/



























































































































































































































































Текст
Я.А.БИРШТЕЙН
ГЕНЕЗИС
ПРЕСНОВОДНОЙ,
ПЕЩЕРНОЙ
И ГЛУБОКОВОДНОЙ
ФАУН
I
АКАДЕМИЯ НАУК СССР
МОСКОВСКОЕ ОБЩЕСТВО ИСПЫТАТЕЛЕЙ ПРИРОДЫ
Я. А. БИРШТЕЙН
ГЕНЕЗИС
ПРЕСНОВОДНОЙ,
ПЕЩЕРНОЙ
И ГЛУБОКОВОДНОЙ
ФАУН
Ответственный редактор
доктор биологических наук Г М. БЕЛЯЕВ
ж
Ji
МОСКВА «НАУКА» 1985
УДК 577.472
Бирштейн Я. А. Генезис пресноводной, пещерной и глубоководной
фаун. — М.: Наука, 1985.
Книга содержит материалы по вопросам географического распро-
странения, происхождения, филогении и эволюции водной фауны конти-
нентальных (наземных и пещерных) водоемов и глубоководной фауны
океана, главным образом на примерах высших ракообразных. В книге
рассматриваются проблема реликтовости, вертикальная биологическая
зональность в океане, вопрос о гигантизме глубоководной фауны, роль
трофического фактора в эволюции этой фауны, обобщены данные по
зоогеографии пещерной фауны Советского Союза, содержатся теорети-
ческие обобщения по эволюции и видообразованию.
Для зоологов, гидробиологов, биоспелеологов, зоогеографов, для
специалистов, разрабатывающих вопросы теории эволюции и видообра-
зования. •
Табл. 9, ил. 17, библиогр. 630 назв.
Рецензенты:
А. П. АНДРИЯШЕВ, М. Е. ВИНОГРАДОВ
2001050100-402
042(02)-85
219-85—IV
© Издательство «Наука», 1985
ПРЕДИСЛОВИЕ
Творческое наследие Якова Авадьевича Бирштейна чрезвычайно велико
и разнообразно: оно охватывает вопросы систематики современных и
ископаемых ракообразных, экологии, этологии, биологии, физиологии,
зоогеографии, эволюции и филогении фауны наземных и подземных (пещер-
ных) пресных водоемов и глубоководной фауны океана. И системати-
ческие работы Я. А. Бирштейна, и его работы, содержащие широкие
теоретические обобщения, в большинстве случаев не утратили своего зна-
чения, а зачастую и научной новизны. Между тем многие публикации
Я. А. Бирштейна, особенно в виде отдельных изданий, уже давно стали
библиографической редкостью, а другие материалы разбросаны по самым
различным периодическим изданиям. Таким образом, назрела настоятельная
необходимость объединить в одном томе по возможности большее число
работ Я. А. Бирштейна, имеющих общебиологический интерес.
Однако выбрать для публикации работы, освещающие все стороны
научного творчества Я. А. Бирштейна, оказалось просто невозможным.
Поэтому пришлось отказаться от публикации чисто систематических статей,
а из работ, посвященных вопросам систематики, в том числе описанию
очень многочисленных новых видов и родов, включить лишь главы, обоб-
щающие изученный материал. В основном же в том включены работы
общего характера, в которых рассматриваются вопросы зоогеографии,
эволюции и филогении пресноводной, пещерной и океанической глубоко-
водной фаун. Из-за ограниченности объема пришлось также отказаться
от повторной публикации двух работ, имеющих несомненный общетеоре-
тический интерес, написанных Я. А. Бирштейном в соавторстве с Л. А. Зен-
кевичем (см. список работ Я. А. Бирштейна за 1955 и 1961 гг.), так как
эти работы вошли в Двухтомное издание избранных трудов Л. А. Зенкевича
(М.: Наука, 1977).
В настоящее издание включены полностью или частично 17 работ
Я. А. Бирштейна, а также приведен по возможности полный список его
научных публикаций. В примечании к названию каждой публикуемой
работы указаны ее первичные выходные данные. В случаях, когда работа
публикуется не полностью, указано, что она дается с сокращениями, или
конкретно указано, какая часть работы помещена.
Литература, на которую имеются ссылки в разных статьях, сведена
в единый список литературы.
В тексты статей внесены исправления замеченных опечаток и погреш-
ностей в литературных ссылках. В тех случаях, когда это удалось установить,
3
внесены уточнения латинских названий животных, которые изменились после
первоначальной публикации той или иной статьи. В списке литературы
уточнены названия работ и их выходные данные, не всегда указанные
в оригиналах статей. В случаях, когда найти ту или иную работу для
внесения таких уточнений не удалось, она приведена в том виде, как
была дана в оригинале статьи. Географические названия даны в современном
написании.
В случаях, когда в статье упоминается впервые описанный в ней
таксон с обозначением sp. n., gen. п. и т. п., эти обозначения заменены
фамилией автора (авторов) таксона в латинской транскрипции и дано
соответствующее примечание. Примечания без пометки были сделаны
автором.
В подборе материала, уточнении литературных ссылок, латинских назва-
ний животных и т. п. принимали участие сотрудники кафедры зоологии
и сравнительной анатомии беспозвоночных животных МГУ Н. А. Заренков
и С. И. Левушкин, а также В. Я. Бирштейн, Е. И. Изюмова и Ю. Г Чиндо-
нова.
4
ОСНОВНЫЕ НАПРАВЛЕНИЯ НАУЧНОЙ ДЕЯТЕЛЬНОСТИ
ЯКОВА АВАДЬЕВИЧА БИРШТЕЙНА
(7 мая 1911—8 июля 1970)
Имя Якова Авадьевича Бирштейна, талантливого советского биолога
с удивительно широким кругом научных интересов, прекрасного человека
и педагога, и сейчас, спустя 15 лет после его кончины, много говорит
ученым разных специальностей. Идеи Якова Авадьевича оказывают влияние
на пути развития определенных направлений науки. Его работы читают
и цитируют океанологи, палеонтологи, зоологи широкого профиля — физио-
логи, систематики, эволюционисты, зоогеографы и экологи.
Я. А. Бирштейн был крупным специалистом по морской, пресноводной
и пещерной фаунам. Сфера его научных интересов была очень обширной:
выявление закономерностей формирования фаун океанических глубин и
подземных водоемов, солоноватоводных морей и пресных водоемов, иссле-
дование морфологических адаптаций животных к чрезвычайно своеобразным
условиям среды, изучение таксономии, зоогеографии, филогении, экологии
и палеонтологии многих групп высших ракообразных, проведение экспе-
риментальных работ по осморегуляции и солеустойчивости водных беспоз-
воночных — это лишь неполный перечень вопросов, которыми Яков Авадье-
вич увлеченно и плодотворно занимался. Он сочетал в себе разносторонние
качества ученого-теоретика, практика, экспериментатора. Его исключи-
тельные одаренность и трудоспособность позволяли ему, помимо научной
деятельности, отдавать много сил и времени педагогической, редакторской
и научно-организационной работе. Он умел работать сам и умел готовить
к научной работе молодую смену.
Яков Авадьевич был интеллигентом в самом лучшем и широком смысле
этого слова. Получив прекрасное воспитание, высококультурный, он тонко
разбирался в литературе и искусстве, обладая незаурядным художественным
вкусом, очень любил поэзию, особенно русскую, хорошо знал ее. Свои
лекции и выступления он умел оживить к месту приведенной стихотворной
строкой. Даже к своим научным работам он нередко брал стихотворные
эпиграфы, которые служили не столько украшением, сколько кратким
и точным введением в развиваемую автором тему.
Культура и знание, эрудиция и оригинальность мышления, а также
многие человеческие качества Якова Авадьевича, среди которых основным
можно считать высокую принципиальность, постоянно привлекали к нему
талантливую молодежь. Вместе с тем он был непримирим к карьеризму
и демагогии в науке. Я. А. Бирштейн обладал яркой творческой инди-
видуальностью. Стиль его научного творчества был неотделим от его
личности и проявлялся не только в различных аспектах интеллектуальной
5
Я. А. Бирштейн
деятельности, но и в характерном для него умении создавать дружескую
атмосферу при работе с коллегами и учениками в повседневной жизни
и в экспедициях. В полевых условиях, как правило, человек познается
наиболее полно. Обаяние Якова Авадьевича способствовало тому, что он
всегда был центром притяжения независимо от возраста, склонностей
и профессиональных интересов его спутников. Те, кому посчастливилось
с ним работать, всегда будут ему благодарны не только за знания, которыми
он щедро делился, но и за радость общения с ним.
Характерной особенностью Якова Авадьевича было ясное, строго логи-
ческое изложение мыслей. Оно давалось ему легко как в устной речи,
так и в стиле его статей. Обладая даром писать четко и понятно о самых
сложных проблемах, заостряя внимание на самом существенном и отодвигая
на второй план второстепенные сведения, он помогал и молодежи учиться
выражать свои мысли строго и логично. Стилю написания научной работы
он всегда уделял много внимания, и его статьи и книги — великолепный
образец стилистики. Якова Авадьевича всегда было интересно слушать
и всегда интересно читать.
Великолепно ориентируясь в нарастающем потоке фактов, гипотез,
вопросов, возникающих в связи с развитием зоологии и смежных дисциплин,
он ясно представлял общее направление развития своей науки и умел
найти те проблемы, заниматься которыми было нужно и интересно. Понимая,
что сложные вопросы быстрее и глубже решаются совместно с другими
учеными, он находил для этих целей научные организации, где такие
проблемные задачи ставились и могли быть решены. Проведение научных
исследований во многих случаях было связано у Якова Авадьевича с боль-
шими или малыми коллективами, в которые он вливался или создавал сам.
Умение работать с соавторами, независимо от их научного ранга,
было характерной чертой Якова Авадьевича. Он всегда участвовал в кол-
6
лективной научной деятельности — от сбора материалов в полевых условиях
до окончательного оформления результатов исследований. Это был надеж-
ный, добрый и внимательный товарищ, ум и эрудиця которого делали
совместную работу с ним интересной и содержательной. Совместные
научные работы с Яковом Авадьевичем были прекрасной школой для
молодежи, а для коллег — тесным дружеским союзом.
Яков Авадьевич не был кабинетным ученым. Он очень любил сам
собирать материал, спускаясь в пещеры или плавая на экспедиционных
судах, и обладал незаурядными качествами полевого работника: был очень
вынослив, мог длительное время спать по три-четыре часа в сутки,
участвовать во всех сторонах экспедиционной деятельности — от усовер-
шенствования методики сбора и обработки материалов до управления
лебедкой и упаковки и погрузки собранных материалов.
Яков Авадьевич родился 7 апреля 1911 года в Киеве в семье врачей —
отец его был хирургом, мать — врачом-окулистом. В 1914 г. семья Бирш-
тейнов переехала в Москву, в которой Яков Авадьевич и прожил до
конца жизни. После окончания средней школы семнадцатилетним юношей
Я. А. Бирштейн поступил на Биологическое отделение Физико-математи-
ческого факультета Московского университета, которое окончил в 1932 г.
После окончания университета Яков Авадьевич в течение шести лет работал
во Всесоюзном научно-исследовательском институте рыбного хозяйства
и океанографии (ВНИРО). За этот период он возглавлял экспедиции
на Каспийское море, участвовал в работах на Дальнем Востоке. С 1936 г.
он начал совмещать работу во ВНИРО с педагогической работой в уни-
верситете, а с 1938 г. полностью перешел на работу в Московский уни-
верситет на кафедру зоологии беспозвоночных, где сначала был ассистен-
том, затем доцентом, профессором, а с 1962 г. — заместителем заведующего
кафедрой.
Уже в первых своих работах Яков Авадьевич не ограничивается одним
описанием фактов, даже интересных и новых, а пытается оценить их
значение для решения крупных проблем. За цикл работ по систематике
и зоогеографии пресноводных, а том числе и подземных, высших ракообраз-
ных ему в 1935 г. была присуждена ученая степень кандидата биологи-
ческих наук без защиты диссертации. Одновременно в составе группы
ученых он начинает большую работу вначале по выявлению причин обед-
нения кормовой базы промысловых рыб Каспийского моря, а затем и по ее
улучшению путем акклиматизации в Каспии некоторых донных беспозво-
ночных Азовского моря. Для этого были предварительно проведены при его
активном участии многочисленные экспериментальные исследования по
осморегулярным способностям беспозвоночных Каспия и других водоемов,
а в 1939 г. Яков Авадьевич непосредственно участвовал в осуществлении
акклиматизационных работ.
В 1948 и 1949 гг. Яков Авадьевич возглавил экспедиции на Каспийское
море, имевшие целью выяснить результаты проведенной до войны аккли-
матизации в нем азовского червя нереис. Результаты этих работ были
поразительными. Нереис стал массовой формой каспийской фауны и основ-
ным компонентом рациона осетровых рыб, что привело к повышению
их упитанности. За книгу «Акклиматизация нереис в Каспийском море»
Якову Авадьевичу вместе с другими ее авторами была присуждена первая
премия Московского общества испытателей природы.
7
В этот же период Яков Авадьевич интенсивно изучает пещерную фауну,
фауну Байкала, водоемов Арктики, Дальнего Востока, Сибири и Кавказа.
Он публикует интереснейшие работы о происхождении фреатической под-
земной фауны, адаптациях пещерных организмов, всесторонне разбирает
проблему реликтов. Эти исследования были обобщены им в докторской
диссертации «Реликты пресных и солоноватых вод СССР», которую он
защитил в 1947 г. В ней высказаны оригинальные мысли о консерватизме
пресноводной фауны и роли пресных вод для сохранения реликтов, о зна-
чении замедления темпа эволюции в происхождении реликтовых форм.
Одной из любимых работ автора была его монография о пресноводных
ракообразных подотряда Asellotta, изданная в 1951 г. в серии «Фауна
СССР» и впоследствии переведенная на английский язык. Он до конца своих
дней сохранил глубокую привязанность к «водяным осликам» и к каспий-
ской фауне, лучшим знатоком которой он был. Каспийское море он начал
изучать сразу же после окончания университета, а незадолго до его смерти
вышел «Атлас беспозвоночных Каспийского моря», одним из редакторов
которого он был и в котором опубликованы написанные им разделы.
С начала 50-х годов Яков Авадьевич включается в исследования глубо-
ководной океанической фауны, сборы которой проводятся советскими
экспедиционными судами в различных районах Мирового океана, и сам
неоднократно принимает участие в тихоокеанских экспедициях на «Витязе».
При этом его занимал широкий круг вопросов — от систематики и зоогео-
графии высших ракообразных до решения проблемы происхождения и
эволюции глубоководной фауны, которой он занимался в течение многих лет.
Сопоставление систематического состава глубоководной фауны с данными
геологии и палеонтологии было одним из подходов к решению вопроса
о геологической древности населения абиссали. За цикл работ по этой
проблема Якову Авадьевичу вместе с Л. А. Зенкевичем была присуждена
Ломоносовская премия Московского университета.
В поисках новых методов оценки темпов эволюции глубоководных
животных Якова Авадьевича привлекают данные по их экологии. Выяв-
ление роли трофического фактора, определяющего различные направления
адаптаций глубоководных животных, позволили ему приблизиться к решению
этой проблемы. Впервые высказав эту мысль на II Международном океано-
графическом конгрессе в 1966 г., он долгое время тщательно подбирал факты
и материалы, прежде чем подготовить к печати совместно с М. Е. Вино-
градовым большую оригинальную работу, которая была опубликована уже
после его смерти.
Сколь бы широки ни были рассматриваемые Яковом Авадьевичем
вопросы, их решение всегда базировалось на изучении громадного мате-
риала по высшим ракообразным. К изучению этой группы он привлекает
многочисленных учеников. Только особенно любимые им равноногие рако-
образные обрабатываются без привлечения коллег. Обработка колоссальных
сборов равноногих ракообразных легла в основу большого числа статей
и монографии «Глубоководные равноногие ракообразные северо-западной
части Тихого океана», за которую Яков Авадьевич в 1963 г. вторично
получил премию Московского общества испытателей природы. В этой
монографии, помимо таксономии, поднимаются вопросы происхождения
и эволюции фауны изопод, их вертикального распределения, питания,
размножения, зоогеографии и многие др.
8
Яков Авадьевич был карцинологом широкого профиля. Он описал
больше сотни новых видов, десятки новых родов, семейства и отряды
ракообразных. В его честь названы многие виды ракообразных, а также
виды и роды во многих других группах животного царства.
Таксономический анализ для Якова Авадьевича всегда был только
первым этапом обработки материала, за которым следовали зоогеографи-
ческие, экологические и эволюционные обобщения. Им написаны разделы
о высших ракообразных в «Основах палеонтологии» и в «Жизни пресных
вод СССР», главы, посвященные ракообразным, в «Курсе зоологии» для
высших учебных заведений и в «Жизни животных».
Особое место в жизни Якова Авадьевича занимала фауна пещер и
подземных вод. Он неоднократно принимал участие в комплексных спелео-
логических экспедициях. Своеобразный мир обитателей пещер изумил его
еще в юности. Едва перешагнув двадцатилетний рубеж, он опубликовал
описание нового вида подземного рачка, открытого им в Закавказье.
В 1935 г. в Грузии, в окрестностях Кутаиси, состоялось первое знакомство
с пещерами. Затем последовали многочисленные поездки в карстовые
районы Крыма и Западного Закавказья. Получив докторскую степень
и звание профессора, он не мог усидеть за столом кабинета, если ему
представлялся случай побывать в любимых пещерах. В 1959 г. он плавает
на резиновой лодке по подземным рекам Абхазии, в начале 60-х годов
спускается в глубокие карстовые колодцы Крыма. И везде с увлечением
собирает обитателей подземных вод. Его богатые пещерные сборы дали
обильный материал для обработки многим специалистам, которых он сумел
заинтересовать проблемами биоспелеологии. Сам же он занимался как
систематик некоторыми группами пещерных ракообразных.
Проблемы пещерной биологии всегда рассматривались Яковом Авадье-
вичем с широких общебиологических позиций. Начав с выяснения состава
малоизученной подземной фауны, он пришел сначала к интересным зоогео-
графическим обобщениям, а затем и к разработке вопросов эволюции
и происхождения этой фауны. Он умел связать пещерную тематику с вопро-
сами сравнительной морфологии, видо- и расообразования. Достойное
место заняли биоспелеологические работы в других его исследованиях,
приведших к широкому освещению проблемы реликтов.
Яков Авадьевич отдал много сил организации биоспелеологических
работ в СССР. Долгие годы, будучи лидером советской биоспелеологии,
он возглавлял и поддерживал любую попытку проведения фаунистического
обследования пещер. По его инициативе в советской зоологической перио-
дике родилась серия публикаций разных авторов под общей рубрикой
«Biospeologica sovietica». Сейчас число публикаций этой серии составляет
уже около 50. Работы Якова Авадьевича по фауне пещер принесли
советской биоспелеологии мировое признание. В качестве одного из ведущих
специалистов мира по пещерной фауне он участвовал в работе между-
народных спелеологических конгрессов, поддерживал тесные деловые связи
с крупнейшими биоспелеологами Франции, Италии, Румынии и других
стран.
В 1956 г. Яков Авадьевич начал обработку большой коллекции иско-
паемых высших ракообразных из отложений самого различного возраста —
от палеозойских до кайнозойских. В работах по ископаемым ракообразным
он также пытался выйти за пределы чистой систематики. В одной из
9
первых своих палеонтологических работ, посвященной десятиногим ракооб-
разным из палеогенового моря Ферганы, он делает попытку восстановить
гидрологические условия, существовавшие в этом водоеме, основываясь
на тщательном изучении ископаемого материала и знании экологии группы.
В других работах прослеживает морфологические изменения в некоторых
группах высших ракообразных во времени, сравнивая современные и иско-
паемые формы, что позволяет ему выявить изменения ареалов и определить
пути расселения этих групп, а также обсудить вопросы их филогении.
Более 30 лет научная деятельность Якова Авадьевича тесно перепле-
талась с педагогической в стенах Московского университетами одновременно
более 10 лет он читал лекции по зоологии беспозвоночных и по гистологии
с основами эмбриологии и руководил аспирантами в Ярославском педа-
гогическом институте. Вместе с сотрудниками и аспирантами этого института
он включился в научно-исследовательские работы по изучению биологии
Рыбинского, а затем и Горьковского водохранилищ.
Яков Авадьевич был блестящим лектором. Изложение материала он
подчинял внутренней логике, фактический материал тщательно отбирал,
включал новейшие данные науки с одной целью — убедительно продемон-
стрировать правоту развиваемых им теоретических концепций. Обладая
великолепной памятью, он не был привязан к конспекту: поступавшие
с места вопросы превращали иной раз его лекцию в увлекательную
беседу, в которой он поражал всех эрудицией и умением вовремя отыскать
нужные факты и доказательства, его всегда слушали с живым интересом,
в любой аудитории его слова находили отклик. Следует особо отметить,
что в беседах или читая лекции Яков Авадьевич никогда не стеснялся
обнаружить свое незнание каких-либо частностей и всегда открыто призна-
вался в этом, обещая уточнить неясное место. Он охотно возвращался
к этому вопросу, когда находил исчерпывающий ответ.
Педагогическая деятельность Якова Авадьевича не ограничивалась чте-
нием лекций, он любил и умел работать с молодежью. Множество учеников
окружало его: студенты, пишущие курсовые и дипломные работы, и аспи-
ранты шли к своему руководителю с надеждами, планами, заботами
и никогда не уходили без умного, дельного совета, всегда чувствовали
неподдельную заинтересованность в своей работе и дружеское участие.
Многие из студентов, начав с курсовых работ, впоследствии защищали
под его руководством диссертационные работы. Среди его учеников,
плодотворно работающих во всех уголках Советского Союза, доктора
и кандидаты биологических наук, имена которых хорошо известны в нашей
стране и за рубежом.
В значительной мере как педагогическую можно рассматривать и ре-
дакторскую деятельность Якова Авадьевича. Эта работа была чрезвычайно
велика по объему и имела громадное значение для формирования молодого
поколения ученых. В своих публичных и журнальных выступлениях он
боролся с недобросовестностью и низким качеством научных публикаций.
Значительное место в его деятельности занимали отзывы, рецензии и раз-
личная редакционная работа. Он умел не только остро и едко критиковать,
когда считал это нужным, но и радоваться чужим успехам и достижениям
в науке, особенно когда они выпадали на долю молодых ученых.
Яков Авадьевич был членом редакционного совета международного
журнала «Crustaceana», где благодаря его инициативе напечатаны многие
10
советские карцинологические работы. В последние годы жизни он был
членом редколлегии биологического отдела «Бюллетеня МОИП».
Нельзя, наконец, не упомянуть о Якове Авадьевиче как о популя-
ризаторе науки. Его перу принадлежат и крупные научно-популярные
очерки, и многочисленные небольшие заметки. Написанные живо и увле-
кательно, они всегда пользовались вниманием широкого круга читателей.
Он предъявлял те же требования к достоверности материала научно-
популярных статей, что и к строго научным публикациям.
Якову Авадьевичу было свойственно высокое чувство долга. В научной
деятельности оно проявлялось прежде всего в исключительной надежности
данных, получаемых при выполнении возложенных на него задач незави-
симо от их важности и степени его личной заинтересованности. Большая
трудоспособность позволила Якову Авадьевичу сделать очень много за свою
жизнь. К сожалению, он не успел подготовить к публикации читанный
им в течение ряда лет в Московском университете и пользовавшийся
неизменным успехом курс «Зоогеография морских и пресных вод», который
он мечтал написать и издать отдельной книгой.
В последние годы жизни Яков Авадьевич совмещал педагогическую
деятельность в МГУ с работой в Лаборатории бентоса ИОАН СССР.
Яков Авадьевич Бирштейн навсегда останется в истории советской
зоологии. Его научное наследие послужит основой для дальнейшего развития
многих отраслей зоологии. Наша обязанность — сохранить в памяти поко-
лений образ замечательного человека — умного и доброго, принципиального
в любом вопросе и в любой ситуации, замечательного наставника молодого
поколения исследователей, видевшего в нем пример служения науке и людям.
Ю. Г Чиндонова, С. И. Левушкин
11
СПИСОК ПЕЧАТНЫХ РАБОТ Я. А. БИРШТЕЙНА
1932
Eine neue Art des Genus Asellus Geoffroy aus Transkaukasien. — Zool.
Anz., Bd. 98, H. 9/10, S. 251—255.
Malacostraca Армении. — Tp. Севан, озер, ст., т. 4, вып. 1/2, с. 139—160.
1933
Malacostraca der Kutais-Hohlen am Rion (Transkaukasien). — Zool. Anz.,
Bd. 104, H. 5/6, S. 143—156.
Die Land- und Siisswasser-Isopoden des arktischen Gebietes. — Fauna
Arctika, Bd. 6, Lief. 5, S. 473—476.
1934
Пресноводные Decapoda СССР и их географическое распространение. —
Зоол. журн., т. 13, вып. 1, с. 39—70. Совм. с Л. Г Виноградовым.
О возможных мероприятиях по повышению производительных свойств
Каспия и Арала. — Рыб. хоз-во СССР, № 3, с. 38—40. Совм. с Л. А. Зен-
кевичем.
1935
К вопросу о происхождении морских ракообразных в реках Понто-
Каспийского бассейна. —Зоол. журн., т. 14, вып. 4, с. 749—761.
Zwei neue Gammariden aus Westasien. — Zool. Anz., Bd. 109, H. 11/12,
S. 293—296.
1936
Рост и распространение Cardium edule заливов Мертвый Култук и
Кайдак в Каспийском море в связи с соленостью. — ДАН СССР, т. 13,
№ 4 (108), с. 187—191.
О водяных осликах Кавказа. — Сб. тр. Зоол. музея МГУ, т. 3, с. 235—242.
12
1937
К вопросу об акклиматизации в Каспийском и Аральском морях новых
видов животных. — Зоол. журн., т. 16, вып. 3, с. 443—447 Совм. с Л. А. Зен-
кевичем.
Распределение биомассы бентоса Северного и Среднего Каспия: Тр. 1-й
Всекасп. науч, рыбохоз. конф. (7—24 января 1935 г.). М.; Л., т. 2,
с. 248—258.
1938
Derzhavinella macrochelata n. gen., n. sp. — новый род и вид Amphipoda
из Северного Каспия. — Зоол. журн., т. 17, вып. 1, с. 180—183.
Заметка о Decapoda из планктона, собранного во время плавания
ледоколов «Садко» и «Литке» в Арктике. — Бюл. МОИП. Отд. биол.,
т. 47, вып. 3, с. 199—244.
Рец. на кн.: Макаров В. В. Anomura. Фауна СССР. Crustacea, т. 10,
вып. 3 (1938). — Зоол. журн., т. 17, вып. 3, с. 554—555.
1939
Зоогеографическая характеристика водяных осликов Байкала. — ДАН
СССР, т. 25, № 3, с. 248—251.
О некоторых особенностях географического распространения пресно-
водных Malacostraca Дальнего Востока. — Зоол. журн., т. 18, вып. 1,
с. 54—59.
О пещерных креветках Абхазии. — Зоол. журн., т. 18, вып. 6, с. 960—975.
Итоги конференции по экологии и физиологии рыб и водных беспозво-
ночных Научно-исследовательского института зоологии МГУ. — Зоол. журн.,
т. 18, вып. 3, с. 461—474. Совм. с Г. Н. Калашниковым.
1940
Исследования фауны пещер СССР в 1935—1939 гг. — Бюл. МОИП.
Отд. биол., т. 49, № 3/4, с. 30—38. Совм. с Г В. Лопашовым.
Пещерные бокоплавы Абхазии с замечаниями о видах Niphargus
Закавказья. — Сб. тр. Зоол. музея МГУ, т. 6, с. 259—269.
К фауне пещерных Amphipoda Абхазии. — Бюл. МОИП. Отд. биол.,
т. 49, № 3/4, с. 47—55.
Высшие раки (Malacostraca). — В кн.: Жизнь пресных вод СССР. М.;
Л.: Изд-во АН СССР, т. 1, с. 405—430.
Жизнь в пещерах. — Успехи соврем, биологии, т. 13, вып. 3, с. 385—402.
Осморегуляторные способности каспийских беспозвоночных. — Зоол.
журн., т. 19, вып. 4, с. 548—565. Совм. с Г. М. Беляевым.
1941
Адаптация и эволюция пещерных животных. — Успехи соврем, био-
логии, т. 14, вып. 3, с. 436—453.
13
1944
Сравнение осморегуляторных способностей волжских и каспийских
Amphipoda. — ДАН СССР, т. 45, № 7, с. 323—326. Совм. с Г М. Беляевым.
1945
Ревизия системы каспийских Gammaridea. — ДАН СССР, т. 50, с. 517—
520.
Заметки о пресноводных высших ракообразных Туркмении и Ирана. —
Учен. зап. МГУ Биология, вып. 83, с. 151 —164.
Годовые изменения бентоса Северного Каспия. — Зоол. журн., т. 24,
вып. 3, с. 133—147.
Первые успехи реконструкции фауны Каспийского моря. — Зоол. журн.,
т. 24, вып. 1, с. 25—31. Совм. с Л. А. Зенкевичем, А. Ф. Карпевич.
Первые успехи планомерной реконструкции фауны Каспийского моря. —
Рыб. хоз-во, № 1, с. 40—44. Совм. с Л. А. Зенкевичем, А. Ф. Карпевич.
1946
Рец. на кн.: Базикалова А. Я. Амфиподы оз. Байкал. — Зоол. журн.,
т. 25, вып. 4, с. 383—384.
География животных. М.: Сов. наука. 455 с. Совм. с Л. А. Зенкевичем,
Н. А. Бобринским.
Осмотическое давление полостной жидкости бокоплавов оз. Байкал.—
ДАН СССР, т. 53, № 3, с. 293—295. Совм. с А. Я. Базикаловой, Д. Н. Талие-
вым.
Осморегуляторные способности бокоплавов оз. Байкал. — ДАН СССР,
т. 53, № 4, с. 381—384. Совм. с А. Я. Базикаловой, Д. Н. Талиевым.
Действие воды озера Балхаш на волго-каспийских беспозвоночных. —
Зоол. журн., т. 25, вып. 3, с. 225—236. Совм. с Г М. Беляевым.
Заметки о географическом распространении понто-каспийских боко-
плавов. — Бюл. МОИП. Отд. биол., т. 51, вып. 3, с. 39—53.
1947
О водяных осликах Сибири. — ДАН СССР, т. 57, № 4, с. 409—412.
Понятие «реликт» в биологии. — Зоол. журн., т. 26 вып. 4, с. 313—330.
1948
Новые взгляды на филогению членистоногих. — Успехи соврем, биологии,
т. 25, вып. 1, с. 143—149.
Третья конференция Научно-исследовательского института зоологии
МГУ по проблемам гидробиологии и ихтиологии. — Зоол. журн., т. 27,
вып. 2, с. 185—192.
Объем и распространение рода Synurella (Crustacea Amphipoda).—
ДАН СССР, т. 60, № 4, с. 701—704.
Нахождение пещерной креветки Troglocaris в грунтовых водах Мацесты
и связанные с этим вопросы. — Бюл. МОИП. Отд. биол., т. 53, вып. 3,
с. 3—10.
14
Новый отряд ракообразных. — Природа, № 2, с. 64—65.
Бокоплавы Таджикистана. — В кн.: Памяти академика С. А. Зернова.
М.; Л.: Изд-во АН СССР, с. 263—273.
Рец. на кн.: Иванов А. В., Петрушевский Г К-, Полянский Ю. И.
и др. Большой практикум по зоологии беспозвоночных. М.: Сов. наука,
1946, ч. 2. 632 с. — Зоол. журн., т. 27, вып. 2, с. 175—181. Совм. с Г Г Абри-
косовым, Ф. А. Лаврехиным.
1949
Новый для СССР род водяных осликов (Stenasellus) из Туркмении
и его значение для зоогеографии Средней Азии. — ДАН СССР, т. 69,
№ 5, с. 691—694. Совм. с И. В. Старостиным.
Некоторые проблемы происхождения и эволюции пресноводной фауны. —
Успехи соврем, биологии, т. 27, вып. 1, с. 119—140.
Рец. на кн.: Павловский Е. Н., Лепнева С. Г Очерки из жизни
пресноводных животных. — Зоол. журн., т. 28, вып. 2, с. 191 —192.
1950
Пещерная фауна Западного Закавказья. — Зоол. журн., т. 29, вып. 4,
с. 354—366.
Жизнь в подземных водах. — В кн.: Жизнь пресных вод СССР. М.;
Л.: Изд-во АН СССР, т. 3, с. 683—706. Совм. с Е. В. Боруцким.
Заметка о бокоплавах Алтая. — Тр. ТГУ. Сер. биол., т. 3, с. 113—117.
1951
Пресноводные ослики (Asellota). — Фауна СССР. М.; Л.: Изд-во
АН СССР, т. 7, вып. 5, с. 1 — 143.
Тонкопанцирные ракообразные (Leptostraca) Берингова и Охотского
морей. — ДАН СССР, т. 78, № 6, с. 1251 — 1254.
Новые и редкие десятиногие ракообразные Охотского моря и Куриль-
ских островов. — ДАН СССР, т. 79, № 2, с. 357—360. Совм. с Л. Г Виногра-
довым.
1952
Подземные бокоплавы района Хоста—Гудаута (Западное Закавказье). —
Бюл. МОИП. Огд. биол., т. 57, вып. 1, с. 26—39.
История одного краба. — Природа, № 9, с. 118.
Новые данные о происхождении фреатической подземной фауны. —
Тр. Всесоюз. гидробиол. о-ва, т. 4, с. 225—229.
Новый вид подземного водяного ослика из бассейна Уссури. —
ДАН СССР, т. 84, № 5, с. 1081 — 1084. Совм. с В. Я. Леванидовым.
Рец. на кн.: Акклиматизация амурских рыб. — Журн. общ. биологии,
т. 13, № 2, с. 162—168. Совм. с Л. А. Зенкевичем.
15
1953
Донная фауна Каспийского моря до и после вселения Nereis succinea. —
В кн.: Акклиматизация нереис в Каспийском море. М.: МОИП, с. 36—114.
Совм. с Н. Н. Спасским.
Изменение размеров и упитанности севрюги Северного Каспия в 1949 г.
по сравнению с 1937—1941 гг. — В кн.: Акклиматизация нереис в Каспий-
ском море. М.: МОИП, с. 233—242. Совм. с Г М. Беляевым.
Питание бентосоядных рыб Каспия (кроме осетровых) в 1948—1949 гг.
и использование ими Nereis succinea. — В кн.: Акклиматизация нереис
в Каспийском море. М.: МОИП, с. 115—144.
Вероятные изменения гидробиологического режима Каспийского моря. —
Тр. Всесоюз. гидробиол. о-ва, т. 5, с. 3—12.
Новые данные по фауне десятиногих ракообразных Берингова моря. —
Зоол. журн., т. 32, вып. 2, с. 215—228. Совм. с Л. Г. Виноградовым.
1954
Изучение фауны Курило-Камчатской впадины. — Природа, № 2, с. 61 —
74. Совм. с Л. А. Зенкевичем, Г М. Беляевым.
Вертикальная зональность планктона Курило-Камчатской впадины. —
ДАН СССР, т. 95, № 2, с. 389—392. Совм. с М. Е. Виноградовым,
Ю. Г Чиндоновой.
По поводу рец. В. В. Кузнецова на книгу Л. А. Зенкевича «Фауна
и биологическая продуктивность моря». — Зоол. журн., т. 33, вып. 1, с. 232—
237. Совм. с Г. М. Беляевым, Л. Г Виноградовым, 3. А. Филатовой.
1955
Заметки о питании глубоководных рыб Курило-Камчатской впадины. —
Зоол. журн., т. 34, вып. 4, с. 842—849. Совм. с М. Е. Виноградовым.
Исследования донной фауны Курило-Камчатской впадины. — Тр.
ИОАН СССР, т. 12, с. 345—381. Совм. с Л. А. Зенкевичем, Г. М. Бе-
ляевым.
Пеллагические гаммариды (Amphipoda—Gammaridea) Курило-Камчат-
ской впадины. — Тр. ИОАН СССР, т. 12, с. 210—287. Совм. с М. Е. Виногра-
довым.
Изучение глубоководной фауны и связанные с ним вопросы. — Вести.
МГУ. Сер. 16. Биология, № 4/5, с. 231—242. Совм. с Л. А. Зенкевичем.
Die Erforschung der Tierwelt des Kurilen-Kamtschatka—Tiefseegraben. —
Urania, Jahrg. 18, H. 1, S. 20—28. Leipzig—Jena. Совм. с Л. А. Зенкевичем,
Г M. Беляевым.
Ракообразные. — БСЭ, изд. 2, т. 36, с. 4—6.
Род Pseudocrangonyx Akatsuka et Komai (Crustacea, Amphipoda)
в СССР. — Бюл. МОИП. Отд. биол., т. 60, вып. 5, с. 77—84.
1956
Результаты акклиматизации Nereis succinea в Каспии и их критика. —
Бюл. МОИП. Отд. биол., т. 61, вып. 1, с. 23—44.
Studies of the deep water fauna and related problems. — Deep-See
16
Res., vol. 4, N 1, p. 54—64. Совм. с Л. А. Зенкевичем.
Методика изучения подземных вод. — В кн.: Жизнь пресных вод СССР.
М.; Л.: Изд-во АН СССР, т. 4, ч. 1, с. 438—452. Совм. с Е. В. Боруцким.
Десятиногие ракообразные палеогена Ферганы. — Бюл. МОИП. Отд.
геол., т. 31, вып. 1, с. 63—75.
Рец. на кн. Иванов А. В. Промысловые водные беспозвоночные. —
М.: Сов. наука, 1955. 356 с. Зоол. журн., т. 35, вып. 1, с. 159—160. Совм.
с Н. И. Тарасовым.
Вертикальное распределение донной фауны. Курило-Камчатской впа-
дины. — Тр. проблем, и темат. совещ. ЗИН АН СССР. Л.: Изд-во АН СССР,
вып. 6, с. 15—16. Совм. с Л. А. Зенкевичем, Г М. Беляевым.
Вертикальное распределение планктона Курило-Камчатской впадины. —
Тр. проблем, и темат. совещ. ЗИН АН СССР. Л.: Изд-во АН СССР,
вып. 6, с. 17—18. Совм. с М. Е. Виноградовым, Ю. Г. Чиндоновой.
1957
О некоторых особенностях ультраабиссальной фауны на примере рода
Storthyngura (Crustacea Isopoda Asellota).—Зоол. журн., т. 36, вып. 7,
с. 961—985.
Новый для СССР род пресноводных креветок. — Зоол. журн., т. 36,
вып. 4, с. 511—514. Совм. с Л. Г Виноградовым.
1958
Новый лангуст (Crustacea, Decapoda) из меловых отложений Запад-
ной Камчатки. — В кн.: Материалы к основам палеонтологии. М.: ПИН
АН СССР, вып. 2, с. 51—55.
Древнейший представитель отряда десятиногих ракообразных из перм-
ских отложений Западной Сибири. — ДАН СССР, т. 122, № 3, с. 477—480.
Пелагические гаммариды (Amphipoda, Gammaridea) северо-западной
части Тихого океана. — Тр. ИОАН СССР, т. 27, с. 219—257. Совм. с М. Е. Ви-
ноградовым.
Глубоководные мизиды северо-западной части Тихого океана. —
Тр. ИОАН СССР, т. 27, с. 258—355. Совм. с Ю. Г Чиндоновой.
Траление на максимальной глубине Мирового океана. — Природа, № 3,
с. 70—71. Совм. с А. И. Савиловым, Г Б. Удинцевым.
Deep-Sea Malacostraca of the North-Western part of the Pacific Ocean,
their distribution and relations. — Proc. XV Internat. Congr. Zool. London,
Sect. Ill, Paper 33, p. 1—4.
1959
Ультраабиссальная фауна Тихого океана. — Internat. Oceanogr. Congr.,
Reprints, Washington, p. 370—372.
Палеотемпературы и проблема происхождения глубоководной фауны. —
Природа, № 5, с. 21—28.
К 70-летию со дня рождения Льва Александровича Зенкевича. — Бюл.
МОИП. Отд. биол., т. 64, вып. 6, с. 141 —146.
Качественная и количественная характеристика глубоководной фауны.—
2 Заказ 333
17
В кн.: Итоги науки. Достижения океанологии. М.: Изд-во АН СССР, № 1,
с. 106—147. Совм. с Л. А. Зенкевичем, Г М. Беляевым, 3. А. Филатовой.
Вертикальная зональность в распределении глубоководной фауны. —
В кн.: Итоги науки. Достижения океанологии. М.: Изд-во АН СССР,
№ 1, с. 166—187. Совм. с Н. Г и М. Е. Виноградовыми.
О схеме вертикальной биологической зональности океана. — ДАН СССР,
т. 129, № 3, с. 658—661. Совм. с Г М. Беляевым и др.
Зоологические исследования в 25-м рейсе экспедиционного судна «Ви-
тязь». — Зоол. журн., т. 38, вып. 2, с. 301—304. Совм. с М. Е. Виноградовым.
I960
Семейство Ischnomesidae (Crustacea, Isopoda, Asellota) в северо-
западной части Тихого океана и проблема амфибореального и биполярного
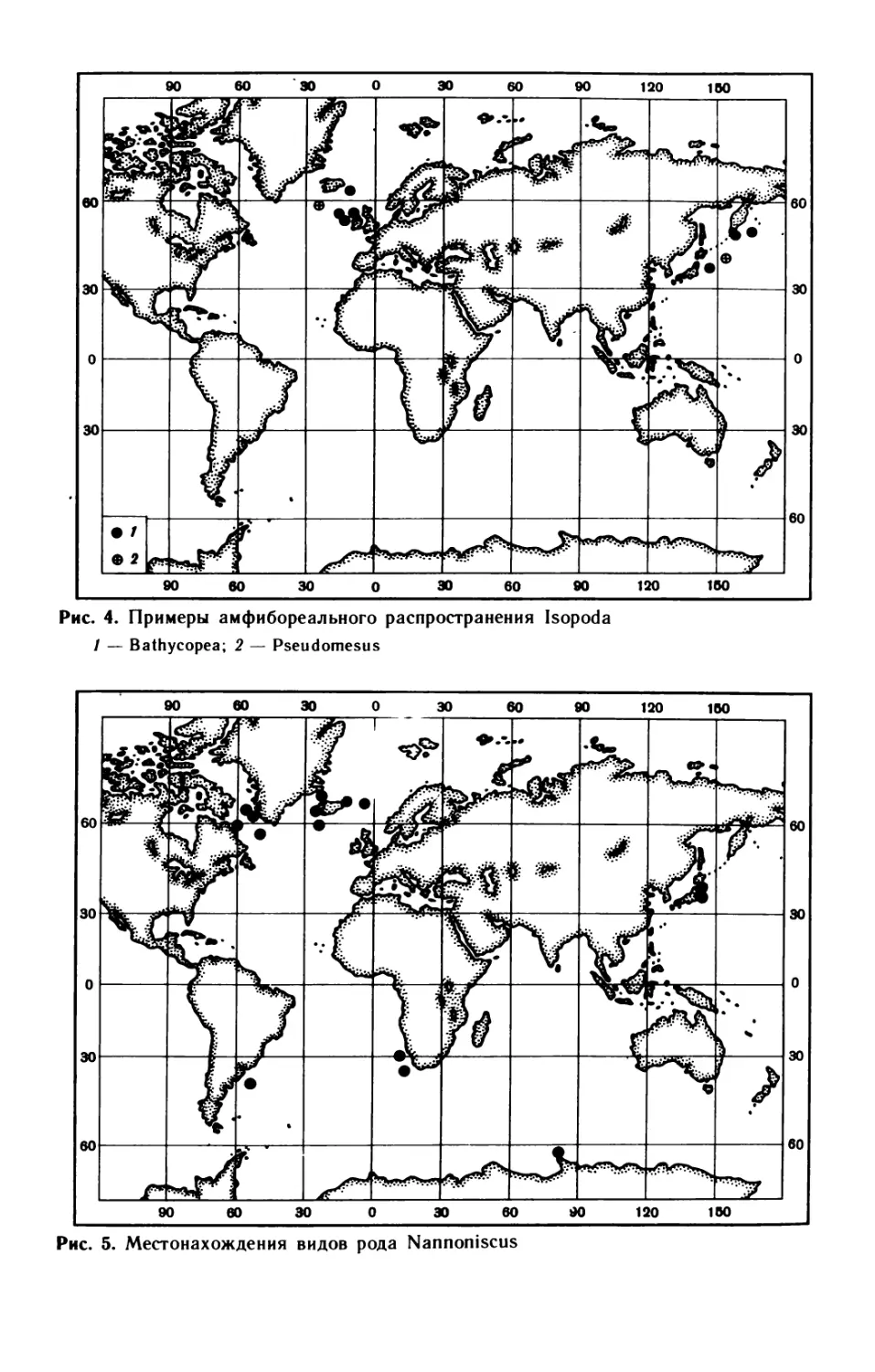
распространения глубоководной фауны. — Зоол. журн., т. 39, вып. 1, с. 3—28.
Класс Crustacea. Ракообразные. — В кн.: Основы палеонтологии: Чле-
нистоногие. М.: Госгеолтехиздат, с. 201—206.
Отряды Mystacocarida (с. 254), Ascothoracida (с. 260—261), подклассы
Cephalocarida (с. 421) и Malacostraca (с. 421—425, 429—460). — В кн.:
Основы палеонтологии: Членистоногие. М.: Госгеолтехиздат.
Донные ультраабиссальные гаммариды северо-западной части Тихого
океана. 1. Семейства Lil 1 jeborgiidae, Astyridae, Lepechinellidae, Gamma-
ridae. — Тр. ИОАН СССР, т. 34, с. 147—164. Совм. с Н. Г. Виноградовой.
Пелагические гаммариды тропической части Тихого океана.—
Тр. ИОАН СССР, т. 34, с. 165—241. Совм. с М. Е. Виноградовым.
Донная фауна Бугенвильской впадины. — Тр. ИОАН СССР, т. 41, с. 128—
131. Совм. с М. Н. Соколовой.
On the problem of the antiquity of the deep-see fauna. — Deep-Sea Res.,
vol. 7, N 1, p. 10—23. Совм. с Л. А. Зенкевичем.
Биоспелеологические исследования в Западном Закавказье осенью
1959 г. — В кн.: Материалы Комис, по изуч. геол, географии карста: Информ,
сб., № 1, с. 174—177. Совм. с С. И. Левушкиным.
1961
Microthambema tenuis n. gen., n.sp. (Isopoda Asellota) and relations of some
Asellote Isopods. — Crustaceana, vol. 2, N 2, p. 132—141.
Подземные бокоплавы Крыма. — Бюл. МОИП. Отд. биол., т. 66, вып. 6,
с. 126—144.
О геологической древности глубоководной фауны. — Океанология, т. 1,
вып. 1, с. 110—124. Совм. с Л. А. Зенкевичем.
Краткий очерк научной, педагогической и общественной деятельности
Л. А. Зенкевича. — В кн.: Лев Александрович Зенкевич: Материалы к био-
библиографии ученых СССР. Сер. геогр. М.: Изд-во АН СССР, вып. 3,
с. 7—21.
1962
Palaeophreatoicus sojanensis gen. et sp. nov. и некоторые вопросы фило-
гении и зоогеографии равноногих ракообразных (Isopoda). — Палеонтол.
журн., № 3, с. 65—80.
18
Uber eine neue Art der Gattung Austroniscus Vanhoffen (Crustacea,
Isopoda, Asellota) aus grossen Tiefen des nordwestlichen Teiles
der Stillen Ozeans. — Izdanie publ. zavoda za ribarstvo NRM — Scopje, t. 3,
N 2, p. 33—38.
Notes on the family Pardaliscidae (Amphipoda) with the description of
a new genus. — Crutaceana, vol. 3, N 4, p. 249—258. Совм.
с M. E. Виноградовым.
Пелагические гаммариды (Amphipoda, Gammaridea), собранные Совет-
ской антарктической экспедицией на д/э «Обь> к югу от 40° ю. ш. — В кн.:
Исследования фауны морей. М.; Л.: Изд-во АН СССР, т. 1 (9), с. 36—57.
Совм. с М. Е. Виноградовым.
Мизиды (Mysidacea), собранные Советской антарктической экспедицией
на д/э «Обь». — В кн.: Исследования фауны морей. М.; Л.: Изд-во АН СССР,
т. 1 (9), с. 58—69. Совм. с Ю. Г Чиндоновой.
1963
Глубоководные равноногие ракообразные (Crustacea, Isopoda) северо-
западной части Тихого океана. М.: Изд-во АН СССР. 214 с.
Равноногие ракообразные (Crustacea, Isopoda) ультраабиссали Буген-
вильской впадины. — Зоол. журн., т. 42, вып. 6, с. 814—834.
Некоторые итоги изучения подземной фауны Крыма. — Тр. Комплекс,
карст, экспедиции АН УССР. Киев: Изд-во АН УССР, вып. 1, с. 123—133.
Глубоководные Amphipoda пелагиали Филиппинской впадины. —
Тр. ИО АН СССР, т. 71, с. 81—93. Совм. с М. Е. Виноградовым.
Исследования подземной фауны Крыма летом 1960 г. — Новости
карстоведения и спелеологии, № 3, с. 69—72.
1964
Третий представитель подземного рода Niphargus (Crustacea, Amphi-
poda) из Крыма. — Бюл. МОИП. Отд. биол., т. 69, вып. 1, с. 119—121.
Нахождение Bathynellacea (Crustacea, Syncarida) в Средней Азии. —
Зоол. журн., т. 43, вып. 1, с. 17—27. Совм. с С. И. Левушкиным.
Новый подвид Bathynella natans Vejd. (Crustacea, Bathynellacea) из
подземных вод Предкавказья. —Зоол. журн., т. 43, вып. 11, с. 1719—1722.
Совм. с С. И. Левушкиным.
Пеллагические гаммариды северной части Индийского океана. — Тр.
ИО АН СССР, т. 65, с. 152—196. Совм. с М. Е. Виноградовым.
Рец. на кн.: Пономарева Л. А.-Эвфаузиевые северной половины Тихого
океана, их распространение и экология массовых видов. М.: Наука, 1963. —
Зоол. журн., т. 43, Bbfh. 12, с. 1879—1884. Совм. с Л. Г Виноградовым.
Итоги изучения подземной фауны СССР. — III Internal. Congr. Spe-
leol., 3, Wien, p. 29—35. Совм. с С. И. Левушкиным.
1965
Распространение Bathynellacea в подземных водах СССР. — В кн.:
Вопросы гидробиологии. М.: Наука, с. 33—34. Совм. с С. И. Левушкиным.
Подземные Paraselloidea (Crustacea, Isopoda) СССР. — Зоол. журн.,
т. 44, вып. 7, с. 997—1013. Совм. с С. И. Левушкиным.
2*
19
Les Malacostraces des eaux souterraines de 1’URSS. — IV Internat. Congr.
Speleol. (Jugoslavia), Summ. of lectures. Ljubljana, p. 74.
Faune des eaux souterraines saumatres de 1’Asie Centrale. — Internat.
J. Speleol., vol. 1, N 3, p. 307—320. Совм. с С. И. Левушкиным.
1966
Заметки о палеозойских высших ракообразных (Malacostraca) в СССР.
Pygocephalomorpha. — Палеонт. журн., № 2, с. 49—56.
IV Международный спелеологический конгресс в Югославии. — Зоол.
журн., т. 45, вып. 5, с. 793—797. Совм. с Г. Я. Бей-Биенко.
Подземная фауна Русской равнины. — В кн.: Вопросы изучения карста
Русской равнины. М.: Изд-во МГУ, с. 115—120. Совм. с С. И. Левушкиным.
Ракообразные. — В кн.: Курс зоологии. М.: Высш, шк., т. 1, с. 335—364.
Смена способа питания как причина таксономической обособленности
глубоководной фауны. — В кн.: Второй Междунар. океаногр. конгр.: (Тез.
докл.). М.: Наука, с. 43—44. Совм. с М. Е. Виноградовым.
1967
Некоторые итоги и задачи изучения подземной фауны СССР. — Зоол.
журн., т. 46, вып. 10, с. 1509—1535. Совм. с С. И. Левушкиным.
Отряд Bathynellacea (Crustacea, Malacostraca) в СССР. 1. Семейство
Bathynellidae. — Бюл. МОИП. Отд. биол, т. 72, вып. 4, с. 51—66. Совм.
с С. И. Левушкиным.
Подземные водяные ослики (Crustacea, Isopoda, Asellota) Закавказья. —
Зоол. журн., т. 46, вып. 6, с. 856—865.
Нахождение Paralomis spectabilis Hansen (Crustacea, Decapoda,
Anomura) в Антарктике. — В кн.: Исследования фауны морей. М.; Л.:
Изд-во АН СССР, т. 4 (12), с. 381—388. Совм. с Л. Г Виноградовым.
1968
Класс ракообразные (Crustacea). — В кн.: Жизнь животных. М.; Л.:
Просвещение, т. 2, с. 377—529.
Класс ракообразные (Crustacea). — В кн.: Атлас беспозвоночных
Каспийского моря. М.: Пищ. пром-сть, с. 118—120.
Бокоплавы (Amphipoda). — В кн.: Атлас беспозвоночных Каспийского
моря. М.: Пищ. пром-сть, с. 241—289. Совм. с Н. Н. Романовой.
Кумовые (Cumacea). — В кн.: Атлас беспозвоночных Каспийского моря.
М.: Пищ. пром-сть, с. 227—241. Совм. с Н. Н. Романовой.
Мизиды (Misidacea). — В кн.: Атлас беспозвоночных Каспийского моря.
М.: Пищ. пром-сть, с. 213—227.
Равноногие (Isopoda). — В кн.: Атлас беспозвоночных Каспийского моря.
М.: Пищ. пром-сть, с. 289—291.
Глубоководные Asellota (Crustacea, Isopoda) из Антарктики и Субан-
тарктики. — В кн.: Исследования фауны морей. М.; Л.: Наука, т. 6 (14),
с. 141 — 152.
Отряд Bathynellacea (Crustacea, Malacostraca) в СССР. II. Семейство
Parabathynellidae и зоогеографические замечания. — Бюл. МОИП. Отд.
биол., т. 73, вып. 6, с. 55—64. Совм. с С. И. Левушкиным.
20
Представитель нового для фауны СССР семейства бокоплавов (Amphi-
poda, Bogidiellidae) в подземных водах Средней Азии. — Зоол. журн., т. 47,
вып. 5, с. 676—683. Совм. с С. И. Левушкиным.
1969
Некоторые вопросы эволюции глубоководной фауны. — Океанология,
т. 9, вып. 5, с. 907—908.
Мизиды (Mysidacea).— В кн.: Тихий океан. Биология Тихого океана.
М.: Наука, кн. 2, с. 69.
Бокоплавы (Amphipoda). — В кн.: Тихий океан. Биология Тихого океана.
М.: Наука, кн. 2, с. 69—72.
Равноногие раки (Isopoda). — В кн.: Тихий океан. Биология Тихого
океана. М.: Наука, кн. 2, с. 75—79.
Равноногие ракообразные (Crustacea, Isopoda) впадины Романш. —
Бюл. МОИП. Отд. биол., т. 74, вып. 3, с. 50—59.
Л. Г Виноградов (1909—1968). — Бюл. МОИП. Отд. биол., т. 74, вып. 5,
с. 150—153.
Malacostraca подземных вод СССР. — Proc. 4th Internat. Congr. Speleol.
in Jugoslavia (12—26.IX 1965), N 4/5, p. 27—33.
1970
Дополнения к фауне равноногих ракообразных (Crustacea, Isopoda)
Курило-Камчатского желоба. — Тр. ИОАН СССР, т. 86, ч. 1, с. 292—340.
О фауне пелагических гаммарид Курило-Камчатского района Тихого
океана. — Тр. ИОАН СССР, т. 86, с. 401—419. Совм. с М. Е. Виноградовым.
О донных десятиногих ракообразных (Crustacea, Isopoda) района Ку-
рило-Камчатского желоба. — Тр. ИОАН СССР, т. 86, с. 420—426. Совм.
с Н. А. Заренковым.
Новые мизиды (Crustacea, Mysidacea) из района Курило-Камчатского
желоба. — Тр. ИОАН СССР, т. 86, с. 277—291. Совм. с Ю. С. Чиндоновой.
Эндемичные для Закавказья роды подземных бокоплавов (Amphipo-
da). — Зоол. журн., т. 49, вып. 10, с. 1471 —1487. Совм. с С. И. Левушкиным.
Caracteristique zoogeographique de la faune souterraine de I’Union Sovie-
tique. — En: Livre du centenaire Emile G. Racovitza, Bucarest, p. 211—221.
1971
Некоторые вопросы эволюции глубоководной фауны. — В кн.: История
Мирового океана. М.: Наука, с. 84—89.
Дополнения к фауне равноногих ракообразных (Crustacea Isopoda)
Курило-Камчатского желоба. Ч. 2. Asellota-2. — Тр. ИОАН СССР, т. 92,
с. 162—238.
Роль трофического фактора в таксономическом обособлении морской
глубоководной фауны. — Бюл. МОИП. Отд. биол., т. 76, вып. 3, с. 59—92.
Совм. с М. Е. Виноградовым.
1972
Une nouvelle espece africaine du genre Stenasellus (Crustacea Isopoda
Asellota) du bassin du Niger. — Intern. J. Speleol., t. 4, p. 9—18.
ФАУНА ПРЕСНЫХ ВОД
ПРЕСНОВОДНЫЕ DECAPODA СССР
И ИХ ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ*
ОБЩАЯ ЧАСТЬ * 1
Вопрос об исторической географии пресноводных десятиногих раков
разработан преимущественно Ортманом (Ortmann, 1902). Нашей задачей
является детализация некоторых положений этого автора по отношению
к фауне СССР, а также их изменения и дополнения на основании новых
геологических данных, и в первую очередь теории Вегенера (1925), значение
которой для биогеографии огромно. В этом отношении мы вполне присоеди-
няемся к мнению Вульфа (1932), пришедшего после анализа географи-
ческого распространения растений к следующему выводу: «Она (теория
Вегенера. — Я. Б. и Л. В.) является наиболее удовлетворяющей современный
уровень знания рабочей гипотезой, на которой может строить свои выводы
историческая биогеография».
Atyidae считаются древнейшими пресноводными Decapoda. Поэтому
нахождение представителя этого семейства Troglocaris в пещерах Закав-
казья является фактом большого зоогеографического интереса. Род Troglo-
caris относится к наиболее примитивным Atyidae, по Бувье (Bouvier, 1925), —
к группе паратиевых («1а serie paratyenne»), объединяющей следующие
роды: Troglocaris (Балканы, Закавказье), Paratya (10 видов в Аннаме,
Японии, Новой Зеландии и Ассаме, с одной стороны, и в Новой Каледонии
и островах Норфолк и Лорд-Хау к востоку от Австралии — с другой),
Syncaris (2 вида в Калифорнии), Palaemonias (Мамонтова пещера в США),
Dugastella (Марокко) и Atyaephyra (Средиземноморские страны).
Ортман (1902) объясняет такое разорванное распространение наличием
материковых мостов, связывавших современные ареалы названных родов.
Садовский (1930) резонно замечает, что к данному случаю вполне прило-
жима теория Вегенера. Действительно, по Вегенеру, до эоцена Австралия,
Индо-Китай и Япония, с одной стороны, и Средиземноморская область,
* Зоол. журн., 1934, т. 13, вып. 1, с. 39—70. (Совм. с Л. Г Виноградовым). Печа-
тается только заключительная «Общая часть» статьи.
1 В первых частях дан обзор систематического состава и данные о распространении
каждого вида пресноводных Decapoda СССР, фауна которых включает 25 видов
и подвидов из 6 семейств: Atyidae (Troglocaris schmidtii kutaissiana), Palaemonidae
(Palaemon superbus, Palaemonetes venerupis и 3 вида Leander), Crangonidae
(Grangon crangon), Potamobiidae (6 видов Astacus, в том числе A. leptodactylus,
представленный пятью подвидами, и 2 вида Cambaroides — С. dauricus с тремя
подвидами и С. schrenckii с двумя), Potamonidae (Potamon potamios) Grapsidae
(Eriocheir japonicus, Helice tridens). — (Прим. ped.).
22
Антильские острова и Восточная Америка — с другой, находились в непо-
средственной близости друг от друга. Таким образом, выявляются два центра
современного распространения Xiphocaridinae: один — на востоке, другой —
на западе.
Возникновение Atyidae Ортман относит к юрскому периоду. В меловом
периоде они достигли своего расцвета. К этому времени, по Бувье, относится
появление рода Paratya, заселившего громадный океан, покрывавший
в то время Южную Европу и Северную Африку и через Северную Индию
соединявшийся с Тихим океаном. Paratya чрезвычайно близка к Troglocaris,
и Бувье считает ее прямым предком последнего. Отделение Troglocaris от
Paratya и его вселение в пещеры, по Бувье, произошло только в четвертичном
периоде. С этим мнением никак нельзя согласиться. Нахождение Troglocaris
в Кутаисской пещере, изолированной от внешнего мира с олигоцена, застав-
ляет отнести вселение этого рода в пещерные воды к гораздо более раннему
времени. Садовский считает, что это событие произошло в меловой г/ёриод,
так как именно к мелу Кавказ впервые поднялся из вод мелового моря в виде
острова и с тех пор больше морем не заливался. Этот срок гораздо более
вероятен. Верхнеюрская—нижнемеловая трансгрессия вытолкнула в прес-
ные воды наряду с Atyidae целый ряд других групп (осетровые, Potamobii-
dae) и явилась, таким образом, чрезвычайно важным этапом в эволюции
гидрофауны.
Два центра распространения современных Xiphocaridinae образовались,
очевидно, в результате постепенного уничтожения соединявшего их прежде
моря, начавшегося в районе Восточной Азии. С обеих сторон увеличивав-
шейся суши возникли разные роды. Сильные колебания солености и появив-
шиеся к тому времени более прогрессивные группы заставили атиид перейти
в пресную воду, где они и сохранились частично до настоящего времени.
Таким образом, эти ракообразные должны считаться реликтами мелового
времени.
Современное распространение семейства Patamobiidae таково: род Asta-
cus занимает Европу и часть Западной Азии, а также западную часть США,
род Cambaroides — бассейн Амура, Японию и п-ов Корея и род Cambarus —
юго-восток Северной Америки (рисунок) ’* Таким образом, имеются четыре
оторванных друг от друга ареала, причем между двумя областями, занятыми
родом Astacus, на востоке находится область, занятая родом Cambaroides,
а на западе — область, занятая родом Cambarus. Это положение вызывало
недоумение ряда авторов. По Гексли (1900), положительный факт, представ-
ляющий затруднение, состоит в том, что сходство, существующее между
амурско-японскими и речными раками (европейскими. — Я. Б. и Л. В.) и
восточноамериканскими видами Cambarus больше, чем между этими по-
следними и западноамериканскими видами Astacus, и сходство между этими
последними и понто-каспийскими раками больше, чем между двумя этими
группами и амурско-японской формой. Если бы факты были обратные и
если бы западно-американские и амурско-японские раки поменялись бы
своими местами, то дело было бы вполне понятно. Мы вполне присоединяемся
к этому пожеланию, но факты — упрямая вещь.
Взгляды на генетические отношения между отдельными родами Potamo-
1 Один из видов р. Cambarus — С. affinis — акклиматизирован во Франции и Гер-
мании.
23
Распространение Potamobiidae (по: Ortmann, 1902; Balss, 1927; Pesta, 1926a, b с изме-
нениями)
/ — Astacus; // — Cambaroides; /// — Cambarus
biidae, высказанные Гексли и Фэксоном (Faxon, 1885), в настоящее время
несколько изменились. Ортман не считает Cambaroides формой, промежу-
точной между Astacus и Cambarus, и сходство между Cambaroides и Camba-
rus объясняет конвергенцией. Доводы, приводимые этим автором, достаточно
убедительны, и с ними приходится согласиться. Однако все же систематика
Potamobiidae требует фундаментальной ревизии. В особенности неясны
отношения между восточно-американскими и европейскими видами Astacus.
Весьма вероятно, здесь мы имеем дело с разными подродами, а может быть,
и родами.
Какова же история расселения и дифференциации на роды Potamobiidae?
На этот счет высказывались разные соображения. Шарф (Scharff, 1911)
считает, что Cambarus произошел от европейского Astacus и проник в Аме-
рику через атлантический мост. По Шимкевичу (1886), колыбель Potamobii-
dae orbis terrarum является Каспий, из которого в первую очередь вышли
на запад A. torrentium, а на восток — «Cambarus sp.?», что доказывается
сходством плевр брюшка и рострума у этих видов. Cambarus прошел всю
Азию (!) и заселил Америку. Берг (1909) называет Cambaroides и европей-
ского Astacus в числе реликтов пресноводного водоема, занимавшего в плио-
цене весь север Евразии и населенного субтропической фауной. Ортман
(1902) выдвинул следующую гипотезу, принятую впоследствии Арльдтом
(Arldt, 1910, 1919—1922): центр распространения семейства Potamobiidae
(и Parastacidae) лежит в Монголии, откуда в верхнемеловое время шло
расселение, с одной стороны, на запад, а с другой — на северо-восток через
северный тихоокеанский материковый мост в Америку. От западноамери-
канского Astacus в древнетретичное время произошел восточноамериканский
Cambarus. Только в позднетретичное время Astacus через всю Азию дошел
до Европы (в противоположном направлении, чем по Шимкевичу).
Учитывая имеющийся, к сожалению, очень скудный палеонтологический
материал, противоречащий большинству изложенных гипотез, мы пришли
к другим выводам.
24
Ископаемые Potamobiidae из пресных вод известны с неокома (нижний
мел). Это Astacus antiquus Horbort из Вестфалии и A. licenti van Straelen
из Монголии (Straelen, 1928с). A politus Marek et Schliiter найден в слоях
того же времени также в Вестфалии, но среди морских или солонова-
товодных организмов (Marek, Schluter, 1885). Таким образом, мы ви-
дим, что уже в нижнем мелу представители рода Astacus жили в местах,
населенных современными Potamobiidae, — Восточной Азии и Европе. В За-
падной Америке, превосходно изученной в палеонтологическом отношении,
род Astacus известен только с миоцена и плиоцена (A. chenoderma Соре),
факт нахождения Astacus в нижнем мелу одновременно и в пресной, и
в морской воде заставляет предполагать вместе с Ортманом и Стреленом,
что примерно в это время (верхняя юра или нижний мел) Potamobiidae
перешли в пресную воду. Стрелен на основании своих палеонтологических
исследований (Straelen, 1928а, Ь) приходит к выводу, что выход Potamo-
biidae из моря в пресные воды должен был иметь место после Оксфорда и
перед портландом, так как именно в это время Северо-Атлантический
материк (Скандис) был соединен с Китайско-Сибирским (Ангарис) и между
ними могло быть пресноводное сообщение. Здесь и встает вопрос: насколько
существенно для всех логических построений это соединение?
Нам кажется, что нет необходимости предполагать, как это делает
Ортман, что Potamobiidae вышли из моря только в одной точке — в Мон-
голии. Гораздо вероятнее более или менее одновременное вхождение этих
животных в пресные воды по всему побережью обитаемого ими моря, как
это наблюдается у других групп в настоящее время. В таком случае можно
обойтись без того, чтобы предполагать прохождение Astacus громадной
территории от Монголии до Каспия по неизвестным гипотетическим путям,
а также превращение его сначала в пресноводное животное, а потом, при
достижении Каспия, опять в морское. Существование Astacus уже в мелу
одновременно в Европе и Монголии подтверждает наше предположение.
Как известно, верхнеюрская—нижнемеловая трансгрессия покрыла Япо-
нию, часть восточного побережья Азии, весь север Азии, почти всю Европу
без Скандинавии и Пиренейского полуострова и юго-восток Северной
Америки (Мексику), по Вегенеру, прилегавшей в это время к Европе.
В этом море должны были водиться Potamobiidae, а в мелу — даже род
Astacus. Затем море постепенно сокращается, исчезая в Сибири и Восточной
Азии, которые уже не были покрыты верхнемеловой трансгрессией. В эоцене
Северная Америка отделилась от Европы, причем Potamobiidae не могли
не остаться в Мексике от верхнеюрской трансгрессии. И действительно,
Мексика, по Ортману, явилась центром заселения видов Cambarus, занявших
обширную область до Скалистых гор. Что касается времени отделения рода
Cambarus от Astacus, то вопрос этот неясен. Следует думать, что оно
произошло до эоцена.
Заселение Западной Америки произошло, по всей вероятности, через
северотихоокеанский мост, как это думал Ортман. Он относил это событие
к верхнему мелу. Между тем в меловой период климатические условия Запад-
ной Америки были настолько суровы, что раки вряд ли могли бы их перенести.
Полюс, по Кёппен и Вегенеру (Кбрреп, Wegener, 1924), находился очень
близко от западного берега Америки, несколько на запад от него. Только
в миоцене полюс отодвинулся на значительное расстояние. К этому времени
и относится наиболее раннее нахождение ископаемого Astacus в Западной
25
Америке — A. chenoderma Соре. Таким образом, есть все основания предпо-
лагать, что переселение раков в Западную Америку произошло в средне-
третичное время. По Арльдту, тогда же происходило переселение через
северотихоокеанский мост ряда пресноводных рыб — Bungia, Aulopyge,
Pseudorasbora и других, известных в Америке в ископаемом состоянии
также с миоцена. Расселение азиатских пресноводных обитателей было,
несомненно, облегчено существованием на северо-западе Северной Америки
в миоценовое время громадных пресных озер. Одним из следов передвижения
Astacus на юго-восток является, как это заметил Ортман, нахождение
A. nigrescens на Аляске.
Западноамериканские Astacus и восточноазиатские Cambaroides пред-
ставляют собой боковые ветви, происшедшие от раков, водившихся в районе
Монголии в дотретичные времена. Некоторое представление об этих жи-
вотных дает описанный Стреленом (Strallen, 1928с) из неокома Монголии
A. licenti. Насколько можно судить по описанию и особенно по таблицам,
этот рак приближаетя больше к Astacus, чем к Cambaroides. Отсюда
можно заключить, что Cambaroides, как это думал Ортман, приобрел свои
конвергентные роду Cambarus черты впоследствии, может быть, даже после
того, как заселил Америку. Все эти вопросы можно разрешить окончательно
только после детального сравнительно-анатомического, эмбриологического и
палеонтологического анализа всех родов Potamobiidae и в особенности
европейских и американских Astacus.
Разрыв между ареалами Potamobiidae Евразии, приходящийся на Си-
бирь, совпадает с разрывом ареалов целого ряда растений и животных.
Берг (1909), сводя большое количество аналогичных случаев, пришел
к убеждению, что охлаждение, имевшее место во время ледникового периода,
уничтожило древнюю флору и фауну Сибири, которая сохранилась в более
благоприятных в отношении климата районах. Это объяснение может быть
принято и для Potamobiidae.
Если принять, что современное распространение Potamobiidae есть след-
ствие верхнеюрской трансгрессии, передвижения материков и миграции
животных через северотихоокеанский материковый мост, станет очевидным,
что те же условия влияли и на другие пресноводные группы, и поэтому
должно найтись некоторое количество аналогичных случаев географического
распространения. Так оно, конечно, и есть. Но мы приведем только один
пример. Это — Acipenseridae. На карте географического распространения
осетровых, данной Миком (Meek, 1916), их ареал почти целиком совпадает
с ареалом Potamobiidae. В Северной Америке есть также два отделенных
друг от друга Скалистыми горами пятна, причем в Европе и Восточной Аме-
рике живет общий вид Acipenser sturio (вегенеровское соединение). Разница
только в том, что у осетровых в Сибири нет типичного для раков разрыва
ареала, что объясняется лучшей приспособленностью первых к арктическим
условиям, а также, может быть, их способностью быстрее передвигаться
и заселять, таким образом, воды раньше раков.
После того как эти идеи сложились у нас, мы познакомились с работой
Колози (Colosi, 1932), в которой распространение Potamobiidae рассматри-
вается как пример, подтверждающий теорию ологенеза. Эта теория предло-
жена Роза (Rosa, 1918) и у нас принята Бергом (1922а), который нашел в ней
много общего с собственными эволюционными идеями. Суть ологенеза заклю-
чается в признании политопного возникновения всех таксономических единиц
26
из примитивных космополитов, населявших Мировой океан. Таким путем
объясняется современный разрыв между ареалами многих групп, для объяс-
нения которого обычно привлекается теория миграций. Здесь не место вхо-
дить в критику ологенеза, тем более что виталистический, резко антидарви-
новский характер этой теории совершенно очевиден. Заметим только, что
теория Вегенера, о которой Колози скромно умалчивает, является чрезвы-
чайно веским аргументом против ологенеза.
Колози, подобно нам, принимает выход Potamobiidae из моря в разных
пунктах, а не их миграцию в широтном направлении по пресным водам (Орт-
ман). Однако ему остались неизвестными многие данные палеонтологии,
и поэтому он относит вселение раков (даже всех Масгига) в пресные воды
к третичному периоду. На самом деле оно произошло гораздо раньше, даже
тогда семейство Potamobiidae уже вполне сформировалось. При вселении
в пресные воды семейство разделилось на роды, а может быть, и на подроды,
а затем прошло разделение на виды. При этом та полная конвергенция, кото-
рую требует теория ологенеза, совершенно не имела места. Принимая
выход раков из моря, а не их длительные миграции как вероятное объяснение
современного распространения Potamobiidae, мы не видим никакого соответ-
ствия между этим фактом и общими виталистическими воззрениями Колози.
Переходя к истории Potamobiidae в третичное и четвертичное время
в Европе, следует отметить громадную роль в эволюции раков внутреннего
моря, чрезвычайно сильно изменявшего как свои границы, так и свои физи-
ческие свойства и сохранившегося до настоящего времени в виде Каспия.
Значение Понто-Арало-Каспийского бассейна в расселении Potamobiidae
было ясно уже старым авторам. Так, Гексли (1900) полагал, что раки
двигались из этого бассейна по Дунаю, причем Astacus torrentium может
представить нам первую группу эмигрантов на запад, A. nobilis (=А. asta-
cus.— Я. Б. и Л. Г.) будет соответствовать второму отряду, a A. lepto-
dactylus со своими вариететами остается в качестве представителя древних
арало-каспийских «раков» (последнее заключение совершенно непонятно;
A. leptodactylus, очевидно, возник позже других, т. е. является самым
молодым). Сходные идеи высказал Шимкевич: в первую очередь из Понто-
Каспия вышли Astacus torrentium и «Cambarus sp.», первый из которых
направился на запад и достиг Англии, а второй — на восток и заселил Аме-
рику; вторая «волна» катится только на восток и дает Cambaroides; третья,
представленная A. kessleri и A. astacus, останавливается на востоке перед
Монгольской пустыней, а на западе перед возникшим Ла-Маншем и, наконец,
четвертая — A. leptodactylus — в наше время вытесняет третью.
На основании изложенного выше ясно, что дальневосточные и американ-
ские раки имели совершенно иную историю и Понто-Арало-Каспийский
бассейн может рассматриваться как центр распространения только евро-
пейских раков, как это думал Гексли. Однако их последовательное расхож-
дение «волнами» совершенно очевидно. Суммируя морфологические и геогра-
фические данные, мы приходим к выводу, что состав этих «волн» таков:
первая «волна» — A. pallipes и A. torrentium, вторая «волна» (широко-
палые раки) — A. astacus, A. pachypus и A. colchicus, третья «волна»
(узкопалые раки) — A. leptodactylus, A. pylzowi, A. kessleri.
Чем объяснить последовательное выхождение групп видов из Понто-
Каспийского бассейна? Нам кажется, что к этому случаю может быть
применена гипотеза, предлагаемая Зенкевичам (1933) для объяснения сту-
27
пенчатости в распространении некоторых животных Ледовитого океана,
и в частности тресковых рыб. Решающую роль в процессе проникновения
морских элементов в пресную воду, согласно этой гипотезе, сыграла «пуль-
сация солености» данного моря. Каждое резкое понижение солености выра-
батывало свою фауну, которая и вгонялась в пресные воды последующим
осолонением. Ряд «пульсаций» дает характерную ступенчатость в распро-
странении многих групп.
Нет сомнения в том, что Понто-Каспийский бассейн за время своей много-
образной истории по нескольку раз испытывал переходы от нормальной
океанической солености до почти полного опреснения и эти изменения втал-
кивали в пресные воды его бассейна пучки видов разнородных групп, как,
например, рыб и ракообразных. Вся трудность заключается в том, чтобы
привязать определенные «волны» к определенным геологическим моментам,
для чего в настоящее время у нас нет достаточных данных. Ясно только, что
образование видов последней «волны» произошло до отделения Каспийского
моря от Черного, так как самый молодой рак — A. leptodactylus — дви-
гался по рекам и Черноморского (Дон, Дунай), и Каспийского (Волга)
бассейнов.
Каждая последующая «волна», если она двигалась по путям предыдущей,
оттесняла эту последнюю от моря в глубь материка. Наряду с этим некоторые
формы приспосабливались к жизни в опресненных участках моря, причем
в случае древности группы образовывали виды (A. pachypus), в случае
относительной молодости — подвиды (A. leptodactylus caspius). Ряд видов
имеет реликтовый характер, населяя участки, занятые когда-то морскими
заливами (A. kessleri, A. colchicus, A. pylzowi). Таким образом, эволюция
европейских раков происходила преимущественно в море, и пресные воды
явились до известной степени убежищем для примитивных форм.
Примером вытеснения более древних примитивных видов более молодыми
может служить межвидовая борьба A. astacus и A. leptodactylus. Побед-
ное шествие последнего происходило на глазах исследователей. Захватив
Волжский бассейн, этот рак по каналам проникает в Северную Двину
(Кесслер, 1875, 1877) и отчасти в Невский бассейн (Шимкевич, 1886).
Искусственно пересаженный человеком в начале XIX в. в р. Исеть, он начи-
нает распространяться по бассейну р. Тобол (70-е годы — Малахов, 1880).
Его ареал вдается в ареал A. astacus. Налегание одних ареалов на другие
сравнительно невелико по площади. Оно имеет место в Белоруссии — в По-
лоцком, Витебском и Могилевском районах. Совместная встречаемость по
водоемам почти отсутствует, и в тех немногих водоемах, где водятся оба
вида, A. astacus быстро идет на убыль и наконец исчезает.
Известны многие примеры, из которых следует, что достаточно A. lepto-
dactylus попасть в озеро, населенное A. astacus, чтобы в 10—15 лет совер-
шенно вытеснить последнего.
Сущность этой борьбы неясна. Оба вида питаются преимущественно
макрофитной растительностью и, казалось бы, не должны испытывать не-
достатка в пище в условиях обычного эвтрофного озера. Мы держали
в аквариуме Зоологического музея МГУ экземпляры обоих видов совместно,
и, хотя драки между ними бывали, но никакого перевеса сил или даже
более частой драки с особями чужого вида, чем с особями своего, нам наблю-
дать не пришлось. Межвидовые спаривания также бывали, но потомства
от них не получено.
28
В настоящее время возможность межвидовой борьбы почти исключена,
так как раки населяют отдельные, часто не сообщающиеся друг с другом
водоемы. В соединяющих отдельные системы крупных реках, как Москва,
Ока, Клязьма, Западная Двина, не говоря уже о Волге, раки встречаются
единичными экземплярами, так как они уничтожены чумой, не затронувшей
только отдельные изолированные водоемы.
Межвидовая борьба имеет место не только между A. astacus и A. lepto-
dactylus. Последний вид вытесняет, по некоторым неопубликованным дан-
ным, также A. pachypus из Одесско-Днепровского залива. На Дальнем Во-
стоке также, по-видимому, имела место борьба между Cambaroides dauricus
и С. schrenckii, как показывает их распространение. Последний водится
в центре системы Амура, а по бокам, в отрезанных друг от друга другим
видом бассейнах притоков Амура, водится С. dauricus.
По отношению к Potamon можно сказать очень немногое. По Ортману,
центр его распространения лежит в Индии и Индо-Китае, откуда он и мигри-
ровал через Персию в Закаспийскую область и на Кавказ и заселил впослед-
ствии берега Средиземного моря. Индия несомненно является центром
распространения целого ряда пресноводных и солоноватоводных форм, и
индийские элементы имеются в фауне южных частей СССР.
В заключение следует сказать несколько слов о креветках бассейна
Амура. Все они представляют собой тропический элемент в фауне Амура,
встречаясь, кроме того, преимущественно в озерах Китая. Амурская фауна
характеризуется целым рядом китайских родов и видов рыб и ракообразных.
Л. С. Берг (1909) считает их древними аборигенами, сохранившимися
с плиоцена и имеющими субтропический характер. Для рода Palaemon
нахождение в оз. Ханка является самым северным. Другие виды этого рода
населяют пресные воды Восточной Африки, Мадагаскара, островов Маври-
кий, Зондских и Филиппинских, Северной Австралии, Индии и т. д. Проник-
новение Palaemon в пресные воды произошло сравнительно недавно — по
Ортману, в неогене. На это указывает наличие солоноватоводных форм,
а также сезонные миграции некоторых видов из пресных вод к морю.
Род Leander распространен еще шире. Он имеет уже чисто морской
характер. Большая часть его видов живет в море, и лишь немногие приспосо-
бились к пресной воде. Нахождение представителя рода Palaemonetes в бас-
сейне Амура чрезвычайно интересно. Как известно, один вид этого рода —
Р. varians — населяет западное побережье Европы и Средиземное и Черное
моря, другой же — Р. exilipes, — к которому приближается Р. venephicus, —
юго-восток США; Р. antrorum Benedict найден в артезианском колодце
в Техасе (США), Р. eigenmanni Науо и Р. calcis Kathbun — в пещерах
о-ва Кубы. Такое распространение требует еще своего объяснения, как и
многие другие вопросы, затронутые в нашей работе.
29
ПРЕСНОВОДНЫЕ ОСЛИКИ (ASELLOTA)*
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
И РОДСТВЕННЫЕ ОТНОШЕНИЯ
Отряд Isopoda объединяет около 2000 видов, обитающих преимущест-
венно в море и на суше. Пресные воды населены весьма незначительным
количеством форм равноногих раков. Однако из 7 подотрядов, на которые
в настоящее время разделяется отряд Isopoda (Asellota, Flabellifera, Val-
vifera, Oniscoidea, Phreatoicidea, Epicaridea, Gnathiidea), только Gnathiidea
не представлены в фауне пресных вод; в то же время Phreatoicidea целиком
являются пресноводными обитателями, живущими главным образом в под-
земных и отчасти в наземных водах Австралии, Новой Зеландии, Тасмании,
Южной Африки. Число представителей пресноводных Flabellifera сравни-
тельно велико: сем. Cymothoidae насчитывает целый ряд свободно живущих
видов (Cirolaninae) в пещерах Европы и Северной Америки и паразитических
видов в Восточной Азии, сем. Sphaeromidae представлено несколькими
видами в водах бассейна северной половины Тихого океана и в пещерах
Европы, сем. Corallanidae — в Китае, Японии и на Зондских островах,
сем. Exocorallanidae — одним видом в Гайане, сем. Anthuridae — одним
видом в грунтовых водах Новой Зеландии.
Пресноводные Valvifera значительно уступают по количеству видов
предыдущему подотряду. Сем. Chetilidae, заключающее всего один род,
известно из Южной Америки; семейство Idotheidae населяет пресные воды
бассейна Северного Ледовитого океана, Южной Америки, Индии и Новой
Зеландии. Oniscoidea, живущие почти исключительно на суше, заходят
иногда в воду, так, например, Ligidium нередко встречается в ручьях Кав-
каза, Desertoniscus birsteini найден в оз. Делили в Туркмении, пещерные
Tithanetes и Spiloniscus входят в водоемы. Наконец, некоторые Epicaridea
паразитируют на тропических и субтропических пресноводных креветках.
Но эти подотряды не характерны для пресноводной фауны СССР; по
существу, только два представителя названных групп (исключая мокриц)
обитают в пресных водах СССР: Mesidothea entomon — в водоемах Поляр-
ного бассейна и Livoneca amurensis -=- в р. Амуре; кроме того, несколько
эвригалинных видов Idotheidae, Sphaeromidae и Anthuridae встречаются
в эстуариях рек, преимущественно на Дальнем Востоке. Подавляющее
большинство пресноводных Isopoda СССР, да и вообще Палеарктики, отно-
сится к подотряду Asellota и к чисто пресноводному надсемейству Aselloidea,
заключающему одно семейство Asellidae.
Семейство Asellidae распространено только в северном полушарии; юж-
ной границей его распространения можно считать 30°, на севере же неко-
торые Asellus известны за полярным кругом — в Западной Гренландии и
в дельте р. Лены. Основной областью распространения Asellidae является
бореальная область. К семейству Asellidae относятся 5 родов: Asellus, Sten-
asellus, Johanella, Synasellus и Lirceus, из которых наиболее обширный
ареал имеет род Asellus, населяющий всю Европу, Северную Африку,
оз. Байкал, восточное побережье Азии, восточную и западную часть Север-
* 1951 г. Ракообразные. М.: Изд-во АН СССР. 144 с. (Фауна СССР; Т. 7; Вып. 5).
Печатается только глава о географическом распространении.
30
ной Америки; род Stenasellus приурочен преимущественно к юго-западной
Европе, и только два из 13 входящих в него видов найдены недавно в эквато-
риальной западной Африке (Remy, 1938; Monod, 1945) и один — в Средней
Азии (Бирштейн, Старостин, 1949); монотипический род Johanella известен
только из одного ключа в Алжире, и, наконец, род Lirceus с 16 видами
занимает восточную часть Северной Америки; близкий к этому роду моноти-
пический род Synasellus обнаружен недавно в подземных водах Португалии
(Braga, 1942, 1944).
Таким образом, общий ареал семейства является разорванным, причем
разорванность ареала присуща не только всему семейству в целом, но и
отдельным его родам и подродам (рис. 1). Это обстоятельство указывает,
разумеется, на значительную древность группы.
Происхождение и время вселения Asellidae в пресные воды совершенно
неясны, поскольку палеонтологические данные по Asellota отсутствуют
(Straelen, 1928а). Морфологические данные также не могут помочь разре-
шить эти вопросы: пресноводное надсемейство Aselloidea со своим един-
ственным семейством Asellidae резко отчленено от преимущественно мор-
ского надсемейства Paraseiloidea; промежуточные формы между обоими
надсемействами не известны. Из морских семейств к надсемейству Aselloidea
несколько приближается только семейство Stenetriidae, населяющее моря
побережий Южной Африки, Австралии, Индо-Китая, Вест-Индии, Южной
и Средней Америки (два вида найдены в Средиземном море). Это семейство
не включается Ганзеном (Hansen, 1905) в группу Paraselloidea (по Ганзену,
семейство Parasellidae), а выделяется в отдельное семейство, равнозначное
Asellidae ( = Aselloidea) и Parasellidae ( = Paraselloidea). По целому ряду
признаков Stenetriidae занимают среднее положение между обоими над-
семействами. Так, например, протоподиты I пары плеоподов 3 у Stenetriidae
сросшиеся, как у Paraselloidea, но их величина незначительная, как у Asel-
loidea. I и II пары плеоподов 3 не соединены вместе, как у Aselloidea; строе-
ние II плеопода 3 среднее между Paraselloidea и Stenasellus. Плеоподы
II пары j сросшиеся, без следов шва, как у Paraselloidea, но operculum
образован III парой плеоподов, как у Aselloidea. IV и V пары плеоподов
также построены по некоторому среднему типу. Однако возможная близость
Asellidae и Stenetriidae пока не помогает разрешить вопрос о происхождении
первого семейства. Тем не менее это обстоятельство представляет значи-
тельный интерес, и оно будет рассмотрено несколько ниже в другом аспекте.
Вселение некоторых Isopoda в пресные воды происходило в очень отда-
ленные времена. Уже упомянутые Phreatoicidea в триасе жили в пресной
воде, как это доказано Чилтоном, описавшим Phreatoicus wianamattensis
из триасовых пресноводных отложений Сиднея (Австралия) (Chilton, 1918).
Пресноводные Sphaeromidae известны из верхней юры (Cyclosphaeroma
woodwardi van Straelen) (Straelen, 1928a). Вероятно, не будет ошибкой
отнести происхождение Asellidae как пресноводной группы к мезозою, когда
формировались и другие группы пресноводных Malacostraca (Decapoda,
Amphipoda).
Географическое распространение современных Asellidae, несмотря на
недостаточную изученность, позволяет наметить некоторые общие законо-
мерности. Весь ареал семейства разбит на 5 отдельных областей, разделен-
ных значительными пространствами суши или моря: 1) Европа и Северная
Африка, 2) оз. Байкал, 3) восточное побережье Азии с островами, 4) запад-
31
Рис. 1. Распространение родов и подродов сем. Asellidae
1 — ареал подрода Asellus s. str.; 2 — ареал подрода Proasellus Dud.; 3 — местонахожде-
ния видов подрода Mesoasellus Birst.; 4 — местонахождения видов подрода Baicaloasellus
Stamm. 5—ареал подрода Conasellus Stamm, и рода Lirceus Raf.; 6—ареал Asellus
tomalensis; 7 — местонахождение рода Johanella Monod.; 8 — местонахождение рода
Synasellus Braga
ная часть Северной Америки и 5) восточная часть Северной Америки.
Кроме того, известны изолированные местонахождения отдельных видов
в Средней Азии, Западной Сибири, Забайкалье и западной экваториальной
Африке. Для европейско-североафриканского ареала характерно присут-
ствие рода Stenasellus (12 видов) и подрода Proasellus (22 вида); кроме
того, вся Европа населена подродом Asellus s. str. (2 вида), отсутствующим
на значительном протяжении Сибири и появляющимся снова на восточных
побережьях Азии (1 вид). В оз. Байкал обитают 4 вида эндемичного
подрода Baicaloasellus и 1 вид подрода Mesoasellus; на восточном побе-
режье Азии, помимо указанного Asellus s. str., из Японии описано 2 вида
Mesoasellus. В западной части Северной Америки известны 1 вид под-
рода Mesoasellus и 2 вида Asellus, систематическое положение которых
не выяснено (A. tomalensis, A. adenta); восточная часть Северной Америки
населена 17 видами подрода Conasellus и 16 видами рода Lirceus (=Мапса-
sellus) (см. рис. 1).
Итак, наиболее богата видами Asellidae Палеарктика, причем палеарк-
тические водяные ослики не представляют собой единой зоогеографической
группировки. Здесь можно отличить 5 основных комплексов: 1) комплекс
кругосредиземноморски^ форм, наиболее характерным представителем
которого является подрод Proasellus; сюда же примыкают роды Stenasellus
и Johanella; 2) комплекс евразиатских форм, к которому принадлежит под-
род Asellus s. str.; 3) комплекс байкальских эндемиков — подрод Baicalo-
asellus; 4) комплекс восточно-азиатских — западноамериканских форм —
подрод Mesoasellus; 5) тяготеющий к восточноамериканским формам род
Synasellus.
На территории Европы происходит смешение кругосредиземноморского
и евразиатского комплексов. Первый, несомненно, древнее и примитивнее
второго и является исконным обитателем кругосредиземноморских стран.
В этом отношении значительный интерес представляет нахождение пред-
ставителей подрода Proasellus на островах Додеканес (A. coxalis, по:
Arcangeli, 1929), Сицилии (A. banyulensis wolfi, по: Dudich, 1925) и,
наконец, Мадейре (A. italicus perarmatus, по: Remy, 1936). Раковица
(Racovitza, 1919) считал, что родиной подрода Proasellus 1 является Малая
Азия, где обитает наиболее примитивный вид подрода A. coxalis; отсюда
произошло заселение северной Африки (A. coxalis peyerimhoffi), а затем
через Гибралтар — Испании (A. banyulensis и др.), далее Южной Франции
и Средней Европы. После работ Раковицы целый ряд видов Proasellus был
открыт главным образом на Аппенинском и Балканском полуостровах, что
1 По терминологии Раковицы: «1а lignee d’Asellus meridianus».
3 Заказ 333
33
заставило Реми (Remy, 1932) пересмотреть гипотезу Раковицы. Реми
считает возможным два объяснения распространения Proasellus.
1. Наличие Эгеиды, существовавшей с начала третичного до начала
четвертичного времени и соединявшей Малую Азию с Балканским полуост-
ровом, причем миграция через Эгеиду должна была осуществиться до
олигоцена, так как в бассейне оз. Охрида, совершенно изолированном с оли-
гоцена, живет несколько видов Proasellus.
2. В соответствии со взглядами Аргана (1935), колоссальный надвиг
северного берега Гондваны на Евразию, при котором Европа была смята
в альпийскую складчатость, после чего Евразия, покрытая в южной части
Гондваной, оторвалась от Гондваны, и на месте разрыва образовалось Среди-
земное море. Оказавшийся таким образом в Европе кусок Гондваны и
явился центром распространения европейских Proasellus.
Следует отметить, что проблема происхождения подрода Proasellus,
по существу, является проблемой происхождения всей древней кругосреди-
земноморской фауны и флоры в целом. Сходное распространение свой-
ственно длинному ряду групп наземных моллюсков, насекомых, паукооб-
разных, пресноводных Malacostraca и т. д. Можно ли приписывать всем им
гондванское происхождение, как это принимают для Proasellus Раковица
и Реми?
Если обратиться к числу видов подрода Proasellus, то окажется, что
в Африке известны 3 вида и 3 подвида, в Сирии и Палестине — 1 вид,
а в Европе с островами — 18 видов и 19 подвидов, т. е. что «массовый центр
ареала* Proasellus лежит в Европе. Нахождение наиболее примитивного
вида — A. coxalis — в Сирии и Палестине не может считаться надежным
доказательством приуроченности центра происхождения подрода к этим
странам, так как нет основания допускать одинаковый темп эволюции
в разных частях ареала. Присутствие подвида A. coxalis в Сицилии — A. co-
xalis cyanophilus — противоречит гипотезе миграции примитивных форм
через Гибралтар в Европу, предложенной Раковицей. Широкое обобщение
Реми также не представляется вполне приемлемым. Положения Аргана,
на которых основывается Реми, далеко не доказаны, а в последнее время
они подверглись серьезной критике. Не входя в подробности этого инте-
ресного вопроса, достаточно привести одно из возражений, собранных
Мушкетовым (1935): мощность пошедших на складкообразование свит не
превосходила 2000—3000 м, тогда как толщина давящей глыбы (Африка)
принимается до 30 000 м. Получается явное несоответствие между, так ска-
зать, грубым инструментом и тонкой, произведенной якобы им работой.
Однако если даже принять построения Аргана, то и в этом случае не полу-
чится соответствия между массой Гондваны, якобы лежащей на Европе, и
ареалом Proasellus. Нахождения A. meridianus и A. cavaticus в Англии и
Германии, A. collingei в Англии, а также A. banyulensis и A. spelaeus в Юж-
ной Франции отстоят весьма далеко от участков Гондваны, как она изобра-
жена Арганом (1935), и переселение туда Proasellus представляется мало-
вероятным. В этом отношении интересно также нахождение единственного
в СССР представителя Proasellus — A. infirmus — около г. Сухуми. Кавказ,
по Аргану (1935), отделен от Аравийского мыса иранскими и анатолийскими
дугами, т. е. и к случаю A. infirmus объяснение Реми не подходит. Таким
образом, резюмируя сказанное, можно утверждать, что нет оснований и для
34
признания гондванского происхождения Proasellus, и для привязывания его
к участкам Гондваны, лежащим на Евразии.
Советские биогеографы, сталкивавшиеся с фактами средиземноморских
разрывов ареалов, довольно единодушно объясняют это явление наличием
прямой континентальной связи между Закавказьем, Крымом, Балканским
полуостровом и Малой Азией, составлявшими до плиоцена и даже до начала
четвертичного периода одно целое (Вульф, 1944; Пузанов, 1927; Цееб, 1947).
В то же время восточная часть Средиземного моря еще была сушей. Эти
данные могут быть приняты и для объяснения современного распростране-
ния Proasellus. Приуроченность ареала Proasellus к южной части Европы
несомненно стоит в связи с великим оледенением. Территория, занятая лед-
ником, лишена Proasellus (рис. 2). При этом у границ максимального оле-
денения, как правило, встречаются подземные или глубоководные слепые
формы. Уже Штаммер (Stammer, 1932а) для Германии показал, что в райо-
нах распространения подземных форм Proasellus отсутствуют наземные
формы, и объяснил это влиянием ледника, высказав предположение о более
широком распространении Proasellus на север в доледниковые времена.
То же установил впоследствии для Бельгии Арканджели (Arcangeli, 1935).
Таким образом, Proasellus может считаться чрезвычайно древним обита-
телем Европы и стран, лежащих вокруг Средиземного моря, занимавшим
в третичное время значительно более широкий ареал, чем сейчас. На древ-
ность подрода указывает, между прочим, нахождение некоторых его предста-
вителей на островах, в частности, на о-ве Мадейре (Remy, 1936), отстоящем,
как известно, на 545 км от Африки и на 800 км от Европы. Мадейра вместе
с Азорскими островами и Островами Зеленого Мыса отделилась от Старого
Света до конца миоцена, и уже в это время A. italicus существовал как вид
и на Мадейре сформировался только его подвид A. italicus perarmatus.
Род Stenasellus также может считаться древним третичным реликтом
(Stammer, 1936с). Большая часть составляющих его видов (10 из 13) оби-
тает в подземных водах Южной Европы, занимая ограниченные, оторванные
друг от друга ареалы. Только три вида зарегистрированы вне этой основной
области распространения рассматриваемого рода — St. chappuisi и St. afri-
canus в западной экваториальной Африке и St. asiaticus — в восточной
Туркмении (рис. 3).
Долготный разрыв ареала (Южная Европа—тропики), помимо Stena-
sellus, свойствен многим подземным группам, как водным, так и сухопутным.
Он показывает, что часть пещерной фауны Кругосредиземноморья имеет
тропическое происхождение. Группы с таким распространением представ-
ляют собой остатки тропической фауны, населявшей Европу до середины
третичного периода. Их сохранению под землей способствовало сходство
условий существования, с одной стороны, под землей, с другой — в тропиках,
прежде всего в отношении постоянства температуры, не подвергающейся
в обоих случаях суточным и сезонным колебаниям. По мере ухудшения
климата температура подземных вод Европы постепенно понижалась, но не
подвергалась резким колебаниям, которые могли оказаться гибельными для
обитателей поверхности земли (Бирштейн, 1950).
Нахождение одного из видов Stenasellus в Средней Азии в отрогах Гис-
сарского хребта с несомненностью подтверждает указания на наличие в со-
ставе среднеазиатской гидрофауны средиземноморских элементов. В своем
з
35
Рис. 2. Распространение подрода Proasellus Dud.
1 — граница максимального оледенения; 2 — Asellus (Proasellus) cavaticus Schiodte;
3 — другие слепые виды Proasellus; 4 — остальные виды Proasellus
анализе фауны моллюсков бассейна Амударьи Жадин (1948) устанавливает
6 групп видов, различающихся по происхождению и характеру распростра-
нения, в том числе группу видов средиземноморского и группу видов тропи-
ческого происхождения. St. asiaticus можно относить с одинаковым основа-
нием к любой из этих групп. В термальном источнике Ходжа-Койнар вместе
с ним обитает моллюск Melanoides tuberculatus, принадлежащий, согласно
Жадину, к тропической группе видов. В то же время подавляющее большин-
ство видов Stenasellus (в том числе виды, морфологически приближающиеся
к St. asiaticus) населяет подземные воды бассейна Средиземного моря.
36
Можно предполагать, что границы между тропическими и средиземномор-
скими элементами в ряде случаев весьма неопределенны, так как многие
виды со средиземноморским распространением имеют тропическое про-
исхождение.
St. asiaticus — древний обитатель Средней Азии. По крайней мере по
отношению к живущему вместе с этим осликом моллюску Melanoides tuber-
culatus палеонтологически доказано присутствие его в Средней Азии еще
в верхнем миоцене (Жадин, 1948). Это подтверждает отмеченную выше
глубокую древность рода Stenasellus, полностью гармонирующую с его при-
митивными морфологическими особенностями.
Поэтому трудно согласиться с Арканджели (Arcangeli, 1939), который
возражает против признания Stenasellus третичным пресноводным реликтом.
Этот автор указывает на существование в середине третичного времени
в Италии моря на том месте, где сейчас найден St. racovitzai, и предполагает,
что виды рода Stenasellus возникли в море, оказались «преадаптирован-
ными» к подземному образу жизни и затем вселились по подземным щелям
в систему грунтовых вод. К какому времени следует относить процесс вселе-
нйя Stenasellus из моря в пресные воды, Арканджели не разъясняет. Шаппюи
(см.: Racovitza, 1950) уже указал, что в Италии несомненно оставались
участки суши, послужившие убежищем целому ряду пресноводных и сухо-
путных третичных форм. К тому же многие современные нахождения видов
рода Stenasellus далеко отстоят от морских берегов; наиболее ярким при-
мером такого случая можно считать нахождение St. asiaticus в Туркмении.
К этому следует добавить, что в составе морской фауны мы не знаем форм,
близких к Stenasellus. Шаппюи, считая Stenasellus пресноводным третичным
реликтом, склонен приписывать этому роду гондванское происхождение,
поскольку в Западной Африке найдено два далеких друг от друга вида Stena-
sellus (chappuisi и africanus), что доказывает давнее обитание Stenasellus
в Гондване. Против мнения Шаппюи говорит существование St. asiati-
cus в Средней Азии. Гораздо естественней признать в Stenasellus остаток
древней тепловодной тропической фауны, захватывавшей в третичное
время огромную территорию — от экваториальной Африки до Средней
Азии.
Подрод Asellus s. str. резко отличается по своей зоогеографической
природе от только что рассмотренных. Один из входящих в него видов —
A. aquaticus — занимает всю Европу, кроме Пиренейского полуострова,
Англию и западную Гренландию; в Африке этот вид отсутствует. Восточная
граница распространения A. aquaticus точно не установлена. По имеющимся
сведениям и материалам, на Урале его нет. Что касается Средней Азии, то
Вальтер (Walter, 1889) считал отсутствие A. aquaticus характерной особен-
ностью фауны ракообразных этой области. Более новые исследователи
(Бирштейн, 1945; Бирштейн, Старостин, 1949) обнаружили в нескольких
озерах западной Туркмении особый подвид A. aquaticus, ареал которого,
вероятно, примыкает к нахождениям типичной формы в Иране. Кроме того,
типичная форма найдена в колодцах Мангышлака. Однако восточнее
A. aquaticus не идет. Другой вид подрода A. monticola живет в Закавказье
и Иране.
На огромном протяжении Сибири и Центральной Азии представители
Asellus s. str. почти не встречаются. Исключение составляют бассейн Оби
и Иртыша, оз. Баунт в системе Ципо-Ципиканских озер (бассейн Витима)
37
и дельта Лены. В каждом из этих водоемов обитает особый вид или подвид
рассматриваемого подрода (Бирштейн, 19476).
Восточная окраина Азии от Китая до побережья Охотского моря включи-
тельно и прилегающие к ней острова (Япония, Сахалин, Рюкю) заселены
A. hilgendorfi. Этот вид проникает на запад до среднего течения Амура и
представлен подвидом martynovi в дельте Лены. В Америке подрод отсут-
ствует (Johansen, 1926), если не считаться с сомнительным указанием на
нахождение A. aquaticus в западной Гренландии.
Таким образом, ареал Asellus s. str. представляется разорванным.
Между двумя областями сплошного распространения подрода (Европа и
Восточная Азия) лежит огромная Сибирь, на протяжении которой встре-
чаются лишь изолированные колонии Asellus.
По вопросу о происхождении Asellus s. str. высказывались разноречивые
мнения. Раковица (Racovitza, 1919) рассматривал A. aquaticus в Европе как
пришельца с севера, Шаппюи (Chappuis, 1927), присоединяясь к этому
мнению, полагал, что A. aquaticus мигрировал в Европу из Сибири и оттеснил
на юг A. meridianus. Арканджели (Arcangeli, 1935) возражал против
арктического происхождения A. aquaticus, считая, что этот вид населял и
в доледниковое время среднюю и восточную Европу, а ледник заставил его
передвинуться к югу.
Все эти гипотезы нельзя считать серьезно обоснованными, поскольку
их авторам оставался неизвестным ареал Asellus s. str. в Азии. И в более
поздних работах нередко можно найти неверные указания на сплошной
палеарктический ареал A. aquaticus (Державин, 1930) или на присутствие /
его на Урале, в Сибири, Китае и Японии (Remy, 1941). '
Анализ имеющихся сейчас более полных и точных данных о видовом
составе и характере ареала Asellus s. str. позволяет ближе подойти к вопросу
об его происхождении.
Разрыв ареала Asellus s. str., приходящийся на Сибирь, совпадает с раз- {
рывом ареалов многих других родов и видов растений и животных, указанных
Бергом (1909). Согласно этому автору, объяснение происхождения разор-
ванных таким образом ранее сплошных ареалов следует искать в условиях
ледникового времени.
По данным Герасимова и Маркова (1939), только Западная Сибирь испы-
тала покровное оледенение. Среднесибирское плато и низины Восточной
Сибири не имели сплошного покрова льда. Здесь шло образование наледей,
превращение промерзших озер в линзы льда, засыхание и погребение
снежных скоплений, глубокое промерзание грунта. Сакс (1947) допускает
значительно более широкое распространение ледника в Западной Сибири и
наличие огромного по площади оледенения Среднесибирского плоскогорья
и Восточной Сибири.
Так или иначе, но суровые условия ледникового периода оказались,
очевидно, гибельными для водяных осликов, и в Сибири они почти повсе-
местно исчезли. Тем интереснее обратить внимание на сохранившиеся в этой
области изолированные колонии представителей подрода Asellus s. str.
Обь и Иртыш, населенные A. latifrons, в ледниковое время текли иначе,
чем сейчас, огибая наступавшие ледники и временами образуя озеровидные
водоемы. Это были независимые друг от друга реки, соединившиеся только
в послеледниковое время (Нагинский, 1949). Поскольку их воды не были
скованы вечным льдом, возможно переживание ледникового периода водя-
38
ними осликами именно в этих реках. В бассейне Лены A. hilgendorfi marty-
novi встречается только в дельте; на всем остальном протяжении Лены обна-
ружить водяных осликов не удалось (устное сообщение П. Л. Пирожникова)
По данным Сакса (1947), ледники с Верхоянского хребта достигали Лены,
но ее нижнее течение с дельтой всегда оставалось свободным ото льда.
Озеро Баунт, в котором обитает третий сибирский представитель Asellus
s. str. — A. epimeralis, населено и другими элементами доледниковой фауны,
в частности байкальскими, и принадлежит к системе древнего байкальского
многоозерья. Замечательно, что не только по присутствию водяных осликов,
но и по наличию ряпушки Coregonus sardinella baunti Much, намечается
связь этого озера с низовьями Лены. Ряпушка живет лишь в нижнем тече-
нии Лены, не поднимаясь выше Жиганска. По мнению М. М. Кожова (1949),
древнее байкальское многоозерье в третичное время могло иметь сток на
северо-восток, в бассейн Лены.
Представление о сибирских водяных осликах как об остатках широко
распространенного в доледниковое время в Сибири Asellus s. str., имевшего
тогда сплошной ареал, подтверждается и морфологическими данными.
Сибирские A. epimeralis и A. latifrons обладают некоторыми общими и притом
примитивными признаками, в частности чрезвычайно крупными по отноше-
нию к протоподиту ветвями второго плеопода самца, удлиненным II плеопо-
дом самки и прямым, лишенным выемки наружным краем экзоподита I плео-
пода самца. Последний признак свойствен также дальневосточному A. hilgen-
dorfi и кавказско-иранскому A. monticola. У A. aquaticus, как было указано
выше, выемка экзоподита I плеопода самца появляется только на определен-
ном этапе постэмбрионального развития. Соотношение ветвей и протопо-
дита II плеопода самца, наблюдающееся у A. latifrons и A. epimeralis, также
характерно для ранних стадий развития A. aquaticus. Таким образом,
последний вид может считаться наиболее далеко зашедшим по пути эволю-
ции передних плеоподов самца, т. е. наиболее молодым. Аналогичное откло-
нение от примитивной формы II плеопода самца обнаруживается у A. hil-
gendorfi и A. manticola, сохраняющих, однако, примитивное строение
I плеопода самца.
Все изложенное позволяет считать Сибирь центром возникновения и
распространения Asellus s. str., a A. aquaticus — северо-восточным при-
шельцем в Европу, как это предполагали Раковица (Racovitza, 1919) и Шап-
пюи (Chappuis, 1927). Сравнительно недавнее проникновение A. aquaticus
в Европу согласуется с отсутствием его на Пиренейском полуострове,
а также в Северной Африке.
В Европе Asellus aquaticus встречается обычно и в большом количестве
в районах, занятых в четвертичное время ледником (север европейской
части СССР, Скандинавия). По всей вероятности, заселение этих районов
произошло после отступания ледника. Весьма интересным в этом отношении
является факт почти полного отсутствия A. aquaticus в Арктике, особенно
на арктических островах, вообще практически лишенных настоящих пресно-
водных Malacostraca (Бирштейн, 19336). Единственное исключение пред-
ставляет собой Гренландия, на западном берегу которой в конце XVIII в.
Фабрициусом и в начале XIX в. Кроером был найден A. aquaticus. Присут-
ствие здесь этого вида совершенно необъяснимо, так как вероятность завоза
его людьми очень незначительна. Возможно, что авторами была допущена
ошибка, подобная той, которая расшифрована Иогансеном в отношении
39
указания A. aquaticus на Лабрадоре (Johansen, 1926). Боваллиус (Boval-
lius, 1886) относит указание Кроера к Jaera.
Нахождение представителей Asellus s. str. на других островах легко
поддается объяснению. Англия и Ирландия были соединены с континентом
еще в среднечетвертичное время (Борисяк, 1934), и ареал A. aquaticus был,
очевидно, тогда сплошным. Точно так же и Япония и Сахалин, населенные
A. hilgendorfi, вступали в четвертичное время в соединение с материком
(Криштофович, 1932).
Для понимания происхождения подрода Asellus s. str. в целом важно
оценить близость его к подроду Mesoasellus. Чрезвычайное сходство в строе-
нии эндоподита II плеопода самца заставляет сближать оба подрода. Этим
признаком они резко отличаются от остальных подродов Asellus. Другие
признаки Mesoasellus напоминают подроды Proasellus, Baicaloasellus и
Conasellus. Mesoasellus, по-видимому, реликт весьма отдаленных времен.
Три (четыре?) несомненно относящиеся к нему вида обнаружены по одному
на больших глубинах Байкала, в пещерах Японии и в пещерах Калифорнии.
Такое распространение свойственно многим архаическим формам, из которых
для примера можно указать пиявку Torix с близким американским родом
Microbdella, Epischura, найденную недавно на Камчатке, а также некоторых
Harpacticoida (Боруцкий, 1948). Все это, несомненно, оттесненные реликты
очень древней фауны.
Близость подродов Mesoasellus и Asellus s. str. позволяет допустить
происхождение второго от первого или, вернее, от сходных с ним форм.
Вероятно, подрод Asellus s. str. зародился в Восточной Сибири, явившейся
очагом возникновения целого ряда пресноводных групп, как, например,
многих ручейников, представляющих собой для Европы также северо-восточ-
ных пришельцев (Мартынов, 1929).
Помимо одного вида Mesoasellus, в Байкале живут 4 вида эндемичного
подрода Baicaloasellus. Это в высокой степени аберрантные формы, резко
отличающиеся от остальных Asellus своеобразным строением эндоподита
II плеопода самца, конец которого вытянут в хитинизированное клювовидное
образование. Однако хитинизация конца эндоподита II плеопода самца
наблюдается и у Mesoasellus, правда, в гораздо меньшей степени. Весьма
вероятно, что Baicaloasellus, так же как и Asellus s. str., является отдален-
ным потомком близких к Mesoasellus форм, причем формирование первого
подрода происходило в специфических условиях Байкала. Такое предполо-
жение вполне соответствует взглядам Берга (19226) на происхождение
байкальской фауны, причем Baicalossellus следует отнести к первой группе
Берга, т. е. к формам, «которые развились в самом Байкале в течение его
долгой геологической жизни», a Mesoasellus — ко второй группе «остатков
верхнетретичной (? — Я. Б.) пресноводной фауны Северной Азии». Сближе-
ние A. angarensis и A. baicalensis с американскими видами, которое допускал
В. В. Алпатов (1923), лишено оснований.
Каковы же в действительности отношения палеарктических Asellidae
к неарктическим? Ван-Нейм (Van Name, 1936) полагает, что американские
Asellidae произошли от форм Старого Света, мигрировавших через перешеек,
существовавший на месте Берингова пролива. С этим можно согласи-ться
только отчасти. Как указывалось выше, в Северной Америке имеются два
разъединенных Скалистыми горами ареала Asellidae — западный и восточ-
ный. В западном ареале констатировано всего два вида — A. tomalensis
40
и A. californicus. Последний вид несомненно относится к подроду Mesoasel-
]us, систематическое положение первого вида неясно из-за совершенно
неудовлетворительного описания, но он определенно не принадлежит к вос-
точно-американскому подроду Conaselliis. Присутствие A. californicus
позволяет сближать фауну западноамериканских Asellidae с восточноазиат-
ской, и, поскольку большая часть видов Mesoasellus известна из Восточной
Азии, можно допустить его азиатское происхождение.
фауна Asellidae восточной части Северной Америки значительно богаче,
и генетические ее отношения гораздо менее ясны. Наибольшее количество
восточноамериканских видов относится к эндемичным — роду Lirceus (16 ви-
дов) и подроду Conasellus (17 видов). Кроме того, известно еще 5 восточно-
американских видов (A. nickajackensis, A. adenta, A. richardsonae, A. smithi
и A. attenuatus), систематическое положение которых остается невыяс-
ненным.
До недавнего времени род *Lirceus со сросшимися с сегментами тела
эпимерами и отсутствием мандибулярного щупика стоял особняком от
других Asellidae. Однако открытие Моно (Monod, 1924) в северной Африке
своеобразной Johanella изменило такое положение. Моно указывает на
чрезвычайную близость этих родов и характеризует свою Johanella как
Lirceus с мандибулярным щупиком. Еще более важным с зоогеографи-
ческой точки зрения фактом приходится считать нахождение в подземных
водах Португалии близкого к Lirceus рода Synasellus (Braga, 1944), что
заставляет внимательнее отнестись к сходству Conasellus с кругосреди-
земноморским Proasellus. Сходство это довольно отдаленное, но еще Рако-
вица (Racovitza, 1919) отметил, что исследованный им Asellus (Conasellus)
communis стоит дальше от Asellus s. str., чем от Proasellus (по современной
терминологии). Итак, одна из возможностей объяснения происхождения
восточноамериканских Asellidae — это сближение их с кругосредиземно-
морской фауной: Johanella и Synasellus — с Lirceus, a Proasellus — с Co-
nasellus. Попытки такого сближения для других групп пресноводных Mala-
costraca уже делались: именно так трактуют Бирштейн и Виноградов
(1934) происхождение восточноамериканского Cambarus. Если обратиться
к другим пресноводным Isopoda, то окажется, что сходное распростра-
нение свойственно преимущественно пещерным Cirolanidae: три рода
(Typhlocirolana с четырьмя видами, Sphaeramides с двумя видами и
Faucheria с одним видом) населяют кругосредиземноморские страны,
а чрезвычайно близкие к ним три рода (Cirolana с двумя видами, Cirolani-
des и Conilera — каждый с одним) известны из Мексики, Кубы и Техаса.
Раковица (Racovitza, 1912) указывает, что среди всех ныне живущих
Cirolanidae ближе всего к Typhlocirolana стоят восточноамериканские роды
Cirolana и Conilera. Близость западноевропейской и восточноамериканской
древней пресноводной фауны (Раковица принимает существование Ciro-
lanidae начиная с мелового или, возможно, юрского периода) может быть
легко объяснена связью обоих материков посредством отрогов погрузивше-
гося впоследствии Атлантического хребта (Берг, 1948). Один из них через
острова Мадейру, Канарские и Азорские тянулся прямо к Пиренейскому
полуострову.
Таким образом, довольно вероятно двустороннее заселение Северной Аме-
рики водяными осликами: с запада через сушу, заменявшую современ-
ный Берингов пролив, и с востока — из Западной Европы через отроги
41
Атлантического хребта. При таком допущении становится понятным и
нахождение Asellus italicus perarmatus на о-ве Мадейра. Реми (Remy,
1936), описавший указанный подвид, держится именно такого мнения.
Не исключается, однако, и менее вероятная возможность происхождения
восточноамериканских Asellidae от западноамериканских; окончательное
решение вопроса, быть может, даст подробное сравнительно-морфоло-
гическое изучение водяных осликов Нового Света, исследованных, к со-
жалению, очень поверхностно.
Обрисованная такими общими чертами картина географического рас-
пространения Asellidae может быть названа закономерной в том отношении,
что аналогичный характер ареала наблюдается и у других групп древних
пресноводных Malacostraca. Если взять, например, карту географического
распространения пресноводных Amphipoda группы Crangonyx, составленную
Шелленбергом (Schellenberg, 1936), то окажется, что в северном полушарии
имеются те же 5 разорванных ареалов — Европа, Байкал, Дальний Восток
и западная и восточная части Северной Америки, которые были намечены
выше для Asellidae. Можно проследить аналогию и для отдельных родов и
подродов. Ареал Synurella совпадает в общем с ареалом Asellus s. str. Совер-
шенно так же, как группа Crangonyx, распространены, по Шелленбергу,
бокоплавы группы Gammarus. Приблизительно тот же характер распро-
странения свойствен и Astacidae (Бирштейн, Виноградов, 1934), с той
только разницей, что речные раки не встречаются в Байкале. Наконец,
все эти группы древних пресноводных Malacostraca сближает между собой
их отсутствие в тропиках 1 В антибореальной области семейство Astacidae
замещается близкими семействами Parastacidae и Austroastacidae, а группа
Crangonyx представлена там несколькими большей частью эндемичными
викарирующими родами. Формы, родственные Asellidae, в пресных водах
южных материков не известны, но преимущественно в антибореальных
морях живет сравнительно близкое семейство Stenetriidae.
Последний список видов единственного рода этого семейства показывает,
что из 25 видов рода Stenetrium 23 обитает в морях южного полушария
и только 2 — в Средиземном море (Nordenstam, 1946).
Трудно предположить, чтобы указанные совпадения оказались случай-
ными; по всей вероятности, распространение всех древних пресноводных
Malacostraca определялось одними и теми же историческими причинами.
Некоторые из них, быть может, удалось подметить, но вся проблема в целом
еще очень далека от своего разрешения.
В заключение этого краткого очерка следует сказать несколько слов
и о пресноводных Paraselloidea—родах Jaera, Microparasellus и Micro-
charon. Последние два рода объединяются в семейство Microparaseilidae,
которое Караман (Karaman, 1934а) сближает с чисто морскими семействами
Macrostylidae и Desmosomatidae. Microparasellidae до сих пор найдены
только в грунтовых водах Югославии и Румынии, но весьма вероятно их
нахождение и в других районах, в частности на Кавказе. Близость Micropa-
rasellidae к морским формам стоит в соответствии с родственными отно-
шениями других высших ракообразных, населяющих грунтовые воды.
В последнее время установлено, что значительный процент подземных
1 Единственное исключение — уже упоминавшиеся Stenasellus chappuisi и St. africa-
nus из экваториальной Африки.
42
Malacostraca имеет близкие родственные формы не в фауне пресных вод,
а среди чисто морских видов. В первую очередь в качестве таких
форм следует назвать род Niphargus, приближающийся, как показал Шел-
ленберг (Schellenberg, 1933), к морским родам Eriopisa и Eriopisella.
Другой представитель подземных Amphipoda — Ingolfiel la acherontis
из грунтовых вод Югославии — принадлежит к подотряду Ingolfiellidea,
заключающему, кроме названного еще 2 вида: I. abyssi, найденную на
глубине 3000 м у входа в Дэвисов пролив, и I. littoralis, найденную среди
кораллов в Сиамском заливе на глубине 2 м (Hertzog, 1935). Из Isopoda за-
мечательное семейство Microcerberidae, представленное одним видом также
в грунтовых водах Югославии, приближается по ряду признаков к чисто
морскому семейству Anthuridae. Такие же связи прослежены для некоторых
пещерных Polychaeta и Archiannelida (Бирштейн, 19406).
Шелленберг (Schellenberg, 1935) и Караман (Karaman, 1935) полагают,
что известная часть фауны грунтовых вод вселилась туда непосредственно
из моря, минуя наземные пресные воды. Такой переход был особенно
легким для форм, которые и в море ведут «подземный» образ жизни,
позволяющий им проникать далеко в глубь грунта. Этим путем они и могли
попасть из системы соленых морских грунтовых вод в систему пресных вод.
Действительно, весь облик Microparasellidae, Microcerberidae, Ingolfiel 1 idae,
имеющих вытянутое червеобразное тело, при очень незначительной вели-
чине животных чрезвычайно характерен для представителей интерстициаль-
ной фауны, и изложенная гипотеза по отношению к Microparasellidae
может быть принята.
Морская природа последнего рассматриваемого здесь рода — Jaera —
не вызывает уже никаких сомнений. В то же время также несомненна
его тенденция к жизни в пресной воде. В настоящее время известно
12 видов и один подвид Jaera, из которых 2 (J. italica и J. schellenbergi)
являются исключительно пресноводными, 2 живут как при нормальной океа-
нической солености, так и в совершенно пресной воде (J. albifrons и J. nord-
manni), один — преимущественно морской, переносящий опреснение до
в %о (J- hopeana), один — солоноватоводный, не заходящий в пресную воду
(J. sarsi caspica) и один — пресноводный, выносящий некоторое осолонение
(J. sarsi), остальные — морские. Этот род населяет северную Атлантику,
Средиземное море и Понто-Каспийский бассейн. Вне Европы достоверно
известен только вдоль североатлантического побережья Америки. Из 12 ви-
дов 11 приурочено к бассейну Средиземного моря, один вид (с подвидом)
эндемичен для Понто-Каспия. Такое распространение заставляет пред-
полагать, что род Jaera сформировался еще в третичное время в бас-
сейне Средиземного моря и Понто-Каспийский бассейн получил предков
J. sarsi вместе с предками других, близких к средиземноморским рако-
образных (Mysidacea, Cumacea) около сарматского времени, как это считал
еще Совинский (1904), принимавший, правда, J. sarsi за J. nordmanni.
Мартынов (1925а) оспаривал мнение Совинского, указывая на то, что
«очень странно, что, несмотря на столь долгий промежуток времени, она
(Jaera) не отклонилась от морских (атлантических и средиземноморских)
представителей этого вида». По мнению этого автора, J. nordmanni попала
в Каспий уже в четвертичное время вместе с Cardium edule. Возражение
Мартынова отпадает, так как каспийская Jaera в действительности отлича-
ется от морских представителей этого рода настолько сильно, что с полным
43
правом выделяется в самостоятельный вид — J. sarsi. Характер ее ареала
(Каспий с притоками, реки, предустьевые пространства и реликтовые озера
Азовско-Черноморского бассейна) с несомненностью указывает на ее принад-
лежность к понто-каспийским эндемикам и заставляет вычеркнуть эту
форму из списка четвертичных средиземноморских вселенцев в Каспий.
Совершенно особую проблему представляет собой вопрос о способе
и времени вселения J. sarsi в реки Понто-Каспийского бассейна. По отно-
шению к другим высшим ракообразным морского происхождения этот
вопрос разрешался по-разному. Державин (1912, 1924), Мартынов (1925а)
и Бенинг (1924а) считали морских ракообразных в реках Понто-Каспийского
бассейна реликтами или псевдореликтами третичных трансгрессий. Саре
(Sars, 1927), Беклемишев (1923) и Бирштейн (1935,1946) рассматривали эти
формы как активных иммигрантов из моря в четвертичное время. За послед-
нее время резкие различия между этими точками зрения в значительной сте-
пени стерлись. В своей недавней работе Державин (1939) относит засе-
ление морскими элементами каспийских рек к четвертичному времени. Во-
прос о том, происходила ли переработка морской фауны в пресноводную
в самом море (Бирштейн) или только в его заливах (Державин), не
имеет принципиального значения. Так или иначе, в четвертичное время
под влиянием значительного опреснения, связанного с притоком талых вод
ледника, Jaera sarsi оказалась способной к обитанию в пресной воде
и попала в реки Понто-Каспийского бассейна. Каким образом она под-
нялась на 1000 км вверх по Волге и на 1800 км вверх по Дунаю —
сказать трудно. С одинаковым правом можно допустить медленное актив-
ное расселение вверх по течению и пассивный перенос на обрастаниях
судов.
Штрохаль (Strouhal, 1939а) обнаружил J. sarsi в одном из искусствен-
ных каналов, соединяющихся с Дунаем, и на этом основании считает воз-
можным активное расселение.
Хотя Державин (1939) и считает, что вряд ли кто-либо из сторон-
ников реликтовой теории будет считать трех понто-каспийских бокоплавов
и Corophium curvispinum, найденных в Дунае под Будапештом, сармат-
скими реликтами, но именно такое мнение высказали Дудич (Dudich, 1930)
и Кессельяк (Kesselyak, 1938) по отношению к J. sarsi в Дунае (и Дудич,
Dudich, 1927, — по отношению к упомянутым Amphipoda). С этим мнением
трудно согласиться, так как сарматская фауна чрезвычайно сильно отли-
чалась от современной, а став на эту точку зрения, приходится допустить
совершенно параллельную эволюцию Jaera в разных частях ее ареала
и в разных условиях, приводящую тем не менее к политопному образо-
ванию одного вида. Гораздо более вероятно недавнее формирование J. sarsi
в едином водоеме (последнее соединение Понта и Каспия относится к рисс-
вюрмскому (Архангельский и др., 1937) или послевюрмскому времени)
и последующее проникновение ее в реки Понто-Каспийского бассейна.
В связи с этим следует отметить присутствие в Каспии эндемичного под-
вида J. sarsi caspica, приуроченного к относительно осолоненным (12—13°/00)
частям моря и, по-видимому, не встречающегося в опресненном северном
Каспии; ареал J. sarsi caspica оказывается, таким образом, оторванным от
ареала типичной J. sarsi, населяющей Волгу и Урал. Можно высказать
предположение, что подвид J. sarsi caspica сформировался уже после
прекращения связи Каспийского и Азовско-Черноморского бассейна при не-
44
давнем осолонении Каспия, последовавшем за послеледниковым опресне-
нием.
Заканчивая обзор распространения Jaera, следует указать на уже отме-
ченную выше для других ракообразных особенность: этот род отсутствует
в тропиках, а в антибореальной области его замещает близкий, недавно
выделенный род Neojaera Nordenstam (Nordenstam, 1933).
К ВОПРОСУ О ПРОИСХОЖДЕНИИ МОРСКИХ РАКООБРАЗНЫХ
В РЕКАХ ПОНТО-КАСПИЙСКОГО БАССЕЙНА*
ВВЕДЕНИЕ
Проникновение морских организмов в пресные воды есть одна из инте-
реснейших зоогеографических проблем. Ставя ее, мы вплотную подходим
к большому общему вопросу о происхождении всей пресноводной фауны
от морской. Немудрено поэтому, что морские организмы, встречающиеся
в пресных водах, издавна привлекали к себе внимание исследователей
и послужили материалом для ряда классических работ. В частности, по
морским ракообразным, найденным в реках Понто-Каспийского бассейна,
существует обширная литература, но вопрос о способе их проникновения
в реки, ставившийся неоднократно, до сих пор не может считаться решенным.
Мне пришлось непосредственно столкнуться с этим вопросом при обра-
ботке фаунистического материала, собранного мной летом 1930 г. на озерах,
расположенных в низовьях Терека. Их история представляет известный
интерес.
По данным Кузнецова (1898), до 1914 г. Терек выше Кизляра рас-
падался на 5 в большинстве копаных рукавов, уровень воды которых был
выше уровня окружающих полей. Аграханский залив, куда впадала часть
рукавов Терека, к концу прошлого столетия был значительно осолонен.
В 1914 г. из-за прорыва плотины у станицы Каргалинской вода хлынула
на лежащие ниже поля, затопила их, и, таким образом, на сухом месте
появилась целая система озер и плавней, имеющих сток в Аграханский
залив и пропускающих через себя 75,3 % воды Терека (Дмитриев, 1929).
С тех пор Аграханский залив стал быстро опресняться.
Озера Нижнего Терека представляют собой обширные, но неглубокие
водоемы, богатые макрофитной растительностью с заиленным дном и боль-
шей частью незаметным течением. Берега сплошь заросли Phragmithes
communis (до 4 м высоты), кусты которого постоянно попадаются и в сере-
дине озер. В собранном мной в этих водоемах материале оказались следую-
щие виды Malacostraca: Dikerogammarus haemobaphes (Eichwald), Ponto-
gammarus robustoides (Grimm), Mesomysis kowalewskyi, Czerniawskyi,
Limnomysis benedeni Czerniawsky, Asellus aquaticus Linne sensu Racovitza.
Первые четыре вида являются морскими. Их нахождение в исследованных
озерах можно объяснить наиболее естественно и просто активным про-
никновением из моря. Действительно, в Аграханском заливе я нашел неда-
леко от протока озер Р. robustoides, М. kowalewckyi и L. benedeni, морфоло-
гически идентичных озерным. Что же касается фауны Терека выше места
* 1935 г. —Зоол. журн., т. 14, вып. 4, с. 749—761.
45
его прорыва, то, к сожалению, исследовать ее я не имел возможности.
Однако характер реки в этом месте позволяет думать, что нахождение там
указанных трех видов маловероятно: Терек в участке ниже г. Кизляра и до
озер представляет собой узкую мелкую речку, довольно быстро текущую
по жесткому грунту и не образующую никаких затонов, заводей и пр.
Как известно из работ Мартынова (1919, 1925а), Р. robustoides, М. kowa-
lewskyi, L. benedeni приурочены к местам с медленным течением, илистым
грунтом и богатой растительностью, и поэтому Терек в районе Кизляра
(и тем более выше) является совершенно неподходящим для них биотопом.
Нахождение их вблизи озер в морском заливе, не отличающемся заметно
по своим физико-химическим свойствам от озер, заставляет предполагать
их активное вселение из моря в пресную воду. Другое возможное предполо-
жение — проникновение морских ракообразных из верхнего течения Терека,
где они могли сохраниться как морские реликты, — в данном случае очень
маловероятно.
Суда из Аграханского залива в Нижне-Терские озера не поднимаются
(с чем, между прочим, можно связать отсутствие в озерах Dreissena
polymorpha), так что возможность пассивной иммиграции исключается.
Если в данном случае вопрос о происхождении морских организмов легко
решается в смысле их активной иммиграции, то для целого ряда водоемов
Понто-Каспийского бассейна он оказывается более сложным и запутанным.
Простое сопоставление уже добытых фаунистических и геологических
данных, не разрешая, может быть, этот вопрос окончательно, приводит
все же к интересным и определенным выводам.
ФАУНИСТИЧЕСКИЕ ДАННЫЕ
Уже Палласу (Pallas, 1773—1776) было известно нахождение в Волге
организма морского происхождения Dreissena polymorpha. Этот моллюск
вслед за Палласом отмечается также для Волги рядом авторов в XIX в.
(Эйхвальд, Бэр, Рулье, Фаренколь и др.). Первые указания морских рако-
образных для Волги принадлежат Зыкову (1903) и Скорикову (1903) и от-
носятся к найденной у Саратова Metamysis strauchi, неправильно опре-
деленной как Mesomysis ulskyi. Та же мизида описывается затем для
верхней Волги Третьяковым (1907) как Mesomysis volgensis. Морские
амфиподы Dikerogammarus haemobaphes, Pontogammarus abbreviatus, Co-
rophium curvispinum фигурируют в списке организмов, найденных Волжской
биологической станцией еще в 1902 г. Затем, главным образом в результате
работ Державина (1912; Державин и др., 1921) и Бенинга (1924а, б, 1925),
этот список был значительно пополнен, и теперь для Волги насчитывается
морских Malacostraca: Isopoda — 1, Amphipoda — 26, Cumacea — 10, Mysi-
dacea — 6 и Decapoda — 1 (Бенинг, 1924a).
Менее подробными данными мы располагаем по другим рекам. 12 видов
морских Malacostraca найдены в дельте Урала, шесть из них поднимаются
до Гурьева (Державин, 1926), два встречаются в среднем течении (Бенинг,
1928а). Для низовьев Куры известно пять видов, часть которых найдена
в девяти мелких реках Южного Каспия (Державин, 1923). Низовья Дона
населены 26 видами (Мартынов, 1925), выше в среднем течении найдено
семь видов (Buchalowa, 1929). В низовьях Днепра констатировано девять
видов (Мартынов, 1925а), у Киева—два (Бенинг, 1915; Шарлеман, 1922),
46
в Днестре — восемь (Фадеев, 1923; Бенинг, 19286; Белинг, 1929). Для
среднего течения Дуная указано четыре (Dudich, 1927), для Вислы — два
(Jarocki, Demianowicz, 1931).
Для объяснения происхождения морских ракообразных в пресных
водах Понто-Каспийского бассейна было предложено две гипотезы. Ско-
риков (1903), Зыков (1903), Саре (1907, Sars, 1927) и Берг (1908) рас-
сматривали морских перакарид, обнаруженных в пресной воде, как активных
иммигрантов из моря в реки. Наиболее четко высказал эту точку зрения
Беклемишев (1923). Речных Peracarida он считает эвригалинными, раз-
новодными в смысле Кесслера животными, активно иммигрировавшими
из моря в порядке совершенно естественного стремления к расширению
ареала.
Державин (1912) выступил с другой гипотезой. Поскольку все извест-
ные в то время местонахождения морских ракообразных не выходили из
области, занятой в акчагыле трансгрессией Каспия, этот автор считает
Peracarida акчагыльскими реликтами, оставшимися на своем коренном ме-
стообитании после отступания моря. Впоследствии, когда Pericarida были
найдены и вне области акчагыльской трансгрессии, Державин признал
за ними способность мигрировать против течения, но только из области
трансгрессии, где, по его мнению, и происходило преобразование морского
животного в пресноводное (псевдореликты, по терминологии Экмана
(Державин и др., 1921)). Помимо реликтов, в реках, по Державину, живут
также пассивные иммигранты, приносимые на судах. В статье, посвященной
этому вопросу, Державин, полемизируя с Беклемишевым, отстаивает
сво.и прежние взгляды (Державин, 1924).
К той же точке зрения происоединилось большинство авторов последнего
времени, в том числе Ильин (1930) и Зернов (1934). Бенинг (19246, 1925)
констатировал усиление некоторых морфологических и биологических от-
личий речных форм от основных морских пропорционально расстоянию от
моря, что он рассматривает как доказательство реликтового происхождения
Peracarida Волги. Следует заметить, что этот факт сам по себе еще ни-
чего не доказывает, так как может быть объяснен и с точки зрения
теории иммиграции. Действительно, степень отличия пресноводной формы
от породившей ее морской пропорциональна времени ее проживания
в пресной воде (Экман), а чем выше по течению она зашла, тем, естественно,
дольше пробыла в пресной воде.
Мартынов в первой своей работе, посвященной морским ракообразным
Дона, рассматривал их как реликты послеледниковой трансгрессии, прибли-
зительно совпадавшей с моментом прорыва Дарданелл (Мартынов, 1919),
а впоследствии — как реликты понтического времени (Мартынов, 1925а).
Что касается морскйх ракообразных Днепра, то, не разбирая вопроса об их
происхождении, Мартынов просто называет их реликтами (Мартынов,
19256).
Белинг (1925) занимает несколько неопределенную позицию: отмечая
более низкое стояние уровня моря в четвертичное время до прорыва
Дарданелл и связанное с этим усиление течения Днепра и размыв и
углубление ложбины, этот автор пришел к такому выводу: «. .вряд ли в то
время фауна понтического бассейна удерживалась в удлиняющей свое
течение и углубляющей свое русло реке. Скорее всего, она заменялась
чисто речной фауной (только как исключение могли сохраниться в реке
единичные легко приспособляющиеся к речному режиму формы, например
47
некоторые виды рыб, ракообразных)» (с. 37). Несогласованность фразы,
стоящей в скобках, с предыдущими бросается в глаза.
В более поздней работе Белинга, посвященной фауне порожистой части
Днепра (Белинг, 1929), также не дается ответа на интересующий нас
вопрос. По Белингу, мизиды не являются сарматскими реликтами, а в таком
случае это же должно относиться и к амфиподам, так как вряд ли они иного
происхождения или древнее мизид. Остается сделать вывод об их активном
проникновении из понтического моря, но прямо об этом Белинг не говорит.
Дудич, изучивший понто-каспийских ракообразных Дуная вблизи Бу-
дапешта, отказывается разрешить вопрос о том, являются) ли они вселен-
цами или реликтами. Он замечает, что если считать их реликтами, то только
реликтами нижнесарматского моря, так как уже со времен среднего
Сармата Венгрия морем не покрывалась (Dudich, 1927).
Фадеев (1930), отмечая понто-каспийские морские виды в фауне Донца,
достаточно определенно высказывается за их активную иммиграцию из моря.
Он указывает на неравномерное распространение этих видов по Донцу,
аналогичное подмеченному Бенингом для Волги. Последний автор пола-
гает, что такое распространение указывает на отдельные периоды транс-
грессий; Фадеев, ссылаясь на мнение Криштофовича и Соболева, указывает
на несовпадение распространения морских животных по Донцу с третичными
трансгрессиями, последняя из которых достигала только устья Донца.
В этой связи интересно нахождение Diamysis mecznikowi nat. pengoi
по всему Донцу. Эта форма средиземноморского происхождения очень
близка к D. bahirensis G. О. S. (Мартынов, 1924). В Черное и Азовское
моря она могла попасть только после прорыва Дарданелл, и ее проникно-
вение в Донец не может быть связано с третичными трансгрессиями.
Что касается представителей малоподвижной морской фауны, то, как
показывают сводки Жадина (1933) и Анненковой (1930), морские моллюски
и полихеты вверх по рекам почти не поднимаются: Hypania invalida (Grube)
отмечена для дельты Волги и нижнего течения Днепра, Hypaniola kowalew-
skyi (Grimm) — для низовьев Днестра и р. Турунчук. Micromelania lincta
и Monodacna colorata известны в нижнем течении Днепра. Правда, не-
сколько видов Theodoxus и Melanopsis населяют значительную часть Ду-
ная, Днестра, Днепра и Буга, но в реках Каспийского бассейна они не
обнаружены. Dreissena polymorpha, по мнению большинства авторов, рас-
селяется пассивно, прикрепляясь к судам и т. д.1
Резюмируя сказанное, можно сделать следующие выводы.
1. Способность к активному продвижению понто-каспийских перакарид
против течения признается всеми* авторами без исключения.
2. В верхнем течении рек находятся наиболее подвижные формы. Мало-
подвижные животные морского происхождения приурочены к устьям рек.
Больше того, зашедшие наиболее далеко виды обладают наибольшей способ-
ностью к активному движению. Dikerogammarus haemobaphes, D. villosus
и Chaetogammarus ischnus, по Мартынову, являются обитателями теку-
чей воды и, очевидно, способны преодолевать силу течения. Именно они
оказываются наиболее распространенными по рекам представителями понто-
каспийских амфипод. Виды рода Pontogammarus — в большинстве обита-
1 Жадин полагает, что Dreissena проникла в Германию еще в третичное время,
но вопрос этот нельзя считать окончательно выясненным.
48
Распространение Dikerogammarus haemobaphes (Eichw.) (/) и Chaetogammariis
ischnus Stebbing (2) и границы: Сарматского моря (по: Мазарович, 1933) (<?),
Понтического (4), Акчагыльско-Куяльницкого моря (5) (по: Архангельский, 1932)
(схематизировано)
тели тихих заиленных водоемов — встречаются недалеко от устьев. Jaera
nordmanni — относительно менее подвижная форма — также не идет так
Далеко, как Dikerogammarus и Chaetogammarus.
Если бы понто-каспийские формы действительно сумели сохраниться
в реке от отступающего моря, то противостоять усиливающемуся течению
могли бы прежде всего сидячие морские формы (Mollusca и Polychaeta,
например), которые фактически в реках не наблюдаются.
4 Заказ 333
49
3. Если стоять на точке зрения сторонников реликтовой природы мор-
ских ракообразных в реках Понто-Каспия, то приходится признать, что
одни и те же виды амфипод являются реликтами разных геологических
эпох: для Дуная они реликты сарматского времени, для Дона — понтиче-
ского, для Волги — акчагыльского. Ясно, что такое положение может
иметь место только в том случае, если на протяжении от сарматского
времени до наших дней: 1) или не происходило никаких морфологических
изменений интересующих нас животных, 2) или эти изменения протекали
в разнородных условиях совершенно одинаково, параллельно. И то и другое,
разумеется, в равной степени чрезвычайно маловероятно. Что касается
первого предположения, то имеющиеся палеонтологические данные ему
решительно противоречат. Ископаемые гаммариды из верхнесарматских
отложений Понто-Каспия описаны Державиным (1927). Все они представ-
лены новыми родами, родственными современным, но резко отличными
от них. Державин пишет, что автохтонные каспийские гаммариды претер-
пели значительную эволюцию со времен верхнего сармата. Из литературы
известно, что уже в акчагыле не существовало также ни одного современного
вида моллюсков и рыб. В сарматском море не было общих видов ни с акча-
гыльским, ни с понтическим. Каждое из них характеризуется специфи-
ческой фауной (это известно для моллюсков и рыб).
Если фауны морей различного времени были неодинаковы, то их реликты
не могут быть представлены одними и теми же видами.
ГЕОЛОГИЧЕСКИЕ ДАННЫЕ
Сторонники гипотезы реликтового происхождения морских ракообразных
в реках Понто-Каспийского бассейна ограничиваются обыкновенно анализом
условий только третичного времени. Они констатируют момент самой поздней
трансгрессии моря, покрывавшей современное местонахождение морских
перакарид или приближавшейся к нему и считают исследуемых животных
реликтами времени этой трансгрессии (Державин, 1912; Державин и др.,
1921; Державин, 1926а; Мартынов, 1925а, б). Послетретичная история
района исследования этими авторами, как правило, не затрагивается.
Между тем в формировании современной пресноводной фауны Палеарк-
тики решающую роль сыграло не третичное, а четвертичное время. Оно
особенно важно также и потому, что, по последним данным, акчагыльская
трансгрессия, реликтами которой Державин и Бенинг считают Peracarida
Волги, относится к послеледниковому времени.
Акчагыльские отложения были впервые установлены и исследованы
Андрусовым, считавшим их первоначально горизонтом, смежным с сар-
матскими слоями (миоцен), так как фауна акчагыла на первый взгляд
кажется близко родственной сарматской (Андрусов, 1902). Впоследствии
Андрусов перенес акчагыл в верхний отдел плиоцена, а вслед за сар-
матскими слоями поместил понтические (Андрусов, 1911).
В начале понтического времени каспийский и азовско-черноморский
бассейны соединял ись друг с другом узким проливом в области современного
Маныча. Северная граница понтического моря в момент наибольшего его
простирания проходила через современный город Запорожье. Вследствие
общего поднятия суши понтическое море довольно быстро сокращалось,
и к концу понтического времени Черное и Азовское моря отделились
от Каспия. В то же время образовались современные долины низовьев
50
Днепра и Дона. Последний процесс Белинг (1925) представляет так: «По
мере отступания моря на юг и понижения базиса эрозии река все энергич-
нее работала над размывом и углублением своего русла». При этом,
помимо резких изменений солевого режима и подвижности воды, изме-
нялся также целый ряд не менее важных факторов, в частности характер
грунта. Река, размывавшая даже твердые породы, безусловно размывала
и уносила ил и мелкий песок, причем ряд форм, приуроченных к этим
грунтам, теперь «не мог найти ни покоя, ни ила» (с. 50).
Каспийское море в конце понтического времени сократилось еще
больше, чем Черное и Азовское. Согласно Батурину (1931), оно занимало
немного больше ’/з площади современного Каспия. Уменьшение Каспия
в течение понтического времени Павловым (1913), Ковалевским (1933)
и Богачевым (1932, 1933) связывается с развитием оледенений в бас-
сейне моря.
Отчлененные друг от друга понтические моря были сильно опреснены
и населены пресноводными и солоноватоводными формами. Поэтому понти-
ческая фауна резко отличается от сарматской, состоявшей из морских
видов.
Последовавшее за понтическим акчагыльское море во всех отношениях
было иным. Оно отличалось многоводностью, и его северный залив просле-
жен далеко по Волге, Каме и Уфе. На западе оно широким проливом
соединялось с Куяльницким морем, занимавшим бассейны Черного и Азов-
ского морей. Соленость акчагыльского моря была высокой, и в нем вновь
появилась настоящая морская фауна, напоминающая сарматскую. Проис-
хождение этой фауны, равно как и внезапное осолонение и обводнение
Каспия, до последнего времени оставались загадкой. Андрусов (1902,
1911), Губкин (1931) и Архангельский (1932) предполагали, что где-то
существовало убежище, в котором сарматская фауна пережила понтическое
время и в акчагыле вернулась в Каспий. Однако детальные геологические
исследования побережий Каспия таких убежищ не обнаружили.
Ковалевский (1933) выступил с другой, чрезвычайно интересной и
изящно изложенной гипотезой. Этот автор находит, что только приток
океанических вод мог превратить маленькое опресненное понтическое море
в мощный соленоводный акчагыльский бассейн. По его мнению, северный
(волжский) залив акчагыльского моря соединился с бореальной трансгрес-
сией Ледовитого океана, относящейся к миндель-рисскому (кромерскому)
веку. Таким образом, вдоль Урала лежал пролив, по которому океани-
ческие воды свободно проникали в Каспий.
Особый интерес представляет мнение Ковалевского о распространении
ледников в бассейне Волги: он считает, что ледники рисского и мин-
дельского оледенений занимали всю долину Волги и спускались до дельты,
и цитирует мнение Голынца о бэровских буграх как о друмлинах одного
из оледенений. Следует отметить, что еще Павлов (1913) принимал рас-
пространение ледника до Волгограда.
Ковалевский подчеркивает, что периодам наиболее сильного развития
ледяного покрова соответствуют фазы наименьшей площади моря, что было
отмечено еще Павловым
1 Связывание воды ледником, на которое указывают Павлов и Ковалевский, противо-
речит гипотезе, выдвигаемой H0gbom (1917; цит. по: Spandl, 1926; Берг, 1928)
для объяснения проникновения арктических форм в Каспий. Согласно этой теории,
ледник, запрудив реки, текущие на север, образовал обширные озера, которые
4*
51
Дальнейшая история Каспия, по Ковалевскому, такова: в голоцене
иольдиево, литориновое и анциловое моря имели сток в Каспий и их воды
оттуда по Манычскому проливу проходили в Азовское и Черное моря. Именно
таким образом можно объяснить проникновение в Каспийское море ком-
плекса арктических форм, вопрос о происхождении которых до сих пор
неясен 1 Комбинируя геологические данные с указаниями древних латинских
и греческих авторов, Ковалевский (1933) показывает, что еще* за 3400 лет
до наших дней «Каспий простирался по древней впадине Волги далеко
на северо-запад и там оканчивался коротким протоком вроде современной
Невы, но много более мощным, питавшим море из „мертвых“ вод ледяного
бассейна» (с. 116). Это соединение прервалось только примерно за 2000 лет
до наших дней * 1 2 При этом следует иметь в виду, что, как указал Яковлев
(1934), «обратное повышение земной коры после прогиба (под давлением
ледника.—Я. Б.) происходило медленнее, чем повышение уровня океана
от таяния ледника» (с. 289).
Благодаря массе талых ледниковых вод и большому количеству мощных
притоков, часть которых в настоящее время уже не существует (Узбой),
Каспий в ранние исторические века (до начала нашей эры) находился
в состоянии почти полного опреснения. Последующее осолонение произошло
совсем недавно.
Из изложенных геологических данных можно сделать следующие два
вывода, интересные с точки зрения вопроса о происхождении морских
элементов в понто-каспийских реках.
1. Понто-Каспийский бассейн пережил резкие изменения солености—
от повышенной в сарматское и акчагыльское время до полного или почти
полного опреснения в понтическое и п ост гл я ци альное время. Таким образом,
процесс превращения морского животного в пресноводное мог происходить
в пределах самого моря.
2. Если действительно южный отрог миндельского ледника по волжской
впадине проникал до моря, то оставшаяся в Волге фауна акчагыльского
моря (если она оставалась), приходившегося на миндель-рисскую межлед-
никовую эпоху, должна была уничтожиться нацело льдами, подобно тому
как это принимает для фауны Белого моря Дерюгин (1928).
Это обстоятельство может послужить, между прочим, ответом на следую-
щее положение Державина: «Для несогласных с высказанным мной пред-
положением (о реликтовой природе волжских Peracarida. — Я. Б.) нет
оснований допускать активную иммиграцию каспийских выходцев в Волгу
из моря в позднейшее время; при подобном допущении останется необъясни-
мым, почему одаренные приспособленностью к пресной воде организмы
сначала покинули область своего распространения под влиянием ее опрес-
нения, а затем заселили ее уже совершенно пресную» (Державин, 1912).
и гнал перед собой, пока не перелил их в бассейн Волги где-то около Рыбинска.
Русские авторы указывают на противоположное явление — обезвоживание районов,
находившихся вблизи ледника.
1 Последними по этому вопросу высказывались Берг (1928) и Гурьянова (1933).
Берг считает, что арктические формы попали в бассейн Волги в послеледниковое
время, когда побережье Балтийского моря, Ладожское и Онежское озера, бассейн
Белого озера и Шексна входили в состав одного обширного озерного бассейна
(Яковлев, 1934). Гурьянова находит, что северные элементы фауны высших рако-
образных Каспийского моря — иммигранты из Сибирского побережья Ледовитого
моря и что проникновение этих форм из Ледовитого океана в Каспий шло где-то
восточнее, ближе к Уралу, вне связи с Балтийским и Белым морями.
2 Л. С. Бергом (19346) данные Ковалевского ставятся под сомнение.
52
ЗАКЛЮЧЕНИЕ
Таким образом, весь комплекс фаунистических и геологических данных,
которыми мы в настоящее время располагаем, заставляет отказаться от
теории реликтового происхождения морских ракообразных в понто-каспий-
ских реках. Я не вижу ни одного факта, ее подтверждающего. Эта теория
создалась на основании случайного совпадения известных до 1914 г. место-
нахождений морских перакарид с границами акчагыльской трансгрессии,
йо теперь, когда ясно, что достаточно полно изученное их распростра-
нение не стоит в связи ни с одной из морских трансгрессий (рис., с. 49), теория
реликтового происхождения должна быть оставлена.
Следует оговориться, что в отдельных случаях мы действительно встре-
чаемся с настоящими морскими реликтами. Так, например, в притоке
Аракса — речке Кара-Су — Державиным (19266) обнаружена колония
каспийских рыб — Rutilus rutilus schelkovnikovi, Blicca bjoerkna transcau-
casica, Alburnus alburnus hohenackeri — и мной (Бирштейн, 1932) — каспий-
ский бокоплав Pontoganimarus robustoides setosus (Schaferna) 1 Харак-
терно, что морские реликты Кара-Су и некоторых других мелких водоемов
отличаются от морских иммигрантов других рек двумя особенностями:
1) их ареал оторван от ареала морских видов, 2) все они морфологически
резко отличаются от морских видов и являются их подвидами. Реликтовая
природа фауны Кара-Су стоит в связи с особенностями геологической
истории этого района. В плиоцене здесь образовались отчленившиеся
от моря озера, фауна которых частично сохранилась в мелких речках
и ручьях (Державин, 19266). Крупные притоки Каспийского, Черного
и Азовского морей находились, разумеется, в совершенно ином положении.
Вслед за вопросом о способе проникновения возникает вопрос о времени
проникновения морских ракообразных в реки. Но прежде чем к нему перейти,
я остановлюсь на одном важном обстоятельстве, на которое до сих пор
не обращалось должного внимания.
Помимо перехода морской фауны в пресные воды, в Понто-Каспий-
ском бассейне наблюдается также противоположный процесс — переход
пресноводной фауны в море. Так, например, Шпандль (Spandl, 1926а)
считает, что своеобразные каспийские Polyphemidae (Cercopagis, Apagis,
Polyphemus exiguus) произошли от представителей родов Bytotrephes и
Polyphemus1 2, населяющих пресные воды Северной Европы и попавших
в Каспий в середине ледникового периода вместе с водой, проталкиваемой
ледником. Но наиболее показательной группой, дающей прекрасный пример
проникновения пресноводных форм в море, могут служить рыбы. В послед-
них списках ихтиофауны Каспийского и Азовского морей, составленных
Книповичем (1921, 1932), из 75 форм в первом 32 и из 82 форм во втором
22 являются по происхождению пресноводными. Интересно отметить, что
паразитирующие на каспийских рыбах ракообразные, по исследованиям
Маркевича (Markewitsch, 1933), представлены главным образом пресновод-
ными видами. Нигде в мире, насколько мне известно, не наблюдается
в таких размерах, как в Понто-Каспийском бассейне, подобное смешение
морской и пресноводной фауны.
1 Синоним Р. robustoides aralychensis Hirstein.
2 Генетическую близость Bytotrephes и Cercopagis признают также Gurney (1908)
и Бенинг (1925).
53
Это интересное явление можно легко объяснить, если вспомнить о после-
ледниковом опреснении наших южных морей, о котором говорилось в обзоре
геологических данных. Грань между морем и пресноводным водоемом в это
время, по всей вероятности, настолько стерлась, что для переработанной
морской (бывшей) фауны река уже не являлась чуждой средой, точно
так же как и пресноводные рыбы без всякого труда расширили свой
ареал за счет недоступного для них раньше в силу своих физико-хими-
ческих условий моря. Последующее постепенное осолонение, по-видимому,
незначительно повлияло на сложившиеся в основном в п ост гл я ци альное
время ареалы пресноводных рыб, может быть несколько сократив область
их распространения в море. Может быть, именно оно рассортировало
рыб на биологические типы — проходных, полупроходных и туводных, уста-
новленные еще Кесслером. На способность некоторых рыб переходить от
жизни в реке к жизни исключительно в море указывают, между прочим,
любопытные данные К. А. Киселевича (1926). Существование в Северном
Каспии отдельных колоний не заходящего в пресную воду «морского»
сазана Киселевич связывает с сокращением дельты Волги, ведущим к изо-
ляции этих колоний от общей массы «ямного» сазана, живущего пре-
имущественно в пресной воде.
На значении изменений солености для проникновения морских организ-
мов в пресную воду остановился Л. А. Зенкевич (1933). Указывая
на вероятное сильное опреснение в четвертичное время окраинных морей
Полярного бассейна, этот автор полагает, что переработанная морская
фауна, пережив резкое понижение солености, стала проникать в пресную
воду, и объясняет, таким образом, распространение ледниковых реликтов.
Как отмечает Зенкевич, между историей окраинных морей Полярного
бассейна, с одной стороны, и Черного, Азовского и Каспийского морей —
с другой, существует полная аналогия, и вселение морских элементов
в реки Понто-Каспийского бассейна может быть объяснено также «солевыми
пульсациями».
Последнее значительное опреснение Каспийского и Азовского морей
имело место, как мы видели, совсем недавно. Если оно действительно
стимулировало проникновение морских элементов в пресную воду, то все
морские животные понто-каспийских рек суть очень молодые недавние
вселенцы. Этот вывод подтверждается некоторыми примерами, иллюстри-
рующими быстрое продвижение ракообразных морского происхождения
вверх по рекам почти на наших глазах. Немалую роль при этом сыграло
развитие искусственных каналов, соединивших отдельные системы. Вунш
(Wundsh, 1915), обнаруживший в 1910 г. в Мюггельском озере Corophium
curvispinum, исходя из вполне достоверных данных предыдущих авторов,
не находивших там этого рака, предполагает, что Corophium распростра-
нился по Германии и дошел до нижнего Одера совсем недавно. Также
в последнее время появился он, согласно Райху и Сцидату (Reich, Szi-
dat) ’, в опреснённой зоне Фришгафена и Куришгафена. Яроцкий и
Демьянович (Jarocki, Demianowicz, 1931), обнаружившие Chaetogammarus
ischnus в Висле, считают, что и в этом случае мы имеем дело с типичной
понто-каспийской автохтонной формой. Очень показательны данные по
недавнему расселению Astacus leptodactylus, на глазах человека двигаю-
1 Цит. по: Яроцкий, Демьянович (Jarocki, Demianowicz, 1931).
54
щегося вверх по рекам Понто-Каспийского бассейна, вытесняя более древний
вид A. astacus (Бирштейн, Виноградов, 1934).
Заселение Средней Европы понто-каспийскими рыбами произошло тоже
недавно. Анализируя происхождение ихтиофауны Венгрии, Ганко (Hanko,
1932) указывает, что понто-каспийские рыбы-иммигранты не встречаются
в Англии, отделение которой от материка относится к литориновому
времени. Отсюда можно сделать вывод о их вселении в Европу только
в около- или послелиториновое время.
Таким образом, проникновение морских элементов в реки Понто-Каспий-
ского бассейна стало возможным после преобразования морской фауны
в условиях резких колебаний солености, и сейчас мы присутствуем при
активном и чрезвычайно интенсивном расселении по системам рек морских
животных, быстро расширяющих свои ареалы.
ПЕЩЕРНАЯ ФАУНА
ЖИЗНЬ В ПЕЩЕРАХ.
ЭКОЛОГО-СИСТЕМАТИЧЕСКИЙ ОЧЕРК*
«Они вступают в море тьмы,
дабы исследовать, что в нем»
(Нубийский географ)
ВВЕДЕНИЕ
Биологическое изучение пещер ставит перед исследователями ряд чрез-
вычайно важных проблем, разрешение которых, несомненно приведет к вы-
яснению многих спорных вопросов общей биологии. Проблемы механизма
приспособления организма к среде, пищевых взаимоотношений, внутри-
видовой дифференциации и т. д. при исследовании населения пещер воз-
никают с особой остротой. Есть основания думать, что пещерная фауна
является весьма благодарным материалом для разработки этих проблем.
Внимание биологов издавна было обращено на эту отрасль работы.
Еще в XVII в. некоторые сведения о подземной фауне сообщил вюрц-
бургский иезуит Кирхер в своей книге «Mundus subterraneus>. Примерна
одновременно Вальвасором был открыт протей. Однако первые, правда,
неполные и приблизительные описания пещерных животных появились
только во второй половине XVIII в. (протей описан Лауренци (Laurenti)
в 1769 г., Oligochaeta — Мейером в 1781 г.). В течение XIX в. было открыто и
описано довольно большое количество подземных животных, и в 80—90-х го-
дах появились первые обобщающие работы, в которых приводились списки
животных для отдельных пещер и пещерных районов (Bilimek, 1867;
Blatchley, 1897; Jurinack, 1888; и др.), а также предлагались гипотезы
для объяснения слепоты пещерных животных и других их специфических
особенностей (Packard, 1888 **; Hamman, 1896). Однако расцвет пещерных
исследований приходится, конечно, на XX век. В начале века начали свои
многолетние исследования Раковица (Racovitza, 1907, 1910, 1912) и Жеан-
нель (Jeannel, 1926, 1931). За 20 лет ими и их сотрудниками было обсле-
довано свыше 1000 пещер (из них 500 во Франции). В то же время на
Балканском полуострове чрезвычайно плодотворно работает Абсолон (АЬ-
solon, Hrabe, 1930; Absolon, Kretochvil, 1932; Absolon, Landrock, 1933).
Штаммер (Stammer, 1932b, 1935a, b, 1936a, b, с) исследует пещеры
северной Италии и Испании, Руффо (Ruffo, 1937) — Италии, Гер-
* Успехи соврем, биологии, 1940, т. 13, вып. 3, с. 385—402.
** В списке литературы к этой статье автор приводит не все упоминаемые в ней
работы, но в примечании к нему указывает, что «полные списки литературы
можно найти в работах Spandl (1926в), Chappuis (1927) и особенно Wolf
(1934—1936)». Наряду с этим в списке литературы приведен ряд работ, посвя-
щенных исследованию фауны пещер разных районов нашей страны (Боруцкий,
1928, 1930а, б; Зайцев, 1940; Лебединский,, 1900—1914; Carl, 1905; Winkler, 1912),
ссылки на которые в тексте статьи отсутствуют. — (Примеч. ред.).
56
цог (Hertzog, 1938) — Эльзаса, Леру (Leruth, 1938, 1939) — Бельгии,
Буреш (Buresch, 1929) — Болгарии, Караман (Кагатап, 1932—1940} —
Югославии, Пирс (Pearse, 1938) —США. Происходит чрезвычайно
быстрое накопление фактов, списки пещерных животных все время по-
полняются новыми видами. Появляются обширные сводки, посвященные
пещерной фауне (Jetfnnel, 1926; Spandl, 1926b; Chappius, 1927, 1930;
Bailey et al., 1933; Wolf, 1934—1936).
Наряду с изучением состава пещерной фауны исследователи работают
над вопросами происхождения приспособительных особенностей подземных
животных (Vire, 1900; Eigenmann, 1909; Kammerer, 1912; Kosswig, 1934—
1939; Kosswig C., Kosswig L., 1936; Vandel, 1938; Lattin, 1939a—c),
а также над исследованием их экологии (Dudich, 1932; Kolosvary, 1934).
Широкий круг поднятых вопросов и большое количество занятых ими
исследователей потребовали создания специальных обществ и научно-
исследовательских учреждений, ставящих своей задачей всестороннее изу-
чение пещерной фауны и условий ее существования, а также издания
специальных спелеологических монографий и журналов. С 1907 г. основ-
ной французский биологический журйал «Archives de Zoologie experimen-
tale et generale» начал публиковать серию «Biospeologica»; в Германии
в 20-х годах начал выходить журнал «Mitteilungen uber Hohlen und Karst-
forschung»; в Австрии — «Spelaologisches Jahrbuch» и серия «Spelaologische
Monographien»; в Румынии — «Bulletin de la Societe de Sciences de Cluj»
и т. д.
В Германии возникает общество по изучению пещер («Gesellschaft fiir
Hohlenforschung und Hohlenkunde»); в Австрии, Румынии и Италии органи-
зуются специальные спелеологические институты, в которых работают перво-
классные специалисты. При некоторых институтах основываются пещерные
станции, позволяющие вести экспериментальные работы непосредственно
в пещерах и оборудованные новейшим инструментарием (например, в пе-
щере Postumia в Италии) (Dudich, 1932, 1933а, Ь).
В СССР изучение пещерной фауны до недавнего времени проводилось
в очень скромных масштабах, и широкие круги биологов остались в сто-
роне от проблематики пещерных исследований. Соответствующей литера-
туры на русском языке пока немного: статья Киршенблатта (1939), в ко-
торой автор, к сожалению, ограничился реферированием только вышедшей
за 12 лет до этого книги Шаппюи (Chappuis, 1927), и статья Щерба-
кова (1909), изложившего работу Раковицы (Racovitza, 1907). Возможности
для работы в этом направлении у нас в стране неограниченные. Очень
большое число пещер имеется на Кавказе, в Крыму, на Урале, Алтае,
Памире, Дальнем Востоке и в Западной Украине; их население остается
до сих пор почти неизвестным. Между тем целый ряд общих вопросов,
возникающих в связи с биологическим изучением пещер, должен быть вклю-
чен во всестороннюю разработку учения Дарвина. Следует отметить, что
многие иностранные исследователи склонны разрешать эти вопросы далеко
не в духе дарвинизма.
Настоящая статья имеет задачей обратить внимание советских иссле-
дователей на новую для них отрасль работы — изучение биологии пещер.
57
УСЛОВИЯ СУЩЕСТВОВАНИЯ В ПЕЩЕРАХ
Подавляющее большинство пещер приурочено к карстовым районам
и обязано своим возникновением действию грунтовых вод карста. По-
токи этих вод размывают известняк, частично растворяя, частично разру-
шая его чисто механически, и в результате эрозионной и коррозионной
деятельности воды внутри известковых массивов образуются пустоты. Рас-
творение известняка водой идет в основном по следующей простой формуле:
СаСО3+ Н2О+ СО2 = СаН2(СО3)2.
Этот процесс протекает различно, в зависимости от того, возникает ли
данная пещера выше или ниже уровня грунтовых вод. Первый случай
наиболее прост; возникающие в этом случае пещеры обычно представляют
собой длинные, узкие, иногда ветвящиеся, но не переплетающиеся между
собой туннели. В другом случае большие массы медленно текущей воды
попадают внутрь горного массива под большим давлением и образуют об-
ширные залы, соединяющиеся между собой переплетающимися тоннелями.
При повышении местности или понижении уровня грунтовых вод пещера
освобождается от воды. Помимо растворения известняка, в пещерах не-
прерывно идет противоположный процесс — выпадение известняка из пере-
сыщенного кальцием водного раствора в виде кристаллов кальцита, образую-
щих сталактиты, сталагмиты, колонны, известковые драпировки, троны,
капеллы и т. д. Скорость образования сталактитов и сталагмитов раз-
лична и зависит прежде всего от количества просачивающейся воды и от
содержания в ней СО2. Измерения скорости прироста сталактитов и сталаг-
митов показали, что в пещере Ingleborough в Англии сталагмит прирастал
за год на 7,46 мм; в пещерах Моравии сталактиты за 10 лет увеличи-
вались на 6—7,5 мм. Подобные данные позволяют вычислить возраст
сталактитов и сталагмитов. Для образования сталактита длиной 20 м
в Агтелекской пещере потребовалось 1100—40 000 лет; для образования
сталагмита высотой 30 м в одной из французских пещер — 150—300 тыс. лет.
Выделение известковых натечных образований постепенно приводит к за-
полнению ими пещеры. Кроме того, в пещерах во многих случаях накапли-
вается глина, образующаяся при растворении известняка.
Известковые пещеры могут достигать значительной длины. Наиболее
крупной из них является Мамонтова пещера (США, штат Кентукки),
общая длина галерей которой составляет около 100 км; Адельсбергская пе-
щера на Балканском полуострове имеет длину 11 км, Агтелекская пещера
в Венгрии — 22 км. Помимо известковых пещер, существуют пещеры в лаве,
искусственные туннели, представляющие в биологическом отношении го-
раздо меньший интерес.
Условия существования внутри известковых пещер резко отличны от
условий существования на поверхности земли. Тинеманн (Thienemann,
1926) указывает на три основных фактора, определяющих своеобразие
подземных биотопов: отсутствие света, относительно постоянную и низкую
температуру и, наконец, бедность пищи. Следует отметить, что, согласно
новым данным, нет достаточных оснований говорить о незначительном коли-
честве питательных веществ в пещерах, и в качестве третьего важного
экологического фактора было бы правильнее назвать высокую влажность.
Отсутствие света не дает возможности существовать зеленым расте-
ниям. Таким образом, пещеры лишены основных продуцентов, способных
58
фотосинтетически создавать из неорганического вещества органическое,
за счет которых на поверхности земли существует весь животный мир.
Однако не следует думать, что зеленые растения исчезают непосредственно
у входа в пещеру; многие из них проникают довольно глубоко внутрь
пещер, довольствуясь тем небольшим количеством света, которое часто
совершенно не воспринимается человеческим глазом. Если принять нормаль-
ную дневную освещенность за единицу, то величина, при которой еще могут
вегетировать зеленые растения, оказывается равной для мхов и водорослей
1 2000, для папоротников— 1 :1700; для цветковых растений — 1 :256, для
лишайников — 1 240. Эти данные объясняют, почему в пещерах зарегистри-
ровано довольно большое количество видов зеленых растений (Morton,
Gams, 1925). Тем не менее в глубинах пещер, куда не доходит солнечный
свет, зеленая растительность отсутствует, а на границе проникновения сол-
нечных лучей растения представлены неспособными к размножению сте-
рильными особями.
Глубина проникновения света в пещеру зависит прежде всего от вели-
чины входного отверстия и поэтому очень сильно варьирует. В превосходно
изученной венгерской пещере Барадла, имеющей три входа, граница про-
никновения света лежит в 60, 70 и 100 м от этих входов (Dudich, 1932).
В глубинах горизонтальных пещер господствует почти постоянная темпе-
ратура, приблизительно соответствующая среднегодовой температуре
местонахождения данной пещеры. Эта закономерность установлена для
большого количества случаев, между прочим, и для некоторых пещер
Абхазии (Балабуев, 1939). Ближе к выходу из пещеры температура
в большей степени зависит от температуры воздуха окружающей мест-
ности: летом она выше, чем в глубине пещеры, а зимой — ниже. Тем-
пература пещерных водоемов очень близка к температуре воздуха,
обычно незначительно ниже. В вертикальных пещерах, весьма много-
численных в Крыму, в нижних частях происходит застаивание охлаж-
денного зимой воздуха, что нередко приводит к образованию пещер-
ного льда. В горизонтальных пещерах, связанных с поверхностью земли
вертикальными шахтами, во время сезонной смены температуры окру-
жающего воздуха возникают сильные ветровые течения, вызывающие
летом интенсивное испарение и образование инея и снега, как, напри-
мер, в Кунгурской пещере на Урале (Крубер, 1915).
Относительно постоянная температура определяет целый ряд биологи-
ческих особенностей пещерных обитателей: апериодичность размножения,
постоянный темп роста, проявляющийся в отсутствии годичных колец на
раковинах моллюсков, и т. д.
Комбинация светового и температурного режимов позволяет различать
в пределах каждой пещеры три основные зоны, характеризующиеся специ-
фическим населением: 1) освещенная зона, в которой произрастают в зна-
чительном количестве зеленые растения и наблюдаются большие коле-
бания температуры; 2) промежуточная зона; 3) глубинная зона, куда не
проникают солнечные лучи и где температурные колебания или ничтожны,
или совсем не наблюдаются.
Пожалуй, не менее характерной, чем температурный и световой режим,
для большинства пещер можно считать чрезвычайно высокую влажность
воздуха. Относительная влажность воздуха, как правило, превышает 90 %.
Так, например, в абхазских пещерах Адзаба, Михайловской и Келасури
59
были отмечены колебания относительной влажности в пределах 85—96 %.
Средняя многолетняя относительная влажность для г. Сухуми, находящегося
вблизи от указанных пещер, для июля, когда были произведены приве-
денные наблюдения, равняется всего 72 % (Балабуев, 1939). Такие же
высокие показатели относительной влажности приводятся для целого ряда
пещер (Jeannel, 1926).
Насыщенность воздуха водяными парами в значительной степени сти-
рает ту резкую грань между водными и сухопутными биотопами, которая
наблюдается на поверхности земли. Сухопутная и водная фауна смеши-
ваются. Водные животные, как, например, некоторые Harpacticoida (Мо-
raria и др.), Asellidae (Stenasellus) и даже Gammaridae (Niphargus),
могут вылезать из воды на сушу и находиться там весьма продолжи-
тельное время. В то же время многие сухопутные формы (мокрицы, много-
ножки, жуки) постоянно заползают в водоемы. Эти удивительные пере-
селения совершаются главным образом в поисках пищи.
Многие наземные пещерные животные в связи с высокой влажностью яв-
ляются формами стеногидробными (Jeannel, 1926). В этом отношении
довольно интересны некоторые морфологические особенности многих пе-
щерных жуков. При тонких удлиненных конечностях и узкой головогруди
они обладают вздутым, каплеобразным или даже шарообразным брюшком.
Между плоской дорсальной поверхностью брюшка и выпуклыми над-
крыльями образуется полость, заполненная насыщенным водяными парами
воздухом. Трахейная система в значительной степени редуцирована, и
дыхание совершается непосредственно дорсальной поверхностью тела,
богатой кровеносными сосудами. У таких жуков (Bathysciinae, Lepto-
dirus, Antroherpon, Aphaenops) отсутствуют защитные приспособления
от высыхания, и они быстро погибают при сравнительно низкой влажности
атмосферы. Зато они свободно переносят длительное погружение в воду,
прекрасно переживают периодические наводнения, гибельные для не-
которых насекомых и могут поэтому называться «полуводными» орга-
низмами (Jeannel, 1926).
Однако, несмотря на постоянные условия температуры, освещения и
влажности атмосферы, в пещерах наблюдаются известные периодические
смены условий существования. По данным Гауэса (Hawes, 1939), в пе-
щерах Югославии зимой происходят наводнения, а летом количество воды
резко уменьшается. На это указывает, между прочим, образ жизни пещер-
ного многощетинкового червя Marifugia cavatica. Летом его известковые
трубки с живыми червями постоянно находятся на берегу водоемов, вне
воды (Remy, 1937), но зимой черви оказываются покрытыми водой. Оче-
видно, Marifugia cavatica, подобно другому представителю семейства
Serpulidae — Mercierella, обитающему вдоль морских побережий, приспосо-
билась к амфибиотическому существованию.
Периодичность в наступлении наводнений определяет периодичность
жизненного цикла некоторых пещерных животных. Так, например, встре-
чающаяся в пещерах Югославии губка Ephydatia miiНеri образует геммулы
зимой, в то время как на поверхности земли образование геммул у этого
вида приходится на весну. Доказано, что образование геммул на поверх-
ности земли стимулируется повышением температуры. Таким же самым
стимулом для подземных губок, по мнению Гауэса, являются зимние навод-
нения. Наводнения несомненно вызывают обильное поступление питатель-
60
ного материала с поверхности земли. Кроме того, они способствуют более
широкому пассивному расселению пещерных животных, хотя, с другой
стороны, известная часть населения может вымываться из пещер и погибать.
Отсутствие света, низкая и постоянная температура и высокая влаж-
ность делают условия существования в пещерах принципиально отлич-
ными от условий существования на поверхности земли. Все остальные фак-
торы среды — грунт, химизм воды и т. д. — уже не являются специфичными
для пещер, и поэтому специально рассматривать их здесь нецелесообразно.
В несколько особом положении находится вопрос об источниках питания
пещерных животных, или, в более широком понимании, проблема дина-
мики органического вещества в пещерах.
ДИНАМИКА ОРГАНИЧЕСКОГО ВЕЩЕСТВА В ПЕЩЕРАХ
Все живые организмы по их роли в превращениях органического ве-
щества, как известно, разделяются на три основные группы: продуценты,
способные синтезировать органическое вещество из неорганического; кон-
сументы, питающиеся органическим веществом продуцентов; редуценты, раз-
лагающие органическое вещество на неорганические соединения. Проду-
центами являются главным образом зеленые растения и отчасти автотроф-
ные бактерии, консументами — все животные, редуцентами — гнилостные
бактерии. Поскольку в глубинной зоне пещер зеленые растения благодаря
отсутствию света не могут произрастать, возникает вопрос об источниках
существования довольно богатого животного населения этой зоны.
До недавнего времени полагали, что органическое вещество попадает
в глубинные зоны пещер только извне. Водные потоки вносят в пещеры
с поверхности земли органический детрит, животных, быстро погибающих
в пещерах, растительные остатки и т. д. Еще большее значение имеют лету-
чие мыши, помет которых часто толстым слоем покрывает пол и стены
пещер. Достаточно сказать, что в абхазской пещере Адзаба мощность
отложений помета летучих мышей превышает 3 м. Пометом летучих мышей
питаются разнообразные животные; многие из них специально приспособи-
лись к жизни в помете летучих мышей. Таковы, например, жуки ста-
филиниды Atheta subcavicola и Phora aptina, для которых характерно
отсутствие обычных признаков пещерных животных; они пигментиро-
ваны и обладают нормально развитыми глазами. Жеаннель полагает, что,
поскольку средой их обитания является именно помет летучих мышей,
они адаптированы к этой среде и условия существования в пещерах не
отразились на их морфологии. Далее, преимущественно, но не исключи-
тельно, пометом летучих мышей питаются почти все жуки, низшие насеко-
мые, мокрицы, многоножки, черви, брюхоногие моллюски, а также все водные
животные. Harpacticoida, как уже указывалось, вылезают из водоемов
на сырой помет летучих мышей и поселяются в нем. Наличие летучих мышей
в значительной степени определяет количественное развитие пещерных жи-
вотных. В этом отношении весьма показательно сравнение двух соседних
абхазских пещер — Ачхше-тыз-гуа и Голова Атапа. В первой летучие
мыши почти совершенно отсутствуют, во второй они встречаются в гро-
мадном количестве. В то же время в Ачхше-тыз-гуа обнаружить каких-
нибудь животных, в частности многоножек, довольно трудно, в Голове
Атапа я насчитал в среднем 25—30 экз. многоножек (Archileucogeorgia
61
abchasica Lohm.) на 1 м2 стен и потолка, не считая других пещерных
обитателей.
Таким образом, вносимое в пещеры извне органическое вещество имеет
колоссальное значение. Но помимо такого аллохтонного органического
вещества, в пещерных водоемах, как показали недавние исследования,
имеется также автохтонное, т. е. образованное внутри пещеры орга-
ническое вещество. В грунте пещерных водоемов были обнаружены авто-
трофные бактерии, способные, окисляя неорганические соединения и исполь-
зуя углекислоту, создавать органическое вещество. Следовательно, эти бак-
терии являются продуцентами, и в глубинных, лишенных света частях
пещеры они заменяют зеленые растения. В венгерской пещере Барадла
были найдены серобактерия Beggiatoa leptomitiformis (Menegh.) Trevis,
окисляющая растворенные в воде сернистые соединения, используя осво-
бождающуюся при этом энергию, и железобактерии Leptothrix ochracea
Kutz., L. crassa Choi., окисляющие гидрокарбонат железа. Химические
анализы воды Агтелекской пещеры показали, что содержание в ней нитра-
тов настолько значительно (0,001—0,0155), что может быть объяснено
только деятельностью автотрофных нитрифицирующих бактерий (Dudich,
1933а).
Все эти данные указывают на то, что продукты разложения орга-
нического вещества (сернистые и азотистые соединения) благодаря деятель-
ности автотрофных бактерий внутри пещеры обращаются вновь в органи-
ческое вещество и включаются в круговорот. Таким образом, цикл орга-
нического вещества в некоторых пещерах частично замкнутый, обратимый.
Учитывая различия в поступлении органического вещества в пещеры
и характер цикла органического вещества внутри пещер, Дудич (Dudich,
1933а) предложил следующую классификацию пещер на биологической
основе.
I. Монотрофные пещеры. Питание из одного источника.
а) экзотрофные пещеры. Продуценты отсутствуют; только один аллохтон-
ный продукционный ряд; цикл органического вещества не замкнут. Зави-
симость от внешней (по отношению к пещерам) среды полная.
б) монотрофно-эндотрофные пещеры. Питание исключительно за счет
пещерных продуцентов. Такие пещеры теоретически могут существовать,
но пока неизвестны.
II. Амфитрофные пещеры.
Питание из двух источников (аллохтонное и автохтонное). Цикл орга-
нического вещества частично обратимый благодаря присутствию продуцен-
тов. Зависимость от внешней среды неполная.
а) фото-эндотрофные пещеры. Два продукционных ряда — один алло-
хтонный, другой автохтонный — за счет фотосинтеза зеленых растений,
приуроченных к участку вблизи входа в пещеру.
б) хемо-эндотрофные пещеры. Два продукционных ряда — аллохтонный
и автохтонный, базирующийся на хемосинтезе органического вещества за
счет автотрофных бактерий.
в) фото-хемо-эндотрофные пещеры. Три продукционных ряда — один
аллохтонный и два автохтонные — за счет фотосинтеза зеленых растений
и за счет хемосинтеза автотрофных бактерий.
Автотрофные бактерии служат пищей многим не только водным, но и
сухопутным пещерным животным. Пещерные креветки Troglocaris schmidti,
62
питающиеся, как показал анализ содержимого их желудков, грунтом, бога-
тым бактериями (Садовский, 1930), обладают специальными приспособ-
лениями, позволяющими захватывать мягкий ил пещерных водоемов.
На пальцах клешней Troglocaris находятся густые пучки длинных щетинок,
при помощи которых креветки набирают ил и подносят его ко рту. Большин-
ство низших ракообразных, а также, по-видимому, все остальные водные
обитатели питаются бактериями. По неопубликованным наблюдениям
Боруцкого, многоножки, жуки и пауки в недоступной ныне пещере Рионгэса
(вблизи г. Кутаиси) заползают в водоем и там едят грунт. Мне также
приходилось неоднократно наблюдать в пещерах Абхазии многоножек, за-
ползших в водоемы и чувствующих себя при этом вполне нормально.
В грунте пещерных водоемов бактерии в некоторых случаях достигают
колоссального количественного развития. Так, например, в упомянутой
пещере Рионгэса грунт водоемов на 10—20 % по весу состоит из бактерий
(Садовский, 1930).
Итак, пещерные обитатели питаются главным образом органическими
остатками, попадающими в пещеры извне, остатками пещерной расти-
тельности и, наконец, автотрофными бактериями. Весьма характерно для
пещерной фауны сравнительно небольшое количество хищников. Главными
являются, во-первых, немногочисленные позвоночные (протей, тритоны,
рыбы, крысы), некоторые жуки, пауки и ложные скорпионы. Если же
говорить о пещерах СССР, в которых пещерные позвоночные отсутствуют,
то в составе их гидрофауны нельзя найти ни одного хищника. В связи
с этим возникает совершенно неразработанный вопрос о характере борьбы
за существование в условиях обилия пищи и при отсутствии хищников,
а также о факторах, ограничивающих в таких условиях рост популяций.
До сих пор остаются загадкой причины ничтожного количественного раз-
вития большинства пещерных гидробионтов.
Отсутствие хищников в пещерных водоемах определяет своеобразие
пищевых отношений гидробионтов. В противоположность сложной сети
длинных пищевых цепей, часто состоящих из большого количества звеньев,
примеры которых для наземных биоценозов можно найти в книгах Зернова
и Кашкарова, схема пищевых цепей для пещерных водоемов, по-видимому,
чрезвычайно проста. Существует какой-то общий пищевой фонд (бактерии,
помет летучих мышей и т. д.), которым пользуются все консументы, не всту-
пающие затем ни в какие дальнейшие пищевые взаимоотношения. Если
изобразить это различие графически, то вместо сложного переплета линий
(для наземных биоценозов) мы получим фигуру, напоминающую сжатое
поле или скошенный луг (для подземных биоценозов).
Очевидно, условия обитания под землей вынуждают некоторых животных
изменить свой первоначально хищнический образ жизни. Так, например,
«полуводные» виды жужелиц Aphaenops (подсемейство Trechinae) перешли
к питанию грунтом и разлагающимся органическим веществом, в связи
с чем утратили способность к быстрому передвижению; их челюстной ап-
парат видоизменился, причем отдельные его части стали тонкими и неж-
ными. Другие виды этого же рода, так же как все остальные представители
подсемейства Trechinae, являются хищниками, быстро настигающими свои
жертвы и убивающие их мощными челюстями.
63
СОСТАВ ПЕЩЕРНОЙ ФАУНЫ
Среди всей массы животных, находимых в пещерах, можно различать
виды, встречающиеся исключительно в пещерах, и виды, обитающие также
на поверхности земли. Еще в 1892 г. Шинер (Schiener) предложил называть
представителей первой группы троглобиями, а второй — троглофилами; слу-
чайно попадающих в пещеры животных он назвал троглоксенами. Эта
терминология была воспринята всеми последующими авторами (Racovitza,
1907; Jeannel, 1926; Chappuis, 1927), но в недавнее время она была не-
сколько детализирована, и современная классификация, которой придержи-
ваются Дудич (Dudich, 1932), Колосвари (Kolosvary, 1934) и Пирс (Pearse,
1938), включает следующие группировки.
1. Эвтроглобионты; обитают исключительно в пещерах и крайне редко
в сходных наземных биотопах. Морфологически приспособлены к под-
земному образу жизни.
2. Гемитроглобионты; встречаются как в пещерах, так и в сходных
с ними биотопах, морфологически не адаптированы к пещерным условиям.
3. Псевдотроглобионты; живут вне пещер, но пользуются пещерами
как местом зимовки, сна, питания и т. д. Размножаются большей частью
вне пещер.
4. Тихотроглобионты; случайно попадают в пещеры и довольно скоро
там погибают.
Здесь уместно остановиться только на представителях первой группы.
Protozoa. Известно только пять эвтроглобионтов: эндопаразит протея
Chloromyxum protei Joseph (Myxosporidia) и эпибионты пещерных раков
Spelaeophrya troglocaridis Stammer (Infusoria Peritricha), Lagenophrys
monolistrae Stammer, Podophrya niphargi Strouhal и Tokophrya stammeri
Strouhal (Suctoria) (Stammer, 1935b; Strouhal, 1939b).
Turbellaria. Целый ряд видов, преимущественно из группы Triclada,
населяет пещерные водоемы. Известна также одна сухопутная пещерная
турбеллярия (Geopaludicola absoloni Komarek).
Rotatoria. Недавно была описана первая и пока единственная пещерная
коловратка — Dissotrocha hertzogi Hauer из грунтовых вод Эльзаса (Hauer,
1939).
Nematoda. Известен также единственный эвтроглобионт — Desmoscolex
aquedulcis Stammer из пещер Словении (Stammer, 1935а).
Archiannelida. В грунтовых водах Швейцарии и Германии водится
Troglochaetus beranecki Delachaux.
Polychaeta. В многочисленных пещерах восточного побережья Адриа-
тического моря встречается Marifugia cavatica Absolon et Hrale (семейство
Serpulidae).
Oligochaeta. 12 видов малощетинковых известны только из пещер,
однако по отношению к большинству из них нет уверенности в том,
что они впоследствии не будут найдены и на поверхности земли (Michael-
sen, 1933).
Hirudinea. Единственный относящийся к этому отряду троглобионт —
Herpobdella (Dina) absoloni Johanson — населяет пещеры Черногории и
отчасти Герцеговины.
Mollusca. Среди двустворчатых моллюсков в пещерах известен только
один вид рода Pisidium (Р. subterraneum Shadin) в пещере Рионгэса близ
Кутаиси (Жадин, 1932) и в пещере в Цебельде (Абхазия), среди брюхоно-
гих — довольно большое количество видов: Bythinella, Iglica, Ра га dhi liopsis,
64
BeIgrandie 1 la, Belgrandia, Horatia, Hauffenia и др. Сухопутные моллюски —
эвтроглобионты — принадлежат к родам Zospeum, Spelaeoconcha и др.
Crustacea. Этот класс представлен в пещерах очень большим количеством
видов, довольно неравномерно'распределенных между отдельными отрядами.
Пещерные Phyllopoda неизвестны. Среди Ostracoda имеется около 20 эвтро-
глобионтов, один из которых — Cryptocandona riongessa Bronstein — найден
в пещере Рионгэса в Грузии. Число видов Copepoda весьма значительно.
Насчитывается около двух десятков Cyclopoida и 120 видов Harpacticoida,
4 пещерных вида относятся к архаическому отряду Anaspidacea, 4 вида —
к Mysidacea.Единственный представитель сравнительно недавно описанного
отряда Thermosbaenacea — Thermosbaena mirabilis Monod — населяет
грунтовые воды Северной Африки (Monod, 1927). Isopoda представлены
большим количеством видов, относящихся к водным семействам: Asellidae,
Janiridae, Microparasellidae, Microcerberidae, Phreatoicidae, Anthuridae,
Cirolanidae, Sphaeromidae и к сухопутным: Trichoniscidae, Oniscidae,
Ligiidae, Armadillidae. He менее многочисленны в пещерах и Amphipoda
(Gammaridae, Talitridae, Calliopidae, Ingolfiel 1 idae). О многообразии этого
отряда можно составить представление, если принять во внимание, что
в настоящее время насчитывается около 80 форм только одного пещерного
рода Niphargus, найденного в пещерах и грунтовых водах Закавказья
(Бирштейн, 1940в) и Крыма. Среди Decapoda уже гораздо меньше эвтрогло-
бионтов. Известны пещерные десятиногие раки из семейств Palaemonidae
(Typhlocaris, Palaemon, Palaemonetes), Atyidae (Troglocaris, Palaemonias),
Hyppolytidae (Barbouria), Astacidae (Cambarus) и Galatheidae (Munidop-
sis).
Arachnoidea. Пещерные скорпионы и сольпуги неизвестны. Зато очень
многочисленны эвтроглобионты — ложные скорпионы, найденные, между
прочим, и в пещерах Закавказья и Крыма. Описано несколько эвтрогло-
бионтов из Palpigradi (Koenenia). Чрезвычайно многочисленные пауки,
представленные несколькими десятками видов, из которых три известны
из пещер Закавказья (Nesticus borutzkyi, N. zaitzewi, N. ponticus). Число
видов клещей гораздо меньше. Описано около 15 видов водяных клещей
(преимущественно из пещер Балканского полуострова) и несколько сухопут-
ных (из группы Oribatidae). Известно также несколько видов сенокосцев,
из них один, лишенный глаз, Nemastoma caecum Grese — из Скельской
пищеры близ Севастополя (Grese, 1911).
Myriapoda. Многоножки, так же как пауки, богато представлены в пе-
щерной фауне. Особенно многочисленны эвтроглобионты среди Diplopoda,
описанных, между прочим, из пещер Закавказья в качестве эндемичных
родов Leucogeorgia и Archileucogeorgia (Verhoeff, 1930; Lohmander, 1936).
Эвтроглобионты Chilopoda сравнительно редки, Pauropoda и Symphyla
неизвестны.
Insecta. Насекомые наряду с другими членистоногими занимают по числу
видов одно из первых мест среди компонентов пещерной фауны. Известно
значительное количество пещерных Apterygota, в особенности Collembola.
Среди прямокрылых в пещерах Закавказья широко распространен Do-
lichopoda euxina Sem.; другие виды этого рода известны из пещер Западной
Европы, Китая. Два вида таракановых, относящихся к роду Nocticola,
описаны из пещер Филиппинских островов.
Ручейники представлены всего одним видом — Wormaldia subterranea —
5 Заказ 333 55
из Югославии (Radovanovic, 1932), бабочки неизвестны. Пожалуй, наиболее
богатым эвтроглобионтами отрядом насекомых являются жуки, особенно
жужелицы из подсемейства Trechinae и Silphidae из подсемейства Bathi-
sciinae. Эвтроглобионты известны в пределах'семейств Pselaphidae и Dyti-
scidae.
Наконец, в последние годы описаны очень любопытные слепые пещерные
мухи — Eccoptomera troglomontana с Балканского полуострова (Absolon,
Landrock, 1933) и Mormotomyiia hirsuta с редуцированными крыльями из
Восточной Африки (Austen, 1936).
Pisces. Пещерные рыбы известны в Африке и Америке. Они относятся
к 6 семействам: Amblyopsidae (бассейн Миссисипи), Siluridae (Техас,
Пенсильвания, Бразилия, Амазонка, Сомали), Cyprinidae (Сомали), Eleo-
tridae (о-в Мадагаскар), Brotulidae (Куба) и Characidae (Мексика).
Amphibia. Пещерные амфибии малочисленней, чем рыбы. В пещерах
Северной Америки обитают Eurycea maculicauda (бассейн Миссисипи),
Е. longicauda (бассейн Миссури), Typhlotriton spelaeus (бассейн Миссури)
и Typhlomolge rathbuni (Техас), в Европе (в Далмации, Герцеговине,
Словении) — Proteus anguineus.
Этот краткий перечень показывает, что из 13 принимаемых в настоящее
время типов 1 шесть — Polifera, Coelenterata, Nemertini, Tentaculata, Ente-
ropneusta и Echinodermata — не включают эвтроглобионтов. Остальные
семь типов чрезвычайно неравномерно представлены в пещерной фауне.
Подавляющее большинство эвтроглобионтов в каждом из других типов
не превышает двух-трех десятков. Следует отметить, что количество видов
эвтроглобионтов в пределах типа непропорционально общему количеству
видов данного типа, хотя членистоногие являются и в пещерах и вообще
наиболее многочисленными животными. Так, например, из 15 000 видов
простейших только 5 являются эвтроглобионтами, в то время как из 5 500
видов плоских червей известно не менее 25 эвтроглобионтов.
Если взять бол*ее мелкие таксономические единицы, то неравномерное
распределение между ними эвтроглобионтов выявится еще более ре^ко.
Так, например, из трех подотрядов, в которые группируются свободно
живущие веслоногие ракообразные, два — Harpacticoida и Cyclopoida —
довольно богато представлены в подземных водоемах, между тем как
третий подотряд — Calanoida — не включает ни одного эвтроглобионта.
В то же время биология Cyclopoida и Calanoida очень сходна. Число
подобных примеров может быть значительно умножено.
На вопрос, почему одни животные оказываются способными существо-
вать в подземных условиях, а другие, подчас и систематически, и биологи-
чески близкие к ним, совершенно не встречаются в пещерах, никто до сих
пор не пытался ответить. И действительно, этот вопрос чрезвычайно труден.
Если для разрешения аналогичной проблемы — происхождения пресновод-
ной фауны — мы располагаем некоторым материалом (осморегуляторные
способности пресноводных форм, значение фосфора для метаморфоза
и т. д.), то в отношении происхождения пещерной фауны в целом
неясны даже подходы к разрешению этой проблемы. В настоящее время
1 См.: Руководство по зоологии / Под ред. Л. А. Зенкевича. М.; Л.: Биомедгиз,
1937. Т. 1. 796 с.
66
делаются попытки выявить родственные отношения отдельных обитающих
в пещерах родов и видов к наземным формам, но от таких частных данных,
основанных на чисто морфолого-систематическом материале, до разрешения
всей проблемы в целом еще очень далеко.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
И ПРОИСХОЖДЕНИЕ ПЕЩЕРНЫХ ЖИВОТНЫХ
Мы не располагаем прямыми указаниями на древность пещерной фауны,
поскольку в ископаемом состоянии пещерные животные до сих пор не были
найдены.
Для разрешения этого вопроса приходится пользоваться косвенными
данными, которые получаются при изучении географического распростране-
ния и родственных отношений пещерных животных, а также немногочислен-
ными случаями сохранения в ископаемом состоянии близких наземных форм.
Так, например, род Phreatoicus (Isopoda), несколько видов которого
обитают в подземных водах Австралии и Новой Зеландии, представлен
в пресноводных триасовых отложениях Нового Южного Уэллса (Австралия)
хорошо сохранившимся видом Ph. wianamattensis, принадлежность которого
к этому роду не вызывает сомнения и у новейших исследователей.
Интересно, что один из видов Phreatoicus — Ph. capensis — встречается
в Южной Африке (все остальные — в Австралии, Тасмании и Новой Зе-
ландии), и Бернар (Bernard, 1914) резонно видит в этом факте лишнее
доказательство древнего соединения Африки и Австралии в единый материк
Гондвану.
Если древность Phreatoicus восходит к началу мезозойской эры, то
Anaspidacea, включающие пещерных Bathynella и Parabathynella, в иско-
паемом состоянии известны еще из палеозойских отложений. Правда,
палеозойские представители этой группы относились к другим родам, но
значительная древность Anaspidacea подтверждается также зоогеографиче-
скими данными: Bathynella natans, В. chappuisi и Parabathynella stygia
известны из грунтовых вод Швейцарии, Германии и Балканского полу-
острова, Parabathynella malaya — из пещеры Малайского полуострова
(Sars, 1929). Наземные пресноводные Anaspidacea — Koonunga, Anaspides
и Paranaspides — живут в Австралии и Тасмании. Такой разорванный
ареал распространения современных Anaspidacea с несомненностью указы-
вает на древность группы. Изложенным материалом, к сожалению, исчерпы-
ваются палеонтологические данные по пещерным животным.
Всех эвтроглобионтов по их происхождению можно разбить на три
группы: 1) виды морского происхождения, 2) виды, происходящие от назем-
ных сухопутных и пресноводных форм и 3) виды, происхождение которых
неясно, так как родственные им формы вообще неизвестны. К первой
группе относится довольно большое количество водных эвтроглобионтов.
Desmoscolex aquedulcis, представитель свободно живущих Nematoda,
найденный в одной из пещер Словении, принадлежит к семейству Desmosco-
lecidae, объединяющему 58 морских видов, и является единственным пре-
сноводным видом этого семейства (Stammer, 1935а). Пещерная архиан-
нелида Troglochaetus чрезвычайно близка к морскому роду Nerillidium
(см.: Remane, 1928) и также должна считаться единственным пресноводным
5* 67
представителем ArchianelIida. Точно так же в пределах семейства Serpulidae
(Polychaeta) не известно ни одного настоящего пресноводного вида, кроме
пещерной Marifugia cavatica. Ближайшие к ней роды Placostegus и Dit-
гира—типично морские животные (Absolon, НгаЬё, 1930).
Довольно много аналогичных примеров дают высшие ракообразные.
Караман (Karaman, 1933) описал из грунтовых вод Югославии любо-
пытного бокоплава Balcanella acherontis, которого он отнес к новому
семейству Balcanellidae. Однако Герцог (Hertzog, 1935) показал, что эта
форма принадлежит к замечательному семейству Ingolfiel 1 idае, выделен-
ному в особый подотряд Ingolfiellidea. До сих пор было известно всего
два представителя этой группы: Ingolfiella abyssi, найденный на глубине
3000 м в Дэвисовом проливе (59 °с. ш., 51 °з. д.), и 1. littoralis, найденный
среди кораллов в Сиамском заливе на глубине 2 м. Оказалось, что третий вид
того же рода I. acherontis (Karaman) живет в грунтовых водах Югославии.
В недавнее время доказано также морское происхождение широко распро-
страненного в Европе пещерного бокоплава Niphargus. Этот род морфологи-
чески близок к родам Eriopisa и Eriopisella. Известно три вида Eriopisa,
один из которых — Е. chilkensis — живет в опресненной зоне побережья
Индии и обладает маленькими глазами, другой, лишенный глаз, Е. philip-
pinensis — в ключах Филиппинских островов и третий, также слепой,
Е. elongata — вдоль атлантического и средиземноморского побережий
Европы. Род Eriopisella объединяет тоже три вида: Е. sechellensis, имеющий
маленькие глаза и живущий вдоль берегов Сейшельских островов, и
слепых Е. capensis (берега Южной Африки) и Е. pusilia (побережье
Бретани, Франция). Последний вид живет в песке, в полостях между
песчинками. Таким образом, морские роды Eriopisa и Eriopisella имеют
тенденцию к редукции глаз, к жизни в условиях значительного опреснения
(Е. chilkensis, Е. phi 1 ippinensis) и к подземному обитанию (Е. pusilia).
Стоит только допустить, что песок Бретани, в котором обитает Е. pusilia,
будет орошаться грунтовыми водами, а рачок вселится в грунтовые воды,
и мы достаточно реально сможем себе представить происхождение Niphargus
(Schellenberg, 1933). Вселившись в грунтовые воды и пещеры из моря,
Niphargus в настоящее время стоит на пути к наземному образу жизни;
целый ряд видов и форм этого рода являются обитателями ключей (Schel-
lenberg, 1935). Надо думать, что переход Niphargus в пресную воду произо-
шел относительно недавно, так как ареал этого рода ограничивается только
Европой и Малой Азией. Если бы Niphargus являлся древним пресноводным
обитателем, мы были бы вправе ожидать его нахождения в Америке,
Северной Африке и на некоторых островах, так или иначе соединявшихся
с Европой и имеющих в составе своей фауны общих с Европой древних
пресноводных животных. Восточноафриканские подземные бокоплавы
(Quadrivisio bengalensis Stebbing и два вида Hyale — Н. jeanneli и
Н. inserta) принадлежат к родам, объединяющим исключительно морские
виды, и несомненно должны считаться формами морского происхождения.
То же относится к новозеландскому Paraleptamphopus subterraneus.
Отряд Isopoda также богат пещерными видами морского происхождения.
Именно к этой группе надо отнести представителей семейств Cirolanidae,
Sphaeromidae, Anthuridae, Microcerberidae, Microparasellidae, Janiridae.
В пределах пещерных Cirolanidae существуют роды, включающие наряду
с пещерными близкие к ним морские формы. Так, например, богатые
68
морскими видами роды Cirolana и Conilera представлены в пещерах Кубы
видом Cirolana cubensis, в пещерах Мексики — видом Conilera stygia.
Другие роды того же семейства объединяют исключительно пещерные
виды, однако их близость к морским родам не подлежит сомнению. Род
Sphaeromides, к которому относятся два вида из пещер Франции и Италии,
очень близок к морскому глубоководному роду Bathynomus и вместе с ним —
к ископаемому роду Palaega, известному из верхнего мела. Род Typhlociro-
lana, состоящий из пещерных видов Северной Америки и о-ва Майорки,•
близок к уже упомянутым родам Cirolana и Conilera (Racovitza, 1912).
Родственные отношения пешерных родов семейства Sphaeromidae (Мопо-
listra, Microlistra, Caecosphaeroma) гораздо менее ясны, но, поскольку все
семейство, за несколькими исключениями, состоит из морских форм, морская
природа пещерных Sphaeromidae не вызывает сомнений (Racovitza, 1910).
То же относится к семействам Anthuridae и Janiridae, единственными пресно-
водными представителями которых являются Crugens fontanus и Janirella
pusilia из грунтовых вод Новой Зеландии и Австралии.
Менее ясно положение в системе описанных недавно из грунтовых вод
Югославии семейств Microparasellidae и Microcerberidae. Караман (Кага-
man, 1935) сближает первое из них с чисто морскими семействами Macrosty-
lidae и Desmosomatidae, второе — с уже упоминавшимися Anthuridae.
Отряд Mysidacea, представленный в пещерах 4 видами, включает всего
368 видов, из которых только 2 являются исключительно пресноводными
обитателями; 3 пещерных вида населяют солоноватые подземные водоемы,
и только один — Troglomysis vjeternicensis — найден в пресном водоеме
(Stammer, 1936). Хотя близкие к нему формы неизвестны, приведенная
характеристика всего отряда исключает сомнения в морском происхожде-
нии Troglomysis.
Среди десятиногих ракообразных в качестве явных вселенцев из моря
следует назвать представителей семейств Galatheidae, Hyppolytidae и Palae-
monidae. Единственный пещерный представитель Galatheidae — Munidopsis
polymorpha — обитает на Канарских островах в лавовой пещере, непосред-
ственно соединенной с морем, и должен считаться по существу морским
животным. Однако, судя по степени редукции его глаз, он весьма продолжи-
тельное время живет под землей. Замечательно, что остальные виды рода
Munidopsis являются глубоководными формами. Точно так же единственный
пещерный представитель Hyppolytidae — Barbouria poeyi — живет на о-ве
Куба, в пещерном солоноватом водоеме, соединяющемся с морем, а все
остальные Hyppolytidae — исключительно морские обитатели. В пределах
семейства Palaemonidae можно указать несколько пресноводных форм, но
выселение их из моря произошло относительно недавно. Род Palaemon объе-
диняет как морские, так и пресноводные виды, в том числе Р. cavernicola
из пещеры в Ассаме. Близкий к предыдущему род Palaemonetes состоит
уже только из пресноводных форм и представлен в пещерах Кубы и Техаса
видами Р eigenmanni, Р calcis и Р antrorum. Наконец, к роду Typhlocaris
относятся только пещерные солоноватоводные и пресноводные виды Т. gali-
laea (Палестина), Т. lethaea (Северная Африка) и Т. salentina (Италия).
Этот род настолько примитивен и своеобразен, что выделяется в особое
подсемейство Typhlocarinae, и родственные ему формы неизвестны — по
всей вероятности, они вымерли. Гораздо менее ясно морское происхожде-
ние семейства Atyidae, представители которого в настоящее время обитают
69
только в пресной воде. Два рода, Atyidae — Troglocaris (Закавказье, Бал-
канский полуостров и Южная Франция) и Palaemonias (Мамонтова пе-
щера), являются эвтроглобионтами. Все современные местонахождения
Atyidae, и в частности той группы родов, к которой относятся Troglocaris и
Palaemonias, приходятся на область, покрытую в мезозое великим средизем-
ным морем Тэтис, и некоторые исследователи (Bouvier, 1925; Бирштейн,
19396) считают Atyidae реликтами этого моря. Ближайшее к Atyidae
семейство Hoplophoridae состоит из исключительно морских, преимущест-
венно глубоководных форм.
Пещерные рыбы морского происхождения относятся к семейству Brotu-
lidae. Два пещерных рода, Brotulidae (каждый с одним видом) — Lucifuga
и Stygicola, населяют грунтовые воды Кубы и являются единственными
пресноводными представителями этого семейства. Все остальные Brotu-
lidae — морские, преимущественно глубоководные формы. Эйгенманн
(Eigenmann, 1909) предполагает, что пещерные Brotulidae произошли от
видов, приспособившихся к жизни в пустотах между коралловыми рифами.
По мере постепенного поднятия коралловых рифов водоемы, заключавшие
предков Lucifuga и Stygicola, опреснялись и эти формы пассивно перехо-
дили к пресноводному образу жизни, оставаясь в то же время подземными
животными. Резюмируя изложенный материал, можно сказать, что довольно
значительная часть эвтроглобионтов, относящихся к различным группам
животного царства, происходит от морских предков. При этом некоторые
из эвтроглобионтов генетически близки к морским животным, обитающим
ныне в море на небольших глубинах (Nematoda, Annelida, Amphipoda,
Palaemonidae, Sphaeromidae, Janiridae, Arthuridae), другие — к глубоковод-
ным формам (Sphaeromides, Munidopsis, Brotulidae). Следует также под-
черкнуть относительное богатство формами морского происхождения грун-
товых вод тропической и субтропической областей. Только там встречаются
десятиногие ракообразные морского происхождения, Brotulidae, Anthuridae,
Janiridae и т. д.
Каким же образом и в какое время происходило заселение пещер
и грунтовых вод морской фауной? Поставленный вопрос осложняется еще
и тем обстоятельством, что вся пресноводная фауна в конечном итоге
произошла от морской, и не всегда удается точно установить, происходит
тот или иной вид непосредственно от морского предка или от пресноводного
животного морского происхождения.
В этой связи важно подчеркнуть тенденции некоторых морских живот-
ных к подземному образу жизни. С одной стороны, наблюдаются приспособ-
ления к жизни в пустотах коралловых рифов, в расщелинах скал и т. д.,
с другой — к жизни в капиллярных ходах между песчинками побережья и
мелководья. Наконец, следует иметь в виду, что целый ряд карстовых рек
имеет подземные русла (особенно в области Адриатики), и возможно
проникновение некоторых форм из моря в реки, минуя наземные пресные
водоемы. Нетрудно подобрать примеры, показывающие вероятность вселения
морской фауны в грунтовые воды именно этими тремя путями.
Переход к подземному образу жизни еще в море с последующим пас-
сивным переходом во внутренние подземные водоемы наглядно демонстри-
рует Munidopsis polymorpha. Если соображения Эйгенманна правильны,
то конечным этапом подобного процесса следует считать пещерных Brotu-
lidae.
70
Отмеченное выше относительное богатство тропических и субтропических
стран пещерными животными морского происхождения может быть связано
с мощным развитием в этих областях коралловых рифов, к которым
приурочена специфическая фауна, частично переходящая к жизни в полостях
между ветвями кораллов, т. е. в условия, аналогичные подземным.
Другой путь — от жизни в капиллярных ходах между песчинками
к жизни в грунтовых водах — не менее реален. Караман полагает, что
Microcerberidae, Microparasellidae и Ingolfiella проникли в систему грунто-
вых вод непосредственно из моря через капиллярные ходы морского по-
бережья. На это указывают незначительная величина названных животных
и их удлиненное и узкое тело, вполне приспособленное к такого рода биото-
пам. Подобное же мнение высказал, как уже указывалось, Шелленберг
в отношении происхождения Niphargus.
Наконец, третий путь, активное внедрение из моря в подземные устья
рек, принимают Абсолон и Грабье (Absolon, НгаЬё, 1930) для пещерного
многощетинкового червя Marifugia cavatica. Эта форма найдена недалеко
от побережья Адриатического моря в нескольких пещерах и ключах, не сое-
диняющихся друг с другом, но имеющих подземный сток в моря. Абсолон
полагает, что таким же путем попала в подземные воды Trogloaega virei,
встречающаяся совместно с Marifugia.
Для некоторых других пещерных форм морского генезиса можно пред-
полагать вселение из моря в наземные пресные водоемы, а оттуда — в под-
земные воды. Вероятно, таким образом надо трактовать происхождение
пещерных Atyidae — Troglocaris и Palaemonias. Единственный вид первого
рода Тг. schmidti населяет пещеры Грузии, Абхазии, Балканского полу-
острова и Южной Франции; единственный вид второго рода Р ganteri —
Мамонтову пещеру (США). Близкие к названным роды Paratya, Асап-
thephyra, Dugastella, Mesocaris, Syncaris известны из наземных вод Новой
Зеландии, Австралии, Экваториальной Африки, Полинезии, Бенгалии, Япо-
нии, п-ова Корея, Калифорнии, Южной Европы. Все эти местонахождения
укладываются в область, занятую в мезозое великим средиземным морем
Тэтис; Бувье (Bouvier, 1925) и Бирштейн (19396) рассматривают Atyidae
(во всяком случае, группу родов, близких к Paratya) как реликты Тэтиса.
При распадении Тэтиса на отдельные постепенно опресняющиеся водоемы
интересующие нас формы оставались в них, а затем в некоторых местах
переходили в пещеры. Раковица (Racovitza, 1912) считает, что Cirolanidae
также сначала сделались пресноводными, а затем вселились в пещеры.
Предок современных Typhlocirolana населял сушу в то время, когда Испания,
Северная Африка и Балеарские острова соединялись воедино, так как именно
в этих странах найдены современные виды Typhlocirolana. Точно так же
современное распространение креветок Typhlocaris (Палестина, Юж-
ная Италия, Северная Африка) заставляет Шаппюи предполагать, что
предок этого рода обитал в водоемах распавшейся в конце третичного
времени Эгейской суши.
Штаммер (Stammer, 1936а) считал, что все пещерные животные морского
происхождения перешли в пресную воду еще до вселения в пещеры.
Иммиграция из моря в пресные воды, по его мнению, может совершаться
только в тропиках. Поскольку с середины мелового периода до неогена
в Европе господствовал тропический климат, именно к этому времени
Штаммер относит происхождение морских элементов пещерной фауны.
71
Однако теперь мы знаем, что внедрение морской фауны в пресные воды
происходит и при умеренном или суровом климате (Понто-Каспийский
бассейн, окраинные моря Полярного бассейна), и поэтому соображения
Штаммера не являются убедительными.
К какому же времени надо отнести проникновение морской фауны в пе-
щеры? Ставя такой вопрос, следует иметь в виду, что переход морской фауны
в пресную воду происходит только при известных условиях и далеко не повсе-
местно. Можно наметить несколько локальных очагов вселения морских
животных в пресные воды, и внимательное изучение каждого из них показы-
вает, что вселению морских организмов в пресную воду предшествовало зна-
чительное опреснение моря или его отдельных участков, часто сопровождав-
шееся последующим осолонением и вызванное геологическими причинами
(таяние великого ледника, вертикальные движения материков и т. д.) 1
По всей вероятности, те же процессы обусловили проникновение морских
обитателей в пещеры. Во многих случаях, как мы видели, источником, из
которого морские животные попали в подземные воды, являлся распадаю-
щийся на отдельные опресняющиеся водоемы Тэтис. Таким образом, проник-
новение морской фауны в пещеры относится к весьма отдаленному времени,
что подтверждается высокой степенью эндемизма большинства эвтрогло-
бионтов морского происхождения. Генетические связи многих пещерных
обитателей с морской глубоководной фауной могут быть поняты только
исходя из предположения, что в отдаленные времена общие предки данной
группы обитали на морском мелководье, откуда они проникли в пещеры,
а впоследствии под давлением более сильных в борьбе за существование
животных они были оттеснены с мелководья на большие глубины, где и
дожили до наших дней. Некоторые группы вымерли в море полностью и
сохранились только в пещерах (креветки Typhlocarinae).
В настоящее время проникновение морской фауны в подземные воды
не наблюдается. Детальные исследования замечательной подземной р. Ти-
маво близ Триеста, имеющей подземное русло, показали, что морская и
пресноводная фауны в этой реке не смешиваются и что современные
морские обитатели не заходят глубоко в грунтовые воды (Stammer, 1939в).
Число видов эвтроглобионтов, происшедших от наземных сухопутных и
пресноводных форм, чрезвычайно велико и значительно превосходит число
видов морского происхождения. К этой группе следует отнести, во-первых,
всех сухопутных пещерных обитателей. Просматривая соответствующие
списки, нетрудно заметить, что подавляющее большинство сухопутных
эвтроглобионтов принадлежит к родам, заключающим также наземные
формы. Роды, состоящие целиком из эвтроглобионтов, сравнительно немно-
гочисленны и в большинстве случаев явно близки к наземным родам.
Исключением из этого правила считалась многоножка из Кутаисской
пещеры Leucogeorgia longipes, выделенная в особое семейство (Verhoeff,
1930), но недавно Ломандер (Lohmander, 1936) показал, что она близка
к обычному наземному роду Unciger и входит в семейство Uncigeridae.
Среди водных пещерных животных также насчитывается большое число
видов, относящихся к обычным наземных родам, но наряду с этим имеется
и довольно значительное количество родов, объединяющих исключительно
пещерные виды.
1 Подробнее см.: Л. А. Зенкевич (1933), Я. А. Бирштейн (1935, 19396).
72
Анализ всей обширной группы видов, происходящих от наземных предков,
показывает, что вселение в пещеры происходило неодновременно и что
в соответствии с этим можно различить недавних и древних пещерных
обитателей.
Совершенно очевидно, например, недавнее отщепление от обычного
водяного ослика Asellus aquaticus лишенной пигмента и глаз пещерной
формы A. aquaticus cavernicola. По мнению Нормана (Normann, 1926) и
Бородина (Borodin, 1927), описанная как самостоятельный род бразильская
пещерная рыба Typhlobagrus kronei на самом деле представляет собой
слепую форму наземной Pimelodella lateristriga. Известна также депигмен-
тированная с редуцированными глазами пещерная форма широко распро-
страненной в юго-восточной Европе Planaria montenegrina.
Если в этих случаях из пещерных форм еще не образовались самостоя-
тельные виды, то при более длительном пребывании в подземных условиях
это имеет место, но возникший вид остается в пределах рода. Известны
роды, объединяющие несколько видов, которые образуют постепенный
переход от типично наземных форм через геми- и псевдотроглобионтов
к эвтроглобионтам.
Так, например, среди пауков из рода Meta известно много видов, не имею-
щих отношения к пещерам, гемитроглобионты М. menardi и М. marianae,
и, наконец, чрезвычайно близкий к ним эвтроглобионт М. bourneti. Число
подобных примеров можно было бы значительно умножить.
Следующий этап — обособление пещерных родов — указывает на чрез-
вычайно продолжительное (в геологическом смысле) пребывание в пещерах.
Часто такие роды сохраняют примитивные признаки и обладают разор-
ванным ареалом, что подтверждает их древность.
Так, жуки рода Geotrechus обитают только в Средней Европе и США.
Балканский пещерный паук Paraleptoneta orientalis очень близок к япон-
ской наземной форме Р. japonica (Absolon, Kratochvil, 1932).
Подрод Mesoasellus представлен тремя видами в пещерах Калифорнии
и Японии, а четвертый вид населяет большие глубины оз. Байкал. Древность
подрода восходит к третичному времени (Бирштейн, 1939а). Род Crangonyx
заключает девять видов из пещер и грунтовых вод Европы, Урала, Даль-
него Востока, Северной Америки и Южной Африки. Единственный близкий
к протею вид — наземный Necturus maculatus — встречается в водоемах
восточного побережья США и Южной Канады.
Некоторые роды, известные из пещер Европы, представлены наземными
видами в тропиках. Таков сенокосец Travunia troglodytes из пещер Балкан-
ского полуострова; другие виды этого рода обитают в Сахаре, Аравии,
Туркестане и Гоби (Absolon, Kratochvil, 1932). Сходное распространение
свойственно родам Harpacticoida — роду Parastenocaris, большая часть
видов которого обитает в грунтовых водах Европы, а близкие к ним
формы — на Яве, Суматре и Сурминаме ’, и роду Elaphoidella, пещерные
европейские представители которого близко родственны видам Индии, Явы
и т. д. Происхождение подобных ареалов еще далеко не ясно. Во многих
случаях названные формы приходится считать оттесненными реликтами.
Говоря о разорванных ареалах, следует отметить чрезвычайно характер-
Не исключено морское происхождение рода Parastenocaris. Другой род семейства
Stenocaridae состоит из морских видов.
73
ное для многих пещерных животных Европы кругосредиземноморское рас-
пространение. Целый ряд родов приурочен к северным побережьям Среди-
земного моря, Западному Закавказью и Северной Африке. Такое распро-
странение свойственно многим жукам, многоножкам, паукам и т. д.
Жеаннель полагает, что центр распространения этих животных лежит
в области ныне погрузившихся под воду Эгейской и Тирренской плит,
связывавших воедино Пиренейские горы, Южную Францию, Италию, Дикар-
ские Альпы, Южные Карпаты и т. д., а также Западное Закавказье.
Связи между этим последним и Балканским полуостровом чрезвычайно
близкие. По существу, пещерная фауна Закавказья представляет собой
обедненную и несколько модифицированную балканскую фауну и может
считаться восточным форпостом единой Тирренской фауны. Однако в послед-
нее время в Закавказье открыты некоторые пещерные эндемичные роды,
придающие закавказской пещерной фауне своеобразный оттенок (Leuco-
georgia, Archileucogeorgia, Zenkevitchia, Caucasonethes).
Довольно ясны также связи южноевропейской и северноафриканской
пещерной фауны. Особенно ярко это явление выражено в Испании, где
отсутствуют обычные в европейских грунтовых водах Niphargus и единст-
венным подземным бокоплавом является Pseudoniphargus africanus, широко
распространенный в грунтовых водах Алжира. Впрочем, аналогичные род-
ственные отношения проявляют и наземные бокоплавы Испании. Таким обра-
зом, по составу фауны Amphipoda Испания оказывается принадлежащей
скорее к Африке, чем к Европе (Schellenberg, 1937а).
В странах, лежащих к северу от областей альпийской складчатости,
состав пещерной фауны резко беднеет. Большая часть пещер Северной Гер-
мании, Северной Франции, Северной Бельгии лишена эвтроглобионтов.
Основной причиной отсутствия эвтроглобионтов в этих районах является
их оледенение. Ледник уничтожил почти всю подземную и наземную третич-
ную фауну, а после его отступления эти области были заселены вновь иной
четвертичной фауной, которая еще не успела приспособиться к подземной
жизни. По-видимому, влияние ледника не ограничилось механическим уни-
чтожением наземного населения и закупориванием пещер. В межледниковые
эпохи потоки талых ледниковых вод промывали пещеры, вынося наружу
их обитателей. Северные границы ареалов целого ряда эвтроглобионтов
совпадают с южными границами гренландского ледника (Niphargus, Asellus
cavaticus, многие жуки). Однако местные оледенения горных хребтов (Альп,
Кавказа) почти не повлияли на пещерную фауну (Holdhaus, 1932).
Наряду с уничтожением пещерной фауны оледенение стимулировало
проникновение некоторых групп в подземные воды. Подземные водяные
ослики из подрода Proasellus приурочены к южной границе ледникового
покрова. Южнее ее встречаются главным образом наземные формы, а се-
вернее подрод совершенно отсутствует (Stammer, 1932). Единственный
подземный бокоплав стран, покрытых ледником, Synurella несомненно под
влиянием оледенения перешел в грунтовые воды и только таким образом
дожил до наших дней.
Проникновение в пещеры эвтроглобионтов, происходящих от наземных
форм, так же мало выяснено, как и форм морского происхождения. И в этом
случае приходится учитывать те тенденции к подземному образу жизни,
которые проявляют наземные обитатели. Прежде всего следует отметить
различных беспозвоночных, закапывающихся в землю, залезающих под
74
камни, в щели и т. д., а также питающихся пометом и живущих в нем.
Кроме того, переход к подземному образу жизни легко совершается у на-
земных гигрофильных видов. Известно довольно большое количество геми-
троглобионтов, встречающихся, помимо пещер, в погребах, шахтах, колодцах
и Других сходных биотопах, характеризующихся повышенной влажностью.
Таковы многие пауки, мокрицы, многоножки и т. д.
По вопросу о происхождении пресноводной пещерной фауны высказы-
валось два мнения: одни авторы считали, что пресноводные животные
пассивно заносились потоками вод под землю, другие настаивали на их
активном внедрении в грунтовые воды. Шпандль (Spandl, 1926b) признает
возможным оба пути; он указывает на постоянно попадающихся в грунто-
вых водах Балканского полуострова обычных наземных рыб, несомненно
заносимых под землю водными потоками. В Закавказье то же самое
известно для форели. Таким же образом попадают под землю многочислен-
ные циклопы, некоторые гаммариды и простейшие. Шаппюи (Chappuis, 1927)
склонен придавать пассивному вселению только второстепенное значение.
Он подчеркивает сходство между ключевой и пещерной фауной и приводит
примеры проникновения ключевых форм далеко в грунтовые воды. Условия
существования в ключах и в грунтовых водах сходны: в обоих
случаях вода имеет сравнительно низкую и более или менее постоянную
температуру. При этом многие ключевые формы отрицательно фототропичны
и предпочитают жить в затемненных местах. Близкие к многочисленным
в ключах и пещерных водоемах виды Harpacticoida на поверхности земли
ведут «полуподземный» образ жизни. Они обитают в моховых болотах, пол-
зая под сплавинами и проникая в капиллярные ходы между веточками мхов.
Итак, приспособления, облегчающие переход к подземному образу
жизни, вырабатываются у животных, обитающих еще на поверхности
земли. Удается проследить ряд постепенных переходов от наземных к под-
земным формам, и на этом основании можно утверждать, что образование
второй, выделенной в начале этой главы группы эвтроглобионтов проис-
ходило постепенно (Asellus aquaticus cavernicola, слепая форма Planaria
montenegrina и др.).
Группа эвтроглобионтов, родственные которым наземные животные неиз-
вестны, чрезвычайно немногочисленна.
К ней относится в первую очередь ракообразное, обитающее в теплых
(44—45 °C) грунтовых водах Северной Африки — Thermosbaena mirabilis.
В первой своей работе Моно (Monod) рассматривал это животное как ли-
чинку, но впоследствии он нашел половозрелые экземпляры и, детально ра-
зобрав строение Thermosbaena, выделил ее в особый отряд Thermosbaenacea,
стоящий в ряде Peracarida между отрядами Mysidacea и Tanaidacea (Monod,
1927). Следовательно, отличия Thermosbaena mirabilis от всех известных
ныне высших ракообразных чрезвычайно значительны. Среди Amphipoda
есть также довольно своеобразные формы, которые, однако, обладают всеми
признаками не только этого отряда, но и семейства Gammaridae. Таковы
род Hadzia и группа родов, близких к Crangonyx. Большая часть этих
родов состоит из подземных видов, распространенных чрезвычайно широко,
но обладающих очень узкими, оторванными друг от друга ареалами
(Евразия, Северная Америка, о-в Мадагаскар). В эту же группу включается
один род эндемичных гаммарид оз. Байкал (Нуа 1 lelopsis Stebbing).
По всей вероятности, вся группа Crangonyx объединяет остатки когда-то
широко распространенной, но впоследствии почти нацело вымершей фауны.
75
АДАПТАЦИЯ И ЭВОЛЮЦИЯ ПЕЩЕРНЫХ ЖИВОТНЫХ*
Изучение биологических особенностей пещерных животных представляет
значительный интерес как для понимания условий развития и жизнедея-
тельности этих животных, так и для освещения некоторых вопросов эволю-
ции.
В биологической литературе имеется описание многих тщательных наблю-
дений за пещерными животными, а также рассуждения о закономерностях
эволюционного развития их. Настоящая статья посвящена критическому
обзору этой литературы и является дополнением эколого-систематического
очерка о пещерных животных (Бирштейн, 19406).
РАСПРЕДЕЛЕНИЕ ЭВТРОГЛОБИОНТОВ ВНУТРИ ПЕЩЕР
И НЕКОТОРЫЕ БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПЕЩЕРНОЙ ФАУНЫ
В пещерах можно различить три основные зоны — освещенную, про-
межуточную и лишенную света глубинную зону. Наибольший интерес
представляет распределение обитателей этой последней зоны. Вопрос о нали-
чии отдельных биоценозов в пределах глубинной зоны разработан очень
слабо. В венгерской пещере Барадла Дудич (Dudich, 1932) различает
несколько биоценозов, но, по его мнению, качественные различия между
ними определяются не средой, чрезвычайно однообразной на всем протяже-
нии пещеры, а той или иной степенью изоляции отдельных биотопов, деятель-
ностью человека и т. д. Жеаннель (Jeannel, 1926), напротив, перечисляет
целый ряд ассоциаций, характеризующихся своим специфическим населе-
нием: ассоциацию обитателей помета летучих мышей, ассоциацию видов,
живущих на сталагмитах, ассоциацию видов, живущих под камнями, ас-
социацию обитателей глинистого грунта, ассоциацию видов периодически
затопляемой зоны. Он указывает на загадочные случаи очень строгой
приуроченности некоторых видов к какому-нибудь определенному месту
в пещере, ничем не отличающемуся от соседних. Так, например, в пещере
Castel-Mouly с 1850 г. многочисленные исследователи находили жука
Aphaenops leshenaulti во все времена года только на правой стене определен-
ного зала. Через некоторое время, после того как жуки собирались очеред-
ным исследователем, они появлялись на том же самом месте вновь. В пещере
Lestelas другой жук — Aphaenops bucephalus — постоянно встречается
только на одном определенном сталагмите. Жеаннель думает, что эти
местонахождения находятся вблизи от глубоких щелей, где проходит весь
половой цикл Aphaenops и где обитают его личинки, однако прямые дока-
зательства правильности такой гипотезы пока отсутствуют. Тот же автор
идет в своих предположениях еще дальше. Он допускает существование
особой недоступной исследователям «фреатической фауны», обитающей
в глубине известковых горных массивов, в щелях, сообщающихся с пеще-
рами. Основанием для этого предположения служит бесспорный факт
отсутствия в пещерах личинок жуков из триб Bathysciinae и Trechinae при
многочисленности взрослых форм. Некоторые пещеры Южной Франции
посещались в течение 40 лет по нескольку раз в месяц, и число собранных
в них жуков колоссально; в то же время ни одна личинка ни разу не была
найдена.
* Успехи соврем, биологии, 1941, т. 14, вып. 3, с. 436—453. (Печатается с сокр.).
76
Эти неизвестные нам личинки, по Жеаннелю, относятся к фреатической
фауне, и их метаморфоз происходит, вероятно, внутри горных массивов,
причем в пещеры попадают только взрослые жуки. Весьма возможно, что
фреатическая фауна состоит не только из личинок Bathysciinae и Trechinae;
может быть, она не менее богата видами, чем собственно пещерная фауна,
но до сих пор она остается совершенно недоступной.
Если о существовании сухопутной фреатической фауны можно только
догадываться, то водная фреатическая фауна благодаря применению насо-
сов, добывающих грунтовые воды из разных горизонтов, уже начинает
изучаться и в ее реальности не приходится сомневаться. Исследуя материал,
полученный при помощи насоса, Караман (Karaman, 1932) установил,
что к разным горизонтам грунтовых вод Югославии приурочены разные
виды бокоплавов. Формы, обитающие в глубокозалегающих грунтовых
водах, не встречаются в пещерах. Аналогичные результаты получил в Эль-
засе Герцог (Hertzog, 1938), которому здесь удалось таким способом
(насосом) обнаружить много неизвестных ранее своеобразных представи-
телей фреатической фауны (коловратка Dissotrocha hertzogi, бокоплав
Bogidiella albertimagni и др.). Еще более полные данные по водной фреати-
ческой фауне добыл Лерю (Leruth, 1938, 1939). Согласно этому автору,
можно говорить о двух самостоятельных водных фаунах — фреатической и
собственно пещерной, причем для Бельгии первая оказывается более богатой
видами и совершенно специфической. Лерю отмечает резкие различия в ка-
чественном и количественном составе населения между отдельными выхо-
дами грунтовых вод, которые он объясняет разными условиями существо-
вания, а также разными биологическими взаимоотношениями. Так, напри-
мер, в случае присутствия в источнике хищных турбеллярий Dendrocoelum
бокоплавы встречаются очень редко, а моллюски отсутствуют, так как и те и
другие истребляются червями.
Распределение животных в пещерных водоемах указывает на существо-
вание водных подземных биоценозов, но данных по этому вбпросу еще мало.
Фауна подземных бокоплавов Югославии, по Караману (Karaman, 1932),
распадается на формы крупных водоемов со слабым течением и илистым
грунтом, формы пещерных ручьев и речек и формы грунтовых вод. Эти же
группировки можно подметить и в других районах. Так, например, в пещерах
Абхазии креветка Troglocaris schmidti и крупный бокоплав Niphargus
longicaudatus magnus приурочены к водоемам со слабым течением и мягким
грунтом, а бокоплавы Niphargus puteanus iniochus, N. ablaskiri и Zenke-
vitchia — к быстротекучей воде и каменистому грунту (Бирштейн, 1940а).
Согласно Шелленбергу (Schellenberg, 1941), обитающий в румынских
пещерах Niphargus longicaudatus maximus распадается на две хорошо
различимые морфологические формы — форму стоячих водоемов (laticau-
data) и форму текучих водоемов (tenuicaudata).
Говоря о приуроченности тех или иных форм к определенным условиям,
следует подчеркнуть почти полное отсутствие сведений о биологии пещерных
обитателей. Между тем эта область несомненно представляет очень большой
интерес. Своеобразие троглобионов сказывается, например, в том, что их
рост и размножение совершаются непрерывно: это обусловлено неизмен-
ностью условий существования под землей, в частности отсутствием
сезонных колебаний температуры.
По вопросу об окончательных размерах пещерных животных существуют
77
довольно противоречивые мнения. Тинеман (Thienemann, 1926) считает
характерным для троглобионтов незначительные размеры, что, по его мне-
нию, стоит в связи с недостатком пищи в пещерах. Однако последнее
соображение, как мы видели, сомнительно. Шаппюи (Chappuis, 1927) в об-
щем присоединяется к мнению Тинемана, указывая, впрочем, на существо-
вание наряду с мелкими также крупных пещерных животных. Незначи-
тельную величину большинства пещерных животных Шаппюи объясняет, на
основании данных Семпера о пропорциональности между размерами водоема
и размерами его обитателей, малой величиной пещерных водоемов. По этому
поводу следует заметить, что в настоящее время результаты опытов Семпера
трактуются не столько как доказательство прямой зависимости между объе-
мом бассейна и величиной обитающих в нем организмов, сколько как след-
ствие тормозящего рост действия продуктов обмена, накопляющихся быстрее
в мелких водоемах. Таким образом, это объяснение неприменимо к обычно
проточным пещерным водоемам.
С другой стороны, указания на малорослость троглобионтов по срав-
нению с близкими наземными формами нельзя считать достаточно обоснован-
ными. Целый ряд групп животных не подчиняется этому правилу. Так,
например, пещерные бокоплавы Niphargus достигают в длину 3—5 см, т. е.
являются наиболее крупными пресноводными представителями этой группы
(не считая байкальских).
Ложноскорпионы, многоножки, жуки, ногохвостки, веслоногие рачки и
многие мокрицы в пещерах не уступают по величине близким им наземным
видам, а в некоторых случаях (Trechus) оказываются более крупными.
Можно думать, что незначительные размеры некоторых троглобионтов,
отмеченные многими авторами, характерны только для представителей
фреатической фауны и являются приспособлением к таким своеобразным
условиям. Увеличение же размеров других троглобионтов, по всей вероят-
ности, стоит в связи с относительно низкой температурой, в подавляющем
большинстве случаев благоприятно отражающейся на предельных размерах
животных.
Образ жизни пещерных обитателей остается до сих пор почти совершенно
неизвестным. Реми (Remy, 1931) показал, что пещерные прямокрылые
Troglophilus и Dolichopoda в поисках пищи совершают миграции. Ночью,
в условиях повышенной влажности и отсутствия света они вылезают из
пещер и охотятся на некоторых насекомых. Днем они или возвращаются
в пещеры, или прячутся под камни, опавшие листья и пр. У самок тех же
прямокрылых имеются на брюшке 2 пары желез, выделяющих секрет,
привлекающий самцов.
Болдырев (1915) произвел ряд интересных наблюдений над закавказским
пещерным прямокрылым Dolichopoda euxina как в естественных условиях,
так и в садках. Просмотр содержимого кишечника этих кузнечиков показал,
что D. euxina, подобно своим западноевропейским родичам, является хищ-
ником и питается преимущественно насекомыми, в частности бабочками-
ночницами, залетающими в пещеры. Однако, кроме остатков насекомых,
среди пищевых масс были обнаружены волосы летучих мышей; по-видимому,
D. euxina, так же как и другие троглобионты, поедает помет летучих мышей,
к которому пристают волосы этих последних. Зимой, как и следовало
ожидать, кузнечики питаются так же интенсивно, как летом. Кузнечики,
помещенные в открытые садки в комнате, становятся активными только
78
с наступлением темноты. Днем они сидят неподвижно, скрываясь в наиболее
темных местах садка. Ночью они быстро передвигаются по садку, чистятся,
питаются и спариваются. Спаривание Dolichopoda происходит весьма своеоб-
разно. При копуляции «самец Dolichopoda вывертывается из-под ног самки
почти под прямым углом в сторону, так как его длинные ноги не умещаются
под телом партнерши». После окончания копуляции оба партнера в течение
1—21 /4 часов не могут разъединиться, хотя и делают соответствующие
попытки. Биологическое значение такого затяжного спаривания, как выяснил
Болдырев, заключается в том, что, пока насекомые остаются соединенными,
сперматофор, введенный в половые пути самки, успевает освободиться
от спермы. Как только партнеры расходятся, самка извлекает сперматофор
и поедает его. Иногда сперматофор уносится в половых частях самца и им
поедается. У других кузнечиковых спаривание гораздо более кратковре-
менно, а защита спермы от преждевременного уничтожения самкой осуще-
ствляется при помощи громадной слизистой массы — сперматофилакса,
которой снабжены сперматофоры. Пока самка поедает сперматофилакс,
сперма успевает излиться в семяприемник самки. Сперматофоры Dolichopoda
лишены сперматофилакса, и самец, помимо своего желания, как бы защи-
щает сперму собственным телом.
Наблюдавший пещерных пауков Troglohyphantes cavernicola Мак-Инду
(Mclndoo, 1910) описывает ловлю пауками низших насекомых, причем
пауки находят добычу, руководствуясь органами чувств, расположенными
на передней паре ног. Любовные игры этого вида значительно упрощены
по сравнению с наземными формами; кладки яиц окружены очень несовер-
шенными коконами. Исследуя экспериментально влияние внешних условий
на Troglohyphantes, Мак-Инду нашел, что наибольшее значение для живот-
ного имеет влажность воздуха, а свет (этот паук отрицательно фототропи-
чен) играет подчиненную роль. Эннендель с соавторами (Annandale, Coggin,
Gravely, 1913) описали замечательный способ защиты от врагов у малай-
ской пещерной многоножки Scutigera. Это многоножка легко отбрасывает
свои длинные конечности, причем автотомия сопровождается скрипучим
шумом, пугающим врага.
Бухман (Buchmann, 1936) изучал различные реакции пещерного реснит-
чатого червя Sphalloplana регсаеса. Оказалось, что в этом отношении
пещерная форма принципиально не отличается от наземных Dendrocoelidae.
Однако ей совершенно несвойствен фотаксис: сильное освещение заставляет
животное судорожно корчиться и довольно быстро приводит его к гибели,
но никаких попыток уйти от света червь не делает. В то же время, по данным
Эйгенмана, слепые пещерные рыбы отрицательно фототропичны, так же как
и личинки Typhlotriton spelaeus, по данным Нобля и Попа (Noble, Pope,
1928). Тем не менее личинки Typhlotriton встречаются преимущественно
вблизи входа в пещеры, что обусловлено температурным режимом.
Аквариальные наблюдения над пещерными рыбами Caecobarbus и
Typhlichtys показали, что главную роль в жизни этих рыб играют чувства
осязания и вкуса. Пользуясь хорошо развитыми органами осязания, рыбы
находят пищу (Petit, Bernard, 1937; Verrier, 1929).
Рассмотрение морофологических особенностей пещерных животных при-
водит всех исследователей к согласному мнению о том, что приспособление
к подземному образу жизни в подавляющем большинстве случаев не вызы-
вает появления новых органов; наблюдается лишь гипертрофия некоторых
79
органов, имеющихся и у наземных форм. В первую очередь у членистоногих
удлиняются конечности, в особенности те из них, на которых находятся
чувствительные щетинки, причем число чувствительных щетинок возрастает.
Примеры этого весьма многочисленны и широко известны. Так, по данным
Лапшова (1940), процентное отношение длины педипальп к длине тела
пещерных ложных скорпионов в среднем равно 198 %, в то время как для
наземных — всего 83 %. Пещерные рыбы Amblyopsidae характеризуются
сильно развитой и своеобразно видоизмененной боковой линией, преобразо-
ванной в ряды чувствительных бугорков, богатых нервными окончаниями.
Обитающие в пещерах плоские черви, моллюски, многие ракообразные
совершенно не отличаются по степени развития органов чувств обоняния,
осязания и химического чувства от близких наземных форм. Зато органы
зрения и пигментация тела в условиях подземной жизни, как правило,
редуцируются, и этот вопрос представляет интерес.
РЕДУКЦИЯ ОРГАНОВ ЗРЕНИЯ
Слепота и отсутствие пигмента считаются чрезвычайно характерными
особенностями пещерных обитателей. Однако эти особенности присущи
далеко не всем эвтроглобионтам. Пещерные пауки Leptonetidae и виды
некоторых других семейств, жуки Pselaphidae, Hydroporus aurasicus,
Atheta subcavicola, Hydraena cavicola, мухи Phora aptina и Limosina
racovitzai, некоторые виды реснитчатых червей, целый ряд Copepoda (Мога-
ria varica, Cyclops unisetiger, C. crinitus, C. kieferi), 9 видов мокриц,
креветка Barbouria poeyi, рыбы Chologaster agassizi и Ch. cornutus, амери-
канские пещерные тритоны Spelerpes maculicaudata и Sp. stejnegeri, будучи
типичными эвтроглобионтами, обладают более или менее нормально разви-
тыми глазами (Lattin, 1939с). Известны также многочисленные случаи
наличия глаз у личинок и молодых особей пещерных видов и последую-
щая редукция органов зрения по мере роста и развития животных. Тем
не менее, поскольку подавляющее большинство троглобионтов слепо и
депигментировано, приходится считать, что эти свойства овязаны с подзем-
ным образом жизни.
Пещерные реснитчатые черви, за исключением нескольких видов с сильно
редуцированными глазами, лишены органов зрения. Следует, впрочем,
иметь в виду, что низко организованные глаза наземных форм реснитчатых
червей довольно легко исчезают в условиях эксперимента. Помещая червей
в темноту и заставляя их голодать, можно вскоре наблюдать полную
атрофию их глаз. Немногочисленные пещерные кольчатые черви абсолютно
слепы.
Среди низших ракообразных известны или лишенные глаз формы (Ostra-
coda, большинство Harpacticoida), или формы, обладающие нормальными
глазами, но утерявшие глазной пигмент (некоторые Cyclopoida). Процесс
редукции пигмента совершается очень быстро у наземных циклопов, слу-
чайно занесенных потоками под землю, причем, по-видимому, даже в первом
поколении. Содержание наземных низших ракообразных (Cladocera) в тем-
ноте (Kapterew, 1910, 1912) довольно быстро приводит к редукции их
глаз. Однако тот же эффект получается и при переводе животных на
голодный режим (Ермаков, 1924), что доказывает неустойчивость несовер-
шенных органов зрения низших ракообразных.
80
Рекукция гораздо более высокоорганизованных глаз высших ракообраз-
ных происходит значительно медленнее. Можно наметить разные стадии
и пут^ этого процесса. Многие пещерные Amphipoda совершенно слепы,
и зачатки их глаз не появляются даже на ранних стадиях эмбрионального
развития. Как показал Сцарский (Szarski, 1935), имеющиеся в литературе
указания на присутствие рудиментов зрительных нервов у Niphargus осно-
ваны на ошибке, и на всех стадиях своего развития Niphargus совершенно
лишен зрительных органов и их рудиментов. Кроме того, он в отличие от
наземных форм не имеет также и статоцистов.
Вольский (Wolsky, 1935), исследовавший другой вид того же рода,
пришел к сходным выводам. Он также не нашел никаких остатков глаз;
что же касается глазного нерва, то Вольский предполагает, что у Niphargus
этот нерв сливается с так называемым nervus pseudofrontalis, образуя
единый nervus optico-pseudofrontalis. Сравнивая различных Amphipoda
с редуцированными органами зрения, Вольский установил, что у этого
отряда в первую очередь разрушается дистальная часть глаза (хрустальный
конус, затем ретинула и, наконец, нерв). Между тем наблюдающиеся
мутации безглазости у Amphipoda сводятся к деградации нерва, а затем
дистальной части глаза. Таким образом, мутационный процесс нельзя
сравнивать с происходившей в естественных условиях редукцией глаз.
Ракообразные, обладающие стебельчатыми глазами, при переходе под
землю сохраняют глазные стебельки, но весь глазной аппарат обычно
подвергается редукции. Среди креветок Barbouria poeyi обладает нормаль-
ными глазами, у Palaemon cavernicolus глаза значительно меньше, чем
у близких наземных видов, но еще функционируют, Typhlocaris, Troglo-
caris и Palaemonias совершенно слепы. При этом, как показало гистологи-
ческое изучение глаза Troglocaris (Babic, 1922), три типичных для рако-
образных глазных ганглия, находящихся в глазном стебельке, не затраги-
ваются редукцией, в то время как роговица, хрустальный конус, рабдом,
сетчатка и пигмент нацело отсутствуют. Кроме того, наблюдается недораз-
витие глазных стебельков Troglocaris, которое является результатом их
замедленного по отношению к телу роста (Юзбашьян, 1940). Длительное
содержание Troglocaris на свету приводит к появлению пигмента в дисталь-
ной части их глаз. Однако далеко не все особи обладают способностью
к образованию пигмента, и большая их часть даже на четвертом году
жизни на свету не обнаруживает никаких изменений. Пигментация глаз
взрослых Troglocaris наблюдалась и на некоторых экземплярах из ключей.
Замечательно, что выведенная на свету личинка Troglocaris schmidti
обладает пигментированным и, по-видимому, нормально построенным гла-
зом; описано его фасеточное строение (Юзбашьян, 1934, 1940).
Процесс редукции глаз у мизид, если судить по хорошо изученной в этом
отношении Troglomysis vjeternicensis (Stammer, 1936), протекает совер-
шенно иначе, чем у Amphipoda и Decapoda. В укороченном глазном стебельке
Troglomysis констатированы только остатки роговицы и хрустального ко-
нуса. Глазные ганглии развиты, но не посылают нервов к поверхности
глазного стебелька. Таким образом, если у Amphipoda и Decapoda редукция
затрагивает в первую очередь дистальную часть глаза, у мизид она начи-
нается с проксимальной части.
Среди Isopoda, обладающих, подобно Amphipoda, сидячими глазами,
как показал Латтин (Lattin, 1939а), можно встретить оба указанных
6 Заказ 333
81
направления редукции (центрипетальный и центрифугальный типы/редук-
ции). Так, например, у рода Androniscus на месте глаз находятся группы
немногочисленных плоских клеток, отличающихся от гиподермальных только
несколько более крупными ядрами и соединенных тонким тяжем неврилеммы
с нормально развитыми зрительными ганглиями. У Asellus, наоборот,
зрительные ганглии сильно редуцированы, но в глазах сохраняются еще
хрустальные конусы. Степень редукции зрительного аппарата у Asellus
cavaticus зависит от возраста животных: у молодых экземпляров глаза
соединяются с мозгом толстым нервным тяжем, а хрустальные конусы
сближены; у более крупных тяж становится очень тонким, а хрустальные
конусы раздвигаются, при этом в строении глаз животных даже одного
возраста наблюдается сильная изменчивость.
Редукция зрительного аппарата Isopoda не идет так далеко, как у других
ракообразных, и у всех пещерных видов существуют рудименты глаз.
Интересно, что степень редукции глаз и глазных нервов и ганглиев не зависит
от высоты их организации у близких наземных форм. У видов, принадле-
жащих к семействам с высокоорганизованным зрительным аппаратом,
редукция может идти дальше, чем у форм, принадлежащих к семействам
с низкоорганизованным зрительным аппаратом.
У пещерных ложноскорпионов, судя по старой работе Гагена (Hagen,
1879) над Blothrus packardi, на месте глаз находятся длинные щетинки
(органы осязания), при основании которых сохраняются светопреломляю-
щие тельца. Зрительный нерв хорошо развит почти на всем протяжении,
за исключением конечной, прилегающей к коже части.
У пещерных моллюсков Lartetia и Vitrei 1а также имеет место интересный,
но недостаточно изученный случай смены функций. Глаз теряет пигмент
и превращается в орган осязания.
Строение глаз пещерных рыб и амфибий известно довольно хорошо
благодаря работам Эйгенмана, Улисса, Жерара, Грессера и Бредера и др.
В пределах семейства Amblyopsidae представители рода Chologaster обла-
дают уменьшенными, но еще функционирующими глазами, причем заме-
чательно, что сетчатка единственного наземного вида этого рода — Ch.
cornutus — редуцирована в большей степени, чем у двух подземных. Глаза
другого рода — Amblyopsis — лежат под кожей, лишены линзы, края радуж-
ной оболочки срослись, пигмент еще есть, сетчатка с колбочками, но лишена
палочек. У Troglichthys глаза также сверху заросли кожей, но по размерам
значительно меньше, чем у предыдущего рода; сетчатка лишена колбочек;
пигментный эпителий развит только в дистальной части глаза. Как
у Amblyopsis, так и у Troglichthys глаза не соединяются с мозгом и, разуме-
ется, не функционируют. У Typhlichthys пигментный эпителий сетчатки
отсутствует, края радужной оболочки не срослись, так как зрачок имеется.
Слои сетчатки в противоположность Amblyopsis и Troglichthys дифференци-
рованы, но ни колбочек, ни палочек нет. Отсутствует также глазной мускул.
Глаз соединяется с мозгом. Следовательно, редукция органов у Amblyopsi-
dae, так же как у ракообразных, идет разными путями и затрагивает
у разных родов разные части глаза.
Глаза единственного изученного в этом отношении пещерного представи-
теля семейства Cyprinidae — Caecobarbus geertsii — также сверху покрыты
кожей. Их глазной бокал замкнут и окружен целиком пигментным эпителием.
Сетчатка представлена утолщением части внутреннего слоя, обращенного
82
к мозгу, имеется сосудистая оболочка, а линза отсутствует. К глазу
прикрепляются три рудимента глазных мышц (Gerard, 1936).
Единственный пещерный представитель семейства Characinidae — Апор-
tichthys jordani — исследован в отношении строения глаз Грессером и
' Бредером (Gresser, Breder, 1940). Глаза этой рыбы также находятся под
кожей, через которую у некоторых экземпляров просвечивает пигментный
эпителий. Эпидермис у одних особей образует толстый эпителиальный
тяж, у других — мешковидное впячивание эпителия; в обоих случаях эпи-
телий прилегает к сгущению мезодермальных клеток, отвечающему роговице.
Гистологическое строение глаза Anoptichthys позволяет утверждать, что
в своем развитии он остановился на стадии первичного глазного бокала,
в котором дифференцировались пигментный эпителий и сетчатка с недораз-
вившимися палочками и колбочками. Мезодермальные элементы — склеро-
тека и сосудистая оболочка — развиты несколько лучше, причем имеется
хориоидальная железа с кровеносными сосудами. Глазной нерв также при-
сутствует, но его клетки имеют эмбриональный характер, а нервные волокна
развиты слабо. Мускулатура глаза представлена шестью, по-видимому,
нефункционирующими мышцами.
Для семейства Brotulidae характерна быстрая редукция глаз в течение
онтогенеза. Постепенно дегенерируют и, наконец, исчезают склеротека,
глазные мышцы, линза, разрушаются хорошо развитая у эмбрионов сетчатка
и пигментный эпителий, при этом, так же как у Amblyopsidae, наблюдается
значительная изменчивость в степени редукции глаз у рыб одного возраста
и даже одного помета (Eigenmann, 1909).
Экспериментальное изучение влияния темноты на глаза рыб производи-
лось на золотых рыбках Огневым (1910). Он нашел, что после трехлетнего
пребывания в темноте животные слепнут. Сетчатка теряет колбочки и
палочки, исчезают слои нервных клеток и волокон, разрушается пигментный
эпителий. Однако наряду с глазами атрофируются и яичники.
Глаза пещерных амфибий построены приблизительно так же, как у рыб.
Еигусеа имеет нормальные глаза, у Typhlotriton дегенерирует сетчатка,
веки срастаются, оставляя свободным небольшое отверстие около центра,
но линза вполне развита. У Proteus и Typhlomolge глаза лежат под кожей,
линза и мышцы отсутствуют, сетчатка или с неотличимыми друг от друга
палочками и колбочками (Proteus), или совсем без них (Typhlomolge),
пигмент не развит, стекловидное тело замещается соединительной тканью.
У молодых животных глаза развиты гораздо лучше. Согласно Ноблю и Попу
(Noble, Pope, 1928), личинки Typhlotriton имеют нормальные глаза. В этом
случае также констатирован широкий диапазон изменчивости степени
редукции глаз у животных одного возраста. Помещение личинок в различные
условия показало, что дегенерация глаз связана с отсутствием света.
У воспитанных на свету животных веки не замыкались, хотя сетчатка
все-таки дегенерировала; в то же время контрольные, воспитанные у темноте
экземпляры прошли через ряд стадий замыкания век. Интересно, что прямое
освещение заставляло расширяться меланофоры взрослых Typhlotriton, но
не влияло на меланофоры личинок, хотя длительное освещение увеличивало
пигментацию личинок, но не сказывалось на пигментации взрослых.
Сходные результаты получил значительно ранее Каммерер (Kammerer,
1912), работавший с протеем. Под действием дневного света у молодых
протеев в глазах под кожей развивался темный пигмент, размеры глаз
6*
83
увеличивались, покрывающая их кожа становилась очень тонкой, дифферен-
цировались отдельные части глаза и в конце концов возникал нормальный
для личинок амфибий орган зрения. Воспитанные при красном свете протеи
обладали лишенными пигмента глазами.
Резюмируя все эти немногочисленные данные, можно сказать, что
широко распространенная среди пещерных обитателей деградация орга-
нов зрения, как правило, не приводит к полному исчезновению последних;
те или иные части зрительного аппарата совершенно не подвергаются
редукции, при этом у молодых экземпляров глаза бывают развиты значи-
тельно лучше, чем у взрослых, а иногда и вполне нормально; таким
образом, редукция органов зрения совершается частично в процессе постэм-
брионального развития. Пути и способы редукции различны даже у близко
родственных форм: иногда в первую очередь разрушается дистальная,
иногда проксимальная часть органов зрения. Свет оказывает сильное
влияние на организацию глаза: помещенные в темноту наземные животные
нередко слепнут; действие света на органы зрения подземных форм часто
задерживает процесс редукции глаз, а иногда вызывает даже их прогрессив-
ное развитие. Все эти выводы имеют большое значение для разрешения
вопроса об эволюции пещерных обитателей.
ЭВОЛЮЦИЯ ПЕЩЕРНЫХ животных
Пути и закономерности эволюции пещерных животных изучались и
обсуждались до сих пор чрезвычайно односторонне. Вся проблема сводилась
к вопросу о причинах редукции глаз и пигмента у троглобионтов, и только
вокруг этого вопроса разгорались горячие споры. Совершенно упускалось
из виду, что пещерные животные обладают бесчисленным количеством
адаптивных признаков и что благодаря этому вся организация каждого
пещерного вида соответствует своеобразным условиям подземного суще-
ствования. Такой односторонний подход к проблеме эволюции пещерной
фауны, вызванный игнорированием значения целостности организма в фило-
генезе, определил целый ряд неверных установок и гипотез.
Вопрос о причинах слепоты пещерных животных разрешался до
недавнего времени в духе учения Ламарка. Даже Дарвин приписывал
потерю троглобионтами органов зрения отсутствию упражнения. Это же
объяснение принимали Спенсер, Паккард, Вире, отчасти Раковица и, нако-
нец, Каммерер. Однако, как указывалось выше, под землей обитает целый
ряд зрячих форм, причем нет оснований приписывать им более короткий
срок жизни в пещерах, чем слепым видам. В этом отношении большой
интерес представляет строение глаз жуков Machaerites subterreneus и
М. mariae, у которых самцы обладают нормальными глазами, а у самок
глаза или редуцированы, или отсутствуют. Довольно веским доводом против
ламаркистского объяснения слепоты пещерных животных являются приве-
денные выше данные о неустойчивой дегенерации глаз многих видов (протея
и др.). Поскольку освещение может привести к развитию нормального
глаза протея, редукция органа зрения этого вида, очевидно, не является
наследственной. Экспериментальная проверка воздействия темноты на ор-
ганы зрения наземных животных также не может служить для подтвержде-
ния правильности ламаркистского объяснения явления. Пэйн (Payne
84
1911*) вырастил в темноте 69 поколений Drosophila и не заметил никаких
следов редукции глаз. Уленгут (Uhlenguth, 1912*, 1914* и 1916*) пере-
саживал глаза молодой саламандры в различные части ее тела, и, несмотря
на то что пересаженные глаза не функционировали, будучи изолированы от
нервной системы, после временной дегенерации они полностью восстанавли-
вались. По более новым данным Гаррисона (Harrison, 1929*), при пере-
садке первичных глазных бокалов от Ambystoma tigrinum другому виду —
A. punctatum, и наоборот, трансплантат не соединяется с мозгом, т. е. не
функционирует, и, несмотря на это, глаз развивается вполне нормально,
только ганглиозные клетки испытывают обратное развитие (Mangold, 1931).
Папаниколау (Papanicolau, 1912*) и Ермаков (1924), Берлингер (Berlinger,
1911) и Графф (Graff, 1917) показали, что редукция глаз ветвистоусых
рачков и реснитчатых червей вызывается голоданием животных. Таким
образом, влияние неупражнения глаз на их строение не является дока-
занным.
Уоллес (1911) считает возможным объяснить редукцию глаз пещерных
животных действием естественного отбора. Он предполагает, что, сделав-
шись в темноте бесполезными, глаза в то же время могли стать и вредными,
так как эти органы легко подвергаются ранениям и заболеваниям. В этом
случае слепые индивидуумы будут обладать известными преимуществами
в борьбе за существование перед зрячими и в конце концов вытеснят этих
последних. Точка зрения Уоллеса, к сожалению, не получила дальнейшего
развития и не подверглась экспериментальной проверке. Впрочем, она не мо-
жет объяснить явления депигментации троглобионтов.
В 1940 г. в нашей литературе появилась попытка совместить взгляды
на эволюцию неоламаркистов и неодарвинистов. Г А. Машталер (1940)
выступил с теорией, сущность которой сводится к признанию прямого
непосредственного действия среды на выработку приспособлений у низших
животных и ведущей роли дарвиновских факторов лишь при выработке
приспособлений у высших животных. Эту теорию, которую автор
ее «скромно» сравнивает с теорией относительности, он распространяет и на
пещерную фауну.
Согласно Машталеру, низшие организмы (черви, низшие ракообразные),
попадая в условия пещер, изменяются непосредственно, адекватно к среде.
Наоборот, развитие троглобионтности у высших организмов совершается
через неопределенную наследственную изменчивость и естественный отбор,
создающий определенное соответствие между организмом и средой. Основа-
нием для такого вывода является совершенно не соответствующее фактам
мнение о том, что в случае низших животных признаки, хотя они и
могут прочно наследоваться в ряде поколений, все же сильно варьируют
в степени своего развития в зависимости от условий вплоть до полного
исчезновения, а в случае высших животных, несмотря на длительное вос-
питание многих поколений в измененных условиях, довольно прочно и по-
стоянно повторяются, т. е. закреплены генотипически. Границу между
высшими и низшими животными Машталер произвольно проводит через
членистоногих, относя ракообразных к низшим, а паукообразных, много-
ножек и насекомых — к высшим организмам. Однако всякому знакомому
с зоологией человеку ясно, что нет совершенно никаких оснований признавать
* Здесь и далее звездочками отмечены работы, не приведенные автором в списке
литературы. — (Примеч. ред.).
85
организацию ракообразных более низкой, чем паукообразных или особенно
многоножек, и приписывать этим группам качественно различные типы
эволюции. Границу между низшими и высшими было бы правдоподобней
проводить между одно- и многоклеточными, двух- и трехслойными, первично-
и вторичноротыми и т. д., но не между ракообразными и паукообразными.
Столь же произвольным является утверждение Машталера о большей
изменчивости и зависимости от среды признаков «низших» троглобионтов
по сравнению с «высшими». Никаких доказательств большей изменчивости
признаков низших троглобионтов автор и не приводит. Что же касается
прямого действия среды на строение низших троглобионтов, то подтвержде-
ние этого Машталер видит в том, что будто бы среди них мы не встречаем
зрячих организмов и, наоборот, среди ближайших их родичей на земле
обычно не находим слепых. На самом же деле, как указывалось в предыду-
щей главе, в пещерах существуют довольно многочисленные зрячие формы
реснитчатых червей, веслоногих и равноногих ракообразных и еще большее
количество слепых представителей групп, относимых Машталером к низшим,
есть среди наземных обитателей. Близкое сходство пещерных и внепе-
щерных форм в родах Asellus, Gammarus, Dendrocoelum, согласно Машта-
леру, наталкивает на мысль, что они в обоих случаях представляют собой
одни и те же виды, изменившиеся в течение ряда поколений под прямым
влиянием среды в сторону редукции зрительного аппарата и утраты пигмента,
и большей частью легко установить непрерывную цепь вариантов, соединяю-
щих два каких-либо вида. В своем предыдущем обзоре (Бирштейн, 1940а)
я приводил в качестве примеров подобных «цепей вариантов», между
прочим, рыб и пауков, т. е. животных, относимых Машталером к высшим.
Что же касается объяснения всего этого явления, то мне по-прежнему
представляется гораздо более вероятным предполагать в таких случаях
относительно недавнее отщепление подземной формы от наземной.
Обращаясь к экспериментальным данным, Машталер находит, что они
также подтверждают его точку зрения на фенотипический характер проис-
хождения приспособлений «низших» представителей троглобионтов. Он при-
водит некоторые из изложенных выше случаев редукции глаз и пигмента
ракообразных и планарий под влиянием темноты. Наоборот, у вышестоящих
организмов, таких, как жуки, пауки, ложноскорпионы и т. д., эксперимен-
тальное получение развития либо редукции зрительного аппарата Машталер
считает невозможным. Последнее утверждение голословно, так как, к сожа-
лению, соответствующие эксперименты ни над жуками, ни над пауками, ни
над ложноскорпионами не ставились. Зато хорошо известны случаи редук-
ции глаз под влиянием темноты у таких высокоорганизованных животных,
как рыбы (Огнев, 1910), и развития глаз под влиянием света у амфибий
(Kammerer, 1912; Noble, Pope, 1928). Опыты Каммерера над протеем зна-
комы Машталеру, который делает из них неожиданный вывод о том, что хотя
протей по организации стоит выше, чем насекомые и пауки, тем не менее
в генетическом отношении его признаки, по-видимому, закреплены менее
прочно. Этот пример показывает, что даже тот скромный фактический
материал, которым располагает Машталер, не укладывается в его «теорию».
В конце своего обсуждения данных по пещерной фауне Машталер
приходит к заключению о различном способе вселения в пещеры низших и
высших животных. Он считает, что для первой группы организмов наиболее
характерный путь возникновения троглобиев лежит через случайные попада-
ния животных в пещеры, где у них постепенно атрофируются глаза и
86
приобретаются прочие характерные признаки. Возникновение же троглобиев
высших форм идет через случайное появление безглазых мутаций, которые
в силу их слабой способности к борьбе за существование в обычных условиях
укрываются в наиболее надежных местах (в пещерах и пр.), где они и
образуют «новые штаммы». Таким образом, в отношении эволюции и проис-
хождения «низших» троглобионов теория Машталера ни в какой мере
не опирается на факты, в отношении же происхождения «высших» трогло-
бионтов Машталер является сторонником теории преадаптации, которой
придерживаются довольно многие иностранные авторы.
Длинный ряд исследователей, начиная с Эйгенмана (Eigenmann, 1909),
Леба (1926) и Кено (Cuenot, 1921*) и кончая Коссвигом (Kosswig, 1934—
1939), Ванделем (Vandel, 1938), Губсом (Hubbs, 1938) и Колосвари (Ко-
losvary, 1936), считают, что редукция органов зрения и пигмента происходит
совершенно независимо от условий существования, но слепые и депигменти-
рованные животные могут выживать только в пещерах, так как на поверхно-
сти земли они погибают в борьбе за существование. Среди советских
авторов того же мнения придерживается Поляков (1937), который пишет:
«. .несомненно, что случайно возникающие как на свету, так и в темноте
безглазые мутанты могут обычно сохраняться в борьбе за существование
только в темноте».
Эйгенман указывает на наличие вне пещер слепых видов рыб Amblyopsi-
dae и Brotulidae; близкие к ним формы являются эвтроглобионтами. Многие
Siluridae находят пищу при помощи органов осязания и не пользуются при
этом органами зрения. По этим признакам указанные рыбы преадаптированы
(Cuenot, 1921*) к жизни под землей. Леб (1926), а в последнее время Коссвиг
и Вандель придают решающее значение мутациям, приводящим к недоразви-
тию глаз. В нескольких работах Коссвиг (Kosswig, 1934, 1937а, б, 1939;
Kosswig С., Kosswig L., 1936) описывает свои наблюдения над пещерной
формой водяного ослика Asellus aquaticus cavernicola из разных пещер
Истрии. Он установил, что каждая популяция Asellus состоит из смеси
особей, в различной степени депигментированных и обладающих в различной
степени редуцированными глазами, причем наряду с нормально окрашен-
ными и вполне зрячими экземплярами попадаются совершенно депигментиро-
ванные и слепые. В противоположность прежним утверждениям не суще-
ствует никакой связи между степенью депигментации и редукции глаз, с од-
ной стороны, и расстоянием от входа в пещеру — с другой, т. е. процент сле-
пых и лишенных пигмента особей по направлению в глубь пещеры не возра-
стает 1 Генетические эксперименты показали, что генотипическая база боль-
шинства типов окраски тела и строения и пигмента глаз очень сложна и что
различные мутации многих генов вызывают сходные фенотипы. Популяция
пещерных Asellus благодаря случайным мутациям гетерозиготна по некото-
рым генам; со временем, при слабой положительной селекции и просто
вследствие флюктуаций, такие немногочисленные популяции могут стать
гомозиготными, что и наблюдается в некоторых пещерах. Если подобные
мутации появляются среди живущих вне пещер водяных осликов, они гибнут
1 Бертосси (Bertossi, 1938) нашел, что в пещерах Италии наряду с пигментированными
особями мокрицы Andronicus boldorii попадаются и депигментированные. На осно-
вании экспериментальных данных автор считает возможным объяснить редукцию
пигмента неблагоприятными условиями существования. К сожалению, генетический
анализ не был проведен.
87
в борьбе за существование и только в условиях подземного обитания имеют
равные шансы с нормальными выжить и размножиться.
Вандель (Vandel, 1938), исследуя наземную мокрицу Trichoniscus
(Spiloniscus) elisabethae, пришел к сходным выводам. Он обнаружил у этой
мокрицы две мутации: alba и pallida. У первой из них пигмент почти нацело
отсутствует, у второй он значительно редуцирован. Обе мутации опреде-
ляются двумя независимыми друг от друга рецессивными генами. Вандель
полагает, что многочисленные пещерные виды Trichoniscus произошли в ре-
зультате аналогичных мутаций, вызывающих, между прочим, изменение тро-
пизма в направлении усиления отрицательного фототропизма.
Логическим завершением этой системы взглядов является статья Коло-
свари (Kolosvary, 1936). Он считает, что эволюция определяется исключи-
тельно внутренними факторами. Заложенные в организме тенденции к редук-
ции осуществляются совершенно независимо от среды. Даже такие мощные
факторы, как циклы горообразований, только стимулировали или задержи-
вали проявление этих тенденций. Псевдотроглобионты, гемитроглобионты
и эвтроглобионты представляют собой отдельные стадии, через которые
проходит по пути редукции организм, направляемый имманентными силами.
На каждой из этик стадий животное активно подыскивает подходящую для
себя среду обитания. Для вполне редуцированных животных подходящим
биотопом являются пещеры. Таким образом, адаптация к пещерным усло-
виям (и вообще к внешней среде) просто не существует, и этот термин
Колосвари берет в кавычки 1
Идеалистический характер взглядов Колосвари достаточно очевиден.
Можно только удивляться, как автор игнорирует факты высокой специали-
зации пещерных животных, рассматривая их только с точки зрения редукции
и не обращая внимания на целый ряд прогрессивных приспособительных
особенностей, которые никак не укладываются в его «теорию».
Постулируемая сторонниками теории преадаптации возможность появ-
ления приспособления вне зависимости от среды, как указал И. И. Шмаль-
гаузен (1938), кажется вероятной только до тех пор, пока мы рассматриваем
развитие какого-нибудь признака изолированно. Так, например, потеря пиг-
мента и органа зрения может, по-видимому, произойти вне пещеры в по-
рядке мутации, и такая мутация может сохраниться в пещерах. Однако
совершенно ясно, что ослепшее и лишенное пигмента животное еще не будет
приспособлено к пещерному образу жизни. Действительно, исследованная
Эйгенманом (Eigenmann, 1909) слепая рыба Chologaster cornutus, несмотря
на отсутствие глаз, не живет в подземных водах. То же относится к много-
численным примерам существования слепых и депигментированных форм вне
пещер. Более того, у нас нет достаточных оснований рассматривать депиг-
ментацию и редукцию органов зрения как приспособительные признаки.
Что же касается таких адаптивных признаков, как удлинение конечностей
и обогащение их органами осязания и обоняния, усовершенствование боко-
вой линии рыб, приспособления к амфибиотическому образу жизни и
питанию бактериальным илом и т. д., то чрезвычайно трудно допустить,
что все они возникли вне пещер в результате случайного совпадения не
направленных и не контролируемых отбором мутаций. Латтин (Lattin, 1939с)
1 Впоследствии Колосвари (Kolosvary, 1938) таким же образом пытался объяснить
особенности организации морских глубоководных животных.
88
в своей последней работе признает, что при образовании компенсаторных
органов чувств (например, органов осязания пещерных рыб) «селекция луч-
ших генотипов вступает в свои права».
И. И* Шмальгаузен (1938) объясняет редукцию органов зрения пещерных
животных как следствие отсутствия отбора по этим биологически беспо-
лезным в условиях пещер признакам. Аналогичные мысли, но не в столь
ясной форме высказал Латтин (Lattin, 1939с). По Шмальгаузену, при выклю-
чении отбора происходит беспорядочное накопление мутаций, большей
частью сдвигающих сроки развития тканей, из которых одни являются индук-
тором, а другие — реактором. Расхождение в периодах деятельности этих
компонентов приводит к недоразвитию или даже полному исчезновению
органа. «При беспорядочном накоплении мутаций, касающихся какого-
либо органа, происходит глубочайшее нарушение процессов его развития,
и это нарушение всегда приводит к недоразвитию. При значительных расхож-
дениях во времени дифференцировки основного индуктора и времени созре-
вания реактора совсем не осуществляется и сама детерминация органа.
В этом случае не образуется даже его зачатка. Орган исчез, не оставив
следа» (Шмальгаузен, 1938, с. 53).
При редукции глаз пещерных животных зачатки органа зрения обычно
сохраняются. В некоторых случаях развитие органа просто останавливается
на известной стадии и дальше не идет, как, например, в случае остановки
развития глаза протея и Anoptichthys на стадии глазного бокала (Каттегег,
1912; Gresser, Breder, 1940). Таким образом, редукция захватывает лишь
поздние стадии онтогенеза. Это обстоятельство находится в соответствии
с развитыми недавно Крушинским (1939) взглядами. Крушинский полагает,
что более поздние стадии развития признака обусловливаются большим
числом последовательно включающихся в процессе онтогенеза факторов,
чем более ранние. Именно благодаря большему числу факторов, опреде-
ляющих конечные этапы онтогенеза, повышается вероятность мутаций,
затрагивающих более поздние стадии развития признака. На беспорядочное
накопление мутаций, нарушающих нормальное развитие глаз пещерных
животных, с несомненностью указывают приведенные выше данные о ши-
роком диапазоне изменчивости в строении органов зрения троглобионтов
на всех возрастных стадиях.
Теория Шмальгаузена несомненно объясняет в основных чертах причины
редукции органов зрения и пигмента у пещерных животных. Однако она
нуждается в экспериментальном подтверждении: в настоящее время мы
еще не располагаем точными наблюдениями над всем ходом процесса
редукции глаз в онтогенезе. Трудно даже сказать, имеет ли здесь место
редукция посредством рудиментации или посредством афанизии (Северцов,
1939). Вероятно, в разных случаях наблюдаются разные способы редукции.
Для протея и Anoptichthys, например, можно считать типичной рудимен-
тацию для Brotulidae, по Эйгенману, и для креветки Troglocaris, судя по
данным Юзбашьяна — афанизию, так как глаз первой личинки Troglo-
caris развит нормально, а в процессе постэмбрионального развития он
испытывает, очевидно, обратное развитие (инволюцию). Далее совершенно
не изучен вопрос о морфогенетической роли функции, которая, несомненно,
имеет известное значение, судя по изложенным выше данным опытов Нобля
и Попа, Каммерера и Юзбашьяна над пещерными амфибиями и креветками.
Следует, наконец, отметить, что теория Шмальгаузена. оставляет невыяс-
89
ненными причины наличия глаз у ряда троглобионтов и замечательный
факт наличия глаз у самцов Machaerites при отсутствии их у самок.
Итак, наиболее вероятно, что редукция глаз и пигмента большинства
троглобионтов произошла после их вселения в пещеры, когда эти признаки
утеряли свое полезное приспособительное значение и были разрушены вслед-
ствие беспорядочного накопления мутаций, не контролируемых отбором.
Этот процесс отнюдь не является адаптацией к условиям существования
в пещерах, а лишь закономерным следствием утраты глазами и пигментом
приспособительного значения, т. е. выведения их из сферы действия есте-
ственного отбора.
Как же обстоит дело с настоящими адаптивными признаками, позволяю-
щими животным постоянно существовать в пещерах? Нет решительно ни-
каких оснований отрицать ведущую роль естественного отбора в формиро-
вании этих признаков. Ряд тихотроглобионт—псевдотроглобионт—гемитро-
глобионт— эвтроглобионт не есть, как думает Колосвари, ряд стадий редук-
ции организма, направляемой имманентными силами, совершенно незави-
симыми от внешней среды. Это — путь приспособлений к реальным условиям
существования, постепенно приводящий к вселению в пещеры. Многие сухо-
путные животные приспособлены к жизни под камнями, в расселинах скал
и в тому подобных биотопах. Их организация и реакции соответствуют
условиям внешней среды, и трудно представить себе, что это соответствие
не является результатом действия естественного отбора. Условия существо-
вания в указанных биотопах, с одной стороны, и в пещерах — с другой,
довольно сходны (высокая влажность, темнота, относительно низкая летом
температура), на что указывает, между прочим, наличие в природе псевдо-
троглобионтов и гемитроглобионтов. Переход к жизни исключительно
в пещерах для таких животных обозначает просто ограничение места
своего пребывания, сопровождающееся дальнейшей специализацией. Этот
переход совершается, очевидно, в результате борьбы за существование,
разыгравшейся в предыдущих занимаемых данным организмом биотопах.
Аналогичные соображения можно высказать и по отношению к водным
животным, так как сходство в составе фауны и во внешних условиях ключей
и пещерных водоемов весьма значительно, и в этом случае также легко
представить себе процесс постепенного перехода от жизни в ключах к жизни
в пещерах.
Эволюцию организма от стадии псевдотроглобионта через стадию ге-
митроглобионта до стадии эвтроглобионта следует рассматривать как после-
довательный переход от более общей среды к более частной. Такое направле-
ние эволюционного процесса Шмальгаузен (1939) предложил называть теле-
морфозом. Телеморфоз характеризуется далеко идущей специализацией,
сопровождающейся часто некоторым упрощением организации, ограниче-
нием пластичности организмов, дифференциацией вида на немногочисленные
изолированные популяции и значительным снижением темпа прогрессивной
эволюции; при этом «специализированный организм может при отсутствии
резких изменений среды существовать неопределенно долго (хотя бы в те-
чение всего ряда геологических периодов от кембрия и доныне)» (Шмаль-
гаузен, 1940, с. 148).
Все указанные особенности телеморфоза при анализе пещерной фауны
выявляются чрезвычайно рельефно. Черты высокой специализации и
частичного регресса подземных обитателей были в достаточной степени
90
описаны выше. Что касается дифференциации вида на отдельные изолиро-
ванные популяции, то и по этому вопросу за последнее время накопился
некоторый материал. Сравнивая между собой популяции Asellus aquaticus
cavernicola из разных истрийских пещер, Коссвиг (Kosswig, 1937а, b, 1939;
Kosswig С., Kosswig L., 1936) обнаружил, что каждая из относительно малых
и изолированных популяций имеет свое типичное соотношение между осо-
бями, характеризующимися определенной степенью депигментации и редук-
ции глаз. Поскольку эти признаки в своем развитии зависят от тех или иных
генов, каждая популяция обладает своей характерной частотой распреде-
ления генов, ответственных за фенотипические особенности составляющих
ее животных. Аналогичное явление Коссвиг описал для бескрылого насеко-
мого Hypogastrura armata (Kosswig, 1937). Правда, соотношения между
различно окрашенными особями Asellus от года к году несколько меняются
(Kosswig, 1939), но это указывает только на существование большого числа
определяющих пигментацию генов, различно комбинирующихся между
собой. В то же время наблюдаются признаки, характерные только для одной
какой-нибудь популяции (100% депигментированных особей, лиловая
окраска глаз).
При изучении креветок Troglocaris schmidti из разных пещер Абхазии
нами (Бирштейн, 1939а) установлено, что в каждой пещере обитает свой
подвид. Отличия между подвидами распространяются на вторичнополовые
признаки. При этом каждый подвид характеризуется комплексом таких
признаков, которые в известной части присущи другим подвидам. Для любого
подвида Troglocaris schmidti специфичны не столько отдельные признаки,
сколько та или иная их комбинация. Сходные данные получил при обработке
многоножек из абхазских пещер Ломандер (Lomander, 1936). Впрочем,
еще в 1850 г. Фитцингер (Fietzinger, 1850) описал несколько узколокализо-
ванных «видов» протея.
Распад видов на микрогеографические расы можно объяснить на основа-
нии соображений Гагедурн (Hagedoorn A. L., Hagedoorn А. С., 1921),
Фишера (Fischer, 1930), Райта (Wright, 1932), Дубинина, Ромашова (1932)
неравномерным накоплением наследственного материала в отдельных изо-
лированных популяциях. Вследствие этого, несмотря на сходные условия
существования, будет наблюдаться расхождение признаков между вновь
образовавшимися формами. Важно подчеркнуть, что изменчивые признаки
не имеют непосредственного отношения к внешней среде — это различные
приспособления для копуляции. Что же касается настоящих адаптивных
признаков, то в различных пещерах у представителей одного и того же вида
они строго константны. Это обстоятельство соответствует представлениям
Шмальгаузена об ограниченности пластичности организмов, эволюциони-
рующих по пути телеморфоза. По-видимому, сам процесс распада вида на
местные формы, отличающиеся друг от друга неадаптивными признаками,
не имеет большого значения для прогрессивной эволюции в целом.
Чрезвычайно интересен вывод Шмальгаузена о наблюдающемся при
телеморфозе замедленном темпе эволюции. Есть все основания предполагать,
что этот вывод полностью приложим к пещерной фауне. Постоянство условий
существования на протяжении длительного в геологическом смысле времени
прежде всего должно было сказаться на замедлении эволюции. Далее, как
отметил еще Дарвин, борьба за существование в пещерах значительно менее
интенсивна, чем на поверхности земли. Численность популяций пещерных
91
животных во многих случаях лимитируется не пищевыми ресурсами или
какими-нибудь другими факторами, из-за которых обычно возникает конку-
ренция, а такими явлениями, как периодические наводнения, вымывающие
пещерных обитателей из их убежищ. Все это делает понятным сохранение
в пещерах древних сравнительно мало изменившихся «остатков прежней
жизни» (Дарвин, 1937). В предыдущем обзоре (Бирштейн, 1940а) были
приведены примеры, иллюстрирующие древность многих эвтроглобионтов.
В дополнение можно указать на интересные данные по бокоплаву (Pseu-
doniphargus africanus). Этот вид найден в грунтовых водах Западной
Африки, Северной Испании, восточного побережья Адриатического моря и,
наконец, о-ва Мадейра. Сравнение экземпляров из разных местонахождений
доказывает их полную тождественность. Поскольку отделение о-ва Мадера
от материка произошло в миоцене, следует считать, что Р. africanus сформи-
ровался как вид еще до миоцена, и с тех пор до наших дней его морфология
не изменилась (Schellenberg, 1939).
Таким образом, пещеры представляют собой палеонтологический зоо-
парк, в значительной степени населенный осколками давно погибших на
поверхности земли фаун. Остается только пожалеть и по примеру Дарвина
изумляться, «что в этих мрачных убежищах не уцелело еще более остатков
прежней жизни благодаря менее суровой конкуренции, которой подвер-
галось их незначительное население» (Дарвин, 1937, с. 209).
НОВЫЕ ДАННЫЕ О ПРОИСХОЖДЕНИИ
ФРЕАТИЧЕСКОЙ ПОДЗЕМНОЙ ФАУНЫ*
Биологические исследования грунтовых вод, произведенные в последние
годы, привели к открытию новой весьма своеобразной фауны. Составляющие
ее виды населяют преимущественно воду, заполняющую пространство между
отдельными частицами грунта. Соответственно такой среде обитания пред-
ставители этой так называемой фреатической фауны отличаются малыми
размерами, укороченными конечностями и удлиненным часто червеобразным
телом, т. е. особенностями, позволяющими существовать в узких капилляр-
ных ходах. Подобно пещерным обитателям, они лишены пигмента и органов
зрения, но снабжены гипертрофированными органами осязания и обоняния.
Из системы грунтовых вод фреатические виды проникают в пещерные
водоемы и глубокие колодцы, питающиеся грунтовыми водами.
При фильтрации через планктонную сетку больших масс грунтовых вод,
получаемых при помощи насосов, а также при обловах планктонной сеткой
глубоких колодцев удалось получить некоторое представление о составе
фреатической фауны. При этом следует иметь в виду слабую степень коли-
чественного развития входящих в нее видов. Многие из них добыты в коли-
честве всего 1 экз.
Основными составляющими эту фауну группами оказались ракообразные
и водяные клещи. Последние представлены в Югославии 11 видами, относя-
щимися к 8 родам, из которых большинство описано впервые и специфично
для фреатической фауны. Почти половина югославских фреатических видов
* Тр. Всесоюз. гидробиол. о-ва, 1952, т. 4, с. 225—229.
92
клещей (5 видов) принадлежит преимущественно к морской группе Halaca-
rida.
Гораздо более многочисленны ракообразные. Не рассматривая здесь
нескольких фреатических видов Ostracoda и Harpacticoida, остановимся
более подробно на весьма своеобразных высших ракообразных. Они пред-
ставлены тремя отрядами — Anaspidacea, Amphipoda и Isopoda.
Все европейские виды Anaspidacea (роды Bathynella и Parabathynella)
принадлежат к фреатической фауне и в капиллярных ходах проходят свое
развитие (Nicholls, 1946). Они вторично вселяются в пещерные водоемы и
колодцы. Недавно найденную в Байкале Bathynella (Базикалова, 1949)
следует рассматривать тоже как вселенца из подземных ключей, бьющих
на дне озера. В водоемах поверхности земли все семейство Bathynellidae
не встречается.
Amphipoda фреатических вод представлены двумя специфическими
семействами Bogidiellidae и Hadziidae, а также двумя семействами, насе-
ляющими и другие типы водоемов, — Ingolfiellidae и Gammaridae. К каж-
дому из первых трех семейств принадлежит по одному роду, к Gammaridae —
два рода: Niphargus со многими видами и Niphargopsis с несколькими видами.
Isopoda фреатической фауны относятся к трем семействам — двум специ-
фическим (Microparasellidae и Microcerberidae) и одному неспецифическому
(Asellidae).
Из этого краткого перечня можно сделать прежде всего заключение
о чрезвычайном своеобразии фреатической фауны, поскольку большинство
составляющих ее видов принадлежит к специфическим родам и даже се-
мействам, не известным ни в море, ни в водоемах поверхности земли.
Совершенно естественным представляется вопрос о том, следует ли
считать фреатическую фауну происходящей от морской или от пресноводной
фауны. Для решения этого вопроса необходимо в первую очередь рассмот-
реть родственные отношения и распространение неспецифических фреати-
ческих родов и семейств.
Шелленберг (Schellenberg, 1933) подробно разобрал положение в си-
стеме и родственные отношения единственных присущих фреатической
фауне родов из сем. Gammaridae — Niphargus, широко распространенного
и в пещерных водоемах, и Niphargopsis. Оказалось, что эти роды весьма
близки к двум морским родам — Eriopisa и Eriopisella. Eriopisella приспо-
соблена к жизни между песчинками морского мелководья. Оба названных
морских рода, подобно подземным, лишены глаз.
Другой неспецифический подземный род — Ingolfiella—был первона-
чально описан в качестве нового рода Balcanella, принадлежащего к новому
семейству Balcanellidae. Герцог (Hertzog, 1935) установил, что Balcanella
тождественна давно известному роду Ingolfiella 1 Этот последний включал
два вида: один (I. littoralis Hansen) был найден среди кораллов в Сиамском
заливе, другой (I. abyssi Hansen) — на глубине 3000 м у ^хода в Дэвисов
пролив (59° с. ш., 51° з. д.). Третий вид, как выяснилось, населяет фреати-
ческие пресные воды Югославии. Род Ingolfiella настолько своеобразен, что
выделяется не только в особое семейство, но даже в особый подотряд (Ingol-
fiellidea) отряда Amphipoda.
1 Этот род изображен в работе В. Н. Беклемишева (1944, с. 204, рис. 107, Д), но оши-
бочно назван Caprella linearis.
93
Рис. 1. Общий вид представителей рода Microcharon
а — Microcharon latus; б — М. teissieri; в — М. stygius (айв по: Кагатап, 1933;
б — по: Levi, 1950)
Для фреатических Isopoda до сих пор не удавалось установить таких же
тесных связей с морской фауной, как для Amphipoda. Правда, фреатические
семейства Microparasellidae и Microcerberidae по ряду признаков прибли-
жаются к морским семействам (Microparasellidae — к Desmosomatidae и
Macrostilidae, Microcerberidae — к Anthuridae), но достаточно четко от-
граничены от них, чтобы считаться самостоятельными (Кагатап, 1940).
В настоящее время возможно с несомненностью доказать тесную связь
сем. Microparasellidae с морской фауной.
В конце 1950 г. появилась небольшая статья «Леви (Levi, 1950), посвя-
щенная описанию нового рода равноногих ракообразных Duslenia, обнару-
женного в море близ Роскова в пробе крупного песка и, очевидно, живущего
в промежутках между песчинками. «Леви затрудняется отнести этот род
к какому-либо известному семейству и лишь указывает на близость Duslenia
к американскому морскому роду Ectias Richardson.
Ознакомление с описанием и рисунками найденной «Леви мелкой (1,5—
2,5 мм) депигментированной и слепой формы не оставляет никаких сомнений
в том, что она принадлежит к сем. Microparasellidae и к одному из двух
родов этого семейства — роду Microcharon Кагатап. Все родовые признаки
Duslenia и Microcharon полностью совпадают, и название первого рода
должно отныне считаться синонимом Microcharon.
Из трех югославских фреатических видов Microcharon [описание четвер-
того фреатического вида из Румынии (Chappuis, 1942) осталось недоступным
для меня] морской вид М. teissieri (Levi) по одним признакам приближается
к М. latus Кагатап, по другим — к М. stygius Каг., занимая между ними
среднее положение. Третий югославский фреатический вид — М. profundalis
Каг. — стоит от них несколько особняком.
К М. latus морской вид близок по своим широким грудным сегментам.
94
Рис. 2. Детали строения конечностей у видов Microcharon
а—г — М. teissieri; д и з — М. stygius; е и ж — М. latus; а и д — I переопод, б и е — I плео-
под ($, в и ж — II плеопод d, г и з — уропод (а—г — по: Levi, 1950; д—з — по: Кагатап,
1933)
Однако различия между этими видами весьма значительны: у М. teissieri
в отличие от М. latus голова вдвое длиннее первого грудного сегмента,
свободный брюшной сегмент хорошо выражен, протоподит уроподов не
короче плеотельсона, а эндоподиты уроподов короче протоподитов. По всем
указанным признакам (кроме ширины грудных сегментов) морской вид пора-
зительно напоминает фреатического М. stygius Каг. (рис. 1,2).
Таким образом, Microparasellidae сначала были найдены в фреатических
водах, а затем под другим названием были описаны по находкам из моря;
Ingolfiellidae, наоборот, сначала стали известны из моря, а затем под другим
названием описаны из фреатических вод. Однако в обоих случаях связь
морской и фреатической фауны выступает чрезвычайно рельефно.
Эта связь достаточно ясна и для Niphargus и Halacaridae. Весьма воз-
можно, что при последующих исследованиях и другие специфические фреати-
ческие группы окажутся близко родственными морским. По всей вероятности,
большинство представителей фреатической фауны имеет морское происхож-
дение.
Это, разумеется, не означает, что в составе фреатической фауны совер-
шенно нет видов пресноводного генезиса. Так, например, сем. Asellidae,
95
представленное в фреатических водах родами Stenasellus и Asellus, в настоя-
щее время совершенно чуждо морю. То же относится к большинству Ostra-
coda и Harpacticoda и к некоторым водяным клещам.
Что касается Bathynellidae, то Шаппюи (цит. по: Nichols, 1946) приписы-
вает им также морское происхождение, указывая на отсутствие этого
семейства в пресных водоемах поверхности земли и на существование близ-
ких морских семейств в пермское время. Однако другие Anaspidacea сейчас
обитают в пресных водах Австралии и Тасмании, и если в какой-то степени
и можно согласиться с морским происхождением Bathynellidae, то переход
этих рачков из моря в пресные грунтовые воды надо относить к весьма
отдаленным временам.
Процесс переселения животных из моря непосредственно в грунтовые
пресные воды, а не через наземные пресноводные водоемы, можно предста-
вить себе следующим образом (Бирштейн, 1940а; Schellenberg, 1935). Еще
в море организмы вырабатывают приспособления к жизни между песчинками
морского мелководья; при этом размеры их уменьшаются, тело удлиняется,
теряются глаза, развиваются органы осязания и обоняния. Все указанные
особенности свойственны морским видам Ingolfiella и Microcharon, а также
родам Eriopisa и Eriopisella. Внедрение из морских грунтовых вод в пресные
несомненно облегчается тем, что при чередовании приливов и отливов моря
морские грунтовые воды то наступают на пресные, то, наоборот, отступают.
Таким образом, под землей создается зона постоянной смены вод разной
солености, а такие условия, как известно, способствуют приобретению
морскими животными широкой эвригалинности. Ставшие эвригалинными и
уже приспособленные к жизни в капиллярных ходах организмы легко
заселяют систему грунтовых пресных вод.
Учитывая слабую степень обособленности части фреатической фауны
от морской (наличие общих родов Ingolfiella и Microcharon), можно допу-
стить, что этот процесс происходит и в наше время или происходил совсем
недавно. Однако он мог протекать и в предшествующие геологические
эпохи, и морские родоначальники некоторых современных фреатических
форм в море могли исчезнуть. Таким образом легко объясняется существо-
вание в фреатических водах своеобразных специфических форм, проявляю-
щих отдаленные родственные связи с морскими видами.
Следует в заключение подчеркнуть явное преобладание среди фреати-
ческих животных морского происхождения высших ракообразных. Как
известно, эта группа благодаря слабой проницаемости своих покровов и
совершенству своего осморегуляторного аппарата легко вырабатывает широ-
кую эвригалинность и в большей степени, чем другие беспозвоночные, спо-
собна к вселению из моря в пресные воды. Поэтому высшие ракообразные
составляют большинство видов морского происхождения как в подземных,
так и в наземных пресноводных водоемах.
96
ЗООГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
ПОДЗЕМНОЙ ФАУНЫ СОВЕТСКОГО СОЮЗА*
Огромная территория СССР до настоящего времени изучена с точки
зрения спелеологии очень неравномерно. Относительно полные сводки, полу-
ченные для Западного Закавказья и Крыма (Бирштейн, 1950, 19636), в по-
следние годы были дополнены данными по другим областям нашей страны:
Средней Азии, Дальнему Востоку, Уралу и Алтаю. В пещерах и подземных
водах было обнаружено около 470 видов животных, из которых по крайней
мере 187 могут считаться троглобионтными (Бирштейн, Левушкин, 1967а, б).
Эти виды чрезвычайно различаются по их распространению и происхож-
дению. Троглобионтные виды, встреченные в СССР, с учетом их связей
и ареалов родов, к которым они относятся, можно отнести к пяти группам:
1) виды, относящиеся к циркумсредиземноморской фауне; 2) виды, отно-
сящиеся к фауне Восточной Азии; 3) виды, относящиеся к комплексу
Северной Америки—Евразии; 4) виды, относящиеся к широко распростра-
ненным подземным родам; 5) виды родов, эндемичных для каждого карсто-
вого района. Некоторые Из этих групп могут быть подразделены на под-
группы.
ЦИРКУМСРЕДИЗЕМНОМОРСКИЙ КОМПЛЕКС
Большинство подземных видов Закавказья и Крыма и многие виды из
подземных вод Средней Азии относятся к родам, характеризующимся
циркумсредиземноморским распространением или обитающим на среди-
земноморском побережье Европы. Эти виды четко распадаются на следую-
щие подгруппы: а) морского происхождения, б) древнепресноводных и
в) наземных.
К первой подгруппе могут быть отнесены еще мало известные форами-
ниферы из интерстициальных вод Средней Азии, обнаруженные в колодцах
пустыни Каракум (Бродский, 1928, Николюк, 1948), в озере пещеры Каптар-
Хана (Бирштейн, Левушкин, 1965а), в котором имеется форма Miliammina
sp., близкая формам, обитающим в подземных водах Сахары (Gauthier-
Lievre, 1935). Из ракообразных в этой подгруппе присутствуют виды родов
Ectinosoma, Nitocra и Nitocrella (Harpacticoida), отмеченных в Закавказье
и Средней Азии, Microcharon (Isopoda) и Niphargus (Amphipoda). Род
Microcharon включает 20 видов, 5 подвидов и 2 формы, до сих пор не опре-
деленные до вида. Они найдены в Югославии, Болгарии, Румынии, юго-
восточной части Франции, на Корсике, в Сирии, в интерстициальных
морских водах Ламанша и средиземноморского побережья Франции и
Италии, в интерстициали побережья озера Иссык-Куль, в подземном соленом
озере пещеры Каптар-Хана и в западном Закавказье, но отсутствуют
в Северной Африке. Богатый видами род Niphargus также отсутствует
в Северной Африке и широко распространен почти во всей Европе, за исклю-
чением Иберийского полуострова. В СССР он обитает на Кавказе, в Крыму,
на Западной Украине, в Молдавии. На востоке зона его распространения
ограничена заливом Кендерли Каспийского моря.
* Опубликована только на французском языке в Румынии: Livre du centenaire
£mile G. Racovitza 1868—1968. Bucarest: Ed. Acad. Rep. Soc. Roum., 1970,
p. 211—221. Пер. О. И. Чибисовой.
7 Заказ 333 97
Морское происхождение может быть приписано креветке Troglocaris
anophthalmias Kollar, широко распространенной в Югославии и Западном
Закавказье и обнаруженной на юге Франции.
К подгруппе пресноводных видов циркумсредиземноморского комплекса
относятся многочисленные представители подземной фауны Закавказья,
Крыма и Средней Азии, из которых следует упомянуть закавказскую
пиявку Erpobdella absoloni ratchaensis Kob. (Кобахидзе, 1958); среди
моллюсков виды родов Horatia, Paladilhiopsis, Belgrandiella и Plagygeyeria
из Западного Закавказья (Старобогатов, 1962) и не определенный до вида
представитель рода Paladilhiopsis из Крыма; среди копепод 4 вида рода
Speocyclops из Закавказья и Крыма (Боруцкий, 1965), Ceuthonectes serbicus
Chappuis из Закавказья; среди изопод 4 вида рода Proasellus из Закавказья
(Бирштейн, 1967) и Stenasellus asiaticus Birst, et Star, из восточной Турк-
мении.
Некоторые из этих видов связаны с тропической фауной. Так, виды рода
Stenasellus населяют подземные воды Южной Европы, Средней Азии и
Экваториальной Африки. Род Speocyclops, поданным Kiefer (1937), близок
роду Bryocyclops Экваториальной Африки, Индии, Явы и Новых Гебридских
островов.
Вопрос о происхождении Bathynellacea до сих пор не имеет однознач-
ного ответа (будем ли мы рассматривать их как очень древнюю пресноводную
группу или припишем им морское происхождение). Три вида из этого отряда,
широко распространенные в Европе — Bathynella natans Wejd., В. stammeri
Jakobi и Parabathynella stygia Chappuis — обнаружены в Западном Закав-
казье (Бирштейн, «Левушкин, 1967а, 19686). Подвид, существенно отличаю-
щийся от В. stammeri, был также обнаружен в Средней Азии в интерсти-
циальных водах на берегу оз. Иссык-Куль (Янковская, 1964).
Наземные виды циркумсредиземноморского комплекса населяют пещеры
Закавказья и Крыма, но полностью отсутствуют в пещерах Средней Азии,
отличаясь в этом отношении от видов циркумсредиземноморских родов
морского и пресноводного происхождения. Сюда можно отнести закавказ-
ских ложных скорпионов из рода Neobisium; пауков-сенокосцев из рода
Nemaspela, близкого роду Nemastoma (Silhavy, 1966), один вид которых
был описан из Закавказья, а другой из Крыма; 5 видов пауков из рода
Nesticus и один вид из рода Troglohyphantes (Пичка, 1965) из Западного
Закавказья и два вида закавказских жужелиц из рода Duvalius (Левушкин,
1963).
Виды этого комплекса таксономически изолированы от видов Западной
Европы в разной степени. В Средней Азии лишь самостоятельные виды.
В Крыму, кроме самостоятельных видов, описан также подвид Niphargus
tatrensis tauricus Birst. (Бирштейн, 1964). В Закавказье были найдены
формы, идентичные обитающим в Западной Европе (Ceuthonectes serbicus
Chappuis, Bryocamptus zchokkei balcanicus Kiefer), многочисленные под-
виды известных в Европе видов (Erpobdella absoloni,Echinocamptus george-
vitschi, Nitocrella hirta, Niphargus puteanus, N. ablaskiri, N. kochianus,
N. longicaudatus, Troglocaris anophthalmus и др.) и большое количество
самостоятельных видов. Пещерный кавказский паук Nesticus borutzkyi
Reimoser недавно был обнаружен в Турции (Wiehle, 1963).
ВОСТОЧНОАЗИАТСКИЙ КОМПЛЕКС
Эта группа включает многие виды из малоизученных пещер и подземных
вод Дальнего Востока. С мейьшей степенью уверенности сюда можно
отнести представителей родов Bathynella и Eobathynella, обитающих на
Дальнем Востоке, в Средней Азии и в оз. Байкал. Как и виды циркумсреди-
земноморского комплекса, они могут быть подразделены на группы видов
морского, пресноводного и наземного происхождения.
Первые представлены единственным видом, Mackinia continentalis Birst,
et Ljov., из пещеры в бассейне р. Партизанская (Бирштейн, Левушкин,
19656). Близкий вид М. japonica Matsumoto был ранее обнаружен в под-
земных водах Хонсю, Сикоку и Кюсю. Недавно подвид М. japonica и еще
два вида этого рода были обнаружены в Южной Корее (Matsumoto, 1967).
Род относится к семейству Janiridae.
Подгруппа пресноводного происхождения в этом комплексе представ-
лена частично родом Pseudocrangonyx, обнаруженным в источниках бас-
сейна Амура, на Камчатке и на Сахалине; другие виды этого рода обитают
в подземных водах Японии и Китая. Кроме того, в источниках бассейна
Уссури обитает Asellus (Mesoasellus) setiferus Birst, et Levan. (Бирштейн,
Леванидов, 1952), близкий группе подземных японских видов, которых
Matsumoto (1962) выделяет в самостоятельный род Nipponasellus.
Среди наземных форм, обнаруженных в пещерах Дальнего Востока,
но еще не определенных, могут быть упомянуты некоторые Trechini (в особен-
ности один вид, по-видимому, из рода Trechiama), относящиеся к японским
троглобионтам.
КОМПЛЕКС СЕВЕРНОЙ АМЕРИКИ —ЕВРАЗИИ
Этот комплекс включает только два рода (один морского происхождения,
а другой наземный), которые встречаются только в СССР и США. Род мок-
риц Caucasonethes был создан для вида С. borutzkyi Verhoeff, обитающего
в одной из пещер в окрестностях Кутаиси (Закавказье). Четыре других
представителя этого рода были обнаружены позже в Виргинии, Теннесси,
Техасе и Орегоне (Vandel, 1965; Боруцкий, 1962а). Второй род, Stygobromus
(Amphipoda) включает 12 видов, в том числе S. pusillus (Mart), обитающего
в глубинах Телецкого озера (Алтай) и еще не описанный вид, который
был найден в источнике в бассейне оз. Байкал. Остальные 10 видов Stygo-
bromus известны из подземных вод разных районов США.
Сходное распространение отмечается и у некоторых водяных осликов.
Зрячие виды, обитающие на небольших глубинах озера Байкал (Baic^la-
sellus), по некоторым признакам сближаются со слепыми видами из пещер
Виргинии (Pseudobaicalasellus). Это сходство представляет собой морфо-
логическую конвергенцию (по: Henry, Magniez, 1968), но пример Caucaso-
nethes и Stygobromus допускает и другое толкование этого явления.
Объяснение подобных связей сопряжено со значительными трудностями.
7*
99
ВИДЫ ШИРОКО РАСПРОСТРАНЕННЫХ РОДОВ
К этой группе относятся виды морского и пресноводного происхождения
и наземные виды. К морским видам, по-видимому, относится Bogidiella
ruffoi Birst, et Ljov. (Amphipoda), описанная нами (Бирштейн, Левушкин,
1968а) из термального источника Хаджа-Кайнар в Восточной Туркмении.
Род Bogidiella включает 15 видов и 3 еще не определенных формы. Он
был обнаружен в Европе, в Мексике и в западной Индии, в Бразилии,
в Алжире, в Израиле и на о-ве Реюньон. Некоторые виды этого рода живут
в морских интерстициальных водах.
К группе пресноводного происхождения относятся два других рода ши-
роко распространенных амфипод, Synurella и Crangonyx, не имеющих связи
с морской фауной. Первый из этих родов представлен пятью видами в евро-
пейской части СССР, тремя видами на Кавказе, двумя видами в дельте
Лены и на Дальнем Востоке, а второй род — двумя видами в Средней
Азии, одним видом на Урале и одним — на Дальнем Востоке. Кроме
Евразии, эти два рода широко представлены в Северной Америке, а кроме
того, Crangonyx обитает и в Южной Африке. Род Parastenocaris (пресно-
водные копеподы) также характеризуется широким ареалом: два вида
этого рода были найдены в Подмосковье и на Кавказе.
Наземное происхождение имеет троглобионт, обнаруженный в пещере
Амир-Темир в Средней Азии — представитель рода Acherontidis (Collem-
bola) (Левушкин, 1972). Остальные виды этого рода обитают в пещерах
Карпат, Афганистана, Японии и Мексики.
ВИДЫ ЭНДЕМИЧНЫХ РОДОВ
Эндемичные роды троглобионтов характерны для Кавказа, Крыма и
Дальнего Востока. Они отсутствуют на Урале и Алтае, во всей равнинной
части СССР и в Средней Азии.
На Кавказе обитает единственный троглобионтный слизень, имеющийся
в мировой фауне, — Troglolestes (Левушкин, Матекин, 1965); встречаются
мокрицы Caucasoligidium, Caucasocyphoniscus, Psachonethes, Pseudobud-
delundiella (Боруцкий, 1950, 1967); амфиподы Zenkevitchia (два вида),
Anopogammarus (один вид) и Lyurella (один вид); жуки Jeannelius (три
вида), Meganophthalmus (один вид) и Cimmerites (один вид) (Курнаков,
1959; Jeannel, 1960; Левушкин, 1963, 1965); диплоподы Leucogeorgia
(один вид) и Archileucogeorgia (два вида). В пещерах Крыма были обна-
ружены: монотипический род копепод Speodiaptomus (Боруцкий, 19626),
три монотипических рода мокриц — Typhloligidium, Tauroiigidium и Tau-
ronethes (Боруцкий, 1950, 1962а), монотипический род псевдоскорпионов
Pseudoplothrus и род жуков Pseudaphaenops с двумя видами. Единственный
эндемичный род, известный в настоящее время для пещер Дальнего Во-
стока, — Epactophanoides (Harpacticoida) (Боруцкий, 1966). И, наконец,
на дне озера Байкал обитает монотипический род Baicalobathynella из
Bathynellacea (Бирштейн, Левушкин, 1967а).
Происхождение всех этих родов остается неясным. Если Jeannelius и
Meganophthalmus сближаются с некоторыми циркумсредиземноморскими
родами, остальные далеко отстоят как от местных родов, обитающих на
поверхности, так и от троглобионтных родов из других стран.
100
ОБСУЖДЕНИЕ
Попытка сгруппировать виды подземной фауны СССР по характеру
ареалов родов, к которым они принадлежат, показала, что распространение
подгрупп различного происхождения может совпадать в одних случаях и
различаться в других. Так, например, наземные роды и роды морского и
пресноводного происхождения имеют сходные ареалы на Кавказе, в Крыму
и, возможно, на Дальнем Востоке, но в Средней Азии подземная фауна
состоит почти исключительно из видов морского происхождения, относя-
щихся к циркумсредиземноморскому комплексу, и не включает ни наземных,
ни пресноводных родов (за исключением Stenasellus).
Очевидно, в некоторых случаях целые фауны, включающие роды раз-
личного происхождения, расширили свое распространение, в то время как
в других случаях такое расширение ареала оказалось возможным только
для видов морского происхождения. Возможно, группы различного проис-
хождения могли и вымирать под влиянием неизвестных нам факторов.
Исходя из сходства ареалов родов различного происхождения в циркум-
средиземноморском регионе и на Дальнем Востоке можно предположить,
что подземные фауны циркумсредиземноморского региона и Средней Азии
сначала формировались как единые, хотя и гетерогенные комплексы, и что
расширение и изменение их ареалов произошло позже.
Принадлежность Закавказья и южной части Крыма к циркумсредиземно-
морской области примечательна, и связи подземной фауны этих районов,
как и всех районов циркумсредиземноморского региона, сходны со связями
всех остальных элементов фауны и флоры наземных биотопов.
Некоторые черты подземных фаун Закавказья и Крыма характерны для
восточной части циркумсредиземноморского региона и соответствуют совре-
менным концепциям геологической истории. Подземная фауна Закавказья
гораздо богаче крымской и более тесно связана с балканской фауной —
настолько, что в ней присутствуют несколько видов, идентичных балканским,
и несколько подвидов балканских видов. Степень эндемизма крымской фауны
значительно выше, и процентное отношение эндемичных родов здесь высоко.
Анализ систематического положения видов рода Niphargus показывает, что
они более близко родственны балканским, чем закавказским, видам (Бир-
штейн, 1961а). Другими словами, связи Крыма и Закавказья с Балканским
полуостровом были независимы друг от друга и, возможно, осуществлялись
в разные эпохи.
В действительности современные геологические данные «заставляют
предположить длительное существование Крымских гор как небольшого
острова, возвышающегося над уровнем понтокаспийского бассейна с ниж-
него миоцена, который периодически соединялся с сушей — со степями
континентальной части Украины и реже — с северной частью Балканского
полуострова (Добруджа)» (Муратов, 1960, с. 178). Бедность подземной
фауны Крыма и высокий процент эндемичных видов объясняются остров-
ным характером этого района.
Что касается Западного Закавказья, трудно сказать, существовала ли
непосредственная связь через сушу с Балканским полуостровом или фау-
нистические связи этих областей осуществлялись через Малую Азию.
Единство подземной фауны восточной Азии находит естественное объяс-
нение в геологической истории этого региона. Существование связей между
Японскими островами и континентом, особенно с п-овом Корея, в конце
101
третичного и даже в начале четвертичного периода допускается большим
количеством геологов и биогеографов. Так, Линдберг (1955, с. 90) считает,
что в начале четвертичного периода «азиатский континент простирался на
восток значительно дальше современных границ Тихого океана, во всяком
случае дальше современной цепи Японских островов, Рюкю и Тайваня>.
Фауна морского происхождения, столь характерная для подземных вод
Средней Азии, представляет собой гораздо более сложную проблему.
Многие формы морского происхождения были обнаружены в солоноватом
озере (11,68 °/00) пещеры Каптар-Хана. Из них следует упомянуть форами-
нифер Milliammina sp., Trochammina sp. и Jodammina zernovi, нематод из
семейства Oncholaimidae, ракообразных Ectinosoma sp., Schizopera para-
doxa, Nitocra sp. и Microcharon halophilus. Недалеко от этой пещеры,
в термальном источнике Ходжа-Кайнар, была найдена амфипода Bogidiella
ruffoi (Бирштейн, Левушкин, 1965а, 1968а). Другой вид Microcharon —
М. kirghisicus — был обнаружен в интерстициальных водах побережья
оз. Иссык-Куль (Янковская, 1964). Кроме того, некоторые виды форамини-
фер были найдены в колодцах пустыни Каракум (Бродский, 1928; Николюк,
1948). Раковинки J. zernovi были обнаружены во многих местах в Средней
Азии, и вполне вероятно, что этот вид широко распространен в подземных
солоноватых водах этого района.
Попытки соотнести места нахождения этих видов с контурами третичных
морей не дали результатов. В палеогене район пещеры Каптар-Хана и
источника Ходжа-Кайнар был занят морем, но в начале неогена он под-
вергался мощным вертикальным колебаниям (достигающим амплитуды
12—14 км), сопровождавшимся резкими (до 60°) изменениями температуры,
что, по-видимому, исключает любую возможность выживания обитателей
подземных вод. Наиболее существенная из последовавших морских транс-
грессий, акчагыльская, остановилась в 300 км от пещеры Каптар-Хана.
Область Иссык-Куля была покрыта отступающим морем уже в каменно-
угольном периоде, и последующие трансгрессии, происходившие в эоцене и
олигоцене, не подходили к озеру ближе 500 км. Следовательно, виды родов
Microcharon и Bogidiella, как и другие виды пещеры Каптар-Хана, не могут
считаться реликтами третичного моря. Следует предположить либо их
распространение подземными водами, либо считать их реликтами гораздо
более древних морей.
Остановимся на распределении эндемичных троглобионтных видов на
территории СССР. Как уже было сказано, большинство этих родов относится
к Закавказью и значительно меньшее количество — к Крыму. Они отсут-
ствуют во всех остальных областях нашей страны, за исключением Baicalo-
bathynella, найденной в оз. Байкал, и Epactophanoides — на Дальнем Во-
стоке. Тот факт, что подземная эндемичная фауна сформировалась лишь
в этих двух местах, представляет большой интерес с точки зрения общей
проблемы происхождения подземной фауны.
По Вандел (Vandel, 1964, s. 551—552), «происхождение реликтов,
пещерных в особенности, следует искать в изменениях климата, происхо-
дивших на поверхности земного шара. Другим фактором «ухода вглубь>
поверхностной фауны является карстообразование, наблюдаемое часто
в областях распространения известняков, что приводит к лишению этих
областей воды и растительного покрова, превращению их в настоящие
пустыни, что вынуждает фауну искать убежище под землей. Что касается
102
дифференциации пещерной фауны в Южной Африке, она, по-видимому,
связана с прогрессирующим иссушением этой зоны земного шара».
Нетрудно понять, что эти выводы не подтверждаются примером нашей
фауны. Закавказье — область, наиболее богатая троглобионтами, и в част-
ности эндемичными родами, была подвергнута резким климатическим изме-
нениям в меньшей степени, чем остальная часть СССР. Согласно мнению
Вульфа (1944, с. 203), «здесь мы имеем в самом строгом смысле этого слова
климат влажных субтропиков, приближающийся к климату Средиземноморья
в первой половине третичного периода». Климатические изменения, которым
был подвергнут Крым, были выражены более резко, но они не могут ни
в какой степени быть сопоставимы с резкими изменениями, имевшими
место в конце третичного и в начале четвертичного периода в более северных
и восточных районах СССР, где тем не менее подземная фауна либо бедна,
либо отсутствует. Процесс карстообразования был более интенсивным
в южной части Крыма, чем в Закавказье, что, однако, не привело к форми-
рованию более богатой подземной фауны. Прогрессирующее иссушение,
столь характерное для Средней Азии, не привело к формированию пещерной
фауны этой области.
Чрезвычайное богатство подземной фауны Балканского полуострова
также не может быть связано с особенно резко выраженными климати-
ческими изменениями.
Таким образом, мы видим, что если различия между подземными фаунами
различных широтных зон Земли в определенной степени могут быть объяс-
нены изменениями климата (Jeanne!, 1959; Vandel, 1964), для более ограни-
ченных районов это объяснение оказывается недостаточным.
Будущие исследования должны выявить закономерности, которым подчи-
няется формирование подземных фаун в разных областях нашей планеты.
ГЛУБОКОВОДНАЯ ФАУНА ОКЕАНА
ИССЛЕДОВАНИЯ ДОННОЙ ФАУНЫ
КУРИЛО-КАМЧАТСКОЙ ВПАДИНЫ*
Летом 1953 г. «Витязь» провел исследование глубоководной фауны
Курило-Камчатской впадины ’, причем из 14 тралений 6 были произведены
на глубинах более 6000 м. Существенно отметить, что при всех тралениях
на глубинах 6000—95002 м в том или ином количестве добывались
донные животные.
Один из наиболее интересных вопросов, встающих при исследовании
фауны больших океанических глубин — выявление закономерностей верти-
кального распределения этой фауны.
Рассмотрение вертикального распространения донной фауны Курило-
Камчатского желоба с различных точек зрения (видовое многообразие,
соотношение различных групп по весу, наличие перерывов во встречаемости
по вертикали, свидетельствующее о смене видового состава и т. д.) позво-
ляет наметить несколько горизонтов, характеризующихся каждый своей,
в той или иной мере специфической донной фауной.
Прежде всего где-то между 1000 и 2000—2500 м проходит граница,
отделяющая зону континентального склона, или, по терминологии Экмана
(Ekman, 1953), архибентальную зону, от нижележащей абиссальной зоны.
К сожалению, у нас не было траловых ловов между 1000 и 2670 м. При
этом Экман справедливо отмечает, что граница эта не может быть точной, так
как в действительности это широкая переходная область, где одна фауна
переходит в другую. Это положение Экмана полностью применимо и к по-
граничным пространствам между другими глубоководными зонами.
Следующим критическим горизонтом является горизонт 5000—7000 м,
к которому приурочен ряд существенных изменений в донной фауне на пере-
ходе к еще большим глубинам — более 7000—8000 м. Таким образом, то,
что принято объединять под названием «абиссаль», следует, по нашему мне-
нию, подразделять по донной фауне на две зоны: верхнюю — абиссальную
зону (от 2000—2500 до 5000—7000 м) и зону наибольших океанических
* Тр. ИОАН СССР, 1955, т. 12, с. 345—381 (Совм. с Л. А. Зенкевичем, Г М. Беляевым).
Печатается со значительными сокр. Включена главным образом заключительная
часть раздела «Вертикальное распространение».
В настоящее время для обозначения таких форм подводного рельефа дна принят
термин «желоб», который и используется в этой и последующих статьях. (Примеч.
ред.).
В статье указана глубина 10 000 м. Впоследствии было установлено, что в действи-
тельности она была около 9500 м (см.: Беляев, 1966, с. 220) (Примеч. ред.).
104
глубин, или «ультраабиссальную» (глубже 7000—8000 м). Между ними
намечается переходная зона с некоторыми характерными элементами.
Приведенные данные о вертикальном распространении донной фауны
Курило-Камчатского желоба позволяют высказать предположение, про-
верка правильности которого будет возможна только в дальнейшем. Можно
предполагать, что какая-то часть современной фауны ложа Тихого океана
способна опускаться на глубины более 5000—7000 м и заселять более глубо-
ководную зону, а может быть, в какой-то степени и дно глубоководных же-
лобов. Однако нельзя исключать и другую возможность — формирование
на дне глубоководных желобов, на глубинах, значительно превышающих
глубину океанического ложа, своей специфической ультраабиссальной
фауны, происходившее, очевидно, с момента возникновения этих желобов.
Вероятнее всего, в природе имеют место оба эти процесса. В таком случае
ультраабиссальная фауна каждого желоба будет в той или иной степени
особой, приуроченной только к нему, так как различные желоба имеют
разный геологический возраст и разделяются значительными простран-
ствами океана, которым свойственны обычные глубины порядка 4000—
6000 м.
Насколько известно, ряд видов, обитающих на средних глубинах ложа
Мирового океана, действительно спускается и глубже — до 7000—7500 м
(Madsen, 1953). В то же время на максимальных глубинах Филиппинского
желоба, как указывает Мадсен, найдены только новые виды иглокожих —
Myriotrochus с глубины 10 200 м и Scotoplanes 1 с глубины 9790 м.
Характерно, что в тех случаях, когда фауну, собранную с максимальных
глубин Курило-Камчатского желоба, уже удалось определить достаточно
точно, она также оказалась принадлежащей к новым видам и даже к новым
родам: несколько новых видов и новый род полихет с глубин более 7000 м
(Ушаков, 1955) и новый род погонофор с глубины 9000 м и более 1 2.
Все это и заставляет предполагать существование на глубинах ниже
7000—8000 м особой ультраабиссальной фауны, состоящей в основном
из видов, способных обитать только на таких огромных глубинах и эндемич-
ных для каждого желоба.
Окончательное доказательство наличия ультраабиссальной фауны в Ку-
рило-Камчатском желобе будет получено лишь после систематической обра-
ботки собранного материала, в том случае, если окажется, что на пре-
дельных глубинах обитают особые виды, не встречающиеся как на меньших
глубинах в этом желобе, так и в других частях океана, в том числе и в других
желобах. Косвенным подтверждением вероятности высказанного предполо-
жения можно считать выводы, вытекающие из сравнения фауны пре-
дельных глубин Курило-Камчатского и Филиппинского (по: Bruun, 1951,
1953) желобов. Если придавать значение этим единичным пробам, то из
этого сравнения следует, что и состав, и соотношение систематических
групп в них различны. Возможно, и виды одних и тех же групп окажутся
разными, как этого следует ожидать в случае независимого формирования
ультраабиссальной фауны того и другого желобов.
1 В дальнейшем этот вид был переведен в род Ellipinion (см.: Hansen, 1975). (Примеч.
ред.).
2 В 1957 г. этот род был описан А. В. Ивановым под названием Zenkevitchiana с един-
ственным видом Z. longissima (Примеч. ред.).
105
Допуская возможность систематической обособленности донной фауны
каждого глубоководного желоба, не следует исключать и возможности
опускания известной части обитателей ложа океана не только на глубины до
7000—8000 м, но и на большие глубины. Весьма вероятно, что и на наиболь-
ших глубинах океанических желобов обитают как специфические ультра-
абиссальные виды, так и некоторые широко распространенные виды океани-
ческого ложа, опускающиеся на предельные глубины желобов.
ВЕРТИКАЛЬНАЯ ЗОНАЛЬНОСТЬ ПЛАНКТОНА
КУРИЛО-КАМЧАТСКОЙ ВПАДИНЫ*
Во время работ Института океанологии Академии наук СССР на экспе-
диционном судне «Витязь» в мае-июне 1953 г. в районе Курило-Камчатского
желоба нами проводились исследования макропланктона. Огромная толща
вод глубоководного Курило-Камчатского желоба позволяла поставить проб-
лему вертикального расчленения состава планктона.
Подобная задача решалась ранее для разных районов Мирового океана.
Были предложены различные схемы вертикального расчленения океана на
зоны по составу планктона. Различия между этими схемами, сведенными
недавно В. Г. Богоровым (1948), касаются слоя до глубины 1000 м.
Вся остальная толща воды, за исключением придонного 50—80-метрового
слоя, относится большинством авторов к единой глубоководной зоне, верхней
границей которой они считают 500 м. Однако еще Ло-Бианко (Lo Bianco,
1903) на основании изучения рыб делил глубины ниже 500 м на две зоны
(500—1500 и 1500—5000 м). Впоследствии это деление было принято Бекке-
ром (Наескег, 1908) для радиолярий. Наше знакомство с глубоководной
пелагической фауной дало возможность несколько иного расчленения глу-
боководной зоны.
Для выяснения неоднородности планктона водной толщи нами были ис-
пользованы сборы, сделанные сетями Богорова-Расса, диаметром 80 и 113 см,
которыми проводились вертикальные ловы с замыканием по следующим
горизонтам: 0—50, 50—100, 100—200, 200—500, 500—1000, 1000—2000,
2000—4000, 4000—6000, 6000—8500 м. Кроме того, использовались серийные
сборы горизонтально буксируемыми на разных глубинах ринг-тралами диа-
метром 160 см. Комбинация данных, полученных при помощи обоих назван-
ных орудий лова, может дать представление о составе фауны каждого
горизонта.
Предварительная обработка собранного материала позволяет предло-
жить следующую схему вертикального расчленения водной толщи Курило-
Камчатского желоба.
0—200 м — ПОВЕРХНОСТНАЯ ЗОНА
Характеризуется развитием фитопланктона и массовыми скоплениями
немногих видов: Calanus cristatus, С. tonsus, Eucalanus bungii, Sagitta ele-
* ДАН СССР, 1954, т. 24, № 2, с. 389—392 (Совм. с М. Е. Виноградовым, Ю. Г. Чиндо-
новой). Печатается с небольшими сокр.
106
gans, нередко Aglantha digitale. Из макропланктона обычны Euphausia paci-
fica, Thisanoessa longipes, Parathemisto japonica, Cyphocaris challenged,
молодь кальмаров, Dimophies arctica. Отсутствуют Mysidae, Gammaridae
(кроме Cyphocaris, challenged), Decapoda, Leptostraca. Ночью в эту зону
поднимаются из более глубоких слоев батипелагические формы: Metridia
pacifica, Candacia columbiae. Racovitzanus antarcticus, Pareuchaeta japonica,
Aetideus pacificus, Heterorhabdus tanned, Pleuromamma scutulata и др.
По числу видов и по биомассе преобладают веслоногие.
200—500 м — ПЕРЕХОДНАЯ ЗОНА
Многообразие фауны резко возрастает, а биомасса падает. Хотя основная
масса планктона состоит из форм, обитающих в поверхностной зоне, но
наряду с ними появляется ряд глубоководных видов, не поднимающихся
в поверхностную зону: молодь Hymenodora frontalis и Gnathophausia gigas,
Euprimno macropa, Scina borealis, S. incerta, larvae Lanceolidae, Pachypti-
lus pacificus, Scaphocalanus magnus, Pseudochirella spinifera, Spinocalanus
stellatus, Haloptilus pseudooxycephalus и полихета Poeobius meseres 1 По
числу видов стоят на первом месте, а по биомассе преобладают веслоногие.
По данным К. А. Бродского (1952), число видов веслоногих рачков в этой
зоне возрастает по сравнению с предыдущей почти втрое.
500—6000 м — ГЛУБОКОВОДНАЯ ЗОНА
По числу видов и по биомассе преобладают представители глубоковод-
ной фауны. Вертикальные миграции захватывают только верхнюю часть
зоны. Основная часть видов распространена в пределах всей зоны.
Глубоководная зона может быть подразделена на две подзоны.
А. 500—2000 м — верхняя глубоководная подзона. По биомассе на пер-
вом месте стоят веслоногие. Только в этой подзоне появляются взрослые
Hymenodora frontalis, молодь Н. glacialis, Gennadas borealis, Eucopia gri-
maldi, Koroga megalops, Paracallisoma alberti, Parandania boecki, Cyclocaris
guilelmi, Eusirella multicaleola, Rhachotropis notator, Scina submarginata,
S. wolterecki, взрослые Lanceola, Scypholanceola, Eukrohnia fowled,
глубоководные Nemertini, Crossota rufobrunnea, Botrynema brucei, Penta-
chogon haeckeli, Halycreas minimum, Atolla bairdii, Periphillia hyascinthina,
глубоководные Dimophiidae. В нижних горизонтах подзоны появляются
Bathycalanus bradyi, молодь Nebaliopsis typica и изредка молодь Benteup-
hausia amblyops.
Б. 2000—6000 м — нижняя глубоководная подзона. По биомассе весло-
ногие уступают щетинкочелюстным, десятиногим и мизидам. Многочисленны,
особенно в верхней части подзоны, Eukrohnia fowled. Появляются Eucopia
australis, взрослые Benteuphausia amblyops, Thysanopoda cornuta 1 2 и их
личинки, два вида Acanthephyra, Cyphocaris richardi, Halice aculeata, два
новых вида Astyra, Ctenoscina brevicaudata, Gigantocypris sp. (?), новые
1 За определение этой формы мы благодарны Л. А. Зенкевичу, который сообщил,
что Р. meseres представляет аберрантную ветвь полихет, близкую к седентарным
2 формам.
По определению Л. А. Пономаревой.
107
виды и роды веслоногих, в том числе Cephalophanes. Многочисленны крупные
Hymenodora glacialis, Scypholanceola и Botrynema brucei. Только в нижних
горизонтах подзоны появляются половозрелые Gnatophausia gigas. Исчезает
Crossota rufobrunnea.
6000 м И НИЖЕ—ЗОНА ВПАДИНЫ
Фауна чрезвычайно бедная как в количественном, так и в качествен-
ном отношении; состоит почти исключительно из Amphipoda, мелких Ostra-
coda и Copepoda. Последние, по-видимому, преобладают по числу видов
и экземпляров, а первые — по биомассе. Окраска всех обитателей этой
зоны грязновато-белая, в то время как многие обитатели глубоководной
зоны окрашены в красные тона. К этой зоне приурочено несколько специ-
фических видов Gammaridae, один из которых относится к новому семейству,
один — к роду Tetronychia (сем. Lysianassidae), один — к роду Andaniexis
(сем. Stegocephalidae) и один — к роду Hyperiopsis (сем. Hyperiopsidae).
К сожалению, характеристика этой зоны основывается главным образом
на единственном лове сетью Богорова—Расса и на одном полноценном и двух
неполноценных ловах ринг-тралами.
Рассматривая распределение отдельных глубоководных видов по перечис-
ленным зонам, можно отметить одну общую для. многих из них закономер-
ность: взрослые крупные особи, как правило, встречаются на больших глу-
бинах, чем молодь, и, следовательно, имеют менее широкое вертикальное
распределение. Таковы, например, Hymenodora glacialis и Н. frontalis из кре-
веток, эвфаузиида Benteuphausia amblyops и тонкопанцирный рачок Neba-
liopsis typica.
По мере увеличения глубины наблюдается изменение средних размеров
планктонных животных. В переходной зоне они больше, чем в поверх-
ностной, где преобладают мелкие копеподы, а в глубинной больше, чем
в переходной, так как здесь преобладают крупные щетинкочелюстные,
мизиды и десятиногие. В нижних горизонтах глубинной зоны и особенно
в зоне впадины средние размеры животных вновь резко уменьшаются.
В предлагаемую нами схему вертикальной зональности планктона хорошо
укладываются данные по вертикальному распределению глубоководных
пелагических рыб, полученные одновременно со сборами планктона и лю-
безно сообщенные нам Е. Б. Куликовой и Т. С. Рассом. В поверхностной
зоне встречаются только личинки Leuroglossus. В переходной зоне обитает
Lampanyctus leucopsarus, опускающийся до 1000 м. Сюда же из более глубо-
ких горизонтов поднимается Chauliodus sloanei. Все остальные виды встре-
чаются только в глубоководной зоне, причем Melamphaes приурочен исклю-
чительно к нижней глубоководной подзоне, a Bathylagus захватывает, по-
мимо нижней глубоководной подзоны, только нижние горизонты верхней
глубоководной подзоны.
При сравнении нашей схемы со схемами предыдущих авторов можно
видеть, что предлагаемая нами нижняя граница поверхностной зоны совпа-
дает с принимаемой Далем и Фоулером (цит. по: Богоров 1948). В. Г Богоров
(1948) проводит ее выше, на глубине 100 м, а слой 100—200 м относит
к верхнему горизонту переходной зоны. Нам кажется, что этот слой правиль-
нее относить к поверхностной зоне, так как в нем не обитают постоянно
108
глубоководные виды, а вегетация еще возможна, хотя и в незначительной
степени.
Разграничение Ло-Бианко и Геккером глубинной зоны на горизонты
выше и ниже 1500 м близко к нашей границе между двумя подзонами глу-
боководной зоны. Такое различие нельзя считать существенным, так как
применявшаяся указанными авторами и нами методика исследований не
позволяет уточнить действительное положение этой границы. То же отно-
сится и к границе между другими принимаемыми нами зонами.
ГЛУБОКОВОДНЫЕ MALACOSTRACA
СЕВЕРО-ЗАПАДНОЙ ЧАСТИ ТИХОГО ОКЕАНА,
ИХ РАСПРЕДЕЛЕНИЕ И ВЗАИМООТНОШЕНИЯ*
Во время исследований больших глубин северо-западной части Тихого
океана, проводившихся в последние годы экспедициями на советском океано-
графическом судне «Витязь», был собран богатый материал по донным
и пелагическим Malacostraca. В настояи/ее время Mysidacea и пелагические
Gammaridea обработаны полностью, Isopoda и Decapoda — в большей мере
и в меньшей степени — донные Gammaridea и Tanaidacea. Фауна глубоко-
водных ракообразных северо-запада Тихого океана характеризуется значи-
тельным разнообразием видов. Из изученного района описаны 47 видов
Mysidacea, 41 вид пелагических Gammaridea, около 30 видов изопод; это
превышает соответствующие данные по другим районам Мирового океана.
Среди исследованных глубоководных Malacostraca новые виды состав-
ляют высокий процент, включая новые роды и даже одно новое семейство
(Vitjazianidае из Gammaridea), причем отношение числа новых видов к коли-
честву уже известных возрастает с глубиной.
Высокий процент новых форм на больших глубинах в океанических
желобах северо-западной части Тихого океана — результат не только недо-
статочного знания фауны этих глубин океанов в целом, но скорее свиде-
тельство присутствия эндемиков на этих глубинах во всех изолированных
океанических желобах.
Исследование вертикального распределения пелагических Malacostraca
в Курило-Камчатском желобе было необходимым для установления границ
вертикальных зон в его водах, основанном на анализе распределения
планктонных и макропланктонных организмов (Бирштейн, и др., 1954). Ос-
новная масса пелагических гаммарид, мизид и декапод обитает в глубоко-
водной зоне (500—6000 м) с ее двумя подзонами (500—2000 м и 2000—
6000 м). Зона желоба (ниже 6000 м) населена редкой, но особой фауной,
состоящей из пелагических Malacostraca: Vitjazianna gurjanovae, Нугоп-
dellea gigas, Hyperiopsis laticarpa, Parargissa arcuata, Andaniexis subabyssi,
Halice quarta.
He все пелагические Malacostraca существуют только в одной верти-
кальной зоне или подзоне. Многие виды встречаются в огромном диапазоне
глубин, что свидетельствует о вертикальных миграциях двух типов — пище-
вых и онтогенетических. Первый тип распределения отмечен для некоторых
* Опубликована только на английском языке: XVth Intern. Congress Zool. London,
1958, Sect. Ill, Paper 33, p. 1—4. Пер. В. Я. Бирштейна.
109
мизид (Boromysis plebeja, В. curtirostris, Hyperamblyops папа); они подни-
маются питаться планктоном на глубину 200—500 м и способны возвра-
щаться на 4000—6000 м. Второй тип распределения, при котором молодые
особи встречаются на меньших, чем крупные половозрелые животные,
глубинах, наблюдался у многих видов, например у Gnathophausia gigas,
Hymenodora glacialis, H. frontalis, Nebaliopsis typica.
Географическое распределение обитателей разных вертикальных зон
сильно различается, поэтому невозможно провести общий зоогеографиче-
ский анализ и каждую вертикальную зону следует рассматривать отдельно.
Фауна ультраабиссали желобов характеризуется значительным эндемиз-
мом. Все 15 видов изопод, 20 видов амфипод и один вид мизид, обнаружен-
ные в северо-западной части Тихого океана ниже 6000 м, — новые, в то время
как для некоторых амфипод установлены новые роды. Так как Курило-
Камчатский желоб соединяется с Японским и Идзу-Бонинским желобами на
глубине 6000 м, неудивительно, что представители некоторых таких
ультраабиссальных видов, как, например, Vitjaziana gurjanovae, Hyperiopsis
laticarpa, распространяются меридионально по всей этой непрерывной цепи
желобов. В Алеутском желобе, изолированном от Курило-Камчатского на
глубине примерно 4000—5000 м, были обнаружены некоторые виды родствен-
ные, но не идентичные формам из Курило-Камчатского желоба. Глубоко-
водные желоба в других частях океана населены особыми ультраабиссаль-
ными видами, во многих случаях относящимися к тем же родам, что и ультра-
абиссальные виды Курило-Камчатского желоба. Так, например, на глубинах
более 6000 м в Бугенвильском желобе летом 1957 г. э/с «Витязь» были соб-
раны виды родов Parargissa, Andaniexis и Hyperiopsis, имеющие отдален-
ное родство с ультраабиссальными видами тех же родов из северо-западной
части Тихого океана.
Эти факты подтверждают ранее высказанное мной предположение, что
каждый изолированный океанический желоб имеет собственную эндемич-
ную ультраабиссальную фауну, которая сформировалась после образова-
ния данного желоба и частично произошла от широко распространенных
абиссальных видов.
Обитатели глубоководной (абиссальной) зоны значительно различаются
по типу географического распределения. Среди них можно выделить: (1) пан-
океанические виды, (2) атлантическо-тихоокеанские и амфибореальные
виды; (3) арктические виды, (4) биполярные виды и (5) эндемичные виды
северной части Тихого океана.
К первой группе относятся многие пелагические гаммариды, мизиды
и декаподы. Среди глубоководных донных Malacostraca нет известных
панокеанических видов. Атлантическо-тихоокеанские и амфибореальные
виды глубоководных Malacostraca очень многочисленны, особенно если к ним
добавляются эндемичные виды Тихого океана, близкородственные атланти-
ческим видам. Среди донных изопод было найдено два атлантических вида,
Haplomesus quadrispinosus и Н. angustus, так же как и несколько новых ви-
дов родов Janirella и Hydroniscus, которые ранее рассматривались как
чисто атлантические. Эти и другие примеры указывают на то, что по фауне
глубоководных Malacostraca северный район Тихого океана ближе к север-
ной части Атлантического, чем любые другие районы Мирового океана.
К арктическим элементам в фауне глубоководных Malacostraca северо-
западной части Тихого океана мы можем отнести лишь единственный вид,
110
Cyclocaris guilelmi, обитающий вдоль азиатского побережья ниже 37° с. ш.
Только одйн вид (Boreomysis inermis) можно рассматривать как биполярный
Особенно многочисленны эндемики северных районов Тихого океана.
Северотихоокеанские эндемики среди пелагических обитателей глубоковод-
ной зоны представлены только видами, тогда как роды, к которым они
относятся, имеют широкие панокеанические ареалы; среди донных глубоко-
водных ракообразных известны и эндемичные северотихоокеанские роды,
например мизиды Scolophthalmus и Holmesiella. Обилие глубоковод-
ных эндемиков в исследованном районе подтверждает существование
центров видового разнообразия и возможного видообразования многих
сублиторальных Malacostraca в северной части Тихого океана (Ekman,
1953; Гурьянова, 1951).
К глубоководным Malacostraca применима классификация, предложен-
ная А. П. Андрияшевым (1935, 1953) для рыб: древние (или истинные)
глубоководные и вторичноглубоководные формы. Среди глубоководных
Malacostraca исследованного района к древним глубоководным формам
можно отнести представителей семейств Oplophoridae (Decapoda), Lopho-
gastridae, Europiidae, Petalophthalmidae и подсемейства Boreomysini
(Mysidacea) и подотряда Asellota (Isopoda). Напротив, виды рода Scle-
rocrangon (Decapoda), трибы Erythropini (Mysidacea), роды Antarcturus
(Isopoda) — вторичноглубоководные формы. Представители обеих групп
встречаются в пределах абиссали и ультраабиссали, но по числу видов
древние глубоководные формы доминируют над вторичными в обеих зонах.
Поскольку первичноглубоководные формы характеризуются большой геоло-
гической древностью (подотряд Lophogastrida известен с карбона, семейство
Oplophoridae — с нижнего мела), то можно допустить, что абиссальная и
ультраабиссальная зоны становятся убежищем для таких примитивных
форм. Следовательно, материал, собранный нашей экспедицией, не подтвер-
ждает мнение Брууна (Bruun, 1956) о недавнем происхождении абиссальной
фауны от батиальных и сублиторальных видов. Эта точка зрения верна
только для вторичноглубоководных видов.
Особенно сложна проблема возраста ультраабиссальной фауны. Сог-
ласно геологическим данным, океанические желоба сформировались в связи
с альпийским орогенезом, т. е. на границе третичного и четвертичного пе-
риодов. Но ультраабиссальная фауна состоит не только из характерных
ультраабиссальных видов, но также из ультраабиссальных родов и даже
семейств, поэтому возраст желобов, указанный геологами, очевидно, недо-
статочен для их образования.
В пределах северо-западной части Тихого океана географическое рас-
пределение пелагических Malacostraca в глубоководной зоне характеризу-
ется резкой сменой видов на 35—40° с. ш. Эти различия очень значительны
в верхней глубоководной подзоне и совсем стираются в ультраабиссали.
Таким образом, проявление широтной зональности в распределении пелаги-
ческих Malacostraca постепенно исчезает с увеличением глубины, что обус-
ловлено гидрологической структурой глубоководных масс воды северной
части Тихого океана (Wfist, 1929).
Границы распределения идентичных пелагических Malacostraca, осо-
бенно Gammaridea, не совпадают в Тихом и Атлантическом океанах. В Тихом
океане Cyphocaris anonyx не встречается севернее 43° с.ш., a Crybelocephalus
megalurus — севернее 38° с. ш., в то время как в Атлантическом океане
111
первый вид поднимается до 65° с. ш., второй — до 57° с. ш. Наоборот, южная
граница распределения Cyclocaris guilelmi в Атлантике расположена при-
мерно на 60° с. ш., граница распределения Koroga megalops — около
63° с. ш. В Тихом океане оба вида распространены на юг до 38° с. ш. Эти раз-
личия, возможно, определяются разницей в температуре глубинных вод
в Тихом и Атлантическом океанах. На 40° с. ш. температура воды на глубине
1000—2000 м равна 2—3 °C в Тихом океане и 4—10 °C в Атлантическом.
Аналогичная разница наблюдается и в более глубинных слоях воды. Это
объясняет сдвиг на юг мест обитания относительно холодноводных видов
в Тихом и на север — относительно тепловодных видов в Атлантическом
океанах.
ГЛУБОКОВОДНЫЕ РАВНОНОГИЕ
РАКООБРАЗНЫЕ (CRUSTACEA, ISOPODA)
СЕВЕРО-ЗАПАДНОЙ ЧАСТИ ТИХОГО ОКЕАНА*
Я изучил морское дно:
Оно пустынно и темно,
И по нему, объят тоской,
Лишь таракан ползет морской.
Леонид Мартынов
ВВЕДЕНИЕ
Изучение глубоководной фауны северо-западной части Тихого океана
экспедициями на э/с «Витязь» в 1949 и в 1953—1958 гг. показало, что
равноногие ракообразные в сравнительно большом количестве экземпляров
и довольно часто встречаются на всех обследованных глубинах, включая
огромные глубины ультраабиссали. Тот же вывод можно сделать и на
основании одновременно проводившихся в других частях океана работ дат-
ской экспедиции на «Галатее». Впоследствии «Витязь» произвел сборы
равноногих ракообразных в нескольких океанических желобах южной части
Тихого океана и в Индийском океане. Этот материал, а также немного-
численные литературные данные позволяют утверждать, что Isopoda по числу
видов и особей представляют собой один из существенных компонентов
абиссальной и ультраабиссальной фауны Мирового океана. Их широкое
вертикальное и горизонтальное распространение позволяет рассчитывать
на возможность использования данных по этой группе для разработки
некоторых общих проблем, возникающих перед исследователями глубоко-
водной фауны. Это прежде всего проблемы ее формирования, особенности
адаптаций, связанных с обитанием на огромных глубинах, районирование
абиссали и ультраабиссали и т. д.
Имея в виду такую возможность, я произвел систематическую обработку
собранных экспедициями «Витязя» глубоководных равноногих ракообразных
северо-западной части Тихого океана. Материал добыт на глубинах от
1000 до 8430 м различными донными и пелагическими орудиями лова.
Немногочисленные в этом материале паразитические виды не обрабаты-
вались.
* М.: Изд-во АН СССР, 1963. 214 с. (Публикуются только введение и обобщающие
заключительные разделы книги).
112
Исследованиями охвачена акватория, ограниченная с севера 57° с. ш.,
с юга 24е с. ш., с запада 138° в. д. и с востока 174° з. д. Следует, однако,
подчеркнуть, что на этом огромном пространстве сделано весьма ограничен-
ное количество станций (Isopoda найдены на 46 станциях), распределенных
к тому же неравномерно как по горизонтали, так и по вертикали. Кроме того,
методика сбора бентоса совершенствовалась от рейса к рейсу. В начале ра-
бот, т. е. во 2-м рейсе в 1949 г. и в 14-м рейсе в 1953 г., промывка грунта из тра-
лов производилась через систему металлических сит, что, как выяснилось
впоследствии, приводило к недоучету мелких видов. В дальнейшем весь при-
носимый тралами и дночерпателем грунт промывался через крупяное сито
№ 140 (14 ячей в 1 см), и непромывавшаяся его часть целиком просматри-
валась под бинокуляром. Таким образом, удалось обнаружить значительное
количество мелких видов изопод (и других животных), к сожалению пропу-
щенных при работах 1949 и 1953 гг. в ультраабиссали Курило-Камчатского
желоба.
Указанные недостатки обработанного материала, так же как и его ограни-
ченный объем, позволяют утверждать, что полученная характеристика
фауны равноногих ракообразных северо-западной части Тихого океана
не может претендовать на исчерпывающую полноту. Эта фауна в действи-
тельности гораздо богаче, чем удалось пока установить. Последующие иссле-
дования несомненно значительно пополнят приводимый ниже список видов,
обнаруженных до сих пор «Витязем».
Это ясно также из того, что в очень многих случаях найденные виды
представлены в сборах каждый одним экземпляром. По-видимому, многие из
них встречаются сравнительно редко. Трудно представить себе, какая часть
действительно существующих видов попала в наши далеко не совершенные
орудия лова, а какая благополучно ускользнула от них или оказалась вне
зоны их действия.
Применяющийся ныне на «Витязе» способ промывки грунта и после-
дующей разборки материала из дночерпателей и тралов дает несомненно
более полное представление о донной фауне, чем практикующаяся обычно
промывка грунта через металлические сита. Преимущества принятой на «Ви-
тязе» методики ясны хотя бы из того, что в некоторых океанических
желобах, обследованных и «Витязем», и «Галатеей», первому кораблю
удалось обнаружить ряд групп мелких животных, пропущенных вторым.
Так, например, в Бугенвильском желобе мы добыли изопод и погонофор,
не найденных там работавшей до нас на тех же глубинах экспедицией
на «Галатее» (Бирштейн, Соколова, 1960).
В то же время специфика этой методики заставляет с большой осто-
рожностью оперировать полученными при ее помощи результатами. Собран-
ный на «Витязе» материал нельзя во всем его объеме сопоставлять с ма-
териалом огромного большинства других морских экспедиций, пользовав-
шихся обычными методами промывки грунта, при которой пропускаются
мелкие виды. Только датская экспедиция на «Ингольфе» обращала специаль-
ное внимание на микробентос.
Поэтому неудивительно явное сходство фаун собранных «Витязем»
мелких изопод северо-западной части Тихого океана и собранных «Ин-
гольфом» мелких изопод Северной Атлантики. Можно предполагать, что при
применении той же методики в каком-либо другом районе океана будет
получена также сходная картина. Следовательно, для зоогеографических
8 Заказ 333
113
выводов пригодны в первую очередь лишь те сравнительно крупные виды
изопод, которые добываются при обычных способах сбора материала,
а мелкие, легко пропускаемые при применении металлических сит формы,
могут привлекаться лишь в качестве дополнительного материала. Впослед-
ствии, при обработке сборов «Витязя» в других частях океана, можно будет
производить полное сопоставление всех обнаруженных видов, как крупных,
так и мелких.
Глубоководные равноногие ракообразные Мирового океана изучены еще
недостаточно. По сборам «Челленджера» описано 37 видов, встреченных
в разных частях океана глубже 914 м (Beddard, 1886). Сборы американского
«Альбатроса» в Атлантическом и Тихом океанах обработаны отчасти Гансе-
ном (Hansen, 1897), отчасти Ричардсон (Richardson, 1908—1910). По Север-
ной Атлантике имеются сведения в работе того же автора (Richardson,
1911), Тэттерсолла (Tattersall, 1906) и, наконец, Гансена (Hansen, 1916),
опубликовавшего превосходную монографию по глубоководным Isopoda,
собранным датской экспедицией на «Ингольфе». В работах Ванхофена
(Vanhoffen, 1914) и отчасти Норденстама (Nordenstam, 1933) освещена
фауна глубоководных Isopoda Антарктики. Е. Ф. Гурьянова (1933, 1935, 1936,
1946, 1955) указала и описала несколько глубоководных видов этих рако-
образных из наших дальневосточных морей и прилежащей части Тихого
океана, а также из Полярного бассейна. В недавних работах Норденстама
(Nordenstam, 1955) и Мензиса (Menzies, 1956а, Ь) содержатся сведения
о глубоководных Isopoda тропической Атлантики. Вольф (Wolff, 1956) посвя-
тил интересное исследование 12 видам, обнаруженным экспедицией на «Гала-
тее» в желобах Филиппинском, Банда и Кермадек на глубине более 6000 м.
Наконец, пишущий эти строки опубликовал данные по семействам Thambe-
matidae и Ischnomesidae, а также по родам Storthyngura и Austroniscus
с больших глубин северо-западной части Тихого океана (Бирштейн, 1957,
1960, 19616, 1962). Интересные материалы недавно опубликованной моно-
графии Мензиса (Menzies, 1962) с описанием 107 новых видов глубоковод-
ных изопод тропической и южной частей Атлантического океана, к сожале-
нию, уже не могли быть полностью использованы в настоящей работе.
По подсчету Вольфа (1956), на глубинах более 3000 м было обнаружено
всего 62 вида равноногих ракообразных (53 вида Asellota, 2 вида Flabelli-
fera, 3 вида Anthuridea и 4 вида Valvifera). На основании результатов
обработки сборов «Витязя» из северо-западной части Тихого океана коли-
чество видов Isopoda с глубин более 3000 м приходится увеличить на
58 видов, т. е. почти вдвое. Кроме того, для тропической и южной Атлантики
Мензис указывает 93 вида с глубин 3000—6000 м. Таким образом, ныне
глубже 3000 м известно уже 213 видов изопод.
Из приводимого ниже перечня собранных экспедициями на «Витязе»
в северо-западной части Тихого океана глубоководных видов равноногих
ракообразных (см. табл. 1) следует, что мне пришлось иметь дело с новой
фауной. Из 86 видов и форм в обработанных сборах только 17, т. е. менее
20 %, были известны ранее, причем в это число входят 2 пелагических вида
и 3 вида, представленных новыми подвидами или формами. Из 15 ранее
известных донных видов 9 найдено на глубинах менее 3000 м и ни один не
найден глубже 5200 м. Для Тихого океана впервые указываются (включая
ранее опубликованные мною данные) 3 семейства и 10 родов равноногих
ракообразных, 2 рода оказались новыми для науки.
114
Выводы настоящей работы частично должны рассматриваться как пред-
варительные, нуждающиеся в проверке и изменениях. На них лежит
печать начального этапа большой и трудоемкой работы по изучению глубоко-
водной фауны северо-западной части Тихого океана.
ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
Для выявления закономерностей вертикального распределения равно-
ногих ракообразных района исследования все пробы, содержавшие этих
животных, были разделены по глубинам на классы, с классовым промежут-
ком в 1000 м, и для каждого класса раздельно составлены списки видов
(табл. 1).
Как уже указывалось выше, использованный материал далеко не безу-
пречен, так как на разные глубины приходится разное число станций,
причем не на всех станциях грунт промывался через мельничный газ для
добывания мелких форм. Учитывая эти обстоятельства, можно смело утвер-
ждать, что для некоторых глубин (например, 3000—4000, 8000—9000 м)
полученные цифры количества видов сильно приуменьшены, хотя, разуме-
ется, и на более детально исследованных глубинах (5000—6000, 6000—
7000 м) в действительности обитает гораздо больше видов, чем их имеется
в наших сборах. Тем не менее, как следует из табл. 1, обработанный мате-
риал позволяет наметить некоторые особенности вертикального распределе-
ния донных Isopoda северо-западной части Тихого океана.
Анализируя эту таблицу, нетрудно заметить, что по мере увеличения глу-
бины происходят довольно закономерные изменения состава фауны: одни
семейства и роды постепенно сходят на нет, другие, наоборот, появляются
им на смену. Такая перестройка фауны зависит от относительной стено-
батности в пределах района исследования подавляющего большинства обна-
руженных родов и даже семейств. Эврибатные роды, встреченные во всем
огромном диапазоне обследованных глубин, представляют собой редкое
исключение (например, род Antarcturus Zur Strassen). Столь же эврибатные
виды в обработанном материале отсутствуют, так как все более или менее
эврибатные роды представлены на различных глубинах особыми относи-
тельно стенобатными видами.
Изменения состава фауны донных равноногих ракообразных по мере
увеличения глубины проявляются прежде всего в постепенном исчезновении
на протяжении верхних четырех километров всех подотрядов Isopoda, кроме
господствующего на больших глубинах подотряда Asellota; лишь этот под-
отряд и единственный род Antarcturus из подотряда Valvifera населяют
все обследованные глубины.
Только выше 4000 м встречены все Cymothoidae, Sphaeromidae, Anthuri-
dae, Idoteidae и Gnathiidae, а также род Astacilla из Arcturidae, причем
нижняя граница их распространения проходит, вероятно, около 3000 м, так
как между 3000 и 4000 м найден лишь один представитель всех этих
семейств (Bathycopea ivanovi). Сходное вертикальное распределение
свойственно также некоторым группам подотряда Asellota, а именно семей-
ству Janiridae и родам Munnopsis, Munnopsurus и Munnopsoides. Перечис-
ленные семейства и роды представляют относительно мелкдводный батиаль-
ный элемент в фауне донных Isopoda района исследования. Наряду
8*
115
Таблица 1. Распределение видов донных равноногих ракообразных по глубинам
Глубина, м Вид
1000—2000 (6 станций) Janiralata tricornis J. serrata * Munnopsurus laevis ' Rocinela angustata Aega plebeia Tecticeps anophthalmus * Bathycopea parallela *
2000—3000 Janiralata rhacuraeformis *
(6 станций) Acanthaspidia hanseni *
Ilyarachna longipes * Munnopsis intermedia * Munnopsoides tattersalli *
3000—4000 Janirella hirsuta *
(5 станций) J. polychaeta * Haploniscus belyaevi * Stylomesus hexaspinosus * Ischnomesus vinogradovi *
4000—5000 Haploniscus belyaevi *
(3 станции) H. hydroniscoides * Mesosignum elegantulum *
Ischnomesus andriashevi* Haplomesus quadrispinosus
5000—6000 Microthambema tenuis*
(14 станций) Janirella ornata * J. quadrituberculata * J. tuberculata *
Haploniscus belyaevi * H. hydroniscoides *
H. menziesi *
Hydroniscus vitjasi * Mesosignum brevispinis * Haplomesus brevispinis*
H. robustus* H. scabriusculus*
. Stylomesus wolffi* S. menziCsi*
S. gracilis* S. inermis pacificus* Pseudomesus similis *
6000—7000 Janirella quadrituberculata ’
(5 станций) J. fusiformis *
Hydroniscus vitjazi * Ischnomesus andriashevi* Haplomesus cornutus*
H. gigas*
7000—8000 Janirella macrura *
(5 станций) Hydroniscus vitjazi * Haplomesus gigas* Ilyarachna kussakini * Eurycope acuticoxalis * E. curticephala *
E. curtirostris *
8000—9000 Janirella spinosa *
(2 станции) Haplomesus gigas*
Calathura brachiata
Leptanthura elegans *
Synidotea pallida
S. neglecta *
Astacilla glabra
Antarcturus hirsutus
A. beddardi
Bathygnathia affinis *
Bathycopea ivanovi *
Synidotea pulchra *
Astacilla anophthalma *
Antarcturus hirsutus
Haplomesus thomsoni
Bathycopea ivanovi *
Antarcturus hirsutus
A. bathybialis *
H. insignis orientals *
Stylomesus wolffi*
S. menziesi*
Micromesus nannoniscoides *
Eurycope linearis *
Macrostylis zenkevitchi *
M. affinis *
M. curticornis *
M. reticulata *
Nannoniscus acanthurus *
N. tenellus *
N. robustus *
Austroniscus karamani*
Desmosoma distinctum *
D. coxale * .
Ilyarachna kussakini *
I. carinata *
Eurycope scabra
E. sp. (cf.~brevirostris)
Storthyngura brachycephala*
S. chelata*
Antarcturus abyssalis *
Ilyarachna kussakini *
Eurycope acuticoxalis *
Storthyngura chelata*
S. herculea*
S. bicornis*
E. magna *
Storthyngura vitjazi*
S. herculea*
S. tenuispinis*
S. tenuispinis kurilica*
Antarcturus ultraabyssalis *
Storthyngura vitjazi*
Eurycope magna *
Виды, отмеченные звездочкой, были описаны в этой работе как новые; виды, отмеченные
знаком 4-, были описаны Я. А. Бирштейном как новые из этой же части Тихого океана в его
более ранних работах 1957, I960, 1962). (Примеч. ред.).
116
с присутствием этих ракообразных для глубин менее 3000 м характерно
отсутствие ряда семейств Asellota, появляющихся только на больших глу-
бинах— Janire11 idae, Haploniscidae, Ischnomesidae, Desmosomatidae, Mac-
rostylidae и родов Eurycope и Storthyngura.
На глубине 3000—4000 м, к сожалению слабо исследованной, по-
видимому, происходит смешение относительно мелководных и относительно
глубоководных элементов. Сюда опускаются найденные и на меньших глуби-
нах Bathycopea ivanovi и Antarcturus hirsutus, но, с другой стороны, здесь
впервые появляются единичные представители глубоководных родов Jani-
rella, Haploniscus, Stylomesus, Ischnomesus, Haplomesus.
Глубины 4000—5000 и особенно 5000—6000 м населены полноценной
богатой видами типично глубоководной фауной, состоящей из представите-
лей семейств Janirellidae, Haploniscidae, Ischnomesidae, Macrostylidae, Nan-
noniscidae, Desmosomatidae, а также некоторых видов родов Ilyarachna,
Eurycope и Storthyngura и одного свойственного только этой зоне вида
Antarcturus (A. abyssalis). В особенности разнообразно население глубин
5000—6000 м, насчитывающее 34 вида.
Глубже 6000 м эта фауна несколько беднеет за счет выпадения некоторых
групп. Так, например, исчезают семейства Munnidae, Macrostylidae, Nan-
noniscidae и Desmosomatidae, из двух родов семейства Haploniscidae оста-
ется только один — Hydroniscus, из трех родов семейства Ischnomesidae —
также только один род Haplomesus. Многие роды, встреченные и выше, на
глубинах более 6000 м представлены другими видами. На глубине 6000—
7000 м взамен Janirella ornata обнаружена J. fusiformis, а на еще больших
глубинах — J. тасгига и J. spinosa, взамен Eurycope scabra — Е. acutico-
xalis, Е. curticephala, Е. curtirostris и Е. magna взамен Haplomesus brevispi-
nis, H. robustus и H. scabriusculus — Н. cornutus, а на еще больших глуби-
нах — Н. gigas. Появляются неизвестные на меньших глубинах виды рода
Storthyngura (S. bicornis, S. tenuspinus, S. herculea, S. vitjazi) и один вид
рода Antarcturus (A. uItraabyssalis).
Переход от глубин менее 6000 м к глубинам более 6000 м происходит, ви-
димо, постепенно. Во всяком случае, существуют виды, общие для обеих зон.
Таковы Janirella quadrituberculata, Hydroniscus vitjazi, Ilyarachna kussakini,
Storthyngura chelata, Ischnomesus andriashevi.
Наибольшая глубина, на которой в северо-западной части Тихого океана
были обнаружены равноногие ракообразные, равняется 8330—8340 м.
В единственном взятом здесь трале (без промывки грунта через мельнич-
ный газ) оказалось три вида, из которых два найдены и на глубине 7000—
8000 м.
Полученные данные по вертикальному распределению изопод хорошо со-
гласуются с предложенной коллективом авторов (Беляев и др., 1959) схе-
мой вертикальной зональности донной фауны Мирового океана. Переходный
горизонт между батиалью и абиссалью приходится, согласно этой схеме, на
глубины от 2500 м до 3500 м, для изопод он приурочен к глубинам 3000—
4000 м. Граница между абиссалью и ультраабиссалью (6000 м), указанная
в схеме и установленная на обработанном материале, совпадает. На равно-
ногих ракообразных можно подтвердить также некоторые различия между
верхнеабиссальной (менее 4500 м) и нижнеабиссальной (глубже 4500 м)
подзонами единой абиссальной зоны. Так, например, представители родов
Storthyngura и Macrostylis появляются только в последней подзоне, видовой
117
состав семейства Ischnomesidae в этих подзонах в основном различен,
фауна изопод нижнеабиссальной подзоны гораздо богаче, чем в верхнеабис-
сальной подзоне.
Полученную картину вертикального распределения изопод в северо-
западной части Тихого океана было бы интересно сопоставить с аналогич-
ными данными для других районов Мирового океана. К сожалению, матери-
алы для такого сравнения в литературе отсутствуют. Только в работе Бед-
дарда имеются списки обнаруженных видов равноногих ракообразных по глу-
бинам, но автор суммировал все данные «Челленджера», относящиеся к са-
мым различным районам его плавания.
В неоднократно цитированной работе Гансена по изоподам из сборов
«Ингольфа» нет специального раздела, посвященного из вертикальному
распределению. Кроме того, в Северной Атлантике, исследованной «Ин-
гольфом», отсутствуют огромные глубины, изученные «Витязем» в Тихом оке-
ане, и все нахождения изопод «Ингольфом» относятся к глубинам менее
3500 м. Тем не менее полнота сборов этой экспедиции и значительная
степень близости родового состава изопод Северной Атлантики и Северной
Пацифики позволяют сравнить между собой вертикальное распределение
одних и тех же родов в этих частях океана.
Это сопоставление можно дополнить гораздо менее многочисленными
данными по вертикальному распределению ряда широко распространенных
родов в Полярном бассейне (Гурьянова, 1946) и в Антарктике (Vanhdffen,
1914; Nordenstam, 1933). В табл. 2 приводится такое сравнение общих
родов, причем преимущественно мелководные в Тихом океане роды из подот-
рядов Flabellifera, Anthuridea и Valvifera из него исключены.
Из табл. 2 следует, что одни и те же роды в Северной Пацифике
приурочены к значительно большим глубинам, чем в Северной Атлантике.
Из 18 сопоставленных родов этому правилу не подчиняется только один
год Munnopsurus, который даже в Тихом океане не принадлежит к глубоко-
водной фауне (в Охотском море М. laevis встречается на глубинах от 80 м).
Верхняя граница обитания остальных 17 родов в Тихом океане располагается
на гораздо большей глубине, чем нижняя граница их обитания в Атланти-
ческом океане.
Этот вывод нельзя объяснить различиями в методике сборов или разной
степенью охвата исследованиями разных глубин экспедициями на «Витязе»
и «Ингольфе». Если нижняя граница обитания некоторых атлантических
видов может быть и определена недостаточно точно, поскольку «Ингольф» не
производил ловы на глубинах более 3500 м, то зато о верхней границе обита-
ния тихоокеанских видов можно судить с большей степенью достоверности,
так как «Витязем» обследованы все глубины начиная с 1000 м. Между тем,
как только что указывалось, весь диапазон вертикального распределения
атлантических представителей 17 родов лежит значительно выше этой верх-
ней границы.
По другим частям Мирового океана аналогичных данных гораздо меньше
и они не столь убедительны. Вертикальное распределение части перечис-
ленных только что родов в Полярном бассейне, судя по работе Е. Ф. Гурьяно-
вой (1946), полностью укладывается в пределы, установленные для Северной
Атлантики.
Для Антарктики можно воспользоваться также весьма фрагментарными
данными немецкой южнополярной (Vanhoffen, 1914) и Шведской антаркти-
118
Таблица 2. Вертикальное распределение некоторых родов изопод в Северной
Пацифике, Северной Атлантике, Полярном бассейне и Антарктике
Род Глубина обитания, м
Северная Пацифика Северная Атлантика Полярный бассейн Антарктика
Acanthaspidia 2940—3042 823—1353 — —
Janirella 3690—8430 873—2583 — —
Haploniscus 2940—5690 667—3321 — 385—3423
Hydroniscus 5803—7286 3388 — —
Ischnomesus 4150—6560 90—3740 — —
Haplomesus 4000—8430 1350—3690 — —
Stylomesus 3920—5690 — 698 2450
Pseudomesus 5430 1372 — —
Macrostylis 5441—6500 36—3350 — —
Nannoniscus 5461—5495 75—3366 — 385
Austroniscus 5461—5495 10—140 — 70—385
Desmosoma 5461—5495 16—3366 40—696 16—2735
Ilyarachna 2940—7230 18—3366 2500 22—3423
Eurycope 4000—7246 1—2800 698—1445 252—3423
Storthyngura 5345—8430 2150—2630 — 400—3423
Munnopsurus 950—1070 70—2583 — 3423
Munnopsis 2915-3015 75—1200 — —
Munnopsoides 2940 840—2580 — —
Bathycopea 1660—4130 364—941 — —
ческой (Nordenstam, 1933) экспедиций, имея в виду, что абиссальные
глубины не были охвачены ими и нижняя граница обитания многих глубоко-
водных родов остается еще не установленной. На основании имеющихся
данных можно заключить, что вертикальное распределение этих родов
в Антарктике и в Северной Атлантике довольно сходно, хотя в Антарктике
для некоторых из них (Haploniscus, Austroniscus, Storthyngura) можно
отметить большую степень эврибатности.
Выявление причин приуроченности одних и тех же родов к большим
глубинам в Северной Пацифике по сравнению с Северной Атлантикой и,
вероятно, с Полярным бассейном и Антарктикой наталкивается на серьезные
затруднения и в настоящее время не представляется возможным.
Для подходов к решению этого сложного вопроса следует учесть особен-
ности температурного режима разных частей океана. Район плавания «Ин-
гольфа» располагается в непосредственной близости от центра формирова-
ния глубинной холодной водной массы, лежащего, согласно Нансену,
в Гренландском море. Соответственно этому значительная часть относи-
тельно глубоководных изопод из сборов «Ингольфа» обитает при низкой
положительной или даже при отрицательной температуре. Так, например,
температурные пределы обитания Haploniscus bicuspis лежат в пределах от
— 1,1 до 0,4°, Haplomesus quadrispinosus — от —1,4 до 2,4° и т. д. Наряду
с такими стенотермными и холодолюбивыми видами в Северной Атлантике
обнаружены и гораздо более эвритермные, а также относительно более
теплолюбивые виды. При этом почти каждый политипический род вклю-
чает виды всех трех категорий, так что ни один из интересующих нас родов
не может считаться целиком холодолюбивым или целиком теплолюбивым.
Температурный режим северо-западной части Тихого океана резко отли-
чен от температурного режима Северной Атлантики. Температура на глубине
119
/ o-------------o z *--------------x J
Рис. 1. Изменение числа видов Asellota и температуры воды в зависимости от
глубины
/ — температура воды; 2 — абсолютное число видов; 3 — среднее число видов на одной
станции
от 1000 до 9000 м изменяется от 1,4 до 2,6°, причем ее минимальные значе-
ния приурочены к горизонту 4000—5000 м, а на меньших и больших глубинах
наблюдается закономерное повынгение (Богоявленский, 1955). Таким обра-
зом, глубины около 5000 м могут считаться наиболее благоприятными для
холодолюбивых видов. Выбирая из табл. 1 данные по числу видов Asellota на
разных глубинах и сопоставляя их с ходом вертикального изменения темпера-
туры, можно отметить тенденцию к обратному ходу кривых изменения темпе-
ратуры и числа видов по мере увеличения глубины в северо-западной
части Тихого океана (рис. 1). Эта зависимость выражена недостаточно четко,
так как, во-первых, максимум количества видов сдвинут на один класс по
отношению к минимуму температуры, и, во-вторых, количество видов зависит
в значительной степени от числа станций, приходящихся на тот или иной
класс глубины, а оно, как уже указывалось, различно.
Для того чтобы элиминировать эти различия, было вычислено среднее
количество видов на станцию путем деления числа видов в пределах
каждого интервала глубины на число приходящихся на данный интервал
станций. Полученная таким способом кривая относительного видового
разнообразия представляет собой довольно точное зеркальное отображение
кривой изменения температуры с глубиной, что как будто подтверждает
зависимость распределения Asellota от температуры. Следует, однако, иметь
120
в виду, что обсуждаемые различия в температуре едва превышают 1° и полу-
ченное совпадение может быть случайным.
Если все же согласиться с тем, что сопоставление рассматриваемых
кривых свидетельствует о приуроченности большинства видов глубоковод-
ных донных изопод к относительно низкой температуре, различия* в их
вертикальном распределении в разных частях океана находят некоторое
объяснение.
В самом деле, и в Северной Атлантике (в районе работ «Ингольфа»),
и в Полярном бассейне, и в Антарктике холодные воды, имеющие темпера-
туру менее 1,8°, располагаются на гораздо меньшей глубине, чем в северо-
западной части Тихого океана, и соответственно этому холодолюбивые
изоподы могут обитать на меньших глубинах, что и наблюдается в действи-
тельности. Против такого заключения можно, однако, возразить, что, судя
по данным Гансена, некоторые виды глубоководных родов Asellota приспо-
собились к существованию при более высокой температуре и, следовательно,
нет оснований считать всех глубоководных Asellota в целом стенотермной
группой, приуроченной исключительно к холодным водам. Для большинства
видов это все же, по-видимому, справедливо. Так, например, по подсчету
Вольфа (Wolff, 1956), из 85 видов пяти родов Asellota, представители
которых опускаются в ультраабиссаль, 66 % найдены при температуре
ниже 3°, из них 15 % — только при отрицательной температуре.
Тем не менее я не склонен считать температурный режим основным
фактором, определяющим особенности вертикального распределения донных
Isopoda разных частей океана. Вполне вероятно, что причины отмеченных
различий кроются в каких-то еще не известных нам факторах, может быть,
и биологического, а не физического или химического характера.
Если сопоставление положения верхней границы обитания глубоковод-
ных донных Isopoda в разных частях океана позволяет высказать некоторые
предположения, то для обсуждения положения нижней границы их обитания
материала оказывается недостаточно. Ультраабиссальные изоподы, помимо
северо-западной части Тихого океана, известны из 4 желобов — Филиппин-
ского (1 вид), Кермадекского (9 видов), Банда (1 вид) и Пуэрто-Рико
(1 вид). В последнем желобе обнаружен эндемичный род Bathyopsurus
(Nordenstam, 1955), в остальных — представители 5 широко распростра-
ненных родов, имеющихся и в северо-западной части Тихого океана (Wolff,
1956).
Род Ischnomesus в Кермадекском желобе достигает глубины 7000 м,
в северо-западной части Тихого океана наибольшая глубина его обитания
сходная — 6560 м. В Кермадекском желобе роды Ilyarachna, Eurycope
и Storthyngura найдены до 7000 м, в желобах северо-западной части
Тихого океана они опускаются несколько глубже. Зато род Macrostylis
добыт в желобе Банда на глубине 7280 м, а в Филиппинском — на глубине
9790 м, между тем как в материалах «Витязя» виды этого рода глубже
6000 м не встречены. Еще больше различия в глубине нахождения видов
рода Leptanthura: в желобе Банда — на глубине 6570 м, в северо-запад-
ной части Тихого океана — на глубине всего 1255 м; правда, в обоих
случаях было обнаружено только по одному экземпляру.
Из этих скудных данных следует, что, по-видимому, некоторые из общих
для разных желобов родов имеют приблизительно одинаковую нижнюю
121
границу обитания, другие в разных желобах доходят до различной пре-
дельной глубины. Таким образом, положение нижней границы обитания
некоторых глубоководных родов в разных частях океана может оказаться
столь же различным, как и положение верхней границы их обитания.
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ГЛУБОКОВОДНЫХ ИЗОПОД
Вопрос о морфологических адаптациях изопод к существованию на боль-
ших океанических глубинах остается почти неосвещенным. Беддард
(Beddard, 1886) не обнаружил никаких особенностей глубоководных видов,
отметив лишь их тенденцию к увеличению размеров тела и его выростов.
Последняя особенность, согласно Беддарду, зависит не столько от глубины
обитания, сколько от низкой температуры, т. е. она наблюдается также
у арктических и антарктических видов.
Врльф (Wolff, 1956), рассматривая морфологию ультраабиссальных
видов, не нашел у них существенных отличий от абиссальных, батиальных
и сублиторальных видов. Он отметил лишь тенденцию наиболее глубоко-
водных видов к редукции выростов тела, к увеличению степени обызвествле-
ния скелета и к увеличению размеров тела. Последняя особенность, ра-
зобранная также мною (1957) на примере рода Storthyngura заслуживает
специального рассмотрения. Здесь нас будет интересовать возможность
выявления на материале «Витязя» других особенностей глубоководных
видов, а также особенностей, подмеченных Беддардом и Вольфом.
Все просмотренные мною особи были депигментированы, и лишь немногие
отличались сероватой или бурой окраской. Последнее относится, в частности,
к пелагической Munneurycope murrayi и связано, быть может, с ее спо-
собностью к вертикальным миграциям огромного диапазона.
По отношению к органам зрения изопод, как заметил еще Беддард,
нельзя говорить об обязательной их редукции в условиях глубоководного
существования. Подавляющее большинство видов обработанной коллекции
полностью лишено глаз, однако отдельные виды с нормально развитыми
глазами существуют на всех глубинах. Таковы виды рода Antarcturus,
обладающие глазами независимо оттого, на какой глубине они обитают.
По строению глаз A. u 1 traabyssalis с глубины 7100—7260 м ничем не
отличается от A. beddardi с глубины 1000 м или от сублиторальных
антарктических представителей этого рода. Еще Беддард обращал внимание
на замечательное и непонятное отсутствие редукции глаз у глубоководных
видов Antarcturus, предполагая, что виды этого рода недавно приспосо-
бились к жизни на больших глубинах и глаза их еще не успели исчезнуть.
Другой пример сохранения глаз глубоководными изоподами — виды рода
Janiralata, не опускающиеся, правда, глубже батиали. Три вида из обра-
ботанной коллекции, найденные на глубине 1060, 1660 и 2280 м, имеют
глаза, развитые приблизительно так же, как у сублиторальных видов этого
рода, хотя невероятно, чтобы на глубине более 1000 м их органы зрения
могли функционировать. Весьма возможно, что эти виды также сравни-
тельно недавно перешли к обитанию на больших глубинах. То же можно
сказать и о видах родов Synidotea, Aega и Rocinela.
В то же время многие слепые виды абиссали и ультраабиссали при-
надлежат к широко эврибатным родам, включающим наряду с глубо-
ководными и мелководные виды, также лишенные органов зрения. Таковы,
122
например, роды Ischnomesus, Nannoniscus, Austroniscus, Macrostylis, Des-
mosoma, Ilyarachna, Eurycope и некоторые другие (см. табл. 2). Приме-
нительно к ним следует предполагать редукцию глаз как следствие при-
способления к жизни в толще грунта, а вовсе не как результат обитания
на больших глубинах.
Отсюда, разумеется, еще не следует, что роды, для которых характерны
глаза, сохраняют их и на больших глубинах, а слепые глубоководные
виды принадлежат к родам, лишенным глаз и на мелководье. На примере
Astacilla сеаса и A. anophthalma — единственных известных слепых видов
рода Astacilla — можно убедиться в существовании, казалось бы, естест-
венного процесса утери глаз при переходе к жизни на больших глубинах.
Этот пример, однако, нисколько не разъясняет непонятного сохранения
глаз видами рода Antarcturus, так как нет оснований считать Astacilla
более давним обитателем больших океанических глубин, чем Antarcturus.
Обращаясь к степени обызвествления покровов, следует прежде всего
отметить отсутствие точных количественных, в том числе и химических
данных поэтому вопросу. Визуальные наблюдения не позволяют согласиться
с мнением Вольфа о большей обызвествленности наружного скелета ультра-
абиссальных видов по сравнению с абиссальными. Можно привести ряд
случаев, противоречащих этому положению. Так, например, ультраабис-
сальная Storthyngura vitjazi обладает более тонкими и мягкими покровами,
чем более мелководные виды рода Storthyngura. Покровы ультраабиссаль-
ной Eurycope magna тоньше, чем абиссальной Е. scabra. Ультраабиссальный
Haplomesus gigas не отличается в этом отношении от абиссальных видов
этого рода. То же справедливо и для видов рода Janirella и Antarcturus.
Случаев, подтверждающих положение Вольфа, при сравнении видов одного
рода из материалов «Витязя» не нашлось. Если же производить сравнение
различных родов, то можно заметить, что некоторые из них, обладающие
особенно тонкими покровами, в ультраабиссаль не опускаются. Таковы,
например, роды Munnopsis, Munnopsoides и Desmosoma. Тем не менее
в составе ультраабиссальной фауны имеются виды с тонким, слабообыз-
вествленным наружным скелетом, в частности виды рода Eurycope.
Трудно также согласиться с Вольфом в том, что для ультраабиссальных
видов характерна тенденция к редукции выростов тела. Этому противоречит,
в частности, большая длина выростов тела Janirella spinosa и Haplomesus
cornutus, чем всех более мелководных видов соответствующих родов.
Адаптивное значение выростов тела изопод может трактоваться двояко.
Можно допускать, что эти выросты помогают животным не погружаться
в мягкий грунт. Возможно также, что они защищают ракообразных от
нападения их врагов, в первую очередь бентосоядных рыб, как это доказано
для «вооруженных» бокоплавов оз. Байкал (Базикалова, Талиев, 1948).
В первом случае мы не вправе ждать закономерных изменений относи-
тельных размеров выростов тела параллельно увеличению глубины, так как
характер грунта по мере увеличения глубины не меняется в каком-либо
определенном направлении. Во втором — возможна редукция выростов
тела у обитателей больших и предельных глубин, т. к. рыбы глубже
7000—7500 м не живут (Wolff, 1961а) и защищающие от них приспособ-
ления становятся там излишними, но, по-видимому, и на глубинах по-
рядка 4000—7000 м количество бентосоядных рыб весьма ограничено
и их воздействие на направление эволюции донных животных должно
сказываться в малой степени.
123
Таким образом, на материалах «Витязя», так же как ранее на материалах
«Челленджера», не удается обнаружить у изопод особенностей строения,
связанных с их существованием на больших океанических глубинах, если
не считать тенденции к увеличению размеров тела. Это отчасти согласуется
с отсутствием или во всяком случае малым количеством исключительно
глубоководных родов рассматриваемого отряда, полностью чуждых малым
глубинам.
Из 32 родов донных изопод, представленных в обработанном материале,
нет ни одного (если не считать трех описанных вновь по мелким особям —
Microthambema, Mesosignum и Micromesus) рода, который был бы известен
только из абиссали или ультраабиссали. Все они обитают и в более
мелководных зонах, причем многие из них широко эврибатны и встречаются
на всех глубинах — от сублиторали до ультраабиссали. Таковы, например,
Ischnomesus, Macrostylis, Ilyarachna, Eurycope, Leptanthura, Antarcturus.
Другие роды распространены в несколько меньшем диапазоне глубин —
от сублиторали до абиссали (Nannoniscus, Austroniscus, Desmosoma) или
в пределах батиали и абиссали (Haploniscus, Stylomesus, Pseudomesus)
и батиали и ультраабиссали (Janirella, Hydroniscus, Storthyngura) (см.
табл. 2).
Таким образом, абиссаль и ультраабиссаль населены представителями
родов, обитающих и на меньших глубинах, в том числе в значительной
части даже в сублиторали. Следовательно, для освоения больших и пре-
дельных океанических глубин изоподам не потребовалась выработка особых
новых признаков. Приспособления, возникшие у обитателей мелководных
зон, оказались пригодными для существования в абиссали и ультраабиссали.
Это в первую очередь приспособления, связанные с жизнью на мягком
грунте и в его толще. Узкое червеобразное тело Microthambema, Pseu-
domesus, Macrostylis, Nannoniscus, Desmosoma и, вероятно, а также всех
родов семейства Ischnomesidae позволяет им передвигаться в промежутках
между частицами грунта. Некоторые роды (Macrostylis, Desmosoma)
обладают копательными переоподами и, как показывают аквариальные
наблюдения Гулта (Hult, 1941) над Desmosoma, роют в грунте ходы.
Pleurogonium spinosissimus, согласно Гулту, также способен закапываться
в мягкий грунт (правда, в горизонтальном положении), несмотря на отсут-
ствие копательных ног и сравнительно короткое и широкое тело. Это
позволяет предполагать такую же способность у Janirella, Haploniscidae,
Austroniscus, Mesosignum и других изопод со сравнительно широким телом.
Другим приспособлением, имеющим, по-видимому, немаловажное зна-
чение в жизни изопод на мягких грунтах, следует считать способность
плавать, связанную со специально модифицированными задними переопо-
дами. Такие переоподы имеются у Pseudomesus, Micromesus, Desmosoma,
некоторых Nannoniscus, а также у всех многочисленных представителей
Munnopsidae. Не менее половины всех изопод абиссали и ультраабиссали
обладает способностью плавать. Лишенные такой способности Janirel 1 idae,
Haploniscidae, Munnidae и Ischnomesidae могут, вероятно, ползать по
поверхности мягкого грунта благодаря расширенному и уплощенному телу
у трех первых семейств и, наоборот, характерному для последнего семейства
удлиненному в виде палочки телу с относительно длинными ногами на
обоих его концах, благодаря чему общая поверхность сопротивления
погружению в грунт оказывается весьма значительной. Таким образом,
124
абиссальные и ультраабиссальные изоподы способны как существовать
в толще грунта, так и передвигаться горизонтально по его поверхности
или даже над ней. Это важно прежде всего потому, что таким образом
обеспечивается широкая возможность использования пищевых ресурсов
грунта.
Указанные приспособления связаны, очевидно, в первую очередь с ха-
рактером грунта, а не с глубиной обитания, поэтому они наблюдаются
и у мелководных видов эврибатных родов. Наличие таких приспособлений
было, вероятно, необходимым условием освоения больших океанических
глубин с господствующими на них мягкими грунтами.
Единственная особенность, присущая только глубоководным видам
Asellota, характерна для чуждого сублиторали семейства Haploniscidae
и заключается в способности видов этого семейства сворачиваться на
брюшную сторону. При этом они не свертываются в правильный шар,
как Armadillidae или Sphaeromidae, а скорее складываются пополам, в связи
с чем их задние грудные сегменты срастаются между собой и с плеотель-
соном. Эта особенность совершенно неизвестна для других групп Asellota,
но ее биологическое значение остается неясным.
Еще более сложным и до сих пор совершенно неясным вопросом прихо-
дится считать вопрос о том, каким образом глубоководные изоподы и другие
глубоководные животные приспособлены к таким специфическим условиям
существования на больших океанических глубинах, как повышенная радио-
активность широко распространенной в абиссали красной глины и. колос-
сальное гидростатическое давление. Согласно Кочи (Koszy, 1956), красные
глины Тихого океана обладают весьма высокой радиоактивностью; надо
полагать, что их обитатели развили какие-то специальные защитные при-
способления, позволяющие им переносить повышенные дозы излучения,
но в чем эти приспособления проявляются, мы пока не знаем. Точно
так же непонятно, как могут существовать глубоководные животные при
гидростатическом давлении, превышающем, по данным Фонтэна (Fontaine,
1930), пределы нормальной жизнедеятельности живой протоплазмы. Неко-
торое отношение к этому вопросу имеет, как мне кажется, разбираемая
ниже проблема глубоководного гигантизма, однако, поскольку он свойствен
далеко не всем глубоководным животным, мы и в данном случае должны
признать свое полное неведение соответствующих адаптаций интересующих
нас организмов.
ГЛУБОКОВОДНЫЙ ГИГАНТИЗМ
Единственной морфологической особенностью глубоководных изопод,
четко проявляющейся у многих родов и обращавшей на себя внимание
ряда авторов (Beddard, 1886; Wolff, 1956, 1960; Бирштейн, 1957), прихо-
дится считать тенденцию к увеличению размеров тела по мере возрастания
глубины. Насколько широко распространено это явление, можно видеть
при сопоставлении глубины обитания и размеров тела 18 родов, общих
для северных частей Тихого и Атлантического океанов (табл. 3).
Из табл. 3 ясно, что прямая пропорциональность л^ежду глубиной
обитания и размерами тела прослеживается у 15 родов и только у 4 родов
(Pseudomesus, Macrostylis, Desmosoma, Nannoniscus) такого соответствия
не наблюдается. Все четыре рода, являющиеся исключением, включают
125
Таблица 3. Глубина обитания и длина тела представителей одинаковых родов
в Северной Пацифике и в Северной Атлантике
Род Северная Пацифика Северная Атлантика
Глубина обитания, м Длина, мм Глубина обитания, м Длина, мм
Acanthaspidia 2940—3042 13—16,5 823—1535 12
Janirella 3690—8430 4,5—10 873—2583 4—5,5
Haploniscus 2940—5690 0,9—4 667—3321 1,5—2,8
Hydroniscus 5803—7286 12 3388 2,8
Haplomesus 4000—8430 5,1 — 15 1350—3690 2—5
Ischnomesus 4150—6560 16 90—3690 3—4,8
Pseudomesus 5430 2,5 . 1372 3,2
Macrostylis 5441—6500 2—4,2 36—3350 2,5—3,2
Nannoniscus 5461—5495 1,8—4 75—3366 1,3—3,3
Austroniscus 5461—5495 2,5—3,75 10—140 2,2
Desmosoma 5461—5495 2,3—3 16—3366 1,6—3,2
Ilyarachna 2940—7230 4—12 18—3360 2,2—10,5
Eurycope 4000— 7246 5—40 1—2800 1,5—11,5
Storthyngura 5345—8430 19—45 2150—2630 4,2
Munnopsurus 950—1070 11 — 16 75—2583 18,2—33,3
Munnopsis 2915—3015 20,5 75—1200 18
Munnopsoides 2940 6,7 840—2580 5,7
Bathycopea 1660—4130 10,3—15 364—941 4,5
мелкие формы с червеобразным телом, и возможно, что их размеры лими-
тируются диаметром капиллярных ходов в грунтах определенного характера.
Судя по данным рассматриваемой таблицы, степень увеличения размеров
с глубиной у разных родов различна. Глубоководные тихоокеанские виды
Hydroniscus, Haplomesus и Storthyngura в 4 раза крупнее более мелковод-
ных атлантических, для родов Haplomesus и Bathycopea этот коэффициент
приближается к двум-трем, для остальных родов он еще меньше. Мате-
риала для окончательных выводов, к сожалению, недостаточно, но все же
имеющиеся данные позволяют допустить, что на увеличение глубины оби-
тания разные роды реагируют по-разному.
Для того чтобы убедиться в закономерности положительной корреляции
между глубиной обитания и размерами тела, следует сопоставить между
собой длину тела половозрелых особей разных видов одного политипи-
ческого эврибатного рода, обитающих на различных глубинах. Поскольку
соответствующих данных в литературе очень мало, приходится доволь-
ствоваться просто максимальными известными размерами каждого вида,
что, разумеется, снижает точность получаемых результатов. Такое сопо-
ставление произведено мною (Бирштейн, 1957) для видов рода Storthyngura,
причем выявлена тесная зависимость между длиной тела и глубиной.
В настоящее время я могу привести еще несколько аналогичных примеров.
Среди широко эврибатных родов семейства Ischnomesidae роды Haplo-
mesus и Ischnomesus показывают строгую параболическую зависимость
размеров составляющих их видов от глубины обитания (рис. 2). Род
Stylomesus не проявляет такой тенденции, но число относящихся к нему
видов весьма ограничено и, может быть, недостаточно для определенных
заключений.
Для видов рода Janirella и Haploniscus ясной картины не получилось,
126
хотя самые крупные виды этих родов приурочены в общем к большим
глубинам, чем самые мелкие. Материал по обоим родам недостаточен
отчасти из-за того, что Ричардсон в своих работах не указывает размеров
описанных ею видов.
Виды рода Macrostylis показывают четкую зависимость длины их тела
от глубины обитания, причем их размеры возрастают с глубиной в гораздо
меньшей степени, чем размеры видов некоторых других родов, в частности
рода Haplomesus (рис. 2).
Для родов Eurycope и Ilyarachna можно отметить тенденцию к увели-
чению размеров по мере увеличения глубины, но вариации длины тела
многих видов, обитающих на сходных глубинах, во многих случаях очень
велики, и разброс точек на графике настолько значителен, что линейная
интерполяция разбираемой закономерности невозможна (рис. 3). Это, веро-
ятно, частично связано с искусственностью рода Eurycope, объединяющего
несколько пока не разделенных естественных родов. Длина тела видов
рода Storthyngura закономерно возрастает с глубиной (Бирштейн, 1957).
Исключением из всех проанализированных указанным способом родов
оказались роды Antarcturus и Leptanthura. Длина относящихся к ним
видов варьирует в широких пределах, но никакой зависимости от глубины
обитания не обнаруживает. Так, например, наиболее крупный вид первого
рода A. oryx Zur Strassen, достигающий длины 60 мм, обитает на глубине
всего 439—450 м (Zur Strassen, 1902).
Тенденция к увеличению размеров тела по мере увеличения глубины
прослеживается не только при сравнении разных видов одного рода, но и при
сопоставлении особей одного эврибатного вида, пойманных на разных
глубинах, как это хорошо показал Вольф (1956). Так, например, длина
тела Ilyarachna antarctica из Антарктики с глубины 3397—3423 м составляет
5,3—6,3 мм, а особей того же вида из желоба Кермадек с глубины
6160—7000 м — 15,2 мм, длина тела Eurycope nodifrons из Северной Атлан-
тики с глубины 2702 м — 5,1 мм, а из желоба Кермадек с глубины 6960—
7000 м — 13—13,5 мм.
Таким образом, явление глубоководного гигантизма обнаруживается
у многих родов и видов глубоководных изопод и выступает в качестве
одной из наиболее характерных для них особенностей. Эта особенность,
однако, не может считаться обязательной для всех родов рассматриваемого
отряда. Есть роды, которым она не свойственна (Antarcturus, Leptanthura),
у других она проявляется в разной степени.
Помимо изопод глубоководный гигантизм отмечен для некоторых мизид
(Бирштейн, Чиндонова, 1958) и моллюсков (Jaeckel, 1955) и, по предва-
рительным данным, присущ некоторым амфиподам. Аналогичное явление
отмечалось для бокоплавов оз. Байкал (Базикалова, 1948). В то же
время некоторые другие группы животных не проявляют подобной тен-
денции, как это показали В. В. Мурина (1958) для сипункулид и Мадсен
(Madsen, 1961а) для Porcellanasteridae.
Последний автор склонен сомневаться в реальности существования
глубоководного гигантизма. «По мнению автора, — пишет он (с. 202),—
более вероятно, что явления глубоководного гигантизма, как его понимают
Бирштейн и Вольф, в действительности не существует (карликовость или
относительное уменьшение размеров, вызванное исключительно неблаго-
приятными условиями существования на больших глубинах в отношении
127
Рис. 2. Зависимость длины тела от глубины обитания у видов родов Haplomesus (/)
и Macrostylus (2)
Длина пг^ла, лг/и
Рис. 3. Зависимость длины тела от глубины обитания у видов рода Eurycope
доступности пищи, есть, напротив, хорошо установленное явление)».
По этому поводу следует заметить, что мне не известны данные об умень-
шении размеров животных с глубиной, в то время как противоположное
явление, судя по представленному здесь материалу, несомненно наблю-
дается, по крайней мере у изопод, а также у некоторых других ракообразных.
Выяснение причин увеличения размеров животных по мере возрастания
глубины их обитания наталкивается пока на непреодолимые трудности,
так как оно должно осуществляться экспериментально. До выработки
соответствующей методики приходится довольствоваться гипотетическими
построениями.
128
Вольф пытался объяснить глубоководный гигантизм изопод низкой
температурой глубинных вод. Он, однако, не учел особенностей изменения
температуры с глубиной, в частности замечательного явления постепенного
повышения температуры глубже 3000—5500 м, происходящего главным
образом за счет адиабатических процессов. Несмотря на повышение темпе-
ратуры с глубиной, размеры ракообразных продолжают увеличиваться.
В связи с этим, как я показал на примере рода Storthyngura, никакой
корреляции между температурой воды и размерами ракообразных не наблю-
дается.
Для бокоплавов оз. Байкал А. Я. Базикалова (1948) также не нашла
зависимости между температурой и размерами их тела. Малые размеры
мелководных видов она связывает с питанием их детритом и растительной
пищей, а также с их способностью прятаться от рыб между камнями
и под ними, а крупные размеры глубоководных видов — с их хищни-
чеством и с незначительным количеством рыб на больших глубинах. Это
объяснение неприменимо к глубоководным океаническим изоподам, так как
характер питания видов одного рода не меняется с глубиной, а коли-
чество питающихся ими рыб в пределах ультраабиссали совершенно
ничтожно, и их воздействие на донных беспозвоночных в диапазоне глубин
5000—10 000 м, по всей вероятности, близко к нулю.
Говоря о возможном воздействии пищевых ресурсов на размеры глубо-
ководных животных, следует иметь в виду, что никакой зависимости между
глубиной и количеством органического вещества в грунте в пределах
абиссали и ультраабиссали не существует. По данным М. Н. Соколовой
(1956а, 1960), в глубоководных желобах Тихого океана наблюдается чере-
дование зон размыва и зон интенсивного осадконакопления, причем грубо
сортирующие детрит животные, к которым относятся, в частности, донные
изоподы, развиваются в большом количестве только на относительно пологих
участках дна, где создаются наиболее благоприятные для них условия
питания. Такие участки в районе Курило-Камчатского желоба распола-
гаются на глубинах около 5000 и 8000 м и на предельных его глубинах.
Условия питания детритоядных животных на промежуточных глубинах
менее благоприятны.
Эти данные заставляют отвергнуть возможное толкование крупных
размеров глубоководных животных как приспособления, позволяющего им
использовать в поисках пищи более обширное пространство. Они заставляют
также усомниться в обоснованности приводимых Мадсеном объяснений
причин отсутствия гигантизма глубоководных животных.
Мадсен (Madsen, 1961) приводит мнение Мосли (Mosley, 1880), связы-
вавшего крупные размеры глубоководных животных с отсутствием на боль-
ших глубинах хищников, добавляя к этому следующее: «Скудные пище-
вые ресурсы могут определять относительно медленный рост абиссальных
животных и соответственно задержку наступления половозрел ост и.
В то же время абиссальные животные могут быть очень долговечными.
Если это так, то можно объяснить некоторые случаи так называемого
гигантизма» (с. 202). Такое построение, в свете данных М. Н. Соколовой,
требует существования чередования на разных глубинах крупных и мелких
видов соответственно чередованию зон с обильными и скудными пище-
выми ресурсами, чего в действительности не наблюдается. Поэтому оно
вряд ли может быть принято.
9 Заказ 333
129
Поскольку размеры тела многих изопод (и некоторых других животных)
меняются пропорционально глубине, а не температуре или количеству пищи,
следует выяснить, какой еще биологически активный фактор внешней
среды столь же прямо, как размеры тела, зависит от глубины, так как
воздействие самой по себе глубины обитания на живые организмы мало-
вероятно. Единственным фактором такого рода приходится считать гидроста-
тическое давление. Его физиологическое влияние на морских обитателей
изучено еще недостаточно, однако, согласно Марсленду (Marsland, 1956),
«здесь (на больших океанических глубинах. — Я. Б.) весьма вероятно прямое
физиологическое воздействие такого огромного давления. Отсюда ясно, что
давление играет активную роль в естественном отборе многих видов. .>
В опытах Фонтена (Fontaine, 1930) и Марсленда (Marsland, 1942, 1951,
1956) было показано, что гидростатическое давление оказывает мощное
воздействие на жизнедеятельность живой протоплазмы. При этом во многих
отношениях повышение давления и понижение температуры влияют на
энергетику протоплазмы в одном направлении и, быть может, сходным
образом обусловливают увеличение размеров тела. Крупные особи, согласно
Фонтену, менее чувствительны к повышению давления, что дает им известные
преимущества перед более мелкими.
Исходя из этих соображений, я предполагал, что глубоководный гиган-
тизм возникает в результате отбора, зависимого от специфического воздей-
ствия гидростатического давления на характер метаболизма (Бирштейн,
1957). Мое объяснение впоследствии было поддержано Вольфом (1960).
Это объяснение не согласуется с многочисленными примерами отсутствия
глубоководного гигантизма у ряда групп. Вопрос о том, почему одни
группы реагируют на повышение гидростатического давления увеличением
размеров тела, а другие — нет, остается открытым, так же как многие
другие вопросы, затронутые в настоящей работе.
ПИТАНИЕ
Питание глубоководных равноногих ракообразных изучено совершенно
недостаточно. Мензис (Menzies, 1956b) обнаружил в кишечнике Eurycope
sp. из желоба Пуэрто-Рико с глубины 2700—2710 м остатки скелетов
фораминифер, кокколитофорид, тинтинноидей, а также мелкие тельца,
напоминающие бактерий. В содержимом кишечника Janirella vemae из того
же лова было много предполагаемых бактерий, а остатки животных и во-
дорослей отсутствовали. Первый вид Мензис считает всеядным детритоедом.
Он допускает существенное значение бактерий в питании обоих видов.
В кишечнике Acanthocope spinosissima из Карибского моря с глубины
1244 м тот же исследователь (Menzies, 1956b) нашел, помимо мелких
телец (бактерий и простейших?), хеты полихет, спикулы губок, раковины
фораминифер и тинтинноидей. По мнению Мензиса, A. spinosissima может
поедать грунт, но также и хищничать, причем способна питаться изби-
рательно. Сходные данные получил Вольф (1956) при исследовании содер-
жимого кишечника Stortyngura pulchra из желоба Кермадек с глубины
6620 м. Были обнаружены скелеты радиолярий, хеты и одна челюсть
полихет, спикулы губок и остатки ракообразных. Вольф подчеркивает
одинаковое строение ротовых придатков глубоководных и мелководных
Isopoda. Те и другие могут быть фильтраторами, грунтоедами, в том
числе потребителями бактерий, а также хищниками.
130
Содержимое кишечников некоторых донных изопод из сборов «Витязя>
изучала М. Н. Соколова (1958). Она вскрыла 10 экз. Eurycope magna
и по 5 экз. Storthyngura herculea и Antarcturus ultraabyssalis с глубины
7140 м, а также 5 экз. A. hirsutus с глубины 1078 м и 5 Haplomesus n. sp.
с глубины 3820 м (последний вид из Берингова моря).
У Eurycope и Storthyngura около 90 % содержимого кишечников
составлял отсортированный детрит с отдельными панцирями диатомей,
остальное приходилось на долю остатков ракообразных и офиур, а также
песка. У Antarcturus ultraabyssalis с той же станции обнаружен только
детрит, притом более тонко отсортированный, чем у двух других родов.
У относительно мелководного A. hirsutus с песчанистого грунта около
половины содержимого кишечника состояло из песка, а около половины —
из особенно тонко отсортированного детрита. Наиболее тонко отсорти-
рованный детрит заполнял целиком кишечник Haplomesus n. sp.
На основании этих данных Соколова относит всех исследованных ею
изопод к группе животных, тонко отсортировывающих детрит с поверхности
грунта, причем Eurycope и Storthyngura, по ее мнению, занимают проме-
жуточное положение между животными, тонко сортирующими детрит,
и животными, грубо сортирующими детрит. В отличие от иностранных
авторов, она объясняет нахождение остатков животных в кишечниках
изопод не их хищничеством, а поеданием ими трупов.
Из двух видов пелагических изопод северо-западной части Тихого
океана только один — Munneurycope murrayi — был изучен со стороны
его питания. По данным Ю. Г Чиндоновой (1959), он представляет сббой
типичного растительноядного фильтратора. В кишечниках 10 экз. М. murrayi
с разных горизонтов оказалось большое количество остатков фитопланктона,
среди которых попадались глобигерины, тинтинноидеи, обрывки медуз и
детрит. Судя по большей степени переваренности пищи у рачков, пой-
манных в глубинных горизонтах, по сравнению с экземплярами, пойманными
ближе к поверхности, М. murrayi совершает пищевые вертикальные ми-
грации в слои, богатые фитопланктоном, а затем, наполнив кишечник,
опускается вниз. Замечательно, что, несмотря на столь своеобразный
способ питания, ротовые придатки М. murraji существенно не отличаются
от ротовых придатков близких родов.
Это обстоятельство заставляет относиться с большой осторожностью
к возможности судить по строению ротового аппарата изопод о характере
их питания. Приведенные выше немногочисленные сведения о составе
содержимого кишечников некоторых глубоководных Asellota и Antarcturus
показывают, что способы захвата пищи у исследованных видов различны:
они в разной степени способны отфильтровывать детрит, могут хватать
крупные пищевые частицы (живых животных или их трупы), могут,
наконец, фильтровать в толще воды. Между тем строение их ротовых
придатков весьма сходно, в частности все они обладают мандибулами
жующего типа с цилиндрическим зубным отростком, снабженным плоской
перетирающей поверхностью. Передний переопод многих из них хвататель-
ный; фильтрационный или сортирующий аппарат в большинстве случаев
морфологически не выражен, если не считать многочисленных щетинок
на II—IV переоподах Antarcturus. Можно предполагать, что способ за-
хвата пищи и ее характер у родов Asellota с редуцированным зубным
отростком мандибул (Macrostylis, Nannoniscus, Austroniscus, Desmosoma,
131
9*
Munnopsurus, некоторые виды Ilyarachna) должны быть особыми, но дан-
ными по этому вопросу мы пока не располагаем.
Говоря о строении ротового аппарата глубоководных Asellota, следует
отметить, что нет возможности целиком согласиться с Вольфом, утвер-
ждающим полное сходство их в этом отношении с мелководными видами.
Исследование мандибул многих глубоководных видов указывает на неко-
торые характерные для них особенности. Перетирающая поверхность
зубного отростка у глубоководных видов часто скошена и снабжена крае-
выми щетинками. У видов рода Janirella по краям этой поверхности
расположены зубцы, дсобенно хорошо развитые у наиболее глубоководных
видов. Возможно, эти приспособления облегчают захват мягкого грунта.
Поднятые вопросы требуют основательного функционально-морфологи-
ческого анализа, который приходится отложить на будущее время.
Учет характера питания равноногих ракообразных может пролить свет
на причины, вызывающие некоторые особенности их вертикального распре-
деления, описанного выше. Из табл. 1 можно видеть, что все Gnathiidae,
Anthuridae и Cymothoidae не опускаются глубже 2000 м, a Idoteidae,
род Astacilla и Sphaeromidae (за исключением Bathycopea ivanovi) —
глубже 3000 м. Судя по литературным данным, для всех перечисленных
семейств и рода Astacilla характерен в основном хищный образ жизни
(среди Idoteidae и Sphaeromidae есть и растительноядные виды), а неко-
торые их представители, например Aega и Rocinella, являются полупара-
зитами. С этими данными хорошо согласуется строение ротового аппарата
перечисленных изопод, столь отличного от ротового аппарата Asellota.
Особый интерес представляет сосущий ротовой аппарат Calathura и Lep-
tanthura, но, к сожалению, остается неизвестным, какие животные служат
их жертвами (Hult, 1941).
Таким образом, по мере увеличения глубины из состава фауны изопод
постепенно выпадают плотоядные формы, и начиная с глубины 3000—4000 м
и кончая предельными глубинами на дне обитают исключительно грунтоеды.
Такая смена способа питания характерна, по-видимому, для многих донных
животных, в то время как для планктона наблюдается, наоборот, увели-
чение количества хищников на больших океанических глубинах (Зенкевич,
Бирштейн, 1955).
РАЗМНОЖЕНИЕ
Данные о размножении глубоководных изопод в литературе отсутствуют.
Обработанные материалы также почти не позволяют осветить этот вопрос.
Из большого количества просмотренных экземпляров только два оказались
самками с яйцами в марсупиальных сумках. У самки Stylomesus inermis
pacifica длиной 4 мм с глубины около 5480 м в марсупиуме было четыре
яйца, у самки Storthyngura tenuispinis kurilica, длиной 19 мм, с глубины
7220 м — 28 яиц. В обоих случаях обращает на себя внимание малая
плодовиость глубоководных видов по сравнению с мелководными. Вольф
констатировал у самки Storthyngura novaezelandiae (Beddard) длиной
14,4 мм с глубины 2102 м 43 эмбриона, причем заметил, что, судя по
размерам марсупиума, их должно было быть вдвое больше. Следовательно,
несмотря на свои более крупные размеры, Storthyngura с глубины 7220 м
132
оказалась значительно менее плодовитой, чем представитель того же рода
с меньшей глубины.
Сокращение числа яиц у глубоководных ракообразных по сравнению
с мелководными отмечалось на примере креветки Sclerocrangon zenkevitchi
Birst, et Win. (Бирштейн, Виноградов, 1953). Возможно, это характерно
и для других глубоководных животных.
ЭПИФИТЫ И ПАРАЗИТЫ
Подавляющее большинство просмотренных особей было свободно от
паразитов и эпифитов. Только у трех видов были обнаружены эпифиты
и паразиты. На поверхности тела Bathycopea ivanovi со ст. 2209 прикреп-
лялись фораминиферы, по определению X. М. Саидовой, принадлежащие
к Haplophragmoides sphaereloculum Cushman и Cubicides ex gr. lobatulus.
Другой представитель рода Cubicides — C. ex. gr. pseudoungerianus — был
найден на дорсальной поверхности плеотельсона Astacilla anophthalma.
Бенедикт (1898) уже указывал фораминифер рода Truncatulina (=Cubi-
cides) на поверхности тела Astacilla caeca с глубины 3336 м. Не менее
интересно нахождение на конечностях и теле Antarcturus bathybialis орга-
низмов, весьма напоминающих Е1 lobiopsidae. Это семейство простейших
паразитирует на каланидах, мизидах, креветках и эвфаузиидах, но для
изопод еще не отмечалось.
Судя по этим скудным данным, эпифиты и паразиты встречаются
только на изоподах батиали и верхних горизонтов абиссали. На больших
и предельных глубинах они исчезают.
ЗООГЕОГРАФИЯ
Слабая изученность глубоководных равноногих ракообразных Мирового
океана затрудняет зоогеографический анализ обработанного материала.
Только для северной половины Атлантического океана и Индоокеанского
сектора Антарктики имеются сравнительно полные данные, да и то до
глубин порядка 3500 м. Кроме того, сравнительно подробно исследована
ультраабиссаль желоба Кермадек. Для остальной огромной акватории
Мирового океана мы располагаем лишь отдельными указаниями на нахож-
дения единичных глубоководных видов Isopoda, но отнюдь не списками
видов, дающими представление о характере фауны этих ракообразных.
Таким образом, полное представление об особенностях географического
распространения равноногих ракообразных на больших глубинах Мирового
океана в настоящее время не может быть получено. Все же можно попы-
таться определить, из каких элементов состоит фауна глубоководных Isopoda
северо-западной части Тихого океана и каковы ее родственные отношения.
Такой анализ приходится проводить на уровне родов, так как огромное
большинство донных видов, обнаруженных в северо-западной части Тихого
океана, пока должно считаться эндемичными для этой акватории. Только
6 видов (Janiralata tricornis, Haplomesus quadrispinosus, H. insignis orien-
talis, Stylomesus inermis pacificus, Eurycope sp. (aff. breviro$tris и Calathura
brachiata) были известны и вне района наших работ, но вопрос о бли-
зости тихоокеанских форм этих видов к типичным требует еще дополни-
тельных исследований.
133
В сборах «Витязя> из северо-западной части Тихого океана оказалось
32 рода равноногих ракообразных с глубины более 1000 м. По характеру
распространения они могут быть разделены на несколько групп.
1. Роды, ограниченные в своем распространении северной половиной
Тихого океана: Microthambema, Micromesus, Tecticeps. Эта группа неодно-
родна: в нее входят, во-первых, один мелководный род, лишь отдельные
представители которого проникают на батиальные глубины (Tecticeps),
и, во-вторых, описанные вновь мелкие монотипические абиссальные роды,
нахождение которых возможно при применении соответствующей методики
сборов и в других частях океана; один из них (Microthambema) близок
к североатлантическому роду Thambema.
2. Роды, общие северным частям Тихого и Атлантического океанов:
Janiralata, Pseudomesus, Bathycopea, Calathura, Bathygnathia (рис. 4).
Из них мелкий род Pseudomesus не имеет существенного значения для
зоогеографического анализа по только что приведенным соображениям.
Следует отметить, что разбираемый тип ареала присущ преимущественно
батиальным (Janiralata, Bathycopea, Calathura, Bathygnathia) родам.
3. Роды, известные, помимо северных частей Пацифики и Атлантики
(включая Полярный бассейн), также из умеренных и высоких широт
южного полушария: Acanthaspidia, Janirella, Hydroniscus, Haploniscus,
Austroniscus, Nannoniscus, Haplomesus, Stylomesus, Desmosoma, Ilyara-
chna, Munnopsurus, Munnopsoides (рис. 5, 6).
4. Роды, приуроченные преимущественно к‘Антарктике, но представлен-
ные ограниченным количеством видов в экваториальных районах Атлан-
тического и Тихого океанов и в Северной Пацифике: Antarcturus (рис. 7).
5. Роды с широким или панокеаническим распространением: Rostylis,
Eurycope, Storthyngura, Rocinela, Aega, Leptanthura, Synidotea и Astacilla.
Из них род Synidotea имеет некоторое сходство с родами первой группы,
поскольку большинство относящихся к нему видов (25) обитает в северной
части Тихого океана (Гурьянова, 1936), а род Astacilla — к родам четвертой
группы, поскольку большинство входящих в него видов приурочено к Антар-
ктике.
В советских работах по географическому распространению глубоковод-
ных животных неоднократно отмечалась необходимость раздельного
зоогеографического анализа по различным вертикальным зонам. На примере
донных равноногих ракообразных также можно видеть, что соотношение
количества родов, принадлежащих к установленным пяти группам, и отно-
сящихся к ним видов в разных зонах различно. Проводя границу между
батиалью и абиссалью по глубине 3000 м, а границу между абиссалью
и ультраабиссалю — по глубине 6000 м и используя данные табл. 1,
получаем следующие соотношения (табл. 4 и 5).
В приведенных в этих таблицах показателях можно усматривать тен-
денцию к расширению ареала родов по мере увеличения глубины, т. е.
явление, ранее отмеченное для видовых ареалов мизид (Бирштейн, Чиндо-
нова, 1958). Все же различия между вертикальными зонами в соотно-
шении групп родов с различным типом распространения не так велики,
как это можно было бы ожидать, судя по ранее опубликованным данным
по другим высшим ракообразным — пелагическим гаммаридам и мизидам
(Бирштейн, Виноградов, 19556, 1958, 1960; Бирштейн, Чиндонова, 1958).
Возможно, здесь имеет значение принадлежность изопод к бентосу, а боль-
134
90 60 30 о ’зо 60 90 120 160
60 30 0 30 60 W У ’С>?> о.-. ЛХ-' **Г. о • ,^'jj 60 30 0 30 во
: £ я* ч< ^9 *'
'я й I •’ 1;
X:
% ’ О
J-
90 60 30 0 30 60 90 120 160
Рис. 6. Местонахождения видов рода Austroniscus
Рис. 7. Местонахождения видов рода Antarcturus (s. str.)
Рис. 8. Местонахождения Haplomesus quadrispinosus (G. О. Sars)
шинства мизид и всех проанализированных с той же целью гаммарид —
к планктону.
С каждым установленным типом ареалов связан ряд важных зоогеогра-
фических вопросов. Наименее сложена характеристика первой группы,
объединяющей северотихоокеанских эндемиков.
Если ограничиться рассмотрением родов, включающих сравнительно
крупные формы, то к этой группе следует относить только Tecticeps.
К этому роду принадлежит пять видов и несколько форм. По мнению
Е. Ф. Гурьяновой (1936), изобилие видов и форм Tecticeps объясняется
недавним интенсивным видо- и формообразованием, явлением «разверты-
вания как бы вееров видов одного и того же рода в каждом из морей» (с. 31).
Кроме Tecticeps, к рассматриваемому типу примыкают роды Janiralata
и Synidotea. Центр видового разнообразия и центр расселения Synidotea
приходится, по Гурьяновой, на северную часть Тихого океана и примы-
кающие к нему дальневосточные моря; из многочисленных видов Janiralata,
обитающих в пределах той же акватории, только один (J. tricornis) распро-
странен и в Северной Атлантике.
Недавнее бурное видообразование многих сублиторальных северотихо-
океанских групп животных достаточно хорошо известно, и изоподы в этом
отношении вполне подобны амфиподам, декаподам, моллюскам, рыбам
и многим другим. Характерно, однако, что лишь немногие возникшие
в северной части Тихого океана роды приспособились к обитанию на
больших глубинах, а изоподы этой группы не проникли глубже батиали.
Среди других глубоководных изопод они представляют собой относительно
молодых вселенцев.
Обращаясь к рассмотрению второго и третьего видов ареалов, нетрудно
137
Таблица 4. Процентные отношения родов разных групп по вертикальным зонам
Зона Группа родов Общее число родов
|-я 2-Я 3-я 4-я 5-я
Батиаль 1 6,2 1 25,0 31,3 31,3 6,2 16
Абиссаль 10,5 15,8 52,6 15,8 5,3 19
Ультраабиссаль — — 62,5 25,0 12,5 .8
Таблица 5. Процентные соотношения видов, относящихся к родам разных групп
по вертикальным зонам
Зона Группа родов Общее число видов
1 -я 2-я 3-я 4-я 5-я
Батиаль 4,3 30,4 21,7 34,8 8,7 23
Абиссаль 4,0 8,0 64,0 18,0 6,0 50
Ультраабиссаль — — 45,0 50,0 5,0 20
заметить, что второй тип может быть приравнен к амфибореальному,
а третий тип — биполярному распространению, хорошо известным для
многих мелководных животных. В одной из своих предыдущих работ
(1960) я указывал на биполярное и амфибореальное распространение
семейства Ischnomesidae в целом, причем род Ischnomesus распространен
так же, как все семейство, род Haplomesus — амфибореален, а род Stylo-
mesus — биполярен. Впоследствии Ischnomesidae были обнаружены в тро-
пиках, в частности в Бугенвильском желобе (Бирштейн, Соколова, 1960).
Они представлены там новым видом рода Ischnomesus. Кроме того, в том же
желобе найдены новые виды родов Janirella, Mesosignum, Haploniscus
и Macrostylis (а также один новый род). Таким образом, распростра-
нение семейства Ischnomesidae и рода Ischnomesus не приходится считать
биполярным, однако ряд приведенных выше случаев распространения
других родов свидетельствует о существовании характерного, приходяще-
гося на тропики разрыва их ареала. Нахождение одного вида Janirella
в Бугенвильском желобе, а также двух описанных Мензисом (Menzies,
1962а) видов в Южной Атлантике отодвигает южную границу ареала
этого рода за экватор, однако пока у нас нет оснований отрицать разобщен-
ность его ареала в области Полярного бассейна. Все же трактовка
распространения родов второй и третьей групп как амфибореального и бипо-
лярного может вызвать сомнения. Возникает вопрос, насколько реален
разрыв их ареала и нельзя ли объяснить этот разрыв недостаточной
изученностью больших океанических глубин, в частности в области Поляр-
ного бассейна и тропиков. Такие сомнения могут подкрепляться уже упоми-
навшимися фактами: наличием сравнительно полных данных по изоподам
Северной Атлантики и Северной Пацифики, добытых экспедициями на
«Ингольфе> и «Витязе>, с одной стороны, а также по изоподам Антарктики,
добытых Немецкой южнополярной и Шведской антарктической экспеди-
циями — с другой.
Только будущие исследования, а также последующая обработка
138
коллекций «Витязя» из других частей океана подтвердят или опровергнут
основательность этого возражения. Следует, однако, иметь в виду, что
некоторые сведения о глубоководных изоподах тропиков в литературе
имеются. Ряд сборов произведен здесь еще «Челленджером», обширные
материалы добыты впоследствии экспедициями на «Зибоге» (Nierstrasz,
1941) и на «Веме» (Menzies, 1962а), тропические желоба исследованы
«Галатеей», некоторые данные получены экспедициями на «Альбатросе» и на
«Дана». Именно благодаря этим сведениям можно допускать панокеани-
ческое, сплошное распространение ряда родов, отнесенных выше к пятой
группе. Трудно представить себе пропуск коллекторами особей сравни-
тельно крупных и довольно часто встречающихся в других частях океана
родов, представляющихся сейчас биполярными и амфибореальными, каковы,
например, Munnopsis, Munnopsurus, Bathycopea и многие др. Вполне веро-
ятно, что впоследствии список биполярных и амфибореальных родов сокра-
тится прежде всего за счет мелких форм, но вряд ли можно сомневаться
в том, что какое-то количество таких родов действительно существует.
Ряд примеров биполярного распространения других глубоководных живот-
ных приводил Л. А. Зенкевич (1960).
Как мне уже приходилось отмечать (Бирштейн, 1960), выявление причин
возникновения разрывов ареалов амфибореальных и биполярных глубоко-
водных животных наталкивается на серьезные затруднения.
По отношению к мелководным морским организмам предложены гипо-
тезы, удовлетворительно объясняющие их амфибореальное и биполярное
распространение (Берг, 1934а, 1947а), но эти построения нельзя перенести
на глубоководную фауну. Согласно Л. С. Бергу, биполярное распростра-
нение организмов возникло в результате похолодания в ледниковое время,
при котором ареалы некоторых бореальных (и субтропических) видов север-
ного полушария сместились на юг, и эти виды получили возможность
пересечь недоступную для них ранее тропическую зону. Последующее
повышение температуры уничтожило их в области тропиков, и, таким
образом, возник характерный для биполярных видов разрыв ареалов.
Глубинные виды в отличие от поверхностных не меняли температуру
своего обитания. На основании изучения соотношения изотопов кислорода
в раковинах фораминифер из донных отложений Эмилиани (Emiliani,
1955) показал, что в течение четвертичных оледенений температура глубин-
ных вод в экваториальной части Тихого океана была такой же, как ныне;
правда, в экваториальной части Атлантического океана она понижалась
на 2,1°
Кроме того, учет температурного режима глубинных вод не только
прошлого, но и настоящего времени ничего не дает для понимания
особенностей распространения глубоководных животных. Температура глу-
бинных вод тропической зоны, как известно, такая же, как и глубинных вод
умеренной зоны. Поэтому нет никаких оснований для признания сущест-
венного значения температуры как фактора, ограничивающего горизон-
тальное распространение донных глубоководных животных. Между тем
среди них имеются не только биполярные формы, но и виды и роды,
строго приуроченные к тропической зоне (Виноградова, 1955; Зенкевич,
1960). В обоих случаях природа преград, препятствующих дальнейшему
расселению этих животных, остается неизвестной. По отношению к циркум-
тропическим животным Н. Г. Виноградова писала: «. . остается непонятным,
139
что не позволяет этим формам проникать в более высокие широты, где
условия существования для них, казалось, благоприятны» (с. 62). Столь же
непонятно, что препятствует биполярным глубоководным животным рассе-
литься в низкие широты и сомкнуть свой разорванный ареал. Таким
образом, на больших глубинах при отсутствии столь характерной для
поверхностных вод поширотной смены температуры, наблюдается все же
широтная зональность в распределении донной фауны, очевидно не свя-
занная с температурным режимом.
Амфибореальное распространение мелководных морских животных,
согласно Л. С. Бергу (1934а), объясняется миграцией бореальных форм
в плиоценовое и послеледниковое время, т. е. при более теплом климате,
чем ныне, из Северной Пацифики в Северную Атлантику (и наоборот)
через Полярный бассейн, где бореальные формы погибли при последующем
похолодании. Этот путь был непроходим для глубоководных видов, так
как в течение всего третичного и четвертичного времени на месте Берингова
пролива существовала суша, на сравнительно короткое время прерывав-
шаяся мелководным проливом (Андрияшев, 1939). Кроме того, глубоко-
водная впадина Полярного бассейна, возникшая только в четвертичное
время, никогда не имела глубоководного соединения с абиссалью Северной
Пацифики и абиссалью Северной Атлантики (Панов, 1961). Следовательно,
глубоководные животные не могли расселятья таким же образом, как
мелководные, и их разорванные ареалы не могут быть обяснены гипотезой
Берга.
Если геоморфология и гидрологический режим океана современной
эпохи и ледникового времени не дают никаких указаний на возможные
причины возникновения биполярного и амфибореального распространения
глубоководных донных животных, то обращение к более древним периодам
оказывается в этом отношении столь же бесперспективным. Геологические
данные не позволяют допускать существования в далеком прошлом резких
температурных различий между глубинными водами тропической и умерен-
ных зон или широкого соединения больших глубин Тихого и Атлантического
океанов через Полярный бассейн. В недавнее время вопрос о климати-
ческой зональности Земли в прошлые геологические эпохи был весьма
основательно и всесторонне рассмотрен Н. М. Страховым (1960). По его
данным, «в течение всего кайнозоя — от начала палеогена до настоящего
времени климатическая зональность на поверхности Земли была ориенти-
рована по одному общему плану, удерживающемуся и доныне; очертания
и размеры одноименных зон от одной эпохи к другой изменялись лишь
в деталях» (с. 168). Принципиально то же положение справедливо и для
значительного отрезка мезозоя: «Юрским периодом кончается весьма дли-
тельный, почти в 130—140x106 лет интервал, в течение которого с пора-
зительным постоянством прослеживается один и тот же план локали-
зации климатических зон на поверхности Земли, и именно тот план,
который мы наблюдаем и в современный геологический момент» (с. 173).
Отрицая вслед за Страховым значительные перемещения полюсов и при-
нимая широтную протяженность климатических зон кайнозоя и большей
части мезозоя, следует допускать относительное постоянство основной
системы перемещений водных масс Мирового океана, в частности долготное
направление глубинных течений, определяющее сравнительное однообразие
пространственного температурного режима абиссали.
140
Невозможность объяснения амфибореального и биполярного распростра-
нения глубоководных животных, в частности изопод, особенностями физико-
географического режима океана в настоящем и прошлом заставляет подойти
к этому трудному вопросу с несколько иных позиций. Разбирая возможные
причины амфибореального и биполярного распространений семейства Isch-
pomesidae и некоторых входящих в него родов, я высказал предположение
д том, что такое распространение было свойственно мелководным предкам
ныне глубоководных форм, причем его возникновение у этих мелководных
форм хорошо объясняется теориями Берга. Мелководные предки могли
впоследствии исчезнуть, а их глубоководные потомки сохранили особенности
их распространения, сложившиеся под воздействием факторов, присущих
поверхностным, а не глубинным водам. Изложенный здесь материал по
другим семействам изопод позволяет привести дополнительные выводы
в пользу этой гипотезы.
Арктическо-бореальный широко эврибатный вид Calathura brachiata,
найденный «Витязем» на глубине 1050—1070 м, имеет почти циркумполярное
распространение, но ареал его в Полярном бассейне все же, вероятно,
прерван, (Hult, 1941). Батиальный род Bathycopea, характеризующийся
четким амфибореальным распространением, в Атлантическом океане
известен начиная с 364 м, в Тихом — с 1660 м. Близкий к нему мелко-
водный род Ancinus распространен также амфибореально. Можно пред-
ставить себе происходившую при иных климатических условиях миграцию
этих или близких к ним форм через Полярный бассейн, как этого требует
гипотеза Берга. Приуроченные к относительно низкой температуре виды
в Тихом океане опустились на большие глубины, чем в Атлантическом,
как это следует из приведенных выше сведений о вертикальном распре-
делении глубоководных изопод в разных частях океана. Вымирание на
мелководьях амфибореальных видов, которое могло вызываться как био-
тическими, так и абиотическими факторами, в ряде случаев не затронуло
их батиальных потомков, которые, в свою очередь, могли дать начало
абиссальным и ультраабиссальным видам. Поскольку в обработанном
материале представлены амфибореальные роды, приуроченные ко всем
вертикальным зонам, от сублиторали и батиали до абиссали и ультра-
абиссали, такое постепенное погружение некогда мелководных форм,
сопровождавшееся, разумеется, видообразованием, представляется вполне
вероятным.
Аналогичные положения легко развить и по отношению к биполярному
распространению. Биполярный род Munnopsurus весьма эврибатен, встре-
чаясь с 75—80 м. Правда, в южном полушарии он был найден на значи-
тельно большей глубине (3423 м), однако вполне вероятно его переселение
по небольшим глубинам из северного полушария в южное через тропи-
ческую/ зону во время одного из похолоданий и последующее опускание
в батиаль в южном полушарии. Еще более эврибатны биполярные роды
Desmosoma, Ishnomesus, Nannoniscus, Austroniscus. Биполярный род Mun-
nopsoides полностью оторван от сублиторали и целиком приурочен к батиали.
Наконец, род Stylomesus — преимущественно абиссальный. Следовательно,
и биполярное распространение прослеживается во всех вертикальных
зонах, причем его происхождение можно истолковать так же, как происхож-
дение амфибореального распространения, т. е. как следствие опускания
потомков биполярных мелководных предков. Если изыскивать для объясне-
141
ния разрывов ареалов глубоководных животных особые факторы, придется
принимать, что разные причины привели к одному следствию, так как
существенных различий между разорванными ареалами родов, обитающих
в разных вертикальных зонах, не наблюдается. Между тем предлагаемая
здесь гипотеза пригодна и для объяснения приводившегося ранее пара-
докса — наличия широтной зональности в распространении глубоководных
животных при ее отсутствии в гидрологическом режиме глубинных вод.
Однако она приемлема при одном непременном условии — при признании
крайне ограниченных способностей к расселению у глубоководных животных.
К сожалению, данными по этому вопросу мы пока не располагаем.
Разбирая биполярное и амфибореальное распространения родов, не сле-
дует забывать о том, что среди глубоководных изопод имеется и некоторое
количество видов с таким же распространением. Среди Ischnomesidae
амфибореальны Haplomesus quadrispinosus (рис. 8, с. 137) и Н. insiggnis,
представленный в Тихом океане подвидом orientalis и биполярен Stylomesus
inermis, представленный в Тихом океане подвидом pacificus. Весьма вероятна
принадлежность к одному виду Eurycope brevirostris особей из Северной Ат-
лантики и Северной Пацифики, что позволяет предполагать амфибореаль-
ность и этого вида. Возможна биполярность Munnopsurus giganteus (отсут-
ствующего в нашем материале), хотя Гансен считает вероятной видовую
самостоятельность описанной Ванхофеном антарктической формы этого вида
М. giganteus forma australis.
Существование не только биполярных и амфибореальных родов, но
наряду с ними и видов с подобным распространением имеет непосредственное
отношение к проблеме темпа эволюции глубоководных животных. Кроме
того, этот факт необходимо иметь в виду при обсуждении пока неразре-
шимого вопроса о геологическом возрасте обсуждаемых разорванных
ареалов.
Амфибореальное и биполярное распространения видов, согласно
Л. С. Бергу, есть следствие ледниковой эпохи. Биполярное распространение
более высоких таксонов он считает доказательством значительной древности
разрыва их ареалов. Мощные материковые оледенения, происходившие
в прошлые геологические периоды, должны были вызвать такие же пере-
селения животных из одного полушария в другое, как и в четвертичное
оледенение. Действительно, древность биполярного распространения дока-
зана на палеонтологическом материале. Н. С. Шатский (1954) для верхнего
мела и эоцена приводит ряд биполярно распространенных морских орга-
низмов. Аналогичные соображения можно высказать и по отношению
к амфибореальному распространению.
К сожалению, пока у нас нет никаких данных для отнесения воз-
никновения разрывов ареалов глубоководных животных к какому-нибудь
определенному отрезку геологического времени. Судя по тому, что в боль-
шинстве случаев приходится иметь дело с разорванными ареалами родов,
а не видов, этот разрыв следует относить к весьма отдаленным от 'нас
периодам. Однако даже в немногочисленных случаях разрывов ареалов
видов легче допустить значительную консервативность последних, чем
представить себе радикальные изменения их экологии и быстрые темпы
их переселения в течение четвертичного периода.
К четвертой группе родов, т. е. к родам, приуроченным преимущественно
к Антарктике, принадлежит единственный род Antarcturus и примыкает,
142
вероятно, род Astacilla. Как видно из рис. 7, центр видового разнообразия
poia Antarcturus sensu lato (включая, согласно Гелю (Hall, 1946), подроды
Microarcturus и Dolichiscus) четко приурочен к Антарктике. Подрод Doli-
scus (два вида) ограничен в своем распространении антарктическими
1ами, подрод Microarcturus (15 видов) не выходит за пределы южного
полушария. Видовое разнообразие наиболее богатого видами подрода
Antarcturus s. str. закономерно уменьшается по направлению с юга на
север. Так, например, у Земли Грейама и у Земли королевы Мод известно
по пять видов, у южной Патагонии, в Магеллановом проливе и у Керге-
лена — по четыре вида, близ берегов южной Африки — два вида. Всего
ch
BQ
в южном полушарии 27 видов Antarcturus. В северном полушарии виды
этого рода распространены только вдоль западных берегов Атлантического
и Тихого океанов. В Карибском море найдено три, и один вид обнаружен у
Флориды. В Тихом океане Antarcturus заходит дальше на север, чем
в Атлантическом, достигая Командорских островов и центральной части
Охотского моря.
Следует отметить, что близ северной границы ареала рода Antarcturus
его представители обитают только на относительно больших глубинах,
между тем как в Антарктике некоторые из них широко эврибатны, а многие
приурочены к сублиторали. Так, например, A. furcatus (Studer), известный
из ряда пунктов Антарктики (Южная Георгия, острова Кергелен, Херд,
Земля Грейама, Земля Виктории и т. д.), найден на глубинах от 10 до 3069 м.
Среди антарктических мелководных видов можно назвать A. brunneus
spinulosus Nordenstam (75—310 м), A. americanus (Bedd.) (40—183 м),
A. granulatus Nord. (160—310 м) и др. Подроды Microarcturus и Doli-
chiscus целиком приурочены к сублиторали. Три вида Antarcturus Кариб-
ского моря найдены между 823 и 1248 м, и только A. floridanus (Rich.) из
прибрежных вод Флориды — на глубине 500 м. Из шести северотихооке-
анских видов наиболее эврибатен A. hirsutus (Rich.) (243—4070 м), далее —
A. beddardi Gurj. (592—1076 м), все остальные обитают на глубинах
более 3000, причем A. ultraabyssalis достигает 7260 м.
Эти данные позволяют предполагать постепенное расселение Antarc-
turus по направлению с юга на север. При таком расселении на разных
его этапах происходило обособление отдельных видов, поскольку ни одного
вида, общего с Антарктикой, в северном полушарии не найдено.
Судя по собранным <Вйтязем> данным, видообразование происходило
и в связи с завоеванием Antarcturus больших океанических глубин. В районе
Курило-Камчатского желоба на разных глубинах обитают близкие между
собой, но самостоятельные виды A. bathybialis на глубине около 4000,
A. abyssalis — 5600—5800 и A. ultraabyssalis — 7100—7260 м.
Обращает на себя внимание подчеркнутое еще Беддардом (Beddard,
1886) отсутствие специфических морфологических особенностей у глубоко-
водных представителей этого рода (по Беддарду, Arcturus). Действительно,
все они ни длиной конечностей, ни характером скульптуры тела, ни степенью
развития глаз принципиально не отличаются от мелководных видов. Даже
живущий на огромной глубине A. ultraabyssalis обладает нормально разви-
тыми пигментированными глазами. Беддард рассматривал эти парадок-
сальные факты как свидетельство сравнительно недавнего проникновения
Antarcturus на большие океанические глубины.
Последняя группа, выделенная по характеру распространения, объеди-
143
няет роды с панокеаническим ареалом. Среди них можно различать, с одЛой
стороны, широкоэврибатные роды (Macrostylis, Eurycope, в меньшей сте-
пени — Storthyngura), с другой — сублиторальные роды, в редких случаях
опускающиеся в батиаль (Aega, Rocinela, Synidotea) и в исключительных
случаях — в ультраабиссаль (Leptanthura).
В заключение этого обзора следует сопоставить количество родов и ви^ов
глубоководных изопод, встречающихся в Северной Пацифике и в других
частях океана, с тем чтобы определить, с какими из них в наибольшей
степени связана северо-западная Пацифика по фауне равноногих ракооб-
разных. Северная Пацифика имеет 26 родов, общих с Северной Атлан-
тикой, и всего 17 родов, общих с морями южного полушария. В Северной
Пацифике обнаружено три или четыре вида, известных также из Северной
Атлантики и один вид, описанный ранее из южного полушария. Отсюда
следует, что широтные связи фаун глубоководных донных изопод теснее, чем
долготные.
Это заключение может служить лишним указанием на проявление
широтной зональности в распространении глубоководной донной фауны.
Оно стоит в явном противоречии с меридиональным расположением основных
материковых массивов и с меридиональным же направлением глубинных
течений. Оба названных фактора должны благоприятствовать долготным,
а не широтным связям глубоководной донной фауны. Остается предполагать,
что наблюдающаяся в действительности картина отражает давние этапы
истории мелководных предков современных глубоководных форм.
НЕКОТОРЫЕ ВОПРОСЫ ПРОИСХОЖДЕНИЯ И ЭВОЛЮЦИИ
ФАУНЫ ГЛУБОКОВОДНЫХ изопод
Сложные проблемы происхождения и эволюции глубоководной фауны
в последнее время привлекают к себе внимание многих исследователей.
Оживленные споры ведутся прежде всего вокруг вопроса о древности
глубоководной фауны, причем одни авторы (Bruun, 1956; Menzies, Imbrie,
1958; Menzies, Imbrie, Heezen, 1961; Madsen, 1961) считают ее относи-
тельно недавно сформировавшейся и бедной архаическими примитивными
формами, другие (Ekman, 1953; Зенкевич, Бирштейн, 1960, 1961) придер-
живаются противоположного мнения. Кроме того, поставлен вопрос об
особенностях процесса видообразования на больших океанических глубинах,
рассматривающийся пока в весьма общей форме (Wieser, 1960; Carter,
1961). Результаты обработки изопод северо-западной части Тихого океана
можно сопоставить с некоторыми суждениями, высказанными по этим
вопросам.
Прежде всего следует отметить малую степень обособленности глубо-
ководной фауны изопод от мелководной. Все обнаруженные на больших
океанических глубинах подотряды и семейства рассматриваемого отряда
обитают и на меньших глубинах, в пределах батиали, и в большинстве
случаев также в пределах сублиторали. Роды, ограниченные в своем верти-
кальном распространении только абиссальной и (или) ультраабиссальной
зонами, крайне немногочисленны. К ним относится только род Bathyopsurus
Nordenstam из желоба Пуэрто-Рико. Родов, не поднимающихся в субли-
тораль и обитающих в пределах батиали, но частично представленных
также в лежащих ниже зонах гораздо больше. Таковы в нашем материале
144
Iicanthaspidia, Janirella, Hydroniscus, Stylomesus, Haplomesus, Pseudo-
lesus, Micromesus, Munnopsoides, Bathygnathia. Однако большинство отме-
енных для больших глубин родов изопод обитает на всех глубинах —
т сублиторали до абиссали или даже до ультраабиссали. Из встреченных
северо-западной части Тихого океана родов к этой группе принадлежат
haploniscus, Ischnomesus, Macrostylis, Nannoniscus, Austroniscus, Desmo-
oma, Ilyarachna, Eurycope, Storthyngura, Bathycopea, Leptanthura, Antarc-
turus.
Пределы вертикального распределения отдельных видов гораздо уже и,
как правило, укладываются в рамки определенных вертикальных зон.
Таким образом, если судить по видовому составу, глубоководная фауна
изопод оказывается вполне специфичной, но таксоны более высокого ранга
ей почти не свойственны. Гораздо более характерны для нее негативные
особенности — отсутствие большого количества семейств и родов.
Из сказанного следует, что заселение изоподами абиссали и ультра-
абиссали почти не сопровождалось формированием крупных таксонов,
а ограничивалось преимущественно видовой дифференциацией. Как уже
указывалось выше, приспособления, выработанные изоподами в пределах
сублиторали, оказались пригодными для освоения больших глубин, а какие-
либо особые адаптации, связанные с жизнью в абиссали и ультраабиссали,
не развились или еще не выявлены исследователями. В этом отношении
изоподы существенно отличаются от многих других групп глубоководных
животных — рыб, иглокожих, десятиногих ракообразных. Как я пытался
выяснить выше, основными приспособлениями эврибатных родов изрпод,
обеспечивающими им возможность существования на различных глубинах,
следует, вероятно, считать приспособления к жизни на поверхности и в толще
мягкого грунта.
Беглый обзор групп глубоководных животных, включающих аберрантные
семейства и роды, резко отличные от обычных мелководных, показывает,
что в большинстве случаев их особенности связаны с хищным образом
жизни и представляют собой приспособления, облегчающие добычу редких
на больших глубинах жертв. Это особенно характерно для рыб и головоногих,
но справедливо также для некоторых десятиногих ракообразных, мизид,
актиний 1 Среди пелагических бокоплавов существуют роды, обладающие
приспособлениями к паразитированию на кишечнополостных и сальпах.
Сестонофаги в условиях больших глубин также должны были сильно
измениться, и мы знаем особые глубоководные крупные таксоны среди
кишечнополостных и погонофор. Грунтоеды, способ захватывания пищи
которых остается почти постоянным на всех глубинах, обычно не подвер-
гаются при переходе к глубоководному обитанию столь резким изменениям.
Меррей (Murray, 1895) отметил тесную связь между фаунами мягких
грунтов мелководья и больших океанических глубин. По его представле-
ниям, по глубинам порядка 50—500 м проходит «линия илов>, соответ-
ствующая зоне интенсивного осаждения детрита и других органических
остатков, сносимых с суши. Из этой зоны происходила миграция обита-
телей мягких грунтов на большие глубины и в сторону открытого моря.
Мадсен принимает это построение и считает «линию илов> зоной форми-
___________ ।
1 Мнение Мадсена (Madsen, 1961) о том, что некоторых Macrouridae следует считать
грунтоедами, не имеет никаких оснований. Эти рыбы — типичные хищники (Бир-
штейн, Виноградов, 1955а).
10 Заказ 333
145
рования морских звезд семейства Porcellanasteridae, мигрировавших в конц^
мезозоя в абиссаль.
Приобретение многими группами приспособлений к существованию
на мягких грунтах в области морского мелководья и последующее исполь-
зование этих приспособлений при заселении больших океанических глубин
представляется весьма вероятным.
Однако реальное существование «линии илов> Меррея и приурочен-
ность именно к ней процесса формирования многих эврибатных родов
•и семейств вызывает некоторые сомнения.
Мягкие грунты, как известно, отлагаются в различных частях Миро-
вого океана в зависимости не столько от глубины, сколько от системы
течений. Зивинг (Slewing, 1958) обратил внимание на сходство условий
существования в интерстициальных водах и в грунтах мелководных заливов,
с одной стороны, и на больших океанических глубинах — с другой, объ-
ясняя этим своеобразное распространение бокоплавов рода Ingolfiella,
виды которого известны из интерстициальных пресных вод Югославии,
морских прибрежных интерстициальных вод Плимута и Перу, из водо-
емов пещер центральной Африки, полостей между кораллами Сиамского
залива и, наконец, с глубины 3521 м из Дэвисова пролива. Сходное
распространение присуще мелким червеобразным изоподам: семейство
Thambemidae обитает в абиссали и нижней батиали, а близкие к нему
семейства Microparasellidae и Microjaniridae — в пресных и прибрежных
морских интерстициальных водах многих стран (Birstein, 1961b). Таким
образом, область возможного формирования приспособлений к условиям
мягких и пористых грунтов была, вероятно, шире и разнообразней, чем
это считали Меррей и Мадсен.
Принимая постепенное опускание на все большие глубины возникших
в области морского мелководья родов, приспособившихся к жизни на мягких
грунтах, мы наталкиваемся на почти неразрешимый вопрос о времени,
к которому следует относить этот процесс, иными словами, на вызыва-
ющую разноречивые суждения проблему геологической древности глубоко-
водной фауны.
Эта проблема в последнее время была рассмотрена Мадсеном, поддер-
жавшим мнение Брууна (Bruun, 1956) об относительной геологической
молодости глубоководной фауны. Мадсен, так же как и Бруун, исходит
из представления о недавнем, относящемся к концу мезозоя или к началу
третичного периода резком понижении температуры (на 8—10°) глубин-
ных вод. Что касается более древних периодов, то Мадсен склонен присо-
единиться к мнению Мосли и Меррея о невозможности жизни на глубинах
океана палеозоя и большей части мезозоя ввиду отсутствия в глубинных
водах кислорода. Большие глубины стали заселяться не ранее мелового
периода, когда резкие климатические изменения создали достаточно интен-
сивную циркуляцию глубинных водных масс. В соответствии с этими
предпосылками и палеонтологическими данными (относящимися к мелковод-
ной фауне) Мадсен различает в составе глубоководной фауны, во-первых,
мезозойские и раннекайнозойские элементы и, во-вторых, позднекайнозой-
ские элементы.
Нетрудно видеть, что если верна основная принимаемая Мадсеном
предпосылка о характере гидрологического режима океана времен мезо-
зоя и палеозоя, то все возражения против предполагаемой геологической
146
молодости глубоководной фауны (Зенкевич, Бирштейн, 1960, 1961) отпадают
с^ми собой. Однако обширный палеоклиматологический материал, сведенный
и (проработанный Н. М. Страховым (1960), как указывалось выше, реши-
тельно противоречит этой предпосылке. Согласно Страхову, современная
климатическая зональность существовала без особых изменений начиная
с мела, а до этого также имелись климатические зоны, но ориентирован-
нее несколько иначе по отношению к современному экватору. Никогда
на земле не было принимаемого Мадсеном вслед за Мерреем равномер-
но теплого климата, и, следовательно, газовый и температурный режимы
океана вряд ли испытывали особенно резкие изменения, поскольку цирку-
ляция водных масс в конечном итоге определяется климатической зональ-
ностью, существовавшей в том или ином виде всегда. Геологическая долго-
вечность последней подтверждена на фаунистическом материале Л. А. Зен-
кевичем (1949). А если условия существования в мезозое на больших
глубинах океана были благоприятными для существования донных живот-
ных и особенно резко не отличались от современных, то нет никаких
оснований отрицать существование раннемезозойской глубоководной фауны,
а также возможность переживания некоторых ее элементов до наших
дней при мало меняющихся условиях абиссали.
Мензис, Имбри и Хизен (Menzies et al., 1961) также считают неверным
представление об относительной стабильности условий существования
на больших океанических глубинах в течение длительных отрезков геоло-
гического времени. Они, правда, не пишут о кислородном дефиците глу-
бинных вод океана вплоть до конца мезозоя, допуская такое положение
только для времени окончания пермского оледенения. Американские авторы
приводят интересные данные об изменениях режима окраинных морей
(Средиземного, Красного, впадины Кариачо у Венесуэлы), однако не утвер-
ждают, что подобные изменения могли происходить также в открытом
океане. Они указывают далее на возможную неравномерность процессов
осадкообразования и выпадения вулканического пепла, интенсивности дея-
тельности мутьевых потоков и т. д., что должно было менять условия
жизни на больших океанических глубинах.
По этому поводу следует заметить, что вряд ли кто-нибудь из сторон-
ников древности глубоководной фауны будет настаивать на абсолютной
стабильности режима абиссали от палеозоя или от начала мезозоя до наших
дней.
Однако, как недавно подчеркнул Картер (Carter, 1961), можно пред-
полагать, что все климатические и связанные с ними изменения, происхо-
дившие на поверхности Земли, должны были затухать с глубиной и воз-
действовать на глубоководную фауну с несравненно меньшей силой, чем
на мелководную. Если такие изменения в отдельных водоемах приобретали
катастрофический характер, как, например, в указываемых Мензисом,
Имбри и Хизеном окраинных морях, глубоководная фануа погибала. Она
действительно отсутствует в Средиземном, Красном и Японском морях,
но ее несомненное присутствие, большое разнообразие и широкое распро-
странение в Мировом океане могут считаться вескими доводами в пользу
длительного существования благоприятного для жизни режима вод абиссали
океана.
Следует, впрочем, отметить, что, по современным представлениям, абис-
сальные глубины возникли только в мезозое. «История развития океанов
ю*
147
и морей показывает, что в геологическом прошлом, до мезозоя, не существо-
вало большой глубины водных пространств. Моря прошлого (до мезозой-
ского времени) были по преимуществу мелководными», — пишет по этому
поводу Д. Г Панов (1961, с. 167). Такое представление, если оно правильно,
позволяет датировать начало формирования абиссальной фауны мезозоем.
Сублиторальная и батиальная фауны должны были возникнуть раньше.
Однако это еще не означает, что абиссальная фауна относительно молодке,
поскольку, как будет разобрано ниже, есть основания допускать замед-
ленный темп ее эволюции.
Что касается целесообразности использования палеонтологических дан-
ных для обсуждения древности глубоководной фауны, то этот вопрос
достаточно подробно разобран недавно нами (Зенкевич, Бирштейн, 1961).
Мы указали на дефектность таких данных, поскольку большинство групп,
составляющих глубоководную фауну, не сохраняется в ископаемом состо-
янии; кроме того, глубоководные отложения не выходят на поверхность
земли, и палеонтологи имеют дело только с мелководными отложениями.
Мадсен упрекает нас в нелогичности, так как в другом месте своей
статьи мы допускаем, что на больших глубинах возникали только таксоны
низкого ранга — роды и виды, а крупные таксономические группы форми-
ровались в области морского мелководья. Следовательно, продолжает нашу
мысль (с которой он, впрочем, не согласен) Мадсен, эти крупные таксоны
могли сохраниться в ископаемом состоянии и с ними можно оперировать.
Мадсен, к сожалению, не учитывает первого приведенного здесь нашего
соображения. Одним из многочисленных примеров важных компонентов
глубоководной фауны, обитающих и на малых глубинах, но вообще
неизвестных в ископаемом состоянии, могут служить объекты настоящей
работы — Asellota. Об их древности на основании палеонтологических дан-
ных нельзя сказать ровно ничего. Это также относится к разбираемым
Мадсеном Porcellanasteridae и Elpidiidae, что не мешает автору датировать
их происхождение и вселение на большие глубины концом мезозоя на осно-
вании весьма спорных палеогидрологических построений. С другой стороны,
в тех сравнительно редких случаях, когда остатки какой-нибудь ныне
глубоководной группы сохранились в ископаемом состоянии, это еще не дает
достаточных оснований уверенно датировать процесс перехода такой группы
к глубоководному существованию. Возражая против общепринятого допуще-
ния обитания Neopilina (или близких к ней форм) на больших глубинах
начиная с конца палеозоя, т. е. со времени исчезновения в геологических
отложениях остатков Monoplacophora, Мадсен указывает на возможность
существования этих моллюсков в течение длительных геологических перио-
дов не в абиссали, а в батиали. Сходное мнение высказывают также
Мензис, Имбри и Хизен. Однако такое же предположение, правда, тоже
без всяких оснований, можно высказать по отношению к любой другой
группе, остатки которой найдены в ископаемом состоянии, что лишний раз
подтверждает защищаемый нами тезис о непредставительности палеонто-
логических данных для решения вопроса о геологическом возрасте глубоко-
водной фауны 1
1 Утверждение Мензиса, Имбри и Хизена (Menzies et al., 1961) об отсутствии данных
по ископаемым Tanaidacea и Cumacea неверно. И тот и другой отряды известны из
юрских отложений (см.: Reiff, 1936 с описанием Palaeotanais quenstedti и Bachmayer,
1961 с описанием Palaeocuma hessi), но это не помогает решению вопроса о времени
заселения ими абиссали.
148
По отношению к преобладающему на больших глубинах подотряду
Изопод Asellota, неизвестному в ископаемом состоянии, невозможно даже
сриентировочно указать, к какому геологическому периоду следует относить
его проникновение в абиссаль и ультраабиссаль (что, по нашему мнению,
нельзя в настоящее время сделать и по отношению к любой другой
группе, входящей в состав глубоководной фауны). Однако можно попы-
таться отметить некоторые вероятные особенности эволюции разбираемых
здесь изопод, протекавшей после их вселения на большие океанические
глубины.
Характеризуя в общих чертах особенности эволюции донных глубоко-
водных животных, Картер (Carter, 1961) обращает внимание на одно-
образие условий существования в пространстве и во времени в пределах
абиссали. По его мнению, пространственное однообразие препятствует
изоляции отдельных популяций, обусловливающей видообразование в усло-
виях сухопутного обитания и морского мелководья. Климатические изме-
нения, связанные с великим оледенением и способствовавшие, как считает
этот автор, видообразованию сухопутных, пресноводных и морских мелко-
водных животных, не могли распространиться в абиссаль и стимулировать
эволюцию ее обитателей. Длительное существование в неизменных условиях
позволило некоторым глубоководным группам пойти по пути узкой спе-
циализации и выработать «фантастические адаптивные модификации». В це-
лом эволюция животных на больших океанических глубинах должна идти
медленнее, чем на малых, а также на поверхности земли.
Однако Визер (Wieser, 1960) рассматривает возможности видообразо-
вания у глубоководных донных беспозвоночных при отсутствии простран-
ственной изоляции отдельных популяций одного вида друг от друга и имми-
грации родственных мелководных форм. Несмотря на однообразие условий
существования на больших океанических глубинах, там обычно наблюда-
ется совместное обитание нескольких близких между собой видов одного
рода, что известно для остракод, амфипод, нематод и, наконец, изопод.
Визер считает такие случаи сосуществования близких видов результатом
симпатрического видообразования. Он допускает возникновение «террито-
риальной фрагментации» обширных популяций, части которых затем начи-
нают дивергировать в силу «эффекта Райта», т. е. случайного различия
концентрации генного материала в изолированных популяциях. Этой фраг-
ментации способствует несомненное отсутствие полной панмиксии в обшир-
ных популяциях сравнительно малоподвижных донных животных.
'С положениями Визера трудно согласиться. Прежде всего нет никаких
оснований к тому, чтобы относить совместное обитание нескольких близ-
ких видов за счет видообразования, происшедшего в данном месте. В дей-
ствительности, как это следует из приводимых автором примеров, в боль-
шинстве случаев каждый из этих близких видов распространен достаточно
широко. Это, в частности, справедливо для упоминаемых Визером видов
изопод Скагеррака, ни один из которых не ограничен в своем распростра-
нении названным проливом, а известен также из ряда пунктов Северной
Атлантики.
Следовательно, несмотря на ограниченные способности к расселению,
упоминаемые Визером виды изопод (а также амфипод, остракод и др.)
успели занять обширные ареалы, причем области их возникновения опре-
149
делить невозможно, но, во всяком случае, нельзя утверждать, что эти
области совпадают с точками их местонахождений.
Что касается значения «эффекта Райта» в процессе видообразования,
то этот вопрос в последнее время подробно разобран Ю. М. Оленовым
(1961), который убедительно показал малую вероятность построений Райта
и их принципиальную ошибочность, связанную с недооценкой творческой
роли естественного отбора в эволюции живых организмов.
Таким образом, построения Визера не разъясняют механизма процесса
видообразования у малоподвижных донных глубоководных животных, в част-
ности у изопод, и не могут быть противопоставлены обычным представ-
лениям, например Картера, о затруденности этого процесса в условиях
существования на больших океанических глубинах. Тем не менее сам
факт видообразования в пределах абиссали и ультраабиссали не вызывает
сомнений, судя по значительному количеству присущих только этим зонам
видов, возникших, вероятно, в их пределах.
Обработанный материал по изоподам позволяет указать по крайней
мере две закономерности возникновения глубоководных видов — дифферен-
циацию различных видов на разных глубинах и обособление особых ультра-
абиссальных видов в разных океанических впадинах, разделенных глуби-
нами менее 6000 м.
В предшествующем изложении не раз упоминались политипические
эврибатные роды, состоящие каждый из цепочки видов, занимающих раз-
ные глубины. Таковы, например, в обработанном материале роды Janirella,
Haplomesus, Eurycope, Storthyngura, Ilyarachna и Antarcturus. Можно
предполагать, что по мере освоения представителями этих родов больших
океанических глубин на определенных горизонтах происходило обособление
особых видов, подобно тому как это принимается для бокоплавов и бычков
оз. Байкал. Анализируя родственные отношения ультраабиссальных видов
изопод ряда южных океанических желобов, Вольф показал родство многих
таких видов с антарктическими батиальными и абиссальными видами
и вероятное заселение исследованных «Галатеей» желобов антарктическими
видами. Последние в процессе освоения предельных океанических глубин
дали начало особым видам. Так, например, известные из желоба Кермадек
ультраабиссальные Storthyngura benti и S. furcata близки к антарктиче-
ским батиальному и абиссальному видам S. robustissima и S. elegans.
Точно так же ультраабиссальные S. chelata из Курило-Камчатского
и Японского желобов и S. bicornis из Японского желоба близки
к S. fragilis, известной с глубины 3429 м близ Японии, и S. inermis
с глубины 5011 м из северо-восточной части Тихого океана, а также
к широко распространенной эврибатной S. pulchra. Эта близость проявля-
ется особенно четко у молодых особей S. bicornis. Один из названных
абиссальных видов был, вероятно, предком обоих ультраабиссальных
видов (Бирштейн, 1957).
Столь же несомненным представляется обособление особых видов
в ультраабиссали изолированных друг от друга океанических желобов.
Среди изопод северо-западной части Тихого океана немногие могут служить
примерами этого явления из-за малого количества произведенных в ультра-
абиссали ловов и разреженности ультраабиссальной фауны. Такова, в част-
ности, S. tenuispinis, собранная в Алеутском и Курило-Камчатском желобах
на сходных глубинах (7246 и 7230 м) и представленная в первом —
150
типичной формой, а во втором — хорошо отличающимся подвидом kurilica.
Если сравнивать желоба северо-западной части Тихого океана с далеко
отстоящими от них, например с относительно хорошо изученным желобом
Кермадек, то при довольно значительном сходстве родового состава
(Eurycope, Storthyngura, Ischnomesus) видовой состав фауны изопод ока-
жется различным. Трудно сомневаться в том, что дальнейшее накопле-
ние сведений об ультраабиссальной фауне других океанических желобов
подтвердит правильность высказанного нами уже давно положения о вероят-
ном эндемизме ультраабиссальной фауны каждого изолированного океани-
ческого желоба (Зенкевич и др., 1954, 1955). Во всяком случае, результаты
обработки ультраабиссальных гаммарид Тихого океана (Бирштейн, Виногра-
дов, 1960), а также данные по некоторым другим группам (Wolff, 1960)
позволяют принять это положение, хотя известно и некоторое количество
ультраабиссальных видов, населяющих не один, а большее число желобов.
Генетический механизм обособления видов как на различных глубинах,
так и в изолированных океанических желобах остается невыясненным,
но в обоих случаях масштаб дивергенции приходится, по-видимому, признать
весьма ограниченным, поскольку этот процесс приводит лишь к образованию
викарирующих видов, не достигающих ранга родов. В этом отношении
эволюция равноногих ракообразных больших океанических глубин отстает
от эволюции бокоплавов оз. Байкал, где процесс освоения больших глу-
бин гаммаридами сопровождался возникновением не только новых видов,
но даже новых родов (Базикалова, 1945).
Скромные масштабы дивергенции глубоководных океанических изопод
несомненно связаны с однообразием условий их существования как на раз-
ных глубинах, так и в разных океанических желобах. Это относится
к характеру населяемого ими грунта, используемого всеми глубоководными
изоподами в качестве пищи (см. выше), температуре, и, вероятно, в какой-
то степени к биоценотическим отношениям, в частности к слабому воздей-
ствию на глубоководных донных животных малочисленных или даже совсем
отсутствующих на больших глубинах бентосоядных рыб.
Согласно И. И. Шмальгаузену (1940), «переход к глубоководной жизни
означает известное упрощение условий существования. Постоянная темпе-
ратура воды, отсутствие света, а следовательно, и тех волн жизни,
которые сопровождают периодические изменения температуры, условий
освещения и т. д. в связи со сменой времен года и случайными колеба-
ниями различных факторов, — все это делает жизнь в глубоких водах
весьма спокойной и однообразной» (с. 145). Переход к существованию
в упрощенной, ограниченной среде обитания ведет эволюцию по пути
специализации. При этом при ограниченности пищевых ресурсов естествен-
ный отбор направляется в сторону создания приспособлений к наиболь-
шей экономии жизненных средств, что вызывает продление жизни особей,
сокращение плодовитости и иногда возрастания размеров тела взрослого
животного, т. е. особенности отмеченные выше для глубоководных изопод.
Такой путь приводит, по Шмальгаузену, к замедлению темпа эволюции
благодаря прежде всего элиминации всех заметно уклоняющихся и со-
хранению лишь узко специализированных особей. «Более длительное раз-
витие, более позднее половое созревание и, наконец, обычное уменьшение
плодовитости ведет к тому, что поколения отделены друг от друга более
значительными промежутками времени и, кроме того, абсолютная числен-
151
ность особей с увеличением размеров тела также снижается. И эти обстоя-
тельства также ведут к замедлению темпа эволюции» (с. 148). «Специали-
зированный организм (вернее, вид.—Я. Б.) может существовать неогра-
ниченно долго при относительно постоянных условиях существования»
(с. 213).
Эти соображения соответствуют представлению об абиссали и ультра-
абиссали как о среде, оказывающей на населяющую их фауну консер-
вирующее воздействие. В своем анализе состава глубоководной фауны
(Зенкевич, Бирштейн, 1960, 1961) мы привели длинный ряд примеров групп
животных, представленных на больших глубинах наиболее примитивными
и, вероятно, архаическими отрядами и семействами. Мы, подобно многим
другим авторам, рассматривали большие океанические глубины как один
из биотопов, играющих роль убежища, в котором отдельные примитивные
архаические группы, вытесненные более прогрессивными и филогенетически
более молодыми группами из благоприятных и разнообразных условий
морского мелководья, способны сохраниться в течение длительного времени,
не подвергаясь существенным морфологическим изменениям.
В качестве одного из таких примеров были проведены и изоподы,
представленные на больших океанических глубинах почти исключительно
видами наиболее примитивного подотряда Asellota. В этом отношении
большие океанические глубины аналогичны пресным водам, также населен-
ным преимущественно представителями того же подотряда, относящимися
к семействам Asellidae и Microparasellidae. Первое из них обладает
некоторыми более примитивными признаками (например, наличие двух
свободных брюшных сегментов перед плеотельсоном), чем морские семей-
ства, свойственные абиссали и ультраабиссали, однако для пресных вод,
как это подчеркивал еще Ч. Дарвин, намечается замедление темпа эволюции
населяющих их организмов по сравнению с темпом эволюции фауны мор-
ского мелководья и сохранение в них многих древних реликтовых форм
(Бирштейн, 1949). Среди господствующего на больших глубинах надсемей-
ства Paraselloidea наиболее примитивной следует считать группу родствен-
ных между собой семейств Microparasellidae, Microjaniridae и Thambemidae,
населяющих, с одной стороны, пресные и морские грунтовые и интерсти-
циальные воды (первые два семейства), а с другой — абиссаль Тихого
и Атлантического океанов (последнее семейство). Аналогичное распро-
странение свойственно архаическому подотряду амфипод Ingolfiellidae,
причем как для них, так и для названных изопод интерстициальные воды
и большие океанические глубины выступают в роли убежищ, в которых
сохранились древние реликты (Бирштейн, 1961). Замедленным темпом
эволюции абиссальных и ультраабиссальных видов можно объяснить
и разорванные ареалы многих глубоководных родов изопод, если принять,
что эти ареалы отражают закономерности распространения мелководных
предков ныне глубоководных форм.
С этих позиций приобретают значительный интерес некоторые особен-
ности вертикального распределения тех групп донных животных, по отно-
шению к которым можно предполагать давность заселения ими абиссали.
В. В. Мурина (1958), Н. Г Виноградова (1958) и мы (Зенкевич, Бирштейн,
1960) обратили внимание на замечательный факт увеличения видового
разнообразия таких групп по мере возрастания глубины (разумеется,
до известных пределов). С нашей точки зрения, это явление может быть
152
объяснено двояко: либо многочисленные на больших глубинах виды пред-
ставляют собой многочисленные остатки некогда господствовавшей на малых
глубинах фауны, либо они возникли на месте своего обитания в результате
происходившего здесь видообразования. В обоих случаях следует допускать
значительную древность группы, так как и для вымирания на мелководье,
и опускания на большие глубины, и для процесса видообразования в небла-
гоприятных для этого условиях абиссали требуется продолжительное время.
Asellota северо-западной части Тихого океана представляют собой именно
такую группу. Выбирая из табл. 1 соответствующие данные, получаем
следующие показатели количества видов этого подотряда, обитающих
на разных глубинах: 1000—2000 м — 3 вида; 2000—3000 м — 5 видов;
3000—4000 м — 6 видов; 4000—5000 м — 10 видов; 5000—6000 м — 33 вида;
6000—7000 м — 11 видов; 7000—8000 м — 12 видов и 8000—9000 — 4 вида.
Отсюда следует, что наибольшее видовое разнообразие Asellota прихо-
дится на нижнеабиссальную подзону, а в сторону как меньших, так и больших
глубин оно закономерно снижается.
Мадсен возражает против нашей трактовки, считая, что увеличение
с глубиной количества видов какой-либо группы говорит лишь о большей
ее приспособленности к жизни на мягких грунтах, а вовсе не о ее древ-
ности. Такие группы сформировались на «линии илов» Меррея и нашли
на больших океанических глубинах наиболее благоприятные для себя
условия существования (Madsen, 1961, р. 191). Однако для некоторых
групп, приуроченных к мягким грунтам, как например, для сипункулид,
наоборот, отмечается закономерное уменьшение числа видов с глубиной
(Мурина, 1958). Поскольку на больших глубинах решительно господствуют
мягкие грунты и все обитающие там донные животные приспособлены
именно к ним, различия в их вертикальном распределении зависят скорее
от древности группы и от ее взаимоотношений с другими группами, чем
от степени приспособленности к мягким грунтам. Если согласиться с гипо-
тезой формирования многих глубоководных животных в пределах «линии
илов» и последующего вселения этих животных в абиссаль, то различный
характер изменения числа видов разных групп по мере увеличения глубины
нельзя объяснить, не считаясь с вероятностью длительности их пребывания
в пределах абиссали.
Если принять наше положение, то появляется возможность разделить
глубоководных изопод на две группы, различные по отношению к глубине
и по длительности обитания в абиссали — древнеглубоководную группу,
число видов которой увеличивается с глубиной, т. е. подотряд Asellota,
и сравнительно недавно заселившую абиссаль группу, число видов которой
уменьшается с глубиной, т. е. подотряды Flabellifera, Anthuridea, Valvi-
fera и Gnathiidea. Никаких оснований к тому, чтобы привязывать к какому-
либо определенному геологическому времени вселение на большие глубины
той или иной группы у нас нет, но вряд ли можно считать случайностью
тот факт, что именно наиболее примитивный подотряд занимает в настоя-
щее время господствующее положение на больших глубинах, между тем
как более совершенные подотряды гораздо более позже начали их осво-
ение. Таким образом, для изопод, так же как и для многих других донных
животных, большие океанические глубины оказались средой, благоприятной
Для переживания медленно эволюционирующих примитивных форм.
153
ДОПОЛНЕНИЯ К ФАУНЕ РАВНОНОГИХ РАКООБРАЗНЫХ
(CRUSTACEA,ISOPODA)
КУРИЛО-КАМЧАТСКОГО ЖЕЛОБА
ЧАСТЬ 2 *
В сборах 39-го рейса «Витязя» на глубинах более 1000 м в Курило-Кам-
чатском желобе и прилегающих к нему участках океана оказалось 76 видов
Asellota. Обработанные ранее материалы из несколько более обширного
района, полностью включающего район работ 39-го рейса, собранные на тех
же глубинах в 8 предшествовавших рейсах «Витязя», содержали 67 видов
этогц подотряда, обнаруженных в пробах с 48 станций (Бирштейн, 1960,
1963а). При проведении 39-го рейса Asellota найдены на 29 станциях
(Бирштейн, 1970).
Из 76 вновь констатированных видов только 32, т. е. менее половины, были
зарегистрированы в прежних сборах, 44 вида оказались новыми. С другой
стороны, 35 обнаруженных ранее видов не были пойманы вновь.
Это сопоставление заставляет сделать весьма пессимистические выводы
в отношении представительности собранного до сих пор материала. Оно
показывает, что взятых только в районе Курило-Камчатского желоба 70 стан-
ций ’, на которых обнаружены Asellota, еще совершенно недостаточно для
суждения о систематическом составе и распределении этих ракообразных.
Мы по-прежнему не можем представить себе, какая часть действительно
существующих видов стала нам известной, а какая благополучно избегла
орудий лова благодаря редкости, малочисленности или в результате чистой
случайности. Невозможно также сказать, сколько же нужно сделать ловов
в Курило-Камчатском желобе для того, чтобы получить более или менее
полную характеристику фауны населяющих его Asellota.
Независимо от представительности обработанного материала он все же
позволяет высказать некоторые суждения, имеющие отношение к общим
вопросам изучения глубоководной фауны.
Несмотря на явную неполноту полученых данных, для всей северо-запад-
ной части Тихого океана стало известно 111, а только для района Курило-
Камчатского и Японского желобов (севернее 35° с. ш.) 103 вида Asellota,
зарегистрированных глубже 1000 м. По данным Вольфа (Wolff, 1962),
с учетом позднее описанных видов (Menzies, Frankenberg, 1968; George,
Menzies, 1968a, b; Menzies, Schultz, 1968; Schultz, 1966; Бирштейн, 1963b,
1968, 1969) во всем Мировом океане, за исключением северо-западной
части Тихого океана, глубже 1000 м известен всего 301 вид Asellota.
Таким образом, на долю северо-западной части Тихого океана приходится
27 %, а на долю района Курило-Камчатского и Японского желобов — около
25 % всех известных глубоководных видов подотряда. Можно не сомневаться
в том, что более детальные исследования других районов океана значи-
тельно увеличат число указанных для каждого из них видов, но это
справедливо и для района наших работ, который пока занимает исклю-
* Тр. ИОАН СССР, 1971, т. 92, с. 162—238. (Печатается только заключительная часть
статьи, дополняющая обобщения, содержащиеся в предыдущей статье этого
сборника).
1 Из подсчета исключены 6 станций, сделанных ранее,за пределами района Курило-
Камчатского желоба, т. е. южнее 35° с. ш.
154
чительное положение в Мировом океане по богатству фауны глубоководных
Asellota.
Все имевшиеся ранее и полученные вновь сведения по составу фауны
и распространению глубоководных Asellota свидетельствуют в пользу уз-
кого эндемизма подавляющего большинства обнаруженных до сих пор
видов. Произведенные в Курило-Камчатском желббе дополнительные сборы
не увеличили, а уменьшили процент известных и за его пределами широко
распространенных видов.
Из 111 видов Asellota северо-западной части Тихого океана только
четыре — Janiralata tricornis (Кгбуег), Haplomesus quadrispinosus
(G. О. Sars), H. insignis Hansen и Eurycope scabra Hansen — обитают
за ее пределами. Первые три распространены также в Северной Атлантике,
причем J. tricornis, кроме того, — в Баренцевом и Чукотском морях.
Н. insignis, описанный из Северной Атлантики, представлен в северо-запад-
ной Пацифике особым подвидом orientalis Birstein. Следует при этом иметь
в виду, что Н. quadrispinosus и Н. i. orientalis были найдены только
на ст. 3520 (28°53',5 с. ш., 137°21' в. д.), расположенной далеко на юго-запад
от границ Курило-Камчатского желоба. Четвертый вид — Е. scabra был
известен у островов Галапагос (Hansen, 1897) и вторично обнаружен
в Японском желобе, причем нет полной уверенности в идентичности
особей из обоих местонахождений.
Если рассматривать только район Курило-Камчатского желоба вместе
с его естественным продолжением — Японским желобом, — ограничив их
с юга 35° с. ш., и соответственно этому исключить 8 видов из общего
списка северо-западнотихоокеанских видов, встреченных только южнее
(Janirella omata, Micromesus nannoniscoides, Eugerdella distincta, Haplo-
mesus quadrispinosus, H. insignis orientalis, Stylomesus wolffi, S. gracilis,
Eurycope linearis), то окажется, что из 103 видов, обнаруженных в ука-
занных желобах и на ложе океана близ их склонов, лишь Janiralata
tricornis и, возможно, Eurycope scabra не эндемичны для района исследо-
вания, причем первый вид нельзя считать глубоководным, поскольку он из-
вестен с глубины от 7 (Кусакин, 1962) до 1610 м. То же относится
и к Munnopsurus laevis (Richardson), широко распространенному, помимо
северо-западной части Тихого океана, в Японском, Охотском и Беринговом
морях на глубинах от 80 до 1100 м. Следовательно, вся глубоководная
фауна Курило-Камчатского и Японского желобов состоит из эндемичных
видов, число которых превосходит сто.
Ограниченность ареалов видов глубоководных изопод характерна, по-
видимому, и для других частей Мирового океана. В этом отношении
показательны выводы, сделанные Вольфом (Wolff, 1962) при пересмотре
систематики некоторых глубоководных видов, считавшихся широко распро-
страненными. Так, например, Storthyngura fragilis (Beddard), указанная
для вод Японии и южной части Индийского океана, оказалась сборным
видом, который пришлось разделить на три различных узко локализованных
вида: S. pulchra (Hansen), указанная для тропической Атлантики и желоба
Кермадек, была разделена на три географических подвида; Ilyarachna
antarctica Vanhoffen из двух антарктических местонахождений и желоба
Кермадек — на три самостоятельных вида.
Ограниченность ареалов глубоководных изопод следует из результатов
обработки материалов по отдельным океаническим бассейнам. Так, напри-
155
мер, все глубоководные виды, описанные Е. Ф. Гурьяновой (1946) из Поляр-
ного бассейна, могут пока считаться для него эндемичными, в обширном
материале из Атлантического океана, как следует из данных Мензиса
(Menzies, 1962а), также не оказалось широко распространенных видов.
Можно утверждать, что такие виды среди донных глубоководных Asel-
lota составляют редкое исключение. Согласно Вольфу, их всего 6: Sty-
lomesus inermis, Ischnomesus armatus, Haploniscus percavix, Hydroniscus
quadrifrons, Eurycope vicarius, Abyssjanira dentifrons. К ним можно доба-
вить Storthyngura triplispinosa Menzies, указанную для южной части
Атлантического и тропической и южной частей Тихого океанов (George,
Menzies, 1968). Однако в этом и подобных случаях, прежде чем уверенно
говорить о широком распространении, необходимо детально сравнить мате-
риал из разных местонахождений, помня о результатах только что цити-
рованной работы Вольфа.
На таксоны более высокого ранга эндемизм не распространяется.
Почти все роды, к которым принадлежат северо-западнотихоокеанские
глубоководные виды, представлены другими иногда близкими видами в иных
районах Мирового океана. Исключение составляют три монотипических рода
(Microthambema, Micromesus и Abyssoniscus), но учитывая малые размеры
относящихся к ним видов, можно предполагать их нахождение в будущем
и за пределами северо-западной части Тихого океана при применении
соответствующих методов сбора материала.
Широкое, во многих случаях панокеаническое распространение большин-
ства глубоководных родов Asellota в некоторых отношениях затрудняет
зоогеографический анализ группы, поскольку такой анализ на уровне родов
теряет смысл, а близость между собой видов одного широко распростра-
ненного рода, обнаруженных в разных частях океана, не всегда может
быть установлена с достаточной степенью достоверности. Это, по-видимому,
удалось сделать по отношению к роду Mesosignum (Menzies, Frankenberg,
1968), а из изопод северо-западной части Тихого океана явные родствен-
ные связи с другими видами проявляют, пожалуй, только Eurycope
affinis и Е. linearis, близкие Е. californiensis из Калифорнии (Schultz,
1966) по такому важному и своеобразному признаку, как редукция щупика
ногочелюсти.
Если нет достаточных оснований допускать формирование эндемичных
родов Asellota на больших глубинах северо-западной части Тихого океана,
то определенные тенденции к возникновению там надвидовых категорий
можно все же отметить. Речь идет об описанных выше внутриродовых
группах близких видов, достаточно, четко отличающихся от видов из других
частей океана. Такие группы обнаружены в составе родов Janirella
(подрод Parjanirella) и Ilyarachna (группа видов без мандибулярного
щупика и с удлиненным 3-м члеником I антенны). Однако более характерным
для рассматриваемой фауны следует считать не формирование надвидовых
таксонов, а интенсивный процесс видообразования, приведший к существо-
ванию в пределах ограниченного участка океана сотни частично близких
между собой видов.
В настоящее время очень трудно представить себе ход этого процесса.
Можно только утверждать, что он затронул разные роды в разной степени.
Так,* например, род Janirella представлен в районе исследования 12 видами,
Haploniscus, Macrostylis и Ilyarachna—9 видами каждый, Haplomesus —
156
8 видами, Storthyngura — 7 видами и 2 подвидами и т. д. С другой стороны,
существуют роды, известные из Курило-Камчатского желоба по одному
виду (Syneurycope, Acanthocope, Xostylus, Acanthaspidia) или вообще вклю-
чающие всего один вид (Microthambema и Abyssoniscus)
При рассмотрении некоторых из богатых видами родов удается разли-
чить внутри каждого из них несколько групп близких между собой видов;
можно предполагать, что каждая такая группа возникла из одного общего
предка, и, следовательно, число предковых форм было значительно меньшим,
чем ныне существующих видов. Такая структура свойственна, например,
как разбиралось выше, родам Haploniscus, в рамках которого намечаются
три группы видов, Macrostylis и Janirella с двумя группами видов в каждом,
Ilyarachna по крайней мере с тремя группами видов и т. д. К этим
группам, очевидно, вполне применим термин «сток», которым широко
пользуются авторы, разбирающие проблему видообразования в крупных
озерах (Brooks, 1950).
Процесс дифференциации видов в рамках каждого стока, так же как
в озере Байкал и других крупных древних озерах, можно представить
себе происходящим как в вертикальной плоскости, что выражается в обо-
соблении близких видов, каждый из которых приурочен к определенной
глубине, так и в горизонтальной плоскости, т. е. в обособлении локально
разграниченных видов в пределах приблизительно одинаковых глубин.
Примерами первого типа видообразования могут служить упоминавшиеся
выше близкие между собой, но приуроченные к разным глубинам виды:
Nannoniscus excavatifrons (1440—1540 м) —N. robustus (5005—6710 м),
Desmosoma coxale (5005—5495 м) — D. tenuipes (6675—6710 м), Eurycope
scabra (5290—5390 м) — E. magna (7210—8345 м), E. affinis (5005—
5495 m) —E. curtirostris (7210—7230 m), Storthyngura paradoxa (2770—
2820 м) —S. brachycephala (5461—5680 m), Janirella polychaeta (3860 м) —
J. spinosa (6435—8430 m), Haploniscus belyaevi (2445—6225 m) —H. pro-
fundicola (6090—7710 m), Hydroniscus minutus (5005—5045 m) — H. vitjazi
(5461—7286 m).
Из этого перечня следует, что в большинстве случаев один из пере-
численных парных видов приурочен к абиссальной, а другой к ультра-
абиссальной зоне или один к батиальной, а другой к абиссальной зоне.
Между глубинами обитания близких между собой видов часто намечается
разрыв, но этому не следует придавать серьезного значения, поскольку
У видов, известных не по одному, а по нескольким или многим место-
нахождениям, области вертикального распределения смыкаются вплотную,
как у Haploniscus belyaevi и Н. profundicola.
Труднее подобрать примеры горизонтальной разобщенности близких
видов, чему особенно сильно препятствует недостаточная изученность района
исследования, исключающая возможность уверенно оконтурить их ареалы.
Из двух близких видов Janirella extenuata и J. ornata первый был обнаружен
на трех станциях, расположенных между 44°48' и 45°22' с. ш. и 158°33'
и 154° 12' в. д., а второй — на единственной станции с координатами
32° 11' с. ш. и 143°09' в. д. на сходных глубинах (5005—5200 м и 5680—
3690 м). Mesosignum brevispinis известен с единственной станции с коор-
динатами 48°51' с. ш., 160°06' в. д.; все три станции, на которых отмечен
близкий вид М. elegantulum, находятся южнее 45°30' с. ш.
Наряду с указанными случаями вертикальной или горизонтальной
157
разобщенности областей распространения близких видов можно отметить
не менее обычные случаи совместной их встречаемости. Так, например,
на одних и тех же станциях неоднократно попадались вместе близкие виды:
р. Macrostylis — М. zenkevitchi и М. affinis, р. Haploniscus — Н. hydro-
niscoides и Н. inermis и р. Ilyarachna — I. scabra и I. venusta.
Следует указать на взаимоисключаемость двух близких видов Storthyn-
gura, обитающих на одинаковой глубине, но ни разу не встреченных
совместно на одной станции. S. vitjazi известна с 6 станций, сделанных
на глубинах 6435—8430 м, S. herculea — с 8 станций, сделанных на глу-
бинах 6475—9345 м, причем районы нахождения обоих видов совпадают.
Никаких отличий в характере грунта и в других физико-географических
характеристиках между этими станциями не отмечено, да они и мало-
вероятны, так что единственным объяснением этой взаимоисключаемости
приходится считать межвидовой антагонизм.
Таким образом, в различных районах Мирового океана на больших
глубинах среди Asellota происходил и происходит более или менее интен-
сивный процесс видообразования. Этот процесс выражен особенно ярко
в районе Курило-Камчатского и Японского желобов.
Уже отмечено (Бирштейн, 1971), что с точки зрения господствующих
ныне представлений о ведущей роли аллопатрического видообразования
механизм этого процесса остается во многом загадочным. Основное затруд-
нение заключается в выявлении барьеров, которые должны преграждать
обмен генами между двумя возникающими видами и, таким образом, гене-
тически изолировать их друг от друга.
Даже в простейшем случае дифференциации видов по разным глубинам
природу такого барьера невозможно себе представить; ничуть не легче
допустить его существование в случае локального разграничения близких
видов в пределах единого горизонта, и вопрос особенно осложняется
в случае совместного обитания близких видов.
С этим затруднением сталкиваются исследователи не только океани-
ческой глубоководной, но и эндемичной фауны крупных озер, как, например,
Байкала, где также, особенно среди бокоплавов, наблюдается значительное
видовое разнообразие, отчасти связанное с дифференциацией близких видов
по разным глубинам. Анализируя это явление применительно к байкалу,
М. М. Кожов (1962) пришел к выводу о вероятности в данном случае
симпатрического видообразования. Применительно к океанической глубоко-
водной фауне, в частности к изоподам, эту же мысль высказывал Визер
(Wieser, 1960), основываясь на совместной встречаемости близких между
собой видов. Он указывал на затрудненность панмиксии в пределах
обширных разреженных популяций мелких малоподвижных донных живот-
ных, включая изопод, и на возможность различной концентрации генов
в разных частях таких популяций. Результатом этого процесса может
быть территориальная «фрагментация» популяции и дивергентная эволюция
обособляющихся ее частей, т. е. симпатрическое видообразование, без
обязательной пространственной изоляции.
Э. Майр (1968) категорически отрицает возможность симпатрического
видообразования вообще и в Байкале и на больших океанических глубинах
в частности. Соображения Визера кажутся ему неубедительными, поскольку,
как это отмечено и нами (Бирштейн, 1963), факт совместного обитания
близких видов еще не доказывает их общего происхождения в данном месте.
158
Что касается присутствия групп видов в Байкале, то Майр вслед за Реншем
(Rensch, 1933) объясняет это явление действием одного (или нескольких)
из следующих факторов: сохранением реликтовых видов, множественной
инвазией, т. е. неоднократным и разновременным вселением предковых
форм из соседних водоемов, слиянием воедино некогда разделенных озер
и наличием географических преград внутри озера.
Значение этих факторов в процессе видообразования на больших океа-
нических глубинах представляется сомнительным. Нет никаких оснований
для предположения о существовании в прошлом изолированных друг
от друга водоемов на месте Курило-Камчатского и Японского желобов,
так же как и каких-нибудь географических преград в их пределах в настоя-
щее время.
Маловероятна и множественная инвазия, которую можно представить
себе как неднократное вселение на большие глубины мелководных форм.
Этому противоречит характер вертикального распределения родов, к которым
принадлежат глубоководные виды и подавляющее большинство которых
чуждо мелководью. Считать эндемичные виды реликтами, конечно, можно,
но это не объясняет их происхождения; кроме того, существование на огра-
ниченном пространстве групп близких между собой видов скорее наводит
на мысль об их недавнем возникновении, чем об их длительном сохра-
нении.
В этой связи представляет некоторый интерес вопрос об изменениях
видового разнообразия Asellota с глубиной, уже освещавшийся на осно-
вании ранее обработанных материалов (Бирштейн, 1963).
Добавление к этим данным обширных материалов 39-го рейса в общем
подтвердило полученные ранее выводы. Полная сводка всех имеющихся
сведений по распределению видового разнообразия Asellota северо-западной
части Тихого океана по глубинам представлена в таблице.
Число видов Asellota по глубинам
Глубина, м Число Индекс Фишера а (50 %-ный доверитель- ный интервал)
станций видов видов на станцию
1000—2000 7 4 0,6 4±4
2000—3000 11 16 1,5 7±3,5
3000—4000 6 9 1,5 3± 1,6
4000—5000 4 14 3,5 14 ±9,8
5000—6000 18 53 2,9 18±3,2
6000—7000 11 35 3,2 10±2,2
7000—8000 9 18 2,0 5± 1,5
8000—9000 6 10 1,7 3± 1,2
9000—10000 3 1 0,3 0
Помимо показателей, аналогичных уже опубликованным, в таблицу
по совету Ю. А. Рудякова 1 включены и вычисленные им на основании
моих подсчетов индексы видового разнообразия Фишера (Fisher, Corbet,
Я очень благодарен Ю. А. Рудякову за помощь в получении этих данных и их
обсуждение. По его мнению, в данном случае лучше пользоваться индексами
разнообразия Фишера, чем примененным им ранее (Рудяков, 1964) индексом
Е/Ещах-
159
Williams, 1943) по различным горизонтам. Эти индексы более четко, чем
показатели общего числа видов и числа видов на станцию, указывают
на приуроченность максимума видового разнообразия к горизонтам 5000—
6000 и 4000—5000 м, как это было уже установлено на основании преж-
них, менее представительных данных.
Если принять, что для таких малоподвижных и узколокализованных
форм, как Asellota, видовое разнообразие отражает интенсивность процесса
видообразования, то можно приурочить наибольшую интенсивность послед-
него к глубинам ложа океана. По всей вероятности, господствующие
там условия наиболее благоприятны для Asellota, в то время как на боль-
ших и меньших глубинах сказывается воздействие различных подавляющих
видообразование факторов как абиотических, так, возможно, и биотических.
Однако и в этих наиболее благоприятных условиях происходит только
образование видов, а не более крупных таксонов, имеющих широкое рас-
пространение и пока не известные нам центры возникновения.
РОЛЬ ТРОФИЧЕСКОГО ФАКТОРА
В ТАКСОНОМИЧЕСКОМ ОБОСОБЛЕНИИ
МОРСКОЙ ГЛУБОКОВОДНОЙ ФАУНЫ *
Проблема эволюции глубоководной фауны послужила в последние годы
предметом оживленной дискусси. Мнения зоологов разделились. Одни (Зен-
кевич, Бирштейн, 1961; Беляев, 1966) защищали традиционные представ-
ления о значительной геологической древности и замедленном темпе эволю-
ции глубоководной фауны, включающей, по их мнению, большое количество
архаических элементов, другие (Bruun, 1956; Menzies, Imbrie, 1958;
Menzies et al., 1961; Madsen, 1961) отстаивали противоположное мнение,
допуская геологически недавнее заселение абиссали мелководными орга-
низмами и вымирание древней глубоководной фауны. Сторонники обоих
взглядов привлекали для их обоснования преимущественно данные по си-
стематическому составу глубоководной фауны, а также некоторые геологи-
ческие и палеонтологические материалы. Мы считаем, что существует
возможность рассмотреть некоторые аспекты этой спорной проблемы
с несколько иных позиций, используя для их освещения имеющиеся скуд-
ные сведения по экологии глубоководных животных.
Естественно, особенности развития, состава и распределения глубоко-
водной фауны определяются сложным комплексом внешних условий и за-
висят от различных биохимических, физиологических и экологических фак-
торов. Однако подавляющее большинство из них пока еще изучены плохо
и не поддаются детальному анализу. Мы располагаем несколько более
подробными сведениями по особенностям питания глубоководных обита-
телей, на которые следует обратить особое внимание, поскольку именно
трофический фактор во многих случаях определяет структуру сообществ,
в частности их видовое разнообразие, и от него зависит их целостность
(Elton, 1927). Направление эволюции также в значительной степени связано
* Бюл. МОИП. Отд. биол., 1971, т. 76, вып. 3, с. 59—92. (Совместно с М. Е. Виногра-
довым). Печатается с сокр.
160
с пищевыми ресурсами: в условиях ограниченной кормовой базы преиму-
щества получают малоплодовитые, узкоспециализированные виды, при оби-
лии пищи — активные, плодовитые виды с широкой амплитудой приспо-
собляемости (Шмальгаузен, 19686). Отсюда следует, что направления
и темпы эволюции в обильных пищей мелководьях были иными, чем в обед-
ненных пищевыми ресурсами глубинных слоях.
ПИЩЕВЫЕ РЕСУРСЫ ГЛУБОКОВОДНОЙ ФАУНЫ
Как известно, пищевые ресурсы океана создаются в поверхностной
эвфотической зоне за счет фотосинтеза растений — пелагических и донных.
Количество органического вещества, сносимого в океан с суши, примерно
на два порядка ниже (Богданов и др., 1971).
Часть созданного органического вещества вновь включается в продукци-
онно-деструкционные циклы сообществ продуцирующей зоны, часть вместе
с мигрирующими животными, отмершими организмами и продуктами жизне-
деятельности опускается в глубинные слои и на дно океана. При этом
органическое вещество в разных формах частично потребляется живот-
ными, обитающими ниже эвфотической зоны, частично растворяется и ми-
нерализуется. Пути, по которым оно попадает из поверхностной зоны
на различные глубины и дно океана, его количество или темп поступле-
ния и форма, в которой оно там присутствует, очевидно, должны накла-
дывать глубокий отпечаток на особенности существования глубоководной
фауны (Виноградов, 1959, 1968; Чиндонова, 1959; Menzies, 1962b; Wickstead,
1962; и др.).
Благодаря исследованиям многих авторов, можно считать установлен-
ным, что общее количество органической взвеси (детрита) заметно умень-
шается в слоях, подстилающих эвфотическую зону, т. е. на глубине 100—
500 м, а затем имеет тенденцию незначительно убывать с увеличением
глубины. Однако биохимический состав взвеси при этом может существенно
меняться. Так, например, белки составляют 70—85 % поверхностного планк-
тона в тропических районах океана. В детрите тех же поверхностных
горизонтов содержание белков ниже в 1,5—2 раза; с увеличением глубины
их доля продолжает уменьшаться. На глубине 6000 м в районе Курило-
Камчатского желоба содержание белков органической взвеси составляет
всего 3—4 % (Богданов и др., 1968), хотя в других случаях на подобных
глубинах оно может быть и значительно выше. Аналогичным образом
изменяется и содержание в детрите нуклеиновых кислот (Агатова, Богда-
нов, 1972). Особенно быстро идет потеря фосфорсодержащих соединений
(Богданов, Шапошникова, 1971; Parsons, 1963).
На глубине более 500 м опустившаяся сюда органическая взвесь состоит
в основном из малоусвояемых стабильных компонентов — гуминовых кислот,
битуминозных и других высокомолекулярных остаточных веществ (Лисицын,
Богданов, Овчинников, 1967; и др.). Даже хитин отчасти разлагается,
а в глубоководном детрите сохраняются лишь его наиболее стойкие ком-
поненты (Krause, 1962). Пищевая ценность такого детрита невелика ’).
1 Во взвешенных органических частицах на всех глубинах сохраняется некоторое
количество белков и углеводов, но они, по-видимому, почти не гидролизуются
ферментной системой глубоководных животных и, следовательно, не ассимилируются
ими (Menzel, Goering, 1966; Gordon, 1970).
11 Заказ 333
161
Таблица 1. Изменение содержания различных компонентов органического веще
'ства в пелитовых илах северо-западной части Тихого океана
Компонент Шельф до 250—300 м Верхняя часть континенталь- ного склона, 300 -400 м Глубоководные желоба (Япон- ский и Курило- Камчатский), 4000— 8000 м Краевая часть ложа океана Центральная часть океани- ческих котло- вин
С °/ ^орг> /О Белки, мкг/г Моно- и олигоса- хариды, мкг/г >2 2,0-1,5 1,5-0,7 0,7—0,3 0,25 и мене
>200 180—130 80 25—30 18 и менее
— 175—40 30 30 25 и менее
Прочие углеводы, мкг/г >5000 5000—3000 2000—1000 1000—500 <500
Липиды, мкг/г >2000 1500 1300 250—150 110 и менее
Гуминовые вещества, мкг/г — -5000 10 000—2000 2000—1000 250—50
Остаточные стабиль- ные компоненты, % — 55 60 70 90
Очевидно, детрит, образовавшийся при отмирании или жизнедеятельное™
самих глубоководных животных, пока он не успел разложиться, обладает
для глубоководных животных гораздо большей пищевой ценностью (Whee
1ег, 1967), но его количество вследствие низкой концентрации постав
щиков весьма мало.
Изменение с глубиной биомассы планктона подчинено совершенно иным
закономерностям, чем распределение детрита, — оно носит экспоненциаль-
ный характер (Виноградов, 1959, 1960, 1968; Яшнов, 1961; Johnston, 1962;
и др.). Это служит одним из оснований (Виноградов, 1955, 1968; и др.),
позволяющих считать, что основной источник питания планктона бати-
пелагиали — не остатки отмерших организмов, опустившихся из поверх-
ностных слоев, а органическое вещество животных, активно мигрирующих
между поверхностной и глубинной зонами.
Приспособления, выработанные глубоководной пелагической фауной
к всемерному сокращению энергетических затрат, позволяют животным
мезо- и батипелагиали существовать при значительно меньшем относи-
тельном (на единицу биомассы) расходе энергии, чем населению эвфоти-
ческой зоны (Marshall, 1960; Виноградов, 1968; и др.).
Рассматривая пищевые ресурсы донного населения, надо иметь в виду,
что если в питании глубоководной фауны пелагиали большую роль играют
животные, активно связанные с продуцирующей зоной, то для глубоководной
донной фауны основой пищевых ресурсов служит органическое вещество
донных осадков, придонная органическая взвесь и развивающаяся здесь
микрофлора, почти нацело отсутствующая в глубоководной пелагиали океана
(Сорокин, 1970). Количество органического вещества, содержащегося
в донных осадках, изменяется не только с глубиной, но обусловлено и рядом
других причин. В некоторых районах и на максимальных глубинах количество
органического вещества в грунте достаточно велико и может быть большим,
чем на промежуточных глубинах. Таковы, например, осадки в прибрежных
глубоководных желобах типа Курило-Камчатского, Яванского и др. Вопрос
осложняется еще и тем, что обычно с увеличением глубины одновре-
менно увеличивается и расстояние до берега. Какой из этих
двух факторов играет более важную роль в обеднении осадков органическим
веществом, не всегда удается решить однозначно. Однако общие тенденции
162
изменения содержания С орг в грунтах сходного гранулометрического состава
с различных глубин и одновременное изменение биохимического состава
органического вещества достаточно ясны (табл. 1, составлена Е. А. Ро-
мановичем).
Можно видеть, что общее содержание органического вещества от
шельфа к дну желобов изменяется всего в 2—3 раза, однако доля в нем
стабильных гуминовых и остаточных веществ возрастает. Другими словами,
с увеличением глубины пищевая ценность органического вещества в осадках
имеет тенденцию уменьшаться. Что же касается абиссальных открытых
районов океана, занимающих подавляющую часть его площади, то там содер-
жание С орг чрезвычайно низко и органическое вещество находится в стадии
глубокого разложения — до 90 % его состава приходится на неиспользу-
емые в пищевых целях остаточные нерастворимые вещества. Максимальное
относительное количество остаточных органических веществ обнаружено
в красных глубоководных глинах тропической области Тихого океана.
Таким образом, очевидно, как в толще воды, так и в донных осадках по
мере увеличения глубины (и удаления от берега для донных осадков) коли-
чество усвояемого органического вещества, которое может быть использо-
вано глубоководными животными, уменьшается.
СРАВНЕНИЕ СОСТАВА ГЛУБОКОВОДНОЙ И МЕЛКОВОДНОЙ ФАУН
Основываясь на сопоставлении глубоководной фауны с мелководной,
некоторые исследователи формулируют чрезвычайно важные выводы. В ка-
честве примера можно привести интересную работу Бернара (Barnard,
1961) по гаммаридам, собранным экспедицией на «Галатее» на глубинах
от 400 до 6000 м. Бернар установил, что большая часть абиссальных
видов (81 вид) донных бокоплавов принадлежит к родам, включающим
преимущественно сублиторальные виды, незначительная часть (21 вид) —
к преимущественно батиальным родам и только 6 видов — к 6 монотипи-
ческим абиссальным родам. По мнению автора, малая степень обособ-
ленности абиссальной фауны гаммарид объясняется недавним заселением
абиссали этими животными. Такие же аргументы используются и некото-
рыми другими авторами. Обсуждая состав глубоководной фауны сипункулид,
В. В. Мурина (1969) специально подчеркивает, что в классе Sipunculida
нет ни одного глубоководного рода или подрода. Отсутствие систематической
обособленности абиссальной фауны сипункулид от мелководной свиде-
тельствует о том, что переход отдельных видов к обитанию на больших
глубинах, вероятно, произошел сравнительно недавно и абиссальная фауна
сипункулид является молодой» (с. 88).
По отношению ко всей донной глубоководной фауне аналогичные
мысли ранее высказал Спэрк (Sparck, 1954) «Ясно. что нет
существенных различий между видами, занимающими область больших
глубин и занимающими литоральную зону... С морфологической точки зрения
нет оснований рассматривать донную фауну больших глубин как очень древ-
нюю фауну. Донная фауна океанического ложа и в особенности глубоко-
водных впадин состоит из тех же групп и большей частью из тех же семейств
и родов, что и фауна мелководная, а в некоторых случаях одни и те же виды
встречаются как в литоральной зоне, так и на больших глубинах» (с. 89).
Однако наряду с группами животных, представленными на больших глу-
н*
163
Таблица 2. Число семейств и родов пелагических животных, известных с глубины
более 500 м, и тех из них, которые не встречены выше 200 м
Г руппа Семейства Роды
Всего Из них не подни- мается выше 200 м Всего Из них не подни- мается выше 200 м
Число | % Число °/о
Siphonophorae 10 0 0 39 1 2,5
Hydromedusae 8 1 12,5 28 10—11 35,9
Scyphomedusae 7 1 14 10 4 40,0
Chaetognatha 1 0 0 4 2 50,0
Cephalopoda 21 8 38,1 105 27 25,7
Copepoda (Calanoida) 18 2 1U 78 34 43,6
Ostracoda — — — 8 1—2 12,5—25
Leptostraca 1 0 0 1 1 100
Mysidacea 4 2 50,0 26 17 65,4
Hyperiidea 7 3 42,9 17 12 70,6
Gammaridea 8 3 37,5 43 34 76,7
Euphausiacea 2 1 50,0 8 1 12,5
Decapoda 7 1 14,0 31 14 45,2
Pisces 51 18 39,1 205 61 30,0
бинах незначительным количеством особых родов или только особыми ви-
дами широко эврибатных родов, существуют группы, включающие не только
глубоководные роды, но даже семейства и отряды, чуждые мелководной
фауне. Можно, конечно, применительно к таким группам допускать давнее
заселение ими абиссали или предполагать более быстрые темпы их эволюции,
хотя никаких оснований для подобных предположений не имеется. С другой
стороны для объяснения этих различий можно попытаться использовать
зависимость между степенью таксономической обособленности фауны глубин
и способом питания разных групп глубоководных животных.
«Беглый обзор групп глубоководных животных, включающих аберран-
тные семейства и роды, резко отличные от обычных мелководных, пока-
зывает, что в большинстве случаев их особенности связаны с хищным обра-
зом жизни и представляют собою приспособления, облегчающие добычу
редких на больших глубинах жертв... Сестонофаги в условиях больших глу-
бин также должны были сильно измениться, и мы знаем особые глубоко-
водные крупные таксоны среди кишечнополостных и погонофор. Грунтоеды,
способ захватывания пищи которых остается почти постоянным на всех глу-
бинах, обычно не подвергаются при переходе к глубоководному обитанию
столь резким изменениям» (Бирштейн, 1963а, с. 196).
Очевидно, фундаментальные различия особенностей добывания пищи
у глубоководных представителей той или иной группы по сравнению с ее
мелководными представителями неизбежно должны вести к резким морфоло-
гическим и таксономическим различиям поверхностной и глубоководной
фаун. У глубоководных пелагических животных «морфологические адап-
тации к глубоководному образу жизни наиболее ярко выражены в тех груп-
пах животных, которые существенно изменили характер питания при пере-
ходе из поверхностных слоев в глубинные, например сменили активное хищ-
ничество на пассивное поджидание добычи» (Виноградов, 1965, с. 661).
Эти соображения высказаны в весьма о(бщей форме и требуют конкрети-
зации. Прежде всего необходимо установить реально существующие разли-
164
Таблица 3. Число семейств и родов донных животных, известных с глубины более
2000 м, и тех из них, которые не встречены выше 500 м
Группа Семейства Роды
Всего Из них не подни- мается выше 500 м Всего Из них не подни- мается выше 500 м
Число % Число %
Calcarea 1 0 0 1 0 0
Demospongia 26 0 0 67 11 16,4
Hydrozoa 10 0 0 26 0 0
Gorgonaria 6 0 0 14 2 14,3
Pennatularia 8 0 0 19 5 26,3
Alcyonaria 6 0 0 8 1 12,5
Actiniaria 9 2 22,2 25 5 20,0
Madreporaria 4 0 0 7 2 28,6
Antipatharia 1 0 0 3 0 0
Zoantharia 1 0 0 1 0 0
Ceriantharia 1 0 0 1 0 0
Bee Coelenterata 46 2 4,3 104 15 14,4
Sipunculoidea 1 0 0 4 0 0
Priapuloidea 1 0 0 1 0 0
Polychaeta 27 0 0 172 2 1,2
Echiuroidea 2 0 0 10 6 60,0
Cirripedia 4 0 0 5 0 0
Mysidacea 2 1 50,0 4 3 75,0
Cumacea 6 0 0 18 0 0
Amphipoda 28 0 0 80 14 17,5
Isopoda 22 4 18,1 57 24 42,1
Tanaidacea 3 1 33,3 13 2 15,4
Deca pod a 14 0 0 25 2 8,0
Pantopoda 6 0 0 13 0 0
Monoplacophora 1 1 100 1 1 100
Amphineura 4 0 0 6 0 0
Prosobranchia 49 1 2,0 128 18 14,1
Opisthobranchia 10 0 0 21 2 9,5
Scaphopoda 2 0 0 4 0 0
Bivalvia 23 1 4,3 50 10 20,0
Cephalopoda 3 0 0 8 3 37,5
Brachiopoda 11 0 0 20 5 25,0
Crinoidea 7 3 41,1 23 17 73,9
Asteroidea 16 2 12,5 64 48 75,0
Ophiuroidea 10 0 0 60 27 44,9
Echinoidea 18 1 5,6 46 19 41,2
Holothurioidea 17 2 11,8 50 30 60,0
Bryozoa 21 1 4,8 39 3 7,4
Ascidia 14 3 21,4 32 20 62,5
Pogonophora 5 2 40,0 14 8 57,1
Pisces 30 6 20,0 101 .65 64,3
чия между отдельными группами по степени таксономической обособлен-
ности глубоководной фауны от мелководной, а затем — с чем эти различия
связаны.
Н. Г Виноградова (1969а; и др.) показала, что роль стенобатных и эври-
батных видов в донной абиссальной фауне у различных таксономических
групп неодинакова. Так, фауна донных кишечнополостных, морских ежей,
усоногих и десятиногих ракообразных почти нацело представлена широкоэв-
рибатными видами, тогда как среди изопод, губок, голотурий-элазипод абис-
165
сальные эндемики составляют значительную и часто доминирующую часть
фауны.
Однако оценка обособленности глубоководной фауны на видовом
уровне в нашем случае не дает достаточно показательных результатов, так
как нас интересуют эволюционные преобразования более крупного масш-
таба, приводящие к формированию высших таксонов, а не процесс узкой
специализации, находящий свое выражение в видообразовании.
Для оценки степени таксономической обособленности глубоководной
фауны от мелководной мы попытались подсчитать для каждой группы живот-
ных общее число известных на больших глубинах родов и семейств, отмечая
при этом, сколько из них встречается также и на малых глубинах (табл. 2 и 3),
Отношение количества исключительно глубоководных родов и семейств
к общему их числу, зарегистрированному на больших океанических глубинах,
должно достаточно наглядно отображать самостоятельность глубоководной
фауны в пределах каждой данной группы.
Все расчеты произведены по классам, и только для ракообразных и дон-
ных кишечнополостных мы сочли возможным привести более детальные
характеристики по отрядам. В данном случае мы считаем возможным
сопоставление разномасштабных показателей.
Соответствущие расчеты для планктона и бентоса целесообразно произ-
водить по-разному, учитывая их различную вертикальную подвижность.
Вертикальные миграции глубоководного планктона происходят на протя-
жении огромного диапазона глубин, что определяет относительную однород-
ность населения всей глубоководной пелагиали глубже фотосинтетической
зоны. Наоборот, глубоководная донная фауна сравнительно стенобатна
и достаточно четко подразделяется на батиальную, абиссальную и ультра-
абиссальную. Поэтому для планктона оказалось удобнее принимать во вни-
мание роды и семейства, указанные для глубин более 500 м, а для бентоса —
более 2000 м. В первом случае для сравнения бралась фауна, обитающая
выше 200 м, во втором — выше 500 м.
РАЗЛИЧИЯ В СТЕПЕНИ ТАКСОНОМИЧЕСКОЙ ОБОСОБЛЕННОСТИ
ГЛУБОКОВОДНОЙ ФАУНЫ В ПРЕДЕЛАХ РАЗНЫХ ГРУПП
И ИЗ ВОЗМОЖНЫЕ ПРИЧИНЫ
Из сводных табл. 2 и 3 следует, что степень обособленности глубоководной
фауны для различных систематических групп весьма различна.
Можно, во-первых, выделить группы, совсем не включающие чисто глу-
боководных родов. К ним относятся Calcarea, Hydrozoa, Antipatharia, Zoan-
tharia, Ceriantharia, Sipunculoidea, Cumacea, Cirripedia, Pantopoda, Scapho-
poda, Amphineura.
Далее следуют группы, включающие не более 25 % исключительно
глубоководных родов. Это Demospongia, Gorgonaria, Alcyonaria, Actiniaria,
Polychaeta, Amphipoda, Tanaidacea (?), Decapoda, Prosobranchia, Opistho-
branchia, Bivalvia, Bryozoa, Brachiopoda, Siphonophora, Euphausiaceae и пе-
лагические Ostracoda.
Более высокая степень таксономической обособленности глубоководной
фауны свойственна группам, исключительно глубоководные роды которых
составляют 25—50 % от всех глубоководных, а процент исключительно
глубоководных семейств иногда значительно превосходит 5 %: Pennatularia,
Madreporaria, пелагические Hydrozoa и Scyphozoa, Chaetognatha, пелаги-
166
ческие Cephalopoda, Isopoda, пелагические Decapoda, Copepoda и рыбы,
Ophiuroidea, Echinoidea.
Наконец, существуют группы, представленные на больших глубинах
50—100 % исключительно глубоководных родов и высоким процентом глубо-
ководных семейств, каковы в бентосе Monoplacophora, Crinoidea, Aste-
roidea, Holothurioidea, Pogonophopa, Ascidia и рыбы, а в планктоне —
Leptostraca, Mysidacea и Amphipoda.
Этот перечень показывает, что степень таксономической обособленности
пелагической глубоководной фауны в целом значительно выше, чем донной,
несмотря на причисление к глубоководным всех животных, встреченных
в пелагиали глубже 500 м, а в бентали — глубже 2000 м. В то же время
в отличие от донной фауны в составе глубоководного планктона отсутствуют
крупные таксономические группы, не представленные на малых глубинах.
Полученные нами данные о степени таксономической обособленности на
уровне родов и семейств глубоководной фауны различных групп очень
сходны с аналогичными данными Н. Г Виноградовой (1969а), рассчитан-
ными на видовом уровне. Таким образом, оказывается, что группы, имеющие
наибольшую степень глубоководного эндемизма на видовом уровне, как
правило, более обособлены от поверхностной фауны и в категориях более
высокого систематического ранга.
Нетрудно заметить, что степень таксономической обособленности глубо-
ководной фауны не связана ни с высотой организации, ни с видовым или
родовым разнообразием, ни с эврибатностью сравниваемых групп. Лопато-
ногие моллюски, несколько групп кишечнополостных, кумовые и усоногие
ракообразные, многоколенчатые, несмотря на различное положение в сис-
теме, разное отношение к глубине и разное общее количество представителей
в равной мере не содержат исключительно глубоководных родов. Точно так
же и группы, включающие значительное количество таких родов, весьма да-
леки друг от друга в систематическом отношении и резко различны по всем
остальным показателям.
Совершенно очевидно также, что малая степень таксономической обособ-
ленности глубоководной фауны никак не может служить указанием на недав-
нее заселение больших глубин группами животных, характеризующимися
низкими показателями такой обособленности, как это считали Спэрк, Бернар
и другие цитированные выше авторы. В самом деле, нельзя представить себе
более позднее вселение в абиссаль полихет или двустворчатых моллюсков,
чем рыб, голотурий или погонофор. Нет далее никаких оснований для призна-
ния более раннего заселения глубоководной пелагиали, чем глубоководной
бентали, как это следует предполагать при таком толковании полученных
результатов.
Степень таксономической обособленности глубоководной фауны может
определяться длумя принципиально разными причинами. Во-первых, при
освоении больших океанических глубин представители той или иной система-
тической группы были вынуждены выработать специальные приспособления,
в результате чего они изменились настолько, что образовали особые роды
и даже семейства. Во-вторых, возможно сохранение на больших океани-
ческих глубинах архаических реликтовых групп, некогда эврибатных, но впо-
следствии вытесненных более сильными конкурентами с малых глубин.
Присутствие Monoplacophora только на больших океанических глубинах
легче всего понять именно таким образом. В организации рецентной Neopilina
167
не удается отметить каких-либо особенностей, которые можно было бы счи-
тать адаптациями к глубоководному существованию. С другой стороны,
несомненна ее близость к силурийским мелководным Pilina, Tryblidium
и другим ископаемым также мелководным родам того же класса.Вымершие
на мелководьях Monoplacophora сохранились только на больших океаниче-
ских глубинах.
Можно допускать аналогичную картину для погонофор, отличающихся
от всех остальных групп приуроченностью наибольшего многообразия
семейств и родов к абиссали. В ископаемом состоянии погонофоры, как
недавно выяснилось (Poulsen, 1963; Соколов, 1965, 1967), подобно монопла-
кофора’м, известны из мелководных отложений палеозоя. Никаких морфоло-
гических особенностей, отличающих исключительно глубоководных предста-
вителей этого класса от эврибатных и мелководных, не указано.
Не исключено, что многообразие и высокая степень таксономической
обособленности глубоководной фауны некоторых других групп животных,
в первую очередь стебельчатых морских лилий и элазипод, также опреде-
ляется преимущественным сохранением их на больших океанических глуби-
нах в результате вытеснения с мелководий.
В частности, единственный исключительно глубоководный род эвфау-
зиид, выделяемый в особое сем. Bentheuphausiidae, характеризуется рядом
примитивных признаков, отличающих его от второго обширного эврибатного
семейства Euphausiidae. Точно так же единственное глубоководное бедное
родами и видами семейство мизид Petalopthalmidae значительно примитив-
нее широко эврибатного богатого родами и видами другого семейства
того же подотряда — Mysidae. Л. А. Зенкевич в ряде своих работ рассматри-
вает глубоководных эхиурид так же, как реликты, некогда более широко
распространенные по вертикали.
Однако подобные построения не применимы к группам, богато представ-
ленным на мелководьях и в то же время характеризующимся весьма различ-
ной степенью таксономической обособленности глубоководной фауны.
Нет сомнения в том, что своеобразие глубоководной фауны в тех случаях,
когда оно четко выражено, определяется как сохранением на больших глуби-
нах архаических вымерших на мелководьях групп, так и морфологическими
изменениями, которым подверглись некоторые животные в процессе приспо-
собления к жизни на больших глубинах. Разграничение этих моментов
представляется весьма трудным делом, между тем иногда мы встречаемся
как раз с их совместным проявлением.
В заключение необходимо отметить еще одно важное обстоятельство.
Мы не находим никакого соответствия между степенью таксономической
обособленности в пределах каждой из рассматриваемых групп, с одной сто-
роны, и ее видовым разнообразием и степенью ее количественного развития
на больших океанических глубинах — с другой. Для донной фауны на-
мечается скорее обратная зависимость. Резко преобладающие по биомассе
на предельных глубинах всех океанических желобов голотурии принадлежат
к широко эврибатному роду Elpidia, весьма многочисленные на больших
глубинах полихеты и моллюски также принадлежат в своем большинстве
к широко эврибатным родам. Наоборот, такие своеобразные формы, как
Monoplacophora, хищные асцидии и многие другие животные, резко отли-
чающиеся от мелководных, представляют собой редкие виды, не образующие
плотных популяций и составляющие ничтожную часть суммарной биомассы
168
глубоководного бентоса. Отсюда следует, что обособление тем или иным
путем глубоководных таксонов высокого ранга нельзя считать необходимым
условием процветания группы на больших глубинах, однако оно несомненно
способствует ее выживанию.
РОЛЬ ТРОФИЧЕСКОГО ФАКТОРА
Изложенные выше данные о закономерном и резком уменьшении пище-
вых ресурсов с глубиной позволяют попытаться выяснить направления
адаптаций глубоководных животных к использованию этих ресурсов
и проверить, не влияет ли трофический фактор на степень таксономи-
ческой обособленности глубоководной фауны от мелководной. В принципе
можно допустить три возможности: одни группы животных по мере
уменьшения пищевых ресурсов принципиально не меняют характер пищи
и способ ее захватывания, другие значительно совершенствуют свой хвата-
тельный аппарат или способ добывания пищи и, наконец, третьи
радикально меняют как характер пищи, так и способ ее захвата. Соответст-
венно этому в разной степени должна происходить перестройка организации
животных, и в обоих последних случаях она может приводить к возможности
особых глубоководных таксонов высокого ранга, что должно отра-
жаться на вычисленных нами показателях таксономической обособленности
глубоководной фауны от мелководной.
Разберем с этой точки зрения наши итоговые данные (табл. 4).
Группы, характеризующиеся малой степенью таксономической обособ-
ленности глубоководной фауны от мелководной (меньше 25% стенобатных
глубоководных родов) и составляющие две первые колонки нашей таблицы,
сохранили неизменным способ питания на всех глубинах. Среди них есть
представители всех основных пищевых группировок. Это, во-первых, сесто-
нофаги, т. е. прикрепленные к субстрату животные, питающиеся органиче-
ской взвесью, приносимой течениями и добываемой при помощи щупалец,
движений мерцательного эпителия или взмахов конечностей — кораллы
Madreporaria, губки Demospongia, часть полихет (Sabellidae, Serpulidae),
мшанки, плеченогие, лопатоногие, усоногие, а также пелагические остракоды.
Вся организация этих групп животных такова, что они не могут изменить
способ захватывания пищи, будучи в этом отношении очень узкоспециа-
лизированными.
Во-вторых, это исключительно или преимущественно грунтоеды: сипун-
кулиды, многие многощетинковые черви, приапулиды. Эти животные рас-
сматриваются как безвыборочно заглатывающие грунт при помощи втяжной
глотки или хобота, причем такой способ захватывания пищи остается
неизменным на всех заселенных ими глубинах (Соколова, 1958). Немного-
численные хищные глубоководные донные полихеты, по данным М. Н. Соко-
ловой (19566), также не изменили характера своего питания по сравнению
с мелководными. Однако, может быть, не случайно оба стенобатных глу-
боководных рода донных полихет — Macellicephaloides и Vitjazia — при-
надлежат как раз к хищникам.
К детритоедам, собирающим пищу с поверхности грунта и не изменив-
шим на больших глубинах такой способ питания, можно отнести большинство
Двустворчатых моллюсков, изученных, к сожалению, в интересующем нас
отношении далеко не достаточно. Иеккель (Jaeckel, 1955) разобрал специ-
ально адаптации глубоководных моллюсков, но никаких особенностей их
Питания не отметил. 3. А. Филатова (1958) указала на преобладание
169
Таблица 4. Группировки классов и отрядов по относительному количеству
в них стенобатных глубоководных родов
0 1-25% 26—50 % 51 — 100 %
Calcarea Demospongia Pennatularia Echiuroidea
Hydrozoa (донные) Gorgonaria Madreporaria Leptostraca
Antipatharia Alcyonaria Hydromedusae Mysidacea (донные и
Zoantharia Actiniaria Scyphomedusae пелагические)
Ceriantharia Siphonophorae Copepoda (пелагиче- Hyperiidea
Sipunculoidea Polychaeta ские) Gammaridea (пелаги-
Priapuloidea Ostracoda (пела- Isopoda ческие)
Cirripedia гические) Decapoda (пелаги- Monoplacophora
Cumacea Amphipoda (дон- ческие) Crinoidea
Pantopoda ные) Cephalopoda Asteroidea
Amphineura Tanaidacea (донные и пелагиче- Holothurioidea
Scaphopoda Euphausiacea Decapoda (дон- ные) Prosobranchia Opisthobranchia Bivalvia Bryozoa Brachiopoda ские Ophiuroidea Echinoidea Chaetognatha Pisces (пелагические) Ascidia Pogonophora Pisces (донные)
на больших глубинах двустворчатых моллюсков — собирателей детрита
с поверхности дна. У абиссальных моллюсков увеличены околоротовые
лопасти, снабженные сократимыми мускулистыми придатками и особым
осязательным жгутом. Эти признаки особенно сильно выражены
у представителей сем. Ma 1 letiidae, включающего наибольшее количество сте-
нобатных глубоководных родов.
Аллен и Сандерс (Allen, Sanders, 1966) подробно изучили адаптации
к глубоководному существованию абиссальной Abra profundorum, относя-
щейся к широко эврибатному роду. По сравнению с более мелководными
видами того же рода A. profundorum обладает в большей степени редуциро-
ванными жабрами, служащими не только для дыхания, но и для транспорти-
ровки пищевых частиц, увеличенными пальпами и удлиненной задней киш-
кой. Все эти особенности связаны с необходимостью захватывать и перераба-
тывать большое количество грунта, бедного органикой по сравнению с мелко-
водными. Таким образом, в данном случае способ и объект питания у глубо-
ководного вида не изменились, но возникли частные приспособления к усво-
ению менее калорийной пищи. К сожалению, мы ничего не знаем об адапта-
циях исключительно глубоководного семейства двустворчатых Nuculanidae
и не располагаем никакими данными по способу питания глубоководных
брюхоногих.
Высокоспециализированные, питающиеся древесиной двустворчатые
моллюски не изменили с глубиной характер своего питания. По отношению
к роду Xylophaga, включающему шесть мелководных, десять батиальных,
пять абиссальных и один ультраабиссальный вид, Кнудсен (Knudsen, 1961,
1967) специально рассмотрел этот вопрос и установил, что на всех глубинах
представители этого ряда питаются одинаково, усваивая целлю’лозу. Донные
бокоплавы используют в пищу преимущественно органические частицы
грунта. Питание глубоководных представителей отряда изучал Бернар
(Barnard, 1962), не обнаруживший различий в этом отношении между ними
170
и мелководными видами. Питанием глубоководных Ostracoda, Cumacea и Та-
naidacea никто специально не занимался, но все известное о способах захвата
пищи этими ракообразными и о ее характере позволяет допустить, что и они
на больших глубинах питаются так же, как на малых. Среди пелаги-
ческих Ostracoda только один глубоководный эндемичный род Gigantocypris
перешел к хищничеству.
Наконец, среди групп хищных беспозвоночных, не изменивших способ
питания с глубиной и характеризующихся малой степенью таксономиче-
ской обособленности, фигурируют актинии, донные десятиногие ракообра-
зные, пантоподы и пелагические сифонофоры.
М. Н. Соколова (1957) тщательно сопоставила питание глубоководных
и мелководных десятиногих ракообразных. Оказалось, что как глубоко-
водные, так и мелководные Crangonidae — стенофаги и питаются одними
и теми же семействами полихет и сходными мелкими ракообразными — кумо-
выми, бокоплавами или изоподами. Раки-отшельники, Galatheidae
и крабы как на малых, так и на больших глубинах выступают в качестве
хищников-эврифагов с весьма сходным набором объектов питания. Таким
образом, донные десятиногие ракообразные представляют собой редкий
пример хищников, сохранивших на больших глубинах тот же способ захвата
пищи и тот же ассортимент жертв, которые свойственны их
мелководным представителям. Можно предполагать, что с такой консерва-
тивностью связано очень быстрое качественное обеднение этой группы, по
мере увеличения глубины обитания и ее отсутствие на глубинах более
5300 м, где разреженность пищевых объектов не позволяет добывать их
в достаточном количестве.
Весьма показателен пример пантопод. Они могут питаться, только выса-
сывая губок и кишечнополостных. При обсуждении эврибатности родов пан-
топод Фаж (Fage,J954) подчеркивает стабильность способа их питания на
всех населенных ими глубинах, объясняя этим отсутствие стенобатных
глубоководных родов. Приблизительно то же можно предполагать и по
отношению к сифонофорам — пассивным хищникам, улавливающим мелких
животных своими арканчиками и, вероятно, не способным выработать
иной способ добывания пищи. Таким образом, между малой степенью
таксономической обособленности глубоководной фауны и неизменностью
способа питания на всех глубинах существует довольно полное соответствие.
Обращаясь к группам, характеризующимся высокой степенью таксономи-
ческой обособленности (больше 25 % стенобатных глубоководных родов)
и составляющим две последние колонки табл. 4, мы сталкиваемся с гораздо
более сложной картиной. Как уже указывалось, высокую степень таксоно-
мической обособленности глубоководной фауны некоторых групп (Pogo-
nophopa, Monoplacophora, стебельчатые Crinoidea, Echiuroidea, Mysidacea)
в какой-то мере можно объяснить их реликтовой природой — сохранением
многих или всех родов только на больших глубинах и исчезновением
на малых. В других случаях (голотурии, офиуры, морские ежи, Pennatularia,
Madreporaria) данных, которыми мы располагаем, оказывается недоста-
точно для объяснения полученных высоких показателей таксономической
обособленности глубоководной фауны. Возможно, что они также частично
связаны с реликтовой природой отдельных родов и семейств, а частично —
с факторами, пока не поддающимися анализу. Однако для многих групп
высокая степень таксономической обособленности глубоководной фауны
171
увязывается с явными усовершенствованиями или даже изменениями спо-
соба питания, возникшими у животных при освоении ими больших глубин.
Среди групп, в пределах которых глубоководная фауна резко обособлена
от мелководной и которые в той или иной степени изменили с глубиной
способ своего питания, также находятся представители всех основных
пищевых группировок, но соотношение их совершенно иное, чем среди групп
с необособленной глубоководной фауной. Так, например, невелико разнооб-
разие безвыборочно заглатывающих грунт детритоедов. К ним можно
отнести только глубоководное семейство морских звезд Porcellanasteridae
и некоторых голотурий из сем. Gephyrothuriidae и роды отряда Molpadonia
(Соколова, 1958). Сравнительно невелико и количество групп животных,
отсортировывающих грунт (большинство голотурий, неправильные морские
ежи), специфические адаптации которых к питанию на больших глубинах
остаются неизвестными.
Сестонофаги также представлены ограниченным количеством стенобат-
ных глубоководных родов, преимущественно асцидий и морских лилий. Среди
них интересен род Situla, приспособленный к фильтрации возможно большего
объема воды. Его вводной сифон настолько расширился, что стал равен
по диаметру туловищу, а жаберный мешок превратился в ловчую сеть,
затягивающую отверстие вводного сифона (Виноградова, 19696).
Нетрудно заметить, что среди животных, глубоководные таксоны которых
резко отграничены от мелководных, велико значение хищников. При этом
в ряде случаев есть основания предполагать, что в от’личие от донных десяти-
ногих ракообразных способ их охоты и набор их жертв на больших
глубинах совершенно иной, чем на малых. Это особенно четко видно на при-
мере пелагических организмов, среди которых в батипелагиали резко пре-
обладают именно плотоядные животные (Виноградов, 1968).
Таким образом, в условиях относительной бедности пищевых ресурсов
средних глубин, т. е. батипелагиали, хищничество оказывается весьма эффек-
тивным способом питания, имеющим явные преимущества перед фильтра-
цией. Действительно, согласно Йоргенсену (Jorgensen, 1962а, b), энергети-
ческие затраты планктонных фильтраторов возмещаются только в том слу-
чае, если количество усвояемого органического детрита в воде превышает
25 мг/л. Фактически на больших глубинах оно может быть значительно
меньше (Кгеу, 1964; Hagmeier, 1964).
Если способ захватывания пищи фильтраторами или пассивными хищни-
ками типа сифонофор принципиально не меняется при переходе животных
к жизни на больших глубинах, то активные хищники вынуждены радикально
перестроить характер охоты за своими жертвами. При крайней разрежен-
ности жертв активный их поиск становится энергетически невыгодным, так
как на преследование жертвы хищник тратит больше энергии, чем приобре-
тает в результате ее поедания. Эволюция пелагических хищников на больших
глубинах пошла но'направлению выработки тонких и совершенных приспо-
соблений, позволяющих предельно сократить энергетические затраты (Вино-
градов, 1968). Это прослеживается на всех системах органов и подробно
разобрано в ряде специальных исследований (Marshall, 1960, 1963);
Walters, 1961, и др.). Добывание пищи такими животными осуществляется
путем пассивного подкарауливания и приманивания жертв, для чего служат
различные фотофоры рыб и головоногих, удлиненные ловчие конеч-
ности некоторых ракообразных и т. п.
172
Способ охоты глубоководных хищных рыб на примере рыб-гадюк (Chau-
liodus) ярко описан Ж. Пересом (цит. по: Marshall, 1960), наблюдавшим
этот процесс из батискафа: «Эти рыбы неподвижно парят в воде, продольная
ось тела наклонена у них под острым углом к горизонту. При этом длинный
второй луч спинного плавника, снабженного светящимся органом, загнут
вперед так, что кончик луча расположен перед ртом». Инертность и почти
полная неподвижность гигантской глубоководной медузы Deepstaria enigma-
tica по сравнению с обычными сцифомедузами поразила исследователей,
наблюдавших за ней из подводного аппарата «Deepstar» (Barham, Pickwell,
1969).
Та же тенденция к переходу от активного к пассивному хищничеству
по мере уменьшения пищевых ресурсов проявляется, но гораздо менее четко,
и у донных животных. Как указывает М. Н. Соколова (1969), плотоядные
морские звезды, которые, по нашему мнению, могут считаться активными
хищниками, ограничены в своем распространении областями с эвтрофными
условиями. В олиготрофных районах среди хищников преобладают актинии
сем. Actinostolidae, т. е. пассивные хищники.
К сожалению, адаптации к пассивному хищничеству на дне морфологи-
чески не столь ясно выражены, как у пелагических животных. Однако
отдельные примеры соответствующих приспособлений известны из литера-
туры. В этом отношении большой интерес представляет донный угорь
Galatheathauma axeli с фотофорами, расположенными внутри огромного
зубастого рта, который закрывается, когда привлеченная светом жертва
окажется в его полости (Wollf, 1961b). Другая донная рыба — Bentho-
saurus — стоит на грунте, опираясь на удлиненные лучи своих грудных
плавников и ожидая в такой позе жертвы.
Другим приспособлением к усвоению редких на больших глубинах жертв
можно считать удлинение лучей у морских звезд, позволяющие облавливать
значительную площадь дна. Своего крайнего выражения эта тенденция дос-
тигает у преимущественно глубоководного семейства Brisingidae.
Следует наконец упомянуть о характерных приспособлениях кишечника
некоторых глубоководных пелагических животных, способствующих их так-
сономической обособленности и связанных с разреженностью их жертв на
больших глубинах. Исключительно глубоководный род и вид тонкопан-
цирного рачка Nebaliopsis typica обладает способным растягиваться «кишеч-
ным мешком»: рачок может поглощать очень большое количество пищи,
высасывая икринки рыб или головоногих. Отряд Leptostraca не имеет
пелагических мелководных представителей, так что особенности строения
Nebaliopsis можно связывать не только с приспособлением к жизни на
больших глубинах, но и с переходом в пелагиаль. Однако способность
единовременно захватывать большое количество пищи свойственно и ряду
Других глубоководных пелагических животных. Так, желудок гиперид Lan-
ceolidae способен настолько раздуваться, что рачок превращается в шар
с торчащими в стороны конечностями, а некоторые батипелагические рыбы,
как хорошо известно, могут заглатывать добычу, мало уступающую им или
Даже значительно превышающую их по размерам.
Наряду с перечисленными случаями усовершенствования глубоковод-
ными животными способов питания, присущих их мелководным родичам,
известны примеры радикальной смены характера питания по мере осво-
ения больших океанических глубин. В таких случаях, естественно, имеет
место и значительная степень таксономической обособленности глубоко-
173
водной фауны от мелководной. Хорошим примером такого явления могут
служить некоторые асцидии.
Представители двух исключительно глубоководных семейств асцидий
(Octacnemidae и Haxacrobylidae, выделяемых одно — в особый подкласс,
а другое — в особый отряд) отличаются от остальных асцидий, как предпо-
лагают занимавшиеся ими исследователи (Ritter, 1906; Madsen, 1947;
Monniot CL, Monniot F., 1968), способностью отрываться от субстрата и пла-
вать в придонных слоях воды, а также хищничеством. Обе эти особенности
несомненно тесно взаимосвязаны и обе развились независимо у неродствен-
ных между собой семейств, объединенных только глубоководностью. Тенден-
ции к развитию хищничества и подвижности совершенно не проявля-
ются у мелководных асцидий, и супруги Монио применительно к асцидиям
макрофагам с полным правом говорят о новом направлении адаптации Pro-
chordata. Очевидно, фильтрация придонного сестона на больших глубинах
оказалась настолько малоэффективным способом захвата пищи, что смени-
лась хищничеством, с чем связано развитие щупалец у Octacnemus, превра-
щение вводного сифона в насос у Gasterascidia и другие изменения
организации, приведшие к возникновению у асцидий не только особых
глубоководных семейств, но даже более высоких таксонов.
У двустворчатых моллюсков по мере увеличения глубины также намети-
лась тенденция к уменьшению роли фильтрации и увеличению роли хищни-
чества, но у них не происходит в связи с этим обособления особых
таксонов, а наблюдается лишь замена одних эврибатных родов другими.
Оккельман (Ockelman, 1958) установил, что в северной Атлантике на глубине
менее 400 м на долю фильтраторов приходится 62—77%, на долю грун-
тоедов — 14 — 28,5 % и на долю хищников — 4 — 9,5 % видов. На глубине
400—1000 м относительное количество фильтраторов понижается до
55,5 %, а грунтоедов и хищников возрастает до 29 и 15,5 % соответственно.
Наконец глубже 1000 м фильтраторов еще меньше (52 %), а грунтоедов
и хищников больше (34,5 % и 13,5 %). По мнению Кнудсена (Knudsen,
1967), относительное значение хищных двустворчатых на больших глубинах
северо-западной части Индийского океана еще выше. Их пищей служат
мелкие ракообразные и полихеты, причем Pectinidae добывают их как со дна,
так и из толщи воды во время плавания. Замечательно, что мелководные
виды Amissium (например, A. pleuronectes) —типичные фильтраторы, в то
время как глубоководные — хищники. В соответствии с этим у них различно
строение жабр.
Можно, однако, привести и притивоположные примеры отказа от хищни-
чества на больших глубинах и переходов к иному способу питания, что, воз-
можно, связано с чрезмерной разреженностью жертв. К сожалению, мы пока
не можем объяснить, почему одни группы на больших глубинах переходят
к хищничеству, а другие, наоборот, отказываются от него, но в обоих случаях
смена способа питания приводит к формированию самостоятельных таксонов.
С этой точки зрения интересен пелагический головоногий моллюск Cirro-
thauma murrayi, пассивно улавливающий взвешенные в воде органические
частицы. Замечательно, что в связи с таким необычным для головоногих
моллюсков способом питания у него в отличие от других головоногих
моллюсков полностью редуцированы глаза. Своеобразие его организации
заставляет выделить его в особый род.
Можно допустить, что исключительно глубоководное семейство морских
174
звезд — Porcellanasteridae — также сформировалось в какой-то степени
в связи с отказом от типичного для всего класса хищничества и переходом
к заглатыванию'грунта.
Помимо перехода на хищное питание или отказа от него, можно наме-
тить еще один тип смены способа питания на больших глубинах, известный
для пелагических животных, — переход к паразитическому существованию.
Высокие показатели степени таксономической обособленности глубоко-
водной фауны, которыми характеризуются гаммариды и гиперииды, связаны
с тем, что большинство представителей этих групп, судя по их строению
и отчасти по содержимому их кишечника, питаются за счет крупных
кишечнополостных и ведут полупаразитический образ жизни. У большин-
ства мелководных представителей гаммарид (донных; мелководные пелаги-
ческие виды единичны) и гипериид способ питания иной.
Сходный путь эволюции прошел из равноногих ракообразных глубоковод-
ный род Anuropus, выделяемый в особое семейство. Как недавно выяснилось
(Barham, Pickwell, 1969), он проникает в гастроваскулярную полость
упоминавшейся выше сцифоидной медузы Deepstaria enigmatica и питается
за ее счет, будучи специально приспособленным к такому образу жизни.
Заканчивая сопоставление групп, характеризующихся высокой степенью
таксономической обособленности глубоководной фауны от мелководной,
с особенностями их питания, следует еще раз подчеркнуть отсутствие
прямой и однозначной зависимости между этими показателями. Высокая сте-
пень таксономической обособленности глубоководной фауны в ряде случаев
связана с реликтовым характером группы, в некоторых случаях просто
не поддается пока объяснению, но во многих случаях явно определяется
трофическим фактором. Его ведущее значение подчеркивается тем фак-
том, что во всех случаях явного усовершенствования или перемены
характера питания в условиях резкого уменьшения пищевых ресурсов
на глубинах наблюдается четкое обособление глубоководных таксонов.
Этот процесс весьма несовершенно отражается полученными нами пока-
зателями, абсолютная величина которых сильно зависит от характера исход-
ного материала, от несовершенства наших функционально-морфологических
представлений, а также от биологии каждой данной группы. Так, например,
поскольку, как разбиралось выше, переход на больших глубинах к пассив-
ному хищничеству произошел в первую очередь у рыб и головоногих,
именно у этих групп можно было бы ожидать наиболее четко выраженную
обособленность глубоководной фауны от мелководной. Получившиеся в дей-
ствительности сравнительно скромные показатели объясняются, помимо не-
точности исходных данных, чрезвычайной вертикальной подвижностью мно-
гих представителей обеих этих групп. Совершая вертикальные миграции
большой амплитуды, они активно питаются в поверхностных слоях, а затем,
опускаясь на глубины, учитываются как глубоководные животные. За счет
этих мигрантов процент стенобатных глубоководных родов головоногих и рыб
несомненно снижается. Тем не менее кардинальные изменения организации
многих из них при переходе от активной в поверхностных слоях к пассивной
на больших глубинах охотно находят свое отражение в сравнительно
высоком абсолютном числе принадлежащих к этим группам эндемичных
глубоководных семейств и родов.
175
ЗАКЛЮЧЕНИЕ
Глубоководную фауну следует рассматривать как сложный, весьма гете-
рогенный комплекс животных, заселивших большие океанические глубины
в различные периоды геологического времени и эволюционировавших при
этом в разных направлениях и с различной скоростью. Прямой зависи-
мости между масштабами морфологических изменений, возникших
у групп при освоении ими больших глубин, и временем их существования
на больших глубинах,по-видимому, нет. Более важное значение приходиться
придавать адаптации к жизни на больших глубинах.
Имеющиеся весьма неполные данные позволяют выявить ведущую роль
трофического фактора, определяющего различные направления адаптации
глубоководных животных. Скудость пищевых ресурсов в значительной сте-
пени обусловливает специфику существования животных на больших океа-
нических глубинах; приспособления именно к этой особенности глубоковод-
ной среды выступают наиболее наглядно и легче поддаются анализу,
чем пока неизвестные приспособления к огромному гидростатическому дав-
лению, повышенной радиоактивности грунта, пониженной концентрации
кальция в воде и т. д.
В соответствии с планом своего строения одни группы животные в про-
цессе заселения бедных пищевыми ресурсами больших океанических глубин
сохранили свойственные их мелководным представителям способы захвата
пищи и ее характер, а другие — решительно их изменили. В первом случае
не потребовалось перестройки организации животных и, за некоторыми
исключениями, не возникли особые глубоководные крупные таксоны (роды,
семейства). Во втором случае, наоборот, приспособление к жизни на больших
глубинах сопровождалось развитием одних и редукцией других органов и
иными морфологическими перестройками, неизбежно приводившими к фор-
мированию новых крупных таксонов.
Высокая степень таксономической обособленности глубоководной фауны
от мелководной, установленная для некоторых животных, определяется не
только перестройкой их организации в процессе освоения ими больших океа-
нических глубин, но иногда и сохранением там представителей архаических
групп, исчезнувших в условиях морского мелководья в результате ожесточен-
ной борьбы за существование с более прогрессивными группами. Не всегда
удается разграничить эти две основные причины высокой степени таксоно-
мической обособленности глубоководной фауны от мелководной, тем более,
что в некоторых случаях они могут проявляться совместно.
У фильтраторов по мере увеличения глубины и оскудения пищевых
ресурсов прежде всего изменяются размеры. На больших глубинах преобла-
дают более мелкие формы. Действительно, более мелкие особи могут удовлет-
вориться меньшим количеством пищи, и поэтому именно они получают на глу-
бинах преимущественные возможности размножения. Грунтоеды, пропуска-
ющие через кишечник большое количество малокалорийной пищи, легко
приспосабливаются к жизни на любой глубине. И те и другие, не меняя
и принципиально не совершенствуя присущий им способ захвата пищи,
обычно не преобразуются на больших глубинах настолько, чтобы дать
начало особым крупным таксонам.
Наиболее интересна судьба хищников. При разреженности жертв обыч-
ная на малых глубинах активная охота оказывается энергетически невыгод-
176
ной и в условиях резкого дефицита пищевых ресурсов заменяется пассивной,
охотой, что требует значительных и разнообразных перестроек организации
и приводит к возникновению новых таксонов. Очевидно, гораздо большая
роль плотоядных форм в глубоководной пелагической фауне, чем в донной,
приводит к ее большей таксономической обособленности, о чем говорилось
выше. И если для пелагиали океана рассматриваемые зависимости могут
считаться более или менее строгой закономерностью, то для более пестрых
условий дна океана можно говорить лишь об общей тенденции, не отрица-
ющей возможности исключений.
Переходя от активной охоты к пассивной, глубоководные хищники сходят
с пути прогрессивной эволюции, по которому следуют активные хищники.
Совершенствование нервной и мышечной систем после перехода к пассивному
хищничеству не должно поддерживаться естественным отбором, направля-
ющим теперь организмы на удлинение ловчих придатков, развитие прима-
ночных органов, совершенствование плавучести и других приспособлений,
позволяющих свести к минимуму энергетические траты, столь трудно
восстанавливаемые при скудости пищевых ресурсов. Таким образом, хищ-
ники, так же как и все остальные питающиеся иным способом животные,
на больших океанических глубинах не имеют перспектив эволюционировать
прогрессивно. Это одна из немногих особенностей, присущих всей глубоко-
водной фауне в целом.
Настоящая работа не могла бы быть выполнена без помощи целого
ряда специалистов, критически просмотревших имеющийся литературный
материал по различным группам, а по ряду разделов давших по нашей
просьбе краткую сводку опубликованных и имеющихся у них данных. Мы
выражаем глубокую благодарность И. И. Акимушкину, Г М. Беляеву,
Н. Г. Виноградовой, О. Н. Зезиной, В. М. Колтуну, Р. Я. Маргулис,
Д. В. Наумову, К. Н Несису, Н. В. Ларину, Ф. А. Пастернаку, Т. С. Рассу,
Е. А. Романкевичу, Ю. А. Рудякову и 3. А. Филатовой.
Авторы чрезвычайно признательны также Л. А. Зенкевичу, Г М. Беляеву,
Н. Г. Виноградовой и М. Н. Соколовой, взявшим на себя труд внимательно
прочитать рукопись и сделать ряд ценных замечаний.
12 Заказ 333
ФИЛОГЕНИЯ и ЭВОЛЮЦИЯ
ПОНЯТИЕ «РЕЛИКТ* В БИОЛОГИИ*
ВВЕДЕНИЕ
Проблема реликтов может считаться одной из основных проблем биогеог-
рафии. Термин «реликт» постоянно употребляется в ботанической и зооло-
гической литературе, им оперируют при построении широких обобщений
(например, «теория оттесненных реликтов»).
Однако далеко не всегда ясно, какой смысл вкладывается различными
авторами в этот термин, и во всяком случае можно с уверенностью утверж-
дать, что единого мнения по вопросу об объеме и содержании понятия
«реликт» не существует. Не давая определения этого понятия, Гандлирш
(Handlirsch, 1909) указывает на возможность рассматривать реликты
с точки зрения: 1) численности составляющих данную систематическую
группу особей (количественные реликты), 2) занимаемого ею пространства
(географические реликты) и 3) времени ее существования (реликты по про-
исхождению). Симпсон (Simpson, 1944) добавляет еще одну категорию,
выделяемую по признаку вариабельности — 4) таксономические реликты,
т. е. группы, ранее вариабельные, а теперь представленные отно-
сительно немногими видами. Оба автора согласны с тем, что две первые
и четвертая категории объединяют близкие и, как правило, взаимно обуслов-
ленные явления, но не пытаются найти основную связывающую их нить.
Филогенетические реликты, по мнению Симпсона, принципиально отличаются
от остальных’, так как им якобы далеко не всегда свойственны сокращенный
ареал, малочисленность и пониженная вариабельность.
Исследователи, пытавшиеся дать определенные понятия «реликт»,
обычно забывали его многогранность, правильно подчеркнутую Гандлиршем,
и подходили к явлению весьма односторонне.
Несмотря на наличие специальных зоологических и ботанических работ,
посвященных проблеме реликтов (Берг, 1916; Ильин, Крашенинников, 1938;
Рылов, 1921; Ekman, 1915), формулировка этого основного в биогеографии
понятия, как я постараюсь показать, еще далека от желательной ясности.
Различия в суждениях по вопросу о признаках реликтов позволяют
наметить три группы исследователей, предлагавших тот или иной основной
критерий реликтовой природы организмов. Границы между этими группами
не всегда достаточно четкие, так как, помимо основного ведущего признака,
для определения понятия нередко привлекаются и другие признаки в качестве
второстепенных.
* Зоол. журн., 1947, т. 26, вып. 4, с. 313—330.
178
ТАКСОНОМИЧЕСКИЙ КРИТЕРИЙ
Первые зоологи, оперировавшие термином «реликт», подходили к явле-
нию прежде всего с позиций систематики, учитывая родственные отношения
изучаемых форм, а затем для объяснения их распространения привлекали
данные палеогеографии. Именно так поступил Ловен (Loven, 1862), впервые
употребивший этот термин в зоологической литературе. Он обнаружил
в озерах Веттерн и Венерн четыре вида ракообразных морского происхожде-
ния, и в том числе мизиду, описанную им в качестве нового вида под назва-
нием Mysis relicta. Для объяснения нахождения в указанных озерах живот-
ных морского происхождения Ловен использовал геологические данные
о морских послеледниковых трансгрессиях, захватывавших озера Ветерн и
Венерн, и рассматривал найденных им ракообразных как остатки фауны
отступившего впоследствии моря. Считая, по всей вероятности, вполне
понятным термин «реликт», Ловен не останавливается на его толковании.
На сходную позицию встал впоследствии Везенберг-Лунд (Wesen-
berg-Lund, 1910), значительно сузив, однако, первоначальное толкование
Ловена. Он отказался от учета географических моментов и ограничился
установлением родственных отношений тех или иных животных. Согласно
Везенберг-Лунду, морскими реликтами в пресных водах следует считать
все формы морского происхождения, независимо от способа и времени
их вселения. Иогансен (Johansen, 1908) предложил подходить к установле-
нию природы организмов гораздо строже и считать реликтами только тех
животных, ближайшие предки которых точно установлены палеонтологи-
чески.
Прямыми последователями Ловена можно считать также тех геологов
и географов, которые имели дело с реликтовыми организмами. Для этих
исследователей реликтовая фауна, понимаемая ими, согласно Ловену,
как остаток морской фауны, представляла интерес не сама по себе, а исклю-
чительно в качестве показателя морского происхождения того или другого
водоема., Термин «реликтовое озеро» был предложен Пешелем (Peschel,
1875) для озер, являющихся остатками морского водоема и обязательно
содержащих морских животных-реликтов.
Так же подходят к реликтовой фауне целый ряд геологов (Кольбрунер,
Рютимейер, Зупан и др.), и в том числе Неймайр. По Неймайру (1904),
эта «реликтовая фауна» позволяет с несомненностью заключить, что упомя-
нутые озера были когда-то частью моря и что население их является остат-
ками первоначальной морской фауны. Такие озера носят, в свою очередь,
название «реликтовых», или «остаточных». Креднер (Credner, 1887) пред-
ложил более широкое понимание реликтовых озер, основываясь исключи-
тельно на геологических данных и не считаясь с присутствием или отсут-
ствием в них морских элементов фауны, поскольку морские животные могут
распространяться активно и попадать в озера, вовсе не связанные в своем
происхождении с морем.
По отношению к озерам взгляды Креднера представляются правильными.
По отношению к животным цитированные выше авторы подчеркивают
лишь их родство или сходство с морскими формами, т. е. учитывают только
одну особенность реликтовых форм. На этом же признаке основывается
определение реликтовой фауны, предложенное выдающимся советским
геологом Н. М. Страховым (1932): реликтовой (или остаточной) фауной
12*
179
вообще называется фауна, которая по составу своему сходна не с господ-
ствующей в данный момент на земле фауной, а с фауной, которая господ-
ствовала некоторое время тому назад и к данному моменту на большинстве
участков земной коры уже вымерла. Применение этого определения для
исследователя современной фауны оказывается почти невозможным из-за
крайней скудности палеонтологических материалов. Кроме того, им трудно
пользоваться в тех случаях, когда объектом исследования являются не целые
фауны, а отдельные виды, особенно если это молодые реликты, как, например,
Mysis oculata relicta и др.
ЭКОЛОГИЧЕСКИЙ КРИТЕРИЙ
Ограниченный систематико-морфологический подход к понятию реликта
вызвал довольно справедливую критику со стороны Экмана (Ekman, 1915).
По мнению Экмана, основным критерием, решающим вопрос о реликтовом
характере того или иного организма, является способ его распространения.
«Вид является в данной области реликтом в том случае, когда его существо-
вание там можно понять только так, что он сам или его предок заселил
данную область при естественных условиях, чуждых современным» (Ekman,
1915). Нетрудно видеть, что это определение является выражением экологи-
ческого подхода к проблеме реликтов, поскольку ударение делается на
условиях существования организма. Следует также отметить, что такое
понимание термина не охватывает огромного количества фактов, имеющих
самое непосредственное отношение к обсуждаемой проблеме. Прежде всего
выпадают многочисленные случаи сохранения древних форм при относи-
тельно неизменных условиях существования — в пещерах, на больших
океанических глубинах, в некоторых биотопах тропиков и пр. Далее, приняв
это определение, приходится предположить, что реликтовый организм
неизбежно должен изменить всю свою экологию, так как иначе он окажется
неприспособленным к современным условиям существования. Иными сло-
вами, приходится приписывать реликтам повышенную экологическую
лабильность, что противоречит многочисленным фактам.
Подходя к проблеме реликтов с экологической точки зрения, Экман
находит нужным изменить трактовку еще одного важного биогеографи-
ческого понятия — «псевдореликта». Под псевдореликтами Натгорст
(Nathorst, 1895) предложил понимать виды, которые, сделавшись реликтами
в другом месте, в данную область попали вторично, уже будучи реликтами;
такое представление весьма прочно укоренилось в фитогеографии. Экман
и к этому понятию применяет экологические критерии, считая псевдоре-
ликтами виды, не являющиеся реликтами в данной местности, а заселившие
ее вновь, но только при иных условиях, чем современные. Реликты и псевдо-
реликты противопоставляются аутоиммигрантам, под которыми Экман
понимает виды, способные заселять данную область при современных
условиях.
В. М. Рылов (1921) уже отметил, что изменение предложенного Натгор-
стом понимания термина «псевдореликт» понадобилось Экману, по-види-
мому, для того, чтобы применить его к формам, длительно существующим
в относительно неизменных условиях, в частности к так называемым леднико-
вым реликтам в высокогорных озерах, которые В. М. Рылов склонен считать
настоящими реликтами.
180
В самом деле, при сопоставлении даваемых Экманом определений
реликтов и псевдореликтов бросается в глаза очень большая близость
этих понятий, объединяемых обязательным для обеих категорий заселением
из ареала при иных, чем сейчас, условиях. Разница сводится лишь к тому,
что псевдореликты, попав в данный водоем при несходных с современными
условиях, могут затем обитать в неизменной среде, а реликты формируются
при изменении среды, что является непременным условием их становления.
Экман возражает против признания реликтами глубоководных обитателей
субальпийских озер, так как в этих водоемах «не происходило никаких
изменений естественных условий, которые могли бы их (этих животных —
Б.) сделать реликтами». С этой точки зрения ни пещерная, ни океани-
ческая глубоководная фауны не могут считаться реликтовыми (да и псевдо-
реликтовыми), несмотря на высказывания занимавшихся ими исследовате-
лей. Так, например, Кларк (Clark, 1913) находит возможным говорить
о глубоководной фауне как о реликтовой, что справедливо также и по отноше-
нию к пещерной фауне (Бирштейн, Виноградов, 1934), да и сам Экман
(Ekman, 1935) называет реликтами биполярные формы, а также считает
тропические виды в Средиземном море и Атлантическом океане реликтами
Тетиса, хотя ни те ни другие не пережили резких изменений среды.
Наряду с явным совершенно искусственным ограничением применения
понятия «реликт», построения Экмана страдают еще одним, не менее
существенным, недостатком. Посвятив много места аргументации целесо-
образности применения экологических показателей для определения реликто-
вости тех или иных видов, Экман не рзъясняет, в чем заключается эта, по его
выражению, «реликтовая природа» (Reliktennatur) организма. Он ограничи-
вается лишь указанием на обязательное ее возникновение при меняющихся
условиях.
Люпобытно, что, вернувшись ровно через 20 лет к формулировке понятия
«реликт», Экман (Ekman, 1935) на первое место ставит географический
признак — обязательное обитание вида изолированно от главной области
его обитания, — только дополняя его цитированным выше экологическим
положением. Однако это не устраняет отмеченных дефектов.
Согласно В. М. Рылову (1921), под реликтом мы подразумеваем остаток,
в зоогеографии — остаток фауны, в прежние геологические эпохи (или
эпоху) имевшей более обширное распространение и сохранившейся в дан-
ное время лишь в некоторых местах. Это определение, как отмечает его
автор, является чисто географическим и чрезвычайно близко к определению,
данному несколько ранее Л. С. Бергом (1916) и Н. Гофштейном (Hofstein,
1912). Реликт в этой формулировке определяется ареалом.
Однако далее В. М. Рылов становится на экологическую точку зрения,
переходя к обсуждению условий сохранения реликтовых форм в изолирован-
ных местонахождениях вне главной области их распространения. По его
мнению, такое сохранение могло произойти вследствие двух принципиально
различных причин: данная форма или приспособилась к изменившимся
Условиям существования и в таком случае должна именоваться адаптивным
реликтом, или условия существования остались неизмененными со времени
предыдущей геологической эпохи и сохранившиеся вместе с ними животные
(и растения) могут быть названы консервативными реликтами.
Отсюда следует, что только «адаптивные реликты» В. М. Рылова отвечают
понятию реликтов в смысле Экмана, так как только они заселили данную
181
область «при естественных условиях, чуждых современным». Однако
В. М. Рылов резонно полемизирует с Экманом, доказывая правомерность
отнесения к категории реликтов и тех форм, которые продолжают существо-
вать при неизменных условиях (консервативных реликтов).
В. М. Рылов отмечает, что между предложенными им категориями
встречаются все переходы и что отнесение какой-либо формы к той или
иной категории зависит от того, какой фактор внешней среды будет подверг-
нут анализу. В конкретном случае ледниковоморских реликтов относимые
к ним формы (Mysis oculata relicta, Limnocalanus grimaldii macrurus
и др.) являются адаптивными реликтами по отношению к солености и
консервативными — по отношению к температуре. Возникает вопрос, какой
же фактор следует считать руководящим при отнесении исследуемого вида
к той или иной категории. В. М. Рылов склонен выбирать те факторы, которые
связаны с происхождением данного организма, например для морских
реликтов в пресноводных водоемах — соленость, а для арктических релик-
тов — температура. Следовательно, первые целиком попадают в категорию
адаптивных, а вторые — в категорию консервативных реликтов. Нетрудно,
однако, заметить, что подобный выбор руководящего фактора является
совершенно произвольным. Мы не знаем, насколько значительную роль
играла температура при формировании ледниковоморских реликтов и не
можем с уверенностью отводить ведущее место в этом процессе солености.
Кроме того, совершенно очевидно, что любой организм обладает способно-
стью переносить известные колебания любого фактора внешней среды
в определенных пределах, и никем не доказано, что эвригалинность ледни-
ковоморских реликтов больше, чем их эвритермность.
Произведенные Е. В. Боруцким (1937) экспериментальные исследования
факторов, определяющих распространение такого типичного чисто пресно-
водного ледникового реликта, как Bryocamptus arcticus (Lili.), показали,
что ведущую роль в этом случае играют особенности химизма воды, а не
температура. Приходится, следовательно, считать его реликтом, адаптивным
по отношению к температуре и консервативным по отношению к химизму
воды, что противоречит установкам В. М. Рылова. Вряд ли можно сомне-
ваться в том, что аналогичные работы по экологии других ледниковых
реликтов выявят самые разнообразные факторы, лимитирующие их распро-
странение, и поставят под вопрос универсальность температурного фактора.
Итак, я не вижу серьезных оснований к противопоставлению друг
другу адаптивных и консервативных реликтов. Любой реликт и, вероятно,
любой организм оказывается в одно и то же время и адаптивным и консер-
вативным в зависимости от того, влияние какого фактора внешней среды
будет подвергнуто исследованию. По отношению к факторам, лимитирующим
распространение, все организмы, очевидно, оказываются консервативными,
по отношению к прочим факторам — более или менее адаптивными, в зави-
симости от экологической валентности вида. Различия между отдельными
видами в этом отношении количественные, зависящие от степени экологи-
ческой валентности. Таким образом, экологический подход В. М. Рылова
к проблеме реликтов не помогает ее разработке.
По Шретеру (Schroeter, 1913), реликтовый ареал характеризуется тем,
что его заселение должно было происходить при ранее существовавших
условиях, иных, чем сейчас. Согласно Штоллеру (Stoller, 1921) и Вангерину
(Wangerin, 1924), напротив, условия существования в реликтовом ареале
182
должны оставаться неизменными с момента проникновения в данный район
реликтового вида.
Ставя во главу угла проблему отношения реликта к окружающим усло-
виям, некоторые исследователи делают из своих определений довольно
странный вывод, противоречащий всем общепринятым представлениям. Они
признают возможность существования неприспособленного к среде орга-
низма. По мнению Шретера (Schroeter, 1913), реликтовое растение должно
находиться в дисгармонии со своими современными условиями обитания.
Е. В. Вульф (1932) считает реликтовым видом остаток древней флоры
или древнего рода, когда-то широко распространенного и находившегося
в полной гармонии со своими условиями обитания, в настоящее же время
ему большей частью не вполне соответствующими. П. П. Сушкин (1938)
полагает, что присутствие организма в экологически чуждой ему обстановке
может служить веским указанием на реликтовый характер этого нахождения
и что «факты распространения, не объясняемые современными условиями
или существующие вопреки им, мы можем объяснить только как реликт».
Завершением этой линии рассуждений можно считать формулировку
И. И. Пузанова (1938): реликтами в биогеографии называются формы
животных и растений, организация и ареалы которых находятся в явном
несоответствии с окружающей их неорганической и органической средой.
Нелегко представить себе реально такие организмы, и, вероятно, Пузанов
испытал бы немалые затруднения, если бы внимательные ученики попросили
его привести конкретные примеры реликтов в его понимании. Ведь приспо-
собленность организмов к среде, иными словами, целесообразность их
строения и отправлений, по справедливому выражению К. А. Тимирязева
(1943), является той основной чертой, которая поражает нас при изучении
органического мира.
Поскольку в высшей степени трудно допустить возможность существова-
ния животных и растений, организация которых находится в явном проти-
воречии со средой, подобные экологические критерии определения реликтов
приходится считать несостоятельными.
ГЕОГРАФИЧЕСКИЙ КРИТЕРИЙ
При использовании географического критерия реликтовой природы
организмов изучение ареалов приобрело в некоторых случаях совершенно
самодовлеющее значение. Ареал оказался оторванным от населяющего его
вида, и проблема реликтов расщепилась на независимые друг от друга
проблему реликтового вида, для решения которой применялись таксономи-
ческий и экологический методы, и проблему реликтового ареала, разре-
шаемую с помощью географического метода. Шретер считает необходимым
отличать реликтовые систематические единицы от реликтовых ареалов,
так как, по его мнению, между этими понятиями может и не быть соответ-
ствия. Он предлагает называть реликтовые систематические единицы
рестанцами, а термин «реликт» применять только по отношению к ареалам.
Для реликтового ареала, по Шретеру (Schroeter, 1913), характерны про-
странственная ограниченность, постепенно идущее сокращение, редкость
занимающего его вида и географическая или экологическая преемственность
с момента появления в нем населяющего ареал вида.
Взгляды Шретера были приняты некоторыми советскими ботаниками,
в частности Е. В. Вульфом. В одной из своих последних работ он также
183
указывает на возможность понимания термина реликт в двух смыслах —
систематическом и географическом, отдавая предпочтение последнему.
Согласно определению Е. В. Вульфа (1941), под реликтовым видом следует
понимать остаток более или менее древней флоры, имеющей реликтовый
ареал, занимаемый им в момент вхождения в состав означенной флоры.
Таким образом, в таком понимании «реликтовый вид» является понятием
географическим, связанным с историей расселения вида и независимым
от того, является ли данный вил систематически изолированным остатком
древнего рода или нет. Для таких систематически изолированных видов
следует применять другой термин, называя их, например, согласно предло-
жению Шретера, «рестанцами». Е. В. Вульф (1941) характеризует реликто-
вый ареал как остаток более обширного ареала, сформировавшегося при
ранее существовавших условиях, часто иных, чем современные. Е. В. Вульф
склонен приписывать реликтам определенный возраст, но, подходя и к этому
вопросу с узко географической точки зрения, он предлагает принимать
возраст реликта не с момента возникновения данного вида, а со времени
вхождения его в состав изучаемой флоры. Продолжая эту мысль, приходится,
очевидно, признать, что какой-либо вид как реликт может быть, например,
ледниковым, а как рестанец — третичным. Далее Е. В. Вульф указывает
на возможность такого положения, когда какой-либо вид оказывается релик-
товым только в части своего ареала. Таким образом, понятия «реликтовый
вид» и «реликтовый ареал» разрываются и во времени, и в пространстве.
Не говоря уже о том, что термин «реликт» впервые был применен к виду,
а не к его ареалу, следует указать на недопустимость такого отрыва. Эволю-
ционное развитие вида неразрывно связано с географическим развитием его
ареала, отображающим те подчас весьма сложные взаимоотношения, кото-
рые возникают у данного вида с другими видами и неорганической средой.
Поэтому формирование ареалов нельзя понять вне эволюции населяющих их
видов, и никаких особых законов, управляющих ареалами, не может суще-
ствовать.
Гроссгейм не ограничивается делением реликтов на адаптантов и эвре-
ликтов. Среди последних он различает еще несколько более мелких катего-
ареалы. Он склонен толковать понятие «реликт» довольно широко, применяя
его не только к видам, принадлежащим к прошлой для данной местности
флоре, но даже к происходящим от таких древних аборигенов формам.
Для расчленения этого общего понятия Гроссгейм обращается к ареалам
и, основываясь на их судьбе, различает среди реликтов виды, сокращающие
свой ареал, которые он именует «эвреликтами», и виды с постоянным
или расширяющимся ареалом, которые он предлагает называть «адаптан-
тами». Реликтам Гроссгейм противопоставляет «совокупность элементов
современной флоры», обозначая их термином «реценты». Среди рецентов
также могут быть виды с сокращающимся ареалом — «реценты-дегра-
данты» — и виды с расширяющимся ареалом — «эвреценты». При этом
Гроссгейм протестует против представления об обязательной индигенно-
сти 1 реликтов и считает ненужным термин «псевдореликт» в понимании Нат-
горста.
Мне кажется, что развиваемые Гроссгеймом соображения в значительной
степени стирают грань между реликтами и нереликтами. Это обстоятельство,
1 От англ, indigenous — туземный, местный. (Примеч. ред.).
184
по-видимому, несколько смущает и автора разбираемого построения. Он
пишет: «Далеко не всегда с полной точностью и определенностью можно
провести границу между реликтами и рецентами данной флоры; это зависит
и от малой изученности флоры. и от той ботанико-географической кон-
вергенции, каковая наблюдается в рядах эвреликты — реценты — дегра-
данты и эвреценты — адаптанты». Однако нетрудно заметить, что последова-
тельное применение терминов Гроссгейма к конкретному материалу застав-
ляет поставить под сомнение возможность существования в природе рецен-
тов. Если, как думает Гроссгейм, реликтами должны считаться все виды
растений, происходящие от древних флор, то нереликтов вообще не может
быть, поскольку преемственность происходящих друг от друга видов неоспо-
рима. Отрицание обязательной индигенности реликтов предоставляет полнук)
возможность относить к этой категории решительно все виды данной флоры
или фауны. В самом деле, в составе флоры или фауны любой местности можно
различать древние консервативные формы, развившиеся от них на месте
молодые виды и пришлые элементы разного возраста, но обязательно
происходящие от каких-то более древних форм. Ко всем этим категориям
можно, очевидно, применять термин «реликт» в расширительном толковании
Гроссгейма. В частности, любой вид, происходящий от древних предков,
для данной местности должен считаться реликтом и не перестает им быть,
переселившись в другую местность.
Гроссгейм не ограничивается делением реликтов на адаптантов и евре-
ликтов. Среди последних он различает еще несколько более мелких катего-
рий, которые отличаются друг от друга по степени реликтовости и совпадают
частично с предложенными в свое время Шретером (Schroeter, 1913) поня-
тиями — реликты эдафические, ценологические, климатические, географиче-
ские. Таким образом, Гроссгейм пользуется всеми намеченными выше
критериями реликтовости: к определению реликтов он применяет таксоно-
мический критерий, разделяет их на эвреликты и адаптанты с помощью
географического критерия и систематизирует эвреликты, используя экологи-
ческие критерии; иными словами, он подходит к категориям разного ранга
с разных точек зрения, но не пытается рассмотреть какую-нибудь категорию
всесторонне.
Несмотря на отмеченные недостатки, в работе Гроссгейма содержится
немало ценных мыслей. Ареал в его представлении является отражением
эволюционного развития вида и не приобретает самодовлеющего значения,
как в построениях Шретера и Вульфа. Формальный подход к изучению
ареала, без учета специфики эволюционного развития вида, может привести,
как следует из определения Гроссгейма, к смешению противоположных
понятий «реликт» и «рецент». Кроме того, Гроссгейм совершенно правильно
подчеркивает относительность понятия «реликт», указывая на реально
существующие в природе различные степени реликтовости.
Попытку сочетать географический и систематический критерии понятия
«реликт» делает также В. В. Алехин (1944). По-видимому, он считает
возможным в одних случаях применять географический, а в других —
систематический метод доказательства реликтовой природы растений.
Сначала он определяет реликты как виды с сократившимся ареалом, но
несколькими строками ниже указывает на существование реликтовых видов,
не имеющих реликтового ареала. Это противоречие объясняется тем, что
в случае невозможности палеонтологически доказать происшедшее сокраще-
185
ние ареала, Алехин рекомендует пользоваться систематическим методом,
считая реликтами виды, занимающие изолированное положение в системе.
Как видно, Алехина не смущает несовпадение выводов, к которым
приводит применение обоих методов; к тому же остается неясным, какой из
них следует считать более доказательным.
Чрезвычайно четкую и довольно удачную формулировку понятия «реликт»
дал Л. С. Берг (1916), основываясь исключительно на характере ареала
как единственном признаке реликтов. «Реликты есть такие подвиды и виды
(и, конечно, более высокие таксономические единицы), которые сохранились
в изолированных местах вне главной области (современного или геологи-
ческого) распространения данного подвида, вида, рода и т. д., причем
относительно промежуточной области можно предположить, что здесь рас-
сматриваемая форма когда-то жила, но затем вымерла. Если данный орга-
низм находится в изолированном местонахождении благодаря активной или
пассивной миграции, то в этом случае мы не говорим о реликте». Весьма
сходное определение понятия «реликт» содержится в работе Гофштёйна
(Hofstein, 1912).
Эти формулировки, предлагая известный признак, позволяющий по
характеру ареала считать тот или иной вид реликтом, не задаются целью
определить существо понятия. Кроме того, их трудно применить к таким
древним консервативным видам, которые продолжают обитать в течение
долгого времени в пределах исконной области своего распространения,
как, например, Lingula и некоторые другие океанические формы. Не совсем
понятно, кроме того, что, собственно, следует разуметь под «главной»
областью распространения, и приводимый Бергом пример не устраняет
возникающего недоумения: «Североамериканская пресноводная рыба Amia
(вид: A. calva) есть реликт в отношении распространения этого рода в тре-
тичное время, когда представители рода Amia были широко распространены
в Европе, Азии и Америке». Мы имеем дело в данном случае с сокращением
ареала, причем нет основания считать Европу и Азию главной областью
распространения Amia, а Америку нет.
ФОРМУЛИРОВКА понятия
Подводя итог этому краткому обзору, я прихожу к заключению об
отсутствии в литературе удовлетворительного определения понятия «реликт».
Разбор отдельных суждений по этому поводу позволяет использовать
некоторые правильно подмеченные цитированными авторами особенности
реликтов и в то же время выясняет требования, которые следует предъявить
к формулировке этого понятия. Надо, очевидно, прежде всего определить
сущность этого понятия, а затем указать признаки, на основании которых
можно отличать реликты при конкретных исследованиях.
В соответствии с буквальным значением этого слова под реликтами
следует понимать остатки прежних фауны и флоры. Отсюда следует, что,
во-первых, реликтовые виды и другие систематические единицы сравнительно
мало изменились с отдаленных времен, т. е. характеризуются замедленным
темпом эволюции, и, во-вторых, близкие им формы исчезли, будучи вытеснены
более прогрессивными видами или уничтожены неблагоприятными условиями
абиотической среды. Факты сохранения в мало измененном состоянии
древних форм были отмечены в свое время Дарвином (1937), указавшим
186
и на условия, благоприятствующие этому. В «Происхождении видов»
сказано: «Но будут ли виды, уступившие свои места другим, измененным
и усовершенствованным видам, принадлежать к тому же самому или к иному
классу, немногие из числа уступивших могут иногда сохраниться на продол-
жительное время благодаря тому, что они приспособились к каким-нибудь
особенным условиям жизни или благодаря тому, что они заняли удаленную
и изолированную область, где они избегли сильной конкуренции. Например,
некоторые виды Trigonia, обширного рода моллюсков, распространенного
во вторичных формациях, сохранились в австралийских морях; немногие
представители большой и почти вымершей группы ганоидных рыб до сих пор
живут в наших пресных водах» (с. 514). Вот эти-то «уступившие», т. е.
побежденные в борьбе за существование, но дожившие при определенных
условиях до наших дней виды и следует, как мне кажется, именовать релик-
тами.
Эти простые установки, базирующиеся на своеобразии эволюции релик-
тов, позволяют без труда наметить основные признаки, характеризующие
реликтовые формы в систематическом, географическом и экологиче-
ском отношении. Исходя из второго тезиса только что приведенного опреде-
ления и указания Дарвина на сохранение «немногих из числа уступивших»,
следует принять, что реликтовые виды (и более высокие систематические
единицы) относятся к родам и семействам, насчитывавшим в прежние
геологические эпохи значительно большее число видов (и родов), чем сейчас.
Ареал реликтовых форм есть остаток или остатки более обширного ареала,
занимавшегося данным видом или родом в прежние геологические эпохи,
поскольку мы имеем дело с формами, «уступившими свои места другим»,
Реликты узко приспособлены к определенным, специфическим условиям
существования, что и является одной из причин сокращения их ареалов и
вообще поражения в борьбе за существование. В отличие от Экмана, я не
вижу никаких оснований к тому, чтобы остерегаться приписывать реликтам
определенный геологический возраст, и считаю, что возраст реликта опреде-
ляется временем формирования данного вида, рода и т. п. Понятие реликта
относительно: разные виды могут быть реликтовыми в разной степени.
Теснейшая взаимосвязь между снижением темпа эволюционного про-
цесса и сокращением ареала не подлежит сомнению. Еще Уолес
(Wallace, 1858) в своей первой работе отметил существование обратной
зависимости между постоянством видов и обширностью занимаемого ими
пространства. Дарвин (1937) также указал на влияние величины ареала
на темп эволюции населяющего его вида. Говоря о значении изоляции,
он, между прочим, утверждал: «Если же изолированная площадь будет очень
мала, .общее количество ее обитателей будет мало и это замедлит образова-
ние новых видов посредством естественного отбора, так как уменьшатся
шансы на появление благоприятных изменений» (с. 160).
«Я прихожу к заключению, — пишет далее Дарвин, — что хотя неболь-
шие изолированные ареалы в некоторых отношениях были крайне благо-
приятными для образования новых видов, но тем не менее в обширных
областях изменения в большинстве случаев совершались быстрее и, что еще
важнее, формы, вновь образовавшиеся на больших территориях и уже
победившие многих соперников, были более способны к широкому рассе-
лению и, следовательно, к образованию наибольшего числа разновидностей
и видов» (с. 162).
187
Дарвин констатировал чрезвычайно важный факт сохранения в пресных
водах «живых ископаемых», объяснив его малой величиной прёсноводных
водоемов по сравнению с океаном и сушей и как следствие этого более
медленным темпом эволюции пресноводных форм благодаря «менее разно-
образной и, следовательно, менее ожесточенной конкуренции». И действи-
тельно, насыщенность реликтами именно пресноводной фауны по сравнению
с морской и сухопутной может считаться твердо установленной.
Теоретические расчеты Фишера (Fisher, 1930) привели его к выводу
о пропорциональности между эффективностью селекции и дисперсией (62)
рассматриваемого вида, откуда следует, что при большей численности
популяции она будет эволюционировать быстрее.
Райт (Wright, 1931, 1940) подверг специальному математическому
анализу вопрос о связи между темпом эволюции и размерами популяции
и выяснил, что в небольших популяциях эволюция должна идти медленнее,
чем в популяциях среднего размера.
Кинси (Kinsey, 1937) исследовал две категории видов галловых ос
(Cynips), из которых одна категория объединяет виды с небольшими (в сред-
нем 4600 миль2), а другая — виды с обширными (в среднем 300 000 миль2)
ареалами. Оказалось, что виды первой категории на 80 % отличаются
высокой степенью константности своих признаков, а виды второй категории,
наоборот, на 80 % весьма вариабельны. По мнению Кинси, «эта большая
разница в размерах видовых популяций является первичной основой разли-
чий их эволюционной истории».
Непосредственные причины замедления эволюционного процесса вслед-
ствие уменьшения ареала, по-видимому, довольно сложны. Возможно, они
связаны с наибольшей мутабильностью в обширных популяциях и низкой —
в небольших изолированных популяциях. В этом отношении интересны
данные А. П. Соколовской и О. С. Стрелковой (1941) по кариологическому
анализу флоры острова Колгуева. Эти авторы показали, что из трех разно-
родных групп растений, произрастающих на о-ве Колгуев, широко распро-
страненные бореальные и арктические виды оказываются в значительной
степени или поголовно полиплоидами. Наоборот, среди аркто-альпийских
видов, обладающих разорванными и ограниченными ареалами, процент
полиплоидов довольно низок (36 %).
Уменьшение ареала того или иного вида, связанное, как правило, с умень-
шением его численности, неизбежно приводит к замедлению темпа его
эволюции. Не менее очевидно, однако, что, выступая в качестве законо-
мерного следствия уменьшения ареала, замедление темпа эволюции может
явиться и причиной этого уменьшения. В самом деле, изменяющиеся медлен-
нее виды в борьбе за существование, согласно принципу Тейлора, будут
вытесняться видами, эволюционирующими более быстрым темпом, и ареал
первых должен начать сокращаться. Это сокращение, по изложенным выше
соображениям, вызовет дальнейшее замедление темпа эволюции, т. е. будет
усугублять реликтовость видов.
Симпсон (Simpson, 1944) возражает против необходимости приписывать
замедленный темп эволюции реликтам как группам с сократившимся
ареалом. По его мнению, каждой «линии» свойствен определенный, характер-
ный темп эволюции. При этом каждая линия проходит начальную фазу
небольшого ареала, затем следует фаза расселения по обширному ареалу и,
наконец, фаза сокращенного ареала. Следовательно, быстро эволюциони-
188
руюшие группы стремятся скорее стать реликтами, чем медленно эволюцио-
нирующие. В то же время «древняя группа скорее будет в фазе сокращенного
ареала, чем молодая и окажется тем самым географическим реликтом
потому, что она старая, а не потому, что она замедленно эволюционирует».
Высказанные Симпсоном взгляды вызывают некоторые сомнения.
Прежде всего остается недоказанной неизменность темпа эволюции «линий».
Гораздо лучше согласуется с фактическим материалом представление об
изменениях темпа эволюции, определяемых переменами интенсивности
естественного отбора в пределах каждой данной линии на протяжении ее
геологической истории. Точно так же и сокращение ареала трудно рас-
сматривать просто как неизбежно наступающую фазу эволюции любой ли-
нии. На самом деле оно происходит, как правило, вследствие борьбы за
существование с другими организмами, причем очевидно, что эволюциониру-
ющие более быстрыми темпами линии окажутся в преимущественном
положении.
Итак, реликтами я предлагаю называть сохранившиеся в малоизменен-
ном состоянии остатки фаун (и флор) прошлых геологических эпох, характе-
ризующиеся замедленным темпом эволюции, принадлежащие к ранее более
многочисленным систематическим группам, населяющие ограниченный ареал
(или ареалы), являющийся частью ранее более обширного ареала, и узко
приспособленные к определенным, большей часть специфическим условиям
существования, мало изменившимся со времени формирования рассматри-
ваемых систематических единиц.
В этом определении не содержится ничего принципиально нового, и
отдельные заключающиеся в нем положения уже указывались предыдущими
авторами. Однако мне кажется, что в предлагаемой трактовке проблема
реликтов выступает прежде всего как эволюционная проблема и как таковая
она требует всестороннего подхода с позиций различных наук — систе-
матики, палеонтологии, экологии и биогеографии. В идеале только синтез
добытых всеми этими науками фактов может разрешить проблему замедлен-
ной эволюции побежденных в борьбе за существование форм, доживших
до нашего времени, т. е. реликтов в понимании Дарвина (не пользовавшегося
этим термином).
МЕТОДЫ ИССЛЕДОВАНИЯ
На практике, особенно в отношении пресноводной и солоноватоводной
фауны, область применения палеонтологии оказывается в высшей степени
ограниченной в связи с ужасающей неполнотой геологической летописи.
Поэтому строгие требования обязательного палеонтологического подтверж-
дения реликтового характера тех или иных форм, предъявленные Иогансе-
ном, Штоллером и Алехиным (Johansen, 1908; Stoller, 1921; Алехин, 1944),
в подавляющем большинстве случаев выполнить невозможно. Задачи
палеонтологии, в частности установление древности разбираемых форм и
вытекающие отсюда соображения о темпе их эволюции, перекладываются
на биогеографию, вынужденную решать,, кроме того, и свои собственные
задачи. Такая двойная нагрузка выдвигает биогеографию на первое место
сРеДи наук, призванных разрабатывать проблему реликтов.
Основным методом, позволяющим биогеографу установить геологический
189
возраст того или другого вида, рода и т. д. при отсутствии палеонтологи-
ческих данных является метод анализа разорванных ареалов. В качестве
примера применения этого метода можно привести хотя бы исследования
Реми (Remy, 1936) и Шелленберга (Schellenberg, 1939), посвященные
высшим ракообразным о-ва Мадейра. Оказалось, что на этом острове
обитает особый подвид итальянского водяного ослика (Asellus italicus
perarmatus) и подземный бокоплав, известный из грунтовых вод Алжира,
северной Испании и восточного побережья Адриатического моря (Pseudonip-
hargus africanus). Отстоящая сейчас на 545 км от Африки и на 800 км от
Европы Мадейра, вместе с Азорскими островами и островами Зеленого
Мыса, отделилась от материка до конца миоцена. Это дает возможность
Реми и Шелленбергу датировать возраст обоих видов по крайней мере
миоценом, причем с тех пор бокоплав не изменился совсем, а водяной ослик
образовал только подвид. С этих же позиций, как мне кажется, следует
подходить и к неоднократно упоминавшейся Mysis oculata relicta. Она
является реликтом постольку, поскольку можно утверждать, что со времени
проникновения в обитаемые ею озера, т. е. с вюрмского оледенения, она
эволюционировала достаточно медленным темпом, мало отклонившись от
родоначальной формы (до степени вариетета).
Отсюда ясно, какое большое значение приобретают для разрешения
проблемы реликтов случаи разрывов ареалов. Не являясь обязательным
признаком реликтовой природы видов, они указывают на возможное сокра-
щение первоначального ареала и иногда позволяют судить о темпе эволюции
населяющих такие ареалы организмов.
Однако исследование конкретных причин снижения темпа эволюции,
сокращения ареала и вообще поражения в борьбе за существование релик-
товых видов не может производиться биографическими методами и отно-
сится к компетенции описательной и экспериментальной экологии. К сожа-
лению, в этом направлении сделано немного. В общей форме значение
некоторых факторов для темпа эволюционного процесса было рассмотрено
И. И. Шмальгаузеном (1946).
Наконец, совершенно очевидно, что базой всей работы над реликтами
должна служить систематика, устанавливающая родственные отношения
изучаемых форм как с ныне живущими, так и с ископаемыми организмами.
ПРЕДЕЛЫ ПРИМЕНЕНИЯ ПОНЯТИЯ
Приложение сформулированных принципов к конкретному материалу
заставляет поставить вопрос о том, насколько глубоко реликтовость прони-
зывает всю эволюцию той или иной группы. Анализ соответствующего
материала показывает, что реликтовый род может включать наряду с релик-
товыми видами и нереликтовые. Более того, реликтовый род или более
крупная таксономическая единица может быть представлен только нерелик-
товыми видами. Очевидно, в этих случаях эволюция родов и видов измеряется
разными масштабами. Длительно задерживаясь на одном уровне высоты
организации, какой-либо род, семейство или отряд в порядке адаптивной
радиации может распадаться на значительное количество сравнительно
быстро изменяющихся нереликтовых видов. В качестве соответствующего
примера можно привести отряд сумчатых млекопитающих.
Этот отряд в целом следует, безусловно, считать реликтовым, поскольку
190
в предшествующие геологические эпохи он имел значительно более широкое
распространение и включал значительно больше представителей, чем
в настоящее время; наряду с этим он сохранил свои древние примитивные
морфологические особенности, т. е. как отряд эволюционировал замедленно.
Тем не менее в пределах своего современного остаточного ареала, в особен-
ности в Австралии, сумчатые за сравнительно короткий срок подверглись
интенсивной адаптивной радиации, причем в течение четвертичного периода
сформировался эндемичный подотряд двурезцовых.
Многие из молодых, быстро эволюционирующих видов сумчатых расши-
рили ареал, вытеснив своих конкурентов. Мне кажется, что нет никаких
оснований считать такие виды или подотряд двурезцовых реликтовыми,
несмотря на несомненную реликтовость всего отряда сумчатых в целом.
Не менее показательным примером является мечехвост Limulus, един-
ственный доживший до наших дней представитель класса Xiphosura,
достигшего своего расцвета в верхнем силуре. Род Limuius в ископаемом
состоянии известен из триаса и, следовательно, может считаться весьма
древним реликтом. Четыре современные вида этого рода населяют мелко-
водья восточного побережья тропической Америки, Молуккских и Зондских
островов и южной Японии. Ареалы отдельных видов сплошные, численность
особей каждого вида весьма значительна, ни один современный вид не был
найден в ископаемом состоянии, так что древность современных видов
остается неизвестной. Все это не позволяет уверенно квалифицировать
виды Limulus как реликтовые; между тем реликтовость рода не вызывает
никаких сомнений.
Можно было бы привести значительное количество аналогичных приме-
ров, но я ограничусь еще только одним, показывающим возможность
объединения в один род реликтовых и нереликтовых видов. Наши речные
раки (род Astacus) относятся к трибе Nephropsidea, включающей четыре
современных семейства: семейство речных раков северного полушария
Astacidae, два семейства речных раков южного полушария — Parastacidae
и Austrastacidae — и, наконец, одно семейство омаров — Nephropsidae,
состоящее главным образом из океанических глубоководных форм. В мезо-
зойское время Nephropsidea были широко распространены по мелководьям
Мирового океана; число семейств, родов и видов трибы было значительно
больше, чем сейчас. Так, например, согласно Стрелену (Straelen, 1936).
из юрских и меловых отложений известно 25 видов омаров, а из третич-
ных — всего 7. С начала кайнозоя, по Стрелену, Nephropsidea находятся
уже в стадии регресса. Они переходят на большие океанические глубины
и несколько ранее — в пресные воды, где и доживают до наших дней.
Род Astacus сформировался еще в юрское время. Центром распростране-
ния европейских видов этого рода явились дериваты сарматского бассейна,
в частности Понто-Каспий, где в разное время возникали разные виды
речных раков. Некоторые из них не расселялись широко, а оставались
в отдельных участках своего коренного ареала. Область их современного
распространения соответствует тому или иному заливу одного из третичных
морей. Другие, наоборот, оказались способными к широкому распростране-
нию. Это относится в особенности к A. leptodactylus, быстро распространяю-
щемуся на глазах человека по рекам Понто-Каспийского бассейна и вытес-
няющему более древний вид A. astacus (Бирштейн, Виноградов, 1934).
Молодой и прогрессивный вид A. leptodactylus обладает рядом биологи-
191
ческих преимуществ перед более древним A. astacus. В частности, плодови-
тость первого вида почти в четыре раза выше, он гораздо менее чувствителен
к дефициту кислорода и т. д. Совершенно очевидно, что нет никаких основа-
ний квалифицировать A. leptodactylus как реликтовый вид, учитывая его
недавнее возникновение и быстрое расширение ареала. Между тем вся триба
Nephropsidea, род Astacus и большая часть европейских видов этого рода
являются реликтами, поскольку темп эволюции трибы и рода весьма медлен-
ный (с середины мезозоя до наших дней не произошло образования новых
родов), количество составляющих трибу родов в настоящее время меньше,
чем в мезозое, триба, род и большая часть его видов ранее были распростра-
нены шире, чем сейчас.
Разобранные примеры показывают, что реликтовая группа при каких-то
неизвестных нам условиях может частично или целиком изменить темп
своей эволюции и свое положение побежденной в борьбе за существование.
Этот вывод подтверждается и практикой. Многие культурные растения и
животные происходят от реликтовых предков, что не мешает им достаточно
быстро эволюционировать и расширять свой ареал в результате творческой
деятельности человечества.
Другие способы воздействия человека на природу заставляют стремиться
к замещению реликтовых видов нереликтовыми. Такая проблема поставлена
и частично разрешена применительно к фауне кормовых (для рыб) беспоз-
воночных Северного Каспия (Зенкевич и др., 1945).
КЛАССИФИКАЦИЯ РЕЛИКТОВ
Представление о реликтах как о замедленно эволюционирующих формах,
побежденных в борьбе за существование и тем не менее доживших до наших
дней, позволяет объединить в эту категорию значительное количество
разнородных во всех отношениях организмов.
Классификация этого материала должна базироваться на тех же призна-
ках, которые определяют само понятие реликта, т. е. на особенностях
эволюции рассматриваемых форм.
Выше уже приводились соображения Дарвина об условиях длительного
сохранения в мало измененном состоянии побежденных в борьбе за существо-
вание видов. Он указывал на возможность такого сохранения: 1) при приспо-
соблении видов к особым условиям существования и 2) при изоляции,
препятствующей сильной конкуренции.
В обоих случаях организмы сохраняются благодаря тому, что они
оказываются вне интенсивной межвидовой борьбы за существование.
Падение интенсивности борьбы за существование влечет за собой умень-
шение эффективности естественного отбора и приводит к замедлению
эволюционного процесса.
Разбирая с этой точки зрения конкретный материал, следует прежде
всего несколько уточнить представление о приспособлении видов к особым
условиям существования. Под таким приспособлением можно понимать два
основных процесса. Организм может перейти к обитанию в малозаселенных
специфических биотопах — на больших океанических глубинах, в подземных
водах и т. д. Наряду с этим он может развить мощные защитные образования,
до известной степени предохраняющие его от врагов (раковины, панцири),
или выработать способность зарываться в грунт и т. д. В таком случае он
192
до некоторой степени выходит из борьбы за существование, оставаясь
в этой же среде обитания, но изменяя свой образ жизни, т. е. условия
существования.
Перечисленные три возможности выхода из интенсивной борьбы за
существование, позволяющие длительно сохраняться эволюционирующим
замедленным темпом организмам, могут быть положены в основу класси-
фикации реликтов. Соответственно этому я предлагаю различать следующие
три основные категории реликтов.
1. Локальные реликты, т. е. формы, сохранившиеся в отдельных,
не изменившихся участках своего первоначального ареала. Сокращение
ареала могло быть вызвано резкими климатическими изменениями, к кото-
рым данная форма не успела приспособиться; она выжила только в участках
с неизменными условиями. Тот же эффект получается при оттеснении
реликтовой формы сильными конкурентами. Замедление темпа эволюции
в этих случаях происходит, как уже указывалось выше, вследствие сокраще-
ния численности популяции и неизбежного ограничения многообразия
биотопических условий, уменьшающего возможные направления адаптации.
К категории локальных реликтов относятся, например, ледниковые реликты,
ксеротермические реликты ботаников, реликты Тетиса в смысле Экмана и
многие другие.
2. Рефугиальные реликты, т. е. формы, сохранившиеся благо-
даря приспособлению к специфической среде обитания. Сюда относятся,
например, многочисленные древние обитатели подземных вод и больших
океанических глубин, консервативности которых способствует, между про-
чим, монотонность и постоянство условий существования. Связи организма
с абиотической средой в таких условиях значительно упрощаются и стано-
вятся однообразнее, что приводит к сокращению возможностей адаптации.
Пресные воды также являются, как заметил Дарвин, рефугием по отношению
к океанической фауне. В пресных водах сохранились многочисленные
архаические формы (например, ганоидные рыбы, ракообразные Anaspida-
сеа и многие другие), уничтоженные в борьбе за существование
в океане.
3. Дефензивные реликты, т. е. формы, обладающие пассивными
средствами защиты, обеспечивающими их длительное существование
в исходной среде обитания. Этот случай подробно разобран И. И. Шмаль-
гаузеном (1946), обратившим внимание на зависимость темпа эволюции
от характера защищенности организма от врагов. Такая защищенность,
как указывает Шмальгаузен, весьма относительна, и существенно, что она
чрезвычайно ограничивает распространение организма определенной обста-
новкой, в которой защитные приспособления оказываются достаточно
эффективными. Снабженные ими организмы на ранних стадиях развития
подвергаются интенсивному истреблению врагами, однако эта форма отбора
имеет характер неизбирательной общей элиминации и не ведет к прогрес-
сивной эволюции. Примерами дефензивных реликтов могут служить многие
Древние океанические формы, в частности упоминавшаяся Дарвином Trigo-
nia, брахиоподы, по не совсем ясным причинам не потребляемые рыбами,
некоторые мшанки, иглокожие и т. д.
Предлагаемые категории, быть может, не исчерпывают всего многообра-
зия реликтов органического мира; кроме того, далеко не всегда удается
провести между ними резкие границы. Так, среди локальных и рефугиальных
* А 13 Заказ 333
реликтов несомненно имеются формы, обладающие пассивными средствами
защиты, т. е. до какой-то степени дефензивные реликты.
Можно сомневаться, например, в том, к какой категории следует отнести
группу характерных обитателей временных водоемов, в частности листоногих
раков Apus и Cyzicus, представляющих собой чрезвычайно древние роды,
в ископаемом состоянии известные из палеозойских отложений. Будучи
по характеру своего обитания рефугиальными реликтами, приспособивши-
мися к специфической и упрощенной среде обитания, они наряду с этим
снабжены крепкими панцирями и, что еще важнее, их яйца имеют весьма
совершенные защитные оболочки, позволяющие свободно переносить
высыхание. В некоторых случаях недостаточно ясной оказывается грань
между локальными и рефугиальными реликтами; это относится, например,
к некоторым реофильным и ключевым формам.
Подобные затруднения неизбежно возникают вследствие нашего незна-
ния хода и движущих сил эволюции той или иной группы, в частности
причин замедления эволюционного процесса на некотором этапе его истори-
ческого развития.
Предлагаемое здесь толкование проблемы реликтов должно способство-
вать разрешению этих основных вопросов.
ПОДКЛАСС MALACOSTRACA. ВЫСШИЕ РАКООБРАЗНЫЕ*.
Высшие ракообразные представляют собою единую естественную группу,
объединяемую строго фиксированным числом сегментов тела и составляю-
щих его отделов, положением половых отверстий и общим планом строения
двуветвистых конечностей.
Палеонтологический материал не дает прямых указаний ни на происхож-
дение высших ракообразных, ни на родственные отношения относящихся
к этому подклассу надотрядов и отрядов. Phyllocarida известны начиная
с нижнего кембрия, Syncarida — с девона, Peracarida — с карбона, Eucari-
da — с пермо-триаса, а может быть, и с кембрия, Hoplocarida — с карбона,
причем во всех случаях древнейшие представители этих надотрядов обла-
дают всеми характерными для них признаками. Очевидно, исходные для
высших ракообразных формы существовали в отдаленные докембрийские
времена, и восстановление их строения, а также выяснение филогенетиче-
ских взаимоотношений групп, составляющих подкласс, возможно главным
образом на основании сравнительно-морфологических данных.
Согласно Зивингу (Slewing, 1956), исходные для всех высших рако-
образных формы «IJrmalacostraca» обладали следующими признаками:
тело их состояло из 20 сегментов (пяти головных, восьми грудных и семи
брюшных) и тельсона, причем грудь была покрыта карапаксом, вытянутым
спереди в рострум; карапакс не срастался с грудными сегментами. Голова
имела свободные простомиум и сегмент первых антенн и несла пару стебель-
чатых фасеточных глаз и непарный науплиальный глаз, а также парный и не-
парный лобные органы; тельсон заканчивался фуркой. Сердце тянулось через
все тело и имело 18 остий и боковых сосудов. Две пары нефридиев открыва-
* Членистоногие трилобитообразные и ракообразные. М.: Госгеолтехиздат, 1960,
с. 422—425. (Основы палеонтологии). Публ. только раздел «Филогенетические
отношения».
194
лись при основании вторых антенн и максилл. Гонады располагались по бо-
кам от перикардия и открывались у самок на шестом, а у самцов на восьмом
грудном сегменте. Конечности (за исключением первых антенн) были дву-
ветвистыми, причем протоподит состоял из трех члеников, снабженных
на грудных ногах наружными придатками. На ротовых придатках эндо-
и экзоподиты были превращены в щупики. К этой гипотетической форме при-
ближается девонский Nahecaris Jaekel, отличающийся от нее, однако,
редукцией задних плеоподов.
Приблизительно так же представляет себе строение предков высших
ракообразных Роу (Raw, 1955). Однако этот автор совершенно иначе
трактует строение головы ракообразных, и принимаемые им гомологии
сегментов ракообразных, многощетинковых червей и трилобитов приводят
к выводам, требующим, впрочем, солидного дополнительного подтверждения.
Голова первичных членистоногих, происшедших от бродячих полихет,
близких к современным Polynoidae, по мнению Роу, состояла из простомиума
с двумя парами глаз, но без придатков, перистомиума с придатками, превра-
тившимися в первую пару антенн, и шести сегментов, несущих двуветвистые
конечности. Из них задние четыре пары образовали головные придатки
ракообразных, а два передних сегмента в связи с развитием верхней губы
втянулись в передний отдел кишечника и редуцировались, дав начало
зубцам желудка. Это позволяет гомологизировать голову высших ракообраз-
ных с головой примитивных трилобитов и просомой примитивных хелицеро-
вых. 15 сегментов груди и брюшка высших ракообразных соответствуют
15 послеголовным сегментам примитивных нижнекембрийских трилобитов,
например Olenellus, а также скорпионов и Eurypteroidea, утерявших тель-
сон ’. При этом у названных животных удается наметить своеобразную
тетерономность сегментации — мероциклию, свойственную полихетам —
предкам членистоногих: послеголовные сегменты группируются в триады,
каждая из которых состоит из двух микросомитов и одного макросомита.
В туловище, таким образом, можно различить пять тройных мероциклов.
У высших ракообразных брюшной отдел начинается и заканчивается макро-
сомитами.
На этом основании Роу считает, что трилобиты и высшие ракообразные
независимо друг от друга произошли от гипотетических <дейтартропод»,
усовершенствовавших мероциклию полихет типа современных Polynoidae.
Следует, однако, подчеркнуть, что его построения не подкрепляются эмбри-
ологическими и сравнительно-морфологическими данными, в частности по
строению и развитию нервной системы, и не могут пока быть приняты безого-
ворочно.
Гораздо более убедительны выводы о филогенетических взаимоотноше-
ниях различных групп высших ракообразных между собой, сделанные
Зивингом из его детальных сравнительно-морфологических исследований
и в общих чертах совпадающие с соображениями Роу.
Наиболее примитивными признаками из всех высших ракообразных
отличается надотряд Phyllocarida, которому свойственны свободные грудные
сегменты, семь брюшных сегментов, тельсон с фуркой, две пары целомодук-
тов, трубкообразное сердце, длинные гонады и т. д. Наряду с этим следует
1 Роу рассматривает хвостовой придаток скорпионов и Eurypteroidea в качестве
гомолога спинного шипа Olenellus.
72 13* 195
PnpAausAa cea
Asapoda
ТолаАРасеа
Pecapoda
PaPompjrP A: и car A da
Шастание карапакса
c e/yfab/Mu сегме///77ами
Afystdacea
Hodompea Ape
Pera car A da /
neped-
кеео ярурнего сег-
мента сголоРои
РоРРилсная /глас-
птиккаяа канди-
AmpAApoda i
ГАегтогРаеласа J I
Pumacea
A ppAos Ar аса
ДАустРорча -
тый карапакс
/Аромор pasPuy
mac
РеРукция карапакса /Арямое
pasPumue. Тано'еяция к слиянию
! лероРнеео яруРнояо сегмента
I г голоРои
РоРатряР Synс ar A da
I ЯреаРраяоРание пероРнис
еруРкысс конечностеАА А
\ ногочелюсти
PnaspAda cea
Ji ат а 6 ара da
\ ААРраяоРание сурнеРраланоАА
\ артерии
\
4 РаРатряР PopAocarAda
РреоРраяоРание послеРней
пари пл напор 0 рропоРи.
ДоРраяРеление рюелуРка
на карриа каленую и пилори-
ческую камери
РреРки AfaAacosAraca
^rdacMirara
Pai АулеAAacea
РстаноРление
/гссюа/нс/аРа
числа сегяех/осА
и лололсенио л ало
Роле отРерстпии.
Схема филогенетических отношений Malacostraca
отметить и черты давней специализации Phyllocarida, например редукцию
задних плеоподов, отсутствие экзоподита второй антенны, двустворчатость
карапакса, снабженного мускулом-замыкателем, строение желудка. Относи-
мый в Phyllocarida девонский Nahecaris лишен большинства этих признаков
высокой специализации и тесно связывает Phyllocarida с другими Malacost-
raca.
Многие примитивные признаки присущи и надотряду Hoplocarida. Из них
следует упомянуть протягивающиеся через все тело сердце и гонады, расчле-
ненность экзоподитов второй антенны и уроподов, присутствие на голове
парного и непарного лобных органов и науплиального глазка. По строению
головы Hoplocarida сближаются с Phyllocarida. Однако Hoplocarida обла-
дают и многими весьма своеобразными особенностями, указывающими на их
раннее обособление от общего ствола Malacostraca, в частности редукцией
первого брюшного сегмента, глубокой специализацией пяти пар грудных
196
конечностей и упрощением строения трех задних пар грудных конечностей,
присутствием субневральной артерии и своеобразным строением карапакса.
Надотряд Syncarida обладает наибольшим количеством признаков, об-
щих с другими надотрядами, и в этом смысле занимает центральное положе-
ние в схеме родственных отношений высших ракообразных. Подобно рас-
смотренным выше надотрядам, Syncarida сохраняют многие примитивные
черты организации (свободные грудные сегменты, у некоторых — 7 брюшных
сегментов, науплиальный глаз у взрослых Paranaspides, фурка у Bathynel-
lacea, протягивающиеся через все тело сердце и гонады, две пары метанефри-
диев). С Eucarida их сближают наличие петазмы, статоциста при основании
первых антенн и нисходящей аорты; с мизидами — наиболее примитивным
отрядом надотряда Peracarida — наличие статоциста в эндоподите уроподов
у пермского Uronectes; с Hoplocarida — некоторые детали строения кишеч-
ника и присутствие науплиального глаза у Paranaspides. Своеобразный орган
внутренней секреции — орган X — имеется у Syncarida, Hoplocarida, Deca-
poda и Mysidacea.
Надотряды Peracarida и Eucarida близко родственны между собой, в осо-
бенности их низкоорганизованные представители — подотряд мизид Lopho-
gastrida и отряд Euphausiacea. Оба надотряда имеют сходное строение мно-
гих внутренних органов, в частности они обладают нисходящей аортой, сое-
диненной с субневральной артерией, и одинаково построенным желудком.
Резкие различия между ними проявляются прежде всего в способе размно-
жения. Для всех Peracarida (за исключением Thermosbaenacea) характерно
образование выводковой сумки (марсупиума) за счет направленных внутрь
отростков грудных конечностей — оостегитов, куда откладываются яйца
и где происходит прямое развите молоди. Такое приспособление в зачатке
имеется только у Phyllocarida, у которых яйца удерживаются под грудью
матери при помощи длинных щетинок грудных ног. Для Eucarida харак-
терно развитие с метаморфозом, причем яйца или откладываются в воду,
или прикрепляются к плеоподам самки.
Низшие представители обоих рассматриваемых надотрядов характеризу-
ются многими весьма примитивными признаками, позволяющими сближать
их с исходными для Malacostraca формами. В особенности интересны в этом
отношении Lophogastrida, обладающие длинным сердцем с большим количе-
ством сегментарных сосудов, двумя парами метанефридиев, эпиподитами
на грудных конечностях и свободным у Gnathophausia седьмым брюшным
сегментом. В качестве примитивных признаков Euphausiacea и низших
Decapoda можно назвать полный метаморфоз и строение жабр.
Все изложенные данные по филогении могут быть резюмированы в виде
следующей схемы (см. рисунок).
197
НЕКОТОРЫЕ ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ И ЭВОЛЮЦИИ
ПРЕСНОВОДНОЙ ФАУНЫ*
Проблемы происхождения и эволюции пресноводной фауны остаются
почти неосвещенными в литературе последнего времени. Первая из них
обсуждалась главным образом во второй половине XIX в. и в самом начале
XX в. (Martens, 1857; Чернай, 1870; Sollas, 1883—1887; Pelseneer, 1905).
Более новые исследователи возвращались к ней лишь попутно, стремясь
увязать результаты своих экспериментов с выводами того или другого
из старых авторов (Schlieper, 1929; Needham, 1930; Panikkar, 1940) или
указывая на возможность использования отдельных зоогеографических дан-
ных для будущего решения всей проблемы происхождения пресноводной
фауны (Державин, 1924; Бирштейн, 1935). Кроме того, высказывались
умозрительные гипотезы о соотношениях пресноводной, морской и почвен-
ной фаун (см.: Берг, 19476). Что касается выяснения особенностей эволю-
ции пресноводной фауны, то после чрезвычайно важных замечаний Дарвина,
отметившего характерные особенности эволюции пресноводных организмов,
этот вопрос почти не привлекал к себе внимания.
1
Наука не располагает еще достаточными данными для категорического
решения вопроса о том, следует ли считать морскую фауну исходной для
фауны пресных вод, как полагает подавляющее большинство авторов, или,
наоборот, как думает Кларк (Clark, 1927), или обе эти фауны происходят
от почвенной, как это считает Берг (19476). Во всяком случае общепризнан-
ным является очень давнее заселение океана организмами, стоявшими на
низких ступенях эволюционного развития. Мы не знаем, в какой среде заро-
дилась жизнь, но мы твердо уверены в том, что развилась она в океане и имен-
но там сформировались почти все ныне существующие крупные систематиче-
ские группы.
Как известно, в кембрии существовали все типы беспозвоночных, причем
все они, за исключением представителей класса Merostomata, обитали в море.
Что касается Merostomata, то даже если принять доводы в пользу пресно-
водного образа жизни древнейших из этих членистоногих, все же следует
иметь в виду их несомненное происхождение от чисто морских форм — три-
лобитов. То же относится и к растениям: в докембрии и кембрии известны
остатки только морских растений (Давиташвили, 1943). Во многих случаях
палеонтология дает прямые указания на исходно морской характер многих
современных пресноводных групп. Так, например, исключительно пресновод-
ные ныне листоногие в кембрийский период обитали только в море; согласно
К. Динеру (1934), для всех пресноводных родов пластинчатожаберных
(кроме Unionidae) мы можем с большой долей вероятности установить как
их отношение к морским группам, так и время их поселения в пресные воды.
По подсчетам Л. А. Зенкевича, половина (31) современных классов
животных обитает исключительно в море, 7 классов живут главным образом
в море и 2 класса представлены как пресноводными, так и морскими фор-
мами; в пресной воде известно всего 2 класса животных, совсем или почти
♦ Успехи соврем, биологии, 1949, т. 27, № 1, с. 119—140.
198
совсем чуждых морю (амфибии и насекомые). Из всей массы пресноводных
первичноводных многоклеточных можно допустить возникновение в пресной
воде только двух классов — коловраток и пиявок. Однако и те и другие пред-
ставлены и в море, где, между прочим, обитает высокоспециализированный
отряд коловраток Seisonidea и многочисленные пиявки из сем. Ichtyobdel-
lidae. Именно это семейство Апати (Apathy, 1888) считал исходным для всего
класса; его мнение не разделяется современными авторами, но морской гене-
зис пресноводных Ichtyobdellidae находит подтверждение во взглядах Лу-
кина (1936а, б), приписывающего морское происхождение наиболее
широко распространенному в пресных водах представителю этого семей-
ства — Piscicola geometra. Поэтому кажется преждевременным категориче-
ски считать родиной коловраток и пиявок пресные воды только на основании
их многочисленности в пресной воде.
Согласно Ромеру и Грову (Roemer, Grove, 1935), в пресной воде возник-
ли рыбы. Но в последней своей работе Ромер (Roemer, 1946) утверждает,
что современные пресноводные рыбы представляют собой реэмигрантов
из моря, куда их предки мигрировали в раннемезозойское время и лишь позже
вернулись в пресные воды.
Таким образом, не предрешая решения вопроса об исходной среде возник-
новения жизни, можно принять, что современная пресноводная первичновод-
ная фауна происходит от морской, поскольку это соответствует всем имею-
щимся палеонтологическим и фаунистическим данным.
II
Для того чтобы проанализировать, при каких условиях, каким образом
и когда происходило формирование пресноводной фауны, следует прежде
всего установить, насколько она самостоятельна и насколько резкая грань
отделяет ее от морской. Соответствующих материалов в литературе нет. Все
авторы, касавшиеся вопроса о происхождении пресноводной фауны, прини-
мали ее полную оторванность от морской как нечто данное и не подлежащее
обсуждению.
Только один из первых исследователей этой проблемы, А. Чернай (1870),
попытался определить степень общности морской и пресноводной фауны,
причем пришел к весьма интересным выводам: «морская вода предшество-
вала при развитии земли пресной и в ней успела развиться многообразная
жизнь из всех главных отделов животных беспозвоночных, прежде чем обра-
зовались на суше пресноводные вместилища. С появлением рек обитатели
моря, в особенности прибрежные, стали входить в устья их. Виды животных,
общие для моря и пресной воды, доказывают, что и ныне подобное переселе-
ние совершается, хотя, может быть, в более ограниченных размерах вслед-
ствие большего преобладания суши от происшедших наносов минеральных
веществ. Родовое сходство многих пресноводных животных с морскими
также говорит в пользу того, что они отдаленные выходцы из моря, изменив-
шиеся впоследствии».
Эти положения, в том числе представление о постепенном замедлении
процесса внедрения морской фауны в пресные воды, оказываются вполне
приемлемыми и с современной точки зрения, если не считаться с наивным
объяснением причины затухания миграции морских организмов в пресные
воды. Однако общая картина, правильно нарисованная Чернаем почти 80 лет
тому назад, сейчас может быть несколько детализирована.
Таблица 1. Степень обособленности пресноводной фауны от морской у животных
различных групп
Группа Общее число заре- гистрированных в пресной воде Из них разноводных
число %
семейств | родов семейств родов семейств родов
Губки 1 2(3?) 10 0 0 0 0
Кишечнополостные 4 8 0 0 0 0
Ресничные черви 18 114 10 17 55 15
Нематоды 7 21 5 2 71 10
Немертины 1 1 1 1 100 100
Гастротрихи 5 13 1 5 20 31
Коловратки 19 100 4 4 21 4
Kamptozoa 1 1 0 0 0 0
Многощетинковые черви 6 11 6 9 100 66
Малощетинковые черви 9 85 3 8 33 9
Пиявки 5 34 2 0 40 0
Мшанки 8 15 1 1 12 7
Брюхоногие моллюски 12 127 7 5 58 4
Пластинчатожаберные 14 108 4 6 28 6
Ветвистоусые ракообразные 8 36 1 2 12 6
Веслоногие ракообразные 15 53 11 17 73 33
Бокоплавы 12 28 10 15 83 53
Равноногие 11 33 8 14 73 43
Мизиды 2 15 1 9 50 60
Кумацеи 4 6 4 6 100 100
Десятиногие ракообразные И 57 6 10 55 17
Используя в качестве исходного материала общеизвестные сводки по
общей зоологии (<Руководство по зоологии», т. I и II; Handbuch der Zoologie)
и некоторые более специальные работы (соответствующие выпуски <Фауны
СССР», определители организмов пресных вод СССР, главы из <Жизни прес-
ных вод СССР», Thiele, 1929—1935) с отдельными дополнениями, взятыми
из периодических изданий, я попытался определить степень обособленности
пресноводной фауны от морской для различных групп свободно живущих
многоклеточных первичноводных беспозвоночных. За меру близости между
пресноводной и морской фаунами я принимаю процент семейств и родов,
объединяющих наряду с пресноводными также морские роды и виды, ко всем
семействам и родам, обитающим в пресной воде. Такие общие морю и пресной
воде семейства и роды я именую для краткости <разноводными», заимствуя
этот термин у Кесслера (1875), употреблявшего его в несколько ином смысле.
Эндемичная каспийская фауна причислена мною к морской, все солоновато-
водные роды, если они не представлены в пресных водах, также сочтены мор-
скими. Результаты соответствующих подсчетов сведены в табл. 1.
Как видно, степень разобщенности морской и пресноводной фаун для
различных групп животных выражается показателями совершенно разного
значения. При этом если не принимать по внимание нехарактерных для пре-
сноводной фауны групп, представленных в пресной воде очень ограниченным
числом видов, к тому же мало распространенных и не достигающих сколько-
нибудь значительного количественного развития (немертины, Kamptozoa,
многощетинковые), то замечается одно общее правило, которое по аналогии
с правилом Мартенса может быть сформулировано следующим образом: сход-
ство между пресноводной и морской фаунами для многоклеточных первич-
200
новодных беспозвоночных тем больше, чем выше систематическое положение
данной группы, т. е. возрастает вверх по системе. В самом деле, пресноводная
и морская фауны низко организованных двуслойных животных — губок
и кишечнополостных — полностью разорваны. Для червей и моллюсков мож-
но отметить от 20 до 50 % разноводных семейств (кроме нематод) и менее
10—15 % разноводных родов (кроме Gastrotricha); к этой же группе примы-
кают примитивные низшие ракообразные — ветвистоусые. Наконец, для
веслоногих и высших ракообразных характерно более 50 % разноводных
семейств и более 15 % разноводных родов. Полученные показатели еще
нуждаются в уточнении, а возможно, и в исправлении при учете более полных
фаунистических списков, но мне кажется, что все вероятные поправки
не опровергнут основного высказанного здесь положения. Оно, разумеется,
вполне гармонирует с общим представлением об известном соответствии
между высотой организации животных и разнообразием условий среды,
в которых они могут обитать.
Анализ полученных данных позволяет сделать еще один общий вывод.
Нетрудно заметить, что в общем наиболее богаты видами те пресноводные
группы (семейства, роды), которые в наибольшей степени оторваны от мор-
ской фауны, что в большинстве случаев приходится относить за счет проис-
шедшего в пресной воде видообразования. Это обстоятельство было в свое
время подмечено А. Чернаем и может с полным правом именоваться «прави-
лом Черная». Для поставленных здесь задач оно имеет значение потому,
что еще резче подчеркивает различия между пресноводной и морской фау-
нами тех групп, которые не могут менять среду обитания.
Как бы ни колебалась степень общности морской и пресноводной фаун
в зависимости, от систематического положения той или другой группы, но
в целом пресноводная фауна оказывается достаточно резко обособленной
от морской, и смешиваются они в ничтожной мере.
Ш
Каковы причины несмешиваемости пресноводной и морской фаун? Один
из первых авторов, ставивших перед собой этот вопрос, — Мартенс (Martens,
1857) обращал внимание на трудности адаптации морских животных к не-
благоприятным условиям существования в пресноводных водоемах, в ча-
стности на их недолговечность (высыхание) и замерзание. Соллэс (Sollas,
1883—1887) считал основным препятствием, не дающим проникать в пресные
воды морским животным, наличие у них пелагических личинок, сносимых
в море течением рек; по его мнению, в пресных водах обитают только формы,
лишенные пелагических личинок.
Указанные этими авторами причины имеют несомненно известное значе-
ние, однако не они должны считаться основными. В самом деле, как указы-
вает и сам Мартенс, в тропиках пресноводные водоемы не замерзают, реки
не высыхают в течение весьма продолжительного времени. Соображениям
Соллэса противоречит существование пресноводного планктона и некоторых
донных животных, имеющих пелагические личинки (креветки, Dreissena
и др.). Если Мартенс преувеличивает значение стоячих водоемов и недооце-
нивает роль текучих вод, то Соллэс поступает как раз наоборот, не принимая
во внимание широкого распространения стоячих пресноводных водоемов.
Основным фактором, разграничивающим пресноводную и морскую фау-
М Заказ 333
201
ны, является, конечно, различие в солености сред их обитания. Вопрос о том,
насколько непреодолима солевая грань между пресной и морской водой, ста-
вился экспериментально еще с начала XIX в., когда несколько исследователей
пытались постепенно приучить морских животных к пониженной солености
и таким образом заставить их в ряду поколений превратиться из морских
в пресноводные организмы (Beudant, 1816).
Полевые наблюдения над зонами смешения пресноводной и морской фаун
привели к выводам о ведущей роли пассивного приобретения эвригалинности
животными при опреснении обширных участков морских прибрежий и отдель-
ных отчленившихся от моря водоемов (Pelseneer, 1905). Полученные резуль-
таты весьма интересны, но они не могут объяснить, почему одни организмы
приобретают способность существовать в пресной воде, а другие, живущие
в том же месте — нет.
Разгадку этого вопроса следует искать в физиологическом воздействии
на морских животных пониженной солености, причем можно предполагать
влияние как суммарной солености, выражающейся в показателях осмотиче-
ского давления, так и отдельных ингредиентов, которыми различа-
ются морская и пресная вода. Так, например, Нидгем (Needham, 1930) отме-
чает значительно меньшее содержание в пресной воде фосфора, потребля-
емого в большом количестве дробящимися яйцами морских животных. Не-
достатком фосфора в пресных водах Нидгем объясняет многочисленные слу-
чаи способности только взрослых животных морского происхождения жить
в пресной воде не размножаясь, так как их яйца не могут развиваться при
столь низком содержании фосфора. Шлипер (Schlieper, 1929, 1931) указы-
вает на затрудненные условия дыхания в пресной воде по сравнению с мор-
ской из-за преобладания в пресной воде бикарбонатов, не связывающих
углекислоту, в то время как в морской воде содержатся преимущественно
монокарбонаты, реагирующие с углекислотой. Отсюда Шлипер делает
вывод о большей легкости перехода в пресную воду форм, способных уско-
рять процесс своего дыхания, к которым он относит рыб и ракообразных.
По этому поводу следует заметить, что и другие организмы, в частности мол-
люски, также могут регулировать интенсивность газообмена, и соображени-
ями Шлипера нельзя объяснить их меньшую приспособленность к обитанию
в пресной воде по сравнению с рыбами и ракообразными. Не помогают здесь
и данные Нидгема, поскольку он не разъясняет, нуждаются ли в одинаковой
степени все животные в фосфоре или рыбы и ракообразные оказываются
в этом отношении в преимущественном положении.
IV
Гораздо более определенные выводы удается сделать при учете реакции
организмов на общую соленость. Особое значение в этом отношении прихо-
дится придавать соотношению осмотического давления полостной жидкости
и среды обитания тех или иных организмов. Способность к активной осморе-
гуляции, т. е. к поддержанию внутреннего осмотического давления на извест-
ном уровне, более или менее независимом от осмотического давления наруж-
ной среды, согласно Шлиперу (Schlieper, 1929, 1933, 1935), является основ-
ным условием, позволяющим морским животным обитать в опресненных
водах. Наряду с этим Шлипер указал на наличие некоторых эвригалинных
морских животных, лишенных органов, осуществляющих процесс осморегу-
ляции, как, например, Asterias rubens. Таким образом Шлипер доказал
202
весьма важный факт: отсутствие резкой грани в отношении осморегуляцион-
ных способностей между пресноводными и морскими беспозвоночными. До
него в литературе господствовало представление об обязательной изотонич-
ности морских беспозвоночных и гипертоничности пресноводных, сформули-
рованное в свое время Ботацци (Botazzi, 1908) и вошедшее во все сводки по
физиологии.
Поднятые Шлипером вопросы породили длинный ряд исследований осмо-
регуляции солоноватоводных и отчасти пресноводных животных. Наличие
хороших сводок результатов этих исследований (Shlieper, 1935; Krogh, 1939;
Beadle, 1943) избавляет меня от необходимости подробно на них останавли-
ваться. Важно отметить, что основное направление, в котором работали мно-
гочисленные иностранные авторы, вело к выяснению механизма осморегуля-
ционного процесса, т. е. преследовало чисто физиологические цели. Осо-
бенно много внимания уделялось проблеме проницаемости покровов тела
(Bethe, 1929; Bethe, Berger, 1931; Beadle, 1937; Robertson, 1937; и др.) и свя-
занной с этим способности регуляции минерального состава крови (Bialasze-
wicz, 1932; Scholles, 1933; Bogucki, 1934; Krogh, 1939; и др.), а также мор-
фологии экскреторных органов в связи с их функцией (Schwabe, 1933; Peters,
1935) и, наконец, вопросам энергетики осморегуляционного процесса (Schli-
ерег, 1930, 1936; Pieh, 1936; Claus, 1937; и др.). При этом основным объектом
исследования служили ракообразные и отчасти насекомые; по другим водным
беспозвоночным сделано чрезвычайно мало.
Для наших целей важно не столько знание деталей физиологии процесса
осморегуляции, сколько самая общая сравнительная характеристика осмо-
регуляторных способностей разных групп беспозвоночных в связи с их раз-
личной способностью переходить из моря в пресную воду.
На более совершенную осморегуляцию ракообразных по сравнению с чер-
вями указывал еще Шлипер. Л. А. Зенкевич (1938) также отметил, что поли-
хеты значительно уступают ракообразным по своим осморегуляторным спо-
собностям. Сходные высказывания можно найти и у других авторов. Но все
они интересовались больше различиями осморегуляторных способностей
в пределах одной группы животных в зависимости от разной степени эврига-
линности тех или иных ее представителей, чем такими различиями между
крупными систематическими группами. На эту сторону вопроса мы (Беляев,
Бирштейн, 1940) впервые обратили внимание во время наших работ на Ка-
спийском море. Мы исследовали соотношение внутреннего и внешнего осмо-
тического давления для трех видов десятиногих, двух видов бокоплавов,
одного вида пластинчатожаберных и одного вида Chironomidae и, сопоставив
полученные данные с имеющимися в литературе указаниями, установили, что
для каждой изученной в этом отношении крупной группы эвригалинных бес-
позвоночных характерен специфический тип осморегуляторного процесса.
Впоследствии мы подтвердили справедливость выясненного нами типа для
бокоплавов на материале из дельты Волги (Беляев, Бирштейн, 1944) и на бо-
гатом материале из оз. Байкал (Базикалова и др., 1946а, б). При работе
на Байкале я имел возможность убедиться в справедливости также типа,
установленного нами для моллюсков.
На основании полученных нами и приведенных другими авторами данных
можно пока установить следующие основные типы осморегуляторного про-
цесса у различных солоноватоводных животных.
1. Декаподный тип. Десятиногие ракообразные пресных и солоноватых
14'
203
вод обладают способностью сохранять более или менее постоянное внутрен-
нее осмотическое давление независимо от солености среды обитания, т. е.
могут считаться гомойосмотичными. На графике соотношения внутреннего
и наружного осмотического давления это выражается линией с более или
менее горизонтальным ходом, без ярко выраженных переломов, направлен-
ной под некоторым характерным для каждого вида углом к диагонали гра-
фика и пересекающейся с последней при высоких показателях солености, что
соответствует моменту перехода животных в состояние гипотонии. Мы (Бе-
ляев, Бирштейн, 1940) получили такую кривую для каспийских Astacus ра-
chypus, A. leptodacty 1 us caspius и Leander adspersus. Л. Зенкевич (1938)
и позже Пора (Рога, 1939) —для черноморских Carcinus maenas и Pachy-
grapsus marmoratus, Дэкин и Эдмондс (Dakin, Edmonds, 1931) —для ав-
стралийского Halaecium cardiformis, Дюваль (Duval, 1927) — для Potamon,
Паниккар (Panikkar, 1941) —для атлантических Palaemonetes varians, Le-
ander serratus и т. д.
2. Амфиподный тип. Организмы более или менее гомойосмотичны и при
сравнительно низкой солености и пойкилосмотичны, но гипертоничны при
более высокой солености. Кривые соотношения внутреннего и внешнего осмо-
тического давления имеют горизонтальный или близкий к горизонтальному
ход только в левой части графика, а при известной солености образуют резкий
перелом, после которого идут параллельно (у понтокаспийских форм) или
под углом (у байкальских форм) к диагонали графика. Мы обнаружили этот
тип осморегуляции у каспийских Dikerogammarus haemobaphes и Pontogam-
marus maeoticus, волжских P. robustoides, P. obesus и Corophium chelicorne
(Беляев, Бирштейн, 1940, 1944), байкальских Eulimnogammarus verrucosus,
E. cyaneus, Carinogammarus, Pallasea cancelloides, Odontogammarus pul-
cherrimus и Gmelinoides fasciatus и широко распространенного в пресных
и отчасти солоноватых водах Gammarus lacustris (Базикалова и др., 19466).
3. Аннелидный тип. Животные пойкилосмотичны и почти изотоничны при
всех соленостях. Лишь на очень небольшом отрезке солевой шкалы наблюда-
ется состояние гипертонии. Малоизученный тип осморегуляции, о котором
можно судить главным образом поданным Л. А. Зенкевича (1938) и отчасти
Бидля (Beadle, 1943). Такой тип осморегуляции обнаружен Зенкевичем у чер-
номорских Nereis diversicolor, N. cultrifera, Nephthys hombergi, Arenicola
grubei, а также нами у байкальского Lamprodrilus wagneri.
4. Моллюсковый тип. Животные полностью пойкилосмотичны и гиперто-
ничны лишь в очень слабой степени, т. е. почти неспособны к активной осмо-
регуляции. Соотношение между внутренним и наружным осмотическим дав-
лением выражается прямой линией, параллельной диагонали графика и ино-
гда сливающейся с ней вблизи верхней солевой границы существования дан-
ного вида. Мы констатировали этот тип у каспийского Cardium edule,
Л. А. Зенкевич (1938) —у черноморских Pecten ponticus и Syndesmya
ovata, Базикалова, Талиев и я — у байкальских Benedictia baicalensis, Baica-
lia herderiana и, наконец, Limnaea auricularia. По нашим с Беляевым пре-
дварительным данным, так же осморегулируют Anodonta.
По всей вероятности, этими типами не исчерпывается все многообразие
способов осморегуляции в животном мире, и возможно, что другие крупные
систематические группы обладают каждая своим специфическим типом осмо-
регуляции. Имеющиеся литературные данные позволяют добавить несколько
дополнительных сведений по другим группам животных.
204
Среди пресноводных простейших наряду с формами, обладающими пуль-
сирующими вакуолями и, следовательно, способными к активной осморегу-
ляции, существуют некоторые виды инфузорий, лишенные органоидов осмо-
регуляции и, очевидно, пойкилосмотичные (Herfs, 1922). Кишечнополостные
пойкилосмотичны, лишены осморегуляторных органов, но в пресной воде
слабо гипертоничны (Palmherst, 1933). Ресничные черви пойкилосмотичны,
если судить по изменению их веса при помещении животных в различную
соленость (Pantin, 1931). Надо думать, что Cladocera также пойкилосмо-
тичны, так как осмотическое давление этих ракообразных в пресной воде, как
показал еще Фриче (Fritsche, 1917) для Daphnia magna, весьма низкое и,
кроме того, подвержено значительным колебаниям. Среди позвоночных круг-
лоротые, судя по экспериментам с Polystotrema stouti (Bond, Сагу, Hutchin-
son, 1932), в море изотоничны, хрящевые рыбы тоже в море почти изото-
ничны, а в пресной воде гипертоничны и, возможно, пойкилосмотичны. На-
конец, костистые рыбы и все вышестоящие группы строго гомойосмотичны.
Таким образом, в пресных и солоноватых водах обитают организмы
с весьма разнообразными осморегуляторными способностями — от строго го-
мойосмотичных до полностью пойкилосмотичных, с многими промежуточ-
ными типами. Тем самым теряют смысл широко распространенные схемы, со-
гласно которым все пресноводные организмы гомойосмотичны, солоновато-
водные также гомойосмотичны или пассивно пойкилосмотичны, а морские,
за исключением хордовых, — пассивно гомойосмотичны (см., например,
Harms, 1934).
Анализируя приведенные данные, можно сделать еще один очень важный
вывод, впервые сформулированный Беляевым и мною в 1940 г. Очевидно,
эвригалинность животных вырабатывается разными путями, из которых уда-
ется пока указать два основных — усовершенствование осморегуляторных
способностей, причем ткани организма продолжают существовать при посто-
янной солености, и приспособление тканей организма к различной солености
омывающей их полостной жидкости, т. е. приобретение эвригалинности са-
мими тканями. По первому пути пошли все гомойосмотичные, по второму —
все пойкилосмотичные в пресной и солоноватой воде животные. К сходным
выводам пришел впоследствии независимо от нас Бидль (Beadle, 1943) 1
При этом следует иметь в виду, что адаптация к существованию в раз-
личной солености не может быть сведена только к осморегуляции. Громадное
значение приобретает и другая сторона той же проблемы, на важности кото-
рой особенно настаивает Крог (Krogh, 1939), а именно способность орга-
низма усваивать соли, растворенные в окружающей среде. Кроме того, надо
учитывать и способности к регуляции минерального состава крови. Не входя
здесь в обсуждение соответствующих данных, отмечу только, что по содер-
жанию калия и магния полостная жидкость гомойосмотичных ракообразных
в большей степени отличается от окружающей среды, чем полостная жид-
кость пойкилосмотичных червей и моллюсков (Bethe, Berger, 1931), и, вообще
говоря, усвоение солей из окружающей среды и регуляция минерального со-
става полостной жидкости тесно связаны с осморегуляторными способ-
ностями.
Еще раньше приблизительно такую же мысль высказал Е. М. Крепе (1929), однако
он предполагал, что у пойкилосмотичных форм, в частности у балянусов, измене-
ние в солевой концентрации внутренней среды ведет только ко временному тормо-
жению функций, а не переносится ими совершенно безболезненно.
205
Таблица 2. Сопоставление степени обособленности пресноводной фауны
от морской и типа осморегуляции у различных групп животных
Г руппа % разноводных Характер осморегуляции
семейств 1 родов
Губки 0 0 Пойкилосмотичны. Неспособны
Кишечнополостные 0 0 к активной осморегуляции
Ресничные черви 10 17 Пойкилосмотичны
Нематоды 71 10 ?
Малощетинковые черви 33 9 Пойкилосмотичны, но в узких
солевых пределах гомойосмо-
тичны
Пиявки 40 0 Вероятно, также пойкилосмо-
Брюхоногие моллюски 58 4 тичны
Пластинчатожаберные 28 6 То же
Ветвистоусые ракообразные 12 6 > >
Веслоногие ракообразные 73 33 ?
Бокоплавы 83 53 В солевых границах нормаль-
Равноногие ракообразные 73 43 ной жизнедеятельности гомой-
осмотичны *
Десятиногие ракообразные 55 17 Гомойосмотичны *
* Эти характеристики в дальнейшем несколько изменились—см. Беляев (1949, 1951).
(Примеч. ред.).
Гораздо важнее остановиться на весьма интересном соответствии, ко-
торое удается установить между степенью разобщенности морской и пресно-
водной фаунЪ и осморегуляторными способностями различных групп живот-
ных. Сводка изложенных выше данных по этим разделам представлена
в табл. 2. Как видно, разрыв между пресноводной и морской фаунами тем
больше, чем менее совершенны осморегуляторные способности той или дру-
гой группы животных. Иными словами, в настоящее время переход пойкил-
осмотичных организмов из моря в пресные воды оказывается весьма затруд-
ненным. Действительно, на примере многих солоноватоводных пойкилосмо-
тичных животных можно показать ограниченность степени их эвригалинно-
сти. Так, лишенная осморегуляторного аппарата Asterias rubens в Балтий-
ском море доходит только до 15 °/00, неспособная к активной осморегуляции
Arenicola не выносит солености ниже 8 %; солоноватоводные моллюски Бал-
тики и Каспия (за исключением Dreissena polymorpha) не заходят в пресную
воду; в Азово-Черноморском бассейне реликтовые Cardidae встречаются
в условиях полного опреснения, но зато не переносят солености выше 5°/00.
Эти и многие другие случаи позволяют признать солевой диапазон существо-
вания пойкилосмотичных животных сравнительно узким.
Среди гомойосмотичных животных можно назвать большое количество
разноводных видов. Здесь прежде всего надо упомянуть о многочисленных
разноводных рыбах, совершающих катадромальные и анадромальные мигра-
ции, далее напомнить о длинном ряде видов высших ракообразных — крабов,
креветок, бокоплавов, мизид, кумацей и т. д., распространенных в пресных
и морских водах. Для всех этих форм постоянство внутреннего осмотического
давления в нормальной для них среде обитания является, по-видимому,
строго обязательным. Чрезвычайно интересно, что оно достигается разными
206
способами. Так, например, у речных раков, креветок и бокоплавов основным
осморегулирующим органом служат определенные участки антеннальных
желез, обеспечивающие гипотонию, а при высоких соленостях — и гиперто-
нию (Беляев, Бирштейн, 1940) мочи по отношению к крови. У крабов и, веро-
ятно, мизид (Vogt, 1933) моча изотонична крови и гомойосмотичность поло-
стной жидкости определяется иными, точно еще не установленными меха-
низмами.
Важно еще отметить, что величина осмотического давления полостной
жидкости гомойосмотичных животных необыкновенно стабильна и мало за-
висит от перемены среды обитания данным видом в течение его филогенеза.
Так, например, Бидль (Beadle, 1943) установил, что осмотическое давление
насекомых и ракообразных, обитающих в гипергалинных водоемах, соответ-
ствует осмотическому давлению полостной жидкости их пресноводных пред-
ков. Мы (Беляев, Бирштейн, 1944) доказали сохранение на одном уровне
осмотического давления полостной жидкости каспийских бокоплавов при их
вселении в Волгу. Подобные факты позволяют использовать величину внут-
реннего осмотического давления тех или иных гомойосмотических живот-
ных в качестве показателя их генезиса. Произведенное нами исследование
40 видов байкальских бокоплавов показало, что между систематическим
положением каждого вида и величиной осмотического давления его полост-
ной жидкости имеется совершенно определенное соотношение: наиболее кон-
сервативные по своим морфологическим признакам формы, близкие к пред-
полагаемым морским исходным предкам байкальских гаммарид (Eulimno-
gammarus), обладают наиболее высоким внутренним осмотическим давле-
нием, вполне сопоставимым с аналогичными показателями морских (каспий-
ских) видов, причем такие близкие к морским предкам байкальские виды со-
хранили способность к поддержанию гомойосмотичности в довольно широких
солевых пределах. Напротив, наиболее специализированным формам, несо-
мненно развившимся в самом Байкале, как глубоководным, вооруженным
шипами (Garjajewia cabanisi, Carinurus reissneri) или узко адаптирован-
ному к жизни на губках Spinacanthus parasiticus, свойственно минимальное
для байкальских гаммарид внутреннее осмотическое давление. Такие формы
утеряли способность к активной осморегуляции и превратились в пойкил-
осмотичных животных. Таким образом, по мере адаптации гаммарид к спе-
цифическим условиям Байкала происходило снижение их внутреннего ос-
мотического давления и ослабление их осморегуляторных способностей (Ба-
зикалова и др., 1946а, б). Приблизительно в таком же духе трактует Паник-
кар (Panikkar, 1941) обнаруженную им незначительную гипотонию полост-
ной жидкости морских креветок Palaemonetes varians, Leander serratus и
L. squilla. По его мнению, они происходят от акклиматизировавшихся в пре-
сных и солоноватых водах предков, вернувшихся затем к существованию
в море. Это мнение не согласуется с зоогеографическими данными и не ка-
жется мне достаточно обоснованным. Гораздо более вероятным представля-
ется сохранение креветками внутреннего осмотического давления, соответст-
вующего несколько пониженной солености океана времени формирования
исследованных Паниккаром видов.
Все это подчеркивает высокое совершенство сложного осморегулятор-
ного механизма гомойосмотичных животных, позволяющее некоторым
из них существовать в чрезвычайно широких пределах колебаний солености
и даже проникать из моря в пресную воду.
207
V
Тем замечательней тот факт, что среди современных первичноводных
животных отнюдь не наблюдается преобладания гомойосмотичных форм над
пойкилосмотичными. Если использовать один из наиболее полных для прес-
ных вод фаунистических списков — каталог водных животных бассейна
Донца А. А. Фадеева (1929) с его же дополнениями (1930), то окажется, что
из 652 видов многоклеточных первичноводных беспозвоночных заведомо
пойкилосмотичны 483, а в той или иной мере гомойосмотичны только 12 (выс-
шие ракообразные), осморегуляторные способности остальных 169 (низшие
ракообразные) неизвестны, хотя ветвистоусые (65 видов), по всей вероятно-
сти, пойкилосмотичны. Подобные же соотношения несомненно свойственны
и другим текучим и стоячим пресным водам с повсеместным господством
в бентосе из первичноводных олигохет и моллюсков, в планктоне — коловра-
ток и ветвистоусых. Следовательно, в современной пресноводной фауне пер-
вичноводных животных, происшедших, согласно общепринятым взглядам,
от морских предков, преобладают те формы, которые в настоящее время не
могут совершить переход из моря в пресную воду. Очевидно, в предшеству-
ющие геологические эпохи имели место условия, допускавшие подобный
переход, т. е. различие в солености между океаном и пресными водами
было не столь значительным, как сейчас. Тогда губки, кишечнополостные,
моллюски и другие пойкилосмотичные животные смогли вселиться из моря
в пресные воды, что в настоящее время для них невозможно.
Таким образом, все изложенное выше приводит к необходимости при-
знать происходившее в течение геологического времени осолонение мирового
океана и присоединиться в этом отношении к мнению Меррэя (Меггеу, 1923),
Макаллума (Macallum, 1907, 1926), Роджерса (Rogers, 1938), Лукина
(1936а,б), Блинова (1945) и других исследователей.
Гидрохимические расчеты, произведенные в недавнее время последним
автором, использовавшим данные по химизму вод нижнепалеозойского Цех-
штейнового моря, показали полную возможность рассматривать солевой
состав современного океана как результат смешения этой части древнего
океана с водами материкового стока. Основываясь на этих выкладках, Бли-
нов определяет соленость океана археозоя или начала палеозоя 10—14 °/00,
т. е. считает ее в 2,5—3,5 раза более низкой, чем соленость современного нам
океана.
К сожалению, палеонтологические доказательства пониженной солено-
сти Мирового океана в предшествующие геологические эпохи крайне немно-
гочисленны, да и трудно было бы надеяться на противоположное. Дело в том,
что, обращаясь за разъяснением по вопросу о солености морей далекого про-
шлого к палеонтологии, мы неминуемо попадаем в заколдованный круг. Па-
леонтологи определяют соленость по фауне и совершенно не считаются с воз-
можностью обитания брахиопод или других современных морских животных
при более низкой солености, чем сейчас. Основываясь на составе фауны, они
определяют соленость и поэтому не в состоянии ответить на вопрос, не
жила ли эта самая ф^на при более низкой солености. Между тем Вальтер
(Walther, 1927) утверждает, что распространение многих руководящих древ-
них брахиопод, пластинчатожаберных, головоногих указывает на их пре-
сноводный образ жизни, а это, как заметил Лукин, может служить доказа-
тельством менее резкого различия в солености между морем и пресными во-
дами в далеком прошлом. Подобный же факт сообщил недавно Трюмэн
208
(Trueman, 1942), обнаруживший на каменноугольных унионидах (в част-
ности, из группы Najas triangulina) трубочки сидячих полихет, очень сходные
с трубками современных Spirorbis, не выдерживающих опреснения. Оче-
видно, в то время или униониды могли выходить в море, или Spirorbis захо-
дить в пресную воду, но, во всяком случае, благодаря меньшему разрыву
в солености между морем и пресной водой создалась невозможная сейчас
фаунистическая комбинация.
Некоторые экологические особенности современных форм также иногда
рассматриваются как показатель повышающегося в течение геологического
времени осолонения Мирового океана. Так, например, Вогэн (Vaughan,
1917) установил, что некоторые кораллы, обитающие сейчас только при
полной океанической солености, в условиях эксперимента выносят
опреснение на 20 %, и объяснил это меньшей соленостью океана в про-
шлом.
В свете приведенных только что материалов по стабильности величины
внутреннего осмотического давления гомойосмотичных животных приобре-
тает большую доказательность мысль Макаллума, Роджерса и Уэрдли (Wa-
rdlaw, 1931) о более низкой солевой концентрации крови позвоночных по
сравнению с морскими беспозвоночными как следствие формирования первых
в древнем опресненном океане.
Таким образом, формирование пресноводной фауны можно представить
себе в следующих са мых общих чертах. При незначительной солености Миро-
вого океана происходило заселение пресных вод многими древними относи-
тельно эвригалинными формами, в частности губками и кишечнополостными.
По мере осолонения океана проникновение в пресные воды затруднялось
прежде всего для пойкилосмотичных животных, и в настоящее время границу
между морем и пресной водой могут преодолеть в основном только
гомойосмотичные организмы. Отсюда ясно, что древность, а следовательно,
и глубина разрыва между морской и пресноводной фаунами той или иной
группы стоит в связи с осморегуляторными ее способностями, и нас не
должна удивлять большая близость морских и пресноводных ракообразных
или рыб по сравнению с губками и кишечнополостными. При большей полноте
геологической летописи можно было бы установить, когда прекратилось мас-
совое внедрение губок, моллюсков и других групп в пресную воду и, быть мо-
жет, увязать такие моменты с процессом осолонения Мирового океана, но
в настоящее время об этом остается только мечтать.
Признание постепенного осолонения Мирового океана, как заметил
Лукин (1936), заставляет сделать один весьма важный вывод применительно
к океанической фауне. Очевидно, ее историческое развитие шло в одном отно-
шении строго направленно по пути приспособления к повышающейся соле-
ности. Отбор благоприятствовал формам, способным существовать при вы-
сокой солености и уничтожал стеногалинных животных, приспособленных
к жизни при низкой солености. Следовательно, никак нельзя судить по со-
временным формам об отношении к солености их далеких предков. Так,
например, единственный современный представитель Merostomata — Limu-
lus — морской обитатель, причем полостная его жидкость изотонична мор-
ской воде, он неспособен к регуляции минерального состава полостной жид-
кости, а также, по-видимому, и к активной осморегуляции (Daley et al.,
1931). Правда, он выдерживает разведение морской воды в 4 раза, но уже
при разведении в 8 раз не живет более 40 ч (Pearse, 1928). Между тем, со-
209
гласно установившимся сейчас взглядам, ближайшие его родичи жили в пре-
сной воде. Очевидно, в палеозое был возможен переход таких пойкилосмо-
тичных форм из пресных вод в море или обратно.
Все эти данные имеют весьма существенное значение для анализа эволю-
ции пресноводной фауны. Из них следует, во-первых, что пресноводная фауна
в части пойкилосмотичных первичноводных животных сложилась очень
давно, когда еще солевые границы между морем и пресной водой были сгла-
жены и могло осуществляться вселение пойкилосмотичных животных из моря
в пресные воды; во-вторых, надо признать, что обогащение пресноводной фа-
уны морскими элементами с течением времени постепенно прекращается
и сейчас возможно только для весьма ограниченного количества групп; на-
конец, в-третьих, в отношении межвидовой конкуренции пресноводные обита-
тели оказываются в весьма благоприятном положении, так как им не угро-
жает борьба с новыми морскими вселенцами.
VI
Каким образом происходила дальнейшая эволюция пресноводной фауны
и можно ли говорить о специфических особенностях эволюции организмов,
населивших пресные воды?
Имеющиеся материалы позволяют утверждать, что пресноводная фауна
развивалась достаточно своеобразно и во многих отношениях принципиально
отлично от океанической. Однако прежде чем привести соответствующие до-
казательства этого положения, необходимо строго ограничить объект нашего
рассмотрения.
Как известно, пресноводная фауна неоднородна и состоит из двух групп
совершенно различного генезиса — первичноводных животных, имеющих
в конечном счете океаническое происхождение, и вторичноводных животных,
происходящих от сухопутных предков. В настоящее время вторичноводные
животные господствуют над первичноводными в бентосе большинства прес-
новодных водоемов, что объясняется решающими преимуществами вторич-
новодных животных в условиях недолговечности пресноводных водоемов
(Лукин, 1936; Hesse, 1924). Тем не менее для темы настоящей работы
вторичноводные животные имеют второстепенное значение, так как, во-пер-
вых, формирование крупных таксономических категорий, к которым они при-
надлежат, происходило в воздушной среде, да и сейчас эволюция многих
из них в значительной своей части связана с условиями воздушного суще-
ствования, и, во-вторых, в геологическом смысле слова они моложе первично-
водных. Самые древние крылатые насекомые известны начиная с карбона,
водные легочные миоллюски появляются только на границе юры и мела. Судя
по палеонтологическим данным, вторичноводные животные начали перехо-
дить к обитанию в пресных водах уже тогда, когда там обитала довольно
богатая фауна первичноводных организмов. Губки, относящиеся к современ-
ному роду Spongilla, известны начиная с юрского времени, переднежаберные
моллюски Valvata и Viviparus — также с юрского времени, a Bithynia и Hyd-
robia — с мелового, пластинчатожаберные Unio — с юры, Sphaerium —
с мела, Anodonta и Pisidium — с эоцена. Среди современных родов низших
ракообразных обнаружены Cyzicus — с девона, Apis — с триаса (Trusheim,
1938), Cypris — с юры, среди высших ракообразных Astacus — с юры (Stra-
elen, 1928b), Phreatoicus—даже с перми (Chilton, 1918).
210
По отношению к другим группам первичноводных животных мы, к сожале-
нию, не располагаем подобными прямыми указаниями на их древность, но,
судя по зоогеографическим данным, по своей консервативности они не усту-
пают уже рассмотренным. Шелленберг (Schellenberg, 1937b) склонен припи-
сывать роду Gammarus палеозойский возраст, Л. А. Зенкевич (1940) относит
формирование пресноводных кишечнополостных к мезозою, сходные взгляды
высказывались Михаельсеном (Michaelsen, 1933) применительно к олигохе-
там и т. д. Следует предполагать, что специфические особенности эволюции
пресноводных организмов выявятся рельефнее именно при анализе первич-
новодных животных, вся геологическая история которых развертывалась
в условиях обитания в пресных водах. Вместе с тем некоторые из общих зако-
номерностей эволюции первичноводных животных могут оказаться справед-
ливыми и для вторичноводных животных, поскольку они все же в значитель-
ной степени связаны с пресноводными водоемами.
Обращаясь к работам, в которых затрагиваются вопросы эволюции пре-
сноводной фауны, нетрудно показать, что пресноводную фауну характери-
зуют следующие основные моменты: 1) пресные воды по сравнению с океаном
представляют собою весьма благоприятную среду для сохранения древних
архаических форм, 2) в пресных водах не происходило формирования круп-
ных таксономических групп и 3) там не происходило вымирания крупных
таксономических групп. Все это может быть объяснено меньшей интенсив-
ностью борьбы за существование и соответственно естественного отбора
в условиях пресноводных водоемов по сравнению с Мировым океаном, о чем
можно судить по замедленному темпу эволюции пресноводной фауны по
сравнению с морской.
VII
Представление о пресных водах как об убежище для древних прими-
тивных форм, вытесненных вследствие ожесточенной борьбы за существова-
ние из океана, высказывалось неоднократно и никем не оспаривалось. Пер-
вым обратил внимание на эту особенность пресноводной фауны Дарвин. «Все
пресноводные бассейны в совокупности — писал он, — по сравнению с морем
и сушей представляют малую область. Следовательно, соперничество между
пресноводными обитателями было менее ожесточенным, чем в какой-либо
другой области; новые формы медленнее образовывались и старые формы
медленнее вымирали. Именно в пресных водах мы встречаем семь родов гано-
идных рыб — остатки когда-то преобладавшего отряда; в пресной же воде мы
встречаем и самые аномальные из существующих на земле форм — Ornitho-
rhynchus и Lepidosiren, которые, подобно ископаемым формам, до некоторой
степени связывают отряды, в настоящее время далеко отстоящие друг от
друга в системе природы. Эти аномальные формы могут быть названы жи-
выми ископаемыми; они уцелели до сих пор, потому что жили в ограниченном
ареале и подвергались менее разнообразной и, следовательно, менее ожесто-
ченной конкуренции» (1937, с. 163). В другом месте он напоминает: «Мы
должны помнить, что многие пресноводные организмы занимают низкое ме-
сто в системе природы, и у нас есть основание думать, что такие существа
изменяются медленнее высших. .» (с. 594).
Сходные мысли высказывал в 1864 г. верный последователь Дарвина
Фриц Мюллер: «Если бы можно было рассматривать филлопод как ближай-
211
ших родственников трилобитов, о чем я не осмеливаюсь судить, то они наряду
с Lepidosteus и Polypterus, Lepidosiren и Ptotopterus служили бы дальней-
шим примером того, что в пресных водах сохраняются формы, давно уже
вымершие в море. Нахождение Artemia в чрезвычайно соленой воде пока-
зало бы при этом, что им удалось избежать гибели не из-за пресной воды,
а в результате более слабого в пресной воде соперничества» (Мюллер,
1932).
И. И. Мечников в 1876 г. (цит. по: Мечников, 1943) подробно обсудил
приведенное выше положение Дарвина о большем постоянстве пресноводных
организмов по сравнению с морскими и сухопутными, подтвердив его на при-
мере пресноводных растений. «Пресноводные растения, — писал Меч-
ников, — вообще бедны числом, видом и, кроме того, они носят на себе отпе-
чаток древнего происхождения» (с. 120). Он подчеркнул невозможность
объяснить это явление с позиций признания ведущего значения внутриви-
довой борьбы, возникающей вследствие перенаселения, так как последнее
постоянно имеет место в пресноводных водоемах. Согласно Мечникову,
«для изменяющего действия борьбы за существование, а следовательно,
и естественного подбора недостаточно простого перенаселения одной формы
и необходима конкуренция по возможности большего количества разнород-
ных существ. .» (с. 122). В этом отношении по изложенным здесь причинам
пресные воды далеко уступают морю и суше, и материал настоящей статьи
вполне подтверждает мнение великого русского ученого.
Гессе (Hesse, 1924) предполагал, что в пресной воде борьба за существо-
вание между животными ослаблена, но зато обострена борьба с абиотиче-
скими факторами: .в таких местах могли сохраниться многие формы,
которые в других местах должны были избегать конкуренции. Так, в пресной
воде переживают последние представители старинных круглоротых и двояко-
дышащих. То же относится к осетровым. . Из костистых рыб в борьбе с моло-
дыми колючеперыми мягкоперые сохранились только на севере, на больших
океанических глубинах и в пресной воде. .» Согласно С. А. Зернову (1934),
«Пресноводные бассейны в истории земли не раз служили последним убежи-
щем для вымирающих морских форм. Можно думать, что в пресной воде
с ее массой изолированных озер и рек они находили возможность укрыться
от вновь явившихся врагов и конкурентов .» (с. 180—181). В. Г Гептнер
(1936) также указывал на то, «. .что в пресной воде в настоящее время
существуют некоторые группы, которые в прежние геологические эпохи жили
в море. Это, по-видимому, примитивные группы, вытесненные из моря и сох-
ранившиеся в менее благоприятных условиях, которые существуют в прес-
ной воде сравнительно с морем» (с. 139).
С этой же точки зрения можно рассматривать и приуроченность наиболее
примитивных современных крылатых насекомых к пресным водам. Я не вижу
никаких поводов к тому, чтобы сомневаться, подобно Е. И. Лукину (1936а),
в правомерности отнесения всех насекомых к вторичноводным на том основа-
нии, что древнекрылые и ручейники в своем развитии связаны с пресной во-
дой. По всей вероятности, из всех примитивных групп крылатых насекомых
до наших дней сохранились преимущественно те, которые приспособились
к прохождению своего длительного личиночного периода в среде с ослаб-
ленной борьбой за существование. Это прослеживается не только на всем
классе насекомых, но и на отдельных его отрядах. Так, например, согласно
212
И. А. Рубцову (1945), наиболее древние и примитивные представители от-
ряда двукрылых имеют водных личинок. В этом случае консервирующее воз-
действие обитания в пресных водах сказалось и на более древних из вторич-
новодных животных.
Не вызывает особых сомнений и чрезвычайно скромная роль пресновод-
ной фауны в формировании новых крупных групп животных. Чернай писал
о том, что пресноводные животные, изменяясь, не развили из себя ни особого
типа, ни класса и уступают в этом отношении сухопутным. Это положение
подтверждается уже приведенными выше современными данными. Что каса-
ется рыб, то, если даже признать вполне убедительным палеонтологические
доказательства их пресноводного происхождения, и в этом случае основной
средой, в которой происходила эволюция этой важнейшей группы, оказы-
вается океан. Уже в девоне существовала хорошо обособленная морская
ихтиофауна (Roemer, 1946), а в дальнейшем многообразие морской ихтио-
фауны быстро возрастало, а пресноводной — падало (Roemer, Grove, 193?),
и современные пресноводные рыбы происходят от морских, а не от древне-
пресноводных (Roemer, 1946).
На другую сторону той же проблемы — на ход вымирания крупных групп
в условиях пресноводного обитания — не было обращено должного внима-
ния, хотя и в этом отношении пресноводная фауна отличается совершенно
определенными особенностями. Если в пресной воде не возникали, за немно-
гими сомнительными исключениями, новые группы, то точно так же там не
происходило, как правило, и столь катастрофического вымирания целых
классов, отрядов и семейств, которое так характерно для бурной истории
океанической фауны. Археоциаты, граптолиты, четырехлучевые кораллы,
Thecoidea, Carpoidca, Blastoidea, рудисты, аммониты, белемниты, трило-
биты и многие другие более мелкие группы зародились, расцвели и вымерли
в море. За это время в пресной воде исчезли только Merostomata, все же
остальные так или иначе оказавшиеся там животные продолжали существо-
вать и в более или менее измененных формах дожили до наших дней. К сожа-
лению, нет достаточных палеонтологических данных для суждения об измене-
ниях в соотношениях различных групп пресноводной фауны в течение геоло-
гического времени, но трудно сомневаться в том, что в пресной воде не имела
места столь кардинальная смена господствующих групп, как'в Мировом оке-
ане. Первые страницы геологической летописи говорят о безраздельном гос-
подстве в океанах кембрия и силура трилобитов,после бурного расцвета вы-
мирающих в конце палеозоя. Та же судьба постигает многочисленных и раз-
нообразных четырехлучевых кораллов, граптолитов, цистоидей, бластоидей
и др. С этого времени на первое место среди морских беспозвоночных высту-
пают аммониты, а несколько позднее — белемниты, с тем чтобы закончить
существование в течение мелового или в самом начале третичного периода.
Известные уже из кембрия панцирные рыбы в силуре достигают значитель-
ного количественного развития, но уже в девоне и карбоне наступает их упа-
док, и в мезозое их сменяют костные ганоиды, в свою очередь отступающие
к концу этой эры на задний план перед бурным расцветом костистых рыб.
Другие типы животного царства Мирового океана пережили такие же резкие
смены господствовавших в разные периоды групп. Насколько можно судить
по скудным палеонтологическим данным, пресноводная фауна не подвер-
галась столь грандиозным перестройкам.
213
Все изложенное выше заставляет присоединиться к мнению Вальтера
(Walther, 1927), характеризующего пресные воды как «биологический ту-
пик» 1
VIII
Перечисленные особенности пресноводной фауны выявляются при самом
общем анализе палеонтологических и зоогеографических данных. Вполне
естественным представляется желание найти более точные показатели раз-
личий в ходе эволюции океанической и пресноводной фаун и прежде всего
сравнить их по степени консервативности. Иными словами, можно поставить
перед собой задачу сопоставления темпа эволюции океанической и пресно-
водной фаун.
С этой целью я произвел некоторые расчеты, основываясь главным обра-
зом на общеизвестной сводке К. Циттеля (1934), переработанной коллекти-
вом советских специалистов 1 2 Впоследствии выяснилось, что совершенно
таким же методом и источником пользовался американский палеонтолог
Симпсон (Simpson, 1944; Симпсон, 1948) для доказательства различного
темпа эволюции разных групп животных (копытных млекопитающих и пла-
стинчатожаберных моллюсков).
Однако выводы, к которым пришел этот исследователь, находятся в рез-
ком противоречии с излагаемым здесь материалом. По мнению Симпсона,
наиболее быстрая «квантовая эволюция» свойственная малым изолирован-
ным популяциям и происходит благодаря случайным мутациям, возникаю-
щим независимо от среды и быстро распространяющимся в популяциях. Ка-
залось бы, что пресноводные организмы, раздробленные на множество мел-
ких популяций, должны были эволюционировать быстрее, чем морские, со-
ставляющие обширные популяции. На самом деле все происходит как раз на-
оборот, и сравнение темпов эволюции морской и пресноводной фаун опровер-
гает формально-генетические построения Симпсона.
Я выбрал для основных групп морских и пресноводных первичноводных
беспозвоночных по отдельности все современные роды, встреченные в ископа-
емом состоянии, а затем, приняв это число для каждой группы за 100 %, вы-
считал процентные отношения родов различной древности. Я вполне отдаю
себе отчет в неточности этого метода, поскольку получившиеся показатели
1 Данные по Decapoda взяты из каталога Глесснера (Glaesner, 1929).
2 В явном противоречии со всем изложенным находится утверждение Г. Верещагина
(1940) о том, что в противоположность морю в большинстве континентальных водо-
емов с исключительно изменчивыми условиями среды мы наблюдаем, как правило,
весьма интенсивный процесс эволюции обитающей в них фауны. Для подтверждения
этого факта достаточно указать на громадное количество рас и подвидов ряда рыб
(сиги, окуни и т. д.), некоторые роды Cladocera (Daphnia, Bosmina) и другие группы.
Распад вида на мелкие изолированные популяции нельзя квалифицировать как
«весьма интенсивный процесс эволюции». Кроме того, для Cladocera характерна не
столько географическая изменчивость, связанная с изоляцией, сколько сезонная,
вызываемая иными факторами. Весьма замечательной особенностью Cladocera
можно считать почти космополитическое распространение отдельных видов и отсут-
ствие или крайне слабую обособленность локальных рас и подвидов. Окуни не распа-
даются на подвиды, а история сигов, связанная с ледниковым временем, не харак-
терна для обычных обитателей пресноводных водоемов.
214
в какой-то степени извращены неполнотой геологической летописи, неоди-
наковой изученностью разных групп и осадков разных горизонтов и стран
и другими случайными и неизбежными обстоятельствами. Тем не менее неко-
торое общее представление о времени формирования основных групп, слага-
ющих по крайней мере современную морскую фауну, а также о степени их
консервативности эти показатели все же дают (табл. 3).
Анализ табл. 3 позволяет сделать следующие выводы. Современная мор-
ская фауна беспозвоночных приблизительно наполовину состоит из сравни-
тельно молодых третичных родов; исключение составляют две группы —
губки и мшанки, сложившиеся в основном в меловое время. Кроме них и голо-
воногих моллюсков, все группы демонстрируют однотипное постепенное, но
довольно быстрое преобразование родового состава по мере приближения
к современности. Архаические роды палеозойского возраста играют во всех
группах весьма незначительную роль, причем их процентное отношение для
всех пяти групп, которым они свойственны, выражается цифрами одного по-
рядка — 5—12 %.
Если расположить проанализированные группы по порядку постепенного
уменьшения степени консервативности, т. е. принять во внимание средний
возраст сохранившихся в ископаемом состоянии родов различных групп, то
получится следующий ряд: мшанки—пластинчатожаберные—лопатоногие—
брюхоногие—иглокожие—брахиоподы—губки—кишечнополостные—усоно-
гие—десятиногие—головоногие, причем для первых четырех групп характе-
рен в среднем юрский возраст, для следующих пяти — меловой и, наконец,
для десятиногих и головоногих — третичный. Такая последовательность
представляется несколько неожиданной. В любом учебнике зоологии в каче-
стве примеров наиболее консервативных групп приводятся в первую очередь
брахиоподы и головоногие моллюски, для чего действительно имеются неко-
торые основания. Противоречия между этим общераспространенным мне-
нием и полученными мною объективными показателями разрешается при
применении сформулированных ранее представлений о реликтах (Бирштейн,
1947а). Мы вправе рассматривать головоногих моллюсков и брахиопод во-
обще как древние, консервативные и реликтовые классы, реликтовые потому,
что число их представителей и свойственные им ареалы в настоящее время
меньше, чем в предыдущие геологические эпохи. Но современная фауна голо-
воногих и брахиопод состоит целиком или в значительной своей части из мо-
лодых третичных родов, по отношению к которым нет оснований применять
термин «реликт>. Реликтовая группа, в данном случае класс, может быть
представлена нереликтовыми родами (головоногие) или включать наряду
с реликтовыми и нереликтовые роды .(брахиоподы).
Примененный здесь простой метод расчета претендует на определение
сроков и темпов формирования той или иной группы только современной
фауны.
Я думаю, что подобный учет древности отдельных родов дает более пра-
вильное представление о степени консервативности разных групп организ-
мов, чем данные о возрасте более крупных таксономических единиц, ко-
торыми оперируют некоторые авторы. Выявившаяся почти для всех групп од-
нотипная картина формирования современной морской фауны, быть может,
не является случайной. Можно допустить, что она в известной мере отражает
некоторый осредненный темп эволюции современной океанической фауны
беспозвоночных.
215
Таблица 3. Степень древности родов морских животных разных групп в % от
современного числа родов, известных в ископаемом состоянии
Г руппа Губки Кишечнополостные Моллюски Мшанки L Плеченогие [Ракообразные
пластинчато- жаберные брюхоногие (V х о X О н са с о головоногие Иглокожие усоногие десятиногие
(14) (68) (ЮЗ) (144) (9) (4) (37) (40) (62) |(9) (65)
Кембрий — — — I — — — — — 1 —
Силур — — 1 3 11 — 8 5 — — —
Девон — — 3 — — — — — — — —
Карбон — — 3 2 — — — — — — —
Пермь — — 4 — — — — — — — —
Триас — 3 14 6 И — 3 — 5 — —
Юра — 13 15 7 — — 16 12 19 11 2
Мел 93 35 16 32 33 — 51 20 22 22 7
Третичный Примем 7 а н и е. 49 44 В табл. 3 и 4 49 цифры 45 100 । в скобках — 22 63 число родов. 53 67 91
Таблица 4. Степень древности родов пресноводных животных разных групп
в % от современного числа родов, известных в ископаемом состоянии
Группа Губки Моллюски Ракообразные
пластин- чатожа- берные брюхо- ногие ракушко- вые листо- ногие десяти- ногие равно- ногие
(1) (6) (Ю) (6) (2) (2) (2)
Силур — — — 67 — — —
Девон — — — — 50 — —
Карбон — — — — — — —
Пермь — — — 17 — — —
Триас — 16 — — 50 — 50
Юра 100 16 30 16 — 50 50
Мел — 34 50 — — — —
Третичный — 34 20 — — 50 —
С этой точки зрения любопытно сравнить с приведенными показателями
океанической фауны аналогичные показатели, полученные для пресновод-
ной, солоноватоводной и сухопутной фаун. К сожалению, палеонтологический
материал по этим последним еще более ограничен, но тем не менее некоторые
характерные различия выявляются достаточно выпукло.
Среди первичноводных пресноводных животных в ископаемом состоянии
известно по нескольку современных родов пластинчатожаберных и брюхоно-
гих моллюсков и ракушковых ракообразных; губки, листоногие ракообразные
и Malacostraca представлены единичными родами. Сводка соответствующих
данных представлена в табл. 4.
Таким образом, если придавать значение приведенным скудным данным,
то можно характеризовать современную морскую фауну как преимущест-
216
венно кайнозойскую, а пресноводной приписывать по крайней мере мезозой-
ский возраст.
Что касается сухопутной и солоноватоводной фаун, то их относительная
молодость не вызывает сомнений. Современные роды таких господствующих
на суше групп беспозвоночных, как насекомые, паукообразные и многоножки
в ископаемом состоянии не найдены. Имеются материалы лишь по брюхо-
ногим моллюскам, из которых 80 % современных родов третичные, а 20 % —
меловые. Сходные соотношения свойственны и солоноватоводным моллю-
скам: пластинчатожаберные на 75 % третичные, на 25 % меловые, брюхоно-
гие — все третичные.
Эти сопоставления могут служить хорошим подтверждением правильно-
сти приведенных выше суждений Дарвина и других авторов о консерватив-
ности пресноводной фауны по сравнению с морской. Если присоединиться
к представлению о происхождении пресноводной фауны (в части первично-
водных животных) от морской, то следует принять, что формирование первой
началось позже, но закончилось раньше, чем второй, причем пресноводная
фауна прошла очень короткий путь эволюционных преобразований и сравни-
тельно мало изменялась. Современные сухопутная и солоноватоводная
фауны, судя по палеонтологическим данным, еще моложе пресноводной.
IX
Консерватизм пресноводной фауны, который отчетливо выражается за-
медленным темпом ее эволюции по сравнению с океанической фауной объяс-
няется несколькими причинами.
1. Недолговечностью (в геологическом смысле слова) подавляющего
большинства пресноводных водоемов;
2. Меньшим разнообразием условий существования в пресноводных во-
доемах по сравнению с океаном, а также меньшей их величиной;
3. Положением пресноводных организмов в пищевых цепях.
Продолжительность существования пресноводных водоемов (примеры
см.: Зернов, 1934; Лукин, 1936а), как правило, оказывается недостаточной
для формирования новых видов. Огромное количество пресноводных организ-
мов выработали специальные приспособления, позволяющие им переносить
высыхание водоемов — цисты, покоящиеся стадии, как, например, геммулы,
статобласты, различные стойкие яйца и другие средства пассивной защиты от
высыхания. Подобные способы пассивной защиты, вероятно, не способствуют
темпам прогрессивной эволюции. В то же время вследствие постоянно совер-
шающегося переноса этих легко транспортабельных стадий из соседних водо-
емов не создается полной изоляции популяции, населяющей каждый данный
водоем, что также препятствует видообразованию. Примером поразительной
консервативности обитателей недолговечных водоемов может служить один
из наиболее характерных для временных водоемов вид — Apus cancriformis.
Как показал Трусгейм (Trusheim, 1938), этот вид существует во временных
весенних лужах с триаса, почти не меняясь морфологически. В немногочис-
ленных геологически древних водоемах, как, например, Байкал, Танганьика,
Охрид и немногие другие, наблюдается интенсивное видообразование, осо-
бенно в случаях, когда этому не препятствуют сильные и активные хищники
(Worthington, 1940). Однако в масштабах геологического времени подавля-
15 Заказ 333
217
ющее большинство пресноводных водоемов, срок жизни которых определя-
ется тысячелетиями, принципиально не отличается от весенних луж.
Разнообразие среды обитания представляется одним из важнейших усло-
вий быстрых темпов прогрессивной эволюции. Только в этом случае обеспе-
чивается возможность разносторонней специализации и, что еще важнее,
только при разнообразных условиях существования могут быть реализованы
естественным отбором те адаптации широкого значения, которые А. Н. Север-
цов (1939) называет ароморфозами.
Нет необходимости доказывать, насколько пресноводные водоемы усту-
пают в указанном отношении Мировому океану. И действительно, именно
в последнем, в частности в наиболее разнообразной биотопически его ча-
сти — в области морского мелководья, происходило формирование основных
прогрессивных групп первичноводной гидрофауны. Гессе называет фауну
этой области матерью всех первичноводных фаун, как пресноводных, так и
морских. Давиташвили (1943), основываясь на палеонтологических данных,
утверждает, что исходные формы всех типов должны были быть обитателями
дна эвфотической полосы мелководья и принадлежали, очевидно, к блужда-
ющему бентосу Экман (Ekman, 1935) прослеживает происхождение осталь-
ных океанических фаун от фауны морского мелководья.
Малочисленность экологических ниш, ограничивающая потенциальные
возможности разносторонней адаптации организмов, накладывается в прес-
ных водоемах на их недолговечность, в свою очередь тормозящую, как мы
видели, ход прогрессивной эволюции. Однако даже в геологически долго-
вечных пресноводных водоемах при монотонных упрощенных условиях суще-
ствования эволюция идет замедленным темпом, что видно на примере гидро-
фауны подземных вод (Бирштейн, 1941).
Наоборот, при значительном разнообразии среды обитания (при условии
относительной долговечности водоема) эволюция пресноводных организмов
совершается весьма быстрыми темпами. В этом отношении особенно ярким
примером можно считать богатую и своеобразную фауну оз. Байкал. Возник-
новение байкальских эндемиков, и в частности бычков, по данным Д. Н. Тали-
ева (1948), произошло в течение четвертичного времени и стало возможным
только после образования больших глубин озера, т. е. после появления огром-
ного количества новых разнообразных экологических ниш.
Биотопическое разнообразие условий существования, естественно вклю-
чается в более широкое понятие величины ареала, на значение которой для
темпа эволюции настойчиво указывал Дарвин. «Я прихожу к заключению, —
пишет он, — что хотя небольшие ограниченные ареалы в некоторых отноше-
ниях были крайне благоприятными для образования новых видов, но тем не
менее в обширных ареалах изменения в большинстве случаев совершались
быстрее и, что еще важнее, новые формы, образовавшиеся на больших ареа-
лах и уже победившие многих соперников, более способны к широкому рас-
селению и, следовательно, к образованию наибольшего числа разновидно-
стей и видов» (Дарвин, 1937, с. 162). Согласно Дарвину, благодаря меньшей
величине пресноводных водоемов по сравнению с океаном «соперничество
между пресноводными обитателями было менее ожесточенным, чем в какой-
либо другой области; новые формы медленнее образовывались и старые фор-
мы медленнее вымирали», (с. 163). Я пытался недавно показать, что тезис
Дарвина о.зависимости темпа эволюции от величины ареала находит себе
подтверждение в ряде современных биогеографических данных (Бирштейн,
218
1947а); точно так же оказывается справедливым и основанное на этом тезисе
его представление о замедленном темпе эволюции пресноводной фауны.
Наконец, одной из причин замедленного темпа эволюции пресноводной
фауны можно считать положение большинства составляющих ее первично-
водных беспозвоночных животных в пищевых цепях водоемов. Почти все
планктонные и многие донные организмы представляют собой низшие звенья
пищевых цепей и подвергаются неизбирательному уничтожению подавляю-
щими по силе хищниками (рыбами).
Немногие первичноводные организмы, не потребляемые рыбами, защи-
щены от них пассивно—крупными размерами (Unionidae), скелетом
(губки) или химическими и механическими средствами обороны (турбелля-
рии, гидра). При этом характерно, что в отличие от океана пресноводные пер-
вичноводные беспозвоночные, за редкими исключениями (ресничные черви),
не занимают положения высших конечных звеньев пищевых цепей и не высту-
пают в роли активных хищников. В пресной воде мы не находим среди первич-
новодных беспозвоночных аналогов многочисленных активных хищных океа-
нических групп — немертин, бродячих полихет, головоногих моллюсков, кра-
бов и т. д., а именно такие формы идут наиболее быстро по пути морфологиче-
ского прогресса.
Все отмеченные моменты определяют своеобразие форм борьбы за суще-
ствование в условиях пресноводных водоемов. В отличие от океана здесь
преобладают пассивные формы защиты от действия биотических и абиоти-
ческих факторов. Активная борьба за существование отступает на задний
план, что приводит к ослаблению процесса избирательной элиминации и бла-
гоприятствует видам, вставшим на путь узкой специализации. Поэтому пре-
сноводные животные никогда не вырабатывали и никогда не выработают
адаптаций широкого значения и всегда будут эволюционировать медленнее,
чем обитатели беспредельно разнообразных океана и суши, где возникали
и продолжают возникать наиболее совершенные представители органиче-
ского мира.
НЕКОТОРЫЕ ВОПРОСЫ ЭВОЛЮЦИИ ГЛУБОКОВОДНОЙ ФАУНЫ*
Особенности эволюции животных на больших океанических гулубинах
издавна привлекали внимание исследователей. Еще знаменитая английская
экспедиция на «Челленджере» была организована в надежде обнаружить на
дне океана древние формы жизни, исчезнувшие из области морского мелко-
водья, поскольку существовало представление о больших океанических глу-
бинах как о среде, способствующей сохранению древних реликтов. Тогда
эти надежды не сбылись.
С тех пор наши сведения о составе, распространении и отчасти биологии
глубоководной фауны обогатились огромным количеством новых данных,
добытых, в частности, советскими экспедициями на «Витязе», датской экспе-
дицией на «Галатее» и американскими экспедициями на «Виме» и других
судах. Однако собранный материал нельзя считать достаточным для форму-
лировки четких выводов об особенностях эволюции глубоководной фауны.
* В кн.: История Мирового океана. М.: Наука, 1971, с. 84—89.
219
Полученные преимущественно фаунистические данные допускают произволь-
ное толкование вопросов эволюции глубоководных животных и нуждаются
в более тщательном анализе и дополнении экспериментальными исследова-
ниями, пока технически невыполнимыми.
Между тем вопрос об эволюции глубоководной фауны и связанный с ним
вопрос об ее геологическом возрасте имеют первостепенное значение для бо-
лее широкой проблемы — истории Мирового океана. Если имеются достаточ-
ные основания для признания длительности существования этой фауны,
следует признать вероятным длительное существование больших океани-
ческих глубин; если, наоборот, глубоководную фауну правильнее рассматри-
вать как недавно возникшую, этот аргумент можно использовать для под-
тверждения представлений о геологически недавнем формировании океани-
ческой абиссали.
Недостаточность материала для суждения об особенностях эволюции глу-
боководной фауны очевидна уже из того, что по этому вопросу высказыва-
ются диаметрально противоположные мнения. С одной стороны, развива-
ются традиционные представления о консервативности и древности глубоко-
водной фауны, насыщенности ее реликтами прошедших эпох и медленных
темпах ее эволюции. Это мнение высказывалось давно, его поддерживал
крупнейший современный морской зоогеограф Свен Экман (Ekman, 1953),
и оно разделяется многими советскими (см., например, Зенкевич, Бирштейн,
1961) и некоторыми зарубежными (Parker, 1961) исследователями.
С другой стороны, большинство современных зарубежных исследовате-
лей, в частности Бруун (Bruun, 1956), Мадсен (Madsen, 1961), Мензис
(Menzies, 1965) и др., считают глубоководную океаническую фауну геологи-
чески молодой и эволюционирующей быстрыми темпами.
Для обоснования обеих гипотез привлекаются аргументы, почерпнутые
из разных областей знания:
1. Используются представления о геологическом прошлом океана.
2. Для трактовки хода видообразования применяются популяционно-
генетические построения.
3. Учитывается положение в системе глубоководных животных и произво-
дится их сравнение с мелководными и ископаемыми, с тем чтобы таким об-
разом получить представление о формировании глубоководной фауны.
Я не буду касаться здесь геологической аргументации, которой пользу-
ются сторонники древности и молодости глубоководной фауны, поскольку
геологической истории океана посвящена вся наша конференция 1 В грубой
схеме сторонники молодости глубоководной фауны опираются на представле-
ния о недавнем возникновении больших океанических глубин или на гипо-
тезу о резких изменениях термического режима океана, якобы происходив-
ших в конце третичного—начале четвертичного времени; их оппоненты при-
нимают древность океанических глубин и относительную стабильность гидро-
логического режима Мирового океана.
Очень коротко следует остановиться на явлениях видообразования на
больших глубинах. Основываясь преимущественно на результатах обработки
глубоководных равноногих ракообразных, я могу отметить три важные
в этом отношении особенности.
1. Некоторые роды на больших глубинах разделяются на группы близко-
' Настоящая статья представляет собой доклад на Совещании по истории Мирового
океана, состоявшемся в Москве 1—4 декабря 1969 г. (Примеч. ред.).
родственных видов — «стоки», другие остаются олиготипическими. В первом
случае получается картина, отмечавшаяся, в частности, Е. Ф. Гурьяновой
(1951) для мелководных дальневосточных амфипод и изопод. Такой тип видо-
образования она назвала «букетным».
2. Часто дифференциация на виды связана с приспособлением каждого
из них к определенной глубине обитания. Границ или разрывов между зонами
обитания, как правило, нет. Наоборот, зоны двух соседних видов перекрыва-
ют одна другую. Бывает, однако, и так, что близкие виды обитают совместно
на одной глубине.
3. При опускании в ультраабиссаль в изолированных друг от друга впа-
динах или желобах обособливаются особые викарирующие виды.
Насколько согласуются эти факты с попытками подойти к проблеме с по-
зиций популяционной генетики?
Немногочисленные высказывания затрагивавших этот вопрос авторов
противоречивы. Картер (Carter, 1961) считает характерной особенностью
глубоководной фауны отсутствие в ней генетически изолированных популя-
ций, разделенных непреодолимыми для потока генов барьерами, что вместе
с относительным постоянством условий существования фауны во времени
должно определять замедленные темпы ее эволюции. Мензис (Menzies,
1965), придерживающийся мнения о более быстром темпе эволюции глубоко-
водных животных по сравнению с мелководными, допускает интенсивный
мутагенез у глубоководных животных, вызываемый повышенной радиоак-
тивностью радиоляриевых илов и, быть может, повышенным гидростатиче-
ским давлением.
Оба эти противоположные допущения не могут быть приняты. Нельзя
представлять себе глубоководную фауну как некий континуум, в пределах
которого господствует полная панмиксия. Этому противоречит достаточно
точно установленный факт ограниченности ареалов очень многих глубоко-
водных видов в горизонтальной плоскости и наличие только что упомянутых
цепочек видов, сменяющих друг друга в вертикальной плоскости. Кроме того,
можно утверждать, что возможность панмиксии у глубоководных донных
животных крайне ограничена их малой подвижностью и отсутствием у них
пелагических личинок, на что указывает, в частности, Торсон (Thorson, 1950).
Допущение повышенной частоты мутаций у глубоководных животных
остается чисто умозрительной гипотезой, причем упускается из виду то несо-
мненное обстоятельство, что за темпы эволюции ответственны не частота
мутаций, а интенсивность и направление естественного отбора, о чем нам
пока, применительно к данному случаю, судить почти невозможно.
При попытке распространить на глубоководную фауну господствующие
в науке представления о видообразовании мы наталкиваемся на серьезные
затруднения. Основное из них — можно ли признать в данном случае веду-
щее значение аллопатрического видообразования, т. е. обязательную полную
генетическую изоляцию непреодолимыми географическими барьерами двух
возникающих видов.
Этому принципу несомненно подчиняется становление ультраабиссальных
видов, эндемичных для каждой изолированной океанической впадины, од-
нако к процессу дифференциации видов по разным глубинам он, очевидно,
неприменим.
Такой процесс вовсе не специфичен для глубоководной океанической
фауны. Аналогичное явление описано для некоторых крупных озер, в част-
221
ности для Байкала. Многие зоологи, например М. М. Кожов (1962),считают,
что в таких случаях происходило симпатрическое видообразование, без про-
странственной генетической изоляции возникавших видов, однако некоторые
теоретики-эволюционисты с этим не соглашаются. Например, крупнейший
современный авторитет Э. Майр (1968) вообще не признает возможности
симпатрического видообразования. Применительно к Байкалу он вслед за
другим крупным эволюционистом Б. Реншем (Rensch, 1933) объясняет при-
сутствие групп близких видов в Байкале действием одйого (или нескольких)
из следующих факторов: сохранение реликтовых видов, множественная ин-
вазия, т. е. неоднократное и разновременное вселение предковых форм из
других водоемов, слияние воедино некогда разделенных озер и наличие геог-
рафических преград внутри озера.
Нетрудно видеть, что в пределах любого ограниченного района океана все
эти факторы, за исключением первого, не могут действовать на больших океа-
нических глубинах.
Из них следует принять во внимание только множественную инвазию,
но и ее вероятность очень мала, поскольку дифференциация видов по глуби-
нам нередко происходит у таких родов, которые полностью отсутствуют
на мелководье.
Поэтому приходится согласиться с авторами, допускающими трактовку
дифференциации видов по глубинам как случай симпатрического видообра-
зования.
Однако генетический механизм этого процесса остается загадочным.
В основе признаваемых случаев симпатрического видообразования на суше
и в пресных водоемах лежит усиливающаяся экологическая изоляция или
расхождение во времени размножения внутривидовых групп. Расхождение
по разным глубинам возникающих видов как в океанической абиссали, так
и в Байкале нельзя связывать с этими факторами. Здесь требуется новое
объяснение, остающееся делом будущего.
Допуская возможность симпатрического видообразования на больших
глубинах, нельзя все же согласиться с Визером (Wieser, 1960), считающим
доказательством этого процесса просто факт совместного нахождения на
больших глубинах близких видов одного рода среди изопод, амфипод, не-
матод и остракод, поскольку нет никаких оснований предполагать возникно-
вение видов в данном месте. В действительности все они достаточно широко
распространены. Вместе с тем соображения Визера об отсутствии полной
панмиксии в обширных разреженных популяциях мелких малоподвижных
донных животных и о вытекающей отсюда возможности создания различной
концентрации разных генов в разных частях каждой популяции достойны
внимания.
Итак, основной вопрос, на который мы сейчас не можем ответить, сво-
дится к природе барьеров, генетически изолирующих возникающие новые
виды глубоководных донных животных. Этот вопрос возникает с еще большей
остротой по отношению к океанической пелагиали. Среди генетиков только
Буциатти-Траверзо (Buzzatti-Traverso, 1960) обратил на него внимание и
предложил модель односторонне направленного по течению дрейфа генов,
и предложил модель односторонне направленного по течению дрейфа генов,
одни из которых в процессе переноса теряются, а другие, наоборот, посте-
пенно концентрируются, но эта модель остается чисто умозрительным по-
строением и не касается проблемы генетического барьера.
222
Из сказанного ясно, что генетический механизм процесса видообразо-
вания на больших глубинах, как на дне, так и в толще воды, остается еще
не изученным. Судя по пониженной плодовитости и растянутому жизнен-
ному циклу обитателей больших глубин, составляющих обширные разрежен-
ные популяции, естественный отбор действует там в своей стабилизирующей,
а не движущей форме (Шмальгаузен, 1968а). Однообразие абиотической
среды и прогрессирующая с глубиной олигомикстность биоценозов упрощают
отношения организмов с внешней средой и между собой и обусловливают
узкую специализацию или способствуют их консервативности.
Итак, данных по проблеме видообразования на больших глубинах совер-
шенно недостаточно для определенных заключений, и обсуждение этой проб-
лемы может вестись лишь в самой общей, малоконкретной форме. Несколько
большим материалом мы можем воспользоваться, переходя к анализу судьбы
не видов, а более крупных таксонов, составляющих глубоководную фауну,
т. е. рассматривая закономерности не микро-, а макроэволюции. При этом
мы можем опираться на некоторые объективные показатели, последовательно
сравнивая между собой состав глубоководной и мелководной фауны. Мы про-
делали такую работу совместно с М. Е. Виноградовым, и я постараюсь сейчас
изложить некоторые сделанные нами выводы.
Первый и основной вопрос, который должен быть решен прежде всего,
сводится к тому, насколько самостоятельна глубоководная фауна, т. е. на-
сколько она обособлена таксономически от мелководной, иными словами, на-
сколько изменилась морская фауна, переходя от жизни на мелководье к жи-
зни на больших глубинах. Для ответа на этот вопрос мы подсчитали по груп-
пам количество родов и семейств, известных для бентоса глубже 2000 м, а для
планктона — глубже 500 м, а затем выделили роды и семейства, не поднима-
ющиеся для бентоса выше 500 м, а для планктона — выше 200 м, т. е. при-
уроченные только к большим глубинам. Процентное отношение исключи-
тельно глубоководных родов и семейств ко всем родам и семействам, зареги-
стрированным на больших глубинах, можно рассматривать как объективную
меру степени таксономической обособленности глубоководной фауны от
мелководной.
Соответствующие расчеты показали, что степень таксономической обо-
собленности глубоководной фауны от мелководной у разных групп весьма
различна.
Можно, во-первых, выделить группы, совсем не включающие ис-
ключительно глубоководные роды (несколько отрядов кишечнополостных,
сипункулиды, многоколенчатые и др.). Далее следуют группы, включающие
менее 20 % таких родов (роговые губки, гидроиды, полихеты, бокоплавы,
десятиногие, двустворчатые моллюски, мшанки и др.). От 20 до 50 % исклю-
чительно глубоководных родов насчитывается среди актиний, гидроидных
и спифоидных медуз, донных мизид и изопод, пелагических десятиногих, вес-
лоногих и рыб и среди большинства классов иглокожих. Наконец, сущест-
вуют группы, представленные на больших глубинах на 50—100 % исключи-
тельно глубоководными родами и высоким процентом глубоководных се-
мейств. В бентосе это моноплакофоры, погонофоры, голотурии и рыбы,
а в планктоне — мизиды и блокоплавы. Степень таксономической обособ-
ленно глубоководной фауны от мелководной в планктоне оказывается в сред-
нем выше, чем в бентосе.
Нетрудно заметить, что степень таксономической обособленности глубо-
223
ководной фауны не связана ни с высотой организации, ни с видовым или ро-
довым разнообразием, ни с эврибатностью сравниваемых групп. Совершенно
ясно также, что малую степень таксономической обособленности глубоковод-
ной фауны нельзя считать показателем недавнего вселения данной группы на
большие глубины, предполагая, что особые роды и семейства просто не ус-
пели сформироваться, как утверждал Сперк (Sparck, 1954) по отношению ко
всей донной фауне, Бернар (Barnard, 1961) — по отношению к гаммаридам
и В. В. Мурина (1958) — по отношению к сицункулидам. В таком случае
пришлось бы признать более раннее вселение в абиссаль рыб, голотурий
и погонофор, чем полихет и двустворчатых моллюсков, а также более раннее
возникновение глубоководного планктона, чем бентоса, что в равной мере
весьма маловероятно.
На наш взгляд, высокая степень таксономической обособленности глубо-
ководной фауны от мелководной может определяться двумя основными при-
чинами, разграничение которых не всегда возможно. Во-первых, на больших
глубинах могут обитать реликтовые представители архаических групп, вы-
тесненных в борьбе за существование из благоприятной для жизни области
морского мелководья, каковы, например, моноплакофоры, погонофоры и не-
которые другие. Во-вторых, некоторые группы в порядке приспособления
к существованию на больших глубинах изменились настолько, что образо-
вали особые глубоководные роды и семейства.
Представляется желательным выяснить, в каком направлении шли эти из-
менения.
Не отрицая возможного воздействия многих специфических для
больших глубин факторов (гидростатического давления, гидро- и биохими-
ческих особенностей), мы попытались сопоставить показатели степени таксо-
номической обособленности глубоководной фауны с характером питания раз-
личных групп животных. Мы исходили из несомненного факта крайнего обе-
днения источников питания на больших океанических глубинах по сравне-
нию с областью морского мелководья и полагали, что одним из основных
направлений адаптации к жизни на больших глубинах Должны быть адапта-
ции к использованию минимальных пищевых ресурсов. Приспособления
именно к этой особенности глубоководной среды обитания выступают более
наглядно и легче поддаются анализу, чем пока неизвестные приспособления
к огромному гидростатическому давлению, повышенной радиоактивности
грунта и т. д.
Для тех групп животных, которые при переходе к жизни на больших глу-
бинах не изменили характера своего питания (грунтоеды, некоторые фильт-
раторы, такие специализированные хищники, как пантоподы), получены
низкие показатели степени таксономической обособленности глубоководной
фауны.
Наоборот, многие хищники при крайней разреженности жертв на
больших глубинах были вынуждены выработать приспособления к пассивной
охоте, обеспечивающей минимум энергетических затрат, что потребовало ра-
дикальной перестройки организации животных и привело к возникновению
особых родов и семейств. То же самое произошло и с группами, решительно
изменившими на больших глубинах характер своего питания — с асцидиями,
перешедшими от фильтрации к хищничеству, с морскими звездами, перешед-
шими от хищничества к потреблению грунта, с пелагическими бокоплавами,
ставшими полупаразитами крупных кишечнополостных, и т. д.
224
Таким образом, эволюционные изменения, которым подверглись разные
группы животных при освоении больших океанических глубин, оказываются
весьма различными.
Одни группы представлены на больших глубинах теми же родами,
что и в области морского мелководья, у других на больших глубинах
возникли особые роды, семейства, а в отдельных случаях даже еще более
высокие таксоны.
Совершенно очевидно, что для формирования таксонов столь высокого
ранга требовалось весьма продолжительное время, особенно если учитывать
вероятные замедленные темпы эволюциии глубоководной фауны, существую-
щей в условиях биологических отношений. Не только присутствие древних
реликтовых форм, вроде моноплакофор и погонофор, но и возникновение
особых таксонов высокого ранга однозначно свидетельствуют в пользу древ-
ности глубоководной фауны, а следовательно, и больших океанических глу-
бин.
ЛИТЕРАТУРА
Агатова А. И., Богданов Ю. А. Биохи-
мический состав взвешенного орга-
нического вещества тропической
части Тихого океана. — Океаноло-
гия, 1972, т. 12, вып. 2, с. 267—
276.
Алехин В. В. География растений. М.:
Сов. наука, 1944. 455 с.
Алпатов JB. В. Новый для оз. Байкал
водяной ослик и сравнение байкаль-
ских видов р. Asellus с европейскими
и американскими. — Рус. гидробиол.
журн., 1923, т. 2, вып. 3/4, с. 64—66.
Алпатов В. В. Среда и рост животных. —
В кн.: Рост животных. М.: Биомедгиз,
1935, с. 326—366.
Андрияшев А. П. Новые данные о глубо-
ководных рыбах Берингова моря. —
ДАН СССР, 1935, т. 4 (II), № 1/2,
с. 105—108.
Андрияшев А. П. Очерк зоогеографии
и происхождения фауны рыб Берин-
гова моря и сопредельных вод. Л.:
Изд-во ЛГУ, 1939. 185 с.
Андрияшев А. П. Древнеглубоководные
и вторичноглубоководные формы рыб
и их значение для зоогеографиче-
ского анализа. — В кн.: Очерки
по общим вопросам ихтиологии. М.;
Л.: Изд-во АН СССР, 1953, с. 58—64.
Андрусов Н. Материалы к познанию
прикаспийского неогена, Акчагыль-
ские пласты. — Тр. Геол, ком., 1902,
т. 15, № 4. 153 с.
Андрусов Н. О возрасте и стратигра-
фическом положении акчагыльских
пластов. — Зап. Минерал, о-ва, 1911,
т. 48, вып. 1, с. 271—296.
Анненкова Н. П. Пресноводные и соло-
новатоводные Polychaeta СССР.
Определители организмов пресных
вод СССР. А. Пресноводная фауна.
Л., 1930, вып. 2. 47 с.
Арган Э. Тектоника Азии. М.; Л.: ОНТИ
НКТП, 1935. 192 с.
Архангельский А. Д. Геологическое стро-
ение СССР: Европейская и Средне-
азиатская части. Л.; М.: Георазвед-
издат, 1932. 425 с
Архангельский А. Д., Шатский Н. С.,
Меннер В. В. и др. Краткий очерк
геологической структуры и геологи-
ческой истории СССР. М.: Изд-во
АН СССР, 1937.
Базикалова А. Я. Амфиподы озера Бай-
кал. — Тр. Байкал, лимнол. ст.
АН СССР, 1945, т. 11, с. 1-140.
Базикалова А. Я. Адаптивное значение
размеров байкальских амфипод. —
ДАН СССР, 1948, т. 61, № 3, с. 569—
572.
Базикалова, А. Я. Находка в оз. Байкал
интересного ракообразного. — При-
рода, 1949, № 7, с. 63—64.
Базикалова А. Я., Бирштейн Я. А., Та-
лиев Д. Н. Осмотическое давление
полостной жидкости бокоплавов оз.
Байкал. — ДАН СССР, 1946а, т. 53,
№ 3, с. 293-295.
Базикалова А. Я., Бирштейн Я. А., Та-
лиев Д. И. Осморегуляторные спо-
собности бокоплавов озера Бай-
кал. — ДАН СССР, 19646, т. 53, № 4,
с. 381—384.
Базикалова А. Я., Талиев Д. Н. О неко-
торых зависимостях дивергентной
эволюции Amphipoda и Cottoidea
в оз. Байкал. — ДАН СССР, 1948,
т. 59, № 3, с. 565—568.
Балабуев А. Г Метеорологические на-
блюдения в пещерах Абхазии в
1935 г. — В кн.: Материалы к фауне
Абхазии. Тбилиси: Груз. фил. АН
СССР, 1939, с. 213—224.
Батурин В. П. 1. Петрография песков
и песчаников продуктивной толщи.
2. Физико-географические условия
века продуктивной толщи. — Тр.
Азерб. нефт. исслед. ин-та, 1931, т. 1.
95 с.
Беклемишев В. Н. К вопросу о речных
Peracarida Понто-Каспийского бас-
сейна. г— Рус. гидробиол. жури.,
1923, т. 2, вып. 11/12, с. 213—218.
226
Беклемишев В. Н. Основы сравнительной
анатомии беспозвоночных. М.: Сов.
наука, 1944. 492 с.
Белинг Д. Е. Материалы по гидрофауне
и ихтиофауне нижнего течения
р. Днепра. — Тр. Всеукр. Черномор.-
Азов. науч.-промысловой опыт, ст.,
1925, т. 1, с. 1—72.
Белинг Д. Е. До ваучування тваринного
населения в порожисты частин! та
в сумежних з нею районах Дне-
пра. — BicTi Н.-Д. инет. води. госп.
Укр., 1929, т. 2, ч. 1, с. 259—276.
Беляев Г М. Осморегуляторные способ-
ности изопод. — ДАН СССР, 1949,
т. 67, № 6, с. 1117—1120.
Беляев Г М. Осмотическое давление
полостной жидкости водных беспоз-
воночных в водоемах различной соле-
ности. — Тр. Всесоюз. гидробиол.
о-ва, 1951, т. 3, с. 92—139.
Беляев Г М. Донная фауна наибольших
глубин (ультраабиссали) Мирового
океана. М.: Наука, 1966. 248 с.
Беляев Г М., Бирштейн Я. А. Осморегу-
ляторные способности каспийских
беспозвоночных. — Зоол. журн.,
1940, т. 19, вып. 4, с. 548—565.
Беляев Г М., Бирштейн Я. А. Сравнение
осморегуляторных способностей
волжских и каспийских Amphi-
poda. — ДАН СССР, 1944, т. 45,
№ 7, с. 323—326.
Беляев Г. М., Бирштейн Я. А., Бого-
ров В. Г и др. Схема вертикаль-
ной биологической зональности
океана. — ДАН СССР, 1959, т. 129,
№ 3, с. 658—661.
Бенинг А. Л. Об амфиподах окрестностей
Киева. — Тр. Днепр, биол. ст., 1915,
№ 2, с. 90—101.
Бенинг А. Л. К изучению придонной
жизни реки Волги. — Монографии
Волж. биол. ст., 1924а, № 1. 398 с.
Бенинг А. Л. О каспийских ракообраз-
ных в бассейне реки Волги. — Рус.
гидробиол. журн., 19246, т. 3,
вып. 3/5, с. 51—54.
Бенинг А. Л. (Benning A. L.). Studien
uber die Malakostraken des Wolga-
basins. 3. — Int. Rev. d. ges. Hydro-
biol. und Hydrogr. 1925, Bd. 13,
S. 46—77.
Бенинг А. Л. Каспийские реликты сред-
него течения Урала. — Рус. гидро-
биол. жури., 1928а, т. 7, вып.
10/12, с. 263.
Бенинг А. Л. Понто-Каспийские элементы
в р. Днестре.— Рус. гидробиол. журн.
19286, т. 7, вып. 10/12, с. 264—265.
Берг Л. С. Аральское море. — Изв.
Туркм. отд-ния Рус. геогр. о-ва,
1908, т. 5, вып. 9. 580 с.
Берг Л. С. Рыбы бассейна Амура. —
Зап. Акад. наук. Физ.-мат. отд-ние,
1909, т. 24, № 9, VII. 270 с.
Берг Л. С. О распространении рыбы Муо-
xocephalus quadricornis и о связан-
ных с этим вопросах. — Изв. Акад,
наук. 1916, т. 10, с. 1343—1360.
Берг Л. С. Номогенез или эволюция
на основе закономерностей. Пг.:
Госиздат, 1922а, VIII. 306 с.
Берг Л. С. Климат и жизнь. М.: Гос-
издат., 19226. 196 с.
Берг Л. С. О прбисхождении северных
элементов в фауне Каспия. — ДАН
СССР, 1928, № 1, с. 107—112.
Берг Л. С. Об амфибореальном распро-
странении морской фауны в север-
ном полушарии. — Изв. геогр. о-ва,
1934а, т. 66, вып. 1, с. 69—78.
Берг Л. С. Уровень Каспийского моря
за историческое время. — Проблемы
физической географии, 19346, вып. 1,
с. 14—64.
Берг Л. С. Биполярное распространение
организмов и ледниковая эпоха. —
В кн.: Климат и жизнь. М.: Гео-
графиздат, 1947а, с. 128—155.
Берг Л. С. Соображения о происхожде-
нии наземной, пресноводной и мор-
ской флоры и фауны. — Бюл.
МОИП. Огд. биол., 19476, т. 52,
№ 5, с. 15—34.
Берг Л. С. О предполагаемом раздви-
жении материков. — Изв. АН СССР.
Сер. геол., 1948, № 3, с. 3—8.
Бирштейн Я. A. Malacostraca Армении. —
Тр. Севан, озер, ст., 1932, т. 4,
№ 1/2, с. 139—160.
Бирштейн Я. A. (Birstein J. A.). Die
Land- und Siisswasser-Isopoden des
artischen Gebites. — Fauna Arctica,
1933b, Bd. 6, N 5/6, S. 473-476.
Бирштейн Я. А. К вопросу о происхожде-
нии морских ракообразных в реках
Понто-Каспийского бассейна. —
Зоол. журн., 1935, т. 14, вып. 4,
с. 749—761.
Бирштейн Я. А. Зоогеографическая ха-
рактеристика водяных осликов оз.
Байкал. — ДАН СССР, 1939а, т. 25,
№ 3, с. 248—251.
Бирштейн Я. А. О пещерных креветках
Абхазии. — Зоол. журн., 19396, т. 18,
вып. 6, с. 960—975.
Бирштейн Я. А. О некоторых особен-
ностях географического распростра-
нения пресноводных Malacostraca
Дальнего Востока. — Зоол. журн.,
1939, т. 18, вып. 1, с. 54—59.
Бирштейн Я. А. К фауне пещерных Am-
phipoda Абхазии. — Бюл. МОИП.
Отд. биол., 1940а, т. 49, вып. 3/4,
с. 47—55.
227
Бирштейн Я. А. Жизнь в пещерах. —
Успехи соврем, биологии, 19406, т. 13,
вып. 3, с. 385—402.
Бирштейн Я. А. Пещерные бокоплавы
Абхазии с замечаниями о видах
Niphargus Закавказья. — Сб. тр.
Зоол. музея МГУ, 1940в, № 6,
с. 259—269.
Бирштейн Я. А. Адаптация и эволюция
пещерных животных. — Успехи со-
врем. биологии, 1941, т. 14, вып. 3,
с. 436—453.
Бирштейн Я. А. Заметка о пресноводных
высших ракообразных Туркмении
и Ирана. — Учен. зап. МГУ, 1945,
№ 83, с. 151 — 164.
Бирштейн Я. А. Заметки о географиче-
ском распространении понтокаспий-
ских бокоплавов. — Бюл. МОИП.
Отд. биол., 1946, т. 51, вып. 3,
с. 39—53.
Бирштейн Я. А. Понятие «реликт» в био-
логии. — Зоол. журн., 1947а, т. 26,
вып. 4, с. 313—330.
Бирштейн Я. А. О водяных осликах Си-
бири. — ДАН СССР, 19476, т. 57,
№ 4, с. 409—412.
Бирштейн Я. А. Некоторые проблемы про-
исхождения и эволюции пресновод-
ной фауны. — Успехи соврем, биоло-
гии, 1949, т. 27, вып. 1, с. 119—140.
Бирштейн Я. А. Пещерная фауна Запад-
ного Закавказья. — Зоол. журн.,
1950, т. 29, вып. 4, с. 354—366.
Бирштейн Я. А. Пресноводные ослики
(Asellota). — Ракообразные. М.; Л.:
Изд-во АН СССР, 1951. 143 с. (Фа-
уна СССР. Н. С.; Т. 7. Вып. 5. № 47).
Бирштейн Я. А. О некоторых особен-
ностях ультраабиссальной фауны
на примере рода Storthyngura (Crus-
tacea Isopoda Asellota). — Зоол.
журн., 1957, т. 36, вып. 7, с. 961—985.
Бирштейн Я. А. Семейство Ischnomesidae
(Crustacea, Isopoda, Asellota) в се-
веро-западной части Тихого океана
и проблема амфипацифического
и биполярного распространения глу-
боководной фауны. — Зоол. журн..,
1960, т. 39, вып. 1, с. 3—28.
Бирштейн Я. А. Подземные бокоплавы
Крыма. — Бюл. МОИП. Отд. биол.,
1961а, т. 66, вып. 6, с. 126—144.
Бирштейн Я. A. (Birstein J. A.). Micro-
thambema tenuis n. gen., n. sp.
(Isopoda Asellota) and relations of
some Asellota Isopods. — Crusta-
ceana, 1961b, vol. 2, N 2, p. 132—141.
Бирштейн Я. A. (Birstein J. A.). Ober
eine neue Art des Gattung Austro-
niscus Vanhoffen (Crustacea, Iso-
poda, Asellota) aus grossen Tiefen
der nordwestlichen Teiles der Stil-
228
len Oceans. — Izdanie publ. zavoda
za ribarstvo NRM-Scopje, 1962, 3,
N 2, p. 33—38.
Бирштейн Я. А. Глубоководные равно-
ногие ракообразные северо-западной
части Тихого океана. М.: Изд-во
АН СССР, 1963а. 214 с.
Бирштейн Я. А. Некоторые итоги изуче-
ния подземной фауны Крыма. — Тр.
Комплекс, карст, экспедиции АН
УССР, 19636, вып. 1, с. 123—133.
Бирштейн Я. А. Равноногие ракообраз-
ные (Crustacea, Isopoda) ультра-
абиссали Бугенвильской впадины. —
Зоол. журн., 1963в, т. 42, вып. 6,
с. 814—834.
Бирштейн Я. А. Третий представитель
подземного рода Niphargus (Crusta-
cea, Amphipoda) из Крыма. — Бюл.
МОИП. Отд. биол., 1964, т. 69, вып. 1,
с. 119—121.
Бирштейн Я. А. Подземные водяные
ослики (Crustacea, Isopoda, Asel-
lota) Закавказья. — Зоол. журн.,
1967, т. 46, вып. 6, с. 856—865.
Бирштейн Я. А. Глубоководные Asellota
(Crustacea Isopoda) из Антарктики
и Субантарктики. — В кн.: Исследо-
вания фауны морей. М.; Л.: Наука,
1968, вып. 6 (14), с. 141 — 152.
Бирштейн Я. А. Равноногие ракообраз-
ные (Crustacea, Isopoda) впадины
Романш. — Бюл. МОИП. Отд. биол.,
1969, т. 74, вып. 3, с. 50—59.
Бирштейн Я. А. Дополнения к фауне
равноногих ракообразных (Crusta-
cea Isopoda) Курило-Камчатского
желоба. Ч. 1. —Тр. ИОАН СССР,
1970, т. 86, с. 292—340.
Бирштейн Я. А. Некоторые вопросы эво-
люции глубоководной фауны. —
В кн.: История Мирового океана.
М.: Наука, 1971, с. 84—89.
Бирштейн Я. А., Виноградов Л. Г Пре-
сноводные Decapoda СССР и их гео-
графическое распространение. —
Зоол. журн., 1934, т. 13, вып. 1,
с. 39—70.
Бирштейн Я. А., Виноградов Л. Г Новые
данные по фауне десятиногих рако-
образных (Decapoda) Берингова
моря. — Зоол. журн., 1953, т. 32,
вып. 2, с. 215—228.
Бирштейн Я. А., Виноградов М. Е. Замет-
ки о питании глубоководных рыб Ку-
рило-Камчатской впадины. — Зоол.
журн., 1955а, т. 34, вып. 4, с. 842—849.
Бирштейн Я. А., Виноградов М. Е. Пела-
гические гаммариды Курило-Камчат-
ской впадины. — Тр. ИОАН СССР,
19556, т. 12, с. 210—287.
Бирштейн Я. А., Виноградов М. Е. Пела-
гические гаммариды (Amphipoda —
Gammaridea) северо-западной части
Тихого океана. — Тр. ИОАН СССР,
1958, т. 27, с. 219-257
Бирштейн Я. А., Виноградов М. Е. Пела-
гические гаммариды тропической
части Тихого океана. — Тр. ИОАН
СССР, 1960, т. 34, с. 165—241.
Бирштейн Я. А., Виноградов М. Е., Чин-
донова Ю. Г Вертикальная зональ-
ность планктона Курило-Камчатской
впадины. — ДАН СССР, 1954, т. 45,
№ 2, с. 389—392.
Бирштейн Я. А., Леванидов В. Я. Новый
вид подземного водяного ослика из
бассейна Уссури. — ДАН СССР,
1952, т. 84, № 5, с. 1081 — 1084.
Бирштейн Я. А., Левушкин С. И. (Bir-
stein J. A., Ljovuskin S. I). Faune
des eaux souterraines saumatres de
1’Asie Centrale. — Intern. J. Speleol.,
1965a, vol. 1, N 3, p. 307—320.
Бирштейн Я. А., Левушкин C.‘ И. Под-
земные Paraselloidea (Crustacea,
Isopoda) СССР. — Зоол. журн.,
19656, т. 44, вып. 7, с. 997—1013.
Бирштейн Я. А., Левушкин С. И. Отряд
Bathynellacea (Crustacea, Malaco-
straca) в СССР. 1. Семейство Ва-
thynellidae. — Бюл. МОИП. Отд.
биол., 1967а, т. 72, вып. 4, с. 51—65.
Бирштейн Я. А., Левушкин С. И. Не-
которые итоги и задачи изучения под-
земной фауны СССР. — Зоол. журн.,
19676, т. 46, вып. 10, с. 1509—1535.
Бирштейн Я. А., Левушкин С. И. Пред-
ставитель нового для фауны СССР
семейства бокоплавов (Amphipoda,
Bogidiel lidae) в подземных водах
Средней Азии. — Зоол. журн., 1968а,
т. 47, вып. 5, с. 676—683.
Бирштейн Я. А., Левушкин С. И. Отряд
Bathynellacea (Crustacea, Mala-
costraca) в СССР. 2. Семейство Ра-
rabathynellidae и зоогеографические
замечания. — Бюл. МОИП. Отд.
биол., 19686, т. 73, вып. 6, с. 55—64.
Бирштейн Я. А., Соколова М. Н. Донная
фауна Бугенвильской впадины. — Тр
ИОАН СССР, 1960, т. 41, с. 128—
131.
Бирштейн Я. А., Старостин И. В. Новый
для СССР род водяных осликов
(Stenasellus) из Туркмении и его
значение для зоогеографии Средней
Азии. — ДАН СССР, 1949, т. 69, № 5,
с. 691—694.
Бирштейн Я. А., Чиндонова Ю. Г Глубо-
ководные мизиды северо-западной
части Тихого океана. — Тр. ИОАН
СССР, 1958, т. 27, с. 258—355.
Блинов Л. К. К вопросу о происхожде-
нии солевого состава морской во-
ды. — Метеорология и гидрология,
1945, № 4.
Богачев В. В. Руководящие окаменелости
разреза Апшеронского полуострова
и прилегающих районов. — Тр.
Азерб. нефт. исслед. ин-та, 1932, 1,
вып. 4. 91 с.
Богачев В. В. Материалы по изучению
третичной ихтиофауны Кавказа. —
Тр. Азерб. нефт. исслед. ин-та, 1933,
вып. 15. 62 с.
Богданов Ю. А., Григорович Ю. А., Ша-
пошникова М. Г Определение белка
в водной взвеси. — Океанология,
1968, т. 8, № 6, с. 1087—1090.
Богданов Ю. А., Лисицын А. П., Роман-
ович Е. А. Органическое вещество
взвеси и донных осадков морей и оке-
анов. — В кн.: Органическое веще-
ство современных и ископаемых осад-
ков. М.: Наука, 1971, с. 35—104.
Богданов Ю. А., Шапошникова М. Г
Содержание взвешенного органи-
ческого вещества в тропических во-
дах Тихого океана. — Океанология,
1971, т. 11, вып. 4, с. 668—673.
Богоров В. Г Вертикальное распределе-
ние зоопланктона и вертикальное
расчленение вод океана. — Тр.
ИОАН СССР, 1948, т. 2, с. 43—59.
Богоявленский А. Н. Химическая харак-
теристика района Курило-Камчат-
ской впадины. — Тр. ИОАН СССР,
1955, т. 12, с. 161 — 176.
Болдырев В. Ф. Материалы к познанию
строения сперматофор и особенности
спаривания у Locustoidea и Gryllo-
idea. — Тр. Рус. энтмол. о-ва, 1915,
т. 41, № 6, с. 1—245.
Борисяк А. А. Курс исторической геоло-
гии. Л.: ОНТИ Горгеонефтеиздат,
1934. 424 с.
Боруцкий Е. В. (Borutzky Е. W.). Ма-
terialen fiber die Fauna der unter-
irdischen Gewasser: Crangonyx chle-
bnikovi sp. n. (Amphipoda) aus den
Hohlen des mittleren Urals. — Zool.
Anz., 1928, Bd. 77, H. 9/10, S. 253—
259.
Боруцкий E. B. (Borutzky E. W.). Unter-
irdische Fauna der Kutais-Hohlen am
Rion (Transcaucasus, Georgien).
Copepoda—Harpacticoida. — Zool.
Anz., 1930a, Bd. 88, H. 5/6, S. 121 —
132.
Боруцкий E. B. (Borutzky E. W.). Zur
Kenntnis der unterirdischen Fauna
der Kutais-Hohlen am Rion (Trans-
caucasus, Georgien). Copepoda—
Cyclopoida. — Zool. Anz., 19306,
Bd. 89, H. 11/12, S. 331—335.
Боруцкий Ё. В. Географическое распро-
странение Bryocamptus (Arctico-
camptus) arcticus Lili, на основе его
экологии. — В кн.: Сб. тр. Зоол.
229
музея МГУ. М.; Л.: Биомедгиз, 1937,
т. 4, с. 135—189.
Боруцкий Е. В. Материалы по фауне.
Copepoda Harpacticoida Байкала.
Род Bryocamptus Chappuis. — ДАН
СССР, 1948, т. 59, № 9, с. 1669—
1672.
Боруцкий Е. В. Наземные Isopoda пещер
Кавказа и Крыма. Сем. Lingiidae. —
Бюл. МОИП. Отд. биол., 1950, т. 55,
вып. 5, с. 69—81.
Боруцкий Е. В. Фауна наземных равно-
ногих раков (Isopoda terrestria)
пещер Крыма в зоогеографическом
аспекте. —Зоол. журн., 1962а, т. 41,
вып. 2, с. 193—202.
Боруцкий Е. В. Первое нахождение тро-
глобионта из Calanoida (Crustacea
Copepoda) в подземных водах. —
ДАН СССР, 19626, т. 147, № 6,
с. 1499—1502.
Боруцкий Е. В. О роде Speocyclops Ki-
efer (Copepoda Cyclopoida). — Зоол.
журн., 1965, т. 44, вып. 6, с. 833—843.
Боруцкий Е. В. Copepoda пещер Примор-
ского края. — Зоол. журн., 1966,
т. 45, вып. 5, с. 77,0—772.
Боруцкий Е. В. Новый род Pseudobud-
delundiella gen. п. и его положение
в надсемействе Trichoniscoidea (на-
земные Isopoda). — Зоол. журн.,
1967, т. 46, вып. 3, с. 338—344.
Бродский А. Л. Foraminifera (Polytha-
lamia) в колодцах пустыни Кара-
кум. — Тр. Сред.-Азиат. ун-та. Сер.
Villa. Зоология, 1928, т. 5, с. 1 —16.
Бродский К. А. Глубоководные весло-
ногие рачки (Calanoida) северо-за-
падной части Тихого океана. —
В кн.: Исследования дальневосточ-
ных морей СССР. Л., 1952, вып. 3,
с. 37—87.
Вегенер А. Происхождение материков
и океанов. М.; Л.: Госиздат, 1925.
145 с.
Верещагин Г Ю. Теоретические вопросы,
связанные с разработкой проблемы
происхождения и истории Бай-
кала. — Тр. Байк, лимнол. ст. АН
СССР, 1940, т. 10, с. 7—72.
Виноградов М. Е. Вертикальные мигра-
ции зоопланктона и их роль в пита-
нии глубоководной пелагической
фауны. — Тр. ИОАН СССР, 1955,
т. 13, с. 71—76.
Виноградов М. Е. Вертикальные мигра-
ции глубоководного зоопланктона. —
В кн.: Итоги науки. Достижения оке-
анологии. М.: Изд-во АН СССР,
1959, т. 1, с. 204—220.
Виноградов М. Е. Количественное рас-
пределение глубоководного планк-
тона в западной и центральной
частях Тихого океана. — Тр. ИОАН
СССР, 1960, т. 41, с. 55—84.
Виноградов М. Е. Вертикальное распре-
деление океанического зоопланктона.
Дис. . . д-ра биол. наук. М.: ИОАН
СССР, 1965.
Виноградов М. Е. Вертикальное распре-
деление океанического зоопланктона.
М.: Наука, 1968. 320 с.
Виноградова Н. Г Некоторые особен-
ности распространения морской глу-
боководной фауны. — Тр. ИОАН
СССР, 1955, т. 13, с. 59— 66.
Виноградова Н. Г Вертикальное распре-
деление глубоководной донной
фауны океана. — Тр. ИОАН СССР,
1958, т. 27, с. 86—122.
Виноградова Н. Г Вертикальное распре-
деление глубоководной донной
фауны. — В кн.: Глубоководная
донная фауна. Плейстон. М.:
Наука, 1969, с. 129—153. (Тихий
океан. Биология Тихого океана;
Кн. 2).
Виноградова Н. Г О нахождении новой
аберрантной асцидии в ультраабис-
сали Курило-Камчатского же-
лоба. — Бюл. МОИП. Отд. биол.,
19696, т. 74, вып. 3, с. 27—43.
Вульф Е. В. Введение в историческую
географию растений. М.: Сельхозгиз,
1932. 356 с.
Вульф Е. В. Понятие о реликте в бота-
нической географии. — В кн.: Мате-
риалы по истории флоры и расти-
тельности СССР. 1941, т. 1, с. 28—60.
Вульф Е. В. Историческая география
растений: История флор земного
шара. М.; Л.: Изд-во АН СССР,
1944, XIX 548 с.
Гексли Г. Г Рак. М.: Изд. бр. Сабаш-
никовых. 1900. VI 272 с.
Гептнер В. Г Общая зоогеография.
М.; Л.: Биомедгиз, 1936. 548 с.
Герасимов И. П., Марков К. К. Леднико-
вый период на территории СССР. —
Тр. Ин-та географии АН СССР, 1939,
т. 33. 462 с.
Гроссгейм А. А. Типы реликтов. — Изв.
Азерб. фил. АН СССР, 1939, № 6,
с. 74— 80.
Губкин И. М. Проблема Акчагыла в свете
новых данных. Л.: Изд-во АН СССР,
1931. 38 с.
Гурьянова Е. Ф. К фауне равноногих
раков (Isopoda) Тихого океана.
2. Новые виды Gnathiidea и Asel-
lota. — Исслед. морей СССР, 1933,
вып. 19, с. 79—91.
Гурьянова Е. Ф. К фауне равноногих
раков (Isopoda) Тихого океана.
3. Новые виды в сборах Тихоокеан-
ской экспедиции Гос. гидрологиче-
230
ского ин-та, 1932. — Исслед. морей
СССР, 1935, вып. 22, с. 25—35.
Гурьянова Е. Ф. Равноногие дальне-
восточных морей. Ракообразные. М.;
Л.: Изд-во АН СССР, 1936. XII
279 с. (Фауна СССР; Т. 7. Вып. 3).
Гурьянова Е. Ф. Новые виды Isopoda
и Amphipoda из Северного Ледови-
того океана. — Тр. Дрейфующей экс-
педиции Главсевморпути на ледо-
кольном пароходе «Г Седов», 1937—
1940 гг., 1946, т. 3, с. 272—297.
Гурьянова Е. Ф. Бокоплавы морей СССР
и сопредельных вод. — Определи-
тели по фауне СССР, издаваемые
ЗИН АН СССР. М.; Л.: Изд-во АН
СССР, 1951, вып. 41. 1032 с.
Гурьянова Е. Ф. К фауне равноногих
раков (Isopoda) Тихого океана.
6. Новые виды Valvifera из Курило-
Сахалинского района. — Тр. ЗИН
АН СССР, 1955, т. 21, с. 208—230.
Давиташвили Л. Ш. Дарвинизм и про-
блема накопления горючих ископа-
емых. — Бюл. Гос. музея Грузии,
1943, т. 12А. НО с.
Дарвин Ч. Происхождение видов. М.;
Л.: Биомедгиз, 1937. 762 с.
Державин А. Н. Каспийские элементы
в фауне бассейна Волги. — Тр.
Ихтиол, лаб. Упр. Касп.-Волж. пром-
сти, 1912, т. 2, № 5, с. 20—44.
Державин А. И. Каспийские ракообраз-
ные в фауне рек Южного Каспия. —
Рус. гидробиол. журн., 1923, т. 2,
вып. 8/10, с. 195—196.
Державин А. Н. Пресноводные Рега-
carida черноморского побережья
Кавказа. — Рус. гидробиол. журн.,
1924, т. 3, вып. 6/7, с. 113—129.
Державин А. Н. К познанию перакарид
р. Урала. — Рус. гидробиол. журн.,
1926а, т. 5, вып. 3/4, с. 48—52.
Державин А. И. Рыбы р. Кара-Су. —
Изв. Бакин, ихтиол, лаб., 19266,
т. 2, вып. 1, с. 161 — 184.
Державин А. Н. К познанию верхне-
сарматских амфипод Понто-Каспий-
ской области. — Бюл. МОИП. Отд.
геол., 1927, т. 35, вып. 2, с. 183—
196.
Державин А. И. Пресноводные Mala-
costraca Дальнего Востока. — Рус.
гидробиол. журн., 1930, т. 9, № 1/3,
с. 1—8.
Державин А. Н. Мизиды Каспия. Баку:
Азерб. фил. АН СССР, 1939. 92 с.
Державин А. И., Дексбах Н. К., Леп-
нева С. Г Каспийские элементы
в фауне верхней Волги. — Тр. Яро-
слав. естеств.-ист. о-ва, 1921, т. 3,
вып. 1, с. 26—43.
Дерюгин К. М. Фауна Белого моря
и условия ее существования. —
В кн.: Исследования морей СССР.
Л.: Гос. гидрол. ин-т, 1928, вып. 7/8.
XII. 511 с.
Динер К. Основы биостратиграфии. М.:
Гос. науч.-техн, горно-геол. нефт.
изд-во, 1934. 271 с.
Дмитриев Н. А. К современному состоя-
нию рыболовства в низовьях реки
Терека. — Изв. Даг. ихтиол, лаб.,
1929, вып. 1, с. 75—96.
Дубинин И. П., Ромашев Д. Д. Генети-
ческие основы строения вида и его
эволюция. — Биол. журн., 1932, № 1,
с. 52—95.
Ермаков Н. В. К вопросу об изменении
глаза у Simocephalus под влиянием
темноты и голода. — Работы Вол ж.
биол. ст., 1924, т. 7, вып. 4/5, с. 149—
173.
Жадин В. И. (Shadin W L). Die Siiss-
wassermollusken aus der Rion-
Hohle bei Kutais (Transkaukasien,
Georgien). — Arch. Molluskenkunde,
1932, Bd. 64, H. I, S. 12—14.
Жадин В. И. Пресноводные моллюски
СССР. Л.: Ленснабтехиздат, 1933.
232 с.
Жадин В. И. Вопросы генезиса фауны
и биоценозов континентальных вод
Советского Союза. — В кн.: Памяти
академика С. А. Зернова. М.; Л.:
Изд-во АН СССР, 1948, с. 56—71.
Зайцев Ф. А. К фауне пещер Абхазии. —
Сообщ. Груз. фил. АН СССР, 1940,
т. 1, вып. 1, с. 63—70.
Зенкевич Л. А. Некоторые моменты зоо-
географии Северного Полярного бас-
сейна в связи с его палеогеографи-
ческим прошлым. — Зоол. журн.,
1933, т. 12, вып. 4, с. 17—34.
Зенкевич Л. А. Действие вод Черного и
Каспийского морей пониженной и по-
вышенной солености на некоторых
черноморских беспозвоночных. Ч. 1,
2. — Зоол. журн., 1938, т. 17, вып. 5,
с. 845—888; вып. 6, с. 976—1002.
Зенкевич Л. А. Распространение пресно-
водных и солоноватоводных ки-
шечнополостных. — Зоол. журн.,
1940, т. 19, вып. 4, с. 580—602.
Зенкевич Л. А. О древности возникно-
вения холодноводной морской фауны
и флоры. — Тр. ИОАН СССР, 1949,
т. 3, с. 191 — 199.
Зенкевич Л. А. Специальная количествен-
ная характеристика глубоководной
жизни в океане. — Изв. АН СССР.
Сер. геогр., 1960, № 2, с. 10—16.
Зенкевич Л. А., Бирштейн Я. А. Изу-
чение глубоководной фауны и свя-
занные с ним вопросы. — Вести.
МГУ, 1955, № 4/5, с. 231—242.
231
Зенкевич Л. А., Бирштейн Я. A. (Zen-
kevitch L. A., >Bir stein J. A.). On
the problem of the antiquity of the
deep-sea fauna. — Deep-Sea Res.,
1960, vol. 7, N 1, p. 10—23.
Зенкевич Л. А., Бирштейн Я. А. О гео-
логической древности глубоководной
фауны. — Океанология, 1961, т. 1,
вып. 1, с. 110—124.
Зенкевич Л. А., Бирштейн Я. А., Бе-
ляев Г М. Фауна Курило-Камчат-
ской впадины. — Природа, 1954,
№ 2, с. 61—74.
Зенкевич Л. А., Бирштейн Я. А., Бе-
ляев Г М. Исследования донной
фауны Курило-Камчатской впа-
дины. — Тр. ИОАН СССР, 1955,
т. 12, с. 345—381.
Зенкевич Л. А., Бирштейн Я. А., Кар-
певич А. Ф. Первые успехи рекон-
струкции фауны Каспийского мо-
ря. — Зоол. журн., 1945, т. 24, вып. 1,
с. 25-31.
Зернов С. А. Общая гидробиология.
М.; Л.: Биомедгиз, 1934. 504 с.
Зыков В. П. Материалы по фауне Волги
и гидрофауне Саратовской губер-
нии. — Bull. Soc. Imp. Nat. Moscou,
1903, t. 17, N 1. 148 c.
Иванов A. B. (Ivanov A. V.). Neue
Pogonophora aus dem nord-westli-
chen Teil des Stillen Ozeans. — Zool.
Jahrb., Abt. Syst., 1957, Bd. 85,
H. 4—5, S. 431—500.
Ильин Б. С. Некоторые данные по рас-
пространению ракообразных (Cirri-
pedia, Peracarida, Decapoda) и быч-
ков (Gobiidae) кубанских лима-
нов. — Тр. Азово-Черномор. науч,
рыбохоз. ст. 1930, вып. 7.
Ильин Н. М. и др. Совещание по исто-
рии флоры и растительности
СССР. — Сов. ботаника, 1938, № 2,
с. 8—87.
Кесслер К. Ф. Русские речные раки. —
Тр. Рус. энтомол. о-ва, 1875, т. 8,
с. 228—320.
Кесслер К. Ф. Сведения о распростра-
нении долгоногого речного рака
(Astacus leptodactylus) в Сибири. —
Тр. Рус. энтомол. о-ва, 1877, т. 10,
с. 1—5.
КиршенбратЯ. Д. Особенности пещерных
животных. — Природа, 1939, № 8,
с. 38—48.
Киселевич К. А. Промысловые рыбы
Волго-Каспийского района, их при-
вычки и особенности. Астрахань,
1926. 49 с.
Книпович И. М. Гидрологические иссле-
дования в Каспийском море в 1914—
1915 гг. — Тр. Касп. экспедиции
1914—1915 гг., 1921, т. 1, П., 943 с.
Книпович И. М. Гидрологические иссле-
дования в Азовском море. — Тр.
Азово-Черномор. науч.-промысловой
экспедиции 1932, вып. 5. 496 с.
Кобахидзе Д. И. Новый подвид пещерной
пиявки из Грузинской ССР (Hirudi-
nea, Herpobdellidae). — Сообщ. АН
ГССР, 1958, т. 21, № 5, с. 519—592.
Кожов М. М. К истории озерных систем
Забайкалья и Прибайкалья и их фа-
уны. — Тр. Всесоюз. гидробиол. о-ва,
1949, т. 1, с. 210—223.
Кожов М. М. Биология озера Байкал.
М.: Изд-во АН СССР, 1962. 315 с.
Крепе Е. М. Исследование над газообме-
ном Balanus crenatus при разной
концентрации солей в окружающей
среде. — Тр. Мурман, биол. ст.,
1929, т. 3, с. 1—32.
Криштофович А. И. Геологический обзор
стран Дальнего Востока. Л.; М.:
Гос. науч.-техн, геол.-развед. изд-во,
1932. 332 с.
Кубер А. А. Карстовая область горного
К>ыма. М., 1915. VIII, 319 с.
Крушинский Л. В. Зародышевое сходство
в свете закономерностей индивиду-
ального развития организма. —
Успехи соврем, биологии, 1939, т. 11,
вып. 2, с. 362—376.
Кузнецов И. Д. Терские речные и при-
терские морские рыбные промыслы.
СПб.: М-во земледелия и гос. иму-
ществ, 1898. 227 с.
Курнаков В. (Kurnakov V.). Les Trechini
de la faune souterraine de 1’Abkha-
zie. — Rev. fran£. entomol., 1959,
t. 26, N 4, p. 231—236.
Кусакин О. Г. К фауне Janiridae (Isopo-
da, Asellota) морей СССР. — Тр.
ЗИН АН СССР, 1962, т. 30, с. 17—65.
Лапшов И. И. Пещерные Pseudoscorpi-
ones Закавказья. — Бюл. МОИП.
Отд. биол., 1940, т. 49, вып. 3/4,
с. 61—74.
Леб Ж. Организм как целое с физико-
химической точки зрения. М.; Л.:
Госиздат, 1926. 290 с.
Лебединский Я. И. К фауне крымских
пещер. — Зап. Новорос. о-ва есте-
ствоиспытателей, 1900, т. 23, вып. 2,
с. 47—64.
Лебединский Я. И. К фауне крымских
пещер. — Зап. Новорос. о-ва есте-
ствоиспытателей, 1904, т. 25, вып. 2,
с. .75—88.
Лебединский Я. И. К фауне крымских
пещер. — Зап. Новорос. о-ва есте-
ствоиспытателей, 1914, т. 40, с. 113—
128.
Левушкин С. И. О фауне Trechini из пе-
щер Западного Закавказья. — Зоол.
журн., 1963, т. 42, вып. 3, с. 451—454.
232
Левушкин С. И. Новый вид троглобиот-
ного кавказского рода Jeannelius
(Coleoptera Trechini) из Анакопей-
ской пещеры вблизи Нового Афо-
на. — Зоол. жури., 1965, т. 44,
вып. 8, с. 1262—1265.
Левушкин С. И. Новый вид Hypogastru-
ridae (Collembola) из пещеры
в окрестностях Самарканда. — Тр.
ЗИН АН СССР, 1972, т. 51, с. 151 —
154.
Левушкин С. И., Матекин П. В. Troglo-
lestes sokolovi sp. n. — первый тро-
глобионтный слизень. — Бюл.
МОИП. Огд. биол., 1965, т. 70, вып. 3,
с. 35—46.
Линберг Г У. Четвертичный период
в свете биогеографических данных.
М.; Л.: Изд-во АН СССР, 1955. 335 с.
Лисицын А. П., Богданов Ю. А., Овчин-
никова Л. И. Некоторые результаты
изучения битуминозных веществ
в водной взвеси Тихого океана. —
Океанология, 1967, т. 7, № 1.
Лукин Е. И. К вопросу о факторах эво-
люции пресноводной фауны. —
Пращ наук.-дослщного зоо-биол.
iH-ту Харьк. ун-та, 1936а, т. 1,
с. 130—143.
Лукин Е. И. О биологических особен-
ностях рыбной пиявки Piscicola geo-
metra. — Прац! наук.-досл!дного
зоо-биол. iH-ту Харьк. ун-та, Г9366,
т. 1, с. 144—161.
Майр Э. Зоологический вид и эволюция.
М.: Мир, 1968. 598 с.
Малахов М. Речные раки в Среднем
и Южном Урале. — Тр. Рус. энтомол.
о-ва, 1880, т. 11, с. 27—44.
Мартынов А. В. О высших ракообраз-
ных окрестностей Ростова-на-До-
ну. — Протоколы заседаний о-ва
естествоисп. при Дон. ун-те, 1919,
т. 1, вып. 3, с. 39—53.
Мартынов А. В. О некоторых инте-
ресных формах Malacostraca из прес-
ных вод Европейской России и их
распространении. — Рус. гидробиол.
журн., 1924, т. 3, вып. 8/10, с. 210—
216..
Мартынов А. В. К познанию реликтовых
ракообразных бассейна нижнего
Дона, их этологии и распростране-
ния. — Ежегодник Зоол. музея Рос.
Акад, наук, 1925а, т. 25 (1924).
115 с.
Мартынов А. В. Gammaridae нижего
Днепра. — Тр. Всеукр. гос. Чер-
номор.-Азов. науч.-промысловой
опыт, ст., 19256, т. 1, с. 133—153.
Мартынов А. В. Экологические пред-
посылки для зоогеографии пресно-
водных бентических животных. —
Рус. зоол. журн., 1929, т. 9, вып. 3,
с. 1—38.
Машталер Г А. Роль фенотипической
и генотипической приспособленности
в эволюции организмов: (Эволюция
приспособлений). Одесса: Одес. ун-т,
1940. 212 с.
Меррей Дж. Океан. Общий очерк науки
о море. Одесса: Госиздат Украины,
1923. 151 с.
Мечников И. И. Очерк вопроса о про-
исхождении видов. — В кн.: Мечни-
ков И. И. О дарвинизме. М.; Л.:
Изд-во АН СССР, 1943, с. 3—213.
Муратов М. В. Краткий очерк геологи-
ческого строения Крымского полу-
острова. М.: Госгеолтехиздат, 1960.
207 с.
Мурина В. В. Систематика и зоогеогра-
фия глубоководных сипункулид:
Дис. . канд. биол. наук. М., 1958.
Мурина В. В. Сипункулиды и приапу-
лиды. — В кн.: Глубоководная фа-
уна. Плейстон. М.: Наука, 1969,
с. 84—89. (Тихий океан. Биология
Тихого океана; Кн. 2).
Мушкетов Д. И. Региональная геотекто-
ника. Л.; М.: ОНТИ НКТП, 1935.
528 с.
Мюллер Ф. За Дарвина. М.: Биомедгиз,
1932. 92 с.
. Ногинский Н. А. Четвертичная история
долины Оби на площади оледене-
ния. — Вопр. географии, 1949, сб. 12.
Неймайр М. История земли. СПб.: Про-
свещение, 1904. Т. 2. 848 с.
Николюк В. Ф. Реликтовые форамини-
феры пустыни Каракум. — Изв. АН
УзССР, 1948, т. 1, с. 79— 88.
Огнев И. Ф. Об изменениях в органах
золотых рыб после трехлетнего пре-
бывания в темноте. — Биол. журн.,
1910, т. 1, № 4, с. 552.
Оденов Ю. М. Некоторые проблемы эво-
люционной генетики и дарвинизма.
Л.: Изд-во АН СССР, 1961. 164 с.
Павлов А. П. О геологической истории
Европейского континента. — Отчет
Моск, ун-та за 1913 г., 1913, ч. 1,
с. 75—124.
Падлас П. С. (Pallas Р. S.). Reise
durch verschiedene Provinzen des
Russischen Reiches. — St. Petersburg
Akad. Wiss., 1773—1776. 747 S.
Панов Д. Г Происхождение материков
и океанов. М.: Географгиз, 1961.
184 с.
Пичка В. Е. О фауне пауков пещер За-
падного Закавказья. — Зоол. журн.,
1965, т. 42, вып. 8, с. 1190—1196.
Плигинский В. Г К. фауне пещер
Крыма. — Рус. энтмол. обозрение?
1912, т. 12, с. 503—506; 1914, т. 14,
16 Заказ 333
233
с. 330—331; 1927, т. 21, с. 171 — 180.
Поляков И. М. Ламарк и ламаркизм. —
В кн.: Ламарк. Философия зоологии.
М.; Л.: Биомедгиз, 1937, т. 2, с. VII —
LXXXVIII.
Пузанов И. И. Материалы к познанию
наземных моллюсков Крыма. Ч. 3.
Состав, распределение и генезис
крымской малокофауны. — Бюл.
МОИП. Огд. биол., 1927, т. 36,
вып. 3/4, с. 221—282.
Пузанов И. И. Зоогеография. М.: Учпед-
гиз, 1938. 359 с.
Рубцов И. А. О неравномерности темпа
эволюции. — Журн. общ. биологии,
1945, т. 6, № 6, с. 411—441.
Рудяков Ю. А. Применение индексов раз-
нообразия в гидробиологических
исследованиях. — Тр. ИОАН СССР,
1964, т. 65, с. 3—15.
Рылов В. М. К вопросу о ледниковых
реликтах в пресноводной фауне. —
Изв. Рос. гидрол. ин-та, 1921, № 1/3,
с. 97—113.
Садовский A. Xiphocardinella kutaissiana
nov. gen. et sp. (Atyidae) из подзем-
ной пещеры под Кутаисом. —
Закавк. краевед, сб., Естеств., 1930.
, Сер. А, 1, № 1, с. 93—104.
Сакс В. Н. Четвертичное оледенение
севера Сибири. — Природа, 1947,
№ 4, с. 16—25.
Саре Г О. Mysidae.—Тр. Касп. экспе-
диции 1904 г., 1907, т. 1. 313 с.
Северцов А. И. Морфологические законо-
мерности эволюции. М.; Л.: Изд-во
АН СССР, 1939. 610 с.
Симпсон Д. Г Темпы и формы эволюции.
М.: Изд-во иностр, лит., 1948. 358 с.
Скориков А. С. Отчет о деятельности
Волжской биологической станции. —
Тр. Сарат. о-ва естествоиспытателей
и любителей естествознания, 1903,
т. 3, вып. 2, с. 3—64.
Совинский В. Н. Введение в изучение
фауны Понто-Каспийско-Аральского
морского бассейна, рассматриваемой
с точки зрения самостоятельной зоо-
географической провинции. — Зап.
Киев, о-ва естествоиспытателей,
1904, т. 18. 497 с.
Соколов Б. С. Древнейшие отложения
раннего кембрия и сабеллитиды. —
Тр. Всесоюз. симпоз. по палеонто-
логии докембрия и раннего кембрия.
Новосибирск, 1965, с. 78—91.
Соколов Б. С. Древнейшие погоно-
форы. — ДАН СССР, 1967, т. 177,
№ 1, с. 201—204.
Соколова М. Н. О закономерностях рас-
пределения глубоководного бентоса:
Влияние микрорельефа и распреде-
ления взвеси в придонных слоях воды
на пищевые группировки глубоко-
водных донных беспозвоночных. —
ДАН СССР, 1956а, т. НО, № 4,
с. 692—695.
Соколова М. Н. Питание глубоководного
бентоса. Питание Laetmonice рго-
ducta v. wyvillei McIntosh. —ДАН
СССР, 19566, т. НО, № 6, с. 1111.
Соколова М. Н. Питание некоторых пло-
тоядных беспозвоночных глубоко-
водного бентоса дальневосточных
морей и северо-западной части Ти-
хого океана. — Тр. ИОАН СССР,
1957, т. 20, с. 279—301.
Соколова М. Н. Питание глубоководных
донных беспозвоночных детрито-
едов. — Тр. ИОАН СССР, 1958, т. 27,
с. 123—153.
Соколова М. Н. Распределение группи-
ровок (биоценозов) донной фауны
глубоководных впадин северо-запад-
ной части Тихого океана.—Тр.
ИОАН СССР, 1960, т. 34, с. 21—59.
Соколова М. Н. Закономерности рас-
пределения глубоководных донных
беспозвоночных в зависимости от
способа и условий их питания.—
В кн.: Глубоководная донная фауна.
Плейстон. М.: Наука, 1969, с. 182—
201. (Тихий океан. Биология Ти-
хого океана; Кн. 2).
Соколовская А. И., Стрелкова О. С. По-
липлоидия и кариологические расы
в условиях Арктики: (Исследование
флоры о-ва Колгуев). — ДАН СССР,
1941, т. 32, № 2, с. 145—147.
Сорокин Ю. И. Численность и продук-
ция бактерий в воде и донных осад-
ках центральной части Тихого оке-
ана. — ДАН СССР, 1970, т. 192, № 3,
с. 655—658.
Старобогатов Я. И. К познанию моллюс-
ков подземных вод Кавказа. — Бюл.
МОИП. Отд. биол., 1962, т. 67, вып. 6,
с. 42—54.
Страхов Н. М. Задачи и методы историче-
ской геологии. М.; Л., 1932. 147 с.
Страхов Н. М. Основы теории литогенеза:
Типы литогенеза и их размещение
на поверхности земли. М.: Изд-во
АН СССР, 1960. Т. 1. 212 с.
Сушкин П. П. Птицы Советского Алтая
й прилежащих частей Северо-Запад-
ной Монголии. М.; Л.: Изд-во АН
СССР, 1938. Т. 2. 434 с.
Талиев Д. Н. К вопросу о темпах и при-
чинах дивергентной эволюции бай-
кальских Cottoidei. — Тр. Байк, лим-
нол. ст. 1948, т. 12, с. 107—158.
Тимирязев К. А. Исторический метод,
в биологии. М.; Л.: Изд-во АН СССР,
1943. 208 с.
Третьяков Д. Mesomysis vol gensis nov.
234
sp.: (Из материала экскурсии 1905 г.
по Волге). — Тр. СПб. о-ва есте-
ствознания, 1907, т. 36, вып. 2. 49 с.
Уоллес А. Р. Дарвинизм: Изложение
теории естественного подбора и неко-
торых из ее приложений. М.: Изд-во
Сабашниковых, 1911. 557 с.
Ушаков П. В. Многощетинковые черви
сем. Aphroditidae Курило-Камчат-
ской впадины. — Тр. ИОАН СССР,
1955, т. 12, с. 311—321.
Фадеев Н. Н. Морские элементы в фауне
бассейна р. Сев. Донец. — Рус.
гидробиол. журн., 1923, т. 2,
вып. 11/12, с. 240—246.
Фадеев Н. Н. Каталог водных животных,
найденных в бассейне р. Донца
и прилежащих местностях за период
работы с 1917 по 1927 г. — Пращ
Харьк. т-ва доел, npip., 1929, т. 52,
с. 7—32.
Фадеев Н. Н. Отчет о работах Донец-
кой научной экспедиции в 1927 г. —
Пращ Донецьк. наук, експ., 1930,
№ 1, с. 5—21.
Филатова 3. А. О некоторых новых видах
двустворчатых моллюсков (Bivalvia)
северо-западной части Тихого оке-
ана. — Тр. ИОАН СССР, 1958, т. 27,
с. 208—218.
Харитонов Д. Е. О пещерных пауках
Абхазии. — В кн.: Материалы к фа-
уне Абхазии. Тбилиси: Груз. фил.
АН СССР, 1939, с. 197—211.
Цееб Я. Я. Зоогеографический очерк
и история крымской гидрофауны. —
Учен. зап. Орлов, пед. ин-та. Сер.
естествознания и химии, 1947, т. 3,
вып. 2, с. 67—112.
Циттель К. Основы палеонтологии: Бес-
позвоночные. Л.; М.: Горнефтеиздат,
1934. Ч. 1. 1056 с.
Чернай А. Животные пресноводные в от-
ношении к морским и сухопутным. —
Тр. О-ва испытателей природы при
Харьк. ун-те, 1870, т. 2, с. 1 —16.
Чиндонова Ю. Г Питание некоторых
групп глубоководного макропланк-
тона в северо-западной части Тихого
океана.— Тр. ИОАН СССР, 1959,
т. 30, с. 166—189.
Шарлеман И. В. Заметка о некоторых
ракообразных (Amphipoda и Clado-
сега) Днепра. — Рус. гидробиол.
журн., 1922, т. 1, с. 322—324.
Шатский Н. С. О зональном и биполярном
размещении глауконитовых форма-
ций в верхнем мелу и эоцене. —
Бюл. МОИП. Отд. геол., 1954, т. 29,
вып. 5, с. 3—19.
Шимкевич В. М. (Schimkewitsch W. М.).
Der turkestanische Flusskrebs. —
Zool. Anz., 1884, Jahrg. 7, N 170,
S. 339—341.
16Ф
Шимкевич В. M. О видовых признаках
и географическом распространении
рода Astacus. — Изв. О-ва любите-
лей естествознания, 1886, т. 50,
вып. 1/2, с. 8—23.
Шмальгаузен И. И. Организм как целое
в индивидуальном и историческом
развитии. М.; Л.: Изд-во АН СССР,
1938. 144 с.
Шмальгаузен И. И. Пути и закономер-
ности эволюционного процесса. М.;
Л.: Изд-во АН СССР, 1940. 232 с.
Шмальгаузен И. И. Факторы эволюции.
М.; Л.: Изд-во АН СССР, 1946.
396 с.
Шмальгаузен И. И. Факторы эволюции.
2-е изд., доп. М.: Наука, 1968а. 451 с.
Шмальгаузен И. И. Контроль и регуля-
ция в эволюции. — В кн.: Киберне-
тические вопросы эволюции. Новоси-
бирск, 19686, с. 34—73.
Щербаков Ф. С. Пещеры как биологи-
ческая среда и их фауна. — Есте-
ствознание и география, 1909, т. 14,
№ 8, с. 36—49.
‘Юбзашьян С. М. Некоторые итоги изу-
чения кутаисской пещерной кре-
ветки. — Тр. Зоол. сектора Груз, отд-
ния Закавк. фил. АН СССР, 1934,
т. 1, с. 175—193.
Юбзашьян С. М. О редукции органов
зрения у пещерных креветок. —
Сообщ. Груз. фил. АН СССР, 1940,
т. 1, вып. 4, с. 293—300.
Яковлев С. А. К вопросу об Иольдиевом
море в Балтике по соединении Бал-
тийского моря с Белым в поздне-
ледниковое время. — Изв. Гос. геогр.
о-ва, 1934, т. 66, вып. 2, с. 277—
272.
Янковская А. И. Реликтовые ракообраз-
ные в грунтовых водах побережья
оз. Иссык-Куль (Северный Тянь-
Шань).— Зоол. журн., 1964, т. 43,
вып. 7, с. 975—986.
Яшнов В. А. Вертикальное распределение
массы зоопланктона тропической
области Атлантического океана. —
ДАН СССР, 1961, т. 136, № 3,
с. 705—708.
Absolon К., Hrabe S. Ober einen neuen
Siisswasser-Polychaeten aus den
Hohlengewassern der Herzegowi-
na. — Zool. Anz., 1930, Bd. 88,
H. 9/10, S. 249—264.
Absolon K., Kratochvil J. Zur Kenntnis der
hohlenbewohnenden Araneae der illy-
rischen Karstgebiete. — Mitt. Hohlen-
und Karstforsch., 1932, Bd. 3, S. 73—
81.
Absolon K., Landrock K. Eine neue
hohlenbewohnende Fliege aus Herze-
govina.— Konowia, 1933, Bd. 11
(1932), S. 266—272.
235
Allen J. A., Sanders A. L. Adaptations
to abyssal life as shown by the bivalve
Abra profundorum. — Deep-Sea Res.,
1966, vol. 13, N 6, p. 1175—1184.
Annandale N., Coggin B. J., Gravely F. H.
The limestone caves of Burma and
the Malay Peninsula. — J. and Proc.
Asiat. Soc. Bengal. N. S., 1913, vol. 9,
p. 391—424.
Apathy S. Siisswasser-Hirudineen. — Zool.
Jb. Abt. Syst. 1888, Bd. 3, H. 5,
S. 725—794.
Arcangeli A. Ricerche faunistiche nelle
isole italiane dell’ Egeo: Isopodi. —
Arch. zool. ital., 1929, vol. 13,
p. 259—268.
Arcangeli A. Asellus delle caverne der
Belgia. — Bill. Mus. roy. hist, natur.
Belg., 1935, vol. 11, N 37, p. 1 — 10.
Arcangeli A. Stenasellus racovitza Razza-
utti, Crostaceo Isopodo aquatico ca-
vernicolo italiano: La famiglia Stena-
sellidae. — Boll. Mus. zool. anat.
comp. Torino, 1939, vol. 46, p. 17—50.
Arldt Th. Die Ausbreitung einiger Deka-
podengruppen. — Arch. Hydrobiol.
und Planktonk., 1910, Bd. 5, S. 257—
282.
Arldt Th. Handbuch der Palaeogeographie.
Leipzig, 1919—1922. Bd. 1, 2.
Austen E. E. A remarcable semi-apterous
fly (Diptera) found in a cave in East
Africa and representing a new family,
genus and species. — Proc. Zool. Soc.
London, 1936, p. 425—431.
Babic K. Ober die drei Atyiden aus
Jugoslavien. — Glasnik Kroatischen
naturwiss. Ges., 1922, Bd. 34, N 3,
S. 300—306.
Bachmayer F. Eine fossile Cumaceenart
(Crustacea, Malacostraca) aus dem
Callovien von La Vonte-Sur-Rhone
(Adreche). — Eclog. geol. helv., 1961,
Bd. 53, S. 422—426.
Bailey V., Bailey F. M., Giovannollia L.
Cave life of Kentucky mainly in the
Mammoth Cave region. — Amer. Mid-
land. Natur., 1933, vol. 14, p. 385—
635.
Balss H. Crustacea Malacostraca—Deca-
poda. — In: Kukenthal—Krumbach:
Handbuch der Zoologie. Berlin; Leip-
zig, 1927. Bd. 3.
Barham E. G., Pickwell G. V The giant
isopod Anuropus: A scyphozoan
symbient. — Deep-Sea Res., 1969,
vol. 16, N 5, p. 525—530.
Barnard J. Gammaridean Amphipoda
from depths of 400 to 6000 meters. —
Galathea Rep., 1961, vol. 5, p. 23—128.
Barnard J. South Atlantic abyssal amphi-
pods collected by R/V «Verna». —
Verna Res. Ser., 1962, N 1. Abyssal
Crustacea, p. 1—78.
Barnard К. H. Contribution to the crusta-
cean fauna of South Africa. — Ann.
S. Afr. Mus., 1914, vol. 10, p. 231—240.
Beadle L. C. Adaptation to changes of
salinity in the polychaetes. 1. — J. Exp.
Biol., 1937, vol. 14, N 1, p. 56—70.
Beadle L. Osmotic regulation and the
faunas of inland waters. — Biol. Rev.,
1943, vol. 18, N 4, p. 172—183.
Beddard F. E. Report of the Isopoda
collected by H. M. S. «Challenger»
during the years 1873—1876. Pt II. —
Challenger Rep., 1886, vol. 17,
p. 1 — 178+25 pl.
Benedict J. E. The Arcturidae in the US
National Museum. — Proc. Biol. Soc.
Wash., 1898, vol. 12, p. 41—51.
Berninger J. Ober die Einwirkung des
Hungers auf Planarien. — Zool. Jb.
Abt. allgem. Zool. 1911, Bd. 30,
S. 181—216.
Bertossi F. Richerche sulla pigmentazione
di Androniscus boldorii Strouhal in
condizioni normali e sperimentali. —
Boll. zool. Torino, 1938, vol. 9,
p. 125—129.
Bethe A. lonendurchlassigkeit der Korpe-
roberflache von wirbellossen Tieren
des Meeres als Ursache der Giftigkeit
von Seewasser abnormer Zusammen-
setzung. — Pfliiger’s Arch. ges. Phy-
siol., 1929, Bd. 221, .S. 344—362.
Bethe A., Berger E. Variationen in Mine-
ralbestand verschiedener Blutarten.—
Pfliiger’s Arch. ges. Physiol., 1931,
Bd. 227, S. 571—584.
Beudant F. S. Memoire sur la possibilite
de faire vivre des mollusques fluviati-
les dans les eaux salees et des
mollusques marins dans les eaux
douces. — Ann. chimie, 1816, t. 2,
p. 32—41.
Bialaszewitz K- Sur la regulation de la
composition minerale de 1’hemo-
lymphe chez le crabe. — Arch. Intern.
Physiol., 1932, vol. 35, p. 98—124.
Bogucki M. Recherches sur la regulation
de la composition minerale du sang
chez 1’ecrevisse (Astacus fluviatilus
L.). — Arch. Intern. Physiol., 1934,
vol. 38, p. 172—179.
Bond R. M., Cary M. K., Hutchinson G. E.
A note on the blood of hag-fish Po-
listotrema stouti. — J. Exp. Biol.,
1932, vol. 9, p. 12—14.
Borodin N. A. A new blind catfish from
Brasil. — Amer. Mus. Novit., 1927,
N 263, p. 1—4.
Botazzi F. Osmotischer Druck und
elektrische Leitf ahigkeit der Flus-
sigkeiten der einzelligen pflanzlichen
und tierischen Organismen. — Er-
geb. Physiol., 1908, Bd. 7, S. 161 —
402.
236
Bouvier E. L. Recherches sur la morpho-
logic des variations, la distribution
geographique des Atyides. — In: En-
cycl. entomoL, 1925, t. 4, p. 370—715.
Bovallius C. Notes on the family Aselli-
dae. — Bidr. Kgl. sven. vetenskapsa-
kad. handl., 1886, bd 11, N 5, s. 1—54.
Braga J. M. Mancasellus mariae, a new
blind isopod inhabiting subterranean
waters in Portugal. — Mem. Mus.
zool. Univ. Coimbra, 1942, vol. 130,
p. 1 — 12.
Braga J. M. Note sur un genre d’Asellides
nouveau du Portugal (Synasellus
n. gen.) et description d’une sou-
sespece de ce genre. — Mem. Mus.
zool. Univ. Coimbra, 1944, vol. 158,
p. 1 — 11.
Brooks J. L. Speciation in ancient lakes. —
Quart. Rev. Biol. 1950, vol. 25, N 2,
p. 30—176.
Bruun, A. F. The Philippine Trench and
its bottom fauna. — Nature, 1951,
vol. 168, N 4277, p. 692.
Bruun A. F. Problems of the life in the
deepest deep-sea. — Geogr. Mag.,
1953, vol. 26, N 5, p. 247—262.
Bruun A. F. The abyssal fauna: Its
ecology, distribution and origin. —
Nature, 1956, vol. 177, N 4520,
p. 1105—1108.
Buchalowa W. J. Die Malakostraken mari-
nen Ursprungs im mittleren Strom-
lauf des Dons. — Zool. Anz., 1929,
Bd. 85, H. 9/10, S. 237—256.
Buchmann J. U7 Notes on an American
cave flatworm, Sphalloplana percae-
ca. — Ecology, 1936, vol. 17, p. 194.
Buresch J. Die Hohlenfauna Bulgariens. —
In: Intern. Congr. Zool. Budapest,
1929, Bd. 10, S. 1427—1437.
Buzzati-Traverso A. A. Discussion on:
Kohn A. J. «Problems of speciation
in marine invertebrates». — In: Per-
spective in marine biology. Berkeley;
Los Angeles: Univ. Cal. press, 1960,
p. 582—583.
Carl J. Materialen zur Hohlenfauna der
Krim. 1. Neue Holencrustaceen. —
Zool. Anz., 1905, Bd. 28, H. 8/9,
S. 322—329.
Carter G. S. Evolution in the deep seas. —
Publ. Amer. Assoc. Adv. Sci., 1961,
N 67, p. 229—237.
Chappuis P. A. Die Tierwelt der unterirdi-
schen gewasser — die Binnengewa-
ser. Stuttgart, 1927. Bd. 3. Schweizer-
bart. 175 S.
Chappuis P. A. Methodik der Erforschung
der subterranean Fauna. — Abder-
haldens Handb. biol. Arbeitsmeth.,
1930, Bd. 9, H. 7, S. 161—226.
Chappuis P. A. Microcharon acherontis
spec, nov., ein neuer subterraner
Isopode. — Fragm. Fauna Hung.,
1942, Bd. 3/4, S. 120—121.
Chilton C. A fossil isopod belonging to the
fresh water genus Phreatoicus. — J.
and Proc. Roy. Soc. N. S. Wales,
1918, vol. 51, p. 365—388.
Chopard L. Biospeologica. 57. Les
Orthopteres cavernicoles de la faune
palearctique. — Arch. zool. exp. et
gen., 1932, t. 74, p. 263—286.
Chopard L. La biologie des Orthopteres. —
In: Encycl. entomoL Ser. A., 1938,
t. 20, p. 1—541.
Clark A. H. On the deep sea and compa-
rable fauna. Pt. 2. — Intern. Rev.
gesamt. Hydrobiol. und Hydrogr.,
1913, Bd. 6, H. 2, S. 133—146.
Clark A. H. Geography and zoology. —
Ann. Assoc. Amer. Geogr., 1927,
vol. 17, N 3, p. 101 — 145.
Claus A. Vergleichende-physiologische
Untersuchungen zur Okologie der
Wasserwanzen mit besonderer Be-
riicksichtigung der Brackwasserwan-
ze Sigara lugubris Fieb. — Zool. Jb.
Abt. allgem. Zool. Physiol. Tiere, 1937,
Bd. 58, H. 3, S. 365—432.
Colosi G. L’Ologenese et la biogeogra-
phic. — Bull. Soc. oceanogr. France,
1932, vol. 68.
Crender R. Die Reliktenseen: Ein physisch-
geographische Monographie. — Peter-
manns Mitt., 1887, Erganzungsheft
N 86, T. 1, S. 1 — 110; 1888, T. 2,
S. 1—50.
Dakin W., Edmonds E. The regulation
of the salt contents of the blood of
aquatic animals. — Austral. J. Exp.
Biol, and Med. Sci., 1931, vol. 8,
N 3, p. 169—187.
Davis W M. The origin of limestone
caverns. — Science. N. S., 1931,
vol. 73, N 1891, p. 327—331.
Dayley M. E., Fremont-Smith F., Car-
roll M. P. The relative composition
of sea water and the blood of Limulus
polyphemus. — J. Biol. Chem., 1931,
vol. 93, p. 17—24.
Dudich E. Systematische Studien an ita-
lienischen Aselliden. — Ann. Mus.
Natur. Hung., 1925, Bd. 22, S. 281 —
301.
Dudich E. Neue Krebstiere in der Fauna
Ungarus. — Arch. Balatonicum, 1927,
Bd. 1, S. 343—387.
Dudich E. A Jaera normanni Rathke, egy
uj viztaseka a magyer faunaban. —
Allat. kozL, 1930, Bd 27. Цит. no:
Kesselyak, 1938.
Dudich E. Biologie (ier Agteleker Tropf-
steinhohle «Baradla» in Ungarn.
Wien, 1932. 246 S. (Spelaolog.
Monogr.; Bd. 13).
237
Dudich E. Die spelaobiologische Station
zu Postumia und ihre Bedeutung
fur die Hohlenkunde. — SpelaoL Jb.,
1933a, H. 13/14, S. 51—65.
Dudich E. Die Klassifikation der Hohlen
auf biologische Grundlage. — Mitt.
Hohlen- und Karstforsch., 1933b,
Bd. 3, S. 35—43.
Duval M. Recherches sur le milieu
interieur de Telphusa fluviatilis
Latr. — Bull. Inst, oceanogr. Monaco,
1927, N 490.
Eigenmann C\ H. Cave vertebrates of
America: A study in degenerative evo-
lution. Wash., 1909. IX, 241 p. (Car-
negie Inst. Wash. Publ.; N 104).
Ekman S. Vorschlage und Erorterungen
zur Reliktenfrage in der Hydrobio-
logie. — Ark. zool., 1915, bd 9 (1914),
N 17, s. 1—36.
Ekman S. Tiergeographie des Meeres.
Leipzig: Akad. VerL, 1935. XII + 542 S
Ekman S. Zoogeography of the sea. L.:
Sidgwick and Jackson, 1953. XIV +
+ 417 S.
Elton C. Animal ecology. N. Y.: MacMillan,
1927.
Emiliani C. Pleistocene temperatures. —
J. GeoL, 1955, vol. 63, N 6, p. 538—578.
Fage L. Biospeologica. 55. Araneae.
Cinquieme serie, precedee d’un essai
sur 1’evolution souterraine et son
determinisme. — Arch. zool. exp. et
gen., 1931, t. 71, p. 99—291.
Fage L. Remarques sur les Pycnogonides
abysseaux. — Union Intern. Sci. Biol.
Ser. B, 1954, p. 49—56.
Faxon UZ A revision of the Astacidae. —
Mem. Mus. Comp. Zool., 1885, vol. 10,
p. 1 — 186.
Fisher R. A. The genetical theory of natural
selection. Oxford: Clarendon press,
1930. 272 p.
Fisher R. A., Corbet A. S., Wiliams С. B.
The relation between the number of
species and the number of individuals
in a random sample of an animal
population. — J. Anim. EcoL, 1943,
vol. 12, p. 42—58.
Fitzinger L. J. Ueber den Proteus angui-
neus. — S.-Вег. Kais. Akad. Wiss.
Math.-naturwiss. KI. Abt. 2, 1850,
S. 291—303.
Fontaine M. Recherches experimentales
sur la reaction des £tres vivantes
aux fortes pressions. — Ann. Inst,
oceanogr., 1930, t. 8, fasc. 1, p. 1—97.
Fritsche H. Studien Ober die Schwankun-
gen des osmotischen Druckes der
Korperflussigkeiten bei D. magnia. —
Intern. Rev. gesamt. Hydrobiol., 1917,
Bd 8, H. 1, S. 22—80, 125—203.
Gauthier-Lievre L. Sur une des singula ri-
tes de 1’oued Rhir: des Foraminiferes
thalassoides vivants dans les eaux
sahariennes. — Bull. Soc. hist, natur.
Afr. Nord, 1935, t. 26, p. 142—147.
George R. Y., Menzies R. J. Species of
Storthyngura (Isopoda) from the
Antarctic with description of six new
species.— Crustaceana, 1968a, vol. 14,
N 3, p. 275—301.
George R. Y.t Menzies R. J. Additions
to the Mediterranean deep-sea isopod
fauna. — Rev. roum. biol. Ser. zool.,
1968b, vol. 13, N 6, p. 367—383.
Gerard P. Sur I’existence des vestiges
oculaires chez Caecobarbus geertsii.—
Mem. Mus. hist, natur. Belg., 1936,
t. 2, N 3, p. 549—552.
Gianferrari L. Elenco dei pesci cavernicoli
e loro distribuzione geografica. —
Atti Soc. ital. Milano, 1933, vol. 71,
N 3/4, p. 217—223.
Glaessner M. F. Crustacea Decapoda. —
Fossil. Catalogus, 1929, ps 41,
p. 1—464.
Gordon D. C. A microscopie study of
organic particles in the North Atlantic
Ocean. — Deep-Sea Res., 1970, vol. 17,
N 1, p. 175—186.
Graff L. von. Tricladida: Bronns Klassen
und Ordnungen des Tierreichs. —
Turbellaria, 1917, Bd. 4, S. 2601 —
3369.
Grese B. S. Ober eine blinde Nemastoma-
Art aus einer Hohle der Krim
(Nemastoma caecum nov. sp.).—
Zool. Anz., 1911, Bd. 37, H. 5, S. 108.
Gresser E. B., Breder С. M. The histology
of the cave Characin, Anoptichthys. —
Zoologica, 1940, vol. 25, N 1, p. 113—
116.
Gurney R. The origin and conditions of
existence of the fauna of freshwater. —
Trans. Norfolk Natur. Soc., 1908,
vol. 9.
Haecker W. Tiefsee-Radiolarien. Allgem.
T. Form und Formbilden bei den
Radiolarien. — Wiss. Ergeb. Tiefsee-
Exped. «Valdivia», 1898—1899, Jena,
1908, Bd. 14, Lfg. 2, S. 477—706.
Hagedoorn A. L., Hagedoorn A. C. The
relative value of the processes causing
evolution. The Hague: Martinus
Nijhoff, 1921.
Hagen H. Hohlen-Chelifer in Nord-Ame-
rica. — Zool. Anz., 1879, Bd. 2, H. 3/4,
S. 399—400.
Hagmeier E. Zum Gehalt an Seston und
Plankton im Indischen Ozean zwi-
schen Australien und Indonesien. —
Kiel. Meeresforsch., 1964, Bd. 20, N 1,
S. 12—17.
238
Hale H. M. Isopoda-Valvifera. — Rep.
В. A. N. Z. Antarct. Res. Exped.,
1929—1931. Ser. B, 1946, vol. 5,
pt 3, p. 162—212.
Handlirsch A. Ober Relikte. — Verh.
ZooL-Bot. Ges. Wien, 1909, Bd. 59,
S. 183—207.
Hanko B. Ursprung und Verbreitung der
Fischfauna Ungarns. — Arch. Hy-
drobiol., 1932, Bd. 23, H. 4, S. 520—
556.
Hansen B. Systematics and biology of the
deep-sea holothurians. Pt 1. Elasi-
poda. — Galathea Rep., 1975, vol. 13,
p. 5—262+ 14 pls.
Hansen H. J. Report on the dredging
«Albatross» during 1891. 22. Iso-
poda. — Bull. Mus. Comp. Zool.,
1897, vol. 31, N 5, p. 93—130 + 6 pls.
Hansen H. J. On the morphology and
classification of the Asellota — group
of Crustaceans, with descriptions of
the genus Stenetrium Hasw. and its
species. — Proc. Zool. Soc. London,
1905, vol. 11, p. 302—331.
Hansen H. J. Crustacea Malacostraca. 3.—
Dan. Ingolf Exped., 1916, vol. 3,
p. 1—262.
Harms J. W Wandlungen des Artgefuges
unter natiirlichen und kiinstlichen
Umweltbedingungen. Tiibingen: Hei-
ne Verl., 1934. 212 S.
Hauer J. Zur Kenntnis subterraner Rota-
torien. — Zool. Anz. 1939, Bd. 126,
H. 1/2, S. 41—44.
Hawes R. S. The flood factor in the ecology
of caves. — J. Anim. Ecol., 1939,
vol. 8, N 1, p. 1—5.
Henry J. P., Magniez G. Sur la systemati-
que et la biogeographic des Aselli-
des. — C. r. Acad, sci., 1968, Ser. D,
t. 267, p. 87—89.
Herfs A. Die pulsierende Vakuole der
Protozoen, ein Schiitzorgan gegen
AussGssung. — Arch. Protistenk.,
1922, Bd. 44, S. 227—260.
Hertzog L. Amphipoden aus dem Grund-
wasser von Scopje. — Zool. Anz.,
1935, Bd. Ill, H. 1/2, S. 50—52.
Hertzog L. Crustaceen aus unterirdischen
Biotopen des Rheintales bei Strass-
burg. 3 Mitt. — Zool. Anz., 1938,
Bd. 123, H. 3, S. 45—56.
Hesse R. Tiergeographie auf okologischer
Grundlage. Jena: G. Fischer, 1924.
613 S.
Hofstein N. Zur Kenntnis der Tiefenfauna
des Bienzer und des Thuner Sees. —
Arch. Hydrobiol. und Planktonk.,
1912, Bd. 7, S. 1—62, 163—229.
Holdhaus K. Die europaische Hohlenfauna
in ihren Beziehungen zur Eiszeit. —
Zoogeographica, 1932, Bd. 1, H. 1,
S. 1—53.
Hubbs C. L. Fishes from the caves of
Jucatan. — Carnegie Inst. Wash.
Publ., 1938, N 491, p. 261—295.
Hubbs C. L., Innes W. T The first kno\\n
blinde fish of the family Characinidae:
A new genus from Mexico. — Occas.
Pap. Mus. Zool. Comp. Univ. Mich.,
1936, N 342, p. 1—7.
Hult J. On the soft-bottom isopods of
the Skager-Rak. — Zool. bidr. Uppsa-
la, 1941, bd. 21, s. 1—234.
Ives J. D. Cave fauna with especial
reference to ecological factors. —
J. E. Mitchell. Sci. Soc., 1927, vol. 43,
p. 84—90.
Jaeckel S. H. Bau und Lebensweise der
Tiefseemollusken. — Neue Brehm-BG-
cherei, 1955, H. 148, S. 1—70.
Jarocki J., Demianowicz A. Ueber das
Vorkommen des ponto-kaspischen
Amphipoden Chaetogammarus te-
nellus (G. O. Sars) in des Wisla
(Weichsel). — Bull. Acad. pol. sci. et
lett. Ser. B, 1931, vol. 2, p. 513—530.
Jeannel R. Fauna cavernicole de la France
avec une etude des conditions d’exi-
stence dans le domaine souterrain.
P.: Lechevalier, 1926. 334 p. (Encycl.
entomol.; T. 7).
Jeannel R. Origine et evolution de la faune
cavernicole du Bihar et des Carpathes
du Banat. — Arch. zool. Torino, 1931,
t. 16, p. 47—59.
Jeannel R. Situation geographique et
peuplement de cavernes. — Ann. Spe-
leol., 1959, t. 14, p. 333—338.
Jeannel R. Revision des «Trechini» du
Caucase (Coleoptera Trechidae).—
Mem. Mus. nat. hist, natur. A, 1960,
t. 17, fasc. 3, p. 1—216.
Johansen A. C. Om Brugen av Betegnelsen'
«Relikt» i Naturhistorien. — Medd.
Dan. geol. foren., 1908, N 14, s. 157—
168.
Johansen F. Asellus aquaticus not found
in Labrador. — Canad. Field-Natur.,
1926, vol. 40. Цит. no: Van Name,
1936.
Johnston R. An equation for the depth
distribution of deep-sea zooplankton
and fishes. — Rapp, et proc.-verb.
reun. Cons. perm, intern, explor. mer.,
1962, vol. 153, N 38, p. 217—219.
Jorgensen В. C. Biology of suspension
feeding. Oxford: Pergamon press,
1962a, XVI, 357 p.
Jorgensen В. C. The food of filter-feeding
organismus. — Rapp, et proc.-verb.
reun. Cons. perm, intern, explor.
mer., 1962b, vol. 153, N 38, p. 99—107.
Kammerer P. Experimente uber Fortpflan-
zung, Farbe, Augen- und Korperre-
duction bei Proteus anguineus
Laur. — Arch. Entwicklung Meeh.,
1912, Bd. 33, S. 350—461.
239
Kapterew P. Experimentelle Untersuchun-
geh fiber die Frage vom Einfluss
der Dunkelheit auf die Gefuhlsorgane
des Daphniden. — Biol. Zentr.-BL,
1910, Bd. 30, S. 239—256.
Kapterew P. Uber den Einfluss des Dun-
kelheit auf das Daphnien-Auge. — Biol
Zentr.-Bl., 1912, Bd 32, S. 233—243.
Кагатап S. 5. Beitrag zur Kenntnis der
Siisswasser-Amphipoden. — Prir.
Raspr., 1932, sv. 2, кн. 1, s. 179—232.
Кагатап S. Uber zwei neue Amphipoden
Balcanella und Jugocrangonyx aus
dem Grundwasser von Skoplje. —
Zool. Anz., 1933, Bd. 103, H. 1/2,
S. 41—47.
Кагатап S. Beitrage zur Kenntniss
der Isopoden-Familia Microparasel-
lidae. — Mitt. Holen- und Karst-
forsch., 1934a, Bd. 2, S. 42—44.
Кагатап S. Weitere subterrane Aselliden
aus Jugoslaviens. — Zool. Anz.,
1934b, Bd. 108, S. 23—26.
Кагатап S. Die Fauna der unterir-
dischen Gewasser Jugoslaviens. —
Verh. Intern. Ver. theor. und angew.
Limnol., 1935, Bd. 7, S. 46—73.
Кагатап S. Die unterirdischen Isopoden
Siidserbiens. — Гласи. Скопского
науч, друштва, 1940, т. 22, с. 19—53.
Kesselyak A. Die Arten der Gattung
Jaera Leach (Isopoda, Asellota).—
Zool. Jb. Abt. Syst., 1938, Bd. 71,
H. 3, S. 219—252.
Kiefer F. Uber Systematik und geographi-
sche Verbreitung einiger Gruppen
stark verkiimmerster Cyclopoiden
(Crustacea Copepoda).—Zool. Jb.
Abt. Syst., 1937, Bd. 70, H. 5/6,
S. 419—442.
Kinsey A. C. An evolutionary analysis
of insular and continental species. —
Proc. Nat. Acad. Sci. US, 1937,
vol. 23, p. 5—11.
Knudsen J. The bathyal and abyssal
Xylophaga (Pholadidae, Bivalvia). —
Galathea Rep., 1961, vol. 5, p. 163—
209.
Knudsen J. The deep-sea Bivalvia. —
Sci. Rep. John Murray Exped.,
1933—1934, 1967, vol. 11, N 3,
p. 237—343.
Kolosvary G. de. Recherches biologique
dans les grottes de pierre a chaux de
la Hongrie. — Folia zool. hydrobiol.,
1934, vol. 6, p. 1 — 12.
Kolosvary G. de. Reflexion sur l’«adapta-
tion> des organismes <lans les grot-
tes. — Acta biotheor., 1936, bd. 2,
biz. 19—22.
Kolosvary G. de. Gedanken zur Kritik
der Anpassung tierischen Tiefseefor-
men. — Acta biotheor., 1938, bd. 4.
Koppen W., Wegener A. Die Klimate
der geologischen Vorzeit. B.: Bornt-
raeger, 1924. 255 S.
Kosswig C. Uber bislang unbekannte
Sinnesorgane bei dem blinden Hohlen-
fisch Stygicola dentatus (Росу). —
Verh. Dt. zool. Ges., 1934, Bd. 36,
S. 185—190.
Kosswig C. Betrachtungen und Experi-
mente fiber die Entstehung von
Hdlentiermerkmalen. — Zfichter,
1937a, Bd. 9, S. 91 — 101.
Kosswig C. Uber Pigmentverlust wahrend
des Hohlenlebens. — Zool. Anz.,
1937b, Bd. 117, H. 1/2, S. 37—43.
Kosswig C. Uber Variability bei Hohlen-
tieren. — Mitt. Hohlen- und Karst-
forsch., 1937c, Bd. 5.
Kosswig C. Zur Farbungsvariabilitat bei
unterirdisch lebenden Wasserasseln,
Asellus aquaticus sensu Racovitza. —
Mitt. Hohlen- und Karstforsch., 1939,
Bd. 7, S. 94—102.
Kosswig C., Kosswig L. Uber Augenrfick-
und -missbildungen bei Asellus
aquaticus cavernicolus. — Verh. Dt.
zool. Ges., 1936, Bd. 38, S. 274—281.
Koszy F. Geochemistry of the radioactive
elements in the ocean. — Deep-Sea
Res., 1956, vol. 3, N 2, p. 93—103.
Krause H. R. Investigation of the decompo-
sition of organic matter in natural
waters. — FAO Fish. Rep., 1962, N 34,
p. 1 — 14.
Krey J. Die mittlere Tiefenverteilung von
Seston, Mikrobiomasse und Detritus
im nordlichen Nordatlantik. — Kiel.
Meeresforsch., 1964, Bd. 20, N 1,
S. 18—29.
Krogh A. Osmotic regulation in aquatic
animals. Cambridge: Univ, press,
1939. 242 p.
Lattin G. de. Untersuchungen an Isopo-
denaugen. — Zool. Jb. Abt. Anat.,
1939a, Bd. 65, H. 3, S. 417—468.
Lattin G. de. Untersuchungen fiber die
Farbvariabilitat der Isopoden. —
Zool*. Anz.. 1939b, Bd. 125, H. 9/10,
S. 309—324.
Lattin G. de. Uber die Evolution der
Hdhientiercharaktere. — S.-Вег. ges.
Naturforsch. Freunde Berlin, 1939c,
N 1, S. 11—41.
Leruth R. Etudes biospSologiques. IX.
La faune de la nappe phreatique du
Gravier de la Meuse a Hermalle-
sous-Argenteau. — Bull. Mus. hist,
natur. Belg., 1938, vol. 14, N 41,
p. 1—37.
Leruth R. La biologie du domaine souter-
rain et la faune cavernicole de la
Belgique. — Mem. Mus. hist, natur.
Belg., 1939, vol. 87, p. 1—506.
240
Levi CL Duslenia teissieri nov. gen.,
n. sp., nouveau Parasellide des cotes
de France. — Arch. zool. exp. et gen.,
1950, t. 87, N 1, p. 42—47.
Lo Bianco S. Lo pesce abissali esequite
du F. A. Kruppcol jacht «Puritan»
nelle adiacenze de Capri ed in altre
localita del Mediterraneo. — Mitt.
Zool. St. Neapel, 1903, vol. 16,
p. 533—1327.
Lohmander H. Ober die Diplopoden des
Kaukasusgebietes. — Goteborgs
KngL vet. vitt.-samh. handl. Ser. B,
1936, Bd. 5, N 1, S. 1—96.
Loven S. Om Crustaceer i Venern och
Vettern. — Ofv. KngL vet.-akad.
fohrhandel., 1862, bd. 18, s. 285—
314.
Macallum A. B. Ancestral features in
blood plasma of vertebrates. —
J. Biol. Chem., 1907, vol. 17.
Macallum A. B. The palaeochemistry of
the fluids and tissues. — Physiol.
Rev., 1926, vol. 6, N 2, p. 316—
357.
Madsen F. J. Octacnemus ingolfi n. sp.,
an Atlantic representative of the
peculiar Tunicate-family Octacne-
midae. — Vid. medd. Dan. na-
turhist. foren., 1947, bd. 110, s. 31 —
44.
Madsen F. J. Some general remarks on the
distribution of the echinoderm fauna
of the deep-sea: Prepr. of XIV Intern.
Zool. Congr. Copenhagen, 1953.
Madsen F. I. On the zoogeography and
origin of the abyssal fauna in view of
the knowledge of the Porcellanasteri-
dae.—Galathea Rep., 1961, vol. 4,
p. 177—218.
Mangold O. Das Wirbeltierauge in der
Entwicklung und Regeneration. —
Ergeb. Biol., 1931, Bd. 7, S. 193—
403.
Marek U7., Schlilter C. Neue Fische und
Krebse aus der Kreide von West-
falen. — Palaeontographica, 1865,
Bd. 15, S. 269—305.
Markewitsch A. P. Les Crustaces para-
sites des poissons de la mer Caspien-
ne. — Bull. Inst, oceanogr. Monaco,
1933, N 658, p. 1—27.
Marshall N. B. Swimbladder structure
of deep-sea fishes in relation to their
systematics and biology. — Discovery
Rep., 1960, N 31, p. 1 — 122.
Marshall N. B. Fishes of the sunless
sea. — New Sci., 1963, vol. 17, N 321,
p. 80—82.
Marsland D. A. Protoplasmic streaming in
plasma gel structure in relation to
form and movement. — In: The
structure ot protoplasm. Iowa: Coll,
press, 1942.
Marsland D. A. The action of hydrostatic
pressure on cell division. — Ann. N. Y.
Acad. Sci., 1951, vol. 51, art. 8.
Marsland D. A. Protoplasmic contractility
in relation to gel structure: tempera-
ture — pressure experiments on cy-
tokinesis and amoeboid move-
ment. — Intern. Rev. CytoL, 1956,
vol. 5.
Martens E. Ober das Vorkommen mariner
Formen in siissen Wasser iiber-
haupt. — Arch. Naturges., 1857, Jg.
23, Bd. 1, S. 149—204.
Matsumoto K. Two new genera and a new
subgenus of the family Asellidae
of Japan. — Annot. Zool. Jap., 1962,
vol. 35, N 3, p. 162—169.
Matsumoto K. Results of the spelacological
survey in South Korea, 1966. 7. Iso-
pod Crustaceans from subterrancen
waters of South Korea. — Bull. Nat.
Sci. Mus., 1967, vol. 10, N 3, p. 259—
283.
Mclndoo N. F. Biology of the Shawnee
cave spiders. — Biol. Bull., 1910,
vol. 19, p. 303—323.
Meek A. The migrations of fish. L.,
1916. 427 p.
Menzel D. W., Goering J. I. The distri-
bution of organic detritus in -the
ocean. — Limnol. and Oceanogr.,
1966, vol. 11, N 3, p. 333—337.
Menzies R. J. New abyssal tropical
atlantic isopods, with observations on
their biology. — Amer. Mus. Novit.,
1956a, N 1798, p. 1 — 16.
Menzies R. J. New bathial Isopoda from
the Caribbean with observations on
their nutrition. — Breviora Mus.
Comp. Zool., 1956b, N 63, p. 1 —
10.
Menzies R. J. The isopods of abyssal
depths in the Atlantic Ocean. —
Verna Res. Ser., 1962a, N 1. Abyssal
Crustacea, p. 79—206.
Menzies R. J. On the food and feeding
habits of abyssal organisms as
examplified by the Isopoda. — Intern.
Rev. Ges. Hydrobiol., 1962b, Bd. 47,
N 3, S. 339—358.
Menzies R. J. Conditions for the existence
of life on the abyssal sea floor. —
Oceanogr. Mar. Biol. Annu. Rev.,
1965, N 3, p. 195—210.
Menzies R. L, Frankenberg D. Systematics
and distribution of the bathyal-
abyssal genus Mesosignum (Crusta-
cea: Isopoda)Antarct. Res. Ser.,
1968, vol. 11, p. 113—140.
Menzies R. J., Imbrie J. On the antiquity
241
of the deep sea bottom fauna. —
Oikos, 1958, vol. 9, N 2, p. 192—210.
Menzies R. J., Imbrie J., Heezen В. C.
Further consideration regarding the
antiquity of the abyssal fauna with
evidence for a changing abyssal envi-
ronment. — Deep-Sea Res., 1961,
vol. 8, N 2, p. 79—94.
Menzies R. J., Schultz G. A. Antarctic
isopod Crustacea. 2. Families, Haplo-
niscidae, Acanthaspidiidae and Jae-
ropsidae, with diagnoses of new
genera and species. — Antarct. Res.
Ser., 1968, vol. 11, p. 141 — 184.
Michaelsen W. Uber Hohlen-Oligochae-
ten. — Mitt. Hohlen- und Karst-
forsch., 1933, Bd. 1, S. 1 — 19.
Monniot CL, Monniot F. Les Ascidides
de grandes profondeurs recoltees par
le navires oceanographique american
Atlantis. 2. — Bull. Inst, oceanogr.
Monaco, 1968, vol. 67, N 1379,
p. 1—48.
Monod Th. Sur quelques Asellides nou-
veaux des eaux douces de 1’Afrique
du Nord. — Bull. Soc. hist, natur.
Afr. Nord, 1924, t. 15, p. 327—336.
Monod Th. Thermosbaena mirabilis Mo-
nod: Remarques sur la morphologic et
sa position systematique. P., 1927.
50 p. (Faune de colonies francaises).
Monod Th. Un nouveau Stenasellus ouest-
african. — Bull. Inst, franc. Afr.
noire, 1945, t. 7.
Morton F., Gams H. Hohlenpflanzen.
Wien, 1925. 227 S.+10 Taf. (Spe-
laolog. Monogr.; Bd. 5).
Mosely N. H. Deep-sea dredging and life
in the deep-sea. — Nature, 1880,
vol. 21, p. 543—552, 569—572, 591 —
593.
Murray J. A summary of the scientific
results obtained at the sounding,
dredging and trawling of H. M. S.
«Challenger». — Challenger Rep.,
1895, vol. 2. Summary of res., p. 797—
1608.
Nathorst A. G. Ett par glaciala «Pseudo-
relikter». — Bot. notis., 1895, s. 29—
34.
Needham J. On the penetration of marine
organisms into freshwater. — Biol.
Zentr.-BL, 1930, Bd. 50, H. 7, S. 504—
509.
Nicholls A. G. Syncarida in relation to the
interstitial habitat. — Nature, 1946,
vol. 158, N 4026, p. 934-936.
Nierstrasz H. F. Die Isopoden der Si-
boga — Expedition. 4. Isopoda Ge-
nuina. 3. Gnathiidea, Anthuridea,
Valvifera, Asellota, Phreatoicoidea. —
Siboga Exped., 1941, Lfvr. 136,
Monogr. 32d, S. 231—308.
Noble G. K., Pope S. H. The effect
of light on the eyes, pigmentation
and behavior of the cave salamander,
Typhlotrition. — Anat. Rec., 1928,
vol. 41, suppl. 1, p. 21.
Nordeustam A. Marine Isopoda of the
families Serolodae, Idotheidae, Pseu-
didotheidae, Arcturidae, Parasellidae
and Stenetriidae mainly from the
South Atlantic. — Fourth Zool. Res.
Swed. Antarct. Exped., 1901 — 1903,
1933, vol. 3, pt 1, p. 1—284.
Nordenstam A. Marine Isopoda from
Professor Dr. Sixten Bock’s Pacific
Expedition, 1917—1918. — Ark. zool.,
1946, bd. 37A, N 7.
Nordenstam A. A new isopod from the
deep sea. — Rep. Swed. Deep-Sea
Exped., 1947—1948, 1955, vol. 2,
fasc. 2, N 16, p. 203—212.
Ockelman IT A. Marine Lamellibran-
chiata. — Medd. Gr0nland, 1958, bd.
122, N 4, s. 1—256.
Ortmann A. E. The geographical distri-
bution of freshwater decapods and
its bearing upon ancient geo-
graphic. — Proc. Amer. Philos. Soc.,
1902, vol. 41, p. 267—400.
Palmherst H. W Beitrage zum Problem
der Osmoregulation einiger Hydropo-
lypen. — Zool. Jb. Abt. allgem. Zool.
Physiol., 1933, Bd. 53, S. 212—260.
Panikkar N. K. Influence of temperature
on osmotic behaviour of some Crusta-
cea and its bearing on problems
of animal distribution. — Nature,
1940, vol. 146, N 3698, p. 366—367.
Panikkar N. K. Osmoregulation in some
palaeomonid prawns. — J. Mar. Biol.
Assoc. U. K., 1941, vol. 25, N 2,
p. 317—359.
Parenzan P. Pesci delle aque sotterranee
della Somalia. — Boll, pesca piscicolt.
e idrobiol., 1938, vol. 14, N 6, p. 752—
754.
Parker R. H. Speculations on the origin
of the invertebrate faunas of the
lower continental slope. — Deep-Sea
Res., 1961, vol. 8, N 3/4, p. 286—293.
Parsons T. R. Suspended organic matter
in sea water. — In: Progress of
oceanography. Oxford: Pergamon
press, 1963, vol. 1, p. 203—239.
Pantin C. F. A. Adaptation of Gunda ulvae
to changes in salinity. Pt 1—3. —
J. Exp. BioL, 1931, vol. 8, N 1,
p. 63—94.
Pearse A. S. Fauna of the caves of
Jucatan. Wash., 1938. 304 p. (Carne-
gie Inst. Wash. Publ.; N 491).
Pelseneer P. L’origine des animaux d’eau
douce. — Bull. Acad. roy. Belg. Cl.
sci., 1905, N 12, p. 699—740.
242
PeschelO. Die Entwicklungsgeschichte des
stehenden Wasser auf der Erde. —
Ausland, 1875, Bd. 48, N 41, S. 805—
808.
Pesta O. Carcinologische Mitteilungen. —
Arch. Hydrobiol., 1926a, Bd. 16,
S. 605—643.
Pesta O. Decapoda. — In: Biologie der
Tiere Deutschlands. B., 1926b, T. 17,
S. 19—58.
Peters H. Der Einfluss der Salzkonzen-
tration des Aussenmediums auf den
Bau und die Funktion der Excretion-
sorgane dekapoder Crustaceen. —
Ztschr. Morphol. 6kol. Tiere, 1935,
Bd. 30, S. 355—381.
Petit G. Un poisson cavernicole aveugle
des eaux douces de Madagascar:
Typhleotris madagascariensis gen.
et sp. nov. — C. r. Acad, sci., 1933,
D, t. 197, p. 347—348.
Petit G., Bernard W. Sur le comportement
en aquarium du Caecobarbus geertsii
Boulenger. — Bull. Mus. hist, natur.,
1937, vol. 9, N 1, p. 50—53.
Pieh S. Uber die Beziehungen zwischen
Atmung, Osmoregulation und Hydra-
tation der Gewebe bei euryhalinen
Meeresevertebraten. — Zool. Jb. Abt.
allgem. Zool. Physiol. Tiere, 1936,
Bd. 56, S. 129—160.
Рога E. A. Sur le comportement des
crustaces brachioures de la Mer Noire
aux variations de salinite du milieu
ambient. — Ann. Sci. Univ. Jassy,
1939, t. 25, p. 286—319.
Poulsen V. Notes on Hyolithellus Billings,
1871, class Pogonophora lohannson,
1937. — Biol. medd. KgL dan. Vid.
selskab., 1963, bd. 23, N 12, s. 1 — 15.
Racovitza E. G. Essai sur les problemes
biospeleologuiques. — Arch. zool. exp.
et gen. Ser. 4, 1907, t. 6, N 7,
p. 371—488.
Racovitza E. G. Spheromiens (l-ere serie)
et revision des Monolistrini (Isopodes
spheromiens). — Arch. zool. exp. et
gen. Ser. 5, 1910, t. 4, p. 625—758.
Racovitza E. G. Cirolanides: Premiere
serie. — Arch. zool. exp. et gen.
Ser. 5, 1912, t. 10, p. 203—329.
Racovitza E. G. Notes sur les Isopodes.
3.—7. — Arch. zool. exp. et gen.,
1919, t. 58, p. 49—77.
Racovitza E. G. Aselides. premiere serie:
Stenasellus. — Arch. zool. exp. et
gen., 1950, t. 70, p. 1—94.
Radovanovic M. Wormaldia subterranea
(n. sp.) eine neue in der Hohlen
Jugoslaviens aufgefundene Trichopte-
ren-Art. — Zool. Anz., 1932, Bd. 100,
H. 3/4, S. 101 — 108.
Raw F. The Malacostraca: Their origin,
relationships and phylogeny. — Ann.
and Mag. Natur. Hist. Ser. 12, 1955,
vol. 8, p. 431—756.
Reiff E. Isopoden aus dem Lias (Amathe-
enschichten) Schwabiens. — Palaon-
toL Ztschr., 1936, Bd. 18, S. 49—90.
Remane A. Nerillidium mediterraneum n.
sp. und seine tiergeographische
Bedeutung. — Zool. Anz., 1928,
Bd. 77, H. 3/6, S. 57—60.
Remy P. Observations sur les moeurs de
quelques orthopteres cavernicoles. —
Ann. Soc. natur. bot. et zool. Ser. 10,
1931, t. 14, p. 263—274.
Remy P. Description d’Asellus (Proasel-
lus) arnautovici n. sp. du lac
d’Ohrid. — Bull. Soc. hist, natur.
Moselle, 1932, t. 33, p. 73—87.
Remy P. Un Aselle nouveau de Madere. —
Broteria. Ser. cienc. natur., 1936,
t. 5 (32), fasc. 1, p. 17—32.
Remy R. Sur Marifugia cavatica Abs.,
Serpulide des eaux douces souterrai-
nes du Karst Adriatique. — Bull.
Mus. nat. hist, natur., 1937, t. 2, N 9,
p. 66—72.
Remy P. Un Stenasellus en Afrique
Occidental Francaise — S. chappuisi
sp. n. de la cote d’Ivoire. — Arch,
zool. exp. et gen., 1938, t. 79, p. 69—74.
Remy P. Asellotes de Jugoslavie et de
Grece. — Arch. zool. exp. et gen.,
1941, L 82, p. 1—25.
Rensch B. Zoologische Systematik und
Artbildungsproblem. — Verh. Dt.
zool. Ges., 1933, Bd. 35, S. 19—83.
Richardson H. Description of a new isopod
of the genus Eurycope from Marthas
Vineyerd. — Bull. US Nat. Mus.,
1908a, N 34, p. 67—69.
Richardson H. Some new isopods of the
superfamily Aselloidea from the At-
lantic coast of North America. —
Bull. US Nat. Mus., 1908b, N 35,
p. 71—86.
Richardson H. Some new isopods of the
family Gnatheidae from the Atlantic
coast of North America. — Bull.
US Nat. Mus., 1909, N 35, p. 483—488.
Richardson H. Isopod collected in the
Northwest Pacific by the US Bureau
of fisheries steamer «Albatross» in
1906.— Bull. US Nat. Mus., 1910,
N 37, p. 75—129.
Richardson H. Les crustaces isopodes du
Travailleur et du Talisman; formes
nouvelles. — Bull. Mus. nat. hist, na-
tur., 1911, t. 7, p. 518—534.
Ritter IF. E. Octacnemus. — Bull. Mus.
Comp. Zool. Harvard Coll., 1906,
vol. 46, N 13, p. 233—252.
243
Robertson J. D. Some feature of the
calcium metabolism of the shore crab
(Carcinus maenas Pennant). — Proc.
Roy. Soc. London B, 1937, vol. 124,
N 835, p. 162—182.
Roemer A. Sh. The early evolution of
fishes. — Quart. Rev. Biol., 1946,
vol. 21, N 1, p. 33—69.
Roemer A. Sh., Grove В. H. Environment
of early vertebrates. — Amer. Mid-
land Natur., 1935, vol. 16, p. 805—856.
Rogers Ch. G. Text book of comparative
physiology. N. Y.; L.: McGraw-Hill,
1938. 715 p.
Rosa R. Ologenesi: Nuova teoria dell’evo-
luzione e della distributione geo-
graphica del virenti. Firenze, 1918.
Ruffo S. Studio sulla fauna cavernicola
della regione Veronese. — Boll. Inst,
entomol. Bologna, 1938, vol. 10
(1937), p. 70—116.
Sars G. O. Notes on the crustacean
fauna of the Caspian Sea. — В кн.:
В честь Н. М. Книповича. М.: Нар-
комзем, 1927, с. 315—329.
Sars G. О. Description of a remarcable
cave-crustacean, Parabathynella ma-
laya G. O. Sars, sp. nov. — J. Fed.
Malay States Mus., 1929, vol. 14,
p. 339—351.
Scharff K. F. Distribution and origin of
life of America. L., 1911. XVI+ 497 p.
Schellenberg A. Niphargus-Probleme. —
Mitt. Zool. Mus. Berlin, 1933, Bd. 19,
S. 406—429.
Schellenberg A. Der Niphargus der Thii-
ringer Waldes und die Glacialrelik-
tenfrage. — Arch. Hydrobiol., 1935,
Bd. 29, S. 274—281.
Schellenberg A. Die Amphipodengattun-
gen um Crangonyx, ihre Verbreitung
und ihre Arten. — Mitt. Zool. Mus.
Berlin, 1936, Bd. 22, H. 1, S. 31-44.
Schellenberg A. Hohlenamphipoden Spa-
niens und ihre Beziehung zu Nordafri-
ca. — Zool. Anz., 1937a, Bd. 118,
H. 7/8, S. 223—224.
Schellenberg A. Kritische Bemerkungen
zur Systematik der Sijsswassergama-
riden. — Zool., Jb. Abt. Syst., 1937b,
Bd. 69, S. 469—516.
Schellenberg A. Verbreitung und Alter der
Amphipoden-Gattung Pseudoniphar-
gus nebst Verbretung der Gattung
Niphargus. — Zool. Anz., 1939,
Bd. 127, H. 11/12, S. 297—304.
Schellenberg A. Subterrane Amphipoden
Osteuropas, ihre Variabilityt und ihre
verwandtschaftlichen Beziehungen. —
Zool. Jb. Abt. Syst., 1941, Bd. 74,
H. 3, S. 243—268.
Schlieper C. Uber die Entwirkung niederer
Salzkonzentration auf marine Orga-
nismen. — Ztschr. vergl. Physiol.,
1929, Bd. 9, H. 2/3, ’ S. 478—
513.
Schlieper C. Die Osmoregulation wasser-
lebender Tiere. — Biol. Rev., 1930,
vol. 5, p. 309—356.
Schlieper C. Uber die Eindringung mari-
ner Tiere in das Siisswasser. — Biol.
Zentr.-BL, 1931, Bd. 51, H. 8, S. 401 —
412.
Schlieper C. Die Brackwasserfauna und
ihre Lebensbedingungen vom physio-
logischen Standpunkt ausbetrach-
tet. — Verh. Intern. Ver. theor. und
angew. LimnoL, 1933, Bd. 6, S. 113—
146.
Schlieper C. Neuere Ergebnisse und
Probleme aus dem Gebiet der Osmore-
gulation wasserlebenden Tiere. —
Biol. Rev., 1935, vol. 10, N 3,
p. 334—360.
Schlieper C. Die Abhangigkeit der At-
mungsintensitat der Organismen vom
Wassergehalt und dem Kolloidalen
Zustand des Protoplasmas. — Biol.
Zentr.-BL, 1936, Bd. 56, S. 87—93.
Scholles U7. Uber die Mineralregulation
wasserlebender Evertebraten. —
Ztschr. vergl. Physiol., 1933, Bd. 19,
S. 522—554.
Schroeter C. Genetische Pflanzengeo-
graphie. — In: Handwdrterbuch der
Naturwissenschaften. Jena, 1913,
Bd. 4, S. 907 -942.
Schultz G. A. Submarine canyons of
Southern California. Pt. 4. Systema-
tics: Isopoda. — Allan Hancock Pacif.
Exped. Rep., 1966, vol. 27, pt 4,
p. 1—56.
Schwabe E. Uber die Osmoregulation ver-
schiedener Krebse (Malacostra-
cen). — Ztschr. vergl. Physiol., 1933,
Bd. 19, S. 183—236.
Slewing R. Untersuchungen zur Morpholo-
gic der Malacostraca (Crustacea). —
Zool. Jb. Abt. Anat., 1956, Bd. 75, H. 1,
S. 39—176.
Slewing R. Ingolfiella ruffoi nov. spec.,
eine neue Ingolfiellidae aus dem
Grundwasser der Peruanischen Ku-
ste. — Kiel. Meeresforsch., 1958, Bd.
14, S. 97—102.
Silhavy V. Uber die Genitalmorphologie
der Nemastomidae (Arach. Opi-
liones). — Senkenberg. biol., 1966,
Bd. 47, N, 1, S. 67—72.
Simpson G. Tempo and mode in evolution.
N. Y.: Columbia Univ, press, 1944.
237 p.
244
Sollas W J. On the origin of freshwater
fauna: A study in evolution. — Sci.
Trans. Roy. Dublin Soc. Ser. 2,
1883—1887, vol. 3, p. 87—118.
Spandl H. Die Bezeiehungen der Wasser-
tierwelt zur Eiszeit. — In: Die Eiszeit.
Leipzig: K. W. Hiersemann, 1926a,
Bd. 2.
Spandl H. Die Tierwelt der unterirdischen
Gewasser. Wien, 1926b. XII, 235 S.
(SpelaoL Monogr.; Bd. 9).
Sparck R. Origin and distribution of the
deep-sea bottom fauna. — Union In-
tern. Sci. Biol. Ser. B, 1954, N 16,
p. 89—90.
Stammer H. J. Zur Kenntnis der Verbrei-
tung und Systematik der Gattung
Asellus, insbesondere der mitteleu-
ropaischen Arten (Isopoda). —Zool.
Anz., 1932a, Bd. 99, H. 5/6, S. 113—
131.
Stammer H. J. Die Fauna des Timavo:
Ein Beitrag zur Kenntnis der Hohlen-
gewasser des Siiss- und Brackwas-
sers im Karst. — Zool. Jb. Abt. Syst.,
1932b, Bd. 63, H. 5/6, S. 521—656.
Stammer H. J. Desmoscolex aquedulcis n.
sp., der erste siisswasserbewohnende
Desmoscolecide aus einer sloweni-
schen Hohle. — Zool. Anz., 1935a,
Bd. 109, H. 11/12, S. 311—318.
Stammer H. J. Zwei neue Protozoen
Spelaeophrya troglocaridis n. sp. von
den Antennen der Hohlengarnele
Troglocaris schmidti Dorm, und Lage-
nophrys monolistrae n. sp. von den
Kiemen (Pleopoden) der Hohlenassel-
gattung Monolistra. — Arch. Proti-
stenk., 1935b, Bd. 84, S. 518—527.
Stammer H. J. Ein neue Hohlenschizopode
Troglomysis vjeternicensis n. gen.,
n. sp. — Zool. Jb. Abt. Syst., 1936a,
Bd. 68, S. 53—104.
Stammer H. J. Alter und Herkunft der
Tierwelt der Hdhiengewasser Euro-
pas. — In: С. г. XII Congr. intern,
zool. Lisbonne, 1936b, p. 1051 —1056.
Stammer H. J. Eine neue Hohlen-
wasserasell aus Spanien, Stenasellus
buchneri und die Verbreitung der
Gattung Stenasellus. — Zool. Anz.,
1936c, Bd. 114, H. 5/6, S. 137—141.
Stoller J. Die Pflanzenwelt des Quar-
ters. — In: Potonie-Gothan. Lehrbuch
der Palaobotanik. 2. AufL B.: Born-
traeger, 1921, S. 1-537.
Stralen V van. Contribution a I’etude des
Isopodes Meso- et Cenozoiques. —
Mem. Acad. Roy. Belg. Cl. sci., 1928a,
t. 9, fasc. 5, p. 1—66.
Stralen V van. Astacus edwardsi Munier-
Chalmas M. S., Astacidae du Paleo-
cene de Sezann (Champagne).—
Bull. Soc. geol. France. Ser. 4, 1928b,
t. 28, fasc. 1/2, p. 3—6.
Stralen V van. On a fossil freshwater
crayfish from eastern Mongolia. —
Bull. Geol. Soc. China, 1928c, vol. 7,
p. 133—138.
Straelen V van. L’anciennete et la regres-
sion du genre Homarus. — Mem.
Mus. Roy. Belg. Mel. Paul. Pelsenner.
Ser. 1-2, 1936, N 3, p. 469—479.
Strouhal H. Einige bemerkenswerte Vor-
komnisse von Wirbellosen, besonders
Isopoden in der Ostmark. — Festschr.
Embrik. Strand., 1939a, Bd. 5.
Strouhal H.^ Die in den Hohlen von
Warmbad Villach, Karnten, fest-
gestellte Tiere. — Folia zool. hydro-
biol., 1939b, vol. 9, N 2, p. 311—313.
Szarski К. О szczatku oka u kielza
studziennego (Niphargus puteanus
Koch).— Kosmos. Ser. A, 1935,
roc. 60, zes. 2, s. 95—111.
Tattersail W. M. The marine fauna of the
coast of Ireland. Pt 5. Isopoda. —
Fish. Ireland. Sci. Invest. (1904),
1906, vol. 2, p. 1—90+2 pls.
Thiele J. Handbuch der systematischen
Weichtierkunde, 1929—19*35. Jena:
G. Fischer. Bd. 1. 1929—1931; Bd 2.
1934—1935, S. 1 — 1154.
Thienemann A. Die Binnengewasser
Mitteleuropas: Ein limnologische Ein-
fuhrung. Stuttgart, 1926. Bd. 1. 255 S.
Thorson G. Reproductive and larval
ecology of marine bottom invertebra-
tes. — Biol. Rev., 1950, vol. 25, p. 1 —
45.
Trueman A. E. Supposed commensalism
of carboniferous Spirorbids on certain
nonmarine Lamellibranchs. — Geol.
Mag., 1942, vol. 77, N 6, p. 312.
Trusheim F. Triopsidae (Crustacea,
Phyllopoda) aus dem Keuper Fran-
kens. — Palaontol. Ztschr., 1938,
Bd. 19, N 3/4, S. 198—216.
Vandel A. Contribution a la genetique
des Isopodes du genre Trichoniscus.
1. Les mutationsalba et pallida
de Trichoniscus (Spiloniscus) elisa-
bethae Herold et I’origine des formes
cavernicoles. — Bull. biol. France
et Belg., 1938, t. 72, p. 121 — 146.
Vandel A. Biospeleologie: La biologie des
animaux cavernicoles. P.: Gauthier-
Villars, 1964. XVIII, 619 p.
Vandel A. Les Trichoniscidae cavernicoles
(Isopoda terrestria: Crustacea) de
1’Amerique du Nord. — Ann. speleoL,
1965, t. 20, N 3, p. 347—389.
Vanhoffen E. Die Isopoden der Deutschen
Sudpolar-Expedition, 1901 — 1903. —
245
Dt. Siidpolar-Exped., 1914, Bd. 15,
H. 7, S. 449—598.
Van, Name IF G. The American land-
and fresh-water isopod Crustacea. —
Bull. Amer. Mus. Natur. Hist., 1936,
vol. 71, p. 1—535.
Vaughan T. IF Corals and the formation
of coral reefs. — Smithsonian Ann.
Rep., 1917, p. 189—238.
Verhoeff K. IF. Eine cavernicole Symphio-
gnathen-Gattung aus Georgien,
Transkaucasien. — Mitt. Hohlen- und
Karstforsch., 1930, H. 2, S. 33—39.
Verrier M. L. Observations sur le compor-
tement d’un poisson cavernicole
(Typhlichthys osborni Eigm). — Bull.
Mus. nat. hist, natur., 1929, t. 1,
N 1, p. 82—84.
Vire A. La faune souterraine de France.
P.: Baillere, 1900. 160 p.
Vogt IF. Uber die Antennendruse von
Mysis relicta. — Zool. Jb. Abt. Anat.
Ontogen., 1933, Bd. 56, S. 373—386.
Wallace A. R. On the tendency of varieties
to depart indefinitely from the original
type. — J. and Proc. Linn. Soc.
London, 1858, vol. 3, N 9, p. 45—62.
Walter A. Transkaspische Binnencrusta-
ceen. 2. Malacostraca. — Zool. Jb.
Abt. Syst., 1889, Bd. 4, S. 1110—1123.
Walters V. A contribution to the biology
of the Giganturidae, with description
of a new genus and species. —
Bull. Mus. Comp. Zool., 1961, vol. 125,
N 10, p. 297—319.
Walther J. Allgemeine Palaeontologie. 3. T.
Die geologische Umwelt der Fossilien.
B., 1927. 809 S.
Wangerin IF Beitrage zur Frage der
pflanzengeographischen Relikte. —
Abh. Naturforsch. Ges. Danzig, 1924,
Bd. 29, H. 1.
Vardlaw H. S. H. Some aspects of the
adaptation of living organisms to their
environment. — Smithsonian Annu.
Rep., 1931, p. 389—411.
Wesenberg-Lund C. Grundziige der Biolo-
gie und Geographie des Siisswasser-
planktons, nebst Bemerkungen uber
Hauptprobleme zukiinftige limnologi-
scher Forschungen. — Intern. Rev.
ges. Hydrobiol. und Hydrogr., 1910,
Bd. 3, Suppl., S. 1—44.
Wheeler E. H. Copepod detritus in the
deep-sea. — Limnol. and Oceanogr.,
1967, vol. 12, N 4, p. 697—701.
Wickstead /. H. Food and feeding in pela-
gic copepods. — Proc. Zool. Soc.
London, 1962, vol. 139, pt 4, p. 545—
556.
Wiehle E. Uber Nesticus borutzkyi Reimo-
ser (Arach. Araneae). — Senkenberg.
bioL, 1963, Bd. 44, N 5, S. 436—439.
Wieser IF. Problems of species formation
in the benthic microfauna of the deep
sea. — In: Perspect. in marine bio-
logy. Berkley; Los Angeles: Univ. Cal.
press, 1960, p. 513—517.
Winkler A. Eine neue blinder Trechus aus
der Krim. — Coleopter. Rdsch., 1912,
Bd. 10, S. 134—135.
Wolf B. Animaliumcavemarum catalogus.
Ps 1—7. B., 1934—1936. 544 S.
Wolff T. Isopoda from depths exceeding
6000 metres. — Galathea Rep., 1956,
vol. 2, p. 85—157.
Wolff T. The hadal community, an intro-
duction. — Deep-Sea Res., 1960, vol. 6,
N 2, p. 95—124.
Wolff T. The deepest recorded fishes. —
Nature, 1961a, vol. 190, N 4772, p. 283.
Wolff T. Animal life from a single
abyssal trawling. — Galathea Rep.,
1961b, vol. 5, p. 129—1624-4 pls.
Wolff T. The systematics and biology
of bathyal and abyssal Isopoda
Asellota. — Galathea Rep., 1962,
vol. 6, p. 1-3204-19 pls.
Wolsky A. Uber eine blinden Hohlengam-
mariden, Niphargus aggtelekiensis
Dudich, mit Bemerkungen fiber die
Riickbildund des Gammaridenauges.
Verh. Intern. Ver. theor. und angew.
Limnol., 1935, Bd. 7, S. 449—463.
Worthington E. IF. Geographical differen-
tiation in freshwaters with special
reference to fish. — In: The new syste-
matics. L.—Oxford: Clarendon press,
1940, p. 287—302.
Wright S. Evolution in Mendelian popula-
tions. — Genetics, 1931, vol. 16,
p. 97—159.
Wright S. The role of mutation, inbreeding,
crossbreeding and selection in evolu-
tion. — Proc. 6th Intern. Congr.
Genet., 1932, vol. 1, p. 356—366.
Wright S. The statistical consequences
of Mendelian heredity in relation
to speciation. — In: The new systema-
tics. London; Oxford: Clarendon
press, 1940, p. 161 — 183.
Wundsch H. Weitere Beitrage zur Frage
der Siisswasserform von Corophium
curvispinum G. O. Sars. — S.-Ber.
Ges. naturforsch. Freunde Berlin,
1915.
Wilst G. Schichtungen und Tiefenzirku-
lation des Pacifischen Ozean. —
Veroff. Inst. Meeresk. Hamburg, 1929,
Bd. A, H. 20.
Zur Strassen O. Uber die Gattung
Arcturus und die Arcturiden der
Deutschen Tiefsee-Expedition. — Zool.
Anz., 1902, Bd. 25, S. 682—689.
246
СОДЕРЖАНИЕ
Предисловие 3
Основные направления научной деятельности Якова Авадьевича Бирштейна
(7 мая 1911—8 июля 1970) 5
Список печатных работ Я. А. Бирштейна 12
ФАУНА ПРЕСНЫХ ВОД
Пресноводные Decapoda СССР и их географическое распространение 22
Пресноводные ослики (Asellota). Географическое распространение и родствен-
ные отношения 30
К вопросу о происхождении морских ракообразных в реках Понто-Каспийского
бассейна 45
ПЕЩЕРНАЯ ФАУНА
Жизнь в пещерах. Эколого-систематический очерк 56
Адаптация и эволюция пещерных животных 7g
Новые данные о происхождении фреатической подземной фауны 92
Зоогеографическая характеристика подземной фауны Советского Союза 97
ГЛУБОКОВОДНАЯ ФАУНА ОКЕАНА
Исследования донной фауны Курило-Камчатской впадины Ю4
Вертикальная зональность планктона Курило-Камчатской впадины . Ю6
Глубоководные Malacostraca северо-западной части Тихого океана, их распре-
деление и взаимоотношения 109
Глубоководные равноногие ракообразные (Crustacea, Isopoda) северо-западной
части Тихого океана 112
Дополнения к фауне равноногих ракообразных (Crustacea, Isopoda)
Курило-Камчатского желоба 154
Роль трофического фактора в таксономическом обособлении морской глубоко-
водной фауны 160
ФИЛОГЕНИЯ И ЭВОЛЮЦИЯ
Понятие «реликт» в биологии 17g
Подкласс Malacostraca. Высшие ракообразные 194
Некоторые проблемы происхождения и эволюции пресноводной фауны 198
Некоторые вопросы эволюции глубоководной фауны 219
Литература 226
247
Яков Авадьевич Бирштейн
ГЕНЕЗИС
ПРЕСНОВОДНОЙ,
ПЕЩЕРНОЙ
И ГЛУБОКОВОДНОЙ
ФАУН
Утверждено к печати
Московским обществом испытателей природы
Редактор издательства
А.М. Гидалевич
Художник
АТ. Кобрин
Художественный редактор
Н.А. Фильчагина
Технический редактор
Н. А. Торгашова
Корректор
В.Л. Крылова
ИБ № 28865
Подписано к печати 11.07.85. Т - 14828
Формат 60 х 90 1/16
Бумага для глубокой печати
Гарнитура Литературная. (Фотонабор)
Печать офсетная Усл. печл. 15,5
Усл.кр.-отт. 15,6. Уч.-издл. 20,9
Тираж 800 экз. Тип. зак. 333
Цена 3 р. 20 к.
Ордена Трудового Красного Знамени.
издательство ’’Наука”
117864 ГСП-7. Москва В-485,
Профсоюзная ул., д. 90
Ордена Трудового Красного Знамени
1-я типография издательства ”Наука”
199034, Ленинград В-34, 9-я линия, 12