/
































































Автор: Заренков Н.А.
Теги: материальные основы жизни биохимия молекулярная биология биофизика систематика теория систематики
Год: 1976




Текст
I i
HA.3APEHKOB
ЛЕКЦИИ
ПО ТЕОРИИ
СИСТЕМАТИКИ
Н. А. ЗАРЕНКОВ
ЛЕКЦИИ
ПО ТЕОРИИ СИСТЕМАТИКИ
ИЗДАТЕЛЬСТВО
МОСКОВСКОГО УНИВЕРСИТЕТА
1976
УДК 577.4
Печатается по постановлению Редакционно-издательского совета Московского университета
В книге рассматриваются таксономический континуум, отношение эмпирической выборки ’видов и их признаков с более обширной биологически возможной их совокупностью, а также структура таксона. Автор считает, что таксономический континуум принципиально невозможен и что структура таксона унаследована от вида-предка, а эмпирическая выборка фактов систематики всегда случайна по отношению к их совокупности. Это подтверждает допустимость разграничения таксонов, реальность их как биологических организаций и обосновывает вероятностную природу таксономического знания. В работе показана возможность предсказания признаков еще неизвестных и потенциально осуществимых в ходе дальнейшей эволюции таксонов на основе теории классической систематики.
Рецензенты:
доктор биол. наук проф. | В. Г. Гентнер | доктор биол. наук проф. С. Э. Шноль
(3) Издательство Московского университета, 1976 г.
^21008—152 077(02)—76
159— 75
СОДЕРЖАНИЕ
Предисловие........................................................ 4
Лекция 1, в которой автор рассказывает об основных понятиях систематики................................................... 7
Лекция 2, в которой автор отвергает некоторые упреки в адрес классической систематики и в свою очередь предъявляет ей новое обвинение....................................................27
Лекция 3, в которой отстаиваются высокие достоинства морфологии и утверждается, что фенотип есть средство осуществления эволюции......................................................... 39
Лекция 4, и которой доказывается, что эмпирическое множество таксонов всегда есть случайная выборка из вообще возможных 56
Лекция 5, в которой автор обсуждает реальность границ таксонов и пытается показать, что они реальны только при условии неосуществимости формального множества событий.....................64
Лекция 6, в которой предлагаются доказательства невозможности континуума таксонов, или, что то же, существование взаимодействий между признаками.................................... . 80
Лекция 7, в которой автор предлагает особую форму ведения протоколов опытов, поставленных природой, для определения взаимодействий между признаками, и называет такие протоколы 4-клеточными решетками в выровненных условиях взаимодействия . 88
Лекция 8, в которой речь пойдет о доказательствах и предсказаниях в систематике ...........................................95
Лекция 9, в которой ранее полученные результаты использованы для рассмотрения некоторых спорных вопросов и служат отправной тонкой для гипотез......................................109
Лекция 10, в которой автор участвует в спорах между систематиками ..................;...................................122
Литература........................................................133
ПРЕДИСЛОВИЕ
Предлагаемая читателю книга представляет собой несколько расширенный курс лекций, читаемых мной на биологическом факультете Московского университета. В лекциях я не стремлюсь к равномерному охвату обширного круга вопросов теории систематики и обращаюсь лишь к тем, которые, как мне кажется, представляют наибольший интерес и вместе с тем недостаточно изучены.
Отношение эмпирической выборки фактов (видов, признаков) к их биологически возможной генеральной совокупности представляется наиважнейшим вопросом. Между тем он был мной увиден едва ли не последним в цепи догадок и доказательств. Решение его, насколько оно мне удалось, получеж> как частный результат и было осознано постепенно.
Вторая по значимости, мне кажется, проблема таксономического континуума, которой не уделяется должного внимания в литературе потому, что многим она кажется несуществующей. Возможно, это происходит прежде всего вследствие смешивания организменной и таксономической организаций. Часто полагают, что очевидная взаимная обусловленность признаков особи в онтогенезе, целостность индивидуума, означает также и взаимную обусловленность признаков видов в эволюции. Отсюда выводится целостность таксонов, прерывистость их множества, невозможность некоторых сочетаний признаков, неосуществимость таксономического континуума. Разбору такого, ошибочного на мой взгляд, хода рассуждений-посвящена почти целая лекция. Она следует после доказательства случайного характера эмпирического множества видов. Таким образом, порядок изложения не отражает действительно имевшей место последовательности умозаключений. Что же касается континуума таксонов, то он, как мне кажется, действительно невозможен, однако по совсем иным причинам, и естественный отбор — лишь одна из них.
Основные усилия были направлены на доказательство этих положений. Их убедительность определяет достоверность конечных выводов, в основном оправдывающих классическую систематику. И наоборот, численные решения задач отнюдь небыли, главной целью. В связи с этим следует подчеркнуть^
4
что решающими в доказательствах я считаю биологические факты, биологические соображения, биологическую аргументацию. Что же касается математических выкладок, то они служат исключительно инструментом, возможности которого, однако, не следует недооценивать. К примеру, количественная оценка границ таксонов позволит не только ликвидировать повод к незаслуженной критике классической систематики со стороны представителей сходственной систематики и «систематики генотипов», но позволит более определенно поставить новые задачи и решить уже имеющиеся. Думаю, что внедрение математических методов в систематику само по себе не шаг вперед. Прогресс систематики связан с математическими методами постольку, поскольку они опираются на разумную биологическую теорию.
Термины «систематика» и «таксономия» в книге использованы как синонимы, но предпочтение отдано первому, ввиду его большей распространенности и принципу приоритета, общепризнанному среди систематиков. Отделение теории и практики классификации (таксономии) от науки о разнообразии видов (систематики) излишне. Такое положение возникло, вероятно, вследствие того, что разные математические методы, призванные решить задачу классификации видов, приобрели большой удельный вес в литературе и породили иллюзию разрешения давних трудностей без привлечения теории эволюции. Поэтому таксономия — это самостоятельная научная дисциплина, имеющая собственный объект, представленный множеством видов, й собственную теорию в лице математических формул. В таком случае за систематикой осталось бы истолкование результатов исследований таксономис-тов в духе эволюционного учения и филогении.
Такой раскол неоправдан прежде всего потому, что математические формулы отнюдь не заменяют теории: они ничего не объясняют, но только констатируют. Стало быть, таксономия не имеет специфической теории и метода и не является самостоятельной научной дисциплиной. Что же касается ее объекта, то он не специфичен для таксономии.
Далее следует иметь в виду, что в систематике давно и естественным образом обособилась практическая систематика, о которой дают представление кодексы н правила номенклатуры, и теория систематики. Последняя должна ответить на вопрос о том, что такое таксон и какова природа таксономического знания, установить место систематики в биологической науке. Наконец, журнал «Systematic Zoology», в котором публикуются статьи по систематике и таксономии (так во всяком случае, если исходить из заглавий статей), уже одним своим названием демонстрирует необходимость термина, которым обозначалось бы все относящееся к изучению органического мира как множества видов. Что же касается рус
5
ского языка, то равноправие двух терминов тем более необходимо, что такие выражения, как «систематическая работа», «систематические замечания», двусмысленны и нуждаются в исправлении на «таксономическая работа», «таксономические замечания».
Считаю своим приятным долгом поблагодарить за помощь и ценные советы В. В. Алпатова, В. Г. Гептнера, Ю. С. Кур-шакову, С. Э. Шноля и М. У. Осипову — за моральную поддержку.
Своим интересом к систематике я обязан моему учителю профессору Московского университета Якову Авадьевичу Бирштейну, систематику высокой биологической культуры. Вкус к количественному подходу во мне воспитал академик Лев Александрович Зенкевич, фактический глава московской школы зоологов в последние десятилетия. Их светлой памяти я посвящаю свой труд.
Лекция I,
В КОТОРОЙ АВТОР РАССКАЗЫВАЕТ ОБ ОСНОВНЫХ ПОНЯТИЯХ СИСТЕМАТИКИ
Систематика изучает множество таксонов. Таксоны суть подмножества генерального множества видов различной мощности, они включают от одного до очень многих видов. Подмножество всех животных есть таксон «царство Животные». Биологический вид с точки зрения систематика — элементарный таксон, единичное подмножество, включающее лишь один вид. Подвиды как зарождающиеся новые виды суть низшие таксоны, пограничные объекты систематики.
Реальность надвидовых таксонов — вопрос первостепенной важности. Ответ на него призвана дать теория систематики. Что же касается видов, то этот вопрос в основном выходит за рамки собственно систематики и изучается эйдологией *. Принимая множество видов как элементарных таксонов в качестве факта, требующего объяснения, систематик обычно не задается вопросом о том, почему вне видов нет жизни. Таким образом, задача автора существенно облегчается.
Положительный ответ на вопрос о реальности надвидовых таксонов во многом предопределяет научный характер таксономического знания. Разногласия по этому поводу носят непримиримый характер. Поводом для них служит специфика таксонов как особой формы биологической организации. Она может быть установлена при сопоставлении таксонов с другими объектами.
В последовательности: особь (индивидуум) — популяция (множество особей одного вида.) — вид (множество популяций особей) — биоценоз (множество популяций особей разных видов) реальность каждой биологической организации обычно не вызывает сомнений, потому что вполне наглядны материальные свидетельства их организованности.
Реальность особи как биологической организации проявляется в наличии онтогенеза, в согласованном развитии признаков (относительный рост), явлении регенерации, в соответ-
1 Эйдос (гр. ei6og то) — вид, рид, «рт, тип.
7
ствиях между органами и тканями, наконец, в явлениях плейотропного действия генов. Непрерывность особи во времени, ее пространственная обособленность, структурированность обычно вполне очевидны.
В популяции особей одного вида также существуют наблюдаемые в природе и опыте вполне реальные отношения, например такие, как у морских котиков. Свидетельства этих отношений доступны наблюдению. Что может быть реальнее ран, полученных секачами в борьбе за. обладание самками? Другой пример — это скопление червей трубочников Tubifex, которые образуют клубок полусферической формы, когда находятся в сосуде с водой; возможно, для создания более сильного тока воды волнообразными изгибаниями тела. Более тесные и вполне наблюдаемые отношения существуют между особями в колониях общественных насекомых (например, у пчел). Они выражаются в полиморфизме, в различном строении маток, рабочих пчел, трутней, особенностях их поведения. Крайний пример — колонии сифонофор, морских животных, обитающих в толще воды. Полиморфизм в таких колониях настолько резок, что отдельные особи совершенно не способны к самостоятельному существованию. Популяцию характеризует пространственное единство и непрерывность во времени, которая достигается посредством размножения, переживания покоящихся стадий (цист, спор) в неактивном состоянии.
Все, что известно о виде 'как организации популяций, — закономерное распределение популяций в пределах видового ареала, колебания численности особей во времени, миграции между популяциями, географическая межпопуляционная изменчивость (часто достаточная для выделения самостоятельных подвидов), — все это обеспечивает существование вида в течение геологически длительного времени и составляет необходимые предпосылки для его эволюции. Единство множества популяций в пределах видового ареала и в то же время их относительная разобщенность характеризуют вид как организацию популяций в пространстве. Непрерывность вида во времени достигается посредством переживания популяций, их способностью восстанавливаться после катастроф благодаря мигрантам из уцелевших популяций.
Биологический вид в эйдологии есть выражение прерывистости органического мира, такая организация особей, которая сделала возможной эволюцию. Эти организации отличаются одна от другой, и в этом смысле существенно различные виды, возможно, эволюционируют по-разному. Например, у некоторых простейших половой процесс отсутствует, размножение осуществляется посредством митотического деления. Поэтому здесь не бывает такой перетасовки наследственных свойств, которая происходит при копуляции гамет у раз-
8
нополых животных. Агамные и разнополые виды, ботанический и зоологический виды, виды у микроорганизмов и других низших форм жизни — таковы объекты эйдологии. Эйдологически различные виды, возможно, имеют неодинаковое значение для построения эволюционного древа (рис. 1). В эйдо-
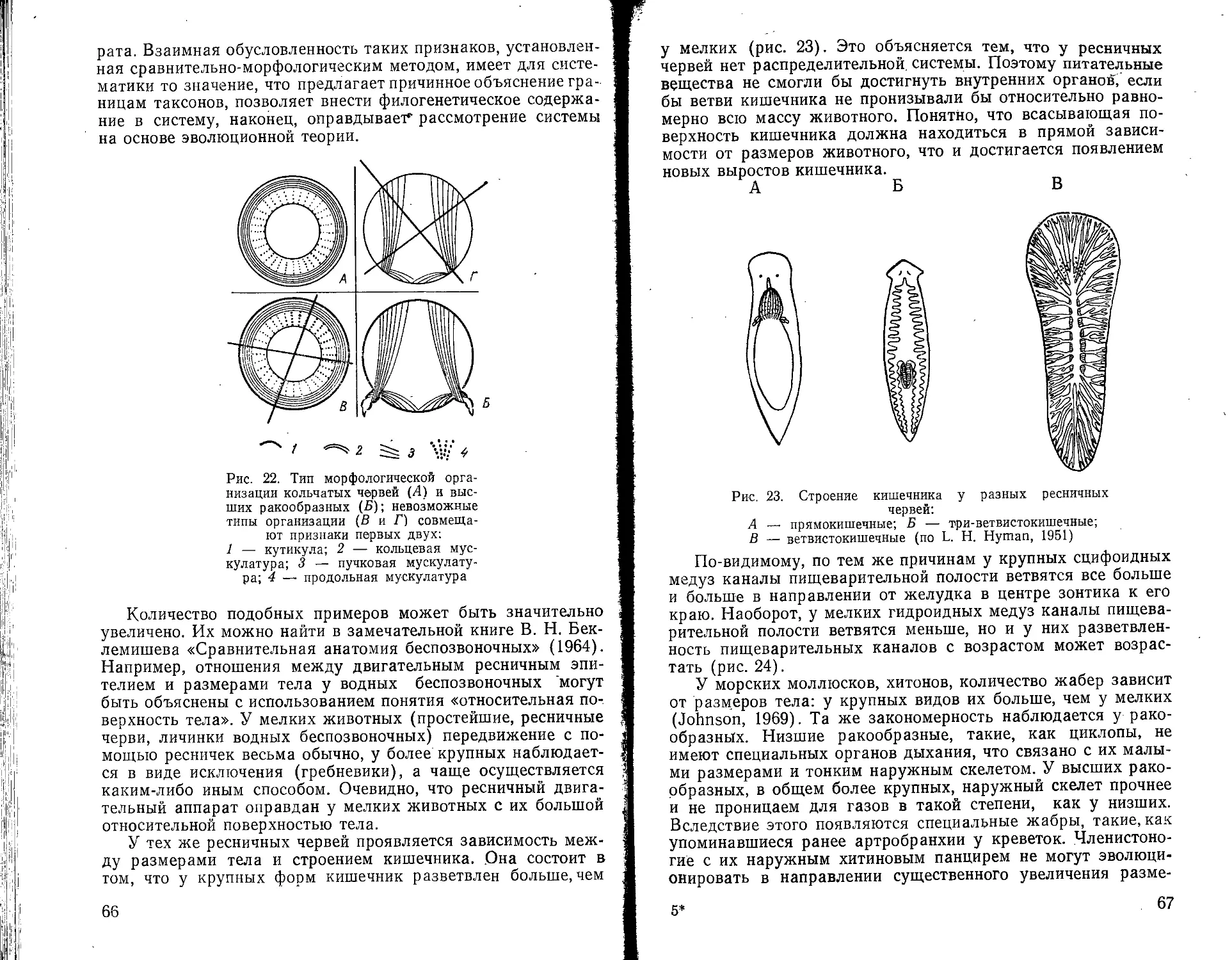
Рис. 1. Эйдологически различные виды растений на филогенетическом древе: жирная линия — обычные диплоидные половые перекрестноопыляемые виды; прерывистая линия — полиплоидные апомиктические и внутриопыляе-мые виды; точки —г гибридные виды (Wagner, 1969)
логии не имеют существенного значения различия, например, между разнополыми -видами; такие виды там равноценны. Наоборот, в систематике различия между эйдологически равноценными видами рассматриваются как существенные. Систематиков занимают не столько «внутренние» общебиологические свойства видов, сколько те их особенности, которые позволяют их различать, распределять по таксонам независимо от эйдологических различий. И. М. Завадский (1968) вполне справедливо отмечает незначительность вклада систематиков в изучение проблемы биологического вида.
Не следовало бы, однако, полагать, что эйдологические различия между видами всецело находятся в сфере интересов популяционной генетики и обусловлены наличием-отсутствием разных полов и способами размножения. Феномен биологического вида имеет исключительное значение для всей биологии и допускает различные подходы к своему изучению (Мошковский, 1969). Например, эйдологически неравноценными можно считать паразитов и их хозяев. Различия между ними очевидны: у паразитов две среды обитания — внешняя
9
среда и хозяин. Сопряженная эволюция паразита и хозяина — явление несколько иного плана, нежели эволюция хищников и их жертв.
Другая биологическая организация, биоценоз, предполагает единство условий обитания и непосредственные контакты между его компонентами во времени и пространстве. Реальность такой организации также не вызывает сомнений. В таксоне как будто бы ничего подобного нет. Обычно к одному таксону принадлежат виды, обитающие в разных уголках Земли, вымершие и ныне живущие. Они могут быть отделены друг от друга громадными расстояниями и геологически длительными разрывами во времени. Их ареалы подвержены крупным изменениям вследствие колебаний климата, дрейфа континентов, морских трансгрессий, межвидовой конкуренции. Поэтому между видами в таксоне непосредственные контакты часто или совсем невозможны, либо воспроизводимы только в опыте; во всяком случае эта особенность не является основанием для объединения видов в таксоне.
Американский речной рак Cambarus limosus Hagen встретился со своими европейскими родичами лишь .в 1890 г., когда после перерыва в 30 миллионов лет (с миоцена) был привезен в Европу. Между тем эти виды принадлежат к одному таксону. Многие морские животные, распространенные в северной части Тихого и Атлантического океанов, не обитают в Северном Ледовитом океане после того, как установился нынешний ледовый режим. Тем не менее они входят в состав таксонов таких, как роды.
Это обстоятельство, по-видимому, послужило поводом к сомнениям в таксонах как реально существующих биологических организациях. Действительно, биологические организации нельзя считать реальностью лишь потому, что они могут быть измерены, взвешены (как особи), подсчитаны (как популяции); это скорее реальность физических объектов, нежели биологических. Точно так же таксон не может считаться реальностью только лишь потому, что биологически реальны составляющие его популяции особей видов. Здесь важнее другое: реальность таксона как биологической организации, как системы взаимоотношений между его элементами. Совершенно необходимо, чтобы относительно организованности его элементов имелись вполне доступные для наблюдения материальные свидетельства.
Отношения между видами в таксоне реальны потому, что вполне реальны и наблюдаемы проявления таких отношений, таксономические признаки. Как, биологическая организация таксон принципиально не отличается от других биологических организаций; доказательство этого положения составляет задачу настоящих лекций. Однако объединение видов в одном таксоне на основе общих признаков — это еще полдела. Не
10
обходимо понять причину этих сходств и различий. Приведем аналогию.
. Известно, что из совершенно равноценных яиц, отложенных пчелиной маткой, развиваются либо матки, либо рабочие пчелы — в зависимости от питания личинок. Можно регистрировать различия между такими пчелами и на этом основывать представление о пчелиной семье как биологической организации популяции. Однако подлинное понимание (не только регистрация) сущности этого объекта приходит только после того, как установлены причины, обусловливающие различия между пчелами, — неодинаковая пища личинок. Точно так же и в систематике сходство между видами становится понятным только после установления причин, его обусловливающих, — общности происхождения. Общие для видов таксона признаки обусловлены общностью, происхождения; различия же свидетельствуют о расхождении видов, их уникальной эволюционной судьбе. Впрочем, такое толкование причин сходства — не единственное, существуют и иные точки зрения. Если все-таки согласиться с ним, то ясно, что кровное родство — достаточное основание для признания реальности таксонов как особого вида биологической организации. Несмотря на то обстоятельство, что между видами одного таксона непосредственные контакты в настоящем необязательны, они безусловно имели место в прошлом через нисходящую цепочку предков-потомков. Реальность филогенетических отношений и, следовательно, реальность таксонов ничуть не уменьшаются от того, что между видами в таксоне сегодня может не быть материальных контактов. Особенность таксона как биологической организации состоит в том, что его непрерывность во времени обеспечивается переживанием, видообразованием — иными, чем у особи и в популяции, эволюционными механизмами. Вследствие медленности процесса эволюции эти механизмы почти недоступны непосредственному наблюдению.
Существует еще одна причина, по которой в реальности таксонов сомневаются чаще, чем, скажем, популяций или биоценозов. Обе последние характеризуются определенной структурой, тогда как таксон как будто бы лишен ее. Например,, в популяции наблюдается определенное соотношение возрастных групп, полов; наконец, имеется кривая изменчивости, которая выражает результаты генетических процессов. Точно так же и в биоценозе наблюдается определенное соотношение организмов с разными потребностями в пище, различным характером питания и отношением к факторам внешней среды. Множество видов таксона как будто бы лишено какой-либо структуры. Известно, например, что между таксономически близкими видами рыб пищевая конкуренция острее, чем между более отдаленными родичами (Шорыгин, 1952). Однако
такие отношения не являются таксономическими. Острота пищевой конкуренции зависит от численности конкурентов, запасов пищи, наличия других конкурентов, совпадения кормовых пастбищ и т. д. Что же касается сходства их признаков как показателя родства, то оно не зависит от указанных факторов. То, что могло бы показаться проявлением структуры таксона (высокая пищевая конкуренция), в действительности не является таксономическим понятием.
Итак, единство таксона проявляется прежде всего в наличии у всех его видов общих таксономических признаков. В соответствии с историческим подходом к природе таксона, и в согласии с установившейся практикой таксономическим признаком следует называть любую особенность, общую всем особям вида (или виду в целом, например половой диморфизм), подверженную межтаксонной изменчивости, действию межвидового 'отбора и закрепленную наследственно. Разнообразие подвидов, клинальная.изменчивость и всякая другая изменчивость есть таксономический видовой признак. Под это определение не подходят общебиологические свойства, такие, например, как механизм считывания генетического кода с ДНК. Этот признак настолько универсален, что, наверное, не подвержен межтаксонной изменчивости. Он принадлежит особям всех видов и, очевидно, относится к организменному уровню биологической организации. Кажется, таких признаков совсем немного.
Под определение таксономического видового признака подходит, например, плодовитость вида, но не его численность и прочие свойства популяции, которые раскрываются при взаимодействии с другими видами и с внешней средой в биоценозах. Численность вида в природе связана с плодовитостью и такими явлениями, как выживание молоди, положение в пищевых цепях, размах изменчивости, темпы эволюции, размеры ареала. Обычно все это неизвестно систематикам, когда они изучают множество видов, и еще не было случая, чтобы их построения были пересмотрены в связи с установлением численности особей видов. Такие сведения интересны систематику a posteriori, но не содержатся в его построениях в явном виде даже в тех редких случаях, когда известны заранее; они не свидетельствуют о родственных отношениях, о происхождении. Наконец, не являются таксономическими отличительные признаки особей, создающие неповторимый облик каждого отдельного индивидуума. Впрочем, как будет показано впоследствии, противопоставление индивидуальных признаков таксономическим не всегда целесообразно.
Области применения разных признаков существенно различны. Организменные признаки всеобщи, они характеризуют собой то, что мы называем живым организмом. Индивидуальные признаки составляют уникальное достояние каждой от-.
12
дельной неповторимой особи. Таксономические признаки в этом смысле занимают промежуточное положение, будучи применимыми в области значительно более обширной, чем индивидуальные, но более узкой, чем организменные.
Таксоны различаются рангом, который возрастает в следующем порядке: вид, род, семейство, отряд, класс, тип, царство. Зоологи и ботаники пользуются несколько различными терминами для обозначения рангов. Число рангов устанавливается в общем пропорционально числу видов изучаемого таксона: чем больше видов, тем пространнее система наименований таксонов. В систематике богатого видами класса насекомых разных рангов больше, чем в типе гребневиков с его 100 видами. В необходимых случаях используются такие наименования, как подрод, надрод, подсемейство, триба и т. д. Ранг таксона обозначает его мощность как подмножества всех вообще видов: чем выше ранг, тем больше мощность подмножества. Поэтому отношения между таксонами разного* ранга выражаются посредством вложений, импликаций.
вид ( род ( семейство ( отряд ( класс ( ... Species genus familia ordo classis
Таксономическое положение вида однозначно определяется последовательностью латинских названий таксонов разного ранга, которая рассматривается как иерархическая. Иерархия таксонов возможна благодаря иерархии признаков. Среди таксономических признаков выделяются такие, которые общи всем видам некоторого рода; это будут признаки родовые. Возможны такие признаки, которые общи всем родам семейства и, стало быть, являются признаками семейства и т. д. Размах изменчивости признака в какой-то степени есть мерило его таксономического значения (ценности): более Изменчивые признаки позволяют различать мелкие таксоны, менее изменчивые — относительно более крупные таксоны. Крупные таксономические признаки — это такие, которые свойственны большому числу видов; более мелкие видовые признаки — свойство отдельных видов. Таксономическое значение первых выше, чем вторых. Следует иметь в виду, что с точки зрения эйдологии каждый раз носителем таких признаков остаются виды. В систематике носителем таких признаков будет таксон.
Таксономическая ценность даже одного и того же признака колеблется в широких пределах. Поэтому иерархия таксономических признаков — понятие относительное. Это обычнейшее явление в системах самых различных таксонов имеет своим следствием несопоставимость (неадекватность) таксономических описаний.
Например, гетеротрофное питание и автотрофное характеризуют собой в целом два царства: первое — животных,
13-
второе — растений. Однако в пределах рода Эвглена — одноклеточных жгутиковых — имеются виды с настоящим автотрофным способом питания и виды с гетеротрофным питанием. В первом случае способ питания есть признак царства, во втором — признак видов. Другой пример, из класса насекомых, относится к отрядам жуков и двукрылых: особенности жилкования крыла у отдельных видов мух весьма различны и часто используются для различения видов. У жуков жилкование крыльев более' однообразно. Крупные таксоны жуков, рангом не ниже семейства, характеризуются каким-то одним типом жилкования, и в пределах таких таксонов изменчивость этого признака не проявляется. У пчелы Andrena africana Muller жилкование крыла подвержено географической изменчивости, будучи различным в отдельных популяциях (Zimmerman, 4933). То же хорошо известно у дрозофилы (Берг, 1960). Число таких примеров весьма велико. В этом легко убедиться, ознакомившись с определительными таблицами': различение соподчиненных таксонов одного и того же ранга в одном более крупном осуществляется по совершенно различным признакам. В этом смысле можно говорить о несопоставимости, неадекватности таксономических описаний. Если же делать ударение на одном признаке, то можно говорить о его непостоянной разрешающей способности: «один и тот же» признак позволяет охарактеризовать в одном случае царства, а в другом — виды. Биологический смысл этого явления станет понятным, если учесть, что таксономические признаки к тому же имеют своих двойников среди популяционных (подвидовых) и индивидуальных; это тот случай, когда противопоставление таксономического признака разным прочим нецелесообразно.
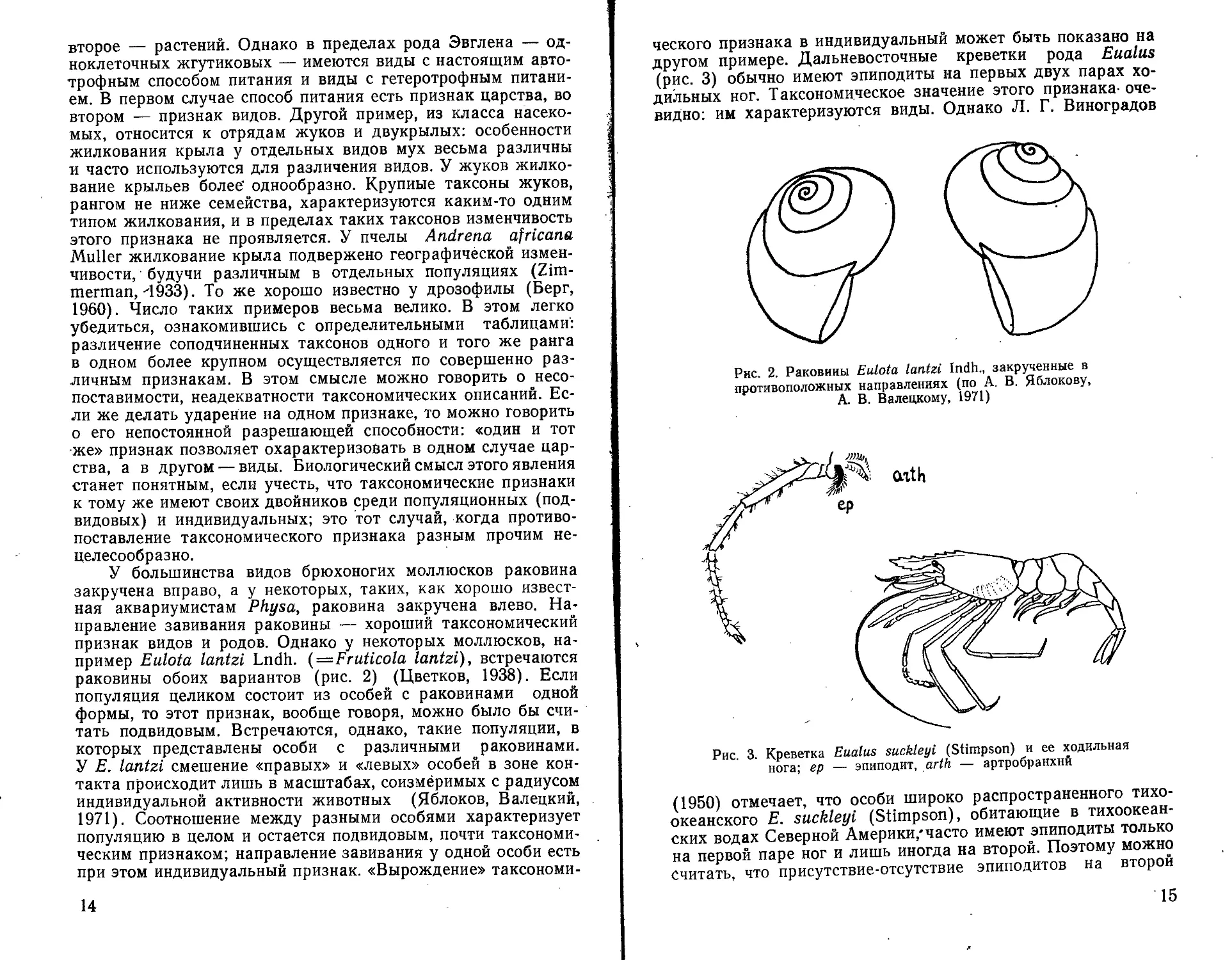
У большинства видов брюхоногих моллюсков раковина закручена вправо, а у некоторых, таких, как хорошо известная аквариумистам Physa, раковина закручена влево. Направление завивания раковины — хороший таксономический признак видов и родов. Однако у некоторых моллюсков, например Eulota lantzi Lndh. (—Fruticola lantzi), встречаются раковины обоих вариантов (рис. 2) (Цветков, 1938). Если популяция целиком состоит из особей с раковинами одной формы, то этот признак, вообще говоря, можно было бы считать подвидовым. Встречаются, однако, такие популяции, в которых представлены особи с различными раковинами. У Е. lantzi смешение «правых» и «левых» особей в зоне контакта происходит лишь в масштабах, соизмеримых с радиусом индивидуальной активности животных (Яблоков, Валецкий, 1971). Соотношение между разными особями характеризует популяцию в целом и остается подвидовым, почти таксономическим признаком; направление завивания у одной особи есть при этом индивидуальный признак. «Вырождение» таксономи-
14
ческого признака в индивидуальный может быть показано на другом примере. Дальневосточные креветки рода Eualus (рис. 3) обычно имеют эпиподиты на первых двух парах ходильных ног. Таксономическое значение этого признака-очевидно: им характеризуются виды. Однако Л. Г. Виноградов
Рис. 2. Раковины Eulota lantzi Indh., закрученные в противоположных направлениях (по А. В. Яблокову, А. В. Валецкому, 1971)
axth
suckleyi (Stimpson) и ее ходильная
Рис. 3. Креветка Eualus нога; ер — эпиподит, arth. — артробранхнй
(1950) отмечает, что особи широко распространенного тихоокеанского Е. suckleyi (Stimpson), обитающие в тихоокеанских водах Северной Америки/часто имеют эпиподиты только на первой паре ног и лишь иногда на второй. Поэтому можно считать, что присутствие-отсутствие эпиподитов на второй
15
паре ног характеризует североамериканскую популяцию вида. У другого близкого вида, Е. fabricii (Кгоуег), первая пара эпиподитов присутствует всегда, что же касается второй пары ног, то она либо несет оба эпиподита (справа и слева), либо имеет его с одной стороны. В данном случае присутствие — отсутствие эпиподита не может быть использовано как таксономический признак, потому что обладатели этого признака не обнаруживают явно приуроченности к определенной части ареала. Очевидно, мы здесь имеем дело с индивидуальным признаком. При этом происходит своего рода «раздвоение» его на «правый» и «левый». Беглое рассмотрение вопроса о неадекватности таксономических описаний, или, что то же, о переменной разрешающей способности одного и того же признака, сразу же привело нас к теории видообразования. В синтетической теории эволюции (Завадский, 1971), в соответствии со взглядами Ч. Дарвина, принимается, что мелкие различия в популяции между особями могут перерасти в таксономические признаки. Обычнейшая практика систематиков, признающих непостоянство разрешающей способности «одного и того же» признака, имеет своим оправданием эту важнейшую эволюционную концепцию.. Индивидуальный признак — прообраз таксономического признака. Эволюция одного и того же таксономического признака проходит через стадии, которые примерно соответствуют рангам таксонов, от вида к роду, далее к, семейству и т. д. В действительности же эта схема значительно сложнее, потому что эволюционируют виды, а не признаки. Несмотря на условность принятого здесь решения, оно, быть может, и не столь умозрительно, как представляется на первый взгляд. Один лишь факт разнообразия органического мира наводил на мысль о изменяемости видов. Так было в. истории эволюционного учения. Переменная разрешающая способность одного и того же признака, быть может, также свидетельствует о его эволюции? Способность признаков эволюционировать вместе с их обладателями причиняет неудобства систематикам, которые вынуждены отыскивать особые признаки для различения видов даже близко родственных таксонов.
Другое затруднение в использовании таксономических признаков возникает вследствие их изменчивости в онтогенезе. Некоторые креветки, близкие к упомянутым, различаются по числу члеников щупика мандибулы (рис. 4). Обычно это родовой признак. Вместе с тем у личинок некоторых видов таких члеников больше, чем у взрослых, т. е. таксономический признак подвержен возрастной изменчивости. Это обстоятельство -причиняет неудобства систематику и он вынужден искать иные признаки для обоснования родов с такими видами. Во всем этом есть, однако, и положительный момент. Благодаря онтогенетической изменчивости систематик может оце
16
нить филогенетическое значение таксономического признака. Дело в том, что имеются основания рассматривать столь своеобразный придаток, как мандибула ракообразных, в качестве производного обычной расчлененной конечности с жевательной лопастью на основании (рис. 4). Своеобразные морские рачки Cephalocarida имеют именно такую примитивную мандибулу. В эволюции произошло усиление основных члеников такой конечности и склеротизация их в связи с жевательной функцией. Прочие членики впоследствии оказались утраченными в той или иной степени. Членистый щупик и является их рудиментом. Можно думать, что наличие этого признака указывает на примитивность его обладателя, причем последний тем примитивнее, чем больше члеников в щупике. Конечно, уровень специализации или степень примитивности вида не следует оценивать по одному лишь признаку. Тем не менее возрастная изменчивость действительно может быть изучена в свете биогенетического закона, и благодаря этой возможности раскрыто эволюционное значение таксономического признака. По-видимому, приведенный пример показателен для обычной практики систематиков. Ясность биологического содержания признаков — такова компенсация за неудобство в их использовании.
Распределение видов по таксонам, разбиение множества видов на подмножества — общеизвестная задача систематики. Однако ею далеко не исчерпывается проблема изучения множества таксонов как единого целого, которое имеет некоторую структуру и требует специального подхода для изучения.
Множество видов, упорядоченное посредством включения их в таксоны надвидового ранга, называется системой. Систематика отнюдь не сводится к систематизации. Описание новых видов, разработка латинских названий для видов и других таксонов — это лишь внешняя, наиболее приметная функция систематики. У большинства людей, далеких от биологии, этим исчерпывается представление о систематике вообще. Вряд ли будет преувеличением утверждать, что и многие биологи видят в системе не более чем каталог. Для того чтобы навести порядок в таком каталоге, казалось бы, требуется всего лишь здравый инженерный подход.
Однако основная функция систематики как научной дисциплины есть объяснение. Объяснение множеству фактов, таксонов, по-видимому, достигается в суждениях относительно положения видов в системе. Классифицируя таксоны, систематик как бы объясняет их. Процесс классификации включается в философское понятие научного объяснения. Только в формально-логическом плане классификация есть всего лишь множество соподчиненных понятий. В материалистической диалектике классификация есть раскрытие внутренней
2
17
Н. А. Заренков
Рис. 4. A — примитивная мандибула с расчлененным двуветвистым щупиком Hutchinsoniella tnacracantha Sanders (Crustacea Cephaloca rida) (no Hessler, 1969);
5, В, Г, — мандибулы разных креветок с 3-, 2- и 1-членистым щупиком и инцизор-ным выростом i. р.; Д — мандибула креветки без щупика и инцизорного выроста
I
необходимой связи между множествами таких объектов (видов и вообще таксонов).
Установив таксономическое положение вида в системе с помощью последовательности латинских названий таксонов различного ранга, систематик как бы объясняет вид в пределах возможностей, методов систематики. Множество таксономических объяснений составляет содержание системы как естественнонаучной теории.
Вид «виноградная улитка» Helix pomatia (Linne) (рис. 5) получает в системе объяснение: «виноградная улитка есть легочный брюхоногий моллюск». Согласно ему, к виноградной улитке как к одному из представителей моллюсков в какой-то степени относятся те сведения, которыми мы располагаем о моллюсках вообще и о легочных брюхоногих моллюсках в частности. Эти сведения (тип организации, физиология, биохимия, происхождение, история расселения, развитие) добыты в ходе сравнительного изучения разных моллюсков и, можно считать, относятся к некоему абстрактному моллюску. Если бы из всех моллюсков нам была известна только одна виноградная улитка, то такое объяснение было бы невозможным; во всяком случае оно не было бы столь обстоятельным. Для объяснения такого изолированного факта пришлось бы обратиться к другим животным, например членистоногим, кольчатым червям, и рассматривать такие признаки (более крупные!), которые допускают сравнение этих столь несхожих животных. Признаки «легкое», «брюхоногость» потеряли бы прежнее таксономическое значение, потому что в пределах таксона «Моллюски» они утратили бы межтаксонную изменчивость. Пришлось бы полагать, что наличие легкого и «брюхоногость» суть признаки всех вообще моллюсков, а не более мелких соподчиненных таксонов. С виноградной улиткой было бы связано представление о моллюсках вообще. Таксономическое суждение сделалось бы более общим. Представление о происхождении этого животного от водных моллюсков было бы менее обоснованным, своим происхождением оно было бы связано с какими-то другими животными.
Единичный факт, вид Н. pomatia, получает в системе объяснение через отнесение его к более широкому классу фактов. Чем богаче наши знания о моллюсках вообще, тем более полное объяснение получает этот факт в системе. Определяя виноградную улитку как представителя легочных моллюсков, мы имеем менее широкое, но более определенное объяснение; относя ее к брюхоногим моллюскам, мы получаем более общее объяснение. Наконец, причисляя ее к моллюскам вообще, мы ограничиваемся тем самым лишь весьма общим объяснением. Благодаря иерархической системе таксонов один и тот же факт (вид как таксон) получает в системе объяснения различной степени определенности. Через таксономическое по-
2*
19
Рис. 5. Таксономическое положение виноградной улитки изображенное с помощью кругов Эйлера. А - по П. В. Матекину (1966), Б - по А. В. Иванову (1940), В - по И М Лихареву и Е С Раммельмейе-ру (1952), Г — по Д. Гейеру (Gayer, 1927)
ложение раскрывается содержание единичного факта — вида в совокупности наших знаний об органическом мире как множестве видов.
• Научная ценность системы как теории зависит от того, в какой мере она считается с особенностями фактов, подлежащих объяснению. При этом способность видов эволюцио-
Рис. 6. Традиционное филогенетическое древо для таксонов, происходящих от единственного предка х. Ветви, поднимающиеся вертикально, — это виды, не изменяющиеся длительное время. Отклоняющиеся ветви обозначают новые виды. Вымершим видам отвечают слепые ветви. Мелкие круги на плоскостях б, k, t3 — надвидовые таксоны
пировать имеет первостепенное значение. Действительно, вопрос об отношениях между систематикой и филогенией вызывает острые разногласия, п споры по этому поводу, в конце концов, отражают противоречия в понимании теории эволюции разными исследователями. Сторонниками сходственной, в частности нумерической систематики, отстаивается
21
Рис. 7. А — система таксонов в момент времени <а, Б — система таксонов в моменты ti и t2, отнесенных к одному моменту U t2
возможность системы, полностью лишенной какого-либо филогенетического содержания. Такая система, по мысли ее приверженцев, представляла бы самостоятельную ценность, могла бы быть рассматриваемой впоследствии с точки зрения филогении и по условиям своего построения была бы полностью свободной от «спекулятивного» духа филогении. Целесообразность такого подхода, казалось бы, можно найти в истории биологии. Действительно, система существовала задолго до утверждения эволюционной теории Дарвина, а в наши дни в систематике вполне плодотворно работают не только дарвинисты, но также их оппоненты. Тем не менее (это будет показано постепенно, по мере получения отдельных результатов) с самого начала в своей работе систематики не были свободны от определенного естественноисторического мировоззрения, и это сказывалось на системе. Вместе с тем (это будет показано впоследствии) содержание основного понятия систематики — «таксон» — полностью определяется взглядами исследователя на основные моменты теории эволюции.
Система, конечно, не есть филогения. Смешивать их — это все равно,
что не отличать следствия от причины. Распределение видов по таксонам есть следствие из их способности эволюционировать. Филогения играет роль объяснения таксономическому объяснению. Филогенетические отношения обычно изображаются в виде древа (рис. 6), система — с помощью кругов Эйлера, графического метода, применяемого в логике (рис. 5, 7). Система на рис. 7, Л получена при повороте на 90° одного из поперечных сечений t2 филогенетического древа рис. 6. Тем не менее и это будет всего лишь констатацией
22
действительного положения вещей, история таксона, филогения, составляет существенное содержание таксономического объяснения в классической систематике.
Обратимся, например, к речным ракам северного полушария — семейству Astacidae. В нем содержится не менее 6 родов и около 100 видов. Географическое распространение этих
Б
Рис. 8. Современное распространение речных раков северного полушария (Л) и распределение суши и моря в нижнемеловое время (В)
ныне исключительно пресноводных животных замечательно. Наши речные раки относятся к двум родам: Astacus, обитающему в Европе и Западной Сибири, и дальневосточному Cambaroid.es. В Северной Америке обитают Cambarus и Paci-fastacus, разделенные Скалистыми горами (рис. 8). Наличие общих признаков, позволяющих включить всех этих животных в один таксон, а следовательно и природу таксона, можно было бы объяснить двояким образом. Во-первых, единым происхождением всех речных раков Astacidae. В нижнемеловое время существовало море Тэтис, ориентированное в ос
23
новном в широтном направлении, и сухопутное соединение Европы с Северной Америкой. От общего предка, обитавшего в Тэтисе, ведут свое происхождение нижнемеловые виды рода Astacus, один из Монголии, другой из Центральной Европы. Поскольку первый был найден в пресноводных отложениях, а второй в морских или солоноватоводных, можно думать, что речные раки действительно были способны перенести смену сред обитания: море на пресные воды. После того как Тэтис прекратил свое существование, а Северная Америка обособилась от Европы, произошел разрыв ареала семейства на три части соответственно дальневосточным, европейским и североамериканским речным ракам, обитающим к востоку от Скалистых гор. Североамериканские речные раки Тихоокеанского побережья, однако, ближе не к своим землякам, а к дальневосточным. Они действительно происходят от последних и возникли в миоцене, когда существовало сухопутное соединение Азии с Аляской. Находка ископаемого речного рака на Аляске подтверждает это. Таксономическое положение родов речных раков Astacidae, их принадлежность к одному семейству получает филогенетическое объяснение. История речных раков, в том виде как она здесь изложена, использует представление об их едином происхождении, едином центре расселения, о происхождении одних видов от других.
Если бы разные речные раки имели разных предков, то все это было бы излишним. Следовало бы, например, полагать, что каждый род имеет собственного предка и что, например, лишь вследствие сходных условий обитания при наличии какой-то внутренней способности все Astacidae приобрели такие признаки, которые сделали возможным таксон «Семейство Речные раки». В этом и заключается второе объяснение. Ясно, что оно имеет в виду не филогенетическое древо, а филогенетический газон, вроде того, который изображен на рис. 9. Система таких таксонов есть прежде всего отражение множества экологических ниш на множество видов.
Первое, классическое обоснование таксона предполагает его монофилетическое происхождение, второе — по-лифилетическое.
Монофилетическая концепция составляет сердцевину исторической биогеографии, в которой современное распространение животных и растений рассматривается в свете данных палеогеографии. История речных раков показывает это со всей очевидностью, и этот пример далеко не единственный. Разорванные ареалы мелководных морских животных, такие, как амфиатлантический, амфипацифический, амфибореаль-ный, во многих случаях не могут быть объяснены действующими ныне факторами, их можно понять только при использовании палеогеографии. Вот еще один пример: тропические воды Тихого и Атлантического океанов соединялись до конца
24
третичного времени, когда возник Панамский перешеек. Сего появлением единая центральноамериканская морская фауна разделилась на тихоокеанскую и атлантическую. Их независимое развитие привело к возникновению новых видов и родов разных животных. Оставшиеся общими для обеих фаун 5% видов и 40% родов крабов — это таксоны, не изменившиеся с конца третичного времени. Сто пар близнецовых ви-
Рис. 9. Полифилетическое происхожде-ние основных групп животных от протестов (Nursall, 1962)
дов крабов (62% от общего их количества) — это слабо разошедшиеся виды. Дивергенция, начавшаяся в конце третичного времени, обусловила таксономическую обособленность фаун. Соответствующие числа характеризуют скорость процесса видообразования. Все это имеет иной смысл, если использовать полифилетическую концепцию. Тогда черты сходства фаун есть следствие сходных условий обитания, а число общих и близнецовых видов характеризует сходство этих условий. Представление о некогда единой фауне оказывается излишним, палеогеография остается не у дел.
Полифилетическая концепция могла бы внести много нового в истолкование находок «живых ископаемых». Когда в 1937 г. у Южной Африки была поймана кистеперая рыба Latimeria, ранее известная из девонских отложений и считавшаяся вымершей, то это рассматривалось как пример исключительной консервативности таксона, не изменившегося за сотни миллионов лет. Наряду с этим находка Latimeria послужила подтверждением неполноты геологической летописи, не сохранившей остатков этой рыбы за громадный отрезок
25
времени от девона до наших дней. Иначе расценили бы это открытие сторонники полифилии. В полном согласии и духом полифилетической концепции они могли бы допустить, что девонские кистеперые и ныне живущая Latimeria не связаны родственными отношениями и развились независимо от разных предков в подходящих условиях обитания. Единство таксона во времени, если следовать полифилетической концепции, совершенно не обязательно: один и тот же таксон, например Кистеперые, может неоднократно возникать заново. Между видами такого таксона нет преемственности ни во времени, ни в пространстве. При таком подходе невозможна датировка отложений разных районов одним возрастом, если даже доказана идентичность их фаун и флор: последние возникли независимо одна от другой благодаря особым, пока неизвестным, законам развития в подходящих условиях обитания.
Ясно, что широкое признание полифилетической концепции эволюции обезоруживает не только историческую биогеографию, но также и другие дисциплины, использующие представление о непрерывности таксона во времени. По сути дела понятие «таксон» утрачивает историческое содержание. Мало того, отрицание непрерывности таксона во времени в сущности есть отрицание его реальности как биологического объекта, потому что между видами такого таксона нет кровного родства, преемственности материальных носителей наследственности. Такой таксон — понятие, но не материальный объект естественноисторической науки.
Надо сказать, что большинство биологов разделяют монофилетическую концепцию, которую обосновал Ч. Дарвин.
Особенность системы как множества таксономических объяснений состоит в том, что они не могут быть выведены одно из другого. Все открытия в систематике, такие, как определение таксономического положения вновь найденных видов, пересмотр объема и границ таксонов, не были предсказаны. Пока не представляется возможным установить, сколько в действительности видов включает данный таксон. До сих пор систематик находится в положении коллекционера, который не ведает, что ему пошлет случай. По-видимому, в этой связи может быть упомянута лишь одна попытка предсказания признаков неизвестных в настоящее время видов, предпринятая Н. И. Вавиловым (1920). Его «гомологические ряды изменчивости» как параллельная изменчивость в близкородственных таксонах дают возможность предсказания видов с такими признаками, которые известны в относительно лучше изученных таксонах. К сожалению, работа Н. И. Вавилова, замечательная уже одной лишь попыткой предсказания (что в биологии бывает не часто), не вызвала должного отклика у систематиков.
26
В целом же предсказательные возможности систематики весьма и весьма невелики. В конце концов, не только систематика вынуждена ограничиваться эмпирическими поисками объяснений отдельных фактов и поисками самих фактов. Развитие других разделов биологии, таких, например, как сравнительная биохимия и физиология, также связано с изучением свойств видов, и в основном (если исключить закономерности физико-химической природы) осуществляется вслепую. Значение формального вывода закономерностей здесь несоизмеримо мало в сравнении с тем, что имеется в математике, физике. Знание тех видов и их свойств, которые нам известны сегодня, как будто бы не дает определенного ответа на вопросы о свойствах неизвестных видов, вымерших, еще неизвестных науке, потенциально осуществимых в ходе эволюции. Поэтому для писателей-фантастов остается слишком широкое поле деятельности, а биология представляется иногда лишь множеством фактов, разрозненных и лишенных объединяющей их теории (Уотермен, 1968).
Систематика, имеющая объектом разнообразие органического мира, которое выражено в свойствах видов, не может отмахиваться от необходимости строгого биологического (без физикохимии) доказательства своих выводов, т. е. таксономических объяснений. Мало того, систематика должна уметь предсказывать, и ее предсказания должны быть биологически обоснованными и объяснимыми.
Лекция 2,
В КОТОРОЙ АВТОР ОТВЕРГАЕТ НЕКОТОРЫЕ УПРЕКИ В АДРЕС КЛАССИЧЕСКОЙ СИСТЕМАТИКИ И В СВОЮ ОЧЕРЕДЬ ПРЕДЪЯВЛЯЕТ Ей НОВОЕ ОБВИНЕНИЕ
Рассмотрение критики в адрес классической систематики позволит полнее усвоить содержание основных понятий систематики. Автору представляется, что эта критика является своеобразным отражением того факта, что множество видов и их таксономических признаков никогда не может быть изучено полностью.
Положение, которое здесь принимается как очевидное и служит отправной точкой для дальнейших рассуждений, состоит в том, что доступное для изучения в настоящее время множество видов составляет лишь часть (и, вероятно, незначительную часть) того генерального множества видов, которое возможно в природе. Генеральное множество видов включает: 1) уже известные вымершие и современные виды, 2) остающиеся неизвестными современные и вымершие виды, 3) виды, потенциально осуществимые в ходе дальнейшей эволюции. Неполнота нашего знания существующих и
27
вымерших видов в какой-то степени доказывается тем, что работы с описаниями новых таксонов, прежде всего видов, занимают видное место в новейшей таксономической литературе.
По-видимому, следует смириться с неполнотой доступного для изучения эмпирического множества видов, а следовательно, и с неполнотой таксономических объяснений. Степень этой неполноты было бы интересно оценить количественно, и в дальнейшем автор попытается решить эту задачу.
Другое важное обстоятельство, которое также дает повод для критики, связано с неполнотой нашего знания признаков видов. Общее число признаков, характеризующих любой вид, весьма велико (хотя никто не берется его подсчитать даже приблизительно), но систематики имеют дело лишь с немногими признаками. Следует думать, что это обстоятельство также каким-то образом сказывается на достоверности таксономических объяснений. Здесь также следует различать г е-неральное множество признаков и эмпирическое м н ож ество признаков соответственных множеств видов. Можно показать, и это будет сделано, что изучение разнообразия видов «вширь» сказывается на достоверности таксономического объяснения примерно таким же образом, что и изучение немногих видов «вглубь».
Критика в адрес классической систематики в значительной степени вызвана неустойчивостью ее построений.
Система постоянно перестраивается в связи с открытием новых таксонов, новых признаков уже известных таксонов и пересмотром таксономического значения ранее известных признаков. Поэтому те объяснения, которые предлагаются в системе отдельным таксонам, также постоянно уточняются и дополняются. Пусть, например, в момент Z2 известны какие-то виды, система которых изображена на рис. 7, А. Если через некоторое время обнаруживаются еще и виды t\, то система может принять такой вид, как на рис. 7, Б. Она отличается от прежней системы не только объемом таксонов (площади кругов), но также иным количеством таксонов (число кругов). Все это отразится на таксономических объяснениях. Внешне для несистематика это выглядит как произвол, перестановка и усложнение системы латинских названий, определяющих положение таксона в системе. В действительности же если оставить в стороне номенклатурные недоразумения (вполне естественные, если иметь в виду почтенный возраст систематики и груз традиций) и временные разногласия в оценке таксономического значения признаков (также вполне понятные, если учесть недостаточную их изученность и громадный объем работы), то кажущиеся формальностью таксономические ревизии вполне оправданы и совершенно необходимы. Каж
28
дый раз в результате ревизий таксоны получают более полное и точное объяснение на основе быстро возрастающей массы сведений о видах и их признаках. Эта кропотливая работа никогда не прекращалась и, к счастью, не прекращается и в наши дни. История изучения видов есть в то же время история их системы. История системы свидетельствует о том, что таксономические ревизии не были волевым актом систематиков, но всегда вызывались необходимостью считаться с новыми фактами (Зенкевич, 1937). Стремление систематиков к возможно более полному охвату громадного разнообразия видов и их признаков, их готовность к пересмотру полученных результатов делают им честь и не могут служить основанием для упреков.
Обращаясь к истории системы, мы видим, что некоторые таксоны сохранились со времени их основания до наших дней, даже несмотря на открытие большого числа новых видов и возросшую их изученность. Многие рода растений, описанные К. Линнеем в 1753 г., сохранили свое значение и в настоящее время. Обширный раздел Членистые Articulata, включающий кольчатых червей и членистоногих, был основан Ж. Кювье в 1817 г. и сохранился в современной системе животного царства. А. Л. Тахтаджян (1947) разделяет мнение М. И. Голен-кина о том, что в ботанике «появление новых систем вовсе не означает разрушение всего старого». С другой стороны, еще в 1809 г. Ж--Б. Ламарк отмечал неустойчивость системы. Перестройки системы даже крупного масштаба не прекращаются и в наши дни. При этом изучение разнообразия органического мира «вширь» действительно имеет то же значение, что и изучение отдельных видов «вглубь».
В последнее время для обоснования быстро развивающейся системы крабов все шире используются особенности строения гениталей самца. Обычно они удобны для различения видов, но использование этого нового признака не сказалось заметно на системе. В систематике довольно трудного семейства креветок Penaeidae было предложено использовать детали строения желудочной мельницы, сложного аппарата из хитиновых зубцов на внутренней поверхности желудка Penaeidae. Однако это новшество не отразилось на системе креветок Penaeidae и не было поддержано более поздними исследователями. Повышенный интерес малокологов к анатомии моллюсков в значительной степени явился реакцией на односторонность таксономических диагнозов ранних авторов, которые ограничивались изучением раковины. Выяснилось, однако, что не у всех моллюсков признаки мягкого тела могут явиться основой для ревизии. Например, у высших переднежаберных моллюсков семейства Buccinidae радула (терка из хитиновых зубцов, используемая для соскребания пищи) устроена одинаково у разных видов, но часто видоспецифична в
29
семействе Trochidae (Галкин, 1955). У пресноводных моллюсков с внутренним оплодотворением детали строения половой системы оказались полезными для диагностики существующих семейств (Старобогатов, 1970), но широкой ревизии на этом основании не состоялось. Имеются, однако, примеры иного рода.
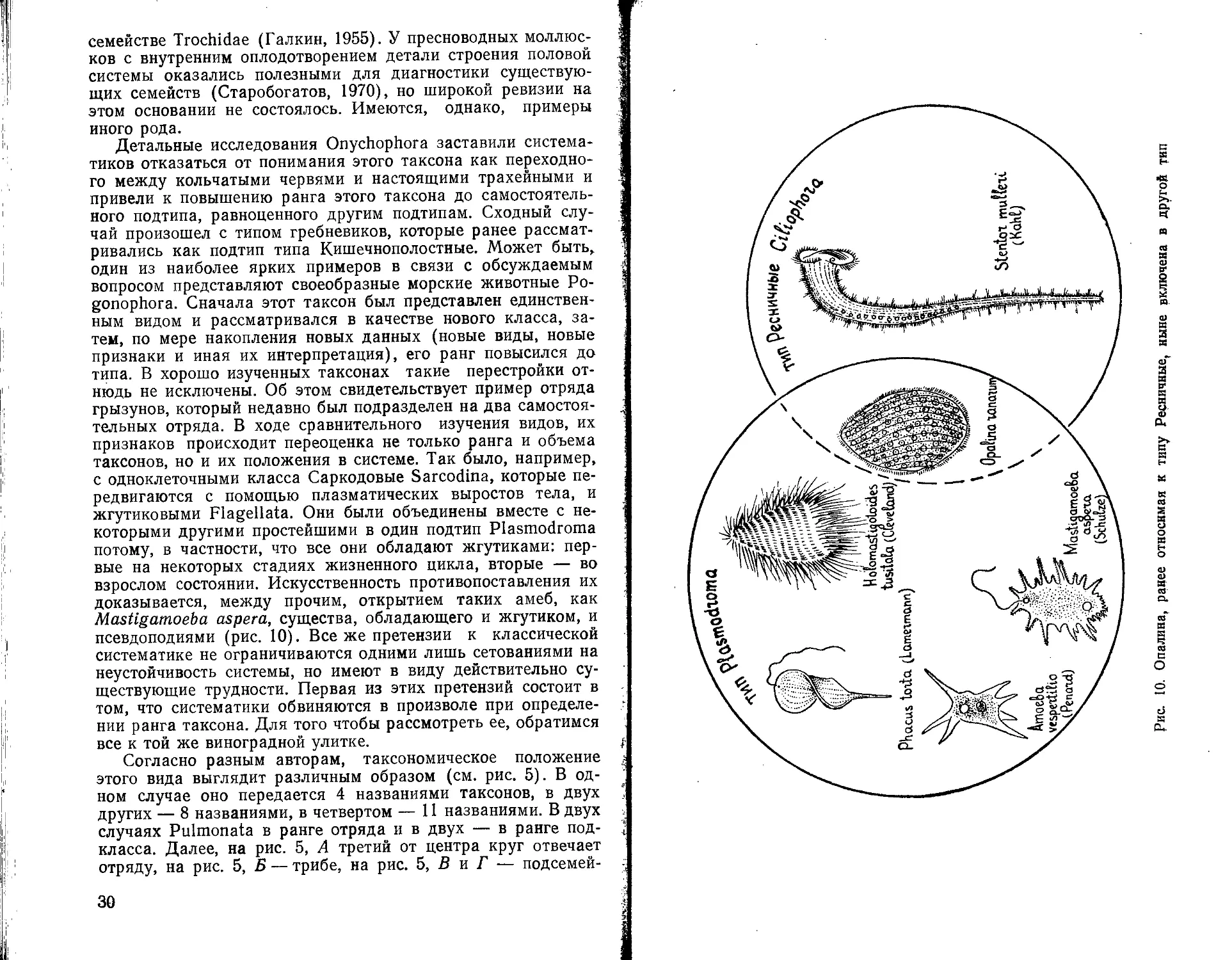
Детальные исследования Onychophora заставили систематиков отказаться от понимания этого таксона как переходного между кольчатыми червями и настоящими трахейными и привели к повышению ранга этого таксона до самостоятельного подтипа, равноценного другим подтипам. Сходный случай произошел с типом гребневиков, которые ранее рассматривались как подтип типа Кишечнополостные. Может быть, один из наиболее ярких примеров в связи с обсуждаемым вопросом представляют своеобразные морские животные Ро-gonophora. Сначала этот таксон был представлен единственным видом и рассматривался в качестве нового класса, затем, по мере накопления новых данных (новые виды, новые признаки и иная их интерпретация), его ранг повысился до типа. В хорошо изученных таксонах такие перестройки отнюдь не исключены. Об этом свидетельствует пример отряда грызунов, который недавно был подразделен на два самостоятельных отряда. В ходе сравнительного изучения видов, их признаков происходит переоценка не только ранга и объема таксонов, но и их положения в системе. Так было, например, с одноклеточными класса Саркодовые Sarcodina, которые передвигаются с помощью плазматических выростов тела, и жгутиковыми Flagellata. Они были объединены вместе с некоторыми другими простейшими в один подтип Plasmodroma потому, в частности, что все они обладают жгутиками: первые на некоторых стадиях жизненного цикла, вторые — во взрослом состоянии. Искусственность противопоставления их доказывается, между прочим, открытием таких амеб, как Mastigamoeba aspera, существа, обладающего и жгутиком, и псевдоподиями (рис. 10). Все же претензии к классической систематике не ограничиваются одними лишь сетованиями на неустойчивость системы, но имеют в виду действительно существующие трудности. Первая из этих претензий состоит в том, что систематики обвиняются в произволе при определении ранга таксона. Для того чтобы рассмотреть ее, обратимся все к той же виноградной улитке.
Согласно разным авторам, таксономическое положение этого вида выглядит различным образом (см. рис. 5). В одном случае оно передается 4 названиями таксонов, в двух других — 8 названиями, в четвертом — 11 названиями. В двух случаях Pulmonata в ранге отряда и в двух — в ранге подкласса. Далее, на рис. 5, А третий от центра круг отвечает отряду, на рис. 5, Б — трибе, на рис. 5, В и Г — подсемей-
30
Рис. 10. Опалина, ранее относимая к типу Ресничные, ныне включена в другой тип
OWOlp0
ству. Эти различия связаны исключительно с детальностью таксономического объяснения и не противоречат одно другому. Везде сохраняется иерархия
Helixcz...eGastropoda с...czMoilusca.
Таксономическое объяснение на рис. 5, А приведено по студенческому учебнику, прочие — по более специальным руководствам. По понятным причинам последние полнее и обстоятельнее первого, которое ограничено рамками учебной программы. Различия примерно такого же порядка обнаруживаются при сравнении ранних таксономических работ с более поздними. Можно надеяться, что дальнейшие исследования мировой фауны моллюсков, углубленное изучение отдельных видов позволят выработать еще более полное объяснение факту «виноградная улитка». Тогда последовательность латинских названий, определяющих положение таксона в системе, усложнится еще больше и возникнет необходимость изменения рангов уже имеющихся таксонов и создания новых рангов и новых таксонов.
Примерно так же обстоит дело и с системой ракообразных. Может показаться, что разные варианты системы низших ракообразных существенно различны и несопоставимы. Вот часть системы этих животных по Я. А. Бирштейну (1960):
superclassis Crustaceomorpha classis Crustacea
subclassis Gnathostraca ordo Anostraca ordo Phyllopoda subordo Notostraca subordo Cladocera
Другой вариант системы этих же видов предлагают Мур и Маккормик (Moore a. McCormick, 1969):
superclassis Crustacea classis Branchiopoda subclassis Calmanostraca ordo Notostraca ordo Cladocera subclassis Sarsostraca ordo Anostraca
Эти системы непротиворечивы, в чем нетрудно убедиться, если их изобразить с помощью кругов Эйлера. Различия между ними в основном относятся к терминологии, чем нисколько не нарушается иерархия таксонов. Однако таксоны одного и того же ранга здесь несопоставимы, например subclassis Gnathostraca и subclassis Calmanostraca. Анализируя положение дел в целом, О. Л. Крыжановский (1969) вполне спра-
32
редливо, на мой взгляд, заключает: «...объективно существует лишь определенные естественные группы видов (т. е. таксоны) и столь же естественна иерархия этих групп. Таксономический же ранг, придаваемый отдельным группам, и ступени иерархии, субъективны».
Примерно такова же позиция в этом вопросе у Б. Б. Ро-дендорфа (1959). На это же обстоятельство указывают и критики классической систематики, когда говорят о «разномасштабное™» существующих таксонов одного и того же ранга, которая состоит в том, что, например, семейство у животных не соответствует семейству растений. Таксоны были бы равномасштабными, если бы, например, степень гомологичности первичных структур ДНК у составляющих их видов была бы одинакова. Такой таксономический признак служил бы для различения таксонов равного ранга у самых различных существ. Показано, однако, что меченая ДНК домашней курицы (класс Птицы) имеет приблизительно 25—38% гомологичных последовательностей ДНК с ДНК птиц других отрядов и 10,5% с ДНК варана (класс Пресмыкающиеся). В то же время между видами разных семейств лилиецветных имеется лишь до 10% гомологичных структур ДНК. На этом основании делается вывод о том, что семейство у однодольных растений по рангу должно быть приравнено к классу хордовых животных и что ранг таксонов установлен произвольно. Иными словами, утверждается, что таксономический признак, в данном случае «10% гомологичных структур ДНК», сохраняет неизменной свою разрешающую способность в ходе эволюции и позволяет различать классы (семейства). «Разномасштабность» распространяется и на таксоны более низкого ранга (Антонов, 1972). Попытки установления универсального критерия ранга таксонов предпринимались неоднократно и раньше. По отношению к бактериям предлагается принять не меньше 70% гомологии ДНК в качестве внутривидового признака «genetic species» и около 45% для видов одного и того же рода (De Ley, 1968).
Более ранняя попытка была предпринята в том же направлении Бойденом (Boyden, 1943). Он нашел, что между рангом таксона и степенью серологического сходства у крабов существует следующая зависимость: члены одного и того же рода — 85%, родственные роды одного и того же семейства — 53, 66, 75%, тесно родственные семейства — 45%, отдаленно родственные семейства — 5%.
В принципе таков же ход мысли у Тарнера (Turner, 1969). В качестве видовых и более мелких признаков растений он предлагает изоферменты, для разрешения задач систематики на уровне родов и ниже — алкалоиды, терпеноиды, флави-ноиды и аминокислоты, а полимеры (ДНК, РНК, белки) — в качестве таксономических признаков более высокого ранга.
3
Н. А. Заренков
33
Для установления равномасштабных таксонов у ж в -них предлагаются следующие градации процента гомологии между их ДНК: 5—15% — классы, 20—45% — отряды, 50— 70% — семейства, 75% — таксоны более мелкие, чем семейства (Медников, Попов, Антонов, 1973).
В классической систематике, наоборот, разрешающая способность одного и того же таксономического признака
Рис. 11. Кривая продолжительности существования родов пластинчатожаберных моллюсков:
жирная линия — вымершие роды; тонкая — процент родов, возникших указанное число лет назад и продолжающих существовать и в настоящее время, определенный на основании возраста ныне существующих родов, известных также и как ископаемые (по Дж. Симпсону с изменениями, 1948)
( = таксономическое значение, таксономическая ценность) принимается переменной, тогда как границы (масштабы) таксонов одного и того же ранга большей частью принимаются равными; во всяком случае в своей практической работе систематики стремятся создать сравнимые таксоны. Нетрудно показать, что часто таксоны одного ранга действительно принимаются как равные, сопоставимые.
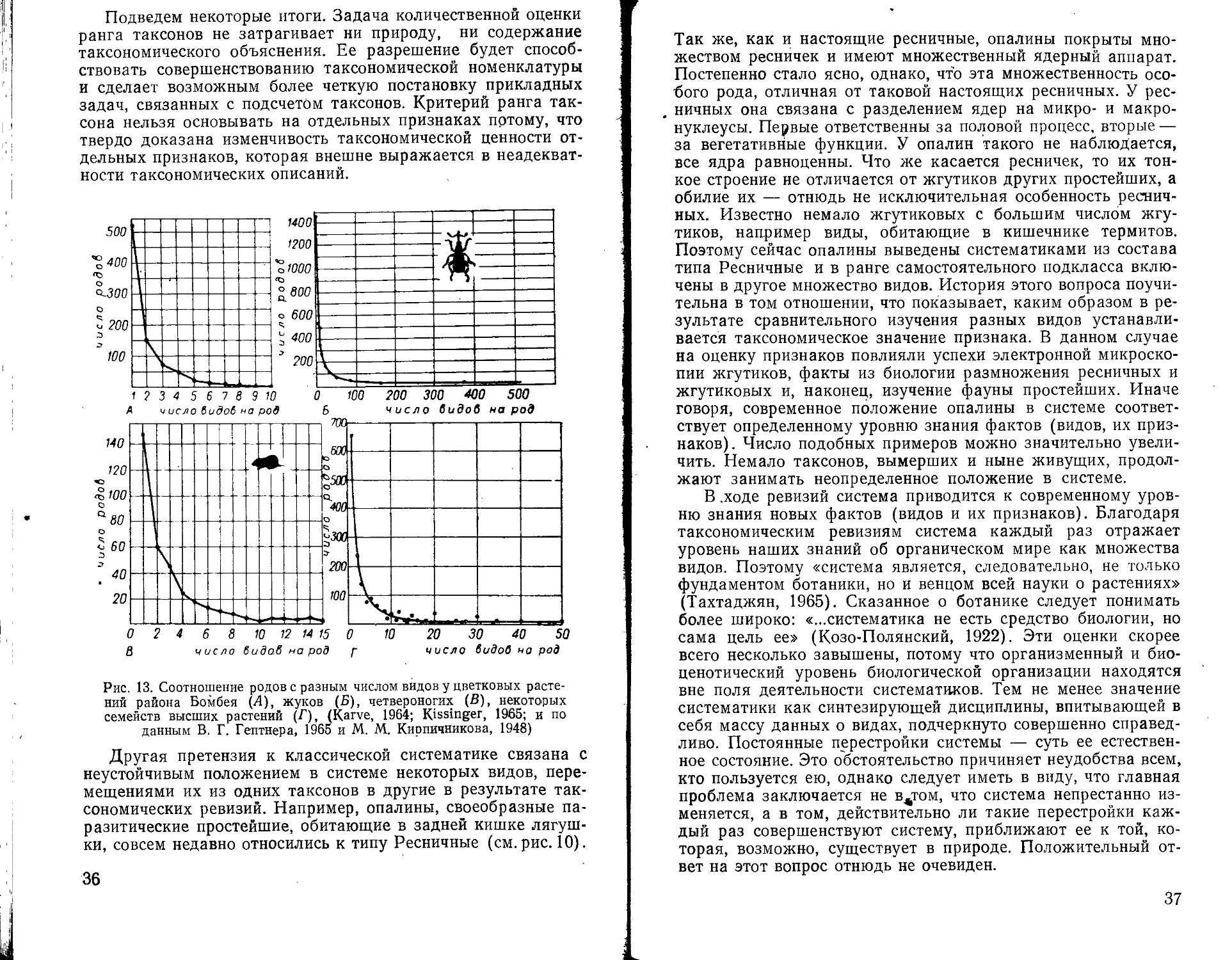
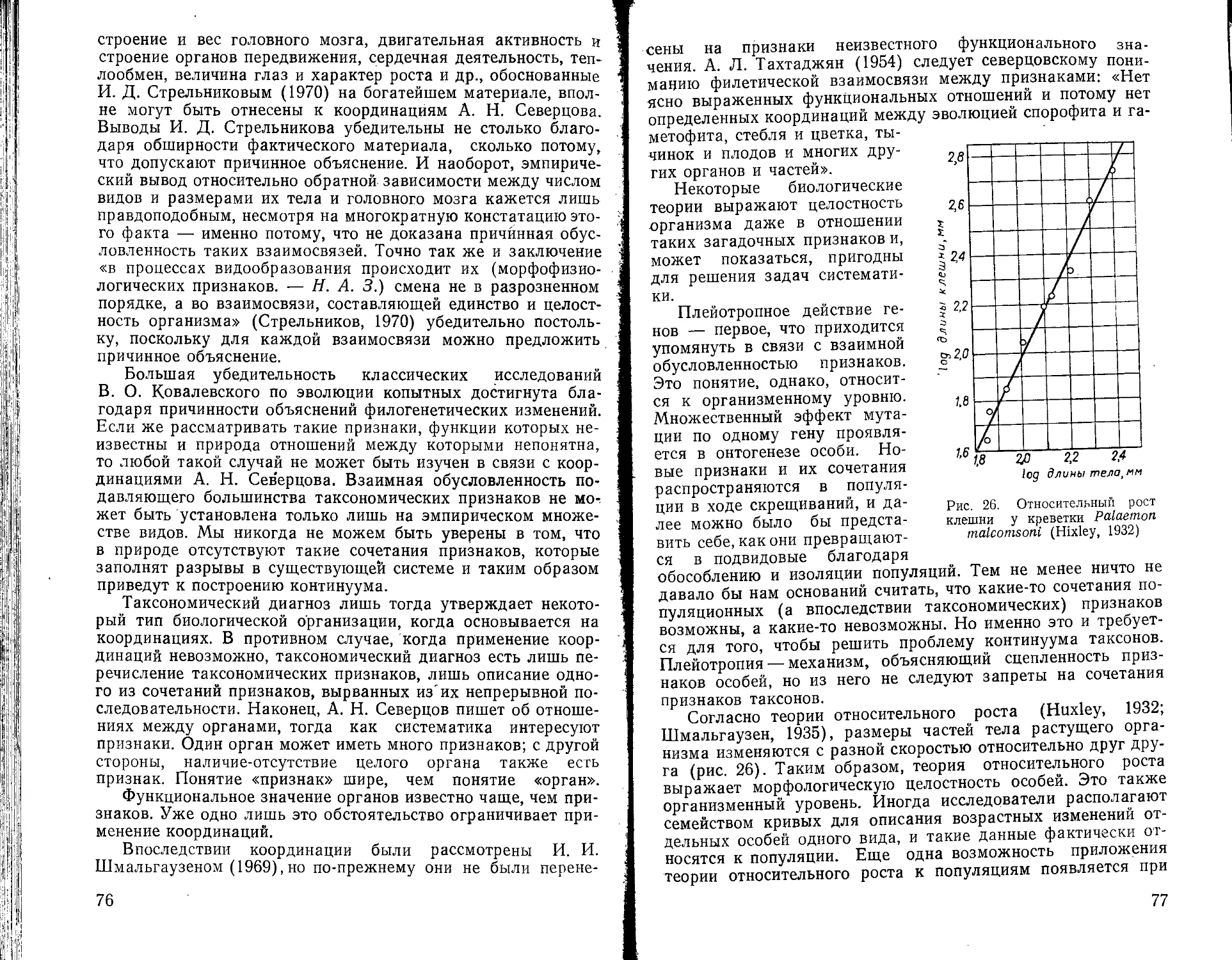
Скорость эволюции таксонов оценивается через подсчет родов, общих для отдельных геологических периодов (рис. 11). Изменение разнообразия крупных групп животных в геологическое время устанавливается по количеству родов для отдельных периодов (рис. 12). Количественное распределение видов по родам составляет предмет специального изучения (Гептнер, 1965; Кирпичников, 1948; Karve, 1964; Kissinger, 1963; Wright, 1941) (рис. 13). В биогеографии практикуется не только подсчет видов, но также таксонов более высокого ранга. Можно видеть, что широкое признание «разномас-штабности» таксонов одного ранга сделало бы эти подсчеты совершенно бессмысленными. Следуя этому принципу, разные, виды эвглен следовало бы отнести к разным царствам.
34
3*
Рис. 12. Изменения во времени числа родов:
А — трилобитов (всего 996 родов), Б — аммонитов (всего 597 родов) (по А. Н. Muller, 1957)
Подведем некоторые итоги. Задача количественной оценки ранга таксонов не затрагивает ни природу, ни содержание таксономического объяснения. Ее разрешение будет способствовать совершенствованию таксономической номенклатуры и сделает возможным более четкую постановку прикладных задач, связанных с подсчетом таксонов. Критерий ранга таксона нельзя основывать на отдельных признаках потому, что твердо доказана изменчивость таксономической ценности отдельных признаков, которая внешне выражается в неадекватности таксономических описаний.
Рис. 13. Соотношение родов с разным числом видов у цветковых растений района Бомбея (Л), жуков (Б), четвероногих (В), некоторых семейств высших растений (Г), (Karve, 1964; Kissinger, 1965; и по данным В. Г. Гептнера, 1965 и М. М. Кирпичникова, 1948)
Другая претензия к классической систематике связана с неустойчивым положением в системе некоторых видов, перемещениями их из одних таксонов в другие в результате таксономических ревизий. Например, опалины, своеобразные паразитические простейшие, обитающие в задней кишке лягушки, совсем недавно относились к типу Ресничные (см. рис. 10).
36
Так же, как и настоящие ресничные, опалины покрыты множеством ресничек и имеют множественный ядерный аппарат. Постепенно стало ясно, однако, что эта множественность особого рода, отличная от таковой настоящих ресничных. У рес-. ничных она связана с разделением ядер на микро- и макронуклеусы. Первые ответственны за половой процесс, вторые — за вегетативные функции. У опалин такого не наблюдается, все ядра равноценны. Что же касается ресничек, то их тонкое строение не отличается от жгутиков других простейших, а обилие их — отнюдь не исключительная особенность ресничных. Известно немало жгутиковых с большим числом жгутиков, например виды, обитающие в кишечнике термитов. Поэтому сейчас опалины выведены систематиками из состава типа Ресничные и в ранге самостоятельного подкласса включены в другое множество видов. История этого вопроса поучительна в том отношении, что показывает, каким образом в результате сравнительного изучения разных видов устанавливается таксономическое значение признака. В данном случае на оценку признаков повлияли успехи электронной микроскопии жгутиков, факты из биологии размножения ресничных и жгутиковых и, наконец, изучение фауны простейших. Иначе говоря, современное положение опалины в системе соответствует определенному уровню знания фактов (видов, их признаков). Число подобных примеров можно значительно увеличить. Немало таксонов, вымерших и ныне живущих, продолжают занимать неопределенное положение в системе.
В .ходе ревизий система приводится к современному уровню знания новых фактов (видов и их признаков). Благодаря таксономическим ревизиям система каждый раз отражает уровень наших знаний об органическом мире как множества видов. Поэтому «система является, следовательно, не только фундаментом ботаники, но и венцом всей науки о растениях» (Тахтаджян, 1965). Сказанное о ботанике следует понимать более широко: «...систематика не есть средство биологии, но сама цель ее» (Козо-Полянский, 1922). Эти оценки скорее всего несколько завышены, потому что организменный и био-ценотический уровень биологической организации находятся вне поля деятельности систематиков. Тем не менее значение систематики как синтезирующей дисциплины, впитывающей в себя массу данных о видах, подчеркнуто совершенно справедливо. Постоянные перестройки системы — суть ее естественное состояние. Это обстоятельство причиняет неудобства всем, кто пользуется ею, однако следует иметь в виду, что главная проблема заключается не в^том, что система непрестанно изменяется, а в том, действительно ли такие перестройки каждый раз совершенствуют систему, приближают ее к той, которая, возможно, существует в природе. Положительный ответ на этот вопрос отнюдь не очевиден.
37
Эмпирическое множество видов, которым располагают биологи, есть выборка из генеральной совокупности видов. Если разнообразие видов изменяемся во времени направленно в отношении существенных для систематики особенностей, то система каждый раз соответствует множеству видов какой-то стадии этого процесса изменения. Такое множество можно было бы изучать с помощью самых тонких методов и рассматривать чрезвычайно важные признаки, но система каждый раз будет отражать только степень изученности именно данного, эмпирического множества видов. Выводы систематиков, вообще говоря, не могут быть распространены на генеральную совокупность видов. Такая систематика регистрирует изученность только эмпирического множества видов. Те изменения, которые возможны в системе вследствие открытия новых видов и их признаков, непредсказуемы. Из отдельных объяснений системы не могут быть выведены другие. Систему можно рассматривать как множество непротиворечивых утверждений, не выводимых одно из другого. Трудность создавшегося положения наглядно проявляется при сопоставлении множества особей (популяции) с множеством видов и вообще таксонов.
Изучение популяции осуществляется на выборке особей. Полученные результаты распространимы на популяцию в целом благодаря случайности выборки. Для оценки ее репрезентативности используется обычный биометрический аппарат. Если выборка не случайна и не репрезентативна, то в большинстве случаев с помощью несложных приемов можно добиться соблюдения этих условий (послойные выборки, увеличение объема выборки).
Систематики, несмотря на услуги палеонтологов, всегда имеют дело лишь с выборкой видов (и вообще таксонов), и поэтому их система остается системой эмпирического множества видов. Ее репрезентативность для системы генерального множества видов — вопрос, игнорируемый в таксономической литературе. Между тем от ответа на него зависит отношение к систематике. Она есть не более чем регистрация с утилитарными целями доступных в настоящее время фактов— если эмпирическое множество видов не представляет генерального их множества; наоборот, система обладает признаками научной теории, если благодаря репрезентативности эмпирического множества по отношению к генеральному множеству можно получить представление о системе генерального множества видов. Если мы вспомним о том, что систематики пользуются лишь выборкой признаков из их обширного множества, то станет ясно, что следует поставить еще одну задачу, аналогичную только что сформулированной: необходимо установить зависимость между устойчивостью системы и количеством использованных при построении системы приз
38
наков. Иначе, требуется определить, в каких отношениях находятся эмпирическое множество признаков с обширным, теоретическим множеством.
Думается, что основной вопрос, с которого следует начинать,— это вопрос о соотношении между эмпирическим множеством видов и теоретически возможным их разнообразием. Необходимо начать именно с этого вопроса. Но какие средства имеются в нашем распоряжении?
Лекция 3,
В КОТОРОЙ ОТСТАИВАЮТСЯ ВЫСОКИЕ’ДОСТОИНСТВА МОРФОЛОГИИ и утверждается, что фенотип ЕСТЬ СРЕДСТВО ОСУЩЕСТВЛЕНИЯ ЭВОЛЮЦИИ
Все доказательства, приводимые в лекциях, осуществлены исключительно на морфологическом материале; тем не менее они претендуют на общее значение. Основанием для этого служат высокие достоинства объектов и методов морфологии. Что же касается ее очевидных недостатков, то они свойственны, вообще говоря, любому другому подходу к проблемам систематики, и вопрос заключается лишь в том, соответствуют ли имеющиеся средства отдельным более частным задачам.
Систематика животных и высших растений продолжает оставаться преимущественно морфологической, несмотря на то что морфология утратила то исключительное положение, которое она когда-то занимала в биологии. Такое состояние систематики могло бы показаться пережитком славного прошлого морфологии. Консерватизм систематиков можно было бы объяснить тем, что биохимические, серологические и другие фенотипические признаки все еще не доказали своих решающих преимуществ и пока не внесли принципиальных изменений в систему, разработанную морфологическими методами. Приверженность систематиков к морфологии можно было бы оправдать большей практичностью морфологических признаков и недостаточной изученностью прочих. Все это в какой-то степени объясняет живучесть морфологической систематики, и некоторые рассматривают такое положение как преходящее. Однако скорее всего глубокое проникновение морфологии в зоологическую и ботаническую систематику объясняется более основательными причинами.
Разнообразие органического мира издавна воспринималось прежде всего как разнообразие морфологических признаков. Прямое доказательство изменяемости животных и растений в крупных масштабах, которые представила палеонтология, почти исключительно относится к морфологии. Таким образом, одна из важнейших предпосылок к появлению и обос-
39
нованию эволюционной теории заключает в себе морфологическое содержание. Эволюционные теории, в том числе и теория Дарвина, в значительной степени основывались на наблюдениях морфологов и в соответствии с преобладавшей в то время морфологической концепцией вида пытались объяснить происхождение морфологических видов. При этом физиология животных не имела решающего значения (Ушаков, 1959). В еще большей степени это относится к биохимии. Сравнительные и эволюционные исследования в биохимии растений почти отсутствовали вплоть до первых десятилетий нынешнего века (Благовещенский, 1966). Лишь впоследствии, благодаря в первую очередь генетике, преимущественно морфологическая теория эволюции сделалась общебиологической. При этом так же, как сегодня системы морфологов подтверждаются работами биохимиков, преимущественно морфологическая теория совпала с общебиологической.
Замечательный взлет генетики вызвал побочные последствия, которые отрицательно сказались на морфологии. Стало казаться, что одни лишь генетические механизмы достаточны для объяснения эволюции, что мутационная изменчивость одна без отбора в состоянии объяснить очевидную направленность процесса эволюции и, стало быть, эволюционная теория, основанная главным образом на морфологии, неверна. В дискуссиях первых двух десятилетий нашего века о соотношении между отбором и мутагенезом ученый мир видел основное: борьбу за материалистическое понимание движущих сил эволюции. Вывод о том, что чисто генетическая теория эволюции невозможна и что естественный отбор остается движущей силой эволюции — основной итог этих споров. Между тем, по сути дела, в них решался также более частный вопрос о научной природе морфологии, о содержательности ее объектов и правомочности методов, о возможности создания эволюционной научной теории на фактах, накопленных морфологией. И если дарвинизм выдержал проверку временем и не вступил в противоречие с современной биологией, то это оказалось возможным, видимо, потому, что с самого начала он опирался на достаточно содержательные факты. Поэтому следует признать, что по крайней мере у высших растений и животных морфологические изменения составляют реальное и существенное содержание эволюционного процесса, а не поверхностную видимость каких-то более важных явлений. Кроме того, отсюда следует, что морфологические методы вполне правомочны, а морфологические признаки заключают в себе богатейшую информацию об организме в целом. По-видимому, в этом и состоит основная причина живучести морфологической систематики.
Биологическое содержание морфологических признаков богаче, чем это представляется на первый взгляд. Пренебре-40
жительное отношение к таким «мелочам», как щетинки и ши-пики членистоногих, совершенно не оправдано. Эти кутикулярные образования формируются у членистоногих в ходе линек и представляют собою конечный результат сложных процессов, протекающих в организме (рис. 14). Даже в таких от-
JNSFCTA
активности желез ^созревание яиц
зо^пог. со^ Icorporo a//g/o| ^
„ > - торможение
неиросекретор- пр ।
„оз го KU ° Нcorpora ggrtf^.
I
^-торножемц^
.^жировое тело ' .
। разбитие
„й л, Л гонад
'-гоР’'оН \
лрот0ракстро™т-*
централь- * пая нервная система
специфическим стимул
ейросекретор=—£ g->|cwyco6oflжелезо}-
>ые клетки j>qa "Г
ные
—------е°.Р.^------>-|У-органП
подавляющий линьку ТI
(X-орган/ ।
CRUSTACEA
торножение
^эпидерм(
созревание яиц
1 hepatopancreas
развитие гонад
Рис. 14. Схема регуляции линьки у ракообразных и насекомых. Пунктиром обозначены гипотетические связи (Passano, 1960)
носительно простых таксономических признаках со сравнительно несложной генетической обусловленностью сконцентрирована богатая информация, относящаяся к организму в целом.
У низших организмов, Prokaryota, таких, как некоторые водоросли и микроорганизмы, разнообразие морфологических признаков невелико и они использовались лишь на первых этапах создания системы. В основном их систематика опирается на биохимические, физиологические и культуральные признаки. Непосредственные палеонтологические доказательства эволюции здесь не очень богаты (или отсутствуют полностью). Тем не менее основные положения дарвинизма распространены и на них. Очевидно, что биохимические и физиологические признаки у низших существ имеют для отбора то же значение, что и морфологические у высших. Действительно, в отношении таких фундаментальных свойств, как, например, тип обмена, низшие организмы представляют поразительное разнообразие в противоположность высшим. Если скудность или отсутствие морфологических признаков у низших организмов указывают на то, что эта особенность фенотипа не имела существенного значения в эволюции, то, наоборот, разнообразие физиологических и биохимических приз
41
наков свидетельствует о том, что именно они послужили точкой приложения движущей силы эволюции, естественного отбора.
Сопоставление систем, разработанных иммунологами и биохимиками, с традиционной системой, как это и ожидалось
Синтез Лизина
Рис. 15. Распространение некоторых биохимических и физиологических признаков среди высших таксонов (по Н. С. Dessauer, 1969) и филогенетическое древо (Whittaker, 1959)
Флоркэном (1947), показывает их вполне удовлетворительное совпадение. Эхо неоднократно отмечалось и в зоологической и в ботанической литературе (Благовещенский, 1966). Пожалуй, сходное положение возникло и в филогении; во всяком случае здесь также пока не наблюдается непримиримых противоречий (рис. 15). Этот своеобразный изоморфизм, кажется, получил всеобщее признание. Он утвердил правиль
42 ’
ность существующей системы и убедил систематиков в правомочности новых признаков. Расхождения, однако, отнюдь не редкость.
Накапливается все больше данных о биохимическом сходстве животных и грибов, и поэтому ставится под сомнение классическое представление о жгутиковых простейших как едином предке высших растений и животных (Белозерский, Антонов, Медников, 1972). У микобактерий «данные серологического анализа... не всегда коррелируют с другими таксономическими признаками» (Головлев, Скрябин, 1972). Имеются и другие примеры подобного же рода среди растений (Хохряков, 1972).
Использование биохимических и серологических признаков затруднено рядом обстоятельств. Считается, что некоторые даже очень сложные вещества могут быть синтезированы в организмах различными способами, и тогда их наличие не будет свидетельствовать о филогенетическом родстве и таксономической близости видов (Erdtman, 1968). Таким образом, помимо гомологичных признаков, имеющих таксономическое значение, в биохимической систематике так же, как в классической, следует различать и аналогичные затрудняющие построение системы. Третично-семантические молекулы белка являются носителями и гомологичных и аналогичных признаков (Margoliash, Fitch, 1969). А. В. Благовещенский (1965) полагает, что присутствие-отсутствие таких веществ, как алкалоиды, терпеноиды, флавиноиды и другие, не может быть полезным в систематике растений, «что все эти вещества вряд ли могут считаться определяющими биохимическую характеристику таксона, все они являются только производными настоящих подлинно специфических веществ». С ним соглашается А. Н. Белозерский (1970). Он считает, что алкалоиды, жиры и жирные кислоты недостаточно связаны со спецификой организма, а ферменты и белки представляют собой весьма сложные смеси, пока еще трудно поддающиеся индивидуализации для их химического изучения. Примерно такие же сомнения высказывает А. Н. Белозерский в отношении серологических тестов.
Существует еще одна особенность, которая роднит «новые» признаки с традиционными: непостоянство их таксономической ценности, внутритаксонная, возрастная и географическая изменчивость.
Установлено, например, что серологические различия между тремя видами рода Магнолия шире, чем между тремя другими родами растений (Johnson, Fairbrothers, 1965). В хемосистематике растений это явление носит всеобщий характер (Федоров, Пименов, 1967). У млекопитающих внутривидовая изменчивость серологических признаков перекрывает межвидовую (Воронцов, Фомичева, Баранов, 1972).
43
Антигенный состав сывороточных белков русского осетра оказался различным в трех аллопатрических популяциях: черноморской, азовской и каспийской (Лукьяненко, Умеров, 1971). Ряд интересных примеров в связи с этим вопросом приводят Ф. И. Страутман, Б. Ф. Сухомлинов, В. А. Кушни-рук (1966). Жабы высокогорья отличаются от обитающих на равнине значительно более высоким содержанием альбуминовых фракций. Высокогорная популяция прыткой ящерицы отличается от популяции низменности наличием дополнительной фракции альбуминов. Биохимический состав и структура белков сыворотки крови лягушки зависит от солености воды. С другой стороны, биохимические признаки так же, как и морфологические, подвержены возрастной и сезонной изменчивости. Помимо особенностей белкового состава крови в период размножения, о чем пишут упомянутые авторы, здесь можно сослаться на выводы И. А. Денисовой (1956) о возрастных изменениях белкового состава крови у малого суслика, Олстона (Alston, 1967)—о качественных и количественных изменениях некоторых веществ (ментол, ментон, эфирные масла) в ходе роста у растений. Немало примеров изменений биохимических параметров в эмбриональном развитии животных, такие, как активность ферментов, синтез новых антигенов, приводит Ж. Браше (1961) в книге «Биохимическая эмбриология». А. В. Благовещенский (1966) считает возможным применять биогенетический закон к объяснению возрастных и филогенетических сопряженных изменений качества ферментов и содержания низкомолекулярных белков у растений.
У ракообразных состав белков сильно меняется в ходе межлинечного цикла и в процессе виттелогенеза у самок. Характеристики белков меняются с возрастом и в течение эмбриогенеза. Саккулина, паразит крабов, влияет на содержание белков в гемолимфе хозяина (Gibert, 1972). Любопытны наблюдения над иммунологическими различиями между кровью здоровых и зараженных паразитом Sacculina крабов Carcinus maenas L. (Bessiere, Fourcade, Ngo-Van-Thien, 1961). В связи с этим обстоятельством ясно, что выводы Бойдена (Boyden, 1943) о серологических взаимоотношениях крабов (рис. 16) могли быть убедительнее, если бы было известно, что крабы не заражены Sacculina.
Такого рода факты иногда рассматриваются как «парадоксальные», препятствующие использованию «новых признаков»; между тем они свидетельствуют прежде всего о том, что «новые признаки» не отличаются принципиально от морфологических.
Возрастная изменчивость утверждает применимость к ним биогенетического закона, без которого биологический смысл морфологических признаков был бы менее понятным.
44
s S га
о a>
»S
О
§ в?
8
с
га я
3
.^са
Географическая изменчивость позволяет рассматривать «новые признаки» в свете современной теории видообразования.
Непостоянство таксономической ценности одного и того же признака свидетельствует о возможности выведения таксономических признаков из индивидуальных, о единой форме эволюционного процесса.
Вместе с тем все эти данные говорят о том, что биохимическая систематика в значительной мере отказалась от типологической концепции, согласно которой можно охарактеризовать вид по одной особи, и переключилась на изучение популяций. Необходимость такой перестройки была указана Расмуссеном (Rasmussen, 1969). Он совершенно справедливо осудил такую практику, когда по сведениям для одной-един-ственной особи многие биохимики считают возможным характеризовать вид, а по данным, относящимся к единственному виду, пытаются характеризовать более крупные таксоны. В целом систематика отказалась от этой практики после классических исследований Гексли (Huxley, 1940). Современные систематики следуют ей не от хорошей жизни: многие таксоны известны только по уникальным экземплярам. Биохимики и иммунологи получили пусть менее удобный в практической работе, но зато биологически более объяснимый материал.
И все-таки положение не так просто. Различное значение морфологических и «новых признаков» в систематике растений, грибов, животных, низших организмов вполне очевидно. Автору представляется, что основной результат из малоуспешных попыток сравнительных биохимиков ревизовать традиционную систему высших организмов и плодотворных исследований в систематике низших заключается в постановке новой проблемы. Это проблема этапности биологической эволюции, которой посвящены работы С. Э. Шноля (1971; 1973).
С. Э. Шноль выделяет три этапа эволюции, из которых нас интересуют первые два. Первый — это достижение предельного биохимического совершенства, второй этап — преодоление диффузионного барьера посредством развития морфологических структур и механизмов перемещения в пространстве. По-видимому, не следует полагать, что первый этап строго отвечает низшим организмам, второй — высшим (животным и высшим растениям). Сейчас хорошо известно, что в эволюции высших организмов также имело место совершенствование биохимической организации, тогда как у низших эволюция в какой-то степени затронула и морфологические признаки. Все же, как мне кажется, различное значение биохимических и морфологических признаков в систематике высших и низших организмов может быть объяснено этапностью биологической эволюции. При переходе от первого этапа ко второму точка приложения движущей силы эволюции естественно
46
го отбора сместилась от биохимических признаков к морфологическим. Эти представления созвучны взглядам Пири (Pi-rie, 1954) («конус Пири») на уменьшение в ходе эволюции химического разнообразия, уникальное возникновение жизни с последующим возрастанием морфологического разнообразия (рис. 17). Показательна также точка зрения Е. М. Крепса
Рис. 17. «Конус Пири», наглядно показывающий, каким образом биохимический отбор («химическая эволюция») приводит к уменьшению химического разнообразия до уникального возникновения жизни, и последующую морфологическую эволюцию, приводящую к морфологической сложности и химическому однообразию (Pirie 1954, из J. R. Nursall, 1962)
(1967): «...если природа нашла удачное химическое решение-биологической задачи, то она сохраняет его в дальнейшей эволюции». Тем не менее полезность серологии и сравнительной биохимии для систематики животных и высших растений очевидна. Благодаря им достигнута большая убедительность таксономических объяснений, таксоны приобрели как бы новое измерение. Приходится сожалеть о том, что некоторая разочарованность, постигшая исследователей в связи с неоправдавшимися надеждами на исключительность «новых признаков», как будто бы препятствует дальнейшему расширению работ в этом направлении.
Благодаря биохимии, генетике, иммунологии арсенал применяемых в систематике методов значительно обогатился, число используемых здесь признаков быстро возрастает. Оценка таксономического значения многих из них возможна на основе схемы классификации содержащихся в организме молекул (Zuckerkandl, Pouling, 1965):
1. Семантические, или сематофорические молекулы, которые несут информацию генов или переносят ее. Сюда относятся гены как первичные смысловые линейные последовательности, транспортная РНК — вторично-семантические молекулы и большинство полипептидов — третично-семантические молекулы.
2. Эписемантические молекулы, которые синтезируются под контролем третично-семантических. Сюда относятся мо-
47
лекулы, построенные при участии ферментов в отсутствие матрицы. Хотя они и не выражают с определенностью информацию семантических молекул, но сами являются выражением этой информации.
АТ-тип
ГЦ-тип
Хордовые
Беспазвоночные
j
Высшие растения
£
Актиномицеты
Грады
J<-
Зеленые водоросли
| Алорофиллсодерзкаирле одноклеточные]^-
Простейшие — 1 1 —
• (возникновение ядер/
Сине-зеленью водоросли
Вирусы (вторичное упрощение/ * 3
Бактерии
20 30 40 50 60 70 80 % Г+Ц
Рис. 18. Распределение частоты встречаемости нуклеотидного состава ДНК в крупных таксонах. На оси абсцисс процентное содержание ГЦ-пар в ДНК (Белозерский, Медников, 1971)
3. Асемантические молекулы, которые не производятся организмом.
До открытия в 1941 г. роли ДНК в явлениях наследственности систематика основывалась исключительно на феноти-
48
пических признаках. В ходе сравнительного изучения ДНК разных видов выяснилось, что особенности ее нуклеотидного состава могут быть использованы в качестве таксономических признаков. Поскольку именно благодаря ДНК сохраняется преемственность в длинном ряду предков-потомков филогенетического древа, следует думать, что в ДНК зафиксированы сведения об историческом развитии вида. Поэтому
Рис. 19. Двухмерное распределение видов бактерий по количеству ДНК на клетку (Л), нуклеотид (5) и нуклеотидному составу (Ашмарин, Харченко, Лойцянская, 1972)
А. Н. Белозерский (1970) считает данные о нуклеотидном составе ДНК наиболее надежными при построении системы. Очевидно, что если посредством изучения структуры ДНК удастся восстановить последовательность предков-потомков, чего еще не сделано, то для изучения филогении откроются новые возможности, таксономические объяснения получат более солидное обоснование. В этом направлении получены интересные результаты. Установлено, что крупные таксоны характеризуются определенным нуклеотидным составом
4
Н. А. Заренков
49
ДНК (рис. 18) и поставлена проблема эволюции этого признака. Показана возможность классификации бактерий на признаках ДНК (рис. 19). Доказано, что между биологическими свойствами бактерий и наиболее общими характеристиками их ДНК несомненно существует корреляция (Ашмарин, Лойцянская, Харченко, 1972). В систематике бактерий широко используется в качестве таксономического признака процент гомологичности ДНК (De Ley, 1968). Сравнивая настоящее положение дел с тем, которое существовало в систематике бактерий раньше, упомянутый автор пишет: «...морфология бактериальной клетки определяется менее чем 100 цистронами. Таким образом, эта морфологическая особенность, используемая уже 30 лет, опирается, самое большее, на несколько сотен цистронов, или около 10% генетической информации, содержащейся в хромосоме».
Перспективы признаков вещества наследственности, ДНК и РНК, оцениваются весьма высоко. ДНК, например, может быть охарактеризована по крайней мере шестью разными признаками, такими, как средний основной состав, длина молекулы, частота последовательностей гуанин-цитозин на участок 20ХЮ6 дальтон и 1ХЮ6 дальтон среднего молекулярного веса и др. (Mandel, 1969). Их преимущества перед традиционными не вызывают сомнений у многих биохимиках, в связи с чем часто пишут о «революции» в систематике. Эти настроения ясно выразил в 1947 г. X. Кихара: «История Земли записана в ее слоях, история живых организмов записана в их хромосомах» (цитировано по Hunziker, 1969). В работах школы А. Н. Белозерского эти принципы получили дальнейшее развитие и привели к появлению «систематики генотипов».
«Систематика генотипов», выражение впервые использованное Э. Майром (1968), не принадлежит к числу удачных высказываний этого маститого зоолога. Во-первых, потому что понятие «генотип» относится к особи; между тем, по мнению Э. Майра, объектом систематики являются популяции, но отнюдь не особи. Следуя логике этого автора, надо было бы говорить о «систематике генофондов». Во-вторых, не следует увлекаться противопоставлением генотипов фенотипам: в природе мы не встречаем их порознь (рис. 20).
Далее, если строить эволюционную систематику, за которую ратует Э. Майр, то следует систематизировать объекты эволюции. Ясно, что фенотипы, объекты непосредственного воздействия движущей силы эволюции, отбора, не могут быть оставлены систематиками. Наконец, этим новшеством подразумевается, что до последнего времени систематика оставалась «систематикой фенотипов», а это- было бы неправильно. «Фенотип — внешнее выражение' генотипа...», «...фенотип вполне закономерно отображает генотип, но только совершен-
50
воздействие срелы, вызывающее отбор фенотипов в популяции
совокупность Фенотипов популяции
4*
но в иной форме» (Шмальгаузен, 1968). Поэтому традиционная система, основанная на фенотипических признаках, есть в то же время система организмов, а правильнее — видов. Думается, что пока не следует вносить раскола в систематику, выделяя «систематику генотипов», тем более что пока ею не выдвинуто каких-либо особых методологических положений. По сути дела пока предлагаются лишь новые средства для решения уже поставленных задач.
Не следует считать, что признаки генотипов явились совершенной новостью для систематики. Такие особенности, как число хромосом и особенности их строения, количество ДНК на клетку, наконец, оценка относительного сродства генотипов посредством межвидовых скрещиваний использовались значительно раньше, до открытия генетического кода. Уже тогда сделалась очевидной относительность таксономической ценности одного и того же генотипического признака. Начнем с растений. Вот что писал по этому поводу Г. А. Левит-ский в 1931 г.: «Хотя таксономическое значение тех или иных кариотипических особенностей, вообще говоря ...изменчиво, все же нельзя не признать, что некоторые из них характеризуют обычно более крупные таксономические единицы, другие — более мелкие. На первом месте в этом отношении следует поставить две особенности, определяющие некоторые о б-щие свойства—всего кариотипа в целом: величину хромосом и их «основное число». «Таким образом, с точки зрения величины хромосом можно говорить о «кариотипе» семейств и даже порядков. Довольно часто этот признак характеризует отдельные части семейств трибы или группы родственных родов... Лишь изредка различия этого типа (достаточно резкие) заходят в пределы рода... Менее резкие отличия в величине хромосом довольно часто харак-’ теризуют как виды... так даже и разновидности». «Различия в одну, две и больше хромосом характеризуют иногда близкие и даже неразличимые расы». «...Кариотипическая характеристика подлежит тому же общему правилу относительности систематических признаков, как и обычных морфологических». Наконец, Г. А. Левитский приходит к весьма знаменательному заключению: «Естественной предпосылкой большой оценки признаков хромосомных является доказанная связь хромосом с наследственными признаками вообще. Но почему хотя бы и этой точке зрения нужно отдавать предпочтение «носителю наследственных признаков» перед самими этими признаками».
Посмотрим, однако, как обстоит дело с животными. Ка-риологические особенности, являющиеся признаками генотипов, приносят несомненную пользу зоологической систематике, и их использование способствует возрастанию неадекватности таксономических диагнозов. Например, у всех 13 изу
52
ченных в этом отношении голожаберных моллюсков отряда Nudibranchia насчитывается по п=13 хромосом, что характерно для отряда в целом (Butot, 1967), а в семействе живородок Viviparidae и=7. Между тем у 37 разных прудовиков семейства Lymnaeidae, моллюсков из отряда Pulmonata, п=10—19, причем у 5 подвидов большого прудовика Lym-паеа stagnalis насчитывается от 10 до 18 хромосом, а у 10 видов близкого рода Stagnicola всегда по п= 18 хромосом (Burch, I960; 1965; Patterson, 1966). У 18 видов упоминавшихся выше моллюсков семейства Helicidae п колеблется от 22 до 30, но все 5 изученных видов рода Helix, к которому относится виноградная улитка, имеют по п—27 хромосом (Burch, 1965). На этих примерах можно убедиться, что изменчивость в таксонах низкого ранга перекрывает изменчивость в более крупных таксонах. Это явление может быть прослежено и у растений в тех случаях, когда обширные полиплоидные ряды сортов (внутривидовая изменчивость) не уступают диапазону изменчивости между разными видами. Есть такие ряды и у простейших. Например, у инфузории туфельки Paramecium caudatum известны формы с макронуклеусом различной степени плоидности, 80п, 50—60 п, 160 п (Райков, (1967). Последний пример свидетельствует в пользу внутривидовой изменчивости еще одного генотипического признака — количества ДНК на клетку. В. Н. Орлов (1970) заключает, что у млекопитающих «не следует любые хромосомные различия форм считать безусловным доказательством их видовой самостоятельности». Становится все более очевидной и географическая изменчивость кариологических признаков. Теперь перейдем к межвидовой гибридизации. Значение этого явления для видообразования обсуждалось неоднократно (Дубинин, 1966; Завадский, 1968). Обзор проблемы в целом и библиографию приводит Н. Г. Рубайлова (1965). Общий вывод состоит в том, что межвидовая гибридизация у растений имела большее значение для ^видообразования, чем у животных. В связи с этим иногда полагают, что у растений филогенетические отношения могут быть изображены с помощью сетки (Stebbins, 1969). Однако существует и другая точка зрения (Wagner, 1969). Случаи отдаленной гибридизации весьма многочисленны, и немногие подобранные мной примеры понадобятся только для того, чтобы привлечь внимание читателя к обстоятельству, которое как будто бы оставалось незамеченным.
Скрещивание между видами есть установление сродства их генотипов. Сопоставление результатов скрещивания одного вида с другими позволяет установить относительность такого сродства. Таксономический признак «сродство генотипов» оказывается относительным, сравнительно независимым от других, и в частности морфологических. Между таксономиче-
53
ски близкими формами скрещивание может оказаться безуспешным, и, наоборот, сравнительно отдаленные виды могут давать плодовитых гибридов. С точки зрения систематики это все та же неодинаковая разрешающая способность одного и того же признака. Обратимся к примерам.
Опыты по гибридизации канарейки с аборигенными вьюрковыми Армении привели к весьма противоречивым результатам. В одних случаях не получено кладок, в других получены кладки неоплодотворенных яиц, затем отмечены случаи, когда оплодотворенные яйца не развиваются. Наконец, от некоторых пар получено потомство (Айрумян, Чилингарян, 1972). Яркую картину разноречивости данных о скрещивании в роде Гравилат дает Гаевски (Gajevski, 1959).
Весьма интересны в связи с рассматриваемым вопросом типы спаривания у некоторых ресничных простейших и, в частности, у видов рода Paramecium. Это явление, генетическая обусловленность которого строго доказана (Сигел, 1969), состоит в том, что между одними особями вида, образующими синген, спаривание возможно, тогда как с особями того же вида, но относящимися к другому сингену, оно невозможно. Приведенные факты имеют отношение к спорам по поводу «биологической» концепции вида, в которой весьма большое значение придается репродуктивной изоляции. Нас, однако, интересует лишь значение этих фактов для систематики. Думается, что из них следует сделать такой вывод: признак генотипа, обратный от сродства, а именно отчужденность, несходство с другими генотипами, относителен и обладает различной разрешающей способностью. У растений он может характеризовать обычно крупные таксоны, у животных — более мелкие, вплоть до внутривидовых форм. Кроме того, кажется почти очевидным, что способность к скрещиванию не связана тесно с морфологическим сходством между гибридизируемыми видамй. Оставляя в стороне эйдологический аспект этого вопроса, значение репродуктивной изоляции для определения вида, систематик не может оказывать предпочтение этому критерию, не вступая при этом в противоречие с морфологическими данными. Кажется более разумным поступить с этим признаком генотипа точно так же, как с другими, и •принять, что он имеет различную разрешающую способность в разных случаях и не может однозначно характеризовать собою какой-то один ранг таксонов, например вид.
Наступление нового периода в систематике, периода «систематики генотипов», о котором достаточно часто сегодня пишут, связано, однако, с другими генотипическими признаками, которые ранее не использовались, с последовательностью нуклеотидов и степенью гомологичности ДНК. Это направление систематики находится лишь в самом начале своего развития и потому, естественно, его вклад в обоснование 54
системы пока сравнительно невелик. Вследствие этого обсуждение его достоинств с точки зрения соответствия вновь полученных результатов ранее достигнутым (история, биогеография таксонов, характер и направление их эволюции) было бы преждевременным. Тем не менее некоторые особенности использования признаков вещества наследственности очевидны.
Прежде всего они не развиваются в онтогенезе так, как это свойственно признакам высших организмов. В соответствии с тем, что вообще известно о явлении наследственности, следует считать, что, например,, соотношение последовательностей нуклеотидов в ДНК зиготы, зародыша и взрослого животного остается без изменений. Поэтому признаки вещества наследственности пока лишены онтогенетического содержания, и их истолкование в связи с биогенетическим законом пока невозможно. Далее, пока ничего не известно о межпопуляционной географической и" индивидуальной, внутрипопуля-ционной изменчивости этих признаков. Вследствие этого пока трудно представить себе, каким образом могло бы происходить превращение их из индивидуальных и популяционных в таксономические.
Наконец, принцип «равномасштабности» препятствует пониманию того, каким образом возникают крупные признаки больших таксонов, и противоречит представлению о единственной форме эволюции. Все это свидетельствует о том, что используемые ныне признаки вещества наследственности, будучи свободными от «недостатков» обычных признаков, действительно могут составить основу для качественно нового этапа в развитии систематики; однако следует не упускать из виду того обстоятельства, что подкупающая ясность и недвусмысленность таксономических решений соседствует здесь с неудовлетворительной изученностью признаков ДНК.
Главное же затруднение при использовании в систематике признаков ДНК, как мне кажется, связано с жесткой однозначностью предлагаемых решений —• в пределах точности методик. Между тем постепенно это делается очевидным, таксономические построения, наоборот, не могут быть жестко однозначными, но лишь вероятностными не вследствие несовершенства методов, а по причине вероятностной сущности объектов систематики. Однозначные решения таксономических задач в принципе недостижимы.
По-видимому, в ближайшем будущем это многообещающее направление с большей очевидностью раскроет свои возможности и более основательно подтвердит их. Современное же состояние молодой «систематики’ генотипов» таково, что не дает оснований считать пройденным этапом классическую систематику; Наоборот, в последней имеются громадные неиспользованные возможности, повышающие ее
55'
научность, — например, возможность предсказания. Не для того природа трудилась сотни миллионов лет, совершенствуя морфологическую, биохимическую (т. е. не только ДНК), физиологическую организацию таксонов, чтобы сегодня мы отвернулись от результатов ее трудов.
В целом же использование признаков ДНК уже принесло систематике большую пользу. Значение таких исследований заключается прежде всего в еще не осознанном биохимиками и осуществленном ими же прямом доказательстве родства между видами таксона. Это означает окончательное крушение типологической и полифилетической концепций таксона.
Поэтому все дальнейшие построения основываются наследующих соображениях. По отношению к понятию «таксономический признак» морфологические признаки у животных играют ту же роль, что и биохимические у низших форм жизни. Вследствие этого возможно рассмотрение общих проблем систематики на зоологическом морфологическом материале и перенесение общих выводов на систематику растений, грибов, низших форм жизни. Тем не менее следует считаться с тем обстоятельством, что по причине морфологической односторонности материала какие-то важные вопросы остались незатронутыми. Поэтому любые иные подходы к теории систематики представляются весьма полезными и необходимыми.
Лекция 4, в которой доказывается, что эмпирическое МНОЖЕСТВО ТАКСОНОВ ВСЕГДА ЕСТЬ СЛУЧАЙНАЯ ВЫБОРКА ИЗ ВООБЩЕ ВОЗМОЖНЫХ
В процессе доказательства этого утверждения используются три положения: 1) представление о стохастическом характере процесса обособления популяций; 2) представление об изолированной популяции как зарождающемся виде; 3) представление об эволюции индивидуальных признаков ДО’ подвидовых и настоящих таксономических признаков.
В изолированных популяциях достаточно низкой численности в результате автогенетических процессов создаются предпосылки для видообразования. Каждая такая популяция может служить источником формирования новых. Ввиду массовости и длительности этого процесса вновь образующиеся популяции чаще всего будут основываться такими особями, которые составляют большинство в исходных уже существующих популяциях. Можно сформулировать это положение в еще более общем виде: обособляющиеся популяции большей частью основываются такими особями, которые составляют большинство всех вообще особей вида, во всех его популяциях вместе взятых.
56
И средняя, и форма кривой распределения в отдельных популяциях зависят от очень многих несогласованно действующих преходящих факторов. К их числу относятся, например, направление и интенсивность отбора, изменения численности и состава популяции. Однако предпосылки к видообразованию складываются в геологическое время, и потому следует полагать, что по истечении достаточно длительного времени в совокупности многочисленных популяций, обитающих в различных меняющихся условиях, осуществятся все мыслимые сочетания некоторых наперед заданных признаков.
Все мыслимое разнообразие четырех альтернативных признаков исчерпывается фенотипами из решетки Пеннета для тетрагибридного скрещивания:
aBCd
AbcD
aBCD aBcD abcD
AbCD AbCd abCd
ABcD abCD aBcd
ABCD ABCd ABcd Abed abed (1)
Здесь различные фенотипы сгруппированы таким образом, что в один и тот же класс попадают особи с одинаковым количеством доминантных признаков, неважно каких. Распределение таких фенотипов оказывается биноминальным и представляет собой разложение бинома Ньютона
(а + а)4 = а4а° + 4а®а^ + 6а2а2 + 4ага3 + а°а4, (2)
если каждый доминантный признак обозначить через а, рецессивный через а.
' По прошествии достаточно длительного времени распределение (1) и формула (2)' смогли бы описать и разнообразие подвидовых признаков в обособленных популяциях. Тогда каждая строчка символов в (1) представляла бы собой диагноз подвида, сочетание «почти таксономических» признаков. Если в согласии с классической теорией видообразования такие признаки имеют шансы превратиться в таксономические, то (1) и (2) смогли бы охарактеризовать разнообразие в таксоне, происходящем от одного предка. Иначе говоря, если исходить из современной теории видообразования, то структура таксона определяется структурой популяции вида-предка.
В действительности же ничего подобного пока не найдено, что и неудивительно. Описания таксонов неадекватны, тогда как (1) по сути дела есть множество адекватных диагнозов. Попробуем представить в адекватной форме разнообразие в пределах конкретного таксона.
В качестве модели рассмотрим морских креветок двух близких семейств Hippolytidae и Pandalidae. Они имеют всесветное распространение, населяя глубины от 0 до 2090 м, и
57
в ископаемом состоянии пока неизвестны. Их видовые диагнозы, к сожалению, часто несовершенны. Признаки, указанные в одних диагнозах, не упоминаются в других. Ввиду неадекватности диагнозов важнейшие сводки по этим семействам (De Man, 1920; Holthuis, 1947) оказались недостаточными (так же, как и довольно многочисленные отдельные статьи), и автору пришлось обратиться к коллекциям, собранным в течение ряда лет советскими экспедициями практически во всех районах Мирового океана. Таким образом, для 336 видов (примерно из 420 известных сейчас видов) со' ставлены такие диагнозы, в которых имеются сведения об 11 одних и тех же альтернативных признаках (табл. 1). Если ог-Таблица 1
Альтернативные таксономические признаки у креветок семейств Hippolytidae, Pandalidae
Положительная форма Отрицательная форма
Концы клешней 1 пары йог с темным пигмеи-
ТОМ . «1 без пигмента .... ai
Артробранхни на I—II ногах имеются . . . «2 отсутствуют а2
Инцизорный вырост мандибулы имеется . • а3 то же а3
Экзоподит на III ногочелюсти имеется . . • ^4 а±
Сочленовная пластинка на заднебоковом углу
VI сегмента брюшка имеется • а5 а*
Эпиподит на III ногочелюсти имеется . . . а» » as
1 супраорбитальный шип имеется .... . а7 at
2 супраорбитальных шипа имеется .... . аа as.
1-члениковый щупик мандибулы имеется . . ая » Лэ
2-члениковый щупик мандибулы имеется . • аю » аю
3-члениковый щупик мандибулы имеется . • ап аю
раничиться меньшим числом признаков, то таких видов оказалось бы больше; если же расширить число видов, то пришлось бы ограничиться меньшим числом признаков из адекватных диагнозов.
Выбор креветок в качестве «модельной группы» совершенно произволен и определен исключительно моими интересами, как карцинолога. Может показаться, чтб сам подход к выбору «модели», каковой являются таксоны, установленные традиционной, исключительно морфологической систематикой, должен ограничивать возможные выводы. В действительности же, и это постепенно сделается очевидным, данное обстоятельство не имеет решительно никакого значения.
Выбор признаков обусловлен двумя обстоятельствами. Во-первых, практикой и традициями систематики этих животных. Рассматриваемые 11 признаков — это те, которыми издавна пользуются для различения видов этих креветок. Во-вторых, доступностью коллекций и литературы. Некоторые приз-
58
даки, быть может, не учтены потому, что в коллекциях не обнаружено соответствующих видов и отдельные литературные источники остались недоступными; кроме того, приходится считаться с несовершенством некоторых диагнозов.
Теперь необходимо ввести следующие обозначения2:
at — признак i без указания на наличие его или отсутствие; '
йг — признак i отсутствует, отрицательная форма;
Of — признак i имеется, положительная форма;
т — число рассмотренных признаков, m=ll, i= 1, 2, 3,..., ..., т.
Первый из признаков, а\, используется для различения семейств: концы пальцев клешни с темным пигментом у Hippo-lytidae и без оного у Pandalidae. .Следующие 5 признаков, йг, йз, Qi, аз, ав, такие же, как и йь т. е. чисто качественные. Из них можно составить 2е сочетаний, и каждое будет отличаться от любого другого по меньшей мере одним символом. Оставшиеся 5 признаков количественные, они не могут сочетаться столь же свободно. Если креветка имеет I супраорбитальный шип, то она не может иметь к тому же и 2 шипа. Это означает, что в эмпирическом материале не может быть сочетания аТа&. Признаки й7 и й8 образуют не 22 сочетания, а лишь 3: а7й8, й7а8, й7й8. Последнее означает полное отсутствие шипов. По этой же причине признаки йэ, йю, йп могут составить не 23 сочетаний, а всего лишь 4 (см. рис. 4): ада10ац, йдОюйц, йэйюйп, йэйюйп. Формально возможное множество сочетаний 11 альтернативных признаков из табл. 1 составляет: 2е-3-4 = = 768.
В дальнейшем такие сочетания будут называться событиями. Любые 2 события различаются по крайней мере одним символом. Их можно рассматривать как таксономические диагнозы, составленные строго адекватно один другому. Относительно любого из 11 использованных признаков в событиях имеется указание, в какой форме он выражен. Событие отличается от таксона формальным содержанием. Обычно в понятие «таксон» вкладывается общность происхождения, особый ареал. В событии же пока зафиксировано лишь состояние признаков.
Множество из 768 формальных событий, при построении которого использовано 11 признаков, будем называть множеством формальных событий и обозначать через А7ц. Пока нет необходимости перечислять все 768 событий, но особая , форма представления N и понадобится уже сейчас. С этой целью введем понятие подобных событий. Подобные события — это такие, которые имеют одинаковое количество положительных
2 Заимствовано у А. С. Кузичева (1968).
59
mit а следовательно, и отрицательных «о признаков; разумеется, всегда m = mi+m0. Например, события
a7asa9aiQan и подобны. В обоих по
«1=5 положительных и по «о=6 отрицательных признаков.
Подобные события различаются лишь самими признаками. Ес-
Рис. 21. Схема порождения событий эмпирического множества Nu при использовании альтернативных признаков
ли положительный признак вообще обозначить через а, отрицательный признак, независимо от индекса I, обозначить как а, то два названных подобных события можно записать как а5а6. Множество Mi =768 событий теперь можно представить как разложение бинома Ньютона (а+а)6- (2аа-\-а2) • (3ad2+ +d3) = a°d11 + llaId10+51a2d9+131a3d8+205a4d7+201a5d6 + -}-121a6d5+41a7a4+6a8d3 (3). Первый сомножитель слева — это формальные сочетания, образуемые первыми 6 качественными признаками. Второй сомножитель — это сочетания
60
йт и а8. Третий сомножитель — все возможные сочетания й9, аю и йп. Здесь не представлены сочетания, вроде а7а8<?э Дыаи, «703619^10^11, — и т. д., бессмысленные в принятой системе кодирования.
Эмпирическое множество событий Ан получим, применив последовательно 11 признаков к 336 видам (рис. 21). Признак 51 обязательно расщепит это множество видов на два подмножества видов — с положительными и отрицательными признаками. Иной исход невозможен, потому что используются лишь такие признаки, которые выражены в разных формах. К двум полученным подмножествам применим признак а9, затем й3 и дальше как в дихотомическом ключе для определения." В конце концов исходное множество видов распределится по Ац=31 событию эмпирического множества. Каждому событию будет отвечать какое-то число видов, от 1 до 44 (табл. 2). Адекватность таксономических диагнозов, сравнимость событий, основанных на одних и тех же признаках, имеет своим следствием очень неравномерное распределение видов по событиям. Используя табл. 2, представим Ап как множество подобных событий:
2а1а10+За2а9+За3а8+8а4а7+11а5а6 +4ава5. (4)
Далее, подсчитаем число видов, приходящихся на каждый член полученного выражения, и запишем результаты как коэффициенты разложения
336 = 9(а1а10)+7(а2а9)+80(а3а8)+77(а4а7)'+ 153(а5а6) + + 10(а6а5). (5)
Теперь предстояло бы проделать основное, ради чего произведены все выкладки: сопоставить внутрипопуляционную изменчивость, как она представлена в (2), с таксономической, выраженной в (4) и (5). По условиям построения это невозможно, потому что в (2) представлены всё формальные сочетания, тогда как из (4) и (5) исключены бессмысленные при данном кодировании формальные сочетания признаков и события. По этой причине моды распределений (4) и (5) смещены вправо, где преобладают отрицательные признаки. Из этого затруднения нас может выручить (3). По сути дела, это модель популяции, в которой все возможные сочетания 11 признаков осуществляются как 768 событий; следует только заменить таксономический признак а, на индивидуальный «j. После этого формальные возражения против сопоставления (3) с (4) и (5) отпадут.
Прежде всего, распределение подобных событий во всех трех случаях сходно. В этом позволяют убедиться расчеты. В основе их положена следующая идея: если распределения трех множеств сходны, то сходны между собой и соотношения Между положительными и отрицательными символами в каждом множестве.
61
03
св
Sf
s
ч
\o св
События эмпирического множества Nn креветок Hippolytidae, Pandalidae
g-OI'rf qiooHiBocfeg O’* b-OcOLO^COLOM^ CD lOOOOO^mtN О ’t4 03 00 1П Ш О CO CO Ш 1П CO OQFHiCb-^ in оз" co ь-Г co о? ь* 0 ст of ь* о of in 03 0 1П intn CO ID «—< ’“• CO СЧ CT
ЭКЭ1 -оно yoHhHiBoatf я Еипчроо dawon in TfH < 03 TfH CD 0 0 0 b-0 -« 03 CD 03 b* in 00 CT 0 0 —<04 co —< in ^inb-CT^cD —< —• 03 03 CO CO CO CO TfH 4* TfH Tt*Tf<^c£)t>t> »—4 •“< • 1—< ^-1 Ч-Н < •—< I »—t »
ЭИ11490Э 10010100001 10010101010 10010110001 10010110010 10100100010 10100100100 10100101010 10100110010 10110000010 10110110001 10110100000 10110100001 10110100010 10110110000 10111100001 11010100001 11110100001 11011100001 а ‘
иоИиа огонь 03 CT —< COCO 03 b- 03 ’-H in ^ООЗСЗ^СО ~ 03 оз 03 CO
CD CD 0
g-OI-d OO 03 03
qiooHiBodag 19 94 CD
ЭКЭ1 CD CO CT
-OHO yOHhHlBOOH1 я вш.14900 dowoH in b-.03 03 CD
0 —«
0 0 О
00 00( 8
ЭИ1Н9ОЭ 00( 001 8
0 0 0
0 0 .
0 p 0
ио1/иа огэиь 1—1 I in co
В #ц (здесь это модель популяции) содержится 3392 положительных признаков и 5056 отрицательных; всего же их 768-11=8448 (3).
• В #ц (множество адекватно описанных таксонов) содержится 132 положительных и 209 отрицательных признаков; в общей сложности 31 • 11 =341 (4).
В диагнозах множества видов содержится 1391 положительный и 2294 отрицательных признаков; всего же их 336-11=3696 (5).
Соотношение между символами в первом множестве 15056 , 209 , _о 2300 ,
-------------- 1,49, во втором = 1,58, в третьем-— 1,65. 3392--------------------------------------------г 132-’ ’ f 1396
Здесь можно было бы применить критерий %2, который позволил бы количественно оценить различие и совпадение резуль-• татов. Однако независимо от него необходимо дать себе отчет, что они означают.
Сходство трех распределений никак не случайно, если иметь в виду хорошо известную систематикам возможность перехода признаков из индивидуальных в таксономические, т. е.
Представим себе предка изучаемых креветок, в популяции которого встречаются все сочетания 11 индивидуальных признаков. В обособившихся случайным образом популяциях эти сочетания превращаются в популяционные подвидовые, «почти таксономические». Сочетания, ранее характеризовавшие особи, теперь характеризуют подвиды. Следующий шаг — это становление таксономических признаков, когда, наконец, сочетания индивидуальных признаков переходят в сочетания таксономических, т. е. события. В силу вероятностного характера возникновения предпосылок к видообразованию (т. е. процесса обособления популяций), таксономические результаты сохранят преемственность с исходным материалом.
Разнообразие в популяции предка оказывается непосредственной причиной разнообразия множества видов таксона., Структура таксона сохраняет черты сходства со структурой популяции вида-предка.
Отсюда ясно, что критерию %2 не следует придавать большого значения. Модель популяции предка, за которую было принято множество #ц, весьма несовершенна. Известно, что-в природе по разным причинам далеко не всегда возможны менделевские популяции и по разным причинам не всегда, симметричны кривые изменчивости. Между тем модель популяции предка #п была построена именно как менделевская Ж популяция с нормальной кривой изменчивости. Даже значи
63-
тельные отклонения распределении (4) и (5) от (3), которые мог бы показать %2, никак не являются основанием для того, чтобы признать ошибочными попытку вывести структуру таксона из структуры популяции. Совсем не обязательно и другое допущение, а именно, что все 11 признаков имелись у вида-предка в качестве индивидуальных. Можно допускать, что они превращались в таксономические последовательно один за другим. Ясно также, что судьба других признаков нисколько не влияет на ход наших рассуждений.
Сходное распределение подобных событий в Nn, Nи и 336 диагнозах множества видов послужило лишь отправной точкой для гипотезы. Статистический критерий мог бы подтвердить ее правильность, но решающее значение при этом имеют биологические соображения, о которых упоминалось в самом начале лекции.
Конечный вывод мог бы быть следующим. Эмпирическое множество подобных событий и адекватным образом описанных видов креветок, представленное в качестве строчек символов, есть случайная выборка из формального их множества. Нет оснований полагать, что факторы становления таксонов были иными в отдаленные от нас геологические эпохи и у других организмов. Поэтому возможна иная, более общая редакция только что сделанного вывода: доступное для изучения множество адекватным образом описанных таксонов в любой момент есть случайная выборка из их формального множества.
Универсальность вывода требует проверки. Следует обратить внимание на то обстоятельство, что в нем не упоминается о репрезентативности. А главное — возможно ли в природе формальное множество событий?
Л е к ц и я 5,
В КОТОРОЙ АВТОР ОБСУЖДАЕТ РЕАЛЬНОСТЬ ГРАНИЦ ТАКСОНОВ И ПЫТАЕТСЯ ПОКАЗАТЬ, ЧТО ОНИ РЕАЛЬНЫ ТОЛЬКО ПРИ УСЛОВИИ НЕОСУЩЕСТВИМОСТИ ФОРМАЛЬНОГО
МНОЖЕСТВА СОБЫТИЙ N*m
Задача, поставленная в заключение лекции 4, имеет один весьма важный для систематики аспект, а именно реальность границ таксонов. Можно думать, что если в природе осуществимо множество событий Nm, где представлены все возможные сочетания признаков, то границы между подмножествами событий, адекватно описанных таксонов, суть условность, навязанная систематиками природе.
Границы таксона определяются таксономическим диагнозом. В нем перечисляются признаки, общие для всех более мелких соподчиненных таксонов. При этом подразумевается,
64
что все они к тому же имеют более крупные признаки того таксона, который включает данный наравне с некоторыми другими, такого же ранга.
Например, диагноз типа кольчатых червей, к которому принадлежит обыкновенный земляной червь, выглядит так: у всех видов кольчатых червей тело разделено на сегменты с гибкими наружными покровами, которые подостланы продольными и кольцевыми мышцами; в каждом сегменте имеется хорошо развитая полость тела, заполненная полостной жидкостью, и т. д. Непосредственное наблюдение за ползущим животным заставляет признать взаимную обусловленность перечисленных признаков. Перистальтический способ движения дождевого червя возможен при наличии сплошного слоя продольных и кольцевых мышц, гибкой кутикулы и упругого внутреннего гидравлического скелета в каждом сегменте. Такой тип организации прослеживается на множестве разных видов, между которыми существуют другие, более мелкие различия.
Другой пример—это высшие ракообразные. Их типичный представитель, речной рак, имеет твердые покровы тела, членистые конечности, мускулатуру пучкового типа. Твердый наружный скелет исключает перистальтический способ движения и делает излишней хорошо развитую полость тела. В связи с этим она не функционирует в качестве опорного органа и заполняется внутренностями. Появляются новые органы передвижения — членистые конечности рычажного типа и соответствующая им пучковая мускулатура.
В обоих примерах причинная, биологическая обусловленность отдельных таксономических признаков не вызывает сомнений. Сочетания признаков не случайны, границы таксона естественны, и в этом смысле сами таксоны (кольчатые черви, высшие ракообразные) кажутся реальными. Систематик уверен в том, что иные комбинации тех же признаков (рис. 22) неосуществимы и эволюция никогда не создаст таких странных существ. Таксономический диагноз представляется не просто перечислением признаков, но также описанием некоторого типа биологической организации. Реальность биологической организации здесь имеет причинное обоснование. Объяснение невозможности одних сочетаний признаков имеет столь же большое значение, что и причинное объяснение существующих.
Тип биологической организации, свойственный ракообразным, выводится из кольчатых червей очень просто, если ограничиться немногими упомянутыми признаками. Для этого достаточно допустить, что у каких-то кольчатых червей произошло упрочение наружных покровов (например, в связи с возрастанием размеров), как следствие—утрата полостью тела опорной функции и появление нового двигательного аппа-
5
Н. А. Заренков
65
рата. Взаимная обусловленность таких признаков, установленная сравнительно-морфологическим методом, имеет для систематики то значение, что предлагает причинное объяснение границам таксонов, позволяет внести филогенетическое содержание в систему, наконец, оправдывает* рассмотрение системы на основе эволюционной теории.
^7 2 3
Рис. 22. Тип морфологической организации кольчатых червей (А) и высших ракообразных (В); невозможные типы организации (В и Г) совмещают признаки первых двух:
1 — кутикула; 2 — кольцевая мускулатура; 3 — пучковая мускулатура; 4 — продольная мускулатура
Количество подобных примеров может быть значительно увеличено. Их можно найти в замечательной книге В. Н. Беклемишева «Сравнительная анатомия беспозвоночных» (1964). Например, отношения между двигательным ресничным эпителием и размерами тела у водных беспозвоночных могут быть объяснены с использованием понятия «относительная поверхность тела». У мелких животных (простейшие, ресничные черви, личинки водных беспозвоночных) передвижение с помощью ресничек весьма обычно, у более крупных наблюдается в виде исключения (гребневики), а чаще осуществляется каким-либо иным способом. Очевидно, что ресничный двигательный аппарат оправдан у мелких животных с их большой относительной поверхностью тела.
У тех же ресничных червей проявляется зависимость между размерами тела и строением кишечника. Она состоит в том, что у крупных форм кишечник разветвлен больше, чем
66
у мелких (рис. 23). Это объясняется тем, что у ресничных червей нет распределительной, системы. Поэтому питательные вещества не смогли бы достигнуть внутренних органов, если бы ветви кишечника не пронизывали бы относительно равномерно всю массу животного. Понятно, что всасывающая поверхность кишечника должна находиться в прямой зависимости от размеров животного, что и достигается появлением новых выростов кишечника.
А Б В
Рис. 23. Строение кишечника у разных ресничных червей:
А — прямокишечные; Б — три-ветвистокишечные;
В — ветвистокишечные (по L. Н. Hyman, 1951)
По-видимому, по тем же причинам у крупных сцифоидных медуз каналы пищеварительной полости ветвятся все больше и больше в направлении от желудка в центре зонтика к его краю. Наоборот, у мелких гидроидных медуз каналы пищеварительной полости ветвятся меньше, но и у них разветвленность пищеварительных каналов с возрастом может возрастать (рис. 24).
У морских моллюсков, хитонов, количество жабер зависит от размеров тела: у крупных видов их больше, чем у мелких (Johnson, 1969). Та же закономерность наблюдается у ракообразных. Низшие ракообразные, такие, как циклопы, не имеют специальных органов дыхания, что связано с их малыми размерами и тонким наружным скелетом. У высших ракообразных, в общем более крупных, наружный скелет прочнее и не проницаем для газов в такой степени, как у низших. Вследствие этого появляются специальные жабры, такие, как упоминавшиеся ранее артробранхии у креветок. Членистоногие с их наружным хитиновым панцирем не могут эволюционировать в направлении существенного увеличения разме
5*
67
ров тела в связи с тем, что при возрастании массы объем увеличивается быстрее, чем поверхность, и нагрузка на панцирь усиливается настолько, что в конце концов может пре-
Рис. 24. А — неразветвленные пищеварительные каналы у молодой гидромедузы Proboscidactyla stellata (Forbes) с зонтиком диаметром 10 мм; Б—то же при диаметре зонтика 45 мм (Russell, 1953); В — пищеварительные каналы у крупной сцифоидной медузы Aurelia aurita L. (Наумов, 1961)
высить разумные пределы. Понятно, что самое крупное водное членистоногое, японский краб Macrocheira kaempferi De Haan, превосходит наиболее крупного из наземных членистоногих — африканского жука Голиафа.
Вообще между размерами тела у животных (относительная поверхность) и различными другими особенностями их ор-68
ганизации существуют самые различные, иногда совершенно неожиданные зависимости.
Актиния и гидра, обитающие на дне, характеризуются радиальной многолучевой симметрией, которая проявляется в наличии того или иного числа щупалец, расположенных вокруг рта. Такой план строения целесообразен, если учесть, что пища и другие раздражители воспринимаются животным как бы с поверхности некой полусферы, опрокинутой над ним.
Рис. 25. А — поперечный срез актинии с радиальными септами пищеварительной полости и щелевидной глоткой; Б — внешний вид актинии; В — поперечный срез и внешний вид восьмилучевого полипа
Однако актиния имеет к тому же щелевидную глотку с сифо-ноглифами (рис. 25) и благодаря ей двулучевую симметрию. Этот признак нельзя объяснить прикрепленным образом жизни животного. Радиальная симметрия второго порядка, наименьшего из всех порядков радиальной симметрии вообще, признак, очевидно, вторичный. Возможно, она связана с крупными размерами этих хищных животных, могущих заглатывать крупную добычу, — ведь для этого надо иметь щелевидный рот, который удобно не только широко раскрывать, но и замыкать. Эта задача была бы более сложной в случае круглого рта, чем щелевидного. Щелевидное ротовое отверстие еще замечательно своими сифоноглифами, углами ротовой
69
щели, снабженными ресничным покровом. Считается, что благодаря сифоноглифам происходит упорядочение тока воды, поступающего в пищеварительную полость и приносящего пищевые частицы и кислород. Такая особенность вполне понятна: у актинии, более крупной, чем гидра, потребность в кислороде значительна. Так что щелевидный рот с обособившимися сифоноглифами также в конце концов объясняется крупными размерами их обладателя.
Все это дало повод Б. Холдейну (1970) заметить, что «сравнительная анатомия есть не что иное, как история борьбы за увеличение поверхности тела живых существ в соответствии с их объемом». Можно, однако, указать возможные исключения из этого правила. Большая относительная поверхность мелких водных беспозвоночных должна облегчать диффузионные физиологические процессы, такие, как выделение. В связи с этим следовало бы ожидать, что у мелких животных будут отсутствовать специальные органы выделения. Действительно, такие органы отсутствуют у мелких кишечнополостных (гидра), однако имеются у коловраток, наимельчайших из многоклеточных (протонефридии). Очевидно, для того чтобы объяснить такие случаи, требуется знать что-то еще.
Сходная проблема существует в систематике сосудистых растений. Например, между признаками древесины разных видов двудольных отмечены корреляции статистического характера (Кедров, 1968). В общем же, однако, причинная обусловленность таксономических признаков проявляется у растений не столь четко, как у животных (Тахтаджян, 1954).
Приведенных примеров, на мой взгляд, вполне достаточно для того, чтобы решительно отвергнуть высказывание В. В. Станчинского (1923) и К. М. Хайлова (1970) о том, что филогенетическая систематика не в состоянии объяснить наблюдаемые факты и выяснить закономерности эволюции отдельных групп животных.
Диагнозы более мелких таксонов, видов, родов, семейств, используют признаки, функциональное значение которых час-' то загадочно и природа отношений между которыми неизвестна. Что можно, например, сказать по поводу функционального значения шипов, покрывающих панцирь и конечности камчатского краба, радующей глаз скульптуры раковин брюхоногих моллюсков, щетинок на насекомых и клещах? Между тем эти особенности используются для различения таксонов. Разнообразие таксонов насекомых можно было бы рассматривать, как множество сочетаний таких признаков, как число члеников лапки и усиков, особенности жилкования крыльев, типов ротового аппарата. Функциональное значение этих признаков, взятых по отдельности, часто известно. Например, уплотненная часть переднего крыла у стрекоз, называемая глазком, служит для предотвращения вредной вибрации крыльев,
70
а жилки являются их каркасом. Но почему такое строение крыльев сопряжено со столь замечательно крупными глазами, можно ли считать, что эти признаки взаимно обусловлены? Здесь нет причинного объяснения невозможности иных сочетаний признаков, даже когда известно функциональное значение каждого в отдельности.
Нет оснований не признавать непрерывность множества сочетаний таких признаков. Такое множество (назовем его континуумом) обладает теми же свойствами, что и формальное множество событий, о котором уже шла речь; в сущности, это синонимы. Существование границ между таксонами кажется лишь следствием нашего незнания мировой фауны и флоры, вымирания промежуточных звеньев и незавершенности эволюционного процесса.
Может показаться, что здесь следует различать таксономический и филогенетический аспекты, как это предлагают, например, А. Кэйн (1958) и Я. И. Старобогатов (1969). В филогенетическом аспекте мы имеем дело с последовательностью связанных преемственностью популяций, и таксономические единицы, если их применять на достаточно полных филогенетических рядах, теряют свои границы и вместе с тем и свою определенность. Употребление таксонов при анализе филогении правомочно только за недостаточной полнотой геологической летописи. В систематическом аспекте мы имеем дело с одновременно существующими группами особей, которые изолированы (и не только пространственно) друг от друга. Они в той или иной степени разграничены разрывами. Необходимо отметить, однако, что на практике систематик имеет дело не только с потомками видов-предков, которые разграничены благодаря вымиранию промежуточных форм, но также и с сосуществующими одновременно предками-потомками, которые, согласно дарвинизму, составляют непрерывную последовательность. Все такие таксоны в равной степени изучаются систематиками, и потому не следовало бы вопрос о возможности их разграничения ставить в зависимость только от неполноты геологической летописи, бесследного вымирания промежуточных форм. Посмотрим, как этот вопрос решается в литературе.
«Филогенетическая реконструкция представляет собой континуум. Это согласуется со взглядами Дарвина, а также с современным дарвинизмом... Если бы эта доктрина стала повсеместно принятой, то она оказала бы сильное и вредное влияние на принципы классификации и понятие вида» (Tom-son, 1966). «...Лишь неполнота палеонтологических данных создает спасительный морфологический hiatus» (Соколов, 1965).
«Дискретность таксонов — следствие вымирания; эволюция непрерывна». Так понимает А. А. Любищев (1966) основ
71
ное положение традиционной систематики, так понимают его и другие исследователи (Синская, 1948; Терентьев, 1966; Тимирязев, 1937; Cain, 1968; Farris, 1954; Michener, 1970).
Непрерывность эволюционного процесса, может показаться, подразумевает безразличные отношения между таксономическими признаками, их способность образовывать все возможные сочетания.
Множество таксонов возникает как множество обособленных популяций, которые различаются сочетаниями мелких признаков. Межпопуляционная изменчивость представляется непрерывной, причем сами признаки лишены способности к избирательному объединению друг с другом (Терентьев, 1966). «Организм состоит из совокупности признаков, которые проделывают эволюцию в значительной степени (иногда совершенно) независимо один от другого» (Берг, 1922). Филогенетический континуум тем 'более вероятен, что множество популяций зарождающихся видов непрерывно возобновляется.
Все было бы вполне ясно, если бы палеонтология располагала полными филогенетическими рядами. К сожалению, это не так. Усилиями палеонтологов было открыто много новых таксонов, благодаря чему спрос на переходные формы может быть даже несколько возрос. Филогенетическое древо украсилось боковыми ветвями, но основания их повсюду остаются висеть в воздухе. Дж. Симпсон (1948) обстоятельно рассмотрел проблему неполноты геологической летописи. Он пришел к выводу, что основная причина этого явления состоит в том, что предковые переходные виды, представленные большей частью особями мелких размеров, состояли из небольших популяций, и потому могли сохраняться только в исключительных случаях. Е. Н. Синская (1948) и А. Н. Криштофович (1950) также полагают, что переходные формы остаются часто неизвестными вследствие своей малочисленности. Одновременно он обращает внимание на трудности распознавания переходных форм. Доводы Дж. Симпсона, основанные на популяционно-генетических соображениях, вызывают к себе интерес и уважение, однако следует иметь в виду, что его построения достаточно гипотетичны. Крупные таксоны формируются геологически длительное время, и этот процесс пока невозможно изучать методами генетики. Принимая дарвиновскую точку зрения на крупные таксоны как результат сильно продвинутой эволюции популяций, следует в то же время считаться со значительно меньшей изученностью макроэволюции по сравнению с микроэволюцией, на что обращает внимание, например, К. М. Завадский (1971).
Отчаявшись найти переходные формы среди вымерших и ныне живущих видов, биологи обратились к воображению. Сравнительное изучение ныне живущих видов, которые находятся на разных стадиях эволюции, а некоторые состоят
72
в родственных отношениях как предки-потомки, до некоторой степени есть изучение эволюции их строения. Отсюда возможность реконструкции их гипотетического предка.
Теория дисимметрии брюхоногих моллюсков, которая объясняет своеобразный план строения этих животных, принадлежит к числу наиболее красивых и обоснованных обобщений сравнительной морфологии. Она опирается на большой материал о разнообразных вымерших и ныне живущих видах. Согласно ей дисимметрия брюхоногих моллюсков выработалась постепенно в связи с регулированием центра тяжести внутренностного мешка; исходно же брюхоногие моллюски были симметричными животными. Поскольку среди известных систематикам видов вполне симметричные формы неизвестны, была предложена реконструкция предка. Подобные гипотетические предки придуманы для рыб и наземных позвоночных, но трудно сказать, насколько такое решение утешительно для систематика, признающего филогенетический континуум и не обнаруживающего такового.
Иногда, однако, разрывы в филогенетической последовательности таксонов объясняют не только вымиранием, но также дискретностью мутационного процесса и временным ускорением темпов эволюции (Darlington, 1940; Ziegler, 1959). Здесь же следует упомянуть о взглядах некоторых палеонтологов, согласно которым новые крупные таксоны могут появляться внезапно, повинуясь еще неизвестной науке особой творческой силе. Однако сам факт внезапного появления новых таксонов не может считаться установленным и для объяснения их происхождения пока не требуется признания иных факторов, чем те, которые известны в настоящее время.
Переходные формы, обязательность которых считается естественным следствием из постепенности эволюции, называются в качестве важнейшего аргумента против целесообразности традиционной системы (Смирнов, Вермель, Кузин, 1924). С действительностью этого аргумента соглашается Е. И. Лукин (1971). Множество таксонов в основном обладает свойствами континуума (Пономаренко, Расницын, 1971). И. А. Наумов (1950) считает, что «каждый из признаков у грибов в очень широкой степени может сочетаться с любым Другим». Отсюда недалеко до отрицания реальности границ таксонов. Действительно, В. Е. Руженцев (1960) замечает, что отрицание объективности таксонов у последователей Ч. Дарвина вызвано тем, что последний в основном признавал только медленный и непрерывный ход эволюции. Например, Дж. Гексли (Huxley, 1940) также считает естественным таксоном только вид и искусственными все прочие таксоны. Эту точку зрения последовательно проводит Э. Майр, ее разделяют многие исследователи (Козо-Полянский, 1922; Аша-don, 1970; Peters, 1970; Simpson, 1961; Thorpe, 1940).
73
Столь же последовательно возражает Э. Майру по данному вопросу В. Г. Гептнер (1965, 1971). Конечно, дело не только в том, что точка зрения Ч. Дарвина зачастую воспринимается излишне упрощенно — Ч. Дарвин полагал, что отбор лишь более резко очерчивает изначала обособленные виды. Главное заключается в том, что на основании громадного опыта биологи относятся к таксонам как к биологически реальным объектам. Если все же таксоны нереальны, то зачем Дж. Симпсон (1948) строит графики вроде того, что изображен на рис. 11, а Э. Майр не только активно их использует в собственных построениях, но также пишет о «жизненных циклах высших [таксономических] категорий»?
Если множество таксонов непрерывно во времени и разрывы во множестве обусловлены неполнотой геологической летописи, то границы между таксонами не отражают сущности объектов систематики. Выделение таксонов из их непрерывного множества возможно лишь благодаря счастливым для систематиков случайным обстоятельствам, неблагоприятным условиям захоронения животных и растительных остатков. Если бы эти обстоятельства сложились иным образом, то мы располагали бы иным набором таксонов, построили бы иную систему и имели бы, вообще говоря, несколько иное представление о филогенетическом древе. Разрывы, возникающие вследствие вымирания, делают возможным лишь распознавание их границ. Сами же таксоны были бы условностью.
Возможно и другое решение. Множество таксонов не образует непрерывной последовательности во времени. Это было бы действительно так, если бы некоторые сочетания признаков оставались бы неосуществленными. Тогда таксоны — реальность, границы между ними — проявление внутренних свойств объекта систематики; все это никак не обусловлено неблагоприятными условиями захоронения. Ясно теперь, что все сводится к вопросу о том, обусловлены ли взаимно таксономические. признаки. Одновременно это есть вопрос о таксоне как типе биологической организации. Л. А. Зенкевич (1939) писал по этому поводу следующее: «...имеются все основания предполагать, что основной набор признаков, характеризующих ту или основную категорию высшего, чем вид, порядка, оказывается чрезвычайно устойчивым, способным сохраниться в течение ряда геологических эпох». Ю. М. Вермель (1931), имея в виду таксономические признаки, считает общеизвестным фактом чрезвычайную распространенность корреляций между ними, и их неравноценность, которая выражается в том, что различные признаки связаны с неодинаковым количеством других признаков.
Зоологи давно освоились с представлением об эволюции видов, как эволюции неких целостностей. Работа И. И. Шмаль-гаузена «Организм как целое в индивидуальном и историче
74
ском развитии» (1942) утверждает это своим названием; в ней он четко различал организменный и видовой уровень. Однако в литературе часто игнорируется это положение.
С представлением о таксоне прежде всего связано понятие об определенном типе биологической организации, свойственной всем видам этого таксона. Поначалу в него вкладывалось преимущественно морфологическое содержание и относилось оно к организму больше, чем к виду. Ж. Кювье изложил свой «принцип корреляции»-таким образом, что можно было бы думать о его эволюционном значении несмотря на то, что это противоречило бы взглядам ученого на неизменяемость видов. Принцип корреляций был применен Ж. Кювье в ходе реконструкции вымерших видов, когда по немногим костям восстанавливался облик животного в целом. Это, быть может, убедительнее всего свидетельствует о том, что Ж- Кювье имел в виду организменную целостность. Н. Н. Воронцов (1967) называет корреляции Ж. Кювье «статическими». В дальнейшем мнение о взаимной обусловленности отдельных признаков в эволюции высказывалось неоднократно. Мастерский обзор этой проблемы, произведенный А. Н. Северцовым (1939), значительно облегчает нашу задачу. Может показаться, что для выражения биологической организации в целях систематики более других подходят координации А. Н. Северцова, которые были определены им так:
«Координациями» или «филетическими координациями» мы будем обозначать постоянное сосуществование двух или нескольких органов одного и того же организма. Критерием координации мы принимаем, таким образом, признак константного сосуществования» (стр. 430).
Для координаций А. Н. Северцова представляется бессодержательным случай, когда у организма имеется лишь один из двух или нескольких рассматриваемых органов или когда отсутствуют оба органа. Примеры координаций, приводимые А. Н. Северцовым, показывают, что, несмотря на неясность определения (говорится об организме, а явление в целом относится к филогенезу), оно заключает в себе эволюционное содержание, а потому могло бы найти применение в систематике. Его примеры относятся к таким органам, функции которых известны и природа отношений между которыми понятна. Таковы примеры со скелетом и мускулатурой, отделами черепа позвоночных. Сюда же относятся разобранные в начале лекции отношения между массой тела, поверхностью и различными особенностями у беспозвоночных. В этом смысле координации А. Н. Северцова могут быть обоснованы на эмпирическом множестве таксонов, они не зависят от полноты знания мировой фауны.
Взаимосвязь между размерами (весом) тела у позвоночных и такими их морфофизиологическими особенностями, как
75
строение и вес головного мозга, двигательная активность и строение органов передвижения, сердечная деятельность, теплообмен, величина глаз и характер роста и др., обоснованные И. Д. Стрельниковым (1970) на богатейшем материале, вполне могут быть отнесены к координациям А. Н. Северцова. Выводы И. Д. Стрельникова убедительны не столько благодаря обширности фактического материала, сколько потому, что допускают причинное объяснение. И наоборот, эмпирический вывод относительно обратной зависимости между числом видов и размерами их тела и головного мозга кажется лишь правдоподобным, несмотря на многократную констатацию этого факта — именно потому, что не доказана причинная обусловленность таких взаимосвязей. Точно так же и заключение «в процессах видообразования происходит их (морфофизиологических признаков. — Н. А. 3.) смена не в разрозненном порядке, а во взаимосвязи, составляющей единство и целостность организма» (Стрельников, 1970) убедительно постольку, поскольку для каждой взаимосвязи можно предложить причинное объяснение.
Большая убедительность классических исследований В. О. Ковалевского по эволюции копытных достигнута благодаря причинности объяснений филогенетических изменений. Если же рассматривать такие признаки, функции которых неизвестны и природа отношений между которыми непонятна, то любой такой случай не может быть изучен в связи с координациями А. Н. Северцова. Взаимная обусловленность подавляющего большинства таксономических признаков не мо-. жет быть установлена только лишь на эмпирическом множестве видов. Мы никогда не можем быть уверены в том, что в природе отсутствуют такие сочетания признаков, которые заполнят разрывы в существующей системе и таким образом приведут к построению континуума.
Таксономический диагноз лишь тогда утверждает некоторый тип биологической организации, когда основывается на координациях. В противном случае, когда применение координаций невозможно, таксономический диагноз есть лишь перечисление таксономических признаков, лишь описание одного из сочетаний признаков, вырванных из'их непрерывной последовательности. Наконец, А. Н. Северцов пишет об отношениях между органами, тогда как систематика интересуют признаки. Один орган может иметь много признаков; с другой стороны, наличие-отсутствие целого органа также есть признак. Понятие «признак» шире, чем понятие «орган».
Функциональное значение органов известно чаще, чем признаков. Уже одно лишь это обстоятельство ограничивает применение координаций.
Впоследствии координации были рассмотрены И. И. Шмальгаузеном (1969), но по-прежнему они не были перене
76
сены на признаки неизвестного функционального значения. А. Л. Тахтаджян (1954) следует северцовскому пониманию филетической взаимосвязи между признаками: «Нет ясно выраженных функциональных отношений и потому нет определенных координаций между эволюцией спорофита и гаметофита, стебля и цветка, ты-
чинок и плодов и многих других органов и частей».
Некоторые биологические теории выражают целостность организма даже в отношении таких загадочных признаков и, может показаться, пригодны для решения задач систематики.
Плейотропное действие генов — первое, что приходится упомянуть в связи с взаимной обусловленностью признаков. Это понятие, однако, относится к организменному уровню. Множественный эффект мутации по одному гену проявляется в онтогенезе особи. Новые признаки и их сочетания распространяются в популяции в ходе скрещиваний, и далее можно было бы представить себе, как они превращаются в подвидовые благодаря
Рис. 26. Относительный рост клешни у креветки Palaemon malcomsoni (Hixley, 1932)
обособлению и изоляции популяций. Тем не менее ничто не
давало бы нам оснований считать, что какие-то сочетания популяционных (а впоследствии таксономических) признаков возможны, а какие-то невозможны. Но именно это и требуется для того, чтобы решить проблему континуума таксонов. Плейотропия — механизм, объясняющий сцепленность признаков особей, но из него не следуют запреты на сочетания
признаков таксонов.
Согласно теории относительного роста (Huxley, 1932; Шмальгаузен, 1935), размеры частей тела растущего организма изменяются с разной скоростью относительно друг друга (рис. 26). Таким образом, теория относительного роста выражает морфологическую целостность особей. Это также организменный уровень. Иногда исследователи располагают семейством кривых для описания возрастных изменений отдельных особей одного вида, и такие данные фактически относятся к популяции. Еще одна возможность приложения теории относительного роста к популяциям появляется при
77
использовании выборки особей разных возрастов. Построен.! ные кривые, несомненно, характеризуют некоторую популяцию! но фактически их относят к организменному уровню, потому] что исследователь не часто может наблюдать' возрастные из-] менения на протяжении индивидуальной жизни одной особи! Такой прием оправдан постольку, поскольку правомочна за-1 мена разных возрастных стадий одной особи особями разных] возрастов. I
Параметры кривых относительного роста (наклон, кри-з визна) могут служить таксономическими признаками. Напри-] мер, разные виды сверчков рода Gryllid.es имеют собственные] показатели относительного роста частей тела (Cousin, 1956).! В качестве показателя степени различий 'между двумя вида-1 ми предлагается отношение числа различающихся параметров.] к общему числу имеющихся (Kurten, 1958). Наконец, суще-] ствует понятие филогенетического относительного1 роста, ког-1 да вместо особей и популяций рассматриваются виды. Возрас-1 тание размеров животных в филогенезе и сопутствующее ему] изменение пропорций устанавливается на выборке видов из! множества вообще возможных, и потому само явление, стро-1 го говоря, не может быть доказано на основе только эмпи-1 рического материала. Решающее значение здесь может иметь] разумная биологическая теория. Если же ее нет, то взаимная] согласованность большинства таксономических признаков не-1 известного биологического значения, которая заключается] в закономерном изменении относительных размеров, не мо-1 жет быть доказана в рамках теории относительного роста.] Исследователь, вообще говоря, ничем не застрахован от того,] что с некоторого момента изучения новые точки не попадут] на эмпирическую кривую графика. 1
По сходным причинам нас не могут выручить и корреляци-1 онные плеяды, усовершенствованные П. В. Терентьевым] (1959). Это понятие относится к популяционному уровню и] определяется как существование среди множества признаков особей одного вида подмножеств, плеяд с более тесными кор- ] реляциями, чем между ними и другими признаками. Так же, ] как в теории относительного роста, здесь практикуется заме-) на разных возрастных стадий одной особи особями разных^ возрастов. Параметры множества коэффициентов корреляции ] между всеми возможными парами принятых во внимание при- 3 знаков и состав плеяд могут служить в качестве таксономиче-1 ских признаков, причем считается, что наиболее тесно скор-] релированные признаки представляют наибольшую ценность] для систематики (Терентьев, 1966). С. В. Канеп (1966), на- ] пример, выявил среди разных особенностей черепа серых по-| левок «признаки-индикаторы», которые, как он полагает, име- ] ют наибольшую таксономическую ценность, поскольку скор-1 релированы с признаками плеяды сильнее, чем с другими |
признаками. Аналогичные исследования выполнены на разных экологических формах зубчатки Odontites serotina (Lamarck) (Scrophulariacea) и других растениях и животных (Шмидт, 1964; Колосова, 1971).
Тем не менее осталось невыясненным, почему наиболее согласованные в онтогенетическом развитии признаки представляют в то же время наибольший интерес для систематики. Это можно рассматривать в качестве гипотезы, но не доказанного положения. В противоположность мнению А. А. Лю-бищева (1971) этот метод пока еще нельзя считать таксономическим. Что же касается проблемы континуума, то для нее из корреляционных плеяд не следует каких-либо выводов. Основываясь на разной скоррелированностй признаков одного вида в популяции ничего нельзя сказать о том, какие корреляционные плеяды, какие сочетания признаков в эволюции осуществимы и какие не осуществимы. Совершенно необязательно, чтобы тесно скоррелированные в онтогенезе признаки были прочно сцеплены и в филогенезе. Совместной встречаемости некоторых двух признаков у двух разных видов могут отвечать разные коэффициенты корреляции, 0,5 и 0,9, например. Они вычисляются для особей, нас же интересуют разные виды. Корреляционные плеяды не могут быть использованы, если один из изучаемых признаков отсутствует у части видов.
Все три понятия: плейотропия, относительный рост, корреляционные плеяды — относятся к организменному и популяционному уровням. Согласованность признаков особей достигается здесь в ходе онтогенеза. Контроль за ходом этих процессов осуществляется в форме команд генотипа. Виды, эволюционирующие объекты, формируются под действием отбора, одних лишь механизмов организменной регуляции здесь недостаточно.
Можно было бы попытаться разрешить проблему континуума на основе биохимии ДНК. Однако имеющиеся здесь трудности настолько велики, что кажутся пока непреодолимыми. Прежде всего, количество возможных сочетаний нуклеотидов выражается астрономически большими числами, тогда как число используемых таксономических признаков сравнительно невелико. Непонятно, каким образом среди сочетаний нуклеотидов можно было бы выбрать те, которые относятся к организменным признакам и какие к таксономическим. Затем, следует иметь в виду неудовлетворительную изученность проблемы «ген—признак», в целом остающейся на уровне гипотез (Медведев, 1968; Тимофеев-Ресовский, 1970). Поэтому пока Невозможно установить соответствие между строением ДНК и структурой таксонов.
В связи с очень возможной согласованностью «запрещений» в генотипе с таковыми в фенотипе представляет инте-
78
79
pec работа И. П. Ашмарина и Е. П. Харченко (1971). Они изучили распространенность разных сочетаний нуклеотидов ДНК у низших организмов (рис. 27) и склоняются к выводу о том, что некоторые сочетания не встречены в эмпирическом материале не потому, что еще не обнаружены, а потому, что, возможно, вообще неосуществимы. При нынешнем положении дел невозможность сочетаний признаков фенотипа, будь она доказана, могла бы оказаться полезной при рассмотрении вопроса о «запрещенных сочетаниях» нуклеотидов. Поступать
наоборот, основывать представление о запрещенности сочетаний признаков фенотипов на предположениях биохимиков, было бы непоследовательным.
В микробиологии, где используются культуральные и физиологические признаки бактерий, представление о «запрещенных комбинациях признаков» предлагается принять как постулат, без доказательства (Старк, 1966; Заварзин, Старк, 1965; Заварзин, палеонтолог Ю. Н. Печер-,
% ГЦ
Рис. 27. Двухмерное распределение вирусов и бактерий по' нуклеотидному составу и содержанию ДНК на геном (Ашмарин, Харченко, 1971)
1969). Точно так же поступает ский (1970).
Совершенно ясно, что проблема континуума таксонов связана с серьезными вопросами эволюционной теории. Важность этой проблемы для систематики очевидна.
Констатация совместной встречаемости таксономических признаков независимо от того, какими методами она осуществляется, никак не может подменить собой доказательства их взаимной обусловленности. Главная сложность состоит в том, что такое доказательство не может быть проведено, по крайней мере в основной своей части (признаки неизвестного функционального значения и не установленной взаимной обусловленности), только на эмпирическом множестве видов.
Лекция 6, В КОТОРОЙ ПРЕДЛАГАЮТСЯ ДОКАЗАТЕЛЬСТВА НЕВОЗМОЖНОСТИ КОНТИНУУМА ТАКСОНОВ, ИЛИ, ЧТО ТО ЖЕ, СУЩЕСТВОВАНИЕ ВЗАИМОДЕЙСТВИЙ МЕЖДУ ПРИЗНАКАМИ
В обоих приводимых здесь доказательствах рассматриваются два множества событий: 1) эмпирическое множество Nu, 2) формальное множество (континуум) Nn (табл. 3).
80
Таблица 3
Формальное множество событий (континуум) N*u, параметры эмпирического JVn и биологически возможного N** множеств
Подобные события множеств
Номер события i 12345.6789 10 11 * "11 *♦ "11
0 1-024 1 1536 9 1792 41 1920 105 1984 233 2076 489 2084 1061 2096 2085 000000 00 000 100000 00 000 606600 об 6.6 i 110000 00 000 000000 01 001 11100000000 000001 01 001 11110000000 0 0 0 0 1 1 6 1 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 1 1 1 0 1 0 0 1 1 1 1 1 1 1 0 0 0 0 0 0 0 1 1 1 1 0 1 0 0 1 111111 0 1 0 0 0 0 111110 1 0 0 1 111111 10 10 0 i i i 1 i 1 6 i 6 6 i а» L11 | Hal a10 51a2 а» 131a3 a8 205a4 a7 201a6 a6 121a* 41a’ a* 6a6 a8 2a1 a10 3a2 a* 3a6 a* 8a* a’ Ila6 a8 4a6 a6 6a1 a10 28a2 a* 69a8 a* 108a4 a’ 105a6 a* 63a* a1 21a’ a* 3a« ~a*
S __ I COoQCOOOCOCO-’-SrsS сосососососоюи^ююю ooooooooooootoiQChOCh
(mW-.mW)i CO £1 P СЧ Ch СТ) о CM QO счсчсч-^счоосчсчсосч^ — т—« СЧ -М
(mW-.mW)** OoQcClOSO-'SOlCOrt СЧ СЧ СЧ СО < со со сч сч сч ’j'Scosocn-’Ocot^ch О — 4t*£tjC4g>ChC4T-.^M<©
6
Н. А. Заренков
81
Кроме того, по ходу доказательства понадобится некое третье множество Mi такое, что Л^цС^пС^ц.
Параметры этого третьего множества нас пока занимать не будут.
Доказательство 1. Возьмем все те же И признаков йц ..., ..., ап, зададим следующие соотношения между положительными a,i и отрицательными й« символами каждого а<:
для ai, .... й6 — по 15 положительных и по 16 отрицательных символов для каждого из трех, неважно каких, признаков; по 16 положительных и 15 отрицательных для трех любых других;
для й7, й8 — по 20,6 отрицательных и 10,3 положительных символов для каждого признака;
для й9, йю, йп — по 23,2 отрицательных и 7,7 положительных символов для каждого признака.
Пусть теперь символы образуют случайным образом линейные последовательности так же, как сталкиваются молекулы газа в' замкнутом пространстве. В результате столкновений и объединений молекул-символов возникают весьма различные события. Ограничимся лишь теми из них, которые-отвечают следующим требованиям:
1) в каждой последовательности имеется точно 11 символов;
2) в каждой последовательности имеется точно один символ с индексом I, либо а,-, либо й,-;
3) невозможно возникновение последовательностей, включающих «бессмысленные» сочетания символов, такие, как а7а8, адйюап, Одаюйц, ада^ац и их комбинаций.
Согласно этим условиям возникнет 31 событие. Равновероятность объединения символов в последовательности есть равновероятность событий и отсутствие взаимодействий между признаками. В действительности же эти вероятности несколько различны вследствие особенностей кодирования, но это нисколько не свидетельствует о взаимодействиях. Подсчитаем эти вероятности, которые представляют собой произведение-только что указанных частот символов.
Для событий, содержащих сочетание й7й8й9йюйц
Р; = (222 . 222 > 20^ . 23^_ = 6>
\ 31 31 ) 31 31
для событий вида (аг.ae)a7asa9a10an, fa ... ав) а7а8а9а10а1г,> («х... ав)
^,2^^=0,9з.10-з,
\ 31 31 ) 31 31 31
для событий вида fa... ae) а,а8а#а10ап, («!•• ae) a,a8a9a10a1
82
D- / 153 163 \ 10,3
^3 ~ ---- --- • -----
\ 31 31 ) 31
для событий вида (ax.
... , (ai-
Pa =
. 121 = 1 38. ю-з, 31 31
. • . GijCZgCZgCZjQCZjj, (^1 • CZijtZgCZgtl^qCZ^, • •
. a6) a?a8a9aluan 153 163 . 20,6
"зГ ’ ~зГ)' 3i
^.2^.^=.0,46.10-з.
31 31 31
Можно вообразить другое множество, в котором символы наделены способностью объединяться неслучайным образом, т. е. «притягиваться» к одним, «отталкиваться от других». * Тогда вероятности возникающих последовательностей будут отличны от только что вычисленных; причем тем больше, чем сильнее взаимодействия. В такой модели, где символы наделены способностью взаимодействовать, вероятности последовательностей будут иными, отличными ОТ Pl, р2, Рз, рл- Этим условиям отвечает Nu с его 31X11 символами. Поскольку вероятности pi, р2, рз, Ра событий Nn достоверно отличаются от pi, р2, Рз, Ра (см. табл. 2), заключаем, что между таксономическими признаками в #ц действительно осуществляются взаимодействия.
Вывод. Эмпирическое множество событий Nlr есть выборка из ЛГп, тогда как из д/п вообще невозможна.
Замечание. Обратное утверждение неверно. Если бы вероятности pi, р2, Рз, Ра не отличались достоверно от Pi, р2, рз, Ра> то с равными основаниями можно было бы представить себе, что есть выборка не только из Уи, но также из такого Л/ц ъ=Л/ц, в котором соотношение,между отрицатель-ными и положительными признаками такое же, как в Л/ц-Такое N**, вообще говоря, возможно. Его легко получить, исключая из попарно «обратные» события такие, как, например, aia2a3a4... и а^азс^.... Тогда соотношение между отрицательными и положительными признаками для каждого (i=l, 2, 3, ..., т) действительно не изменится, и Pj=p1', ... , р4 =р'4 на множестве N**x меньшем Af*i (Ni *^N*i).
Доказательство 2. Расположим 31 строчку символов (события Мн) в колонку таким образом, чтобы символы с номером i разных строчек составляли бы отдельные столбцы (табл. 2). То же проделаем с 768 строчками символов Л/ц. Заменим символы а, на «1», символы а{ на «0» (табл. 3). Подсчитаем количество единиц и нулей в каждом столбце, т. е. найдем (/«<’); zz?(°))£ и (т^>: щ<0))ь Сравним эти показатели одних и тех же столбцов разных множеств.
6
83
Поскольку JVn есть случайная- выборка из возможных- в природе событий, то различия между (т<9: т<0)); и (т(1) ' mV))* должны быть статистически недостоверными для каждого столбца i. В действительности же они достоверно различаются (в 95-процентном доверительном интервале) для всех столбцов, кроме i=7. Значит, Л7ц получена не из Nu, а из какого-то другого множества. Любое другое множество, пусть это будет Mi, обязательно входит в Mi, MicMi- Следовательно, некоторые события вообще невозможны, т. е. формальное множество Mi неосуществимо.
Вывод. Рассмотренные признаки объединены в события не случайным образом, между признаками осуществляются взаимодействия.
Замечание. Доказательство 2 также проходит лишь в одну сторону. Если бы между (mV): и (mV): zzz<0))i достоверные
различия не были бы найдены, то заключение о том, что Mi невозможна, не было бы правомочным, взаимная обусловленность не была бы показана.
Рассмотрим йодель из 4 признаков:
* "4 ** "4 ** *4
0000 0000
0001 0001 0001
0010 0010
0100 0100 0100
1000 1000 1000 1000 1000
ООН ООП
1100 1100 1100 1100
0101 0101 0101 . 0101 0101
1010 1010 1010 1010 1010
оно оно оно оно
1001 1001 1001 1001 1001
1110
1101 1101 1101 1101
1011 1011 1011 1011 1011
0111 0111 0111
1111 1111
Все 5 множеств событий включают поровну символы «1» и «О» и биномиальные. Формальное множество М состоит из 24 событий и в каждом столбце i— 1, 2, 3, 4 содержит поровну символы «4» и «О». Если бы существовало эмпирическое множество, такое, как М, с равным числом' символов «1» и «О» в каждом столбце, то его с равными основаниями можно было бы считать полученным как из М, так и из некоторого Ni . В этом случае нельзя было бы отдать предпочтения
84
какому-либо из двух множеств, нельзя делать какие-либо заключения относительно неосуществимости некоторых сочетаний признаков, т. е. взаимодействий между ними.
В действительности же наше эмпирическое множество ЛГц походит на в котором столбцы содержат неравное число единиц и нулей. N4 может принадлежать лишь к такому множеству, в каждом столбце символов которого примерно такое же, неравное, соотношение между разными символами. Этому условию отвечает Mi •
Здесь же, на модели из 4 признаков, можно получить ответ еще на один вопрос по поводу обоснованности сделанных выводов. В сущности оба доказательства используют то обстоятельство, что (т(б : существенно отлично от (т(1):
Однако значимость этих отличий сильно колеблется' от одного столбца символов к другому. Спрашивается, сколько требуется столбцов символов, таких, что (mW : mW) , существенно неравно (m(1):/п(0))ь для того, чтобы признать неслучайный характер выборки Nn из Xi? Ответ на этот вопрос дает упомянутая модель: достаточно одного столбца с существенно различающимися (nW : nW)L и (nW : nW)i для того, чтобы признать принадлежность случайной выборки Мц к 2V11, а не к Мь
После проведенных доказательств полезно вернуться к лекции 4. Когда в ней говорилось о том, что эмпирическое множество событий (видов) есть выборка из некоторого другого множества, то под этим последним имелось в виду формальное множество Na- Теперь следует внести поправку в этот вывод.
Эмпирическое множество адекватным образом описанных таксонов (событий) всегда есть случайная выборка из биологически возможного множества адекватным образом описанных таксонов (событий), которое, в свою очередь, составляет часть формального и не может достигнуть мощности по-следнего.
Множество Nu биологически возможных событий включает: эмпирические ныне живущие и вымершие события, остающиеся неизвестными современные и вымершие события, потенциально осуществимые в ходе дальнейшей эволюции события.
Отношения между множествами выглядят так: с Nm, причем Nm неосуществимо (рис. 28).
В какой мере выводы из доказательств 1 и 2 приложимы к другим таксонам, не являются ли они достоянием одних лишь креветок Hyppolytidae и Pandalidae?
85
Думается, что он имеет универсальное значение. Действительно, в обоих доказательствах используется то обстоятельство, что в столбцах содержится неравное количество символов «1».и «О». Причина этого явления состоит в том, что любой признак расщепляет множество видов на неравные части. На рис. 21, где представлена схема порождения событий, это наблюдается на всех стадиях ветвящегося процесса. По сути дела это обычнейшая практика систематиков, когда любой таксономический признак, любое сочетание признаков позво-
Рис. 28. Внутрипопуляционная изменчивость (4) как непосредственная причина межтаксонной изменчивости (Б). Множества событий: Nm — эмпирическое; Nm** — биологически возможное; Nm* — формальное; заштриховано — Nm** П Nm
ляет распределить множество видов по таксонам лишь неравномерно. Всеобщность неравномерного распределения видов по таксонам (которая сохраняется даже при неадекватности таксономических диагнозов) означает универсальность взаимодействий между признаками. Правильнее было бы считать, что виды потому неравномерно распределены по таксонам, что между таксономическими признаками осуществляется взаимодействие.
86
В какой мере, однако, вывод о взаимодействии обусловлен спецификой изучаемых признаков, следует ли предполагать, что, например, на других признаках даже тех же креветок взаимодействие показать нельзя? В сущности это часть более общего вопроса о выборке таксономических признаков. Ответ, на него был получен еще раньше, в решении-задачи об эмпирической выборке таксонов. Этот ответ кажется очевидным: выборка признаков задается выборкой видов. Генеральное биологически возможное множество видов обладает генеральным множеством признаков, случайная выборка видов характеризуется случайной выборкой признаков. Не-исключено, что множество тех значений разрешающей способности одного и того же признака, которое проявляется на выборке видов, распространимо в какой-то степени на свойства генерального множества значений того же признака в генеральном множестве видов.
Систематики, таким образом, отнюдь не свободны в выборе признаков. Эти ограничения, однако, не только объективного характера. Они носят также «субъективный»3 характер в том смысле, что зависят от характера человеческой деятельности, не обусловлены объективными законами живой природы.
Известно, например, сколь сильно изменилась система микроорганизмов вследствие совершенствования биохимических и физиологических методов исследования по сравнению с ранней системой, в которой по понятным причинам («субъективного» характера) большое место занимали морфологические признаки. Здесь «субъективные» причины перевешивали значение объективных. Иное положение в зоологической систематике. Морфологические изменения — основное содержание эволюции животных. Здесь объективное ограничение в выборе признаков не было перекрыто ограничениями «субъективного» свойства. Что же касается самих морфологических признаков, то степень их доступности неодинакова: ультрамикро--скопические структуры, например, сделались доступными сравнительно недавно, тогда как примечательные особенности внешнего строения и анатомии (число пальцев, строение позвонков) были известны издавна. В’этом смысле использование различных морфологических признаков также обусловлено «субъективными» обстоятельствами. Однако пусть мы имеем выборку животных с очень доступными признаками и потому стеснены в выборе признаков исключительно объектив-
3 Систематик зачастую субъективен в своей деятельности в том смысле, что отдает предпочтение одним признакам перед другими, основываясь на личном опыте и традициях школы. Здесь, однако, говорится не об отдельных личностях, а о систематиках вообще, точнее, о репрезентативной выборке систематиков из их генеральной совокупности.
87
ними обстоятельствами. Каким образом выборка признаков задается выборкой видов? На этот вопрос я попытаюсь ответить в лекции 8, когда сделается более конкретным понятие о генеральном множестве признаков генерального множества видов.
Далее возникает вопрос о «силе», характере взаимодействий' между признаками. Например, можно ли считать, что признак йь для которого имеем (тО): т^) =8 : 23, «сильнее» взаимодействует с другими, чем, к примеру, а3, (ml1): /n(°))3= = 20: 11. Наконец, следует выяснить, что могут дать выводы из проведенных доказательств для понимания природы таксонов. Этим вопросам будут посвящены следующие лекции.
; Лекция?,
В КОТОРОЙ АВТОР ПРЕДЛАГАЕТ ОСОБУЮ ФОРМУ ВЕДЕНИЯ ПРОТОКОЛОВ ОПЫТОВ, ПОСТАВЛЕННЫХ ПРИРОДОЙ, ДЛЯ ОПРЕДЕЛЕНИЯ ВЗАИМОДЕЙСТВИЙ МЕЖДУ ПРИЗНАКАМИ, И НАЗЫВАЕТ ТАКИЕ ПРОТОКОЛЫ 4-КЛЕТОЧНЫМИ РЕШЕТКАМИ В ВЫРОВНЕННЫХ УСЛОВИЯХ - ВЗАИМОДЕЙСТВИЯ
Представим себе один из опытов, которые беспрестанно осуществляет эволюция и который пока невозможно воспроизвести искусственно.
Пусть у бабочки имеется шип на членике ноги ах и цветное пятно на крыле ау. Спрашивается, что произойдет с первым признаком, если возникнет новый вид без цветного пятна на крыле, т. е. с признаком ау. Возможен и другой опыт. У бабочки оба признака в отрицательной форме ахау, т. е. нет ни шипа, ни цветного пятна. Следует установить, появится ли ах, если в ходе эволюции возникнет новый вид с ау.
Обобщая, можно вообразить четыре результата таких опытов: 1) оба признака в положительной форме, ахау; 2) оба признака в отрицательной форме, ахау; 3) первый признак положительный, второй отрицательный, ахау; 4) первый признак отрицательный, второй положительный, ахау. Результаты этих опытов, которые мы наблюдаем в разнообразии видов, можно было бы представить в виде 4-клеточной решетки ах1ау, изображенной на рис. 29. Можно воспользоваться несколько иными обозначениями, записывая символ «1», если данное сочетание представлено в эмпирическом множестве, и «0» если оно не найдено. Пусть, однако, из четырех всевозможных сочетаний в эмпирическом множестве видов представлены лишь виды с признаками ахау и ахау и отсутствуют виды с ахау и ахау. Тогда получим иную решетку. В тексте первую
88
из решеток будем обозначать ах/ау 1111, вторую ах/ау 1010,
перечисляя символы в клетках решетки последовательно по часовой стрелке, начиная с верхней левой клетки.
Размерные признаки, такие, как форма и величина того же шипа, разные индексы, абсолютные размеры тела и отдельных органов также могут быть представлены в альтер-
нативной форме, и взаимодействие между ними также представимо с помощью 4-клеточной решетки. Фактически это уже было проделано при кодировании члеников щупика мандибулы у креветок в лекции 4. При
рой записаны ~все четыре сочетания
признаков ах, av, возможные в совокупности видов
кодировании размерных
признаков следует подразделить весь диапазон
изменчивости на отдельные классы и рассматривать ее как дискретную. Классы изменчивости могут быть выделены на основании чисто практических соображений, например точности измерений.
Между такой 4-клеточной решеткой и корреляционным полем существует очевидное соответствие. В частности, решетке ах1ау 1111 отвечает корреляционное поле с rx/j/~0,5, решетке ах/ау 1010 — корреляционное поле с гх/и~1.
Если бы мы попытались изобразить 4-клеточные решетки для любых двух признаков на событиях из табл. 3, то получили бы в основном решетки типа 1111—материал явно неинтересный для дальнейшего анализа. Главное же заключается в том, что, строго говоря, взаимодействие между двумя
лишь таксономическими признаками, по-видимому, никогда не имеет места. Взаимодействие между любыми двумя признаками в общем случае отражает всевозможные взаимодействия между всеми признаками вида. Следует допускать, что в каком-то смысле эти взаимодействия различны и некоторые из них несущественны, тогда как другие имеют большее значение. Тем не менее до тех пор, пока это не установлено, следует исходить из того соображения, что морфологическая организация суть организация, каждый элемент которой (таксономический признак) связан со всеми прочими элементами.
Представление о целостности морфологической организации можно было бы конкретизировать следующим образом. Пусть между ах и ау у видов одного таксона осуществляется взаимодействие по типу 1010. Тогда у видов другого таксона между теми же ах и ау может осуществляться взаимодействие другого типа, например 0101. Различия между решетками
89
следует отнести за счет других признаков, по которым разли- 1 чаются изучаемые виды.
Признаки а{, ..., йц— это выборка из множества таксономических признаков. Запишем их следующим образом. Слева от Й1 поместим символы, обозначайэщие признаки всех вообще креветок, отряда десятиногих ракообразных, класса Ракообразные и т. д. — эти признаки выражены одинаково у ; изучаемых креветок и очевидно, что не ими обусловлены раз- ' личия между решетками для одних и тех же признаков. Справа от йц запишем признаки, выраженные по-разному у различных видов тех же_ креветок; это признаки родов, видов, подвидов.
pjB иных условиях между теми же Й1 и й2 осуществляются со-I. вершенно отличные взаимодействия:
а2
о
о
Я1
1
О
... 100000000
ч-256
1314
«1
о
1
1
о
Можно думать, что последние, относительно мелкие таксоно- мические признаки, являются причиной возможных различий ' между решетками, построенными для одних и тех же призна- ' ков, но разных таксонов. Все же задача о взаимодействии : между двумя лишь признаками совсем не безнадежна. «Искажающее» влияние относительно мелких признаков «справа» на взаимодействие любых двух признаков можно свести к минимуму посредством выравнивания условий взаимодейст- < вия.
Взаимодействием между таксономическими признаками й» и й,- из т рассматриваемых будем называть наличие' 1, 2, 3 сочетаний разных форм признаков йг и й; из 4 воз- 1 можных (соответственно отсутствие 3, 2, 1 сочетаний форм тех же признаков) в решетках, построенных на событиях, неотличимых между собою ни по одному из прочих (т—2) признаков.
Например, для того чтобы построить 4-клеточную решетку 1 для Й1 и й2 в выровненных условиях ..010100001, следует отоб- 1 рать из эмпирического множества Nu такие события, кото- | рые неразличимы по , 9 прочим признакам ..010100001. з В табл. 3 имеется два таких события: 1185, 1697. Тогда меж- 1 ду Й1 и й2 осуществляется в условиях ..010100001 взаимодей-ствие 4
1697-►
а2
1
1185
1
di
0
0
... 100100010
834
стороны, в иных условиях ..110100001 взаимодейст-______________— е£ ЛЛ ттП1ГЛТТАППТ1'
| С другой ( t
I- вие Й1 с а.2 сохраняется без изменений:
------е-929
о
1
Я1
о
1
1441
Взаимодействие между и й2 не было бы установлено, если бы не были выровнены условия взаимодействия между ними:
«1
1697
1441
Разнообразие решеток, построенных для взаимодействий в выровненных условиях, значительно превосходит то, которое возможно, если такого выравнивания не производить, и потому возникает' задача их перечисления.
91
90
Для каждой пары признаков вообще возможны 16 раза решеток:
оно
1001
0001 0101 1110
0010 1010 1101
0100 ООН 1011
0000 1000 1100 0111 1111
Тип решетки сохраняется независимо от порядка заполня ния клеток, т. е. места символов at и йф Поэтому для рещд ток всех типов имеет место тождество НапримеЯ
а{/а3 1011 = аг7й3- 1110. 1|
Некоторые решетки допускают наглядное истолкование тя взаимодействий, которые они описывают. Решетка 1010 преД ставляет такое взаимодействие, когда оба признака либо соД местно встречаются, либо совместно отсутствуют. Решетки 0101 свидетельствует об «избегании» признаками друг другД
Было бы интересно рассмотреть подробно эмпирическое множество решеток, построенных для 11 признаков на эмпия рических множествах событий и видов. Такая задача имели бы смысл постольку, поскольку последние репрезентативна по отношению к их более обширному биологически возможной му множеству. Если эмпирические множества репрезентатиЛ ны по отношению к биологически возможным, то в таком случаи множество эмпирических решеток следует считать репрезеД тативным по отношению к биологически возможному. Болев важной, однако, представляется задача определения достовер! ности отдельных решеток. Она в сущности имеет в виду ни только определение достоверности границ таксонов, но также определение достоверности отдельных эмпирических заключе! ний о взаимной обусловленности признаков в эволюции, т. е| выводы из сравнительной морфологии. I
До сих пор мы ограничивались лишь 11, однако их чис! ло можно значительно увеличить. Рассматривались лишь 2 се| мейства, 39 родов и 336 видов, между тем как известны И другие; кроме того, какое-то число видов пока не открыто зо« ологами, но будет описано впоследствии. Наконец, следует! считаться с потенциальными возможностями этой группы жич вотных как в отношении видообразования, так и в отношении новых признаков. Необходимо иметь уверенность в том, чт(Я пополнение списка событий в табл. 3 не приведет к полному! отказу от полученных результатов. Увеличение числа исполь-) зуемых признаков (уточнение условий взаимодействия) при-| ведет к расширению табл. 3 вправо и одновременно к удли-j нению ее. Действительно, новый признак может оказаться положительной форме у видов одного события и в отрица-1 тельной у других видов того же события, и тогда произойдем
расщепление ранее существовавшего события новым призна-
Г В качестве примера рассмотрим, что произойдет с решеткой Ю10 aja, в условиях С, построенной на эмпирическом множестве видов с использованием т признаков. Допустим, что каждому из двух событий, а^С и (ца3С, отвечает больше одного вида.
Если в дополнение к уже использованным т признакам двести некоторый новый признак am+i (рис. 30), то может произойти следующее:
а о. С
а НС
I случай
1О1о(алСЛт*1
ЮОО-а^Са^ mio(ajaJcra-'n*1 Xa,ajCam,i ioio<ail’ii:.a^ at a:Cam^ 0010~a} ajCam+1 ООЮ^аГа^
1000 aiaJCam.} 0010 aiaJCamt.
1000 a^Ca^ 00Ю ata^Cam^ ioio<a-‘ajcram'
Ш случай
10Ю
Л случай
Рис. 30. Схема испытаний решетки adas 1010, построенной на т признаках, новым признаком
ЭДт+1
1) в каждом из двух событий новый признак выражен положительно у одних видов и отрицательно у других. 1огда каждое из ранее имевшихся событий претерпит расщепление на два новых; _
2) у всех видов одного события признак am+i представлен в одной лишь форме, тогда как у видов другого события тот же признак в положительной форме у одних и в отрицательной у других. Тогда расщеплению подвергнется только одно событие из двух. Понятно, что здесь возможны 2 подслучая, по числу расщепившихся событий;
92
93
3) у всех видов каждого события новый признак выраже^ только в одной из двух форм. ;
В общей сложности из 16 возможных исходов испытаний в 8 взаимодействие 1010 в новых условиях сохранится, в 8’ других решетка 1010 перейдет в 1000 и 0010. Таким образом, может получиться, что в уточненных условиях между а{ и а; осуществляются иные взаимодействия, отличные от ранее наблюдавшихся.
Все мыслимое разнообразие вновь построенных решеток изображено на рис. 30. Результаты испытаний решеток прочих типов приведены в табл. 4. Становится очевидным, что
Таблица 4 :
Все возможные исходы испытаний решеток - различных типов, построенных на т признаках в условиях С, новым признаком ат^ в новых
условиях
Исходный тип решетки в условиях С т. е. число единиц в клетках решетки Частота решеток различных типов (с разным числом единиц после испытания новым признаком ат^.|
1 единица 2 единицы 3 единицы 4 единицы
1 4 0 0 0
2 8 8 0 0
3 12 24 16 0
4 16 48 52 32
Таблица 5
Параметры, характеризующие процесс порождения событий на формальной модели с 11-ю признаками и эмпирическом множестве событий
Формальная модель Эмпирические данные
Я g » •' я сдые k поддают ст по собы- р аспр еделение резул ь-тативностей Л признаков ИЗ И1==11 средняя ^±д °k ффициент 1 мметрии
И II 5 а и « Ф s
JJ» g и а О’ Е- я «
1 2 11 22 2-11=22 2,00
2 4 55 220 3-26+4-29=194 3,53±0,3 0,52 —
3 8 165 1320 4-'23+5 -60+6 -46+ +7-22+8-14=934 5,66±0,06 1,13 0,47 2,02
4 16 330 5280 4-4+5-28+6-44+ +7-62+8-64+9 X X60+10-34+11 X Х19+12-13+13Х 8,23±0,04 2,02
х9+14-2=2706
94
. проверка предположения об определяющем значении условий взаимодействия на тип решетки сводится к определению ис-хода серии последовательных испытаний признаками ат+1, Ят+2, Om+s, —> йт+ь. Далее, очевидно, что исход таких испытаний в значительной степени зависит от того, сколько видов отвечает каждому эмпирическому событию. Чем больше видов, в событии, тем выше вероятность того, что новый признак окажется в положительной форме у одних видов и в отрицательной у других; иначе говоря, чем больше подмножество видов,, тем они разнообразнее. Для события с единственным видом вероятность расщепления следует принять за нуль. До сих пор эти рассуждения относились только к эмпирическому множеству видов. В действительности же их необходимо перене-, сти на биологически возможное множество видов и событий.
Только тогда наши выводы могут претендовать на большее,, чем на одну лишь констатацию фактов. Что же касается чис-; ла признаков, то их следует использовать столько, чтобы чис-ло возникающих событий поравнялось с числом видов в биологически возможном множестве событий. Тогда каждому i событию такого множества будет отвечать один вид. Уточне-- ние условий взаимодействия за этим пределом приведет к ‘ использованию нетаксономических индивидуальных призна-, ков, а это выходит за пределы нашей задачи.
Поскольку между таксономическими признаками, осущест-" вляется взаимодействие, общее число событий в табл. 3 никог-да не достигнет 7.68, мощности множества формально возмож-ных событий. Следовательно, пополнение списка событий АГц до Nn не приведет к вырождению разных решеток в решетку одного типа—1111. При использовании новых признаков опять же табл. 2 пополнится новыми событиями, но не настолько, чтобы ПО числу событий Nm+k сравнялось С Ntn+k-Поэтому эмпирическое множество решеток в какой-то степени сохранится независимо от полноты изученности видового-состава мировой фауны и изученности отдельных видов.
В связи с введением новых понятий требуются дополнительные .обозначения: S — число видов эмпирического множества, S** — число видов биологически возможного множества, М — число признаков, достаточных для того, чтобы в каждом событии биологически возможного множества Nm содержался бы один вид, т. е. чтобы имело место равенство-Nm = S’*.
Лекция 8, В КОТОРОЙ РЕЧЬ ПОЙДЕТ О ДОКАЗАТЕЛЬСТВАХ И ПРЕДСКАЗАНИЯХ В СИСТЕМАТИКЕ
Поставленная в предыдущей лекции задача определения достоверности эмпирических обобщений о взаимодействиях . между признаками и границами таксонов состоит из несколь-
95-
>
ких более частных задач, которые автор попытается разрешить последовательно.
Прежде всего следует установить число событий в биологически возможном их множестве Мь показать единственность решения и перечислить эти события, дополнить Уц до Мь События М1ПМ1 — сегодня неизвестные и потенциально осуществимые в ходе эволюции.
Далее, необходимо определить число морфологически различимых видов S** биологически возможного множества событий Mi • Затем потребуется установить число признаков М, необходимых для того, чтобы S** видов распределить по событиям таким образом, чтобы в каждом содержался точно •: один вид, NM = S**.
Наконец, следует разобраться в вопросе о выборе признаков. Каким образом выбор признаков мог бы повлиять на ; все проведенные до сих пор рассуждения? Каким образом, на- : пример, выбор признаков мог бы отразиться на структуре множества Nu, на разнообразии эмпирического множества решеток?
Задача 1. Построение множества биологически возможных событий. Структура искомого Mi должна отвечать следующим условиям. Во-первых, она должна быть выводимой из структуры популяции вида-предка, т. е. допускать объяснение на основе теории видообразования. Это означает, что распре- деление подобных событий в искомом Mi должно подчи- ; няться биномиальному закону. Поэтому частоты подобных со- ' бытий искомого Mi есть уменьшенные частоты' теоретиче- • ского множества Мь Они не могут быть увеличенными частотами Nn, в котором распределение явно не биномиальное и репрезентативность которого по отношению к искомому множеству сомнительна. Поэтому в данном случае построение искомого Mi следует вести от теоретического, но не эмпирического множества. В пользу такого подхода говорит и еще i одно соображение. В сущности задача состоит в том, чтобы построить такую кривую, которая на графике располагается между кривыми для Mi и Мь Ее можно получить либо под- -1 нимая кривую Мь либо опуская кривую Мь В первом слу- | чае в искомую Mi не попадут такие события, как а10ан, 1 а7а4, записанные справа и слева от крайних членов раз- 3 ложения (4); между тем нет оснований считать, что такие со- .; бытия невозможны. Наоборот, построение искомого Mi от тео-ретического Mi не исключает присутствия таких событий в Nn. Второе условие, которое следует соблюсти при по- ? строении, — это сохранение в столбцах i такого соотношения | между символами «1» и «О», которое наблюдается в эмпири- 3
96 1
ческом множестве Мц. В самом деле, если учесть, что видообразование связано не только с появлением новых признаков (новые единицы и нули), но также и с повышением таксономической ценности одного отдельного признака, то эмпирическую выборку видов следует рассматривать, так же и как выборку значений этого показателя, из более обширного их множества. Можно ли считать, что если выборка видов случайна, то тогда и выборка значений показателя также случайна?
Вообще говоря, достаточных оснований для такого утверждения нет. В то же время как будто бы нет другой возможности построить Nu, не используя при этом сведений о соотношении символов в столбцах Nu, и не следует не учитывать того обстоятельства, что выборка признаков задается выборкой видов. Искомая совокупность, к которой мы сейчас пе-; рейдем, остается в связи с вышесказанным достаточно гипо-' тетической.
i Может быть, больший интерес представляет не столько са-г ма искомая совокупность, сколько те возможности ее построе-[ ния, которые, как мне кажется, заложены в рассмотрении эм-лирического материала систематиков в связи с классическими представлениями о видообразовании. Таким образом, в дальнейшем допускается, чтоМц походит на Мп распределением подобных событий и на Мп соотношением между символами в столбцах. Последнее условие означает, что х-(т!М: /п(0))£ = (яМ*:/п(0))£ , где х>1. Если мы обратимся к столбцу i=l в
384
( табл. 4, то найдем, что х = — = 48. Ясно, что этого не мог
!' жет быть. Если допустить, что х=48, то число нулей в Мц / в том же столбце составило бы 23-48=1104, т. е. больше имеющихся в Мц 384 нулей. Следовательно, х<48. Перебирая таким же образом один за другим все столбцы, мы в конце концов придем к столбцу i=5, для которого имеем х =
384 „
=-----13.Очевидно, что это значение х — наименьшее из
29
всех. Его можно использовать для каждого столбца i, и каж-**
Дый раз «расход» символов для построения Мц не превысит их числа, допустимого объемом Мц. Например, в столбце i=l число нулей т<°>=23-13=299, единиц т<1)=8-13 = == 104. Теперь нетрудно найти число событий в Мц. Оно составит 31-13=403. Каждый член биномиального разложения Мц
ба1^10 + 28а2а9 -} 69а3а8 + 108а4а7 + 105а8а® + 63а6о® +
+ 21aW + 3a8a3 (6)
^/4 Н. А. Заренков
403 -Л
меньше соответствующего члена Л)ц в — . Я
Множество Л^п = 403 событий есть максимально воз- ’ можное из числа тех, которые отвечают обоим условиям задачи. Не существует другого множества такой же мощности, чтобы оно не отличалось от Мп по числу событий, но отличалось бы самими событиями. Пусть, однако, такое множество существует, оно получено из Nn после того, как поменяли местами два символа, «1» и «0», в столбце i. В таком случае _ сохранится исходное (m(1): ; но нарушится соотношение
между подобными событиями. Если же поменять местами в Мп любые два символа «1» и «0» в одном событии, то на- Я рушится соотношение между разными символами в двух Я столбцах. Всякий раз соотношение между нулями и единица- ,Я ми во множестве в целом останется без изменений. Будь наша Я выборка репрезентативной, можно было бы говорить о пред- Я сказании неизвестных видов с той степенью уверенности, ко- И торая определяется репрезентативностью. Поскольку Мц — Я единственное, задача перечисления составляющих его собы- Я тий разрешима, например, непосредственным перебором мно- Я жеств, удовлетворяющих условиям «по столбцам» и «по строч- Я кам» символов. Задача эта, возможно, несколько громоздка, Я а перечисление всех строчек символов, событий, здесь явно Я неуместно. К тому же полученные выводы уже представле- Я ны в явной форме и допускают эмпирическую проверку непо- Я средственно' систематиками. Автор утверждает, что будущие Я фаунистические открытия будут сделаны за счет (403—31) со- Я бытий. Вновь найденные виды должны обладать сочетания- Я ми признаков, свойственных этим последним событиям; при- Я чем будут обнаружены прежде всего такие виды, которые от- Я носятся к моде биномиального распределения подобных со- : бытий Nn. Вероятность обнаружения видов с тем или иным соотношением положительных и отрицательных признаков пропорциональна частоте подобных событий в биномиальном распределении (6). Например, вероятность обнаружения а4а7 есть 108/403. Следует лишь обратить внимание на существенное уточнение, которое вносится в эти утверждения случайным характером выборки эмпирического множества таксонов по отношению к биологически возможному.
Для оценки репрезентативности коэффициентов биномиального разложения Nn по отношению к таковым Mi необходим обычный статистический аппарат. Точно так же следует оценивать репрезентативность (mt1): т<°)) j по отноше-нию к (mt1) : mt°))/ • Все утверждения относительно обнаружения в природе креветок с заданными сочетаниями признаков носят вероятностный характер ввиду случайности эмпириче-98
ской выборки. Вероятностный характер процесса видообразования — причина, лежащая совсем в иной плоскости. Первое обстоятельство следует из случайности самого процесса изучения мировой фауны и носит, так сказать, «субъективный» характер (в том смысле, что относится к человеческой деятельности). Второе следует из природы эволюционного процесса, объективного характера. Естественно, они неразличимы. Чтобы не перегружать текста, автор в дальнейшем не будет приводить обычные оценки репрезентативности, которые легко могут быть осуществлены самим читателем.
Задача 2. Построение множества биологически возможных видов S**, отвечающих множеству биологически возможных СобЫТИЙ jVii-
Число видов в эмпирических событиях Л^ц, как в этом нетрудно убедиться, не связано с числом положительных или отрицательных признаков событий (табл. 2). Биологический смысл биномиального распределения числа видов по подобным событиям обсуждался в лекции 4. Этот феномен объясняется случайным характером процесса видообразования и способностью подвидовых признаков превращаться в таксономические. В таком случае и общее число видов в совокупности событий будет прямо пропорционально общему числу событий и распределено по подобным событиям согласно закону биномиального распределения.
В 31 событии, построенном с использованием 11 признаков, содержится 336 видов. Очевидно, что в 403 событиях, основанных на тех же 11 признаках, может содержаться 336.403 ,
——— = 4355 видов как элементарных морфологически различимых таксонов, описания которых строго адекватны одно другому. В подобных событиях эти £**=4355 видов распределены приблизительно следующим образом:
65 (a+10) + 303 (а2а9) + 746 (я3а8) +_1167 (а№) +
+ 1135 (а5я6) + 652 (ава6) +217 (+а4) + 32 (++)• (7)
Задача 3. Требуется определить то число морфологических альтернативных признаков, которое потребуется для распределения £**=4355 видов по Nm = 4355 событиям так, чтобы в каждом событии содержался бы один вид. Для упрощения расчетов примем, что среди множества таксономических признаков можно подобрать М таких, что признаки, сходные по кодированию с (а<, ..., ц6), (й7, й8), (й9, йю, йп), содержатся в М в том же соотношении, что и в т. Это означает, что среди М признаков имеется -уу- Л4 таких, как
2 3
(йь ..., й6), -уу- М таких, как (й7, й8), -yj- М таких, как (йд, дю, йп).
71/г Н. А. Заренков ^9
По аналогии с рис. 21, на котором изображен процесс порождения событий Mi признаками т=11 на множестве видов 3 = 336 (табл. 2), следует представить себе, как будет протекать процесс порождения событий Nm на множестве видов 3** = 4355 с использованием Л4 признаков. Очевидно, что любой признак из т, использованный в качестве первого, обязательно приведет к возникновению 2 событий. Это действительно так, потому что используются лишь такие признаки, которые выражены по-разному у разных видов 3. Возникшие события будут включать различное число видов. Любой признак из оставшихся (т—1), использованный в качестве второго, вероятнее всего вызовет расщепление каждого из двух уже имеющихся событий надвое, и тогда появятся 4 события. Другой вариант, когда появятся 3 события, менее вероятен, потому что 2 события содержат много видов и среди них почти наверняка найдутся с разными формами одного и того же признака. Это второе испытание может быть осуществлено С2щ разными способами. Точно такие же рассуждения приложимы ко всем прочим признакам: третьим признаком из (т—3) оставшихся расщепление уже имеющихся 3— 4 событий может быть осуществлено Ст способами на 4—8 новых, четвертым признаком из (т—4) оставшихся признаков 4—8 событий будут расщеплены на 6—16 событий и т. д. На каком-то этапе этого процесса появятся слепые ветви. В процессе для Mi они отвечают только биологически невозможным событиям. В этом отличие схемы порождения событий от аналогичной схемы для множества Mi (табл. 2), где слепые ветви соответствуют как биологически невозможным событиям, так и неизвестным, но биологически осуществимым. На схеме для множества Nm биологически невозможные события появятся, вообще говоря, начиная со второй стадии процесса, и каждое из них приводит к невозможности появления 2, 4, 8, ... на последующих стадиях. Поэтому скорость процесса порождения событий будет быстро затухать. Если в начале процесса соотношение между аргументом (число используемых признаков k=l, 2, 3, ..., т, ... М) и функцией (число событий Nk,k->~M) приблизительно представимо показательной функцией, то в дальнейшем приращение функции на приращение аргумента уменьшится. Ход таких процессов описывает логистическая функция (рис. 30)
М =------------- + 1> k = 1, 2, 3, ... , т, ... , М,
1 4. e^-c-k
Ь, с — const. (8>
Поэтому те данные из табл. 5, которые относятся к модели с 11 признаками, никогда не могут быть наблюдаемы в дейст
100
вительности,- Например, далеко не каждая пара признаков из Cii = 55 возможных пар способна породить 4 события.
Среди 55 пар найдутся такие, которые породят только по 3 события каждая.
В связи с некоторыми другими особенностями процесса формула (8) также должна быть несколько видоизменена. Дело в том, что замедление процесса порождения собы-
тии вызывается не только невозможностью некоторых сочетаний признаков, но также появлением событий с одним-единственным видом, которые не могут быть расщепл'ены по условиям задачи, относящейся к области систематики, но не к популяционному уровню. Процесс будет затухать
по мере приближения Nk к =3 и возраста-
ния числа событий с един-
Рис. 31. График функции (8) (штриховая линия) и (9) (жирная линия)
ственным видом. В конце концов в какой-то момент процесса будет наблюдаться много событий с единственным видом и немного с большим числом видов. Применение к
ним нового признака лишь в исключительных случаях может привести к расщеплению сразу всех последних. В практической систематике это явление известно в качестве тезиса о неравноценности таксономических признаков. Для подразделения разных крупных таксонов на более мелкие не могут, быть использованы одни и те же признаки. Система разных таксонов использует разные признаки. По этим соображениям в (8) следует внести уточнение:
л,Г = _^
1 _I_ J-c-k1
t< 1.
(9)
График функции (8) симметричен относительно точки перегиба, кривая функции (9) состоит из двух участков: короткий вогнутый до точки перегиба (где приращение функции превышает приращение аргумента) и более длинный выпуклый после точки перегиба (где приращение аргумента превышает приращение функции) (см. рис. 32). Еще одно и последнее замечание по поводу процесса порождения событий следует сделать в связи с особенностями кодирования признаков. Среди признаков Qi ... йц есть такие, сочетания которых бессмысленны (одна и та же креветка не может иметь одновременно и 3- и 2-члениковый щупик мандибулы). Это еще одна
7^2*
101
дополнительная причина, по которой некоторые, пары, тройки, четверки и т. д. признаков не приведут к порождению событий. Например, в эмпирических Си — 55 парах признаков имеется 26 таких, что каждая порождает 3 события, и 29 пар, порождающих каждая 4 события. В среднем же одна пара 26-3 + 29-4 0
порождает -----—------= 3,53
55
зультаты для троек и четверок
Рис. 32. График уравнения (9) в полосе значений результативностей различных сочетаний k признаков из общего их числа М
события. Аналогичные ре-
приведены в табл. 4. Асимметрия наблюдаемых распределений есть следствие из особенностей кодирования признаков.
В связи со всем вы-график функции следует
в
шесказанным логистической (9) на рис. 31 внести такое уточнение: кривая функции проходит через средние распределений результативности сочетаний признаков, т. е. является наиболее вероятным значением. Кривая находится в полосе значений этого показателя, ши
рина которой отвечает лимитам их распределений (рис. 32). Поскольку распределения асимметричны, кривая смещена к яижней кромке полосы (рис. 33, 34).
Рис. 33. Участок кривой функции (9), описывающей процесс порождения событий. По эмпирическим данным табл. 5
102
Ширина полосы, если ее оценивать посредством ak как функции от k (при постоянных S**, М), определится из следующих соображений. Начало полосы всегда есть точка [1, 2] согласно определению таксономического признака. Любой ai из М обязательно приведет к порождению точно 2 собы-
Рис. 34. Схема, поясняющая соотношение между кривой порождения событий функций (9) и результативностью k признаков из общего их числа М
тий. Конец полосы есть точка [М, 4355] согласно условию задачи. Любые М признаков из М, что, естественно, может быть осуществлено единственным способом С^, обязательно породят 4355 событий.
Наибольшее количество сочетаний признаков возможно при
— См т. е. возле точки перегиба логистической кривой. Уже по одной лишь этой причине здесь будет максимальный разброс результативностей сочетаний признаков, максимальная щ. Если выпрямить одну из сторон полосы, то получится биномиальное распределение для аА = /(См). На рис. 34 и табл. 5 вертикальное сечение полосы для отдельных значений k на абсциссе есть диапазон разброса результативностей сочетаний k признаков из общего числа признаков М(См)- На эмпирическом материале эти распределения оказались биномиальными. Они и не могли оказаться иными, потому что по сути дела это есть распределение частот эмпирических подобных событий. Тогда ввиду случайности эмпирической выборки распределения биологически возможных подобных событий ( = результативностей сочетаний признаков) — также биномиальные. Отсюда следует важный вывод: средняя для результативностей эмпирических может считаться средней для искомых М признаков. Иными словами, точки [2; 3, 53], [3; 5, 66], [4; 8, 23] лежат не только на эмпирической кри
103
вой, но и на искомой кривой для биологически возможного S** = Nm- Думаю, что это относится ко всем точкам восходящего участка эмпирической кривой до точки ее перегиба.
В таком случае, параметры уравнения (9) могут быть по- ' лучены из
5,66 =
4355
3,53 = —4335-Л7
,b-c-2'
,6—сУ
8,23 = -----——7
Подставляя найденные значения b, с, t в уравнение
4355 =
4355
1 + еь-с-м(
+ 1,
находим М. Таким образом, для того чтобы 4335 биологически возможных событий (элементарных таксонов, описанных адекватным образом, морфологически различимых видов) распределить по 4355 событиям, требуется применить М признаков. На этом множестве событий могут быть построены совершенные решетки, которые описывают взаимодействия между признаками такие, что не изменятся ни в ходе фаунистических исследований (потому что новые находки невозможны), ни в случае дальнейшего углубленного изучения признаков видов (потому что любой новый признак не будет таксономическим) .
Каждая эмпирическая решетка для т=11 признаков и 5 = 335 видов должна подвергнуться (М—11) испытаниям прежде, чем ее можно будет признать достоверной.
Теперь ясно, какое именно значение в вышеприведенных рассуждениях занимает представление о случайном характере выборки значений результативностей признаков из их генерального множества. Некоторые подтверждения в пользу этого положения были получены в ходе вычисления параметров логистической кривой. Эта гипотеза заслуживает того, чтобы на ней остановиться специально, тем более что в математических методах систематики необходимое число признаков определяется чисто произвольно (Sokal a. Sneath, 1963), и систематиками, которые пользуются такими методами, хорошо показана зависимость результатов от выборки признаков (Вайнштейн, 1968, Soule, 1967).
Итак, имея в виду случайный характер выборки видов и, следовательно, их признаков, можно ли быть безразличным к выбору признаков, сохранится ли система неизменной независимо от выборки признаков? Ответ на этот вопрос должен быть дан независимо от весьма противоречивых резуль
104
татов практиков. При этом довольно очевидные ^субъективные» причины выбора признаков должны быть опущены. Следует установить, каким образом систематик объективно ограничен в выборе признаков. Именно так этот вопрос был поставлен в лекции 6. Здесь необходимо различать до некоторой степени два различных обстоятельства: общее число признаков и их качество. Последнее также действительно имеет смысл: для построения системы одних и тех же видов, вообще говоря, можно воспользоваться одним и тем же числом, но разных признаков.
В отношении только лишь числа признаков однозначный ответ, очевидно, невозможен: он зависит от того, в какой степени случайная выборка видов (и, следовательно, их признаков) репрезентативна по отношению к биологически осуществимому множеству видов и их признаков. Если ввиду своей малости эмпирическая выборка видов не репрезентативна по отношению к биологически осуществимому множеству, то такие перестройки системы в ходе таксономических ревизий неизбежны. Пусть, однако, условие репрезентативности соблюдено. Как будет вести себя система в ходе углубленного изучения видов и их признаков? В общей форме ответ на этот вопрос содержится в неявном виде в логистической кривой на рис. 36. Он тривиален: устойчивость системы, конечно же, зависит от степени изученности видов и их признаков. По мере приближения т к М, Nm к S** вероятность построения различных систем на одном и том же множестве видов, но с использованием разных выборок т из М уменьшается, что выражается в сужении полосы. Что же касается выбора самих признаков (не числа — об этом только что было сказано) , то этот вопрос заслуживает того, чтобы на нем остановиться более подробно. Таксономическое исследование начинается с некоторой выборки видов, и объем ее постепенно возрастает. В любой момент такая выборка есть случайная по отношению к множеству биологически возможных видов; так же, как, по-видимому, и признаки, точнее их параметры, по отношению к биологически возможным. Если это действительно так, то тем самым предопределен выбор признаков исследователями, использующими обычные приемы классической систематики.
Пусть сначала известно только три вида одного рода с признаками
^'Х^'2^'3^'4
^'Х^'2^'3^'4
(10)
105.
Такое предположение более естественно, чем если бы было допущено существование
а1а2а3а4
^'Х^'2^'3^'4
(Юа)
потому что (10) состоит из средних членов биномиального разложения, а не крайних, как (Юа). Признаки вновь обнаруженных видов чаще всего будут такими, что распределение подобных событий будет приближаться к биномиальному. Поэтому более вероятно, что вместе со вновь открытыми видами (отмечены знаком *) совокупность (10) примет вид
—* — — — — *
^'i^'2^'3^'4 ^'1^'2^'3^'4
^1^'2^'3^'4
(И)
нежели
^'i^'2^'3^'4
ага2а^
ах а2 а3 а\
a1a2asai
(Па)
Когда число видов в эмпирической выборке превысит некоторый предел, то обязательно обнаружится какой-то, ранее не замечавшийся признак й0; в противном случае вновь обнаруженные виды составили бы в конце концов вместе с ранее известными континуум Л/< =16, в котором все виды были бы неразличимы по одной и той же форме некоторого признака а0. Между тем чем больше видов, тем вероятнее обнаружение у них нового свойства. Мало того, поскольку признаки взаимодействуют, новый признак разделит имеющуюся совокупность лишь на неравные половины. Например, может возникнуть такая картина
Яо ^1 ^2 ^3 ^4
аоах а2а3а4
(WWk а3аха2 а3а4
(12)
106
Совокупность (12) более вероятна, нежели (12а), в которой наблюдается равномерное расщепление не только в i=2, но также еще и в i=0 столбце:
айахй2агсй a,flAa2a.iai а0ага2аъа\
а0ага2а^а4 afjala^ii
(12а)
В (10) форма «о не имела таксономического значения, не проявлялась потому, что присутствовала у всех трех видов. Начиная с (12) она могла бы послужить в качестве признака нового подрода в пределах прежнего рода. В (12) два подрода очерчены пунктиром.
Диагнозы таких подродов основаны на «одном и том же» признаке й0, они сопоставимы, адекватны. Несопоставимость диагнозов может появиться уже в силу того, что признак йо выражен по-разному в двух родах. С формой его обязательно, в силу взаимодействий между признаками, окажутся связанными какие-то другие признаки, например а*\ отрицательная форма Й5 допускает внутривидовую изменчивость (сг-|-й)5 по тому же признаку
а5 айага^л^ (а + а)а авсгх а2 а3«4 (а + в)5
аоа1а2«з а4 а5 айага2а^ (а + а)5
a0WA (? + а) 5
С этого момента диагнозы можно считать неадекватными.
Для того чтобы эти рассуждения не показались умозрительными и допускающими любую другую историю изучения, следует иметь в виду, что эмпирическая выборка событий каждый раз должна отвечать двум условиям: во-первых, распределение подобных событий приближается к биномиальному, во-вторых, в столбцах содержится большей частью неравное количество отрицательных и положительных символов. При соблюдении этих условий количество возможных историй изучения не столь уж велико, как это может показаться сначала. Главное же состоит в том, что вероятности таких историй были бы различными. Вероятность той или иной формы некоторого нового признака у уже известного события есть функция от (mi—m0); когда разность равна нулю, то обе формы, вообще говоря, равновероятны, когда mi <?то, то вероятнее, что новый признак окажется положительным, когда m0>mi, то новый признак скорее всего будет отрицательным.
107
Необходимо подчеркнуть, что конечный результат, система видов, репрезентативных по отношению к биологически возможному множеству видов, не может зависеть от очередности изучения видов, от «траектории» исследования. Действительно, если множество (12) удовлетворяет обоим условиям, то любая случайная выборка из него, например аоа^а^, а0«1й2«за4, также будет им удовлетворять. Однако исследователем такие виды одного рода будут восприниматься как а0а2, а0а2 потому, что неразличимы по а3, а±. Конечно, дело случая, какие виды сделаются известными науке вначале и какие впоследствии. В биологическом смысле — дело случая, что изучение фауны Европы предшествовало изучению Антарктики. Но биологически возможное множество видов, доступное для изучения, — объективная реальность. Точно так же объективной реальностью является множество их признаков. Поэтому на одном и том же множестве видов могут быть использованы только одни и те же признаки. При соблюдении условия репрезентативности решение задачи систематизации такого множества — единственное. Что же касается устойчивости системы, то она может быть достигнута только на множестве Nm — S**; в противном случае возможны перестройки системы в связи с новыми открытиями видов и признаков. Думается, что то обстоятельство, что Nm(]NM состоит как из неизвестных, так и из потенциально осуществимых в эволюции видов, не должно настраивать исследователя на скептическое отношение к Nm(\NM- зачем в систему вводить потенциально осуществимые виды, которые, возможно, не осуществятся. Дело в том, что, во-первых, пока невозможно отличить такие виды от уже осуществленных (вымерших и ныне живущих), во-вторых, кажется совершенно неразрешимой задача о том, какие потенциально' осуществимые в эволюции виды возникнут и какие не возникнут по причине гибели предка, осуществившегося, но вымершего бесследно. Иначе результаты действия отбора непредсказуемы; здесь же имела место попытка предсказать свойства объектов его воздействия.
Таким образом, выяснилось, что в действительности систематик не волен в выборе признаков. Его построения стохастически предопределены биномиальным распределением подобных событий. Границы таксонов вероятностны, каждое таксономическое объяснение суть вероятностное объяснение. Сам вопрос о возможности произвольного выбора признаков неправомочен. Выбор признаков исследователем часто ограничивается техническими возможностями (микроскопирование, анатомирование, гистологические, биохимические и цитологические методики) — доступностью признаков, но это совсем иная плоскость, на которой нет места объективным закономерностям живой природы.
108
Лекция 9,
В КОТОРОЙ РАНЕЕ ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ ИСПОЛЬЗОВАНЫ ДЛЯ РАССМОТРЕНИЯ НЕКОТОРЫХ СПОРНЫХ ВОПРОСОВ И СЛУЖАТ ОТПРАВНОЙ ТОЧКОЙ ДЛЯ ГИПОТЕЗ
Основные результаты уже прочитанных лекций сводятся к следующему. Во-первых, показано, что изучение объективных закономерностей органического мира как множества таксонов возможно на их эмпирической выборке. Полученные та-|ким образом обобщения носят вероятностный характер. Вто-|рым результатом таксон утверждается в качестве особой биологической организации близко родственных соподчиненных (таксонов. Она унаследована от множества популяций вида-|предка и представляет собой биномиальное распределение адекватно описанных соподчиненных таксонов. В ходе дока-'зательства этих двух положений пришлось выйти за пределы Собственно систематики и прибегнуть к помощи теории видообразования. Третий результат, возможность объективного разграничения реально существующих таксонов, приведен в лекциях как тезис о взаимодействии между признаками. Причины этого явления оставались пока не рассмотренными, их следует искать на организменном уровне биологической организации.
Начнем, однако, с первого. Опираясь на него, можно показать, что вероятностные представления о границах таксонов позволяют считать недоказанным полифилетическое развитие; во всяком случае теми способами, которыми при этом пользуются.
Иногда полагают, что конвергентное развитие, чрезвычайно широко распространенное в живой природе (Светлов, 1962), может в конце концов привести к идентичному набору признаков у разных не родственных между собой видов. При этом ссылаются также на многочисленные случаи параллелизмов, якобы ограничивающие и направляющие изменчивость, на гомологические ряды Н. И. Вавилова, якобы противоречащие представлению о неопределенной изменчивости. Примеры якобы полифилии использованы А. А. Любищевым (1968) в качестве важнейшего аргумента против дарвиновских представлений о видообразовании расхождением видов, против классической систематики. Связь полифилетической концепции с номогенезом Л. С. Берга (1922) вполне очевидна. Несмотря на то что номогенез был подвергнут глубокой критике К- М. Завадским и М. Т. Ермоленко (1966), актуальность критики полифилии сохраняется, потому что ее сторонники взывают к фактам.
Различают полифилию на разных уровнях. Полифилия на уровне отряда — это независимое происхождение соподчинен
8 Н. А. Заренков
109
ных таксонов отряда таких, что каждый имеет признаки дан- J ного отряда. «Установление полифилии стало, пожалуй, мо- я дой» (Воронцов, 1967). Разделены на самостоятельные таксо- . " ны и на этом основании объявлены полифилетическими кито-образные, ластоногие, грызуны. Постоянно обсуждается по-лифилия членистоногих, четвероногих. Несмотря на то что . представление о полифилетическом происхождении млекопитающих и наземных позвоночных не принадлежит к числу доказанных (Матвеев, 1969; Шмальгаузен, 1959) и изучение палеонтологических свидетельств эволюции этих животных еще не завершено (Hopson, Crompton, 1969), возникшие споры произвели большое впечатление на ученый мир ввиду широкой популярности и казавшейся удовлетворительной изучен-ности млекопитающих. При этом, однако, преданы забвению куда более резкие потрясения системы беспозвоночных, по-следовавшие после К. Линнея, Ж.-Б. Ламарка и Ж- Кювье. |
Отряд «Полипы с полипняком» Ж--Б. Ламарка включал Я мшанок, губок, оболочников и некоторых полипов. Первые Д сейчас выделены в самостоятельный тип, вторые представля- Ц ют раздел Parazoa (ранг, средний между царством и под- Д царством), третьи возведены в ранг типа Хордовые, а послед- Я ние оказались кишечнополостными. Подобные катаклизмы Я системы принято объяснять низким уровнем науки XVIII— Я XIX веков. Считается, что современная зоология застрахована Я от подобных неприятностей. Я
Своеобразный эгоцентризм некоторых современных систе- Я матиков выражается в том, что нынешний уровень знаний Я объявляется абсолютным, а претензии предъявляются той Я теории эволюции, которая не признает полифилии. Между тем Я сегодня так же, как в XVIII в., таксоны остаются вероятност- я ным понятием. Наши знания о видах и их признаках замеча-тельно выросли после К- Линнея, но эмпирическое множест- 5' во видов по-прежнему остается (и останется таковым всегда) лишь выборкой из биологически возможной их совокупности. > : Ввиду этого следует считаться с возможностью разделения даже привычных хорошо изученных таксонов благодаря открытию новых признаков и новых видов, пересмотру значения уже известных признаков. Если говорить более конкретно, то, например, у разных млекопитающих, которые, как полагают сторонники полифилии, выводятся независимыми корнями от разных пресмыкающихся и сегодня неразличимы на уровне класса, могут быть найдены такие признаки, которые позволят разделить млекопитающих на самостоятельные таксоны независимо от палеонтологических данных о пресмыкающихся; возможно, даже совсем иным образом, не так как в современной системе. С другой стороны, рептилии, рассматриваемые в качестве предков млекопитающих, при дальнейшем изучении могут оказаться обладателями таких признаков, что
ПО ?
г
выведение именно из них млекопитающих сделается невозможным. Оба решения вероятны ввиду случайного характера эмпирической выборки признаков. То же допустимо и для видов. Среди пресмыкающихся возможны такие пока неизвестные виды, которые могли бы- послужить в качестве еще более правдоподобного предка млекопитающих, чем известные ныне рептилии. Среди неизвестных нам родичей млекопитающих отнюдь не исключены потомки тех рептилий, которые послужили поводом для провозглашения полифилии млекопитающих. Оба этих решения вероятны ввиду случайного характера эмпирической выборки видов. Сравнительное изучение видов и признаков позволяет высказать вероятностное суждение о преемственности планов строения, но такое суждение лишь одно из прочих, претендующих на истинность. Оно отвечает определенному уровню знаний, но никак не абсолютно. Бесспорно, что одни виды происходят от других — это доказана теорией видообразования. Ретроспективно, пользуясь только сравнительными методами, можно высказать лишь вероятностное суждение по поводу происхождения одного конкретного вида от другого. Вопрос о полифилии принципиально неразрешим сравнительным методом, которым пользуются при его обсуждении; это не вопрос сравнительной морфологии, но только теории видообразования. Пользуясь сравнительным методом, полифилию невозможно ни доказать, ни опровергнуть. Есть недостаточно изученные таксоны, нет таксонов полифи-летических. Что же касается теории видообразования, то она пока не дает оснований для полифилии. Таксон есть не просто множество сходных видов, но прежде всего биологическая организация, унаследованная*от популяции вида-предка. Гомологические ряды Н. И. Вавилова и многочисленные случаи конвергенции относятся к проблеме происхождения «одних и тех же» признаков, но не организаций. Недавно В. Н. Тихомиров (1972) еще раз напомнил о том, что схождение признаков не есть полифилия.
То обстоятельство, что гомологические ряды Н. И. Вавилова занимают столь видное место в спорах вокруг полифилии — отнюдь не заслуга самого Н. И. Вавилова. Причины параллельной изменчивости в родственных таксонах четко объяснены Н. И. Вавиловым. Они заключаются в сходстве наследственности таких таксонов. В этом объяснении нет ничего сверх того, что присутствует в дарвиновских представлениях об эволюции. Именно так идеи Н. И. Вавилова восприняты Н. В. Тимофеевым-Ресовским, Н. Н. Воронцовым и А. В. Яблоковым (1969), когда они допускают возможность сходной реакции близких таксонов на сходные воздействия внешней среды. Параллельная изменчивость была объявлена свидетельством новых, еще неизвестных факторов эволюции сторонниками полифилии (Любищев, 1968; Махотин, 1959;
8*
111
Мейен, 1972). Укладываются ли гомологические ряды в развиваемые здесь положения, которые, как можно видеть, оправдывают классическую систематику? Нетрудно показать, что противоречий здесь не обнаруживается.
Возьмем два близких таксона. Если исследователь заинтересуется изменчивостью по 4 признакам йь й2, й3, й4, то в каждом из таксонов он найдет следующую изменчивость:
0101
1010 1010
1110 0101 1000 1011 ОНО 0001
0111 1001 0001 1101 ООН 0100
Оба таксона характеризуются: биномиальным распределением подобных событий, неравным соотношением между символами «1» и «0» в каждом столбце, принадлежностью к одному и тому же Nt (см. стр. 84). Если бы исследователя заинтересовала изменчивость по признакам йь й2, то в каждом таксоне был бы увиден ряд
11.., 01.., 10.., 00...
Любые пары из тех же 4 признаков также составляют более или менее полные ряды, например
.11., .01., .10., .00.,
но, заинтересовавшись 3 признаками, исследователь не получит столь полных рядов, некоторых сочетаний будет недоставать. Они, однако, могут быть получены на более обширном множестве биологически возможных событий, отдельно для каждого из рассматриваемых таксонов. Поэтому могло бы показаться, что гомологические ряды — явление универсальное, т. е. распространимое на все признаки. Это, конечно, не так, оно сделалось бы универсальным только на таксономическом континууме, который принципиально неосуществим. Предложенная модель, как только что было показано, допускает возможность построения гомологических рядов изменчивости и одновременно — возможность всех тех выводов, которые уже получены на креветках. Таким образом, гомологические ряды изменчивости никак не противоречат принципам классической систематики. Наконец, следует иметь в виду, что, несмотря на внешнюю одинаковость, например И., в первой из упомянутой пары рядов, такие сочетания не взаимозаменяемы: это сочетания «одних и тех же» признаков. Это сделается более очевидным в ходе рассмотрения третьего результата. Он был получен как факт. Причины, по которым существуют «запрещенные» сочетания признаков, оставались нерассмотренными. Мне кажется, что эти причины могут быть поняты при 'использовании понятий организменного
112
уровня биологической организации и связаны с давней проблемой макромутаций.
С ’самого начала необходимо оговориться, что естественный отбор, в какой-то степени ответственный за разрывы между таксонами, предлагает тем самым свое объяснение взаимодействиям через уничтожение тех или иных сочетаний признаков. Отбор не исключает закономерностей, присущих таксону как биологической организации, взаимодействия между признаками допускают объяснение на основе внутренних свойств объектов систематики. Для начала попробуем представить себе, каким образом здесь проявляется действие отбора. •
Ограничившись фиксированным числом признаков, мы в общем случае можем найти в популяции особь с наперед заданным их сочетанием. Если такая особь погибнет, то в результате скрещивания мы можем получить равноценную ей по тем же признакам новую особь. Уничтожение отбором отдель-j ных популяций — носителей сочетаний полутаксономических , признаков, по-видимому, также поправимая беда. Можно вообразить, как те же самые сочетания признаков проявятся .
• в других популяциях, занявших место погибших. Эти надежды тем более обоснованы, чем меньшее число признаков принято во внимание. Вымирание видов—носителей сочетаний таксоно-‘ мических признаков, по-видимому, оставляет более глубокий след на потенциальном разнообразии таксонов-потомков, и последствия этой потери будут нарастать со временем. Так произойдет потому, что сочетания, исчезнувшие вместе с их носителем, не могут возродиться у видов с другими признака-i ми ввиду взаимодействий между признаками. В этом смысле можно говорить о последействии отбора. Такие потери могут быть восполнены благодаря ресинтезу гибридных видов, своего рода аналога скрещиваний между особями; но с повышением таксономического ранга такие возможности сокращаются, причем быстрее у животных, чем у растений. Поэтому следует еще раз подчеркнуть, что биологически возможная совокупность событий N*m включает потенциально осуществимые события, которые могут не состояться по внешней причине — прямому действию и последействию отбора. В лекциях сделана попытка предсказать свойства материала, с которым имеет дело отбор, но не ожидаемые результаты его воздействия. Мало того, из всего сказанного ясно, что параметры Nm обусловлены не только свойствами таксона как биологической организации, но также и воздействием отбора в прошлом и настоящем. Пока не . представляется возможным установить, в какой мере оригинал Nm и его увеличенное изображение Nm сформировались под воздействием отбора и в какой степени обусловлены внут
113 '
ренними свойствами таксона как биологической организации. В обеих совокупностях, безусловно, присутствует исторический момент, но это не означает возврата к обычному толкованию разрывов в таксономическом континууме.
Прямое действие и последействие отбора выражается в уменьшении частот подобных событий, не влияет на форму кривой их распределения. Оно не нарушает равенства
(m(’): т<0)); = (mt1): m^))?, потому что осуществляется по строчкам символов, точнее — по видам, реально существующим в природе, а не по столбцам, изобретению автора. Взаимодействие между признаками приводит к исключению некоторых сочетаний и, следовательно, к уменьшению частот подобных событий; но, кроме того, оно еще приводит к тому, что (т<‘): т(0)); ф (tv№ : т(0))г.
Разнообразие видов следовало бы рассматривать как результаты опытов, поставленных природой для изучения взаимодействий между признаками. Иногда результаты ее опытов наглядны и объяснимы — это в случае координаций А. Н. Северцова. Чаще, однако, результаты не столь наглядны, и исследователь воспринимает межтаксонную изменчивость как комбинаторную, когда, вообще говоря, допустимы все исходы опытов; если же некоторых пока не найдено, то нет причин, почему бы им не найтись впоследствии. Следует допускать все же, что многие из таких взаимодействий в действительности относятся к координациям и допускают причинное объяснение вроде того, которое предлагается на основе физиологического значения относительной поверхности тела. Тем не менее интуитивно оно кажется недостаточным для основной массы таксономических признаков. Здесь напрашивается совсем иное объяснение.
В классе Ресничные черви ротовое отверстие может быть расположено в различных точках брюшной поверхности от переднего конца до заднего. Этот признак не превышает ранга «отрядного». В то же время целые типы животных имеют строго определенное положение ротового отверстия на переднем конце тела, и этот признак в данном случае имеет значительно более высокую таксономическую ценность. Анализируя организацию ресничных червей, В. Н. Беклемишев (1964) замечает, что перемещение ротового отверстия мало отражается на общем плане строения этих животных вследствие их низкой организации в целом. Перемещение ротового отверстия у ресничных червей не влечет за собой коренной перестройки связанных с ротовым отверстием относительно простых органов и систем органов. Попробуем представить себе что-либо подобное, например, у членистоногих. В случае смещения рта изменения сказались бы у них по крайней мере на: 1) органах чувств (вкуса, обоняния, зрения), 2) ротовом ап
114
парате, 3) железах и мышцах, связанных с передним отделом пищеварительного тракта, 4) передних отделах центральной нервной системы. В связи с этим интересен один момент в эволюции ракообразных, который представляют усоногие раки, единственный отряд класса, перешедший к прикрепленному образу жизни. Подвижные личинки этих животных прикрепляются к подводным предметам посредством предротового придатка. Возникающая вследствие такого явно неудобного положения тела животного задача перемещения наверх ротового отверстия не может быть разрешена, каку ресничных червей, по причине общей сложности организации. Ротовое отверстие здесь не сместилось — произошло сильное искривление продольной оси тела, как это показано на рис. 35. А вот другой пример: личинка иглокожих, прикрепившаяся ко дну сходным же образом (рис. 35). Здесь также осуществилось смещение рта, и последствия этой замечательной операции налицо: кишечник приобрел петлеобразную форму. Может быть, наиболее удивительна судьба губок. Личинки этих весьма примитивных животных, оседающие на дно, имеют двигательные жгутиковые клетки снаружи. Между тем у взрослой губки те же клетки выстилают внутреннюю полость и создают ток воды, приносящей кислород и пищевые частицы. Для того чтобы разрешить эту относительно несложную функцию, губкам пришлось буквально вывернуться наизнанку (см. рис. 35). В. Н. Беклемишев (1964) замечает по этому поводу следующее: «...чем ниже организация какой-либо группы животных, тем более глубокие изменения вызывает в ней всякое приспособление».
Тип симметрии — иногда весьма важный признак крупных таксонов животных. Его изменчивость находится в обратной зависимости от сложности всего животного в целом. Обычные гидры, простейшие из кишечнополостных, характеризуются непостоянным числом щупалец и соответственно этому неопределенным порядком лучевой симметрии. Здесь это индивидуальный признак, не имеющий таксономического значения. Два класса коралловых полипов, Шестилучевые и Восьмилучевые, характеризуются установившимся порядком лучевой симметрии, что и отражено в их названиях. Порядок симметрии определяется здесь, прежде всего, числом септ пищеварительной полости. Сложное строение септ, включающих разнообразные клетки, их связанность с глоткой (отсутствовавшей у гидры), наконец, относительно более сложные щупальца — все это имеет своим следствием сравнительную стабильность порядка симметрии. И все же различного рода уклонения не представляются исключительной редкостью. У иглокожих порядок лучевой симметрии определяется количеством антимер, секторов тела с примерно равноценным содержимым, и обычно характеризует собой надвидовые таксоны. Например, все
115
Б
Рис. 35. А — личинка губки с расположенными снаружи жгутиковыми клетками; Б — взрослая губка, у которой жгутиковые клетки выстилают внутреннюю полость; В, Г, Д—последовательные стадии искривления продольной оси тела в развитии морской уточки (Crustacea Cirripedia);
Е. /К—искривление оси тела у некоторых иглокожих
116
морские ежи имеют 5 антимер и соответственно 5-лучевую симметрию; в данном случае это признак класса. У морских звезд и офиур даже виды одного семейства могут иметь различное число антимер. Здесь этот признак характеризует виды и роды.
Понятие «один и тот же» признак оказывается, следовательно, двусмысленным: в нем подчеркнута внешняя тождественность некоторого признака в разных таксонах и не отражено неодинаковое его положение в системе морфофизиологических связей в организменной организации. В общем случае «один и тот же» признак не может быть таксономически равноценным даже в двух близких таксонах, в двух членах гомологических рядов Н. И. Вавилова. В ходе эволюции один и тот же признак различных подвидов по-разному обрастает морфофизиологическими связями с другими вновь появляющимися. Такие различия нарастают вследствие расхождения видов в процессе видообразования. Возрастание сложности приводит к тому, что «один и тот же» признак приобретает неодинаковое таксономическое значение даже в близких таксонах. Вследствие возрастающей сложности такой признак мало-помалу утрачивает былую способность образовывать сочетания с другими, делается элементом сложного каркаса признаков, который называется таксономическим диагнозом — но уже более высокого ранга. Виды, объединенные в одном таксоне на основе такого диагноза, обладают другими, более мелкими признаками, сочетающимися относительно более свободно. Поэтому они не сказываются на комбинаторных возможностях более крупных признаков. Так, однако, в пределах одного таксона; в другом же они могут оказывать такое влияние. Чтобы эти рассуждения не показались умозрительными, вернемся к лекции 7. Там были приведены именно такие два противоположных случая: 1) когда решетка для «одних и тех же» признаков сохраняется без изменений в разных условиях взаимодействия — «1/Й2 0101 в условиях ..100100010 и ...110100001, 2) когда взаимодействие «одних и тех же» признаков зависит от условий взаимодействия — Й1/Й2 И00 в условиях ..010100001 и 0010 условиях ..100000000. Отсюда, вероятно, следует возможность количественной оценки таксономического значения признаков: более крупные должны характеризоваться высокой инвариантностью решеток с малым числом единица них. Пока не выполнено специальных исследований, эти соображения остаются гипотезой. Для дальнейшего рассмотрения понадобится лишь более очевидное соображение о том, что способность признаков сочетаться с другими ограничивается по мере их усложнения. По-видимому, для характеристики таксономического аспекта этого явления целесообразно ввести понятие прообраза признака: индивидуальный признак суть прообраз популяционного подвидового, по
117
следний суть прообраз видового. В свою очередь видовой признак — прообраз родового и т. д.
Необходимость по возможности более четкого различения таксономических признаков от их прообразов обнаруживается при рассмотрении давней проблемы макромутаций. Она весьма полно разобрана в трудах И. И. Шмальгаузена (1969), Дж. Симпсона (1948), Э. Майра (1968), Б. Ренша (Rensh, 1960). В последнее время к ней обращались И. М. Оленов (1961), Н. В. Тимофеев-Ресовский, Н. Н. Воронцов, А. В. Яблоков (1969). Общее в их понимании этой проблемы состоит, как мне кажется, в том, что, оставаясь на позициях дарвинизма и отрицая особую форму эволюции, приводящую к внезапному возникновению крупных таксонов, назвднные авторы считают возможным внезапное возникновение отдельных крупных признаков таксонов. Используемая при этом терминология дает, однако, повод для недоумений. Думается, что здесь в действительности речь идет не о крупных таксономических признаках, а об их прообразах.
В качестве примеров макромутаций ранга отряда приводятся, в частности, мутация дрозофилы по числу сегментов брюшка, мутация aristopedia, приводящая к появлению ножки на месте антенны и уменьшающая число члеников лапки с 5 до 4, мутация tetraptera, которая выражается в появлении задней пары крыльев на месте жужжалец. Эти примеры в какой-то степени характерны для других, приводимых разными авторами в подтверждение чисто мутационного происхождения крупных таксономических признаков.
Обратимся для начала к мутации, увеличивающей число сегментов брюшка. В ходе сравнительного изучения разных видов зоологи установили, что конструктивный элемент, называемый сегментом тела насекомого, включает по меньшей мере следующее: 1) сочленение сегмента с соседним, чем обеспечивается их подвижность, 2) систему мышц, 3) необходимую иннервацию и соответствующий нервный ганглий в составе центральной нервной системы, 4) элементы дыхательной трахейной системы. Когда такого содержимого нет, то данное образование не есть сегмент, не есть признак отряда. К сожалению, сегменты мутантов в этом отношении как будто бы не изучены; но в таком случае преждевременно говорить о признаке отряда. Такие же сомнения вызывают и другие примеры. Антенна у насекомых есть предротовой придаток, иннервируемый от головного мозга, и его строение довольно разнообразно в пределах отрядов; ходильные конечности иннервируются от брюшной нервной цепочки. Первая есть чувствительный придаток, вторые служат для передвижения. Пока не доказана иннервация вновь возникшего придатка от брюшной нервной цепочки, нет оснований рассматривать его в качестве ножки. В понятие «крыло» у насеко
118
мых вкладывается, в частности, особое сочленение с телом, допускающее его вибрацию, а также мышцы и их иннервация, У. двукрылых насекомых совершенно иная манера полета, не такая, как у четырехкрылых. Превращение лапки из 5- в 4-члениковую означает исчезновение не только соответствующих элементов наружного скелета, но также и мышц. Если все эти условия не выполняются, то нет оснований говорить о новом крупном таксономическом признаке.
Одним словом, в понятие «признак» следует вкладывать не только какое-то элементарное свойство вида, но также определенный уровень его сложности, взаимосвязь с другими признаками, устанавливаемая сравнительным методом на множестве видов. Если же показано, что некоторые два признака существенно различной степени сложности занимают неодинаковое положение во взаимосвязях с другими признаками, то нет оснований считать их одними и теми же признаками в таксономическом понимании. В таких случаях, как мне кажется, можно говорить лишь о прообразах признаков.
Примеры крупных мутаций расширяют представление о внутривидовой изменчивости, подтверждают возможность выведения крупных признаков от мелких, таксономических от индивидуальных через популяционные. Такие мутанты являются носителями материала для формирования таксономических признаков, но не являются представителями нового таксона. Проблема происхождения крупных таксономических признаков относится не только к организменному уровню, на котором проявляются мутации, но также к более высокому, популяционному. Для того чтобы возник новый вид, с новым признаком, необходимо, чтобы обособилась популяция такая, что все особи имеют 4-члениковую лапку. Усложнение такого признака, становление его таксономической ценности предстает перед систематиками, как эволюционная гетерохрония признаков в смысле А. Л. Тахтаджяна (1947). Таксономические признаки формируются исторически длительное время под воздействием не только мутагенеза, но также и отбора. Закономерности этого процесса выявляются в ходе сравнительного изучения видов в историческом аспекте. Таким образом, в согласии с вышеназванными авторами я полагаю, что пока нет надобности в особой форме эволюции, макроэволюции и что, кроме того, возникновение крупных признаков не следует сводить исключительно к одному лишь мутагенезу.
Тянитолкай, персонаж сказки К. Чуковского, может быть и допустим как чудо на организменном уровне, как редчайший мутант. Находки двухголовых змей в принципе допускают возможность такого события на организменном уровне. Значительно труднее допустить существование популяции тянитолкаев — и не только вйиду малой частоты подобных мутаций. Не следует упускать из виду, что помимо контроля
119
на уровне организменной организации существует еще контроль на уровне организации популяций, видовом уровне. Строгости, проявляемые здесь отбором, исключительно суровы: при поступательном движении в трехмерном пространстве возможны исключительно организмы с гетерополярной осью.
Теперь применим полученные результаты к другой весьма почтенного возраста проблеме — неполноте геологической летописи. Отсутствие переходных форм в филогенетических рядах рассматривается дарвинистами, как следствие неполноты геологической летописи, и в то же время используется для обоснования особой творческой взрывной эволюции их оппонентами.
Прежде всего, филогенетический континуум суть иное понятие, нежели континуум таксономический: первый подразумевает лишь непрерывность переходов между тем или иным числом членов, которые, будучи взятыми совместно, в совокупности, не обязательно составляют континуум таксономический. Вот пример таксономического континуума, на котором обозначены стрелками полные филогенетические ряды филогенетического континуума:
077 707. 770
001 010 юо i / ООО
Полные филогенетические ряды возможны и на неполном множестве событий, меньшем континуума:
оо)х
Ш-~7О7^
^000
Наконец, при условии осуществимости континуума возможна такая ситуация, когда филогенетические ряды будут неполными, прерывистыми. Так может получиться, например, если некоторые события неизвестны науке по каким-либо чисто случайным причинам, например по причине последействия вымирания. Одним словом, филогенетический и таксономический континуумы не связаны между собой. Принципиальная неполнота множества биологически возможных таксонов никак не означает неполноты филогенетических рядов: они могут быть и полными, и неполными.
Большую пользу здесь принесет представление о взаимодействии между таксономическими признаками. Принявши это
120
положение, мы должны вслед за тем признать, что различия между членами рядов не могут быть исчезающе малыми. Изменение даже одного признака должно сказаться на тех или иных других признаках, и потому соседние члены рядов не могут не различаться набсГром признаков. Тем не менее переходы в рядах постепенны. Противоречий в этих высказываниях нет, все становится на место, если использовать представление о прообразе таксономических признаков.
Пусть в филогенетическом ряду имеется два вида с признаками 0^2 и oio2. Они различаются двумя формами признаков, и потому здесь имеется разрыв в таксономическом понимании. На популяционном же уровне такой разрыв не обязателен и переход существует. Его составляют популяционные прообразы таксономических признаков. Дело в том, что отрицательные формы di и а2 отнюдь не предполагают отрицательной же формы признаков индивидуальных — популяционных. В популяциях первого вида может наблюдаться изменчивость (a+a)i, у второго вида — (а+а)2. Это положение о прообразе таксономических признаков ранее доказывалось в связи с вопросом о неадекватности таксономических диагнозов и при обсуждении таксономической неравноценности «одного и того же» признака.
Таким образом, неполнота палеонтологической летописи возникает не только вследствие бесследного вымирания промежуточных звеньев. В какой-то степени она кажущаяся. Филетический ряд в общем случае, может быть полным даже тогда, когда между его членами, видами, обнаруживаются резкие разрывы. Ошибочное впечатление о неполноте такого ряда складывается за недостаточной изученностью популяционной изменчивости таких видов. С другой стороны, кажущиеся достаточно полными филогенетические ряды в действительности могут скрывать разрывы, незаметные из-за недостаточной изученности таксономических признаков их обладателей.
В любом случае сравнительное изучение видов не может ни подтвердить, ни опровергнуть представления об особой форме эволюции. Здесь точно так же, как и при обсуждении полифилии, в качестве верховного судьи выступает теория эволюции. Последняя же утверждает, что эволюция — это эволюция видов, а превращения видов на основе межпопуляционной изменчивости совершаются постепенно; с точки зрения систематики такие изменения прерывисты. Палеонтологическим материалом преимущественно морфологического характера чрезвычайно рискованно пользоваться для подкрепления возможности особой формы эволюции, называемой макроэволюцией. При воссоздании филогенетического древа следует искать не только переходные виды, но и переходные популяции четко обособленных видов.
121
Лекция 10,
В КОТОРОЙ АВТОР УЧАСТВУЕТ В СПОРАХ МЕЖДУ СИСТЕМАТИКАМИ
Существует своеобразная система биологических систематик (Brundin, 1972; Любищев, 1972), Ее возникновение в некоторой степени вызвано разнообразием методов классификации. Чаще других упоминаются *сходственная (фонетическая) и филогенетическая классическая систематики. Можно, однако, говорить лишь о двух направлениях в систематике.
В сходственной систематике таксон есть множество сходных видов. Сходство здесь часто оценивается количественно, что дает повод говорить о математической таксономии. Математические методы, обзор которых произведен Н. Бейли (1964), Р. Блэкит (1968), используются все шире и потому ее часто отождествляют с количественной систематикой вообще. Вряд ли это правильно: количественные методы применимы, например, к вычислению родства и построению системы на иных принципах.
В филогенетической систематике таксон есть множество родственных видов. Родственные отношения между видами устанавливаются в ходе сравнительного разностороннего изучения взрослых организмов, а также с привлечением данных эмбриологии и палеонтологии. Так, однако, было бы в идеале. В действительности же исследователи большей частью ограничиваются изучением взрослых особей и к одному таксону относят сходные виды. Возможность ошибок при этом, естественно, не исключена, история систематики полна ими. Сторонники классического направления, как мне кажется, обычно полагают, что более детальными исследованиями такие ошибки можно свести к минимуму; принципиальных же противопоказаний к использованию сходства для определения родства как будто бы нет. Схождение диагнозов в независимых линиях филогенетического развития послужило бы наиболее действенным возражением против такого понимания причин сходства и, следовательно, природы таксона; но в теории видообразования нет места полифилии.
Термин «фенетический» дал повод к недоразумениям, так как он имеет общий корень со словом «фенотип». Оба, по-видимому, происходят от латинизированного phaenon (сияющий) греческого слова tpaecvos (сияющий, ясный, внятный), и целесообразность их использования очевидна, хотя первое обозначает наглядные свойства в понимании систематика, а второе — генетика. Кажется, впервые слово «фенетический» было употреблено в следующей цитате:
«Таксономические отношения между таксонами должны быть оценены только на основе сходств, существующих те
122
перь в доступном материале. Такие отношения являются статическими (Michener, 1957) или ф енетическим и, как мы теперь предпочитаем их называть» (Sokal a. Sneath, 1963). О независимости таксономического понимания от генетического свидетельствуют и другие цитаты: «...относительное сходство среди организмов (phenetic)» (Michener, 1970). «Таксономические отношения (affinities) вполне определены сходствами признаков таксонов (фенетическим сходством)» (Sokal, 1964). «Когда мы ввели термин «фенетический», мы приложили его к классификации, основанной на полном сходстве» (Harrison, 1964). «Полное сходство (или близость (affinity) ) между двумя объектами есть функция сходства многих признаков, по которым объекты сравниваются» (Sokal a. Sne-ath, 1963).
Филогенетическая систематика отличается от сходственной не используемыми признаками (в обоих они в основном фенотипические), но иной их трактовкой. В первой они служат показателем родства, во втором — только таксономических отношений. Е. С. Смирнов (1969), автор одного из сходственных методов, пишет: «...вовсе не очевидно, что эти отношения (сходственные. — Н. 3.) можно одновременно трактовать и как родственные, т. е. свидетельствующие об общности исторического развития...». Позиция Е. С. Смирнова, к которой необходимо будет вернуться впоследствии, изложена им и в следующем высказывании: «...сходство вовсе не должно выражать родства, ...систематические единицы, основанные на учете сходства, не могут быть выражением кровных связей. Другими словами, мы приходим к признанию полифилии систематических категорий...» (Смирнов, Вермель, Кузин, 1924). В понимании таксономических категорий названные авторы имеют в виду определение Ю. М.-Вермеля, предложенное им в 1931 г.: «Всякое скопление индивидов или их групп, в котором все члены сходны друг с другом больше, чем с любым членом другого скопления, мы будем называть систематической категорией». Таким образом, не следует смешивать фенетическую систематику с фенотипической, повод говорить о которой появился только совсем недавно в связи с «систематикой генотипов». Довольно широкое распространение математических методов классификации является реакцией на ходячее представление о субъективности систематиков, отсталости традиционных методов работы. Надо признаться, что систематики классического направления дают тому повод. Затруднение, однако, состоит в том, что разные математические методы, будучи примененными к одним и тем же видам, приводят к неодинаковым результатам (Пономаренко, Расницын, 1971; Тамарин, 1971; Шаталкин, 1968; Heiser, Soria, Burton, 1965; Воусе, 1964). Возникла проблема субъективного выбора объективных методов исследования.
123
«Методологическая ошибка, связанная с использованием математических методов в таксономии, заключается в том, что были смешаны два понятия объективности: объективность как соответствие результата наблюдаемым в природе отношениям и объективность как соответствие результата независимому от исследователя методу» (Шаталкин, 1973).
Снова натуралисты убеждаются в том, что применение любого метода предполагает биологическое обоснование и понимание биологического смысла полученных результатов. Математические методы сами по себе не есть вклад в теорию науки о разнообразии видов, так как предназначены констатировать, но не объяснять. Вместе с тем обсуждение правомочности того или иного метода объективно полезно для систематики. Дело в том, что независимо от измерений авторов предлагаемые ими формулы обычно выражают определенное понимание природы таксона и задач систематики. В свою очередь, эти разногласия основываются на противоречиях в понимании принципиальных вопросов теорий видообразования. Поэтому не следует думать, что не будь многочисленных трудностей при установлении родства сравнительными методами, все предпочли бы филогенетическую систематику. На первый взгляд, однако, все обстоит иным образом.
Систематики как будто бы решают свои задачи в отрыве ст эволюционистов. Внешне развитие систематики связано исключительно с количественным ростом наблюдений — видов, их признаков и совершенствованием правил номенклатуры. Подход систематиков к объекту, основные приемы классификации как будто бы не претерпели существенных изменений за последние двести лет. Мнение Ф. Добжанского (Dobzhan-sky, I960) о том, что эволюционная теория не повлияла на классификацию, еще не встретило серьезных возражений. Все же с этой точкой зрения согласиться невозможно.
К. Линней, Ж--Б. Ламарк, Ж- Кювье и Ч. Дарвин.— каждый из этих замечательных биологов занимался практической систематикой, и вклад каждого в систему как упорядоченное множество видов действительно никак не связан с признанием эволюции в той или иной форме или с отрицанием ее. Способность создавать классы объектов, сходных по каким-либо признакам, есть общее свойство ума. Оно подвержено индивидуальной изменчивости и не зависит от общебиологических воззрений. Но система есть, кроме того, научная теория. Ее объяснительная функция не может не зависеть от того, каковы взгляды автора на происхождение видов. В некоторых случаях это может быть установлено с достаточной ясностью.
Общеизвестна система растений К. Линнея, в которой растения классифицируются по числу тычинок цветка. В дальнейшем она не получила подтверждения, и теория систематики может объяснить, почему это произошло: потому что этот
124
признак эволюционирует так же, как и все другие, и не может характеризовать собой разные таксоны одного ранга. Переменная разрешающая способность признака, в свою очередь, следует из признания изменяемости видов, из признания единой формы эволюционного процесса; но это уже относится к теории эволюции. Объективно, даже независимо от общих взглядов К. Линнея, его система растений основана на отрицании способности видов эволюционировать. Общие взгляды К. Линнея сказались на его системе растений.
Теория типов Ж. Кювье, в которой утверждается разобщенность, несводимость типов организации, никак не может явиться той почвой, на которой выросло бы филогенетическое древо. Объяснение его системы через филогению невозможно, привлечение теории эволюции к истолкованию результатов исследования — излишне. Например, тип Radiata Ж- Кювье, включавший, в частности, иглокожих, не имеет ничего общего с другими типами, включающими организмы с двубокой симметрией. Сегодня зоологи не признают несводимость этих типов организаций, потому что в эмбриональном' развитии иглокожих имеется личинка диплеврула двусторонне-симметричного плана строения. Признак типа Ж. Кювье оказался подверженным возрастной изменчивости; кроме того, известны иглокожие с двухлучевой симметрией весьма близкой к двубокой. Благодаря возрастной и межтаксонной изменчивости можно понять, каким образом могла быть выведена радиальная симметрия иглокожих. В сущности, системе Ж. Кювье свойственна та же особенность, что и системе К. Лин-, нея, — неизменность таксономической ценности признака, в данном случае типа симметрии. Следуя Ж. Кювье, мы должны были бы отделить иглокожих от других двусторонне-симметричных вторичноротых, разрушить раздел Deuterostomia. Иными словами, общие взгляды Ж. Кювье действительно отражаются на системе.
Тем не менее было бы несправедливостью упрекать К. Линнея и Ж. Кювье в том, что они видели задачи систематики в формальной каталогизации. Система в их понимании не была самоцелью, она допускала объяснение на теологическом принципе. Основная объяснительная функция системы сохраняла свое значение. Задачу систематики оба видели в раскрытии плана Творца. С точки зрения современного материалистически настроенного натуралиста их система представляется формальностью, но это именно потому, что им отвергнуто теологическое объяснение. С самого своего зарождения систематика претендовала на нечто большее, чем просто регистрацию фактов. Она низведена до такого уровня трудами сторонников сходственной систематики. Особенно показательна нумерическая таксономия, может быть, наиболее заметное сейчас течение в области сходственной систематики с
9 Н. А. Заренков
125
весьма характерным для нее признанием равного веса любых признаков. Идейный багаж ее небогат. Стремление к точной регистрации биологически не проанализированных фактов довлеет здесь над всеми иными соображениями. «Таксономия, как представляется нам, есть строго эмпирическая наука» (Sokal a. Sneath, 1963) — этот постулат нумерической таксономии в сущности отрицает ее научность. Думается, что нечто не есть наука, если оно ограничивается лишь регистрацией фактов. Отказ от попыток объяснить полученные результаты в свете теории эволюции — другая примечательная особенность этого направления, на что раньше обратил внимание И. И. Канаев (1966). Он же подметил преемственность идей основателей нумерической таксономии с типологической школой. Вследствие этого претензии нумерической таксономии на революцию в систематике (Сокал, 1968) кажутся чрезмерными. В свое время идея эволюции оказала революционизирующее воздействие на все разделы биологии. Поэтому отказ от эволюционизма есть скорее контрреволюция.
Далее, обращает на себя внимание то обстоятельство, что методы нумерической таксономии, так же как и других направлений сходственной систематики, игнорируют биологическую природу объектов и в принципе применимы к классификации самых различных предметов. Р. Сокал (1967) усматривает в этом достоинство нумерических методов. А. А. Люби-щев (1967) обсуждает методы классификации видов наравне с методами классификации химических элементов Д. И. Менделеева, кристаллов Е. С. Федорова и галактик. Разумеется, можно говорить о общих проблемах классификации и о категории сходства вообще, когда речь идет о внешней функции систематики; но не следует упускать из виду особенности.биологических объектов, когда мы обращаемся к основной, объяснительной функции системы.
Соседство элементов в клетках таблицы Д. И. Менделеева не означает с необходимостью генетической связи между ними. Известно, например, что при распаде урана-235 образуются цезий и рубидий, которые не соседствуют ни друг с другом, ни с ураном. Между тем соседство таксонов в системе есть свидетельство их общего происхождения, во всяком случае в классической систематике. Свойства химических элементов обусловлены структурой атомов и молекул, и здесь мы не встречаем ничего подобного возрастной или географической изменчивости животных и растений; таксономические признаки есть результат действия отбора на основе внутривидовой изменчивости, виды есть объект исторический. Место элемента в таблице Д. И. Менделеева определяется двумя признаками: атомным весом и валентностью. В биологической системе нет таких признаков, которые обладали бы такой стойкой разрешающей способностью. Каким образом отсюда
126
следует специфика биологической классификации?
Рассмотрим в связи с этим поле деятельности сходственных систематиков. Уже отмечалось (Родендорф, 1965; Sharov, 1965), что в первую очередь их внимание привлекают сравнительно небольшие таксоны; ревизия таксонов высокого ранга пока откладывается на будущее (Sokal, 1964). Исключение составляет микробиология, где исследователями охвачены крупные таксоны. Однако повсюду разнообразие воспринимается как континуум, разрывы в котором обусловлены внешними причинами и не следуют из. внутренних свойств видов. В таком континууме невозможно установить преемственность между разными типами биологических организаций методом сравнения, и исследователи вынуждены прибегать к сходственным методам. Чем более приближающимся к континууму представляется множество таксонов, тем более оправданными кажутся сходственные методы. Наоборот, крупные таксоны, такие, как классы и типы, разграничены столь резко, что взаимная обусловленность признаков представляется очевидной; и хотя причины этого явления не всегда понятны, континуум представляется невозможным. Здесь понятие сходства теряет свое значение, уступая преемственности, вы-, водимости биологических организаций. К одному таксону относят соподчиненные, если последние выводимы от общего предка. Сходственные методы — это вынужденная замена сравнительно-морфологическим, сравнительно-биохимическим, сравнительно-физиологическим. Она осуществляется по двум причинам: во-первых, множество мелких таксонов воспринимается как почти континуум, во-вторых, (и отчасти вследствие), отсутствует такое причинное объяснение разрывов в эмпирическом разнообразии, которое основывалось бы на внутренних свойствах таксона как биологической организации, не сводилось бы только к внешней причине, вымиранию. Математический бум в сходственной таксономии, как мне кажется, вызван, в частности, кажущейся/бесперспективностью филогенетического объяснения на кажущемся весьма вероятным таксономическом континууме. Отношение сходства симметрично: вид А походит на вид Б ровно столько, сколько Б походит на А. Отношение родства, напротив, не симметрично: если А есть предок Б, то нельзя поменять их местами, не меняя отношение. Используя одно лишь сходство, в одном таксоне можно объединить потомков некоего предка, но нельзя отобразить отношений между предками и потомками. Сходственную систематику вполне устроил бы даже крайний полифилетизм, потому что в таком случае не приходится устанавливать родственных отношений. Не случайно, что энтузиазм по поводу сходственной таксономии соседствует с благожелательным отношением к полифилии. Это заметно в работах А. А. Любищева (1967, 1972).
9*
127
Другой путь к оправданию континуума — ламарковская концепция.
Определяющая роль внешних факторов в формировании видов при наличии у последних способности целесообразно изменяться означает, что по прошествии достаточно длительного времени разнообразные условия существования приведут к возникновению видов с любыми наперед заданными признаками. В таком случае объяснение системы следует искать не в родственных отношениях между таксонами, а во внешних факторах. Сознательное проведение ламарковской концепции в систематику сказывается не только на объяснительной функции системы, но даже на практических приемах ее построения. В связи с этим следует остановиться на творчестве Е. С. Смирнова. К сожалению, в капитальном сочинении «Таксономический анализ» (1969) его общие взгляды изложены весьма кратко и, лишь пользуясь другими работами, можно составить более или менее полную картину. Прежде всего, объект систематики, по Е. С. Смирнову, ограничен во времени. Система существует только для определенного отрезка времени, для отдельных геологических периодов, не существует единой системы. По этой причине энтомолог, изучающий современных насекомых, не должен помещать в’нее вымершие таксоны. Отношения между системами разных геологических эпох — задача филогении. Если это так, то в системе может не оказаться предков и потомков, а тогда, стало быть, в общем случае нельзя говорить о филогенетических отношениях между таксонами одной системы. Действительно, Е. С. Смирнов говорит о «таксономических отношениях», противопоставляет системе филогению, протестует против филогенетического подхода в систематике. Задача систематики, по Е. С. Смирнову, заключается в четкой регистрации таксонов, причем последние полностью основаны на сходстве-разли^ чии. В филогенетической систематике сходство есть показатель родства, а что означает сходство в таксономии Е. С. Смирнова? Ответ на этот вопрос дан в работах 1923, 1924 гг.: обитание в сходных условиях. Виды потому сходны и относятся к одному таксону, что обитают в сходных условиях. Тогда система для отдельного геологического периода приобретает такой смысл: система каждый раз есть отражение множества существующих в данное время экологических ниш на множество видов. В структуре таксона отражено разнообразие тех условий, в которых обитают составляющие таксон виды. Перед нами последовательно проведенное в теорию и практику систематики ламарковское понимание эволюции.
Установленное Е. С. Смирновым (1924) нормальное распределение коэффициентов корреляции для видовых признаков (средних, полученных на многих особях) автор склонен рассматривать «как отражение соответствующей группировки 128
экологических факторов». Понятно, что вследствие особого понимания задач систематики Е. С. Смирнов не ставит вопроса о том, насколько распространим полученный им результат на все биологически возможное множество таксонов. Точно так же регистрация корреляций между таксономическими признаками представляется достоверным фактом, хотя они вычислены только на эмпирическом множестве видов.
Автор же, напротив, исходил из того, что подобные выводы нельзя делать только на эмпирическом материале и что отношение последнего к биологически возможному — вопрос вопросов. Продуманность отдельных взаимосвязанных положений таксономии Е. С. Смирнова представляет контраст с бездумностью «революционеров» от систематики. Целесообразность метода Е. С. Смирнова может оспариваться за рамками собственно систематики, в области эволюционной теории. Однако, принявши ламарковскую модель эволюции, мы найдем его метод вполне оправданным. Единственную непоследовательность Во взглядах Е. С. Смирнова я нахожу в признании им реальности таксона. Таксоны существуют геологически длительное время. Поэтому, ограничиваясь, например, изучением только ныне живущих видов, мы искусственно разрываем таксон, делаем его условностью. Это еще один пример, когда общие взгляды систематика сказываются на результатах его исследований.
Отказ от представлений К. Линнея и Ж- Кювье о неизменности таксономического значения одного и того же признака (тычинок цветка, геометрического плана строения) по сути дела означал признание изменяемости видов и единой формы эволюционного процесса. Этот отказ в основном совершался стихийно, по мере того, как прежние методы классификации вступали в противоречие с фактами. Исход этого столкновения — крушение классов К. Линнея, несостоятельность типа Radiata Cuvier.
Теперь ясно, что вышеприведенное утверждение Ф. Доб-жанского о независимости принципов систематики от теории эволюции ошибочно. В этих происшествиях’’повинна, прежде всего, ошибочная теологическая теория. С принятием другой теории, пусть даже стихийным, неосознанным, система растений и животных существенно изменилась. Новый метод — неадекватные описания — получил обоснование несколько позже в замечательном труде Ч. Дарвина. В этом смысле дарвинизм, безусловно, вызвал революцию в систематике. Это напоминает положение, сложившееся в животноводстве и растениеводстве в новое время. Хотя выведение пород животных и сортов растений осуществлялось издавна, научное обоснование селекционно-генетических методов все же вызвало революцию в этом виде человеческой деятельности.
Некоторые современные, ведущие свое происхождение от
129
Ч. Дарвина представления о видообразовании сказались даже на практических приемах работы систематиков. Правило 50% (75%), о котором уже упоминалось, отражает дарвиновские представления о постепенности превращения видов. Рекомендуемые Э. Майром, Э. Линсли и Р. Юзингером (1956) географические и экологические названия для подвидов подчеркивают значение пространственной изоляции для видообразования. Итак, практическая систематика отнюдь не безучастна к событиям в области теории. Классификация «вообще» невозможна, она обязательно опирается на некоторую биологическую теорию. Это вполне естественно. Более общий вопрос о соотношении между методом и методологией давно уже нашёл положительное разрешение в философии. Пренебрежение этой истиной привело к необоснованному, на мой взгляд, пересмотру структуры систематики. Этому немало способствовали Дж. Симпсон (Simpson, 1961) и Э. Майр (1971). В той или иной степени им следуют и другие биологи. Из систематики в обычном понимании они выделяют таксономию как науку о теории и практике классификации и собственно систематику как науку о разнообразии видов. Дж. Симпсон считает, что «объектом классификации служат организмы, а объектом таксономии являются классификации». А. Л. Тахтаджян (1973) рассматривает таксономию лишь как часть систематики, определяя ее как «грамматику систематики», и занимает, таким образом, более умеренную позицию. В классической систематике термин «таксономический» почти полностью перекрывается теми понятиями, с которыми имеет дело, например Кодекс зоологической номенклатуры (1966), т. е. с чисто номенклатурными, процедурными и юридическими вопросами, не имеющими никакого отношения к принципиальным теоретическим вопросам классификации, обоснования таксонов, их реальности.
Если оставить в стороне явно ошибочное утверждение Дж. Симпсона об организмах как объектах систематики (кстати, это заблуждение разделяют и другие исследователи), то тогда останется более основательный повод для критики. Возведение таксономии в ранг научной дисциплины означает, что объективную научную ценность могут представлять разные, даже взаимоисключающие классификации, что в природе много разных систем и каждая из них — реальность. Иначе получается, что объекты научной дисциплины таксономии есть биологически нереальные объекты. Если же мы примем положение, что реально существует только одна система организмов, то таксономия окажется лишенной существенного признака естественнонаучной дисциплины — специфического объекта. В противном случае приходится обделить систематику—науку о разнообразии видов. Однако к одному объекту возможны различные подходы, и поэтому последние доводы
130
против самостоятельности таксономии как науки вследствие неспецифичности объекта, вообще говоря, недостаточны. Любая научная дисциплина характеризуется, кроме того, еще собственным методом, теорией. Здесь мы как будто находим оправдание самостоятельности таксономии. Литература, посвященная методам классификации, быстро возрастает. В связи с'наплывом разных методов построения системы, действительно, казалось бы, возникает потребность в особом «хозяине», который навел бы порядок в этом множестве формул и методов. Но уже упоминалось, что претендующие на объективность новейшие методы приводят к существенно различающимся системам. Причины этого явления были прекрасно разъяснены А. И. Шаталкиным (1973). Поэтому систематика нуждается в таком методе, который основывался бы на разумной биологической теории, не только на общем понятии сходства. Мне представляется, что сегодня 'такую основу может составить только дарвинизм, дополненный открытиями современной биологии, синтетическая теория эволюции. Однако объяснение многообразия видов на такой основе есть функция систематики, даже в узком смысле слова. Выходит, что таксономия не обладает и собственной теорией. Не может же явиться таковой теория корреляций, широко используемая при вычислении сходства... Биометрические методы используются повсюду в биологии и никак не специфичны именно для таксономий.
К этому следует добавить, что специфические биологические методы упорядочения множества видов давно и успешно используются. Это сравнительно-морфологический, сравнительно-физиологический, сравнительно-биохимический. Создание таксономии с ее методами вычисления сходства означает вредное умаление и заслуг и значения этих методов. В действительности же, наоборот, область их применения может быть расширена за счет мелких таксономических признаков неизвестного функционального значения. Думается, что недалеко то время, когда из одних сочетаний мелких признаков мы сможем выводить другие, повторяя ход эволюционного процесса так же успешно, как в случае с крупными, явно взаимно обусловленными признаками. Для этого надо установить вероятностные правила, по которым такие признаки сочетаются на множестве биологически возможных событий.
В систематике как будто бы нет ничего, что оправдывало бы выделение таксономии из систематики в качестве самостоятельной научной дисциплины. Тем не менее объективные причины раскола налицо, но они не связаны со спецификой таксономического знания.
Внешнее давление на систематику все время возрастает. Все биологи, и не только биологи, заинтересованы в добротном каталоге, в котором виды были бы расставлены навсег-
131
да каждый на своем месте. Большой удельный вес исследований на организменном уровне (энергетика клетки, ДНК, биологические мембраны), где исторический момент пока дей-. ствительно не проступает с такой ясностью, как, например, в зоогеографии, приводит к тому, что главная объяснительная функция системы оказывается позади ее второстепенной внешней функции упорядочения. Между тем в действительности классическая систематика дает меньше оснований для скептического к себе отношения, чем это кажется на первый взгляд. Во-первых, таксономические ревизии далеко не всегда затрагивают сущность таксономического объяснения. Во-вторых, таксономические решения не могут быть жестко однозначными по причине вероятностной сущности объекта. Соблазн здравого инженерного решения в данном случае весьма велик, объективность математических методов представляется избавлением от «произвола», которому не видно конца. Отсюда пренебрежение к традиционным сравнительным методам исследования, настойчивые поиски объективных сходственных методов. Традиционные таксономические ^систематические) методы упорядочения, конечно, нуждаются в совершенствовании, это составляет предмет практической систематики; но любые математические приемы обязательно должны основываться на биологической теории.
Оценка достоверности таксономических построений — может быть, наиболее актуальная задача. Другая задача — достижение равномасштабности таксонов одного ранга :— также требует разрешения. Не следует думать, что здесь речь идет в сущности только о количественном измерении очевидных объектов, таксонов. Внедрение количественных методов означает не просто методическое усовершенствование. Оно не только повысит научность систематики, но также сделает возможными разные подсчеты, например, близости современной или ископаемой фауны, поможет установить закономерности эволюции таксонов, будет способствовать постановке новых задач и решению старых. Следует иметь при этом в виду, что количественная оценка ранга таксона не может быть однозначной, но только вероятной. Вероятность таких оценок следует не столько из тех погрешностей измерения, которые неизбежны при измерении вообще, но прежде всего из сущности объекта систематики. Любые самые солидные методы определения родства и границ таксонов несостоятельны, если они обещают жестко однозначные решения.
Наконец, мне представляется полезным построение такой системы, которая включала бы и неизвестные науке таксоны. В такой системе перестройки были бы следствием открытия новых признаков уже известных видов, если исключить просто ошибочные определения; но и в этом случае достоверность каждого таксономического объяснения могла бы быть уста-132
новлена. Генеральная система биологически возможных таксонов способствовала бы повышению престижа систематики, стимулировала бы развитие исследований в разных других направлениях. Построение такой системы целесообразно по достижении необходимой репрезентативности эмпирического множества фактов по отношению к биологически возможному. В этом, как мне кажется, и заключается смысл благородной деятельности систематиков, неустанно открывающих новые виды и новые признаки.
ЛИТЕРАТУРА
Айр ум ян К. А., ЧилингарянА. А. Гибридизация канарейки с аборигенными вьюрковыми (Passeriformes, Fringillidае) Армении. «Зоол. журн.», 1972, т. 56, № 2.
Антонов А. С. О специфичности первичных структур ДНК хордовых животных. Сб. «Строение ДНК и положение организмов в системе». Изд-во МГУ, 1972.
Ашмарин И. П., ЛойцянскаяМ. С., Харченко Е. П. Насколько тесно связан ряд биологических особенностей бактерий с составом и содержанием ДНК? Сб. «Строение ДНК и положение организмов в системе». Изд-во МГУ, 1972.
Ашмарин И. П., Харченко Е. П. «Запрещенные» сочетания нуклеотидного состава и количество ДНК в геноме микроорганизмов. «Журн. эвол. биохим. и физиол.», 1971, т. 7, № 5.
Бейли Н. Статистические методы в биологии. М., «Мир», 1964.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных, т. 1, т. 2. М., «Наука», 1964.
Белозерский А. Н. Молекулярная биология — новая ступень познания природы. М., «Советская Россия», 1970.
Белозерский А. Н., Медников Б. М. Нуклеиновые кислоты и систематика организмов. «Природа», 1971, № 8.
Берг Л. С. Номогенез или эволюция на основе закономерностей. «Тр. Геогр. ин-та», 1922, вып. 1.
Берг Р. Л. Межвидовая и внутривидовая изменчивость жилкования крыла в семействе дрозофилид (Drosophilidae). Сб. «Применение математики в биологии», вып. 2. Изд-во ЛГУ, 1960.
Б ир штейн Я. А. Класс Crustacea Ракообразные. «Основы палеонтологии: Членистоногие, трилобитообразные, ракообразные». М., Изд-во АН СССР, 1960.
Благовещенский А. В. Биохимические критерии таксономии цветковых растений. Сб. «Проблемы современной ботаники», вып. 1. М.—Л., «Наука», 1965.
Благовещенский А. В. Биохимическая эволюция цветковых растений. М., «Наука», 1966.
Б л э к и т Р. Э. Морфометрический анализ. Сб. «Теоретическая и математическая биология». М., «Мир», 1968.
Браше Ж. Биохимическая эмбриология. М., ИЛ, 1951.
Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. М.—Л., «Наука», 1967.
Вайнштейн Б. А. О числовой таксономии. «Журн. общ. биол.», 1968,. т. 29, вып. 2.
Вермель Ю. М. Эскизы о факторах, направляющих эволюцию. «Тр. НИИ зоологии», 1931, т. 4, вып. 3.'
Виноградов Л. Г. Определитель креветок, раков и крабов Дальнего Востока. «Изв. ТИНРО», 1950, т. 32.
Воронцов Н. Н. Эволюция пищеварительной системы грызунов (Мышеобразные). Новосибирск, 1967.
133-
Воронцов Н. Н., Фомичева И. И., Баранов О. К. Перспективы и границы применения электрофоретических и иммунологических методов в таксономии млекопитающих. «З.оол. журн.», 1972, т. 51, № 12. Г а л к и н Ю. И. Брюхоногие моллюски трохиды дальневосточных и северных морей СССР (сем. Trochidae). «Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР», 57. М,—Л.,- Изд-во АН СССР, 1955.
Гептнер В. Г. Предисловие к русскому изданию книги Э. Майра, Э. Лиисли, Р. Юзингера «Методы и принципы зоологической систематики». М., ИЛ, 1956.
Гептнер В. Г. Структура систематических групп и биологический прогресс. «Зоол. журн.», 1965, т. 44, № 9.
Гептнер В. Г. Предисловие к русскому изданию книги Э. Майра «Принципы зоологической систематики». М., «Мир», 1971.
Головлев Е. Д., Скрябин Г. К. Систематика микобактерий и родственных организмов. «Изв. АН СССР», сер. биол., 1972, № 5.
Даревский И. С., Даниелян Ф. Д. Диплоидные и тетраплодные особи в потомстве партеногенетических самок скалистых ящериц, естественно спаривающихся с самцами близких бисексуальных видов. ДАН СССР, 1969, т. 184, № 3.
Денисова И. А. Электрофоретическое исследование возрастных изменений в соотношении белковых фракций сыворотки крови малого суслика. Сб. «Внутривидовая изменчивость наземных позвоночных животных и мироэволюция». Свердловск, 1956.
Дубинин Н. П. Эволюция популяций и радиация. М., Атомиздат, 1966. Завадский К. М. Вид и видообразование. Л., Изд-во АН СССР, 1968. Завадский К. М. Синтетическая теория эволюции и диалектический материализм. «Философские проблемы эволюционной теории», вып. 2. М, Изд-во АН СССР, 1971.
Завадский К. М., Ермоленко М. Т. К критике неономогенеза. «Философские проблемы современной биологии». Л., «Наука», 1966.
Заварзин Г. А. Несовместимость признаков и теория биологической системы. «Журн. общ. биол.», 1969, т. 30, № 1.
3 а в а р з и н Г. А., Старк Ю. С. Анализ запрещенных вариантов в систематике микроорганизмов. «Изв. АН СССР», сер. биол., 1965, № 5. Зенкевич Л. А. История системы беспозвоночных. «Руководство по зоологии», т. 1. М.—Л., «Советская наука», 1937.
Зенкевич Л. А. Система и филогения. «Зоол. журн.», 1939, т. 18, № 4. Иванов А. В. Класс Брюхоногих моллюсков (Gastropoda). Руководство по зоологии, т. 2. М.—Л., «Советская наука», 1940.
Канаев И. И. Очерки из истории проблемы морфологического типа от Дарвина до наших дней. М,—Л., «Наука», 1966.
К а н е п С. В. Корреляционная структура черепа некоторых серых полевок. Сб. «Внутривидовая изменчивость наземных позвоночных животных и микросистематика». Свердловск, 1966.
Кедров Г. Б. Взаимосвязь некоторых признаков древесины двудольных и ее эволюционное значение. Сб. «Морфология высших растений». М., «Наука», 1968.
Кирпичников М. М. К вопросу о количественном анализе в систематике растений. «Бот. журн.», 1948, т. 33, № 3.
Козо-Полянский Б. М. Введение в филогенетическую систематику высших растений. Воронеж, 1922.
Колосова Л. Д. К вопросу об эволюции корреляционных плеяд (на примере двух видов льнянок). «Журн. общ. биол.», 1971, т. 32, № 4. Крепе Е. М. Фосфолипиды клеточных мембран нервной системы в развитии животного мира. «Баховские чтения», вып. 22. Л., «Наука», 1967. Криштофович А. Н. Эволюция растений по данным палеоботаники. «Проблемы эволюции», вып. 1. Новосибирск, 1950.
Крыжаиовский О. Л. Объективность и реальность высших таксономических категорий. «Зоол. журн.», 1969, т. 47, № 6.
134
Кузичев А. С. Диаграммы Венна. М., «Наука», 1968.
КэйнА. Вид и его эволюция. М., ИЛ, 1958.
Ламарк Ж--Б. Философия зоологии. Избранные сочинения в двух томах, т. 1. М., Изд-во АН СССР, 1955.
Левитский Г. А. Морфология хромосом и понятие кариотипа в систематике. «Тр. по прикл. бот., ген. и селекции», 1931, т. 27, № 1.
Лихарев И. М., Р амм ел ьменер Е. С. Наземные моллюски фауны СССР. Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР. Л., «Наука», 1952.
Лукин Е. И. Неравномерность темпов органической эволюции. Сб. «Философские проблемы эволюционной теории», вып. 2. М., Изд-во АН СССР, 1971.
Лукьяненко В. И., У мер о в Ж. Г. Особенности антигенного состава сывороточных белков трех аллопатрических популяций русского осетра. «Журн. общ. биол.», 1971, т. 32, № 4.
Л ю б и щ е в А. А. О некоторых новых направлениях в математической таксономии. «Журн. общ. биол.», 1966, т. 27, № 6.
Любищев А. А. Проблемы систематики. Сб. «Проблемы эволюции», вып. 1. Новосибирск, 1968.
Любищев А. А. Рецензия на книгу Е. С. Смирнова «Таксономический анализ». «Энтомол. обозрение», 1971, т. 50, № 2.
Любищев А. А. К логике систематики. Сб. «Проблемы эволюции», вып. 2. Новосибирск, 1972.
Майр Э. Систематика и происхождение видов с точки зрения зоолога. М., ИЛ, 1947.
Майр Э. Зоологический вид и эволюция. М., «Мир», 1968.
.Майр Э. Принципы зоологической систематики. М., «Наука», 1971.
.МайрЭ., ЛинслиЭ., ЮзингерР. Методы и принципы зоологической систематики. М., ИЛ, 1956.
Матвеев Б. С. О задачах эволюционной морфологии. «Вести, зоол.», 1969, вып. 4.
Матекин П. В. Тип 10. Моллюски (Mollusca). «Курс зоологии. 1. Зоология беспозвоночных». Под ред. Б. С. Матвеева и П. В. Матекина, изд. 7. М., «Высшая школа», 1966.
М а хоти» А. А. О принципах составления филогенетических схем. «Тр. Ин-та морфологии животных им. А. Н. Северцова», 1959, вып. 27.
Медведев Ж- А. Молекулярно-генетические механизмы развития. М., «Медицина», 1968.
Медников Б. М., П о п о в Л. С., А н т о н о в А. С. Характеристики первичной структуры ДНК как критерий для построения естественной системы рыб. «Журн. общ. биол.», 1973, т. 34, № 4. /
Международный кодекс зоологической номенклатуры, принятый XV Международным зоологическим конгрессом. М.—Л., «Наука», 1966.
М е й е н С. В. Путь к новому синтезу, или куда ведут гомологические ряды. «Знание — сила», 1972, № 8.
Мошковский Ш. Д. Проблема вида и протозоология. Успехи протозоологии. «Тез. докл. III Междунар. конгр. протозоол.». Л., 1969.
Наумов Н. А. О некоторых актуальных вопросах микологии. «Проблемы ботаники», вып. 1. М.—Л., Изд-во АН СССР, 1950.
Наумов Д. В. Гидроиды и гидромедузы морских', солоноватых и пресноводных бассейнов СССР. Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР, вып. 70. М.—Л., Изд-во АН СССР, 1960.
О л ено в Ю. М. Некоторые проблемы эволюционной генетики и дарвинизма. М.—Л., Изд-во АН СССР, 1961.
Орлов В. Н. Эволюционные аспекты хромосомной дифференциации млекопитающих. «Зоол. журн.», 1970, т. 49, № 6.
Пономаренко А. Г., РасницынА. П. О фенетической и филогенетической системах. «Зоол. журн.», 1971, т. 50, № 1.
135
Печерский Ю. Н. О распознавании зависимостей -между признаками палеонтологических объектов. «Применение математических методов в палеонтологии». Кишинев, 1970.
Райков И. Б. Кариология простейших. Л., «Наука», 1967.
Родендорф Б. Б. Вопросы палеозоологической систематики. «Палеон-тол. журн.», 1959, № 3.
Родендорф Б. Б. Многообразие органического мира и его историческое развитие. «Журн. общ. биол.», 1965, т. 26, № 4.
Рубайлова Н. Г. Развитие взглядов Ч. Дарвина по вопросам отдаленной гибридизации. Сб. «Идея развития в биологии». М., «Наука», 1965.
Руженцев В. Е. Принципы систематики, система и филогения палеозойских аммоноидей. М.—Л., Изд-во АН СССР, 1960.
Светлов П. Г. -О целостном и элементаристском методах в эмбриологии. «Арх. ант., гист. и эмбриол.», 1964, т. 46, № 4.
Северцов А. Н. Морфологические закономерности Эволюции. М.— Л., Изд-во АН СССР, 1939.
Сигел Р. У. Генетика типов спаривания — взгляд в прошлое и будущее. Успехи протозоологии. «Тез. докл. III Междунар. конгр. прото-зоол.». Л., 1969.
Симпсон Дж. Г. Темпы и формы эволюции. М., ИЛ, 1948.
Синская Е. Н. Динамика вида. М.—Л., Сельхозгиз, 1948.
Смирнов Е. С. Теория типа и естественная система. «Тр. I Всеро.с. съезда зоологов, анатомов и гистологов», 1923.
СмирновЕ. С. О строении систематических категорий. «Русск. зоол. журн.», 1923, 3, 3—4.
Смирнов Е. С. Анализ распределения и соотношения признаков в систематических категориях. «Докл. Российск. АН», 1924, IV—VI.
Смирнов Е. С. Систематика живого. Сб. «Структура и формы материи. Диалектический материализм и современное естествознание». М., «Наука», 1967.
Смирнов Е. С. Таксономический анализ. Изд-во МГУ, 1969.
СмирновЕ. С., Вермель Ю. М., Кузин Б. С. Очерки по теории эволюции. М., 1924.
Соколов И. И. О некоторых методах и принципах систематики. «Тр. ЗИН», 1965, вып. 35.
С о к э л Р. Р. Современные представления о теории систематики. «Журн-общ. биол.», 1967, т. 28, № 6.
Сокэл Р. Р. Нумерическая таксономия: методы и современное развитие. «Журн. общ. биол.», 1968, т. 29, № 3.
Станчинский В. В. Биологический принцип в классификации. «Тр. I Всерос. съезда зоологов, анатомов и гистологов», 1923.
С т а р к Ю. С. Принцип запрещения в систематике микроорганизмов. «Изв. АН СССР», сер. биол., 1966, т. 5.
Старобогатов Я- И. Фауна моллюсков и зоогеографическое районирование континентальных водоемов. Л., «Наука», 1970.
Старобогатов Я- И. Практические приемы систематики и вопрос о критерии вида. «Зоол. журн.», 1969, т. 47, № 6.
Страутман Ф. И., Сухомлинов Б. Ф., Кушнирук В. А. Применение биохимических методов для выяснения формообразующей роли среды. Сб. «Внутривидовая изменчивость позвоночных животных и микроэволюция». Свердловск, 1966.
Стрельников И. Д. Анатомо-физиологические основы видообразования позвоночных. Л., «Наука», 1970.
Тамарин П. В. О противоположном характере результатов, получаемых с помощью весового и адансоновского принципов в таксономии. «Журн. общ. биол.», 1971, т. 32, № 4.
Тахтаджян А. Л. О принципах, методах и символах филогенетических построений в ботанике. «Бюлл. МОИП», биология, 1947, т. 52, № 5..
136
Тахтаджян А. Л. Вопросы эволюционной морфологии растений. Изд-во ЛГУ, 1954.
Тахтаджян А. Л. Насущные задачи систематики высших растений. Сб. «Проблемы современной ботаники», 1965.
Тахтаджян А. Л. Наука о разнообразии живой природы. «Природа», 1973, № 6.
Терентьев П. В. Метод корреляционных плеяд. «Вести. Моск, ун-та», 1959, т. 9, № 2.
Терентьев П. В. Методические соображения по изучению внутривидовой географической изменчивости. Сб. «Внутривидовая изменчивость позвоночных животных и микроэволюция». Свердловск, 1966.
Тимирязев К. А. Чарлз Дарвин и его учение. М., 1937.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М., «Наука», 1969.
Тихомиров В. Н. Об отражении некоторых особенностей эволюции покрытосеменных в филогенетической системе. «Бюлл. МОИП», биология, 1972, т. 77, № 3.
У от ер м ен Т. Проблема. Сб. «Теоретическая и математическая биология». М., «Мир», 1968.
Ушаков Б. П. Физиология клетки и проблема вида в зоологии. «Цитология», 1959, т. 1, № 5.
Федоров А. А., Пименов М. Г. Хемосистематика, ее проблемы' и практическое значение. «Растительные ресурсы», 1967, т. 3, № 1.
‘Ф л о р к э н М. Биохимическая эволюция. М., ИЛ, 1947.
Хайлов К. М. Системы и систематизация в биологии. Сб. «Проблемы методологии системного исследования». М., «Мысль», 1970.
Холдейн Б. С. О целесообразности размера. «Природа», 1970, №9. Хохряков А. П. Роль А. В. Благовещенского в развитии теории эволюции и систематики растений. «Биохимия и филогения растений». М., «Наука», 1972.
Цветков Б. Н. Правые и левые формы моллюска Fruticola lantzi Lndh. и их географическое распространение. «Бюлл. МОИП», биология, 1938, т. 47, № 5—6.
III а т а л к и н А. И. Коэффициенты сходства в таксономии. «Науч. докл. высш, школы», биол. науки, 1968, вып. 7.
III ат ал кин А. И. Математические методы в таксономии и их использование при построении системы мух-журчалок (Syrphidae Diptera). Автореф. канд. дисс. М., 1973.
Шмальгаузен И. И. Рост и дифференцировка. Сб. «Рост животных». М,—Л., 1935.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом развитии. М.—Л., 1942.
III мальгаузен И. И. Вопрос о монофилии и полифилии в проблеме происхождения наземных позвоночных. «Бюлл. МОИП», биология, 1959, т. 69, № 4.
Ш мальгаузен И. И. Факторы эволюции. Теория стабилизирующего отбора, изд. 2. М., «Наука», 1968.
Ш мальгаузен И. 'И. Контроль и регуляция в эволюции. Сб. «Кибернетические вопросы биологии». М., «Наука», 1968.
Ш мальгаузен И. И. Проблемы дарвинизма, изд. 2. М., «Наука», 1969. Ш н о л ь С. Э. Возможная детерминированность биологической эволюции физическими и химическими свойствами эволюционирующей системы. Сб. «Философские проблемы эволюционной теории», ч. 1. М., «Наука», 1971.
Ш н о л ь С. Э. Физико-химический аспект процесса биологической эволюции. Возможная детерминированность основных этапов эволюции физическими и химическими свойствами эволюционирующей системы. «Журн. общ. биол.», 1973, т. 34, № 3.
Шор ы г и н А. А. Питание и пищевые взаимоотношения .рыб Каспийского моря. М., «Пищепромиздат», 1952.
137
Яблоков А. В., Валецкий В. В. Об изменении ареала, левозакру-ченных форм Eulota lantzi в Заилийском Алатау за последние десятилетия. «Зоол. журн.», 1971, т. 50, № 3.
Alston R. Е. Biochemical Systematics. In: «Evolutionary biology», vol. 1. N.Y., 1967.
Am a don D. Taxonomic categories below the level of genus: theoretical and practical aspects. «J. Bombay Natur. Hist. Soc.», vol. 67, No. 1. 1970.
BessiereC., FourcadeS., Ngo-Van-Hien. Etude serologique de I’hemolymphe de quelques crustaces decapodes (Fixation du complements et immuno—electrophorese). «Trav. Soc. Pharm. Montpelier», 1961, t. 20, N 2.
Boyce A. J. The value of some methods of numerical taxonomy with reference to Hominoid Classification. In: «Phenetic and Phylogenetic classification». Systematics Assoc.», vol. 6, 1964.
Boyden A. Serology and animal systematics. «Amer. Natur.», 1943, vol. 77.
В r u n d i n L. Evolution, Causal biology, and classification. «Zool. Scripta», 1972, vol. 1, No. 3—4.
Burch J. B. Chromosome studies of aquatic Pulmonate snails. «Nucley», 1960, vol. 3, No. 2.
Burch J. B. Chromosome numbers and systematics in Euthyneuran snails. «Proc. First. Europ. Malacol. Congr.», 1965.
В u t о t L. J. M. Phylogenetic position of Heterurethra (Gastropoda Euthy-neura) in the light of cytotaxonomy. «Genen en phaenen», 1967, vol. 11, No. 4.
Cain A. J. The assessment of new types of character in taxonomy. In: «Chemotaxonomy and serotaxonomy. «The Syst. Assoc.», 1968, Spec, vol. 2.
Cousin G. Biometrie et definitions de morphologie quantitative des espe-ces et de leurs hybrides. «Bull. soc. zool. France», 1956, vol. 81.
Darlington C. D. Taxonomic species and genetic systems. «The New Systematics». Oxford, 1940.
Dessauer H. C. Molecular data in animal systematics. In: «Systematic Biology. Proc. Int. Conf. Nat. Acad. Sci.», publ. 1692. Washington, 1969.
Dobzhansky Th. Genetics and the origin of species. N. Y., 1960.
Erdtman H. The assessment of biochemical techniques in plant taxonomy. In: «Chemotaxonomy and Serotaxonomy». The System. Assoc., 1968, spec. vol. 2.
Farris J. S. Categorical ranks and evolutionary taxa in numerical taxonomy. «Syst. Zool.», 1968, vol. 17, No. 2.
G a j e v s k i W. Evolution in the genus Geum L. «Evolution», 1959, vol. 13, No. 3.
Geyer D. Unsere Land- and Susswasser-Mollusken. «Einfiihrung in die Molluskenfauna Deutschlands.» Stuttgart, 1927.
G i b e r t J. Synthese bibliographique des recherches electrophoretique sur les proteines des Crustaces. «Anne biol.», 1972, vol. 11, No. 7—8.
Harrison G. A. Discussion. 2. In: «Phenetic and Phylogenetic classification.» The System. Assoc. 1964, vol. 6.
H e i s e г С. B., S о r i a Jr. J., В u r t о n D. L. A numerical study of Sola-num species and hybrids. «Amer. Natur.», 1965, vol. 99.
Hessler R. R. Cephalocarida. In: «Treatise on Invertebrate Paleontology», I pt. (R) Artropoda 4, «Univ. Kansas, Geol. Soc. Am.», 1969.
H о 11 h u i s L. B. The Decapoda of the Siboga—Expedition. 9. The Hippo-lytidae & Rhynchocinetidae collected by the Siboga and Snellius Expeditions with remarks on the species. «Siboga Expedition», t. 140, mon. (39 a8), Leiden, 1947.
H о p s о n J. А., С г о m p t о n A. W. Origin of Mammals. In: «Evolutionary biology», vol. 3, 1969.
138
Hunziker J. H. Molecular data in plant systematics. In: «Systematic biology». Proc. Int. Conf. Nat. Acad. Sci., publ. 1692. Washington, 1969.
Huxley J. S. Problems of relative growth. London, 1932.
Huxley J. S. Toward a new systematics. «The New Systematics». Oxford, 1940.
Hyman L. H. The Invertebrates: Plathelminthes and Rhynchocoela. «The Acoelomata Bilateria», 1951, vol. 2.
Johnson К. M. Quantitative relationships between gill number, respiratory surtace and cavity shape in chitons. «Veliger», 1969, vol. 11, No. 3.
Johnson M. A., Fairbrothers D. E. Comparison and interpretation of serological data in the Magnoliaceae. «Bot. Gas.», 1965, vol. 126, No. 4.
К a r v e A. D. A. A theoretical study of the distribution of genera according to the numbers of species. Indian. «J. Genet. Plant Breed.», 1964, vol. 24, No. 3.
Kissinger D. G. The frequency of genera of a particular size in three families of phytophagous beetles. «Syst. Zool.», 1963, vol. 12.
Kurten B. A differentiation index, a new measure of evolutionary races. «Evolution», 1958, vol. 12, No. 2.
Ley De J. Molecular biology and bacterial phylogeny. In: «Evolutionary biology», vol. 2. N. Y., 1968.
Man De J. G. Families Pasiphaeidae, Stylodactilidae, Hoplophoridae, Ne-matocarcinidae, Thalassocaridae, Pandalidae, Psalidopodidae, Gnatho-phyllidae, Processidae, Glyphocrangonidae and Crangonidae. The Deca-poda of the Siboga—Expedition. «Siboga Expedition», mon. 39 a3, Leiden, 1920.
Mandel M. Roundtable discussion. In: «Systematic biology». Proc. Inst. Conf. Nat. Acad. Sci. publ. 1692, Washington, 1969.
M a r g о 1 i a s h E., Fitch W. M. Roundtable discussion. In: «Systematic biology». Proc. Inst. Conf. Nat. Acad. Sci., publ. 1692. Washington, 1969.
Michener Ch. D. Diverse Approaches to Systematics. In: «Evolutionary biology», vol. 4, 1970.
Moore R. C., McCormick L. General features of Crustacea. «Treatise on Invertebrate palaeontology», vol. 1 pt. (R) Arthropoda. «Univ. Kansas Geol. Soc. Am.», 1969.
M ii 11 e r A. H. Lehrbuch der Palaozoologie. «Allgemeine Grundlagen», Bd. 1. Jena, 1957.
N u r s a 11 I. R. On the origin of the major groups of animals. «Evolution», 1962, vol. 16, No. 1.
P ass ano L. M. Moulting and its control. In: «Physiology of Crustacea», vol. I. Metabolism and growth, 1960.
Patterson С. M. Chromosome numbers and systematics in Streptoneu-ran snails. «Malacologia», 1966, vol. 5, No. 1.
Peters D. S. Uber den Zusammenhang von biologischen Artbegriff und phylogenetischer Systematik. «Aufsatze und Reden Senckenberg. Natur-forsch. Ges. 1970, Bd. 18.
Pirie N. W. On making and recognising life. «New Biol.», 1954, vol. 16. Rasmussen D. I. Molecular taxonomy and typology. «Bioscience», 1969, vol. 19, No. 5.
R e n s c h B. Evolution above the species level. N. Y„ 1960.
Sharov A. G. Evolution and taxonomy. «Zeitschr. zool. Syst. Evolution-forsch.», 1965, vol. 3, No. 3—4.
Simpson G. G. Principles of animal taxonomy. N. Y„ 1961.
Sokal R. The future Systematics. In: «Taxonomic Biochemistry and Serology». N. Y., 1964.
S о к a 1 R„ S h e a t h P. H. A. Principles of numerical taxonomy. San Francisco, 1963.
Soule M. Phenetics of natural populations. I. Phenetic Relationships of insular populations of the Side—Blotched Lizard. «Evolution», 1967, vol. 21, No. 3.
139
Stebbins G. L. The significance of hybridisation for plant taxonomy. «Taxon», 1969, vol. 18.
T omson W. The status of species. In: «Philosophical problems in biology»,
Thorpe W. N. Ecology and the future of systematics. In: «The New Systematics». Oxford, 1940.
Turner B. L. Chemosystematics: recent developments. «Taxon», 1969, vol. 18.
W a g n e r W. H. Jr. The construction of a classification. In: «System, biology». Proc. Int. Conf. Nat. Acad. Sci. publ. 1692. Washington, 1969.
Wright S. The «Age and Area» concept Extended. Reviews. «Ecology», 1941, vol. 22.
Ziegler B. Evolution in upper Jurassic ammonites. «Evolution», 1959, vol. 13, No. 2.
Zimmerman K. Uber Mutationen in wilden Populationen. 1. Zur Varia-bilitat des Geaders in Bienenfliigel. «Mitt. Zool. Mus. Berlin», 1933, Bd. 19.
Zuckerkandl P., Pauling L. Evolutionary divergence and convergence in proteins. In: «Evolving genes and proteins». N. Y., 1965.
НИКОЛАЙ АЛЕКСЕЕВИЧ ЗАРЕНКОВ
ЛЕКЦИИ ПО ТЕОРИИ СИСТЕМАТИКИ
Тематический план 1975 г. № 159
Редактор И. Г. Комлева. Художник Н. А. Князьков Технический редактор И. А. Лебедева Корректоры И. А. Большакова, Н. И. Коновалова
Сдано в набор 28/Ш 1975 г. Подписано к печати 12/11 1976 г. Л-54498 Формат
60X90'/ie Бумага тип. № 3 Физ. печ. л. 8,75 Уч.-изд. л. 8,92 Изд. № 2519
Заказ 621 Тираж 3160 экз. Цена 72 коп.
Издательство Московского университета.
• Москва. К.-9, ул. Герцена, 5/7.
Типография Изд-ва МГУ. Москва, Ленинские горы