/


























































































































































































































































































































Автор: Москул Г.А..
Теги: pisces рыбы ихтиология систематика животных специальные зоологические науки рыбное хозяйство
ISBN: 978-5 -8209-1869-8
Год: 2021




Текст
(ОПРЕДЕЛИТЕЛЬ)
1
Министерство науки и высшего образования
Российской Федерации
КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Г. А. МОСКУЛ
РЫБЫ
ВОДОЁМОВ
БАССЕЙНА КУБАНИ
Определитель
Второе издание, исправленное и дополненное
Краснодар
2021
2
УДК 597.2/.5.081.3(083)(470.62)
ББК 28.693.32(2Рос-4Кра)я73
М82
Рецензенты:
Доктор биологических наук, профессор
М. С. Чебанов
Доктор биологических наук, доцент
В. В. Тюрин
Изложены сведения о распространении, биологии и рыбохозяй-
ственном значении каждого вида. Приводится полная классификация
рыбообразных и рыб бассейна Кубани и прилегающих к нему азово-
кубанских и закубанских рек с указанием латинского и русского назва-
ний.
Адресуется студентам биологических и сельскохозяйственных на-
правлений университетов, специалистам в области ихтиологии, зооло-
гии позвоночных, работникам рыбного хозяйства, учителям-биологам,
а также всем любителям природы и любителям рыбной ловли.
Ил. 143. Библиогр. 133 назв.
УДК 597.2/.5.081.3(083)(470.62)
ББК 28.693.32(2Рос-4Кра)я73
Москул, Г. А.
Рыбы водоёмов бассейна Кубани: определитель / Г. А. Мос-
кул. — 2 -е изд., испр. и доп. — Краснодар: Кубанский гос. ун-т,
2021. — 312 с. — 500 экз.
ISBN 978-5 -8209-1869-8
М82
© Кубанский государственный
университет, 2021
© Москул Г. А., 2021
ISBN 978-5-8209-1869-8
3
ПАМЯТИ
доктора биологических наук,
заведующего кафедрой зоологии
Кубанского госуниверситета
АБАЕВА
ЮРИЯ ИВАНОВИЧА
(1935–1997)
ПРЕДИСЛОВИЕ
Фауна рыб бассейна Кубани довольно разнообразна и представля-
ет значительный интерес как в научном плане, так и в рыбохозяйствен-
ном отношении. Кроме того, рыбы служат предметом рыбной ловли —
любимого спорта многих тысяч людей.
Задача определителя — установить научное название рыбы, пой-
манной в водоёмах бассейна Кубани и прилегающих к нему азово-
кубанских и закубанских рек. Кроме того, в определитель включены
проходные, полупроходные и некоторые морские рыбы, которые вхо-
дят иногда в пресные и солоноватые воды. Описаны также некоторые
рыбы, обитающие в прудах и озёрах Краснодарского края и Республи-
ки Адыгея.
Всего в определителе описаны 115 видов и подвидов рыб и ры-
бообразных, относящихся к 30 семействам, 19 отрядам, 2 классам и 1
типу (приложение 1).
Однако нельзя считать, что фауна рыбообразных и рыб бассейна
Кубани изучена полностью и приведённые сведения являются оконча-
тельными. В процессе дальнейшего исследования могут быть обнару-
жены новые виды и сведения о рыбах, которые будут дополнять дан-
ную работу.
Настоящий определитель рыб основан не только на собственных
наблюдениях автора, проводившего исследования на водоёмах бассей-
на Кубани и прилегающих к нему азово-кубанских и закубанских рек
в 1970—2019 гг. в рамках НИР КрасНИИРХ (1970—2011 гг.) и Кубан-
ского госуниверситета (1997—2019 гг.), но и на многочисленных лите-
ратурных данных (Данилевский, 1871; Книпович, 1923; Александров,
1927; Троицкий, 1948; Крыжановский, Троицкий,1954; Чижов, Аба-
ев,1968; Троицкий, Позняк, 1980; Ходячий, Москул, Никитина, 1982;
4
Троицкий, Цуникова, 1988; Абаев, 1996; Емтыль, Иваненко, 2002; Аб-
рамчук, Иваненко, 2018, 2019 и др.)
Надеемся, что определитель облегчит процесс определения рыб
как ихтиологам, зоологам, работникам рыбного хозяйства, студентам
биологических факультетов, так и широкому кругу любителей приро-
ды. Если определить какую-либо рыбу при помощи данного определи-
теля не предоставляется возможным, то необходимо воспользоваться
классическим трёхтомным определителем Л. С. Берга (1948—1949) для
пресноводных рыб и работой А. Н. Световидова (1964) для морских
рыб. Поскольку современный список видов рыб Чёрного моря претер-
пел ряд существенных изменений, то за современными взглядами сто-
ит обратиться к работе Е. Д. Васильевой (2007).
В определителе возможны различные неточности и пробелы, если
читатели, обнаружившие те или иные упущения, неточности, сооб-
щат о них, автор будет очень признателен. Все критические замеча-
ния и пожелания просим направлять по адресу: 350040, г. Краснодар,
ул. Ставропольская, 149, Кубанский государственный университет, ка-
федра водных биоресурсов и аквакультуры.
Автор благодарен заведующему кафедрой водных биоресур-
сов и аквакультуры, кандидату сельскохозяйственных наук, доценту
А. В. Абрамчуку и доценту той же кафедры, кандидату биологических
наук Н. Г. Пашиновой за ценные советы, полученные в процессе рабо-
ты над вторым изданием определителя.
Особая благодарность — А. М. Иваненко за научное редактирова-
ние и подготовку рукописи к печати.
5
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ
ХАРАКТЕРИСТИКА БАССЕЙНА РЕКИ КУБАНИ
Кубань — одна из крупнейших рек Кавказа. Среднее и нижнее её
течение, а также бóльшая часть притоков находятся в пределах Крас-
нодарского края, для которого Кубань является главной водной маги-
стралью. Значение этой реки для края в прошлом и в настоящее время
так велико, что по исторически сложившейся традиции Краснодарский
край часто называют просто Кубанью (Нагалевский, Чистяков, 2001).
Зарождаясь на склонах Эльбруса, за пределами края, Кубань рас-
секает его пополам и впадает у г. Темрюка в Азовское море. Кубань
и её притоки собирают воду с обширного бассейна, природные осо-
бенности которого предопределяют облик реки и характер её водного
режима.
Водосборный бассейн Кубани располагается в основном в запад-
ной части северного склона Большого Кавказа, захватывая часть Став-
ропольской возвышенности, Западно-Кубанской низменности, Азово-
Кубанской равнины и Таманского полуострова. По своей величине и
водности бассейн р. Кубани является самым крупным на Северном
Кавказе. Он протягивается от Таманского полуострова на западе до
Эльбруса на востоке. За исток р. Кубани принимается место слияния
рек Учкулан и Уллукам. При площади водосбора 57 900 км2 суммар-
ная длина рек бассейна составляет 38 325 км, а общее число рек равно
13 569. Длина самой Кубани — 870 км (Борисов, 2005).
Бассейн р. Кубани односторонний, несимметричный, так как река
принимает притоки в основном слева. Конфигурация его почти гру-
шевидная (рис. 1). В высотном отношении бассейн делится на четыре
основные зоны: равнинную высотой до 200 м над уровнем моря, пред-
горную — от 200 до 500 м, горную — от 500 до 1 000 м, высокогор-
ную — свыше 1 000 м над уровнем моря (Лотышев, 2000; Нагалевский,
Чистяков, 2001).
По водному режиму реки средней и нижней части бассейна р. Ку-
бани можно разделить на следующие три типа:
а) верховья рек Кубани, Малой и Большой Лабы относятся к типу
рек с преимущественно снежно-ледниковым питанием и стоком в ос-
новном летом;
б) реки Лаба с её притоками Фарсом, Чамлыком и другими, Уруп,
Белая, среднее и нижнее течение Кубани принадлежат к типу рек со
6
смешанным питанием; преобладает дождевое, сток во все сезоны мень-
ше 50 %, преобладает летний сток;
в) все притоки, впадающие в Кубань западнее р. Белой, относятся
к типу рек со смешанным питанием и преобладанием дождевого; сток
во все сезоны меньше 50 %, преобладает зимне-весенний (Нагалевский,
Чистяков, 2001).
Гидрографическая сеть бассейна р. Кубани включает в себя не
только реки, число которых превышает 13,5 тыс., но и 617 озёр об-
щей площадью 1 771 км2, в том числе: горных и предгорных — 291 с
Рис. 1. Бассейн р. Кубани (выделен цветом), закубанские и приазовские реки
7
площадью больше 20 км2; плавневых и дельтовых — 326 с площадью
1 751 км2. Основная часть горных озёр расположена в пределах Главно-
го и Бокового хребтов (Комлев, Мельникова, 2008).
Сток р. Кубани и её притоков зарегулирован 20 водохранилищами
с суммарным объёмом 4 963,7 млн м3 и площадью 751,2 км2 (табл. 1).
Таблица 1
Морфометрическая характеристика водохранилищ бассейна р. Кубани
Название
водохранилища
Год
соору-
жения
Параметры
Объём,
млн м3
Площадь,
км2
Длина,
км
Ширина,
км
Краснодарское (Кк и РА) 1973 2 350,0 420,00
40,00
15,00
Кубанское (Большое) (КЧР) 1967 587,0
50,00
—
—
Сенгилеевское (Ск)
1958 810,0
42,0
10,50
5,70
Усть-Джегутинское (КЧР) 1962 475,0
49,80
—
—
Варнавинское (Кк)
1964 160,0
42,50
11,00
4,23
Шапсугское (РА)
1952 130,0
46,00
9,00
8,00
Новотроицкое (Ск)
1952 130,0
18,00
11,00
—
Крюковское (Кк)
1969 111,0
40,20
8,80
6,50
Егорлыкское (Ск)
1952 111,4
16,80
7,00
1,40
Головное (КЧР)
—
36,0
2,67
—
—
Октябрьское (РА)
1964 18,0
1,04
3,40
0,31
Шенджийское (РА)
1965 15,0
7,80
4,00
3,6
Ганженское (Кк)
1954
9,0
4,40
5,00
2,0
Эшкаконское (КЧР)
1989
8,5
1,20
2,60
0,44
Белореченское (Кк)
1954
6,0
5,80
4,30
3,00
Кужорское (РА)
1978
2,1
0,50
1,30
2,20
Четукское (РА)
1963
1,5
1,50
2,60
2,50
Аксаутское (КЧР)
1999
1,2
0,30
0,84
0,75
Марухское (КЧР)
2002
1,0
0,21
0,58
0,57
Майкопское (РА)
1950
1,0
0,50
0,50
0,40
Примечание — Кк — Краснодарский край; Ск — Ставропольский край;
РА — Республика Адыгея; КЧР — Карачаево-Черкесская Республика
Из них 11 находятся в Краснодарском крае и Республике Адыгея
и 9 в Ставропольском крае и Карачаево-Черкесской Республике. Наи-
более крупными в Краснодарском крае и Республике Адыгея являют-
ся: Краснодарское — на р. Кубани, Шапсугское — в устье р. Афипс,
Варнавинское — на р. Абин и Крюковское, вбирающее в себя воды рек
8
Хабль, Бугай, Иль, Песчанка, Ахтырь; в Ставропольском крае и Кара-
чаево-Черкесской Республике: Большое, Сенгилеевское, Новотроицкое
и Егорлыкское (Веселовский, Данилова, Емельянова, 1973; Канонни-
ков, 1977; Коровин В. И., Коровин А. В., 1981; Ефремов, 1991).
Краснодарское водохранилище — один из крупнейших искусст-
венных водоёмов Северного Кавказа, внёсшее значительные изменения
в режим уровней воды в его нижнем бьефе. Оно создавалось в народно-
хозяйственных целях: противопаводковое назначение — срезка пиков
паводков, защита населения и устранение угрозы разрушительных на-
воднений для 600 тыс. га пойменных земель в низовьях Кубани; обес-
печение водой рисовых оросительных систем на площади 215 тыс. га;
осуществление мероприятий по воспроизводству ценных пород рыбы;
улучшение условий судоходства (Огурцов, Поляков, Гольднер, 1975;
Ефремов, 1991; Москул, 1994; География Республики Адыгея, 2001;
Комлев, Мельникова, 2008).
Кроме того, река Кубань через магистральные каналы соединяется
с реками Азово-Кубанской равнины (Еей, Челбасом, Кирпилями, Бей-
сугом, Понурой и др.), а в Ставропольском крае воды Кубани по Не-
винномысскому каналу поступают в реку [Большой] Егорлык (приток
Маныча) и по Егорлыку в Сенгилеевское, Новотроицкое и Егорлыкское
водохранилища.
9
ПРАВИЛА НАУЧНОЙ СИСТЕМАТИКИ
Основоположником зоологической системы является шведский
учёный-естествоиспытатель Карл Линней (1707–1778). В основу отече-
ственной систематики рыб положена система Л. С. Берга (1876–1950).
Однако в последние годы в эту систему внесены некоторые изменения
и дополнения, основные из них были нами учтены при составлении
данного определителя.
В систематике животных существует определённая иерархия сис-
тематических категорий. Виды объединяют в роды, роды — в семейст-
ва, семейства — в отряды, отряды — в классы, а классы — в типы. Но не
всегда эти основные систематические группы объединяют все органи-
ческие формы, существующие в природе. Поэтому применяют некото-
рые промежуточные категории (подтип, надкласс, подкласс, надотряд,
подотряд, надсемейство, подсемейство, подрод, подвид). Исходя из это-
го полная современная иерархия систематических групп выглядит сле-
дующим образом: тип → подтип → надкласс → класс → подкласс →
→ надотряд → отряд → подотряд → надсемейство → семейство →
→ подсемейство → род → подрод → вид → подвид → форма.
Основной систематической единицей является вид (species). Для
обозначения вида принята система двойных названий (бинарная, или
биномиальная номенклатура), например, серебряный карась — Car-
assius gibelio (Bloch, 1782). Согласно пункту 5.1. статьи 5 Междуна-
родного кодекса зоологической номенклатуры (4-е издание), научное
название вида, но не таксона любого другого ранга есть сочетание двух
названий (биномен), из которых первое — родовое название, а вто-
рое — видовое название. Родовое название должно в с е г д а н а ч и -
наться с прописной буквы, а видовое — со строчной
б у к в ы. Также согласно пункту 5.2. того же Кодекса научное название
подвида есть сочетание трёх названий (триномен, т. е. биномен, со-
провождаемый подвидовым названием). Подвидовое название должно
всегда начинаться со строчной буквы. После названия
вида указывается фамилия автора, который впервые описал этот вид и
год описания. Если вид был впервые описан в пределах другого рода,
то фамилия автора заключается в круглые скобки. В современной но-
менклатуре родовые и видовые названия обычно выделяют курсивом,
а фамилию автора-описателя набирают капителью (малыми пропис-
ными).
10
Вид характеризуется относительной морфобиологической ста-
бильностью, являющейся результатом приспособления к определённой
среде, в которой вид формировался и живёт. Вид есть свободно скре-
щивающееся сообщество организмов, всё время обновляющееся в ре-
зультате смерти старых особей и нарождения молодых. Вид занимает
определённую область распространения — а р е а л, в пределах кото-
рого условия существования соответствуют его морфобиологическим
потребностям.
Вид относительно стабилен во времени, он сохраняет морфобио-
логическую специфику в течение всей своей истории.
В отечественной систематике рыб широко представлены внутри-
видовые таксономические категории — п о д в и д (subspecies), п л е м я
(natio), р а с а (infraspecies) и м о р ф а (morpha).
Как уже отмечалось, для обозначения подвидов применяют трино-
минальную номенклатуру (к названию вида добавляют название под-
вида), например, вырезуб (Rutilus frisii (Nordmann, 1840) в водоёмах
Кубани кроме номинативного, представлен подвидом кутум — Rutilus
frisii kutum (Kamensky, 1901).
П л е м я (natio) — как бы подвид подвида, или подвид второго по-
рядка. В обозначении применяются четыре наименования (родовое, ви-
довое, подвидовое и племенное), причём перед племенным названием
ставится слово natio, например, ранее русскую быстрянку — Alburnoi-
des rossicus Berg, 1924, обитающую на Кубани, называли кубанской
быстрянкой — Alburnoides bipunctatus rossicus natio kubanicus Berg,
1932, считая кубанским племенем обыкновенной быстрянки.
Р а с о й, или э к о т и п о м (infraspecies), называют стойкие вариа-
ции, которые отличаются от типичной формы определённой комбина-
цией признаков, например, одомашненная форма сазана — карп. Раса
обычно передаёт свои признаки по наследству и при изменении внеш-
них условий.
М о р ф а — слабо закреплённое наследственное видоизменение,
при соответствующей перемене внешних условий легко возвращается
в исходное состояние, например форель. Проходная рыба лосось кум-
жа — Salmo trutta Linnaeus, 1758, оставшаяся после нереста в реке, даёт
начало ручьевой форели — Salmo trutta morpha fario Linnaeus, 1758.
Морфа, как и раса, не имеет определённого ареала, может встре-
чаться вместе с основной формой. В отличие от расы морфа — наслед-
ственно неустойчивая категория и существует у многих видов рыб.
11
П о м е с и (гибриды) обозначаются посредством соединения на-
званий производителей знаком ×; например, помесь плотвы и леща
обозначают: Rutilus rutilus × Abramis brama.
В ихтиологии принято, что единицы выше рода до отряда включи-
тельно носят названия по первоописанному роду и имеют определён-
ные окончания:
подсемейства оканчиваются на -inae (-ы или -и) — Cyprininae —
Карпы; Clupeinae — Сельди;
семейства на -idae (-овые, или -евые) — Cyprinidae — Карповые;
Clupeidae — Сельдевые;
надсемейства на -oidea ( -подобные) — Cyprinoidea — Карпоподоб-
ные;
подотряды на -oidei (-видные) — Cyprinoidei — Карповидные;
отряды на -formes (-образные) — Cypriniformes — Карпообразные.
Единицы выше отряда не имеют определённых окончаний.
При написании на русском языке видовое название, данное в честь
какого-либо учёного, исследователя и т. д., пишется с заглавной буквы,
например: Knipowitschia longicaudata (Kessler,1877) — длиннохвостый
бычок Книповича.
Далее приводятся некоторые правила чтения латинских названий,
которые часто встречаются в данном определителе (табл. 2).
Таблица 2
Правила чтения латинских названий (из География рыб, 2020)
Латынь
Транс-
крипция
Правило
Пример
1
2
3
4
с
[к]
перед «а», «о», согласными carpio, Percoidei, pectoralis
c
[ц]
перед «al», «ol», «i», «у»
Perciformes, Cyprinus
y,i
[и]
—
Perciformes, Cyprinus
z
[з]
—
Schizothorax
s
[з]
между гласными
Alosa
s
[с]
начало, конец, перед со-
гласными
Salmo, Cyprinus, Carassius
sch
[ш]
в немецких словах
schrenki (нем.)
sch
[cx]
в латинских словах
Schizothorax
x
[кc]
—
Schizothorax
rh, th [p], [т] «h» — не читается
Schizothorax
12
1
2
3
4
h, ch [г], [х]
—
hypohyale, chorda
f, ph
[ф]
—
Fluviatilis, Phoxinus
gu
[гв]
перед гласными
Anguilla
gu
[гу] перед согласными
Gulaphallus
qu
[кв] дифтонг
Squalus
ae
[э]
дифтонг
Percidae
oe
[е]
дифтонг
Capoeta
tio
[цио]
—
natio
tia
[ция]
—
Botia
u
[ю]
после мягких согласных
fluviatilis
u
[у] после твёрдых согласных
fasciatus
При составлении данного определителя за основу была принята
«Система рыб» Л. С. Берга (1948—1949), работы Г. В. Никольского
(1971), Ю. С. Решетникова с соавторами (Пятиязычный... , 1989; Спи-
сок..., 1997; Аннотированный каталог..., 1998; Атлас..., 2002, 2003;
Рыбы в заповедниках..., 2010; Рыбы в заповедниках..., 2013), А. В. Аб-
рамчука, А. М. Иваненко (2018, 2019) и определители Н. М. Книповича
(1923), А. Н. Световидова (1964), Б. А. Кузнецова (1974), Е. А. Весело-
ва (1977), С. К. Троицкого, Е. П. Цуниковой (1988) и др.
Окончание табл. 2
13
КАК ПОЛЬЗОВАТЬСЯ ОПРЕДЕЛИТЕЛЕМ
Задача определителя — установление научного названия вида
рыбы. Но если не известно, к какому классу относится животное, то
сначала надо определить класс, затем отряд, далее семейство, потом
род и наконец вид. Иногда определение ведётся не только до вида, но
и до подвида, племени, расы или морфы данной рыбы. С помощью
таблиц определителя можно начинать определение животного с над-
класса.
Определительные таблицы составлены по дихотомическому (ди-
хотомия — раздвоенность, последовательное деление на две части)
принципу, при этом имеем дело с выбором двух противоположных по-
ложений, обозначенных цифрами — т е з ы (цифра слева без скобок) и
антитезы (цифрасправавскобках).
Все таблицы следует рассматривать с первой тезы. Если признаки
рыбы совпадают с указанными в тезе, можно переходить к следующей
по порядку тезе. При несоответствии признаков данной тезе необхо-
димо обратиться к антитезе, обозначенной в скобках. Так, следуя шаг
за шагом, доходим до названия семейства, потом рода, затем вида и
подвида.
Для уточнения правильности определения семейства или вида сле-
дует прочесть их характеристику, в которой даются дополнительные
морфологические признаки, приводятся сведения о водоёмах, в кото-
рых эти рыбы встречаются, их предельные размеры и сведения по био-
логии, характеризующие данный вид. Почти все виды рыб изображены
на рисунках, которые помогут ориентироваться в признаках и прове-
рить правильность определения.
Если определить вид не удаётся, это может быть вызвано, во-пер-
вых, ошибкой определяющего, во-вторых, тем, что данная рыба ещё
не отмечена в водоёмах бассейна Кубани, или тем, что определяемый
экземпляр является помесью двух видов рыб.
В рассматриваемых водоёмах помеси могут встречаться у осетро-
вых и особенно у карповых рыб. При хорошем знании видового состава
рыб можно определить, какие рыбы дали помеси.
Определение рыб связано с установлением признаков, характер-
ных для данного вида (число чешуй в боковой линии, количество вет-
вистых и неветвистых лучей в плавниках, число позвонков и ряд других
морфологических признаков рыбы). Другие признаки (длина головы,
14
длина хвостового стебля, высота тела и др.), необходимые для опреде-
ления рыбы, устанавливают путём промеров (Правдин, 1966).
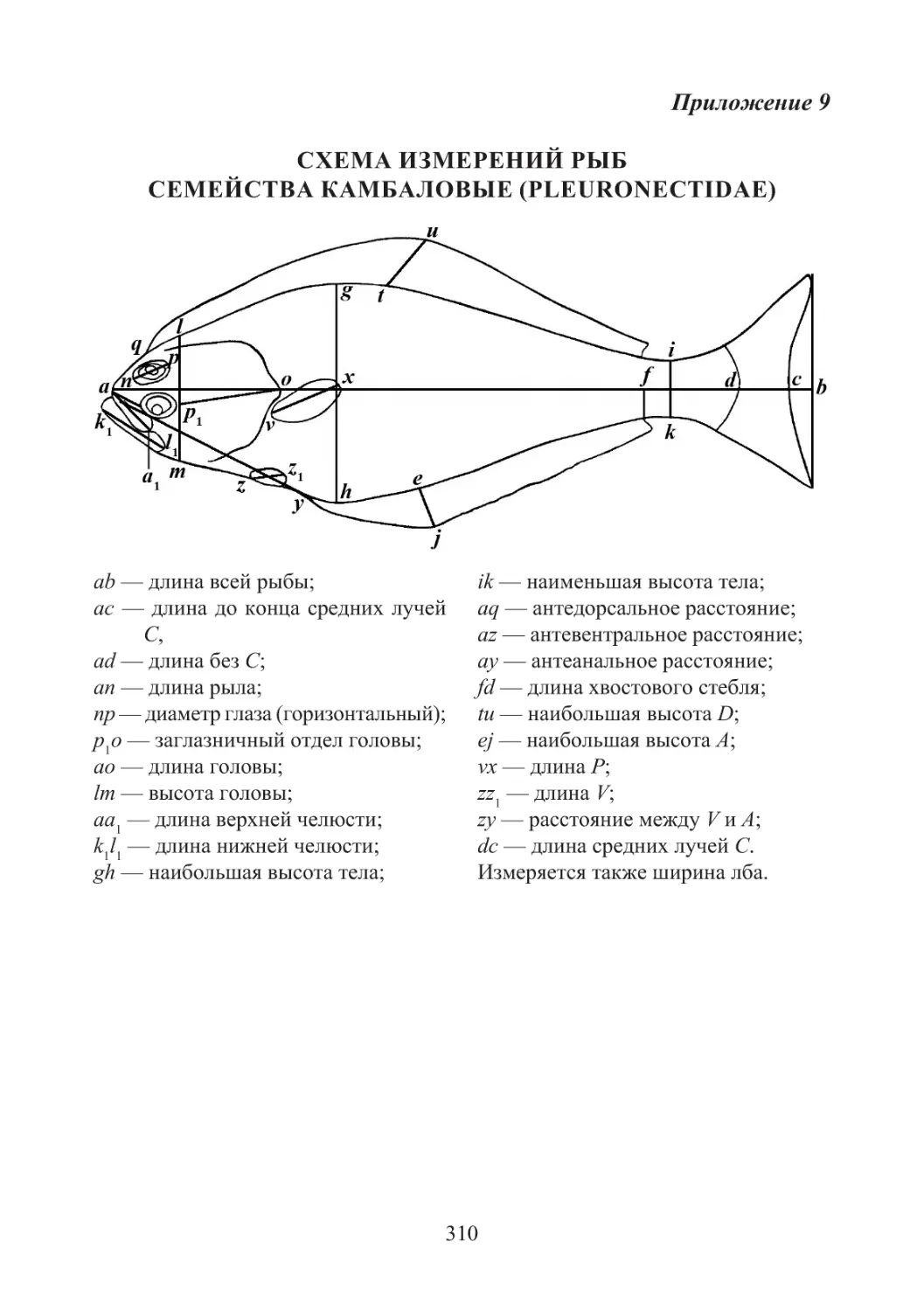
Цветные рисунки описываемых видов помещены в прил. 2, а табли-
цы и рисунки промеров рыбообразных и рыб приведены в прил. 3—10.
15
МАТЕРИАЛЫ И ИНСТРУМЕНТАРИЙ
Определяют рыбу в свежем виде, когда у неё хорошо выражены
все признаки и окраска. В тех случаях, когда необходимо сохранить
рыбу, её фиксируют 70%-м спиртом. В исключительных случаях при
отсутствии спирта рыб фиксируют формалином. Формалин должен
быть 4%-м (1 часть аптекарского 40%-го формалина на 9 частей воды).
У крупных экземпляров надо делать небольшие надрезы на брюш-
ке, чтобы фиксирующая жидкость могла проникнуть во внутренние ор-
ганы рыбы.
Чтобы сохранить у рыб их естественную окраску, пользуются ре-
цептом В. Ч. Дорогостайского: 20 г азотнокислого калия, 10 г уксусно-
кислого натрия и 10 г сернокислого натрия растворяют в 100 мл горячей
воды. После охлаждения раствора к нему прибавляют 30 мл формалина
и 20 мл глицерина, а затем фильтруют. В полученном растворе рыба в
течение нескольких лет сохраняет эластичность и естественную окра-
ску.
Для определения рыб необходимы следующие материалы и препа-
ровальные инструменты: кювета; кусок клеёнки, на которую кладётся
рыба; сантиметровая лента длиной 150 см для измерения крупных рыб;
штангенциркуль длиной 15—25 см; простой измерительный циркуль;
ручная лупа; пинцет; скальпель; препаровальные иглы и ножницы.
Если рыба фиксирована формалином, то перед началом определе-
ния следует 2—3 ч держать её под проточной водой для полного рас-
творения формалина, чтобы избежать ожогов кожи рук.
16
ОБЪЯСНЕНИЕ ОСНОВНЫХ
ПРИЗНАКОВ, ПРОМЕРОВ И ТЕРМИНОВ,
УПОТРЕБЛЯЕМЫХ В ОПРЕДЕЛИТЕЛЕ
Аборигены — виды рыб, издавна живущие в данном водоёме; ко-
ренное население водоёма.
Абсолютная длина тела — см. Длина тела.
Абсолютная плодовитость — число зрелых икринок в рыбе.
Абсолютная потенциальная плодовитость — число всех икри-
нок в рыбе.
Адаптация — приспособление организма или отдельных его ор-
ганов к определённым условиям среды.
Аквакультура — разведение и выращивание водных организмов.
Акклиматизация — 1) вселение новых видов рыб из других бас-
сейнов; 2) приспособление организма к новым условиям существова-
ния.
Аксиллярная лопасть — увеличенная
чешуйка в основании брюшного плавника,
имеется у некоторых сельдей, барабульки
(рис. 2).
Аля (лат. alae) — крыловидные чешуи
у основания лопастей хвостового плавника,
выполняющие функцию обтекателей (рис. 3);
имеются у некоторых сельдей и сардин.
Анадромная миграция — см. Миграция
рыб.
Анальное отверстие — заднепроход-
ное отверстие.
Анальный плавник (лат. analis — А) —
непарный плавник, расположенный за аналь-
ным отверстием. Пример: формула плавника
А III 8 означает, что в анальном плавнике три
неветвистых луча и 8 ветвистых.
Антеанальное расстояние — расстоя-
ние от вершины рыла до начала основания
анального плавника.
Антевентральное расстояние — рас-
стояние от вершины рыла (или от переднего
Рис. 2. Аксиллярная ло-
пасть (а) у основания
брюшного плавника
Рис. 3. Крыловидные че-
шуи (а) у основания лопа-
стей хвостового плавника
17
края верхнечелюстной кости) до начала основания брюшного плавника
по прямой линии.
Антедорсальное расстояние — расстояние от вершины рыла до
основания первых лучей спинного плавника по прямой линии.
Ареал — область земного шара, где встречается данный вид.
Бентос — организмы, живущие на дне водоёма; если это живот-
ные, то говорят зообентос, если растения — фитобентос.
Бентофаги (бентосоеды) — гидробионты, питающиеся донными
организмами.
Бестер — гибрид белуги со стерлядью Б × С (БС).
Биомасса (ихтиомасса) — масса гидробионта, пересчитанная на
единицу площади или объёма воды.
Бластула — стадия развития зародыша.
Боковая, или латеральная, линия (лат. linea lateralis — l.l.)
—
число чешуй, прободённых отверстиями бокового органа, служащего
у рыб для восприятия колебаний водной среды. Боковая линия обычно
тянется вдоль всего туловища, но у верховки и горчака она пронизыва-
ет всего несколько чешуй, т. е. неполная.
Брачный наряд рыб — изменение внешнего вида рыбы в нерес-
товый период, происходит преимущественно у самцов к моменту раз-
множения и проявляется в смене окраски (бычки, лососёвые и др.), по-
явлении белых ярких роговых бугорков на голове (осетровые) и чешуе
(карповые); это признак готовности рыбы к нересту.
Брачный период — наступает у рыб к моменту размножения и
характеризуется появлением брачного наряда.
Брызгальце (лат. spiraculum) — отверстие, расположенное позади
глаза (например у осетровых), через которое вода может поступать в
ротовую полость.
Брюшина — оболочка, выстилающая брюш-
ную полость. Окраска брюшины — систематиче-
ский признак, имеется у подуста и серебряного ка-
рася.
Брюшная присоска (у бычков и некоторых
других рыб) — видоизменение брюшных плавни-
ков; образуется путём срастания брюшных плавни-
ков или их мускулистых оснований (рис. 4).
Брюшные плавники — парные плавники, рас-
положенные у разных рыб по-разному: далеко за
Рис. 4. Брюшная
присоска бычка
18
грудными на брюхе, недалеко за грудными, под грудными или впереди
грудных плавников.
Веберов аппарат — ряд косточек, соединяющих плавательный
пузырь с органом слуха.
Веретеновидная форма тела рыб — сжатое с боков тело с слегка
заострённой головой (плотва, окунь, сельди и др.).
Веко жировое — прозрачные плёнки, налегающие на глаза и ос-
тавляющие посередине свободную щель, имеется у кефалей и некото-
рых сельдей (рис. 5).
Рис. 5. Жировое веко:
а — сельдь (Alosa fallax); б — лобан (Mugil cephalus)
Верхнечелюстная кость — см. Челюсти.
Верхнечелюстная пластинка (имеется у миног) — см. Зубы.
Верхний рот — см. Рот.
Вершина рыла — наиболее подвинутая вперёд точка верхней (у
подуста) или нижней (у сельдей рода Caspialosa) челюсти.
Весенненерестующие рыбы — рыбы, нерест которых происходит
весной.
Ветвистый луч — см. Лучи плавников.
Вид (лат. species) — группа организмов с общими морфофизио-
логическими, биохимическими и поведенческими признаками, способ-
ная к свободному взаимному скрещиванию (панмиксия), которое даёт в
ряду поколений плодовитое потомство; закономерно распространяется
в пределах определённого ареала и сходно изменяется под влиянием
факторов внешней среды.
Видовая плодовитость — сумма икринок, откладываемых сам-
кой в продолжение всей жизни.
19
Водоросли (лат. Algae) — низшие талломные растения, обитаю-
щие в водоёмах, а также во влажных местах на суше; взвешенные в
толще воды составляют фитопланктон, живущие на субстрате — фи-
тобентос.
Водохранилище — водоём, образованный водоподпорным соору-
жением и предназначенный для регулирования стока воды.
Возрастная группа — рыбы, объединённые в группы с одинако-
вым возрастом.
Ворсинки — конические выросты кожи.
Выдвижной рот — см. Рот.
Выемка межчелюстная [медиальная вырез-
ка] — выемка в верхней челюсти, имеется у некото-
рых сельдей (рис. 6).
Выклев зародышей рыб — освобождение от
оболочки яйца.
Вылупление личинок — выклев зародышей
рыб.
Высшие водные растения — см. Макрофиты.
Высота головы — высота у затылка (над жа-
берными щелями); измеряют над местом прикреп-
ления первого позвонка к черепу.
Высота плавника — длина наибольшего (са-
мого длинного) луча.
Высота тела (наибольшая, наименьшая): наибольшая — расстоя-
ние в самом высоком месте тела; наименьшая — расстояние между са-
мыми близкими точками спинного и брюшного краёв хвостового стебля.
Высота спинного или анального плавников — длина их наиболь-
шего луча.
Высота хвостового стебля — то же, что наименьшая высота
тела.
Галобионты — гидробионты, населяющие солёные воды.
Гаметы — см. Половые клетки.
Ганоидная чешуя — см. Чешуя.
Генерация — см. Поколение.
Генипоры (щёчные поры) — простейшая форма органов бокового
чувства в виде мелких отверстий, находящихся на голове. Расположе-
ние этих пор, как и чувствительных (слизевых) каналов и их пор, имеет
значение при определении некоторых видов бычков (рис. 7).
Рис. 6. Межчелю-
стная выемка (а)
20
Генитальное отверстие — см. Поло-
вое отверстие.
Гетероцеркальный хвостовой плав-
ник — см. Плавники.
Гидробионты — водные организмы
(растения, бактерии, животные).
Гиногенез — частный случай партено-
генеза, при котором рождаются только самки
(серебряный карась).
Гипофиз — железа внутренней секреции, расположенная у осно-
вания головного мозга и выделяющая гормоны, стимулирующие рост
тела и развитие половых органов.
Годовик — перезимовавший сеголеток.
Годовое кольцо — откладывается на чешуе, плоских костях чере-
па, отолитах, костных лучах плавников; используется для определения
возраста рыб и обратного расчисления темпов роста.
Гомоцеркальный хвостовой плавник — см. Плавники.
Гонадотропный гормон — вырабатывается у осетровых рыб пе-
редней, у костистых — промежуточной долей гипофиза.
Гоноподий — подвижный непарный
совокупительный орган некоторых живоро-
дящих рыб (подсемейство Poeciliinae), яв-
ляющийся видоизменённым первым лучом
анального плавника (рис. 8).
Глоточные зубы — зубы, расположен-
ные на глоточных костях; у карповых рыб
они находятся на нижнеглоточных костях, на
пятой жаберной дуге и располагаются в один,
два или три ряда (рис. 9). Чтобы их рассмот-
реть, необходимо, перерезав мышцы, извлечь
через жаберное отверстие пятую жаберную дугу.
Горло — пространство между местом прикрепления жаберных пе-
репонок и основанием грудных плавников.
Градусо-дни — произведение среднесуточной температуры воды
на число дней с этой температурой.
Грудные плавники (лат. pectoralia — P) — парные плавники, рас-
положенные позади жаберных отверстий, у некоторых рыб под жабер-
ными отверстиями или даже впереди них.
Рис. 7. Генипоры (а) на го-
лове бычка
Рис. 8. Гоноподий (а) у
самца гамбузии
21
Грудь — пространство на брюшной стороне тела рыбы, непосред-
ственно следующее за основанием грудных плавников.
Губы — мягкие складки вокруг рта; различают сложные губы,
двухлопастные, мясистые.
Детрит — органический ил и остатки организмов в водной среде,
на которых концентрируются бактерии.
Детритофаги (детритоеды) —гидробионты, питающиеся дет-
ритом.
Диаметр глаза — диаметр роговицы; различают продольный (го-
ризонтальный), или длину глаза, и поперечный (вертикальный) диамет-
ры глаза.
Длина анального плавника — длина основания анального плав-
ника от первого луча до основания последнего луча.
Длина брюшного плавника — расстояние от переднего края
брюшного плавника до его вершины (но не длина наибольшего луча).
Длина головы — расстояние от вершины рыла (при закрытом рте)
до наиболее удалённой точки крышечной кости (в расчёт не принима-
ют перепонку, окаймляющую задний край жаберной крышки).
Длина грудного плавника — расстояние от переднего края груд-
ного плавника до его вершины (ни в коем случае не длина наибольшего
луча!).
Длина рыла (предглазничный отдел головы) — расстояние от вер-
шины рыла до переднего края глаза.
Длина спинного плавника — длина основания спинного плавника
от переднего (хотя бы и зачаточного) луча до последнего луча (или до
конца перепонки, если она есть).
Рис. 9. Однорядные (а, б), двухрядные (в, г) и трёхрядные (д, е) глоточные зубы
карповых рыб:
а — плотва; б — карась; в — жерех; г — краснопёрка; д — сазан; е — маринка
22
Длина тела — различается в зависимости от способа измерения:
1) абсолютная, или общая длина тела (вся длина тела,
полная длина) — расстояние от вершины рыла до перпендикуляра, вос-
становленного от конца самой длинной лопасти хвостового плавника
(L);
2)длина тела —расстояниеотконцарыладоконцачешуй-
ного покрова у основания хвостового плавника, а если чешуи нет, до
основания лучей хвостового плавника (l);
3)длина тела по Смитту—улососёвыхинекоторыхсель-
девых рыб — длина до конца средних лучей хвостового плавника (ls).
Длина хвостового стебля — расстояние от вертикали конца ос-
нования анального плавника до конца чешуйного покрова (или до ос-
нования хвостового плавника, считая посередине тела).
Донные организмы — см. Бентос.
Донные рыбы — бентические рыбы, живущие у дна (камбалы,
бычки и др.).
Жабры — система органов
внешнего дыхания у первично-
водных животных; у костистых
рыб жабры представляют собой
складчатые выросты — жабер-
ные лепестки, размещённые на
жаберных дугах (рис. 10).
Жаберная дуга (лат. arcus
branchialis) — дуга, на которой
расположены с одной стороны
жаберные тычинки, а с другой —
жаберные лепестки.
Жаберная крышка — ко-
стная крышка, закрывающая жа-
берную полость.
Жаберные лепестки — палочковидные или пластинчатые вырос-
ты, расположенные на внешней стороне жаберной дуги (образуют соб-
ственно жабры).
Жаберные лучи — лучи жаберной перепонки, поддерживающие
жаберную перепонку; они прикреплены к первой дуге жаберного ап-
парата.
Жаберные отверстия — жаберные щели.
Рис. 10. Строение жаберной дуги (жаб-
ры):
а—б
—
длина нижней части дуги;
б—в
—
длина верхней части дуги;
а—б + б—в
—
длина всей дуги
23
Жаберные перепонки — окаймляют сзади жаберные крышки и
служат для более плотного закрывания жаберных отверстий.
Жаберные тычинки (лат. spinae branchialis — Sp. br.) — костяные
или хрящевые палочки на передней стороне жаберной дуги; число жа-
берных тычинок — важный систематический признак, считают их на
первой жаберной дуге.
Жевательные зубы — см. Зубы.
Жерновок — твёрдое роговое образование, заменяющее у карпо-
вых верхние глоточные зубы, служащее совместно с нижнеглоточными
зубами для перетирания пищи.
Жёсткий луч — см. Лучи плавников.
Жилые рыбы — рыбы, постоянно живущие и размножающиеся в
одном и том же водоёме.
Жировой плавник — небольшой мягкий, не поддерживаемый лу-
чами плавник, расположенный позади спинного плавника, имеется у
лососёвых.
Жировые веки — см. Веко жировое.
Жучки — костные образования у осетровых, имеющие кониче-
скую форму и расположенные на теле продольными рядами.
Заглазничное пространство — расстояние от заднего края глаза
до конца жаберной крышки (без перепонки).
Заглазничные кости — окологлазничные кости за глазами.
Затылок — место над прикреплением позвоночника к черепу
(обычно над жаберной крышкой).
Затылочные бугры — округлые возвышения с шипами на голове.
Затылочные гребни — удлинённые возвышения с шипами на го-
лове.
Зимненерестующие рыбы — рыбы, размножающиеся зимой (на-
лим, лососёвые и др.).
Зообентос — см. Бентос.
Зоопланктон — группа мельчайших организмов, обитающих в
толще воды.
Зрелые производители — рыбы, у которых икра и сперма пригод-
ны для оплодотворения.
Зубы — различны у разных видов рыб по расположению и фор-
ме. Могут располагаться на челюстях, сошнике, нёбных костях, языке,
губах, глоточных костях; различают резцевидные, стреловидные, ще-
тинковидные, или волосовидные, трёхвершинные зубы. Бывают зубы
24
крупные — клыки. У рыб некоторых видов зубы жевательного типа или
дробящие пищу. Роговые зубы миног расположены на верхнечелюст-
ной и нижнечелюстной пластинках, на поверхности ротовой воронки и
на языке. Срединные губные зубы могут быть двух- и трёхвершинными.
Инбридинг — близкородственное разведение.
Индекс наполнения кишечника — определяется отношением
массы содержимого пищеварительного тракта к массе тела; при умно-
жении полученного индекса (числа) на 10 000 получают коэффициент
наполнения в продецимилле (‱).
Индивидуальная плодовитость — общее количество икринок,
вымётываемых самкой за один нерестовый период.
Индивидуальная абсолютная плодовитость рыбы — абсолют-
ное количество зрелых икринок, вымётываемых одной самкой за один
нерестовый сезон.
Индивидуальная относительная плодовитость — число зре-
лых икринок, вымётываемых одной самкой за один нерестовый сезон
в пересчёте на 1 г массы рыбы без внутренностей; известны пересчёты
на10г,100г,1кг.
Индифферентные рыбы — рыбы, откладывающие икру на раз-
ные субстраты (окунь, ёрш, судак).
Инкубационный период — сроки развития от оплодотворения до
выклева эмбриона.
Истмус (лат. isthmus) — промежуток между жаберными щелями.
Каннибализм — поедание особей своего же вида.
Капсула — хрящевая или костная оболочка, в которую, в частно-
сти, заключён плавательный пузырь рыб из сем. Вьюновых.
Катадромная миграция — см. Миграция рыб.
Килевые чешуйки — см. Киль.
Киль — острый край тела (брюха, спины или боков хвостового
стебля); может быть покрыт чешуёй или голый, кожистый. У некото-
рых видов сельдевых и карповых киль покрыт особой формой крыше-
образных килевых чешуек.
Клейкость икры — свойство, при котором оболочка оплодотво-
рённой икры с помощью выделяемой жидкости приклеивается к суб-
страту или другим икринкам; известна у рыб фитофилов, псаммофилов
и литофилов.
Кожистый киль — киль, не покрытый чешуёй, голый.
Конечный рот — см. Рот.
25
Кормовые ресурсы — совокупность растительных и животных
организмов автохтонного и аллохтонного происхождения и их продук-
тов распада, находящихся в водоёме.
Крыловидные чешуи —
вытянутые чешуи, попарно
расположенные на хвостовом
плавнике сельдей (см. рис. 3),
а также за грудными плавни-
ками у кефалей (рис. 11).
Ктеноидная чешуя —
см. Чешуя.
Летненерестующие
рыбы — условное деление рыб,
у которых сезон нереста — лето
(линь, краснопёрка и др.).
Лимнофильные рыбы —
обитатели стоячих вод.
Литофилы — экологи-
ческая группа рыб, откладывающих икру при нересте на каменистый
грунт, чаще в реках на течении (осетровые, лососёвые, подуст, усач,
рыбец, шемая и др.).
Личинка — стадия развития рыбы от момента исчезновения жел-
точного мешка до приобретения общей формы тела, характерной для
данного вида.
Личиночный период — часть жизненного цикла рыбы, когда пи-
тание происходит за счёт внешнего корма, а рыба ещё не приняла об-
лика и внутреннего строения
взрослого организма.
Лоб — промежуток ме-
жду глазами (межглазничный
промежуток).
Лопастинка — удли-
нённая
крыловидная
че-
шуйка, например у кефалей
(см. рис. 11), или боковые
выросты кожи на брюшной
присоске, например у бычков
(рис. 12).
Рис. 11. Удлинённая лопастинка (а) над
основанием грудного плавника кефали
(Mugil hospes)
Рис. 12. Брюшная присоска у бычка:
а — воротник без лопастинок; б — ворот-
ник с лопастинками (1)
26
Лучи плавников — бывают неветвистые и ветвистые; неветвистые
лучи не разветвляются и, в свою очередь, подразделяются на нечле-
нистые (обычно твёрдые, жёсткие и колючие) и членистые (обычно
мягкие на вершине, гибкие); ветвистые лучи ветвятся или от самого
основания, или в своей верхней части (рис. 13).
Рис. 13. Лучи плавников:
1 — неветвистый, нечленистый, мягкий, гибкий; 2 — неветвистый, члени-
стый; 3 — ветвистый; 4 — колючий, гладкий; 5 — колючий, зазубренный;
6 — неветвистый, нечленистый, твёрдый, жёсткий
Макрофиты — крупные низшие и высшие водные растения, об-
разующие ряд экологических группировок в водоёме: плавающие (кув-
шинка, кубышка, водокрас, рдест, ряска и др.), надводные (тростник,
рогоз и др.), подводные (элодея, роголистник, уруть и др.).
Малёк — стадия развития рыбы после личинки с вполне сформи-
ровавшимися лучами плавников и слабо выраженным чешуйчатым по-
кровом.
Малоценные рыбы — рыбы не использующиеся человеком в
пищу, а также рыбы, потребляемые в пищу, но плохо использующие
кормовые ресурсы водоёма на прирост, конкурирующие с более цен-
ными рыбами.
Марикультура — разведение и выращивание морских организ-
мов.
Медиальная вырезка — см. Выемка межчелюстная.
Межглазничное пространство, межглазничный промежу-
ток — то же, что лоб.
Межжаберный промежуток — перегородка, разделяющая жа-
берные полости; у некоторых рыб (карповые) жаберные перепонки
приращены к межжаберному промежутку (лат. isthmus) — участку ме-
жду жаберными щелями.
27
Мера промысловая — размеры, меньше которых запрещается ло-
вить рыбу.
Метаморфоз — преобразование организма в период постэмбрио-
нального развития (например, превращение пескоройки в миногу).
Миграция рыб — наследственно закреплённое массовое переме-
щение рыб от одного местообитания к другому; различают нерестовую,
кормовую и зимовальную миграцию рыб; большинство рыб совершают
регулярные анадромные миграции из моря в реки и реже — катадром-
ные — из рек в море (речной угорь).
Микропиле — микропилярные канальцы в количестве до 10 в обо-
лочке анимального полюса яйца рыб (икринки), через которые прони-
кает сперматозоид в процессе оплодотворения.
Молодой — лат. juveniles (juv.)
Молодь — условная возрастная и размерная категория, характери-
зующая состояние рыбы до наступления половой зрелости.
Монофаги — рыбы и другие животные, питающиеся однотипной
пищей.
Морские рыбы — рыбы, живущие в солёной воде морей.
Мягкий луч — см. Лучи плавников.
Наибольшая высота тела — см. Высота тела.
Наименьшая высота тела — см. Высота тела.
Нёбная кость — парная кость в верхней части рта рыбы.
Нёбные зубы — зубы, расположенные на нёбных костях.
Нёбо — верхний свод ротовой полости.
Неветвистый луч — см. Лучи плавников.
Непарные плавники — см. Плавники.
Неполная боковая линия — см. Боковая линия.
Неполовозрелые рыбы — все возрастные группы рыб до первого
нереста.
Нерест — процесс размножения рыб.
Нижнечелюстная кость — см. Челюсти.
Нижнечелюстная пластинка — см. Зубы.
Нижний рот — см. Рот.
Общая длина тела — см. Длина тела.
Овуляция — процесс выхода яйца из яичника самки.
Окологлазничные кости — кости, расположенные вокруг глаз.
Остракофилы — экологическая группа рыб, откладывающих
икру при нересте в мантийную полость моллюсков (горчак).
28
Относительная плодовитость — количество икры, приходя-
щееся на единицу массы самки.
Отолиты, или слуховые камешки — костные образования во
внутреннем ухе рыб; используют для определения возраста рыб.
Панцирь — различные костные образования на теле и голове рыб;
панцирь на теле может состоять из кольцевых тонких костных пласти-
нок, костных поясков (сем. Морские иглы).
Паракаудальный орган — ряд кожных
листочков, сидящих на лучах хвостового
плавника (например у хамсы) (рис. 14); рас-
сматривается как образование, гомологичное
alae.
Парапофизы — боковые отростки туло-
вищных позвонков рыб.
Парные плавники — см. Плавники.
Пелагические рыбы — рыбы, населяю-
щие толщу воды.
Пелагофилы — экологическая группа
рыб, вымётывающих икру в толщу воды (ке-
фали, толстолобики, сельди).
Пилорические придатки —
отростки пищеварительного тракта
пальцевидной формы, расположен-
ные перед желудком или за ним
(рис. 15).
Пищевод — передняя часть
пищеварительного тракта, соеди-
няющая ротовую полость с желуд-
ком.
Плавательный пузырь — на-
полненный газом мешок, располо-
женный в дорзальной части полос-
ти тела большинства костистых рыб.
Плавники — бывают парные (грудные и брюшные) и непарные
(спинной, анальный, жировой и хвостовой). У некоторых рыб бывают
дополнительные плавники, расположенные за спинным и анальным
плавниками. Плавники, кроме жирового, состоят из лучей и натяну-
той между ними плавниковой перепонки. У низкоорганизованных рыб
Рис. 14. Паракаудаль-
ный орган (а) у хамсы
(по В. А. Водяницкому и
З. М. Пчелиной, 1955)
Рис. 15. Желудок и пилорические
придатки (а)
29
(сельдеобразные, карпообразные) брюшные плавники расположены на
брюхе за груднымиплавниками, занимая абдоминальное поло-
жение. У высокоорганизованных рыб (окунеобразные и др.) брюшные
плавникинаходятсявпереднейчаститела—торакальное поло-
жение. Если брюшные плавники располагаются впереди грудных — на
горле, то такое расположение называется ю г у л я р н ы м (трескооб-
разные, окунеобразные и др.). Если в плавниковой формуле цифра по-
мещена в скобки, значит величины, проставленные в скобках, встрече-
ны в единичных случаях. Это относится как к формулам плавников, так
и к формулам боковой линии.
Плакоидная чешуя — см. Чешуя.
Планктон — гидробионты, живущие в толще воды в состоянии
парения; различают фитопланктон и зоопланктон.
Планктофаги — животные, потребляющие планктонные орга-
низмы (из рыб — толстолобики, молодь многих рыб).
Плодовитость рыб — количество икринок, выбрасываемое ры-
бой в один период икрометания.
Подбородок — пространство на брюшной стороне головы между
нижней челюстью и местом прикрепления жаберных перепонок.
Позвонки — кости, составляющие позвоночник рыб.
Позвоночные (лат. Vertebrata) — наиболее организованные хордо-
вые животные, имеющие позвоночный столб и череп.
Половозрелые, или зрелые, рыбы — все рыбы, принимавшие уча-
стие в первом нересте.
Половой диморфизм — различия признаков мужских и женских
особей раздельнополых видов; развивается к периоду половой зрело-
сти.
Половой сосочек — см. Урогенитальный сосочек.
Половые железы — гонады, у самцов рыб — семенники, у са-
мок — яичники.
Полупроходные рыбы — экологическая группа солоноватовод-
ных рыб, заходящих для размножения в низовья рек.
Поперечные ряды чешуй (лат. squama — Squ.)
—
ряды чешуй,
проходящие через весь бок рыбы от спины до брюшка. Эти ряды обыч-
но просчитывают у тех рыб, у которых нет боковой линии (у некоторых
бычков).
Постдорсальное расстояние (или постдорсальное пространст-
во) — расстояние от вертикали конца спинного плавника до конца че-
30
шуйчатого покрова (или до основания хвостового плавника, если че-
шуй нет), считая посередине тела.
Предглазничная кость — окологлазничная кость впереди глаза.
Предглазничное пространство — см. Рыло.
Предличинка, или свободный эмбрион, — личинка с момента вы-
хода её из яйца до исчезновения желточного мешка.
Пресные воды — природные воды с минерализацией воды до
0,5 г/л.
Пресноводные рыбы — группа рыб, обитающая в пресных водо-
ёмах.
Придонные рыбы — рыбы, обитающие в придонных слоях водо-
ёма.
Присоска — см. Брюшная присоска.
Продецимилле (‱) — см. Индекс наполнения кишечника.
Производители — половозрелые особи обоего пола.
Промежуток межжаберный, или истмус (перешеек) (лат. isth-
mus) — пространство, разделяющее жаберные полости.
Промилле (‰) — см. Солёность воды.
Промысловая мера — см. Мера промысловая.
Проходные рыбы — рыбы, заходящие для размножения из морей
в реки или из реки в море.
Псаммофилы — экологическая группа рыб, часто откладываю-
щих икру при нересте на песчаный грунт (пескари, бычки, гольцы и
др.)
Птеригоподии — видоизменённые брюшные плавники самцов
акул и скатов, приспособленные для внутреннего оплодотворения.
Рабочая плодовитость — количество икры, идущее для целей
искусственного оплодотворения.
Радужина (лат. iris) — окрашенная часть сосудистой оболочки
глаза, окружающая зрачок (пиленгас).
Развилка — выемка между лопастями
хвостового плавника.
Растительноядные рыбы — рыбы-фи-
тофаги, питающиеся преимущественно фи-
топланктоном или макрофитами.
Расщеп — ряд расширенных чешуй,
окаймляющих анальное отверстие и основа-
ние анального плавника (рис. 16).
Рис. 16. Расщеп (а)
31
Реофильные рыбы — рыбы, развитие которых приурочено к во-
доёмам или участкам водоёмов, имеющим высокую скорость течения
воды.
Роговые зубы — зубы, расположенные на верхнечелюстной и
нижнечелюстной пластинках (у миног).
Рострум — см. Рыло.
Рот — его расположение — систематический признак; различают
следующие виды рта (рис. 17):
Рис. 17. Различные формы рта у рыб:
а — верхний; б — конечный; в — полунижний; г — нижний; д — выдвижной;
е — воронкообразный (круглый); ж — нижний полулунный; з — нижний по-
перечный
1)верхний рот —нижняячелюстьсильновыступаетвперёд;
разрез рта направлен вверх;
2)конечный рот —челюстивыдаютсяодинаковоиразрез
рта направлен по длине тела;
3)полунижний рот —верхняячелюстьвыдаётсявперёдне-
сколько больше нижней;
4)нижний рот —рыловыдаётсянаднижнейчелюстью;
5)выдвижной рот —образуетвоткрытомсостояниитрубку,
которая при закрывании рта складывается;
6)воронкообразный(круглый)—уминог.
32
Различают также по форме поперечный, косой и полулунный рот.
Рыло — часть головы, расположенная впереди глаза, предглаз-
ничное пространство.
Рыльная площадка — часть рыла между передними концами
верхнечелюстных костей.
Свободный эмбрион — зародыш со дня выклева до перехода на
смешанное питание.
Сеголеток (от слов «сего лета») — вполне сформировавшаяся
рыбка; обозначается значком 0+.
Сенсорные каналы — чувствительные клетки, воспринимающие
колебания водной среды.
Сестон — совокупность взвешенных в воде органо-минеральных
частиц и планктонных организмов.
Симфизис — место соединения костей нижней челюсти (в ниж-
ней части головы).
Скат рыб — наследственно закреплённое перемещение рыб вниз
по течению, происходящее в определённые периоды их жизненного
цикла.
Солёность воды — суммарное количество растворённых в воде
минеральных солей; солёность воды выражается в промилле, обозна-
чаемых символом «‰». 1 ‰ отвечает содержанию в 1 л воды 1 г солей
(пресными считаются воды до 0,5 ‰; миксогалинными, или солонова-
тыми, — 0,5—30 ‰; эугалинными, или морскими, — 30 —40 ‰; ги-
пергалинными, или пересоленными, — более 40 ‰); миксогалинные
воды подразделяются на олигогалинные (0,5—5 ‰), мезогалинные
(5—18 ‰) и полигалинные (18—30 ‰).
Солоноватые воды — см. Солёность воды.
Солоноватоводные рыбы — рыбы, населяющие опреснённые
участки морей.
Сорные рыбы — см. Малоценные рыбы.
Сошник (лат. vomer) — непарная кость, расположенная в передней
части основания черепной коробки рыб.
Спектр питания — состав пищи животного, характеризующийся
набором потребляемых кормов и их количественным соотношением.
Спинной плавник (один или два) (лат. pinna dorsalis — D) — плав-
ник, расположенный на спине рыбы. Например, D III 8—9 означает: в
спинном плавнике три неветвистых луча и от 8 до 9 ветвистых. Если
два спинных плавника, то лучи первого плавника обозначают рим-
33
скими цифрами, ветвистые лучи второго — арабскими. Если спинные
плавники не слиты вместе, а отделены один от другого, то между циф-
рами, характеризующими первый плавник и второй плавник, ставят
запятую. Например, XIII—XVI, I—III 13—15 (окунь) означает: два
спинных плавника, не слитых вместе; в первом плавнике от 13 до 16
колючих лучей, во втором — от 1 до 3 неветвистых лучей и от 13 до 15
ветвистых лучей.
Спиральный клапан — расположенная по спирали слизистая
складка внутренней поверхности кишечника осетровых, способствую-
щая лучшему всасыванию перевариваемой пищи.
Стеногалинные организмы — животные, не выдерживающие
больших изменений концентрации солей.
Стенотермные рыбы — рыбы, существующие лишь при опре-
делённой или меняющейся в очень узких пределах температуре воды.
Стенофаги — рыбы, питающиеся небольшим набором кормовых
организмов.
Субстрат — подстилающий слой, место, на котором развиваются
и живут организмы.
Таксон — классификационная единица в систематике растений и
животных (вид, род, семейство, отряд, класс, тип).
Теплолюбивые рыбы — группа рыб, основные жизненные функ-
ции которых (питание, рост, размножение) происходят весной и летом
при температуре воды 15—20 °С и выше.
Толщина головы — самое широкое место головы в области жа-
берных крышек.
Туводные рыбы — пресноводные рыбы, не заходящие из реки в
море.
Урогенитальный сосочек — мясистый отросток, который пред-
ставляет собой плотное выпячивание ткани, расположенное позади
анального отверстия; появляется во время полового созревания у сам-
цов миноги и канального сома.
Фаунистический комплекс рыб — группа видов рыб, связанных
общностью географического происхождения.
Фитобентос — растительность дна водоёма.
Фитопланктон — совокупность микроскопических растений,
находящихся во взвешенном состоянии в толще воды.
Фитофилы — экологическая группа рыб, откладывающих клей-
кую икру при нересте обычно на растительность.
34
Фулькры — рудиментарная ганоид-
ная чешуя у осетровых рыб, представляет
собой треугольные чешуи на верхнем крае
основания хвостового плавника осетровых
рыб, построенные по типу ганоидной чешуи
(рис. 18).
Хвостовой плавник (лат. caudalis —
С) — по форме и расположению выделяют
несколько типов хвостового плавника: г е -
тероцеркальный (несимметричный,
неравнолопастной)—уакул,осетровыхидр.; гомоцеркальный
(ложносимметричный) — у большинства костистых рыб (сельдевые,
карповые идр.); дифицеркальный (симметричный)—усарга-
новых, угрёвых, тресковых). По соотношению размеров верхней и ниж-
ней лопастей хвостовые плавники могут быть эпи-, гипо- и изобатными
(церкальными). При э п и б а т н о м (эпицеркальном) типе верхняя ло-
пасть длиннее (акулы, осетровые), при г и п о б а т н о м (гипоцеркаль-
ном) верхняя лопасть короче (летучие рыбы, лещ), при и з о б а т н о м
(изоцеркальном) обе лопасти имеют одинаковую длину (сазан, рыбец,
судак и др.).
Хвостовой стебель — часть тела, расположенная позади аналь-
ного плавника или заднепроходного отверстия.
Хирономиды (тендипедиды) — некровососущие комары-толкун-
цы сем. Chironomidae, личинки которых известны под названием мо-
тыль.
Хищные рыбы — группа рыб, основным источником питания ко-
торых является рыба.
Хоаны — внутренние ноздри.
Холодолюбивые, или холодноводные, рыбы — группа рыб, кото-
рые всю жизнь проводят в холодной воде.
Хорда — эластичный стержень, служащий осевым скелетом хор-
довых животных, к которым относятся рыбы; у большинства рыб хорда
имеется только у зародышей, но у круглоротых, осетровых и некото-
рых других рыб она сохраняется на протяжении всей жизни.
Циклоидная чешуя — см. Чешуя.
Челюсти — верхняя и нижняя.
Чешуя — наружный покров рыб (рис. 19):
Рис. 18. Фулькры (а) осёт-
ра
35
Рис. 19. Типы чешуи рыб:
1 — плакоидная, а — чешуйка в разрезе; 2 — ганоидная; 3 — циклоидная;
4 — ктеноидная
1)плакоидная чешуя — пластинкисзубчиками(уакул,
скатов);
2)ганоидная чешуя —ромбовидныепластинки,покрытые
эмалеподобным веществом ганоином; остатки ганоидной чешуи сохра-
нились у осетровых на верхней лопасти хвоста, фулькры и жучки этих
рыб представляют собой видоизменённые ганоидные чешуи;
3)циклоидная чешуя — тонкиеокруглые пластинки(са-
зан, лещ);
4)ктеноидная чешуя —болееплотныепластинкисзубчи-
ками на свободном (заднем) крае (судак, окунь).
Членистый луч — см. Лучи плавников.
Чувствительные (слизевые) каналы — органы кожного чувства
у рыб (см. Генипоры).
Ширина лба — межглазничное пространство.
Щёки — пространство между глазом и задним краем предкрыш-
ки.
Эвригалинные организмы — гидробионты, приспособленные к
существованию в условиях значительных изменений солёности воды.
Эвритермные рыбы — рыбы, приспособленные к жизни в усло-
виях значительных изменений температуры окружающей среды.
Эврифаги — рыбы, питающиеся разнообразной пищей.
Эмбриональный период — зародышевый период, часть жизнен-
ного цикла рыб от момента оплодотворения до перехода на внешнее
питание.
Эфемерные рыбы — рыбы, живущие один год и отмирающие по-
сле нереста.
36
ТАБЛИЦЫ
ДЛЯ ОПРЕДЕЛЕНИЯ РЫБООБРАЗНЫХ И РЫБ
Рыбообразные и рыбы принадлежат к типу Позвоночные — Verte-
brata, подтипу Черепные — Craniata.
Подтип Черепные — Craniata объединяет представителей трёх
надклассов: Миксиноморфы — Myxinimorphi, Петромизонтоморфы —
Petromyzontomorphi и Челюстноротые — Gnathostomata.
В водоёмах бассейна Кубани и прилегающих к нему реках и озё-
рах обитают представители двух надклассов: Петромизонтоморфы —
Petromyzontomorphi и Челюстноротые — Gnathostomata.
Надкласс Петромизонтоморфы — Petromyzontomorphi объединяет
представителей одного класса — Миноги — Petromyzontida Bonaparte, 1832
с одним отрядом Миногообразные — Petromyzontiformes Bonaparte, 1832.
Отряд Миногообразные — Petromyzontiformes Bonaparte, 1832
Черепные хордовые животные, не имеющие обособленных челю-
стей. Ротовое отверстие имеет вид округлой присоски, усаженной внутри
многими роговыми зубами. Хорда сохраняется в течение всей жизни. Ске-
лет неокостеневающий. Населяют как пресные, так и морские водоёмы.
Тело голое, угреобразное, с каждой стороны тела по семь жаберных
отверстий. Нет парных плавников. Есть два спинных плавника и хвосто-
вой. Глаза хорошо развиты. Рот присасывательный. Миноги испытывают
превращение (метаморфоз). Личинка миноги — пескоройка (Ammocoetes)
отличается от взрослой миноги ртом (в виде треугольной щели), лишённым
зубов, плохо развитыми плавниками, жаберными отверстиями, лежащими
в бороздке. Пескоройки долгое время обитают в иле, питаясь детритом.
У взрослых рот круглый, присасывательный, усаженный по внешнему
краю кожистой бахромой. Роговые зубы сидят на верхнечелюстной и ниж-
нечелюстной пластинках, на поверхности ротовой воронки — губные зубы.
В водах бассейна Кубани отряд представлен одним семейством —
Миноговые — Petromyzontidae Bonaparte, 1831.
Семейство Миноговые — Petromyzontidae Bonaparte, 1831
Тело удлинённое, угреобразное, голое. Грудных и брюшных плав-
ников нет. Два спинных плавника разделены небольшим промежутком:
37
задний спинной плавник переходит в хвостовой. В период икромета-
ния тело миноги укорачивается, спинные плавники становятся выше и
сближаются, у самцов около анального отверстия образуется половой
сосочек. После нереста самцы и самки миноги погибают.
Семейство Миноговые объединяет представителей 8 родов с 34 ви-
дами. В водах России встречается 10 видов. В водоёмах Кубани обита-
ют представители рода Зубатые миноги — Eudontomyzon Regan, 1911.
Род Зубатые миноги — Eudontomyzon Regan, 1911
Нижнегубные зубы обычно расположены в несколько рядов. На
верхнечелюстной пластинке, как правило, 2 зуба, но бывает 3 и более,
на нижнечелюстной 5—10.
Род Зубатые миноги включает как проходные, так и жилые виды.
В бассейне Кубани обитает один вид — украинская минога — Eudonto-
myzon mariae (Berg, 1931).
Украинская минога — Eudontomyzon mariae (Berg, 1931) (рис. 20, 2.1*)
Рис. 20. Самец (вверху) и самка (внизу) украинской миноги — Eudontomyzon
mariae (Berg, 1931)
Тело цилиндрическое, удлинённое. Кожа голая, слизистая, без
парных плавников. Челюсти отсутствуют, а в ротовой воронке разме-
щены роговые зубы. Ротовая присоска поддерживается кольцевым хря-
щом. Носовое отверстие одно. Череп хрящевой. Сердце двухкамерное,
один круг кровообращения.
Нижнегубные зубы в несколько рядов, но обычно не образу-
ют сплошной полоски. На нижнечелюстной пластинке 5—10 зубов
*Здесь и далее в двойной нумерации, например 2.1, цифра 2 означает но-
мер приложения, а следующая после точки цифра — номер рисунка в прило-
жении.
38
(рис. 21, 8). На верхнечелюстной — 2 зуба (рис. 21, 3), но иногда ме-
жду ними бывает маленький зуб, изредка между крупными краевыми
зубами 3 маленьких зубчика. Боковых губных зубов с каждой стороны
3 или 4 (рис. 21, 5).
Рис. 21. Ротовая присоска (а) украинской миноги и её схема (б):
1 — ротовое отверстие; 2 — верхнечелюстной зуб; 3 — верхнечелюстная
пластинка; 4 — верхнегубные зубы; 5 — боковые зубы; 6 — краевые губные
зубы; 7 — нижнегубные зубы; 8 — нижнечелюстная пластинка
Зубы у миноги во время нереста могут быть как тупыми, так и ост-
рыми. Миноги имеют признаки полового диморфизма. У половозрелых
особей спинные плавники увеличиваются и просвет между ними со-
кращается, у самки вырастает анальный плавник, а у самца — половой
сосочек (см. рис. 20, 2.1).
Нерест миноги происходит в апреле—мае. Плодовитость
2—3 тыс. икринок. Икра крупная, диаметром 0,9—1,3 мм. После не-
реста взрослые особи обычно погибают. Личинки выклёвываются на
10—12 -й день после оплодотворения. Личиночный период у миноги
продолжается 3—4 года. Затем около полугода происходит процесс
метаморфоза. Молодые миноги держатся на участках реки с камени-
стыми, галечными и крупнопесочными грунтами, предпочитая места
затенённые.
Длина половозрелых особей колеблется от 14 до 20 см.
39
В водоёмах бассейна Кубани минога не имеет рыбохозяйственного
значения. В настоящее время из-за значительного загрязнения водоёмов
численность миноги резко сократилась. Изредка встречаются единич-
ные экземпляры в реках бассейна Кубани (Афипс, Псекупс, Белая, Лаба,
Уруп, Зеленчук) и их притоках, а также в причерноморских, приазов-
ских и закубанских реках (Москул, 1988, 1994, 1995, 1998, 2007; Троиц-
кий, Цуникова, 1988; Абаев, 1996; Лужняк, 2003; Решетников, Пашков,
2009; Емтыль, Иваненко, 2002; Абрамчук, Иваненко, 2018 и др.).
Биология миноги изучена недостаточно. В литературе очень мало
данных по биологии украинской миноги водоёмов бассейна Кубани.
Многие вопросы биологии как пескоройки, так и взрослой миноги ос-
таются неизученными.
Украинская минога занесена в Красную книгу РФ (2001), катего-
рия 2 — сокращающийся в численности вид, в Красную книгу Красно-
дарского края (2007, 2017).
Надкласс Челюстноротые — Gnathostomata
Объединяет представителей, характеризующихся преимуществен-
но водным дыханием при помощи жабр, расположенных на жаберных
дугах. Хорда сохраняется в течение всей жизни или заменяется хряще-
выми либо костными позвонками. Есть челюсти. Имеются парные но-
совые отверстия и парные конечности в виде парных плавников. Внут-
ренний скелет более или менее окостеневший.
Класс Лучепёрые рыбы — Actinopterygii Klein, 1885
Первичноводные позвоночные животные, вся организация кото-
рых приспособлена к жизни в воде. Тело у большинства рыб обтекае-
мой формы. Органами движения служат парные и непарные плавники.
Кожа рыб слизистая, обычно покрыта чешуёй различного строения.
Дыхание жаберное. Жабры образованы многочисленными тонкими ле-
пестками.
У низших рыб хорда сохраняется всю жизнь, у высших она во
взрослом состоянии остаётся только между позвонками. У хрящевых
рыб (акулы, скаты) скелет хрящевой, у костно-хрящевых (осетровые)
внутренний скелет в основном хрящевой, но череп покрыт накладными
костями.
40
Класс Лучепёрые рыбы объединяет три подкласса: Кладистии
(Cladistia), Хрящекостные рыбы, или Хрящевые ганоиды (Chondrostei),
и Новопёрые рыбы (Neopterygii).
Лучепёрые — группа ныне живущих рыб, включающая 3 подкласса,
44 отряда, 453 семейства, 4 289 родов и 26 891 вид. Около 44 % видов из-
вестны только из пресных вод (Пономарев, Баканева, Федоровых, 2016).
В водоёмах бассейна Кубани и прилегающих к нему закубанских
рек и рек Азово-Кубанской равнины обитают представители двух под-
классов: Хрящекостные рыбы, или Хрящевые ганоиды (Chondrostei), и
Новопёрые рыбы (Neopterygii).
Таблица
для определения подклассов класса
Лучепёрые рыбы — Actinopterygii Klein, 1885
1 (2). Верхняя лопасть хвоста значительно больше нижней. Тело
или голое, или покрыто 5 рядами ромбических костных щитков (жу-
чек). Ганоидная чешуя только на хвосте. Внутренний скелет хрящевой.
Подкласс Хрящекостные рыбы, или Хрящевые ганоиды, —
Chondrostei Müller, 1844 (с. 40)
2 (1). Верхняя лопасть хвоста примерно равна нижней. Тело покры-
то костными тонкими чешуями или голое. Внутренний скелет костный.
Подкласс Новопёрые рыбы — Neopterygii Cope,1871 (с. 56)
Подкласс Хрящекостные рыбы,
или Хрящевые ганоиды, — Chondrostei Müller, 1844
Подкласс характеризуется небольшим количеством хондриальных
(хрящевого происхождения) костей. Осевой скелет представлен в тече-
ние всей жизни хордой и сегментально расположенными хрящевыми
верхними и нижними дугами позвонков. Череп в основе хрящевой, но
снаружи покрыт накладными костями.
Голова заканчивается вытянутым рылом, на нижней стороне кото-
рого расположен рот, имеющий вид поперечной щели. Зубов нет. Тело
удлинённое, хвостовой плавник гетероцеркальный (асимметричный).
На верхней лопасти хвостового плавника есть ганоидная чешуя и фуль-
кры. Тело или голое, или покрыто пятью рядами жучек. Рот нижний,
есть более или менее сильно развитый рострум.
41
Подкласс Хрящекостные рыбы, или Хрящевые ганоиды, включа-
ет представителей одного отряда — Осетрообразные (Acipenseriformes
Berg, 1940).
Отряд Осетрообразные — Acipenseriformes Berg, 1940
Признаки отряда совпадают с указанными признаками подкласса
Хрящекостные рыбы, или Хрящевые ганоиды (Chondrostei). Это древние
рыбы. В водоёмах бассейна Кубани обитают представители двух семейств.
Таблица
для определения семейств отряда
Осетрообразные — Acipenseriformes Berg, 1940
1 (2). Тело покрыто пятью рядами костных жучек. Рот нижний, по-
перечный, впереди него четыре усика.
Семейство Осетровые — Acipenseridae Bonaparte, 1831 (с. 41)
2 (1). Тело голое. Рот нижний, есть сильно развитый рострум.
Семейство Веслоносые — Polyodontidae Bonaparte, 1838 (с. 55)
Семейство Осетровые — Acipenseridae Bonaparte, 1831
Осетровые — проходные и пресноводные рыбы. Тело вытянутое в
длину и суживающееся к хвосту. Рыло удлинённое, коническое или ло-
патовидное. Рот нижний, поперечный, выдвижной. Имеется две пары
усиков (рис. 22).
Рис. 22. Головы различных осетровых рыб снизу:
а — белуги; б — шипа; в — осётра; г — стерляди; д — севрюги
42
Ромбическая чешуя ганоидного типа покрывает только верхнюю
лопасть хвостового плавника, по верхнему краю основания этого плав-
ника расположен ряд видоизменённых чешуй треугольной формы
(фулькры). На теле имеется пять продольных рядов костных образова-
ний — жучек; между ними по спинной стороне тела разбросаны кост-
ные пластинки и шипики.
Осетровые — теплолюбивые рыбы. Они отличаются продолжи-
тельным периодом жизни. Половозрелыми становятся поздно. Нерест
не ежегодный. Интервалы между нерестами составляют в среднем 2—4
года. Нерест происходит в пресной воде. У проходных осетровых (бе-
луга, русский осётр, севрюга) выражены экологические расы. Рыбы
озимой расы входят в реки осенью, зимуют в них и, отнерестившись
весной, скатываются в море. Рыбы яровой расы входят в реки весной,
нерестятся в мае—июне и уходят в море. Икра донная, клейкая. Личин-
ки и мальки осетровых не задерживаются в реке, а быстро скатываются
в море, за исключением стерляди.
Рыбы этого семейства относятся к наиболее ценным в промысло-
вом отношении. Уловы осетровых в недалёком прошлом (1986 г.) со-
ставляли 1 091 т, а через 10 лет (1995 г.) из-за сокращения естественно-
го воспроизводства уловы сократились и составили 639 т. В настоящее
время промысел осетровых в Азовском море запрещён. Осетровые ши-
роко используются в пресноводной пастбищной и товарной аквакуль-
турах.
В бассейне Кубани обитают представители 2 родов.
Таблица
для определения родов семейства
Осетровые — Acipenseridae Bonaparte, 1831
1 (2). Жаберные перепонки соединены между собой, образуя под
межжаберным промежутком свободную складку. Усики длинные, уп-
лощённые, рот большой полулунный.
Род Белуги — Huso Brandt, 1869 (с. 43)
2 (1). Жаберные перепонки приращены к межжаберному проме-
жутку, не образуя под ним складку. Усики гладкие, неуплощённые, рот
относительно небольшой, имеет вид поперечной щели.
Род Осётры — Acipenser Linnaeus, 1758 (с. 45)
43
Род Белуги — Huso Brandt, 1869
Массивное веретенообразное тело, суживающееся к хвосту. Боль-
шая и высокая голова с коротким рылом. Жаберные перепонки сра-
щены между собой и образуют свободную складку. Рот полулунный,
очень большой, занимает всю нижнюю поверхность рыла. Усики сплю-
щены с боков. Род представлен двумя видами.
Таблица
для определения видов рода
Белуги — Huso Brandt, 1869
1 (2). Первая спинная жучка меньше последующих в ряду. В спин-
ном плавнике более 60 лучей. Усики с листовидными придатками.
Белуга — Huso huso (Linnaeus, 1758) (с. 43)
2 (1). Первая спинная жучка больше последующих в ряду. В спин-
ном плавнике менее 60 лучей. Усики без листовидных придатков.
Калуга — Huso dauricus (Georgi, 1775) (с. 45)
Краткая биологическая характеристика видов рода
Белуги — Huso Brandt, 1869
Белуга — Huso huso (Linnaeus, 1758) (рис. 23, 2.2)
Рис. 23. Белуга — Huso huso (Linnaeus, 1758): вверху неполовозрелая рыба
длиной 110 см, внизу взрослый самец длиной 180 см
44
В Азовском море образует подвид Huso huso maeoticus ( Sal’nikov &
Maliatsky, 1934) — азовская белуга. Спинных жучек 11—14, боко-
вых — 41 —52, брюшных — 7 —14. У азовского подвида окраска свет-
ло-серая, у черноморского — более тёмная.
D 62—73, A 28—41. Спинных жучек 9—17, боковых — 37—53,
брюшных — 7—14. Первая спинная жучка меньше последующих в
ряду. Усики длинные, достигают верхней губы. Большая и высокая го-
лова с коротким рылом. Рот полулунной формы, очень большой, за-
нимающий всю нижнюю поверхность головы. Жаберные перепонки
образуют сплошную складку.
Белуга — самая крупная рыба не только в Азово-Черноморском
бассейне, но и среди всех проходных и пресноводных рыб. Масса бе-
луг, вылавливаемых в прибрежных водах Кубани, колеблется от 10 до
150 кг. Отдельные особи достигали массы 750 кг при длине 4,6 м (Му-
сатова, 1973). Продолжительность жизни белуги — более 50 лет.
Белуга — проходная рыба, для нереста заходит в реки. Половозре-
лой становится в возрасте 12—16 лет. Плодовитость в зависимости от
размера самки колеблется от 210 до 1 600 тыс. икринок. Икра отклады-
вается на камни, к которым приклеивается. Икрометание происходит с
апреля по июнь при температуре воды от 12 до 20 °С. Развитие опло-
дотворённых икринок — 6 —8 сут. Личинки скатываются по течению
воды в море. Питается молодь беспозвоночными животными (гамма-
риды, мизиды, личинки хирономид и др.), а старшие возраста — рыбой
(бычки, кильки, сельди и др.).
Белуга — ценная промысловая рыба. Уловы белуги очень неве-
лики — от 0,2 т (1995 г.) до 329 т (1978 г.). Резкое снижение запасов
белуги связано со строительством плотин на реках, что привело к пре-
граждению хода белуги, да и всех осетровых к нерестилищам.
Начиная с 1994 г. стадо белуги пополняется молодью, полученной
искусственным путём. Так, в 1994 г. выпущено 0,1 млн экз., в 1996—
1998гг. — по0,1,в1999г. — 0,2,в2004г. — 0,124млнэкз.
В 2001 г. впервые в р. Кубань была выпущена молодь белуги, полу-
ченная от производителей, выращенных в Южном филиале Федераль-
ного селекционно-генетического центра рыбоводства (ЮФ ФСГЦР)
(Чебанов, Галич, Чмырь, 2001; Чебанов, Галич, 2003). Начиная с 2013 г.
небольшое количество разновозрастной молоди белуги массой от 3 до
300 г выпускает ГБУ КК «Кубаньбиоресурсы» (Чебанов, Галич, Мер-
кулов, 2017).
45
Внесена в Красную книгу МСОП, Красную книгу РФ (2001) и
Краснодарского края (2007, 2017). Поддержание численности белуги
возможно при заводском воспроизводстве на рыборазводных заводах.
Является объектом пастбищной и товарной аквакультуры.
Калуга — Huso dauricus (Georgi, 1775) (рис. 24, 2.3)
Рис. 24. Калуга — Huso dauricus (Georgi, 1775)
D 43—57, A 26—35. Спинных жучек 10—16, боковых — 32—46,
брюшных — 8 —12. Тело между жучками покрыто мелкими зёрныш-
ками, среди которых иногда у крупных экземпляров попадаются более
крупные округлые пластинки. Первая спинная жучка — самая большая
в ряду. Рот очень большой. Усики длинные, достигают верхней губы,
без листовидного придатка.
Калуга является эндемиком бассейна р. Амур. Молодые особи в
летние месяцы мигрируют в Охотское и Японское моря. Растёт калуга
медленнее белуги, половозрелость наступает поздно — в 17—20 лет.
Нерест весенний (май—июнь). Нерестится на песчаном и галечнико-
вом грунте при температуре воды 12—14 °С. Плодовитость колеблется
от 0,5 до 4 млн икринок. Молодь питается беспозвоночными, а взрос-
лые особи — рыбой.
Внесена в Красную книгу МСОП, Красную книгу РФ (2001). Име-
ется небольшое по масштабам искусственное воспроизводство на пред-
приятиях Государственного казённого учреждения Краснодарского
края «Кубаньбиоресурсы».
Род Осётры — Acipenser Linnaeus, 1758
Жаберные перепонки прикреплены к межжаберному промежутку,
не образуя свободной складки, как у рода белуг.
Пресноводные и проходные рыбы. В бассейне Кубани, а также в
46
Чёрном и Азовском морях в пределах Краснодарского края встречают-
ся представители 6 видов.
Таблица
для определения видов рода
Осётры — Acipenser Linnaeus, 1758
1 (2). Нижняя губа сплошная, не прервана посредине (см. рис. 22, б).
Первая спинная жучка крупнее остальных. Усики бахромчатые, почти
достигают конца рыла.
Шип — Acipenser nudiventris Lovetsky, 1828 (с. 47)
2 (1). Нижняя губа посередине прервана. Первая спинная жучка не
выделяется своими размерами.
3 (4). Рыло острое, очень длинное, мечевидное: длина его состав-
ляет более 60 % длины головы. Боковых жучек 26—43.
Севрюга — Acipenser stellatus Pallas, 1771 (с. 50)
4 (3). Рыло более короткое, длина его менее 60 % длины головы.
5 (6). Жучек в боковых рядах более 55 (56—71). Усики бахромча-
тые. Речная рыба.
Стерлядь — Acipenser ruthenus Linnaeus, 1758 (с. 48)
6 (5). Жучек в боковых рядах менее 50. Усики обычно небахром-
чатые.
7 (8). Между спинным и боковыми рядами жучек расположены
правильными рядами мелкие ромбические пластинки.
Атлантический осётр — Acipenser sturio Linnaeus, 1758 (с. 52)
8 (7). Между спинным и боковыми рядами жучек беспорядочно
разбросаны звёздчатые пластинки и/или твёрдые зёрнышки.
9 (10). Жаберные тычинки на первой жаберной дуге веерные, так
как каждая из них состоит из трёх двойных рожков. Тело между рядами
жучек усеяно довольно большими зёрнами или мелкими звёздчатыми
пластинками, разбросанными в беспорядке.
Сибирский осётр — Acipenser baerii Brandt, 1869 (с. 51)
10 (9). Жаберные тычинки на первой жаберной дуге невеерные:
они заканчиваются одним остриём каждая. Боковых жучек менее 50.
Усики обычно небахромчатые. Рыло короткое, более или менее закруг-
лённое, туповатое. На теле у взрослых особей между спинным и боко-
выми рядами жучек расположены звёздчатые пластинки.
Русский осётр — Acipenser gueldenstaedtii Brandt, 1833 (с. 49)
47
Краткая биологическая характеристика видов рода
Осётры — Acipenser Linnaeus, 1758
Шип — Acipenser nudiventris Lovetsky, 1828 (рис. 25, 2.4)
Рис. 25. Шип — Acipenser nudiventris Lovetsky, 1828
D 40—57, A 23—37. Спинных жучек 11—17, боковых — 49—72,
брюшных — 12 —17. Жаберных тычинок 24—36.
Тело удлинённое, веретенообразной формы. Рыло сравнительно
короткое, но заострённое. Жаберные перепонки приращены к межжа-
берному промежутку. Нижняя губа сплошная, не прервана (см. рис. 22,
б), что чётко отличает его от других видов осётров. Усики бахромча-
тые. Первая спинная жучка самая крупная. Брюшные жучки с возрас-
том часто стираются и бывают почти незаметны (отсюда видовое на-
звание nudiventris, что в переводе с латинского означает голобрюхий).
Пластинок на теле между рядами жучек нет. Спина серовато-зелёная,
бока светлые, брюхо желтовато-белое, плавники сероватые. Шип —
крупная рыба. Длина тела половозрелых особей колеблется от 97 до
220см.Массасамцов—от5до30кг,асамок—от8до50кг.Половая
зрелость наступает у самок в возрасте 12—14 лет, у самцов — в 9 лет.
Шип — ценная, но очень редкая рыба. В 1930-е гг. шип встречал-
ся как в Чёрном, так и единично в Азовском море. В настоящее время
ведутся работы по искусственному разведению шипа и реакклиматиза-
ции его в водоёмах Краснодарского края (Чебанов, 2010).
В 2005 г. от искусственно сформированного маточного стада шипа
был произведён выпуск 300 тыс. экз. молоди этого вида в Краснодар-
ское водохранилище (Чебанов, Козырицкая, 2007; Чебанов, Галич,
2013). Повторный выпуск разноразмерной молоди шипа массой от 3
до 15 г в этот же водоём был осуществлён в 2013 г. (Чебанов, Галич,
Меркулов, 2017).
Вид внесён в Красную книгу РФ (2001) и Краснодарского края
(2007, 2017).
48
Стерлядь — Acipenser ruthenus Linnaeus, 1758 (рис. 26, 2.5)
Рис. 26. Стерлядь — Acipenser ruthenus Linnaeus, 1758
D 37—54, A 19—31. Спинных жучек 12—17, боковых — 56—71,
брюшных 10—19. Жаберных тычинок на первой жаберной дуге 16—26,
тычинки простые невеерообразные. Рот небольшой, нижний. Нижняя
губа посредине прервана (рис. 22, г). Усики бахромчатые, достигают
рта. От других видов рода Acipenser чётко отличается большим числом
боковых жучек (до 71). Окраска спины от тёмно-серой до серовато-ко-
ричневой, брюхо белое.
Стерлядь — речная рыба, но в небольшом количестве встречается
и в опреснённых участках моря. Длина половозрелых самок колеблется
от 37 до 50 см, самцов — от 32 до 45 см. Масса самок — 1,8—2,5 кг,
самцов — 1,5—2,0 кг. Половозрелости достигает на 3—5 -м году жизни.
Плодовитость колеблется от 3 до 30 тыс. икринок. Нерест происходит
в мае при температуре воды 14—17 °С. Инкубационный период длится
4—5 сут.
Стерлядь — самая мелкая из азовских осетровых рыб. До строи-
тельства Фёдоровского гидроузла и плотины Краснодарского водохра-
нилища стерлядь встречалась в Кубани от устья до ст-цы Тбилисской.
Последний случай поимки стерляди в р. Кубань от естественного не-
реста относится к 1940-м гг. (Мусатова, 1973). В настоящее время ве-
дутся работы по реакклиматизации стерляди в р. Кубань (Березовская,
Чебанов, 1998). В 1999 г. начата реакклиматизация стерляди в Крас-
нодарском водохранилище и продолжается в реках Кубань и Протока.
За период с 1999 по 2015 г. было выпущено более 50 млн экз. молоди
(Чебанов, Галич, Меркулов, 2017).
Вид внесён в Приложение II Конвенции о международной торговле
видами дикой фауны и флоры, находящимися под угрозой исчезнове-
ния (CITES), как «уязвимый вид», а также в Красные книги РФ (2001)
и Краснодарского края (2017). Вылов в России запрещён во всём Волж-
ско-Каспийском и Азово-Черноморском рыбохозяйственных бассейнах.
49
Русский осётр — Acipenser gueldenstaedtii Brandt, 1833 (рис. 27, 2.6)
Рис. 27. Русский осётр — Acipenser gueldenstaedtii Brandt, 1833
D 29—44, A 18—25. Спинных жучек 9—14, боковых — 25 —37,
брюшных — 6 —13. Жаберных тычинок 16—26.
Над линией боковых жучек беспорядочно разбросаны крупные ко-
стные пластинки. Рот поперечный. Нижняя губа прервана. Усики без
бахромок.
Жаберные перепонки приращены к межжаберному промежутку.
Голова и предглазничное пространство (рыло) относительно короткое,
как бы обрубленное.
Населяет бассейны Чёрного и Азовского морей. Проходная рыба:
нагуливается в море, а нерестится в реке. Половозрелым становит-
ся на 8—14 -м году жизни. Самцы, как и у других осетровых, созре-
вают раньше самок. Длина половозрелых самок — 90 —130 см, сам-
цов — 70—110 см, масса — от 8 до 30 кг. Плодовитость колеблется
от 30 до 450 тыс. икринок. Нерест происходит при температуре воды
12—15 °С. Инкубационный период 5—10 сут. Скат личинок и мальков
происходит в мае—июле. Осётр питается как донными беспозвоноч-
ными, так и рыбой. Молодь отдаёт предпочтение личинкам хирономид,
гаммаридам и ручейникам. Переход на питание рыбой происходит у
осётра в возрасте 2 лет при длине около 30 см. В пище преобладают
килька, атерина и бычки.
Осётр — ценная промысловая рыба. Ранее уловы осётра занима-
ли первое место среди осетровых. В 1986 г. уловы осётра составляли
605 т, в 1995 г. — 334 т, в 1999 г. — 65 т. Снижение уловов осетровых
в Азово-Кубанском районе связано с антропогенным воздействием на
экосистему и значительным загрязнением водоёмов. В настоящее вре-
мя промысел осётра в Азовском море запрещён. В последние годы за-
пасы осетровых поддерживаются путём искусственного разведения на
рыборазводных заводах.
В период 1986—1990 гг. в Азовское море в среднем выпускалось
50
по 8,4 млн экз. молоди осётра в год, в период с 1991 по 1995 г. — в сред-
нем по 13,7 млн экз. Затем началось снижение объёмов выпуска молоди
русского осётра, а в период с 2010 по 2013 г. количество выпускаемой
рыбы сократилось до 1,2—1,5 млн экз. в год (Чебанов, Галич, Мерку-
лов, 2017).
Вид внесён в Красный список МСОП, Красную книгу РФ (2001) и
Краснодарского края (2007, 2017).
Севрюга — Acipenser stellatus Pallas, 1771 (рис. 28, 2.7)
Рис. 28. Севрюга — Acipenser stellatus Pallas, 1771
D 40—46, A 24—29. Спинных жучек 11—14, боковых — 30 —36,
брюшных — 9 —14. Жаберных тычинок 24—26.
Жаберные перепонки приращены к межжаберному промежутку.
Нижняя губа прервана (см. рис. 22, д). Спина и бока окрашены в тём-
ный цвет, брюхо светлое (см. рис. 2.7). Севрюга — проходная рыба:
нагуливается в море, а для икрометания заходит в р. Кубань. Длина
тела половозрелых самок достигает 180 см, масса — 18—20 кг, сам-
цов — 120 см и 12—15 кг. Самцы созревают в возрасте 5—7 лет, сам-
ки — 10—12 лет. Максимальная зарегистрированная длина севрюги —
220см,амасса—80кг.
Ранее севрюга являлась важным объектом промысла. По уловам
занимала среди осетровых второе место после осётра. Уловы севрюги
в1985г.составляли438т,в1995г. — 303т,в1999г. — 38т.Впро-
шлом ценная промысловая рыба, с начала 2000-х гг. вылов севрюги в
России запрещён из-за перепромысла и резкого сокращения численно-
сти. Запасы севрюги поддерживаются путём искусственного разведе-
ния на осетровых заводах.
В период с 1986 по 1990 г. выпуск молоди средней навеской 1,5 г
составил 15,3 млн экз. в год, с 1991 по 1995 г.
—
по 11,4 млн экз. В
последующие годы количество выпускаемой молоди снизилось и в
51
2004 г. составило 1,46 млн экз., а к 2014 г. — 0,5 млн экз. в год (Чебанов,
Галич, Меркулов, 2017).
Вид внесён в Красный список МСОП, Красную книгу РФ (2001) и
Краснодарского края (2007, 2017).
Сибирский осётр — Acipenser baerii Brandt, 1869 (рис. 29, 2.8)
Рис. 29. Сибирский осётр — Acipenser baerii Brandt, 1869
Сибирский осётр имеет ряд общих признаков с русским, от кото-
рого отличается веерообразными жаберными тычинками.
D 38—52, A 20—31. Спинных жучек 10—19, боковых — 32—59,
брюшных — 7 —16. Жаберных тычинок 28—45. Форма рыла подвер-
жена вариациям; обычно оно короткое, уплощённое и умеренно закруг-
лённое, иногда более или менее удлинённое и заострённое. Тело между
рядами жучек усеяно довольно большими зёрнами или мелкими звёзд-
чатыми пластинками, разбросанными в беспорядке. Жаберные тычин-
ки веерообразные, каждая с тремя двойными рожками. Выделяют две
формы: тупорылую и острорылую.
Проходная и пресноводная рыба. Достигает в длину 2 м.
Сибирский осётр обитает в реках Сибири — от Оби до Колымы.
Этот вид считают монотипичным или выделяют до трёх—четы-
рёх подвидов: Acipenser baerii baicalensis Nikolsky, 1896 (байкальский
осётр), Acipenser baerii chatys Drjagin, 1948 (якутский осётр, или якут-
ский стерлядевидный осётр), Acipenser baerii stenorrhynchus Nikolsky,
1896 (ленский осётр, или восточносибирский осётр, или длиннорылый
осётр, или острорылый осётр), Acipenser baerii baerii Brandt, 1869 (об-
ский осётр, или западносибирский осётр). Однако в конце 1990-х гг.
показано отсутствие различий подвидового уровня у популяций, отно-
симых ранее к разным подвидам. В крупных речных бассейнах осётр
образует популяционные континуумы, характеризующиеся клиналь-
ной изменчивостью ряда морфологических признаков. При этом разли-
52
чия между северными и южными популяциями превышают различия
между популяциями разных речных бассейнов, что делает выделение
отдельных подвидов необоснованным (Рубан, 1998, 1999).
В Краснодарский край завезён в 2005 г., является объектом товар-
ного рыбоводства; скрещивают с другими осётрами.
Атлантический осётр — Acipenser sturio Linnaeus, 1758
(рис. 30, 2.9)
Рис. 30. Атлантический осётр — Acipenser sturio Linnaeus, 1758
D 31—43, A 22—26. Спинных жучек 9—13, боковых — 24 —33.
Жаберных тычинок 18—25. Рыло удлинённое, заострённое. Усики рас-
положены ближе ко рту, чем к концу рыла. Нижняя губа прервана. Тело
между спинным и боковыми рядами жучек покрыто косыми рядами
ромбических, густо сидящих пластинок (см. рис. 2.9).
Проходная рыба. Плодовитость от 200 тыс. до 5,7 млн икринок.
Питается беспозвоночными и рыбой.
Распространён по Европейскому побережью Атлантики от Се-
верной Норвегии до Балтийского, Средиземного и Чёрного морей.
В России атлантический осётр ранее считался балтийским осётром.
Встречался в Ладожском и Онежском озёрах. Исчезающий редкий вид.
Достигал длины 3 м, массы более 200 кг.
Вид внесён в Красный список МСОП, Красные книги РФ (2001) и
Краснодарского края (2007, 2017).
Большинство видов осетровых образует в природе помеси, назы-
ваемые обычно шипами. Для бассейна Чёрного и Азовского морей из-
вестны приведённые далее помеси (Световидов, 1964).
Белуги и севрюги — Huso huso (L.) × Acipenser stellatus Pall.
Спинных жучек 13—14, боковых — 38 —42, брюшных — 9 —12. Рот,
губы, усики и межжаберный промежуток у одних, как у белуги, у дру-
гих, как у севрюги.
Белуги и осётра — Huso huso (L.) × Acipenser gueldenstaedtii Br.
53
Спинных жучек 11—13, боковых — 34—51, брюшных — 8 —12. Рот
больше, чем у осётра, но меньше, чем у белуги. Складки под межжа-
берным промежутком нет. Усики, как у осётра. Между рядами жучек
мелкие зёрнышки, пластинок нет.
Стерляди и севрюги — Acipenser ruthenus L. × Acipenser stella-
tus Pall. Спинных жучек 13—16, боковых — 43—61. Число и форма
жучек, как у стерляди. Рыло удлинённое, как у севрюги, или немного
короче, уплощённое или в форме трёхгранной пирамиды. Между ря-
дами жучек обычно гребенчатые зёрнышки, под спинными иногда ряд
пластинок.
Существует гибрид, полученный путём искусственного скрещи-
вания белуги со стерлядью — б е с т е р (по первым слогам слов бе-
луга и стерлядь) — Huso huso × Acipenser ruthenus. Впервые получен
в 1952 г. в СССР профессорами Н. И. Николюкиным и Н. А. Тимофее-
вой в Тепловском рыбопитомнике Саратовской области. Бестер соче-
тает быстрый рост белуги и раннее созревание стерляди. Длина — до
1,8 м, масса — до 30 кг. Плодовит. В аквакультуре гибриды первого
поколения за 2 года выращивания в садках и бассейнах достигают
массы в 1 кг и даже больше. Родственные формы: белужий бестер и
стерляжий бестер.
Гибрид, полученный при скрещивании самки белуги и самца стер-
ляди, кратко называемый БС — бестер, обладает высоким темпом рос-
та и скороспелостью.
При скрещивании самки белуги и самца бестера (Б × БС) получа-
ется возвратный гибрид ББС — белшип, который растёт ещё быстрее,
чем бестер. Сеголетки достигают 500, трёхлетки — 3 500 г.
Эти гибриды являются перспективными для товарного осетровод-
ства.
Отличительные признаки бестера и белшипа.
Бестер. Голова менее широкая, чем у белуги; форма рта полулун-
ная, как у белуги, но размеры значительно меньше. Первая жучка боль-
ше, чем две последующие; жаберные перепонки приращены к межжа-
берному промежутку.
Белшип. Высота головы больше, чем у бестера, рыло более широ-
кое и тупое; рот меньше, чем у белуги, но больше, чем у бестера. Пер-
вая жучка — самая массивная в ряду. Часто отсутствует перегородка,
разделяющая ноздри.
На рыборазводных осетровых заводах получены и зарегистриро-
54
ваны три породы бестера: аксайская, бурцевская и внировская. Все эти
породы впервые получены профессором Н. И. Николюкиным в Теплов-
ском рыбопитомнике Саратовской области и получили широкое при-
знание как в России, так и в других странах (Польша, Латвия, Италия,
Япония, Венгрия, США, Китай, Корея и др.).
Аксайский бестер,или бестер стерляжий,выведенв
1958 г. путём осеменения икры стерляди спермой бестера — методом
возвратного скрещивания стерляди самцами F1 бестера (СБС). В 1969 и
1973 гг. возвратные гибриды «стерлядь × бестер» были получены также
в Аксайском рыбхозе Ростовской области. По внешнему виду напоми-
нает стерлядь, но имеет более крупный размер и бóльшую массу тела.
У породы следующие биологические показатели: строение межжа-
берного промежутка без складки; число боковых жучек 55—59, лучей
в спинном плавнике 42—46, в анальном — 24 —27; половой зрелости
достигают самцы в возрасте 3—4 года при массе 2 кг, самки — в 6—8
лет при массе 3 кг; плодовитость самок — 40 тыс. икринок. Выход то-
варной рыбы от одной самки — 4,7 т.
Бурцевский бестер —межродовойгибридотскрещивания
самки белуги и самца стерляди (БС), впервые получен в 1952 г., а в
1999 г. внесён в Государственный реестр селекционных достижений
и допущен к использованию. По внешнему виду очень похож на стер-
лядь.
У породы следующие биологические показатели: строение меж-
жаберного промежутка с небольшой кожной складкой; число боковых
жучек 49—54, лучей в спинном плавнике 51—52, в анальном — 28—
30; половой зрелости достигают самцы в возрасте 4 года при массе 4 кг,
самки — в 8 лет при массе 8 кг; плодовитость самок — 120 тыс. икри-
нок. Выход товарной рыбы от одной самки — 16,9 т.
Внировский бестер, или бестер белужий — межро-
довой гибрид, получен в 1958 г. от скрещивания самок белуги с самца-
ми F1 бестера (ББС). По своим внешним признакам бестер внировский
приближается к белуге.
Биологические показатели гибрида следующие: строение межжа-
берного промежутка с кожной складкой; число боковых жучек 49—53,
лучей в спинном плавнике 55—58, в анальном — 29—31; половой зре-
лости достигают самцы в возрасте 8 лет, самки — в 14 лет при массе
30 кг; плодовитость самок — 300 тыс. икринок. Выход товарной рыбы
от одной самки — 25,7 т.
55
Все три породы используются для товарного осетроводства и по-
лучения пищевой икры.
В племенных хозяйствах проводятся опыты по разведению и выра-
щиванию других гибридов осетровых, например белуга × шип, шип ×
× севрюга, осётр × шип, осётр × белуга (Пономарев, Лагуткина, 2008).
Семейство Веслоносые — Polyodontidae Bonaparte, 1838
Веслоносы отличаются от осетровых отсутствием жучек на теле,
ганоидная чешуя есть только на верхней лопасти хвостового плавника.
Рыло очень длинное, имеет форму весла. На рыле 2 коротких усика. На
челюстях есть очень мелкие зубы. Рот большой, выдвижной. На жабер-
ных дугах длинные и тонкие жаберные тычинки.
Семейство представлено двумя видами: Polyodon spathula (Wal-
baum, 1792) — веслонос, обитающий в восточной части США, и Pse-
phurus gladius (Marten, 1862) — псефур (китайский веслонос), энде-
мик р. Янцзы и её притоков.
В водоёмах бассейна Кубани обитает один вид — веслонос.
Веслонос — Polyodon spathula (Walbaum, 1792) (рис. 31, 2.10)
Рис. 31. Веслонос — Polyodon spathula (Walbaum, 1792)
Пресноводная рыба Северной Америки. Достигает длины более
2 м и массы свыше 70 кг. Половое созревание наступает у самцов в воз-
расте 6—8, у самок — 7 —10 лет. Нерестится весной при температуре
воды 13—16 °С на песчаном или каменистом грунте. Плодовитость —
от 80 до 200 тыс. икринок. Питается главным образом зоопланктоном.
Американский веслонос завезён в Россию (Краснодарский край) в
1974 г. и выпущен в водоёмах рыборазводного завода Горячий Ключ с
целью акклиматизации (Виноградов, Ерохина, 1976; Мельченков, 1988).
Веслонос — крупная быстрорастущая рыба. При благоприятных усло-
56
виях прирост массы тела за сезон составляет 3—4 кг. В Краснодарском
водохранилище сеголетки достигали массы 50—150 г, двухлетки —
1 500—1 900 г, трёхлетки — 4 100 г (Москул, 1994).
Веслонос как объект пастбищной аквакультуры является перспек-
тивным видом для выращивания в прудах, озёрах и водохранилищах.
Подкласс Новопёрые рыбы — Neopterygii Cope, 1871
Подкласс объединяет группу современных рыб. Тело различной
формы. Хвост равнолопастный. Тело покрыто чешуёй, имеющей фор-
му тонких костных пластинок, или голое. Скелет полностью или поч-
ти полностью окостеневает. Осевой скелет в виде костных позвонков.
Хорда у взрослых особей сохраняется только между позвонками.
Отдел Костистые рыбы — Teleostei Müller, 1845
Костистые включают наибольшее число видов и являются наибо-
лее разнообразной группой из всех позвоночных животных. Костистые
насчитывают 26 840 современных видов (96 % всех современных видов
рыб), объединяющихся в 40 отрядов, 448 семейств и 4 278 родов (Поно-
марев, Баканева, Федоровых, 2016; Абрамчук, Иваненко, 2019).
В водоёмах бассейна Кубани обитают представители, принадле-
жащие к 17 отрядам.
Таблица
для определения отрядов отдела
Костистые рыбы — Teleostei Müller, 1845
1 (15). Спинной плавник один, в нём нет жёстких лучей или их не
более двух.
2 (4). Жирового плавника нет.
3 (12). Брюшные плавники расположены позади грудных. Боковая
линия отсутствует. Килевые чешуйки на брюхе обычно имеются.
Отряд 1. Сельдеобразные — Clupeiformes Bleeker, 1859 (с. 58)
4 (2). Жировой плавник имеется. Чешуя циклоидная, плотно сидя-
щая на теле. Боковая линия полная.
Отряд 2. Лососеобразные — Salmoniformes Bleeker, 1859 (с. 68)
5 (14). Брюшные плавники отсутствуют.
57
6 (18). Спинной и анальный плавники очень длинные и сливаются
с хвостовым. Тело змеевидное.
Отряд 3. Угреобразные — Anguilliformes Goodrich, 1909 (с. 75)
7 (8). Зубы на челюстях отсутствуют. Имеются глоточные зубы.
Рот обычно выдвижной. Тело покрыто циклоидной чешуёй, у некото-
рых голое. Жирового плавника нет. Спинной плавник один.
Отряд 4. Карпообразные — Cypriniformes Rafinesque, 1810
(с. 77)
8 (7). Зубы на челюстях имеются. Рот невыдвижной с усиками.
Тело голое.
Отряд 5. Сомообразные — Siluriformes Cuvier, 1816 (с. 158)
9 (10). Боковой линии нет или она слабо развита. Спинной плавник
один. В плавниках колючих лучей нет. В основном живородящие рыбы.
Отряд 6. Карпозубообразные — Cyprinodontiformes Berg, 1940
(с. 163)
10 (9). Боковая линия проходит на теле ближе к брюху. Рот боль-
шой в виде клюва, с мелкими зубами. Спинной плавник расположен
далеко сзади над анальным.
Отряд 7. Сарганообразные — Beloniformes Berg, 1940 (с. 165)
11 (13). Брюшные плавники обычно расположены на брюхе. Грудные
плавники посажены высоко. Боковой линии нет. Два спинных плавника,
разделённых хорошо заметным промежутком. На верхней части головы
чешуй нет. Вдоль бока тела проходит тёмная или серебристая полоса.
Отряд 8. Атеринообразные — Atheriniformes Rosen, 1966
(с. 168)
12 (3). Брюшные плавники впереди грудных, почти на горле. На
подбородке обычно один усик. Спинных плавников обычно три, ино-
гда два или один, анальных — два или один.
Отряд 9. Трескообразные — Gadiformes Goodrich, 1909 (с. 170)
13 (11). Брюшные плавники на груди или их нет, если они рас-
положены на груди, то имеют вид колючки. Тело голое или покрыто
костными пластинками. Рыло имеет трубковидную форму.
Отряд 10. Колюшкообразные — Gasterosteiformes Greenwood,
1966 (с. 171)
14 (5). Брюшные плавники заметно отодвинуты назад от груди. Спин-
ных плавников два; они короткие и разделены значительным промежутком;
первый состоит из 4 колючих неветвистых лучей, второй — из мягких ветви-
стых лучей. Боковой линии нет. Тело и голова покрыты циклоидной чешуёй.
58
Отряд 11. Кефалеобразные — Mugiliformes Günther, 1880 (с. 177)
15 (1). Два спинных плавника соединены перепонкой. Первый со-
стоит из колючих или гибких неветвистых лучей. Если плавники сли-
ты, то передняя часть имеет колючие или гибкие неветвистые лучи.
16 (17). Брюшные плавники расположены под основанием груд-
ных или впереди них — на горле, не срастаются между собой.
Отряд 12. Окунеобразные — Perciformes Bleeker, 1859 (с. 187)
17 (16). Брюшные плавники расположены под основанием груд-
ных или впереди них — на горле, срастаются между собой, образуя
воронку (присасывательный диск).
Отряд 13. Бычкообразные — Gobiiformes Günther, 1880 (с. 202)
18 (6). Спинной и анальный плавники длинные. Спинной заходит
на голову. Тело сильно сжато с боков, высокое, несимметричное. Глаза
расположены на одной стороне.
Отряд 14. Камбалообразные — Pleuronectiformes Bleeker, 1859
(с. 220)
19 (20). Спинной плавник сдвинут далеко назад, расположен над
анальным плавником. Тело стреловидной формы, покрытое мелкой че-
шуёй. Рот большой, с крупными крепкими зубами.
Отряд 15. Щукообразные — Esociformes Bleeker, 1859 (с. 73)
20 (19). Тело покрыто костными кольцами. Рыло удлинённое в
виде трубки. Рот маленький, обращён вверх, беззубый.
Отряд 16. Иглообразные — Syngnathiformes Helfman, Col-
lette & Facey, 1997 (с. 174)
21 (22). Тело высокое сжатое с боков. Рот большой, губы вздутые.
22 (21). Спинной плавник один, состоит из передних жёстких и
задних мягких лучей.
Отряд 17. Цихлидообразные — Cichliformes Betancur-R et al.,
2013 (с. 183)
Отряд Сельдеобразные — Clupeiformes Bleeker, 1959
Рыбы среднего и мелкого размера, с сжатым с боков телом. Хвосто-
вой плавник гомоцеркальный. Фулькр нет. Чешуя циклоидная, легко спа-
дает. В плавниках нет жёстких лучей. Это примитивные костистые рыбы.
Плавательный пузырь у них соединён каналом с пищеводом. Нет Веберо-
ва аппарата. Обитатели пресных и морских вод. Есть и проходные рыбы.
В водоёмах бассейна Кубани встречаются представители двух семейств.
59
Таблица
для определения семейств отряда
Сельдеобразные — Clupeiformes Bleeker, 1959
1 (2). Брюхо, сжатое с боков, снабжено шипообразными чешуй-
ками от горла до анального отверстия, хорошо заметны в виде пилки.
Киль выражен сильно или слабо, но всегда имеется.
Семейство Сельдевые — Clupeidae Cuvier, 1816 (с. 59)
2 (1). Брюхо закруглённое, костных щитков вдоль его края нет.
Киля на брюхе нет. Рот нижний, очень большой, задний конец верхне-
челюстной кости заходит далеко за вертикаль заднего края глаза, дос-
тигая предкрышечной кости.
Семейство Анчоусовые — Engraulidae Gill, 1861 [Engraulidi-
dae] (с. 67)
Семейство Сельдевые — Clupeidae Cuvier, 1816
Брюхо с килевыми чешуйками. Небольшой спинной плавник распо-
ложен над брюшными плавниками. Грудные плавники расположены за
жаберной крышкой. Хвостовой плавник сильно выемчатый. Голова не по-
крыта чешуёй. Боковой линии нет. Усиков нет. Зубы мелкие, нередко отсут-
ствуют. Рот невыдвижной, конечный или верхний, большой или маленький.
Жирового плавника нет. Сельдевые в основном морские рыбы, отдельные
виды постоянно живут в пресных водоёмах, а некоторые виды — проход-
ные. Сельдевые питаются планктонными беспозвоночными. Некоторые во
взрослом состоянии питаются мелкой рыбой. У большинства рыб икра пе-
лагическая, но отдельные виды откладывают донную икру.
Сельдевые — важнейшие объекты промысла. В бассейне Кубани
семейство представлено двумя родами.
Таблица
для определения родов семейства
Сельдевые — Clupeidae Cuvier, 1816
1 (2). При основании лопастей хвостового плавника есть удлинённые
крыловидные чешуйки (alae) (см. рис. 3). На верхней челюсти есть медиаль-
ная вырезка. Удлинённых последних двух лучей анального плавника нет.
Род Алозы — Alosa Linck, 1790 (с. 60)
60
2 (1). Удлинённых чешуй (alae) при ос-
новании лопастей хвостового плавника нет.
Верхняя челюсть без медиальной вырезки.
Последние два луча анального плавника уд-
линены (рис. 32).
Род Тюльки, или Каспийские киль-
ки, — Clupeonella Kessler, 1877 (с. 64)
Род Алозы — Alosa Linck, 1790
Крупные и средних размеров сельди, отличающиеся от близких
родов большим ртом и зубами на сошнике. На верхней челюсти есть
медиальная вырезка. На глазах хорошо развито жировое веко. На зад-
нем крае хвостового стебля с каждой стороны по две удлинённые че-
шуйки (alae), налегающие на основание хвостового плавника. Брюхо
сжатое с боков; на всём протяжении от горла до начала анального плав-
ника покрыто килевыми чешуйками. Размеры взрослых рыб — более
30 см. Алозы — проходные рыбы, в реке не питаются. Часть особей
после нереста гибнет. Нерест порционный. Икра у всех сельдей этого
рода — пелагическая. По характеру питания алозы весьма разнообраз-
ны. Одни виды ведут хищный образ жизни, другие питаются пелагиче-
скими беспозвоночными.
Алозы распространены в прибрежных водах Атлантического океа-
на, а также в Средиземном, Чёрном, Азовском и Каспийском морях.
Алозы имеют большое хозяйственное значение.
Род представлен 10 видами, среди них имеются морские, проход-
ные и полупроходные сельди.
В водоёмах бассейна Кубани встречаются представители трёх видов.
Таблица
для определения видов рода
Алозы — Alosa Linck, 1790
1 (4). Зубы на челюстях развиты хорошо.
2 (3). Жаберных тычинок на первой дуге 27—39, они недлинные,
короче жаберных лепестков (рис. 33).
Черноморско-азовская морская, или керченская, сельдь — Alosa
maeotica (Grimm, 1901) (с. 61)
Рис. 32. Анальный плавник
Sardinella
aurata
с
удлинёнными лучами
61
3 (2). Жаберных тычинок на первой дуге
40—73. Глаза большие, тело прогонистое. На
челюстях есть зубы.
Черноморско-азовская
проходная
сельдь — Alosa immaculata (Bennett, 1835)
[Alosa pontica (Eichwald, 1838)] (с. 62)
4 (1). На нижней челюсти нет зубов. Жа-
берных тычинок на первой дуге 65—78; они
длинные, тонкие (длиннее жаберных лепест-
ков). На глазах имеется жировое веко.
Азово-черноморский пузанок — Alosa
tanaica (Grimm, 1901) (с. 63)
Краткая биологическая характеристика видов рода
Алозы — Alosa Linck, 1790
Черноморско-азовская морская, или керченская, сельдь — Alosa
maeotica (Grimm, 1901) (рис. 34, 2.11)
Рис. 34. Черноморско-азовская морская, или керченская, сельдь — Alosa
maeotica (Grimm, 1901)
D III—IV 13—15, A II—III 16—20. Брюшных шипов 30—36. По-
звонков 48—52. На челюстях есть зубы. Длина тела — до 40 см. Про-
должительность жизни — до 6 лет. Боковой линии нет.
Крупная сельдь с толстым и мясистым телом. Спина и голова ок-
рашены в тёмно-фиолетовый цвет. Грудные плавники короткие, зубы
хорошо развиты. Жаберных тычинок 27—39, они недлинные, короче
жаберных лепестков, крепкие, толстые.
Рис. 33. Жаберные тычинки
(а) и жаберные лепестки
(б) первой жаберной дуги
Alosa maeotica
62
Размножается в море. Нагуливается как в море, так и в ку-
банских лиманах. По Кубани черноморская сельдь в небольшом
количестве поднимается до Краснодарского водохранилища. По-
ловое созревание наступает не ранее трёхлетнего возраста. Пло-
довитость от 20 до 250 тыс. икринок. Икра крупная, полупела-
гическая. Развитие икринок в зависимости от температуры воды
длится 42—56 ч.
Черноморско-азовская морская сельдь — хищник. Питается мел-
кой рыбой (килька, атерина, бычки). Очень жирная — 11 —18 % жира.
Морская стайная рыба, ведёт пелагический образ жизни. На зиму (но-
ябрь—декабрь) уходит из Азовского в Чёрное море, возвращается об-
ратно в марте.
Черноморско-азовская проходная сельдь — Alosa immaculata (Ben-
nett, 1835) [син. Alosa pontica (Eichwald, 1838)] (рис. 35, 2.12)
Рис. 35. Черноморско-азовская проходная сельдь — Alosa immaculata
(Bennett, 1835)
D III 13—14, A II—III 16—20. Брюшных шипов 31—35, позвонков
48—53. Жаберных тычинок на первой дуге 40—73, чаще всего 46—55.
Глаза большие, тело прогонистое. На челюстях есть зубы.
Чешуя слабая, легко спадает. Тело окрашено слабо, спина светло-
зеленоватая. Бока серебристые. Голова относительно длинная, в сред-
нем 25 % длины тела. Рот крупный, верхняя челюсть протягивается за
вертикаль середины глаза. На обеих челюстях зубы хорошо развиты и
заметны на ощупь. За жаберными крышками имеется по одному тёмно-
му пятну. Азовская сельдь достигает 27—30 см. Максимальная зареги-
стрированная длина 39 см.
63
Проходная рыба, для икрометания заходит в реки. Нагуливается
как в море, так и в кубанских лиманах, иногда заходит в Кубань и Крас-
нодарское водохранилище. Питается мелкой рыбой и беспозвоночны-
ми. Имеет промысловое значение. Однако уловы невелики.
В Чёрном и Азовском морях черноморско-азовская проходная
сельдьобитаетмежду50°с.ш.и41°с.ш.имежду27°в.д.и44°в.д.
Здесь данный вид является объектом коммерческого промысла.
Встречается на глубине до 90 м.
Азово-черноморский пузанок — Alosa tanaica (Grimm, 1901)
(рис. 36, 2.13)
Рис. 36. Азово-черноморский пузанок — Alosa tanaica (Grimm, 1901)
D III—IV 12—15, A III 16—19. Позвонков 47—51. Тело высокое,
укороченное в хвостовой области. Грудные плавники длинные. На
нижней челюсти нет зубов. Жаберных тычинок на первой дуге 65—78;
они длинные, тонкие (длиннее жаберных лепестков). На глазах имеет-
ся жировое веко. За жаберной крышкой имеются (до 8) тёмные пятна
вдоль тела (см. рис. 2.13).
Азово-черноморский пузанок не достигает больших размеров. Его
длина не превышает 20 см, обычно 14—16 см. Питается планктоном,
ракообразными (мизиды, гаммариды).
Полупроходная рыба, размножающаяся в низовьях рек. Пузанок
в апреле—мае для икрометания заходит как в лиманы, так и в Кубань,
достигая Краснодарского водохранилища. После нереста пузанок ска-
тывается в море, нагуливается и ранней осенью (сентябрь) мигрирует
64
на зимовку в Чёрное море. Половозрелым становится в два года, иногда
к концу первого года жизни. Нерестовое стадо состоит из 2—4 -годова-
лых особей. Нерест проходит в апреле—июне. Плодовитость — 10—
40 тыс. икринок.
Вид может рассматриваться как подвид черноморско-каспийского
пузанка (Alosa caspia tanaica (Grimm, 1901)) (Парин, Евсеенко, Василь-
ева, 2014).
Род Тюльки,
или Каспийские кильки, — Clupeonella Kessler, 1877
Рот верхний, беззубый, небольшой. Верхняя челюсть доходит до
вертикали переднего края глаза. На середине верхней челюсти меди-
альной вырезки нет. Последние два луча анального плавника удлине-
ны. Удлинённые чешуи у основания хвостового плавника отсутствуют.
Есть киль от горла до анального отверстия. Тюльки — эвригалинные и
эвритермные рыбы, живущие как в морской, солоноватой, так и в пре-
сной воде при температуре от 0 до 30 °С. Нерест порционный. Икра пе-
лагическая. Распространены в Чёрном, Азовском и Каспийском морях
и их бассейнах. Род содержит 4 вида, 2 из них встречаются в водоёмах
бассейна Кубани и причерноморских реках.
Представители рода Clupeonella — объекты питания хищных
сельдей, судака, осетровых и других рыб, а также являются объектами
промысла.
Таблица
для определения видов рода
Тюльки, или Каспийские кильки, — Clupeonella Kessler, 1877
1 (2). Высота тела — 20,5—24,7 % его длины. Жаберных тычинок
52—64.
Черноморско-азовская тюлька — Clupeonella cultriventris (Nord-
mann, 1840) (с. 65)
2 (1). Высота тела — 14 —20,5 % его длины. Брюшных шипов 24—
27. Жаберных тычинок 38—50. Эндемик оз. Абрау.
Абрауская тюлька [сарделька] — Clupeonella abrau (Maliatsky,
1930) (с. 66)
65
Краткая биологическая характеристика видов рода
Тюльки, или Кильки, — Clupeonella Kessler, 1877
Черноморско-азовская тюлька — Clupeonella cultriventris
(Nordmann, 1840) (рис. 37, 2.14)
Рис. 37. Черноморско-азовская тюлька — Clupeonella cultriventris (Nordmann,
1840)
D III—IV 11—13, А III 14—18. Брюшных шипов 24—30. Тело
сравнительно высокое, наибольшая высота его составляет 20,5—24,7 %
длины тела. Голова небольшая. Жаберных тычинок на первой дуге от
52 до 64, в среднем — 57,8. Глаза небольшие. Рот верхний, нижняя
челюсть выдаётся вперёд. Чешуя крупная, легко опадающая, на брю-
хе образует хорошо выраженный киль из килевых чешуек (24—30).
Окраска типично пелагическая. Спина и верхняя часть головы более
тёмные, светло-зелёного или голубоватого оттенков, бока и брюшко
серебристо-белые. Радужина глаз чёрная.
Рыба стайная, держится в толще воды. Пелагическая, эвригалин-
ная рыба: встречается как в опреснённых участках, так и в солёных;
входит в дельты Кубани, Протоки, встречается в кубанских лиманах,
отмечена в Краснодарском водохранилище (Москул, 1994).
Половой зрелости достигает в возрасте одного года при достиже-
нии длины 4,5—5,0 см. Икра пелагическая, мелкая. Инкубационный
период при температуре воды 9 °С — 4 сут. Икрометание порционное.
Общая плодовитость колеблется от 4 до 20 тыс. икринок, в среднем —
11 тыс. Нерестится как в море, так и в лиманах и в пресной воде.
Питается в основном пелагическими ракообразными. Тюлька яв-
ляется объектом промысла в Азовском и Чёрном морях. Уловы значи-
66
тельно колеблются по годам: в 1986 г. — 15 тыс. т; 1990 г. — 133 т;
2009г. — 1,6тыс.т;2012г. — 468тыс.т.
Абрауская тюлька [сарделька] — Clupeonella abrau (Maliatsky,
1930) (рис. 38, 2.15)
Рис. 38. Абрауская тюлька [сарделька] — Clupeonella abrau (Maliatsky, 1930)
D III 11—12, А II—III 17—19. Брюшных шипов 24—27. Жаберных
тычинок 38—50, позвонков 41—42.
Абрауская тюлька отличается низким телом: его высота в среднем
составляет 16,3 % от длины (до конца средних лучей С), длина головы к
длине тела — 24,3 %, высота головы — 15,9 %. Диаметр глаза составля-
ет в среднем 29,8 % (25,8—33,6 %) длины головы. Длина тела до конца
средних лучей хвостового плавника — 8,5 см, масса — до 10 г.
Абрауская тюлька — эндемик оз. Абрау (недалеко от г. Ново-
российска). Пелагический пресноводный вид, питающийся планктон-
ными рачками и водорослями. Ведёт преимущественно ночной образ
жизни.
Половая зрелость наступает на втором году жизни при длине
3,5—4,5 см. Икрометание с мая по октябрь. Икра пелагическая. Нерест
происходит вечером при температуре воды 21—22 °С, выклев личи-
нок наблюдается уже утром. Инкубационный период длится 10—12 ч.
Выклюнувшиеся личинки опускаются на дно, избегая поверхностного
волнения воды. После рассасывания желточного мешка они переходят
на внешнее питание и держатся в 2—3 м от поверхности воды.
Внесена в Красный список МСОП, Красную книгу РФ (2001) (ка-
тегория 4 — неопределённый по статусу узкоареальный вид), Красную
книгу Краснодарского края (2007, 2017).
67
Семейство Анчоусовые — Engraulidae Gill, 1861
[Engraulididae]
Брюшко закруглённое. Рот нижний, большой. Задний конец верх-
ней челюсти заходит далеко за глаз. Рыло выступает над нижней че-
люстью. Стайные пелагические, морские рыбы. К семейству относят-
ся представители 16 родов и 139 видов. В водах бассейнов Чёрного и
Азовского морей встречаются представители рода Анчоусы — Engrau-
lis Cuvier, 1816.
Род Анчоусы — Engraulis Cuvier, 1816
Тело удлинённое, не сжатое с боков. Брюхо закруглённое, брюш-
ные килевые чешуи развиты слабо. Род включает 6 видов. В водах бас-
сейнов Чёрного и Азовского морей встречается один вид — европей-
ский анчоус, или хамса (Engraulis encrasicolus (Linnaeus, 1758)).
Европейский анчоус, или хамса, — Engraulis encrasicolus
(Linnaeus, 1758) (рис. 39, 2.16)
Рис. 39. Европейский анчоус, или хамса, — Engraulis encrasicolus (Linnaeus,
1758)
Европейский анчоус в пределах ареала образует ряд подвидов
(географических рас), отличающихся между собой в основном длиной
тела.
Черноморский анчоус [черноморскаяхамса]—Engraulis
encrasicolus ponticus (Aleksandrov, 1927). Морская стайная пелагиче-
ская рыба. Населяет Чёрное море, длина тела — 12 —14,5 см.
68
Азовский анчоус [азовскаяхамса]—Engraulisencrasicolus
maeoticus (Pusanov, 1926). Населяет Азовское море и восточную часть
Чёрного, длина тела — 10—12 см.
Как черноморский, так и азовский анчоус зимует в Чёрном море в
местах с температурой воды около 8 °С. В начале весны начинается ми-
грационный путь к местам нагула. Черноморская хамса подходит к за-
падному и северному побережью Чёрного моря. Азовская хамса подхо-
дит к восточному побережью, а затем движется к Керченскому проливу.
Войдя в Азовское море, хамса усиленно питается до начала икрометания.
Половой зрелости хамса достигает в возрасте одного года. Нерест
азовской хамсы очень растянут. Икрометание порционное. В Азовском
море нерест происходит почти по всей площади моря с мая по август.
Плодовитость азовской хамсы составляет 10—13 тыс. икринок. Икрин-
ки пелагические. Инкубационный период при температуре воды 15 °С —
65 ч. Растёт хамса быстро. Годовики имеют длину 8,5 см, двухлетки —
9,5 см, трёхгодовики — 10—12 см. Продолжительность жизни не более
3—4 лет. Питается хамса беспозвоночными. Азовская хамса отличается
высокой жирностью. Осенью содержание жира колеблется от 20 до 28 %.
Хамса является также объектом питания хищных видов рыб.
Как в Чёрном, так и в Азовском море хамса является важнейшим
объектом промысла. Уловы значительно колеблются по годам. Так в
1987 г. было выловлено 17,3 тыс. т, а в 1991 г. лишь 6 т, в 1995 г. —
25,2тыс.т,в2009г. — 325тыс.т,в2010г. — 525тыс.т,в2012г. —
82 тыс. т.
Отряд Лососеобразные — Salmoniformes Bleeker, 1859
Лососеобразные — открытопузырные, мягкопёрые рыбы различ-
ных размеров. Это проходные и пресноводные рыбы умеренных вод.
Распространены в северных и умеренных зонах Европы, Азии и Се-
верной Америки. Спинной плавник один, но позади его расположен
жировой плавничок. Боковая линия есть. Чешуя циклоидная, крепко
сидящая на теле. Брюшные плавники расположены абдоминально (по-
середине брюха). В плавниках нет колючих лучей. Являются важными
объектами аквакультуры, спортивного и промышленного рыболовства.
Отряд включает три семейства. В водоёмах бассейна Кубани и
причерноморских реках обитают представители одного семейства —
Лососёвые (Salmonidae).
69
Семейство Лососёвые — Salmonidae Cuvier, 1816
Рыбы крупного и среднего размера. Тело сжато с боков. Наиболее
высокий участок тела лежит перед спинным плавником. Боковая линия
полная. Она тянется от головы до основания хвостового плавника. Че-
шуя относительно мелкая, прочно сидящая в коже. Спинной плавник
короткий, содержит не более 17 лучей. За спинным плавником на спине
имеется небольшой жировой плавник. В анальном плавнике 7—19 мяг-
ких лучей. Брюшные плавники лежат под спинным. Морские, проход-
ные и пресноводные рыбы, населяют в основном холодные водоёмы.
Имеют большое промысловое значение. Являются важными объектами
аквакультуры и спортивного рыболовства.
Семейство включает 10 родов и 223 вида. В водах бассейна Кубани
встречаются представители двух родов: (Настоящие) Лососи — Salmo
Linnaeus, 1758 и Тихоокеанские лососи — Oncorhynchus Suckley, 1861.
Таблица
для определения родов семейства
Лососёвые — Salmonidae Cuvier, 1816
1 (2). Чешуя мелкая, в боковой линии 110—120 чешуй. В анальном
плавнике 7—10 мягких ветвистых лучей.
Род (Настоящие) Лососи — Salmo Linnaeus, 1758 (с. 69)
2 (1). Чешуя мелкая, в боковой линии 135—150 чешуй. В анальном
плавнике 10—16 мягких ветвистых лучей.
Род Тихоокеанские лососи — Oncorhynchus Suckley, 1861 (с. 72)
Род (Настоящие) Лососи — Salmo Linnaeus, 1758
Проходные и пресноводные рыбы, отличающиеся от других родов
семейства лососёвых большим ртом, сравнительно мелкой чешуёй, ко-
ротким анальным плавником, длинным сошником.
Рыбы различных размеров с вальковатым или уплощённым с бо-
ков телом. Спинной плавник один, но позади его расположен жировой
плавник. Брюшные плавники лежат посередине брюха. В плавниках
нет колючих лучей. Морские и пресноводные рыбы населяют в основ-
ном холодные водоёмы. Жилые формы этого рода служат объектом ис-
кусственного разведения в холодных прудах и бассейнах.
70
Образует несколько подвидов, в России встречается 4, и они со-
гласно международной таксономии (Нельсон, 2009) выделены в от-
дельные виды, из которых в причерноморских реках Краснодарского
края встречается один вид — Salmo labrax Pallas, 1814 [Salmo trutta
labrax Pallas, 1814] — черноморский лосось [черноморская кумжа].
Черноморский лосось [черноморская кумжа] — Salmo labrax
[Salmo trutta labrax] Pallas, 1814 (рис. 40, 2.17)
Рис. 40. Черноморский лосось [черноморская кумжа] — Salmo labrax [Salmo
trutta labrax] Pallas, 1814
D IV 9—11, А III 7—9, l.l. 115—132. Жаберных тычинок 10—12.
Верхняя челюсть заходит за вертикаль заднего края глаза. Хвостовой
плавник имеет слабую выемку. На боках головы крупные тёмные пятна.
Черноморский лосось относится к проходным рыбам, обитает в
море, на нерест заходит в черноморские реки (Мзымта, Шахе, Псоу и др.).
В Азовском море встречаются единичные экземпляры. С. К. Троицкий
(1948) отмечает, что в 1920—1930-е гг. лосось встречался в прибрежных
участках Азовского моря в районе Ачуево. А. Н. Световидов (1964) отме-
чает также, что лосось обитал в Кубани и Лабе. М. Х. Емтыль, Г. К. Плот-
ников, Ю. И. Абаев (1988) приводят сведения о присутствии черноморско-
го лосося в среднем и нижнем течении Кубани и её притоках. В результате
многолетних исследований (1970—2012 гг.), проведённых нами в рамках
научно-исследовательских работ Краснодарского НИИ рыбного хозяйст-
ва (КрасНИИРХ) на водоёмах бассейна Кубани (р. Кубань с левобереж-
ными притоками, водохранилища, озёра, лиманы и др.), данный вид не
был обнаружен. Но несмотря на это, мы приводим краткую биологиче-
скую характеристику этого вида по литературным данным.
71
Половозрелым лосось становится на четвёртом году жизни. Пло-
довитость небольшая — 4 —7 тыс. икринок. Икра крупная (3,5—5,1 мм
в диаметре), жёлтого или розового цвета, прозрачная. Преднерестовый
ход в реки начинается весной (апрель—май), лето проводит в реках, не-
рестится зимой. Икру, подобно всем лососёвым, закапывает в галечни-
ковый грунт, гнездо выкапывает самец. Продолжительность развития
икры — 50—120 дней. Личинки некоторое время находятся в гнёздах,
затем покидают свои убежища и живут в реках около двух лет, потом
скатываются в море. Питается в реках личинками комаров, водными
и воздушными насекомыми, а в море — рыбой. Растёт лосось доволь-
но быстро. К концу третьего года жизни достигает длины 30—40 см
,
массы — 1 —3 кг. Половозрелые особи весят 3—5 кг, имея длину 50—
70 см. Лосось — ценная промысловая рыба. Вследствие малочислен-
ности лов лосося запрещён. Вид внесён в Красную книгу МСОП, РФ и
Краснодарского края (2017).
Черноморский лосось [черноморская кумжа], попав в реки и не
имея возможности вернуться в море, остаётся в пресной воде и превра-
щается в форель, которая никогда не достигает такой величины, как
кумжа.
Пресноводные формы лосося — форели постоянно живут в пре-
сной воде, предпочитают горные реки с холодной водой и быстрым те-
чением.
В водоёмах бассейна Кубани встречается одна форма форелей.
Ручьевая форель — Salmo labrax morpha fario Linnaeus, 1758
(рис. 41, 2.18)
Рис. 41. Ручьевая форель — Salmo labrax morpha fario Linnaeus, 1758
72
D III—V 9—11, A II—IV 7—9, l.l. 115—132. Чешуя мелкая. На
теле и на спинном плавнике имеются чёрные и красные пятна (послед-
ние со светлым ободком). Верхняя челюсть у взрослых рыб заходит
за вертикаль заднего края глаз. Живут в ручьях. Часто её называют
пеструшкой, потому что тело её покрыто красными, чёрными и белыми
пятнышками. Форель — мелкая рыба. Её обычные размеры находятся
в пределах 25—35 см и 200—500 г. Но известны и более крупные эк-
земпляры. Л. П. Сабанеев пишет, что в притоках Кубани встречалась
форель массой до 2 кг. Ручьевая форель очень требовательна к высоко-
му содержанию в воде кислорода, снижение до 3,5—4,0 мг/л является
критическим. Питается она в течение всего года, но наиболее усиленно
при температуре воды 15—17 °С воздушными насекомыми и их личин-
ками, жуками и рыбой.
Встречается форель во всех горных реках бассейна Кубани и их
притоках. Нерестится в осенне-зимний период. Икра крупная, оран-
жевого цвета. Биология ручьевой форели близка к лососю и от него
отличается замедленным темпом роста, меньшими размерами и пло-
довитостью. Промыслового значения в наших реках не имеет, в основ-
ном — объект любительского рыболовства.
Род Тихоокеанские лососи — Oncorhynchus Suckley, 1861
У всех представителей рода Тихоокеанские лососи — Oncorhyn-
chus в анальном плавнике 10—16 ветвистых лучей. Чешуя мелкая в
боковой линии 135—150 чешуй. Рот большой конечный. Зубы на че-
люстях хорошо развиты. Все виды проходные, нагуливаются в море,
а нерестится возвращаются в родные реки. Хоминг развит хорошо.
Нерест происходит осенью, литофилы закапывают икру в грунт. Не-
рестятся один раз в жизни, после нереста погибают. Питаются мелкой
рыбой и ракообразными. Род включает 6 видов. В Краснодарском крае
акклиматизирован один вид — микижа, или радужная форель, — Onco-
rhynchus mykiss (Walbaum, 1792) [= Parasalmo gairdnerii Richardson,
1836]. Радужная форель является пресноводной формой стальноголо-
вого лосося (Parasalmo [Salmo] gairdnerii Richardson, 1836).
Стальноголовый лосось — обитатель побережья Тихого океана от
Аляски до Калифорнии. Акклиматизирован в бассейне Чёрного моря.
По форме тела напоминает форель. Верхняя челюсть заходит за верти-
каль заднего края глаза. На теле многочисленные тёмные пятна. Голо-
73
ва небольшая. В спинном плавнике 11—12 лучей. Жаберных тычинок
16—22. Достигает длины более 100 см. Проходная рыба, образует жи-
лые формы. Нерестится в конце зимы, весной. Питается ракообразны-
ми и мелкой рыбой.
Радужная форель — Oncorhynchus mykiss (Walbaum, 1792) (рис. 42,
2.19)
Рис. 42. Радужная форель — Oncorhynchus mykiss (Walbaum, 1792)
D III—IV 9—12, A II—IV 10—16, l.l. 135—150. Жаберных тычи-
нок 16—22, позвонков 60—65. На боках тела вдоль боковой линии тя-
нется широкая радужная полоса. На теле выше и ниже боковой линии,
на спинном и анальном плавниках много тёмных пятен. Они есть на
спинном, жировом и хвостовом плавниках (см. рис. 2.19).
Радужная форель является ценным объектом прудового и садково-
го рыбоводства. Разработана методика выращивания радужной форели
в прудах и садках, получения икры и молоди в искусственных услови-
ях.
В Краснодарском крае большую известность получило специали-
зированное Адлерское форелевое хозяйство, которое расположено в
15 км от г. Адлера на р. Мзымте. Производственная мощность — 160 т
форели в год.
Отряд Щукообразные — Esociformes Bleeker, 1859
Тело удлинённое, голова большая с вытянутым и сплющенным
рылом. Рот большой. Брюшные плавники расположены далеко поза-
ди грудных, на середине брюха. Спинной плавник сильно отодвинут
74
назад и находится над анальным. Жирового плавника нет. Боковой ли-
нии нет. Рыло заострённое, челюсти большие, на нижней челюсти ряд
крупных зубов.
В водоёмах бассейна Кубани обитают представители одного се-
мейства.
Семейство Щуковые — Esocidae Cuvier, 1816
Рыло вытянутое, нижняя челюсть немного выдаётся вперёд. Тело
стреловидной формы, покрытое мелкой плотно сидящей чешуёй. Голо-
ва большая, с длинным сплющенным рылом и большим количеством
конусообразных острых зубов. Жаберные перепонки не приращены к
межжаберному промежутку и не сращены между собой. Окраска тела
очень изменчива по цвету в зависимости от среды обитания. Основной
цвет серо-зеленоватый или серо-бурый. Бока покрыты крупными пят-
нами бурого, оливкового или черноватого цвета, иногда образующи-
ми поперечные полосы. На спинном, хвостовом и анальном плавниках
тёмные пятна.
Семейство представлено одним видом.
Обыкновенная щука — Esox lucius Linnaeus, 1758 (рис. 43, 2.20)
Рис. 43. Обыкновенная щука — Esox lucius Linnaeus, 1758
D IV—X 13—16, A IV—VII 10—13, l.l. 105—144. Жаберные ты-
чинки короткие и толстые, их обычно 29—45. Рыло вытянутое, упло-
щённое. Длина рта примерно равна 1⁄2 длины головы. Хвост выемча-
тый. Рот очень велик, челюсти усажены длинными и острыми зубами.
На теле обычно светлые пятна. Щёки покрыты чешуёй, крышечная
кость — лишь в верхней части (см. рис. 2.20).
Щука пресноводная рыба, не совершает больших передвижений.
75
Держится обычно среди зарослей, предпочитая стоячие или малопро-
точные воды. Встречается практически во всех водоёмах бассейна
Кубани, кроме горных рек и озёр. Растёт щука быстро при богатой
кормовой базе. Годовалые щуки весят 150—200 г, двухгодовалые —
800—1 000 г. Это одна из самых долгоживущих рыб: некоторые эк-
земпляры доживают до 40—50 лет и имеют массу более 35 кг, но чаще
всего встречаются особи в возрасте 10—12 лет массой 8—10 кг при
длине 80—90 см.
Половое созревание наступает на 2—3-м году жизни. Нерестится
ранней весной при температуре воды 5—7 °С на глубине 25—60 см. Н е -
рест шумный. Нерестовый субстрат — прошлогодняя растительность.
Икра крупная (2,1 мм), клейкая, желтоватого цвета. Плодовитость —
50—320 тыс. икринок. Период инкубации продолжается 8—14 дней.
Личинки питаются зоопланктоном, по достижении 1,2—1,5 см пере-
ходят на хищный образ жизни. Взрослая щука ведёт хищный образ
жизни. Питается в основном массовыми видами (плотва, карась, лещ,
окунь и др.). Подкарауливает добычу в зарослях. Щука — промысловая
рыба, а также объект любительского рыболовства.
Отряд Угреобразные — Anguilliformes Goodrich, 1909
К этому отряду относятся рыбы с очень характерной змеевидной
формой тела. Жаберные щели маленькие. Тело покрыто циклоидной
чешуёй. В боковой линии до 110 отверстий. Брюшных плавников нет.
Спинной и анальный плавники длинные и сливаются с хвостовым. По-
звонков у некоторых видов до 260.
Развитие происходит с метаморфозом.
Отряд Угреобразные объединяет представителей 15 семейств, 141
род и 791 вид. Угреобразные обитают преимущественно в тёплых мо-
рях. В пресных водах встречаются представители только одного семей-
ства.
Семейство Угрёвые, или Пресноводные угри, —
Anguillidae rafinesque, 1810
Тело покрыто мелкой, погружённой в кожу чешуёй. Спинной
плавник начинается далеко позади грудных плавников. Грудные плав-
ники хорошо развиты. В семействе один род и 15 видов. В водоёмах
76
бассейна Кубани данное семейство представлено одним видом из рода
Речные угри — Anguilla Schrank, 1798.
Речной угорь — Anguilla anguilla (Linnaeus, 1758) (рис. 44, 2.21)
Рис. 44. Речной угорь — Anguilla anguilla (Linnaeus, 1758)
Тело почти цилиндрическое, в задней части сжатое с боков. Окра-
ска без пятен. Спинной и анальный плавники слиты с хвостовым. Спин-
ной плавник начинается далеко позади грудных плавников. Грудные
плавники хорошо развиты. Брюшных плавников нет. Глаза маленькие,
расположены над задним концом рта. Рот конечный. Зубы небольшие.
Позвонков 110—120. Колючек в плавниках нет. Чешуя мелкая, цикло-
идная, скрытая в коже. В боковой линии до 110 отверстий. Достигает
2,0 м длины и 6 кг массы. Единственный вид в фауне бассейна Куба-
ни, который нагуливается в пресной воде, а для икрометания уходит в
море, т. е. совершает катадромные миграции.
Угорь нерестится в 8 000 км от мест нагула в центральной части
Атлантического океана в Саргассовом море между Бермудскими и Ба-
гамскими островами на глубине 300—400 м (иногда до 1 000 м) при
температуре воды не менее 7 °С и солёности не менее 35 ‰, после чего
погибает. Выклюнувшиеся личинки (лептоцефалы) поднимаются в по-
верхностные слои воды и пассивно переносятся к берегам Западной
Европы. Миграция длится до 2—3 лет. За это время молодь достига-
ет длины 7,5 см. Затем молодь приобретает цилиндрическую форму и
входит в устья рек и озёр. Питается молодь в основном зоопланктоном
и фитопланктоном. На 3—4 -м году жизни, в основном при обитании в
пресных водах, угри начинают хищничать. В это время угорь наиболее
интенсивно растёт, достигая массы 4—6 кг при длине тела до 2 м.
77
Угорь ведёт ночной образ жизни, днём он находится в убежище или
зарывается в ил. Сезон нагула длится с апреля по ноябрь, зимой не пита-
ется. В море угорь скатывается в возрасте 6—9 лет и мигрирует на места
нереста для икрометания. В это время у угрей меняется окраска, рыло
вытягивается, глаза заметно увеличиваются; весь этот метаморфоз длит-
ся до года и более. Уходит из реки угорь очень упитанным, но в море пи-
тание прекращается и рыба теряет массу. После нереста угорь погибает.
Ценный промысловый вид. Промысловая масса угрей обычно ко-
леблется от 500 до 2 000 г.
В водоёмах бассейна Кубани речной угорь встречается изредка в еди-
ничных экземплярах в Кубани, на рисовых чеках, в приазовских лиманах.
Вид внесён в Красную книгу Краснодарского края (2017).
Отряд Карпообразные — Cypriniformes Rafinesque, 1810
В отряде 13 семейств, 489 родов и около 4 205 видов (Абрамчук,
Иваненко, 2019). На основе исследований ДНК восстановлен статус се-
мейств Nemacheilidae и Vaillantellidae, а также описаны новые Serpenti-
cobitidae, Ellopostomatidae и Barbuccidae (Романов, 2019). Внешний вид
рыб этого отряда очень разнообразен. Но у всех представителей отряда
имеются общие признаки. Все они имеют Веберов аппарат (модифи-
кация первых четырёх передних позвонков, соединяющих плаватель-
ный пузырь с внутренним ухом для передачи звука), который служит
в качестве органа восприятия давления окружающей среды. Брюшные
плавники расположены далеко за грудными, посередине брюха, зани-
мают абдоминальное положение. В спинном и анальном плавниках
имеется по 2—4 неветвистых (в том числе 1—2 колючих) и различное
число ветвистых лучей. В брюшных плавниках обычно 1—2 неветви-
стых и ряд ветвистых лучей. Спинной плавник всегда один. Жировой
плавник отсутствует. Боковая линия обычно хорошо развита. Тело по-
крыто циклоидной чешуёй, у некоторых голое. Голова голая, без че-
шуи. Рот более или менее выдвижной, у некоторых снабжён усиками.
Плавательный пузырь подразделён на два или больше отделов. Зубы на
челюстях отсутствуют, но имеются сильные зубы на нижних глоточ-
ных костях жаберного аппарата. Населяют преимущественно пресные
воды Европы, Азии, Северной Америки и Африки. Многие виды этого
отряда являются ценными промысловыми рыбами и имеют важное зна-
чение в промысле.
78
В фауне вод бассейна Кубани и прилегающих к нему азово-ку-
банских и закубанских рек встречаются представители четырёх се-
мейств: Карповые — Cyprinidae, Вьюновые — Cobitidae, Гольцовые —
Nemacheilidae (включая Balitoridae) и Чукучановые — Catostomidae.
Таблица
для определения семейств отряда
Карпообразные — Cypriniformes Rafinesque, 1810
1 (3). Рот не окружён сплошной широкой губой.
2 (4). Жерновок имеется, усиков или нет, или, если они есть, то
обычно не более двух пар. Глоточных зубов в одном ряду не более 8.
Семейство Карповые — Cyprinidae Rafinesque, 1810 (с. 78)
3 (1). Рот окружён широкой сплошной губой. Усиков нет. Тело по-
крыто чешуёй. Жерновок отсутствует. Имеются многочисленные гло-
точные зубы, сидящие в один ряд.
Семейство Чукучановые — Catostomidae Agassiz, 1855 (с. 142)
4 (2). Усиков 3—5 пар.
5 (6) Имеется выдвижной шип под глазом.
Семейство Вьюновые — Cobitidae Swainson, 1838 (с. 149)
6 (5) Выдвижной шип под глазом отсутствует.
Семейство Гольцовые — Nemacheilidae (включая Balitoridae
Swainson, 1839) Regan, 1911 (с. 147)
Семейство Карповые — Cyprinidae Rafinesque, 1810
Это самое большое семейство пресноводных рыб из числа позво-
ночных. Всего в нём насчитывается около 367 родов и около 3 006 ви-
дов (Абрамчук, Иваненко, 2019). Особенностью семейства является на-
личие глоточных зубов. Зубы немногочисленны, расположены в 1—3
ряда на нижних глоточных костях жаберного аппарата, в ряду не более
8 зубов, есть жерновок (роговая подушечка над глоточными зубами).
Рот окаймлён только межчелюстными костями. Одна группа рыб имеет
одно- и двухрядные зубы и не имеет усиков, другая — двух- и трёхряд-
ные зубы и в углах рта одну или две пары усиков.
Плавники состоят из ветвистых, но первые 2—3 или 4 луча спинно-
го и анального плавников неветвистые, самые передние из неветвистых
лучей часто зачаточны, едва заметны. При счёте лучей все неветвистые
79
лучи, даже зачаточные, принимаются и обозначаются римскими циф-
рами, в отличие от ветвистых, обозначаемых арабскими. Последний
или последние два неветвистые луча спинного, а нередко и анально-
го плавника часто бывают утолщены, окостеневают и превращаются в
шип, иногда сзади зазубренный.
Для семейства характерна ярко-серебристая или золотистая окра-
ска, у более взрослых рыб она значительно темнее (оливково-бурая).
Форма тела рыб разнообразна: есть высокотелые со сплющенными бо-
ками (лещ, густера, белоглазка и др.); уплощённое в спиннобрюшном
направлении (пескари); у некоторых на брюшке имеется ярко выражен-
ный киль (чехонь, верховка, белый толстолобик и др.).
Семейство очень разнообразно и включает рыб-хищников и мир-
ных, имеющих различное строение рта и кишечника. В период раз-
множения у карповых появляется брачный наряд. Самцы несколько
меньше самок. Часть видов относится к проходным, полупроходным
и пресноводным рыбам. Живут как в холодных быстро текущих водах,
так и в сильно прогреваемых, с недостаточным количеством кислоро-
да. Большинство карповых — фитофилы и используют в качестве не-
рестового субстрата свежезалитую и прошлогоднюю растительность,
но есть и литофилы, псаммофилы, пелагофилы, остракофилы и другие
экологические группировки. Карповые имеют большое промысловое
значение. К наиболее ценным видам относятся рыбец и шемая, к цен-
ным промысловым — сазан, лещ, тарань, плотва, толстолобики и др., к
малоценным — чехонь, караси, линь и др.
В бассейне Кубани семейство представлено 27 родами и 36 вида-
ми и подвидами рыб.
Таблица
для определения родов семейства
Карповые — Cyprinidae Rafinesque, 1810
1 (6). Спинной плавник начинается над брюшными. Чешуя срав-
нительно мелкая, в боковой линии 56—61 чешуя. Голова небольшая,
лоб выпуклый. Спинной плавник усечённый, анальный — выемчатый.
Окраска тела серебристо-желтоватая. Все плавники красноватые.
Род Ельцы — Leuciscus (Cuvier, 1816) (с. 83)
2 (4). В боковой линии 36—40 чешуй.
3 (8). Рот конечный. Чешуя крупная, в боковой линии 33—40 чешуй.
80
По бокам тела тянется тёмная полоса, которая иногда доходит до основа-
ния хвостового плавника. Спина тёмная, на верхней части боков при ос-
новании каждой чешуйки бурое пятно. Плавники серые. Анальный плав-
ник закруглённый, реже усечённый, но никогда не бывает выемчатым.
Род Бобырцы — Petroleuciscus Bogutskaya, 2002 (с. 85)
4 (2). Чешуя сравнительно крупная, в боковой линии от 41 до 47
чешуй.
5 (7). Боковая линия дугообразная, расположена ближе к брюху. В
боковой линии 43—47 чешуй. По заднему краю каждой чешуйки прохо-
дит полоска из тёмных пигментных точек. Рот большой, конечный, рыло
удлинённое. Окраска серо-серебристая. Спина тёмная, бока серебристые.
Грудные плавники оранжевые, брюшные и анальный — красноватые.
Род Голавли — Squalius Bonaparte, 1837 (с. 86)
6 (1). Спинной плавник начинается позади вертикали заднего кон-
ца основания брюшных. В боковой линии более 70 чешуй. Мелкие (до
20 см) рыбки — речные и озёрные.
Род Гольяны — Phoxinus Rafinesque, 1820 (с. 89)
7 (5). Боковая линия прямая, проходит посреди тела. Чешуя круп-
ная, в боковой линии 35—39 чешуй. Рот верхний, поперечный, малень-
кий. Окраска спины желтовато-серая. Брюшные, грудные и анальный
плавники светло-жёлтые.
Род Псевдорасборы (Амурские чебачки) — Pseudorasbora
Bleeker, 1860 (с. 90)
8 (3). Рот небольшой, полунижний. Боковая линия неполная, в ней
всего 5—11чешуй. По бокам тела проходит узкая зеленовато-синяя
продольная полоска, расширяющаяся к задней части тела. Анальный
плавник слегка выемчатый. Во время нереста самцы приобретают ра-
дужную окраску, а у самок вырастает длинный яйцеклад.
Род Горчаки — Rhodeus Agassiz, 1832 (с. 92)
9 (10). Рот конечный или полунижний. Глоточные зубы одноряд-
ные, обычно 6—5 или 5—5. Чешуя серебристо-белая, плотно сидящая.
В боковой линии 41—68 чешуй. Жилые и полупроходные рыбы.
Род Плотвы — Rutilus Rafinesque, 1820 (с. 93)
10 (9). Рот верхний или полуверхний. Тело высокое, покрытое до-
вольно крупной чешуёй. В боковой линии от 38 до 43 чешуй. Глоточ-
ные зубы двухрядные, зазубренные. Окраска очень яркая, бока золоти-
стые, плавники ярко-красные.
Род Краснопёрки — Scardinius Bonaparte, 1837 (с. 98)
81
11 (12). Тело и плавники чёрные. В боковой линии от 39 до 42
чешуй. По краю каждой чешуйки светлая полоска. Глоточные зубы с
широкими жевательными поверхностями, сильные, одно- или двухряд-
ные. Анальный и спинной плавники с малым числом лучей — 7 —8.
Род Чёрные амуры — Mylopharyngodon Peters, 1881 (с. 99)
12 (11). Тело и плавники не чёрные. В боковой линии более 42 че-
шуй. По краю каждой чешуйки тёмная полоска. Глоточные зубы двух-
рядные, острые, зазубренные.
Род Белые амуры — Ctenopharyngodon Steindachner, 1866 (с. 100)
13 (15). Боковая линия полная.
14 (17). За брюшными плавниками есть киль, покрытый чешуёй.
Нижняя челюсть снабжена бугорком, входящим в выемку на верхней че-
люсти. Рот большой конечный или полуверхний. Глоточные зубы силь-
ные, крючкообразные, двухрядные. В боковой линии от 62 до 105 чешуй.
Род Жерехи — Aspius Agassiz, 1832 (с. 101)
15 (13). Боковая линия неполная, прекращается на первых 2—13
чешуях, начинаясь от жаберной крышки. Рот полуверхний. Глоточные
зубы двухрядные, зазубренные, или однорядные. Маленькие рыбки
длиной до 9 см. Вдоль боков тянется слегка голубоватая полоска.
Род Верховки — Leucaspius Heckel & Kner, 1858 (с. 103)
16 (36). Кожистый киль между брюшным и анальным плавниками
не покрыт чешуёй. Спинной плавник отнесён далеко назад, конец его
не достигает вертикали начала анального плавника (шемая) или дос-
тигает и заходит за неё (уклейка). В спинном плавнике 9—12, в аналь-
ном — 17 —23 луча. В боковой линии от 45 до 82 чешуй. Чешуя у одних
видов легко спадающая (уклейка), у других плотно сидящая (шемая).
Пресноводные и проходные рыбы.
Род Уклейки — Alburnus Rafinesque, 1820 (с. 104)
17 (14). Киль на брюхе на протяжении от брюшных плавников до
анального отверстия не покрыт чешуёй. Глоточные зубы незазубрен-
ные. Жаберные тычинки короткие и редкие. А III 9—18.
18 (23). На спине впереди спинного плавника нет киля.
Род Быстрянки — Alburnoides Jeitteles, 1861 (с. 108)
19 (22). Глоточные зубы однорядные. В боковой линии более 50
чешуй. Киля позади спинного плавника нет.
20 (21). В анальном плавнике 24—31 луч. В боковой линии 51—57
чешуй.
Род Лещи — Abramis Cuvier, 1816 (с. 109)
82
21 (20). В анальном плавнике 36—42 луча. В боковой линии 48—
53 чешуи.
Род Синцы — Ballerus Heckel, 1843 (с. 110)
22 (19). Глоточные зубы двурядные. В боковой линии не более 50
(43—50) чешуй. Впереди спинного плавника имеется бороздка, не по-
крытая чешуёй. В анальном плавнике 19—23 луча.
Род Густеры — Blicca Heckel, 1843 (с. 112)
23 (18). Между затылком и началом спинного плавника борозд-
ка без чешуи. За спинным плавником есть киль, покрытый чешуёй, а
позади брюшных плавников — киль, не покрытый чешуёй. Рыло вы-
двигается вперёд. Рот нижний, полулунной формы. Глоточные зубы
однорядные.
Род Рыбцы — Vimba Fitzinger, 1873 (с. 113)
24 (25). Рот верхний. Тело удлинённое, сплющенное с боков.
Спинной плавник отнесён назад и расположен над анальным. Груд-
ные плавники длинные, конец их доходит до основания брюшных.
На брюхе острый киль, не покрытый чешуёй. Чешуя мелкая, легко
спадающая. Боковая линия изогнута зигзагообразно, в ней от 90 до
115 чешуй.
Род Чехони — Pelecus Agassiz, 1835 (с. 117)
25 (24). Рот конечный, небольшой. Тело толстое закруглённое, по-
крыто очень мелкой чешуёй, спинной и анальный плавники короткие.
В углах рта имеется по одному небольшому усику. Все плавники мяг-
кие, закруглённые, хвостовой — с очень слабой выемкой. В боковой
линии 97—120 чешуй. Глоточные зубы однорядные.
Род Лини — Tinca Cuvier, 1816 (с. 118)
26 (27). Рот нижний, поперечный и слабодугообразный. Брюшина
чёрная. Боковая линия делает слабый изгиб книзу, в ней от 56 до 63
чешуй. Брюшного киля нет. Усиков нет. Глоточные зубы однорядные
6—5 или 6—6.
Род Подусты — Chondrostoma Agassiz, 1835 (с. 119)
27 (26). Рот нижний, полулунной формы. Нижняя губа трёхраз-
дельная. Боковая линия прямая, в ней 52—68 чешуй. Глоточные зубы
трёхрядные. В спинном плавнике 8—10 ветвистых лучей, как правило,
зазубренная колючка.
28 (29). Усиков две пары: одна из них расположена на конце рыла,
а другая — в углах рта. Рыло удлинённое.
Род Усачи — Barbus Cuvier, 1816 (с. 122)
83
29 (28). Усиков одна пара, расположена в углах рта.
30 (32). Усики сравнительно короткие: не достигают заднего края
предкрышки. Вдоль боков тела 6—14 тёмных пятен.
31 (33). Рот нижний или конечный. Нижняя губа прервана. Мелкие
рыбки.
Род Пескари — Gobio Cuvier, 1816 (с. 124)
32 (30). Усики длинные, достигают заднего края предкрышки.
Род Румынские пескари — Romanogobio Banarescu, 1961 (с. 126)
33 (31). Рот нижний, сильно выдвижной, с образованием хобот-
ка. Тело покрыто крупной чешуёй. Спинной плавник очень длинный в
нём насчитывается от 15 до 22 ветвистых лучей. В спинном и анальном
плавниках имеется по одному зазубренному лучу.
34 (35). Около рта расположены две пары коротких усиков. Гло-
точные зубы трёхрядные, с мощно развитой жевательной площадкой.
Род Сазаны, или Карпы, — Cyprinus Linnaeus, 1758 (с. 129)
35 (34). Усики отсутствуют. Глоточные зубы однорядные. Тело
высокое, обычно золотистого цвета, но встречаются особи с тёмной
или почти серебристой окраской. В спинном и анальном плавниках
есть зазубренный костяной луч.
Род Караси — Carassius Jarocki, 1822 (с. 132)
36 (16). На брюхе имеется киль от горла до анального отверстия
или только между брюшными плавниками и анальным отверстием. Гла-
за расположены низко. Чешуя мелкая, в боковой линии 92—125 чешуй.
Род Толстолобы [Толстолобики] — Hypophthalmichthys Bleek-
er, 1859 (с. 138)
Согласно последним данным по систематике рыб [Дж. Нельсон
с соавторами (Nelson, Grande, Wilson, 2016), В. И. Романов (2019),
А. В. Абрамчук, А. М. Иваненко (2018, 2019)], восстановлен статус
некоторых семейств, родов и видов, а также описаны новые отряды,
семейства, рода и виды, в частности, восстановлен статус семейства
Гольцовые — Nemacheilidae Regan, 1911, но упразднено семейство Ба-
литоровые — Balitoridae Swainson, 1839.
Род Ельцы — Leuciscus Cuvier, 1816
Продолговатые рыбки с полной боковой линией, сравнительно
коротким анальным плавником, двухрядными глоточными зубами,
отсутствием киля за брюшными плавниками. Голова небольшая, с бо-
84
ков несколько сжата. Рот конечный. Спинной плавник расположен над
брюшными. Речные и озёрные рыбы. Род содержит 5 видов, распро-
странённых в Европе и Азии.
В водоёмах бассейна Кубани можно встретить один вид.
Язь — Leuciscus idus (Linnaeus, 1758) (рис. 45, 2.22)
Рис. 45. Язь — Leuciscus idus (Linnaeus, 1758)
D III 8 (7—9), A III 9—12, l.l. 56—61. Жаберные тычинки короткие
и редкие, на первой дуге 10—14 штук. Спинной плавник усечённый.
Рыба средних размеров с довольно толстым телом, небольшой
головой с выпуклым лбом. Анальный плавник с небольшой, а хвосто-
вой — с глубокой выемкой. Верхняя часть туловища тёмная, нижняя —
светлая. Плавники красноватые (см. рис. 2.22). Глоточные зубы двух-
рядные, 3.5—5.3, гладкие, на конце загнуты в крючок. Рот конечный,
косой, небольшой, челюсти равной длины или верхняя чуть длиннее.
Язь — пресноводная рыба, в море встречается очень редко. Еди-
ничные экземпляры встречаются в дельте Кубани и в приазовских ли-
манах. Длина тела — 30 —40 см, масса — 0,5—2,6 кг. Размножается в
марте—апреле при температуре воды 10,5 °С. Плодовитость — от 30 до
90 тыс. икринок. Питается беспозвоночными. Язь в бассейне Кубани
не имеет промыслового значения, но представляет интерес для люби-
телей-рыболовов.
Возможны помеси язя и жереха.
Leuciscus idus (L.) × Aspius aspius (L.) — Краснопёр
D III 8, A III 11—12, l.l. 58—63 (Берг, 1949). Спинной плавник усе-
85
чённый, анальный — выемчатый. Окраска тела серебристо-желтоватая.
Все плавники красноватые.
Род Бобырцы — Petroleuciscus Bogutskaya, 2002
Род Petroleuciscus включает в себя 6 видов, ранее помещавшихся в
род Leuciscus (Bogutskaya, 2002). Они встречаются в бассейне Чёрного
моря и восточных стоках Эгейского, на восток до Ирана и Узбекистана.
Один вид известен из вод континентальной Европы, а другой обитает
в районе греческого о. Лесбос, расположенного у побережья Турции.
По бокам тела тянется тёмная полоса, в задней части иногда дохо-
дит до основания хвостового плавника, причём бывает довольно ши-
рокой. Спина тёмная, на верхней части боков при основании каждой
чешуйки бурое пятно. Плавники серые. Анальный плавник закруглён-
ный, реже усечённый, но никогда не бывает выемчатым.
В водоёмах бассейна Кубани и прилегающих к нему рек обитает
один вид.
Калинка, или бобырец, — Petroleuciscus borysthenicus (Kessler,
1859) (рис. 46, 2.25)
Рис. 46. Калинка, или бобырец, — Petroleuciscus borysthenicus (Kessler, 1859)
D III 8—9, A III 9—10, l.l. 36—40. Бобырец — небольшая рыбка,
длина взрослых особей — 8 —14 см. Рот конечный, зубы двухрядные,
гладкие, чаще 2.5—5.2 или 2.5—4.2. Спина тёмная, во время нереста
брюшные плавники окрашены в оранжевый цвет. От молоди красно-
86
пёрки, тарани и плотвы отличается анальным плавником, который у
бобырца слегка закруглённый и реже усечённый, но никогда не бывает
выемчатым. Спина тёмная; в верхней части боков при основании каж-
дой чешуи бурое пятно. Вдоль боков в задней части тела тянется тём-
ная полоса.
Нерестится в мае—июне. Плодовитость — 2,5—3,5 тыс. икринок.
Питается личинками насекомых и беспозвоночными.
Бобырец обитает в кубанских лиманах, степных реках и других
водоёмах бассейна Кубани. Промыслового значения не имеет.
Род Голавли — Squalius Bonaparte, 1837
Виды рода Squalius — это мелкие и крупные рыбы, обитающие в
горных ручьях, озёрах и медленно текущих больших реках. Они харак-
теризуются отсутствием чешуйчатого киля перед анусом, полной боко-
вой линией, обычно конечным или субконечным ртом, спинным плав-
ником позади основания брюшных, двухрядными глоточными зубами,
отсутствием заметной тёмной боковой полосы, 8—11 разветвлёнными
лучами в анальном и 7—9 разветвлёнными лучами в спинном плавни-
ке. Систематика рода Squalius до сих пор не закончена.
Чешуя сравнительно крупная, по заднему краю каждой чешуйки
имеется полоска тёмных пигментных точек. Боковая линия дугообраз-
ная, расположена ближе к брюху, в ней от 40 до 46 чешуй. Окраска
тела серо-серебристая. Спина тёмная, бока серебристые. Спинной,
анальный и брюшные плавники слегка выпуклые или усечённые.
В водоёмах бассейна Кубани и прилегающих к нему азово-кубан-
ских и закубанских рек обитают две формы (вид и подвид).
Таблица
для определения видов и подвидов рода
Голавли — Squalius Bonaparte, 1837
1 (2). Чешуя крупная, в боковой линии от 41 до 46 чешуй. D III (7) 8,
А III 8—9. По заднему краю каждой чешуйки имеется полоска тёмных
пигментных точек. Спина тёмная. Позади жаберной крышки есть тём-
ное пятно.
Кавказский голавль — Squalius cephalus orientalis Nordmann,
1840 (с. 87)
87
2 (1). В боковой линии 40—43 чешуи. D III 8, А III 9. Тёмная
полоска в задней части тела отсутствует. Окраска серо-серебристая,
без пятен на боках. Позади жаберной крышки тёмное пятно отсут-
ствует.
Афипский елец [голавль], или калинка, — Squalius aphipsi Alek-
sandrov, 1927 (с. 88)
Краткая биологическая характеристика видов и подвидов рода
Голавли — Squalius Bonaparte, 1837
Кавказский голавль — Squalius cephalus orientalis (Nordmann,
1840) (рис. 47, 2.23)
Рис. 47. Кавказский голавль — Squalius cephalus orientalis (Nordmann, 1840)
D III (7) 8, A III 8—9, l.l. 41—46. Глоточные зубы двухрядные
(2.5—5.2). Кавказский голавль является подвидом обыкновенного го-
лавля — Squalius cephalus (Linnaeus, 1758). Рыба небольших размеров,
длина тела обычно не превышает 45 см. Чешуя довольно крупная, в
боковой линии от 41 до 46 чешуй. Тело невысокое. Длина головы мень-
ше или равна высоте тела. Спина тёмно-зелёная, бока желтоватого, а
брюшко серебристого цвета. Грудные, брюшные и анальный плавники
оранжевого и красного цвета.
Голавль — пресноводная рыба, в море никогда не встречается.
Предпочитает небольшие, но чистые и без растительности реки. Ик-
рометание весеннее. Икра мелкая, откладывается на коряги и камни.
Нерест порционный с апреля по май.
Голавль питается насекомыми и их личинками, во взрослом со-
стоянии — мелкой рыбой (пескарём). Промыслового значения не име-
88
ет. Широко распространён по всему бассейну Кубани и причерномор-
ских рек. Является объектом любительского рыболовства.
Возможны помеси кавказского голавля и шемаи.
Squalius cephalus orientalis (Nordmann) × Alburnus mento (Heckel)
D III 8, A III 11, l.l. 47—47. Глоточные зубы 2.5—5.2, слегка зазуб-
ренные. Киля, не покрытого чешуёй, впереди анального отверстия нет
(р. Псекупс (Берг, 1949)).
Афипсская калинка, или афипсский голавль [елец],
или бекас — Squalius aphipsi (Aleksandrov, 1927) (рис. 48, 2.24)
Рис. 48. Афипский голавль [елец] — Squalius aphipsi (Aleksandrov, 1927)
D III 8, A III 9, l.l. 40—43. Афипский елец — небольшая рыбка,
достигающая в длину 15 см и массы 42 г. Обитает в р. Кубани и лево-
бережных притоках (Афипс, Псекупс, Пшиш, Белая, Лаба и др.). Не-
рест порционный с апреля по июнь. Нерест происходит на перекатах с
быстрым течением, на глубине 10—20 см. Икру откладывает на галеч-
ный грунт. Питается в основном насекомыми и их личинками, а также
поедает икру других рыб (рыбца, шемаи). Промыслового значения не
имеет. Эндемик бассейна Кубани. Занесён в Красную книгу Краснодар-
ского края (2007, 2017).
Размещение этого вида в роде Squalius носит предположительный
характер (Богуцкая, Позняк, 1994). Он не имеет нескольких остеоло-
гических признаков, диагностических для рода, и согласно предвари-
тельным молекулярным данным его помещают близко к роду Бобырцы
(Petroleuciscus) (Bogutskaya, 2002).
89
Род Гольяны — Phoxinus Rafinesque, 1820
Гольяны — мелкие (до 20 см) рыбки, речные и озёрные. Зубы
двухрядные. Анальный плавник короткий, киль отсутствует. Чешуя
мелкая. Окраска тёмная с мелкими тёмными пятнышками. В боковой
линии более 70 чешуй. Спинной плавник начинается позади вертикали
заднего конца основания брюшных. Верхний край спинного плавника
слегка выпуклый. Рот конечный или полунижний.
В роде 10 видов, в водоёмах бассейна Кубани водится один вид.
Обыкновенный гольян [красавка] — Phoxinus phoxinus (Linnaeus,
1758) (рис. 49, 2.28)
Рис. 49. Обыкновенный гольян — Phoxinus phoxinus (Linnaeus, 1758)
D III 7, А III 6—7, l.l. 80—92.
Гольян — типичный реофил, живёт в быстрых неглубоких реках
с песчаным или каменистым дном и хорошим насыщением воды ки-
слородом. Обнаружен в реках бассейна Кубани, а также в закубанских
(Хабль, Абин, Псебес и др.) и черноморских реках. Тело удлинённое,
прогонистое, веретенообразное. Рот маленький, полунижний.
Глаза расположены высоко. Имеет тёмную окраску, с мелкими
пятнышками на спине и большие тёмные пятна по бокам. Боковая ли-
ния прерывистая. Грудные и брюшные плавники закруглённые.
Гольян — типичный литофил, откладывает икру на течении меж-
ду камнями, личинки до всасывания желточного мешка прячутся меж-
ду камнями. Плодовитость — от 200 до 600 икринок. В период нереста
окраска самцов резко отличается от окраски самок. У самцов спина ста-
90
новится очень тёмной, парные плавники — жёлтыми, брюхо — крас-
ным, углы рта — малиновыми, брюшные и анальный плавники — яр-
ко-красными. У самок брачный наряд не выражен.
Питается гольян водными беспозвоночными. Сам является объек-
том питания хищных видов рыб.
Род Псевдорасборы (Амурские чебачки) — Pseudorasbora Bleeker,
1860
Тело умеренно удлинённое. Рот очень маленький, верхний, попе-
речный. Усиков нет. Чешуя крупная. Горло покрыто чешуёй. Боковая
линия прямая, проходит посередине тела. Жаберные тычинки зачаточ-
ные. Спинной плавник короткий (7 ветвистых лучей), без колючки.
Анальный плавник тоже короткий (6 ветвистых лучей). Брюхо без киля.
В бассейне Кубани обитает один вид.
Амурский чебачок — Pseudorasbora parva (Temminck & Schlegel,
1846) (рис. 50, 2.26)
Рис. 50. Амурский чебачок — Pseudorasbora parva (Temminck & Schlegel,
1846)
D III 7, A III 6, l.l. 35—39. Длина тела — до 8—10 см (редко до
11 см). Тело окрашено в бронзово-серебристый цвет, его высота не-
много больше длины головы или равна ей. Вдоль тела от глаза до ос-
нования хвостового плавника тянется тёмная полоска, у половозрелых
особей она становится малозаметной. Радужина глаз светлая, над зрач-
91
ком тёмное пятно. Тело покрыто относительно крупной чешуёй. По
внешнему краю каждой чешуйки проходят тёмные пятна, образующие
тёмный ободок и формирующие на чешуйном покрове полулунные
тёмные пятнышки. Плавники закруглены, на спинном и анальном име-
ются мелкие тёмные пятна. Парные плавники и анальный светло-жёл-
тые. Рот верхний, поперечный, маленький. Нижняя челюсть заходит за
верхнюю. Глоточные зубы однорядные, их формула 5—5, сжатые. У
половозрелых самцов на нижней и верхней челюстях по 2—4 роговых
шипика, на щеке под глазом 4—6 шипиков. Усики отсутствуют. Брюхо
без киля.
Половозрелость у амурского чебачка в водоёмах бассейна Кубани
наступает на 1—2 -м году жизни при длине 4—5 см. Самцы крупнее
самок. Во время нереста на голове впереди глаз появляются острые бу-
горки (шипики).
В яичниках самок амурского чебачка отчётливо видны три порции
икры. Индивидуальная абсолютная плодовитость (с учётом всех трёх
генераций) колеблется от 374 ± 0,21 (1+) до 1 374 ± 0,38 (4+) икринок.
Нерест амурского чебачка в зависимости от температурных и дру-
гих условий проходит с третьей декады апреля до конца июня. Опти-
мальная температура воды, при которой происходит нерест, колеблется
от16до28°С.
В основном нерестится амурский чебачок на глубине от 0,4 до
1,5 м. Нерест порционный, в мае—июне. Икра откладывается тонким
слоем в одну икринку в затишных местах. Икра приклеивается к любо-
му субстрату (растительность, затонувшие ветки, палки, коряги, камни,
куски дерева, пустые раковины моллюсков и др.). Икра эллипсовид-
ная. Инкубационный период — от 6 до 14 дней: при температуре воды
16,5—18,5 °С он длится 12—14 дней, а при 22—28 °С — 6 —8 дней.
Основная пища чебачка — зоопланктон и личинки хирономид,
вследствие чего при большой численности становится конкурентом в
питании молоди ценных промысловых рыб (сазана, леща, рыбца, ше-
маи и др.). Злостный икроед. В некоторых водоёмах достигает значи-
тельной численности.
Естественный ареал амурского чебачка был ограничен реками
Китая, Кореи, Японии и бассейном Амура. В 1960-е гг. при интродук-
ции рыб амурского комплекса (белый и пёстрый толстолобики, белый
и чёрный амуры) вид случайно попал в водоёмы республик бывшего
СССР (Киргизия) и стал компонентом ихтиофауны многочисленных
92
рек и водохранилищ, в том числе и водоёмов бассейна Кубани (реки,
озёра, водохранилища, лиманы и пруды). Впервые в крае обнаружен в
1988 г. близ ст-цы Казанской. В дальнейшем неоднократно отмечался
как в Ростовской области, так и в Краснодарском крае (рисовые чеки
близ ст-цы Фёдоровской, озеро на Сазальницкой косе и др.).
Амурский чебачок — тугорослая рыба, не имеет хозяйственного
значения и является нежелательным видом в водоёмах с интенсивным
рыбоводством. Прекрасный живец, используется в качестве наживки в
любительском рыболовстве.
Род Горчаки — Rhodeus Agassiz, 1832
Тело высокое, сжатое с боков. Чешуя крупная (32—40 попереч-
ных рядов). Боковая линия неполная. Рот маленький, слегка косой,
полунижний. Усиков нет. Глоточные зубы однорядные, 5—5, сжатые
с боков. Спинной плавник удлинённый (8—11 ветвистых лучей), без
колючки, начинается немного позади оснований брюшных. Анальный
плавник без колючки с 8—10 ветвистыми лучами, начинается впереди
конца спинного плавника. Брюшина чёрная.
В бассейне Кубани встречается один подвид.
Обыкновенный горчак — Rhodeus sericeus amarus (Bloch, 1782)
(рис. 51, 2.27)
Рис. 51. Обыкновенный горчак — Rhodeus sericeus amarus (Bloch, 1782)
93
D III 9—10, A III 8—9 (10), l.l. 4—6 (7), 34—40 поперечных ря-
дов чешуй. Усиков нет. Длина тела — 4,3—7,5 см (до 9,5 см), масса
тела — до 3,5 г. У горчака высокая спина, сжатое с боков тело, малень-
кий конечный рот. Однорядные глоточные зубы, их формула 5—5. У
самок в период нереста появляется длинный яйцеклад, через который
она откладывает икру в мантийную полость двустворчатых моллюсков.
Предпочитает медленно текущие и стоящие воды.
Род Плотвы — Rutilus Rafinesque, 1820
Тело покрыто крепкосидящей чешуёй. В боковой линии 33—67 че-
шуй. Рот конечный или полунижний. Ротовая щель полулунная. Спин-
ной плавник расположен над основанием брюшных. Глоточные зубы
однорядные, кососрезанные, иногда со слабым крючком. Жаберные ты-
чинки редкие и короткие. Жилые, полупроходные и проходные рыбы.
Род включает 7—8 видов и несколько подвидов. В пределах рассматри-
ваемого бассейна этот род представлен двумя видами и одним подвидом.
Таблица
для определения видов рода
Плотвы — Rutilus Rafinesque, 1820
1 (4). В боковой линии менее 50 чешуй. Плавательный пузырь на зад-
нем конце округлый, без отростка. За брюшными плавниками киль, покры-
тый чешуёй. В спинном и анальном плавниках обычно 10 ветвистых лучей.
2 (3). Рот конечный или полунижний. Высота тела обычно более 1⁄4 его
длины. В боковой линии чаще всего 43—45 чешуй. Спинной и хвостовой
плавники серые, остальные оранжевые или красные. Непроходная рыба.
Обыкновенная плотва — Rutilus rutilus (Linnaeus, 1758) (с. 94)
3 (2). Рот нижний. Высота тела — более 1⁄3 его длины. В боковой
линии обычно 43 чешуи. Парные плавники имеют черноватые края.
Полупроходная рыба.
Тарань — Rutilus heckelii (Nordmann, 1840) (с. 95)
4 (1). В боковой линии более 50 чешуй. Высота тела обычно менее
1⁄4 его длины.
Вырезуб — Rutilus frisii (Nordmann, 1840) (с. 96)
Образует подвиды, один из них обитает в бассейне Кубани.
Кутум — Rutilus frisii kutum (Kamensky, 1901) (с. 97)
94
Краткая биологическая характеристика видов и подвидов рода
Плотвы — Rutilus Rafinesque, 1820
Обыкновенная плотва — Rutilus rutilus Linnaeus, 1758 (рис. 52,
2.29)
Рис. 52. Обыкновенная плотва — Rutilus rutilus Linnaeus, 1758
D III 10, А III 10, l.l. 41—48. Глоточные зубы однорядные, причём
с правой стороны имеется 5, а с левой — 6 зубов. Жаберных тычинок
на первой жаберной дуге — 9 —10.
Оседлая рыба, предпочитает жить в медленно текучих реках, озё-
рах, лиманах, водохранилищах и других водоёмах. По внешним при-
знакам типичная плотва приближается к краснопёрке, но тело её имеет
более серебристый цвет боков и более прогонистое тело, а рот полу-
нижний, тогда как у краснопёрки он полуверхний. Спинной и хвосто-
вой плавники бледно-жёлтые, брюшные и анальный — красноватые.
Радужная оболочка глаза оранжевая, вверху с красным пятном. Боль-
ших размеров плотва не достигает, в заросших малопроточных водо-
ёмах она не превышает 6—8 см, масса — 30 —40 г. Но при благоприят-
ных условиях обитания, например, в Краснодарском водохранилище,
обычны особи длиной 25—28 см и массой 250—280 г. Некоторые дос-
тигают 350—600 г и являются объектом промысла.
Половое созревание наступает в 3—4 года (тугорослая форма со-
зревает быстрее), нерест происходит в апреле при температуре 10—
95
12 °С. Для нереста плотва использует заводи, затоны, заливы и другие
мелководные участки. Нерестовый субстрат — прошлогодняя расти-
тельность. Плодовитость плотвы Краснодарского водохранилища —
20—128 тыс. икринок, а других водоёмов — 5 —25 тыс. икринок. Ин-
кубационный период — 4 —5 дней. Молодь питается зоопланктоном,
а начиная со второго года жизни — водными растениями, личинками
насекомых, мелкими моллюсками.
Плотва является объектом любительского рыболовства.
В естественных условиях плотва образует помеси: плотва × лещ,
плотва × густера, плотва × уклея, плотва × краснопёрка и др. (Берг, 1949).
Помеси плотвы и леща — Rutilus rutilus (L.) × Abramis brama (L.)
D III (8) 9—10 (11), А III—IV (14) 15—20, l.l. 42—54. Киль за
брюшными плавниками резко выражен, но покрыт чешуёй.
Помеси плотвы и густеры — Rutilus rutilus (L.) × Blicca bjoerkna (L.)
D III 8—10, А III 14—16, l.l. 43—46. Впереди D нет бороздки, по-
крытой чешуёй. Киль впереди анального отверстия покрыт чешуёй.
Помеси плотвы и уклеи — Rutilus rutilus (L.) × Alburnus alburnus (L.)
D III 8—11, А III 13—15, l.l. 45—52. Встречается очень редко.
Тарань — Rutilus heckelii (Nordmann, 1840) (рис. 53, 2.30)
Рис. 53. Тарань — Rutilus heckelii (Nordmann, 1840)
D III 9—11, А III 9—10 (11), l.l. 41—48 (43). Тарань — полупроход-
ная рыба, бо
́
льшую часть своей жизни проводит в Азовском море, а на
96
нерест заходит в лиманы и реки. Внешне очень похожа на плотву. От-
личается более высоким телом (наибольшая высота тела — 34—36 %
егодлины,ауплотвы—менее30%)ибо
́
льшим количеством жабер-
ных тычинок (14), глоточными зубами, их формула 6—5. Растёт тарань
быстрее плотвы. Средний размер промысловой тарани — 14 —22 см,
масса — 200—400 г. Максимальная длина тела — до 35 см, масса — до
1,8 кг. Весенний ход кубанской тарани начинается в феврале—марте
и продолжается до апреля. Нерест происходит с середины апреля до
середины мая. Самцы покрываются «жемчужной сыпью», т. е. брач-
ным нарядом. Основные места нереста тарани — кубанские лиманы
и пойменные водоёмы приазовских рек (Ея, Челбас, Бейсуг и др.). В
Кубань тарань поднимается до Фёдоровского гидроузла. Половозрелой
становится на 3—4 -м году жизни. Плодовитость — от 6 до 200 тыс.
икринок. Икра, как и у плотвы, откладывается на растения. После не-
реста тарань скатывается в море и нагуливается там до глубокой осени.
Питается в основном моллюсками и ракообразными.
Ценный объект промысла; ныне уловы колеблются по годам от
207 до 1 445 т, а в 1933—1973 гг. среднегодовые уловы были 5 500 т.
Резкое сокращение запасов тарани объясняется ухудшением условий
размножения и в лиманах, и в реках из-за сильного загрязнения.
Ранее считалась подвидом обыкновенной плотвы (Rutilus rutilus
heckelii (Nordmann, 1840)), но современные молекулярно-генетиче-
ские исследования (Matrilinear phylogeography..., 2009; Phylogeny and
phylogeography..., 2017) позволяют повысить статус подвида до вида.
Вырезуб — Rutilus frisii (Nordmann, 1840) (рис. 54, 2.31)
Рис. 54. Вырезуб — Rutilus frisii (Nordmann, 1840)
97
D II—IV 8—10, А III—IV 9—12, l.l. 53—68. Жаберных тычинок
7—12. Вырезуб — крупная рыба, достигающая длины 70 см и массы
более 6 кг. Распространён в бассейне Чёрного и Азовского морей. По-
ловозрелым становится в 5-летнем возрасте. Для нереста входит в реки,
ход начинается в конце марта — апреле, икрометание — в конце апреля
и в мае на каменистых участках реки. Плодовитость колеблется от 80
до 250 тыс. икринок. После нереста скатывается вниз по течению и
кормится в опреснённых участках моря и в кубанских лиманах. Пита-
ется в основном моллюсками. Глоточные зубы крепкие, мощные, спо-
собны раздавливать крупные раковины моллюсков.
Вырезуб — ценная рыба. Мясо обладает высокими вкусовыми
качествами. Промыслового значения не имеет из-за малочисленности.
Единичные экземпляры встречаются в р. Кубань и кубанских лиманах.
Вырезуб (Rutilus frisii frisii (Nordmann, 1840)) внесён в Красную книгу
Краснодарского края (2017).
Вырезуб образует подвиды, к таким подвидам относится кутум.
Кутум — Rutilus frisii kutum (Kamensky, 1901) (рис. 55, 2.32)
Рис. 55. Кутум — Rutilus frisii kutum (Kamensky, 1901)
D III 9, А III 10, l.l. 55—58. Тело кутума прогонистое, высота тела
меньше длины головы, по окраске близок к тарани. Высота спинного
плавника равна его длине, анального — больше его длины. Хвостовой
плавник выемчатый.
Естественный ареал распространения кутума — бассейн Каспий-
ского моря. В 1957—1961 гг. из Дагестана в рыбоводные хозяйства
Кубани было завезено 75,3 млн оплодотворённой икры кутума. Икра
инкубировалась в искусственных условиях. Личинок выпускали в ку-
банские лиманы. Через четыре года половозрелые особи кутума были
98
обнаружены на побережье Азовского моря от Темрюка до Ачуево (Тро-
ицкий, Цуникова, 1988). Длина самок — 39 —52 см, масса — 800 —
950 г, самцов — соответственно 33—45 см и 570—920 г. Нерестовый
ход производителей в Кубань и Протоку начинается в марте—апреле
при температуре воды 12—14 °С. Самцы в этот период покрываются
брачным нарядом. Нерест кутума происходит при температуре воды
15—16 °С, инкубационный период длится 10—12 дней. С. К. Троиц-
кий, Е. П. Цуникова (1988) отмечают, что имеются сведения о нересте
кутума в гирлах лиманов и в самих лиманах.
Кутум — ценная промысловая рыба. Из-за малочисленности на
Кубани промыслового значения не имеет. Внесён в Красную книгу РФ
(2001) (категория 2 — сокращающийся в численности подвид вырезуба).
Род Краснопёрки — Scardinius Bonaparte, 1837
Чешуя плотная, налегающая друг на друга. Боковая линия пол-
ная, в ней 37—44 чешуи. Глоточные зубы двухрядные — 3.5—5.3, пи-
лообразно-зазубренные. Рот конечный, обращённый вверх. Жаберные
тычинки короткие, редкие, малочисленные, около 12 на первой дуге.
В водоёмах бассейна Кубани встречается один вид.
Краснопёрка — Scardinius erythrophthalmus (Linnaeus, 1758)
(рис. 56, 2.33)
Рис. 56. Краснопёрка — Scardinius erythrophthalmus (Linnaeus, 1758)
99
D III 8—10, A III 9—12, l.l. 37—44. Жаберных тычинок 10—12.
Тело высокое, покрыто довольно крупной чешуёй. Окраска очень яр-
кая, бока серебристые или золотистые, плавники ярко красные, глаза
оранжевые с красным пятном (см. рис. 2.33). Голова маленькая, рот ко-
нечный, обращённый вверх, спинной плавник расположен за вертика-
лью брюшных плавников и заканчивается чуть не доходя до вертикали
от анального плавника. Брюшко за брюшными плавниками сильно сжа-
то с боков, имеется киль, покрытый чешуёй. Боковая линия смещена
ближе к брюшку. Жаберных тычинок 10—12.
По внешнему виду краснопёрка похожа на плотву, но её легко раз-
личить по обращённому вверх рту, сильно отодвинутому кзади спинно-
му плавнику и килю за брюшными плавниками. Наиболее простой спо-
соб отличия — золотистый цвет чешуи и огненно-красные плавники,
а также цвет глаз: у краснопёрки глаза оранжевые, с красным пятном
вверху, тогда как у плотвы — кроваво-красные.
Типичный стайный вид рыб, обитает в озёрах и реках с медленно
текущими водами, любит заросли, хорошо прогретую воду.
Созревает краснопёрка на 2—3 -м году жизни. Нерестится в мае—
июне на свежезалитой растительности. Плодовитость колеблется от 10
до 100 тыс. икринок. Инкубационный период при температуре воды
20°С —4—5сут.
Больших размеров краснопёрка не достигает, длина — обычно
17—20 см, масса — 50—120 г. Но встречаются в лиманах и более круп-
ные экземпляры — до 35 см и массой 560 г. Питается в основном на-
секомыми и их личинками, растительностью, фитопланктоном и зоо-
планктоном, а также икрой рыб. Промыслового значения практически
не имеет. Является объектом любительского рыболовства.
В естественных условиях возможны помеси: краснопёрка × уклея,
краснопёрка × густера, краснопёрка × плотва и др. (Берг, 1949).
Род Чёрные амуры — Mylopharyngodon Peters, 1881
Род включает один вид — чёрный амур — Mylopharyngodon piceus
(Richardson, 1846), интродуцированный (вселённый) в бассейне Куба-
ни в 1960 г. Естественный ареал — бассейн р. Амур (ниже г. Хабаров-
ска), р. Уссури, оз. Ханка, широко распространён в реках Китая, где
является объектом промысла. С 1972 г. в российской части бассейна
Амура установлен запрет на вылов чёрного амура.
100
Чёрный амур — Mylopharyngodon piceus (Richardson, 1846)
(рис. 57, 2.34)
Рис. 57. Чёрный амур — Mylopharyngodon piceus (Richardson, 1846)
D III 7—8, А III 8, l.l. 39—43. Тело покрыто крупной чешуёй, ок-
раска тёмная, почти чёрная, брюшко светлее. По краю каждой чешуй-
ки светлая полоска. Лоб широкий, глаза средней величины. Глоточные
зубы мощные, двухрядные (1.4—4.1) незазубренные, с широкой же-
вательной поверхностью. Рот конечный. Все плавники почти чёрные.
Спинной плавник выдвинут несколько вперёд от вертикали брюшных
плавников.
Половая зрелость наступает в 7—9 лет при длине 70 см. Нерест
происходит в начале лета на быстром течении, при подъёме воды, в
нижних слоях, при температуре воды 26—28 °С. Икра пелагическая.
Растёт довольно интенсивно: годовики имеют массу 35—60 г, в 13—14
лет чёрный амур весит 15 кг.
Питается исключительно моллюсками. Перспективный вид пру-
дового рыбоводства.
Род Белые амуры — Ctenopharyngodon Steindachner, 1866
Род включает один вид — белый амур — Ctenopharyngodon idella
(Valenciennes, 1844). Широко распространённый вид в пресноводных
водоёмах от р. Амур до Южного Китая. Интродуцирован в водоёмах
Краснодарского края в 1960 г. и используется как биологический ме-
лиоратор в борьбе с высшей растительностью.
101
Белый амур — Ctenopharyngodon idella (Valenciennes, 1844)
(рис. 58, 2.35)
Рис. 58. Белый амур — Ctenopharyngodon idella (Valenciennes, 1844)
D III 7, А III 8, l.l. 43—45. Жаберные тычинки короткие, редкие,
их 12—18. Голова составляет 1⁄4 длины тела. Тело прогонистое, относи-
тельно невысокое, длиной до 1,5 м. Чешуя средней величины. По краю
каждой чешуи (кроме расположенных на брюхе) тёмный ободок. Ок-
раска спины зеленоватая или желтовато-серая, бока тёмно-золотистые.
Спинной и анальный плавники тёмно-серые, остальные более светлые.
На жаберной крышке имеются радиальные полосы (см. рис. 2.35).
Питается белый амур исключительно высшей растительностью.
Глоточные зубы двухрядные (2.5—4.2 или 2.4—4.2), имеют боль-
шую жевательную поверхность. Кишечник очень объёмистый, в 6—8
раз превышает длину тела. Растёт быстро. Средняя масса составляет
5—6 кг. Но встречаются особи массой 15—18 кг.
Белый амур является объектом прудового рыбоводства, хорошо
себя чувствует в кубанских лиманах, где много высшей водной расти-
тельности.
Род Жерехи — Aspius Agassiz, 1832
Голова и рыло длинные. Верхняя челюсть короче нижней. Тело
удлинённое, сжатое с боков. Чешуя плотно сидящая, средней величи-
ны. В водоёмах бассейна Кубани род Жерехи представлен одним ви-
дом — обыкновенным жерехом — Aspius aspius (Linnaeus, 1758).
102
Обыкновенный жерех — Aspius aspius (Linnaeus, 1758) (рис. 59, 2.36)
Рис. 59. Обыкновенный жерех — Aspius aspius (Linnaeus, 1758)
D III 8, А III 12—14, l.l. 65—76. Жаберных тычинок 7—11. Тело
удлинённое, рот большой, глоточные зубы хватательные, двухрядные
(3.5—5.3). Жаберные тычинки редкие и короткие, на первой жабер-
ной дуге их не более 11. Чешуя плотно сидящая, средней величины. За
брюшными плавниками есть киль, покрытый чешуёй.
Рот большой, конечный. Нижняя челюсть выдвигается вперёд и
имеет бугорок, входящий в выемку на верхней губе.
Окраска жереха неяркая: спина зеленоватая, бока и брюшко сереб-
ристые. Спинной и хвостовой плавники серые, брюшные и анальный
плавники имеют желтоватый оттенок, непарные плавники выемчатые
(см. рис. 2.36).
В бассейне Кубани ведёт полупроходной образ жизни, но больших
миграций не совершает. Предпочитает обитать в реках с небольшим те-
чением, встречается и в замкнутых водоёмах, но для нереста выбирает
участки с галечниковым грунтом и более сильным течением.
Половое созревание наступает в 3—4 -летнем возрасте. Самцы в
период нереста становятся более ярко окрашены и покрываются бугор-
ками. Плодовитость самок — 50—500 тыс. икринок. Нерест происхо-
дит в марте—апреле при температуре воды 8—14 °С, но возможен и
при более низкой температуре. Икра слабоклейкая, с нерестилищ сно-
сится водой под гальку и там развивается. Длительность развития икры
зависит от температуры воды: чем она выше, тем короче инкубацион-
ный период. Так, например, при температуре воды 8,5 °С — 17, при
12,5 °С — 10,5 сут.
103
Обычная длина взрослых рыб составляет 35—40 см, масса —
1,2—1,5 кг. Но встречаются более крупные — до 50 см и 2 кг и даже до
5—6 кг. Первое время молодь питается зоопланктоном, но по достиже-
нии 6—8 см переходит на питание личинками других рыб. Несмотря на
то что жерех типичный хищник, обособленного желудка у него нет. В
связи с этим основу его рациона составляют мелкие рыбы (уклея, вер-
ховка, плотва и молодь промысловых рыб — сазана, леща и др.).
Существенной роли в промысловых уловах жерех не играет. Го-
довой улов на Кубани составляет 200—400 кг. В основном это объект
спортивного рыболовства.
Род Верховки — Leucaspius Heckel & Kner, 1858
Тело умеренно-удлинённое. Чешуя легко спадающая. Боковая ли-
ния неполная. Спинной плавник короткий, анальный — удлинённый,
начинается под задним концом спинного. Рот конечный, полуверхний.
В водоёмах бассейна Кубани род представлен одним видом — вер-
ховкой — Leucaspius delineatus Heckel, 1843.
Верховка — Leucaspius delineatus Heckel, 1843 (рис. 60, 2.37)
Рис. 60. Верховка — Leucaspius delineatus Heckel, 1843
D III 7—9, А III 10—14. Мелкая рыбка с крупной чешуёй, непол-
ной боковой линией из 2—13 чешуй. Чешуя легко спадающая. Вдоль
боков, особенно в задней части тела, тянется слегка голубоватая полос-
ка. Глоточные зубы двухрядные (1.5—5.1 или 1.5—4.1), иногда с одной
стороны однорядные. Жаберные тычинки густосидящие, 14—16.
Верховка встречается во всех водоёмах (лиманы, озёра, водохра-
нилища, пруды, реки). Предпочитает заросшие мелководные участки
104
водоёмов, держась в верхних слоях воды. Наибольшая длина тела —
6—7 см, масса — 2 —3 г. Нерест порционный. Икринки приклеивают-
ся к нижней поверхности плавающих растений (рдест, стрелолист, ку-
бышки и др.), а иногда к гладким палкам, доскам и другим предметам.
Кладки имеют характерный вид строчек в 1—2 икринки шириной. В
каждой кладке бывает от 10 до 30 икринок. Икрометание происходит в
мае—июне. Питается в основном зоопланктоном и воздушными насе-
комыми. Верховка промыслового значения не имеет. Является объек-
том питания хищных видов рыб.
Род Уклейки — Alburnus Rafinesque, 1820
Туводные и проходные стайные рыбы с удлинённым телом, сжа-
тым с боков. Рот конечный. Глоточные зубы двухрядные. Чешуя тон-
кая, блестящая, у одних видов легко спадающая (уклейка), у других
плотно сидящая (шемая). В боковой линии от 40 до 82 чешуй. Пресно-
водные и проходные рыбы. Род насчитывает 4 вида. В водоёмах бас-
сейна Кубани обитают представители двух видов.
По современной системе (Пономарев, Баканева, Федоровых, 2016)
род Шемаи (Chalcalburnus Berg, 1932) входит в состав рода Уклей-
ки (Alburnus Rafinesque, 1820). Дж. Нельсон с соавторами (Nelson,
Grande, Wilson, 2016) сводит Chalcalburnus в синоним Alburnus, хотя
ранее (Nelson, 2006) он признавал этот род самостоятельным.
Как пишут М. Котла и Дж. Фрейхоф (Kottelat, Freyhof, 2007),
нынешний род Alburnus включает виды, которые традиционно были
помещены в два различных рода: Alburnus (уклейки) и Chalcalburnus
(шемаи). Более раннее различие между этими двумя родами было ос-
новано на различии в вентральном киле, который не был покрыт или
частично покрыт чешуёй, в количестве и размере жаберных тычинок
и степени зазубренности глоточных зубов. Эти признаки проявляют
изменчивость и перекрываются у нескольких видов из двух групп, а
также внутри некоторых видов и не имеют диагностической ценности
на уровне рода. Длина бесчешуйной части брюшного киля измеряет-
ся как количество поперечных рядов чешуй вдоль обнажённой части
киля, отсчитываемых от анального отверстия вперёд. Виды Alburnus
alburnus и Alburnus chalcoides ранее относились к ряду подвидов. Изу-
чение фактического материала показало, что некоторые из них являют-
ся валидными видами.
105
Таблица
для определения видов рода
Уклейки — Alburnus Heckel, 1843
1 (2). В боковой линии 45—55 чешуй. Чешуя легко спадающая.
Обитает преимущественно в пресных водоёмах.
Обыкновенная уклейка — Alburnus alburnus (Linnaeus, 1758)
(с. 105)
2 (1). В боковой линии 60—75 чешуй. Чешуя плотно сидящая.
Проходная рыба.
Черноморско-азовская шемая — Alburnus mento (Heckel, 1836)
(с. 106)
Краткая биологическая характеристика видов рода
Уклейки — Alburnus Heckel, 1843
Обыкновенная уклейка — Alburnus alburnus (Linnaeus, 1758)
(рис. 61, 2.39)
Рис. 61. Обыкновенная уклейка — Alburnus alburnus (Linnaeus, 1758)
D III—IV (7) 8 (9), A III (14) 16—19 (20), l.l. (45) 40—52 (55), чаще
всего 46—52. Киль за брюшными плавниками на всём протяжении не
покрыт чешуёй. Глоточные зубы двухрядные, обычно 2.5—5.2. Чешуя
тонкая, легко спадает. Жаберные тычинки длинные, густосидящие, их
13—15, изредка больше.
Уклейка — пресноводная, стайная рыба, держится у поверхности
воды. Имеет пелагическую окраску — спинка тёмная, серовато-голу-
бая с зеленоватым отливом, а брюшко и бока серебристые, со светлым
106
отблеском. Спинной и хвостовой плавники тёмные, а остальные желто-
ватые или красноватые (см. рис. 2.39). Встречается практически повсе-
местно: в реках, озёрах, лиманах, водохранилищах, каналах. Предпочи-
тает чистую воду. Отсутствует в горных озёрах и горных участках рек,
где температура воды низкая.
Половая зрелость наступает на 2—3 -м году жизни при длине тела
8—10 см и массе 5—10 г. Плодовитость колеблется от 2 до 10 тыс.
икринок. Икринки липкие, откладываются на растительность. Икроме-
тание порционное. Нерест происходит в мае—июне. Инкубационный
период при температуре 15—18 °С длится 3—4 сут.
Питается уклейка ракообразными — планктонными организмами,
иногда икрой и личинками рыб, а также насекомыми.
Представляет интерес для рыболовов-любителей, как наживка.
Уклейка иногда образует помеси с плотвой, краснопёркой и гус-
терой.
Черноморско-азовская шемая — Alburnus mento (Heckel, 1836)
(рис. 62, 2.38)
Рис. 62. Черноморско-азовская шемая — Alburnus mento (Heckel, 1836)
D III 8 (9), A III 14—16, l.l. 56—74 (66). Небольшая рыбка с уд-
линённым телом, покрытым чешуёй средних размеров. За брюшными
плавниками имеется киль, в задней части не покрытый чешуёй. Жабер-
ные тычинки частые и длинные, на первой жаберной дуге их 19—25.
Глоточные зубы двухрядные (2.5—5.2), слабо зазубренные. Голова не-
большая, рот полуверхний.
107
Шемая — обитатель пелагиали (толщи воды). В соответствии с
этим окраска спины и головы тёмная с зеленоватым оттенком, бока и
брюшко — серебристые. Хвостовой плавник серый, все остальные —
розовые, а по краям — слегка сероватые (см. рис. 2.38). Годовики ше-
маи сходны с уклейкой по внешнему виду, но у шемаи, в отличие от
уклеи, плотно сидящая чешуя, больше чешуй в боковой линии и цвет
спины имеет голубоватую окраску.
Шемая — проходная рыба, для икрометания из мест нагула (Азов-
ское море) заходит в р. Кубань, её притоки: Лабу, Псекупс, Пшиш,
Белую, а также закубанские реки: Адагум, Абин, Псебес, Шебш и др.
Заход шемаи в реки начинается осенью, поднявшись до среднего тече-
ния Кубани, она зимует в условиях реки, а весной поднимается в вер-
ховья. Для нереста шемая выбирает галечниковый грунт в местах, где
образуются перекаты (скорость течения воды — до 1 м/с).
Половозрелость наступает на 3—4 -м году жизни. Нерест происхо-
дит в мае при температуре воды 18 °С. Плодовитость шемаи в среднем
составляет 15—20 тыс. икринок. Икрометание происходит в ночное
время в неглубоких местах с быстрым течением. Икра сначала приклеи-
вается к камням, а потом смывается с них и забивается между галькой,
где происходит её развитие. Инкубационный период при температуре
18—20 °С длится около 3 сут. После всасывания желточного мешка
личинки выходят из своих убежищ и скатываются в Азовское море.
Взрослые особи после нереста также скатываются в море.
Крупных размеров шемая не достигает: длина колеблется от 20 до
30 см, масса от 150 до 300 г. Максимальная масса — 400 г. Питается
шемая в основном беспозвоночными и насекомыми. Шемая — одна из
самых ценных рыб Кубано-Азовского бассейна, обладающая высокими
пищевыми качествами, особенно в вяленном виде. Мясо очень вкус-
ное и нежное. Шемая переводится с персидского «царская рыба» (шах-
маи).
Ранее годовые промысловые уловы шемаи колебались по годам
от 1 до 60 т. В последние годы из-за неблагоприятных условий естест-
венного размножения запасы шемаи резко снизились и вид включён в
Красную книгу Краснодарского края (2017).
Строительство плотин на реках отрезало путь производителям к
нерестилищам и скату молоди в море. После строительства Краснодар-
ского водохранилища образовалась жилая форма шемаи, которая нагу-
ливается в водохранилище, а для нереста поднимается в притоки Куба-
108
ни (Псекупс, Пшиш, Белую, Лабу и др.) (Москул, Никитина, Гришкин,
1990; Москул, 1995).
Род Быстрянки — Alburnoides Jeitteles, 1861
Род Быстрянки мало чем отличается от рода Уклейки. Быстрянки
отличаются более высоким телом, незазубренными глоточными зубами
и обычно более короткими и редкими тычинками. Род включает пять
видов (Романов, 2014). В фауне бассейна Кубани обитает один вид —
русская быстрянка — Alburnoides rossicus Berg, 1924.
Русская быстрянка — Alburnoides rossicus Berg, 1924 (рис. 63, 2.40)
Рис. 63. Русская быстрянка — Alburnoides rossicus Berg, 1924
D III 7—9, чаще III 8; A III 13—16 (обычно 14—15), l.l. (45) 47—53
(56). Тело высокое, рот конечный. Отверстия боковой линии сверху и
снизу окаймлены чёрными точками, поэтому вдоль боковой линии тя-
нется пунктирная двойная полоска. Выше боковой линии широкая тём-
ная полоса тянется от верхнего края жаберной крышки до основания
хвостового плавника (см. рис. 2.40).
Зубы двухрядные, гладкие, 2.5—5.2. Длина тела достигает
9—11 см, обитает в верхних участках реки. Половозрелой становится
на 2-м году жизни. Икрометание происходит в мае—июне на галеч-
никовом грунте, на перекатах, где глубина не превышает 15—20 см.
109
Плодовитость — около 1 тыс. икринок. Питается насекомыми и их
личинками, а также растительностью и икрой рыб. Населяет водоёмы
бассейна Кубани (левобережные притоки с быстрым течением).
Род Лещи — Abramis Cuvier, 1816
Высокотелые рыбы с длинным анальным плавником (24—30 вет-
вистых луча). Нижняя лопасть хвостового плавника длиннее верхней.
За брюшными плавниками имеется киль, не покрытый чешуёй. За за-
тылком — бороздка без чешуи. Боковая линия полная. Глоточные зубы
однорядные. Род включает 1 вид (лещ), который встречается в фауне
бассейна Кубани.
Лещ — Abramis brama (Linnaeus, 1758) (рис. 64, 2.41)
Рис. 64. Лещ — Abramis brama (Linnaeus, 1758)
D III 9—10, A III 24—30, l.l. 51—57. Широко распространённый
вид, водится в реках, лиманах, озёрах и водохранилищах. По строению
тела лещ легко отличается от других видов, оно широкое, сильно сплю-
щено с боков. Рот полунижний, маленький, выдвижной. Глоточные
зубы однорядные, с каждой стороны по 5—6 зубов. Жаберных тычи-
нок в первой жаберной дуге от 19 до 24 шт. Окраска плавников и тела
леща зависит от возраста, местообитания, грунта и др. Грудные плав-
ники немного не доходят до основания брюшных, а брюшные доходят
до анального отверстия (см. рис. 2.41).
110
Питается лещ во взрослом состоянии бентосом, но не отказывает-
ся и от различных насекомых, растительности и семян растений. Мо-
лодь предпочитает зоопланктонные организмы.
В водоёмах бассейна Кубани встречаются две формы леща: тувод-
ная и полупроходная. Полупроходной лещ кормится в море, а для икро-
метания заходит в лиманы и реки. Туводный лещ всю жизнь проводит
в пресной воде.
Половое созревание у леща наступает на 2—4-м году жизни. Сам-
цы созревают на год раньше самок. На голове и чешуе появляются бу-
горки из рогового эпителия — брачный наряд. Нерест очень шумный:
на мелководье самцы устраивают брачные игры, выпрыгивают из воды
на 50 см и плашмя падают в воду. Продолжительность нереста длится
40—50 дней. Начинается, как правило, в апреле при температуре воды
16 °С и заканчивается в июне. В отношении нерестилищ лещ неприхот-
лив. Нерестовым субстратом может служить как свежезалитая, так и
прошлогодняя растительность. При недостатке таких нерестилищ лещ
успешно нерестится на подводные предметы на глубине 1,5—2,0 м.
Плодовитость леща колеблется от 40 до 430 тыс. икринок. Инкубаци-
онный период длится около трёх дней. Подобно всем карповым, личин-
ки леща проходят стадию покоя, а затем начинают активно питаться и
двигаться.
Темп роста леща относительно высокий: двухлетки достигают
массы 120 г и длины 18 см, трёхлетки — 250 г и 22 см, четырёхлетки и
пятилетки — соответственно 450 г и 28 см, 670 г и 30 см. Максималь-
ный возраст — 10—12 лет. Но обычно в промысловых уловах не попа-
даются особи старше 10-летнего возраста и массы 1,5—2,2 кг.
Лещ — ценная промысловая рыба. До зарегулирования речного
стока шёл интенсивный промысел полупроходного леща до 1 500—
2 000 т в год. В настоящее время уловы леща заметно снизились и
колеблются от 120 до 346 т в год. Лещ представляет большой интерес
для спортивного рыболовства.
Род Синцы — Ballerus Heckel, 1843
Высокотелые рыбы с очень длинным анальным плавником, на-
считывающим от 35 до 76 ветвистых лучей. Тело более вытянуто в
длину, чем у леща. Непарные плавники сероватые с тёмными краями,
парные желтоватые, на концах тёмные. В боковой линии от 46 до 75
111
чешуй. Глоточные зубы однорядные, их формула 5—5. Род включает
2 вида, один из которых обитает в рассматриваемых водоёмах (бело-
глазка).
Белоглазка — Ballerus sapa (Pallas, 1814) (рис. 65, 2.42)
Рис. 65. Белоглазка — Ballerus sapa (Pallas, 1814)
D III 7—8, A III 35—42, l.l. 48—53. Отличается от леща длинным
анальным плавником. Тело, как у леща, сжато с боков. Рыло толстое,
выпуклое, рот полунижний. Чешуя плотно сидящая. Глоточные зубы
однорядные, их формула 5—5, реже 5—6.
В отличие от леща, больше привязана к пресной воде. Для нереста
поднимается вверх по течению. Нерестится в русле рек. Плодовитость
составляет 10—50 тыс. икринок. Икрометание происходит в апреле—
мае. Питается бентическими организмами.
Рыбохозяйственного значения из-за малочисленности практиче-
ски не имеет. Внесена в Красную книгу Краснодарского края (2017).
Согласно современным взглядам (Kottelat, Freyhof, 2007; Nelson,
Grande, Wilson, 2016), род Ballerus Heckel, 1843 состоит из двух ви-
дов (Ballerus ballerus (Linnaeus, 1758) — синец и Ballerus sapa (Pal-
las, 1814) — белоглазка) с очень длинным анальным плавником. Оба
они были традиционно включены в род Abramis, но молекулярные и
морфологические данные показывают, что они представляют собой
112
отдельный род. Впрочем, молекулярные данные свидетельствуют и о
том, что рода Abramis, Ballerus, Blicca, Vimba и западноазиатские рода
Acanthalburnus и Acanthobrama тесно связаны.
Род Густеры — Blicca Heckel, 1843
От рода Лещи — Abramis отличается двухрядными глоточными
зубами, а также более коротким анальным плавником. Единственный
представитель рода в ихтиофауне бассейна р. Кубани — густера (Blicca
bjoerkna (Linnaeus, 1758)).
Густера — Blicca bjoerkna (Linnaeus, 1758) (рис. 66, 2.43)
Рис. 66. Густера — Blicca bjoerkna (Linnaeus, 1758)
D III 8 (9), A III 19—23, l.l. 43—51. Жаберных тычинок 14—21.
Густера по внешнему виду напоминает леща: тело высокое, плоское,
с заметным горбом, небольшой полунижний рот. Но она имеет более
крупную чешую, двухрядные глоточные зубы, непарные плавники ко-
роткие. Первые жёсткие лучи не зазубренны. На спине за затылком
имеется бороздка, не покрытая чешуёй. Лопасти хвостового плавника
одинаковой длины, у леща нижняя лопасть длиннее верхней.
113
Обитает густера в медленно текучих или стоячих, хорошо прогре-
ваемых водах, реже встречается в осолонённых лиманах. В горных, бы-
стротекучих и холодных речках не встречается. Предпочитает обитать
в местах с глинисто-илистым дном вблизи небольших зарослей макро-
фитов. Темп роста значительно медленнее, чем у леща, и таких разме-
ров, как лещ, никогда не достигает. Лишь редкие особи имеют массу
1 кг и длину 35—40 см, чаще всего она весит 300—400 г, а длина не
превышает 25—28 см. В малопроточных водоёмах, почти полностью
заросших, обитает тугорослая форма, максимальные размеры которой
не превышают 15—17 см, а масса — 40—80 г.
Созревает густера рано, уже на второй год жизни при длине 10—
13 см, некоторые особи участвуют в нересте. Нерестится густера позже
леща при более высокой температуре воды (15—17 °С), в мае—июне.
Икра откладывается на растениях. Икрометание порционное, порций
обычно три. Плодовитость составляет 12—120 тыс. икринок.
Питается густера во взрослом состоянии бентическими организ-
мами, а также растительностью и семенами растений.
Густера является объектом любительского рыболовства.
В природе густера может образовывать помеси с плотвой, красно-
пёркой, уклейкой, лещом, рыбцом и, возможно, с другими видами.
Род Рыбцы — Vimba Fitzinger, 1873
Тело умеренно высокое. Рыло выдаётся вперёд, рот нижний по-
лулунный. Глоточные зубы однорядные, их формула 5—5. Анальный
плавник начинается позади вертикали конца спинного. Лучей в спин-
ном плавнике III 7—9, в анальном — III 15—23. В боковой линии 48—
64 чешуи. На спине между спинным и хвостовым плавником имеется
хорошо заметный киль, покрытый чешуёй, за головой и до спинного
плавника свободная от чешуи бороздка. На брюхе между брюшными и
анальным плавниками имеется киль, не покрытый чешуёй.
Согласно сводке «Промысловые рыбы России» (2006), род Vimba
содержит один вид. В составе вида, кроме номинативного подвида Vim-
ba vimba vimba (Linnaeus, 1758) — обыкновенный рыбец, или сырть,
обитающего в бассейнах Северного и Балтийского морей, различают
ещё три подвида: Vimba vimba persa (Pallas, 1814) — каспийский ры-
бец (обитает в бассейне Каспийского моря); Vimba vimba carinata (Pal-
las, 1814) — черноморско-азовский рыбец (бассейн Чёрного и Азов-
114
ского морей); Vimba vimba tenella (Nordmann, 1840) — малый рыбец
(Лужняк, 2003; Туниев, 2005; Решетников, Пашков, 2017). Два послед-
них подвида обитают в бассейне Кубани и причерноморских рек.
Таблица
для определения подвидов рода
Рыбцы — Vimba Fitzinger, 1873
1 (2). В боковой линии 56—58 чешуй. В анальном плавнике 18—
21 луч. Чёрное и Азовское моря, бассейн Кубани.
Черноморско-азовский рыбец — Vimba vimba carinata (Pallas,
1814) (с. 114)
2 (1). В боковой линии 54—55 чешуй. В анальном плавнике 15—
17 лучей. Реки Черноморского побережья и закубанские реки.
Малый рыбец — Vimba vimba tenella (Nordmann, 1840) (с. 116)
Краткая характеристика подвидов рода
Рыбцы — Vimba Fitzinger, 1873
Черноморско-азовский рыбец — Vimba vimba carinata (Pallas,
1814) (рис. 67, 2.44)
Рис. 67. Черноморско-азовский рыбец — Vimba vimba carinata (Pallas, 1814)
D III 8 (9), А III (17) 18—21, l.l. 56—58. Жаберных тычинок 16—
20. Проходные, полупроходные и пресноводные рыбы. От других рыб
рыбец отличается наличием бороздки, не покрытой чешуёй между за-
115
тылком и началом спинного плавника, между концом спинного и на-
чалом хвостового плавника — ясно заметный киль, покрытый чешуёй.
Позади брюшных плавников — киль, не покрытый чешуёй.
Рот нижний, полулунной формы, выдвигается вперёд и вниз при
питании бентосными организмами. Глоточные зубы однорядные. Жа-
берные тычинки короткие, редкие, на первой жаберной дуге их насчи-
тывается 16—20.
Окраска рыбца изменчива и в период нереста наиболее красива:
спина тёмно-серого цвета, бока краснеют, брюшные и анальный плав-
ники ярко-оранжевые. У самцов появляется брачный наряд. После не-
реста спина становится голубовато-серой, брюшко — серебристо-бе-
лое, брюшные и анальный плавники — желтоватого цвета.
Обычно в период нагула рыбец больших косяков не образует, но
перед миграцией собирается в стаи. Существует две расы проходного
рыбца: озимая, которая начинает подниматься по рекам осенью (сен-
тябрь—декабрь) с незрелыми половыми продуктами, зимует здесь и
нерестится весной, и яровая, которая идёт на нерест в марте—апре-
ле с почти зрелой икрой. Нерест происходит при температуре воды
18—25 °С на глубине 0,5—0,8 м. Скорость течения должна быть
0,7—0,9 м/с. Нерест порционный, чаще всего 3 раза с интервалом ме-
жду икрометанием 10—12 дней. У самок, нерестующих первый раз,
бывает 2 порции икры, а у старых самок — 1 —2 порции. Для нереста
выбирает места с галечниковым грунтом, перекатами. Основные не-
рестилища находятся в притоках Кубани: Лабе, Белой, Афипсе, Псе-
купсе, Пшише и др. В Кубани нерест происходит в среднем её течении
(район ст-цы Казанской). Нерест рыбца наиболее активно протекает в
утренние и вечерние часы. На нерестилищах самцов значительно боль-
ше, чем самок. Икра слабоклейкая, течением сносится под камни и там
происходит развитие эмбрионов (2—3 сут.). После выклева личинки
некоторое время находятся под камнями, затем начинают скатывать-
ся вниз по течению реки, но в море скатываются через 5—12 месяцев.
Средняя плодовитость колеблется от 80 до 90 тыс. икринок.
Больших размеров рыбец не достигает, максимальная длина его
40—41 см, масса — 700—800 г, предельный возраст — 10—12 лет. В
годовалом возрасте рыбец имеет длину 4—6 см, массу — 1 —3 г.
Обитает в бассейнах Чёрного и Азовского морей.
Питание рыбца в различных условиях имеет некоторые особенно-
сти: молодь потребляет зоопланктон, личинки комаров и других насе-
116
комых, взрослые особи — детрит, частично растительность, бентосные
организмы.
Черноморско-азовский рыбец образует две формы: проходную и
туводную. Проходной рыбец до перекрытия Кубани гидроузлами (Фё-
доровским и Краснодарским) нагуливался в Азовском море, а на нерест
заходил в Кубань и левобережные притоки (Афипс, Псекупс, Пшиш,
Белую, Лабу, Зеленчук и др.). В результате гидростроительства на
р. Кубани был преграждён миграционный путь рыбца к естественным
нерестилищам.
Включён в Приложение 3 «Перечень таксонов животных, нуж-
дающихся в особом внимании к их состоянию в природной среде Крас-
нодарского края» к Красной книге Краснодарского края (2017).
Многолетние исследования, проведённые автором, показали, что
в Краснодарском водохранилище образовалась туводная форма (мест-
ное стадо) рыбца, которое нагуливается в водохранилище, а нерестится
в реках, впадающих в него (Псекупсе, Пшише, Белой, Лабе и др.). По
морфологическим признакам не отличается от проходного рыбца. Но
данная группа требует дополнительных исследований и уточнения сис-
тематики (Москул, 1994).
Малый рыбец — Vimba vimba tenella (Nordmann, 1840) (рис. 68,
2.45)
Рис. 68. Малый рыбец — Vimba vimba tenella (Nordmann, 1840)
D III (7) 8, А III 15—17, l.l. 54—55. Отличается от проходного
117
меньшими размерами, а также морфологическими признаками: мень-
шим количеством лучей в плавниках, меньшим количеством чешуй в
боковой линии и тычинок на первой жаберной дуге. На спинном и хво-
стовом плавниках имеются чёрные пятнышки. Максимальная длина —
18—20 см, причём при длине 12,5 см особи становятся половозрелыми.
Биология мало изучена. Обитает в притоках Кубани, закубанских ре-
ках (Афипсе, Убине и др.), а также в реках Черноморского побережья
(Мзымте, Туапсе, Пшаде и др.). Систематика малого рыбца требует
уточнения.
Малый рыбец внесён в Красную книгу Краснодарского края (2017).
Род Чехони — Pelecus Agassiz, 1835
В водоёмах бассейна Кубани род представлен одним видом.
Чехонь — Pelecus cultratus (Linnaeus, 1758) (рис. 69, 2.46)
Рис. 69. Чехонь — Pelecus cultratus (Linnaeus, 1758)
D II—III 6—7, A II—III 24—29, l.l. 90—115. Жаберных тычинок
15—25. Тело удлинённое, сильно сжатое с боков, саблевидное. Спи-
на почти прямая. Голова небольшая, рот верхний, довольно крупный.
Спинной плавник отнесён назад и расположен над анальным. Грудные
плавники длинные, конец их доходит до основания брюшных. Тело по-
крыто мелкой чешуёй, легко спадающей. На брюхе имеется киль, не по-
крытый чешуёй, тянущийся от горла до анального отверстия. Боковая
линия проходит вблизи брюха, зигзагообразная с сильным изгибом у
грудного плавника. Глоточные зубы двухрядные, их формула 2.5—5.2.
Окраска пелагическая, бока серебристые, спина зеленоватого цве-
та, чешуя с ярко-серебристым блеском (см. рис. 2.46).
118
Чехонь живёт как в пресноводных, так и в солоноватых водах. Пи-
тается водными личинками насекомых, рачками, молодью рыб, а также
насекомыми, падающими в воду.
Растёт чехонь довольно медленно. Масса трёхлетних особей дос-
тигает 150—200 г. В промысловых уловах встречаются в основном
особи длиной 25—35 см при массе 200—350 г. Максимальная длина —
45—50 см, масса — 500—600 г.
Половозрелой чехонь становится на 3-м году жизни. Плодови-
тость колеблется от 10 до 60 тыс. икринок. Нерест происходит в пре-
сной воде в русле рек. Икринки крупные, полупелагические. Нерест в
Кубани, Псекупсе, Пшише, Белой, Лабе и других реках происходит в
мае—июне при температуре воды 16—18 °С. Инкубационный период
короткий — от 3 до 3,5 сут.
Чехонь является объектом промысла как в море, так и в водохра-
нилищах. Годовые уловы колеблются от 45 до 184 т.
Чехонь является также объектом любительского рыболовства.
Род Лини — Tinca Cuvier, 1816
В фауне бассейна Кубани обитает один вид.
Линь — Tinca tinca (Linnaeus, 1758) (рис. 70, 2.47)
Рис. 70. Линь — Tinca tinca (Linnaeus, 1758)
119
D III—IV 7—9, A III—IV 6—8, l.l. 87—115. Жаберные тычинки
длинные, их 10—16. Тело толстое закруглённое. Рот конечный, неболь-
шой. В углах рта имеется по одному небольшому усику. Все плавники
мягкие, закруглённые, хвостовой — с очень слабой выемкой. У самок
брюшные плавники короче грудных, у самцов — длиннее, у половозре-
лых самцов второй неветвистый луч брюшных плавников заметно утол-
щён и широк, что служит хорошим признаком для определения пола.
Спина окрашена в тёмно-зелёный цвет, переходящий по бокам в зеле-
новато-бурый и зеленовато-жёлтый, плавники тёмные. Окраска линя
меняется иногда до чёрного цвета. Радужная оболочка глаз красная
(см. рис. 2.47). Тело покрыто мелкой, плотно сидящей чешуёй и всегда
обильно покрыто слизью. Это защитная функция организма. Глоточные
зубы однорядные, чаще всего 4—5, обычно слева 4, а справа 5.
Линь — пресноводная и оседлая рыба, обитает во всех водоёмах со
слабым течением, а также в водоёмах заиленных, с зарослями водной
мягкой растительности. Держится линь преимущественно у дна. Очень
нетребователен к содержанию кислорода в воде и может жить при очень
малом проценте насыщения. Далёких миграций не совершает.
Растёт медленно. Средняя масса — 400—600 г. В кубанских ли-
манах и водоёмах, расположенных на малых степных реках, линь дос-
тигает массы 1,5—1,8 кг и длины 38—44 см. Самцы растут несколько
медленнее самок. Половозрелым линь становится на 3—4-м году жиз-
ни. Икрометание порционное, икра мелкая и липкая, приклеивающаяся
к растениям. Плодовитость колеблется от 200 до 800 тыс. икринок. Не-
рест происходит в мае—июне при температуре воды 19—20 °С. Инку-
бационный период длится 2,5—3 сут. В первое время молодь питается
зоопланктоном, а взрослые — донными беспозвоночными: личинками
хирономид, мелкими моллюсками и ракообразными. Кишечник корот-
кий — 100 % длины тела.
Промысловое значение линь имеет только в лиманах и некоторых
водоёмах, расположенных на степных реках. Линь является объектом
спортивного рыболовства.
Род Подусты — Chondrostoma Agassiz, 1835
Род представлен 5 видами. В реках бассейна Кубани и Черномор-
ского побережья Краснодарского края обитают 2 вида. Ранее выделяли
два подвида, в том числе кубанский подуст, но, по данным последней
120
ревизии (Пономарев, Баканева, Федоровых, 2016), кубанский подвид
признан видом.
Таблица
для определения видов рода
Подусты — Chondrostoma Agassiz, 1835
1 (2). Вдоль боковой линии тянется двойная тёмная полоска из
чёрных пигментных точек. Брюшные плавники доходят до ануса.
Колхидский подуст — Chondrostoma colchicum Derjugin, 1899
(с. 120)
2 (1). Продольная тёмная полоска из пигментных точек отсутству-
ет. Брюшные плавники не доходят до ануса.
Кубанский подуст — Chondrostoma kubanicum ( Berg, 1914) (с. 121)
Краткая характеристика видов рода
Подусты — Chondrostoma Agassiz, 1835
Колхидский подуст — Chondrostoma colchicum Derjugin, 1899
(рис. 71, 2.48)
Рис. 71. Колхидский подуст — Chondrostoma colchicum Derjugin, 1899
D III 8 (9), A III 9 (10), l.l. 57—64. Жаберных тычинок 22—30. По-
звонков 43—47. Глоточные зубы однорядные, формула 6—5 или 6—6.
Вдоль боковой линии тянется двойная тёмная полоска из чёрных пиг-
ментных точек (см. рис. 2.48).
121
Тело высокое, сжатое с боков, удлинённое. Начало спинного плав-
ника расположено над началом брюшных. Брюшного киля нет. Рыло
заметно выдаётся вперёд. Рот поперечный и слабодугообразный. Ниж-
няя челюсть покрыта роговым чехликом. Брюшина чёрная. Брюшные
плавники почти достигают анального отверстия. Живёт в русле рек,
обычно на песчаном и каменистом грунте. Больших размеров подуст не
достигает, средние его размеры — 22 —26 см, но встречаются и более
крупные экземпляры — 30 —32 см.
Созревает подуст на 3—4-м году жизни, плодовитость неболь-
шая — 2,1—13,2 тыс. икринок. Икрометание происходит в русле
реки в апреле—мае. Икра сносится течением под гальку и там раз-
вивается. Растёт молодь подуста быстро, сеголетки достигают длины
6—7 см, двухлетки — 12 —15 см. Молодь питается зоопланктоном,
а со второго года жизни исключительно растительной пищей (водо-
рослями, обрастаниями), поэтому кишечник в 2—3 раза превышает
длину тела.
Промыслового значения не имеет, а часто это трофей рыболова-
любителя.
Кубанский подуст — Chondrostoma kubanicum (Berg, 1914) (рис. 72,
2.49)
Рис. 72. Кубанский подуст — Chondrostoma kubanicum (Berg, 1914)
D III 8—9 (10) (чаще 8), A III 9—11 (чаще всего 10), l.l. 54—62.
Позвонков 42—47. Грудные и брюшные плавники короткие. Брюшные
122
не доходят до ануса (у колхидского почти достигают). Ротовая щель
почти прямая. Рыло заострённое. Формула глоточных зубов чаще всего
6—5, иногда 5—5, редко 6—6 (у колхидского всегда 6—5). Продоль-
ная тёмная полоса из пигментных точек отсутствует (у колхидского она
есть). Встречаются вытянутые в длину экземпляры, у которых высота
тела составляет менее 24 % длины тела (без С), и широкие, у которых
соответственное соотношение более 26 % (см. рис. 2.49).
Эндемик р. Кубани. Биология этого вида практически не изуче-
на. Систематика требует уточнения. Промыслового значения не имеет,
часто это трофей рыболова-любителя.
Род Усачи — Barbus Cuvier, 1816
Средние и мелкие рыбы. Характерны 2 пары усиков — одна в уг-
лах рта, другая на конце рыла, короткие анальный и спинной плавники,
трёхрядные глоточные зубы. Рыло удлинённое. Губы развиты хорошо.
Спинной плавник короткий, расположен над брюшными. Последний
неветвистый луч спинного плавника утолщён и сзади с зубчиками. К
этому роду относится большое число видов, населяющих воды Сред-
ней Азии и Европы. В водоёмах бассейна Кубани и прилегающих к
нему рек род представлен двумя видами: колхидский усач — Barbus
escherichii Steindachner, 1892 (обитает в реках Черноморского побе-
режья Краснодарского края: Мзымте, Пшаде, Туапсе, Вулане и др.) и
кубанский усач — Barbus kubanicus Berg, 1912 (населяет водоёмы бас-
сейна Кубани и прилегающих к нему рек). Ранее считались подвидами
крымского усача — Barbus tauricus Kessler, 1877.
Таблица
для определения видов рода
Усачи — Barbus Cuvier, 1816
1 (2). Жаберных тычинок снаружи 12, изнутри 17. На теле и плав-
никах тёмные пятнышки. Обитает в реках Черноморского побережья
Краснодарского края (Мзымте, Пшаде. Туапсе, Вулане и др.).
Колхидский усач — Barbus escherichii Steindachner, 1897 (с. 123)
2 (1). Жаберных тычинок снаружи 10, изнутри 16. Нижняя губа
трёхраздельная. Населяет водоёмы бассейна Кубани и закубанских рек.
Кубанский усач — Barbus kubanicus Berg, 1912 (с. 123)
123
Краткая характеристика видов рода
Усачи — Barbus Cuvier, 1816
Колхидский усач — Barbus escherichii Steindachner, 1897 (рис. 73,
2.50)
Рис. 73. Колхидский усач — Barbus escherichii Steindachner, 1897
D IV 8, A III 5, l.l. 52—60 (в среднем 56,2). Жаберных тычи-
нок снаружи 12, изнутри 17. На теле и плавниках тёмные пятнышки
(см. рис. 2.50). Тело вальковатое, покрытое мелкой чешуёй. Утолщён-
ный луч спинного плавника с острыми зубчиками. Длина половозрелых
особей — 25 —30 см. Является объектом любительского рыболовства.
Икра ядовита, употреблять в пищу не рекомендуется.
Обитает в реках Черноморского побережья Краснодарского края
(Мзымте, Пшаде, Туапсе, Вулане и др.).
Кубанский усач — Barbus kubanicus Berg, 1912 (рис. 74, 2.51)
Рис. 74. Кубанский усач — Barbus kubanicus Berg, 1912
124
D IV 8, A III 5, l.l. 57—68 (в среднем 62,5). Тело невысокое, вытя-
нутое, с утолщением в области спины. Рыло тупое с большой мясистой
верхней губой. Нижняя губа трёхраздельная. Рот нижний полулунной
формы. Две пары усиков, причём задняя пара, находящаяся в углах рта,
длиннее передней. Спинной плавник расположен над брюшным. Гло-
точные зубы трёхрядные, их формула 2.3.5—5.3.2. Хвостовой плавник
выемчатый, нижняя лопасть несколько больше верхней. Чешуя мелкая,
плотно прилегает к телу, на каждой чешуе имеется один заметный про-
дольный гребень. Жаберных тычинок на первой жаберной дуге снару-
жи 10, изнутри 16.
Окраска: спина от серо-коричневого цвета до тёмно-коричневого
с зеленоватым отливом. Брюшко белое. Спина и бока с мелкими тём-
ными пятнами.
В реках бассейна Кубани усач не достигает больших размеров.
Длина его — 35—36 см. Максимальная длина 45 см, масса — 2,5 кг. По
некоторым данным вырастает до 70 см (Богуцкая, Насека, 2001).
Нерест усача происходит в мае—июне на песчано-галечниковом
грунте, на перекатах. Икра усача ядовита. Личинки, после выклева,
сносятся течением реки в нижние её участки. Подросшая молодь под-
нимается вверх по течению.
Питается усач бентосными организмами, поедает также икру рыб.
Этим самым наносит ущерб рыбному хозяйству: собираясь на нерести-
лищах осетровых рыб усачи поедают их икру.
Усач является объектом любительского рыболовства. Икра слабо
ядовита, употреблять в пищу не рекомендуется.
Обитает в водоёмах бассейна Кубани (реках, водохранилищах),
преимущественно в среднем и верхнем течении Кубани и основных её
притоках. Встречается усач и в Краснодарском водохранилище (Мос-
кул, 1996).
Род Пескари — Gobio Cuvier, 1816
Небольшие рыбки. Тело умеренно удлинённое цилиндрической
формы. Вдоль боков тела 6—14 тёмных пятен. Рот нижний или конеч-
ный. Нижняя губа прервана. Боковая линия полная, расположена по-
середине тела. Чешуя крупная, в боковой линии 34—46 чешуй. Спин-
ной плавник немного впереди основания брюшных. Глоточные зубы
двухрядные. В углах рта имеется по одному усику. Пресноводные
125
рыбы. Род включает около 40 видов, обитающих главным образом
в Восточной Европе, преимущественно на территории современной
России и смежных государств. Наиболее широко распространённый
представитель рода — обыкновенный пескарь — Gobio gobio (Lin-
naeus, 1758), объект любительского и спортивного рыболовства. Об-
разует до 20 подвидов (Атлас..., 2002, 2003). На основании сравни-
тельного морфологического и кариологического анализа (Васильева,
Васильев, Куга, 2004) обыкновенному пескарю Gobio gobio gobio
(Linnaeus, 1758) из бассейна Кубани предложен видовой статус —
Gobio kubanicus Vasil’eva, 2004 — кубанский пескарь.
Кубанский пескарь — Gobio kubanicus Vasil’eva, 2004 (рис. 75,
2.52)
Рис. 75. Кубанский пескарь — Gobio kubanicus Vasil’eva, 2004
D III 7, А II—III 6, l.l. 40—43. Глоточные зубы загнуты крючком,
их формула 3.5—5.3. Усики сравнительно короткие и не достают зад-
него края предкрышки. Хвостовой стебель короткий, окраска тела пё-
страя с различными оттенками, имеются пятна (см. рис. 2.52). На горле
имеется чешуйный покров. Держится в придонных слоях, предпочитает
проточные воды. Небольшие рыбки, длина тела — не более 15—20 см.
Икрометание в конце апреля и мае, икринки откладываются на дне
реки. Питается донными организмами: личинками комаров, подёнок, а
также мелкими моллюсками и икрой промысловых рыб, чем наносит
определённый вред.
126
В бассейне Кубани кубанский пескарь обитает преимущественно в
притоках и частично в русле Кубани, а также в степных реках. Промы-
слового значения не имеет.
Род Румынские пескари — Romanogobio Bănărescu, 1961
Тело удлинённое, слегка сжатое с боков. Хвостовой стебель очень
низкий. Голова конусовидная, широкая. Рот нижний. Верхняя губа
сплошная, нижняя широко прервана посередине. Усиков одна пара, они
длинные, заходят за вертикаль заднего края глаза. Хвостовой плавник
глубоко вырезан, его лопасти заострены. Окраска тела светло-жёлтая.
Верх головы и туловища темнее, чем бока и брюхо. У одних окраска
тела бледная, без пятен, у других вдоль боков тела ряд тёмных округ-
лых пятен (6—12). Спинной, хвостовой, иногда и грудные плавники
имеют до 4 рядов мелких пятнышек.
В водоёмах бассейна Кубани род представлен двумя видами: севе-
рокавказский длинноусый пескарь — Romanogobio ciscaucasicus (Berg,
1932) и кубанский длинноусый пескарь — Romanogobio pentatrichus
Naseka & Bogutskaya, 1998. Некоторые авторы (Naseka, Freyhof, 2004)
описывают для бассейна Кубани ещё один вид — малый длинноусый
пескарь — Romanogobio parvus Naseka & Freyhof, 2004. По внешним
признакам этот пескарь практически не отличается от кубанского длин-
ноусого пескаря.
Таблица
для определения видов рода
Румынские пескари — Romanogobio Bănărescu, 1961
1 (2). Окраска тела бледная, без пятен. Усики достигают заднего
края предкрышки и в среднем равняются половине длины головы.
Северокавказский длинноусый пескарь — Romanogobio ciscauca-
sicus (Berg, 1932) (с. 127)
2 (1). Вдоль боков тела расположено (6) 7—11 (12) тёмных пятен.
Усиков одна пара, они длинные, заходят за вертикаль заднего края гла-
за.
Кубанский длинноусый пескарь — Romanogobio pentatrichus
Naseka & Bogutskaya, 1998 (с. 128)
127
Краткая характеристика видов рода
Румынские пескари — Romanogobio Bănărescu, 1961
Северокавказский длинноусый пескарь — Romanogobio
ciscaucasicus (Berg, 1932) (рис. 76, 2.53)
Рис. 76. Северокавказский длинноусый пескарь — Romanogobio ciscaucasicus
(Berg, 1932)
D III 7, А III 5—7, l.l. 41—48. Глоточные зубы двухрядные, их
формула обычно 3.5—5.3. Тело удлинённое, вальковатое, несколько
более высокое, чем у кубанского пескаря. Одна пара длинных усиков,
достигающих заднего края предкрышки, и в среднем равняются по-
ловине длины головы. Рот нижний. Хвостовой стебель сравнительно
длинный — 19,1—27,0, в среднем 24,0 % длины тела. Горло от межжа-
берного промежутка и до линии, соединяющей задние края оснований
грудных плавников, не покрыто чешуёй (голое). Окраска тела бледная,
без пятен. Общий фон окраски сверху до боковой линии зеленовато-
бурый, а ниже неё светло-жёлтый. Отверстия боковой линии сверху и
снизу окаймлены мелкими пятнышками (см. рис. 2.53). Предпочитает
текучие воды. Длина тела половозрелых особей — от 6,6 до 13,7 см.
Обитает в Кубани как в самом русле реки, так и в лиманах, в зоне
выноса речных вод, а также в водоёмах на степных реках. Образ жизни
практически не изучен.
Северокавказский длинноусый пескарь — Romanogobio ciscauca-
sicus (Berg, 1932) — был изначально описан С. Н. Каменским (1901)
как кавказская вариация (Gobio uranoscopus var. caucasica) дунайского
длинноусого пескаря — Romanogobio uranoscopus (Agassiz, 1828). Поз-
128
же Л. С. Берг (1910, 1914) описал его под названием Gobio uranoscopus
caucasica. В дальнейшем Л. С. Берг (1932) обосновал видовой статус
этого пескаря и дал ему новое замещающее название Gobio ciscauca-
sicus Berg, 1932. Под этим названием длинноусые пескари из рек Се-
верного Кавказа (Терека, Кумы, Кубани и др.) фигурировали вплоть до
таксономической ревизии пескарей региона (Naseka, 1996; Насека, Бо-
гуцкая, 1998), в ходе которой они были отнесены к роду Romanogobio
Bănărescu, 1961 и разделены на 2 вида: северокавказский длинноусый
пескарь — Romanogobio ciscaucasicus (Berg, 1932) и кубанский длинно-
усый пескарь — Romanogobio pentatrichus Naseka & Bogutskaya, 1998.
В среднем и верхнем течении Кубани (у г. Армавира и с. Успенского)
эти виды имеют симпатричное распространение (Насека, Позняк, 2000).
Кубанский длинноусый пескарь — Romanogobio pentatrichus
Naseka & Bogutskaya, 1998 (рис. 77, 2.54)
Рис. 77. Кубанский длинноусый пескарь — Romanogobio pentatrichus Naseka &
Bogutskaya, 1998
D III 7, А III 5, l.l. 42—46. Горло голое. Жаберные тычинки корот-
кие, редкие, их 2—5. Глоточные зубы двухрядные, их формула обычно
3.5—5.3, реже 2.5—5.3 и другие сочетания, вершина зуба крючкообраз-
ная (Насека, Богуцкая, 1998; Naseka, Bogutskaya, Poznjak, 2002).
Тело удлинённое, слегка сжатое с боков. Хвостовой стебель очень
низкий. Голова конусовидная, широкая. Рот нижний. Верхняя губа
сплошная, нижняя широко прервана посередине. Усиков одна пара, они
длинные, заходят за вертикаль заднего края глаза. Хвостовой плавник
129
глубоко вырезан, его лопасти заострены. Окраска тела светло-жёлтая.
Верх головы и туловища темнее, чем бока и брюхо. Вдоль боков тела про-
ходит ряд тёмных округлых пятен (7—11). Спинной, хвостовой, иногда
и грудные плавники имеют до 4 рядов мелких пятнышек (см. рис. 2.54).
Обитает только в Кубани и её левом притоке Лабе (Атлас..., 2003).
Достигает длины 12 см. Образ жизни не изучен. Вероятно, являет-
ся эндемиком бассейна Кубани (Атлас..., 2002, 2003).
Род Сазаны, или Карпы, — Cyprinus Linnaeus, 1758
Род Сазаны характеризуется телом, покрытым крупной чешуёй,
наличием двух пар усиков. Глоточные зубы трёхрядные, с мощнораз-
витой жевательной площадкой. Род включает три вида, один из кото-
рых встречается в водоёмах бассейна Кубани.
Сазан, или карп, — Cyprinus carpio Linnaeus, 1758 (рис. 78, 2.55)
Рис. 78. Сазан, или карп, — Cyprinus carpio Linnaeus, 1758
D III—IV (15) 16—21 (22), A III 5 (6), l.l. 33—40. Сазан — широко
распространённая и хорошо известная рыба. Обитает практически во
всех водоёмах бассейна Кубани, а также в опреснённых участках Азов-
ского моря. Нет сазана в горных притоках Кубани и горных озёрах с
холодной водой.
Сазан имеет прогонистое тело, покрытое крупной чешуёй, у рта
расположены две пары усиков, причём верхние короче нижних. Окра-
130
ска сазана изменчива и зависит от места его обитания, но в целом имеет
тона от золотистого до тёмно-оливкового. Цвет спинки более тёмный,
на боках ярко-золотистый, плавники коричневого цвета, хвост с крас-
новатым оттенком (см. рис. 2.55).
Спинной плавник длинный, имеет небольшую выемку. Брюшные
плавники расположены чуть дальше вертикали, проведённой от спин-
ного плавника. Первые жёсткие лучи плавников окостенелые и зазуб-
рены. На первой жаберной дуге от 23 до 26 жаберных тычинок.
Рот нижний, выдвижной. Глоточные зубы массивные, трёхрядные
1.1.3.—3.1.1, иногда 1.2.3—3.2.1. Питается сазан разнообразной пищей,
в пищевой рацион его входят бентосные организмы, зоопланктон, рас-
тительность (бо
́
льшей частью прошлогодняя, полуразложившаяся), ли-
чинки и взрослые формы воздушных и водных насекомых, жуки и др.
Половое созревание у сазана наступает рано: у самцов в три—че-
тыре (иногда в два года), у самок — в четыре—пять (иногда в три года).
Плодовитость самок большая и достигает 2—3 млн икринок, но в сред-
нем — 700—800 тыс. Нерест сазана растянутый — с конца апреля до
конца июня. Массовый нерест происходит в начале мая. Икрометание
порционное.
Нерестится он на глубине от 25 до 40 см, когда температура воды
поднимается до 18—20 °С, причём полупроходная форма нерестится
несколько позже.
Икрометание начинается рано утром, на восходе солнца, и продолжа-
ется до 10—11 ч утра. Обычно одну самку сопровождают два—три самца.
Рост сазана зависит от условий обитания и поэтому сильно отлича-
ется в различных водоёмах. В среднем к концу первого года сазан дос-
тигает 10—15 см, к концу второго — 20 —25 см, третьего — 30 —36 см,
четвёртого — 41 —46 см. Продолжительность жизни сазана большая —
15—20 лет, но такие особи очень редки. В водоёмах Краснодарского
края встречаются экземпляры в возрасте 10—12 лет, массой 11—15 кг.
Сазан является промысловой рыбой. Уловы составляют 100—
500 т в год. В последние годы уловы сазана заметно упали и составля-
ют 50—100 т (1996 г.). Сазан является также объектом любительского
рыболовства.
В результате селекционной работы выведены новые расы сазана,
обладающие теми свойствами, которые нужны человеку (рис. 79): от-
сутствием или малым количеством чешуи на теле (рис. 79, г), более
высоким телом и высокой скороспелостью. Обычно породы карпа
131
оцениваются по характеру чешуйного покрова (см. рис. 2.56). Разли-
чают карпов чешуйчатых (рис. 79, а), у которых тело сплошь покрыто
чешуёй; карпов с неполным чешуйным покровом (рис. 79, б, в, д), у
которых чешуя имеется вдоль боковой линии и редко разбросана по
другим участкам тела — зеркальные карпы; карпов, лишённых чешуи,
называемых голыми, и др.
Рис. 79. Карп и его формы:
а — чешуйчатый; б — зеркальный; в — линейный; г — голый; д — голый, или
кожистый (без чешуи); е — рамчатый
Карп — один из самых распространённых объектов товарного
рыбоводства. Это объясняется его высокими вкусовыми качествами, а
также освоенной технологией воспроизводства и выращивания его как
в прудах и озёрах, так и на тёплых водах ТЭЦ.
Ещё на рассматриваемой территории можно встретить карпов кои
(японского парчового карпа) — декоративных одомашненных рыб,
выведенных из амурского подвида Cyprinus carpio haematopterus (Lin-
132
naeus, 1758) сазана (Cyprinus carpio). История разведения парчовых
карпов в прудах насчитывает около трёх тысяч лет. В настоящее время
наиболее продвинутая страна в селекции карпов кои — Япония. Кар-
пом кои считается рыба, прошедшая 6 селекционных отборов, после
чего ей присваивается определённая категория. В настоящее время в
Японии существует много разновидностей кои, но за стандарт принято
считать всего четырнадцать цветных форм и раскрасок.
Род Караси — Carassius Jarocki, 1822
Караси — крайне выносливые и малотребовательные к условиям
среды рыбы (кислородному режиму, температуре воды, её качеству,
характеру водоёма).
Это обстоятельство обусловливает широкий ареал их распро-
странения, но особенно они многочисленны в малопроточных, сильно
заросших водоёмах. Очень часто караси встречаются в таких водо-
ёмах, где ни одна другая рыба обитать не может. От сазана караси от-
личаются отсутствием усиков вокруг рта и однорядными глоточными
зубами.
В водоёмах бассейна Кубани обитают два вида: обыкновенный,
или золотой, карась — Carassius carassius (Linnaeus, 1758) и серебря-
ный карась — Carassius gibelio (Bloch, 1782). Данное латинское би-
номинальное название принято с 2003 г., ранее — Carassius auratus
gibelio (Bloch, 1782). К последнему виду ранее относили китайского
карася, или золотую рыбку. Её предок был одомашнен человеком ещё
в VII в. н. э. Является одной из самых популярных аквариумных рыб и
представлена целой группой пород домашних аквариумных животных,
полученной в результате многовековой направленной селекции.
Таблица
для определения видов рода
Караси — Carassius Jarocki, 1822
1 (2). В боковой линии 31—36 чешуй. На первой жаберной дуге
23—33 тычинки. Свободный край спинного плавника выпуклый (за-
круглён); последний неразветвлённый луч анального и лучи спинного
плавников слабо зазубрены (30 мелких зубчиков). Брюшина светлая.
Золотой карась — Carassius carassius (Linnaeus, 1758) (с. 133)
133
2 (1). В боковой линии 26—33 чешуйки. На первой жаберной дуге
38—52 тычинки. Свободный край спинного плавника вогнутый или
прямой; последний неразветвлённый луч спинного плавника сильно
зазубрен (10—15 крупных зубчиков). Брюшина обычно чёрная.
3 (4). Окраска золотисто-бурого или бронзового цвета, l.l. 26—31.
Китайский карась — Carassius auratus (Linnaeus, 1758) (с. 134)
4 (3). Окраска серебристо-бурого цвета, l.l. 29—33.
Серебряный карась — Carassius gibelio (Bloch, 1782) (с. 137)
Краткая характеристика видов рода
Караси — Carassius Jarocki, 1822
Золотой карась — Carassius carassius (Linnaeus, 1758) (рис. 80, 2.56)
Рис. 80. Золотой карась — Carassius carassius (Linnaeus, 1758)
D III—IV 14—21, A II—III 6—8, l.l. 31—36. Тело широкое, сжатое
с боков. Высота его составляет более 2/5 его длины. Жаберных тычинок
на первой жаберной дуге от 23 до 33.
134
Окраска золотого карася зависит от места обитания: в заросших с
тёмным грунтом водоёмах она тёмно-бурая, но в более просторных со
светлым дном бока светло-золотистые (могут быть медно-золотистые),
парные плавники красные с тёмными концами (см. рис. 2.57). Спинной
плавник длинный, слегка закруглён, хвостовой — выемчатый, окраска
их тёмно-серая. Первые жёсткие лучи в спинном, анальном и брюшных
плавниках зазубренные.
Несмотря на неприхотливость к среде обитания, при недостаточно
благоприятных гидрологических, гидробиологических условиях золо-
той карась превращается в тугорослую форму со значительными внеш-
ними изменениями: тело становится прогонистым, голова сильно уве-
личивается. В водоёмах с благоприятными условиями золотой карась
растёт относительно быстро: на третий год жизни он достигает длины
20—25 см и веса 250—300 г.
Половой зрелости золотой карась достигает на 3—4 -м году жизни,
как и все карповые, самцы созревают раньше самок. Нерестится при
температуре воды 14—15 °С. По характеру размножения — фитофил,
икрометание порционное; чаще всего 3 раза, но известны случаи пяти
подходов на нерест. Среднепопуляционная плодовитость — 200—
300 тыс. икринок. Инкубационный период — 5 —7 дней. Стадия покоя
личинок длится 4—6 дней.
В отношении пищи золотой карась неприхотлив: пищей служат
различные бентосные организмы, личинки насекомых, низшие и выс-
шие водоросли. Вместе с бентосом заглатывает много ила. Является
объектом промысла в кубанских лиманах и в водоёмах на степных ре-
ках.
Китайский карась, или золотая рыбка, — Carassius auratus
(Linnaeus, 1758) [Carassius auratus auratus (Linnaeus, 1758)] (рис. 81,
2.59)
Естественным ареалом этого карася является Китай, Япония,
острова Тайвань и Хайнань. Благодаря искусственному разведению
распространился по всему миру. В России китайский карась и его по-
роды (золотые рыбки) впервые появились в XVII в. и содержались
в царских прудах. Разводили его также в рыбоводных хозяйствах
Курской и Белгородской областей и в Краснодарском крае (Атлас...,
2002).
135
Рис. 81. Китайский карась — Carassius auratus (Linnaeus, 1758)
Хромист (окрашенная форма) золотая рыбка — одомашненная
форма типичного китайского карася (Carassius auratus (Linnaeus,
1758) [Carassius auratus auratus (Linnaeus, 1758)]). Более 1 000 лет
назад в Китае, а затем в Японии путём селекции и отбора аквариуми-
стами были выведены различные породы золотой рыбки (около 200
разновидностей): вуалехвосты, телескопы, кометы, львиноголовки и
др. (рис. 82, 2.59), служащие излюбленными объектами аквариумно-
го рыбоводства. Они представляют собой наследственно закреплённые
уродства, связанные главным образом с нарушением деятельности же-
лёз внутренней секреции. Эти формы — результат длительной селек-
ции (Никольский, 1971). В настоящее время золотые рыбки — одни из
самых популярных аквариумных и прудовых рыбок. Широкому рас-
пространению способствует их неприхотливость и чрезвычайное раз-
нообразие. В Китае, Корее и Японии выведено несколько сотен пород
этой декоративной рыбы (Жизнь животных, 1971).
Китайская золотая рыбка — Carassius auratus (Linnaeus, 1758)
[Carassius auratus auratus (Linnaeus, 1758)] — в естественных ус-
ловиях встречается только на востоке России и является объектом
декоративного рыбоводства (Васильева, 1990; Список..., 1997; Ан-
нотированный каталог..., 1998).
136
Рис. 82. Некоторые породы золотой рыбки (Carassius auratus (Linnaeus, 1758)):
а — вуалехвост; б — комета; в — небесное око; г — телескоп; д — шубункин;
е — львиноголовка
В естественных условиях китайский карась встречается в южных
регионах России, в частности в водоёмах Краснодарского и Ставро-
польского краёв. В отдельные годы единичные экземпляры встречают-
ся в азово-кубанских лиманах, водоёмах, расположенных на азово-ку-
банских равнинных реках (Ея, Бейсуг, Кирпили и др.). В Карасунских
озёрах бассейна Кубани встречаются караси-мутанты.
Систематика не изучена.
137
Серебряный карась — Carassius gibelio (Bloch, 1782) (рис. 83, 2.58)
Рис. 83. Серебряный карась — Carassius gibelio (Bloch, 1782)
D III—IV 15—19, A II—III 5—6, l.l. 29—33. В отличие от золотого,
серебряный карась не живёт в сильно заросших водоёмах, он очень ред-
ко встречается в илистых водоёмах. Серебряный карась обитает в р. Ку-
бани и её притоках, а также в пойменных водоёмах и водохранилищах.
Серебряный карась отличается от золотого светлой окраской
(спинка зеленоватая, бока и брюшко серебристо-серые), более про-
долговатой формой тела, спина у него менее выпуклая, высота тела не
превышает 1⁄3 длины тела, хвостовой плавник более вырезан. Спинной
плавник несколько выемчатый (см. рис. 2.58). По внутренним анато-
мическим признакам отличается более длинным кишечником и чёрной
брюшиной. Жаберных тычинок на первой жаберной дуге 38—52.
Темп роста несколько выше, чем у золотого карася: к моменту созре-
вания (3—4 года) достигает 300—400 г веса и длины 25 см. В Краснодар-
ском крае и во всей южной зоне России популяции серебряного карася
состоят преимущественно из самок. Однако нерест происходит ежегодно
при участии самцов других видов (золотой карась, линь, сазан, плотва),
в потомстве вновь получаются одни самки. Такое явление, называемое
гиногенезом (рождение самок), вызвано тем, что при оплодотворении
икры молоками слияние ядер самки и самца в яйце не происходит, муж-
138
ские клетки лишь стимулируют развитие эмбриона. Развитие эмбрионов
происходит нормально, но в потомстве получаются только самки. Такое
состояние популяций серебряного карася носит приспособительный ха-
рактер. Однако при резком ухудшении условий, в популяции появляются
самцы, причём воспроизводительная способность самцов высокая.
Нерест в мае—июне. Плодовитость самки средних размеров ко-
леблется в пределах 160—280 тыс., в среднем — 170 тыс. икринок.
Нерест порционный. Икра клейкая, время от оплодотворения до выкле-
ва личинок при температуре воды 20 °С — 3 —4 сут.
Питается карась разнообразно: в рацион входит животная и рас-
тительная пища. Большое значение в питании играет фитопланктон и
зоопланктон. Максимальная интенсивность питания — после нереста,
особенно в июле, прекращение питания наблюдается с понижением
температуры воды до 14—16 °С.
Очень часто серебряный карась обитает в неспускных или полу-
спускных прудах. При потреблении различных видов кормов темп рос-
та его значительно выше, чем в естественных водоёмах.
В связи с падением запасов ценных видов рыб промысловое зна-
чение карасей в последние годы увеличивается как в лиманах, прудах,
озёрах, так и в реках и водохранилищах. Кроме того, они являются объ-
ектами любительского рыболовства.
Род Толстолобы (Толстолобики) — Hypophthalmichthys
Valenciennes, 1844
Чешуя мелкая, в боковой линии 110—125 чешуй. Спинной плав-
ник короткий (7 ветвистых лучей), без костного луча, анальный удли-
нённый. Усиков нет, рот косой, смотрит вверх. Лоб намного шире, чем
у других карповых (отсюда название). Глаза сидят очень низко, ниж-
ний край их ниже уровня углов рта, поэтому лоб выглядит ещё больше.
Жаберные перепонки не приращены к межжаберному промежутку, а
сращены между собой. Населяют воды Восточной Азии (бассейн Аму-
ра, реки Китая и др.). Акклиматизированы в Средней Азии и южных
европейских странах. Раньше он подразделялся на два рода: Белые
толстолобики — Hypophthalmichthys Valenciennes, 1844 и Пёстрые
толстолобики — Aristichthys Ōshima, 1919. Сейчас, вслед за G. J. How-
es (1981), род Aristichthys рассматривается как синоним Hypophthal-
michthys. Они определённо родственны, но некоторые исследователи
139
считают, что они слишком непохожи, чтобы их можно было отнести к
одному роду. Синонимизация требует дальнейшего изучения (Kottelat,
2013). В водоёмах бассейна Кубани обитают представители двух видов.
Таблица
для определения видов рода
Толстолобы, или Толстолобики, — Hypophthalmichthys
Valenciennes, 1844
1 (2). На всём протяжении брюха имеется острый киль. Жаберные
тычинки длинные, соединённые между собой.
Белый толстолобик — Hypophthalmichthys molitrix (Valenciennes,
1844) (с. 139)
2(1). Киль тянется от брюшных плавников до анального отверстия.
Жаберные тычинки не срастаются между собой.
Пёстрый толстолобик — Hypophthalmichthys [Aristichthys] nobilis
(Richardson, 1846) (с. 141)
Краткая характеристика видов
рода Толстолобы, или Толстолобики, — Hypophthalmichthys
Valenciennes, 1844
Белый толстолобик — Hypophthalmichthys molitrix (Valenciennes,
1844) (рис. 84, 2.60)
Рис. 84. Белый толстолобик — Hypophthalmichthys molitrix (Valenciennes,
1844)
140
D II 7, A II—III 11—14, l.l. 109—125. Родина белого толстолоби-
ка — бассейн Амура и реки Китая. Интродуцирован в бассейне Куба-
ни в 1960 г. вместе с пёстрым толстолобиком, белым и чёрным аму-
рами. Вселённых рыб амурского, или дальневосточного, комплекса
часто называют растительноядными рыбами из-за способности белого
толстолобика и белого амура потреблять растительные корма — фи-
топланктон и макрофиты. Окраска пелагическая: спинка зеленоватого
цвета, бока и брюшко серебристые (см. рис. 2.60). Жаберные перепон-
ки сращены и образуют большую складку поперёк межжаберного про-
межутка, к которому они не приращены. Глоточные зубы однорядные,
их формула 4—4.
Питается белый толстолобик микроскопическими водорослями —
фитопланктоном, поэтому эта рыба является прекрасным мелиорато-
ром водоёмов. При помощи своего цедильного ротового аппарата бе-
лый толстолобик профильтровывает цветущую и мутную от детрита
воду.
Биология растительноядных рыб имеет много общих черт. В ес-
тественных условиях нерест происходит в русле рек при скорости
течения воды 0,3—1,7 м/с в период подъёма уровня воды, во время
паводков. Нерестовая температура колеблется от 19 до 26 °С. Для ик-
рометания выбираются участки реки, имеющие турбулентное тече-
ние, на перекатах или в местах слияния основной реки с протоками.
В р. Кубани нерест происходит на перекатах выше г. Усть-Лабинска
до г. Армавира. Нерест белого толстолобика и белого амура проис-
ходит в верхних слоях воды, пёстрого — в нижних. На одну самку
приходится два—три самца. Нерестовая миграция очень шумная:
рыбы выпрыгивают из воды, преодолевая препятствия. Нерестятся
растительноядные рыбы рано утром. Икра полупелагическая, мел-
кая, в плавучем состоянии поддерживается турбулентными потоками
воды, в стоячих водах икра быстро опускается на дно и гибнет. Та-
кое явление наблюдается ежегодно в Краснодарском водохранилище
(Москул, Никитина, 1986). Икрометание единовременное. Сроки не-
реста растянуты от одного до двух месяцев. Половое созревание у бе-
лого толстолобика наступает в 3—4 -летнем возрасте. Половозрелых
самцов можно определить по наличию роговых зубчиков на лучах
внутренней стороны грудных плавников. На всём протяжении брюха
имеется острый киль. Кишечник очень длинный, более чем в 10 раз
превосходит длину тела.
141
Белый толстолобик — стайная пелагическая рыба средних размеров.
Растёт он очень быстро: сеголетки достигают массы 30—50 г, двухлет-
ки — 500—600 г. Свои высокие генетические способности роста белый
толстолобик начинает реализовывать с третьего года жизни: в Красно-
дарском водохранилище четырёхлетние особи достигают 3,5—4,5 кг,
пятилетки — 4,5—5,5 кг, шестилетки — 5,5—6,8 кг, в десятилетнем воз-
расте встречаются особи длиной 95 см и массой 17,7 кг (Москул, 1995).
Абсолютная плодовитость самок белого толстолобика длиной
56,1—86,1 см и массой 4 100—15 800 г колеблется от 381 до 2 125 тыс.
икринок, в среднем 1 213 тыс. икринок. Относительная плодовитость
варьирует от 93 до 134 икр./г, в среднем — 125 икр./г.
Белый толстолобик является объектом промысла как в реках, водо-
хранилищах, лиманах, так и в опреснённых участках Азовского моря.
Ежегодный вылов колеблется от 100 до 500 т.
Пёстрый толстолобик — Hypophthalmichthys [Aristichthys] nobilis
(Richardson, 1846) (рис. 85, 2.61)
Рис. 85. Пёстрый толстолобик — Hypophthalmichthys [Aristichthys] nobilis
(Richardson, 1846)
D III 7, A III 11—14, l.l. 92—115. Ареал естественного распростра-
нения близок к белому толстолобику. От последнего отличается тем,
что жаберные тычинки свободны, киль тянется от брюшных плавников
до анального отверстия, более тёмной окраской, длинными грудными
плавниками, а также большой головой. Питается в основном зоопланк-
тоном, частично фитопланктоном и детритом. Кишечник пёстрого
142
толстолобика значительно короче и жаберные тычинки не образуют
фильтрующего сетчатого аппарата.
Пёстрый толстолобик обладает быстрым ростом, в десятилетнем
возрасте достигает массы 35 кг, а отдельные экземпляры — 42 кг при
длине 105 см (Москул, 1995).
Половозрелым становится в 4—5 -летнем возрасте при длине 55—
75 см и массе 5,8—6,5 кг. Плодовитость колеблется от 910 до 2 275 тыс.
икринок.
Пёстрый толстолобик имеет промысловое значение как в естест-
венных водоёмах, так и в прудах. Мясо очень вкусное, нежное, не ко-
стлявое. Содержание жира колеблется от 12 до 23 %. Крупные экземп-
ляры от 5 кг и выше идут на изготовление балычных изделий, которые
отличаются высокими вкусовыми качествами.
Семейство Чукучановые — Catostomidae Cope, 1871
Рыбы, относящиеся к этому семейству, распространены от Канады
до Мексики, на Атлантическом побережье США (в Великих озёрах), в
Китае (бассейн р. Янцзы) и Восточной Сибири. Всего насчитывается
13 родов и около 70 видов. В 1971 г. из США в Краснодарский край
(рыбопитомник «Горячий Ключ») были завезены три вида рыб рода
Буффало — Ictiobus Rafinesque, 1820 (большеротый, или буйвол-рыба,
малоротый и чёрный буффало).
Род Буффало, или Иктиобусы, — Ictiobus Rafinesque, 1820
От карпа виды буффало отличаются однорядными многочислен-
ными глоточными зубами, более длинными и тонкими жаберными
тычинками, отсутствием жерновка над глоточными зубами, большим
ртом и толстыми губами. Кроме того у них нет усиков, но губы по-
крыты многочисленными ворсинками и расстояние от начала рыла до
анального плавника в 2,5 раза больше, чем расстояние от начала аналь-
ного до хвостового плавника (у карпа оно меньше). Все три вида буф-
фало предпочитают водоёмы с небольшим течением, хорошо прогре-
ваемые мелководные озёра и водохранилища. Все они имеют высокое
тело и крупную чешую. Рот выдвижной, нижний. Спинной плавник
длинный. Все виды буффало — стайные рыбы, что очень важно при
вылове их из озёр, лиманов и водохранилищ.
143
Таблица
для определения видов рода
Буффало, или Иктиобусы, — Ictiobus Rafinesque, 1820
1 (2 ). Рот большой, полуверхний. Жаберных тычинок 59—68.
Большеротый буффало — Ictiobus cyprinellus (Valenciennes, 1844)
(с. 143)
2 (1). Рот небольшой, нижний. Жаберных тычинок 29—39.
3 (4). Тело высокое, по форме напоминает леща. Голова маленькая.
Чёрный буффало — Ictiobus niger (Rafinesque, 1820) (с. 146)
4 (3). Тело прогонистое, по форме напоминает сазана. Жаберных
тычинок 29—33. Рот нижний небольшой.
Малоротый буффало — Ictiobus bubalis (Rafinesque, 1819) (с. 145)
Краткая биологическая характеристика видов рода
Буффало, или Иктиобусы, — Ictiobus Rafinesque, 1820
Большеротый буффало — Ictiobus cyprinellus (Valenciennes, 1844)
(рис. 86, 2.63)
Рис. 86. Большеротый буффало — Ictiobus cyprinellus (Valenciennes, 1844)
D III 23—30, A II 7—10, V 10—11, P 14—15. Длина тела составляет
в среднем 35 см, при идеальных условиях местообитания максимальная
масса рыбы может достигать 45 кг при длине 120 см. Голова крупная. Жа-
берная крышка высокая, широкая, радиально исчерчена. Глаз небольшой
и расположен ближе к концу рыла. Рот большой, конечный, невсасываю-
144
щий, направлен косо вверх. Глоточные зубы однорядные, тонкие и сла-
бые. Спинной и анальный плавники сдвинуты назад, хвостовой — очень
широкий, слегка выемчатый; спинной длинный, серповидной формы.
Окраска тела тёмно-коричневая, спинка тёмно-серая или оливково-брон-
зовая, бока более светлые, брюхо жёлтое или беловатое, плавники одно-
тонные, обычно коричневато-серые, тёмные, спинной плавник длинный.
Большеротый буффало обитает в пресных мелководных озёрах
и водохранилищах, хорошо переносит мутную воду, в течение всего
лета ведёт стайный образ жизни. Относится к быстрорастущим видам.
В Канаде и Америке — популярный объект спортивного рыболовства.
Большое место занимает в прудовом рыбоводстве, особенно практи-
куется выращивание большеротого буффало на рисовых полях, где на
второй год жизни он достигает 1—1,5 кг.
Половой зрелости большеротый буффало достигает в трёхлетнем
возрасте: в период нереста голова и тело покрывается «жемчужной
сыпью» — брачным нарядом. Икра донная, мелкая, клейкая. Нерес-
товым субстратом служат прошлогодняя растительность, корневища
растений, камни, т. е. он менее прихотлив, чем сазан, которому необ-
ходима свежезалитая вегетирующая растительность. Нерест длится
50—80 дней. В Канаде самки нерестятся не каждый год. Для размноже-
ния буффало заходит в мелководные заливы, когда температура воды
поднимается до 16—17 °С, но может нереститься и при температуре
25—27 °С. Инкубационный период длится 9—10 сут. и значительно
сокращается при высокой температуре воды. Выклюнувшиеся личинки
проходят стадию покоя на дне, под корневищами и камнями, а затем
поднимаются в верхние слои воды.
Сначала питается в основном мельчайшими организмами, затем
более крупными ракообразными. Во взрослом состоянии большеротый
буффало — типичный планктофаг, бентические организмы составляют
в пище небольшой процент.
Для прудового рыбоводства большеротый буффало — перспектив-
ный объект, так как помимо зоопланктона охотно потребляет комбикор-
ма. Опыт его выращивания в прудах Краснодарского края дал положи-
тельный результат: сеголетки достигают 200—400 г, некоторые — до
500—550 г, а двухлетки — до 1,5 кг, трёхлетки — 2,5 кг, четырёхлет-
ки — 3,5 кг. Мясо большеротого буффало вкусное, нежное, но костля-
вее, чем у карпа. В естественных водоёмах не прижился. В настоящее
время и в прудовых хозяйствах буффало практически отсутствует.
145
Малоротый буффало — Ictiobus bubalis (Rafinesque, 1819) (рис. 87,
2.64)
Рис. 87. Малоротый буффало — Ictiobus bubalis (Rafinesque, 1819)
Тело умеренно удлинённое, сжатое с боков, высокое. Его окраска
более светлая, чем у большеротого буффало. У молодых особей спин-
ка тёмно-серо-бронзовая с синеватым отливом, бока более светлые с
золотистым отливом, брюхо белое, плавники однотонные, коричнева-
то-серые. По мере увеличения размеров рыб наблюдается некоторое
осветление окраски. Спина отчётливо дугообразная, перед спинным
плавником сильно сжата с боков в виде киля. Глаза крупные. Рот ма-
ленький, почти горизонтальный. Спинной плавник длинный, его пер-
вые 6—7 лучей значительно длиннее всех остальных, профиль плавни-
ка почти серповидный.
Малоротый буффало предпочитает текучие воды и от большеро-
того отличается медленным темпом роста, расположением рта (у мало-
ротого нижний, у большеротого — полуверхний), короткими и редки-
ми жаберными тычинками, несколько поздним половым созреванием
(основная масса рыб созревает на четвёртый год жизни), а также харак-
тером питания: сеголетки в двухмесячном возрасте почти полностью
переходят на питание зообентосом и при искусственной подкормке
начинают поедать комбикорма. К концу первого года жизни молодь
достигает 70—150 г, на второй год — до 1 кг, на третий — 1,5—2 кг,
146
на четвёртый — до 2,5 кг. Максимальный вес — 15 —18 кг. Пищевые
качества мяса малоротого буффало выше, чем большеротого.
Чёрный буффало — Ictiobus niger (Rafinesque, 1820) (рис. 88, 2.62)
Рис. 88. Чёрный буффало — Ictiobus niger (Rafinesque, 1820)
Тело стройное (высота его три раза укладывается в длине тела), но
более толстое, чем у малоротого буффало. Передняя часть спины закруг-
лена или со слабым килем. Рот маленький, нижний, губы толстые, силь-
но исчерченные. Передняя часть верхней губы расположена много ниже
глаза. Окраска тела тёмная или тёмно-бронзовая, на спине с зеленоватым
отливом. Плавники однотонные, темно-серые. Несколько светлее окра-
ска бывает лишь у рыб, постоянно обитающих в очень мутных водах.
Места обитания чёрного буффало часто совпадают с местами оби-
тания большеротого буффало, но он более предпочитает мутные воды с
сильно заиленным дном, мелкие заиленные пруды и болота. В отличие
от двух предыдущих видов, держится в придонных слоях и по характе-
ру поведения напоминает карпа.
Чёрный буффало — бентофаг, растёт быстрее, чем малоротый.
Созревает на 4—5 -й год жизни. Масса сеголеток составляет 50—70,
двухлеток — 700—1 200, трёхлеток — 2 200—3 000, четырёхлеток —
2 800—5 300 г. При совместном выращивании с карпом интенсивно
потребляет задаваемые карпу комбикорма.
Все три вида завезены в СССР в 1971 г. в рыбопитомник «Горячий
Ключ», затем в 1974 г. в другие прудовые хозяйства европейской части
России и водоёмы Алтая. В настоящее время все виды буффало практи-
чески не встречаются в водоёмах бассейна Кубани.
147
Семейство Гольцовые — Nemacheilidae reGan, 1911
Тело удлинённое, покрыто мелкой чешуёй или голое. Рот нижний
или полунижний, маленький, окружённый 6—8 усиками (3—4 пары).
Выдвижного шипа под глазом нет. Зубы однорядные. Пресноводные,
придонные рыбы. Литофилы. Питаются донными беспозвоночными
и водорослями. В водоёмах бассейна Кубани семейство представлено
двумя родами, в каждом по одному виду.
Таблица
для определения родов семейства
Гольцовые — Nemacheilidae Regan, 1911
1 (2). Задний край хвостового плавника прямой или слегка вы-
пуклый. В спинном плавнике 3—4 жёстких и 7 ветвистых лучей (всего
10—11), в анальном — 3 —4 жёстких и 5 ветвистых (всего 8—9).
Род Усатые гольцы — Barbatula Linck, 1790 (с. 147)
2 (1). Задний край хвостового плавника с выемкой. В спинном
плавнике 2 жёстких и 7 ветвистых лучей (всего 9), в анальном — 2 жё-
стких и 5 ветвистых (всего 7).
Род Южноевропейские гольцы — Oxynoemacheilus Bănărescu &
Nalbant, 1966 (с. 148)
Краткая биологическая характеристика видов рода
Усатые гольцы — Barbatula Linck, 1790
Усатый, или обыкновенный, голец — Barbatula barbatula
(Linnaeus, 1758) (рис. 89, 2.65)
Рис. 89. Усатый, или обыкновенный, голец — Barbatula barbatula (Linnaeus,
1758)
148
D III—IV 7, A III—IV 5, V I—II 6—7, P I 10—12. Под глазом нет
складного шипа. Голова не сжата с боков. Усиков 6: четыре на конце
рыла и два в углах рта. Спинной плавник короткий или умеренной дли-
ны, с 7 ветвистыми лучами; начало его немного впереди или немного
позади начала брюшных. Хвостовой плавник прямой, по краям слег-
ка закруглённый. Тело на жёлтом фоне покрыто буроватыми пятнами.
Плавники, особенно спинной и хвостовой, с рядами тёмных пятнышек.
Бока тела покрыты мелкими чешуйками, не налегающими друг на дру-
га (см. рис. 2.65).
Встречается в реках, ручьях, озёрах и прудах. Держится у дна. Не-
рест в мае—июне. Питается донными беспозвоночными. Максималь-
ная длина тела — 10—15 см.
Промыслового значения не имеет.
Краткая биологическая характеристика видов рода
Южноевропейские гольцы — Oxynoemacheilus Bănărescu & Nal-
bant, 1966
Голец Крыницкого — Oxynoemacheilus merga (Krynicki, 1840)
(рис. 90, 2.66)
Рис. 90. Голец Крыницкого — Oxynoemacheilus merga (Krynicki, 1840)
D II 7, A II 5. Хвостовой плавник слегка выемчатый. Тело покры-
то бурыми пятнышками. За спинным плавником на спине находятся
4—6 крупных бурых пятен. Спинной и хвостовой, а иногда и грудные
плавники с рядами тёмных пятнышек (см. рис. 2.66). Длина — до 9 см.
Промыслового значения не имеет.
Гольцы имеют короткий жизненный цикл. Становятся половозре-
лыми обычно уже на 2—3 -м году жизни. Плодовитость гольцов невели-
ка — 250—500 икринок. Откладывается икра на камни и песок. Нерест
149
происходит в мае при температуре воды 18—20 °С. Вылупившиеся ли-
чинки лежат на дне, опираясь на большие грудные плавники. Питаются
микроскопическими организмами, живущими в песке. Взрослые голь-
цы питаются беспозвоночными.
Семейство Вьюновые — Cobitidae swainson, 1838
Небольшие рыбки обычно с длинным вальковатым телом, покры-
тым мелкой чешуёй или голым. Имеется выдвижной шип под глазом.
Рот почти у всех нижний, маленький, без зубов. Вокруг рта от 6 до 12
усиков (3—6 пар). Глоточные зубы малочисленные, однорядные, жерно-
вок отсутствует. Окраска пёстрая: тело покрыто чёрными и бурыми пят-
нами. Ведут придонный образ жизни, питаются обрастаниями, личин-
ками ручейников, комаров и подёнок. Весьма активны в ночное время
суток. Переносят недостаток кислорода в воде, могут заглатывать воз-
дух с поверхности. В большинстве случаев это рыбы с коротким циклом
жизни, не достигающие больших размеров (максимум до 20 см). Поло-
вая зрелость наступает в 2—3 года. Икра вымётывается на различный
субстрат (камни, песок, растительность). Промыслового значения почти
не имеет, используется в качестве наживки при ловле хищных рыб.
В фауне бассейна Кубани вьюновые представлены 3 родами.
Таблица
для определения родов семейства
Вьюновые — Cobitidae Swainson, 1838
1 (4). Усиков три пары (две пары на конце рыла и одна в углах
рта). Голова слегка уплощена. Под глазами имеется шип с двумя ветвя-
ми, иногда скрытый в коже. Задний край хвостового плавника прямой
или слегка выпуклый.
2 (3). У самцов на самом длинном луче грудного плавника есть ко-
стная пластинка (орган Канестрини). У основания хвостового плавника
нет тёмной поперечной полосы.
Род Щиповки — Cobitis Linnaeus, 1758 (с. 150)
3 (2). У самцов на самом длинном луче грудного плавника нет ко-
стной пластинки (орган Канестрини). У основания хвостового плавни-
ка имеется тёмная поперечная полоса.
Род Щиповки Сабанеева — Sabanejewia Vladykov, 1929 (с. 154)
150
4 (1). Усиков пять пар (две пары на верхней, две пары на нижней
челюсти, одна пара в углах рта). Шипа под глазом нет. Хвостовой плав-
ник закруглённый.
Род Вьюны — Misgurnus Lacépède, 1803 (с. 156)
Род Щиповки — Cobitis Linnaeus, 1758
Тело удлинённое, сжатое с боков, покрытое очень мелкой чешуёй.
Голова сжата с боков, голая. Рот маленький, 6 усиков (3 пары), распо-
ложенных на конце рыла (4) и в углах рта (2). Под глазами имеется шип
с двумя ветвями, иногда скрытый в коже, при опасности выдвигается
наружу и ранит обидчика.
Все виды этого рода имеют орган Канестрини — маленькую кост-
ную пластинку на самом длинном луче грудного плавника, покрытую
кожей. У разных видов пластинка имеет и различную форму: тонкую и
длинную, широкую, с зазубринами и без них. Размеры, форма и распо-
ложение органа Канестрини являются систематическими признаками.
Обитают в водоёмах Европы и Северной Азии, а также Переднего
Востока. Встречаются в различных биотопах — реках, ручьях, озёрах и
прудах. Предпочитают мягкий грунт (ил, песок), куда любят зарываться,
но могут просто прятаться под камнями. Ведут ночной образ жизни. Пи-
таются донными беспозвоночными и детритом. Вырастают от 6 до 20 см.
В бассейне Кубани обитают четыре вида: обыкновенная (Cobitis
taenia Linnaeus, 1758), сибирская (Cobitis melanoleuca Nichols, 1925),
азовская (Cobitis tanaitica Băcescu & Maier, 1969) и южнорусская (Co-
bitis rossomeridionalis Vasil’eva & Vasil’ev, 1998) щиповки (Васильева,
Васильев, 1998).
Систематика двух последних щиповок требует дополнительного
изучения с привлечением новых методов исследования (молекулярной
генетики) и определением чётких видовых критериев.
Таблица
для определения видов рода
Щиповки — Cobitis Linnaeus, 1758
1 (2). Края спинного плавника округлены. У основания хвостового
плавника нет удлинённых тёмных пятен.
Обыкновенная щиповка — Cobitis taenia Linnaeus, 1758 (с. 151)
151
2 (1). Спинной плавник усечённый. У основания хвостового плав-
ника имеются два удлинённых тёмных пятна.
Сибирская щиповка — Cobitis melanoleuca Nichols, 1925 (с. 152)
3 (4). Спинной плавник расположен чуть впереди начала брюш-
ных плавников.
Южнорусская щиповка — Cobitis rossomeridionalis Vasil’eva &
Vasil’ev, 1998 (с. 153)
4 (3). Спинной плавник сдвинут вперёд и расположен впереди на-
чала брюшных плавников.
Азовская щиповка — Cobitis tanaitica Băcescu & Maier, 1969
(с. 153)
Краткая биологическая характеристика видов рода
Щиповки — Cobitis Linnaeus, 1758
Обыкновенная щиповка — Cobitis taenia Linnaeus, 1758 (рис. 91,
2.67)
Рис. 91. Обыкновенная щиповка — Cobitis taenia Linnaeus, 1758
D II—III 6—7, A II—III 5—7, P I 6—8, V II 5—7, C 16. Тело свет-
ло-жёлтой окраски; по бокам тела тянется ряд бурых полос. Усики ко-
роткие: самые длинные из них достигают только вертикали переднего
края или середины глаза. Хвостовой стебель без кожистой складки.
Подглазничный шип иногда скрыт в коже. Орган Канестрини имеет
вид широкого топорика. Основной цвет тёмно-бурый, по бокам тела от
10 до 18 тёмных бурых округлых или четырёхугольных пятен, иногда
сливающихся и тогда принимающих вид широкой полоски. Тёмная по-
лоска тянется от конца рыла к глазу. Вдоль боковой линии ряд крупных
тёмных пятен, а над ними многочисленные тёмные пятнышки, распо-
ложенные неясными рядами. Хвостовой стебель без кожистой складки
(см. рис. 2.67).
152
Щиповка обычно держится на мелководьях со слабым течением и
среди растительности. Придерживается дна, иногда зарывается в грунт
или скрывается среди растительности. Наибольшая длина тела — 9 см.
Питается донными беспозвоночными, моллюсками, семенами расте-
ний и др.
Нерестится в мае—июне при температуре воды 14—20 °С. Икрин-
ки крупные, диаметром 1,8—2,6 мм, откладываются на растительность.
Нерест порционный, длится 1—2 месяца. Плодовитость — от 450 до
2 300 икринок.
Промыслового значения не имеет, однако, играет роль в питании
хищных рыб. Встречается практически во всех реках, водохранилищах
и лиманах бассейна Кубани.
Сибирская щиповка — Cobitis melanoleuca Nichols, 1925 (рис. 92,
2.68)
Рис. 92. Сибирская щиповка — Cobitis melanoleuca Nichols, 1925
D II—III 6—7, A II—III 5—6, P I 7—8, V I—II 5—6, C 16. Жабер-
ных тычинок 10—12. Позвонков 42—46. Длинное веретенообразное
тело. Глаза маленькие, посажены высоко и близко друг к другу. Вокруг
рта шесть усиков: четыре усика спереди и два по углам рта. Под глазом
небольшой острый складной шип.
Отличается от обыкновенной наличием двух тёмных пятен у ос-
нования хвостового плавника и узкой бутылкообразной формой органа
Канестрини, который у самцов C. taenia по форме похож на округлый
топорик.
Населяет русла Кубани и её левобережных притоков. Встречает-
ся также в лиманах, озёрах, прудах и солоноватых заливах Азовского
моря.
153
Азовская щиповка — Cobitis tanaitica Băcescu & Maier, 1969
(рис. 93, 2.69)
Рис. 93. Азовская щиповка — Cobitis tanaitica Băcescu & Maier, 1969
D II—III 8—9, A II—III 5—6, P I 7—8, V I—II 5—6. Тело удлинён-
ное, покрыто мелкой чешуёй. Голова голая, без чешуи. Спинной плав-
ник сдвинут вперёд и расположен впереди начала брюшных плавников.
На верхнем крае основания хвостового плавника расположено чёрное
пятно. Рыло тупое, 6 коротких усиков на верхней челюсти (4 спере-
ди и 2 над углами рта). Рот маленький, нижний. Под глазом имеется
подвижный шип. Спина желтоватая, брюхо и бока светлые с полосами
чёрно-бурых пятен. На голове, спинном, хвостовом плавниках пятна
(см. рис. 2.69). Длина — до 10 см. Обитает в водоёмах бассейна Кубани
и прилегающих к нему реках (Ее, Челбасе, Бейсуге и др.).
Южнорусская щиповка — Cobitis rossomeridionalis Vasil’eva &
Vasil’ev, 1998 (рис. 94)
Рис. 94. Южнорусская щиповка — Cobitis rossomeridionalis Vasil’eva &
Vasil’ev, 1998
154
D II—III (6) 7—8, A I—III 5—7, P I 7—9 (10). Чешуя мелкая, ок-
руглой формы. Спинной плавник сдвинут вперёд и располагается не-
сколько впереди начала брюшных плавников. В верхней части основа-
ния хвостового плавника имеется одно тёмное пятно. Усики короткие.
Подглазничный шип хорошо развит. Жировые гребни на хвостовом
стебле хорошо выражены. Орган Канестрини у самцов широкий, по
форме напоминает топорик и доходит до конца 3—6 -го члеников при-
креплённого к нему луча грудного плавника.
Обитает в реках, мелких ручьях и лиманах. В Кубани придержива-
ется прибрежных мелководий, предпочитает места с медленным течени-
ем, с зарослями водной растительности и илистым или песчаным дном.
Вид выделен из состава обыкновенной щиповки (Cobitis taenia) и
фактически является её видом-двойником (Васильева, Васильев, 1998).
Биология мало изучена.
Род Щиповки Сабанеева — Sabanejewia Vladykov, 1929
Представители рода Щиповки Сабанеева [Южноевропейские щи-
повки] — Sabanejewia Vladykov, 1929 отличаются от рода Щипов-
ки — Cobitis Linnaeus, 1758 отсутствием у самцов органа Канестрини.
Чешуя мельче, не налегающая друг на друга. В водоёмах бассейна Ку-
бани род представлен двумя видами: предкавказская щиповка — Sa-
banejewia caucasica (Berg, 1906) и кубанская щиповка — Sabanejewia
kubanica Vasil’eva & Vasil’ev, 1988
Таблица
для определения видов рода
Щиповки Сабанеева — Sabanejewia Vladykov, 1929
1 (2). Вдоль боков тела ряд (10—15) бурых пятен. Усики короткие,
не достигают заднего края глаза.
Кубанская щиповка — Sabanejewia kubanica Vasil’eva & Vasil’ev,
1988 (с. 155)
2 (1). Вдоль боковой линии ряда больших пятен нет. Усики длин-
ные, доходят до заднего края глаза. Тело покрыто мелкими тёмными
пятнами.
Предкавказская щиповка — Sabanejewia caucasica (Berg, 1906)
(с. 156)
155
Краткая биологическая характеристика видов рода
Щиповки Сабанеева — Sabanejewia Vladykov, 1929
Кубанская щиповка — Sabanejewia kubanica Vasil’eva & Vasil’ev,
1988 (рис. 95, 2.70)
Рис. 95. Кубанская щиповка — Sabanejewia kubanica Vasil’eva & Vasil’ev,
1988
D II—III 6—7, А II—III 5—6, P I 6—8, V II 5—6. Тело удлинённое.
Голова довольно крупная. У основания хвостового плавника имеется
тёмное пятно в виде полоски, прерванное посередине. Вдоль боковой
линии (по центру) проходит ряд больших овальных тёмных пятен (10—
15), над которыми разбросано много тёмных пятнышек, не имеющих
тенденции образовывать продольные ряды. Промежутки между боко-
выми пятнами с очень небольшим количеством или вообще без тём-
ного пигмента, узкое беловатое седло позади головы с тёмными пят-
нами. Хвостовой и спинной плавники в мелких пятнах. Вентральный
жировой кожистый гребень на хвостовом стебле отсутствует или менее
развит, чем дорсальный. Чешуя очень мелкая, поперечных рядов от 170
до 200 чешуй. У половозрелых самцов по бокам тела впереди спинного
плавника имеется вздутие.
Тело окрашено в желтоватый цвет; по бокам имеются коричневые
пятна (см. рис. 2.70).
Предпочитает проточные водоёмы с твёрдым грунтом и переката-
ми. Встречается в Кубани от низовьев до Армавира. Длина тела — до
10 см, обычно 6—8 см, масса — 3—5 г. Живёт до 4—5 лет. Икроме-
тание порционное в мае—июне. Плодовитость — до 2 500 икринок.
Питается личинками комаров, водорослями.
Промыслового значения не имеет. Редкий малоизученный вид.
156
Предкавказская щиповка — Sabanejewia caucasica (Berg, 1906)
(рис. 96, 2.71)
Рис. 96. Предкавказская щиповка — Sabanejewia caucasica (Berg, 1906)
D I—II 6—7 (8), А II 5, P I 7—8, V I 5—6. Тело покрыто многочис-
ленными бурыми пятнышками. По бокам тела нет больших тёмных пя-
тен, но есть полоска золотистого цвета. Усики длинные, задние дости-
гают вертикали заднего края глаза. На спине перед спинным плавником
и за ним расположены тёмные пятна, иногда сливающиеся в сплошную
бурую полоску. У основания хвостового плавника нет тёмного пятна,
но имеется тёмная поперечная полоска, иногда прерванная посередине
в виде скобок (см. рис. 2.71). Подглазной шип сильный. Чешуя сравни-
тельно крупная, налегающая.
Обитает в реках, предпочитает верхнее течение Кубани. Нерестит-
ся в мае—июне. Длина — до 10 см. Биология практически не изучена.
Включена в Красную книгу РФ (2001) как редкий вид.
В целом систематика щиповок требует дополнительного изучения
с привлечением новых методов исследования (молекулярной генетики)
и определением чётких видовых критериев.
Вьюновые имеют довольно существенное значение как одно из
основных звеньев пищевых цепей во многих водоёмах. Потребляя мел-
ких бентических беспозвоночных, многие из них являются в свою оче-
редь важным объектом питания хищных рыб.
Род Вьюны — Misgurnus Lacépède, 1803
Обычная длина вьюнов — 15 —18 см, некоторые виды достига-
ют 30 см. Под глазом нет складного шипа. Удлинённое тело покрыто
157
мелкой, но хорошо заметной чешуёй. Спинной плавник короткий, рас-
положен над брюшными плавниками. Рот окаймлён 10—12 усиками,
хвостовой плавник закруглён.
В бассейне Кубани обитает один вид.
Вьюн — Misgurnus fossilis (Linnaeus, 1758) (рис. 97, 2.72)
Рис. 97. Вьюн — Misgurnus fossilis (Linnaeus, 1758)
DII—IV5—7,AIII—V5(6),PI8—11,VII5—6,CI13—14I.
Тело удлинённое, вальковатое, покрытое очень мелкой чешуёй. Бо-
ковая линия не видна. Спинной плавник небольшой, отнесён назад и
расположен над брюшными. Голова небольшая, вытянутая вперёд. Рот
обращён книзу и окружён десятью усиками, из них четыре на верхней
челюсти на конце рыла, два — в углах рта, четыре — на нижней че-
люсти. Чешуя мелкая, 135—175 поперечных рядов. Длина тела — до
20—25 см.
Цвет тела желтоватый, на боках тела имеется небольшая тёмная
полоса и отдельные тёмные пятна (см. рис. 2.72).
Обитает в нижнем течении Кубани, кубанских лиманах и в водо-
ёмах на степных реках. Держится в основном в заросших и заиленных
местах. Нерестится у дна среди растительности в мелких водоёмах со
слабым течением. Питается личинками комаров и рачками. Способен
переносить недостаток кислорода в воде. У него хорошо развит допол-
нительный орган воздушного дыхания — небольшой участок задней
кишки, обильно снабжённый кровеносными сосудами. При нехватке
кислорода в воде он поднимается на поверхность и глотает воздух, ко-
торый проходит по кишечнику, обогащая кровь кислородом. Нерест в
мае—июне в местах обычного обитания у берегов, в густых зарослях.
Питается личинками насекомых и детритом. Промыслового значения
не имеет. В водоёмах бассейна Кубани и прилегающих к нему рек, озёр,
прудов малочислен, очень редко встречается.
158
Отряд Сомообразные — Siluriformes Cuvier, 1816
Обычно пресноводные рыбы, но имеются и морские виды. Тело го-
лое или покрыто костными пластинками. Рот невыдвижной, большой, на
челюстях есть зубы. Усиков около рта несколько пар. Есть предчелюст-
ные и челюстные кости. Глоточных зубов нет. В грудных, а иногда и в
других плавниках у сомов развиваются сильные колючки. Сомообразные
очень разнообразны по величине, достигают длины от 1,2 см до 2,5 м, а
по некоторым источникам, до 5 м при массе до 300 кг (европейский сом).
В фауне бассейна Кубани обитают или могут быть встречены
представители трёх семейств.
Таблица
для определения семейств отряда
Сомообразные — Siluriformes Cuvier, 1816
1 (4). Анальный плавник очень длинный — от 36 до 92 лучей, кон-
чается у хвостового или переходит в него. За спинным нет жирового
плавника.
2 (3). Спинной плавник короткий (не более 7 мягких лучей).
Семейство Сомовые — Siluridae Cuvier, 1816 (с. 158)
3 (2). Спинной плавник длинный (51—80 мягких лучей).
Семейство Клариевые — Clariidae Bonaparte, 1846 (с. 160)
4 (1). В анальном плавнике от 17 до 24 мягких лучей. На спине за
спинным плавником имеется жировой плавник.
Семейство Икталуровые, или Кошачьи сомы, — Ictaluridae
Gill, 1861 (с. 162)
Семейство Сомовые — Siluridae Cuvier, 1816
Тело голое, чешуя и костные пластины отсутствуют, хвост обыч-
но намного длиннее туловища и сжат с боков. Голова большая сплю-
щенная. Жаберные перепонки не соединены между собой. Спинной
плавник маленький (у некоторых видов отсутствует) без колючего
луча, мягких лучей не более 7, расположен на передней части спины.
Брюшные плавники расположены в средней части брюха, у многих
видов редуцированы в размерах или отсутствуют. Жирового плавника
нет. Анальный плавник очень длинный, составлен 41—110 лучами. Рот
159
большой. Нижняя челюсть выдаётся вперёд. На челюстях и сошнике
имеются многочисленные мелкие зубы. В грудных плавниках есть по
одному костному лучу, иногда со слабой зазубренностью. На верхней
челюсти имеется одна пара длинных усов, на нижней — одна—две
пары более коротких. Спина, голова и бока тёмные, брюхо светлое.
В бассейне Кубани один род c одним видом.
Обыкновенный сом — Silurus glanis Linnaeus, 1758 (рис. 98, 2.73)
Рис. 98. Обыкновенный, или европейский, сом — Silurus glanis Linnaeus, 1758
D3—5,A77—92,PI14—17,VI(9)11—13.Сом—однаизнаи-
более крупных пресноводных рыб, весьма распространённая и обычная
для всего бассейна Кубани. Особенно многочислен сом в устьях рек,
встречается в опреснённых участках моря.
Для сома характерны миграции сравнительно небольшой протя-
жённости. Нерестилища и места откорма расположены на небольшом
расстоянии друг от друга. Длина сома, выловленного в Краснодарском
водохранилище в 1998 г., составила 210 см при массе 95 кг. Однако в
промысловых уловах преобладают 5—10 кг. Икрометание происходит
в мае. Отложенная икра охраняется самцом до её выклева. Темп роста
сома достаточно высок. Годовики достигают длины 25—28 см и массы
180—200 г. Половое созревание наступает на третьем или четвёртом
году жизни. Икрометание происходит в мае—июне при температуре
23—27 °С. Перед откладыванием икры сом устраивает «гнездо», раз-
двигая растительность, корневища камыша, рогоза, тростника, ивняка
и др. Откладываемая икра прилипает к растениям. Икринки крупные
имеют в диаметре около 3 мм. Личинки вылупляются из икринок обыч-
но через 3—5 сут.
160
Плодовитость сома в зависимости от размеров и возраста силь-
но колеблется. У самки Краснодарского водохранилища длиной тела
65 см и массой 1,8 кг было обнаружено 5,2 тыс. икринок, а у самки дли-
ной 120 см и массой 25 кг — 430 тыс. икринок. Средняя плодовитость
сома — 90 тыс. икринок.
Сом — типичный хищник. При достижении 10—15 см основной
рацион сома составляет рыба, а до этого он питается головастиками и
беспозвоночными. Взрослые сомы питаются также лягушками, речны-
ми раками и др. В желудках сома из Краснодарского водохранилища
обнаружено 27 видов рыб, но среди них преобладали лещ, плотва, гус-
тера, сазан, бычки и др., часто встречались осетровые. Сом в недалёком
прошлом являлся промысловой рыбой, а в настоящее время из-за его
малочисленности вылавливается как прилов. Мясо сома очень ценится
как пищевой продукт. Является объектом любительского лова.
Семейство Клариевые — Clariidae Bonaparte, 1846
Клариевые имеют гладкое, удлинённое, цилиндрическое тело с
длинными анальными и спинными плавниками, доходящими до хво-
стового и состоящими только из мягких лучей. Жирового плавника
нет. Наружный луч грудного плавника зазубрен. В брюшном плавнике
в норме шесть мягких лучей. На узкой голове четыре пары усов. Кла-
риевые сомы имеют различную расцветку, среди цветов возможны со-
четания розового, чёрного, белого и серого.
В бассейне Кубани один род c одним видом.
Африканский клариевый сом — Clarias gariepinus (Burchell,
1822) (рис. 99, 2.74)
Рис. 99. Африканский клариевый сом — Clarias gariepinus (Burchell, 1822)
161
D 55—75, A 36—59, P I 6—11, V 4—6. На первой жаберной дуге
24—55 жаберных тычинок. Передний край первого луча грудного
плавника с зазубринами.
Африканский клариевый сом, или африканский лабиринтовый
сом, или мраморный клариевый сом, или нильский клариас, или кла-
риевый сом — теплолюбивая пресноводная рыба, но легко переносит
засоление воды до 10 ‰. Встречается по всей Африке, включая водо-
ёмы Сахары, в бассейне р. Иордан, в Южной и в Юго-Восточной Азии.
В последнее время вид получил широкое распространение в аква-
культуре многих государств, в том числе в России. В Россию был заве-
зён из Нидерландов в 1993 г. в рыбоводное хозяйство Новолипецкого
металлургического комбината, где была освоена биотехника разведе-
ния и выращивания вида.
Клариевый сом имеет гладкое удлинённое тело, лишённое че-
шуи. Спинной и анальный плавники длинные, достигающие хвостово-
го плавника. Хвостовой стебель сильно сжат с боков. Голова широкая
и плоская, несёт четыре пары неразветвлённых усов: одна назальная,
одна максиллярная (самая длинная и наиболее подвижная) на сошнике
и две мандибулярные — внутренняя и наружная. Зубы имеются на че-
люстях и сошнике. Плавательный пузырь маленький, состоит из двух
долей и заключён в капсулу, образованную поперечными выростами
парапофизов четвёртого и пятого позвонков.
Окраска спины сома меняется в зависимости от освещённости и
варьирует от песочно-жёлтого до серого с оливковыми и зеленовато-
коричневыми отметинами и даже до синевато-тёмного цвета, брюхо
светлое (рис. 2.74).
Благодаря наличию специального наджаберного органа (на второй
и четвёртой жаберных дугах) для дыхания атмосферным воздухом, по
строению напоминающего лёгкие наземных позвоночных, клариевый
сом нетребователен к кислородному режиму водоёма, поэтому его мож-
но выращивать при очень высоких плотностях посадки. Эта же биологи-
ческая особенность позволяет ему оставаться живым без воды в течение
длительного времени (около 48 ч).
Клариевый сом — хищник, однако охотно питается водными на-
секомыми, моллюсками, водной растительностью. Оптимальная темпе-
ратура для выращивания — 25 —30 °С. Массы 1 кг достигает за первый
год выращивания (за 8—12 мес.). При максимальных размерах достига-
ет 170 см длины и веса 60 кг. Продолжительность жизни — около 8 лет.
162
Созревает клариевый сом в возрасте 1—2 года. Плодовитость —
от 5 до 100 тыс. икринок.
Семейство Икталуровые,
или Кошачьи сомы, — Ictaluridae Gill, 1861
В фауне бассейна Кубани обитает один вид этого семейства аккли-
матизированный в 1972 г.
Канальный сом — Ictalurus punctatus (Rafinesque, 1818) (рис. 100,
2.75)
Рис. 100. Канальный сом — Ictalurus punctatus (Rafinesque, 1818)
D I 6, A 23—25, P I 8—9. Жаберных тычинок 14—18. Позвонков
42—44. Родина — Средняя и Центральная Америка. В естественных
условиях канальный сомик обитает в пресных водах, но выдерживает
солёность воды до 8—10 ‰. Хищник, но потребляет и бентические ор-
ганизмы.
Тело веретенообразной формы, голова и тело покрыты толстой
кожей без чешуи. Окраска тела от тёмной до чёрной, иногда пятнистая
(см. рис. 2.75). Усиков 4 пары: 2 пары на верхней части головы и 2
пары на подбородке. Спинной плавник находится между грудными и
брюшными плавниками. Грудной плавник снабжён сильными шипами.
В анальном плавнике от 23 до 25 мягких лучей. В грудном плавнике
первый неветвистый луч мощный, спереди слегка зазубренный, а сзади
зазубрины острые и многочисленные (12—17). На спине имеется жиро-
вой плавник, который не соединяется со спинным. Хвостовой плавник
выемчатый.
163
Канальный сом — теплолюбивая рыба. Наилучшая для него тем-
пература воды 24—28 °С. Половое созревание наступает на 3—4 -м году
жизни. Икрометание в мае—июне при температуре воды 18—22 °С.
Плодовитость — 7 —20 тыс. довольно крупных (3—4 мм) икринок. Во
время нереста сомики разбиваются на пары.
Самец устраивает гнездо на глубине до 120 см, где самка отклады-
вает икру. После оплодотворения самец охраняет икру, а затем и потом-
ство. Икра клейкая, светло-оранжевого цвета. Инкубационный период
при температуре воды 25 °С длится 6—10 сут., при 30 °С — 4,5 сут.
Мясо сома очень нежное и вкусное, напоминает мясо форели.
В последнее время канальный сом стал объектом любительского
рыболовства. Встречается в среднем и нижнем течении Кубани, в во-
доёмах на степных реках, единичные экземпляры — в Краснодарском
водохранилище, а также в сбросном канале Краснодарской ТЭЦ, где
и нерестится. В уловах преобладают особи массой 1—2 кг, достигают
массы 5 кг, а на родине — 25—35кг.
Отряд Карпозубообразные — Cyprinodontiformes Berg, 1940
Небольшие рыбки с телом, покрытым циклоидной чешуёй. Плав-
ники без колючих лучей. Боковой линии нет, или она слабо выражена.
Усики отсутствуют. Во рту имеются зубы. Спинной плавник один, рас-
положен над анальным.
Тропические и субтропические рыбы, обитатели рек и озёр. В во-
доёмах бассейна Кубани акклиматизирован один вид из семейства Гам-
бузиевые, или Пецилиевые, — Poeciliidae Garman, 1895.
Семейство Гамбузиевые,
или Пецилиевые, — Poeciliidae Garman, 1895
В семействе более 300 видов небольших живородящих рыбок,
обитающих в пресных, частично солоноватых и морских водах Амери-
ки — от Калифорнии и Филадельфии в Северной Америке до Эквадо-
ра и Уругвая в Южной. Анальный плавник выдвинут вперёд, и первые
лучи его у самцов превращены в совокупительный орган (гоноподий).
Семейство объединяет более 40 родов. Из них род Гамбузии — Gambu-
sia Poey, 1854 искусственно завезён на Кубань для борьбы с малярий-
ными комарами.
164
Род Гамбузии — Gambusia Poey, 1854
Родина — южные и восточные штаты США, Мексика, Куба, где
род Гамбузии встречается в мелких озёрах, каналах и лагунах. Боль-
шинство видов пресноводные, некоторые могут обитать в солоноватой
или солёной воде. Ряд видов акклиматизирован в различных регионах
мира для борьбы с малярией и другими инфекционными заболевания-
ми — рыбки активно поедают личинок комаров. В водоёмах бассейна
Кубани акклиматизирован один вид — хольбрукская гамбузия — Gam-
busia holbrooki (Girard, 1859).
Хольбрукская гамбузия — Gambusia holbrooki (Girard, 1859)
(рис. 101, 2.76)
Рис. 101. Самка (вверху) и самец (внизу) хольбрукской гамбузии — Gambusia
holbrooki (Girard, 1859)
165
D 7—8, A 8—11, P 11—14. Небольшие рыбки. Длина тела самок
от 3 до 6 см, самцов — до 3,5 см. Самцы встречаются реже самок; у них
анальный плавник приближен к брюшным, а три луча его превращены
в совокупительный орган — гоноподий, позволяющий вводить сперму
в организм самки. Половое созревание наступает рано: через 30—40
дней после рождения. Живородящая рыба. Период размножения — с
апреля по ноябрь при температуре воды 15 °С и выше. За это время ка-
ждая зрелая самка производит 5—6 поколений. Первый помёт — через
месяц после осеменения. Последующие помёты могут происходить без
повторного осеменения, оплодотворение обеспечивается за счёт спер-
мы, сохраняющейся в яйцеводах самки. Число вымётываемых мальков
колеблется от 15 до 120. Малёк вымётывается уже достаточно сформи-
ровавшимся и скоро приступает к активному питанию.
Пища молоди на 90 % состоит из коловраток. Взрослые особи пи-
таются зоопланктонными и бентическими организмами, в том числе
личинками малярийного комара.
Хольбрукская гамбузия завезена в СССР из Италии в 1925 г. и вы-
пущена в болотистые водоёмы Абхазии. В последующем широко рас-
пространилась по южным водоёмам СССР. Обитает гамбузия в мед-
ленно текучих водоёмах, а также в реке Кубани (среднее течение), в
Карасунских озёрах, в водоёмах на Старой Кубани и сбросных водах
Краснодарской ТЭЦ.
Отряд Сарганообразные — Beloniformes Berg, 1940
Плавники без колючих лучей. Спинной расположен далеко по-
зади над анальным. Брюшные плавники на брюхе. Отряд представ-
лен 6 семействами, 34 родами и 283 видами. В рассматриваемом
бассейне обитают представители двух семейств, в каждом по одно-
му виду.
Таблица
для определения семейств отряда
Сарганообразные — Beloniformes Berg, 1940
1 (2). Хвостовой плавник закруглённый. Длина тела — до 5 см.
Семейство Адрианихтовые — Adrianichthyidae Weber, 1913
(с. 166)
166
2 (1). Хвостовой плавник выемчатый (дифицеркальный), обе лопа-
сти имеют одинаковую длину. Длина тела — до 60 см.
Семейство Саргановые — Belonidae Bonaparte, 1832 (с. 167)
Семейство Адрианихтовые — Adrianichthyidae weBer,
1913
Пресноводные и солоноватоводные маленькие рыбки (до 5 см).
Распространены от Индии до Японии и Индо-Австралийского архипе-
лага. Обитают в мелких речках, озёрах, рисовых чеках Китая, Кореи,
Японии.
В водоёмах бассейна Кубани обитает один вид: китайская медака
(оризия), или китайская рисовая рыбка — Oryzias sinensis Chen, Uwa &
Chu, 1989.
Вид завезён в Краснодарский край из Казахстана в 1970-х гг. в це-
лях борьбы с малярийным комаром.
Китайская медака — Oryzias sinensis Chen, Uwa & Chu, 1989
(рис. 102, 2.77)
Рис. 102. Китайская медака, или китайская рисовая рыбка — Oryzias sinensis
Chen, Uwa & Chu, 1989
D 5—8, A 14—22, P 7—11, V 5—7. Поперечных рядов чешуй 24—
36, жаберных тычинок 10—16. Тело сжатое с боков, голова уплоще-
на сверху, спинной плавник сдвинут назад у хвоста. Чешуя крупная,
циклоидная. Чёрная продольная полоса на спине тянется от затылка до
начала спинного плавника, по бокам тела — слабая темноватая полоска
вдоль средней линии. Хвостовой плавник усечённый или закруглённый
по заднему краю. Спина желтоватая или слабо-коричневая, брюшко
167
светлое. Небольшой рот и большие глаза. Рыло меньше диаметра глаза.
Длина тела — до 5 см. Продолжительность жизни — 3 —4 года.
Половая зрелость наступает на первом—втором году жизни. Ик-
рометание порционное, в каждой порции 15—45 икринок. В среднем
за сезон самка вымётывает около 2 тыс. икринок. Размножение про-
исходит ночью или под утро. Сначала икра повисает за анальным от-
верстием самки в виде грозди, где она оплодотворяется. Затем самка
рассеивает икринки по водным растениям. Развитие икры при темпера-
туре воды 24—28 °С продолжается 10—15 сут. Вылупившиеся личин-
ки через некоторое время приступают к внешнему питанию. Основу
питания составляют коловратки, личинки комаров, а также мелкие зоо-
планктонные ракообразные.
Морфологическое изучение медаки, интродуцированной в бас-
сейне Азовского моря (Насека, Дирипаско, 2005), показало, что она
должна быть идентифицирована как Oryzias sinensis, а не Oryzias latipes
Temminck & Schlegel, 1846, как считалось ранее (Атлас пресноводных
рыб..., 2002, 2003). Китайская медака не входит в состав аборигенной
фауны, а натурализовалась после преднамеренных и непреднамерен-
ных интродукций в водоёмы Краснодарского края (Емтыль, Иваненко,
2002; Богуцкая, Насека, 2004).
Семейство Саргановые — Belonidae Bonaparte, 1832
Саргановые имеют длинное и тонкое тело, покрытое очень мелкой
чешуёй. Челюсти снабжены острыми клыковидными зубами. Обе че-
люсти удлинённые в виде клюва.
В семействе 10 родов, около 47 видов. У всех саргановых кости
имеют ярко-зелёный цвет. В Чёрном и Азовском морях обитает под-
вид — черноморский сарган — Belone belone euxini (Günther, 1866),
который изредка заходит в гирла и кубанские лиманы.
Черноморский сарган — Belone belone euxini (Günther, 1866)
(рис. 103, 2.78)
Рис. 103. Черноморский сарган — Belone belone euxini (Günther, 1866)
168
D II 14—17, A II 17—20. Тело сильно вытянутое, верхняя и нижняя
челюсти образуют длинный клюв. Зубы мелкие. Вдоль боков проходит
фиолетово-бурая линия.
Половозрелым сарган становится в возрасте 3—4 лет при длине
тела 30—40 см и массе 35—50 г. Нерест происходит в мае—июле при
температуре воды 18—20 °С. Плодовитость саргана колеблется от 1 до
30 тыс. икринок. Икрометание порционное. Инкубационный период
длится 15—25 дней. Молодь питается беспозвоночными, а взрослые
особи в основном рыбой — хамсой, атериной, молодью кефали и дру-
гих рыб. Сарган имеет некоторое промысловое значение, но в основ-
ном служит объектом любительского рыболовства.
Черноморский сарган обитает в Чёрном и Азовском морях, изред-
ка заходит в гирла и кубанские лиманы.
По данным базы FishBase (www.fishbase.org), подвид, обитаю-
щий в Чёрном, Азовском и Мраморном морях, недавно выделен в от-
дельный вид — Belone euxini Günther, 1866 (Fricke, Bilecenoglu, Sari,
2007).
Отряд Атеринообразные — Atheriniformes Rosen, 1966
Два спинных плавника, разделённых хорошо заметным промежут-
ком. Первый плавник состоит из гибких неветвистых лучей. Брюшные
плавники расположены на брюхе. Боковая линия отсутствует. Вдоль
бока тела проходит тёмная или серебристая полоса.
Атеринообразные обитают у берегов морей, а также в солонова-
тых и пресных водоёмах.
В бассейне Кубани и прилегающих к нему лиманах отряд пред-
ставлен одним семейством.
Семейство Атериновые — Atherinidae Günther, 1961
Небольшие прибрежные морские рыбки. Тело удлинённое. На
верхней части головы чешуй нет. Грудные плавники посажены высо-
ко. Рот конечный. Боковой линии нет. Имеется два спинных плавни-
ка. Вдоль боков тела проходит тёмная или серебристая полоса. Пило-
рических придатков нет. В фауне бассейна Кубани встречается один
подвид.
169
Черноморская атерина — Atherina boyeri pontica Eichwald, 1831
(рис. 104, 2.79)
Рис. 104. Черноморская атерина — Atherina boyeri pontica Eichwald, 1831
D1 VII—IX, D2 I—II 10—12, A II 13—15. Жаберных тычинок на
1-й дуге 26—31. Поперечных рядов чешуй впереди спинного плавника
18—22. Два спинных плавника. Первый плавник с 7—9 гибкими невет-
вистыми лучами, второй плавник с 1—2 неветвистыми и 10—12 ветви-
стыми лучами, анальный плавник с двумя неветвистыми и 13—15 вет-
вистыми лучами. Рыба пелагическая, стайная. Обитает как в море, так и
в устьях рек и лиманах. Рот конечный, большой. Глаза большие. Чешуя
циклоидная. Нижняя челюсть входит в выемку верхней челюсти. Се-
ребристая полоска на боках тела в средней части занимает в ширину
один ряд чешуй (см. рис. 2.79).
Длина тела — от 6 до 13 см, масса — от 2,5 до 13,5 г. Предельный
возраст — 4 —5 лет.
Половое созревание у атерины наступает уже на втором году жиз-
ни. Средняя плодовитость составляет 590 икринок. Икрометание пор-
ционное.
Икринки с помощью нитевидных отростков прикрепляются к рас-
тительности. Нерест в мае—июне.
Питается атерина беспозвоночными организмами, насекомыми и
молодью рыб. Сама атерина входит в состав пищи осетровых, судака
и других хищных видов рыб. Весной заходит из Чёрного моря в Азов-
ское, осенью выходит обратно. Нагуливается в кубанских лиманах до
конца ноября. Промысловое значение небольшое, так как промысел не
развит вследствие малой ценности этой рыбы.
170
Отряд Трескообразные — Gadiformes Goodrich, 1909
Трескообразные — морские, преимущественно холодноводные и
придонные рыбы. Отличительные признаки трескообразных: спинные,
анальные и брюшные плавники без колючих лучей; если есть брюшные
плавники, то они обычно расположены перед грудными; межчелюст-
ные и верхнечелюстные кости подвижны; нижние глоточные отделены
друг от друга; жабры гребёнчатые; плавательный пузырь не имеет про-
тока, он есть не всегда. Чешуя циклоидная, редко ктеноидная.
В пресной воде встречается один вид — Lota lota (Linnaeus, 1758)
из семейства Налимовые — Lotidae Bonaparte, 1832. Однако не все
учёные, особенно зарубежные, признают самостоятельность семейства
Налимовые, они относят его в ранге подсемейства Lotinae в семейство
Тресковые — Gadidae Rafinesque, 1810.
Семейство Налимовые — Lotidae Bonaparte, 1832
Тело удлинённое с двумя спинными плавниками. Второй спинной
и анальный плавники длинные, задние их концы либо отделены не-
большим промежутком от хвостового, либо соприкасаются или слиты
с хвостовым плавником, имеющим закруглённую форму. Колючек в
плавниках нет. Брюшные плавники впереди грудных. Чешуя циклоид-
ная. Очень характерен для многих трескообразных усик на подбородке.
Семейство представлено одним видом — обыкновенный налим —
Lota lota (Linnaeus, 1758), который обитает в пресных водах.
Обыкновенный налим — Lota lota (Linnaeus, 1758) (рис. 105, 2.80)
Рис. 105. Обыкновенный налим — Lota lota (Linnaeus, 1758)
D1 10—15, D2 71—88, A 69—85, P 16—23, V 6—8. Жаберных ты-
чинок 4—10. Единственный вид из тресковых, перешедший из морских
171
вод в пресные. Голова несколько приплюснутая. Глаза маленькие. Рот
большой, полунижний, верхняя челюсть выдаётся вперёд. На подбо-
родке хорошо развитый усик, а у переднего края ноздрей два коротких
усика. Челюсти и сошник вооружены щетинковидными зубами. Тело
покрыто мелкой циклоидной чешуёй.
Налим сохранил холодолюбивость, свойственную семейству трес-
ковых. Он предпочитает чистые и холодные воды. Размножается зи-
мой. Растёт налим медленно. В 6—7 лет он достигает длины 60—70 см
и массы 1,5—2,6 кг. Налим — хищная рыба, питается в основном мел-
кой рыбой.
В бассейне Кубани данный вид практически не встречается. Име-
ется лишь одно сообщение С. К. Троицкого и Е. П. Цуниковой (1988),
что пойман налим в р. Протоке у Ачуева в 1929 г. Ни до, ни после этого
сообщений о поимке обыкновенного налима в водоёмах бассейна Ку-
бани не поступало. Нами за весь период исследований (1970—2019 гг.)
водоёмов бассейна Кубани налим не был встречен.
Отряд Колюшкообразные — Gasterosteiformes Greenwood,
1966
Мелкие рыбки. Впереди спинного плавника имеются две или боль-
ше не связанных перепонкой отдельных колючек. Спинной и анальный
плавники содержат ветвистые лучи. Брюшные плавники находятся не-
далеко позади от грудных, концы которых заходят за их основание. В
брюшных плавниках всегда есть колючий луч. Тело голое или покры-
тое костными пластинками. Рот выдвижной. Рыло коническое, иногда
слегка трубковидное.
В водоёмах бассейна Кубани встречаются представители одного
семейства.
Семейство Колюшковые — Gasterosteidae Bonaparte,
1831
Жёсткие лучи передней части спинного плавника не соединены
друг с другом перепонкой, они превратились в ряд очень острых твёр-
дых колючек. Тело голое или покрыто костными пластинками.
В фауне бассейна Кубани обитают представители двух родов, в
каждом из которых по одному виду.
172
Таблица
для определения родов семейства
Колюшковые — Gasterosteidae Bonaparte, 1831
1 (2). Перед спинным плавником 3 (как исключение 2 или 4) ко-
лючки.
Род Трёхиглые колюшки — Gasterosteus Linnaeus, 1758
Трёхиглая колюшка — Gasterosteus aculeatus Linnaeus, 1758
(с. 172)
2 (1). Перед спинным плавником 8—11 колючек.
Род Девятииглые, или Многоиглые, колюшки — Pungitius
Costa, 1848
Малая южная колюшка — Pungitius platygaster Kessler, 1859
(с. 173)
Краткая биологическая характеристика видов семейства
Колюшковые — Gasterosteidae Bonaparte, 1831
Трёхиглая колюшка — Gasterosteus aculeatus Linnaeus, 1758
(рис. 106, 2.81)
Рис. 106. Трёхиглая колюшка — Gasterosteus aculeatus Linnaeus, 1758
D (II) III (IV) (9) 10—14, A I (7) 8—10, P 10—11, V I 1. Тело по-
крыто не чешуёй, а костными пластинками. На спине перед плавником
обычно три колючки, причём первые две из них длиннее последней.
Живёт как в морской солёной, так и в совершенно пресной воде, иногда
поднимается вверх по рекам. Держится преимущественно в неглубо-
ких стоячих и полустоячих водоёмах. Маленькая рыбка — максимум
173
5—6 см. Самцы мельче самок. В нерестовый период у самцов красное
брюхо и голубые глаза (см. рис. 2.81).
Половозрелой становится в возрасте одного года. Нерест очень
растянут — с апреля по июнь. Для нереста колюшки подходят в при-
брежную зону (кубанские лиманы, озёра, водохранилища, пруды), так
как для размножения необходим растительный субстрат, из которого
самцы устраивают гнездо. Самка откладывает от 60 до 500 икринок, по-
сле их оплодотворения самец охраняет отложенную икру. Икрометание
порционное. Икра вымётывается до четырёх раз в сезон. Инкубацион-
ный период длится в зависимости от температуры воды от 4 до 15 сут.
Молодь питается зоопланктоном, взрослые — переходят на дон-
ный корм. Во время нереста других рыб (судак, тарань, лещ и др.) ко-
люшка охотно поедает их икру и личинок. Живёт до трёх лет. Промы-
слового значения не имеет.
Малая южная колюшка — Pungitius platygaster Kessler, 1859
(рис. 107, 2.82)
Рис. 107. Малая южная колюшка — Pungitius platygaster Kessler, 1859
D VIII—XI 7—10, A I 7—9, P 10—11. Длина тела половозрелой
самки — 2,5—4,6 см, масса — 0,3—0,9 г. Половозрелой становится уже
к концу первого года жизни. Нерестится среди растений, однако гнездо
устраивает не на грунте, как трёхиглая, а на некотором расстоянии от
дна.
Нерест происходит в апреле—мае. Питается главным образом
беспозвоночными. Встречается как в кубанских лиманах, так и в усть-
ях рек и водохранилищах бассейна Кубани. Не является объектом про-
мысла.
174
Отряд Иглообразные — Syngnathiformes Helfman,
Collette & Facey, 1997
Рыло в виде трубки, удлинённое, рот маленький, беззубый. Тело
удлинённое, тонкое, сплошь покрыто костными щитками. Брюшных
плавников нет. Спинной плавник без колючих лучей. Хвостовой плав-
ник небольшой или его нет вовсе. Морские и пресноводные рыбы. Жи-
вут в прибрежной зоне, реже в пелагиали.
В фауне бассейна Кубани обитают два вида одного семейства.
Семейство Игловые — Syngnathidae rafinesque, 1810
Тело удлинённое, сплошь покрытое костными кольцами (щитками,
пластинками), опоясывающими всё тело. Рыло удлинённое трубкообраз-
ное, рот маленький, беззубый. Жабры не в виде листков, а в виде пучков
(отсюда другое название отряда — пучкожаберные). Самцы проявляют
заботу о потомстве, вынашивая икру в выводковой камере, расположен-
ной на нижней стороне хвостовой части тела или на брюхе. Самка откла-
дывает в сумку икру небольшими порциями. В процессе откладывания
икра оплодотворяется. У игл-рыб выводковая сумка длинная, продоль-
ная, с центральной продольной щелью и двумя боковыми створками,
которые у многих видов в период вынашивания икры могут полностью
закрываться, изолируя развивающихся эмбрионов от внешней среды.
Питаются мелкими планктонными организмами и икрой рыб.
В описываемом регионе семейство представлено двумя видами:
змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758) и черно-
морская игла-рыба — Syngnathus nigrolineatus Eichwald, 1831.
Таблица
для определения видов семейства
Игловые — Syngnathidae Rafinesque, 1810
1 (2). Хоботок сильно сжат с боков. Обе половины грудного пояса
внизу подвижны, не сращены друг с другом. Выводковая камера у сам-
ца на брюшной стороне туловища. Хвостового плавника нет.
Змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758)
(с. 175)
2 (1). Хоботок почти цилиндрический. Обе половины грудно-
175
го пояса внизу сращены
друг с другом (рис. 108).
Выводковая камера у
самца на хвостовом отде-
ле тела. Хвостовой плав-
ник маленький.
Черноморская игла-
рыба — Syngnathus nigro-
lineatus Eichwald, 1831 (с. 176)
Краткая биологическая характеристика видов семейства
Игловые — Syngnathidae Rafinesque, 1810
Змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758)
(рис. 109, 2.83)
Рис. 109. Змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758)
D 32—38. Туловищных колец 28—30, хвостовых — 60 —74, чаще
всего 68—70.
Тело тонкое, покрыто костными кольцами. Спинной плавник
один. Брюшных плавников нет. Выводковая камера у самцов на брюш-
ной стороне туловища, открытая, не защищённая кожными складками
и пластинками, икринки прикрепляются к брюху самца. Рыло трубча-
тое, сверху вогнутое. Окраска желтовато-серая или зеленовато-жёлтая
с бурыми пятнышками на спине (см. рис. 2.83).
Небольшие рыбки удлинённой формы. Длина — до 22 см, самцы
мельче самок. Размножение происходит в мае—июне. Самка отклады-
Рис. 108. Передний конец сросшихся половин
грудного пояса с расположенными на них щит-
ками на теле снизу у Syngnathus nigrolineatus
176
вает икру в выводковую камеру самца. Икра крупная. Плодовитость —
30—60 икринок. Во время развития в выводковой камере икра получа-
ет кислород из крови, протекающей по сосудам слизистой оболочки
выводковой камеры самца. После вылупления из икры молодь некото-
рое время прячется в выводковую камеру самца.
Питается морская игла беспозвоночными организмами и молодью
рыб. В свою очередь служит пищей хищным рыбам. Промыслового
значения не имеет. Обитает как в пресных водоёмах, так и в водоёмах
с солёностью, близкой к морской. Встречается в лиманах, Кубани и из-
редка в Краснодарском водохранилище.
Черноморская игла-рыба — Syngnathus nigrolineatus Eichwald,
1831 (рис. 110, 2.84)
Рис. 110. Черноморская [пухлощёкая] игла-рыба — Syngnathus nigrolineatus
Eichwald, 1831: а — самка; б — самец; в — самец, вид снизу (половина вы-
водковой камеры открыта)
D 32—40, A 3, P 11—14, C 10. Туловищных колец 15—17, хво-
стовых — 36 —41. Передний конец сросшихся внизу половин грудного
пояса заострён (см. рис. 108). Тело шестигранное и удлинённое, покры-
тое костными пластинками. Рыло в виде трубки. Жаберное отверстие
очень маленькое. Брюшных плавников нет. Хвостовой плавник неболь-
шой, анальный зачаточный. Тело зеленовато- или красновато-бурое,
брюхо беловатое (см. рис. 2.84).
Обитает как в солоноватых водах, так и в совершенно пресных.
Встречается в большом количестве в Краснодарском и Шапсугском во-
дохранилищах, в среднем течении Кубани и кубанских лиманах. Длина
тела — до 21 см, обычно — 10—15 см. Самцы мельче самок. Размно-
жается в мае—июне. Плодовитость — 20—35 икринок. Икринки от-
кладываются самкой в выводковую камеру самца, где он и вынашивает
их до вылупления личинок; иногда молодь некоторое время остаётся
177
в камере. Икринки в выводковой камере у мелких (молодых) самцов
расположены в два ряда, у крупных — спереди в три ряда, сзади — в
два. Растёт молодь быстро, достигая к концу первого года жизни длины
тела 6—8 см. Пищей служат беспозвоночные организмы. Сама игла-
рыба является объектом питания хищных видов рыб.
Промыслового значения не имеет.
Сейчас данный вид считают синонимом Syngnathus abaster Risso,
1827 — итальянская рыба-игла (Решетников, 2013).
Отряд Кефалеобразные — Mugiliformes Günther, 1880
Преимущественно морские прибрежные или пелагические рыбы.
Чешуя циклоидная (у некоторых ктеноидная). Некоторые представи-
тели приспособились к жизни в пресной воде. Имеются два спинных
плавника, первый с четырьмя колючими лучами, второй, располо-
женный далеко от первого, с мягкими лучами. Грудные плавники
расположены высоко, а брюшные приближены к ним и находятся на
брюхе.
В бассейне Кубани обитают представители семейства Кефале-
вые — Mugilidae Cuvier, 1829.
Семейство Кефалевые — Mugilidae Cuvier, 1829
Это небольшая группа рыб с замкнутым плавательным пузырём
и без боковой линии. Тело удлинённое с большой сплющенной свер-
ху головой. Ротовая щель поперечная. Циклоидная чешуя покрывает
не только тело, но и голову. Живут при различной солёности: от со-
вершенно пресной воды до солёности в 40 ‰. Два спинных плавника,
первый с четырьмя колючими лучами, второй, расположенный далеко
от первого, с мягкими лучами. В анальном плавнике 2—3 колючки и
7—12 ветвистых лучей.
Кефалевые — стадные кочующие рыбы. Всего в семействе кефа-
левых 17 родов и 72 вида рыб. В бассейне Кубани встречаются пред-
ставители двух родов: Серые кефали — Mugil Linnaeus, 1758 с одним
видом (лобан — Mugil cephalus Linnaeus, 1758) и Кефали-лизы — Liza
(Jordan & Swain, 1884) с тремя видами: сингиль — Liza aurata (Risso,
1810), остронос — Liza saliens (Risso, 1810), пиленгас — Liza haemato-
cheilus (Temminck & Schlegel, 1845).
178
Таблица
для определения видов семейства
Кефалевые — Mugilidae Cuvier, 1829
1 (2). На чешуях спины и верхней части головы сзади желобки
системы боковой линии ординарные. Чешуйный покров на конце рыла
доходит только до задней пары ноздрей и оканчивается спереди не
более чем одним рядом мелких чешуй. Жировое веко не доходит до
зрачка. Пилорических придатков 8, все равной длины (иногда 7 или 9)
(рис. 111, г).
Сингиль — Liza aurata (Risso, 1810) (с. 179)
Рис. 111. Дорсальный вид головы Liza aurata (а), Liza saliens (б) и пилориче-
ские придатки Liza saliens (в), Liza aurata (г)
2 (1). На чешуях спины и в верхней части головы сзади по не-
скольку (2—3) желобков
системы боковой линии.
Чешуйный покров на конце
рыла доходит до передней
пары ноздрей и оканчива-
ется многими рядами мел-
ких чешуй. Пилорических
придатков 8, из них 3 длин-
ные, остальные короткие
(рис. 111, в). Жаберных ты-
чинок 60—65.
Остронос — Liza sa-
liens (Risso, 1810) (с. 180)
Рис. 112. Головы Liza haematocheilus (а)
и Mugil cephalus (б) сверху
179
3 (4). Чешуя на голове по размеру такая же, как на спине, мелких
чешуек на конце рыла нет (рис. 112, а). Радужина глаз оранжевая. Пи-
лорических придатков 6. Хвостовой плавник слабо выемчатый.
Пиленгас — Liza haematocheilus (Temminck & Schlegel, 1845)
(с. 181)
4 (3). Жировое веко покрывает глаз вплоть до зрачка. Чешуйный по-
кров на голове мельчает и доходит почти до конца рыла (рис. 112, б). Пи-
лорических придатков 2, один удлинённый. Жаберных тычинок до 140.
Лобан — Mugil cephalus Linnaeus, 1758 (с. 182)
Краткая биологическая характеристика видов семейства
Кефалевые — Mugilidae Cuvier, 1829
Сингиль — Liza aurata (Risso, 1810) (рис. 113, 2.85)
Рис. 113. Сингиль — Liza aurata (Risso, 1810)
D IV, I 9, A III 9. Голова широкая. Рыло покрыто чешуёй только
до задней пары ноздрей. Чешуя на спине и голове имеет посередине по
одному каналу. Желудок удлинённой формы с короткими и равными
по длине пилорическими придатками. Жировое веко вокруг глаза зача-
точное. Сверху рыло голое, вплоть до задних ноздрей. Сингиль растёт
довольно быстро, особенно до наступления половой зрелости. Половое
созревание наступает у самцов не раньше чем в трёхлетнем возрасте,
у самок — в пятилетнем возрасте. Плодовитость самок очень велика
и колеблется от 740 до 2 800 тыс. икринок. Икра мелкая, пелагиче-
ская. Икрометание происходит в июле—августе при температуре воды
20—22 °С. Личинки и сеголетки нагуливаются вблизи берегов. Зимует
180
сингиль в Чёрном море на глубине. После зимовки молодь заходит в
причерноморские лиманы (Кизилташская группа) и приазовские, где
она быстро растёт, питаясь беспозвоночными и детритом. Двухлет-
ки достигают массы 100—120 г, длины 18—20 см. Взрослый сингиль
практически в приазовских лиманах не встречается. В кизилташских
лиманах нагуливается как молодь, так и взрослые особи. Масса — до
1 200 г. Продолжительность жизни сингиля — 10—11 лет.
В Чёрном море имеет промысловое значение.
Некоторые современные ихтиологи переносят данный вид в род
Толстогубые кефали — Chelon Artedi, 1793 (Chelon aurata (Risso,
1810)), однако У. Эшмайер с коллегами (Eschmeyer, Fricke, van der
Laan, 2019а) поддерживает валидность вида в форме Liza aurata (Ris-
so, 1810).
Остронос — Liza saliens (Risso, 1810) (рис. 114, 2.86)
Рис. 114. Остронос — Liza saliens (Risso, 1810)
D IV, (I) 9, A III (8) 9. Голова более узкая, чем у сингиля, профиль
прямой. Чешуя на голове и ближе к концу рыла становится мельче. Че-
шуя, расположенная на голове и передней части спины, имеет по 2—3
канала. Желудок расширенной формы с неровными по длине пилори-
ческими придатками, их у остроноса 8, из них 3 длинные, остальные
короткие. Жировые веки около глаз зачаточные.
Остронос становится половозрелым в возрасте 3—4 лет. Плодови-
тость колеблется от 500 до 2 000 тыс. икринок. Икрометание происхо-
дит при температуре воды 22 °С. Нерест происходит в открытом море.
181
Личинки и мальки остроноса держатся вблизи берега. Молодь заходит
в Азовское море и приазовские лиманы. Остронос более устойчив к
низким температурам, чем сингиль. Отмечены случаи пребывания мо-
лоди в пресной воде (устье Кубани и Протоки).
Растёт остронос медленнее, чем сингиль. В кизилташских лиманах
отмечены особи длиной 33—36 см и массой 700—800 г. Наиболее ста-
рые особи были в возрасте 8+. Вид имеет промысловое значение и как
объект любительского рыболовства.
Остроноса, как и сингиля, предложено перенести в род Толстогу-
бые кефали — Chelon Artedi, 1793, и в данном случае У. Эшмайер с
коллегами (Eschmeyer, Fricke, van der Laan, 2019б) поддерживают ва-
лидность вида Chelon saliens (Risso, 1810).
Пиленгас — Liza haematocheilus (Temminck & Schlegel, 1845)
(рис. 115, 2.87)
Рис. 115. Пиленгас — Liza haematocheilus (Temminck & Schlegel, 1845)
D IV, I 8—9, A III 8—9. Жировое веко развито очень слабо. Пи-
лорических придатков шесть. Верхняя губа умеренно утолщена, без
придатков. Канальцы на чешуях спины однорядные. Мелких чешуек на
конце рыла нет. От конца рыла до первого спинного плавника 23—28
чешуй. Хвостовой плавник слабо выемчатый (см. рис. 2.87).
Нативным ареалом является залив Петра Великого, Амурский за-
лив до Амурского лимана. В 1971 г. пиленгас акклиматизирован в Чёр-
ном, а позднее и в Азовском море. Пиленгас весьма эврибионтный вид,
обитающий в водах с широким диапазоном солёности, от пресной до
океанической, выдерживает сезонные колебания температуры воды от
182
отрицательной до +34 °С. Нерестится в солёной воде, а зимует в пре-
сной. Является объектом прудового рыбоводства (Москул, 1995; Аб-
рамчук, Иваненко, 2018).
Уловы пиленгаса предприятиями Краснодарского края в Азовском
мореснизилисьот3239тв2006г.до483тв2012г.
Ранее в отечественной литературе данный вид определялся как
Mugil soiuy Basilewsky, 1855, сейчас, согласно работе Т. Л. Чесали-
ной и М. В. Чесалина (2002), предложено вернуть пиленгасу его пер-
воначальное латинское название — Liza haematochila (Temminck &
Schlegel, 1845). По данным базы (http://researcharhive.calacademy.org/
research/ichthyology/catalog/fishcat-main.asp), куратором которой явля-
ется доктор У. Эшмайер (W. N. Eschmeyer), ранее вид был включён в
род Mugil (как Mugil soiuy), но теперь считается членом рода Planiliza
Whitley, 1945. Таким образом, валидным является название — Planili-
za haematocheila (Temminck & Schlegel, 1845) (Eschmeyer, Fricke, van
der Laan, 2019в).
Лобан — Mugil cephalus Linnaeus, 1758 (рис. 116, 2.88)
Рис. 116. Лобан — Mugil cephalus Linnaeus, 1758
D IV, I 8—9, A III 8. У лобана на глазах имеются широкие жиро-
вые веки, достигающие зрачка. Пилорических придатков 2, один из них
удлинённый. Спинных плавников два, причём один расположен далеко
от другого.
Лобан, как сингиль и остронос, — рыба теплолюбивая, зимует в
море у берегов Крыма, где в это время служит объектом промысла. Вес-
ной кефаль начинает мигрировать на север и входит в Азовское море
и приазовские лиманы. Ход в Азовское море происходит в апреле, где
183
до июня—июля нагуливается, а в конце июля взрослые половозрелые
особи уходят из лиманов в море для икрометания. После нереста воз-
вращаются обратно в кизилташские и кубанские (приазовские) лиманы
для нагула. Осенью выходят из лиманов и перемещаются к местам зи-
мовки.
Половозрелым лобан становится в возрасте 6—7 лет. Плодови-
тость лобана — 2 500—6 200 тыс. икринок. Икра пелагическая. Инку-
бационный период длится 30—40 ч. Икринки имеют низкий удельный
вес, большую жировую каплю и поэтому плавают у самой поверхно-
сти. Выклюнувшиеся личинки для совершения своей первой миграции
(из открытого моря к берегу) имеют ряд приспособлений: в области
спинных плавников спинка их не смачивается и во время движения
образуются пузырьки воздуха, а при волнении мальки изменяют ок-
раску с типично тёмной (зеленовато-синяя спинка, серебристые бока)
на частично серебристую. Первое приспособление позволяет малькам
держаться в поверхностных слоях воды и облегчает движение, второе
выполняет защитную функцию: маскируясь под белые гребешки волн,
они малозаметны для многочисленных хищников. Подошедшая к бе-
регам молодь кефали заходит в лиманы и находится там до осени. К
концу первого года длина кефали достигает 5—7 см. Осенью вместе с
взрослыми рыбами сеголетки совершают миграции вдоль Кавказского
побережья на юг, в сторону Батуми. На второй год длина увеличивается
до 16—18 см, масса — до 40—60 г. Молодь кефали длиной 15—20 см
называют чуларой, ларич.
Уловы кефалей (без пиленгаса) в Азовском море колеблются от
0,2до3,25твгод.
Отряд Цихлидообразные — Cichliformes Betancur-R et al.,
2013
Ранее входил в отряд Окунеобразных (Perciformes), но по данным
последней ревизии подотряд поднят в ранг отряда. Отряд состоит из
двух семейств: Pholidichthyidae Jordan, 1896 — это небольшое семей-
ство, включающее один род с двумя видами, и Цихловые, или Цихли-
ды, — Cichlidae Bonaparte, 1835, — одно из самых крупных семейств
позвоночных, насчитывающее более 202 родов и свыше 1 700 видов.
Именно молекулярные данные позволили поместить эти два семейства
в один и тот же таксон.
184
В бассейне Кубани обитают представители семейства Цихловые,
или Цихлиды, — Cichlidae Bonaparte, 1835.
Семейство Цихловые, или Цихлиды, — Cichlidae
Bonaparte, 1835
В основном это пресноводные рыбы, обитатели тропических вод. Тело
довольно высокое, спинной плавник один, окраска яркая. Распространены
в Южной и Центральной Америке, Африке, Юго-Западной Азии и Индии.
Семейство насчитывает в общей сложности около 1 762 видов, от-
носящихся к более чем 202 родам. Большинство — это хищные рыбы,
но среди них имеются всеядные.
В 1970-х гг. в большой род Тиляпии (Tilapia Smith, 1840) включа-
ли более ста видов, распространённых в основном в тропических во-
дах. Впоследствии из старого рода Tilapia Smith, 1840 было выделено
несколько самостоятельных родов: Oreochromis Günther, 1889 и Sa-
rotherodon Rüppell, 1852. В качестве напоминания о тиляпии осталось
несколько небольших родов семейства Цихловые, виды которых до сих
пор именуются тиляпиями.
Пищевой рацион тиляпий состоит из зоопланктонных и бентиче-
ских организмов, личинок насекомых, водорослей и высшей водной
растительности. В отношении условий жизни тиляпия неприхотлива:
обитает как в пресной, так и в солёной воде, выдерживает температуру
воды до 30 °С. Быстро созревает и уже в 5-месячном возрасте может
давать потомство. Плодовитость — до 1 тыс. икринок. В год может
давать от 6 до 11 генераций. Самцы крупнее самок. Икра откладывается
в гнездо, которое имеет вид ямки, после оплодотворения самки одних
видов забирают икру в ротовую полость, где происходит инкубация,
до появления личинок. Некоторое время самка охраняет личинок и при
опасности прячет их в ротовую полость. Другие виды приклеивают
икру к предметам и самцы охраняют икру от поедания другими рыба-
ми. Тиляпии — рыбки небольших размеров. Обычно длина составляет
18—20 см, масса — 130—150 г. Но встречаются особи массой до 850 г
и даже до 1 200 г (мозамбикская тиляпия). Тиляпии ярко окрашены, на
боках имеются две боковые линии — верхняя и нижняя, на рыле два
носовых отверстия, тогда как у других костистых рыб их четыре.
Тиляпии являются одним из основных объектов рыбоводства
в странах Ближнего Востока и Африки. В настоящее время тиляпии
185
культивируются в Юго-Восточной Азии, Центральной и Южной Аме-
рике, а также в Японии, Бельгии, Голландии, США и других странах. В
нашу страну в 1960-е гг. были завезены два вида рода Тиляпии: мозам-
бикская тиляпия — Oreochromis mossambicus Peters, 1852 и голубая
(золотая) тиляпия — Oreochromis aureus Steindachner, 1864.
Таблица
для определения видов семейства
Цихлиды — Cichlidae Bonaparte, 1835
1 (2). В окраске тела преобладают серые и чёрные цвета. На пер-
вой жаберной дуге 16—18 тычинок. 3—4 ряда щёчных чешуек. Есть
тёмное пятно на заднем конце жаберной крышки.
Мозамбикская тиляпия — Oreochromis mossambicus (Peters, 1852)
(с. 185)
2 (1). Окраска тела — светло-голубая и светло-серая. На первой
жаберной дуге 28—30 тычинок. 2 ряда щёчных чешуек. Тёмное пятно
на заднем конце жаберной крышки отсутствует.
Голубая (золотая) тиляпия — Oreochromis aureus (Steindachner,
1864) (с. 186)
Мозамбикская тиляпия — Oreochromis mossambicus (Peters, 1852)
(рис. 117, 2.89)
Рис. 117. Мозамбикская тиляпия — Oreochromis mossambicus (Peters, 1852)
186
D XI—XXIII 9—16, A III—IV 5—6. Позвонков 25—33. В Крас-
нодарском крае тиляпии выращиваются в прудах, снабжаемых геотер-
мальной водой овощеводческого объединения «Мостовский». Тиляпии
за один вегетационный сезон достигают товарной массы 200—250 г.
Самцы растут быстрее самок. Половая зрелость у мозамбикской тиля-
пии наступает в возрасте 3—6 месяцев. Число икрометаний у них до-
ходит до 16 в год. Плодовитость самки массой 800—1 000 г достигает
2,5 тыс. икринок. Общее производство тиляпии составляет ежегодно
около 100 т. В последние годы тиляпии выращивались в садках на тё-
плых водах Краснодарской ТЭЦ, откуда проникли в сбросной канал
ТЭЦ, где они нерестятся и являются объектом любительского рыболов-
ства. Отдельные экземпляры достигают массы более 1 кг.
Голубая (золотая) тиляпия — Oreochromis aureus (Steindachner,
1864) (рис. 118, 2.90)
Рис. 118. Голубая (золотая) тиляпия — Oreochromis aureus (Steindachner,
1864)
D XIV—XVII 11—15, A III 8—11. Позвонков 28—31. Пресновод-
ная рыба, хорошо приспособленная к жизни в солоноватой воде. Дли-
на взрослых особей обычно составляет 13—18 см, вес — 2,3—2,7 кг.
Самый крупный зарегистрированный экземпляр вида достигал 53 см и
187
весил 4,5 кг. Питается в основном растительной пищей, однако от слу-
чая к случаю может употреблять и зоопланктон, а молодь ест мелких
беспозвоночных.
Производители голубой тиляпии вынашивают икру во рту, при-
чём преимущественно самки. Плодовитость в диапазоне от 160 до
1 600 икринок на одну самку. Самка голубой тиляпии откладывает
около 9—10 икринок на 1 г массы тела. Икра имеет жёлто-коричне-
вый цвет и вытянутую форму, словно яйцо. Весь процесс, от опло-
дотворения до прекращения охраны выводка, занимает примерно 3
недели. Самцы тиляпий растут быстрее самок и имеют почти одина-
ковую длину тела. Половая зрелость наступает в 5—6 месяцев. Спо-
собна регулярно размножаться с интервалом 25—80 сут.
Тиляпии легко размножаются по сравнению с другими рыбами,
что в ряде случаев ведёт к перенаселению водоёмов, снижению про-
дуктивности и является одной из сложнейших проблем при её культи-
вировании.
Так как самцы у большинства видов растут значительно быстрее
смок, то выращивание одних самцов позволяет значительно увеличить вы-
ход продукции. Однако сортировка и отбор однополых особей трудоёмки.
Перспективен способ межвидовой гибридизации, позволяющий
получать преобладающее количество самцов в потомстве. Так, при
скрещивании самки Oreochromis mossambicus с самцом Oreochromis
hornorum количество самцов в потомстве равно 100 %, а при скрещива-
нии самки Oreochromis niloticus с самцом Oreochromis aureus — 80 —
90 % самцов.
Представляет интерес способ получения однополого потомства
путём искусственной реверсии (изменения) пола производителей. Так,
скармливание личинкам с пищей половых гормонов, например тесто-
стерона, в течение первых недель после выклева позволяет увеличить
выход самцов.
Отряд Окунеобразные — Perciformes Bleeker, 1859
Наиболее многочисленный отряд ныне живущих костистых рыб.
В отряде 20 подотрядов, 160 семейств, 1 539 родов и более 10 тыс.
видов рыб (точнее 10 033 вида) (Пономарев, Баканева, Федоровых,
2016). Преимущественно хищные рыбы, обитающие как в море, так и в
пресных водоёмах. Спинных плавников у подавляющего большинства
188
видов два. Передний из них имеет жёсткие, колючие нерасчленённые
лучи. Задний плавник состоит из мягких расчленённых лучей, лишь
два первых луча имеют вид колючек. У некоторых видов задний край
переднего и передний край заднего спинного плавника соединяются
перепонкой, в результате чего образуется один плавник. В анальном
плавнике 1—3 колючки, остальные лучи мягкие. Брюшные плавники
расположены под грудными или немного позади их. Первый луч колю-
чий. Чешуя, если есть, обычно ктеноидная, реже циклоидная. Предста-
вители отряда Окунеобразные широко распространены как в морских,
так и в пресных водах.
В водах бассейна Кубани обитают представители 4 семейств.
Таблица
для определения семейств отряда
Окунеобразные — Perciformes Bleeker, 1859
1 (2). В анальном плавнике 1—2 колючих, жёстких луча. Первая
колючая часть спинного плавника выше задней.
Семейство Окунёвые — Percidae Rafinesque, 1815 (с. 188)
2 (1). В анальном плавнике 3 колючих жёстких луча. Первая колю-
чая часть спинного плавника ниже задней. Окраска тела зеленоватая,
иногда по бокам тянется продольная полоса.
Семейство Центрарховые, или Ушастые окуни, — Centrarchi-
dae Bleeker, 1859 (с. 197)
3 (4). На подбородке 2 усика. Два спинных плавника, расположен-
ных на довольном расстоянии друг от друга. Окраска тела с красными,
жёлтыми и серебристыми пятнами.
Семейство Барабулевые, или Султанковые, — Mullidae Rafin-
esque, 1815 (с. 199)
4 (3). Усиков нет. Окраска серебристая, на туловище с каждой сто-
роны тела по восемь рядов продольных полос.
Семейство Лавраковые, или Перцихтовые, — Percichthyidae
Jordan & Eigenmann, 1890 (с. 200)
Семейство Окунёвые — Percidae rafinesque, 1815
Хищные рыбы. Верхнечелюстные кости не соединены плотно с
предчелюстными. Спинных плавников обычно два, но иногда они со-
189
единяются в один. В анальном плавнике 1—2 колючих луча. В брюш-
ных плавниках 1 колючий и 5 ветвистых мягких лучей. На челюстях
имеются зубы, а у некоторых видов и клыки. Тело покрыто ктеноидной
чешуёй.
В фауне бассейна реки Кубани обитают представители четырёх
родов.
Таблица
для определения родов семейства
Окунёвые — Percidae Rafinesque, 1815
1 (2). Зубы мелкие, расположены на челюстях широкими полоса-
ми. Клыков нет. На конце первого спинного плавника тёмное пятно. По
бокам тела 5—9 тёмных поперечных полос. Боковая линия не продол-
жается на хвостовой плавник.
Род Окуни [Пресноводные окуни] — Perca Linnaeus, 1758
(с. 189)
2 (1). Зубы на челюстях расположены узкими полосами. Клыки
обычно есть. Боковая линия продолжается на хвостовой плавник.
Род Судаки — Sander Oken, 1817 (с. 191)
3 (4). Спинные плавники разделены заметным промежутком.
Верхнечелюстная кость сзади свободная.
Род Перкарины — Percarina Nordmann, 1840 (с. 194)
4 (3). Спинные плавники слиты друг с другом в один. Верхнечелю-
стная кость покрыта предглазничной.
Род Ерши — Gymnocephalus Bloch, 1793 (с. 195)
Род Окуни [Пресноводные окуни] — Perca Linnaeus, 1758
Тело сжатое с боков, овальное, покрытое мелкой, крепкоси-
дящей ктеноидной чешуёй. Щёки сплошь покрыты чешуёй, голова
сверху голая. Щетинковидные зубы расположены полосами во много
рядов на челюстях и сошнике. Клыков нет. Жаберных лучей 7. Жа-
берные перепонки не сращены между собою. Два спинных плавника,
соприкасающихся или немного раздвинутых. Анальный плавник ко-
роткий с двумя передними колючими лучами, хвостовой выемчатый.
Боковая линия не продолжается на хвостовой плавник. Позвонков
более 24.
190
Род насчитывают три вида, которые водятся в пресных и отчасти
солоноватых водах умеренного пояса северного полушария. В водо-
ёмах бассейна Кубани обитает один вид этого рода — обыкновенный
[речной] окунь — Perca fluviatilis Linnaeus, 1758.
Обыкновенный, или речной, окунь — Perca fluviatilis Linnaeus,
1758 (рис. 119, 2.91)
Рис. 119. Обыкновенный окунь — Perca fluviatilis Linnaeus, 1758
D1 XIII—XVII, D2 I—III 13—15, A II (7) 8—9 (10), l.l. 57—77. Окунь
имеет сжатое с боков тело, которое покрыто плотной мелкой ктеноид-
ной чешуёй. Крышечная кость покрыта чешуёй только вверху. Верх-
няя челюсть доходит до вертикали середины глаза. Брюшные плавники
немного длиннее грудных. Тело зеленовато-жёлтое, на боках 5—9 по-
перечных чёрных полос. В конце первого спинного плавника чёрное
пятно. Первый спинной плавник серого цвета, второй спинной — зеле-
новато-жёлтого, анальный и грудные плавники — жёлтые, иногда крас-
ные, брюшные плавники — светлые с ярко-красной каймой. Хвостовой
плавник тёмного цвета у основания и красного по бокам и у окончания
(см. рис. 2.91).
Обыкновенный окунь — одна из самых распространённых пре-
сноводных рыб. Встречается практически во всех водоёмах бассейна
Кубани, избегая мутную воду и отсутствует в горных участках рек и
горных озёрах.
191
Половозрелым окунь становится в возрасте 2—3 лет. Нерест начи-
нается в конце марта и продолжается до конца апреля при температуре
воды 7—8 °С. Икра откладывается в виде студенистых лент главным
образом на прошлогодние растения. Плодовитость колеблется от 10 до
280 тыс. икринок. Икра не охраняется родителями. Личинки выводят-
ся на 10—12 -й день. В первое время питаются зоопланктоном, а молодь
длиной 4 см переходит на питание рыбой.
Растёт окунь медленно. Сеголетки достигают 5—8 см и 7—10 г,
двухлетки — 10—12 см и 15—20 г. Обычная длина взрослого окуня —
30—50 см, масса — 0,8—1,2 кг, изредка в Краснодарском водохранили-
ще и кубанских лиманах встречаются особи до 2,0—2,5 кг.
Окунь — популярный объект любительского рыболовства, в от-
дельных водоёмах имеет важное промысловое значение. Ежегодный
вылов в кубанских лиманах колеблется от 21 до 60 т.
Род Судаки — Sander Oken, 1817
Пресноводные, солоноватоводные и полупроходные рыбы. Тело
удлинённое, сжатое с боков. Зубы расположены узкими рядами на
челюстях, сошнике и нёбных костях. На челюстях и нёбных костях
бывают клыки. Жаберные тычинки у взрослых в виде бугорков. Два
спинных плавника, соприкасающихся или слегка раздвинутых. Боко-
вая линия продолжается на хвостовой плавник.
В бассейне Кубани встречаются представители двух видов.
Таблица
для определения видов рода
Судаки — Sander Oken, 1817
1 (2). В анальном плавнике 11—14 ветвистых лучей. Щёки (пред-
крышки) голые или лишь частично покрыты чешуёй. На челюстях
большие острые зубы — клыки.
Судак — Sander lucioperca (Linnaeus, 1758) (с. 192)
2 (1). В анальном плавнике 9—10 ветвистых лучей. Щёки (пред-
крышки) сплошь покрыты чешуёй. На нижней челюсти больших зубов
(клыков) обычно нет.
Бёрш — Sander volgensis (Gmelin, 1789) (с. 193)
192
Краткая биологическая характеристика видов рода
Судаки — Sander Oken, 1817
Судак — Sander lucioperca (Linnaeus, 1758) (рис. 120, 2.92)
Рис. 120. Судак — Sander lucioperca (Linnaeus, 1758)
D1 XIII—XVII, D2 I—III 19—24, A II—III 11—14, l.l. 80—97. Су-
дак — теплолюбивая рыба, обитатель толщи воды, очень требователен
к большому содержанию кислорода в воде и её чистоте. Обитатель пе-
лагиали. Окраска неяркая — спинка зеленовато-серая, бока серебри-
стые, имеются 8—10 поперечных полос (см. рис. 2.92).
Тело удлинённое, сжатое с боков. Плавники светло-серые. Спин-
ной плавник состоит из двух частей: мягкой и колючей. Чешуя мелкая.
Голова средней величины, рот большой, на челюстных костях имеются
клыки. Верхняя челюсть заходит на задний край глаза. На первой жа-
берной дуге 13—15 жаберных тычинок.
Половозрелым судак становится на 3—4-м году жизни. Плодо-
витость кубанского судака при массе 2—3 кг составляет в среднем
480 тыс. икринок. Преднерестовый ход судака начинается в марте. По
Кубани и Протоке высоко не поднимается, заходит сразу в лиманы. Не-
рест происходит в апреле при температуре воды от 8 до 12 °С. Икра
липкая, мелкая, нерест единовременный, вымётывание икры у одной
самки продолжается 1—2 ч. Место для кладки икры выбирает самец,
расчищает его, устраивает небольшое гнездо, затем отложенную икру
оплодотворяет и охраняет до появления личинок. Продолжительность
инкубации — от 3—4 до 10—11 дней.
Первый месяц личинки потребляют ракообразных, а затем пере-
ходят на питание молодью более мелких рыб (уклеи, сазана, леща).
193
Максимальная интенсивность питания отмечена в весенний и осенний
периоды, в мае и сентябре потребляют 20 %, в июле — 18 % годового
рациона. В течение суток судак наиболее интенсивно питается в часы
восхода и захода солнца. Темп роста судака в старшем возрасте уве-
личивается: средний годовой прирост массы тела в дельте Кубани со-
ставляет 500—600 г, в Краснодарском водохранилище — 400—800 г. В
четырёхлетнем возрасте достигает 1,5—1,8 кг, в семи- и восьмилетнем
возрасте — соответственно 4 и 5 кг.
Судак относится к ценным промысловым рыбам. Вылов его со-
ставляет до 2 тыс. т в год.
Бёрш, или берш, или волжский судак, — Sander volgensis (Gmelin,
1789) (рис. 121, 2.93)
Рис. 121. Бёрш — Sander volgensis (Gmelin, 1789)
D1 XII—XIV, D2 I—II 20—22, A II 9—10, l.l. 70—83. Отличается
от судака прежде всего отсутствием клыков и меньшими размерами.
Спинные плавники бёрша выше, чем у судака, и соприкасаются. Щёки
покрыты чешуёй. Верхняя челюсть доходит до вертикали середины
глаза. Окраска тела светлее, чем у судака; тёмные поперечные полосы
выделяются резче (см. рис. 2.93).
Рыба пресных вод, только изредка в единичных экземплярах встре-
чается в опреснённых участках моря. Половое созревание наступает на
3—4 -м году жизни. Икрометание происходит с конца апреля по май.
Икра мелкая. Инкубационный период длится 6—8 сут. Молодь питается
беспозвоночными, а со второго года жизни в его рационе преобладают
мелкие рыбы. Держатся стаями, подобно обыкновенным судакам.
194
Бёрш встречается в реках Каспийского, Азовского и Чёрного мо-
рей, выходит в Каспийское море. В водоёмах бассейна Кубани, в том
числе в Краснодарском водохранилище, встречается с 1995 г. До этого
лишь единичные экземпляры поднимались по Кубани до Краснодар-
ского водохранилища (Москул, 2001, 2003).
Род Перкарины — Percarina Nordmann, 1840
Тело сжатое с боков, чешуя тонкая, легко спадает. Рот выдвиж-
ной. Голова голая или часть покрыта чешуёй. Предкрышка снабже-
на шипами по краю. Род состоит из 2 видов. В Азовском море и ку-
банских лиманах обитает один вид: азовская перкарина — Percarina
demidoffii Nordmann, 1840.
Азовская перкарина — Percarina demidoffii Nordmann, 1840
(рис. 122, 2.94)
Рис. 122. Азовская перкарина — Percarina demidoffii Nordmann, 1840
D1 VIII—X, D2 II—III 9—12, A II 7—11, l.l. 33—37. Маленькая
рыбка: самки достигают длины 10—11 см, самцы — 7 —8 см. Тело уд-
линённое, довольно высокое, сжатое с боков, покрытое тонкой чешу-
ёй, которая при дотрагивании легко спадает с тела. Основание грудных
плавников находится над основанием брюшных. Боковая линия полная.
195
Рот большой, вооружённый мелкими зубами. На боках головы большие
слизеотделительные полости, тело покрыто слоем слизи. Голова, в том
числе и жаберные крышки, и грудь не покрыты чешуёй. Диаметр гла-
за равен длине рыла. Половозрелой становится на втором году жизни.
Нерест происходит в июне—июле. Плодовитость — от 1,8 до 3,2 тыс.
икринок. Икра донная, инкубационный период около 2 сут. Продол-
жительность жизни — 3 —4 года. Питается донными беспозвоночными
животными. Служит пищей судака.
Промыслового значения не имеет (почти непригодна для употреб-
ления в пищу). Выделяет слизь, снижая ценность основного улова; ис-
пользуется на тук и кормовую муку. Обитает в Азовском море, но часто
встречается как в осолонённых, так и опреснённых лиманах.
Род Ерши — Gymnocephalus Bloch, 1793
Тело сжатое с боков. Рот небольшой, выдвижной. На обеих челю-
стях имеются щетинковидные зубы. На сошнике и нёбных костях зубов
мало или совсем нет. Голова голая, предкрышка с шипами. Спинные
плавники слиты вместе. Хвостовой плавник выемчатый.
В водоёмах бассейна Кубани встречаются представители двух
видов: обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758)
и донской ёрш, или бирючок, или носарь, — Gymnocephalus acerinus
(Gmelin, 1774).
Таблица
для определения видов рода
Ерши — Gymnocephalus Bloch, 1793
1 (2). В спинном плавнике 11—16 колючих и 11—15 мягких лучей,
в анальном 2 колючих и 5—6 мягких лучей. В боковой линии не более
40 чешуй.
Обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758)
(с. 196)
2 (1). В спинном плавнике 17—19 колючих и 12—14 мягких лучей,
в анальном 2 колючих и 5—6 мягких лучей. В боковой линии 50—55
чешуй.
Донской ёрш, или бирючок, — Gymnocephalus acerinus (Gmelin,
1774) (с. 197)
196
Краткая биологическая характеристика видов рода
Ерши — Gymnocephalus Bloch, 1793
Обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758)
(рис. 123, 2.95)
Рис. 123. Обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758)
D XI—XVI 11—15, A II 5—6, l.l. 35—40. На предкрышечной и
крышечной костях есть острые шипы. Во рту имеются мелкие зубы.
Тело окрашено в серовато-зелёный цвет, на спине есть ряды тёмных
пятнышек, бока желтоватые, пузо светло-серое или белое. Спинной и
хвостовой плавники с чёрными точками. Общий окрас этой рыбы зави-
сит от окружающей среды: ерши светлее в реках и озёрах с песчаным
дном и темнее в водоёмах, где дно илистое. Глаза мутно-розовые, ино-
гда с синей радужкой (см. рис 2.95).
Обитает в основном в пресноводных водоёмах. Небольшая рыба:
длина — до 12 см, масса — 30 г. Самки крупнее самцов. Живёт в во-
доёмах с медленно текучей и стоячей водой, предпочитает заилен-
ные, мягкие грунты. Нерест порционный, с апреля по июнь. Плодови-
тость — от 1 до 5 тыс. икринок. Откладывает мелкую икру на дно или
растительность. Развитие икры продолжается в течение 10—15 дней.
Ёрш питается донными организмами, а также поедает икру других рыб
и молодь. Промыслового значения не имеет, но является объектом лю-
бительского рыболовства.
197
Донской ёрш, или бирючок, — Gymnocephalus acerinus (Gmelin,
1774) (рис. 124, 2.96)
Рис. 124. Донской ёрш — Gymnocephalus acerinus (Gmelin, 1774)
D XVII—XIX 12—14, A II 5—6, l.l. 50—55. Рыло удлинённое, на
спине, боках тела и на колючей части спинного плавника резко выра-
жены чёрные пятнышки. Длина тела — до 15 см.
Бирючок — пресноводная речная рыба. Живёт в русле рек на до-
вольно быстром течении, предпочитает плотные, песчаные грунты. Не-
рестится в апреле—мае в реке на быстром течении, на чистом песчаном
и каменистом грунте. Икра донная, клейкая, с большой жировой кап-
лей. Питается в основном донными беспозвоночными. Является объек-
том любительского рыболовства. Мясо у бирючка нежное и вкусное.
Особенно ценят рыбаки уху из бирючка.
Семейство Центрарховые,
или Ушастые окуни, — Centrarchidae Bleeker, 1859
Представители этого семейства отличаются наличием одного
спинного плавника, передняя колючая часть которого ниже задней.
Населяют пресные воды Северной Америки от Канады до Мексики.
Многие виды акклиматизированы в водоёмах западной Европы. Один
вид — большеротый окунь, или форелеокунь, акклиматизирован в
оз. Абрау и Лиманчик у Новороссийска и в некоторых водоёмах Мос-
ковской области.
198
Большеротый окунь, или форелеокунь, — Micropterus salmoides
(Lacépède, 1802) (рис. 125, 2.97)
Рис. 125. Большеротый окунь, или форелеокунь, — Micropterus salmoides
(Lacépède, 1802)
D X 12—13, A III 10—11, l.l. 62—68. Жаберные тычинки длинные,
их 8—10 штук. Позвонков 30—32. Спинной плавник разделён глубо-
кой выемкой почти на две части. Верхняя челюсть доходит до заднего
края глаза и иногда заходит за задний край глаза.
Окраска сверху зелёная, иногда с небольшими тёмными пятнами;
на щеках две косые тёмные полосы с каждой стороны (см. рис. 2.95).
Большеротый окунь достигает половой зрелости в возрасте
2—3 лет. Плодовитость колеблется от 30 до 180 тыс. икринок. Нерест
происходит в мае—июне. Икра откладывается в гнездо, устраиваемое
самцом в виде ложбинки, выстланной гравием. Самец охраняет не
только икру, но и личинок несколько дней после выклева. Длительность
развития икринок — от 3 до 14 дней.
Большеротый окунь — отличный биологический мелиоратор, по-
едает мелкую сорную рыбу, головастиков, лягушек, но предпочитает
питаться мелкими рыбами. В первые три месяца мальки поедают бес-
позвоночных, а потом питаются рыбой и головастиками. Растёт боль-
шеротый окунь сравнительно быстро. Так, двухлетние особи достигают
массы 300—500 г, трёхлетние — 600 —800 г. Отдельные экземпляры в
семилетнем возрасте имеют массу тела 3,5 кг. Мясо отличается высо-
кими вкусовыми качествами, поэтому его разводят во многих водоёмах
Америки и Европы.
199
В 1905 г. из Германии завезли восемь экземпляров большеротого
окуня и выпустили в оз. Абрау, где они успешно акклиматизировались
и быстро увеличивали свою численность. В 1937—1939 гг. молодь из
оз. Абрау выпустили в оз. Лиманчик, расположенное в 4 км от Абрау, а
в 1939 г. — в водоём Фастовецкого рыбхоза Краснодарского края (Чи-
жов, Абаев, 1868; Ходячий, Москул, Никитина, 1982; Москул, 1998).
Интенсивный бесконтрольный промысел как браконьерами, так и ры-
боловами-любителями привёл к истреблению этого ценного вида. В на-
стоящее время большеротый окунь исчез из состава ихтиофауны этих
озёр (Скляров, Корнаухов, 2012).
Семейство Барабулевые, или Султанковые, — Mullidae
rafinesque, 1815
Небольшие рыбки со стройным, уплощённым с боков телом, ту-
пым рылом. На подбородке 2 усика. Зубы на челюстях мелкие. Глаза
посажены высоко. Спинных плавников два, первый из них состоит
из колючих лучей. Чешуя крупная. В состав семейства включают 6
родов с 85 видами. В Чёрном море водится один вид и несколько
подвидов. Встречается в Азовском море и единично в кубанских ли-
манах.
Черноморская барабуля (султанка) — Mullus barbatus ponticus
Essipov, 1927 (рис. 126, 2.98)
Рис. 126. Черноморская барабуля (султанка) — Mullus barbatus ponticus
Essipov, 1927
200
D VIII, I 8, A II 6, l.l. 37—41. Жаберных тычинок 21—23. Тело
удлинённое и сжатое с боков, длиной до 30 см, обычно 10—20 см.
Спинные и анальный плавники короткие, хвостовой — вильчатый. Это
ярко окрашенные рыбки, на теле имеются поперечные красные полосы,
брюшко серебристое, светло-жёлтые плавники (см. рис. 2.98). В Чёр-
ном море образуют несколько местных стад, одно из которых для нагу-
ла мигрирует в Азовское море, единичные экземпляры встречаются в
кубанских лиманах.
Половозрелой становится уже в двухлетнем возрасте. Икромета-
ние длится с мая по сентябрь при температуре воды от 19 до 23 °С.
Икрометание порционное, происходит почти каждую ночь в прибреж-
ной зоне. За сезон вымётывается до 100 порций икры. Общая плодови-
тость — до 1 млн икринок. Вымётывание икры начинается вечером и
длится всю ночь. Икра мелкая с жировой каплей. Развитие икры длится
при температуре воды 18 °С около 2 сут. Молодь живёт в пелагиали,
вдали от берегов, а по достижении 3—3,5 мм длины молодь подходит к
берегам и опускается на дно.
Обитает барабулька у морских берегов, обыкновенно на неболь-
ших глубинах — 15 —30 м, хотя попадается и на глубине в 100—300 м.
Питается в основном донными беспозвоночными. Основную роль в ра-
зыскивании пищи играют усики. Держится стаями и в местах, где она
кормится, обычно довольно сильно взмучивает грунт, поэтому рыбаки
легко обнаруживают её скопления.
Барабуля — ценная рыба. Мясо очень вкусное и высоко ценится.
Является объектом промысла.
Семейство Лавраковые, или Перцихтиевые, —
Percichthyidae Jordan & eiGenmann, 1890
Голова крупная, треугольной формы. Рот большой, верхняя че-
люсть заходит за середину глаза, нижняя челюсть слегка выдвинута
вперёд. Крупная ктеноидная чешуя покрывает тело, жаберные крышки
и щёки. На теле 7—8 горизонтальных тёмных полос, совпадающих с
рядами чешуи, причём 5 верхних идут до хвоста, а 2 нижних — только
до середины анального плавника.
Морские и пресноводные рыбы. В водоёмах бассейна Кубани
встречается один вид — полосатый окунь (лаврак), который проник из
Азовского моря.
201
Полосатый окунь, или лаврак, — Morone saxatilis (Walbaum, 1792)
(рис. 127, 2.99)
Рис. 127. Полосатый окунь, или лаврак, — Morone saxatilis (Walbaum, 1792)
D1 VII—XI, D2 I 8—14, A III 7—12, l.l. 58—64. Жаберных тычинок
20—29. Позвонков 22—25. Это один из наиболее популярных и цен-
ных видов рыб Атлантического и Тихоокеанского побережий Северной
Америки. Отличается высокой эвригалинностью: живёт и созревает в
воде любой солёности — от пресной до океанической. Для размно-
жения обычно заходит в реки, но может нереститься и в лиманах при
солёности до 3,5 ‰ и температуре воды 12—23 °С. Икра развивается
нормально при солёности от 0 до 16,5 ‰.
Полосатый окунь — эвритермный вид, нагуливается при темпера-
туре воды 5—25 °С. Растёт интенсивно. Встречаются экземпляры мас-
сой 20—30 кг.
В период нереста одну самку сопровождают несколько самцов.
Нерест происходит у поверхности воды. Самка вымётывает зеленова-
тую неклейкую икру, которая немного тяжелее пресной воды. Плодо-
витость зависит от размера самки и колеблется от 20 тыс. икринок у
самок массой 1,5 кг до 5 млн икринок у самок массой 22 кг. Самки со-
зревают в 4—5 -летнем возрасте, самцы — в 2—3-летнем возрасте.
В 1965 г. из США молодь полосатого окуня завезена в рыбопитом-
ник «Горячий Ключ» для искусственного разведения. В 1973 г. было
пересажено более 7,5 тыс. сеголеток в Шапсугское водохранилище.
В настоящее время встречается в небольшом количестве в кубанских
лиманах. Питается в основном рыбой. Молодь питается зоопланкто-
202
ном, а при длине 8,0—8,5 см сеголетки переходят на питание мелкой
рыбой.
Отряд Бычкообразные — Gobiiformes Günther, 1880
Небольшие придонные рыбы с большой широкой головой и за-
круглённым хвостом. Брюшные плавники сращены в присасыватель-
ный диск (воронку) расположенный на горле. Спинных плавников два
или один. Колючие лучи в первом спинном плавнике слабые. Плава-
тельный пузырь обычно отсутствует. Зубы небольшие. Тело покрыто
мелкой чешуёй, а иногда голое. В анальном плавнике один колючий
луч или колючек нет совсем. Морские и пресноводные рыбы. Отряд
представлен 9 семействами, в которых насчитывается 2 211 видов (По-
номарев, Баканева, Федоровых, 2016).
В фауне бассейна Кубани встречается одно семейство — Бычко-
вые — Gobiidae Cuvier, 1816.
Семейство Бычковые — Gobiidae Cuvier, 1816
Тело покрыто циклоидной или ктеноидной чешуёй, а у некоторых
видов — шипиками или совершенно голое. Спинных плавников обыч-
но два, передний развит слабее заднего и представлен неветвистыми
лучами. Брюшные плавники расположены на груди под грудными,
они слиты в присоску (воронку). Жаберные отверстия довольно узкие,
жаберные перепонки прикреплены к межжаберному промежутку. Бо-
ковой линии на теле нет. Органы системы боковой линии (генипоры)
имеются на голове, где также расположены чувствительные каналы и
поры. Плавательный пузырь у одних видов имеется, у других его нет.
Икринки двух типов: мелкие — полиплазматические, т. е. содержащие
одинаковое количество плазмы и желтка, личинки ведут пелагический
образ жизни; крупные — олигоплазматические, содержащие мало
плазмы и много желтка, личинки ведут донный образ жизни. У многих
видов имеется половой диморфизм: самцы крупнее самок и отличают-
ся окраской, которая усиливается во время нереста. У самцов окраска
темнее (некоторые становятся совсем чёрные), плавники удлиняются,
голова изменяет свою форму. Все бычки откладывают донную икру.
Самцы устраивают гнезда и охраняют отложенную самкой икру вплоть
до выхода молоди. Икринки прикрепляются к нижней стороне нересто-
203
вого субстрата (камни, ракушки и др.). Питаются бычки донными бес-
позвоночными, иногда мелкой рыбой.
Многие виды имеют промысловое значение (кругляк, сирман,
мартовик, песочник и др.). Уловы бычков в Азовском море колеблются
от 367 до 1 500 т в год. Все бычки являются существенной частью
пищи многих ценных рыб (осетровых, судака и др.).
В бассейне Кубани семейство Бычковые (Gobiidae Cuvier, 1816)
представлено 9 родами.
Таблица
для определения родов семейства
Бычковые — Gobiidae Cuvier, 1816
1 (10). Тело покрыто чешуёй. На голове есть чувствительные ка-
налы и поры. Во втором спинном плавнике не более 10—11 лучей, в
анальном — один жёсткий и 7—10 мягких лучей.
2 (3). Наиболее длинные лучи расположены в средней части хво-
стового плавника, поэтому хвостовой плавник имеет правильную сим-
метричную форму.
Род Бычки-лысуны, или Бычки-бубыри, — Pomatoschistus
Gill, 1864 (с. 204)
3 (2). Хвостовой плавник несимметричен или закруглён. Наиболее
длинные лучи хвостового плавника расположены в его верхней части,
поэтому его задний край косо срезан. Углы рта находятся на уровне
середины глаз.
Род Бычки Книповича — Knipowitschia Iljin, 1927 (с. 205)
4 (5). Жаберные крышки голые или покрыты чешуёй только свер-
ху. Плавательный пузырь есть. Во втором спинном и в анальном плав-
никах есть по одному жёсткому лучу. Икра мелкая — 1,2—1,8 мм в
диаметре.
Род Бычки — Gobius Linnaeus, 1758 (с. 207)
5 (4). Темя, затылок, горло и брюшко покрыты чешуёй. Плаватель-
ного пузыря нет. Икра крупная, около 3 мм в диаметре.
Род Черноморско-каспийские бычки — Neogobius Iljin, 1927
(с. 208)
6 (7). Передние ноздри вытянуты в усиковидные трубочки, свеши-
вающиеся над верхней губой. Жаберные крышки голые.
Род Тупоносые бычки — Proterorhinus Smitt, 1900 (с. 210)
204
7 (6). Передние носовые отверстия не вытянуты в трубочки. Хво-
стовой плавник закруглённый. Между глазами нет генипор. Тело голое.
Плавательного пузыря нет.
Род Каспиосомы — Caspiosoma Iljin, 1927 (с. 212)
8 (9). Верхняя губа к углам рта не расширена. На первом спинном
плавнике обычно 3 тёмные полоски. Воротник присоски брюшного
диска без лопастинок.
Род Бабки — Babka Iljin, 1927 (с. 212)
9 (8). Верхняя губа к углам рта расширяется. На первом спинном
плавнике 3 светлых и 2 тёмных поперечных полосы.
Род Понтиколы — Ponticola Iljin, 1927 (с. 213)
10 (1). Тело покрыто костными пластиночками, чешуи нет. Чув-
ствительных каналов и пор нет. Во втором спинном плавнике 1 жёст-
кий и 6—11 мягких лучей, в анальном — 1 жёсткий и 6—10 мягких
лучей.
Род Пуголовки — Benthophilus Eichwald, 1831 (с. 217)
Род Бычки-лысуны, или Бычки-бубыри, — Pomatoschistus Gill,
1864
Голова, горло, брюхо и основания грудных плавников голые. Бока
тела покрыты ктеноидной чешуёй. На голове хорошо развита сейсмосен-
сорная система: есть чувствительные каналы, поры и генипоры. Меж-
глазничный промежуток узкий, глаза направлены вверх или в стороны.
Ноздри не вытянуты в виде трубочки. Плавательный пузырь имеется.
Небольшие рыбы. В водоёмах бассейна Кубани встречается один вид.
Леопардовый лысун [бычок-бубырь] — Pomatoschistus marmoratus
(Risso, 1810) (рис. 128, 2.100)
Рис. 128. Леопардовый лысун — Pomatoschistus marmoratus (Risso, 1810)
205
D1 VI, D2 I 8—10, A I 7—10. Поперечных рядов чешуй 38—45.
Позвонков 31. Встречается как в солёных, так и в пресных водоёмах.
Небольшая рыбка, достигающая длины около 8,0 см, но обычно не бо-
лее 5 см и массы 1,8—2,2 г.
Обладает резким половым диморфизмом. Самцы темнее самок.
Самки обычно с немногими резкими неправильными пятнами; спин-
ной плавник с многочисленными тёмными пятнышками. На основании
хвостового плавника имеется тёмное пятно.
Продолжительность жизни — около двух лет. После нереста сна-
чала отмирают самки, затем самцы, которые охраняют, очищают и
аэрируют икру, откладываемую самкой в створки мёртвых моллюсков.
Нерестится в мае—июле на глубине до 50—60 см. Плодовитость — от
1 250 до 2 340 икринок, икрометание порционное. Икра мелкая (0,75—
0,95 мм в диаметре), полиплазматическая. Обитает в Азовском море,
кубанских лиманах, в устьях Кубани, Протоки и приазовских рек (Бей-
суг, Кирпили, Ея и др.), изредка, единично в Краснодарском водохра-
нилище у хут. Ленина на песчаном грунте (Москул, 1982, 1986). Явля-
ется объектом питания осетровых, судака, окуня и других хищных рыб.
Род Бычки Книповича — Knipowitschia Iljin, 1927
Голова, передняя часть спины до начала второго спинного плавни-
ка и низ тела до анального плавника голые, бока покрыты ктеноидной
чешуёй. Межглазничной поры нет. Глаза смотрят вверх.
В водоёмах бассейна Кубани встречаются два вида.
Таблица
для определения видов рода
Бычки Книповича — Knipowitschia Iljin, 1927
1 (2). Глаза расположены близко друг к другу и направлены вверх.
Хвостовой плавник несимметричный, у его основания большое непра-
вильной формы тёмное пятно.
Длиннохвостый бычок Книповича — Knipowitschia longicaudata
(Kessler, 1877) (с. 206)
2 (1). Глаза направлены в стороны, а не вверх. Хвостовой плавник
закруглённый, у его основания длинное прямоугольное тёмное пятно.
Бычок-бубырь — Knipowitschia caucasica (Berg, 1916) (с. 207)
206
Краткая биологическая характеристика видов рода
Бычки Книповича — Knipowitschia Iljin, 1927
Длиннохвостый бычок Книповича — Knipowitschia longicaudata
(Kessler, 1877) (рис. 129, 2.101)
Рис. 129. Длиннохвостый бычок Книповича — Knipowitschia longicaudata
(Kessler, 1877)
D1 (V) VI (VII), D2 I 7—9, A I 8—9. Поперечных рядов чешуй
36—45. Самцы крупнее самок. Наибольшая длина самок — 35—42 мм,
самцов — 50 мм, масса — 0,7—0,8 г. Хвостовой плавник несиммет-
ричный. Спинные плавники разделены значительным промежутком.
Голова, передняя часть спины до второго спинного плавника и низ
тела до анального плавника голые, на боках тела ктеноидная чешуя.
Глаза расположены близко друг к другу и направлены вверх. Окраска
желтовато-белая, на спине мелкие бурые пятнышки. Вдоль оснований
спинных и анального плавников ряды тёмных пятнышек. У основа-
ния хвостового плавника большое неправильной формы тёмное пятно
(см. рис. 2.101). Есть плавательный пузырь. У самца во время нереста
непарные плавники удлиняются и становятся тёмными, на боках обра-
зуются буроватые поперечные полосы.
Продолжительность жизни — один год. Половая зрелость наступа-
ет в возрасте менее 1 года при длине 20 мм. В размножении этого быч-
ка наблюдаются две генерации: в мае размножаются перезимовавшие
особи, а в июле и августе — сеголетки, достигшие половой зрелости.
Широко распространён в Каспийском, Азовском и Черноморском
бассейнах. Обитает как в морской воде, так и в пресноводных лима-
нах. Встречается в реках и водохранилищах. Длиннохвостый бычок
Книповича является объектом питания молоди судака в кубанских
лиманах.
207
Бычок-бубырь — Knipowitschia caucasica (Berg, 1916) (рис. 130,
2.102)
Рис. 130. Бычок-бубырь — Knipowitschia caucasica (Berg, 1916)
D1 VI, D2 I 6—9, A I 7—9. Поперечных рядов чешуй 30—37. Тело
веретеновидное, невысокое, относительно толстое, уплощённое, не-
сколько более вытянутое у самок, покрытое средней по размерам че-
шуёй, которая отсутствует на голове, на спине до окончания основания
второго спинного плавника и на груди. Глаза направлены в стороны, а
не вверх. Лоб выпуклый. Грудные плавники широкие и длинные, поч-
ти достигают анального плавника. Хвостовой плавник обычно закруг-
лён. Есть плавательный пузырь. Окраска тёмно-серая с желтоватым
оттенком или буровато-коричневатого цвета. Самцы отличаются от
самок более тёмной окраской тела, по бокам у них тёмные пятна. У
них на заднем конце первого спинного плавника выраженное тёмное
пятно (см. рис. 2.102), а у самок интенсивного чёрного цвета подборо-
док.
Обитает как в морской воде, так и в пресноводных лиманах.
Продолжительность жизни — около 2 лет. Наибольшая длина
тела — 45 —50 мм. Половой зрелости достигает в возрасте 8—11 меся-
цев при длине тела 2,0—2,4 см. Размножение с конца марта до августа.
Является объектом питания молоди судака в кубанских лиманах.
Род Бычки — Gobius Linnaeus, 1758
Есть плавательный пузырь. Икринки мелкие, содержат сравни-
тельно мало желтка. Личинки пелагические. В бассейне Кубани оби-
тает один вид.
208
Бычок-травяник — Gobius ophiocephalus Pallas, 1814 (рис. 131, 2.103)
Рис. 131. Бычок-травяник — Gobius ophiocephalus Pallas, 1814
D VI, I (13) 14—15 (16), А I (II) 12—15 (16). Поперечных рядов
чешуи 57—63. Жаберные крышки или совсем голые или покрыты че-
шуёй лишь по верхнему краю. Темя, затылок, горло, брюхо и стебли
грудных плавников покрыты циклоидной чешуёй. По бокам имеются
извилистые бурые поперечные полосы (см. рис. 2.103). Достигает в
длину 15—17 см.
Размножается в апреле—июле в двухгодовалом возрасте, икру
откладывает в два приёма. Гнездо устраивает самец из остатков водо-
рослей, икру откладывают от 5 до 10 самок. Плодовитость — от 15 до
30 тыс. икринок.
Хищник, питается мелкой рыбой, ракообразными и моллюсками.
Является объектом промысла, но численность очень мала.
Род Черноморско-каспийские бычки — Neogobius Iljin, 1927
Плавательного пузыря у взрослых особей нет. Тело удлинённое,
сзади сжатое, покрытое чешуёй. Рот не заходит за глаза. Щёки и жа-
берные крышки голые. Затылок покрыт чешуёй. Многие виды имеют
большое промысловое значение. В бассейне Кубани обитают 2 вида.
Таблица
для определения видов рода
Черноморско-каспийские бычки — Neogobius Iljin, 1927
1 (2). Чешуя крупная, менее 55 поперечных рядов. На переднем
спинном плавнике есть большое чёрное пятно.
Бычок-кругляк — Neogobius melanostomus (Pallas, 1814) (с. 209)
209
2 (1). Чешуя мелкая, более 55 поперечных рядов. Чёрного пятна на
переднем спинном плавнике нет.
Речной бычок-песочник — Neogobius fluviatilis (Pallas, 1814)
(с. 210)
Краткая биологическая характеристика видов рода
Черноморско-каспийские бычки — Neogobius Iljin, 1927
Бычок-кругляк — Neogobius melanostomus Pallas, 1814 (рис. 132,
2.104)
Рис. 132. Бычок-кругляк — Neogobius melanostomus Pallas, 1814
D VI, I 14—16, A I 12—13. Поперечных рядов чешуи 47—54. Не-
большая рыбка, достигающая длины 18—20 см. Самки мельче самцов.
Плавательного пузыря нет. Высота второго спинного плавника на всём
протяжении равномерна. На конце первого спинного плавника есть
чёрное продолговатое пятно (см. рис. 2.104). Самцы при достижении
половой зрелости полностью чернеют, а их непарные плавники силь-
но разрастаются в высоту. Самки становятся половозрелыми на втором
году жизни. Плодовитость колеблется от 650 до 1 200 икринок. Икро-
метание происходит с мая по июль. Икринки приклеиваются к донным
предметам и охраняются самцом. Питается бычок моллюсками и рако-
образными.
Бычок-кругляк является объектом промысла как в кубанских ли-
манах, так и в прибрежных участках Азовского моря. Общие уловы
бычков в Азовском море до 1985 г. колебались от 18 до 375 тыс. ц, из
них 80—90 % приходилось на кругляка. В настоящее время уловы рез-
ко сократились из-за ухудшения условий размножения.
210
Речной бычок-песочник — Neogobius fluviatilis (Pallas, 1814)
(рис. 133, 2.105)
Рис. 133. Речной бычок-песочник — Neogobius fluviatilis (Pallas, 1814)
D1 VI, D2 I 15—17, A I 13—15. Поперечных рядов чешуи 58—65.
Второй спинной плавник к заднему концу заметно понижается. Темя,
затылок, спина, жаберные крышки (на треть), основания грудных плав-
ников, задняя половина горла и брюхо покрыты циклоидной чешуёй.
Ширина головы равна её высоте или чуть больше. Рот конечный или
верхний, рыло заострённое. На челюстях имеются мелкие конические
зубы. Нижняя челюсть выдаётся вперёд, верхняя губа не утолщена по
бокам. Самцы во время нереста становятся чёрными с белыми каёмка-
ми на непарных плавниках.
Половозрелым становится на втором году жизни. Нерест проис-
ходит с конца апреля по июнь. Икрометание на донные предметы и
отмершую растительность. Плодовитость — 800 —1 200 икринок. Пи-
тается ракообразными, личинками насекомых, червями и др.
Является объектом промысла. В уловах занимал третье место
после кругляка и мартовика (морской, эвригалинный вид). Обитает в
Азовском море, кубанских лиманах, устьях Кубани и других рек, встре-
чается также в Шапсугском и Краснодарском водохранилищах.
Род Тупоносые бычки — Proterorhinus Smitt, 1900
Передние носовые отверстия вытянуты в усиковидные трубочки,
свешивающиеся вниз над верхней губой, что является характерным
211
признаком представителей рода. Жаберные крышки голые или покры-
ты чешуёй только вдоль верхнего края или на четверть. В бассейне Ку-
бани обитает один вид.
Бычок-цуцик, или мраморный тупоносый бычок, — Proterorhinus
marmoratus (Pallas, 1814) (рис. 134, 2.106)
Рис. 134. Бычок-цуцик, или мраморный тупоносый бычок, — Proterorhinus
marmoratus (Pallas, 1814)
D1 VI (VII), D2 I 14—17 (19), A I 11—15 (16). Поперечных рядов
чешуи обычно 36—48, жаберных тычинок 6—9. Толщина головы за-
метно меньше её высоты. Жаберные крышки покрыты немногочислен-
ными чешуйками только вдоль своего верхнего края. Темя, затылок,
основания грудных плавников, брюхо и задняя часть горла покрыты
чешуёй. Чешуя довольно большая, циклоидная. Основной цвет бурова-
то-серый, чаще желтовато-серый с 4—5 тёмными полосами на спине,
переходящими ниже середины тела в пятна. На рыле с каждой стороны
по тёмному пятну. Плавники обычно полосатые. На основании хвосто-
вого плавника имеется треугольное чёрное пятно, окаймлённое двумя
белыми пятнами (см. рис. 2.106). Плавательного пузыря нет. Длина
тела половозрелых особей — 5 —10 см. Продолжительность жизни —
2—3 года. Нерест происходит в мае—июне. Питается донными орга-
низмами.
Обитает как в солоноватых лиманах, так и в пресных водохрани-
лищах. Встречается в Азовском море, кубанских лиманах, устьях Ку-
бани и других рек, изредка в Шапсугском и Краснодарском водохра-
нилищах.
Промыслового значения не имеет, является объектом питания
хищных видов рыб.
212
Род Каспиосомы — Caspiosoma Iljin, 1927
Передние носовые отверстия не вытянуты в трубочки. Тело голое.
Чувствительных каналов и пор нет. Плавательного пузыря нет. Хвосто-
вой плавник закруглённый.
В бассейне Кубани обитает один вид.
Каспиосома — Caspiosoma caspium (Kessler, 1877) (рис. 135, 2.107)
Рис. 135. Каспиосома — Caspiosoma caspium (Kessler, 1877)
D1 (V) VI (VII), D2 I (II) (11) 12 (13), A I (8) 9—10. Тело голое, уме-
ренно удлинённое, лишь немного сжатое с боков, уплощённое. Плава-
тельного пузыря нет. Перед вторым спинным плавником имеется полу-
лунное бурое пятно и такое же у конца этого плавника (см. рис. 2.107).
Глаза находятся по верхним краям головы. Передние носовые отвер-
стия не вытянуты в трубочки. Чувствительных каналов и пор нет. Хво-
стовой стебель удлинённый, невысокий, несколько сжат по бокам. Хво-
стовой плавник закруглённый. Небольшая рыба длиной до 45 мм. Икра
крупная.
Обитает в основном в кубанских лиманах.
Род Бабки — Babka (Neogobius) Iljin, 1927
Верхняя губа к углам рта не расширена. На первом спинном плав-
213
нике обычно 3 тёмных полоски. Воротник присоски брюшного диска
без лопастинок. В бассейне Кубани обитает один вид.
Бычок-гонец — Babka (Neogobius) gymnotrachelus (Kessler, 1857)
(рис. 136, 2.108)
Рис. 136. Бычок-гонец — Babka (Neogobius) gymnotrachelus (Kessler, 1857)
D1 VI—VII, D2 I 15—18, A I 12—15. Поперечных рядов чешуи 56—
68. Плавательного пузыря нет, икра крупная. Нижняя челюсть не выда-
ётся вперёд. Верхняя губа к углам рта не расширена. Затылок, спина до
спинного плавника, жаберные крышки, горло и стебли грудных плав-
ников голые. Второй спинной плавник равномерной высоты. Окраска
желтовато-серая. На боках имеются бурые пятна, иногда образующие
косые поперечные полоски. На голове сверху и на боках извилистые
тёмные полоски. На первом спинном плавнике обычно 3 тёмные полос-
ки. Второй спинной, хвостовой и грудные плавники имеют узкие ряды
тёмных точек. Воротник присоски брюшного диска без лопастинок.
Обитает как в солёной, так и в пресной воде. Встречается в кубан-
ских лиманах, изредка в левобережных притоках Кубани. Отмечен в
Краснодарском и Шапсугском водохранилищах (Москул, 1994). Дли-
на тела — 12 —16 см. Икрометание в мае—июне. Плодовитость — до
1 500 икринок. Является объектом питания судака, окуня и других ви-
дов хищных рыб. Промысловое значение невелико.
Род Понтиколы — Ponticola (Neogobius) Iljin, 1927
Изначально род Ponticola был описан Б. С. Ильиным в 1927 г. как
подрод рода Neogobius, но потом повысили его статус до рода. В бас-
сейне Кубани обитают четыре вида.
214
Таблица
для определения видов рода
Понтиколы — Ponticola (Neogobius) Iljin, 1927
1 (2). Второй спинной плавник выше в задней части. Длина и высота
хвостового стебля почти равны. Верхняя губа к углам рта расширяется.
Бычок-рыжик — Ponticola (Neogobius) eurycephalus (Kessler,
1874) (с. 214)
2 (1). Второй спинной плавник сзади не повышается, наибольшей
высоты достигает посередине.
3 (4). Нижняя челюсть ясно выдаётся вперёд. На боках тела круп-
ные светло-бурые пятна, расположенные в шахматном порядке.
Бычок cирман (ширман) — Ponticola (Neogobius) syrman (Nord-
mann, 1840) (с. 217)
4 (3). Нижняя челюсть не выдаётся вперёд. Верхняя губа к углам
рта не расширена.
5 (6). Второй спинной плавник наиболее высок в задней части.
Кавказский речной бычок — Ponticola (Neogobius) constructor
(Nordmann, 1840) (с. 215)
6 (5). Второй спинной плавник равномерной высоты.
Речной бычок Родиона — Ponticola (Neogobius) rhodioni (Va-
sil’eva & Vasil’ev, 1994) (с. 216)
Краткая биологическая характеристика видов рода
Понтиколы — Ponticola (Neogobius) Iljin, 1927
Бычок-рыжик — Ponticola (Neogobius) eurycephalus (Kessler, 1874)
(рис. 137, 2.109)
Рис. 137. Бычок-рыжик — Ponticola (Neogobius) eurycephalus (Kessler, 1874)
215
D1 VI, D2 I 15—19, A I 12—15. Поперечных рядов чешуи 55—67.
Темя, затылок, верхняя часть жаберных крышек, горло, брюхо покрыты
циклоидной чешуёй. Тело удлинённое, не слишком высокое, несколь-
ко сжатое с боков. Второй спинной плавник повышается кзади. Голова
относительно короткая, её высота меньше ширины. Рот большой. Рыло
сверху совсем или слегка уплощённое. Верхняя губа расширяется на-
зад, иногда не доходя до углов рта. Хвостовой стебель сильно сжат с
боков. Нижняя челюсть несколько выдаётся вперёд. Лопастинки ворот-
ника брюшной присоски остроконечные. Окраска буроватая или охри-
сто-буроватая с буроватыми неправильной формы пятнами на боках;
вдоль края второго спинного плавника светлая кайма (см. рис. 2.109).
Самцы во время нереста приобретают чёрную окраску.
Встречается в горных притоках Кубани, а также в гирлах, приле-
гающих к ним лиманах с каменистым грунтом.
Кавказский речной бычок — Ponticola (Neogobius) constructor
(Nordmann, 1840) (рис. 138, 2.110)
Рис. 138. Кавказский речной бычок — Ponticola constructor (Nordmann, 1840)
D1 VI—VII, D2 I 16—18, A I 12—14. Поперечных рядов чешуи
56—73. Ширина лба вдвое меньше диаметра глаза. Нижняя челюсть
выдаётся за верхнюю. Губы очень мясистые, особенно верхняя, по бо-
кам расширенная. Первый спинной плавник заметно ниже второго, ко-
торый равномерной высоты и лишь посередине немного выше; аналь-
ный повышается к заднему концу. Окраска коричнево-бурая. Самцы во
время нереста становятся чёрными с белой каёмкой по верхнему краю
первого спинного плавника.
216
Встречается в Кубани и левобережных притоках (Афипсе, Псе-
купсе, Пшише, Белой), а также в Краснодарском и Шапсугском водо-
хранилищах. Держится в основном на участках с галечным грунтом и
прозрачной водой.
Речной бычок Родиона — Ponticola (Neogobius) rhodioni
(Vasil’eva & Vasil’ev, 1994) (рис. 139, 2.111)
Рис. 139. Речной бычок Родиона — Ponticola (Neogobius) rhodioni (Vasil’eva &
Vasil’ev, 1994)
D1 VI—VII, D2 I 17—19 (20), A I (12) 13—15 (16). Поперечных
рядов чешуи 56—84. Широкая уплощённая голова, её ширина заметно
больше высоты. Верхняя губа к углам рта то расширена, то не расши-
рена. Нижняя челюсть чуть выдаётся вперёд. Тело и затылок покрыты
циклоидной чешуёй. Окраска коричнево-бурая (см. рис. 2.111). Спина
перед первым спинным плавником однотонная, бока однотонные или
мраморные. Первый спинной плавник равномерно покрыт мелкими
крапинками или полосами мелких пятнышек. Самцы во время нерес-
та чёрные с белой каёмкой по верхнему краю первого спинного плав-
ника. Хвостовой плавник закруглённый. Брюшная присоска округлой
формы, реже продолговатая, не доходит до анального отверстия. Лопа-
стинки воротника остроконечные.
Встречается в Кубани и левобережных притоках (Афипсе, Псе-
купсе, Пшише, Белой), а также в Краснодарском и Шапсугском водо-
хранилищах. Держится в основном на участках с галечным грунтом и
прозрачной водой.
Достигает длины 12—16 см и массы 18—24 г.
217
Бычок сирман (ширман) — Ponticola (Neogobius) syrman
(Nordmann, 1840) (рис. 140, 2.112)
Рис. 140. Бычок сирман — Ponticola (Neogobius) syrman (Nordmann, 1840)
D1 VI, D2 I 16—18, A I 12—14. Поперечных рядов чешуи 58—71.
Окраска серая, на боках крупные светло-бурые пятна; от глаза к
верхней губе тёмная полоска; на первом спинном плавнике 3 светлые и
2 тёмные поперечные полосы; на втором спинном, анальном и хвосто-
вом плавниках имеются голубые полоски вдоль лучей (см. рис. 2.112).
Голова вальковатая, её высота немного больше ширины. Рот большой,
несколько скошенный, с выступающей вперёд нижней челюстью. Рыло
тупое, верхняя губа длинная, тонкая. Уголки рта под передней частью
зрачка глаза. Длина — до 24 см, обычно 16—18 см. Масса — до 120 г,
обычно 90—100 г. Продолжительность жизни — до 4—5 лет.
Встречается в лиманах, реках с галечниковым и ракушечным грун-
том. Длина тела достигает 18—20 см, масса — 50—75 г. Нерестится в
мае—июне. Питается в основном молодью рыб и ракообразными.
Имеет небольшое промысловое значение в Азовском море.
Род Пуголовки — Benthophilus Eichwald, 1831
Тело покрыто костными пластиночками или зёрнышками, но по-
ловозрелые самцы теряют их. Поэтому вид пуголовок лучше опреде-
лять на самках. Голова широкая, плоская. Чувствительных каналов и
пор нет. Плавательного пузыря нет. Передние ноздри вытянуты в не-
большие трубочки, прилегающие к верхней губе.
В бассейне Кубани обитают два вида.
218
Таблица
для определения видов рода
Пуголовки — Benthophilus Eichwald, 1831
1 (2). Тело густо покрыто костными зёрнышками. В спинном ряду
22—27 бугров. На спине нет бурых перевязок.
Азовская пуголовка — Benthophilus magistri Iljin, 1927 (с. 218)
2 (1). На теле преобладают бугры, костных зёрнышек мало. Спин-
ных бугров обычно 27—30. На спине 3 бурые перевязки.
Черноморская (звёздчатая) пуголовка — Benthophilus stellatus
(Sauvage, 1874) (с. 219)
Краткая биологическая характеристика видов рода
Пуголовки — Benthophilus Eichwald, 1831
Азовская пуголовка — Benthophilus magistri Iljin, 1927 (рис. 141, 2.114)
Рис. 141. Азовская пуголовка — Benthophilus magistri Iljin, 1927:
а — вид сбоку; б — вид сверху
219
D III—IV, I 9—11, A I 8—10. Затылок и темя покрыты зёрнышка-
ми, пластинок нет. В спинном ряду пластинок обычно 22—27, в брюш-
ном — чаще всего 18—24. Наибольшая длина тела равна 10 см.
Обитает азовская пуголовка в опреснённых частях Азовского
моря, кубанских лиманах и гирлах. В реках встречаются единичные эк-
земпляры. Азовская пуголовка относится к эфемерным рыбам. Нерест
проходит в мае—июне. Плодовитость — от 500 до 2 000 икринок. Сам-
цы крупнее самок.
Промыслового значения не имеет.
Черноморская (звёздчатая) пуголовка — Benthophilus stellatus
(Sauvage, 1874) (рис. 142, 2.113)
Рис. 142. Черноморская (звёздчатая) пуголовка — Benthophilus stellatus
(Sauvage, 1874):
а — вид сбоку; б — вид сверху
D III—IV, I 8—9, A I (7) 8 (9). Костяные пластинки на теле и голове
крупные, с острыми шипами. Среди пластинок выдаются более круп-
ные по бокам головы и три ряда по бокам туловища. В спинном ряду
обычно 27—30 пластинок, в брюшном — 22—25. На спине три бурых
пятна. У самца во время нереста грудные плавники длиннее брюшных.
Длина половозрелых самцов — до 11 см, самок — до 9 см.
220
Обитает как в море, так и в лиманах и реках. В море предпочитает
опреснённые места с песчаным или ракушечным грунтом, в реках —
илистые места. Нерест происходит в мае—июне. Плодовитость колеб-
лется от 600 до 2 000 икринок. После нереста все отнерестившиеся осо-
би погибают. Звёздчатая пуголовка является эфемерным однолетним
позвоночным. После нереста производители погибают.
Промыслового значения не имеет.
Внутривидовой статус кубанских бычков требует уточнения.
Отряд Камбалообразные — Pleuronectiformes Bleeker, 1859
Близки к окунеобразным, но отличаются от них несимметричным
во взрослом состоянии телом. Глаза на одной стороне, другая (слепая)
сторона обычно не пигментирована. Колючих лучей в плавниках нет.
Почти все камбалы откладывают пелагическую икру, которая проходит
развитие, плавая в толще воды. Личинки симметричные, один глаз —
на правой, другой — на левой стороне. Некоторое время они живут в
толще воды и питаются зоопланктоном. Позднее оседают на дно, ло-
жатся на один бок и переходят к донному образу жизни. Нижняя сто-
рона тела остаётся непигментированной, а другая приобретает окраску
под цвет дна. Камбалообразные обладают удивительной особенностью
менять окраску глазной стороны тела под цвет грунта, очень быстро
«перекрашиваются» при попадании на грунт иной окраски.
Камбалообразные — важнейшие промысловые рыбы. Начаты ра-
боты по разведению камбал в рыбоводных хозяйствах и выпуску моло-
ди в естественные водоёмы.
В отряде начитывают 14 семейств,134 рода и 678 видов, 10 из ко-
торых обитают в пресной воде. В водоёмах бассейна Кубани встреча-
ются представители одного семейства — Камбаловые (Pleuronectidae
Cuvier, 1816).
Семейство Камбаловые — Pleuronectidae Cuvier, 1816
Глаза у представителей семейства расположены на правой сто-
роне головы, это так называемые правосторонние камбалы. Однако у
некоторых видов встречаются и левосторонние (реверсивные) формы.
Брюшные плавники симметричны, с узким основанием. Икринки не со-
держат жировой капли, плавающие, развиваются в верхних слоях или
221
в толще воды. У некоторых видов икра придонная. Семейство в водо-
ёмах бассейна Кубани представлено одним подвидом рода Platichthys
Girard, 1854.
Род Речные камбалы — Platichthys Girard, 1854
Речные камбалы отличаются от других камбал костными бугор-
ками у основания лучей непарных плавников. Заходят в пресную воду,
поднимаясь по рекам иногда на значительное расстояние. В пределах
своей области распространения речные камбалы образуют подвиды.
Черноморская камбала, или глосса, — Platichthys flesus luscus
(Pallas, 1811) (рис. 143, 2.115)
Рис. 143. Черноморская камбала, или глосса, — Platichthys flesus luscus (Pallas,
1811)
D 52—65, обычно 57—62; A 36—45, обычно 39—44, l.l. 74—82.
Глосса не достигает больших размеров, как правило, длина составляет
до 25 см, изредка до 29 см, масса — 290 г. Как и у других камбал, сам-
цы несколько мельче самок. Тело с обеих сторон покрыто циклоидной
чешуёй. Шиповатых костяных пластинок на слепой стороне тела нет,
есть в небольшом количестве на глазной стороне вдоль передней части
боковой линии и на голове. Окраска грязно-зелёная с бурыми звёзд-
чатыми пятнами, окаймлёнными светлым ободком на теле и круглы-
222
ми бурыми пятнами на непарных плавниках; иногда тело бурого цвета
(см. рис. 2.115). Изредка встречаются экземпляры с пигментированной
слепой стороной тела, особенно у особей, живущих на илистом грунте.
Половозрелой становятся на 3—4 -м году жизни. Икрометание
происходит в воде, где солёность достигает 10—12 ‰. Икра пела-
гическая и при более низкой солёности опускается на дно и гибнет.
Нерестится глосса в феврале—марте при температуре воды 6—10 °С.
Инкубационный период длится 6—7 дней. Плодовитость составляет в
среднем 270 тыс. икринок. Личинки прозрачные и симметричные. По
достижении длины 7—10 мм они переходят к донному образу жизни, и
в это время глаза перемещаются на одну сторону. Молодь нагуливается
в лиманах и опреснённых участках, а также в гирлах и устьях Кубани и
Протоки. Встречается глосса и в руслах рек.
Питается глосса ракообразными организмами, моллюсками, чер-
вями и рыбой. Растёт глосса сравнительно быстро и к концу первого
года жизни достигает массы 4—5 г и длины 5—7 см, к трём годам —
120—150 г и длины 18—20 см. Глосса является объектом промысла.
223
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
Абаев Ю. И. Эколого-зоогеографический анализ и рыбохозяйст-
венная оценка современной ихтиофауны бассейна реки Кубани: авто-
реф. дис. ... д-ра биол. наук. — М., 1996. — 60 с.
Абрамчук А. В., Иваненко А. М. Ихтиофауна бассейна Кубани:
учеб. пособие. — Краснодар: Кубанский гос. ун-т, 2018. — 195 с.
Абрамчук А. В., Иваненко А. М. Система ныне живущих рыбо-
образных и рыб мировой фауны: учеб. пособие: в 2 ч.
—
Краснодар:
Кубанский гос. ун-, 2019а. — Ч. 1. — 409 с.
Абрамчук А. В., Иваненко А. М. Система ныне живущих рыбо-
образных и рыб мировой фауны: учеб. пособие: в 2 ч.
—
Краснодар:
Кубанский гос. ун-т, 2019б. — Ч. 2. — 385 с.
Абрамчук А. В., Пашинова Н. Г., Москул Г. А. Биоразнообразие
ихтиофауны Закубанских рек (бассейн реки Кубань) // Рыбное хозяйст-
во. — 2018. — No 5. — С.64—67.
Абрамчук А. В., Пашинова Н. Г., Москул Г. А. Естественное вос-
производство и состояние популяций основных промысловых видов
рыб водохранилищ бассейна Кубани // Естественные и технические
науки. — 2018. — No 8 (122). — С. 28—31.
Александров А. И. Материалы по ихтиофауне бассейна р. Куба-
ни // Тр. Керченск. науч. рыбохоз. ст.
—
1927.
—
Т. 1, вып. 2—3.
—
С. 151—178.
Аннотированный каталог круглоротых и рыб континентальных
вод России / под ред. Ю. С. Решетникова. — М.: Наука, 1998. — 218 с.
Атлас пресноводных рыб России = Atlas of Russian Freshwater
Fishes: в 2 т. / Ю. С. Решетников [и др.]; под общ. ред. Ю. С. Решетни-
кова. — М.: Наука, 2002. — Т. 1. — 378 с.
Атлас пресноводных рыб России = Atlas of Russian Freshwater
Fishes: в 2 т. / Ю. С. Решетников [и др.]; под общ. ред. Ю. С. Решетни-
кова. — М.: Наука, 2002. — Т. 2. — 251 с.
Атлас пресноводных рыб России = Atlas of Russian Freshwater
Fishes: в 2 т. / Ю. С. Решетников [и др.]; под общ. ред. Ю. С. Решетни-
кова. — 2-еизд. — М.:Наука,2003. — Т.1. — 379с.
Атлас пресноводных рыб России = Atlas of Russian Freshwater
Fishes: в 2 т. / Ю. С. Решетников [и др.]; под общ. ред. Ю. С. Решетни-
кова. — 2-еизд. — М.:Наука,2003. — Т.2. — 253с.
Барсуков Д. У., Позняк В. Г. О таксономии и расширении ареала
224
северокавказского длинноусого пескаря Romanogobio ciscaucasicus //
Проблемы сохранения и рационального использования биоразнооб-
разия Прикаспия и сопредельных регионов: материалы IV Междунар.
заоч. науч. конф. — Элиста: Изд-во КалмГУ, 2006. — С. 60—63.
Берг Л. С. Отчёт о командировке на Кавказ в 1909 году // Ежегод-
ник Зоол. музея Имп. акад. наук. — 1910. — Т. XV. — С. 153—170.
Берг Л. С. Рыбы бассейна Кубани // Ежегодник Зоол. музея Имп.
акад. наук. — 1912. — Т. XVII. — С. 116—122.
Берг Л. С. Фауна России и сопредельных стран. Рыбы (Marsipo-
branchii и Pisces). Т. III: Ostariophysi, вып. 2. — Петроград: Изд-во Имп.
Акад. Наук, 1914. — С. 337—846.
Берг Л. С. Рыбы пресных вод Российской империи = Les poissons
des eaux douces de la Russie.
—
М.: Типография Т-ва Рябушинских,
1916. — XXVII. — 563 с.
Берг Л. С. Рыбы пресных вод России. — 2 -е изд. — М.: Гос. изд-во,
1923. — XXIX. — 536 с.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран: в
2 ч. — 3-е изд., испр. и доп. — Л.: Изд-во Леноблисполкома и Ленсове-
та,1932. — Ч.1. — 543с.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран: в
2 ч. — 3-е изд., испр. и доп. — Л.: Изд-во Леноблисполкома и Ленсове-
та,1933. — Ч.2. — 899с.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран: в
3 ч. — 4-е изд., испр. и доп. — М.; Л.: Изд-во АН СССР, 1948—1949. —
Ч. 1—3. — 1384 с.
Березовская В. И., Савельева Э. А., Чебанов М. С. Реакклиматиза-
ция стерляди и шипа в бассейне р. Кубань // Ресурсосберегающие тех-
нологии в аквакультуре: материалы II Междунар. симп.; г. Адлер, 4—7
октября 1999 г. — Краснодар: КрасНИИРХ, 1999. — С. 16.
Богуцкая Н. Г., Насека А. М. О восстановлении видового статуса
описанных Л. С. Бергом корейского пескаря и кубанского усача (Pisces:
Cyprinidae) // Академику Л. С. Бергу — 125 лет: сб. науч. статей.
—
Бендеры, 2001. — С. 54—58.
Богуцкая Н. Г., Насека А. М. Каталог бесчелюстных и рыб пре-
сных и солоноватых вод России с номенклатурными и таксономиче-
скими комментариями.
—
M.: Товарищество научных изданий KMK,
2004. — 389 с.
Богуцкая Н. Г., Позняк В. Г. Переописание афипсского ельца Leu-
225
ciscus aphipsi Aleksandrov (Leuciscinae, Cyprinidae) // Вопр. ихтиоло-
гии. — 1994. — Т. 34, вып. 3. — С. 308—315.
Борисов В. И. Реки Кубани.
—
Краснодар: Кубан. кн. изд-во,
2005. — 120 с.
Борисов В. И. Занимательное краеведение. — Краснодар: Кубан.
кн. изд-во, 2006. — 160 с.
Борисов П. Г., Овсянников Н. С. Определитель промысловых рыб
СССР.
—
4-е изд., доп. и перераб.
—
М.: Пищевая пром-ть, 1964.
—
319 с.
Васильева Е. Д. Морфология черепа бычка Gobius niger L. в связи
с его положением в роде Gobius sensu lato (Gobiidae, Pisces) // Вестн.
Моск. ун-та. Сер. 16. Биология. — 1990. — No 2. — С. 67—73.
Васильева Е. Д. Рыбы Чёрного моря. Определитель морских, со-
лоноватоводных, эвригалинных и проходных видов с цветными ил-
люстрациями, собранными С.В. Богородским. —
М.: Изд-во ВНИРО,
2007. — 238 с.
Васильева Е. Д., Васильев В. П. Виды-двойники в роде Cobitis (Co-
bitidae). 1. Южнорусская щиповка Cobitis rossomeridionalis sp. nova //
Вопр. ихтиологии. — 1998. — Т. 38, No 5. — С. 604—614.
Васильева Е. Д., Васильев В. П., Куга Т. И. К таксономии пескарей
рода Gobio (Gobioninae, Cyprinidae) Европы: новый вид пескаря Gobio
kubanicus sp. nova из бассейна реки Кубань // Вопр. ихтиологии.
—
2004. — Т. 44, No 6. — С. 766—782.
Васильева Е. Д., Лужняк В. А. Рыбы бассейна Азовского моря. —
Ростов н/Д: Изд-во ЮНЦ РАНЕ, 2013. — 272 с.
Веселов Е. А. Определитель пресноводных рыб фауны СССР: по-
собие для учителей. — М.: Просвещение, 1977. — 238 с.
Веселовский Н. В., Данилова Г. Н., Емельянова В. П. Изменение
гидрохимического режима рек Кубани и Терека под влиянием заре-
гулирования стока и изъятия воды на орошение // Географические
проблемы изучения охраны и рационального использования природ-
ных условий и ресурсов Северного Кавказа. — Ставрополь, 1973.
—
С. 82.
Вилер А. Определитель рыб морских и пресных вод Северо-Евро-
пейского бассейна / пер. с англ. Т. И. Смольяновой. — М.: Лёгк. и пищ.
пром-сть, 1983. — 430 с.
Виноградов В. К., Ерохина Л. В. Новые объекты рыбоводства и
акклиматизации // Рыбное хозяйство. — 1976. No 10. — С. 10—13.
226
География Республики Адыгея / А. Ш. Бузаров [и др.].
—
2-е
изд. — Майкоп: Адыг. респ. кн. изд-во, 2001. — 199 с.
География рыб: учеб. пособие для ВО / Н. А. Абросимова [и
др.]. — 2-е изд., испр. и доп. — СПб.: Лань, 2020. — 120 с.
Данилевский Н. Я. Описание рыболовства на Чёрном и Азовском
морях // Исследования о состоянии рыболовства в России. — СПб.: Ти-
пография В. Безобразова и комп., 1871. — Т. 8. — 316 с.
Емтыль М. Х., Иваненко А. М. Рыбы юго-запада России: учеб. по-
собие. — Краснодар: Кубанский гос. ун-т, 2002. — 340 с.
Емтыль М. Х., Плотников Г. К., Абаев Ю. И. Современное состоя-
ние ихтиофауны бассейна реки Кубань // Актуальные вопросы изуче-
ния экосистемы бассейна Кубани: материалы науч.-практ. конф.
—
Краснодар, 1988. — С. 98—108.
Ефремов Ю. В. В стране горных озёр. — Краснодар: Кн. изд-во,
1991. — 190 с.
Жизнь животных: в 6 т. — Т. 4, ч. 1: Рыбы / под ред. проф. Т. С. Рас-
са. — М.: Просвещение, 1971. — 655 с.
Казанчеев Е. Н. Рыбы Каспийского моря: определитель.
—
2-е
изд. — М.: Лёгк. и пищ. пром-сть, 1981. — 167 с.
Каменский С. Н. Карповые (Cyprinidae) Кавказа и Закавказья. —
Тифлис: Типография канцелярии Главноночальствующего граждан-
ской частью на Кавказе, 1901. — Вып. 2. — 192 с.
Канонников А. М. Природа Кубани и Причерноморья. — Красно-
дар: Кн. изд-во, 1977. — 112 с.
Кирюхина Н. А. Морфологическая изменчивость пухлощёкой иг-
лы-рыбы Syngnathus nigrolineatus в связи с её инвазией в водоёмы бас-
сейна Волги // Российский журнал биологических инвазий. — 2013. —
No2. — С.2—10.
Книпович Н. М. Определитель рыб Чёрного и Азовского морей. —
М.: 40-я типография М. С. Н. Х., 1923. — 130 с.
Комлев А. М., Мельникова Т. Н. Водный режим рек Северо-Запад-
ного Кавказа: закономерности формирования и методы расчёта: моно-
графия. — Пермь: Пресстайм, 2008. — 111 с.
Коровин В. И., Коровин А. В. Гидрографическая сеть Северного
Кавказа: учеб. пособие. — Краснодар: Кубанский гос. ун-т, 1981. — 78 с.
Красная книга Краснодарского края (животные) / отв. ред. А. С. За-
мотайлов.
—
2-е изд.
—
Краснодар: Центр развития ПТР Краснодар.
края, 2007. — 477 с.
227
Красная книга Краснодарского края. Животные / отв. ред. А. С. За-
мотайлов, Ю. В. Лохман, Б. И. Вольфов. — 3 -е изд. — Краснодар: Адм.
Краснодар. края, 2017. — 720 с.
Красная книга Российской Федерации (животные) / РАН; гл. ред-
кол.: В. И. Данилов-Данильян [и др.].
—
М.: АСТ; Астрель, 2001.
—
862 с.
Крыжановский С. Г., Троицкий С. К. Материалы об ихтиофау-
не рек Черноморского побережья // Вопр. ихтиологии.
—
1954.
—
Вып. 2. — С. 144—159.
Кудерский Л. А. Ещё о саморасселении рыб // Рыбохозяйственное
изучение внутренних водоёмов. — 1971. — Сб. 6. — С. 22—24.
Кузнецов Б. А. Определитель позвоночных животных фауны
СССР:в3ч.
—
Ч. 1: Круглоротые, рыбы, земноводные, пресмыкаю-
щиеся. — М.: Просвещение, 1974. — 190 с.
Лотышев И. П. География Краснодарского края.
—
5-е изд.,
испр. — Краснодар: Печат. двор Кубани, 2000. — 135 с.
Лужняк В. А. Ихтиофауна рек и лиманов Черноморского побере-
жья России // Вопр. ихтиологии. — 2003. — Т. 43. — С. 457—463.
Мельченков Е. А. Питание сеголеток веслоноса // Растительнояд-
ные рыбы и новые объекты рыбоводства и акклиматизации: сб. науч.
тр. — М.: ВНИИПРХ, 1988. — Вып. 54. — С. 20—30.
Мельченков Е. А. Биологические основы разведения и выращи-
вания веслоноса (Polyodon spathula (Walbaum)): автореф. дис. ... д-ра
биол. наук: 03.00.10 / ВНИИ пресноводного рыбного хоз-ва.
—
М.,
2001. — 63 с.
Москул Г. А. Видовой состав рыб и условия их размножения в
Краснодарском водохранилище // Актуальные вопросы изучения эко-
системы бассейна Кубани. — Краснодар, 1988. — С. 108—113.
Москул Г. А. Рыбохозяйственное освоение Краснодарского водо-
хранилища. — СПб.; Краснодар: КрасНИИРХ, 1994. — 136 с.
Москул Г. А. Биологические основы рыбохозяйственного освое-
ния внутренних водоёмов Северного Кавказа: автореф. дис. ... д-ра
биол. наук. — М., 1995. — 51 с.
Москул Г. А.Рыбы водоёмов бассейна Кубани: определитель.
—
Краснодар: Краснодарский НИИ рыбного хозяйства, 1998. — 177 с.
Москул Г. А., Москул Н. Г. Современная ихтиофауна водоёмов
Краснодарского края // Проблемы литодинамики и экосистем Азовско-
го моря и Керченского пролива = Problems of lithodynamics and ecosys-
228
tems of the Azov sea and Kerch strait: тез. докл. Междунар. науч.-практ.
конф.; г. Ростов-на-Дону, 8—9 июня 2004 г.
—
Ростов н/Д, 2004.
—
С. 58—59.
Москул Г. А., Москул Н. Г. Ихтиофауна водоёмов бассейна Куба-
ни и прилегающих к нему рек Азово-Кубанской равнины и Закубанских
рек // Исследования по ихтиологии и смежным дисциплинам на внут-
ренних водоёмах в начале XXI века (к 80-летию профессора Л. А. Ку-
дерского): сб. науч. тр. ГосНИОРХ.
—
СПб.: Товарищество научных
изданий КМК, 2007. — Вып. 337. — С. 258—269.
Москул Г. А., Пашинова Н. Г. Современная ихтиофауна бассейна
реки Кубани //Академику Л. С. Бергу — 140 лет: сб. науч. статей. —
Бендеры, 2016. — С. 438—441.
Москул Г. А., Никитина Н. К., Гаврикова Е. Г. Современное со-
стояние и пути развития рыбного хозяйства на водохранилищах Крас-
нодарского и Ставропольского краёв // Тр. ГосНИОРХ. — Л., 1982. —
Т. 186 — С. 43—143.
Москул Г. А., Никитина Н. К., Гришкин С. А. Некоторые данные
о состоянии популяций рыбца и шемаи Краснодарского водохранили-
ща // Актуальные вопросы экологии и охраны природы Азовского моря
и Восточного Приазовья: материалы науч.-практ. конф. — Краснодар,
1990. — С. 130—133.
Москул Н. Г. Берш — новый вид для водоёмов Кубани // Актуаль-
ные вопросы экологии и охраны природы экосистем южных регионов
России и сопредельных территорий: материалы науч.-практ. конф. —
Краснодар, 2001. — С. 299—304.
Москул Н. Г. Морфобиологическая характеристика берша Stizoste-
dion volgense (Gmelin) и его роль в экосистеме водоёмов бассейна Ку-
бани: автореф. дис. ... канд. биол. наук: 03.00.10 / Азов. науч.-исслед.
ин-т рыб. хоз-ва (АзНИИ РХ). — Ростов н/Д, 2003. — 23 с.
Мусатова Г. Н. Осетровые рыбы реки Кубани и их воспроизводст-
во. — Краснодар : Кн. изд-во, 1973. — 110 с.
Нагалевский Ю. Я., Чистяков В. И. Физическая география Крас-
нодарского края: учеб. пособие. — 2 -е изд., испр. и доп. — Краснодар:
Кубанский гос. ун-т, 2001. — 187 с.
Насека А. М., Богуцкая Н. Г. Новый вид пескаря Romanogobio pen-
tatrichus (Gobioninae, Cyprinidae) из бассейна реки Кубань // Вопр. их-
тиологии. — 1998. — Т. 38, No 2. — С. 173—181.
Насека А. М., Дирипаско О. А. Новые рыбы-вселенцы в водоёмах
229
Северного Приазовья // Вестник зоологии.
—
2005. —
Т.39,No4.
—
С. 89—94.
Насека A. M., Позняк В. Г. Северокавказский длинноусый пескарь
Romanogobio ciscaucasicus в бассейне Кубани (Gobioninae, Cyprini-
dae) // Вопр. ихтиологии. — 2000. — Т. 40, No 3. — С. 406—410.
Нельсон Дж. С. Рыбы мировой фауны: перевод четвёртого пере-
работанного английского издания Н. Г. Богуцкой. — М.: ЛИБРОКОМ,
2009. — 876 с.
Никольский Г. В. Частная ихтиология. — 3 -е изд. испр. и доп. —
М.: Высш. школа, 1971. — 471 с.
Огурцов Н. А., Поляков Ю. Н., Гольднер Р. М. Краснодарское во-
дохранилище. — Краснодар: Кн. изд-во, 1975. — 64 с.
Парин Н. В., Евсеенко С. Л., Васильева Е. Д. Рыбы морей России:
аннотированный каталог. — М.: Товарищество научных изданий КМК,
2014. — 733 с.
Плотников Г. К., Емтыль М. Х., Абаев Ю. И. Современное состоя-
ние ихтиофауны азовских и черноморских лиманов Краснодарского
края // Актуальные вопросы экологии и охраны природы Азовского
моря и Восточного Приазовья: материалы науч.-практ. конф. — Крас-
нодар, 1989. — С. 117—124.
Позняк В. Г. К характеристике ихтиофауны верхнего течения реки
Большой Егорлык // Ихтиологические и рыбохозяйственные исследо-
вания на реках и водохранилищах. — Воронеж: Изд-во ВГУ, 2001. —
С. 79—84.
Пономарев С. В., Баканева Ю. М., Федоровых Ю. В. Ихтиология:
учеб. для студентов вузов, обучающихся по направлению подготовки
«Водные биоресурсы и аквакультура».
—
2-е изд., доп.
—
СПб.; М.;
Краснодар: Лань, 2016. — 558 с.
Пономарев С. В., Лагуткина Л. Ю. Фермерское рыболовство: учеб-
ник. — М.: Колос, 2008. — 347 с.
Попова М. С. Изменения в ихтиофауне бассейна реки Егорлы-
ка после его обводнения // Материалы по изучению Ставропольского
края. — Ставрополь, 1955. — Вып. 7. — С. 195—199.
Правдин И. Ф. Руководство по изучению рыб (преимуществен-
но пресноводных) / под ред. проф. П. А. Дрягина и канд. биол. наук
В. В. Покровского. — 4 -е изд., перераб. и доп. — М.: Пищ. пром-сть,
1966. — 376 с.
Промысловые рыбы России: в 2 т. / под ред. О. Ф. Гриценко,
230
А. Н. Котляра, Б. Н. Котенёва. — М.: Изд-во ВНИРО, 2006. — 1280 с.
Пятиязычный словарь названий животных. Рыбы / Ю. С. Решет-
ников, А. Н. Котляр, Т. С. Расс, М. И. Шатуновский.
—
М.: Рус. яз.,
1989. — 733 с.
Решетников С. И., Пашков А. Н. 365. РЫБЕЦ МАЛЫЙ Vimba vim-
ba tenella (Nordmann, 1840) // Красная книга Краснодарского края. Жи-
вотные / отв. ред. А. С. Замотайлов, Ю. В. Лохман, Б. И. Вольфов. — 3 -е
изд. — Краснодар: Адм. Краснодар. края, 2017. — С. 469—470.
Решетников С. И., Пашков А. Н. Экосистемы малых рек Черно-
морского побережья Северо-Западного Кавказа. — Краснодар: Кубан-
ский гос. ун-т, 2009. — 152 с.
Решетников Ю. С. 36. Syngnathus abaster Risso, 1827 — черномор-
ская игла (малая игла-рыба) // Рыбы в заповедниках России: в 2 т. /под
ред. Ю. С. Решетникова. — Т. 2: Морские рыбы. — М.: Товарищество
научных изданий КМК, 2013. — С. 116—119.
Романов В. И. Современные представления о системе рыбообраз-
ных и рыб мировой фауны: учеб. пособие. — Томск: Издательский Дом
Томского гос. ун-та, 2019. — 310 c.
Рубан Г. И. О структуре вида сибирского осётра Acipenser baerii
Brandt // Вопр. ихтиологии. — 1998. —Т. 38, вып. 3. — С. 307—327.
Рубан Г. И. Сибирский осётр Acipenser baerii Brandt: структура
вида и экология. — М.: ГЕОС, 1999. — 235 с.
Рыбы в заповедниках России: в 2 т. / под ред. Ю. С. Решетнико-
ва. — Т. 1: Пресноводные рыбы. — М.: Товарищество научных изданий
КМК, 2010. — 627 с.
Рыбы в заповедниках России: в 2 т. / под ред. Ю. С. Решетни-
кова.
—
Т. 2: Морские рыбы.
—
М.: Товарищество научных изданий
КМК, 2013. — 673 с.
Световидов А. Н. Рыбы Чёрного моря. — М.; Л.: Наука, 1964. —
550 с.
Современное состояние и перспективы рыбохозяйственного ис-
пользования Азово-Кубанских лиманов / Г. А. Москул, Ю. И. Ковален-
ко, Н. Г. Пашинова, О. А. Болкунов // Современные рыбохозяйственные
и экологические проблемы Азово-Черноморского региона: материалы
7 Междунар. конф. — Керчь, 2012. — Т. 1. — С. 68—75.
Список рыбообразных и рыб пресных вод России / Ю. С. Решет-
ников [и др.] // Вопр. ихтиологии. — 1997. — Т. 37, No 6. — С. 723—
771.
231
Троицкий С. К. Рыбы Краснодарского края. — Краснодар: Крае-
вое книгоиздательство, 1948. — 80 с.
Троицкий С. К., Позняк В. Г. Общая характеристика ихтиофауны
Северного Кавказа // Ресурсы живой фауны: монография: в 3 ч. / отв.
ред. А. К. Темботов. — Ч. 1: Водные животные. — Ростов н/Д: Изд-во
Ростовского ун-та, 1980. — С. 82—110.
Троицкий С. К., Цуникова Е. П. Рыбы бассейнов нижнего Дона и
Кубани: руководство по определению видов. — Ростов н/Д: Кн. изд-во,
1988. — 112 с.
Туниев С. Б. Современное состояние и перспективы изучения их-
тиофауны Сочинского национального парка // Проблемы устойчивого
развития регионов рекреационной специализации / Сочинский научно-
исследовательский центр РАН. — Сочи, 2005. — С. 163—174.
Ходячий Н. П., Москул Г. А., Никитина Н. К. Рыбоводство в водо-
ёмах Кубани. — Краснодар: Кн. изд-во, 1982. — 94 с.
Чебанов М. С., Галич Е. В. Руководство по искусственному вос-
производству осетровых рыб: технический доклад ФАО по рыбному
хозяйству No 558. — Анкара: ФАО, 2013. — 325 с.
Чебанов М. С., Галич Е. В., Меркулов Я. Г. 353. БЕЛУГА АЗОВ-
СКАЯ Huso huso maeoticus Salnikov et Malyatskij, 1934; 354. ШИП
Acipenser nudiventris Lovetsky, 1823; 355. ОСЁТР РУССКИЙ Acipenser
gueldenstaedtii Brandt et Ratzeburg, 1833; 356. СЕВРЮГА Acipens-
er stellatus Pallas, 1771; 357. СТЕРЛЯДЬ Acipenser ruthenus (Linnae-
us, 1758) // Красная книга Краснодарского края. Животные / отв. ред.
А. С. Замотайлов, Ю. В. Лохман, Б. И. Вольфов. — 3 -е изд. — Красно-
дар: Адм. Краснодар. края, 2017. — С. 454—461.
Чебанов М. С., Галич Е. В., Чмырь Ю. Н. Созревание произво-
дителей белуги в условиях аквакультуры и рыбоводно-биологическая
оценка полученного потомства // Проблемы и перспективы развития
аквакультуры в России: материалы докл. науч.-практ. конф.; г. Адлер,
24—27 сентября 2001 г. — Краснодар; КрасНИИРХ, 2001. — С. 118—
119.
Чебанов М. С., Савельева Э. А., Колос А. А. Современное состоя-
ние производителей осетровых рыб в Азово-Кубанском районе и эф-
фективность их использования в промышленном воспроизводстве //
Осетровые на рубеже XXI века: тез. докл. Междунар. конф.; г. Аст-
рахань, 11—15 сентября 2000 г. — Астрахань: КаспНИИРХ, 2000. —
С. 280—281.
232
Чебанов М. С., Козырицкая Ю. Е. 230. ШИП Acipenser nudiventris
Lovetsky, 1828 // Красная книга Краснодарского края (животные) / отв.
ред. А. С. Замотайлов. — 2 -е изд. — Краснодар: Центр развития ПТР
Краснодар. края, 2007. — С. 316—317.
Чесалина Т. Л., Чесалин М. В. Liza haematochila (Temminck &
Schlegel, 1845) — правильное латинское название кефали-пиленгаса
Mugil soiuy Basilewsky, 1855 (Pisces: Mugilidae) // Экология моря.
—
2002. — Вып. 62. — С. 41—45.
Чижов Н. И., Абаев Ю. И. Рыбы водоёмов Краснодарского края. —
Краснодар: Кн. изд-во, 1968. — 94 с.
Behnke R. J. Genus Oncorhynchus // Trout and Salmon of North Amer-
ica. — New York; London; Toronto: The Free Press, 2002a. — P. 10—21.
Behnke R. J. Rainbow and Redband Trout // Trout and Salmon of
North America. — New York; London; Toronto: The Free Press, 2002b. —
P. 67.
Behnke R. J. Relationships of the Far Eastern Trout, Salmo mykiss
Walbaum // Copeia. — 1966. — No. 2. — P. 346—348.
Bogutskaya N. G. Petroleuciscus, a new genus for the Leuciscus bo-
rysthenicus species group (Teleostei: Cyprinidae) // Zoosystematica Rossi-
ca. — 2002. — Vol. 11 (1). — P. 235—237.
Eschmeyer W. N., Fricke R., van der Laan R. Mugil auratus // Catalog
of Fishes / California Academy of Sciences. — URL: http://researcharchive.
calacademy.org/research/ichthyology/catalog/fishcatmain.asp (дата обра-
щения: 22.12.2019а).
Eschmeyer W. N., Fricke R., van der Laan R. Mugil saliens // Catalog
of Fishes / California Academy of Sciences. — URL: http://researcharchive.
calacademy.org/research/ichthyology/catalog/fishcatmain.asp (дата обра-
щения: 22.12.2019б).
Eschmeyer W. N., Fricke R., van der Laan R. Mugil haematochei-
lus // Catalog of Fishes / California Academy of Sciences. — URL: http://
researcharchive.calacademy.org/research/ichthyology/catalog/fishcatget.as -
p?spid=16926 (дата обращения: 22.12.2019в).
Fricke R., Bilecenoglu M., Sari H. M. Annotated checklist of fish and
lamprey species (Gnathostoma and Petromyzontomorphi) of Turkey, includ-
ing a Red List of threatened and declining species // Stuttgarter Beitr. Naturk.
Serie A (Biologie). — 2007. — No 706. — S. 1—172.
Howes G. J. Anatomy and phylogeny of the Chinese major carps
Ctenopharyngodon Steind., 1866 and Hypophthalmichthys Blkr., 1860 //
233
Bulletin of the British Museum (Natural History), Zoology.
—
1981.
—
Vol. 41 (1). — P. 1—52.
Kottelat M. The Fishes of the Inland Waters of Southeast Asia: A Cata-
logue and Core Bibliography of the Fishes Known to Occur in Freshwaters,
Mangroves and Estuaries // The Raffles Bulletin of Zoology.
—
2013.
—
Sup. 27. — P. 1—663.
Kottelat M., Freyhof J. Handbook of European freshwater fishes.
—
Delémont: Imprimerie du Démocrate SA, 2007. — 660 p.
Matrilinear phylogeography and demographical patterns of Rutilus ru-
tilus: implications for taxonomy and conservation / M. H. D. Larmuseau,
J. Freyhof, F. A. M. Volckaert, J. K. J. Van Houdt // Journal of Fish Biolo-
gy. — 2009. — Vol. 75. — P. 332—353.
Moskul G. A., Abramchuk A. V. Pashinova N. G. The role of the silver
carp in increase fish productivity and the reducing the level of eutrophication
of reservoirs of Kuban basin // IOP Conference Series: Earth and Environ-
mental Science: Water Power Energy Forum 2018. 29 October to 2 Novem-
ber 2018, Kazan, Russian Federation. — 2019a. — Vol. 288. — P. 1—5.
Moskul G. A., Abramchuk A. V., Pashinova N. G. Biological, ecological
and morphological characteristics of the silver carp (Hypophthalmichthys mo-
litrix (Val., 1844) Kuban basin // Scientific research of the SCO countries: syn-
ergy and integration. Part 2: Participants reports in English: Materials of the In-
ternational Conference. March 26, 2019. — Beijing, 2019b. — P. 108—116.
Moskul G. A., Pashinova N. G., Abramchuk A. V. Hypophthalmichthys
nobilis (Richardson,1846) Kuban basin (biological, ecological and mor-
phological) // Scientific research of the SCO countries: synergy and integra-
tion. Part 2: Participants reports in English: Materials of the International
Conference. April 29, 2019. — Beijing, 2019b. — P. 108—116.
Naseka A. M. Comparative study on the vertebral column in the Gobi-
oninae (Cyprinidae, Pisces) with special reference to its systematics // Publ.
Spec. Inst. Esp. Oceanogr. —1996. — No 21. — P. 149—167.
Naseka A. M., Bogutskaya N. G., Poznjak V. G. Romanogobio penta-
trichus Naseka et Bogutskaya, 1998 // The Freshwater Fishes of Europe. —
Vol. 5: Cyprinidae — 2. Pt. 3. Carassius to Cyprinus: Gasterosteidae /
P. M. Banarescu, H.-J. Paepke (Editors). —
Wiebelsheim: AULA-Verlag,
2002. — P. 187—200.
Naseka A. M., Freyhof J. Romanogobio parvus, a new gudgeon from
River Kuban, southern Russia (Cyprinidae, Gobioninae) // Ichthyol. Explor.
Freshwaters. — 2004. — Vol. 15, No 1. — P. 17—23.
234
Nelson J. S., Grande T. C., Wilson M. V. H. Fishes of the World. —
5-th ed. — Hoboken, 2016. — 750 p.
Phylogeny and phylogeography of the roaches, genus Rutilus (Cyprin-
idae), at the Eastern part of its range as inferred from mtDNA analysis /
B. A. Levin [et al.] // Hydrobiologia. — 2017. — Vol. 788, Iss. 1. — P. 33—
46.
Smith G. R., Stearley R. F. The Classification and Scientific Names
of Rainbow and Cutthroat Trouts // Fisheries. — 1989. — Vol. 14, No 1. —
P. 4—10.
235
Указатель русских названий рыб
А
Адрианихтовые 165, 166
Алозы 59, 60, 61
Амурские чебачки 80, 90
Амур
–
белый 100, 101, 101*1
–
чёрный 100, 100*
Амуры
–
белые 81, 100
–
чёрные 81, 99
Анчоус
–
азовский 68
–
европейский 67
–
черноморский 67
Анчоусовые 59, 67
Анчоусы 67
Атерина
–
черноморская 169, 169*
Атериновые 168
Атеринообразные 57, 168
Б
Бабки 204, 212
Балиторовые 83
Барабуля 200
–
черноморская 199, 199*
Барабулевые 188, 199
Бекас 88
Белоглазка 111, 111*
Белуга 43, 43*, 44
–
азовская 44
Белуги 42, 43
Белшип 53
Берш 193
Бёрш 191, 193, 193*
Бестер 53
–
аксайский 54
–
белужий 54
–
бурцевский 54
–
внировский 54
–
стерляжий 54
Бирючок 195, 197
Бобырец 85, 85*, 86
Бобырцы 80, 85, 88
Буйвол-рыба 142
Буффало 142, 143
–
большеротый 142, 143, 143*
–
малоротый 142, 143, 145, 145*
–
чёрный 142, 143, 146, 146*
Быстрянка
–
русская 108, 108*
Быстрянки 81, 108
Бычки 203, 207
–
бубыри 203, 204
–
Книповича 203, 206
–
лысуны 203, 204
–
тупоносые 203, 210
–
черноморско-каспийские
203, 208, 209
Бычковые 202, 203
Бычкообразные 58, 202
Бычок
–
бубырь 204, 204*, 205, 207,
207*
–
гонец 213, 213*
–
Книповича
–
–
длиннохвостый 205, 206,
206*
1
Звёздочкой отмечены страницы, на которых размещены рисунки.
236
–
кругляк 208, 209, 209*
–
песочник
–
–
речной 209, 210, 210*
–
речной
–
–
кавказский 214, 215, 215*
–
–
Родиона 214, 216, 216*
–
рыжик 214, 214*
–
сирман 214, 217, 217*
–
травяник 208, 208*
–
тупоносый мраморный 211,
211*
–
цуцик 211, 211*
–
ширман 214, 217
В
Верховка 103, 103*
Верховки 81, 103
Веслонос 55, 55*
–
китайский 55
Веслоносые 41, 55
Вуалехвост 136*
Вьюн 157, 157*
Вьюновые 78, 149
Вьюны 156
Вырезуб 93, 96, 96*, 97
Г
Гамбузиевые 163
Гамбузии 163, 164
Гамбузия
–
хольбрукская 164, 164*
Глосса 221, 221*, 222
Голавли 80, 86, 87
Голавль 87
–
афипсский 87, 88, 88*
–
кавказский 86, 87, 87*
–
обыкновенный 87
Голец
–
Крыницкого 148, 148*
–
обыкновенный 147, 147*
–
усатый 147, 147*
Гольцовые 78, 83, 147
Гольцы 148
–
усатые 147
–
южноевропейские 147, 148
Гольян 89
–
обыкновенный 89, 89*
Гольяны 80, 89
Горчак 20
–
обыкновенный 92, 92*
Горчаки 80, 92
Густера 112, 112*, 113
Густеры 82, 112
Е
Елец
–
афипсский 87, 88, 88*
Ельцы 79, 83
Ёрш
–
донской 195, 197, 197*
–
обыкновенный 195, 196, 196*
Ерши 189, 195, 196
Ж
Жерех
–
обыкновенный 102, 102*
Жерехи 81, 101
И
Игла-рыба
–
змеевидная 174, 175, 175*
–
черноморская 174, 175, 176,
176*
Игловые 174, 175
Иглообразные 58, 174
Иктиобусы 142, 143
237
Икталуровые 158, 162
К
Калинка 85, 85*, 87, 88
Калуга 43, 45, 45*
Камбала
–
черноморская 221, 221*
Камбаловые 220
Камбалообразные 58, 220
Камбалы
–
речные 221
Караси 83, 132, 133
Карась
–
золотой 132, 133, 133*
–
китайский 133, 134, 135, 135*
–
обыкновенный 132
–
серебряный 132, 133, 137,
137*
Карп 129, 129*, 131, 131*
–
голый 131, 131*
–
зеркальный 131, 131*
–
кожистый 131, 131*
–
линейный 131, 131*
–
рамчатый 131, 131*
–
чешуйчатый 131, 131*
–
японский парчовый 131
Карповые 78, 79
Карпозубообразные 57, 163
Карпообразные 57, 77, 78
Карпы 83, 129
Каспиосома 212, 212*
Каспиосомы 204, 212
Кефалевые 177, 178, 179
Кефалеобразные 58, 177
Кефали
–
лизы 177
–
серые 177
–
толстогубые 180, 181
Кильки 65
–
каспийские 60, 64
Клариевые 158, 160
Клариевый сом 161
–
африканский 160, 160*, 161
Колюшка
–
малая южная 172, 173*
–
трёхиглая 172, 172*
Колюшки
–
девятииглые 172
–
многоиглые 172
–
трёхиглые 172
Колюшковые 171, 172
Колюшкообразные 57, 171
Комета 136*
Красавка 89
Краснопёр 84
Краснопёрка 98, 98*, 99
Краснопёрки 81, 98
Кумжа
–
черноморская 70, 70*, 71
Кутум 93, 97, 97*, 98
Л
Лаврак 200, 201, 201*
Лавраковые 188, 200
Лещ 109, 109*, 110
Лещи 82, 109, 112
Лини 82, 118
Линь 118, 118*, 119
Лобан 177, 179, 182, 182*
Лососеобразные 56, 68
Лососёвые 68, 69
Лососи (Настоящие) 69
–
тихоокеанские 69, 72
Лосось 71
–
стальноголовый 72
–
черноморский 70, 70*, 71
238
Лучепёрые рыбы 39
Львиноголовка 136*
Лысун
–
леопардовый 204, 204*
М
Медака
–
китайская 166, 166*, 167
Микижа 72
Минога
–
украинская 37, 37*, 38, 38*
Миноги
–
зубатые 37
Миноговые 36
Миногообразные 36
Н
Налим 171
–
обыкновенный 170, 170*
Налимовые 170
Небесное око 136*
Носарь 195
О
Окунёвые 188, 189
Окунеобразные 58, 183, 187, 188
Окуни 189
–
пресноводные 189
–
ушастые 188, 197
Окунь 191
–
большеротый 198, 198*
–
обыкновенный 190, 190*
–
полосатый 200, 201, 201*
–
речной 190
Оризия 166
Осётр
–
атлантический 46, 52, 52*
–
байкальский 51
–
восточносибирский 51
–
длиннорылый 51
–
западносибирский 51
–
ленский 51
–
обский 51
–
острорылый 51
–
сибирский 46, 51, 51*
–
русский 46, 49, 49*
–
якутский 51
–
–
стерлядевидный 51
Осетровые 41, 42
Осетрообразные 41
Осётры 42, 45, 46, 47
Остронос 177, 178, 180, 180*
П
Перкарина
–
азовская 194, 194*
Перкарины 189, 194
Перцихтовые 188, 200
Пескари 83, 124
–
румынские 83, 126, 127
Пескарь
–
кубанский 125, 125*, 126
–
кубанский длинноусый 126,
128, 128*
–
малый длинноусый 126
–
обыкновенный 125
–
северокавказский длинно-
усый 126, 127, 127*, 128
Пескоройка 36
Пецилиевые 163
Пиленгас 177, 179, 181, 181*
Плотва 95
–
обыкновенная 93, 94, 94*
Плотвы 80, 93, 94
Подуст
–
колхидский 120, 120*
239
–
кубанский 120, 121, 121*
Подусты 82, 119, 120
Позвоночные 36
Понтиколы 204, 213, 214
Псевдорасборы 80, 90
Псефур 55
Пуголовка
–
азовская 218, 218*
–
звёздчатая 218, 219, 219*
–
черноморская 218, 219,
219*
Пуголовки 204, 217, 218
Пузанок
–
азово-черноморский 61, 63,
63*
–
черноморско-каспийский 64
Р
Рыба-игла
–
итальянская 177
Рыбец
–
каспийский 113
–
обыкновенный 113
–
малый 114, 116, 116*, 117
–
черноморско-азовский 113,
114, 114*, 116
Рыбка
–
золотая 134, 136*
–
–
китайская 135
–
рисовая китайская 166, 166*
Рыбообразные 36
Рыбцы 82, 113, 114
Рыбы
–
костистые 56
–
лучепёрые 39, 40
–
новопёрые 40, 56
–
хрящекостные 40
С
Сазан 129, 129*, 130, 132
Сазаны 83, 129
Сарган
–
черноморский 167, 167*
Саргановые 166, 167
Сарганообразные 57, 165
Сарделька 64, 66, 66*
Севрюга 46, 50, 50*
Сельдевые 59
Сельдеобразные 56, 58, 59
Сельдь
–
керченская 60, 61, 61*
–
черноморско-азовская мор-
ская 60, 61, 61*, 62
–
черноморско-азовская про-
ходная 61, 62, 62*, 63
Сингиль 177, 178, 179, 179*
Синец 111
Синцы 82, 110
Сом 160
–
европейский 159, 159*
–
канальный 163
–
обыкновенный 159, 159*
Сомик канальный 162, 162*
Сомовые 158
Сомообразные 57, 158
Сомы кошачьи 158, 162
Стерлядь 46, 48, 48*
Судак 191, 192, 192*
–
волжский 193
Судаки 189, 191, 192
Султанка 199, 199*
Султанковые 188, 199
Т
Тарань 93, 95, 95*, 96
Телескоп 136*
240
Тиляпии 184, 185
Тиляпия
–
голубая 185, 186, 186*
–
золотая 185, 186, 186*
–
мозамбикская 185, 185*
Толстолобик
–
белый 139, 139*, 141
–
пёстрый 139, 141, 141*, 142
Толстолобики 83, 138, 139
–
белые 138
–
пёстрые 139
Толстолобы 83, 138, 139
Тресковые 170
Трескообразные 57, 170
Тюлька
–
абрауская 64, 66, 66*
–
черноморско-азовская 64, 65,
65*
Тюльки 60, 64, 65
У
Угри
–
пресноводные 75
–
речные 76
Угорь 76, 77
–
речной 76, 76*
Угрёвые 75
Угреобразные 57, 75
Уклейки 81, 104, 105
Уклейка 105, 106
–
обыкновенная 105, 105*
Украинская минога 37, 37*, 38,
38*
Усач 124
–
колхидский 122, 123, 123*
–
кубанский 122, 123, 123*
–
крымский 122
Усачи 82, 122, 123
Ф
Форелеокунь 198, 198*
Форель 71, 72
–
озёрная
–
радужная 72, 73, 73*
–
ручьевая 71, 71*, 72
Х
Хамса 67, 68
–
азовская 68
–
черноморская 67
Хрящевые ганоиды 40, 41
Хрящекостные рыбы 40, 41
Ц
Центрарховые 188, 197
Цихлидообразные 58, 183
Цихлиды 183, 184, 185
Цихловые 183, 184
Ч
Чебачок
–
амурский 90, 90*, 92
Челюстноротые 39
Черепные 36
Чехони 82, 117
Чехонь 117, 117*, 118
Чукучановые 78, 142
Ш
Шемаи 104
Шемая 107
–
черноморско-азовская 105,
106, 106*
Шип 46, 47, 47*
Шубункин 136*
241
Щ
Щиповка
–
азовская 150, 151, 153, 153*
–
обыкновенная 150, 151, 151*
–
кубанская 154, 155, 155*
–
предкавказская 154, 156,
156*
–
сибирская 150, 151, 152, 152*
–
южнорусская 150, 151, 153,
153*
Щиповки 149, 150, 151
–
Сабанеева 149, 154, 155
Щука 74, 75
–
обыкновенная 74, 74*
Щуковые 74
Щукообразные 58, 73
Я
Язь 84, 84*
242
Указатель латинских названий рыб
A
Abramis 82, 109, 111, 112
–
brama 11, 95, 109, 109*
Acanthalburnus 112
Acanthobrama 112
Acipenser 42, 45, 46, 48
–
baerii 46, 51, 51*
–
–
baerii 51
–
–
baicalensis 51
–
–
chatys 51
–
–
stenorrhynchus 51
–
gueldenstaedtii 46, 49, 49*, 52
–
nudiventris 46, 47, 47*
–
ruthenus 46, 48, 48*, 53
–
stellatus 46, 50, 50*, 52, 53
–
sturio 46, 52, 52*
Acipenseridae 41, 42
Acipenseriformes 41
Actinopterygii 39
Adrianichthyidae 165, 166
Alburnoides 81, 108
–
bipunctatus 10
–
–
rossicus 10
–
–
–
natio kubanicus 10
–
rossicus 108, 108*
Alburnus 81, 104, 105
–
alburnus 95, 104, 105, 105*
–
chalcoides 104
–
mento 105, 106, 106*
Alosa 59, 60, 61
–
caspia tanaica 64
–
immaculata 61, 62, 62*
–
maeotica 60, 61, 61*
–
pontica 61, 62
–
tanaica 61, 63, 63*
Ammocoetes 36
Anguilla 76
–
anguilla 76, 76*
Anguillidae 75
Anguilliformes 57, 75
Aristichthys 139
–
nobilis 139, 141, 141*
Aspius 81, 101
–
aspius 84, 101, 102, 102*
Atherina
–
boyeri pontica 169, 169*
Atherinidae 168
Atheriniformes 57, 168
B
Babka 204, 212
–
gymnotrachelus 213, 213*
Balitoridae 78, 83
Ballerus 82, 110, 111, 112
–
ballerus 111
–
sapa 111, 111*
Barbatula 147
–
barbatula 147, 147*
Barbuccidae 77
Barbus 82, 122, 123
–
escherichii 122, 123, 123*
–
kubanicus 122, 123, 123*
–
tauricus 122
Belone
–
belone euxini 167, 167*
–
euxini 168
Belonidae 166, 167
Beloniformes 57, 165
1
Звёздочкой отмечены страницы, на которых размещены рисунки.
243
Benthophilus 204, 217, 218
–
magistri 218, 218*
–
stellatus 218, 219, 219*
Blicca 82, 112
–
bjoerkna 95, 112, 112*
C
Carassius 83, 132, 133
–
auratus 132, 133, 134, 135*,
135, 136*
–
–
auratus 134, 135
–
–
gibelio 132
–
carassius 132, 133, 133*
–
gibelio 9, 133, 137, 137*
Caspiosoma 204, 212
–
caspium 212, 212*
Catostomidae 78
Centrarchidae 188, 197
Chalcalburnus 104
–
chalcoides 88
Chelon 180, 181
–
aurata 180
–
saliens 181
Chondrostei 40, 41
Chondrostoma 82, 119, 120
–
colchicum 120, 120*
–
kubanicum 120, 121, 121*
Cichlidae 183, 184, 185
Cichliformes 58, 183
Cladistia 40
Clariidae 158, 160
Clarias
–
gariepinus 160
Clupeidae 11, 59
Clupeiformes 56, 58, 59
Clupeinae 11
Clupeonella 60, 64, 65
–
abrau 64, 66, 66*
–
cultriventris 64, 65, 65*
Cobitidae 78, 149
Cobitis 149, 150, 151, 154
–
melanoleuca 150, 151, 152,
152*
–
rossomeridionalis 150, 151,
153, 153*
–
taenia 150, 151, 151*, 152,
154
–
tanaitica 150, 151, 153, 153*
Craniata 36
Ctenopharyngodon 81, 100
–
idella 100, 101, 101*
Cyprinidae 11, 78, 79
Cypriniformes 11, 57, 77, 78
Cyprininae 11
Cyprinodontiformes 57, 163
Cyprinoidea 11
Cyprinoidei 11
Cyprinus 83, 129
–
carpio 129, 129*, 132
–
–
haematopterus 131
E
Ellopostomatidae 77
Engraulidae 59, 67
Engraulididae 59, 67
Engraulis 67
–
encrasicolus 67, 67*
–
–
ponticus 67
–
–
maeoticus 68
Esocidae 74
Esociformes 58, 73
Esox
–
lucius 74, 74*
Eudontomyzon 37
–
mariae 37, 37*
244
G
Gadidae 170
Gadiformes 57, 170
Gambusia 163, 164
–
holbrooki 164, 164*
Gasterosteidae 171, 172
Gasterosteiformes 57, 171
Gasterosteus 172
–
aculeatus 172, 172*
Gnathostomata 36, 39
Gobiidae 202, 203
Gobiiformes 58, 202
Gobio 83, 124
–
ciscaucasicus 128
–
gobio 125
–
gobio gobio 125
–
kubanicus 125, 125*
–
uranoscopus 127
–
–
caucasica 128
–
–
var. caucasica 127
Gobius 204, 207
–
ophiocephalus 208, 208*
Gymnocephalus 189, 195, 196
–
acerinus 195, 197, 197*
–
cernuus 195, 196, 196*
H
Huso 42, 43
–
dauricus 43, 45, 45*
–
huso 43, 43*, 52, 53
–
–
maeoticus 44
Hypophthalmichthys 83, 138, 139
–
molitrix 139, 139*
–
nobilis 139, 141, 141*
I
Ictaluridae 158, 162
Ictalurus
–
punctatus 162, 162*
Ictiobus 142, 143
–
bubalis 143, 145, 145*
–
cyprinellus 143, 143*
–
niger 143, 146, 146*
K
Knipowitschia 203, 205, 206
–
caucasica 205, 207, 207*
–
longicaudata 11, 205, 206,
206*
L
Leucaspius 81, 103
–
delineatus 103, 103*
Leuciscus 79, 83, 85
–
idus 84, 84*
Liza 177
–
aurata 177, 178, 178*, 179,
179*, 180
–
haematocheilus 177, 178*,
179, 181, 181*
–
haematocheila 182
–
saliens 177, 178*, 178, 180,
180*
Lota
–
lota 170, 170*
Lotidae 170
M
Micropterus
–
salmoides 198, 198*
Misgurnus 150, 156
–
fossilis 157, 157*
Morone
–
saxatilis 201, 201*
Mugil 177, 182
–
cephalus 177, 178*, 179, 182
245
–
soiuy 182
Mugilidae 177, 178
Mugiliformes 58, 177
Mullidae 177, 188, 199
Mullus
–
barbatus ponticus 199, 199*
Mylopharyngodon 81, 99
–
piceus 99, 100, 100*
Myxinimorphi 36
N
Nemacheilidae 77, 78, 83, 147
Neogobius 203, 208, 209, 212, 213,
214
–
constructor 214, 215, 215*
–
eurycephalus 214, 214*
–
fluviatilis 209, 210, 210*
–
gymnotrachelus 213, 213*
–
melanostomus 208, 209, 209*
–
rhodioni 214, 216, 216*
–
syrman 214, 217, 217*
Neopterygii 40, 56
Nerophis
–
ophidion 174, 175, 175*
O
Oncorhynchus 69, 72
–
mykiss 72, 73, 73*
Oreochromis 184
–
aureus 185, 186, 186*, 187
–
hornorum 187
–
mossambicus 185, 185*, 187
–
niloticus 187
Oryzias
–
latipes 167
–
sinensis 166, 166*, 167
Oxynoemacheilus 147, 148
–
merga 148, 148*
P
Parasalmo gairdnerii 72
Pelecus 82, 117
–
cultratus 117, 117*
Perca 189
–
fluviatilis 190, 190*
Percarina 189, 194
–
demidoffii 194, 194*
Percichthyidae 188, 200
Percidae 188, 189
Perciformes 58, 187, 188
Petroleuciscus 80, 85, 88
–
borysthenicus 85, 85*
Petromyzontida 36
Petromyzontidae 36
Petromyzontiformes 36
Petromyzontomorphi 36
Pholidichthyidae 183
Phoxinus 80, 89
–
phoxinus 89, 89*
Planiliza 182
–
haematocheila 182
Platichthys 221
–
flesus luscus 221, 221*
Pleuronectidae 220
Pleuronectiformes 58, 220
Poeciliidae 163
Polyodon
–
spathula 55, 55*
Polyodontidae 41, 55
Pomatoschistus 203, 204
–
marmoratus 204, 204*
Ponticola 204, 213, 214
–
constructor 214, 215, 215*
–
eurycephalus 214, 214*
–
rhodioni 214, 216, 216*
–
syrman 214, 217, 217*
Proterorhinus 203, 210
246
–
marmoratus 211, 211*
Psephurus gladius 55
Pseudorasbora 80, 90
–
parva 90, 90*
Pungitius 172
–
platygaster 172, 173, 173*
R
Rhodeus 80, 92
–
sericeus amarus 92, 92*
Romanogobio 83, 126, 127, 128
–
ciscaucasicus 126, 127, 127*,
128
–
parvus 126
–
pentatrichus 126, 128, 128*
–
uranoscopus
Rutilus 80, 93, 94
–
frisii 10, 93, 96, 96*
–
–
frisii 97
–
–
kutum 10, 93, 97, 97*
–
rutilus 11, 93, 94, 94*, 95
–
–
heckelii 93, 95, 95*
–
heckelii 93, 95, 95*
S
Sabanejewia 149, 154, 155
–
caucasica 154, 156, 156*
–
kubanica 154, 155, 155*
Salmo 69
–
gairdnerii 72
–
labrax 70, 70*
–
–
morpha fario 71, 71*
–
trutta 10
–
trutta labrax 70, 70*
–
–
morpha fario 10
Salmonidae 68, 69
Salmoniformes 56, 68
Sander 189, 191, 192
–
lucioperca 191, 192, 192*
–
volgensis 191, 193, 193*
Sardinella aurata 60*
Sarotherodon 184
Scardinius 81, 98
–
erythrophthalmus 98, 98*
Serpenticobitidae 77
Siluridae 158
Siluriformes 57, 158
Silurus
–
glanis 159, 159*
Squalius 80, 86, 86, 87, 88
–
aphipsi 87, 88, 88*
–
cephalus orientalis 86, 87,
87*, 88
Syngnathidae 174, 175
Syngnathiformes 58, 174
Syngnathus
–
abaster 177
–
nigrolineatus 174, 175, 175*,
176, 176*
T
Teleostei 56
Tilapia 184
Tinca 82, 118
–
tinca 118, 118*
V
Vaillantellidae 77
Vertebrata 36
Vimba 82, 112, 113, 114
–
vimba carinata 113, 114, 114*
–
–
persa 113
–
–
tenella 114, 116, 116*
–
–
vimba 113
247
ПРИЛОЖЕНИЯ
248
Приложение 1
СИСТЕМАТИЧЕСКИЙ СПИСОК РЫБООБРАЗНЫХ И РЫБ,
ВСТРЕЧАЮЩИХСЯ В ОПРЕДЕЛИТЕЛЕ
Тип Позвоночные — Vertebrata
Подтип Черепные — Craniata
Надкласс Петромизонтоморфы — Petromyzontomorphi
Класс Миноги — Petromyzontida Bonaparte, 1832
Отряд I. Миногообразные — Petromyzontiformes Bonaparte, 1832
Семейство 1. Миноговые — Petromyzontidae Bonaparte, 1831
Род Зубатые миноги — Eudontomyzon Regan, 1911
1. Украинская минога — Eudontomyzon mariae (Berg, 1931) (с. 37)
Надкласс Челюстноротые — Gnathostomata
Класс Лучепёрые рыбы — Actinopterygii Klein, 1885
Подкласс Хрящекостные рыбы, или Хрящевые ганоиды, — Chondrostei
Müller, 1844
Отряд II. Осетрообразные — Acipenseriformes Berg, 1940
Семейство 2. Осетровые — Acipenseridae Bonaparte, 1831
Род Белуги — Huso Brandt, 1869
2. Белуга — Huso huso (Linnaeus, 1758) (с. 43)
3. Калуга — Huso dauricus (Georgi, 1775) (с. 45)
Род Осётры — Acipenser Linnaeus, 1758)
4. Шип — Acipenser nudiventris Lovetsky, 1828 (с. 47)
5. Стерлядь — Acipenser ruthenus Linnaeus, 1758 (с. 48)
6. Русский осётр — Acipenser gueldenstaedtii (Brandt, 1833) (с. 49)
7. Севрюга — Acipenser stellatus Pallas, 1771 (с. 50)
8. Сибирский осётр — Acipenser baerii (Brandt, 1869) (с. 51)
9. Атлантический осётр — Acipenser sturio (Linnaeus, 1758) (с. 52)
Семейство 3. Веслоносые — Polyodontidae Bonaparte, 1838
Род Веслоносы — Polyodon Lacépède, 1797
10. Веслонос — Polyodon spathula (Walbaum, 1792) (с. 55)
Подкласс Новопёрые рыбы — Neopterygii Cope, 1871
Отдел Костистые рыбы — Teleostei Müller, 1845
Отряд III. Сельдеобразные — Clupeiformes Bleeker, 1959
Семейство 4. Сельдевые — Clupeidae Cuvier, 1816
Род Алозы — Alosa Linck, 1790
11. Черноморско-азовская морская, или керченская, сельдь — Alosa maeotica
(Grimm, 1901) (с. 61)
12. Черноморско-азовская проходная сельдь — Alosa immaculata (Bennett,
1835) [син. Alosa pontica (Eichwald, 1838)] (с. 62)
249
13. Азово-черноморский пузанок — Alosa tanaica (Grimm, 1901) (с. 63)
Род Тюльки, или Каспийские кильки, — Clupeonella Kessler, 1877
14. Черноморско-азовская тюлька — Clupeonella cultriventris (Nordmann,
1840) (с. 65)
15. Абрауская тюлька [сарделька] — Clupeonella abrau (Maliatsky, 1930)
(с. 66)
Семейство 5. Анчоусовые — Engraulidae Gill, 1861 [Engraulididae]
Род Анчоусы — Engraulis Cuvier, 1816
16. Европейский анчоус, или хамса, — Engraulis encrasicolus (Linnaeus, 1758)
(с. 67)
Отряд IV. Лососеобразные — Salmoniformes Bleeker, 1859
Семейство 6. Лососёвые — Salmonidae Cuvier, 1816
Род (Настоящие) Лососи — Salmo Linnaeus, 1758
17. Черноморский лосось [черноморская кумжа] — Salmo labrax [Salmo trutta
labrax] Pallas, 1814 (с. 70)
18. Ручьевая форель — Salmo labrax morpha fario Linnaeus, 1758 (с. 71)
Род Тихоокеанские лососи — Oncorhynchus Suckley, 1861
19. Радужная форель — Oncorhynchus mykiss (Walbaum, 1792) (с. 73)
Отряд V. Щукообразные — Esociformes Bleeker, 1859
Семейство 7. Щуковые — Esocidae Cuvier, 1816
Род Щуки — Esox Linnaeus, 1758
20. Обыкновенная щука — Esox lucius Linnaeus, 1758 (с. 74)
Отряд VI. Угреобразные — Anguilliformes Goodrich, 1909
Семейство 8. Угрёвые, или Пресноводные угри, — Anguillidae
rafinesque, 1810
Род Речные угри — Anguilla Schrank, 1798
21. Речной угорь — Anguilla anguilla (Linnaeus, 1758) (с. 76)
Отряд VII. Карпообразные — Cypriniformes Rafinesque, 1810
Семейство 9. Карповые — Cyprinidae rafinesque, 1810
Род Ельцы — Leuciscus Cuvier, 1816
22. Язь — Leuciscus idus (Linnaeus, 1758) (с. 84)
Род Бобырцы — Petroleuciscus Bogutskaya, 2002
23. Калинка, или бобырец, — Petroleuciscus borysthenicus (Kessler, 1859)
(с. 85)
Род Голавли — Squalius Bonaparte, 1837
24. Кавказский голавль — Squalius cephalus orientalis (Nordmann, 1840) (с. 87)
25. Афипсская калинка, или афипсский голавль [елец], или бекас, — Squalius
aphipsi (Aleksandrov, 1927) (с. 88)
Род Гольяны — Phoxinus Rafinesque, 1820
26. Обыкновенный гольян [красавка] — Phoxinus phoxinus (Linnaeus, 1758)
(с. 89)
250
Род Псевдорасборы (Амурские чебачки) — Pseudorasbora Bleeker, 1860
27. Амурский чебачок — Pseudorasbora parva (Temminck & Schlegel, 1846)
(с. 90)
Род Горчаки — Rhodeus Agassiz, 1832
28. Обыкновенный горчак — Rhodeus sericeus amarus (Bloch, 1782) (с. 92)
Род Плотвы — Rutilus Rafinesque, 1820
29. Обыкновенная плотва — Rutilus rutilus Linnaeus, 1758 (с. 94)
30. Тарань — Rutilus heckelii (Nordmann, 1840) (с. 95)
31. Вырезуб — Rutilus frisii (Nordmann, 1840) (с. 96)
32. Кутум — Rutilus frisii kutum (Kamensky, 1901) (с. 97)
Род Краснопёрки — Scardinius Bonaparte, 1837
33. Краснопёрка — Scardinius erythrophthalmus (Linnaeus, 1758) (с. 98)
Род Чёрные амуры — Mylopharyngodon Peters, 1881
34. Чёрный амур — Mylopharyngodon piceus (Richardson, 1846) (с. 100)
Род Белые амуры — Ctenopharyngodon Steindachner, 1866
35. Белый амур — Ctenopharyngodon idella (Valenciennes, 1844) (с. 101)
Род Жерехи — Aspius Agassiz, 1832
36. Обыкновенный жерех — Aspius aspius (Linnaeus, 1758) (с. 102)
Род Верховки — Leucaspius Heckel & Kner, 1858
37. Верховка — Leucaspius delineatus Heckel, 1843 (с. 103)
Род Уклейки — Alburnus Rafinesque, 1820
38. Обыкновенная уклейка — Alburnus alburnus (Linnaeus, 1758) (с. 105)
39. Черноморско-азовская шемая — Alburnus mento (Heckel, 1836) (с. 106)
Род Быстрянки — Alburnoides Jeitteles, 1861
40. Русская быстрянка — Alburnoides rossicus Berg, 1924 (с. 108)
Род Лещи — Abramis Cuvier, 1816
41. Лещ — Abramis brama (Linnaeus, 1758) (с. 109)
Род Синцы — Ballerus Heckel, 1843
42. Белоглазка — Ballerus sapa (Pallas, 1814) (с. 111)
Род Густеры — Blicca Heckel, 1843
43. Густера — Blicca bjoerkna (Linnaeus, 1758) (с. 112)
Род Рыбцы — Vimba Fitzinger, 1873
44. Черноморско-азовский рыбец — Vimba vimba carinata (Pallas, 1814)
(с. 114)
45. Малый рыбец — Vimba vimba tenella (Nordmann, 1840) (с. 116)
Род Чехони — Pelecus Agassiz, 1835
46. Чехонь — Pelecus cultratus (Linnaeus, 1758) (с. 117)
Род Лини — Tinca Cuvier, 1816
47. Линь — Tinca tinca (Linnaeus, 1758) (с. 118)
Род Подусты — Chondrostoma Agassiz, 1835
48. Колхидский подуст — Chondrostoma colchicum Derjugin, 1899 (с. 120)
49. Кубанский подуст — Chondrostoma kubanicum (Berg, 1914) (с. 121)
251
Род Усачи — Barbus Cuvier, 1816
50. Колхидский усач — Barbus escherichii Steindachner, 1892 (с. 123)
51. Кубанский усач — Barbus kubanicus Berg, 1912 (с. 123)
Род Пескари — Gobio Cuvier, 1816
52. Кубанский пескарь — Gobio kubanicus Vasil’eva, 2004 (с. 125)
Род Румынские пескари — Romanogobio Bănărescu, 1961
53. Северокавказский длинноусый пескарь — Romanogobio ciscaucasicus
(Berg, 1932) (с. 127)
54. Кубанский длинноусый пескарь — Romanogobio pentatrichus Naseka &
Bogutskaya, 1998 (с. 128)
Род Сазаны, или Карпы, — Cyprinus Linnaeus, 1758
55. Сазан, или карп, — Cyprinus carpio Linnaeus, 1758 (с. 129)
Род Караси — Carassius Jarocki, 1822
56. Золотой карась — Carassius carassius (Linnaeus, 1758) (с. 133)
57. Китайский карась, или золотая рыбка — Carassius auratus (Linnaeus, 1758)
[Carassius auratus auratus (Linnaeus, 1758)] (с. 134)
58. Серебряный карась — Carassius gibelio (Bloch, 1782) (с. 137)
Род Толстолобы (Толстолобики) — Hypophthalmichthys Valenciennes,
1844
59. Белый толстолобик — Hypophthalmichthys molitrix (Valenciennes, 1844)
(с. 139)
60. Пёстрый толстолобик — Hypophthalmichthys [Aristichthys] nobilis
(Richardson, 1846) (с. 141)
Семейство 10. Чукучановые — Catostomidae Cope, 1871
Род Буффало, или Иктиобусы, — Ictiobus Rafinesque, 1820
61. Большеротый буффало — Ictiobus cyprinellus (Valenciennes, 1844) (с. 143)
62. Малоротый буффало — Ictiobus bubalis (Rafinesque, 1819) (с. 145)
63. Чёрный буффало — Ictiobus niger (Rafinesque, 1820) (с. 146)
Семейство 11. Гольцовые — Nemacheilidae reGan, 1911
Род Усатые гольцы — Barbatula Linck, 1790
64. Усатый, или обыкновенный, голец — Barbatula barbatula (Linnaeus, 1758)
(с. 147)
Род Южноевропейские гольцы — Oxynoemacheilus Bănărescu & Nal-
bant, 1966
65. Голец Крыницкого — Oxynoemacheilus merga (Krynicki, 1840) (с. 148)
Семейство 12. Вьюновые — Cobitidae swainson, 1838
Род Щиповки — Cobitis Linnaeus, 1758
66. Обыкновенная щиповка — Cobitis taenia Linnaeus, 1758 (с. 151)
67. Сибирская щиповка — Cobitis melanoleuca Nichols, 1925 (с. 152)
68. Азовская щиповка — Cobitis tanaitica Băcescu & Maier, 1969 (с. 153)
69. Южнорусская щиповка — Cobitis rossomeridionalis Vasil’eva & Vasil’ev,
1998 (с. 153)
252
Род Щиповки Сабанеева — Sabanejewia Vladykov, 1929
70. Кубанская щиповка — Sabanejewia kubanica Vasil’eva & Vasil’ev, 1988
(с. 155)
71. Предкавказская щиповка — Sabanejewia caucasica (Berg, 1906) (с. 156)
Род Вьюны — Misgurnus Lacépède, 1803
72. Вьюн — Misgurnus fossilis (Linnaeus, 1758) (с. 157)
Отряд VIII. Сомообразные — Siluriformes Cuvier, 1816
Семейство 13. Сомовые — Siluridae Cuvier, 1816
Род Сомы — Silurus Linnaeus, 1758
73. Обыкновенный, или европейский, сом — Silurus glanis Linnaeus, 1758
(с. 159)
Семейство 14. Клариевые — Clariidae Bonaparte, 1846
Род Кларии — Clarias Scopoli, 1777
74. Африканский клариевый сом — Clarias gariepinus ( Burchell, 1822) (с. 160)
Семейство 15. Икталуровые, или Кошачьи сомы, — Ictaluridae Gill,
1861
Род Американские сомики — Ictalurus Rafinesque, 1820
75. Канальный сомик — Ictalurus punctatus (Rafinesque, 1818) (с. 162)
Отряд IX. Карпозубообразные — Cyprinodontiformes Berg, 1940
Семейство 16. Гамбузиевые, или Пецилиевые, — Poeciliidae Garman,
1895
Род Гамбузии — Gambusia Poey, 1854
76. Хольбрукская гамбузия — Gambusia holbrooki (Girard, 1859) (с. 164)
Отряд X. Сарганообразные — Beloniformes Berg, 1940
Семейство 17. Адрианихтовые — Adrianichthyidae weBer, 1913
Род Оризии — Oryzias Jordan & Snyder, 1906
77. Китайская медака — Oryzias sinensis Chen, Uwa & Chu, 1989 (с. 166)
Семейство 18. Саргановые — Belonidae Bonaparte, 1832
Род Сарганы — Belone Cuvier, 1816
78. Черноморский сарган — Belone belone euxini (Günther, 1866) (с. 167)
Отряд XI. Атеринообразные — Atheriniformes Rosen, 1966
Семейство 19. Атериновые — Atherinidae Günther, 1961
Род Атерины — Atherina Linnaeus, 1758
79. Черноморская атерина — Atherina boyeri pontica Eichwald, 1831 (с. 169)
Отряд XII. Трескообразные — Gadiformes Goodrich, 1909
Семейство 20. Налимовые — Lotidae Bonaparte, 1832
Род Налимы — Lota Oken, 1817
80. Обыкновенный налим — Lota lota (Linnaeus, 1758) (с. 170)
Отряд XIII. Колюшкообразные — Gasterosteiformes Greenwood, 1966
Семейство 21. Колюшковые — Gasterosteidae Bonaparte, 1831
Род Трёхиглые колюшки — Gasterosteus Linnaeus, 1758
81. Трёхиглая колюшка — Gasterosteus aculeatus Linnaeus, 1758 (с. 172)
253
Род Девятииглые, или Многоиглые, колюшки — Pungitius Costa, 1848
82. Малая южная колюшка — Pungitius platygaster Kessler, 1859 (с. 173)
Отряд XIV. Иглообразные — Syngnathiformes Helfman, Collette &
Facey, 1997
Семейство 22. Игловые — Syngnathidae rafinesque, 1810
Род Змеевидные морские иглы — Nerophis Rafinesque, 1810
83. Змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758) (с. 175)
Род Обыкновенные морские иглы — Syngnathus Linnaeus, 1758
84. Черноморская игла-рыба — Syngnathus nigrolineatus Eichwald, 1831 (с. 176)
Отряд XV. Кефалеобразные — Mugiliformes Günther, 1880
Семейство 23. Кефалевые — Mugilidae Cuvier, 1829
Род Кефали-лизы — Liza (Jordan & Swain, 1884)
85. Сингиль — Liza aurata (Risso, 1810) (с. 179)
86. Остронос — Liza saliens (Risso, 1810) (с. 180)
87. Пиленгас — Liza haematocheilus (Temminck & Schlegel, 1845) (с. 181)
Род Серые кефали — Mugil Linnaeus, 1758
88. Лобан — Mugil cephalus Linnaeus, 1758 (с. 182)
Отряд XVI. Цихлидообразные — Cichliformes Betancur-R et al., 2013
Семейство 24. Цихловые, или Цихлиды, — Cichlidae Bonaparte, 1835
Род Ореохромис (Тиляпия) — Oreochromis Günther, 1889
89. Мозамбикская тиляпия — Oreochromis mossambicus (Peters, 1852) (с. 185)
90. Голубая (золотая) тиляпия — Oreochromis aureus (Steindachner, 1864)
(с. 186)
Отряд XVII. Окунеобразные — Perciformes Bleeker, 1859
Семейство 25. Окунёвые — Percidae rafinesque, 1815
Род Окуни [Пресноводные окуни] — Perca Linnaeus, 1758
91. Обыкновенный, или речной, окунь — Perca fluviatilis Linnaeus, 1758 (с. 190)
Род Судаки — Sander Oken, 1817
92. Судак — Sander lucioperca (Linnaeus, 1758) (с. 192)
93. Бёрш, или берш, или волжский судак — Sander volgensis (Gmelin, 1789)
(с. 193)
Род Перкарины — Percarina Nordmann, 1840
94. Азовская перкарина — Percarina demidoffii Nordmann, 1840 (с. 194)
Род Ерши — Gymnocephalus Bloch, 1793
95. Обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758) (с. 196)
96. Донской ёрш, или бирючок, — Gymnocephalus acerinus (Gmelin, 1774)
(с. 197)
Семейство 26. Центрарховые, или Ушастые окуни, — Centrarchidae
Bleeker, 1859
Род Чёрные окуни — Micropterus Lacépède, 1802
97. Большеротый окунь, или форелеокунь, — Micropterus salmoides (Lacépède,
1802) (с. 198)
254
Семейство 27. Барабулевые, или Султанковые, — Mullidae
rafinesque, 1815
Род Барабули — Mullus Linnaeus, 1758
98. Черноморская барабуля (султанка) — Mullus barbatus ponticus Essipov,
1927 (с. 199)
Семейство 28. Лавраковые, или Перцихтиевые, — Percichthyidae
Jordan & eiGenmann, 1890
Род Лавраки — Morone Mitchill, 1814
99. Полосатый окунь, или лаврак, — Morone saxatilis (Walbaum, 1792) (с. 201)
Отряд XVIII. Бычкообразные — Gobiiformes Günther, 1880
Семейство 29. Бычковые — Gobiidae Cuvier, 1816
Род Бычки-лысуны, или Бычки-бубыри, — Pomatoschistus Gill, 1864
100. Леопардовый лысун [бычок-бубырь] — Pomatoschistus marmoratus (Risso,
1810) (с. 204)
Род Бычки Книповича — Knipowitschia Iljin, 1927
101. Длиннохвостый бычок Книповича — Knipowitschia longicaudata (Kessler,
1877) (с. 206)
102. Бычок-бубырь — Knipowitschia caucasica (Berg, 1916) (с. 207)
Род Бычки — Gobius Linnaeus, 1758
103. Бычок-травяник — Gobius ophiocephalus Pallas, 1814 (с. 208)
Род Черноморско-каспийские бычки — Neogobius Iljin, 1927
104. Бычок-кругляк — Neogobius melanostomus Pallas, 1814 (с. 209)
105. Речной бычок-песочник — Neogobius fluviatilis (Pallas, 1814) (с. 210)
Род Тупоносые бычки — Proterorhinus Smitt, 1900
106. Бычок-цуцик, или мраморный тупоносый бычок, — Proterorhinus
marmoratus (Pallas, 1814) (с. 211)
Род Каспиосомы — Caspiosoma Iljin, 1927
107. Каспиосома — Caspiosoma caspium (Kessler, 1877) (с. 212)
Род Бабки — Babka (Neogobius) Iljin, 1927
108. Бычок-гонец — Babka (Neogobius) gymnotrachelus (Kessler, 1857) (с. 213)
Род Понтиколы — Ponticola (Neogobius) Iljin, 1927
109. Бычок-рыжик — Ponticola (Neogobius) eurycephalus (Kessler, 1874) (с. 214)
110. Кавказский речной бычок — Ponticola ( Neogobius) constructor ( Nordmann,
1840) (с. 215)
111. Речной бычок Родиона — Ponticola (Neogobius) rhodioni (Vasil’eva &
Vasil’ev, 1994) (с. 216)
112. Бычок сирман (ширман) — Ponticola (Neogobius) syrman (Nordmann,
1840) (с. 217)
Род Пуголовки — Benthophilus Eichwald, 1831
113. Азовская пуголовка — Benthophilus magistri Iljin, 1927 (с. 218)
114. Черноморская (звёздчатая) пуголовка — Benthophilus stellatus (Sauvage,
1874) (с. 219)
255
Отряд XIX. Камбалообразные — Pleuronectiformes Bleeker, 1859
Семейство 30. Камбаловые — Pleuronectidae Cuvier, 1816
Род Речные камбалы — Platichthys Girard, 1854
115. Черноморская камбала, или глосса, — Platichthys flesus luscus (Pallas,
1811) (с. 221)
256
Приложение 2
ЦВЕТНЫЕ ИЛЛЮСТРАЦИИ РЫБООБРАЗНЫХ И РЫБ
БАССЕЙНА КУБАНИ
Рис. 2.1. Украинская минога — Eudontomyzon mariae (Berg, 1931) (вверху
самка, а внизу — самец)
Рис. 2.2. Белуга — Huso huso (Linnaeus, 1758)
Рис. 2.3. Калуга — Huso dauricus (Georgi, 1775)
257
Рис. 2.4. Шип — Acipenser nudiventris Lovetsky, 1828
Рис. 2.5. Стерлядь — Acipenser ruthenus Linnaeus, 1758
Рис. 2.6. Русский осётр — Acipenser gueldenstaedtii Brandt, 1833
Рис. 2.7. Севрюга — Acipenser stellatus Pallas, 1771
258
Рис. 2.8. Сибирский осётр — Acipenser baerii Brandt, 1869
Рис. 2.9. Атлантический осётр — Acipenser sturio Linnaeus, 1758
Рис. 2.10. Веслонос — Polyodon spathula (Walbaum, 1792)
Рис. 2.11. Черноморско-азовская морская сельдь — Alosa maeotica (Grimm, 1901)
259
Рис. 2.12. Черноморско-азовская проходная сельдь — Alosa immaculata (Ben-
nett, 1835)
Рис. 2.13. Азово-черноморский пузанок — Alosa tanaica (Grimm, 1901)
Рис. 2.14. Черноморско-азовская тюлька — Clupeonella cultriventris (Nord-
mann, 1840)
260
Рис. 2.15. Абрауская тюлька — Clupeonella abrau (Maliatsky, 1930)
Рис. 2.16. Европейский анчоус — Engraulis encrasicolus (Linnaeus, 1758)
Рис. 2.17. Черноморский лосось — Salmo labrax Pallas, 1814
Рис. 2.18. Ручьевая форель — Salmo labrax morpha fario Linnaeus, 1758
261
Рис. 2.19. Радужная форель — Oncorhynchus mykiss (Walbaum, 1792)
Рис. 2.20. Обыкновенная щука — Esox lucius Linnaeus, 1758
Рис. 2.21. Речной угорь — Anguilla anguilla (Linnaeus, 1758)
262
Рис. 2.22. Язь — Leuciscus idus (Linnaeus, 1758)
Рис. 2.23. Кавказский голавль — Squalius cephalus orientalis (Nordmann, 1840)
Рис. 2.24. Афипсский голавль — Squalius aphipsi (Aleksandrov, 1927)
263
Рис. 2.25. Бобырец — Petroleuciscus borysthenicus (Kessler, 1859)
Рис. 2.26. Амурский чебачок — Pseudorasbora parva (Temminck & Schlegel, 1846)
Рис. 2.27. Обыкновенный горчак — Rhodeus sericeus amarus (Bloch, 1782)
264
Рис. 2.28. Обыкновенный гольян — Phoxinus phoxinus (Linnaeus, 1758)
Рис. 2.29. Обыкновенная плотва — Rutilus rutilus Linnaeus, 1758
Рис. 2.30. Тарань — Rutilus heckelii (Nordmann, 1840)
265
Рис. 2.31. Вырезуб — Rutilus frisii (Nordmann, 1840)
Рис. 2.32. Кутум — Rutilus frisii kutum (Kamensky, 1901)
Рис. 2.33. Краснопёрка — Scardinius erythrophthalmus (Linnaeus, 1758)
266
Рис. 2.34. Чёрный амур — Mylopharyngodon piceus (Richardson, 1846)
Рис. 2.35. Белый амур — Ctenopharyngodon idella (Valenciennes, 1844)
Рис. 2.36. Обыкновенный жерех — Aspius aspius (Linnaeus, 1758)
267
Рис. 2.37. Верховка — Leucaspius delineatus Heckel, 1843
Рис. 2.38. Черноморско-азовская шемая — Alburnus mento (Heckel, 1836)
Рис. 2.39. Обыкновенная уклейка — Alburnus alburnus (Linnaeus, 1758)
268
Рис. 2.40. Русская быстрянка — Alburnoides rossicus Berg, 1924
Рис. 2.41. Лещ — Abramis brama (Linnaeus, 1758)
Рис. 2.42. Белоглазка — Ballerus sapa (Pallas, 1814)
269
Рис. 2.43. Густера — Blicca bjoerkna (Linnaeus, 1758)
Рис. 2.44. Черноморско-азовский рыбец — Vimba vimba carinata (Pallas, 1814)
Рис. 2.45. Малый рыбец — Vimba vimba tenella (Nordmann, 1840)
270
Рис. 2.46. Чехонь — Pelecus cultratus (Linnaeus, 1758)
Рис. 2.47. Линь — Tinca tinca (Linnaeus, 1758)
Рис. 2.48. Колхидский подуст — Chondrostoma colchicum Derjugin, 1899
271
Рис. 2.49. Кубанский подуст — Chondrostoma kubanicum (Berg, 1914)
Рис. 2.50. Колхидский усач — Barbus escherichii Steindachner, 1897
Рис. 2.51. Кубанский усач — Barbus kubanicus Berg, 1912
272
Рис. 2.52. Кубанский пескарь — Gobio kubanicus Vasil’eva, 2004
Рис. 2.53. Северокавказский длинноусый пескарь — Romanogobio
ciscaucasicus (Berg, 1932)
Рис. 2.54. Кубанский длинноусый пескарь — Romanogobio pentatrichus
Naseka & Bogutskaya, 1998
273
Рис. 2.55. Сазан, или Карп, — Cyprinus carpio Linnaeus, 1758
Рис. 2.56. Породы карпа:
а — зеркальный; б — голый; в — чешуйчатый
274
Рис. 2.57. Золотой карась — Carassius carassius (Linnaeus, 1758)
Рис. 2.58. Серебряный карась — Carassius gibelio (Bloch, 1782)
275
Рис. 2.59. Породы золотой рыбки (Carassius auratus (Linnaeus, 1758)):
а — вуалехвост; б — оранда; в — шубункин; г — чёрный телескоп; д — золо-
тая рыбка
Рис. 2.60. Белый толстолобик — Hypophthalmichthys molitrix (Valenciennes, 1844)
276
Рис. 2.61. Пёстрый толстолобик — Hypophthalmichthys nobilis (Richardson, 1846)
Рис. 2.62. Чёрный буффало — Ictiobus niger (Rafinesque, 1820)
Рис. 2.63. Большеротый буффало — Ictiobus cyprinellus (Valenciennes, 1844)
277
Рис. 2.64. Малоротый буффало — Ictiobus bubalis (Rafinesque, 1819)
Рис. 2.65. Усатый, или обыкновенный, голец — Barbatula barbatula
(Linnaeus, 1758)
Рис. 2.66. Голец Крыницкого — Oxynoemacheilus merga (Krynicki, 1840)
278
Рис. 2.67. Обыкновенная щиповка — Cobitis taenia Linnaeus, 1758
Рис. 2.68. Сибирская щиповка — Cobitis melanoleuca Nichols, 1925
Рис. 2.69. Азовская щиповка — Cobitis tanaitica Băcescu & Maier, 1969
Рис. 2.70. Кубанская щиповка — Sabanejewia kubanica Vasil’eva & Vasil’ev,
1988
279
Рис. 2.71. Предкавказская щиповка — Sabanejewia caucasica (Berg, 1906)
Рис. 2.72. Вьюн — Misgurnus fossilis (Linnaeus, 1758)
Рис. 2.73. Обыкновенный сом — Silurus glanis Linnaeus, 1758
280
Рис. 2.74. Молодой африканский клариевый сом — Clarias gariepinus
(Burchell, 1822)
Рис. 2.75. Канальный сомик — Ictalurus punctatus (Rafinesque, 1818)
Рис. 2.76. Хольбрукская гамбузия — Gambusia holbrooki (Girard, 1859)
Рис. 2.77. Китайская медака — Oryzias sinensis Chen, Uwa & Chu, 1989
281
Рис. 2.78. Черноморский сарган — Belone belone euxini (Günther, 1866)
Рис. 2.79. Черноморская атерина — Atherina boyeri pontica Eichwald, 1831
Рис. 2.80. Обыкновенный налим — Lota lota (Linnaeus, 1758)
Рис. 2.81. Трёхиглая колюшка — Gasterosteus aculeatus Linnaeus, 1758
282
Рис. 2.82. Малая южная колюшка — Pungitius platygaster Kessler, 1859
Рис. 2.83. Змеевидная игла-рыба — Nerophis ophidion (Linnaeus, 1758)
Рис. 2.84. Черноморская игла-рыба — Syngnathus nigrolineatus Eichwald,
1831
Рис. 2.85. Сингиль — Liza aurata (Risso, 1810)
283
Рис. 2.86. Остронос — Liza saliens (Risso, 1810)
Рис. 2.87. Пиленгас — Liza haematocheilus (Temminck & Schlegel, 1845)
Рис. 2.88. Лобан — Mugil cephalus Linnaeus, 1758
284
Рис. 2.89. Мозамбикская тиляпия — Oreochromis mossambicus (Peters, 1852)
Рис. 2.90. Голубая тиляпия — Oreochromis aureus (Steindachner, 1864)
285
Рис. 2.91. Обыкновенный окунь — Perca fluviatilis Linnaeus, 1758
Рис. 2.92. Судак — Sander lucioperca (Linnaeus, 1758)
Рис. 2.93. Бёрш — Sander volgensis (Gmelin, 1789)
286
Рис. 2.94. Азовская перкарина — Percarina demidoffii Nordmann, 1840
Рис. 2.95. Обыкновенный ёрш — Gymnocephalus cernuus (Linnaeus, 1758)
Рис. 2.96. Донской ёрш — Gymnocephalus acerinus (Gmelin, 1774)
287
Рис. 2.97. Большеротый окунь, или форелеокунь — Micropterus salmoides
(Lacépède, 1802)
Рис. 2.98. Черноморская барабуля, или султанка, — Mullus barbatus ponticus
Essipov, 1927 (две цветовые вариации)
288
Рис. 2.99. Полосатый окунь, или лаврак, — Morone saxatilis (Walbaum, 1792)
Рис. 2.100. Леопардовый лысун [бычок-бубырь] — Pomatoschistus marmoratus
(Risso, 1810)
Рис. 2.101. Длиннохвостый бычок Книповича — Knipowitschia longicaudata
(Kessler, 1877)
289
Рис. 2.102. Бычок-бубырь — Knipowitschia caucasica (Berg, 1916)
Рис. 2.103. Бычок-травяник — Gobius ophiocephalus Pallas, 1814
Рис. 2.104. Бычок-кругляк — Neogobius melanostomus Pallas, 1814
290
Рис. 2.105. Речной бычок-песочник — Neogobius fluviatilis (Pallas, 1814)
Рис. 2.106. Бычок-цуцик, или мраморный тупоносый бычок — Proterorhinus
marmoratus (Pallas, 1814)
Рис. 2.107. Каспиосома — Caspiosoma caspium (Kessler, 1877)
291
Рис. 2.108. Бычок-гонец — Babka (Neogobius) gymnotrachelus (Kessler, 1857)
Рис. 2.109. Бычок-рыжик — Ponticola (Neogobius) eurycephalus (Kessler,
1874)
Рис. 2.110. Кавказский речной бычок — Ponticola (Neogobius) constructor
(Nordmann, 1840)
292
Рис. 2.111. Речной бычок Родиона — Ponticola (Neogobius) rhodioni
(Vasil’eva & Vasil’ev, 1994)
Рис. 2.112. Бычок сирман (ширман) — Ponticola (Neogobius) syrman
(Nordmann, 1840)
Рис. 2.113. Черноморская (звёздчатая) пуголовка — Benthophilus stellatus
(Sauvage, 1874)
293
Рис. 2.114. Азовская пуголовка — Benthophilus magistri Iljin, 1927
Рис. 2.115. Черноморская камбала, или глосса, — Platichthys flesus luscus
(Pallas, 1811)
294
Приложение 3
СХЕМА ИЗМЕРЕНИЙ ПРЕДСТАВИТЕЛЕЙ
СЕМЕЙСТВА МИНОГОВЫЕ (PETROMYZONTIDAE)
ab — вся длина;
gh — наибольшая высота тела;
lm — высота головы;
ae — расстояние от конца рыла до
первого жаберного отверстия;
an — длина рыла;
nр — диаметр глаза (горизонталь-
ный);
ре — промежуток между глазом и
жаберным аппаратом;
аА — расстояние от конца рыла до
ануса;
ik — диаметр ротового диска;
ао — длина головы с жаберным ап-
паратом;
aq — антедорсальное расстояние;
sq1
—
промежуток между ID и IID;
qs — длина основания ID;
tu — наибольшая высота ID;
q1s
1
—
длина основания IID;
t1u
1
—
наибольшая высота IID;
s1b — длина спинной части С;
Аb — расстояние от ануса до конца С.
Таблица промеров миноговых (Petromyzontidae)
Признак
Номер рыбы
Вид рыбы
Масса, г
Вся длина (ab или L), мм
Наибольшая высота тела (qh)
Наибольший обхват тела
Высота головы (lm)
Расстояние от конца рыла до пер-
вого жаберного отверстия (ae)
Длина жаберного аппарата (co)
Длина рыла (an)
Диаметр глаза (np)
Промежуток между глазом и жа-
берным аппаратом (pe)
295
Признак
Номер рыбы
Диаметр ротового диска (ik)
Длина головы с жаберным аппара-
том (ao)
Ширина лба
Антедорсальное расстояние (aq)
Расстояние от конца рыла до ануса
(aА)
Промежуток между ID и IID (sq1)
Длина основания ID (qs)
Наибольшая высота ID (tu)
Длина основания IID (q1s
1
)
Наибольшая высота IID (t1u
1
)
Длина спинной части С (s1b)
Расстояние от ануса до конца С
(Аb)
Зубов на верхнечелюстной пла-
стинке
Зубов на нижнечелюстной пла-
стинке
Зубов на язычной пластинке
Пол и стадия зрелости
296
Приложение 4
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА ОСЕТРОВЫЕ (ACIPENSERIDAE)
А — вид рыбы сбоку:
ab — длина всей рыбы;
aс — длина до конца средних лучей
С;
ad — длина до корней средних лучей
С;
od — длина туловища;
an — длина рыла;
nр — диаметр глаза (горизонталь-
ный);
ро — заглазничный отдел головы;
ас — длина головы;
lm — высота головы у затылка;
gh — наибольшая высота тела;
ik — наименьшая высота тела;
fd — длина хвостового стебля;
aq — антедорсальное расстояние;
аz — антевентральное расстояние;
ау — антеанальное расстояние;
qs — длина основания D;
tu — наибольшая высота D;
yy1
—
длина основания A;
ej — наибольшая высота А;
vх — длина Р;
zz1
—
длина V;
vz — расстояние между Р и V;
zу — расстояние между V и А.
Б — голова снизу:
1—2
—
расстояние от конца рыла до
хрящевого свода рта;
1—3
—
расстояние от конца рыла до
средних усиков;
4—5
—
длина наибольшего усика;
6—7
—
ширина рта.
Таблица промеров рыб семейства Осетровые (Acipenseridae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина всей рыбы (ab или L)
297
Признак
Номер рыбы
Длина рыбы до конца средних лу-
чей С (ac)
Длина до корней средних лучей С
(ad или l)
Длина туловища (od)
Спинных жучек
Боковых жучек
Брюшных жучек
Лучей в D
Лучей в А
Тычинок на первой жаберной дуге
Длина рыла (an)
Ширина рыла
Расстояние от конца рыла до губы
Расстояние от конца рыла до сред-
них усиков (1—3)
Длина наибольшего усика (4—5)
Диаметр глаза (горизонтальный)
(np)
Заглазничный отдел головы (po)
Длина головы (ao)
Высота головы у затылка (lm)
Наибольшая ширина головы
Ширина лба
Высота лба
Наибольшая высота тела (qh)
Наибольшая толщина тела
Наибольший обхват тела
Наименьшая высота тела (ik)
Длина хвостового стебля (fd)
Антедорсальное расстояние (aq)
Антевентральное расстояние (az)
Антеанальное расстояние (aу)
Длина основания D
298
Признак
Номер рыбы
Наибольшая высота D (tu)
Длина основания А (уу1)
Наибольшая высота А (ej)
Длина P (vx)
Длина V (zz1)
Расстояние между P и V (vz)
Расстояние между V и А (zу)
Пол и стадия зрелости
299
Приложение 5
СХЕМА ИЗМЕРЕНИЯ РЫБ
СЕМЕЙСТВА КАРПОВЫЕ (CYPRINIDAE)
аb — длина всей рыбы;
ас — длина по Смитту;
ad — длина без С;
оd — длина туловища;
an — длина рыла;
np — диаметра глаза (горизонтальный);
po — заглазничный отдел головы;
ao — длина головы;
1m — высота головы у затылка;
gh — наибольшая высота тела;
ik — наименьшая высота тела;
aq — антедорсальное расстояние;
rd — постдорсальное расстояние;
fd — длина хвостового стебля;
qs — длина основания D;
tu — наибольшая высота D;
yy1
—
длина основания A;
ej — наибольшая высота A;
vx — длина Р;
zz1
—
длина V;
vz — расстояние между Р и V;
zy — расстояние между V и А;
w — промер высоты лба;
w1
—
промер ширины лба.
Заштрихованные ряды поясняют просчёты чешуй в боковой линии (l.l. — linia
lateralis), над и под ней (формула боковой линии имеет примерно такой вид:
11
..
48 52,
8
ll=
где левое число обозначает наименьшее, а правое — наибольшее число чешуй,
над чертой — число чешуй над боковой линией, под чертой — число чешуй
под боковой линией).
300
Таблица промеров рыб семейства Карповые (Cyprinidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина всей рыбы (аb или L)
Длина рыбы по Смитту (ac)
Длина рыбы без С (ad или l)
Длина туловища (od)
Чешуй в боковой линии (l.l.)
Чешуй над боковой линией
Чешуй под боковой линией
Лучей в D
Лучей в А
Тычинок на первой жаберный дуге
Длина рыла (an)
Диаметр глаза (горизонтальный)
(nр)
Заглазничный отдел головы (ро)
Длина головы (ао)
Высота головы у затылка (lm)
Ширина лба
Наибольшая высота тела (gh)
Наименьшая высота тела (ik)
Антедорсальное расстояние (aq)
Постдорсальное расстояние (rd)
Длина хвостового стебля (fd)
Длина основания D (qs)
Наибольшая высота D (tu)
Длина основания А (yy1)
Наибольшая высота A (ej)
Длина Р (vx)
Длина V (zz1)
Расстояние между Р и V (vz)
Расстояние между V и A (zy)
Формула глоточных зубов
Позвонков (туловищных и хвосто-
вых)
Возраст
Пол и стадия зрелости
301
Таблица признаков рыб семейства Карповые (Cyprinidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина рыбы без С (ad или l)
Чешуй в боковой линии
Лучей в D
Лучей в А
Тычинок на первой жаберной дуге
Позвонков (туловищных и хвосто-
вых)
Возраст
Пол и стадия зрелости
В % длины тела
Длина рыла
Диаметр глаза
Заглазничный отдел головы
Длина головы
Высота головы у затылка
Ширина лба
Наибольшая высота тела
Наименьшая высота тела
Антедорсальное расстояние
Постдорсальное расстояние
Длина хвостового стебля
Длина основания D
Наибольшая высота D
Длина основания А
Наибольшая высота А
Длина P
Длина V
Расстояние между P и V
Расстояние между V и А
В % длины головы
Длина рыла
Диаметр глаза
Ширина лба
302
Приложение 6
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА СЕЛЬДЕВЫЕ (CLUPEIDAE)
А — вид рыбы сбоку:
аb — длина всей рыбы;
ас — длина до конца средних лучей
С;
ad — длина без С;
an — длина рыла;
n1р
1
—
диаметр глаза (вертикальный);
ро — заглазничный отдел головы;
aо — длина головы;
lm — высота головы у затылка;
w1
—
ширина лба (измеряется как у
карповых);
a1a
6
—
длина верхнечелюстной кости;
al1
—
длина нижней челюсти;
qh — наибольшая высота тела;
ik — наименьшая высота тела;
aq — антедорсальное расстояние;
rd — постдорсальное расстояние;
аz — антевентральное расстояние;
ау — антеанальное расстояние;
fd — длина хвостового стебля;
qs — длина основания D;
tu — наибольшая высота D;
уу1
—
длина основания A;
ej — наибольшая высота A;
vx — длина P;
zz1
—
длина V;
vz — расстояние между Р и V;
zy — расстояние между V и A;
dc — длина средних лучей С.
Б — жаберная дужка: верхний ряд —
тычинки, нижний — лепестки.
Таблица промеров рыб семейства Сельдевые (Clupeidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина всей рыбы (ab или L)
Длина рыбы до конца средних лу-
чей С (ac)
303
Признак
Номер рыбы
Длина рыбы без С (ad или l)
Поперечных рядов чешуй
Лучей в D
Лучей в А
Тычинок на первой жаберной дуге
Пилорических придатков
Позвонков (отдельно туловищных
и хвостовых)
Брюшных шипиков впереди ануса
Брюшных шипиков позади ануса
Зубов на верхней челюсти
Зубов на нижней челюсти
Длина рыла (an)
Диаметр глаза (вертикальный)
(n1p
1
)
Заглазничный отдел головы (po)
Длина головы (ao)
Высота головы у затылка (lm)
Ширина лба
Длина нижней челюсти (a1l
1
)
Наибольшая высота тела (qh)
Наименьшая высота тела (ik)
Антедорсальное расстояние (aq)
Постдорсальное расстояние (rd)
Антевентральное расстояние (az)
Антеанальное расстояние (aу)
Длина хвостового стебля (fd)
Длина основания D (qs)
Наибольшая высота D (tu)
Длина основания А (уу1)
Наибольшая высота А (ej)
Длина P (vx)
Длина V (zz1)
Расстояние между P и V (vz)
Расстояние между V и А (zу)
Длина средних лучей С (dc)
Возраст
Пол и стадия зрелости
304
Приложение 7
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА ЛОСОСЁВЫЕ (SALMONIDAE)
аb — длина всей рыбы;
ас — длина по Смитту;
ad — длина без С;
od — длина туловища;
an — длина рыла;
nр — диаметр глаза (горизонталь-
ный);
аа5
—
длина средней части головы;
ао — длина головы;
ро — заглазничный отдел головы;
lm — высота головы у затылка;
w1
—
ширина лба (как у карповых);
aa6
—
длина верхнечелюстной кости;
k1l
1
—
длина нижней челюсти;
qh — наибольшая высота тела;
ik — наименьшая высота тела;
аq — антедорсальное расстояние;
rd — постдорсальное расстояние;
аz — антевентральное расстояние;
ау — антеанальное расстояние;
fd — длина хвостового стебля;
qs — длина основания D;
tu — наибольшая высота D;
уу1
—
длина основания A;
ej — наибольшая высота А;
vx — длина Р;
zz1
—
длина V;
vz — расстояние между Р и V;
zу — расстояние между V и А.
Таблица промеров рыб семейства Лососёвые (Salmonidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина всей рыбы (ab или L)
Длина тела по Смитту (ac)
Длина тела без С (ad или l)
Длина туловища (od)
305
Признак
Номер рыбы
Лучей в D
Лучей в А
Тычинок на первой жаберной дуге
Длина наибольшей жаберной ты-
чинки
Длина жаберной дуги
Пилорических придатков
Позвонков (отдельно туловищных
и хвостовых)
Длина рыла (an)
Диаметр глаза (горизонтальный)
(np)
Заглазничный отдел головы (po)
Длина средней части головы (aa5)
Длина головы (ao)
Высота головы у затылка (lm)
Ширина лба
Длина верхнечелюстной кости
(aa6)
Ширина верхнечелюстной кости
(nm1)
Длина нижней челюсти (k1l
1
)
Наибольшая высота тела (qh)
Наименьшая высота тела (ik)
Антедорсальное расстояние (aq)
Постдорсальное расстояние (rd)
Антевентральное расстояние (az)
Антеанальное расстояние (aу)
Длина хвостового стебля (fd)
Длина основания D (qs)
Наибольшая высота D (tu)
Длина основания А (уу1)
Наибольшая высота А (ej)
Длина P (vx)
Длина V (zz1)
Расстояние между P и V (vz)
Расстояние между V и А (zу)
Возраст
Пол и стадия зрелости
306
Приложение 8
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА ОКУНЁВЫХ (PERCIDAE)
ab — длина всей рыбы;
ас — длина по Смитту;
ad — длина без С;
od — длина туловища;
an — длина рыла;
nр — диаметр глаза (горизонтальный);
ро — заглазничный отдел головы;
ао — длина головы;
w1
—
ширина лба (как у карповых);
lm — высота головы у затылка;
a1a
2
—
длина верхнечелюстной кости;
n1m
1
—
ширина верхнечелюстной кости;
k1l
1
—
длина нижней челюсти;
gh — наибольшая высота тела;
ik — наименьшая высота тела;
aq — антедорсальное расстояние;
rd — постдорсальное расстояние;
az — антевентральное расстояние;
ay — антеанальное расстояние;
fd — длина хвостового стебля;
qs — длина основания ID;
q1s
1
—
длина основания IID;
tu — наибольшая высота ID;
t1u
1
—
наибольшая высота IID;
yy1
—
длина основания A;
ej — наибольшая высота A;
vx — длина Р;
vx1
—
ширина основания Р;
zz1
—
длина V;
vy — расстояние между Р и А;
zу — расстояние между V и А;
Aу — расстояние между анусом и А.
Таблица промеров рыб семейства Окунёвые (Percidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
307
Признак
Номер рыбы
Длина всей рыбы (ab)
Длина рыбы без С (ad)
Чешуй в боковой линии (l.l.)
Лучей в ID
Лучей в IID
Лучей в А
Тычинок на первой жаберной дуге
Длина первой жаберной дуги
Длина наибольшей жаберной ты-
чинки
Длина головы (ao)
Высота головы (lm)
Длина рыла (an)
Диаметр глаза (горизонтальный)
(np)
Заглазничный отдел головы (po)
Длина верхнечелюстной кости (al)
Длина нижнечелюстной кости (k1l1)
Ширина лба
Наибольшая высота тела (qh)
Наименьшая высота тела (ik)
Длина хвостового стебля (fd)
Антедорсальное расстояние (aq)
Расстояние от ануса до А (Ау)
Длина основания ID (qs)
Длина основания IID (q1s
1
)
Расстояние между ID и IID
Высота ID (tu)
Высота IID (t1u
1
)
Длина P (vx)
Ширина P (vx1)
Длина V (zz1)
Длина основания А (уу1)
Высота А (ej)
Хвостовая выемка (cb)
Позвонков (туловищных и хвостовых)
Возраст
Пол и стадия зрелости
308
Таблица признаков рыб семейства Окунёвые (Percidae)
Признак
Номер рыбы
Вид рыбы
Масса рыбы, г
Длина тела
Длина головы
Пол и стадия зрелости
В % длины тела
Длина головы
Высота головы у затылка
Наибольшая высота тела
Наименьшая высота тела
Длина хвостового стебля
Антедорсальное расстояние
Постдорсальное расстояние
Расстояние от ануса до А
Длина основания ID
Длина основания IID
Промежуток между ID и IID
Высота ID
Высота IID
Длина основания А
Высота А
В % длины головы
Длина рыла
Диаметр глаза горизонтальный
Заглазничный отдел головы
Длина верхнечелюстной кости
Длина нижнечелюстной кости
Ширина лба
Длина головы в % наибольшей вы-
соты тела
Длина жаберной тычинки в % дли-
ны жаберной дуги
Ширина основания P в % длины P
Хвостовая выемка
309
Приложение 9
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА ТРЕСКОВЫЕ (GADIDAE)
аb — длина всей рыбы;
ad — длина без С;
o1d — длина туловища;
an — длина рыла;
nр — диаметр глаза (горизонталь-
ный);
ро — заглазничный отдел головы;
ао — длина головы;
lm — высота головы у затылка;
aa1
—
длина верхней челюсти;
k1l
1
—
длина нижней челюсти;
qh — наибольшая высота тела;
ik — наименьшая высота тела;
aq — антедорсальное расстояние;
ау — антеанальное расстояние;
fd — длина хвостового стебля;
qs — длина основания ID;
q1s
1
—
длина основания IID;
q2s
2
—
длина основания IIID;
tu — наибольшая высота ID;
yy1
—
длина основания IA;
у2у
3
—
длина основания IIA;
ej — наибольшая высота IА;
vx — длина Р;
zz1
—длина V;
zу — расстояние между V и IА.
310
Приложение 9
СХЕМА ИЗМЕРЕНИЙ РЫБ
СЕМЕЙСТВА КАМБАЛОВЫЕ (PLEURONECTIDAE)
ab — длина всей рыбы;
ас — длина до конца средних лучей
С,
ad — длина без С;
an — длина рыла;
nр — диаметр глаза (горизонтальный);
р1о — заглазничный отдел головы;
ао — длина головы;
lm — высота головы;
аа1
—
длина верхней челюсти;
k1l
1
—
длина нижней челюсти;
gh — наибольшая высота тела;
ik — наименьшая высота тела;
aq — антедорсальное расстояние;
аz — антевентральное расстояние;
ау — антеанальное расстояние;
fd — длина хвостового стебля;
tu — наибольшая высота D;
ej — наибольшая высота А;
vx — длина Р;
zz1
—
длина V;
zy — расстояние между V и A;
dc — длина средних лучей С.
Измеряется также ширина лба.
311
Предисловие..................................................
Краткая физико-географическая характеристика бассейна реки Ку-
бани......................................................
Правиланаучнойсистематики................................
Какпользоватьсяопределителем..............................
Материалыиинструментарий................................
Объяснение основных признаков, промеров и терминов, употреб-
ляемыхвопределителе........................................
Таблицыдляопределениярыбообразныхирыб.................
Отряд Миногообразные — Petromyzontiformes Bonaparte,
1832......................................................
Семейство Миноговые — Petromyzontidae Bonaparte, 1831 . . . . . . .
Класс Лучепёрые рыбы — Actinopterygii Klein, 1885 . . . . . . . . . . . .
Отряд Осетрообразные — Acipenseriformes Berg, 1940 . . . . . . .
Семейство Осетровые — Acipenseridae Bonaparte, 1831 . . . . . . . . . .
Семейство Веслоносые — Polyodontidae Bonaparte, 1838 . . . . . . . .
Отряд Сельдеобразные — Clupeiformes Bleeker, 1959 . . . . . . .
СемействоСельдевые—ClupeidaeCuvier,1816.................
Семейство Анчоусовые — Engraulidae Gill, 1861 [Engraulididae] . . .
Отряд Лососеобразные — Salmoniformes Bleeker, 1859 . . . . . .
СемействоЛососёвые—SalmonidaeCuvier,1816...............
Отряд Щукообразные — Esociformes Bleeker, 1859 . . . . . . . . . .
СемействоЩуковые—EsocidaeCuvier,1816....................
Отряд Угреобразные — Anguilliformes Goodrich, 1909 . . . . . . . .
Семейство Угрёвые, или Пресноводные угри, — Anguillidae
Rafinesque,1810...........................................
Отряд Карпообразные — Cypriniformes Rafinesque, 1810 . . . . .
СемействоКарповые—CyprinidaeRafinesque,1810..............
СемействоЧукучановые—CatostomidaeCope,1871..............
Семейство Гольцовые — Nemacheilidae Regan, 1911 . . . . . . . . . . . . .
СемействоВьюновые—CobitidaeSwainson,1838...............
Отряд Сомообразные — Siluriformes Cuvier, 1816 . . . . . . . . . . . .
СемействоСомовые—SiluridaeCuvier,1816....................
СемействоКлариевые—ClariidaeBonaparte,1846...............
Семейство Икталуровые, или Кошачьи сомы, — Ictaluridae Gill,
1861......................................................
3
5
9
13
15
16
36
36
36
39
41
41
55
58
59
67
68
69
73
74
75
75
77
78
142
147
149
158
158
160
162
ОГЛАВЛЕНИЕ
312
Отряд Карпозубообразные — Cyprinodontiformes Berg, 1940
Семейство Гамбузиевые, или Пецилиевые, — Poeciliidae Garman,
1895......................................................
Отряд Сарганообразные — Beloniformes Berg, 1940 . . . . . . . . . .
Семейство Адрианихтовые — Adrianichthyidae Weber, 1913 . . . . . . .
Семейство Саргановые — Belonidae Bonaparte, 1832 . . . . . . . . . . . . .
Отряд Атеринообразные — Atheriniformes Rosen, 1966 . . . . . . .
Семейство Атериновые — Atherinidae Günther, 1961 . . . . . . . . . . . .
Отряд Трескообразные — Gadiformes Goodrich, 1909 . . . . . . . .
СемействоНалимовые—LotidaeBonaparte,1832...............
Отряд Колюшкообразные — Gasterosteiformes Greenwood,
1966......................................................
Семейство Колюшковые — Gasterosteidae Bonaparte, 1831 . . . . . . .
Отряд Иглообразные — Syngnathiformes Helfman, Collette
&Facey,1997...............................................
Семейство Игловые — Syngnathidae Rafinesque, 1810 . . . . . . . . . . . .
Отряд Кефалеобразные — Mugiliformes Günther, 1880 . . . . . . .
СемействоКефалевые—MugilidaeCuvier,1829.................
Отряд Цихлидообразные — Cichliformes Betancur-R et al.,
2013......................................................
Семейство Цихловые, или Цихлиды — Cichlidae Bonaparte, 1835 . .
Отряд Окунеобразные — Perciformes Bleeker, 1859 . . . . . . . . . .
СемействоОкунёвые—PercidaeRafinesque,1815................
Семейство Центрарховые, или Ушастые окуни, — Centrarchidae
Bleeker,1859..................................................
Семейство Барабулевые, или Султанковые, — Mullidae Rafinesque,
1815......................................................
Семейство Лавраковые, или Перцихтиевые, — Percichthyidae
Jordan&Eigenmann,1890...................................
Отряд Бычкообразные — Gobiiformes Günther, 1880 . . . . . . . .
СемействоБычковые—GobiidaeCuvier,1816..................
Отряд Камбалообразные — Pleuronectiformes Bleeker, 1859
Семейство Камбаловые — Pleuronectidae Cuvier, 1816 . . . . . . . . . . .
Списокиспользованнойлитературы............................
Указательрусскихназванийрыб..............................
Указательлатинскихназванийрыб..............................
Приложения..................................................
163
163
165
166
167
168
168
170
170
171
171
174
174
177
177
183
184
187
188
197
199
200
202
202
220
220
223
235
242
247
313
Научное издание
Москул Георгий Алексеевич
РЫБЫ ВОДОЁМОВ БАССЕЙНА КУБАНИ
Определитель
Второе издание, исправленное и дополненное
Подписано в печать 15.03.21. Выход в свет 30.04.21
Формат 60×84 1/
16
. Гарнитура «Таймс». Печать цифровая.
Уч.-изд. л. 19,5. Тираж 500 экз. Заказ No
.
Кубанский государственный университет
350040, г. Краснодар, ул. Ставропольская, 149.
Издательско-полиграфический центр КубГУ
350040, г. Краснодар, ул. Ставропольская, 149.