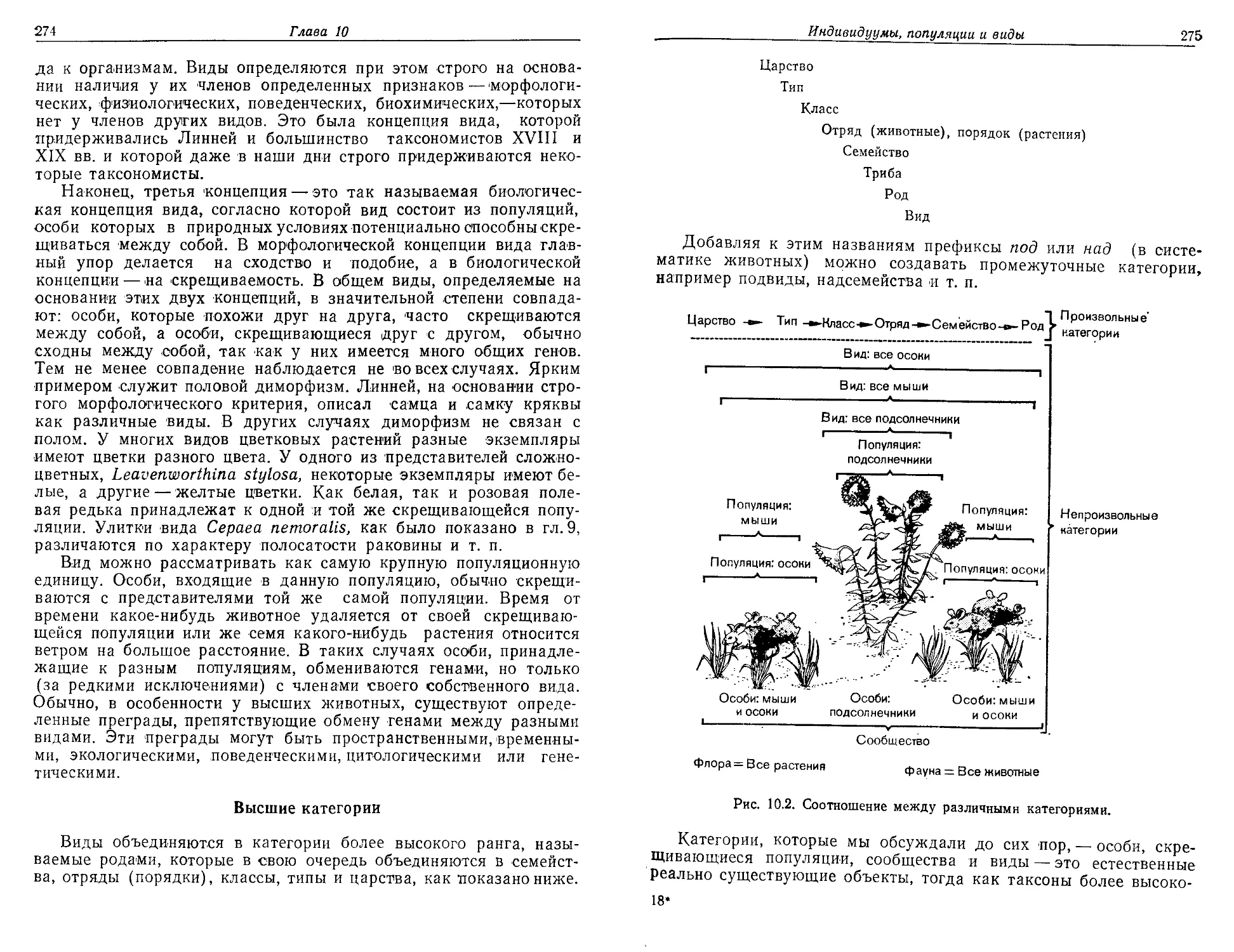
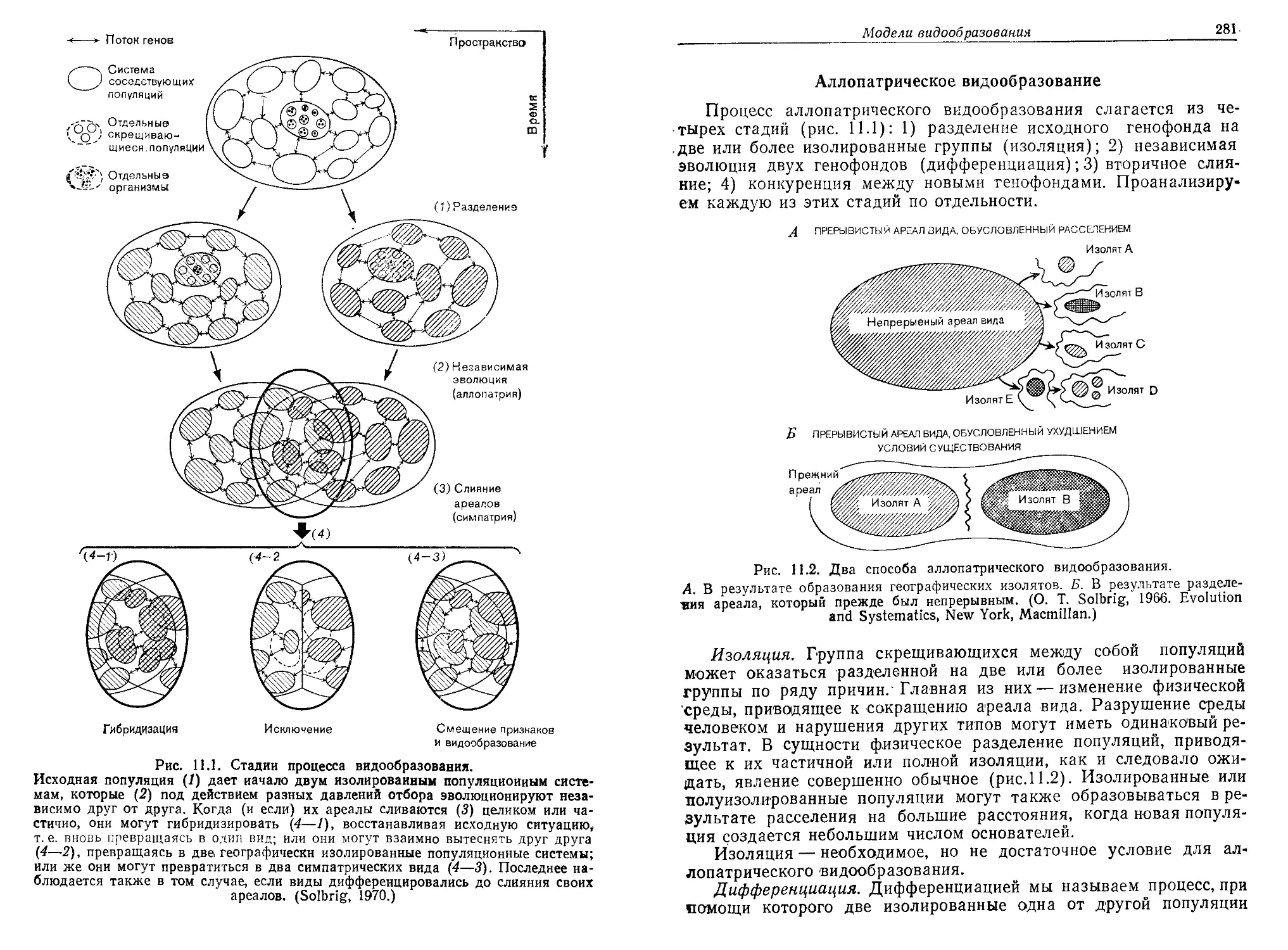
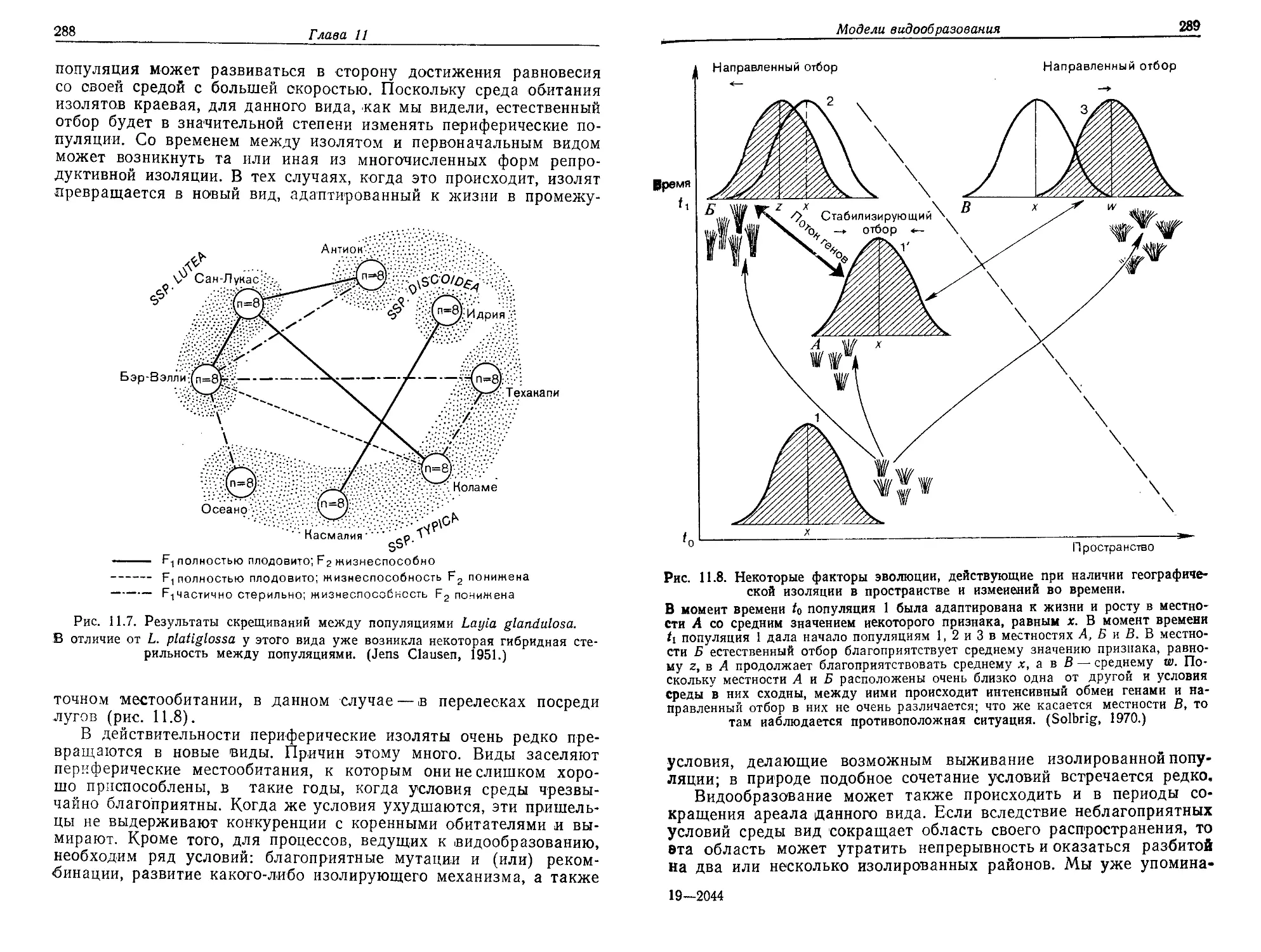
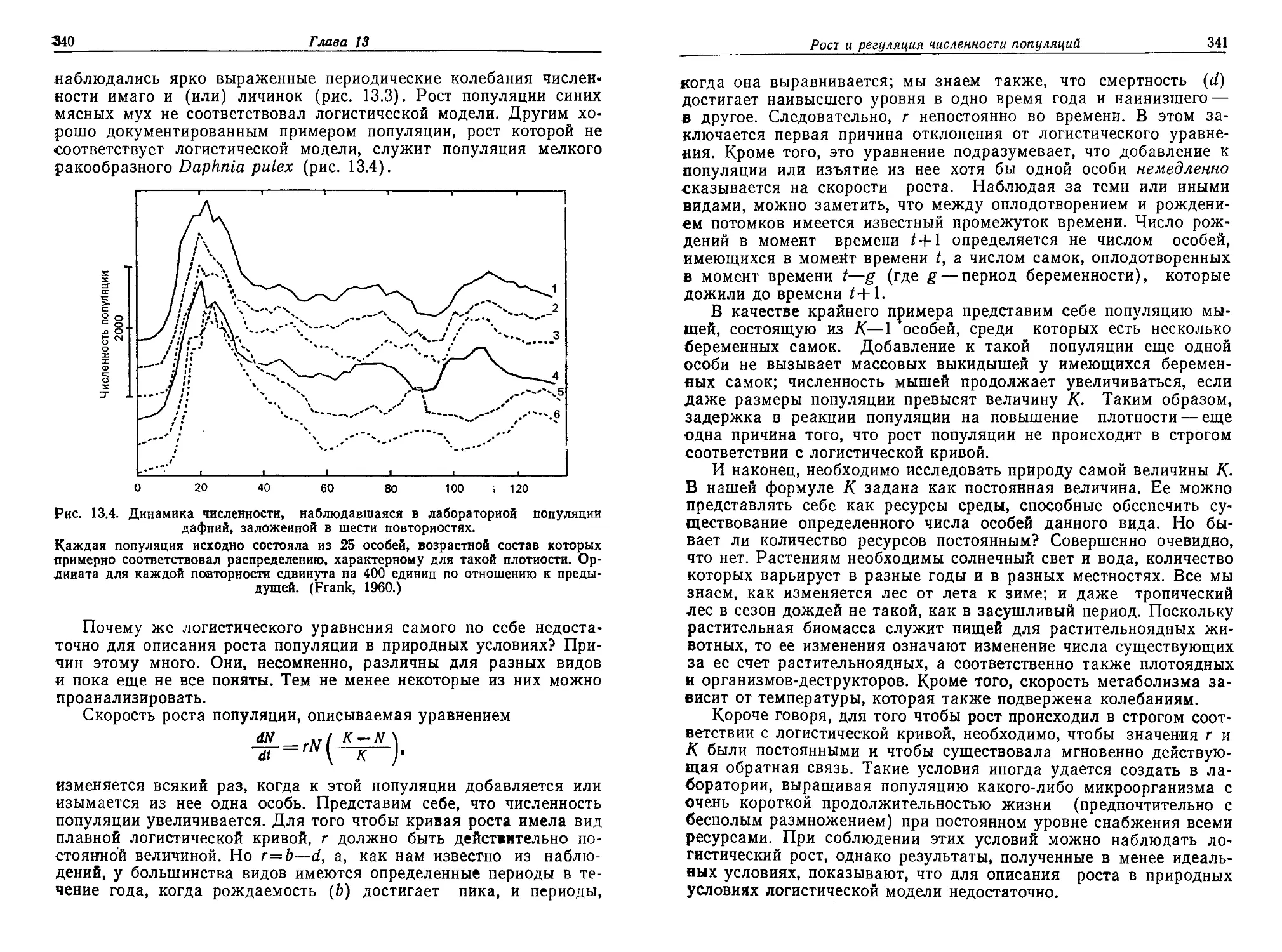
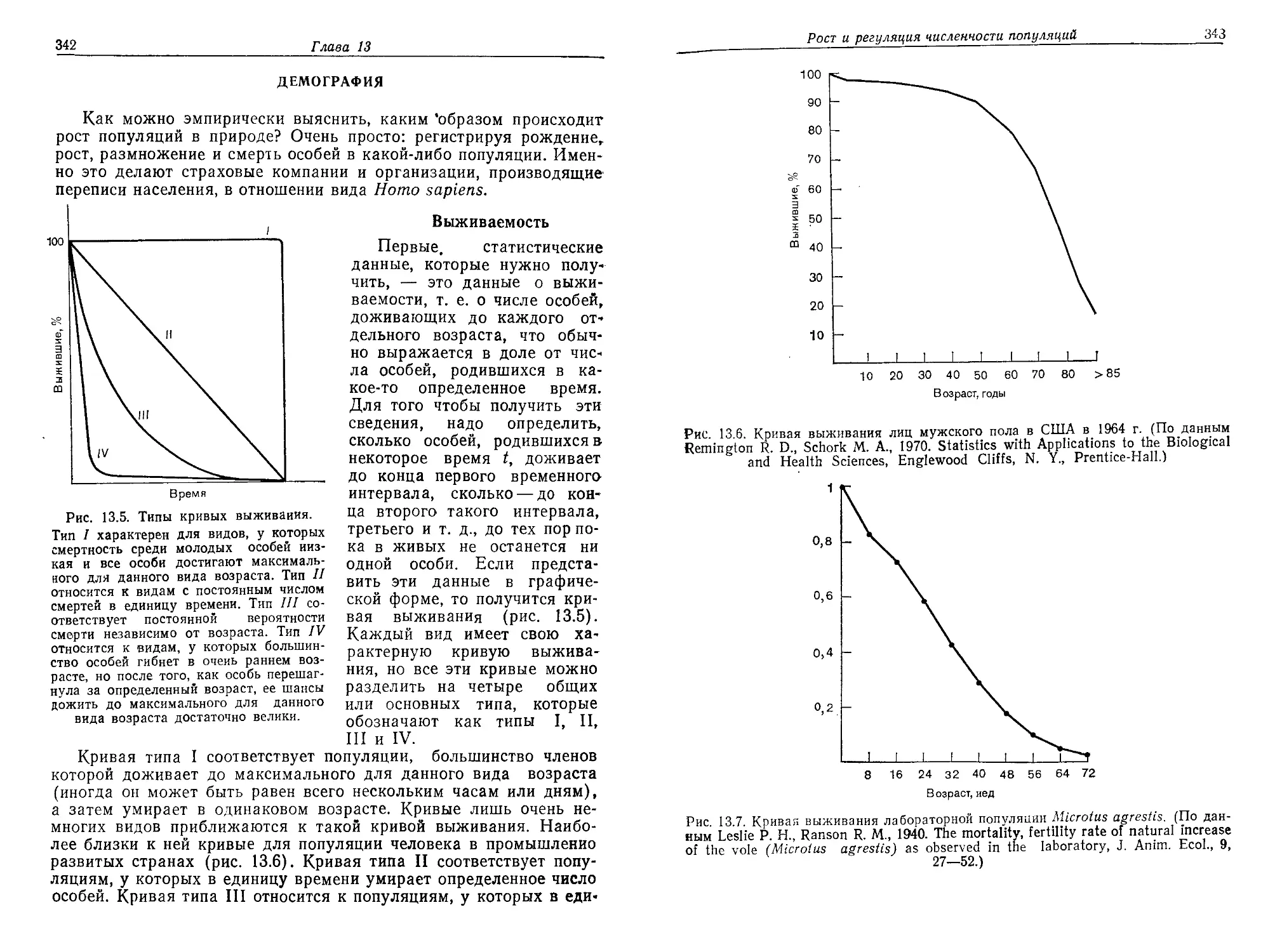
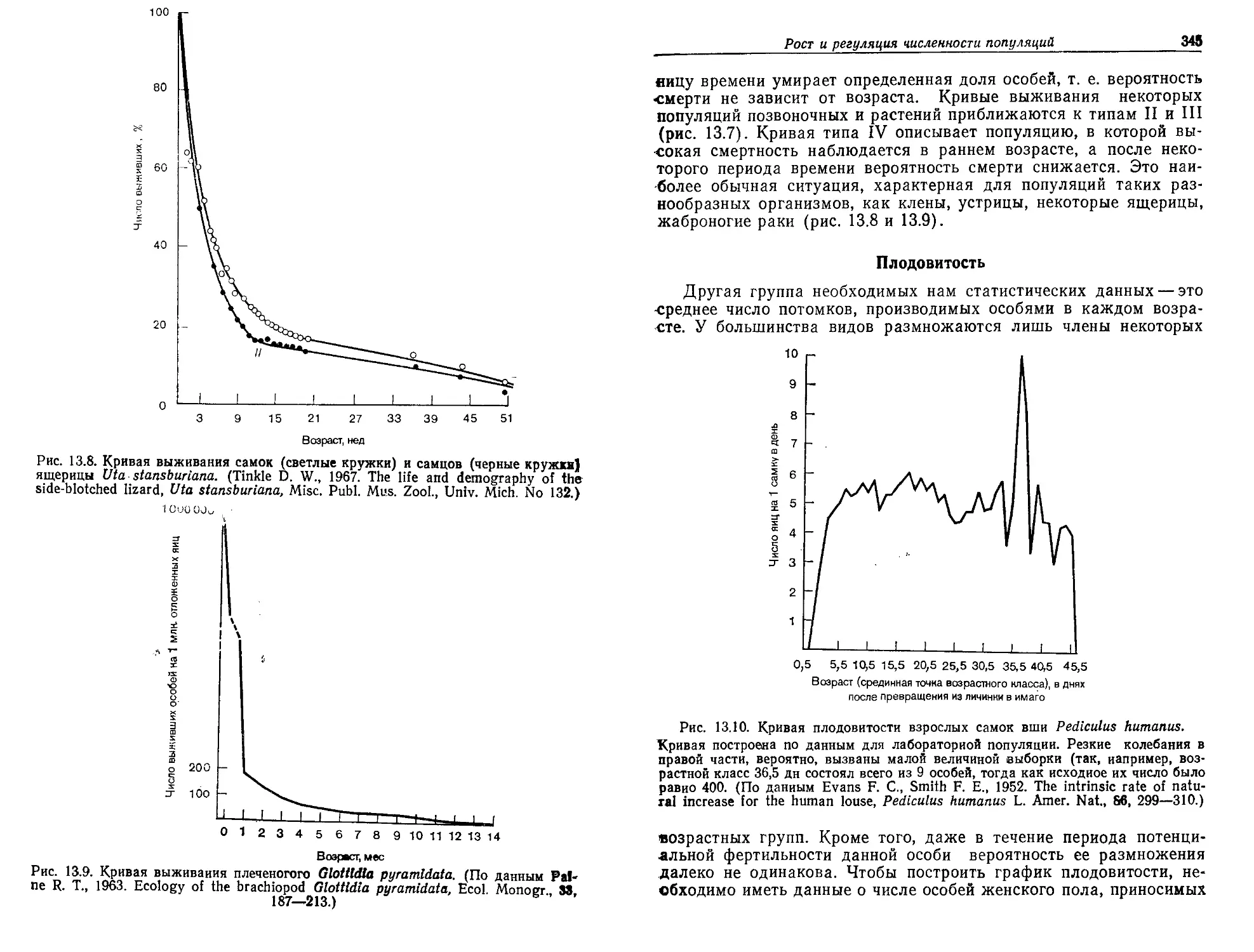
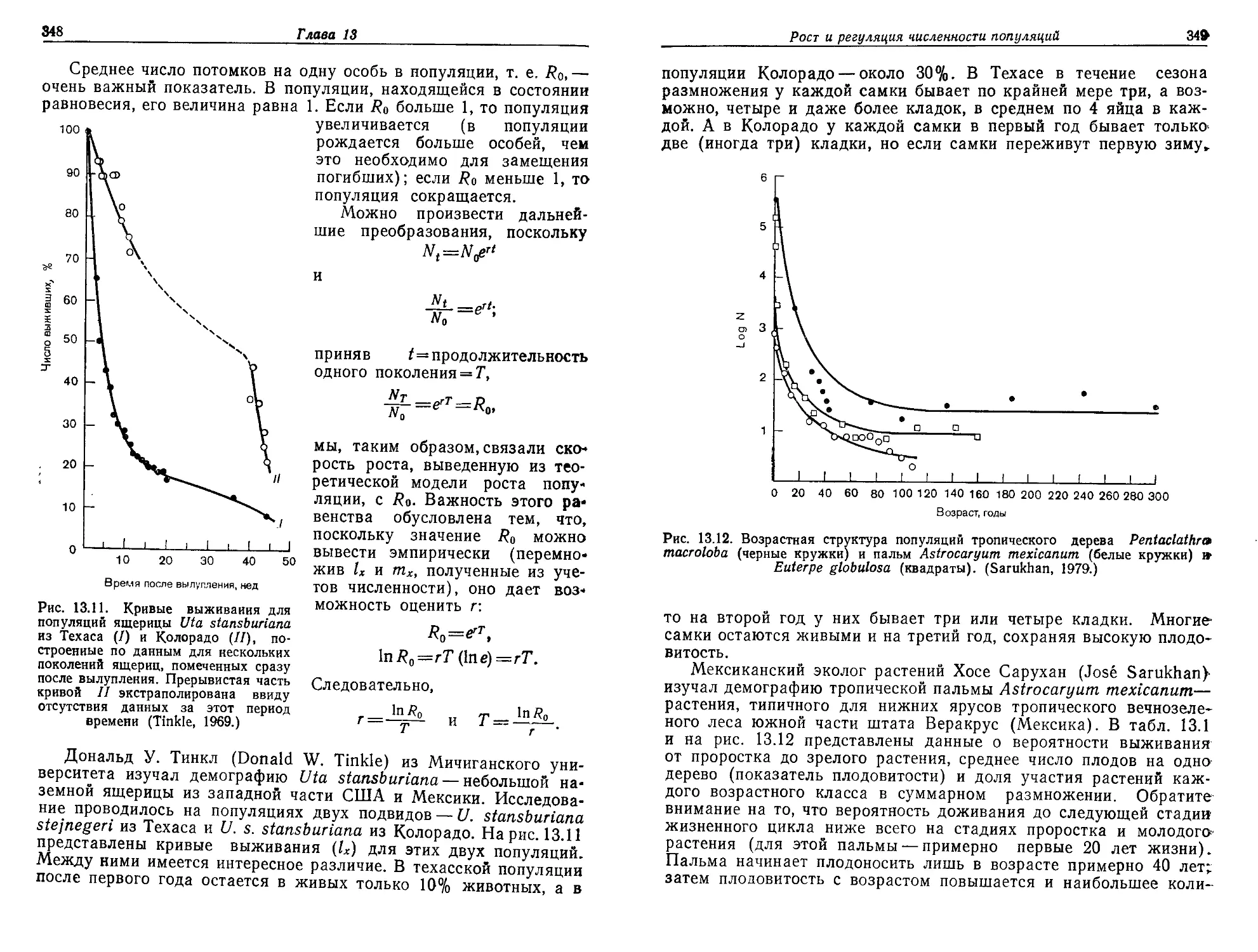
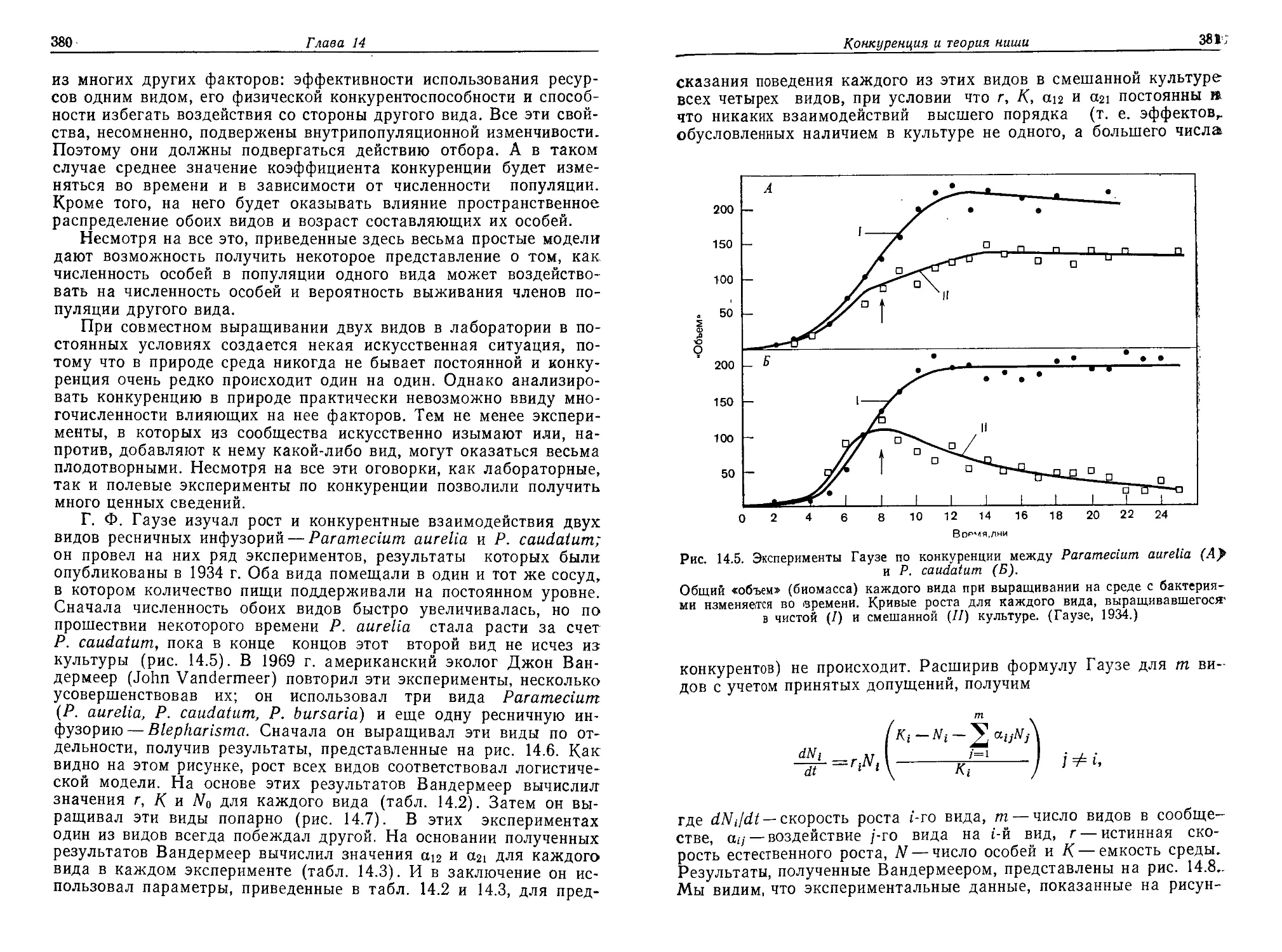
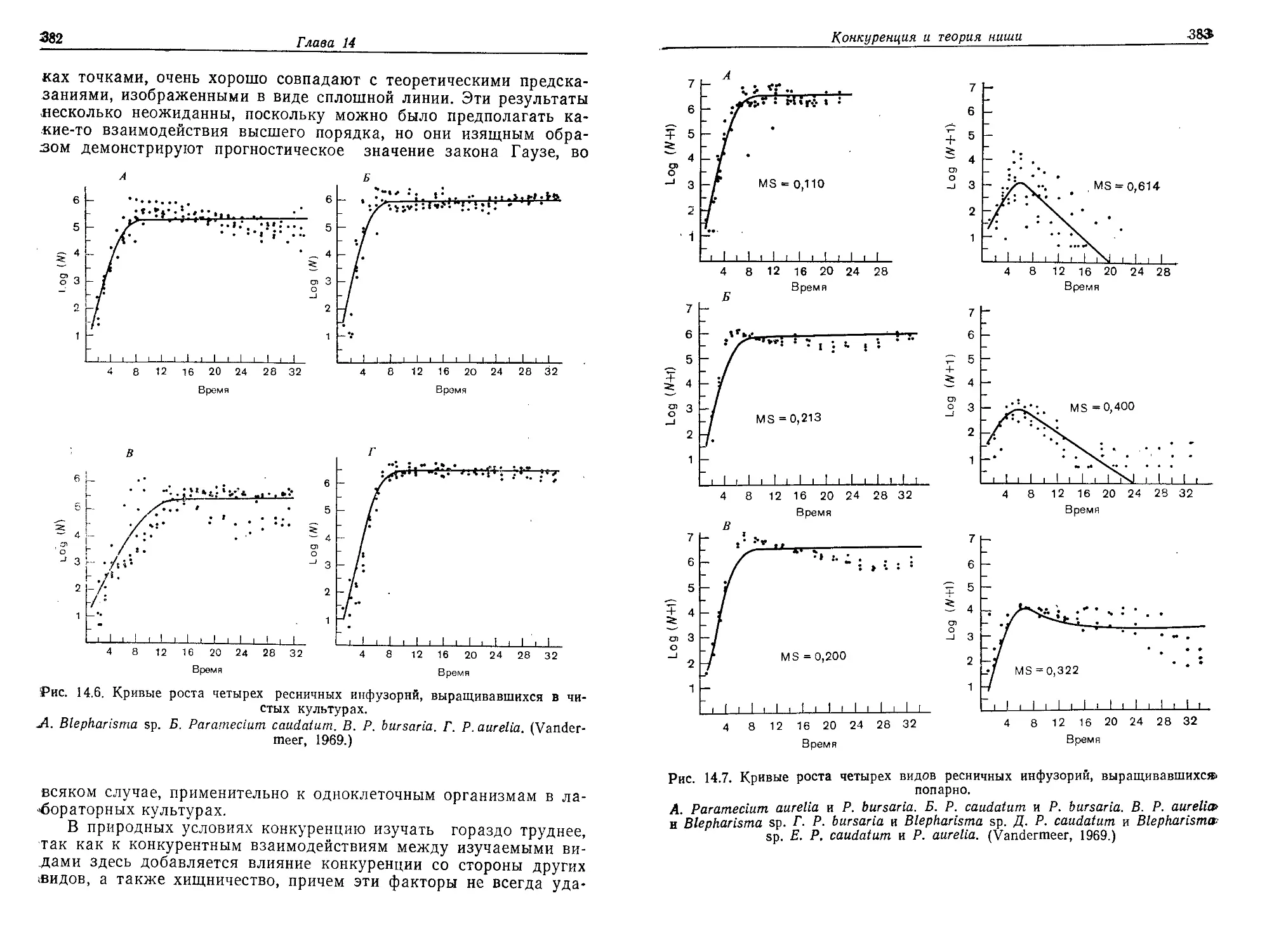
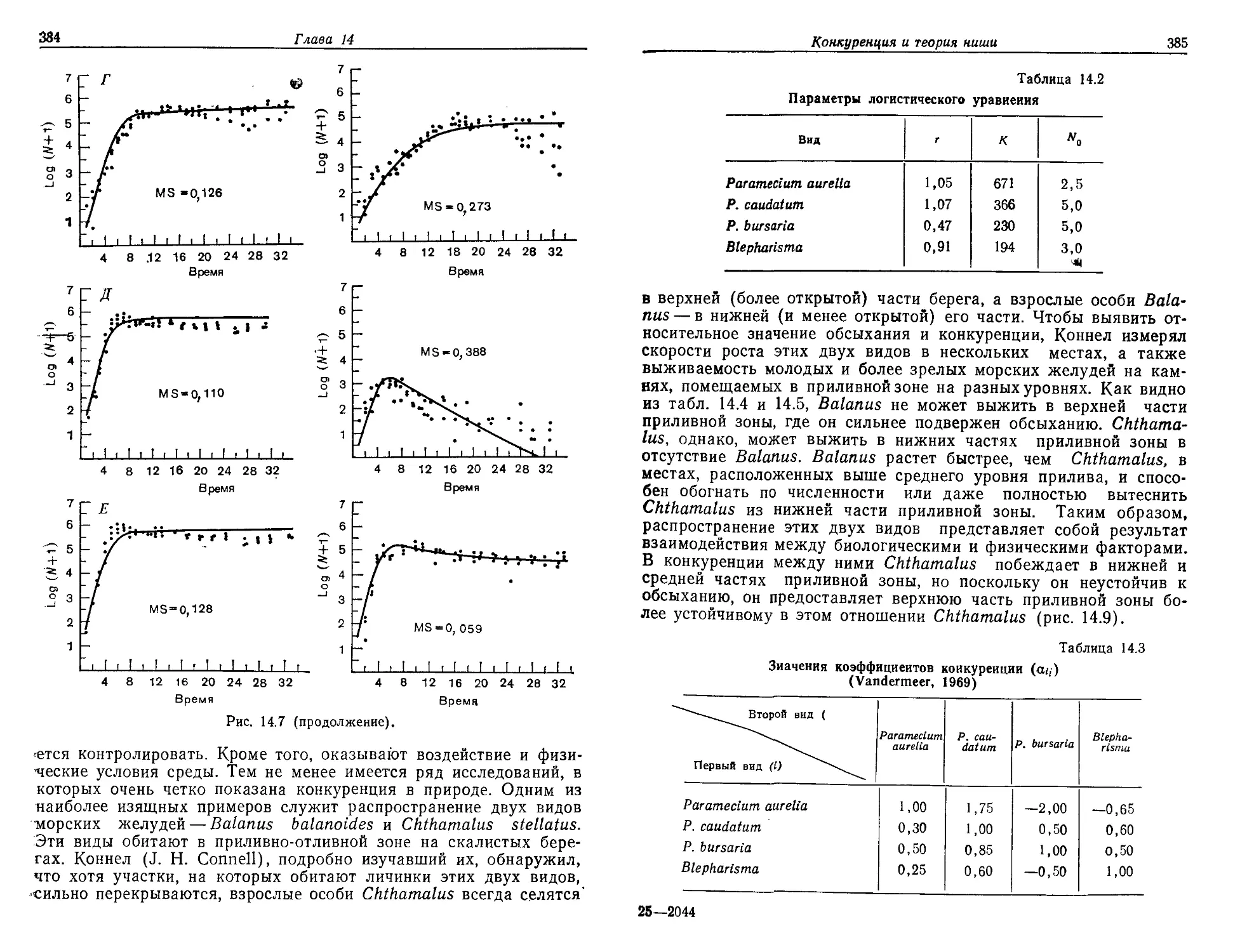
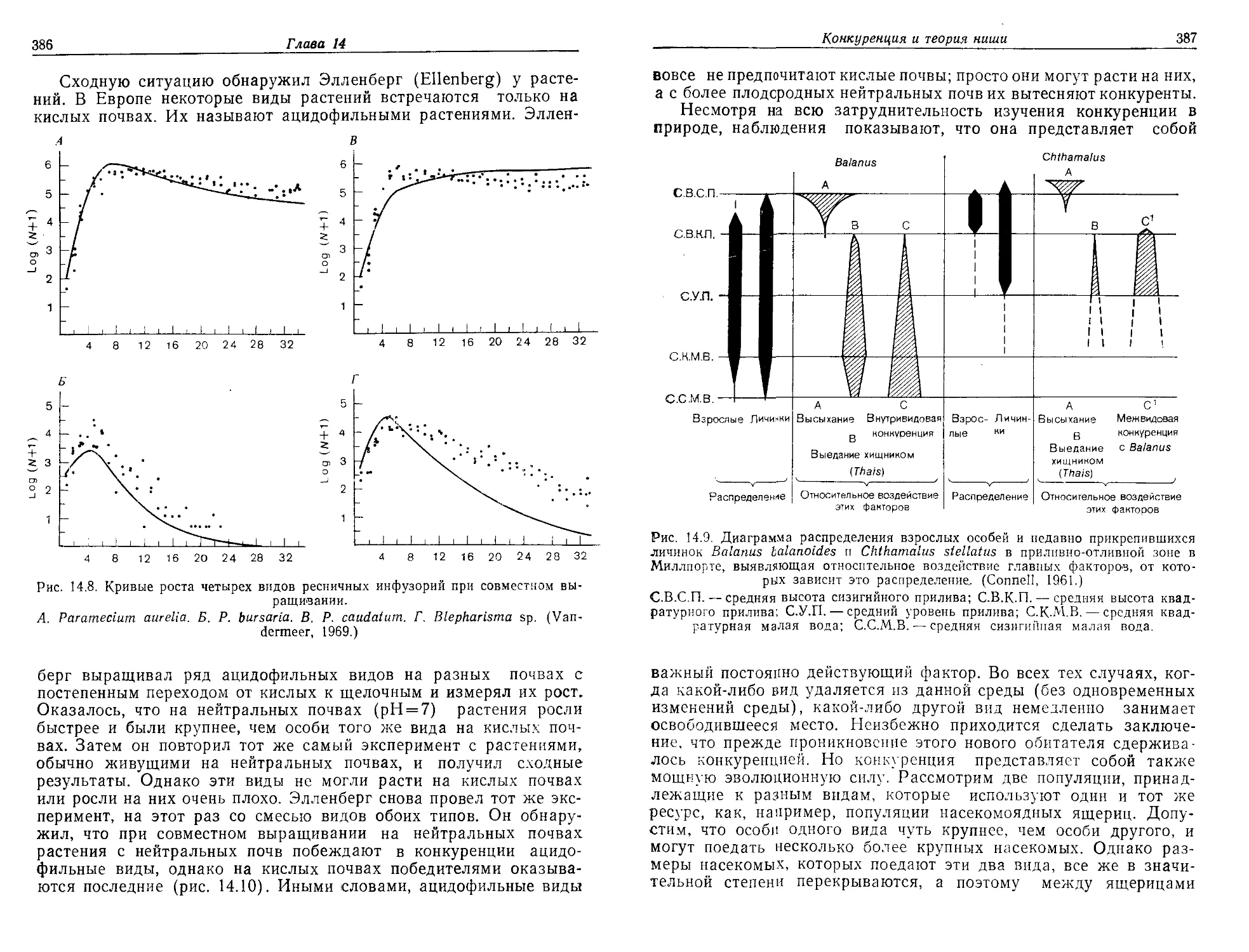
/


































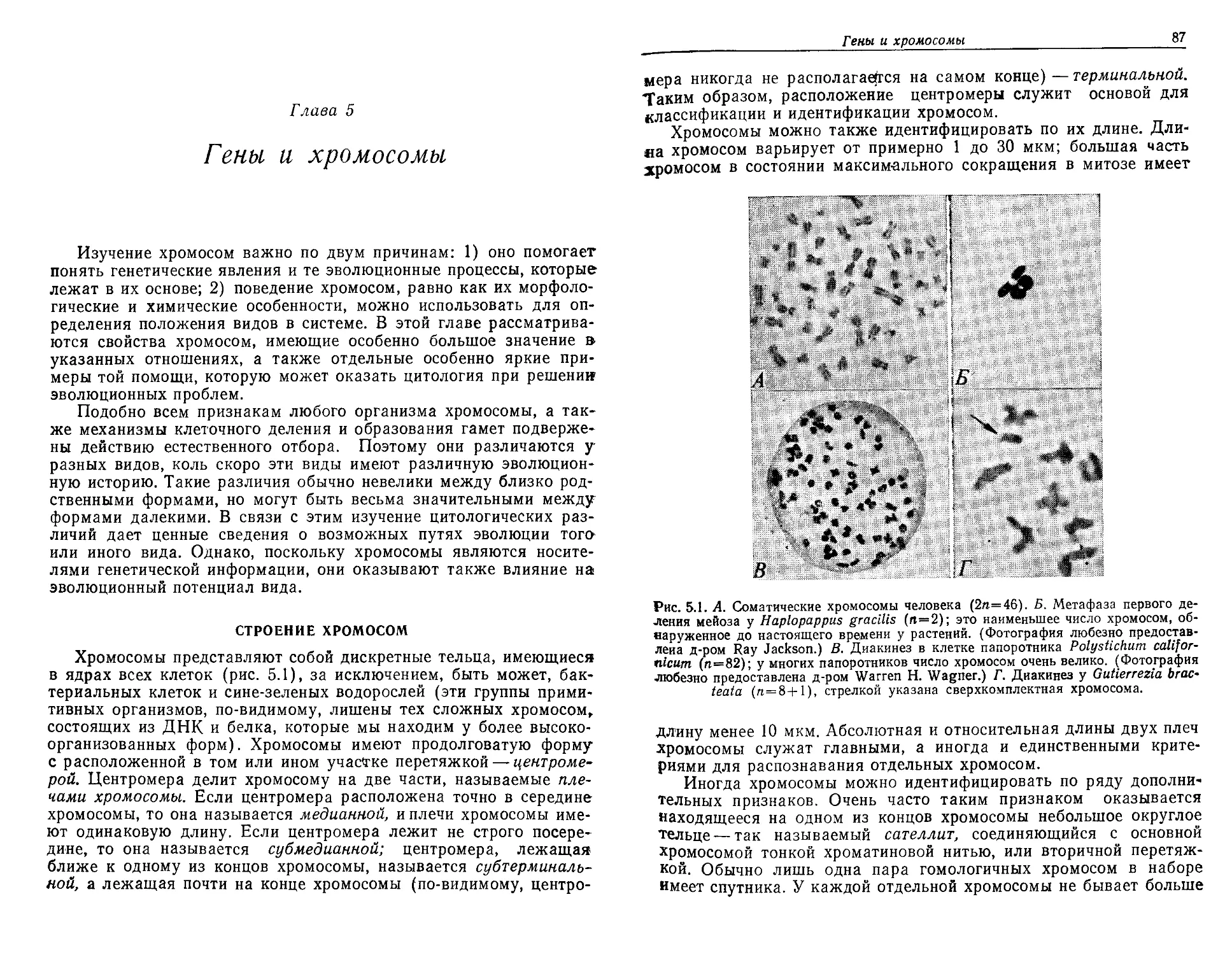
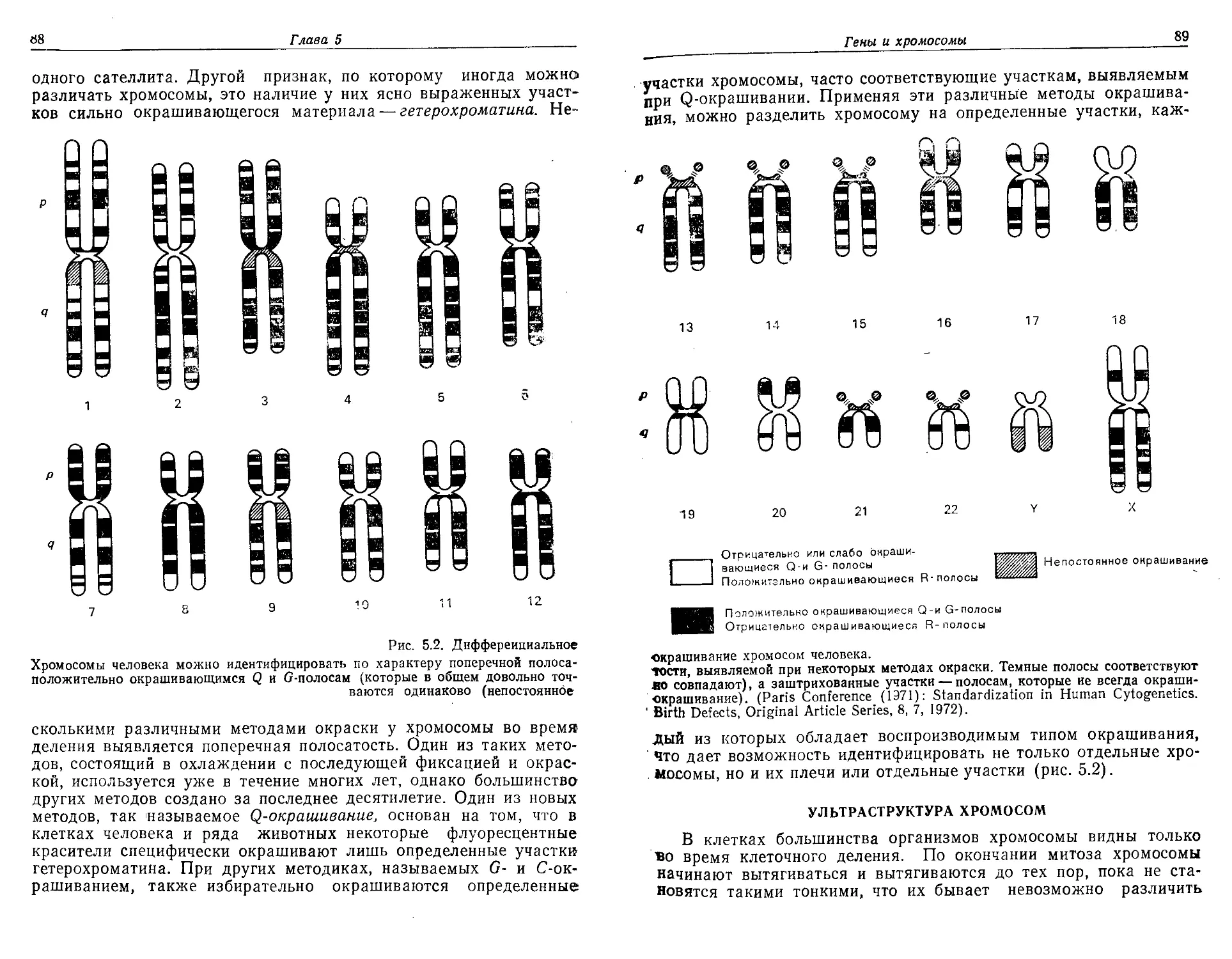
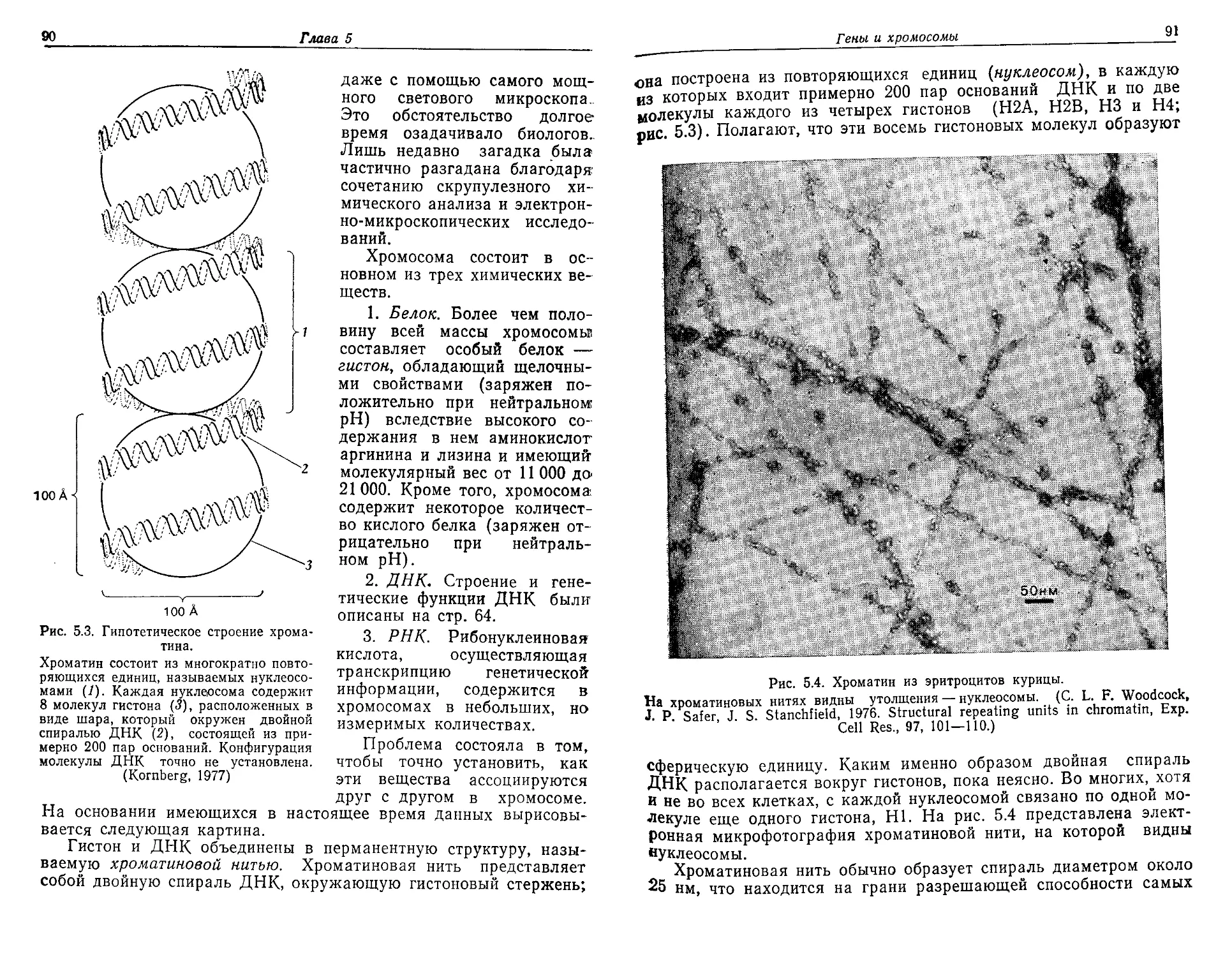
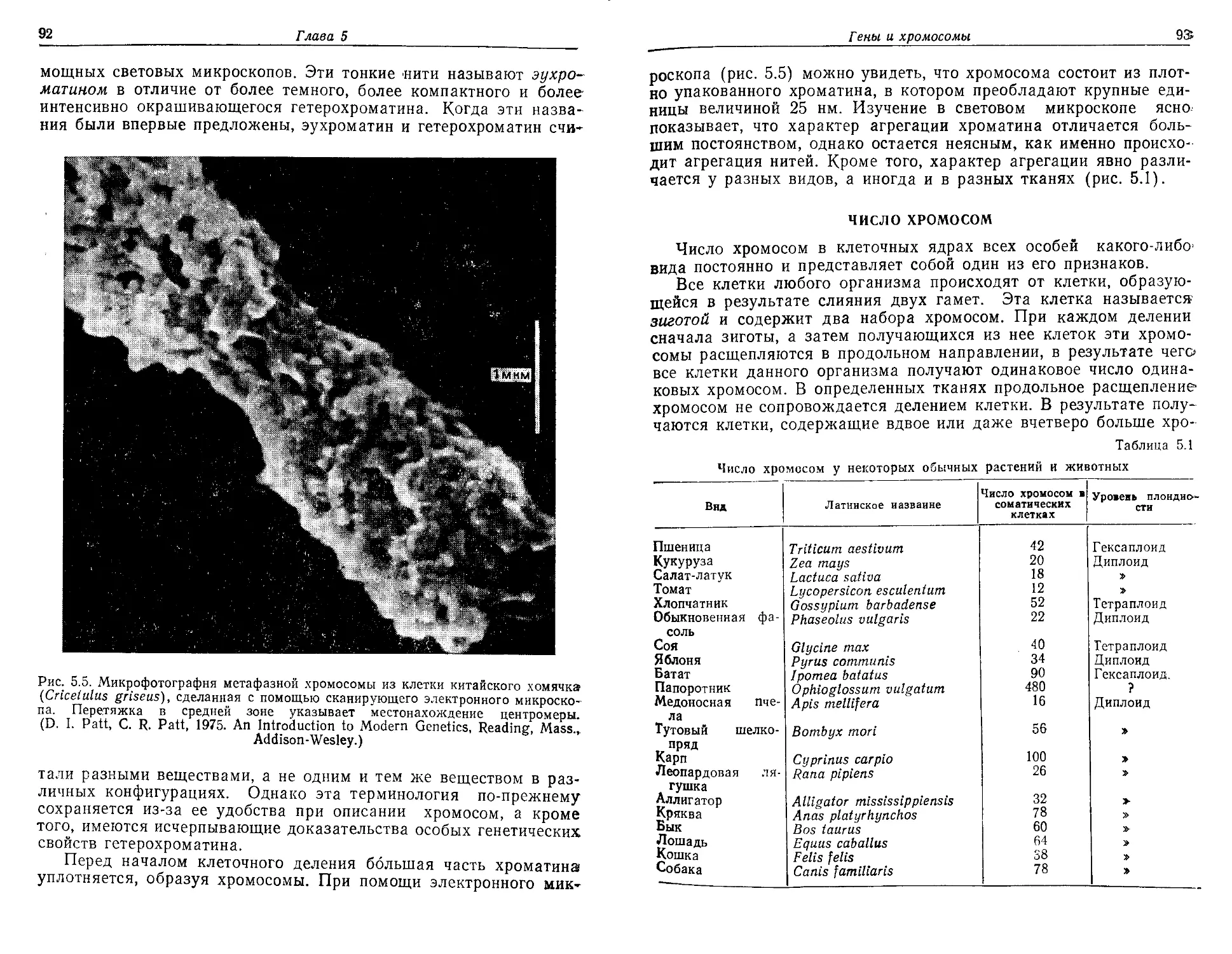










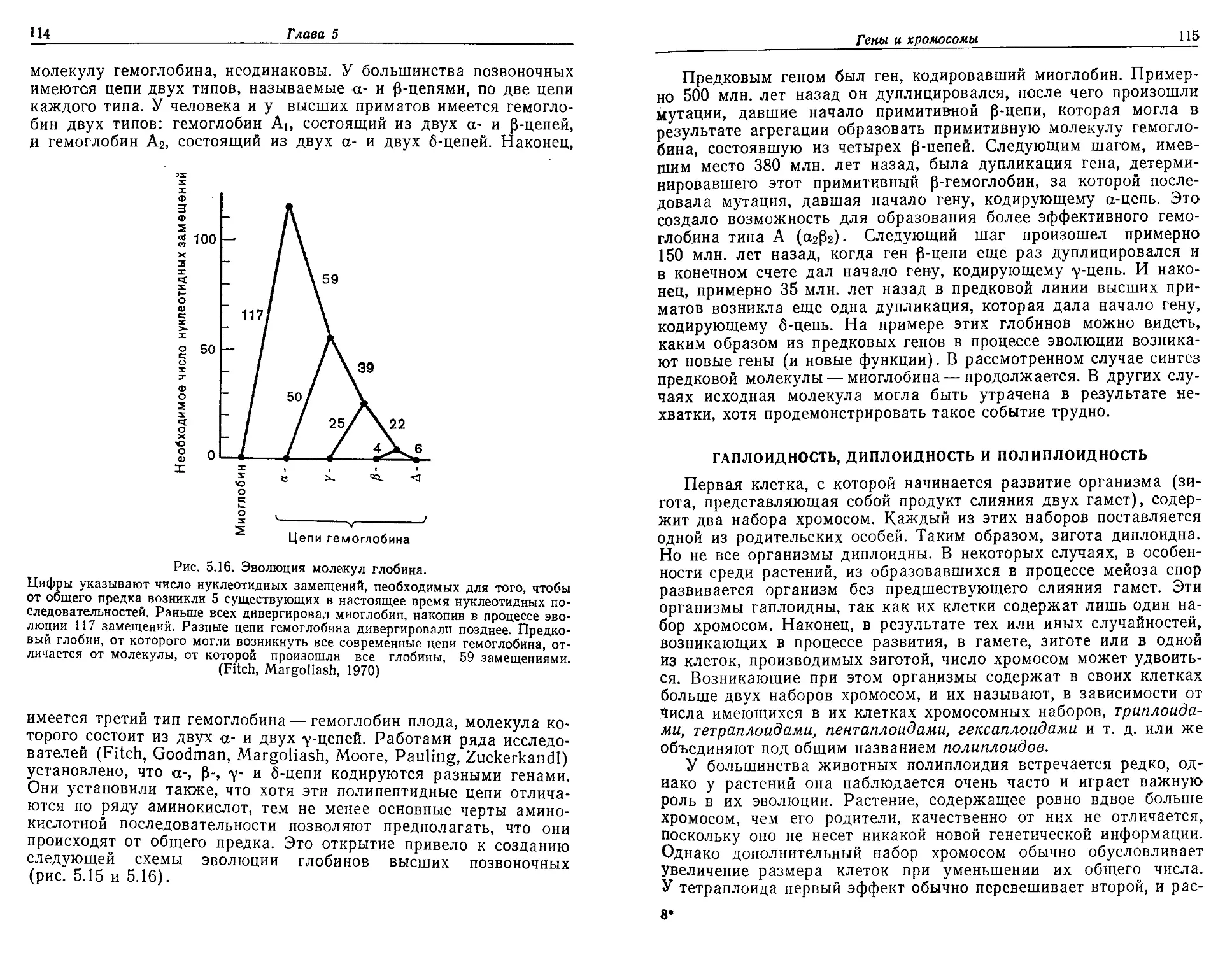

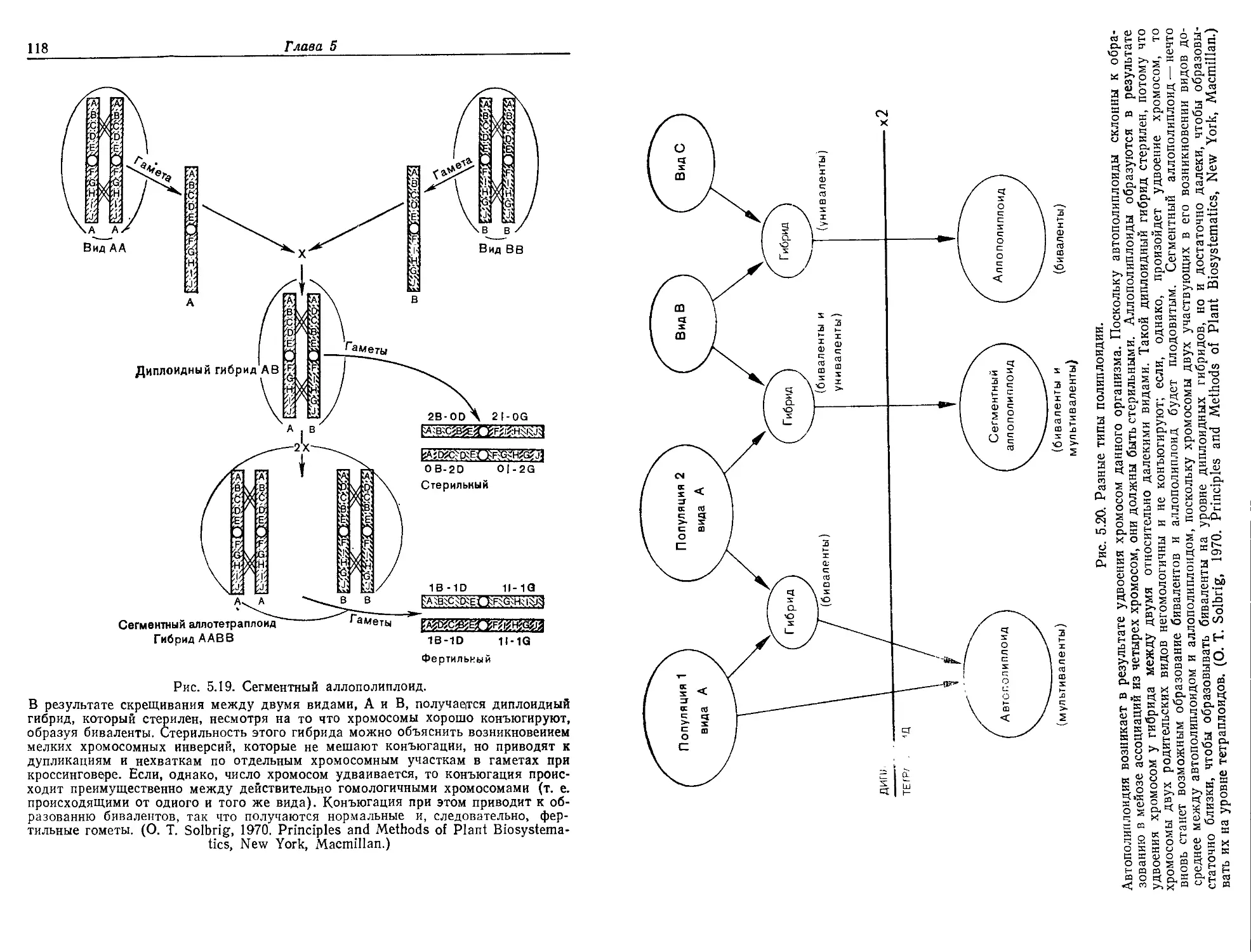

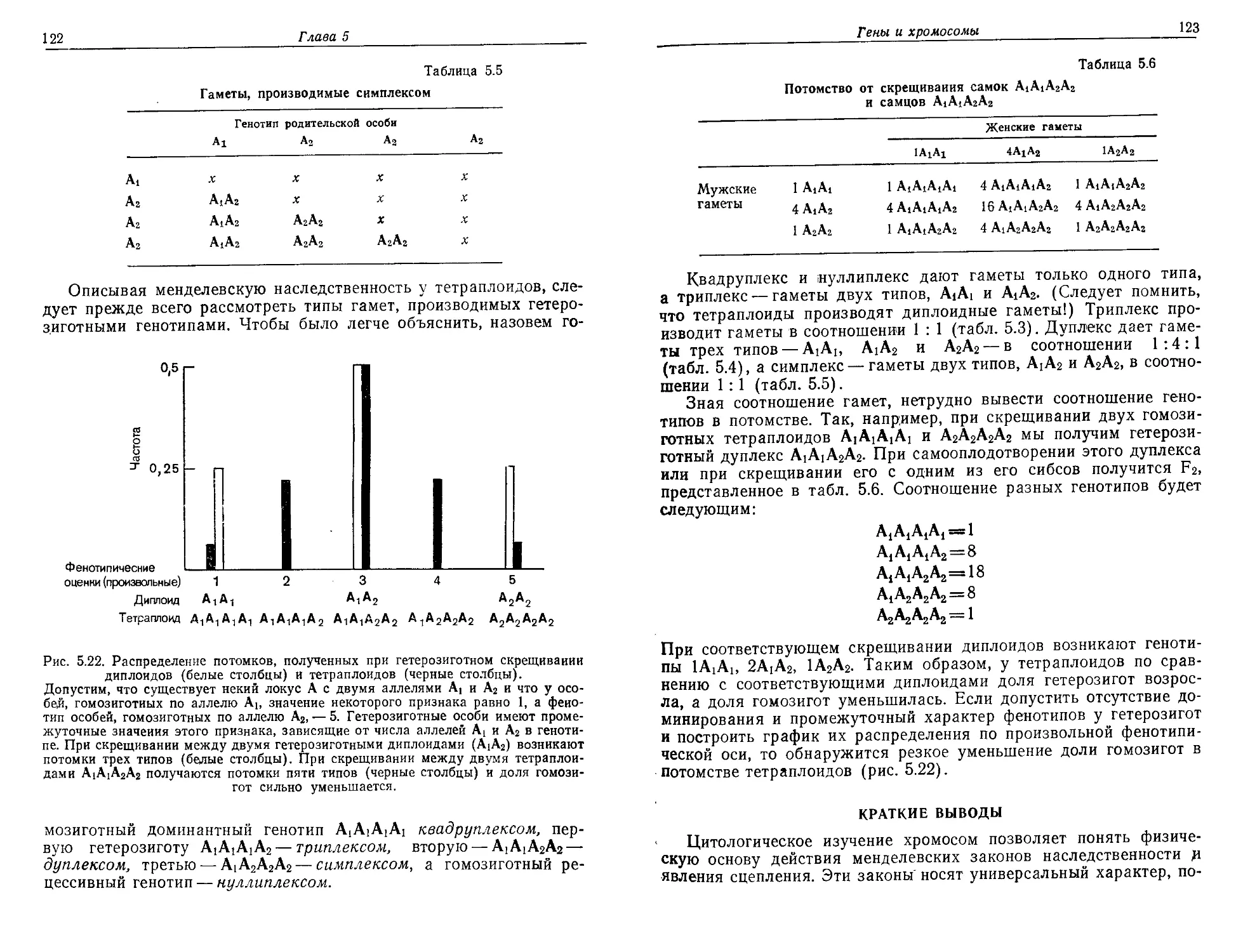




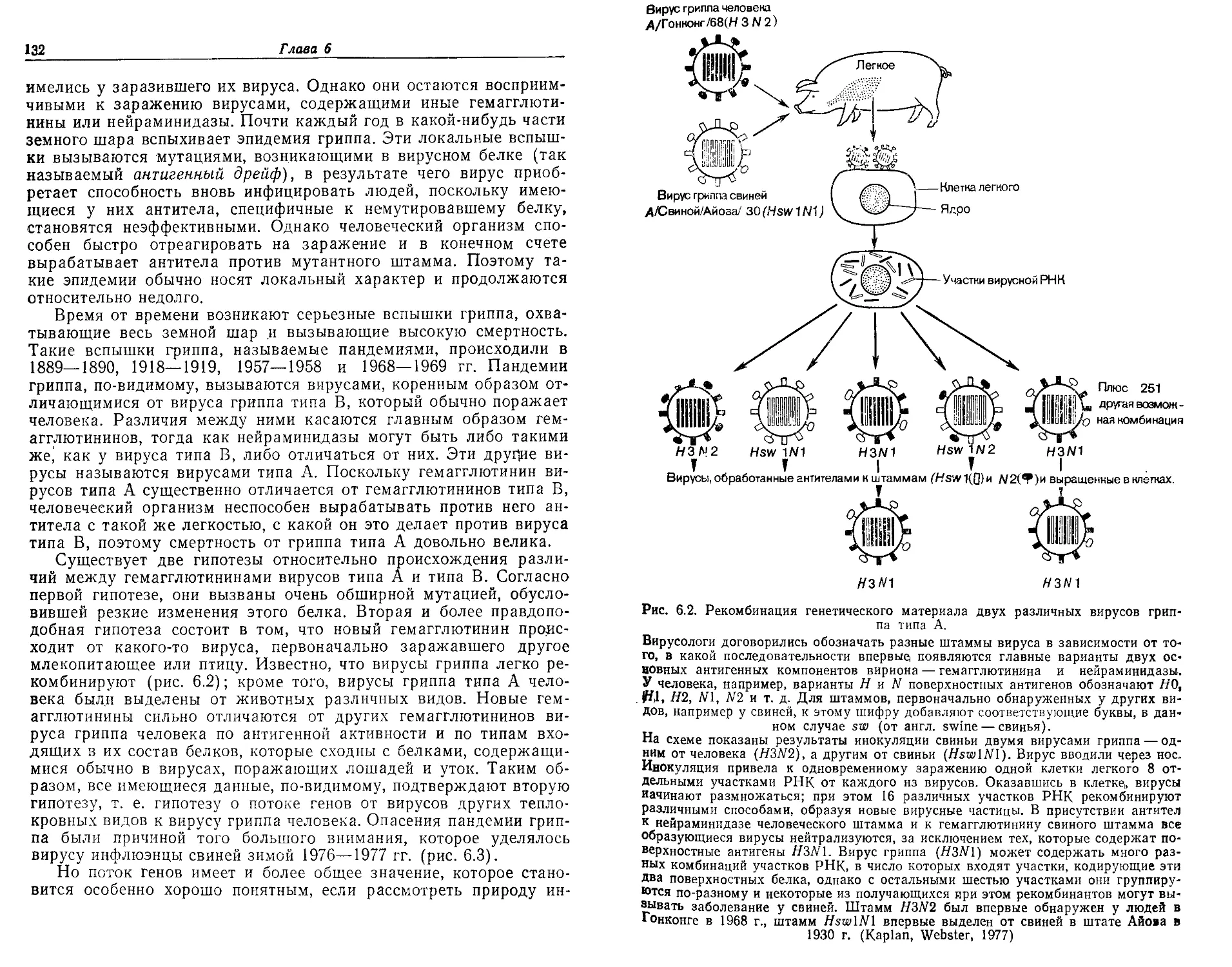





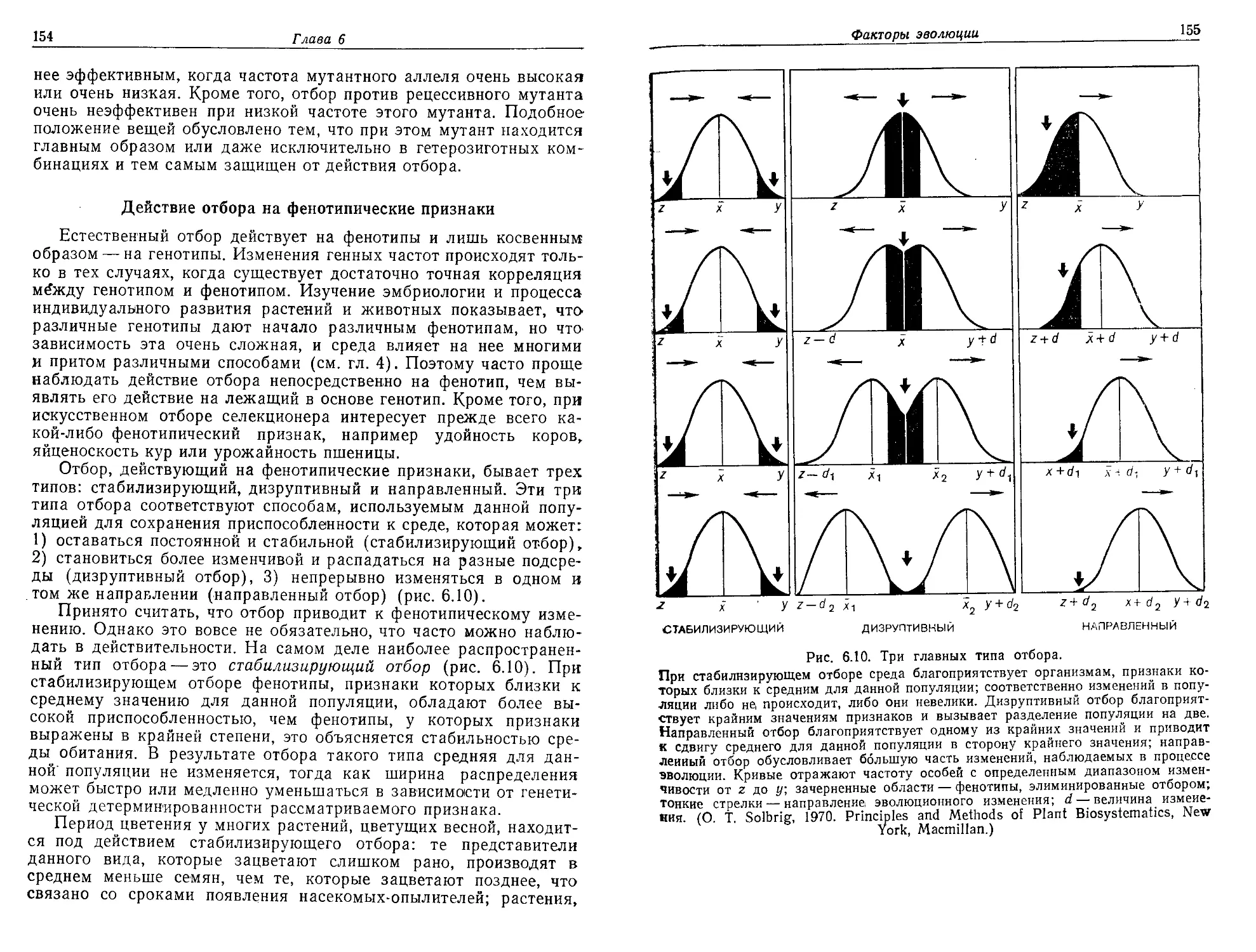






























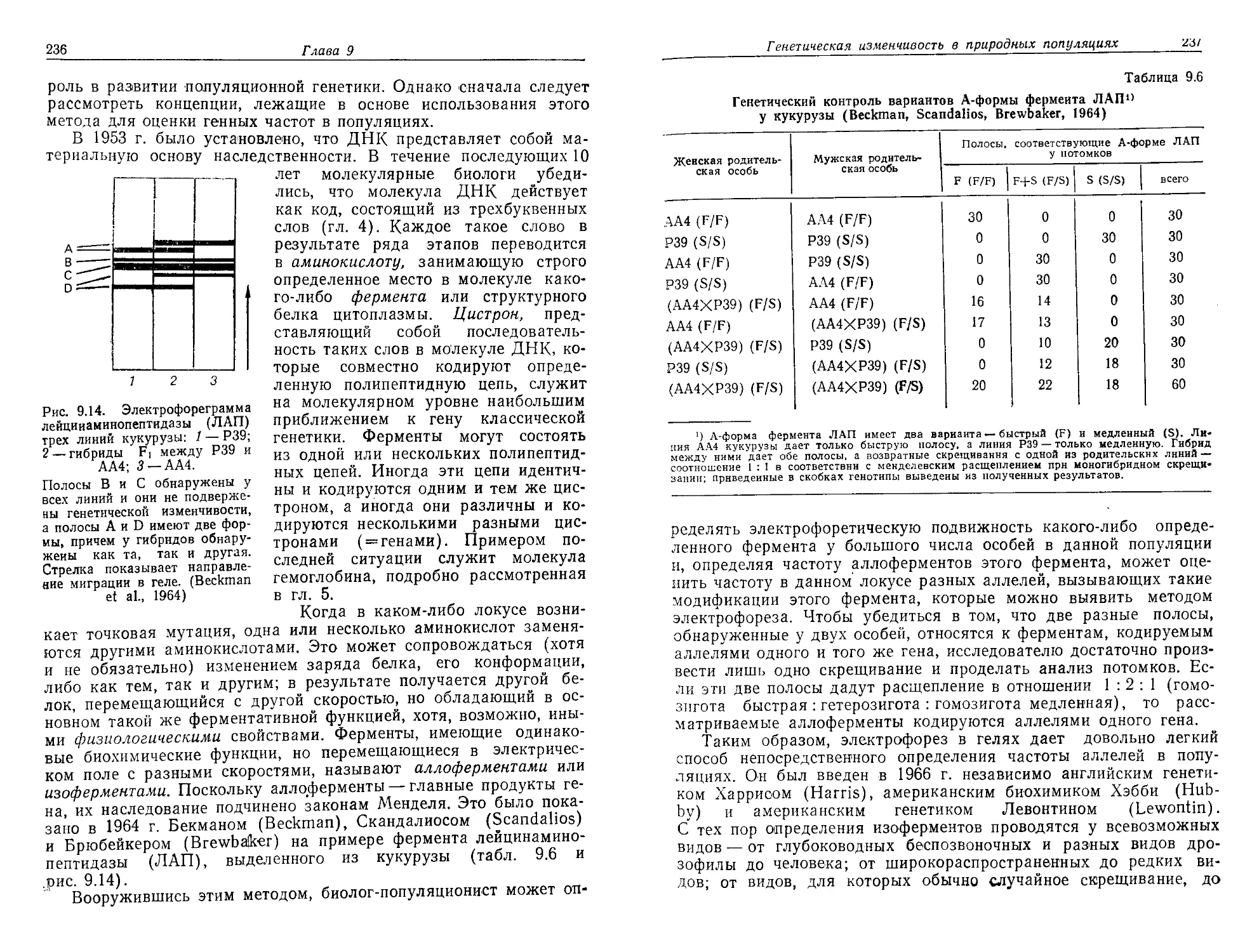
















































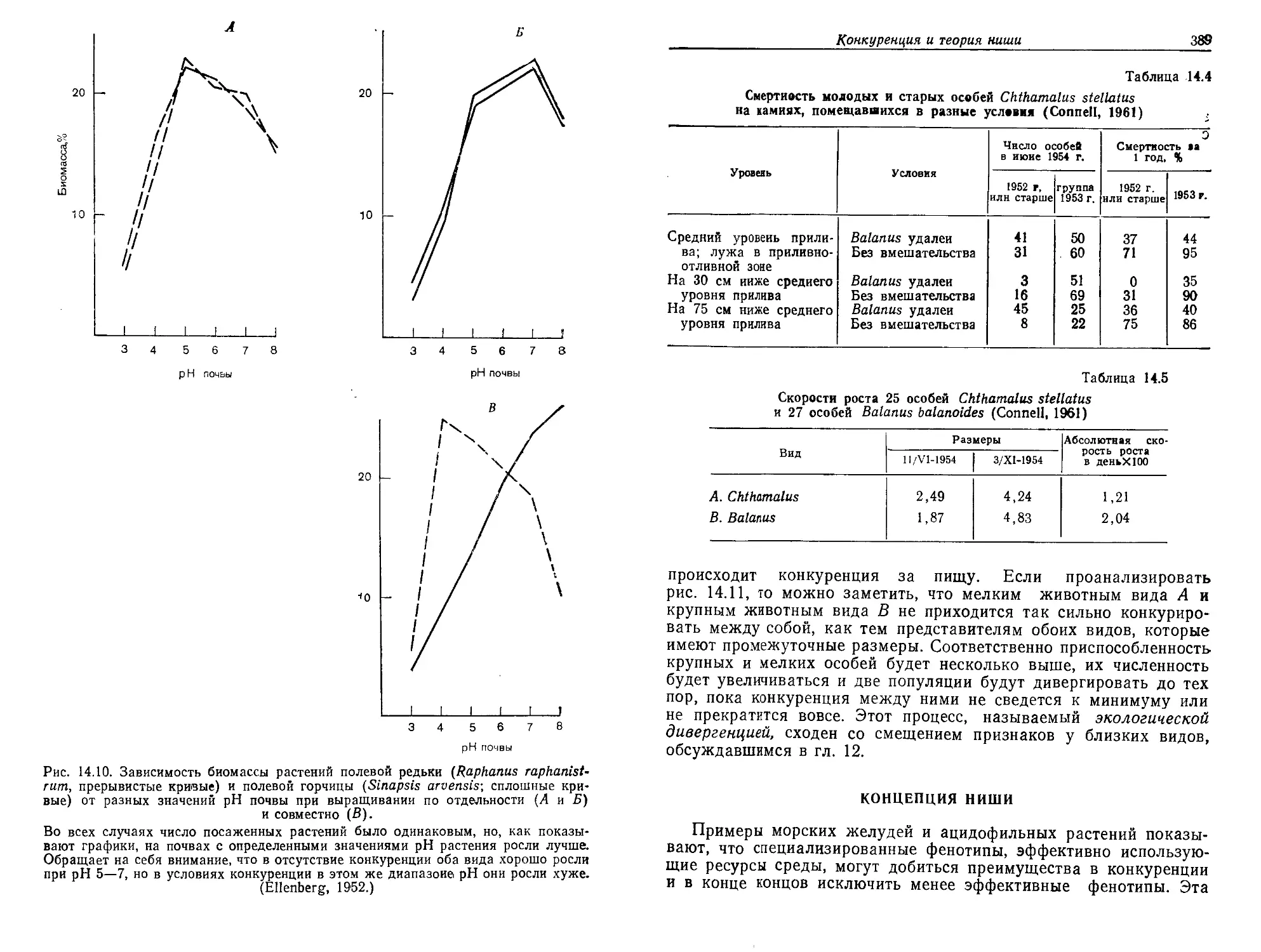
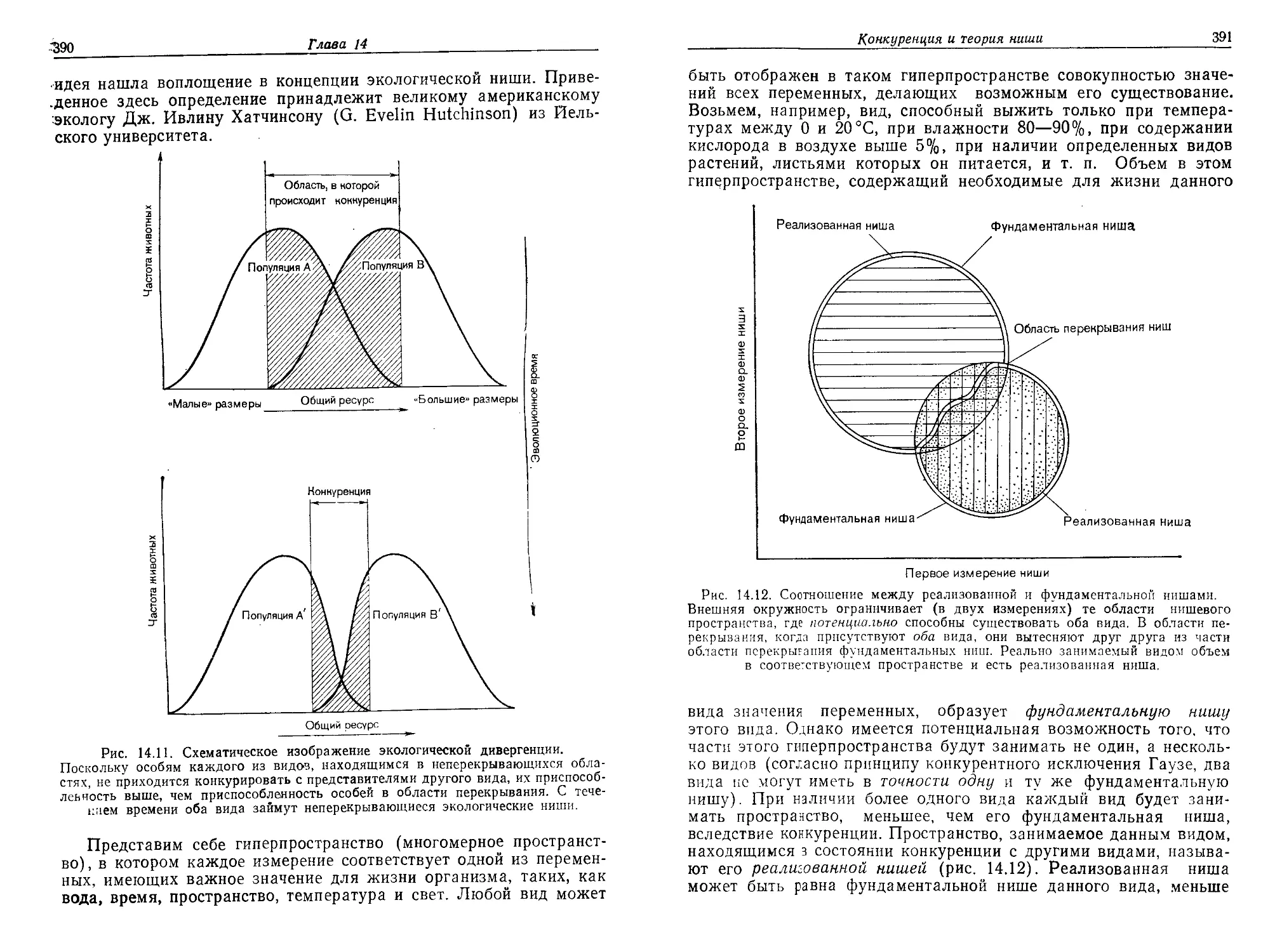


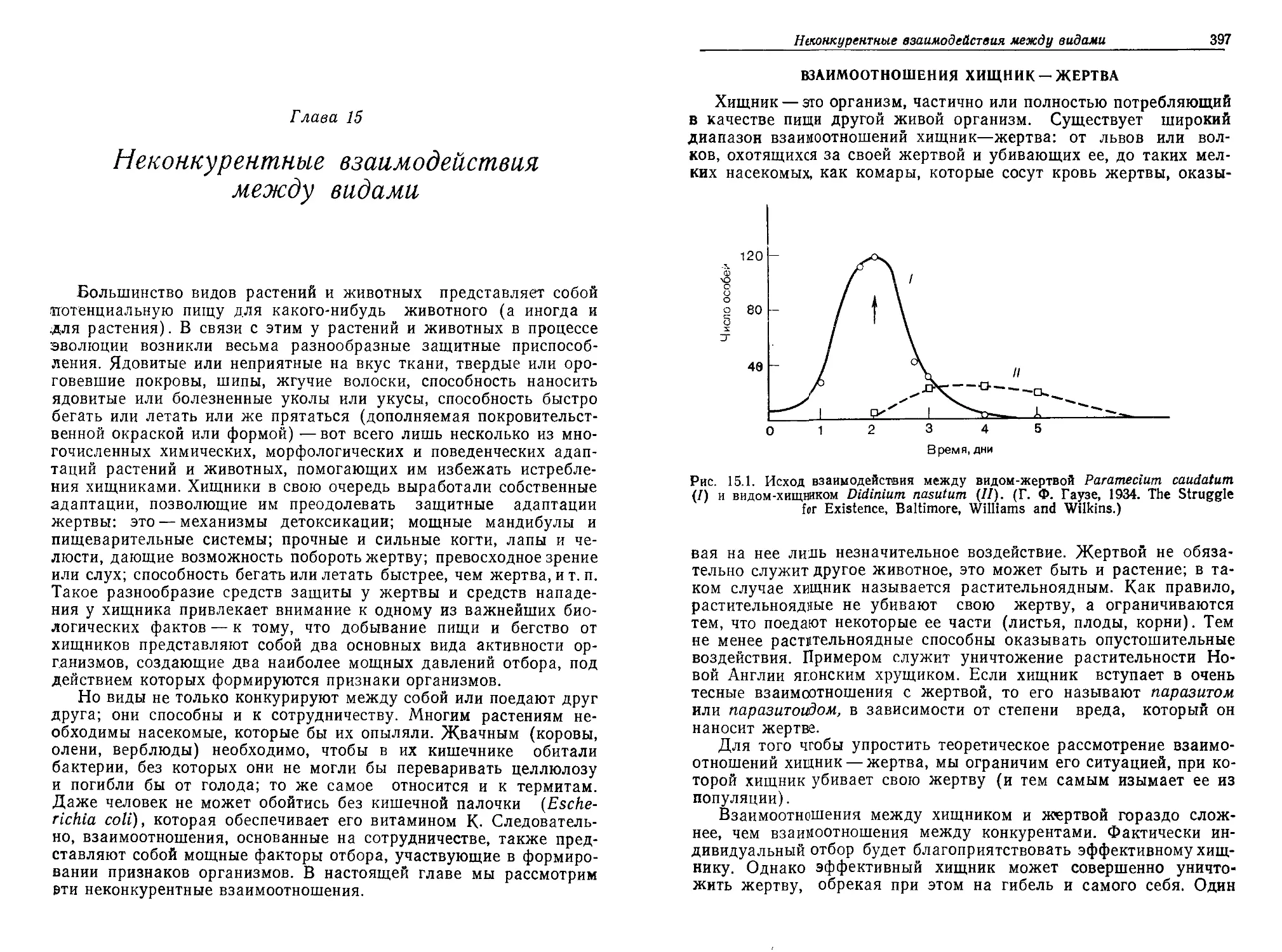
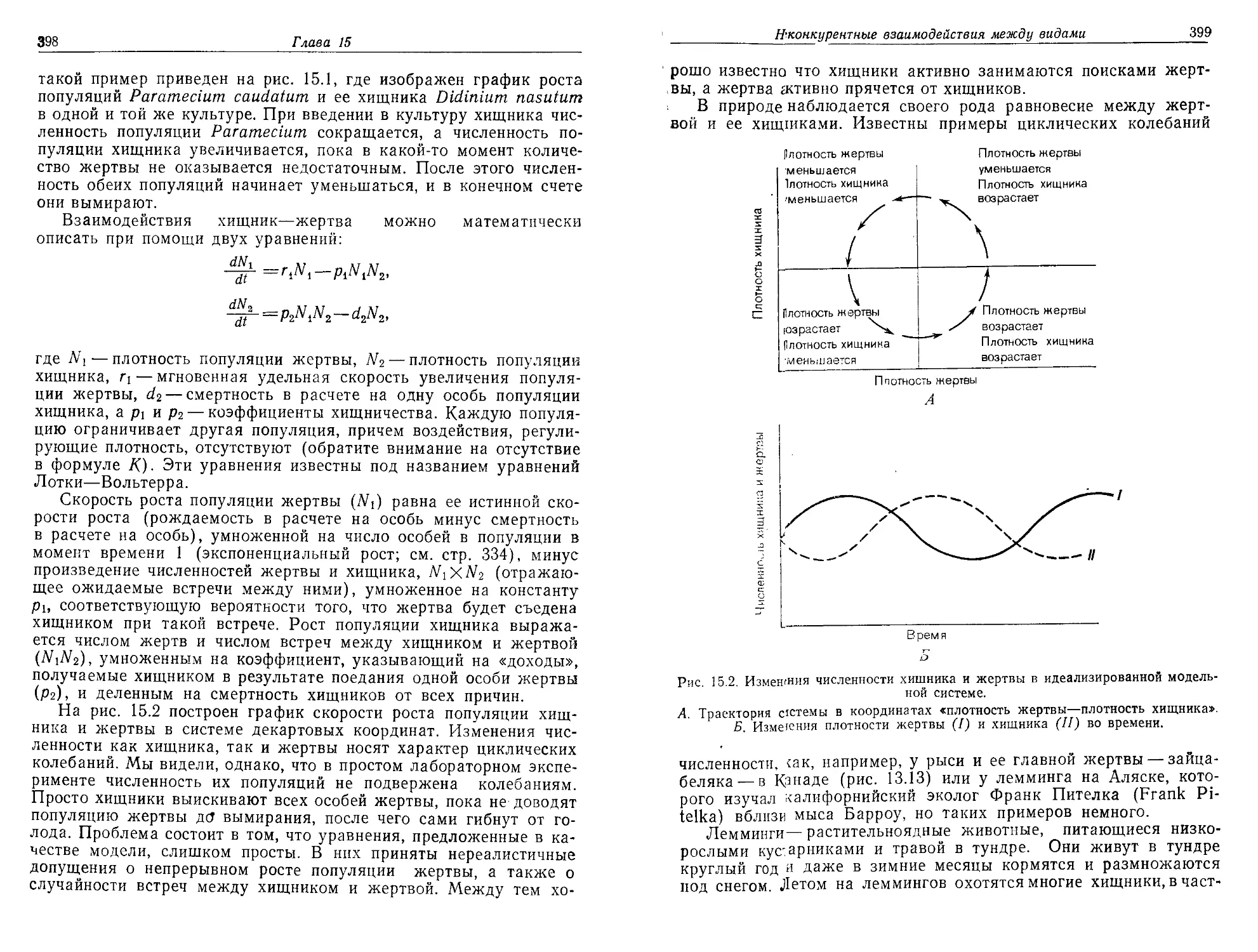
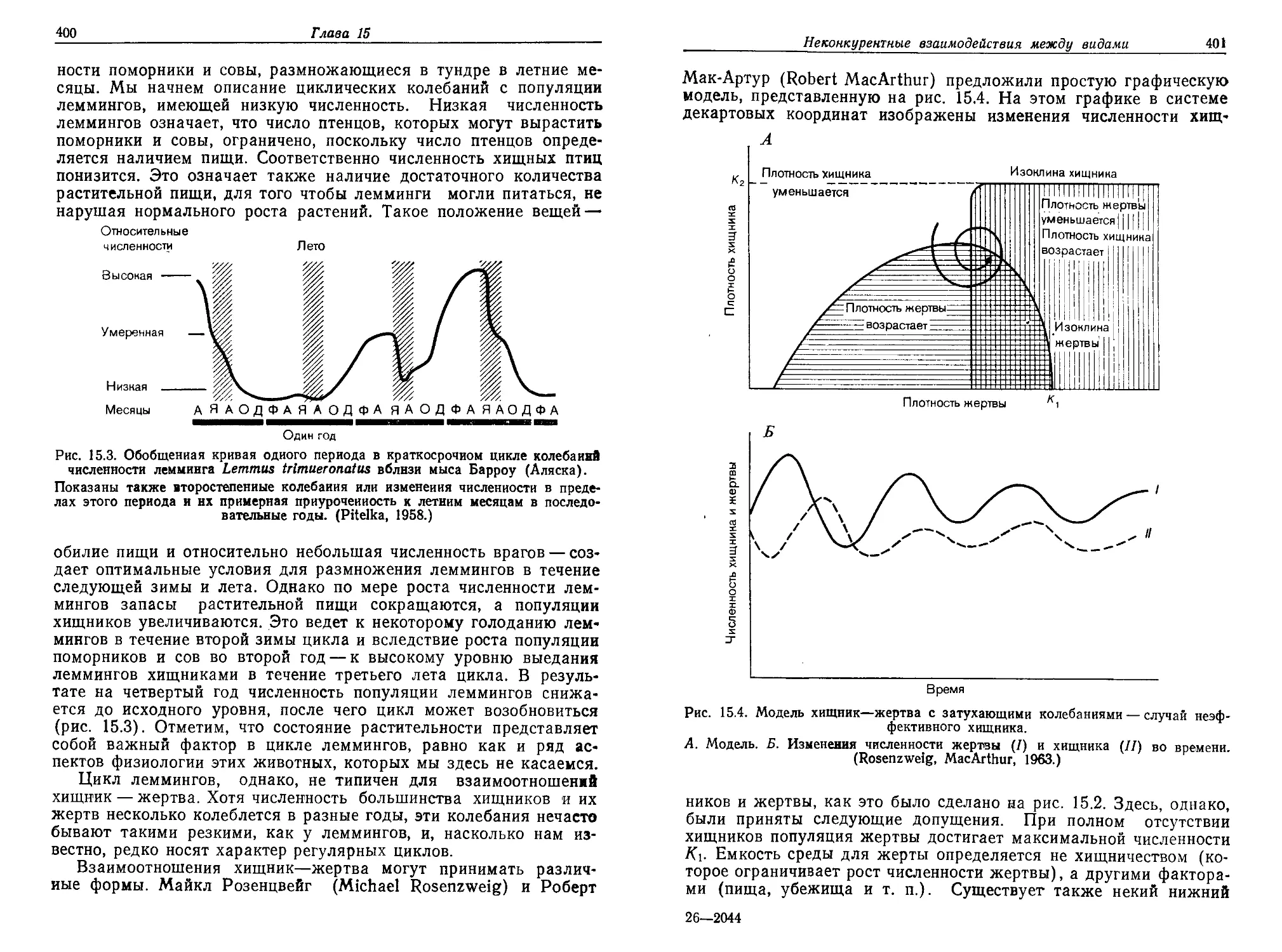

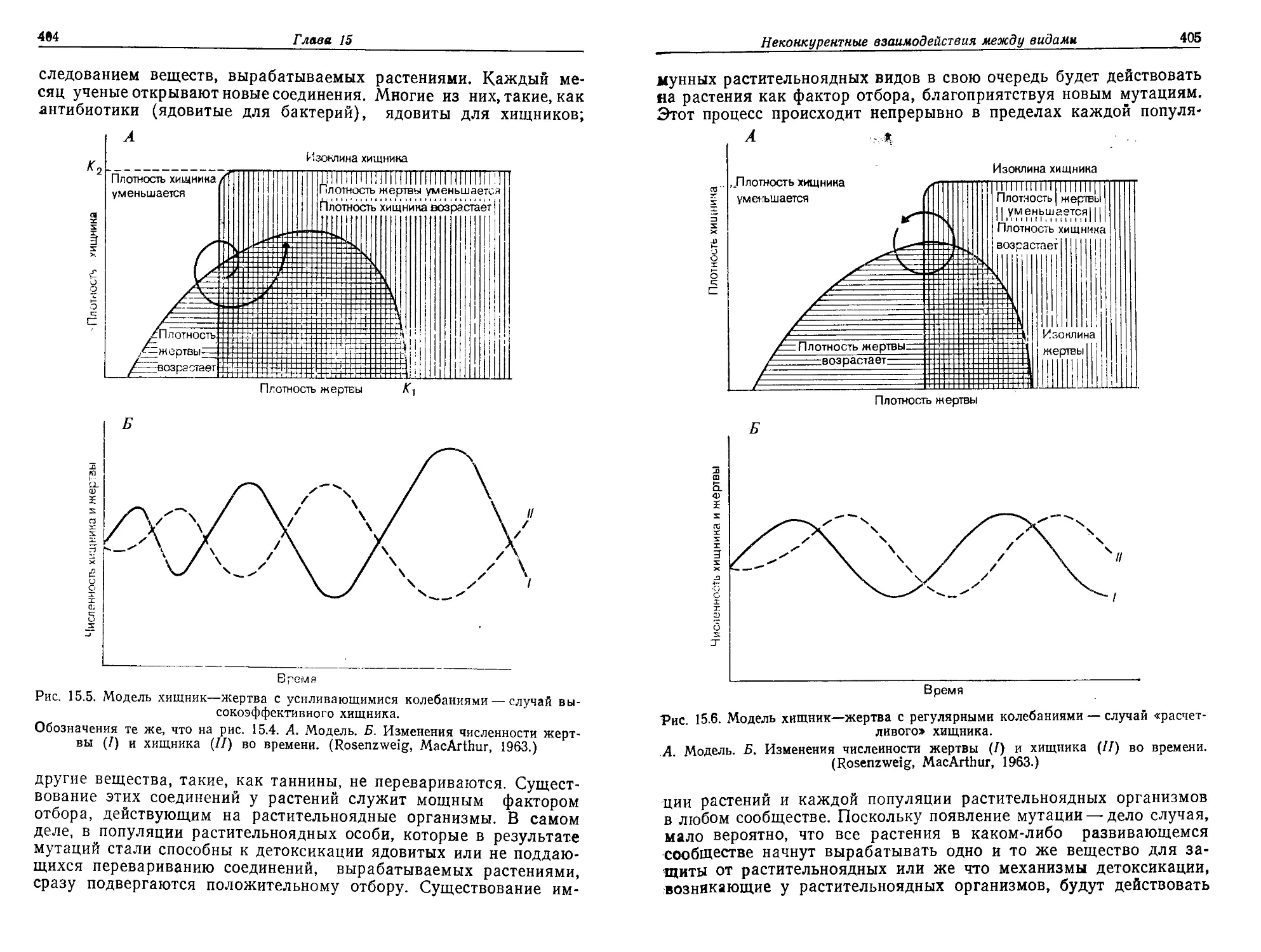
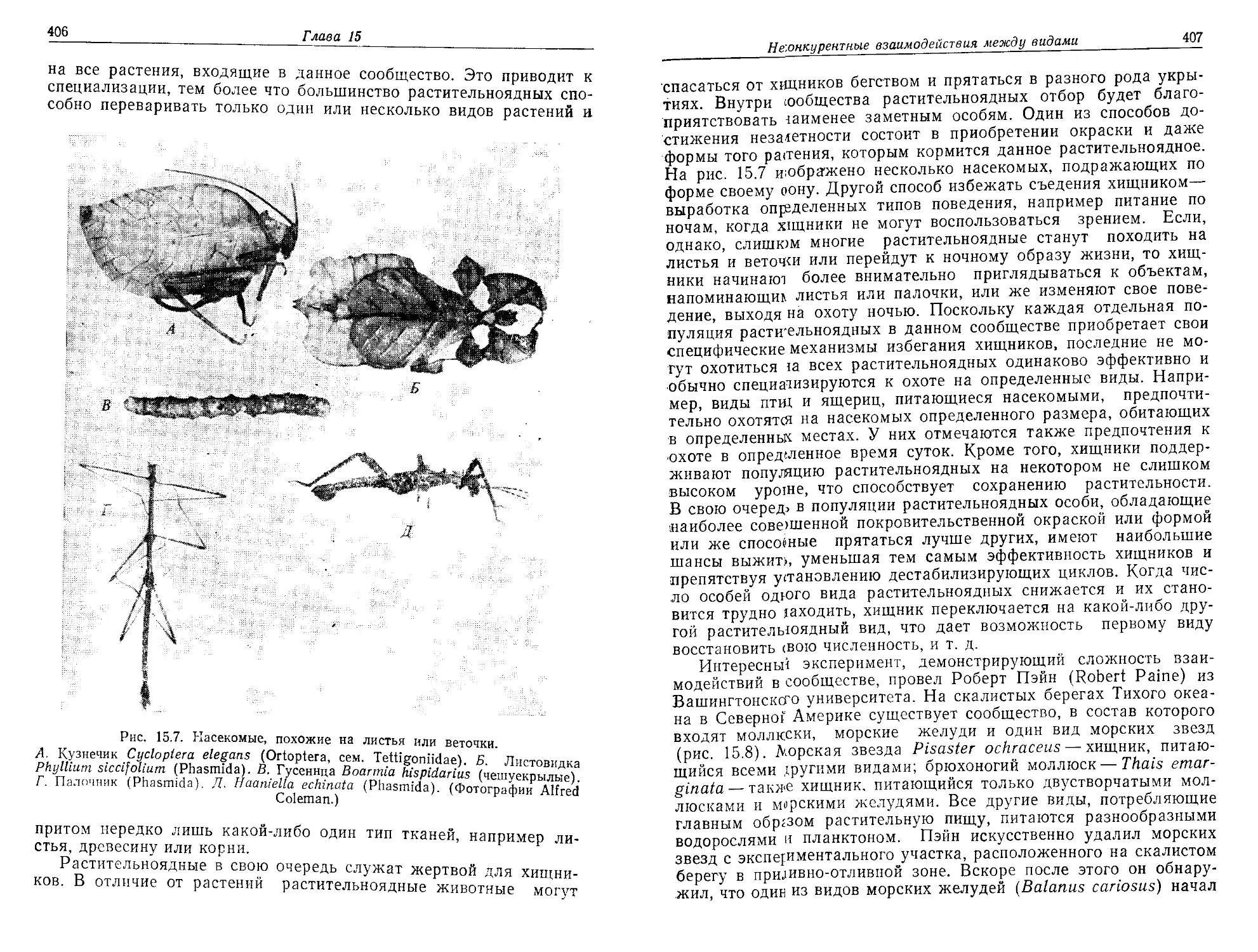
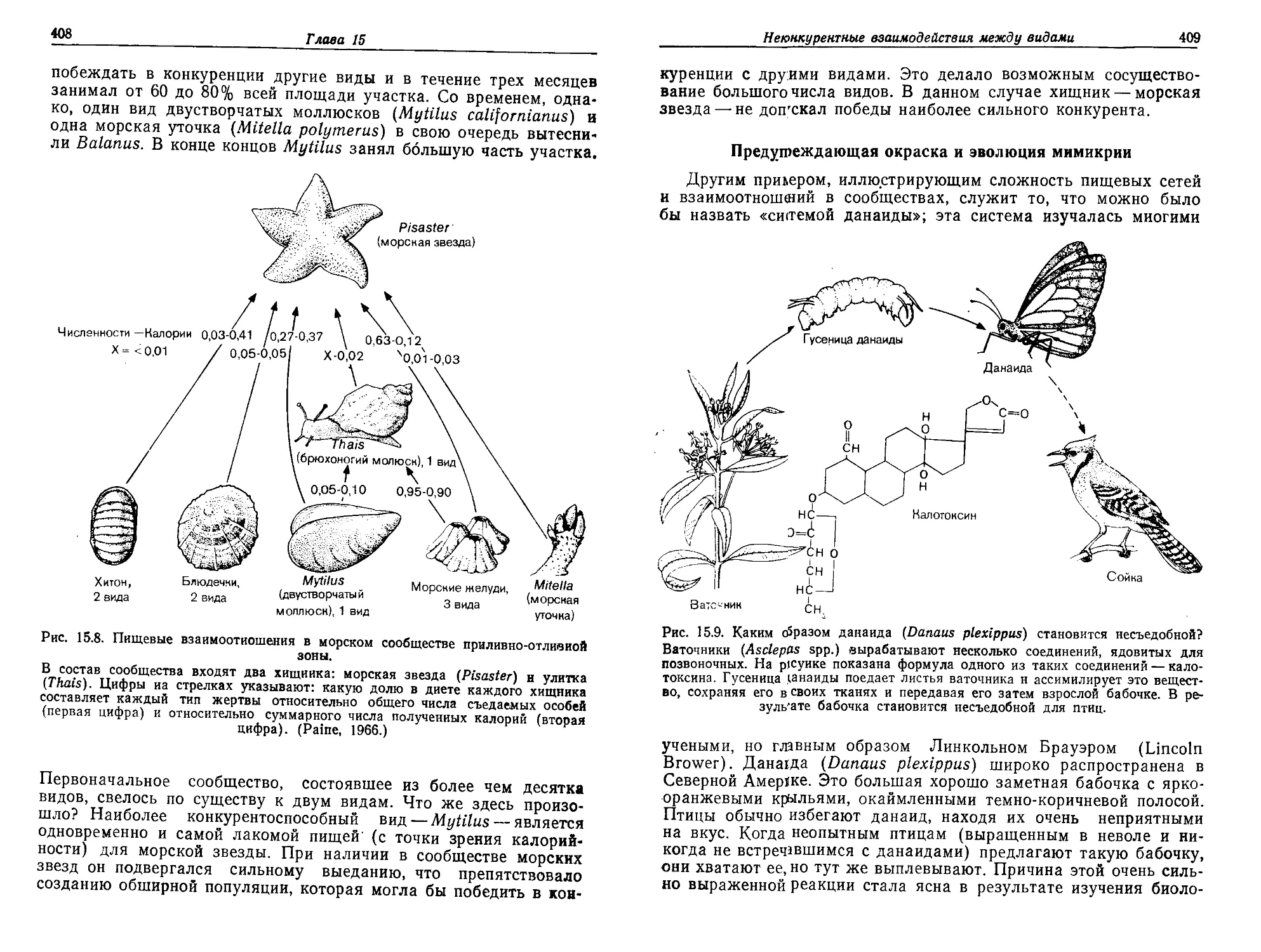























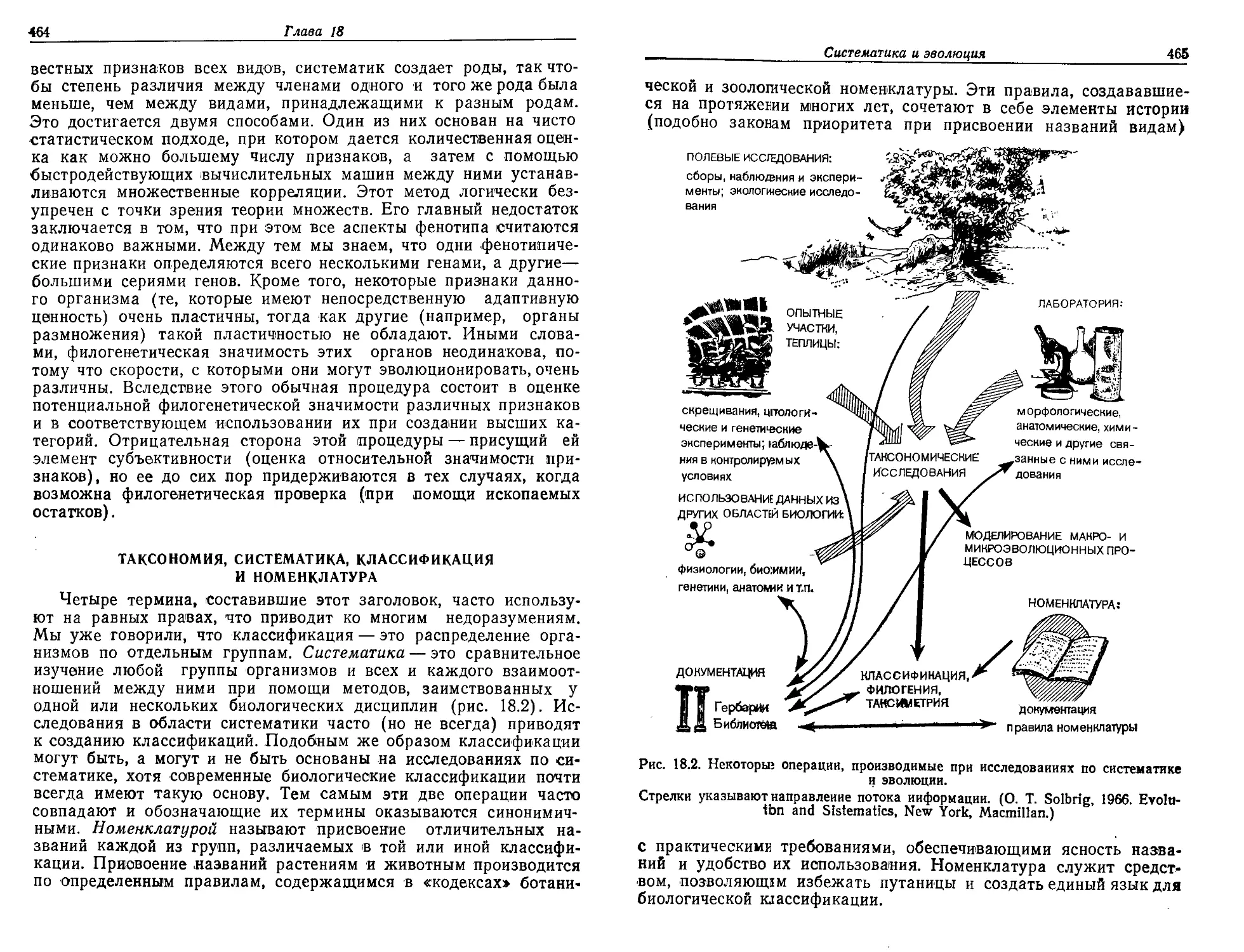


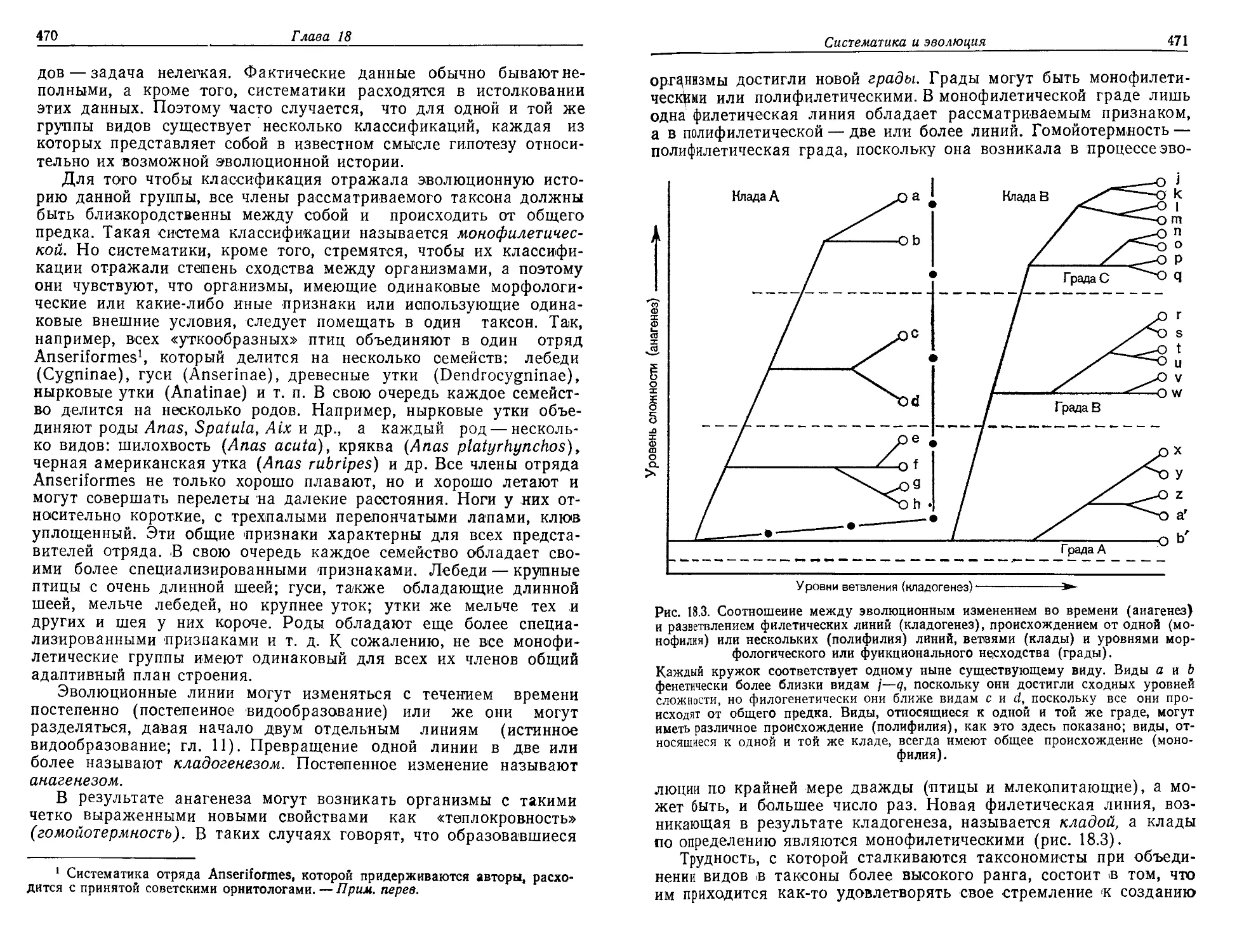
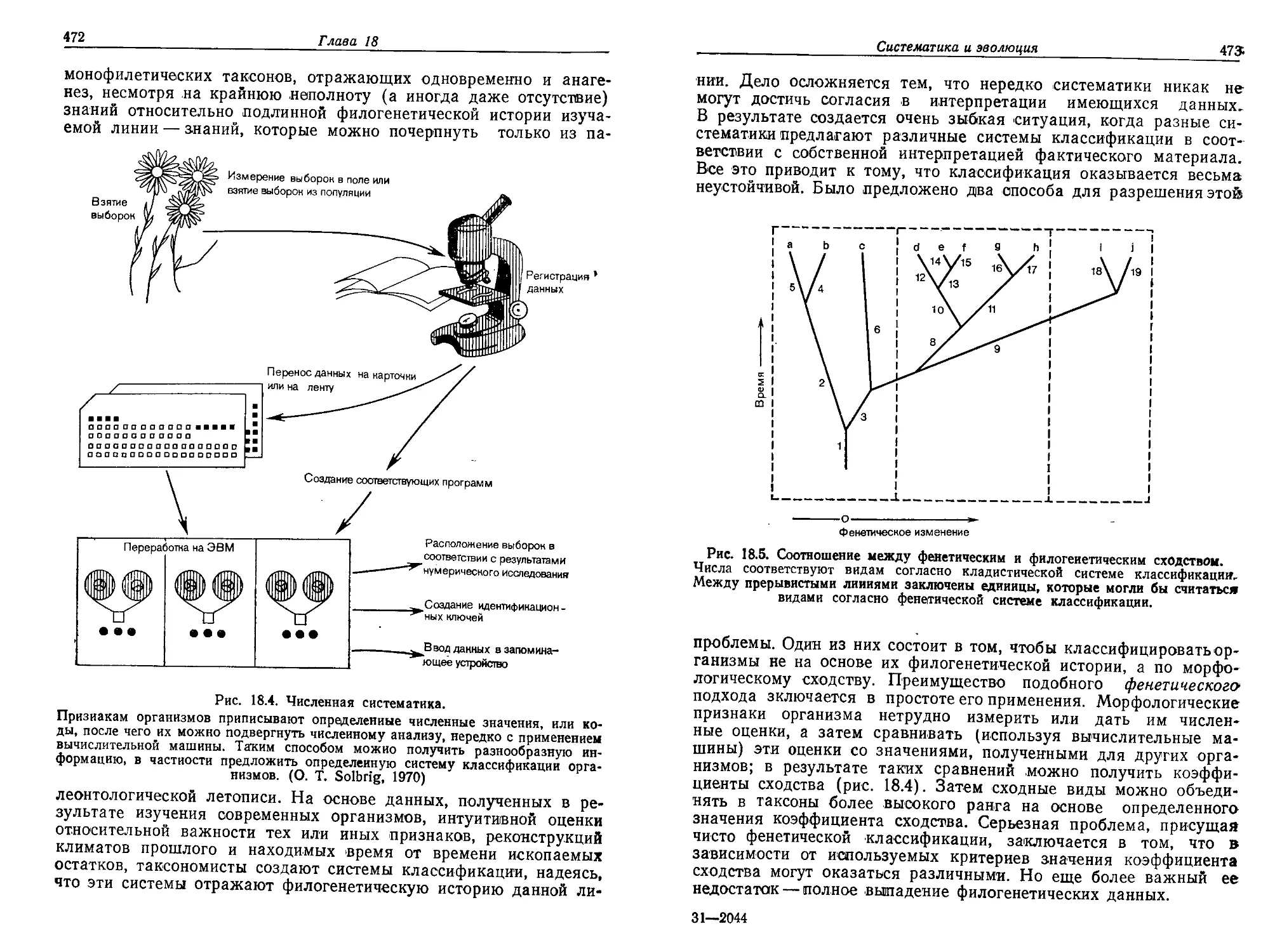












Текст
Introduction
to Population
Biology
and Evolution
Otio T. Solbrig
Dorothy J. Solbrig
Addison-Wesley Publishing Company
Reading, Massachusetts. Menlo Park, California
London. Amsterdam
Don Mills, Ontario. Sydney
О. Солбриг
Д.Солбриг
Популяционная
биология
и эволюция
Перевод с английского
канд. физ.-мат. наук Т. И. Штилькинда
под редакцией
канд. физ.-мат. наук А. Д. Базыкина
Москва
«Мир»
1982
ББК 28.04
С 60
УДК 575.3/.7
$ 60 Солбриг О., Солбриг Д.
Популяционная биология и эволюция: Пер. с англ./Пере-
вод Штилькинда Т. И.; Под ред. и с предисл. А. Д. Базыки-
на. — М.: Мир, 1982. — 488 с., с ил.
В книге американских ученых О. и Д. Солбригов изложены основы эволюцион-
ного учения, теория Дарвина, основы генетики, рассмотрены факторы эволюции, фор-
мы естественного отбора и пути видообразования, гибридизация, адаптация, рост
популяций и его регуляция, теория ниш и конкуренция, эволюция поведения, связь
между палеонтологией и теорией эволюции, основы таксономии. Приведены в до-
ступной форме математические модели рассматриваемых процессов.
Для преподавателей, аспирантов и студентов университетов и педагогически»
институтов, а также для биологов всех специальностей.
C’04i(0iT=82 l03~82’ ч« ’> 2001000000 БьК 28.04
Редакция литературы по биологии
Отто Солбриг, Дороти Солбриг
ПОПУЛЯЦИОННАЯ БИОЛОГИЯ И ЭВОЛЮЦИЯ
Научный редактор М. Б. Николаева. Мл. редактор 3. Е. Кожанова. Художник Д. А. Аникеев.
Художественный редактор А. В. Проценко. Технический редактор Н, Д. Толстяков».
Корректор Т. И. Стифеева
ИБ № 2659
Сдано в набор i5.09.81. Подписано к печати 07.01.82. Формат 60Х90*/ы. Бумага типографская
М I. Гарнитура литературная. Печать высокая. Объем 15,25 бум. л. Усл. печ. л. 30.50. Усл.
кр.-отт. 30,50. Уч.-изд. л. 32.42. Изд. № 4/1257. Тираж 15 000 экз. Зак. 2044. Цена 2 р. 60 к.
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2.
Московская типография № 11 Союаполиграфпрома при Государственном комитете СССР
по делам издательств, полиграфии н книжной торговли. Москва, 113105, Нагатинская. 1.
© 1979 by Addison-Wesley Publishing Compa-
ny, Inc.
© Перевод на русский язык <Мир», 1982
Предисловие редактора перевода
Эволюционное учение составляет фундамент современной био-
логии и одновременно венчает собой все ее здание. Нет такой кон-
кретной отрасли биологии, которая бы не была пронизана эволю-
ционными идеями и не входила бы составной частью в систему'
эволюционных представлений о живом. Более того, эволюционная
идеология является связующим звеном между биологией и иными
науками, как предбиологического, так и социального циклов. Ре-
шение проблем возникновения жизни, ранних стадий формирова-
ния и развития биосферы, геологической истории Земли немыслимо
вне рамок общих представлений о биологической эволюции. Вме-
сте с тем вопросы происхождения человека, эволюции обществен-
ного поведения и коммуникаций у животных лежат на стыке об-
ластей компетенции биологических и социальных наук и носят,
несомненно, эволюционный характер.
Такое уникальное положение эволюционного учения в системе
наших представлений о мироздании создает специфические труд-
ности для преподавания теории эволюции. Для понимания идей
эволюционного учения, правильной постановки проблемы, соот-
ветствующей оценки результатов экспериментов и наблюдений от
студента требуются значительные знания в области конкретных
биологических дисциплин. В то же время из предыдущего должно
быть ясно, сколь желательно, чтобы преподаванию конкретных
биологических дисциплин предшествовало основательное знаком-
ство с современными представлениями об эволюции. Ситуация
усугубляется тем, что, вопреки распространенному мнению, теория
эволюции не является к настоящему времени чем-то окончатель-
но сложившимся и полностью завершенным. Многие фундамен-
тальные проблемы эволюции, например соотношение между слу-
чайным и детерминированным в эволюции, сравнительная роль
моно- и полифилии (т. е. происхождение тех или иных групп от
общего или различных корней), взаимосвязь между закономерно-
стями микро- и макроэволюции и многие другие, являются остро
дискуссионными. Не следует забывать также, что в наши дни тео-
рия эволюции перестает быть предметом чисто академическим и
обретает важное практическое значение: вмешательство человека
в окружающую его среду достигло масштабов, когда он начинает
оказывать заметное воздействие на эволюцию больших групп жи-
вых существ (например, болезнетворных микроорганизмов) и са-
мой биосферы. В обозримой перспективе маячит вопрос об управ-
лении процессом эволюции. Очевидно, однако, что искусственное
б
Предисловие редактора перевода
управление эволюцией без понимания закономерностей эволюции
естественной не только не может быть эффективным, но и чревато
катастрофическими последствиями.
Настоящая книга фактически является учебником по эволю-
ционному учению. Соответствующих современных пособий доволь-
но много, но большинство из них носит отпечаток той биологиче-
ской дисциплины, профессиональный опыт в которой имеет автор
книги, будь то зоология или ботаника, генетика или биохимия и
т. п. Солбригам удалось, на мой взгляд, избежать такого крена.
В соответствии со своим замыслом они создали общую, довольно
элементарную — рассчитанную на студентов младших курсов, но
достаточно полную и содержательную сводку по современному
эволюционному учению.
Специального упоминания заслуживают несколько обстоя-
тельств. Во-первых, — это постоянное на протяжении всей книги
внимание авторов к методологическим аспектам науки. Авторы
последовательно акцентируют важность таких этапов научной
работы, как постановка проблемы, формирование альтернативных
гипотез, замысел ключевых проверочных экспериментов, позво-
ляющих сделать выбор между гипотезами, и т. д. Во-вторых, пра-
вильное общее понимание авторами' роли математического моде-
лирования в биологии: они не склонны ни к свойственному многим
биологам-натуралистам пренебрежительному отношению к мате-
матическому моделированию как инструменту исследований, ни
к фетишизации результатов исследования математических моделей,
характерной для некоторых ученых, пришедших в биологическую
проблематику из математики и физики. В-третьих, авторы не за-
тушевывают открытых проблем и трудных мест в современном
учении об эволюции, знакомя читателя, например, с явлением не-
ожиданно высокой полиморфности, обнаруженной в природных
популяциях в последние 10—15 лет, и другими нерешенными за-
дачами.
Живой характер изложения довольно сложных идей, четкость
в постановке проблем, обилие нового интересного эксперименталь-
ного материала, привлекаемого авторами для доказательства и
иллюстрации тех или иных теоретических положений, делают кни-
гу не только полезной для студентов, на которых она в первую
очередь рассчитана, но и интересной для биологов-профессиона-
лов, так же как и для всех интересующихся современными проб-
лемами эволюции.
А. Базыкин
Предисловие
Эта книга по замыслу авторов должна удовлетворить сущест-
вующую потребность в современном и исчерпывающем учебнике
по биологии популяций и эволюции для студентов первого или
второго курсов. Она задумана главным' образом как учебное по-
собие по вводному курсу, рассчитанному на один семестр.
В учебнике строго излагаются главные проблемы генетики,
экологии популяций и эволюционной теории, однако авторы исхо-
дят из того, что читатель совершенно незнаком с этими пробле-
мами. Своеобразие книги состоит также в том, что она начинается
с подробного обсуждения научного метода и процесса создания
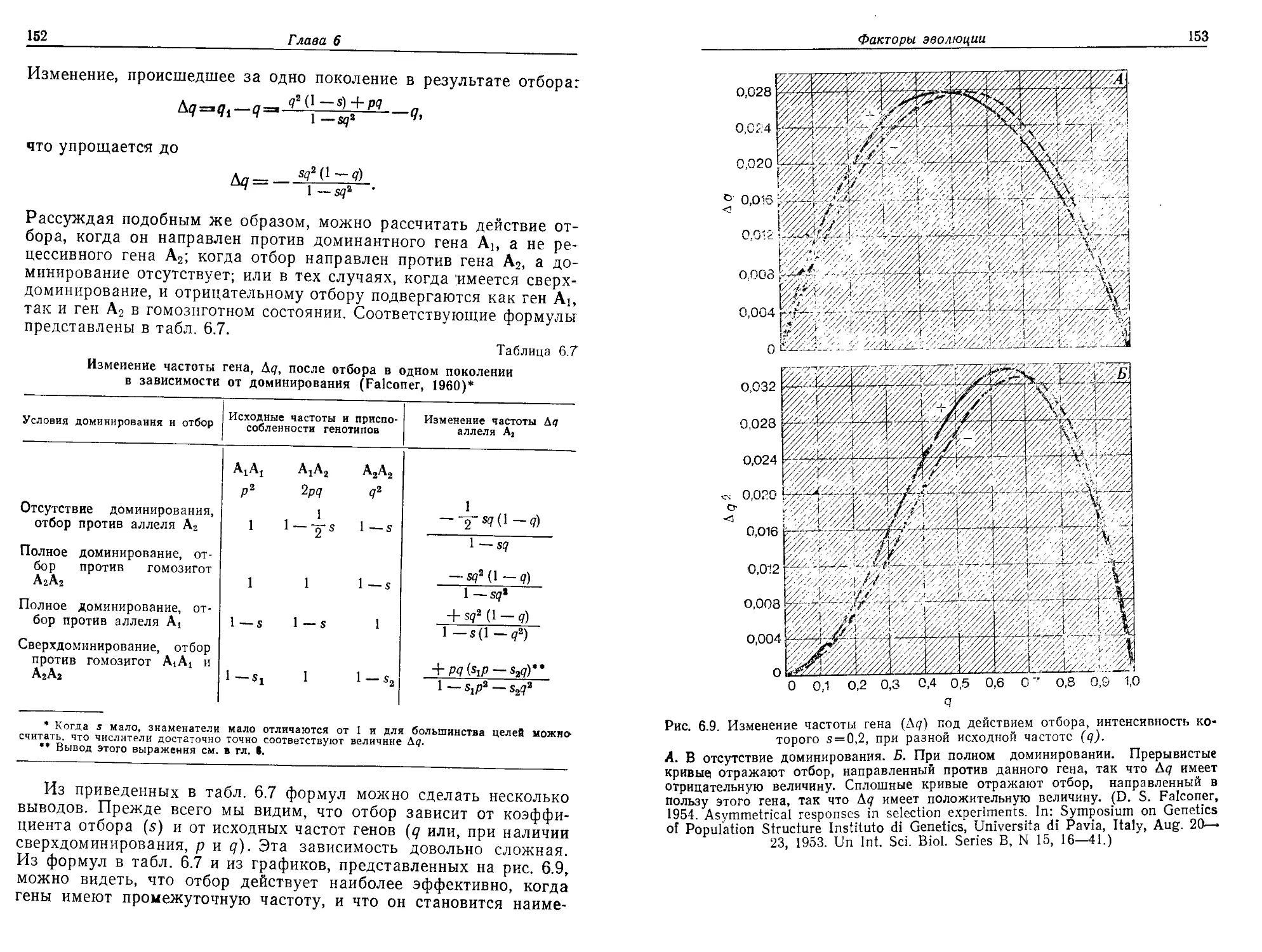
гипотез и их проверки, рассматриваемых непосредственно в при-
ложении к теории эволюции. Авторы вновь и вновь возвращаются
к этой общей теории при изложении более узких и частных ги-
потез.
Главное внимание в книге уделяется описанию и разбору раз-
личных теоретических представлений в сочетании с необходимыми
эмпирическими данными. На протяжении всей книги авторы ставят
четыре вопроса, связывая их с каждой очередной темой: 1) В чем
состоит рассматриваемая проблема? 2) Каковы относящиеся к ней
данные — эмпирические «факты»? 3) Какие были выдвинуты ги-
потезы для того, чтобы объяснить эти факты? 4) Какие существу-
ют эксперименты или наблюдения, позволяющие сделать выбор
между альтернативными гипотезами? Мы надеемся, что примене-
ние метода построения гипотез в сочетании с дедуктивным мето-
дом убедит учащихся в важности строгой проверки гипотез и по-
кажет им, что подобный подход имеет для популяционной биоло-
гии такое же большое значение, как и для биологии молекулярной.
Другая особенность книги — использование математических моде-
лей во всех тех случаях, когда они уместны. Мы считаем, что ма-
тематика при разумном применении может дисциплинировать по-
пуляционную биологию и способствовать расширению ее горизон-
тов. Если, однако, математические модели основаны на нереали-
стических допущениях, то они могут нанести науке большой вред.
Поэтому мы особенно подробно указываем для каждой исполь-
зуемой модели принятые при ее построении допущения, а также ее
недостатки. Кроме того, каждая формула выводится последова-
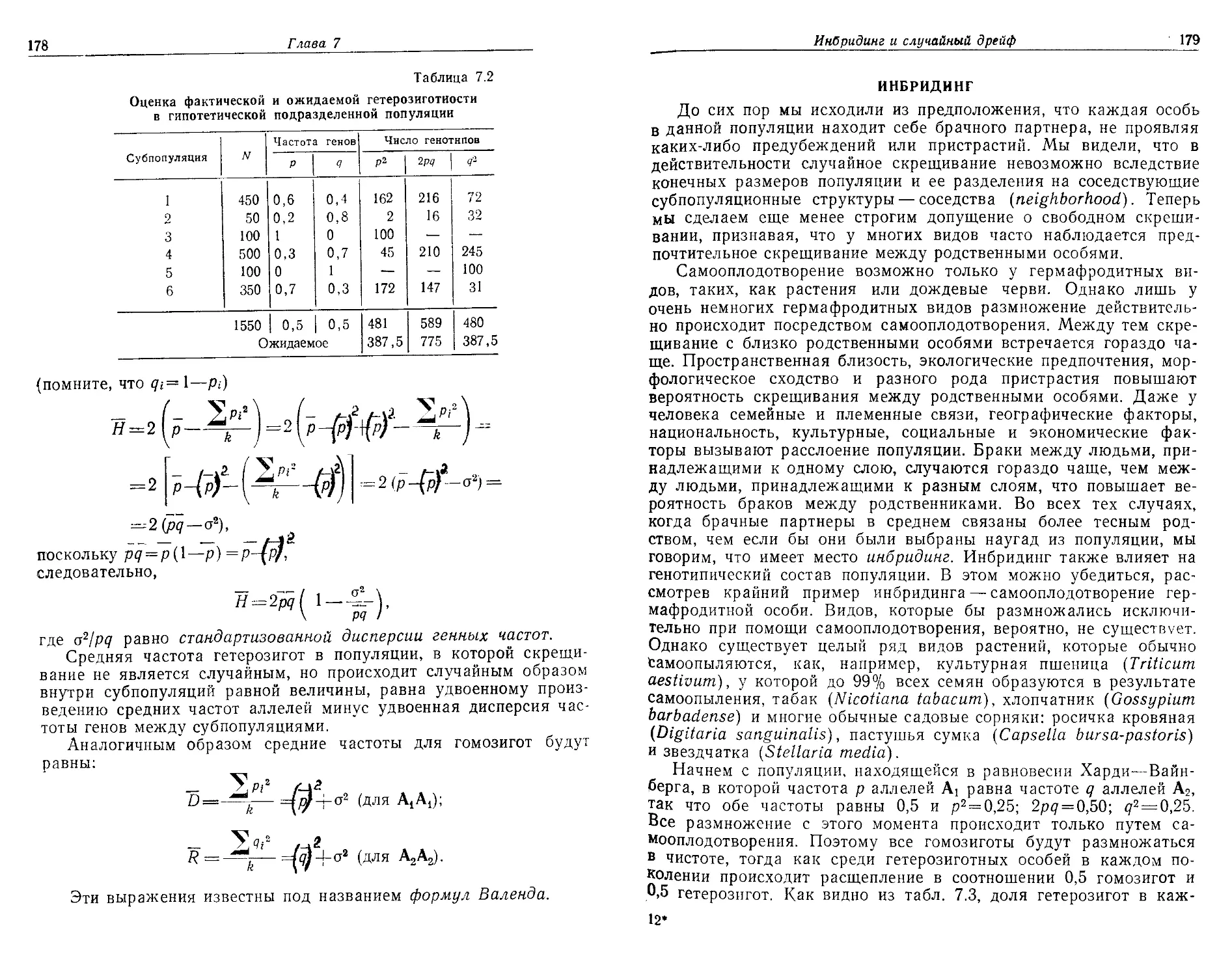
тельно, шаг за шагом, с тем чтобы сделать ее понятной даже сту-
дентам, математические познания которых ограничиваются школь-
ным курсом алгебры. И, наконец, мы старались привести как
в
Предисловие
можно больше примеров и результатов конкретных исследований,
причем единственным ограничивающим фактором в этом отношении
были соображения объема.
Популяционная биология создает основу для изучения разнооб-
разия природы; изучение этого разнообразия необходимо для
того, чтобы понять и попытаться разрешить такие проблемы, как
загрязнение среды, охрана природы и рациональное использование
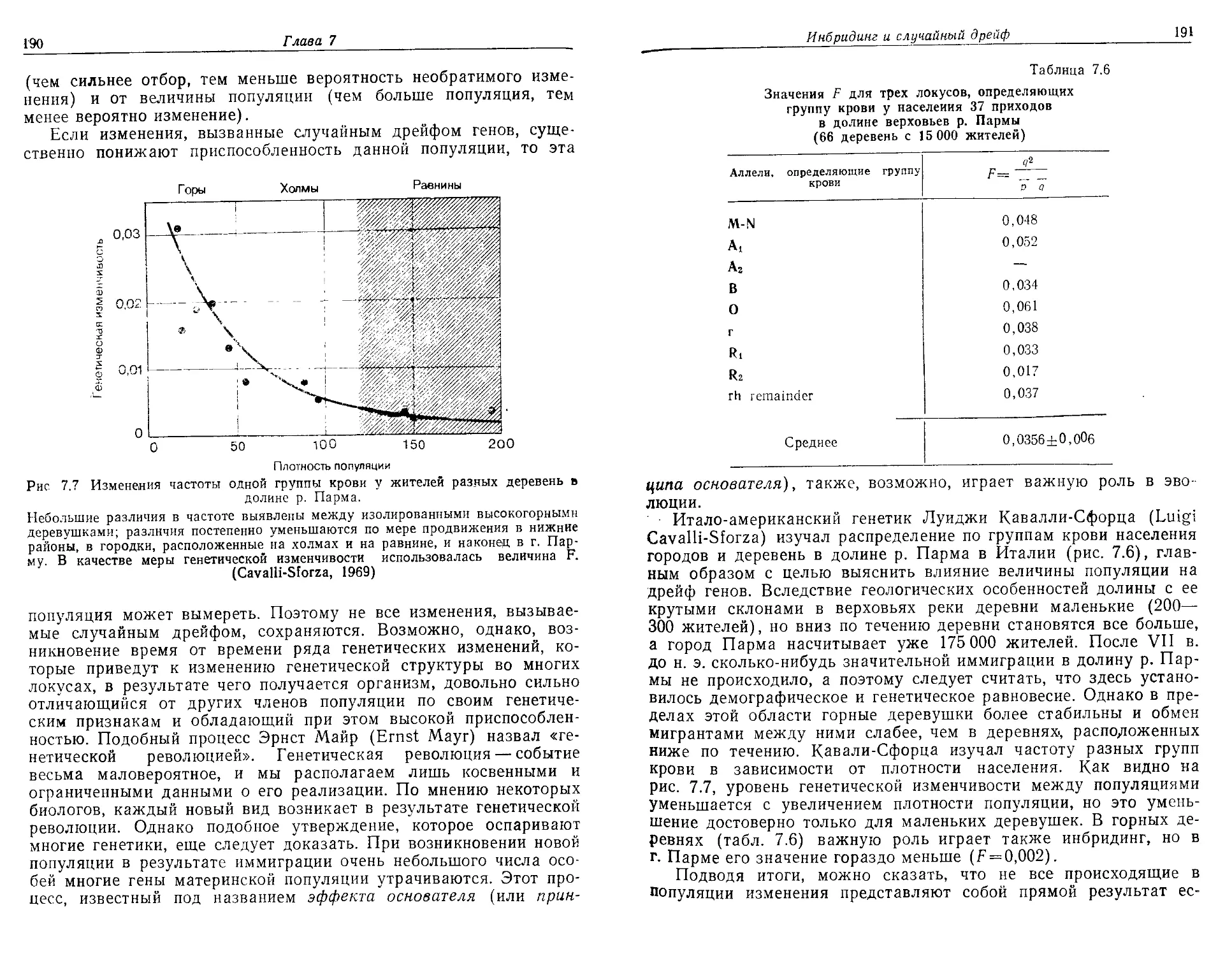
природных ресурсов. Это весьма динамичная быстро развиваю-
щаяся область. Авторы надеются, что им удалось воплотить в
своей книге живой дух и важное значение популяционной биоло-
гии и что они смогут передать свое восприятие читателю.
В подготовке книги принимали участие многие лица. Мы при-
знательны студентам Гарвардского университета, для которых в
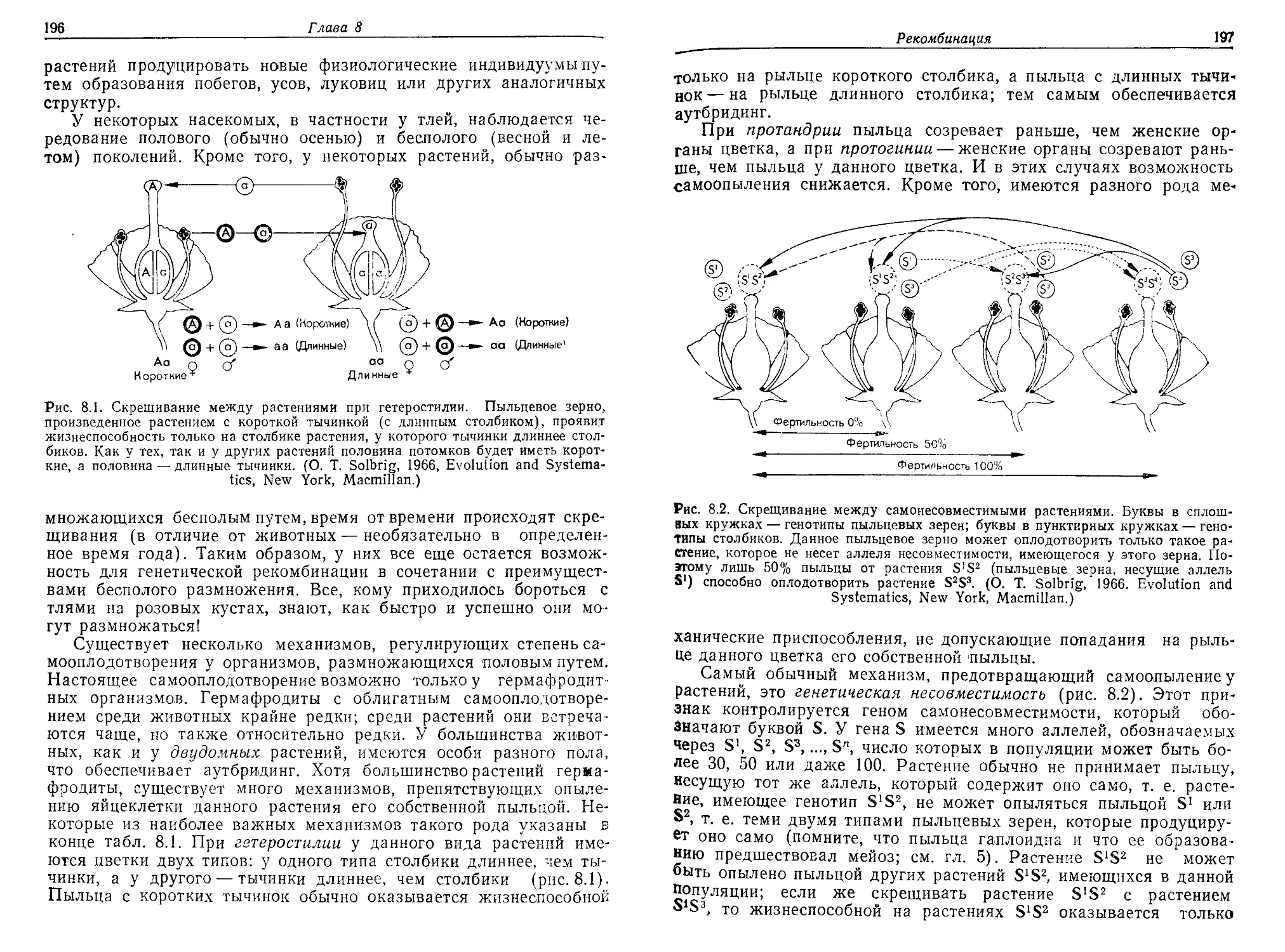
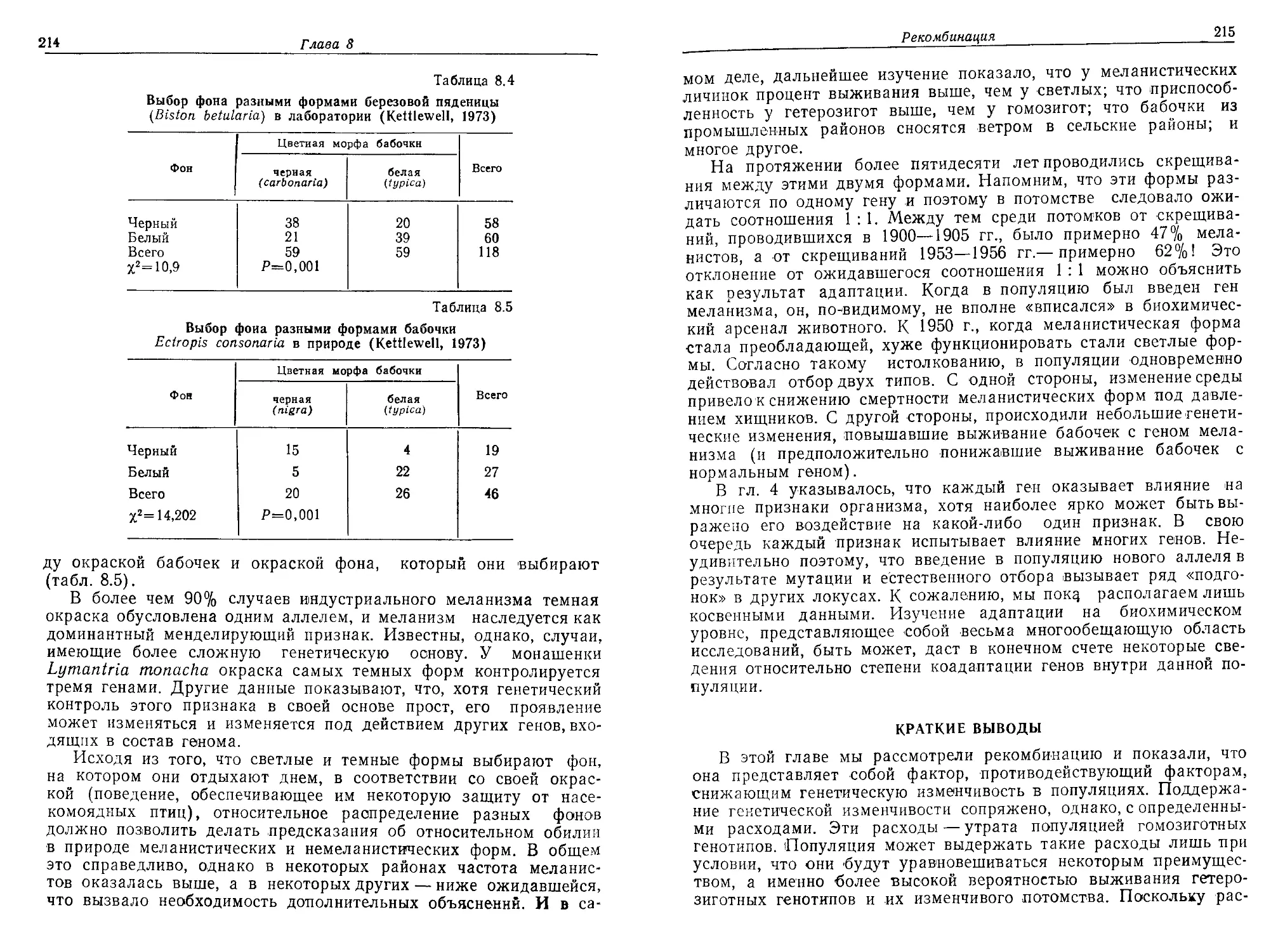
первую очередь предназначалась книга. Мы очень обязаны д-рам
Уильяму Брауну из Корнелльского университета, Дж. Мерриту
Имлену из университета штата Индиана, Уильяму Хили из кол-
леджа «Хоули Крое» и в особенности Генри Уилбуру из универ-
ситета Дьюка, которые внимательно прочитали рукопись и сделали
много полезных замечаний. Разумеется, ответственность за любые
ошибки, которые могут обнаружиться в книге, несут авторы. Мы
благодарны г-ну Джеймсу Фенстону из издательства «Эддисон-
Уэсли» за поддержку, оказанную им нашему начинанию. Неизмен-
ная преданность делу г-жи Маргарет Кассиди, ее аккуратность и
точность сыграли очень большую роль в создании этой книги, и мы
ей глубоко признательны. И в заключение мы хотим поблагода-
рить г-жу Аниту Фэхи за помощь в технической подготовке руко-
писи, далеко выходящую за рамки ее обязанностей.
О. Солбриг,
Д. Солбриг
Часть первая
Глава 1
Эволюция, популяции
и естественный отбор
На сегодняшний день обнаружено и описано примерно пол-
миллиона видов ныне живущих растений и около миллиона видов
животных, а число еще не описанных видов оценивается по мень-
шей мере в один или два миллиона. Кроме того, как полагают
ученые, число вымерших видов растений и животных, когда-либо
живших на земле, в 5—9 раз больше числа современных видов.
Возникшее сто с лишним лет назад представление о том, что виды
появлялись на протяжении последних 3 млрд, лет в результате
эволюции путем естественного отбора, было одним из самых зна-
чительных открытий в биологии.
ВИДЫ И ПОПУЛЯЦИИ
Чисто интуитивно мы считаем, что вид — это группа очень
сходных между собой организмов, например людей, лошадей или
морковок. Научное определение вида с течением времени неод-
нократно изменялось (гл. 10), и в настоящее время наиболее ши-
роко распространено следующее определение: «Вид — это группа
морфологически сходных организмов, имеющих общее происхож-
дение и потенциально способных к скрещиванию между собой в
естественных условиях». Это определение порождает ряд практи-
ческих проблем, таких, как установление общего происхождения
и разъяснение смысла, вкладываемого в выражения «сходные» и
«потенциально способные к скрещиванию». В дальнейшем мы об-
судим эти проблемы, а пока приведенное определение должно дать
некоторое представление о том, что такое вид.
Особи, из которых слагается данный вид, не всегда живут в
непосредственном соседстве друг с другом. Обычно они образуют
более или менее изолированные группы, называемые популяция-
ми. Изучая процесс эволюции, мы рассматриваем составляющие
вид популяции. Популяция — это сообщество скрещивающихся
между собой организмов, поскольку особи в пределах данной по-
пуляции обычно скрещиваются друг с другом, хотя иногда неко-
торые особи могут покидать свою популяцию и скрещиваться с
особями, принадлежащими другой популяции. Определенную та-
ким образом популяцию, называемую скрещивающейся или мен,-
делевской популяцией, следует отличать от популяции, изучаемой
10
Глава 1
статистиками или демографами. И статистики, и демографы так-
же определяют популяцию как группу особей, объединяемых, од-
нако, на другой основе — на основе «взаимодействия в простран-
стве» или «обитания в данном географическом районе».
Скрещивающиеся популяции не обязательно представляют со-
бой дискретные единицы; нередко границы между ними провести
трудно. Хорошим примером служат леса бальзамической пихты
(Abies balsamea), которые простираются через всю Канаду — от
Ньюфаундленда до Альберты, а в США — от Новой Англии до
Айовы, иногда на сотни километров без видимых перерывов. Как
разграничить популяции в этом случае? Не обесценивается ли
сформулированная концепция невозможностью провести границы
между популяциями? На этот вопрос следует ответить: «Нет».
Невозможность дифференцировать разделенные пространственно
скрещивающиеся популяции в одних случаях и их перекрывание
в других не обязательно означает, что границ не существует. Проб-
лему разделения скрещивающейся популяции на обособленные
единицы можно сравнить с проблемой разделения группы орга-
низмов на молодых и старых. Несмотря на отсутствие четких кри-
териев, разделение на «молодых» и «старых» полезно и осмыс-
ленно.
У организмов с бесполым размножением (многие бактерии и
некоторые растения) нет скрещивающихся популяций, потому что
каждая особь размножается независимо от остальных. Однако
большинство организмов, размножающихся бесполым путем, время
от времени прибегают к половому размножению. Например, мно-
гие виды боярышника (Crataegus) размножаются семенами, кото-
рые образуются без оплодотворения. И все же иногда эти растения
образуют некоторое количество семян обычным половым путем,
т. е. в результате слияния двух гамет — яйцеклетки и спермия.
Популяция организмов, размножающихся бесполым путем, пред-
ставляет собой скрещивающуюся популяцию в той мере, в какой
у нее происходит половое размножение.
Для того чтобы понять, каким образом протекает эволюция,
необходимо понимать процессы, происходящие в пределах популя-
ции. Изучение этих процессов составляет предмет популяционной
биологии. Ее главные составные части — это популяционная гене-
тика, т. е. изучение процессов наследования в пределах популяции,
и популяционная экология, т. е. изучение взаимодействий членов
данной популяции друг с другом, а также с окружающей их фи-
зической и биологической средой. Хотя популяционного биолога
интересуют главным образом эволюционные явления на уровне
популяций, он должен быть хорошо осведомлен о том, какие
именно виды возникают в результате процесса эволюции. Разно-
образие современных и вымерших видов, составляющее область
исследования систематики и палеонтологии, вкратце рассматри-
вается в конце книги.
Эволюция, популяции и естественный отбор
11
ВОЗНИКНОВЕНИЕ ЖИЗНИ НА ЗЕМЛЕ
Постепенное осознание того, что все живые существа состоят
исключительно из химических соединений, оказало огромное влия-
ние на биологию. Прежде полагали, что «неживое» и «живое»
имеют различную природу, но сегодня мы знаем, что это не так.
Поскольку живая материя сложнее и обладает более высоким
уровнем организации, чем материя неживая, организмы, очевидно,
возникли в процессе эволюции из более простых, неживых компо-
нентов. Попытки проследить происхождение живых форм позво-
ляют заглянуть в самую сущность жизни.
Совершенно очевидно, что современные представления о воз-
никновении жизни далеко не полны. Многие из них спекулятивны,
поскольку основаны на свойствах современных организмов и на
данных, заимствованных у физиков, химиков, геологов и астроно-
мов. Поэтому в лучшем случае мы имеем лишь общее представле-
ние о том, как живое могло возникнуть из неживого, и это пред-
ставление служит не более чем фундаментом для исследования
природы жизни и только приоткрывает завесу над ней.
Происхождение жизни и ее природа интересуют человека с тех
самых пор, как он стал мыслящим существом. Одно из первых
объяснений состояло в том, что жизнь возникла из неживой ма-
терии в результате акта самопроизвольного зарождения. В основе
этой идеи, несомненно, лежали наблюдения, свидетельствующие о
разложении всех мертвых тел на более простые элементы, о воз-
вращении всего на свете в землю, а также невежество во всем, что
касается жизни личинок и плесеней, появляющихся как бы ниот-
куда. Когда в конце XIX в. Пастер доказал, что в современных
условиях самопроизвольного возникновения живого на Земле не
происходит, эта теория происхождения жизни из неживого стала
казаться несостоятельной с научной точки зрения и на некоторое
время была оставлена. Самопроизвольное зарождение жизни в
настоящее время невозможно ввиду существования на Земле жи-
вых организмов, потребляющих все имеющиеся в наличии богатые
энергией органические соединения. Это обстоятельство препятст-
вует превращению простых органических соединений в более
сложные — процесс, протекающий крайне медленно.
Однако когда-то условия на Земле были иными. На заре су-
ществования нашей планеты никаких организмов на ней не было,
а поэтому низкая скорость процессов, в результате которых про-
стые химические соединения превращались в более сложные, роли
не играла. Предполагается, что в таких условиях самопроизволь-
ное зарождение происходило путем медленного превращения ато-
мов и молекул в простые химические соединения, которые затем
превращались во все более и более сложные вещества, и в конеч-
ном счете это привело к появлению живых организмов. Для того
чтобы такие последовательные события могли происходить, были
12
Глава 1
необходимы известные условия: 1) колебания температуры не
должны были выходить за узкие пределы между точкой замерза-
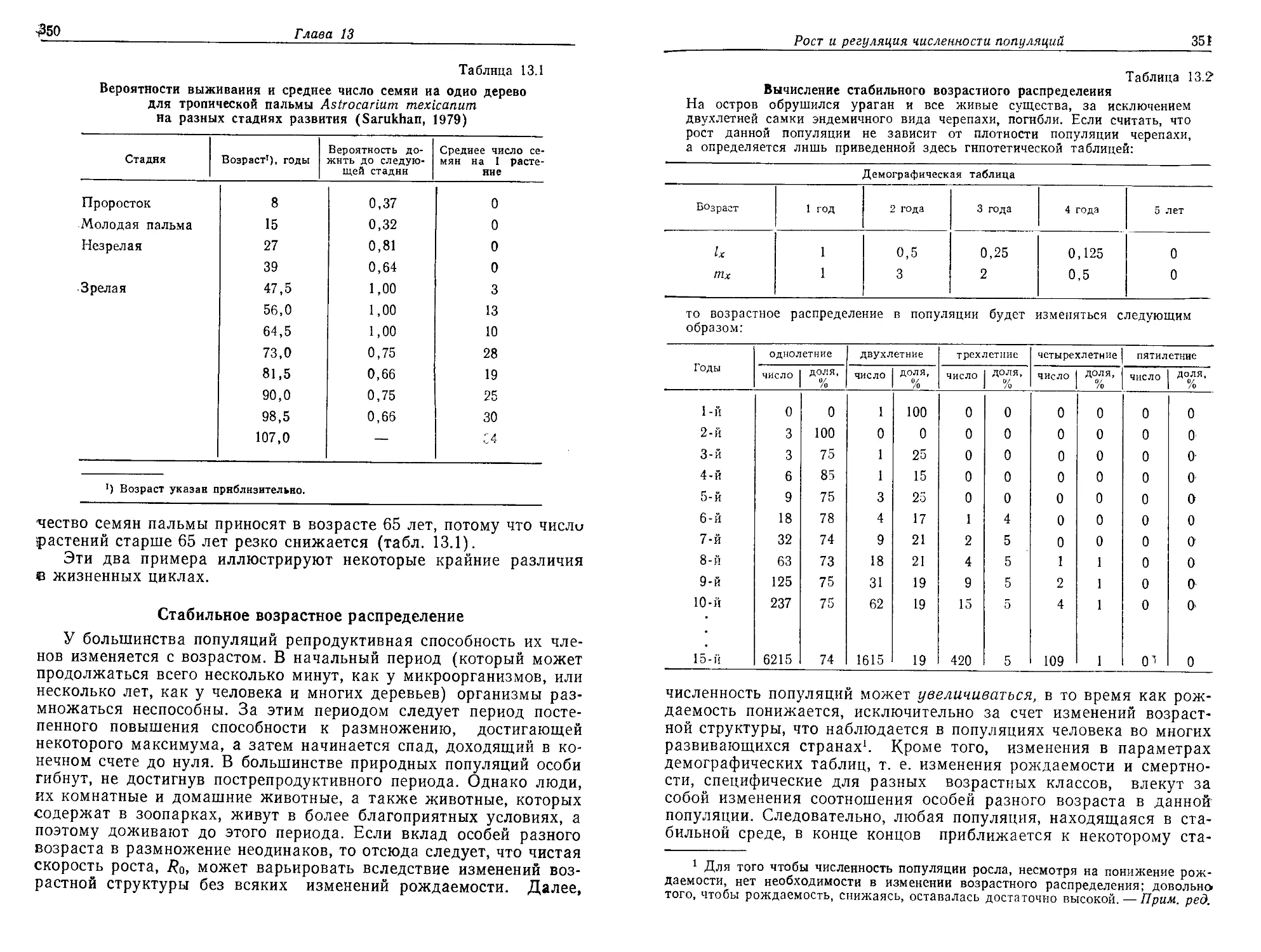
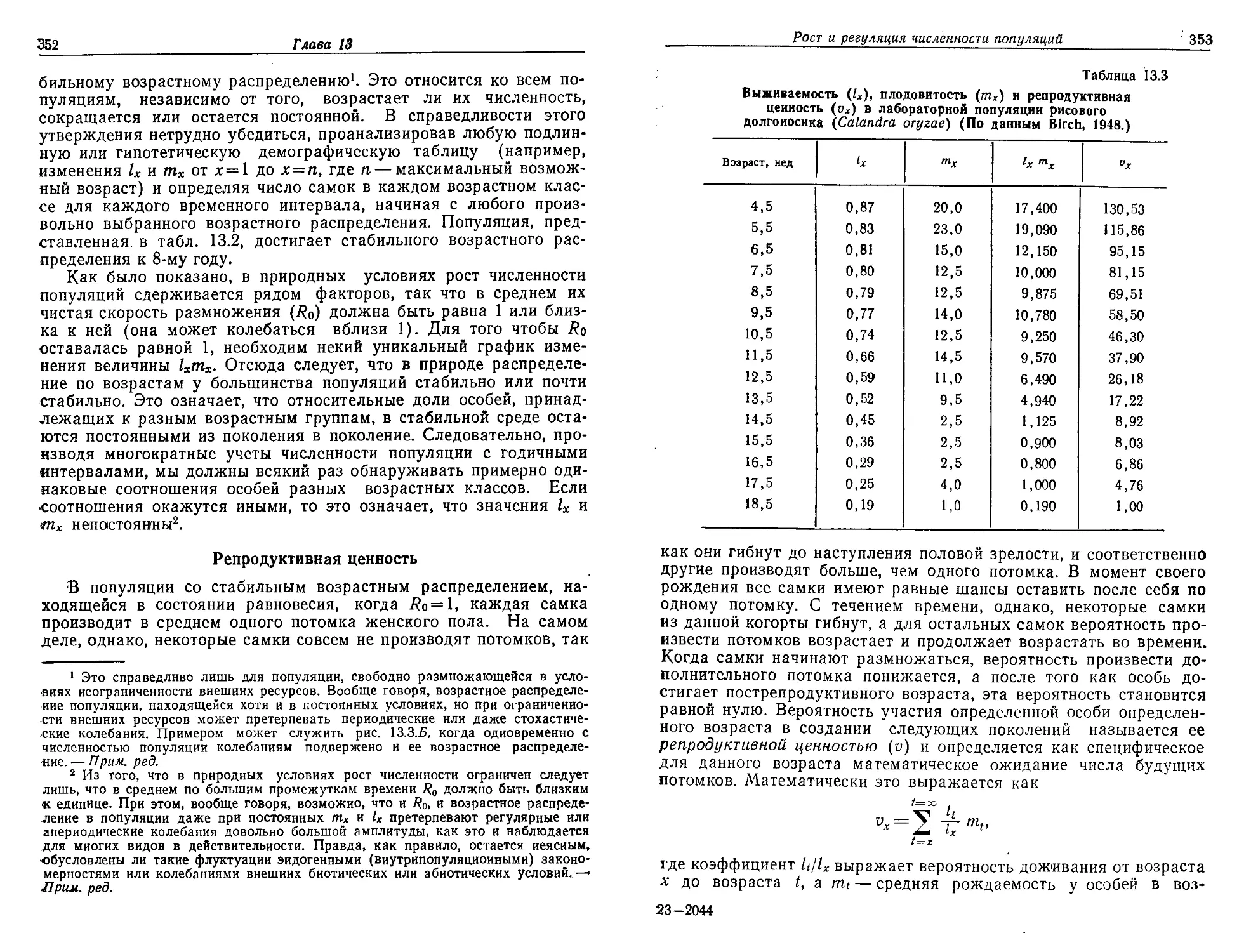
ния и точкой кипения воды; 2) требовалось наличие источника
энергии — в данном случае солнечного излучения. В какой-то мо-
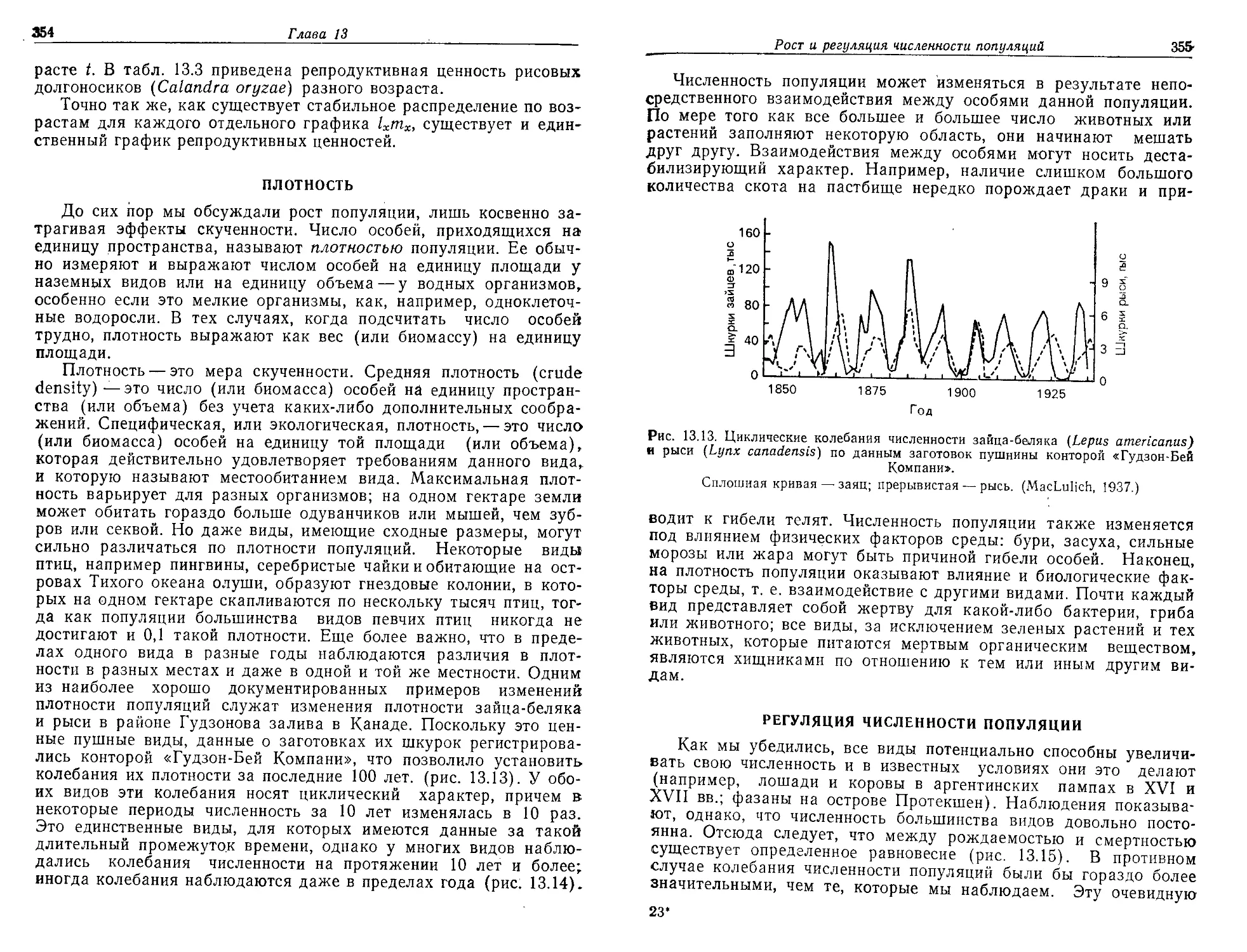
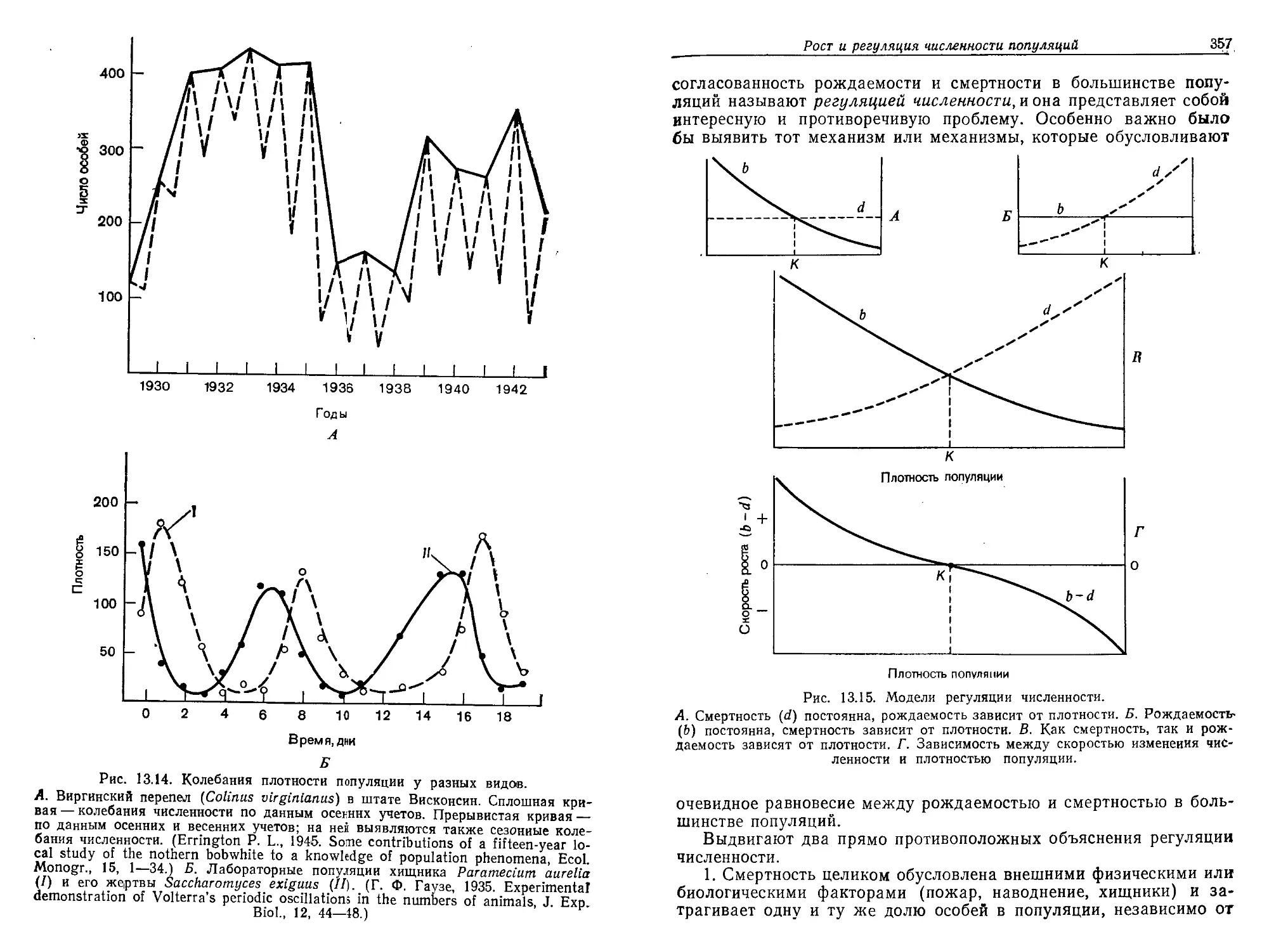
мент на Земле начала скапливаться вода, и образовались океаны,
в которых растворились различные химические соединения. На-
копление воды и растворенных в ней химических веществ облег-
чало протекание реакций между разными веществами и образо-
вание еще более сложных веществ.
Многие из возникших таким путем соединений распадались на
более простые, а затем и на отдельные молекулы. Благодаря этим
процессам концентрация сложных веществ оставалась низкой.
Однако при некоторых особых условиях концентрация химических
веществ, по-видимому, могла возрастать в несколько раз, посколь-
ку для того, чтобы могли происходить все более сложные реакции,
такие высокие концентрации необходимы. Считается, что одним
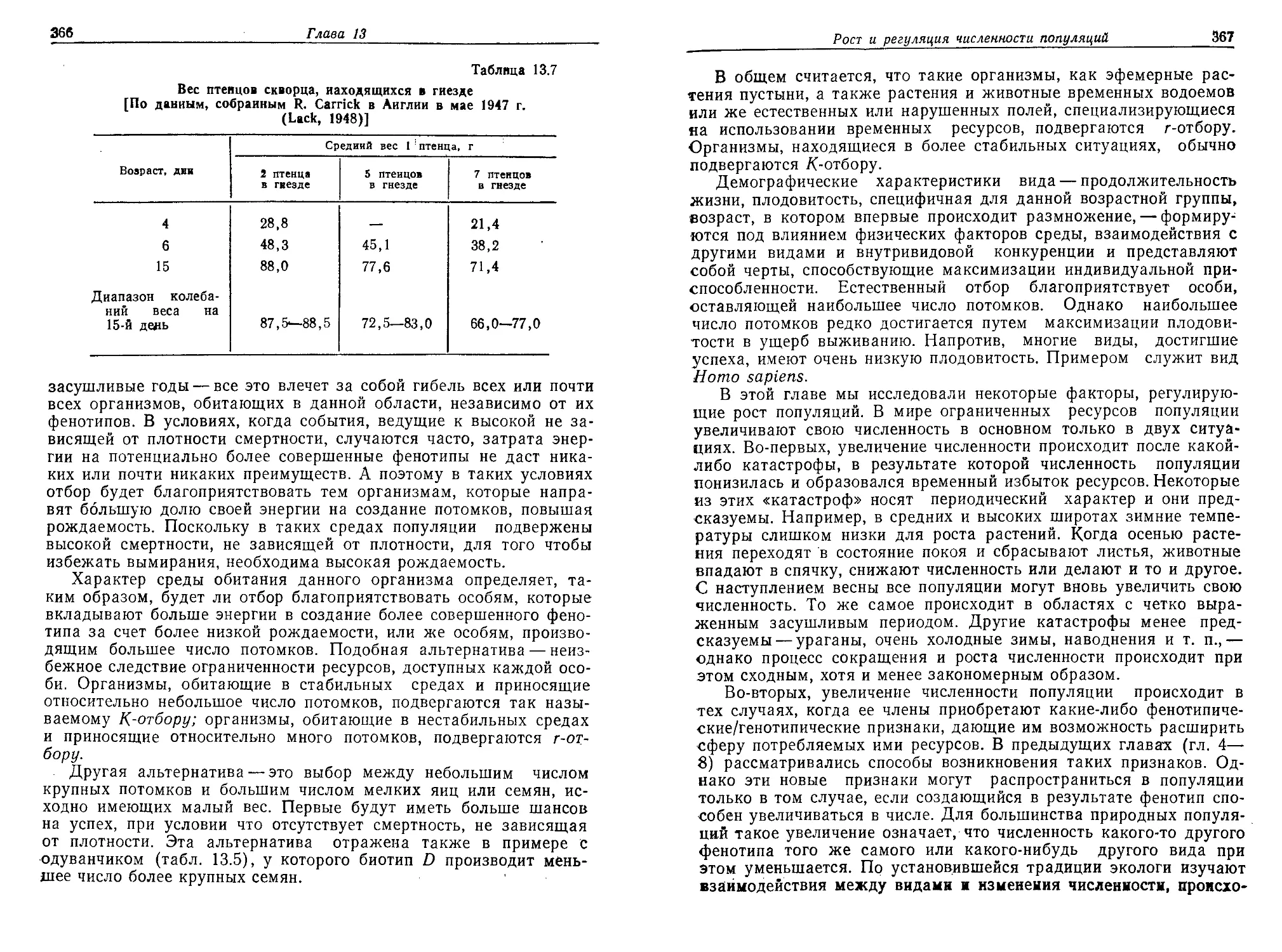
из важных механизмов повышения концентрации было поглощение
органического вещества растворимыми коллоидными частицами —
так называемыми коацерватами. Каждая из этих частиц обладала
индивидуальностью, отличаясь от всех других, а поэтому некото-
рые из них добывали энергию и делились быстрее, чем другие.
Это привело в конечном счете к возникновению соединений, спо-
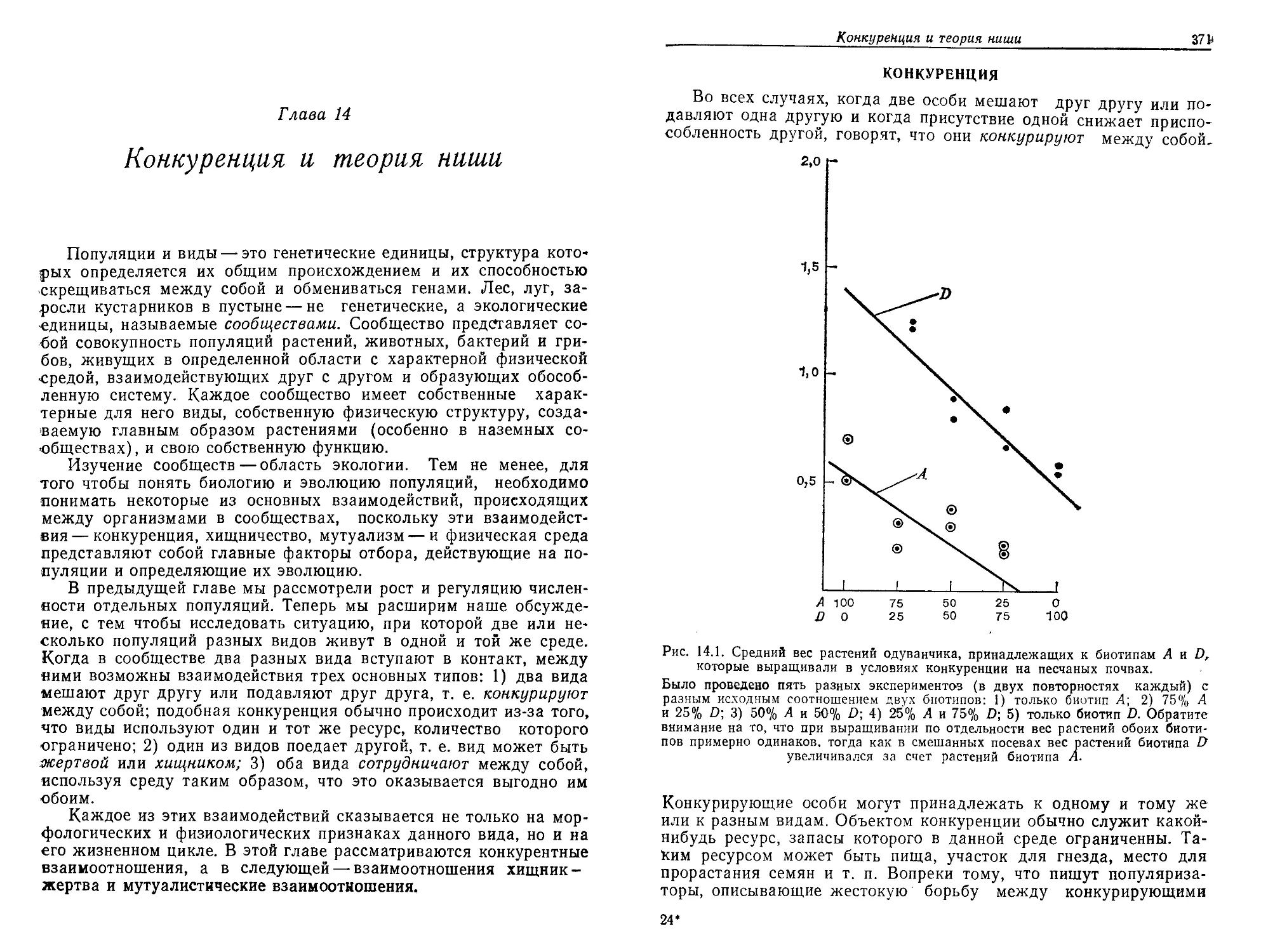
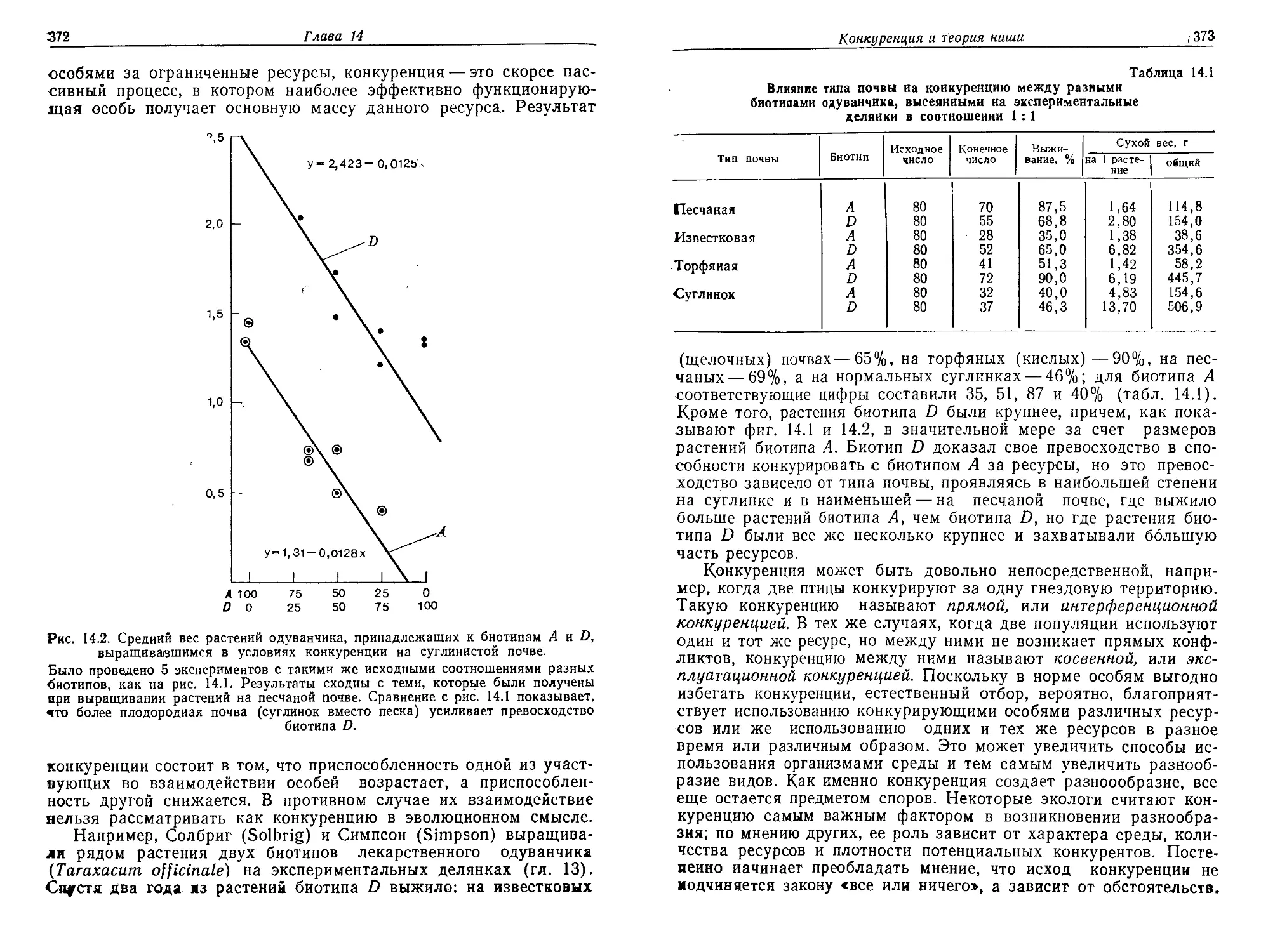
собных воспроизводить себе подобные соединения путем дуплика-
ции. Дупликация, или способность к размножению, представляет
собой отличительное свойство жизни. Когда появилось вещество,
обладавшее способностью к самовоспроизведению, каким бы про-
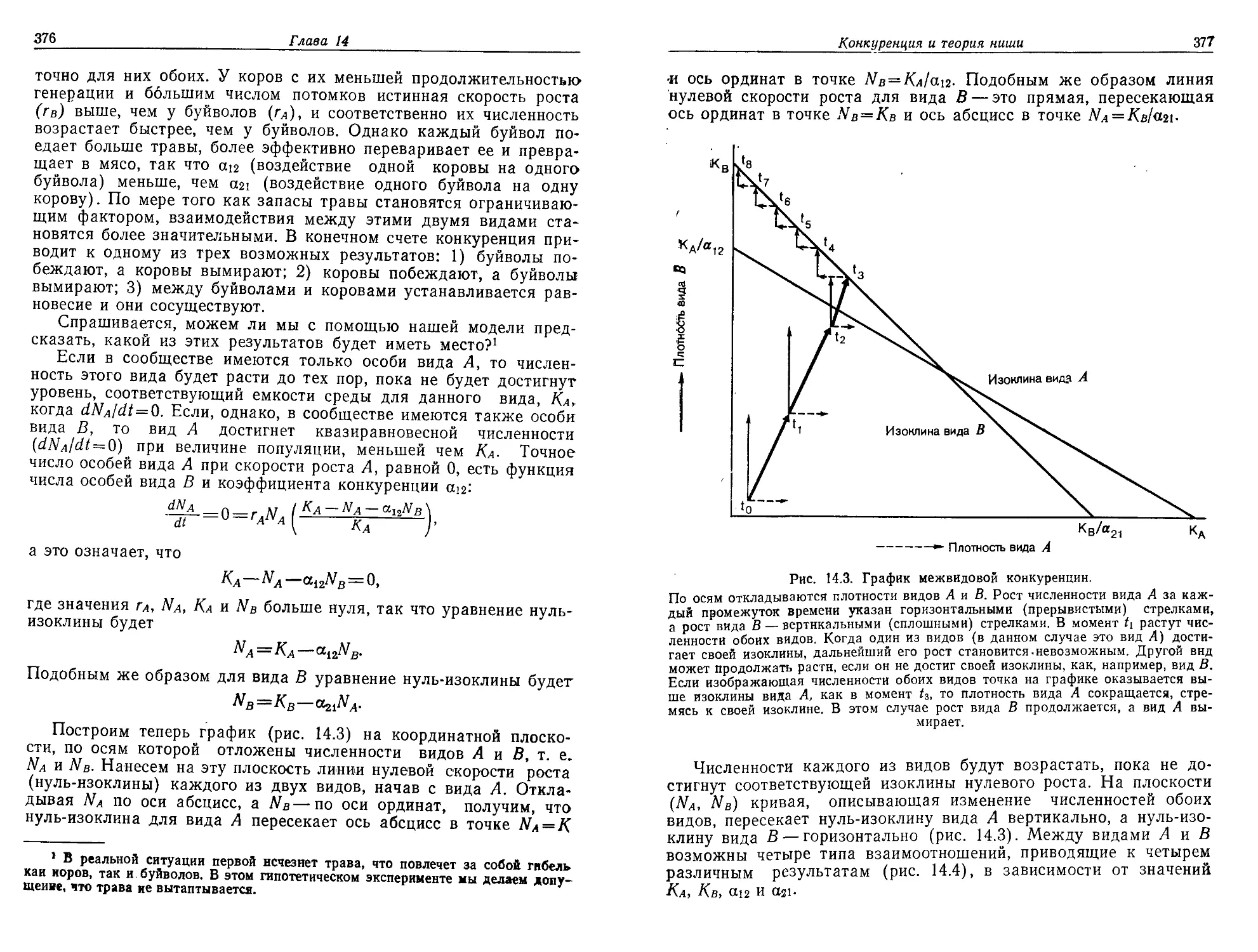
стым это вещество не было, возникло то, что мы называем жизнью.
Начиная с этого времени в результате процесса, который мы на-
зываем эволюцией путем естественного отбора, простые организмы
постепенно становились все более сложными, приобретая черты и
функции, присущие современным растениям и животным.
Природа всех конкр>етных процессов, приведших к возникнове-
нию жизни, неизвестна. Тем не менее было высказано несколько
правдоподобных предп сложений о возможных путях этого • про-
цесса. Мы не располагаем методами, позволяющими точно выяс-
нить, какими были условия на Земле в то время, поэтому в этих
вопросах очень большое место занимают догадки. Кроме того,
ввиду изменившихся условий, существующих на нашей планете
в настоящее время, и медленности рассматриваемого процесса,
экспериментальная проверка предположений о путях, приведших
к возникновению живого вещества, невозможна. Но несмотря на
все эти затруднения, в 1953 г. Стэнли Л. Миллер (Stanley L. Mil-
ler) из Чикагского университета провел простой эксперимент,
подтверждающий правильность основной идеи. В этом экспери-
менте в герметически закрытом приборе циркулировала смесь га-
зов, которые, по общему мнению, наиболее вероятно содержались
в ранней атмосфере Земли: метан, аммиак, водород и водяные
Эволюция, популяции и естественный отбор
13
пары. В определенном месте через эту смесь пропускали электри*
ческий разр'яд, имитирующий молнию (рис. 1.1). Миллер проводил
этот эксперимент в течение недели, а затем подверг анализу об-
разовавшиеся продукты и обнаружил среди них небольшие ко-
личества аминокислот — основных компонентов белков и одних
Рис. 1.1. Схема эксперимента, проведенного С. Л. Миллером.
По этой замкнутой системе в течение примерно недели циркулировала смесь ме-
тана, аммиака, водорода и водяных паров. В каждом цикле через газовую смесь
пропускали электрический разряд. Аминокислоты накапливались в нижией части
прибора. (О. Т. Solbrig, 1966. Evolution and Systematics, New York, Macmillan.)
из главных органических веществ, содержащихся в любом живом
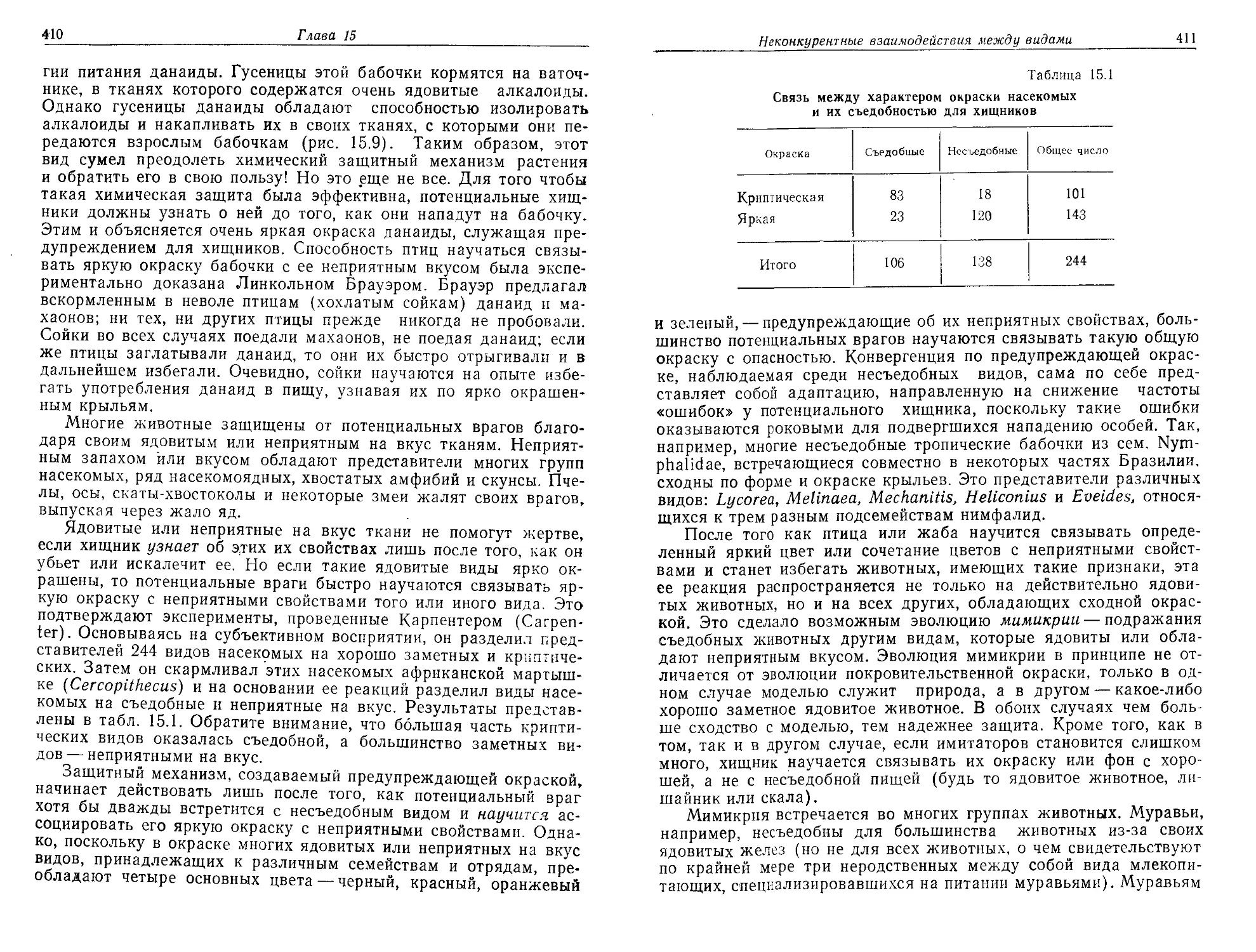
существе.
Одна из наиболее привлекательных особенностей рассматри-
ваемой гипотезы происхождения жизни состоит в том, что, в соот-
ветствии с ней, после того как на Земле создались соответствую-
щие условия, образование сложных органических веществ и жиз-
ни стало неизбежным. Вероятность каждого из необходимых для
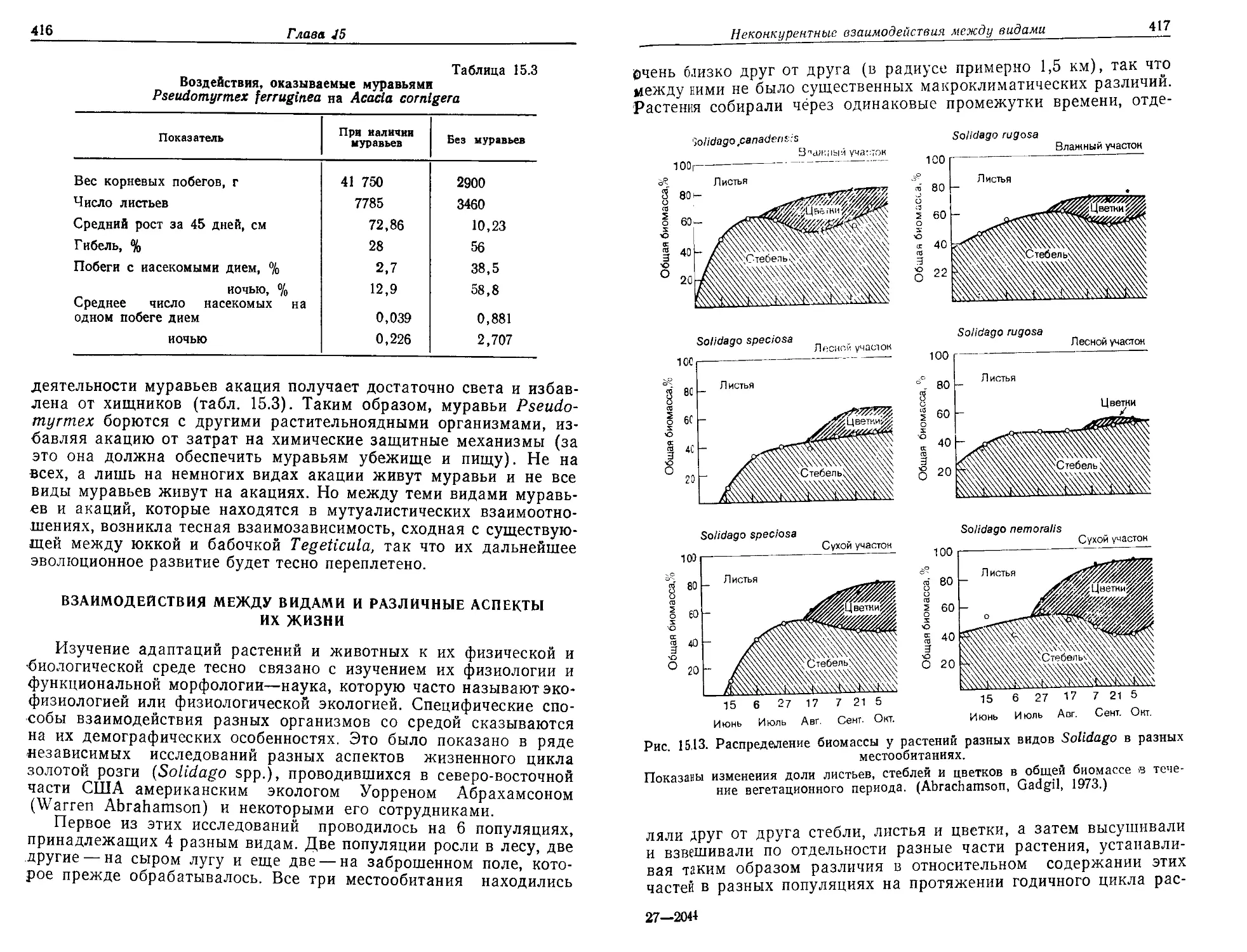
этого событий в единицу времени была крайне мала, однако сум-
марное время было колоссально велико. Кроме того, положение
Земли в Солнечной системе, вероятно, обусловило неизбежность
возникновения соответствующих условий. Но Солнце — не един-
ственная в своем роде звезда во Вселенной; только в нашей Га»
14
Глава 1
лактике насчитываются миллионы таких звезд, а число их во всем
космосе исчисляется миллиардами. По данным покойного астро-
нома Харлоу Шепли (Harlow Shapley) из Гарвардского универ-
ситета, из примерно 1020 звезд, существующих во Вселенной, 20%
по величине, яркости и химическому составу идентичны нашему
Солнцу. Если хотя бы 1% этих солнц имеет планетные системы
и если хотя бы 1 % планет расположен по отношению к своим
солнцам так, как Земля расположена относительно нашего Солнца,
то можно считать вероятным наличие той или иной формы жизни
по крайней мере на 2-Ю15 планетах. А в таком случае жизнь мо-
жет оказаться в общем-то довольно распространенным явлением!
Кроме того, Шепли полагал, что условия, в которых возможно
возникновение жизни, существуют также и на потухших звездах,
имеющих твердую кору и горячие недра. Читатель должен ясно
понимать, что «жизнь» определяется здесь как некая самовоспро-
изводящаяся система, не обязательно представленная такими фор-
мами, которые населяют Землю.
КОНЦЕПЦИЯ ЕСТЕСТВЕННОГО ОТБОРА
Эволюцию от атомов и молекул к простым, а затем и сложным
соединениям и далее к еще более сложным, способным к само-
воспроизведению, называют химической эволюцией, чтобы отли-
чать ее от эволюции организмов, называемой органической эволю-
цией. Различие между этими двумя типами эволюции обусловлено
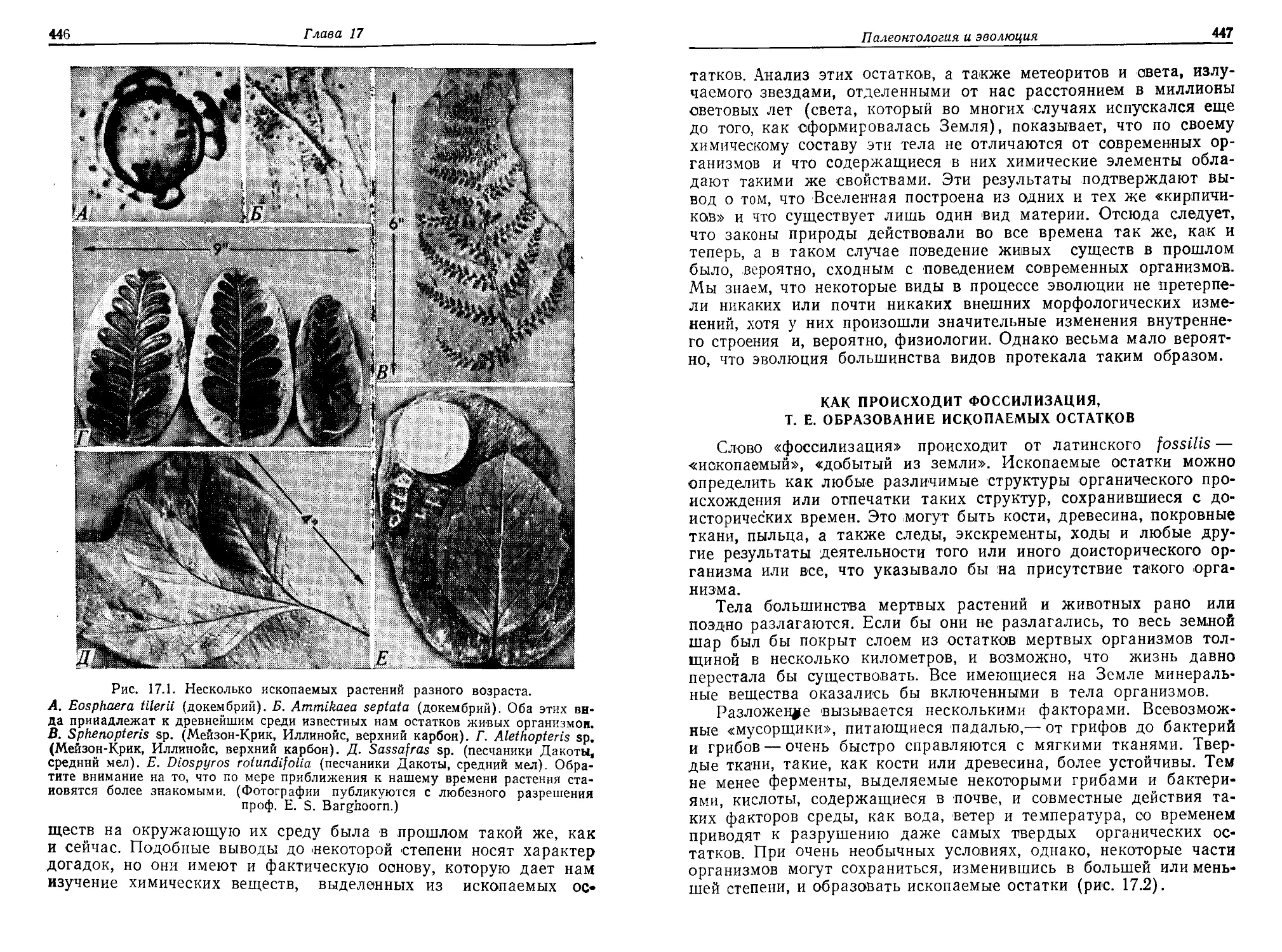
основным свойством живой материи — ее способностью к само-
воспроизведению. На последовательных стадиях, приведших к
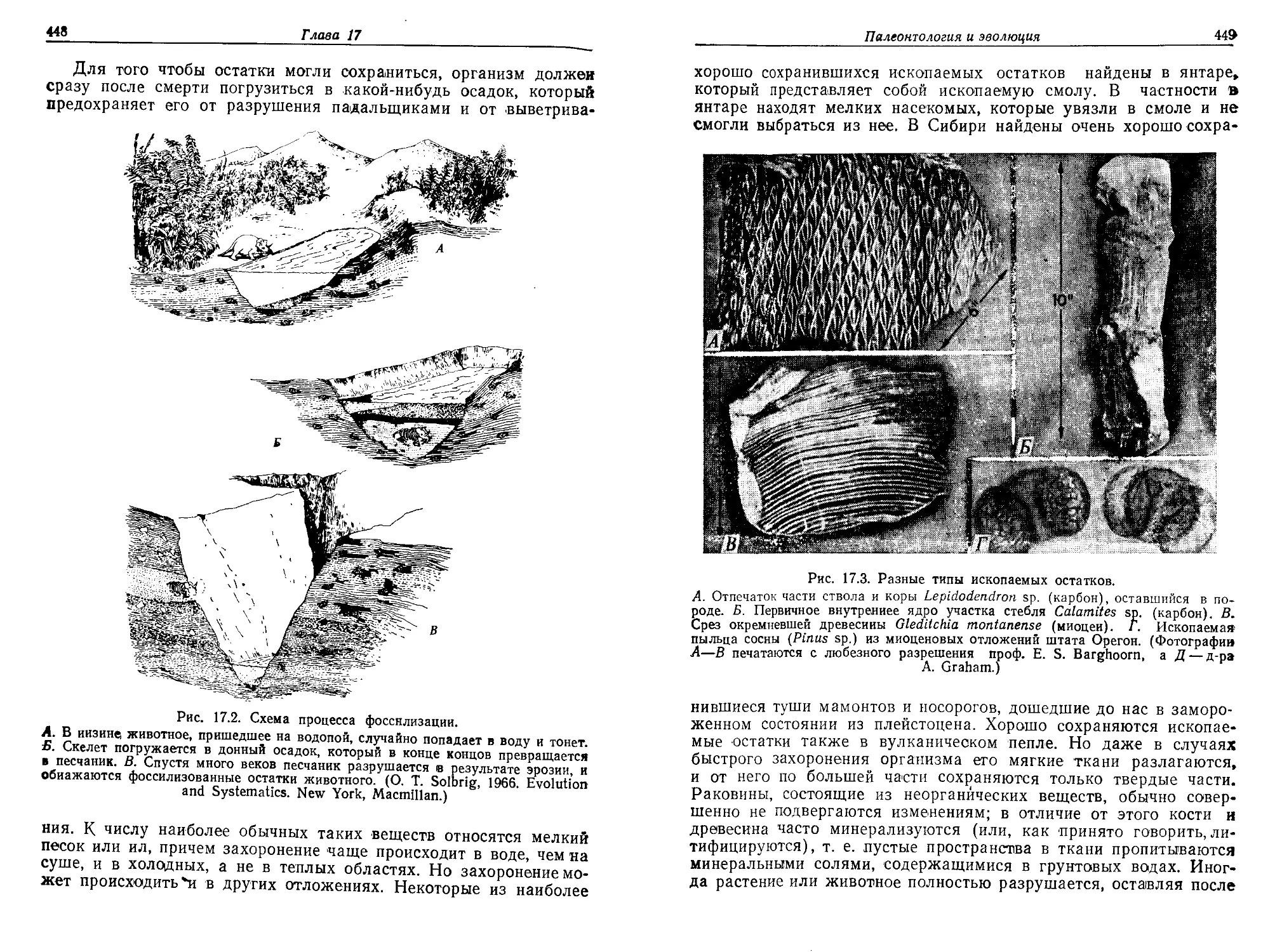
возникновению жизни, происходило образование все более и более
сложных веществ во все больших количествах по мере связывания
все больших количеств солнечной энергии и увеличения числа
химических веществ, способных вступать в реакции друг с другом.
Эти вещества обладали также способностью к росту, сходному с
ростом кристалла, и могли случайным образом распадаться на
части. Однако подобный процесс еще нельзя считать размноже-
нием. В тот момент, когда такое вещество —по всей вероятности,
нуклеиновая кислота, сходная с ДНК, содержащейся в хромосо-
мах современных растений и животных, или идентичная ей, — обре-
ло способность к самоудвоению, оно смогло начать образовывать
все новые и новые количества подобного себе вещества за счет
других, возможно, более простых соединений. Иными словами, оно
могло расти и репродуцироваться. Если два химических вещества
или два «штамма» одного и того же вещества обладают одинако-
выми свойствами, то из них, которое продуцирует большее число
выживающих «потомков», станет более обильным. В этом состоит
сущность процесса, называемого естественным отбором и прису-
щего исключительно органической эволюции.
Эволюция, популяции и естественный отбор
15
путем
Концепция естественного отбора была впервые доложена Чарл-
зом Дарвином и Альфредом Уоллесом на одном и том же засе-
дании Линнеевского общества в Лондоне 1 июля 1858 г. Годом
позже Дарвин (рис. 1.2) развил свои идеи в теперь широко изве-
стной книге «Происхождение видов
Принцип естественного отбора
основан на трех главных наблюде-
ниях и на двух сделанных из них
выводах. Во-первых, Дарвин обра-
тил внимание на то, что все орга-
низмы потенциально способны к
увеличению своей численности в
геометрической прогрессии. Во-вто-
рых, он заметил, что численность
организмов каждого вида обычно
остается постоянной из поколения
в поколение. Из этих двух наблю-
дений Дарвин сделал вывод, что
должна происходить «борьба за су-
ществование». Поскольку потомков
производится больше, чем может
выжить, они должны конкуриро-
вать между собой за ресурсы, необ-
ходимые для выживания. Дарвин
обратил также внимание на то, что
ни у одного вида нельзя найти двух
особей, которые были бы совершен-
естественного отбора».
Рис.
фии,
1.2. Чарлз Дарвин (с фотогра-
находящейся в коллекции
де Кандолля в Женеве).
но идентичны друг другу, или, вы-
ражаясь иначе, что изменчивость представляет собой одно из яв-
лений природы. Если изменчивость наблюдается среди взрослых
особей, то она должна наблюдаться и среди молодых. Когда мо-
лодые особи конкурируют за ресурсы, то выживать будут пре-
имущественно те из них, которые обладают признаками, способ-
ствующими победе в конкуренции (благоприятные вариации), а
гибнуть в большей степени особи с неблагоприятными вариация-
ми. Это явление было названо дифференциальной смертностью.
В результате в каждом следующем поколении популяция будет
содержать относительно большую долю особей с благоприятными
вариациями, чем в предыдущем. Многократное повторение этого
процесса с течением времени приводит к элиминации неблаго-
приятных вариаций, и к отбору благоприятных вариаций, обеспе-
чивающих победу в борьбе за существование (рис. 1.3).
Один из признаков, по которым различаются особи, — это число
производимых ими потомков. Вероятность того, что какой-либо
потомок данной родительской пары доживет до репродуктивного
возраста при прочих равных условиях, тем больше, чем большее
Второе поколение
Новорож- .Полово-
денные зрелые
Родительские
особи -
Третье поколение
Новорожденные Половозрелые
Двадцать Восемь
60%
гибнет
Десять
Четыре
О о о о 40% О О О О
о о гибнет О О
О О
Десять Шесть
О О О О О О О О
О О О О О О О О
О О О О О 40% 0-0 О
О О О О О гибнет О О О
О О О О О О О О
О О О О о О О о
Тридцать Восемнадцать
© 0 0 С
© О 1о% © ®
® © гибнет © ©
О 0 О О
0 0 ©
©©©•©© ©0©©®
© © © © © © © © © 0
0 Э О О О О О О О О
о © © о о 10% © © © © ©
© ® © ® © гибнет ® о © © ©
9 0 0 0 0 © ® © © ©
© ® '© © О © © © © ©
© ® © © © ©ООО©
© ® © © ©
Десять Девять Сорок пять
Рис. 1.3. Дифференциальное выживание.
Сорок
Три гипотетические пары кроликов различаются только по окраске. Каждая пара
приносит по 10 крольчат. Черные и белые кролики более заметны, чем серые, а по-
этому соответственно 60 и 40% их потомков гибнет до достижения половозрело-
сти, тогда как у серых кроликов гибнет только 10% крольчат. По прошествии трех
поколений черные кролики, составлявшие 33’/з% исходной популяции, составляют
всего лишь 16/250=6,4%, несмотря иа увеличение их абсолютного-числа. Относи-
тельная численность белых кроликов также снизилась до 55/250= 21,6%. Числен-
ность же серых кроликов, напротив, возросла с ЗЗ’/з до 180/250= 72%.
Эволюция, популяции и естественный отбор 17
Четвертое поколение
Новорожденные • • • • • • • Половозрелые
• • • • • • •
• • • • • • •
• • • • • 60% _ • •
• • • • • гибнет • •
• • • • • • •
© • • • • • •
• • • • э • •
Сорок Шестнадцать
О О о о о о о о о о О о О о о о
О О о о о о о о о о О о о о о о
О о о о о о о. о о о О о о о о о
О о о о о о о о о о о о о о о о
О о о о о о о о о о 40% о о о о о о
о о о о о о о о о о гибнет о о о о о о
о о о о о о о о о о о о о о о о
о о о о о о о о о о о о о о о о
о о о О' о о о о
о о о о о о о о
Девяносто Пятьдесят четыре
© © © © © о © © © © © о © 0 © ® © о о
© © ©. о © © © © © © © © © о о © © © ©
© © © © © © © о © © © о о о 0 о о 0 ©
0 © © © © © ® © о © © © © © © © © © ©
© © © © © © о о © © © © © G о о о о © ©
© © © © © © © © © © о © © © © © о © о
© © © © © © © © © © о © 0 о 0 © © © ©
© © © © © © © © о © © © © © 0 © © © © <s>
© © © © © © о © © 0 э. О © © 0 © о о
о © © © © © © © 0 © 10% © © © 0 © © о © ©
© о © © © © © © © © гибнет © © о © о 0 © © © о
© о © © © © © © © © © о 0 о © © © © © ©
© © © © © © © © © © © © о о © © © © © ©
© © © © О © © © © © © © © © © о © 0 © ©
о © о о © © © © © © © © © © © © © © © ©
© о о © © © © © © © © © © © © © © © © 0
© © © © © © о © © © о о о © ©
о © 0 © © © о © © © © © о о
Q © © © © © © © © © © © 'о о ©
© © о © .© о © е о © © © о © ©
Двести Сто восемьдесят
е—2С44
18
Глава 1
Родительские особи Второе поколение Новорожденные Половозрелые Третье поколение
Новорожденные Половозрелые 9 9 О 9
• в 9 9 9 9 9 9 9 9
9 9 9 • • • _ - Л 9 9 сева 9 9 9 9 9 9 9 9
в • • 60% _ • • * • 9 9 9 9 60% 9 9
~ ~ ~ гибнет Л Л Л • 9 9 9 9 гибнет ® *
Л Л Л • • • • • 9 9
• 9 9 9 9 9 О
• • • • • 9 9
Пятнадцать Шесть Сорок пять Восемнадцать
о о о о о о о о о о о о о
о о ООО
о о о о
о 40% Г О О о о ооо 40% о о о
о о гибнет о о о о ООО гибнет о о о
о О' о о ООО о о о
о о ООО о о о
Десять
Шесть .
Тридцать
Восемнадцать
О' © о о о © © © о © о © 0.0 ®
о о © © о
о о о © о о о о о © о ООО
° ° 10% ® 0 © © о о © 10% @ о ООО
о © — о © © о о © о - о о ООО
© © ™бнет о © © о © О о гибнет © о ООО
о о © © о о о © о о О О О'
© о © о © о © © о ©
© о © © ©
Десять
Девять
Сорок пять
Сорок
Рис. 1.4. Дифференциальное размножение.
Допустим, что черные кролики (см. рис. 1.3) производят в результате каждого
спаривания ие 10, а 15 крольчат и что все прочие условия сохраняются без изме-
нений. В таком случае через три поколения численность черных кроликов в популя-
ции составит 54/288=18,75% (в отличие от 6,4% в случае, изображенном на рис.
1.3), численность белых—18,75% и численность серых — 180/288—62,5%.
Эволюция, популяции и естественный отбор
19
Четвертое поколение
Половозрелые
• • • Ф
Пятьдесят четыре
ООО о о о о о о о о о ООО ООО ООО ООО
о о о о о ООО
о о о о о о о ООО ООО ООО
о о о о о о о ООО ООО ООО
о о о о о о о ООО 40% ООО ООО
о о о о о о о ООО гибнет ~ о о о ООО
о о о о о о о ООО ООО ООО
о о о о о о о ООО ООО ООО
о о о о о ООО
о о о о о ООО
Девяносто Пятьдесят четыре
о © о © © © о О О © 0 © 0 О © 0 © © ©
о о © о о о о ООО © о © о ООО®©
о О о о 0 о © ООО 0 © о 0 О О О О о
© о 0 о © © © © © о 0 о о о о о о о о
© с о © © о о О 0 © о о о о О 0 о о о
о о 0 О 0 © о ООО О 0. 0 о ООО©©
© 0 © © о о о ООО о о о о о © о 0 ©
© © 0 0 © о о ООО о о © о ©ООО©
© 0 0 О 0 0 о © 0'0 о о О 0 О О О о о
о © 0 © о © о ООО 10% t о о О О О О О О О
о © о о о о ООО гибнет о о о о © © О О ©
© 0 0 о о о о ООО ©ООО о о © о о
© © о о о О 0 ООО О 0 о о о о о о о
о 0 о 0 0 о о ООО о с о о ©ООО©
0 © 0 О © о © GO © О О © 0 о о о о о
о о 0 © о © © ©о о о о о о о © о о о
© о о 0 © 0 о ООО 0 0 0-0 о
0 о © © © о о ООО 0 о © 0 о
© 0 © О о о © о о © © о о о О
0 © © 0 О о © © © 0 © о © о о
Двести Сто восемьдесят
0@©©©©<0©©©©©(Э©©Ф
ЙО
Глава 1
число потомков произведет эта пара. Поэтому в популяции, со-
стоящей из особей, сходных во всех других отношениях, особи,
производящие наибольшее число потомков, будут иметь больше
шансов на то, что некоторые из их потомков доживут до взрослого
состояния. Этот процесс, назы-
ваемый дифференциальным раз-
множением, вместе с дифферен-
циальной смертностью лежит в
основе естественного отбора
(рис. 1.4).
Какие же признаки приводят
к повышению выживаемости по-
томков и тем самым оказывают
влияние на дифференциальную
смертность? Таких признаков,
как это уже ясно понял Дарвин,
много и они чрезвычайно разно-
образны. В сущности отбор бу-
дет благоприятствовать сохране-
нию любого свойства, повышаю-
щего вероятность выживания
данной особи и ее (или его) по-
томков. Любой унаследованный
признак, увеличивающий способ-
ность обладающей им особи вы-
носить суровые условия среды,
называется адаптивным. Адап-
тивные признаки помогают орга-
Рис. 1.5. Грегор Мендель (Dorfler, Во- низму справляться с неблаго-
taniker Portrats, 1907). приятными условиями среды, до-
бывать больше пищи и притом
лучшего качества, избегать хищников и тому подобное. Эти приз-
наки обеспечивают выживание большего числа потомков. Адап-
тивные признаки могут быть морфологическими, как, например,
наличие густого меха и подкожного жирового слоя у белых мед-
ведей, помогающих им переносить жестокие морозы. Они могут
быть физиологическими, как, например, у яблони, которой для
хорошего цветения и плодоношения необходимо подвергнуться
действию низких температур — адаптация к тому, чтобы цветение
не могло начаться осенью, во время «бабьего лета». Адаптивными
могут стать и поведенческие признаки, как, например, сложные и
чрезвычайно специфические брачные церемонии у птиц, которые
гарантируют, что спаривание произойдет только между особями,
принадлежащими к одному и тому же виду, предотвращая тем
самым возникновение плохо приспособленных гибридов. Наконец,
очень важная адаптивная черта — это способность производить
как можно больше потомков, но не в ущерб их способности к вы-
Эволюция, популяции и естественный отбор 21
живанию. Все главные признаки растений и животных несомнен-
но адаптивны или же представляют собой проявления признаков,
бывших адаптивными прежде. Некогда считалось, что всякий да-
же самый незначительный наследуемый фенотипический признак
живых организмов носит приспособительный характер, однако,
как полагают в настоящее время, это, по-видимому, не так.
Самым слабым местом в дарвиновской теории естественного
отбора было отсутствие адекватного объяснения механизма на-
следственности. Поскольку естественный отбор не мог бы рабо-
тать, если бы потомки не наследовали признаки своих родителей,
действие естественного отбора определяется механизмом наслед-
ственной передачи этих признаков. Теория наследственности, обще-
принятая в настоящее время, была впервые предложена Грегором
Менделем (рис. 1.5) в 1865 г., но приобрела широкую известность
только в начале XX в. Проводя эксперименты с разными сортами
гороха, Мендель получил данные, позволяющие предположить, что
наследование обусловлено некими частицами, передаваемыми от
родителей потомкам. Теперь мы называем эти частицы генами.
Законы, которым подчиняется передача генов от одного поколения
другому, рассматриваются в гл. 4. Идея корпускулярной наслед-
ственности имеет огромное значение для понимания того, каким
образом естественный отбор действует в популяциях. Все выте-
кающие из этой идеи следствия подробно описаны в гл. 5—9.
эволюция
Эволюцию можно рассматривать как изменения любого свой-
ства данной популяции с течением времени. В некоем общефи-
лософском смысле в этом и заключается суть эволюции. Однако
для исследователя, желающего количественно оценить эволюцион-
ное изменение и изучить механизмы, с помощью которых оно про-
исходит, такое определение слишком туманно. Безусловно, не
всякое изменение есть эволюция, а, кроме того, приведенное опре-
деление не дает никакого представления о масштабах времени,
в котором она происходит.
В одном из весьма популярных определений эволюция рассмат-
ривается как изменение частоты отдельных генов из поколения
в поколение. Это очень точное определение. Теоретически частоту
генов можно измерить; можно также регистрировать изменения
частоты каждого гена с течением времени. Однако и это опреде-
ление порождает ряд проблем. Хотя частоты генов в некоторых
случаях удается установить, в настоящее время такое измерение
возможно лишь для относительно небольшого числа генов, а спо-
собов, которые позволяли бы решить, что именно эти гены обу-
словливают изменения, повышающие выживаемость рассматривае-
мых организмов, у нас нет.
Третье определение пытается преодолеть трудности, порождае-
мые двумя первыми, определяя эволюцию как изменение адаптив-
Глава 1
них признаков популяции и лежащих в их основе генов с течением
времени. Хотя это последнее определение кажется более точным,
поскольку в нем рассматриваются только те изменения, которые
повышают выживаемость популяции, оно не содержит указаний
на то, о каких именно изменениях идет речь. А поэтому такое
определение неоперационально и создает порочный круг.
Разнообразие существующих определений свидетельствует о
том, как трудно оценить эволюцию количественно и предложить
меру эволюционных изменений. Большинство ученых согласны с
тем, что эволюция представляет собой изменение, происходящее
во времени, и что это изменение должно быть связано с генетикой
организмов. Однако эволюция — процесс очень сложный, слагаю-
щийся из многих явлений. Поскольку далеко не все эти явления
достаточно хорошо поняты, дать безукоризненно строгое определе-
ние эволюции пока еще. невозможно. Перейдем теперь к рассмот-
рению некоторых факторов, имеющих важное значение для эво-
люции.
Первый и важнейший из них — это механизм наследственности.
Способ, которым гены определяют проявление признаков, и спо-
соб передачи генов от родителей потомкам формируют эволюцион-
ную картину в целом. В свою очередь на механизмы наследствен-
ности, а тем самым и на эволюцию значительное влияние оказы-
вают организация генов в хромосомах и поведение хромосом во
время клеточного деления. А поэтому эволюцию нередко опреде-
ляют как изменение частоты генов в популяции.
Хотя, как известно, эволюция может происходить и в отсутст-
вие полового процесса, это нельзя считать нормой. Половой про-
цесс представляет собой механизм, объединяющий генетический
материал отдельных особей и создающий новые, ранее неизвест-
ные сочетания родительских генов. В результате создается огром-
ный запас изменчивости, а это, по-видимому, дает такие большие
преимущества, что половой процесс стал преобладающим способом
размножения у всех растений и животных. Половое размножение,
очевидно, возникло на очень ранней стадии эволюции организ-
мов1.
В отсутствие полового процесса и скрещивания виды в той
форме, в какой они сейчас нам известны, не могли бы существо-
вать. Однако столь же важную роль в эволюции и, в частности,
в увеличении числа видов играло развитие преград — географиче-
ских, экологических, поведенческих или генетических, — препят-
ствующих свободному обмену генами. Первые очень простые ор-
ганизмы, возможно, обладали способностью обмениваться генами
с другими живыми существами, находящимися на том же уровне
организации, однако обмен генами между различными современ-
1 Этот вопрос подробно рассматривается в монографии Мэйнарда Смита
«Эволюция полового размножения», М., «Мир», 1981. — Прим. ред.
Эволюция, популяции и естественный отбор 23
ными формами с их очень специализированными и сложными пу-
тями индивидуального развития невозможен. В тех случаях, когда
такой обмен происходит, это приводит к гибели потомков или в
лучшем случае к их стерильности. Селективная ценность преград,
препятствующих обмену генами в таких случаях, очевидна. Раз-
мышляя и рассуждая об эволюции, мы почти всегда имеем в виду
эволюцию видов. Это ясно свидетельствует о важной роли в эво-
люции полового процесса и изоляции.
В несколько ином аспекте можно также говорить о том, что
эволюция отчасти определяется внешними условиями. Дифферен-
циальное выживание всегда частично обусловлено способностью
адаптироваться к среде, в частности к физической. Химическая
эволюция могла начаться только после того, как наша планета
из первоначального «огненного шара» превратилась в тело, где
могло происходить накопление воды. Органическая эволюция,
приведшая к появлению огромного числа различных организмов,
частично представляет собой следствие адаптации этих организ-
мов к бесконечному разнообразию условий, имеющихся на земле.
ОГРАНИЧЕНИЯ, СОПРЯЖЕННЫЕ С ЭВОЛЮЦИЕЙ
Живые организмы — это сложные системы, которые потребля-
ют энергию п простые химические соединения, используя их на
построение собственного тела и производство потомства. Однако
количество необходимых организмам энергии и химических ве-
ществ не безгранично и, кроме того, они распределены на земном
шаре неравномерно.
Организмы получают энергию из двух источников. Одним ис-
точником служит Солнце, излучающее доступную организмам
энергию в форме света. Другой источник энергии — это опреде-
ленные богатые химической энергией неорганические вещества,
такие, как нитраты и сульфаты, но главным образом — богатые
энергией органические соединения, содержащиеся в телах живых
и мертвых организмов. Неорганические молекулы, необходимые
организмам в качестве строительных блоков, также распределены
неравномерно: они содержатся в низких концентрациях в виде
ионов в водах морей и рек и в почвенных растворах, а в высоких
концентрациях — в составе живых организмов.
Энергию света зеленые растения поглощают при фотосинтезе,
связывая ее в богатых энергией молекулах углеводов. Однако
растения способны использовать менее 5% падающей лучистой
энергии; остальная энергия переизлучается или рассеивается в
форме тепла. С помощью химической энергии, содержащейся в са-
харах, образованных в процессе фотосинтеза, растение строит свои
ткани из атомов и молекул, которые оно получает из воды (если
это водное растение) или из почвенного раствора. Для поглощения
света необходимы специализированные структуры («солнечные
24
Глава 1
батареи»), которые мы называем листьями (а иногда это зеленые
стебли). Листья должны располагаться таким образом, чтобы не
затенять друг друга; они должны быть достаточно прочными, что-
РАСТЕНИЕ
- Вещества (Н,0,
биогенные элементы)
'ПЛОТОЯДНОЕ
ЖИВОТНОЕ
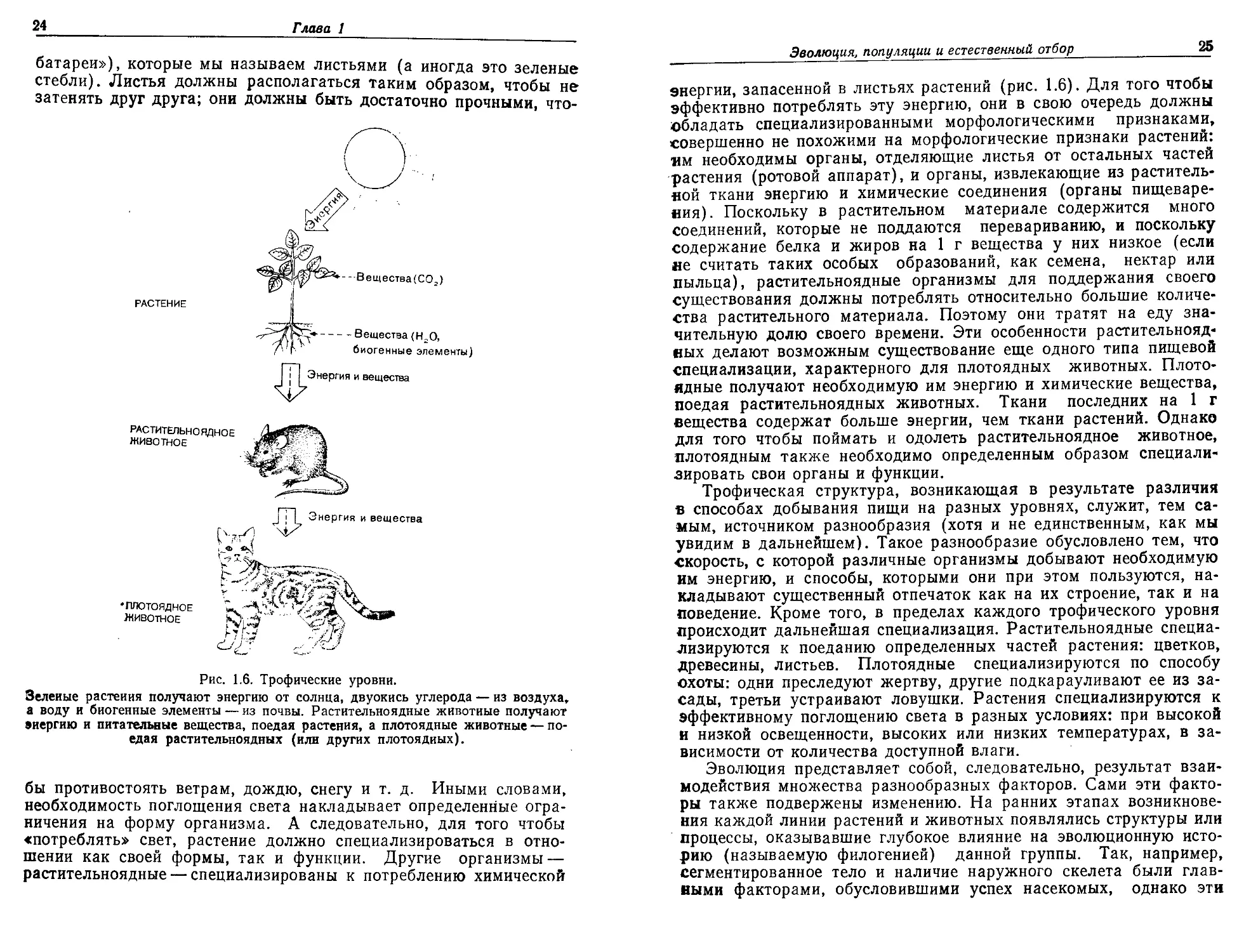
Рис. 1.6. Трофические уровни.
Зеленые растения получают энергию от солнца, двуокись углерода — из воздуха,
а воду и биогенные элементы — из почвы. Растительноядные животные получают
энергию и питательные вещества, поедая растения, а плотоядные животные — по-
едая растительноядных (или других плотоядных).
бы противостоять ветрам, дождю, снегу и т. д. Иными словами,
необходимость поглощения света накладывает определенные огра-
ничения на форму организма. А следовательно, для того чтобы
«потреблять» свет, растение должно специализироваться в отно-
шении как своей формы, так и функции. Другие организмы —
растительноядные — специализированы к потреблению химической
Эволюция, популяции и естественный отбор 25
энергии, запасенной в листьях растений (рис. 1.6). Для того чтобы
эффективно потреблять эту энергию, они в свою очередь должны
обладать специализированными морфологическими признаками,
совершенно не похожими на морфологические признаки растений:
мм необходимы органы, отделяющие листья от остальных частей
растения (ротовой аппарат), и органы, извлекающие из раститель-
ной ткани энергию и химические соединения (органы пищеваре-
ния). Поскольку в растительном материале содержится много
соединений, которые не поддаются перевариванию, и поскольку
содержание белка и жиров на 1 г вещества у них низкое (если
не считать таких особых образований, как семена, нектар или
пыльца), растительноядные организмы для поддержания своего
существования должны потреблять относительно большие количе-
ства растительного материала. Поэтому они тратят на еду зна-
чительную долю своего времени. Эти особенности растительнояд-
ных делают возможным существование еще одного типа пищевой
специализации, характерного для плотоядных животных. Плото-
ядные получают необходимую им энергию и химические вещества,
поедая растительноядных животных. Ткани последних на 1 г
вещества содержат больше энергии, чем ткани растений. Однако
для того чтобы поймать и одолеть растительноядное животное,
плотоядным также необходимо определенным образом специали-
зировать свои органы и функции.
Трофическая структура, возникающая в результате различия
в способах добывания пищи на разных уровнях, служит, тем са-
мым, источником разнообразия (хотя и не единственным, как мы
увидим в дальнейшем). Такое разнообразие обусловлено тем, что
скорость, с которой различные организмы добывают необходимую
им энергию, и способы, которыми они при этом пользуются, на-
кладывают существенный отпечаток как на их строение, так и на
поведение. Кроме того, в пределах каждого трофического уровня
происходит дальнейшая специализация. Растительноядные специа-
лизируются к поеданию определенных частей растения: цветков,
древесины, листьев. Плотоядные специализируются по способу
охоты: одни преследуют жертву, другие подкарауливают ее из за-
сады, третьи устраивают ловушки. Растения специализируются к
эффективному поглощению света в разных условиях: при высокой
и низкой освещенности, высоких или низких температурах, в за-
висимости от количества доступной влаги.
Эволюция представляет собой, следовательно, результат взаи-
модействия множества разнообразных факторов. Сами эти факто-
ры также подвержены изменению. На ранних этапах возникнове-
ния каждой линии растений и животных появлялись структуры или
процессы, оказывавшие глубокое влияние на эволюционную исто-
рию (называемую филогенией) данной группы. Так, например,
сегментированное тело и наличие наружного скелета были глав-
ными факторами, обусловившими успех насекомых, однако эти
26
Глава 1
факторы в свою очередь ограничили размеры и образ жизни чле-
нов этой группы.
Именно многогранность эволюционных процессов делает их
изучение столь увлекательным. А наши знания еще далеко не
полны. Многое предстоит открыть, многое изучить. Применение
такой новой техники, как быстродействующие вычислительные
машины, или методов сравнительной биохимии для выяснения пу-
тей развития или эволюционных взаимоотношений на химическом
уровне открывает широчайшие перспективы.
В последующих главах подробно описываются некоторые ос-
новные факторы, обусловливающие эволюцию, анализируется их
роль и относительная важность и исследуется их становление.
Мы попытаемся также связать эти факторы с проблемами эколо-
гии. Везде, где только возможно, мы старались показать, в какой
мере методология и средства научного исследования, которыми мы
пользуемся, накладывают ограничения на возможности познания
природы и в какой мере они ему способствуют. Однако прежде
всего следует развеять некоторые широко распространенные оши-
бочные представления относительно эволюции.
ОШИБОЧНЫЕ ПРЕДСТАВЛЕНИЯ ОБ ЭВОЛЮЦИИ
Ошибочные концепции относительно эволюции довольно мно-
гочисленны. Внимательное чтение этой книги должно рассеять
большую часть предубеждений такого рода, однако существуют
два представления, настолько прочно обосновавшиеся в умах ши-
рокой публики, что их следует рассмотреть особо. Первое из них —
отождествление эволюции с борьбой. Выражение «выживание
наиболее приспособленных» принадлежит Дарвину, но из его про-
изведений ясно следует, что он вкладывал в него не тот смысл,
который ему часто приписывают. Популяризация идеи о борьбе
принадлежит английскому философу Герберту Спенсеру (1820—
1903), который использовал ее в своей концепции социальной
эволюции. В ту историческую эпоху идея о борьбе за существо-
вание между индивидуумами, в которой наиболее приспособлен-
ный побеждает и пользуется плодами победы, была привлекатель-
ной для известных социальных слоев. Вторая половина XIX в. со-
ответствует расцвету идей свободного предпринимательства и
вседозволенности, эпохе наивысшего расцвета Британской импе-
рии. Проповедь Спенсера, якобы основывавшаяся на достижениях
естественных наук, служила кажущимся научным (а для некото-
рых— и нравственным) оправданием всей царившей в мире не-
справедливости. Перенося закономерности органической эволюции
в социальную сферу, Спенсер неверно истолковал основную идею
Дарвина — дифференциальное выживание и производство потом-
ства. Он заменил ее одним из элементов теории естественного от-
бора — дифференциальной смертностью, обусловленной большей
Эволюция, популяции и естественный отбор 27
•приспособленностью к среде одних особей по сравнению с други-
ми, и выдвинул идею прямой конкуренции как причины диффе-
ренциальной смертности. В такой форме эти идеи целиком при-
надлежат самому Спенсеру и не имеют под собой никакой науч-
ной основы. Мы должны постоянно помнить, что под «наиболее
приспособленными» Дарвин имел в виду особей, оставляющих
наибольшее число выживающих потомков. Это могут быть сравни-
тельно слабые особи, и нередко так оно и бывает в действитель-
ности.
Спустя почти 100 лет после Дарвина советский генетик Лысен-
ко во имя дарвинизма отверг ряд положений современной генетики
и цитологии и принял теорию наследования приобретенных при-
знаков. В этом случае опять-таки пренебрежение научной стро-
гостью создало путаницу, и хотя все мы хорошо знаем ошибки
и заблуждения Лысенко, мы не всегда так же ясно отдаем себе
отчет в ошибочности взглядов Спенсера.
Эволюция неспособна к предвидению. Одно время полагали,
что эволюции присуща некая «внутренняя направляющая сила»
н, следовательно, эволюция целенаправленна. Эти взгляды носят
название теории «ортогенеза». Эволюционное изменение представ-
ляет собой, однако, результат действия генетических и экологи-
ческих факторов, складывающихся в каждый данный момент в
определенном месте на земном шаре. Природе этих факторов и их
взаимодействию друг с другом посвящена эта книга.
Другой важный момент, который следует усвоить, заключается
в недопустимости давать результатам эволюции какие-либо мо-
рально-этические оценки. Высокоразвитые в эволюционном отно-
шении организмы не стали «лучше» в нравственном отношении; они
просто лучше адаптированы к своей среде обитания по сравнению
со своими вымершими предками. Следует также помнить, что все
современные организмы в силу самого факта своего существова-
ния, по-видимому, приспособлены к занимаемой ими среде весьма
эффективно, независимо от своего положения на филогенетическом
древе. Эволюция столь же беспристрастна, как, полагают, должно
быть беспристрастно правосудие: будут увеличиваться в числе
лишь те организмы, которые производят наибольшее число жиз-
неспособных потомков, безотносительно к тому, «плохими» или
«хорошими» их считают люди.
В нескольких последующих главах закладываются основы, не-
обходимые для понимания эволюции. В гл. 2 рассматривается на-
учный метод как таковой, а в гл. 3 — его приложение к теории
естественного отбора. Первые три главы части II (гл. 4—6) по-
священы основам генетики. Сначала излагаются основные законы
генетики, затем описываются хромосомы — физические структуры,
в которых заключен генетический материал, и, наконец, подробно
рассматриваются главные факторы, обусловливающие генетические
изменения в популяциях. В гл. 7 разбирается роль случайности в
28
Глава 1
эволюции, а в гл. 8 — процесс размножения. Рассмотрение основ
популяционной генетики завершает гл. 9, в которой дан подроб-
ный обзор известных типов генетической изменчивости в популя-
циях, чтобы выяснить, соответствуют ли они сделанным ранее-
предсказаниям.
Часть III посвящена формам эволюции на уровне популяции,,
вида и сообщества. Что такое эволюционирующие единицы и ка-
ким образом генетическая структура, описанная в ч. II, вопло-
щается в особях, популяциях и видах, составляет предмет обсуж-
дения гл. 10. В двух следующих главах рассматривается процесс
видообразования, а в гл. 13 — рост популяций. В гл. 14 и 15 раз-
бирается структура сообществ. Гл. 16 посвящена поведению — его-
сущности, его влиянию на эволюцию, степени, в которой оно пе-
редается по наследству. Завершается книга двумя главами —о ро-
ли палеонтологии и таксономии в понимании эволюции.
ЛИТЕРАТУРА
Anfinsen С. В. 1965. The Molecular Basis of Evolution, New York, Wiley. [Имеется
перевод 1-ro изд.: Анфннсеи К- Молекулярные основы эволюции. — М.: ИЛ.
1962.]
Bell Р. R. 1959. Darwin’s Biological Work, New York, Wiley.
Commoner B. 1964. DNA and chemistry of inheritance, American Scientist, 52,
365—388.
Darwin C. 1964. On the Origin of Species by Means of Natural Selection. Facsimile
of the first (1869) with introduction by E. Mayr, Cambridge, Mass., Harvard
University Press.
Dobzhansky T. 1956. What is an adaptive trait? Am. Nat., 90, 337—347.
Ghiselin M. T. 1969. The Triumph of the Darwinian Method, Berkeley, University
of California Press.
Irvine W. 1959. Apes, Angels and Victorians, New York, Meridian Books.
Lewontin R. C. 1970. The units of selection, Ann. Rev. Ecol. Syst., 1, 1—18.
Miller S. L., Orgel L. E. 1974. The Origins of Life on the Earth, Englewood Cliffs,
N. J., Prentice-Hall.
Глава 2
Научный метод
На протяжении всей своей истории человек наблюдал природу
и пытался постичь ее. Но за несколько последних веков наши1
знания о природе стали во много раз глубже и шире, чем за все
предшествующие тысячелетия. Причиной этого было развитие на-
учного метода, позволившего сделать много достоверных и полез-
ных обобщений. В этой главе мы рассмотрим все те черты, кото-
рыми научный метод отличается от других способов приобретения
знаний.
ПРИОБРЕТЕНИЕ ЗНАНИЙ
Формальная логика изучает возможности установления истин-
ности или ложности суждений, независимо от тех конкретных
объектов, к которым эти суждения относятся. Логика формирует
необходимые условия получения правильных выводов, с тем чтобы
избежать неверных умозаключений. Однако формальной логик»’
недостаточно для приобретения новых фактических знаний в той
или иной конкретной области. Какими же способами мы приобре-
таем знания?
Наши представления в большинстве случаев бывают основаны,
на принятии существующих взглядов или на наших собственных
не вызывающих сомнений допущениях. Большинство из нас время'
от времени сталкиваются с необходимостью отстаивать свои взгля-
ды и убеждения или изменять их. При этом мы прибегаем к раз-
личным методам. Любой метод представляет собой некую про-
цедуру, применяемую для решения определенной группы проб-
лем— в данном случае для отстаивания определенных убеждений.
По мнению американского философа М. Коэна (М. Cohen), чаще-
всего используются следующие четыре метода.
1. Предубежденность. Этот метод обусловлен привычкой или
инерцией. Мы защищаем некое положение просто потому, что мы-
издавна в него верили. Именно к этому методу чаще всего при-
бегают в вопросах, касающихся отношения к различным странам,.,
расам, языкам, спортивным командам и т. п. Совершенно очевид-
но, что таким способом вряд ли можно приобрести новые знания»
Тем не менее очень многие споры между людьми, в том числе №
самые жаркие, касаются утверждений, отстаиваемых этим мето-
дом.
30
Глава 2
2. Ссылка на авторитеты. Ссылка на высокие авторитеты —
другой способ, к которому прибегают люди для защиты своих
взглядов. Эти авторитеты бывают различны в зависимости от
убеждений данного лица и от обсуждаемого предмета: люди цити-
руют Маркса, конституцию, Че Гевару, Красную книжечку Мао,
Библию, Коран, Эми Вандербильт, Винса Ломбарди и т. п.. счи-
тая это достаточным доказательством правильности своих убеж-
дений в области политики, религии, правил хорошего тона или
спорта.
К авторитетам обычно прибегают в двух случаях. В одном
случае человек, который сам не располагает прямыми фактиче-
скими данными, может обратиться к какому-либо источнику. Так
мы поступаем, когда пользуемся словарем, ищем нужную нам дату
в учебнике истории или формулу какого-нибудь соединения в учеб-
нике химии. Подобное обращение к авторитетам неизбежно и оп-
равдано, поскольку ни у кого нет достаточно времени, чтобы до-
сконально изучить каждый вопрос, с которым ему приходится
сталкиваться. При этом человек доверяет конкретным сведениям,
почерпнутым из источника, однако оставляет за собой право не
согласиться с мнением его автора.
В другом случае ссылаются на авторитет, который сам себя
наделил или был наделен другими некой непогрешимостью. Осо-
бенно часто такая ситуация возникает в связи с политическими,
социальными или религиозными проблемами. Сам же «авторитет»
объявляется глашатаем истины в последней инстанции, что обеспе-
чивает ему право на не подлежащие сомнению суждения. Индиви-
дуум, подвергающий сомнению эти суждения, объявляется ерити-
ком или отступником.
Хотя метод ссылки на авторитеты преследует цель достижения
единомыслия и стабильности во взглядах, цель эта недостижима,
потому что «авторитеты» обычно расходятся во мнениях, а число
«еретиков» всегда бывает довольно значительным.
3. Интуиция. Некоторые оценки так прочно укореняются в об-
ществе, а некоторые явления так воспринимаются нашими орга-
нами чувств, что представляются «самоочевидными». Земля пло-
ская, а Солнце вращается вокруг нее, плесень самозарождается
в гниющих продуктах, профессора всегда правы — все эти концеп-
ции в течение долгого времени принимались как «самоочевидные».
Впоследствии оказалось, однако, что многие «самоочевидные»
истины ложны: Земля имеет форму шара, Солнце не вращается
вокруг Земли и бывает, что даже профессора ошибаются. Иными
словами, наши органы чувств нельзя считать непогрешимыми, и
если какое-либо суждение не вызывает сомнений лишь на том
основании, что оно представляется «самоочевидным», то это еще
не гарантирует его истинности.
4. Научный метод или рефлективное исследование. Все три
описанных выше метода претендуют на абсолютные и окончатель-
Научный метод
31
ные ответы на любые вопросы. Однако все они могут привести к
неверным выводам, поскольку человеку свойственно ошибаться и
проявлять необъективность. Кроме того, ни один из этих методов
не содержит в себе способа обнаружения и исправления ошибок.
Научный метод не дает окончательных ответов, но он лучше всех
других методов сочетается с наблюдением и экспериментом.
В основе научного метода лежат наблюдения в природе, по-
строение гипотез, объясняющих эти наблюдения, и эксперимен-
тальная проверка выводов, вытекающих из сделанных гипотез.
Новые наблюдения постоянно сравниваются с прежними наблю-
дениями и существующими теориями. Это позволяет выявлять
ошибки и модифицировать теории по мере появления новой инфор-
мации. Вследствие этого научные представления об окружающем
мире не бывают окончательными и всеобъемлющими. Такая не-
определенность не исключает научного прогресса, а скорее под-
разумевает его неизбежность, поскольку при этом неверные кон-
цепции, которых, учитывая подверженность человека ошибкам,
избежать нельзя, постоянно устраняются и уточняются. Научные
выводы, подобно заключениям, которые делаются на основе здра-
вого смысла, представляют собой мнения, но мнения, основанные
на наблюдениях, а не произвольные высказывания. Рассмотрим
теперь вкратце, что такое научный метод.
СТРУКТУРА НАУЧНОГО МЕТОДА
Между философами и логиками нет полного согласия относи-
тельно структуры научного метода. Ученые, даже наиболее пре-
успевшие, вообще уделяют мало внимания этому вопросу и обыч-
но могут высказать лишь очень туманные и неточные мнения от-
носительно логических основ своей работы. Ниже (рис. 2.1) дана
общая схема научного метода, нарисованная широкими мазками.
Более подробное обсуждение проблемы можно найти в ряде книг
(Bunge, 1967; Kuhn, 1957; Cohen, Nagel, 1934; Popper, 1959; Mad-
den, 1960).
Исходная причина любого научного исследования — это осо-
знание недостаточности имеющихся знаний для удовлетворитель-
ного объяснения того или иного наблюдаемого в природе явления.
Почему, например, потомки, хотя они и похожи на своих родите-
лей, не бывают полностью идентичны одному или другому родите-
лю или не носят строго промежуточный характер? Эту первую
стадию ученые называют «постановкой проблемы». Проблем, ко-
торые можно изучать, существует бесчисленное множество. Неко-
торые из них тривиальны, другие — нет. Некоторые никак не влия-
ют на развитие науки или на благополучие людей; другие же
имеют огромное значение для человека. При выборе проблемы
для исследования ученый должен решить, какое значение она
может иметь как для развития науки, так и для человечества.
32
Глава 2
Иногда кажется, что данная проблема несущественна для прак-
тики, одиако в конечном итоге она оказывается чрезвычайно важ-
вой. Примером служит сделанное Рентгеном наблюдение, что не-
Рис. 2.1. Одна из возможных схем научного метода.
-которые типы излучения не отклоняются магнитным полем, при-
ведшее к открытию носящих его имя лучей со всеми проистекши-
ми отсюда последствиями.
После того как подлежащая исследованию проблема выбрана,
юна должна быть ясно и точно сформулирована. Ученый должен
очень четко изложить, в чем именно заключается данная проб-
лема.
Следующий этап научного исследования — сбор всех фактов,
относящихся к изучаемой проблеме. Под фактами имеются в виду
результаты наблюдений и измерений, образцы или экземпляры,
.прежние представления и тому подобное; все это исследователь
черпает из книг, из разговоров с коллегами, однако главный источ-
ник фактов — собственное изучение природы. На этой стадии вся
«сложность реального мира упрощается и сводится к тем его чер-
пай, которые имеют отношение к изучаемой проблеме и могут
•быть охвачены ученым.
Научный метод
33
После того как проблема сформулирована и все имеющиеся
данные собраны, ученый пытается найти объяснение, т. е. пред-
ложить такое решение изучаемой проблемы, которое не противо-
речит «фактам», собранным им самим и другими исследователя-
ми. Это связано с созданием одной или нескольких гипотез или
моделей и с их проверкой, для того чтобы выяснить, какая из них
ближе всего к истине.
Гипотеза создается в результате индуктивного мышления1: на
основании ряда отдельных частных «фактов» ученый предлагает
общее объяснение, т. е. объяснение, выходящее за пределы имею-
щихся фактов и сохраняющее значимость для таких ситуаций и
объектов, которые еще не удавалось наблюдать или исследовать.
Философы неистово спорят о том, можно ли логически оправдать
индуктивный метод, и каковы его рациональные основы. Между
тем использование индуктивного метода при построении новых ги-
потез, по-видимому, ведет к новым успехам в науке.
Согласно классическим представлениям, при проведении науч-
ных исследований создание гипотез следует за сбором фактических
данных. На самом же деле оба эти процесса обычно идут рука об
руку.
Гипотеза должна не только давать объяснение явлениям при-
роды, но и делать некоторые предсказания — предсказания, под-
дающиеся проверке. Например, говорить, что потомок похож на
своих родителей потому, что он связан с ними родственными уза-
ми,— это еще не значит дать настоящее объяснение, хотя такое
утверждение безусловно справедливо. Если же, однако, сказать,
что потомок похож на своих родителей потому, что они передали
ему некоторое число генов определенного типа и в соответствии
с определенными правилами, то это будет достоверным научным
объяснением, поскольку существование генов, факт их передачи
и правила, по которым происходит эта передача, могут быть про-
верены при помощи экспериментов и наблюдений.
Проверка той или иной гипотезы нередко принимает форму
заранее спланированного лабораторного эксперимента. Такие экс-
перименты служат обычным способом проверки гипотез в физике,
химии и по большей части в биологии, однако они не являются
обязательными, а в некоторых областях науки, например в геоло-
гии, лабораторная проверка невозможна. При проверке гипотез в
области эволюции и популяционной генетики лабораторный экспе-
римент нередко заменяется наблюдениями в природе. Другими
способами проверки служат эксперименты на опытных делянках
(например, в сельском хозяйстве), изменения естественной среды
(например, в экологии), создание реальных физических моделей,
1 По вопросу о том, как именно происходит создание гипотез, существуют
разногласия (см. Popper, 1959; Chiselin, 1974).
3-2044
34
Глава 2
имитирующих естественную ситуацию, создание математических
моделей с помощью ЭВМ. Еще один очень важный способ про-
верки правильности гипотез состоит в сравнении равноценных или
почти равноценных ситуаций: структуры, органы или процессы
сравниваются в разных ситуациях, у разных органов и (или) у
организмов, взятых в разных местах, если есть основания пред-
полагать, что они подвергались действию сходных факторов, на-
пример одинаковых давлений отбора. Обнаруживаемые при этом
сходства и различия позволяют получить представление о том,
как протекают некие общие процессы. Этот «сравнительный
метод» играет очень важную роль в эволюционных исследова-
ниях.
Как именно проводится проверка правильности данной гипо-
тезы, не имеет решающего значения. Самое главное, чтобы объяс-
нение, данное изучаемой проблеме, поддавалось проверке через
предсказания, соответствующие выдвинутой гипотезе. Эти пред-
сказания представляют собой обычно общие выводы, логически
вытекающие из предложенной гипотезы, а любая гипотеза должна
быть сформулирована таким образом, чтобы из нее следовали
предсказания или общие положения, поддающиеся проверке. На-
пример, одно время было принято объяснять многие исключитель-
ные свойства живых организмов существованием некоего «жизнен-
ного порыва» или «жизненной силы». Поскольку эта жизненная
сила считалась нематериальной, никакого способа проверить пред-
положение о ее существовании не было. Поэтому постулирование
«жизненного порыва» было псевдообъяснением, которое в конце
концов отвергли и заменили вполне материалистическими взгля-
дами.
Если предсказания, выведенные из данной гипотезы, оправды-
ваются, такой результат принимается за временное доказательство
справедливости этой гипотезы. Ответ на поставленный вопрос по-
лучен, и исследователь может перейти к изучению в таком же
порядке какого-либо нового вопроса.
Описанная здесь картина соответствует классическим представ-
лениям о применении научного метода. При этом предполагается,
что: 1) ученый совершенно беспристрастен; 2) для каждого случая
известны все относящиеся к нему данные; 3) эксперименты или
наблюдения, предназначенные для проверки гипотезы, всегда дают
однозначные результаты. На самом деле все эти допущения почти
никогда не реализуются. В действительности гипотезы строятся
людьми более или менее пристрастными, основываются обычно
на неполном знании природы, а приводимые в качестве доказа-
тельств их справедливости наблюдения и эксперименты нельзя
считать дающими однозначный ответ на поставленные вопросы.
Такие условия приводят к тому, что ученому-автору приходится
защищать свою гипотезу с помощью приведенных выше «ненауч-
ных» способов. Ученый, особенно если он пользуется широкой из-
Научный метод
35
вестностью, для защиты своей гипотезы может призвать на помощь
свой авторитет. Хороший пример этого можно найти в ранней
истории генетики. Опубликовав свою работу по генетике гороха,
Мендель послал один экземпляр статьи немецкому ботанику Кар-
лу Негели (Carl Nageli), который в то время считался крупнейшим
авторитетом в вопросах наследственности. Он был профессором
ботаники в весьма уважаемом Берлинском университете. Негели
отнесся к работе Менделя скептически, не оценив всей простоты
и изящества его экспериментов. В своем письме Менделю он
указал, что поверит в его открытие лишь в том случае, если Мен-
дель сумеет повторить свои эксперименты на ястребинке (Hiera-
ciutn), на которой Негели работал в то время, и получит анало-
гичные результаты. Мендель попытался сделать это, но результаты
©казались отрицательными. Ни Негели, ни Мендель (равно как
и никто другой в то время) не знали, что у ястребинки семена
Образуются бесполым путем. Таким образом, Негели, воспользо-
вавшись своим авторитетом, поставил Менделя в условия, в ко-
торых его идеи оказались совершенно дискредитированы и не
могли быть приняты. Хотя то, что Негели настаивал на необходи-
мости проверки теорий Менделя, было совершенно закономерным,
справедливость его требования о проведении Менделем экспери-
ментов именно на ястребинке весьма сомнительна.
Еще одна проблема состоит в том, что теории, предложенные
для решения одного вопроса, не независимы от теорий, предло-
женных для решения различных других вопросов; напротив, все
эти теории составляют отдельные части общего представления об
окружающем нас мире. Поэтому теории на самом деле объединя-
ются в некие совокупности и обычно, рассматривая какую-либо
новую проблему, исследователь старается предложить объяснение,
согласующееся с другими принятыми теориями. Иными словами,
выдвигаемая теория обычно представляет собой «субтеорию», впи-
сывающуюся в рамки одной из уже принятых теорий, а не что-то
совершенно новое. Так, например, в настоящее время любая не-
эволюционная теория, предложенная для объяснения строения
какого-либо организма, была бы сочтена неприемлемой, даже если
бы она соответствовала фактам и выдерживала проверку. Только
тогда, когда все здание из субтеорий, составляющих основную
теорию, становится слишком громоздким и сложным, сомнению
подвергается сама основная теория. При этом в научном мире
обычно возникает раскол. Как правило, многие ученые старшего
поколения отказываются принимать новые общие гипотезы, тогда
как другие, в особенности молодые ученые, восторженно одобряют
новые концепции. Это различие во взглядах приводит к бурным
Дискуссиям. Одно из таких столкновений в XVI в. было вызвано
переходом от системы Птолемея к системе Коперника. Другим
примером служит переход от ньютоновской к релятивистской кос-
мологии, происшедший в начале XX в. Примером из области био-
3*
36
Глава 2
логии служит переход в 1860-х гг. от креационистских к эволюци-
онным взглядам на мир. В наши дни крупное изменение произо-
шло в геологии: на смену прежнему представлению о том, что рас-
положение континентов друг относительно друга было всегда
одинаковым, пришла теория «дрейфа континентов».
В идеальном случае научный метод представляет собой .чрез-
вычайно эффективную процедуру для получения информации от-
носительно окружающей нас природы. Однако он вовсе не обяза-
тельно свободен от ошибок и даже может быть использован для
обоснования ошибочных представлений о природе. Большинство
ученых стараются использовать научный метод в его наиболее
строгой или идеальной форме, но многие, а может быть и все, в
той или иной степени обречены на неудачу по причинам, изложен-
ным выше. В более глубоком смысле эти неудачи обусловлены
тем, что ученые живут в определенной культурной среде и не мо-
гут полностью выйти за рамки системы взглядов, принятых в том
обществе, к которому они принадлежат. Их мышление и их подход
к своей работе формируются под влиянием этой системы взглядов
и ценностей.
СОСТАВНЫЕ ЧАСТИ,
КОТОРЫЕ ДОЛЖНА СОДЕРЖАТЬ ЛЮБАЯ ГИПОТЕЗА
Ученый не только открывает, но и создает, а поэтому вообра-
жение ему так же необходимо, как призвание и упорная работа.
Вместе с тем научная работа сопряжена с большими ограничения-
ми, чем искусство, и гипотезы или модели должны удовлетворять
определенным правилам.
Прежде всего гипотеза должна быть сформулирована таким
образом, чтобы из нее можно было вывести поддающиеся про-
верке заключения, что позволило бы решать, объясняет ли данная
гипотеза факты или нет. Ученые и философы вели ожесточенные
споры о том, каково должно быть происхождение концепции или
гипотезы. Некоторые философы, так называемые эмпирики, счи-
тают, что все концепции или гипотезы в науке должны возникать
на основе проведенных ранее исследований конкретных объектов
или событий и их взаимоотношений. Однако такой крайний эмпи-
ризм не мог бы объяснить возникновение многих концепций, на-
пример концепций гена или естественного отбора, поскольку ни
Мендель, ни Дарвин с этими понятиями никогда не встречались.
В настоящее время философы и логики соглашаются с тем, что
гипотеза может иметь любой из целого ряда альтернативных
источников: интуиция, метод проб и ошибок, прошлый опыт, слу-
чай или воображение. Главное, чтобы она была плодотворной и
могла быть подтверждена. Часто сама гипотеза не может быть
проверена, но поддаются проверке вытекающие из нее следст-
Научный метод
37
Еия (например, мутационная теория, см. гл. 6). Следует всегда
омнить, что гипотеза, которую нельзя проверить, не имеет
даысла.
Другой важный аспект гипотезы — ее логическая форма. Ги-
потеза состоит в основном из утверждений трех типов. Во-первых,.
SSO ряд утверждений, указывающих, в чем состоит явление, кото-
Ш предстоит дать объяснение. Такие утверждения называют
посылками данной гипотезы. Например, для классических
[цсспериментов Менделя (см. гл. 1 и 3) предпосылками могут быть
утверждения о существовании сортов гороха с высоким и низким
стеблем, с гладкими и морщинистыми семенами, белыми и крае-
выми цветками и т. п., которые размножаются в чистоте; возмож-
ность скрещивания между любыми двумя сортами без снижения
плодовитости; принадлежность всех сортов к одному и тому же
вдду. Предпосылки — это описание некоторых фактов, которые
реализуются до подлежащего объяснению явления или одновре-
менно с ним. Предпосылки — это формализованная «проблема»,
стоящая перед ученым. Во-вторых, это ряд формализованных ут-
верждений, имеющих всеобщую или универсальную значимость,
Т. е. законы природы, приложимые к данному явлению. В рассмат-
риваемом примере это законы Менделя. Предпосылки в совокуп-
ности с соответствующим общим законом объясняют (или пыта-
ются объяснить) явление, о котором идет речь, и в результате
ведут к утверждению третьего типа, или предсказанию, которое
Может быть проверено путем наблюдения или эксперимента. Из
утверждений этих трех типов только вторые, т. е. формулировка
общих положений, рассматриваются как собственно гипотеза. Но
без предпосылок и без поддающихся проверке предсказаний гипо-
теза превращается в догматическое утверждение, от которого мало
ПОЛЬЗЫ.
При создании гипотез и их проверке следует принимать во вни-
мание все конкурирующие с ними гипотезы. Процесс элиминации
состоит в том, чтобы установить, способны ли эти другие гипотезы
объяснить все имеющиеся факты. В конечном итоге из всех аль-
тернативных гипотез остается одна, поскольку лишь она согласу-
ется со всеми фактами. Существует, однако, некое осложняющее
Дело обстоятельство. Ученый, проверяющий альтернативные гипо-
тезы, может делать выбор только среди известных гипотез. Но
вполне возможно, что существует еще одна, пока несформулиро-
ванная гипотеза, которая так же хорошо соответствует всем имею-
щимся фактам, но более проста и экономична и обладает большей
поэтому, посколь-
рассмотрели все
вероятной, а не
Это ограничение
достоверность его
ЩИМСЯ фактам, но более проста и экономична и
ценностью в качестве всеобъемлющей теории. А
ку мы не можем быть уверены в том, что мы
возможные альтернативы, следует говорить о
безусловной справедливости данной гипотезы,
касается, в сущности, любого научного знания:
лишь вероятна, но не безусловна.
38
Глава 2
Имеются еще и другие осложнения. Иногда не удается выбрать
такую гипотезу, которая объясняла бы все факты, относящиеся к
данной проблеме, но имеются две или несколько гипотез, позво-
ляющих объяснить многие, хотя и не все эти факты. В таком слу-
чае ни одна из гипотез не может быть безоговорочно принята.
Если такие гипотезы нельзя объединить в одну, поскольку Они со-
держат противоречащие друг другу утверждения, то ученые ока-
зываются в затруднительном положении. Возможна также проб-
лема противоположного характера. Существует две или даже
несколько гипотез, объясняющих все имеющиеся факты, но нет
способа, позволяющего сделать между ними выбор. В таких слу-
чаях следует, как правило, выбирать логически самую простую
гипотезу. Мы выбираем ту гипотезу, которая объясняет факты
проще всего, т. е. с привлечением наименьшего числа новых по-
нятий и произвольных допущений. Это правило называют «брит-
вой Оккама» в честь Уильяма Оккама — английского философа-
схоласта начала XIV в. Сформулированный Оккамом принцип
гласит: «Сущности не следует умножать сверх необходимости».
Еще один важный компонент модели — это ее дедуктивное раз-
витие. Под этим мы имеем в виду, что гипотеза должна допускать
дальнейшую разработку, с тем чтобы можно было выявить все ее
следствия. Многие гипотезы формулируются словесно, однако к
разработке гипотез все больше и больше привлекается математи-
ка. Хорошим примером из области популяционной биологии слу-
жит теория увеличения численности популяции и ее регуляции,
которая рассматривается в гл. 13. Поскольку математика — наука
дедуктивная, она служит идеальным орудием для дедуктивной
разработки новой гипотезы, разумеется, при условии что состав-
ляющие эту гипотезу концепции можно выразить количественно.
Однако математика — это лишь средство, а не цель. Гипотеза,
сформулированная в математических терминах, по сути своей не
более точна, чем та, которая выражена словами. Правда, когда
гипотезы сформулированы математически, в них легче выявить
несоответствия, а также вывести из них предсказания, которые
раньше не приходили в голову. В этом заключается сила мате-
матики. Но в популяционной биологии часто приходится иметь
дело со сложными и не до конца понятными фактами (предпо-
сылками). Для математической разработки гипотез, относящихся
к этим фактам, нередко приходится делать упрощающие допуще-
ния, которые могут оказаться нереалистичными. Короче говоря,
хотя математика может быть очень полезна для формулирования
и разработки гипотез, она никак не заменяет воображение и био-
логическую прозорливость.
Последний этап в создании и развитии модели или гипотезы —
это ее проверка при помощи наблюдений и экспериментов, с тем
чтобы выяснить, совпадают ли предсказания с действительностью.
Эксперименты должны быть универсальными и хорошо воспроиз-
Научный метод
39
'родимыми. Эксперименты, которые не удается повторить, или та-
jjjje, которые могут быть воспроизведены только в одной лабора-
тории, в одной стране, одним лицом или одной группой лиц, не-
приемлемы. Проверочные эксперименты должны проводиться па-
раллельно с контрольными; иначе говоря, в них должны учиты-
ваться все относящиеся к делу переменные факторы. Поскольку'
переменные факторы легче контролировать в искусственных усло--
виях, создаваемых экспериментатором, лабораторные эксперимен-
ты с этой точки зрения предпочтительнее наблюдений в природе.
Вместе с тем искусственно воссоздать природу невозможно, а по-
этому контролируемые условия, в которых проводятся лаборатор-
ные эксперименты, могут оказаться настолько далекими от есте-
ственных, что проверка гипотезы будет обесценена. Это особенно
Справедливо в отношении эволюционных гипотез.
В следующей главе мы подвергнем анализу теорию эволюции
путем естественного отбора в свете правил, налагаемых научным
методом.
ЛИТЕРАТУРА
Bunge М. 1967. Scientific Research, 2 vol., Berlin, Springer-Verlag.
Cohen M. R., Nagel E. 1934. An Introduction to Logic and the Scientific Method,
New York, Harcourt, Brace.
Ghiselln M. T. 1974. The Economy of Nature and the Evolution of Sex, Berkeley,
University of California Press.
Kuhn T. S. 1957. The Copernican Revolution, Cambridge, Mass., Harvard Univer-
sity Press.
Kuhn T. S. 1962. The Structure of Scientific Revolutions, Chicago, University of
Chicago Press.
Madden E. H. 1960. The Structure of Scientific Thought, Boston, Houghton Mifflin.
Popper K. R. 1969. The Logic of Scientific Discovery, New York, Harper Torchbooks.
Глава 3
Теория эволюции
путем естественного отбора
В идеале теория эволюции путем естественного отбора должна
была бы предсказывать состояние земной биоты в будущем. Од-
нако для того чтобы это было возможно, необходимо располагать
полным описанием современного состояния биоты, а также доско-
нальным знанием правил, или законов, по которым происходит
изменение организмов во времени. Подобных сведений у нас нет
и они вряд ли когда-либо появятся. Число отдельных организмов
так велико и каждый из них так сложен, что описать их все со-
вершенно невозможно. В настоящее время мы располагаем лишь
неполным и отрывочным описанием главных признаков большин-
ства видов растений и животных (гл. 18).
У нас не только нет подробного описания организмов, населяю-
щих земной шар; мы все еще не понимаем истинную природу эво-
люционных изменений, происходящих в отдельных линиях. Кроме
того, для предсказания течения эволюции в будущем необходимы
точные сведения о тех изменениях, которые будут происходить в
среде. Из всего этого следует, что создание действительно прогно-
стической теории эволюции невозможно.
Но даже если бы мы располагали всеми необходимыми сведе-
ниями, т. е. исчерпывающими знаниями о современном состоянии
биоты и достаточными знаниями о правилах, определяющих эво-
люционные изменения, а также умели предсказывать изменения,
происходящие в среде, то все же полностью предсказать ход эво-
люции было бы невозможно. Причина того, что эволюционные из-
менения могут быть в конечном счете непредсказуемыми, кроется
в стохастической природе некоторых явлений. Мы имеем в виду,
что некоторые эволюционные явления носят случайный характер,
а следовательно, они предсказуемы лишь в статистическом смысле
(гл. 7).
В этой главе излагается в общем виде теория естественного от-
бора, область ее приложения и недостатки, что закладывает ос-
новы для более подробного ее рассмотрения в последующих гла-
вах.
Создавая «Происхождение видов», Дарвин ставил себе цель
показать, что имеющейся в природных популяциях изменчивости
Теория эволюции путем естественного отбора
41
«остаточно, для того чтобы при наличии соответствующих условий
Дать начало новым видам. Таким образом, его целью было не
предсказывать исход эволюции в будущем, а построить чисто ма-
териалистическую гипотезу, объясняющую происхождение разно-
образия живых организмов. Взамен преобладающих в то время
идеалистических концепций Дарвин предложил для объяснения
происхождения видов механизм, поддающийся проверке, — есте-
ственный отбор. Хотя эта теория не утверждает, что естественный
отбор — единственный механизм эволюции, Дарвин подразумевал,
что эволюция происходила только таким образом. В этом смысле
его теория предсказывает, что в будущем виды будут возникать
только в результате естественного отбора.
Теперь, используя концепции, развитые в гл. 2, мы изложим
теорию эволюции путем естественного отбора более строгим и точ-
ным образом, чем это было сделано в гл. 1. Мы разделим наше
изложение на три части. Сначала мы рассмотрим основные наблю-
дения, положенные в основу теории; иными словами, мы изучим
ту «проблему» и те «факты», которым теория эволюции путем
естественного отбора пытается дать объяснение. Затем мы рас-
смотрим саму теорию под заголовком «Минимальные условия,
необходимые для эволюции путем естественного отбора». Завер-
шается глава обсуждением того, представляет ли собой эволюци-
онная теория общую теорию, гипотезу или мировоззрение и может
ли она быть безоговорочно опровергнута при помощи эксперимента
или наблюдения.
ЭВОЛЮЦИЯ ПУТЕМ ЕСТЕСТВЕННОГО ОТБОРА.
ПРОБЛЕМА И ФАКТЫ
Куда бы мы ни бросили взгляд — в небесную ли высь или глу-
бины океана, от тропиков до полярных областей — мы везде на-
ходим живые организмы. Число их огромно, а их форма, облик и
образ жизни столь разнообразны, что они часто кажутся нам при-
чудливыми и непостижимыми. Все это разнообразие представляет
собой не что иное, как выражение высокой степени химической
организации, которую мы называем жизнью. Дошедшие до нас
сведения указывают на то, что люди с древнейших времен пыта-
лись объяснить разнообразие живых существ и создавали для
этого подробные мифы, сказки, системы взглядов и религиозные
учения.
Начиная с XVI в. многих европейских мыслителей, среди кото-
рых наиболее известны Бэкон, Декарт и Галилей, перестали удов-
летворять существовавшие в то время объяснения устройства
природы. Они все яснее сознавали, что природа часто ведет себя
совсем не так, как подсказывает интуиция, т. е. что многое обстоит
не всегда так, как кажется на первый взгляд; этому способство-
вало появление в ту пору новых приборов, в особенности телеско-
42
Глава 3
па, а позже микроскопа, а также таких новых разделов матема-
тики, как разработанное Ньютоном дифференциальное исчисление.
Все это привело к тому, что ученые стали требовать доказательств
интуитивным утверждениям о природе вещей. Так зародилось то,
что мы теперь называем научным методом, который был рассмот-
рен в гл. 2.
Когда естествоиспытатели стали применять новую методологию
к органическому миру, они сделали четыре важных открытия:
1. Между организмами существует известное сходство в от-
ношении анатомии, морфологии и поведения.
2. Растения и животных на основе таких сходных черт
можно объединить в группы, а эти группы можно в свою оче-
редь объединить в ряды, начиная от простейших и до самых
сложных.
3. «Окаменевшие» (фоссилизованные) кости вымерших жи-
вотных также четко распределяются по группам, которые мож-
но отнести к различным упорядоченным рядам современных
организмов. (До XVI в. считалось, что окаменевшие кости при-
надлежали животным, погибшим во время всемирного потопа.)
4. Подобное рождает подобное, но иногда потомки отлича-
ются от своих родителей в незначительной, а подчас и в до-
вольно существенной степени. (Такие изменения получили на-
звания «мутаций», но их не следует путать с тем, что мы на-
зываем мутациями в настоящее время.)
Все эти наблюдения логически привели к мысли о том, что раз-
нообразие живых организмов могло возникнуть в результате про-
цесса «эволюции». Сперва существовали простые примитивные
организмы, которые в результате возникавших время от времени
«мутаций» давали начало более развитым или «прогрессивным»
формам, а те в свою очередь давали начало еще более «прогрес-
сивным» и т. д., что в конечном счете должно было привести к
появлению человека. Так идея об эволюции в XVII и XVIII вв.
проникла в естественную историю через труды Эразма Дарвина
(деда Чарлза Дарвина), немецкого поэта Иоганна Вольфганга
Гёте и в особенности великого французского натуралиста Жана
Батиста Пьера Антуана де Монэ, больше известного как шевалье
де Ламарк (1744—1829). Ламарк был автором одного из первых
трудов, в котором виды рассматривались не как неизменные, а
как изменяющиеся во времени.
Однако ни Ламарк, ни кто-либо другой из эволюционистов до-
дарвиновского периода не предлагали ни убедительных доводов
в пользу существования эволюции, ни правдоподобных поддаю-
щихся проверке механизмов, которые были бы ответственны за
эволюционные изменения. Их объяснение относилось лишь к су-
ществованию сходных признаков у разных организмов, но, по-
скольку оно не содержало никаких поддающихся проверке пред-
Теория эволюции путем естественного отбора 43
сказаний, его нельзя было считать строгой научной теорией. Идея
эволюции не пользовалась в то время популярностью.
Дарвин внес в эволюционную гипотезу недостающий компо-
нент. Он обратил внимание на то, что потомков производится го-
раздо больше, чем может выжить. Следовательно, между орга-
низмами существует конкуренция или, пользуясь выражением
Дарвина, «борьба за существование». Любой признак, рассуждал
Дарвин, повышающий вероятность выживания его обладателей
или их потомков, будет постепенно встречаться все чаще, пока,
наконец, им не окажутся наделены все особи данной группы
(рис. 1.3). Этот процесс Дарвин назвал естественным отбором,
потому что он близок к процессу отбирания лучших семян для
посева или лучших производителей при разведении скота. Отбор,
производимый человеком, Дарвин назвал «искусственным отбо-
ром».
С добавлением к наблюдениям натуралистов додарвиновского
периода концепции естественного отбора как движущей силы эво-
люции возникла научная гипотеза, объяснявшая разнообразие
живых организмов. Эта гипотеза излагает конкретные необходи-
мые условия, каждое из которых следует подтвердить (или отбро-
сить) путем наблюдения и (или) эксперимента. Эти условия от-
носятся к размножению, наследственности, корреляции между
выживанием и признаками особей и происхождению изменчивости.
Рассмотрим каждое из этих условий более подробно.
МИНИМАЛЬНЫЕ УСЛОВИЯ, НЕОБХОДИМЫЕ ДЛЯ ЭВОЛЮЦИИ
ПУТЕМ ЕСТЕСТВЕННОГО ОТБОРА
Размножение
Для того чтобы мог происходить естественный отбор, организ-
мы должны производить на свет больше потомков, чем это необ-
ходимо только для замещения родительских особей. Необходимость
соблюдения этого условия очевидна. Если каждая особь или каж-
дая пара особей в данной популяции будет производить ровно
столько особей, сколько необходимо, чтобы заменить эту пару, т. е.
ровно по одной на каждую особь, и если каждый потомок дожи-
вет до половозрелости и произведет в свою очередь по одному
потомку и т. д., то в результате создается идеальное демографиче-
ское равновесие. В таких условиях эволюция путем естественного
отбора, в той форме, в какой она здесь определена, происходить
не может, поскольку при этом ни дифференциального размноже-
ния, ни дифференциальной смертности не будет. Если же на каж-
дого индивидуума будет приходиться меньше одного потомка, то
популяция окажется обреченной на вымирание.
Обратите внимание на то, что здесь подчеркивается факт про-
дуцирования на каждую особь точно одного потомка, который
44
Глава 3
всегда доживает до половозрелости. Однако такая ситуация не-
реальна. Всегда возможны какие-либо случайности, не говоря уже
о том, что некоторые виды существуют, питаясь другими, зачастую
главным образом более беззащитной молодью. Если какая-то часть
особей погибнет до того, как у нее появится потомство, то вид, в
котором каждая взрослая особь производит только одного 'потом-
ка, окажется на пути к вымиранию. Поскольку в общем и целом
(на протяжении относительно коротких промежутков времени)
численность видов не снижается, большинство видов производит
больше потомков, чем это строго необходимо для замещения есте-
ственной убыли.
Это утверждение можно проверить путем наблюдений в при-
роде. В табл. 3.1 и 3.2 указано число потомков, производимых
самками разных видов. Здесь следует обратить внимание на два
момента. Во-первых, можно видеть, что это условие соблюдается
у всех видов. Не известно ни одного организма, который бы про-
изводил лишь одного потомка на каждую особь. Все организмы
производят больше потомков. Во-вторых, виды очень сильно раз-
личаются по числу производимых потомков. Некоторые виды, как,
например, человек, производят сравнительно немного потомков,
тогда как у других, например у пресловутой устрицы и у многих
Таблица 3.1
Число потомков, производимых разными видами животных
за один сезон размножения1)
Латинское название Общепринятое название Число потомков
Homo sapiens Canis familiaris Sturnus vulgaris Terrapene Carolina Rana catesbiana Человек Собака Скворец Коробчатая черепаха Лягушка-бык 1 7(1—22) 4—6 яиц в кладке 2—7 яиц в кладке 6000—20 000 икрииок в
Salmo .trutta Ручьевая форель кладке 200—6000 икрииок в
Salmo aquabonita Лосось кладке 1380—2280 икрииок в
Homarus americanus Agropecten irradians (Pecten irradians) Crassostrea virginica Омар Гребешок Дальневосточная уст- кладке 8500 яиц в кладке 2 000 000 яиц в кладке 500 000—1 000 000 яиц в
Drosophila melanogaster Vespula maculata рица Плодовая мушка Пятнистая оса кладке 100 яиц иа 1 самку 25 000—35 000 яиц на
Musca domestica Комнатная муха 1 матку 75—200 яиц на 1 самку
) По данным Altman Р. L„ Dittmer D. S. (eds.), Biology Data Book, ▼. I, 2nd ed.,
1972.
Теория эволюции путем естественного отбора
45
Таблица 3.2
Число семян иа 1 растение в год для некоторых видов
травянистых растений, произрастающих на поле,
вспаханном год назад, поле, заросшем сорняками
(ие пахавшемся 10 лет) и в лесу1»
Вид Число растений Среднее число семян иа 1 растение Пределы колеба- ний числа семяи иа 1 растение
Поле, вспаханное год назад Ambrosia artemlsiifolia 12 1190 110—2690
Chenopodium album 57 4820 90—50 100
Поле, заросшее сорняками Solidago altissima и S. canadensis 11 3070 960—5330
Лес Dentaria laciniata 9 24 9—42
Cardamine bulbosa 10 33 5—90
Sanicula gregaria 27 26 3—50
Prenanthes alba 7 118 95—170
0 Newell, Tramer, Ecology, 59, 228—234, 1978.
растений, потомки исчисляются тысячами. Кроме того, у различных
видов очень сильно варьируют энергетические затраты на произ-
водство потомства. Некоторые виды растений вкладывают в созда-
ние потомства более 30% всей имеющейся у них энергии, а другие
затрачивают на это менее 1 % энергии.
Корреляция между вероятностью выживания и фенотипом
Если принять, что у каждого вида число потомков больше чис-
ла родителей, то в отношении вероятности выживания отдельного
потомка возможна следующая альтернатива: 1) выживание це-
ликом зависит от случая; 2) выживание данного потомка по край-
ней мере частично зависит от его признаков (морфологических,
физиологических, биохимических). И в той и в другой ситуации
в популяции будут наблюдаться изменения (некоторые признаки
могут быть закреплены или утрачены чисто случайно), однако в
первой ситуации наблюдаемые изменения будут носить случайный
характер и будут подвержены колебаниям во времени. Поскольку
в палеонтологических рядах некоторые эволюционные изменения
носят несомненно адаптивный характер (например, развитие орга-
нов зрения), гипотезу о том, что эволюция представляет собой
чисто случайное явление, следует отбросить. Однако вопрос, какая
именно доля эволюционных изменений носит адаптивный характер,
остается открытым.
Наследственность
Если признаки, способствующие выживанию некоторых особей,
не будут передаваться потомкам этих особей, то все потомки могут
погибнуть ввиду отсутствия необходимых приспособительных при-
46 Глава3
знаков. Наблюдения и эксперименты показывают, что большинство
признаков организма передается по наследству, хотя они могут
быть выражены у потомков в измененной форме вследствие воз-
действия факторов среды. Мы знаем также, что в число этих при-
Рис. 3.1. Типичная и меланистическая (carbonaria) формы березовой пяденицы на
покрытом лишайниками древесном стволе (А) в незагрязненной сельской местно-
сти (Дорсет, Англия) и на стволе дуба, свободном от лишайников, близ Бирмин-
гема, Англия (Б). (С фотографии, любезно предоставленной д-ром Н. В. D. Kett-
lewell, Оксфордский университет.)
знаков входят и те, которые имеют непосредственное значение для
повышения выживаемости потомков.
Одним из примеров этого служит березовая пяденица {Biston
betularia) в Англии. Эта бабочка, которая, как и многие другие,,
активна ночью, днем сидит, раскрыв крылья, на стволах берез.
Обычно стволы берез бывают покрыты лишайниками, белыми с
серыми пятнышками. Крылья березовой пяденицы также белые с
серыми пятнами, так что она хорошо сливается с фоном (рис. 3.1).
Днем на этих бабочек охотятся птицы. Чем лучше бабочка слива-
ется с фоном, тем меньше вероятность того, что птицы ее заметят
и съедят (табл. 3.3). Примерно 100 лет назад коллекционеры, ло-
вившие бабочек в окрестностях Бирмингема (Англия), обнаружили
другую форму березовой пяденицы, которую они назвали carbona-
ria, потому что крылья у нее были серо-черные. Промежуточная
форма insularia, которую вначале объединяли с carbonaria, была
выявлена позднее. С течением времени численность формы carbo-
naria в промышленных районах Англии и вокруг них все более
Теория эволюции путем естественного отбора
47
Таблица 3.3
Зависимость выедания птицами разных форм бабочек от фона,
на котором они сидят (Kettlewell, 1973)
А. Число бабочек, съеденных птицами за два дня в загрязненном лесу, после
того как было выпущено одинаковое число особей обеих форм
Форма typica 43
Форма carbonaria 15
Всего 58
Б. Количество бабочек, вторично отлавливавшихся к концу дня на протяжении
семидневного периода в загрязненном лесу (в % к числу выпущенных)
Форма typica 45,79
Форма carbonaria 62,57
Возрастала и в настоящее время стала преобладающей. Что же
случилось? С наступлением промышленной революции лишайники
вымерли в результате загрязнения воздуха, так что стволы и вет-
ви деревьев стали темными. Вследствие этого окраска пядениц,
бывшая ранее покровительственной, перестала служить им защи-
той и они стали более легкой добычей для птиц. Что же касается
формы carbonaria, то она, напротив, стала более многочисленной,
так как ее темная окраска хорошо сливалась с темной корой не-
покрытых лишайниками деревьев, число которых также все время
возрастало. Результаты исследований Кеттлуэлла (Kettlewell) по-
казали, что изменения возникли также и в генетической структуре
популяции.
Следовательно, эволюция происходит потому, что в популяциях
различных организмов, каждый из которых производит избыточ-
ное число потомков и передает им свои признаки, вероятность
выживания некоторых особей и их потомков выше, поскольку они
производят больше потомков или обладают некоторыми фенотипи-
ческими признаками, повышающими выживаемость, или же по
обеим этим причинам одновременно. Такие признаки называются
приспособительными, а соответствующие процессы — дифференци-
альным размножением и дифференциальным выживанием.
Важно помнить, что естественный отбор действует только на те
признаки, которые передаются по наследству. Если семени оду-
ванчика посчастливится упасть на плодородную почву, то вырос-
шее из него растение произведет больше семян, чем растение, ока-
завшееся на более бедной почве. Поскольку, однако, причина избы-
точного производства семян — плодородная почва — не представля-
ет собой наследуемой особенности, никакого эволюционного изме-
нения не последует.
Неизменность внешних условий
Для того чтобы процесс дифференциального размножения
и (или) дифференциального выживания вел к систематическим из-
менениям в популяции, условия среды должны благоприятствовать
48
Глава 3
одним и тем же фенотипам более чем в одном поколении. Это
означает, что естественному отбору необходимо сохранение извест-
ного постоянства условий во времени. Между тем этому требова-
нию иногда не уделяют внимания. Поскольку среда никогда не
остается совершенно неизменной для последовательных поколений
или в различных местообитаниях, оценить степень постоянства
среды, необходимую для того, чтобы естественный отбор мог дей-
ствовать, очень трудно. Некоторые условия среды, например по-
года, очень изменчивы, тогда как другие ее характеристики (дол-
гота дня, интенсивность света, смена времен года) очень постоян-
ны. Соответственно наличие у организмов структур, непосредствен-
но связанных с этими неизменными характеристиками, таких, как
глаза у животных или листья у растений (функции которых свя-
заны со светом), более вероятно, чем наличие у них структур для
защиты от таких непредсказуемых явлений, как град. В среде,
сохраняющей однообразие по времени, процессы дифференциаль-
ного размножения и дифференциального выживания привели бы
к возникновению популяции одинаковых особей, поскольку особи,
обладающие менее адаптивными признаками и (или) производя-
щие меньше потомков, элиминируются. После этого выживание
зависело бы исключительно от случая, и мы не встречали бы в
природе того разнообразия, которое мы в ней наблюдаем. Следует
ли считать такое несоответствие противоречащим эволюции? Для
того чтобы объяснить причину столь большого разнообразия, нам
надо рассмотреть источники изменчивости в популяциях.
ИСТОЧНИКИ ИЗМЕНЧИВОСТИ
Существует два источника изменчивости. Один из этих источни-
ков внутренний и обусловливает возникновение новых признаков.
Другой — внешний, и хотя он не «создает» новых признаков, также
вносит свой вклад в поддержание наблюдаемой изменчивости. Как
будет подробно объяснено в гл. 4—6, родители передают своим
потомкам химические вещества, которые направляют развитие и
рост их потомков по родительским или очень сходным путям. При
этом изменение химических свойств или количества передаваемого
генетического вещества может привести к возникновению у потом-
ков новых признаков. Такие изменения, которые можно объеди-
нить под общим названием мутаций и рекомбинаций (более под-
робно они рассматриваются в гл. 6), служат причиной возникнове-
ния новых наследственных признаков. В результате этих процес-
сов появляются особи, обладающие признаками, отличными от
признаков их предков. Некоторые из таких новых признаков ока-
жутся адаптивными и будут распространяться в популяции, за-
мещая признаки, имевшиеся в ней раньше. Иными словами, если
в результате какого-либо внутреннего изменения в генах возника-
ет фенотип, который может произвести большее число выживаю-
Теория эволюции путем, естественного отбора 4S>
щих потомков путем дифференциального размножения и (или)
дифференциального выживания, то этот фенотип распространится
в популяции. В строгом смысле слова это и есть подлинный источ-
ник новой изменчивости. Однако большая часть наблюдаемой нами
изменчивости обусловлена в популяции изменениями внешних ус-
ловий.
В действительности тот или иной признак создает для данного-
организма преимущество только в определенных фиксированных
условиях. Biston betularia со светлыми крыльями обладает преи-
муществом в незагрязненной среде, форма carbonaria — в загряз-
ненной. Следовательно, как та, так и другая форма существуют в-
природе лишь постольку, поскольку существуют как те, так »
другие условия. Исчезновение какого-либо из этих условий, воз-
можно, привело бы к вымиранию формы, связанной с ними.
’ Среда представляет собой сумму всего того, что окружает дан-
ный организм. В нее входят такие факторы, как почва, рельеф*
местности, погода и все другие организмы. Из этих факторо»
наиболее значительным колебаниям подвержена погода, измене-
ния которой связаны с рядом независимых причин (вращением
Земли вокруг своей оси, ее вращением вокруг Солнца, вихревыми
потоками в атмосфере и т. п.). В свою очередь погода, в особен-
ности атмосферные осадки, влияют на процесс образования почвы.
А следовательно, различные режимы выпадения осадков с тече-
нием времени приводят к образованию почв разного типа. Рельеф*
местности в значительной степени обусловлен характером горо-
образовательных процессов и эрозией, происходящими в резуль-
тате дрейфа континентов и под действием климатических условий..
Растения и животные, обитающие в определенном месте, также
зависят от всех этих трех факторов. Исходя из того, что «успех»,,
т. е. выживание данной особи и ее потомков, коррелирует с опре-
деленной средой и что сами условия среды подвержены измене-
ниям, «преимущество» каждого данного признака — явление пре-
ходящее. Кроме того, среда изменяется как во времени, так и в
пространстве, а поэтому следует ожидать, что наблюдаемые в раз-
ных местах и в разные моменты времени организмы должны раз-
личаться.
В популяции, в которой все особи одинаковы, никакого естест-
венного отбора происходить не может. Однако вследствие измене-
ний генетического материала (этот вопрос подробно рассматри-
вается в гл. 4—6) и вследствие реакций организмов на временные
и пространственные изменения в среде (гл. 13—15) все известные
нам популяции растений и животных изменчивы по своим призна-
кам и поэтому подвержены действию естественного отбора.
Итак, для того чтобы происходил естественный отбор, необхо-
димы следующие условия:
1. Избыточное число потомков по сравнению с тем их числом,
которое может выжить при нормальных условиях.
4—2044
50
Глава 3
2. Передача признаков от родителей потомкам.
3. Наличие в популяции различных типов особей, отличаю-
щихся по одному или нескольким признакам.
4. Более высокая вероятность выживания потомков, обладаю-
щих определенным фенотипическим признаком.
5. Достаточное постоянство среды во времени.
При наличии этих условий число особей, имеющих благоприят-
ные признаки, будет возрастать во времени, а в этом и заключа]
ется сущность эволюции путем естественного отбора.
Естественный отбор часто определяют как процесс, при кото-
ром организмы, наиболее хорошо приспособленные к данной cpef-
де, оставляют наибольшее число потомков. Если при этом крите-
рием для определения наиболее приспособленных организмов слу-
жит выживание потомков, такое определение будет тавтологией,
поскольку по существу утверждается, что организмы, производя-
щие наибольшее число потомков, будут производить наибольшее
число потомков. А нам нужно иметь возможность предсказывать
для данной среды те признаки, которые позволят обладающим ими
организмам оставлять наибольшее число выживающих потомков.
Каждое из перечисленных выше условий действия естественного
отбора может быть подтверждено или отброшено в результате на-
блюдений или экспериментов, как это и должно быть возможно
для любой научной теории.
Теория эволюции путем естественного отбора представляет со-
бой, следовательно, общую схему, с которой должны согласовы-
ваться детализированные и частные объяснения для более узких
проблем. Поскольку возможность проверить общую теорию в ши-
роком масштабе биоты в целом никогда не представится, эволю-
ционные биологи обращаются к более конкретным проблемам, пы-
таясь исследовать отдельные системы во всех подробностях, в
надежде, что находимые ими решения окажутся приложимыми
также и к другим организмам.
О ХАРАКТЕРЕ ТЕОРИИ ЕСТЕСТВЕННОГО ОТБОРА
Прошло более ста лет с тех пор, как Дарвин сформулировал
свою теорию. Это были годы тщательных наблюдений, анализа и
экспериментирования в области естественных наук. Было накопле-
но множество подробных сведений, важнейшие особенности кото-
рых будут рассмотрены на последующих страницах. Эти исследо-
вания обнаружили, что сформулированные в предыдущем разделе
пять условий, необходимых для действенности теории естествен-
ного отбора, не просты и многогранны. Эти исследования показа-
ли также, что существуют закономерности, определяющие харак-
тер взаимоотношений между взрослыми особями, и то, как они
производят гаметы и как размножаются. Мы относительно хорошо
знаем те правила, по которым гаметы объединяются для образо-
Теория эволюции путем естественного отбора 51
вания зигот (см. гл. 4—6). Мы начали до некоторой степени разби-
раться во взаимодействиях между взрослыми особями (гл. 12—15}
и в законах, по которым происходит их спаривание и образование
гамет (гл. 8), однако мы почти ничего не знаем о законах, касаю-
щихся развития организма от зиготы до взрослой особи. Между тем,
Бесполое
размножение
Рис. 3.2. Обобщенная схема жизненного цикла.
Не все стадии этой общей схемы имеются у каждого организма. Например, у жи-
вотных гаплоидная фаза не развивается до взрослого организма, а непосредствен-
но переходит от мейоза к оплодотворению. Мы хотим подчеркнуть, что естествен-
ный отбор действует на все стадии жизненного цикла, а не только на взрослую»
стадию.
для того чтобы делать правдоподобные предсказания относительно-
эволюционных событий, необходимо понимать жизненный цикл в-
целом (рис. 3.2). Следовательно, теория естественного отбора слу-
жит в настоящее время путеводной нитью и стимулом для созда-
ния более частных гипотез. Только после того, как различные ас-
пекты поведения отдельных особей и популяций будут поняты и
обобщены, теория естественного отбора станет подлинно прогно-
стической теорией. Это время еще далеко, и, принимая во внима-
ние сложность мира живого, оно, возможно, никогда не настанет.
Открывать новые направления, идти неизведанными путями —
в этом и состоит привлекательность популяционной биологии.
ЛИТЕРАТУРА
Darwin С. 1964. On the Origin of Species by Means of Natural Selection. Facsimile
of the first edition (1859) with introduction by E. Mayr, Cambridge, Mass.,
Harvard University Press.
4
52 Глава 3
Darwin С. 1975. Charles Darwin’s Natural Selection. Edited from manuscript by
R. C. Stauffer, Cambridge University Press, pp. 692f.
Ghiselin M. T. 1969. The Triumph of the Darwinian Method, Berkeley, University of
California Press, pp. 287f.
Ghiselin M. T. 1974. The Economy of Nature and the Evolution of Sex, Chapter 1
and 2, pp. 1—48, Berkeley, University of California Press.
Horn H. 1979. On the concept of optimality. In: О. T. Solbrig et al. (eds.), Topics in
Plant Population Biology, New York, Columbia University Press.
Kettlewell B. 1973. The Evolution of Melanism, Oxford, Clarendon Press, pp. 423f.
Mayr E. 1976. Evolution and the Diversity of Life. Chapter 1, Basic concepts of
evolutionary biology, pp. 9—16, Cambridge, Mass., Belknap Press of Harvard
University Press.
Solbrig О. T. 1976. Plant population biology: an overview, Systematic Botany 1,
202—208.
Часть вторая
Глава 4
Наследственность
и источники изменчивости
Эволюция невозможна без передачи потомкам признаков их
родителей. Только при этом условии признаки могут распростра-
няться в популяции в результате дифференциального выживания
и (или) дифференциального размножения. Дарвин понимал, что
наследственность играет важную роль в эволюции. Его очень ин-
тересовали механизмы, лежащие в основе наследственности и раз-
множения, и он провел много экспериментов для того, чтобы по-
нять их, однако ему не удалось выявить фундаментальные законы
генетики. Проблема наследственности была слабым местом в про-
возглашенной Дарвином теории эволюции.
Мендель открыл законы наследственности в 1865 г., но они не
оказали никакого влияния на биологию до тех пор, пока их не от-
крыли вторично в конце XIX в. С момента их вторичного открытия
наши знания о наследственности стали бурно развиваться. В этой
и следующих трех главах разбираются главные концепции гене-
тики и представления о механизмах наследственности, а также то,
как они влияют на эволюцию путем естественного отбора и какие
накладывают на нее ограничения.
КОРПУСКУЛЯРНАЯ и СЛИТНАЯ НАСЛЕДСТВЕННОСТЬ
Очень часто особь не представляет собой точной копии своих
родителей; иногда она больше похожа на одного родителя, чем на
другого, а иногда — на кого-либо из более отдаленных предков.
Случается, что у некоторых потомков появляются и совершенно
новые признаки. На протяжении многих лет выдвигались тщатель-
но разработанные теории для объяснения этих наблюдений. Так,
например, высказывалось мнение, что признаки, относящиеся к
одним частям организма растения или животного, наследуются
только по материнской линии, а относящиеся к другим частям —
только по отцовской. Высказывались предположения, что на неко-
торые признаки потомков оказывает влияние время года. Предпо-
лагалось также, что пища, поедаемая матерью, обусловливает не-
которые особенности потомства и тому подобное. В основе всех
этих идей лежит концепция, 'называемая теорией слитной наслед-
ственности; согласно этой концепции, наследственность каким-то
образом определяется «смешением кровей» предковых форм. Та-
54
Глава 4
кое представление все еще сохраняется среди широкой публики.
Оно проявляется в таких выражениях, как «чистокровный» приме-
нительно к чистопородным животным или же «кровное» родство —
о людях. Согласно теории слитной наследственности, совокупность
всех признаков каждого из родителей передается в виде некоего
целого потомку, у которого эти совокупности смешиваются и утра-
КОРПУСКУЛЯРНАЯ
НАСЛЕДСТВЕННОСТЬ
Рис. 4.1. Изменчивость при слитной и корпускулярной наследственности.
При скрещивании двух крайних форм в случае слитной наследственности все по-
томки в последующих поколениях носят промежуточный характер; в случае кор-
пускулярной наследственности после первого поколения будет происходить рас-
щепление. То, что мы наблюдаем в природе, легче поддается объяснению на основе
теории корпускулярной наследственности. (О. Т. Solbrig, 1966. Evolution and Sys-
tematics, New York, Macmillan.)
чивают свою индивидуальность. Тот факт, что потомок отличается
как от одного, так и от другого из своих родителей, приписывают
этому смешению. Различия между сестрами и братьями объясняют
тем, что «сила крови» того и другого родителя подвержена измен-
чивости. В качестве довода в пользу слитной наследственности
часто ссылаются также на то, что признаки потомков в общем
представляют собой нечто среднее между признаками их родите-
лей.
Последнее заключение служит одновременно и тестом для про-
верки правильности этой теории. Природные популяции растений
и животных состоят из особей, которые не вполне сходны друг с
другом. Их изменчивость поставляет сырье для естественного от-
бора. Однако если исходить из теории слитной наследственности,
то эта необходимая изменчивость должна была бы уменьшаться
вдвое в каждом поколении и всего за несколько поколений члены
данной популяции достигли бы полного единообразия по всем
своим признакам (рис. 4.1). Дальнейшая эволюция была бы не-
возможна без введения какого-либо нового источника изменчиво-
Наследственность и источники изменчивости
55
сти. Сторонники теории слитной наследственности постулировали,
что новая изменчивость вносится в популяцию со скоростью, экви-
валентной скорости утраты изменчивости, обусловленной слиянием
«аследственных задатков. Предполагалось, что механизмом, при
помощи которого вносится изменчивость, служит наследование
приобретенных признаков.
ТЕОРИЯ ПАНГЕНЕЗА
Чарлз Дарвин понимал, что для действия естественного отбо-
ра и для происходящей в результате этого эволюции необходимо
наличие в популяции известного запаса изменчивости. В популя-
ции, все члены которой совершенно одинаковы, из поколения в по-
коление не возникает никаких изменений, даже если имеет место
дифференциальное размножение и (или) дифференциальная смерт-
ность. Наблюдения над флорой и фауной, сделанные во время
кругосветного путешествия, а также у себя на родине в Англии,
навели Дарвина на мысль о том, что популяции животных и расте-
ний не однородны, а чрезвычайно изменчивы. Кроме того, его
обширные эксперименты по скрещиванию растений показали, что
эта изменчивость носит наследственный характер, а не обуслов-
лена случайными событиями в среде. Эволюция путем естествен-
ного отбора требует наличия большого запаса изменчивости, од-
нако если исходить из теории слитной наследственности, которая
в то время принималась почти всеми, то изменчивость в скрещи-
вающейся популяции должна практически отсутствовать. Дарвин
пытался примирить это противоречие, выдвинув теорию пангенеза.
Согласно этой теории, каждая часть тела вырабатывает особые
частицы — пангены, несущие информацию относительно наследст-
венных и приобретенных признаков; эти частицы включаются в
гаметы и передаются потомству (рис. 4.2). Такую сложную теорию
никак нельзя было проверить экспериментально, а кроме того, она
не давала исчерпывающего объяснения всем наблюдавшимся фак- :
там. Теория эта никогда не получала полного признания. j
ЗАРОДЫШЕВАЯ ЛИНИЯ И СОМАТИЧЕСКАЯ ЛИНИЯ |
Август Вейсман (August Weismann, 1834—1914), профессор 1
зоологии во Фрейбургском университете (Германия), сформули-
ровал в 1883 г. теорию зародышевой плазмы, пытаясь примирить
теоретическое требование большого запаса изменчивости в попу-
ляциях со своими наблюдениями, свидетельствовавшими о том, что ।
приобретенные признаки не наследуются. Вейсман провел экспе- i
римент, в котором он на протяжении 22 поколений отрезал мышам
хвосты и изучал влияние этого на длину хвостов у потомков этих
мышей. Он установил, что ампутация хвоста у родительских осо-
бей не оказывала никакого влияния на длину хвоста у их потом-
56
Глава 4
ков. Из этого Вейсман сделал вывод, что приобретенные признаки
обычно не передаются по наследству. Вейсман отметил также, что
наиболее примитивные организмы — одноклеточные — размножа»
Рис. 4.2. Теория пангенеза.
Согласно этой теории, каждая часть тела вырабатывает особые частицы, называе-
мые пангенами, которые несут в себе информацию о всех признаках организма.
Пангеиы перемещаются к половым органам, где оии включаются в гаметы и, та-
ким образом, передаются потомкам. Поскольку пангены несут информацию о всех
признаках организма, теория паигеиеза подразумевает, что признаки, приобретен-
ные организмом в течение его жизни, могут быть переданы его потомкам.
ются путем деления надвое, передавая своим потомкам все содер-
жимое своего тела. Что же касается более сложных многоклеточ-
ных организмов, то они передают потомкам только содержимое
своих половых клеток, или гамет, тогда как остальные части их
тела умирают и разлагаются. Вейсман пришел к заключению, что
каждый организм состоит из двух частей: зародышевой плазмы,
Наследственность и источники изменчивости
57
которая живет вечно, и развивающейся из нее сомы, которая
смертна (рис. 4.3). При помощи теории зародышевой плазмы Вейс-
ман смог примирить результаты своих экспериментов по' отреза-
нию хвостов с наблюдаемой в природе изменчивостью. Только те
изменения, которые затрагивают зародышевую плазму, передаются
потомкам; изменения, затрагивающие сому (например, разного рода
Смерть
/
Молодая особь
(дифференцировка
половых органов)
Рис. 4.3. Зародышевая линия и соматическая линия.
Гаметы и продуцирующие нх структуры составляют зародышевую линию организ-
ма (сплошные стрелки), которая ие прерывается, проходя через последовательные
поколения. Тело животного и растения, составляющее соматическую линию (пре-
рывистые стрелки), представляет собой временную вспомогательную структуру.
травмы), по наследству не передаются. Таким образом, хотя из-
менчивость в популяциях не может поддерживаться за счет на-
следования приобретенных признаков в том смысле, в каком это
обычно понимают, теория Вейсмана допускает возможность изме-
нений в зародышевой плазме, могущих служить источником измен-
чивости.
Теория Вейсмана пользовалась большим влиянием в свое время
и была важным этапом в развитии биологии. Наши современные
взгляды по-прежнему представляют собой модифицированную
концепцию зародышевой плазмы и сомы. Важным вкладом, вне-
сенным в науку теорией Вейсмана, была идея о том, что не все
признаки, приобретенные родителями, передаются потомству. Од-
нако, несмотря на то что Вейсман смог объяснить, почему многие
приобретенные признаки не передаются потомкам, его теория не
объясняла происхождения необходимой для эволюции изменчи-
вости и механизмов, поддерживающих ее в популяции.
58
Глава 4
КОРПУСКУЛЯРНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
Австрийский монах Грегор Мендель в 1865 г. сформулировал
корпускулярную теорию наследственности. Согласно его теории,,
родительские особи передают своим потомкам частицы, которые
теперь называют генами и которые несут информацию, необходи-
мую для воспроизведения родительских признаков. Гены служат
калькой, по которой создаются признаки.
В настоящее время нам известно, что гены находятся в субкле-
точных органеллах, называемых хромосомами. Каждый ген обычно'
занимает в хромосоме одно и то же определенное положение, на-
зываемое локусом (в латинском языке locus означает «место»)
данного гена. Большинство растений и животных диплоидны; это
означает, что их клетки содержат два набора хромосом — по одно-
му набору от каждого из родителей. Таким образом, у диплоидных
организмов имеется по два гена каждого типа, причем эти гены
занимают одинаковые локусы в гомологичных хромосомах. Если
два гена, занимающие одинаковые локусы, оказывают различное
влияние на признаки данного организма, то их называют разными
аллелями одного гена.
Число аллелей у того или иного гена может варьировать от од-
ного до многих, но обычно оно не превышает 10. Каждый дипло-
идный организм одновременно может содержать не более двух
аллелей, один из которых он получает от матери, а другой — от
отца. Если данная особь получит от обоих родителей одинаковые
аллели, то ее называют гомозиготной; если же она получит разные
аллели, то она будет гетерозиготной. Гетерозиготная особь может
быть похожа на особь, гомозиготную по одному из ее аллелей;
в таком случае этот аллель называют доминантным. Гетерозигот-
ная особь может также быть промежуточной между особями, го-
мозиготными по тем двум аллелям, которые она несет. Сочетание
доминантности по одним локусам и промежуточного наследования
по другим частично обусловливает черты сходства и различия
между родителями и их потомками. Если в популяции каждый из
генов, определяющих некий признак, представлен лишь одним ал-
лелем, то все особи будут одинаковыми по этому признаку.
Теория корпускулярной, или, как ее часто называют, менде-
левской, наследственности содержит три главных концепции.
1. Наследственность корпускулярна (дискретна), и каждая из
родительских особей передает своему потомку одинаковое число
генов, за исключением тех животных и растений, у которых имеют-
ся половые хромосомы1. У этих организмов вклады родителей
1 Половые хромосомы определяют пол данной особи. У некоторых видов осо-
би одного пола несут две половые хромосомы (XX), а особи другого пола — одну
(ХО), как у клопа-ромбовика Anasa tristis. У многих видов, в том числе у чело-
века и у дрозофилы, особи мужского пола имеют хромосому (Y), которой нет у
особей женского пола и которая образует пару (конъюгирует) с хромосомой X,
представленной у особей женского пола в двух экземплярах (XX).
Наследственность и источники изменчивости
59
по тем генам, которые локализованы в половых хромосомах, не-
одинаковы. Вывод о существовании генов был сделан теоретически,
однако впоследствии гены были идентифицированы биохимически.
2. Хотя некоторые гены могут подавлять или изменять дейст-
вие других генов, наследственные факторы непосредственно не
Розовы й
Рис. 4.4. Менделевский закон расщепления.
Среди генов, детерминирующих окраску цветков у львиного зева, нет доминиро-
вания. .В Fj цветки розовые, т. е. имеют промежуточную окраску между окраской
цветков родительских особей. (О. Т. Solbrig, 1966. Evolution and Systematics, New
York, Macmillan.)
влияют друг на друга. Иными словами, фундаментальная струк-
тура генов не изменяется при их ассоциации с другими генами и
они передаются следующему поколению в той же форме, в какой
они были получены предыдущим.
3. Гены исключительно стабильны по своему составу и функ-
ции. Тем не менее они могут изменяться, или мутировать, что со-
провождается изменением их функции; изменившийся ген называ-
ется мутантным. Гены могут мутировать под действием постоянно
существующих естественных причин; в этом случае процесс их
изменения называется спонтанной мутацией. Они могут также
мутировать в результате искусственного воздействия посторонних
факторов, например некоторых химических веществ или некоторых
60
Глава 4
видов излучения, в частности рентгеновских лучей (и атмосферного
излучения, испускаемого радиоактивными осадками!). Вещества,
способные индуцировать мутации, называют мутагенами, а воз-
Рис. 4.5. Менделерский закон расщепления.
У растений гороха высокий стебель представляет собой доминирующий признак.
Все гибриды Fi имеют высокий стебель. В F2 наблюдается расщепление в отноше-
нии 3:1. (О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystematics, New
York, Macmillan.)
никающие под их действием мутации — индуцированными мута-
циями.
Мендель сформулировал два главных следствия, вытекающие
из его теории, которые известны под названием «законов Менде-
ля»; первый из них — закон расщепления признаков (рис. 4.4, 4.5).
В своей современной форме он гласит, что если сперва скрестить
между собой двух особей, называемых родительским поколением
(Р), которые гомозиготны по разным аллелям одного и того же
Наследственность и источники изменчивости
61
гена, и затем скрестить между собой их потомков, или гибридоо
первого поколения (Fi), то потомство от этого второго скрещива-
ния (гибриды второго поколения F2) будет состоять из особей трех
Рис. 4.6. Менделевский закон независимого распределения.
Обратите внимание на то, что расщепление по признаку окраски семян у гороха
не зависит от расщепления по признаку морщинистых или гладких семян. В дан-
ном случае наблюдается доминирование: все растения Fi сходны с одним из роди-
телв|й и имеют желтые гладкие семена. (О. Т. Solbrig, 1966. Evolution and Syste-
matics, New York, Macmillan.)
типов в следующем соотношении: одна четверть их будет сходна
с одной из особей исходного родительского поколения (Р), поло-
вина— с гибридными родителями (Fi) и одна четверть — с другой
особью исходного родительского поколения (Р) (рис. 4.4). В тех
случаях, когда в первом гибридном поколении (FJ наблюдается
<62
Глава 4
полное доминирование, т. е. когда проявляется только один из
аллелей, класс особей в F2, сходных с Fb морфологически не будет
отличаться от класса, соответствующего доминантной особи исход-
ного родительского поколения (Р) (рис. 4.5). В таких случаях в
F2 соотношение особей составит 3:1, причем три четверти потом-
ков будут похожи на доминантную родительскую особь, а - одна
четверть — на другую родительскую особь Р, которая несет аллель,
называемый рецессивным. Закон расщепления, действие которого
было продемонстрировано многократно, представляет собой логи-
ческое следствие стабильности генов, обсуждавшейся выше. Закон
расщепления помогает также объяснить тот факт, что нередко
потомки сходны по какому-нибудь признаку не с отцом или ма-
терью, а с дедом или бабкой.
Второй закон Менделя — закон независимого распределения —
касается наследования по двум локусам. Этот закон гласит, что
.аллели одного гена будут расщепляться в соответствии с первым
законом, независимо от того, с каким аллелем второго гена он был
связан в родительском поколении. Возьмем, например, двух особей,
каждая из которых гомозиготна по двум локусам А и В, причем
по разным аллелям (1 и 2). Обозначим их генотипы AiAiB^ и
А2А2В2В2. Скрещивание между ними даст в Fi гибридов AiA2BiB2.
Если скрестить между собой особей из Fi, то в F2 получится неко-
торое число особей с генотипами исходной родительской пары (Р);
часть из них будет нести оба аллеля одного гена от одного из ис-
ходных родителей (Р) и оба аллеля другого гена — от другого
(например, А1А1В2В2), а часть будет нести иные сочетания алле-
лей (рис. 4.6). При любом сочетании аллелей одного гена аллели
другого гена расщепляются в соответствии с первым законом Мен-
деля. Так, например, среди особей, содержащих аллели AjA2, чет-
верть составляют AiA2BiBi, половину — А^гВ^г и четверть —
А1А2В2В2.
Во многих случаях независимого распределения не происходит.
Причина этого кроется в том, что гены не представляют собой фи-
зически независимые тела, а составляют часть хромосом. Если
два гена находятся в одной и той же хромосоме, то они передают-
ся вместе и не могут распределяться независимо друг от друга.
Юднако, как мы увидим в гл. 5, существует ряд специальных ме-
ханизмов, делающих возможным обмен аллелями между гомо-
логичными хромосомами в процессе формирования гамет (см. разд.
«Сцепление» и «Кроссинговер»).
ЕДИНИЧНЫЕ ГЕНЫ И СИСТЕМЫ МНОЖЕСТВЕННЫХ ГЕНОВ
Согласно первоначальной формулировке теории корпускулярной
наследственности, предложенной Менделем, каждый признак оп-
ределяется парой аллелей одного гена. Хотя известны случаи, ког-
да признак контролируется лишь одним геном, как правило, все
Наследственность и источники изменчивости
63
же большинство признаков контролируется многими генами. Мен-
дель считал также, что каждый ген оказывает влияние только на
один признак Однако теперь известно, что большинство генов вли-
Гуанин
Цитозин
Рис. 4.7. Компоненты молекулы ДНК.
Скелет каждой двухцепочечной молекулы ДНК построен из чередующихся фос-
фатных остатков и дезоксирибозы. Две цепи удерживаются вместе водородными-
связями, соединяющими соответственно молекулы гуанина и тимина одной цепи
с молекулами цитозина и аденина другой. Никакие другие сочетания оснований
невозможны по пространственным соображениям. (О. Т. Solbrig, 1970. Principles-
and Methods of Plant Biosystematics, New York, Macmillan.)
нет одновременно на много признаков, хотя их главное действие-
может явно сказываться только на какой-либо определенной струк-
туре или в какой-то один период развития. Может показаться, что
это утверждение противоречит корпускулярной наследственности.
Если мы полагаем, что ген хранит в себе определенную информа-
цию, то каким образом он может влиять на столь многие аспекты
<64
Глава 4
жизни данного организма? Ответ на это дает изучение биохимии
гена и процесса развития признака.
Гены расположены в хромосомах, содержащихся в ядрах всех
жлеток организма. По своему химическому составу гены пред-
ставляют собой дезоксирибонуклеиновую кислоту, или ДНК (у не-
которых вирусов гены состоят из РНК). Сложная молекула'ДНК
.(рис. 4.7) построена из шести более простых молекул: дезоксири-
А
Б -----------------------------------------------
Рис. 4.8. Репликация ДНК.
Л. При репликации ДНК цепи двойной спирали расходятся и подле каждой из
«их синтезируются отдельные, участки (которые позже соединяются) дочерних це-
пей, причем материнская цепь служит матрицей. Б. В эукариотических клетках
репликация ДНК начинается во многих точках молекулы.
бозы (сахар), фосфорной кислоты и четырех азотистых оснований
(двух пуринов — аденина и гуанина, и двух пиримидинов — цито-
зина и тимина). Полимеризуясь, эти шесть молекул образуют
ДНК. Каждая молекула сахара соединяется с остатком фосфорной
кислоты и с азотистым основанием (одним из четырех), образуя
нуклеотид. Нуклеотиды в свою очередь соединяются друг с другом
за счет образования связи между дезоксирибозой одного нуклео-
тида и фосфатом другого, в результате чего получаются длинные
цепи, закрученные в спираль таким образом, что основания ока-
зываются обращенными внутрь. Две такие цепи могут далее соеди-
ниться за счет образования водородных связей между их азоти-
стыми основаниями, в результате чего возникает двойная спираль
(нормальная или стабильная форма ДНК, содержащаяся в хро-
Наследственность и источники изменчивости
65
росомах). Образование двойной спирали возможно при соблюде-
нии следующего условия: против каждого тимина в одной цепи в
яругой цепи должен располагаться аденин, а против каждого ци-
тозина— гуанин и наоборот. Таким образом, последовательность
Азотистых оснований в одной цепи определяет их последователь-
ность в другой. При репликации молекулы ДНК (рис. 4.8) водо-
иодные связи между основаниями разрываются, обе цепи раскру-
чиваются и расходятся. Затем каждая цепь выступает в роли мат-
рицы для синтеза новой комплементарной ей цепи, что приводит
;к образованию двух новых двойных цепей с точно такой же по-
.следовательностью азотистых оснований.
Порядок расположения пар оснований в молекуле ДНК служит
«одом. Место тройки расположенных рядом пар нуклеотидов, так
называемого «триплета», в молекулах ДНК определяет — через
ряд промежуточных реакций — место соответствующей аминокис-
лоты в молекуле белка (сборка которой происходит в цитоплазме).
.Аминокислоты — это блоки, из которых построены белки. Фермен-
ты представляют собой важный класс белков, обладающих катали-
тическим действием; в качестве катализаторов они контролируют
специфические химические реакции в клетке. Для образования
таких структур, как цветок растения, конечность животного или
даже волос, для возникновения окраски цветочного лепестка не-
обходим целый ряд сложных химических реакций. Моменты «вклю-
чения» и «выключения» каждой из этих реакций и их скорости
контролируются определенным ферментом, синтез которого зави-
сит от определенного гена или набора генов. Однако большая
часть химических реакций, протекающих в клетках в процессе
развития, например крыла насекомого, связана не только с этой
•структурой. Структуру задают, вероятно, лишь последовательность,
•относительная важность и скорости. А следовательно, нетрудно
понять, что в развитии какого-либо одного признака могут участ-
вовать не один, а несколько генов (рис. 4.9) и что один ген может
действовать не на один, а на несколько признаков1.
Сроки и скорости синтеза того или иного фермента обычно ре-
гулируются по меньшей мере двумя генами. Один из них, называе-
мый структурным геном, содержит код, определяющий последова-
тельность аминокислот в полипептидной цепи данного фермента.
Другой — ген-регулятор — продуцирует фермент (или какое-либо
Другое вещество), которое воздействует на фермент, продуцируе-
!Мый структурным геном (или на сам этот структурный ген), тем
самым., регулируя активность структурного гена (т. е. управляя
'«го действием). Как именно осуществляется эта регуляция, более
1 Природа единиц наследственности (цистронов, генов-регуляторов и т. п.) и
Яо^еИСТВИе на су®клетОчнОМ и клеточном уровнях в настоящее время достаточно
тио Ш° ИЗуЧенЫ- ^ем не менее при объяснении генетических явлений на популя-
, “ном уР?вне правомерно пользоваться общим термином «ген», поскольку наши
ания о действии гена на этом уровне все еще неполны.
5—2044
66
Глава 4
НОРМАЛЬНЫЕ ГЕНЫ
МУТАНТНЫЕ ГЕНЫ
Мухи, несущие
доминантный
ген у +
ПРЕДШЕСТВЕННИК ТРИПТОФАНА
Накапливают триптофан
Мухи, несущие
доминантный
ген сп -ь
Мухи, несущие
доминантные
reHbist+ и cd+
СО—СН2—CHNHj—соон
вместе
ОН
(Г идроксикинуренин)
Рис. 4.9. Некоторые этапы образования коричневого глазного пигмента у дрозо-
филы и гены, контролирующие эти этапы.
Слева — нормальная ситуация, справа — действие мутантных генов. Гены дейст-
вуют на специфические химические реакции в соответствующих клетках, хотя
внешне их действие проявляется в виде морфологического изменения. (О. Т. Sol-
brig, 1966. Evolution and Systematics, New York, Macmillan.)
Наследственность и источники изменчивости
«Г
«ли менее точно установлено только для бактерий, у которых гены-
регуляторы способны подавлять действие структурных генов. У бо-
лее высокоорганизованных растений и животных такого механизма
регуляции, по-видимому, нет. Хотя точный механизм регуляции,
действующий у этих организмов, остается неясным, некоторые
эволюционисты считают, что изменения генов-регуляторов, воз-
можно, важнее, чем изменения структурных генов. Мы вернемся
к этому вопросу в гл. 9.
Если действие, производимое данным аллелем на некий при-
знак, невелико, то наличие или отсутствие такого аллеля окажет
лишь незначительное влияние на химические реакции, ведущие к
формированию этого признака. Если измерить этот признак у ор-
ганизмов, содержащих и не содержащих соответствующий аллель,
то различие окажется несущественным. То же самое наблюдается
в тех случаях, когда на какой-либо признак действует ряд раз-
личных неаллельных генов. В таких случаях в популяциях наблю-
дается непрерывная изменчивость между двумя крайними форма-
ми. Системы генов, которые поодиночке обладают небольшим ку-
мулятивным действием, а совместно контролируют непрерывную
изменчивость, называют системами множественных генов, или
полигенами. Гены, обладающие резко выраженным самостоятель-
яым действием и обусловливающие прерывистую изменчивость,
называют главными генами, олигогенами или, иногда, менделее-
вскими генами.
Между системами множественных генов и главными генами
нет абсолютного различия, поскольку известны гены, дающие весь
диапазон промежуточных эффектов. Кроме того, один и тот же
ген может оказывать резко выраженное действие на какой-либо
один признак, на основании чего его следует отнести к разряду
-«главных» генов, и вместе с тем обладать слабым действием на
какой-нибудь другой признак, что позволяет считать его частью
•системы множественных генов; тем не менее, несмотря на отсут-
ствие четкого различия между главными генами и системами
множественных генов, концепция системы генов полезна и вполне
применима для многих целей.
Можно думать, что такие признаки, как высота тела, под-
верженная в популяции непрерывной изменчивости, или интенсив-
ность окраски, которая также образует непрерывный ряд — от сов-
сем светлой до очень темной, — не могут контролироваться глав-
ными генами, обладающими четким и определенным действием.
Аллели главных генов, конечно, не могут обусловить такую непре-
рывную изменчивость, однако большое число генов, каждый из ко-
торых обладает небольшим кумулятивным действием, способно к
этому. Впервые кумулятивное действие генов было показано швед-
ским генетиком Нильссоном-Эле в 1911 г. Работая с пшеницей, ои
обнаружил, что окраска зерен зависит от трех неаллельных генов.
У растений, которые были гомозиготными рецессивами по всем трем
5*
68
Глава 4
генам, зерна были белые; у растений, содержавших доминантные
аллели всех трех генов, зерна были темно-красные, а у растений,
имевших некоторые, но не все, доминантные аллели, зерна был»
а 2 а о ® з ’Ч
а1а 1
Рис. 4.10. Генетическая основа окраски зерен у пшеницы.
А. Потомки растений, гомозиготных по рецессивным аллелям в двух локусах к
гетерозиготных по одному и тому же третьему локусу, расщепляются в соотноше-
нии 3 красных : 1 белый. Б. У растений, рецессивных по одному локусу и гетеро-
зиготных по двум другим, потомки расщепляются в отношении 15 красных: 1 бе-
лый. В. В случае гетерозиготности по всем трем локусам наблюдается расщепле-
ние в отношении 63 красных : 1 белый. Красные зерна различаются по интенсив-
ности красной окраски, которая пропорциональна числу имеющихся у растений
доминантных аллелей.
также красные, но не такие темные. Все эти гены обладали сход-
ным действием. Когда каждое растение изучали по отдельности,
то растения, бывшие гомозиготными рецессивами по двум .локусам и
гетерозиготами по одному, расщеплялись в соотношении 3 красных
зерна: 1 белое зерно (рис. 4.10,Д) Растения, гетерозиготные по
двум генам, расщеплялись в соотношении 15 красных:! белое.*
Наследственность и источники изменчивости
69
Рис. 4.11. Результаты экспериментов Иста (East) на Nicotiana longlflora, иллюст-
рирующие гипотезу множественных факторов.
Скрещивали растения с короткими (40—46 см) и длинными (88—97 см) цветками.
У растений Fi цветки были промежуточной (55—70 см) длины, а в Fj наблюда-
лось расщепление по этому признаку (52—88 см). Растения Fa, относившиеся к
разным классам по длине цветка (52, 61, 73 и 82 см), подвергали самоопылению.
Как и ожидалось, в их потомстве происходило расщепление, ио в F3 длина цветков
в общем была близка к длине цветков родителей. Такие же результаты были по-
лучены в F< и Fs. Если бы изменчивость была обусловлена факторами среды, а не
генетической конституцией, то во всех поколениях длина цветка была бы пример-
но одинакова; если же изменчивость определялась бы лишь одной парой расщеп-
ляющихся аллелей, то после второго дочернего поколения в некоторых линиях
расщепления не должно было бы происходить. (Числа в таблице соответствуют
числу растений с определенной длиной цветка в данном поколении. Обратите вни-
мание на распределение по этому признаку в пределах каждого поколения.)
(О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystematics, New York,
Macmillan.)
(рис. 4.10,5). Растения, гетерозиготные по всем трем генам, давали
соотношение 63 красных: 1 белое (рис. 4.10,5). При выращивании
красных зерен от растений F2, полученных от скрещивания особей,
гетерозиготных по всем трем генам, иногда получали расщепление
в отношении 3:1, иногда 15:1, а иногда 63:1, как того и ожидали.
Ясно, что в этом случае каждый ген ведет себя как настоящий
менделевский фактор и что все три подчиняются первому и второ-
му законам Менделя. Аддитивность эффектов этих трех генов
подтверждается также изменчивостью интенсивности окраски крас*
70
Глава 4
них зерен. Серии таких генов, каждый из которых обладает малым
эффектом, обусловливают наследование количественных призна-
ков, внешне давая картину непрерывной изменчивости. Существо-
вание множественных генов было окончательно продемонстриро-
вано в 1916 г. гарвардским генетиком Истом (East Е. М.), рабо-
тавшим на табаке (рис. 4.11).
Но даже и в тех случаях, когда признак определяется не од-
ним геном, а большим их числом, между генетически различными
группами должна существовать известная прерывистость, хотя
разграничение генотипических классов может оказаться при этом
нелегким делом. Границу провести не всегда возможно из-за не-
наследуемого компонента изменчивости, т. е. из-за действия таких
факторов среды, как пища, температура, свет, убежища. Эти фак-
торы создают изменчивость среди генетически идентичных особей.
Так, например, хорошо питающееся животное будет иметь больший
вес, а возможно, и большие размеры, чем истощенное животное
одинаковой с ним генетической конституции. Правда, этот компо-
нент не играет сколько-нибудь существенной роли в эволюции, по-
тому что, если потомкам той и другой особей давать одинаковое
количество пищи, они будут иметь в общем одинаковый вес. Од-
нако этот ненаследственный компонент создает непрерывную фе-
нотипическую изменчивость, которая накладывается на генотипи-
ческую дискретность. Для того чтобы избежать такого воздействия
среды, генетики стараются выращивать растения и животных в
одинаковых условиях, хотя в поле почти невозможно избежать
небольших различий по таким факторам, как количество осадков,
дренаж и минеральный состав почвы, точно так же, как трудно
выдавать животным совершенно одинаковые количества корма.
Эффекты нескольких дискретных генов, каждый из которых оказы-
вает на данный признак небольшое действие, сочетаются с нена-
следуемыми изменениями, обусловливаемыми средой, в результате
чего создается действительно непрерывная изменчивость
(рис. 4.12).
ЭПИСТАЗ И ПЛЕЙОТРОПИЯ
Иногда один из аллелей одного гена маскирует или модифи-
цирует фенотипический эффект одного из аллелей в другом ло-
кусе. Такое взаимодействие называют эпистазом. Этот множест-
венный эффект аллеля обусловлен тем, что главные генные про-
дукты— ферменты — катализируют определенную химическую ре-
акцию в метаболизме клетки. Когда на одном и том же метаболи-
ческом пути действуют два разных фермента, это приводит к эпи-
стазу.
Два независимых гена могут воздействовать на один и тот же
метаболический путь (см. рис. 4.9). В таких случаях скорость и
сроки действия одного гена должны соответствовать скорости и
Рис. 4.12. Сочетание множественных генов и пластичности приводит к непрерыв-
ному (штрихпунктирная кривая), а не к прерывистому (вертикальные линии) рас-
пределению.
В самом деле, если допустить, что каждый доминантный аллель на изображенной
здесь шахматной доске оказывает на величину гипотетического признака х влия-
ние, равное 7 единицам, а каждый рецессивный аллель — влияние, равное 2 еди-
ницам, то мы получим семь дискретных размерных классов: 12, 17, 22, 27, 32, 37
и 42 единицы с частотами соответственно 1,6; 9,4; 23,4; 31,3; 23,4; Э,4 и 1,6%, от-
вечающие наличию у особей 0, 1, 2, 3, 4, 5 и 6 доминантных аллелей. Одиако если
вследствие влияний среды отклонение признака у отдельной особи от величины,
ожидаемой для ее генотипа, может доходить до 3 единиц, то возникает серия фе-
нотипов со значениями данного признака, непрерывно изменяющимися от 9 до
45 единиц при указанных на графике ожидаемых частотах. Чем больше число ге-
нов, действующих на данный признак, тем более гладким будет его распределение.
(О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystematics, New York,
Macmillan.)
72
Глава 4
срокам действия другого гена. Строго говоря, эпистатическое взаи-
модействие между аллелями разных генов выражается в том, что
один ген маскирует эффект другого, в результате чего фенотип
определяется первым, а не вторым геном. Ген, маскирующий дру-
гой ген или препятствующий его проявлению, называется эписта-
тическим то отношению к последнему, который при этом называ-
ется гипостатическим.
Рис. 4.13. Наследование окраски у обыкновенной тыквы.
Плоды тыквы обычно бывают белыми, желтыми или зелеными. При скрещивани-
ях между растеииими с белыми и желтыми и между растениями с белыми и зе-
леными плодами во всех случаях доминирует белая окраска. В скрещиваниях
между растениями с желтыми и зелеными плодами всегда доминирует желтая
окраска. Таким образом, желтая окраска — рецессивный признак по отношению
к белой окраске, ио доминантный — по отношению к зеленой окраске. У растений
тыквы есть ген W, эпистатичный по отношению к гену Y. Во всех случаях, когда
ген W присутствует в доминантной форме, плод будет белым. В отсутствие алле-
ля W (при генотипе ww) плоды будут желтыми при наличии аллеля Y и зелены-
ми, если Он отсутствует. Аллель Y блокирует образование зеленого пигмента, но
не блокирует синтез желтых каротиноидов, а аллель W блокирует синтез обоих
пигментов.
Пример эпистаза обнаружен у дикорастущей фиалки Viola
tricolor, изучавшейся датским генетиком Йенсом Клаузеном (Jens
Clausen) в 1926 г. Обычно цветки у этого вида фиолетового цвета.
Окраска цветков зависит от пяти генов, обозначаемых Мь М2, М3,
М4 и М5. Если первый из этих генов (Mi) представлен доминант-
ным аллелем, то растение будет иметь фиолетовые цветки, неза-
висимо от того, будут ли остальные четыре аллеля представлены
доминантным или рецессивным аллелем. Если же первый ген пред-
ставлен в форме гомозиготного рецессива (пили), то цветки будут
светло-пурпурными, при условии, что второй ген представлен в до-
минантной форме (mimi, М2М2, ... или mimi, М2ш2, ...). Если первые
два гена рецессивны, а третий доминантен (пинц, т2т2, М3...),
то цветки будут пурпурными; а если первые три рецессивны, а
Наследственность и источники изменчивости
73
четвертый доминантен (mimb т2т2, т3т3, М4 ...), то темно-пур-
пурными; если первые четыре рецессивны, а пятый доминантен
(ГП1ГП1, m2m2, т3гпз, т4т4, М5...), то бархатисто-черными; а если
рецессивны все пять генов (tnjtnj, m2m2,
m3m3, гщгщ, ГП5ГП5), то блестяще-черными.
Другим хорошо известным примером
эпистаза служит наследование окраски
плодов у тыквы (рис. 4.13).
Термином плейотропия обозначают
детерминирование нескольких фенотипи-
ческих эффектов одним аллелем какого-
ss
3S
либо гена (рис. 4.14). Пример плейот-
ропного действия наблюдается у гена
S растений табака. У растения, несуще-
го доминантный аллель S, листья имеют
длинные черешки и заостренные кончи-
ки, чашечки с длинными острыми зубца-
ми, венчики со слабовыраженными ло-
пастями и слегка выступающими кончи-
ками, длинные пыльники и продолгова-
тые плоды. В отличие от этого у расте-
ний, гомозиготных по рецессивному ал-
лелю, листья сидячие, без заостренных
кончиков, чашечки с короткими зубца-
ми, венчики округлые, пыльники корот-
кие и плоды округлые. Плейотропия
представляет собой результат действия
гена, осуществляемого на одной из ран-
них стадий развития и затрагивающего
поэтому многие структуры. Большинст-
во, а может быть, и все гены обладают
плейотропным действием, но лишь в не-
многих случаях их эффекты проявляют-
ся так резко, как в приведенном приме-
ре. А поэтому термин плейотропия
обычно применяется для обозначения
именно таких крайних случаев.
Рис. 4.14. «Черешковый»
ген S у растений табака
(Nicotiana tabacum) облада-
ет рядом плейотропных эф-
фектов, обусловливая такие
признаки, как длинные че-
решки и заостренные кончи-
ки листьев, чашечки с длин-
ными тонкими зубцами, вен-
чики со слегка выраженны-
ми кончиками, длинные
пыльники и продолговатые
плоды.
ГЕНЫ, ГЕНОТИПЫ И ФЕНОТИПЫ
До сих пор мы описывали наследование признаков у организ-
мов так, как если бы между наследственным признаком и теми
генами, которые его в конечном счете определяют, существовала
простая зависимость. В некоторых случаях, особенно в экспери-
ментальных условиях, когда обеспечивается постоянство среды и
аллелей других генов, замещение в каком-либо организме одного
74
Глава 4
аллеля другим приводит к простым изменениям признаков, как при
проводившихся Менделем скрещиваниях растений гороха с длин-
ным и коротким стеблем. Известны тысячи примеров, когда заме-
щение одного аллеля другим приводило к изменениям признаков.
У растений такими признаками могут быть общие размеры, форма
листьев, окраска цветков, содержание крахмала в семенах. У’ жи-
вотных это может быть окраска шерсти, телосложение, группа
А Б
Рис. 4.15. Влияние гена Curly иа форму крыльев у Drosophila melanogaster.
А. Мухи с нормальными крыльями во время отдыха складывают крылья над
брюшком. Б. Мухи, несущие доминантный ген Curly (Су), не могут сложить
крылья таким образом, потому что у них крылья загнуты вверх.
крови, восприимчивость к болезням, способность ощущать вкус
некоторых веществ и т. п. Однако зависимость между генами и
признаками не является простой, поскольку проявление гена за-
висит от ряда других факторов, прежде всего от среды, в которой
развивается организм, и от других его генов, т. е. от так назы-
ваемого генетического фона.
Влияние среды на проявление гена можно проиллюстрировать
на примере гена Curly (Су) плодовой мушки (Drosophyla melano-
gaster) (рис. 4.15). Этот доминантный ген обусловливает развитие
у мух загнутых крыльев при условии, что куколок содержат при
температуре 25 °C. Если же куколок, имеющих в точности такую
же генетическую конституцию, содержать при 19 °C, то у многих
мух крылья будут нормальными, а у остальных наблюдаются раз-
нообразные отклонения в форме крыльев — от слегка до сильно
загнутых. По-видимому, ген Curly обусловливает развитие крыль-
ев, необычайно чувствительных к теплу в то время, когда муха
вылетает из влажной оболочки, покрывающей куколку. При высо-
кой температуре верхняя и нижняя поверхности крыльев обсыха-
ют с разной скоростью, в результате чего и получаются загнутые
крылья; при более низкой температуре скорость обсыхания ниже
и крылья загибаются у меньшего числа мух.
Влияние генетического фона исследовать труднее. На рис. 4.16
изображена частичная родословная пяти поколений одной семьи,
А. Руки девочки с полидактилией. Эта девочка не принадлежит к семье, родо-
словная которой представлена на схеме Б. Б. Некоторые представители семьи,
в которой была полидактилия. В каждой группе из четырех цифр две верхние
цифры соответствуют числу пальцев на левой и правой руках, а две нижние —
числу пальцев на левой и правой ногах. Звездочкой отмечены те случаи, когда в
первоисточнике тип полидактилии не был указан достаточно точно. (Lucas, Guy’s
Hosp. Rpts., 3rd sen, 25, 1881.)
члены которой несли доминантный ген полидактилии [избыточное
число пальцев на руках и (или) ногах]. Как показывает эта ро-
дословная, проявление гена сильно варьирует. У некоторых членов
семьи число пальцев на руках было нормальным, а на ногах было1
по 6 пальцев; у других были шестипалые кисти, но нормальные
ступни, у одного члена семьи было по 6 пальцев на руках и на
76
Глава 4
ногах, у одного было больше пальцев на правых, чем на левых
конечностях; а у двоих и на руках и на ногах число пальцев было
нормальным. В этом случае невозможно установить, были ли на-
блюдаемые различия вызваны влияниями среды или генетического
фона, однако, по всей вероятности, имело значение как то, так и
Другое.
Естественный отбор — это процесс дифференциального выжи-
вания и (или) дифференциального размножения особей. Если дан-
ный организм обладает каким-либо признаком, способствующим
его выживанию и (или) размножению, аллели, которые в конеч-
ном счете определяют этот признак, также сохраняются отбором.
Если, однако, эти аллели находят фенотипическое выражение толь-
ко у некоторых особей и только при некоторых условиях среды,
как, например, полидактилия у человека или загнутые крылья у
дрозофилы, то отбор будет действовать на них только в том слу-
чае, когда они выражены. Рассмотрим признак, понижающий вы-
живаемость и (или) размножение особей, которые им обладают.
У особей, несущих такой признак, число потомков будет ниже
среднего для данной популяции. Частота аллеля или аллелей, де-
терминирующих этот признак, будет понижаться, и в конце концов
они элиминируются. Однако скорость их исчезновения зависит от
того, как часто эти признаки оказываются выраженными. Для
того чтобы у особей, несущих данный аллель, проявился соответ-
ствующий признак, необходимо наличие каких-то особых условий
среды; а этот аллель, даже будучи вредным, может сохраняться в
популяции, поскольку он остается фенотипически невыраженным.
Теперь представим себе иную ситуацию. При каком-то одном
сочетании условий среды данный признак благоприятен для орга-
низма, а при другом — гибелен. Примером служит величина листь-
ев у растений. Во влажной среде растению лучше иметь большие
листья, а в засушливой — мелкие. Следовательно, растение, способ-
ное образовывать крупные листья в сезон дождей и мелкие —
в сезон засухи, будет обладать преимуществом. Одно из таких
растений — кустарник Encelia farinosa, растущий в пустынных об-
ластях на юго-востоке США (рис. 4.17). При высоком содержании
влаги в почве клетки растущего листа увеличиваются, в результате
чего лист достигает больших размеров. Если же влаги в почве
мало, то клетки не увеличиваются и листья получаются мелкими.
Точный генетический механизм, обусловливающий конкретное про-
явление данного признака в одной среде, а не в другой, и создаю-
щий тем самым два благоприятных признака, неизвестен. Очевид-
но, отбор благоприятствует аллелям, контролирующим такое из-
менчивое проявление признака.
Совокупность всех генов данного организма называют его гено-
типом; совокупность всех его признаков — фенотипом. Естествен-
ный отбор действует на фенотипы и лишь косвенно — на генотипы.
Если выражение данного признака, например величины листьев у
Наследственность и источники изменчивости
77
Encelia farinosa, зависит от факторов среды, то такой признак на-
зывается пластичным. Если данный ген детерминирует данный
признак только при определенных условиях, а в других условиях
этот признак не проявляется, то говорят, что этот ген обладает
неполной пенетрантностью; если степень выраженности признака,
Рис. 4.17. Растущий в пустыне кустарник Encelia farinosa дает мелкие, покрытые
волосками листья в засушливое время года и более крупные, менее густо опушеи-
«ые листья в дождливый период. (Фотография публикуется с любезного разре-
шения д-ра James Ehrelinger.)
детерминируемого данным геном, варьирует у различных особей,
как в случае гена Curly, то говорят, что этот ген обладает варьи-
рующей экспрессивностью. Следовательно, пластичный признак —
это результат фенотипического проявления генов с неполной пене-
трантностью и (или) варьирующей экспрессивностью.
Явления доминирования, эпистаза, неполной пенетрантности и
варьирующей экспрессивности приводят к тому, что несходные
генотипы могут формировать сходные фенотипы. Эти явления от-
ветственны также за то, что организмы, которые фенотипически
сходны (но генотипически различны) в одной среде, могут оказать-
ся фенотипически различными в другой среде. А следовательно,
Для того чтобы понять, как наследуются признаки в любых ситуа-
циях, недостаточно знать, какие аллели определяют их. Необхо-
димо также понять процессы роста и развития, т. е. этапы пути
от молекулы ДНК до фенотипических признаков, а также правил
78
Глава 4
ла, определяющие прохождение этих этапов. Они до сих пор не
получили объяснения, и пройдет еще некоторое время, прежде чем
мы обретем соответствующие знания. А поэтому возможности эво-
люциониста и популяционного генетика, пытающихся установить
эволюционное значение генетической структуры популяций, чрез-
вычайно ограничены. Чтобы решить эту проблему, при моделиро-
вании эволюционного процесса принимают ряд упрощающих допу-
щений, например, допущение о том, что отбор действует на гены,
допущение о постоянстве коэффициентов отбора или же о том, что
признаки определяются исключительно генотипом. Эти допущения
рассматриваются в нашей книге, но читатель должен помнить
том, что, поскольку истинная зависимость между генотипом и фе-
нотипом в настоящее время неизвестна, современная эволюционная
теория носит в значительной мере временный и условный характер.
Таким образом, мы должны уяснить себе природную ситуацию,
т. е. факты, и довольствоваться тем, что наша теория соответствует
этим фактам.
ЗАКОН ХАРДИ-ВАЙНБЕРГА И ПОДДЕРЖАНИЕ
ИЗМЕНЧИВОСТИ
Значение изменчивости в популяциях едва ли возможно пере-
оценить. Для того чтобы естественный отбор мог действовать, чле-
ны популяций должны отличаться друг от друга. Выше уже гово-
рилось, что существуют два источника изменчивости: генетический
и зависящий от среды, или негенетический. В эволюции главную
роль играет первый источник. Как непрерывная, так и прерыви-
стая изменчивость, как мелкие, так и крупные различия имеют
одну и ту же — генетическую основу, т. е. определяются генами..
В соответствии с теорией слитной наследственности генетическая
изменчивость утрачивается в результате гибридизации. Тот факт,
что генетическая изменчивость существует в природе, был одной
из причин отказа от этой теории. Теория же корпускулярной на-
следственности позволяет объяснить передачу отдельных призна-
ков от родителей потомкам без утраты изменчивости. Английский
математик Дж. Харди и немецкий генетик Г. Вайнберг независимо-
друг от друга установили это в 1908 г. Они показали, что в отсут-
ствие факторов, способных повысить или понизить изменчивость,
имеющаяся в данной популяции изменчивость будет оставаться на
исходном уровне. Эта концепция известна как формула (или за-
кон) Харди — Вайнберга.
Рассмотрим популяцию, состоящую из очень большого числа
особей (теоретически — из бесконечно большого числа), в которой
скрещивание происходит совершенно случайным образом. Допу-
стим, что ген А представлен в популяции двумя аллелями, Aj и А2,
с частотами paq соответственно, так что p+q=l. Гаметы (особые
клетки, содержащие один из двух имеющихся в организме наборов
Наследственность и источники изменчивости
79
г^нов), несущие аллель Аь встречаются с частотой р. Если их со-
единение с другими гаметами происходит случайным образом, то
дбля, соединяющаяся с гаметами Аь будет равна р, а доля, соеди-
няющаяся с гаметами Аг, будет равна q. Зиготы, получающиеся
в результате такого слияния гамет, будут иметь генотипы AiAi с
частотой р2 и генотипы А1А2 с частотой pq (рис. 4.18). Аналогии-
А,
Частота- р
Гамета
А2
Ч астота q
Гамета
а2
Частота р Частота q
АтА, Р2 А) А2 pq
а2а1 pq А 2 2 <?2
Рис. 4.18. Частота зигот в популяции.
При наличии гамет двух типов — Ai и А2 — с частотами, соответственно равными
и q, возможно образование зигот трех типов: А1А1 с частотой р2, AiA2 с частотой
2pq и А2А2 с частотой q2.
ным образом гаметы, несущие аллель А2, будут сочетаться с А2-
или Аггаметами, давая начало новым особям с генотипами А2А2
с частотой q2 и A2Aj — с частотой qp. Поскольку генетически гено*
типы AiA2 и A2Aj идентичны, их суммарная частота составляет
*2.pq. Таким образом, частоты различных генотипов в рассматривае-
мой популяции составят:
Генотип Частота
A4Ai р*
А1А3 2pq
А2А2 q2
В следующем поколении гомозиготные генотипы дадут гаметы
только одного типа, а гетерозиготные — равное число гамет одного
и другого типа. Если каждый организм в данной популяции про-
80
Глава 4
изводит в среднем одинаковое число гамет, то доля гамет каждого
типа будет зависеть от частоты в популяции разных фенотипов,
так что
доля А1 -гамет: p2+l/3(2pq) = p(p+q) =р,
доля Аг-гамет: q2 + 42(2pq) =q(q+p) =q.
Частота Аг и А2-гамет, производимых популяцией после сво-
бодного скрещивания в одном поколении при отсутствии отбора
или мутаций, будет такой же, как среди гамет, давших начало
этой популяции. Отсюда следует, что соотношение генотипов не
изменится и в последующих поколениях, а это означает, что в по-
пуляции поддерживается имевшаяся в ней исходная изменчивость.
Формула Харди—Вайнберга показывает, что при постоянных
условиях в популяции сохраняются первоначальные частоты генов
и что любые изменения этих частот возникают лишь под действием
внешних факторов. Для того чтобы объяснить наблюдаемое сохра-
нение изменчивости, нет нужды выдвигать какие-либо иные гипо-
тезы, например гипотезу о наследовании приобретенных признаков..
По существу, формула Харди—Вайнберга демонстрирует совме-
стимость менделевской теории наследственности и дарвиновской
теории эволюции. Поскольку в большинстве случаев естественный
отбор снижает изменчивость в популяции, необходимым условием
для непрерывной эволюции остается наличие источника новой из-
менчивости, даже при корпускулярной природе наследственности.
Однако источник этой новой изменчивости может быть значительно
скромнее того, который был бы необходим в рамках гипотезы слит-
ной наследственности.
ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И НАСЛЕДУЕМОСТЬ
Для того чтобы могла происходить эволюция путем естествен-
ного отбора, необходимо наличие в популяции изменчивости фено-
типических признаков, детерминированной генетически; иными
словами, образующие популяцию особи не могут быть генетически
идентичными (см. гл. 3). Поскольку в настоящее время мы обычно
не можем определить эту изменчивость непосредственно исследуя
генотип, приходится ограничиваться анализом фенотипической из-
менчивости с помощью статистических методов. Этими методами
пользуются в селекции растений и животных, где они оказались
весьма полезными. Иногда, однако, применение статистических
методов оказывалось совершенно неправильным, как, например, в
тех случаях, когда их пытались использовать для оценки различий
в умственных способностях между представителями разных рас
или между бедными и богатыми. Поэтому необходимо иметь не-
которое представление о способах получения статистических оце-
нок.
Наследственность и источники изменчивости 8В
Задача состоит в том, чтобы оценить, какую часть изменчиво-
сти, наблюдаемой в данной популяции, обитающей в определенной:
среде, можно отнести за счет генетических факторов, а какую — за
счет совокупного воздействия факторов среды.
Генетические свойства популяции нередко описывают, исполь-
зуя частоты генов. Однако при изучении популяций мы реально
наблюдаем лишь количественные различия между особями по ка-
кому-либо фенотипическому признаку или признакам. Любые на-
блюдения и измерения, производимые на составляющих популя-
цию особях, относятся к их фенотипам. Имея дело с тем или иным:
количественным признаком, мы обычно вычисляем его среднее:
значение и дисперсию. Среднее значение какого-либо призна-
ка (X) для данной популяции равно сумме его значений для от-
дельных особей, деленной на число измерений, т. е.
i=N
2*.
V 1=1
а дисперсия (s2) равна сумме квадратов отклонений от среднего^,
деленной на число измерений минус 1:
1-N
2(Хг-X)2
(Стандартное отклонение, или среднее квадратичное отклонение,,
равно квадратному корню из дисперсии.)
Среднее значение соответствует, грубо говоря, диапазону из-
менчивости данного признака, тогда как дисперсия характеризует
разброс значений вокруг среднего для данной популяции. Стоящая
перед нами задача заключается в том, чтобы выделить в разбросе
фенотипических значений составляющие, обусловленные различны-
ми генетическими факторами и факторами среды. В какой степени,
например, телосложение составляющих популяцию особей обу-
словлено средой, а в какой — генетическими факторами? Рассмот-
рим вкратце основные черты этой операции.
Генетика метрического (непрерывно изменяющегося) признака
основывается на изучении его изменчивости. Это объясняется тем,
что когда генетику приходится иметь дело с популяцией, в которой
имеется один или несколько непрерывно изменяющихся признаков,
он не может описывать отдельные классы, как он это делает, изу-
чая признаки, характеризующиеся прерывистой изменчивостью. Он
может определить лишь среднее значение, диапазон изменчивости
и дисперсию для данной популяции. После того как относительные
величины разных компонентов изменчивости в данной популяции
6-2044
«2
Глава 4
известны, генетик может на их основе делать выводы о генетиче-
ской структуре популяции.
Степень изменчивости измеряется и выражается через диспер-
сию. Если за значения признака принимаются отклонения от сред-
него для данной популяции, то дисперсия — это просто среднее
квадратов этих значений. (В данном случае дисперсия равйа
—X)2/N.) Полная дисперсия — это реально измеряемая фено-
типическая дисперсия, и она равна сумме генетической дисперсии
и дисперсии, обусловленной факторами среды, поскольку наблю-
даемое значение того или иного признака зависит от тех генов,
которые его детерминируют, и от среды обитания данного орга-
низма.
Для того чтобы непосредственно определить генетическую дис-
персию, генетик должен был бы выращивать растения или жи-
вотных в абсолютно одинаковых условиях и, допустив, что в таких
.условиях все отклонения негенетического происхождения равны
нулю, отождествлять наблюдаемую дисперсию с генетической.
К такой процедуре прибегают, однако, редко, так как создать аб-
солютно одинаковые условия практически невозможно.
Одно из свойств метрических признаков, определяемых систе-
мами множественных генов, — это сходство между родственными
особями, которое легко измерить и которое дает возможность оце-
нить генетическую дисперсию. Для проведения такого измерения
генетик делит популяцию на группы особей, связанных между со-
бой одинаковой степенью родства, называемые генетическими семь-
ями (например, сибсы, полусибсы, потомки и один или оба роди-
теля и т. п.). При помощи математического метода, известного
под названием дисперсионного анализа, наблюдаемую в популя-
ции изменчивость можно разделить на две компоненты; 1) внутри-
семейная дисперсия, т. е. отклонения особей от среднего для от-
дельных семей; 2) межсемейная дисперсия, т. е. отклонения сред-
них для семей от среднего для популяции. Если внутри семей все
особи идентичны, то вся изменчивость данной популяции — это
межсемейная изменчивость. Если же особи, составляющие отдель-
ные семьи, различаются между собой, и во всех семьях наблюда-
ется одинаковая изменчивость, то различия между семьями будут
минимальными, а изменчивость внутри семей — максимальной.
Мерой сходства между особями при данном разбиении популяции
на семьи может служить отношение «межсемейной» дисперсии к
сумме «внутри-» и «межсемейной» дисперсий:
/==..._g.a
В» 4- W3 ’
где В2 —дисперсия между семьями, IF2 — дисперсия в пределах
отдельных семей, a t—коэффициент корреляции между классами.
Чем больше t, тем больше сходство между членами данной семьи,
т. е. тем ближе родство между ними. Этот общий подход в соче-
Наследственность и источники изменчивости
83.
тании с рядом сложных статистических методов дает возможность
оценить генетическую дисперсию в популяции.
Зная общую фенотипическую дисперсию и генетическую дис-
персию, можно вычислить другой показатель, называемый насле-
дуемостью-.
где № — наследуемость (Но не квадрат наследуемости), —
генетическая дисперсия, a Vp— полная фенотипическая дисперсия..
Поскольку Vp всегда больше, чем Vg, значение й2 всегда будет
меньше единицы (табл. 4.1). Чем ближе h2 к 1, тем большую роль,
играет генетическая компонента в определении соответствующего'
признака. Заметим, однако, что, поскольку VG получают из корре-
ляции родственных особей в пределах одной популяции, сравни-
вать между собой популяции по наследуемости неправомерно1..
Наследуемость — это мера того, в какой степени изменчивость дан-
ного признака в пределах данной популяции обусловлена генети-
ческими факторами. Как таковая она дает селекционеру растений?
или животных представление о наличии или отсутствии запаса
генетической изменчивости, которую можно использовать для се-
лекции, и о величине этого запаса. Продолжив рассуждения, мож-
но сказать, что естественный отбор будет действовать только на те
Таблица 4.1
Приблизительные значения наследуемости различных признаков
у домашних и лабораторных животных (Falconer, 1960)
Крупный рогатый скот
Количество белых пятен у коров фризской породы 0,95-
Содержание жира в молоке 0,60'
Удойность 0,30
Частота зачатия при первой случке 0,01
Свиньи
Толщина хребтового сала 0,55
Длина тела 0,50
Вес в возрасте 180 дней 0,30
Величина помета 0,15
Куры
Вес яйца 0,60
Возраст при откладке 1-го яйца 0,50
Годовая продукция яиц 0,30
Вес тела 0,20
Выживаемость 0,10
Мыши
Длина хвоста в 6 недель 0,60
Вес тела в 6 недель 0,35
Величина помета 0,15
1 Имеется в виду сравнение интеллектуальных и других способностей у людей:,
принадлежащих к различным расовым и социальным группам. — Прим. ред.
«4
Глава 4
фенотипические признаки, которые наследуются (т. е. для которых
/г2>0), независимо от того, насколько они изменчивы в природе.
Мы рассмотрели концепцию наследуемости довольно подробно,
потому что эта концепция трудна для понимания и часто исполь-
зуется неправильно. Следует обратить внимание на то, что насле-
дуемость— это отношение генетической дисперсии к полной фено-
типической дисперсии. Ни одна из этих величин не является абсо-
лютным свойством конкретной особи или конкретного признака;
обе они — статистические характеристики, справедливые лишь в
условиях данной среды и данного генетического состава конкрет-
ной популяции. Поэтому мы не можем говорить о наследуемости.
того или иного признака безотносительно к той среде и той популя-
ции, для которой были получены рассматриваемые данные. Осо-
бенно опасны утверждения, затрагивающие наследуемость при-
знаков человека, в частности при сравнениях различных групп
людей. Такие утверждения лишены смысла и их следует избегать1.
Концепция наследуемости также важна для проблемы естест-
венного отбора, как это станет ясно в гл. 6, при рассмотрении ос-
новной теоремы Фишера о естественном отборе, поскольку чем
выше наследуемость данного признака, тем больше потенциальная
возможность его изменения под действием естественного отбора.
ЛИТЕРАТУРА
Общие вопросы
Falconer D. S. 1960. Introduction to Quantitative Genetics, New York, Ronald
Press.
Goodenough U., Levine R. P. 1978. Genetics, 2nd ed., New York, Holt, Rinehart and
Winston.
Spiess E. B. 1977. Genes in Populations, New York, Wiley.
Strickberger M. U7. 1976. Genetics, 2nd ed., New York, Macmillan.
Watson J. D. 1976. Molecular Biology of the Gene, 3rd ed., Menlo Park, Cal., Benja-
min. [Имеется перевод: Уотсон Дж. Молекулярная биология гена. — М.: Мир,
1978.]
•Специальные статьи и обзоры
Britten R. J., Davidson Е. Н. 1969. Gene regulation for higher cells: a theory, Scien-
ce, 165, 349—357.
Calvo J. M., Fink G. R. 1971. Regulation of biosynthetic pathways in bacteria and
fungi, Ann. Rev. Biochem., 40, 943—968.
Clausen J. 1926. Genetical and cytological investigations on Viola tricolor L. and
V. arvensis Murr, Hereditas 8, 1—156.
East E. M. 1916. Studies on size inheritance in Nicotiana, Genetics 1, 164—176.
Ehrelinger J. R., Bjorkman O., Mooney H. A. 1976. Leaf pubescence: effects on ab-
sorbance and photosynthesis in a desert species, Science, 192, 376—377.
Hardy G. H. 1908. Mendelian proportions in a mixed population, Science, 28, 49—
1 Такое сравнение ничего не дает, поскольку популяции различаются как по
генетической структуре, так и по условиям обитания, а наследуемость определя-
ется обоими этими факторами. — Прим. ред.
Наследственность и источники изменчивости
85
jNilsson-Ehle Н. 1909. Krenzungsuntersuchungen an Hafer und Weizen, Lunds
Univ. Arsskr. Afd., 2, 7 (2), 1—84.
Nirenberg M. IF. 1963. The genetic code: II. Scientific American 190 (March),
80—94.
Nirenberg AL W., Leder P. 1964. RNA codewords and protein synthesis: the effect
of trinucleotides upon the binding of sRNA to ribosomes, Science, 145, 1399—
1407.
Stebbins G. L. 1959. Genes, chromosomes and evolution. In: W. Turrill (ed.), Vis-
tas in Botany, London, Pergamon.
Stern C. 1973. Principles of Human Genetics, 3rd ed., San Francisco, Freeman.
Watson J. D., Crick F. H. C. 1953. A structure for deoxyribose nucleic acids, Natu-
re, 171, 737—738.
История вопроса
Brink R. A. (ed.) 1967. Heritage from Mendel, Madison, University of Wisconsin
Press.
Carlson E. A. 1966. The Gene: A Critical History, Philadelphia, Saunders.
Sturtevant A. H. 1965. A History of Genetics, New York, Harper and Row.
Watson J. D. 1968. The Double Helix, New York, Atheneum. Also 1969, New Yorki
The New American Library.
Глава 5
Гены и хромосомы
Изучение хромосом важно по двум причинам: 1) оно помогает
понять генетические явления и те эволюционные процессы, которые
лежат в их основе; 2) поведение хромосом, равно как их морфоло-
гические и химические особенности, можно использовать для оп-
ределения положения видов в системе. В этой главе рассматрива-
ются свойства хромосом, имеющие особенно большое значение в
указанных отношениях, а также отдельные особенно яркие при-
меры той помощи, которую может оказать цитология при решении
эволюционных проблем.
Подобно всем признакам любого организма хромосомы, а так-
же механизмы клеточного деления и образования гамет подверже-
ны действию естественного отбора. Поэтому они различаются у
разных видов, коль скоро эти виды имеют различную эволюцион-
ную историю. Такие различия обычно невелики между близко род-
ственными формами, но могут быть весьма значительными между
формами далекими. В связи с этим изучение цитологических раз-
личий дает ценные сведения о возможных путях эволюции тога
или иного вида. Однако, поскольку хромосомы являются носите-
лями генетической информации, они оказывают также влияние на
эволюционный потенциал вида.
СТРОЕНИЕ ХРОМОСОМ
Хромосомы представляют собой дискретные тельца, имеющиеся
в ядрах всех клеток (рис. 5.1), за исключением, быть может, бак-
териальных клеток и сине-зеленых водорослей (эти группы прими-
тивных организмов, по-видимому, лишены тех сложных хромосом,
состоящих из ДНК и белка, которые мы находим у более высоко-
организованных форм). Хромосомы имеют продолговатую форму
с расположенной в том или ином участке перетяжкой — центроме-
рой. Центромера делит хромосому на две части, называемые пле-
чами хромосомы. Если центромера расположена точно в середине
хромосомы, то она называется медианной, и плечи хромосомы име-
ют одинаковую длину. Если центромера лежит не строго посере-
дине, то она называется субмедианной; центромера, лежащая
ближе к одному из концов хромосомы, называется субтерминаль-
ной, а лежащая почти на конце хромосомы (по-видимому, центро-
Гены и хромосомы
87
мера никогда не располагаемся на самом конце) — терминальной.
“Таким образом, расположение центромеры служит основой для
классификации и идентификации хромосом.
Хромосомы можно также идентифицировать по их длине. Дли-
на хромосом варьирует от примерно 1 до 30 мкм; большая часть
хромосом в состоянии максим-ального сокращения в митозе имеет
Рис. 5.1. А. Соматические хромосомы человека (2п=46). Б. Метафаза первого де-
ления мейоза у Haplopappus gracilis (п=2); это наименьшее число хромосом, об-
наруженное до настоящего времени у растений. (Фотография любезно предостав-
лена д-ром Ray Jackson.) В. Диакинез в клетке папоротника Polystichum califor-
nicum (п=82); у многих папоротников число хромосом очень велико. (Фотография
любезно предоставлена д-ром Warren Н. Wagner.) Г. Диакинез у Gutierrezia brae-
teata (ге=8+1), стрелкой указана сверхкомплектная хромосома.
длину менее 10 мкм. Абсолютная и относительная длины двух плеч
хромосомы служат главными, а иногда и единственными крите-
риями для распознавания отдельных хромосом.
Иногда хромосомы можно идентифицировать по ряду дополни-
тельных признаков. Очень часто таким признаком оказывается
находящееся на одном из концов хромосомы небольшое округлое
тельце — так называемый сателлит, соединяющийся с основной
хромосомой тонкой хроматиновой нитью, или вторичной перетяж-
кой. Обычно лишь одна пара гомологичных хромосом в наборе
имеет спутника. У каждой отдельной хромосомы не бывает больше
68
Глава 5
одного сателлита. Другой признак, по которому иногда можно
различать хромосомы, это наличие у них ясно выраженных участ-
ков сильно окрашивающегося материала — гетерохроматина. Не-
Рис. 5.2. Дифференциальное
Хромосомы человека можно идентифицировать по характеру поперечной полоса-
положительно окрашивающимся Q и G-полосам (которые в общем довольно точ-
ваются одинаково (непостоянное
сколькими различными методами окраски у хромосомы во время
деления выявляется поперечная полосатость. Один из таких мето-
дов, состоящий в охлаждении с последующей фиксацией и окрас-
кой, используется уже в течение многих лет, однако большинство
других методов создано за последнее десятилетие. Один из новых
методов, так называемое Q-окрашивание, основан на том, что в
клетках человека и ряда животных некоторые флуоресцентные
красители специфически окрашивают лишь определенные участки
гетерохроматина. При других методиках, называемых G- и С-ок-
рашиванием, также избирательно окрашиваются определенные
Гены и хромосомы
89
участки хромосомы, часто соответствующие участкам, выявляемым
при Q-окрашивании. Применяя эти различные методы окрашива-
ния, можно разделить хромосому на определенные участки, каж-
Отрицательно или слабо окраши-
вающиеся Q-и G- полосы
Положительно окрашивающиеся R-полосы
Непостоянное окрашивание
Положительно окрашивающиеся Q-и G-полосы
Отрицательно окрашивающиеся R-полосы
окрашивание хромосом человека.
тости, выявляемой при некоторых методах окраски. Темные полосы соответствуют
яо совпадают), а заштрихованные участки — полосам, которые ие всегда окраши-
юкрашивание). (Paris Conference (1Э71): Standardization in Human Cytogenetics.
Birth Defects, Original Article Series, 8, 7, 1972).
Дый из которых обладает воспроизводимым типом окрашивания,
Что дает возможность идентифицировать не только отдельные хро-
мосомы, но и их плечи или отдельные участки (рис. 5.2).
УЛЬТРА СТРУКТУРА ХРОМОСОМ
В клетках большинства организмов хромосомы видны только
"Во время клеточного деления. По окончании митоза хромосомы
начинают вытягиваться и вытягиваются до тех пор, пока не ста-
новятся такими тонкими, что их бывает невозможно различить
so
Глава 5
wo A
Рис. 5.3. Гипотетическое строение хрома-
тина.
Хроматин состоит из многократно повто-
ряющихся единиц, называемых нуклеосо-
мами (/). Каждая нуклеосома содержит
8 молекул гистона (3), расположенных в
виде шара, который окружен двойной
спиралью ДНК (2), состоящей из при-
мерно 200 пар оснований. Конфигурация
молекулы ДНК точно не установлена.
(Kornberg, 1977)
даже с помощью самого мощ-
ного светового микроскопа.
Это обстоятельство долгое-
время озадачивало биологов..
Лишь недавно загадка была
частично разгадана благодаря
сочетанию скрупулезного хи-
мического анализа и электрон-
но-микроскопических исследо-
ваний.
Хромосома состоит в ос-
новном из трех химических ве-
ществ.
I. Белок. Более чем поло-
вину всей массы хромосомы
составляет особый белок —
гистон, обладающий щелочны-
ми свойствами (заряжен по-
ложительно при нейтральном
pH) вследствие высокого со-
держания в нем аминокислот
аргинина и лизина и имеющий
молекулярный вес от 11 000 ДО'
21 000. Кроме того, хромосома
содержит некоторое количест-
во кислого белка (заряжен от-
рицательно при нейтраль-
ном pH).
2. ДНК. Строение и гене-
тические функции ДНК были
описаны на стр. 64.
3. РНК. Рибонуклеиновая
кислота, осуществляющая
транскрипцию генетической
информации, содержится в
хромосомах в небольших, но
измеримых количествах.
Проблема состояла в том,
чтобы точно установить, как
эти вещества ассоциируются
друг с другом в хромосоме.
На основании имеющихся в настоящее время данных вырисовы-
вается следующая картина.
Гистон и ДНК объединены в перманентную структуру, назы-
ваемую хроматиновой нитью. Хроматиновая нить представляет
собой двойную спираль ДНК, окружающую гистоновый стержень;
Гены и хромосомы
91
она построена из повторяющихся единиц (нуклеосом), в каждую
из которых входит примерно 200 пар оснований ДНК и по две
молекулы каждого из четырех гистонов (Н2А, Н2В, НЗ и Н4;
рис. 5.3). Полагают, что эти восемь гистоновых молекул образуют
Рис. 5.4. Хроматин из эритроцитов курицы.
На хроматиновых нитях видны утолщения — нуклеосомы. (С. L. F. Woodcock,
J. Р. Safer, J. S. Stanchfield, 1976. Structural repeating units in chromatin, Exp.
Cell Res., 97, 101—110.)
сферическую единицу. Каким именно образом двойная спираль
ДНК располагается вокруг гистонов, пока неясно. Во многих, хотя
и не во всех клетках, с каждой нуклеосомой связано по одной мо-
лекуле еще одного гистона, Н1. На рис. 5.4 представлена элект-
ронная микрофотография хроматиновой нити, на которой видны
нуклеосомы.
Хроматиновая нить обычно образует спираль диаметром около
25 нм, что находится на грани разрешающей способности самых
92
Глава 5
мощных световых микроскопов. Эти тонкие нити называют эухро-
матином в отличие от более темного, более компактного и более-
интенсивно окрашивающегося гетерохроматина. Когда эти назва-
ния были впервые предложены, эухроматин и гетерохроматин счи-
Рис. 5.5. Микрофотография метафазной хромосомы из клетки китайского хомячка
(Cricelulus griseus), сделанная с помощью сканирующего электронного микроско-
па. Перетяжка в средней зоне указывает местонахождение центромеры.
(D. I. Patt, С. R. Patt, 1975. An Introduction to Modern Genetics, Reading, Mass.,.
Addison-Wesley.)
тали разными веществами, а не одним и тем же веществом в раз-
личных конфигурациях. Однако эта терминология по-прежнему
сохраняется из-за ее удобства при описании хромосом, а кроме
того, имеются исчерпывающие доказательства особых генетических
свойств гетерохроматина.
Перед началом клеточного деления большая часть хроматина
уплотняется, образуя хромосомы. При помощи электронного мик-
Гены и хромосомы
93
роскопа (рис. 5.5) можно увидеть, что хромосома состоит из плот-
но упакованного хроматина, в котором преобладают крупные еди-
ницы величиной 25 нм. Изучение в световом микроскопе ясно
показывает, что характер агрегации хроматина отличается боль-
шим постоянством, однако остается неясным, как именно происхо-
дит агрегация нитей. Кроме того, характер агрегации явно разли-
чается у разных видов, а иногда и в разных тканях (рис. 5.1).
ЧИСЛО ХРОМОСОМ
Число хромосом в клеточных ядрах всех особей какого-либо1
вида постоянно и представляет собой один из его признаков.
Все клетки любого организма происходят от клетки, образую-
щейся в результате слияния двух гамет. Эта клетка называется
зиготой и содержит два набора хромосом. При каждом делении
сначала зиготы, а затем получающихся из нее клеток эти хромо-
сомы расщепляются в продольном направлении, в результате чего
все клетки данного организма получают одинаковое число одина-
ковых хромосом. В определенных тканях продольное расщепление-
хромосом не сопровождается делением клетки. В результате полу-
чаются клетки, содержащие вдвое или даже вчетверо больше хро-
Таблица 5.1
Число хромосом у некоторых обычных растений и животных
Вид Латинское название Число хромосом в соматических клетках Уровеиь плондио~ сти
Пшеница Triticum aestivum 42 Гексаплоид
Кукуруза Zea mays 20 Диплоид
Салат-латук Lactuca sativa 18
Томат Lycopersicon esculentum 12
Хлопчатник Gossypium barbadense 52 Тетраплоид
Обыкновенная фа- соль Phaseolus vulgaris 22 Диплоид
Соя Glycine max 40 Тетраплоид
Яблоня Pyrus communis 34 Диплоид
Батат Ipomea batatas 90 Гексаплоид.
Папоротник Ophioglossum vulgatum 480 ?
Медоносная пче- ла Apis mellifera 16 Диплоид
Тутовый шелко- пряд Bombyx mori 56 >
Карп Cyprinus carpio 100 >
Леопардовая ля- гушка Rana pipiens 26
Аллигатор Alligator mississippiensis 32 >
Кряква Anas platyrhynchos 78 »
Бык Bos taurus 60
Лошадь Equus caballus 64 »
Кошка Felis felis 38
Собака Canis familiaris 78 1 >
Э4
Глава 5
мосом, чем зигота. При этом процессе, носящем название эндо-
митоза, изменяется число хромосом, но не их морфология. Эндо-
митоз имеет физиологическое, а не генетическое значение, так как
он в норме не происходит в клетках тканей, продуцирующих га-
меты. Число хромосом в соматических клетках варьирует от ми-
Рис. 5.6. Редукция числа хромосом.
Две хромосомы, АА' и ВВ', у каждой из которых имеется терминальная центро-
мера, сливаются, образуя одну хромосому АВ с медианной центромерой и струк-
туру, представляющую собой по существу единичную центромеру с двумя корот-
кими плечами А'В'. Утрата этой центромеры не окажет никакого генетического
эффекта на организм. Описанный механизм требует одновременного возникнове-
ния разрывов в точках 1 н 2. (О. Т. Solbrig, 1970. Principles and Methods of
Plant Biosystematics, New York, Macmillan.)
-нимум двух пар — в клетках представителя семейства сложно-
цветных Haplopappus gracilis (рис. 5.1) до нескольких сотен —
у ряда насекомых и низших животных (табл. 5.1). У большинства
видов растений и животных число хромосом составляет от 5 до
30 пар. У человека их 23 пары (рис. 5.1). У аскариды (Ascaris
megalocephala var. univalens) мейотические клетки и клетки заро-
дышевого пути содержат по одной хромосоме, но в соматических
клетках она распадается на несколько хромосом.
Хотя число хромосом у каждого вида постоянно, в гаметах
или в тканях, которые их продуцируют, иногда возникают откло-
нения. Такие отклонения возникают в основном двумя путями:
Гены и хромосомы
95»
1) при делении хромосом в клетке, из которой должны образо-
ваться гаметы, когда сама клетка при этом не делится; в резуль-
тате получается клетка, содержащая ровно вдвое больше обычного
Б
А
С. kashmirica
л = 6
hA A A i с. типь
л=6
в • * 11
||1“
38
С. leontodontoides
л = 5
С. conyzaefolia
п=4
21
С. suffreniand
п=4
С. capillaris
п = 3
38! Л
1'1
22
С. fuliginosa
л = 3
Рис. 5.7. Эволюция хромосом в двух линиях Crepis: с крупными (X) и мелкими
(5) хромосомами.
Наиболее примитивный вид каждой линии помещен ввеоху, а наиболее продвину-
тый— внизу. Числа указывают общую длину всех хромосом у каждого вида по
отношению к их длине у Crepis kashmirica, принятой за 100. (Babcock, 1947)
числа хромосом; 2) при слияниях или разрывах отдельных хромо-
сом, в результате чего получаются клетки с избыточным или умень-
шенным числом хромосом (рис. 5.6). Для того чтобы клетка оста-
валась жизнеспособной, увеличение или уменьшение числа хро-
мосом не должно сопровождаться сколько-нибудь существенного
потерей генетического материала. Поэтому утрата целых хромосом
"96
Глава 5
будет гибельной, если только у данной особи не произошло ранее
удвоения числа хромосом. При слиянии двух хромосом одна из
двух центромер должна быть утрачена. Иначе во время клеточного
деления хромосома оказалась бы прикрепленной к двум нитям ве-
ретена (нити веретена участвуют в перемещении хромосом при
делении клетки), а это имело бы гибельные последствия. Чаще
всего при слиянии происходит утрата чрезвычайно коротких плеч
и частей терминальной центромеры двух хромосом, которые затем
соединяются при помощи оставшихся частей центромеры и дают
начало хромосоме с медианной или субмедианной центромерой.
.Длина этой хромосомы примерно равна суммарной длине двух
исходных хромосом. Виды с низким хромосомным числом часто
содержат хромосомы с медианной или субмедианной центромерой,
указывающие на то, что у предков этих видов, имевших большее
число хромосом, произошла редукция их числа. Хорошим приме-
ром служат растения рода Crepis. Для этой группы растений ока-
залось возможным восстановить эволюцию кариотипа; кариотипом
называют основной набор хромосом у данного вида. Те виды рода
Crepis, которые с эволюционной точки зрения примитивны, содер-
жат основное число хромосом (и = 6), а у родственных им более
•специализированных форм хромосом меньше—5, 4 и 3 (рис. 5.7
и табл. 5.2).
Таблица 5.2
Число хромосом у некоторых видов
(по данным Babcock, 1947)
Вид Число хромосом в соматических клетках Уровень плоидно- сти Тип размножения Продолжитель- ность жизни
С. elegans 14 Днплоид Половое Многолетник
>С. paludosa 12 » >
С. sibirica 10 > >
С. chrysantha 8 >
С. polytrlcha 16 Тетраплоид » »
С. clliata 40 Октоплонд Двулетник
•С. alpina 10 Днплоид Однолетник
•С. patula 8 >
С. capillaris 6 >
С. monticola 22—88 Аллополиплоид Бесполое Многолетник
С. intermedia 33—88 В »
С. barbigera 44—88 » >
МИТОЗ И МЕЙОЗ
Все клетки имеют примерно одинаковые очень небольшие раз-
меры; организм животного такого размера, как лошадь, или та-
кого растения, как дуб, состоит из миллионов и миллиардов кле-
ток. Продолжительность жизни многих клеток относительно неве-
лика, а поэтому они должны непрерывно замещаться. Делению
Гены и хромосомы
97
рлетки предшествует сложный процесс деления ядра, называемый
яитозом (рис. 5.8).
Деление клеток у разных видов различается во многих отно-
шениях; между делением растительных и животных клеток суще-
ствует ряд резких различий по второстепенным чертам этого про-
Рис. 5.8. Митоз — процесс, обеспечивающий распределение удвоенного генетическо-
го материала по дочерним клеткам.
На ранних стадиях (профаза, I) хроматин собирается в глыбки и становится вид-
ным в световой микроскоп; к концу профазы каждая хромосома состоит из двух
сестринских хроматид. В метафазе (II) у большинства клеток ядериая мембрана
исчезает; появляется веретено, идущее от одного полюса клетки к другому; хро-
мосомы выстраиваются в плоскости экватора. Во время аиафазы (III) центроме-
ры делятся, сестринские хроматиды отделяются одна от другой и перемещаются к
двум противоположным полюсам клетки. В телофазе (IV) вокруг каждого набо-
ра дочерних хромосом образуется ядерная мембрана, веретено исчезает и хромо-
сомы- постепенно деспирализуются. У большинства клеток вслед за митозом про-
исходит деление самих клеток. А. Митоз в животной клетке. Б. Митоз в раститель-
ной клетке. 1 — клеточная мембрана, 2 — центриоли, 3 — веретено, 4 — звезда, 5 —
ядрышко, 6 — клеточная стенка, 7 — клеточная пластинка. (D. I. Patt, G. R. Patt,
1975. An Introduction to Modern Genetics, Reading, Mass., Addison-Wesley.)
цесса. Что касается митоза, то он протекает в основном одинаково
У всех организмов. В делении ядра должен участвовать механизм,
обеспечивающий строго равномерное распределение наследствен-
ных частиц — генов — между дочерними клетками. И хотя некото-
рые особенности митоза иногда используют в качестве диагности-
ческих признаков для идентификации различных групп животных
Или растений (например, у видов с диффузной центромерой), такие
случаи редки.
С генетической точки зрения наиболее важная черта митоза —
это точное копирование хромосомы и ее основных составляющих
7-2044
98
Глава 5
частей, с тем чтобы оба дочерних ядра получили одинаковое ко-
личество генетической информации. Это необходимо для сохране-
ния генетической непрерывности вида. Физико-химические свойст-
ва ДНК обеспечивают точное копирование генетического содержа-
ния хромосом.
Профаза I
Интеркинез
Tелофаза I
Метафаза I Анафаза I
Рис. 5.9. Стадии мейоза в схематизированной животной клетке.
Физические состояния хромосом и их хромонем на каждой стадии профазы I изо-
бражены по отдельности, чтобы более четко показать явления конъюгации, ре-
комбинации и образования хиазмы. Стрелки указывают точки разрыва и обмена
участками. (D. I. Patt, G. R. Patt, 1975. An Introduction to Modern Genetics, Rea-
ding, Mass., Addison — Wesley.)
Клеточное деление необходимо для роста и поддержания жиз-
недеятельности организма. У одноклеточных организмов оно также
служит механизмом увеличения числа особей. У многоклеточных
организмов имеются специальные клетки — гаметы, которые при
оплодотворении сливаются, образуя зиготу. В процессе последо-
вательных делений зигота дает начало новой особи того же вида.
Поскольку при слиянии двух гамет их хромосомы объединяются,
Гены и хромосомы
99
т. е. число их удваивается, образование гамет у следующего по-
коления должно сопровождаться уменьшением числа хромосом
вдвое. В противном случае всего за несколько поколений число
хромосом увеличилось бы так сильно, что их объем превысил бы
объем не только ядра, но и всей клетки. Процесс, сводящий число
хромосом к исходному, называется мейозом.
Мейоз слагается из двух последовательных делений, в резуль-
тате которых образуется 4 дочерних ядра (рис. 5.9). Однако удвое-
ние хромосом при этом происходит лишь один раз. У большинства
организмов деление ядра сопровождается делением цитоплазмы.
Мейоз — это процесс, имеющий огромное значение как для наслед-
ственности, так и для эволюции. С генетической точки зрения
самыми важными чертами мейоза представляются уменьшение
числа хромосом ровно вдвое, независимое распределение гомоло-
гичных хромосом по дочерним ядрам и обмен участками гомоло-
гичных хромосом в результате кроссинговера. Гомологичными на-
зывают хромосомы, несущие аллели одних и тех же генов. При
слиянии двух гамет для каждой хромосомы из одной гаметы нахо-
дится гомолог из другой гаметы.
Профаза мейоза делится на несколько стадий (рис. 5.9), пото-
му что она протекает дольше и сложнее, чем профаза митоза. Пер-
вое важное различие между двумя профазами состоит в том, что
когда в профазе мейоза хромосомы становятся видимыми, они
представляют собой целостные структуры, т. е. не разделены про-
дольно на две хроматиды, как в профазе митоза (рис. 5.9). Вто-
рое и очень важное отличие профазы мейоза — происходящая весь-
ма специфическим образом конъюгация гомологичных хромосом,
в результате которой центромеры и гомологичные участки хромо-
сом располагаются друг против друга. Это приводит к тому, что
вместо диплоидного числа хромосом получается гаплоидное число
пар гомологичных хромосом, называемых бивалентами. В процес-
се конъюгации хромосомы обвиваются одна вокруг другой и ста-
новятся короче и толще.
Спустя некоторое время после завершения процесса конъюга-
ции гомологичные члены каждого бивалента начинают расходить-
ся. В участках расхождения можно видеть, что каждый гомолог
теперь продольно разделен на две хроматиды. Однако в одном или
нескольких местах по длине бивалента одна хроматида от каж-
дого гомолога обменивается участками со своим партнером; при
этом две хроматиды от разных членов бивалента перекрещиваются
Друг с другом (рис. 5.10). Каждая из этих выявляемых цитологи-
ческими методами точек перекреста называется хиазмой. Хиазмы
Удерживают гомологичные хромосомы вместе и, по мере того как
расхождение хромосом продолжается, между последовательными
хиазмами образуются петли, а между концом хромосомы и бли-
жайшей к ней хиазмой — полупетли. Процесс расхождения вынуж-
дает хиазмы перемещаться к концам хромосомы, чему способству-
7*
100
Глава 5
ют также продолжающаяся спирализация и сокращение хромосом.
По мере приближения профазы к концу центромеры гомологичных
хромосом каждого бивалента располагаются по обе стороны эква-
тора веретена на равных расстояниях от него (рис. 5.8). В какой-
то момент связь между сестринскими хроматидами разрушается,
хиазмы распадаются и каждая половинка бивалента отходит к
одному из полюсов, куда ее тянет соответствующая центромера.
Поскольку от каждого бивалента к каждому полюсу отходит
только по одной хромосоме, у каждого полюса собирается гапло-
Рис. 5.10. Конъюгировавшие хромосомы в мейозе.
Часто во время профазы одна из хроматид перекручивается с хроматидой гомо-
логичной хромосомы. Точки, в которых хроматиды соприкасаются друг с другом,
называются хиазмами.
идное число хромосом. Они, однако, отличаются от хромосом в
анафазе митоза тем, что каждая из них состоит из двух хроматид.
Вскоре они проделывают второе деление, механически сходное с
митотическим делением. Перед вторым делением мейоза хромосо-
мы могут на короткое время образовать интерфазное ядро. Иногда
после первого деления мейоза происходит также деление цито-
плазмы, а у растений может даже образоваться клеточная стенка.
В других же случаях второе деление мейоза непосредственно
следует за первым без промежуточной интерфазы и может иногда
сопровождаться делением цитоплазмы.
Второе деление мейоза начинается с того, что все хромосомы
выстраиваются по экватору второго веретена деления. Вслед за
этим происходит одновременное разделение хроматид каждой хро-
мосомы. Таким образом, в результате мейоза образуется четыре
ядра, каждое из которых содержит гаплоидное число единичных
хроматид.
Мейотическое деление описано здесь так подробно, потому что
менделевские законы расщепления и независимого распределения
непосредственно вытекают из поведения хромосом вовремя мейоза.
Рассмотрим организм с генетической конституцией А^г, уна-
следовавший каждый из этих аллелей от гомозиготных родителей
AiAi и А2А2. Эти два аллеля занимают совершенно одинаковое
положение в гомологичных хромосомах. При мейотическом деле-
нии клетки гомологичные хромосомы расходятся во время первого
деления мейоза и делятся во время второго деления мейоза, в ре-
Гены и хромосомы
101
зультате чего возникают четыре диплоидные клетки, из которых
две несут аллель А1 и две—аллель Аг, в соответствии с менделев-
ским законом расщепления.
Переходя далее к рассмотрению двух пар аллелей А1А2 и В^г-
для двух генов, расположенных в разных хромосомах, можно ви-
деть, что поскольку разные биваленты располагаются на эквато-
риальной пластинке метафазы I независимо друг от друга, по-
стольку негомологичные хромосомы при мейозе распределяются
независимо; иными словами, каждая из образующихся четырех
клеток с равной вероятностью может получить отцовский или ма-
теринский гомолог. Это служит физической основой второго зако-
на Менделя — закона независимого распределения.
Однако не все гены данного генома (т. е. полного набора генов
данного индивидуума) распределяются независимо, потому что ге-
ны, лежащие в одной хромосоме, физически связаны между собой.
Эта связь, правда, не абсолютна, что обусловлено явлением, по-
лучившим название кроссинговера.
СЦЕПЛЕНИЕ И КРОССИНГОВЕР
Очень важно, чтобы при каждом делении ядра дочерние ядра
получали точные копии генетического набора материнского ядра.
То обстоятельство, что гены собраны в хромосомы, значительно
облегчает эту задачу. В самом деле, если бы каждый из более
чем 10 000 содержащихся в клетке генов должен был самостоя-
тельно делиться, а затем каждая из двух его половин направля-
лась бы к своему полюсу, то на их пути неизбежно возникала бы
«пробка», которая привела бы к образованию дочерних ядер раз-
ной величины. В таких условиях вряд ли было бы возможно раз-
витие сложных многоклеточных организмов. Объединение генов в
хромосомы сделало возможным равномерное распределение генов,
но одновременно ограничило независимость этого распределения.
В сущности, при мейозе гены, находящиеся в одной хромосоме,
должны все вместе отойти к одному полюсу. Это явление, извест-
ное под названием сцепления, имеет большое генетическое зна-
чение.
Гены, лежащие в разных хромосомах, не сцеплены между со-
бой, в отличие от генов, локализованных в одной хромосоме, ко-
торые сцеплены друг с другом. Для каждого вида характерно
определенное число генов; число групп сцепленных между собой
генов соответствует числу хромосом у данного вида. Чем меньше
число групп сцепления при постоянном числе генов, тем больше
число генов в каждой группе сцепления и тем формально менее
справедлив закон независимого распределения. Это явление было
впервые отмечено двумя английскими генетиками — У. Бэтсоном
(W. Bateson) и Р. К. Пеннетом (R. С. Punnett) —в 1906 г. Когда
эти авторы скрещивали растения душистого горошка, имевшие
102
Глава 5
пурпурные цветки и продолговатые («длинные») пыльцевые зерна,
с растениями, имевшими красные цветки и округлые пыльцевые
зерна, то они получали только растения с пурпурными цветками
и длинными пыльцевыми зернами; это показывает, что пурпурные
цветки и длинные пыльцевые зерна — доминантные признаки. В F2
они получили четыре комбинации, которых и следовало ожидать в
соответствии с независимым распределением генов (пурпурные/
длинные, пурпурные/округлые, красные/длинные, красные/округ-
лые), но не в ожидавшемся соотношении 9:3:3: 1, а в соотноше-
нии, в котором число двух родительских комбинаций (пурпурные/
длинные и красные/округлые) было выше ожидаемого. Соотноше-
ние четырех классов составляло примерно 11:1:1:3. Этот не-
ожиданный результат противоречил законам Менделя.
Ответ на загадку, порожденную экспериментами Бэтсона и
Пеннета, удалось получить только после исследований на плодовой
мушке — дрозофиле (1910—1915), проведенных лауреатом Нобе-
левской премии американским генетиком Томасом Г. Морганом
(Thomas Н. Morgan) и его учениками и близкими сотрудниками:
Альфредом Стертевантом (Alfred Stertevant), К- Б. Бриджесом
(С. В. Bridges) и Германом Мюллером (Hermann Muller). Бэтсон
и Пеннет показали, что закон независимого распределения непри-
меним к наследованию окраски цветков и формы пыльцевых зерен
у душистого горошка. Если бы, однако, гены, обусловливающие
эти признаки, лежали в одной неразрывной хромосоме, то все по-
томство должно было бы состоять только из родительских типов.
Наличие среди потомков растений с пурпурными цветками и
округлыми пыльцевыми зернами, а также растений с красными
цветками и продолговатыми пыльцевыми зернами обусловлено об-
меном соответствующими участками между двумя хроматидами
гомологичных хромосом во время мейоза. Это явление называют
кроссинговером (рис. 5.11), а цитологически оно проявляется в
образовании хиазм.
На рис. 5.11 показана схема кроссинговера. Обратите особое
внимание на то, что кроссинговер происходит между двумя хро-
матидами на четырехцепочечной стадии профазы мейоза и при-
водит к тому, что в телофазе мейоза имеется по одной хромосоме
с каждой из родительских генетических комбинаций (Ах Bt и А2В2)
и две рекомбинантные хромосомы (AiB2 и AzBt). Следовательно,
участки, лежащие по обе стороны от перекреста, распределяются
независимо.
Число хиазм (и соответствующих перекрестов), происходящих
в каждом хромосомном плече, ограничено. Оно зависит от ряда
факторов, таких, как длина данной хромосомы, генетическая кон-
ституция вида и особи, наличие других хиазм, но обычно колеб-
лется от 1 до 3 на плечо. В некоторых случаях, например у самцов
дрозофилы, хиазмы не образуются вовсе, но такие случаи редки.
Кроме того, точное положение хиазм во время мейоза варьирует
Гены и хромосомы
103
Рис. 5.11. Модель процессов разрыва, синтеза и репарации, предложенная для объ-
яснения реципрокного и нереципрокного кроссинговера.
А. Спирали ДНК от двух родительских особей на стадии образования тетрады.
На всех последующих схемах, кроме последней, изображены только две средние
спирали. Стрелками показана полярность цепи ДНК в каждом ее участке. Б. Раз-
рыв двух цепей ДНК. В. Вдоль двух н^разорванных цепей синтезируются новые
нуклеотидные последовательности. Новообразованная ДНК показана прерывистой
линией. Г. Отделение новообразованных цепей ДНК от их матриц. Д. Новообра-
зованная ДНК соединяется с одной из цепей другой молекулы. Е. Завершение
кроссинговера. Ж- Неконъюгировавшие участки наружной родительской молекулы
удалены. Штриховкой отмечен тот участок ДНК («гетерозиготный» участок), в ко-
тором произошло неправильное спаривание оснований. 3. На этой схеме вновь по-
казаны исходные наружные родительские молекулы ДНК, так что в результате
получается картина тетрады после того, как произошла рекомбинация. В «гетеро-
зиготном» участке произошла репарация. Если в качестве! .матрицы используется
(+)-цепь, то наблюдается реципрокная рекомбинация; если же матрицей для ре-
парации служит a-цепь, то наблюдается нереципрокная рекомбинация. (D. I. Patt,
G. R. Patt, 1975. An Introduction to Modern Genetics, Reading, Mass., Addison-
Wesley.)
от клетки к клетке. Вероятность возникновения хиазмы между
Двумя генами прямо пропорциональна физическому расстоянию
между ними по длине хромосомы и частоте образования перекрес-
тов для каждой хромосомы.
Рекомбинанты
Родительские (нереком-
бинировавшие) типы
(0,20 + 0.30)-(0,2ХО,3)
0,50-0,06=0, 44
Ожидаемые
фенотипы
без перекреста= А1 В1С1 и А2В2С2
< 1 пере к ре ст = А А ВА С 2 и А2В2Ст или А-|В2С2и А 2В.| С-|
2перекреста= АаВ2С, и А2В1С2
2
Б
Рис. 5.12. А. Единичный и двойной перекресты, затрагивающие два локуса, нахо-
дящиеся на большом расстоянии друг от друга.
Единичный обмен приведет к иному расположению генов, которое поддается вы-
явлению, тогда как расположение генов, получающееся в результате двойного об-
мена, идентично исходному. Допустим, что гены А( и А2 различаются фенотипиче-
ски; гены С] и С2 также различны по своему действию. (D. I. Patt, G. R. Patt,
1975. An Introduction to Modern Genetics, Reading, Mass., Addison-Wesley.)
5. Установление местоположения генов в хромосоме. Гетерозиготную особь скре-
щивают с гомозиготной по рецессивному гену. Вероятность перекреста возрастает
с увеличением расстояния. Если частота перекрестов между генами А и В равна
20%, а между генами В и С — 30%, то частота перекрестов между генами А и С
при расположении генов в последовательности АВС составит 50%, а при последо-
вательности АСВ — 10%. На самом же деле частота перекрестов между концевы-
ми генами меньше суммы расстояний от А до В и от В до С вследствие двойных
перекрестов (в данном случае она равна 6%).
Гены и хромосомы
105
Это обстоятельство можно использовать для определения по-
рядка расположения генов в хромосоме. В самом деле, если часто-
та перекрестов между любыми двумя генами пропорциональна
физическому расстоянию между ними, то в скрещивании между
особями, гетерозиготными по трем сцепленным генам, число ре-
комбинантов между ними можно использовать для графического
Изображения (составления генетической карты) их относительно-
го местоположения в хромосоме (рис. 5.12).
Сцепление представляет собой механизм, предотвращающий
независимое распределение генов, локализованных в одной и той
Же хромосоме. Однако благодаря кроссинговеру доля в популяции
хромосом, содержащих различные сочетания аллелей по двум ло-
кусам (А1ВЬ AiB2, A2Bi и А2В2), в конечном счете совпадает с
ожидаемым соотношением в отсутствие сцепления. Но если для
несцепленных генов такое равновесие достигается за одно поко-
ление случайного скрещивания, у сцепленных генов приближение
к равновесию происходит медленно. Это обстоятельство может
играть важную роль в эволюции.
Неравновесность по сцеплению
Гены, лежащие в одной и той же хромосоме, не могут распре-
деляться при мейозе независимо, так как они физически связаны
друг с другом. Однако кроссинговер разделяет эти физически сцеп-
ленные гены, так что в популяции, в которой не происходит отбо-
ра, частоты разных комбинаций генов в конечном счете соответ-
ствуют тем, которые можно было ожидать при независимом рас-
пределении. Рассмотрим подробнее причины этого явления, а также
роль сцепления.
Представим себе популяцию организмов, различающихся по
двум локусам, в каждом из которых имеется по два аллеля, Аь
А2 и Вь В2. Допустим далее, что эти два локуса лежат в одной и
той же хромосоме. В такой популяции может быть четыре генети-
чески различные хромосомы:
А^:
AiB2: Х2
А2В.: Х3
Wr Х<,
где Xlt Х2, Х3 и Х4— частоты этих хромосом в популяции. Если
обозначить частоты каждого аллеля через р (АО, 9(А2), s(Bx) и
‘(Вг), то
р=Л'14-Х2, д=Х3-]-Х4,
s=Xi~\-X3, 1=Х2-\-Х4.
106
Глава 5
Зная частоты четырех хромосом, мы можем оценить частоты
разных аллелей. Однако обычно мы располагаем сведениями о
частоте аллельных генов, а не о частоте хромосом (что в данном
случае эквивалентно сведениям о частоте генотипов, поскольку
Xi, Х2, Х3, и Х4 представляют четыре возможных генотипа). Обыч-
ный способ оценки частоты генотипов на основании частот ал-
лелей состоит в том, что частоту первого аллеля первого гена
умножают на частоту первого аллеля второго гена, получая таким
образом частоту первого генотипа; и так далее, аллель за аллелем.
При этом, однако, предполагается, что частоты аллелей в первом
локусе независимы от частот аллелей во втором локусе. Представим
себе, например, популяцию гамет, в которой гаметы, встречающие-
ся с частотой р, несут аллель Аь с частотой q — аллель А2, с час-
тотой s — аллель Вь а с частотой t— аллель В2. Чтобы установить,
какая доля гамет несет аллели А1В1, надо умножить р на $. Это
всегда будет верно при независимом распределении. Из этого не-
посредственно вытекает, что произведение частот хромосом ХД4
минус произведение частот Х2Х3 должно быть равно нулю (Х]Х4=
= ps-qt-, Х2Х3 = qs-pt).
Однако при наличии сцепления XtXt—Х2Х3, вообще говоря, не
будет равно нулю, потому что при этом некоторые генные комби-
нации встречаются в свободно скрещивающейся популяции чаще,
чем можно ожидать при независимом распределении. А следова-
тельно, при наличии сцепления частоту хромосом в популяции
нельзя вычислить путем простого перемножения частот имеющих-
ся в них генов (т. е. X^ps). Так, например, если в данной попу-
ляции аллели А] и В] полностью сцеплены, то гаметы, несущие
аллели Аь будут все нести также и ген Вь а не только часть этих
гамет (s). Поэтому в случаях сцепления Х1Х4—Х2Х3 уже не будет
равно нулю, так как Xi уже не равно произведению ps; так же
обстоит дело и с хромосомами других типов. Частота Xt будет
равна ps плюс некоторый избыток или нехватка, которые мы на-
зовем неравновесностью по сцеплению и обозначим буквой D. Зна-
чение D дается выражением ХгХ4—Х2Х3. Соответственно частоты
хромосом в популяции при наличии сцепления можно выразить в
зависимости от частот генов как
X1=ps+(X1X4-^3)=Ps+^
х2=pt - (ХЛ4 - Х2Х3) = pt - D,
X3=qs—(XlXi—X2X3')—qs—D,
Х4 = р/+(Х1Х4-Х2Х3) =qt^D.
Когда Al и Bi (равно как А2 и В2) сцеплены, состояние, известное
под названием «притяжения», D имеет положительное значение.
Если сцеплены Ai и В2 (равно как А2 и Bi), что соответствует «от-
талкиванию» аллелей At и Вь то D отрицательно. Чтобы проде-
Гены и хромосомы
107
монстрировать это, заменим в уравнениях ps и D их хромосомны-
ми эквивалентами. Например,
ps-}-D — (Xl~\-X2) (Х4-]-Х3) +(ХЛ-ВД =
=x*+xlx3+xix2+x2x3+xlxi-x2x3,
где + Х2Х3 и —Х2Х3 взаимно уничтожаются, так что после преоб-
разования получим
ps+D = Х42+Х,Х2+Х,Х3+Х.Х, =
^Х^Хх + Х2+Х3 + Х,).
Величина в скобках представляет собой сумму частот всех воз-
можных хромосом, равную 1, так что
ps+r>=Xj,
что дает нам первое уравнение. Остальные три получают анало-
гично.
В диплоидной популяции, особи которой различаются по двум
локусам с двумя аллелями в каждом, может быть всего девять
генотипов. Если эти локусы сцеплены, то частоты генотипов нельзя
предсказать на основании генных частот в соответствии с форму-
лой Харди — Вайнберга. В этом случае приходится вычислять час-
тоту каждой хромосомы (или гаметы) с учетом возникновения
перекрестов.
Для того чтобы предсказать частоту каждой хромосомы в сле-
дующем поколении, мы сначала перечислим разные способы воз-
никновения в популяции четырех типов хромосом: AiBb AtB2, A2Bi
и А2В2. Начнем с хромосомы AtBi. Она может возникнуть двумя
разными способами. Во-первых, ее могут поставлять генотипы, име-
ющие состав AjBi/** (где * обозначает любой произвольный ал-
лель), при условии что между локусами А[ и Bi нет перекреста.
Если обозначить вероятность рекомбинации (вероятность перекре-
ста) через г, то вероятность получения хромосомы Xi из генотипа
А]В]/** равна 1—г. Во-вторых, хромосому Xt можно получить из
генотипа A‘/*Bi при условии перекреста. Частота генотипов
AiB,/** в популяции равна частоте хромосомы AiBb т. е.
Хь Частота генотипов А1/*ВЬ очевидно, равна произведению ген-
ных частот p-s. Следовательно, частота Xt хромосомы А^ в по-
колении t+1
Х1Ж=Х1Д1—r)+psr, (1)
И аналогичным образом
-Г) >
^3t+l=^3t(l~ г)+^г,
Х4/+1 = Х4/ (1 —r)-\-qtr.
108
Глава 5
Но нам известно, что p = Xi + X2, a s = X1+X3; следовательно,
ps=(Х4+х2) (Х1+Х3) = X?+Х,Х2+XtX3+Х2Х3=
=Х1(Х1+Х2+Х3)+Х2Х3 =
= Х1(1-Х4) + Х2Х3.
Подставив полученное значение в уравнение (1), получим
Xlz+I=Xu(l-r)+r =
= Xlt-rXit^rXlt-rXltXit^rX2(X3t =
= Xlt rt (Xltxit x2tx3ly
Но, как мы видели, XiX4—X2X3 = Z); поэтому
^i/4-i —Хц rDt. (2)
и аналогичным образом
^2/+l =^2/ + r^)t>
^J/44 = X3(-]-rDt,
-^4/4-1 ^4/ rDf.
Иными словами, частота данной хромосомы в поколении /+1 рав-
на ее частоте в предыдущем поколении минус коэффициент реком-
бинации между двумя локусами, умноженный на неравновесность
по сцеплению в поколении t.
Поскольку Xi=ps + Dl и X4+=ps + Dt+i, то, подставляя их зна-
чения в уравнение (2), получим
Ps 4"^t+l= (Ps-h^i) r^z>
Dt+l=ps—ps+Dt—rDt=Dt (1 — r)
(1 —r) (1 -r) (1 —r)2,
так что
DM = D0(l-r)‘,
где Do — исходное значение неравновесности по сцеплению в дан-
ной популяции. Соответственно неравновесность по сцеплению в
популяции понижается со скоростью г при условии, что отбор от-
сутствует, а скрещивание происходит случайно. Поэтому следует
ожидать, что в обычных условиях неравновесность по сцеплению
не представляет собой важной эволюционной силы. Для того что-
бы неравновесность по сцеплению стала существенным фактором,
необходим эпистатический отбор, направленный на сохранение
данной генной комбинации.
Гены и хромосомы
109
Доказательства существования в природе неравновесности по
сцеплению были получены при изучении популяций разных видов
дрозофилы, главным образом Drosophila pseudoobscura. Так, на-
пример, Пракаш и Левонтин (Prakash, Lewontin) показали, что
в третьей хромосоме этого вида аллель 1,04 гена Pt-Ю всегда
встречается вместе с так называемой инверсией standard, тогда
как три другие аллеля (0,94; 1,02; 1,06) этого же самого гена ни-
когда не бывают ассоциированы с ней. Т. Престон Уэбстер (Т. Pre-
ston Webster) получил данные, свидетельствующие о существо-
вании неравновесности по сцеплению у пепельной саламандры
(Plethodon cinereus), обитающей в лесах на востоке США. В по-
пуляциях этого животного аллели двух эстеразных локусов на-
ходятся в состоянии неравновесности по сцеплению. Имеются так-
же данные, свидетельствующие о наличии неравновесности по сцеп-
лению у некоторых растений, например у ячменя и овса. Кроме
того, об этом явлении известно у человека (резус-фактор) и у
брюхоногих моллюсков. Немногочисленность примеров неравновес-
ности генов в природе не обязательно означает, что оно встреча-
ется редко; скорее это свидетельствует о технических трудностях,
связанных с его изучением. Неравновесность по сцеплению, по
всей вероятности, наиболее часто возникает между соседними ге-
нами какой-либо хромосомы. В настоящее время, однако, не су-
ществует другого метода определения всех генов в данном участке
хромосомы, кроме процесса нанесения на карту всех генов данно-
го вида. Однако этим способом не удается идентифицировать ни
гены, по которым гомозиготны все исследованные особи, ни гены,
оказывающие небольшое влияние на фенотип. В результате мно-
гие гены остаются неучтенными и до сих пор еще не составлено
полной генетической карты ни для одного вида, причем малове-
роятно, что это когда-нибудь удастся сделать. Точную оценку от-
носительной частоты неравновесности по сцеплению в природе при-
ходится отложить до тех пор, пока не будут разработаны новые
методы исследования.
Сцепление — это механизм, регулирующий частоту, с которой
неаллельные гены, лежащие в одной и той же хромосоме, будут
смешиваться с другими такими генами, локализованными в гомо-
логичных хромосомах, происходящих от разных родителей. При
полном сцеплении возникают только родительские типы, т. е. не
появляется никаких новых комбинаций. Если же сцепление отсут-
ствует, то происходит независимое расхождение генов в мейозе и
вероятность сохранения родительских комбинаций очень мала.
В тех случаях, когда родительские комбинации очень благоприят-
ны, естественный отбор будет способствовать развитию механиз-
мов, которые усиливают сцепление. И напротив, если изменчи-
вость потомков выгодна популяции, то естественный отбор будет
благоприятствовать ослаблению сцепления и повышению числа
рекомбинантных типов. К числу механизмов, усиливающих сцеп-
по
Глава 5
ление, относятся снижение частоты кросоинговера и уменьше-
ние числа хромосом. Последнее достигается в результате слу-
чайных поперечных разрывов и воссоединения оборванных концов
или нескольких хромосом по-новому, утраты центромер и отбора
в пользу этих новых структур. Каким образом возникают те или
другие конкретные изменения, точно неизвестно. Но если они про-
исходят, то естественный отбор им благоприятствует (и сохраняет
их) в тех популяциях, в которых преимущество дает низкая час-
тота рекомбинации, и элиминирует их в популяциях, в которых
низкая частота рекомбинации нежелательна. Различные механиз-
мы отбора и соответствующие примеры рассматриваются в гл. 8.
ИЗМЕНЕНИЯ СТРУКТУРЫ ХРОМОСОМ
Положение генов в хромосоме может измениться вследствие
изменений в структуре хромосомы. Такие изменения возникают в
результате изменения числа хромосом, происходящего в тех слу-
чаях, когда структурное изменение приводит к появлению новых
центромер иди к их утрате, но они могут и не сопровождаться
изменением числа хромосом. В обоих случаях характер сцепления
генов в новых хромосомах меняется. Некоторые изменения струк-
туры хромосом приводят также к увеличению числа генов или к
их утрате.
Морфологические изменения хромосом отражают изменения их
внутренней структуры. Все эти изменения известны под общим на-
званием хромосомных перестроек или аберраций, хотя, строго го-
воря, это не аберрации в обычном смысле слова, а скорее явления,
происходящие с низкой частотой в больщинстве популяций. Суще-
ствуют следующие основные типы перестроек (рис. 5.13).
Дупликации. Какой-либо участок данной хромосомы удваива-
ется. Удвоение происходит, вероятно, в результате ошибок репли-
кации.
Нехватки. Какой-нибудь участок хромосомы утрачивается. Не-
хватки могут возникать под действием рентгеновского, ультрафио-
летового или какого-либо другого излучения высокой энергии. Все
нехватки, за исключением самых мелких, детальны.
Транспозиции (внутренняя транслокация). Участок данной хро-
мосомы перемещается в ее пределах на другое место; это проис-
ходит в результате разрывов, за которыми следует воссоединение
фрагментов хромосомы в порядке, отличающемся от первоначаль-
ного.
Инверсии. Поворот участка внутри данной хромосомы на 180°.
Инверсию называют перицентрической, если инвертированный уча-
сток содержит центромеру, и парацентрической, если центромера
в него не входит.
Транслокации. Участок одной хромосомы перемещается в дру-
гую хромосому. Если какой-либо участок хромосомы А перемеща-
Гены и хромосомы
111
ется в хромосому В, а участок хромосомы В — в хромосому А, то
транслокация называется реципрокной, при этом перемещающиеся
участки не обязательно должны быть одинаковой длины.
I }’EjFj£?DCBAl
Перицентрическая
OfeIFba]
Транслокация Нереципрокная
Рис. 5.13. Разные типы хромосомных перестроек и нх влияние на морфологию хро-
мосом.
Степень изменения зависит от величины участка, затронутого перестройкой.
(О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystematics, New York,
Macmillan.)
Хромосомные перестройки, в частности транслокации, могут
привести к изменению длины хромосомы и ее вида. В таких слу-
чаях они обнаруживаются в митозе.
Однако обычно о существовании перестроек судят на основа-
нии поведения хромосом в мейозе гибридов. На рис. 5.14 показа-
но, как выглядят соответствующие хромосомы на стадии пахине-
112
Глава 5
мы. Хромосомные перестройки играют важную роль в эволюции.
Одним из примеров этого служат дупликации, давшие начало раз-
личным генам гемоглобина у высших приматов.
Рис. 5.14. Влияние различных хромосомных перестроек на конъюгацию хромосом
в мейозе в случае гетерозиготности по данной перестройке. (О. Т. Solbrig, 1970.
Principles and Methods of Plant Biosystematics, New York, Macmillan.)
Дупликации можно разделить на три группы. Первую группу
составляют дупликации единичных генов, при которых данный ген
имеется во многих копиях, что, по-видимому, дает клетке возмож-
ность продуцировать одновременно большое количество опреде-
ленного вещества, например рибосомной РНК. Ко второй группе
Гены и хромосомы
113
относятся повторения в сто, тысячи и даже миллионы раз довольно
длинных участков ДНК, обычно порядка 400—2000 нуклеотидных
пар. По-видимому, эти повторяющиеся последовательности, иногда
называемые также избыточной ДНК., участвуют в регуляции ак-
тивности генов. Наконец, иногда возникают дупликации, приводя-
щие к эволюции новых биохимических функций. Наиболее извест-
ный пример таких дупликаций дает эволюция глобинов — белков,
Многие аминокислотные последовательности ван 0-цепях гемоглобина одинако-
вы. Кроме того, в «ранней» части a-цепи имеются аминокислотные последователь-
ности, которые сильно коррелируют с последовательностями, расположенными в
0-цепи на 66 аминокислот дальше, и наоборот. Отсюда делается вывод, чго уча-
сток длиной в 66 аминокислот дуплицировался в предковой молекуле и от этой
предковой молекулы произошли как а-, так и 0-цепи.
связанных с переносом кислорода в мышечной ткани (миоглобин)
и в крови (гемоглобин).
Миоглобин — относительно простой белок, состоящий из еди-
ничной полипептидной цепи и одной группы гема (в данном слу-
чае это протопорфириновое кольцо, содержащее один атом желе-
за), с молекулярным весом примерно 17 000. У всех позвоночных,
за .исключением очень примитивных бесчелюстных, молекулы мио-
глобина очень сходны. Поэтому считается, что у всех позвоночных
гены, определяющие синтез миоглобинов, представляют собой мо-
дифицированных потомков одного предкового гена. Молекула ге-
моглобина имеет более сложное строение. Она состоит из четырех
полипептидных цепей, каждая из которых содержит гем, и имеет
молекулярный вес 67 000, т. е. примерно в четыре раза выше, чем
у миоглобина. Гемоглобины более эффективно отдают свой кис-
лород, чем миоглобины, благодаря взаимодействию между четырь-
мя гемами в их молекуле. Кроме того, четыре цепи, составляющие
8—2044
114
Глава 5
молекулу гемоглобина, неодинаковы. У большинства позвоночных
имеются цепи двух типов, называемые а- и ^-цепями, по две цепи
каждого типа. У человека и у высших приматов имеется гемогло-
бин двух типов: гемоглобин Аь состоящий из двух а- и р-цепей,
и гемоглобин Аг, состоящий из двух а- и двух б-цепей. Наконец,
Цепи гемоглобина
Рис. 5.16. Эволюция молекул глобина.
Цифры указывают число нуклеотидных замещений, необходимых для того, чтобы
от общего предка возникли 5 существующих в настоящее время нуклеотидных по-
следовательностей. Раньше всех дивергировал миоглобин, накопив в процессе эво-
люции 117 замещений. Разные цепи гемоглобина дивергировалн позднее. Предко-
вый глобин, от которого могли возникнуть все современные цепи гемоглобина, от-
личается от молекулы, от которой произошли все глобины, 59 замещениями.
(Fitch, Margoliash, 1970)
имеется третий тип гемоглобина — гемоглобин плода, молекула ко-
торого состоит из двух а- и двух у-цепей. Работами ряда исследо-
вателей (Fitch, Goodman, Margoliash, Moore, Pauling, Zuckerkandl)
установлено, что a-, 0-, у- и б-цепи кодируются разными генами.
Они установили также, что хотя эти полипептидные цепи отлича-
ются по ряду аминокислот, тем не менее основные черты амино-
кислотной последовательности позволяют предполагать, что они
происходят от общего предка. Это открытие привело к созданию
следующей схемы эволюции глобинов высших позвоночных
(рис. 5.15 и 5.16).
Гены и хромосомы
115
Предковым геном был ген, кодировавший миоглобин. Пример-
но 500 млн. лет назад он дуплицировался, после чего произошли
мутации, давшие начало примитивной p-цепи, которая могла в
результате агрегации образовать примитивную молекулу гемогло-
бина, состоявшую из четырех p-цепей. Следующим шагом, имев-
шим место 380 млн. лет назад, была дупликация гена, детерми-
нировавшего этот примитивный р-гемоглобин, за которой после-
довала мутация, давшая начало гену, кодирующему a-цепь. Это
создало возможность для образования более эффективного гемо-
глобина типа А (а2р2)- Следующий шаг произошел примерно
150 млн. лет назад, когда ген p-цепи еще раз дуплицировался и
в конечном счете дал начало гену, кодирующему у-цепь. И нако-
нец, примерно 35 млн. лет назад в предковой линии высших при-
матов возникла еще одна дупликация, которая дала начало гену,
кодирующему 6-цепь. На примере этих глобинов можно видеть,
каким образом из предковых генов в процессе эволюции возника-
ют новые гены (и новые функции). В рассмотренном случае синтез
предковой молекулы — миоглобина — продолжается. В других слу-
чаях исходная молекула могла быть утрачена в результате не-
хватки, хотя продемонстрировать такое событие трудно.
ГАПЛОИДНОСТЬ, диплоидность и полиплоидность
Первая клетка, с которой начинается развитие организма (зи-
гота, представляющая собой продукт слияния двух гамет), содер-
жит два набора хромосом. Каждый из этих наборов поставляется
одной из родительских особей. Таким образом, зигота диплоидна.
Но не все организмы диплоидны. В некоторых случаях, в особен-
ности среди растений, из образовавшихся в процессе мейоза спор
развивается организм без предшествующего слияния гамет. Эти
организмы гаплоидны, так как их клетки содержат лишь один на-
бор хромосом. Наконец, в результате тех или иных случайностей,
возникающих в процессе развития, в гамете, зиготе или в одной
из клеток, производимых зиготой, число хромосом может удвоить-
ся. Возникающие при этом организмы содержат в своих клетках
больше двух наборов хромосом, и их называют, в зависимости от
Числа имеющихся в их клетках хромосомных наборов, триплоида-
ми, тетраплоид ами, пент аплоид ами, гексаплоидами и т. д. или же
объединяют под общим названием полиплоидов.
У большинства животных полиплоидия встречается редко, од-
нако у растений она наблюдается очень часто и играет важную
роль в их эволюции. Растение, содержащее ровно вдвое больше
хромосом, чем его родители, качественно от них не отличается,
поскольку оно не несет никакой новой генетической информации.
Однако дополнительный набор хромосом обычно обусловливает
увеличение размера клеток при уменьшении их общего числа.
У тетраплоида первый эффект обычно перевешивает второй, и рас-
8*
116
Глава 5
тения, как правило, бывают крупнее (результат, непосредственно
используемый в плодоводстве, например при разведении земляни-
ки). Дальнейшее удвоение числа хромосом нередко приводит в
конечном итоге к уменьшению величины растений.
Полиплоиды, получаемые искусственно путем химической об-
работки, нередко бывают стерильны. Поскольку клетки тетраплои-
дов содержат по четыре гомологичных хромосомы, при конъюгации
Рис. 5.17. Мейоз у тетраплоидов.
Клетки тетраплоидов содержат по четыре хромосомы каждого типа, которые при
ме,йозе обычно собираются вместе (А). При расхождении сестринские хроматиды
не направляются по две к каждому из полюсов, как следовало бы ожидать (5);
вместо этого три хроматиды могут мигрировать к одному полюсу, а одна — к дру-
гому (В).
этих хромосом в мейозе вместо бивалентов образуются тетрава-
ленты. При расхождении тетравалентов к каждому из двух полю-
сов не всегда направляется по две хромосомы, в результате чего
образуются гаметы, в которых наблюдается нехватка одних хро-
мосом и избыток других (рис. 5.17).
Тем не менее существует особый тип полиплоидов, которые пло-
довиты. При скрещивании двух видов, связанных далеким родст-
вом, их хромосомы при мейозе иногда не образуют бивалентов.
Вместо бивалентов получаются униваленты (неконъюгировавшие
хромосомы, которые не претерпевают нормального мейоза) и соот-
ветственно гибридные растения оказываются стерильными. Если
у такого растения произойдет удвоение хромосом, то плодовитость
восстанавливается, поскольку при этом хромосомы каждого типа
будут представлены в двух экземплярах и будут конъюгировать
друг с другом в профазе мейоза. Такой гибрид называют аллопо-
липлоидом, в отличие от предыдущего типа, который называют
автополиплоидом. Примером аллополиплоида служит растение та-
Гены и хромосомы
117
бака, Nicotiana tabacum. Это растение возникло в результате удвое-
ния хромосом у естественного гибрида между N. otophora и N. syl-
vestris. Скрещивание между этими видами произошло в природе
несколько сотен, а может быть, и тысяч лет назад, но затем его
удалось успешно повторить в искусственных условиях (рис. 5.18).
Существуют полиплоиды третьего типа, промежуточного между
Рис. 5.18. Возможное происхождение табака (Nicoiiana tabacum) в результате
гибридизации между N. otophora и N. sylvestris, за которой следовало удвоение
хромосом. (Т. Н. Goodspeed, 1954. The Genus Nicotiana, Waltham, Mass., Chronica
Botanica.)
двумя описанными выше. Иногда некоторые из хромосом двух ро-
дительских типов достаточно близки, так что они могут конъюги-
ровать друг с другом, тогда как между другим;и такой близости
нет. В этих случаях возникают сегментные аллополиплоиды
(рис. 5.19). Дальнейшая судьба сегментного аллополиплоида за-
висит главным образом от того, преобладают ли в нем алло- или
автокомпоненты (рис. 5.20). Большинство природных полиплоидов
представляют собой либо аллополиплоиды, либо сегментные ал-
лополиплоиды. Известно также несколько природных автополи-
плоидов.
Полиплоидия — это один из механизмов, посредством которых
могут возникать новые виды. Мы вернемся к этому вопросу в
гл. 11. Полиплоидия оказывает также влияние на характер на-
следования.
Многие сельскохозяйственные культуры являются полиплоида-
ми (например, пшеница, хлопчатник, табак, представители семей-
ства крестоцветных). Происхождение геномов этих растений мож-
но проследить с помощью специальных генетических и цитологи-
118
Глава 5
Фертильный
Рис. 5.19. Сегментный аллополиплоид.
В результате скрещивания между двумя видами, А и В, получается диплоидный
гибрид, который стерилен, несмотря на то что хромосомы хорошо конъюгируют,
образуя биваленты. Стерильность этого гибрида можно объяснить возникновением
мелких хромосомных инверсий, которые не мешают конъюгации, но приводят к
дупликациям и нехваткам по отдельным хромосомным участкам в гаметах при
кроссинговере. Если, однако, число хромосом удваивается, то конъюгация проис-
ходит преимущественно между действительно гомологичными хромосомами (т. е.
происходящими от одного и того же вида). Конъюгация при этом приводит к об-
разованию бивалентов, так что получаются нормальные и, следовательно, фер-
тильные гометы. (О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystema-
tics, New York, Macmillan.)
Популяция 1 \ I Популяция 2 1 I Вид В ) 1 Вид С
вида А / к вида д
120
Глава 5
ческих методик. На основании полученных при этом данных се-
лекционер может вносить в культурный сорт гены от родственного
дикорастущего вида. Примером полиплоида с улучшенными при-
знаками служит садовая земляника.
Диплоидный гид
putchetla
511-л'а'
Фертильность = 99%
Оегментно-аллотетраплоидный гибрид
(pulchells х peruviana)
1011 - a’a’.bb
Фертильность > 70%
Х2
Диплоидный гибрид
(puionei'a х peruviana)
511-д'в
Фертильности < 30%
Автстетрагг.оид
'peruvianas
5 IV- ВЗ,ЗВ
Фертильност» < 5 %
Х2
Диплоидный зид
peruviana
5 II BS
Фертильность = 99%
Тетсатлоидный гибрид
(elegans X pulcnelia)
1011- A1 A2 CD
Фертильность < 10 %
Тетраплоидный гибрид
(elegans X peruviana)
5IV - Д2В CD
Фертильность <5?р
Сегмекгно-а>т._й0-<-ог1г>сидный
ГИбриД
(elegans х peruviana)
2OII-А2А2 ВВ СС 00
Сегментно-аллогенсаллоидный вид Фертильность >70%
elegans
15Н - а2А? СС DD
Фертильность -- 0’^%
Рис. 5.21. Полиплоидия у Glandularia.
Скрещивали два диплоидных и один гексаплоидный (шесть хромосомных наборов)
вид Glandularia. Обрабатывая некоторые растения колхицином, получили искус-
ственные полиплоиды. На схеме указаны: фертильность пыльцы (в % %), конъюга-
ция хромосом и предполагаемые геномы. Обратите внимание на то, что фертиль-
ность восстанавливается при образовании сегментного аллополиплоида, но снижа-
ется при образовании автополиплоида. [О. Т. Solbrig, С. Passani, R. Glass, 1968.
Artificial hybridization between different polyploid levels in Glandularia (Verbena-
ceae), Amer. J. Bot., 55, 1235—1239.]
В Европе земляника известна, вероятно, с тех самых пор, как
существует человек. Европейский вид, Fragaria vesca, — диплоид
с очень мелкими плодами. После открытия Америки в Европу
ввезли виды Нового Света — Fragaria chiloensis (из Чили) и F. vir-
giniana (из восточных областей Северной Америки). Эти виды —
октоплоиды (восемь хромосомных наборов) с несколько более
Гены и хромосомы
121
крупными плодами. В результате скрещиваний этих двух видов
(а возможно, и других) были выведены сорта с более крупными
плодами, что в конце концов привело к созданию сорта F. апа-
nassa, плоды которого более чем в 10 раз крупнее, чем у роди-
тельских видов, и который обладает также другими желательными
с хозяйственной точки зрения признаками. Другим примером,
представленным на рис. 5.21, служат дикие родичи Glandularia
(сем. Verbenaceae).
МЕНДЕЛЕВСКАЯ НАСЛЕДСТВЕННОСТЬ У ТЕТРАПЛОИДОВ
Наследственность у тетраплоидов осложняется тем, что у них
в ядрах соматических клеток каждый локус представлен не два,
а четыре раза. Следовательно, у тетраплоида при рассмотрении
ситуации с одним локусом, для которого имеется два аллеля, при-
ходится иметь дело с пятью возможными генотипами (AiAjAiAi,
А1А1А1А2, AiAiA2A2, AiA2A2A2, АгАгАгАг), а не с тремя, как в ана-
логичной ситуации у диплоида. То же самое относится и к доми-
нированию. Вместо одной гетерозиготы у тетраплоида их три, они
могут быть идентичны одной из гомозигот — при наличии полного
доминирования, или же — в случае количественного эффекта — мо-
жет наблюдаться градиент фенотипического выражения, а не пять
четко выраженных типов.
Таблица 5.3
Гаметы, производимые триплексом
Генотип родительской особи Аг
А1 А1 А1
А, X X X X
А, A4Ai X X X
а4 а4а4 AiAi X X
Аг А4Аг A1A2 А1А2 X
Таблица 5.4
Гаметы, производимые дуплексом
Генотип родительской особи Аг
At А1 Аг
А4 X X X X
а4 AiAi X X X
Аг А1А2 А4Аг X X
Аг А4Аг A1A2 АгАг X
122
Глава 5
Таблица 5.5 Гаметы, производимые симплексом
Генотип родительской особи
А1 Аз а2 а2
А, X X X X
А2 AfA2 X X X
А2 AiA2 а2а2 X X
Аг А]А2 а2а2 А2А2 X
Описывая менделевскую наследственность у тетраплоидов, сле-
дует прежде всего рассмотреть типы гамет, производимых гетеро-
зиготными генотипами. Чтобы было легче объяснить, назовем го-
Рис. 5.22. Распределение потомков, полученных при гетерозиготном скрещивании
диплоидов (белые столбцы) и тетраплоидов (черные столбцы).
Допустим, что существует некий локус А с двумя аллелями А] и А2 и что у осо-
бей, гомозиготных по аллелю А], значение некоторого признака равно 1, а фено-
тип особей, гомозиготных по аллелю А2,— 5. Гетерозиготные особи имеют проме-
жуточные значения этого признака, зависящие от числа аллелей Aj и А2 в геноти-
пе. При скрещивании между двумя гетерозиготными диплоидами (AiA2) возникают
потомки трех типов (белые столбцы). При скрещивании между двумя тетраплои-
дами AiAiA2A2 получаются потомки пяти типов (черные столбцы) и доля гомози-
гот сильно уменьшается.
мозиготный доминантный генотип квадруплексом, пер-
вую гетерозиготу AiAiA]A2— триплексом, вторую — A]AiA2A2—
дуплексом, третью — AiA2A2A2 — симплексом, а гомозиготный ре-
цессивный генотип — нуллиплексом.
Гены и хромосомы
123
Таблица 5.6
Потомство от скрещивания самок AjAtAjAz
и самцов А1А1АгА2
Женские гаметы
lAiAt 4А1А2 IA2A2
Мужские 1 AtA4 1 AjAiAiAi 4 А1А1А4Аг 1 А1А1А2А2
гаметы 4 А1Аг 4 А1А1А1А2 16 AiAi А2А2 4 А1А2А2А2
1 А2А2 1 А1А1А2А2 4 А1А2А2А2 1 А2А2А2А2
Квадруплекс и нуллиплекс дают гаметы только одного типа,
а триплекс — гаметы двух типов, AiAi и AjA2. (Следует помнить,
что тетраплоиды производят диплоидные гаметы!) Триплекс про-
изводит гаметы в соотношении 1 : 1 (табл. 5.3). Дуплекс дает гаме-
ты трех типов — AiAb А[А2 и А2А2— в соотношении 1:4:1
(табл. 5.4), а симплекс — гаметы двух типов, AtA2 и А2А2, в соотно-
шении 1:1 (табл. 5.5).
Зная соотношение гамет, нетрудно вывести соотношение гено-
типов в потомстве. Так, например, при скрещивании двух гомози-
готных тетраплоидов AjAjA^i и А2А2А2А2 мы получим гетерози-
готный дуплекс A!AiA2A2. При самооплодотворении этого дуплекса
или при скрещивании его с одним из его сибсов получится F2,
представленное в табл. 5.6. Соотношение разных генотипов будет
следующим:
А| A|AjA| 1
Aj А1А1А2=-8
Aj Al А2 Аг =18
At А2А2А2 = 8
А2 A2 А2 A2 = 1
При соответствующем скрещивании диплоидов возникают геноти-
пы IAjAi, 2AtA2, 1А2А2. Таким образом, у тетраплоидов по срав-
нению с соответствующими диплоидами доля гетерозигот возрос-
ла, а доля гомозигот уменьшилась. Если допустить отсутствие до-
минирования и промежуточный характер фенотипов у гетерозигот
и построить график их распределения по произвольной фенотипи-
ческой оси, то обнаружится резкое уменьшение доли гомозигот в
потомстве тетраплоидов (рис. 5.22).
КРАТКИЕ ВЫВОДЫ
Цитологическое изучение хромосом позволяет понять физиче-
скую основу действия менделевских законов наследственности и
явления сцепления. Эти законы носят универсальный характер, по-
124
Глава 5
зволяя делать широкие предсказания. Однако по второстепенным
хромосомным характеристикам разные виды сильно различаются.
Даже близко родственные виды могут значительно различаться
по длине хромосом, а число хромосом может различаться в два-
три раза (если один из видов — полиплоид). Эти различия по хро-
мосомным признакам означают существование различий в харак-
тере наследственности, подобных тем, которые описаны здесь меж-
ду полиплоидами и диплоидами, а также в степени сцепления.
Все присущие отдельным видам особенности имеют свои преиму-
щества и свои недостатки, и все они подвержены действию естест-
венного отбора, которое рассматривается в следующей главе.
ЛИТЕРАТУРА
Общие вопросы
Dawson G. W. Р. 1962. An Introduction to the Cytogenetics of Polyploids, Oxford,
Blackwell Scientific Publications.
Dyson R. D. 1978. Cell Biology: A Molecular Approach, 2nd ed., Boston, Allyn and
Bacon.
Lewis K. R., John B. 1963. Chromosome Marker, London, Churchill.
Swanson С. P. 1957. Cytology and Cytogenetics, Englewood Cliffs, N. J., Prentice-
Hall.
White M. J. D. 1973. Animal Cytology and Evolution, 3rd ed., Cambridge, Cambrid-
ge University Press.
Специальные статьи и обзоры
Babcock Е. В. 1947. The Genus Crepis, Part 1, Berkeley, University of California
Press, Botany series, vol. 21.
Bateson W., Punnett R. C. 1906. Experimental studies in the physiology of heredity.
Reports to the Evolution Committee of the Royal Society (London).
Casperson T., Zech L. (eds.) 1973. Chromosome identification: technique and appli-
cation in biology and medicine (Nobel Symposium 23), Stockholm, Nobel Foun-
dation.
Fitch W. M. 1966. Evidence suggesting a partial, internal duplication in the ancest-
ral gene for heme-contining globins, J. Mol. Biol., 16, 17—27.
Fitch W. M. 1973. Aspects of molecular evolution, Ann. Rev. Genetics, 7, 343—380.
Fitch W. M., Margoliash E. 1970. The usefulness of amino acid and nucleotide se-
quences in evolutionary studies, Evol. Biol., 4, 67—109.
Gillies С. B. 1975. Synaptonemal complex and chromosome structure, Ann. Rev. Ge-
netics, 9, 91—110.
Goodspeed T. H. 1954. The genus Nicotiana, Waltham, Mass., Chronica Botanica.
Jackson R. C. 1971. The karyotype in systematics, Ann. Rev. Ecol. Syst., 2, 327—
368.
Kornberg R. D. 1977. Structure of chromatin, Ann. Rev. Biochem., 46, 931—954.
Lewis H. 1957. Genetics and cytology in relation to taxonomy, Taxon, 6, 42—46.
Paris Conference. 1972. Standardization in human cytogenetics, Cytogenetics, 11,
Renwick J. H. 1971. The mapping of human chromosomes, Ann. Rev. Genetics, 5.
81—120.
Ris H., Kubai D. F. 1970. Chromosome structure, Ann. Rev. Genetics, 4, 263—294.
Stebbins G. L. 1947. Types of polyploids: their classification and significance, Adv.
in Genetics, 1, 403—430.
Stebbins G. L. 1960. The comparative evolution of genetic systems. In: S. Tax (ed.).
Evolution after Darwin, vol. 1, Chicago, University of Chicago Press.
Глава 6
Факторы эволюции
Предмет популяционной генетики составляют явления наслед-
ственности в группах организмов в целом. Ее (интересует не меха-
низм наследственности как таковой, а его роль в эволюции попу-
ляций. Популяционная генетика занимается изучением влияния,
оказываемого на наследственность мутационным процессом, ре-
комбинациями и отбором, а также значениями при этом величины
популяции, типа скрещивания, продолжительности жизни отдель-
ных особей и т. п. Кроме того, она рассматривает воздействия на
генетический состав популяции таких явлений, как сцепление, мно-
жественный аллелизм, сцепле.нность с полом, и других подобных
процессов. Поскольку все эти' явления могут вызывать изменения
в популяции, изучение органической эволюций неизбежно связано
с изучением генетики популяций.
С генетической точки зрения эволюция — это изменение часто-
ты генов в данной популяции. Частота какого-либо гена (а точ-
нее— аллеля) зависит от ряда различных факторов (рис. 6.1),
среди которых есть несколько главных.
1. Мутационный процесс. Мутации — это изменения, затрагива-
ющие последовательность нуклеотидов или их число в молекуле
ДНК и создающие тем самым новые аллели. Мутации могут
изменять отдельные нуклеотиды или целые хромосомы.
2. Поток генов. Частота генов в данной популяции может из-
меняться в результате эмиграции или иммиграции особей. Это
явление называется потоком генов.
3. Рекомбинацией называют процесс, при помощи которого
имеющиеся в популяции аллели перемешиваются (рекомбинируют)
в результате скрещиваний между особями. Хотя рекомбинация
сама по себе не влечет изменения генных частот в данной попу-
ляции, она оказывает влияние на действие естественного отбора
и ее поэтому следует рассматривать как важный фактор эво-
люции.
4. Естественный отбор, или дифференциальное размножение и
выживание фенотипов, представляет собой главный способ измене-
ния генных частот.
5. Дрейф генов. Генные частоты подвержены также чисто слу-
чайным изменениям. Дрейф генов может играть важную роль в
126
Глава 6
возникновении эволюционных изменений в небольших популяциях
(численность которых не превышает 100 особей).
В настоящей и следующей главах дается подробный обзор всех
этих факторов и делается попытка оценить их относительное зна-
Родительская популяция
Новая величина популяции потомков
с новыми частотами генов
Рис. 6.1. Факторы эволюции.
Такие факторы, как мутационный процесс, рекомбинация, поток генов и наруше-
ния расщепления, повышают изменчивость популяции. Другие факторы — отбор и
дрейф генов — снижают изменчивость. Взаимодействие между этими факторами
мажет привести к тому, что в новом поколении генные частоты будут иными.
чение. Мы начнем с изменения частот в одном локусе с двумя ал-
лелями. В последующих главах будут рассмотрены изменения во
многих локусах.
МУТАЦИОННЫЙ ПРОЦЕСС
Мутация — это изменение в последовательности, типе или чис-
ле нуклеотидов в молекуле ДНК, составляющей основную часть
хромосомы. Изменение может быть очень небольшим, затрагива-
ющим лишь одну пару нуклеотидов, или же очень крупным, и:
меняющим целый участок хромосомы или даже всю хромосому.
В предыдущей главе под названием хромосомных аберраций мы
описали мутации, связанные с изменениями участков хромосомы.
Факторы эволюции
127
В этом разделе наше внимание будет обращено на те мутации,
которые затрагивают один генный локус. Такие мутации обычно
называют точковыми мутациями.
Некоторые точковые мутации возникают так редко, что явля-
ются буквально единственными в своем роде. Они не оказывают
сколько-нибудь долговременного влияния на популяцию, поскольку
их шансы на сохранение в большой популяции крайне малы, если
только они не дают своим носителям какого-либо очень большого
и немедленного сказывающегося преимущества, что весьма мало-
вероятно. Другие мутации, называемые повторными мутациями,
возникают с частотой, поддающейся измерению. Их частота варь-
ирует для разных генов ц у разных видов (табл. 6.1), однако
обычно она составляет 10-4—10-8 на один ген на одно поколение
(т. е. от одной мутации в данном локусе на 10 000 генов в одном
поколении до всего лишь одной на 100 000 000 генов в одном по-
колении). Повторные мутации оказывают очень большое влияние
на генные частоты в популяции, причем это влияние можно пред-
сказать математически.
Рассмотрим популяцию организмов, все особи которой несут в
данном локусе лишь один аллель Аь Аллель At мутирует в ал-
' Таблица 6.1
Частоты спонтанных мутаций в некоторых локусах различных
организмов (Strickberger, Genetics, 1976, New York)
Организм Признак Частота мутацив на гамету (или клетку)
Escherichia coli (кишечная па- Устойчивость к стрептомицину 4-10-10
лочка) Устойчивость к фагу Т1 3-10-8
Salmonella typhimurium (палоч- Устойчивость к треонину 4,1 10-6
ка крысиного тифа) Способность к синтезу трипто- фана 5-Ю-8
Diplococcus pneumoniae (мико- бактерия туберкулеза) Устойчивость к пенициллину 1 10-7
Neurospora crassa (хлебная Способность к синтезу аденина 3-Ю-8
плесень) Способность к синтезу инозита 5-Ю-8
Drosophila melanogaster (пло- Желтое тело 1,2-10-4
довая мушка) Коричневые глаза Черное тело Безглазость ЗЮ-5 2-Ю-5 6-Ю-5
Zea mays (кукуруза) Сморщенные семена Бесцветный алейрон Сахаристый эндосперм 1,2-10-® ЗЮ-5 2,4-10-®
Mus musculus (домовая мышь) Коричневая шерсть Розовые глаза Пегая окраска 8,5-10-® 8,5-10-® 1,7-10-5
Homo sapiens (человек) Эпиболия Ретинобластома Врожденное отсутствие радуж- ной оболочки Хорея Гентингтона 6-ю-« (1,2—2,3) -10-® 5-Ю-5 5-10-»
128
Глава 6
лель А2 с постоянной частотой, которую мы обозначим через и.
Если исходная частота аллеля At равна р0, то частота qx аллеля А2
в следующем поколении составит
Если представить себе, что имеется 1 000 000 гамет, каждая из
которых содержит аллель Ai, и что частота, с которой они мути-
руют к аллелю А2 составляет и = 10-4 (0,0001), то можно вычис-
лить, что:
р0=1 (все аллели AJ;
u-p0 = 1 • 10'4 = 0,0001;
а новая частота А[
Р1= р0-(и-р0) = 1 —0,0001 —0,9999.
По прошествии одного поколения 999 900 гамет будут нести ал-
лель Ai и 100 гамет — аллель А2.
Но новый аллель А2 может вновь мутировать к аллелю At —
явление, называемое обратной мутацией. Если обозначить через
v частоту мутирования от А2 к А] и рассуждать таким же обра-
зом, как при вычислении изменений, сопровождающих мутирова-
ние от А] к А2, то получимз
Исходная частота А4 н Аз.’ ро и qo
и
Частоты мутаций: А1ч==Аз;
о
Новые частоты: pi—po—(«-ро) +(о •?(>),
?1 = <7о— (v •?») + («-ро).
С учетом прямых и обратных мутаций суммарное изменение
частот р и q составит:
Др=Р1 — Ро = (v • q0) — (и р0),
ро = (и.ро)—(и-^о).
По мере увеличения частоты одного аллеля частота другого
аллеля уменьшается, а тем самым уменьшается и возможность
мутирования в этом направлении, но зато становится больше ал-
лелей, способных мутировать в обратном направлении, т. е. к ис-
ходному типу. В конечном счете будет достигнуто равновесие, при
котором никаких дальнейших изменений частоты аллелей в ре-
зультате мутаций происходить не будет. Точку равновесия нахо-
дят, приравнивая изменение частот q и р к нулю (когда эти две
частоты находятся в состоянии равновесия, никаких изменений не
происходит). Таким образом, при равновесии
Д<7=Др = 0,
p-u=q-v Или _£_ = _Е_
q и ’
Факторы эволюции 129
И
Прибавляя q к обеим частям уравнения и решая его, получим
p+<7=v9+<7.
। vq + uq __ / уи \
и \ и I
и
q~~- .—.
7 и + V
Частота аллеля At зависит исключительно (в отсутствие отбо-
ра) от частоты, с которой он мутирует к аллелю Аг, и от частоты,
с которой аллель А2 мутирует обратно к аллелю Аь Поскольку эти
частоты очень низки (частота обратных мутаций обычно составля-
ет всего 0,1 частоты прямых мутаций), изменения, вносимые в
популяцию одним только мутационным процессом (при равнове-
сии Харди—Вайнберга), очень незначительны. При нормальной
частоте мутаций (10-4—!0~9), для того чтобы в данной популяции
половина аллелей At была заменена аллелями А2, Потребуется от
минимум 5000 до более чем 50 000 000 поколений. Эффект мута-
ционного процесса тем больше, чем короче продолжительность
жизни каждого поколения. Для человека 50 000 поколений соот-
ветствуют 1—2 млн. лет, а у бактерий с их быстрой сменой поко-
лений (порядка получаса на каждое) на столько же поколений
потребуется менее трех лет. Во всех тех случаях, когда частота
прямых мутаций выше частоты обратных, мутанты становятся бо-
лее многочисленными, чем исходный ген, если только они не эли-
минируются отбором. Можно также сделать еще один вывод. По-
вышение частоты мутаций не вызовет никаких изменений, если
только оно одинаково изменяет частоту прямых и обратных мута-
ций. В этом случае исходное равновесие, определяемое формулой
Харди—Вайнберга, не изменится.
Описанный выше пример относится к некоторой заданной си-
туации, при которой точно 10~4 аллелей At мутируют к аллелю А2,
и возникающие в результате этого изменения частот точно отра-
жаются в потомстве. Однако в природе все обстоит иначе. Число
10~4 соответствует лишь средней частоте мутирования аллеля Ai
к аллелю А2. В любом конкретном поколении истинная частота
мутаций может быть выше или ниже этого значения. Поскольку
мутантов в любом случае мало, важно выяснить следующее: если
некая гамета содержит мутантный аллель, то какова вероятность
развития из нее взрослого организма?
Австралийский генетик и статистик Рональд А. Фишер (Ronald
A. Fisher) исследовал эту проблему в 1930 г. Он поставил сле-
9—2044
130
Глава 6
дующий вопрос: какова вероятность сохранения данного гена в
популяции, если этот ген селективно нейтрален? Используя мате-
матический аппарат теории ветвящихся процессов (в ее детерми-
нистическом варианте), Фишер показал, что вероятность сохране-
ния такого гена близка к нулю (табл. 6.2). Если этот ген обладает
селективным преимуществом, равным 1%, то вероятность его со-
хранения равна всего примерно 2%. Следовательно, для того что-
бы мутация могла закрепиться, она должна обладать не только
селективным преимуществом, но и достаточной исходной частотой?
Таблица 6.2
Вероятность сохранения и элиминации нейтральной мутации (Fisher, 1958)
Число поколений Вероятность элиминации Разность Вероятность сохранения
не дает преимущества 1 %-ное преимущество не дает преимуще ства 1%-ное преимущество
1 0,3679 0,3642 0,0037 0,6321 0,6358
3 0,6259 0,6197 0,0062 0,3741 0,3803
7 0,7905 0,7825 0,0080 0,2095 0,2175
15 0,8873 0,8783 0,0090 0,1127 0,1217
31 0,9411 0,9313 0,0098 0,0589 0,0687
63 0,9698 0,9591 0,0107 0,0302 0,0409
127 0,9847 0,9729 0,0118 0,0153 0,0271
Предел 1,000 0,9803 0,0197 0,0000 0,0197
ПОТОК ГЕНОВ
Поток генов — это обмен генами между популяциями, осущест-
вляющийся в форме происходящих время от времени спариваний
между особями, принадлежащими к различным скрещивающимся
популяциям, проникновения в данную популяцию животного, ро-
дившегося в другом месте, или же внесения чужих семян или
пыльцы. Во всех этих случаях в популяцию вводятся гены, возник-
шие в результате мутации в какой-либо иной среде и сохраненные
отбором. Непосредственный эффект такого события сходен с эф-
фектом мутаций: в популяцию вносится новый источник изменчи-
вости. Если у иммигранта имеется аллель, совершенно новый для
данной популяции, то эффект в точности аналогичен эффекту му-
тации. Чтобы вычислить эффект иммиграции особей, каждая из
которых несет новый аллель Сг, на популяцию, в которой прежде
имелся только аллель Сь мы используем те же самые уравнения,
которые были выведены для мутационного процесса, заменив и
(частота мутаций) на i (частота иммиграции), v (частота обрат-
ных мутаций) —на е, или частоту эмиграции аллелей Сг из попу-
Факторы эволюции
131
ляции. Если частоту аллеля Ci обозначить через р0, а частоту ал-
леля С2— через qQ, то после обмена генами в одном поколении
получим
Др=е-<70—1-р0 (изменение частоты CJ
и
A<7 = i-p0—e-q0 (изменение частоты С2),
а при равновесии
Может оказаться, однако, что иммигранты содержат те же са-
мые аллели Ci и С2, что и изучаемая популяция, но частоты этих
аллелей в двух популяциях различны. В таком случае новые час-
тоты вычисляют следующим образом. Рассмотрим популяцию, осо-
би которой несут в некотором данном локусе аллель С2 с частотой
q0. В эту популяцию в каждом поколении начинает поступать не-
которое число иммигрантов, у которых аллель С2 встречается с
иной частотой, qm. После обмена генами в одном поколении доля
иммигрантов в этой популяции составит tn, а доля особей неимми-
грантов 1—tn. Частота гена С2 будет теперь равна сумме доли
неиммигрантов, умноженной на частоту q0 у них гена С2, плюс доля
иммигрантов, умноженная на частоту qm у них гена С2.
qi—mqm-\-(l — m)qa=
=m(qm—q0)+q0.
Изменение Aq частоты аллеля С2, вызванное потоком генов, — это
разность между его исходной частотой qQ и частотой qi после при-
тока новых генов:
<7о =
=m(qm—?0) + q0—q0==m (qm—q0).
Таким образом, изменение генных частот, вызванное потоком ге-
нов, зависит от доли иммигрантов (т), исходной частоты q0 ал-
леля С2 и частоты С2 у иммигрантов.
Примером действия мутационного процесса и потока генов мо-
жет служить эволюция вируса гриппа, поражающего человека.
Этот вирус содержит два типа белков, действующих как антигены,
стимулирующие вырабатывание организмом человека антител к
этим белкам. Один из этих белков — гемагглютинин — содержится
в оболочке вируса и образует выступы, способные связывать эрит-
роциты, другой — нейраминидаза—фермент, вызывающий отделе-
ние вирусных частиц от эритроцитов. Люди, переболевшие грип-
пом, приобретают иммунитет к повторным заражениям вирусом,
содержащим такие же гемагглютинины и нейраминидазы, какие
9*
132
Глава 6
имелись у заразившего их вируса. Однако они остаются восприим-
чивыми к заражению вирусами, содержащими иные гемагглюти-
нины или нейраминидазы. Почти каждый год в какой-нибудь части
земного шара вспыхивает эпидемия гриппа. Эти локальные вспыш-
ки вызываются мутациями, возникающими в вирусном белке (так
называемый антигенный дрейф), в результате чего вирус приоб-
ретает способность вновь инфицировать людей, поскольку имею-
щиеся у них антитела, специфичные к немутировавшему белку,
становятся неэффективными. Однако человеческий организм спо-
собен быстро отреагировать на заражение и в конечном счете
вырабатывает антитела против мутантного штамма. Поэтому та-
кие эпидемии обычно носят локальный характер и продолжаются
относительно недолго.
Время от времени возникают серьезные вспышки гриппа, охва-
тывающие весь земной шар и вызывающие высокую смертность.
Такие вспышки гриппа, называемые пандемиями, происходили в
1889—1890, 1918—1919, 1957—1958 и 1968—1969 гг. Пандемии
гриппа, по-видимому, вызываются вирусами, коренным образом от-
личающимися от вируса гриппа типа В, который обычно поражает
человека. Различия между ними касаются главным образом гем-
агглютининов, тогда как нейраминидазы могут быть либо такими
же^ как у вируса типа В, либо отличаться от них. Эти другие ви-
русы называются вирусами типа А. Поскольку гемагглютинин ви-
русов типа А существенно отличается от гемагглютининов типа В,
человеческий организм неспособен вырабатывать против него ан-
титела с такой же легкостью, с какой он это делает против вируса
типа В, поэтому смертность от гриппа типа А довольно велика.
Существует две гипотезы относительно происхождения разли-
чий между гемагглютининами вирусов типа А и типа В. Согласно
первой гипотезе, они вызваны очень обширной мутацией, обусло-
вившей резкие изменения этого белка. Вторая и более правдопо-
добная гипотеза состоит в том, что новый гемагглютинин проис-
ходит от какого-то вируса, первоначально заражавшего другое
млекопитающее или птицу. Известно, что вирусы гриппа легко ре-
комбинируют (рис. 6.2); кроме того, вирусы гриппа типа А чело-
века были выделены от животных различных видов. Новые гем-
агглютинины сильно отличаются от других гемагглютининов ви-
руса гриппа человека по антигенной активности и по типам вхо-
дящих в их состав белков, которые сходны с белками, содержащи-
мися обычно в вирусах, поражающих лошадей и уток. Таким об-
разом, все имеющиеся данные, по-видимому, подтверждают вторую
гипотезу, т. е. гипотезу о потоке генов от вирусов других тепло-
кровных видов к вирусу гриппа человека. Опасения пандемии грип-
па были причиной того большого внимания, которое уделялось
вирусу инфлюэнцы свиней зимой 1976—1977 гг. (рис. 6.3).
Но поток генов имеет и более общее значение, которое стано-
вится особенно хорошо понятным, если рассмотреть природу ин-
вирус гриппа человека
Д/Гонконг/68(Н 3 N 2)
//ЗЛИ
H3N1
Рис. 6.2. Рекомбинация генетического материала двух различных вирусов грип-
па типа А.
Вирусологи договорились обозначать разные штаммы вируса в зависимости от то-
го, в какой последовательности впервые появляются главные варианты двух ос-
новных антигенных компонентов вириона — гемагглютинина и нейраминидазы.
У человека, например, варианты Н и N поверхностных антигенов обозначают /70,
Н2, ЛИ, N2 и т. д. Для штаммов, первоначально обнаруженных у других ви-
дов, например у свиней, к этому шифру добавляют соответствующие буквы, в дан-
ном случае sw (от англ, swine — свинья).
На схеме показаны результаты инокуляции свиньи двумя вирусами гриппа — од-
ним от человека (H3N2), а другим от свиньи (HswlNl). Вирус вводили через нос.
Инокуляция привела к одновременному заражению одной клетки легкого 8 от-
дельными участками РНК от каждого нз вирусов. Оказавшись в клетке,, вирусы
начинают размножаться; при этом 16 различных участков РНК рекомбинируют
различными способами, образуя новые вирусные частицы. В присутствии антител
к нейраминидазе человеческого штамма и к гемагглютинину свиного штамма все
образующиеся вирусы нейтрализуются, за исключением тех, которые содержат по-
верхностные антигены H3N1. Вирус гриппа (7/ЗЛП) может содержать много раз-
ных комбинаций участков РНК, в число которых входят участки, кодирующие эти
Два поверхностных белка, однако с остальными шестью участками они группиру-
ются по-разному и некоторые из получающихся при этом рекомбинантов могут вы-
зывать заболевание у свиней. Штамм H3N2 был впервые обнаружен у людей в
Гонконге в 1968 г., штамм HswlNl впервые выделен от свиней в штате Айова в
1930 г. (Kaplan, Webster, 1977)
t34
Глава 6
Животный штамм(Н2Л/2)
Вирус гриппа человека
/3 'Гонконг' 68 1Н 3 N 2 )
Рис. 6.3. Предполагаемое происхождение штамма Гонконг/1968 вируса гриппа че-
ловека.
Согласно этой гипотезе, некий предковый вирус гриппа (возможно, птичий) дал
начало ряду родственных между собой вирусов животных, в том числе одному
вирусу, выделенному от лошадей в Майами в 1963 г., и другому, выделенному в
том же году от уток на Украине. Спустя пять лет еще один вирус, принадлежащий
к этой группе и родственный как лошадиному, так и утиному штаммам, реком-
бинировал с широко распространившимся в то время азиатским, или H2N2, штам-
мом вируса гриппа человека, дав начало штамму Гонконг, или H3N2. Этот но-
вый штамм вируса человека получил ген гемагглютинина (ИЗ) от предполагаемо-
го промежуточного животного штамма, а от азиатского штамма сохранил как ген
нейраминидазы (N2), так и гены, необходимые для того, чтобы возбуждать забо-
левание гриппом у людей. (Kaplan, Webster, 1977)
тродуцируемых генов. Поскольку эти иммигранты представляют
собой случайную выборку из своих популяций, они будут нести с
высокой частотой те аллели, которым в тех прежних условиях бла-
гоприятствовал отбор, и очень редко будут привносить гибельные
аллели, частота которых удерживается отбором на низком уровне.
Факторы эволюции
135
Хотя аллели, повышающие приспособленность особей популяции
А, не обязательно будут повышать приспособленность особей по-
пуляции В вследствие разного генетического фона и различных
давлений отбора в этих двух популяциях, тем не менее аллели,
внесенные потоком генов, обычно элиминируются медленно, так
как условия среды в соседствующих популяциях часто отличаются
очень незначительно. Кроме того, по сравнению с аллелями, воз-
никающими в результате мутаций, среди аллелей, приобретенных
благодаря потоку генов, оказывается относительно высокая доля
благоприятных, которые обычно закрепляются в популяции. В не-
которых случаях эти аллели играют очень важную роль, особен-
но, как мы увидим в дальнейшем, когда возможно образование
межвидовых гибридов.
В конечном итоге поток генов действует как некая связующая
сила, которая может несколько нивелировать действие отбора, не
влияя в сколько-нибудь значительной степени на приспособлен-
ность, но препятствуя возникновению слишком сильных различий
между популяциями одного и того же вида. Поскольку эффект
потока генов прямо зависит от числа иммигрантов, чем дальше
друг от друга расположены две популяции, тем меньше ощущает-
ся эта связующая сила. Если две популяции становятся совершен-
но изолированными, то отбор может толкнуть их к эволюции в
разных направлениях, но как только между ними устанавливается
контакт, поток генов будет вновь стремиться объединить их.
Решающие данные, необходимые для оценки роли иммигра-
ции— это скорость распространения гамет в зависимости от рас-
стояния, расстояние, на которое животные отходят от своей попу-
ляции, и расстояние, на которое семена разносятся ветром или
другими факторами от места своего возникновения. Мы не распо-
лагаем достаточными данными, чтобы дать общую оценку этой
роли. Однако в общем скорость потока генов, по-видимому, экспо-
ненциально уменьшается с увеличением расстояния, хотя она зна-
чительно различается у разных видов. Одни виды, например чело-
век, очень подвижны, другие, как некоторые бабочки (рис. 6.4),
довольно ограничены в своих передвижениях.
РЕКОМБИНАЦИЯ
Рекомбинацией называют смешение у потомков генов и хро-
мосом их родителей. Строго говоря, рекомбинация не относится к
числу факторов, изменяющих частоту генов в популяции. Однако
она имеет важное значение главным образом по двум причинам.
Во-первых, разные гены растения или животного взаимодействуют
между собой, и некоторые сочетания генов оказываются при этом
лучше, чем другие. Во-вторых, число рекомбинантов неизмеримо
выше возможного числа мутаций. Хотя в конечном счете источни-
ком новой генетической изменчивости служат мутации, возникно-
Рис. 6.4. Карта плотности популяции бабочки Euphydryas edit ha в 1960—1963 гг.
Особей из разных популяций из года в год метили и наблюдали за ними. Каждая
точка обозначает первую поимку одной особи. Обратите внимание на то, что чис-
ленность популяции Н возросла, популяции G значительно уменьшилась, а попу-
ляции С колебалась. Особи довольно свободно перемещались в пределах той об-
ласти, где они были впервые пойманы, но за ее пределами меченых особей находи-
ли редко. Популяции располагались на расстоянии не более чем 1900 м друг от
друга, что не превышает расстояния, на которое эти бабочки способны летать,
и тем не менее они оставались обособленными. (Р. R. Ehrlich, 1965. The popula-
tion biology of the butterfly Euphydryas editha. II. The structure of the Jasper Rid-
ge colony, Evolution, 19, 327—336.)
Факторы эволюции
137
вение в популяции новых типов по большей части бывает обуслов-
лено рекомбинацией. Этот процесс может создавать большее число
более разнообразных форм и с большей скоростью, чем мутацион-
ный процесс, и непосредственным источником изменчивости в по-
пуляции служат рекомбинации, а не мутации. Раздельнополость
представляет собой внешнее выражение модификаций, происшед-
ших в фенотипе, для того чтобы обеспечить рекомбинацию. Ре-
комбинация имеет место в тех случаях, когда ядра и хромосомы
мужской и женской гамет сливаются, образуя ядро зиготы (см.
гл. 5).
Если гетерозиготный организм, имеющий генетическую консти-
туцию А^гВ^г, скрестить с другим организмом, несущим такое
же сочетание генов, то их потомки (при отсутствии сцепления меж-
ду двумя локусами) будут иметь (в соответствии со вторым зако-
ном Менделя) следующие генотипы: AiAiBjBj; AiAiBjF^; AiA[B2B2;
A^BiBf, A1A2B1B2I А1А2В2В2; А2А2В|В15 АгАгВ^г; А2А2В2В2.
Всего девять разных типов. Каждый из этих типов может возник-
нуть в результате мутации, но вероятность возникновения всех
девяти типов путем мутаций в одном поколении очень мала — так
мала, что это практически невозможно. (Если принять частоту
мутаций равной 10~4, то вероятность возникновения этих девяти
типов в популяции, состоящей из особей А1А2В1В2, за одно поколе-
ние в результате мутаций равна 10~24- Л/, где — число особей в
данной популяции.)
Число диплоидных генотипов, которые можно скомпоновать из
любого числа аллелей какого-либо гена, описывается выражением
где g — число диплоидных генотипов, а г — число аллелей в каж-
дом локусе.
Если рассматривать два локуса, расположенных в разных хро-
мосомах так, что они могут свободно рекомбинировать, общее
число возможных диплоидных генотипов с участием этих двух ге-
нов составит
а а — f 1) г (г + 1) _ Г г(г + 1) ]а
SA sb 2 ' 2 [ 2 J ’
Если, например, принять, что г = 2, т. е. в каждом из двух ло-
кусов имеется по два аллеля, то
2 (2 + 1) 1» _
8а'8ъ— §-- “9’
что, как мы видели, соответствует числу возможных сочетаний ал-
лелей при рекомбинации у двух организмов, гетерозиготных по
Двум локусам, в каждом из которых имеется по два аллеля.
138
Глава 6
Если вместо двух рассматривать три гена, то число рекомби-
наций будет равно произведению gA-gB-gc и т. д.; таким образом,
для п генов (локализованных в разных хромосомах) число реком-
бинантов составит
„ „ п г(г+ 1) Г (г+ 1)
gА'§В'§С •• •'ёп— 2 ' 2 •••• 2 И 2 I '
Число возможных рекомбинаций зависит, следовательно, от
числа аллелей в данном локусе (г) и от числа независимых локу-
сов (л); оно довольно велико, если г и п больше трех, как это
часто бывает. Например, при п=10 и г = 3 число рекомбинаций
равно 610 = 60 466 176. Ясно, что изменчивость, получающаяся за
счет рекомбинации, очень велика, так велика, что в процессе эво-
люции возник целый ряд механизмов для регуляции и ограничения
рекомбинации (см. гл. 8).
Интересным примером, демонстрирующим важную роль реком-
бинации, служит гриб, вызывающий ржавчину пшеницы (Puccinia
graminis). Диплоидные (дикариотические, т. е. содержащие по два
гаплоидных ядра на клетку) споры этого гриба заражают пшеницу
(а также овес, рожь, ячмень и многие другие злаки) весной. Внеш-
ние признаки заражения хорошо заметны: это вытянутые в верти-
кальном направлении красновато-коричневые или черноватые зер-
нистые образования на стебле и листьях, по которым заболевание
и получило название «ржавчины». На растениях пшеницы гриб про-
дуцирует споры двух типов. Споры одного типа, называемые уре-
доспорами, диплоидные, они продуцируются в течение всего лета
и могут заражать другие растения пшеницы. К концу лета гриб
производит вместо уредоспор споры другого типа — телейтоспоры.
Каждая телейтоспора претерпевает мейоз и производит четыре га-
плоидные споры, называемые базидиоспорами. Мейоз и образова-
ние базидиоспор нередко происходят только следующей весной,
а до тех пор телейтоспоры продолжают оставаться на отмерших
растениях пшеницы или падают на землю. Базидиоспора неспособ-
на заражать пшеницу и может дать начало мицелию (вегетативное
тело гриба, грибница) лишь в том случае, если она попадет на
растение барбариса. Оказавшись на листе или побеге барбариса,
базидиоспора развивается в гаплоидный мицелий. В конце концов
два гаплоидных мицелия разного пола (или разного типа скрещи-
ваемости, как это обычно называется у грибов) объединяются, об-
разуя диплоидный мицелий, все еще на барбарисе. А затем этот
диплоидный мицелий дает начало диплоидным спорам — так на-
зываемым эцидиоспорам, заражающим растения пшеницы (они
не способны заражать барбарис), и весь цикл возобновляется
(рис. 6.5).
Переход с одного вида хозяина на другой в жизненном цикле
ржавчины пшеницы имеет интересные эволюционные и экономиче-
ские последствия. Рекомбинация не может происходить в отсутст-
Факторы эволюции
139
вие второго растения-хозяина—барбариса. Кроме того, в областях
с суровой зимой все уредоспоры погибают от холода, и без расте-
ний барбариса ржавчина не может выжить. Поэтому в таких об-
ластях (например, на пшеничных полях Дакоты и центральных
Рис. 6.5. Жизненный цикл ржавчины (Puccinia gramltiis).
Споры (эцидиоспоры) прорастают на пшенице, образуя мицелий, состоящий нз
двуядерных клеток. Этот мицелий дает начало уредоспорам, которые могут зара-
жать другие растения пшеницы. В конце сезона продуцируются телейтоспоры. Два
содержащихся в телейтоспоре ядра сливаются и в таком состоянии гриб перезимо-
вывает. Весной телейтоспоры претерпевают мейоз, в результате чего образуются
гаплоидные базидиоспоры, которые способны прорастать только на барбарисе.
Из базидиоспор развивается гаплоидный мицелий, дающий начало спермогониям
и рецептивным гифам. Клетки из спермогониев переносятся на эти гифы, где об-
разуются двуядерные эцидиоспоры, которые могут прорастать только на пшенице.
провинций Канады) искоренение кустов барбариса задерживает
весной заражение пшеницы ржавчиной. Эта мера не может со-
вершенно ликвидировать заражение, потому что уредоспоры при-
носятся ветром из областей с более мягким климатом, где перези-
мовавшие уредоспоры уже успели заразить растения пшеницы и
начали продуцировать новые споры. Очень важно, однако, что ис-
коренение барбариса элиминирует рекомбинацию, происходящую
при половом процессе у ржавчины. Современные сорта пшеницы
иммунны, или высоко устойчивы, к заражению ржавчиной. Но в
но
Глава 6
процессе эволюции непрерывно возникают новые генотипы ржав-
чины, устойчивостью к которым пшеница не обладает, что застав-
ляет заниматься созданием все более устойчивых сортов. Генети-
ческая изменчивость в популяции ржавчины возникает в резуль-
тате мутаций и рекомбинации. Устранение возможности половой
рекомбинации, достигаемое путем искоренения барбариса, замед-
лит процесс эволюционного приспособления у ржавчины. Этот
пример немногим отличается от ситуации с вирусом гриппа у Ното
sapiens. Мы в состоянии справиться с вирусом, который живет в
популяции человека и может преодолеть его защитные механизмы
только при помощи мутации; однако вирусы, возникающие в ре-
зультате рекомбинации вирусов человека с вирусами других видов,
например обезьян или свиньи, представляют серьезную проблему.
НАРУШЕНИЕ РЕКОМБИНАЦИИ
В гл. 4 упоминалось о том, что второй закон Менделя (закон
независимого распределения генов) приложим только к генам, ле-
жащим в разных хромосомах. На рекомбинацию генов, лежащих
в одной и той же хромосоме, влияют сцепление и кроссинговер, что
подробно рассмотрено в гл. 5. Другое явление, нарушающее про-
цесс рекомбинации, — мейотический драйв.
Мейотический драйв
Два американских генетика Л. Сандлер (L. Sandler) и Е. Но-
вицкий (Е. Novitski) сообщили в 1957 г., что в некоторых линиях
Drosophila melanogaster, гетерозиготных по рецессивным генам,
определяющим киноварный (cinnabar, сп) и коричневый (brown,
bw) цвет глаз и локализованным в правом плече второй хромосо-
мы, наблюдались нехватка гамет с генами cnbw и соответствую-
щий избыток хромосом дикого типа. Когда этих мух скрещивали
с рецессивными гомозиготами сп bw/cn bw (у которых глаза бе-
лые), то вместо соотношения 1:1 красноглазых (дикий тип) и
белоглазых мух получали соотношение 25 красноглазых: 1 бело-
глазая. Аналогичным примером среди растений служат томаты —
культурный сорт Lycopersicon esculentum, а также его дикорасту-
щий родич и предполагаемый предок L. pimpinellifolium. ЧарлзМ.
Рик (Charles М. Rick) из Калифорнийского университета в Дей-
висе обнаружил в четвертой хромосоме томатов ген, который он
назвал «элиминатором гамет» (Ge). В локусе этого гена было най-
дено три аллеля, Ge”, Gec и Gep, но нарушение расщепления на-
блюдалось только у гетерозигот Gec/Ger, образующих гаметы в
соотношении примерно 5 Gec: 95 GeP вместо ожидавшегося соот-
ношения 1:1.
Мейотический драйв играет, по-видимому, важную роль, влияя
на сохранение определенного гена или сочетания генов. Не вполне
Факторы эволюции
141
ясно, однако, распространено ли это явление широко, или встре-
чается в порядке исключения. В любом случае мейотический драйв
представляет собой дополнительный фактор, влияющий на расщеп-
ление. Как и при сцеплении, это может приводить к тому, что
хромосомы, благоприятные с точки зрения отбора, продуцируются
в избытке, а гибельные — в гораздо меньшем числе, в результате
чего снижается число зигот с низкой приспособленностью, и умень-
шается цена, которую надо платить за поддержание в популяции
благоприятных сочетаний генов.
Рекомбинация сама по себе не создает никаких изменений в
генетической информации, но она выявляет сочетания генов, на ко-
торые может действовать отбор, сохраняя одни из них и элими-
нируя другие. Рассмотрим теперь, каким способом отбор действует
на популяцию, чтобы вызвать эти изменения.
ОТБОР
Как мы уже видели, естественный отбор — это процесс, опреде-
ляющий дифференциальный вклад особей в следующее поколение.
Этот относительный вклад в создание следующего поколения на-
зывают приспособленностью или адаптивной ценностью. Число по-
томков, которое данный организм вносит в следующее поколение,
зависит от ряда факторов. В некоторых популяциях растений и
животных наблюдаются циклические колебания численности, вы-
ражающиеся в периодических вспышках и спадах числа особей.
В период увеличения численности популяции большинство особей,
по всей вероятности, оставит некоторое число выживающих потом-
ков, в периоды спада потомство производят лишь немногие члены
популяции. Однако в каждый момент времени некоторые особи
будут производить относительно большее число выживающих по-
томков, чем другие. Чтобы упростить изучение отбора на всех ста-
диях цикла численности и на протяжении нескольких поколений,
генетик оперирует не абсолютными численностями, а частотами.
Абсолютная приспособленность — это участие особи в создании
следующего поколения, относительная приспособленность — это
вклад в следующее поколение по сравнению с вкладом какого-либо
другого генотипа, имеющегося в данной популяции, обычно такого,
который обладает более высокой абсолютной приспособленностью.
Обозначим абсолютную приспособленность данной особи через пг,
а относительную приспособленность — через w, М и W— средняя
абсолютная и средняя относительная приспособленности для дан-
ной популяции. Таким образом, строчными буквами обозначается
индивидуальная приспособленность, а прописными — соответству-
ющие значения для популяции в целом. Далее, при сравнении дей-
ствия отбора на разных членов популяции, всегда исходят из того,
что отбор направлен скорее против потомков определенного орга-
низма, чем в их пользу.
142
Глава 6
У растений и животных, размножающихся половым путем, по-
томки отличаются по своему генному составу как от одной, так и
от другой родительской особи. А поэтому проще изучать действие
отбора на гены, чем на организмы. Приспособленность генотипа,
передающегося следующему поколению с наивысшей частотой,
обычно принимается равной единице, независимо от действитель-
ного числа выживающих потомков. Однако следует ясно понимать,
что отбор действует на отдельные организмы и лишь через них —
на генотипы и гены. Только когда различия в приспособленности
между отдельными особями связаны с наличием или отсутствием
определенного аллеля или группы аллелей в генотипе особи, отбор
действует на этот аллель и на генотип, в состав которого он вхо-
дит. Следовательно, результат действия отбора, определяющий на-
правление эволюции, задается совместным влиянием всех генов(
на приспособленность организма.
Большинство, если не все новые мутации, понижают приспо-
собленность особи, которая гомозиготна по этой мутации. Многие
мутантные гены в гомозиготном состоянии обладают летальным
действием, т. е. особи, несущие двойную дозу мутантного гена,
гибнут. Другие мутации полулегальны, т. е. большинство гомози-
готных по ним особей гибнет, хотя некоторые выживают. Наконец,
существуют субвитальные мутации, понижающие приспособлен-
ность гомозиготных по ним особей, но не слишком резко. Однако-
в гетерозиготном состоянии летальные, полулегальные и субви-
тальные аллели могут как снижать приспособленность своих носи-
телей, так и быть совершенно нейтральными (рецессивы) или даже
повышать приспособленность. В первом случае мутация не обла-
дает доминантностью по приспособленности (строго говоря, доми-
нантность промежуточная). Во втором она полностью рецессивна,
поскольку в гетерозиготе действие мутантного гена полностью за-
маскировано. В третьем случае мутантный ген обладает сверхдо-
минантностью или гетерозисным эффектом (рис. 6.6). Очевидно,
поведение аллеля как в гомозиготном, так и в гетерозиготном со-
стоянии определяет его селективное преимущество или неблаго-
приятный эффект в данной популяции. Напомним, что при своем:
первом появлении мутантный аллель всегда находится в гетеро-
зиготном состоянии, его судьба на ранних этапах зависит от того,
каким он обладает действием в этом состоянии. Только после того-
как частота мутантного гена повышается (в результате отбора),
он становится у некоторых особей гомозиготным.
Прежде чем перейти к подробному изучению этих примеров,
следует также напомнить, что гены в хромосомах данной особи
взаимодействуют друг с другом и что приспособленность каждого-
отдельного гена в известной степени зависит от других генов. На-
пример, некоторые гены полностью рецессивны в одних генотипах,
но обладают неблагоприятным действием даже в гетерозиготном
состоянии в других генотипах. Соответственно, когда мы приписы-
Факторы эволюции
143
ваем определенному аллелю некоторую приспособленность, мы
имеем в виду среднюю приспособленность этого аллеля по попу-
ляции в целом.
Отбор имеет место во всех тех случаях, когда определенный
фенотип подвержен более высокой смертности, чем другие фено-
Состояние 1(—)
ПРИСПОСОБЛЕННОСТЬ
Состояние 2 (0) Состояние 3 (4-)
АА
Аа
-•
АА
•-
аа
Рис. 6.6. Доминирование по приспособленности.
Гетерозиготы могут иметь фенотипы: I — промежуточные между фенотипами двух
гомозиготных родительских особей (кодоминирование); II — сходные с доминант-
ным родительским фенотипом, но не идентичные ему (неполное доминирование);
III— неотличимые от гомозиготного родительского доминантного фенотипа (пол-
ное доминирование); IV—выходящие за пределы изменчивости гомозиготных ро-
дительских фенотипов (гетерозис или сверхдоминирование). Под фенотипом по-
нимается любой признак организма — морфологический, физиологический или ре-
продуктивный. (О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystema-
tics, New York, Macmillan.)
типы в данной популяции, когда он производит меньше потомков
или когда суммарный результат смертности и скорости размноже-
ния выражается в уменьшении числа выживающих потомков.
•С точки зрения конечного результата не имеет значения, когда и
как происходят эти явления, если только организм гибнет до того,
как он произвел потомство. Смерть, наступающая после заверше-
ния размножения, не оказывает влияния на отбор, за исключением
тех случаев, когда родители заботятся о подрастающем потомстве,
как у человека и у многих других позвоночных. Высокая доля в
популяции особей, уже завершивших размножение, может также
сказывать косвенное влияние на отбор, поглощая ресурсы, необ-
ходимые молоди. Это последнее обстоятельство, однако, представ-
ляет особый случай и наблюдается главным образом у человека
144
Глава 6
и домашних животных. В природе особи большинства, если не всех,
видов гибнут в том возрасте, когда они еще сохраняют способ-
ность к половой активности.
Эволюция устойчивости к пестицидам у насекомых
Очень интересным примером естественного отбора, который от-
ражает многие проблемы, обсуждавшиеся в предыдущих разделах
этой книги, служит устойчивость к пестицидам, возникшая за по-
следние 30 лет у более чем 200 видов насекомых, наносящих ущерб
здоровью и хозяйственной деятельности человека.
Во время Второй мировой войны был создан инсектицид ДДТ,
обладающий остаточным действием; это означает, что после при-
менения он не улетучивается и не разрушается до конца. Солдаты,
одежда которых была обработана ДДТ, долгое время не заража-
лись вшами и блохами. В послевоенный период было создано боль-
шое число химических соединений с остаточным действием, на-
шедших широкое применение в сельском хозяйстве, медицине и
промышленности. Однако уже в 1947 г. появилось сообщение о
том, что в Швеции мухи не гибнут от ДДТ. Сначала заподозрили,
что в этом повинно качество препарата, но после того, как за
первым сообщением последовал ряд других, стало ясно, что у на-
секомых вырабатывается устойчивость к инсектициду. Для пре-
одоления этой устойчивости стали увеличивать дозу пестицида и
разрабатывать новые виды инсектицидов. Так, например, в 1950 г.
для борьбы с ореховой тлей в Калифорнии рекомендовалось при-
менять инсектицид паратион в разведении 25 г на 100 л воды,
а спустя 7 лет доза инсектицида возросла в 6 раз (150 г на 100 л
воды). Соответственно возросла, конечно, и стоимость борьбы с
вредителем.
Это, однако, еще не самое худшее. Поскольку применяемые
инсектициды часто токсичны для всех насекомых, они убивают
также других насекомых и насекомоядных птиц, обычно поедаю-
щих тех вредителей, с которыми человек старается бороться. Все
это вызывает нарушение существующего экологического равнове-
сия. Уничтожение полезных насекомых и птиц потребует еще бо-
лее частого применения еще более мощных инсектицидов, посколь-
ку теперь на растительные культуры нападает большее число ви-
дов вредителей, в том числе ряд вредителей, считавшихся прежде
второстепенными. В крайних, но не столь уж редких случаях для
защиты посевов их приходится обрабатывать инсектицидами еже-
недельно, а это сильно повышает стоимость борьбы с насекомы-
ми. Джордж Джорджью (George Georghiou) сообщает, что в
Центральной Америке для защиты хлопковых полей приходится
проводить до 30 обработок пестицидами за один вегетационный
сезон, продолжающийся 6 мес. В результате разведение хлопка
может стать нерентабельным, что заставит отказаться от выращи-
Факторы эволюции
145*
Рис. 6.7. Действие инсектицида мала-
оксона на активность ацетилхолин-
эстеразы чувствительных (кружки) и
устойчивых (квадраты) мух.
Фермент выделяли от мух и испыты-
вали in vitro в присутствии малаок-
сона. Видно, что активность чувстви-
тельного фермента подавлялась край-
не резко, тогда как активность устой-
чивого подавлялась незначительно.
(Devonshire, 1975)
вания этой культуры, особенно в странах «третьего мира», как это
произошло с хлопководством в долине реки Каньете в Перу.
Устойчивость к пестицидам усиленно изучалась, и нет ничего
удивительного в том, что, как оказалось, ее возникновение сопря-
жено с генетическим изменением
ряда признаков, в том числе
биохимических, физиологических
и поведенческих. Эволюция ус-
тойчивости к пестициду особен-
но хорошо исследована у комнат-
ной мухи (Musca domestica). По-
этому мы сосредоточили внима-
ние на этом виде.
Действие пестицидов. К со-
жалению, точный биохимический
механизм действия инсектицида
не всегда известен. Однако из-
вестно, что инсектициды, принад-
лежащие к классу фосфорорга-
нических соединений (малатион,
паратион и др.), убивают насеко-
мых, связываясь с ферментом
ацетилхолинэстеразой (AChE),
участвующей в синаптической пе-
редаче импульсов. Когда инсек-
тицид «отравляет» этот фермент,
последний перестает выполнять
свою обычную функцию — рас-
щеплять ацетилхолин на холин
и уксусную кислоту. В результа-
те при раздражении двигатель-
ного нерва возникают множест-
венные мышечные сокращения,
при этом «сжигаются» все ре-
зервные вещества организма п,
в конце концов, животное гибнет
от истощения. Биохимические
исследования Девоншира (Devonshire) и других авторов показа-
ли, что у мух, устойчивых к инсектициду, возникают изменения
в холинэстеразе, которые значительно снижают скорость ее инги-
бирования фосфорорганическими соединениями (табл. 6.3). По-
томки устойчивых и чувствительных к инсектициду штаммов ком-
натной мухи содержат ацетилхолинэстеразу, которая обладает
свойствами смеси чувствительных и устойчивых ферментов, дей-
ствующих независимо друг от друга с соотношением активностей
2:1 (рис. 6.7). Эти данные можно объяснить, допустив, что гиб-
риды содержат устойчивый и чувствительный ферменты в равных
Количествах и что чувствительный фермент вдвое более чувствите-
Ю—2044
846
Глава 6
Таблица 6.3
Константы скорости (А,) реакций ингибирования
ацетилхолииэстеразы, выделенной из голов устойчивых
и чувствительных мух; измерения проводились
в отсутствие субстрата (Devonshire, 1975)
Инсектицид 10“’-^ (М-'-мин-1)
устойчивые чувствительные
Штамм 608 Штамм Cooper
О-метоат 1,77±0,04 19,9±0,32 20,5±0,53
Метилпараоксон 21,6± 1,0 — 90,1± 4,9
Параоксон 139±6,5 578±22 —
Изопропилпараоксон 9,23±0,43 76,3±3,5 80,6±6,0
Малаоксон 47,3±3,6 867 ±35 601 ±21
Этилмалаоксон 94,3±3,5 932± 12 —
Тетрахлорвннфос 24,7±0,2 — 172±26
лен к инсектициду, чем устойчивый. Правильность этой модели
была подтверждена, когда установили, что устойчивые мухи не-
сут мутантный аллель (обозначаемый AChE-R) одного гена, лока-
лизованного во второй хромосоме (укомнатной мухи 5хромосом).
Хотя аллель AChE-R повышает устойчивость мух к фосфорор-
ганическим инсектицидам, он не делает мух совершенно иммунны-
ми к этому яду. Вскоре ученые обнаружили, что мухи, кроме того,
обладают биохимическими механизмами, которые производят де-
токсикацию инсектицида; иными словами, муха изменяет химиче-
ский состав инсектицида, обезвреживая его. К настоящему време-
ни у комнатной мухи в шести разных локусах обнаружены аллели,
обеспечивающие способность к детоксикации различных инсекти-
цидов (табл. 6.4). Некоторые из этих аллелей, как, например,
Ох-2 и Ses, сообщают мухе устойчивость не к одному, а к несколь-
ким инсектицидам. Поэтому применение какого-либо одного ин-
сектицида может непроизвольно вызвать отбор на устойчивость к
другому инсектициду. Возможно также, что существуют и другие,
еще не выявленные гены, поскольку разные устойчивые штаммы
содержат лишь один или несколько, но не все соответствующие
аллели.
Помимо этих биохимических механизмов исследователи обна-
ружили у комнатной мухи физиологический механизм, который
создает некоторую устойчивость к инсектициду, снижая проницае-
мость наружных покровов насекомого. Значение этого механизма
устойчивости состоит в том, что даже небольшое снижение скоро-
Факторы эволюции
147
Таблица 6.4
Некоторые из генов, обусловливающих устойчивость
комнатной мухи (Musca domestica) к пестицидам
Ген Хромосома Действие
AChE-R II Синтез модифицированной ацетилхолинэстеразы- устойчивой к фосфорорганическим соединениям.
Ох-2 II Детоксикация различных инсектицидов
Deh II Синтез ДДТ — дегидрохлориназы
Ох-2 II Сообщает устойчивость к фосфорорганические соединениям
ру-ех II Сообщает устойчивость к пиретруму
У II Стимуляция интенсивных глутатион-зависимых. превращений фосфорорганических соединений
Реп III Снижает поглощение инсектицида
kdr III Полная устойчивость. Механизм неизвестен
Dld-4 IV Сообщает устойчивость к циклодиенам
Ses v ] Сообщает устойчивость к ряду различных инсек-
DDT-md v } тицидов. Может быть многоаллельным
Ox-5 V J
py-ses V J Сообщает устойчивость к пиретруму
сти проникновения инсектицида в организм насекомого может со-
здать условия, которые позволят механизмам детоксикации справ-
ляться с инсектицидом по мере его проникновения в метаболиче-
скую систему мухи. Были идентифицированы два гена, влияющие'
йа скорость проникновения инсектицида. В отношении гена Реп,
локализованного в хромосоме III, было непосредственно показано,
что он снижает скорость проникновения ДДТ. Что касается гена
kdr (от англ, (knock down resistance), также локализованного в
хромосоме III, то он создает устойчивость к пиретруму и ДДТ,
однако подлинный способ его действия неизвестен.
Другой путь, позволяющий животному избежать гибели от ин-
сектицида остаточного действия, состоит в таком изменении пове-
дения, которое предотвращает контакт с инсектицидом. Избегание
инсектицида может быть вызвано повышенной чувствительностью
к его раздражающему действию, вследствие чего насекомое уда-
ляется или прячется, почуяв близость препарата. Или же оно мо-
жет выработаться под действием отбора, сохраняющего только
тех особей, которые вели себя определенным образом до примене-
ния инсектицида. Известно одно такое изменение, возникшее у
малярийного комара Anopheles gambiae в Родезии. До обработки
Жилищ человека ДДТ комары встречались с одинаковой частотой
снаружи и внутри помещений. Однако после обработки домов и
хижин ДДТ на протяжении 8 лет комары практически перестали
встречаться как в обработанных, так и в необработанных жили-
щах, и такое положение вещей заметно не изменилось даже после
10*
148
Глава 6
Таблица 6,5
Число самок Anopheles gambiae, пойманных вручную
во время их дневного отдыха в Шамве
(Южная Родезия) (Mulrhead-Thomson, 1960)
Место поимки 1926—1928 гг. (до обработки) 1958 г. (после обработки гексахлорциклогек- саном в течение 8 лет)
Внутри помещений 269 II*
Вне помещений 205 619
• 96 необработанных домов.
прекращения обработок (табл. 6.5). Отбор сохранял комаров, ко-
торые залетают в жилище человека, кусают свою жертву, после
чего сразу же вылетают наружу, никогда не садясь на стены для
отдыха.
ИНСЕКТИЦИД
ПОВЕДЕНИЕ
Избегание контакта
МЕХАНИЧЕСКАЯ
УСТОЙЧИВОСТЬ
Устойчивость к проникновению
РАЗРУШЕНИЕ
Механизмы детоксикации
ЗАЩИТНЫЕ
Г МЕХАНИЗМЫ
БИОХИМИЧЕСКАЯ
УСТОЙЧИВОСТЬ
Ферментагионые изменения в [
точках приложения действия j
Рис. 6.8. Устойчивость к инсектицидам, возникающая в результате сочетания ря-
да различных механизмов — поведенческих, морфологических, биохимических и
физиологических.
Ни один из этих механизмов сам по себе ие обеспечивает полного иммунитета, но
все вместе они сводят потенциально летальное воздействие (толстая верхняя
стрелка) к воздействию довольно незначительному (тонкая нижняя стрелка).
В общей сложности за 30 лет у комнатной мухи возникло не
менее 9 мутаций (в разных штаммах и в разных частях земного
шара), обеспечивающих своим носителям известную устойчивость
к новым пестицидам. Большая часть этих мутаций (особенно му-
Факторы эволюции
149
тации, определяющие способность к детоксикации ферментов) об-
условливают неполную устойчивость, так что в присутствии лишь
одного такого аллеля смертность от инсектицидов в популяции мух
понижается, но не ликвидируется полностью. Однако одновремен-
ное присутствие двух или нескольких таких генов приводит к муль-
типликативному повышению устойчивости (рис. 6.8). За очень ко-
роткий период времени многие виды насекомых по всему земному
шару в результате совместного действия мутаций, рекомбинации,
отбора и, возможно, потока генов приобрели способность прино-
равливаться к крупным изменениям в среде. Применение инсекти-
цидов грозило сделать среду обитания и источник пищи — сельско-
хозяйственные культуры — непригодными для них. Как это будет
подробнее рассмотрено в гл. 15, по крайней мере некоторые из
этих насекомых были, возможно, преадаптированы к тому, чтобы
справиться с этой новой напастью — химическими ядами, потому
что растения сами вырабатывают в листьях и других органах боль-
шое число ядовитых веществ. Изменения устойчивости, свидете-
лями которых мы были за последние 30 лет, весьма примечательны
как по простоте лежащего в их основе механизма, так и по слож-
ности затрагиваемых при этом взаимодействий. В целом этот про-
цесс служит нам хорошим уроком. В своих попытках нацело унич-
тожить вредных насекомых мы, возможно, добились лишь того,
что уничтожили некоторых полезных насекомых и птиц и нанесли
вред самим себе, поскольку большинство созданных нами соеди-
нений ядовиты для человека. К сожалению, мы неспособны выра-
ботать устойчивость к этим веществам с такой же скоростью, с ка-
кой это удается мухам, так как возраст достижения половозрело-
сти у нас слишком велик, а скорость размножения слишком низ-
кая.
ВЫЧИСЛЕНИЕ ПРИСПОСОБЛЕННОСТИ ПОПУЛЯЦИИ
Выживаемость. Выживаемость % для каждого генотипа — это
просто число имеющихся в данной популяции особей с таким ге-
нотипом после действия отбора, деленное на исходное число осо-
бей этого генотипа. Возьмем, например, популяцию, состоявшую
дзначально из 10000 особей, в которой частота р аллеля А! со-
ставляла 0,6, а частота q аллеля А2—0,4, исходные численности и
возможная смертность, обусловленная отбором, представлены в
табл. 6.6.
Выживаемость X трех генотипов вычисляют следующим об-
разом:
Число после отбора 3300
AiAi’ Число до отбора 3600 ’ ’
Число после отбора 2880
AiAa’ Число до отбора 4800 ’ ’
Число после отбора 800
АаАг' Число до отбора ~ 1600 ’
150
Глава 6
Хотя погибло почти 2000 гетерозиготных особей и всего 800 го-
мозиготных рецессивов, выживаемость последних была ниже вы-
живаемости первых. Пользуясь понятием «выживаемость», а не
абсолютными числами, можно производить достоверные сравнения
достоинств разных фенотипов и генотипов.
Таблица 6.6
Вычисление выживаемости трех генотипов в гипотетической
популяции
Показатель А,А, А,А, А2А2 Всего
Численность исходной популяции /A.V 2pq-N q*-N N
До отбора 0,62-10 000 = =3600 2 06-0,4х X Ю 000 = 4800 0,42 10 000 = = 1600 10 000
После отбора 3300 2880 800 6980'
Потери —300 —1920 —800 —3020
Выживаемость 0,9 0,6 0,5 0,7
Относительная приспособленность. В любой популяции смерт-
ность не остается постоянной во времени. Например, процент ги-
бели у многих видов растений в засуху будет выше, чем в год с
нормальным количеством осадков. Для того чтобы выяснить по-
ведение данного фенотипа и генотипа во времени, следует устра-
нить годичные колебания смертности путем обычного математиче-
ского приема — деления выживаемости каждого генотипа на вы-
живаемость того генотипа, у которого эта величина максимальна.
Получаемое при этом число и есть относительная приспособлен-
ность (w) данного генотипа. В нашем случае
0,9 < пп
“'AiA2=-^9 =1,00,
0,6
аУА1А2=-о^- = О,66,
0,5 АСК
®А,А2=^д-=0,55.
На относительной приспособленности не сказываются факторы,
одинаково изменяющие смертность всех генотипов; она отражает
только дифференциальную выживаемость разных генотипов. Обра-
тите внимание, что у генотипа, характеризующегося самой низкой
смертностью, приспособленность равна I. Это не означает, что осо-
би с этим генотипом до размножения не гибнут вовсе. Это лишь
означает, что по сравнению со всеми другими генотипами смерт-
ность у него самая низкая.
Факторы эволюции
151
Коэффициент отбора. Теперь мы можем определить еще одну
величину — коэффициент отбора s. Коэффициент отбора находят
простым вычитанием из 1 относительной приспособленности, так
что
Sa1a1 = 1—№a1Aj = 1 —1,000 = 0,
Sa1a3 — 1—®a1a2 = 1—0,66 = 0,34,
Sa,a2 = 1 —®a2a2 = 1 — 0,55 = 0,45.
Отбор против гомозиготных рецессивов
Сначала мы рассмотрим изменения частоты аллеля А2, вызы-
ваемые отбором в тех случаях, когда имеется полное доминирова-
ние, т. е. когда мутантный ген подвергается отрицательному отбо-
ру только в гомозиготном состоянии, потому что, находясь в гете-
розиготном состоянии, он не оказывает никакого воздействия на
приспособленность (полное доминирование). Из гл. 4 нам извест-
ны (закон Харди—Вайнберга) частоты трех возможных сочета-
ний двух аллелзй:
А1Л1=р2, А1А2 = 2р^, А2А2=^2
и
Р2+2р?+<?2 = 1.
Относительная приспособленность этих комбинаций в данном
случае составляет по определению
^AiAi^l,
даА1А2 = 1,
W2a2 = 1—s,
где s — коэффициент отбора, направленного против гомозиготных
рецессивов. Вклад, вносимый гаметами разных типов в создание
следующего поколения, выразится в следующих долях:
для AiAi — р2-1; для AiA2— 2рд-1; для А2А2 — q2-(l—s),
поскольку вклад гамет равен произведению частот генотипов на
относительную приспособленность. Суммарный вклад гамет всех
трех типов уже не будет равен единице, он будет равен 1—sq2. Для
того чтобы найти частоту аллеля А2 в следующем поколении, надо
сложить половину вклада гамет AjA2 и вклад гамет А2А2, а затем
разделить полученную сумму на новый общий вклад; таким обра-
зом, частота аллеля А2 в первом поколении составит:
____<72(1— s) + pq
41 ~~ 1 — St?2
152
Глава 6
Изменение, происшедшее за одно поколение в результате отбора:
что упрощается до
sg2 (I — д)
1—S?2 •
Рассуждая подобным же образом, можно рассчитать действие от-
бора, когда он направлен против доминантного гена Аь а не ре-
цессивного гена А2; когда отбор направлен против гена А2, а до-
минирование отсутствует; или в тех случаях, когда .имеется сверх-
доминирование, и отрицательному отбору подвергаются как ген Аь
так и ген А2 в гомозиготном состоянии. Соответствующие формулы
представлены в табл. 6.7.
Таблица 6.7
Изменение частоты гена, &.q, после отбора в одном поколении
в зависимости от доминирования (Falconer, I960)*
Условия доминирования н отбор Исходные частоты и приспо- собленности генотипов Изменение частоты Д? аллеля As
АЛ AjA2 ^2^2
Р2 2р? <72
Отсутствие доминирования, отбор против аллеля Д2 1 1 1 1 —s I — у s<7 (1 — <7)
Полное доминирование, от- бор против гомозигот А2А2 1 1 —s 1 — sq — sq2 (1 — q)
1 —sq* + $q2 (1 — q)
Полное доминирование, от- бор против аллеля At 1 — S 1 — s 1
1 -5(1-^)
Сверхдоминирование, отбор против гомозигот AiAi и А2А2 1 —Si 1 1 + pg (hP-stf)** 1 — siP2 — s2g2
* Когда s мало, знаменатели мало отличаются от 1 и для большинства целей можно
считать, что числители достаточно точно соответствуют величине
Вывод этого выражения см. в гл. I.
Из приведенных в табл. 6.7 формул можно сделать несколько
выводов. Прежде всего мы видим, что отбор зависит от коэффи-
циента отбора (s) и от исходных частот генов (q или, при наличии
сверхдоминирования, р и q). Эта зависимость довольно сложная.
Из формул в табл. 6.7 и из графиков, представленных на рис. 6.9,
можно видеть, что отбор действует наиболее эффективно, когда
гены имеют промежуточную частоту, и что он становится найме-
Факторы эволюции
153
0,032
0,024
0,016
0,003
0,028
, 0,020
О 0,1 0,2 0,3 0,4 0,5 0,6 0 ’’ 0,8 0,0 1,0
о
Рис. 6.9. Изменение частоты гена (Дд) под действием отбора, интенсивность ко-
торого s=0,2, при разной исходной частоте (q).
А. В отсутствие доминирования. Б. При полном доминировании. Прерывистые
кривые отражают отбор, направленный против данного гена, так что Ад имеет
отрицательную величину. Сплошные кривые отражают отбор, направленный в
пользу этого гена, так что Ад имеет положительную величину. (D. S. Falconer,
1954. Asymmetrical responses in selection experiments. In: Symposium on Genetics
of Population Structure Institute di Genetics, Universita di Pavia, Italy, Aug. 20—
23, 1953. Un Int. Sci. Biol. Series B, N 15, 16—41.)
154
Глава 6
нее эффективным, когда частота мутантного аллеля очень высокая
или очень низкая. Кроме того, отбор против рецессивного мутанта
очень неэффективен при низкой частоте этого мутанта. Подобное
положение вещей обусловлено тем, что при этом мутант находится
главным образом или даже исключительно в гетерозиготных ком-
бинациях и тем самым защищен от действия отбора.
Действие отбора на фенотипические признаки
Естественный отбор действует на фенотипы и лишь косвенным
образом — на генотипы. Изменения генных частот происходят толь-
ко в тех случаях, когда существует достаточно точная корреляция
мёжду генотипом и фенотипом. Изучение эмбриологии и процесса
индивидуального развития растений и животных показывает, что
различные генотипы дают начало различным фенотипам, но что'
зависимость эта очень сложная, и среда влияет на нее многими
И притом различными способами (см. гл. 4). Поэтому часто проще
наблюдать действие отбора непосредственно на фенотип, чем вы-
являть его действие на лежащий в основе генотип. Кроме того, при
искусственном отборе селекционера интересует прежде всего ка-
кой-либо фенотипический признак, например удойность коров,
яйценоскость кур или урожайность пшеницы.
Отбор, действующий на фенотипические признаки, бывает трех
типов: стабилизирующий, дизруптивный и направленный. Эти три
типа отбора соответствуют способам, используемым данной попу-
ляцией для сохранения приспособленности к среде, которая может:
1) оставаться постоянной и стабильной (стабилизирующий отбор),
2) становиться более изменчивой и распадаться на разные подсре-
ды (дизруптивный отбор), 3) непрерывно изменяться в одном и
том же направлении (направленный отбор) (рис. 6.10).
Принято считать, что отбор приводит к фенотипическому изме-
нению. Однако это вовсе не обязательно, что часто можно наблю-
дать в действительности. На самом деле наиболее распространен-
ный тип отбора — это стабилизирующий отбор (рис. 6.10). При
стабилизирующем отборе фенотипы, признаки которых близки к
среднему значению для данной популяции, обладают более вы-
сокой приспособленностью, чем фенотипы, у которых признаки
выражены в крайней степени, это объясняется стабильностью сре-
ды обитания. В результате отбора такого типа средняя для дан-
ной' популяции не изменяется, тогда как ширина распределения
может быстро или медленно уменьшаться в зависимости от генети-
ческой детерминированности рассматриваемого признака.
Период цветения у многих растений, цветущих весной, находит-
ся под действием стабилизирующего отбора: те представители
данного вида, которые зацветают слишком рано, производят в
среднем меньше семян, чем те, которые зацветают позднее, что
связано со сроками появления насекомых-опылителей; растения,
Факторы эволюции
155
/-0*2 Х1
У + d% z+d2 x±d2 У + d%
СТАБИЛИЗИРУЮЩИЙ ДИЗРУПТИВНЫЙ
НАПРАВЛЕННЫЙ
Рис. 6.10. Три главных типа отбора.
При стабилизирующем отборе среда благоприятствует организмам, признаки ко-
торых близки к средним для данной популяции; соответственно изменений в попу-
ляции либо не, происходит, либо они невелики. Дизруптивный отбор благоприят-
ствует крайним значениям признаков и вызывает разделение популяции на две.
Направленный отбор благоприятствует одному из крайних значений и приводит
к сдвигу среднего для данной популяции в сторону крайнего значения; направ-
ленный отбор обусловливает большую часть изменений, наблюдаемых в процессе
эволюции. Кривые отражают частоту особей с определенным диапазоном измен-
чивости от z до у, зачерненные области — фенотипы, элиминированные отбором;
тонкие стрелки — направление эволюционного изменения; d — величина измене-
ния. (О. Т. Solbrig, 1970. Principles and Methods of Plant Biosystematics, New
York, Macmillan.)
156
Глава 6
зацветающие слишком поздно, также производят в среднем мень-
ше семян, чем те, которые зацветают раньше, вследствие недостат-
ка влаги или же более сильной конкуренции со стороны других
растений.
Другой возможный тип отбора — это отбор, благоприятствую-
щий сохранению крайних, а не средних значений признака в связи
с разделением среды на два или несколько разных типов
(рис. 6.10). Однако, поскольку особи с такими крайними значе-
ниями скрещиваются между собой, в каждом поколении будет
появляться большое число особей, имеющих промежуточные при-
знаки. В среде, благоприятствующей крайним значениям, любые
механизмы, препятствующие скрещиванию между особями с про-
тивоположными крайними значениями признака, будут создавать
преимущество своим носителям, приводя ко второму типу отбо-
ра— дизруптивному. Дизруптивному отбору необходим какой-либо
механизм, обеспечивающий преимущественное скрещивание меж-
ду сходными особями, — иными словами, какой-либо изолирующий
механизм.1
Механизм отбора, благоприятствующего крайним значениям,
т. е. дизруптивного отбора, недостаточно ясен. Английские генети-
ки Тодей (Thodey) и Боам (Воат) провели в 1959 г. интересный
эксперимент на Drosophila melanogaster. В начале эксперимента
8 самкам и 8 самцам дрозофилы была предоставлена возможность
свободно скрещиваться между собой, после чего Тодей и Боам
отбирали из каждого поколения по 4 самца и 4 самки с наимень-
шим числом стерноплевральных щетинок и по 4 самца и 4 самки
с их наибольшим числом и снова давали им возможность свободно
скрещиваться. Поскольку при этом обоим крайним типам оказы-
валось одинаковое предпочтение, отбор вначале был стабилизиру-
ющим, благоприятствуя крайним типам. Но к концу эксперимента
в популяции не осталось промежуточных типов, очевидно, потому,
что скрещивания между мухами с большим и с малым числом
щетинок давали стерильных потомков. Отбор из стабилизирующе-
го стал дизруптивным.2
Если ситуации, подобные созданным Тодеем и Боамом в экс-
перименте, существуют в природе, то дизруптивный отбор мог бы
служить механизмом, ведущим к видообразованию.
Третий тип отбора — направленный отбор — благоприятствует
одному из крайних фенотипических выражений данного признака,
как это случается в природе, когда среда непрерывно изменяется
1 Это не точно. Дизруптивный отбор — это ситуация, когда особи с крайними
значениями признака обладают преимуществом по сравнению с особями, характе-
ризующимися промежуточным значением того же признака. Предпочтение при
скрещивании — одно из возможных следствий действия дизруптивного отбора,
а не его необходимое условие. — Прим. ред.
2 Правильнее, следуя авторам цитируемой работы, считать, что отбор с самого
начала эксперимента был дизруптивным. — Прим. ред.
Факторы эволюции
157
в одном направлении (рис.
6.10). Направленный отбор
вызывает фенотипические и
генотипические изменения
быстрее, чем отбор любого
другого типа. Искусствен-
ный отбор по большей час-
ти бывает направленным.
За последние 60 лет на
Иллинойсской эксперимен-
тальной сельскохозяйствен-
ной станции было проведе-
но несколько селекционных
экспериментов на кукурузе
(Zea mays). В одном из
этих экспериментов в каж-
дом поколении отбирались
растения с низко располо-
женным початком. В исход-
ной популяции средняя вы-
сота расположения початка
колебалась у разных особей
от 109 до 142 см от поверх-
ности почвы. После отбора
в 24 поколениях у всех рас-
тений початок располагал-
ся на высоте не более 20,3
см, причем продолжение от-
бора не приводило к даль-
нейшему изменению, по-ви-
димому, вследствие истоще-
ния запаса генетической из-
менчивости по этому приз-
наку. Одновременно в дру-
гой линии, происходящей
от той же исходной популя-
ции, проводился отбор рас-
тений, у которых початок
располагается на большой
высоте. После 24 поколе-
ний были получены расте-
ния, у которых початки рас-
полагались в среднем на
высоте 3 м от поверхности
земли!
В другой серии экспери-
Поколение
£
Рис. 6.11. Результаты направленного отбо-
ра у Zea mays, проводившегося на протяже-
нии 76 поколений.
А. Отбор на высокое и низкое содержание
масла в зернеи Б. Отбор на высокое и низ-
кое содержание белка в зерне. Ввиду того
что содержание белка варьирует в зависи-
мости от погоды, в данные по содержанию
белка за каждый год вносилась поправка
путем вычитания среднего за данный год
содержания белка для линий с высоким и
низким содержанием масла. (J. W. Dudley,
1977.. 76 generations of selection for oil and
protein percentage in maize, In: E. Pollak
and others (eds.), Proc, of the Intern. Conf,
on Quant. Genetics, Aug. 16—21, 1976, Amesr
Yowa State Univ. Press.)
i5'
Глава 6
ментов производился отбор растений на высокое и низкое содер-
жание белка в зернах. В «высокой» линии после 76 поколений от-
бора содержание белка возросло на 15% по сравнению с исход-
ной линией, а в «низкой» линии содержание белка уменьшилось
примерно на 9%. В третьей серии экспериментов проводился от-
бор на высокое и низкое содержание масла. Если в исходной ли-
нии растения давали примерно 5% масла, то в «высокой» селек-
тированной линии содержание масла в зернах составляло почти
19%, а в «низкой» — менее 1% (рис. 6.11).
Возможно, большая часть фенотипических изменений, возни-
кающих в процессе эволюции, обусловлена направленным отбо-
ром. Однако по определению отбор любого типа вызывает геноти-
пические изменения. Если фенотипические и (или) генотипические
изменения отсутствуют или если подобные изменения хотя и име-
ют место, но являются совершенно случайными, то это свидетель-
ствует об отсутствии естественного отбора.
МУТАЦИОННЫЙ ПРОЦЕСС И ОТБОР
Как мы уже видели, мутационный процесс наиболее эффекти-
вен, когда частота исходного гена высокая (а частота мутантного
гена низкая), поскольку при этом имеется больше генов, способ-
ных мутировать (p-и). Это совпадает с условиями, при которых
•отбор оказывается наименее эффективным. Но с повышением час-
тоты мутантного аллеля эффективность отбора повышается, а эф-
фективность мутационного процесса понижается до тех пор, пока
не установится равновесие, после чего дальнейших изменений час-
тоты генов не происходит.
Рассмотрим случай, когда мутантный аллель подвергается от-
рицательному отбору только в гомозиготном состоянии. При рав-
новесии изменения, вызванные в одном поколении мутационным
процессом, должны быть равны изменениям, вызванным отбором,
так что
/1 \ W2 (1 — q)
u(\—q)—v.q= 4l^sq24’-.
Это уравнение можно упростить, если незначительно поступиться
точностью, для того чтобы зависимости стали более понятными.
Во-первых, его можно упростить, если пренебречь обратными му-
тациями (—v-q), которые практически отсутствуют при низкой
частоте аллеля А2, а во-вторых, можно отбросить знаменатель
1—sq2, поскольку при низкой частоте аллеля А2 его значение близ-
Факторы эволюции
159
ко к 1- При этом получим
и (1 — <?) =s<?2 (1 —-</),
u=sq2,
Это приблизительное выражение, описывающее частоту мутант-
ного аллеля А2 при равновесии между мутационным процессом и
отбором, показывает, что равновесная частота аллеля А2 должна
зависеть от относительных значений частоты мутаций и коэффици-
ента отбора и, по крайней мере теоретически, может принимать
самые различные значения. Но при обычной низкой частоте мута-
ций (порядка 10~4—10-8) частота мутантного аллеля будет оста-
ваться низкой, даже если на него не будет действовать слишком
сильный отрицательный отбор. Поэтому в результате совместного
действия отбора и мутационного процесса частота вредных мута-
ций, не обладающих гетерозисным эффектом, будет оставаться на
низком уровне. Этот вывод подтверждается результатами наблю-
дений над природными популяциями. Следующий вывод состоит в
том, что один лишь мутационный процесс вряд ли может вызвать-
эволюционное изменение и что главную роль в определении того,
удастся ли данному гену распространиться в популяции или же он
останется редким, играет отбор.
Прекрасным примером взаимодействия между отбором и му-
тационным процессом служит эволюция миксоматоза у кроликов-
в Австралии. В 1859 г. в Австралию из Европы завезли кроликов.
Вскоре они сильно размножились и превратились в форменное
бедствие. В 1950 г. для борьбы с кроликами их заразили вирусом
миксомы, вызывающим у них миксоматоз — острое заболевание,
от которого животное гибнет вскоре после заражения. Этот способ-
оказался очень эффективным: в первый же год от миксоматоза в
Австралии погибло свыше 99% кроликов.
Вирус миксоматоза переносится комарами. Летом комаров очень-
много и болезнь распространяется быстро. Однако зимой полово-
зрелых комаров нет, так что вирус (не имеющий промежуточного
хозяина) зимой также погибает. Поэтому каждую весну фермеры
Должны были заново интродуцировать вирус. Однако в самой ви-
русной популяции отбор благоприятствует мутантным штаммам,
которые, будучи менее вирулентными, не убивают своего хозяина-
кролика, переживают вместе с ним зиму и могут распространяться
Из года в год естественным путем. Вирусы исходного интродуциро-
ванного штамма быстро убивали своих хозяев и поэтому не могли
распространяться на следующее лето (если не считать тех мест,
куда их интродуцировали вторично). В отличие от них менее ви-
рулентный штамм оказывается доступным комарам на следующее-
лето после интродукции и, следовательно, может распространяться
160
Глава 6
естественном путем. В самом деле, штаммы, выделенные в Авст-
ралии от .иких кроликов через 3—4 года после интродукции, были
менее вирулентны, чем исходный штамм.
В то >е время среди кроликов происходил сильный отбор на
устойчиво'ть к миксоматозу. В популяциях, продолжающих под-
вергаться заражению вирусом, генетическая устойчивость к нему
возрастал;, вероятно, в результате мутаций и последующего отбо-
ра. В одн>й популяции кроликов гибель от одного штамма вируса
за 7 лет снизилась от 30 до 25%.
Таким образом между вирусом и кроликами происходил про-
цесс коэв'люции. Изменения в популяции кроликов влияют на ха-
рактер отюра, действовавшего на комаров, и наоборот. В высоко-
чувствптежных популяциях кроликов отбор, действующий на ви-
русы, бла'оприятствует сохранению менее вирулентных штаммов.
В отношении самих кроликов отбор благоприятствует сохранению
устопчивьх особей. По мере возрастания устойчивости в популя-
ции кролжов отбор благоприятствует все более вирулентным
штаммам вируса. В обеих популяциях отбор действует независи-
мо в нащавлении максимизации их приспособленности. Конечным
результатам будет обширная популяция кроликов, служащих хо-
зяевами бширной популяции вирусов.
ПОЛНАЯ ЭЛИМИНАЦИЯ РЕЦЕССИВОВ
Мнопе мутации, будучи летальными в гомозиготном состоянии,
в гетерозтготном состоянии проявляют лишь незначительное дей-
ствие ил! не проявляют его вовсе. Вычисление изменения частоты
генов в таких случаях представляет собой сравнительно простую
операции и дает возможность глубже понять действие отбора.
Исхощая частота генных комбинаций в рассматриваемой по-
пуляции шовь принимается равной p2 + 2pq + q2 = 1. Относительная
приспосо!ленность этих комбинаций
®AjAj = K Wa!A2 = 1 И Ша2а2 = 0.
Соответственно после отбора разные генотипы будут иметь
следуюпце частоты:
AiAp p2-wA1A1=^pQ2,
A4A2: 2pq-wAlAi=2p0q0,
A2A2: <72-ayA2A2 = 0.
Итого
Po2H“2/7o<7o=po (Po+^o+^o) — Po 0 +?o)>
а частота
n __ PuPo ______ 4o
1 Ро (1 + 9o) 1 + Po
Факторы эволюции
161
После одного поколения размножения равновесие Харди—
Вайнберга будет восстановлено (во всяком случае, в популяции
зигот), отбор будет снова действовать, и, рассуждая таким же
образом, мы находим
?0
я _ 41 __ 1 Ф) ____ Ф)
’2 - 1+?I - qt - 1 +2?о ’
* + Я»
и т. д.; следовательно,
qn = -r~г— (гармонический ряд),
а изменение Ад равно
1 ™ l+<7. 1 Н-<7о Ч-<?9
Для полной элиминации рецессивов отбор должен быть очень
интенсивным. Однако эффективность действия отбора зависит от
частоты аллеля, на который он действует. При очень высокой
(близкой к 1) частоте аллеля значение числителя будет близким
к 1, а значение знаменателя приблизится к 2. Поэтому почти -по-
ловина генов будет элиминирована. Если частота вредного гена
равна 0,5, то числитель в приведенном выражении будет равен 0,25,
а знаменатель— 1,5 и примерно лишь */6 аллелей А2 будет элими-
нирована из популяции. По мере дальнейшего уменьшения часто-
ты генов А2 числитель будет уменьшаться пропорционально д2,
а знаменатель — пропорционально 1 + д0, так что Ад будет стано-
виться все меньше и меньше. Таким образом, отбор против ре-
цессивов очень интенсивен при их высокой частоте в популяции,
Ио очень неэффективен при их низкой частоте.
Из формулы для частоты аллеля А2 в n-м поколении
д =________
Чя 1 + nq,
можно вычислить число поколений, необходимых для достижения
данного изменения частоты гена:
Яп Яп,
д— я>-Яп __1________1_
ЯпЯа Яп Я»
Отсюда следует, что, для того чтобы понизить частоту аллеля с
0,99 до 0,5, потребуется
п = ----= 2 —1 — 1 поколение (приблизительно),
и,о и,УУ
И—2044
162
Глава 6
но для того, чтобы понизить частоту с 0,5 до 0,25, потребуется
п—--------—!-=-=4 — 2 — 2 поколения,
U , V , D
а с 0,25 до 0,125 —
Л=”о;Г2У--0J25 ^8”4 "4 поколения-
Полная элиминация представляет собой предельный случай
действия крайне интенсивного отбора. Если отбор менее интенси-
вен, изменение будет происходить не так быстро, но процесс в
общем будет носить такой же характер. Иначе говоря, отбор бу-
дет очень эффективным, когда частота гена высокая, и очень не-
эффективным при низкой частоте гена. Кроме того, поскольку за-
пасы вредного аллеля в популяции постоянно пополняются за счет
повторных мутаций, рецессивный ген никогда не может быть эли-
минирован, каким бы интенсивным не было его вредное действие.
Примером служит наследственное заболевание человека, назы-
ваемое фенилкетонурией. Эта болезнь вызывается рецессивным ал-
лелем, нарушающим превращение аминокислоты фенилаланина в
тирозин. Люди, страдающие этой болезнью, выделяют с мочой
аномально большие количества фенилпировиноградной кислоты.
Почти у всех людей, которые систематически выделяют с мочой
фенилпировиноградную кислоту, наблюдается та или иная степень
умственной отсталости. Обычно эта отсталость выражена доволь-
но сильно, приближаясь к идиотии или имбецильности, так что до
размножения дело доходит редко.
Фенилкетонурия встречается у 2—10 детей на каждые 100 000
(или 0,2—1 на 10 000). Для дальнейших вычислений мы примем
наибольшую частоту: 1 на 10 000= 10-4;
<72 = 10-\
(?=j/TOLi = ю-2^0,01.
Это означает, что в среднем один человек из 100 несет рецессив-
ный ген фенилкетонурии. Если мужчина и женщина, являющиеся
носителями этого рецессивного гена, вступают в брак, то следует
ожидать, что четвертая часть их детей будет страдать фенилкетон-
урией. Частота этой болезни определяется частотой мутаций.
Недавно было обнаружено, что если у новорожденного ребенка
удается диагносцировать фенилкетонурию (путем анализа мочи),
то вредные последствия болезни удается предотвратить, посадив
ребенка на диету, не содержащую фенилаланина; при этом чело-
век нормально развивается и может иметь потомство. Следователь-
но, в каждом поколении частота дефектного аллеля будет повы-
шаться со скоростью, соответствующей частоте мутаций (если
Факторы эволюции
163
удастся выявить все случаи). Такое повышение можно рассчитать.
<70 = 0,0!,
<71 = <7о + (Ро ‘ = 0,01 + [ 0,99 • (2 • 10’4) ] =
=0,014-0,000199 = 0,010199,
q2=Я14- (^. и) = 0,010199 + (0,9898 • 2 • 10-4) = 0,010395,
<73 =<72 • (р2 «) = 0,010395+(0,9896 4- 2 • 10’4) = 0,010594.
Таким образом, по прошествии трех поколений, или приблизитель-
но 100 лет, частота появления людей с этим геном возрастет от
примерно 100 до примерно 106 на каждые 10 000, а число людей,
больных фенилкетонурией, — от 10 до 11 на 10 000. Если же в
дальнейшем частота болезни возрастет еще больше, достигнув, на-
пример, 10% (0,1), то больные с фенилкетонурией будут встречать-
ся гораздо чаще (один на каждые 100 человек); это создаст серь-
езные затруднения, поскольку фенилаланин содержится в боль-
шинстве пищевых продуктов и исключить его из диеты сложно и
дорого. Предотвращение создания такой популяции, в которой поч-
ти каждый страдает каким-нибудь серьезным генетическим забо-
леванием, требующим повседневного медицинского вмешательства,
представляет собой важную моральную и социальную проблему.
ОСНОВНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА
Основная концепция теории эволюции путем естественного от-
бора состоит в том, что генотипы с более высокой приспособлен-
ностью (да) оставляют пропорционально большее число потомков
и что соответственно их гены будут представлены в следующем
поколении с большей частотой. A priori ясно, что для любого из-
менения необходимо наличие в популяции некоторой генетической
изменчивости, причем чем выше эта изменчивость, тем больше воз-
можностей для изменения. Но существует ли какая-либо строгая
зависимость между генетической изменчивостью и максимально
возможной скоростью отбора?
Существование подобной зависимости было показано в 1930 г.
Рональдом Фишером (Ronald Fisher), который назвал ее «основной
теоремой естественного отбора». Эта теорема гласит: «Скорость
повышения приспособленности в любой популяции в любой момент
равна ее генетической дисперсии по приспособленности в этот мо-
мент». Смысл этой теоремы можно проиллюстрировать на примере
одного локуса с двумя или более аллелями при свободном скре-
щивании. Если локусов больше одного и скрещивание не свобод-
ное, то она может оказаться справедливой не во всех случаях.
Рассмотрим популяцию, в которой приспособленность опреде-
ляется одним локусом, причем в этом локусе имеется только два
пллеля, Ai и А2, с частотами р и q (q— 1—р), соответственно. Обо-
4.1’
164
Глава 6
значим приспособленность гомозигот AtA[ через гетерозигот
AjA2 — через оцг и гомозигот А2А2— через да22. Частота аллеля At
после отбора в одном поколении
п а + witPo4o __ “Wo2 + “Wo?o
“Wo2 + 2“W<tf9 + “Wo* ’
где
=^иР2+2аУ12Р<7+®22^.
что соответствует средней приспособленности популяции.
Рассмотрим теперь, как изменится W после отбора в одном по-
колении.
Л№ =
= (®11Р12 + 2Ш12Р1<71 Ч-^гг?!2) — (а'иРо2 +
4-2Ш12р09о+®22<7о2) =^и (Pi2—Ро2) +
+ 2ш12 (Р1<71—РоРо) +®22 (Q12—<7о2)-
Это выражение можно упростить при помощи нескольких ал-
гебраических преобразований.
ДГ = Wi j [ (Р14- Ро) (р4—р0) J 4- 2да1г [Р1 (1 — Р1) — р0 (1 — р0) ] +
+аъ [ (ft + <7оЖ—%) 1 =
= I (Pi + Ро) (Pi—Ро) 1 + 2ш12 [ (Pi —р^ — (р0—Ро®) 1 +
Pi) + (I— Ро)П(1— Pi)~ (1— Ро)П =
= 1 [ (Pi +Ро) (pi - Ро) ] + 2^12 [ (Pi — Ро) + (Ро2 — Pl2) J +
4-W22 1(1 —Pi + 1 —р0) (1 —pi — 1 - J Ро» =
= ®h[(Pi+Po) (Pi -Ро))+2®12[(Р1—ро) + (Ро —Pi) (Ро+Р1)1 +
+ ®22 (2 — Р1 — Ро) (—Pi + Ро) =
=^н1(Р1—Ро) (Р1+Ро))+2ш12[(р,—р0) (1— р0 — Pi)]4-
+®22[(Pi—Ро) (Pi+Po—2)].
Вынеся за скобки (р1 — р0), получим
ДГ = (Р1 — р0) [Юн (р4 4-р0) 4- 2w12 (1 — Ро —Pi) +
+ да22 (Р14~Ро — 2)J.
Но
Pi—р0 = Др и
Факторы эволюции
165
После соответствующей подстановки и алгебраического преобра-
зования получим
ДЙ? = (Др)2 2®12+ie.'224--^y-) =
\ /'ОИо /
= 2ро9о [“’про+®12 (1 — 2р0) — ®22<7о1 2 =
=2ро<7о [“’про+®127о—W 12р0—ю22р0]2.
Если теперь ввести обозначения Ег и £2:
Et=™iiP0+w12qn—W,
Е2=wl2p0 4- w22qQ—W,
то приведенное выше уравнение можно записать как
&W—2p0q0(Ei—E2')i.
Положительная часть выражения для Еь Wup0 + wl2qQ соответ-
ствует приспособленностям генотипов AiAj и половины генотипов
А1А2, содержащих аллель Аь а положительная часть выражения
для Е2, wl2po + w22qo— приспособленностям половины генотипов
А]А2, содержащих аллель А2, плюс приспособленность генотипов
А2А2. В таком случае £i и Е2 представляют отклонения приспособ-
ленностей аллелей Ai и А2 соответственно от средней для популя-
ции приспособленности. Величину Et—Е2 Фишер назвал «средним
избытком» аллелей Аь
Если изложить все это словесно, то мы показали, что измене-
ние приспособленности (Д№) равно произведению частот двух ал-
лелей на квадрат разности между средними для популяции при-
способленностями этих двух аллелей. По определению дисперсии —
это аддитивная дисперсия приспособленности двух рассматривае-
мых аллелей (см. гл. 4). Во всех случаях, когда р = 0 или р=1,
значение W будет равно нулю; это легко показать путем подста-
новки в соответствующие места только что выведенных уравнений.
В случае превосходства гетерозигот, когда щ12>ю22 и tth2>®n,
провести доказательство сложнее; однако, применяя методы регрес-
сионной статистики, можно показать, что независимо от значений
®и, ®12 и w22, аддитивная генетическая дисперсия равна 0, когда
№=0.
Теорема Фишера имеет двоякий смысл. Во-первых, она фор-
мально доказывает невозможность отбора в однородной популяции,
а во-вторых, эта теорема показывает, что скорость изменения опре-
деляется не значением приспособленности, которое создает данный
аллель, а степенью изменчивости по приспособленности, создавае-
мой в популяции всеми аллелями данного локуса. Например, по-
пуляция, в которой ши = 0,9, р = 0,5, ш,2=1 и 9=0,5, изменится в
166
Глава 6
следующем поколении сильнее, чем популяция, в которой щц=0,1,
р=0,01, w22— I и 47 = 0,99, потому что в первой популяции суммар-
ная генетическая дисперсия по приспособленности больше.
СЛУЧАЙНЫЕ ЯВЛЕНИЯ
До сих пор мы исходили из допущения, что имеем дело с очень
большой (теоретически — бесконечно большой) популяцией и что
скрещивание в ней происходит совершенно случайным образом.
В природе ни одно из этих двух условий никогда не выполняется.
Все популяции имеют конечные, в большинстве случаев средние
(102—103 особей) или малые (менее 102 особей), размеры и про-
ходят через периоды резкого уменьшения численности, особенно
в периоды ухудшения окружающих условий (недостаток пищи,
экстремальные температуры, засуха и т. п.). Кроме того, скрещи-
вание не бывает случайным; такие простые факторы, как физиче-
ская близость в популяциях растений или предпочтения при спа-
ривании у животных, делают случайное скрещивание невозмож-
ным.
Конечные размеры популяции (гл. 7) и неслучайность скрещи-
вания (гл. 8) приводят к тому, что каждое поколение представля-
ет собой до некоторой степени случайную выборку из предыдущего.
Это означает, что элиминация или сохранение генов в данной по-
пуляции носит отчасти случайный характер. Вероятность того, что
данный ген сохранится в популяции или будет элиминирован, за-
висит от его частоты, которая определяется мутационным процес-
сом, отбором и потоком генов. Но в популяциях в разные годы
могут происходить случайные флуктуации, которые не представ-
ляют собой результата отбора. Эти флуктуации редко вызывают
длительные эволюционные изменения, но они не позволяют попу-
ляции достигнуть ее максимальной приспособленности. Случайные
явления могут иметь эволюционное значение лишь при условии,
что некоторое число важных генных комбинаций будет полностью
элиминировано из популяции. Вероятность того, что это произой-
дет, даже при такой небольшой численности популяции, как 100
особей, очень мала. Сомнительно, чтобы случайный дрейф генов
(так называется это явление) был когда-либо важным фактором
эволюции. Мы вернемся к этим вопросам в следующей главе.1
КРАТКИЕ ВЫВОДЫ
В этой главе мы кратко проанализировали простейшую ситуа-
цию, при которой особи в популяции различаются по одному ло-
кусу с двумя аутосомными аллелями. Подобная ситуация, возмож-
но, никогда не наблюдается в природе. Организмы обычно разли-
1 Вопрос о роли случайного дрейфа в эволюции в высшей степени дискуссио-
нен. — Прим. ред.
Факторы эволюции
167
чаются по многим локусам. Кроме того, лишь в редких случаях на
приспособленность влияет только один аллель, независимо от дру-
гих генов данного организма. Однако для того чтобы понять более
сложные ситуации, встречающиеся в природе, следует сначала ра-
зобраться в простых случаях. Поэтому вполне оправдано и целе-
сообразно изучать ситуацию, которая сама по себе довольно аб-
страктна.
Популяционный генетик изучает изменения генных частот в
популяциях и явления, вызывающие эти изменения. Орудиями тру-
да ему служат статистика и математика, а также эксперименты в
лабораториях и в природных условиях. За последние два десяти-
летия к этому арсеналу добавилось два новых метода — моделиро-
вание на вычислительных машинах и электрофорез в гелях. На
основе своих теоретических и экспериментальных исследований по-
пуляционный генетик создает модели для объяснения общих эво-
люционных явлений. Процессы эволюции при заданных предполо-
жениях можно имитировать на цифровых вычислительных маши-
нах, проверяя логику и внутреннюю согласованность заложенных
в модель предпосылок. Однако истинной проверкой модели служат
эксперимент и наблюдения, проводимые в полевых и лаборатор-
ных условиях на природных популяциях растений, животных и
микроорганизмов. Определяя методом электрофореза, созданным
на основе знаний, полученных молекулярной биологией за послед-
ние два десятилетия, частоты ферментов, популяционный генетик
оценивает частоты аллелей в природе. Таким способом можно ре-
гистрировать изменения генных частот, действительно происходя-
щие в природе. Результаты некоторых из этих исследований и
порожденные ими разногласия обсуждаются в гл. 9.
ЛИТЕРАТУРА
Общие вопросы
Crow J. F., Kimura М. 1970. An Introduction to Population Genetics Theory, New-
York, Harper and Row.
Dobzhansky T. 1970. Genetics of the Evolutionary Process, New York, Columbia
University Press.
Falconer D. S. 1960. Introduction to Quantitative Genetics, New York, Ronald Press.
Grant V. 1963. The Origin of Adaptations, New York, Columbia University Press.
Grant V. 1975. Genetics of Flowering Plants, New York, Columbia University Press.
Li С. C. 1976. First Course in Population Genetics, Pacific Grove, Cal., Boxwood
Press. [Имеется перевод: Ли Ч. Введение в популяционную генетику. — М.:
. Мир, 1978.]
Spiess Е. В. 1977. Genes in Populations, New York, Wiley.
Специальные статьи и обзоры
Beckwith I„ Rossow P. 1974. Analysis of genetic regulatory mechanisms, Ann. Rev.
Genetics, 8, 1—14.
Brussard p. F.. Ehrlich P. R., Singer M. C. 1974. Adult movements and population
structure in Euphydryas editha, Evolution, 28. 408—415.
168 Глава 6
Devonshire A. L. 1975. Studies of the acetylcholinesterase from houseflies (Musca
domestica L.) resistant and susceptible to organophosphorus insecticides, Bio-
chem. J., 149, 463—469.
Ehrlich P. R., Raven P. H. 1969. Differentiation of populations, Science, 165, 1228—
1232.
Fenner F. J. 1974. The Biology of Animal Viruses, 2nd ed., New York, Academic
Press.
Fisher R. A. 1958. The Genetic Theory of Natural Selection, New York, Dover.
Georghiou G. P. 1972. The evolution of resistance to pesticides, Ann. Rev. Ecol.
Syst, 3, 133—168.
Kaplan M. M., Webster R. G. 1977. The epidemiology of influenza, Scientific Ameri-
can, 237 (6), 88—106.
Levin D. A., Kerster H. W. 1974. Gene flow in seed plants, Evol. Biol., 7, 139—220.
Muirhead-Thomson R. C. 1960. The significance of irritability, behavioristic avoidan-
ce and allied phenomena in malarial eradication, Bull. WHO, 22, 721—734.
Thoday J. M., Boam T. B. 1959. Effects of disruptive selection: 111. Polymorphism
and divergence without isolation, Heredity, 13, 205—218.
Woodworth С. M., Leng E. R., Jugenheimer R. W. 1952. Fifty generations of selec-
tion for protein and oil in corn, Agron. J., 44, 60—65.
Wright S. 1931. Evolution in Mendelian populations, Genetics, 16, 97—159.
Глава 7
Инбридинг и случайный дрейф генов
Обсуждение факторов эволюции в предыдущей главе (гл. 6)
было основано на допущении, что популяция очень велика и что
скрещивание в ней происходит случайным образом. Но популяций
с такими характеристиками в природе, вероятно, не существует.
Все популяции имеют ограниченные размеры и, что особенно важ-
но, скрещивание обычно не бывает случайным. В этой главе мы
рассмотрим влияние неслучайного скрещивания и конечных раз-
меров популяции на ее генетическую структуру, о чем в гл. 6 упо-
миналось лишь мимоходом.
СЛУЧАЙНОЕ СКРЕЩИВАНИЕ
Что именно означает выражение «случайное скрещивание»?
Ойо означает, что в данной популяции вероятность скрещивания
особи а с особью b равна вероятности скрещивания особи а с
особью с, d, х и т. д. Проанализируем это утверждение в практи-
ческих терминах. Допустим, что мы имеем дело с видом, пред-
ставленным четко выраженными дискретными популяциями, кото-
рые можно ясно разграничить в пространстве (что само по себе
является исключением). Если допустить далее, что это популяция
растений, то, для того чтобы скрещивание было совершенно слу-
чайным, вероятность опыления пыльцевым зерном от растения а
яйцеклетки соседнего растения b должна быть в точности равна
вероятности опыления этим самым пыльцевым зерном растения s,
находящегося на противоположном конце поля. Однако выполне-
ние этого условия нереально. Вероятность опыления пыльцевым
зерном от растения а яйцеклетки растения b больше вероятности
опыления им яйцеклетки растения s. То же самое относится и к
популяции животных. Вероятность встреч (а следовательно, и спа-
риваний) между особями, живущими по соседству, больше, чем
Между особями, удаленными друг от друга.
Если построить график вероятности скрещивания между осо-
бями в зависимости от расстояния между ними, то получится
кривая, изображенная на рис. 7.L Вероятность спаривания снижа-
ется очень резко с увеличением расстояния, но не достигает нуля;
всегда остается небольшая вероятность того, что между двумя осо-
бями, находящимися на большом расстоянии друг от друга, про-
170
Глава 7
изойдет спаривание. Подобную кривую называют лептокуртиче-
ской кривой (она несколько «уже» колоколообразной кривой нор-
мального распределения, или иными словами, ее среднее квадра-
тичное отклонение меньше, чем у соответствующей нормальной
кривой).
Представим себе теперь трехмерное распределение вероятнос-
тей. Оно будет иметь вид конуса (рис. 7.2); если же изобразить ве-
Рис. 7.1. Лептокуртическая и нормальная кривые.
Лептокуртическая кривая (/) имеет больше точек, располагающихся вблизи от
среднего значения, чем нормальная кривая (//). Зависимость вероятности спари-
вания (ось ординат) от расстояния между особями (ось абсцисс) описывается леп-
токуртической кривой.
роятности спаривания для всех составляющих данную популяцию
организмов в трех измерениях, то получится, по выражению Брюса
Уоллеса (Bruce Wallace), нечто вроде «ложа, утыканного гвоздя-
ми» (рис. 7.3).
Помимо этих пространственных ограничений, которые накла-
дываются на свободное спаривание, существуют ограничения, об-
условленные конечными размерами популяции и разного рода осо-
бенностями механизма размножения.
До сих пор, обсуждая генетический механизм, мы исходили из
допущения, что частоты генов в популяции гамет можно точно
предсказать на основании генотипов родительских особей. Мы до-
пускали, например, что гетерозигота А|А2 продуцирует гаметы
двух типов А] иА2в отношении 1:1. Мы допускали, далее, что в
оезультате объединения этих гамет образуется три типа зигот:
А1АЬ AjA2 и А2А2 в отношении 1:2:1. Ниже мы рассмотрим это
допущение и покажем, что приведенные отношения представляют
собой лишь вероятности и что вряд ли именно такие отношение
будут когда-либо обнаружены на самом деле.
В норме отдельные организмы производят большое число га-
мет. Количество пыльцевых зерен и сперматозоидов может исчис-
Инбридинг и случайный дрейф
171
ляться тысячами; число яйцеклеток у некоторых животных и рас-
тений может быть ниже, но даже и у таких видов оно обычно
больше 100 на одну особь. При нормальном течении мейоза, от-
Рис. 7.2. Трехмерная лептокуртическая кривая дисперсии.
Рис. 7.3. Вероятности спаривания между особями в популяции.
Каждый конус соответствует вероятности спаривания отдельного растения или
животного, ведущего сидячий образ жизни, в двумерном пространстве. Расстоя-
ния между конусами отражают распределение особей в популяции. Чем ближе
друг от друга находятся две особи, тем больше шансов на то, что между ними
произойдет скрещивание.
сутствии систематических нарушений расщепления (гл. 5) и жиз-
неспособности всех гамет аллельный состав гамет и их частоты
должны быть предсказуемы на основании генотипов родителей.
Однако некоторые пыльцевые зерна или сперматозоиды бывают
неполноценными. Поскольку вероятность того, что такие гаметы
распределятся пропорционально среди гамет разных типов, крайне
мала, состав гамет и их частота уже не будут точно соответство-
вать таковым у родительских особей. Имеются и другие факторы,
способные нарушить эти соотношения в поколении потомков. При
172
Глава 7
образовании яйцеклеток как у высших растений, так и у живот-
ных только одна из четырех возникающих в результате мейоза
клеток превращается в гамету. Кроме того, не все члены данной
популяции производят в точности одинаковое число гамет. Из
большого числа гамет, продуцируемых особями данной популяции,
лишь немногие участвуют в успешных скрещиваниях, в результате
которых появляются потомки. А следовательно, фонд зигот, воз-
никших в данной популяции,—-это всего лишь выборка из того
фонда зигот, который могла бы произвести эта популяция, если
бы результатом каждого мейоза были 4 жизнеспособные гаметы и
каждая гамета в конечном итоге приняла бы участие в скрещива-
нии. Будучи выборкой из теоретически возможной популяции, ре-
альная популяция подчиняется всем вероятностным законам, при-
ложимым к выборкам. Иными словами, процесс образования зигот
в ряде отношений сходен с процедурой взятия выборки красных
и черных шаров из мешка, в котором находится множество таких
красных и черных шаров. Доля красных шаров в выборке будет
близка к их доле в мешке, но вероятность того, что соотношение
красных и черных шаров в выборке будет точно таким же, как
в мешке, очень мала. Согласно теории вероятности, чем больше
выборка (т. е. в нашем случае — чем больше популяция), тем мень-
ше эти отклонения; чем меньше выборка (т. е. чем меньше попу-
ляция), тем больше отклонения.
Идеальное соотношение вряд ли достижимо, как это видно из
табл. 7.1, в которой представлено расщепление по семи признакам,
Таблица 7.1
Отношения при расщеплении, наблюдавшиеся Менделем
в F2 от скрещивания разных сортов гороха
Признак Число изученных особей Fs Отношение
Гладкие семена Морщинистые семена 5474 1850 2,96:1
Желтые семядоли Зеленые семядоли 6022 2001 3,01:1
Серовато-коричневая кожура семян Белая кожура семян 705 224 3,15:1
Уплощенный боб Сужающийся боб 882 299 2,95:1
Зеленый боб Желтый боб 428 152 2,82:1
Пазушные цветки Верхушечные цветки 651 207 3,14:1
Длинные стебли Короткие стебли 787 277 2,84:1
Инбридинг и случайный дрейф
173
полученное Менделем в F2 от скрещивания разных сортов гороха
(Pisum sativum). Ожидаемым отношением во всех случаях было
3:1, и хотя наблюдаемые отношения были очень близки к ожи-
даемому, они никогда не совпадали с ним в точности. Самое близ-
кое соответствие было получено для самых больших популяций.
Конечные размеры популяции и отклонения от случайного ха-
рактера скрещивания — свойства, присущие всем популяциям. Они
функционируют как ограничения, накладываемые природой. Эти
два свойства вносят в эволюцию вероятностный элемент. Под этим
мы подразумеваем, что конечный результат взаимодействия основ-
ных факторов эволюции — мутационного процесса, рекомбинации
и отбора — нельзя предсказать с абсолютной уверенностью; его
можно предсказать лишь в статистическом смысле. Это означает,
что если бы одно и то же эволюционное событие повторялось мно-
гократно, мы могли бы предсказать лишь наиболее вероятный его
результат. Это означает также, что если бы в точности одно и
то же эволюционное событие повторялось многократно, результат
его всякий раз был бы несколько иным, чем во всех других слу-
чаях. Но каждое эволюционное событие происходит лишь однаж-
ды. В большинстве случаев оно приводит к тому, чего скорее всего
можно было ожидать, но время от времени результат оказывается
совершенно неожиданным. Это очень важный момент, который сле-
дует твердо усвоить.
СЛУЧАЙНАЯ УТРАТА АЛЛЕЛЕЙ В ПОПУЛЯЦИИ
Выше было подчеркнуто, что уравнение равновесия Харди—
Вайнберга p2+2pq + q2 = 1 выполняется только: 1) в бесконечно
больших популяциях и 2) при случайном скрещивании. Как мы
теперь убедились, ни то, ни другое допущение, строго говоря, не
вполне оправдывается. Как это влияет на равновесие Харди —
Вайнберга?
Допустим, что мы начинаем с популяции, имеющей конечную
численность х. Допустим также, что в локусе А одна половина
особей несет аллель А], а другая половина — аллель А2; иными
словами, аллель At имеет частоту р = 0,5, а аллель А2— частоту
<7 = 0,5. Допустим далее, что локус А не подвержен ни мутациям,
ни нарушениям расщепления, ни отбору. Можно ожидать, что при
этом одна половина гамет будет нести аллель Аь а другая—ал-
лель А2. Однако по причинам, приведенным выше, отношение гамет
будет несколько отклоняться от отношения 1:1. Кроме того, в ре-
зультате не вполне случайного скрещивания и конечной величины
популяций зигот в процессе оплодотворения произойдут дальней-
шие статистические отклонения. Направления этих отклонений мо-
гут быть противоположны тем, которые имели место в процессе
формирования гамет (например, в популяции гамет может быть
избыток аллелей А2, но тем не менее в образовании зигот примет
174
Глава 7
участие большая доля аллелей AJ, так что они могут до некото-
рой степени погашать друг друга; иногда же они происходят в.
одном и том же направлении, усиливая друг друга. Во всяком
случае в следующем поколении частота р аллеля Аь по всей ве-
роятности, не будет равна в точности 0,5, как это предсказывает
теорема Харди—Вайнберга, а установится некое новое равнове-
сие. Допустим, что новые частоты составляют: р = 0,55 для алле-
ля А! и 9 = 0,45 для аллеля А2. В следующем поколении произойдет
Рис. 7.4. Случайность выборки может быть причиной значительных колебаний ча-
стоты данного гена в разных популяциях.
В этом вычислении, проведенном со случайными числами, аллель А! появляется
у 50% членов обеих популяций. Всего лишь после 27 поколений аллель Ai «за-
крепился» в одной популяции (/), тогда как в другой его частота колеблется от
40 до 60% (//). (Cavalli-Sforza, 1969)
аналогичного рода ошибка выборки, но при этом выборка будет
браться из популяции, где аллелей А! чуть больше, чем аллелей А2.
Вероятность нового небольшого уменьшения или увеличения час-
тоты аллелей Ai составляет ровно 0,5. Иными словами, то обстоя-
тельство, что доля аллелей А! в одном поколении случайно уве-
личилась, не означает, что в следующем поколении она непремен-
но уменьшится. Значения частот могут быть: р = 0,6 и 9 = 0,4 или
же р = 0,51 и 9 = 0,49. Допустим, что новая частота аллеля Ai рав-
на р = 0,6. Мы вновь ожидаем отклонения от генной частоты в
родительской популяции в сторону либо уменьшения, либо уве-
личения р, причем эти два события равновероятны. Новое значе-
ние может оказаться равным р = 0,7. Если эти отклонения приво-
дят к попеременному повышению и понижению частоты аллеля Аь
Инбридинг и случайный дрейф 175
тФ в популяции будут происходить колебания между значениями,
слегка превышающими исходное р = 0,5 или несколько не дости-
гающими его. Возможно, однако, что в ряде последовательных
поколений большинство изменений будет происходить в одном и
том же направлении. В таком случае один из имевшихся в по-
пуляции аллелей может быть в конце концов утрачен (рис. 7.4).
Вероятнцсть такого события зависит от величины популяции и
от времени. При наличии достаточного времени в популяции ко-
нечной величины один из гетерозиготной пары аллелей в конеч-
ном счете утратится в результате только лишь случайных собы-
тий, если ни отбора, ни мутирования происходить не будет. Это
весьма важный момент.
Подразделение популяции и формулы Валенда
Перейдем теперь к рассмотрению другого фактора, оказываю-
щего стохастическое воздействие на частоту генов — к простран-
ственному распределению особей.
На рис. 7.5 представлена карта распределения растений вида
Clematis fremontii var. riehlii в шт. Миссури. Подобное распреде-
ление встречается довольно часто, хотя его нельзя считать уни-
версальным. Главное, на что следует обратить внимание, это то,
что плотность особей не одинакова по всей территории; на неко-
торых лужайках растения тесно примыкают друг к другу, тогда
как на других они разделены большими промежутками. Посколь-
ку вероятность скрещивания между растениями, находящимися в
тесной близости, больше, чем между растениями, удаленными
друг от друга, можно считать, что популяция Clematis fremontii,
изображенная на рис. 7.5, эффективно подразделена на субпопу-
ляции.
Хотя скрещивания возможны между всеми растениями, расту-
щими на данной лужайке, в большинстве случаев они происходят
в пределах одной субпопуляции и притом чаще всего — между
соседними растениями. Попробуем проследить за изменениями
частоты генов в такой популяции.
Допустим еще раз, что в нулевом поколении частота аллеля
А! равна частоте аллеля А2, т. е. p = q = 0,5 по всей популяции,
причем отбора не происходит и мутаций не возникает. В 1-м по-
колении возникнут отклонения от этой исходной частоты, вызван-
ные случайными факторами. Однако, поскольку события, проис-
ходящие в разных субпопуляциях, независимы, вызванные ими
отклонения будут в каждой из этих популяций различными: в не-
которых частота аллеля Ai будет больше 0,5, а в других — меньше.
Наиболее вероятно, что примерно в половине популяций она по-
высится, а в другой — понизится. То же самое относится к сле-
дующему поколению и к каждому из дальнейших. Однако, по-
скольку отклонения взаимно погашаются, в целом частота генов
178
Глава 7
Рис. 7.5. Распространение кустарника Clematis fremontii var. riehlii.
Распространение этой разновидности ограничено прогалинами в области, занима-
ющей в целом около 1000 км2 на востоке центральной части штата Миссури.
В пределах этой области (А) можно выделить целую иерархию естественных под-
разделений: области с большим количеством прогалин (В), скопления прогалин
(В), колонии растений, примерно соответствующие прогалинам (Г), скопления
растений (от нескольких до, возможно, тысячи особей) на каждой прогалине (Д).
(Erickson, 1945)
Инбридинг и случайный дрейф 177
будет оставаться примерно одинаковой, в данном случае равной
0,5. Такое положение вещей сохранится даже после того, как в
каждой из субпопуляций один аллель будет утрачен. В самом
деле, вероятность закрепления аллеля Aj в одной из субпопуля-
ций равна его исходной частоте. Если исходные частоты аллелей
p=^q=0,5, то в конце концов примерно в половине субпопуляций
закрепится аллель Аь а в другой половине—аллель А2, и тем
самым частоты аллелей Ai и А2 в популяции в целом не изменятся
(табл. 7.2).
Однако, несмотря на то что частоты генов остались неизмен-
ными, частоты генотипов изменились. Ведь если в каждой из суб-
популяций закрепится какой-либо один аллель, либо Аь либо А2,
то популяция в целом будет состоять исключительно из двух го-
мозиготных генотипов. Отсюда можно сделать вывод, что главные
результаты, обусловленные неслучайностью скрещивания и конеч-
ными размерами популяции, это: 1) подразделение популяции на
более мелкие субпопуляции; 2) повышение вероятности закреп-
ления в субпопуляции одного аллеля; 3) повышение частоты го-
мозиготных генотипов, не обязательно сопровождающееся изме-
нением частоты аллелей в популяции в целом. Эти результаты
представляют собой отклонение от равновесия Харди—Вайн-
берга.
Представим теперь ту же самую концепцию в математических
терминах.
Допустим, что имеется k субпопуляций с разными частотами
аллелей: р для аллеля А! и q для аллеля А2; каждая субпопу-
ляция находится в состоянии равновесия Харди—Вайнберга. До-
пустим также, что все субпопуляции состоят из одинакового числа
особей. Частоты генов и генотипов представлены в табл. 7.2.
Средняя частота генов в популяции в целом будет равна
а дисперсия (см. гл. 4) генных частот между субпопуляциями со-
ставит
Pi* _з -— -2
0Г*:=------р ={^ — р ;
т. е. дисперсия равна среднему квадратов генных частот минус
квадрат средней генной частоты.
Средняя частота гетерозигот AjA2 в популяции в целом со-
ставит
22Р| 22р*’
~ k 1 I ’
12 - 2044
178
Глава 7
Таблица 7.2
Оценка фактической и ожидаемой гетерозиготиости
в гипотетической подразделенной популяции
Субпопуляция Частота генов Число генотипов
р <7 р2 2р<7
1 450 0,6 0,4 162 216 72
2 50 0,2 0,8 2 16 32
3 100 1 0 100 — —
4 500 0,3 0,7 45 210 245
5 100 0 1 — — 100
6 350 0,7 0,3 172 147 31
1550 0,5 0,5 481 589 480
Ожидаемое 387,5 775 387,5
(помните, что ^,= 1—р<)
=-2(р<7—о2),
поскольку р<7 = р(1—р) = р-£р^
следовательно,
Н=2^1 1 — 4!-),
\ pq /
где a2lpq равно стандартизованной дисперсии генных частот.
Средняя частота гетерозигот в популяции, в которой скрещи-
вание не является случайным, но происходит случайным образом
внутри субпопуляций равной величины, равна удвоенному произ-
ведению средних частот аллелей минус удвоенная дисперсия час-
тоты генов между субпопуляциями.
Аналогичным образом средние частоты для гомозигот будут
равны:
— /Ч2
(для АЛО;
- /Л
Я =—=(v+ct2 (для АаА2)'
Эти выражения известны под названием формул Валенда.
Инбридинг и случайный дрейф
179
ИНБРИДИНГ
До сих пор мы исходили из предположения, что каждая особь
в данной популяции находит себе брачного партнера, не проявляя
каких-либо предубеждений или пристрастий. Мы видели, что в
действительности случайное скрещивание невозможно вследствие
конечных размеров популяции и ее разделения на соседствующие
субпопуляционные структуры — соседства (neighborhood). Теперь
мы сделаем еще менее строгим допущение о свободном скрещи-
вании, признавая, что у многих видов часто наблюдается пред-
почтительное скрещивание между родственными особями.
Самооплодотворение возможно только у гермафродитных ви-
дов, таких, как растения или дождевые черви. Однако лишь у
очень немногих гермафродитных видов размножение действитель-
но происходит посредством самооплодотворения. Между тем скре-
щивание с близко родственными особями встречается гораздо ча-
ще. Пространственная близость, экологические предпочтения, мор-
фологическое сходство и разного рода пристрастия повышают
вероятность скрещивания между родственными особями. Даже у
человека семейные и племенные связи, географические факторы,
национальность, культурные, социальные и экономические фак-
торы вызывают расслоение популяции. Браки между людьми, при-
надлежащими к одному слою, случаются гораздо чаще, чем меж-
ду людьми, принадлежащими к разным слоям, что повышает ве-
роятность браков между родственниками. Во всех тех случаях,
когда брачные партнеры в среднем связаны более тесным род-
ством, чем если бы они были выбраны наугад из популяции, мы
говорим, что имеет место инбридинг. Инбридинг также влияет на
генотипический состав популяции. В этом можно убедиться, рас-
смотрев крайний пример инбридинга — самооплодотворение гер-
мафродитной особи. Видов, которые бы размножались исключи-
тельно при помощи самооплодотворения, вероятно, не существует.
Однако существует целый ряд видов растений, которые обычно
йамоопыляются, как, например, культурная пшеница (Triticum
aestivum), у которой до 99% всех семян образуются в результате
самоопыления, табак (Nicotiana tabacum), хлопчатник (Gossypium
barbadense) и многие обычные садовые сорняки: росичка кровяная
(Digitaria sanguinalis), пастушья сумка (Capsella bursa-pastoris)
и звездчатка (Stellaria media).
Начнем с популяции, находящейся в равновесии Харди—Вайн-
берга, в которой частота р аллелей Ai равна частоте q аллелей А2,
так что обе частоты равны 0,5 и р2 = 0,25; 2р^ = 0,50; ^2 = 0,25.
Все размножение с этого момента происходит только путем са-
мооплодотворения. Поэтому все гомозиготы будут размножаться
в чистоте, тогда как среди гетерозиготных особей в каждом по-
колении происходит расщепление в соотношении 0,5 гомозигот и
0,5 гетерозигот. Как видно из табл. 7.3, доля гетерозигот в каж-
12*
180
Глава 7
Таблица 7.3
Соотношение гомозиготных «и гетерозиготных
генотипов в популяции, размножающейся
путем самооплодотворения
Поколение Генотипы
AjAj AjA3 А3А2
0 — 1 —
1 1 2 1
2 3 <> 3
3 7 2 7
4 15 2 15
5 31 2 31
10 1023 2 1023
Л 2п—1 2 2 л— 1
дом поколении уменьшается наполовину. В конце концов такая
популяция будет состоять сплошь из гомозиготных особей.
Такие же результаты получатся при иных исходных частотах
генов, например при частоте р = 0,3 аллеля Aj и частоте 9 = 0,7
аллеля Аг. Доля гетерозигот в каждом поколении уменьшается
вдвое, а доли обеих гомозигот увеличиваются на равное число
особей, пока вся популяция не будет состоять из гомозигот AiAb
имеющих частоту р, и гомозигот А2Аг с частотой q. В общем ожи-
даемая частота гетерозигот в популяции, размножающейся путем
самооплодотворения, в любом поколении п составляет 2pq-(xlz)n,
где 2pq — начальная частота гетерозигот в нулевом поколении,
а п-—число последовательных поколений самооплодотворения.
Если лишь некоторые особи в данной популяции размножают-
ся путем самооплодотворения, а другие скрещиваются случайным
образом, доля гомозигот будет выше, а доля гетерозигот ниже,
чем предсказывает равновесие Харди—Вайнберга, но зависимость
эта перестает быть простой. Здесь следует ввести новое понятие—
коэффициент инбридинга F.
Коэффициент инбридинга
Этот очень полезный и широко используемый показатель был
впервые введен в 1923 г. одним из создателей популяционной ге-
нетики — американским ученым Сьюэллом Райтом, который на-
звал его индексом «фиксации».
Прежде чем дать определение показателя F, следует опреде-
лить, что означают термины «идентичны по происхождению» и
«одинаковы по состоянию». Мы говорим, что некая гомозиготная
Инбридинг и случайный дрейф 181
особь имеет два гена, которые идентичны по происхождению, ес-
ли можно проследить происхождение этих генов от одного общего
предка. Такого рода гомозиготы называют также аутозиготными.
Однако если у гомозиготной особи имеются два идентичных ал-
леля, возникших в результате двух независимых мутаций, то их
называют одинаковыми по состоянию, а особь — аллозиготной.
Это различие может показаться слишком тонким, и оно в самом
деле тонкое, однако проводить его необходимо, потому что, как
мы увидим, инбридинг повышает частоту аутозиготных особей.
Коэффициент инбридинга F служит для измерения идентичности
по происхождению на основании степени родства между брачны-
ми партнерами. Мы определим коэффициент инбридинга как «ве-
роятность того, что данный индивидуум получит от родителей в
данном локусе два гена, идентичных по происхождению».
Допустим на данном этапе наших рассуждений, что мы имеем
дело с некоторой идеализированной популяцией самосовместимых
растений, в которой скрещивание происходит строго случайным
•образом. Допустим далее, что в этой исходной популяции каждая
из особей содержит в изучаемом локусе два аллеля, отличающих-
ся друг от друга и от любого другого аллеля в этой популяции.
Будучи гетерозиготной, каждая особь продуцирует гаметы двух
типов (одну — с одним аллелем и другую —с другим). Поскольку
популяция не содержит двух одинаковых аллелей, общее число
типов производимых в ней гамет составит 2N0. Когда эти гаметы
объединяются (что происходит, как было указано, строго случай-
ным образом), чтобы сформировать зиготы следующего поколе-
ния, вероятность объединения двух идентичных гамет с образо-
ванием гомозиготы равна l/2.V0. Поскольку в нашей популяции в
нулевом поколении все гаметы были разными, вероятность того,
что в первом поколении потомков у какой-либо особи два его
аллеля окажутся идентичными по происхождению, равна веро-
ятности наличия у нее двух идентичных аллелей, как было опре-
делено выше, а именно 1/2Л\>. Это и есть величина коэффициента
инбридинга F для первого поколения.
В следующем поколении число особей будет равно Nx. Одни
из них будут гомозиготными, другие —гетерозиготными. Допу-
стим, однако, что все аллели различны, как это было в предыду-
щем поколении. В таком случае вероятность того, что две иден-
тичные гаметы объединятся с образованием гомозиготы, была бы
равна 1/2Л^1. Хотя во втором поколении образуются и другие го-
мозиготы, лишь эти гомозиготы будут в данном поколении ре-
зультатом объединения двух гамет, идентичных по происхождению
от одного аллеля в поколении 1. Но не все аллели различны, не-
которые из них идентичны, поскольку они происходят от общего
«предка» в поколении 0. Поэтому помимо гомозигот, идентичных
по происхождению от одного из аллелей поколения 1, будут по-
являться и другие гомозиготы, например, гомозиготы, образую-
Г82
Глава 7
щиеся в результате объединения двух гамет от гомозиготной осо-
би, или гомозиготы, образующиеся в результате объединения двух
гамет от двух разных особей Fb которые, однако, происходят от
общего предка из поколения 0. Какова вероятность возникнове-
ния аутозигот второго типа? Если доля зигот, аутозиготных вслед-
ствие того, что они образуются в результате объединения двух
гамет, идентичных по происхождению от одного аллеля поколе-
ния 1, составляет 1/2М, то, очевидно, остается (1—1/2Л^) зигот,
не относящихся к этому типу. Вероятность того, что среди этого
остатка возникнет гомозигота путем случайного скрещивания двух
идентичных гамет, происходящих от предкового аллеля поколе-
ния 0, уже была вычислена как F для первого поколения
( = 1/2 No)-, она соответствует вероятности того, что любые два ал-
леля в одной зиготе окажутся копиями одного и того же аллеля
из поколения 0. Следовательно, коэффициенты инбридинга F для
р ___ 1
?(i)— 2ЛГ0 ’
F(3>=w+( 1—^г)F(2)’
разных поколении равны:
для первого поколения
для второго поколения
для третьего поколения
для л-го поколения
Таким образом, мы видим, что коэффициент инбридинга со-
стоит из двух частей: части 1/2 Nn-i, относящейся к инбридингу в
данном поколении, и части, которую можно отнести за счет пред-
шествующего инбридинга и в которую входит коэффициент ин-
бридинга для предыдущего поколения.
Важное следствие из этого состоит в том, что если величина
популяции внезапно резко возрастает, так что новый вклад в
коэффициент инбридинга окажется пренебрежимо мал, поскольку
l/2A^n-i станет очень маленьким числом, то в целом коэффициент
инбридинга не будет уменьшаться, а останется на том же уровне,
на каком он был до увеличения размеров популяции.
Величина l/2Nn-i относится, конечно, только к идеализирован-
ной популяции. Это ограничение можно устранить, назвав 1/2Л7п—х
«дельтой F» (т. е. AF) — изменением коэффициента инбридинга.
Тогда
F„ = AF +(\-\F)F{n_l} =
= ^-F—(AF)
1 — Г(П-1)
Инбридинг и случайный дрейф 183
где Л/7 — мера увеличения инбредности и равна отношению той
доли популяции, которая участвовала в инбридинге, к той ее до-
ле, которая в нем не участвовала. Когда коэффициент инбридинга
выражается в терминах AF, уравнение для Fn становится прило-
жимым к любой размножающейся системе, а не только к исход-
ной популяции.
Вернемся теперь к исходной популяции и допустим, что все
особи размножаются путем самооплодотворения. Допустим снова,
что в исходной популяции каждая особь несет в изучаемом локу-
се два аллеля, отличающиеся друг от друга и от любого другого
аллеля в этой популяции. Половину потомков каждой гетерози-
готы составляют гомозиготы, идентичные по происхождению в
каждом поколении. Соответственно АА=!/2 для каждого поко-
ления.
Таким образом:
для первого поколения
для второго поколения К(2) — l/2+(l — х/2) А(1) —3/4)
для третьего поколения ^(3) =-1/2-f-(1 *—1/2) ^(2) = 7/8.
для л-го поколения F(n} = J/2+(1 —1/2) F(n_l}.
Число гетерозигот (Я) в л-м поколении вычисляется следую-
щим образом:
в первом поколении = 112Н0,
во втором поколении —
и после подстановки
Н2 = но - 1/2Я0 -1/2 (Но —х/2Я0) =
= Яо-3/Ло=^о(1-3/4),
или, иными словами, число гетерозигот равно первоначальному
числу гетерозигот, умноженному на разность между единицей и
долей гетерозигот, превратившихся в гомозиготы в результате
многократного повторяющегося самооплодотворения, т. е. F.
А поэтому
в л-м поколении Н,, ~Н0 (1 — F,,).
Такая зависимость между числом гетерозигот и коэффициентом
инбридинга F справедлива при любых значениях F.
Равновесие при сочетании самооплодотворения со случайным
скрещиванием. При непрерывном самооплодотворении доля гете-
розиготных генотипов в каждом поколении уменьшается вдвое.
Однако если самооплодотворение прерывается одним поколением
184
Глава 7
совершенно свободного скрещивания, то первоначальное равнове-
сие генотипов (p2 + 2pq + q2) по Харди—Вайнбергу восстанавли-
вается (гл. 4). В природе не происходит ни непрерывного само-
оплодотворения, ни совершенно случайного скрещивания; в попу-
ляциях у одних особей наблюдается самооплодотворение (или
инбридинг с близкими родичами), а у других — более или менее
случайное скрещивание. Можно считать, что в популяциях имеет-
ся некая доля ($) самооплодотворяющихся особей и доля (1—$)
свободно скрещивающихся особей. Значение F в популяциях, на-
ходящихся в состоянии равновесия при сочетании самооплодотво-
рения и случайного скрещивания, равно1
Генетики Д. Р. Маршалл (D. R. Marshall) и Р. У. Аллард
(R. W. Allard) установили, что средняя степень ауткроссинга (сво-
бодного скрещивания) в одной калифорнийской популяции овса
бородатого (Avena barbata) составляет всего 0,014; т. е. 98,6%
растений в этой популяции производили семена в результате са-
моопыления. Коэффициент инбридинга в этом случае
р__ s ____ 0.986 __„ gyn
г—2_s— 1014 —u,y/z.
Эти авторы определили затем частоту генотипов для шести
локусов в той же популяции (табл. 7.4). Из приведенных значе-
ний можно получить независимую оценку F:
Hn = H0(l-F), (стр. 183)
а поэтому
1—Р=—
Яо ’
где Но получено из ожидаемого числа гетерозигот при свободном
скрещивании (2pq).
Для четырех гетерозиготных локусов величины F, вычислен-
ные на основании числа гетерозигот в популяции, были устойчиво
ниже (табл. 7.3) оценок, сделанных на основании степени аут-
кроссинга. Следовательно, можно предположить, что либо уровень
ауткроссинга выше, либо происходит отбор в пользу гетерозигот-
ных генотипов. Маршалл и Аллард приняли вторую гипотезу. Мы
вернемся к ней в гл. 9.
1 Вывод этой формулы см. Е. В. Spiess, Genes in populations, p. 246.
Инбридинг и случайный дрейф
185
Таблица 7.4
Частоты генотипов и значения F для шести генных
локусов в популяции Avena barbata
из одной местности в округе Напа, Калифорния.
Генотипы 11 и 22 — гомозиготы, генотип 12 — гетерозигота.
(Marshall D. R., Allard R. W., 1970. Maintenance of isozyme
polymorphisms in natural populations of Avena barbata.
Genetics, 66, 393—399)
Ген Генотип Частота Величина выборки F
Е1 11 0
12 0 54 1,00
22 1
Е< И 0,30
12 0,11 54 0,76
22 0,59
е7 11 0
12 0 54 1,00
22 1
Ею 11 0,46
12 0,13 85 0,70
22 0,41
р, 11 0,40
12 0,15 86 0,70
22 0,45
арх5 11 0,48
12 0,11 86 0,78
22 0,41
Инбридинг в популяциях человека
У каждого из нас двое родителей, четверо бабок и дедов,
восемь прабабок и прадедов и т. д.; таким образом, если про-
следовать на п поколений назад, каждый из нас имеет 2п пред-
ков. Это означает, что за 20 последних поколений у каждого из
нас было более 700 000 предков. Если исходить из того, что на
каждое столетие приходится по четыре поколения, то на 20 по-
колений требуется пять столетий. Если бы все жители США,
не связанные непосредственным родством (допустим, один на
каждые 20 человек), имели совершенно независимые наборы пред-
186
Глава 7
ков, то для этого было бы необходимо, чтобы в XV веке жили
3 триллиона человек, которые были бы их предками. Это число
на несколько порядков выше, чем число людей населяющих весь
земной шар в настоящее время. По всей вероятности, родствен-
ные связи между людьми гораздо сильнее, чем это подозревает
большинство из нас.
В популяциях человека принято измерять среднюю идентич-
ность по происхождению (называемую теперь кровным родством)
для данной популяции как средний коэффициент инбридинга вхо-
дящих в нее особей. Эта величина, обозначаемая через а, опре-
деляется как
где pt — частота особей в популяции с коэффициентами инбри-
динга Fi.
Из приведенных в табл. 7.5 оценок можно видеть, что для
разных групп населения величина а варьирует более чем на два
порядка. В крупных социальных или религиозных группировках,
в которых браки между близкими родственниками строго запре-
щены, как, например, у католиков, значения а низкие, тогда как
в небольших группах (например, у самаритян) или в группах,
в которых разрешаются браки между двоюродными братьями и
сестрами (как у некоторых индийских племен), значения а гораз-
до выше.
Инбридинг может иметь несколько последствий: 1) он ведет
к повышению гомозиготности; 2) приводит к проявлению рецес-
Таблица 7.5
Некоторые оценки инбридинга а (По Cavalli-Sforza,
Bodmer, 1971.)_______________________________________________
Популяция а
США, католики 0,00009
Аргентина, католики 0,00058
США, мормоны 0,00038
Япония 0,0046
Гвинея 0,026
Индия, Андхра-Прадеш 0,032
Израиль и Иордания, самаритяне 0,0434
Браки между четвероюродными братьями и сестрами 0,0019
Браки между троюродными братьями и сестрами 0,0078
Браки между двоюродными братьями и сестрами 0,0625
Браки между дядей и племянницей 0,125
Инбридинг и случайный дрейф 187
сивных генотипов; 3) поскольку многие рецессивы обладают вред-
ным эффектом, инбридинг обычно ведет к уменьшению размеров,
плодовитости, мощности, урожайности и приспособленности (Од-
нако степень выраженности этих эффектов пропорциональна чис-
лу вредных гомозиготных генов в популяции. У видов, для кото-
рых инбридинг является нормой, таких, как пшеница и многие
другие культурные растения, число вредных генов в популяции
очень невелико.); 4) отмечается повышение фенотипической из-
менчивости, поскольку сходные гены имеют тенденцию накапли-
ваться у одной и той же особи, которая при этом будет отличать-
ся от других особей.
ЭФФЕКТИВНАЯ ЧИСЛЕННОСТЬ ПОПУЛЯЦИИ
До сих пор в этой главе мы допускали, что популяция имеет
конечные и постоянные размеры и состоит из N особей, образую-
щихся в каждом поколении в результате соединения мужских и
женских гамет, взятых случайным образом из предыдущего по-
коления. Мы показали, что в такой популяции гетерозиготность
имеет тенденцию к снижению, а гомозиготность — к повышению.
Размеры реальных популяций редко остаются постоянными из
года в год, поскольку в одни годы обстоятельства благоприятст-
вуют росту и размножению, а в другие — нет. В популяциях, за-
нимающих постоянные местообитания, годичные колебания чис-
ленности выражены менее резко, чем в популяциях, использую-
щих временные местообитания, например, у сорняков или видов,
обитающих в периодически пересыхающих водоемах. Кроме того,
популяция представляет собой смесь размножающихся и нераз-
множающихся особей, причем среди размножающихся особей ус-
пех размножения варьирует в зависимости от возраста и разме-
ров, а также от других факторов.
Как мы видели (см. стр. 182), в идеализированной популяции
самосовместимых растений, в которых скрещивание происходит
строго случайным образом, существует вероятность, равная 1/2 No,
что две идентичные гаметы объединятся с образованием гомози-
готы, и что вообще в л-м поколении вероятность такого события
составляет 1/2 Nn-i.
Если мы имеем дело не с популяцией гермафродитных само-
совместимых растений, а с популяцией животных, состоящей из
Nm самцов и Nf самок, то эти формулы придется несколько из-
менить, но ход рассуждений останется прежним. Поскольку каж-
дая зигота — результат объединения одной мужской и одной жен-
ской гаметы, вероятность происхождения двух генов, содержа-
щихся у разных особей поколения п, от самцов из поколения п—1
равна !/4 (Уг-'/г), а вероятность их происхождения от одного и
того же самца равна 1/4 Nm. Подобным же образом вероятность их
происхождения от одной и той же самки равна 1/4 Nf.
188
Глава 7
Во всех случаях, когда число самцов в данной популяции рав-
но числу самок (Nm=Nf),
1 . 1 _ 1 1 1 _ 2 _ 1 _ 1
4Nm + 4N, ~ 4Nm “Г 4Л'т — 4Nm ~ 2Nm ~ N '
Если, однако, Nm Ф Nf, то
1 д_ 1 — 1 1
~4N^ +“4Л7 ~ Ne
где Ne — эффективная численность популяции, равная величине
идеальной популяции, гомозиготность которой возрастает так же,
как в наблюдаемой нами популяции с численностью самцов Nm
и самок Nt.
Любое отклонение от идеальной популяции (где Nm=Nf) при-
ведет к тому, что эффективная численность популяции будет ни-
же ее фактической численности. Мы уже упоминали о двух фак-
торах, понижающих эффективную численность популяции: 1) раз-
личная численность самцов и самок; 2) колебания численности
популяции от поколения к поколению. К числу других таких фак-
торов относятся неравномерный вклад разных особей в фонд га-
мет популяции, неслучайное скрещивание и подразделение попу-
ляции на субпопуляции. Поскольку все эти факторы присутствуют
в реальных популяциях, эффективная численность популяции всег-
да ниже действительного числа имеющихся в ней особей. Точных
исследований по эффективной численности популяции проводи-
лось мало, однако в работах на дрозофиле она варьировала от
0,48 до 0,71 -N, у человека — от 0,69 до 0,95 -N, а у улитки Lym-
паеа составила 0,75-N. У растений она, по-видимому, еще ниже.
Эффективная величина популяции служит мерой степени откло-
нения реальной популяции от популяции идеальной. Чем меньше
отношение Ne/N, тем больше в данной популяции тенденция к
инбридингу и к повышению уровня гомозиготности.
СЛУЧАЙНЫЙ ДРЕЙФ ГЕНОВ
Генофонд каждого последующего поколения данной популя-
ции представляет собой выборку из родительского поколения,
давшего ему начало. А поэтому он подвержен изменчивости, об-
условленной ошибкой выборки. Эти колебания тем больше, чем
меньше выборка, т. е. чем меньше эффективная численность по-
пуляции. Все популяции по крайней мере иногда претерпевают
неблагоприятные периоды, в которые их численность резко сокра-
щается. Следовательно, в популяциях происходят изменения ген-
ных частот, не являющиеся прямым результатом естественного
отбора. Этот процесс известен под названием случайного дрейфа
генов.
Значение случайного дрейфа генов в качестве фактора эволю-
ции было предметом многочисленных дискуссий. Когда популяция
Инбридинг и случайный дрейф
189
подвергается отбору, любое случайное изменение, происходящее
в том же направлении, в каком действует отбор, приводит к уси-
лению, или ускорению, отбора. И наоборот, любое случайное из-
Ьедаию /Парма;-
Смньятико вУ - Z ’
1М /
s<
... V
fe/
у-4 a
\ Петриньяиола дУ 'к|
- -^/нароббио
уЖч
• f.j’-.' •/л. <•
, . . ...... .. at- у-х
) лУк. :уВиллула'''*—А’
./.' ^»...ак.,У?.”/-.;й.-У<УУ27 А5'у. ' '
1 Корнильо t W -< >Г' • ;---
'./У- \ \
Марра и Канетоло .ру, пру.,
’ Й1М*. ' : ; .'У V Баллоне ' У"-
./ 133
У t, Моссале У \У,-’.УУ^
>130 "АГрамматика.>.• . *“*
:у2а&- •</
И!
Вестана
’/.32-
У.. ..МРенОиж./ '' '4/ ’ - ... .
- .’fe235. fe/fe У.’--у ’
( -'Г. А-Карпането - ,
Тиццано 1 t-
?'.Т32- J /
Т/г/™»"»,, Мораньяно
I •< . .. 193 ;•
Музиара
г' ЗЬБ- у2 •
- ; fe Мадурера / fe
Тревиньяно У
у -59
.'А yJ' '
V Ранцано
'• fey" 316
Пратопьяно!-<ХЦ •• .
:',/’.^7Лл'Л ;
.’Насароло Седа;
,,.fe.J43^'< M(Wko ./лЛуганьяно ..Ваиро»'..
; Пьянадетго 272.*;^манья /Валь^на ’{*/’ / •
Вальдитанна^ - A/J6 ’ >- •У/н>фОне•-<’
-у. л-. ^у-А.Трефиуми )sy'“'".y.’,.•
"ч \ • .... Л' У'Ьр Ригозо У >г/ У \
<АУу .... J".;-А'У,, .;^М..Тоскана-\<4'!';'''’ f (
* ;404 .. "
Паланцано
• Вестано -
. /133. ~!
....
1П
^уБосю^
Рис. 7.6. Карта долины верхнего течения р. Парма (Северная Италия) с обозна-
чением деревень, главных дорог между ними и численности населения в 1950 г.
(Cavalli-Sforza, Bodmer, 1971)
менение в противоположном направлении замедляет отбор. В этом
смысле случайный дрейф генов эквивалентен «статическому шу-
му» в системе. Однако иногда такие пертурбации могут привести
к утрате наиболее приспособленного аллеля и к закреплению ме-
нее приспособленного. В таких случаях наступает необратимое
изменение (если только утраченный аллель не восстанавливается
в результате мутации). Вероятность возникновения таких изме-
нений находится в обратной зависимости от интенсивности отбора
190
Глава 7
(чем сильнее отбор, тем меньше вероятность необратимого изме-
нения) и от величины популяции (чем больше популяция, тем
менее вероятно изменение).
Если изменения, вызванные случайным дрейфом генов, суще-
ственно понижают приспособленность данной популяции, то эта
Рис 7.7 Изменения частоты одной группы крови у жителей разных деревень в
долине р. Парма.
Небольшие различия в частоте выявлены между изолированными высокогорными
деревушками; различия постепенно уменьшаются по мере продвижения в нижние
районы, в городки, расположенные на холмах и на равнине, и наконец в г. Пар-
му. В качестве меры генетической изменчивости использовалась величина F.
(Cavalli-Sforza, 1969)
популяция может вымереть. Поэтому не все изменения, вызывае-
мые случайным дрейфом, сохраняются. Возможно, однако, воз-
никновение время от времени ряда генетических изменений, ко-
торые приведут к изменению генетической структуры во многих
локусах, в результате чего получается организм, довольно сильно
отличающийся от других членов популяции по своим генетиче-
ским признакам и обладающий при этом высокой приспособлен-
ностью. Подобный процесс Эрнст Майр (Ernst Мауг) назвал «ге-
нетической революцией». Генетическая революция — событие
весьма маловероятное, и мы располагаем лишь косвенными и
ограниченными данными о его реализации. По мнению некоторых
биологов, каждый новый вид возникает в результате генетической
революции. Однако подобное утверждение, которое оспаривают
многие генетики, еще следует доказать. При возникновении новой
популяции в результате иммиграции очень небольшого числа осо-
бей многие гены материнской популяции утрачиваются. Этот про-
цесс, известный под названием эффекта основателя (или прин-
Инбридинг и случайный дрейф
191
Таблица 7.6
Значения F для трех локусов, определяющих
группу крови у населения 37 приходов
в долине верховьев р. Пармы
(66 деревень с 15 000 жителей)
Аллели, определяющие группу
крови
Р Q
M-N
At
Аг
В
О
г
Ri
R2
rh remainder
0,048
0,052
0,034
0,061
0,038
0,033
0,017
0,037
Среднее 0,0356±0,00б
ципа основателя), также, возможно, играет важную роль в эво-
люции.
Итало-американский генетик Луиджи Кавалли-Сфорца (Luigi
Cavalli-Sforza) изучал распределение по группам крови населения
городов и деревень в долине р. Парма в Италии (рис. 7.6), глав-
ным образом с целью выяснить влияние величины популяции на
дрейф генов. Вследствие геологических особенностей долины с ее
крутыми склонами в верховьях реки деревни маленькие (200—
300 жителей), но вниз по течению деревни становятся все больше,
а город Парма насчитывает уже 175 000 жителей. После VII в.
до н. э. сколько-нибудь значительной иммиграции в долину р. Пар-
мы не происходило, а поэтому следует считать, что здесь устано-
вилось демографическое и генетическое равновесие. Однако в пре-
делах этой области горные деревушки более стабильны и обмен
мигрантами между ними слабее, чем в деревнях, расположенных
ниже по течению. Кдвали-Сфорца изучал частоту разных групп
крови в зависимости от плотности населения. Как видно на
рис. 7.7, уровень генетической изменчивости между популяциями
уменьшается с увеличением плотности популяции, но это умень-
шение достоверно только для маленьких деревушек. В горных де-
ревнях (табл. 7.6) важную роль играет также инбридинг, но в
г. Парме его значение гораздо меньше (F = 0,002).
Подводя итоги, можно сказать, что не все происходящие в
популяции изменения представляют собой прямой результат ес-
192
Глава 7
тественного отбора. Некоторые благоприятные изменения могут
возникать чисто случайно. Чем меньше популяция, тем сильнее
эффекты случайности выборки и тем чаще изменения носят слу-
чайный характер. Такие случайные эффекты порой могут оказать-
ся важными событиями в эволюции какой-либо линии, однако
это бывает редко. Вместе с тем некоторое изменение частоты ге-
нов, вызванное случайными эффектами, представляет собой один
из многократно наблюдавшихся аспектов эволюционного про-
цесса.
Тенденции к инбридингу и к повышению уровня гомозиготно-
сти могут противодействовать мутационный процесс, рекомбина-
ция и отбор, и это чревато определенными генетическими послед-
ствиями, которые будут рассмотрены в гл. 8.
ЛИТЕРАТУРА
Общие вопросы
Cavalli-Sforza L. L., Bodmer HZ. F. 1971. The Genetics of Human Populations, San
Francisco, Freeman.
Crow J. F., Kimura M. 1970. An Introduction to Population Genetics Theory, New
York, Harper and Row.
Nei M. 1975. Molecular Population Genetics and Evolution, Amsterdam, North-Hol-
land.
Wallace B. 1968. Topics in Population Genetics, New York, Norton.
Специальные статьи и обзоры
Buri Р. 1956. Gene frequency in small populations of mutant Drosophila, Evolution,
10, 367—402.
Carson H. L. 1967. Inbreeding and gene fixation in natural populations. In:
R. A. Brink (ed.), Heritage from Mendel, Madison, University of Wisconsin
Press.
Cavalli-Sforza L. L. 1969. Genetic drift in an Italian population, Scientific American,
221 (2), 30—37.
Crow I. F. 1954. Breeding structure of populations, II. Effective population number.
In: 0. Kempthorne, T. A. Bancroft, J. W. Gowen and J. L. Lush (eds.), Statistics
and Mathematics, Ames, Iowa, Iowa State College Press.
Crow J. F., Maruyama T. 1971. The number of neutral alleles maintained in a finite
geographically structured population, Theoret. Pop. Biol., 2, 437—453.
Erickson R. O. 1945. The Clematis fremontii var. richlii population in the Ozarks,
Ann. Mo. Bot. Gard., 32, 413—460.
Felsenstein J. 1971. Inbreeding and variance effective numbers in populations with
overlapping generations, Genetics, 68, 581—597.
Greenwood J. J. D. 1974. Effective population numbers in the snail (Cepaea nemora-
Us), Evolution, 24, 513—526.
Kimura M. 1962. On the probability of fixation of mutant genes in a population
Genetics, 47, 713—719.
Kimura M., Crow J. F. 1964. The number of alleles that can be maintained in a finite
population, Genetics, 49. 725—738.
Kimura M., Ohta T. 1969. The average number of generations until fixation of a mu-
tant gene in a finite population, Genetics, 61, 763—771.
Nei M., Feldman M. W. 1972. Identity of genes by descent within and between popu-
lations under mutation and migration pressures, Theoret. Pop. Biol., 3 , 460—
465.
Wright S. 1923. Mendelian analysis of the pure breeds of livestock, I. The measu-
rement of inbreeding and relationship, J. Heredity, 14, 339—348.
Wright S. 1951. The genetical structure of populations, Ann. Eugenics, 15, 323—
354.
Глава 8
Рекомбинация
В гл. 7 было показано, что неслучайное скрещивание и конеч-
ные размеры популяции оказывают глубокое влияние на ее гене-
тическую структуру, вызывая главным образом снижение доли
гетерозиготных генотипов и утрату аллелей. Поэтому если бы в
условиях относительно однородной среды этим факторам не про-
тиводействовали другие факторы, то рано или поздно популяции
становились бы полностью гомозиготными по всем локусам. Су-
ществуют, однако, силы, противодействующие утрате аллелей (гл.
6). Главная из них — мутационный процесс, но рекомбинация и
некоторые типы отбора также играют очень важную роль. Кроме
того, среда редко бывает совершенно однородной. В этой главе
мы более подробно, чем это было сделано в гл. 6, рассмотрим ре-
комбинацию.
ФАКТОРЫ, ОКАЗЫВАЮЩИЕ ВЛИЯНИЕ НА РЕКОМБИНАЦИЮ
Ниже перечислены главные факторы, регулирующие процесс
рекомбинации (Grant V., 1958).
1. Число хромосом.
2. Частота кроссинговера.
3. Поток генов.
4. Продолжительность поколения.
5. Величина популяции.
6. Система скрещивания.
7. Несовместимость.
8. Преграды, препятствующие скрещиванию, и внешние изо-
лирующие механизмы.
Влияние на рекомбинацию числа хромосом и потока генов об-
суждалось в гл. 5 и 6.
Продолжительность поколения. Влияние продолжительности
поколения можно сформулировать очень просто. Если число ре-
комбинаций выражается формулой (стр. 138)
то число рекомбинаций в единицу времени (например, за год) по-
лучают, умножая число рекомбинаций на число поколений (N) в
год
13—2044
194
Глава 8
При прочих равных условиях у животного с двумя поколениями в
год число рекомбинантов за этот период будет вдвое больше, чем
у животного с одним поколением в год.
Продолжительность поколения имеет и другое важное значе-
ние. Не все потомки доживают до стадии половой зрелости. В ес-
тественных условиях в большинстве случаев число выживающих
потомков в среднем примерно равно числу умирающих взрослых
особей. Если продолжительность жизни особей данного вида ве-
лика, то популяция обновляется медленно. Но при одинаковой
плодовитости организмы, живущие долго, производят в течение
своей жизни больше потомков, чем недолгоживущие организмы.
Отсюда следует, что у долгоживущего организма гораздо меньшая
доля потомков доживает ежегодно до половой зрелости, чем у
короткоживущего. В качестве крайнего примера приведем сек-
войю. Если допустить, что в области, занимаемой в настоящее
время этими деревьями, растет максимально возможное их число,
скорость замещения будет равна примерно одному потомку на
зрелое дерево каждые 100—500 лет, хотя число семян, произво-
димых одним деревом за это время, достигает нескольких милли-
онов. И напротив, в популяции однолетних растений, например
салата, каждое растение должно замещаться ежегодно, и все же
число семян, производимых каждой особью, гораздо меньше, чем
у секвойи. У большинства животных продолжительность жизни
обычно очень небольшая и превышает два года лишь у очень не-
большого числа видов. Среди растений известны виды, живу-
щие по очень многу лет, причем это не только древесные формы,
но и многие травянистые виды, размножающиеся вегетативным
путем.
Величина популяции. Чем больше популяция, тем больше у
нее возможностей для запасания генетической изменчивости. Если
взять экстремальный случай — популяцию, состоящую из двух
особей, — то в ней может быть самое большее четыре разных ал-
леля для каждого гена; потенциально эти аллели в результате
полового размножения могут образовать в последующих поколе-
ниях 10 различных комбинаций с участием каждого из генов. В по-
пуляции, состоящей из трех особей, для каждого гена может быть
максимум по 6 аллелей и потенциальная возможность образовать
21 различную комбинацию; при четырех особях — 8 аллелей и
36 комбинаций; при пяти особях—10 аллелей и 55 комбинаций;
при десяти — 20 и 210, а при п особей — ‘максимум 2п аллелей и
потенциальная возможность образовать п(2л-|-1) рекомбинантов
(вывод этой формулы приведен в гл. 6). Другие аспекты влияния
величины популяции на ее генетическую структуру были рассмот-
рены в гл. 7.
Неизвестно ни одной популяции, которая была бы гетерозигот-
на по каждому генному локусу; однако чем больше популяция,
тем, при прочих равных условиях, больше вероятность образова-
Рекомбинация
195
дня новых комбинаций и тем выше возможный в ней уровень из-
менчивости.
Система размножения и система несовместимости. Не все ор-
ганизмы размножаются одним и тем же способом. Различные ис-
пользуемые при этом механизмы называются системами размно-
жения.
В табл. 8.1 перечислены главные системы размножения у рас-
тений и животных. Системы бесполого размножения — это те, при
которых не происходит рекомбинации. Поскольку при этом типе
размножения каждое растение или животное производит себе по-
добных совершенно независимо от других особей, механизм раз-
множения сам по себе очень эффективен. У облигатно апомикти-
тических видов (растения, продуцирующие семена без оплодотво-
рения) и у партеногенетических животных единственным источни-
ком новой изменчивости служат генные мутации. Облигатная
бесполость встречается очень редко, возможно, потому что образую-
щиеся в результате такого размножения очень однородные попу-
ляции с течением времени элиминируются при изменении условий
среды. Вегетативным размножением называют способность многих
Таблица 8.1
Главные системы размножения у растений и животных
А. Бесполое
Партеногенез и апомик-
сис
Вегетативное размноже-
ние
Сезонные (животные)
Факультативные (растения)
Псевдогамия (по большей части растения, неко-
торые животные)
Облигатные (растения и животные)
С половым размножением (в большинстве слу-
чаев растения, некоторые животные)
Без полового размножения (некоторые животные
и растения)
Б. Половое
Полный инбридинг (растения и несколько видов животных)
Тесный инбридинг (растения н животные)
Животные
Моногамия
Полигамия
Полигиния
Промискуитет
При численном
равенстве или
неравенстве осо-
бей разного по-
ла
Аутбридинг
Растения
Двудомность
Гетеростилия
Протандрия, протогиния
Механическое предотвращение самоопыле-
ния
Мультиаллельная генетическая самонесов-
местимость
13*
196
Глава 8
растений продуцировать новые физиологические индивидуумы пу-
тем образования побегов, усов, луковиц или других аналогичных
структур.
У некоторых насекомых, в частности у тлей, наблюдается че-
редование полового (обычно осенью) и бесполого (весной и ле-
том) поколений. Кроме того, у некоторых растений, обычно раз-
Рис. 8.1. Скрещивание между растениями при гетеростилии. Пыльцевое зерно,
произведенное растением с короткой тычинкой (с длинным столбиком), проявит
жизнеспособность только на столбике растения, у которого тычинки длиннее стол-
биков. Как у тех, так и у других растений половина потомков будет иметь корот-
кие, а половина — длинные тычинки. (О. Т. Solbrig, 1966, Evolution and Systema-
tics, New York, Macmillan.)
множающихся бесполым путем, время от времени происходят скре-
щивания (в отличие от животных — необязательно в определен-
ное время года). Таким образом, у них все еще остается возмож-
ность для генетической рекомбинации в сочетании с преимущест-
вами бесполого размножения. Все, кому приходилось бороться с
тлями на розовых кустах, знают, как быстро и успешно они мо-
гут размножаться!
Существует несколько механизмов, регулирующих степень са-
мооплодотворения у организмов, размножающихся половым путем.
Настоящее самооплодотворение возможно только у гермафродит-
ных организмов. Гермафродиты с облигатным самооплодотворе-
нием среди животных крайне редки; среди растений они встреча-
ются чаще, но также относительно редки. У большинства живот-
ных, как и у двудомных растений, имеются особи разного пола,
что обеспечивает аутбридинг. Хотя большинство растений герма-
фродиты, существует много механизмов, препятствующих опыле-
нию яйцеклетки данного растения его собственной пыльцой. Не-
которые из наиболее важных механизмов такого рода указаны в
конце табл. 8.1. При гетеростилии у данного вида растений име-
ются цветки двух типов: у одного типа столбики длиннее, чем ты-
чинки, а у другого — тычинки длиннее, чем столбики (рис. 8.1).
Пыльца с коротких тычинок обычно оказывается жизнеспособной
Рекомбинация
197
только на рыльце короткого столбика, а пыльца с длинных тычи-
нок— на рыльце длинного столбика; тем самым обеспечивается
аутбридинг.
При протандрии пыльца созревает раньше, чем женские ор-
ганы цветка, а при протогинии — женские органы созревают рань-
ше, чем пыльца у данного цветка. И в этих случаях возможность
самоопыления снижается. Кроме того, имеются разного рода ме-
Фертильность 100%
Рис. 8.2. Скрещивание между самонесовместимыми растениями. Буквы в сплош-
ных кружках — генотипы пыльцевых зерен; буквы в пунктирных кружках — гено-
типы столбиков. Данное пыльцевое зерно может оплодотворить только такое ра-
стение, которое не несет аллеля несовместимости, имеющегося у этого зерна. По-
этому лишь 50% пыльцы от растения S'S2 (пыльцевые зерна, несущие аллель
S1) способно оплодотворить растение S2S3. (О. Т. Solbrig, 1966. Evolution and
Systematics, New York, Macmillan.)
ханические приспособления, не допускающие попадания на рыль-
це данного цветка его собственной пыльцы.
Самый обычный механизм, предотвращающий самоопыление у
растений, это генетическая несовместимость (рис. 8.2). Этот при-
знак контролируется геном самонесовместимости, который обо-
значают буквой S. У гена S имеется много аллелей, обозначаемых
Через S1, S2, S3,..., S", число которых в популяции может быть бо-
лее 30, 50 или даже 100. Растение обычно не принимает пыльцу,
несущую тот же аллель, который содержит оно само, т. е. расте-
ние, имеющее генотип S'S2, не может опыляться пыльцой S1 или
S2, т. е. теми двумя типами пыльцевых зерен, которые продуциру-
ет оно само (помните, что пыльца гаплоидна и что ее образова-
нию предшествовал мейоз; см. гл. 5). Растение S'S2 не может
быть опылено пыльцой других растений S'S2, имеющихся в данной
Популяции; если же скрещивать растение S'S2 с растением
S'S3, то жизнеспособной на растениях S'S2 оказывается только
198
Глава 8
пыльца S3. Действие аллелей, определяющих самонесовмести-
мость, по-видимому, состоит в регуляции скорости роста пыльце-
вой трубки или даже прорастания пыльцы. Ткани столбика вы-
рабатывают какое-то вещество (возможно, антигенного характе-
ра), которое подавляет или резко замедляет рост пыльцевых тру-
бок, имеющих ту же самую генетическую конституцию.
Преграды скрещиванию и внешние изолирующие механизмы
рассматриваются в гл. 11 и 12.
Таким образом, рекомбинация представляет собой цепь про-
цессов, начинающихся с генной мутации и продолжающихся во
время полового цикла. Гены объединены в хромосомы, и это ог-
раничивает рекомбинацию. Отдельные особи объединены в скре-
щивающиеся популяции и виды, что также устанавливает грани-
цы рекомбинации. Гены кита не могут рекомбинировать с генами
моржа. А поэтому лишь некоторая доля потенциально возможной
рекомбинации реализуется в природе.
КЛАССИФИКАЦИЯ РЕКОМБИНАЦИОННЫХ СИСТЕМ
Доля достижимой рекомбинации варьирует главным образом
в зависимости от системы размножения, а также от таких факто-
ров, как величина популяции, частота кроссинговера и число хро-
мосом. В соответствии с классификацией, предложенной амери-
канским генетиком Хамптоном Карсоном (Hampton Carson), ре-
комбинационные системы делятся на относительно открытые, ог-
раниченные и закрытые.
Закрытые рекомбинационные системы, примером которых слу-
жат организмы с облигатным партеногенезом или апомиктичес-
кие виды, такие, как общеизвестный одуванчик лекарственный
(Taraxacum officinale) —это системы с полным отсутствием ре-
комбинации. Ограниченные рекомбинационные системы создают
мощные преграды рекомбинации; к ним относятся факультатив-
ные апомикты (растения, в норме размножающиеся бесполым
путем, но тем не менее способные к половому размножению) или
виды, для которых характерен тесный инбридинг (например, виды
тлей, обычно размножающиеся бесполым путем, но дающие
осенью половое поколение). Открытые рекомбинационные системы
существуют у форм, которые в норме дают большое число по-
тенциальных рекомбинантов, как большинство позвоночных.
Возникает вопрос, почему существуют эти различия в реком-
бинационных системах? Почему некоторые виды имеют закрытые
рекомбинационные системы, тогда как у других, иногда даже
близко родственных, эти системы открытые? На этот вопрос еще
не получено полного ответа. Один из подходов к его решению со-
стоит в том, чтобы рассматривать его как экономическую проб-
лему. Рекомбинационная система каждого типа предоставляет ор-
ганизму известные выгоды и сопряжена с различными «расходами».
Рекомбинация
199
Выгоды минус «расходы» дают (повышение (или понижение) при-
способленности, которое мы называем «доходом». Естественный
отбор в пределах каждой популяции будет благоприятствовать
особям с такой рекомбинационной системой, которая приносит
наибольший «доход», т. е. наивысшую приспособленность.
Открытые рекомбинационные системы требуют больших «рас-
ходов» и, по всей вероятности, приносят больше «выгод». Рас-
ходы, связанные с открытой рекомбинационной системой, можно
произвольно разделить на три части: 1) генетический груз;2) энер-
гетические расходы на аутбридинг; 3) расходы на мейоз. Выгоды
предположительно состоят в повышении приспособленности, до-
стигаемой организмом при производстве потомка, у которого ве-
роятность дожить до зрелого возраста выше, потому что он обла-
дает новой генетической конституцией, возникшей в результате
скрещивания. Рассмотрим теперь каждый вид таких «расходов» и
«выгод» более подробно.
ГЕНЕТИЧЕСКИЕ РАСХОДЫ
И КОНЦЕПЦИЯ ГЕНЕТИЧЕСКОГО ГРУЗА
В предыдущей главе было показано, что все популяции склон-
ны к инбридингу; но при этом число гомозиготных генотипов по-
вышается и происходит утрата аллелей. Мы увидели, что все
это — следствие конечных размеров популяции, неслучайного скре-
щивания и полустохастической природы системы размножения.
Противодействие этой тенденции означает благоприятствование
гетерозиготам в ущерб гомозиготам и редким аллелям — в ущерб
широко распространенным. Следовательно, при этом происходит
снижение суммарной приспособленности, или «генетический рас-
ход», который называют генетическим грузом.
Термин «генетический груз» был впервые введен в 1950 г. из-
вестным американским генетиком, лауреатом Нобелевской премии
Германом Мёллером (Hermann J .Miiller). Он использовал этот
термин в очень специфическом и гораздо более ограниченном
Смысле, чем его применяют в настоящее время. Мёллер называл
генетическим грузом накапливание в популяциях человека в ге-
терозиготных комбинациях аллелей, возникающих в результате
повторных мутаций и обладающих в гомозиготном состоянии
вредным или даже летальным действием. По мере повышения
частоты этих аллелей начинают продуцироваться гомозиготные
зародыши, которые либо гибнут на разных стадиях развития
(спонтанные выкидыши), либо приводят к появлению неполноцен-
ных индивидуумов.
Но выдвинутая Мёллером концепция имеет гораздо более ши-
рокое приложение. Представим себе популяцию, состоящую из
Генетически различных организмов, живущих в совершенно одно-
родной среде, и допустим, что мы можем точно измерить приспособ-
200
Глава 8
ленность каждой особи. Некоторые особи непременно окажутся
при этом более приспособленными, чем другие, а одна будет обла-
дать наивысшей приспособленностью IFmax. Очевидно, что средняя
приспособленность такой популяции ниже, чем популяции, состоя-
щей из генетически однородных особей, каждая из которых обла-
дает приспособленностью W'max. Проигрыш в приспособленности L
у первой популяции можно представить как
т _ ^тах —
^тах ’
где IF —средняя приспособленность популяции. Проигрыш в
приспособленности L называют генетическим грузом. Обратите
внимание на то, что этот груз относится к популяции, а не к
особи. Если допустить, что особь с приспособленностью 1Етах ге-
терозиготна, то L может служить мерой расходов, связанных с
противодействием тем силам, которые снижают гетерозиготность.
Любой фактор, который понижает приспособленность той или
иной особи, увеличивает генетический груз популяции. Однако
самую важную роль в этом играют два явления — мутационный
процесс и расщепление.
Мутационный груз. В гл. 6 было показано, что равновесная
частота рецессивного аллеля при отборе, направленном против
него, составляет
<7
где и — частота мутаций, as — коэффициент отбора против гомо-
зигот по данному аллелю. В случае доминантного аллеля равно-
весная частота равна
где h — степень доминирования.
В популяции, состоящей из трех генотипов, AiAi, AjA2 и
А2А2, где А2 — полностью рецессивный вредный аллель, средняя
приспособленность популяции равна сумме произведений частот
генотипов на их индивидуальные приспособленности, как показано
ниже (см. также гл. 6).
Частота Приспособ-
ленность (W)
A1A1 — p2 X 1
A[A2= 2pq X 1
A2A2“ q2 X 1 — s
Средняя приспо-
собленность
= р2
= 2р</
= q2—sq2
=р2+2pq 4- q2—sq2
= 1—sq2
Рекомбинация
201
Таким образом, в этом случае величина генетического груза
равна sqz. Подставляя ^u/s вместо q, в состоянии равновесия
можно вычислить ту долю генетического груза, которая обуслов-
лена повторными мутациями.
, 2 Л1/ « \2 ( U \
L~sq2—s I у — ) =sl — ]=и.
Иными словами, мутационный груз при равновесии, обусловлен-
ный полностью рецессивным аллелем, равен частоте, с которой
этот аллель вносится в популяцию в результате мутаций, и не
зависит от его влияния на приспособленность. Это объясняется
тем, что равновесная частота такого аллеля обратно пропорцио-
нальна его вредному действию.
Рассуждая подобным же образом, можно показать, что для
полудоминантных аллелей
W = 1 —2pqhs—sq2 1 — 2hsq,
поскольку р в случае популяций с вредными генами близко и
единице и значение sq2 очень мало.
Подставив u/hs вместо q, получим
L=hsq=-^-=2u.
7 hs
Таким образом, мутационный груз, создаваемый каким-либо вред-
ным аллелем в данной популяции, лежит между частотой, с ко-
торой он мутирует, и ее удвоенным значением — в зависимости
от степени доминантности этого аллеля и независимо от степени
его вредоносности.
Сегрегационный груз. В гл. 6 было показано, что иногда ге-
терозигота превосходит по приспособленности обе гомозиготы.
Здесь эта ситуация будет рассмотрена более подробно. Предста-
вим себе популяцию, особи которой различаются только по одно-
му локусу с двумя аллелями. А! и А2, с частотами р и q соответ-
ственно, при следующих исходных частотах и относительных при-
способлсгпостях трех генотипов:
генотип AtAj AtA2 А2Аг,
исходная частота ро2 2рс,гд ф2,
приспособленность 1—s 1 I—t,
где s и t— коэффициенты отбора, направленного соответственно
против генотипов AjAj и А2А2; эти генотипы имеют следующие ча-
стоты:
A±A±—р2; AiA2—2pq; А2А2 —q2.
202
Глава 8
Поэтому вклад разных генотипов в создание гамет составит
АЛ—р2-(1 — $),
А4А2—2pq-1,
А2А2—<z2-(l —/),
поскольку вклад в создание гамет равен произведению частоты
каждого генотипа на коэффициент отбора для этого генотипа.
Суммарный вклад популяции в создание гамет составит
р2 (1 — s) 4- 2pq q2 (1 — t) = p2—sp2 -}- 2pq p2 — tq2 =
= 1— sp2~ tq2.
Для того чтобы найти частоту генов А2 в следующем поколе-
нии, надо прибавить половину вклада генотипа AiA2 к вкладу
генотипа А2А2 и разделить на полученный выше суммарный вклад
гамет, так что частота аллеля А2 в поколении 1 будет равна
_ <72(1- 0+Р?
41 1 — sq2 _ fq2 '
а изменение, возникающее за одно поколение в результате отбо-
ра,
Л„_„ п— () + pq q2 — tq2 + pq-q + sP2q-}-tqs
— 71 — 9 — i „ sp2 _ tq2 J _sp2_tq2 ,
что после упрощения сводится к
Д/,-_ pq(sp — tq)
4 — 1 — sp2 — /р2
Во всех случаях, когда произведение s-p больше, чем t-q,
разность между ними будет больше нуля и величина Др будет
положительной, т. е. частота А2 будет возрастать. Когда s-p
меньше, чем t-q, то числитель и Ар будут отрицательны и частота
А2 будет уменьшаться. Когда sp—tq—O, никаких дальнейших из-
менений генных частот в популяции не происходит. Таким обра-
зом, в состоянии равновесия
sp=tq,
и поскольку,
1—9-
то s(l—р) =tq и s—sq = tq, или
s=sp-Hp=p(s-H),
и после преобразования
S
7
Рекомбинация
203
Аналогичным образом
Следовательно, когда приспособленность гетерозигот выше
приспособленности как одной, так и другой гомозиготы, ни один
из генотипов не закрепляется в ущерб другому, а оба аллеля
сохраняются в популяции с промежуточными частотами. Это яв-
ление называется сбалансированным полиморфизмом или одноло-
кусным гетерозисом.
Средняя приспособленность популяции в состоянии равнове-
сия, когда никаких дальнейших изменений генных частот не про-
исходит и А<7 = Ар = 0, равна
- / / \2 / $ \2
B/=l-sp>_^=l-s(7±T) -'(ттт) =
Поэтому генетический груз, возникающий в результате сверх-
доминирования, составляет
Такого рода груз называют сегрегационным грузом, и он доволь-
но значителен, когда величины $ и t выше 0,1. В экстремальном
случае, когда обе гомозиготы детальны и s = / = l, L = 0,5, т. е.
половина потомков ежегодно гибнет.
Другие виды груза. Рассуждая подобным же образом, можно
вычислить потери приспособленности, испытываемые популяцией,
когда некоторые скрещивания в ней не удаются вследствие не-
совместимости между партнерами (груз, обусловленный несов-
местимостью) или же когда в популяции появляется мутация,
обладающая селективным преимуществом (субституционный груз).
Общезначима ли концепция генетического груза?
Произведенный выше анализ показывает, что такие повышаю-
щие генетическое разнообразие явления, как мутационный процесс
и преимущество гетерозигот, сопряжены с определенными «рас-
ходами». Однако эти расходы легко преувеличить. Читателю сле-
дует напомнить об ограничивающем допущении, принятом для
описанной модели, а именно — допущении об однородности среды
во времени для всех членов рассматриваемой популяции. Только
при этом условии Ц/шах будет иметь фиксированное значение и
только при этом каждому генотипу можно приписать определен-
ную приспособленность. Кроме того, концепция генетического гру-
204
Глава 8
за применима к популяции, а не к особи. Рассмотрим более
обычную ситуацию, когда физическая среда изменяется как в про-
странстве, так и во времени.
При введении концепции относительной приспособленности
было показано, что приспособленность какого-либо конкретного
генотипа измеряется относительно приспособленностей всех дру-
гих генотипов данной популяции. Поэтому в изменчивой среде
приспособленность какого-либо генотипа по сравнению с другими
в той же популяции изменяется с изменением среды. Представим
себе популяцию, в которой имеется два фенотипа, причем каждый
из них соответствует исключительно одному генотипу. Один фено-
тип приспособлен к засухе, а другой функционирует наилучшим
образом при высокой влажности. Соответственно приспособлен-
ность каждого из этих генотипов будет зависеть от среды, в ко-
торой он находится. Болотистые места благоприятны для второго
генотипа, а сухие — для первого. Кроме того, в годы с очень боль-
шим количеством осадков приспособленный к влажным усло-
виям фенотип будет функционировать лучше, чем приспособлен-
ный к засухе, и наоборот. Естественный отбор будет непрерывно
элиминировать из популяции определенные фенотипы, но отрица-
тельному отбору будут подвержены оба фенотипа, в зависимости
от места и от условий в данном году. Если воспользоваться соот-
ветствующими формулами, то окажется, что средняя приспособ-
ленность такой популяции ниже теоретического максимального
значения для однородной популяции, обитающей либо только во
влажной, либо только в засушливой среде.
Допустим теперь, что популяция обитает в аналогичных физи-
ческих условиях, но содержит лишь один фенотип, например,
приспособленный к высокой влажности. В этом случае приспособ-
ленность всех генотипов будет довольно однородной и, применив
имеющиеся формулы, мы убедимся, что приспособленность попу-
ляции соответствует 1Етах или близка к ней. Однако такая попу-
ляция сможет существовать только на болотистых участках, а в
засушливые годы ей будет угрожать опасность элиминации.
Этот пример показывает, что относительная приспособленность,
равная 1, т. е. 1Етах> при нулевой величине груза не есть мера ус-
пеха популяции в изменчивой среде. Существование груза указы-
вает на то, что разнообразие влечет за собой генетические «рас-
ходы», а именно возникновение генотипов, которые оказываются
менее приспособленными в определенные моменты времени и в
данных участках ареала (см. также гл. 9).
Но отбор действует прежде всего на уровне особей. Если дан-
ная особь может произвести большее число выживающих потом-
ков, то ее гены будут сохранены отбором, независимо от судьбы
тех ее потомков, которые не смогли выжить. Погибли ли они
вследствие неприспособленности своих генотипов или в результа-
те конкуренция, не играет большой роли. Однако в некоторых
Рекомбинация
205
ситуациях расходы, связанные с утратой зигот, обусловленной
высоким генетическим грузом, могут оказаться гибельными, a
поэтому концепция «груза» имеет известное практическое значе-
ние.
Энергетические расходы, связанные с аутбридингом
Всегда и везде любая особь, вынужденная искать себе потен-
циального брачного партнера, затрачивает на это некоторое ко-
личество энергии. Организмы, выделяющие свои гаметы в воду,
не несут этого расхода; он минимален также у видов, которые
живут стадами, состоящими из самцов и самок. Энергетические
расходы достигают наибольшей величины у .сидячих видов, на-
пример у растений, у которых перенос гамет от одной особи к дру-
гой осуществляется внешними факторами. Кроме того, аутбри-
динг всегда сопряжен с некоторой потерей гамет.
Хороших работ по изучению энергетических расходов на аут-
бридинг, в сущности, мало. Для ряда различных видов растений
установлено, что число пыльцевых зерен, необходимых для опло-
дотворения одной яйцеклетки, у перекрестноопыляющихся расте-
ний на несколько порядков больше, чем у самоопыляющихся
(табл. 8.2). Подобным же образом у перекрестноопыляющихся
видов цветки крупнее и ярче и они вырабатывают большие коли-
чества нектара, чтобы привлекать насекомых. Затраты энергии на
эти структуры компенсируются уменьшением числа цветков или
размеров растения, а иногда и того и другого.
Таблица 8.2
Соотношение числа пыльцевых зерен и яйцеклеток
в цветках растений с различными системами
размножения (Cruden, 1977. Pollen — ovule ratios,
Evolution, 31, 32—66)
Система размножения
Число
изученных
видов
Соотношение
пыльца/яйцеклетка
(среднее)
Клейстогамия
Облигатная автогамия
Факультативная автогамия
Факультативный ауткроссинг
Облигатный ауткроссинг
4,7
27,7
168,5
796,6
5859,2
Мейотические расходы
При размножении путем самооплодотворения потомки насле-
дуют все свои гены от одной родительской особи, а при перекрес-
тном оплодотворении потомки наследуют одну половину своих
206
Глава 8
генов от одного, а другую — от другого родителя. Поэтому при
ауткроссинге родительская особь передает своим потомкам вдвое
меньше генов, чем при самооплодотворении. Разницу в количестве
передаваемых генов Дж. К- Уильямс (G. С. Williams) назвал
мейотическими расходами. Эти расходы составляют 50%, если
две скрещивающиеся между собой особи не несут общих генов,
Возможные результаты мейо-
Рис. 8.3. Расходы на мейоз.
При оогенезе половина аллелей материнских яйцеклеток остается «неиспользован-
ной». Дж. К. Уильямс (G. С. Williams) назвал это «мейотическими расходами».
Поскольку только одна из четырех потенциальных гамет превращается в яйце-
клетку, половина аллелей утрачивается. (Если, например, яйцеклетка имеет гено-
тип А2Вь то аллели А, и В2 утрачиваются. Конечно, в других яйцеклетках, про-
дуцируемых той же особью, «утраченными» могут оказаться другие аллели.)
идентичных по происхождению (Г = 0; см. гл. 7), но они ниже, ес-
ли, как это часто бывает, между особями существует известная
степень родства (рис. 8.3).
Выгоды, которые дает ауткроссинг
Поскольку огромное большинство организмов размножается
при помощи перекрестного оплодотворения и поскольку аутбри-
динг требует больших расходов, чем инбридинг, следовательно,
продукция жизнеспособного потомства дает явное преимущество,
и это преимущество компенсирует расходы, порождаемые генети-
ческим грузом, энергетические расходы на ауткроссинг и мейоти-
ческие расходы.
Мейотические расходы зависят от степени инбридинга; они
равны нулю в совершенно гомозиготной и гомогенной популяции
и максимальны, когда в популяции нет двух генетически родст-
венных между собой особей (Г = 0; см. гл. 7). Генетический груз,
Рекомбинация
207
создаваемый мутациями, зависит, как мы видели, от частоты му-
таций; сегрегационный же груз зависит от уровня гетерозиготнос-
ти и будет поэтому возрастать с увеличением аутбридинга. На-
конец, расходы, связанные со случайным скрещиванием, т. е. с
поисками брачного партнера, зависят от величины популяции, от
ее плотности и от того, в какой степени каждая особь ищет себе
партнера вне круга родственных и соседних особей. Поэтому рас-
ходы в общем возрастают при открытых рекомбинационных си-
стемах.
Если относительно расходов, связанных с ауткроссингом, су-
ществует единство во взглядах, то вопрос о преимуществах, ко-
торые дает генетически разнообразное потомство, вызывает раз-
ногласия. Можно, однако, указать на два благоприятных обстоя-
тельства, не исключающих одно другое.
Во-первых, явные преимущества, которые дает создание гете-
розиготных потомков; эти преимущества будут обсуждаться® гл.
9. Во-вторых, преимущество, которое связано с условиями среды,
изменяющейся в пространстве и во времени. Каждый отдельный
организм существует в среде, в которой обитают многие другие
организмы. Одни из них относятся к тому же виду, что и данный
организм, и могут быть потенциальными конкурентами; другие
принадлежат к иным видам и могут также конкурировать с ними
за ресурсы, но могут оказаться потенциальными хищниками или
потенциальной жертвой. Эта «биологическая среда» очень сложна
и непрерывно изменяется, часто довольно быстро. Кроме того,
эта биологическая среда отзывается на любое адаптивное измене-
ние у любого вида. Мы видели, например, что у мух в ответ на
введение в среду ДДТ выработалась устойчивость к нему. Ана-
логичным образом у растительноядных насекомых в ответ на по-
явление у какого-либо вида растений токсичного для этих насеко-
мых вещества нередко развивается механизм детоксикации. В та-
ких случаях непосредственные выгоды создания путем рекомбина-
ции разнообразного потомства ясны. Организм, который в своих
взаимодействиях с окружающей средой всегда полагается на один
и тот же способ защиты, нападения или конкуренции, рискует
потерять всех своих потомков, если (и когда) какой-либо хищник
или патоген «прорвет линию обороны», у потенциальной жерт-
вы появится способ защиты или у одного из конкурентов разовь-
ется новый механизм конкуренции. Следовательно, для данного
организма вероятность создания жизнеспособного потомства по-
высится, если у каждого из его потомков морфологические, фи-
зиологические, химические и поведенческие механизмы взаимо-
действия со средой слегка различаются как в качественном, так
и в количественном отношении. Гомозиготность может привести
к гибели всего потомства, тогда как гетерозиготность повышает
вероятность его выживания. Поскольку возможность произвести
потомство, доживающее до репродуктивного возраста, частично
208
Глава 8
определяется случайными факторами, вероятность выигрыша (т. е.
создания такого потомства) обычно выше при наличии разнооб-
разных, а не гомозиготных и одинаковых потомков, подобно тому
как вероятность выиграть в лотерее выше при наличии большого
числа билетов с разными номерами, чем такого же их числа с
одинаковыми номерами.
Как показало предшествующее обсуждение, и открытые и за-
крытые рекомбинационные системы имеют не только свои досто
инства, но и недостатки. Поэтому весьма вероятно, что в каждой
среде селективным преимуществом обладает та рекомбинацион-
ная система, которая дает наибольшую «выгоду» (доходы — рас-
ходы). Однако нет ни одной работы, в которой были бы проана-
лизированы все расходы и доходы, связанные с рекомбинацион-
ной системой одного конкретного вида. Дело не только в том, что
такое исследование требует много труда и времени, но и в том,
что в настоящее время не существует какого-либо общего способа
перевода генетических расходов (т. е. мейотических расходов или
мутационного груза) в расходы, выражающиеся в снижении при-
способленности (т. е. в потере потомков). Результаты некоторых
частных исследований этой проблемы показывают, что такой пе-
ресчет, вероятно, возможен.
НАРУШЕНИЕ РЕКОМБИНАЦИИ
В гл. 4 говорилось о том, что второй закон Менделя (закон
независимого распределения) применим только к генам, лежащим
в разных хромосомах. Неприменимость второго закона к генам,
лежащим в одной хромосоме, обусловлена явлениями сцепления
(см. гл. 5) и мейотического драйва (см. гл. 6). Теперь мы пока-
жем, что нарушение рекомбинации в некоторых случаях может
быть благоприятным и поэтому может поддерживаться отбором.
Благоприятное влияние сцепления может быть связано с тем,
что сцепление служит одним из способов сохранения в популяции
некоторых генных комбинаций без потери гамет. Допустим, что в
некой популяции генотипы AiAiBjBj и A2A2B2B2 имеют одинако-
вую высокую приспособленность и что генотипы, содержащие ал-
лели А; и В2 или А2 и Вь имеют одинаковую низкую приспособ-
ленность. Если А и В — два локуса, лежащие в разных хромосо-
мах, и если допустить, что частоты аллелей А! и А2 равны p — q =
= 0,5, а частоты аллелей Bi и В2 равны m = n = 0,5, то генотипы
будут иметь следующие частоты:
Генотип А(А1В1В1-. р2-т2 = (0,5)2-(0,5)2 = 0,0625
Генотип А2А2В2В2: q2-n2 = (0,5)2-(0,5)2 = 0,0625
Итого 0,125
Рекомбинация
209*
Таким образом, лишь Vs всей популяции будет обладать геноти-
пом, определяющим высокую приспособленность.
Если, однако, локусы А и В сцеплены и лежат в одной хро-
мосоме, причем исходная частота хромосом AjBi равна Xi — 0,5, а
хромосом АгВ2 равна Х4 = 0,5 с коэффициентом рекомбинации
г=0,01, то тогда, в соответствии с уравнением, приведенным на
стр. 107,
Xj,+I =Xlr(l — r)+pm (г) =
=0,5 (1 —0,01) + (0,5) (0,5) • (0,01) =
=0,495 + 0,0025 = 0,4975
и аналогичным образом
Х4г+1 = 0,4975
Итого 0,995 ,
тогда как частота Х2 хромосом AtB2 составит
==*г< (1 -г)+рп (Г) = 0 (1 -0,01) + (0,5). (0,5) • (0,01) =
= 0+0,0025
и аналогичным образом для хромосом A2Bi
*3(+1= °-0025
Итого 0,005 ,
а генотипический состав будет следующим:
АЛВ.В^АД.АД: (*1Ж) • (Х1/+1) = (0,4975)2 = 0,2475
А2А2В2В2 = АА'.АА-- (Х4/+1) 0++1) = (0,4975)2 = 0,2475
Итого 0,495,
где АА указывает, что аллели At и Bi лежат в одной и той же
хромосоме. Таким образом, 49,5%, или почти половина, популяции
имеет генотип, обусловливающий высокую приспособленность, тог-
да как в отсутствие сцепления такой генотип имеет лишь Vs осо-
бей. Поскольку потеря половины потомков менее катастрофична,
Чем потеря 7/в (87,5%), сцепление является механизмом, который
Может понизить расходы, связанные с сохранением в популяции
сочетаний генов, обусловливающих высокую приспособленность.
КОАДАПТЛЦИЯ
Нарушение расщепления приводит к повышению частоты не-
которых генных комбинаций в гаметах данной особи и у ее по-
томков. В некотором смысле этот эффект сходен с действием ин-
14—2044
210
Глава 8
бридинга, 'поскольку при этом частота одних аллелей возрастает,
а другие аллели утрачиваются и характер этих процессов не со-
ответствует предсказаниям формулы Харди—Вайнберга. Однако
если инбридинг действует на все аллели и притом случайным об-
разом, а кроме того, снижает уровень гетерозигот в популяции,
то нарушение расщепления оказывает влияние только на опре-
Рис. 8.4. Изменения частоты хромосомных перестроек в экспериментальных усло-
виях.
Была создана популяция Drosophila pseudoobscura, содержащая особей с двумя
различными последовательностями генов ST и СН. Популяцию содержали при
16 °C в течение некоторого времени. Соотношение частот разных последователь-
ностей достигло равновесия на уровне 86% ST и 14% СН. (По данным S. Wright,
Dobzhansky, 1946.)
деленные генные комбинации и может в некоторых случаях (на-
пример, в случае инверсий у Drosophila pseudoobscura) повысить
уровень гетерозиготности.
Мы можем задать себе вопрос, благоприятны ли такие явле-
ния, как мейотический драйв и сцепление для образующих попу-
ляцию особей и благоприятствует ли им поэтому отбор. Многие
генетики склонны считать эти явления благоприятными, потому
что они обеспечивают сохранение в хромосоме или в отдельной ее
части сочетаний генов, которые особенно хорошо функционируют
вместе. Такие группы неаллельных генов называют коадаптиро-
ванными генными комплексами.
В настоящее время у нас нет неопровержимых доказательств
справедливости концепции биохимической природы действия не-
адаптированных генных комплексов, но в пользу этой концепции
свидетельствует множество косвенных данных.
Наиболее хорошо известны данные, полученные в результате
изучения хромосомных инверсий у Drosophila pseudoobscura (см.
Рекомбинация
211
также гл. 9). Когда в лаборатории создают искусственные попу-
ляции мух этого вида, смешивая в разных соотношениях особей с
последовательностью Чирикахуа (СН) и со стандартной последо-
вательностью (ST), взятых из одной и той же местности, то при-
мерно через год популяция достигает равновесия, причем частота
Рис. 8.5. Изменение частоты хромосом со стандартной последовательностью ге-
нов (ST) в популяции Drosophila pseudoobscura в эксперименте с четырьмя по-
вторностями. Исходно во всех четырех экспериментальных популяциях было 20%
хромосом ST из Пинои-Флэтс (Калифорния) и 80% хромосом СН из Чихуахуа
(Мексика). Обратите внимание на то, что частота ST возрастает и закрепляется
на почти одинаковом уровне в трех популяциях из четырех. Ср. с рис. 8.4. (По
данным Dobzhansky, Pavlovsky, 1953.)
хромосом с инверсией ST составляет примерно 0,86, а с инверси-
ей СН — около 0,14 (рис. 8.4). Райт (Wright) и Добржанский
(Dobzhansky) вычислили относительную приспособленность (И7)
трех возможных генотипов, которая оказалась равной 0,90 для ST/
/ST, 1,00 для ST/СН и 0,41 для СН/СН, а коэффициенты отбора
составляют s = 0,10 для СН и ( = 0,59 для ST (см. гл. 9). Из этих
величин можно вычислить ожидаемые частоты хромосом (стр.
202—203):
( 0,58 п осс
р~~ s + t ~ 0,69 —
и
14*
212
Глава 8
Если, однако, при создании искусственных популяций мух с
хромосомой СН и с хромосомой ST берут из разных местностей,
то примерно через год популяция становится гомозиготной по
ST/ST (рис. 8.5). В этом случае приспособленности оказываются
иными. Например, если хромосома ST происходит из Пинон-
Флэтс, а хромосома СН—либо из Пинон-Флэтс, либо из Матера
(обе местности находятся в горах Сьерра-Невада в Калифорнии),
либо из Мексики, то получаются приспособленности, представ-
ленные в табл. 8.3. Чтобы облегчить сравнение, приспособленность
гетерозигот во всех случаях принята за 1,00 (№=1,00). Можно
видеть, что при скрещивании мух из разных местностей наиболь-
шей приспособленностью обладают гомозиготы ST/ST. Считается,
что причина этого превосходства гетерозигот в случаях, когда
хромосомы происходят из одной местности, состоит в «коадапти-
рованности» хромосом к совместному функционированию.
Таблица 8.3
Относительная приспособленность особей Drosophila pseudoobscura, полученных
в результате скрещиваний между особями с хромосомами ST и СН,
имеющими различное географическое происхождение
(Dobzhansky, Pavlovsky, 1953. Indeterminate outcome
of certain experiments on Drosophila
populations, Evolution, 7, 198—210.)
ST СН Приспособленность (W)
ST/ST ST/CH сн/сн
Пинон-Флэтс (Кал.) Пинон Флэте (Кал.) 0,90 1,00 0,41
Пинон-Флэтс (Кал.) Матер (Кал.) 1,38 1,00 0,43
Пинон-Флэтс (Кал.) Мексика 1,26 1,00 0,87
Другим примером служит Biston betularia, о которой кратко
упоминалось в гл. 3; у этой бабочки новая меланистическая фор-
ма carbonaria за последние 150 лет вытеснила доминировавшую
прежде форму с серыми пятнами. Это наиболее хорошо изученный
пример эволюционного возникновения индустриального меланиз-
ма.
Эволюция индустриального меланизма
За последние сто лет ученые имели возможность наблюдать
и тщательно документировать процесс замещения светлой окрас-
ки у многих различных видов бабочек темной или черной. Это
изменение окраски, получившее название индустриального мела-
низма, считается одним из интереснейших эволюционных явле-
ний. В результате полевых наблюдений и экспериментов биологам
удалось выяснить преимущества и недостатки темной окраски в
Рекомбинация
213
природе, а при помощи лабораторных экспериментов они выяви-
ли различные механизмы наследования этого признака.
Темная окраска может обеспечить животному три совершенно
различных преимущества. Во-первых, темную особь труднее за-
метить на темном фоне —так называемая критическая, или по-
кровительственная, окраска. Во-вторых, черная поверхность лучше
поглощает тепло, а поэтому темно окрашенное насекомое утром
согревается быстрее, чем светлое. Наконец, в-третьих, меланисти-
ческие формы лучше защищены от ультрафиолетового излучения.
С середины XIX в. и до наших дней в промышленных районах
северного полушария постоянно отмечается появление черных
форм бабочек, принадлежащих к самым различным видам и ро-
дам. Во всех таких случаях меланистические формы принадлежат
к криптическим видам, т. е. к видам, которые пытаются избежать
поимки хищниками, стараясь слиться с фоном, создаваемым их
средой обитания. Наиболее обширные исследования таких форм
проведены на бабочках Британских островов, где появление ме-
ланистических форм совпало с периодом увлечения коллекцио-
нированием бабочек.
На Британских островах обитает около 780 видов бабочек и
примерно у 100 видов налюдается один и тот же процесс замеще-
ния их сложной и специализированной окраски более темной.
Протекает это во всех случаях в основном одинаково. Промышлен-
ная революция XIX в. вызвала изменения естественной среды
обитания в крупных промышленных городах и вокруг них. Глав-
ным изменением было общее потемнение среды, обусловленное
частично отложением копоти, а частично — исчезновением свет-
лых лишайников, покрывавших стволы деревьев. Светлоокрашен-
ные насекомые, спасавшиеся от хищников благодаря тому, что
они сливались со светлым фоном, на темном фоне стали замет-
ными, в отличие от темных мутантов. А поэтому птицы, которые
прежде находили и выедали темных особей, теперь стали нахо-
дить и выедать главным образом светлых особей, так что коэф-
фициенты отбора для темных и светлых бабочек обменялись значе-
ниями.
Для того чтобы это объяснение оказалось правильным, необ-
ходимы два условия: 1) бабочки должны действительно выбирать
тот фон, который соответствует их окраске; 2) различие между
светлой и темной формами должно иметь простую генетическую
основу. Кеттлуэлл (Kettlewell) помещал темных и светлых осо-
бей березовой пяденицы (Biston betularia) в бочонок, обитый из-
нутри материей в черную и белую полоску, и наблюдал за тем,
куда они садились. Как видно из табл. 8.4, темные бабочки выби-
рали темный фон, а светлые — светлый, чаще, чем можно было
ожидать по случайным причинам. В природе Кеттлуэлл тщательно
регистрировал места отдыха темной и светлой форм бабочки
Ectropis consonaria и также обнаружил сильную корреляцию меж-
214
Глава 8
Таблица 8.4
Выбор фона разными формами березовой пяденицы (Biston betularia) в лаборатории (Kettlewell, 1973)
Фон Цветная морфа бабочки Всего
черная (carbonaria) белая (typica)
Черный 38 20 58
Белый 21 39 60
Всего 59 59 118
Х2= 10,9 Р=0,001
Таблица 8.5
Выбор фона разными формами бабочки
Ectropis consonaria в природе (Kettlewell, 1973)
Фон Цветная морфа бабочки Всего
черная (nigra) белая (typical
Черный 15 4 19
Белый 5 22 27
Всего Хг= 14,202 20 Р=0,001 26 46
ду окраской бабочек и окраской фона, который они выбирают
(табл. 8.5).
В более чем 90% случаев индустриального меланизма темная
окраска обусловлена одним аллелем, и меланизм наследуется как
доминантный менделирующий признак. Известны, однако, случаи,
имеющие более сложную генетическую основу. У монашенки
Lymantria monacha окраска самых темных форм контролируется
тремя генами. Другие данные показывают, что, хотя генетический
контроль этого признака в своей основе прост, его проявление
может изменяться и изменяется под действием других генов, вхо-
дящих в состав генома.
Исходя из того, что светлые и темные формы выбирают фон,
на котором они отдыхают днем, в соответствии со своей окрас-
кой (поведение, обеспечивающее им некоторую защиту от насе-
комоядных птиц), относительное распределение разных фонов
должно позволить делать предсказания об относительном обилии
в природе меланистических и немеланистических форм. В общем
это справедливо, однако в некоторых районах частота меланис-
тов оказалась выше, а в некоторых других — ниже ожидавшейся,
что вызвало необходимость дополнительных объяснений. И в са-
Рекомбинация
215
мом деле, дальнейшее изучение показало, что у меланистических
личинок процент выживания выше, чем у светлых; что приспособ-
ленность у гетерозигот выше, чем у гомозигот; что бабочки из
промышленных районов сносятся ветром в сельские районы; и
многое другое.
На протяжении более пятидесяти лет проводились скрещива-
ния между этими двумя формами. Напомним, что эти формы раз-
личаются по одному гену и поэтому в потомстве следовало ожи-
дать соотношения 1:1. Между тем среди потомков от скрещива-
ний, проводившихся в 1900—1905 гг., было примерно 47% мела-
нистов, а от скрещиваний 1953—1956 гг.— примерно 62%! Это
отклонение от ожидавшегося соотношения 1 : 1 можно объяснить
как результат адаптации. Когда в популяцию был введен ген
меланизма, он, по-видимому, не вполне «вписался» в биохимичес-
кий арсенал животного. К 1950 г., когда меланистическая форма
стала преобладающей, хуже функционировать стали светлые фор-
мы. Согласно такому истолкованию, в популяции одновременно
действовал отбор двух типов. С одной стороны, изменение среды
привело к снижению смертности меланистических форм под давле-
нием хищников. С другой стороны, происходили небольшие генети-
ческие изменения, повышавшие выживание бабочек с геном мела-
низма (и предположительно понижавшие выживание бабочек с
нормальным геном).
В гл. 4 указывалось, что каждый ген оказывает влияние на
многие признаки организма, хотя наиболее ярко может быть вы-
ражено его воздействие на какой-либо один признак. В свою
очередь каждый признак испытывает влияние многих генов. Не-
удивительно поэтому, что введение в популяцию нового аллеля в
результате мутации и естественного отбора вызывает ряд «подго-
нок» в других локусах. К сожалению, мы покз располагаем лишь
косвенными данными. Изучение адаптации на биохимическом
уровне, представляющее собой весьма многообещающую область
исследований, быть может, даст в конечном счете некоторые све-
дения относительно степени коадаптации генов внутри данной по-
пуляции.
КРАТКИЕ ВЫВОДЫ
В этой главе мы рассмотрели рекомбинацию и показали, что
она представляет собой фактор, противодействующий факторам,
снижающим генетическую изменчивость в популяциях. Поддержа-
ние генетической изменчивости сопряжено, однако, с определенны-
ми расходами. Эти расходы — утрата популяцией гомозиготных
генотипов. Популяция может выдержать такие расходы лишь при
условии, что они 'будут уравновешиваться некоторым преимущес-
твом, а именно более высокой вероятностью выживания гетеро-
зиготных генотипов и их изменчивого потомства. Поскольку рас-
216
Глава 8
ходы и преимущества у разных видов различны, следует ожидать,
что у разных видов степень генетической изменчивости будет раз-
личной. В следующей главе рассматриваются имеющиеся в на-
стоящее время данные относительно генетической изменчивости в
природных популяциях растений и животных.
ЛИТЕРАТУРА
Общие вопросы
Frankel R., Galan Е. 1977. Pollination Mechanisms, Reproduction and Plant
Breeding, Berlin, Springer-Verlag.
Hawkes J. G. (ed.) 1966. Reproductive Biology and Taxonomy of Vascular Plants,
Oxford, Pergamon Press.
Jain S. K. 1976. The evolution of inbreeding in plants, Ann. Rev. Ecol. Syst., 7,
469—496.
Kettlewell B. 1973. The Evolution of Melanism: The Study of a Recurring Necessity,
Oxford, Clarendon Press.
Wallace B. 1970. Genetic Load: Its Biological and Conceptual Aspects, Englewood
Cliffs, N. J., Prentice-Hall.
Zimmering S., Sandler L., Nicoletti B. 1970. Mechanisms of meiotic drive, Ann. Rev.
Genetics, 4, 409—436.
Специальные статьи и обзоры
Carson Н. L. 1957. The species as a field for gene recombination. In: E. Mayr (ed.).
The Species Problem, Washington, American Association for the Advancement
of Science.
Dobzhansku T., Pavlovsky 0. 1953. Ideterminate outcome of certain experiments on
Drosophila populations, Evolution, 7, 198—210.
Grant V. 1958. The regulation of recombination in plants, Cold Spring Harbor Sym-
posia on Quantitative Biology, 23, 337—363.
Maguire M. P. 1963. High transmission of a Tripsacum chromosome in corn, Gene-
tics, 48, 1185—1194.
Muller H. J. 1950. Our load of mutations, Am. J. Hum. Gen., 2, 111—176.
Prakash S., Lewontin R. C. 1968. A molecular approach to the study of genic hete-
rozygosity in natural populations, III. Direct evidence of coadaptation in gene
arrangements of Drosophila. Proc. Nat. Acad. Sci., 59, 398—405.
Prakash S., Lewontin R. C. 1971. A molecular approach to the study of genic hete-
rozygosity in natural populations, V. Further direct evidence of coadaptation in
inversions of Drosophila, Genetics, 69, 405—408.
Rick С. M. 1959. Non-random gene distribution among tomato chromosomes, Proc.
Nat. Acad. Sci., 45, 1515—1519.
Sandler L., Novitscki E. 1957. Meiotic drive as an evolutionary force, Am. Nat., 91,
105—110.
Solbrig О. T. 1977. On the relative advantage of cross- and self-fertilization, Ann.
Mo. Bot. Carden, 63, 262—276.
Williams G. C. 1975. Sex and Evolution, Princeton, N. J., Princeton University
Press.
Wright S., Dobzhansky T. 1946. Genetics of natural populations. XII. Experimental
reproduction of some of the changes caused by natural selection in certain po-
pulations of Drosophila pseudoobscura, Genetics, 31, 125—156.
Глава 9
Генетическая изменчивость
в природных популяциях
Для того чтобы мог происходить естественный отбор, в попу-
ляции должен иметься запас наследуемой изменчивости. В пре-
дыдущих главах мы показали, что теория Менделя дает такую
модель наследственности, которая удовлетворительным образом
объясняет возникновение изменчивости и ее поддержание. Мы по-
казали также, что зависимости между фенотипом и генотипом не
являются однозначными, и что в настоящее время обычно нет воз-
можности точно определить связь между ними. Однако мы при-
вели несколько примеров (эволюция меланистических форм у ба-
бочки Biston betularia, развитие устойчивости к пестицидам у
комнатной мухи и к штаммам вируса гриппа у человека), в кото-
рых связь между генетическим изменением и отбором на феноти-
пическом уровне достаточно ясна и однозначна. Но не всегда воз-
можно объяснить наблюдаемую изменчивость так четко, как в
приводившихся до сих пор примерах. В данной главе рассматри-
ваются несколько глубже некоторые типы генетической изменчи-
вости, наблюдаемой в природных популяциях, в связи с чем изла-
гаются и обсуждаются некоторые гипотезы, выдвинутые за послед-
нее время для объяснения этой изменчивости.
Отдельные особи, составляющие природные популяции, отли-
чаются одна от другой как по количественным, так и по качес-
твенным признакам. Часть этой изменчивости носит генетический
характер, а часть обусловлена влиянием среды. В тех случаях,
когда какой-либо количественный признак подвергается сильному
отбору, следует ожидать, что генетическая компонента дисперсии
(см. гл. 4) мала, и изменчивость обусловлена главным образом
влиянием среды. Например, у коров фризской породы наследуе-
мость [/i2= Vg/(Vg+ Ve)] количества белых пятен очень велика
(0,95). Это признак, по которому животноводы не ведут активной
селекции. Однако наследуемость удойности составляет 0,3, а час-
тоты зачатия при первой случке — только 0,01. Любую корову, не
зачавшую при первой случке, обычно удаляют из стада, т. е. се-
лекция по этому признаку ведется с максимально возможной ин-
тенсивностью. Удойности также уделяется крайне большое внима-
ние, однако, ввиду того что селекцию можно начать лишь после
того как корова давала молоко в течение одного-двух лет, отбор
приходится вести только посредством отбора самцов; т. е. в каче-
218
Глава 9
стве производителей в селекционных программах используются
лишь бычки, рожденные высокоудойными коровами. Вследствие
этого в стадах сохраняется более высокая генетическая изменчи-
вость по удойности, чем по частоте зачатия. По этой же причине
в природных популяциях следует ожидать низкой генетической из-
менчивости по признакам, подверженным сильному давлению со
стороны отбора. Это находит формальное выражение в фишеров-
ской основной теореме естественного отбора (см. гл. 6). Подобные
рассуждения исходят из допущения, что среда довольно однород-
на как в пространстве, так и во времени и что интенсивность от-
бора во времени более или менее постоянна. Эти условия не обя-
зательно выполняются, и в настоящее время принято считать, что
большинство организмов живет в среде, изменяющейся и во вре-
мени и в пространстве. Дополнительное осложнение состоит в
том, что когда наиболее приспособленным генотипом оказывается
гетерозигота, популяция всегда будет обладать генетической из-
менчивостью. Все это делает невозможным предсказание уровня
генетической изменчивости в популяциях. В таких условиях даже
интерпретировать наблюдаемую генетическую изменчивость очень
трудно.
ТИПЫ ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
В ПОПУЛЯЦИЯХ РАСТЕНИЙ И ЖИВОТНЫХ
Уровень генетической изменчивости по большинству признаков
в популяциях большинства организмов до сих пор остается неиз-
вестным, хотя за последние 10 лет достигнуты значительные успе-
хи в разработке методов изучения типов генетической изменчи-
вости в природных популяциях.
Поскольку отбор действует на фенотипическом уровне, на
первый взгляд может показаться, что для выяснения степени из-
менчивости в данной популяции достаточно измерить фенотипи-
ческую изменчивость. Однако при таком подходе нельзя устано-
вить, какая часть наблюдаемой изменчивости является генетичес-
кой, а именно это нас интересует. Произведя соответствующие
скрещивания и статистическую обработку полученных результатов,
можно оценить наследуемость данного признака, как было пока-
зано в гл. 4. Этот громоздкий метод, поглощающий к тому же
массу времени, применяется почти исключительно в программах
по искусственному разведению. В природных ситуациях генетики
старались заменить его изучением признаков с простой наслед-
ственной основой (предпочтительно контролируемых одним геном),
у которых альтернативные аллели обусловливают ярко выражен-
ные фенотипические эффекты, легко поддающиеся измерению или
подсчету (например, разные цветовые формы).
Русский генетик Н. В. Тимофеев-Ресовский в 1940 г. опублико-
вал статью о цветовых формах («морфах») берлинской популяции
божьей коровки Adalia bipunctata. В общей сложности он обна-
Генетическая изменчивость в природных популяциях
219
Рис. 9.1. Морфы божьей коровки Adalia bipunctata. А—В. «Красные» морфы: А —
typica; Б и В — annulata. Г—3. Черные 'морфы: Г — sublunata; Д — 2-maculata;
Е—4-maculata; Ж — б-postulata; 3 — lunigera. (Тимофеев-Ресовский, 1940)
Рис. 9.2. Процент черных морф Adalia bipunctata при весенних (апрель) и осен-
них (октябрь) учетах. Два последних столбца — частота аллелей, определяющих
черную морфу. (Тимофеев-Ресовский, 1940)
ружил в этой популяции 8 разных типов окраски (рис. 9.1), прав-
да, 3 из них встречались гораздо чаще, чем 5 остальных. Для
трех морф характерны черные пятна на красном фоне («красные»
формы), а для пяти — красные пятна на черном фоне («черные»
формы). Путем соответствующих экспериментов было установле-
но, что цвет фона определяется простым доминированием гена
220
Глава 9
Таблица 9.1
Весенняя и осенняя численности красных и черных морф в популяциях
божьей коровки Adalia bipunctata (Тимофеев-Ресовский, 1940)
Весна Осень
Год Морфы число | % число 0 ' ,0
1930 Черные 176 29,1 783 62,9
Красные 428 70,9 461 37,1
Итого 604 100,0 1244 100,0
1931 Черные 334 43,0 622 55,7
Красные 443 57,0 494 44,3
Итого 777 100,0 1116 100,0
1933 Черные 213 37,8 675 66,8
Красные 351 62,2 334 33,2
Итого 564 100,0 1011 100,0
1934 Черные 149 34,5 708 57,2
Красные 283 65,5 529 42,8
Итого 432 100,0 1237 100,0
1938 Черные 192 40,8 432 49,1
Красные 279 59,2 448 50,9
Итого 471 100,0 880 100,0
Всего Черные 1064 37,4 3220 58,7
Красные 1784 62,6 2268 41,3
Итого 2448 100,0 5488 100,0
красной окраски над геном черной, а точный характер пятен —
системой множественных аллелей. Тимофеев-Ресовский изучал эту
популяцию на протяжении 8 лет, уделяя главное внимание соот-
ношению красных и черных форм. Ему удалось установить, что
красные формы более многочисленны в зимние месяцы, тогда как
численность черных форм увеличивается летом (табл. 9.1 и рис.
9.2). Он установил также (табл. 9.2), что выживаемость красных
форм зимой выше. Очевидно, черные формы либо быстрее размно-
жаются, либо лучше выживают в летние месяцы.
Другим примером фенотипической изменчивости, на этот раз
у растения, служит изменчивость габитуса полевицы Agrostis
stolonifera в Англии, изучавшаяся Дж. Л. Эстоном (J. L. Aston)
и А. Д. Брэдшоу (A. D. Bradshaw). В местообитаниях, удаленных
от моря, это растение сильно разрастается при помощи длинных
подземных побегов — столонов (рис. 9.3), тогда как в прибрежных
Генетическая изменчивость в природных популяциях 2'21
районах оно нередко имеет вид маленького плотного пучка с
множеством коротких столонов. Эти различия сохраняются, если
пересадить растения в одинаковые ус-
ловия на садовые делянки; раз-
личия имеются также у растений, выра-
щенных из семян, что свидетельствует об
их наследственном характере. Эстон
и Брэдшоу изучали растения из раз-
ных местностей с различной длиной
столонов. Полученные ими результаты
подытожены в табл. 9.3 и на рис. 9.4.
Обратите внимание на то, что длина
столонов зависит от места, на котором
растет растение, и что различия по это-
му признаку между популяциями могут
быть статистически значимыми. Еще бо-
лее интересен тот факт, что при нали-
чии резкого градиента физических усло-
вий наблюдается соответствующее рез-
кое различие в длине столонов; если же
Рис. 9.3. Схематическое изо-
бражение Agrostis stolonife-
га из внутренних районов.
Обратите внимание на со-
единяющие растения столо-
ны, по которым оно получи-
ло свое название.
условия изменяются постепенно, то из-
менения морфологии растения от одной
популяции к другой также происходят
постепенно. По-видимому, этому расте-
нию выгодно иметь короткие столоны на
прибрежных скалах и длинные — в ме-
стах, удаленных от моря.
ПОЛИМОРФИЗМ ОКРАСКИ В ПРИРОДНЫХ популяциях
САДОВОЙ УЛИТКИ СЕРАЕА NEMORALIS
Одним из классических примеров изменчивости служат узоры
окраски раковин у двух близкородственных видов садовых улиток—
Сераеа hortensis и С. nemoralis. Необычайная изменчивость окрас-
ки раковин у этих улиток привлекает внимание европейских гене-
тиков и эволюционистов уже на протяжении более чем 50 лет.
Особи вида С. nemoralis, изученного более детально, чем дру-
гой вид, различаются по цвету раковины (бурая, розовая пли
желтая), по наличию или отсутствию продольных полос и по чис-
лу этих полос (1—5); полосы могут быть темными или светлыми
наконец, перистом раковины может быть окрашенным или белым
(рис. 9.5). Генетические исследования показали, что цвет ракови-
ны определяется одним геном с тремя аллелями, причем аллель
бурой окраски (YB) доминирует над аллелем розовой (Y) и оба
они доминируют над аллелем желтой окраски (у). Независимый,
но тесно связанный с геном окраски ген, определяющий наличие
или отсутствие полос, эпистатичен по отношению к генам, опре-
222
Глава 9
Рис. 9.4. Длина столонов в популяциях Agrostis stolonifera.
Выборки из популяции отбирали по трансектам, проведенным через склоны с по-
степенными (В, Г) и резкими (А, Б) изменениями высоты. Представлены средние
для популяции длины столона; для четырех популяций показана длина столонов
у отдельных растений из этих популяций. (Aston, Bradshaw, 1966)
£ Пастбище
I защищенное(9)
15 30 45 60
Вершина
берегового
обрыва (8)
J
60
Пастбище
открытое(5)-
15 30 45 60
Длина столонов в см
1 ст, с чл. ала от -
дельных значений1-4
Генетическая изменчивость в природных популяциях
223
деляющим характер полосатости. Так, улитки, несущие доминант-
ный аллель U, не имеют полос, тогда как у гомозиготных рецессивов
(UU) раковина полосатая. Число полос у улитки с генотипом uu
определяется отдельным геном, а еще один ген определяет нали-
чие или отсутствие пигмента в полосах. Таким образом цвет и
характер полосатости раковины у Сераеа nemoralis контролиру-
ются по крайней мере четырьмя главными генами, но возможно,
что число их еще больше.
Рис. 9.5. Раковины шести особей Сераеа nemoralis, собранные в парке Версаль-
ского дворца под Парижем.
Обратите внимание на различия в полосатости и в окраске перистома. Раковина,
изображенная справа, лишена полос.
Вид Сераеа nemoralis широко распространен в Западной Ев-
ропе— от северной части Испании и Италии до Англии. Недавно
он был завезен также в восточную часть ОША. Он образует обо-
собленные популяции, в которых насчитывается от нескольких
До двухсот особей и которые находятся на расстоянии от несколь-
ких метров до нескольких километров друг от друга. Обитают эти
улитки в живых изгородях, высокой траве, кустах и в лесной под-
стилке. Определение факторов эволюции, обусловливающих на-
блюдаемую изменчивость фенотипа раковины у улиток рода Се-
раеа, оказалось нелегкой задачей и вместе с тем послужило пре-
красной иллюстрацией сложности эволюционного процесса. Изла-
гаемые ниже данные взяты по большей части из работ англий-
ских эволюционистов А. Дж. Кэйна (A. J. Cain) и П. М. Шеппар-
224
Глава 9
Таблица 9.2
Переживание зимы красными и черными морфами божьей коровки
Adalia bipunctata (Тимофеев-Ресовский, 1940)
Год Морфы Исходное число Число выживших Выживание, %
1934 Черные 739 23 3,11
Красные 334 21 6,29
Итого 1073 44 4,10
1937 Черные 528 31 5,87
Красные 305 41 13,44
Итого 833 72 8,64
1938 Черные 578 24 4,15
Красные 405 54 13,33
Итого 983 78 7,93
Всего Черные 1845 78 4,23
Красные 1044 116 11,11
Итого 2889 194 6,72
да (Р. М. Sheppard), а также французского ученого Максима Ла-
мота (М. Lamotte).
Биолог-популяционист, изучающий тип изменчивости какого-
либо фенотипического признака, пытается сначала установить, на-
следуется ли этот признак. После того как это установлено, он
тщательно изучает характер изменчивости признака в природе,
стараясь найти корреляции между этим признаком и каким-либо
фактором среды, который мог бы играть роль фактора отбора.
Одновременно с полевыми исследованиями популяционист про-
водит лабораторные эксперименты, для того чтобы точно выяс-
нить, как реагирует популяция на тот или иной физический или
биологический фактор.
Мы уже упоминали о том, что, по данным генетических иссле-
дований, цвет и характер полосатости раковины Сераеа петога-
lis контролируются несколькими главными генами, а кроме того
на них действуют другие генные системы. Что касается корреля-
ции этих признаков с факторами среды, то тщательные наблюде-
ния с применением сложных статистических методов обработки
данных выявили несколько таких корреляций, однако этих резуль-
татов недостаточно, чтобы полностью объяснить существующие
типы изменчивости. Например, улитки с белым перистомом встре-
чаются только в некоторых популяциях Пиренеев и Ирландии,
т. е. на границе ареала вида, где они существуют с особями, име-
Генетическая изменчивость в природных популяциях
225
Таблица 9.3
Изменчивость средней длины столонов у разных популяций
Agrostis stolonifera в прибрежных местообитаниях
(Aston, Bradshaw, 1966)
Популяция Характер местообитания Длина столона, см
1 Основание берегового склона; открытое 14,5
15 Грейт-Орме; открытое 15,9
8 Бровка берегового склона; открытое 16,5
3 Береговой склон; открытое и сухое 17,0
6 Крутой утес; открытое и сухое 18,0
7 Осиоваиие берегового склона, открытое и влаж-
иое 24,1
16 Литтл-Орме; пастбище на бровке склона 32,2
2 Ручей вблизи моря; открытое 33,6
5 Пастбище; открытое 35,5
18 Нэш-Волли (Ь) 41,9
11 Пастбище; умеренно открытое 42,3
14 Береговой склон Бомерис 44,0
17 Нэш-Волли (а) 45,4
10 Пастбище; обычное 46,7
4 Ручей 47,5
9 Пастбище; защищенное 47,9
12 Ручей 48,7
20 Нэш-Волли (d) 49,8
19 Нэш-Волли (с) 51,2
13 Ручей 57,0
Ющими бурый перистом, которые преобладают по всему ареалу.
Что общего между условиями среды в Ирландии и на Пиренеях,
остается неясным.
Что касается бурых раковин, то с ними ситуация яснее. Осо-
бей с бурой раковиной нет почти ни в одной из популяций, оби-
тающих на юге Франции и в Средиземноморье; они редко встре-
чаются также в центральных и северных районах Франции, хотя
в буковых лесах этих районов частота их несколько выше. На
земле Рейнальд-Пфальц (ФРГ) и на Британских островах они ста-
новятся довольно многочисленными, составляя 10% всех особей.
Создается впечатление, что частота бурой окраски раковины свя-
зана с климатическими и микроклиматическими факторами. В
15—2044
226
Глава 9
пользу такой точки зрения свидетельствует также наблюдение о
том, что улиток с бурыми раковинами никогда не находят в мес-
тах, ярко освещаемых солнцем; они предпочитают затененные мес-
тообитания. Вместе с тем число полос не обнаруживает такой
строгой корреляции с факторами среды. Шеппард выявил извест-
ию -
80 -
-.О
°з 70 -
m 60 f-
О
8. 50 -
ф
£ 40 -
ф
£ 30 -
ф- Ьукосыелеса
э - Дубравы
Q-Смешанные листопадные леса
© - Живые изгороди
© - Г рубая растительность
О - Низкая трава
10 20 30 40 50 60 70 80 90 100
Раковины, в значительной степени лишенные полос,%
Рис. 9.6. График корреляции между процентом желтых раковин, процентом рако-
вин, в значительной мере лишенных полос (отсутствуют по крайней мере две верх-
ние полосы), и местообитанием колоний Сераеа nemoralis.
График показывает, каким образом естественный отбор воздействует на частоту
желтой окраски и тип полосатости. Так, в лесах с однородной подстилкой из бу-
рых опавших листьев (особенно в буковых лесах) велика доля менее заметных,
в значительной мере лишенных полос бурых или розовых раковин. В отличие от
этого в местообитаниях с грубой растительностью (в том числе в живых изгоро-
дях) велика доля желтых полосатых раковин, которые здесь менее заметны. Не-
сколько колоний, составляющих, казалось бы, исключение, также удовлетворяют
этому объяснению; так, популяция смешанного листопадного леса, в котором у
80% особей были желтые раковины, обитала в лесу с особыми условиями, а имен-
но: вся почва в этом лесу была покрыта не бурыми листьями, а низкой зеленой
травой; отсюда и высокая частота желтых, почти бесполосых раковин. (Sheppard,
1958)
ную степень корреляции (рис. 9.6) между частотой разных типов
полосатости и микроместообитанием. Он показал, что в лесах, где
поверхность земли бурая от гниющих листьев, и особенно там, где
она достаточно однородна на вид, наиболее часто встречаются
бесполосые бурые и розовые раковины, а также розовые раковины
с одной полосой, тогда как в живых изгородях и в грубой зеленой
растительности чаще встречаются желтые раковины с пятью по-
лосами. Кроме того, он установил, что чем меньше зеленого в
фоне, тем ниже доля желтых раковин, и чем более однороден фон,
тем чаще встречаются бесполосые формы и формы с одной поло-
Генетическая изменчивость в природных популяциях 227
сой. Как показали работы Ламота, аналогичные корреляции су-
ществуют во Франции, но, по-видимому, с многочисленными иск-
лючениями. Возможной причиной различий в распределении ули-
ток по биотопам во Франции и на Британских островах служит
то, что во Франции популяции Сераеа nemoralis часто встречают-
ся вдоль изгородей, где сосуществуют разные виды растений, или
в окультуренных лесах, или на лугах с разбросанными кое-где
деревьями и кустами, тогда как на Британских островах они встре-
чаются в густых лесах и на лугах.
Фактором отбора, обусловливающим такое распределение, оче-
видно, служат птицы, питающиеся этими моллюсками. В Англии
поедающий улиток певчий дрозд (Tardus ericetorum) разбивает
раковины о камни. Подсчитывая разбитые раковины разного цве-
та и с разным числом полос и сопоставляя полученные числа с
соотношением раковин разного типа в популяции живых улиток,
удалось установить, что в местообитаниях с пестрым фоном, таких,
как живые изгороди и грубая травянистая растительность, поло-
сатые особи лучше защищены от хищников, на однородном же
фоне, например в лесах, лучшую защиту создает отсутствие по-
лос. Можно также показать, что особи с желтой раковиной (име-
ющей зеленоватый оттенок, когда в ней находится улитка) реже
оказываются жертвами хищников, на зеленом фоне, а на буром
фоне, например на голой земле на сухих листьях, преимущество
имеют особи с бурой или розовой раковиной. Сходные результа-
ты получил Ламот во Франции. Таким образом, выедание птицами
представляет собой важный фактор в поддержании наблюдаемого
полиморфизма. Однако, как показывает далеко не абсолютная
корреляция между характером полосатости и типом местообита-
ния, это не единственный фактор. Другим фактором отбора слу-
жат, возможно, физические условия среды, поскольку разные
формы улиток обладают различной устойчивостью к разнооб-
разным факторам среды. Например, бесполосые особи, особенно
те, у которых раковина желтая, по-видимому, более устойчивы к
Таблица 9.4
Смертность среди улиток Сераеа nemoralis при содержании ,
в течение 40 ч при 35 °C (по данным Lamotte, 1960)
Характер полосатости Желтые Розовые Всего
ЧИСЛО изученных смертность, % число изученных смертность, % число изученных смертность %
Бесполосые 66 13,6 15 33,3 81 17,2
Одна полоса в поло- жении 3 45 26,6 26 42,3 71 32,3
Пять полос 39 38,4 22 36,3 61 37,7
15
228
Глава 9
высоким температурам, чем полосатые (табл. 9.4). Кроме того,
особи с желтыми раковинами более устойчивы к холоду, чем ро-
зовые, причем бесполосые более холодоустойчивы, чем полосатые.
Это обстоятельство, вероятно, имеет более важное значение, чем
устойчивость к жаре, потому что улитки обычно не имеют воз-
можности укрываться на зиму в убежищах. Корреляция, наблюда-
емая между частотой бесполосых раковин и июльскими темпера-
турами, а также между частотой желтых раковин и январскими
температурами, свидетельствует о том, что высокие и низкие тем-
пературы среды в самом деле могут играть роль селективного
фактора (табл. 9.5).
Таблица 9.5
Частота «бесполосых» и «желтых» раковин
у Сераеа nemoralis в зависимости от средних
температур июля и января соответственно
(Lamotte, 1960)
Средняя температура
июля, °C
16—18 18—19 19—20 20—21
21
Средняя частота бес-
полосых, % 22
24 26 29
30
Средняя температура
января, °C 4
4—2 2—0 0
Средняя частота жел-
тых, °/о 57,9 61,0
73,4
78,8
Итак, у Сераеа nemoralis было выявлено несколько корреля-
ций между фенотипической изменчивостью и факторами среды, что
частично объясняет наблюдаемые в природе типы изменчивости.
Тем не менее, даже если учесть кумулятивное действие этих фак-
торов, всегда остается некоторое не получившее объяснения оста-
точное разнообразие.
Число известных случаев полиморфизма по фенотипическим
признакам очень велико как у растений, так и у животных. Лишь
немногие организмы были изучены так тщательно, как Сераеа ne-
moralis, но подробное обследование показывает, что полиморфный
признак обычно коррелирует с изменчивыми физическими и (или)
биотическими факторами среды, хотя в большинстве случаев эта
корреляция далеко не абсолютна. Изменчивы, однако, не только
фенотипические, но также цитологические и молекулярные при-
знаки.
ПОЛИМОРФИЗМЫ ПО ХРОМОСОМНЫМ ИНВЕРСИЯМ У ДРОЗОФИЛЫ
Любопытная генетическая изменчивость обнаружена у разных
видов плодовой мушки (Drosophila). За немногими исключениями,
Генетическая изменчивость в природных популяциях
229
в популяциях всех видов дрозофилы имеются особи с одной или
несколькими хромосомными инверсиями (гл. 5) в одной, двух или
трех хромосомах. Эти инверсии были обнаружены еще в 1926 г.
Стертевантом (Sturtevant), но интенсивно изучать их стали лишь
в 30-х годах, после того как в слюнных железах плодовой мушки
были открыты гигантские политенные хромосомы (рис. 9.7). Это
Рис. 9.7. Политениая хромосома из слюнной железы Drosophila melanogaster.
(С любезного разрешения S. Elgin; прежде была опубликована в статье Sil-
ver L. М., Elgin S. С. R., 1976. A method for determination of the in situ distribu-
tion of chromosomal proteins, Proc. Nat. Acad. Sci. USA, 73, 423—427.)
очень облегчило изучение инверсий в природе. Первым генетиком,
занявшимся исследованием этого признака, был Феодосий Доб-
ржанский (Theodosius Dobzhansky).
Представим себе дрозофилу с генными последовательностями
ABCDEFGHI и AEDCBFGHI в двух гомологичных хромосомах.
Для того чтобы эти хромосомы могли конъюгировать в мейозе, одна
из них должна образовать петлю (рис. 9.8, А). Со временем в той
же хромосоме может возникнуть вторая инверсия. Если эта ин-
версия не захватывает участок, который уже был инвертирован,
то она называется независимой, и при конъюгации такой хромо-
сомы с нормальным гомологом можно будет наблюдать две петли
(рис. 9.8,5). Однако если вторая инверсия происходит внутри пер-
вой, то во время мейоза гетерозиготы по этой инверсии образуют
петлю внутри первой петли (рис. 9.8,В). Такие инверсии называют
вставочными. Наконец, вторая инверсия может одним своим концом
находиться внутри первой инверсии, а другим — вне ее. Такие ин-
версии называют перекрывающимися (рис. 9.8, Г), и они во время
мейоза образуют восьмерку. Поскольку в хромосомах слюнных
желез дрозофилы наблюдается однозначно упорядоченное чередо-
вание темных и светлых полос, можно точно определить, какой
именно участок хромосомы инвертирован. Тщательно выяснив
230
Глава 9
порядок возникновения инверсий, можно установить их вероят-
ную филогению, или эволюционное происхождение, при условии
что все они связаны между собой перекрывающимися инверсия-
ми, как это изображено на рис. 9.9.
ABCDEFGHI
▲ ▲
AEDCBFGHI
ABCDEFGHI
AEDCBFGHI
ABCDEFGHI
AEDCBFGHI
AEDCBFHGI
ABCDEFGHI
AEDCBFHGI
ABCDEFGHI
AEDCBFGHI
AECDBFGHI
ABCDEFGHI
AECDBFGHI
ABCDEFGHI
AEDCBFGHI
AEHGFBCDI
ABCDEFGHI
AEHGFBCDI
Рис. 9.8. Конъюгация хромосом у особей дрозофилы, гетерозиготных по инвер-
сиям.
А. Одна инверсия. Б. Независимые инверсии. В. Вставочные инверсии. Г. Пере-
крывающиеся инверсии. (Dobzhansky Т„ 1970. Genetics of the Evolutionary Pro-
cess, New York, Columbia Univ. Press.)
Допустим, что имеются три перекрывающиеся инверсии с по-
следовательностями полос 1 ABCDEFGHI, AEDCBFGHI и
AEHGFBCDI. Мы хотим выяснить возможный и притом кратчай-
ший путь их эволюции, т. е. путь, состоящий из минимального
Генетическая изменчивость в природных популяциях 231
числа этапов. Первая инверсия может возникнуть из второй (или
наоборот) за один этап; таким же образом вторая может возник-
нуть из третьей; однако, для того чтобы третья произошла из
первой (или первая из третьей), необходимо по крайней мере
два этапа. Иными словами, наиболее экономными последователь-
ностями следует считать 1->2->3 или 3->2->1, или 1+-2->3.
Пользуясь таким способом, можно расположить все известные
инверсии в данной хромосоме у данного вида в филогенетическую
последовательность. На рис. 9.10
изображено полученное таким
образом филогенетическое древо
для трех очень близко родствен-
ных видов — Drosophila pseudo-
obscura, D. persimilis и D. rtiiran-
da. Обратите внимание на то,
что у двух видов (D. pseudoob-
scura и D. persimilis) имеется
общая хромосомная последова-
тельность (Стандарт), а одна
важная последовательность (Ги-
потетическая) никогда не наблю-
дается. Существование предска-
занных ранее последовательнос-
тей было подтверждено данны-
ми, собранными позднее в при-
роде Добржанским, что придало
достоверность этой филогенети-
ческой схеме.
Изменчивость инверсий в
природных популяциях наиболее
ное древо.
Последовательность Б может дать на-
чало последовательностям А и В в
результате простой инверсии. Для то-
го чтобы последовательность В могла
произойти из А, понадобилось бы две
инверсии, и наоборот.
подробно изучалась у вида
D. pseudoobscura. Было установлено, что все популяции поли-
морфны по инверсиям в третьей хромосоме; что нет таких после-
довательностей, которые встречались бы на всем протяжении
ареала распространения этого вида; что некоторые последователь-
ности (Стандарт, Чирикахуа, Эрроухед) встречаются довольно
часто, тогда как другие были обнаружены лишь в какой-либо од-
ной популяции; и что поэтому ни одна популяция не содержит
полного комплекта всех последовательностей, хотя в некоторых
местностях было обнаружено до 8 разных последовательностей.
В природе обнаружены как структурные гомозиготы (мухи,
обе гомологичные хромосомы которых содержат одинаковые по-
следовательности), так и структурные гетерозиготы. Еще больший
интерес представляет тот факт, что частота данной инверсии
варьирует в различных местностях, а также в пределах одной
местности как в течение одного года, так и на протяжении не-
скольких лет (рис. 9.11 и 9.12). Однако, несмотря на то что эти
232
Глава 9
ГИПОТЕТИЧЕСКАЯ
Рис. 9.10. Филогенетические связи Drosophila pseudoobscura, D. persimilis и
D. miranda, устанавливаемые на основе последовательностей генов в третьей хро-
мосоме. (Dobzhansky, 1970)
Генетическая изменчивость в природных популяциях 233
хромосомные инверсии изучаются уже в течение примерно 40 лет
и в природе, и в лабораторных условиях, мы все еще не можем
точно сказать, какой именно фактор или факторы обусловливают
их сохранение или изменение частоты, наблюдаемые в природ-
ных популяциях. В предыдущей главе мы (привели данные, сви-
детельствующие о некоторых возможных преимуществах, создава-
емых этими инверсиями.
Рис. 9.11. Частоты хромосом ST в выборках из популяций Drosophila pseud»-
obscura, отбиравшихся около 1940 г. (черные столбцы), в 1957 г. (столбцы, по-
крытые точками) и в 1963—1965 гг. (белые столбцы); белый ромбик означает, что
в данный период выборки не отбирались. На шкале в левом нижнем углу показа-
ны эти частоты в процентах. (Dobzhansky et al., 1966)
ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Сложная наследственная основа фенотипической изменчивости
затрудняет понимание участвующих в этом генетических и эволю-
ционных факторов. Если бы статистические оценки фенотипичес-
234
Глава 9
ких признаков можно было заменить частотами аллелей, опреде-
ляющих данный признак, то эволюционная картина стала бы, воз-
можно, яснее. Поэтому генетики стараются оценить генные часто-
ты непосредственно в популяциях. Новый метод, впервые исполь-
зованный для этой цели в 1966 г., позволяет легко оценивать ген-
ные частоты в популяции.
Март Апр. Май Июнь Июль Авг. Сент. Окт.
Рис 9.12. Сезонные изменения частоты хромосомных инверсий у Drosophila pseu-
doobscura.
Регулярные сезонные колебания частоты хромосом ST и СН наблюдались в од-
ной местности на горах Сан-Джасинто (Калифорния) в 1939—1946 гг. (Т. Dob-
zhansky, 1970. Genetics of the Evolutionary Process, New York, Columbia Univ.
Press.)
Метод электрофореза в геле
Если смесь ферментов или других белков, нанесенных на ка-
кую-либо подложку — полоску бумаги, ацетатцеллюлозу или же
крахмальный гель,— поместить в электрическое поле, то отдель-
ные ферменты и другие белки начинают перемещаться к катоду
или аноду, в зависимости от того, несут ли они положительный
или отрицательный заряд; если же белки незаряжены, то они ос-
таются на месте (рис. 9.13). Скорость движения к полюсам за-
висит от величины заряда молекулы (чем больше заряд, тем вы-
ше скорость) и от сопротивления, оказываемого подложкой пере-
мещению фермента. Грубо говоря, сопротивление пропорциональ-
но величине молекул фермента. Если по прошествии некоторого
времени выключить ток, а подложку (например, гель) зафикси-
ровать и окрасить, то на ней появляется ряд окрашенных полос,
каждая из которых соответствует какому-либо одному ферменту
(а иногда двум или нескольким ферментам, перемещающимся с
одинаковой скоростью). Вместо фиксации и окраски можно инку-
бировать подложку с подходящим субстратом и каким-либо рас-
творимым красителем, который выпадает в осадок только в том
случае, если тот или иной из содержащихся в смеси ферментов
взаимодействует с субстратом. Например, исследователь, желаю-
Генетическая изменчивость в природных популяциях
235
щий установить, содержит ли исходная смесь белков фермент
эстеразу, инкубирует подложку с а-нафтилацетатом и раствори-
мым красителем — прочным синим. Эстераза разлагает а-нафти-
Рис. 9.13. Электрофореграмма белков, выделенных из семян Agropyron scabrigla-
me и A. tilcarense, собранных в разных частях Аргентины.
Электрофорез проводился в акриламидном геле. (С любезного разрешения
J. Н. Hunziker.)
лацетат на уксусную кислоту и а-нафтил, используя в качестве ис-
точника электронов краситель прочный синий,который при окисле-
нии становится нерастворимым и.выпадает в осадок. Через неко-
торое время подложку фиксируют и исследователь может уста-
новить скорость передвижения белков, обладающих той или иной
специфической ферментативной активностью. Каждая данная по-
лоса может соответствовать одному ферменту или же нескольким
ферментам, перемещающимся с одинаковыми скоростями. Этот
биохимический метод, как мы сейчас убедимся, сыграл огромную
236
Глава 9
роль в развитии популяционной генетики. Однако сначала следует
рассмотреть концепции, лежащие в основе использования этого
метода для оценки генных частот в популяциях.
В 1953 г. было установлено, что ДНК представляет собой ма-
териальную основу наследственности. В течение последующих 10
Рис. 9.14. Электрофореграмма
лейцииаминопептидазы (ЛАП)
трех линий кукурузы: 1 — Р39;
2— гибриды F, между Р39 и
АА4; 3 —АА4.
Полосы В и С обнаружены у
всех линий и они не подверже-
ны генетической изменчивости,
а полосы А и D имеют две фор-
мы, причем у гибридов обнару-
жены как та, так и другая.
Стрелка показывает направле-
ние миграции в геле. (Beckman
et al., 1964)
лет молекулярные биологи убеди-
лись, что молекула ДНК действует
как код, состоящий из трехбуквенных
слов (гл. 4). Каждое такое слово в
результате ряда этапов переводится
в аминокислоту, занимающую строго
определенное место в молекуле како-
го-либо фермента или структурного
белка цитоплазмы. Цистрон, пред-
ставляющий собой последователь-
ность таких слов в молекуле ДНК, ко-
торые совместно кодируют опреде-
ленную полипептидную цепь, служит
на молекулярном уровне наибольшим
приближением к гену классической
генетики. Ферменты могут состоять
из одной или нескольких полипептид-
ных цепей. Иногда эти цепи идентич-
ны и кодируются одним и тем же цис-
троном, а иногда они различны и ко-
дируются несколькими разными цис-
тронами ( = генами). Примером по-
следней ситуации служит молекула
гемоглобина, подробно рассмотренная
в гл. 5.
Когда в каком-либо локусе возни-
кает точковая мутация, одна или несколько аминокислот заменя-
ются другими аминокислотами. Это может сопровождаться (хотя
и не обязательно) изменением заряда белка, его конформации,
либо как тем, так и другим; в результате получается другой бе-
лок, перемещающийся с другой скоростью, но обладающий в ос-
новном такой же ферментативной функцией, хотя, возможно, ины-
ми физиологическими свойствами. Ферменты, имеющие одинако-
вые биохимические функции, но перемещающиеся в электричес-
ком поле с разными скоростями, называют аллоферментами или
изоферментами. Поскольку аллоферменты — главные продукты ге-
на, их наследование подчинено законам Менделя. Это было пока-
зано в 1964 г. Бекманом (Beckman), Скандалиосом (Scandalios)
и Брюбейкером (Brewbafker) на примере фермента лейцинамино-
пептидазы (ЛАП), выделенного из кукурузы (табл. 9.6 и
.рис. 9.14).
Вооружившись этим методом, биолог-популяционист может оп-
Генетическая изменчивость в природных популяциях
Таблица 9.6
Генетический контроль вариантов A-формы фермента ЛАП1'
у кукурузы (Beckman, Scandalios, Brewbaker, 1964)
Женская родитель- ская особь Мужская родитель- ская особь Полосы, соответствующие A-форме ЛАП у потомков
F (F/F) F4-S (F/S) S (S/S) всего
АА4 (F/F) АА4 (F/F) 30 0 0 30
Р39 (S/S) Р39 (S/S) 0 0 30 30
АА4 (F/F) Р39 (S/S) 0 30 0 30
Р39 (S/S) АА4 (F/F) 0 30 0 30
(АА4ХР39) (F/S) АА4 (F/F) 16 14 0 30
АА4 (F/F) (АА4ХР39) (F/S) 17 13 0 30
(АА4ХР39) (F/S) Р39 (S/S) 0 10 20 30
Р39 (S/S) (АА4ХР39) (F/S) 0 12 18 30
(АА4ХР39) (F/S) (АА4ХР39) (F/S) 20 22 18 60
*) A-форма фермента ЛАП имеет два варианта — быстрый (F) и медленный (S). Ли-
ния АА4 кукурузы дает только быструю полосу, а линия Р39 — только медленную. Гибрид
между ними дает обе полосы, а возвратные скрещивания с одной из родительских линий —
соотношение 1 : 1 в соответствии с менделевским расщеплением прн моногибридном скрещи-
вании; приведенные в скобках генотипы выведены из полученных результатов.
ределять электрофоретическую подвижность какого-либо опреде-
ленного фермента у большого числа особей в данной популяции
и, определяя частоту аллоферментов этого фермента, может оце-
нить частоту в данном локусе разных аллелей, вызывающих такие
модификации этого фермента, которые можно выявить методом
электрофореза. Чтобы убедиться в том, что две разные полосы,
обнаруженные у двух особей, относятся к ферментам, кодируемым
аллелями одного и того же гена, исследователю достаточно произ-
вести лишь одно скрещивание и проделать анализ потомков. Ес-
ли эти две полосы дадут расщепление в отношении 1:2:1 (гомо-
зигота быстрая : гетерозигота : гомозигота медленная), то расс-
матриваемые аллоферменты кодируются аллелями одного гена.
Таким образом, электрофорез в гелях дает довольно легкий
способ непосредственного определения частоты аллелей в попу-
ляциях. Он был введен в 1966 г. независимо английским генети-
ком Харрисом (Harris), американским биохимиком Хэбби (Hub-
by) и американским генетиком Левонтином (Lewontin).
С тех пор определения изоферментов проводятся у всевозможных
видов — от глубоководных беспозвоночных и разных видов дро-
зофилы до человека; от широкораспространенных до редких ви-
дов; от видов, для которых обычно случайное скрещивание, до
238
Глава 9
Генетическая изменчивость в природных популяциях 239
самооплодотворяющихся видов и т. д. Собрано огромное количе-
ство данных, но интерпретировать их очень трудно.
Одна из трудностей сопряжена с тем, что не все изоферменты
данного фермента, обнаруженные в изучаемой популяции, непре-
менно обусловлены изменчивостью аллелей в локусе, кодирующем
этот фермент. Альтернативная гипотеза состоит в том, что разные
изоферменты могут быть результатом широких модификаций од-
ного фермента под действием продуктов активности дру-
гих генов. Это явление называют посттрансляционной мо-
дификацией (рис. 9.15). Такая посттрансляционная модификация
обладает многими чертами, присущими единичному локусу со мно-
гими аллелями: 1) она наследуется; 2) при наличии нескольких
активных модифицирующих локусов может образоваться очень
много вариантных форм фермента; 3) если модифицирующие ло-
кусы действуют независимо (т. е. вызывают изменение различных
свойств фермента), то их эффект будет мультипликативным, что
приведет к асимметричному распределению по частоте, при кото-
ром обычных вариантов будет мало, а редких — много. Именно
такая картина наблюдается чаще всего.
Как измерить изменчивость аллоферментов в популяциях
Для того чтобы сравнивать популяции, принадлежащие к од-
ному и тому же или к разным видам, необходимо выразить час-
тоты аллелей в ряде локусов некоторым числом, которое служит
показателем суммарной или средней генной изменчивости в дан-
ной популяции.
Если какой-либо ген представлен в популяции более чем одним
аллелем, его называют полиморфным геном. Полиморфный ген
может иметь два или более аллелей, причем каждый из них мо-
жет быть либо редким (иметь низкую частоту), либо распростра-
Рис. 9.15. Источники наследственной изменчивости фенотипов, выявляемой мето-
дом гель-электрофореза.
Исследовалась система ксантиндегидрогеназы (XdH) у Drosophila melanogaster.
Альтернативные аллели помечены звездочками. Локус ту обусловливает синтез
субъединиц XdH, которые изменяются под действием гена-модификатора та-1.
Субъединицы объединяются попарно, образуя димеры. Димеры, состоящие из
разных субъединиц, имеют различную подвижность в геле и дают различные по-
лосы. Изменчивость, выявляемая методом гель-электрофореза, может быть вызва-
на изменениями как гена-модификатора, так и структурного гена. А. Изменчи-
вость всецело обусловлена полиморфизмом структурного гена (гу). Б. Изменчи-
вость обусловлена полиморфизмом гена-модификатора (та-1). В. Несмотря на по-
лиморфизм гена-модификатора, методом гель-электрофореза не удается выявить
изменчивости вследствие наличия доминирования. (Finnerty, Johnson, 1979)
24®
Глава 9
ненным (иметь высокую частоту). Возведя в квадрат частоту
каждого имеющегося в популяции аллеля и сложив эти величины,
мы получим число, которое называют гомозиготнсстью в данном
локусе и обозначают буквой j:
где х, — частота i-ro аллеля. Единица минус гомозиготность дает
гетерозиготность в этом локусе, обозначаемую буквой h:
й = 1—/s=l—
Если усреднить j или h по всем локусам, то мы получим среднюю
гомозиготность (J) или среднюю гетерозиготность (Н) для дан-
ной популяции:
Н 2*' J 2*
п =------. </=------.
n п
где hi и /; — соответственно гетеро- и гомозиготность i-ro локуса,
ап — число локусов.
Концепция средней гомозиготности и средней гетерозиготности
была развита применительно к случайно скрещивающейся попу-
ляции. Пусть, например, в популяции, где изучалась изменчи-
вость изоферментов, детерминируемых одним локусом, и было
обнаружено, что в этом локусе имеется два аллеля с частотами
р = 9=0,5; гомозиготность и гетерозиготность для этого локуса со-
ответственно равны
/ =2xi2==(0’5)2+(°.5)2= °>25 + 0’25 = 0>5>
h= 1 —/=г0,5.
Таким образом, половина этих локусов гетерозиготна, а поло-
вина гомозиготна, что и следовало ожидать для популяции со
случайным скрещиванием, находящейся в равновесии Харди—
Вайнберга (гл. 4). Если, однако, скрещивание не является слу-
чайным, приведенная формула неприменима для измерения гомо-
и гетерозиготности в популяции. Тем не менее она служит мерой
генного единообразия (J) и разнообразия (Н) в популяции и как
таковая может быть использована. Поскольку все популяции в
той или иной мере отклоняются от модели случайного скрещива-
ния (гл. 7), предпочтительнее рассматривать J и Н как меры со-
ответственно единообразия и разнообразия генов, а для опреде-
ления уровней гетерозиготности использовать более прямые изме-
рения.
Генетическая изменчивость в природных популяциях 241
генетическая изменчивость
У РАЗНЫХ ВИДОВ РАСТЕНИЙ И ЖИВОТНЫХ
Хотя число видов, у которых была изучена изменчивость ал-
лоферментов, значительно выше 100, они составляют слишком ма-
ленькую выборку по сравнению с общим числом известных видов
растений и животных, которое уже превысило 2 млн. Тем не ме-
нее результаты изучения этих 100 видов, несмотря на их некото-
рую разноречивость, дают кое-какие предварительные оценки.
Самым удивительным результатом этих исследований было то,
что количество обнаруженной изменчивости оказалось очень боль-
Таблица 9.7
Сравнение генетической изменчивости у животных и растений
(Hamrick J. L., 1979. Genetic variation and longevity.
In: Solbrig et al., eds., Topics
in Plant Population Biology, N. Y.)
Группа Число видов или ферм Среднее число ло- кусов на 1 вид Средняя доля локусов
полиморфных на I популяцию (С.О.) гетерозиготных в» 1 особь (С.О.)
Насекомые Дрозофила 28 24 0,529(0,030) 0,150(0,010)
Другие виды 4 18 0,531 0,151
Гапло-диплоидные осы 6 15 0,243(0,039) 0,062(0,007)
Морские беспозвоночные 9 26 0,587(0.084) 0,147(0,019)
Брюхоногие Наземные 5 18 0,437 0,150
Морские 5 17 0,175 0,083
Все беспозвоночные 57 Среднее=21,8 Среднее=0,469 Среднее=0,135
Рыбы 14 21 0,306(0,047) 0,078(0,012)
Амфибии И 22 0,336(0,034) 0,082(0,008)
Рептилии 9 21 0,231(0,032) 0,047(0,008)
Птнцы 4 19 0,145 0,042
Грызуны 26 26 0,202(0,015) 0,043(0,005)
Крупные млекопитающие 4 40 0,233 0,037
Все позвоночные 68 Среднее=24,1 Среднее = 0,247 Среднее=0,061
Однолетние растения 36/33 11 0,446(0,296) 0,142(0,107)
Двулетние растения 13/11 18 0,220(0,250) 0,079(0,081)
Травянистые многолет- ники 17/14 12 0,250(0,312) 0,116(0,118)
Деревья и кустарники 10/7 12 0,652(0,314) 0,359(0,102)
Все растения 76/65 Среднее= 12,6 Среднее = 0,391 Среднее=0,149
16—2044
242
Глава 9
шим. Например, в популяциях многих видов доля полиморфных
локусов достигает 25—50%, а у составляющих популяцию особей
от 5 до 15% всех локусов гетерозиготны.
В табл. 9.7 приведены некоторые оценки гетерозиготности и
полиморфизма для разных видов, принадлежащих к различным
типам растений и животных. Поскольку выборки малы, любые
общие заключения были бы преждевременными. Максимальная
cin
0,0
Idx
ma-l
—"I e
64,8
гу aidox
a-
52,35 56,8
Рис. 9.16. 1ене,тические элементы системы ксантиндегидрогеназы (XdH) у Droso-
phila melanogaster.
Числа указывают место, занимаемое геном на карте хромосомы. Ген гу —струк-
турный ген для XdH, a aidox — структурный ген для альдегидоксидазы (АО). Ну-
левые мутанты в локусе гу лишены XdH-активности, но нормальны в отношении
АО-активности, тогда как aldox-отрицательные мутанты лишены АО-активности,
но нормальны в отношении XdH-активности. Мутанты та-1 совершенно лишены
как XdH-, так и АО-активности. У мутантов Ixd как XdH-, так и АО-активность
значительно понижена по сравнению с активностью этих ферментов у дикого ти-
па. У различных мутантов cin и XdH-, н АО-активности гораздо ниже, чем у ди-
кого типа. (Finnerty, Johnson, 1979)
изменчивость среди позвоночных, по-видимому, меньше, чем в не-
которых других группах, таких, как растения и некоторые беспоз-
воночные, однако необходимо накопить гораздо больше данных,
прежде чем делать какие-либо определенные выводы.
Другой интересный факт состоит в том, что генное разнообра-
зие (J) варьирует у различных генов: одни гены очень мономорф-
ны, а другие чрезвычайно полиморфны. Из всех известных в на-
стоящее время примеров генетической изменчивости наиболее яр-
ко выражена изменчивость фермента ксантиндегидрогеназы у
дрозофилы. У 60 самок Drosophila persimilis, которых искусствен-
но сделали гомозиготными в лабораторных условиях, Койн (Coy-
ne) обнаружил 23 аллеля. Эти мухи происходили из трех разных
популяций. У 146 гомозиготных самок близкородственного вида
Drosophila pseudoobscura, происходивших из 12 различных попу-
ляций, было найдено 37 аллелей. Ввиду проводившихся над этими
Генетическая изменчивость в природных популяциях 243
мухами лабораторных манипуляций установить, насколько рас-
пространены эти мутантные аллели в природе, невозможно, но
многочисленность аллелей удивительна.
Фермент ксантиндегидрогеназа участвует в метаболизме нор-
мального красного пигмента глаз дрозофилы. Локус розовых глаз
(гу) — структурный локус, но есть еще три локуса — каштановых
глаз (та-1), низкого содержания ксантиндегидрогеназы (Ixd) и
киноварных глаз (cin), — которые также влияют на уровни ксан-
тиндегидрогеназы, а следовательно, и на цвет глаз (рис. 9.16). Эти
другие гены называют генами-модификаторами; принято считать,
что они оказывают действие путем посттрансляционной модифика-
ции, хотя точная природа этой модификации неизвестна. Посколь-
ку ген гу (считающийся структурным геном ксантиндегидрогена-
зы) лежит в третьей хромосоме Drosophila melanogaster,
эффект посттрансляционной модификации на изоферменты можно
изучить, используя мух с идентичными третьими хромосомами
(т. е. происходящими от одной и той же родительской особи), но
разными Х-хромосомами, каждая из которых содержит свой ал-
лель гена та-1. Если аллели та-1 оказывают влияние на фермент
ксантиндегидрогеназу, то в таком эксперименте будут обнаружены
разные изоферменты. Когда Финнерти (Finnerty) и Джонсон
(Johnson) провели такой эксперимент, они обнаружили, что в
каждом случае происходило достоверное изменение формы моле-
кулы белка, приводившее к изменению электрофоретической под-
вижности. Финнерти и Джонсон экспериментировали не с
D. persimilis, как Койн (Coyne), а с D. melanogaster-, поэтому
нельзя утверждать, что изменчивость, выявленная Койном, час-
тично или целиком обусловлена посттрансляционной модифика-
цией. Однако их эксперимент показывает, что понимание генети-
ческой изменчивости признаков под действием естественного от-
бора остается не простой задачей, которая в результате открытия
изменчивости аллоферментов не облегчилась, а, пожалуй, ослож-
нилась еще более.
МЕХАНИЗМЫ ПОДДЕРЖАНИЯ ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
Выяснение причин высокой генной изменчивости в природных
популяциях и расшифровка механизмов ее поддержания состав-
ляют в настоящее время центральные проблемы популяционной
генетики.
Полиморфизм в данном локусе может быть переходным или
устойчивым. Переходный полиморфизм возникает в популяции,
когда происходит замещение аллеля, бывшего некогда обычным,
другим аллелем, придающим своим носителям более высокую
приспособленность. Состояния переходного полиморфизма имеют
важное значение; они вызывают специфические генетические эф-
фекты, уже обсуждавшиеся в гл. 8 (генетический груз). Некото-
16*
244
Глава 9
рые из них будут обсуждаться также и здесь. Устойчивые (или
сбалансированные) полиморфизмы поддерживаются той или иной
формой уравновешивающего отбора (гл. 6). Их можно искусствен-
но разделить на случаи однолокусного гетерозиса, отбора в поль-
зу гетерозигот и отбора в изменчивой среде.
Однолокусный гетерозис
В гл. 6 был приведен вывод формулы для изменений генных
частот в популяции, особи которой различаются лишь по аллелям
одного локуса и на которую действует направленный отбор (стр.
151), а в гл. 8 были выведены генные частоты для случаев, когда
относительная приспособленность (w) гетерозиготного генотипа
выше приспособленности как одной, так и другой гомозигот, и
было показано, что превосходство гетерозигот накладывает на
популяцию генетический груз в результате непрерывного выщеп-
ления гомозиготных генотипов с более низкой приспособленно-
стью. Чем больше преимущество гетерозигот, тем больше груз.
Наилучший и до настоящего времени единственный несомнен-
ный пример однолокусного гетерозиса дает серповидноклеточная
анемия — заболевание, встречающееся в Африке.
Серповидноклеточная анемия представляет собой генетическое
заболевание человека, при котором нарушается способность 13-
цепи гемоглобина к переносу кислорода. Оно обусловлено единич-
ной мутацией, вызывающей замену в нормальной молекуле гемо-
глобина глутаминовой кислоты в положении 6 аминокислотой ва-
лином. Глутаминовая кислота кодируется триплетами ГАА и ГАГ
(Г — гуанин, А — аденин, см. гл. 4), а валин — триплетами ГУА
и ГУГ (У — урацил); таким образом, изменение лишь в одной
молекуле, т. е. замещение аденина тимином в молекуле ДНК, вы-
зывает такие изменения в процессах роста и развития, что че-
ловек, унаследовавший эту молекулу от обоих родителей, умирает
в раннем возрасте от серповидноклеточной анемии.
Казалось бы, мутация, которая в гомозиготном состоянии ле-
гальна, должна была бы встречаться в популяции с очень низ-
кой частотой (q =У и/s, гл. 6). Однако ген серповидноклеточной ане-
мии встречается с высокой частотой в некоторых областях Афри-
ки (рис. 9.17) и с частотой, выше ожидаемой, среди представите-
лей черной расы в Новом Свете. Эта проблема изучалась в
1955—1965 гг. А. К- Эллисоном (А. С. Allison), который совершен-
но твердо установил, что человек, гетерозиготный по гену серпо-
видноклеточности, устойчив к тропической малярии, вызываемой
Plasmodium falciparum. Таким образом, в этом случае относи-
тельная приспособленность гетерозиготы равна единице, а отно-
сительная приспособленность гомозиготы по рецессивному гену
равна нулю (или близка к этому), тогда как приспособленность
гомозиготы по доминантному гену зависит от условий среды. В об-
Генетическая изменчивость в природных популяциях
245
ластях с высокой частотой малярии приспособленность доминант-
ной гомозиготы будет низкой, так как многие люди с таким гено-
типом будут умирать от малярии (табл. 9.8). В тех областях, где
малярия не представляет серьезной проблемы или же не сущест-
вует вовсе, нормальные гомозиготы превосходят гетерозиготы по
приспособленности, поскольку система переноса кислорода у таких
Рис. 9.17. Частота генов серповидноклеточиости в разных частях Африки, Аравии,
Индии и юга Европы.
В нижнем левом углу указаны доли носителей этого гена в разных популяциях
в процентах. (Allison, 1961)
людей функционирует лучше, чем у гетерозиготных индивиду-
умов. Мероприятия по борьбе с малярией, проводившиеся в раз-
ных частях земного шара, а также эмиграция африканских жите-
лей в течение последних 100 лет как в незараженные, так и в
зараженные малярией области Нового Света создаютпринципиаль-
ную возможность проверки этой гипотезы.
В период между 1520 и 1800 гг. африканцев, принадлежащих
к черной расе, привозили в качестве рабов из разных частей за-
падного побережья Африки в Новый Свет. Согласно историческим
сведениям, наибольшее число людей привозили из мест, располо-
женных по соседству с Ганой, где частота серповидноклеточности,
как и на большей части западного побережья Африки, достигает
246
Глава 9
Таблица 9.8
Число смертей от малярии в зависимости от наличия
гена серповидиоклеточности у африканских детей
(Allison, 1965)
Местность Число смертей Число носителей гена серповидио- клеточностн (гете- рознгот) Частота гена сер* повидноклеточ- нод в популяции. %
Кампала (Уганда) 16 0 0,20
Киншаса (Заир) 23 0 0,235
Кананга (Заир) 23 1 0,25
Аккра (Гана) 13 0 0,08
Ибадан (Нигерия) 29 0 0,24
20%. У нас нет, однако, возможности точно определить, с какой
частотой этот ген встречался в популяции эмигрантов. По всей
вероятности, его частота варьировала в разное время и в зависи-
мости от географического происхождения рабов. В Новом Свете
популяция черной расы смешивалась с другими расами, причем
степень этого смешения не всегда легко установить. Следователь-
но, существует два источника неясности относительно ожидаемого
снижения частоты гена серповидиоклеточности: 1) пониженная ча-
стота гена в популяциях Нового Света может быть обусловлена
его изначальной низкой частотой в популяции эмигрантов; 2) она
может быть вызвана смешением с популяциями, в которых этот
ген отсутствует. Имеются, однако, методы исследования, позволя-
ющие частично обойти эти затруднения.
В табл. 9.9 приведены частоты гена серповидиоклеточности сре-
ди населения различных мест Карибского бассейна. Обратите вни-
мание на то, что она варьирует от 23% (что равно частоте гена в
Африке) до 7%. Интересно, что высокие частоты гена обнаружены
в Гондурасе и Суринаме — областях, где малярия очень распро-
странена, а низкие — на островах Кюрасао и Барбадос, где маля-
рии нет! В США путем изучения 15 удобных генетических марке-
ров в популяциях черной расы было установлено, что миграция
генов белой расы в эти популяции была порядка т — 0,1—0,2.
Однако вычисленное значение т для гена серповидиоклеточности
составило 0,46—0,69. Поскольку это значение слишком велико по
сравнению со значениями для других генов, был сделан вывод, что
среди американских популяций черной расы происходил значи-
тельный отрицательный отбор против гена серповидноклеточности.
Помимо гена серповидноклеточности в популяциях человека
есть ряд других полиморфных генов, которые, как предполагается,
вызывают однолокусный гетерозис; это, в частности, гены, опре-
Генетическая изменчивость в природных популяциях
247
Таблица 9.9
Частота генов серповидноклеточности в популяциях
Карибского бассейна (по Allison,
1965, с изменениями)
Популяция Число обследован- ных Носители гена серповидиоклеточ- ностн, %
Гондурас (черные караибы) 705 23,3
Кюрасао 1502 7,2
Суринам, Кейбл 519 16,8
Суринам, Менго 172 20,3
Суринам, Стельманс-Айленд 275 11,3
Суринам (джьюки) 343 15,0
Ямайка 1018 10,9
Сент-Винсент 748 8,7
Доминика 664 9,5
Барбадос 912 7,0
Сент-Люсия 825 14,0
деляющие другие варианты гемоглобина. Однако ни в одном дру-
гом случае участвующие в этом факторы отбора не получили
столь изящного объяснения, как в случае серповидноклеточной
анемии.
Интересный пример полиморфизма у растений описан изра-
ильскими ботаниками Апирионом (Apirion) и Зохари (Zohary)
у диплоидной (2л=14) формы ежи сборной (Dactylis glomerata
var. judaica}—вида, у которого имеются также полиплоидные
разновидности. Этот полиморфизм интересен тем, что в нем участ-
вуют аллели, обусловливающие недостаточность хлорофилла и ле-
тальные в гомозиготном состоянии, причем все это можно объяс-
нить, допустив, -что в данном случае имеет место однолокусный
гетерозис.
Подвид judaica ежи сборной встречается в Израиле главным
образом в трех местах: на холмах в окрестностях Иерусалима, в
районе Назарета и в окрестностях Цфата в Верхней Галилее. В
этих районах ежа входит в состав сообщества низкорослых кус-
тарничков и травянистых растений, обычных для Средиземноморс-
кой области. Эти растения не способны к самоопылению (см. гл.
8). Сотню семян от разных растений из разных местностей высея-
ли на одинаковых делянках и подсчитали процент проростков-
альбиносов (гомозиготных по гену недостаточности хлорофилла).
Результаты представлены в табл. 9.10. При помощи соответству-
ющих статистических методов можно оценить процент гетерози-
248
Глава 9
готных растений в популяции, а также процент мутантов с недо-
статочностью хлорофилла. Обратите внимание на то, что частота
мутантов очень высокая — от 2% в одних популяциях до 15% в
других. Поскольку такие высокие частоты не могут быть резуль-
татом одних только мутаций и в гомозиготном состоянии мутации
детальны, единственным возможным объяснением служит наличие
сильного отбора в пользу гетерозигот. Простой расчет подтверж-
дает справедливость такого предположения. Если частота рецес-
сивного гена q = 0,15, а частота доминантного р = 1—0,15=0,85
(популяция в Кирьят-Анавим), то в поколении потомков можно
предсказать следующие частоты генотипов:
для ДА: р2 = 0,852 = 0,7225,
для Аа: 2pq = 0,85 -0,15- 2 = 0,255,
для аа: <72 = 0,152 = 0,0225.
Но поскольку гомозиготы аа детальны, предсказанная доля
гомозигот составит 0,739^74%, а гетерозигот — 0,26=26%. Одна-
ко фактическая их доля (допуская, что она остается постоянной
из года в год) составляет соответственно 66% и 30% • Иными сло-
вами, гетерозигота должна производить больше семян или иметь
более низкую смертность, чем гомозигота, или же обладать од-
новременно обеими этими особенностями.
Хотя мы не знаем, каковы преимущества гетерозигот перед го-
мозиготами, эти результаты показывают, что, по всей вероятности,
какое-то преимущество у них имеется. Вся эта ситуация напоми-
нает то, что наблюдается в случае серповидноклеточной анемии.
Отбор в пользу гетерозигот
При серповидноклеточной анемии наблюдается сильный урав-
новешивающий отбор против обеих гомозигот: доминантный ген
в гомозиготном состоянии обусловливает восприимчивость к маля-
рии, а рецессивный ген в гомозиготном состоянии детален. Имеет-
ся бесчисленное множество примеров гибридов, мощность кото-
рых выше, чем у обеих родительских форм; наиболее известные из
них — гибридная кукуруза и другие культурные злаки. Однако тот
факт, что искусственный гибрид обладает большей мощностью, не-
обязательно означает, что в природных условиях он оставляет
больше потомков, а именно это служит истинным критерием пре-
восходства гибридов, или гетерозиса. Тем не менее многие гене-
тики и эволюционисты полагают, что гетерозиготы обычно более
приспособлены, чем гомозиготы. Они исходят из того, что при на-
личии двух разных аллелей организм имеет больше возможностей
в процессе развития соответствующим образом отладить свои би-
охимические механизмы, но способ реализации такого преимуще-
ства пока неизвестен. В качестве примера, подтверждающего эту
Генетическая изменчивость в природных популяциях
249
Таблица 9.10
Популяции Dactylis glomerata, содержащие мутантов
с недостаточностью хлорофилла (Apirion, Zohary, 1961)
Местность н тип почвы Общее число обследо- ванных растений Число нор- мальных гомозигот- ных растений Число гетеро- зиготных растений Количество гетерозигот в выборке, % Оценка частоты гамет с геном albino, %
Холмы в окрестностях Иеруса- лима Долина Мацлевы краснозем . 47 35 12 26 13
Ора рендзина 79 65 14 18 9
Миинадав рендзииа 94 81 13 14 7
Шнеллер-Камп краснозем 78 55 23 29 14,5
Университетский городок краснозем 64 58 6 9 4,5
Кирьят-Анавим краснозем 80 66 24 30 15
Кирьят-Анавим краснозем 94 66 28 30 15
Кастель краснозем 100 72 28 28 14
Назаретские холмы Кефар-Аореш рендзина 82 79 3 4 2
Назарет рендзина 95 90 5 5 2,5
Верхняя Галилея, Эйнзэйтим рендзина 79 76 3 4 2
Цафат рендзина 88 77 11 13 6,5
Бирийа смешанная почва 86 76 10 12 6
Миар рендзина 97 82 15 15 7,5
точку зрения, часто приводят хлебную плесень Neurospora crassa.
Штаммы, гетерозиготные по гену, регулирующему синтез пара-
аминобензойной кислоты, растут лучше, чем штаммы, гомозигот-
ные по одному или другому из этих двух аллелей. Как показали
биохимические исследования, плюс-гомозиготы синтезируют очень
большое, а минус-гомозиготы — слишком малое количество пара-
аминобензойной кислоты, причем как избыток, так и недостаток
этой кислоты подавляет рост.
Имеются многочисленные данные, свидетельствующие о том,
что гетерозиготы (по одному или нескольким локусам) обладают
более высокой приспособленностью, чем гомозиготы. Добржанский
250
Глава 9
(Dobzhansky) и Спасский (Spassky) в 1963 г. опубликовали ре-
зультаты ряда исследований, проведенных на Drosophila pseudo-
obscura, пойманных в природе. При помощи особого метода эти
ученые создали мух, гомозиготных по одной из их собственных
хромосом, а затем сравнивали жизнеспособность этих особей с
жизнеспособностью особей, гетерозиготных по этой же хромосоме
Жизнеспособность гомозигот
Рис. 9.18. Жизнеспособность дрозофил, гетерозиготных или гомозиготных по ка-
кой-либо из хромосом дикого типа.
Мух, гомозиготных по одной из хромосом дикого типа, сравнивали с мухами, ге-
терозиготными по этой же хромосоме. За единицу жизнеспособности принята
средняя жизнеспособность гетерозигот в популяции. По оси абсцисс отложена
жизнеспособность каждой гомозиготы, а по оси ординат — средняя жизнеспособ-
ность тех гетерозигот, у которых имеется эта хромосома. (Dobzhansky, Spassky,
1963)
(рис. 9.18), Вывод из этих экспериментов очень прост: нет ника-
кой корреляции между жизнеспособностью гетерозиготной мухи,
содержащей пару хромосом из природной популяции, и жизне-
способностью мухи, гомозиготной по тем же самым хромосомам.
Знание того, что данная хромосома легальна или полулегальна
(потому что она несет летальные или полулегальные гены) в гомо-
зиготном состоянии, не дает нам возможности предсказать ее по-
ведение в гетерозиготном состоянии. То же самое относится к
хромосомам, обладающим в гомозиготном состоянии средней или
низкой приспособленностью. Гетерозигота как в том, так и дру-
гом случае может быть нормальной или же обладать приспособ-
ленностью выше или ниже нормальной. Иными словами, резуль-
таты этого эксперимента наводят на мысль, что гетерозис необя-
Генетическая изменчивость в природных популяциях 251
зательно определяется поведением генов в гомозиготном состоянии;
скорее он обусловлен самим фактором гетерозиготности.
Брюс Уоллес (Bruce Wallace) из Корнеллского университета
провел ряд экспериментов, в которых он подвергал особей Droso-
phila melanogaster рентгеновскому облучению в дозах, достаточно
высоких, чтобы вызвать мутации. При помощи чрезвычайно тон-
ких манипуляций ему удалось установить, что некоторые гетеро-
зиготы, содержавшие облученную хромосому и гомологичную ей
Мухи дикого типа
--------+/+
I
I
I
Облученные
Рт/+ б/сГ ХСу1_/+£ф Pm/Ф cTc/xCyL/+ фф
/------------74------------\ г—————к
Pm/CyL Рт/+ CyL/+ +/-U Pm/CyL Рт/-Р CyL/9) +/Ф
(Стандарт) (Стандарт)
Рис. 9.19. Эксперименты Брюса Уоллеса с облучением.
Мухи дикого типа при помощи соответствующих приемов делят на группы с иден-
тичными хромосомами, несущими гены-маркеры Pm (сливовый цвет глаз). Одну
группу подвергают облучению, а другую нет. Затем мух скрещивают с гомозигот-
ными аналитическими линиями, несущими гены Су (загнутые крылья) и L (лопа-
стевидные глаза). Таким путем создаются стандарты, с которыми можно сопо-
ставлять гомозиготных особей дикого типа (+/+) и облученных гетерозигот
(+/©)•
нормальную хромосому, обладали более высокой приспособленно-
стью, чем гомозиготы, содержавшие две нормальные хромосомы
(рис. 9.19). Если же гомозиготная особь содержала две идентич-
ные облученные хромосомы, то это во всех случаях приводило к
летальному исходу. Здесь вновь создается впечатление, что имен-
но гетерозиготность как таковая, а не качество содержащихся в
гетерозиготе генов обусловливает более высокую приспособлен-
ность.
Если гетерозиготы обладают более высокой приспособленно-
стью, чем гомозиготы, но выщепляющиеся гомозиготы нормальны
и жизнеспособны, то проблема генетического груза оказывается
несерьезной. В самом деле, в такой популяции в каждом поко-
лении будут элиминироваться только те особи, которые гомози-
готны по крайне вредным аллелям. Число таких особей должно
быть относительно небольшим. Остальная популяция будет состо-
ять из смеси гетерозигот и гомозигот, причем приспособленность
252
Глава 9
гетерозигот должна быть несколько выше. Особи должны конку-
рировать между собой за жизненные ресурсы и за брачных парт-
неров; исход конкуренции будет определяться совместным дейст-
вием случайности и наследственности, при этом гетерозиготные ге-
нотипы будут обладать небольшим преимуществом. Уоллес назвал
Гомозиготны по
большинству ___________________________________—»-
локусов
Высоко
гетерозиготны
Рис. 9.20. Графическое изображение мягкого и жесткого отбора.
Некоторые генотипы элиминируются из популяции вследствие их летальности
(заштрихованная область). Это — жесткий отбор. Среди нелетальных генотипов
некоторое число не сможет дожить до репродуктивного возраста и произвести
потомство вследствие недостатка ресурсов, причем число это зависит от количе-
ства ресурсов и от других условий (область, покрытая точками). Чем ниже гете-
розиготность, тем выше вероятность элиминации.
элиминацию особей, несущих вредные аллели, жестким отбором,
а элиминацию нормальных особей, обусловленную их поражени-
ем в конкуренции за ресурсы, мягким отбором (рис. 9.20).
Отбор в изменчивой среде
Представьте себе снова популяцию, различающуюся только по
одному локусу с двумя аллелями. Мы видели (гл. 4), что боль-
шинство, а возможно, и все гены проявляют в разных условиях
среды различную экспрессивность и различную пенетрантность.
Большая часть аллелей также оказывает на приспособленность
своих носителей различное влияние в разных средах. Поэтому
приспособленность данного генотипа представляет собой не ка-
кую-то постоянную величину, а зависит от той конкретной среды,
в которой он обитает. Например, представим себе такое положе-
ние, когда какой-либо организм живет в четырех слегка разли-
чающихся средах, которые можно расположить в определенной
Генетическая изменчивость в природных популяциях 253-
последовательности на некой произвольной оси. Допустим, далее,
что фенотип, создаваемый гомозиготным генотипом А1АЬ обладает
наибольшими преимуществами в среде 1 и наименьшими в среде
4, а фенотип, создаваемый генотипом А2А2 — наоборот. Гетерози-
гота всегда обладает такой же приспособленностью, как наиболее
приспособленная гомозигота, но никогда не превосходит ее. Соот-
ветствующие приспособленности приведены в табл. 9.11.
Таблица 9.11
Приспособленности, создаваемые гомозиготными и гетерозиготными генотипами в разных средах
Среда Приспособленность генотипа
Ai At Ai A2 Аз Аз
1 1 1 1—2/
2 1 . 1 1—t
3 1—S 1 1
4 1—2s 1 1
Среднее 1—3/4s 1 I-3/?
Таким образом, наибольшей приспособленностью во всех
четырех средах обладает гетерозигота, хотя в каждой среде в;
конце концов произойдет закрепление одной из гомозигот в ре-
зультате отбора,направленного против другой гомозиготы. Однако
при наличии потока генов из одной среды в другую устанавлива-
ется динамическое равновесие, приводящее к полиморфизму. Име-
ем ли мы дело со средами, различающимися в пространстве, или
же с последовательными изменениями одной среды во времени,
как, например, при смене времен года, значения не имеет.
Примером полиморфизма, который может быть результатом
изменения давлений отбора во времени, служит инверсионный по-
лиморфизм у Drosophila pseudoobscura, уже обсуждавшийся в
этой главе. Добржанский (Dobzhansky) тщательно изучал попу-
ляцию этого вида с гор Сан-Джасинто (Калифорния) с 1939' по
1946 г. (рис. 9.12). В этой популяции в хромосомах преобладают
три последовательности: Стандарт (ST), Эрроухед (AR) и Чири-
кахуа (CH). С марта по июнь частота хромосом ST уменьшается,
а частота СН возрастает, тогда как между июнем и августом на-
блюдается обратная картина. Сходные изменения обнаружены и
в других популяциях. В 1972 г. Добржанский и Айала сообщили-
об аналогичном исследовании, проведенном на ранчо Мак-До-
нальда в округе Напа (Калифорния). Ежемесячные выборки, брав-
шиеся с апреля 1971 по октябрь 1972 г., подтвердили наличие се-
зонных изменений в хромосомном полиморфизме (рис. 9.21), но
выявили также значительные изменения в полиморфизме по фер-
ментам.
254
Глава 9
Вместе с тем Райт (Wright) и Добржанский (Dobzhansky) по-
казали, что в популяции, содержащей смесь мух с последователь-
ностями ST и СН и выращенной в популяционном ящике в лабо-
ратории, после достижения равновесия соотношение хромосом со-
ставляет 70% ST и 30% СН; отсюда можно сделать вывод, что в
этих условиях относительная приспособленность гетерозигот
Рис. 9.21. Изменение частоты некоторых хромосомных транслокаций у Drosophila
pseudoobscura.
Частота последовательностей Стандарт (ST), Чирикахуа (СН) и Эрроухед (AR)
в одной популяции в Калифорнии изменялась в разное время года на протяжении
1971 (слева) и 1972 (справа) гг. (По данным Dobzhansky, Ayala, 1973.)
ST/СН равна 1, а двух гомозигот — 0,7 (ST/ST) и 0,3 (СН/СН).
Таким образом, в данном случае происходит как отбор в пользу
гетерозигот, так и изменение коэффициентов отбора в результате
изменений среды.
Во многих случаях существует связь между параметрами
среды и генотипами, как, например, у обсуждавшихся уже Сераеа,
Drosophila pseudoobscura, Adalia bipunctata, Biston betularia. Эти
и другие примеры составляют довольно внушительную совокуп-
ность данных, позволяющих считать, что разные среды обитания
создают различные давления отбора и что изменчивость среды в
пространстве и во времени может быть причиной значительной
части наблюдаемой генетической изменчивости. Однако на осно-
вании имеющихся данных нельзя совершенно определенно заклю-
чить, что давления отбора, оказываемые разными средами обита-
ния, служат единственной причиной наблюдаемых картин измен-
чивости. Необходимо еще экспериментально установить, какой
Генетическая изменчивость в природных популяциях 255
именно фактор (или факторы) среды регулирует частоту генов.
Из всех известных случаев лишь серповидноклеточная анемия по-
падает в эту категорию.
НЕЙТРАЛЬНАЯ ИЗМЕНЧИВОСТЬ
В качестве еще одной причины наличия в природных популя-
циях растений и животных большой изменчивости, не получившей
до сих пор объяснения, выдвигалось мнение о том, что эта измен-
чивость нейтральна. Иными словами, если в результате мутации
возникает новый аллель с приспособленностью не выше и не ниже
той, которая имеется в популяции, он мог бы закрепиться случай-
ным образом и сохраняться (по крайней мере в течение некоторо-
го времени) по чистой случайности.
Это объяснение, носящее название гипотезы случайного дрей-
фа, было предложено японскими генетиками Кимурой и Отой
(Kimura, Ohta) в 1971 г. Согласно этой гипотезе, большая часть
новых мутаций оказывается вредной и поэтому сразу элиминиру-
ется естественным отбором. Небольшое число мутаций, оказавше-
еся благоприятным, может широко распространяться. Однако
очень небольшое, но статистически значимое число мутаций будет
нейтральным в селективном отношении (ни вредным, ни полез-
ным) и их судьба определяется случайным дрейфом генов.
Существует несколько возможных способов возникновения ге-
нов, нейтральных с точки зрения отбора. Генетический код обла-
дает избыточностью (см. гл. 4), поскольку для кодирования всего
лишь 26 аминокислот имеется 64 различных триплета. Вследствие
этого некоторые генные мутации не влияют на белок, кодируемый
данным геном, так как мутировавший триплет все еще кодирует
ту же самую аминокислоту. Кроме того, считается, что некоторые
мутации (например, сопровождающиеся заменой триплета ГУУ,
кодирующего валин, триплетом ГЦУ, кодирующим аланин) приво-
дят к синтезу аминокислот с очень сходными свойствами, не влия-
ющими на свойства молекулы белка, а поэтому могут быть ней-
тральными в селективном отношении. Есть и такие мутации, ко-
торые детерминируют замены аминокислот, оказывающие более
сильное влияние на синтезируемый фермент, однако, по мнению
некоторых генетиков, и эти мутации также могут быть селективно
нейтральны.
Нейтральные аллели не обязательно идентичны в своем воз-
действии на приспособленность популяции. Просто аллели счита-
ются «нейтральными», если их воздействие на приспособленность
так мало, что их элиминация или закрепление в данной популя-
ции зависит главным образом от случая. Допустим, что в популя-
ции имеется два аллеля, Ai и А2, и что аллель Ai имеет приспо-
собленность, равную 1, а аллель А2—1+«, где s может быть как
положительной, так и отрицательной величиной, т. е. А2 может
256
Глава 9
быть приспособлен либо лучше, либо хуже, чем Аь Эти два алле-
ля считаются нейтральными, если абсолютная величина произве-
дения $ и эффективной численности популяции (см. гл. 7) намного
меньше 1:
WeXs< 1.
Таким образом, будет ли данный аллель считаться нейтраль-
ным, зависит от того, в какой популяции он появится. Чем боль-
ше эффективная численность популяции, тем меньше должна быть
s, чтобы аллель оказался действительно нейтральным, поскольку
чем меньше эффективная численность популяции, тем больше ве-
роятность закрепления аллеля в результате случайного дрейфа.
Вопрос состоит в том, существуют ли такие мутации и сколь часто
они возникают. Одни авторы считают, что такие мутации очень
часты, а другие — что они редки. Решение этой проблемы затруд-
няется тем, что, хотя у нас и есть метод (гель-электрофорез), по-
зволяющий установить наличие у данного вида многих форм бел-
ков, мы не располагаем методикой, которая бы дала возможность
непосредственно устанавливать (если не считать таких примеча-
тельных исключений, как гемоглобин) воздействие этих ферментов
и других белков на приспособленность. В отсутствие таких данных
можно выдвинуть два альтернативных положения: 1) поскольку
не обнаружено никакого воздействия на приспособленность, фер-
мент должен быть нейтральным; 2) хотя известен ряд случаев,
когда ферменты влияют на приспособленность, ни в одном случае
не была доказана их нейтральность, а поэтому все мутации или
•большая их часть должны воздействовать на приспособленность.
Есть еще и третья альтернатива, а именно что большинство изо-
ферментных вариантов обусловлено посттрансляционной модифи-
кацией под действием генов в других локусах, а не однолокусны-
ми мутациями. В таком случае необходимый уровен гетерозигот-
•ности в одном локусе может оказаться на целый порядок величи-
ны ниже, чем мы теперь считаем. А поэтому его можно было бы
-объяснить в свете классической селекционистской гипотезы.
КРАТКОЕ РЕЗЮМЕ И ЗАКЛЮЧЕНИЕ
Популяциям растений и животных, по-видимому, свойственна
высокая генетическая изменчивость — условие, необходимое для
эволюции путем естественного отбора. Уровень генетической из-
менчивости, выявляемый методом гель-электрофореза, гораздо вы-
ше, чем предполагалось первоначально. Насколько именно он
выше — до сих пор остается неясным, потому что, во-первых, не-
обходимо еще решить ряд методических и теоретических проблем
(в частности, выяснить, какая часть наблюдаемой изменчивости
•обусловлена аллельной изменчивостью структурного гена, а ка-
кая— посттрансляционной модификацией) и, во-вторых, число ис-
Генетическая изменчивость в природных популяциях
257
следованных видов и число изученных у этих видов генов очень
мало. Нам уже известно, что между видами и даже между груп-
пами видов существуют значительные различия в этом отношении
(табл. 9.7), Найти в каждом случае механизмы, поддерживающие
эту изменчивость,— задача чрезвычайно трудная.
Генотипы, гомозиготные по Различные генотипы, гетерсзигот-
большинству локусоз. неболь- —------------------------ нь:е по большинству локусов
дрейфа
Генетический Генетическая Генетическая
груз нору а ’ ' элита
Рис. 9.22. Графическое изображение некоторых гипотез, предложенных для объ-
яснения высокой частоты гетерозиготных генотипов, наблюдающейся в природных
популяциях.
Штриховая кривая соответствует теоретически ожидаемому распределению при
случайном скрещивании и отсутствии отбора. Сплошная линия аппроксимирует
реально наблюдаемое распределение. Жесткий отбор — это процесс элиминации
летальных и субвитальных гомозиготных генотипов. Возможен также прямой от-
бор гетерозигот либо вследствие их превосходства, либо вследствие их большей
способности к выживанию в изменчивой среде. Наблюдаемый избыток гетерози-
гот может быть также обусловлен потоком генов и случайным дрейфом. Геноти-
пы, гомозиготные по летальным аллелям, образуют «генетический груз»; «более
высококачественные» (в смысле приспособленности) генотипы получили название
«генетической элиты». Промежуточные генотипы составляют «генетическую
норму».
Существует несколько различных общих теорий, пытающихся
дать объяснение существованию столь большой изменчивости, од-
нако единого общего объяснения в настоящее время нет. На рис.
9.22 в графической форме представлена как сама эта проблема,
так и различные объяснения наблюдаемых явлений. Показано
ожидаемое распределение генотипов (прерывистая линия) в боль-
шой свободно скрещивающейся популяции. Несколько особей бу-
дут гомозиготными, а несколько — гетерозиготными по большин-
ству своих локусов. У большей же части особей примерно поло-
вина локусов будет гомозиготна, а другая половина — гетерози-
готна. Описанные выше наблюдения над природными популяция-
17—2044
258
Глава 9
ми свидетельствуют об избытке гетерозигот (сплошная линия) и
о том, что число аллелей на 1 локус гораздо выше, чем это ожи-
далось. Вот некоторые из предложенных объяснений. 1) Жесткий
отбор, т. е. элиминация вредных гомозиготных генотипов, позво-
лил бы объяснить элиминацию большинства гомозиготных гено-
типов. Эта концепция связана с представлениями о мутационном
грузе, обусловленном неблагоприятными мутациями, и о сегрега-
ционном грузе — в случаях однолокусного гетерозиса (например,
серповидноклеточная анемия). 2) Отбор в пользу гетерозигот по
одному или нескольким локусам. Добржанский назвал таких осо-
бей, гетерозиготных по многим локусам, генетической элитой. Эта
гипотеза, хотя она и приложима в некоторых случаях, неприемле-
ма в качестве общего объяснения, потому что она не может объ-
яснить элиминацию выщепляющихся гомозиготных генотипов.
3) Для того чтобы объяснить некоторую нехватку гомозигот,
Уоллес (Wallace) предложил концепцию мягкого отбора. Отбор
благоприятствует гетерозиготам по многим локусам (т. е. они об-
ладают преимуществом при конкуренции и размножении), но при
этом особи, гомозиготные по этим локусам (то, что Добржанский
назвал генетической нормой), необязательно должны элиминиро-
ваться. 4) Еще одно объяснение состоит в том, что коэффициенты
отбора подвержены изменениям вследствие изменяющейся [в
пространстве и (или) во времени] среды. Гетерозигота необяза-
тельно обладает большей приспособленностью, чем любая из го-
мозигот, но в изменяющихся условиях в среднем превосходит их.
5) Некоторые из имеющихся в популяции аллелей могут вноситься
в нее потоком генов из соседних популяций. 6) Многие из имею-
щихся в популяции аллелей подлинно «нейтральны» по своему
воздействию на приспособленность и присутствуют в популяции
вследствие случайного дрейфа генов. Поскольку эти различные
гипотезы не обязательно исключают друг друга, нынешняя задача
состоит в оценке относительной роли каждого из приведенных
факторов >в генетической структуре популяции.
Мутационный процесс, рекомбинация, поток генов и отбор
представляют собой основные факторы, определяющие эволюцию
путем естественного отбора. В гл. 6 мы описали эти факторы, а
также простые модели их действия на генные частоты в локусе с
двумя аллелями в бесконечно большой популяции, обитающей в
однородной среде. Читатель должен был понять, что эти модели
полезны для учебных целей, но совершенно нереалистичны при-
менительно к настоящим популяциям, развивающимся в природных
условиях. Мы показали, что природные популяции имеют конеч-
ные размеры, что они живут в изменчивых средах, что гены взаи-
модействуют между собой при определении фенотипа и что они
объединены в хромосомы. Кроме того, мы привели достаточно
примеров, чтобы убедить читателя в наличии в популяциях очень
больших запасов генетической изменчивости, которые трудно или
Генетическая изменчивость в природных популяциях 259
даже невозможно объяснить и с учетом всех известных фак-
торов. Предстоит еще многое узнать, многое исследовать и, несом-
ненно, придется выдвигать и доказывать новые теории, прежде
чем будет создана такая действительно всеобъемлющая теория
эволюции путем естественного отбора, о которой говорилось в
гл. 3.
Теперь мы обратимся к другому аспекту эволюции, а именно к
происхождению видов и их взаимодействиям.
ЛИТЕРАТУРА
Общие вопросы
Ayala F. J. (ed.). 1976. Molecular Evolution, Sunderland, Mass., Sinauer Associa-
tes.
Ciblett E. R. 1977. Genetic polymorphisms in human blood, Ann. Rev. Genetics, 11,
13—28.
Giesel J. T. 1976. Reproductive strategies as adaptations to life in temporally hete-
rogeneous environments, Ann. Rev. Ecol. Syst., 7, 57—80.
Hedrick P. W., Ginevan M. E., Ewing E. P. 1976. Genetic polymorphism in hetero-
geneous environments, Ann. Rev. Ecol. Syst., 7, 1—32.
Johnson G. B. 1977. Assessing electrophoretic similarity, Ann. Rev. Ecol. Syst., 8,
309—328.
Kimura M., Ohta T. 1971. Theoretical Aspects of Population Genetics, Princeton,
N. J., Princeton University Press.
Powell J. R. 1975. Protein variation in natural populations of animals, Evol. Biol.,
8, 79—120.
Sheppard P. M. 1958. Natural selection and heredity, London, Hutchinson.
Selander R. K. 1973. Genetic variation among vertebrate species, Ann. Rev. Ecol.
Syst., 4, 75—92.
Специальные статьи и обзоры
Allison А. С. 1961. Genetic factors in resistance to malaria, Ann. N. Y. Acad. Sci.,
91 (3), 710—729.
Allison A. C. 1965. Polymorphism and natural selection in human populations, Cold
Spring Harbor Symp. Quant. Biol., 29, 137—149.
Anderson W., Dobzhansky T., Pavlovsky 0., Powell J., Yarkley D. 1975. Genetics
of natural populations. XLII. Three decades of genetic change in Drosophila
pseudoobscura, Evolution, 29, 24—36.
Apirion D., Zohary D. 1961. Chlorophyll lethals in natural populations of the orchard
grass (Dactylis glomerata L.), Genetics, 46, 393—399.
Aston J. L., Bradshaw A. D. 1966. Evolution in closely adjacent plant populations,
HAgrostis stolonifera in maritime habitats, Heredity, 21, 649—664.
Beckman L., Scandalios J. G., Brewbaker J. L. 1964. Genetics of leucine amino pep-
tidase isozymes in maize, Genetics, 50, 899—904.
Brussard P. F. 1975. Geographic variation in North American colonies of Сераеа
nemoralis, Evolution, 29, 402—410.
Cain A. J., Sheppard P. M. 1950. Selection in the polymorphic land snail Capaea
nemoralis, Heredity, 4, 275—294.
Coyne J. A., Felton A. A. 1977. Genic heterogeneity at two alcohol dehydrogenase
loci in Drosophila pseudoobscura and Drosophila persimilis, Genetics, 87, 285—
304.
Dobzhansky T., Anderson W., Pavlovsky O. 1966. Genetics of natural populations.
XXXVH1. Continuity and change of populations of Drosophila pseudoobscura
in western United States, Evolution, 20, 418—427.
17
260
Глава 9
Dobzhansky Т., Ayala F. J. 1973. Temporal frequency changes of enzyme and chro-
mosomal polymorphisms in natural populations of Drosophila, Proc. Nat. Acad.
Sci. USA, 70, 680—683.
Dobzhansky T., Spassky B. 1963. Genetics of natural populations. XXXIV. Adaptive
norm, genetic load and genetic elite in Drosophila pseudoobscura, Genetics, 48,
1467—1485.
Finnerty V., Johnson G. 1979. Post-translational modification as a potential explana-
tion of high levels of enzyme polymorphism: xanthine dehydrogenase and alde-
hyde oxidase in Drosophila melanogaster, Genetics 91 (in press).
Harris H. 1969. Genes and isozymes, Proc. Roy. Soc. Ser. B, 174, 1—31.
Hubby J. L., Lewontin R. C. 1966. A molecular approach to the study of genic hete-
rozygosity in natural populations. I. The number of alleles at different loci in
Drosophila pseudoobscura, Genetics, 54, 577—594.
Jones J. S., Leith В. H., Rawlings P. 1977. Polymorphism in Cepaea, Ann. Rev. Ecol.
Syst., 8, 109—146.
Lamotte M. 1960. Polymorphism of natural populations of Cepaea nemoralis, Cold
Spring Harbor Symp. Quant. Biol., 24, 65—84.
Livingstone F. B. 1971. Malaria and human polymorphisms, Ann. Rev. Genetics, 5,
33—64.
Markert C. L. 1968. The molecular basis for isozymes, Ann. B. Y. Acad. Sci., 151,
14—40.
Markert C. L., Moller F. 1959. Multiple forms of enzymes: tissue, ontogenetic and
species specific patterns, Proc. Nat. Acad. Sci., 45, 753.
Ohta T., Kimura M. 1975. Theoretical analysis of electrophoretically detectable po-
lymorphisms: models of very slightly deleterious mutations, Am. Nat., 109,
137—145.
Wallace B. 1958. The average effect of radiation induced mutations on viability in
Drosophila melanogaster, Evolution, 12, 532—552.
Wallace B., 1975. Hard and soft selection revisited, Evolution, 29, 465—473.
Тимофеев-Ресовский H. B. 1940. Zur Analyse des Polymorphismus bei Adalia Ы-
punctata, Biol. Zentral., 60, 130—137.
Часть третья
Глава 10
Индивидуумы, популяции и виды
В гл. 3 были изложены основные аксиомы эволюционной тео-
рии. Для того чтобы могла происходить эволюция, необходимо,
чтобы организмы были способны производить себе подобных;
чтобы они производили избыточное число потомков; чтобы выжи-
вание потомков зависело от их признаков (биохимических, физио-
логических, морфологических и т. п.) и чтобы существовал меха-
низм, с помощью которого эти признаки передавались бы после-
дующим поколениям. В гл. 4—9 все эти условия были подробно
рассмотрены. Мы показали, что организмы не передают потомкам
свои признаки как таковые, а передают им только потенциальную
способность к развитию этих признаков при наличии соответству-
ющей внешней и внутренней среды. Кроме того, мы показали, что
преимущества, которые дают организму обладание данным при-
знаком, не абсолютны, а относительны и зависят от свойств сре-
ды, от потенциальных соседей и конкурентов. Была показана так-
же важная роль случайности в эволюции.
Мутационный процесс, рекомбинация и поток генов определя-
ют генотип особей, составляющих популяцию. В результате согла-
сованного действия этих генотипов в клетках данного организма и
в рамках условий данной среды обитания в каждой популяции
создается некоторый набор фенотипов. Суть естественного отбора
заключается в дифференциальном выживании этих фенотипов в
популяции, т. е. в относительной успешности, с которой отдельные
организмы преодолевают неблагоприятные условия -среды, кон-
курируют с другими организмами за ресурсы и, наконец, произ-
водят потомков. Поэтому наилучший способ изучения и описания
мутационного процесса и рекомбинации состоит в том, чтобы про-
водить их на уровне генов и генотипов, т. е. выражать их посредст-
вом изменений генных частот во времени и факторов, вызываю-
щих изменения. Естественный отбор лучше всего изучать на уров-
не фенотипов, т. е. изучать изменения численности особей во вре-
мени.
ГЕНОТИП И ФЕНОТИП
Согласно общепринятым представлениям, фенотипический со-
став данной популяции в данной среде определяется исключи-
тельно ее генотипическим составом. Однако способа, позволяюще-
262
Глава 10
го экспериментально подтвердить справедливость этих представ-
лений, не существует. Известно немало исследований, в которых
показано, что некоторые фенотипические различия имеют генетиче-
скую основу. Например, различия в окраске и форме плодов тыквы
определяются генами, содержащимися в клетках этих растений; раз-
личия в окраске зерен у кукурузы или в цвете глаз у дрозофилы
также зависят от определенных генов. -Но вместе с тем разные ге-
нотипы могут давать начало одинаковым фенотипам (как в слу-
чае рецессивных мутаций), а идентичные генотипы, оказавшись в
Рис. 10.1. Крыло нормальной (Л) и мутантной (Б) особей Drosophila melanogas-
ler.
Мутация crossveinless (cv) обусловливает отсутствие поперечных жилок на
крыльях.
разных средах обитания,— разным фенотипам. Мы знаем также,
что разные генотипы могут детерминировать развитие идентичных
фенотипов в одной среде и различных фенотипов в другой. В гл.
6 было приведено два примера этого: полидактилия у человека и
загнутые крылья у дрозофилы. Еще одним примером служит му-
тация crossveinless у Drosophila melanogaster, обусловливающая
отсутствие на крыльях поперечных жилок (рис. 10.1). Если куко-
лок из линий мух с нормальными крыльями в возрасте между
21-м и 23-м часами подвергнуть в течение 4 ч действию темпера-
туры 40 °C, то примерно у 40% вылетающих из этих куколок мух
поперечные жилки на крыльях отсутствуют. Различие между нор-
мальными крыльями и крыльями, лишенными поперечных жилок,
обусловлено генетически; в результате совместного разведения
мух, реагирующих на воздействие высокой температуры, в конеч-
ном счете получается линия мух, крылья которых лишены по-
перечных жилок даже в отсутствие теплового воздействия.
Поэтому даже если в конце концов удастся доказать, что каж-
дый набор генотипов создает в данной среде один и только один
набор фенотипов, то правила превращения генотипов в фенотипы
окажутся чрезвычайно сложными. В настоящее время эта область,
известная под названием генетики развития, еще очень мало раз-
работана.
Отсутствие в настоящее время точных сведений о том, как ге-
нотипические изменения влияют на фенотипический состав популя-
Индивидуумы, популяции и виды 263
ции, и наоборот, затрудняет доскональное понимание эволюции.
Имеется два альтернативных способа временно обойти эту труд-
ную проблему: 1) допустить, что естественный отбор изменяет
частоты генов и изучать эволюцию на генетическом уровне,
2) допустить, что наблюдаемые в популяции различные фенотипы
создаются исключительно в результате взаимодействия мутацион-
ного процесса, рекомбинации, потока генов и рекомбинационных
нарушений и изучать эволюцию прежде всего на фенотипическом
уровне. Оба эти допущения резонны. Изменения частоты феноти-
пов в популяции, вызванные естественным отбором, приводят к
изменениям в популяции гамет и в аллелях, которые несут эти га-
меты, а изменения частот генов и генотипов приводят к фенотипи-
ческим изменениям. Тем не менее, когда генам и генотипам при-
писываются определенные приспособленности, как это было сдела-
но в гл. 4—9, или когда генотипический состав выводится из фе-
нотипических признаков, принимается два сильно упрощающих
допущения, которые обычно нельзя проверить экспериментально.
В гл. 4—9 мы изучили основные концепции того, что можно
назвать «генотипической» формой теории эволюции. Иначе гово-
ря, мы изложили наиболее яркие аспекты принятой теории, каса-
ющиеся процессов, которые обусловливают возникновение, под-
держание и трансформацию генотипической изменчивости в по-
пуляциях растений и животных. В следующих шести главах рас-
смотрим некоторые стороны того, что можно назвать «фенотипи-
ческой» формой теории эволюции. Иначе говоря, сосредоточим
внимание на разнообразии популяций организмов в пространстве
и во времени и исследуем их взаимодействия. Следует, однако,
помнить, что это лишь дидактический прием и что эволюционное
изменение затрагивает одновременно все уровни — от генов до
сообществ.
УРОВНИ ОРГАНИЗАЦИИ ЖИВОГО
Живые организмы населяют почти все уголки земного шара —
от тропиков до полярных широт и от глубин океана до больших
высот в атмосфере. Число их огромно, а разнообразие по типу,
форме и образу жизни кажется беспредельным. Все это много-
образие представляет собой выражение того высокого уровня
химической организации, которое мы называем жизнью. Нукле-
иновые кислоты, белки, липиды, вода, некоторые минеральные ве-
щества и углеводное «топливо», собранные в маленьких пузырь-
ках с полупроницаемыми мембранами, имеются у всех организ-
мов — от мельчайших сине-зеленых водорослей и бактерий до ги-
гантских китов.
Куда бы ни устремился наш взор — в небесную высь или в
глубины морей, в недра материи или в просторы Вселенной,—
везде находим множество систем организации, которые относятся
264
Глава 10
к разным наслаивающимся друг на друга уровням, взаимосвязан-
ным и невероятно сложным. Наблюдая и изучая это разнообразие
форм, мы невольно принимаем тот уровень организации, к кото-
рому принадлежим сами как индивидуумы, т. е. уровень отдель-
ного организма, за единицу измерения. Любая система ниже
этого уровня считается «маленькой» или «простой», а все, что вы-
ше — «большой» и «сложной». Однако индивидуальный орга-
низм — это такой уровень организации, который иногда бывает
трудно определить. Представление о том, что индивидуум не мо-
жет быть разделен без того, чтобы не утратить своих свойств «жи-
вого», применимо к большинству высших животных. Но большин-
ство растений и многих так называемых низших животных можно
разбить на более мелкие единицы, каждая из которых способна
регенерировать недостающие участки,— явление, используемое
при размножении растений с помощью черенков.
Далее, в некоторых случаях «индивидуумы» неспособны к
самостоятельному существованию, но должны агрегировать с дру-
гими индивидуумами своего или чужого вида. Наиболее ярким при-
мером этого служат лишайники. Лишайник состоит из водоросли
и гриба, живущих в тесном симбиозе. Водоросли, входящие в со-
став лишайников, обычно встречаются и сами по себе, но почти
все грибы, образующие с водорослями лишайники, встречаются
только в составе последних. Других примеров взаимозависимости,
выраженной в столь же резкой форме, как у лишайников, неиз-
вестно, однако можно составить длинный перечень паразитов и
симбионтов, существование которых прямо зависит от других ин-
дивидуумов, хотя подобная связь не всегда бывает взаимной.
Так, например, человеку для того, чтобы жить, необходимы не
только растения и животные, служащие ему пищей, но и кишечная
палочка {Escherichia coli) — бактерия, синтезирующая витамин К,
без которого человек не может существовать. В свою очередь ки-
шечная палочка не может жить в естественных условиях вне тол-
стого кишечника человека.
Примером организации другого типа служат колониальные
формы. Индивидуумы таких различных видов, как зеленые водо-
росли Volvox u Pandorina, пчелы и муравьи, существуют в тесной
связи с другими членами своего вида, причем каждый член коло-
нии выполняет определенную роль, жизненно важную для всех
остальных. У таких форм отдельные члены колонии уже не спо-
собны к самостоятельному существованию. Единицей служит ко-
лония, а не отдельные ее члены. Но ведь в сущности, если не счи-
тать гермафродитных самоопыляющихся растений и нескольких
партеногенетических растений и животных, каждый организм в от-
дельности неполон, поскольку он не может размножаться в оди-
ночку. Для размножения необходимы минимум два индивидуума
противоположного пола. Однако организмы обычно живут не изо-
лированными парами, а группами, состоящими из варьирующего
Индивидуумы, популяции и виды
265
числа особей разного пола и возраста, обитающих в каждой ме-
стности. Эти группы представляют собой скрещивающиеся попу-
ляции. Популяция ограничена в пространстве и является некой
реальностью только у видов, размножающихся половым путем.
Следующий тип организации — это сообщество. Популяция
представляет собой генетическую единицу, единство которой во
времени поддерживается происхождением составляющих ее осо-
бей и их размножением. Сообщество же — единица экологичес-
кая. Оно состоит из одной или нескольких скрещивающихся попу-
ляций, принадлежащих к одному или нескольким видам, приспо-
собленным к одной и той же среде. В сообщества входят расте-
ния и животные как одноклеточные, так и многоклеточные. В той
мере, в какой всякий организм изменяет среду, в которой он оби-
тает, он представляет собой один из факторов этой среды, а сле-
довательно, между членами данного сообщества существует оп-
ределенная взаимозависимость. Эта зависимость носит довольно
расплывчатый характер, так же как и физические границы сооб-
щества. Тем не менее сообщества — это экологические единицы,
соответствующие известным критериям и обладающие известной
непрерывностью во времени, и в качестве таковых они несомнен-
но существуют.
Одно из главных свойств материи, в том числе живой мате-
рии,— ее организованность. Индивидуумы, симбиотические и коло-
ниальные формы, скрещивающиеся популяции не являются ре-
зультатом случайности, а представляют собой продукт высокоин-
тегрированной системы организации. Но в то же время не всегда
легко определить все эти уровни организации. Причин этому мно-
го, и в каждом отдельном случае они различны. Можно, однако,
выдвинуть одну общую причину: в эволюции нет двух одинаковых
ситуаций. Качество и частота мутаций, система рекомбинации,
экологическая обстановка, к которой организмы должны при-
спосабливаться, различны для каждой популяции, каждой коло-
нии и каждого сообщества. Лжетсута в Северной Америке сплошь
покрывает площадь в тысячи гектаров, и выделить дискретные
размножающиеся популяции невозможно. В отличие от этого се-
ребристый клен ограничен влажными участками, где он образует
густые древостои. Кролики разбросаны парами по всей террито-
рии прерий и Большого Бассейна, тогда как луговые собачки, так-
же обитающие в этих областях, образуют колонии, состоящие из
сотен особей.
ЕДИНИЦЫ ОТБОРА
До сих пор обсуждение естественного отбора касалось того,
что обычно называют «индивидуальным отбором». Мы описывали
отбор в терминах альтернативных аллелей одного локуса или аль-
тернативных признаков особей, принадлежащих к одной популя-
266
Глава JO
ции; иначе говоря, мы рассматривали двух или более особей, раз-
личающихся ,по своей генетической конституции. Поскольку имен-
но так представлял себе естественный отбор Дарвин, этот тип от-
бора называют «дарвиновским отбором».
Естественный отбор может, однако, происходить не только на
уровне особей, но и на уровне других биологических единиц, при
условии что эти единицы удовлетворяют следующим критериям:
1) способны реплицироваться; 2) производят избыток единиц по
сравнению с тем их количеством, которое необходимо для заме-
щения родительского поколения, 3) их выживаемость коррелиру-
ет с их признаками; 4) у них имеется механизм, обеспечивающий
передачу этих признаков потомкам (см. гл. 3). Ниже рассматри-
ваются четыре единицы отбора, отличные от особей, и дается
оценка эволюционным последствиям отбора на этих уровнях. К
этим единицам относятся: 1) гаметы (гаметный отбор); 2) семей-
ные группы (отбор родичей); 3) популяции (групповой или
междемовый отбор) и 4) половой отбор.
Гаметный отбор
По своей генетической конституции гаметы отличаются от то-
го диплоидного организма, в котором они возникли; они продуци-
руются в избыточном количестве; обладают своими особыми при-
знаками; передают эти признаки (через зиготы) другим гаметам.
Следовательно, они удовлетворяют условиям, необходимым для
того, чтобы естественный отбор мог действовать на популяцию
гамет, созданных индивидуальным диплоидным организмом, не-
зависимо от отбора, действующего на диплоидных родительских
особей. Признаками гамет, которые должны подвергаться силь-
ному отбору, должны быть такие их черты, как подвижность спер-
матозоидов, жизнеспособность различных гаплоидных генотипов
гамет (например, их способность выжить в различных условиях
среды и продолжительность их жизни) и способность соединять-
ся с другой гаметой, чтобы образовать зиготу. Если, например,
у растений пыльцевые зерна одного типа могут дать начало пыль-
цевым трубкам, которые растут быстрее, чем пыльцевые трубки
других зерен, то пыльцевые зерна такого типа смогут оплодотво-
рить большее число яйцеклеток. Поскольку гаметы гаплоидны, от-
бору подвергаются все их гены, и, таким образом, гаметный от-
бор очень эффективен. Следовательно, можно ожидать, что любая
вредная мутация будет элиминироваться очень быстро, а любая
благоприятная мутация так же быстро распространится. Если,
однако, какой-либо аллель обладает плейотропным действием и,
давая преимущество гамете, в то же время снижает приспособ-
ленность диплоидного организма, то частота такого аллеля будет
поддерживаться на некотором промежуточном уровне.
Индивидуумы, популяции и виды
2&-г
Отбор родичей
Действие естественного отбора состоит в изменении частоты
генов в популяции. Частота данного аллеля повышается, когда
несущие его особи способны пережить неблагоприятные условия
среды и произвести большее .число потомков, чем те члены попу-
ляции, которые не содержат этого гена. Если, однако, особь, при-
надлежащая к данному виду, способна повысить выживаемость
родственных особей, несущих общий с ней аллель, то при этом
повысится также частота генов, которые содержит эта особь,
даже в случаях, когда выживаемость самой этой особи или число
ее потомков не изменились. Представим, например, трех кормя-
щихся на лугу луговых собачек из одного помета (сибсов), гете-
розиготных по одному и тому же аллелю, Представим далее,
что к ним бесшумно приближается хищник, который может
схватить любую из трех особей, причем остальные две ничего не
заметят. Но если одна из луговых собачек, заметив хищника, по-
даст сигнал тревоги, то она отвлечет этим внимание хищника на
себя и поэтому: 1) будет почти наверное съедена хищником и
2) даст возможность своим сибсам спрятаться в норку. Совершен-
но йсно, что подача сигнала в такой ситуации гибельна для дан-
ной особи (поскольку, привлекая к себе внимание хищника, она
повышает вероятность своей гибели), однако это повышает веро-
ятность выживания данного аллеля, так как гибнет лишь один из
трех его носителей. Если рассматриваемая черта (подача сигнала
тревоги) определяется именно этим аллелем, то можно предста-
вить себе, каким образом он мог бы распространяться в популя-
ции. Итак, мы видим, что некоторые черты, хотя они и гибельны
для своего носителя, могут тем не менее сохраняться отбором, ес-
ли они дают достаточное преимущество родственным ему особям.
Такие черты называют альтруистическими, а соответствующий тип
отбора — отбором родичей (kin selection).
Когда будет происходить отбор в пользу гена альтруизма? Он
будет происходить в тех случаях, когда повышение приспособлен-
ности родственных особей (поскольку вероятность наличия этого
гена для них выше, чем для других членов популяции) достаточно
велико, чтобы компенсировать снижение приспособленности альт-
руистической особи. Если же оно недостаточно велико, то данный
ген не подвергнется положительному отбору.
Таким образом, чтобы предсказать существование альтруизма,
важно знать степень родства между членами группы. Коэффици-
ентом родства между двумя особями называют вероятность того,
что обе они несут в любом локусе аллели, идентичные по проис-
хождению. Для диплоидной родительской особи и ее потомков
коэффициент родства равен у2, поскольку она передает потомку
половину своих хромосом. По этой же причине коэффициент род-
ства между потомками одних и тех же родителей также равен 72.
268
Глава 10
Для любого аллеля в любом локусе одного сибса шансы на то,
что родительская особь, от которой этот аллель был получен, пе-
редала его и другому сибсу, составляют 50%. Гены альтруизма,
определяющие альтруистический акт, который благоприятствует
одному из сибоов, будут сохраняться отбором в том случае, если
благодаря этому акту приспособленность этого сибса возрастет
на величину, вдвое большую той, на которую понизится приспо-
собленность альтруиста.
Наиболее яркие примеры отбора родичей наблюдаются у об-
щественных насекомых. Например, у медоносной пчелы (Apis
mellifera) имеется каста стерильных самок — рабочих особей. Ка-
ким образом естественный отбор, в основе действия которого ле-
жит передача приспособительных признаков из поколения в поко-
ление, мог бы создать целую касту стерильных особей? У медонос-
ной пчелы самец гаплоиден, а самка диплоидна. Коэффициент
родства между двумя рабочими особями равен 3/4. так как наборы
генов, полученные каждой из них от отцовской особи (половина
их генома), идентичны, а из генов, полученных от материнской
особи, идентична в среднем половина. В отличие от этого коэф-
фициент родства между материнской и дочерней особями равен
всего */2- Таким образом, выращивание сестринских особей может
быть более выгодным, чем выращивание собственных потомков.
В какой мере отбор родичей действует у человека? Здесь по-
ложение гораздо сложнее. Вследствие способности человека к
приспособительному поведению, трудно определить, ib какой мере
данный тип поведения определяется генотипом, а не культурным
наследием. Для всех цивилизаций характерно широкое распрост-
ранение и поощрение таких актов, которые направлены на благо
других людей, но могут дорого обойтись тому, кто их совершает.
Преобладание у человека таких типов поведения, непосредственные
результаты которых приносят больше пользы другим людям, чем
ему самому, заставляет предполагать существование для этого
какой-то общей основы — генетического предрасположения к не-
которому альтруизму. Тип альтруизма и его степень сильно варьи-
руют, а его конкретные формы, по-видимому, различны в каждой
цивилизации.
Принято считать, что на протяжении большей части ранней
истории человечества основной социальной группой была семья.
В таком случае альтруистические акты должны были благоприят-
ствовать родственным особям и, очевидно, действовал отбор ро-
дичей. С развитием умственных способностей и языка, сопровож-
давшего эволюцию вида Homo sapiens, увеличились возможности
для более сложных взаимоотношений. Одним из них является
реципрокный альтруизм — концепция, введенная Р. Триверсом
(R. Trivers). Реципрокный альтруизм означает, что какой-либо
альтруистический акт, который может оказаться выгодным для
неродственной особи, совершается в расчете на известное возна-
Индивидуумы, популяции и виды 269
граждение. Таким вознаграждением могло бы быть ожидание по-
мощи в будущем; им могла быть также признательность со сто-
роны остального сообщества. Поэтому бескорыстные действия не
всегда в самом деле бескорыстны. Будучи сопряжена с опреде-
ленной выгодой, способность к альтруизму может в соответствую-
щих условиях подвергаться отбору.
Групповой отбор
Представим себе популяцию животных, нуждающихся в каком-
либо возобновляемом ресурсе среды, запасы которого в каждый
отрезок времени ограничены, как, например, трава или какой-ни-
будь вид жертвы. Составляющие популяцию особи должны расти
и размножаться, а естественный отбор, как всегда, будет благо-
приятствовать тем фенотипам, которые производят наибольшее
число выживающих потомков. Может случиться, что популяция
израсходует все имеющиеся ресурсы, прежде чем они будут во-
зобновлены. В таком случае эта популяция вымрет. Представим
себе теперь другую популяцию, в которой вследствие отсутствия
соответствующих мутаций не возникло фенотипов, способных до-
чиста выесть весь ресурс. Такая популяция не только доживет до
того времени, когда этот ресурс возобновится, но и сможет засе-
лить области, которые прежде были заняты быстро размножив-
шимися, но ныне вымершими популяциями. Такой отбор, действу-
ющий на уровне групп и противостоящий индивидуальному отбо-
ру, называют групповым или междемовым отбором.
Для того чтобы групповой отбор мог действовать, должно
иметь место дифференциальное вымирание групп, состоящих из
«эгоистичных» членов, вслед за которым происходила бы реколо-
низация занимавшейся ими местности группами альтруистов. В то
же самое время поток генов не может быть слишком обширным,
так как в противном случае гены «эгоизма» наводнили бы коло-
нии «альтруистов», где им благоприятствовал бы индивидуальный
естественный отбор.
Групповой отбор — проблема, по которой существует много
разногласий. Нет никаких прямых данных, свидетельствующих о
том, что он действительно происходит; эволюцию большинства, а
может быть, и всех признаков организмов можно объяснить на
основании индивидуального отбора и (или) отбора роди-
чей. Однако отсутствие прямых данных еще не опровергает су-
ществования группового отбора. На самом деле некоторые при-
знаки, в особенности поведенческие, легче всего объяснить именно
с помощью группового отбора. Может ли групповой отбор дейст-
вовать в противовес индивидуальному естественному отбору —
вопрос иного рода, который в настоящее время еще не разрешен,
но вызывает горячие дискуссии.
270
Глава 10
Половой отбор
Если на каждую самку в данной популяции приходится по не-
скольку самцов, самка может выбирать самца по своему усмотре-
нию. Но если один из самцов способен отгонять других самцов,
прибегая либо к угрозам, либо к настоящей драке, в результате
чего ему удается оплодотворить одну или несколько самок, то
такой самец произведет больше потомков, чем те самцы, которых
он прогнал. (В экстремальных случаях другие самцы вообще не
будут участвовать в размножении.) Те признаки, которые делают
самца или самку более привлекательными для особей противопо-
ложного пола или более угрожающими для соперников одного с
ними пола и которые участвуют в обеспечивании ее или его ус-
пеха, будут иметь приспособительное значение и будут сохранять-
ся отбором. В этом состоит суть теории полового отбора, впервые
выдвинутой Чарлзом Дарвином в 1871 г. в его книге «Происхож-
дение человека» для объяснения различий между самцами и сам-
ками, наблюдаемых у многих видов животных, например причуд-
ливых перьев или огромных рогов, не имеющих, очевидно, общего
приспособительного значения.
В первоначальном изложении этой теории Дарвин подчеркивал
непосредственное столкновение как очень важный фактор отбора,
который может обусловить эволюцию таких структур, как рога у
оленей, шпоры на крыльях и на ногах у многих птиц, рога у
жуков-рогачей. Такого рода половой отбор, возникающий в ре-
зультате конкуренции между особями одного и того же пола, на-
зывается внутриполовым отбором. Во многих случаях между жи-
вотными происходят настоящие драки, однако возможно, что угро-
зы, выражающиеся в выставлении напоказ таких органов нападе-
ния, как рога или клыки, или же во взъерошивании шерсти и
перьев, с тем чтобы создать впечатление более крупных размеров,
играют более важную роль, чем настоящее нападение. Сверхаг-
рессивное поведение может понизить, а не повысить приспособлен-
ность, как у многих видов лосей, у которых настоящая драка мо-
жет привести к гибели обоих противников, или же дать возмож-
ность другим самцам увести самок за то время, пока происходит
драка. А поэтому структуры, которые можно использовать в дра-
ке, могут развиваться и в том случае, если драки как таковые
возникают редко, по крайней мере между представителями одного
и того же вида.
Внутриполовой отбор будет наиболее интенсивным у тех видов,
у которых конкуренция между представителями одного и того же
пола достигает наибольшей остроты. Дарвин заметил этой потому
придавал большое значение системе скрещивания того или иного
вида. Строгая моногамия при численном равенстве особей того и
другого пола — единственная система, для которой внутриполовой
отбор окажется неэффективным, при условии что данная пара со-
Индивидуумы, популяции и виды 271
храняется по крайней мере в течение всего сезона размножения.
Все другие системы скрещивания — полигамия, промискуитет и
численное неравенство полов — создают возможность по меньшей
мере для некоторого внутриполового отбора. Поскольку наиболее
распространенные системы скрещивания — это моногамия с из-
бытком самцов и полигиния (при которой один самец оплодотво-
ряет многих самок), отбор среди самцов должен происходить ча-
ще, чем отбор среди самок, и так оно и есть на самом деле.
Теория полового отбора, предложенная Дарвином, не получила
всеобщего признания. По мнению некоторых авторов, нахождение
брачных партнеров — это просто еще один аспект естественного
отбора. Если, например, какой-нибудь стерильный самец добьется
особенно больших успехов в отстранении других самцов и в спа-
ривании со всеми самками, он тем не менее не будет иметь потом-
ков и его гены не будут воспроизводиться. Успеха в размножении
может добиться также самец, имеющий яркую окраску (у птиц
или у насекомых), однако его потомки вследствие своей заметно-
сти будут, вероятно, в большей мере уничтожаться хищниками,
так что через одно поколение в популяции может оказаться мень-
ше, а не больше ярко окрашенных особей. Очевидно, половой от-
бор действует на одну из составных частей общей приспособленно-
сти, и эволюция полового диморфизма не может продолжаться,
если он оказывается гибельным для особи. Остается, однако, от-
крытым вопрос о том, возможна ли эволюция признаков, которые
создают преимущество только или главным образом для нахож-
дения брачных партнеров.
У дрозофилы известны мутанты с желтым телом вместо обыч-
ного серо-желтого. Когда «желтых» и серо-желтых самцов вводят
в популяцию нормальных серо-желтых самок, то серо-желтые
самцы оставляют гораздо больше потомков, потому что самки
редко подпускают к себе желтых самцов. Желтые же самки не
делают различия между желтыми и серо-желтыми самцами. Эти
данные показывают, что выбор брачных партнеров в самом деле
может оказать влияние на судьбу признака, который в остальном
нейтрален.
К проблеме полового отбора, играющего очень важную роль
в эволюции поведения и общественного образа жизни, мы вернем-
ся в гл. 16.
Культурная эволюция
До сих пор естественный отбор рассматривался применитель-
но к признакам, передающимся по наследству. Некоторые приз-
наки, однако, распространяются в популяции негенетическими спо-
собами. Мы будем называть это явление культурой, хотя этот тер-
мин часто используется в менее общем смысле для обозначения
лишь определенных видов деятельности в человеческом обществе
272
Глава 10
или даже в качестве признака, отличающего человека от других
животных.
Если принять наше определение, то культурными признаками
обладают многие виды животных. Перелетные птицы из года в
год и из поколения в поколение следуют по одним и тем же
маршрутам. Молодые луговые собачки учатся у старых членов
колонии различать границы своей колонии. Некоторые виды птиц
научаются разным диалектам основной песни своего вида от дру-
гих птиц, живущих по соседству.
Интересный пример распространения культурного признака
наблюдался в одном стаде японских макак (Масаса fuscata). Это
стадо приохотилось к новой пище — бататам. Одна полуторагодо-
валая самка, прежде чем съесть батат, стала не просто соскребать
с него песок, а отмывать его в море. Такое поведение переняли
другие молодые особи. Матери научились этому у своих подрас-
тающих детенышей и стали в свою очередь обучать такому пове-
дению более молодых потомков. В конце концов все стадо, за ис-
ключением старых самцов, которые мало общаются с остальным
стадом, стало мыть бататы, прежде чем съесть их.
Та же самая самка в возрасте 4 лет придумала еще одну
процедуру. Обезьян кормили пшеницей, которую бросали им на
песок, откуда они ее выбирали по зернышку. Самка же прибегла
к способу, которым пользуются старатели для отмывания золото-
го песка: она бросала горсть зерен, перемешанных с песком, в
воду и собирала всплывающее зерно, после того как песок опус-
кался на дно. Этот навык также распространился в стаде.
Совершенно очевидно, что на культурную эволюцию оказыва-
ют влияние генетические признаки. Способность к обучению име-
ет важное значение для культурной эволюции, а эта способность,
по крайней мере частично, обусловлена генетически. Кроме того,
между генетической и культурной эволюцией существует, воз-
можно, обратная связь. Такая обратная связь играет особенно
большую роль в эволюции человека. Многие в настоящее время
считают, что развитие различных видов активности, передаваемых
культурным путем (например, использование орудий или язык),
после того как оно стало возможным благодаря наличию опреде-
ленной структуры головного мозга, обусловило давление отбора,
способствовавшее дальнейшему развитию мозга, что в свою оче-
редь создавало условия для новых успехов в области культурного
развития. Прежде считалось, что все поведение человека целиком
определяется культурой, без всякого участия генетических фак-
торов. Теперь же, однако, представляется более вероятным, что
мы наследуем не только способность к культурной эволюции, нои
тенденцию или даже потребность в развитии определенных черт
культуры, таких, как брак, танцы или язык. Во всех известных
человеческих обществах наблюдаются эти черты, хотя их конкрет-
ные формы весьма разнообразны.
Индивидуумы, популяции и виды
273
Культурная эволюция отличается от эволюции генетической
рядом особенностей. Она носит ламаркистский характер, поскольку
приобретенные признаки могут передаваться по наследству. Кроме
того, эти признаки могут передаваться не только потомкам, но и
другим особям. Наконец, культурные признаки могут распростра-
няться в популяции гораздо быстрее, чем генетические признаки.
Между культурной и генетической эволюцией существует так-
же ряд сходных черт. Популяции различаются по своим культур-
ным признакам и эти различия могут, подобно потоку генов, пе-
редаваться от одной популяции к другой и со сходными резуль-
татами. Может также происходить культурный дрейф, при кото-
ром признаки изменяются случайным образом. И, разумеется, ес-
тественный отбор может действовать на культурные признаки
так же, как он действует на генетические признаки.
вид
До сих пор в этой главе говорилось о том, что живая материя
высокоорганизованна и что некоторые единицы ее организации —
гаметы, особи, семейные группы, а возможно, даже и популя-
ции — служат или могут служить независимыми единицами отбо-
ра, поскольку они обладают необходимыми для этого качествами
(гл. 3). Однако наиболее важная биологическая единица — это,
вероятно, вид. Вид представляет собой основную единицу класси-
фикации. Даже человек, не имеющий отношения к науке, без тру-
да поймет, что различные представители семейства кошачьих —
львы, тигры, пумы — явно отличаются друг от друга. Однако бо-
лее детальное изучение показывает, что определение и истолкова-
ние природы вида дело нелегкое.
Концепция вида
Группы организмов, называемые видами, выделяются уже дав-
но, однако в вопросе о том, что такое на самом деле вид, все еще
остается много неясного.
Существуют три главные концепции вида. Первая, философ-
ская и логическая концепция вида, восходит к Аристотелю. Вид —
Это категория мышления, точнее — некий класс (в логическом
смысле), к которому принадлежат все объекты (живые или нежи-
вые), обладающие определенными общими для всех них свойства-
ми. Так, например, «вилка» — это некий вид, поскольку все вилки
обладают рядом фундаментальных общих свойств — имеют зубцы
и ручку, устроенные таким образом, что с их помощью можно
протыкать и поднимать предметы. В остальном вилки могут раз-
личаться по форме и размерам и быть изготовленными из самых
разных материалов.
Вторая концепция, так называемый морфологический крите-
рий, представляет собой приложение логического определения ви-
18—2044
274
Глава 10
да к организмам. Виды определяются при этом строго на основа-
нии наличия у их членов определенных признаков— морфологи-
ческих, физиологических, поведенческих, биохимических,—которых
нет у членов других видов. Это была концепция вида, которой
придерживались Линней и большинство таксономистов XVIII и
XIX вв. и которой даже в наши дни строго придерживаются неко-
торые таксономисты.
Наконец, третья концепция — это так называемая биологичес-
кая концепция вида, согласно которой вид состоит из популяций,
особи которых в природных условиях потенциально способны скре-
щиваться между собой. В морфологической концепции вида глав-
ный упор делается на сходство и подобие, а в биологической
концепции — на скрещиваемость. В общем виды, определяемые на
основании этих двух концепций, в значительной степени совпада-
ют: особи, которые похожи друг на друга, часто скрещиваются
между собой, а особи, скрещивающиеся друг с другом, обычно
сходны между собой, так как у них имеется много общих генов.
Тем не менее совпадение наблюдается не во всех случаях. Ярким
примером служит половой диморфизм. Линней, на основании стро-
гого морфологического критерия, описал самца и самку кряквы
как различные виды. В других случаях диморфизм не связан с
полом. У многих видов цветковых растений разные экземпляры
имеют цветки разного цвета. У одного из представителей сложно-
цветных, Leavenworthina stylosa, некоторые экземпляры имеют бе-
лые, а другие — желтые цветки. Как белая, так и розовая поле-
вая редька принадлежат к одной и той же скрещивающейся попу-
ляции. Улитки вида Сераеа nemoralis, как было показано в гл. 9,
различаются по характеру полосатости раковины и т. п.
Вид можно рассматривать как самую крупную популяционную
единицу. Особи, входящие в данную популяцию, обычно скрещи-
ваются с представителями той же самой популяции. Время от
времени какое-нибудь животное удаляется от своей скрещиваю-
щейся популяции или же семя какого-нибудь растения относится
ветром на большое расстояние. В таких случаях особи, принадле-
жащие к разным популяциям, обмениваются генами, но только
(за редкими исключениями) с членами своего собственного вида.
Обычно, в особенности у высших животных, существуют опреде-
ленные преграды, препятствующие обмену генами между разными
видами. Эти преграды могут быть пространственными, временны-
ми, экологическими, поведенческими, цитологическими или гене-
тическими.
Высшие категории
Виды объединяются в категории более высокого ранга, назы-
ваемые родами, которые в свою очередь объединяются в семейст-
ва, отряды (порядки), классы, типы и царства, как показано ниже.
Индивидуумы, популяции и виды
275
Царство
Тип
Класс
Отряд (животные), порядок (растения)
Семейство
Триба
Род
Вид
Добавляя к этим названиям префиксы под или над (в систе-
матике животных) можно создавать промежуточные категории,
например подвиды, надсемейства и т. п.
_ 1 Произвольные’
Царство -в- Тип -»-Класс-»-Отряд-»-СемейстВО-«»-Род >
rid I Cl ирИп
Вид: все осоки
!--------------------------*----------------------1
Вид: все мыши
1------------------------*----------------------|
Вид: все подсолнечники
Популяция:
подсолнечники
Особи: мыши Особи: Особи: мыши
и осоки подсолнечники и осоки
1--------------------------,---------------------
Сообщество
Непроизвольные
категории
флора = Все растения
фауна = Все животные
Рис. 10.2. Соотношение между различными категориями.
Категории, которые мы обсуждали до сих пор, — особи, скре-
щивающиеся популяции, сообщества и виды — это естественные
реально существующие объекты, тогда как таксоны более высоко-
18*
276
Глава 10
го ранга представляют собой абстрактные логические категории
(рис. 10.2). Каждая из них — это класс объектов, в который вхо-
дят все категории более низкого ранга, имеющие определенные
общие признаки. Например, все виды млекопитающих, имеющие
когти и зубы определенного типа, относятся к роду Felis, в том
числе кошки, львы, тигры. Обратите внимание на то, что хотя род
Felis — удобная концепция, помогающая нам выразить одним сло-
вом определенные связи, и хотя, возможно, все виды Felis имеют
общее происхождение, род Felis лишен той осязаемой реальности,
которая присуща виду Felis leo — льву. Разные виды этого рода
обычно не могут скрещиваться друг с другом, они не испытывают
одних и тех же давлений отбора, не обитают в одной и то же сре-
де и, хотя, возможно, имели общее происхождение, не имеют об-
щего настоящего или будущего.
Иерархическая система категорий, в которой группы на каж-
дом уровне образуются путем объединения групп, стоящих на од-
ну ступень ниже, легла в основу методики таксономии. Она была
в значительной мере создана Линнеем в XVIII в. и сохраняется до
сих пор. Эта система дала возможность выявлять родственные
связи, но она часто оказывалась неадекватной для того, чтобы
формулировать некоторые заключения относительно эволюцион-
ных взаимоотношений организмов. Такая неадекватность привела
к тому, что некоторые ученые, в частности цитологи, генетики и
экологи, отказались от классической системы классификации и
создали свою собственную систему категорий. С другой стороны,
некоторые из особенно ортодоксальных систематиков в своих
таксономических исследованиях пренебрегали генетическими дан-
ными и данными о скрещивании или же не придавали им серьез-
ного значения. Однако конфликт этот надуманный. Цель система-
тика — навести порядок и создать способ систематизации и име-
нования организмов. Цель эволюциониста — объяснить разнооб-
разие организмов и их прошлую историю, а также разработать
общую теорию, позволяющую делать предсказания. Эволюционис-
ты не могли бы изучать разнообразие животного мира, если бы он
не был как-то систематизирован такоономистом, а таксономист
зависит от других специалистов, без данных которых он не может
понять свойства организмов. Мы вернемся к этому в гл. 18, при
обсуждении принципов и методов классификации.
В следующей главе рассматриваются некоторые пути возник-
новения новых видов и при этом обсуждаются более подробно
трудности, порождаемые применением морфологической концеп-
ции вида, а также так называемой биологической концепции вида.
ЛИТЕРАТУРА
Общие вопросы
Darwin С. 1871. The Descent of Man and Selection in Relation to Sex, London,
Murray.
Fisher R. A. 1958. The Genetical Theory of Natural Selection, New York, Dover.
Индивидуумы, популяции и виды
277
Fox R. 1971. The cultural animal. In: J. F. Eisenberg and W. S. Dillon (eds.), Man
and Beast: Comaparative Social Behavior (Smithsonian Annual III), pp. 273—
311.
Grant V. 1971. Plant Speciation, New York, Columbia University Press.
Maynard-Smith J. 1978. The Evolution of Sex, Cambridge, Cambridge University
Press. [Имеется перевод: Мэйнард Смит Дж. Эволюция полового размноже-
ния.— М.: Мир, 1981.]
Специальные статьи и обзоры
Hamilton W. D. 1964. The genetical evolution of social behavior, I, II, J. Theor.
Biol., 7, 1—52.
Kawai M. 1965. Newly acquired pre-cultural behavior of the natural troop of Japa-
nese monkeys on Koshima inlet, Primates, 6(1), 1—30.
King J. L., Jukes T. H. 1969. Non-Darwinian evolution, Science, 164, 788—798.
Kawamura S. 1963. The process of subculture propagation among Japanese maca-
ques. In: С. H. Southwick (ed.), Primate Social Behavior: An Enduring Prob-
lem, Princeton, N. J., Van Nostrand.
Lewontin R. C. 1970. The units of selection, Ann. Rev. Evol. Syst, 1, 1—18.
Maynard-Smith J. 1964. Kin selection and group selection, Nature, 201, 1145—1147.
Sammeta К- P. V., Levins R. 1970. Genetics and ecology, Ann. Rev. Genetics, 4,
469—488.
Глава 11
Модели видообразования
Одна из центральных проблем популяционной биологии —это
проблема происхождения видов. Дарвин дал своей знаменитой
книге название «О происхождении видов» (хотя он не разрешил
эту проблему), и с тех пор вопрос о том, каким образом возника-
ют новые виды, служит предметом многих научных дискуссий и
горячих споров. В наши дни профессор Гарвардского университета
Эрнст Майр (Ernst Мауг) достиг больших успехов в выяснении
этой проблемы, однако ряд вопросов все еще остается нерешен-
ным. Причина такого большого интереса к проблеме образования
новых видов заключается в том, что этот процесс представляет
собой один из решающих этапов эволюции.
Когда мы говорим о происхождении видов, мы имеем в виду
появление группы особей, которую мы называем видом и отли-
чаем от других таких групп. Однако принято считать, что вид воз-
никает в результате отбора особей, которые обладают определен-
ными новыми признаками и, размножаясь, дают начало виду. При
этом не обязательно исключается групповой отбор, но действие
его доказать трудно. В предыдущей главе мы определили вид как
группу потенциально скрещивающихся между собой популяций,
которые изолированы в репродуктивном отношении от других та-
ких групп. Это так называемая биологическая концепция вида.
Такое определение нельзя считать хорошим в абсолютном смысле,
поскольку выражение «изолированы в репродуктивном отноше-
нии» можно понимать по-разному и оно поэтому неоднозначно.
Кроме того, практически установить наличие или отсутствие у
двух данных популяций потенциальной способности к скрещива-
нию— занятие нелегкое и трудоемкое. Однако альтернативные
объяснения сопряжены с еще большими проблемами (гл. 12).
В конечном счете видообразование — это возникновение преград,
препятствующих обмену генами между близкородственными попу-
ляциями. Предполагается, что две популяции или группы популя-
ций, которые стали изолированными в репродуктивном отноше-
нии, приспособлены исключительно к своим локальным средам, и
внесение генов из одной такой популяции в другую будет снижать
приспособленность участвующих в этом особей.
Модели видообразования
279
СПОСОБЫ ВИДООБРАЗОВАНИЯ
Существует два главных способа возникновения новых видов
(табл. 11.1). Прежде всего — это постепенное превращение во
времени одного вида в другой, или так называемое филетическое
видообразование. Кроме того, возможно превращение одних видов
в другие в результате гибридизации и в различной степени слия-
ния видов. Ни одна из этих двух систем не связана ни с развити-
ем изолирующих механизмов, ни с увеличением числа видов.
Таблица 11.1
Возможные способы происхождения видов
I. Превращение видов (филетическое видообразование)
1. Аутогенная трансформация (в результате мутаций, отбора и т. п.)
2. Аллогенная трансформация (в результате интрогрессии от других видов)
II. Увеличение числа видов (истинное видообразование)
1. Внезапное видообразование (отдельные особи)
а. Хромосомные перестройки
б. Полиплоидия
2. Постепенное видообразование (популяции)
а. Симпатрическое видообразование
б. Парапатрическое видообразование
в. Аллопатрическое видообразование (географическое видообразование)
Второй главный способ видообразования—это процесс, с по-
мощью которого один предковый вид дает начало одному или не-
скольким видам, необязательно утрачивая при этом свою само-
стоятельность. Такой процесс называют увеличением числа видов,
или истинным видообразованием. В этой главе основное внимание
уделено именно второму процессу. Различают мгновенное и посте-
пенное видообразование, причем последнее можно разделить на
три главных типа: 1) аллопатрическое, или географическое видо-
образование; 2) парапатрическое видообразование; 3) симпатриче-
ское видообразование.
Среди гипотез, выдвигавшихся для объяснения процесса уве-
личения числа видов, или истинного видообразования, наибольшее
распространение получила так называемая географическая теория
видообразования, впервые сформулированная немецким биологом
Морицем Вагнером (Moritz Wagner) в 1868 г. В ней категоричес-
ки утверждается, что первый шаг в процессе видообразования —
репродуктивная изоляция, создаваемая физическим (географи-
ческим) разделением. Второй шаг — независимая эволюция -этих
изолированных в репродуктивном отношении популяций. Если
ареалы таких популяций затем вновь сливаются и если репродук-
тивная изоляция сохраняется благодаря развитию 'какого-либо
другого изолирующего механизма, например генетической несов-
местимости, то процесс видообразования считается завершенным.
Далее будут рассмотрены три главных типа постепенного видо-
образования, начиная с аллопатрического видообразования.
Гибридизация
Исключение
Смещение признаков
и видообразование
Рис. 11.1. Стадии процесса видообразования.
Исходная популяция (2) дает начало двум изолированным популяционным систе-
мам, которые (2) под действием разных давлений отбора эволюционируют неза-
висимо друг от друга. Когда (и если) их ареалы сливаются (3) целиком или ча-
стично, они могут гибридизировать {4—1), восстанавливая исходную ситуацию,
т. е. вновь превращаясь в один вид; или они могут взаимно вытеснять друг друга
(4—2), превращаясь в две географически изолированные популяционные системы;
или же они могут превратиться в два симпатрических вида (4—3). Последнее на-
блюдается также в том случае, если виды дифференцировались до слияния своих
ареалов. (Solbrig, 1970.)
Модели видообразования
281
Аллопатрическое видообразование
Процесс аллопатрического видообразования слагается из че-
тырех стадий (рис. 11.1): 1) разделение исходного генофонда на
две или более изолированные группы (изоляция); 2) независимая
эволюция двух генофондов (дифференциация); 3) вторичное слия-
ние; 4) конкуренция между новыми генофондами. Проанализиру-
ем каждую из этих стадий по отдельности.
А ПРЕРЫВИСТЫЙ АРЕАЛ ВИДА, ОБУСЛОВЛЕННЫЙ РАССЕЛЕНИЕМ
Изолят А
Б ПРЕРЫВИСТЫЙ АРЕАЛ ВИДА, ОБУСЛОВЛЕННЫЙ УХУДШЕНИЕМ
УСЛОВИЙ СУЩЕСТВОВАНИЯ
Рис. 11.2. Два способа аллопатрического видообразования.
А. В результате образования географических изолятов. Б. В результате разделе-
ния ареала, который прежде был непрерывным. (О. Т. Solbrig, 1966. Evolution
and Systematics, New York, Macmillan.)
Изоляция. Группа скрещивающихся между собой популяций
может оказаться разделенной на две или более изолированные
группы по ряду причин. Главная из них — изменение физической
среды, приводящее к сокращению ареала вида. Разрушение среды
человеком и нарушения других типов могут иметь одинаковый ре-
зультат. В сущности физическое разделение популяций, приводя-
щее к их частичной или полной изоляции, как и следовало ожи-
дать, явление совершенно обычное (рис.11.2). Изолированные или
полуизолированные популяции могут также образовываться в ре-
зультате расселения на большие расстояния, когда новая популя-
ция создается небольшим числом основателей.
Изоляция — необходимое, но не достаточное условие для ал-
лопатрического видообразования.
Дифференциация. Дифференциацией мы называем процесс, при
помощи которого две изолированные одна от другой популяции
282
Глава 11
эволюционируют в разных направлениях, так что они становятся
морфологически и (или) физиологически различными. Изоляция
необязательно влечет за собой дифференциацию, хотя часто дело
обстоит именно так. Факторы, регулирующие процесс дифферен-
циации, все еще нуждаются в более подробном исследовании.
Ч
Рис. 11.3. Силуэты гербарных экземпляров растений Potentilla glandulosa из четы-
рех выборок, выращенных в Стенфорде в одинаковых условиях.
Внизу изображен поперечный разрез через центральную часть Калифорнии при-
мерно на 38° с. ш. с указанием природных местообитаний популяций, из которых
были взяты выборки. Обратите внимание на изменение морфологии растений в на-
правлении с востока на запад, а также на частичное перекрывание ареалов под-
видов reflexa и hanseni. На поперечном разрезе показано также местонахождение
трех экспериментальных станций в Стенфорде, Матере и Тимберлайне. (Clausen
et al., 1940.)
Имеется, однако, много данных в пользу концепции о том, что
популяции довольно хорошо адаптируются в локальной среде. В гл.
4—9 были проанализированы динамика скрещивающихся популя-
ций и факторы, вызывающие их изменения, а также факторы, ко-
торые противодействуют изменению. Эти факторы—мутационный
процесс, рекомбинация и отбор,— оказывая независимо друг
от друга влияние на особей, 'принадлежащих к различным скре-
щивающимся популяциям, обусловливают несколько различные
приспособления к среде. Насколько различны эти приспособления,
Модели видообразования
283
зависит от степени различия между средами обитания разных
скрещивающихся популяций, от степени обмена генами между по-
пуляциями и в меньшей степени — от случайных явлений. Все это
приводит к тому, что разные скрещивающиеся популяции в пре-
делах одного вида слегка различаются генетически, физиологичес-
ки и морфологически. Чем больше обмен генами между двумя по-
пуляциями, тем теснее их близость, и наоборот. Однако обмен ге-
нами обычно очень невелик. Наиболее интенсивно типы изменчи-
вости популяций изучались на растениях.
Одно из наиболее подробно исследованных растений—это лап-
чатка Potentilla glandulosa, изучением которой занимались Йенс
Клаузен (Jens Clausen), Дэвид Кек (David Keck) и Уильям Хиси
(William Hiesey) более 30 лет, начиная с 1930-х гг. Potentilla
glandulosa — травянистое растение, распространенное по всей за-
падной части Северной Америки, где она образует ряд локальных
популяций. Клаузен, Кек и Хиси взяли выборки из большого чис-
ла калифорнийских популяций. По внешнему облику этот вид
можно разделить на четыре подвида (рис. 11.3): подвид
glandulosa, встречающийся на склонах Берегового хребта в Ка-
лифорнии и на изолированных участках в предгорьях Сьерра-Не-
вада; подвид reflexa, встречающийся на теплых солнечных скло-
нах, в предгорьях и на средних высотах (от 300 до 2000 м) в го-
рах Сьерра-Невада; подвид hanseni — луговая форма, встречаю-
щаяся на высоте от 1300 до 3000 м; и, наконец, подвид nevaden-
sis — высокогорная форма, встречающаяся на лугах и на склонах
гор на высоте от 3000 до 3600 м. Преград к скрещиванию между
этими подвидами Potentilla
glandulosa не существует,
если не считать преград,
создаваемых их простран-
ственным экологическим •
Выборки из нескольких популяций каждого из этих подвидов
были пересажены на три экспериментальные станции в Калифор-
нии примерно на 38° северной широты. Первая станция находится
в Стенфорде вблизи залива Сан-Франциско. Климат здесь мягкий,
морской, а станция расположена на высоте всего 30 м. Хотя зи-
мой случаются небольшие морозы, условия в общем благоприятны
для пышного роста, даже в зимнее время, потому что минималь-
ная температура остается довольно высокой. На Стенфордской
станции росту растения скорее могут повредить летняя жара и
засуха, чем зимние холода. Вторая станция находится в Матере, на
высоте 1400 м на западном склоне гор Сьерра-Невада. Климат
Матера благоприятствует активному росту растений только в пе-
риод с мая по октябрь. Весна обычно наступает в начале апреля,
хотя заморозки бывают до самого начала июня. Лето мягкое, с
теплыми днями и прохладными ночами, дождей выпадает мало.
Осень прохладная и сухая. Снегопады случаются с октября по
апрель; толщина снежного покрова редко превышает 60—90 см.
284
Глава ]1
Третья станция расположена в Тимберлайне, в горах Сьерра-Не-
вада на высоте 3050 м. Здесь климат очень суровый, морозы пре-
кращаются лишь на 3—бнед в году, причем и в этот период могут
ударить в любой момент. Снежный покров достигает толщины
3—6 м.
Этот эксперимент дал следующие результаты. Прибрежная
форма подвида glandulosa достигла наибольшей высоты в Стен-
форде, в Матере росла хуже и не выдерживала зимы Тимбер-
лайна (рис. 11.4). Подвид reflexa лучше всего рос в Матере, т. е.
на своей родине, был менее мощным в Стенфорде, а в Тимбер-
лайне также погиб. Подвид hanseni в Стенфорде зимой впадал в
состояние глубокого покоя, несмотря на мягкую зиму, лучше все-
го рос в Матере и выжил в Тимберлайне, хотя на больших вы-
сотах семена его редко вызревали. Наконец, подвид nevadensis
едва выжил в Стенфорде, где он зимой находился в состоянии
покоя, достигал наибольшей мощности в Матере, а в Тимберлай-
не рос хорошо и систематически завязывал семена. Таким обра-
зом, четыре морфологических подвида этого растения представ-
ляют одновременно и четыре различных физиологических типа.
Кроме того, каждый подвид функционировал — в отношении рос-
та и продукции семян — наилучшим образом на той станции, ус-
ловия которой были наиболее близки к условиям его родного
местообитания, за исключением подвида nevadensis, функциони-
ровавшего в Матере лучше, чем в Тимберлайне. Однако подвид
nevadensis был единственным, способным нормально существовать
в крайне суровых условиях на станции в Тимберлайне.
Дифференциация популяций может также происходить при
очень небольших расстояниях между ними, как показали английс-
кий эколог Энтони Брэдшоу (Anthony Bradshaw) и его сотрудни-
ки на примере Agrostis tenuis. Этот широко распространенный мно-
голетний пастбищный злак растет в разнообразных местообитани-
ях в умеренных областях. В Великобритании он растет на отвалах
в районе заброшенных шахт и на соседних с ними лугах. Почва
на некоторых из этих отвалов содержит много свинца, меди или
цинка и токсична для большинства растений. Поэтому здесь мо-
гут расти лишь немногие виды растений, в том числе Agrostis
tenuis, у которого возникли популяции, толерантные к этим ме-
таллам. Эта толерантность, очевидно, не индуцирована условиями
выращивания, так как она не утрачивается при выращивании в
отсутствие высокого содержания металлов и передается по на-
следству. В результате подробного изучения популяций в районе
Гогинанокого свинцового рудника в Уэльсе Брэдшоу показал, что
ширина переходной зоны между толерантными к свинцу и нето-
лерантными популяциями составляет менее 20 м. Брэдшоу и его
сотрудники показали также, что растения по обе стороны от пе-
реходной зоны обладают такими же уровнями толерантности и
нетолерантности, как и растения горадо более удаленные от
Модели видообразования
285
В СТЕНФОРДЕ, В МАТЕРЕ, В ТИМБЕРЛАЙНЕ,
высота 30м высота~1400м высота 3000м
Рис. 11.4. Поведение четырех экотипных подвидов Potentilla glandulosa в трея
резко различающихся средах на экспериментальных станциях института Кариег»
в Стенфорде, Матере и Тимберлайне.
Выборки четырех подвидов были взяты: nevadensis — из местности, расположен-
ной на высоте 3000 м; hanseni — на высоте 1380 м; reflexa—из предгорий Сьер-
ра-Невады на высоте 750 м; glandulosa — с внешних склонов Берегового хребта
в Калифорнии на высоте 270 м. (Clausen et al., 1940.)
286
Глава 11
этой зоны (рис. 11.5). Анализ экологических и исторических фак-
торов показывает, что толерантные к свинцу формы должны бы-
ли возникнуть недавно и появились буквально на расстоянии
нескольких метров от нетолерантных форм, в пределах зоны рас-
А
Рис. 11.5. Гистограммы, показывающие распределение по толерантности к метал-
лу в двух рядах популяций Agrostis tenuis, расположенных по границе областей
с высоким содержанием в почве свинца (Л) или меди (5). (Bradshaw et al., 1965.)
„ .Расстояние между
участками, м
Показатель выносливости
пространения пыльцы последних. Однако давления отбора, по-ви-
димому, так сильны, что гибридам нигде не удается обосноваться.
Они не могут выдержать высокое содержание в почве свинца по
одну сторону границы, но не могут выдержать и конкуренцию с
растениями, растущими по другую ее сторону. Невозможность вы-
живания гибридов создает в данном случае абсолютную или поч-
ти абсолютную преграду обмену генами.
Можно привести много других примеров локального приспо-
собления популяций как у растений, так и у животных. Следует
Модели видообразования
287
ожидать, что возникновение различных сочетаний генов, определя-
ющих различные адаптивные комбинации, приведет в качестве
побочного эффекта к снижению возможностей скрещивания меж-
ду популяциями. Иными словами, следует ожидать, что у попу-
ляций, развивающихся в изоляции, в конечном счете возникнет та
или иная репродуктивная изоляция. Рис. 11.6 и 11.7 иллюстриру-
ют возможности скрещивания между двумя представителями Не-
Полностью плодовитые гибоиды
Частично стерильныр гибриды
Дженнер
Пт. Хоэ
Те кате
Мюир-Бич
(стелющийся)
; •'У’ - у •Сан-Бернардино
Этиванда (хлопьевидно-
::\7.-.';'опушеннь1й)
Пала (хлопьевидно-.’
опушенный) Анца (желтые.-ул-’
язычковые цветки)
Рис. 11.6. Скрещивания между популяциями в пределах вида Layia platiglossa-
Обратите внимание на то, что некоторые популяции приобрели новые морфоло-
гические признаки. Тем не менее репродуктивная изоляция между ними (стериль-
ность гибридов) еще только зарождается. (Jens Clausen, 1951.)
п=7). Пачеио Пасс
Холон (желтые
:Мл==7}:- язычновые цветки)
;:Северная Калифорния
Южная Калифорния
liantheae (сем. сложноцветные), изучавшимися Йенсом Клаузе-
ном (Jens Clausen), у которых проявляется некоторая стериль-
ность между географически разделенными популяциями.
Популяции из противоположных краевых местообитаний ареа-
ла данного вида (как в географическом, так и в экологическом
смысле) будут различаться в наибольшей степени. Благодаря
обмену генами между промежуточными популяциями вид сохра-
няет известную связность. Однако по краям ареала распростране-
ния вида процесс дифференциации ускоряется. Подходящих место-
обитаний здесь часто оказывается мало, и, кроме того, они бы-
вают изолированы. Например, там, где лес переходит в луга,
обычно можно встретить небольшие перелески посреди луга. То
же самое можно наблюдать у верхней границы леса на горах.
Популяция какого-либо лесного вида, обитающая в одном из пе-
релесков в степи, оказывается изолированной от других скрещи-
вающихся популяций. Такая популяция называется географичес-
ким или периферическим изолятом. В периферических изолятах
обмен генами перестает действовать в качестве связующей силы и
288
Глава 11
популяция может развиваться в сторону достижения равновесия
со своей средой с большей скоростью. Поскольку среда обитания
изолятов краевая, для данного вида, как мы видели, естественный
отбор будет в значительной степени изменять периферические по-
пуляции. Со временем между изолятом и первоначальным видом
может возникнуть та или иная из многочисленных форм репро-
дуктивной изоляции. В тех случаях, когда это происходит, изолят
превращается в новый вид, адаптированный к жизни в промежу-
----- F-i полностью плодовито; F2 жизнеспособно
----- F, полностью плодовито; жизнеспособность F2 понижена
-----F-|4acTH4HO стерильно; жизнеспособность F2 понижена
Рис. 11.7. Результаты скрещиваний между популяциями Layla glandulosa.
В отличие от L. platiglossa у этого вида уже возникла некоторая гибридная сте-
рильность между популяциями. (Jens Clausen, 1951.)
точном местообитании, в данном случае — в перелесках посреди
лугов (рис. 11.8).
В действительности периферические изоляты очень редко пре-
вращаются в новые виды. Причин этому много. Виды заселяют
периферические местообитания, к которым они не слишком хоро-
шо приспособлены, в такие годы, когда условия среды чрезвы-
чайно благоприятны. Когда же условия ухудшаются, эти пришель-
цы не выдерживают конкуренции с коренными обитателями и вы-
мирают. Кроме того, для процессов, ведущих к видообразованию,
необходим ряд условий: благоприятные мутации и (или) реком-
бинации, развитие какого-либо изолирующего механизма, а также
Модели видообразования
289
Рис. 11.8. Некоторые факторы эволюции, действующие при наличии географиче-
ской изоляции в пространстве и изменений во времени.
В момент времени t0 популяция 1 была адаптирована к жизни и росту в местно-
сти А со средним значением некоторого признака, равным х. В момент времени
<1 популяция 1 дала начало популяциям 1, 2 и 3 в местностях А, Б и В. В местно-
сти Б естественный отбор благоприятствует среднему значению признака, равно-
му z, в А продолжает благоприятствовать среднему х, а в В — среднему w. По-
скольку местности А и Б расположены очень близко одна от другой и условия
среды в них сходны, между ними происходит интенсивный обмен генами и на-
правленный отбор в них не очень различается; что же касается местности В, то
там наблюдается противоположная ситуация. (Solbrig, 1970.)
условия, делающие возможным выживание изолированной попу-
ляции; в природе подобное сочетание условий встречается редко,
Видообразование может также происходить и в периоды со-
кращения ареала данного вида. Если вследствие неблагоприятных
условий среды вид сокращает область своего распространения, то
эта область может утратить непрерывность и оказаться разбитой
на два или несколько изолированных районов. Мы уже упомина-
19-2044
290
Глава 11
ли о том, что популяции, занимающие местообитания у противо-
положных границ ареала, обычно довольно сильно различаются;
иногда эти различия достигают такой степени, что при скрещива-
ниях между особями из таких популяций наблюдается некоторое
понижение плодовитости (см. рис. 11.6). Поскольку обмен генами
может при этом происходить через промежуточные популяции,
целостность вида не нарушается. Однако в случае внезапного ис-
чезновения этих промежуточных популяций различия между кра-
евыми популяциями приобретают огромное значение, и, если им
удается выжить, они нередко оказываются на пути к превраще-
нию в новые виды (ом. рис. 11.7).
Факторы, вызывающие слияние ареалов. Факторы, ведущие к
перекрыванию ареалов двух прежде изолированных популяций,
сходны с теми, которые ведут к их разделению. В основном это
изменения среды, вызывающие расширение ареала или миграцию
популяций в область, которая прежде была незанятой. Эволюция
популяций может также привести к физиологическим и морфоло-
гическим изменениям (например, к развитию более совершенного
механизма распространения семян), которые прямо или косвенно
ответственны за расселение. Этот этап не представляет собой не-
пременного условия в процессе видообразования. Однако, когда
он имеет место, он ведет к важным взаимодействиям между дву-
мя единицами, которые прежде были разделены.
Наложение друг на друга ареалов популяций, при которых их
члены сближаются настолько, что могут скрещиваться, и возника-
ющие в результате этого взаимодействия составляют последнюю
ступень процесса видообразования. Две встречающиеся таким об-
разом популяции могли дифференцироваться физиологически и
генетически, так что они будут обладать разными экологическими
требованиями и их гаметы уже не смогут сливаться и давать жиз-
неспособные зиготы. В таких случаях процесс видообразования
можно считать завершенным.
Однако если встретившиеся популяции имеют одинаковые эко-
логические требования и, кроме того, способны к гибридизации,
то они вступают во взаимодействие друг с другом. Это взаимодей-
ствие может привести к одному из трех результатов:
1) один из видов возьмет верх над другим, и восстановятся
прежние неперекрывающиеся ареалы;
2) в результате отбора два вида так поделят между собой
среду, что конкуренция между ними прекращается ив то же
время возникает та или иная преграда, препятствующая скрещи-
ванию между ними, поэтому гибридов образуется мало или не
образуется вовсе;
3) две популяции гибридизируют и образуют одну интерфер-
тильную популяцию.
Конкурентное исключение. Когда две популяции, бывшие до
того разделенными, вступают в контакт друг с другом, члены
Модели видообразования
291
разных популяций начинают взаимодействовать между собой. Да-
же при наличии поведенческой и (или) генетической изоляции
особи из разных популяций часто нуждаются в одних и тех же
или сходных ресурсах, таких, как пища, пространство, места для
гнездования и т. п. Если между этими популяциями еще не воз-
никла репродуктивная изоляция, то их особи могут также делать
попытки к скрещиванию друг с другом. Одна из популяций может
обладать столь большим преимуществом в конкуренции, что со-
существование популяций окажется невозможным.
Каждая скрещивающаяся популяция обладает своим особым
набором требований к таким условиям среды, как тип местооби-
тания, пища, наличие воды и т. п. Во всех случаях, когда две
популяции предъявляют одинаковые требования к некоторым или
ко всем факторам среды, что приводит к столкновению их инте-
ресов, между популяциями имеет место конкуренция за эти ре-
сурсы. Две популяции, находящиеся в состоянии конкуренции,
стремятся вытеснить одна другую, если только требования одной
из них или обеих не изменяются таким образом, что конкуренция
ослабевает. В этом состоит сущность так называемого принципа
исключения, сформулированного советским биологом Г. Ф. Гаузе
в 1934 г.: «Две формы, предъявляющие совершенно одинаковые
требования к среде, не могут сосуществовать в течение неопреде-
ленно долгого времени; в конечном счете одна форма вытеснит
другую».
Принято считать, что большинство видов, изначально входящих
в состав каждой данной биоты, находится в состоянии динамичес-
кого равновесия. Это означает, что они поделили между собой
среду таким образом, чтобы наносить наименьший ущерб друг
другу. Конечно, какая-то конкуренция за факторы среды всегда
существует, потому что ресурсы среды ограниченны. Например,—
мы берем крайний случай — всем растениям необходимо некото-
рое пространство для роста, и совершенно очевидно, что на том
месте, где растет одно растение, никакое другое растение расти не
может. Свет также необходим всем растениям. В лесу компоненты
различных растительных ярусов — деревьев, образующих лесной
полог, кустарников и травянистых растений — так подогнали свои
потребности в необходимом им для фотосинтеза солнечном свете,
что все они могут сосуществовать. Большинство деревьев не мо-
жет расти в тени, тогда как кустарники и травянистые растения
способны к этому; причем травы часто обходятся меньшим коли-
чеством света, чем кустарники. Точно так же всем растениям не-
обходима вода. Однако некоторые из них могут жить при скудном
снабжении водой, а другие нет; причем нередко приспособления,
делающие возможным сосуществование, основаны на относитель-
но небольших различиях в потребностях в воде. Например, в се-
верном дубово-буковом лесу серебристый клен всегда встречается
в участках с наиболее влажной почвой.
19*
292
Глава 11
Однако конкуренция особенно хорошо выявляется там, где
равновесие отсутствует. Например, любое возделываемое поле
или сад, если их перестать обрабатывать, очень быстро изменяют
свой флористический состав. Выращивавшиеся на них культуры
обычно оказываются неспособными к размножению и их вытес-
няют однолетние растения, которые адаптировались к нарушен-
ным условиям, возникшим под влиянием обработки. Но спустя
несколько лет, по мере того как состав и структура почвы стаби-
лизируются, однолетняя растительность постепенно замещается
растительностью, типичной для данной области. Растения, непред-
намеренно завезенные человеком с других материков, — это, как
правило, сорняки сельскохозяйственных культур, неспособные
конкурировать с местной растительностью в естественных усло-
виях. Их распространение ограничено полями и садами, обочи-
нами дорог и другими такого рода местообитаниями, созданными
человеком. Местная растительность, адаптированная к другому на-
бору условий, не может расти в этих нарушенных местообитаниях.
Исключение, т. е. элиминация одной популяции,— один из воз-
можных результатов перекрывания ареалов. Вытесняющей конку-
рента популяцией может быть как предковая, так и популяция,
происходящая от нее. Другой результат перекрывания ареалов —
это смещение признаков.
Смещение признаков. Только в тех случаях, когда области, в
пределах которых расселяются два сходных вида животных или
происходит эффективное опыление двух сходных видов растений,
перекрываются, можно с абсолютной уверенностью считать, что
две самостоятельные, но близко родственные скрещивающиеся по-
пуляции действительно представляют собой два отдельных вида.
В таких случаях сомнений быть не может: если между ними воз-
никла репродуктивная изоляция, они не будут смешиваться; если
же они смешиваются, значит репродуктивная изоляция еще не
установилась.
Когда два таких близко родственных вида животных или рас-
тений, возникших аллопатрически, вновь встречаются в пределах
своих ареалов, некоторые особи могут ошибочно принять предста-
вителей других видов за своих и скрещиваться с ними; потомки
от таких скрещиваний, если они получатся, стерильны или почти
стерильны. Поскольку эти особи производят очень мало потомков
или не производят их вовсе, они и их генотипы элиминируются
естественным отбором. В пределах каждой популяции существу-
ет фенотипическая изменчивость. Ошибки в опознавании скорее
всего случаются среди тех особей, которые наиболее сходны с
другим видом. Поэтому те особи каждого вида, которые меньше
похожи на членов другого вида, имеют больше шансов произвести
жизнеспособное потомство. В результате с течением времени два
рассматриваемых вида будут все сильнее отличаться один от дру-
гого. Это явление получило название смещения признаков. По-
Модели видообразования
293
скольку смещение признаков возможно только в случае совмест-
ного существования двух видов, популяции, живущие раздельно,
различаются меньше, чем популяции, живущие вместе. Кроме то-
го, поскольку животные в отличие от растений активно выбирают
себе партнеров, смещение признаков более заметно проявляется у
животных, хотя у растений смещение признаков может привести
к развитию структур, препятствующих оплодотворению чужим ви-
дом.
Явление смещения признаков можно проиллюстрировать на
примере двух видов поползней — Sitta пеутауег и Sitta tephrono-
ta. Первый из этих видов занимает область от Балканского полу-
острова и доходит на востоке до западной половины Ирана, а
второй — от Туркменской до Армянской ССР. В нескольких рай-
онах Ирана ареалы этих двух видов перекрываются. В тех частях
ареалов, где виды встречаются по отдельности, они очень сходны
друг с другом, но в зонах перекрывания у S. пеутауег отмечает-
ся явное уменьшение общих размеров, длины клюва и выражен-
ности полос по бокам головы, тогда как у 5. tephronota эти при-
знаки не изменяются.
Гибридизация. Третьим результатом слияния ареалов может
быть гибридизация, т. е. объединение генофондов двух популяций.
Этот вопрос подробно рассматривается в гл. 12.
Парапатрическое видообразование
У малоподвижных или неподвижных организмов, таких, как
растения, роющие (т. е. обитающие в подземных норах) грызу-
ны или нелетающие насекомые, видообразование может происхо-
дить путем возникновения уникальных в генетическом отношении
организмов, которые: 1) способны использовать слегка отличные,
но физически смежные местообитания и 2) репродуктивно изоли-
рованы от остального вида. Парапатрическое видообразование,
возможно, встречается не так часто, как аллопатрическое, но, в
частности у растений, оно может быть более распространенным,
чем это принято считать. Для иллюстрации парапатрического ви-
дообразования можно привести два примера — один из царства
животных, а другой из царства растений.
Spalax ehrenbergi — слепыш, обитающий в Старом Свете. День
он проводит в вырытых им норах. Самцы охраняют свои норы и
окружающую местность. Однажды устроив себе нору, животное
остается в ней в течение большей части, если не всей своей жиз-
ни. Когда молодые особи достигают определенного возраста, они
покидают гнездо и устраивают собственные норы. Израильский би-
олог Эвиатар Нево (Eviatar Nevo) и его сотрудники, очень под-
робно изучавшие вид S. ehrenbergi, установили, что он представ-
294
Глава 11
Негев
Рис. 11.9. Распространение разных хромосомных форм Spalax ehrenbergi. (Nevo,
Shkolnik 1974).
ляет собой комплекс из нескольких близкородственных видов,
различающихся по числу хромосом.
В этом регионе встречаются четыре хромосомные формы
(рис. 11.9): 1) в горах Верхней Галилеи, во влажном и субгумидном
климате обитает форма с 52 хромосомами; 2) вид-двойник с 54 хро-
мосомами предпочитает области с климатом от субгумидного до
влажного, но с более низкими температурами, в окрестностях го-
Модели видообразования
295
•ры Мейрон и Голанских высот; 3) форма с 58 хромосомами в го-
рах Нижней Галилеи и в Центральном Израиле—область с кли-
матом от субгумидного до полузасушливого; 4) вид-двойник с
60 хромосомами в области с климатом от полузасушливого до
засушливого — в Самарии на Иудейских горах. Эти четыре вида-
двойника интенсивно изучались Нево и его сотрудниками в отно-
шении ряда морфологических, поведенческих и биохимических при-
знаков. Результаты показывают, что эти четыре хромосомные фор-
мы действительно очень сходны. Так, например, помимо большого-
морфологического сходства и одинаковых приспособлений к под-
земному образу жизни они, по-видимому, почти идентичны по-
своим генам, о чем свидетельствуют данные электрофоретического
исследования белков (96% исследованных белков, соответствую-
щие 25 локусам, идентичны). Но вместе с тем между этими че-
тырьмя формами обнаружены физиологические различия: ско-
рость метаболизма у них понижается при переходе от обитателей
влажных северных к обитателям засушливых южных районов.
Как возникли эти формы и существует ли между ними репродук-
тивная изоляция? По счастью, на оба эти вопроса можно дать
ответ. Spalax ehrenbergi, вероятно, произошел от предковой формы
S. mimtus, которая обитала в этой области в ледниковую эпоху и
ископаемые остатки которой найдены в отложениях, относящихся
ко времени, отделенному от настоящего 0,5 млн. лет. Ископаемые
формы, неотличимые от современных S. ehrenbergi, найдены в отло-
жениях вблизи Иерусалима, датируемых 250 000 лет до настоящего
времени. К сожалению, у ископаемых форм невозможно опреде-
лить число хромосом. Морфологические различия, наблюдаемые
между предковым видом S. mimtus и происходящим от него
S. ehrenbergi, — результат постепенного видообразования. Что ка-
сается хромосомных рас, то они, по-видимому, возникли в более
недавнем прошлом путем парапатрического видообразования. Ка-
кая из этих рас была предковой, сказать нельзя.
Хотя хромосомные расы различаются по своим физиологиче-
ским предпочтениям, в зоне контакта они живут «на виду» друг
у друга. Поэтому представители разных хромосомных рас имеют
возможность скрещиваться между собой. Такие скрещивания мож-
но проводить в лаборатории. Гибриды, содержащие промежуточ-
ное число хромосом, обладают пониженной плодовитостью и мо-
гут скрещиваться с любой из родительских форм. В природе гиб-
риды встречаются в зонах контакта, но лишь изредка и только
в очень узкой полосе. Нево показал, что самки умеют отличать
самцов с таким же, как у них, числом хромосом от самцов с иным
числом хромосом. Сочетание пониженной плодовитости гибридов
и умения самок различать разные хромосомные расы приводит к
тому, что обмена генами практически не происходит. Все это сви-
детельствует о парапатрическом видообразовании, когда вид рас-
селялся на новые территории, которые экологически слегка отли-
296
Глава 11
I____________I___________i________
40 50 60
Длина внутреннего листочка обертки
17 20 23 26 29 32 4 5 6
Ширина язычка
Число цветков
Рис. 11.10. Различия между Stephanomeria exigua ssp. coronaria и S. tnalhearensis.
Показаны диапазоны изменчивости, средние значения и стандартные отклонения
для длины цветка, длины внутренних листочков обертки, длины семянки и шири-
ны язычка (за единицу принята 1/6 мм). Показано также распределение по чис-
лу цветков. (Gottlieb, 1973.)
чались от его родной территории, после чего возникали хромосом-
ные изменения, возможно, повлекшие за собой также мелкие фи-
зиологические изменения. Репродуктивная изоляция, порождаемая
хромосомными и экологическими различиями, затем усиливалась
в результате развития способности к распознаванию разных хро-
мосомных рас.
Модели видообразования
297
Примером парапатрического видообразования у растений слу-
жит пара очень близко родственных видов сложноцветных Stepha-
nomeria exigua ssp. coronaria и S. malheurensis, усиленно изучав-
шаяся американским эволюционистом Лесли Готлибом (Leslie
Gottlieb).
S. malheurensis произрастает только в одной местности в запад-
ной части штата Орегон, где в 1973 г. он был представлен менее
чем 250 особями. Этот вид отличается, хотя и очень незначитель-
но, от своего предка S. exigua ssp. coronaria по пяти морфологиче-
ским признакам (рис. 11.10). Кроме того, примерно у 98% расте-
ний S. exigua ssp. coronaria цветки розовые, а у 2% цветки белые,
тогда как у S. malheurensis 80% растений имеют розовые, а 20% —
белые цветки. Изучение сходства по генному составу, проводив-
шееся методом электрофореза белков, показало, что все или боль-
шинство аллелей, имеющихся у S. malheurensis, имеются также у
S. exigua ssp. coronaria, но не наоборот. Между этими двумя ви-
дами существуют некоторые физиологические различия в скорости
роста и условиях, необходимых для прорастания, но не ясно, име-
ют ли эти различия какое-нибудь значение, однако указанные
виды репродуктивно изолированны. Для S. malheurensis характер-
на высокая степень самоопыления, а это означает, что обмен пыль-
цой между ним и генетически самонесовместимым и перекрестно-
опыляющимся S. exigua ssp. coronaria резко ограничен. Кроме
того, когда растение одного вида опыляется пыльцой другого ви-
да, оно производит только половину нормального количества
семян. Наконец, у гибридов Fb если они и образуются, жизнеспо-
собно только 25% пыльцевых зерен, потому что среди различий,
существующих между этими видами по структуре хромосом, есть
одна реципрокная транслокация.
Все эти данные указывают на то, что процесс видообразования
в двух рассмотренных случаях протекает очень сходно. По краям
ареала распространения вида, но не в изоляции, появляются особи,
обладающие как новыми физиологическими признаками, которые
впоследствии дают им возможность занять новую экологическую
нишу, так и известной степенью репродуктивной изоляции. Во
всяком случае, у слепышей репродуктивная изоляция была в даль-
нейшем усилена отбором. В то время как при аллопатрическом
видообразовании дифференциация предшествует развитию изоля-
ции, при парапатрическом видообразовании оба процесса происхо-
дят одновременно или почти одновременно.
Симпатрическое видообразование
Когда репродуктивная изоляция предшествует дифференциации
и когда она возникает в пределах одной популяции или в преде-
лах области распространения популяции, то имеет место симпатри-
ческое видообразование.
298
Глава 11
Симпатрическое видообразование — явление редкое. Кроме
того, его не всегда можно отличить от парапатрического видооб-
разования, поскольку лишь в редких случаях удается установить,
предшествовала ли изоляция дифференциации, или же они про-
исходили одновременно.
Наиболее хорошо известные примеры симпатрического видооб-
разования относятся к насекомым, паразитирующим на растениях.
Техасский биолог Ги Баш (Guy Bush), подробно изучавший эту
проблему, считает, что симпатрическое видообразование, возмож-
но, происходит только у паразитических и паразитоидных видов
животных, потому что они обычно спариваются на хозяине. Одна-
ко число таких видов превышает 500000. Репродуктивная изоля-
ция может возникнуть в результате одного лишь перехода на но-
вый вид хозяина. Популяции, оказавшейся на новом хозяине, надо
теперь будет приспосабливаться к нему, что обычно влечет за
собой морфологические и физиологические изменения.
Лучше всего изучены в этом отношении новые расы по хозяи-
ну, возникшие у пестрокрылки Rhagoletis pomonella. У этого вида,
паразитирующего на плодах северо-американских видов боярыш-
ника (Crataegus), примерно в 1864 г. образовалась раса, парази-
тирующая на некоторых зимних сортах яблок, которые были вве-
зены из Старого Света. Эта раса впервые появилась в Нью-Йорке,
а затем распространилась в другие места. В 1960 г. новая
раса Rh. pomonella была обнаружена в вишневом саду в Вискон-
сине.
Переход на нового хозяина сопровождается, по данным Баша,
мелкими генетическими изменениями. Главную роль играют, по-
видимому, гены двух типов: 1) связанные с распознаванием и вы-
бором хозяина и 2) связанные с выживанием на хозяине. Число
изменений, связанных с распознаванием и выбором нового хозяи-
на, может быть небольшим и, по крайней мере у насекомых, по-
видимому, затрагивает хеморецепцию, поскольку распознавание
хозяина, вероятно, основано главным образом на химических, а не
на физических признаках. При переходе на нового хозяина первый
шаг состоит в обеспечении передачи потомкам способности узна-
вать этого нового хозяина. Конечно, животное должно быть спо-
собным выжить на новом хозяине. В зависимости от того, в какой
степени новый хозяин отличается химически и физически от ста-
рого, переход будет сопровождаться большей или меньшей диффе-
ренциацией.
Подводя итоги, можно сказать, что аллопатрическое, парапат-
рическое и симпатрическое видообразования—это разные способы,
с помощью которых популяция или группа скрещивающихся меж-
ду собой популяций, адаптированных к данному набору физиче-
ских и биологических факторов среды, дают начало новой популя-
ции или группе популяций, приспособленных к другому набору
условий и репродуктивно изолированных от своих предков.
Модели видообразования
299
Мгновенное видообразование
Дальнейшие осложнения связаны с тем, что иногда видообра-
зование происходит не более или менее постепенно, а внезапно.
Это обычно случается в результате удвоения числа хромосом — яв-
ления, называемого полиплоидией, — или же при помощи особых
эволюционных механизмов, приводящих к видообразованию в ре-
зультате так называемого катастрофического отбора.
Полиплоидия. Проблема полиплоидии подробно разбиралась
в гл. 5. Полиплоидию можно определить как явление, возникающее
в результате удвоения числа хромосом у данного организма. По-
лучающийся при этом организм не способен давать плодовитое по-
томство при скрещивании с членами диплоидной предковой попу-
ляции. Таким образом, этот организм сразу становится репродук-
тивно изолированным, а если он окажется способным размножать-
ся и найдет такую экологическую нишу, которую он может ис-
пользовать, то можно сказать, что возник новый вид. Полиплоидия
часто встречается у растений, но редко у животных. Среди цвет-
ковых растений почти 40% всех видов, по-видимому, являются
полиплоидами, так что полиплоидия в самом деле представляет
собой очень важное явление.
Полиплоиды бывают в основном двух типов (см. рис. 5.20):
1) аутополиплоиды, возникающие в результате удвоения хромосом
у нормальной особи, и 2) аллополиплоиды, представляющие собой
результат удвоения числа хромосом у гибридной особи. Второй
тип, по-видимому, встречается чаще. Вероятность возникновения
аллополиплоидов выше по цитологическим соображениям (у них
скорее можно ожидать нормальное течение мейоза), а также по-
тому, что у гибрида могут оказаться новые признаки, которые по-
зволят ему использовать местообитания, не доступные ни для од-
ной из родительских особей. Поэтому полиплоидию можно рас-
сматривать как один из случаев парапатрического или симпатри-
ческого видообразования.
Видообразование в результате катастрофического отбора. Оче-
видно, в некоторых случаях может происходить очень быстрое
видообразование, ведущее к генетической изоляции при неболь-
шой морфологической дифференциации или без нее и в отсутствие
полиплоидии. Это явление известно под названием катастрофиче-
ского отбора. Наилучшим его примером служат растения рода
Clarkia из сем. Onagraceae.
Clarkia — группа растений, произрастающих на западе США,
в частности в Калифорнии. В течение последних 20 лет системати-
ку этой группы усиленно изучали калифорнийский эволюционист
Харлан Льюис (Harlan Lewis) и его ученики. В процессе своих
исследований они проводили межвидовые скрещивания между
большинством видов этого растения. Цитологический анализ полу-
ченных гибридов показал, что многие виды резко различаются по
300
Глава It
последовательности генов в хромосомах. Это различие особенно
четко выражено у С. franciscana. Этот вид обнаружен только в
одной местности — на серпентинитовом склоне, обращенном к Зо-
лотым Воротам, на восточной оконечности форта в Сан-Франциско
Рис. 11.11. Распространение Clarkia атоепа, С. franciscana и С. rubricunda в Ка-
лифорнии.
Обратите внимание на большое морфологическое сходство этих трех видов. Наи-
лучшим диагностическим признаком для различения этих видов служит лепесток,
в частности имеющееся на нем пятно. У С. атоепа это пятно ярко-красного цвета
и расположено примерно в середине; у С. franciscana оно четко выражено, крас-
новато-пурпурное и расположено в основании лепестка; у С. rubricunda оно не
столь хорошо выражено и границы его несколько размыты. (Н. Lewis, 1958.)
(рис. 11.11). Он морфологически сходен с двумя другими видами
С. rubricunda и С. атоепа, которые произрастают поблизости.
Первый из этих видов встречается по побережью и в долинах Бе-
регового хребта, от северной части округа Сан-Луис-Обиспо до
северной части залива Сан-Франциско в округе Марин. Вид
С. атоепа замещает С. rubricunda в центральной части округа
Марин и идет дальше на север, по побережью Британской Колум-
бии и острова Ванкувер. Льюис (Lewis) и Равен (Raven) полу-
Модели видообразования
301
чали межвидовые гибриды между С. franciscana, С. атоепа и
С. rubricunda во всех возможных сочетаниях, а затем изучали их
плодовитость и различия в структуре хромосом, выявлявшиеся при
конъюгации хромосом в мейозе. Фертильность пыльцы ни в одном
случае не превышала 2%. Конъюгация хромосом у гибридов по-
казала, что хромосомы этих трех видов резко различаются по рас-
положению гомологичных участков. С. franciscana отличается от
С. rubricunda не менее чем по трем крупным транслокациям и
четырем парацентрическим инверсиям, а от С. атоепа — не менее
чем по двум транслокациям и двум или более парацентрическим
инверсиям и, кроме того, вероятно, по ряду более мелких пере-
строек. С. атоепа в свою очередь отличается от С. rubricunda по
трем транслокациям и двум инверсиям. Эти хромосомные измене-
ния имеют совершенно иные масштабы, чем у большинства других
видов, например у Glandularia, упоминавшейся в гл. 5 (см. также
рис. 5.9).
Когда два близкородственных вида или две близкие популяции
различаются в отношении порядка расположения генов в хромосо-
мах, совершенно очевидно, что в одной или в обеих этих группах
произошли изменения структуры хромосом, так как у общего пред-
ка этих видов или популяций должна была быть последователь-
ность генов только одного типа (во всяком случае, когда-то).
Как происходит такое изменение в строении хромосом? Обычно
предполагается, что у какой-либо одной особи возникает неболь-
шое изменение, придающее ей более высокую приспособленность.
Со временем этим новым качеством будут обладать все члены
данной популяции. В результате повторения этого процесса рас-
сматриваемые популяции могут стать совершенно различными в
цитологическом отношении. Предполагается, что указанные изме-
нения сопровождаются морфологическими и физиологическими
изменениями, возникающими по мере того как популяции стано-
вятся приспособленными к различным экологическим условиям.
Означают ли большие различия в последовательности генов в
хромосомах Clarkia franciscana, С. атоепа и С. rubricunda при
большом морфологическом сходстве этих видов, что они возникли
не путем постепенного накопления мелких изменений, а каким-то
иным путем? Допустим, что имеется одна особь, в одной из хро-
мосом которой гены расположены в несколько ином, чем обычно,
порядке. Если эта особь гетерозиготна, то перекрест в изменен-
ном участке хромосомы приведет к тому, что из четырех образую-
щихся при нормальном мейозе гамет две окажутся стерильными
вследствие нехватки генов. Если измененный участок невелик, то
вероятность возникновения в нем перекреста очень мала; поэтому
в большинстве клеток, претерпевающих мейоз, перекреста в изме-
ненном участке не произойдет и новая последовательность закре-
пится в популяции. Но если измененный участок велик, то дело
обстоит совсем иначе. Таким образом, вероятность того, что после-
302
Глава И
довательность генов в хромосойах С. franciscana сложилась в ре-
зультате накопления мелких изменений, в самом деле очень мала,
потому что многократное возникновение мелких хромосомных из-
менений без соответствующих изменений морфологии — событие
маловероятное. В связи с этим Харлан Льюис выдвинул гипотезу
о существовании механизма, который он называет «катастрофиче-
ским отбором».
Согласно этой гипотезе, при некоторых неизвестных нам усло-
виях (к ним относятся, возможно, стрессовые условия среды, как,
например, засуха, или «мутаторные генотипы», т .е. генотипы, даю-
щие высокую частоту мутаций) происходят глубокие хромосомные
перестройки. Если особь с такой глубокой хромосомной перестрой-
кой отделится от своей популяции, она может основать новую
популяцию, содержащую новую хромосомную перестройку и с са-
мого своего возникновения изолированную от предковой попу-
ляции.
Силы, вызывающие такие резкие изменения в одних случаях
и такие небольшие изменения — в других, остаются неизвестными.
Так, у некоторых видов, изолированных на протяжении миллионов
лет, не возникло никаких генетических или хромосомных различий.
Одним из примеров длительной изоляции без генетического измене-
ния служат два вида платана Platanus orientalis и Р. occidentalis.
Первый вид — уроженец Среднего Востока, второй — Северной
Америки. Эти виды очень ясно различаются как морфологически,
так и физиологически, потому что они произрастают в разных кли-
матических областях. Установить сколько-нибудь достоверно, как
долго эти два вида были изолированы друг от друга, невозможно,
однако, по всей вероятности, они были изолированы по крайней
мере с конца третичного периода, т. е. в течение миллиона лет или
более того. Искусственный гибрид между этими видами, назван-
ный Р. acerifolia, широко выращивается как декоративное дерево;
у этого жизнеспособного и высоко плодовитого растения мейоз
протекает нормально, что свидетельствует об отсутствии каких-
либо резких изменений в последовательности генов.
Видообразование, вызванное катастрофическим отбором, пред-
ставляет собой, так же как и большинство случаев полиплоидии,
особый случай парапатрического видообразования. Итак, аллопат-
рическое видообразование всегда происходит постепенно, тогда как
парапатрическое и симпатрическое видообразование может быть
мгновенным.
ЛИТЕРАТУРА
Общие вопросы
Clausen J. 1951. Stages in the Evolution of Plant Species, Ithaca, N. Y., Cornell
University Press.
Гаузе Г. Ф. 1935. The Struggle for Existence, Baltimore, Williams and Wilkins,
Модели видообразования
303
Grant V. 1971. Plant Speciation, New York, Columbia University Press.
Mayr E. 1963. Animal Species and Evolution, Cambridge, Mass., Belknap Press of
Harvard University Press.
Solbrig О. T. 1970. Principles and Methods of Plant Biosystematics, New York,
Macmillan.
Stebbins G. L. 1974. Flowering Plants: Evolution Above the Species Level, Cambrid-
ge, Mass., Belknap Press of Harvard University Press.
Специальные статьи и обзоры
Ayala F. J. 1975. Genetic differentiation during the speciation process, Evol. Biol.,
8, 1—78.
Ayala F. J., Tracy M. L., Hedgecock D., Richmond R. C. 1974. Genetic differentia-
tion during the speciation process in Drosophila, Evolution, 28, 576—592.
Bradshaw A. D. 1959. Population differentiation in Agrostis tenuis Sibth. I. Morpho-
logical differences, New Phytol., 58, 208—227.
Bradshaw A. D. 1960. Population differentiation in Agrostis tenuis Sibth. III. Po-
pulations in varied environments, New Phytol., 59, 92—103.
Bradshaw A. D., McNeilly T. S-, Gregory R. P. 1965. Industrialization, evolution and
the development of heavy metal tolerance in plants, Brit. Ecol. Symp., 6, 327—
343.
Bush G. L. 1975. Modes of animal speciation, Ann. Rev. Ecol. Syst., 6, 339—364.
Clausen J., Keck D. D., Hiesey W. 1940. Experimental studies on the nature of spe-
cies. I. Effects of varied environments on western North American plants, Car-
negie Inst. Wash., PubL, 520.
Clausen J., Hiesey W. 1958. Experimental studies on the nature of species. IV. Gene-
tic structure of ecological races, Carnegie Inst. Wash. Publ., 615.
. Ehrlich P. R., Raven P. H. 1969. Differentiation of populations, Science, 165, 1228—
1232.
Gottlieb L. D. 1973. Genetic differentiation, sympatric speciation and the origin of
a diploid species of Stephanomeria, Am. J. Bot., 60, 545—553.
Jain S. K., Bradshaw A. D. 1966. Evolutionary divergence among adjacent plant
populations. I. The evidence and its theoretical analysis, Heredity, 21, 407—
442.
Lewis H. 1958. Rapid evolution in Clarkia, Evolution, 12, 319—336.
Nevo E., Bar-El H. 1’976. Hybridization and speciation in fossorial mole rats, Evolu-
tion, 30, 831—840.
Nevo E., Shkolnik A. 1974. Adaptive metabolic variation of chromosome forms in
mole rats, Spalax, Experientia, 30, 724—726.
Wilson E. O. 1965. The challenge from related species. In: H. G. Baker and
G. L. Stebbins (eds.), The Genetics of Colonizing Species, New York, Academic
Press.
Глава 12
Гибридизация,
преадаптация и адаптация
В предыдущей главе рассматривались три главных способа воз-
никновения видов: 1) аллопатрическое, или географическое, видо-
образование, которое многие считают наиболее обычным способом
возникновения видов; 2) парапатрическое видообразование и
3) симпатрическое видообразование. Видообразование — это про-
цесс дивергенции: одна популяция или группа популяций, члены
которых имеют одинаковые гены и свободно скрещиваются между
собой (во всяком случае, в пределах данной популяции), превра-
щаются в две отдельные группы популяций, уже различающиеся
по генному составу. Эта глава посвящена гибридизации между
видами — процессу, в результате которого два самостоятельных,
но близкородственных вида иногда получают общие гены. В ней
рассматриваются также процессы, с помощью которых виды при-
обретают совершенно новые признаки.
Видообразование сопровождается разделением генофонда, но
сам по себе этот процесс не приводит к возникновению новых при-
знаков. Просто изменчивость, существовавшая внутри вида, ста-
новится изменчивостью между новыми видами. Но возникновение
видов, происходящее во времени, сопряжено с появлением новых
признаков — того, что иногда называют «эволюционными новшест-
вами»; об этом свидетельствует палеонтологическая летопись.
В конце главы мы рассмотрим значение адаптаций в свете того,
что нам известно относительно действия естественного отбора.
ГИБРИДИЗАЦИЯ
Гибридизация — это третий возможный результат встречи меж-
ду двумя частично дифференцированными и до того географически
изолированными популяциями (два других результата — взаимное
исключение и смещение признаков — были рассмотрены в преды-
дущей главе). Строго говоря, гибридизация — синоним аутбридин-
га. А поэтому во всех тех случаях, когда в результате слияния
яйцеклетки одной особи со сперматозоидом другой получается зи-
гота, возникает гибрид. Сперматозоид может происходить от осо-
би, принадлежащей к той же популяции, что и яйцеклетка, от осо-
би из другой популяции того же вида или же от особи другого
Гибридизация, преадаптация и адаптация 305
вида. Чтобы различать эти разные случаи, мы ограничим значение
термина гибридизация, применяя его лишь к скрещиваниям меж-
ду особями, которые предположительно относятся к разным ви-
дам. Для скрещиваний в пределах одного вида будет использовать-
ся термин рекомбинация.
Ранее мы обычно считали, что если особи, относящиеся к двум
разным популяциям, скрещиваются между собой, то в конечном
счете генофонды популяций полностью перемешиваются, так что-
выживает единая интегрированная размножающаяся популяция.
Однако теперь мы знаем, что существуют механизмы, которые
дают возможность популяциям, принадлежащим к разным видам,,
время от времени гибридизировать, но не настолько интенсивно,
чтобы популяции оказались затопленными потоком чужих генов.
Всякий, кому покажется удивительным, почему такие две изредка
гибридизирующие популяции рассматриваются как самостоятель-
ные виды, найдет много единомышленников, поскольку вопрос а
том, какого уровня должна достигнуть степень гибридизации меж-
ду двумя популяциями, чтобы их можно было считать принадле-
жащими одному, а не двум разным видам, остается одним из са-
мых запутанных в биологии.
У высших растений гибридизация происходит довольно часто,
тогда как у позвоночных животных она наблюдается редко. Сле-
дуя за Эрнстом Майром (Ernst Мауг), мы различаем пять основ-
ных типов гибридизации. Следует, однако, помнить, что между эти-
ми типами нет резких границ и что существуют промежуточные
ситуации.
1. Происходящие время от времени скрещивания между сим-
патрическими видами, приводящие к появлению гибрид-
ных особей, которые нежизнеспособны или стерильны, что
исключает возможность возвратных скрещиваний с роди-
тельскими видами.
2. Появление более или менее жизнеспособных гибридов
между симпатрическими видами, причем отдельные такие
гибриды способны к возвратным скрещиваниям с одним
или с обоими родительскими видами.
3. Образование вторичной зоны контакта и частичного скре-
щивания между двумя прежде изолированными популя-
циями, которые не достигли полной репродуктивной изо-
ляции в течение предшествовавшего периода географиче-
ской изоляции.
4. Полное локальное нарушение репродуктивной изоляции
между двумя симпатрическими видами, приводящее к об-
разованию скоплений гибридов, в которых может быть
представлен весь диапазон изменчивости родительских
видов.
5. Образование нового вида в результате гибридизации и
последующего удвоения хромосом (аллополиплоидия).
20—2044
.'306
Глава 12
Первая из этих ситуаций не имеет серьезных последствий, по-
скольку гибриды будут элиминированы естественным отбором.
Последняя ситуация — аллополиплоидия — уже рассматривалась,
а в четвертом из описанных случаев происходит полное растворе-
ние родительских типов, поскольку возникают все промежуточные
комбинации родительских типов. Здесь будут рассмотрены второй
Рис. 12.1. Области распространения Salvia mellifera н S. apiana в Калифорнии и
зона перекрывания между ними. (Epling, 1947.)
и третий типы гибридизации, известные под названием интрогрес-
сия.
Иногда образуются межвидовые гибриды, которые почти пол-
ностью стерильны. У растений такие гибриды часто совершенно
не производят жизнеспособных семян, но дают некоторое количе-
ство фертильной пыльцы, которая может оплодотворять особей
родительских видов. В других случаях гибриды плодовиты, но они
плохо адаптированы. Эплинг (Epling) изучал два вида шалфея,
Salvia apiana и Salvia mellifera, которые произрастают совместно
на многих тысячах квадратных километров в прибрежных районах
южной части Калифорнии. Между этими видами существует ре-
продуктивная изоляция, но там, где их местообитания были на-
рушены человеком, они образуют скопления гибридов, которые
способны к возвратным скрещиваниям с родительскими видами
(рис. 12.1). Андерсон (Anderson) изучал два вида традесканции,
Гибридизация, преадаптация и адаптация 307
Tradescantia caniculata и Т. subaspera, которые растут вместе на
плато Озарк: первый на скалистых склонах, ярко освещаемых
солнцем, а второй на богатой почве в сильно затененных местах..
Но в лощинах и на пологих склонах имеются местообитания про-
межуточных типов. Здесь растут гибриды между двумя видами,
способные также к возвратным скрещиваниям с родительскими:
видами. Для того чтобы понять, каким образом эти два вида со-
храняют свою самостоятельность, несмотря на гибридизацию, не-
обходимо учитывать действие естественного отбора. Очевидно,,
гибриды плохо адаптированы как к одному, так и к другому из
родительских местообитаний и продолжительность их жизни неве-
лика. Особи, получающиеся в результате возвратных скрещиваний,
не так хорошо адаптированы, как чистые исходные виды, и отбор'
будет направлен против них. Таким образом, целостность вида
поддерживается отбором. Но если численность гибридов сильно
увеличится — например, при разрушении их местообитаний в ре-
зультате деятельности человека, — вероятность опыления растений
родительских видов пыльцой гибридов повышается, и в конце кон-
цов большинство, а может быть, и все растения могут оказаться
гибридами или беккроссами. Таким образом, судьба двух видов,
между которыми происходит гибридизация, частично зависит от
интенсивности гибридизации между ними и от уровня генетической
и экологической стерильности гибридов и беккроссов. Однако
эволюционная роль гибридизации не обязательно зависит от часто-
ты образования гибридов, а определяется влиянием гибридизации
на генетический состав родительских популяций. Иначе говоря, в
большинстве случаев эволюционное значение имеют не сами гиб-
риды, а продукты возвратных скрещиваний гибридов с родитель-
скими формами. Этот процесс называют интрогрессивной гибриди-
зацией. В результате интрогрессивной гибридизации гены одного-
вида могут вводиться в популяции другого вида. Ныне покойный,
блестящий эволюционист Эдгар Андерсон (Edgar Anderson) неод-
нократно высказывал мнение, что в случаях интрогрессивной
гибридизации естественный отбор благоприятствует беккроссам
из-за тех новых генов, которые они содержат.
Отдельные организмы, образующие скрещивающуюся популя-
цию, более или менее хорошо адаптированы к среде, в которой
они растут. В каждом поколении в результате мутаций и реком-
бинаций возникают как особи, генотипически очень сходные с ро-
дительскими формами, так и особи, в той или иной степени отли-
чающиеся от них. В стабильной среде большинство потомков, до-
живающих до зрелого возраста, будут и генотипически, и феноти-
пически сходны с родительским поколением; в изменяющейся
среде это сходство будет меньше. Степень, в которой поколение
потомков отличается от родительского поколения, зависит от сте-
пени рекомбинации (и от всех факторов, которые ее контролиру-
20
308
Глава 12
ют) и от интенсивности отбора, как уже говорилось в гл. 4—9.
Чем больше генетическая рекомбинация, тем выше вероятность
того, что потомки не будут сходны со своими родителями; чем ин-
тенсивнее отбор, тем сильнее будут генетические изменения. При
стабилизирующем отборе изменения будут минимальными и глав-
ным образом генотипическими; при направленном же отборе из-
менения будут более значительными и отразятся как на генотипе,
так и на фенотипе (см. гл. 6, разд. «Действие отбора на фенотипи-
ческие признаки»).
При гибридизации в популяцию вносятся новые гены и тем
самым возрастает вероятность генетического изменения. Этот про-
цесс действует как фактор, увеличивающий изменчивость.
Действие гибридизации и интрогрессии на примере подсолнечника
Прекрасным примером роли гибридизации в эволюции служат
подсолнечники, изучавшиеся Чарлзом Хейзером (Charles Heiser)
в Университете штата Индиана, в частности однолетний подсол-
нечник (Melianthus annuus). Этот вид распространен по всем
центральным и западным областям Северной Америки — от юж-
ной части Канады до северной части Мексики. Он отличается
значительной морфологической изменчивостью, не только внутри-,
но и в особенности межпопуляционной. Хейзер разбил этот вид
на три подвида и одну разновидность: подвид lenticularis, подвид
texanus, подвид annuus (к которому относится культурный под-
солнечник— разновидность macrocarpus). Подвид annuus — сор-
няк, широко распространенный на Среднем Западе США, в осо-
бенности вблизи железных дорог и на городских пустырях. Под-
вид lenticularis — дикорастущий подсолнечник, встречающийся на
обочинах дорог и на заброшенных полях на западе и юго-западе
США. Наконец, распространение подвида texanus ограничено во-
сточной частью штата Техас. Гипотетическая история происхожде-
ния этих трех форм служит прекрасным примером роли гибриди-
зации в эволюции.
Предковая форма подсолнечника точно не установлена. Одна-
ко на основании своих обширных исследований Хейзер постули-
ровал, что подвид lenticularis из западных областей Северной Аме-
рики представляет собой более вероятную предковую форму этого
вида, чем любой другой из ныне живущих подвидов. Этот подвид,
согласно Хейзеру, под косвенным влиянием деятельности человека,
дал начало подвиду annuus. Ряд индейских племен собирали се-
мена подвида lenticularis, используя их в пищу, в результате чего
подсолнечник, возможно, стал одним из первых сорняков, приспо-
собившихся к росту вокруг бивуаков и поселений индейцев, где
природные условия были нарушены. С течением времени в резуль-
тате либо сознательного, либо бессознательного отбора со стороны
человека подвид lenticularis мог дать начало несколько более
Гибридизация, преадаптация и адаптация
309
крупным формам, из которых получился подвид annuus. Подвид
annuus var. macrocarpa— культивируемый подсолнечник — отлича-
ется от annuus var. annuus главным образом одним мутантным
геном, который ограничивает ветвление стебля и обусловливает
образование одной или очень немногих крупных корзинок. Разу-
меется, кроме того, человек путем селекции создал более урожай-
ные формы, особенно за последнее время.
История третьего подвида, Н. annuus texanus, более запутан-
ная. По-видимому, он возник после того, как в Техас был занесен
естецтвенным путем или интродуцирован человеком либо подвид
lenticularis, либо подвид annuus. В настоящее время Н. annuus
широко распространен по обочинам дорог и на заброшенных по-
лях, особенно в центральных частях Техаса. В восточных районах
этого штата растет в качестве сорняка на обочинах и в нетрону-
тых природных местообитаниях другой однолетний вид подсолнеч-
ника— Н. debilis var. cucumerifolius. В тех местах, где Н. debilis
var. cucumerifolius и Н. annuus texanus растут вместе, иногда мож-
но обнаружить отдельные гибридные растения. Их рост подавлен
по сравнению с родительскими видами, которые растут поблизо-
сти, и они в большинстве случаев стерильны. Однако у некоторых
из них небольшая доля пыльцы иногда оказывается жизнеспособ-
ной.
Изменчивость Н. annuus в восточной части Техаса (табл. 12.1)
явно смещена в сторону Н. debilis, о чем свидетельствует большая
доля листьев с глубоко пильчатым краем, пятнистый стебель, не-
большой диаметр корзинок, мелкие семянки, небольшое число
язычковых цветков, малая ширина листочков обертки и тенденция
к ветвлению от самого основания стебля. На основании всего этого
Хейзер постулировал, что подвид texanus возник в результате гиб-
ридизации и интрогрессии между Н. debilis var. cucumerifolius
с Н. annuus lenticularis или Н. annuus annuus, когда тот или дру-
гой из двух последних подвидов был интродуцирован в Техас. По-
мимо морфологических признаков, которые Н. annuus получил от
Н. debilis, он, возможно, приобрел также физиологические при-
знаки, которые сделали его более приспособленным к климату и
другим условиям Техаса, — признаки, выработавшиеся у Н. debi-
lis за многие годы существования в Техасе.
На другом конце ареала Н. annuus — в Калифорнии — можно
наблюдать сходное явление гибридизации. Подсолнечник появился
в Калифорнии сравнительно недавно. Он, вероятно, был интроду-
цирован индейцами, которые употребляли его семена в пищу, а
цветки использовали при религиозных обрядах. Сначала его рас-
пространение ограничивалось нарушенными местообитаниями вок-
руг бивуаков индейцев. Примерно за последние сто лет в резуль-
тате массового переселения людей на запад США, количество
разрушенных природных местообитаний в Калифорнии возросло
в огромной степени; соответственно увеличилось и количество
310
Глава 12
Таблица 12.1
Сравнение числа язычковых цветков и диаметра корзинки*
у Helianthus annuus и Н. debilis var. cucumerifolius
(По данным Heiser, 1951.)
Вид Местность Среднее число язычковых цвет- ков Диаметр корзин* ки, см
Н. debilis var. си- Округ Накогдочес, Техас 13,7 1,62
cumerifolius Округ Фрио, Техас 15,7 1,84
Округ Хьюстон, Техас 19,0 2,24
Н. annuus Округ Галвестон, Техас 17,3 2,30
Округ Фрио, Техас 20,4 2,58
Округ Галвестон, Техас 22,0 3,28
Округ Коллин, Техас** 23,2 3,60
Округ Питтсбург, Оклахо-
ма** 25,9 3,54
* Обратите внимание на то, что в популяциях Н. debilis число язычковых цветков и
диаметр корзннкн увеличиваются по мере продвижения на север н на восток (область рас-
пространения Н. annuus). а у Н. annuus число цветков и диаметр корзинки уменьшаются
по мере продвижения на юг и на восток (область Н. debilis).
** Эти популяции обитают за пределами ареала Н. debilis.
подсолнечника, который теперь очень широко распространен по
обочинам дорог и на полосах отчуждения вдоль железнодорож-
ного полотна, а также как сорняк на возделываемых полях.
Н. bolanderi— уроженец Калифорнии и Орегона. Он произрастает
в местообитаниях двух типов. Одна его раса встречается на сер-
пентиновых пустошах Берегового хребта. В этих местах почвы
отличаются крайне низким содержанием кальция и высоким со-
держанием магния и тяжелых металлов. На них произрастает
чрезвычайно своеобразная флора. Кроме того, Н. bolanderi встре-
чается на пустошах в долинах центральной и северной части Ка-
лифорнии, в тех же местах, что и Н. annuus. В своем подробном
исследовании Хейзер показал, что между этими двумя видами
иногда возникают гибриды. Хотя эти гибриды в значительной сте-
пени стерильны, они иногда производят некоторое количество жиз-
неспособной пыльцы. По мнению Хейзера, Н. annuus, когда он
был интродуцирован в Калифорнию, гибридизировал с Н. bolande-
ri, который в то время, вероятно, произрастал только на серпенти-
новых обнажениях. В результате повторных и возвратных скрещи-
ваний между этими двумя формами появилась некая форма
Гибридизация, преадаптация и адаптация
311
Листочки
обертки
Рис. 12.2. Влияние гибридизации на морфологические признаки Helianthus annuus
и Н. bolanderi. Bl—Н. bolanderi — серпентиновая раса; В2 — Н. bolanderi —
яесерпентиновая раса, предположительно интрогрессировавшая с Н. annuus; Н —
естественный гибрид между Н. annuus и Н. bolanderi; Al —Н. annuus из Кали-
форнии, предположительно интрогрессировавший с Н. bolanderi; А2 — Н. annuus
из Сент-Луиса (штат Миссури), вне ареала Н. bolanderi и, следовательно, не под-
верженный интрогрессии со стороны последнего. Обратите внимание на то, что
у предположительно интрогрессировавших типов сходство с гибридами сильнее,
чем у любого из «чистых» видов. (С. Heiser, 1949. Study in the evolution of the
sunflower species Helianthus annuus and H. bolanderi. Univ. Calif. Pub. Bot. 23,
157—208.)
312
Глава 12
Таблица 12.2
Сравнение числа язычковых цветков и диаметра корзинок
у двух видов подсолнечника Helianihus annuus
и Н. bolanderi (По данным Heiser, 1949.
Study in the evolution of the sunflower species Heliantus annuus
and H. bolanderi. Univ. Calif. Pabl. Bot., 23, 157—208.)
Вид Место Среднее число язычковых цвет- ков Диаметр корзин- ки, см
Н. bolanderi Серпентин 12,7* 1,70*
Н. bolanderi Долина 14,8* 2,07*
Гибрид Н. bolande- ri X И. annuus — 14—20** 2—3**
Н. annuus Калифорния 18,2* 2,41*
Н. annuus Не из Калифорнии 20,7* 2,98’
* Среднее значение для растений, выращенных в саду (измерения проводили в разных
популяциях на 29—61 растениях).
’* Диапазон значений для 6 растений.
Н. annuus, которая была сходна с Н. bolanderi (табл. 12.2;
рис. 12.2) и хорошо адаптировалась к условиям среды в Калифор-
нии, возможно, благодаря заимствованию генов у последнего вида.
В результате интрогрессии генов от Н. annuus и Н. bolanderi по-
явилась та форма Н. bolanderi, которая способна расти в нару-
шенных местообитаниях.
Помимо Н. bolanderi и Н. debilis var. cucumerifolius есть и дру-
гие однолетние виды подсолнечника, ареалы которых перекрыва-
ются с ареалом Н. annuus. Один из них — Н. petiolaris — вид, ши-
роко распространенный на западе США и иногда заходящий на
восток, ареал которого сходен с ареалом Н. annuus, хотя послед-
ний вид имеет более широкое распространение. Оба вида во мно-
гих местах растут вместе, однако они, по-видимому, несколько
различаются по своим экологическим предпочтениям. В общем,
Н. annuus, очевидно, больше приурочен к тяжелым, а Н. petiola-
ris— к песчаным почвам. Кроме того, Н. petiolaris зацветает в
июне и цветет в течение всего июля и части августа, а Н. annuus
обычно зацветает не раньше июля и продолжает цвести еще в
сентябре; это несколько ограничивает возможности обмена гена-
ми. Скопления гибридов между этими двумя видами образуются
не часто, однако они были обнаружены в ряде случаев. Эти гиб-
риды опять-таки в значительной степени стерильны, но производят
некоторое количество жизнеспособной пыльцы, что создает воз-
можность для интрогрессии. У интрогрессировавших популяций
Н. petiolaris листья шире, с более выраженными пильчатыми края-
ми, листочки обертки опушены и вообще все части растения круп*
Рис. 12.3. Вероятный путь эволюции Helianthus annuus в США по данным Хейзера.
Подвид lenticularis (или происходящий от него подвид annuus) распространился
из юго-западных областей США в Калифорнию, где он гибридизировал с Н. bo-
landeri. Подвид lenticularis (или подвид annuus) распространился также на Сред-
ний Запад, где он гибридизировал с Н. petiolaris, и в Техас, где он гибридизиро-
вал с Н. debilis var. cucumerifolius, дав начало подвиду texanus. Одновременно
происходила также дифференциация в результате отбора, о чем свидетельствуют
различия в размерах корзинок, окраске пыльников и числе язычковых цветков.
Размеры схематических изображений корзинок пропорциональны их размерам в
природных популяциях. Каждый «лепесток» соответствует двум язычковым цвет-
кам. Черный цвет означает пурпурные, точки — красные, а белый — черные пыль-
ники; сочетание этих обозначений — наличие в популяции цветков с пыльниками
разного цвета. Контуры листьев: А—Н. bolanderi; Б — Н. annuusXH. bolanderi;
В — Н. annuus ssp. annuus; Г — Н. petiolaris; Д— Н. petiolaris'xH. annuus; Е —
Н. annuus ssp. annuus; Ж — Н. annuus ssp. lenticularis; 3 — H. annuus ssp. texa-
nus; И — H. annuus'X.H. debilis var. cucumerifolius; К — H. debilis var. cucumeri-
folius. (Solbrig, 1970.)
Цифрами на карте обозначены штаты: 1 — Вашингтон; 2— Монтана; 3— Север-
ная Дакота; 4 — Миннесота; 5 — Орегон; б — Айдахо; 6—Вайоминг; 8— Южная
Дакота; 9 — Небраска; 10— Айова; 11 — Висконсин; 12 — Мичиган; 13— Невада;
14— Юта; 15—Колорадо; 16— Канзас; 17 — Миссури; 18— Иллинойс; 19 —
Индиана; 20— Огайо; 21— Пенсильвания; 22 — Нью-Йорк; 23—Нью-Гемпшир;
24 — Мэн; 25 — Вермонт; 26 — Массачусетс; 27 — Род-Айленд; 28 — Коннектикут;
29 — Нью-Джерси; 30 — Делавэр; 31—Мэриленд; 32—Западная Виргиния; 33—
Виргиния; 34 — Северная Каролина; 35 — Южная Каролина; 36—Кентукки; 37—
Калифорния; 38 — Аризона; 39—Нью-Мексико; 40— Техас; 41 — Оклахома; 42—
Арканзас; 43 — Теннесси; 44 — Луизиана; 45—Миссисипи; 46 — Алабама; 47 —
Джорджия; 48 — Флорида.
314
Глава 12
нее. Популяции Н. annuus, подверженные интрогрессии со сто-
роны Н. petiolaris, напротив, имеют более узкие листья, более
узкие листочки обертки, более мелкие корзинки, более густо опу-
шенные чешуйки и меньшее число язычковых цветков (табл. 12.3;
рис. 12.3).
Таблица 12.3
Сравнение числа язычковых цветков и диаметра
корзинки у двух видов подсолнечника,
Helianthus annuus и Н. petiolaris
(По данным Heiser, 1947.)
Вяд
Число язычковых
цветков*
Диаметр корзни-
ки**, см
Н. petiolaris
Н. petiolaris X Н. an-
nuus
Н. annuus
13—16
15—21
21—30
2,3
2,9
3,3
* Диапазон значений для нескольких растений, выращивав-
шнхся в саду.
* * Средние значения для популяций из Ист-Сент-Луиса.
На основании этих трех примеров гибридизации и интрогрес-
сии вырисовывается определенная картина, которая наблюдается
и у ряда других видов. Очевидно, четыре рассмотренных нами вида
подсолнечника развивались в изоляции один от другого. В этих
условиях они дифференцировались и адаптировались к специфиче-
ским экологическим условиям. Тем не менее между ними не воз-
никло полной генетической изоляции. Когда ареал И. annuus уве-
личился (возможно, в результате деятельности человека), этот
вид пришел в контакт с тремя другими видами. Образовались гиб-
риды, которые в свою очередь вступали в возвратные скрещива-
ния с родительскими формами, что приводило к передаче генов-
от одного вида к другому. Некоторые из переданных таким обра-
зом генов сообщили получившим их видам признаки, благодаря
которым эти виды оказались лучше адаптированы к новой среде
обитания. Поскольку ареал Н. annuus расширялся, он, по всей
вероятности, приобрел физиологические признаки, давшие ему
возможность лучше адаптироваться к условиям среды в Техасе,
Калифорнии и на Среднем Западе. Этот вид в свою очередь пере-
дал трем другим видам признаки, позволившие им произрастать на
почвах с нарушенной структурой. Харлан (Harlan) и Де Уэт (De
Wet) назвали виды, подобные Н. annuus «компиляторами» или
«видами-грабителями», потому что они, «заимствуя» посредством
гибридизации гены у местных видов, адаптируются в результате
к более широкому диапазону условий среды.
Гибридизация, преадаптация и адаптация 315
Таким образом, эволюционная роль интрогрессивной гибриди-
зации состоит, вероятно, в том, чтобы дать возможность некоторым
видам приобретать гены у других видов и тем самым повышать
свою приспособленность и адаптацию к новым местообитаниям.
ИЗОЛИРУЮЩИЕ МЕХАНИЗМЫ
Теперь читателю уже должно быть ясно, что главное свойство
вида — это его репродуктивная изоляция от всех других сущест-
вующих в природе видов. Наличие репродуктивной изоляции не
означает, что данный вид абсолютно лишен возможности скрещи-
ваться с другими видами или же что подобное скрещивание, если
оно произойдет, будет стерильным. Многие виды могут скрещи-
ваться, а иногда и действительно скрещиваются в природе и дают
плодовитых гибридов. Гибридизация иногда может даже привести
к приобретению новых генов, как в случае подсолнечника. Репро-
дуктивная изоляция означает, что обычно большинство членов
данного вида не скрещивается с членами других видов. Проиллю-
стрируем это на двух примерах —одном из царства животных и
одном из царства растений.
Кряква Anas platyrhynchos и шилохвость A. acuta — два вида
уток, вероятно, наиболее широко распространенных в пресновод-
ных водоемах северного полушария. Согласно Эрнсту Майру
(Ernst Мауг), численность этих двух видов на всем земном шаре
превышает 100 000 000 и они гнездятся бок о бок на миллионах
озер, прудов и рек в Азии, Европе и Северной Америке. В неволе
эти два вида скрещиваются друг с другом, давая нормальные и
вполне плодовитые гибриды в Fi, F2 и последующих поколениях.
Но в природе гибриды встречаются реже, чем в одном случае на
тысячу, и нет никаких данных, свидетельствующих о наличии воз-
вратных скрещиваний или интрогрессии.
Salvia apiana и S. mellifera— два вида шалфея, произрастаю-
щих в прибрежных шалфейных сообществах южной и центральной
Калифорнии (рис. 12.1). Их ареалы перекрываются в области
между Санта-Барбарой и мексиканской границей. S. apiana отдает
некоторое предпочтение более сухим почвам, хотя оба вида неред-
ко образуют смешанный травостой. S. mellifera цветет раньше,
примерно с апреля до конца мая, a S. apiana — с середины мая до
конца июня. Таким образом, их периоды цветения несколько пере-
крываются. У S. mellifera цветки мелкие, а у S. apiana они гораздо
крупнее. Наконец, первый вид обычно опыляется мелкими пчела-
ми, мухами и бабочками, тогда как S. apiana опыляется крупными
пчелами-плотниками (рис. 12.4). Такие экологические и механиче-
ские различия совместно обеспечивают репродуктивную изоляцию
этих двух видов. Тем не менее иногда образуются гибриды, чаще
всего в нарушенных местообитаниях. В некоторых случаях гиб-
риды достигают зрелости, но они обладают лишь частичной плодо-
Рис. 12.4. Цветки Salvia mellifera, S. apiana, гибридов между ними и беккроссов,
а также некоторых из специфических опылителей.
A. Salvia mellifera. Б. Chloralictus sp. — небольшая одиночная пчела, опыляющая
S. mellifera. В. Anthophora sp. — средних размеров пчела, опыляющая 3. mellife-
ra. Г. Гибрид Fi между 3. mellifera и S. apiana. Д—Ж- Беккроссы от скрещива-
ния гибридов с обеими родительскими особями. 3—И. Цветки S. apiana в обычном
и «опрокинутом» положениях. К. Xylocopa brasilianorum — опылитель 3. apiana.
Нетрудно увидеть, что различия в величине цветков у этих двух видов и в раз-
мерах опыляющих их видов пчел действуют как изолирующие механизмы. (Ри-
сунки цветков взяты из Epling, 1947; пчел — из К. Grant, V. Grant, 1964, Mecha-
nical isolation of Salvia apiana and S. mellifera [Labiata], Evolution, 18, 196—242.)
Гибридизация, преадаптация и адаптация 317
витостью. Хотя в таких случаях происходит некоторая интрогрес-
сия, она, по-видимому, распространяется не слишком широко.
Эти два вида уток и два вида шалфея служат иллюстрациям»
того, каким образом виды, не разделенные абсолютными генетиче-
скими преградами, могут жить в тесном соседстве без всякой или
почти без всякой гибридизации. Пример с шалфеем показывает
также, что существует не одна абсолютная преграда скрещива-
нию, а множество частичных преград, дополняющих друг друга »
создающих в результате почти полную репродуктивную изоляцию.
В табл. 12.4 систематизированы основные типы изолирующих
механизмов. Среди них единственные две преграды, обеспечиваю-
щие действительно абсолютную изоляцию (и, возможно, необрати-
мые),— это: 1) несовместимость гамет, т. е. неспособность спер-
матозоида одного вида образовать жизнеспособную зиготу с яйце-
клеткой другого вида; 2) абсолютная стерильность гибридов, т. е.
неспособность гибридов производить жизнеспособные гаметы. Все-
другие преграды время от времени нарушаются и их можно на-
звать «дырявыми» изолирующими преградами. Географически изо-
лированные виды могут с течением времени встретиться; некоторые
пыльцевые зерна могут иногда прорастать на рыльцах видов, счи-
тающихся несовместимыми с ними; у видов, размножающихся
путем инбридинга, и у апомиктических видов иногда происходит
аутбридинг и гибридизация; окончание сезона размножения одного
вида может приходиться на начало сезона размножения другого;
гибриды могут обитать в областях с условиями среды, промежуточ-
ными между условиями среды обитания двух родительских видов,
причем даже и в тех случаях, когда гибриды не отличаются особой
мощностью. Как мы уже видели, после того как гибриды обосно-
вались, они способны скрещиваться с родительскими видами, что
приводит к образованию беккроссов и к обмену генами между по-
пуляциями.
Если среди изолирующих преград так много «дырявых», позво-
ляющих многим видам гибридизировать друг с другом, то не сви-
детельствует ли это о наличии какой-то ошибки в нашей концепции
вида? В самом деле, строгое применение биологической концепции
вида привело бы к выделению в качестве самостоятельных лишь
тех таксонов, которые разделены абсолютными преградами. Это
означает, что многочисленные виды дуба рассматривались бы как
один вид, так же как практически все березы и ивы, многие насе-
комые, рыбы, папоротники, орхидеи, злаки и т. п. Очевидно, одна-
ко, что в каждом из этих случаев мы имеем дело не с одним, а с
многими видами. Поэтому даже при отсутствии механизмов, соз-
дающих абсолютную изоляцию, возможно образование и сохране-
ние новых видов.
Рассматривая процесс видообразования, всегда следует прини-
мать во внимание не только репродуктивную изоляцию, но также
отбор и поток генов. Биологические явления всегда многогранны.
318
Глава 12
Таблица 12,4
Изолирующие механизмы
Снижение частоты
контактов
Действующие до
оплодотворения
Снижение частоты
скрещиваний
.Действующие пос-
ле оплодотворе-
ния
Затруднение в фор-
мировании зигот
(презиготические
механизмы)
Уменьшение обмена
генами путем гиб-
ридизации (постзи-
готические меха-
низмы)
1. Географическое разделе-
ние*
2. Экологическое разделе-
ние
3. Несовместимость га-
мет** (гаметическая
изоляция)
4. Инбридинг и бесполое
размножение
5. Этологическая изоляция
6. Разное время размно-
жения (аллохроническая
изоляция)
7. Несовместимость га-
мет**
8. Стерильность гибри-
дов**
9. Неполноценность или
разрушение гибридов**
10. Неспособность гибридов
обосноваться
* Не всегда считается настоящим изолирующим механизмом.
Может быть абсолютным н необратимым.
Чтобы легче разобраться в этих явлениях, мы изучаем все участ-
вующие в них факторы поочередно, однако во многих случаях это
может привести к ошибкам.
ПРОЦЕСС ВИДООБРАЗОВАНИЯ И РЕГУЛИРУЮЩИЕ
ЕГО ФАКТОРЫ
В эволюционном плане основной единицей принято считать
генофонд, который приравнивается к локальной скрещивающейся
популяции. Но даже концепция генофонда представляет собой
некоторую абстракцию, потому что понятие генофонда нельзя
определить в строгих терминах. Кроме того, как мы уже видели
в гл. 8, механизм скрещивания оказывает большое влияние на ве-
личину генофонда и его изменчивость. У некоторых аутбредных
видов генофонд может быть довольно большим; в противополож-
ность этому в популяциях некоторых бесполых организмов из по-
коления в поколение обмен генами может почти отсутствовать.
Любой механизм, ограничивающий поток генов, благоприятст-
вует видообразованию. Обратное положение, т. е. что видообра-
зование будет происходить во всех тех случаях, когда поток генов
Гибридизация, преадаптация и адаптация
319
ограничен, неверно. Когда на две популяции действуют разные се-
лективные силы, то популяции обычно дивергируют генетически,
а тем самым также физиологически и морфологически. Обмен
генами замедляет процесс дивергенции вследствие постоянного
образования промежуточных форм. Если отбор недостаточно ин-
тенсивен, то образование двух обособленных популяций невозмож-
но. Однако при интенсивном отборе образование двух резко раз-
граниченных популяций возможно и оно действительно происходит.
Хорошим примером служит эволюция популяций, адаптированных
к экстремальным эдафическим условиям, например, популяций
Agrostis tenuis, изучавшихся Брэдшоу, о которых уже упоминалось
в гл. 11.
Вместе с тем теперь известен ряд случаев, когда, несмотря на
наличие абсолютных преград, препятствующих обмену генами,
морфологическая дифференциация очень незначительна или не-
происходит вовсе. Одним из примеров служит группа видов Gilia
inconspicua, изучавшаяся американским эволюционистом В. Гран-
том (Verne Grant). Эта группа однолетних растений встречается
во всех засушливых областях в западной части Северной Америки,
и южной части Южной Америки, где они растут на открытых пес-
чаных участках в различных местообитаниях, таких, как русла
пересыхающих ручьев в пустыне или песчаные поймы рек, прога-
лины в зарослях пустынных кустарников и склоны гор. Особи
этих видов самосовместимы, и пыльца автоматически попадает на
лопасти рыльца. Как показал Грант в своих экспериментах, расте-
ния, получающиеся в результате самоопыления, сохраняют жиз-
неспособность на протяжении многих поколений. По достижении
зрелости у растений, принадлежащих к комплексу видов Gilia
inconspicua, на верхушках стеблей появляются мелкие невзрачные
цветки. Эти цветки сходны по общим признакам, но различаются
по незначительным морфологическим особенностям. В результате
тщательных морфологических и цитологических исследований и
экспериментов по скрещиванию удалось показать, что эта группа,
считавшаяся одним видом, на самом деле состоит из 6 независимо
возникших групп видов, в которые входят в общей сложности не
менее чем 25 видов (рис. 12.5). За очень немногими исключения-
ми, все эти 25 видов генетически совершенно изолированы друг
от друга и, несмотря на многократные попытки скрещивать их
между собой, гибридные семена получить не удалось. В тех не-
многих случаях, когда гибриды все же были получены, они ока-
зались стерильными или же обладали низкой фертильностью.
Другим примером служит Lasthenia (сем. сложноцветных).
При изучении этой группы мелких озимых однолетних травянистых;
растений, встречающихся в весенних лужах в Калифорнии, Орндаф*
(Ornduff) столкнулся со сложной картиной. Так, например, у
Lasthenia chrysostoma (секция Baeria) были обнаружены диплоид-
ные и тетраплоидные расы, а у другого вида, L. macrantha, —
320
Глава 12
диплоидные и гексаплоидные расы. В результате скрещивания
популяций L. chrysostoma с разным числом хромосом образовы-
вались гибриды, у которых пыльца обладала пониженной фертиль-
Рис. 12.5. Преграды, создаваемые неспособностью к скрещиванию и стерильно-
стью гибридов, между 23 видами-двойниками из комплекса Gilia inconspicua.
Прерывистые линии — полная невозможность скрещиваний; тонкие линии — вы-
сокая стерильность гибридов (0—6% фертильной пыльцы); жирные линии — по-
лустерильность гибридов (12—18% фертильной пыльцы). (Grant, 1964.)
ностью, хотя иногда фертильность пыльцы достигала 50%. Гибри-
ды между диплоидными популяциями также обладали понижен-
ной фертильностью, которая в одном случае была ниже 20%
(рис. 12.6). Скрещивания внутри вида L. macrantha, напротив,
давали гибриды с высокой фертильностью даже в тех случаях,
когда скрещиваемые виды обладали разным числом хромосом.
Наконец, межвидовые гибриды были относительно фертильными
Гибридизация, преадаптация и адаптация
321
или стерильными в зависимости от того, между какими популяция-
ми производились скрещивания, и от числа хромосом в этих по-
пуляциях, но фертильность таких гибридов обычно была выше
50% (рис. 12.7).
Рис. 12.6. Диаграмма, на которой показаны средние значения для жизнеспособ-
ности пыльцы у гибридов, полученных в результате искусственных межвидовых
скрещиваний у Lasthenia, секция Baeria. (Ornduff, 1966.)
Прекрасный пример из животного мира дали интенсивные ис-
следования, проводившиеся калифорнийским генетиком Айалой
(Ayala) и его сотрудниками на шести видах так называемой груп-
пы Drosophila willistoni, распространенной в Западном полушарии
(рис. 12.8). Четыре из этих видов (D. willistoni, D. paulistorum,
D. equinoxialis и D. tropicalis) довольно широко распространены
в Центральной и Южной Америке; один вид (D. insularis) встре-
21—2044
322
Глава 12
чается только на некоторых из Малых Антильских островов, а
D. pavlovskiana — только в Гайане. В каждой отдельной местности
в пределах ареала группы D. willistoni могут симпатрически оби-
Жизнеспособность
пыль,.м % .......0-19--------20-39 ------ 40-59 ------ 60-79 ---—80-100
Рис. 12.7. Диаграмма, на которой показаны средние значения для жизнеспособ-
ности пыльцы гибридов, полученных в результате внутривидовых скрещиваний
между диплоидными и тетраплоидными расами Lasthenia chrysostoma. (Ornduff,
1966.)
тать два или более вида этой группы. Названные шесть видов в
морфологическом отношении практически идентичны; единственное
различие между ними касается половых органов самца. При содер-
жании в лабораторных условиях между этими видами наблюдается
сильная этологическая изоляция, которая, возможно, действует
также и в природе.
Если в лаборатории иногда возникают гибриды, то самцы всег-
да бывают стерильны; гибридные же самки иногда бывают фер-
Гибридизация, преадаптация и адаптация 323
тильными. В природе гибридов не обнаруживали ни разу. Айале
и его сотрудникам удалось также выявить в пределах вида D. ра-
ulistorum шесть групп, проявлявших четко выраженные предпоч-
тения к спариванию с членами своей группы, несмотря на то что
эти группы были совершенно неразличимы морфологически. Одна-
Рис. 12.8. Распространение четырех видов-двойников, принадлежащих к группе
Drosophila willistoni. (Ayala, 1975.)
ко этологическая изоляция между этими группами не так сильна,
как изоляция между видами. При образовании гибридов между
разными группами самцы опять-таки стерильны, но самки всегда
•фертильны. Каждая из этих групп имет свой ареал, отличный от
.других (рис. 12.9), хотя все же эти ареалы слегка перекрываются.
В природе гибридов не обнаружено. Поскольку группы, выявляе-
мые в пределах вида D. paulistorum, обладают многими признака-
ми видов, их стали называть полувидами.
Таким образом, как мы видим, существуют виды, не разделен-
ные абсолютными изолирующими преградами, и, что столь же
важно, существуют виды, популяции которых не вполне интерфер-
дильны. Кроме того, следует помнить, что большинство популяций
21*
324
Глава 12
данного вида изолировано от других его популяций вследствие
разделенности в пространстве и во времени. Ведь между популя-
циями полевок из Нашвилла (штат Теннесси) и из Гейнсвилла
(штат Флорида) значительный обмен генами просто невозможен.
Между этими полувидами нет морфологических различий, однако некоторые из
них разделены репродуктивными преградами. Большинство гибридов между по-
лувидами дают плодовитых самок, но стерильных самцов. (Ayala, 1975.)
Поскольку вероятность прямого обмена генами между этими по-
пуляциями так мала, что практически почти равна нулю, любой
обмен происходит через географически промежуточные популяции.
Для такого процесса понадобится в лучшем случае несколько де-
сятков или сотен поколений. Однако, как показали эксперименты,
между популяциями полевок из Нашвилла и Гейнсвилла репродук-
тивной изоляции не возникло.
ОПРЕДЕЛЕНИЕ ВИДА
Как можно определить вид? Согласно биологической концеп-
ции, вид — это «группа скрещивающихся популяций, репродуктив-
но изолированных от любой другой группы популяций». Имеется
Гибридизация, преадаптация и адаптация 325
слишком много случаев, к которым эта концепция в ее абсолютно
строгом смысле неприменима. Однако она по-прежнему остается
единственной попыткой определить вид в содержательных терми-
нах—содержательных потому, что для определения вида исполь-
зуется такое вполне конкретное биологическое свойство, как спо-
собность к скрещиванию. Тем не менее остается фактом, что у нас
нет возможности дать такое содержательное определение вида,
которое бы отражало все аспекты эволюционной ситуации. Если
выбрать в качестве критерия скрещиваемость, то окажется, что
многие морфологически обособленные группы интерфертильны;
если же критерием будет служить морфологическое сходство, то
вскоре станет ясно, что созданные таким образом группы представ-
ляют собой совокупности взаимно изолированных или частично
изолированных популяций. Избрав физиологические или экологи-
ческие критерии, можно обнаружить подобное же несоответствие.
Провозглашение более четверти века назад биологической кон-
цепции вида было большим шагом вперед в истории биологии.
Эта концепция привлекла внимание к роли изолирующих механиз-
мов и к важности содержательного определения вида. Однако за
прошедшие с тех пор годы мы узнали много нового о структуре
и генетике популяций и о механизмах видообразования. Говорят,
что наука слагается из ряда последовательных шагов, все больше
приближающих нас к истине. Концепция вида служит превосход-
ной иллюстрацией к этому высказыванию.
ПРЕАДАПТАЦИЯ И ПРОИСХОЖДЕНИЕ ЭВОЛЮЦИОННЫХ НОВШЕСТВ
На примере эволюции подсолнечника мы увидели, что вид при-
обретает новые признаки в результате происходящей время от
времени гибридизации с другими видами. В гл. 11 было показано,
что смещение признаков в процессе дифференциации видов ведет
к изменению какого-либо признака или, путем усиления данного
признака, к развитию нового признака (например, иной брачной
песни у лягушек, возникшей в результате усиления какой-либо
части исходной песни). Однако иногда в процессе эволюции виды
приобретают признаки, которые отличаются от исходных признаков
не только количественно. Особенно важное значение имеет приоб-
ретение организмами в процессе эволюции структур или свойств,
которые дают им возможность освоить новую функцию, как это
было, например, когда животные вышли из воды на сушу и при-
обрели легкие и ходильные конечности, или же когда они научи-
лись летать и приобрели крылья. Если бы первые наземные жи-
вотные не могли каким-то образом вдыхать воздух, они не смогли
бы выжить на суше. Точно так же первые животные, поднявшиеся
в воздух, должны были каким-то образом удержаться в нем. Но
как же может животное приобрести путем накопления благоприят-
ных мутаций способность дышать воздухом до того, как оно по-
селилось на суше?
326
Глава 12
Совершенно очевидно, что организм не может начать выполнять
какую-то новую функцию, например дышать воздухом, до того,
как он приобрел соответствующую новую структуру. А следова-
тельно, такая структура уже должна иметься у него заранее. Иначе
говоря, организм должен обладать структурой, которая^в резуль-
тате положительного отбора в некой среде и для некой функции
достигла такого состояния, что стала способна выполнять новую
функцию, применительно к которой она не подвергалась отбору.
Мы называем такие структуры преадаптированными. Хорошей
иллюстрацией этой концепции служит история первых амфибий.
Водные животные обитают в среде (вода), которая обеспечи-
вает им сохранение влажности тела, снабжение кислородом и пла-
вучесть. Выходя из воды на сушу, они сталкиваются с необходи-
мостью преодолевать силу тяжести в менее плотной среде, извле-
кать кислород из воздуха и поддерживать свои клетки и ткани
во влажном состоянии. Впервые примитивные легкие появились у
некоторых рыб в виде полых выростов в передней части пищевода.
Такие легкие первоначально давали возможность этим рыбам жить
в водоемах с низким содержанием кислорода; примитивные легкие
сохранились у ряда костных рыб до сих пор. У этих же групп
развились также мускулистые грудные плавники, с помощью ко-
торых они, вероятно, могли передвигаться по скалам и по дну.
Животные с неким подобием легких и с возможностью переме-
щаться по дну водоемов впервые возникли среди рыб в ответ на
давление со стороны своей среды обитания — мелководных при-
брежных участков, стоячих водоемов с низким содержанием кис-
лорода. Приобретая такие признаки, эти рыбы получили возмож-
ность покидать на время водную среду либо для того, чтобы спас-
тись от хищника, либо для того, чтобы уйти из пересыхающего
водоема. Такие выходы на сушу, вероятно, могли иметь место
только во влажные ночи или в дождливые дни, подобно тому как
это делают некоторые рыбы (например, интродуцированный во
Флориду сомик-кошка) в настоящее время. Новый тип поведения,
давший животным непосредственное преимущество, Дут же привел
в действие мощный ряд новых давлений отбора, которые стимули-
ровали быструю эволюцию и специализацию преадаптированных
структур, в данном случае — легких и конечностей. Давления отбо-
ра, изначально обусловившие возникновение легких, необходимых
для дыхания, и конечностей, необходимых для поддержания тела
и передвижения в воздушной среде, были связаны с жизнью в
мелких, бедных кислородом водоемах, так что, когда понадоби-
лись новые функции (вдыхание воздуха, передвижение по суше),
эти структуры уже существовали. Новые давления отбора усилили
и усовершенствовали легкие и передние конечности, а также ока-
зали сильное влияние на развитие грудной клетки и головы, но все
это не могло бы произойти, если бы примитивные легкие и плав-
ники не были преадаптированы к жизни на суше.
Гибридизация, преадаптация и адаптация 327
Концепция преадаптации чрезвычайно важна, потому что она
позволяет объяснить, каким образом в изменяющейся среде могут
одновременно происходить и эволюция и адаптация. Неадаптиро-
ванные особи, популяции и виды обречены на вымирание. Многие
специализированные и процветающие организмы оказывались не
в состоянии справиться с проблемами, возникшими в результате
изменений условий среды, и вымирали. Тысячи видов, которые
когда-то господствовали на нашей планете, вымерли, иногда после
долгих периодов процветания. Примерами служат динозавры и
южноамериканские Notoungulata. Из растений можно назвать
гигантские каламиты и семенные папоротники, которые некогда
были господствующими формами в растительности Земли. В сов-
сем недавнее время мы явились и продолжаем быть свидетелями
вымирания многих животных и растений, таких, как странствую-
щий голубь, американский бизон, красивый кустарник Franklinia
(ныне сохранившийся лишь в качестве культурного растения), и
многих других организмов, не способных существовать в среде, в
которой непрерывно возникают резкие изменения, создаваемые
человеком. О том, что вымирание отнюдь не неизбежная судьба
всех организмов, свидетельствуют такие разнообразные примеры,
как ржавчина пшеницы, тараканы, японский хрущик, крысы и
скворцы, которые процветают, несмотря на вмешательство челове-
ка в природу, и в сущности в значительной степени именно благо-
даря этому вмешательству.
АДАПТАЦИЯ
Слово адаптация использовалось эволюционистами и эколога-
ми уже очень давно, причем в различных значениях. Оно исполь-
зовалось главным образом в трех разных смыслах: 1) в сочетании
с каким-либо признаком («длинный мех служит адаптацией к хо-
лоду»); 2) для обозначения состояния («кактусы адаптированы к
жизни в пустынях»); 3) для обозначения процесса («растения
адаптируются к засухе, закрывая устьица»). Эти различные ис-
пользования слова создают трудности для понимания. Мы будем
использовать слово «адаптация» главным образом для обозначе-
ния состояния адаптации.
Ученых давно интересуют очень тщательно сбалансированные
и вызывающие восхищение приспособительные признаки, вырабо-
танные организмами для высокоэффективного выполнения опреде-
ленных функций. Тонкие длинные ноги скаковых лошадей, их
стройное обтекаемое тело, длинная шея и вытянутая голова — все
эти признаки созданы путем искусственного отбора с целью по-
высить скорость бега. У ломовых лошадей ноги короткие, массив-
ные и сильные, грудь широкая, а шея короткая — селекция ведется
по признакам, увеличивающим силу. Точно так же у гепардов,
преследующих свою жертву, тело тонкое и длинное и длинная
328
Глава 12
шея, а у львов и тигров, скрадывающих добычу, тело сильное и
широкое. Следовательно, не без оснований можно заключить, что
естественный отбор формирует признаки организмов таким обра-
зом, чтобы они могли наиболее успешно выполнять определенные
задачи. Такие структуры называют адаптированными.
Однако при более глубоком и строгом рассмотрении этого пред-
мета возникает много вопросов. Все ли признаки организмов адап-
тивны или только некоторые и какие именно? Адаптивны ли толь-
ко морфологические признаки, или также поведенческие, биохими-
ческие и признаки, связанные с размножением? Каким именно
способом организм, приспособленный к одной среде, становится
приспособленным к новой среде? И можно ли объяснить эволю-
цию таких сложнейших органов, как глаз млекопитающих, одним
лишь накоплением мелких мутаций под действием естественного
отбора?
Адаптацию можно рассматривать как явление, подчиняющееся
закону «все или ничего». В этом случае единственным критерием
адаптивности служит сам факт существования. Популяция адап-
тирована к определенной среде, если она может жить и размно-
жаться в этой среде. Но если считать адаптацию лишь явлением
типа «все или ничего», то применимость этой концепции окажется
очень ограниченной. Можем ли мы, так сказать, количественно
оценивать адаптацию?
В повседневной жизни, называя организм (или человека)
«адаптировавшимся», мы имеем в виду, что данный организм
справляется со стоящей перед ним задачей «надлежащим обра-
зом». Продолжая в том же духе, можно назвать ту или иную
структуру адаптивной, если она подходит для выполнения опреде-
ленной задачи. Но некоторые структуры оказываются более «под-
ходящими», чем другие. А следовательно, имеет смысл говорить
о степени адаптации. Проблема состоит в том, чтобы установить
критерии, которые помогали бы решать, в какой мере данный
признак «подходит» для данного организма, или, иначе говоря,
определять степень создаваемого этим признаком преимущества.
Процесс естественного отбора, рассмотренный в гл. 3, дает
нам критерий, позволяющий определять степень «адаптивности»
того или иного признака. В сущности естественный отбор ведет к
увеличению частоты в популяции одних вариантов данного при-
знака и уменьшению частоты других. Те варианты, которых стано-
вится больше, следует считать более «подходящими» в том смыс-
ле, что они повышают выживаемость обладающих ими организмов,
т. е. они более адаптивны. Иными словами, адаптивен такой
признак, которому благоприятствует отбор, сохраняя особей, об-
ладающих этим признаком, и элиминируя особей, лишенных его,
причем более адаптивным признакам отдается предпочтение по
сравнению с менее адаптивными. Так, например, темная окраска
у Biston betularia адаптивна по сравнению со светлой в загрязнен-
Гибридизация, преадаптация и адаптация 329
ной среде, потому что ей благоприятствует естественный отбор.
Заметьте, что мы не высказываем никакого суждения относительно
того, что темная окраска лучше, эффективнее или обладает каким-
либо иным достоинством. Обратите также внимание на то, что о
степени адаптивности данного состояния признака можно судить
только по отношению к другим состояниям этого признака, таким,
как темная или светлая окраска у бабочек, съедобные или несъе-
добные насекомые (гл. 15), устойчивые илн чувствительные к ДДТ
мухи (гл. 6). Что касается признаков, закрепившихся в популяции
и не подверженных изменчивости, то любые высказывания относи-
тельно степени их адаптивности носят сомнительный и неопреде-
ленный характер.
Являются ли адаптивные признаки оптимальными?
Нередко утверждают, что эволюция njfreM естественного отбо-
ра ведет к развитию оптимальных фенотипов, т. е. таких феноти-
пов, которые теоретически с наибольшей эффективностью способ-
ствуют выживанию и размножению организмов.
При этом возникает кардинальная проблема: как можно ска-
зать, что такое наиболее эффективный генотип? При построении
оптимизационных моделей исследователь никогда не может быть
уверен в том, что он перебрал все имеющиеся альтернативы. Мо-
жет оказаться, что какой-то совершенно иной вариант сработал бы
лучше, чем вариант, принятый за оптимальный. Аналогичным об-
разом естественный отбор может действовать только на те при-
знаки или сочетания признаков, которые реально существуют.
Резко отличающийся от имевшихся ранее признак (или сочетание
признаков) дает возможность организму использовать новую си-
туацию в среде обитания таким образом, каким ее никогда не
смогли бы использовать его предки, хотя при этом он может функ-
ционировать в этой среде весьма неэффективно по сравнению с тео-
ретическим оптимумом. Примером служат самые первые наземные
позвоночные. Они были лучше, чем рыбы, адаптированы к жизни
на суше, но при том мало эффективны в абсолютном смысле.
Факт возникновения в процессе эволюции организмов, отличаю-
щихся от своих предков, даже когда среда остается неизменной,
служит указанием на то, что оптимум еще не достигнут.
Естественный отбор — это процесс, благоприятствующий наиме-
нее ущербным из имеющихся фенотипов. Но этот процесс необя-
зательно создает оптимальные фенотипы. Отбираемые фенотипы
удовлетворяют лишь двум основным требованиям: 1) они адапти-
рованы в абсолютном смысле; 2) они сохраняются из поколения в
поколение.
Имеются дополнительные соображения, главным образом гене-
тического характера, которые препятствуют эволюции оптималь-
ных фенотипов. Это, во-первых, случайная природа мутационного
33'- Глава 12
процесса. Большинство мутаций гибельны, а поскольку каждая
особь в данной популяции несет несколько вредных аллелей, ни
один фенотип никогда не будет действительно оптимальным. Во-
вторых, большинство генов обладают плейотропным действием
(т. е. оказывают множественные эффекты) и хотя некоторые из
этих эффектов бывают благоприятными, другие обычно неблаго-
приятны. В-третьих, существует такое явление, как сцепление ге-
нов. Частота в популяции аллелей со слабым вредным действием
может возрасти, если они тесно сцеплены с благоприятными ге-
нами. При наличии достаточного времени такое неравновесие по
сцеплению (гл. 8) может быть нарушено, однако процесс этот,
вероятно, протекает медленно. В-четвертых, у гетерозигот происхо-
дит выщепление менее приспособленных гомозиготных генотипов.
Хотя каждый из этих генетических факторов сам по себе может
быть тривиальным, их аддитивные эффекты могут оказаться до-
вольно значительными.
Итак, естественный отбор элиминирует худшие (в смысле вы-
живания и плодовитости) фенотипы в популяции, но выживающие
фенотипы, хотя они и лучше приспособлены, чем фенотипы элими-
нируемые, ни в коем случае нельзя считать оптимальными; т. е.,
иначе говоря, это не обязательно наилучшие возможные фенотипы.
Теперь мы перейдем к более подробному изучению взаимодей-
ствий между особями, популяциями и видами и влияния этих про-
цессов на эволюцию путем естественного отбора.
ЛИТЕРАТУРА
Общие вопросы
Anderson Е. 1949. Introgressive Hybridization, New York, Wiley.
Dawson G. W. P. 1962. An Introduction to the Cytogenetics of Polyploids, Oxford,
Blackwell Scientific Publications.
Solbrig О. T. 1970. Principles and Methods of Plant Biosystematics, New York,
Macmillan.
Stebbins G. L. 1971. Chromosomal Evolution in Higher Plants, London, Edward
Arnold.
Специальные статьи и обзоры
Ayala F. J. 1975. Genetic differentiation during the speciation process, Evlo. Biol.,
8, 1—78.
Epling C. 1947. Natural hybridization of Salvia apiana and S. mellifera, Evolution,
1, 69—78.
Grant V. 1964. The biological composition of a taxonomic spesies in Gilia, Adv. in
Genetics, 12, 281—327.
Heiser С. 1951. Hybridization in the annual sunflowers: Helianthus annuusXH. debi-
lis var. cucumerifolius, Evolution, 5, 42—51.
Heiser C. 1961. Natural hybridization and introgression with particular reference to
Helianthus, Recent Adv. in Botany, 1, 874—877.
Jacksan R. C. 1976. Evolution and systematic significance of polyploidy, Ann. Rev.
Ecol. Syst, 7, 209—234.
Lewis H. 1962. Catastrophic selection as a factor in speciation, Evolution, 16, 257—
271.
Гибридизация, преадаптация и адаптация
331
Ornduff R. 1966. A biosystematic survey of the goldfield genus Lasthenia, Univ.
Calif. Publ. Botany, 40, 1—92.
Packard G. C. 1974. The evolution of air-breathing in paleozoic gnathostome fishes,
Evolution, 28, 320—325.
Remington C. L. 1968. Suture-zones of hybrid interaction between recently joined
biotas, Evol. Biol., 2, 321—428.
Roose M. L., Gottlieb L. D. 1976. Genetic and biochemical consequences of polyploi-
dy in Tragopogon, Evolution, 30, 818—830.
Stebbins G. L. 1947. Types of polyploids: their classification and significance, Adv.
in Genetics, 1, 403—429.
Глава 13
Рост и регуляция численности
популяций
Генетик, изучающий эволюцию, старается выявить изменения
в частоте или в соотношении генов в популяции. Генетик иссле-
дует частоты, а не абсолютные численности, потому что числен-
ность особей в популяциях растений и животных изменяется из
года в год или даже в пределах одного года. Изменения эти иног-
да бывают очень резкими. Когда популяция растет, увеличивается
также и число большинства генов, а когда она сокращается, то
это число уменьшается. Однако генные частоты представляют со-
бой статистические абстракции. Для того чтобы понять взаимоот-
ношения организма со средой, нам необходимо знат-ь число отдель-
ных особей каждого вида, годичные колебания их численностей,
число ежегодно погибших и рожденных, а также сроки гибели и
рождения, поскольку все эти параметры могут оказывать влияние
на течение эволюции.
Число особей данного вида в области может изменяться, при-
чем иногда очень резко. Например, в 1536 г. испанский аделан-
тадо Педро де Мендоза, закладывая город Буэнос-Айрес, привез
в аргентинские пампы 20 коров и 72 лошади. Спустя три года по-
селение было сожжено дотла индейцами, и испанцы его покинули.
Лошади и коровы оказались предоставленными самим себе. Они
размножились в пампах и к 1700 г. численность популяции коров
и популяции лошадей достигла миллиона голов каждая. Испанские
мореплаватели XVI и XVII вв. систематически завозили также на
океанические острова коз, чтобы обеспечить себе источник пищи
на случай кораблекрушения. Один такой путешественник Хуан
Фернандес завез пару коз на острова Тихого океана вблизи побе-
режья Чили, которые затем были названы его именем. В 1704 г.,
когда Александр Селкирк (послуживший Даниэлю Дефо прототи-
пом Робинзона Крузо) был оставлен на этих островах капитаном
его корабля, численность стада коз, которым дала начало эта
пара, превышала 10 000, и стадо существует до сих пор. Ежегодно
возникают новые штаммы вируса гриппа (гл. 6), которые за не-
сколько месяцев распространяются по всему земному шару. Ржав-
чина пшеницы ежегодно поражает посевы в северных районах
пшеничного пояса США, где за несколько месяцев ее количество
увеличивается в несколько раз (гл. 6). На органических тканях за
Рост а регуляция численности популяций
333
несколько часов с удивительной быстротой размножаются бакте-
рии; при хлебопечении или пивоварении число дрожжевых клеток,
содержащихся в закваске, быстро увеличивается в несколько раз.
Способность организмов увеличиваться в числе при надлежащих
условиях поистине колоссальна.
Но виды способны и вымирать, иногда очень быстро. В 1919 г.
в США был случайно завезен сумчатый гриб, вызывающий рак
каштанов. Этот гриб поселяется на каштановых деревьях, в том
числе и на американских каштанах Castanea dentata, и приводит
их к гибели. Через 10ле(г после интродукции гриба популяция этого
величественного дерева, ранее преобладавшего в лесах, так сокра-
тилась, что в настоящее время оно практически вымерло. До
XIX в. странствующий голубь был одним из. наиболее многочис-
ленных видов птиц в США. Однако охота на этих голубей при-
вела к их исчезновению. Вместе с тем никакой отстрел не дает
возможности сдержать распространение в США скворцов и черных
дроздов.
Хотя примеры резких колебаний численности популяций в сто-
рону как увеличения, так и сокращения найти нетрудно, большин-
ство видов находится в состоянии своего рода равновесия, во
всяком случае на протяжении сравнительно коротких отрезков
времени. В численности большинства видов растений и животных
явных изменений не отмечается.
Изучение изменений численности особей в популяции, называе-
мое демографией, представляет собой высокоспециализированную
область популяционной биологии. Основной толчок к развитию
демографических исследований дали страховые компании, которым
нужны были сведения о вероятности смерти людей, желавших
приобрести страховой полис. В результате демография человека
изучена довольно хорошо, тогда как о демографии популяций
растений и животных известно относительно мало.
Как показывают наблюдения, разные виды сильно различают-
ся по демографическим показателям. Продолжительность жизни
некоторых видов деревьев исчисляется десятками или даже сотня-
ми лет, тогда как жизнь большинства бактерий продолжается
несколько часов или даже минут. Большинство видов животных
живут менее года, а большинство видов сосудистых растений —
более года. Вместе с тем некоторые эфемерные растения живут
всего один-два месяца, а лишь несколько видов животных — более
50 лет. Продолжительность жизни большинства микроорганизмов
коротка (несколько часов или дней), но и среди них наблюдается
в э|том отношении довольно значительная изменчивость.
Демографические характеристики того или иного вида зависят
от той среды, в которой он обитает, и представляют собой адап-
тивный признак вида, выработавшийся в процессе естественного
•отбора. В данной и двух следующих главах мы изучим этот аспект
эволюции более подробно.
334
Глава 13
РОСТ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ
Каждый организм рождается и умирает1. Если он проживет
достаточно долго, он успевает также размножиться. Продолжи-
тельность жизни каждой особи и число ее потомков индивидуаль-
ны, и их нельзя точно предсказать. Однако популяция, слагающая-
ся из отдельных особей, обладает статистическими свойствами,
зависящими от доли особей со сходными характеристиками. Эти
свойства можно анализировать теоретически в математических тер-
минах. Они касаются главным образом роста, размножения и ги-
бели.
Мы опишем сначала некоторые демографические свойства по-
пуляций, а затем обсудим ряд проблем, с которыми приходится
сталкиваться при изучении реальных популяций, а также рассмот-
рим несколько конкретных примеров.
Экспоненциальный рост
Популяциями с дискретными или неперекрывающимися поко-
лениями называют такие популяции, в которых потомки начинают
размножаться лишь после смерти родительских особей. Например,
в популяциях таких однолетних растений, как полевая горчица,
растения гибнут после того, как они произведут семена, а эти но-
вые семена прорастают лишь на следующий год. Популяциями с
непрерывными или перекрывающимися поколениями называют та-
кие, в которых взрослые особи разного возраста размножаются
одновременно (например, популяции человека, многих видов де-
ревьев и многих животных). Математические методы, необходи-
мые для анализа этих двух ситуаций, несколько различны.
Дискретные поколения. Начнем с простейшего случая, а именно
с популяции, состоящей из особей, которые рождаются, размно-
жаются один раз, а затем умирают. Допустим, что ни одна особь
не гибнет до тех пор, пока она не размножится. Число особей в
каждом поколении будет зависеть от скорости размножения, Ьо,
которую мы определим как среднее число потомков, оставленное
после себя каждой особью. Если Ьо больше 1, то численность попу-
ляции увеличивается; если Ьо равно в точности 1, то численность
популяции остается постоянной; если же Ьо меньше 1, то числен-
ность популяции сокращается и она в конечном счете элиминиру-
ется. У видов с соотношением полов 1 : 1 скорость размножения
обычно определяется как среднее число потомков женского пола,
производимых каждой самкой. У большинства видов репродуктив-
ная способность сильно превышает ту, которая необходима для
1 У организмов, размножающихся делением (как бактерии), почкованием (как.
гидра) или бесполым путем (как многие растения), вопрос о том, что именно сле-
дует считать «рождением» или «смертью», решить довольно трудно.
Рост и регуляция численности популяций 335
замещения родительского поколения, а поэтому численность боль-
шинства популяций обычно увеличивается, если только их рост
не сдерживается каким-либо внешним фактором. Например, было
установлено, что самка комнатной мухи откладывает за одно по-
коление примерно 120 яиц, среди которых в среднем из 60 вылуп-
ляются самки. У мухи бывает примерно по 7 поколений в год.
Если допустить, что мухи погибают лишь после того, как отложат
яйца, то популяция, состоявшая в начале года из одного самца
и одной самки, к концу года возрастет до 5 598720 000 000 особей,
или примерно до 5,6-1012! Считая, что каждая муха весит 0,01 г,
вес популяции увеличился бы при этом от 0,02 г до 28 000 тонн!
Выразив это соотношение в виде уравнения, получим
где Nt — число особей в поколении t; b0 — чистая скорость размно-
жения самок; No— число особей в 0-м поколении. Иными слова-
ми, число особей в любой момент времени (измеряемый в поколе-
ниях) равно их начальному числу, умноженному на скорость раз-
множения в степени t. В рассматриваемом нами случае
N1 = 2 607 = 5 598 720 000 000.
Такой рост называется экспоненциальным.
Совершенно очевидно, что численность видов может увеличи-
ваться таким образом лишь в особых обстоятельствах и лишь в
течение очень коротких периодов времени.
Непрерывнее размножение. Многие виды размножаются по-
стоянно и у них не удается вычленить отдельные поколения. Для
того чтобы подсчитать число особей у такого вида в данный мо-
мент времени, необходимо знать число особей, имевшихся у него
в какой-то момент времени в прошлом, и скорость, с которой про-
исходил рост популяции в промежутке между этими двумя момен-
тами. Скорость роста популяции описывается уравнением
^-=bN—dN^rN,
at
где N— число особей в популяции в некоторый данный момент;
b — рождаемость; d — смертность; г — константа, специфичная для
определенного вида и его среды, называемая истинной (intrinsic)
скоростью увеличения численности популяции и равная b — d,
т. е. рождаемость минус смертность. Таким образом, скорость ро-
ста популяции равна произведению константы г на число особей
в этой популяции, N. Число особей в данной популяции в данный
момент времени находят следующим образом. Уравнение
336
Глава 13
можно представить в виде
1 dN __
N' dt ~Г
ИЛИ
dinW _ .
dt ~Г'
интегрируя это выражение, получим
In Nt—\nN0<=rt.
Освободившись от логарифмов, имеем
е(1п Л/(-1п Nt) _grl
или
где No — число особей в начальный момент; Nt— число особей в
момент времени t, а е — основание натуральных логарифмов.
Логистический рост
Экспоненциальный рост может происходить лишь на протяже-
нии очень короткого периода времени вследствие ограниченности
ресурсов. Затем наступает недостаток пищи, а в конечном счете
и пространства. Если бы каждую весну каждая пара мух, раз-
множаясь, давала 28 тонн мух, то на земном шаре не осталось бы
места ни для чего другого, кроме мух. При низкой численности
популяции, например если она состоит всего из двух мух, рост
может быть примерно экспоненциальным в течение одного-двух
поколений. Однако по мере увеличения численности популяции
она приблизится к некоторому верхнему пределу, когда число гиб-
нущих особей равно числу появляющихся на свет. Размер популя-
ции, при котором скорость ее роста равна нулю, называется ем-
костью среды и обозначается буквой К. Емкость среды опреде-
ляется наличными ресурсами.
Таким образом, рост популяции регулируется двумя фактора-
ми. Первый, который можно рассматривать как врожденную ха-
рактеристику данного вида, это его способность размножаться с
некоторой максимальной скоростью. Второй, который можно рас-
сматривать как сопротивление среды, — это ограниченность ресур-
сов среды, приводящая к тому, что с увеличением числа особей
в популяции возрастает смертность и снижается рождаемость.
Приводившиеся ранее формулы следует модифицировать, с тем
чтобы в них учитывался тот факт, что число новых особей, сущест-
вование которых может обеспечить среда, зависит от числа особей,
которые в ней уже обитают. Проще всего это сделать, допустив,
что данная зависимость является линейной и что имеется некая
Рост и регуляция численности популяций 337
предельная численность, по достижении которой добавление к по-
пуляции новых особей возможно лишь при условии, что некоторые
из уже имеющихся в ней особей погибнут. Эта предельная чис-
ленность и есть К—емкость среды.
Рис. 13.1. Экспоненциальная (7) и логистическая (//) кривые роста.
Для более общей модели непрерывного роста уравнение ско-
рости роста принимает вид
dN _rNfK~N\
Эту формулу, которую часто называют логистическим уравнением,
или уравнением Вольтерра в честь предложившего его итальян-
ского математика, очень легко проанализировать. Обратите вни-
мание на то, что когда число особей в популяции (N) мало по
сравнению с емкостью среды (К), отношение (К — N)/K близко
к 1 и рост популяции почти экспоненциален. Однако по мере того
как N приближается к К, отношение (К — N)/K приближается
к 0, достигая его при N=K. Если построить график роста числен-
ности популяции в зависимости от времени, то получится S-образ-
ная кривая (рис. 13.1). Рост такого типа называется логистическим
ростом.
Как показали эксперименты, рост некоторых микроорганизмов
(например, Paramecium, рис. 14.6, или дрожжей) в искусственно
регулируемых условиях соответствует логистической кривой. Слу-
жит ли, однако, логистическая модель точным описанием роста
популяций всех или хотя бы большинства организмов?
В 1937 г. на остров Протекшей в Пьюджет-Саунд у побережья
штата Вашингтон завезли двух самцов и шесть самок обыкновен-
ного фазана (Phasianus colchicus). За пять лет численность попу-
ляции фазана достигла 1325 размножающихся особей, т. е. воз-
22—2044
338
Глава 13
росла в 166 раз. Судя по некоторым признакам, рост этой популя-
ции происходит по логистической кривой (рис. 13.2). От двух сам-
цов и шести самок, привезенных на остров в 1937 г., она возросла
до 30 особей к весне 1938 г., до 81 —в 1939 г., до 282 — в 1940 г.,
641—в 1941 г., 1194 — в 1942 г. и до 1898 — в 1943 г., когда на
остров прибыли военные части и солдаты начали охотиться на
Рис. 13.2. Рост популяции фазанов на острове Протекшей.
Исходно популяция состояла из двух самцов и шести самок. (Einarsen, 1945.)
фазанов. Однако создается впечатление, что к 1943 г. численность
популяции начала стабилизироваться. За предшествующие 6 лет
учеты численности проводили весной, до начала размножения, и
осенью, после того как размножение заканчивалось и молодые
особи почти завершали рост. При этом обнаружилось, что хотя
число яиц, приходящихся на одну самку, за изучавшийся период
не изменялось, ежегодный прирост неуклонно снижался, потому
что доля успешных гнездований все больше уменьшалась, а смерт-
ность среди птенцов повышалась. В результате наблюдались коле-
бания численности между весной и осенью. В зимние месяцы про-
исходила только гибель птиц, а не размножение.
Результаты других исследований показывают, однако, что ло-
гистическая модель малопригодна для описания роста популяции.
Рост и регуляция численности популяций 339
Австралийский эколог А. Дж. Николсон (A. J. Nicholson) провел-
ряд экспериментов на лабораторных популяциях зеленой мясной’
мухи (Lucilia cuprina). В большинстве экспериментов ограничи-
П сололжительногть эксперимента, дни
Рис. 13.3. Динамика численности лабораторной популяции зеленой мясной мухи-
(Lucilia cuprina).
Взрослых мух ежедневно кормили измельченной печенью, причем количество кор-
ма в одних опытах (А и В) периодически изменялось, а в других (Б) оставалось
одинаковым (0,4 г); ритм дачи корма показан пунктирной кривой в верхней ча-
сти каждого графика. (Nicholson, 1958.)
вающим рост фактором служило количество корма, ежедневно
вносившегося в экспериментальную культуру. Личинкам мясной
мухи необходим определенный уровень питания для того, чтобы
окуклиться, а затем превратиться во взрослых особей. Чем больше
количество корма, достающегося каждой личинке, тем больших
размеров достигает личинка, прежде чем она окукливается. Чем
крупнее личинка, тем крупнее получающаяся из нее взрослая особь
и тем большее число яиц она может отложить. Был проведен ряд
экспериментов, в которых количество корма, дававшегося взрос-
лым особям и личинкам, варьировали независимо одно от другого.
При этом оказалось, что даже при самых неблагоприятных усло-
виях популяции удавалось сохраниться. В большинстве случаев-
2 •
340
Глава 13
наблюдались ярко выраженные периодические колебания числен-
ности имаго и (или) личинок (рис. 13.3). Рост популяции синих
мясных мух не соответствовал логистической модели. Другим хо-
рошо документированным примером популяции, рост которой не
соответствует логистической модели, служит популяция мелкого
ракообразного Daphnia pulex (рис. 13.4).
Рис. 13.4. Динамика численности, наблюдавшаяся в лабораторной популяции
дафний, заложенной в шести повторностях.
Каждая популяция исходно состояла из 25 особей, возрастной состав которых
примерно соответствовал распределению, характерному для такой плотности. Ор-
дината для каждой повторности сдвинута на 400 единиц по отношению к преды-
дущей. (Frank, 1960.)
Почему же логистического уравнения самого по себе недоста-
точно для описания роста популяции в природных условиях? При-
чин этому много. Они, несомненно, различны для разных видов
и пока еще не все поняты. Тем не менее некоторые из них можно
проанализировать.
Скорость роста популяции, описываемая уравнением
dN кт/ К —N \
sr=rN(—K—J-
изменяется всякий раз, когда к этой популяции добавляется или
изымается из нее одна особь. Представим себе, что численность
популяции увеличивается. Для того чтобы кривая роста имела вид
плавной логистической кривой, г должно быть действительно по-
стоянной величиной. Но г=Ь—d, а, как нам известно из наблю-
дений, у большинства видов имеются определенные периоды в те-
чение года, когда рождаемость (6) достигает пика, и периоды,
Рост и регуляция численности популяций 341
когда она выравнивается; мы знаем также, что смертность (d)
достигает наивысшего уровня в одно время года и наинизшего —
в другое. Следовательно, г непостоянно во времени. В этом за-
ключается первая причина отклонения от логистического уравне-
ния. Кроме того, это уравнение подразумевает, что добавление к
популяции или изъятие из нее хотя бы одной особи немедленно
сказывается на скорости роста. Наблюдая за теми или иными
видами, можно заметить, что между оплодотворением и рождени-
ем потомков имеется известный промежуток времени. Число рож-
дений в момент времени /4-1 определяется не числом особей,
имеющихся в момейт времени t, а числом самок, оплодотворенных
в момент времени t—g (где g— период беременности), которые
дожили до времени /4-1.
В качестве крайнего примера представим себе популяцию мы-
шей, состоящую из К—1 особей, среди которых есть несколько
беременных самок. Добавление к такой популяции еще одной
особи не вызывает массовых выкидышей у имеющихся беремен-
ных самок; численность мышей продолжает увеличиваться, если
даже размеры популяции превысят величину К. Таким образом,
задержка в реакции популяции на повышение плотности — еще
одна причина того, что рост популяции не происходит в строгом
соответствии с логистической кривой.
И наконец, необходимо исследовать природу самой величины К.
В нашей формуле К задана как постоянная величина. Ее можно
представлять себе как ресурсы среды, способные обеспечить су-
ществование определенного числа особей данного вида. Но бы-
вает ли количество ресурсов постоянным? Совершенно очевидно,
что нет. Растениям необходимы солнечный свет и вода, количество
которых варьирует в разные годы и в разных местностях. Все мы
знаем, как изменяется лес от лета к зиме; и даже тропический
лес в сезон дождей не такой, как в засушливый период. Поскольку
растительная биомасса служит пищей для растительноядных жи-
вотных, то ее изменения означают изменение числа существующих
за ее счет растительноядных, а соответственно также плотоядных
и организмов-деструкторов. Кроме того, скорость метаболизма за-
висит от температуры, которая также подвержена колебаниям.
Короче говоря, для того чтобы рост происходил в строгом соот-
ветствии с логистической кривой, необходимо, чтобы значения г и
К были постоянными и чтобы существовала мгновенно действую-
щая обратная связь. Такие условия иногда удается создать в ла-
боратории, выращивая популяцию какого-либо микроорганизма с
очень короткой продолжительностью жизни (предпочтительно с
бесполым размножением) при постоянном уровне снабжения всеми
ресурсами. При соблюдении этих условий можно наблюдать ло-
гистический рост, однако результаты, полученные в менее идеаль-
ных условиях, показывают, что для описания роста в природных
условиях логистической модели недостаточно.
342
Глава 13____________________________
ДЕМОГРАФИЯ
Как можно эмпирически выяснить, каким ’образом происходит
рост популяций в природе? Очень просто: регистрируя рождение,
рост, размножение и смерть особей в какой-либо популяции. Имен-
но это делают страховые компании и организации, производящие
переписи населения, в отношении вида Homo sapiens.
Рис. 13.5. Типы кривых выживания.
Тип / характерен для видов, у которых
смертность среди молодых особей низ-
кая и все особи достигают максималь-
ного для данного вида возраста. Тип //
относится к видам с постоянным числом
смертей в единицу времени. Тип III со-
ответствует постоянной вероятности
смерти независимо от возраста. Тип IV
относится к видам, у которых большин-
ство особей гибнет в очень раннем воз-
расте, но после того, как особь перешаг-
нула за определенный возраст, ее шансы
дожить до максимального для данного
вида возраста достаточно велики.
Выживаемость
Первые, статистические
данные, которые нужно полу-
чить, — это данные о выжи-
ваемости, т. е. о числе особей,
доживающих до каждого от-
дельного возраста, что обыч-
но выражается в доле от чис-
ла особей, родившихся в ка-
кое-то определенное время.
Для того чтобы получить эти
сведения, надо определить,
сколько особей, родившихся в
некоторое время t, доживает
до конца первого временного
интервала, сколько — до кон-
ца второго такого интервала,
третьего и т. д., до тех пор по-
ка в живых не останется ни
одной особи. Если предста-
вить эти данные в графиче-
ской форме, то получится кри-
вая выживания (рис. 13.5).
Каждый вид имеет свою ха-
рактерную кривую выжива-
ния, но все эти кривые можно
разделить на четыре общих
или основных типа, которые
обозначают как типы I, II,
III и IV.
Кривая типа I соответствует популяции, большинство членов
которой доживает до максимального для данного вида возраста
(иногда он может быть равен всего нескольким часам или дням),
а затем умирает в одинаковом возрасте. Кривые лишь очень не-
многих видов приближаются к такой кривой выживания. Наибо-
лее близки к ней кривые для популяции человека в промышленно
развитых странах (рис. 13.6). Кривая типа II соответствует попу-
ляциям, у которых в единицу времени умирает определенное число
особей. Кривая типа III относится к популяциям, у которых в еди-
Рост и регуляция численности популяций
343
Возраст, годы
Рис. 13.6. Кривая выживания лиц мужского пола в США в 1964 г. (По данным
Remington R. D., Schork М. А., 1970. Statistics with Applications to the Biological
and Health Sciences, Englewood Cliffs, N. Y., Prentice-Hall.)
Возраст, иед
Рис. 13.7. Кривая выживания лабораторной популяции Microtus agrestis. (По дан-
ным Leslie Р. Н., Ranson R. М., 1940. The mortality, fertility rate of natural increase
of the vole (Microtus agrestis) as observed in the laboratory, J. Anim. Ecol., 9,
27—52.)
100
60
40
Возраст, нед
Рис. 13.8. Кривая выживания самок (светлые кружки) и самцов (черные кружки]
ящерицы Uta stansburlana. (Tinkle D. W., 1967. The life and demography of the
side-blotched lizard, Uta stansburlana, Misc. Publ. Mus. Zool., Univ. Mich. No 132.)
Возраст, мес
Рис. 13.9. Кривая выживания плеченогого Glottldla pyramldata. (По данным Pai-
ne R. Т., 1963. Ecology of the brachiopod Glottldla pyramldata, Ecol. Monogr., S3,
187—213.)
Рост и регуляция численности популяций 345
яйцу времени умирает определенная доля особей, т. е. вероятность
смерти не зависит от возраста. Кривые выживания некоторых
популяций позвоночных и растений приближаются к типам II и III
(рис. 13.7). Кривая типа IV описывает популяцию, в которой вы-
сокая смертность наблюдается в раннем возрасте, а после неко-
торого периода времени вероятность смерти снижается. Это наи-
более обычная ситуация, характерная для популяций таких раз-
нообразных организмов, как клены, устрицы, некоторые ящерицы,
жаброногие раки (рис. 13.8 и 13.9).
Плодовитость
Другая группа необходимых нам статистических данных — это
среднее число потомков, производимых особями в каждом возра-
сте. У большинства видов размножаются лишь члены некоторых
Возраст (срединная точка возрастного класса), в днях
после превращения из личинки в имаго
Рис. 13.10. Кривая плодовитости взрослых самок вши Pediculus humanus.
Кривая построена по данным для лабораторной популяции. Резкие колебания в
правой части, вероятно, вызваны малой величиной выборки (так, например, воз-
растной класс 36,5 дн состоял всего из 9 особей, тогда как исходное их число было
равно 400. (По данным Evans F. С., Smith F. Е., 1952. The intrinsic rate of natu-
ral increase for the human louse, Pediculus humanus L. Amer. Nat, S6, 299—310.)
возрастных групп. Кроме того, даже в течение периода потенци-
альной фертильности данной особи вероятность ее размножения
далеко не одинакова. Чтобы построить график плодовитости, не-
обходимо иметь данные о числе особей женского пола, приносимых
346
Глава 13
в каждом помете, и о возрасте матери. Располагая такими с в еде*
киями, можно построить путем вычисления для каждого возраст*
ного класса среднего числа потомков, приносимых его членами,
график плодовитости, соответствующий рождаемости, специфич-
ной для каждого возрастного класса (рис. 13.10).
Чистая скорость размножения
Построив кривую выживания и график плодовитости для дан-
ной популяции, можно определить ее чистую скорость размноже-
ния, называемую также скоростью замещения* 1. Эта величина по-
казывает (если допустить, что скорости выживания и размножения
постоянны), растет ли численность популяции, остается ли она
постоянной или же сокращается.
Для вычисления чистой скорости размножения необходимо
знать: 1) число особей в данной популяции, рожденных в опреде-
ленное время (такую группу особей называют когортой), ту их
долю, которая будет успешно размножаться, и если они размно-
жаются не один, а несколько раз, то ту их долю, которая доживет
до каждого из последовательных актов размножения; 2) среднее
число потомков женского пола, которое в среднем производит
самка при каждом акте размножения.
Используя зависимость выживаемости от возраста, можно вы-
числить вероятность выживания после первого временного интер-
вала (он обычно равен одному году, но выбор его зависит от
изучаемого вида). Эту вероятность получают, разделив число осо-
бей, доживших до времени 1, на число всех особей, рожденных в
момент времени 0:
M = J£- = 1—
'vo
где d — смертность, которая равна
_N0-Nt
1~~ No
Вероятность того, что некая особь, пережившая первый интервал,
переживет и второй, дается выражением
X2 = -^- = l— (L,
л Nt
того, что особь, рожденная в момент времени t0,
моменту /г (обозначим эту вероятность через 1г},
l^X^-^x^-^.
1 Последний термин малоупотребителен и в отечественной литературе прак-
тически не встречается. — Прим. ред.
а вероятность
будет жива к
равна
Рост и регуляция численности популяций
347
Соответственно доля особей, доживающих от рождения до возра-
ста х, определяется по формуле
/JC = X1xX2X- • • хХА. = (1 — dj) X(l— d2)X • • • X(l—dj.
Таким образом, для того чтобы вычислить вероятность доживания
до определенного возраста, достаточно располагать статистиче-
скими сведениями о смертности, специфической для данного воз-
раста.
Рассмотрим теперь когорту, состоящую из No особей, родивших-
ся в данном году. Число особей, доживших до возраста х, будет
равно Noxlx, т. е. числу рожденных особей, умноженному на ве-
роятность того, что они доживут до возраста х. Число потомков,
которое произведет эта когорта в возрасте х, определяют следую-
щим образом.
Если b — число рождений, то в момент времени х число рож-
дений в данной когорте будет равно числу особей, доживших до
времени х, умноженному на рождаемость, специфическую для дан-
ного возраста (тх):
&л.=Л^хХтл.=
Для возраста х—1: —NoxlxXtnx. bx-i^N^xl^Xm^ bx-2 Nд X Iх^2 х tnx^2
Для возраста х—2:
Для возраста 1: bi=Noxli хт{
Общее число рождений *=*max 6= 2 NOlxmx
Таким образом, зная число особей, родившихся в определенное
время, мы можем вычислить наиболее вероятное число потомков,
которое они произведут; для этого достаточно лишь располагать
сведениями о зависимостях выживаемости и плодовитости в дан-
ной популяции от возраста.
Чтобы получить чистую скорость размножения Ro, т. е. среднее
число потомков на одну особь в популяции, достаточно разделить
полученное выше выражение на No-
х==хтах
N 1)1 хтХ
No
х=0
No
xmax
348
Глава 13
Среднее число потомков на одну особь в популяции, т. е. /?0, —
очень важный показатель. В популяции, находящейся в состоянии
равновесия, его величина равна
Время после вылупления, нед
Рис. 13.11. Кривые выживания для
популяций ящерицы Uta stansburiana
из Техаса (/) и Колорадо (II), по-
строенные по данным для нескольких
поколений ящериц, помеченных сразу
после вылупления. Прерывистая часть
кривой II экстраполирована ввиду
отсутствия данных за этот период
времени (Tinkle, 1969.)
1. Если Ro больше 1, то популяция
увеличивается (в популяции
рождается больше особей, чем
это необходимо для замещения
погибших); если Ro меньше 1, то
популяция сокращается.
Можно произвести дальней-
шие преобразования, поскольку
Nt=N^‘
и
приняв ^продолжительность
одного поколения = Т,
-^L—erT=R
No е
мы, таким образом, связали ско-
рость роста, выведенную из тео-
ретической модели роста попу-
ляции, с Ro. Важность этого ра-
венства обусловлена тем, что,
поскольку значение Ro можно
вывести эмпирически (перемно-
жив 1Х и тх, полученные из уче-
тов численности), оно дает воз-
можность оценить г:
Ro=<?T,
In R0—rT (1пе)=гТ.
Следовательно,
г_______ In/?о .. 1п/?(
Т — —= И 1 --—
Дональд У. Тинкл (Donald W. Tinkle) из Мичиганского уни-
верситета изучал демографию Uta stansburiana — небольшой на-
земной ящерицы из западной части США и Мексики. Исследова-
ние проводилось на популяциях двух подвидов — U. stansburiana
stejnegeri из Техаса и U. s. stansburiana из Колорадо. На рис. 13.11
представлены кривые выживания (1Х) для этих двух популяций.
Между ними имеется интересное различие. В техасской популяции
после первого года остается в живых только 10% животных, а в
Рост и регуляция численности популяций 34fr
популяции Колорадо — около 30%. В Техасе в течение сезона
размножения у каждой самки бывает по крайней мере три, а воз-
можно, четыре и даже более кладок, в среднем по 4 яйца в каж-
дой. А в Колорадо у каждой самки в первый год бывает только-
две (иногда три) кладки, но если самки переживут первую зиму»
Возраст, годы
Рис. 13.12. Возрастная структура популяций тропического дерева Pentaclathra
macroloba (черные кружки) и пальм Astrocaryum mexicanum (белые кружки) »
Euterpe globulosa (квадраты). (Sarukhan, 1979.)
то на второй год у них бывает три или четыре кладки. Многие*
самки остаются живыми и на третий год, сохраняя высокую плодо-
витость.
Мексиканский эколог растений Хосе Сарухан (Jose Sarukhan)
изучал демографию тропической пальмы Astrocaryum mexicanum—
растения, типичного для нижних ярусов тропического вечнозеле-
ного леса южной части штата Веракрус (Мексика). В табл. 13.1
и на рис. 13.12 представлены данные о вероятности выживания
от проростка до зрелого растения, среднее число плодов на одна
дерево (показатель плодовитости) и доля участия растений каж-
дого возрастного класса в суммарном размножении. Обратите
внимание на то, что вероятность доживания до следующей стадии
жизненного цикла ниже всего на стадиях проростка и молодого-
растения (для этой пальмы — примерно первые 20 лет жизни).
Пальма начинает плодоносить лишь в возрасте примерно 40 лет;
затем плодовитость с возрастом повышается и наибольшее коли-
-350
Глава 13
Таблица 13.1
Вероятности выживания и среднее число семяи иа одно дерево
для тропической пальмы Astrocarium mexicanutn
на разных стадиях развития (Sarukhan, 1979)
Стадия Возраст1), годы Вероятность до- жить до следую- щей стадии Среднее число се- мян на 1 расте- ние
Проросток 8 0,37 0
Молодая пальма 15 0,32 0
Незрелая 27 0,81 0
39 0,64 0
Зрелая 47,5 1,00 3
56,0 1,00 13
64,5 1,00 10
73,0 0,75 28
81,5 0,66 19
90,0 0,75 25
98,5 0,66 30
107,0 — С 4
) Возраст указан приблизительно.
чество семян пальмы приносят в возрасте 65 лет, потому что числи
растений старше 65 лет резко снижается (табл. 13.1).
Эти два примера иллюстрируют некоторые крайние различия
в жизненных циклах.
Стабильное возрастное распределение
У большинства популяций репродуктивная способность их чле-
нов изменяется с возрастом. В начальный период (который может
продолжаться всего несколько минут, как у микроорганизмов, или
несколько лет, как у человека и многих деревьев) организмы раз-
множаться неспособны. За этим периодом следует период посте-
пенного повышения способности к размножению, достигающей
некоторого максимума, а затем начинается спад, доходящий в ко-
нечном счете до нуля. В большинстве природных популяций особи
гибнут, не достигнув пострепродуктивного периода. Однако люди,
их комнатные и домашние животные, а также животные, которых
содержат в зоопарках, живут в более благоприятных условиях, а
поэтому доживают до этого периода. Если вклад особей разного
возраста в размножение неодинаков, то отсюда следует, что чистая
скорость роста, Ro, может варьировать вследствие изменений воз-
растной структуры без всяких изменений рождаемости. Далее,
Рост и регуляция численности популяций
351
Таблица 13.2
Вычисление стабильного возрастного распределения
На остров обрушился ураган и все живые существа, за исключением
двухлетней самки эндемичного вида черепахи, погибли. Если считать, что
рост данной популяции не зависит от плотности популяции черепахи,
а определяется лишь приведенной здесь гипотетической таблицей:
Демографическая таблица
Возраст 1 год 2 года 3 года 4 года 5 лет
1х 1 0,5 0,25 0,125 0
tnx 1 3 2 0,5 0
то возрастное распределение в популяции будет изменяться следующим
образом:
Годы однолетние двухлетние трехлетние четырехлетние пятилетние
число ДОЛЯ, % ЧИСЛО доля, % число ДОЛЯ, % число ДОЛЯ, % число ДОЛЯ, %
1-й 0 0 1 100 0 0 0 0 0 0
2-й 3 100 0 0 0 0 0 0 0 0
3-й 3 75 1 25 0 0 0 0 0 о-
4-й 6 85 1 15 0 0 0 0 0 0
5-й 9 75 3 25 0 0 0 0 0 0
6-й 18 78 4 17 1 4 0 0 0 0
7-й 32 74 9 21 2 5 0 0 0 0
8-й 63 73 18 21 4 5 1 1 0 0
9-й 125 75 31 19 9 5 2 1 0 0
10-й 237 75 62 19 15 5 4 1 0 0.
15 - й 6215 74 1615 19 420 5 109 1 от 0
численность популяций может увеличиваться, в то время как рож-
даемость понижается, исключительно за счет изменений возраст-
ной структуры, что наблюдается в популяциях человека во многих
развивающихся странах1. Кроме того, изменения в параметрах
демографических таблиц, т. е. изменения рождаемости и смертно-
сти, специфические для разных возрастных классов, влекут за
собой изменения соотношения особей разного возраста в данной
популяции. Следовательно, любая популяция, находящаяся в ста-
бильной среде, в конце концов приближается к некоторому ста-
1 Для того чтобы численность популяции росла, несмотря на понижение рож-
даемости, нет необходимости в изменении возрастного распределения; довольно
того, чтобы рождаемость, снижаясь, оставалась достаточно высокой. — Прим. ред.
352
Глава 13
бильному возрастному распределению1. Это относится ко всем по-
пуляциям, независимо от того, возрастает ли их численность,
сокращается или остается постоянной. В справедливости этого
утверждения нетрудно убедиться, проанализировав любую подлин-
ную или гипотетическую демографическую таблицу (например,
изменения 1Х и тх от х=1 до х=п, где п — максимальный возмож-
ный возраст) и определяя число самок в каждом возрастном клас-
се для каждого временного интервала, начиная с любого произ-
вольно выбранного возрастного распределения. Популяция, пред-
ставленная. в табл. 13.2, достигает стабильного возрастного рас-
пределения к 8-му году.
Как было показано, в природных условиях рост численности
популяций сдерживается рядом факторов, так что в среднем их
чистая скорость размножения (/?0) должна быть равна 1 или близ-
ка к ней (она может колебаться вблизи 1). Для того чтобы
оставалась равной 1, необходим некий уникальный график изме-
нения величины 1хтх. Отсюда следует, что в природе распределе-
ние по возрастам у большинства популяций стабильно или почти
стабильно. Это означает, что относительные доли особей, принад-
лежащих к разным возрастным группам, в стабильной среде оста-
ются постоянными из поколения в поколение. Следовательно, про-
изводя многократные учеты численности популяции с годичными
интервалами, мы должны всякий раз обнаруживать примерно оди-
наковые соотношения особей разных возрастных классов. Если
соотношения окажутся иными, то это означает, что значения 1Х и
тх непостоянны2.
Репродуктивная ценность
В популяции со стабильным возрастным распределением, на-
ходящейся в состоянии равновесия, когда /?о = 1, каждая самка
производит в среднем одного потомка женского пола. На самом
деле, однако, некоторые самки совсем не производят потомков, так
1 Это справедливо лишь для популяции, свободно размножающейся в усло-
виях неограниченности внешних ресурсов. Вообще говоря, возрастное распределе-
ние популяции, находящейся хотя и в постоянных условиях, но при ограниченно-
сти внешних ресурсов может претерпевать периодические нли даже стохастиче-
ские колебания. Примером может служить рис. 13.3.5, когда одновременно с
численностью популяции колебаниям подвержено и ее возрастное распределе-
ние. — Прим. ред.
2 Из того, что в природных условиях рост численности ограничен следует
лишь, что в среднем по большим промежуткам времени /?0 должно быть близким
к единице. При этом, вообще говоря, возможно, что и Ro, и возрастное распреде-
ление в популяции даже при постоянных тх и 1Х претерпевают регулярные или
апериодические колебания довольно большой амплитуды, как это и наблюдается
для многих видов в действительности. Правда, как правило, остается неясным,
•обусловлены ли такие флуктуации эндогенными (виутрипопуляциоиными) законо-
мерностями или колебаниями внешних биотических или абиотических условий,—•
Прим. ред.
Рост и регуляция численности популяций
353
Таблица 13.3
Выживаемость (1х), плодовитость (тх) и репродуктивная
ценность (vx) в лабораторной популяции рисового
долгоносика (Calandra огугае) (По данным Birch, 1948.)
Возраст, нед 1х тх тх °х
4,5 0,87 20,0 17,400 130,53
5,5 0,83 23,0 19,090 115,86
6,5 0,81 15,0 12,150 95,15
7,5 0,80 12,5 10,000 81,15
8,5 0,79 12,5 9,875 69,51
9,5 0,77 14,0 10,780 58,50
10,5 0,74 12,5 9,250 46,30
11,5 0,66 14,5 9,570 37,90
12,5 0,59 11,0 6,490 26,18
13,5 0,52 9,5 4,940 17,22
14,5 0,45 2,5 1,125 8,92
15,5 0,36 2,5 0,900 8,03
16,5 0,29 2,5 0,800 6,86
17,5 0,25 4,0 1,000 4,76
18,5 0,19 1,0 0,190 1,00
как они гибнут до наступления половой зрелости, и соответственно
другие производят больше, чем одного потомка. В момент своего
рождения все самки имеют равные шансы оставить после себя по
одному потомку. С течением времени, однако, некоторые самки
из данной когорты гибнут, а для остальных самок вероятность про-
извести потомков возрастает и продолжает возрастать во времени.
Когда самки начинают размножаться, вероятность произвести до-
полнительного потомка понижается, а после того как особь до-
стигает пострепродуктивного возраста, эта вероятность становится
равной нулю. Вероятность участия определенной особи определен-
ного возраста в создании следующих поколений называется ее
репродуктивной ценностью (и) и определяется как специфическое
для данного возраста математическое ожидание числа будущих
потомков. Математически это выражается как
t=x
где коэффициент Щ1Х выражает вероятность доживания от возраста
х до возраста t, a mt — средняя рождаемость у особей в воз-
23-2044
354
Глава 13
расте t. В табл. 13.3 приведена репродуктивная ценность рисовых
долгоносиков (Calandra oryzae) разного возраста.
Точно так же, как существует стабильное распределение по воз-
растам для каждого отдельного графика 1хтх, существует и един-
ственный график репродуктивных ценностей.
ПЛОТНОСТЬ
До сих пор мы обсуждали рост популяции, лишь косвенно за-
трагивая эффекты скученности. Число особей, приходящихся на
единицу пространства, называют плотностью популяции. Ее обыч-
но измеряют и выражают числом особей на единицу площади у
наземных видов или на единицу объема — у водных организмов,
особенно если это мелкие организмы, как, например, одноклеточ-
ные водоросли. В тех случаях, когда подсчитать число особей
трудно, плотность выражают как вес (или биомассу) на единицу
площади.
Плотность — это мера скученности. Средняя плотность (crude
density)—это число (или биомасса) особей на единицу простран-
ства (или объема) без учета каких-либо дополнительных сообра-
жений. Специфическая, или экологическая, плотность, — это число
(или биомасса) особей на единицу той площади (или объема),
которая действительно удовлетворяет требованиям данного вида,
и которую называют местообитанием вида. Максимальная плот-
ность варьирует для разных организмов; на одном гектаре земли
может обитать гораздо больше одуванчиков или мышей, чем зуб-
ров или секвой. Но даже виды, имеющие сходные размеры, могут
сильно различаться по плотности популяций. Некоторые виды
птиц, например пингвины, серебристые чайки и обитающие на ост-
ровах Тихого океана олуши, образуют гнездовые колонии, в кото-
рых на одном гектаре скапливаются по нескольку тысяч птиц, тог-
да как популяции большинства видов певчих птиц никогда не
достигают и 0,1 такой плотности. Еще более важно, что в преде-
лах одного вида в разные годы наблюдаются различия в плот-
ности в разных местах и даже в одной и той же местности. Одним
из наиболее хорошо документированных примеров изменений
плотности популяций служат изменения плотности зайца-беляка
и рыси в районе Гудзонова залива в Канаде. Поскольку это цен-
ные пушные виды, данные о заготовках их шкурок регистрирова-
лись конторой «Гудзон-Бей Компани», что позволило установить
колебания их плотности за последние 100 лет. (рис. 13.13). У обо-
их видов эти колебания носят циклический характер, причем в
некоторые периоды численность за 10 лет изменялась в 10 раз.
Это единственные виды, для которых имеются данные за такой
длительный промежуток времени, однако у многих видов наблю-
дались колебания численности на протяжении 10 лет и более;
иногда колебания наблюдаются даже в пределах года (рис. 13.14).
Рост и регуляция численности популяций 355-
Численность популяции может изменяться в результате непо-
средственного взаимодействия между особями данной популяции.
По мере того как все большее и большее число животных или
растений заполняют некоторую область, они начинают мешать
друг другу. Взаимодействия между особями могут носить деста-
билизирующий характер. Например, наличие слишком большого
количества скота на пастбище нередко порождает драки и при-
160
1850 1875 1900 1925
Год
Рис. 13.13. Циклические колебания численности зайца-беляка (Lepus americanus)
и рыси (Lynx canadensis) по данным заготовок пушнины конторой «Гудзон-Бей
Компани».
Сплошная кривая — заяц; прерывистая — рысь. (MacLulich, 1937.)
водит к гибели телят. Численность популяции также изменяется
под влиянием физических факторов среды: бури, засуха, сильные
морозы или жара могут быть причиной гибели особей. Наконец,
на плотность популяции оказывают влияние и биологические фак-
торы среды, т. е. взаимодействие с другими видами. Почти каждый
вид представляет собой жертву для какой-либо бактерии, гриба
или животного; все виды, за исключением зеленых растений и тех
животных, которые питаются мертвым органическим веществом,
являются хищниками по отношению к тем или иным другим ви-
дам.
РЕГУЛЯЦИЯ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ
Как мы убедились, все виды потенциально способны увеличи-
вать свою численность и в известных условиях они это делают
(например, лошади и коровы в аргентинских пампах в XVI и
XVII вв.; фазаны на острове Протекшей). Наблюдения показыва-
ют, однако, что численность большинства видов довольно посто-
янна. Отсюда следует, что между рождаемостью и смертностью
существует определенное равновесие (рис. 13.15). В противном
случае колебания численности популяций были бы гораздо более
значительными, чем те, которые мы наблюдаем. Эту очевидную
23*
Годы
Б
Рис. 13.14. Колебания плотности популяции у разных видов.
Л. Виргинский перепел (Colinus virginianus) в штате Висконсин. Сплошная кри-
вая — колебания численности по данным осегннх учетов. Прерывистая кривая —
по данным осенних и весенних учетов; на ней выявляются также сезонные коле-
бания численности. (Errington Р. L., 1946. Some contributions of a fifteen-year lo-
cal study of the nothern bobwhite to a knowledge of population phenomena, Ecol.
Monogr., 15, 1—34.) Б. Лабораторные популяции хищника Paramecium aurelia
(I) и его жертвы Saccharomyces exiguus (II). (Г. Ф. Гаузе, 1935. Experimental
demonstration of Volterra’s periodic oscillations in the numbers of animals, J. Exp.
Biol., 12, 44—48.)
Рост и регуляция численности популяций
357.
согласованность рождаемости и смертности в большинстве попу-
ляций называют регуляцией численности, и она представляет собой
интересную и противоречивую проблему. Особенно важно было
бы выявить тот механизм или механизмы, которые обусловливают
Рис. 13.15. Модели регуляции численности.
А. Смертность (d) постоянна, рождаемость зависит от плотности. Б. Рождаемость-
(&) постоянна, смертность зависит от плотности. В. Как смертность, так и рож-
даемость зависят от плотности. Г. Зависимость между скоростью изменения чис-
ленности и плотностью популяции.
очевидное равновесие между рождаемостью и смертностью в боль-
шинстве популяций.
Выдвигают два прямо противоположных объяснения регуляции
численности.
1. Смертность целиком обусловлена внешними физическими или
биологическими факторами (пожар, наводнение, хищники) и за-
трагивает одну и ту же долю особей в популяции, независимо от
'358
Глава 13
Численности последней. Иначе говоря, смертность не зависит от
плотности (рис. 13.15,Л). С этой точки зрения популяции регу-
лируют свою численность, подгоняя рождаемость под смертность.
2. Смертность всегда зависит от числа входящих в популяцию
особей: чем больше число особей, тем выше риск гибели. Согласно
этой точке зрения, особи способны найти убежище или же выра-
ботатъ какие-то способы защиты от наводнения, пожара или хищ-
ника, но число убежищ или путей спасения ограниченно. Поэтому
с увеличением численности популяции начинает ощущаться недо-
статок ресурсов — пищи, убежищ, путей к бегству, так что все
большее число особей гибнет от хищников, наводнений или от го-
лода, и в конце концов смертность становится равной рождаемо-
сти. С этой точки зрения регуляция численности популяции всегда
достигается за счет смертности, зависящей от плотности
(рис. 13.15,5). ,
Ни ту, ни другую точку зрения нельзя принять целиком или
полностью отвергнуть. Некоторые причины смертности не зависят
от плотности популяции (в качестве крайнего примера можно при-
вести извержение вулкана на острове Кракатау в 1883 г.), а мно-
гие другие от нее зависят (наиболее очевидный пример — недоста-
ток пищи). Проблема состоит в том, чтобы установить относитель-
ную важность этих двух, казалось бы, различных типов смертно-
сти и выяснить, способны ли виды регулировать свою рождаемость,
а если да, то при помощи какого механизма и при каких условиях.
На все виды оказывают воздействие как зависящие, так и не
зависящие от плотности факторы смертности, однако не у всех
видов демографические реакции одинаковы. Численность некото-
рых видов довольно постоянна. Плотность других видов резко ко-
леблется под действием внешних факторов, причем колебания эти
кажутся нерегулярными. Наконец, плотность таких видов, как,
например, заяц-беляк, подвержена резким, но циклическим коле-
баниям. Чем объясняются эти глубокие различия между видами?
Весьма вероятно, что существуют различные способы, при помощи
которых тот или иной вид справляется с проблемами, создаваемы-
ми средой и другими видами, и что эти различные способы пред-
ставляют собой альтернативные эволюционные решения, называе-
мые адаптивными стратегиями.
СМЕРТНОСТЬ, ЗАВИСЯЩАЯ И НЕ ЗАВИСЯЩАЯ
от плотности
Когда в какой-либо области случается опустошительный пожар
или наводнение, уничтожаются все организмы, независимо от их
числа. В случае же ограниченного количества доступной пищи
число организмов, которые погибнут голодной смертью (если это
вообще произойдет), зависит от их первоначальной численности:
если их немного, то они, возможно, выживут все; если же их мно-
Рост и регуляция численности популяций 359*
го, то число смертей, вызванных недостатком пищи, может быть
весьма значительным. Факторы среды, «убивающие» независимо
от наличного числа особей, обусловливают так называемую не
зависящую от плотности смертность; те факторы, на действие ко-
торых влияет плотность популяции, вызывают смертность, завися-
щую от плотности. Климатические и физические факторы (земле-
трясения, наводнения, резкие колебания температуры и т. п.) от-
носятся к факторам, не зависящим от плотности. Ресурсы среды
(пища, убежища) считают обычно факторами, зависящими от
плотности. На самом деле эти два класса факторов не обособле-
ны, а образуют непрерывный ряд — от факторов, в основном неза-
висящих от плотности (например, всепоглощающий пожар), до
факторов, в основном зависящих от плотности (пища). Зависящие
и не зависящие от плотности факторы воздействуют на параметры,
входящие в демографические таблицы, по-разному.
Представим себе стабильную среду, в которой не происходит
никаких катастроф. Численность популяций организмов, живущих
в такой среде, будет по большей части соответствовать емкости
этой среды или приближаться к ней. Большая часть производимых
потомков не будет доживать до репродуктивного возраста. В такой
среде особь, повышающая число производимых ею потомков, ока-
зывает лишь незначительное влияние на их приспособленность,
потому что вероятность выживания каждого потомка очень мала.
Отбор будет благоприятствовать мутациям, повышающим вероят-
ность выживания в возрасте от t до f+1, поскольку такие особи,
благодаря своей большей продолжительности жизни, могут раз-
множаться в течение нескольких сезонов и оставлять гораздо
больше потомков. В отличие от этого в нестабильной среде, где
часто происходят катастрофы, вероятность выживания в возрасте
от t до ^+1 понижена, независимо от фенотипа, но при этом ве-
роятность сохранения потомков повышается. Это очень несложно
доказать математически.
Допустим, что у нас имеется смесь двух фенотипов: 1) корот-
коживущего, который размножается один раз и гибнет, и 2) жи-
вущего дольше и размножающегося дважды. Это фенотипическое
различие детерминировано генетически. Мы хотим выяснить, в ка-
ких условиях среды отбор будет благоприятствовать одному или
другому из этих фенотипов. По определению отбор будет благо-
приятствовать фенотипу, который в момент времени /+1 имеет
наибольшее число потомков:
Число особей в момент времени /-1-1 равно произведению их числа
в поколении t на скорость роста популяции (%) (см. стр. 346).
Для фенотипа с короткой продолжительностью жизни число
особей в момент времени f+1 равно числу потомков, производи-
мых на одну особь, умноженному на вероятность того, что они
3360
Глава 13
доживут до репродуктивного возраста, и умноженному на число
родительских особей:
^<Ж)Л =Nma '$А ,
\A=N0[fA-SA,
где NottA — среднее число потомков, a SA— вероятность выжива-
ния. Для фенотипа с большей продолжительностью жизни к этому
расчету следует добавить число особей, переживших первый год:
^<Ж>а =^3 Гонв-Зв)+*].
%s = (AroffB-SB)+*.
где R— выживаемость (1Х) особей, имеющих фенотип В в период
от t до £+1. Если мы теперь приравняем эти две скорости роста
XB = V,
(^онв-3в)+/?=^о[[л -S\
и допустим далее, что выживаемости потомков (Ss и 5Л) для обо-
их фенотипов одинаковы, то сможем предсказывать, в каких слу-
чаях отбор будет отдавать предпочтение фенотипу с большей про-
должительностью жизни по сравнению с короткоживущим феноти-
пом и наоборот:
(Ko[iB-SB) + R=NoiiA.SA,
-----------=JVoffA ,
^offB Н s’ ~N0[[A.
Будет ли отдано предпочтение долгоживущему фенотипу, зависит
ют отношения вероятности его выживания к вероятности выжи-
вания его потомков. Отношение R/S выражается большим числом
во всех тех случаях, когда выживаемость взрослых особей высо-
кая, а новорожденных потомков — низкая; оно выражается малым
числом, если их выживаемости примерно равны; если же S на-
много больше R, то отношение это пренебрежимо мало.
Мы исходили из допущения, что V равно V. Теперь мы хотим
предсказать, какие факторы среды благоприятствуют каждому из
.фенотипов, обеспечивая им одинаковую степень успеха. В стабиль-
ных средах, как мы видели, обоснованию молодых особей может
препятствовать недостаток ресурсов, поскольку превосходящие их
по размерам взрослые особи более конкурентоспособны. В таких
средах S будет мало, a R/S — велико. Согласно нашей формуле,
для поддержания одинаковой скорости роста Nott а должно быть
Рост и регуляция численности популяций
361
гораздо больше NottB- Чем больше R/S, тем больше вероятность
того, что предпочтение будет отдаваться долгоживущему фенотипу.
В отличие от этого в нестабильной среде вероятность выжива-
ния молоди равна вероятности выживания взрослых особей. В та-
ком случае потенциальная возможность прожить еще один год не
дает существенного преимущества, а поэтому величина R/S пре-
Рис. 13.16. Смертность как функция времени для двух биотипов одуванчика, вы-
ращивавшихся по отдельности в чистых культурах, но при одинаковых условиях.
Обратите внимание, на большое сходство обеих кривых. (По данным Solbrig О. ,Т.»
Simpson В. В., 1974. Components of regulation of a population of dandelionsHim
Michigan. J. Ecol., 62, 473—486.) , l
небрежимо мала. Возвращаясь к нашему уравнению, мы видим",,
что значение NotfB близко к значению NottA- Иными словами, в
нестабильной среде отбор благоприятствует фенотипу А. Экспери-:
ментальное подтверждение сделанного выше предсказания было;
получено Солбригом (Solbrig) и Симпсоном (Simpson) с различ-
ными фенотипами лекарственного одуванчика.
Растения одуванчика были собраны с трех смежных полей к
штате Мичиган и пересажены на садовые делянки с одинаковыми
условиями: их генотипический состав установили методом опреде-
ления изоферментов (табл. 13.4). У 300 изученных растений было,
выявлено минимум четыре различных генотипа. Чаще всего встре-
чались два генотипа — так называемые биотипы А и D. Однако»
они не были равномерно распределены по трем полям: биотип А
был наиболее обилен на двух первых полях, а биотип D — на'
третьем поле. Поля слегка различались по влажности, а также по»
количеству получаемых ими солнечных лучей; однако главный
362
Глава 13
различием между ними была степень нарушенности природных ус-
ловий— одна из причин смертности, не зависящей от плотности,—
которая была наибольшей для первого поля (где доминировал
биотип Л) и наименьшей для третьего поля (где доминировал
биотип D). Соответственно можно было ожидать, что у биотипов
на третьем поле выживаемость окажется выше, а продукция семян
ниже, а у обитателей первого поля картина будет обратная. Как
Ряс. 13.17. Смертность двух биотипов одуванчика, выращивавшихся вместе в сме-
шанной культуре, в которой исходно оба биотипа были представлены поровну.
(По данным Solbrig О. Т., Simpson В. В., 1974. Components of regulation of a po-
pulation of dandelions in Michigan. J. Ecol., 62, 473—486.)
показало экспериментальное исследование, именно так повели се-
бя биотипы А и D (рис. 13.16, 13.17 и табл. 13.5).
Истинная скорость роста (г) равна рождаемости минус
смертность. Рождаемость в известных пределах видоспецифична.
У некоторых организмов, например у дрозофилы, одна самка от-
кладывает в среднем 300 яиц; другие, например дуб, за всю свою
жизнь производят много тысяч семян. Наконец, у ряда видов,
например у человека и слонов, рождаемость очень низкая —
в среднем менее 20 потомков. Поскольку все эти виды достигли
одинакового успеха, в том смысле, что все они существуют уже в
течение достаточно длительного времени, напрашивается вывод,
что эволюционный успех того или иного вида определяется не од-
ной только рождаемостью. Чем же объяснить такие значительные
различия в рождаемости?
Рост и регуляция численности популяций
•363
Таблица 13.4
Распределение четырех биотипов одуванчика
в трех смежных популяциях
(Solbrig О. Т., Simpson В. В., 1974. Components
of regulation of a population of Dandelions
in Michigan, J. Ecol., 62, 473—486.)
Местообитание Число выборок Биотипы» %
А В с D
1. Сухое; ярко ос- вещается солн- цем; очень силь- но нарушенное 94 73 13 14 0
2. Сухое; затенен- ное; средняя степень наруше- ния 96 53 32 14 1.
3. Сырое; полуза- теиениое; нена- рушенное 94 17 8 11 ОС
Таблица 13.5
Сравнение двух биотипов одуванчика, выращенных в теплице
Показатель Биотип А (г—отбор*) БиЬтнп D (К—отбор*)
Среднее число соцветий на 1 рас-
тение 0,24 0,04
Среднее число семян на 1 соцве-
тие 105,5 204,3
Средний вес семян, мг 0,315 0,437
Общее число семян на 1 растение
(в среднем) 25,3 8,2
* См. стр. 366.
ДЕМОГРАФИЯ И ЕСТЕСТВЕННЫЙ ОТБОР
Каждый отдельный организм располагает лишь ограниченным
количеством пищи, воды и других необходимых ресурсов. За счет
этих ресурсов он создает собственные ткани и производит потом-
ков (яйца, споры, семена или живых детенышей). Если особи круп-
ные, сильные, выносливые, хорошо адаптированные к среде и спо-
собны защитить себя от врагов, то их смертность будет ниже, чем
у других. Поэтому потенциальная продолжительность жизни у них
будет выше, чем у других особей данной популяции, не обладаю-
щих указанными признаками. Но ресурсы, необходимые для созда-
ния таких свойств, ограниченны. Если крупная особь не может
384
Глава 13
получить доступа к каким-либо новым ресурсам, то увеличение
размеров тела может нанести ущерб ее плодовитости, а это приве-
дет к снижению приспособленности. Здесь возможны две альтер-
нативы.
1. Большая продолжительность жизни компенсирует снижение
специфической для данной возрастной группы плодовитости, так
что суммарная плодовитость данной особи за всю ее жизнь не
уменьшается. Примерами такого положения вещей служат попу-
ляции Uta stansburiana и биотипы D одуванчика.
2. Большая продолжительность жизни не компенсирует сниже-
ния специфичной для данной возрастной группы плодовитости, но
производимые потомки имеют более крупные размеры (или боль-
ше весят, или родители кормят их больше или же дольше). Вслед-
ствие своих больших размеров, такой потомок имеет больше шан-
•сов выжить, что может компенсировать уменьшение числа потом-
ков; в результате его вклад в создание следующего поколения
-окажется больше, чем вклад какой-либо другой особи из данной
популяции, которая производит больше потомков, но потомки ко-
торой имеют меньше шансов выжить.
Рассмотрим, например, популяцию растений, в которой в ре-
зультате мутаций появились экземпляры, вырабатывающие в своих
листьях новое химическое вещество, ядовитое для какого-либо
хищника. Однако на вырабатывание этого вещества расходуется
некоторая энергия, которая могла бы быть вложена в создание
потомков, так что плодовитость при этом снижается. А следова-
тельно, такая мутация распространится в популяции лишь в том
случае, если более высокая вероятность выживания потомков ком-
пенсирует уменьшение их числа.
Это лишь некоторые примеры альтернативы, возникающей при
распределении энергии между вегетативным ростом (повышающим
выживаемость и способность запасать ресурсы), защитными при-
способлениями (снижающими смертность от хищников и болезней)
и размножением. Отбор будет сохранять тот организм, который
вносит наибольшее число потомков в следующее поколение. Для
того чтобы обеспечить этот вклад, необходимо максимизировать
произведение выживаемости на размножение, т. е. Zx-mx. Прекрас-
ный пример решения этой проблемы дает величина кладки у птиц,
изученная английским орнитологом Девидом Лэком (David Lack).
Лэк изучал величину кладки у ряда видов птиц. Он хотел вы-
яснить, оставляют ли особи, откладывающие наибольшее число
яиц, наибольшее число потомков или же число яиц и вероятность
выживания каким-то образом компенсируют друг друга. Среди
прочих видов Лэк изучал обыкновенного скворца. Он окольцевал
несколько тысяч птенцов в гнездах и зарегистрировал число птен-
цов в каждом гнезде. Оценка успешности размножения произво-
дилась на основе числа молодых особей старше 3 мес, обнаружен-
ных повторно (наибольшая смертность наблюдается в течение
Рост и регуляция численности популяций
365
Таблица 13.6
Выживание окольцованных птенцов обыкновенного скворца
(Sturnus vulgaris) после вылета из гнезда
в зависимости от величины кладки (Lack, 1948)
Величина кладки Число гнезд Число птенцов Пойманы спустя более чем 3 мес после вылета Относительная продуктивность, %-ное отношение числа отловлен- ных слетков к числу гнезд
число %
1 65 65 0 — —
2 164 328 6 1,8 3,7
3 426 1278 26 2,0 6,1
4 989 3956 82 2,1 8,3
5 1235 6175 128 2,1 10,4
6 526 3156 53 1,7 10,1
7 93 651 10 1,5 10,2
8 15 120 1 0,8 6,4
9 2 18 0 — —
Гв 1 10 0 — —
первых 3 мес). Результаты исследований Лэка приведены в
табл. 13.6. Обращают на себя внимание несколько особенностей:
1) величина кладки варьирует от 1 до 10 яиц; 2) распределение
кладок по величине неравномерно (примерно треть всех гнезд со-
держит по 5 яиц, а число отклонений в обе стороны тем меньше,
чем они больше отличаются от пяти); 3) вероятность выживания
(оцениваемая по числу молодых птиц, вторично отловленных спу-
стя 3 мес) по'существу одинакова для птенцов из гнезд с 3, 4 или
5 птенцами, но уменьшается, если птенцов больше чем 5; 4) если
умножить число птенцов в гнезде на вероятность выживания, то
скажется, что наибольшее число слетков дают гнезда, в которых
было по 5 птенцов. Судя по результатам взвешивания птенцов из
гнезд, содержащих разное число птенцов (табл. 13.7), из больших
кладок не получается большего числа потомков вследствие недо-
статочного питания птенцов. Очевидно, пять птенцов — это опти-
мальный вариант; птенцы при этом имеют несколько меньший вес,
чем в том случае, когда число их меньше, но вес этот не настолько
мал, чтобы снизить вероятность их выживания. Следовательно,
при числе птенцов, равном 5, приспособленность максимизируется.
В природе встречаются, однако, такие обстоятельства, когда
гибель неизбежна, какими бы ни были фенотипические признаки.
Обвалы, наводнения, пожары, землетрясения, разрушение среды
человеком, крайне и необычно холодные, дождливые, жаркие или
366
Глава 13
Таблица 13.7
Вес птенцов скворца, находящихся в гнезде
[По данным, собранным R. Carrick в Англии в мае 1947 г.
(Lack, 1948)]
Возраст, дви Средний вес 1'птенца, г
2 птенца в гнезде 5 птенцов в гнезде 7 птенцов в гнезде
4 28,8 — 21,4
6 48,3 45,1 38,2
15 88,0 77,6 71,4
Диапазон колеба- ний веса на 15-й день 87,5—88,5 72,5—83,0 66,0—77,0
засушливые годы — все это влечет за собой гибель всех или почти
всех организмов, обитающих в данной области, независимо от их
фенотипов. В условиях, когда события, ведущие к высокой не за-
висящей от плотности смертности, случаются часто, затрата энер-
гии на потенциально более совершенные фенотипы не даст ника-
ких или почти никаких преимуществ. А поэтому в таких условиях
отбор будет благоприятствовать тем организмам, которые напра-
вят большую долю своей энергии на создание потомков, повышая
рождаемость. Поскольку в таких средах популяции подвержены
высокой смертности, не зависящей от плотности, для того чтобы
избежать вымирания, необходима высокая рождаемость.
Характер среды обитания данного организма определяет, та-
ким образом, будет ли отбор благоприятствовать особям, которые
вкладывают больше энергии в создание более совершенного фено-
типа за счет более низкой рождаемости, или же особям, произво-
дящим большее число потомков. Подобная альтернатива — неиз-
бежное следствие ограниченности ресурсов, доступных каждой осо-
би. Организмы, обитающие в стабильных средах и приносящие
относительно небольшое число потомков, подвергаются так назы-
ваемому К-отбору; организмы, обитающие в нестабильных средах
и приносящие относительно много потомков, подвергаются г-от-
бору.
Другая альтернатива — это выбор между небольшим числом
крупных потомков и большим числом мелких яиц или семян, ис-
ходно имеющих малый вес. Первые будут иметь больше шансов
на успех, при условии что отсутствует смертность, не зависящая
от плотности. Эта альтернатива отражена также в примере с
одуванчиком (табл. 13.5), у которого биотип D производит мень-
шее число более крупных семян.
Рост и регуляция численности популяций 367
В общем считается, что такие организмы, как эфемерные рас-
тения пустыни, а также растения и животные временных водоемов
или же естественных или нарушенных полей, специализирующиеся
на использовании временных ресурсов, подвергаются г-отбору.
Организмы, находящиеся в более стабильных ситуациях, обычно
подвергаются /(-отбору.
Демографические характеристики вида — продолжительность
жизни, плодовитость, специфичная для данной возрастной группы,
возраст, в котором впервые происходит размножение, — формиру-
ются под влиянием физических факторов среды, взаимодействия с
другими видами и внутривидовой конкуренции и представляют
собой черты, способствующие максимизации индивидуальной при-
способленности. Естественный отбор благоприятствует особи,
оставляющей наибольшее число потомков. Однако наибольшее
число потомков редко достигается путем максимизации плодови-
тости в ущерб выживанию. Напротив, многие виды, достигшие
успеха, имеют очень низкую плодовитость. Примером служит вид
Homo sapiens.
В этой главе мы исследовали некоторые факторы, регулирую-
щие рост популяций. В мире ограниченных ресурсов популяции
увеличивают свою численность в основном только в двух ситуа-
циях. Во-первых, увеличение численности происходит после какой-
либо катастрофы, в результате которой численность популяции
понизилась и образовался временный избыток ресурсов. Некоторые
из этих «катастроф» носят периодический характер и они пред-
сказуемы. Например, в средних и высоких широтах зимние темпе-
ратуры слишком низки для роста растений. Когда осенью расте-
ния переходят в состояние покоя и сбрасывают листья, животные
впадают в спячку, снижают численность или делают и то и другое.
С наступлением весны все популяции могут вновь увеличить свою
численность. То же самое происходит в областях с четко выра-
женным засушливым периодом. Другие катастрофы менее пред-
сказуемы— ураганы, очень холодные зимы, наводнения и т. п.,—
однако процесс сокращения и роста численности происходит при
этом сходным, хотя и менее закономерным образом.
Во-вторых, увеличение численности популяции происходит в
тех случаях, когда ее члены приобретают какие-либо фенотипиче-
ские/генотипические признаки, дающие им возможность расширить
сферу потребляемых ими ресурсов. В предыдущих главах (гл. 4—
8) рассматривались способы возникновения таких признаков. Од-
нако эти новые признаки могут распространиться в популяции
только в том случае, если создающийся в результате фенотип спо-
собен увеличиваться в числе. Для большинства природных популя-
ций такое увеличение означает, что численность какого-то другого
фенотипа того же самого или какого-нибудь другого вида при
этом уменьшается. По установившейся традиции экологи изучают
взаимодействия между видами и изменения численности, происхо-
369
Глава 13
дящие в популяциях, тогда как популяционные генетики уделяют
все свое внимание динамике генетических изменений. Следует,
однако, понимать, что все это составляет единый процесс и что
генетические и демографические изменения происходят одновре-
менно.
Виды различаются по ряду характеристик жизненного цикла,
главными из которых являются: 1) число и величина семян, яиц
или новорожденных особей; 2) средняя продолжительность жизни;
3) форма кривой выживания (т. е. смертность, специфическая для
каждого возрастного класса); 4) возраст, в котором особь впервые
участвует в размножении; 5) плодовитость, специфическая для
каждого возрастного класса (т. е. число потомков, производимых
особью в каждом возрасте, если она участвует в размножении не
один, а несколько раз). Из всех этих аспектов слагается жизнен-
ный цикл организма. Естественный отбор формирует жизненный
цикл таким же образом, как он формирует фенотипические при-
знаки.
Изучение жизненных циклов и концепция жизненной стратегии
тесно связаны с изучением демографии. Демографические иссле-
дования предоставляют данные, позволяющие нам охарактеризо-
вать жизненный цикл. Интерпретация жизненного цикла в терми-
нах факторов отбора, действующих со стороны окружающей среды,
дает представление о «стратегии». Таким образом, жизненная
стратегия — это концепция, позволяющая объяснить происхожде-
ние того или иного комплекса демографических характеристик.
А в таком случае она должна делать предсказания, поддающиеся
проверке. В данном контексте слово «стратегия» не имеет ника-
кого телеологического оттенка. Хотя «стратегия» организма дает
ему возможность успешно справляться со всем разнообразием,
факторов среды, а также с конкурентами и хищниками, организм
не планирует свою «стратегию» подобно тому, как военачальник
разрабатывает общий план кампании. Однако только те особи,
которые обладают соответствующим комплексом морфологических
и поведенческих признаков, побеждают в борьбе за существование
и способны передать эти признаки своим потомкам. В следующей
главе рассматривается вопрос о том, каким образом взаимодейст-
вия с другими видами способствуют формированию особенностей,
характеризующих жизненный цикл различных организмов.
ЛИТЕРАТУРА
Общие вопросы
Boughey A. S. 1973. Ecology of Populations, 2nd ed., New York, Macmillan.
den Boer P, J., Gradwell G. R. (eds.). 1971. Dynamics of Populations, Waagenin-
gen, Centre for Agricultural Publishing and Documentation.
Harper J. 1977. Population Biology of Plants, London, Academic Press.
MacArthur R. H. 1972. Geographical Ecology: Patterns in the Distribution of Spe-
cies, New York, Harper and Row.
Рост и регуляция численности популяций 369*
May R. М. (ed.). 1976. Theoretical Ecology: Principles and Applications, Oxford,.
Blackwell Scientific Publications.
Solbrig О. T. (ed.). 1979. Demography and Evolution of Plant Populations, Oxford,.
Blackwell Scientific Publications.
Whittaker R. H. 1975. Communities and Ecosystems 2nd ed., New York, Macmil-
lan.
Специальные статьи и обзоры
Elnarsen A. S. 1945. Some factors affecting ring-necied pheasant population densi-
ty, Murrelet, 26, 39—44.
Fox S. F. 1975. Natural selection on morphological phenotypes of the lizard Uta
stansburiana, Evolution, 29, 95—107.
Frank P. W. 1960. Prediction of population growth form in Daphnia pulex cultures,
Am. Nat., 94, 357—372.
Harper J. L., White J. 1971. The dynamics of plant populations. In: P. J. den Boer
and G. R. Gradwell (eds.), Dynamics of Populations, Waageningen, Centre for
Agricultural Publishing and Documentation.
Harper J. L„ White J. 1974. The demography of plants, Ann. Rev. Ecol. Syst., 5,
419—464.
Lack D. 1948. Natural selection and family size in the starling, Evolution, 2, 95—
110.
MacLulich. 1937. Fluctuations in the numbers of the varying hare (Lepus america-
nus), Univ. Toronto Studies, Biol. Ser., 43.
McNaughton S. J. 1975. r- and А-selection in Typha, Am. Nat., 109, 251—261.
Mertz D. B. 1972. The Tribolium model and the mathematics of population growth,
Arm. Rev. Ecol. Syst., 3, 51—78.
Nicholson A. J. 1948. Competition for food among LuCilia cuprina larvae, Proc. 8th'
Intern. Cong. Entom., pp. 277—281.
Nicholson A. J. 1958. The self-adjustment of populations to change, Cold Spring-
Harbor Symp. Quant. Biol., 22, 153—173.
Sarukhan J. 1979. Demographic problems in tropical systems. In: О. T. Solbrig (ed.),
Demography and Evolution of Plant Populations, Oxford, Blackwell Scientific
Publications.
Tinkle D. W. 1969. Evolutionary implications of comparative population studies ire
the lizard Uta stansburiana. In: Systematic Biology: Proceedings of an Inter-
national Conference, Nat. Acad. Sci. Publ., 1692, pp. 133—154.
24—2044
Глава 14
Конкуренция и теория ниши
Популяции и виды — это генетические единицы, структура кото-
рых определяется их общим происхождением и их способностью
скрещиваться между собой и обмениваться генами. Лес, луг, за-
росли кустарников в пустыне — не генетические, а экологические
-единицы, называемые сообществами. Сообщество представляет со-
бой совокупность популяций растений, животных, бактерий и гри-
бов, живущих в определенной области с характерной физической
-средой, взаимодействующих друг с другом и образующих обособ-
ленную систему. Каждое сообщество имеет собственные харак-
терные для него виды, собственную физическую структуру, созда-
ваемую главным образом растениями (особенно в наземных со-
обществах) , и свою собственную функцию.
Изучение сообществ — область экологии. Тем не менее, для
того чтобы понять биологию и эволюцию популяций, необходимо
понимать некоторые из основных взаимодействий, происходящих
между организмами в сообществах, поскольку эти взаимодейст-
вия — конкуренция, хищничество, мутуализм — и физическая среда
представляют собой главные факторы отбора, действующие на по-
пуляции и определяющие их эволюцию.
В предыдущей главе мы рассмотрели рост и регуляцию числен-
ности отдельных популяций. Теперь мы расширим наше обсужде-
ние, с тем чтобы исследовать ситуацию, при которой две или не-
сколько популяций разных видов живут в одной и той же среде.
Когда в сообществе два разных вида вступают в контакт, между
ними возможны взаимодействия трех основных типов: 1) два вида
мешают друг другу или подавляют друг друга, т. е. конкурируют
между собой; подобная конкуренция обычно происходит из-за того,
что виды используют один и тот же ресурс, количество которого
ограничено; 2) один из видов поедает другой, т. е. вид может быть
жертвой или хищником; 3) оба вида сотрудничают между собой,
используя среду таким образом, что это оказывается выгодно им
обоим.
Каждое из этих взаимодействий сказывается не только на мор-
фологических и физиологических признаках данного вида, но и на
его жизненном цикле. В этой главе рассматриваются конкурентные
взаимоотношения, а в следующей — взаимоотношения хищник -
жертва и мутуалистические взаимоотношения.
Конкуренция и теория ниши
37 Ь
КОНКУРЕНЦИЯ
Во всех случаях, когда две особи мешают друг другу или по-
давляют одна другую и когда присутствие одной снижает приспо-
собленность другой, говорят, что они конкурируют между собой.
Рис. 14.1. Средний вес растений одуванчика, принадлежащих к биотипам А и D,
которые выращивали в условиях конкуренции на песчаных почвах.
Было проведено пять разных экспериментов (в двух повторностях каждый) с
разным исходным соотношением двух биотипов: 1) только биотип А; 2) 75% А
и 25% £>; 3) 50% А и 50% £>; 4) 25% А и 75% £>; 5) только биотип D. Обратите
внимание на то, что при выращивании по отдельности вес растений обоих биоти-
пов примерно одинаков, тогда как в смешанных посевах вес растений биотипа D
увеличивался за счет растений биотипа А.
Конкурирующие особи могут принадлежать к одному и тому же
или к разным видам. Объектом конкуренции обычно служит какой-
нибудь ресурс, запасы которого в данной среде ограниченны. Та-
ким ресурсом может быть пища, участок для гнезда, место для
прорастания семян и т. п. Вопреки тому, что пишут популяриза-
торы, описывающие жестокую борьбу между конкурирующими
24
372
Глава 14
особями за ограниченные ресурсы, конкуренция — это скорее пас-
сивный процесс, в котором наиболее эффективно функционирую-
щая особь получает основную массу данного ресурса. Результат
Рис. 14.2. Средний вес растений одуванчика, принадлежащих к биотипам А и D,
выращивавшимся в условиях конкуренции на суглинистой почве.
Было проведено 5 экспериментов с такими же исходными соотношениями разных
биотипов, как на рис. 14.1. Результаты сходны с теми, которые были получены
при выращивании растений на песчаной почве. Сравнение с рис. 14.1 показывает,
что более плодородная почва (суглинок вместо песка) усиливает превосходство
биотипа D.
конкуренции состоит в том, что приспособленность одной из участ-
вующих во взаимодействии особей возрастает, а приспособлен-
ность другой снижается. В противном случае их взаимодействие
нельзя рассматривать как конкуренцию в эволюционном смысле.
Например, Солбриг (Solbrig) и Симпсон (Simpson) выращива-
ли рядом растения двух биотипов лекарственного одуванчика
(Taraxacum officinale) на экспериментальных делянках (гл. 13).
Спустя два года из растений биотипа D выжило: на известковых
Конкуренция и т'еория ниши
,373
Таблица 14.1
Влияние типа почвы иа конкуренцию между разными
биотипами одуванчика, высеянными иа экспериментальные
делянки в соотношении 1 : 1
Тип почвы Биотип Исходное число Конечное число Выжи- вание, % Сухой вес, г
на 1 расте- ние общий
Песчаная А 80 70 87,5 1,64 114,8
D 80 55 68,8 2,80 154,0
Известковая А 80 28 35,0 1,38 38,6
D 80 52 65,0 6,82 354,6
Торфяная А 80 41 51,3 1,42 58,2
D 80 72 90,0 6,19 445,7
Суглинок А 80 32 40,0 4,83 154,6
D 80 37 46,3 13,70 506,9
(щелочных) почвах — 65%, на торфяных (кислых)—90%, на пес-
чаных— 69%, а на нормальных суглинках — 46%; для биотипа А
соответствующие цифры составили 35, 51, 87 и 40% (табл. 14.1).
Кроме того, растения биотипа D были крупнее, причем, как пока-
зывают фиг. 14.1 и 14.2, в значительной мере за счет размеров
растений биотипа А. Биотип D доказал свое превосходство в спо-
собности конкурировать с биотипом А за ресурсы, но это превос-
ходство зависело от типа почвы, проявляясь в наибольшей степени
на суглинке и в наименьшей — на песчаной почве, где выжило
больше растений биотипа А, чем биотипа D, но где растения био-
типа D были все же несколько крупнее и захватывали большую
часть ресурсов.
Конкуренция может быть довольно непосредственной, напри-
мер, когда две птицы конкурируют за одну гнездовую территорию.
Такую конкуренцию называют прямой, или интерференционной
конкуренцией. В тех же случаях, когда две популяции используют
один и тот же ресурс, но между ними не возникает прямых конф-
ликтов, конкуренцию между ними называют косвенной, или экс-
плуатационной конкуренцией. Поскольку в норме особям выгодно
избегать конкуренции, естественный отбор, вероятно, благоприят-
ствует использованию конкурирующими особями различных ресур-
сов или же использованию одних и тех же ресурсов в разное
время или различным образом. Это может увеличить способы ис-
пользования организмами среды и тем самым увеличить разнооб-
разие видов. Как именно конкуренция создает разноообразие, все
еще остается предметом споров. Некоторые экологи считают кон-
куренцию самым важным фактором в возникновении разнообра-
зия; по мнению других, ее роль зависит от характера среды, коли-
чества ресурсов и плотности потенциальных конкурентов. Посте-
пенно начинает преобладать мнение, что исход конкуренции не
подчиняется закону «все или ничего», а зависит от обстоятельств.
374
Глава 14
К изучению эффекта конкуренции можно подходить с двух
сторон. Если мы имеем дело с конкуренцией за какой-либо ресурс
между членами одного и того же вида, то можно определить по-
терю приспособленности каждым генотипом, а затем заняться вы-
числениями, описанными в гл. 6. Таким образом мы установим
влияние конкуренции на частоту генов и генотипов в изучаемой
популяции. Примером такого подхода служит вычисление значе-
ний s и t для Drosophila pseudoobscura, гомозиготных по инвер-
сиям СН/СН и ST/ST (см. гл. 8).
Другой подход к изучению конкуренции состоит в том, чтобы
исследовать ее влияние на рост и размножение конкурирующих
групп. В этом случае изучается воздействие конкуренции на чис-
ленность в популяции особей каждого фенотипа. Это обычный»
а в случае принадлежности конкурирующих особей к разным ви-
дам и единственный, подход. Однако при этом не удается получить,
прямых данных о частоте генов и генотипов.
Модели конкуренции и роста двух видов, сосуществующих
в одной среде
При рассмотрении логистической модели роста (гл. 13) дела-
лось допущение, что рост популяции регулируется исключительно-
доступными ресурсами. Обычно же за имеющиеся ресурсы конку-
рируют многие виды данного сообщества, так что на скорость ро-
ста того или иного вида влияет также число особей других, вхо-
дящих в сообщество видов, которые используют этот ресурс. Кро-
ме того, хотя влияние это взаимно, оно редко бывает одинаковым:
некоторые виды оказывают на рост других сильное, а некоторые —
лишь незначительное воздействие; присутствие на лугу одной ко-
ровы влияет на популяцию кроликов (в результате выедания тра-
вы) сильнее, чем присутствие одного кролика — на популяцию
коров.
Логистическую модель роста можно усовершенствовать, с тем
чтобы в ней в явном виде учитывалось наличие других видов. Это
впервые сделал американский ученый А. Дж. Лотка (A. J. Lotka)
в 1925 г., а в 1926 г. независимо от него и более подробно — италь-
янский ученый В. Вольтерра (V. Volterra). Начнем с двух конку-
рирующих между собой видов—А и В. В гл. 13 было показано,
что когда каждый вид растет отдельно от другого, скорости их
роста описываются уравнениями:
v dNA ,, ( к A — -VA
вид А. = rANA - Ка j;
„ dNB / Кв — Мв\
вид В: = rBNB --------у
Однако при совместном обитании этих двух видов скорость ро-
ста вида А зависит от числа имеющихся в наличии особей вида В
Конкуренция и теория ниши
375
и наоборот. Птребности особей, принадлежащих видам А и В,
могут совпасть но вероятность такого положения вещей невелика;
поэтому влияне особи вида А на особь вида В будет либо боль-
ше, либо менте, чем обратное. Обозначим через 012 величину
влияния особи вида В на особь вида А и через 021 — особи вида А
на особь видаЗ. Тогда скорости роста видов А и В при совмест-
ном обитании южно записать следующим образом:
dNA „ ! КА — NA — a12NB\
идЛ; = ------J:
„ ,, (Kb—Nb—o.2ina\
пд5: ~^Г=гвКв{----------------)
Мы видим, 1то скорость роста каждого вида изменяется в за-
висимости от оеднего влияния каждой особи другого вида, умно-
женного на обдую численность этого другого вида. Хотя присут-
ствие на пастбнце одного кролика оказывает минимальное воз-
действие на пшуляцию коров, большое число кроликов оказывает
очень существиное влияние, что хорошо известно любому фер-
меру.
Обобщая мидель на п конкурирующих видов, получим
п
I Ki-Ni-Ум
r.tf I ___i*1
ri‘ \ K.
dNt _
dt
где n — число шдов, rt — истинная скорость роста вида i, Ki —
емкость средыдля r-го вида, Ni— плотность популяции i-ro вида
и ац — коэффициент, описывающий конкурентное воздействие ви-
да j на вид г.
В состояни! равновесия все dNfdt должны быть равны 0; ины-
ми словами, Н( происходит ни увеличения, ни уменьшения числен-
ности ни одною из видов. Плотности популяции будут при этом
равны
п
N^Kt-^jN?,
/¥=*
где звездочка юответствует равновесной плотности.
Такая мод<ль конкуренции очень сильно упрощает процесс
межвидовой конкуренции. Поясним сущность этого процесса на
гипотетическое примере. Представим себе пастбище, на котором
пасется пара буйволов и пара коров. Те и другие питаются травой,
и мы допустим что количество доступной им травы остается посто-
янным круглы! год. В начале эксперимента численность обоих ви-
дов будет рас'И, потому что имеющихся ресурсов вполне доста-
376
Глава 14
точно для них обоих. У коров с их меньшей продолжительностью
генерации и большим числом потомков истинная скорость роста
(гв) выше, чем у буйволов (га), и соответственно их численность
возрастает быстрее, чем у буйволов. Однако каждый буйвол по-
едает больше травы, более эффективно переваривает ее и превра-
щает в мясо, так что сиг (воздействие одной коровы на одного
буйвола) меньше, чем 021 (воздействие одного буйвола на одну
корову). По мере того как запасы травы становятся ограничиваю-
щим фактором, взаимодействия между этими двумя видами ста-
новятся более значительными. В конечном счете конкуренция при-
водит к одному из трех возможных результатов: 1) буйволы по-
беждают, а коровы вымирают; 2) коровы побеждают, а буйволы
вымирают; 3) между буйволами и коровами устанавливается рав-
новесие и они сосуществуют.
Спрашивается, можем ли мы с помощью нашей модели пред-
сказать, какой из этих результатов будет иметь место?1
Если в сообществе имеются только особи вида А, то числен-
ность этого вида будет расти до тех пор, пока не будет достигнут
уровень, соответствующий емкости среды для данного вида, Na,
когда dNAldt=G. Если, однако, в сообществе имеются также особи
вида В, то вид А достигнет квазиравновесной численности
(dNA/dt = Q) при величине популяции, меньшей чем Ка- Точное
число особей вида А при скорости роста А, равной 0, есть функция
числа особей вида В и коэффициента конкуренции 012-'
dNA _А_г м I KA — NA — a12NB\
~dT -U - rANA J,
а это означает, что
Кд NA—al2NB=Q,
где значения rA, NA, Ка и Nb больше нуля, так что уравнение нуль-
изоклины будет
Кд = Кд — СС12Кв.
Подобным же образом для вида В уравнение нуль-изоклины будет
Кв = Кв — ^21Кд.
Построим теперь график (рис. 14.3) на координатной плоско-
сти, по осям которой отложены численности видов А и В, т. е.
Na и Nb. Нанесем на эту плоскость линии нулевой скорости роста
(нуль-нзоклины) каждого из двух видов, начав с вида А. Откла-
дывая Na по оси абсцисс, a NB— по оси ординат, получим, что
нуль-изоклина для вида А пересекает ось абсцисс в точке NA = K.
* В реальной ситуации первой исчезнет трава, что повлечет за собой гибель
каи норов, так и буйволов. В этом гипотетическом эксперименте мы делаем допу-
щеиие, что трава не вытаптывается.
Конкуренция и теория ниши
377
и ось ординат в точке Мв — Кл/ап. Подобным же образом линия
нулевой скорости роста для вида В — это прямая, пересекающая
ось ординат в точке Nb = Kb и ось абсцисс в точке Na = Kb/u21-
Рис. 14.3. График межвидовой конкуренции.
По осям откладываются плотности видов Л и В. Рост численности вида А за каж-
дый промежуток времени указан горизонтальными (прерывистыми) стрелками,
а рост вида В — вертикальными (сплошными) стрелками. В момент Л растут чис-
ленности обоих видов. Когда один из видов (в данном случае это вид Л) дости-
гает своей изоклины, дальнейший его рост становится.невозможным. Другой внд
может продолжать расти, если он не достиг своей изоклины, как, например, вид В.
Если изображающая численности обоих видов точка на графике оказывается вы-
ше изоклины вида А, как в момент /з, то плотность вида А сокращается, стре-
мясь к своей изоклине. В этом случае рост вида В продолжается, а вид А вы-
мирает.
Численности каждого из видов будут возрастать, пока не до-
стигнут соответствующей изоклины нулевого роста. На плоскости
(NA, Nb) кривая, описывающая изменение численностей обоих
видов, пересекает нуль-изоклину вида А вертикально, а нуль-изо-
клину вида В — горизонтально (рис. 14.3). Между видами А и В
возможны четыре типа взаимоотношений, приводящие к четырем
различным результатам (рис. 14.4), в зависимости от значений
Na, Кв, ящ и а21-
378
Глава 14
Случай 1. Емкость среды для вида А (Ка) больше, чем Кв1аг\г
а емкость среды для вида В (Кв) больше, чем Кд/аы- Поэтому
а^^Кл/Кв и ац>КвКА.
Если провести нулевые изоклины, то они пересекаются. В точ-
Число особей вида А
Рис. 14.4. Графики конкуренции между двумя видами.
Случай 1: Лл>К&/а2, и Кв> Ка/чц. Соответствующее сосуществование видов
неустойчиво и либо вид А, либо вид В в конечном счете вымирает. Случай 2:
Кл<Лв/а21 и К.в<Кл1а\2> достигается устойчивое равновесие, соответствующее
сосуществованию обоих видов. Случай 3: Ка> Кв/a2i и Лв<Лл/а12; выживает
только вид А. Случай 4: Лл<Лв/а2| и ЛвЖл/а12; выживает только вид В.
сосуществовать. Однако любое возмущение приведет к возраста-
нию численности одного вида и сокращению другого. Таким обра-
зом, случай 1 представляет собой неустойчивое равновесие, кото-
рое приведет к вымиранию одного или другого вида — в зависимо-
сти от того, какой из них первым выйдет за нулевую изоклину
другого. Это зависит от конкретной ситуации, в результате кото-
рой виды впервые появились в данном месте.
Случай 2. В этом случае Ка меньше, чем Кв!ам и Кв<Ка!ол2,
а следовательно, а^СКлЖв и Ъ2\<.Кв1Ка.
В этом случае изоклины также пересекаются, но равновесие
является устойчивым. Если вид В достигает своей нулевой изо-
клины раньше, чем вид А, слева от точки равновесия, то числен-
Конкуренция и теория ниши 379
ность вида А будет продолжать расти. При высоких численностях
вида А численность вида В будет сокращаться до тех пор, пока
не будет достигнута точка равновесия, после чего дальнейшего
увеличения численности вида А происходить не может. Подобным
же образом, если вид А первым достигнет своей изоклины, то
численность вида В продолжает увеличиваться, вынуждая вид А
к сокращению численности вплоть до достижения этой точки рав-
новесия.
Случай 3.
п Ка //г / Ад \ Ад /дг Кв \
“12 < АГ“21 > Ад
В этом случае независимо от начальных условий выживает только
вид А.
Случай 4.
„ Ад (к Лд\ КВ (к Кв \
12 > А? Р3 > а1г ) И а2‘ < КА < а21 )•
В этом случае независимо от начальных условий выживает только
вид В.
Эти модели двух видов, рост которых происходит в условиях
конкуренции, предсказывают, что два вида, использующие один
и тот же ресурс, могут сосуществовать только в том случае, если
вид, численность которого ниже, всегда способен расти быстрее
преобладающего вида1. Это означает, что на рост каждого вида
его собственные члены оказывают в каком-то отношении более
сильное влияние, чем особи другого вида. Подобные обстоятель-
ства могут возникнуть, вероятно, лишь в том случае, если каждый
из этих видов потребляет какие-то ресурсы, не используемые дру-
гим видом. Во всех других случаях сосуществование невозможно.
Впервые эту проблему проанализировал советский биолог
Г. Ф. Гаузе. Он сформулировал «принцип взаимного исключения»,
иногда называемый принципом Гаузе. Согласно этому принципу,
два вида не могут сосуществовать, если они используют в точности
одни и те же ресурсы (гл. 11).
Описанная выше графическая модель позволяет получить пред-
ставление о взаимодействиях двух видов. Однако, так же как и
в случае моделей роста одного вида, ее ценность зависит от того,
насколько реалистичны лежащие в ее основе допущения. В этой
модели га, гв, Ка и Кв снова приняты постоянными, а кроме того,
к ним добавлены еще две постоянные — ац и a2i, коэффициенты
конкуренции или влияние особи вида А на особь вида В и особи
вида В на особь вида А. Коэффициенты конкуренции слагаются
1 Как ясно из предыдущего текста, условия устойчивого сосуществования за-
даются равновесными численностями видов в отсутствие конкурентов (Ка и Кв)
и коэффициентами конкуренции. Скорости роста в это условие не входят. — Прим,
ред.
380
Глава 14
из многих других факторов: эффективности использования ресур-
сов одним видом, его физической конкурентоспособности и способ-
ности избегать воздействия со стороны другого вида. Все эти свой-
ства, несомненно, подвержены внутрипопуляционной изменчивости.
Поэтому они должны подвергаться действию отбора. А в таком
случае среднее значение коэффициента конкуренции будет изме-
няться во времени и в зависимости от численности популяции.
Кроме того, на него будет оказывать влияние пространственное
распределение обоих видов и возраст составляющих их особей.
Несмотря на все это, приведенные здесь весьма простые модели
дают возможность получить некоторое представление о том, как
численность особей в популяции одного вида может воздейство-
вать на численность особей и вероятность выживания членов по-
пуляции другого вида.
При совместном выращивании двух видов в лаборатории в по-
стоянных условиях создается некая искусственная ситуация, по-
тому что в природе среда никогда не бывает постоянной и конку-
ренция очень редко происходит один на один. Однако анализиро-
вать конкуренцию в природе практически невозможно ввиду мно-
гочисленности влияющих на нее факторов. Тем не менее экспери-
менты, в которых из сообщества искусственно изымают или, на-
против, добавляют к нему какой-либо вид, могут оказаться весьма
плодотворными. Несмотря на все эти оговорки, как лабораторные,
так и полевые эксперименты по конкуренции позволили получить
много ценных сведений.
Г. Ф. Гаузе изучал рост и конкурентные взаимодействия двух
видов ресничных инфузорий — Paramecium aurelia и Р. caudatum;
он провел на них ряд экспериментов, результаты которых были
опубликованы в 1934 г. Оба вида помещали в один и тот же сосуд,
в котором количество пищи поддерживали на постоянном уровне.
Сначала численность обоих видов быстро увеличивалась, но по
прошествии некоторого времени Р. aurelia стала расти за счет
Р. caudatum, пока в конце концов этот второй вид не исчез из
культуры (рис. 14.5). В 1969 г. американский эколог Джон Ван-
дермеер (John Vandermeer) повторил эти эксперименты, несколько
усовершенствовав их; он использовал три вида Paramecium
(Р. aurelia, Р. caudatum, Р. bursaria) и еще одну ресничную ин-
фузорию— Blepliarisma. Сначала он выращивал эти виды по от-
дельности, получив результаты, представленные на рис. 14.6. Как
видно на этом рисунке, рост всех видов соответствовал логистиче-
ской модели. На основе этих результатов Вандермеер вычислил-
значения г, К. и No для каждого вида (табл. 14.2). Затем он вы-
ращивал эти виды попарно (рис. 14.7). В этих экспериментах
один из видов всегда побеждал другой. На основании полученных
результатов Вандермеер вычислил значения ai2 и 021 для каждого
вида в каждом эксперименте (табл. 14.3). И в заключение он ис-
пользовал параметры, приведенные в табл. 14.2 и 14.3, для пред-
Конкуренция и теория ниши 381;
сказания поведения каждого из этих видов в смешанной культуре
всех четырех видов, при условии что г, К, ап и 021 постоянны №
что никаких взаимодействий высшего порядка (т, е. эффектов,,
обусловленных наличием в культуре не одного, а большего числа
Общий «объем» (биомасса) каждого вида при выращивании на среде с бактерия-
ми изменяется во времени. Кривые роста для каждого вида, выращивавшегося"
в чистой (/) и смешанной (II) культуре. (Гаузе, 1934.)
конкурентов) не происходит. Расширив формулу Гаузе для m ви-
дов с учетом принятых допущений, получим
m
где dNildt — скорость роста i-го вида, m — число видов в сообще-
стве, а,/— воздействие /-го вида на i-й вид, г — истинная ско-
рость естественного роста, N — число особей и К — емкость среды.
Результаты, полученные Вандермеером, представлены на рис. 14.8..
Мы видим, что экспериментальные данные, показанные на рисун-
382
Глава 14
ках точками, очень хорошо совпадают с теоретическими предска-
заниями, изображенными в виде сплошной линии. Эти результаты
.несколько неожиданны, поскольку можно было предполагать ка-
кие-то взаимодействия высшего порядка, но они изящным обра-
зом демонстрируют прогностическое значение закона Гаузе, во
Рис. 14.6. Кривые роста четырех ресничных инфузорий, выращивавшихся в чи-
стых культурах.
Л. Blepharisma sp. Б. Paramecium caudatum. В. Р. bursaria. Г. Р. aurelia. (Vander-
meer, 1969.)
всяком случае, применительно к одноклеточным организмам в ла-
бораторных культурах.
В природных условиях конкуренцию изучать гораздо труднее,
так как к конкурентным взаимодействиям между изучаемыми ви-
дами здесь добавляется влияние конкуренции со стороны других
жидов, а также хищничество, причем эти факторы не всегда уда-
Конкуренция и теория ниши
383
Log (Л/+1) Log (W+1) Log (Л/+1)
Время
Время
Время
Время Время
Рис. 14.7. Кривые роста четырех видов ресничных инфузорий, выращивавшихся»
попарно.
A. Paramecium aurelia и Р. bursaria. Б. Р. caudatum и Р. bursaria. В. Р. aurelia»
н Blepharisma sp. Г. Р. bursaria и Blepharisma sp. Д. Р. caudatum a Blepharisma
sp. Е. Р. caudatum и Р. aurelia. (Vandermeer, 1969.)
384
Глава 14
Время Время
Рис. 14.7 (продолжение).
ется контролировать. Кроме того, оказывают воздействие и физи-
ческие условия среды. Тем не менее имеется ряд исследований, в
которых очень четко показана конкуренция в природе. Одним из
наиболее изящных примеров служит распространение двух видов
•морских желудей — Balanus balanoides и Chthamalus stellatus.
Эти виды обитают в приливно-отливной зоне на скалистых бере-
гах. Коннел (J. Н. Connell), подробно изучавший их, обнаружил,
что хотя участки, на которых обитают личинки этих двух видов,
сильно перекрываются, взрослые особи Chthamalus всегда селятся’
Конкуренция и теория ниши
385
Таблица 14.2
Параметры логистического уравнения
Вид Г к. "о
Paramecium aurelia 1,05 671 2,5
Р. caudatum 1,07 366 5,0
Р. bursaria 0,47 230 5,0
Blepharisma 0,91 194 3,0
в верхней (более открытой) части берега, а взрослые особи Bala-
nus — в нижней (и менее открытой) его части. Чтобы выявить от-
носительное значение обсыхания и конкуренции, Коннел измерял
скорости роста этих двух видов в нескольких местах, а также
выживаемость молодых и более зрелых морских желудей на кам-
нях, помещаемых в приливной зоне на разных уровнях. Как видно
из табл. 14.4 и 14.5, Balanus не может выжить в верхней части
приливной зоны, где он сильнее подвержен обсыханию. Chthama-
lus, однако, может выжить в нижних частях приливной зоны в
отсутствие Balanus. Balanus растет быстрее, чем Chthamalus, в
местах, расположенных выше среднего уровня прилива, и спосо-
бен обогнать по численности или даже полностью вытеснить
Chthamalus из нижней части приливной зоны. Таким образом,
распространение этих двух видов представляет собой результат
взаимодействия между биологическими и физическими факторами.
В конкуренции между ними Chthamalus побеждает в нижней и
средней частях приливной зоны, но поскольку он неустойчив к
обсыханию, он предоставляет верхнюю часть приливной зоны бо-
лее устойчивому в этом отношении Chthamalus (рис. 14.9).
Таблица 14.3
Значения коэффициентов конкуренции (<х</)
(Vandermeer, 1969)
Второй вид ( Первый вид (1) Paramecium aurelia P. cau- datum p, bursaria Blepha- risma
Paramecium aurelia 1,00 1,75 —2,00 —0,65
Р. caudatum 0,30 1,00 0,50 0,60
Р. bursaria 0,50 0,85 1,00 0,50
Blepharisma 0,25 0,60 —0,50 1,00
25—2044
386
Глава 14
Сходную ситуацию обнаружил Элленберг (Ellenberg) у расте-
ний. В Европе некоторые виды растений встречаются только на
кислых почвах. Их называют ацидофильными растениями. Эллен-
Рис. 14.8. Кривые роста четырех видов ресничных инфузорий при совместном вы-
ращивании.
A. Paramecium aurelia. Б. Р. bursaria. В. Р. caudatum. Г. Blepharisma sp. (Van-
dermeer, 1969.)
берг выращивал ряд ацидофильных видов на разных почвах с
постепенным переходом от кислых к щелочным и измерял их рост.
Оказалось, что на нейтральных почвах (рН = 7) растения росли
быстрее и были крупнее, чем особи того же вида на кислых поч-
вах. Затем он повторил тот же самый эксперимент с растениями,
обычно живущими на нейтральных почвах, и получил сходные
результаты. Однако эти виды не могли расти на кислых почвах
или росли на них очень плохо. Элленберг снова провел тот же экс-
перимент, на этот раз со смесью видов обоих типов. Он обнару-
жил, что при совместном выращивании на нейтральных почвах
растения с нейтральных почв побеждают в конкуренции ацидо-
фильные виды, однако на кислых почвах победителями оказыва-
ются последние (рис. 14.10). Иными словами, ацидофильные виды
Конкуренция и теория ниши
387
вовсе не предпочитают кислые почвы; просто они могут расти на них,
а с более плодородных нейтральных почв их вытесняют конкуренты.
Несмотря на всю затруднительность изучения конкуренции в
природе, наблюдения показывают, что она представляет собой
Chthamalus
А
Baianus
Распределение
Выедание хищником
(Thais)
Относительное воздействие
этих факторов
С1
Межвидовая
конкуренция
с Baianus
А
Взрос- Личин- Высыхание
лые ки в
Выедание
хищником
_____________t (Thais)
Распределение Относительное
воздействие
этих факторов
Рис. 14.9. Диаграмма распределения взрослых особей и недавно прикрепившихся
личинок Baianus balanoides п Chthamalus stellatus в приливно-отливной зоне в
Миллпорте, выявляющая относительное воздействие главных факторов, от кото-
рых зависит это распределение.. (Connell, 1961.)
С.В.С.П. — средняя высота сизигийного прилива; С.В.К.П. — средняя высота квад-
ратурного прилива; С.У.П. — средний уровень прилива; С.К-М.В. — средняя квад-
ратурная малая вода; С.С.М.В. — средняя сизигийная малая вода.
важный постоянно действующий фактор. Во всех тех случаях, ког-
да какой-либо вид удаляется из данной среды (без одновременных
изменений среды), какой-либо другой вид немедленно занимает
освободившееся место. Неизбежно приходится сделать заключе-
ние, что прежде проникновение этого нового обитателя сдержива-
лось конкуренцией. Но конкуренция представляет собой также
мощную эволюционную силу. Рассмотрим две популяции, принад-
лежащие к разным видам, которые используют один и тот же
ресурс, как, например, популяции насекомоядных ящериц. Допу-
стим, что особи одного вида чуть крупнее, чем особи другого, и
могут поедать несколько более крупных насекомых. Однако раз-
меры насекомых, которых поедают эти два вида, все же в значи-
тельной степени перекрываются, а поэтому между ящерицами
A
3 4 5 6 7 8
p H почьы
20
3 4 5 6 7 8
pH почвы
Рис. 14.10. Зависимость биомассы растений полевой редьки (Raphanus raphanist-
гит, прерывистые кривые) и полевой горчицы (Sinapsis arvensis; сплошные кри-
вые) от разных значений pH почвы при выращивании по отдельности (Л и Б)
и совместно (В).
Во всех случаях число посаженных растений было одинаковым, но, как показы-
вают графики, на почвах с определенными значениями pH растения росли лучше.
Обращает на себя внимание, что в отсутствие конкуренции оба вида хорошо росли
при pH 5—7, но в условиях конкуренции в этом же диапазоне pH они росли хуже.
(Ellenberg, 1952.)
Конкуренция и теория ниши
389
Таблица 14.4
Смертность молодых и старых особей Chthamalus stellatus
на камнях, помещавшихся в разные условия (Connell, 1961)
Уровень Условия Число особей в июне 1954 г. э Смертность (а 1 год, %
1952 г, или старше группа 1953 г. 1952 г. или старше 1953 г.
Средний уровень прили- Balanus удалей 41 50 37 44
ва; лужа в приливно- Без вмешательства 31 60 71 95
отливной зоне
На 30 см ниже среднего Balanus удалей 3 51 0 35
уровня прилива Без вмешательства 16 69 31 90
На 75 см ниже среднего Balanus удалей 45 25 36 40
уровня прилива Без вмешательства 8 22 75 86
Таблица 14.5
Скорости роста 25 особей Chthamalus stellatus
и 27 особей Balanus balanoides (Connell, 1961)
Вид Размеры Абсолютная ско* рость роста в деньХЮО
1I/V1-1954 3/Х1-1954
A. Chthamalus 2,49 4,24 1,21
В. Balanus 1,87 4,83 2,04
происходит конкуренция за пищу. Если проанализировать
рис. 14.11, то можно заметить, что мелким животным вида А и
крупным животным вида В не приходится так сильно конкуриро-
вать между собой, как тем представителям обоих видов, которые
имеют промежуточные размеры. Соответственно приспособленность
крупных и мелких особей будет несколько выше, их численность
будет увеличиваться и две популяции будут дивергировать до тех
пор, пока конкуренция между ними не сведется к минимуму или
не прекратится вовсе. Этот процесс, называемый экологической
дивергенцией, сходен со смещением признаков у близких видов,
обсуждавшимся в гл. 12.
КОНЦЕПЦИЯ ниши
Примеры морских желудей и ацидофильных растений показы-
вают, что специализированные фенотипы, эффективно использую-
щие ресурсы среды, могут добиться преимущества в конкуренции
и в конце концов исключить менее эффективные фенотипы. Эта
390
Глава 14
•идея нашла воплощение в концепции экологической ниши. Приве-
.денное здесь определение принадлежит великому американскому
экологу Дж. Ивлину Хатчинсону (G. Evelin Hutchinson) из Йель-
ского университета.
Область, в которой
происходит конкуренция
«Малые» размеры
Общий ресурс
«Большие» размеры
Общий ресурс
Рис. 14.11. Схематическое изображение экологической дивергенции.
Поскольку особям каждого из видов, находящимся в неперекрывающихся обла-
стях, не приходится конкурировать с представителями другого вида, их приспособ-
ленность выше, чем приспособленность особей в области перекрывания. С тече-
нием времени оба вида займут неперекрывающиеся экологические ниши.
Представим себе гиперпространство (многомерное пространст-
во), в котором каждое измерение соответствует одной из перемен-
ных, имеющих важное значение для жизни организма, таких, как
вода, время, пространство, температура и свет. Любой вид может
Конкуренция и теория ниши
391
быть отображен в таком гиперпространстве совокупностью значе-
ний всех переменных, делающих возможным его существование.
Возьмем, например, вид, способный выжить только при темпера-
турах между 0 и 20°C, при влажности 80—90%, при содержании
кислорода в воздухе выше 5%, при наличии определенных видов
растений, листьями которых он питается, и т. п. Объем в этом
гиперпространстве, содержащий необходимые для жизни данного
Первое измерение ниши
Рис. 14.12. Соотношение между реализованной и фундаментальной нишами.
Внешняя окружность ограничивает (в двух измерениях) те области нишевого
пространства, где потенциально способны существовать оба вида. В области пе-
рекрывания, когда присутствуют оба вида, они вытесняют друг друга из части
области псрекрыгапия фундаментальных ниш. Реально занимаемый видом объем
в соответствующем пространстве и есть реализованная ниша.
вида значения переменных, образует фундаментальную нишу
этого вида. Однако имеется потенциальная возможность того, что
части этого гиперпространства будут занимать не один, а несколь-
ко видов (согласно принципу конкурентного исключения Гаузе, два
вида нс могут иметь в точности одну и ту же фундаментальную
нишу). При наличии более одного вида каждый вид будет зани-
мать пространство, меньшее, чем его фундаментальная ниша,
вследствие конкуренции. Пространство, занимаемое данным видом,
находящимся з состоянии конкуренции с другими видами, называ-
ют его реализованной нишей (рис. 14.12). Реализованная ниша
может быть равна фундаментальной нише данного вида, меньше
'392
Глава 14
нее, но никогда не бывает больше нее. Фундаментальная ниша име-
ет абсолютные границы, границы же реализованной ниши могут
«изменяться в зависимости от того, сколько имеется конкурентов
и хищников и какова их численность.
Концепция ниши—абстракция, дающая нам возможность лучше
представлять себе эволюционные и экологические факторы, воздей-
ствующие на биологию того или иного вида, и лучше определять
оси, соответствующие отдельным ресурсам. Если известны фун-
даментальные ниши видов, входящих в данное сообщество, то
можно делать предсказания относительно интенсивности конку-
ренции и ее возможных исходов. Однако такая стадия еще не до-
стигнута. Тем не менее экологи уже производят точные определе-
ния осей, соответствующих отдельным ресурсам, для многих орга-
низмов. Хорошим примером разделения ресурсов между разными
видами, иллюстрирующим полезность концепции ниши, служит
исследование, проведенное Томасом У. Шенером (Thomas W. Schoe-
пег) из Вашингтонского университета на ящерицах.
Четыре вида ящериц рода Anolis живут на маленьком (8 км2)
островке Саут-Бимини (Багамские острова). Эти четыре вида
обитают совместно на островке с очень пологим рельефом и до-
вольно однообразной растительностью. Что же делает возможным
сосуществование этих близких видов со сходными пищевыми по-
требностями? Шенер провел количественное исследование ниш
этих четырех видов по нескольким параметрам: 1) предпочитаемый
тип растительности, 2) диаметр веток, на которых отдыхает яще-
рица, и их высота над землей; 3) окраска коры этих веток; 4) до-
ля времени, проводимого среди листвы; 5) размеры жертвы;
6) виды жертвы. Он обнаружил между всеми видами некоторое пе-
рекрывание по каждому из этих параметров. Не было ни одного
параметра, который бы использовался только одним видом. Но
если рассматривать все параметры одновременно, то оказывается,
что каждый вид занимает вполне определенную и обособленную
часть гиперпространства ниши. Например, Anolis sagrei — частично
наземный вид и чаще отдыхает на маленьких, а не на больших
ветках вблизи поверхности почвы; A. distichus предпочитает отды-
хать на стволах и крупных ветвях деревьев, имеющих средние или
большие размеры; A. angusticeps предпочитает небольшие веточки,
расположенные высоко над землей; A. carolinensis чаще всего
встречается на листьях или прилегающих к ним побегах или вет-
ках. Хотя все четыре вида имеют примерно одинаковые размеры,
они несколько различаются по максимальной длине головы, рав-
ной 19,7 мм для A. carolinensis (средняя длина 18 мм), 16,4 мм для
A. angusticeps (средняя—15,7мм), 15,5 мм для A. sagrei (средняя—
14,4) и 13,5 мм для .4. distichus (средняя—12,6). Длина головы имеет
важное значение, поскольку от нее зависит максимальная величина
жертвы, которую может заглотить ящерица. Эти ящерицы питают-
ся разнообразными насекомыми и пауками, а также поедают не-
Конкуренция и теория ниши
393
которую растительную пищу, например мелкие плоды. Однако,
хотя в максимальных размерах, которых достигают самцы, между
этими видами имеются некоторые различия, размеры более мел-
ких особей (а также самок и молодых особей) в значительной
мере перекрываются. Анализируя содержимое желудков большого
числа ящериц, Шенер установил, что в пределах одного вида более
крупные особи обычно поедают небольшое число сравнительно
Рис. 14.13. Видовые ниши, изображенные в виде «облаков» в иишевом простран-
стве.
Каждая точка представляет собой популяцию какого-либо вида о трехмерном ни-
шевом пространстве. Обратите внимание, что между популяциями разных видов
иет четких границ; видовые ниши перекрывают друг друга вследствие разнооб-
разия результатов локальной конкуренции, зависящих от того, какой вид занял
данную нишу первым, от изменений числеииости, колебаний факторов внешней
среды и случайных событий.
крупных пищевых объектов и что диапазон этих объектов по ве-
личине шире. Он обнаружил также, что A. distichus предпочитает
всем другим насекомым муравьев, которые составляют 75—90% его
рациона, а Л. sagrei поедает гораздо больше плодов, чем три дру-
гих вида. Диеты A. carolinensis и A. angusticeps очень сходны.
Однако, когда Шенер определял относительное обилие этих яще-
риц в зависимости от различных типов растительности (табл. 14.6),
он обнаружил, что виды, которые встречаются очень часто в расти-
тельности одного типа {angusticeps — distichus; carolinensis — sag-
rei), особенно сильно различаются по своей диете и что те два
вида, диета которых наиболее сходна {angusticeps — carolinensis),
предпочитают различные типы растительности. Эти последние
данные представляют особый интерес, поскольку они показывают,
что два вида могут использовать ресурс одного и того же типа,
394
Глава 14
Таблица 14.6
Обилие видов Anolis в разных типах растительности
иа острове Саут-Бимини (Schoener, 1968)
Местообитание Обилие*
A. angu- siiceps А. саго- 1 inert sis A. dlstichus A. sagrei
Черная земля (во внутренних частях
острова) о р О О
'Coccothrinax (кустарник) н О Р—О О
•Мангры (Rhizophora) н н н р
Маигры (Avicennia) н р н р
Salicornia на приливно-отливной отмели н н н р
'Uniola на морском берегу н р н р
Прибрежные скалы н н н н
Черная земля (начало образования) Число местообитаний, в которых обиа- О О О О
ружен данный вид 2 5 3 7
• О — обилен по сравнению с другими местами, где данный
Р — имеется, но обнаружить удается редко; Н — не обнаружен.
вид был обнаружен;
если они разделены физически или во времени, как это можно
было предсказать на основании концепции многомерной ниши, где
координаты времени и пространства отложены по разным осям.
Концепция многомерной ниши дает возможность количествен-
но описать активность отдельного вида (добывание пищи, забота
о потомстве, размножение, сон) в пространстве и во времени и
точно определить, как данный вид взаимодействует в сообществе.
Каждый вид можно представить в' виде облака в многомерном про-
странстве сообщества (рис. 14.13). Располагая этой информацией,
можно задать вопрос: сколько видов может теоретически сосуще-
ствовать в данной области? Число и относительное обилие видов
в какой-либо области называют видовым разнообразием. Возвра-
щаясь к многомерной нише, можно видеть, что видовое разнооб-
разие зависит от числа осей ресурсов, а также от разнообразия
и обилия каждого ресурса, определяющих длину оси ресурса. На-
пример, если пищевых ресурсов в какой-либо области мало и при
этом они однообразны, то эти ресурсы не могут быть поделены
столь разными способами, как в том случае, когда область богата
пищевыми ресурсами. Видовое разнообразие зависит также от
ширины ниши, т. е. от разнообразия ресурсов, используемых дан-
ным видом, а это в свою очередь зависит от относительного обилия
одного конкретного ресурса во времени и пространстве. Теория
ниши — область экологии, интенсивно разрабатываемая как мате-
матически, так и экспериментально, а это позволяет надеяться, что
теоретические и эмпирические результаты исследований в популя-
ционной биологии удастся объединить с более классическими
аспектами экологии сообществ.
Конкуренция и теория ниши
395
ЛИТЕРАТУРА
Общие вопросы
Emlen I. М. 1973. Ecology; An Evolutionary Approach, Reading, Mass., Addison-
Wesley.
Гаузе Г. Ф. 1934. The Struggle for Existence, Baltimore, Williams and Wilkins.
Hutchinson G. E. 1965. The Ecological Theater and the Evolutionary Play, New Ha-
ven, Yale University Press.
McR aughton 5. J., Wolf L. L. 1973. General Ecology, New York, Holt, Rinehart and
Winston.
May R. M. (ed.). 1976. Theoretical Ecology: Principles and Applications, Oxford,
Blackwell Scientific Publications.
Ricklefs R. E. 1973. Ecology, Portland, Ore, Chiron Press.
Slobodkin L. B. 1966. Growth and Regulation of Animal Populations, New York,
Holt, Rineliart and Winston.
Специальные статьи и обзоры
Colwell R. К-, Fuentes E. 1975. Experimental studies of the niche, Ann. Rev. Ecol.
Syst, 6, 281-310.
Connell J. H. 1961. The influence of interspecific competition and other factors on
the distribution of the barnacle Chthamalus stellatus, Ecology, 42, 710—723.
Ellenberg H. 1952. Physiologisches und okologisches Verhalten derselben Pflan-
zenarten, Ber. Deut. Bot. Ges., 65, 350—361.
MacArthur R. H„ Levins R. 1967. The limiting similarity, convergence and divergen-
ce of co-existing species, Am. Nat., 101, 377—385.
Roughgarden J. 1974. Niche width: biogeographic patterns among Anolis lizard
populations. Am. Nat., 108, 533—540.
Schoener T. W. 1968. The Anolis lizards of Bimini: resource partitioning in a com-
plex fauna. Ecology, 49, 704—726.
Solbrig О. T.. Simpson В. B. 1974. Components of regulation of a population of
dandelions in Michigan, J. Ecol., 62, 473—486.
Solbrig О. T., Simpson, В. B. 1977. A garden experiment on competition between
biotypes o: the common dandelion (Taraxacum afficinale), J. Ecol., 65, 427—
430.
Stearns S. C. 1977. The evolution of life history traits, Ann. Rev. Ecol. Syst., 8,
145—172.
Steiner W. M. 1977. Niche width and genetic variation in Hawaiian Drosophila, Am.
Nat., Ill, 1037—1045.
Vandermeer J. H. 1969. The competitive structure of communities: an experimental
approach with Protozoa, Ecology, 50, 362—371.
Vandermeer J. H. 1972. Niche theory, Ann. Rev. Ecol. Syst., 3, 107—132.
Wilbur H. M. 1977. Propagule size, number and dispersion pattern in Ambystoma
and Asclepias, Am. Nat., Ill, 43—68.
Yoshiyame R. M., Roughgarden J. 1977. Species packing in two dimensions, Am.
Nat. Ill, 107—121.
Глава 15
Неконкурентные взаимодействия
между видами
Большинство видов растений и животных представляет собой
потенциальную пищу для какого-нибудь животного (а иногда и
для растения). В связи с этим у растений и животных в процессе
эволюции возникли весьма разнообразные защитные приспособ-
ления. Ядовитые или неприятные на вкус ткани, твердые или оро-
говевшие покровы, шипы, жгучие волоски, способность наносить
ядовитые или болезненные уколы или укусы, способность быстро
бегать или летать или же прятаться (дополняемая покровительст-
венной окраской или формой) —вот всего лишь несколько из мно-
гочисленных химических, морфологических и поведенческих адап-
таций растений и животных, помогающих им избежать истребле-
ния хищниками. Хищники в свою очередь выработали собственные
адаптации, позволющие им преодолевать защитные адаптации
жертвы: это — механизмы детоксикации; мощные мандибулы и
пищеварительные системы; прочные и сильные когти, лапы и че-
люсти, дающие возможность побороть жертву; превосходное зрение
или слух; способность бегать или летать быстрее, чем жертва, и т. п.
Такое разнообразие средств защиты у жертвы и средств нападе-
ния у хищника привлекает внимание к одному из важнейших био-
логических фактов — к тому, что добывание пищи и бегство от
хищников представляют собой два основных вида активности ор-
ганизмов, создающие два наиболее мощных давлений отбора, под
действием которых формируются признаки организмов.
Но виды не только конкурируют между собой или поедают друг
друга; они способны и к сотрудничеству. Многим растениям не-
обходимы насекомые, которые бы их опыляли. Жвачным (коровы,
олени, верблюды) необходимо, чтобы в их кишечнике обитали
бактерии, без которых они не могли бы переваривать целлюлозу
и погибли бы от голода; то же самое относится и к термитам.
Даже человек не может обойтись без кишечной палочки (Esche-
richia coli), которая обеспечивает его витамином К. Следователь-
но, взаимоотношения, основанные на сотрудничестве, также пред-
ставляют собой мощные факторы отбора, участвующие в формиро-
вании признаков организмов. В настоящей главе мы рассмотрим
эти неконкурентные взаимоотношения.
Неконкурентные взаимодействия между видами
397
ВЗАИМООТНОШЕНИЯ ХИЩНИК — ЖЕРТВА
Хищник — это организм, частично или полностью потребляющий
в качестве пищи другой живой организм. Существует широкий
диапазон взаимоотношений хищник—жертва: от львов или вол-
ков, охотящихся за своей жертвой и убивающих ее, до таких мел-
ких насекомых, как комары, которые сосут кровь жертвы, оказы-
Рис. 15.1. Исход взаимодействия между видом-жертвой Paramecium caudatum
(I) и видом-хищником Didinium nasutum (II). (Г. Ф. Гаузе, 1934. The Struggle
for Existence, Baltimore, Williams and Wilkins.)
вая на нее лить незначительное воздействие. Жертвой не обяза-
тельно служит другое животное, это может быть и растение; в та-
ком случае хищник называется растительноядным. Как правило,
растительноядные не убивают свою жертву, а ограничиваются
тем, что поедают некоторые ее части (листья, плоды, корни). Тем
не менее растительноядные способны оказывать опустошительные
воздействия. Примером служит уничтожение растительности Но-
вой Англии яг.онским хрущиком. Если хищник вступает в очень
тесные взаимоотношения с жертвой, то его называют паразитом
или паразитоидом, в зависимости от степени вреда, который он
наносит жертве.
Для того чтобы упростить теоретическое рассмотрение взаимо-
отношений хищник — жертва, мы ограничим его ситуацией, при ко-
торой хищник убивает свою жертву (и тем самым изымает ее из
популяции).
Взаимоотношения между хищником и жертвой гораздо слож-
нее, чем взаимоотношения между конкурентами. Фактически ин-
дивидуальный отбор будет благоприятствовать эффективному хищ-
нику. Однако эффективный хищник может совершенно уничто-
жить жертву, обрекая при этом на гибель и самого себя. Один
398
Глава ]5
такой пример приведен на рис. 15.1, где изображен график роста
популяций Paramecium caudatum и ее хищника Didinium nasutum
в одной и той же культуре. При введении в культуру хищника чис-
ленность популяции Paramecium сокращается, а численность по-
пуляции хищника увеличивается, пока в какой-то момент количе-
ство жертвы не оказывается недостаточным. После этого числен-
ность обеих популяций начинает уменьшаться, и в конечном счете
они вымирают.
Взаимодействия хищник—жертва можно математически
описать при помощи двух уравнений:
где JVj — плотность популяции жертвы, N2 — плотность популяции
хищника, Г1 — мгновенная удельная скорость увеличения популя-
ции жертвы, d2— смертность в расчете на одну особь популяции
хищника, a pi и р2— коэффициенты хищничества. Каждую популя-
цию ограничивает другая популяция, причем воздействия, регули-
рующие плотность, отсутствуют (обратите внимание на отсутствие
в формуле К). Эти уравнения известны под названием уравнений
Лотки—Вольтерра.
Скорость роста популяции жертвы (А^) равна ее истинной ско-
рости роста (рождаемость в расчете на особь минус смертность
в расчете на особь), умноженной на число особей в популяции в
момент времени 1 (экспоненциальный рост; см. стр. 334), минус
произведение численностей жертвы и хищника, Ni%N2 (отражаю-
щее ожидаемые встречи между ними), умноженное на константу
Pi, соответствующую вероятности того, что жертва будет съедена
хищником при такой встрече. Рост популяции хищника выража-
ется числом жертв и числом встреч между хищником и жертвой
(IV1IV2), умноженным на коэффициент, указывающий на «доходы»,
получаемые хищником в результате поедания одной особи жертвы
(р2), и деленным на смертность хищников от всех причин.
На рис. 15.2 построен график скорости роста популяции хищ-
ника и жертвы в системе декартовых координат. Изменения чис-
ленности как хищника, так и жертвы носят характер циклических
колебаний. Мы видели, однако, что в простом лабораторном экспе-
рименте численность их популяций не подвержена колебаниям.
Просто хищники выискивают всех особей жертвы, пока не доводят
популяцию жертвы да вымирания, после чего сами гибнут от го-
лода. Проблема состоит в том, что уравнения, предложенные в ка-
честве модели, слишком просты. В них приняты нереалистичные
допущения о непрерывном росте популяции жертвы, а также о
случайности встреч между хищником и жертвой. Между тем хо-
Неконкурентные взаимодействия между видами
399
рошо известно что хищники активно занимаются поисками жерт-
вы, а жертва гктивно прячется от хищников.
В природе наблюдается своего рода равновесие между жерт-
вой и ее хищшками. Известны примеры циклических колебаний
Плотность жертвы
меньшается
1лотность хищника
'меньшается
Плотность жертвы
уменьшается
Плотность хищника
возрастает
Плотность жертвы
взрастает
Плотность хищника
уменьшается
Плотность жертвы
возрастает
Плотность хищника
возрастает
Плотность жертвы
А
Рис. 15.2. Изменения численности хишника и жертвы в идеализированной модель-
ной системе.
А. Траектория сютемы в координатах «плотность жертвы—плотность хищника».
Б. Изме(ения плотности жертвы (I) и хищника (II) во времени.
численности, <ак, например, у рыси и ее главной жертвы — зайца-
беляка-— в Канаде (рис. 13.13) или у лемминга на Аляске, кото-
рого изучал калифорнийский эколог Франк Пителка (Frank Pi-
telka) вблизи мыса Барроу, но таких примеров немного.
Лемминги— растительноядные животные, питающиеся низко-
рослыми кустарниками и травой в тундре. Они живут в тундре
круглый год и даже в зимние месяцы кормятся и размножаются
под снегом. Летом на леммингов охотятся многие хищники, в част-
400
Глава 15
ности поморники и совы, размножающиеся в тундре в летние ме-
сяцы. Мы начнем описание циклических колебаний с популяции
леммингов, имеющей низкую численность. Низкая численность
леммингов означает, что число птенцов, которых могут вырастить
поморники и совы, ограничено, поскольку число птенцов опреде-
ляется наличием пищи. Соответственно численность хищных птиц
понизится. Это означает также наличие достаточного количества
растительной пищи, для того чтобы лемминги могли питаться, не
нарушая нормального роста растений. Такое положение вещей —
Один год
Рис. 15.3. Обобщенная кривая одного периода в краткосрочном цикле колебаний
численности лемминга Lemmus trimueronatus вблизи мыса Барроу (Аляска).
Показаны также второстепенные колебания или изменения численности в преде-
лах этого периода и их примерная приуроченность к летним месяцам в последо-
вательные годы. (Pitelka, 1958.)
обилие пищи и относительно небольшая численность врагов — соз-
дает оптимальные условия для размножения леммингов в течение
следующей зимы и лета. Однако по мере роста численности лем-
мингов запасы растительной пищи сокращаются, а популяции
хищников увеличиваются. Это ведет к некоторому голоданию лем-
мингов в течение второй зимы цикла и вследствие роста популяции
поморников и сов во второй год — к высокому уровню выедания
леммингов хищниками в течение третьего лета цикла. В резуль-
тате на четвертый год численность популяции леммингов снижа-
ется до исходного уровня, после чего цикл может возобновиться
(рис. 15.3). Отметим, что состояние растительности представляет
собой важный фактор в цикле леммингов, равно как и ряд ас-
пектов физиологии этих животных, которых мы здесь не касаемся.
Цикл леммингов, однако, не типичен для взаимоотношений
хищник — жертва. Хотя численность большинства хищников и их
жертв несколько колеблется в разные годы, эти колебания нечасто
бывают такими резкими, как у леммингов, и, насколько нам из-
вестно, редко носят характер регулярных циклов.
Взаимоотношения хищник—жертва могут принимать различ-
ные формы. Майкл Розенцвейг (Michael Rosenzweig) и Роберт
Неконкурентные взаимодействия между видами
401
Мак-Артур (Robert MacArthur) предложили простую графическую
модель, представленную на рис. 15.4. На этом графике в системе
декартовых координат изображены изменения численности хищ-
Рис. 15.4. Модель хищник—жертва с затухающими колебаниями — случай неэф-
фективного хищника.
А. Модель. Б. Изменения численности жертвы (/) и хищника (II) во времени.
(Rosenzweig, MacArthur, 1963.)
ников и жертвы, как это было сделано на рис. 15.2. Здесь, однако,
были приняты следующие допущения. При полном отсутствии
хищников популяция жертвы достигает максимальной численности
Ki- Емкость среды для жерты определяется не хищничеством (ко-
торое ограничивает рост численности жертвы), а другими фактора-
ми (пища, убежища и т. п.). Существует также некий нижний
26—2044
40?
Глава 15
предел численности, перейдя который популяция жертвы не может
продолжать свое существование ввиду отсутствия достаточных
возможностей для спаривания. Следовательно, численность попу-
ляции жертвы будет увеличиваться лишь в том случае, если она
ниже кривой ее роста, представленной на рис. 15.4. Аналогичным
образом существует некая пороговая численность и для хищников,
ниже которой они не могут найти достаточно пищи для поддер-
жания своего существования, а также верхний предел, или емкость
среды для хищников, выше которого рост популяции хищника
становится невозможным из-за других факторов (например, недо-
статка мест для гнездования). Если хищники не конкурируют меж-
ду собой, то кривая их роста будет близка к прямой линии вплоть
до того, как будет достигнута емкость среды для хищника, Кг-
При наличии же взаимодействия между хищниками кривая нуле-
вого роста будет отклоняться вправо с уменьшением численности
популяции хищника (рис. 15.4).
Разберем теперь три гипотетических примера, которые мы на-
зовем случаями неэффективного хищника, высокоэффективного
хищника и «расчетливого» хищника, и проанализируем их при
помощи графического метода Розенцвейга и Мак-Артура. Неэф-
фективному хищнику, для того чтобы выжить, необходима высо-
кая плотность жертвы. Поэтому нижнему пределу его численности
соответствует высокая плотность жертвы (рис. 15.4). Популяция
жертвы будет расти, пока не достигнет уровня, соответствующего
емкости среды. Когда ее численность достигает пороговой для хищ-
ника, последний начинает истреблять ее, в результате чего повы-
шается его собственная численность. По мере роста популяции
хищника численность жертвы начинает снижаться. В конце концов
численность жертвы оказывается недостаточной, чтобы обеспечи-
вать существование большого числа хищников, и популяция по-
следних также начинает сокращаться. Если проследить за общей
траекторией обеих популяций (стрелка), то можно видеть, что она
образует скручивающуюся спираль, которая заканчивается на пе-
ресечении обеих кривых. Неэффективный хищник может благопо-
лучно существовать, находясь в равновесии со своей жертвой. При
нарушении этого равновесия возникают затухающие колебания,
продолжающиеся до тех пор, пока не восстановится равновесие.
В случае высокоэффективного хищника наблюдается прямо про-
тивоположная ситуация. В результате его взаимодействий с жерт-
вой создается очень неустойчивый цикл, и в конечном счете хищ-
ник истребляет свою жертву и гибнет сам (рис. 15.5). Именно это
в течение десятилетия произошло с возбудителем рака каштанов
в США, куда этот вирус был завезен в 1915 г. Популяция возбу-
дителя заболевания росла до тех пор, пока зараженными не ока-
зались все деревья, но с гибелью деревьев погиб и возбудитель
болезни. Наконец, в случае «расчетливого» хищника создается
устойчивый цикл. При этом две кривые, соответствующие нуль-
____________Неконкурентные взаимодействия между видами 403
изоклинам, должны пересекаться именно в той точке, в которой
изоклина жертвы достигает максимума (рис. 15.6)
Модель Розенцвейга—Мак-Артура гораздо более реалистич-
на, чем уравнения Лотки—Вольтерра. В ней учитывается, что на
рост как хищника, так и жертвы оказывают влияние факторы, не
относящиеся к взаимодействиям между ними. Кроме того, эта
модель позволяет получить более реалистичные результаты и по-
казывает, что сосуществование хищника и жертвы возможно лишь
в том случае, если естественный отбор не допускает чрезмерного
повышения эффективности хищника. Парадоксально, но сверхэф-
фективный хищник сам обрекает себя на гибель, если только жерт-
ва не выработает механизмы, позволяющие ей избегать хищника.
Возможно, что при этом действует какая-либо форма группового
отбора: выживают только популяции «расчетливых» или неэффек-
тивных хищников. К другим факторам относятся: наличие убежищ
для жертвы; иммиграция и эмиграция как хищников, так и жерт-
вы; способность некоторых видов хищников переживать периоды
отсутствия жертвы, переходя в то или иное покоящееся состояние
(цисты, яйца).
Конкуренция и хищничество — важные факторы отбора, кото-
рые в значительной степени определяют эволюцию фенотипиче-
ских признаков и жизненные циклы организмов. Проиллюстриру-
ем это на нескольких примерах.
Растительные ткани служат пищей очень многим и разнообраз-
ным животным и грибам и даже нескольким видам растений, на-
пример повилике (Cuscuta spp). Растения не могут спастись бег-
ством или укрыться от своих хищников (известных под названием
растительноядных). Однако отдельные растения, ткани которых в
результате мутаций становятся несъедобными или ядовитыми,
меньше подвержены уничтожению, и поэтому им будет благо-
приятствовать естественный отбор. Таким образом, не должно
вызывать удивления, что растения вырабатывают фантастическое
разнообразие химических веществ. Целая отрасль химии, извест-
ная под названием химии природных соединений, занимается ис-
1 Автор неточно описывает ситуацию и неправильно употребляет термины
устойчивый и неустойчивый предельный циклы. За вычетом некоторых деталей,
как правило, дело обстоит следующим образом. Если вертикальный участок нмль-
изоклины хищника лежит правее максимума нуль-изоклины жертвы, то равнове-
сие устойчиво. Если он лежит немного левее максимума, то равновесие неустой-
чиво и возникает устойчивый предельный цикл, т. е. незатухающие колебания чис-
ленностей обеих популяций с определенной амплитудой и формой, продолжающие-
ся сколь угодно долго. Если же он лежит сильно левее максимума, то цикл раз-
рушается и в конечном счете обе популяции вымирают. Подробно эта ситуация
разобрана в работе: А. Д. Базыкин, Ф. С. Березовская «Эффект О.: : нижняя
критическая численность популяции и динамика системы хищник—жертва». В сб.
«Проблемы экологического мониторинга и моделирования экосистем» т. 2. Л.. Гид-
рометеоиздат 1979. — Прим. ред.
26s
404
Глава 15
следованием веществ, вырабатываемых растениями. Каждый ме-
сяц ученые открывают новые соединения. Многие из них, такие, как
антибиотики (ядовитые для бактерий), ядовиты для хищников;
Рис. 15.5. Модель хищник—жертва с усиливающимися колебаниями — случай вы-
сокоэффективного хищника.
Обозначения те же, что на рис. 15.4. А. Модель. Б. Изменения численности жерт-
вы (/) и хищника (II) во времени. (Rosenzweig, MacArthur, 1963.)
другие вещества, такие, как таннины, не перевариваются. Сущест-
вование этих соединений у растений служит мощным фактором
отбора, действующим на растительноядные организмы. В самом
деле, в популяции растительноядных особи, которые в результате
мутаций стали способны к детоксикации ядовитых или не поддаю-
щихся перевариванию соединений, вырабатываемых растениями,
сразу подвергаются положительному отбору. Существование им-
Неконкурентные взаимодействия между видами
405
мунных растительноядных видов в свою очередь будет действовать
яа растения как фактор отбора, благоприятствуя новым мутациям.
Этот процесс происходит непрерывно в пределах каждой популя-
А
Изоклина хищника
Рис. 15.6. Модель хищник—жертва с регулярными колебаниями — случай «расчет-
ливого» хищника.
А. Модель. Б. Изменения численности жертвы (/) и хищника (II) во времени.
(Rosenzweig, MacArthur, 1963.)
ции растений и каждой популяции растительноядных организмов
в любом сообществе. Поскольку появление мутации — дело случая,
мало вероятно, что все растения в каком-либо развивающемся
сообществе начнут вырабатывать одно и то же вещество для за-
щиты от растительноядных или же что механизмы детоксикации,
возникающие у растительноядных организмов, будут действовать
406
Глава 15
на все растения, входящие в данное сообщество. Это приводит к
специализации, тем более что большинство растительноядных спо-
собно переваривать только один или несколько видов растений и
Рис. 15.7. Насекомые, похожие на листья или веточки.
А. Кузнечик Cycloptera elegans (Ortoptera, сем. Tettigoniidae). Б. Листовидка
Phy Ilium siccifolium (Phasmida). В. Гусеница Boarmia hispidarius (чешуекрылые).
Г. Палочник (Phasmida). Д. Haaniella echinata (Phasmida). (Фотографии Alfred
Coleman.)
притом нередко лишь какой-либо один тип тканей, например ли-
стья, древесину или корни.
Растительноядные в свою очередь служат жертвой для хищни-
ков. В отличие от растений растительноядные животные могут
Неконкурентные взаимодействия между видами
407
спасаться от хвдников бегством и прятаться в разного рода укры-
тиях. Внутри ^общества растительноядных отбор будет благо-
приятствовать гаименее заметным особям. Один из способов до-
стижения незаметности состоит в приобретении окраски и даже
формы того растения, которым кормится данное растительноядное.
На рис. 15.7 июбражено несколько насекомых, подражающих по
форме своему оону. Другой способ избежать съедения хищником—
выработка определенных типов поведения, например питание по
ночам, когда хпцники не могут воспользоваться зрением. Если,
однако, слишюм многие растительноядные станут походить на
листья и веточги или перейдут к ночному образу жизни, то хищ-
ники начинают более внимательно приглядываться к объектам,
напоминающик листья или палочки, или же изменяют свое пове-
дение, выходя на охоту ночью. Поскольку каждая отдельная по-
пуляция расти'ельноядных в данном сообществе приобретает свои
специфические механизмы избегания хищников, последние не мо-
гут охотиться ia всех растительноядных одинаково эффективно и
обычно специатизируются к охоте на определенные виды. Напри-
мер, виды птиг и ящериц, питающиеся насекомыми, предпочти-
тельно охотятся на насекомых определенного размера, обитающих
в определенным местах. У них отмечаются также предпочтения к
охоте в определенное время суток. Кроме того, хищники поддер-
живают популяцию растительноядных на некотором не слишком
высоком уроше, что способствует сохранению растительности.
В свою очеред» в популяции растительноядных особи, обладающие
наиболее сове>шенной покровительственной окраской или формой
или же способные прятаться лучше других, имеют наибольшие
шансы выжит», уменьшая тем самым эффективность хищников и
препятствуя установлению дестабилизирующих циклов. Когда чис-
ло особей одюго вида растительноядных снижается и их стано-
вится трудно даходить, хищник переключается на какой-либо дру-
гой растителыоядный вид, что дает возможность первому виду
восстановить свою численность, и т. д.
Интересным эксперимент, демонстрирующий сложность взаи-
модействий в сообществе, провел Роберт Пэйн (Robert Paine) из
Вашингтонского университета. На скалистых берегах Тихого океа-
на в Северно! Америке существует сообщество, в состав которого
входят моллкски, морские желуди и один вид морских звезд
(рис. 15.8). Аорская звезда Pisaster ochraceus — хищник, питаю-
щийся всеми другими видами; брюхоногий моллюск — Thais emar-
ginata — также хищник, питающийся только двустворчатыми мол-
люсками и морскими желудями. Все другие виды, потребляющие
главным образом растительную пищу, питаются разнообразными
водорослями и планктоном. Пэйн искусственно удалил морских
звезд с экспериментального участка, расположенного на скалистом
берегу в приливно-отливной зоне. Вскоре после этого он обнару-
жил, что один из видов морских желудей (Balanus cariosus) начал
408
Глава 15
побеждать в конкуренции другие виды и в течение трех месяцев
занимал от 60 до 80% всей площади участка. Со временем, одна-
ко, один вид двустворчатых моллюсков (Mytilus californianus) и
одна морская уточка (Mitella polymerus) в свою очередь вытесни-
ли Balanus. В конце концов Mytilus занял большую часть участка.
Хитон, Блюдечки, Mytilus ~ Морские желуди, Mitella
2 вида 2 вида (двустворчатый 3 вида (морская
моллюск), 1 вид уточка)
Рис. 15.8. Пищевые взаимоотношения в морском сообществе приливно-отливной
зоны.
В состав сообщества входят два хищника: морская звезда (Pisaster) и улитка
(Thais). Цифры иа стрелках указывают: какую долю в диете каждого хищника
составляет каждый тип жертвы относительно общего числа съедаемых особей
(первая цифра) и относительно суммарного числа полученных калорий (вторая
цифра). (Paine, 1966.)
Первоначальное сообщество, состоявшее из более чем десятка
видов, свелось по существу к двум видам. Что же здесь произо-
шло? Наиболее конкурентоспособный вид — Mytilus — является
одновременно и самой лакомой пищей' (с точки зрения калорий-
ности) для морской звезды. При наличии в сообществе морских
звезд он подвергался сильному выеданию, что препятствовало
созданию обширной популяции, которая могла бы победить в кон-
Неконкурентные взаимодействия между видами
409
куренции с друими видами. Это делало возможным сосущество-
вание большого числа видов. В данном случае хищник — морская
звезда — не доп’скал победы наиболее сильного конкурента.
Предутеждающая окраска и эволюция мимикрии
Другим призером, иллюстрирующим сложность пищевых сетей
и взаимоотношений в сообществах, служит то, что можно было
бы назвать «системой данаиды»; эта система изучалась многими
Рис. 15.9. Каким сбразом данаида (Danaus plexippus) становится несъедобной?
Ваточники (Asclepas spp.) вырабатывают несколько соединений, ядовитых для
позвоночных. На р|суике показана формула одного из таких соединений — кало-
токсина. Гусеница еанаиды поедает листья ваточника н ассимилирует это вещест-
во, сохраняя его в своих тканях и передавая его затем взрослой бабочке. В ре-
зультате бабочка становится несъедобной для птиц.
учеными, но главным образом Линкольном Брауэром (Lincoln
Brower). Данагда (Danaus plexippus) широко распространена в
Северной Амертке. Это большая хорошо заметная бабочка с ярко-
оранжевыми крыльями, окаймленными темно-коричневой полосой.
Птицы обычно избегают данаид, находя их очень неприятными
на вкус. Когда неопытным птицам (выращенным в неволе и ни-
когда не встречавшимся с данаидами) предлагают такую бабочку,
они хватают ее, но тут же выплевывают. Причина этой очень силь-
но выраженной реакции стала ясна в результате изучения биоло-
410
Глава 15
гии питания данаиды. Гусеницы этой бабочки кормятся на ваточ-
нике, в тканях которого содержатся очень ядовитые алкалоиды.
Однако гусеницы данаиды обладают способностью изолировать
алкалоиды и накапливать их в своих тканях, с которыми они пе-
редаются взрослым бабочкам (рис. 15.9). Таким образом, этот
вид сумел преодолеть химический защитный механизм растения
и обратить его в свою пользу! Но это еще не все. Для того чтобы
такая химическая защита была эффективна, потенциальные хищ-
ники должны узнать о ней до того, как они нападут на бабочку.
Этим и объясняется очень яркая окраска данаиды, служащая пре-
дупреждением для хищников. Способность птиц научаться связы-
вать яркую окраску бабочки с ее неприятным вкусом была экспе-
риментально доказана Линкольном Брауэром. Брауэр предлагал
вскормленным в неволе птицам (хохлатым сойкам) данаид и ма-
хаонов; ни тех, ни других птицы прежде никогда не пробовали.
Сойки во всех случаях поедали махаонов, не поедая данаид; если
же птицы заглатывали данаид, то они их быстро отрыгивали и в
дальнейшем избегали. Очевидно, сойки научаются на опыте избе-
гать употребления данаид в пищу, узнавая их по ярко окрашен-
ным крыльям.
Многие животные защищены от потенциальных врагов благо-
даря своим ядовитым или неприятным на вкус тканям. Неприят-
ным запахом или вкусом обладают представители многих групп
насекомых, ряд насекомоядных, хвостатых амфибий и скунсы. Пче-
лы, осы, скаты-хвостоколы и некоторые змеи жалят своих врагов,
выпуская через жало яд.
Ядовитые или неприятные на вкус ткани не помогут жертве,
если хищник узнает об этих их свойствах лишь после того, как он
убьет или искалечит ее. Но если такие ядовитые виды ярко ок-
рашены, то потенциальные враги быстро научаются связывать яр-
кую окраску с неприятными свойствами того или иного вида. Это
подтверждают эксперименты, проведенные Карпентером (Carpen-
ter). Основываясь на субъективном восприятии, он разделил пред-
ставителей 244 видов насекомых на хорошо заметных и криптиче-
ских. Затем он скармливал этих насекомых африканской мартыш-
ке (Cercopithecus) и на основании ее реакций разделил виды насе-
комых на съедобные и неприятные на вкус. Результаты представ-
лены в табл. 15.1. Обратите внимание, что большая часть крипти-
ческих видов оказалась съедобной, а большинство заметных ви-
дов — неприятными на вкус.
Защитный механизм, создаваемый предупреждающей окраской,
начинает действовать лишь после того, как потенциальный враг
хотя бы дважды встретится с несъедобным видом и научится ас-
социировать его яркую окраску с неприятными свойствами. Одна-
ко, поскольку в окраске многих ядовитых или неприятных на вкус
видов, принадлежащих к различным семействам и отрядам, пре-
обладают четыре основных цвета — черный, красный, оранжевый
Неконкурентные взаимодействия между видами
411
Таблица 15.1
Связь между характером окраски насекомых
и их съедобностью для хищников
Окраска Съедобные Несъедобные Общее число
Критическая 83 18 101
Яркая 23 120 143
Итого 106 138 244
и зеленый, — предупреждающие об их неприятных свойствах, боль-
шинство потенциальных врагов научаются связывать такую общую
окраску с опасностью. Конвергенция по предупреждающей окрас-
ке, наблюдаемая среди несъедобных видов, сама по себе пред-
ставляет собой адаптацию, направленную на снижение частоты
«ошибок» у потенциального хищника, поскольку такие ошибки
оказываются роковыми для подвергшихся нападению особей. Так,
например, многие несъедобные тропические бабочки из сем. Nym-
phalidae, встречающиеся совместно в некоторых частях Бразилии,
сходны по форме и окраске крыльев. Это представители различных
видов: Lycorea, Melinaea, Mechanitis, Heliconius и Eveides, относя-
щихся к трем разным подсемействам нимфалид.
После того как птица или жаба научится связывать опреде-
ленный яркий цвет или сочетание цветов с неприятными свойст-
вами и станет избегать животных, имеющих такие признаки, эта
ее реакция распространяется не только на действительно ядови-
тых животных, но и на всех других, обладающих сходной окрас-
кой. Это сделало возможным эволюцию мимикрии — подражания
съедобных животных другим видам, которые ядовиты или обла-
дают неприятным вкусом. Эволюция мимикрии в принципе не от-
личается от эволюции покровительственной окраски, только в од-
ном случае моделью служит природа, а в другом — какое-либо
хорошо заметное ядовитое животное. В обоих случаях чем боль-
ше сходство с моделью, тем надежнее защита. Кроме того, как в
том, так и в другом случае, если имитаторов становится слишком
много, хищник научается связывать их окраску или фон с хоро-
шей, а не с несъедобной пищей (будь то ядовитое животное, ли-
шайник или скала).
Мимикрия встречается во многих группах животных. Муравьи,
например, несъедобны для большинства животных из-за своих
ядовитых желез (но не для всех животных, о чем свидетельствуют
по крайней мере три неродственных между собой вида млекопи-
тающих, специализировавшихся на питании муравьями). Муравьям
412
Глава 15
подражают в разных частях земного шара пауки, кузнечики, жуки,
мухи и другие насекомые. То же самое относится к шмелям, ко-
торым подражают разные виды мух и бабочек. Но наиболее ши-
роко известные и хорошо изученные примеры мимикрии относятся
к бабочкам, подражающим данаиде.
В восточной части Северной Америки распространена бабочка
Limenitis archippus (рис. 15.10). По внешним признакам, особенно
Рис. 15.10. Бэйтсовская мимикрия.
А. Несъедобная модель — данаида (Danaus plexippus). Б. Подражающая ей ба-
бочка Limenitis archippus.
по окраске крыльев и узору на них, эта бабочка похожа на данаи-
ду. Поскольку L. archippus вполне съедобна, уже давно высказы-
вались предположения, что она подражает данаиде. Чтобы прове-
рить это, Брауэр (Brower) кормил бабочками L. archippus группу
соек, которые прежде никогда не сталкивались с этими бабочка-
ми. Большинство птиц поедало бабочек, не проявляя никаких от-
рицательных реакций. Другой группе соек были предложены да-
наиды, и, как уже говорилось выше, они быстро научились отка-
зываться от них. Когда затем этим сойкам предложили бабочек
L. archippus, большинство птиц отказалось и от них. Так было
установлено, что бабочки L. archippus в самом деле спасаются от
хищников благодаря своему сходству с несъедобными данаидами.
Поскольку защита, создаваемая мимикрией, зависит от того,
научится ли потенциальный хищник связывать окраску или форму
данной бабочки с неприятными свойствами той модели, которой
она подражает, судьба вида-имитатора тесно связана с модельным
видом. Если имитаторы гораздо более многочисленны, чем модель,
то хищник может научиться ассоциировать их окраску со съедоб-
ностью имитатора и, учитывая заметность их окраски, они могли
бы оказаться в весьма опасном положении. Подобная ситуация
была бы вредна не только для имитатора, но и для его модели.
Кроме того, если в процессе обучения хищник будет часто ловить
имитаторов, то обучение замедлится. Наконец, вследствие своей
Неконкурентные взаимодействия между видами 413
заметности имитатор должен жить только в том же месте и в то
же время, что и его модель.
Брауэр попытался выразить взаимоотношения между имита-
торами и моделями в количественной форме. Он кормил скворцов
(Sturnus vulgaris) личинками мучного хрущака (Tenebrio molitor),.
часть которых предварительно окунал в раствор хинина, что де-
лало их очень неприятными на вкус. Вместе с этими несъедобны-
ми личинками («моделями») Брауэр давал скворцам съедобных
личинок, которых он окунал просто в воду («имитаторы»). Сквор-
цы быстро научились ассоциировать личинок хрущака с неприят-
ным вкусом и избегали их. При соотношении моделей и имитато-
ров примерно 9: 1 птицы отвергали 80% всех съедобных личинок;
при соотношении 2:3, т. е. когда число имитаторов было чуть вы-
ше числа моделей, птицы все еще отвергали примерно те же 80%
съедобных личинок. При соотношении 1 модель на 9 имитаторов
птицы поедали большое число имитаторов, но отвергали примерно
17% съедобных личинок, принимая их за модели. По-видимому,
даже в тех случаях, когда численность имитаторов значительно
выше численности моделей, мимикрия все-таки обеспечивает сво-
им носителям некоторую защиту. Однако вряд ли какой-либо вид
может перенести истребление хищниками 83% его особей.
Обычную предупреждающую окраску называют мюллеровской
мимикрией в честь Фрица Мюллера (Fritz Miller) — немецкого
натуралиста, правильно объяснившего это явление в 1879 г. Сход-
ство животного, лишенного других защитных свойств, с неродст-
венным ему несъедобным или опасным видом называется бэйтсов-
ской мимикрией по имени английского натуралиста Генри Уолтера
Бэйтса (Henry Walter Bates), который в 1862 г. дал объяснение
мимикрии этого типа.
Число съедобных видов, подражающих какой-либо несъедобной
модели, не должно быть слишком большим; в противном случае
хищник научится ассоциировать данную окраску со съедобной
пищей и оба вида окажутся лишенными защиты. Эволюция ми-
микрии представляет собой прямой результат взаимоотношений
хищник—жертва. В настоящее время проводится подробное изу-
чение многих примеров таких взаимоотношений, лежащих на гра-
ни между эволюционными и экологическими исследованиями.
СИМБИОЗ
Не все межвидовые взаимоотношения гибельны для одного из
участвующих в них видов. Во многих случаях они выгодны обоим
видам; хорошим примером служат цветковые растения и опыляю-
щие их насекомые. Опылитель получает необходимую ему пищу
в виде нектара и (или) пыльцы, а растение благодаря посещению
насекомого оказывается опыленным. В отсутствие какого-либо
альтернативного источника пищи насекомое без растения погибло
414
Глава 15
Таблица 15.2
Типы взаимоотношений, выгодные и (или) безвредные для обоих видов
А. В пределах одного вида.
1. Спаривание.
2. Некоторые социальные взаимодействия (например, образование косяков
у рыб или стай у птиц).
3. Разделение труда между членами колонии (например, у общественных
насекомых или у вольвоксовых).
Б. Между видами.
1. Комменсализм — один вид выигрывает, а другой ничего не выигрывает,
но и не теряет (например, пауки-комменсалы, живущие в паутине другого
паука и питающиеся остатками его пищи; комары и растения сарраценип).
2. Мутуализм — выигрывают оба вида.
а. Необлигатный (например, неспециализированные цветки и их опылите-
ли; коровы и египетская цапля).
б. Облигатный (например, юкка и ее опылитель Tegeticula spp.)
3. Симбиоз — крайние случаи облигатного мутуализма, когда оба вида нахо-
дятся в теснейшей ассоциации. Классический пример — лишайники.
бы от голода; точно так же без опылителя и растение вымерло бы.
Подобные взаимоотношения известны под общим названием сим-
биоза или мутуализма. Некоторые авторы придерживаются весьма
дробной классификации мутуалистических взаимоотношений
(табл. 15.2).
Выгода, которую извлекают из симбиотических взаимоотноше-
ний участвующие в них виды, различна. Одну крайность пред-
ставляют виды, которые не могут существовать друг без друга,
как, например, растения юкки и ее облигатный опылитель — чешуе-
крылое Tegeticula (рис. 15.11). Другая крайность — опылители,
посещающие многие различные растения, цветки которых опыля-
ются многими из таких насекомых. В последнем случае взаимоот-
ношения между любыми двумя видами довольно случайны.
Интересным примером симбиоза служат взаимоотношения меж-
ду муравьями и некоторыми видами растений, среди которых наи-
более хорошо известна центрально-американская Acacia cornigera,
изучавшаяся американским экологом Даниэлем Джензеном (Da-
niel Janzen). У этой акации имеются крупные роговидные полые
колючки (рис. 15.12), на одной стороне которых имеется полоска
тонкой ткани. Мелкие жалящие муравьи рода Pseudomyrmex, от-
личающиеся очень агрессивным поведением, пробуравливают эту
тонкую ткань и используют полость колючки в качестве гнезда.
Кроме того, у основания листьев этой акации имеются нектарники,
а на концах листьев — особые узелки, называемые пищевыми тель-
цами, которые богаты белком и другими питательными вещества-
ми. Муравьи питаются как нектаром, так и этими пищевыми тель-
цами, что позволяет им совершать весь свой жизненный цикл на
акации. За это муравьи нападают на всех насекомых, которые
пытаются поедать листья акации. Кроме того, они перегрызают
побеги растений, растущих по соседству с акацией. Благодаря
Неконкурентные взаимодействия между видами
415
Рис. 15.11- Опыление цветков Yucca spp. бабочкой Tegeticula.
А. Цветки юкки, с пыльников которых насекомое собирает пыльцу. Б. Пыльник с
сидящей на нем бабочкой. В. Разрез через завязь и примыкающий к ней пыльник.
Собрав пыльцу, б1бочка откладывает в завязь яйца. Развивающиеся из них гу-
сеницы питаются .'емязачатками юкки, уничтожая таким образом несколько се-
мян. Г. После отмалывания яиц бабочка переползает на рыльце и засовывает в
него комок пыльна которую она собрала для кормления будущих гусениц; на
самом же деле oia опыляет при этом цветок. (Фотографии стеклянных моделей
цветка из Гарвар.ского университета публикуются с любезного разрешения
С. Е. Wood.)
Рис. 15.12. Колючки Acacia cornigera.
416
Глава J5
Таблица 15.3
Воздействия, оказываемые муравьями
Pseudomyrmex ferruginea на Acacia cornigera
Показатель При наличии муравьев Без муравьев
Вес корневых побегов, г 41 750 2900
Число листьев 7785 3460
Средний рост за 45 дней, см 72,86 10,23
Гибель, % 28 56
Побеги с насекомыми днем, % 2,7 38,5
ночью, % 12,9 58,8
Среднее число насекомых на одном побеге днем 0,039 0,881
НОЧЬЮ 0,226 2,707
деятельности муравьев акация получает достаточно света и избав-
лена от хищников (табл. 15.3). Таким образом, муравьи Pseudo-
myrmex борются с другими растительноядными организмами, из-
бавляя акацию от затрат на химические защитные механизмы (за
это она должна обеспечить муравьям убежище и пищу). Не на
всех, а лишь на немногих видах акации живут муравьи и не все
виды муравьев живут на акациях. Но между теми видами муравь-
ев и акаций, которые находятся в мутуалистических взаимоотно-
шениях, возникла тесная взаимозависимость, сходная с существую-
щей между юккой и бабочкой Tegeticula, так что их дальнейшее
эволюционное развитие будет тесно переплетено.
ВЗАИМОДЕЙСТВИЯ МЕЖДУ ВИДАМИ И РАЗЛИЧНЫЕ АСПЕКТЫ
ИХ ЖИЗНИ
Изучение адаптаций растений и животных к их физической и
•биологической среде тесно связано с изучением их физиологии и
функциональной морфологии—наука, которую часто называют эко-
физиологией или физиологической экологией. Специфические спо-
собы взаимодействия разных организмов со средой сказываются
на их демографических особенностях. Это было показано в ряде
независимых исследований разных аспектов жизненного цикла
золотой розги (Solidago spp.), проводившихся в северо-восточной
части США американским экологом Уорреном Абрахамсоном
(Warren Abrahamson) и некоторыми его сотрудниками.
Первое из этих исследований проводилось на 6 популяциях,
принадлежащих 4 разным видам. Две популяции росли в лесу, две
другие — на сыром лугу и еще две — на заброшенном поле, кото-
рое прежде обрабатывалось. Все три местообитания находились
Неконкурентные взаимодействия между видами
417
Очень близко друг от друга (в радиусе примерно 1,5 км), так что
между ними не было существенных макроклиматических различий.
Растения собирали чёрез одинаковые промежутки времени, отде-
Solitiago rugosa
Лесной участок
Soddago,canadensis
Влажный участок
ТОО,—-----------------------------
Santiago speciosa
1001------------------
участок
100
80
60
40
20
Л истин
Цветки
Июнь Июль Авг. Сент. Окт.
Рис. 15.13. Распределение биомассы у растений разных видов Solidago в разных
местообитаниях.
Показаны изменения доли листьев, стеблей и цветков в общей биомассе в тече-
ние вегетационного периода. (Abrachamson, Gadgil, 1973.)
ляли друг от друга стебли, листья и цветки, а затем высушивали
и взвешивали по отдельности разные части растения, устанавли-
вая таким образом различия в относительном содержании этих
частей в разных популяциях на протяжении годичного цикла рас-
27— 2044
418
Глава 15
тения. У растений с заброшенного поля наибольшая часть био-
массы приходилась на цветки, затем шли популяции сырого луга,
а у растений лесной популяции цветки содержали наименьшую
долю энергии, запасенной растением. В отличие от этого у расте-
ний с сырого луга большую часть биомассы составляли стебли, а у
лесных растений — листья (рис. 15.13). Эти различия (передавае-
мые по наследству) можно легко объяснить как реакции на среду,
в которой обитают эти популяции. Заброшенное поле — наименее
надежная среда, а поэтому (см. гл. 13) можно предсказать, что
растущие здесь растения затрачивают большую долю энергии на
производство семян. Сырой луг в смысле нарушения природной
среды занимает промежуточное положение. Он хорошо освещает-
ся солнцем, но растительность здесь очень густая и растения могут
легко оказаться затененными более высокими растениями. Конку-
рируя за свет, растения золотой розги растут вверх, чтобы избе-
жать затенения. Поэтому значительная доля энергии идет на соз-
дание тканей стебля. Лесная подстилка, напротив, освещена хуже,
чем поле или луг, так как деревья создают тень. Однако в лесной
подстилке растительность не такая густая и составляющие ее тра-
вянистые растения не так сильно конкурируют между собой за
свет. Поскольку золотая розга не может перерасти по высоте де-
ревья, с тем чтобы забрать у них солнечный свет, и поскольку
растения самой золотой розги не очень мешают друг другу, они
в лесу обычно не бывают высокими, так что основная часть энер-
гии может вкладываться в развитие листьев. Таким образом, мор-
фология и образ жизни растений золотой розги в этих трех место-
обитаниях непосредственно связаны с условиями их среды. Кроме
того, поскольку эти местообитания находятся так близко друг от
друга, что семена из одного местообитания часто заносятся ветром
в другое, отсутствие лесной формы на полях и лугах и наоборот
означает, что различия в их морфологии и образе жизни имеют
непосредственную адаптивную ценность.
В другой работе Абрахамсон (Abrahamson) и его сотрудники
повторили это исследование на одном из прежних видов {Solidago
canadensis), использовав популяции, обитающие на заброшенных
полях. На этот раз они изучали также и подземные части растений
(корни и корневища) и определяли не только вес разных тканей,
но и содержание в них калорий. В этой работе они брали выборки
трех типов: 1) растения, совершенно свободные от паразитов;
2) растения, зараженные галлообразующими растительноядными
насекомыми; 3) растения, зараженные галлобразующими насеко-
мыми, на которых в свою очередь паразитирует их естественный
враг — одно из перепончатокрылых. Растения, свободные от пара-
зитов, были крупнее, чем те, на которых паразитировали раститель-
ноядные насекомые, поскольку значительную часть энергии расте-
ний золотой розги паразиты расходовали на собственные нужды
и на образование галлов. Когда же сами галлообразователи ста-
Неконкурентные взаимодействия между видами
419
ровятся объектом паразитизма, они уже не в состоянии забирать у
золотой розги так много калорий, как прежде (рис. 15.14), и ча-
стью этих калорий завладевает перепончатокрылое (рис. 15.15).
Таким образом, это перепончатокрылое оказывает благоприятное
действие на золотую розгу и представляет собой природное био-
логическое средство борьбы с ее врагами-галлообразователями.
Абрахамсон не только убедился, что насекомые вызывали из-
менение общего количества доступной растению энергии, но и об-
Рис. 15.14. Кривые роста галлообразующих насекомых на растениях золотой роз-
ги (Solidago canadensis), свободных от галлообразователей (I) и зараженных
ими (II).
А. При заражении золотарниковой молью Gnorimoshema gallaesolidaginis. Б. При
заражении пестрокрылкой Eurosta solidaginis. На самих галлообразователях па-
разитируют другие насекомые. (Stinner, Abrahamson, 1979.)
наружил, что растение отвечает на это изменением распределения
энергии между разными своими тканями. При нападении расти-
тельноядных насекомых растения золотой розги снижают количе-
ство репродуктивных тканей (цветки и корневища) пропорцио-
нально сильнее, чем количество ткани вегетативных органов. Это
объяснили адаптивной реакцией растений, обеспечивающей выжи-
вание золотой розги, поскольку для более мелких экземпляров
(с менее развитыми стеблем и корнем и меньшим числом листьев)
вероятность выживания ниже.
В исследованиях Абрахамсона была также выявлена опреде-
ленная последовательность событий, которые происходят на за-
брошенном поле и в которых участвуют растения и насекомые. На
возделывавшееся прежде поле, оставшееся незасеянным, или же
на вырубленный лесной участок, с которого были вывезены де-
ревья, ветер заносит семена золотой розги; эти семена в конце
концов прорастают и создают популяцию. Вначале новая попули-
ст*
420
Г/aea 15
ция обычно бывает свободна от галлообразователей. Однако рано
или поздно взрослая пестрокрылка или золотарниковая моль (гал-
лообразующие насекомые, чаще всего нападающие на золотую
розгу) находят этот участок и откладывают яйца на растущие
верхушки растений. Вылупляющиеся личинки вбуравливаются в
растительные ткани, оказывая на них химическое воздействие и
заставляя растение образовывать галлы. Внутри каждого галла из
личинки развивается взрослое насекомое. С течением времени
Рис. 15.15. Поток энергии (в джоулях на 1 м2) по цепи «золотая розга — галлооб-
разователь—паразит».
Проставленные в «каналах» модели проценты указывают долю участия каждого
компонента в создании растительной продукции на 1 м2. Обратите внимание на
то, что на паразитов насекомых растение «овсе не расходует никакой энергии.
(Stinner, Abrahamson, 1979.)
популяция растительноядных распространяется в популяции зо-
лотой розги, но в конце концов популяцию галлообразователей
находят паразитические осы, которые в свою очередь откладывают
яйца в галлы, с тем чтобы их личинки могли развиваться на расти-
тельноядных насекомых. Это ведет к сокращению популяции гал-
лообразующих насекомых и соответственно к восстановлению по-
популяции золотой розги.
Приведенный пример ясно показывает сложность взаимоотно-
шений между организмом и средой и их взаимосвязь с демографи-
ческими характеристиками данного организма. Он позволяет оце-
нить пластичность реакций золотой розги на происходящие в среде
изменения. Популяции золотой розги, сложившиеся в каждой из
физически различных сред, по-разному распределяют имеющуюся
у них энергию между листьями, стеблем и репродуктивными тканя-
ми, по-видимому, для того, чтобы максимизировать свою общую
приспособленность на протяжении всей жизни. Как уже было по-
казано в гл. 13, это не всегда выражается в продукции наиболь-
шего числа семян в единицу времени. Это могут быть изменения
Неконкурентные взаимодействия между видами
421
продолжительности жизни или изменения в распределении ресур-
сов, позволяющие понизить смертность. Кроме того, в пределах
каждой популяции происходит отбор растений, способных изменять
•характер распределения энергии в особых условиях — например,
при выёдании травоядными. Подобная реакция, конечно, совер-
шенно бессознательна, однако не исключено, что она происходит
при участии гормонов, хотя способ этого участия пока неясен.
До сих пор мы рассматривали организмы с ростом открытого
типа (к ним относятся растения, многие беспозвоночные, рыбы,
рептилии и амфибии). Но большинство животных не обладает спо-
собностью изменять скорость своего роста или пропорции тела.
Однако и у этих животных также имеют место «товарообмены»,
хотя они затрагивают другие аспекты жизненного цикла. Приме-
• ром служит эволюция заботы о потомстве.
Адаптивные преимущества, создаваемые заботой о потомстве
Забота о потомстве состоит в том, что взрослые особи обеспе-
чивают своих детенышей пищей, а также затрачивают время и под-
вергают себя опасности, для того чтобы защитить их от хищников.
Взрослые особи могут непосредственно приносить детенышам со-
бранную ими пищу, как это делают многие птицы; обеспечение
пищей может происходить и косвенным образом — путем снабже-
ния яйца большим запасом желтка. Забота о потомстве требует
передачи некоторой части ресурсов, потенциально принадлежащих
родителям, потомкам, подобно тому как при цветении часть по-
тенциальных ресурсов материнского растения переходит к потом-
ству. Поскольку количество этих ресурсов (энергия, время) всегда
бывает ограничено, чем больше заботы уделяют родители своим
потомкам, тем меньше число потомков, о которых они могут за-
ботиться. Существование такой обратной зависимости было дока-
зано для многих морских беспозвоночных (морские ежи, морские
звезды, черви, моллюски, ракообразные), для рыб и для птиц. Но
вместе с тем забота о потомстве повышает вероятность его выжи-
вания. Поэтому, хотя у вида, проявляющего заботу о потомстве,
число яиц может быть меньше, число выживших потомков может
оказаться больше, чем у вида, не заботящегося о своем потомстве.
Кроме того, если родители кормят и охраняют своих детенышей,
то последним не нужно обладать при рождении всеми чертами, не-
обходимыми для самостоятельной жизни. Это делает возможным
более раннее появление на свет и более быстрый последующий
рост, как показало сравнительное изучение двух видов птиц, про-
веденное американским экологом Робертом Риклефсом (Robert
Ricklefs).
Одним из видов, изучавшихся Риклефсом, был обыкновенный
скворец (Sturnus vulgaris). У этого вида период насиживания
длится всего лишь 12 дней. Вылупившиеся птенцы лишены перьев,
28—2044
422
Глава 15
глаза у них закрыты и они совершенно беспомощны. Для того что-
бы они выжили, родители должны кормить их и ухаживать за
ними в гнезде в течение по крайней мере следующих трех недель.
Но при этом птенцы растут очень быстро, поскольку большую
часть их тела составляет пищеварительный аппарат. Другим ви-
дом был обыкновенный перепел (Coturnix coturnix). У этого вида
период насиживания яиц 18 дней; вылупившиеся птенцы покрыты
пухом, глаза у них открыты, и они вполне способны бегать и са-
мостоятельно кормиться. Родители все же заботятся о птенцах,
показывая им, какую пищу следует брать, и обогревая их в холод-
ную погоду, но заботы эти не достигают такой степени, как у
скворцов. У перепелов число яиц в кладке больше, но смертность
среди птенцов выше, так как они развиваются медленнее и дольше
находятся на таких стадиях, на которых им угрожают разного
рода опасности. К этому следует добавить, что перепелу прихо-
дится снабжать яйцо большим количеством питательного материа-
ла, чем скворцу.
Можно привести много других примеров повышения выживае-
мости растений и животных в данной среде благодаря наличию
у них различных признаков, позволяющих им забрать себе боль-
шую долю ресурсов, победить в конкуренции с другими видами,
снизить опасность, ожидаемую со стороны хищников, или, напро-
тив, самим более эффективно вылавливать жертву, или же стать
более эффективными симбионтами. Всем адаптациям присущи
некоторые общие черты: все они так или иначе повышают веро-
ятность выживания данной особи или ее потомков, все они в из-
вестных пределах увеличивают эффективность использования ре-
сурсов и все они связаны с неким «товарообменом»; что-то дела-
ется лучше, но за это всегда приходится поступиться чем-то дру-
гим. Однако одно лишь строгое описание отдельных адаптаций
само по себе не объясняет, как они возникли, и не помогает пред-
сказывать поведение видов или популяций в будущем. Для этого
необходимы теории, позволяющие делать прогнозы и поддающие-
ся проверке.
На первых страницах этой книги уже говорилось, что в настоя-
щее время описано свыше полумиллиона ныне существующих ви-
дов растений и свыше миллиона видов животных. Сколь ни велики
эти числа, они, возможно, соответствуют лишь половине действи-
тельно существующих на земле видов организмов. Мы говорили
также, что число видов, которые когда-то существовали, а ныне
вымерли, вероятно, на порядок выше. В пяти последних главах
обсуждались некоторые процессы и механизмы, участвующие в
создании и поддержании этого огромного разнообразия организ-
мов, однако недостаток места заставил нас опустить многие важ-
ные темы. Мы не касались вопроса о потоке энергии в экосистеме,
не пытались выяснить, представляет ли собой разнообразие посто-
янное или преходящее явление. Точно так же мы не рассматри-
Неконкурентные взаимодействия между видами 423
вали проблему экологической сукцессии, теории относительно оп-
тимального сбора урожая ресурсов или числа видов, способных
сосуществовать в данной области. Это всего лишь некоторые из
многих интересных и увлекательных проблем, которыми занимают-
ся сегодня популяционные биологи и экологи.
В мире, находящемся под угрозой разрушения в результате
деятельности человека, важно как можно быстрее понять механиз-
мы взаимодействия видов между собой и их реакции на создавае-
мые нами возмущения, ибо это единственный способ избежать
необратимых экологических нарушений.
ЛИТЕРАТУРА
'Общие вопросы
Cott Н. В. 1940. Adaptive Coloration in Animals, New York, Oxford University
Press.
Gilbert L. E., Raven P. H. (eds.). 1975. Coevolution of Animals and Plants, Austin,
University of Texas Press.
Mey R. M. 1973. Stability and Complexity in Model Ecosystems, Princeton, N. J.,
Princeton University Press.
Maynard Smith J. 1974. Models in Ecology, Cambridge, Cambridge University Press.
[Имеется перевод: Мэйнард Смит Дж. Модели в экологии.—М.: Мир, 1976.]
Специальные статьи и обзоры
Abrahamson W. G., Gadgil М. 1973. Growth form and reproductive effort in golden-
rods (Solidago, Compositae), Am. Nat., 107, 651—661.
Brower I. V. Z. 1958. Experimental studies of mimicry in some North American but-
terflies. Part 1. The monarch, Danaus plexippus and viceroy, Limenitis archip-
pus archippus, Evolution, 12, 32—47.
Brower L. P. 1969. Ecological chemistry, Scientific American, 220 (2), 22—29.
Brower L. P., Brower J. V. Z. 1964. Birds, butterflies and plant poisons: A study in
ecological chemistry, Zoologica, 49, 137—159.
Carpenter G. D. H. 1921. Experiments on the relative edibility of insects, with special
reference to their coloration, Trans. Ent. Soc. London.
Connell J. H. 1972. Community interactions on marine rocky intertidal shores, Ann.
Rev. Ecol. Syst., 3, 169—192.
Ehrlich P. R., Raven P. H. 1967. Butterflies and plants, Scientific American, 216
(6), 104—113.
Janzen D. H. 1966. Coevolution of mutualism between ants and acacias in Central
America, Evolution, 20, 249—275.
Oaten A., Murdoch Я7. W7. 1975. Functional response and stability in predator-prey
systems, Am. Nat., 109, 289—298.
Paine R. T. 1966. Food web complexity and species diversity, Am. Nat., 100, 65—75.
Paine R. T. 1974. Intertidal community structure: Experimental Studies on the re-
lationship between a dominant competitor and its principal predator. Oecolo-
gia, 15, 93—120.
Pitelka F. A. 1958. Some aspects of population structure in the short-term cycle of
the brown lemming in northern Alaska, Cold Spring Harbor Symp. Quant. Biol.,
22 237______251.
Ricklefs R. E. 1968. Patterns of growth in birds, Ibis, 110, 419—451.
Rosenzweig M. L., MacArthur R. H. 1963. Graphical representation and stability
conditions of predator-prey interactions, Am. Nat, 97, 209—223.
Stinner B. R., Abrahamson W. G. 1979. Energetics of the Solidago canadensis-stem
gall insect-parasitoid guild interaction, Ecology (in press).
28'
Глава 16
Поведение и эволюция
В предыдущих главах рассматривались гены и детерминиру-
емые ими морфологические признаки на популяционном уровне.
Теперь мы займемся обсуждением поведенческих признаков.
В той или иной ситуации животное может вести себя по-раз-
ному. Разумеется, не все реакции животного благоприятны для
него. Некоторые из них могут сразу погубить его, тогда как другие
понижают его приспособленность в далекой перспективе. Есть и
такие типы поведения, которые повышают вероятность выжива-
ния и размножения. В этой главе мы попытаемся ответить на два
вопроса: подвержено ли поведение естественному отбору и если да,
то каким образом под действием естественного отбора вырабаты-
ваются различные типы поведения?
Типы поведения,’ на которые способно данное животное, огра-
ничены его морфологией и физиологией в трех отношениях: по
движениям, которые оно может совершать, по информации, кото-
рую оно может получать из окружающей его среды, и по тому, в
какой мере оно может регулировать свои движения в ответ на
полученную информацию. В животном мире наблюдается широкое
разнообразие по всем этим трем факторам.
Нетрудно понять, что поведение ограничено физической фор-
мой животного: возможности амебы в смысле поведения лежат в
очень узких пределах; для того чтобы летать, животному необхо-
димы какие-то крылья; для того, чтобы жить на суше, а не вво-
де,— легкие или какие-либо другие специальные структуры; для
того, чтобы вскармливать детенышей молоком— специальные же-
лезы.
Чтобы поведение было эффективным, животное должно уметь
различать объекты окружающей среды и происходящие в ней со-
бытия. Эту информацию оно получает при помощи органов чувств.
Тип информации и ее сложность весьма разнообразны. Животные
способны распознавать множество различных внешних сигналов»
(табл. 16.1). У них имеются органы, реагирующие на температу-
ру, свет, давление, электрический ток, силу тяжести и различные
химические вещества. Информация, получаемая животным, зави-
сит от тех органов чувств, которыми оно обладает. Органы чувств
собаки сходны с нашими, но она в большей степени полагается
на обоняние. Глаз пчелы реагирует на свет, но в отличие от чело-
Поведение и эволюция
425
века пчела воспринимает ультрафиолетовые лучи и не восприни-
мает дальний красный свет. Кроме того, пчела воспринимает по-
ляризованный свет. Эти способности пчел оказывают влияние на
их поведение.
' Таблица 16.1
Типы внешних стимулов, распознаваемых животными
Тип стимула Стимул Пример животного Рецептор животного
Температурный Световой Поляризованный свет Цвет Ямкоголовая змея Пчела Птицы Лицевые ямки Сложный глаз Глаз (с колбочками в сетчатке)
Звуковой Воздушные коле- бания То же » » Колебания суб- страта Бабочки Насекомые Человек Пчела Тимпанальный орган Джонстонов орган Ухо Субгенуальиый орган
Химический Запах Феромоны Вкус (контакт ис- точника стимула с рецептором) Вкус Человек Бабочки Падальные мухи Собака Нос Рецепторы усиков Хемосенсорные ще- тинки Вкусовые почки
Механический Гравитационный Прикосновение Давление Млекопитающие Кошка Ракообразные Тельца Мейснера Тельца Пачини Статоциты
Для того чтобы животное могло выжить, оно должно быть спо-
собно использовать информацию, воспринимаемую его органами
чувств, и соответствующим образом перерабатывать ее. Регуля-
цию поведения осуществляют главным образом две системы. Пер-
вая из них — нервная система — передает информацию от органов
чувств в головной мозг и двигательным мышцам. Для осуществ-
ления сложных форм поведения животному необходима высоко-
развитая нервная система.
Вторая система регуляции — гормональная—имеет химическую
природу. Некоторые гормоны вызывают быстрые изменения пове-
дения, как, например, гормон вылупления у бабочки Cecropia,
стимулирующий подготовку к вылету имаго из кокона. Другие гор-
моны обладают более длительным действием. Например, у пере-
летных птиц увеличение длины дня ранней весной стимулирует
выделение гормонов, вызывающих физиологические изменения, ко-
торые ведут к накоплению жировой ткани, необходимой птице для
длительных перелетов. Аналогичным образом гормоны координи-
руют длинный ряд физиологических и поведенческих изменений,
служащих подготовкой к размножению. Взаимодействие между
426
Глава 16
гормонами и поведением очень сложное. В общем гормоны влия-
ют на поведение животного, и в свою очередь многие типы пове-
дения могут вызывать гормональные изменения.
АДАПТИВНОСТЬ ПОВЕДЕНИЯ
Мы уже говорили об ограничениях, налагаемых на поведение
животного строением его тела. Совершенно ясно, что морфологи-
ческие и физиологические признаки животного подвержены есте-
ственному отбору. А как обстоит дело с формами поведения? Если
поведение подвергается естественному отбору, то оно должно
удовлетворять двум критериям: оно должно быть адаптивным и
должно наследоваться. Мы сначала рассмотрим адаптированность
поведения.
Есть много способов, с помощью которых характер поведения
может повысить приспособленность организма. В одну из общих
категорий, относящихся к поддержанию существования организма,
входят формы поведения, повышающие эффективность особи и
увеличивающие продолжительность ее жизни. Различные типы по-
веденческих реакций позволяют животным использовать благо-
приятные аспекты среды и до некоторой степени защитить себя от
ее неблагоприятных воздействий. В качестве примера можно при-
вести ящерицу, которая регулирует температуру своего тела, вы-
ползая на солнце или прячась от него. Разные формы поведения
могут также обеспечить защиту от хищников. Некоторые способы
защиты от хищников состоят в том, чтобы устроить надежное
убежище (например, глубокую нору), спасаться бегством при виде
хищника или притворяться мертвым. Другая важная категория
форм поведения, связанных с поддержанием существования
животного, касается питания — как поисков пищи, так и обраще-
ния с уже добытой пищей. Некоторые из этих форм поведения
простые, как, например, хемотаксическая реакция многих простей-
ших на определенные химические вещества, а некоторые другие—
довольно сложные, например согласованные действия прайда львов
во время охоты.
Приспособленность повышается также в результате форм по-
ведения, объединяемых под общей рубрикой типов поведения,
связанных с размножением. Входящие в эту категорию формы
поведения можно подразделить на привлечение самцов и самок
друг к другу, ритуал ухаживания и заботу о потомстве.
Основная функция всех видов репродуктивного поведения со-
стоит в том, чтобы обеспечить оплодотворение яйцеклеток спер-
мой. Первый шаг к осуществлению этой функции—встреча по-
тенциальных брачных партнеров. У разных видов имеются раз-
нообразные механизмы для привлечения особей противоположного
пола друг к другу: некоторые виды насекомых выделяют спе-
циальные химические аттрактанты — феромоны, светляки произ-
427
Поведение и эволюция
водят световые вспышки, многие виды лягушек квакают. Эти сиг-
налы часто видоопецифичны (т. е. производятся только представи-
телями данного вида), и особь одного вида не реагирует на сиг-
Рис. 16.1. Ритуал ухаживания у Drosophila melanogaster.
А. Ориентация — самец приближается к самке и следует за ней. Б. «Ножницы» —
самец сначала складывает крылья на спинке параллельно одно другому, а затем
разводит их в стороны (у D. melanogaster этот тип ухаживания наблюдается ред-
ко, но у других видов дрозофилы он встречается часто). В. Трепетание — одно
крыло располагается перпендикулярно к телу и быстро вибрирует. Г. Вылизыва-
ние — самец вылизывает брюшко самки. Д. Попытка к копуляции. Е. Копулирую-
щая пара. (Manning, 1965.)
нал, подаваемый особью, принадлежащей к другому виду. Напри-
мер, два близкородственных вида квакш, относящихся к комплек-
су видов Hyla versicolor, морфологически очень сходны, но разли-
чаются по брачным крикам самцов: самцы одной группы выводят
быструю трель, а самцы другой — более медленную. Эти крики
428
Глава 16
можно записать, а затем воспроизвести при помощи двух громко-
говорителей. Если самку одного из этих видов поместить на рав-
ных расстояниях от обоих громкоговорителей, то она направится
либо к одному, либо к другому из них. Если затем повторить экс-
перимент несколько раз, то самка всякий раз направляется на
один и тот же сигнал. Таким образом, самки способны различать
разные сигналы.
Поведение животных с момента встречи самца и самки и до
выделения гамет называют ухаживанием. Простейшая форма уха-
живания состоит в узнавании представителя противоположного
пола (того же самого вида) и одновременном выделении гамет.
Подобного рода поведение характерно для многих беспозвоноч-
ных. У многих видов наблюдаются более сложные церемонии уха-
живания, а у некоторых животных они очень сложны, так что
каждая особь данного вида (или, во всяком случае, одного из по-
лов этого вида) выполняет чрезвычайно замысловатые действия.
Примеры таких сложных церемоний можно видеть у многих птиц.
У одних видов в ухаживании участвуют оба пола, реагирующие
друг на друга, а у других весь ритуал совершается представите-
лем одного из полов. Например, у плодовой мушки Drosophila
melanogaster ритуал ухаживания исполняет главным образом
самец (рис. 16.1). Самец начинает с того, что приближается к
самке, поворачивается к ней головой и следует за ней, когда она
удаляется. Затем он ставит одно крыло перпендикулярно к своему
телу и начинает вибрировать им. После этого самец заходит сза-
ди и пытается взобраться на самку. Несколько первых попыток
к спариванию самка обычно отвергает, но самец продолжает свои
действия до тех пор, пока не достигает успеха или же отказыва-
ется вовсе от своих намерений.
Довольно обычное явление при ухаживании (так же как и
при других формах поведения) представляет собой ритуализация.
Ритуализация состоит в том, что в процессе эволюции первона-
чальная функция какой-либо формы поведения модифицируется и
становится коммуникационной. Ритуализация агрессивного пове-
дения обычно дает возможность получать информацию о вероят-
ности того, вступит ли противник в бой или обратится в бегство.
При ухаживании ритуализованные формы поведения сигнализиру-
ют о готовности к спариванию; часто они указывают на пол осо-
би; и поскольку формы поведения обычно видоспецифичны, они
дают информацию о том, принадлежат ли две участвующие в це-
ремонии ухаживания особи к одному и тому же виду. В некото-
рых случаях удается проследить за эволюцией той или иной фор-
мы поведения путем сравнительного изучения близкородственных
видов. Сигналами могут служить очень многие формы поведения,
такие, как приливы крови к определенным участкам тела, корм-
ление, дефекация и т. п. В процессе ритуализации первоначальное
поведение обычно упрощается и одновременно усиливается, пре-
Поведение и эволюция
429
вращаясь тем самым в более эффективный сигнал. Иногда это со-
провождается морфологическими изменениями, делающими сигнал
еще более заметным. Пример ритуализованного поведения при
Рис. 16.2. Десять поз, наблюдаемых у кряквы и у других уток, которые добывают
корм на поверхности воды.
А — начальное покачивание клюва: 5 — вскидывание головы; В — помахивание
хвостом; Г — свистящее бормотание; Д—«голова вверх—хвост вверх»; Е—по-
ворот в сторону самки; Ж—поклоны при плавании; 3 — поворот головы затыл-
ком к самке; И — выпячивание груди; К— «вверх—вниз». (Lorenz, 1958.)
ухаживании можно наблюдать у настоящих уток, таких, как кряк-
вы. На рис. 16.2 изображены 10 поз, наблюдаемых при ухажива-
нии у разных видов этих уток. У кряквы яркое оперение самца
особенно хорошо заметно при позе «голова вверх—хвост вверх»
(рис. 16.2 Д). Такие же движения можно наблюдать у родствен-
ных видов с другой расцветкой. Последовательность этих поз у
птиц, принадлежащих к разным видам, может быть различной.
430
Глава 16
Церемония ухаживания выполняет несколько функций. Она
обеспечивает одновременное половое возбуждение у самца и сам-
ки, так что оба они приходят в состояние готовности к спарива-
нию в одно и то же время. Она подавляет возможное агрессивное
поведение между брачными партнерами и дает возможность осо-
бям, принадлежащим к одному и тому же виду, узнавать друг
друга. Это очень важно, особенно в областях, где обитают два
близкородственных вида. Если бы особи, относящиеся к разным
видам, предприняли попытку к спариванию, то, вернее всего, они
не смогли бы произвести потомство, а если бы и смогли, то их
потомки скорее всего оказались бы стерильными или нежизнеспо-
собными. Следовательно, отбор должен благоприятствовать тем
особям, которые не спариваются с особями, принадлежащими к
другим видам.
Что же касается заботы о потомстве, то, согласно современным
теориям, ее эволюция может вызываться рядом факторов внешней
среды. Уилсон (Е. О. Wilson) приводит четыре таких фактора:
1) высокая активность хищников, поскольку при этом, защищая
своих детенышей, родительская особь повышает свою приспособ-
ленность; 2) специализация к труднодобываемой пище, поскольку
при этом детеныши не в состоянии сами прокормить себя до тех
пор, пока они не достигнут определенных размеров или не приоб-
ретут достаточных навыков. Волкам и крупным кошкам, которые
охотятся на крупную добычу, для успешной охоты необходимо
достигнуть определенных пороговых размеров и научиться под-
крадываться к добыче; эти животные заботятся о своих детены-
шах и обучают их до тех пор, пока они не становятся способными
сами обеспечивать свое существование; 3) если вид обитает в
среде с необычайно суровыми физическими условиями, то молодь
может оказаться неспособной выжить без помощи родителей; 4) у
видов, адаптированных к предсказуемым условиям среды, которые
ведут к /(-отбору (см. гл. 13), обычно также сильнее развита за-
бота о потомстве, /(-отбор благоприятствует большим размерам,
большей продолжительности жизни и многократому размножению
в течение жизни. Эти условия благоприятствуют пометам, состоя-
щим из относительно небольшого числа потомков, вероятность вы-
живания которых повышается благодаря заботе о них со стороны
родителей.
НАСЛЕДУЕМОСТЬ ПОВЕДЕНИЯ
Мы рассмотрели адаптивную ценность различных форм поведе-
ния, в том числе поведения, обеспечивающего поддержание жизни
особи, а также репродуктивного поведения. Однако, для того что-
бы естественный отбор мог действовать на поведение, поведенчес-
кие признаки должны не только обладать адаптивной ценностью,
но и передаваться от родителей потомкам.
Поведение и эволюция
431
До какой степени возможна генетическая передача поведения?
Приобретенные признаки не могут наследоваться, а поэтому и не
подвергаются очбору, однако возможно, что существует отбор на
генетические признаки, которые облегчают определенные формы
'поведения или предрасполагают к ним животное. Некоторые из
имеющихся данных свидетельствуют о том, что поведение нахо-
дится под генетическим контролем. Определенные типы поведения
появляются внезапно, до того как выполняющая их особь имела
случай наблюдать такое поведение у других особей, принадлежа-
щих к тому же виду; к таким формам относится поведение при
вылуплении у гтиц. Поведение при вылуплении, наблюдаемое у
большинства птиц, начинается с того, что птенец принимает оп-
ределенную позу, засовывая голову под правое крыло и направляя
клюв к воздушной камере, расположенной на тупом конце яйца.
Затем птенец прорывает клювом оболочку, отделяющую воздуш-
ную камеру, и начинает дышать легкими. После этого птенец над-
калывает скорлупу изнутри, образуя первую щель в яйце. За этим
следует ряд движений, с помощью которых птенец с силой ударя-
ет клювом о скорлупу, а одновременные движения ног вызывают
поворот тела птенца. Все это приводит к постепенному расшире-
нию щели в скорлупе. Таким образом, вылупление связано с ря-
дом довольно сложных стереотипных движений, которым, однако,
птенец не мог научиться.
Другие данные, указывающие на то, что поведение может пе-
редаваться по наследству, получены в результате изучения межви-
довых гибридов, в частности у птиц. Если формы поведения роди-
тельских видов идентичны, то аналогичное поведение наблюдается
у гибрида. Если поведение родительских видов различается коли-
чественно, то поведение гибридов обычно бывает промежуточным.
Если же в поведении родителей наблюдаются качественные раз-
личия, то поведение гибридов может сильно варьировать. Иногда
оно бывает промежуточным (приближаясь по мере накопления
животным опыта к поведению одного из родителей). В других слу-
чаях черты поведения одного из родительских видов у потомков
бывают подавлены или же в их поведении появляются новые чер-
ты. Так, Лоренд (Lorenz), скрещивая чирка и багамскую шило-
хвость (Anas bahamensis), получил интересный гибрид: ни тот, ни
другой из родительских видов во время ухаживания не принима-
ет позу «голова вверх—хвост вверх», характерную для некоторых
родственных им видов уток (рис. 16.2), но у гибрида эта поза на-
блюдается. Судя по результатам этих экспериментов, в поведении
имеется генетический компонент.
Для поведения многих видов характерны определенные стерео-
типы, одинаковые у всех представителей данного вида (или по
крайней мере у всех представителей одного из полов) и повторя-
ющиеся данной особью всякий раз одинаково. Некоторые дейст-
вия совершаются животными только в совершенно определенных
432
Глава IS
ситуациях. Многие из таких поведенческих стереотипов видоспе-
цифичны. Это, однако, не обязательно означает, что подобное по-
ведение наследуется. Оно может быть также обусловлено сходным
опытом, приобретенным всеми членами данного вида в сходных
условиях.
Установлено также, что некоторые различия в поведении конт-
ролируются единичными генами. Одним примером служит мутант-
ный аллель «yellow» у плодовой мушки, обусловливающий у го-
мозиготных по этому гену самцов желтую окраску тела и более
медленную вибрацию крыла при ухаживании. Успешность спари-
вания этих самцов существенно ниже, чем у самцов с нормальной
окраской, потому что самки чаще отвергают их.
Все эти данные подтверждают представление о том, что гены
оказывают сильное влияние на поведение. Но на поведение вли-
яют не только гены. Животные изменяют свое поведение на ос-
нове опыта, а иногда приобретенную таким образом форму по-
ведения родители передают потомкам путем обучения (см. гл. 10).
Особи могут научиться соответствующему той или иной ситуации
поведению от своих родителей или от других представителей дан-
ного вида. Луговые собачки узнают о границах своей колонии
от более старых ее членов.
Песни, по-видимому, наследуются у разных видов птиц в раз-
ной степени. Некоторые виды, как, например, канарейки, достиг-
нув половой зрелости, начинают петь, даже если их выращивали
изолированно от других особей их вида. У других видов, например
у щеглов, при выращивании в изоляции песня несколько упроща-
ется. Вариациям основного трафарета песни и дополнениям, в том
числе характерной концовке, отсутствующей в пении большинства
выращенных в неволе птиц, щеглы, по-видимому, научаются в ес-
тественных условиях от других щеглов.
Было много споров о том, какие типы поведения передаются
генетически и какие приобретаются в результате обучения. В на-
стоящее время все признают, что граница, проводившаяся между
наследственным и благоприобретенным, была излишне строгой.
Гены и среда сильно взаимодействуют, и таких форм поведения,
которые можно было бы отнести исключительно к одной из этих
категорий, очень немного. Более уместно задаться вопросом, в ка-
кой степени и каким образом генетическая природа организма и
среда его обитания участвуют в создании наблюдаемой формы
поведения.
ОБЩЕСТВЕННОЕ ПОВЕДЕНИЕ
Мы обсуждали поведение отдельных животных с точки зрения
их адаптации к физическим условиям среды и к воспроизведению.
Перейдем теперь к вопросу об их взаимодействиях с другими осо-
бями. Большинство животных проводят часть своего жизненного
цикла или весь цикл в контакте с другими представителями того
Поведение и эволюция
433
же вида. У некоторых видов этот контакт ограничивается брач-
ным периодом; другие же проводят всю свою жизнь в качестве
членов какой-либо группы.
Общественный образ жизни характерен для многих таксонов
животного царства и притом в чрезвычайно разнообразных фор-
мах. Общественные группы различаются по размерам, то степени
взаимосвязанности их членов, соотношению разных демографичес-
ких классов и распределению функций внутри группы. Каковы те
факторы, которые приводят к возникновению разного рода груп-
пировок?
Формы общественной структуры, возможной для данной попу-
ляции, ограничиваются ее историей, определившей ее нынешний
генетический состав. Скорость эволюции этой структуры и ее
формы не могут выходить за пределы, создаваемые степенью гене-
тической пластичности популяции. Эти пределы, однако, могут
•быть достаточно широкими, особенно если рассматривать их на
протяжении длительного периода времени. В диапазоне генети-
ческих возможностей данного вида уровень его общественной ор-
ганизации и формы сообщества определяются рядом факторов
среды, создающих главные давления отбора. Ниже приведен в
несколько измененной форме перечень преимуществ, которые да-
ет общественный образ жизни, составленный социобиологом
Уилсоном (Е. О. Wilson).
1. Защита от хищников. Когда животные образуют скопления,
хищнику труднее приблизиться к ним, оставаясь незамеченным.
Поэтому особь, входящая в состав группы, имеет больше шансов
спастись от хищника, чем одиночная особь. Отсюда следует так-
же, что, будучи членом сообщества, данная особь может тратить
меньше времени на проявление бдительности по отношению к
хищникам, уделяя его другим видам активности, например добы-
ванию пищи. Хищники, со своей стороны, могут счесть нападение
на группу особей более опасным, чем нападение на одну особь.
У многих птиц стая «окрикивает» нападающего хищника. «Окри-
кивание» наблюдается также у некоторых общественных млекопи-
тающих, таких, как павианы.
2. Повышение способности к конкуренции. Группа обществен-
ных животных уже одной своей величиной может подавить потен-
циальных конкурентов, принадлежащих к другим видам. Способ-
ность к конкуренции повышается также в результате сотрудниче-
ства. Например, бродячие муравьи, которые фуражируют больши-
ми группами, набрасываются на свою жертву всем скопом, а
поэтому добывают пищу более эффективно, чем те муравьи, кото-
рые фуражируют поодиночке. Кроме того, бродячие муравьи мо-
гут обычно рассчитывать на более крупную добычу. (Этот пример
подходит также под следующую рубрику.)
3. Повышение эффективности добывания пищи. Общественный
образ жизни позволяет также животным добывать пищу более
434
Глава 16
Рис. 16.3. Регуляция температуры и вентиляция в гнезде термитов (Macroterm.es).
Термитам необходимы тепло и высокая влажность. Гнезда Macrotermes могут до-
стигать почти 5 м в высоту и такого же диаметра. Центральная камера опирает-
ся иа столбы, поднимающиеся из подземной части гнезда (е), и содержит ячейки
для вскармливания личинок (б), окруженные «грибными садами» (а). Теплый
воздух из центральной камеры поднимается вверх под самую крышу гнезда (в).
Затем он проходит по ряду каналов, расположенных в имеющихся на поверхно-
сти гнезда гребиях (г и д). Здесь СО2 выделяется в атмосферу, и воздух в гнезде
обогащается кислородом, (ж) —камера матки. Верхняя цифра у каждой буквы —
температура (°C), а нижняя — содержание СО2 (%). Влажность равна 98—99%.
(Luscher, 1961.)
эффективно благодаря тому, что он дает им возможность наблю-
дать за соответствующими формами поведения у других особей
и использовать их опыт. Кроме того, некоторые животные кормят-
ся более эффективно, охотясь группой, а не поодиночке. Так, стая
насекомоядных птиц поднимает в воздух больше насекомых, чем
Поведение и эволюция
435
такое же число птиц, летящих поодиночке, в результате чего
каждая особь иьеет больше шансов поймать добычу.
Характер распределения пищи представляет собой теоретичес-
ки один из важшх факторов, определяющих тип организации со-
общества. Если пища распределена в среде довольно равномерно,
то животные затимают каждый свою территорию и охраняют ее
от других особе! того же вида (при условии что эта территория
достаточно богага пищей, а следовательно, охрана ее имеет
смысл). Если тща сосредоточена на отдельных участках и до-
ступна либо все время, либо через определенные промежутки, то
животные поселяются как можно ближе к ней и собирают ее
каждый для себя. Если же пища распределена в среде неравно-
мерно и встречается совершенно нерегулярно, то наибольшая эф-
фективность добывания пищи достигается при групповом образе
жизни.
4. Повышение эффективности размножения. Для многих групп
насекомых характерно образование роев, состоящих из особей,
которые проделывают довольно далекий путь для того, чтобы при-
нять участие в церемониях ухаживания и в спаривании. По всей
вероятности, рошие имеет особенно важное значение для редких
видов, членам юторых иначе трудно было бы найти себе брачного
партнера. Друпе возможные функции роения состоят в умень-
шении инбридинга и предотвращении гибридизации с другими
видами (докопуляционные изолирующие механизмы).
5. Изменена? среды. Объединившись в группы, животные мо-
гут изменять среду в благоприятную для них сторону. Простей-
ший пример этого дают грызуны, которые зимой сбиваются в ку-
чу, чтобы согрегься. А наиболее сложные изменения среды дости-
гаются в гнездах таких высокоорганизованных общественных на-
секомых, как пчелы и термиты, в которых температура и влаж-
ность чрезвычайно строго регулируются (рис. 16.3).
ОРГАНИЗАЦИЯ СООБЩЕСТВ
Рассмотрим теперь некоторые специфические черты органи-
зации сообществ и, в тех случаях, когда это возможно, проана-
лизируем порождающие их экологические и эволюционные фак-
торы и их адаптивное значение. Мы изучим здесь такие явления,
как территориальность, иерархическая организация и системы
скрещивания.
Территория
Уилсон (Е. О. Wilson) определяет территорию данного живот-
ного или группы животных, как «область, занимаемую более или
менее исключительно этим животным или группой животных, ко-
торые не допускают на нее посторонних особей, либо непосредст-
венно защищая ее, либо предупреждая о том, что она занята».
436
Глава 16
Концепция территории впервые возникла в результате иссле-
дований, проводившихся на птицах, и именно в этой группе мож-
но наблюдать наиболее четко выраженные классические приме-
ры. У многих видов птиц, например у зарянки, самец весной за-
нимает гнездовой участок (или территорию). Он заявляет о сво-
ем присутствии пением, которое предупреждает других самцов,
что данная территория занята, и одновременно служит для при-
влечения самки. Самец охраняет свой участок и после того, как
образуется пара, продолжая делать это в течение всего периода
размножения. У некоторых видов самка также принимает участие
в защите территории.
Охрана территории сопряжена со стычками с вторгающимися
в нее чужими особями. Иногда при этом возникают настоящие
драки; но чаще дело ограничивается проявлениями агрессивного
поведения со стороны владельца территории и чужака. Обычно
хозяин выгоняет пришельца, причем вероятность его победы тем
выше, чем ближе он находится к центру своей территории.
Размеры территории зависят от наличных ресурсов и от плот-
ности популяции. Площадь территории больше, когда лимитирую-
щего ресурса мало, и меньше — когда его достаточно. Когда чис-
ло представителей данного вида в данной местности невелико,то
их территории, вероятно, несколько больше, чем когда их числен-
ность велика. Однако существуют некоторые минимальные разме-
ры территории, зависящие от ресурсов, и если число особей слиш-
ком велико, так что не все они могут найти себе участки в хоро-
ших местах, некоторые особи занимают территории в краевых
областях, где успешное размножение менее вероятно; другие же
образуют не участвующую в размножении бродячую популя-
цию, члены которой вовсе не имеют территорий. Такие птицы
могут замещать владельцев территорий в случае гибели послед-
них.
«Территория» несет весьма разнообразные функции (табл.
16.2). Охраняемая область — это обычно место для гнездования,
как у многих птиц, которые гнездятся группами, причем каждая
охраняет свое собственное гнездо; но эта область может быть
также местом кормежки или одновременно и тем и другим. Или
же она может быть токовищем — местом, где собираются самцы
для брачных демонстраций и игр; каждый самец занимает на
токовище определенный участок, а самки прохаживаются рядом и
выбирают себе партнера. Особь может охранять некую ограничен-
ную территорию, совершая регулярные обходы, чтобы уберечь ее
от вторжений, или же охранять лишь тот участок, на котором она
находится в данный момент (в тех случаях, когда вся область
слишком велика, чтобы можно было обеспечить ее охрану). За-
щитниками территории могут быть отдельные особи или группы
особей, самцы (чаще) или самки. Некоторые животные ведут тер-
риториальный образ жизни на одной стадии своего жизненного
Поведение и эволюция
437
Таблица 16.2
Функции территории
Охраняемый ресурс Примеры
Пища Макаки-резусы, американ- ские певчие воробьи
Пространство для брач- ных церемоний Гнездовые участки Убежище Лягушка-бык, луговой тете- рев Игуаны Лангусты
цикла или в какое-то одно время года и одиночный или колони-
альный образ жизни в другие периоды.
Преимущество, которое дает территориальность отдельной осо-
би, состоит, по-видимому, в том, что она гарантирует доступ к
желаемому ресурсу, будь то место для гнездования, убежище,
пища, брачный партнер или любое сочетание этих факторов. Один
из возможных недостатков территориальности заключается в том,
что при этом возрастает опасность нападения со стороны хищни-
ков. Но главный ее недостаток — это расход энергии на охрану
территории. Если животное затрачивает время и энергию на то,
чтобы обходить свою территорию и прогонять чужих особей, то
тем самым оно не может направить эту энергию на выполнение
функций, более непосредственно повышающих приспособленность,
т. е. на поиски пищи или заботу о потомстве. Учитывая такую
утечку энергии, территориальному животному выгоднее узнавать
своих соседей и не тратить времени на конфликты с ними. Многие
птицы и в самом деле узнают своих соседей по их пению и не об-
ращают на них внимания, но агрессивно реагируют на появление
чужаков.
Таким образом, следует ожидать, что территориальность будет
наблюдаться в тех местах, где ресурсы имеются на протяжении
достаточно длительного периода, чтобы животному имело смысл
оставаться на одном месте. Другое необходимое условие— доволь-
но равномерное распределение ресурсов. При пятнистом распре-
делении ресурсов другие животные также будут стараться посе-
литься вблизи скопления ресурсов и территориальные конфликты
станут слишком частыми — такими частыми, что будут занимать
слишком большую долю времени животного. В таких условиях
более выгодна другая стратегия, когда каждая особь собирает
ресурсы для удовлетворения собственных потребностей.
Иерархическая структура
Территориальное поведение представляет собой взаимодействие
двух особей (или двух групп особей), тогда как иерархическая
структура отражает взаимоотношения в пределах одной группы.
29—2044
438
Глава 16
Иерархическая структура проявляется в комплексе постоянно под-
держиваемых отношений доминирования и подчинения в группе
животных, обитающих совместно на данной территории. Сущест-
вуют разные формы иерархической структуры (рис. 16.4). В од-
них случаях какое-либо
,О
'/I
4
4
О
Различные струк-
Рис. 16.4.
туры доминирования в со-
обществе.
Стрелки направлены в сто-
рону доминирующей особи.
Взаимоотношения в преде-
лах сообщества могут стро-
иться по одной из этих схем
или по их различным соче-
таниям.
одно животное доминирует над всеми
остальными, между которыми нет раз-
личий по рангу. Иерархия может быть
линейной: при этом одна особь домини-
рует над всеми остальными, следующая
подчиняется первой, но доминирует над
прочими, и т. д. Возможны и другие бо-
лее сложные системы соподчинения, со-
держащие кольцевые элементы.
Иерархия устанавливается в резуль-
тате агрессивных действий, иногда до-
ходящих до настоящей драки. Какие
именно качества позволяют особи за-
нять доминирующее положение — не-
известно. К числу некоторых таких ка-
честв относятся: пол (самцы обычно
доминируют над самками); возраст
(взрослые особи доминируют над моло-
дыми; возраст, по-видимому, служит од-
ним из решающих факторов при уста-
новлении соподчинения среди половоз-
релых самцов у разных видов копыт-
ных, птиц и рыб); размеры; степень аг-
рессивности; материнство (у некоторых
обезьян); история поражений и побед
в прошлом.
Для начального периода формирования той или иной группы
животных характерны значительные проявления агрессии, сопро-
вождающие установление иерархической структуры. Но после то-
го как иерархия установлена, наступает относительное спокойст-
вие. Животные мирно занимаются своими делами, если не счи-
тать возникающих время от времени мелких стычек из-за пищи
или чего-нибудь в этом роде. Изменение ранга отдельных особей
возможно, но после того как иерархия уже установилась, такие
изменения сопряжены с гораздо большими трудностями, чем вна-
чале. Во многих случаях при удалении особи, доминирующей «ад
всеми остальными, проявления агрессивности в сообществе уси-
ливаются, поскольку между особями, занимавшими ранее одина-
ковое подчиненное положение, начинается борьба за первое место.
Селективное преимущество, которое дает доминирующее поло-
жение, очевидно: животное, занимающее такое положение, имеет
первоочередной доступ к пище, местам для устройства гнезда и
брачным партнерам. Кроме того, особь, занимающая «место на-
Поведение и эволюция
439
верху», меньше подвержена стрессовым воздействиям, а следова-
тельно, и эндокринным нарушениям.
Преимущества иерархической системы для особей, занимаю-
щих подчиненное положение, не столь очевидны. В некоторых
случаях, в частности у общественных насекомых, возможно, име-
ет место отбор родичей (см. гл. 10). Если шансы данного живот-
ного на то, чтобы выжить и вырастить собственных потомков, ма-
лы, то оно может повысить приспособленность, помогая вместо
этого своим родичам. Например, у Polistes группы фертильных
самок (многие из них — сестры) совместно основывают гнездо.
Одна из этих самок занимает доминирующее положение и откла-
дывает яйца, тогда как другие выполняют функции рабочих осо-
бей. Было высказано предположение, что, поскольку далеко не
все новые гнезда обеспечивают успешное размножение и по-
скольку у подчиненных особей в общем яичники развиты слабее,
чем у доминирующей особи, эти подчиненные особи, по всей веро-
ятности, окажутся неспособными вырастить собственное потомст-
во. Поэтому для них, может быть, более целесообразно помогать
одной из своих сестер.
В большинстве сообществ животные, находящиеся на низшей
ступени иерархии, имеют возможность эмигрировать. Эта возмож-
ность дает им шансы выжить, особенно в условиях перенаселен-
ности. В случае недостатка какого-либо ресурса вероятность вы-
живания особи низшего ранга вне данного сообщества может при-
мерно соответствовать вероятности ее выживания в сообществе.
У видов, живущих во временных местообитаниях, эмиграция —
частое явление. Примером служит полевка Microtus pensilvanicus.
Она часто живет на сырых лужках, вроде тех, которые образуют-
ся после того, как бобры покидают свою плотину, и которые бы-
стро зарастают лесными деревьями. (Эта полевка в настоящее
время часто встречается также на обрабатываемых землях).Чис-
ленность популяции полевки колеблется в зависимости от време-
ни года. Чем выше численность, тем сильнее давление, испытыва-
емое подчиненными особями и вынуждающее их эмигрировать. Их
изгоняют из сообщества доминантные особи в процессе прямых
конфликтов. Шансы на выживание у эмигрантов невелики, но
некоторым из них удается найти новые местообитания с благо-
приятными условиями среды и занять в них доминирующее поло-
жение. Кроме того, как считает Дж. Кристиан (J. J. Christian),
подчиненные животные, вынужденные расселяться, представляют
собой важный фактор в эволюции млекопитающих. Многие из
таких животных эмигрируют в области, менее соответствующие
потребностям их вида. При этом значительная часть их гибнет,
но среди них может оказаться несколько особей с мутациями,
дающими им преимущество в этих новых местообитаниях. При
подходящих условиях такие особи могут давать начало новым
видам.
29*
440
Глава It
У некоторых других видов эмиграция наблюдается редко. Опа-
сности, подстерегающие особь за пределами группы, так велики,
что перспектива остаться в своем сообществе, пусть даже в под-
чиненном положении, оказывается для нее достаточно привлека-
тельной. Примером служат павианы. В их сообществах особи низ-
шего ранга хорошо питаются и время от времени спариваются,
тогда как одинокие особи обычно живут недолго. Кроме того, не
исключена возможность изменения их положения в иерархии в
случае гибели доминантных животных. Во многих системах ранг
особи зависит, по крайней мере частично, от ее возраста; поэтому
чем дольше живет особь, тем больше у нее шансов достигнуть
высокого ранга.
Иерархические системы сходны с территориальными системами
в том отношении, что в основе как тех, так и других лежат аг-
рессивные взаимодействия. Иерархическое сообщество может од-
новременно быть территориальным. В некоторых группах наблю-
дается смешанная иерархически-территориальная структура. Для
некоторых ящериц характерна территориальность, но вместе с
тем среди них имеются доминирующие особи, которым дозволя-
ется нарушать границы территории. У некоторых кошек сущест-
вует иерархическая структура, но особи низшего ранга защи-
щают свои логовища от особей более высокого ранга. Многие ви-
ды, для которых в нормальных условиях характерна территори-
альность, при перенаселенности переходят к иерархической струк-
туре.
ПОЛОВОЙ ОТБОР И СИСТЕМЫ СКРЕЩИВАНИЯ
Ёстественный отбор теоретически можно рассматривать в двух
аспектах: отбор на выживание, максимизирующий способность
организма к конкуренции, а следовательно, продолжительность
его жизни (1Х популяции), и отбор на размножение, действующий
на другую компоненту приспособленности — на успех размноже-
ния (тх популяции). Отбор на размножение, или репродуктивный
отбор, действует на все признаки, имеющие отношение к разм-
ножению,— от выбора брачного партнера до выращивания потом-
ства; однако наибольшее внимание исследователей привлекала
дифференциальная способность особей различных генетических
типов к нахождению брачных партнеров — тот аспект отбора, ко-
торый Дарвин назвал половым отбором (гл. 10).
Различают два типа полового отбора: эпигамный отбор, свя-
занный с выбором, производимым самцами между самками и на-
оборот, и интрасексуалъный отбор, обусловленный конкуренцией
между представителями одного и того же пола, обычно между
самцами. Дак полагал Дарвин, оба эти типа отбора, по-видимому,
обусловили эволюцию вторичных половых признаков (т. е. тех
различий между особями разного пола, которые не относятся к
органам, непосредственно участвующим в размножении). Элитам-
Поведение и эволюция
441
ный отбор мог привести к развитию разного рода украшений, за-
пахов, демонстративного поведения и т. п., которые привлекают
особей противоположного пола. Интрасексуальный отбор, веро-
ятно, привел к развитию органов нападения и защиты, агрессив-
ности и т. п., необходимых особи, которая конкурирует с 'другими
особями своего же пола. В природе существуют вторичные поло-
вые признаки как того, так и другого типа. В некоторых случаях
такие признаки (например, пышное оперение райской птицы илн
внушительные рога некоторых оленей) достигают чрезмерного
развития и могут оказаться гибельными для их обладателей. Так,
хотя более броское, чем у других самцов, оперение повышает
привлекательность данного самца для самок, оно вместе с тем
делает его более заметным для хищников. Согласно современной
теории, действие отбора направлено на то, чтобы максимизиро-
вать приспособленность с учетом как полового отбора, так и от-
бора на выживание. Половой отбор может вести к развитию чрез-
мерно выраженных признаков лишь до тех пор, пока они повыша-
ют общую приспособленность.
Развитие полового диморфизма в результате полового отбора
свидетельствует о том, что отбор может действовать по-разному
на самцов и самок. Какой-либо признак или какая-либо стратегия
необязательно должны быть одинаково выгодны для обоих полов.
Это потенциальное различие в значительной мере обусловливает
наблюдаемое в природе разнообразие систем скрещивания.
Назовем родительским вкладом любое поведение по отноше-
нию к потомку, которое повышает шансы данного потомка на вы-
живание за счет способности данной родительской особи обеспе-
чивать выживание другого потомка. Самки обычно продуцируют
гаметы, содержащие значительный запас энергии, тогда как га-
меты, продуцируемые самцами, содержат генетические инструк-
ции, но очень небольшой запас энергии. .Поэтому число яйцекле-
ток ограничено по сравнению с числом спермиев, а родительский
вклад самки в одну гамету больше, чем вклад самца. Самец мо-
жет увеличить число своих потомков путем многократных спари-
ваний; все яйца самки обычно оплодотворяются в данном сезоне
размножения в результате одного акта спаривания. Поэтому ошиб-
ки в выборе брачного партнера для самки гораздо серьезнее, чем
для самца. Спаривание самки с представителем другого вида или
с нежелательной особью собственного вида может означать, что
в данном сезоне она не произведет жизнеспособного потомства;
для самца же это лишь означает потерю некоторого времени и
небольшого количества спермы. Поэтому возможность распозна-
вания при выборе брачного партнера имеет очень большое зна-
чение для самки, тогда как для самца она менее важна.
Гордон Ориенс (Gordon Orians) разработал теорию систем
скрещивания, основанную на том, что выбор брачного партнера,
•осуществляемый самкой, максимизирует индивидуальную приспо-
442
Глава 16
собленность. Он считает, что самка руководствуется при выборе
двумя критериями.
1. Генетическое качество самца. Чем выше приспособленность
самца, тем выше приспособленность потомков самки. Поэтому сам-
ка должна реагировать на те признаки самца, которые отражают
его приспособленность.
2. Вероятность помощи со стороны самца в размножении и за-
боте о потомстве. У некоторых видов самец обеспечивает охрану
территории с имеющимися на ней ресурсами, корм для молоди,
насиживание яиц и т. п. У территориальных видов выбор самки
может зависеть от качества территории.
В тех случаях, когда самка основывает свой выбор на гене-
тическом качестве партнера, следует ожидать полигинии (один
самец и несколько самок), поскольку при этом для самки не име-
ет значения то обстоятельство, что самец, возможно, ранее спари-
вался с другими самками. В таких случаях часто наблюдается
резко выраженный половой диморфизм и сильное развитие у сам-
ца признаков, способствующих привлечению самки и полезных в
конкуренции с другими самцами.
В тех же случаях, когда вклад самца не ограничивается его
генами, часто наблюдается моногамия (один самец и одна сам-
ка). При этом самка, по всей вероятности, предпочтет избрать
партнера несколько более низкого качества, но зато еще не уча-
ствовавшего в спаривании, а не самца, который уже спаривался
с другой самкой и на помощь которого она поэтому может рас-
считывать лишь частично. Полигиния может иметь место в тех
случаях, когда приспособленность самки выше, если она делит
самца с другими самками на высококачественной территории, чем
если она принимает ухаживания не участвовавшего в спаривании
самца на территории более низкого качества.
При каких же условиях следует ожидать, что самец будет по-
могать самке в выращивании потомства? Иногда условия среды
бывают настолько суровыми, что самка просто не может одна
вырастить потомство. В таких случаях приспособленность самца
будет выше, если он помогает добывать пищу, убежище, обеспе-
чить защиту от хищников и т. п., вместо того чтобы стараться
произвести на свет больше потомков. Например, может оказаться
необходимым, чтобы гнездовые участки или источники пищи, ко-
личество которых недостаточно, охраняли оба родителя. Другой
фактор, способствующий моногамии, имеет значение для видов, у
которых самец и самка образуют пару, сохраняющуюся на всю
жизнь, как, например, у моевки — чайки, гнездящейся на скалах.
У этого вида размножение может начинаться раньше, потому что
самцы и самки уже имеют партнеров и им нет необходимости
проделывать всю церемонию ухаживания. У моевки особи, уже
образовавшие пары, в среднем выращивают больше потомков,
чем те, которым еще предстоит найти себе партнера.
Повед ение и эволюция
443
Заключение
Итак, мы убедились, что поведение подвержено действию ес-
тественного отбора. Поводе знческие признаки проистекают из ана-
томии и физиологии живо-гного и неотделимы от них. В плане
организма как целого формы поведения обычно адаптивны и часто
они могут, по крайней мере до некоторой степени, передаваться
либо генетически, либо в результате обучения.
У многих видов существуют определенные специфические фор-
мы поведения. Одна из главных задач текущих исследований по
эволюционным аспектам поведения состоит в том, чтобы точно
выяснить те экологические факторы, которые обусловливают эти
формы поведения. В насто ящей главе были рассмотрены неко-
торые из таких исследований.
ЛИТЕРАТУРА
Общие вопросы
Brown J. L. 1975. The Evolution of Behavior, New York, Norton.
Hinde R. A. 1970. Animal Behavior: A Synthesis of Ethology and Comparative Psy-
chology, 2nd ed., New York, M. cGraw-Hill. [Имеется перевод: Хайнд P. Поведе-
ние животных. — М.: Мир, 19Т5.]
Thorpe W. Н. 1961. Bird Song: Tfie Biology of Vocal Communication and Expres-
sion in Birds, Cambridge, Carnibridge University Press.
Wilson E. 0. 1975. Sociobiology: The New Synthesis, Cambridge, Mass., Harvard
University Press.
Wilson E. 0. 1978. On Human N ature, Cambridge, Mass., Harvard University
Press.
Специальные статьи и обзоры
Alexander R. D. 1974. The evolution of social behavior, Ann. Rev. Ecol. Syst., 5,
325—384.
Christian J. J. 1970. Social subordination, population density and mammalian evo-
lution, Science, 168, 84—90.
Hamilton W. D. 1972. Altriusm and related phenomena, Ann. Rev. Ecol. Syst., 3,
192—232.
Littlejohn M. J., Fouquette M. I., Johnson C. 1960. Call discrimination by female
frogs of the Hyla versicolor complex, Copeia, 1960, 47—49.
Lorenz R. Z. 1958. The evolution of behavior, Scientific American 199 (Dec.),
67—78.
Luscher M. 1961. Air-conditioned termite nests, Scientific American, 205 (July),
138—145.
Manning A. 1965. Drosophila and the evolution of behavior. In: J. D. McCarthy
and C. L. Duddington (eds.), Viewpoints in Biology, 4, London, Butterworths.
Orians G. H. 1969. On the evolution of mating systems in birds and mammals, Am.
Nat., 103, 589—603.
Глава 17
Палеонтология и эволюция
До сих пор мы рассматривали три основных компонента тео-
рии эволюции. Сначала мы обсуждали генетическую изменчивость,,
поставляющую сырье для эволюции. Существование генетической
изменчивости обусловлено в конечном счете физическими и хи-
мическими свойствами тех веществ, из которых состоят организ-
мы, в частности свойствами нуклеиновых кислот (ДНК и РНК) и
белков. Эти вещества, образующие гены и хромосомы, обеспечи-
вают преемственность генетической информации из поколения в
поколение. В то же время характер деления хромосом делает
возможным возникновение ошибок при дупликации, называемых
мутациями, а также перераспределение генов, в результате чего
синтез генетической информации происходит по-новому. Второй
компонент эволюции — это отбор. Ввиду того что на свет рож-
дается слишком много особей по сравнению с имеющимися запа-
сами пищи, отбор будет благоприятствовать тем особям, которые
способны использовать ресурсы наиболее эффективным образом
и оставляют в результате этого наибольшее число потомков. Тре-
тий компонент обусловлен разнообразием среды, благодаря кото-
рому в данной области может происходить отбор организмов бо-
лее чем одного типа. Но помимо этих трех компонентов сущест-
вует еще четвертый — время, на фоне которого разыгрывается
взаимодействие между генетической изменчивостью, отбором и
изменчивой средой. В каждый данный момент на поверхности зем-
ного шара существует множество различных сред. При наличии
достаточного времени в каждом данном месте различные среды
последовательно сменяют одна другую. Эти непрерывные измене-
ния среды в пространстве и во времени придают ей пестроту, по-
зволяющую сравнивать ее с лоскутным одеялом. Организмы, су-
ществующие в каждой ее точке, отражают в своей генетической
конституции и морфологических особенностях адаптированность к
имеющимся в данный момент условиям, а также к условиям, в
которых жили их непосредственные предки и, в меньшей степени,
предки более далекие. Поэтому, для того чтобы получить истин-
ное представление об эволюции, следует уделить должное внима-
ние этому четвертому ее измерению.
Палеонтология и эволюция
445
ВАЖНОЕ ЗНАЧЕНИЕ ИСКОПАЕМЫХ ОСТАТКОВ
ДЛЯ ЭВОЛЮЦИОННЫХ ИССЛЕДОВАНИЙ
Великое разнообразие растений и животных как в прошлом,
так и в настоящем отражает реакцию организмов на колебания
условий среды на протяжении тысячелетий. Само возникно-
вение живых форм из более простых неживых соединений —
ответ на изменяющиеся условия среды. В самом деле, если бы
Земля не начала остывать и если бы различные элементы не
распределились в ней так, как это случилось, или же если бы
температурные и энергетические условия не сделали возможным
образование воды, то жизнь — во всяком случае в той ее форме,
в какой она нам известна,— не могла бы возникнуть. Исходные
причины, обусловившие изменения среды, являются внеземными
и недостаточно поняты. Тем не менее мы имеем известное пред-
ставление о характере изменений, происходивших на Земле со
времени ее образования, в частности о тех, которые имели место
в сравнительно недавнее время. Сведения об этом дают остатки
растений и животных, некогда обитавших на поверхности Земли
и в морских водах. Эти остатки называют ископаемыми остатка-
ми (рис. 17.1).
Область биологии, занимающуюся изучением ископаемых ос-
татков и условий, существовавших на Земле в прошлом, называ-
ют палеонтологией. Зная типы и формы организмов, населявших
определенную часть земного шара в определенный период време-
ни, можно по аналогии с тем, что нам известно о потребностях
современных живых существ, делать выводы относительно клима-
та, господствовавшего в тех местах в тот период. Мы знаем, на-
пример, что распространение обезьян ограничено тропическими
областями и что они не переносят холодные зимы. Ископаемые
остатки обезьян были найдены в третичных отложениях в
Патагонии, на юге Аргентины — в области, которая в на-
стоящее время отличается очень холодными зимами, так что
обезьяны жить в ней не могут. Возможны два объяснения: либо
обезьяны, жившие в третичный период, были адаптированы к
холодам, либо климат Патагонии в третичный период был гораздо
теплее, чем сейчас. Обнаружение в тех же слоях ископаемых ос-
татков других тропических животных, таких, как муравьеды, а
также тропических деревьев свидетельствует о справедливости
второго предположения. В тех же отложениях найдены остатки
других ныне вымерших животных — сумчатых саблезубых тигров
и гигантских ленивцев. Путем аналогий и экстраполяции мы мо-
жем сделать вывод, что эти животные также предпочитали теплый
климат. Подобным же образом тип растений, с которыми ассоци-
ированы эти живвтные, может дать некоторые сведения о харак-
тере их питания и среде обитания.
Таким образом, палеонтолог старается восстановить жизнь и
среду прошлого, исходя из допущения, что реакция живых су-
446
Глава 17
Рис. 17.1. Несколько ископаемых растений разного возраста.
A. Eosphaera tilerii (докембрий). Б. Ammikaea septata (докембрий). Оба этих ви-
да принадлежат к древнейшим среди известных нам остатков живых организмов.
В. Sphenopteris sp. (Мейзон-Крик, Иллинойс, верхний карбон). Г. Alethopteris sp.
(Мейзон-Крик, Иллинойс, верхний карбон). Д. Sassafras sp. (песчаники Дакоты,
средний мел). Е. Diospyros rotundifolia (песчаники Дакоты, средний мел). Обра-
тите внимание на то, что по мере приближения к нашему времени растения ста-
новятся более знакомыми. (Фотографии публикуются с любезного разрешения
проф. Е. S. Barghoorn.)
ществ на окружающую их среду была в прошлом такой же, как
и сейчас. Подобные выводы до некоторой степени носят характер
догадок, но они имеют и фактическую основу, которую дает нам
изучение химических веществ, выделенных из ископаемых ос-
Палеонтология и эволюция 447
татков. Анализ этих остатков, а также метеоритов и света, излу-
чаемого звездами, отделенными от нас расстоянием в миллионы
световых лет (света, который во многих случаях испускался еще
до того, как сформировалась Земля), показывает, что по своему
химическому составу эти тела не отличаются от современных ор-
ганизмов и что содержащиеся в них химические элементы обла-
дают такими же свойствами. Эти результаты подтверждают вы-
вод о том, что Вселенная построена из одних и тех же «кирпичи-
ков» и что существует лишь один вид материи. Отсюда следует,
что законы природы действовали во все времена так же, как и
теперь, а в таком случае поведение живых существ в прошлом
было, вероятно, сходным с поведением современных организмов.
Мы знаем, что некоторые виды в процессе эволюции не претерпе-
ли никаких или почти никаких внешних морфологических изме-
нений, хотя у них произошли значительные изменения внутренне-
го строения и, вероятно, физиологии. Однако весьма мало вероят-
но, что эволюция большинства видов протекала таким образом.
КАК ПРОИСХОДИТ ФОССИЛИЗАЦИЯ,
Т. Е. ОБРАЗОВАНИЕ ИСКОПАЕМЫХ ОСТАТКОВ
Слово «фоссилизация» происходит от латинского fossilis —
«ископаемый», «добытый из земли». Ископаемые остатки можно
определить как любые различимые структуры органического про-
исхождения или отпечатки таких структур, сохранившиеся с до-
исторических времен. Это могут быть кости, древесина, покровные
ткани, пыльца, а также следы, экскременты, ходы и любые дру-
гие результаты деятельности того или иного доисторического ор-
ганизма или все, что указывало бы на присутствие такого орга-
низма.
Тела большинства мертвых растений и животных рано или
поздно разлагаются. Если бы они не разлагались, то весь земной
шар был бы покрыт слоем из остатков мертвых организмов тол-
щиной в несколько километров, и возможно, что жизнь давно
перестала бы существовать. Все имеющиеся на Земле минераль-
ные вещества оказались бы включенными в тела организмов.
Разложение вызывается несколькими факторами. Всевозмож-
ные «мусорщики», питающиеся падалью,— от грифов до бактерий
и грибов — очень быстро справляются с мягкими тканями. Твер-
дые ткани, такие, как кости или древесина, более устойчивы. Тем
не менее ферменты, выделяемые некоторыми грибами и бактери-
ями, кислоты, содержащиеся в почве, и совместные действия та-
ких факторов среды, как вода, ветер и температура, со временем
приводят к разрушению даже самых твердых органических ос-
татков. При очень необычных условиях, однако, некоторые части
организмов могут сохраниться, изменившись в большей или мень-
шей степени, и образовать ископаемые остатки (рис. 17.2).
448
Глава 17
Для того чтобы остатки могли сохраниться, организм должен
сразу после смерти погрузиться в какой-нибудь осадок, который
предохраняет его от разрушения падальщиками и от выветрива-
Рис. 17.2. Схема процесса фосснлизации.
А. В низине животное, пришедшее на водопой, случайно попадает в воду и тонет.
5. Скелет погружается в донный осадок, который в конце концов превращается
в песчаник. В. Спустя много веков песчаник разрушается в результате эрозии, и
обнажаются фоссилизованные остатки животного. (О. Т. Solbrig, 1966. Evolution
and Systematics. New York, Macmillan.)
ния. К числу наиболее обычных таких веществ относятся мелкий
песок или ил, причем захоронение чаще происходит в воде, чем на
суше, и в холодных, а не в теплых областях. Но захоронение мо-
жет происходить '"и в других отложениях. Некоторые из наиболее
Палеонтология и эволюция
хорошо сохранившихся ископаемых остатков найдены в янтаре»
который представляет собой ископаемую смолу. В частности в
янтаре находят мелких насекомых, которые увязли в смоле и не
смогли выбраться из нее. В Сибири найдены очень хорошо сохра-
Рис. 17.3. Разные типы ископаемых остатков.
А. Отпечаток части ствола и коры Lepidodendron sp. (карбон), оставшийся в по-
роде. Б. Первичное внутреннее ядро участка стебля Catamites sp. (карбон). В.
Срез окремневшей древесины Gleditchia montanen.se (миоцен). Г. Ископаемая
пыльца сосны (Pinus sp.) из миоценовых отложений штата Орегон. (Фотографии
А—В печатаются с любезного разрешения проф. Е. S. Barghoorn, а Д — д-ра
A. Graham.)
вившиеся туши мамонтов и носорогов, дошедшие до нас в заморо-
женном состоянии из плейстоцена. Хорошо сохраняются ископае-
мые остатки также в вулканическом пепле. Но даже в случаях
быстрого захоронения организма его мягкие ткани разлагаются,
и от него по большей части сохраняются только твердые части.
Раковины, состоящие из неорганических веществ, обычно совер-
шенно не подвергаются изменениям; в отличие от этого кости и
древесина часто минерализуются (или, как принято говорить, ли-
тифицируются), т. е. пустые пространства в ткани пропитываются
минеральными солями, содержащимися в грунтовых водах. Иног-
да растение или животное полностью разрушается, оставляя после
450
Глава 17
себя лишь пустую полость, которая тем не менее точно отражает
внешние признаки мертвого организма; такие ископаемые остатки
называют отпечатками. Впоследствии эта полость может запол-
ниться каким-либо посторонним веществом, в результате чего об-
разуется слепок (рис. 17.3).
Ископаемые остатки после захоронения могут претерпевать
очень сильные изменения. Чаще всего они уплощаются под дав-
лением осадков, накапливавшихся над содержащим их слоем.
Этот слой может также подвергнуться изменениям в результате
складкообразования и горизонтальных движений блоков земной
коры. Для того чтобы мы могли обнаружить ископаемые остатки,
они должны быть выведены на дневную поверхность в результате
эрозии, в процессе которой они могут быть разрушены. Некото-
рые ископаемые остатки, вышедшие на поверхность земли, впос-
ледствии иногда бывают снова погребены. Таким образом, веро-
ятность нахождения неповрежденных экземпляров, особенно
древних, очень мала.
ПРОБЛЕМА НЕДОСТАЮЩИХ ЧАСТЕЙ
Палеонтолог в большинстве случаев, а возможно и всегда, име-
ет дело с ископаемым материалом, представляющим собой силь-
но измененный и обычно неполный остаток организма, жившего в
далеком прошлом. Цель его состоит в том, чтобы узнать как мож-
но больше об этом организме и об условиях, в которых он жил.
Начинать следует с определения систематической принадлежности
ископаемого организма и, если это возможно,— установления его
родственных связей с какой-либо известной группой. Это совсем
непростая проблема, когда речь идет о фрагментарных и изменен-
ных ископаемых остатках вымершего организма. Иногда такие ос-
татки содержат ключевые структуры — зубы, челюсти или че-
реп, но во многих случаях находят лишь бедренную кость, ребро
или предплюсневые кости. Поскольку по таким структурам можно
только приблизительно установить, каким было вымершее живот-
ное, палеонтолог прибегает в формальным видам. Каждому иско-
паемому остатку присваивается видовое название, и он регистри-
руется как отдельный вид; при этом делается научно обоснован-
ное предположение о его возможных родственных связях с теми
или иными группами. Если впоследствии удается найти более
полный скелет и показать, что два или несколько формальных
видов представляют собой части одного и того же организма,
названия этих формальных видов отбрасывают, заменяя их пра-
вильным названием животного, положение которого в системе
теперь можно установить более точно. Применение концепции
формальных видов позволяет палеонтологу производить научную
обработку неполных ископаемых остатков, которые иначе оказа-
лись бы бесполезными. Особенно большую помощь эта концепция
Палеонтология и эволюция
451
оказывает палеоботаникам. В самом деле, позвоночных животных
сплошь и рядом удается определять по остаткам их костей и по
зубам — структурам, вероятность сохранения которых наиболее
велика. Совсем иное дело растения. Отпечатков древесных стволов
и листьев, в виде которых чаще всего предстают ископаемые ос-
татки растений, нередко оказывается недостаточно, чтобы уве-
ренно определить принадлежность к той или иной группе.
ДАТИРОВАНИЕ ИСКОПАЕМЫХ ОСТАТКОВ.
ОТНОСИТЕЛЬНЫЙ И АБСОЛЮТНЫЙ ВОЗРАСТ
Ископаемые остатки представляют ценность лишь в том слу-
чае, если известен их возраст. А возраст слоев, содержащих фау-
ну, палеонтологи определяют на основании найденных в них ис-
копаемых остатков! Это звучит парадоксально, хотя на самом де-
ле парадокса здесь нет. Отец современной геологии Уильям Смит
(William Smith) еще в 1790 г. обратил внимание на то, что для
каждого геологического слоя характерно свое особое сочетание
ископаемых остатков и что поэтому различные слои можно опре-
делять по содержащимся в них остаткам, даже если физические
признаки этих слоев изменились. В областях, не подвергавшихся
сильным нарушениям, можно установить, в какой последователь-
ности происходило отложение слоев, поскольку очевидно, что вы-
шележащие слои должны были откладываться позднее. К тому
времени, сопоставляя ископаемые остатки, найденные в пластах из
разных областей, палеонтологи смогли создать геохронологичес-
кую шкалу, известную под названием геологической колонки
(табл. 17.1). Выразить временные интервалы в годах оказалось
более трудной задачей, и лишь сравнительно недавно появилась
возможность делать это с достаточной точностью. Начиная с
1950 г. стали разрабатываться методы более точного определения
возраста пород, основанные на скорости радиоактивного распада
содержащихся в них элементов. Как показали эти методы, оцен-
ка возраста Земли, производившаяся прежними методами, была
занижена. Кроме того, при помощи этих методов была подтверж-
дена правильность относительных возрастов, которыми пользова-
лись палеонтологи.
Для определения возраста костеносного слоя палеонтологи
стараются найти ископаемые остатки, ограниченные в своем рас-
пространении какими-то определенными слоями, положение ко-
торых в геохронологической шкале известно. Всем неизвестным
ранее ископаемым остаткам из данного слоя приписывают тот же
возраст. Чрезвычайно важно при этом точно установить местопо-
ложение ископаемых остатков в породе и знать вышележащие и
нижележащие отложения, поскольку с течением геологического
времени эти остатки нередко в силу разных геологических про-
цессов изменяют свое первичное положение. В настоящее время
452
Глава 17
Геологическая колонка
Таблица 17.1
бремя е окойяа- ння периода, млн. лет ; Примерная про- должительность, i млн. лет 1 Эра Период Эпоха Некоторые важные эволюционные собы- тия
1 Четвертичный Современная Плейстоцен Появление чело- века
1 62 Кайнозой Третичный Плиоцен Миоцен Олигоцен Эоцен Палеоцен Развитие совре- менных семейств птиц, млекопитаю- щих и цветковых растений
63 135 181 72 46 59 Мезозой Мел Юра Триас Первые цветковые растения. Эпоха рептилий, голосе- менных
230 280 50 65 Пермь Карбон Радиация амфи- бий; первые реп- тилии
345 405 60 20 Палеозой Девон Снлур Первые наземные растения и живот- ные
425 500 600 75 100 —3000 Докембрий Ордовик Кембрнй Бесчелюстные; первые акулы
~3500 Первые живые су- щества, 2500 млн. лет назад
в тех случаях, когда это возможно, отбирают также образцы
горных пород для определения их возраста радиоактивными ме-
тодами.
МОЛЕКУЛЯРНЫЕ ЧАСЫ ЭВОЛЮЦИИ
Если окажется, что теория эволюции белков, основанная и?
нейтральности аллелей, верна для достаточно большого числа
белков, то в таком случае скорость замены аминокислот можно
будет использовать для определения времени тех или иных фи-
логенетических событий. В самом деле, согласно нейтралистской
теории, скорость аллельных замещений равна скорости возникно-
вения аллелей в результате мутаций. Если частота мутаций дан-
ного гена остается довольно постоянной на протяжении длитель-
ных периодов времени, то значит аллельные замещения происходят
Палеонтология и эволюция
453
с постоянной ia-стотой. В таком случае, установив число амино-
кислотных затеи в данном гомологичном белке у двух или не-
скольких оргашзмов, можно точно определить время, когда они
отделились от какого-то общего предка. Иными словами, если
наши допущешя верны, то белки обладают своего -рода встроен-
Рис. 17.4. Нуклеоидные замещения на протяжении палеонтологического времени.
По оси ординат (тложено число нуклеотидных замещений по семи белкам для
парных видов млекопитающих, а по оси абсцисс — время дифференцировки этих
видов от общего ередка. Прямая линия проведена из начала координат до самой
удаленной точки. (Fitch, 1976)
ными в них «часами», которые датируют филогенетические собы-
тия. Это, безуловно, стохастические часы (подобно радиоактив-
ному распаду) но в масштабах длительных периодов .времени эти
часы довольно точны, а кроме того, их можно сделать еще более
точными, если юпользовать несколько белков.
Мы не знае1, конечно, правильна ли нейтралистская теория эво-
люции. Современные исследования эволюции белков показали, од-
нако, что общ;я скорость аминокислотных замен непостоянна во
времени. А слщовательно, скорость эволюции белков нельзя счи-
тать стохастически постоянной. Тем не менее среднюю скорость
этой эволюции для длительных периодов времени и для многих
белков можно использовать в качестве приблизительных эволю-
ционных часов (рис. 17.4). Эти данные очень важны и перспектив-
ны для устансвления времени 'некоторых эволюционных событий.
КОНЦЕ1ЦИЯ ВИДА ВО ВРЕМЕНИ И В ПРОСТРАНСТВЕ
Мы определяли вид как группу -потенциально скрещивающихся
между собой гопуляций. Мы показали также, что вид — это един-
454
Глава 17
ственная непроизвольная категория, в отличие от всех категорий
более высокого ранга, которые создаются искусственно. Но если
учитывать четвертое измерение эволюции, то наша концепция
скрещиваемости становится неприменимой. Современная лошадь
не может скрещиваться с лошадью, жившей в средние века, и
даже с лошадью, жившей в прошлом веке, не говоря уже о тех
лошадях, которые жили в плейстоцене. Следует ли считать, что
все эти лошади принадлежат к разным видам? Очевидно, что
нельзя. Но вид во времени не имеет тех границ, которыми наде-
ляют современные виды на основании критерия скрещиваемости.
Кроме того, с течением времени популяции эволюционируют и
незаметно переходят одна в другую.
Проблему эту еще больше осложняет неполнота палеонтологи-
ческих данных. В очень немногих случаях палеонтолог обладает
достаточно обширным материалом, позволяющим судить об из-
менчивости того или иного вида на данном временном уровне.
Границы между видами обычно (и слишком часто) палеонтологи
устанавливают в местах разрывов в палеонтологической летопи-
си. Эволюция более ранней формы достигает к этому времени та-
кого уровня, что на основании морфологических признаков ее
можно признать самостоятельным видом (здесь опять-таки при-
бегают к аналогии с соответствующими современными видами,
разделяя ископаемые виды в тех случаях, когда различия между
ними сравнимы с различиями между современными видами). Од-
нако при этом нельзя исключить, что при наличии известных
промежуточных форм получился бы непрерывный ряд. Хорошим
примером того, к чему приводят такие непрерывные ряды, служит
запутанная терминология предков человека; было время, когда
практически каждому найденному черепу присваивали свое родо-
вое название. Наличие непрерывного ряда во времени, конечно, не-
обходимо палеонтологу, для того чтобы установить, действительно
ли более ранняя форма была предком более поздней формы; в
этом отношении очень показательно изучение эволюции лошадей.
ЭВОЛЮЦИЯ СЕМЕЙСТВА ЛОШАДЕЙ (EQUIDAE)
В настоящее время семейство Equidae представлено одним ро-
дом с шестью видами: лошадь (Equus caballus), осел (Е. asinus),
кулан (Е. hemionus) и три вида зебры (Е. zebra, Е. grevyinE. bur-
chellt). Род Equus возник в конце плиоцена, но семейство лоша-
дей существовало еще 60 млн. лет назад, в эоцене. Предками
Equidae были давно вымершие примитивные млекопитающие, при-
надлежавшие к отряду Condylarthra и жившие в начале третичного
периода. Первым возникшим от них представителем Equidae был
Hyracotherium; это животное с небольшой головой и длинным хвос-
том напоминало по форме и размерам собаку—одни виды достига-
ли в высоту 25 см, а другие 50 см. При ходьбе оно опиралось на по-
Палеонтология и эволюция
455
Рис. 17.5. Эволюцюнное древо лошадей. Все предки лошади изображены в оди-
наковом масштабе.
456
Глава 17
душечки пальцев, как это делают и современные собаки. У него бы-
ло по четыре пальца на передних и по три на задних ногах, причем
каждый палец был одет своим маленьким копытом. Судя по строе-
нию зубов, имевших низкую коронку со слаборазвитыми низкими
складками, Hyracotherium питался сочными листьями, мелкими
плодами и мягкими семенами. Это было, по-видимому, животное,
объедавшее листву деревьев и обитавшее в лесу. Хотя Hyracothe-
rium был распространен и в Новом и в Старом Свете, эволюция
семейства лошадей происходила главным образом в Северной Аме-
рике с многократными инвазиями в Старый Свет (рис. 17.5).
К началу олигоцена в результате эволюции Hyracotherium воз-
ник род Mesohippus. Это животное было несколько крупнее, с
подушечками на трехпалых конечностях; голова у него была
больше похожа на лошадиную, со сдвинутыми назад глазами
и вытянутой мордой. Головной мозг также увеличился, а зубы,
хотя в общем сходные с зубами Hyracotherium, имели более вы-
раженные складки. В миоцене появляется Merychippus — гораздо
более крупное животное, высотой до 1 м. У наиболее далеко
продвинувшегося вида этого рода подушечки исчезли и, хотя он
опирался на три пальца, основная нагрузка приходилась на ко-
пыто среднего пальца. Зубы имели высокие коронки со сложной
складчатой поверхностью, а кроме того, у них появился зубной
цемент, отсутствовавший у их предков. Следовательно, животное
жило уже не в лесу, а в местообитаниях типа саванны и пита-
лось главным образом травой. Следующее звено —Pliohippus —
представлено однопалой щиплющей траву лошадью, населявшей
североамериканские равнины в плиоцене; от нее к концу этой
эпохи произошел род Equus. По сравнению со своими предками
Equus — крупное животное с однопалыми конечностями и боль-
шими копытами; зубы у него имеют высокие коронки со сложной
складчатостью и приспособлены к питанию травой. В Новом Све-
те этот род вымер, но сохранился в Старом Свете. (Так называе-
мые дикие лошади, живущие в западных областях Северной
Америки, — это одичавшие европейские домашние лошади, ввезен-
ные в Америку человеком.)
Помимо этого главного ствола в эволюции лошадей было не-
сколько различных боковых ветвей, приведших к Anchitherium,
Archaeohippus и Hypohippus в эоцене и к Hipparion, Nannippus и
Neohipparion в плиоцене (рис. 17.5). Семейство лошадей пред-
ставляет собой группу, по которой мы располагаем наиболее пол-
ными и хорошо изученными ископаемыми остатками. В тех об-
ластях, где палеонтологическая летопись по этому семейству осо-
бенно богата, один род постепенно переходит в другой и поэтому
границы между родами произвольны.
На примере палеонтологической летописи Equidae можно про-
следить за тем, как возникают и в конечном счете закрепляются
адаптивные признаки; особенно показательна в этом отношении
Палеонтология и эволюция 457
эволюция зубов. На одном из верхних коренных зубов у Equus
имеется невольной выступ, известный под названием кроше. У
большинства экземпляров Mesohippus и у других лошадей из оли-
гоцена эта складка отсутствует. Но у некоторых особей, несом-
ненно представляющих собой мутантов, имеются маленькие кро-
ше. У Miohippus число экземпляров, имеющих кроше, больше, но
все же есть осоСи, у которых оно отсутствует. У миоценового Pa-
rahippus кроше имелось во всех популяциях, хотя и оставалось
маленьким. Позднее у предков Equus размеры его увеличились.
Аналогичным обэазом обстояло дело с зубным цементом, появив-
шимся в миоцен! у Parahippus и у последовавшего за ним Meryc-
hippus. Сначала зубной цемент встречался лишь у некоторых осо-
бей, но в конце концов получил всеобщее распространение, после
чего толщина ею стала увеличиваться. Эта последовательность
событий совпадгет по времени с переходом лошадей от питания
древесной листвэй к питанию травой. Более жесткая трава тре-
бовала более п {очных зубов. Цемент, повышающий прочность зу-
бов и делающий их более удобными для перетирания пищи, явно
создавал селективное преимущество. Такое же преимущество да-
вало более силшое развитие складчатости на жевательной по-
верхности корежых зубов и увеличение размеров -коронки. Инте-
ресно отметить быстрое развитие всех этих признаков в миоцене
с появлением лопадей, щиплющих траву.
СКОРОСТИ эволюции
Эволюция семейства лошадей прекрасно иллюстрирует также
тот факт, что скорость эволюционного процесса непостоянна и что
направление его может изменяться. Это уже должно было стать
очевидным на основании всего того, что было сказано о способах
действия естественного отбора. Мы видели, как у Equidae переход
от обитавших в лесу форм, питавшихся древесной листвой, к рав-
нинным формам щипавшим траву, повлек за собой ускорение
эволюции тех структур, которые были в наибольшей степени за-
тронуты происшедшими изменениями, в частности зубов. Кроме
того, сильно увегичились общие размеры животных и изменилась
механика их конечности: из ноги, опирающейся на подушечки паль-
цев, она превратилась в пружинящую ногу современной Equus.
В других линиях в течение того времени, когда происходила эво-
люция лошадей, изменений было мало или не было вовсе. Приме-
ром служит опоссум, очень мало изменившийся за последние
80 млн. лет, т. е. с позднемелового времени.
В сущности шт никаких причин, по которым эволюция долж-
на была бы всепа идти в одном направлении. Хотя современная
лошадь гораздо крупнее, чем Hyracotherium, она меньше некото-
рых предков видев Equus из плиоцена. Кроме того, некоторые вы-
мершие плиоценовые лошади, не являвшиеся предками Equus (на-
30-2044
458
Глава 17
пример, Megahippus и Hypohippus), были еще крупнее, а, напри-
мер, Calippus и Nannippus значительно мельче.
Различия в скоростях эволюции можно объяснить главным об-
разом изменениями среды или их отсутствием. В однородной сре-
де та или иная линия, достигшая высокого адаптивного уровня,
в дальнейшем изменяется мало. В отличие от этого в быстро из-
меняющейся среде непрерывно возникают новые давления отбора,
к которым организму приходится адаптироваться. Кроме факторов
среды определенную роль играют и внутренние факторы. Все те
признаки, которые повышают изменчивость (высокая численность
популяций, большое хромосомное число, малая продолжитель-
ность генерации и т. п.), в периоды флуктуаций среды оказывают-
ся выгодными и обладающие ими организмы эволюционируют с
большей скоростью.
Последний вывод, который можно сделать из нашего краткого
анализа палеонтологической летописи и который также хорошо ил-
люстрирует история семейства Equidae, состоит в том, что боль-
шинству, если не всем, видам и родам суждено вымирание. Не-
которые— их меньшинство — преобразуются в более продвинутые
в эволюционном отношении типы, но большинство в конце концов
становится не в состоянии адаптироваться к постоянно возникаю-
щим новым условиям среды или конкурировать с более адапти-
рованными организмами и вымирает. Такая судьба постигла 17
из 18 родов Equidae (рис. 17.5) и 9/ю всех их видов!
В заключение следует сказать, что палеонтологическая летопись
дает весьма существенные доказательства в пользу справедливос-
ти синтетической теории эволюции. Тем не менее в большой мере
из-за неполноты палеонтологической летописи и ее неслучайнос-
ти, палеонтологические данные сами по себе не могут служить
доказательством эволюции, несмотря на широко распространенное
мнение, что они таким доказательством служат. Только при разум-
ном использовании всей совокупности данных, предоставляемых
палеонтологической летописью, современными организмами и
экспериментами, можно получить подтверждение теории Дарвина.
ЛИТЕРАТУРА
Общие вопросы
Calvin М. 1969. Chemical Evolution: Molecular Evolution Towards the Origin of
Living Systems on the Earth and Elsewhere, Oxford, Oxford University
Press. [Имеется перевод: Кальвин M. Химическая эволюция. — М.: Мир,
1971.]
Kummel В. 1961. History of the Earth, San Francisco, Freeman.
Simpson G. G. 1949. The Meaning of Evolution, New Haven; Yale University
Press.
Simpson G. G. 1951. Horses, New York, Oxford University Press.
Палеонтология и эволюция
459
Специальные статьи и обзоры
Axelrod D. I. 1970. Mesozoic paleogeography and early angiosperm history, Bot.
Rev., 36, 277—319.
Bock W. I. \9T2. Species interactions and macroevolution, Evol. Biol., 5, 1—24.
Fitch W. Л1. 1976. Molecular evolutionary clocks. In: F. J. Ayala (ed.), Molecular
Evolution, Sunderland, Mass., Sinauer Associates.
Simpson G. G. 1950. History of the fauna of Latin America, Am. Scient., 38, 361—
389.
Wolfe J. A., Barghoorn E. S. 1960. Genetic change in Tertiary floras in relation to
age, Am. J. Sci., 258A, 388—399.
30
Глава 18
Систематика и эволюция
Для изучения и понимания окружающего нас мира организ-
мов необходимо привести все множество входящих в него форм
в какую-либо рациональную систему. А поэтому систематика так
же стара, как стремление человека к познанию, и усилия, при-
кладываемые им к тому, чтобы классифицировать животных и
растения и понять сущность их разнообразия, влекут за собой
развитие всех других разделов биологии. В свою очередь данные,
получаемые в этих других более специализированных разделах,
оказывают влияние на систематику и вызывают в ней глубокие
изменения, в результате чего из простой попытки к классификации
она становится наукой, задача которой состоит в том, чтобы вы-
явить наблюдаемую в природе упорядоченность и понять ее при-
чины.
СОВРЕМЕННАЯ СИСТЕМАТИКА
Процесс превращения систематики в науку протекал медленно,
хотя скорость его постоянно возрастала. Совершенно очевидно, что
лишь после установления предварительных границ каждой группы
организмов и распределения этих групп по более крупным кате-
гориям, т. е. родам, семействам, отрядам и классам, можно пред-
принимать сколько-нибудь серьезные попытки изучения причин их
существования. До настоящего времени описано и классифициро-
вано примерно 4 500 000 видов. Более или менее полные обзоры
существуют лишь по двум из всех крупных групп организмов: сре-
ди животных — это птицы, насчитывающие примерно 8650 видов,
а среди растений — голосеменные, насчитывающие примерно 650
видов. Наши знания о видах, принадлежащих к другим группам,
и об их числе неполны, и хотя любые предсказания о числе еще
неописанных видов очень проблематичны, принято считать, что
существует свыше 100 000 неописанных видов цветковых растений,
около 20 000 видов рыб и более 1 000 000 видов насекомых! Таким
образом, конца работы по описанию и классификации видов ор-
ганизмов пока не предвидится.
Однако работа систематиков не ограничивается описанием ви-
дов и их классификацией. Они хотят знать генетические взаимо-
отношения и историю видов, которые они исследуют, а также ме-
Систематика и эволюция
461
ханизмы, с помощью которых эти виды возникли. Они хотят так-
же открыть законы, управляющие поведением всех организмов.
По мере того как биологи, работающие в других областях биоло-
гии, узнают все больше и больше о химии, физиологии, анатомии
Рис. 18.1. Хемосистематика.
Химия — одна из отраслей науки, которая в настоящее время служит источником
данных для систематики. Разные группы организмов нередко содержат характер-
ные химические соединения или классы соединений, помогающие систематику
установить их филогению. (О. Т. Solbrig, 1970)
и поведении растений и животных, на систематика обрушивается
лавина данных, которые ему следует учитывать при создании сис-
тем классификации и эволюционных гипотез (рис. 18.1). Быть мо-
жет, именно небольшая когорта практических систематиков наи-
более остро ощущает взрыв научной информации, свидетелями
которого мы являемся! В то же время возросшее понимание всех
аспектов функций и поведения организмов сделали систематику
занятием более увлекательным и интересным, чем она была ког-
462
Глава 18
да-либо прежде, и эта старейшая из всех биологических дисцип-
лин переживает сейчас небывалый подъем.
В этой книге мы рассмотрели некоторые стороны ряда сравни-
тельно новых областей биологии — цитологии, генетики, науки о
поведении, а также таких более классических областей, как эко-
логия и палеонтология. Следует помнить, что толчок к развитию
всех этих наук дала систематика и что изобилие сведений, кото-
рые она сейчас получает от них, это плоды того вклада, который
они в свое время получили от систематики. Поэтому представля-
ется полезным дать краткий обзор некоторых моментов в истории
развития систематики, а также ее методологии и методов.
ОСНОВЫ КЛАССИФИКАЦИИ
Классификацией называют распределение организмов (или лю-
бых других объектов) по классам. Организмы, принадлежащие к
каждому классу, объединяет та или иная система связей между
ними. Язык представляет собой простейшую систему классифика-
ции, в которой каждый класс «вещей» получает обозначение (имя).
Всех членов данного класса объединяет наличие у них общих
признаков или родственные связи. Первый шаг в любой системе
классификации состоит в разграничении классов; второй шаг —
установление отношений между классами и создание иерархии.
Отношения могут быть установлены двумя путями: во-первых, на
основании перекрывания или совпадения неидентичных классов;
во-вторых, путем подчинения одних классов другим или включе-
ния одного класса в другой. В качестве примера перекрывания
можно привести следующий: можно выделить класс длинношер-
стных собак и класс коричневошерстных собак, неидентичные один
другому. Класс длинношерстных собак коричневого цвета перекры-
вается с обоими этими классами и имеет одинаковую протяжен-
ность в пространстве и <во времени с перекрыванием между ними.
Классификация путем соподчинения классов и их включения
один в другой используется в языке наряду с перекрыванием.
«Рыжие лисицы», «серые лисицы» и «черно-бурые лисицы» — все
принадлежат к более обширному и включающему их всех клас-
су «лисица». В биологической классификации обычно пользуются
методом соподчинения и включения одних классов в другие, и
метод этот восходит к аристотелевой логике. Все классы включа-
ются в более крупные классы. Так, например, виды
Fells fells (кошка), Fells concolor (пума), Fells tigris (тигр) и
Fells опса (ягуар) все включаются в род Fells. Род Fells в свою
очередь включается в семейство Felidae, в отряд Carnivora и в
класс Mammalia. Подобное распределение представляет собой
иерархическую систему категорий.
Согласно этому методу, вид обладает всеми характерными чер-
тами своего рода плюс собственные видовые отличия. Роц харак-
Систематика и эволюция
463
теризуется всеми чертами, общими для всех входящих в него видов.
«Видовые отличия» — это те признаки, некоторым мы отличаем
один вид от другого в пределах данного рода. Аналогичным образом
семейство определяется по признакам, общим для всех входящих^
в данное семейство родов, и т. д. для каждой категории более вы-
ского ранга. Эгот метод классификации полезен, если нас инте-
ресует установление системы, основанной только на логических
отношениях между родом и входящими в него видами и между
самими этими зидами. Так, если бы мы определяли принадлеж-
ность к роду Falls по наличию хвоста и строго придерживались
формальной логики, то нам пришлось бы исключить из этого ро-
да (а также из вида Fells fells — кошка) всех кошек с обрезан-
ными хвостами. Но на самом деле кошки с обрезанными хвоста-
ми принадлежат и к виду Fells felis и к роду Fells.
На практике положение данного организма в системе опреде-
ляют на основе всей совокупности его признаков, не придавая
главного значения логическому соответствию. Если у животного
отсутствуют некоторые типичные для данной группы признаки, но
имеются все другие, его относят к той группе, к которой бы оно
принадлежало при наличии у него и этих недостающих призна-
ков. Научное обоснование подобной практики дает эволюционная
теория. В процессе адаптации к различным средам популяции
могут подвергаться любым изменениям, в результате которых
изменяются как «родовые» признаки, так и «видовые отличия».
Мы определяем виды, когда это только возможно, прежде всего
на основе их генетических отношений и их скрещиваемости и
только во вторую очередь — по их морфологическим признакам.
Однако при этом возникает мекая проблема. Если родственные (в
филогенетическом смысле) классы не имеют никаких общих при-
знаков, то как можно определить ту категорию более высокого
ранга, в которую они входят? Частичное решение этой проблемы
дает теория множеств. В теории множеств класс определяется по
нескольким признакам (чем их больше, тем лучше) и каждый вид
должен обладать по крайней мере неким минимальным числом
этих признаков, чтобы его можно было включить в данный класс.
Эта процедура была бы вполне удовлетворительной, если бы бы-
ли известны филогенетические взаимоотношения. Но они извест-
ны редко. Обычно сначала устанавливают классификацию, а за-
тем выводят из нее филогению.
В практической систематике используется эмпирический пра-
гматический подход, сочетающий в себе элементы формальной
логики, теории множеств и эволюционной теории. Полевые наблю-
дения и эксперименты, дополняемые изучением музейных коллек-
ций и гербариев, дают сведения об изменчивости популяций и их
скрещиваемости. На основе этих данных устанавливается эле-
ментарный класс — вид. Затем, используя всю совокупность из-
464
Глава 18
вестных признаков всех видов, систематик создает роды, так что-
бы степень различия между членами одного и того же рода была
меньше, чем между видами, принадлежащими к разным родам.
Это достигается двумя способами. Один из них основан на чисто
статистическом подходе, при котором дается количественная оцен-
ка как можно большему числу признаков, а затем с помощью
быстродействующих вычислительных машин между ними устанав-
ливаются множественные корреляции. Этот метод логически без-
упречен с точки зрения теории множеств. Его главный недостаток
заключается в том, что при этом все аспекты фенотипа считаются
одинаково важными. Между тем мы знаем, что одни фенотипиче-
ские признаки определяются всего несколькими генами, а другие—
большими сериями генов. Кроме того, некоторые признаки данно-
го организма (те, которые имеют непосредственную адаптивную
ценность) очень пластичны, тогда как другие (например, органы
размножения) такой пластичностью не обладают. Иными слова-
ми, филогенетическая значимость этих органов неодинакова, по-
тому что скорости, с которыми они могут эволюционировать, очень
различны. Вследствие этого обычная процедура состоит в оценке
потенциальной филогенетической значимости различных признаков
и в соответствующем использовании их при создании высших ка-
тегорий. Отрицательная сторона этой процедуры — присущий ей
элемент субъективности (оценка относительной значимости при-
знаков), но ее до сих пор придерживаются в тех случаях, когда
возможна филогенетическая проверка (при помощи ископаемых
остатков).
ТАКСОНОМИЯ, СИСТЕМАТИКА, КЛАССИФИКАЦИЯ
И НОМЕНКЛАТУРА
Четыре термина, составившие этот заголовок, часто использу-
ют на равных правах, что приводит ко многим недоразумениям.
Мы уже говорили, что классификация — это распределение орга-
низмов по отдельным группам. Систематика — это сравнительное
изучение любой группы организмов и всех и каждого взаимоот-
ношений между ними при помощи методов, заимствованных у
одной или нескольких биологических дисциплин (рис. 18.2). Ис-
следования в области систематики часто (но не всегда) приводят
к созданию классификаций. Подобным же образом классификации
могут быть, а могут и не быть основаны на исследованиях по си-
стематике, хотя современные биологические классификации почти
всегда имеют такую основу. Тем самым эти две операции часто
совпадают и обозначающие их термины оказываются синонимич-
ными. Номенклатурой называют присвоение отличительных на-
званий каждой из групп, различаемых в той или иной классифи-
кации. Присвоение названий растениям и животным производится
по определенным правилам, содержащимся в «кодексах» ботани-
Систематика и эволюция
465
ческой и зоологической номенклатуры. Эти правила, создававшие-
ся на протяжении многих лет, сочетают в себе элементы истории
(подобно законам приоритета при присвоении названий видам)
скрещивания, щтологи
ческие и генетидеские
эксперименты; (аблюде-
ния в нонтролируэмых
условиях
ПОЛЕВЫЕ ИССЛЕДОВАНИЯ:
сборы, наблюдения и экспери-
менты; экологжеские исследо-
вания
НОМЕНКЛАТУРА:
ДОКУМЕНТАЦИЯ
КЛАССИФИКАЦИЯ,
ФИЛОГЕНИЯ,
ТАНСИМЕТРИЯ
МОДЕЛИРОВАНИЕ МАНРО- И
МИКРОЭВОЛЮЦИОННЫХ ПРО-
ЦЕССОВ
документация
правила номенклатуры
м орфологические,
анатомические, хими-
ческие и другие свя-
занные с ними иссле-
дования
ОПЫТНЫЕ
УЧАСТКИ,
ТЕПЛИЦЫ:
ЛАБОРАТОРИЯ:
ТАКСОНОМИЧЕСКИЕ
ИССЛЕДОВАНИЯ
ИС ПОЛЬЗО ВАНИЕ ДАННЫХ ИЗ
ДРУГИХ ОБЛАСТЕЙ БИОЛОГИИ:
физиологии, биаимии,
генетики, анатомти и Т.П
Библиотека
Рис. 18.2. Некоторье операции, производимые при исследованиях по систематике
и эволюции.
Стрелки указывают направление потока информации. (О. Т. Solbrig, 1966. Evolu-
tbn and Sistematics, New York, Macmillan.)
с практическими требованиями, обеспечивающими ясность назва-
ний и удобство их использования. Номенклатура служит средст-
вом, позволяющим избежать путаницы и создать единый язык для
биологической классификации.
466
Глава 18
Таксономии давали различные определения: «теоретическое
изучение классификации, рассматривающее ее основы, принципы,
процедуры и правила» (Simpson, 1961); «концепция, представляю-
щая собой синтез всех фактических данных [об организмах], и
выражение взаимоотношений [организмов] (Mason, 1950); «изу-
чение принципов и практических приемов классификации,... о
частности... методов, принципов и даже частично результатов био-
логической классификации» (Heslop-Harrison, 1953). Соотношение
между таксономией и систематикой несколько сходно с соотноше-
нием между теоретической физикой и всей физикой в целом. Так-
сономия включает в себя классификацию и номенклатуру, нов
своих концепциях сильно опирается на систематику. Эти две об-
ласти часто ошибочно приравнивают друг другу.
ИСКУССТВЕННЫЕ, ЕСТЕСТВЕННЫЕ
И ФИЛОГЕНЕТИЧЕСКИЕ СИСТЕМЫ КЛАССИФИКАЦИИ
Самые ранние системы классификации, которые нам известны,
были откровенно практическими, преследовавшими лишь одну
цель — создать систему, дающую возможность как-то упорядочить
элементы окружающей среды. Так, например, в трудах древних
греков и римлян животные делились на морских, наземных и
воздушных, а растения — на травы, кустарники и деревья. Эти
системы классификации античной эпохи, известные нам главным
образом из трудов Аристотеля и Теофраста, постепенно разраба-
тывались и совершенствовались, особенно после эпохи средневе-
ковья. Усовершенствование системы классификации достигло своей
кульминационной точки в середине XVIII в., в работе великого
шведского естествоиспытателя Карла Линнея, которому принадле-
жит первая попытка создания всеобщей системы классификации
для всех организмов.
Система Линнея обладает рядом черт, снискавших ей в свое
время заслуженную славу. Так, например, в эту систему можно
включать вновь открываемые единицы, не нарушая ее. Основные
таксоны — роды и виды — были в общем достаточно хорошо раз-
граничены, и Линней ввел для обозначения видов очень удобную
бинарную номенклатуру, сохранившуюся до сих пор. Однако при
всем том концепция вида Линнея мало чем отличалась от аристо-
телевой, будучи основана исключительно на морфологическом
сходстве, а его высшие категории, хотя логически приемлемые,
нередко устанавливались на основе случайных признаков. Линней
был очень наблюдательным человеком и обладал пытливым умом,
побудившим его заняться, помимо всего прочего, экспериментами
по гибридизации и наследственности у растений. Но, несмотря на
эти свои занятия, Линней, как и большинство натуралистов того
времени, считал, что виды неизменяемы и что они, очевидно, были
когда-то созданы творцом в той форме и в том числе, в каком они
существовали в его время. А поэтому Линней, бывший очень на-
Систематика и эволюция
467
божным человеком, не стремился к тому, чтобы открыть всеобщую
систему упорядоченности во Вселенной. Его профессиональный и ре-
лигиозный обр;з мыслей, возможно, не позволял ему заняться этим.
К открытии такой всеобщей системы стремилось следующее
поколение естествоиспытателей, в особенности французские биоло-
ги. Корни этою движения восходят к началу революции в на-
уке. Открытие Ньютоном законов тяготения и движения небесных
тел, а также щей и 'методы Бэкона и Декарта повлекли за собой
стремление к гознанию законов природы, которое в конечном счете
привело к созданию новых взглядов на биологический мир. Боль-
шинство ученьк все еще верило в то, что виды были созданы не-
ким высшим существом, но допускали, что этот творец создавал
их в соответстши с неким великим замыслом или системой и что
задача систематика — открыть эту систему. Начались поиски так
называемых естественных родственных связей между организма-
ми, в результате чего было проведено много хороших исследова-
ний, в которые были выявлены многие эволюционные отношения,
хотя в то вреья их эволюционная природа не была понята. Зна-
чительный стилул получило также изучение сравнительной ана-
томии, в этот «е период появились первые серьезные физиологи-
ческие исследования.
Среди первях французских естествоиспытателей, разделявших
эти новые взгляды, были Бюффон (Buffon) и Ламарк (Lamarck),
которому Бюффон покровительствовал; их работы относятся ко
второй половше XVIII века. Ламарк был автором одной из первых
систем, в которой виды рассматривались не как неизменные, а
как продукт вакого-то развития. Исследования, ставившие себе
целью выявлеше великого замысла творца, процветали во Фран-
ции и в несколько меньшей степени — в Германии и Англии. По-
мимо Бюффонаи Ламарка здесь следует привести имена Джеффри
(Geoffrey), Де Кандолля (De Candolle), Окена (Oken) и Эразма
Дарвина (Erasnus Darwin).
«Естественняе системы» не означали полного отрыва от искус-
ственной систем Линнея. Формальная логика все еще оставалась,
во всяком слутае в теории, основой классификаций. Однако за-
трачивалось больше усилий на то, чтобы изучать всю совокуп-
ность признаксв данного организма и на то, чтобы классифика-
ции отражали ютественные родственные связи. В результате под-
ход к классифигации стал более эмпирическим, с применением ло-
гических концетций, которые в настоящее время естественно было
бы ассоциировать с теорией множеств.
Натуралистя первой половины XIX в. очень близко подошли
к открытию теэрии эволюции. В их работах можно найти много
современно зв/чащих рассуждений относительно взаимоотно-
шений между растениями и животными. По-видимому, они не
смогли дойти до того, чтобы высказать мысль об эволюционирую-
щей биоте лишь потому, что не знали механизма, с помощью ко-
468
Глава 18
торого можно было бы объяснить происходящие в ней изменения.
Таким образом, величайшим вкладом Чарлза Дарвина была не
сама идея об эволюции, а его теория естественного отбора, пред-
ложившая механизм эволюции. Ламарк, а после него и другие
предлагали системы, по которым происходит трансформация ви-
дов, но эти системы неприемлемы, так как их авторы не могли
представить рационального объяснения того, каким образом про-
исходила эта трансформация.
Не сразу после ©формулирования и принятия теории естест-
венного отбора проявилось ее влияние на систему классификации.
Главная причина этого состояла в великолепной работе система-
тиков того времени, которые в поисках «естественных родствен-
ных связей» открывали эволюционное родство. Сам Дарвин при-
знает влияние своего четырехтомного труда по классификации морс-
ких желудей на создание теории эволюции. Таксономисты поеледар-
виновского периода сменили выражение «естественное» родство на
«филогенетическое» родство, но в течение некоторого времени ни-
каких резких изменений в манере их работы не происходило.
Вторичное открытие в 1900 г. законов Менделя сначала вызва-
ло отход от Дарвина. Корпускулярная наследственность на пер-
вых порах считалась противоречащей идее естественного отбора.
Как можно объяснять эволюцию накоплением мелких признаков,
если признаки наследуются как дискретные единицы? Реакция
против дарвинизма была частично обусловлена тем, что некото-
рые из обнаруженных в числе первых мутации обладали резко
выраженными эффектами. Она была обусловлена также неудач-
ным выбором Гуго де Фриза (один из тех, кто вторично открыл
законы Менделя и был ярым противником теории естественного
отбора), проводившим исследования на растениях рода Oenothera.
У видов, принадлежащих к этому роду, встречаются морфологи-
ческие изменения, ошибочно принимавшиеся в то время за на-
следуемые точковые мутации. Несмотря на все эти осложнения,
менделизм и дарвинизм довольно скоро объединились. Этому
объединению особенно способствовали расширение и углубление
изучения генетических явлений и развитие популяционной гене-
тики, приведшие в конечном итоге к созданию синтетической те-
ории эволюции.
Выдающаяся черта синтетической теории состоит в переходе
от типологического мышления к популяционному. Одна из осо-
бенностей линнеевской системы, которая еще сохраняла свое зна-
чение к концу прошлого века, была концепция Платона об эйдос
(идее, типе, сущности). Этот принцип нашел воплощение в кон-
цепции типа, т. е. некой идеализированной особи, являющейся
моделью всех других особей данного вида. Представление о типе
было непосредственным порождением того периода, когда вид счи
тался неизменяемым. Единственная особь — тип — могла адекват-
но представлять данный вид. Дарвин понимал важность измен-
Систематика и эволюция
469
чивости, но в еп время типологическое мышление еще оставалось
в силе, отчасти потому, что динамика эволюции популяций не
была известна, "олько в результате развития генетики и понима-
ния того, что единицей эволюции служит не отдельная особь, a
популяция, в образе мышления систематиков произошел сдвиг и
они перешли от изучения отдельных экземтляров к изучению се-
рий и обширны: выборок, покидая, когда это было возможным,
стены музеев ддя полевых исследований. Концепция типа, как во-
площения эйдос данного вида, была отброшена, хотя типовые эк-
земпляры используются до сих пор для номенклатурных целей.
Другим важном результатом воздействия синтетической тео-
рии эволюции fa систематику было изменение концепции вида.
Линней определи виды в соответствии с предписаниями аристо-
телевой логики. В его определении подчеркивались «видовые от-
личия», поскольку признаки рода были общими для целой группы
видов. Из-за тсго что в его концепции главное внимание уделя-
лось различиям а не сходству, она затемняла родственные свя-
зи. Во времена Линнея сведений о генетике и размножении было
мало, что дела;о невозможным использование каких-либо призна-
ков, за исключшием чисто морфологических. А отсутствие хоро-
ших оптических приборов ограничивало возможности использова-
ния морфологических признаков организмов для выявления ес-
тественных родственных связей, и при классификации организмов
учитывались ка: их сходные черты, так и различия между ними.
Развитие физислогии, анатомии, эмбриологии и (к концу века)
цитологии и гететики сильно увеличило число используемых кри-
териев. Исследования по систематике в большинстве случаев ста-
ли основой биологических классификаций, и такое положение
сохраняется до сих пор. Дарвиновская теория создала для нее
твердую теоретическую базу. Это заставило ученых придавать
значение не тооько различиям, но и сходству между видами. Од-
нако, несмотря на все это, линнеевская концепция вида не пре-
терпела серьезных изменений.
Только бурщчй рост генетики в первой половине XX в. и раз-
витие синтетической теории эволюции привели к коренному изме-
нению концепции вида. Репродуктивные взаимоотношения стали
главным критещем при определении видов. Это новое определе-
ние вида тесно связано с важным значением, которое придается
в синтетической теории скрещивающимся популяциям.
СОПОСТАВЛЕНИЕ ФЕНЕТИЧЕСКОЙ
И ФИЛОГЕНЕТИЧЕСКОЙ КЛАССИФИКАЦИЙ
Конечная цель систематика состоит в том, чтобы создать си-
стему классификации, которая отражала бы эволюционную ис-
торию входящих в нее видов. Однако установление эволюционной
истории и филогенетического родства той или иной группы ви-
470
Глава 18
дов — задача нелегкая. Фактические данные обычно бывают не-
полными, а кроме того, систематики расходятся в истолковании
этих данных. Поэтому часто случается, что для одной и той же
группы видов существует несколько классификаций, каждая из
которых представляет собой в известном смысле гипотезу относи-
тельно их возможной эволюционной истории.
Для того чтобы классификация отражала эволюционную исто-
рию данной группы, все члены рассматриваемого таксона должны
быть близкородственны между собой и происходить от общего
предка. Такая система классификации называется монофилетичес-
кой. Но систематики, кроме того, стремятся, чтобы их классифи-
кации отражали степень сходства между организмами, а поэтому
они чувствуют, что организмы, имеющие одинаковые морфологи-
ческие или какие-либо иные признаки или использующие одина-
ковые внешние условия, следует помещать в один таксон. Так,
например, всех «уткообразных» птиц объединяют в один отряд
Anseriformes1, который делится на несколько семейств: лебеди
(Cygninae), гуси (Anserinae), древесные утки (Dendrocygninae),
нырковые утки (Anatinae) и т. п. В свою очередь каждое семейст-
во делится на несколько родов. Например, нырковые утки объе-
диняют роды Anas, Spatula, Aix и др., а каждый род — несколь-
ко видов: шилохвость (Anas acuta), кряква (Anas platyrhynchos),
черная американская утка (Anas rubripes) и др. Все члены отряда
Anseriformes не только хорошо плавают, но и хорошо летают и
могут совершать перелеты на далекие расстояния. Ноги у них от-
носительно короткие, с трехпалыми перепончатыми лапами, клюв
уплощенный. Эти общие признаки характерны для всех предста-
вителей отряда. В свою очередь каждое семейство обладает сво-
ими более специализированными признаками. Лебеди — крупные
птицы с очень длинной шеей; гуси, также обладающие длинной
шеей, мельче лебедей, но крупнее уток; утки же мельче тех и
других и шея у них короче. Роды обладают еще более специа-
лизированными признаками и т. д. К сожалению, не все монофи-
летические группы имеют одинаковый для всех их членов общий
адаптивный план строения.
Эволюционные линии могут изменяться с течением времени
постепенно (постепенное видообразование) или же они могут
разделяться, давая начало двум отдельным линиям (истинное
видообразование; гл. 11). Превращение одной линии в две или
более называют кладогенезом. Постепенное изменение называют
анагенезом.
В результате анагенеза могут возникать организмы с такими
четко выраженными новыми свойствами как «теплокровность»
(гомойотермность). В таких случаях говорят, что образовавшиеся
1 Систематика отряда Anseriformes, которой придерживаются авторы, расхо-
дится с принятой советскими орнитологами. — Прим, перев.
Систематика и эволюция
471
организмы достигли новой грады. Грады могут быть монофилети-
ческими или полифилетическими. В монофилетической граде лишь
одна филетическая линия обладает рассматриваемым признаком,
а в полифилетической — две или более линий. Гомойотермность —
полифилетическая града, поскольку она возникала в процессе эво-
Уровень сложности (анагенез)
Рис. 18.3. Соотношение между эволюционным изменением во времени (анагенез)
и разветвлением филетических линий (кладогенез), происхождением от одной (мо-
нофилия) или нескольких (полифилия) линий, ветвями (клады) и уровнями мор-
фологического или функционального несходства (грады).
Каждый кружок соответствует одному ныне существующему виду. Виды а и Ь
фенетически более близки видам j—q, поскольку онн достигли сходных уровней
сложности, но филогенетически они ближе видам cud, поскольку все они про-
исходят от общего предка. Виды, относящиеся к одной и той же граде, могут
иметь различное происхождение (полифилия), как это здесь показано; виды, от-
носящиеся к одной и той же кладе, всегда имеют общее происхождение (моно-
филия).
люции по крайней мере дважды (птицы и млекопитающие), а мо-
жет быть, и большее число раз. Новая филетическая линия, воз-
никающая в результате кладогенеза, называется кладой, а клады
по определению являются монофилетическими (рис. 18.3).
Трудность, с которой сталкиваются таксономисты при объеди-
нении видов в таксоны более высокого ранга, состоит в том, что
им приходится как-то удовлетворять свое стремление « созданию
472
Глава 18
монофилетических таксонов, отражающих одновременно и анаге-
нез, несмотря на крайнюю неполноту (а иногда даже отсутствие)
знаний относительно подлинной филогенетической истории изуча-
емой линии — знаний, которые можно почерпнуть только из па-
Рис. 18.4. Численная систематика.
Признакам организмов приписывают определенные численные значения, или ко-
ды, после чего их можно подвергнуть численному анализу, нередко с применением
вычислительной машины. Таким способом можно получить разнообразную ин-
формацию, в частности предложить определенную систему классификации орга-
низмов. (О. Т. Solbrig, 1970)
леонтологической летописи. На основе данных, полученных в ре-
зультате изучения современных организмов, интуитивной оценки
относительной важности тех или иных признаков, реконструкций
климатов прошлого и находимых время от времени ископаемых
остатков, таксономисты создают системы классификации, надеясь,
что эти системы отражают филогенетическую историю данной ли-
Систематика и эволюция
473.
нии. Дело осложняется тем, что нередко систематики никак не
могут достичь согласия в интерпретации имеющихся данных.
В результате создается очень зыбкая ситуация, когда разные си-
стематики предлагают различные системы классификации в соот-
ветствии с собственной интерпретацией фактического материала.
Все это приводит к тому, что классификация оказывается весьма
неустойчивой. Было предложено два способа для разрешения этой
Фенетическое изменение
Рис. 18.5. Соотношение между фенетическим и филогенетическим сходством.
Числа соответствуют видам согласно кладистической системе классификации.
Между прерывистыми линиями заключены единицы, которые могли бы считаться
видами согласно фонетической системе классификации.
проблемы. Один из них состоит в том, чтобы классифицировать ор-
ганизмы не на основе их филогенетической истории, а по морфо-
логическому сходству. Преимущество подобного фенетического
подхода зключается в простоте его применения. Морфологические
признаки организма нетрудно измерить или дать им числен-
ные оценки, а затем сравнивать (используя вычислительные ма-
шины) эти оценки со значениями, полученными для других орга-
низмов; в результате таких сравнений можно получить коэффи-
циенты сходства (рис. 18.4). Затем сходные виды можно объеди-
нять в таксоны более высокого ранга на основе определенного
значения коэффициента сходства. Серьезная проблема, присущая
чисто фенетической классификации, заключается в том, что в
зависимости от используемых критериев значения коэффициента
сходства могут оказаться различными. Но еще более важный ее
недостаток — полное выпадение филогенетических данных.
31—2044
474
Глава 18
При другом подходе, известном под названием кладистичес-
кого, таксоны классифицируются исключительно на основе числа
ветвлений данной филогенетической линии, имевших место на
протяжении ее эволюции. Сторонники такого подхода настаивают
на том, что ветвление филогенетических линий служит объектив-
ным кртерием для разделения их на разные таксоны; в тех же
случаях, когда эволюция филогенетической линии происходит пу-
тем анагенеза, такой критерий отсутствует. Согласно кладисти-
ческому подходу, при каждом ветвлении возникают сестринские
таксоны однакового ранга, таксономически обособленные от своих
предков (рис. 18.5). Преимущество кладистического метода состо-
ит в том, что он придает системам классификации известное еди-
нообразие и простоту, устраняя субъективность многих классифи-
каций. Использование молекулярных часов (гл. 17) может по-
мочь точно установить, когда именно происходили кладистические
события. Как и при фенетическом подходе, в жертву этой просто-
те приходится приносить эволюционную и экологическую инфор-
мацию.
Невозможно создать систему, которая отражала бы эволюци-
онную историю видов, поскольку виды классифицируются при на-
личии весьма смутных (ввиду недостатка данных) представлений
об этой истории. В таких условиях любая система классификации
будет в значительной степени субъективной. Строгая привержен-
ность фонетическим или кладистическим критериям устранит
субъективность. Но субъективные классификации — это гипотезы,
подлежащие проверке и видоизменению, и в этом смысле они
служат орудиями, помогающими эволюционисту в его поисках
истины.
ЗАДАЧИ НА БУДУЩЕЕ
В начале этой главы мы указали на грандиозность стоящей
перед систематиками задачи по присвоению названий организмам
и их описанию. Но систематика становится теперь более точной
наукой, чем она была во времена Линнея; она обладает рядом
основополагающих принципов и известной теоретической базой.
Современные систематики часто могут строить предположения и
ставить эксперименты для их проверки. Создаваемые таким обра-
зом классификации гораздо богаче по содержащейся в них ин-
формации. Во времена Линнея видовое название было всего лишь
средством для обозначения того или иного вида растений или
животных и ничем более. В современной классификации видовое
название — это ключ ко всему тому, что известно о данном виде,
о его прошлой истории и взаимоотношениях с другими организма-
ми, потому что наши знания об организме, которому присвоено
данное название, гораздо обширнее.
Тем не менее возможности систематики не безграничны. Глав-
Систематика и эволюция
475
ный камень преткновения— отсутствие данных относительно вы-
мерших организмов, о направлениях эволюции и ее скоростях.
Этот недост.ток знаний создает серьезные трудности, поскольку
без таких дашых количественная оценка признаков, используемых
в классифиюции, всегда будет оставаться в значительной степе-
ни субъектишой и построить подлинно филогенетическую класси-
фикацию неюзможно.
Скептики могут поэтому сомневаться в том, стоит ли пытаться
классифипир>вать организмы так, чтобы классификация отража-
ла их прошлою историю, когда история эта неизвестна. Однако, как
было показаю в этой книге, внимательное изучение генетики, цито-
логии, строешя, функции, поведения и экологических взаимоотно-
шений може: дать нам много сведений о прошлом. Разумное ис-
пользование этих сведений позволит создать классификацию, ко-
торая будет [учше соответствовать прошлому органического мира,
чем искусствшные классификации, строившиеся двести лет назад.
Мы должны юегда помнить, что наука может в лучшем случае
дать статистшеское приближение к реальной действительности и
самое большее, чего мы 'можем добиться—это выявления относи-
тельной истшы.
ЛИТЕРАТУРА
Общие вопросы
Davis Р. Н., Hetuoood V. Н. 1963. Principles of Angiosperm Taxonomy, Edinburgh,
Oliver and Joyd.
Heslop-Harrison J. 1953. New Concepts in Flowering Plant Taxonomy, London,
W. Heinemai.
Simpson G. G. D61. Principles of Animal Taxonomy, New York, Columbia Univer-
sity Press.
Solbrig О. T. 1170. Principles and Methods of Plant Biosystematics, New York,
Macmillan.
Специальные ста'ьи и обзоры
Ashlock Р. D. 194. The uses of cladistics, Ann. Rev. Evol. Syst., 5, 81—100.
Estabrook G. F. 1972. Cladistic methodology) a discussion of the theoretical bases
for the induction of evolutionary history, Ann. Rev. Evol. Syst., 3, 427—456.
Hull D. L. 1970. Contemporary systematic philosophies, Ann. Rev. Ecol. Syst., 1,
19-54.
Mason H. L. 195). Taxonomy, systematic botany and biosystematics, Madrono, 10,
William W. T. 1(71. Principles of clustering, Ann. Rev. Evol. Syst., 2, 303—326.
31
Предметный указатель
Автополиплоид 116, 299
Адаптация 327—330
Адаптивная стратегия 358
— ценность см. Приспособленность
Аллели 58
— доминантные 58
— рецессивные 62, 151—152, 154
---элиминация 160—163
Аллозиготная особь 181
Аллополиплоид 116, 117, 299
— сегментный 117, 118
Аллофермеиты 236—239
Альтруизм реципрокный 268
Альтруистические черты 287
Анагенез 470
Антибиотики 404
Антигенный дрейф 132
Антигены вируса гриппа 131
Аутозиготная особь 181
Ацетилхолииэстераза 145, 146, 147
Базидиоспоры 138—139
Бесполое размножение 195, 196
Биваленты 99
Вегетативное размножение 195
Взаимоотношения растение—опыли-
тель 410—416
Видовое разнообразие 394
Вид, биологическая концепция 9,
273—274, 278, 324—325, 453—454,
469
— морфологическая концепция 273—
274
— философская концепция 273
— формальный 450
Видообразование 278—279, 318—324,
470
— аллопатрическое 279, 281—293
— гибридизация 279, 293, 304—315
— дифференциация 281—290
— изоляция 281
— катастрофическое 299—302
— конкурентное исключение 290—293
— мгновенное 299, 302
— парапатрическое 279, 293—297
— симпатрическое 297—298
— слияние ареалов 290
— у паразитов 298
— филетическое 279
Виды апомиктические 197
— ацидофильные 386, 388
Виды-компиляторы 314
Вирус гриппа, эволюция 132, 133,
134
Витамин К 264, 381
Вклад родительский 441
Внутриполовой отбор 270, 440
Возрастное распределение стабиль-
ное 350—352
Выживаемость 342—345, 353
Галлы 418—420
Гаметный отбор 266
Гаметы 56, 57, 78
Гаплоидиость 115—121
Гемагглютинин 131—133
Гемоглобин 113—115
Генетическая норма 258
— революция 190
— элита 258
Генетический груз 199—208, 258
— — мутационный 200—201
---обусловленный несовместимо-
стью 203
---сегрегационный 201—203
— — субституционный 203
— фон 74
Генное единообразие 240
— разнообразие 240
Генные частоты, стандартизованная
дисперсия 178
Геном 101
Генотип 76
— и фенотип 262—263
Гены 21, 58, 59, 65—78
— гипостатические 72
— варьирующая экспрессивность 77
— главные (менделевские, олигоге-
ны) 67
— коадаптированные комплексы 210
— множественные 67, 71
— пенетрантность- неполная 77
— полиморфные 239
— регуляторные (гены-регуляторы)
65
— структурные 65
— эпистатические 72, 221
Гены-модификаторы 243
Геологическая колонка 451—452
Гетерозиготность в локусе 240
Гетерозиготы у тетраплоидов 121—
123
Гетерозис одиолокусиый 142, 203,
244, 252, 258
Гетеростилия 196—197
Гетерохроматин 88, 92
Предметный указатель
477
Гибридизация иипрогрессивная 307
Гипотеза, создали; и проверка 33—39
Гистоны 90, 91
Глобины, эволюция 113—115. См.
также Миоглозин, Гомоглобин
Гомознготность в локусе 240
Гомозиготы у тетраплоидов 121—123
Града 470
Групповой отбор 269
Демография 333, 342—354, 363—368
Дивергенция эколзгическая 389
Дисперсия 81—83
— внутрисемейнат 82
— межсемейная 82
ДНК 14, 63—65 90, 103, ИЗ, 236
— избыточная 1В
— репликация 64—65
Дрейф генов 125, 166, 188—192, 258
Дрожжи, увеличений численности по-
пуляции 333, <37
Дрозофила, инве}сии 210, 213, 228—
234
— эффективная тисленность популя-
ции 188
Дуплекс 122
Единицы отбора 265—271
Емкость среды (Л) 336, 337, 341, 359,
374—381, 398, 401, 402
Естественный отбор 125
Жизненная стратегия, концепция 368
Жизненный цикт, обобщенная схе-
ма 51
----характеригики 368
Жизнь, возиикнотение 11—14
Закон независимого распределения
62, 100
— расщепления признаков 60—62,
100
Зародышевая лития 55—57
— плазма 55—57
Зигота 79, 93
Идентичность по происхождению 181
Изменчивость нейтральная и отбор
255—256
<— фенотипическая у божьей коровки
219—220, 224
—-------полевицы 220—221, 222, 225
—-------садовыо улиток 221, 223—
228
Изолирующие механизмы 315—318,
435
Изоферменты 236—239
Инбридинг 179—187
Инверсии. вставочные 229
— независимые 229
— перекрывающиеся 229
— хромосомные, филогения 229—233
Инсектициды фосфорорганические
144—149
Интрогрессия 306—315
Ископаемые остатки, датирование
451
Кариотип 96
Квадруплекс 122
Клада 471
Кладогенез 470
Классификация 462—475
— искусственная 466
— кладистический подход 474
— монофилетическая 470
— фенетический подход 473
Коадаптация 209—215
Коацерваты 12
Когорта 346
Комменсализм 414
Конкуренция 373—389, 433
— интерференционная 373
— коэффициент (а) 375—381
— эксплуатационная 373
Коэволюция 160
Коэффициент инбридинга (F) ISO-
185
— отбора 151
— родства 287
Кровное родство в популяциях чело-
века 186
Кроссинговер 101—ПО
Кряква, изолирующие механизмы 315,
317
— ритуализованное поведение 429
Ксантиндегидрогеназа 238, 239, 240,
242, 243
Кукуруза, изоферменты 236—237
— направленный отбор 157—158
— частота спонтанных мутаций 127
— число хромосом 93
Культурная эволюция 271—273
К-отбор 366, 367, 430
Лемминги, колебания численности
399—400
Лейцииаминопептидаза 236—237
Лептокуртическая кривая 170
Лишайники 264, 414
Логика аристотелева 462, 469
478
Предметный указатель
Малаоксон 145, 146
Междемовый отбор 269
Мейоз 96—101
Мейотические расходы 205—206
Мейотический драйв 140
Меланнзм индустриальный, эволюция
212—215
Менделевская наследственность см.
Наследственность корпускулярная,
Закон расщепления признаков, За-
кон независимого распределения
Метод научный 30—39
Миксомы вирус, эволюция 159—160
Мимикрия 409—413
— бэйтсовская 413
— мюллеровская 413
Миоглобин ИЗ, 115
Митоз 96—101
Модель Розенцвейга — Мак-Артура
402—403
Моногамия 271, 442
Муравьи 411—412
— взаимоотношения с Acacia 414—
416
Мутации 59, 60, 125—130
— индуцированные 60
— и отбор 158—160
— летальные 142
— обратные 128
— повторные 127
— полулегальные 142
— спонтанные 59
— субвнтальные 142
— точковые 127
Мутуализм см. Симбиоз
Наследственность корпускулярная
53—55, 58, 78
— слитная 53—55
Наследуемость 80—84
— у коров фризской породы 218
Нейраминидаза 131—133
Несовместимости система 195—198
Ниша 389—394
— реализованная 391
— фундаментальная 391
Номенклатура 464
Нуклеосомы 91
Нуклеотиды 64
Нуллиплекс 122
Ортогенез 27
Отбор естественный 14—21, 40—51,
68, 141—149, 151—167, 265—271.
См. также Направленный отбор,
Дизруптивный отбор, Групповой
отбор, Отбор родичей, Половой от-
бор, Стабилизирующий отбор
— жесткий 252, 258
— мягкий 252, 258
— родичей 267—269, 439
Пангенез, теория 55
Пангены 55
Паразитоид 397
Паразиты 397
— галлы Solidago 418—420
Паратион 144—146
Партеногенез 195
Пихта бальзамическая 10
Плейотропня 70—73, 330
Плодовитость 345—346, 347, 353
Плотоядные 24—25
Поведение, адаптивность 426—430
— генетический компонент 431—432
— забота о потомстве 421—422, 430
— наследуемость' 430—432
— общественное 432—435
-----защита от хищников 433
-----изменение среды 435
-----повышение способности к кон-
куренции 433
--------эффективности добывания
пищи 433—434
----------размножения 435
— регуляция нервная 425
----- гормональная 425
— репродуктивное 426—430
Поколение родительское 60
Полигамия 271
Полигиния 442
Полидактилия 75
Полиморфизм 243—248
— переходной 243
— устойчивый (сбалансированный)
203, 243—255
Полиплоидность (полиплоидия) 115—
121. См. также Автополиплоид,
Аллополиплоид
Половой отбор 270—271, 440—442
— процесс 22
Полувиды 323
Популяционая биология 10
— генетика 126
Популяция 9. См. также Скрещиваю-
щиеся популяции
— истинная скорость роста (г) 335,
341, 362, 374—381, 398
— плотность 354—355
— подразделение 175—178
— размеры (величина) 166, 194
— рост численности 334—341
-------- логистический 336—341, 374
--------экспоненциальный 334—336,
337
Предметный указатель
479
Посттрансляциошая модификация
239 256
Поток генов 125, 130—132, 134—135,
193, 258. См. также Интрогрессия,
Репродуктивная изоляция
— энергии 420
Преадаптация 315—327
Превосходство 'преимущество) гете-
розигот 248—251
Признаки приобретенные, наследова-
ние 55—57, 273
Принцип конкуэентного исключения
291, 379, 391
— Оккама 38
Приспособленность (адаптивная цен-
ность) 141, 149—154
— абсолютная 141
— относительна! 141, 151, 212
Продолжительнссть поколения 193—
194
Протандрия 197
Протогиния 197
Птицы, величин! кладки 364—366
Пчела медоносная, коэффициент род-
ства 268
— — число хромосом 93
Пшеница, генепческая основа окрас-
ки зерен 68—70
Растительноядные 24—25, 397, 403—
407, 419—420
Рекомбинациошые системы 198—199
Рекомбинация 25, 133, 135—140
— нарушение 140—141, 208—209.
См. также Кроссинговер, Сцепле-
ние, Мейотигеский драйв
Репродуктивная изоляция 278—282
— ценность 352—354
Ритуализация 428
Род, логическа! категория 462—463
РНК 64, 90, 112
г-Отбор 366, 367
Самонесовместимость 197—198
Сателлит 87
Сверхдоминиро>ание см. Гетерозис
Семьи генетические 82
Серповидноклеточная анемия 244—
247
Симбиоз 264, 413—416
Симплекс 122
Систематика 460—462, 464—466
Системы размножения 195—198
— скрещивани! 270—271, 440—442
Скрещивающиеся популяции 265
Случайного дрейфа гипотеза 255, 258
Случайное скрещивание 169—173,
183—184
Случайные явления 166
Смертность, зависящая от плотности
358—362
— не зависящая от плотности 358—
362, 366
Сообщество 265, 370, 407—408
— иерархическая структура 437—440
— организация 435—440
Стабилизирующий отбор 154, 155
Стандартное отклонение 81
Сцепление 101—ПО, 330
— неравновесность 105—ПО, 330
Таксономия 276, 456
Таннины 404
Телейтоспоры 138—139
Термиты 396, 434
Территория 435—437, 440, 442
Тетраплоиды, гаметы 116, 121—123
— менделевская наследственность
121—123
Тли, чередование поколений 196
Трофическая структура сообщества
23—25
Уравнение Вольтерра 337
— Лотки—Вольтерра 398
Уредоспоры 138—139
Устойчивость к ДДТ 144, 147, 207
— — инсектицидам 144—149
----малярии 244—246
Фенилкетонурия 162—163
Фенотип 76
— оптимальный 329—330
Филогения 25, 230, 463
Формула Валенда 178
Харди—Вайнберга закон 78—80, 151
— ----- отклонения 173—180
Хиазма 99, 100, 102
Хищничество 397—413
Хлебная плесень, отбор в пользу ге-
терозигот 249
-----частота спонтанных мутаций
127
Хроматиды 99
Хроматиновая нить 90, 91
Хромосомные аберрации ПО, 210—
212
-----у Clarkia 299—302
Хромосомы 58, 62
— гомологичные 99
— дифференциальное окрашивание
88—89
480 Предметный указатель
Хромосомы
— плечи 87
— половые 58
— слюнных желез дрозофилы 229
— строение 86—89
— человека, число 87, 94
— число у разных видов 93—96
Центромера 86, 94, 96
Цистрон 236
Человек, кривая выживания 343
— инбридинг в популяциях 185—187,
191
— отбор родичей 268
— частота спонтанных мутаций 127
— число потомков 44
Численность популяции эффективная
187—188
Чистая скорость размножения 346—
348, 352
Шалфей, изолирующие механизмы
315—317
Шилохвость, изолирующие механиз-
мы 315
Шмели 412
Эволюция 21—28
— молекулярные часы 452—453
— органическая 14
— химическая 14
Эмиграция см. Поток генов
Эндомитоз 94
Эпигамный отбор 440
Эпистаз 70—73
Эпистатический отбор 108
Эстераза, определение методом элек-
трофореза в геле 235—236
Эухроматин 92
Эффект (принцип) основателя 190—
191
Эцидиоспоры 138—139
Указатель
латинских
названии
Acacia cornigera, симбиотические взаи-
моотношения 414—416
Adalia bipunctata, цветовые морфы
219, 220, 224
Agropecten irradians (Pecten irra-
dians), число потомков 44
Agropyron scabriglutne, электрофоре-
грамма белков 235
— tilcarense, электрофореграмма бел-
ков 235
Agrostis stolonifera, фенотипическая
изменчивость 220, 221, 222, 225
— tenuis, дифференциация популяций
284, 286
Alethopterls 446
Alligator mississippiensis, число хро-
мосом 93
Ambrosia artemisiifolia, число семян
45
Ammikaea septata 446
Anas acuta 315
— platyrhynchos 315
-----число хромосом 93
Anasa tristis 58
Anatinae 470
Anchitherium 456
Anolis, обилие видов 394
— angusticeps 392, 393, 394
— carolinensis 392—394
— distichus, разделение ниш 392, 393,
394
•— sagrei, разделение ниш 392, 393,
394
Anopheles gambiae, избегание инсек-
тицида 147
Anseriformes 470
Anserinae 470
Anthophora 316
Apis mellifera, число хромосом 93
Ascaris megalocephala N&t. univalence,
число хромосом 94
Asclepias 409
Astrocaryum mexicanum, 350
— — возрастная структура популя-
ций 349
Avena barbata, степень ауткроссинга
184
— — частота генотипов 185
В alarms balanoides 384, 385, 387, 389
— cariosus, взаимодействия в сообще-
стве 407
Biston betularia 328
— критическая окраска 46, 49
эволюция индустриального ме-
ланизма 212—214
Blepharisma 380, 382, 383, 385, 386
Doarmia hispidarius, взаимоотношение!
хищник—жертва, мимикрия 406
Bombyx той, число хромосом 93
Bos taurus, число хромосом 93
Calandra oryzae, выживаемость 353
-----плодовитость 353
репродуктивная ценность 353
Calippus 458
Canis familiaris, число потомков 44
--------хромосом 93
Capsella bursa-pastoris, самоопыление
179
Cardamine bulfosa, число семян 45
Castanea dentata 333
Cecropia 425
Cepaea hortensis, фенотипическая из-
менчивость 221
— nemoralis, фенотипическая измен-
чивость 221, 223, 226—228
Cercopithecus 410
Chenopodium album, число семян 45
Chloralictus 316
Chthamalus stellatus 384, 385, 387,
389
Clarkia, мгновенное видообразование
299—302
Clematis fremontii, подразделение, по-
пуляции 175, 176
Colinus virginianus, колебания плот-
ности популяции 356
Condylarthra 454
Coturnix coturnix, заботы о потомст-
ве 422
Crassostrea virginica, число потомков
44
Crataegus 10
Crepis, эволюция хромосом 95, 96
Cricetulus griseus 92
Cycloptera elegans, взаимоотношения
хищник—жертва 406
Cygninae 470
Cyprinus carpio, число хромосом 93
Dactylis glomerata, отбор в пользу ге-
терозигот 247, 249
-----полиморфизм 247
Danaus plexippus 409, 412
Daphnia pulex, динамика численности
340
Dendrocygninae 470
Dentaria laciniata, число семян 45
482
Указатель латинских названий
Didinium nasutum, взаимоотношения
хищник—жертва 397, 398
Digitaria sanguinalis, число хромосом
179
Diplococcus pneumoniae, частота му-
таций 127
Drosophila equinoxiales 321
— insularis 321
— melanogaster, генетическая измен-
чивость 239, 242, 243
----дизруптивный отбор 156
----нарушение рекомбинации 140
---- отбор в пользу гетерозигот 251
----политенные хромосомы 229
----проявление геиов 74
----ритуал ухаживания 427, 428
---- связь генотипа и фенотипа 262
---- частота мутаций 127
----число потомков 44
— miranda, филогенетические связи
231, 232
— paulistorum 321, 323
•--- распространение полувидов 324
— pavlovskiana 322
— persimilis, генетическая изменчи-
вость 242
---- филогенетические связи 231,
232
— pseudoobscura, изменчивость по ин-
версиям 231
----неравновесность по сцеплению
109
---- отбор в пользу гетерозигот 250
---- относительная приспособлен-
ность 212
----полиморфизм 253, 254
------- филогенетические связи 232
----хромосомные перестройки 210
— — частота хромосом 211
-----------ST 233, 243
— tropicalis 321
— willistoni 321, 322, 323
Ectropis consonaria, эволюция индуст-
риального меланизма 213, 214
Encelia farinosa 76
Eosphaeria tilerii 446
Equidae эволюция 454—458
Equus caballus 454
— — число хромосом 93
Escherichia coli 264
— — частота мутаций 127
Euphydryas editha, плотность попу-
ляции 136
Eurosta solidaginis 419
Euterpe globulosa, возрастная струк-
тура популяций 349
Felis 276, 463
— concolor 462
— felis 462
----число хромосом 93
— leo ZU
— onca 462
— tigris 443
Fragaria ananassa 121
— chiloensis 120
— vesca 120
— virginiana 120
G'ilia inconspicua, группа видов, гене-
тическая изоляция 319, 320
Glandularia, полиплоидия 120, 121
Gleditchia montanense 449
Glottidia pyramldata, кривые выжива-
ния 344
Glycine max, число хромосом 93
Gossipium barbadense 179
----число хромосом 93
Gnorimeshema gallaesolidaginis 419
Gutierrezia bracteata 87
Haaniella echinata, мимикрия 406
Haplopappus gracilis 87
----число хромосом 94
Helianthus annuus ssp. annuus 308—
314
----annuus var. macrocarpus 308,
309
--------lenticularis 308—314
--------texanus 308—314
— bolandery 310—313
— debilis 309
----var. cucumerifolius 309, 310,
312, 313
— petiolaris 312—314
Heliconius 411
Hieracium и законы Менделя 35
Hipparion 456
Homarus americanus, число потомков
44
Homo sapiens, частота мутаций 127
---- число потомков 44
Hyla versicolor, репродуктивное пове-
дение 427
Hypohippus 456
Ipomea batatus, число хромосом 93
Lactuca saliva, число хромосом 73
Lasthenia chrysostoma, жизнеспособ-
ность пыльцы гибридов 322
— — репродуктивная изоляция 319,
320 321
— macrantha 319, 320, 321
Layia glandulosa, дифференциация
популяций 288
— platiglossa, дифференциация попу-
ляций 287
Указатель латинских названий
483
Leavenworthina :ylosa 274
Lemmus trimueraatus, колебания чис-
ленности 400
Lepidodendron 44
Limenitis archipps, бэйтсовская ми-
микрия 412
Lucilia cuprina, динамика численно-
сти 339
Lycopersicon esulentum, нарушение
рекомбинации 40
---- число хроисом 93
— pimpinellifolitn 140
Lycorea 411
Lymantra monaca, эволюция индуст-
риального мелнизма 214
Lynx canadensis.KoneQawiH численно-
сти 355
Macaca fuscata, развитие культурных
признаков 272
Macrotermes, изенение среды 434
Mechanitis 411
Melineae 411
Merychippus 456-457
Mesohippus 456-157
Microtus agresti, кривая выживания
лабораторной опуляции 343
— pensilvanicuspMmpauHa 439
Miohippus 457
Mitella polymere, взаимодействие в
сообществе 40
Musca domestic, гены устойчивости
к пестицидам 47
----устойчивось к инсектицидам
145—147, 148
— — число потиков 44
Mus musculus, чстота мутаций 127
Mytilus californmus, взаимодействия
в сообществе 08
Nannippus 456
Neohipparion 45(
Neurospora crasi, частота мутаций
127
Nicotiana longifma, гипотеза множе-
ственных фактров 69
— otophora 117
• — sylvestris 117
— - tabacum, аллполиплоидия 117
•---плейотропя 73
----самоопы.тше 179
----система можественных генов
Notoungulata 32
Ophioglossum vlgatum, число хро-
мосом 93
Ortoptera 406
Pandorina 264
Parahippus 457
Paramecium 337
— aurelia, колебания плотности по-
пуляции 356
----модели конкуренции 380—383,
385, 386
— bursaria 380, 382, 383, 385, 386
— caudatum, взаимоотношения хищ-
ник—жертва 397, 398
----модели конкуренции 380—383,
386
Pediculus humanus, кривая плодови-
тости 345
Pentaclathra macroloba, возрастная
структура популяции 349
Phaseolus vulgaris, число хромосом
93
Phasianus colchicus, рост численности
популяции 337
Phyllium sissifolium, мимикрия 406
Pisaster ochraceus, взаимодействия в
сообществе 407
Pisum sativum, расщепление призна-
ков 173
Plasmodium falciparum 244
Platanus acerifolia 302
— occidentalis 302
— orientalis 302
Pletodon cinereus, неравновесность no
сцеплению 109
Pliohippus 456
Polistes, иерархическая система 439
Polystichum californicum 87
Potentilla glandulosa, дифференциа-
ция популяций 282—285
Prenanthes alba, число семян 45
Pseudomyrmex ferruginea, симбиоти-
ческие взаимоотношения 414, 416
Puccinia graminis, жизненный цикл
139
Pyrus communis, число хромосом 93
Rana catesbiana, число потомков 44
— pipiens, число хромосом 93
Raphanus raphanistrum, конкуренция
388
Rhagoletis pomonella, симпатрическое
видообразование 298
Saccharomyces exiguus, колебания
плотности популяции 357
Salmo aquabonita, число потомков 44
— trutta, число потомков 44
Salmonella typhimurium, частота му-
таций 127
Salvia apiana, интрогрессивная гибри-
дизация 306
— — репродуктивная изоляция 31о,
316
— mellifera, интрогрессивная гибриди-
зация 306
484
Указатель латинских названий
Salvia mellifera
----репродуктивная изоляция 315,
316
Sanicula gregaria, число семян 45
Sassafras 445
Sinapsis arvensis, конкуренция 388
Sitta neymayer, смещение признаков
296
— tephronota, смещение признаков
296
Solidago 416
— распределение биомассы в различ-
ных местообитаниях 417
— altissima, число семян 45
— canadensis 418
----кривые роста 419
----число семян 45
Spalax ehrenbergi, парапатрическое
видообразование 293—295
— mimtus 295
Sphenopteris 446
Stellaria media, самоопыление 179
Stephanomeria exigua ssp. coronaria,
парапатрическое видообразование
296, 297
— malheurensis, парапатрическое ви-
дообразование 296, 297
Sturnus vulgaris, выживаемость птен-
цов 365
— — забота о потомстве 421
----число потомков 44
Taraxacum officinale, закрытые ре-
комбинационные системы 198
-----конкуренция между биотипам»
373
Tegeticula, симбиотические взаимоот-
ношения 414, 415
Tenebrio moliior, соотношение между
имитаторами и моделями 413
Terrapene Carolina, число потомков 44
Тettigoniidae 406
Thais emarginata, взаимодействия в
сообществе 407
Tradescantia caniculata, интрогрессив-
ная гибридизация 307
— subaspera, интрогрессивная гибри-
дизация 307
Triticum aestivum 179
-----число хромосом 93
Тurdus ericetorum 227
Uta stansburiana, кривая выживания
344, 348
-----плодовитость 364
Vespula maculata, число потомков 44
Viola tricolor, эпистаз 72
Volvox 264
Xylocopa brasilianorum 316
Zea mays, направленный отбор 157
-----частота мутаций 127
-----число хромосом 93
Yucca, симбиотические взаимоотноше-
ния 415
Оглавление
Предисловие редктора перевода
5
Предисловие.......................................................... Т
ЧАСТЬ ПЕРВАЯ
Глава 1. Эволюия, популяции н естественный отбор.................... 9>
Виды и популяции................................................
Возникиовене жизни иа Земле.........................................11
Концепция етественного отбора...................................14-
Эволюция .......................................................21
Ограничения сопряженные с эволюцией................................23-
Ошибочные редставления об эволюции.................................26-
Литератра.......................................................28-
Глава 2. Научиь4 метод..............................................29
Приобретеии знаний..................................................29
Структура нучиого метода .......................................... 31
Составные чети, которые должна содержать любая гипотеза . . 3&
Литератра ...................................................39
Глава 3. Теория эволюции путем естественного отбора....................40-
Эволюция птем естественного отбора. Проблема и факты ... 41
Минимальны условия, необходимые для эволюции путем естественно-
го отбора ......................................................43
Размножеие....................................................43
Корреляции между вероятностью выживания и фенотипом 45-
Наследствнность...............................................45-
Неизмеииост внешних условий.....................................47
Источники именчивости...........................................43
О характеретеории естественного отбора..........................50»
Литератра....................................................51
ЧАСТЬ ВТОРАЯ
Глава 4. Наследтвеиность и источники изменчивости......................53
Корпускулярая и слитная наследственность...........................53
Теория паигнеза...................................................55-
Зародышева линия и соматическая линия.............................55-
Корпускулярая теория наследственности..................... . . 58
Единичные ены и системы множественных генов ... 62
Эпистаз и пейотропия.......................................... . 79
Гены, генотпы и фенотипы.................................. . . 73
Закон Хардг—Вайнберга и поддержание изменчивости . , . 78
Фенотипичесия изменчивость и наследуемость ... , . 89
Литератра.............................................. . 84
Глава 5. Гены и хромосомы................................... . 88
Строение хрмосом..................................................88
Ультраструкура хромосом..........................................89-
Число хромсом.....................................................93
Митоз и мевз.................................................... 96-
Сцепление и кроссииговер...................................... . Ю1
Неравиовеность по сцеплению.......................................105-
486
Оглавление
Изменения структуры хромосом....................................110
Гаплоидность, диплоидность и полиплоидность.....................115
Менделевская наследственность у тетраплоидов....................121
Краткие выводы..................................................123
Литература..................................................124
Глава 6. Факторы эволюции..........................................125
Мутационный процесс ............................................126
Поток генов.....................................................130
Рекомбинация . . ........................................135
Нарушение рекомбинации..........................................140
Мейотический драйв............................................140
Отбор...........................................................141
Эволюция устойчивости к пестицидам у насекомых................144
Вычисление приспособленности популяции..........................149
Отбор против гомозиготных рецессивов..........................151
Действие отбора на фенотипические признаки....................154
Мутационный процесс и отбор.....................................158
Полная элиминация рецессивов....................................160
Основная теорема естественного отбора...........................163
Случайные явления .............................................. 166
Краткие выводы..................................................166
Литература..................................................167
Глава 7. Иибридииг и случайный дрейф генов.........................169
Случайное скрещивание...........................................169
Случайная утрата аллелей в популяции............................173
Подразделение популяции и формулы Валенда.....................175
Инбридинг.......................................................179
Коэффициент инбридинга........................................180
Инбридинг в популяциях человека...............................185
Эффективная численность популяции...............................187
Случайный дрейф генов...........................................188
Литература..................................................192
Глава 8. Рекомбинация.................................- .... 193
Факторы, оказывающие влияние на рекомбинацию...................193
Классификация рекомбинационных систем...........................198
Генетические расходы и концепция генетического груза . . . . 199
Общезначима ли концепция генетического груза?.................203
Энергетические расходы, связанные с аутбридингом..............205
Мейотические расходы.............................................205
Выгоды, которые дает ауткроссинг..............................206
Нарушение рекомбинации.........................................208
Коадаптация.......................................................209
Эволюция индустриального меланизма............................212
Краткие выводы.................................................215
Литература.................................................216
Глава 9. Генетическая изменчивость в природных популяциях .... 217
Типы генетической изменчивости и популяциях растений и животных 218
Полиморфизм по окраске в природных популяциях садовой улитки Ce-
paea nemoralis....................................................221
Полиморфизмы по хромосомным инверсиям у дрозофилы .... 228
Генотипическая изменчивость....................................233
Метод электрофореза в геле....................................234
Оглавление
487
Как измерить изменчивость аллоферментог з популяциях
Генетическая изменчивость у разных_видов рутений и животных
Механизмы поддержания генетической измендвости . . . .
Однолокусный гетерозис....................................
Отбор в пользу гетерозигот • ................
Отбор в изменчивой среде .................................
Нейтральная изменчивость . ................
Краткое резюме и заключение ................
Литература........................ .....................
ЧАСТЬ ТРЕТЬЯ
Глава 10. Индивидуумы, популяции и виды.......................
Генотип н фенотип.................... .....................
Уровни организации живого..................................
Единицы отбора.............................................
Гаметный отбор...........................................
Отбор родичей............................................
Групповой отбор ................... .....................
Половой отбор............................................
Культурная эволюция................ .....................
Вид........................................................
Концепция вида ..........................................
Высшие категории.........................................
Литература....................... .....................
Глава 11. Модели видообразования
239
241
243
244
248
252
255
256
259
261
261
263
265
266
268
269
270
271
273
273
274
276
278
Способы видообразования ........................................ 279
Аллопатрическое видообразование................................281
Парапатрическое видообразование................................293
Симпатрическое видообразование.................................297
Мгновенное видообразование.....................................299
Литература...................................................302
Глава 12. Гибридизация, преадаптация и адаптация
Гибридизация .............................................
Действие гибридизации и интрогрессии на примере подсолнечника
Изолирующие механизмы......................................
Процесс видообразования и регулирующие до факторы
Определение вида..........................................
Преадаптация и происхождение эволюционное новшеств
Адаптация ................................................
Являются ли адаптивные признаки оптимиьными? . . . .
Литература.............................................
Глава 13. Рост и регуляция численности популяшй
Рост численности популяции................................
Экспоненциальный рост...................................
Логистический рост......................................
Демография................................................
Выживаемость............................................
Плодовитость............................................
Чистая скорость размножения.............................
Стабильное возрастное распределение ....................
Репродуктивная ценность ................................
Плотность.................................................
304
304
308
315
318
324
325
327
329
330
332
334
334
336
342
342
345
346
350
352
354
488
Оглавление
Регуляция численности популяции . 355
Смертность, зависящая и не зависящая от плотности....................358
Демография и естественный отбор......................................363
Литература . 368
Глава 14. Конкуренция и теория ниши
370
Конкуренция....................................................371
Модели конкуренции и роста двух видов, сосуществующих в одной
среде........................................................374
Концепция ниши.................................................389
Литература..................................................395
Глава 15. Неконкурентные взаимодействия между видами
396
Взаимоотношения хищник—жертва..................................397
Предупреждающая окраска и эволюция мимикрии..................409
Симбиоз........................................................413
Взаимодействия между видами и различные аспекты их жизни . 416
Адаптивные преимущества, создаваемые заботой о потомстве . . 421
Литература..................................................423
Глава 16. Поведение и эволюция.......................................424
Адаптивность поведения...........................................426
Наследуемость поведения..........................................430
Общественное поведение...........................................432
Организация сообществ............................................435
Территория.....................................................435
Иерархическая структура........................................437
Половой отбор и система скрещивания..............................440
Заключение.......................................................443
Литература...................................................443
Глава 17. Палеонтология н эволюция
444
Важное значение ископаемых остатков для эволюционных исследований 445
Как происходит фоссилизация, т. е. образование ископаемых остатков 447
Проблема недостающих частей ... -...........................450
Датирование ископаемых остатков. Относительный и абсолютный воз-
раст ....’..................................................451
Молекулярные часы эволюции..................................452
Концепция вида во времени и в пространстве ....................... 453
Эволюция семейства лошадей (Equidae)........................454
Скорость эволюции...........................................457
Литература..............................................458
Глава 18. Систематика и эволюция
Современная систематика......................................
Основы классификации.........................................
Таксономия, систематика, классификация и иомеиклатура
Искусственные, естественные и филогенетические системы классифика
ции..........................................................
Сопоставление феиетической и филогенетической классификации
Задача иа будущее............................................
Литература...............................................
'Предметный указатель............................................
Указатель латинских названий.....................................
460
462
464
466
469
474
475
476
481