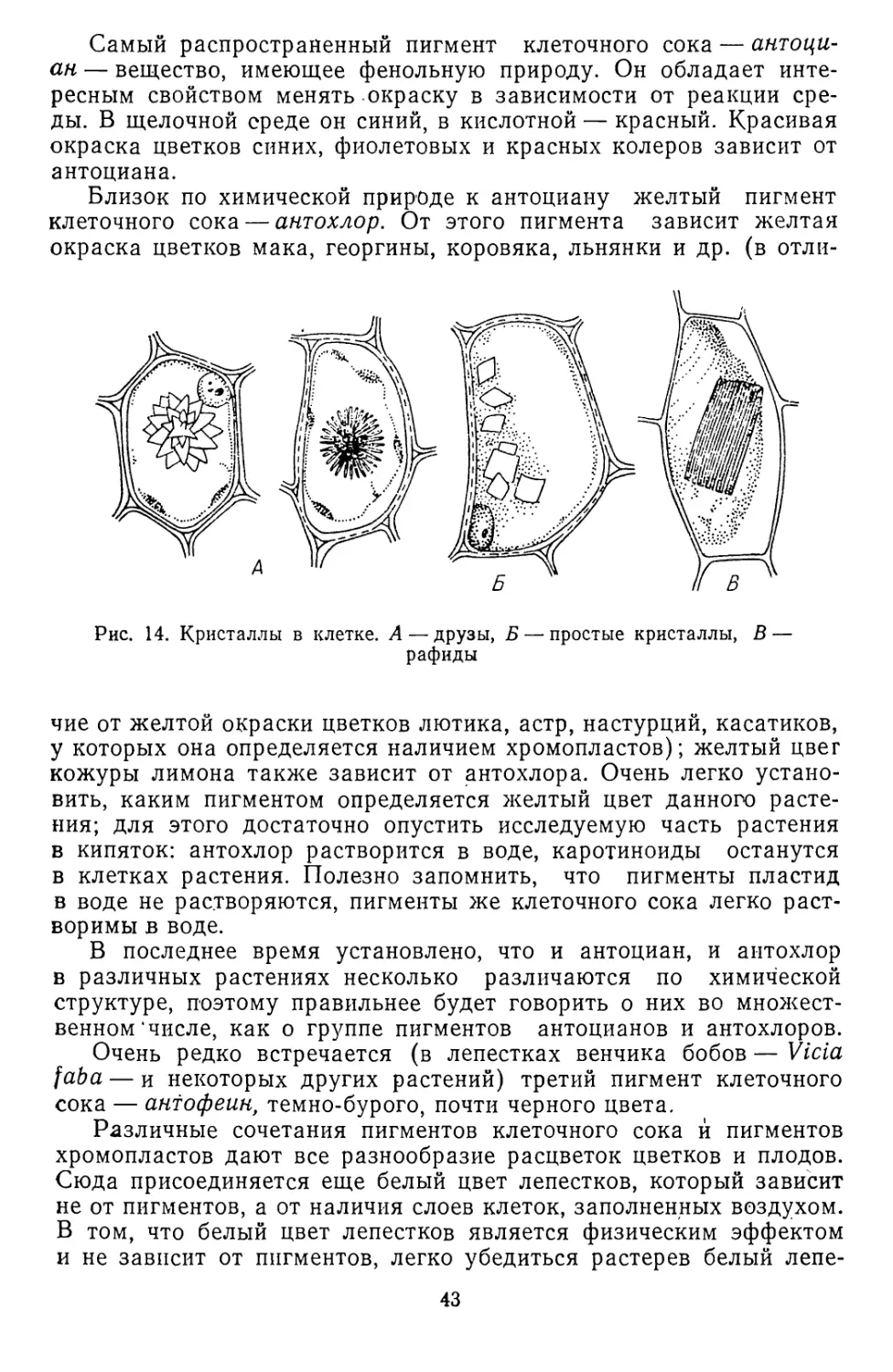
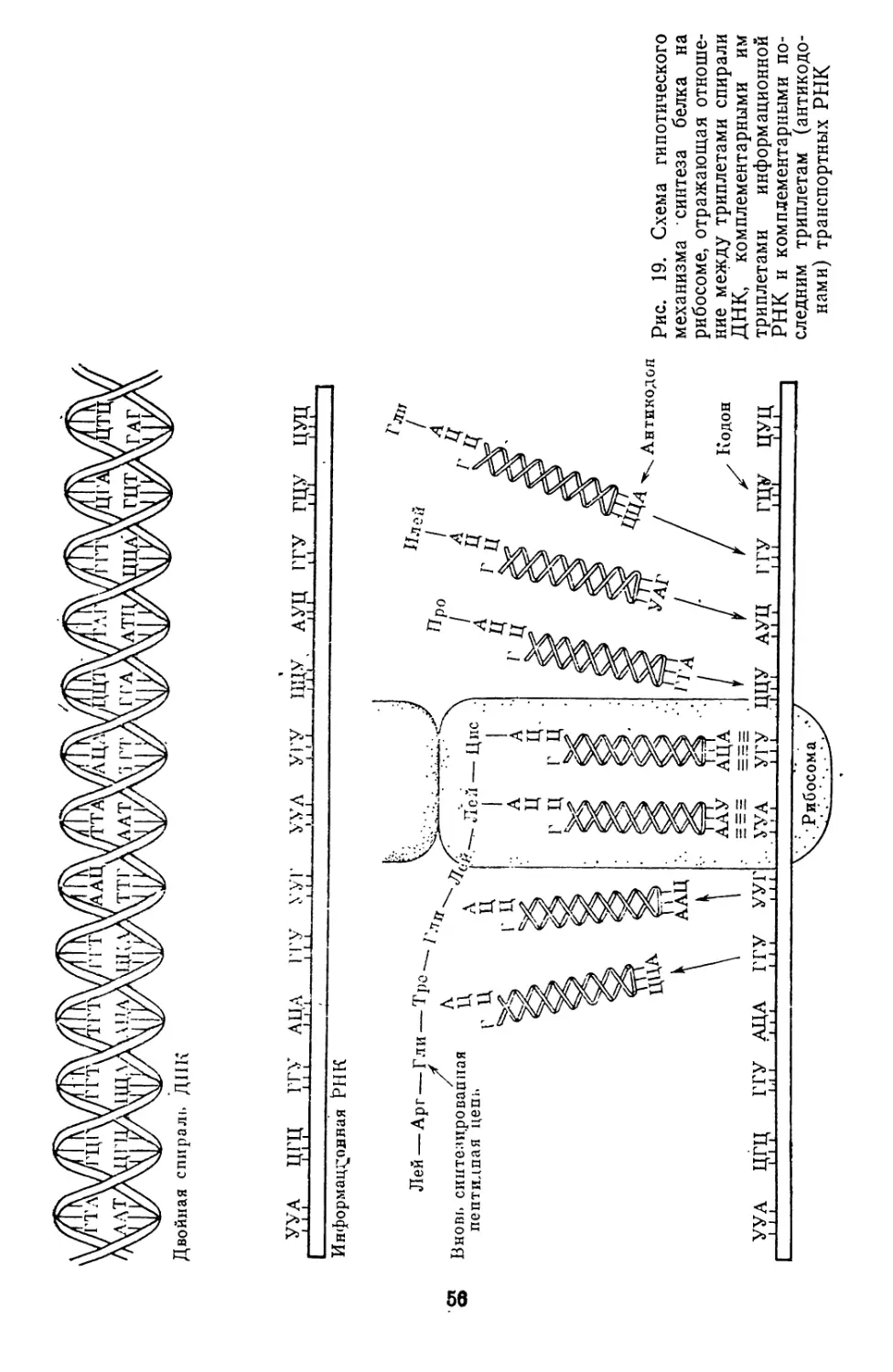
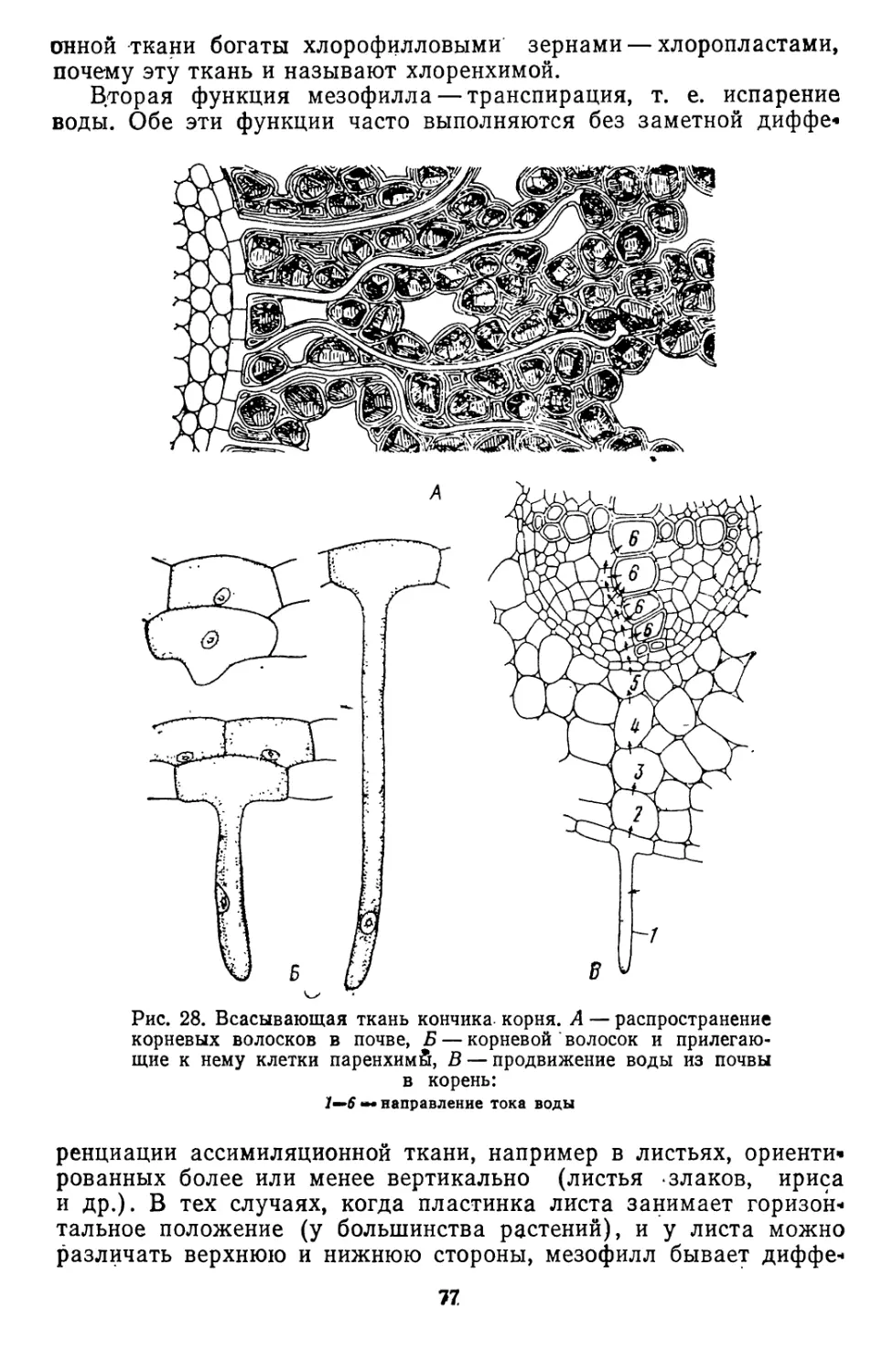
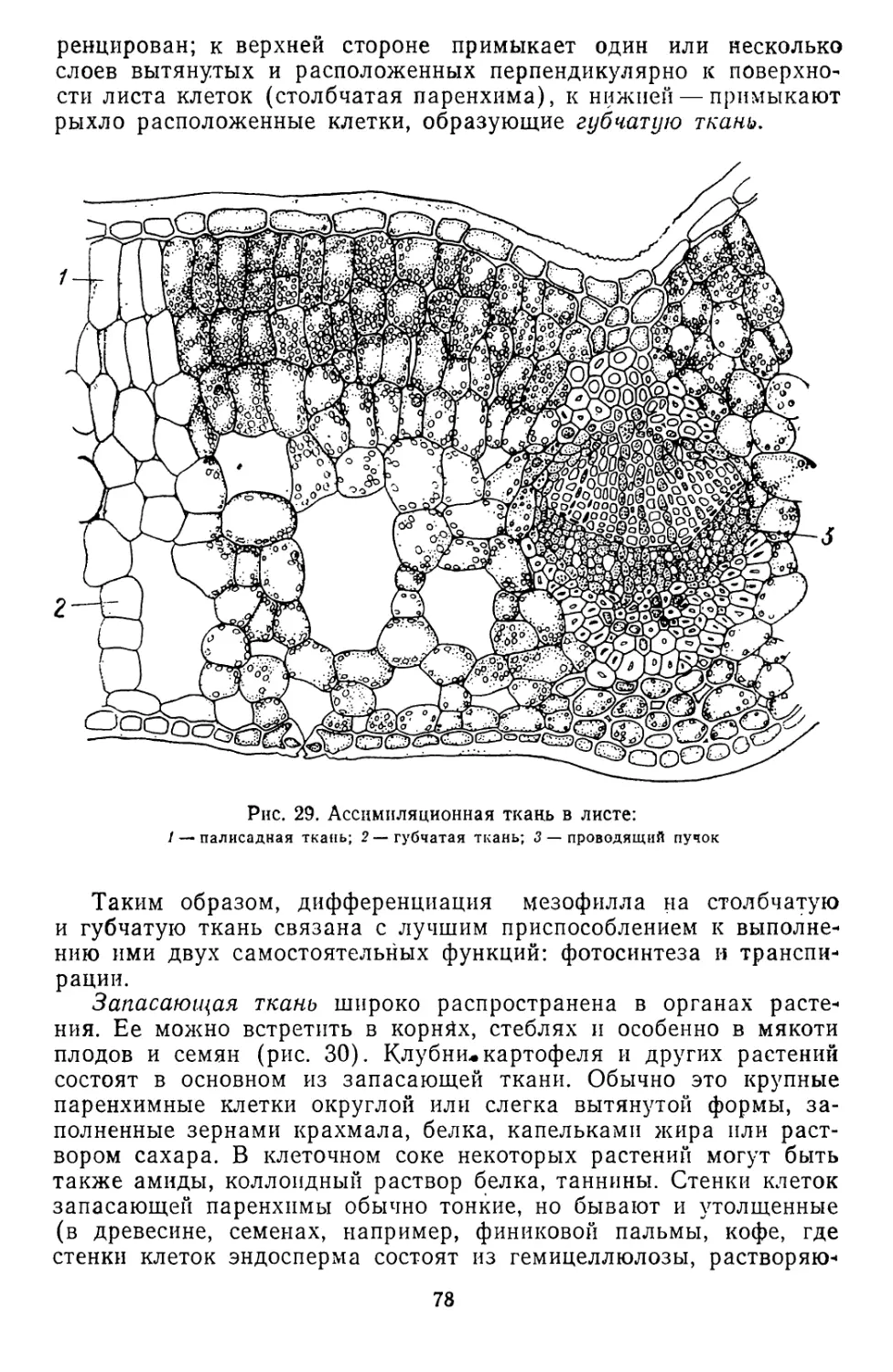
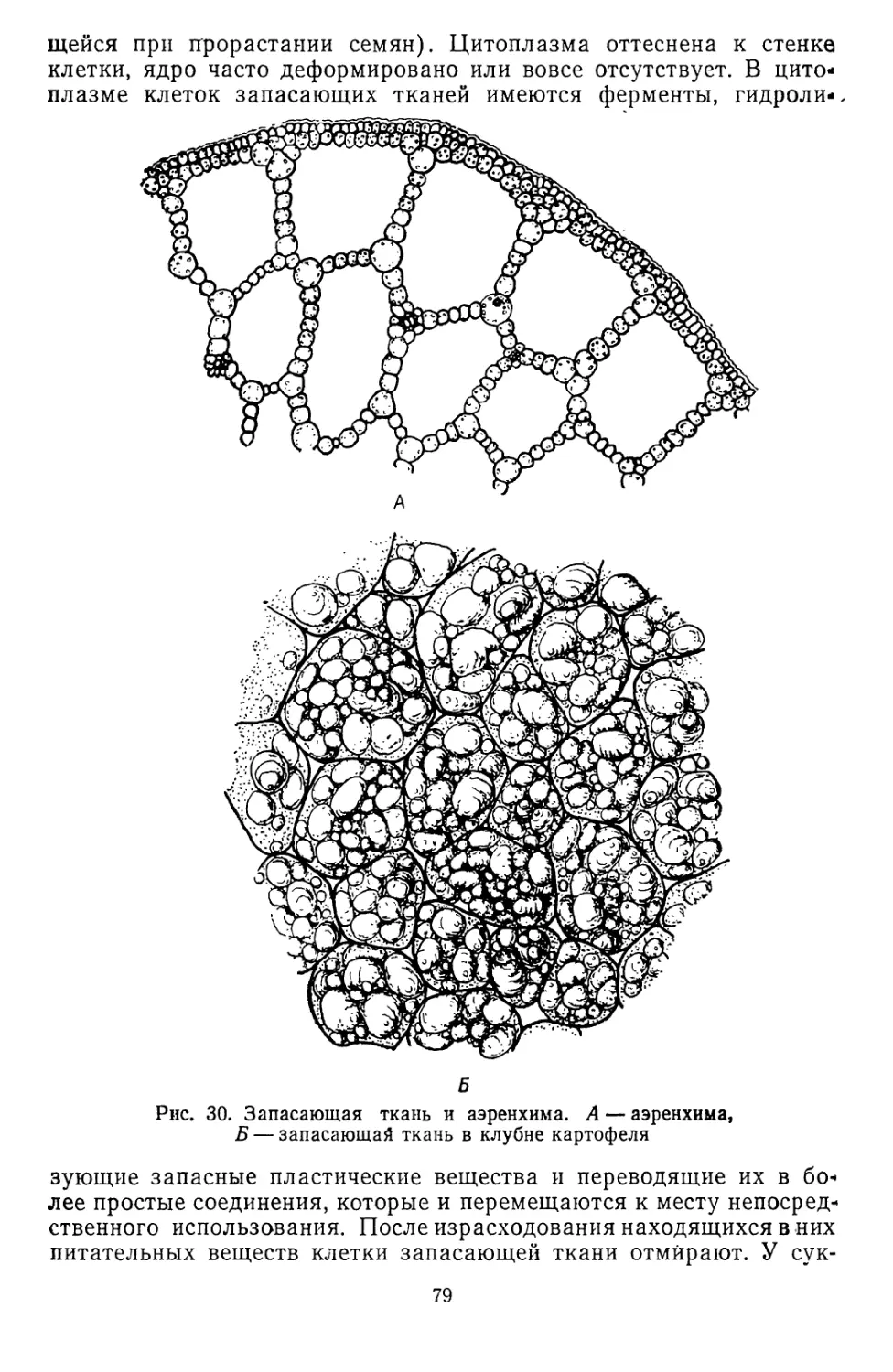
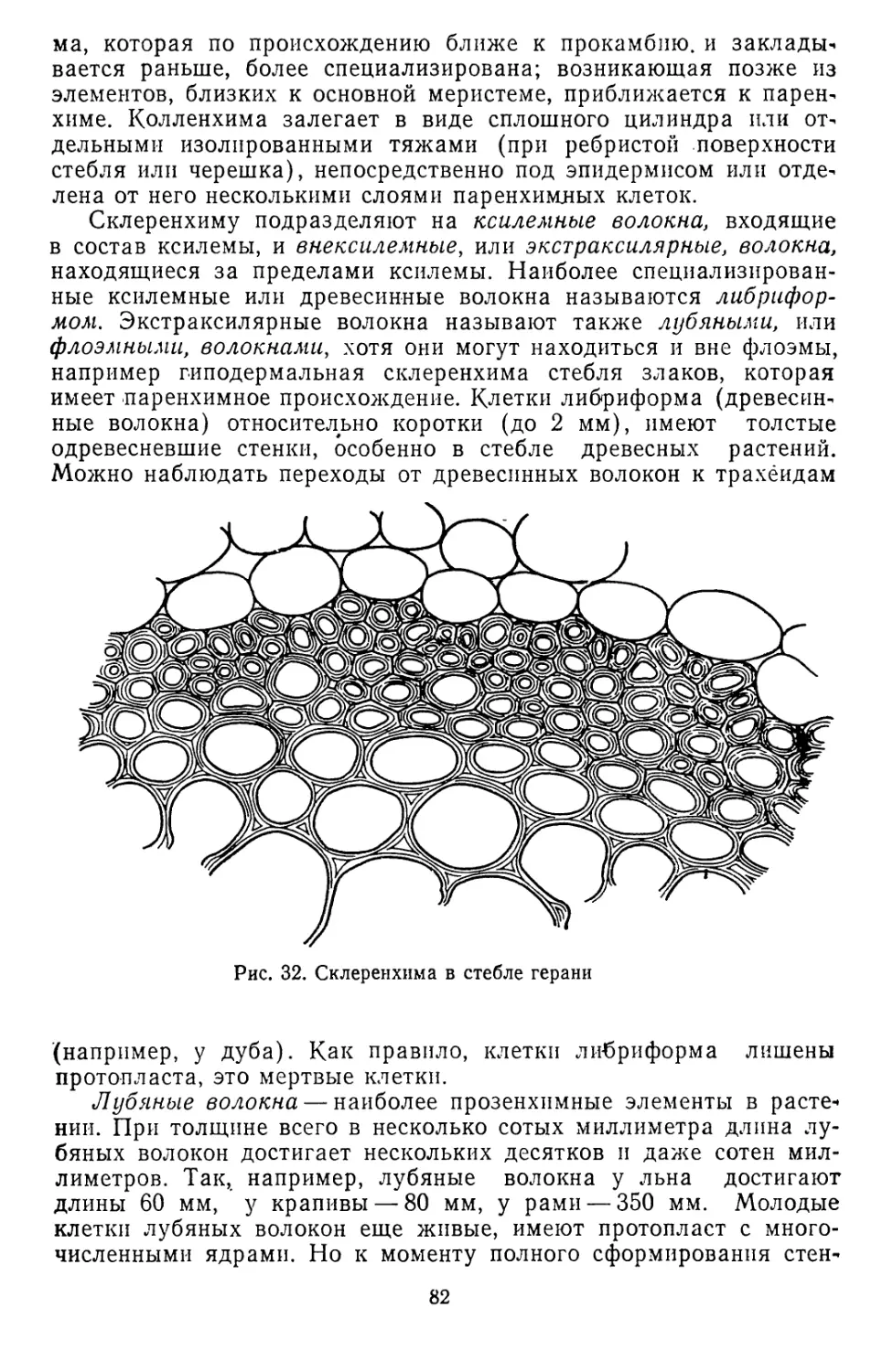
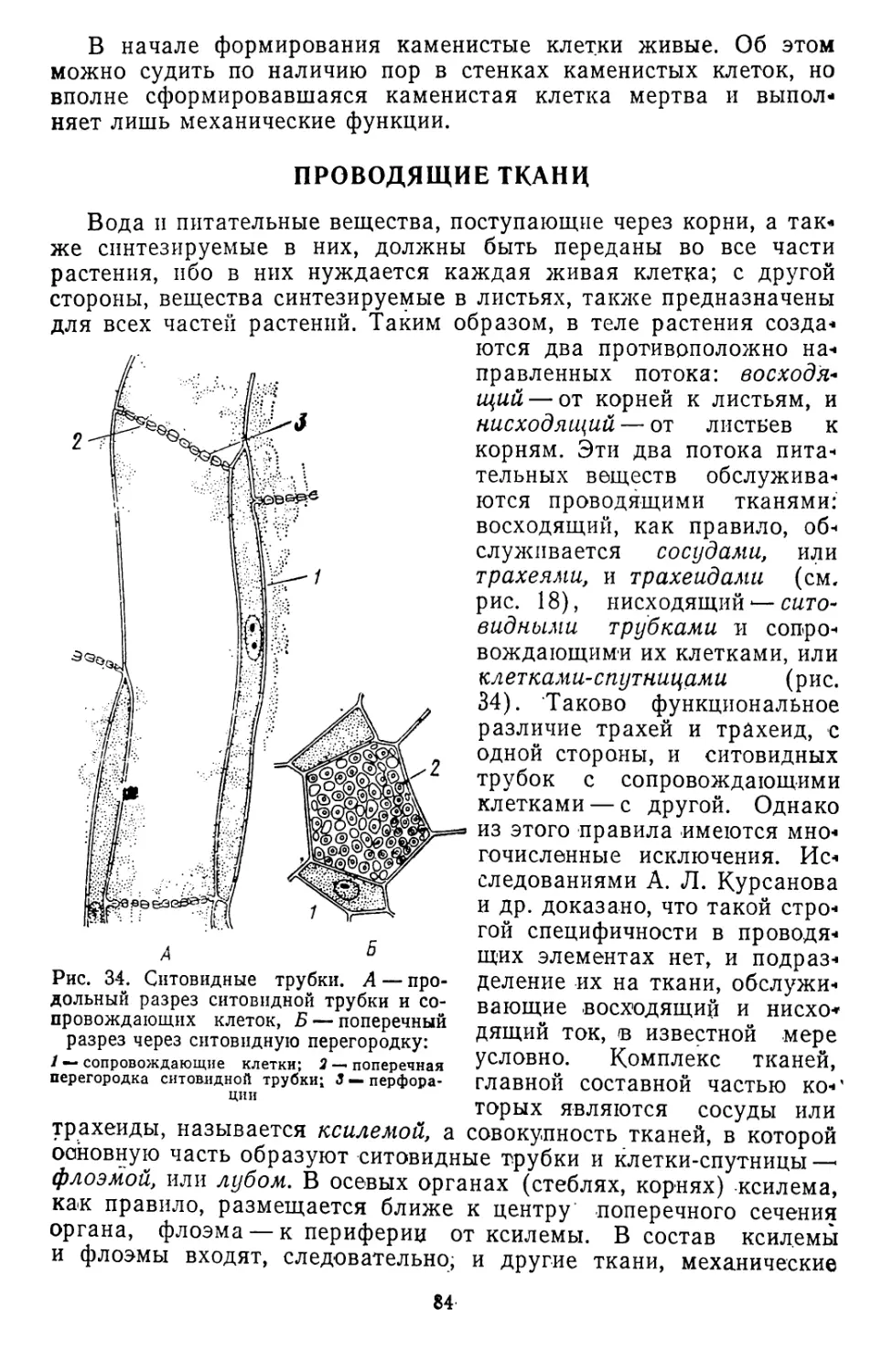
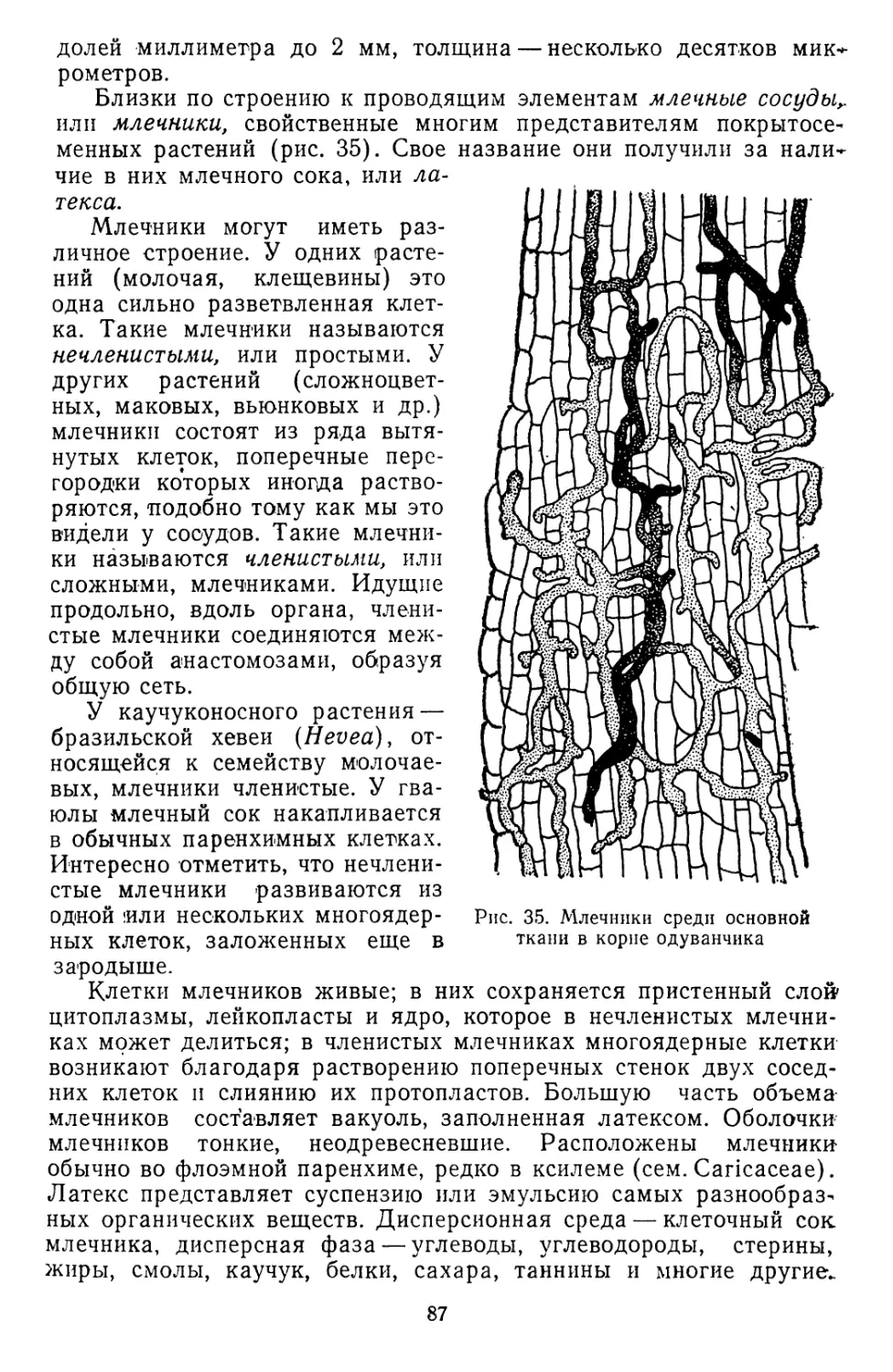
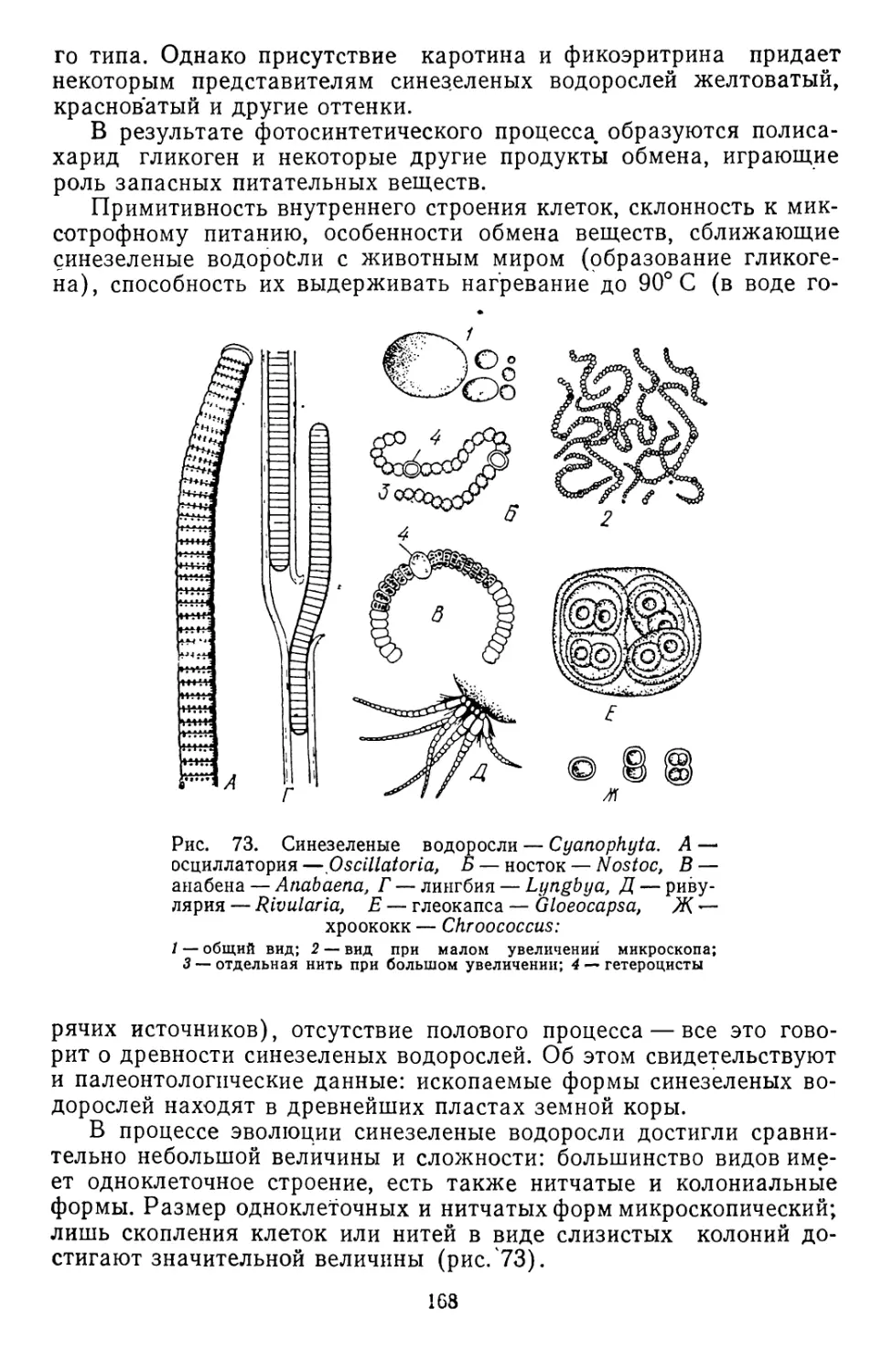
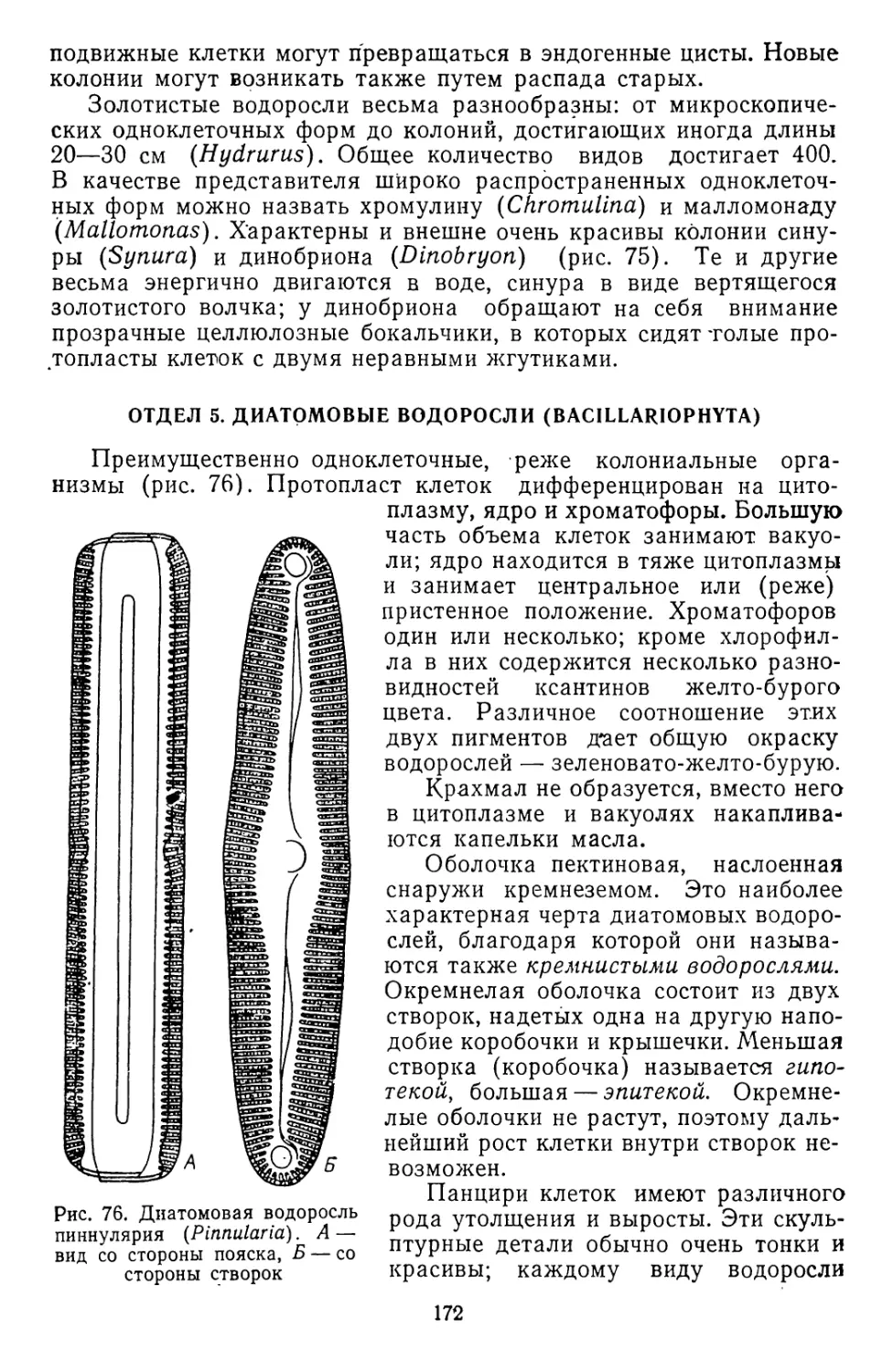
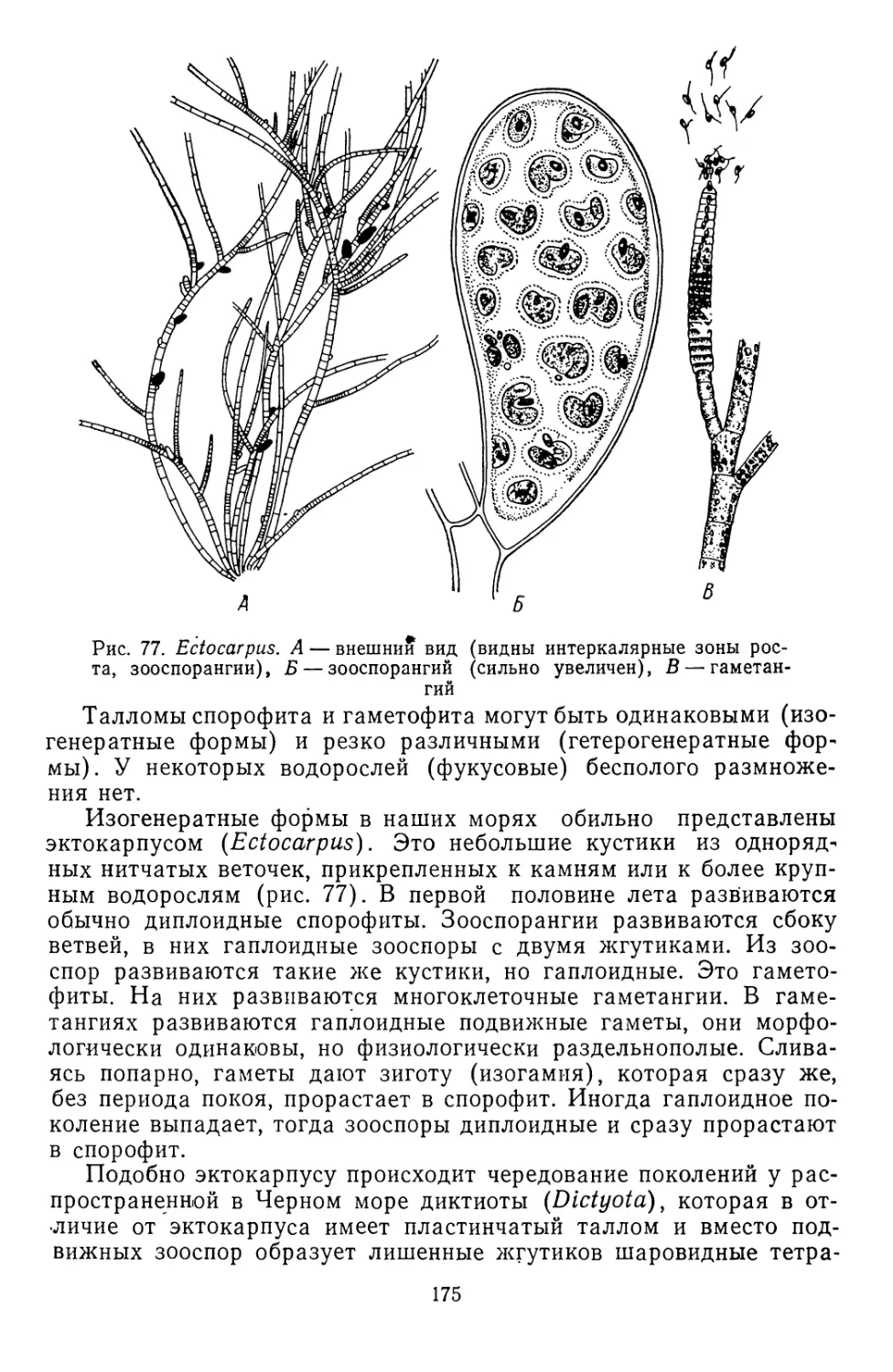
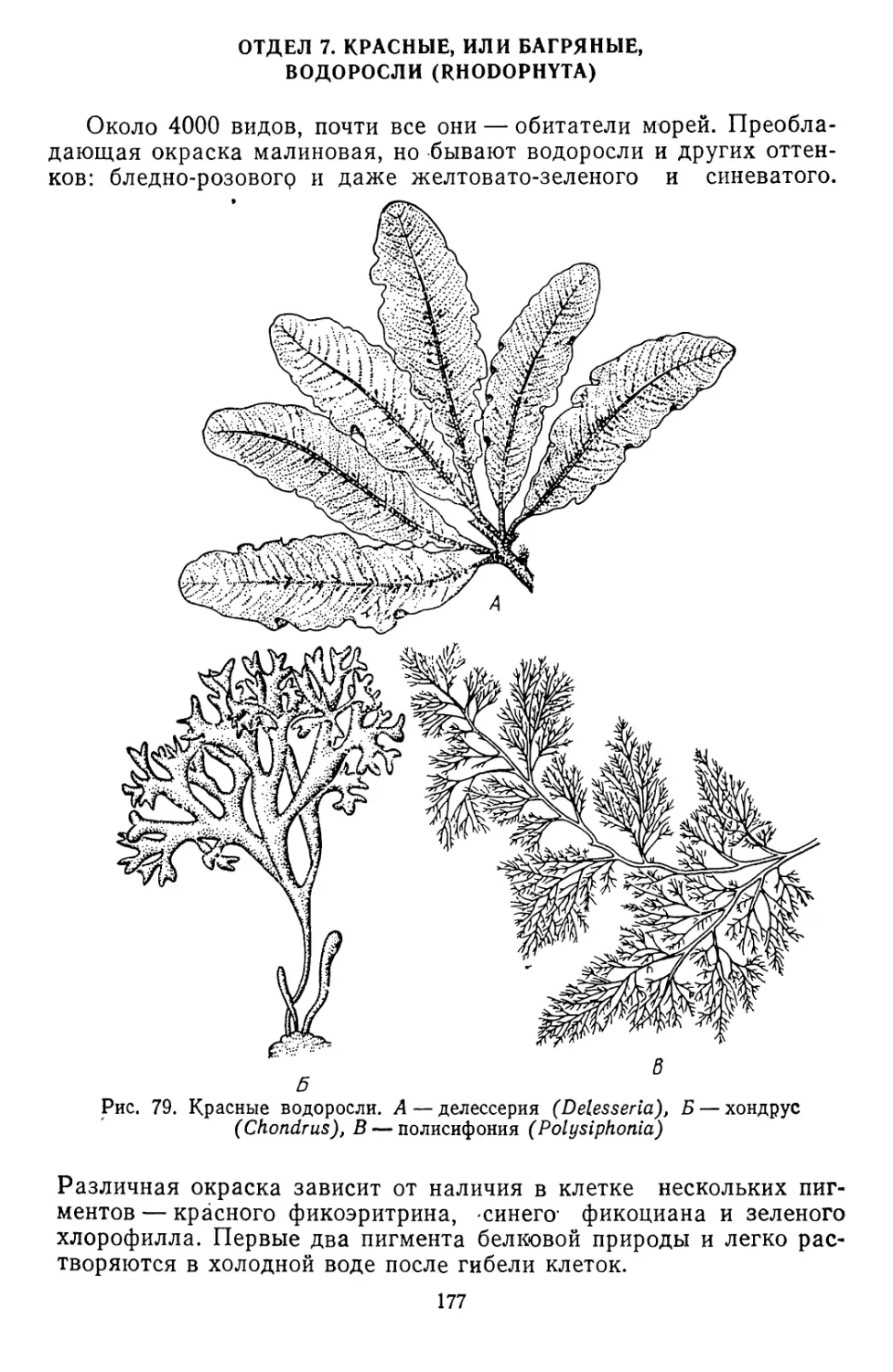
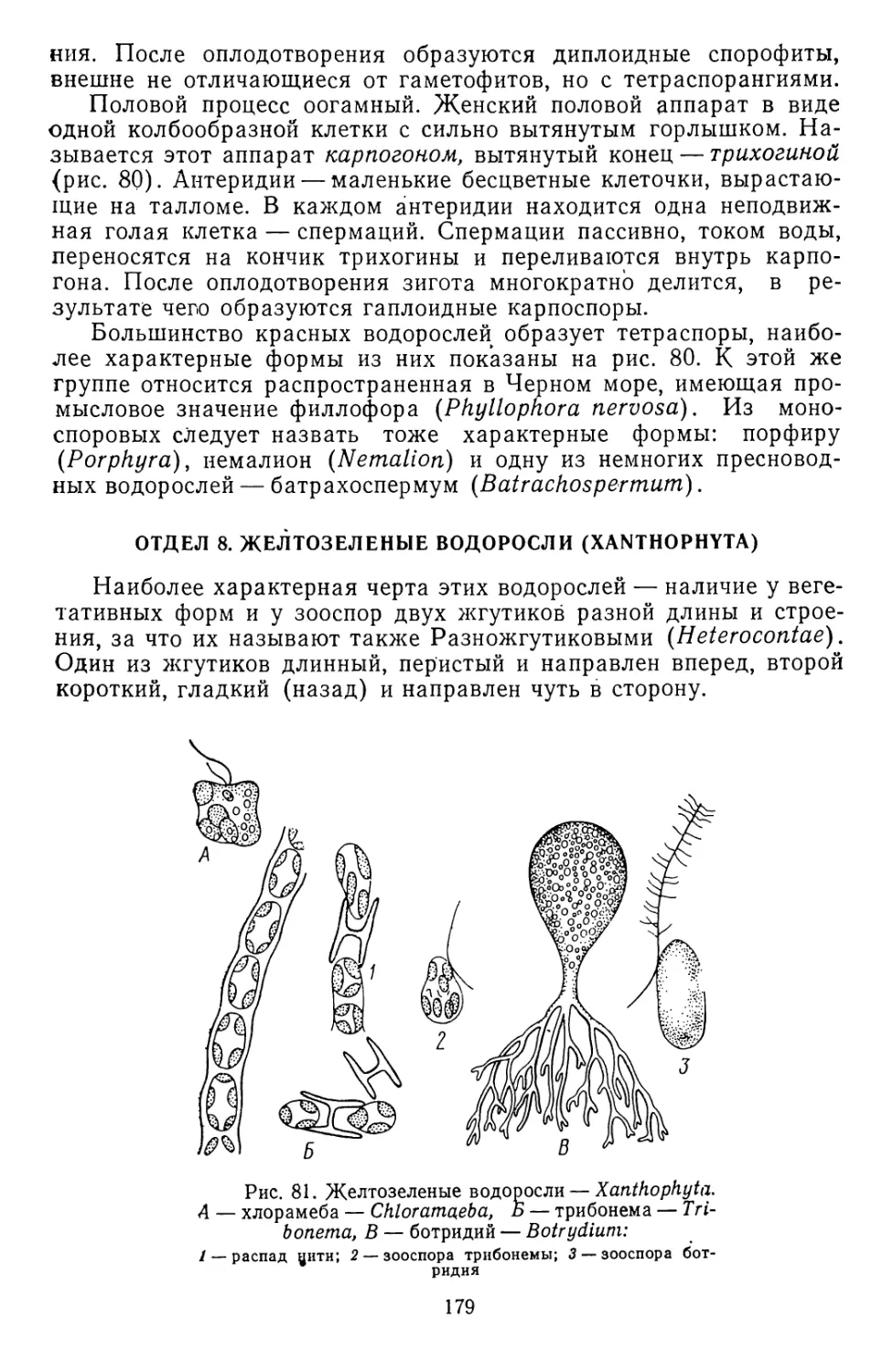
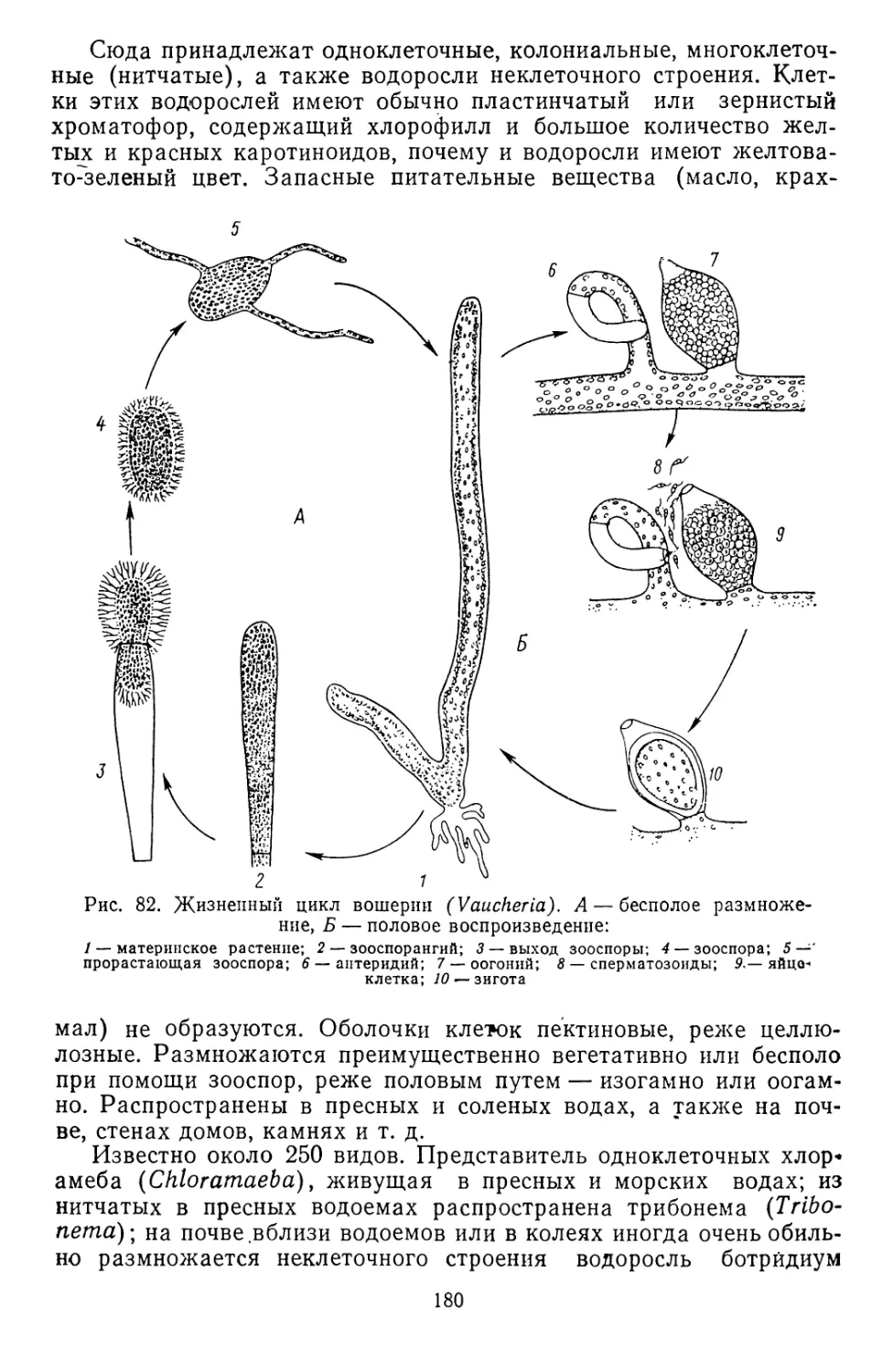
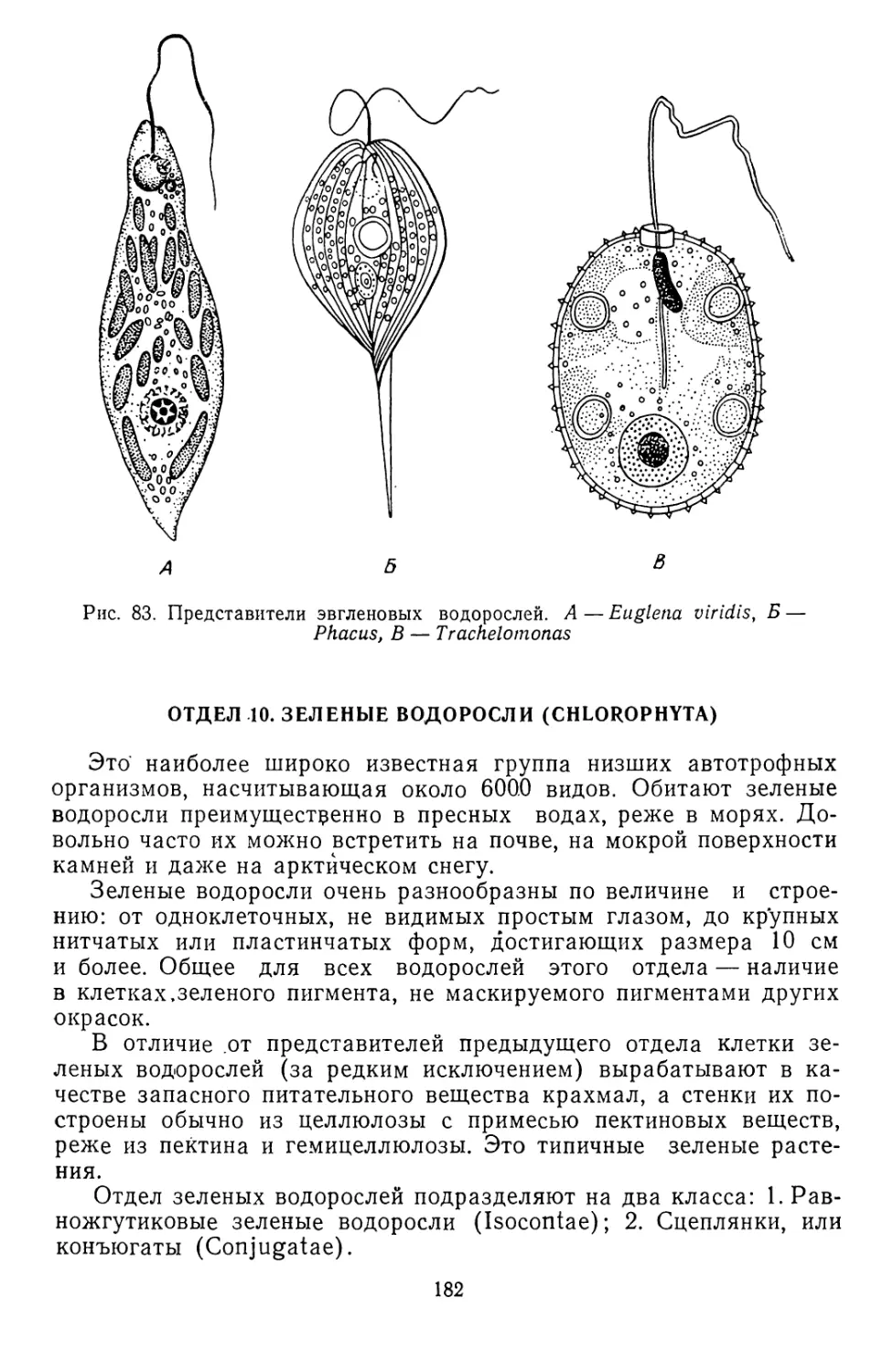
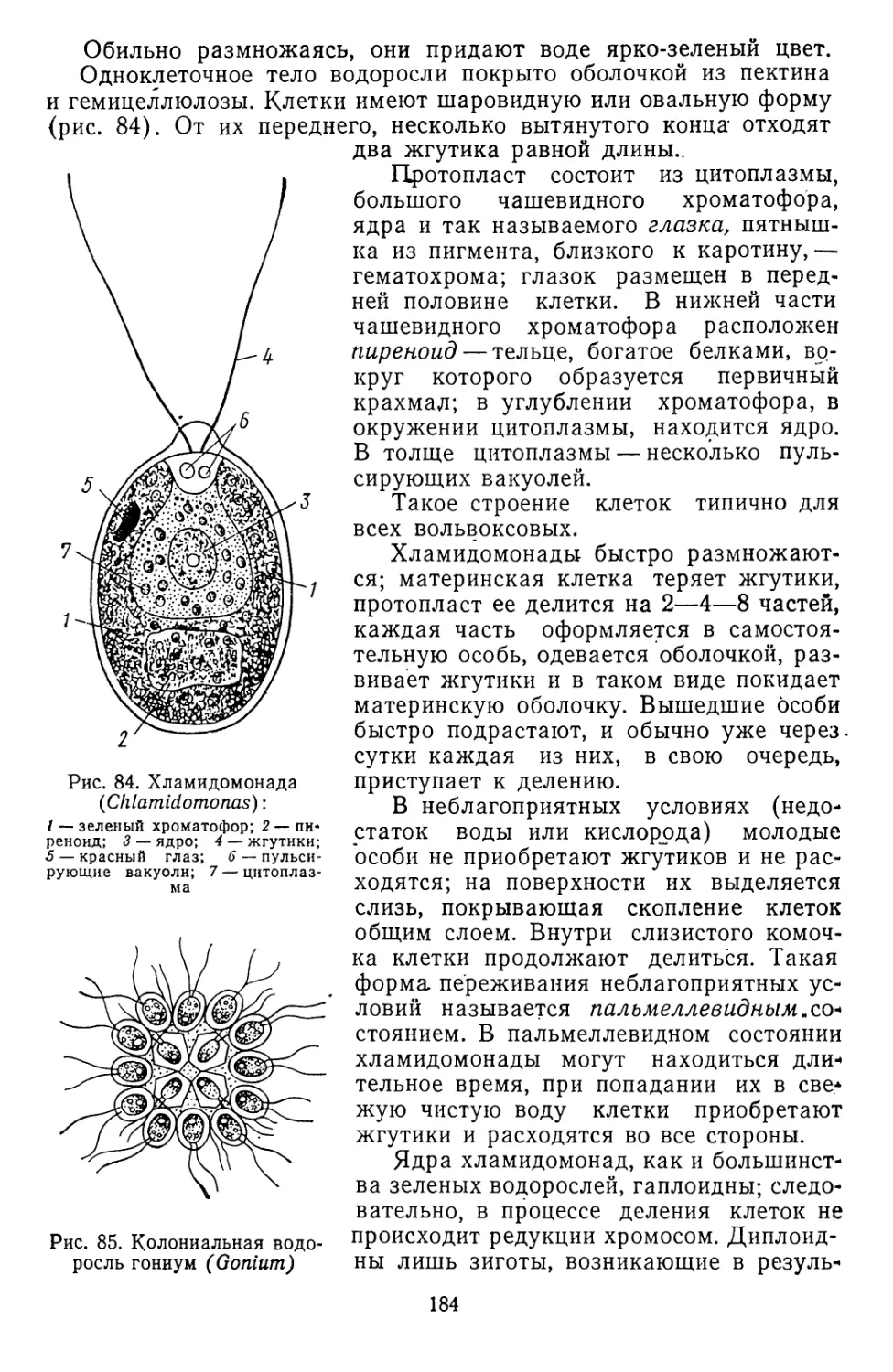
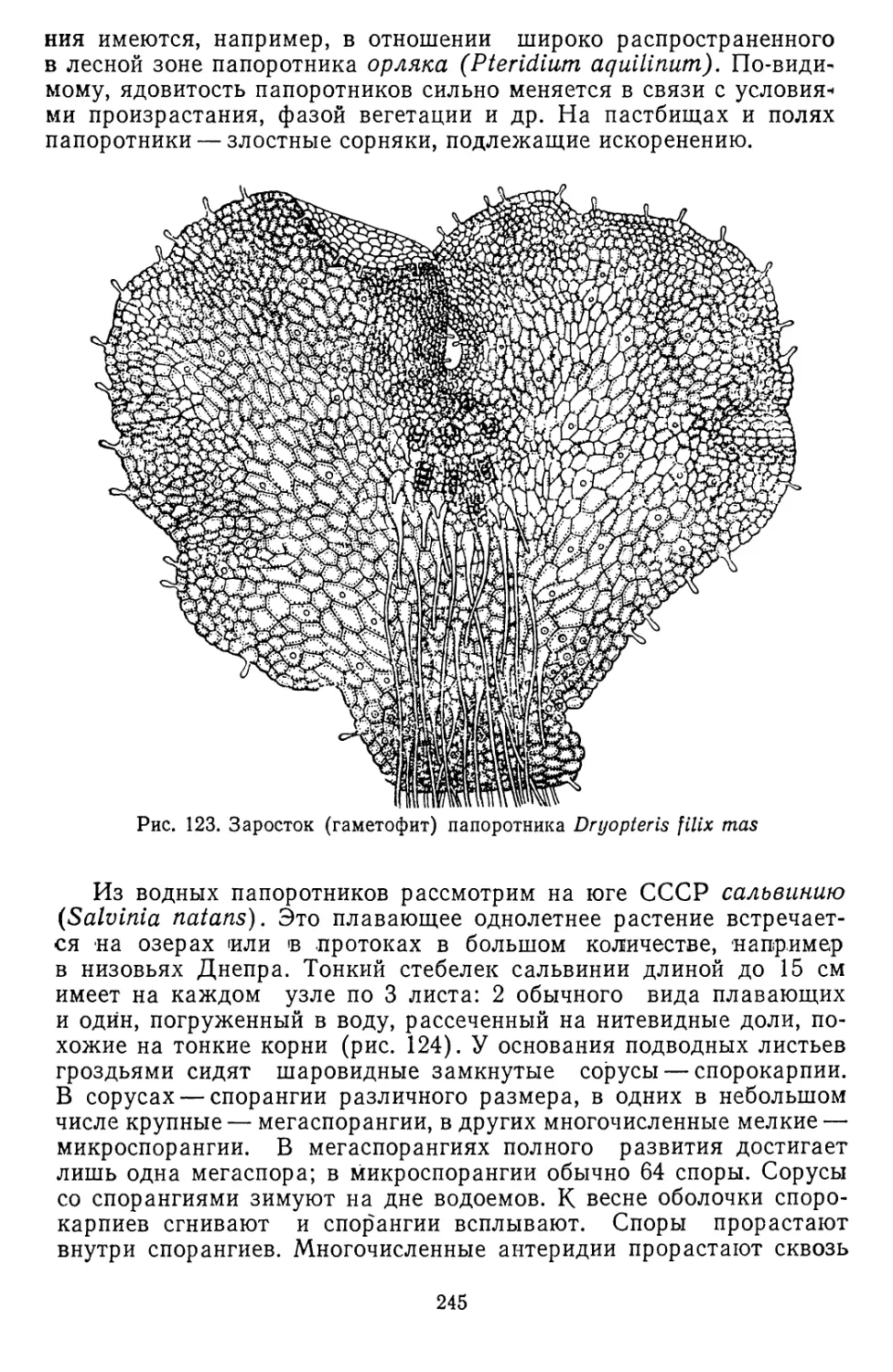
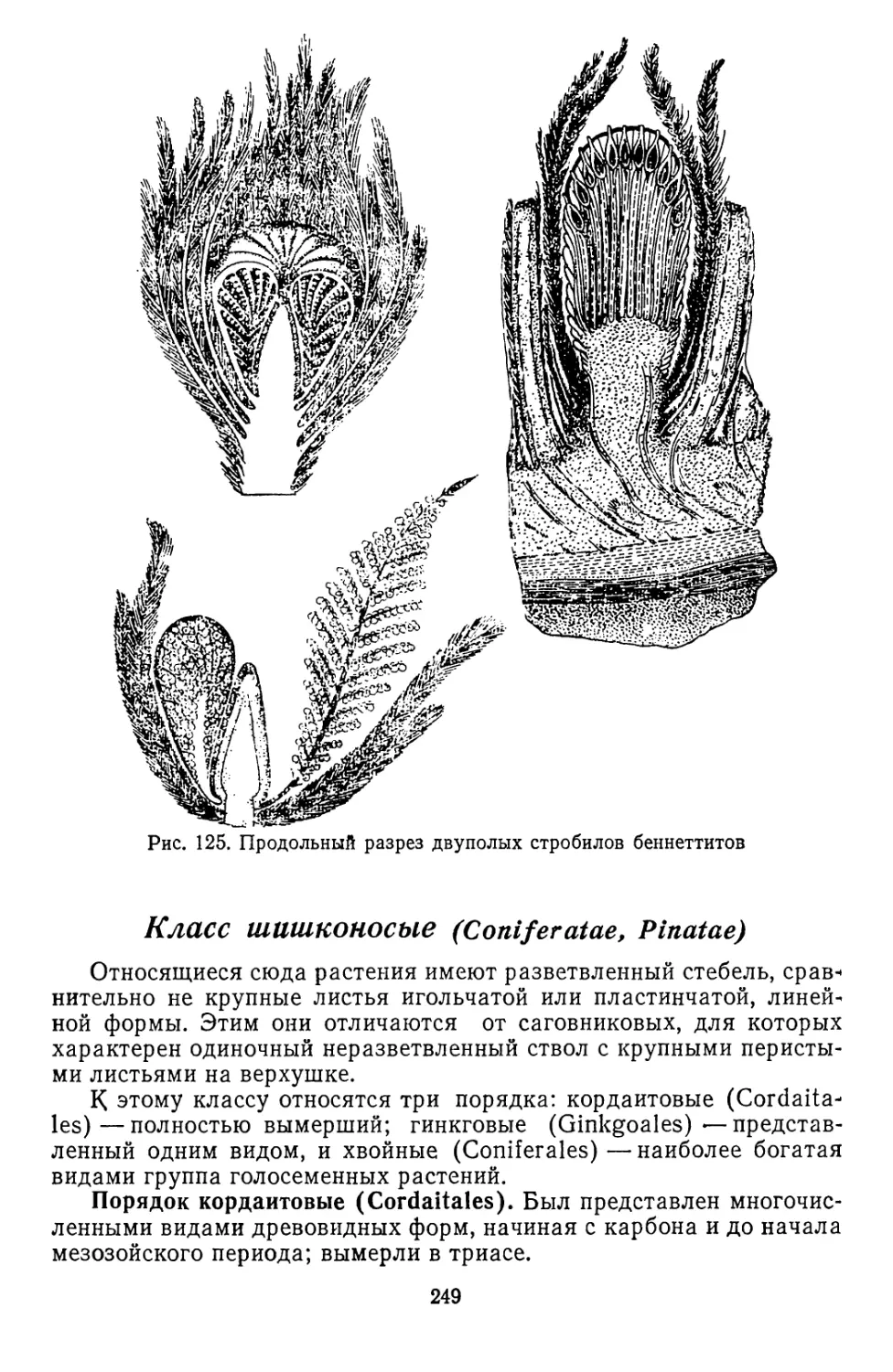
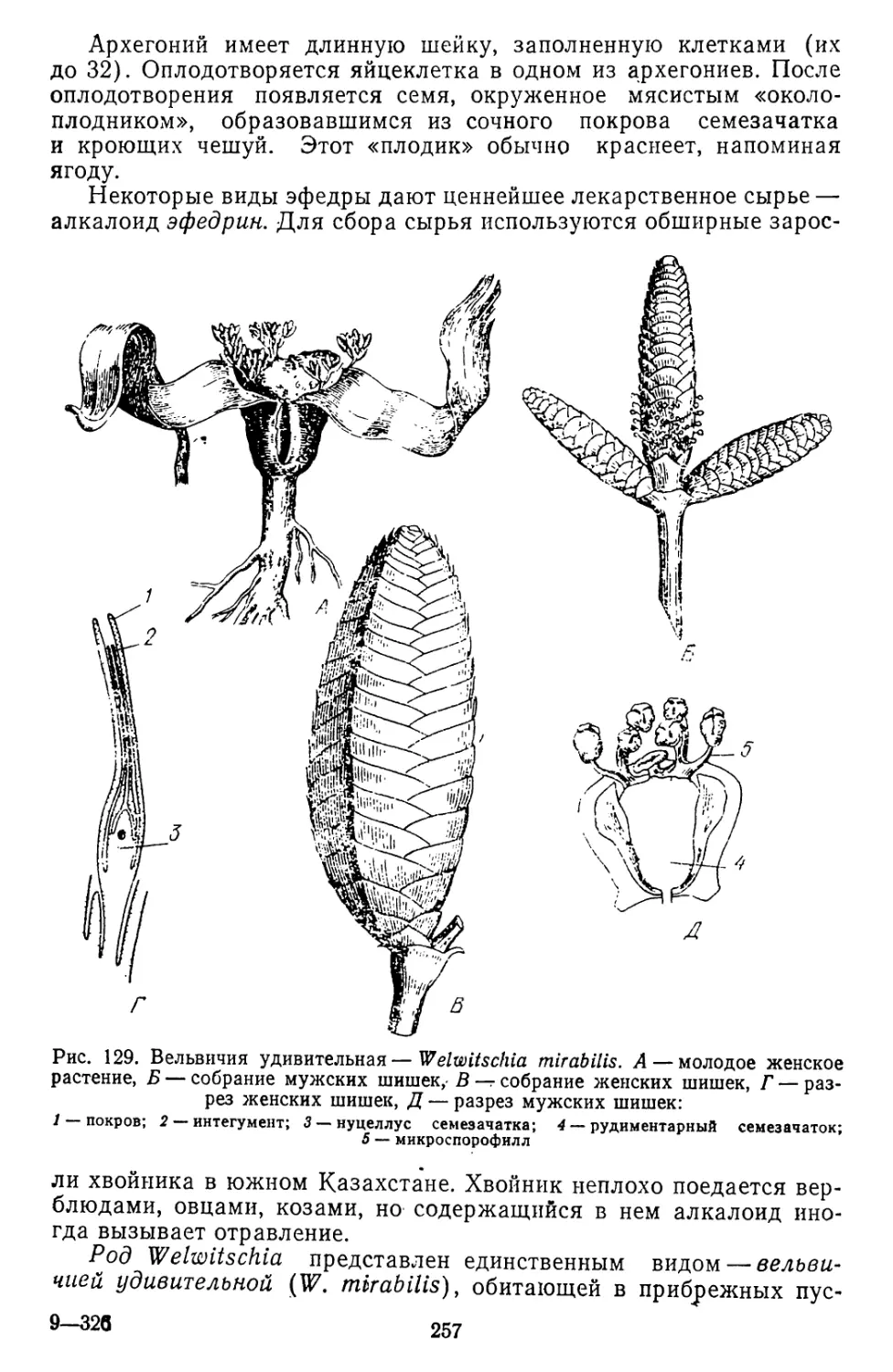
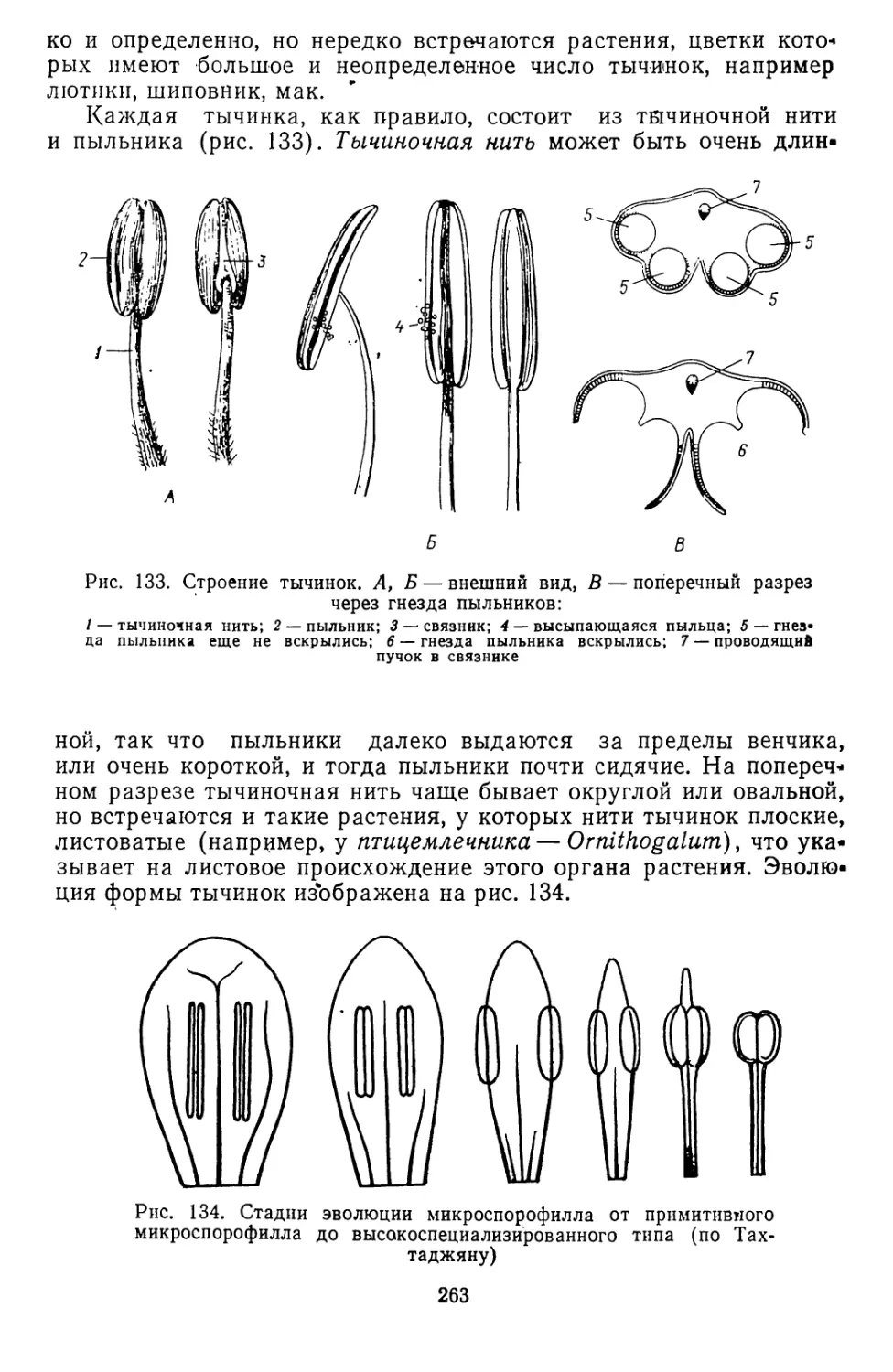
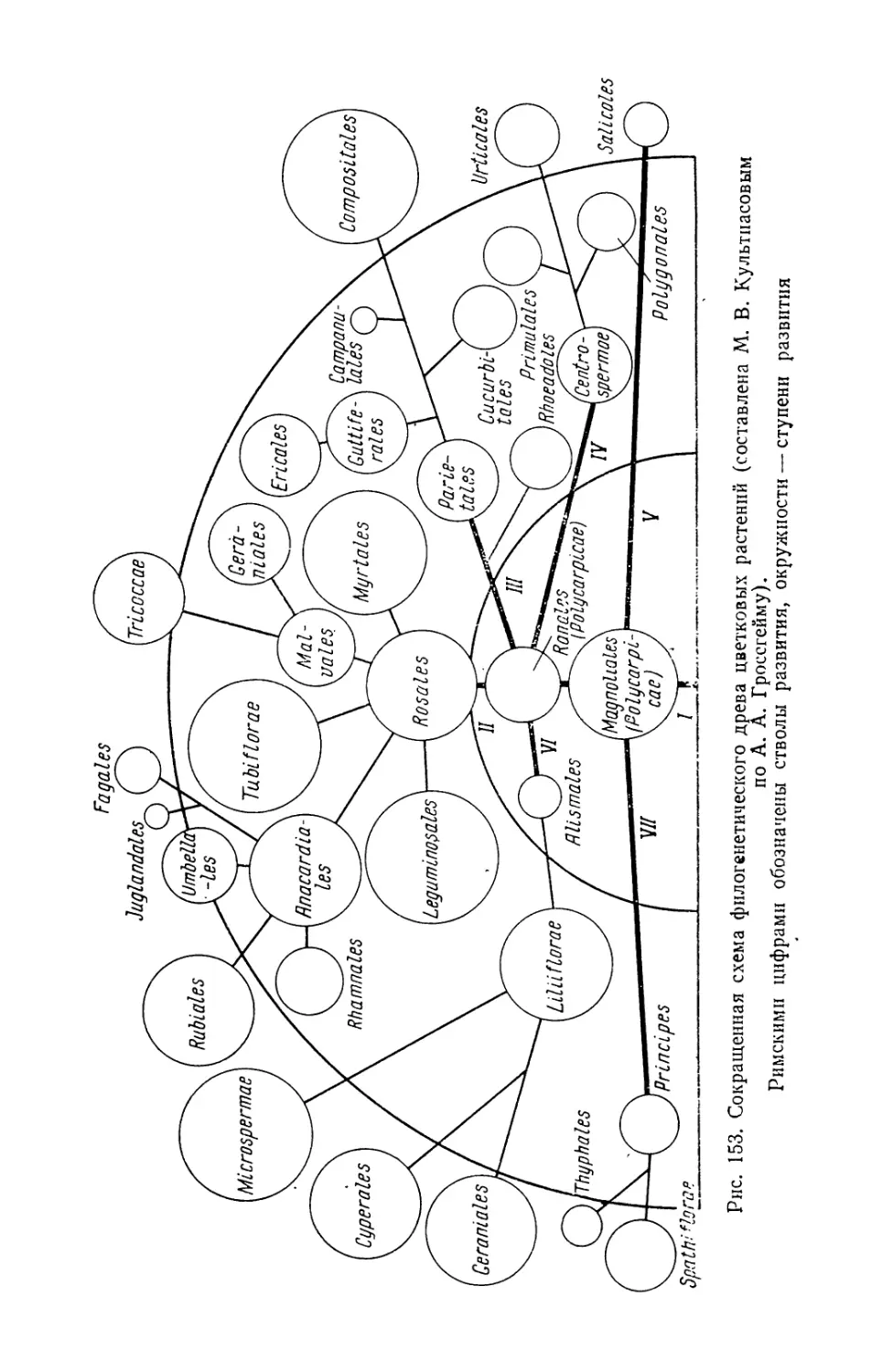
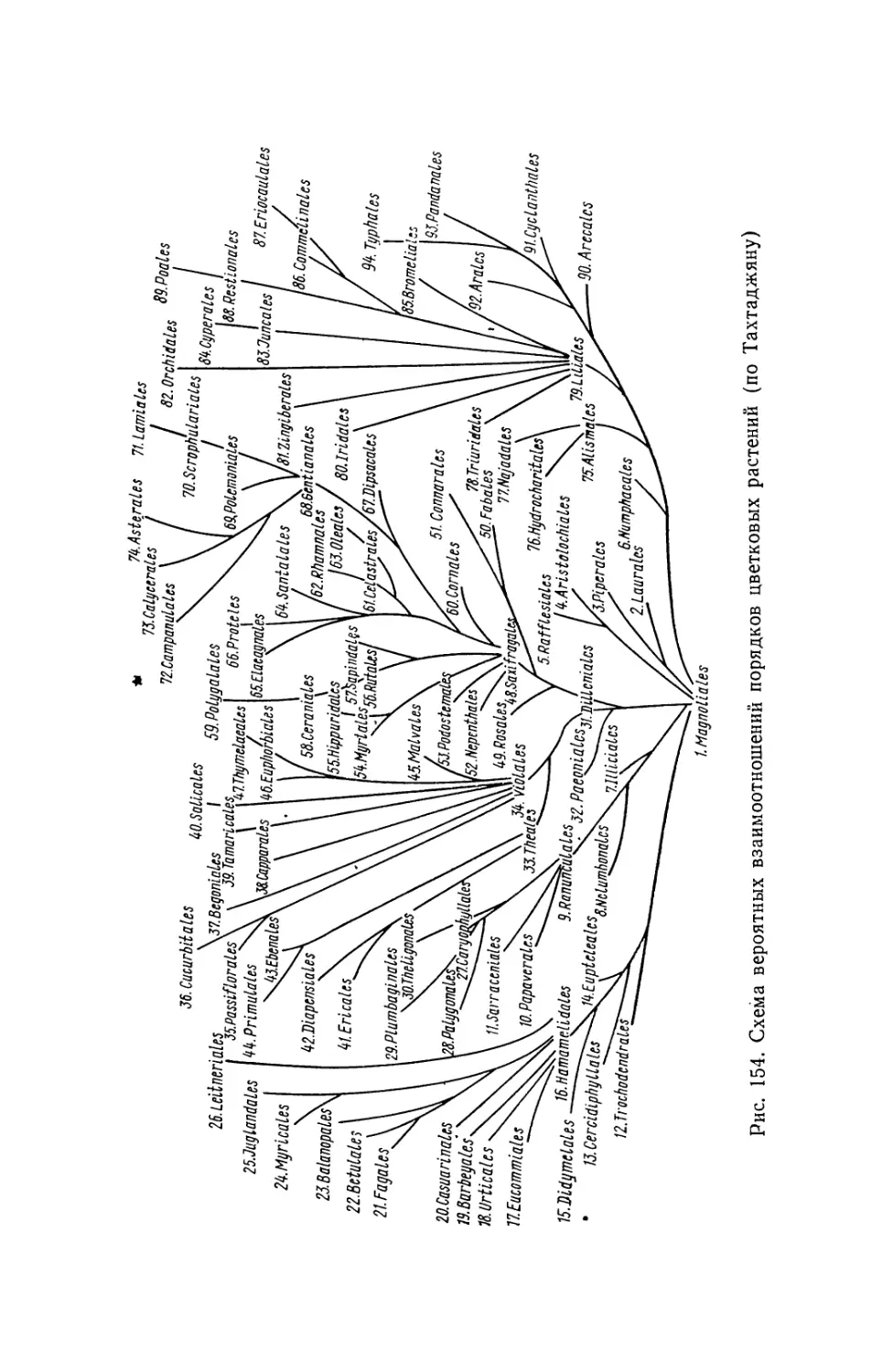
/













































































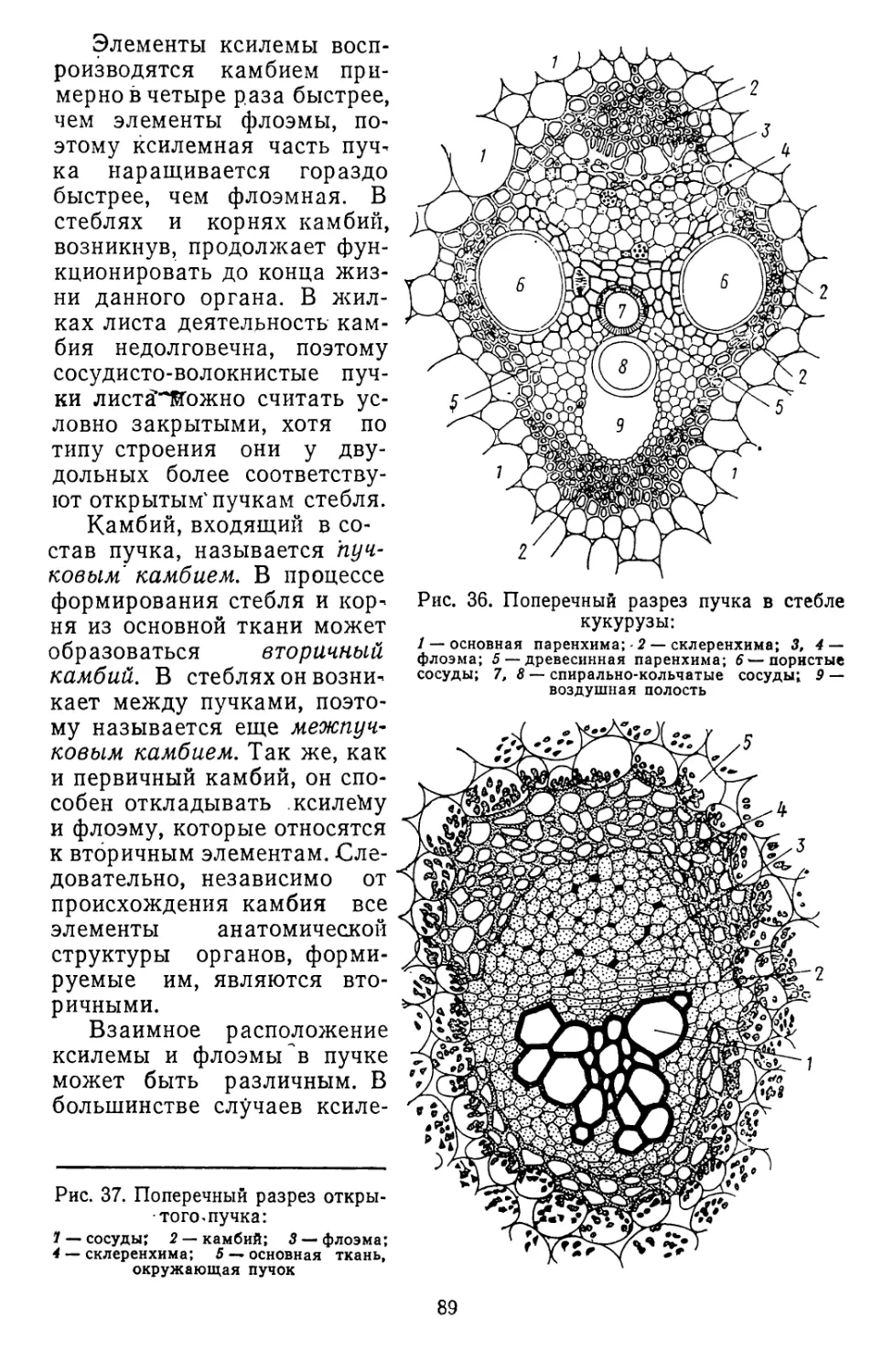
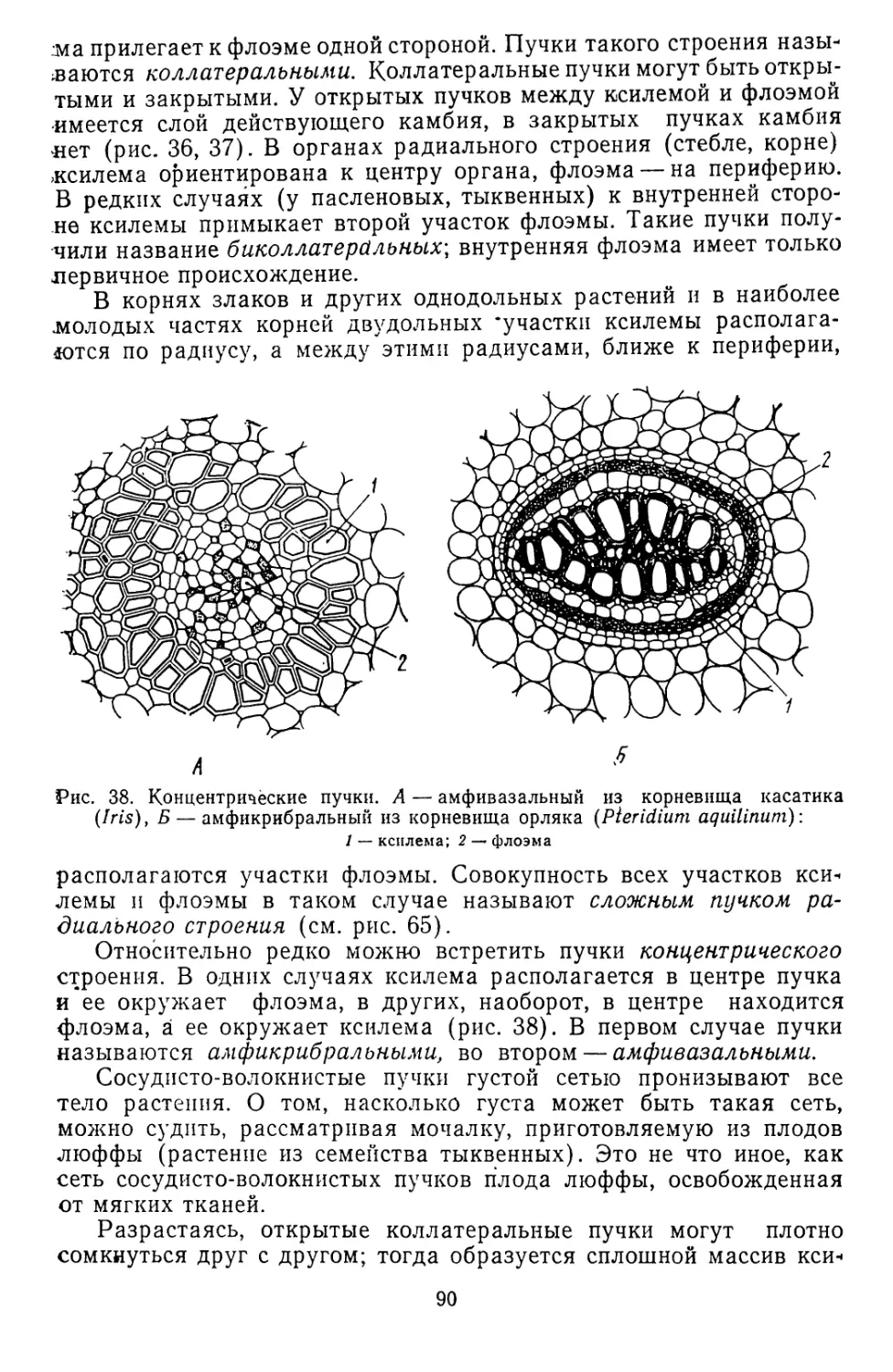

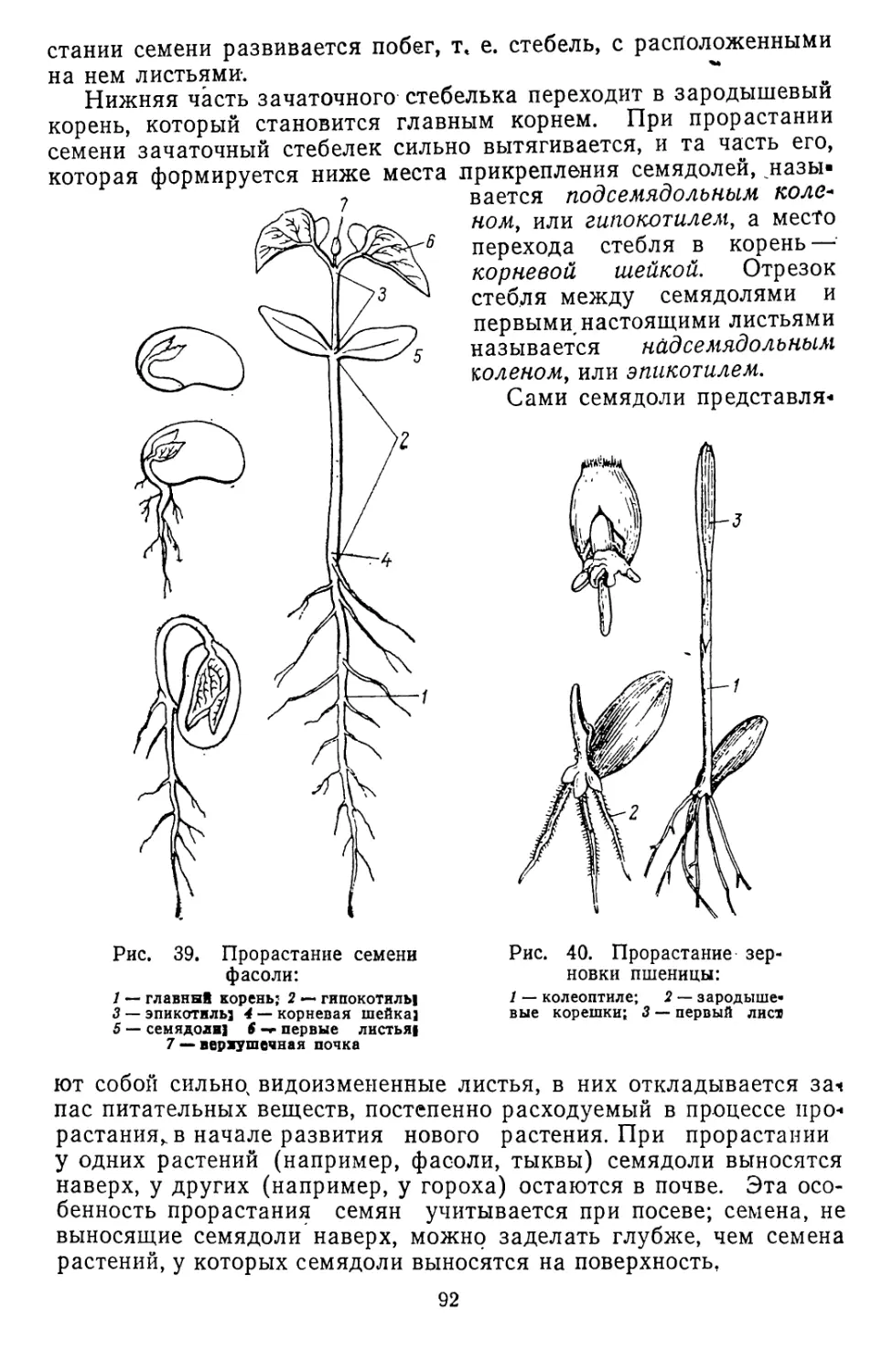

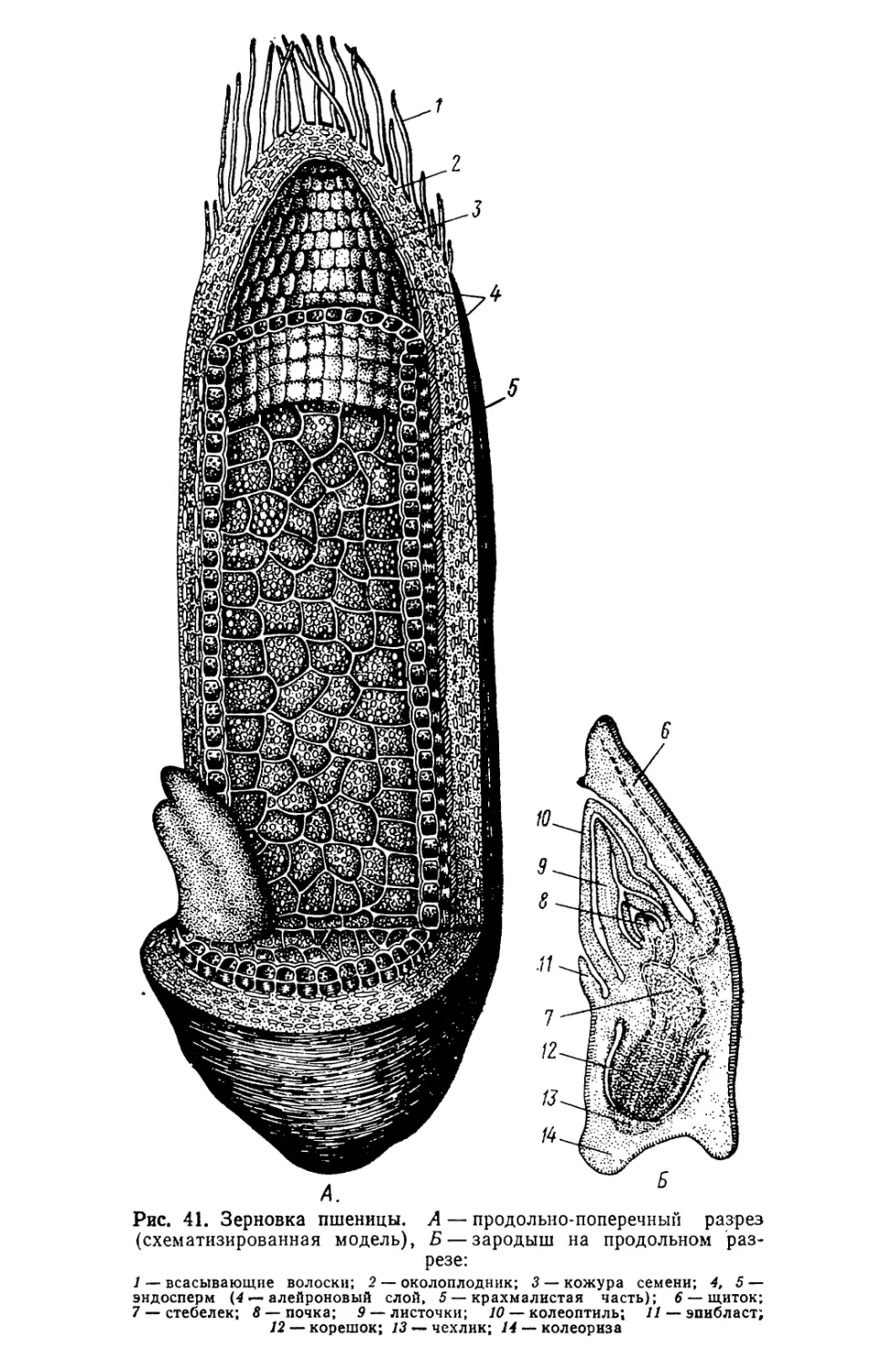











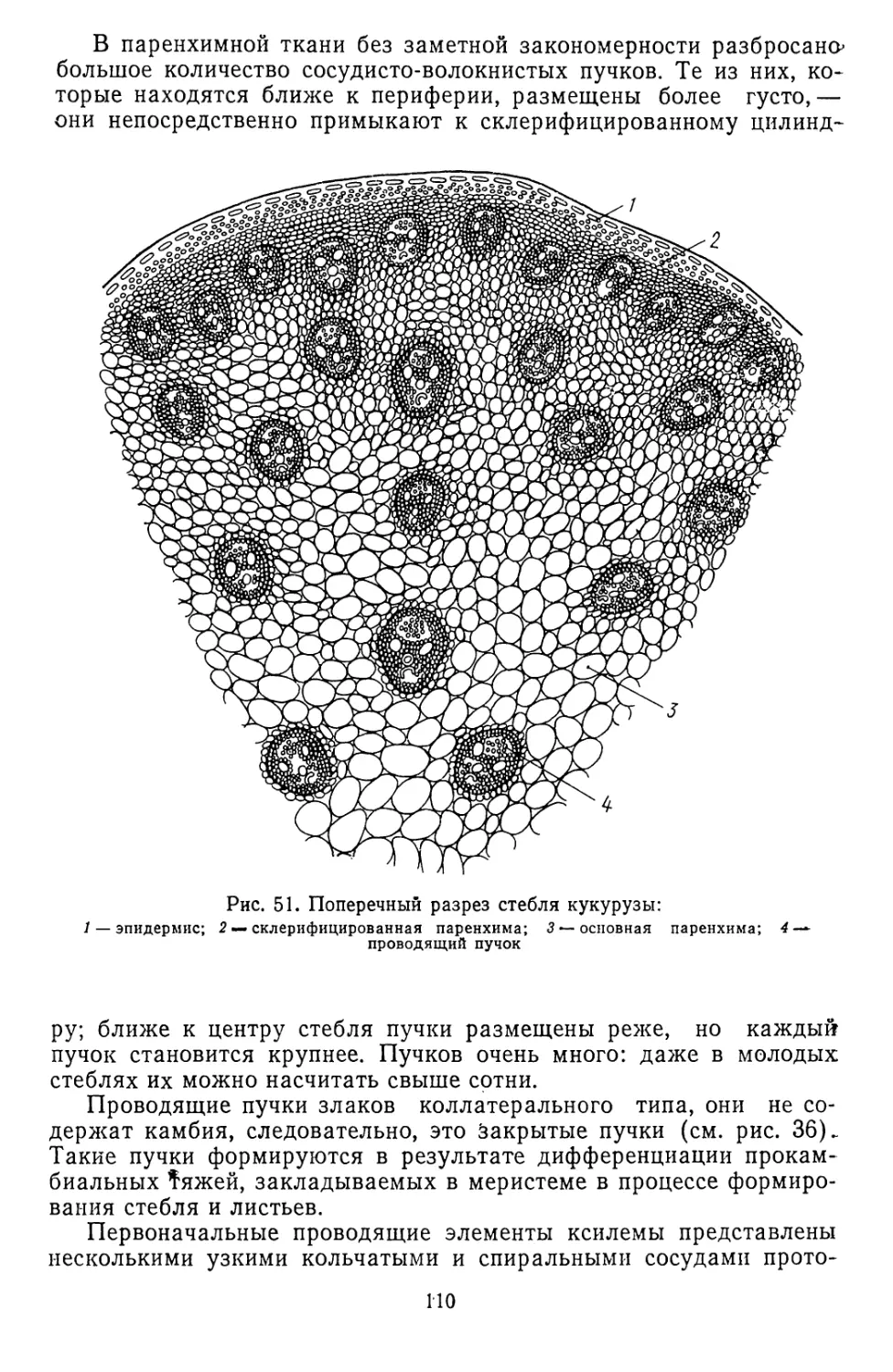
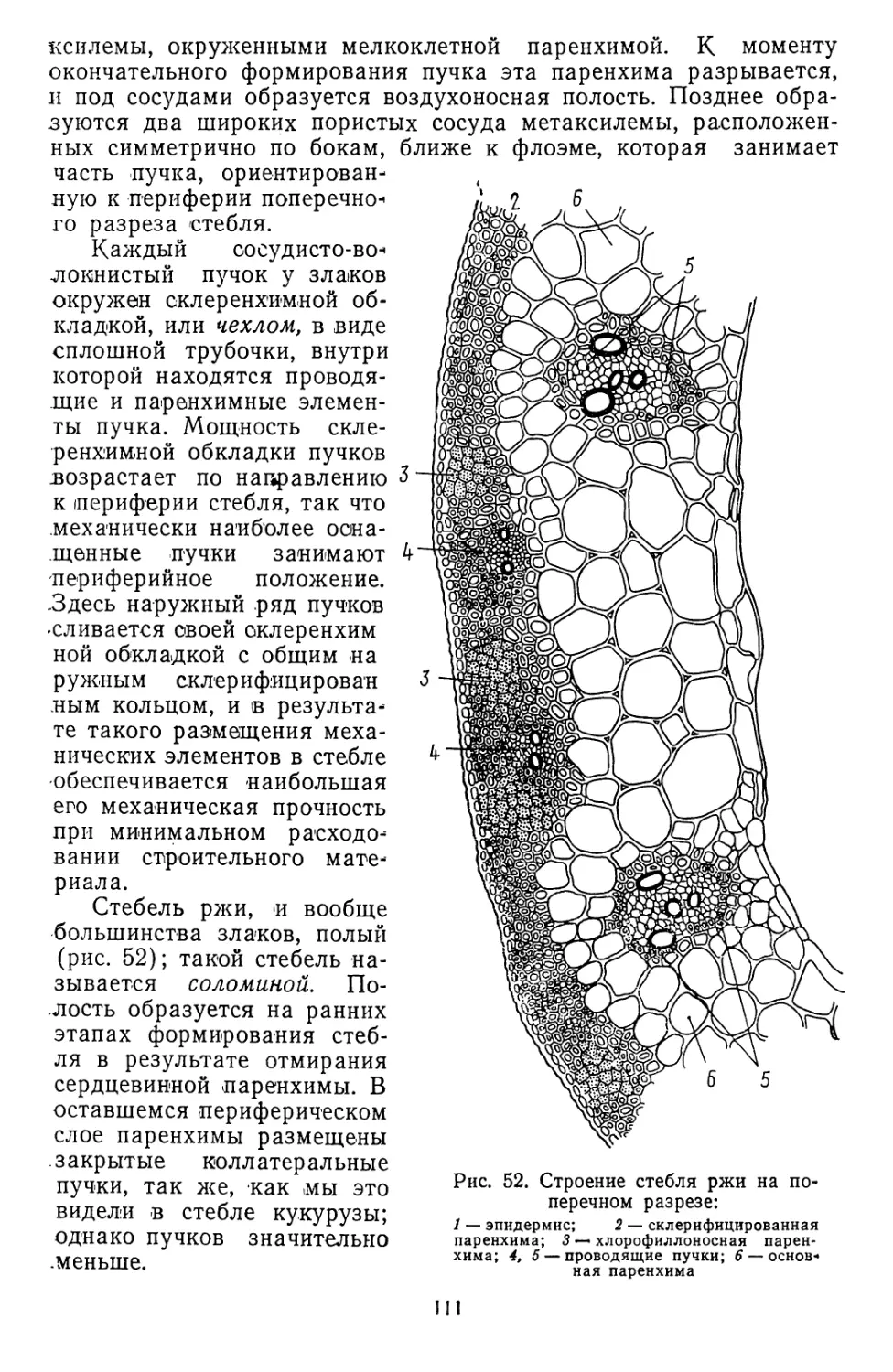
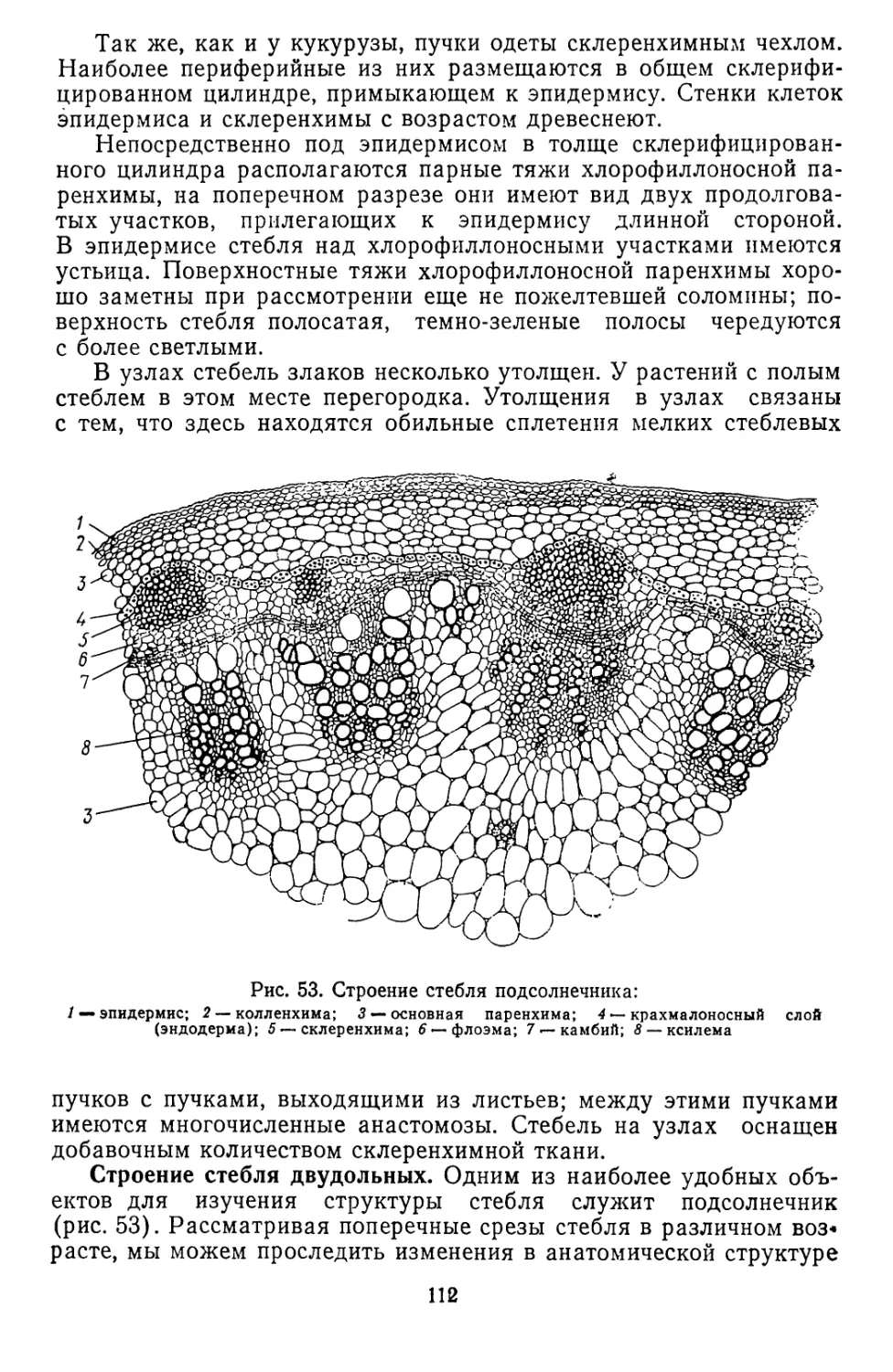



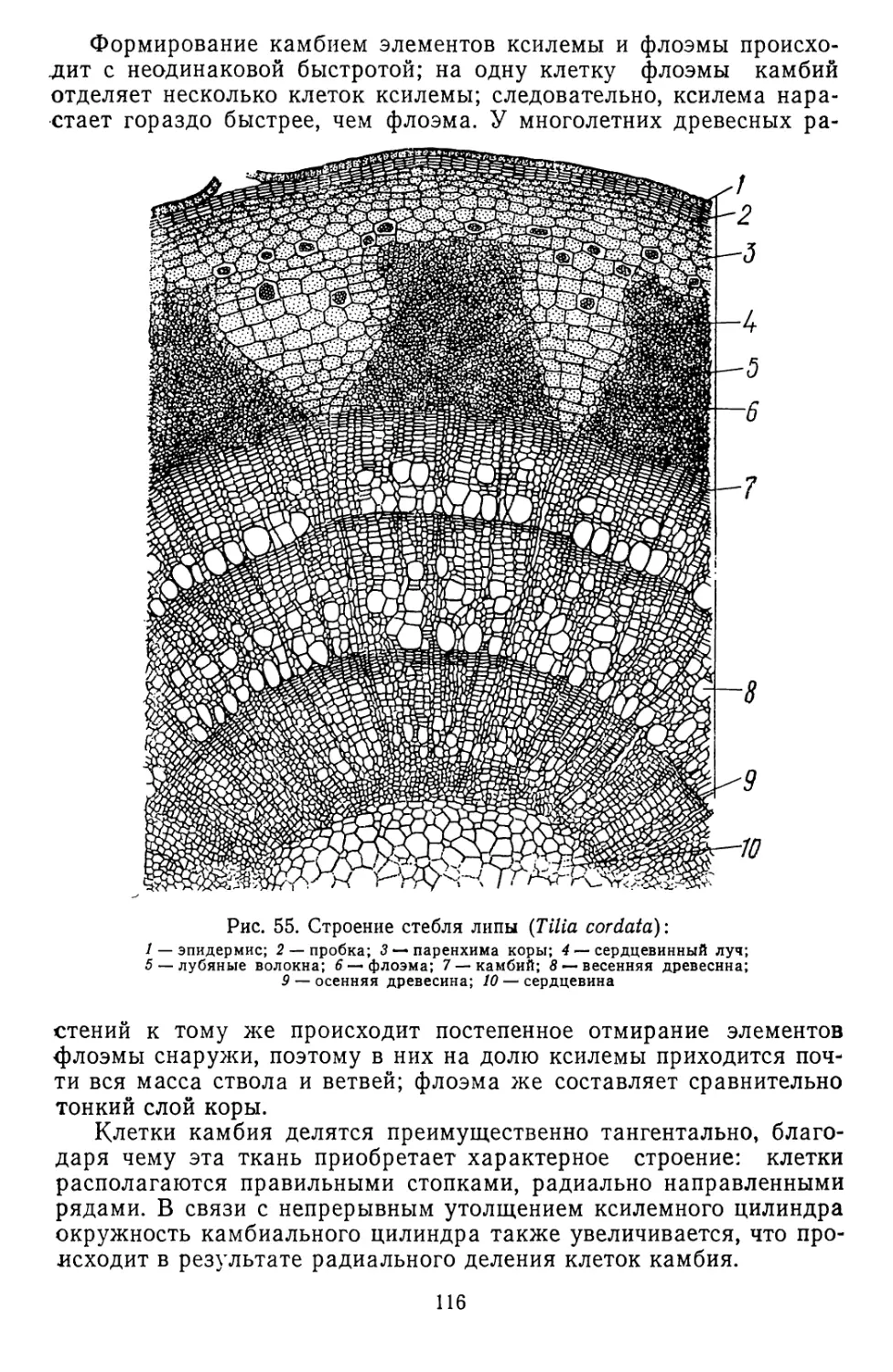






















































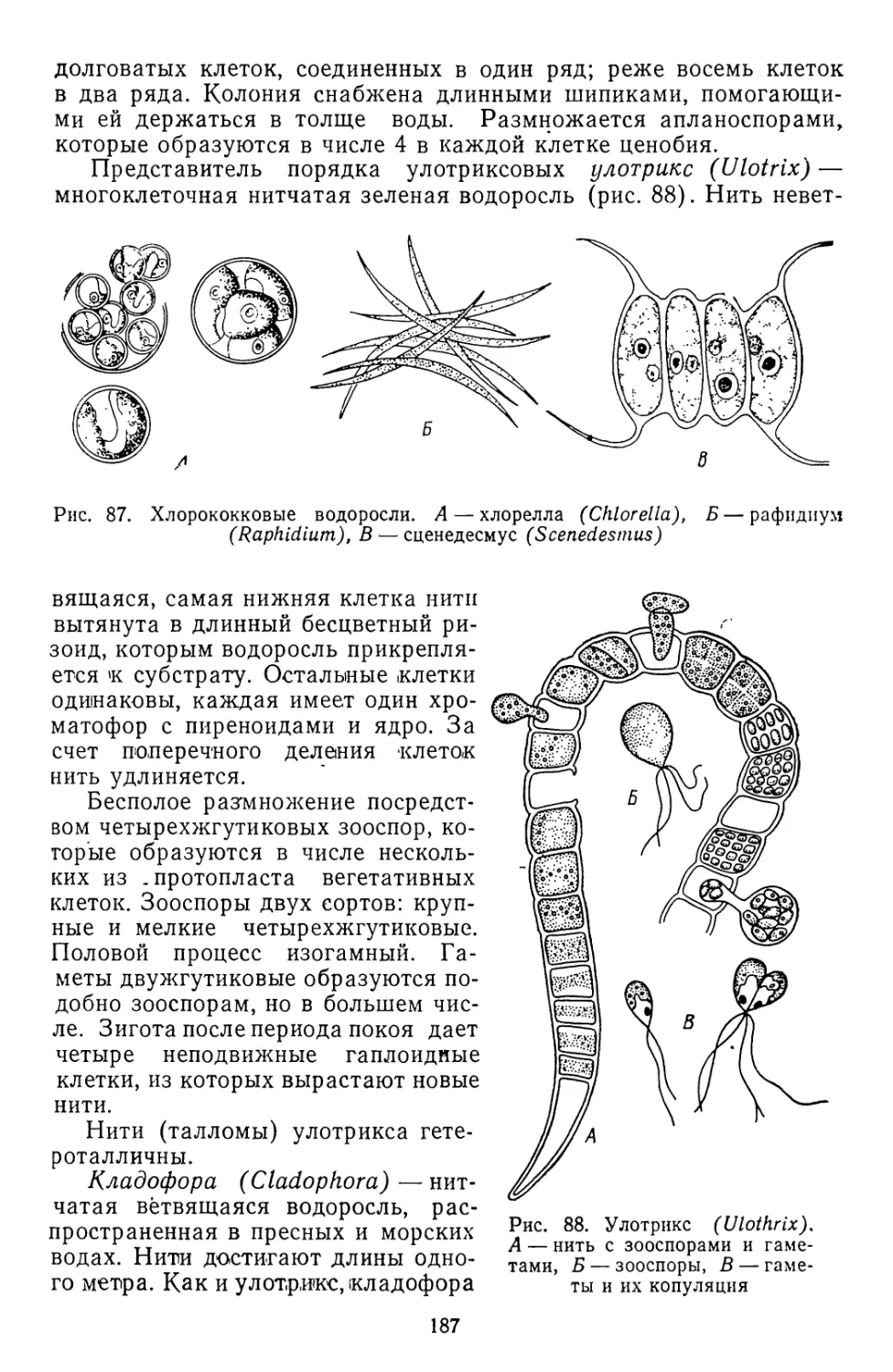


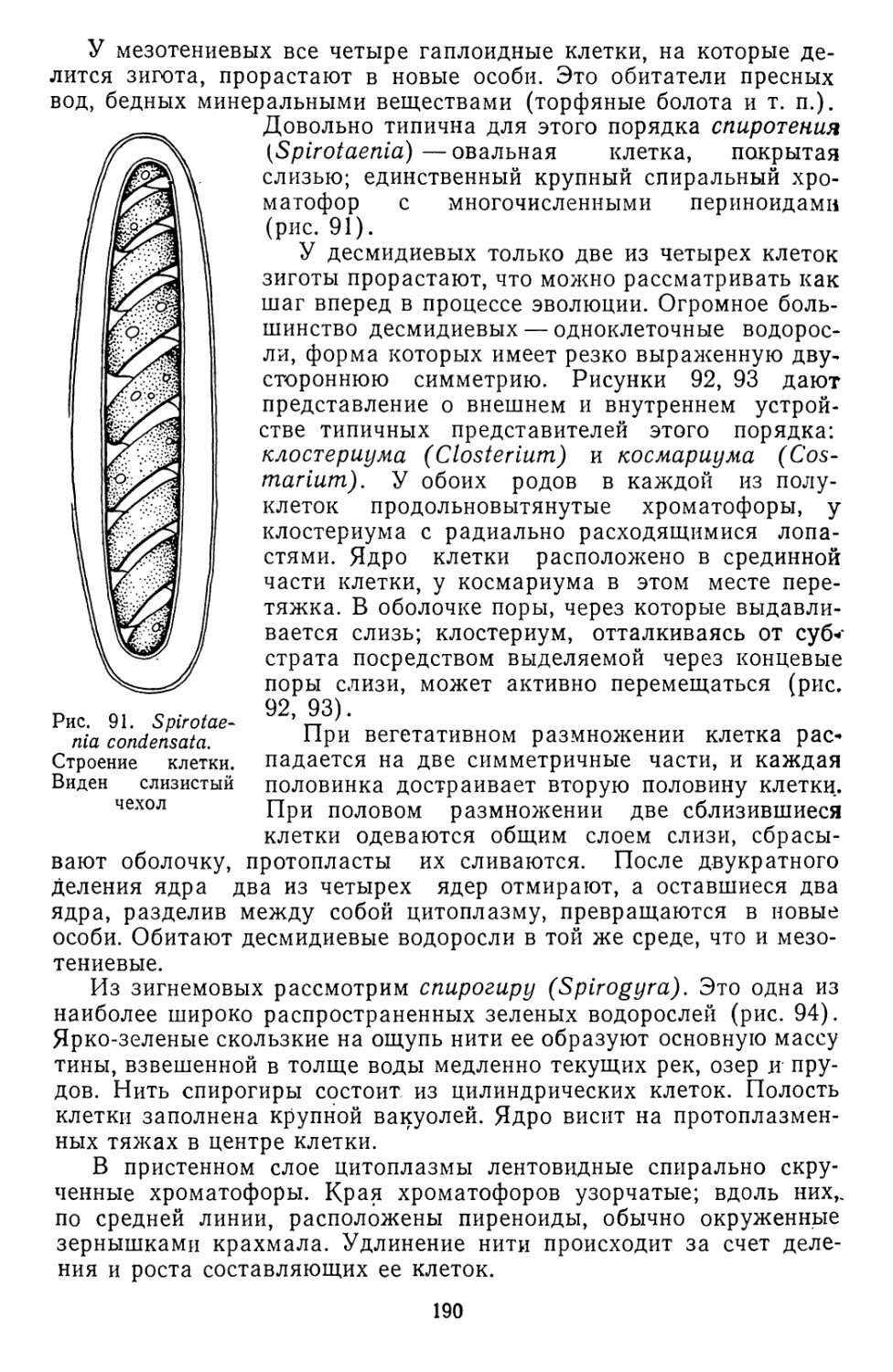
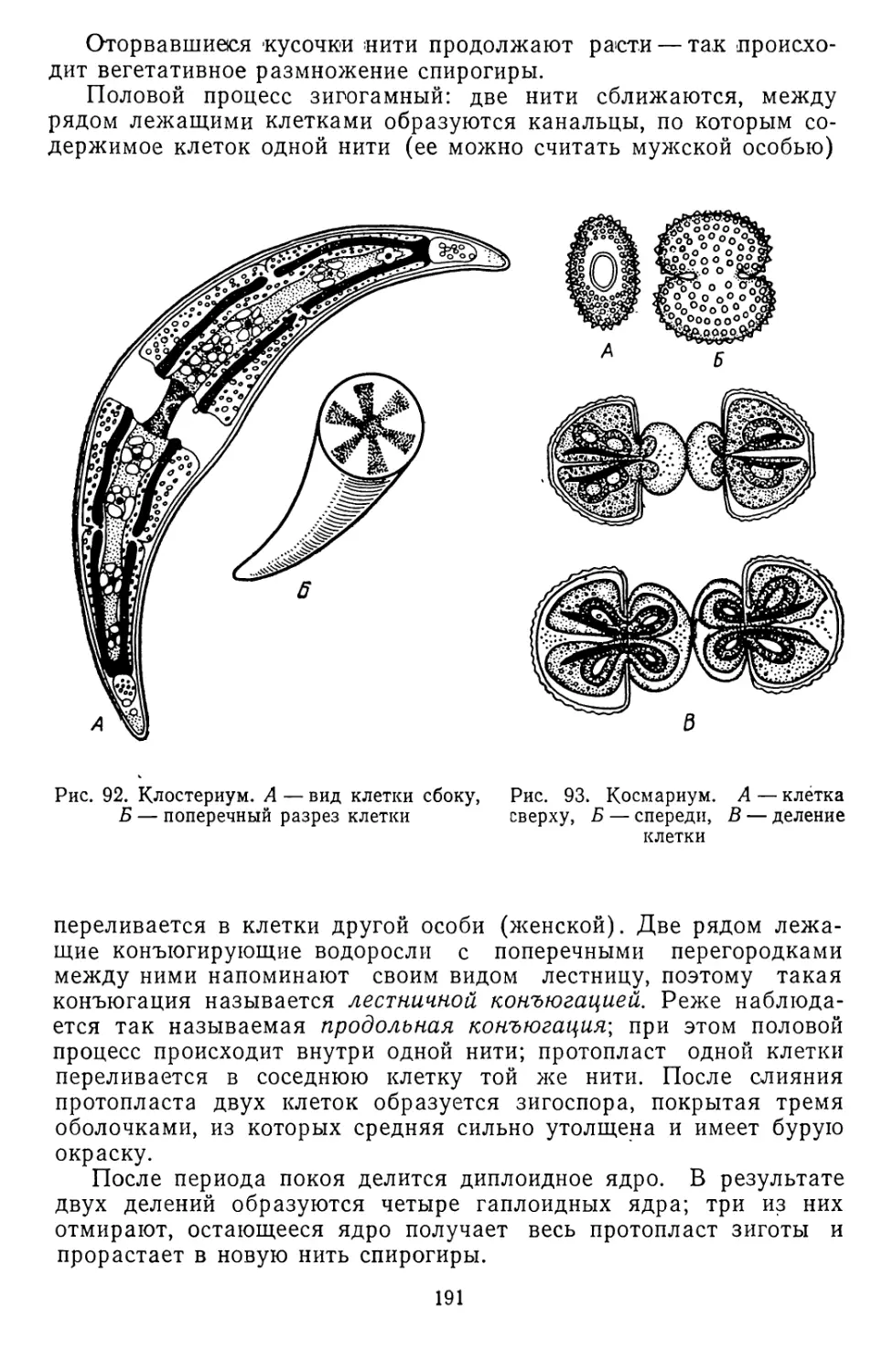





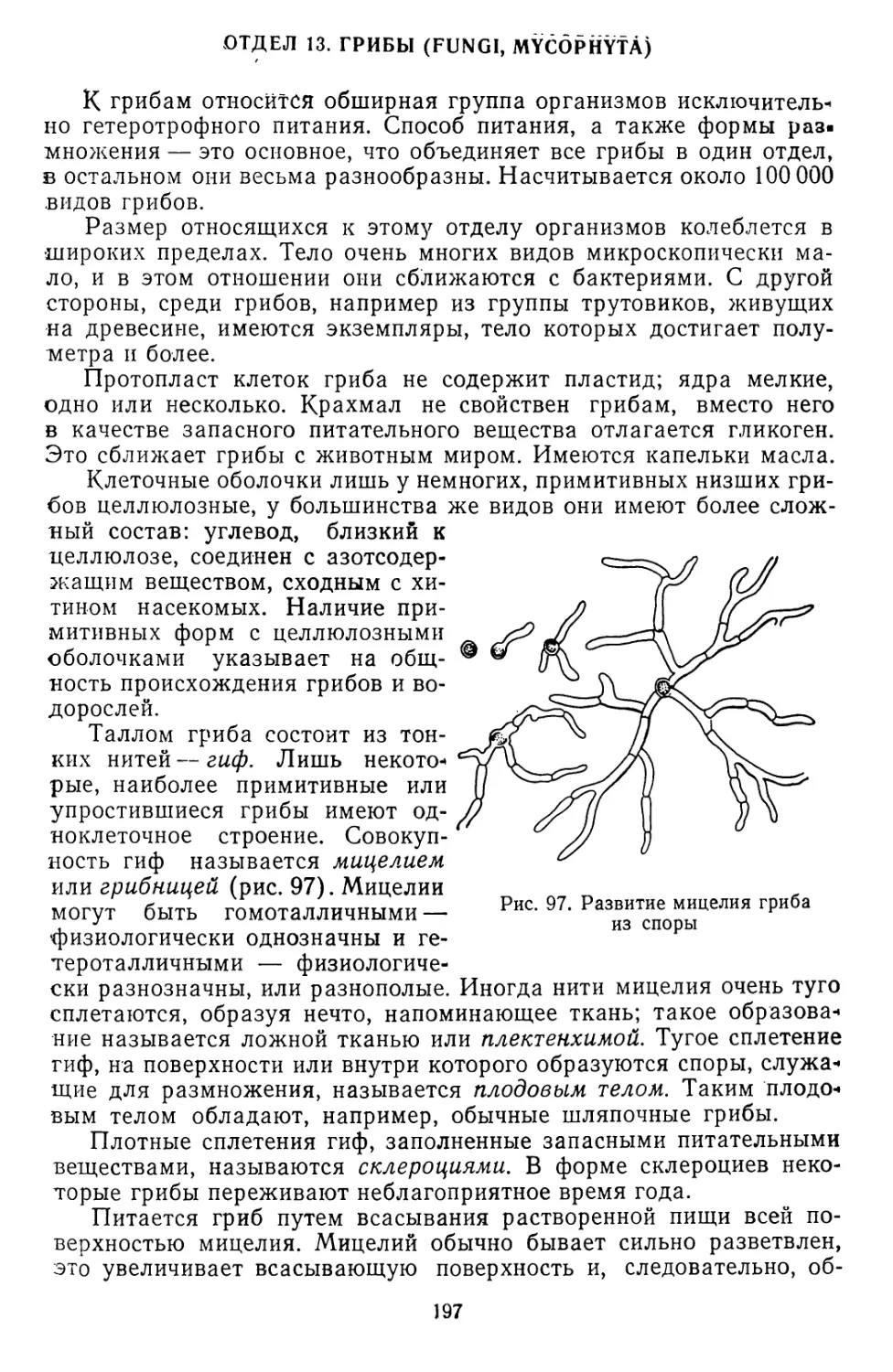




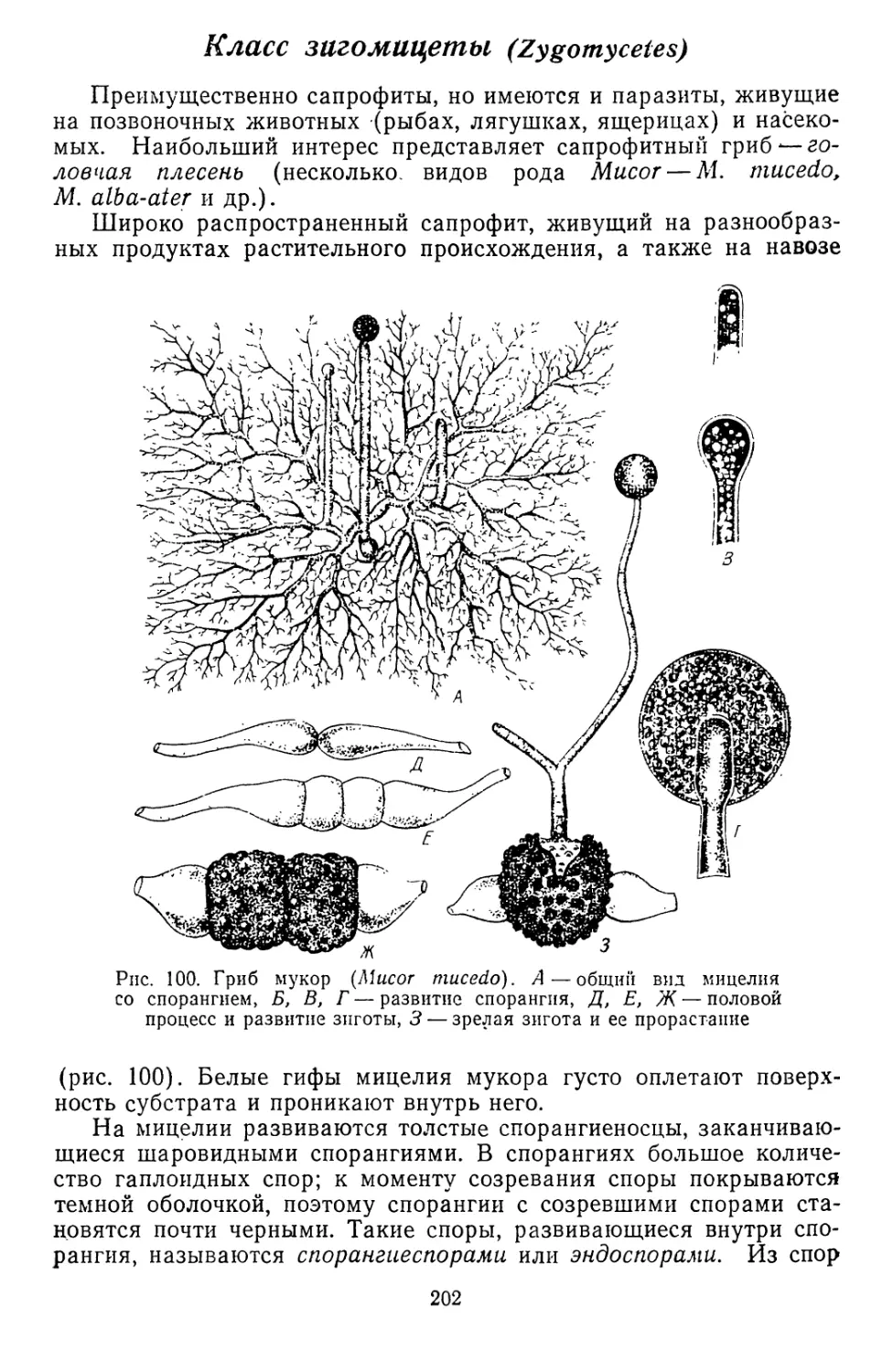

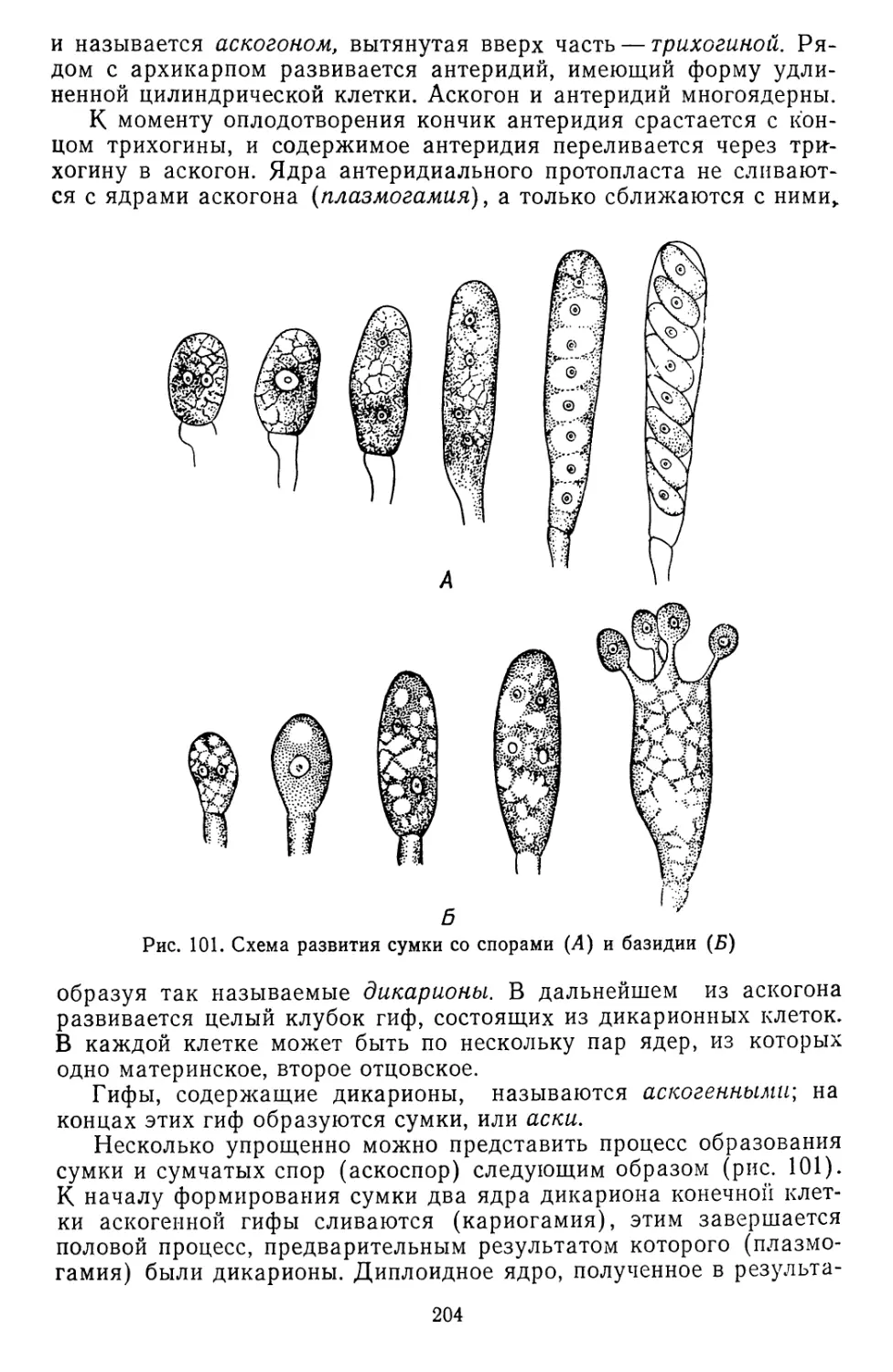

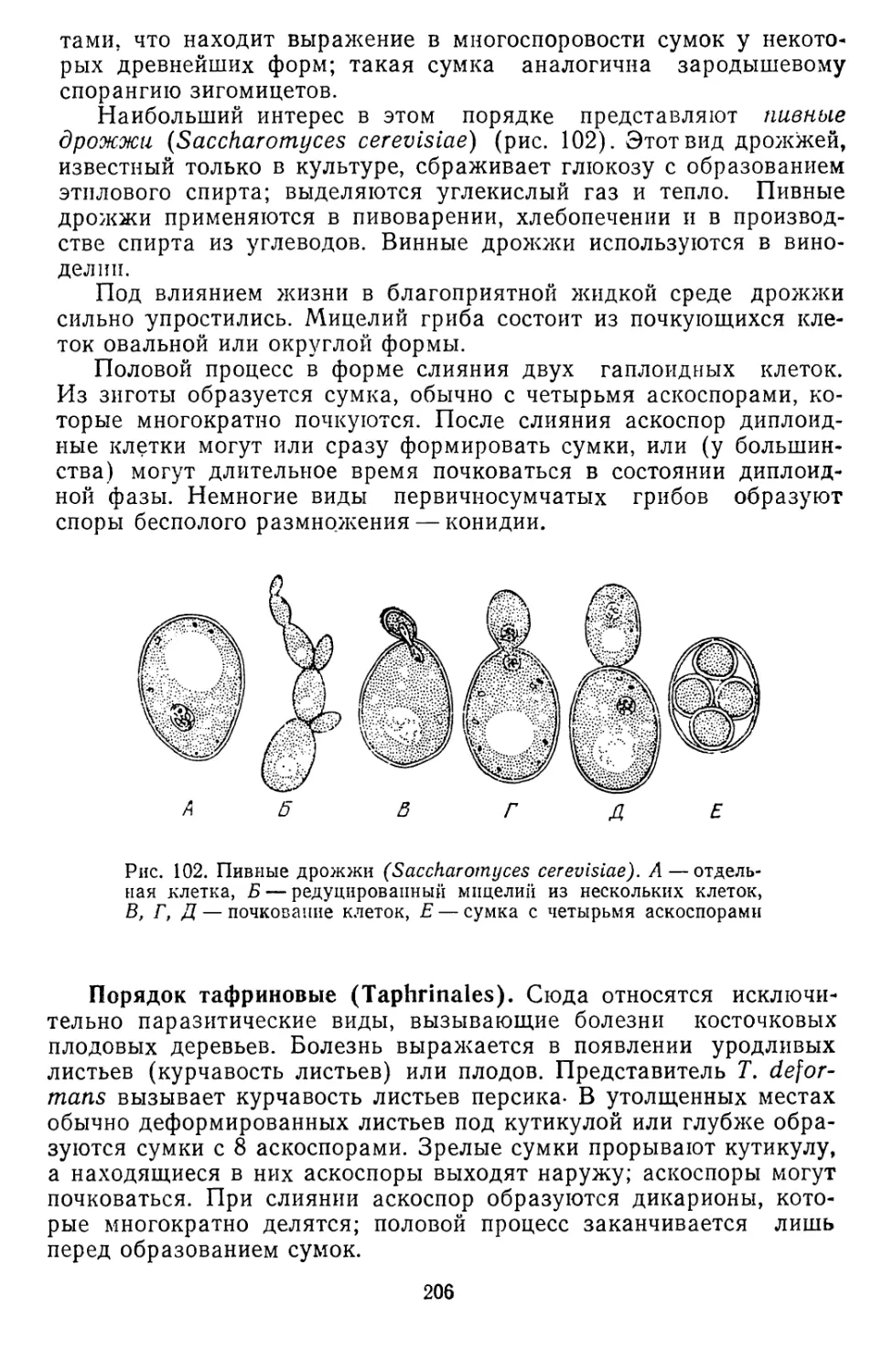














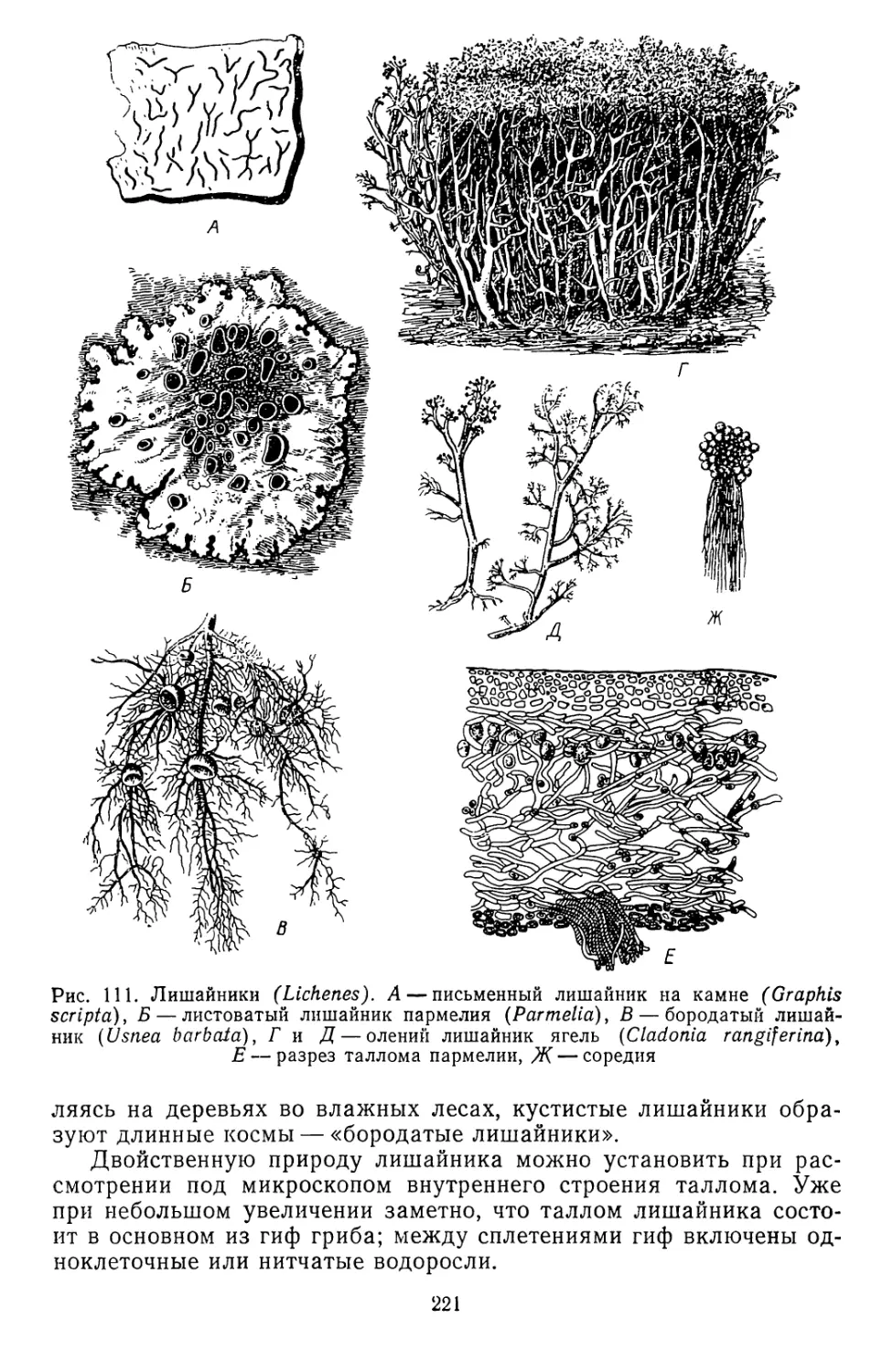



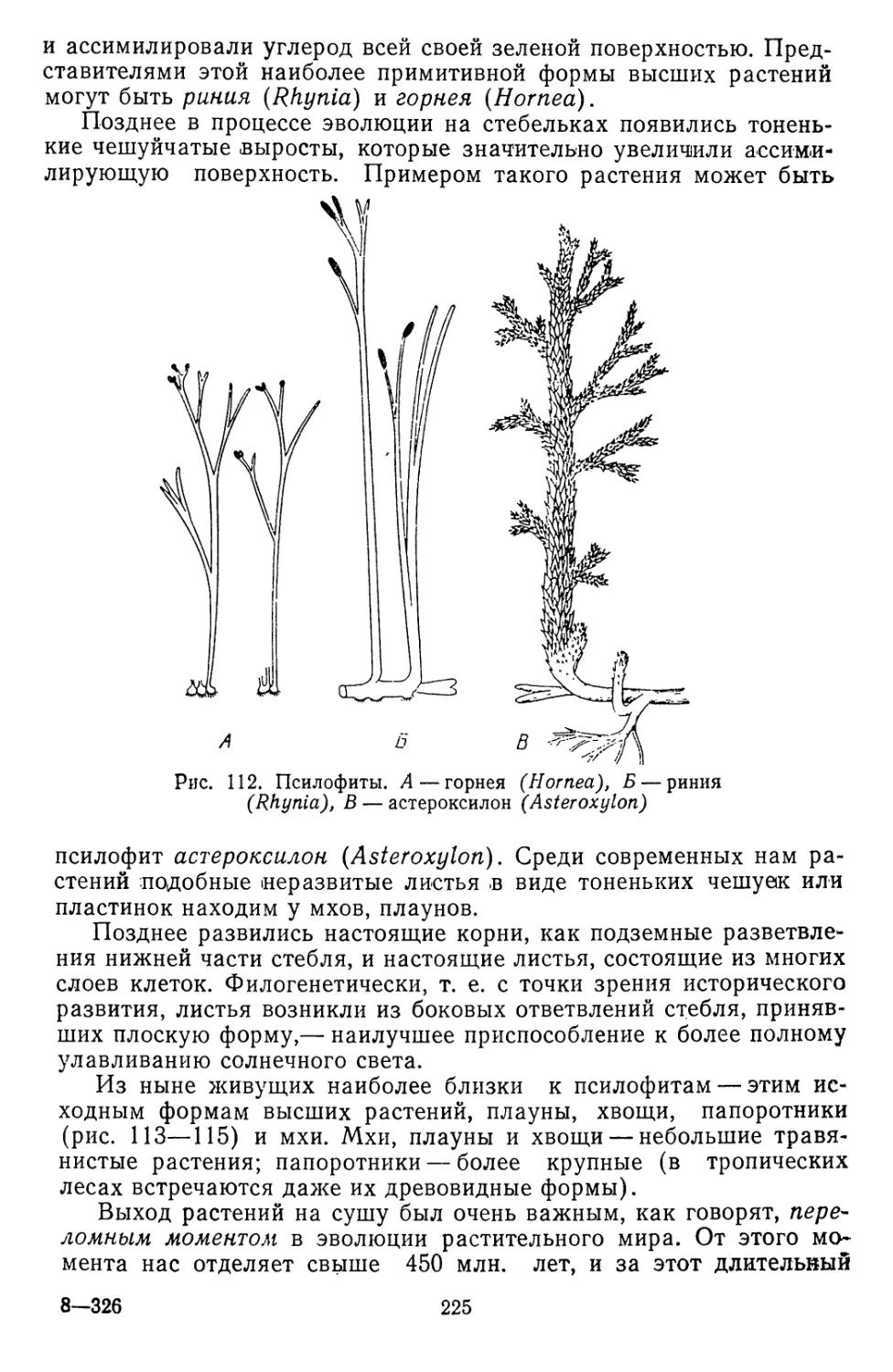



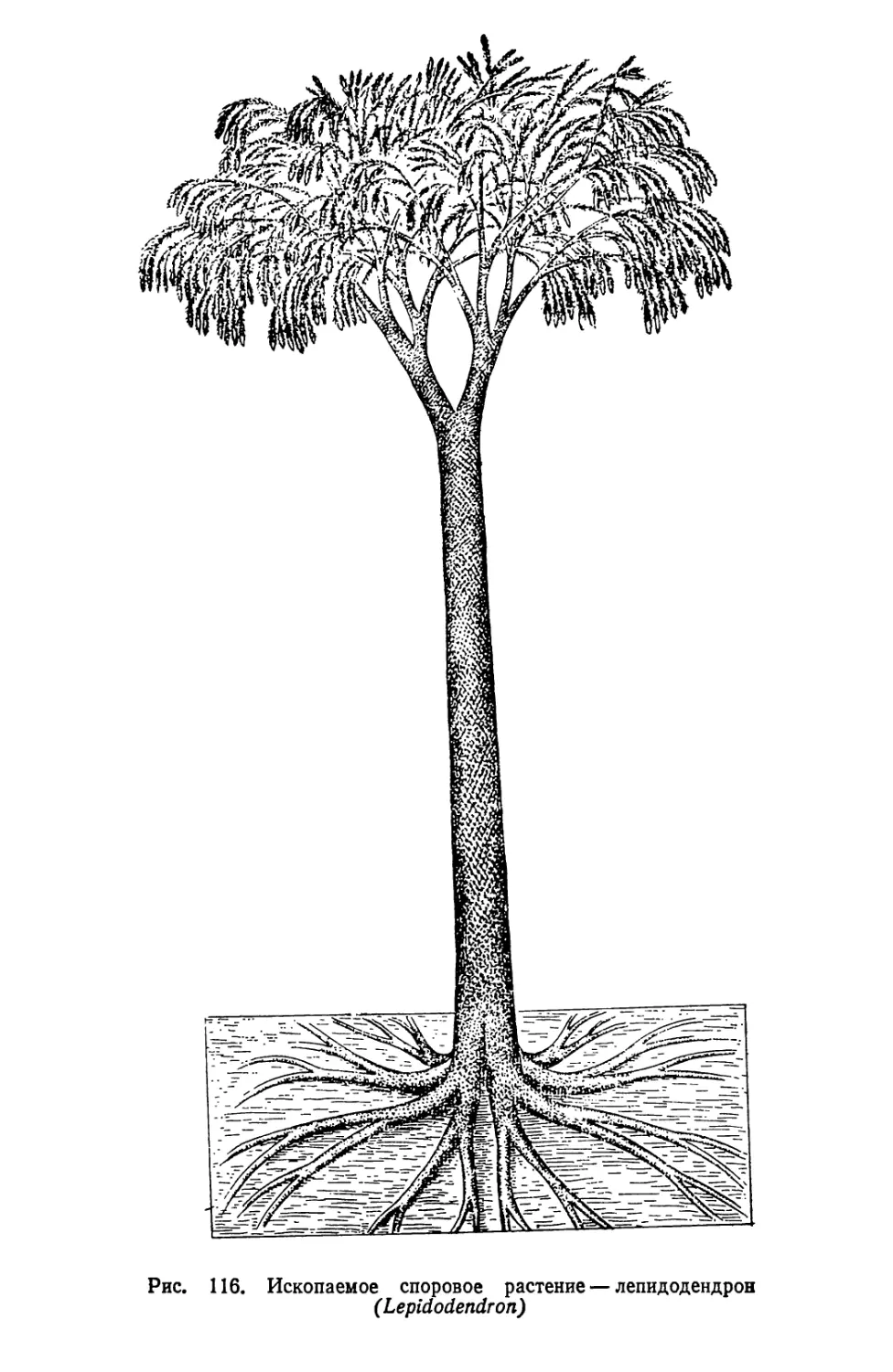



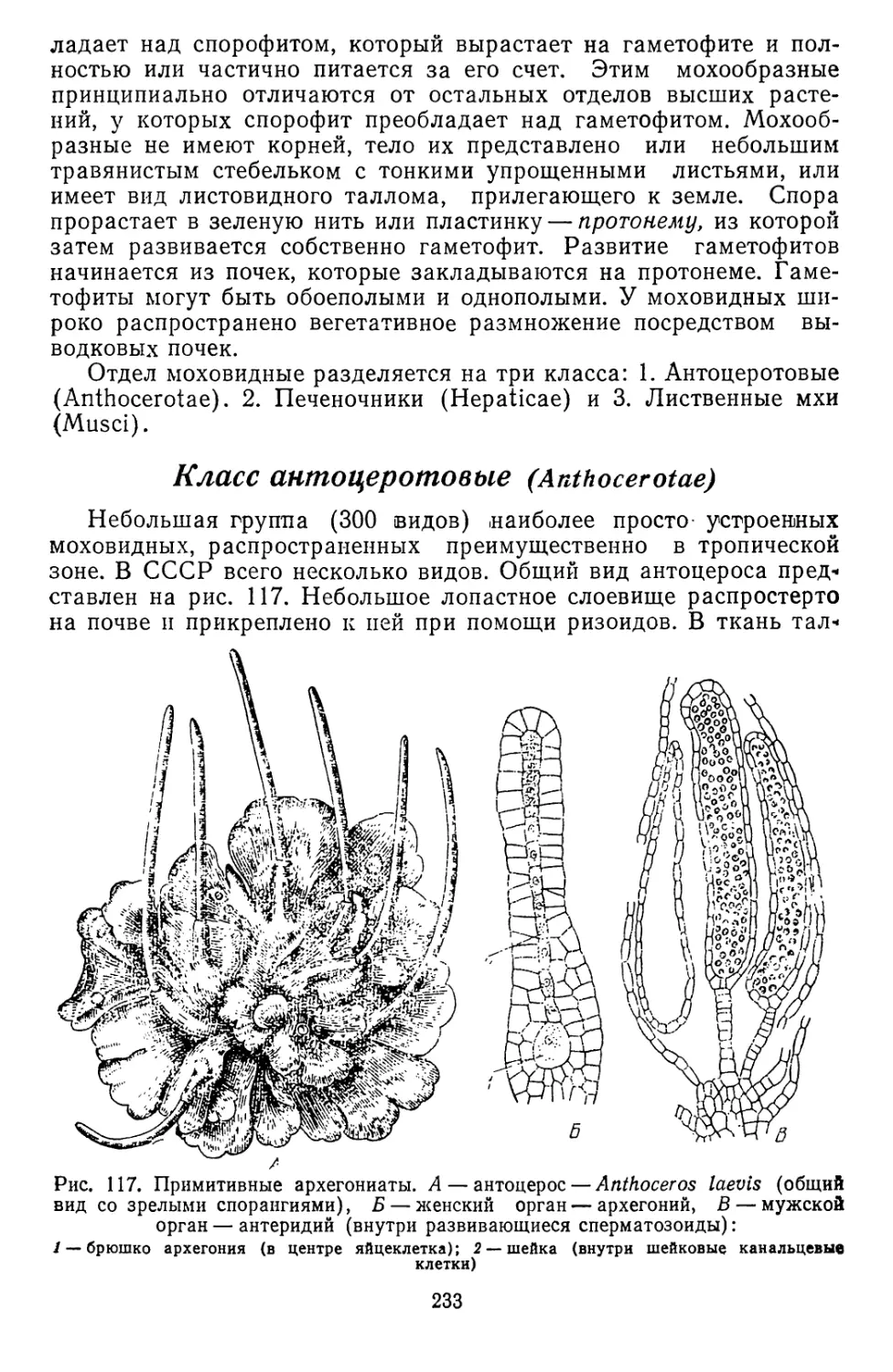


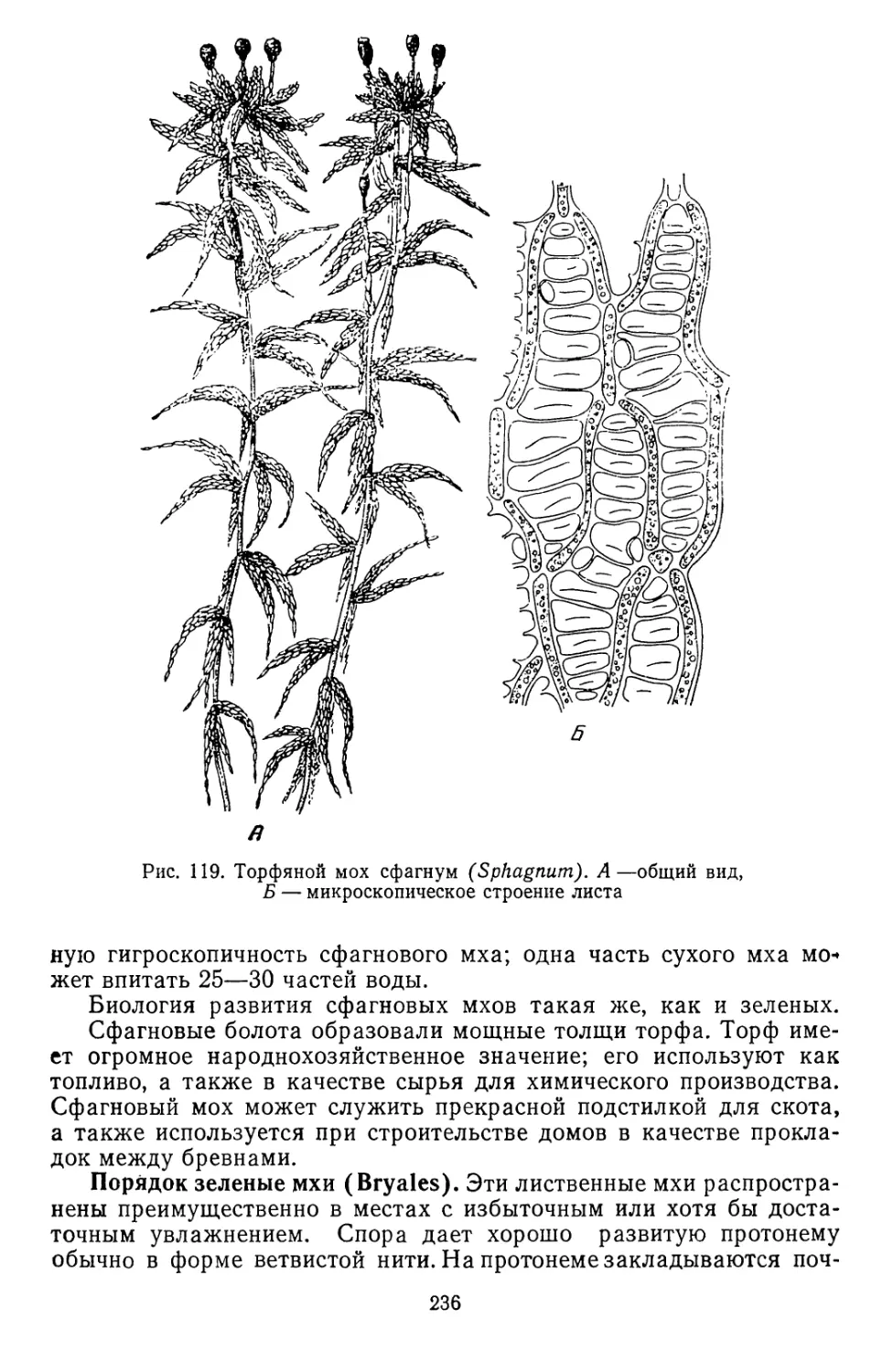























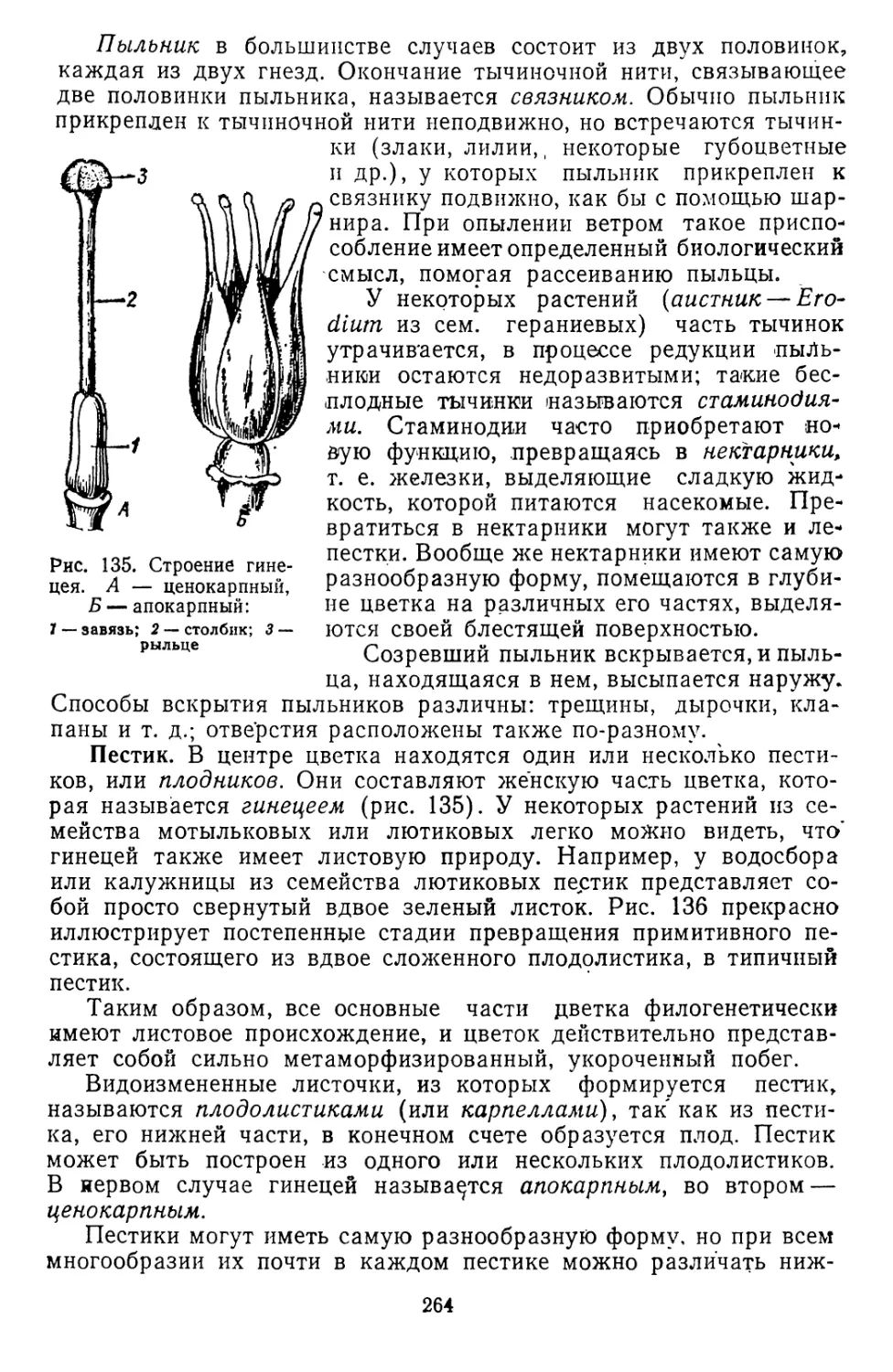
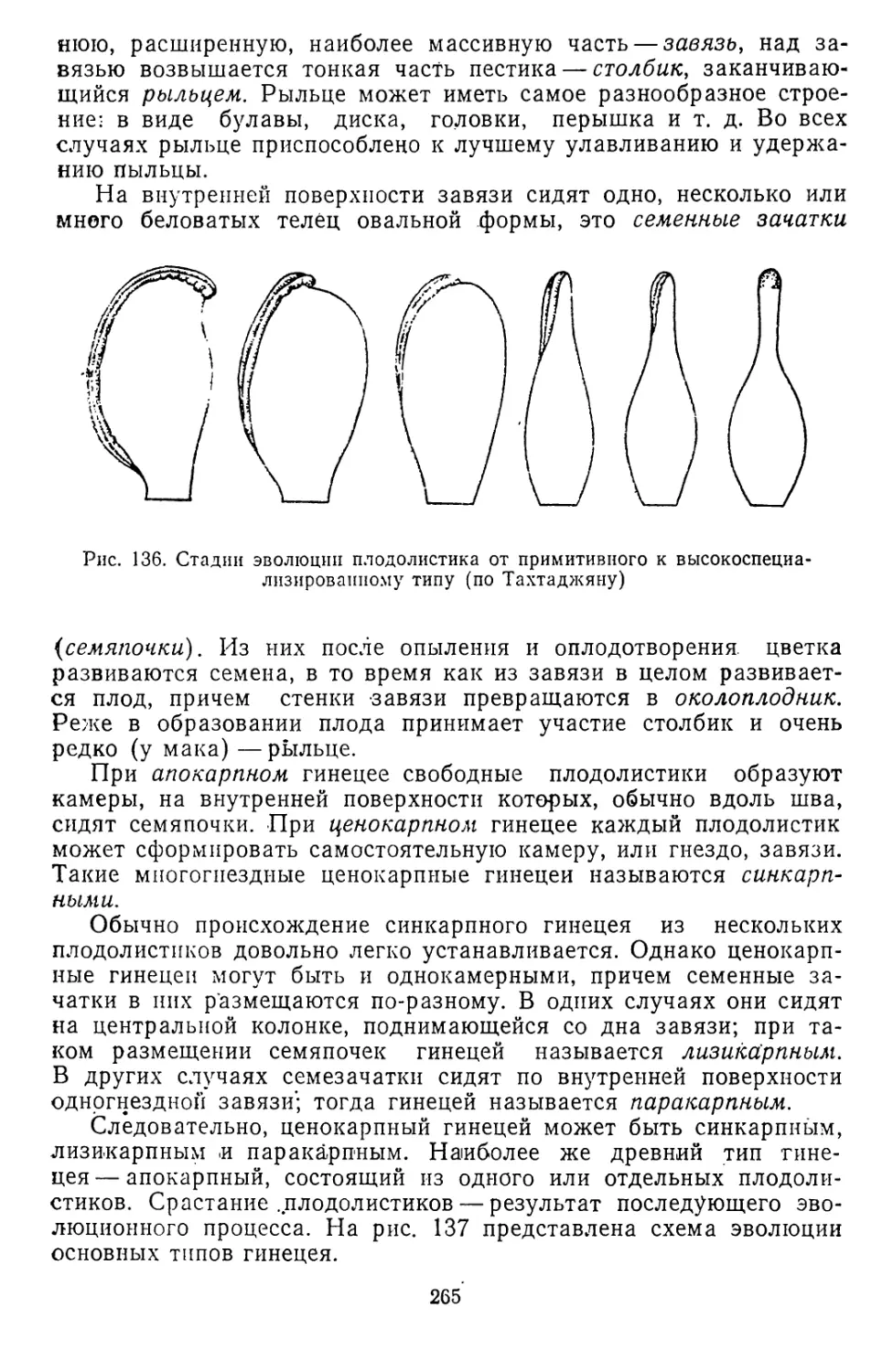
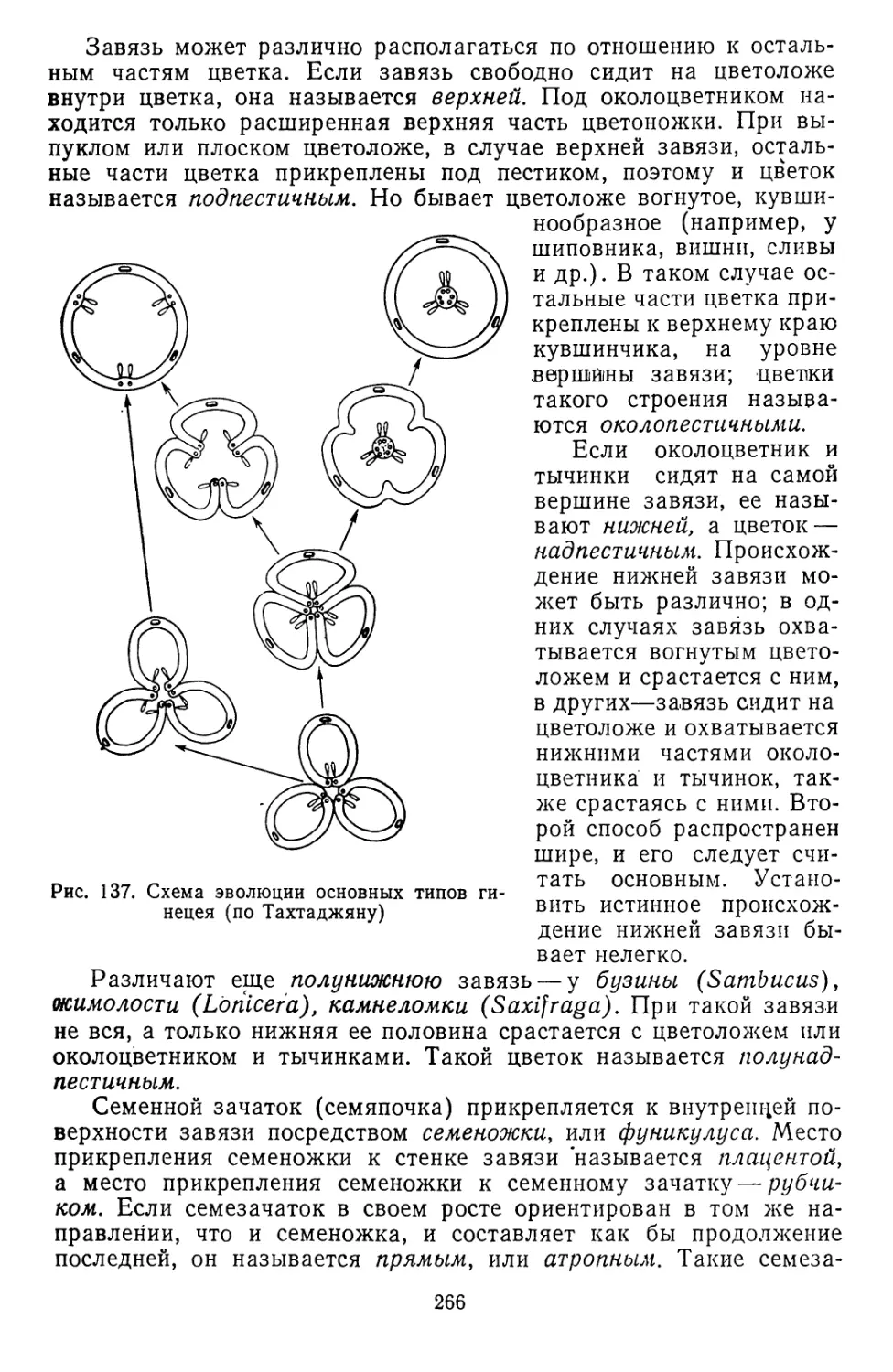
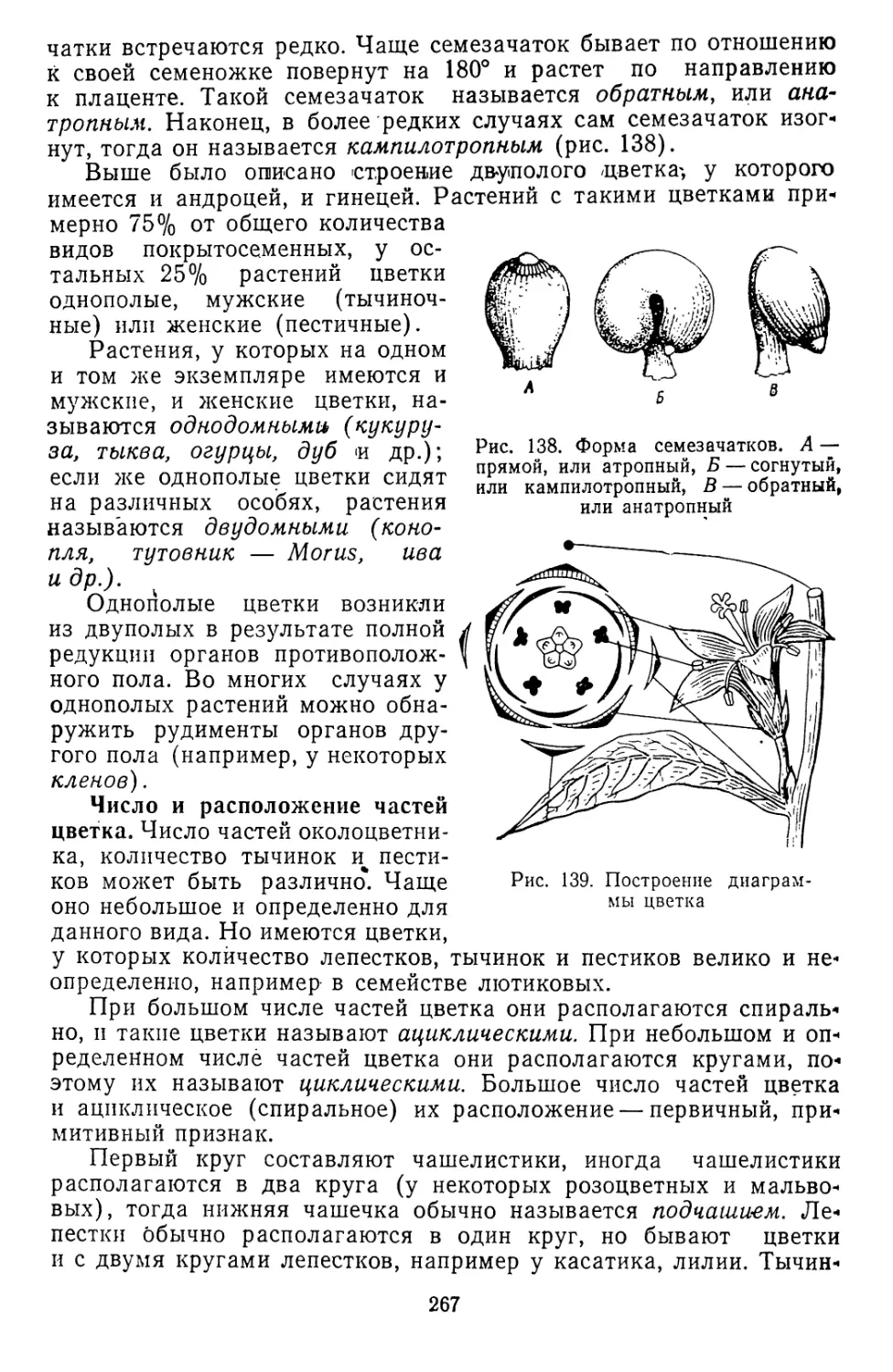






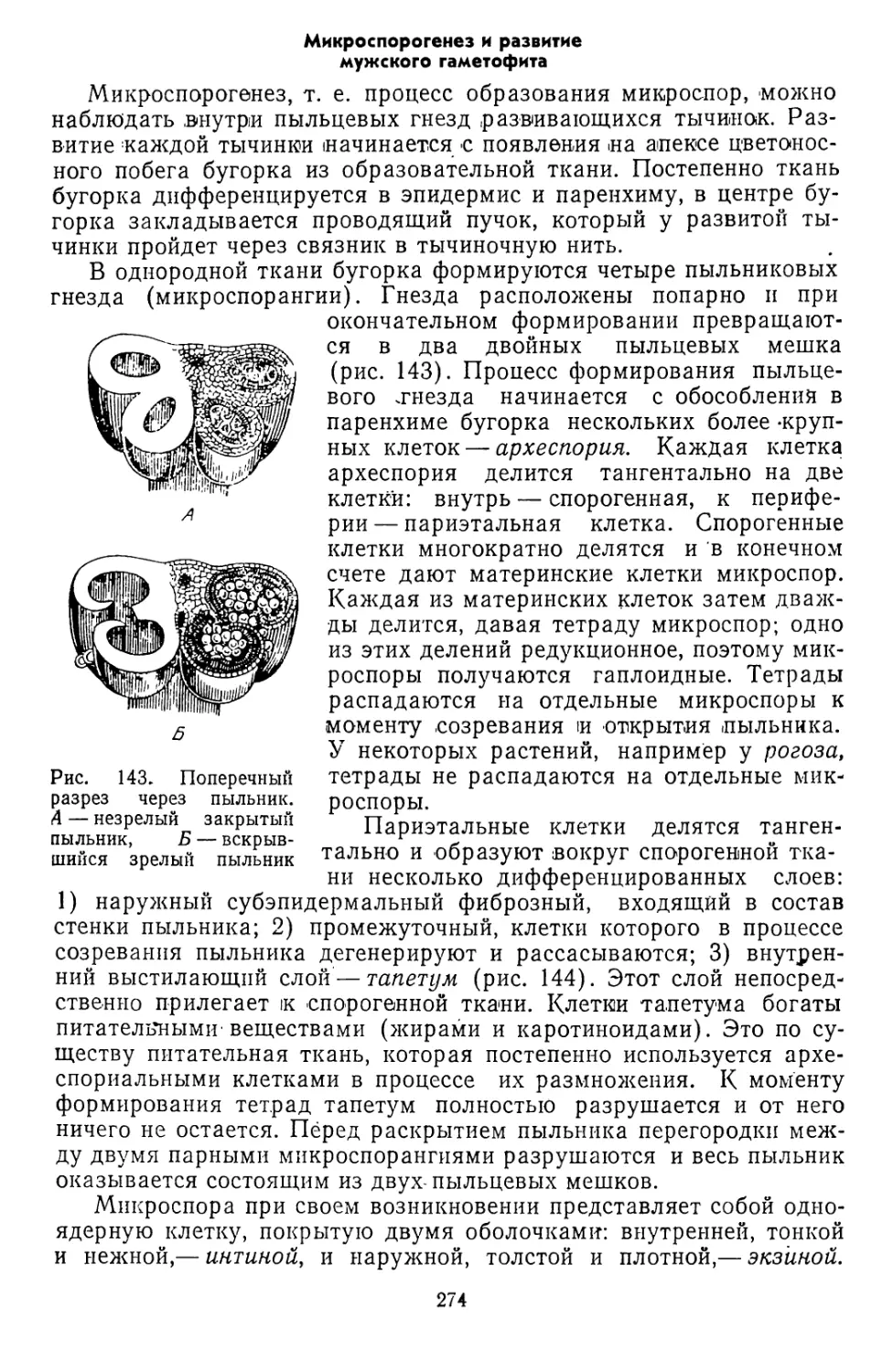



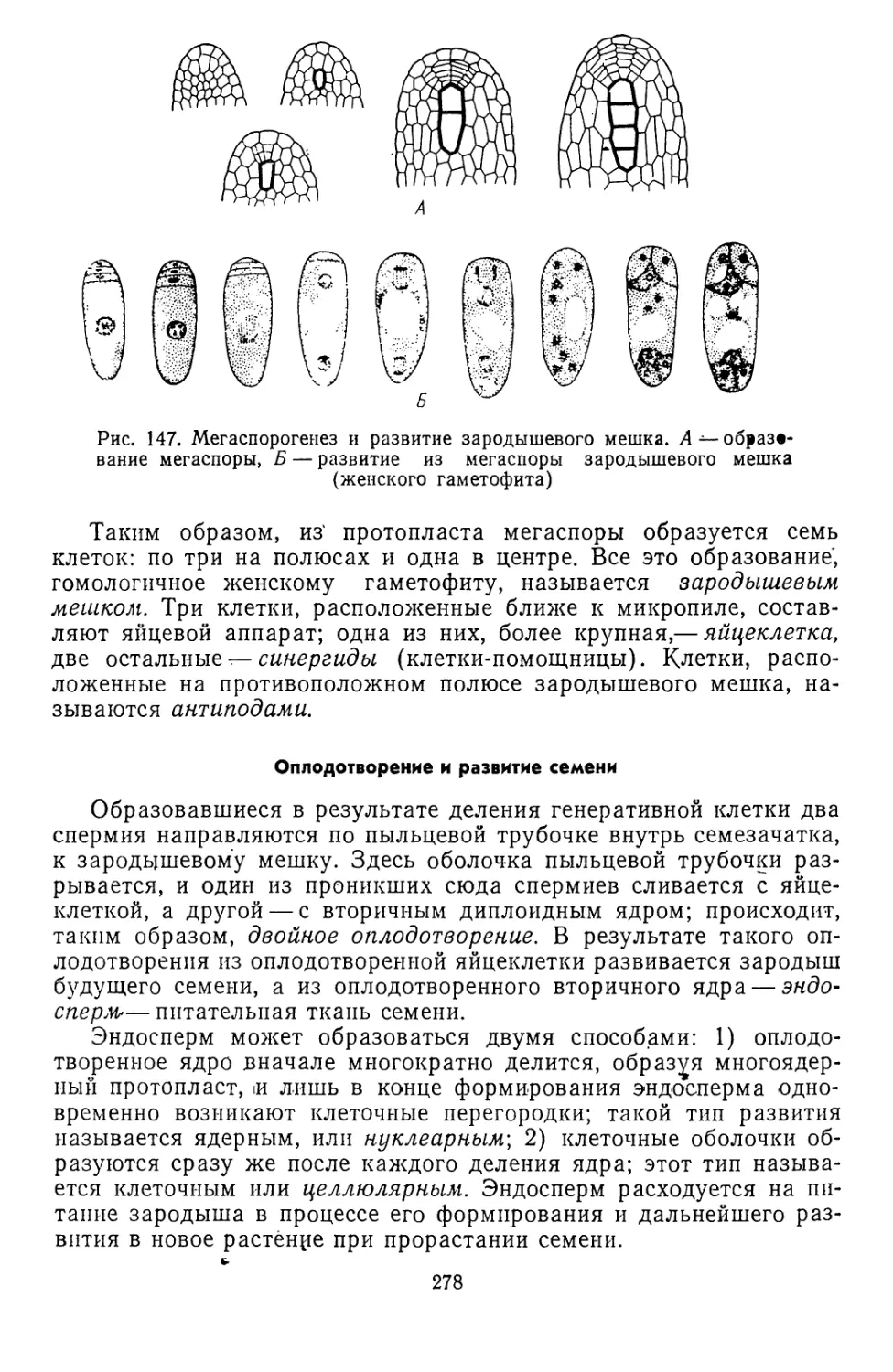





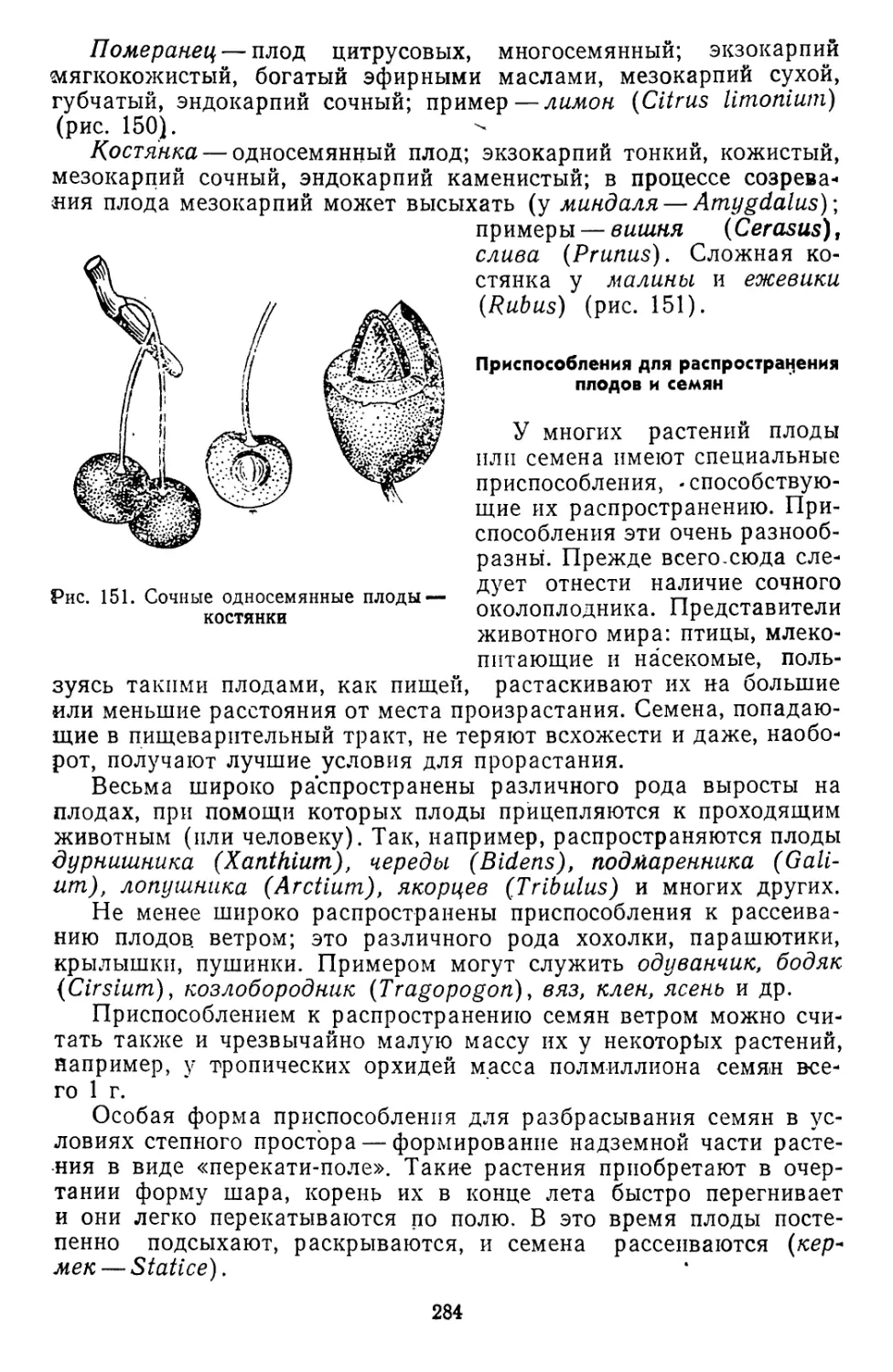





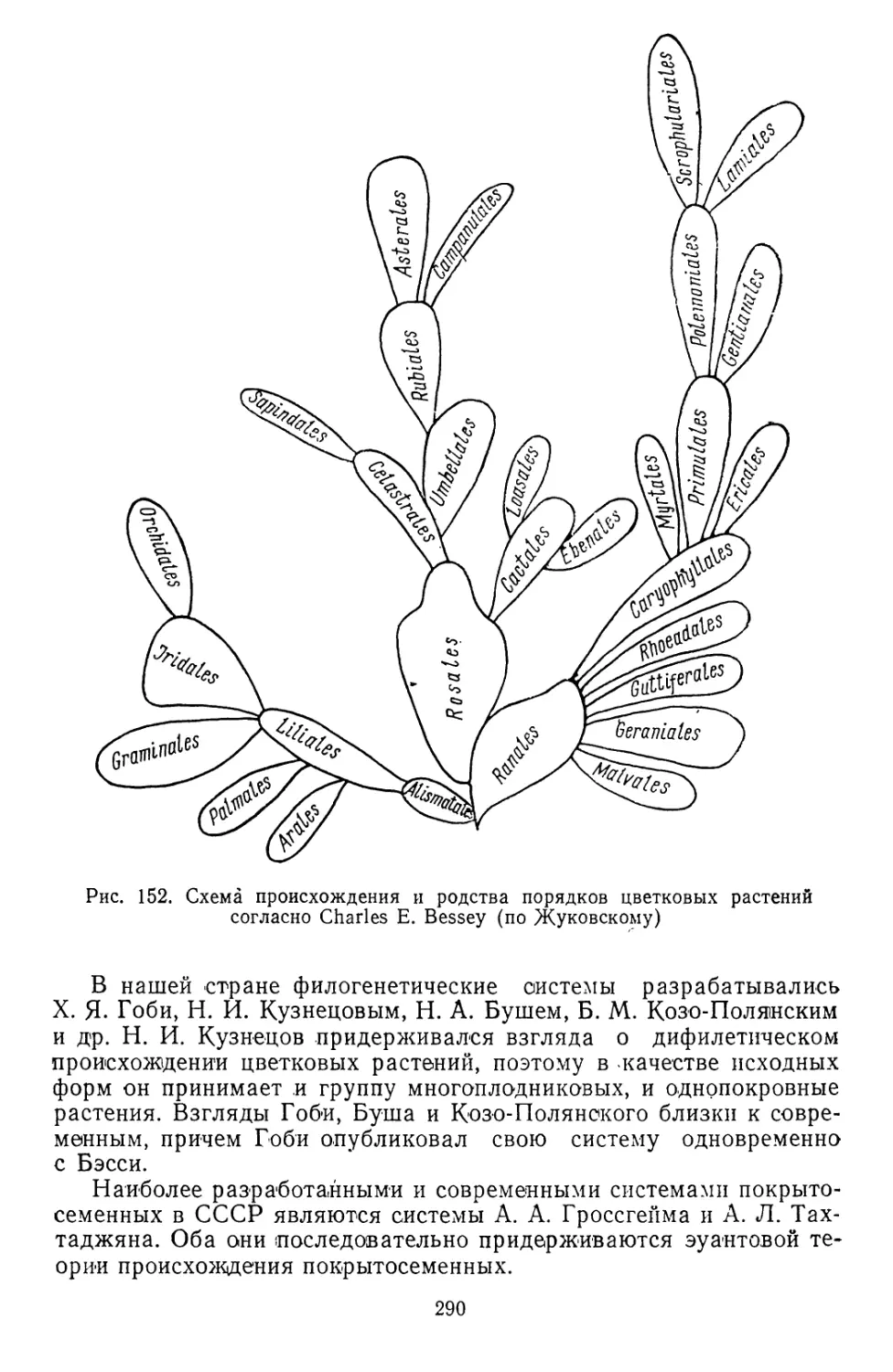

















































































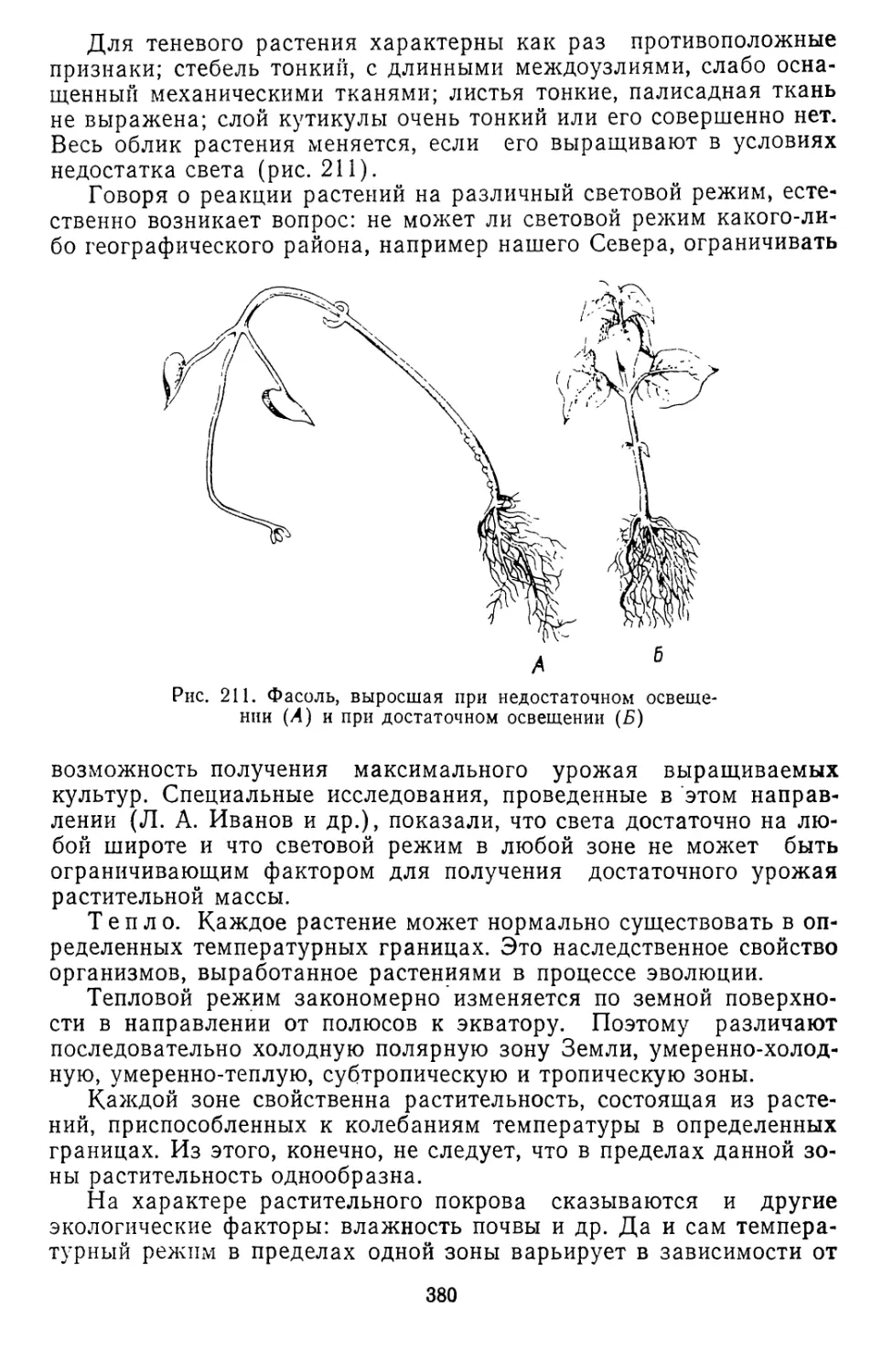
































































Текст
<Л К. ТИХОМИРОВ
ТЮТАТТТТКД
JL> v_z A21JOLJrJLJlVxjl
Ф. к. ТИХОМИРОВ
БОТАНИКА
ИЗДАНИЕ 4-е, ДОПОЛНЕННОЕ
Допущено
Главным управлением
высшего и среднего
сельскохозяйственного образования
Министерства сельского хозяйства ССЖР
в качестве учебника для студенто
высших сельскохозяйственных
учебнж «ведений
по сжЕпяальности
«жэжехния»
МОСКВА «ВЫСШАЯ ШКОЛА» 1978
ББК 28.5
Т 46
Рецензент:
кафедра ботаники Ленинградского сельскохозяйственного
института (зав. кафедрой проф. В. К. Пельменев)
Тихомиров Ф. К.
Т46 Ботаника: Учебник для с.-х. вузов.—4-е изд., доп.— М.:
Высш, школа, 1978. —439 с., ил. и табл.
В пер.: 1 р. 40 к.
В учебнике в сжатой форме изложены основы ботаники: внешнее и внутреннее
строение высших и низших растений. Более подробно описаны растения, имеющие
практическое значение.
Предназначается для студентов зооинженерных факультетов сельскохозяйствен-
ных вузов, а также с успехом может быть использован практиками сельского хо-
зяйства.
Т 21006~470 . БЗ—102—24—77 ББК 28.5
001(01)—78 58
© Издательство «Высшая школа», 197£
ВВЕДЕНИЕ
ОПРЕДЕЛЕНИЕ НАУКИ И ОБЪЕКТ
ЕЕ ИЗУЧЕНИЯ
Ботаника — наука о растениях. Она изучает строение и жизнь
растений в связи с условиями их обитания; классифицирует расте-
ния и устанавливает систему растительного мира, отражающую
историю его развития; исследует растительный покров земной по-
верхности и закономерности сочетания в нем отдельных растений.
Ботаника — биологическая наука, имеющая огромное практи-
ческое значение. В настоящее время огромное внимание уделяется
широкому использованию ботаники, приобретающей все большее
значение для поднятия плодородия почвы, выработки пищевых
и кормовых средств, изыскания новых приемов агротехники.
Ботанические знания широко используются человеком для удов-
летворения его материальных и духовных потребностей. Она
является биологической основой агрономических наук, изучающих
растения с прикладными, производственными целями. В системе
подготовки зоотехников ботанические знания необходимы для
усвоения таких дисциплин, как растениеводство, луговодство,
кормление сельскохозяйственных животных.
Жизнь человека тесно связана с окружающим его раститель-
ным миром, и мы не всегда отчетливо и полно представляем себе
все многообразие этих связей. Растительный мир гораздо богаче
и разнообразнее, чем нам обычно представляется. Кроме крупных
растений, видимых простым, невооруженным глазом, существует
богатейший мир растений микроскопических, населяющих и воды,
и сушу.
В практической жизни мы чаще имеем дело с зелеными расте-
ниями: деревьями, кустарниками, травами. Однако каждому из-
вестны и такие организмы, как грибы, например съедобные шля-
почные грибы. Они совсем не похожи на обычные зеленые расте-
ния, гораздо проще устроены и совершенно иначе питаются.
Зеленые растения из неорганических веществ создают вещества
органические. Можно, например, положить семя какого-нибудь
растения на прокаленный песок, лишенный органического вещест-
ва, поливать этот песок растворами минеральных солей, необходи-
мых растению, и из этого семени вырастет нормальное растение.
Наука, занимающаяся изучением жизни растений,— физиология —
3
давно уже установила, что при этом растение берет воду и мине-
ральные вещества из почвы, углекислый газ — из воздуха и из
этих неорганических веществ строит различные органические ве-
щества; при этом выделяется свободный кислород.
Такой процесс протекает с участием света, и световая энергия
при этом накапливается в растении в виде скрытой энергии орга-
нических веществ. Поэтому органическое вещество, например ве-
щество растения, при сжигании может выделять энергию. Это как
раз и есть та световая энергия, которая была раньше усвоена ра-
стением в процессе создания органического вещества.
Растения, которые сами создают из неорганических веществ
необходимые им органические соединения, называются автотроф-
ными, что означает самопитающиеся растения. Этим названием
подчеркивается, что такие растения могут сами создавать органи-
ческое вещество из элементов неживой природы. Кроме зеленых
растений, населяющих сушу, к автотрофным растениям относятся)
и многочисленные водоросли, обитающие в основном в воде. *|
Совершенно иначе питаются грибы: им обязательно необходи-i
мо готовое органическое вещество. Там, где такого вещества нет,
нет и грибов. Одни грибы поселяются на отмерших организмах,
это грибы-сапрофиты. К ним относятся, например, большинство
съедобных грибов и различные плесени. Но многие грибы поселя-
ются на живых растениях и даже на животных, это грибы-парази-
ты. К ним относятся, например, грибы, вызывающие заболевания
растений: головня, ржавчина, мучнистая роса и др.
В противоположность зеленым растениям грибы называются
гетеротрофными растениями, что означает инакопитающиеся.
Этим названием подчеркивается, что грибы питаются иначе, чем
обычные зеленые растения: они не могут создавать органические
вещества из неорганических. Подобно грибам питается большин-
ство бактерий — мельчайших организмов, невидимых простым, не-
вооруженным глазом.
Роль гетеротрофных растений в природе противоположна роли
зеленых, автотрофных растений. Последние создают органическое
вещество, гетеротрофные же растения, грибы и бактерии, разлага-
ют, разрушают органическое вещество; при этом освобождаются
и возвращаются в почву или воду минеральные вещества, необхо-
димые для питания зеленых, автотрофных растений. Так, посред-
ством этих двух групп растений, а также и животных совершается
беспрерывный круговорот веществ в природе.
Животные питаются подобно гетеротрофным растениям: они
также не могут заново создавать органическое вещество и нужда-
ются в органической пище. Однако в отличие от растений живот-
ные переваривают пищу внутри своего тела, в то время как расте-
ния способны всасывать пищу лишь в растворенном состоянии.
Наиболее низкоорганизованные, первичные растения и животные
бывают настолько сходны между собой, что указанное различие
в способах поглощения и переваривания пищи становится основ-
ным критерием для их разделения.
4
Разделяя растительный мир на автотрофов и гетеротрофов
и подчеркивая противоположную роль этих групп в круговороте
веществ в природе, следует указать, что разделение это не носит
абсолютного характера. Некоторые растения, например синезеле-
ные водоросли, питаются смешанно, их можно назвать миксотрофа-
ми. С другой стороны, в группе бактерий, где преобладает гетеро-
трофное питание, имеются и автотрофные организмы.
возникновение и эволюция
РАСТИТЕЛЬНОГО МИРА
Современное состояние естественных наук дает нам возмож-
ность представить картину возникновения и эволюции жизни на
Земле. Возраст нашей планеты около четырех с половиной мил-
лиардов лет. Первые живые организмы на Земле появились, по-ви-
димому, около трех миллиардов лет назад. В течение полутора
миллиардов лет на поверхности Земли постепенно складывались
условия, необходимые для возникновения простейших живых орга-
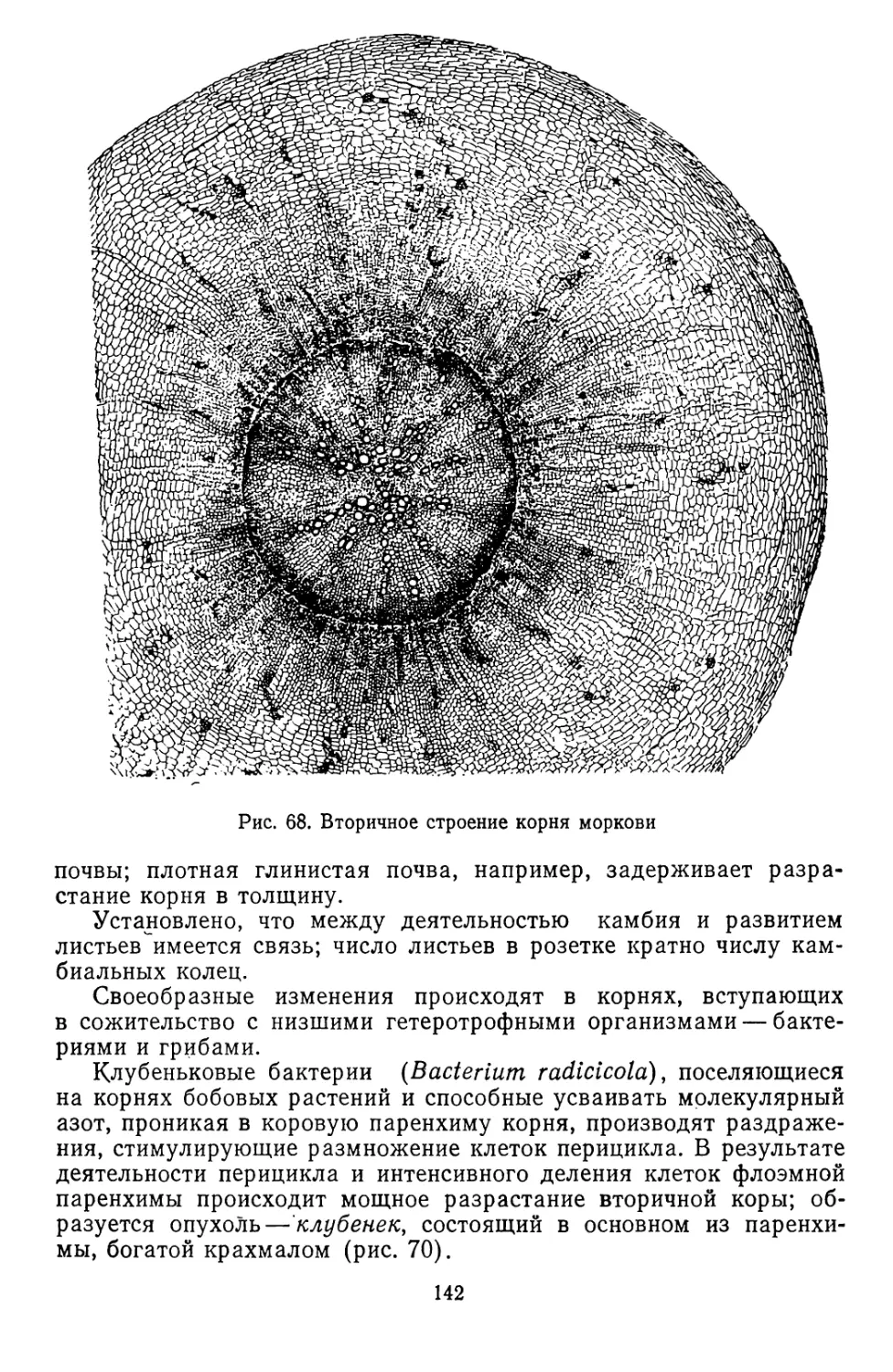
низмов. После того как Земля достаточно остыла, впадины ее по-
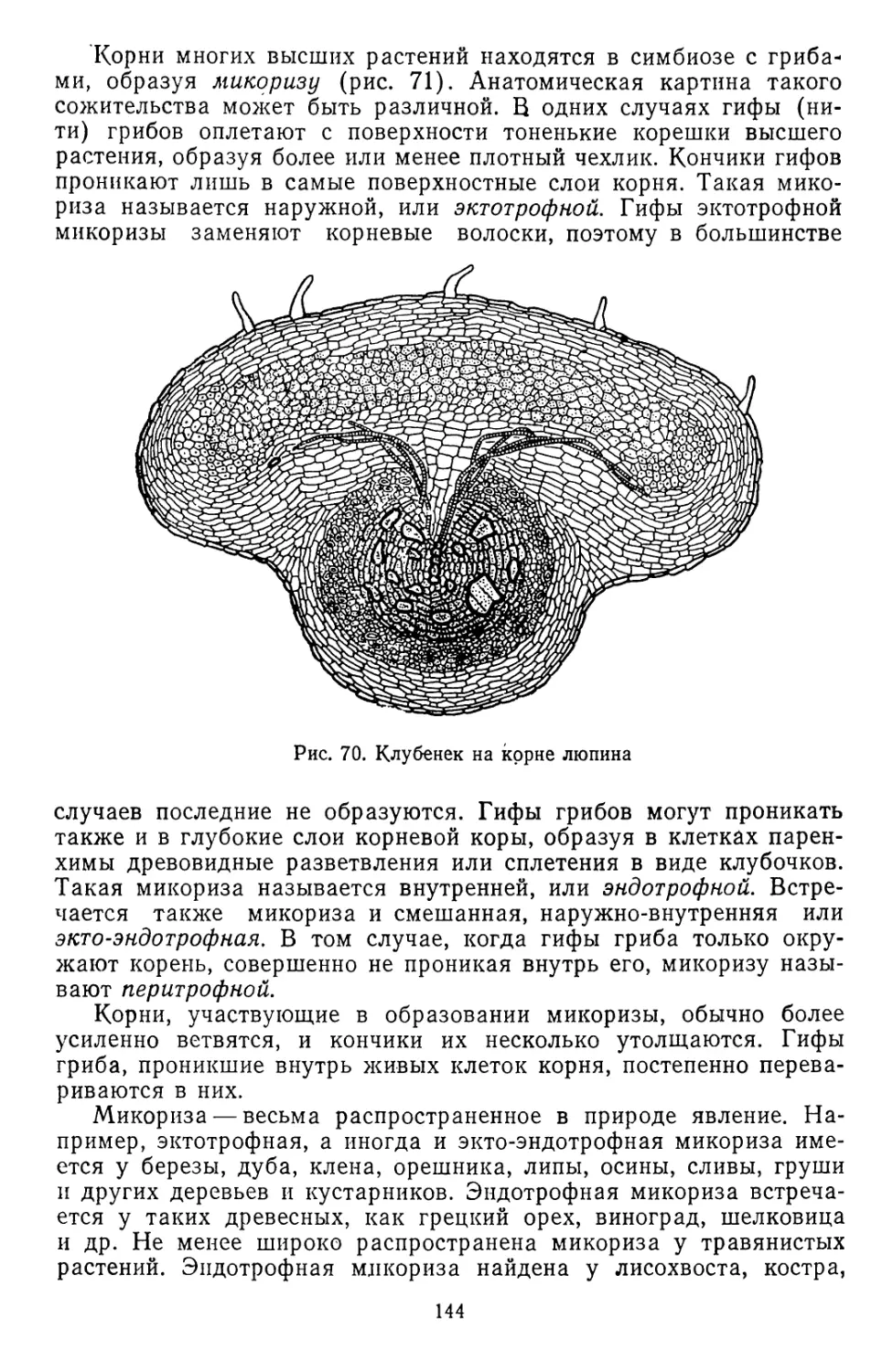
верхности заполнились водой, возник Мировой океан — колыбель
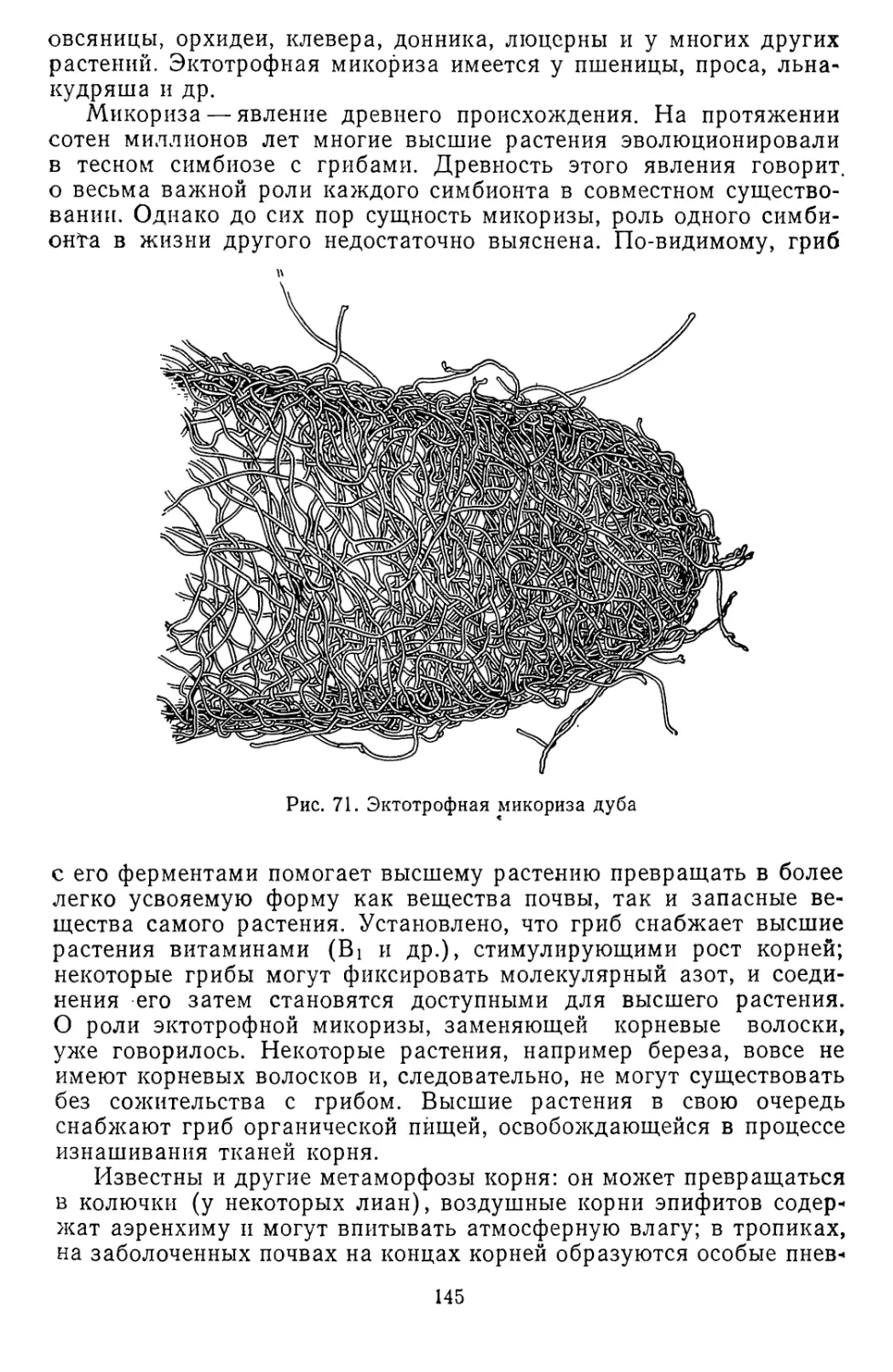
жизни. В первичной атмосфере, составлявшей наружную газовую
оболочку Земли, содержались пары воды, водород, азот, метан,
аммиак, окись углерода. Протекавшие в недрах Земли интенсивные
процессы радиоактивного распада были причиной бурной вулка-
нической деятельности планеты. В составе солнечных лучей к по-
верхности Земли свободно проникало коротковолновое ультрафио-
летовое излучение — мощный источник энергии для разнообразных
синтетических процессов. Неравномерность нагрева земной поверх-
ности способствовала возникновению мощных грозовых разрядов.
Как показали специально организованные опыты, под действи-
ем высоких температур, электрических разрядов и коротковолно-
вого ультрафиолетового излучения из смеси названных веществ,
свойственных первичной атмосфере, в водной среде могут синтези-
роваться более сложные органические соединения: сахар, амино-
кислоты, азотистые основания; из этих простейших органических
веществ первичного синтеза в благоприятных условиях водной
среды могут быть построены и те вещества, которые необходимы
для возникновения жизни: белки, нуклеиновые кислоты, углеводы,
липиды и др. Подобные реакции, по-видимому, протекали и в пер-
вичной атмосфере Земли, и в Мировом океане, прежде чем могли
сформироваться первые живые самовоспроизводящиеся системы.
Первые живые организмы нельзя было назвать ни животными,
ни растениями, это могли быть просто капельки живой материи,
способные к самообновлению. Они, очевидно, имели более простое
строение, чем простейшие клеточные организмы, например бакте-
рии. Более того, в свете современных представлений свойствами
живого мог обладать уже молекулярный белок в соединении с мо-
лекулами нуклеиновых кислот: дезоксирибонуклеиновой (ДНК)
5
или рибонуклеиновой (РНК). Комплекс из этих двух компонентов
составляет как бы материальный субстрат жизни дарке на самом
низком уровне ее организации: науке не известны биологические
системы, способные к самовоспроизведению, в которых не присут-
ствовали бы оба названных компонента: белок и нуклеиновая
кислота.
Первичные живые организмы были гетеротрофные, они не со-
держали хлорофилла и питались органическими веществами абио-
тического синтеза и, очевидно, продуктами распада отмирающих
организмов. Такая гетеротрофная стадия жизни длилась очень
долго, около миллиарда лет. По всей вероятности это была очень
скудная жизнь, не приобщенная еще к энергии Солнца. Современ-
ные наши бактерии могут дать представление о степени сложности
этих древних организмов; хотя они и имели нуклеиновые кислоты,
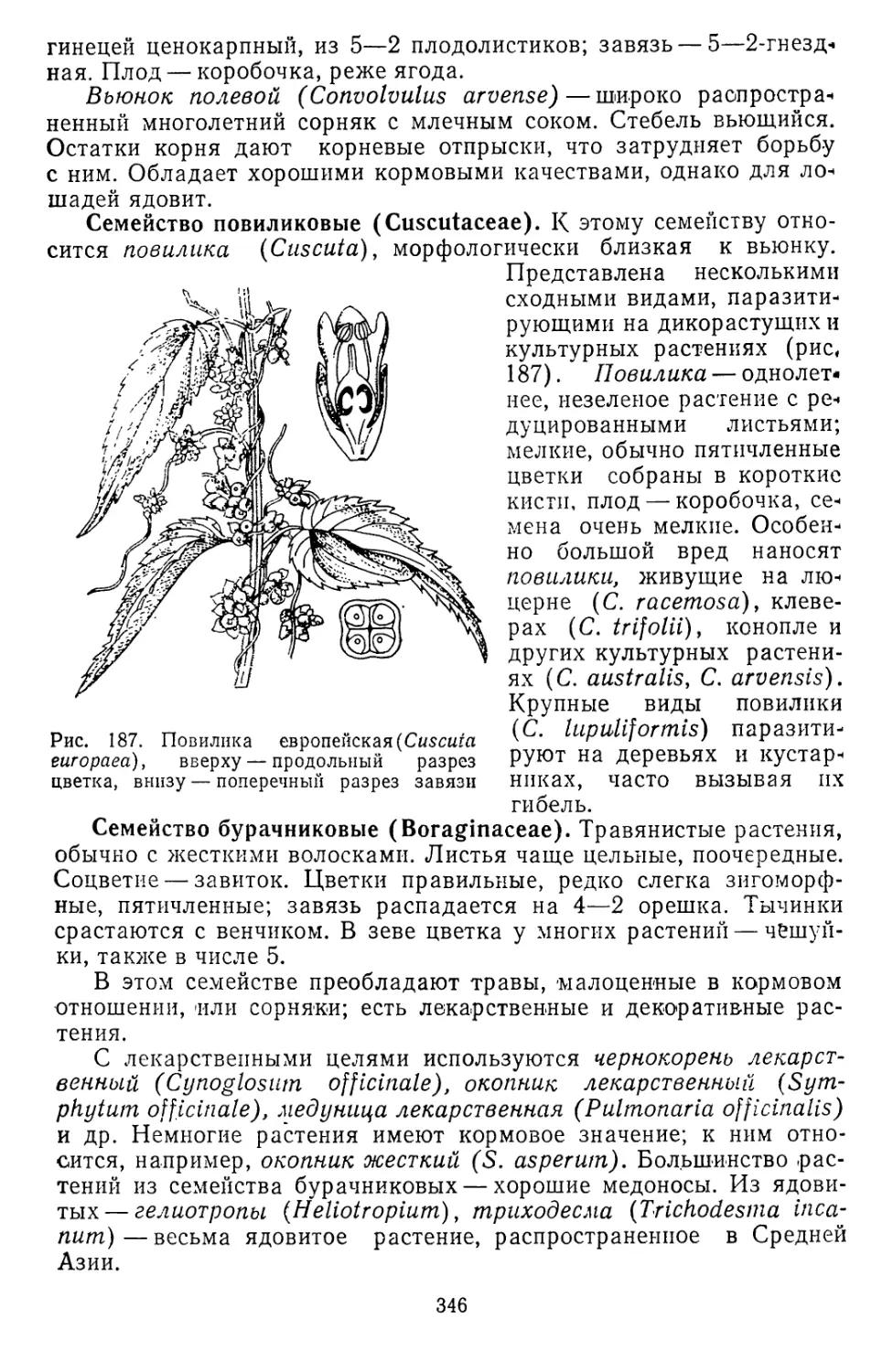
но морфологически оформленных ядер еще не было. Такие орга-
низмы называются прокариотными. В состоянии прокариотных
клеток жизнь существовала около двух миллиардов лет. Первые
фотосинтезирующие и азотфиксирующие организмы — синезеленые
водоросли были также еще прокариотными. Возникновение этих
водорослей около 2 миллиардов лет назад можно рассматривать
как переломный момент в развитии жизни на Земле. Это были пер-
вые автотрофные растения. Значительно позже, около миллиарда
лет назад, возникли эукариотные растения: зеленые, бурые, крас-
ные и другие водоросли; в клетках этих водорослей основная мас-
са нуклеиновых кислот сконцентрировалась в морфологически
оформленном ядре, а красящие вещества — пигменты, в том числе
и хлорофилл,— в особых органоидах: хроматофорах и пластидах.
Возникновение автотрофных растений связано с образованием
особого вещества — хлорофилла, от которого зависит зеленый цвет
растений. При помощи хлорофилла организмы приобрели способ-
ность использовать солнечную энергию для создания органического
вещества из неорганических. Поглощенная солнечная энергия на-
капливается в создаваемом органическом веществе. Тем самым
жизнь на Земле приобретает источник энергии для своего разви-
тия.
Именно через хлорофилл беспредельный поток солнечной энер-
гии переливается в живые организмы на Земле, и в этом отношении
не только зеленые растения, но и всех животных и людей можно
назвать детьми Солнца.
Процесс создания органического вещества с участием световой
энергии называется фотосинтезом.
Способность зеленых растений создавать органическое вещест-
во и в нем аккумулировать энергию Солнца явилась мощным фак-
тором эволюции растительного и животного мира. Зеленые расте-
ния достигли высокой степени совершенства. В итоге эволюции,,
длившейся сотни миллионов лет, возникли многочисленные груп-
пы зеленых растений.
Низшие зеленые растения — водоросли, обитающие, как прави-
ло, в воде, дали начало высшим растениям, приспособившимся
6
жить на суше. Венцом эволюции явились цветковые, или покрыто-
семенные, растения, составляющие на большей части суши основу
растительного покрова.
Совершенство строения высших растений связано с их приспо-
соблением к жизни в условиях суши.. Выйдя из водной среды, от-
даленные предки современных высших растений стали приспосаб-
ливаться к гораздо более сложной жизненной обстановке. В ходе
этого приспособления и возникла большая сложность внешнего
и внутреннего строения тела высших растений.
Процесс постепенного приспособления к новым условиям жизни
длился многие миллионы лет. Ныне живущие растения, а также
ископаемые остатки когда-то живших растений позволяют нам
в общих чертах восстановить картину эволюции растительного ми-
ра с момента первоначального освоения суши и до наших дней.
Остатки древнейших высших растений, непосредственно произо-
шедших от водорослей, не сохранились. Поэтому трудно решить,
какая именно группа водорослей дала начало высшим растениям.
Скорее всего это были зеленые водоросли, однако не исключена
возможность, что начало высшим растениям деЛи бурые водоросли.
Массовый выход растений на сушу начался примерно около
450 млн. лет назад. До этого жизнь свыше двух миллиардов лет
развивалась в водной среде.
Время появления и существования той иди иной группы расте-
ний ботаническая наука исчисляет, пользуясь хронологией, приня-
той в исторической геологии, изучающей историю и закономерно-
сти развития Земли и земной коры. Историческая геология делит
историю земной коры на эры, а эры — на периоды. В дальнейшем
изложении мы будем пользоваться терминам# геологического ле-
тоисчисления. Чтобы легче было ориентироваться в этих терминах,
приводим схематическую таблицу 1 геологической хронологии
(данные Комиссии АН СССР по определению абсолютного возра-
ста геологических формаций, Т. Д. Афанасьевой др., 1964).
Подпериоды: палеоген и неоген. В палеогене три эпохи: палео-
цен (7 млн. лет), эоцен (23 млн. лет), олигоцен (12 млн. лет).
В неогене две эпохи: миоцен (13 млн. лет) и плиоцен (10—10,5 млн.
лет).
Четвертичный период делят на две эпохи: плейстоцен продол-
жительностью 1,5—2 млн. лет и современного эпоху — голоцен —
11 000 лет.
На протяжении сотен миллионов лет лик Земли неоднократно
менялся. Целые материки и части их то уходили под воду, то снова
появлялись. Поднявшиеся, участки суши разрешались, и продукты
разрушения в виде ила, глины и песка накапливались на дне мо-
рей и океанов. Слои осадочных пород — это та летопись, которую
ученые читают, чтобы узнать историю Земли и возраст живших
на ней организмов.
По данным палеонтологии, за всю историю Земли на ней оби-
тало около 500 млн. видов различных организмов, но ко времени
появления человека (не менее 2 млн. лет назад) их было около
7
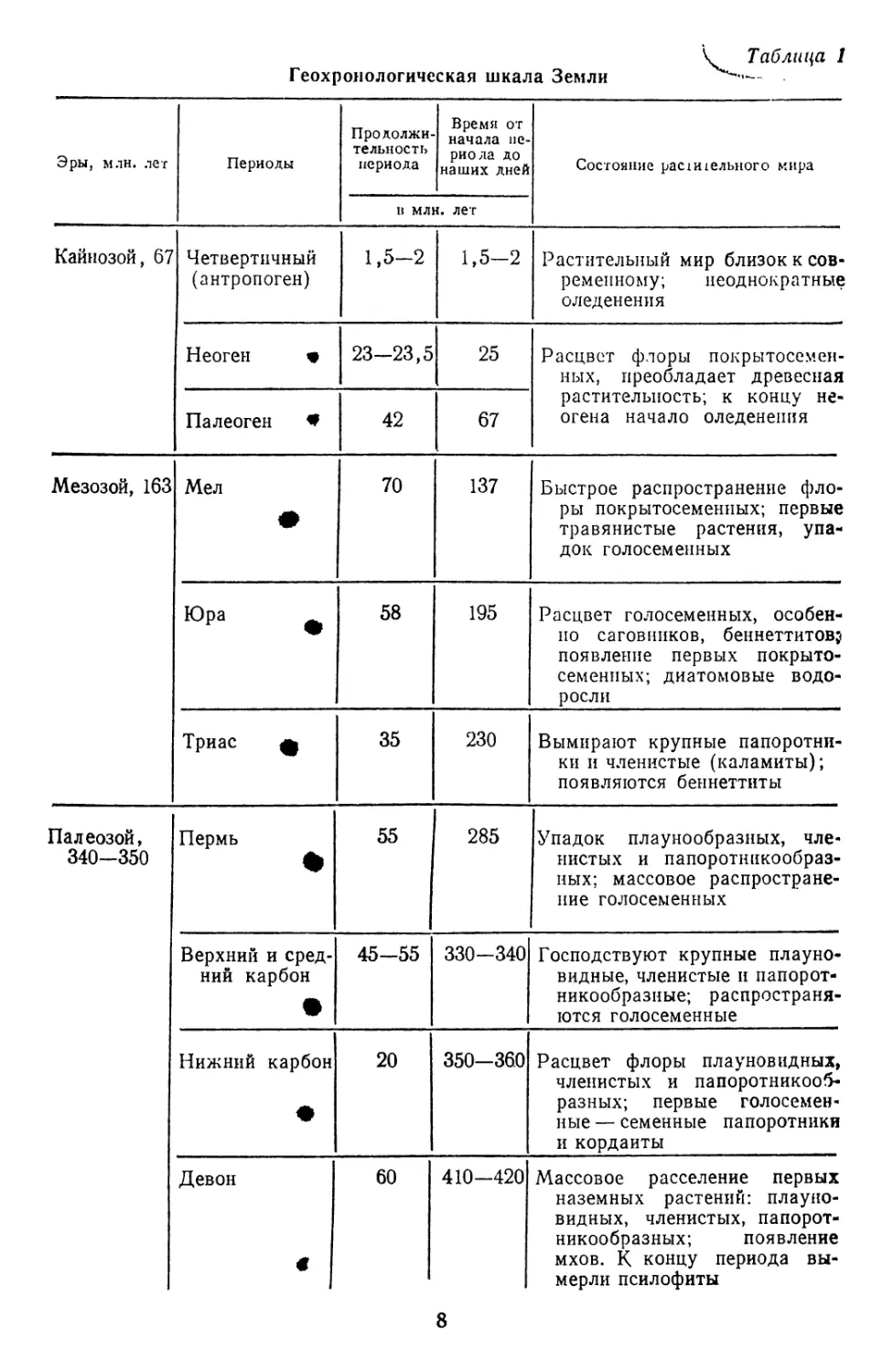
Таблица 1
Геохронологическая шкала Земли
Эры, млн. лег Периоды Продолжи- тельность периода Время от начала пе- риода до наших дней Состояние рас1И1ельного мира
в млн. лет
Кайнозой, 67 Четвертичный (антропоген) 1,5-2 1,5-2 Растительный мир близок к сов- ременному; неоднократные оледенения
Неоген « 23-23,5 25 Расцвет флоры покрытосемен- ных, преобладает древесная растительность; к концу не- огена начало оледенения
Палеоген W 42 67
Мезозой, 163 Мел • 70 137 Быстрое распространение фло- ры покрытосеменных; первые травянистые растения, упа- док голосеменных
Юра 58 195 Расцвет голосеменных, особен- но саговников, беннеттитов? появление первых покрыто- семенных; диатомовые водо- росли
Триас ф 35 230 Вымирают крупные папоротни- ки и членистые (каламиты); появляются беннеттиты
Палеозой, 340-350 Пермь • 55 285 Упадок плаунообразных, чле- нистых и папоротникообраз- ных; массовое распростране- ние голосеменных
Верхний и сред- ний карбон • 45-55 330-340 Господствуют крупные плауно- видные, членистые и папорот- никообразные; распространя- ются голосеменные
Нижний карбон • 20 350-360 Расцвет флоры плауновидных, членистых и папоротникооб- разных; первые голосемен- ные — семенные папоротники и кордаиты
Девон < 60 410—420 Массовое расселение первых наземных растений: плауно- видных, членистых, папорот- никообразных; появление мхов. К концу периода вы- мерли псилофиты
8
Продолжение табл. 1
Эры, млн. лег Периоды Продолжи- тельное! ь периода в млн Время от начала пе- риода до наших дней . лег Состояние растительного мира
Палеозой, 340-350 Силур * 30 440—450 Выход растений (псилофитов) на сушу. Господствуют водо- росли
Ордовик 60 500-510 Вероятное появление первых наземных растений. Господ- ство водорослей
Кембрий * 70 570-580 Господство водорослей; воз- можны единичные выходы растений на сушу
Докембрий, 3000 Протерозой, 2000 * 2580 Жизнь протекала в воде; зеле- ные, бурые, красные и сине- зеленые водоросли; возмож- ны грибы
Археи, 1000 ♦ >3500 Зарождение жизни в воде; бак- терии и синезеленые водорос- ли
2 млн., в том числе до 500 тыс. видов растений и около 1,5 млн.
видов животных. Следовательно, история жизни на Земле — это
история возникновения и отмирания все новых и новых видов ра-
стений и животных, сменяющих друг друга в ходе приспособления
к изменяющимся условиям обитания. Те организмы, которые в на-
стоящее время обитают на нашей планете,— это в основном виды,
наиболее приспособленные к существованию в современных усло-
виях.
РАСТЕНИЯ —ОСНОВНОЙ КОМПОНЕНТ
БИОСФЕРЫ
Биосфера — это оболочка Земли, состав, структура и энергети-*
ка которой в существенных чертах обусловлены прошлой или со-
временной деятельностью живых организмов. Они — создатели
первичного органического вещества, за счет которого существует,
практически, весь остальной живой мир; деятельностью их атмо-
сфера насыщена кислородом, необходимым для дыхания живых
организмов и, наконец, за счет кислорода, освобожденного зелены-
ми растениями, в верхних слоях атмосферы, на высоте 10—50 км
9
создан слой из озона, как экран, защищающий поверхность Земли
от ультрафиолетового излучения короче 290 нм. Излучение это-
смертельно для всего живого. Поэтому лишь после возникновения
озонового экрана растения и животные, до того обитавшие в воде,
могли выйти на сушу. Как уже сказано, это произошло около
450 млн. лет назад. Кислород, насытивший к этому времени атмо-
сферу и создавший озоновый экран, был освобожден из воды
фотосинтетической деятельностью водорослей, населявших Миро-
вой океан. Не легко точно определить гранты биосферы. Как по-
казали исследования, споры бактерий и грибов встречаются на
высоте нескольких десятков километров (40—80) над поверхно-
стью Земли, однако совершенно очевидно, ^то на такой высоте
отсутствуют условия для обмена веществ. В связи с этим возникает
необходимость введения в науку понятия парабиосферной, пере-
ходной зоны, где хотя и встречаются диаспоры (зачатки) живых
организмов, но нет активного метаболизма.^С этой точки зрения
к парабиосферной зоне следует, по-видимому, отнести и глубин-
ные слои коры земного шара, где могут быть сосредоточены горные
породы биогенного происхождения, но отсутствует в настоящее
время активный метаболизм.
Зеленые растения и в наше время играют ведущую роль в био-
сфере. Достаточно сказать, что на их долю приходится не менее
99% всей живой биомассы Земного шара, на долю же животных
и гетеротрофных растений — всего 1%- Однако и гетеротрофные
растения, грибы и бактерии, также являются необходимым компо-
нентом биосферы: без их участия круговорот жизни на Земле был
бы невозможен: для того чтобы круговорот веществ в природе про-
должался, необходимо, чтобы вся созданная зелеными растениями
биомасса (не менее 200 млрд, т в год) была минерализована; эту
огромную работу в конечном счете выполняют гетеротрофные орга-
низмы: животные, грибы и бактерии. Фотосинтезирующая деятель-
ность зеленых растений в большей части сосредоточена на суше:
лишь одна четверть всей создаваемой ежегодно биомассы прихо-
дится на Мировой океан, и здесь находится лишь около 0,1% всей
накопленной живой биомассы, которая в целом на планете состав-
ляет огромную величину — свыше двух триллионов тонн.
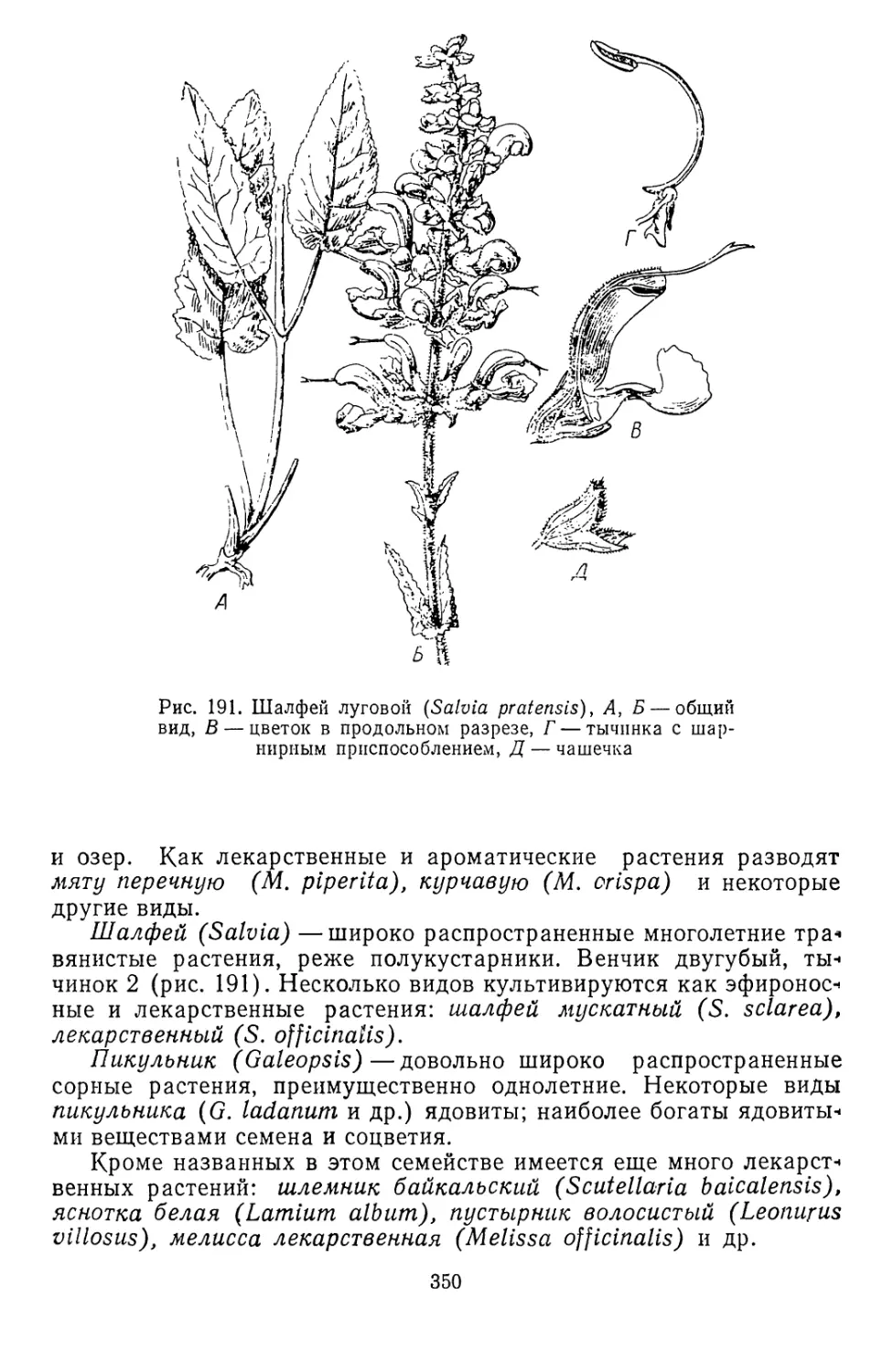
Зеленые растения, будучи первосоздателями биосферы, являют-
ся также и хранителями ее. Охранная роль зеленых растений за-
ключается в том, что они поддерживают биосферу в состоянии,
пригодном для продолжения жизни в ее современных формах, свя-
зывая энергию Солнца с жизнью на Земле, поддерживая в атмо-
сфере и биосфере оптимальное содержание кислорода и двуокиси
углерода. При непосредственном и ведущем участии зеленого ра-
стения создана почва — материальная основа человеческой цивили-
зации; они же и охраняют почву от разрушения и уничтожения.
Начиная с середины двадцатого столетия, биосфера подверга-
ется серьезной опасности со стороны негативных последствий науч-
но-технического прогресса. Заметно изменяется обычный состав
атмосферного воздуха: за последние 100 лет количество СО2 в ат-
10
мосфере увеличилось на 10%; этот процесс в наше время в быст-
рых темпах ускоряется, так что до конца XX в., как предполагает-
ся, количество СО2 в атмосфере может возрасти еще на 25%.
Уменьшилось на несколько процентов (3—4%) содержание кисло-
рода в атмосфере, а в Мировом океане даже более чем на 12%.
Атмосфера прогрессирующе загрязняется пылью и ядовитыми про-
дуктами неполного сгорания, это представляет прямую угрозу
жизни на Земле. Охрана растительного мира — основа всех меро-
приятий, направленных на предотвращение этой угрозы.
РАЗДЕЛЫ БОТАНИКИ
На протяжении многовековой истории ботанической науки
в связи с потребностями человеческого общества и уровнем техни-
ки складывались различные направления в изучении растений.
Накапливались знания, с различных сторон освещавшие строение
и жизнь растений, все более совершенствовались методы изучения
растений.
К настоящему времени ботаническая наука, имеющая единым
объектом своего исследования растение, растительный мир, разде-
лилась на ряд хотя и связанных между собой, но все же более или
менее самостоятельных ботанических дисциплин: морфологию, ана-
томию, физиологию, систематику и экологию растений, фитоцено-
логию, географию растений, палеоботанику и др. Каждая из этих
наук имеет свою область исследования, применяет свойственные ей
методы изучения.
Морфология изучает внешние формы растений. Прослеживая
изменения, которые происходят в формах растений на протяжении
длительной эволюции растительного мира и в течение индивиду-
альной жизни растения, эта наука выявляет общие закономерности
их образования, зависимость формы растений от изменяющихся
условий среды.
Анатомия изучает внутреннее строение растений. Внутреннюю
структуру растений можно хорошо рассмотреть только с помощью
увеличительных приборов, поэтому развитие анатомии тесно свя-
зано с усовершенствованием микроскопа. В анатомию растений
обычно в качестве самостоятельного раздела включают учение
о клетке. Этот раздел давно уже выделился в самостоятельную нау-
ку— цитологию, переживающую сейчас эпоху бурного развития.
Знание внутреннего строения растений необходимо для понима-
ния протекающих в них процессов. Этим занимается следующая
отрасль ботаники — физиология растений, которая, исследуя про-
цессы, протекающие внутри организма, помогает понять все тонко-
сти строения растений, установить закономерную связь между
формой растений, их структурой и условиями существования. Зна-
ние сущности физиологических процессов, протекающих в растени-
ях, дает нам ключ к управлению этими процессами; изменяя же
течение жизненных процессов, мы можем в нужную нам сторону
11
изменять и формы растений. В конечном счете знание физиологии
растений помогает нам решать практические задачи: повышать
урожай растений, больше получать растительной массы нужного
нам качества. Наряду с агрохимией физиология растений является
основой рационального земледелия (К. А. Тимирязев).
К физиологии растений очень тесно примыкает биохимия, ис-
следующая химические процессы, лежащие в основе жизненных
явлений. С другой стороны, биохимия примыкает к микробиоло-
гии— науке о микроорганизмах. Исследование биохимической
структуры живого вещества микробов и процессов, связанных с их
деятельностью, помогает нам раскрыть более общие закономерно-
сти явления жизни.
В последние десятилетия бурно развивается биофизика — наука,
изучающая физические и физико-химические процессы, протекаю-
щие в живых организмах. Объектом исследования этой науки яв-
ляется также структура организмов, биологические системы на
всех уровнях: от молекулярного до организменного.
Систематика растений ставит своей задачей классифицировать
все разнообразие растительных организмов, привести их в систему
возможно точнее отображающую процесс эволюции растительного
мира. Используя данные морфологии и других ботанических дис-
циплин, систематика объединяет в группы сходные растения, выяв-
ляет взаимную близость их в происхождении. Располагая эти
группы в систему, систематика пытается нарисовать общую карти-
ну эволюции растительного мира от древнейших времен и до наших
дней, от простейших и самых древних растений до наиболее высо-
коразвитых, возникших в наиболее близкие к нам периоды сущест-
вования Земли.
Систематика растений имеет большое практическое значение,
так как без классификации растений было бы невозможно разоб-
раться в огромном разнообразии видов и планомерно использовать
богатства растительного мира для нужд человека.
Экология изучает растения в связи с условиями их обитания,
выявляет закономерности приспособления растений к условиям
среды; исследует роль морфологических и анатомических струк-
тур растения в процессе использования ими таких факторов жизни,
как свет, тепло, вода, элементы пищи и др.
Фитоценология, или геоботаника, изучает закономерности сло-
жения растительного покрова Земли, процессы формирования
и эволюции растительных группировок, изменения их состава и со-
стояния в связи с условиями внешней среды и взаимовлияния ра-
стений, слагающих группировку.
География растений изучает закономерности распространения,
расселения растений по земной поверхности в прошлом и настоя-
щем.
Палеоботаника изучает ископаемые остатки растений, живших
в прошлое геологическое время. Изучение этих остатков позволяет
восстановить историю растительного мира, выяснить связи между
группами ныне живущих растений. Тем самым палеоботаника тес-
12
но соприкасается с систематикой, и данные этой науки привлека-
ются для построения системы растительного мира наравне с дан-
ными о ныне живущих растениях.
РАЗВИТИЕ БОТАНИЧЕСКОЙ НАУКИ
Истоки ботанической науки находятся в глубокой древности.
Зачатки науки о растениях возникли из практических потребностей
человека в пище, одежде, крове, лекарственных средствах.
Первые письменные упоминания о растениях находят в древ-
нейших надписях на египетских гробницах, относимых к третьему
тысячелетию до нашей эры.
Зачатки ботанических знаний мы находим в трудах великого
греческого философа Аристотеля, жившего в IV в. до н. э. Много
ценных сведений о растениях собрал ближайший ученик Аристо-
теля Феофраст (370—286 гг. до н. э.). Сохранилось шесть книг его
сочинения «Причины растений» и девять книг большого ботаниче-
ского труда «Исследования о растениях».
В эпоху средневековья, наступившую после падения Римской
империи, естественные науки находились в состоянии застоя, шло
главным образом накопление фактов. После этого длительного
периода в XVI в. начинается быстрый расцвет ботаники как науки.
Этому расцвету способствовали великие географические открытия,
которые дали доступ ученым к растительным богатствам тропиче-
ских стран Азии и Америки. Быстро накапливаются гербарии —
коллекции засушенных растений; увеличивается количество описа-
ний вновь открытых и изученных растений; к концу XVI в. число
их достигает уже 6000; делаются первые попытки классификации
растений (Чезальпино, 1519—1603).
Изобретение микроскопа позволило углубиться во внутреннее
строение растений, и таким образом закладываются основы ана-
томии, основанной на микроскопическом изучении строения расте-
ний. В 1665 г. Роберт Гук (1635—1703) впервые установил ячеи-
стое строение растительной ткани и дал этим ячейкам название
«клетка», которое сохранилось до наших дней, хотя и приобрело
иное содержание. В 1675—1679 гг. была опубликована капиталь-
ная работа Мальпиги (1628—1694) под названием «Анатомия ра-
стений», а в 1682 г. под тем же названием вышла книга, написан-
ная Грю (1641 —1712). Эти два научных труда положили начало
новой отрасли ботаники — анатомии растений.
Выдающуюся роль в развитии ботаники сыграл шведский нату-
ралист Карл Линней (1707—1778). Он привел в порядок накопив-
шийся к тому времени огромный описательный ботанический мате-
риал, создал искусственную систему растений, сыгравшую в то
время большую положительную роль. Линней ввел систему двой-
ных названий для растений и животных и по определенной схеме
описал большинство известных в то время растений- В системе
13
Линнея все известные в то время растения были распределены
между 24 классами.
В XVIII в. все большее участие в развитии ботанической науки
начинают принимать наши отечественные ученые. В начале этого
столетия в Москве, а затем в Петербурге закладываются первые
в России ботанические сады. Научным центром в России в то вре-
мя была Петербургская академия наук, основанная Петром I
и открытая в 1725 г. Выдающуюся роль в развитии естественных
наук сыграл первый русский академик М. В. Ломоносов (1711 —
1765).
В конце XVIII в. закладываются основы физиологии растений.
Постепенно раскрываются тайны питания зеленого растения.
В 1771 г. Д. Пристли устанавливает, что воздух, «испорченный»
дыханием животного, может быть «исправлен» растением. Позднее
(в 1779 г.) Ингенгауз уточняет, что способностью исправлять ис-
порченный животными воздух могут обладать только зеленые ра-
стения, поглощая при этом углекислый газ, и что процесс этот
может идти только на свету. Сущность этого процесса была рас-
крыта только в обстоятельных опытах Сенебье (1742—1809), ко-
торый установил, что зеленые растения питаются углекислотой,
выделяя при этом в окружающую среду кислород. В этом и заклю-
чается очищающая роль зеленых растений.
В XIX в. исследование фотосинтеза было продолжено Н. Соссю-
ром (1767—1845), Ж. Б. Буссенго (1802—1887) и К. А. Тимиря-
зевым (1843—1920). Швейцарский ученый Н. Соссюр к исследова-
нию фотосинтеза применил точные методы количественного анали-
за и экспериментально доказал, что зеленое растение в
фотосинтезе выделяет в окружающую среду кислород, усваивая
углерод из углекислоты, а в процессе дыхания подобно животным
поглощают кислород из окружающей среды, выделяя в нее угле-
кислоту. Французский исследователь Ж. Б. Буссенго по праву счи-
тается основателем современной агрономической химии. Наряду
с фотосинтезом он исследовал все стороны питания растений, ши-
роко используя при этом и вегетационный метод исследования.
Великий русский ученый — дарвинист К- А. Тимирязев свыше 40 лет
своей жизни посвятил изучению фотосинтеза. Тимирязев исследо-
вал зависимость фотосинтеза от интенсивности и качественного
состава света, роль частей светового спектра в фотосинтетическом
процессе; при этом он применил точные количественные методы
исследования. Заслугой Тимирязева было так же и то, что он впер-
вые исследование фотосинтеза перенес в природную обстановку.
Развитие физиологии растений тесно связано с достижениями
химии и физики. О тесной связи физиологии растений с химией
говорит существование науки биохимии растений, изучающей
химические превращения в растениях и опирающейся на законо-
мерности, открытые общей и органической химией. Развивается
также и такой раздел биологии, как биологическая физика, или
биофизика, изучающая физические закономерности в биологиче-
ских явлениях.
14
В физиологии растений широко используются новые методы ис-
следования, основанные на новейших открытиях физики; так, на-
пример, в последнее время для изучения процессов, совершающих-
ся в растениях, все шире применяются радиоактивные изотопы
элементов — так называемый метод «меченых» атомов.
Огромную положительную роль в развитии всех биологических
наук, в том числе и ботаники, сыграли два великих открытия сере-
дины XIX столетия: клеточная теория и эволюционное учение Дар-
вина. Эти два научных достижения были высоко оценены Марксом
и Энгельсом и поставлены ими наравне с открытием закона со-
хранения энергии.
Успехи в развитии учения о клетке в XVIII—XIX вв. были тесно
связаны с усовершенствованием микроскопа и микроскопической
техники. Создателями клеточной теории наука по праву считает
немецкого ботаника Шлейдена (1804—1881) и зоолога Шванна
(1810—1882). В трудах Шлейдена (1838) и Шванна (1839), а затем
и А. Вирхова (1855) клеточная теория превратилась в универсаль-
ное биологическое учение. Дальнейшее исследование клетки связа-
но с именами выдающихся ученых — Флемминга, Бючли, Левитско-
го, Данжара, Насонова и др.
Биологическая наука всегда была ареной острой идеологиче-
ской борьбы материалистического и идеалистического взглядов на
возникновение и развитие органического мира. До XIX в. в науке
господствовал идеалистический взгляд на природу; признавали
в той или иной форме «сотворение» мира, постоянство и неизмен-
ность видов. В начале XIX в. идея эволюции была последовательно
развита французским натуралистом Ламарком (1744—1829).
Однако честь создания эволюционной теории принадлежит вели-
кому английскому натуралисту Чарльзу Дарвину, именем
которого эта теория и названа. Его книга «Происхождение видов
путем естественного отбора, или сохранение благоприятствуемых
пород в борьбе за жизнь» (1859) открыла новую эру в развитии
биологических наук и окончательно утвердила в умах прогрессив-
ного человечества материалистическую идею развития органиче-
ского мира. Учение Дарвина об эволюции, или дарвинизм, является
естественнонаучной основой всех биологических наук, на основе
этой теории разрабатываются научные системы растительного
и животного мира.
Успеху идеи единства и развития растительного мира во многом
способствовали еще додарвиновские работы немецкого ботаника
В. Гофмейстера (1824—1877), установившего понятие чередования
поколений и на основе изучения этого явления у высших споровых
и семенных растений выдвинувшего идею генетического единства
всех групп высших растений. Позднее эта идея была окончательно
утверждена работами польского ботаника Э. Страсбургера (1844—
1912).
Большую роль в пропаганде ботанических знаний и дарвинист-
ских взглядов на природу сыграли известные циклы лекций
К. А. Тимирязева, опубликованные в книгах «Жизнь растений»
15
и «Исторический метод в биологии». В этих книгах К. А. Тимирязев
выступает как выдающийся популяризатор науки, защитник и про-
пагандист идей Дарвина в биологии.
Пропагандируя и развивая дарвинизм, К. А. Тимирязев борол-
ся против различных проявлений антидарвинизма, отечественного
и зарубежного. Талантливая пропаганда идей Дарвина сыграла
большую роль в развитии дарвинизма в нашей стране.
К. А. Тимирязев создал большую школу ученых, работавших
в области различных вопросов физиологии растений; среди них
наиболее известны В. И. Палладии и Д. Н. Прянишников.
В. И. Палладии (1859—1922) изучал химизм дыхания и тесно свя-
занные с ним процессы спиртового брожения. Д. Н. Прянишников
(1865—1948)—крупнейший ученый в области физиологии расте-
ний и агрохимии. Его работы по азотистому обмену растений со-
ставляют основу современных научных представлений по этому
вопросу.
С. Г Навашин сделал открытие величайшего биологического
значения. Он установил, что процесс оплодотворения у покрытосе-
менных растений принципиально отличается от процесса оплодот-
ворения у голосеменных растений тем, что у покрытосеменных
в оплодотворении участвуют два спермия, один из которых слива-
ется с яйцеклеткой и дает начало зародышу, второй сливается
с одной из клеток зародышевого мешка и дает начало эндосперму.
Следовательно, в отличие от голосеменных эндосперм покрытосе-
менных растений является продуктом полового процесса. Процесс,
открытый акад. С. Г Навашиным, получил в науке название двой-
ного оплодотворения.
Во второй половине XIX в. и в первой половине XX в. много-
численная армия отечественных ботаников изучает растительный
покров нашей страны и сопредельных стран. Особенно усилились
и стали планомерными исследования растительности нашей страны
после Великой Октябрьской социалистической революции.
Выдающийся советский ботаник В. Л. Комаров (1869—1945),
много лет возглавлявший Академию наук СССР, изучал флору
Средней Азии, Дальнего Востока, п-вов Камчатки и Кореи, Северо-
Западного Китая. Изучению флоры и растительности Средней Азии
много лет отдали О. А. Федченко (1845—1921) и Б. А. Федченко
(1872—1947). Крупнейшими исследователями флоры и раститель-
ности Сибири были П. Н. Крылов (1850—1931) и В. В. Сапожни-
ков (1861 —1924), Кавказа — Н. И. Кузнецов (1864—1932) и
А. А. Гроссгейм (1888—1948).
В итоге огромной работы, проделанной нашими ботаниками,
в настоящее время имеется достаточно полное представление
о флоре и растительности нашей страны. Создана многотомная
«Флора СССР», а также флоры союзных республик и некоторых
естественных районов Союза. Составлена сводка по растительности
СССР.
В области изучения культурных растений и исследования зако-
номерностей наследственной изменчивости большие заслуги имеет
16
акад. Н. И. Вавилов (1887—1943), руководивший Всесоюзным на-
учно-исследовательским институтом растениеводства. Им много
сделано для теоретического* обоснования работы по улучшению
и созданию новых сортов культурных растений. Н. И. Вавиловым
и его сотрудниками собраны богатые коллекции культурных ра-
стений всех стран мира, изучены вопросы происхождения и рассе-
ления культурных растений, установлены главнейшие очаги проис-
хождения культурных растений. Коллекции семян растений,
созданные Н. И. Вавиловым и его учениками и постоянно обнов-
ляемые, и в наше время служат незаменимым источником для
создания новых сортов растений.
И. В. Мичурин (1855—1935) практически доказал значение гиб-
ридизации для создания новых форм растений, разработал методы
воспитания гибридных растений. Им были разработаны новые ме-
тоды акклиматизации растений, выдвинута и широко реализована
идея использования мирового богатства культурных и дикорасту-
щих форм для создания новых полезных растений.
ГЛАВА I
РАСТИТЕЛЬНАЯ КЛЕТКА
За редким исключением любой растительный организм состоит
из клеток. Тело бактерии, как правило, состоит из одной клетки;
очень многие водоросли и некоторые грибы также имеют однокле-
точное «строение или состоят всего из нескольких клеток. Тело боль-
шинства растений имеет многоклеточное строение, причем у круп-
ных растений количество клеток исчисляется многими миллионами
и миллиардами. Лишь у некоторых грибов и немногих водорослей
тело не расчленено на клетки, т. е. имеет неклеточное строение, но
это как раз служит признаком несовершенства -строения. Всеобщ-
ность клеточного строения растительных и животных организмов —
одно из доказательств единства происхождения всего органическо-
го мира.
Мы будем рассматривать строение наиболее «совершенно устро-
енной растительной клетки, свойственной высшим растениям. Од-
нако надо иметь в виду, что такого сложного устройства клетка дос-
тигла в результате длительного пути эволюционного развития на
протяжении более двух миллиардов лет. Первые, примитивные
клетки были устроены гораздо проще. В частности, они не имели
ядер, нуклеиновые кислоты, обязательные для клетки и сконцентри-
рованные у высокоорганизованных клеток в ядрах, у первобытных
клеток находились непосредственно в цитоплазме, ядро не было
морфологически выделено. Такие простейшие безъядерные клет-
ки называются прокариотными в отличие от клеток с ядрами — эу-
кариотных, свойственных большинству растений. Прокариотные
клетки у бактерий и синезеленых водорослей — наиболее древних
из известных клеточных организмов. Совершенствование и специа-
лизация клеток, входящих в состав многоклеточных организмов,
шли одновременно с эволюцией растений, в процессе которой груп-
пы клеток приспо'Сабливались к выполнению той или иной функции.
ФОРМА И ВЕЛИЧИНА КЛЕТОК
Клетки могут иметь весьма разнообразную форму. У однокле-
точных организмов это шар, эллипсоид, прямая или изогнутая па-
лочка, нередко извилистая ниточка. У многих водорослей и у боль-
шинства грибов клетки имеют форму более или менее вытянутых
нитей. У высших растений, имеющих сложное внутреннее строение,
форма клеток зависит от месторасположения их в теле растения
18
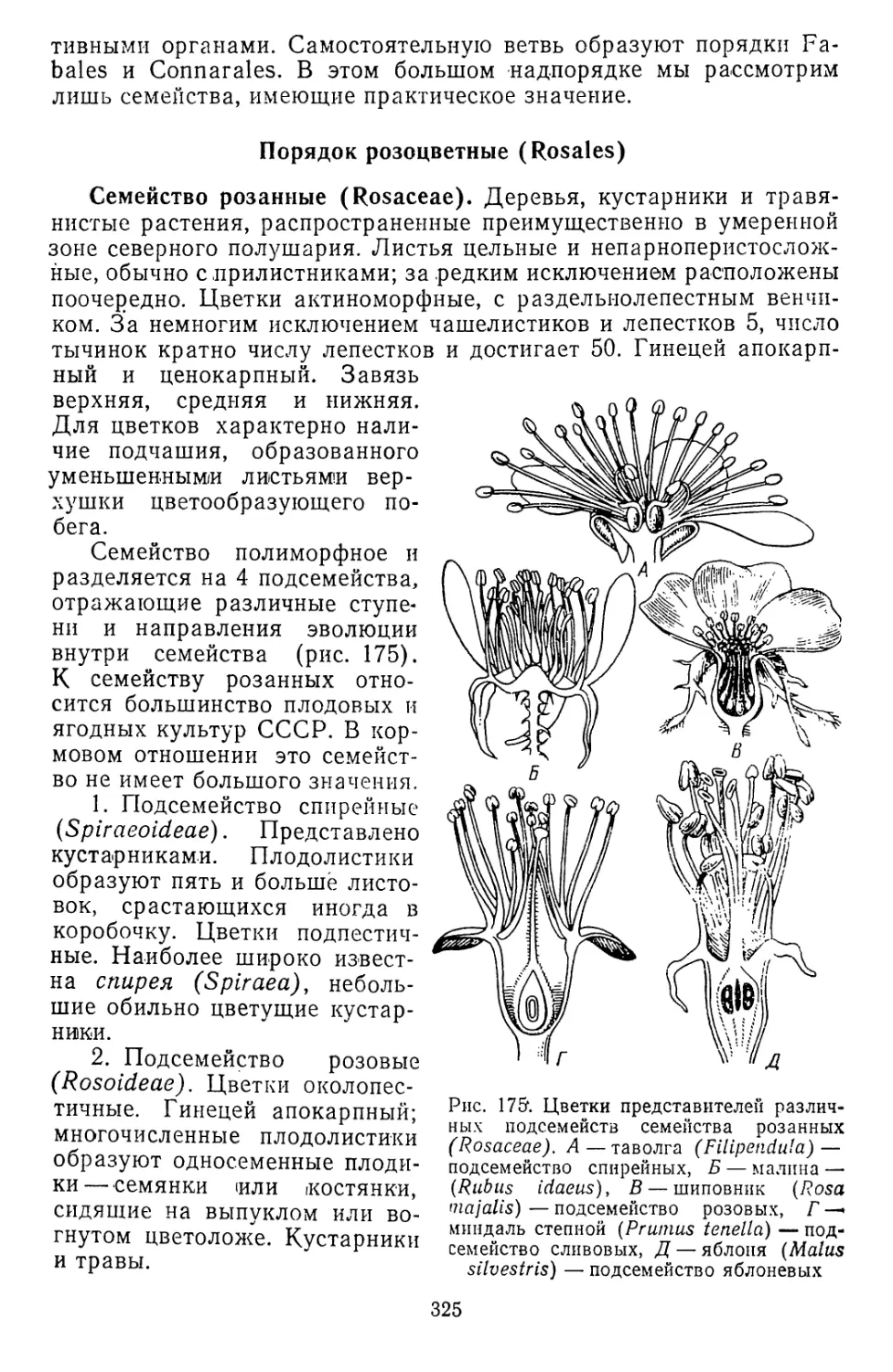
Рис. 1. Разнообразие клеток высших растений. А, Б — меристема-
тические клетки, В — крахмалоносные клетки запасающей парен-
химы, Г — клетки эпидермиса, Д — двухъядерная клетка одного
из слоев пыльцевого гнезда, Е — клетка ассимиляционной ткани
листа, Ж — членик ситовидной трубки с клеткой-спутницей, 3 —
каменистая клетка, И — членик сосуда
и выполнения ими тех или иных функций. Например, поверхност-
ные клетки плоские; клетки механических волокон тонкие и сильно
вытянутые; клетки, образующие мякоть плодов, более или менее
округлые и т. д. (рис. 1). В морфологическом отношении клетки выс-
ших растений принято подразделять на два типа: паренхимные,
по форме близкие к шару, кубу, эллипсоиду, параллелепипеду или
цилиндру, и прозенхимные, сильно вытянутые, у которых длина во
много раз превышает ширину.
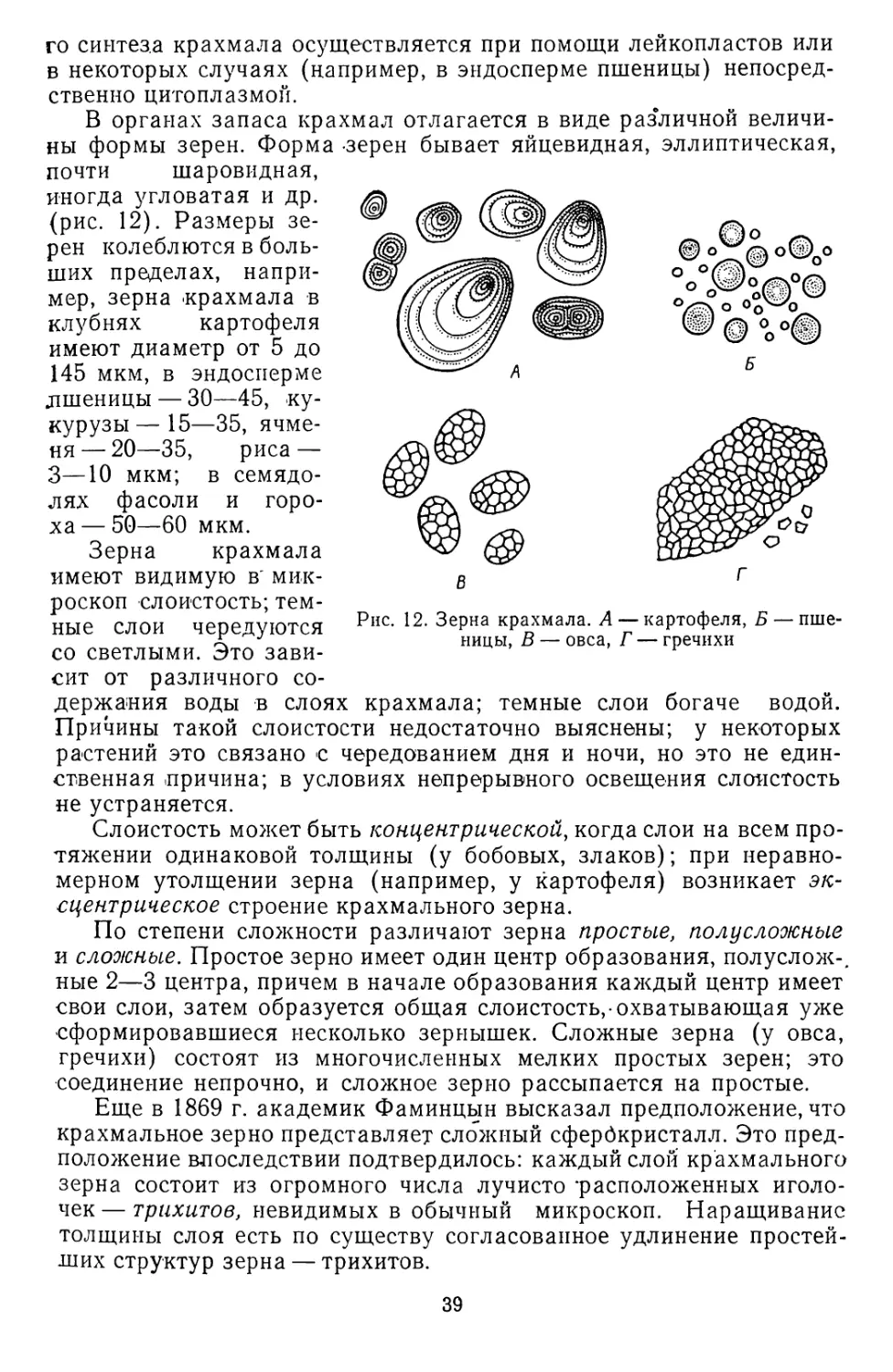
Размер клеток обычно микроскопический, измеряемый тысяч-
ными долями миллиметра — микрометрами, но сильно вытянутые
клетки достигают длины нескольких миллиметров и даже сантимет-
ров. Например, волоконца на семенах хлопчатника имеют длину до
65 мм, лубяное волокно у конопли и льна — 20—60 мм, а у крапи-
вы — даже 80 мм и т. д. В мякоти плодов, например яблока, тома-
та, лимона, арбуза, клетки крупные, округлые, достигают в диамет-
ре 1 мм; такие клетки вполне различимы и без микроскопа.
Тип строения высшего растения, при котором большое тело раз-
делено на огромное количество мельчайших единиц — клеток,—
результат длительного процесса эволюции; такое построение орга-
низма имеет глубокий биологический смысл; создаются условия для
сложной анатомо-физиологической дифференциации единого орга-
19
низма, в связи с чем облегчается выполнение его многообразных
функций; неизмеримо увеличивается прочность и способность его
противостоять вредным воздействиям окружающей среды; в связи
со сложной дифференциацией частей организма совершенствуются
органы воспроизведения и обеспечивается больший успех размно-<
жения в условиях изменяющейся внешней среды.
МОЛЕКУЛЯРНЫЕ КОМПОНЕНТЫ КЛЕТКИ
Большую часть массы живой действующей клетки составляет
вода; ее количество в растительных клетках обычно в пределах
75—85%, но в некоторых случаях, например в клетках .сочных пло-
дов, молодых корней, достигает 90—95%. Около 95% всей воды
составляет свободная вода, служащая растворителем или диспер-
сионной средой коллоидной системы протоплазмы. На долю, при-
мерно, остальных 5% приходится связанная вода, настолько проч-
но соединенная с макромолекулами органических веществ, что от-
нять ее можно, только разрушив структуру макромолекул, что
влечет за собой гибель клетки.
В воде растворены соли. В клетке они диссоциированы на кати-
оны и анионы, необходимые для поддержания осмотического давле-
ния и кислотно-щелочного равновесия. В некоторых случаях, нап-
ример, в клетках растений, произрастающих на солончаках, осмо-
тическое давление в клетках может достигать нескольких десятков
атмосфер. Присутствие многих неорганических катионов и анионов
необходимо для активации ферментов и протекания различных про-
цессов обмена веществ в клетке.
В растительных клетках всегда присутствуют углеводы: моно-
сахариды, дисахариды и полисахариды. Такие полисахариды, как
крахмал, гликоген, целлюлоза, представлены крупными молекулами
полимерного строения и вместе с нуклеиновыми кислотами и белка-
ми, также имеющими полимерную структуру, относятся к категории
макромолекул.
В состав протоплазмы клеток обязательно входят липиды — жи-
ры и жироподобные вещества. К простым липидам относятся жиры,
представляющие собой сложные эфиры глицерина и жирных кис-
лот; воска — также относятся к простым липидам; однако в их сос-
тав кроме глицерина входят и другие спирты.
Сложные липиды — соединения простых липидов с другими ве-
ществами, например с фосфорной кислотой, фосфолипиды, или фос-
фаты.
Нуклеиновые кислоты — РНК (рибонуклеиновая) и ДНК (де-
зоксирибонуклеиновая) — обязательный компонент клетки — пред-
ставляют собой сложные полимеры, звенья которых называются
нуклеотидами. Каждый нуклеотид состоит из трех компонентов: уг-
левода рибозы (РНК) или дезоксирибозы (ДНК), остатка фосфор-
ной кислоты и одного из пяти азотистых оснований, двух пуриновых
(аденин и гуанин) и трех пиримидиновых (тимин, цитозин и ура-*
20
цил); урацил входит в состав только РНК, тимин — только в ДНК;
аденин, гуанин и цитозин — в состав нуклеотидов обеих нуклеино-
вых кислот. Молекулы белков состоят из остатков примерно 20 ами-
нокислот, соединенных пептидными связями. Такое количество эле-
ментов обеспечивает возможность безгранично большого числа
сочетаний, поэтому количество белков, встречающихся в клетках
растений, практически бесконечно. Это могут быть структурные
белки и белки-ферменты, а также запасные белки.
Аминокислоты, вошедшие в состав белковой молекулы, могут
быть различной величины и сложности, и сама молекула может
иметь то сравнительно несложную структуру в виде простой или
разветвленной цепи, то очень сложную, представляющую клубок из
многочисленных разветвленных цепей.
Для удобства обозначения каждой из двадцати аминокислот,
входящих в состав белковых молекул, присвоен определенный сим-
вол, состоящий из трех букв:
Глицин — Гли
Аланин — Ала
Валин — Вал
Лейцин — Лей
Изолейцин — Иле
Глутаминовая кислота—Глу
Аспарагиновая кислота — Асп
Аргинин — Apr
Лизин — Лиз
Оксилизин — Оли
Треонин — Тре
Серин — Сер
Цистин — Цис
Метионин — Мет
Фенилаланин — Фен
Тирозин — Тир
Триптофан — Три
Пролин — Про
Оксипролин — Опр
Гистидин — Гис
СТРОЕНИЕ КЛЕТКИ, ЕЕ СОСТАВНЫЕ
ЧАСТИ
Клетка высшего растения, несмотря на микроскопически малую
величину, имеет очень сложное строение, которое, как уже сказано,
явилось результатом длительной эволюции. Однако высокая степень
организации клеток выражается не только в сложности их строения,
но и в бесконечно большом разнообразии процессов, совершающих-
ся в клетках, результатом чего является огромное количество хими-
ческих веществ, добываемых из продуктов растительного происхож-
дения. Первичной лабораторией, где вырабатываются эти вещества,
служит растительная клетка.
Принято начинать изучение клетки с ее наиболее существенных
частей — органоидов. Совокупность органоидов клетки составляет
протопласт. Это активное, живое начало клетки. В процессе жизне-
деятельности клетки в ней накапливаются различного рода вклю-
чения в протопласт; они могут иметь жидкую и твердую консистен-
цию; по своей биологической природе это запасные питательные
вещества, продукты промежуточного обмена или отбросы обмена,
уже не нужные растению. Стенка, или оболочка клетки,— также
производное протопласта. Такое подразделение составных частей
клетки в некоторой мере условно, тем не менее всегда надо иметь
21
в виду, что наиболее существенная часть клетки — ее протопласт;
нет протопласта — нет и клетки как живой элементарной единицы
растительного организма.
ПРОТОПЛАСТ
Протопласт состоит из цитоплазмы (протоплазмы) с митохон-
дриями и одного или нескольких ядер с ядрышками; в протоплас-
те автотрофных организмов, водорослей и высших растений имеют-
ся также хроматофоры или пластиды. Однако синезеленые водорос-
ли не содержат оформленных хроматофоров или пластид п ядра.
Внутри цитоплазмы обычно можно обнаружить пространства, за-
полненные жидкостями. Такие пространства называются вакуоля-
ми. Вакуоли заполнены так называемым клеточным соком — раст-
вором самых разнообразных органических и неорганических ве-
ществ. Иногда в вакуолях скопляются жидкие масла. Содержимое
вакуолей, а также гранулы веществ, имеющие плотную консистен-
цию, называются включениями. Это или запасные питательные ве-
щества, или же различного рода отбросы, не используемые уже рас-
тениями. Снаружи у растений протопласт обычно покрыт твердой
оболочкой, придающей клетке определенную форму.
Такова структура растительной клетки, обнаруживаемая при
помощи обычного оптического микроскопа. Она представляется
весьма несложной. Однако это лишь кажущаяся простота: обычный
Рис. 2. Современная обобщенная схема растительной клетки:
/ — аппарат Гольджи; 2 — свободно расположенные рибосомы: 3 — хлоропласты; 4 — меж-
клеточные пространства; 5 — полирибосомы; 6 — митохондрии; 7 — лизосомы; 8 — грану-
лярная эндоплазматическая сеть; 9 — агранулярная эндоплазматическая сеть; 10—микро-
трубочки; // — пластиды; 12 — плазмодесмы; 13 — клеточная оболочка; 14 — ядрышко;
J5—18 — ядерная оболочка; 16 — поры в ядерной оболочке; 17 — плазмалемма; 19 — гиало-
плазма (матрикс); 20 — тонопласт; 21 — вакуоль; 22 — ядро
22
оптический микроскоп не дает возможности проникнуть в тонкую
структуру протопласта. Современные методы исследования с при-
менением электронного микроскопа, увеличивающего в десятки
и сотни тысяч раз, позволили проникнуть в ультраструктуру, в суб-
микроскопическую и молекулярную организаций) клетки. Обнару-
жилась чрезвычайно сложная структура внутренней организации
клетки. Схемы, помещенные на рис. 2 и 3, дают представление о
сложности строения протопласта клетки. В дальнейшем изложении
мы будем неоднократно, обращаться к деталям этих схем.
Кроме форменных, морфологических элементов клетки в состав
протопласта и клеточного сока входят в ничтожно малых количес-
твах особые органические вещества, обладающие сильным физиоло-
гическим действием: энзимы (ферменты), гормоны, витамины, анти-
биотики. Все эти вещества, так же как и включения и оболочка, —
продукты жизнедеятельности протопласта.
Цитоплазма. В клетках, находящихся в состоянии активной жиз-
ни, цитоплазма представляет собой полупрозрачную тягучую жид-
кость, сходную с яичным белком. Она бесцветна, но в воде хорошо-
заметна, так как сильнее преломляет свет. Плотность цитоплазмы
выше плотности воды, в пределах 1,01 —1,06, поэтому в воде она то-
нет. Более чем на 90% цитоплазма состоит из воды.
Цитоплазма может в большой степени обезвоживаться, напри-
мер в клетках семян и спор, не теряя способности вернуться в дея-
тельное состояние; в таких случаях консистенция ее меняется до
состояния твердого студня (геля).
Лишь в редких случаях в клетках образовательной, зародышевой
ткани цитоплазма и ядро занимают почти весь объем клетки. Обыч-
но же в клетках незародышевых тканей большая часть объема за-
нята вакуолями или твердыми продуктами обмена, а цитоплазма
ограничивает эти вакуоли
или запасные питательные
вещества со всех сторон.
Однако как бы ни была
вакуолизирована клетка,
между оболочкой и вакуо-
лями (или единственной ва-
куолью) всегда находится
слой цитоплазмы.
Цитоплазма клеток весь-
ма чутко реагирует на из-
меняющиеся условия внеш-
ней среды, но вместе с тем
обладает и достаточной
стойкостью, позволяющей
растительным организмам
сохраняться в живом состоя-
нии даже при очень резких
колебаниях факторов внеш-
ней среды. Характер реак-
Рис. 3. Схема пространственного изображе-
ния участка клетки с эндоплазматической
сетью при очень сильном увеличении:
1 — стенка; 2 — плазмалемма; 3 — мембрана эндо-
плазматической сети; 4 — цистерна эндоплазмати-
ческой сети (элемент диктиосомы); 5 — плазмо*
десма; 6 — гиалоплазма (матрикс); 7 — вакуоль
23
ций цитоплазмы на воздействия внешней среды во многом зависит
от ес физико-химической структуры.
Цитоплазма — это гетерогенная система, подобная коллоиду.
В этой системе дисперсионной средой является вода с растворен-
ными в ней веществами (минеральные соли, углеводы, аминокисло-
ты и др.), а дисперсной фазой — комплексы белковых молекул, сое-
динённых с молекулами воды. Такие системы называются гидрозо-
лями, поскольку здесь жидкая фаза диспергирована в водной среде.
При температуре 50—60° С белковые вещества цитоплазмы боль-
шинства растений дегидратизируются, т. е. комплексы белковых мо-
лекул утрачивают воду и оседают; происходит необратимый процесс
коагуляции (свертывания), и клетка погибает. Лишь некоторые
Рис. 4. Плазмолиз клетки (справа — плазмолиз клетки
при большой вязкости цитоплазмы)
простейшие водоросли из типа синезеленых, приспособившиеся жить
в горячих источниках, способны выдерживать температуру до 90° С
без разрушения структуры цитоплазмы. Однако в сильно обезво-
женном состоянии (например, в спорах) цитоплазма может выдер-
живать в течение некоторого времени нагревание даже свыше
100° С. Поэтому, например, чтобы освободить инфицированный ма-
териал от спор патогенных бактерий, его подвергают длительному
нагреванию при температуре 102—105° С. Цитоплазма клеток мо-
жет активно двигаться, причем это движение может стимулировать-
ся пли, наоборот, тормозиться внешними условиями. В клетках выс-
ших растений наблюдается двоякого рода движение цитоплазмы:
круговое и струйчатое. Вязкость цитоплазмы и способность ее сок-
ращаться хорошо наблюдаются в явлениях плазмолиза и деплазмо-
лиза (рис. 4). В клетках, не испытывающих недостатка в воде,
плазма плотно прилегает к внутренней стенке клеточной оболочки.
Как уже было сказано, обычно в клетке имеются вакуоли, за-
полненные клеточным соком. В нормальных условиях существова-
ния растений концентрация клеточного сока бывает выше, чем кон-
центрация окружающего, например, почвенного раствора. Поэто-
му, согласно закону диффузии, вода стремится извне проникнуть
в клетки (где ее относительно меньше) до тех пор, пока концентра-
ции растворов не выравняются. Создается внутреннее давление кле-
24
точного сока на слой цитоплазмы, прилегающий к клеточной обо-
лочке. Такое состояние внутреннего напряжения клетки называется
тургором. Благодаря тургору в клетках, составляющих тело рас-
тения, органы его находятся в упругом, не увядшем состоянии. Это
обычное, нормальное состояние растения.
Однако в состоянии тургора клетка может находиться лишь до
того момента, пока концентрация клеточного сока не ниже, чем
в окружающей среде. Стоит только поместить клетку в раствор, где
концентрация растворенных веществ выше, чем в клеточном соке,
и напряженное состояние клетки, тургор, исчезает.
Для искусственного создания таких условий клетки помещают
в какой-либо нейтральный раствор, в 8%-ный раствор поваренной
соли, или 10%-ный раствор калийной селитры, или в 30%-ный
раствор тростникового сахара и др. В качестве объекта берут ко-
жицу с чешуи лука, кусочек мякоти корня свеклы или любую дру-
гую ткань с вакуолизированными клетками. Спустя уже несколько
минут можно наблюдать в микроскоп, как цитоплазма постепенно
отстает от клеточных стенок, пока не сожмется в округлый комо-
чек.
Если клеточный сок окрашен, легко можно заметить, что интен-
сивность окраски в меру сжимания цитоплазмы повышается, при-
чем кажется, что окрашена цитоплазма; на самом деле это клеточ-
ный сок просвечивает сквозь слой цитоплазмы. Такое явление от-
ставания цитоплазмы от стенок и сжимания ее называется плазмо-
лизом. Нетрудно найти объяснение этому явлению: концентрация
воды в растворе, окружающем клетку, ниже, чем в клеточном соке;
по закону диффузии вода устремляется из вакуоли (где концентра-
ция ее выше) сквозь слой цитоплазмы, объем вакуоли уменьшает-
ся, а цитоплазма — вязкое и упругое вещество — сжимается.
В результате сжатия объем вакуоли может сократиться во многс
раз.
Если такие плазмолизированные клетки положить в чистую воду,
быстро начнется обратный процесс: вода проникает внутрь вакуоли,
объем последней увеличивается, и цитоплазма постепенно расправ-
ляется, пока опять наружный слой ее не будет прижат к клеточной
оболочке. Этот обратный процесс называется деплазмолизом. В ре-
зультате деплазмолиза клетка возвращается в свое нормальное со-
стояние.
Явления плазмолиза и деплазмолиза возможны потому, что по-
верхностные слои цитоплазмы обладают особыми биофизическими
свойствами полупроницаемости.
Слои цитоплазмы, непосредственно граничащие с оболочкой или
вакуолью, физически отличаются от остальной массы протоплазмы
благодаря более развитой тут силе поверхностного натяжения. Эти
слои получили следующие названия: слой, прилегающий к оболоч-
ке, называется плазмалеммой; слой прилегающий к вакуолям, —
тонопластом; между ними находится мезоплазма.
Цитоплазма каждой клетки посредством тончайших протоплаз-
менных нитей 1соединяется с протоплазмой всех смежных клеток
25
поэтому протопласты всех клеток растения образуют живое един-
ство, единый организм.
Основное вещество цитоплазмы, в котором размещены все ее
структуры, называется гиалоплазмой (матрикс). Это относительно
однородная среда, состоящая из белковых молекул в соединении с
большим количеством молекул воды. В гиалоплазме находится
большое количество различных ферментов, с помощью которых
и протекают разнообразные процессы обмена веществ. Физико-хи-
мические свойства цитоплазмы, описанные выше, — это свойства
прежде всего гиалоплазмы. Предполагают, что вещество гиалоплаз-
мы может иметь гранулярную и фибриллярную структуру.
Пограничный слой гиалоплазмы представляет из себя тончай-
шую пленку — плазматическую элементарную мембрану толщиной
около 10 нм. С помощью электронного микроскопа удалось устано-
вить, что плазматическая мембрана состоит из трех слоев (рис. 5):
Рис. 5. Схема молекулярной архитектуры плазматической
мембраны:
/ — молекулы белка; 2 — гидрофильная часть молекулы; 3 — угле-
водородные цепи; 4 — двойной ряд фосфолипидных молекул
между двумя наружными мономолекулярными слоями белка на-
ходится бимолекулярный слой фосфолипидных молекул. Такой трех-
слойной плазматической мембраной являются упоминавшаяся уже
плазмалемма, прилегающая к оболочке клетки, и тонопласт, приле-
гающий к вакуолям. В обоих случаях плазматическая мембрана об-
ладает избирательной способностью; вещества сквозь нее проника-
ют активно, обычно против градиента концентрации.
Внутрицитоплазменные структуры также ограничиваются плаз-
матической мембраной. Плазматическая мембрана весьма подвиж-
на, активна. Она может образовывать различного рода складки,
впячивания. В случае ранения она быстро регенерирует и закрывает
отверстия, если они не слишком велики. При больших разрывах
цитоплазма может вытечь через образовавшиеся отверстия. Возни-
кает плазматическая мембрана, по-видимому, спонтанно, как ре-
зультат биохимических процессов, протекающих в матриксе цито-
плазмы.
Плазматическая мембрана на поверхности цитоплазмы играет
важную роль в процессе поступления питательных веществ внутрь
клетки. Так, например, путем впячивания наружной мембраны
26
внутрь цитоплазмы происходит как бы заглатывание капелек жид-
кости, окружающей клетку. Такое явление, по-видимому, широко
распространенное в природе, получило название пиноцитоз. По
средством пиноцитоза «растения могут поглощать высокомолекуляр-
ные соединения, проникновение которых в клетку иными способами
затруднено.
Как видно из приведенных схем (см. рис. 2, 3), гиалоплазма
пронизана многочисленными канальцами, которые образуют эн-
доплазматическую сеть, или эндоплазматический ретикулум,— ор-
ганоид клетки, свойственный всем растениям. Стенки канальцев эн-
доплазматической сети состоят из описанной выше плазматичес-
кой мембраны. Диаметр канальцев сети 30—50 нм.
С эндоплазматической сетью функционально связаны особые
органы цитоплазмы — диктиосомы, или так называемые аппараты
Гольджи (см. рис. 2 и 3). Это пакеты плоских цистерн толщиной до
1 мкм, стенки которых состоят из той же плазматической мембра-
ны. Возле цистерн можно видеть, по-видимому, отпочковывающиеся
от них пузырьки, которые, вероятно, увеличиваясь, превращаются
в вакуоли. Количество диктиосом в одной клетке может достигать
1000 и более.
Цистерны непосредственно соединены с канальцами эндоплаз-
матической сети. В диктиосомах конденсируются и временно накап-
ливаются органические вещества (белки и др.), подлежащие в даль-
нейшем транспортировке в другие участки протопласта. Диктносо-
мам свойственна и секреторная деятельность, они секретируют
целлюлозу, идущую на построение клеточных стенок, а также и дру-
гие углеводы. В диктиосомах образуются также эфирные масла, а у
диатомовых водорослей — поликремневая кислота. Очень важно
подчеркнуть, что канальцы эндоплазматической сети связывают,
с одной стороны, цитоплазму данной клетки с ее ядром и, с другой
стороны, через плазмодесмы канальцы связывают в единое целое
протопласты соседних клеток. Важность такой всепроникающей се-
ти канальцев для функционирования организмов трудно переоце-
нить. Эндоплазматическая сеть не только способна воспринимать
извне и транспортировать различного рода продукты, но и прини-
мает непосредственное участие в процессах синтеза. Синтезирующая
роль эндоплазматической сети подчеркивается еще тем обстоятельст-
вом, что на наружной поверхности канальцев, составляющих эту
сеть, мы обнаруживаем особые тельца — рибосомы, функция кото-
рых — синтез белка.
Рибосомы имеют форму гранул, поэтому эндоплазматическая
сеть, канальцы которой снаружи покрыты рибосомами, называет-
ся гранулярной эндоплазматической сетью; участки эндоплазмати-
ческой сети, примыкающие к наружной мембране (плазмалемме),
не имеют снаружи рибосом и поэтому называются агранулярной.
Соединенные между собой канальцами эндоплазматической сети по-
лости цистерн диктиосом, перинуклеарное пространство между дву-
мя мембранами, окружающими ядро и вакуоли, составляют вакуо-
лярную систему клетки.
27
Рибосомы имеют форму сплющенных сфероидов размером око*
ло 25X15 нм. Установлено, что тело рибосомы делится желобком
на две субъединицы, большую и малую; к поверхности канальца
эндоплазматической сети рибосомы прикрепляются своей большей
субъединицей. Молекулы РНК, входящие в состав рибосом, обра-
зуют одинарные и двойные спирали, которые располагаются бли-
же к поверхности рибосом; промежутки между спиралями РНК за-
полнены молекулами белка и небольшим количеством липидов, пос-
ледние могут и отсутствовать в рибосомах. Предполагают, что в
каждой рибосоме находится всего несколько молекул РНК. Коли-
чество рибосом в одной клетке различно и измеряется десятками
тысяч. Образуются рибосомы в виде субъединиц в ядрышках, и окон-
нательная сборка их происходит в цитоплазме, куда переходят из
ядра субъединицы в форме нуклеопротеидов. Рибосомы также най-
дены в митохондриях и хлоропластах. Рибосомы могут соединять-
ся в комплексы по нескольку, цепочкой или спиралью, образуя по-
лирибосомы, или полисомы.
Как уже сказано, функция рибосом — синтез белка. Установле-
но, что рибосомы, связанные с мембранами эндоплазматической
сети, синтезируют белок более энергично, чем свободно располо-
женные в цитоплазме. Взрослые и старые клетки почти совсем или
совсем не содержат рибосом.
Молекулы РНК, входящие в состав рибосом, обладают лишь по-
тенциальной способностью синтезировать белок. Эти молекулы ак-
тивизируются особыми молекулами информационной РНК, которые
синтезируются на молекулах дезоксирибонуклеиновой кислоты
(ДНК), находящихся в ядре. Закодированные в молекулах ДНК
(генах) специфические свойства белков данного растения воспроиз-
водятся рибосомами благодаря тому, что соответствующая инфор-
мация переносится молекулами информационной РНК. Эти молеку-
лы называются еще РНК-посредниками. Следовательно, информа-
ционная РНК не только активирует молекулы РНК рибосом, но
и сообщает им способность синтезировать белки определенного со-
става, свойственного данному растению.
Аминокислоты, необходимые для создания молекулы белка, пе-
реносятся к месту синтеза молекулами особой, транспортной РНК.
Более детально о процессе синтеза белка будет сказано в одном из
последующих разделов учебника. Вопрос о происхождении рибосом
недостаточно исследован. По-видимому, основная масса рибосомной
РНК вырабатывается в ядрах, при непосредственном участии хро-
матинового вещества, однако не исключена возможность возникно-
вения рибосом непосредственно на месте, в цитоплазме. Не решен
вопрос о возможности самовоспроизведения рибосом.
Используя ультрацентрифугу со скоростями 200—300 тыс.
об/мин, удалось отделить рибосомы от гиалоплазмы. При этом ри-
босомы отделяются вместе с обрывками канальцев эндоплазмати-
ческой сети, к которым они были прикреплены. Эта фракция по-
лучила в свое время название микросом. Было установлено, что
фракция микросом составляет 15—20% сухой массы цитоплазмы.
28
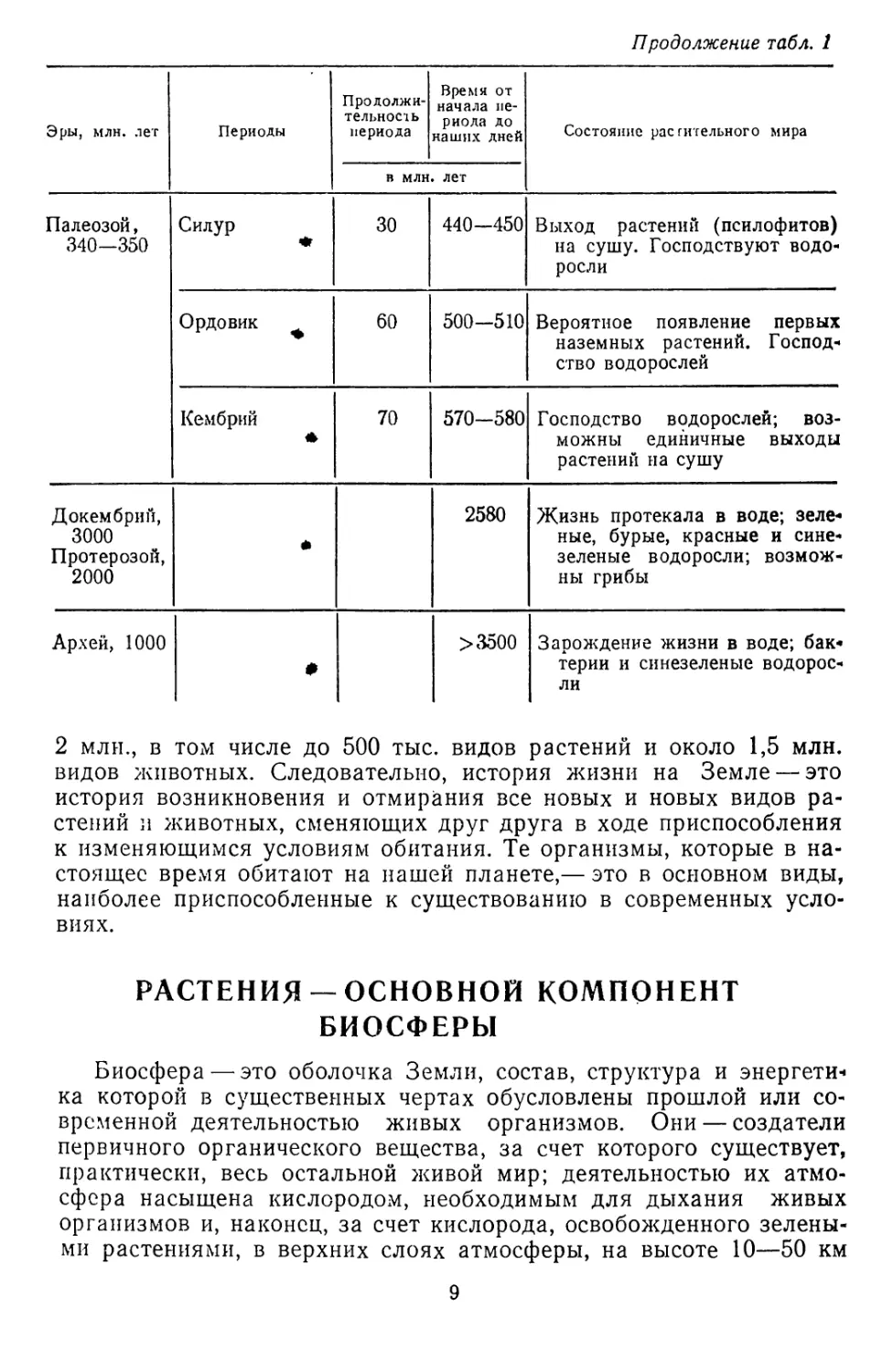
J
Рис. 6. Строение митохондрии:
/ — наружная мембрана; 2 —внутренняя мембоа-
на; 3 —кристы; 4—матрикс
Митохондрии (хондриосомы), встречающиеся в цитоплазме всех
растений, за исключением бактерий, имеют форму овальных телец
длиной до 1 мкм; располагаясь рядом, они могут образовать види-
мую под микроскопом нить (см. рис. 2, 3, 6). Плотность митохонд-
рий несколько выше, чем гиалоплазмы, поэтому их можно отделить
центрифугированием. Количество митохондрий в клетке весьма из-
менчиво и измеряется сот-
нями и тысячами. Продол-
жительность жизни мито-
хондрии около недели, по-
этому они непрерывно во-
зобновляются.
Митохондрии — органои-
ды довольно сложного стро-
ения. Содержимое этих те-
лец отграничивается от гиа-
лоплазмы двойной плазмати-
ческой мембраной; наруж-
ная гладкая, внутренняя об-
разует впячивания внутрь митохондрии в виде гребней — крист,
трубочек и вздутий, мешочков (рис. 6). Такие впячивания увели-
чивают внутреннюю поверхность митохондрий и повышают их
прочность. Поверхность внутренней мембраны не гладкая, на ней
шаровидные выросты на ножках — оксисомых. Внутренняя полость
митохондрий заполнена матриксом, или хондриоплазмой, в кото-
рой ферменты, липиды и молекулы РНК и ДНК. На долю белков
приходится 45—50% сухого вещества митохондрий. Остальное со-
ставляют липиды, РНК и ДНК и неорганические соединения.
Роль митохондрий огромна; это как бы энергетические центры
клетки: здесь сосредоточена основная масса дыхательных фермен-
тов, обеспечивающих энергией многочисленные процессы, проте-
кающие в клетке. Процессы эти совершаются при участии адено-
зпнтрифосфорной кислоты (АТФ), которая синтезируется в мито-
хондриях.
В вопросе о возникновении митохондрий в онтогенезе нет обще-
го мнения. Существует несколько точек зрения; митохондрии обра-
зуются: 1) каждый раз заново из инициальных частиц, обнаружи-
ваемых в цитоплазме; 2) из старых митохондрий путем деления,
почкования; 3) в результате отшнуровывания двойной мембраны
ядра.
В клетках растений обнаружены также особые шаровидные тель-
ца размером до 1мкм — сферосомы, функции их — синтез жира, об-
разуются они, по-видимому, за счет отшнуровки участков эндоплаз-
матического ретикулума; мембрана у сферосом одинарная.
В клетках животных организмов и грибов обнаружены тельца,
похожие на митохондрии, — лизосомы. В отличие от митохондрий
мембрана их одинарна и не образует внутренних выростов. В лизо-
1 Размер оксисом 7—9 нм.
29
матофоров или пластид. Хроматофоры
Рис. 7. Различные формы пластид у растений,
хроматофоры у зеленых водорослей:
1 — спирогиры; 2 — хламидомонады; 3 — улотрикса;
4 _ зигнемы; 5 — хлоропласт цветкового растения;
6 — хромопласты
сомах содержатся гидролитические ферменты, расщепляющие про-
теины, нуклеиновые кислоты, полисахариды. В растительных клет-
ках высших растений лизосомы пока не обнаружены.
Сравнительно недавно в клетках высших растений обнаружены
еще особые микротельца — пероксисомы. Подобно сферосомам и
лизосомам они образуются из канальцев эндоплазматической сети
и имеют одинарную мембрану. Пероксисомы контактируют с мито-
хондриями и хлоропластами, в них находятся ферменты, катализи-.
рующие восстановление молекул кислорода путем окисления амино-
кислот и других органических кислот; их главная функция — защи-
та клеток от активного кислорода.
Пластиды. Функция фотосинтеза у автотрофных растений связа-
на с наличием особых пигментов: хлорофилла и др. Лишь у наиболее
примитивных синезеленых водорослей пигменты рассеяны не-
посредственно в цитоплазме. У всех же остальных автотрофных рас-
тений хлорофилл и другие пигменты, принимающие участие в фото-
синтезе, находятся внутри особых телец, органелл клетки — хро-
свойственны большинству
водорослей, они относи-
тельно крупны, находятся
в клетке в числе одного
или нескольких, могут
иметь различную форму:
пластинки, ленты, чаши
и др. (рис. 7). У высших
растений и у наиболее вы-
сокоорганизованных водо-
рослей пластиды мелкие,
величина их колеблется в
пределах 1 —12 мкм, и в
каждой клетке их до 100
штук. В большинстве слу-
чаев пластиды имеют
форму несколько вытяну-
того диска, иногда они
могут быть иной формы:
веретеновидной, полулун-
ной, звездчатой и др.
Превращение в процессе
эволюции крупных хро-
матофоров в большое количество мелких пластид с гораздо боль-
шей суммарной поверхностью — признак прогресса, поэтому круп-
ные одиночные хроматофоры мы встречаем только у водорослей,
да и то не у всех: наиболее высокоорганизованные водоросли (ча-
сто зеленые, бурые и красные) имеют мелкие зернистые пластиды.
В зависимости от окраски и функции, выполняемой пластидами,
их делят на три группы: хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют зеленую окраску, хромопласты желтую, оран-
30
жевую или красную, лейкопласты бесцветны. Наиболее важную
роль в жизни растения играют хлоропласты. От них, собственно, и
зависит зеленый цвет растений.
В развивающихся органах растения пластиды образуются из
пропластид, возникающих еще в зародыше растения. Это тельца,
сходные с митохондриями, но несколько крупнее их. Снаружи про-
пластиды одеты двойной плазматической мембраной, причем внут-
ренняя мембрана может образовать впячивания в глубь пропласти-
ды. В зависимости от того, какая формируется ткань, пропластиды
могут превращаться в любой из трех типов пластид, хлоропласт,
хромопласт или лейкопласт. Внутренняя структура пропластиды
при этом превращается в структуру соответствующей пластиды.
Размножаются пластиды путем деления. Наиболее хорошо исследо-
вано строение хлоропластов.
Хлоропласты более или менее округлой дисковидной формы. Те-
ло пластиды одето двойной мембраной и состоит из бесцветной гид-
рофильной белковой основы — стромы (матрикса), имеющей тон-
козернистую структуру, ламелл и из красящих веществ — пигмен-
тов. Хлоропласт может утратить пигменты, и тогда останутся одна
Рис. 8. Строение хлоропласта. А — продольный разрез через хлоропласт (участок
внизу показан в увеличенном виде), Б — трехмерная схема расположения и взаи-
мосвязи ламелл и гран внутри хлоропласта:
/ — граны; 2—оболочка; 3 — матрикс (строма); 4 — ламеллы; 5 — капли жира, образовав-
шегося в хлоропласте
строма и ламеллы; при этом он сохранит свою форму. Ламеллы
состоят из парных мембран, отходящих, по-видимому, от внутрен-
них мембран оболочки хлоропласта; ламеллы протягиваются через
весь хлоропласт параллельно его оси. В определенных местах меж-
ду мембранами ламелл располагаются в числе нескольких упло-
щенные пузырьки дисковидной формы — тилакоиды, также состоя-
щие из элементарной мембраны; тилакоиды располагаются стопка-
ми, напоминающими стопки монет, и называются гранами. В каж-
31
дой стопке может быть до 100 таких дисковидной формы тилакоидов,
а количество стопок (гран) в одном хлоропласте до 40—60 (рис. 8).
Предполагают, и для этого есть достаточно оснований, что мономо-
лекулярный слой хлорофилла размещается в мембранах тилакоидов
между белковым и фосфолипидным слоем, а каротиноиды в липид-
ном слое мембраны. Хлорофилл, как известно, зеленого цвета, каро-
тиноиды подразделяются на группу каротинов (оранжево-красного
цвета) и ксантофиллов (желтого цвета).
В хлоропластах высших растений два зеленых пигмента: хло-
рофилл а, имеющий синеватый оттенок, и хлорофилл b — с желтова-
тым. оттенком. Каротины и ксантофиллы также представлены нес-
колькими изомерами. К изомерам каротина относится красящий
пигмент томатов — ликопин. Пластиды, имеющие только кароти-
ноиды, называются хромопластами, лейкопласты совсем не имеют
пигментов.
Три вида пластид — хлоропласты, хромопласты и лейкопласты,
по-видимому, могут взаимно переходить друг в друга: позеленение
клубней картофеля (превращение лейкопластов в хлоропласты),
покраснение плодов шиповника (превращение хлоропластов в хро-
мопласты), позеленение головки корнеплода моркови (превращение
хромопластов в хлоропласты) и т. д.
С утратой пигментов пластиды лишаются функции фотосинтеза.
Однако лейкопласты играют важную роль в жизни растения: они
синтезируют из первичных продуктов фотосинтеза более сложные
соединения: крахмал, белки и др. Лейкопласты, синтезирующие из
глюкозы вторичный крахмал, называются амилопластами, синтези-
рующие белок — протеопластами. Наиболее широко распростране-
ны лейкопласты, синтезирующие вторичный крахмал, поэтому их
нередко называют крахмалообразователями. Способность лейко-
пластов синтезировать белки и другие вещества (кроме крахмала)
в последнее время подвергается сомнению.
Физиологическая роль хромопластов пока недостаточно выясне-
на. Некоторые современные исследования указывают на участие
каротиноидов в окислительно-восстановительных процессах фото-
синтеза; установлено также, что каротин является фактором роста,
а также играет какую-то важную роль в половом процессе; однако
роль этих пигментов еще подлежит исследованию.
У водорослей специфическую роль в процессе отложения пер-
вичных продуктов фотосинтеза играют особые тельца белковой
природы — пиреноиды, являющиеся структурными элементами
хроматофоров. Первичный крахмал или заменяющие его поли-
сахариды в клетках водорослей накапливаются вокруг пирено-
идов.
Химическая природа хлорофилла изучается уже свыше 100 лет,
и лишь сравнительно недавно установлена химическая природа это-
го вещества. Оба хлорофилла высших растений — сложные эфиры
двухосновных кислот — хлорофиллина’а и b и спиртов — фитола
и метилового. Хлорофиллины — сложные многоатомные соедине-
ния, содержащие азот и магний.
32
Эмпирические формулы обоих хлорофиллов сходны:’хлорофилла
a— C55H72O5N4Mg; хлорофилла b --- CssHyoOeNWg. Химическая
природа каротиноидов проще: каротин — углеводород эмпиричес-
кой формулы С40Н56, ксантофилл — двухатомный спирт формулы
С4оН5б02. Смесь пигментов, так называемый сырой хлорофилл, лег-
ко извлекается из растертой массы листьев этиловым спиртом. По-
лучается темно-зеленая спиртовая вытяжка пигментов.
Русский ботаник М. С. Цвет в начале XX в. (1901 —1910), поль-
зуясь разработанным им хроматографическим методом, отделил хло-
рофилл от каротина и разделил смесь хлорофиллов па хлорофиллы
а и Ь. Разделение хлорофиллов основано па различной адсорбцион-
ной способности этих веществ. В хлоропластах обнаружена в не-
большом количестве РНК, следы ДНК й рибосомы, поэтому в хло-
ропластах может происходить синтез белка.
Хлорофилл играет основную роль в процессе фотосинтеза. В при-
сутствии хлорофиллов в этом процессе, по-видимому, активное уча-
стие принимают и каротиноиды. Явление фотосинтеза и физиологи-
ческую роль хлорофилла изучают давно. Великий русский ученый
К. А. Тимирязев посвятил изучению этого вопроса большую часть
-своей жизни. Этот важнейший для жизни растений и всего органи-
ческого мира процесс и до наших дней полностью еще не раскрыт,
однако применение новейших, методов, в том числе и так называе-
мого’метода меченых атомов, привело к крупным открытиям в этой
области. Большие заслуги в изучении фотосинтеза имеют ученые
Байер, Бах, Любименко, Вилынтеттер, Виноградов, Рубен и Камен,
Арион и др.
Размножение пластид проходит путем деления их посредством
простой перетяжки; при делении клеток пластиды материнской клет-
ки распределяются между дочерними..
В процессе приспособления к использованию света различной
интенсивности выработалась способность хло’ропластов к активному
перемещению внутри клеток, в толще цитоплазмы. При ярком ос-
вещении, которое действует разрушающе на хлорофилл, хлороплас-
ты располагаются преимущественно у клеточных стенок, направ-
ленных параллельно солнечным лучам. Происходит взаимное
затенение пластид, и этим смягчается резкое действие света. При за-
тенении, когда количество света становится недостаточным для мак-
симальной производительности фотосинтеза, хлоропласты распола-
гаются преимущественно вдоль стенок, расположенных перпенди-
кулярно к падающим лучам. При этом максимально увеличивается
освещенность пластид. Такие перемещения пластид называются
ориентированными движениями.
Ядро. Как и цитоплазма, ядро — обязательная составная часть
живой клетки растения. Клетка без ядер у высших растений встре-
чается лишь как исключение, и это носит дегенеративный характер.
У большинства бактерий и у синезеленых водорослей ядерное ве-
щество морфологически не обособлено.
Ядро всегда находится в окружении цитоплазмы. Даже в тех
случая^, когда клетка сильно вакуолизирована и цитоплазма при-
2—326
33
жата в виде тонкой пленки к клеточной стенке, ядро все же нахо-
дится в окружении цитоплазмы, слой которой в этом месте обра-
зует заметный бугорок. Это обстоятельство подчеркивает тесную,
органическую связь этих двух основных компонентов протопласта.
Связь тут, конечно, не только морфологическая, но и физиологичес-
кая: цитоплазма и ядро выступают как единое целое в сложных
процессах обмена веществ, протекающих в клетке; это, однако, не
исключает своеобразия функций ядра как органоида клетки.
Размер ядра связан с размерами клетки. То же можно сказать
и о его форме. Чащ.е ядра имеют форму шара или эллипсоида, ред-
ко они вытянуты или имеют другую, более сложную форму. Диаметр
ядра обычно 5—20 мкм, у грибов же ядра могут быть мелкими —
до 1 мкм. С ростом клетки и ее вакуолизацией размер ядра заметно
не увеличивается, поэтому ядра у клеток, выросших и сильно ваку-
олизированных, занимают относительно меньший объем.
В клетках высших растений обычно одно ядро, у низших — бы-
вает и несколько, а в клетках некоторых грибов можно наблюдать
•много ядер, но они очень мелкие. В состоянии относительного по-
коя, в момент, когда клетка не делится, ядро представляется одно-
родным и лишь заметны ядрышки — мелкие круглые тельца в ко-
личестве 1—4. Однако если ядра разрушить и подвергнуть центри-
фугированию, то вещество их разделится на две фракции: плотный
осадок — хроматин и ядерный сок — нуклеоплазму, или кариоплаз-
му, представляющую собой белковый коллоид подобно гиалоплаз-
ме, таким же способом можно отделить и ядрышки, которые име-
ют плотность больше, чем все остальные органоиды клетки.
Биохимический анализ показывает, что основную массу ядра —
70—96% сухого вещества — составляют белки, среди которых пре-
обладают нуклеопротеиды, т; е. белки, связанные с нуклеиновыми
кислотами; 10—20% сухой массы ядра составляют липиды, как и в
цитоплазме, представленные в основном фосфолипидами, содержа-
щими фосфорную кислоту.
С поверхности ядро одето оболочкой, состоящей из двух слоев
плазматической мембраны типичного строения. Между мембранами
находится перинуклеарное пространство. Общая толщина всей обо-
лочки около 40 нм. Перинуклеарное пространство непосредственно
соединяется с каналами эндоплазматической сети таким образом,
что наружная мембрана ядерной оболочки переходит в стенки ка-
нальцев эндоплазматической сети. В двойной оболочке ядра имеют-
ся отверстия, диаметр которых достигает 60 нм,— поры. Тот факт,
что каналы пор заполнены плазмой, биохимически отличающейся
и от цитоплазмы, и от нуклеоплазмы, говорит о том, что проникно-
вение веществ из цитоплазмы в ядро — не простой механический
процесс перемещения частиц. Оболочка ядра отличается от плаз-
малеммы иной проницаемостью и неспособностью восстанавливать-
ся после механических повреждений.
Хроматиновое вещество, получившее свое название по способ-
ности более интенсивно окрашиваться, лишь при рассмотрении в
оптический микроскоп кажется бесструктурным, аморфным. При ис-
34
следовании с помощью электронного микроскопа с высокой разре-
тающей способностью хроматиновое вещество оказывается состоя-
щим из тонких извитых нитей (протофибрилл) диаметром 2—4 нм.
Как показывают более тонкие исследования, основу нитей состав-
ляют двойные спирали. Эти двойные спирали представляют моле-
кулы дезоксирибонуклеиновой кислоты (ДНК). В хроматиновых
нитях эти молекулы соединены с белками, образуй дезоксирибо-
нуклеопротеиды (ДНП). Протофибриллы
в пучки. Длина молекулы ДНК в яд-
рах клеток высшего растения может
достигать нескольких сантиметров, в
то время (ка'к толщина ее составляет
всего несколько нм.
ДНК конденсируются
хромосомы из про-
Рис. 10. Схема, показы-
вающая двухцепочное
строение молекулы ДНК
Рис. 9. Модель строения хромосомы из про-
тофибрилл ДНК. А и Б — интерфазная хро-
мосома, В — метафазная хромосома (по де
Робертису и др.)
Особенно отчетливо хроматиновое вещество выделяется в период
деления клетки. В этот период протофибриллы ДНП соединяются
в особые тельца, имеющие форму комочков, палочек, крючков, —
хромосомы. В хромосомах протофибриллы ДНК (точнее ДНП)
свернуты в тугую спираль, толщина которой достигает 20—30 нм.
Эта спираль в свою очередь складывается в сложные тугие петли,
в результате чего образуется более или менее нитевидное тельце,
видимое в оптический микроскоп, — хромосома. Не исключена воз-
можность, что хромосома представляет собой одну единственную
2*
35
огромной длины молекулу ДНК (соединенную с белком), дважды
«утолщенную» за счет скручивания в спираль с последующей уклад-
кой этой спирали плотными петлями (риб. 9). Следовательно, как
хромосомы делящейся клетки,’ так и хроматин покоящейся клетки
состоят из одних и тех же элементов — протофибрилл ДНП, наибо-
Полинуклеотидная
цепь А
Полинуклеотидная
цепь В
Сахар — Аденин
Фосфат
Тимин — Сахар
Сахар — Цитозин
Фосф^
Сахар — Тимин
Фосфат
Гуанин — Сахар
Фосфат
Аденин —- Сахар^
Фосфат^
Сахар— Цитозин
Фосфат
Гуанин — Сахар
Фосфат^
Сахар— Аденин
Фосфат
Тимин — Сахар
Фосфат
^Сахар— Гуанин
Фосфат
Фосфат
Цитозин — Саха^^
Фосфат
Рис. 11. Схема участка молекулы ДНК с двумя полинукле-
отидными цепями, соединенными водородными связями
лее существенной частью которых являются двойные спирали дезо-
ксирибонуклеиновой кислоты (ДНК).. Схематически строение моле-
кулы ДНК изображено на рис. 10.
Клеткам каждого вида растений свойственно определенное ко-
личество хромосом; причем в вегетативных клетках, т. е. клетках,
не служащих для размножения, число хромосом обычно бывает чёт-
ным и хромосомы по своей форме парные. К хромосомам мы еще
вернемся при рассмотрении процесса деления клеток.
Не вдаваясь в подробности строения молекулы ДНК, что явля-
ется предметом изучения в биохимии и генетике, здесь можно ска-
зать, что молекула дезоксирибонуклеиновой кислоты состоит из
молекул сахара (дезоксирибозы), фосфорной кислоты и четырех
азотистых оснований: двух пуриновых (аденин и гуанин) и двух пи-
римидиновых (цитозин и тимин). Посредством молекул фосфата ос-
татки сахаров соединены в спиральные цепи, на внешней стороне
которых располагаются сахарофосфатные связи. На внутренней сто-
36
роне спиралей к остатку сахара присоединены основания. Посред-
ством водородных связей пуриновые.основания одной цепи соединя-
ются с пиримидиновым основанием другой цепи (рис. 11). Звено мо-
лекул ДНК, состоящее из сахара, фосфорной кислоты и азотистого
основания, называется нуклеотидом.
В состав хроматинового вещества входят также рибонуклеино-
вая кислота (РНК), аминокислоты и липиды; однако специфичес-
ким веществом хроматина является ДНК. По современным пред-
ставлениям в молекулах ДНК «закодирована» вся наследственная
информация клетки, а следовательно, и организма, развивающего-
ся из репродуктивной клетки. Рибонуклеиновая кислота специфична
для следующего органоида клетки — ядрышка.
Из всех органоидов клетки ядрышко обладает наибольшей плот-
ностью, нередко оно имеет полутвердую консистенцию. Биохимичес-
ки, как уже сказано, ядрышки характеризуются высокой концент-
рацией рибонуклеиновой кислоты (РНК), которая здесь связана
с фосфопротеидами. Как и ядро, ядрышки обычно имеют сферичес-
кую форму, хотя из этого правила также бывают исключения: встре-
чаются ядрышки вытянутые, лентовидные. Размер ядрышек зависит
от функционального состояния клетки; в частности, замечено, что
у быстрорастущей клетки, в которой идут интенсивные процессы син-
теза белка, ядрышки увеличиваются в размерах. Поскольку синтез
белка осуществляется при помощи рибосом, было высказано пред-
положение, что местом образования рибосом являются ядрышки,
откуда рибосомы поступают в цитоплазму.
Применение электронного микроскопа позволило обнаружить
тонкую, субмйкроскопическую структуру ядрышка. Оказалось, что,
в отличие от ядра и другйх органоидов клетки, на поверхности ядры-
шек нет плазматической мембраны; само вещество ядрышек гетеро-
генно, оно состоит из нуклеонемы — пучков фибрилл, весьма сход-
ных с протофибриллами хроматинового вещества, и из аморфного
матрикса, имеющего гранулярную структуру. Эти факты указывают
па теснейшую связь ядрышка с остальными элементами ядра,
в частности и с хромосомами.
Ядерный сок, или кариоплазма (нуклеоплазма), оптически пред-
ставляется гомогенной жидкостью; биохимически — это, подобно
гиалоплазме, матрикс — гидрозоль различных белков, в том числе
ферментов; электронномикроскопические исследования позволяют
обнаружить в кариоплазме гранулы, состоящие из РНК. По-види-
мому, это РНК, образованная в ядрышке и находящаяся на пути
в протоплазму. Это еще раз подчеркивает теснейшую функциональ-
ную связь ядрышка, ядра и цитоплазмы. Рибосомы, продуцируемые
ядрышком, синтезируют белок в цитоплазме.
Установленная тесная связь ядра и цитоплазмы в процессах син-
теза и обмена веществ делает понятным, почему клетки, лишенные
ядра, утрачивают жизнеспособность и в конечном счете довольно
быстро погибают. Однако следует отметить, что клетки автотроф-
ных растений могут, по сравнению с животными клетками, значи-
тельно дольше существовать в безъядерном состоянии, что, очевид-
37
но, обусловлено автотрофным способом питания и наличием в пла-
стидах ДНК.
О роли ядра в процессе размножения и в явлениях наследствен-
ности будет 'Сказано при рассмотрении вопроса деления клетки.
ВКЛЮЧЕНИЯ В ПРОТОПЛАСТ
В процессе жизнедеятельности протопласта в нем образуются са-
мые разнообразные продукты. Это или питательные вещества, ко-
торые далее используются организмом, или отбросы. Водные раст-
воры многих веществ собираются в толще цитоплазмы в виде все
увеличивающихся капель клеточного сока, заполняющего вакуоли.
Нередко вакуоли или одна вакуоля занимают почти весь объем
клетки, а цитоплазма со всеми органоидами клетки располагается
тонким слоем под оболочкой.
Первоначально вакуоли возникают в результате накопления в ка-
нальцах эндоплазматической сети растворимых в воде низкомоле-
кулярных продуктов синтеза. В этих местах диаметр канальца уве-
личивается и стенка его, представляющая плазматическую мембра-
ну, становится тонопластом, ограничивающим вакуоль. Вакуоли
свойственны всем клеткам растений, в том числе и зародышевым
клеткам высших растений, в которых они плохо заметны из-за не-
большой величины.
При формировании эндосперма семян или других органов запа-
са размер вакуолей может уменьшаться за счет заполнения их за-
пасными питательными веществами. В качестве запасных питатель-
ных веществ в клетках откладываются, часто в больших количест-
вах, крахмал, белок и жиры. Крахмал отлагается в виде зерен
различного размера и строения, запасной белок вначале в жидком
состоянии образует капельные включения, которые затем, в меру
обезвоживания цитоплазмы, превращаются в твердые алейроновые
зерна. Жиры накапливаются в цитоплазме или в виде крупных ка-
пель, или в виде тонкой эмульсии. Подобно жирным маслам в клет-
ках отлагаются также эфирные масла.
Крахмал. Это самое распространенное питательное вещество в
растениях. Различают первичный или ассимиляционный, крахмал,
отлагающийся в хлоропластах (зернышки этого крахмала очень
мелкие), и вторичный, или запасной, крахмал, синтезируемый с по-
мощью лейкопластов или непосредственно цитоплазмой и накоп-
ляющийся преимущественно в органах запаса — семенах, плодах,
клубнях, корнях и др. Основная масса первичного крахмала синте-
зируется в листьях. Естественно, что в этих органах не может отло-
житься значительное количество крахмала. Под действием фермен-
та диастазы крахмал, гидролизуясь превращается в дисахарид —
мальтозу, которая под действием фермента мальтазы — в глюкозу.
Крахмал не растворим в воде, глюкоза, наоборот, легко раство-
рима; в состоянии раствора глюкоза транспортируется из листьев
в органы запаса, и здесь происходит ресинтез ее обратно в крахмал,
который называется v^e вторичным, запасным. Процесс вторично-
38
го синтеза крахмала осуществляется при помощи лейкопластов или
в некоторых случаях (например, в эндосперме пшеницы) непосред-
ственно цитоплазмой.
В органах запаса крахмал отлагается в виде различной величи-
ны формы зерен. Форма -зерен бывает яйцевидная, эллиптическая,
почти шаровидная,
иногда угловатая и др.
(рис. 12). Размеры зе-
рен колеблются в боль-
ших пределах, напри-
мер, зерна ^крахмала в
клубнях картофеля
имеют диаметр от 5 до
145 мкм, в эндосперме
лшеницы — 30—45, ку-
курузы— 15—35, ячме-
ня— 20—35, риса —
3—10 мкм; в семядо-
лях фасоли и горо-
ха — 50—60 мкм.
Зерна крахмала
имеют видимую в' мик-
роскоп слоистость; тем-
ные СЛОИ чередуются Рнс- 12- 3еРна крахмала. А - картофеля, Б - пше-
со светлыми. Это зави- г
сит от различного со-
держания воды в слоях крахмала; темные слои богаче водой.
Причины такой слоистости недостаточно выяснены; у некоторых
растений это связано с чередованием дня и ночи, но это не един-
ственная причина; в условиях непрерывного освещения слоистость
не устраняется.
Слоистость может быть концентрической, когда слои на всем про-
тяжении одинаковой толщины (у бобовых, злаков); при неравно-
мерном утолщении зерна (например, у картофеля) возникает эк-
сцентрическое строение крахмального зерна.
По степени сложности различают зерна простые, полу сложные
и сложные. Простое зерно имеет один центр образования, полуслож-,
ные 2—3 центра, причем в начале образования каждый центр имеет
свои слои, затем образуется общая слоистость,-охватывающая уже
сформировавшиеся несколько зернышек. Сложные зерна (у овса,
гречихи) состоят из многочисленных мелких простых зерен; это
соединение непрочно, и сложное зерно рассыпается на простые.
Еще в 1869 г. академик Фаминцын высказал предположение, что
крахмальное зерно представляет сложный сфербкристалл. Это пред-
положение впоследствии подтвердилось: каждый слой крахмального
зерна состоит из огромного числа лучисто ‘расположенных иголо-
чек— трихитов, невидимых в обычный микроскоп. Наращивание
толщины слоя есть по существу согласованное удлинение простей-
ших структур зерна — трихитов.
39
Крахмал — наиболее распространенное питательное вещество,
входящее в состав пищевых и кормовых средств, а также исполь-
зуемое как сырье для производства спирта и других продуктов.
В клетках низших растений (бактерии, грибы) накапливается
полисахарид, близкий к крахмалу, — гликоген, однако, в отличие
от крахмала, он представляет гидрофильный коллоид, а не твер-
дое вещество.
Алейроновые зерна. Запасной белок можно наблюдать в семе-
нах (рис. 13). Очень мелкие алейроновые зерна заполняют прост-
Рис. 13. Алейроновый слой и зер-
на крахмала в зерновке пшеницы:
1 — сухой околоплодник; 2 — кожура
семени; 3 — клетки с алейроновыми
зернами (алейроновый слой); 4 — клет*
ки с зернами крахмала-
ранства между крупными, оваль-
ной формы, крахмальными зер-
нами в клетках семядолей фасо-
ли или гороха. Алейроновые зер-
на сплошь заполняют полости
одного или нескольких наружные
слоев клеток эндосперма в зер^
новках хлебных злаков, поэтому
и сам слой этих клеток называет-
ся алейроновым. Белок может
отлагаться также в виде кристал*
лов, например в семенах клеще*
вины.
Запасной белок — важная со-
ставная часть пищевых и кормо*
вых продуктов, получаемых из
плодов и семян растений. Семена
бобовых, например, горох, бобы,
фасоль, чечевица и др., ценятся
как раз за высокое содержание
в них белков.
Жирные масла. До 90% всех
видов растений содержат в качестве запасного питательного веще-
ства жиры. Наиболее широко распространены жиры в семемах.
Это имеет определенный* биологический смысл, ведь жиры очень
калорийны: 1 г жира при сжигании дает 39 кДж, в то время как
1 г крахмала — всего 17 кДж. Таким образом, при меньшем объе-
ме и массе достигается большая энергообеопеченность семян.
Жирные масла обычно соединены с другими жироподобными
веществами, сообщающими жирам окраску и различный запах.
Группа жироподобных веществ, включая и жиры, называется ли-
пидами. Липиды обладают общими физическими свойствами: не
растворяются в воде, но растворяются в спирте, эфире, бензине
и других жирорастворителях, оставляют на бумаге масляное пят-
но, при мелком раздроблении в воде образуют эмульсию и т. д.
Многие растения, объединяемые в группу масличных культур,
высеваются специально для получения масла: подсолнечник, лен,
клещевина, рапс и пр. Масла широко используются как пищевой
продукт, а также в технике для производства олифы, лаков, мас-
ляных красок, смазочных веществ. После извлечения масла из се*
40
мян подсолнечника, льна и некоторых других растений остается
жмых, или шрот, — ценный корм, богатый белком.
Эфирные масла. Эфирные масла получили свое название по спо-
собности легко улетучиваться и, подобно жирным маслам, остав-
лять на бумаге жирные пятна, а также растворяться в жирораст-
ворителях. Это сборная группа продуктов обмена, .обладающих
хорошо различимыми запахами; сюда относятся спирты, альдеги-
ды, кетоны, углеводороды, органические кислоты и другие органи-
ческие соединения. Среди них есть много лекарственных веществ,
а также веществ, обладающих приятным запахом; поэтому эфир-
ные масла находят применение в медицине, парфюмерии, конди-
терском производстве. Некоторые растения выращивают специаль-
но для получения эфирных масел: розы, лаванду {Lavandula
vera), мускатный шалфей {Salvia sclarea), розовую герань {Pe-
largonium roseum), мяту {Mentha piperita) и др.
Растения, богатые эфирными маслами, неохотно поедаются жи-
вотными.
Клеточный сок. От клеточного сока зависит вкус растений и в
большинстве случаев окраска плодов, цветков, а иногда и других
органов растений. В' клеточном соке могут быть растворены самые
разнообразнее вещества: сахара, органические кислоты, гликози-
ды, дубильные вещества, алкалоиды, пектиновые вещества, мине-
ральные соли и красящие вещества — пигменты. Некоторые веще-
ства могут выкристаллизовываться из клеточного сока, образуя
кристаллические включения в клетку. На долю сухого вещества
клеточного сока приходится от 5 до 30%, остальное составляет
вода.
Каждому растению специфичен определенный состав клеточного
сока, поэтому мы легко различаем на вкус сочные плоды раз-
личных растений. Но у одного и того же растения состав клеточ-
ного сока изменяется. Например, при созревании яблок, слив и ви-
шен в них уменьшается количество кислот и увеличивается коли-
чество сахара; меняется окраска цветков в процессе цвете-
ния и т. д.
Из сахаров наиболее часто встречаются в клеточном соке мо-
носахариды— глюкоза и фруктоза. Глюкозу называют также ви-
ноградным сахаром, фруктозу — плодовым; фруктоза слаще глю-
козы. В большом количестве эти сахара находятся в плодах
винограда, яблок, груш, слив, персика, арбуза, в стеблях сорго
и кукурузы. Вместе с моносахаридами обычно находится и диса-
харид — сахароза, тростниковый сахар, но особенно много саха-
розы в стеблях сахарного тростника и в корнеплодах сахарной
свеклы.
В клеточном соке растений из семейства сложноцветных, коло-
кольчиковых и некоторых других семейств широко распространен
высокомолекулярный полисахарид инулин. Действием спирта ину-
лин может быть осажден в виде крупных слоистых сферокристал-
лов. Такие сферокристаллы легко получить, например, в клетках
клубней георгины после длительного вымачивания их в спирте.
41
Органические кислоты в клеточном соке находятся в свободном
состоянии или в виде солей. Наиболее часто встречаются в кле-
точном соке щавелевая, яблочная, винокаменная и лимонная кис-
лоты. Кислый сок листьев и плодов многих растений зависит в ос-
новном от присутствия одной или нескольких из этих кислот.
Кроме обычных органических кислот, в клеточном соке часто нахо-
дятся аминокислоты, содержащие азот. Из этих кислот растение
синтезирует важнейшее органическое вещество — белок.
Гликозиды — безазотистые органические соединения, производ-
ные глюкозы. Некоторые из них используются как лекарственные
и ароматические вещества. Так, например, к гликозидам относятся
лекарственные вещества, добываемые из цветков ландыша, листь-
ев наперстянки {Digitalis'), побегов горицвета {Adonis vernalis);
от гликозидов зависит аромат чая, кофе, миндаля, ванили, запах
горчицы и др. Некоторые гликозиды в больших количествах ядо-
виты; при гидролизе иногда образуют синильную кислоту.
Близки по своей природе к гликозидам дубильные вещества
(танниды), обладающие вяжущим вкусом. Дубильные вещества
имеют также лекарственное значение, но в основном они идут на
дубление кожи, а также используются в текстильном производстве.
Наилучшие дубители у нас — дуб, ива, лиственница {Larix), скум-
пия {Cotinus) и др. В коре дуба содержится 10—20% дубильных
веществ, в коре ивы — 9—13%, в коре скумпии еще больше.
Алкалоиды — азотсодержащие щелочные вещества — обладают
сильным физиологическим действием, поэтому многие из них ис-
пользуются как лекарственные средства, а в больших количествах
являются ядами. Из лекарственных веществ алкалоидной природы
наиболее известны морфин, кокаин, кофеин, хинин, атропин, эфед-
рин. К алкалоидам относятся сильнейшие яды: стрихнин, бруцин,
кураре, добываемые из растений тропического семейства логание-
вых (чилибуха и Др.).
Пектиновые вещества по своей природе близки к углеводам;
при кипячении с сахаром они желатинизируются, дают студень,
желе. Этими веществами богаты плоды цитрусовых (лимон,
апельсин), айвы, яблони, сливы и других растений из семейства
розоцветных.
В клеточном соке встречаются также соли азотной, фосфорной,
соляной и других кислот. Нитратами богаты растения сорных
мест — крапива, паслен, лебеда, щирица и др.; хлоридами — расте-
ния морских побережий — солончаков и солонцов; фосфаты широ-
ко распространены в клеточном соке многих растений.
Из кристаллических включений наиболее широко распростра-
нены кристаллы щавелевокислого кальция; встречаются простые
кристаллы, сложные кристаллы — друзы, образующиеся в результа-
те сращения нескольких или многих кристаллов, и рафиды в виде
тонких иголочек (рис. 14). Гораздо реже встречаются кристал-
лы щавелевокислого магния и*гипса. Довольно широко распрост-
ранены отложения кремнезема, например в кожице некоторых зла-
ков и осок.
42
Самый распространенный пигмент клеточного сока — антоци-
ан — вещество, имеющее фенольную природу. Он обладает инте-
ресным свойством менять -окраску в зависимости от реакции сре-
ды. В щелочной среде он синий, в кислотной — красный. Красивая
окраска цветков синих, фиолетовых и красных колеров зависит от
антоциана.
Близок по химической природе к антоциану желтый пигмент
клеточного сока — антохлор. От этого пигмента зависит желтая
окраска цветков мака, георгины, коровяка, льнянки и др. (в отли-
Рис. 14. Кристаллы в клетке. А — друзы, Б — простые кристаллы, В—
рафиды
чие от желтой окраски цветков лютика, астр, настурций, касатиков,
у которых она определяется наличием хромопластов); желтый цвет
кожуры лимона также зависит от антохлора. Очень легко устано-
вить, каким пигментом определяется желтый цвет данного расте-
ния; для этого достаточно опустить исследуемую часть растения
в кипяток: антохлор растворится в воде, каротиноиды останутся
в клетках растения. Полезно запомнить, что пигменты пластид
в воде не растворяются, пигменты же клеточного сока легко раст-
воримы в воде.
В последнее время установлено, что и антоциан, и антохлор
в различных растениях несколько различаются по химической
структуре, поэтому правильнее будет говорить о них во множест-
венном‘числе, как о группе пигментов антоцианов и антохлоров.
Очень редко встречается (в лепестках венчика бобов — Vicia
faba — и некоторых других растений) третий пигмент клеточного
сока — антофеин, темно-бурого, почти черного цвета.
Различные сочетания пигментов клеточного сока й пигментов
хромопластов дают все разнообразие расцветок цветков и плодов.
Сюда присоединяется еще белый цвет лепестков, который зависит
не от пигментов, а от наличия слоев клеток, заполненных воздухом.
В том, что белый цвет лепестков является физическим эффектом
и не зависит от пигментов, легко убедиться растерев белый лепе-
43
сток между пальцами, при этом он обесцветится, сделается про-
зрачным.
Млечный сок. В клетках и особых вместилищах многих расте-
ний содержится так называемьГй млечный сок. Это непрозрачная
жидкость белого, желтоватого, оранжевого или красноватого цвета.
Растения с млечным соком встречаются в семействах молочайных,
маковых, сложноцветных, вьюнковых и др. Широко известны такие
растения с млечным соком, как одуванчик, молочай, мак, вьюнок
полевой.
Состав млечного сока разнообразен: здесь могут быть и запас-
ные питательные вещества — белки, сахара, жиры; и продукты
обмена, более не используемые растениями. Млечный сок богат
водой (50—80%)’; диспергированные в воде вещества могут обра-
зовать и эмульсию, и суспензию, т. е. находиться и в жидком, и в
твердом состоянии; сахара, соли и алкалоиды образуют настоящие
молекулярные растворы.
Млечный сок некоторых растений представляет большую цен-
ность и широко используется. В млечном соке некоторых растений
содержится каучук, получаемый в огромных количествах, главным
образом, для производства технической резины. Почти вся мировая
продукция натурального каучука добывается из млечного сока
тропического дерева хевеи (Hevea). Не менее важное значение
имеет 'гуттаперча, получаемая у нас из коры бересклета бородав-
чатого. (Evonimus verrucosa) или листьев гуттаперчевого дерева —
эвкоммии (Eucommia). Млечный сок опийного мака, застывая, да-
ет опиум, из которого получают ценные лекарственные средства.
Ферменты, витамины, ауксины, антибиотики. Все названные ве-
щества не относятся к форменным элементам содержимого клеток,
т. е. их нельзя, подобно уже рассмотренным составным частям
клетки, увидеть в микроскоп; для обнаружения этих веществ поль-
зуются методами химии, биохимии, физиологии и др. Ферменты,
витамины, ауксины и антибиотики обладают некоторыми общими
свойствами; все они — производные протопласта; находясь в нич-
тожных количествах, они способны оказывать сильное физиологи-
ческое действие, поэтому играют большую роль как в жизни расте-
ния, в котором они вырабатываются, так и в жизни животных, по-
требляющих растения.
Ферменты, называемые еще энзимами, играют роль катализа-
торов. Находясь в клетках (в протопласте и в клеточном соке)
в ничтожно малых количествах, они способны возбуждать, ускорять
или, наоборот, замедлять реакции, протекающие в клетке. Можно
сказать, что все процессы обмена веществ в живых организмах
протекают при участии ферментов, которых насчитывается очень
много. Каждый фермент обладает специфичностью: Так, например,
липазы действуют на жиры, протеазы на белки, диастаза осахари-
вает крахмал, инвертаза гидролизует тростниковый сахар на глю-
козу и фруктозу. Есть группа ферментов, регулирующих окисли-
тельно-восстановительные процессы (дыхание), — пероксидазы,
каталазы. Будучи отделены от протопласта клетки, ферменты сох-
44
раняют активность. Изменения условий внешней среды могут из-
менять активность ферментов или вовсе прекращать их действие.
Близки по своему действию к ферментам витамины, Основопо-
ложник учения о витаминах — К. Функ (1911), хотя влияния вита-
минной недостаточности наблюдались Н. И. Луниным еще в 1880 г.
Однако наиболее бурно учение о витаминах развивается лишь
в последние десятилетия. В настоящее время хорошо изучено свы-
ше 30 витаминов, и примерно такое же количество открыто и изу-
чается. Витамины обозначаются буквами латинского алфавита:
А, В, С, D, Е, К, G и др.; их обычно подразделяют на жирораст*
воримые (A, D, Е, К) и водорастворимые (остальные).
Роль витаминов в жизни растений еще недостаточно изучена.
Гораздо лучше изучена роль витаминов в жизни животных и чело-
века. При недостатка в организме большинства витаминов живот-
ные заболевают, а при полном их отсутствии заболевание приво-
дит к* смерти. Набор разнообразных кормов в рационе животных,
включая и зеленые корма, обычно полностью удовлетворяет их по-
требность в витаминах.
Ауксины, или гормоны роста, регулируют рост клеток и расте-
ния в целом. Предположение о существовании ростовых веществ
высказал еще в 1880 г. Ч. Дарвин в связи с изучением роста
и движения растений. В науку о ростовых веществах внес большой
вклад советский ученый Н. Г. Холодный.
Сходны по своему действию с ауксинами недавно открытые ве-
щества гиббереллины, являющиеся продуктом жизнедеятельности
микроскопического грибка, обычно поражающего рис. Гибберелли-
ны стимулируют удлинение побегов и ускоряют развитие высших
растений. В целом действие гиббереллинов более сложно и раз-
носторонне, чем ауксинов.
Антибиотики — вещества, вырабатываемые низшими раститель-
ными организмами и' способные -убивать микроорганизмы. Широ-
кое применение получили антибиотические вещества, выделяемые
некоторыми видами дктиномицетов и грибов. Наибольшей4 извест-
ностью пользуется антибиотик пенициллин, открытый в 1929 г.
английским ученым А. Флеммингом. Пенициллин вырабатывается
грибком, относящимся к группе зеленых плесёней, он подавляет
жизнедеятельность возбудителей многих инфекционных заболева-
ний. Продукт жизнедеятельности одного из актиномицетов — стреп-
томицин — помогает излечивать такие тяжелые болезни, как ту-
беркулез, чума, туляремия и др. В настоящее время известно свы-
ше 3000 антибиотиков; свыше 30 из них нашли применение
в лечебной практике.
Антибиотическим, бактерицидным действием обладают также
й летучие вещества, выделяемые некоторыми высшими растениями:
луком, чесноком, хреном, редькой, горчицей, черемухой и многими
другими. Советский ученый Б. П. Токин впервые обратил внимание
на это явление. Такие летучие вещества получили название фитон-
цидов, Давно известное целебное свойство чеснока, лука объясня-
ется наличием в них фитонцидов.
45
КЛЕТОЧНАЯ ОБОЛОЧКА
Клетки высших растений имеют целлюлозную оболочку; этим
они отличаются от клеток животных организмов, которые за един-
Рис. 15. Схема строения клеточной оболоч-
ки волоска хлопчатника. А — трехмерная
модель, 5 — поперечный разрез, В — деталь
модели при большом увеличении
Рис. 16. Схема строения клеточной оболоч-
ки. А — общий вид, Б — при большом уве-
личении в профиль, В — с поверхности:
1 — первичная оболочка; 2, 3, 4 — соответственно
внешний, средний и внутренний слои вторичной
оболочки; 5 — пора; 6 — слепая пора; 7 — плазмо-
десменные канальцы
ственным исключением
(асцидии) вообще пе име-
ют в составе своего тела
целлюлозы, или клетчат-
ки. У многих низших ра-
стений также нет целлю-
лозы. У большинства гри-
бов клеточные оболочки
из хитина, у многих водо-
рослей —щз пектинового
вещества, а оболочки бак-
терий— липидные, *из ге-
мицеллюлозы или хити-
на; однако и в этих слу-
чаях клетки одеты жест-
кой ригидной оболочкой,
и это отличает их от жи-
вотных клеток.
Целлюлоза по химиче-
ской природе близка к
сахарам и крахмалу. Эм-
пирическая формула цел-
люлозы такая же, как и
у крахмала (C6Hio05)n,
только число п здесь
значительно больше, чем
у крахмала. Частица
СбН 10О5 — глюкозный ос-
таток: СбН12О5—Н2О =
= СбНюРб.
В отличие от плазма-
леммы, которая полупро-
ницаема, клеточная обо-
лочка, состоящая из по-
лисахаридов, характери-
зуется полной проницае-
мостью.
Молекулы целлюлозы
образуют тончайшие фиб-
риллы с диаметром 3—
4 нм — элементарные фиб-
риллы, которые объеди-
няются в тяжи обычно
сильно уплощенной фор-
мы шириной 10—25 нм,
46
это микрофибриллы, -видимые в электронный микроскоп. Микро-
фибриллы погружены в основное вещество клеточной оболочки —
матрикс, состоящий в основном из гемицеллюлозы с большей или
меньшей примесью пектиновых веществ. На долю матрикса прихо-
дится до 60% сухого вещества оболочек, а на долю скелетного ве-
щества— целлюлозы — всего 30%, остальные 10% составляют бел-
ки, липиды и другие вещества. При соприкосновении с водой
клеточные стенки разбухают
в основном за счет матрик-
са, который несравненно
гидрофильней фибрилл цел-
люлозы.
Микрофибриллы, по-раз-
ному ориентируясь по отно-
шению к оси клетки, обра-
зуют различную ультра-
структуру клеточных оболо-
чек. В матриксе оболочек
изодиаметричеоких клеток
фибриллы располагаются в
беспорядке, образуя дис-
персную текстуру, в удли-
ненных клетках фибриллы
могут быть ориентированы
совершенно или почти пер-
пендикулярно длинной оси
клетки, в таком случае тек-
стура кольчатая, или труб-
чатая; при наклонной ори-
ентации фибрилл ультратек-
стура спиральная и, нако-
нец, если микрофибриллы
направлены строго или поч-
ти параллельно к продоль-
ной оси клетки, говорят о
волокнистой или волокнопо-
добной структуре. Клеткам,
которые в процессе форми-
рования вытягиваются, свой-
ственна кольчатая или труб-
чатая ультратекстура. В
оболочках прозенхимных
клеток микрофибриллы с
одинаковой текстурой рас-
полагаются слоями, образуя
перекрестные текстуры (рис.
15). В некоторых случаях,
например в семенах фини-
ковой пальмы или лютика,
Рис. 17. Различные пары пор. А — простая,
Б — окаймленная, В — полуокаймленная:
1 ~ замыкающая пленка; 2 — внутреннее отверстие,
поры; 3 — полость поры; 4 — торус
Рис. 18. Типы утолщений клеточных оболо-
чек в трахеях. А — кольчатые утолщения,
Б, В, Г — спиральные, Д — сетчатые
47
стенки клеток образуются без целлюлозного скелета, -из гемицел-
люлозы. В процессе прорастания семени гемицеллюлоза гидроли-
зуется до сахара, который и используется растущим проростком.
Следовательно, гемицеллюлоза в данных случаях выступает как
запасное питательное вещество и вместе с тем, отличаясь боль-
шой твердостью, играет и механическую роль.
Возникновение, рост и утолщение оболочек. Там, где должна
возникнуть клеточная оболочка, между двумя вновь образующи-
мися клетками, вначале наблюдается густое сплетение из,каналь-
цев эндоплазматической сети и цистерны диктиосом. Затем в этом
месте появляются пузырьки, заполненные пектиновым веществом;
пузырьки эти отделяются от цистерн аппарата Гольджи (диктио-
сом). Пузырьки увеличиваются в размере, сливаются и образуют
сплошную пектиновую пленку — срединную пластинку. По обе сто-
роны этой пластинки, за счет деятельности цитоплазмы, начинают
выкристаллизовываться молекулы целлюлозы, которые, полимери-
зуясь в фибриллы, формируют первичную оболочку клетки. Обо-
лочка эта не сплошная, в результате раздвигания пучков фибрилл
образуются Просветы — перфорации, сквозь которые из одной клет-
ки в другую протягиваются тончайшие тяжи цитоплазмы — плаз-
модесмы. Как уже сказано, внутри плазмодесмы проходят каналь-
цы эндоплазматической сети.
- В меру роста клетки фибриллы первичной оболочки растягива-
ются и скользят одна относительно другой, что облегчается про-
слойками матрикса; между растянутыми старыми фибриллами
синтезируются новые. Так происходит увеличение, рост поверхно-
сти оболочки. Этот рост стимулируется и регулируется гормонами
роста — ауксинами. Первичной оболочке свойственна дисперсная
ультраструктура.
Рост оболочки в толщину происходит за счет наложения новых
слоев целлюлозы, причем в каждом новом слое фибриллы бывают
ориентированы под различным углом к продольной оси клетки, что
обеспечивает большую прочность стенки (рис. 15). Наслоения-но-
вых фибрилл целлюлозы образуют вторичную оболочку клетки, ко-
торая, следовательно, имеет многослойное строение (рис. 16). Если
клетка входит в состав ткани и окружена со всех сторон другими
клетками, вторичные наслоения формируются внутрь от первичной
оболочки. Но в клетках поверхностных (эпидермис) вторичная
оболочка может наслаиваться и кнаружи от первичной оболочки.
Вторичное утолщение никогда не бывает сплошным, равномер-
ным; наоборот, всегда наряду .с утолщенными местами оболочки
сохраняются участки, где имеется только первичная оболочка. Эти
иеутолщенпые места называются порами. Поры имеют различное
строение: они бывают простыми, окаймленными, полуокаймленцы-
ми, причем, естественно, пора одной клетки располагается напро-
тив поры в стенке другой клетки, поэтому правильнее говорить
о паре пор (рис. 17).
Утолщения клеточной оболочки могут быть самые разнообраз-
ные. Наряду с массивными утолщениями, оставляющими свобод-
48
ними только поры, у клеток прозенхимных широко распростране-
ны утолщения кольчатые, спиральные или сетчатые (рис. 18).
Кольчатые или спиральные утолщения представляют собой кольца
или спирали круглого сечения, расположенные внутри клетки, име-
ющей форму цилиндрической трубки. Благодаря круглому сечению
этих утолщений, они соединяются с внутренней поверхностью пер-
вичной оболочки лишь узкой спайкой. Поэтому при повреждении
клеток, например при разрезе, кольчатые и спиральные утолщения
легко выпадают из клеток в виде, колец и пружинок.
Кольчатые и. спиральные утолщения не препятствуют удлине-
нию клеток, росту их в длину, это очень важно, ибо вытянутые,
цилиндрические клетки, соединяясь, образуют,общие трубки -г- со-
суды, и в молодых,-растущих частях растений такие сосуды могут
удлиняться вслед за ростом данного органа растения, например
корня или стебля. Утолщения могут быть и в виде сетки, находя-
щейся на внутренней поверхности клеточной оболочки, и в виде
ступенек, вдающихся внутрь клетки, и, наконец, как уже сказано,
утолщение может быть' почти сплошное, так что -неутолщенными
остаются лишь узкие пространства округлой или щелевидной фор-
мы. При значительном утолщении вторичной оболочки в таком
случае образуются каналы, идущие в толще клеточной оболочки,
такие каналы двух соседних клеток совпадают, перегораживает
их лишь тонкая пленка первичной оболочки. Наличие неутолщен-
ных первичных оболочек в виде тонких пленок, через которые про-
ходят плазмодесмы с-канальцами эндоплазматической сети внутри,
обеспечивает непрерывную связь протопластов всех клеток данного
организма в единое целое.
Инкрустация и адкрустация клеточных оболочек. С возрастом
оболочки растительных клеток могут приобретать новые физико-
механические свойства в связи с пропитыванием их пли наслоением
особых веществ. В первом случае процесс пропитывания назы-
вается инкрустацией, а пропитывающие вещества — инкрустирую-
щими— лигнин, таннин, кремнезем, углекислый кальций. Во вто-
ром случае, процесс наслаивания вещества называется адкруста-
цией, а наслаивающиеся вещества — адкрустирующими — суберин,
кутин, воск, спорополленин.
Лигнин — вещество фенольного ряда с кислыми свойствами.
Элементарно он, так же как и целлюлоза, состоит из углерода,
кислорода и водорода, но углерода в нем относительно больше,
чем в клетчатке, и калорийность лигнина значительно выше по
сравнению с клетчаткой. Оболочки клеток древесины содержат
20—30% лигнина.
В процессе инкрустации молекулы лигнина проникают в' проме-
жутки между фибриллами, заменяя матрикс и не образуя какой-
либо особой структуры. Заменяя матрикс, лигнин придает стенкам
клетки высокую прочность на сжатие. Инкрустация клеточных
ьстенок лигнином приводит к одревеснению их, при этом надо
иметь в виду, что вторичная оболочка одревесневает в большей
степени по сравнению с первичной.
49
Одревеснение клеточных оболочек почти не изменяет их прони-
цаемости, и клетки с одревесневшими стенками могут оставаться
живыми, в них могут накапливаться, а затем расходоваться пита-
тельные вещества, .а протопласт таких клеток может даже делить-
ся. Вместе с тем одревеснение клеточных оболочек делает их более
устойчивыми против разрушения микроорганизмами и грибами,,
причем эта стойкость еще более усиливается одновременным про-
питыванием оболочек дубильными веществами — таннинами. По-
следнее можно наблюдать в центральной части ствола многих де-
ревьев (так называемая ядровая древесина,).
Одревеснение — обратимый процесс. Посредством ферментов,
вырабатываемых протопластом клетки, оболочка, пропитанная лиг-
нином, может полностью раствориться и, есЛи при этом отомрет
и протопласт, от клетки ничего не останется, она исчезнет бесслед-
но. Процесс раздревеснения клеточных стенок можно наблюдать,
например, при созревании плодов айвы, они становятся значитель-
но мягче.
Оболочки некоторых водорослей пропитываются углекислой из-
вестью (СаСОз), а у диатомовых водорослей кремнеземом (ЗЮг).
Интересно отметить, что в клеточных оболочках диатомовых водо-
рослей отсутствуют микрофибриллы, и роль скелетного вещества
играет, следовательно, -кремнезем, молекулы которого образуют
трехмерные аморфные структуры. Кремнезем откладывается так-
же в клеточных оболочках эпидермиса многих осок и некоторых
злаков, сообщая листьям этих растений особую твердость, благо-
даря чему листьями таких растений легко обрезаться.
Суберин, кутин, а также воск и спорополленин по своей приро-
де близки к жирам, поэтому клеточные оболочки, содержащие эти
вещества, не смачиваются водой и почти непроницаемы для воды
и газов. В отличие от лигнина, эти вещества наслаиваются на кле-
точную оболочку. Суберин наслаивается на первичную оболочку
изнутри, делая ее непроницаемой. В результате этого протопласт
отмирает и клетка заполняется воздухом. Этот процесс называется
опробковением. Иногда клетки с опробковевшими стенками в про-
цессе адкрустации заполняются твердым экскретом, например,
у березы белым веществом бетулином.
Кутин и воск наслаиваются снаружи внешней поверхности
клеточных оболочек эпидермиса. Кутин при этом образует проч-
ную пленку, неотделимую от оболочки, — кутикулу, воск — легко
стираемый налет, хотя частично может и пропитывать наружный
слой клеточной стенки. Спорополленин образует наружную обо-
лочку микроспоры — экзину, которая характеризуется большим
морфологическим разнообразием и структурной сложностью. Ку-
тин, воск и спорополленин.— продукты протопласта, которые в по-
лужидком состоянии проникают на поверхность оболочки, надо
полагать, при помощи эктодесм. Это — плазмодесмы, проникающие
в наружные оболочки клеток эпидермиса вплоть до ее поверхно-*
сти. Роль эктодесм значительно шире, они служат не только для.
выведения наружу продуктов протопласта, но и для проникнове-’
50
ния растворов из внешней среды внутрь, на чем основано примене-
ние внекорневой подкормки и гербицидов..
Ослизнение клеточных оболочек. Этот процесс широко распро-
странен в природе и в большинстве случаев имеет положительное
биологическое значение. Ослизнение клеточных оболочек может
быть нормальное, полезное для растений, и патологическое, вызы-
ваемое бактериями. Набухает и ослизняется матрикс — вся вто-
ричная оболочка, или только наружный ее слой. Разбухший мат--
рикс может заполнить всю полость клетки, при этом клетка
лопается. К нормальным явлениям относятся случаи ослизнения
целлюлозных оболочек поверхностного слоя клеток семян некото-
рых растений. Так, например, при намачивании ослизняются семе-
на льна, айвы, тыквы, некоторых видов ромашки. К семени, окру-
женном слоем липкой слизи, легко прилипают частички почвы, что
способствует закреплению семени в почве и создает лучшие условия
для прорастания. Ослизнение семян полупаразита омелы способст-
вует перенесению их на лапках и клюве птиц, питающихся плода-
ми этого растения.
В патологических случаях могут ослизняться и стенки клеток
не поверхностных, а глубинных. Этот процесс вызывается специ-
альными бактериями и является, собственно, болезнью растения.
Такая болезнь, называемая гуммозом, часто поражает плодовые
деревья, особенно вишни, из коры дерева обильно вытекает слизь,
так называемый вишневый клей. Гуммоз развивается медленно и в
конечном .счете приводит к гибели дерева.
ОСНОВНЫЕ ФУНКЦИИ КЛЕТКИ
Обмен между клеткой и окружающей средой. Живая клетка
находится в состоянии постоянного обмена с внешней средой; для
одноклеточных растений такой средой являются прилегающие слои
гидросферы или атмосферы, для многоклеточных — также и сосед-
ние клетки. Основную роль в этом процессе обмена играет плазма-
тическая мембрана, одевающая снаружи протопласт клетки; как
уже сказано, мембрана полупроницаема и обладает в силу этого
свойства избирательной способностью. Будучи полупроницаемой,
плазматическая мембрана обусловливает осмотические свойства
клетки и, следовательно, играет первостепенную роль в таком важ-
ном процессе, как поступление молекул воды в клетку. Вода про-
никает по закону диффузии: со стороны большей ее (воды) кон-
центрации в сторону меньшей концентрации. При этом вода будет
проникать в клетку до тех пор, пока давление этого потока не
уравновесится давлением растягивающихся клеточных стенок (со-
стояние тургора), или до тех пор, пока с обоих сторон концентра-
ция воды не станет одинаковой.
Гораздо сложнее в клетку проникают или, наоборот, из клетки
выходят ионизированные в воде частицы растворенного вещества
или макромолекулы, образующие коллоидный раствор. Для осуще-
51
ствления этих процессов нужна энергия, источником ее служат за-
пасы АТФ, и, следовательно, процесс поглощения клеткой элемен-
тов питания неразрывно связан с процессом дыхания. Благодаря
затрате энергии ионы и молекулы растворенного вещества могут
перемещаться в клетку (или из клетки) против градиента концен-
трации, т. е. со стороны меньшей концентрации. Так, например,
необходимый растению катион К+ может проникать в клетки кор-
ня даже в том случае, когда концентрация его в почвенном раст-
воре ниже, чем в клеточном соке; наоборот, ненужный растению
катион Na+ клетки корня удаляют в окружающую среду даже при
более высокой концентрации в ней этого элемента. Плазматическая
мембрана является механизмом, обеспечивающим такой перенос
ионов.
Крупные молекулы полимеров (макромолекулы) проникают
внутрь клетки при помощи механизма пиноцитоза; при этом .на-
ружная плазматическая мембрана впячивается внутрь цитоплазмы,
это впячивание затем отшнуровывается в виде пузырька, который,
оказавшись внутри матрикса цитоплазмы, отдает ей свое содержи-
мое; при этом мембранная стенка пузырька- распадается. Пиноци-
тоз — это тоже активный процесс, на осуществление которого рас-
ходуется энергия, освобождаемая из АТФ.
Фотосинтез.' Фотосинтез — процесс углеродного питания расте-
ний. В ходе этого процесса растение из неорганических веществ,
воды и углекислого газа создает первичное органическое вещество.
В создаваемом веществе аккумулируется солнечная энергия, пог-
лощаемая зеленым пигментом — хлорофиллом, а также другими
фотосинтезирующими пигментами. Суммарная формула фотосин-
теза может быть представлена так:
6СО2Ц-6Н2О-|-2864 кДж->СбН12О6+6О2
В процессе аккумуляции солнечной энергии, создания первично-
го органического вещества совершаются многочисленные реак-
ции, одни из которых требуют .света и называются световыми реак-
циями, другие не требуют света и называются темновыми реак-
циями.
Наиболее существенными реакциями световой фазы фотосинте-
за являются реакции фотолиза (разложения) воды и реакция фо-
тосинтетического фосфорилирования.
Схематически световая фаза фотосинтеза может быть выраже-
на следующими формулами:
4Chl-|-4hv—*4Chl*
4СЫ* + 4Н2О -> 2 (Н2О2) + 4СЫН
2 (Н2О2) —*2Н2ОЦ-О2
4 СЫН->4Chl + 4H
4СЫ + 4h v+4Н2О 2Н2О+02+4СЫ+4Н
52
Поглощая кванты света hv, хлорофилл Chi становится актив*
ным: поглощенная им энергия световых лучей превращается
в энергию возбужденных электронов, которая идет на разложение
воды, в результате чего в окружающую среду выделяется кисло*
род и освобождается активный водород. В темновой фазе фото-
синтеза через систему катализаторов этот водород передается на
восстановление двуокиси углерода: 4Н + СО2->СН2О4-Н2О, в ре-
зультате чего получается первичный промежуточный продукт фо-
тосинтеза (СН2О) и вода. Полимеризируясь, первичный продукт
может дать глюкозу, а затем крахмал и другие органические ве-
щества:
6СН2О —> С6Н12Об
пС6Н12Об —► (СбН10О5) п-\- пН2О
Эти реакции после освобождения активного водорода не требу-
ют света и относятся к темновым реакциям фотосинтеза.
Реакции фотосинтетического фосфорилирования, в процессе
которых энергия солнца переходит в химическую' энергию АТФ,
заключаются в присоединении к молекулам аденозиндифосфата
(аденинрибонуклеотид + 2 остатка фосфорной кислоты) молекул
фосфорной кислоты:
л. АДФ + лН3РО4 — л. АТФ
В результате этой реакции образуется аденозинтрифосфат
(АТФ), в котором связь между второй и третьей фосфатными
группами нуклеотида особенно богата химической энергией. Это
так называемая макроэргическая связь. Химическая энергия ее,
являющаяся в данном случае аккумулированной световой энерги-
ей, затем легко может быть передана другим химическим соедине-
ниям в ходе внутриклеточного обмена веществ. Таким образом,
энергия макроэргической связи АТФ служит как бы «разменной
энергетической монетой» в различного рода биохимических превра-
щениях. При переходе АТФ в АДФ отщепляется одна .молекула
фосфорной кислоты и за счет утраты макроэргической связи (обо-
значена значком ~) освобождается 41,9 кДж энергии. Реакция
перехода АТФ в АДФ может быть представлена в виде следующе-
го уравнения:
ООО
II II II
Аденин — Рибоза — Р—О — Р—О — Р—ОН -|- Н2О —>
I III
ОН ОН | ОН
Аденинрибозонуклеотид
О О
II II
—> Аденин — Рибоза—Р—О ~ Р—ОН -J- Н3РО4
I I
ОН ОН
53
При усвоении одной грамм-молекулы СО2 в продуктах фотосин-
теза аккумулируется в виде химической энергии органического
вещества в среднем 477 кДж. У многих растений в качестве конеч-
ного продукта фотосинтеза может быть дисахарид сахароза, моле-
кула которого синтезируется из двух молекул моносахаридов глю-
козы и фруктозы. Темновые реакции протекают вне гран, в мат-
риксе хлоропластов, где с помощью соответствующих ферментов
могут образоваться жиры, а в присутствии азотистых веществ,
растворимых в воде, — аминокислоты и белки. Необходимая для
этих сложных процессов синтеза энергия берется из молекул АТФ.
Дыхание. Дыхание клетки — сложнейший процесс, во всех де-
талях еще не изученный. В исследовании его принимали участие
виднейшие ботаники: Л. П. Бородин, В. И. Палладии, А. Н. Бах,
С. П. Костычев и др. В основе дыхания, как известно, лежит окис-
лительный распад органического вещества, в первую очередь угле-
водов, до конечных продуктов с выделением СО2 и Н2О.
Суммарная формула процесса дыхания очень проста; она
обратна формуле фотосинтеза: СбН12О6 + 6О2->6СО2 + 6Н2О +
+ 2864 кДж. При этом освобождается та энергия, которая была
-аккумулирована в процессе фотосинтеза. Непрерывный приток этой
энергии — обязательное условие жизни.
Сложная цепь превращений соединяет исходные продукты, ис-
пользуемые в дыхании, с простейшими, конечными продуктами
окислительного распада. В. ходе этих превращений расщепляются
и синтезируются многочисленные органические соединения. Обра-
зуется масса побочных продуктов, вовлекаемых в дальнейшие
синтетические и аналитические процессы. Таким образом, процесс
дыхания тесно связан с процессом обмена веществ, является частью
общего процесса метаболизма. В этом едином процессе может пре-
обладать то анаболизм, то катаболизм. В первом случае идет на-
копление вещества и аккумулированной в нем энергии, во вто-
ром— расход преобладает над приходом и происходит потеря ор-
ганического вещества.
Суммарное направление единого процесса метаболизма за-
висит от многих факторов; например, в клетках запасающей ткани
прорастающего семени преобладает процесс катаболизма, в клет-
ках эндосперма созревающих семян или в образовательной ткани,
наоборот, преобладают процессы анаболизма.
Местом, где протекает единый процесс метаболизма, является
все содержимое клетки: его протопласт, включения, клеточный сок,
а в молодых клетках и оболочка. Объем этого процесса зависит
от состояния растения, условий, в которых оно находится, индиви-
дуальных качеств организма и других факторов. Количественно
процесс обмена веществ может быть выражен в интенсивности ды-
хательного процесса, которую выражают дыхательным коэффици-
ентом: отношением СО2 О2; чем меньше этот коэффициент, тем вы-
ше тепловой эффект дыхания. Чаще всего величина коэффициента
дыхания бывает близка к единице. Это объясняется тем, что ос-
новным дыхательным веществом у растений являются углеводы,
54
а при полном окислении углеводов объем поглощенного кислоро-
да равен объему выделенной углекислоты:
СбО12Об + 6О2-> 6СО2 + 6Н2О+2864 кДж
СО2:02= 1:1
В том случае, когда окисляется вещество, относительно бедное
кислородом, например жиры или белки, для полного его окисле-
ния потребуется больше кислорода, соответственно чему дыха-
тельный коэффициент станет меньше единицы. Так, для жиров он
будет равен 0,7.
Процесс дыхания в нормальных условиях распадается на два-
этапа. Первый этап, называемый гликолизом, анаэробный, заклю-
чается в распаде исходного вещества (сахаров) на более простые-
вещества без присоединения кислорода воздуха. Гликолиз приво-
дит к образованию пировиноградной кислоты, он протекает вне-
митохондрий. Второй этап, называемый циклом Кребса, — аэроб-
ный, когда с присоединением кислорода извне происходит окисле-
ние до образования конечных продуктов: СО2 и Н2О. Этот этап
протекает в митохондриях. Первый этап осуществляется под дей-
ствием группы ферментов анаэробных дегидраз; на втором — дей-
ствует группа аэробных дегидраз и многочисленных оксидаз (пе-
роксидаза, каталаза, фенолоксидаза и др.).
Если бы окисление глюкозы, а затем пировиноградной кислоты
до конечных продуктов происходило бы сразу, одноступенчато, ос-
вободилось бы слишком много энергии подобно тому, как это про-
исходит при сжигании топлива. На самом деле такие бывает, окис-
ление идет многоступенчато, через образование все более бедных
энергией и более простых соединений. При этом с участием кислоро-
да происходит окислительное фосфорилирование с образованием
АТФ, куда и переходит большая часть энергии пировиноградной
кислоты. Затем энергия АТФ используется в самых разнообраз-
ных эндергонических реакциях, при этом АТФ переходит в АДФ.
Вполне основательно поэтому митохондрии, где сосредоточена
основная масса АТФ, называют «силовыми станциями клеток».
В условиях недостатка кислорода, например при прорастании
семян в воде, окисление идет почти без поглощения кислорода из-
вне. При таком анаэробном дыхании дыхательный коэффициент
будет больше единицы.
Синтез белка. Белковая молекула собирается из более или ме-
нее сложных структурных единиц — аминокислот. Известно 20 ами-
нокислот, которые используются для конструирования белковых
молекул. Соединяясь, аминокислоты образуют полипептидную цепь,,
которая может состоять из десятков и сотен аминокислотных ос-
татков. Свойство белков зависит от набора и взаимного располо-
жения аминокислот в белковой молекуле.
Сборка полипептидной цепи происходит на рибосоме (рРНК),
которая играет организующую роль. Специфическую же роль
в этом процессе играют ДНК, информационная, или матричная,.
55
Двойная спираль ДНК
УУА ЦГЦ ГГУ АЦА ГГУ УУГ
1 III Г I I* 111 I I1 1_I I 1_I I I
Информационная 1?HK
У'.'У < А.УЧ ГГУ W ЦУЦ
Рис. 19. Схема гипотического
механизма синтеза белка на
рибосоме, отражающая отноше-
ние между триплетами спирали
ДНК, комплементарными им
триплетами информационной
РНК и комплементарными по-
следним триплетам (антикодо-
нами) транспортных РНК
мРНК и транспортная тРНК. Для объяснения процесса синтеза
белка воспользуемся рис. 19. Двойная спираль ДНК состоит, из
соединенных поперечными (водородными) связями цепей монону-
клеотидов, число которых колеблется от 77 до нескольких миллио-
нов. Каждый нуклеотид состоит из фосфорной кислоты, сахара
(у ДНК — дезоксирибрза, у РНК — рибоза) и азотистого основа-
ния (см. рис. 11). В состав нуклеотидов ДНК входят два пурино-
вых основания — аденин (А) и гуанин (Г) и два пиримидиновых'—
цитозин (Ц) и тимин (Т). У РНК первые три основания те же, но
вместо тимина — пиримидиновое основание урацил (У). Следова-
тельно, нуклеотиды отличаются только основаниями, которых
у ДНК и РНК по четыре, и поэтому может существовать только
четыре категории нуклеотидов, которые мы назовем по начальной
букве названия азотистого основания: у ДНК — А, Г, Т, Ц;
у РНК —А, Г, У, Ц.
Каждые три следующих друг за другом нуклеотида образуют
триплет, управляющий включением в белок определенной амино-
кислоты. Количество различающихся триплетов из четырех раз-
личных нуклеотидов будет равно 43 = 64. Однако аминокислот 20г
поэтому одной и той же аминокислотой может управлять несколь-
ко триплетов. Молекула РНК имеет одинарную и более короткую
цепь нуклеотидов, чем ДНК.
Расположение нуклеотидов в двойной цепи и попарное соеди-
нение их всегда комплементарно; это означает, что данный нукле-
отид может соединяться только с одним из остальных трех.
ДНК РНК
А—Т А —У
Ц —Г Ц—Г
Г —Ц Г —Ц
Т—А У —А
Пуриновое азотистое основание, входящее в состав одного ну-
клеотида, всегда соединяется только с пиримидиновым основанием
другого нуклеотида комплементарной цепи, причем аденин всегда
с тимином (в ДНК) или с урацилом (в РНК), гуанин всегда с ци-
тозином.
Триплеты в двойной цепи ДНК называются кодогенами, в оди-
нарной цепи мРНК— кодонами. По принципу комплементарности
для каждого кодогена существует комплементарный ему кодон.
Передача первичной информации с ДНК осуществляется путем
сборки молекулы мРНК на разъединившихся цепях молекулы
ДНК, причем эта сборка выполняется комплементарно, т. е. на
нуклеотиде А ДНК собирается нуклеотид У РНК, на нуклеотиде
Ц — соответственно Г, на Г — Ц и на Т — А. Следовательно, на
триплете ДНК (кодогене) состава ТТА собирается триплет РНК
(кодон) ААУ, на кодогене ГЦГ — соответственно кодон ЦГЦит. д.
В верхней части рис. 19 указанная закономерность хорошо просле-
живается. Каждая молекула мРНК управляет сборкой определен-
ной полипептидной цепи, и количество, набор кодонов и последо-
57
©ательность расположения их в молекуле мРНК точно соответству-
ют количеству, набору и последовательности, размещения в цепи
-аминокислот, входящих в синтезируемый белок, иначе можно ска-
зать — в каждой молекуле мРНК записан шифр для соответствую-
щей молекулы белка.
Транспортная РНК состоит примерно из 80 нуклеотидов. На
рис. 19 она имеет форму спирали, на одном из концов которой
всегда располагается триплет ЦЦА, а на другом нуклеотид Г.
Данная молекула тРНК может транспортировать только одну, оп-
ределенную аминокислоту.
В операции сборки полипептидной цепи роли между мРНК и
тРНК распределяются следующим образом: мРНК, выйдя из ядра
в цитоплазму, приближается к рибосоме и размещается на ней
таким образом, чтобы с поверхностью рибосомы контактировали
два первых кодона. По мере сборки мРНК перемещается слева
направо, подавая на «сборочную площадку» все новые кодоны до
тех пор, пока не будет пропущена вся цепь кодонов данной моле-
кулы мРНК. В это время молекулы тРНК выбирают из цитоплазм
мы каждая «свою» аминокислоту, прицепляют ее к своему ЦЦА —
концу и подтягивают к месту сборки молекулы белка, т. е. к рибо-
соме. Как только на рибосоме появляется кодон, соответствующий
антикодону данной молекулы тРНК, она тотчас же к нему присо-
единяется и некоторое время задерживается, пока не подойдет
следующая тРНК со «своей» аминокислотой. Как только это про-
изойдет, две оказавшиеся рядом аминокислоты соединяются, обра-
зуя пептидную связь. Тем временем мРНК перемещается на один
кодон влево и на «сборочную площадку» выдвигается следующий
кодон, к которому присоединяется антикодон следующей тРНК до
тех пор, пока все кодоны информационной РНК не будут пропу-
щены через рибосому. Могут ли тРНК и мРНК выполнять свои
функции повторно — неизвестно; но зато известно, что одна рибо-
сома может обеспечить сборку полипептидной цепи лишь опреде-
ленной длины; в случае необходимости собрать белок с более длин-
ной цепью аминокислот потребуется несколько рибосом, рас-
положенных рядом. Такие комплексы рибосом, расположенные
цепочкой, действительно в изобилии встречаются в клетках и на-
зываются полирибосомы.
Новейшие исследования показывают, что субъединицы рибосо-
мы, продуцируемые в ядрышке и выходящие через поры в ядер-
нбй оболочке в цитоплазму, соединяются лишь к началу процесса
синтеза белковой молекулы; по окончании сборки белка субъеди-
ницы разъединяются и существуют раздельно до начала следую-
щего рибосомного цикла. Молекула матричной РНК «протягивает-
ся» между соединившимися субъединицами рибосомы, а в полири-
босомах через все «ожерелье» рибосом.
Сборка белковых молекул идет с огромной быстротой, за одну
минуту образуется 5—6 тыс. пептидных связей (между аминокис-
лотами). Для построения одной средней по размеру молекулы бел-
ка требуется всего четверть секунды.
58
Раздражимость. Раздражимость свойственна всем живьш
клеткам, но у растений проявляется слабее, чем у животных, хотя
у некоторых растений, например у стыдливой мимозы (Mumosa
pudica), это свойство выражено очень хорошо. В передаче раздра-
жений непосредственное участие принимает цитоплазма клеток.
Проявлением свойства раздражимости можно считать воздействие
на клетки растений гормонов. Действие гормонов весьма разносто-
ронне. К гормонам, регулирующим рост, относятся* ауксины, ока-
зывающие влияние на большинство ростовых процессов, кинины,
стимулирующие деление клеток, и гиббереллины, действующие на
растительную клетку более разносторонне: они стимулируют рост
молодых клеток в длину, ускоряют развитие растения, усиливают
клеточное деление. Явления тропизмов также связаны с воздейст-
вием гормонов на рост клеток. Факторы, побуждающие растения
к формированию генеративных органов (цветение), также по-ви-
димому, гормональной природы, и действие их связано с раздра-
жимостью клеток, это же свойство клеток лежит в основе настиче-
ских движений частей растения. В отличие от тропизмов, которые
обусловлены односторонним воздействием раздражителя, настии
вызываются равномерно действующими раздражителями, напри-
мер, влияние влажности, света, тепла. И в том, и в другом случае
эффект воздействия раздражителя проявляется в движении расте-
ний. Никтинастические движения связаны со сменой дня и ночи,
например, закрывание и открывание цветков или соцветий (табак,
одуванчик,* подсолнечник и др.), складывание на ночь сложных
листьев (клевер, горох, фасоль и др.).
Движение сложных листьёв стыдливой мимозы — проявление
сейсмонастии, вызываемой механическим воздействием, то же са-
мое раскрытие плодов у недотроги (Impatiens) в результате при-
косновения.
РАЗМНОЖЕНИЕ КЛЕТОК
Клеткам свойственно размножаться, в результате чего увели-
чивается их общая масса и происходит рост растения. Расти клет-
ки могут лишь до известного, в общем ограниченного, предела.
Обычный способ воспроизведения клеток — деление на две. Также
размножаются одноклеточные организмы.
Большой вклад в изучение сложного процесса деления клеток
внесли польский ботаник Э. Страсбургер (1878), немецкий гисто-
лог В. Флемминг (1882), русский ботаник И. Д. Чистяков, своими
открытиями заложившие основу для дальнейших исследований
в этой области.
Различают два типа деления клеток: прямое, или амитоз, и не-
прямое — митоз и мейоз. Митоз называется еще кариокинезом,
или эквационным делением; мейоз называют также редукционным
делением.
59
Локоящееся ядро
Профаза
Профаза
Профаза
Анафаза
Телофаза
Рис. 20. Митоз
Прямое деление клетки, или амитоз. Этот тип деления редко
удается наблюдать у высших растений; высказывали даже пред-
положение, что он свойствен лишь клеткам, утрачивающим способ-
ность к делению, а также клеткам больного растения.
При прямом делении ядро перетяжкой делится на две более
или менее равные части, вслед за ядром таким же способом де-
лится и все содержимое клетки; иногда ядро делится на несколько
частей, и тогда образуются многоядерные клетки.-При этом не на-
блюдается каких-либо сложных структур ни в ядре, ни в цитоплаз-
ме клетки, Отсюда и название способа деления — амитоз, в проти-
воположность митозу, что по-гречески означает «нить», при кото-
ром в цитоплазме наблюдаются особые нитчатые структуры.
Митоз, или кариокинез. При этом типе деления происходят
сложные перемещения составных частей ядра, которое по-гречески
называется «карион», вторая часть слова происходит от греческо-
го слова «кпнезис» — движение. Непрерывный процесс деления
клетки условно подразделяется на 4 ф’азы: 1) профаза, 2) мета-
фаза, 3) анафаза, 4) телофаза (рис. 20). Однако подготовка к де-
60
А—Т
Рис. 21. Схема редупликации молекулы ДНК. А — исходная
молекула ДНК, 5 — процесс достройки дочерних цепей,
В — две дочерние молекулы ДНК, возникшие в результате
репликации
а
лению начинается еще в период между двумя кариокинетическими
делениями, этот период называется интеркинезом или интерфазой.
Проследим изменения, происходящие в делящейся клетке в про-
должение каждой из этих фаз, начиная с интерфазы.
Интерфаза. Происходит важнейший процесс удвоения хромати-
нового вещества за счет редупликации молекул* ДНК. Вначале
происходит синтез РНК и белка. Затем хроматиновое вещество де-
конденсируется, в результате чего хромосомы оказываются нераз-
личимыми; в конце концов все хроматиновое вещество распадается
на отдельные молекулы ДНК, а'последние дисперализируются;
двойные спирали распадаются и каждая из одинарных цепей до-
страивает себе вторую комплементарную цепь; в результате полу-
чаются две совершенно идентичные молекулы ДНК, которые за-
тем при реконструкции хромосом расходятся, входя в состав двух
идентичных хроматид (рис. 21). Перед началом профазы в ядре
опять усиливается синтез РНК и белков.
Профаз'а. Реконструируются хромосомы, которые имеют форму
палочек, петель, крючков. Видно, что хромосомы уже удвоены,
каждая состоит из двух хроматид, между которыми к концу про-
фазы заметна щель.
Каждому виду растений свойственно определенное число хро-
мосом, причем в соматических клетках это число четное. Напри-
мер, у твердой пшеницы 28 хромосом, мягкой — 42, подсолнечни-
ка— 34, кукурузы — 20, гороха—14 и т. д. Это количество
хромосом, наблюдаемое в клетках в начале деления, называется
диплоидным числом хромосом; ядрышки к этому моменту обычно
61
исчезают. Таким образом, все вещество ядра к концу профазы раз-
деляется на хромосомы ц ядерный сок, или кариоплазму. Ядерная
оболочка исчезает, и кариоплазма смешивается с цитоплазмой,
образуя миксоплазму: обозначаются ахроматиновые нити.
Метафаза. Хромосомы располагаются по экватору клетки вог-
нутыми частями внутрь, образуя так называемую экваториальную
пластинку. В это время при взгляде с полюса клетки хорошо вид-
на форма хромосом; установлено, что каждая пара хромосом име-
ет определенную форму, повторяющуюся при каждом делении
клетки; таким образом, каждая хромосома индивидуальна, отли-
чима от других.
От экваториальной пластинки к полюсам протягиваются бес-
цветные нити, образующие ахроматиновые конуса; в целом вся
эта фигура из бесцветных нитей, простирающаяся от одного полю-
са клетки к другому, называется ахроматиновым веретеном L Уста-
новлено, что ахроматиновые нити имеют белковую природу с не-
большой примесью РНК, консистенция их плотнее, чем окружаю-
щая цитоплазма.
Тщательные электронно-микроскопические исследования пока-
зали, что ахроматиновые нити на самом деле представляют тон-
чайшие (диаметром около 25 нм) трубочки, микротрубочки, стен-
ки которых состоят из белковых молекул. Такие же микротрубоч-
ки обнаружены в жгутиках подвижных клеток. Имеются также
наблюдения, устанавливающие связь движения цитоплазмы (цик-
лоз) и наличия в ней большого количества микротрубочек.
К концу метафазы каждая хромосома окончательно расщепля-
ется надвое, так что число хромосом удваивается и дочерние хро-
мосомы начинают расходиться.
Анафаза. Дочерние хромосомы расходятся к противоположным
концам ахроматинового веретена, причем каждая пара дочерних
хромосом, образовавшаяся в результате расщепления, распределя-
ется между двумя полюсами, и, таким образом, на полюсах ока-
зываются одинаковые наборы хромосом, повторяющие набор
материнских хромосом в начале деления клетки. Ахроматиновые
нити в процессе расхождения дочерних хромосом играют какую-то
организующую роль, направляя хромосомы к полюсам клетки.
Телофаза. Хромосомы, отошедшие к полюсам, спутываются, ут-
рачивают видимую индивидуальность. Восстанавливаются оболоч-
ка ядра и ядрышки; ахроматиновые нити постепенно исчезают,
а по экватору клетки возникает первичная оболочка. Образование
перегородки идет от центра клетки к ее боковым стенкам.
Так происходит реконструкция Двух новых клеток. Телофаза —
процесс, обратный профазе. В процессе разделения цитоплазмы на
две дочерние клетки — цитокинезе — происходит распределение
пластид и митохондрий между двумя дочерними клетками.
1 У некоторых (подвижных) водорослей и грибов на полюсах ахроматиновых
конусов обнаружены (как и у животных) округлые тельца —1 центросомы, имею-
щие трубчатую структуру и содержащие двуспиральную ДНК, упакованную в
клубок.
62
Весь процесс митоза занимает 1—2 ч. причем наиболее продол-
жительны профаза и телофаза — 30—60 мин, в то время как мета-
фаза проходит за несколько минут, а анафаза — лишь немного
длиннее метафазы. Частота митозов различна, в клетках образо-
вательной ткани в момент интенсивного роста они могут протекать
почти без перерывов.
Редукционное деление, или мейоз. В мейозе два деления быстро
следуют одно за другим. В начале профазы первого деления длин-
ные и тонкие хромосомные нити располагаются в ядре без опреде-
ленного порядка. Затем начинается попарное сближение гомоло-
гических (парных) хромосом, которые при этом утолщаются
и становятся короче. Парные хромосомы настолько тесно сближа-
ются, что ядро кажется гаплоидным, т. е. содержащим простое,
одинарное число хромосом. Такие пары хромосом называются би-
валентами, а стадия — диакинезом. Далее становится заметным,
что каждая из сблизившихся парных хромосом расщеплена вдоль
на две хроматиды (редупликация).
Следовательно, каждый бивалент состоит из четырех хроматид.
В это время происходит важнейший процесс обмена гомологичны-
ми участками между двумя сблизившимися, парными хромосома-
ми. Диакинезом заканчивается профаза.
В метафазе биваленты располагаются по экватору клетки,
и начинается расщепление парных хромосом; в анафазе гомоло-
гичные хромосомы, каждая из которых состоит из двух хроматид,
расходятся к полюсам (рис. 22).
В отлидие от митоза на полюсах теперь оказываются гаплоидные
наборы хромосом; следовательно, это первое деление — редукци-
онное. Второе деление в мейозе эквационное.
После непродолжительной телофазы первого деления сразу же
начинается метафаза второго деления; каждая хромосома гапло-
идного набора делится надвое, и одиночные хроматиды расходятся
к полюсам, составляя ядерный набор новых клеток.
Таким образом, в результате мейоза из одной исходной мате-
ринской клетки с диплоидным набором хромосом образуется четы-
ре, каждая из которых имеет гаплоидный набор. Редуцируется
число хромосом вдвое, отсюда и название — редукционное деление.
Биологический смысл редукционного деления для нас станет яс-
ным, если мы проследим дальнейшую судьбу клеток с гаплоидным
ядром. У высших растений редукционное деление предшествует
образованию половых клеток, или гамет и спор. Образовавшись,
гаметы рано или поздно попарно сливаются, давая зиготу, из ко-
торой развивается зародыш нового организма. В процессе оплодот-
ворения происходит слияние гаплоидных ядер и восстанавливается
диплоидный набор хромосом, свойственный обычным клеткам выс-
шего растения. Следовательно, редукция числа хромосом в момент
образования половых клеток, гамет, предупреждает увеличение
числа хромосом цри оплодотворении против обычного, свойствен-
ного данному виду. При отсутствии такой редукции каждый акт
оплодотворения был бы связан с удвоением числа хромосом, и та-
63
ким образом в последующих
поколениях число хромосом
увеличивалось бы в геометрии
ческой прогрессии.
Из спор высших растений
вырастает поколение (напри-
мер, у папоротников, хвощей),
у которого все клетки гаплоид*
ные. Это поколение без после-
дующей редукции числа хро-
мосом образует половые клет-
ки, которые, сливаясь, дают
поколение растений с дипло-
идными клетками. Таким об-
разом, и здесь редукционное
деление связано с процессом
воспроизводства, в ходе кото-
рого совершается половой про-
цесс, восстанавливающий дип-
лоидный набор хромосом. Од-
нако в природе, нередко без
видимой внешней причины,
спонтанно, может происходить
удвоение, утроение и т. д. чис-
ла хромосом. Это явление на-
зывается полиплоидией, В от-
личие от обычных, диплоид-
ных организмов с двойным на-
бором хромосом, возникают
полиплоиды с числом хромо-
сом, кратным 4, 6, 8 и больше.
Такие формы в результате от-
бора и размножения .могут
стать самостоятельными вида-
Рис. 22. Схематическое сопоставление
митоза (Л) и мейоза (Б)
ми. Под действием химических и физических реагентов можно
вызвать искусственную полиплоидию. Этот путь все шире исполь-
зуют для получения форм, нужных человеку.
Давно уже установлено, что сложная картина изменения в хро-
мосомном аппарате клеток, наблюдаемая в момент их деления,
связана с явлениями наследственности, т. е. передачей потомству
признаков и свойств организма. Наука, занимающаяся изучением
явлений наследственности ш изменчивости, называется генетикой,
а раздел ее, исследующий явления наследственности и изменчи-
вости в связи с превращениями, наблюдаемыми в' хромосомном ап-
парате клеток, — цитогенетикой,
В современной цитогенетике огромное внимание уделяется спе-
цифическому веществу хромосом — дезоксирибонуклеиновой кисло-
те (ДНК). Своеобразная структура этого вещества, раскрытая
наиболее совершенными методами исследования, дает основание
G1
считать дезоксирибонуклеиновую кислоту наиболее ответственной
за сложные явления наследственности и наследственной изменчи-
вости организмов.
Согласно современным представлениям, наследственность, т. е*
способность организмов передавать в потомство свои признаки
и особенности развития, определяется набором молекул ДНК, вхо-
дящих в состав хромосом репродуктивных клеток, и структурой,
составом этих молекул. Совокупность всех наследственных задат-
ков, факторов наследственности (генов) называется генотипом,
а фактическое проявление этих задатков в свойствах, признаках
•и особенностях развития организма — его фенотипом. Генотип бу-
дущего организма, следовательно, «закодирован» в молекулах
ДНК хромосом, входящих в состав клетки или клеток (в случае
полового воспроизводства), из которых развивается этот организм.
Как же этот код реализуется в свойствах и признаках организма,
в особенностях его развития? Схематически это можно представить
себе так: молекулы ДНК продуцируют молекулы информационной
РНК специфического состава; молекулы информационной РНК
служат матрицей, на которой синтезируются молекулы ферментов
и других белков, специфичных для данного типа развития, детали
этого процесса мы рассмотрели, когда говорили о синтезе белка;
деятельность определенного набора и состава ферментов определя-
ют пути морфогенеза, т. е. формообразовательных процессов, при-
водящих, наконец, к созданию организмов с определенными свой-
ствами и признаками.
Потребность в генах, определяющих развитие тех или иных
признаков, неодинакова; например гены, ответственные за форми-
рование и функционирование генеративных органов, необходимы
только в относительно короткий период жизни растений, в то вре-
мя как гены, от которых зависит формирование вегетативных орга-
нов, необходимы на протяжении всего вегетационного периода, не
говоря уже о генах, создающих рибосомы. Исследование этих во-
просов привело к открытию явления «размножения» генов. Этс
явЛение заключается в том, что молекулы ДНК хромосом проду-
цируют не только молекулы информационной РНК, но и неболь-
шие отрезки ДНК, комплементарные информационной РНК, и этот
процесс совершается многократно, в меру потребности клеток
именно в данном гене. Небольшие молекулы ДНК воспроизводят
соответствующие молекулы информационной РНК в соотношениях
нужных для обеспечения формообразовательного процесса. Саме
собой разумеется, «то успех или неуспех этого процесса, конкрет-
ное выражение признаков и свойств нового организма будет зави
сеть от условий среды, в которой протекает развитие организма
Такова прямая связь между генотипом и формирующимся фе
нотипом: развитие фенотипа контролируется генотипом. Но одно!
прямой связью нельзя объяснить всех явлений морфогенеза. По
видимому, существует и обратная связь, т. е. влияние протопласта
клеток формирующегося организма на совокупную деятельность ге
нов, на интенсивность и направленность этой деятельности.
ГЛАВА II
ТКАНИ И ВЕГЕТАТИВНЫЕ ОРГАНЫ РАСТЕНИЙ
Совершенствование организмов неразрывно связано с диффе-
ренциацией его тела на части, специализированные для выполнения
определенных функций. Такая дифференциация возникла уже на
клеточном уровне, и в первой главе учебника мы видели, какой
высокой степени расчленения достигает даже микроскопического
размера клетка. С возникновением многоклеточных организмов
создались предпосылки и для межклеточной дифференциации,
причем первые признаки дифференциации мы наблюдаем даже
у некоторых колониальных форм, например у вольвокса.
Простейшие многоклеточные организмы — синезеленые и мно-
гие зеленые водоросли,— как правило, представляют из себя це-
почку клеток, образующую более или менее прочную нить тол-
щиной в одну клетку. Уже у свободно плавающих нитей мы можем
наблюдать явления дифференциации, связанные с процессом раз-
множения, например гетероцисты у синезеленых водорослей. При-
крепление нити к субстрату «потребовало» специального приспо-
собления одной или нескольких нижних клеток: они утратили
хлорофилл и превратились в ризоиды, выполняющие механиче-
скую функцию прикрепления водоросли к субстрату. Часть клеток
таких нитчатых водорослей приобретает специальную функцию
бесполого или полового размножения.
Увеличение размера водорослей было связако с дальнейшим
усложнением структуры их тела. Так, например, у наиболее круп-
ных бурых водорослей можно обнаружить группы клеток, приспо-
собленных к выполнению механических функций; выделяются по
своей форме и строению покровные клетки, выполняющие защит-
ную функцию; внутри таллома различаются также группы прово-
дящих запасающих, ассимиляционных клеток. Так в процессе эво-
люции возникли, как составные части единого организма, группы
клеток общего анатомо-физиологического назначения — ткани.
Зачатки тканей, следовательно, имелись уже у Низших растений —
водорослей.
Однако наибольшей степени внутреннего и внешнего расчлене-
ния достигли высшие растения. Это было связано с приспособле-
нием к жизни з условиях суши, в гораздо более сложной обста-
новке, чем водная среда, в которой обитали водоросли. В ходе
этого приспособления, а точнее в результате естественного отбора,
сохраняющего в организмах только то, что наиболее соответствует
66
условиям жизни, у высших растений развились не только настоя-
щие ткани, но и специализированные органы — внизу корни, ввер-
ху листья. Стебель выполняет, в основном, связующие функции;
как мы дальше увидим, этот орган был исходным для формиро-
вания и листьев и корней. Рассмотрение общих элементов струк-
гуры тела высших растений мы начнем с тканей.
ТКАНИ
Для того чтобы лучше составить первое представление о расти-
гельных тканях, можно рассмотреть поперечный разрез одного
из основных органов растения, например стебля. На таком разрезе
мы увидим, что все поле зрения занято клетками различной фор-
мы и величины, с разнообразным характером* оболочек и содержи-
мого.
'Сходные клетки, как правило, располагаются группами, слоя-
ми в определенной закономерности, а не хаотически. Каждая
группа клеток выполняет в жизни растения определенную функ-
цию. Такие группы внешне однородных клеток, связанных между
собой и выполняющих общую функцию, называют тканями.
Как мы увидим дальше, каждая группа однородных клеток
имеет и общее происхождение. Связь между клетками может быть
различная. Если клетки живые, связь осуществляется посредством
плазмодесм; при отмирании клеток между ними остается только
механическая связь, и такое механическое соединение клеток так-
же продолжает оставаться тканью.
Все ткани растений можно разделить на две не равные по объ-
ему группы: 1) недифференцированные, ^образовательные ткани,
или меристемы, 2) дифференцированные, или постоянные, ткани.
Дифференцированные ткани подразделяются на покровные, основ-
ные, проводящие, механические и выделительные. В основу клас-
сификации тканей положены функции тканей, а также их анато-
мическое строение. Поэтому такую классификацию можно назвать
физиолого-анатомической.
Проводящие и механические ткани обычно располагаются в ор-
ганах растений во взаимной связи, образуя сосудисто-волокнистые,
или проводящие, пучки. Такие пучки выполняют двоякую функ-
цию: механическую, сообщая в комплексе со всеми остальными
тканями необходимую прочность органам растения, и проводящую,
обеспечивая перемещёние по растению воды и растворенных в во-
де питательных веществ. Все тело растения, от корней до кон-
чиков листьев, пронизано системой сосудисто-волокнистых пучков,
связывающих органы высшего растения в единую систему.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ — МЕРИСТЕМЫ
Первоисточником образовательной ткани высших растений яв-
ляется эмбриональная ткань зародыша. В процессе развития за-
родыша ткань его дифференцируется, но на кончиках корней и по-
3*
67
«Ж
‘ Is*
?о5Ф)т~
Рис. 23. Образовательная
ткань в конусе нарастания
(апексе)
бегов сохраняется образовательная ткань, постоянно возобновляе-
мая за счет деления верхушечных клеток.
Верхушечная часть побега и корня называется апексом, или
апикальной меристемой, которая возникает здесь за счет деления
одной или нескольких инициальных клеток. Та часть апикальной
меристемы, которая прилегает непосредственно к инициальным
клеткам и не обнаруживает абсолютно никаких признаков диф-
ференциации, называется промеристемой. В зависимости от того,
в какие постоянные ткани будет диффе-
ренцироваться апикальная меристема, ее
подразделяют на протодерму, которая
дает эпидермальные и субэпидермальные
ткани, прокамбий, который дает начало
проводящим .и механическим тканям, а
также камбию, и основную меристему,
образующую основные паренхимные тка-
ни (рис. 23).
Клетки протодермы в процессе даль-
нейшей дифференциации .становятся все
более плоскими, растут в поверхность и
превращаются в наружный слой специа-
лизированной ткани, одевающей моло-
дой побег или корень.
Клетки прокамбия прозенхимные,
длина их в несколько раз превышает ши-
рину. Тяжи из таких клеток закладываются в глубину формирую-
щихся органов в окружении -основной ткани, клетки которой де-
лятся в разных направлениях.
В процессе развития органов клетки прокамбия или нацело
превращаются в элементы проводящей й механической ткани, или
часть прокамбиальных клеток не дифференцируется; они .сохраня-
ют способность деления и становятся таким образом клетками
камбия — длительно функционирующей образовательной ткани.
Клетки камбия сходны с клетками прокамбия, на поперечном-
разрезе они имеют форму четырехгранников, реже многогранни-
ков. Отличается камбий от прокамбия более правильным разме-
щением клеток, имеющих на поперечном разрезе форму как бы
сложенных в стопки кирпичиков. Однако камбий может состоять
и из одного слоя клеток.
Камбий закладывается параллельно боковой поверхности орга-
на, в котором он находится. Поэтому он относится не к апикаль-
ной, а боковой или латеральной меристеме; к латеральной же
меристеме относится и пробковый камбий, или феллоген, который
образуется из уже дифференцированных тканей основной парен-
химы или эпидермиса. Камбий также может возникать из-слабо-
дифференцированной основной ткани. Меристемы, возникающие
из дифференцированных тканей, принято называть вторичными,
поэтому и камбий, образованный клетками основной паренхимы,
мы называем вторичным камбием; феллоген, следовательно, тоже
68
относится к вторичным меристемам. Структуры, возникающие из
камбия и феллогена,- называются вторичными.
гУчастки меристемы могут также образовать в растении особые
зоны роста, например, в нижней части междоузлия злаков, в ба»
зальной (нижней) части, молодого листа. Такая меристема
называется вставочной или интеркалярной. Соответственно и рост
органов за счет этих зон роста называется вставочным, или интер-
калярным. Цитоплазма меристемных клеток слабо вакуолизиро-
вана. В них отсутствуют эргастические вещества и менее развита
эндоплазматическая сеть; митохондрии й пластиды только форми-
руются ядро занимает относительно больший объем по сравнению
с клетками дифференцированных тканей. Оболочки клеток сильно
гидратизированы и состоят из пектиновых веществ, гемицеллюло-
зы с небольшой примесью целлюлозы. 'Процессу дифференциации
меристематических клеток предшествует увеличение объема кле-
ток, связанное с их вакуолизацией, поэтому клетки дифференци-
рованных тканей отличаются от клеток меристемы более крупными
размерами и относительно меньшим ядром.
ПОКРОВНЫЕ ТКАНИ
Поверхность тела высших растений защищена от неблагопри-
ятных воздействий внешней среды покровной тканью. Такая защи-
та необходима прежде всего для предохранения органов растения
от высыхания. Не меньшее значение имеют покровные ткани в пре-
дохранений более нежных внутрилежащих тканей от различного
рода механических повреждений, а также и от повреждений на-
секомыми и другими вредителями растений.
На листьях и однолетних зеленых, побегах покровная ткань
имеет вид тонкой 'прозрачной пленки и называется кожицей или
эпидермисом. Обычно эпидермис однослойный, но бывает и много-
слойный. Многолетние побеги приобретают темную, буроватых
оттенков поверхность. Это объясняется тем, что под эпидермисом
закладывается многослойная ткань — пробка, или феллема. В свя-
зи с происхождением эпидермис называют первичной покровной
тканью, пробку — вторичной покровной тканью.
На многолетних деревянистых побегах и стволах деревьев об-
разуется мощный защитный слой — корка. Она возникает-вслед-
ствие отмирания и высыхания поверхностных слоев различных
тканей: покровных, основных, механических и проводящих; отми-
рание происходит в результате образования внутренних слоев
пробки. Ввиду того, что этот ссохшийся поверхностный слой вы-
полняет ту же функцию, что и остальные покровные ткани; его
также относят к категории тканей; учитывая своеобразие проис-
хождения корки, ее называют третичной покровной тканью.
Первичная покров.ная ткань, или эпидермис. Как уже было
сказано, в процессе отногенеза эпидермис образуется из апикаль-
ной меристемной ткани — протодермы. Клетки эпидермиса обычно
плоские, боковые стенки их обычно извилистые, выступы стенки
69
одной клетки заходят в вогнутости соседней клетки. Благодаря
такому скреплению клеток обеспечивается прочность и эластич-
ность эпидермиса. В некоторых случаях форма клеток эпидермиса
отличается от обычной, например, в кожуре многих семян клетки
вытянуты вертикально, высота их больше ширины. Толщина кле-
точных стенок неодинакова, наиболее толсты наружные стенки,
Рис. 24. Эпидермис листа ириса (/) и кукурузы (//). А — общий
вид с поверхности, Б — одно из устьиц, В — устьице на поперечном
разрезе:
1 — замыкающие клетки; 2 — обычные клетки эпидермиса; 3 — клетки мезо-
филла с хлоропластами; 4— воздухоносная полость; 5 —побочная клетка
мецее толсты боковые, перпендикулярные к поверхности органа,
и наиболее тонки внутренние стенки клеток, которыми эпидермис
граничит с нижележащими тканями.
Большую часть объема клетки эпидермиса занимает крупная
вакуоль, заполненная клеточным соком, иногда окрашенным. Ци«
топлазма тонким слоем прилегает к внутренней стенке клетки,
в ней можно видеть ядро и лейкопласты:, хлоропласты бывают не
во всех клетках эпидермиса, а обычно лишь в особых клетках, за-
мыкающих устьица,— щелевидные отверстия 'в эпидермисе, через
которые происходит газообмен между внешней средой и тканями
растения. Под устьицами находятся подустьичные полости, соеди-
няющиеся с межклетниками губчатого мезофилла,— ткани из рых-
ло расположенных клеток.
Форма клеток эпидермиса, если смотреть на него сверху, силь-
но варьирует и в большой степени зависит от формы пластинки
листа (рис. 24). У листа с широкой пластинкой эпидермальные
70
Рис. 25. Схема, объясняющая
механизм работы устьиц
клетки также широкие, с крупноизвилистыми краями; наоборот,
на листьях вытянутых, линейных, например, у злаков, или на стеб-
лях клетки эпидермиса вытянуты, извилистость их краев обычно
более мелкая.
Особую форму имеют клетки, замыкающие устьица. У боль-
шинства растений таких клеток одна пара и, когда устьице от-
крыто, они имеют полулунную форму. У злаков каждое устьице
окружено двумя парами клеток. Само устьице имеет форму узкой
длинной щели. Щель замыкается
двумя удлиненными клетками своеоб-
разного строения. Устьица у злаков
расположены рядами, подустьичные
полости при этом соединяются в об-
щий канал. К замыкающим клеткам
примыкают так называемые побочные
клетки. Устьица служат для газообме-
на: через них удаляются наружу во-
дяные пары и воздух, состав которого
изменяется в процессе дыхания и фото-
синтеза растения. При наступлении сухой и жаркой погоды из-
быточное испарение с поверхности листьев растения может быть
вредно для них. Открытое состояние устьиц в таких условиях мог-
ло бы оказаться гибельным для растения.
В процессе эволюции у растения выработалось приспособление
в работе устьиц, разрешающее это противоречие: устьица облада-
ют свойством автоматически закрываться и открываться по мере
надобности растения. Механизм этого процесса можно объяснить
в связи со строением клеток, замыкающих устьица (рис. 24, 25)«
Оболочка замыкающих клеток устьиц утолщена неравномер-
но: стенки, параллельные поверхности листа, толстые; стенки, об-
ращённые к устьицам и противоположные им, тонкие. В состоянии
тургора между стенками, охватывающими устьичную щель, оста-
ется зазор, сами стенки туго натянуты. Как только вследствие
недостатка воды в растении тургор в клетках падает, клетка спа-
дается, и при этом, естественно, выпячиваются наиболее тонкие,
обращенные к отверстию стенки; встречные выпячивания клеточ-
ных стенок сближаются, и устьице замыкается. При заполнении
замыкающих клеток водой тургор восстанавливается, и происхо-
дит обратный процесс — устьице открывается.
Работа устьиц связана также и с чередованием дня и ночи.
Днем, благодаря деятельности пластид, всегда имеющихся в за-
мыкающих клетках, в клеточном соке этих клеток накапливается
сахар; в силу разности концентраций вода устремляется в замы-
кающие клетки и тургор в них возрастает; устьица открываются.
Ночью количество сахара уменьшается, тургор падает и устьица
закрываются.
От описанного выше устьичного аппарата бывают, конечно,
и отклонения, связанные с разнообразным строением устьиц у раз-
личных растений, но принцип их работы такой же. Например,
71
удлиненные замыкающие клетки у злаков имеют на концах рас-
ширения, вздутия, которые в состоянии тургора раздуваются,
в силу чего между параллельными сторонами замыкающих клеток
образуется щель. Тугор падает—и параллельные стенки смы-
каются.
У большинства растений, листья которых расположены более
или менее горизонтально, устьица размещены преимущественно
на нижней стороне листа; это можно рассматривать как одно из
приспособлений к более экономному испарению влаги. У листьев,
ориентированных более или менее вертикально, устьица располо-
жены почти одинаково густо на обеих сторонах листа. Так бывает,
например, у злаков. У листьев, плавающих на поверхности воды, на-
пример у белых кувшинок, устьица расположены на верхней сто-
роне листа, обращенной в атмосферу. Количество устьиц на еди-
ницу поверхности листа сильно варьирует у различных растений
одного и того же вида в зависимости от внешних условий.-В сред-
нем количество устьиц на 1 мм2 колеблется от нескольких десят-
ков до нескольких сотен. Устьица образуются из клеток протодер-
мы; в большинстве случаев две замыкающие клетки возникают
путем деления одной клетки протодермы; между двумя дочерними
клетками образуется щель — устьице. Реже замыкающие клетки
имеют независимое друг от друга происхождение.
Кроме обычных устьиц, служащих для газообмена, у многих
растений имеются приспособления, выделяющие капельно-жидкую
воду; такие образования расположены по краям листа, имеют до-
вольно разнообразное строение и называются гидатодами; процесс
выделения жидкой воды называется гуттацией. Явления гуттации
можно наблюдать у земляники, шиповника, гороха, злаков и мно-
гих других травянистых и древесных растений.
Выделение капельно-жидкой воды растением наблюдается
обычно, когда затруднено испарение; при тихой погоде и избытке
влаги в атмосфере, ночью или ранним утром.
На поверхности эпидермиса можно видеть различные образо-
вания, усиливающие защитную роль покровной ткани или выпол-
няющие иную функцию (рис. 26). Обычно это волоски, представ-
ляющие собой одноклеточные выросты эпидермиса. Образовав-
шись, волоски вскоре отмирают, утрачивая протопласт, и запол-
няются воздухом; вследствие этого они кажутся белыми, а когда
волосков много, то и вся поверхность, покрытая ими, кажется
седой или совсем белой. Волоски могут быть простыми или развет-
вленными, в последнем случае они имеют внутренние перегородки;
неразветвленные волоски также бывают с перегородками; волоски
могут состоять также из двух и нескольких рядов клеток. Оболочки
волосков иногда древеснеют. Густо расположенные волоски обра-
зуют на поверхности листа ветрозащитный слой, задерживающий
испарение; отражая солнечные лучи, волоски также уменьшают
нагревание листьев. Выросты эпидермиса иногда имеют форму
красивых звездочек или чешуек, например у лоха. Следует также
иметь в виду, что и простой волосок может быть прикреплен
72
к эпидермису не концом, а серединой, в виде двуплечего рычаж-
ка.'Реже встречаются не мертвые, а живые волоски, содержащие
в себе жидкость. Такие волоски называются железистыми и слу-
жат часто органами выделения: в них накапливаются эфирные
масла и другие выделения, обычно распространяющие запах при
испарении. Головка железистых волосков может быть многокле-
Рис. 26. Волоски и чешуйки на поверхности эпидермиса. Л,
Б, В — мертвые волоски, Г — чешуйки на листе'лоха; Д —
железистые волоски, Е — жалящие волоски крапивы
точная. Еще реже встречаются жгучие волоски, например у кра-
пивы. Волосок крапивы содержит муравьиную кислоту; стенки его
пропитаны углекислой известью, а конец — кремнеземом. Такой
волосок, если к нему прикоснуться, легко обламывается и цара-
пает поверхность кожи; в царапину попадает жгучая жидкость,
вызывающая у человека и животных покраснение кожи и зуд.
Следовательно, жгучие или жалящие волоски можно рассматри-
73
вать как приспособление к защите растения от повреждения
и поедания.
В клетках многослойного эпидермиса некоторых растений, на-
пример у фикуса, развиваются гроздевидные образования из угле-
кислой извести — цистолиты; при этом, в меру роста цистолита,
клетка, в которой он образовался, сильно увеличивается в раз-»
мере.
Выросты на эпидермисе могут формироваться и из основной
ткани. Такие многоклеточные волоски древеснеют и приобретают
прочность и значительную толщину, превращаясь в шипы, прицеп*
ки (на плодах) и другие выросты. Им дают общее название —
эмергенцы. Весьма разнообразны по величине, форме и окраске
бывают выросты, возникающие под воздействием различных пара-
зитов,— галлы. Многоклеточные выросты в форме чешуй, малень-
ких подушечек могут быть и органами выделения, подобно желе-
зистым волоскам. Эти выделительные приспособления называются
наружными выделительными органами.
Поверхностная стенка клеток эпидермиса бывает покрыта осо-
бым жироподобным веществом — кутином. Кутин — продукт жиз-
недеятельности протопласта, выделяется через оболочку наружу
и образует сплошной наружный слой — кутикулу, которая не про-
пускает воду и почти непроницаема для газов. Слой кутикулы
предохраняет растение от испарения воды с поверхности листьев
или других органов. У некоторых осок кутикула пропитывается
кремнеземом и становится очень твердой, поэтому листьями осоки
легко порезаться. Кутикула может быть пропитана воском, что
еще более усиливает защитную функцию эпидермиса. Воск может
выделяться и на поверхности эпидермиса, обусловливая несмачи-
ваемость его.
Вторичная покровная ткань, или пробка. Поверхность стеблей
большинства древесных растений и кустарников уже в первый год
их существования начинает приобретать буроватую окраску; такое
изменение свидетельствует о том, что на смену эпидермису обра-
зовалась новая покровная ткань — пробка, или феллема
(рис. 27).
Появление пробки — результат деятельности образовательной
ткани — пробкового камбия, или феллогена. Слой этой ткани ча-
ще образуется сразу же под кожицей в результате деления клеток
эпидермиса (например, у яблони, груши, ивы) или клеток основ-
ной паренхимы, прилегающей к эпидермису (у березы, дуба, клена
и др.). У некоторых растений первый слой пробки образуется на
глубине нескольких слоев субэпидермальной паренхимы (напри-
мер, у так называемой белой акации, у гледичии) или на большой
глубине, вблизи проводящих тканей (у барбариса, винограда
и др*.); последующие прослойки закладываются под первой на еще
большей глубине. Клетки феллогена делятся в тангентальном на-
правлении, откладывая кнаружи правильные радиальные слои
клеток, постепенно превращающиеся в слои пробки. Стенки клеток
адкрустируются суберином и воском, который образует ультратон-
74
кие прослойки, становятся непроницаемыми для воды, что и вызы-
вает гибель клеток.
В течение года может отложиться 24-20 слоев пробки. При
этом, в связи с утолщением побега, наряду с периклинальным де-
лением (в тангентальном направлении) возникают п антиклиналь-
ные (радиально направленные) деления.
Внутрь от себя феллоген отделяет клетки, богатые хлорофил-
ловыми зернами. Располагаются клетки рыхло, слой этих клеток
А
Б
Рис. 27. Вторичная покровная ткань ветки бузины. А — чечевичка, Б —
строение перидермы:
/ — пробка (феллема); 2 — феллоген; 3 — феллодерма; 4 — эпидермис
называется феллодермой, а совокупность пробки, феллогена и фел-
лодермы — перидермой.
Выполняя защитную роль, пробка образуется и на подземных
частях растения: корнях, корневищах и клубнях. Защитная роль
пробки хорошо заметна* при ранении растений; раны довольно
быстро затягиваются слоем пробки, например, царапины на по-
верхности плодов арбуза, яблок, на стеблях кактуса, на стволах
деревьев. То же происходит при опадании листьев и побегов: ос-
тающийся рубец быстро закрывается защитным слоем пробки.
Покровные ткани, защищая растение от неблагоприятных воз-
действий внешней среды, вместе с тем изолируют его от этой сре-
ды. Это противоречие разрешается образованием в эпидермисе
устьиц, через которые и совершается газообмен между растения-
ми и внешней средой. Такую же роль играют так называемые че-
75
чевички, образующиеся в перидерме вначале на месте некоторой
части устьиц, а потом вне связи с расположением устьиц (рис, 27)
Чечевички — это разрывы в покровной ткани, заполненные слоем
рыхлых клеток, не препятствующих газообмену. В месте разрыва
образуется бугорок, по форме напоминающий семя чечевицы, от-
сюда и название. Бугорок из клеток образуется в результате дея-
тельности феллогена. Эти клетки называются выполняющими.
Оболочки их могут опробковевать и тогда образуется рыхлый слой
клеток, сквозь который свободно проходит воздух. Если же обо-
лочки выполняющих клеток не пробковеют, на зиму образуется
сплошной слой из опробковевших клеток, закрывающих чечевич-
ку. Весной в результате деятельности феллогена этот слой разры-
вается.
ОСНОВНЫЕ, ИЛИ ПИТАЮЩИЕ, ТКАНИ
Рассмотрев под микроскопом срез листа, молодого стебля или
корня, мы можем легко установить, что любой из этих органов
состоит в основном из паренхимных, обычно тонкостенных живых
клеток. Поскольку эта ткань составляет как бы основу органа,
а в ней размещены другие ткани, ее называют основной тканью.
Главная функция этой ткани — функция питания, поэтому основ-
ные ткани называют также питающими тканями. Клетки парен-
химной ткани слабо дифференцированы, способны к возобновлению
меристематической деятельности. Паренхимная ткань пер-
вичной коры, сердцевины и мезофилла дифференцируется иа ос-
новной меристемы, паренхима же, связанная с проводящими тка-
нями,— из прокамбия и камбия.
В зависимости от того, в какой части растения находится -ос-
новная ткань и какую функцию выполняет, различают три вида
ее: 1) поглощающая, или всасывающая, паренхима; 2) ассимиля-
ционная паренхима, или хлоренхима, и 3) запасающая паренхима.
Поглощающая, или всасывающая, паренхима находится в кон-
чикйх корней (рис. 28). Корневые волоски и поверхностные клетки
всасывающей части корня, непосредственно контактирующие
с почвой, получают из нее элементы почвенного питания и пере-
дают их в расположенные- ниже слои паренхимы клеток. Отсюда
вода и растворенные в ней вещества проникают вглубь и затем
поступают в проводящие элементы корня. Клетки поглощающей
ткани сильно вакуолизированы, стенки их очень тонки и нежны.
Ассимиляционная ткань, или хлоренхима, составляет основу
листа, а также находится’в поверхностных слоях молодых стеблей
и зеленых плодов (рис. 29). Клетки ассимиляционной ткани обыч-
но сильно вакуолизированы, одеты тонкой первичной оболочкой.
По положению в листе между двумя слоями эпидермиса хлорен-
химу называют мезофиллом.
Основная функция ассимиляционной ткани, как указывает уже
само название,— фотосинтез, В связи с этим клетки ассимил'яци-
76
онной ткани богаты хлорофилловыми зернами — хлоропластами,
почему эту ткань и называют хлоренхимой.
Вторая функция мезофилла — транспирация, т. е. испарение
воды. Обе эти функции часто выполняются без заметной диффе*
Рис. 28. Всасывающая ткань кончика корня. А — распространение
корневых волосков в почве, Б — корневой ' волосок и прилегаю-
щие к нему клетки паренхимй, В — продвижение воды из почвы
в корень:
1—6 направление тока воды
ренциации ассимиляционной ткани, например в листьях, ориенти*
рованных более или менее вертикально (листья -злаков, ириса
и др.). В тех случаях, когда пластинка листа занимает горизон*
тальное положение (у большинства растений), и у листа можно
различать верхнюю и нижнюю стороны, мезофилл бывает диффе-
77
ренцирован; к верхней стороне примыкает один или несколько
слоев вытянутых и расположенных перпендикулярно к поверхно-
сти листа клеток (столбчатая паренхима), к нижней — примыкают
рыхло расположенные клетки, образующие губчатую ткань.
Рнс. 29. Ассимиляционная ткань в листе:
1 — палисадная ткань; 2 — губчатая ткань; 3 — проводящий пучок
Таким образом, дифференциация мезофилла на столбчатую
и губчатую ткань связана с лучшим приспособлением к выполне-
нию ими двух самостоятельных функций: фотосинтеза и транспи-
рации.
Запасающая ткань широко распространена в органах расте-
ния. Ее можно встретить в корнйх, стеблях и особенно в мякоти
плодов и семян (рис. 30). Клубни* картофеля и других растений
состоят в основном из запасающей ткани. Обычно это крупные
паренхимные клетки округлой или слегка вытянутой формы, за-
полненные зернами крахмала, белка, капельками жира или раст-
вором сахара. В клеточном соке некоторых растений могут быть
также амиды, коллоидный раствор белка, таннины. Стенки клеток
запасающей паренхимы обычно тонкие, но бывают и утолщенные
(в древесине, семенах, например, финиковой пальмы, кофе, где
стенки клеток эндосперма состоят из гемицеллюлозы, растворяю-
78
щейся при прорастании семян). Цитоплазма оттеснена к стенке
клетки, ядро часто деформировано или вовсе отсутствует. В цито-
плазме клеток запасающих тканей имеются ферменты, гидроли-
Б
Рис. 30. Запасающая ткань и аэренхима. А — аэренхима,
Б — запасающая ткань в клубне картофеля
зующие запасные пластические вещества и переводящие их в бо-*
лее простые соединения, которые и перемещаются к месту непосред*
ственного использования. После израсходования находящихся в них
питательных веществ клетки запасающей ткани отмирают. У сук-
79
кулентных растений в клетках паренхимной ткани накапливается
вода (например, у кактусов, алоэ, агав). Это водоносная парен-
хима.
Клетки паренхимной ткани в органах некоторых,растений об-
разуют широкие воздухоносные ходы, посредством которых воз-
дух доставляется к тем частям растения, связь которых с атмос-
ферой затруднена. Такая воздухоносная паренхима называется
аэренхимой (рис. 30). Аэренхимой богаты стебли и корни водных
растений. Бывает она также и в корнях растений, произрастающих
в сильно уплотненной почве целинных степей.
ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
В толще паренхимной ткани образуются также вместилища
выделений. Они могут образоваться двумя путями: в результате
разъединения клеток и скопления секрета, в межклеточном прост-
ранстве или в результате растворения оболочек нескольких смеж-
ных клеток. В первом случае вместилища называются схизоген-
ные, во втором — лизигенные. В паренхиме встречаются одиночные,
клетки, заполненные различными секретами, иногда кристаллами;
они могут иметь различную форму и называются секреторными
идиобластами. В результате слияния ряда клеток могут образо-
ваться длинные ходы; таковы, например, смоляные ходы у хвой-
ных и некоторых растений семейства зонтичных (Ferula и др.).
Скопления клеток, в которых находятся продукты выделения,
включая сюда железки и железистые волоски на поверхности
эпидермиса, ввиду особой их функцйи классифицируют как осо-
бую ткань — выделительную. Клетки выделительной'ткани недол-
говечны, после заполнения их секретом протопласт обычно от-
мирает.
К выделительным ткайям следует отнести различного устрой-
ства нектарники, которые, выделяя жидкости, богатые сахаром,
играют важную биологическую роль в процессе перекрестного
опыления при помощи насекомых. Однако, роль нектарников не
ограничивается только привлечением насекомых —опылителей.
Нектар обладает антибиотическими свойствами, он может обрат-
но всасываться растениями и содержащиеся в нем питательные
вещества использоваться в процессе обмена веществ растения,
особенно в метаболизме самого цветка.
МЕХАНИЧЕСКИЕ ТКАНИ
Органы растений обладают известной прочностью, иногда очень
большой. Прочность растения определяется всей совокупностью
тканей, их взаимным расположением, однако основную роль здесь
играют специальные механические ткани. Общее свойство всех
механических тканей — утолщенность клеточных оболочек. Утол-
щения могут быть равномерными и неравномерными.
83
При всем значительном разнообразии, которое наблюдается
в строении механических тканей, их можно разделить* на три типа:
колленхиму, склеренхиму и каменистые клетки, или склереиды.
Каменистые клетки и клетки склеренхимы при окончательном
оформлении ткани становятся мертвыми; колленхима — живая
ткань.
Колленхиму можно обнаружить в периферической части расту-
щих молодых стеблей, черешков, плодоножек, листовых жилок
и др. Клетки колленхимы вакуолизированы, живой протопласт их
содержит ядро и хлорофилловые зерна. Оболочки колленхимы не
древеснеют и на анатомическом срезе выделяются красивым сере-
бряным блеском; они состоят из целлюлозы и пектина с большим
содержанием воды, что дает им возможность свободно растяги-
ваться в связи с ростом клетки. Эти свойства дают основание
Рис. 31. Уголковая колленхима в черешке листа свеклы
считать оболочки клеток колленхимы первичными, пластичность
колленхимы — наиболее характерная ее черта. Клетки колленхи-
мы не утратили способность к делению и даже меристематической
активности? и в этом отношении, а также по форме колленхима
весьма близка паренхиме; клетки колленхимы меняют свою форму
от явно прозепхимной до паренхимной. Можно наблюдать и посте-
пенный переход от паренхимы к колленхиме.
Стенки клеток утолщаются по-разному. В одних случаях утол-
щены тангентальные стенки. На поперечном разрезе они предста-
вляются в виде выемчатых пластинок, благодаря чему этот вид
колленхимы получил название пластинчатой. В других случаях
четырехугольные или многоугольные на поперечном разрезе клет-
ки имеют утолщения в уголках; такая колленхима называется
уголковой (рис. 31). Но часто можно встретить почти равномер-
ные утолщения. Выделяют еще рыхлую колленхиму с развитыми
межклеточниками, с которыми граничат наиболее утолщенные
стенки.
Возникает колленхима из апикальной меристемы в начале ее
дифференциации на прокамбий и основную паренхиму. Колленхи-
81
ма, которая по происхождению ближе к прокамбию, и заклады-
вается раньше, более специализирована; возникающая позже из
элементов, близких к основной меристеме, приближается к парен-
химе. Колленхима залегает в виде сплошного цилиндра пли от-
дельными изолированными тяжами (при ребристой поверхности
стебля или черешка), непосредственно под эпидермисом или отде-
лена от него несколькими слоями паренхимных клеток.
Склеренхиму подразделяют на ксилемные волокна, входящие
в состав ксилемы, и внексилемные, или экстраксилярные, волокна,
находящиеся за пределами ксилемы. Наиболее специализирован-
ные ксилемные или древесинные волокна называются либрифор-
мом. Экстраксилярные волокна называют также лубяными, или
флоэмными, волокнами, хотя они могут находиться и вне флоэмы,
например гиподермальная склеренхима стебля злаков, которая
имеет паренхимное происхождение. Клетки либриформа (древесин-
ные волокна) относительно коротки (до 2 мм), имеют толстые
одревесневшие стенки, особенно в стебле древесных растений.
Можно наблюдать переходы от древесинных волокон к трахеидам
Рис. 32. Склеренхима в стебле герани
(например, у дуба). Как правило, клетки либриформа лишены
протопласта, это мертвые клетки.
Лубяные волокна — наиболее прозенхимные элементы в расте-
нии. При толщине всего в несколько сотых миллиметра длина лу-
бяных волокон достигает нескольких десятков и даже сотен мил-
лиметров. Так, например, лубяные волокна у льна достигают
длины 60 мм, у крапивы — 80 мм, у рами — 350 мм. Молодые
клетки лубяных волокон еще живые, имеют протопласт с много-
численными ядрами. Но к моменту полного сформирования стен-
82
ки клеток сильно утолщаются, и протопласт их обычно отмирает
(рис. 32). Оболочки лубяных волокон обычно в большей или мень-
шей степени древеснеют, но у некоторых растений, например,
у льна, конопли и рами, они остаются чисто целлюлозными, что
определяет их высокие текстильные качества. Лубяные волокна
объединяются в пучки, прочность которых зависит также и от ха-
рактера соединения волокон; подобно трахеидам концы одних во-
локон вклиниваются между концами смежных. Прочность самого
лубяного волокна определяется также тончайшим строением его
фибрилл, из которых построено волокно.
Волокна ксилемы (либриформ) и волокна флоэмы образуют-
ся из прокамбия и камбия. Экстраксилярные волокна, располо-
женные за пределами флоэмы, образуются из основной меристемы,
а у некоторых осоковых даже из протодермы.о
В связи с ростом органов первичного происхождения волокна,
входящие в состав 'их, бывают длиннее, чем волокна вторичного
происхождения, возникающие из камбия.
Волокна ксилемы и флоэмы могут септироваться, т. е. в них
могут возникать поперечные перегородки; такие волокна называ*
ются септированными, или перегородчатыми волокнами.
Склереиды (рис. 33) встречаются в различных органах: в стеб-
лях, листьях, корнях, плодах. Форма склереид более или менее
изодиаметрическая, но имеют-
ся склереиды ветвистые и вы-
тянутые. Стенки клеток сильно
утолщены и обильно пропита-
ны лигнином, благодаря чему
они обладают большой проч-
ностью, твердостью. Отсюда
другое название — каменистые
клетки. Твердость незрелых
плодов груши и айвы зависит
от каменистых клеток; из по-
добных же клеток состоит ко-
сточка сливы, абрикоса, перси-
ка и др. Каменистые клетки,
разбросанные в одиночку сре-
ди мякоти листа, стебля или
другого органа, придают им
повышенную твердость.
Склереиды имеют различ-
ное происхождение. Чаще все-
го это склерофицированные
разросшиеся паренхимные
клетки, но склереиды мезо-
филла листьев возникают из
Рис. 33. Склереиды, или каменистые
клетки, в мякоти плода груши. А —
при слабом увеличении (к группе ка-
менистых клеток прилегают клетки
паренхимы), Б — при сильном увели-
чении
клеток губчатой или палисадной хлоренхимы; склереиды, находя-
щиеся вблизи проводящих элементов, образуются из прокамбия
и камбия, а склереиды семенной кожуры — из клеток протодермы.
83
В начале формирования каменистые клетки живые. Об этом
можно судить по наличию пор в стенках каменистых клеток, но
вполне сформировавшаяся каменистая клетка мертва и выпол-
няет лишь механические функции.
ПРОВОДЯЩИЕ ТКАНИ
Вода и питательные вещества, поступающие через корни, а так-
же синтезируемые в них, должны быть переданы во все части
растения, ибо в них нуждается каждая живая клетка; с другой
стороны, вещества синтезируемые в листьях, также предназначены
для всех частей растений. Таким образом, в теле растения созда-
ются два противоположно на-
правленных потока: восходя*
щий — от корней к листьям, и
нисходящий — от листьев к
корням. Эти два потока пита-
тельных веществ обслужива-
ются проводящими тканями:
восходящий, как правило, об-
служивается сосудами, или
трахеями, и трахеидами (см.
рис. 18), нисходящий — сито-
видными трубками и сопро-
вождающими их клетками, или
клетками-спутницами (рис.
34). Таково функциональное
различие трахей и трахеид, с
одной стороны, и ситовидных
трубок с сопровождающими
клетками — с другой. Однако
из этого правила имеются мно-
гочисленные исключения. Ис-
следованиями А. Л. Курсанова
и др. доказано, что такой стро-
гой специфичности в проводя-
щих элементах нет, и подраз-
деление их на ткани, обслужи-
вающие восходящий и нисхо-
дящий ток, в известной мере
условно. Комплекс тканей,
главной составной частью ко-’
торых являются сосуды или
совокупность тканей, в которой
Рис. 34. Ситовидные трубки. А — про-
дольный разрез ситовидной трубки и со-
провождающих клеток, Б — поперечный
разрез через ситовидную перегородку:
1 — сопровождающие клетки; 2 — поперечная
перегородка ситовидной трубки; 3 — перфора-
ции
трахеиды, называется ксилемой, а ______________ _______,
основную часть образуют ситовидные трубки и клетки-спутницы —
флоэмой, или лубом. В осевых органах (стеблях, корнях) ксилема,
как правило, размещается ближе к центру поперечного сечения
органа, флоэма — к периферии от ксилемы. В состав ксилемы
и флоэмы входят, следовательно; и другие ткани, механические
84
и основные, но наиболее характерными анатомическими элёмен»
там,и ксилемы и флоэмы являются проводящие ткани.
Вполне сформировавшиеся сосуды представляют собой различи
ной толщины трубки, состоящие из члеников; каждый членик
сформировался из одной вытянутой клетки. Поперечные стенки
у толстых сосудов ’бывают более или менее горизонтальными,
в узких сосудах поперечные перегородки бывают скошенными,
часто под острым углом. Сосуды очень редко располагаются
в одиночку, обычно рядом располагается целая пачка сосудов,
примыкающих друг к другу. Диаметр сосуда —0,1—0,2 мм и лишь
в очень редких случаях (у лиан) достигает 0,3—0,7 мм. Длина
сосудов достигает нескольких метров, а в редких случаях (у лиан
и некоторых деревьев) —нескольких десятков метров.
Одновременно с формированием сосудов на внутренней поверх*
ности их образуются целлюлозные утолщения. Без таких утолще*
ний сосуды не могли бы выполнять свою роль, так как разраста*
ющиеся смежные ткани легко бы смяли их тонкую стенку.
Прежде всего образуются кольчатые и спиральные утолщения;
они филогенетически более древние, следовательно, это наиболее
примитивные утолщения. Формирование кольчатых и спиральных
утолщений в молодых сосудах не препятствует их удлинению.
Позже возникают более широкие сосуды с лестничными и сетча*
тыми утолщениями, занимающими большую поверхность стенок.
Наконец, наибольшая площадь утолщения у точечных или пори*
стых сосудов, у которых не утолщены лишь поры.
Сосуды свойственны наиболее высокоразвитым покрытосемен*
ным растениям, хотя в • единичных случаях встречались уже
у высших споровых и голосеменных; это высшая ступень в эволю-
ции проводящих элементов ксилемы. До возникновения сосудов
их функцию выполняли трахеиды, свойственные голосеменным
и высшим споровым растениям, но сохранившиеся и у больщин*
ства покрытосеменных растений.
Трахеиды — это отдельные прозенхимные клетки со скошенны-
ми концами, которыми они и причленяются друг к другу, образуя
водопроводящую сеть. Между соседними трахеидами нет сплош-
ных отверстий, какие мы видели у сосудов (трахей); связь между
соседними трахеидами осуществляется только посредством круг*
лых окаймленных пор. Трахеидам свойственны такие же утолще-
ния, как и сосудам. В трахеидах, которые уже вполне сфор*
мировались, протопласт отмирает; следовательно, и эти про-
водящие элементы, подобно сосудам, относятся к мертвым
тканям.
Только в фазе формирования стенки трахеид и сосудов состоят
из чистой целлюлозы, вскоре же начинается процесс одревеснения,
вначале пропитываются лигнином утолщения, а затем и вся обо-
лочка клеток. Это в большей степени увеличивает прочность
и твердость ксилемы. Достаточно сказать, что древесина хвойных
растений (а у некоторых, например у тисса, она очень твердая)
состоит в основном из массы одревесневших трахеид.
S5
Проводящие элементы ксилемы лишь для удобства изучения
рассматриваются нами изолировано от других тканей. На самом
деле этого не бывает. Сосуды и трахеиды всегда контактируют
с прослойками паренхимной ткани; это живая ткань, клетки ее
обычно содержат запасные питательные вещества, которые, раст-
воряясь при помощи энзим, проникают в сосуды или трахеиды
!И транспортируются ими по телу растения.
Клетки паренхимы, примыкающие к сосудам, могут врастать
'через поры внутрь сосуда и даже размножаться там, образуя
тиллы. С возрастом у древесных растений все большая масса со-
судов заполняется тпллами, при этом сосуды утрачивают проводят
щие функции и превращаются по существу в механическую ткань.
Оболочки пузыреобразных выростов в сосудах — тиллов посте-
пенно утолщаются и сильно древеснеют, в результате чего ксиле-
ма становится очень плотной, твердой. Прочность ксилемы увели-
чивается еще больше в результате заполнения тиллов дубильными
и смолистыми веществами, имеющими темную окраску. Благодаря
этому ксилема темнеет, а иногда принимает красивую окраску
красноватых оттенков.
В отличие от проводящих элементов ксилемы, ситовидные труб-
ки флоэмы состоят из живых клеток. Свое название ситовидные
трубки получили за строение поперечных перегородок, разделяю-
щих смежные клетки, из которых построена трубка. Эти. перего-
родки пронизаны большим количеством сквозных отверстий, обра-
зующих как бы решето или сито, сквозь которые проходят плаз-
модесмы. соединяющие протопласты соседних клеток. Поверхность
ситовидных пластинок и канальцев в них покрыта слоем углево-
да — каллозы.
Стенки ситовидных трубок и клеток-спутниц состоят из чистой
щеллюлозы и лишь к концу вегетации иногда у некоторых трубок
•слегка древеснеют.
Ядро и тонопласт в ситовидных трубках разрушаются, распа-
даются мембраны канальцев эндоплазматической сети и диктио-
сом; исчезают внутренние мембраны в митохондриях; клеточный
сок перемешивается с миксоплазмой.
Размещаются ситовидные трубки в одиночку, в окружении па-
5ренхимных и механических тканей или пачками; в последнем Слу-
чае трубки могут тесно соприкасаться, и тогда в местах соприкос-
новения образуются такие же ситовидные поверхности, как и на
поперечных перегородках. Ситовидными порами трубки сообща-
ются также и с клетками-спутницами; с прилегающими паренхим-
ными клетками ситовидные трубки сообщаются простыми порами.
Клетки-спутницы заметно отличаются от клеток, из которых
слагаются ситовидные трубки; они тоньше и короче, менее вакуо-
лизированы, сохранили ядро, митохондрии, дпктиосомы и эндо-
плазматическую сеть.
Клетки-спутницы имеются не у всех высших растений, их нет
у хвойных и некоторых покрытосеменных (например, у картофе-
ля). Длина клеток у ситовидных трубок небольшая, от десятых
86
долей миллиметра до 2 мм, толщина — несколько десятков мик*
рометров.
Близки по строению к проводящим элементам млечные сосуды».
или млечники, свойственные многим представителям покрытосе-
менных растений (рис. 35). Свое название они получили за нали-
чие в них млечного сока, или ла-
текса.
Млечники могут иметь раз-
личное строение. У одних расте-
ний (молочая, клещевины) это
одна сильно разветвленная клет-
ка. Такне млечники называются
нечленистыми, или простыми. У
других растений (сложноцвет-
ных, маковых, вьюнковых и др.)
млечники состоят из ряда вытя-
нутых клеток, поперечные пере-
городки которых иногда раство-
ряются, подобно тому как мы это
видели у сосудов. Такие млечни-
ки называются членистыми, или
сложными, млечниками. Идущие
продольно, вдоль органа, члени-
стые млечники соединяются меж-
ду собой анастомозами, образуя
общую сеть.
У каучуконосного растения —
бразильской хевеи (Hevea), от-
носящейся к семейству молочае-
вых, млечники членистые. У гва-
юлы млечный сок накапливается
в обычных паренхимных клетках.
Интересно отметить, что нечлени-
стые млечники развиваются из
одной или нескольких многоядер-
ных клеток, заложенных еще в
Рис. 35. Млечники среди основной
ткани в корне одуванчика
зародыше.
Клетки млечников живые; в них сохраняется пристенный слой
цитоплазмы, лейкопласты и ядро, которое в нечленистых млечни-
ках может делиться; в членистых млечниках многоядерные клетки
возникают благодаря растворению поперечных стенок двух сосед-
них клеток и слиянию их протопластов. Большую часть объема
млечников составляет вакуоль, заполненная латексом. Оболочки
млечников тонкие, неодревесневшие. Расположены млечники
обычно во флоэмной паренхиме, редко в ксилеме (сем. Caricaceae).
Латекс представляет суспензию или эмульсию самых разнообраз-
ных органических веществ. Дисперсионная среда — клеточный сок
млечника, дисперсная фаза — углеводы, углеводороды, стерины,
жиры, смолы, каучук, белки, сахара, таннины и многие другие.
87
В основном это секрет, однако питательные вещества, находящие-
ся в латексе, по-видимому, могут быть использованы вторично,
в процессе метаболизма.
СОСУДИСТО-ВОЛОКНИСТЫЕ ПРОВОДЯЩИЕ
ПУЧКИ
Элементы ксилемы и флоэмы в органах растения, как правило,
закономерно сочетаются в пучки. К проводящим элементам кси-
лемы и флоэмы обычно присоединяются механические элементы —
склеренхима, поэтому пучки и называются сосудисто-волокнисты^
ми. Тонкие разветвления пупков, например, в листьях постепенно
утрачивают механические элементы и выполняют только проводя-
щую функцию; такие пучки называются просто проводящими.
В начале формирования структуры органов, близ апекса, сосу-
дисто-волокнистые пучки образуются из прокамбия. У однодоль-
ных растений, по израсходовании этой первичной образовательной
ткани, дальнейшее разрастание пучка прекращается; такие пучки,
не содержащие уже образовательной ткани, называются закры*
тыми.
В стеблях двудольных и голосеменных растений весь прокам-
бий не расходуется в процессе формирования пучка, часть его
в виде узенькой прослойки между ксилемой и флоэмой остается;
клетки этой прослойки прокамбия приобретают свойство непре-
рывно делиться в тангентальном и радиальном направлениях. За
счет образовательной ткани — камбия происходит разрастание
пучка в тангентальном и радиальном направлениях. Такие пучки
с действующей прослойкой камбия называются открытыми*
И прокамбий и камбий образуют проводящие и механические эле-
менты ксилемы и флоэмы.
Элементы ксилемы и флоэмы, сформированные деятельностью
прокамбия, называются первичными; камбий дает вторичные кси-
лему и флоэму. Первичная ксилема, формирующаяся в начале
дифференциации меристемы, называется протоксилемой. Прово-
дящие элементы протоксилемы представлены, как правило, только
кольчатыми и спиральными сосудами. К моменту завершения ро-
ста органа протоксилема утрачивает проводящую функцию и за-
меняется метаксилемой, у которой сосуды со ступенчатыми, сет-
чатыми и сплошными утолщениями. В процессе формирования
вторичной ксилемы сосуды метаксилемы обычно перестают функ-
ционировать. Соответственно ксилеме в первичной флоэме* также
различают протофлоэму и метафлоэму. Ситовидные трубки прото-
флоэмы, возникнув еще в апикальной меристеме, функционируют
короткое время и в процессе роста органа быстро разрушаются
и исчезают (облитерируются), заменяясь метафлоэмой. Ситовид-
ные трубки протофлоэмы уже, чем у метафлоэмы, и у однодоль-
ных не сопровождаются клетками-спутницами. При вторичном
утолщении проводящие элементы метафлоэмы обычно сжимаются
И полностью облитерируются.
88
Элементы ксилемы восп-
роизводятся камбием при-
мерно в четыре раза быстрее,
чем элементы флоэмы, по-
этому ксилемная часть пуч-
ка наращивается гораздо
быстрее, чем флоэмная. В
стеблях и корнях камбий,
возникнув, продолжает фун-
кционировать до конца жиз-
ни данного органа. В жил-
ках листа деятельность кам-
бия недолговечна, поэтому
сосудисто-волокнистые пуч-
ки листа^ожно считать ус-
ловно закрытыми, хотя по
типу строения они у дву-
дольных более соответству-
ют открытым'пучкам стебля.
Камбий, входящий в со-
став пучка, называется пуч-
ковым' камбием. В процессе
формирования стебля и кор-
ня из основной ткани может
образоваться вторичный
камбий. В стеблях он возни-
кает между пучками, поэто-
му называется еще межпуч-
ковым камбием. Так же, как
и первичный камбий, он спо-
собен откладывать ксилему
и флоэму, которые относятся
к вторичным элементам. Сле-
довательно, независимо от
происхождения камбия все
элементы анатомической
структуры органов, форми-
руемые им, являются вто-
ричными.
Взаимное расположение
ксилемы и флоэмы "в пучке
может быть различным. В
большинстве случаев ксиле-
Рис. 37. Поперечный разрез откры-
того . пучка:
7 —сосуды; 2 — камбий; 3 —флоэма;
4 — склеренхима; 5 — основная ткань,
окружающая пучок
Рис. 36. Поперечный разрез пучка в стебле
кукурузы:
1 — основная паренхима; - 2 — склеренхима; 3, 4 —
флоэма; 5 — древесинная паренхима; 5—пористые
сосуды; 7,8 — спирально-кольчатые сосуды; 9 —
воздушная полость
89
гм а прилегает к флоэме одной стороной. Пучки такого строения назы-
ваются коллатеральными. Коллатеральные пучки могут быть откры-
тыми и закрытыми. У открытых пучков между ксилемой и флоэмой
имеется слой действующего камбия, в закрытых пучках камбия
нет (рис. 36, 37). В органах радиального строения (стебле, корне)
>ксилема ориентирована к центру органа, флоэма — на периферию.
В редких случаях (у пасленовых, тыквенных) к внутренней сторо-
не ксилемы примыкает второй участок флоэмы. Такие пучки полу-
чили название биколлатеральных\ внутренняя флоэма имеет только
первичное происхождение.
В корнях злаков и других однодольных растений и в наиболее
.молодых частях корней двудольных -участки ксилемы располага-
ются по радиусу, а между этими радиусами, ближе к периферии,
Рис. 38. Концентрические пучки. А — амфивазальный из корневища касатика
(Iris), Б — амфикрибральный из корневища орляка (Pieridium aquilinum):
1 — ксилема; 2 — флоэма
располагаются участки флоэмы. Совокупность всех участков кси-
лемы и флоэмы в таком случае называют сложным пучком ра-
диального строения (см. рис. 65).
Относительно редко можно встретить пучки концентрического
строения. В одних случаях ксилема располагается в центре пучка
и ее окружает флоэма, в других, наоборот, в центре находится
флоэма, а ее окружает ксилема (рис. 38). В первом случае пучки
называются амфикрибральными, во втором — амфивазальными.
Сосудисто-волокнистые пучки густой сетью пронизывают все
тело растения. О том, насколько густа может быть такая сеть,
можно судить, рассматривая мочалку, приготовляемую из плодов
люффы (растение из семейства тыквенных). Это не что иное, как
сеть сосудисто-волокнистых пучков плода люффы, освобожденная
от мягких тканей.
Разрастаясь, открытые коллатеральные пучки могут плотно
сомкнуться друг с другом; тогда образуется сплошной массив кси-
90
лемы— древесина и сплошной слой флоэмы — луб. Так, например*,,
бывает в^стеблях древесных растений. Между разросшимися пуч-
ками сохраняются лишь тонкие прослойки паренхимы — радиаль-
ные лучи, а между древесиной и лубом — цилиндр из нескольких;
слоев клеток камбия. Однако в стеблях некоторых растений (на-
пример, у мыльнянки, подмаренника) прокамбий (а затем и кам-
бий) сразу закладывается сплошным кольцом, при этом форми-
руется сплошной цилиндр ксилемы и флоэмы, минуя пучковое
строение.
ВЕГЕТАТИВНЫЕ ОРГАНЫ ВЫСШИХ РАСТЕНИЙ,
ИХ ОНТОГЕНЕЗ И ФУНКЦИИ
К вегетативным органам высшего растения относят стебель,
корень и лист; это — органы, необходимые для существования дан-
ной особи. Органы, приспособленные специально для размноже-
ния, называют генеративными. Строение генеративных органов’
специфично для каждой группы высших растений, поэтому рас-
смотрение их удобней отложить до систематического изучения
этих растений. Здесь мы рассмотрим строение только вегетатив*
ных органов, сходных у всех высших растений.
У семенных растений зачатки вегетативных органов заклады-
ваются, как правило, в процессе формирования семени и составля-
ют наиболее существенную часть зародыша. Поэтому изучение
вегетативных органов мы начинаем с рассмотрения семени и его
прорастания.
РАЗВИТИЕ ВЕГЕТАТИВНЫХ ОРГАНОВ
ИЗ ЗАРОДЫША СЕМЕНИ
В крупном семени зародыш будущего растения можно рас-
смотреть невооруженным глазом пли при помощи лупы. Обычно
для первоначального знакомства принято рассматривать крупное
двудольное семя фасоли (рис. 39) и однодольное семя какого-либо
злака, например пшеницы (рис. 40). Мы также -рассмотрим се-
мена этих двух растений. Чтобы легче было препарировать и рас-
сматривать семя, его предварительно намачивают. С намоченного
и набухшего семени фасоли легко снимается кожура — плотная
оболочка, предохраняющая внутренние нежные части семени от
неблагоприятных воздействий внешней среды. Основная масса
вынутого из кожуры крупного зародыша семени состоит из двух
мясистых семядолей, заключающих в себе запас питательных ве-
ществ.
Семядоли прикреплены к укороченному зародышевому сте-
бельку, который кверху заканчивается почечкой. При рассмотре-
нии в лупу можно видеть, чтсг почечка состоит из нескольких за-
родышевых листочков, прикрывающих удлиненный бугорок, так
называемый конус нарастания, или апекс. Из почечки при прора-
91
станин семени развивается побег, т< е. стебель, с расположенными
на нем листьями.
Нижняя часть зачаточного стебелька переходит в зародышевый
корень, который становится главным корнем. При прорастании
семени зачаточный стебелек сильно вытягивается, и та часть его,
которая формируется ниже места
Рис, 39. Прорастание семени
фасоли:
1 — главны! корень; 2 —* гипокотиль!
3 — эпикотильз 4 — корневая шейка]
5 — семядоли] б -*• первые листья!
7 — верхушечная почка
прикрепления семядолей, назы-
вается подсемядольным коле-
ном, или гипокотилем, а место
перехода стебля в корень —
корневой шейкой. Отрезок
стебля между семядолями и
первыми настоящими листьями
называется надсемядолъным
коленом, или эпикотилем.
Сами семядоли представля-
Рис. 40. Прорастание зер-
новки пшеницы:
1 — колеоптиле; 2 — зародыше-
вые корешки; 3 — первый лист
ют собой сильно, видоизмененные листья, в них откладывается за<
пас питательных веществ, постепенно расходуемый в процессе про-*
растания^в начале развития нового растения. При прорастании
у одних растений (например, фасоли, тыквы) семядоли выносятся
наверх, у других (например, у гороха) остаются в почве. Эта осо-
бенность прорастания семян учитывается при посеве; семена, не
выносящие семядоли наверх, можно заделать глубже, чем семена
растений, у которых семядоли выносятся на поверхность,
92
Семена однодольных устроены иначе (рис. 41). У пшецицы, так
как и у фасоли, семя покрыто сверху плотной кожурой, но она име-
ет здесь несколько иное происхождение. В нее входят слои, относя-
щиеся не только к самому семени, но и плодовая оболочка, которая
прочно срастается с семенной. Это объясняется тем, что у пшени-
цы, как и других злаков, плод односемянный, и околоплодник при
созревании плода остается тонким и плотно облегает семя. Такие
плоды называются зерновками; почти весь объем зерновки состоит
из семени.
Кожура зерновки пшеницы не сдирается так легко, как кожу-
ра семени фасоли, поэтому для рассмотрения частей семени его
следует разрезать вдоль через хорошо заметную бороздку. Разре-
зав таким образом зерновку, мы увидим, что большая часть ее
объема занята запасающей тканью, которая называется эндоспер-
мом. К эндосперму у нижнего конца зерновки сбоку плотно при-
легает зародыш. В хорошую лупу или при слабом увеличении
микроскопа можно рассмотреть, что зародыш состоит из щитка,
плотно прилегающего непосредственно к эндосперму,— это единст-
венная семядоля семени пшеницы — и из зачаточного, сильно уко-
роченного стебелька, заканчивающегося с одного конца почечкой,
а с другого конца переходящего в зародышевый корешок. Эти
части зародыша состоят из образовательной ткани. Корешок окру-
жен особой сумкой — колеоризой. При прорастании семени зача-
точный корешок разрывает колеоризу и кожуру и выходит нару-
жу, образуя главный корень. Кончик корня прикрыт особым чех-
ликом, защищающим нежную точку роста корня от повреждений;
одновременно, ослизняясь, наружные клетки чехлика помогают
проникновению корня в глубь почвы. На некотором расстоянии
от кончика корня его поверхность покрыта тонкими корневыми
волосками, которые представляют собой выросты поверхностных
клеток корня.
Зачаточный стебелек при прорастании сильно разрастается
и выносит почечку поближе к поверхности почвы. В момент всхо-
дов почечка имеет вид толстой иголки, состоящей из нескольких
плотно свернутых в трубочку листбчков, окружающих конус на-
растания, из которого затем будут развиваться побеги растения.
Самый наружный листочек, имеющий форму сильно удлиненного
колпачка, называется колеоптиле. Часть стебелька прорастающего
растения между щитком и местом прикрепления колеоптиле назы-
вается мезокотилем. Длина мезокотиля в большей степени зависит
от глубины заделки семян. При слишком глубокой заделке семени
большая часть питательных веществ эндосперма расходуется на
формирование длинного мезокотиля, и тогда на развитие почечки
остается мало питательных веществ; всходы получаются сильно
ослабленными, и взошедшие растения при неблагоприятных усло-
виях легко погибают. Слишком мелкая заделка семян вредна тем,
что прорастающая почечка оказывается совсем близко к поверх-
ности почвы, и при неблагоприятных условиях растение также мо-
жет погибнуть.
Рис. 41. Зерновка пшеницы. А — продольно-поперечный разрез
(схематизированная модель), Б — зародыш на продольном раз-
резе:
1 — всасывающие волоски; 2 — околоплодник; 3 — кожура семени; 4, 5 —
эндосперм (4 — алейроновый слой, 5 — крахмалистая часть); 6 — щиток;
7 — стебелек; 8— почка; 9 — листочки; 10 — колеоптиль; 11— эпнбласт;
12 — корешок; 13 — чехлик; 14 — колеориза
Для успешного прорастания семени необходимы вода, кисло-
род воздуха и определенная температура, благоприятная для дан-
ного вида растения. Вода вызывает набухание семени и затем
принимает участие во всех процессах, связанных с ростом и раз-
витием растения, кислород необходим для дыхания прорастающего
растения, причем в этот момент его расходуется особенно много;
процессы, протекающие в растении, начинаются и получают даль-
нейшее развитие лишь в определенных температурных границах.
Во время прорастания семени запасные питательные вещества
эндосперма или семядолей переходят в растворимое состояние
й расходуются на построение проростка и добывание энергии, не-
обходимой для обеспечения ростовых процессов. Проростки бога-
ты водой. Объем их благодаря этому во много раз больше тех ча-
стей семени, за счет которых они образуются. Количество же сухого
вещества в прорастающем семени уменьшается до тех пор, пока
расход его не начнет возмещаться в результате фотосинтеза, про-
текающего в зеленых частях молодого растения. С этого момента
растение начинает усваивать солнечную энергию и аккумулиро-
вать ее в форме скрытой, химической энергии органических ве-
ществ.
Созревшие семена обычно не сразу прорастают, чаще проходит
некоторый период покоя, после которого семена приобретают
способность к активной жизни. Отсутствие необходимых для про-
растания условий может продлить период покоя неопределенно
долго. В отличие от глубокого покоя, необходимого растению, та-
кой покой называется вынужденным покоем. Чтобы прекратить
вынужденный покой, достаточно поместить семена в определенные
условия внешней среды. Для большинства растений такими усло-
виями; как уже сказано, являются влага, доступ воздуха и опре-
деленная температура; лишь некоторым, очень немногим растени-
ям для прорастания семян необходим свет (например, для мят-
лика).
ПОБЕГ И ЕГО ВИДОИЗМЕНЕНИЯ
Из почечки зародыша семени развивается побег, т. е. стебель
с расположенными на нем листьями. То место стебля, где прикреп-
ляется лист (или несколько листьев), называется стеблевым
узлом, а отрезок стебля между двумя соседними узлами — междо-
узлием. Междоузлия могут быть длинными или короткими, поэто-
му и побег бывает удлиненным или укороченным. Угол, образуе-
мый стеблем и отходящим от него листом, называется листовой
пазухой.
По существу побег — единый орган; единство стебля и листьев
в этом органе, хорошо прослеживаемое в онтогенезе, отражает их
общее происхождение: филогенетически, т. е. с точки зрения исто-
рического развития, листья возникли из боковых ответвлений стеб-
ля, принявших плоскую форму,— наилучшее приспособление к бо-
лее полному улавливанию солнечного света.
95
Рис. 42. Продольный раз-
рез через верхушечную
почку
Почки. Побег заканчивается верхушечной почкой. В пазухах
листьев часто сидят пазушные почки. Если почку разрезать вдоль,
то с помощью лупы можно рассмотреть ее строение (рис. 42).
Центр почки занимают клетки, за счет размножения которых
идет нарастание побега; эта часть почки, имеющая вид конуса,
называется конус нарастания, или апекс, а
ткань, из которой эта часть состоит, назы-
вается апикальной меристемой. В нижней
части конуса можно заметить выросты —
зачатки будущих листьев. Ниже распола-
гаются все более крупные выросты, посте-
пенно приближающиеся по своему виду к
маленьким нежным светло-зеленым листоч-
кам, которые и составляют основную мас-
су почек. Листочки, из которых состоит
почка, плотно прилегают друг к другу, об-
разуя черепитчатую поверхность. В пазухах
зачаточных листьев можно обнаружить бу-
горки— здчатки будущих боковых побегов.
Заложенные почки могут сразу же тро-
нуться в рост, что мы и можем часто наб-
людать в течение лета у деревьев и кустар-
ников. Однако далеко не все они трогают-
ся в рост в этом же году. В странах, где
в течение года бывают периоды, неблаго-
приятные для жизни -растения (у нас — зи-
ма, в жарких странах — сухое лето), у ра-
стений образуются спящие почки, длитель-
ное время пребывающие в состоянии покоя.
Такие почки имеют особые приспособления,
предохраняющие их от высыхания. Наруж-
ные листочки у них превращаются в плотные чешуи, не пропускаю-
щие или почти не пропускающие ни газов, ни воды. Защитная роль
этих чешуй усиливается еще тем, что они с внутренней стороны
выделяют особое смолистые вещества, бальзамы, а снаружи бы-
вают покрыты густым пушком. Иногда в пазухе листа располага-
ется несколько почек. В.таком случае при вертикальном располо-
жении, вдоль побега, они называются сериальными; при горизон-
тальном расположении, поперек побега,— коллатеральными.
Заложенные в пазухах листа почки могут не распускаться не-
сколько' и даже много лет. По мере роста ствола или ветки эти
почки подрастают за счет .удлинения своей оси, так что верхушка
их остается вблизи поверхности коры. Просыпаются такие спящие
почки обычно тогда, когда бывает повреждена верхушка побега
выше спящей почки. Пробуждение спящих почек происходит так-
же и в том случае, если будет удалена или отомрет верхушечная
почка. Это явление объясняется тем, что верхушечная почка вы-
деляет особые вещества — ауксины, которые тормозят развитие
спящих почек.
96
Помимо обычных верхушечных и пазушных почек, растения
могут закладывать почки на любой части стебля, на корнях и да-
же на листьях; такие почки называются придаточными. Заклады-
ваются они на большей или меньшей глубине от поверхности ор-
гана. Способность растений закладывать придаточные почки имеет
Рис. 43. Типы ветвления побегов (схема). А — моноподи-
альное, Б — симподиальное, В — дихотомическое
большое биологическое значение: расширяются возможности вос-
становления растений при повреждении; это свойство использует-
ся в вегетативном размножении растений при помощи частей по-
бега, корня или листа.
Есть еще особые выводковые почки; они закладываются в па-
зухах листьев (например, у лилии) или в соцветиях (у лука, мят-
лика живородящего и др.). Выводковые почки свойственны также
и споровым растениям — мхам. Отпадая, выводковые почки дают
начало новому растению, отсюда такое их название.
Ветвление побегов. Сравнительно редко надземная часть выс-
шего растения состоит из одного побега; чаще побегов бывает не-
сколько или очень много, отходящих один от другого, например,
у древесных растений. Такое своеобразное явление, широко рас-
пространенное в растительном мире, связано с ветвлением побегов
(рис. 43).
Это свойство имеет большое биологическое значение, так как
оно в огромной степени расширяет возможности растения увеличи-
вать массу листьев — основных ассимилирующих органов.
Филогенетически наиболее древний тип ветвления — дихотомиче-
ское, или вильчатое, ветвление; оно свойственно низшим растениям,
а среди высших встречается у наиболее древних форм: ископаемых
псилофитов, плаунов, мохообразных. При дихотомическом ветвле-
нии верхушечная почка главного побега разделяется в процессе
своего формирования на две почки, которые и дают два новых по-
бега следующего порядка.
4—326 97
Принято главный побег растения, отходящий от корня, назы-
вать осью первого порядка; возникающие из него или на нем —
осями второго порядка, на осях второго порядка — оси третьего
порядка и т. д. Следовательно, при дихотомическом ветвлении, раз-
дваиваясь, побег дает оси второго порядка, те в свою очередь
и таким же путем дают оси третьего порядка и т. д.; каждый побег
низшего порядка может дать только два побега следующего по-
рядка и то лишь взамен себя.
Значительно шире представлено у высших растений моноподи-
альное ветвление. При этом типе ветвления верхушечная почка на
протяжении всей жизни растения продолжает наращивать длину
растения, и поэтому главный побег образует как бы стержень всего
растения, занимает главенствующее положение. Растение обычно
принимает стройную вытянутую форму, свойственную ели, пихте
и др. При моноподиальном ветвлении на главном побеге (ось пер-
вого порядка) за счет пазушных почек развиваются боковые побе-
ги (оси второго порядка), на осях второго порядка развиваются
оси третьего порядка, и таким образом общее количество побегов
растет в геометрической прогрессии. Побеги высших порядков
обычно несколько короче побегов предыдущего порядка, в резуль-
тате чего растение приобретает характерную форму «елочки».
Наиболее прогрессивно симподиальное ветвление- При этом
типе ветвления один, два или несколько боковых побегов, образую-
щихся на главном побеге, быстро обгоняют его в росте, происходит
так называемое перевершинивание. Главный побег или замедляет
рост, или вовсе его прекращает. Оси второго порядка, обогнав ось
первого порядка, затем отстают, уступая первенство побегам сле-
дующего порядка, и т. д. Нередко на главном стволе образуется
несколько почти одинаковых крупных боковых стволов. Это созда-
ет возможность развития обширной развесистой кроны, что очень
ценится в садоводстве. Симподиально ветвятся многие наши пло-
довые и декоративные деревья: яблоня, груша, слива, абрикос,
цитрусовые, липа, вяз, клен, грецкий орех и многие другие. Особен-
но характерно симподиальное ветвление для кустарников. Из тра-
вянистых растений симподиальное ветвление свойственно картофе-
лю, томатам, тыквенным и др.
Сравнительно редко встречается тип ветвления, сходный с дихо-
томическим,— ложная дихотомия (например, у сирени, конского
каштана). При этом типе ветвления верхушечная почка замирает.
Усиленно развиваются две пазушные (супротивные) почки, бли-
жайшие к верхушечной. Образуется характерная вилка; верхушеч-
ная почка отмирает и от нее остается лишь едва заметный след.
Создается подобие дихотомического ветвления.
В зависимости от типа ветвления, направления роста побегов
и степени их развития создается определенный внешний облик,
или габитус растения, характерный для того или иного вида, раз-
новидности или сорта.
Побег неоднороден по длине; от основания к его верхушке дли-
на междоузлий уменьшается, и побег становится тоньше. По этому
98
признаку обычно легко различать верхний и нижний конец отре-
занного побега-
Физиологические различия по длине побега можно установить,
если отрезок побега, так называемый черенок, используется для
вегетативного размножения. Если такой черенок, способный к ве-
гетативному восстановлению целого растения, положить горизон-
тально, например на влажный песок, то можно заметить, что при-
даточные корни всегда образуются на морфологически нижнем
конце побега, а почки распускаются на морфологически верхнем
конце его. Это свойство морфологи-
ческого и физиологического разли-
чия по длине побега называется
полярностью; оно присуще не толь-
ко побегам, но и другим органам
растения, а у низших -растений —
всему таллому.
Особый способ симподиального
ветвления свойствен злакам. У них
формируется куст из большего или
меньшего количества побегов, осно-
вания которых сильно сближены и
придвинуты к поверхности почвы.
Такой способ ветвления называется
кущением. Свойством куститься об-
ладают и наши хлебные злаки, осо-
бенно озимые формы. У них про-
цесс кущения начинается с того, что
из нескольких сильно сближенных
подземных узлов главного побега
выходят первые боковые побеги и Рис- 44. Схема кущения рыхло-
одновременно пучки придаточных кустового злака
корней.
На боковых побегах, а также
на сильно сближенных узлах, в свою очередь могут образоваться
боковые побеги следующего порядка; причем каждый побег раз-
вивает собственную мочку придаточных корней. При благоприят-
ных условиях этот процесс может повториться несколько раз, так
что в конечном счете из группы сильно сближенных узлов куще-
ния кверху одним пучком отходит несколько или много побегов,
книзу несколько пучков мочковатых корней, образующих общую
корневую систему злака. Обычно весь этот комплекс сильно сбли-
женных, почти слитых узлов носит общее название — узел куще-
ния злака (рис. 44).
Злаки, у которых побеги из узла кущения отходят друг от друга
под некоторым углом, называются рыхлокустовыми. К ним отно-
сится большинство злаков. У некоторых злаков боковые побеги
сильно сближены, плотно прилегают друг к другу, их называют
плотнокустовыми (например, ковыль — Stipa, типчак — Festuca
sulcata). Злаки (например, пырей), у которых от подземного узла
4*
99
Рис. 45. Схема кущения корневищного злака
кущения отходят не только надземные, но и горизонтально идущие
подземные стебли, называются корневищными (рис. 45)-
У многолетних злаков, произрастающих в умеренной и холод-
ной зонах, надземные побеги к зиме отмирают, а весной из узлов
кущения развиваются новые побеги. В результате ежегодного от-
мирания в течение нескольких лет над узлом кущения скапливает-
ся целый пучок оснований отмерших побегов, защищающих узел
кущения от высыхания и вымерзания в неблагоприятные периоды
года. Узел кущения, таким образом, играет очень важную роль
в жизни злаков; у хлебных злаков удаление или отмирание его
ведет к гибели растения в целом. Поэтому важно, чтобы в суровую
зиму узел кущения озимых хлебов был хорошо защищен.
Листорасположение. Листья расположены на стебле в опреде-
ленной закономерности, обеспечивающей наименьшую взаимную
затеняемость. Такая закономерность — результат естественного от-
бора форм, обеспечивающих наилучшее пользование солнечным
светом.
Различают следующие три типа размещения листьев: 1) очеред-
ное, или спиральное, 2) супротивное и 3) мутовчатое (рис. 46).
При очередном размещении на каждом узле прикреплен один
лист; если места прикрепления листьев в порядке их размещения
соединить линией, то получится спираль. Идя по такой спирали от
какого-либо места, мы найдем лист, сидящий над тем, с которого
начат счет. Очередное расположение листьев свойственно большин-
ству цветковых растений.
При супротивном расположении листья сидят на каждом узле
попарно, один против другого; каждая такая пара листьев ориен-
тирована по отношению к другой под прямым углом. При таком
100
Рис. 46. Типы расположения листьев. А — очередное, или спираль-
ное, Б — супротивное, В — мутовчатое
расположении достигается наименьшая затеняемость листьев друг
другом. Супротивное расположение листьев встречается реже, чем
очередное, оно свойственно представителям семейства губоцвет-
ных, маслинных (сирень, ясень), кленовых и др.
Еще реже встречается мутовчатое расположение листьев, при
котором на одном узле размещается три и большее число листьев.
Такое листорасположение встречается, например, у олеандра (Ne-
rium oleander), вороньего глаза (Paris quadrifolia) и др-
При любом расположении листья на растениях размещаются
так, что обеспечивается, в пределах возможного, наименьшая вза-
имная затеняемость. Такое размещение листьев достигается не
только определенной закономерностью в расположении точек при-
крепления листьев к стеблю, но и неравномерным ростом листьев
и побегов, связанным с интенсивностью освещения.
При расположении листьев одного или нескольких побегов бо-
лее или менее в одной плоскости, если смотреть по направлению
падающего света, создается впечатление ковра, сложенного из от-
дельных листьев. Такое сочетание листьев получило название
листовой мозаики (рис. 47). Листовая мозаика служит подтверж-
дением большой пластичности вегетативных органов.
Для травянистых растений, используемых в корм животным,
большое значение имеет степень и характер олиственности побе-
гов. Чем больше листьев на побегах, тем выше кормовая ценность
данного растения. Это объясняется тем, что листья почти вдвое
питательнее стеблей, в них гораздо больше протеина и значительно
меньше клетчатки.
Травы, у которых листья сосредоточены преимущественно в
нижней части побегов, называются низовыми. Их выгоднее исполь-
зовать в пастбище. Травы с равномерным размещением листьев
по длине побега называются верховыми. Они более подходят для
сенокосного использования. Бывают и промежуточные формы. Для
низовых злаков характерно большое количество укороченных веге-
101
тативных побегов, за счет которых и создается основная масса
листьев. У верховых злаков в кусте преобладают генеративные
и удлиненные вегетативные побеги-
Видоизменения побегов. Стебель, как правило, несет на себе
листья и генеративные органы; общая масса этих органов в боль-
Рис. 47. Листовая мозаика (розетка листьев герани)
шой степени зависит от развития стебля. Стебель может выполнять
также и другие функции; в нем очень часто отлагаются запасные
питательные вещества; нередко он выполняет функцию вегетатив-
ного размножения. В процессе эволюции стебель приобретал и не-
которые другие функции.
В связи с новыми функциями стебель (а вместе с ним обычно
и листья) более или менее резко изменяет свою внешнюю и внут-
реннюю структуру, образуя видоизменения, или метаморфозы,
стеблей и побегов: корневища, клубни, луковицы и др. (рис. 48).
Корневище — сильно измененный подземный побег, свое назва-
ние он получил по сходству с корнем. Развиваясь под землей, по-
бег утрачивает зеленый цвет, а листья превращаются в бесцветные
чешуи. В зависимости от количества отложенных внутри стебля
питательных веществ корневища могут быть то очень толстые
(у касатика — Iris, купены — Polygonatum и др.), то более тонкие,
например у пырея (Elytrigia repens).
В условиях рыхлой почвы такие корневища сильно разрастают-
ся, от узлов их время от времени отходят обычные надземные
побеги, а книзу придаточные корни, и в конечном счете от неболь-
шого одиночного куста пырей может распространиться по всему
полю. Такие свойства делают его очень опасным и трудно искоре-
нимым сорняком.
W2
Клубни — сильно изменившиеся формы подземных стеблей. Они
образуются на конце длинных подземных стеблей — столонов.
Листья на поверхности клубней редуцированы в^ бесцветные чешуи,
которые, опадая в начале формирования клубней, оставляют на них
следы в виде листовых рубцов.
Луковица — своеобразные изменения побега- Она состоит из
сильно укороченного стебля, так называемого донца, от которого
книзу отходит пучок придаточных корней; донце кверху переходит
в конус нарастания, окруженный сильно измененными толстыми
Рис. 48. Подземные видоизменения побегов. А — луковица, Б — клубни
картофеля
листьями, которые, собственно, и образуют мякоть луковицы: Ли-
стья эти толстые, богатые питательными веществами; наружные
листья луковицы превращаются в тонкие омертвевшие, темноокра-
шенные чешуи, предохраняющие луковицу от высыхания и вообще
от вредных воздействий извне.
Луковица формируется к концу вегетацци растения, а весной
от центра ее выходит цветоносный побег, олиственный или окру-
женный несколькими листьями. На смену отмершей материнской
луковицы образуется одна или несколько новых.
Луковичные и клубнелуковичные растения наиболее широко
распространены в степях, полупустынях и пустынях. Наличие ор-
ганов запаса позволяет растениям, используя короткий период
вегетации, быстро выгнать цветоносный побег, зацвести и дать
плоды; тем самым растение как бы убегает от неблагоприятного
периода года, в данном случае — жаркого и сухого лета. Многолет-
103
ние .растения, имеющие такие биологические особенности, называ-
ются эфемероидами. К эфемероидам относятся также и некоторые
корневищные растения, например пустынная осока — Carex phy-
sodes.
Своеобразными метаморфозами являются изменения побегов
суккулентов (сочных, мясистых растений). У них стебли покрыты
колючками, или сильно редуцированными листьями, имеющими
вид коротких чешуй. К суккулентам относятся кактусы, произра-
стающие в естественном состоянии в Мексике, преимущественно
в районах с сухим и жарким климатом. У нас, в умеренной зоне,
суккулентные растения встречаются в условиях физиологической
сухости, вызванной избытком солей в почве, на мокрых солончаках
вблизи морей и соленых озер; они относятся к группе солянок.
Наиболее типичен как стеблевой суккулент солерос (Salicornia
herbacea).
К стеблевым метаморфозам относятся также филлокладии, или
кладодии,— стебли, имеющие форму листьев и выполняющие их
функции. Филлокладии вместо листьев имеет произрастающая
в Крыму и на Кавказе иглица (Ruscus).
Наконец, побег может метаморфизироваться в усики и колюч-
ки. Усики стеблевого происхождения — у тыквы, огурца, дыни, ви-
нограда и др.; колючки — у терна, боярышника, дикой груши, лоха
(Elaeagnus) и др. У лоха можно найти постепенный ряд переход-
ных форм — от настоящего нормального стебля до типичной колюч-
ки. Усики и колючки бывают и листового происхождения (усики
у гороха, вики).
Классификация цветковых растений в связи с типами побегов
и продолжительностью жизни. Соответственно типам побегов и их
возрасту высшие цветковые растения подразделяют на несколько
биоморфологических групп. Сравнительное изучение ископаемых
остатков и современных высших растений показывает, что наибо-
лее древние цветковые растения были представлены деревьями.
В дальнейшем, в процессе приспособления к крайним условиям су-
ществования, древесные формы на различных линиях эволюции
дали начало травянистым формам. Так возникли цветковые расте-
ния, обитающие в воде (например, белая кувшинка — Nytnphaea
alba); во влажных тропических лесах возникла интересная форма
травянистых растений, живущих на стволах крупных деревьев,—
эпифиты. В условиях затенения в этих же лесах широко представ-
лены растения-паразиты.
Мощным фактором формирования травянистых форм явилось
похолодание климата в приледниковой зоне. В тех же условиях
холодного климата формировалась травянистая растительность
высокогорного пояса. Такую же роль сыграло иссушение клима-
та— в пустынях и полупустынях также преобладают травянистые
формы. Промежуточными формами между деревьями и травами
являются кустарники и полукустарники.
Большую роль в эволюционном превращении древесных форм
в травянистые сыграло широко распространенное в живом мире
104
явление неотении-, оно заключается в преждевременном заверше-
нии онтогенеза, в результате чего еще юношеские (ювенильные)
фазы развития быстро сменяются окончательной (дефинитивной)
фазой, характеризующейся способностью к генеративному (поло-
вому) возобновлению. Травянистые растения — это наследственно
закрепленные неотенические формы древесных растений.
Деревья — деревянистые растения, имеющие главный ствол
с более или менее развитой кроной боковых ветвей. В большинстве
случаев деревья достигают размера нескольких метров или десят-
ков метров, но бывают и маленькие деревца.
Кустарники — деревянистые растения, но главный ствол у них
начинает симподиально ветвиться почти у самой поверхности поч-
вы, отчего и образуется куст. Кустарники бывают различной вы-
соты— от 1 до 6 м и их иногда в свою очередь подразделяют на
крупные, средние и мелкие. К кустарниковым формам относятся
желтая акация (Caragana arborescens), шиповник, смородина,
крыжовник и др. Кустарники в общем лучше приспособлены к пе-
реживанию неблагоприятных времен года, так как у них почки
расположены ближе к поверхности почвы. Еще лучше приспособ-
лены к суровым условиям зимы или жаркого и сухого лета полу-
кустарники.
Полукустарники — отличаются от кустарников тем, что у них
стебли одревесневают только в нижней части, которая и становит-
ся многолетней. На многолетних стеблях ежегодно отрастают и от-
мирают однолетние травянистые побеги. Примерами полукустар-
ников могут быть многие виды полыни, астрагала, терескен (Euro-
tia ceratoides) и др.
Травянистые формы — растения в общем более приспособлен-
ные к переживанию неблагоприятных времен года. Наиболее древ-
ние среди травянистых многолетние травы; у них в неблагоприят-
ный период года надземная часть полностью отмирает. Почки
находятся у самой поверхности или под поверхностью почвы и,
следовательно, хорошо защищены от неблагоприятных условий
внешней среды.
Многолетние травы могут существовать десятки лет, ежегодно
возобновляя надземную часть от узла кущения или корневой шей-
ки, где закладываются все новые и новые побеги. К этой группе
относятся многие кормовые травы: люцерна, клевер, эспарцет из
семейства бобовых; тимофеевка, житняки, мятлики и др.— из се-
мейства злаков.
Из многолетних форм в процессе естественного отбора возникли
однолетние и двулетние травы.
Однолетние травянистые растения ежегодно возобновляются из
семян: вырастают, цветут, плодоносят и отмирают. Продолжитель-
ность жизни однолетних трав различна: от нескольких недель до
нескольких месяцев. Неблагоприятный период года эти растения
переживают в форме семян. У большинства дикорастущих видов
семена могут сохраняться в почве несколько лет — это очень ценное
приспособительное свойство. К однолетним травам относится боль-
105
шинство культурных растений: хлебные злаки, лен, конопля, под-
солнечник, горох и другие. Сюда также относится много сорных
растений.
Двулетние растения в первый год жизни развивают только ве-
гетативные органы — обычно это прикорневая розетка листьев. На
следующий год отрастает цветоносный стебель, который приносит
плоды, после чего все растение отмирает. К двулетним относятся
такие широко известные растения, как морковь, свекла, капуста,
белена, многие сорняки из семейства крестоцветных.
Различают еще яровые и озимые растения. Яровые растения
прорастают из семян весной и не требуют для своего полного раз-
вития действия низких температур. Озимые растения для заверше-
ния своего развития должны после прорастания подвергнуться
действию низких температур, иначе они не дают цветоносных по-
бегов. Обычно такие растения прорастают из семени осенью, а
весной продолжают свое развитие.
Хлебные злаки — пшеница, рожь, ячмень — выращиваются как
в яровых, так и в озимых формах. В зависимости от особенностей
климатических условий в отдельных районах нашей страны выра-
щиваются исключительно или преимущественно то озимые, то яро-
вые формы хлебных культур.
МОРФОЛОГИЯ СТЕБЛЯ
Формы стебля. По направлению и способу роста наиболее ча-
сто встречаются прямостоячие стебли; они свойственны большин-
ству растений. Приподнимающиеся стебли вначале растут горизон-
тально или несколько наискось к поверхности почвы, затем прини-
мают вертикальное положение.
Нередко встречаются стебли ползучие, или стелющиеся,— при-
легающие к почве на большей части своей длины. В узлах такие
стебли обычно образуют придаточные корни, которыми закрепля-
ются в почве. Ползучие стебли с очень длинными междоузлиями
называют усами, а с укороченными — плетями. Стебли ползучих
побегов мягкие, механически слабо оснащены и не способны дер-
жаться самостоятельно. Такими же свойствами обладают стебли
у растений вьющихся и цепляющихся (рис. 49).
Вьющиеся или цепляющиеся растения с деревянистым стеблем
называются лианами. В тропических лесах лианы достигают огром-
ной длины — 200—300 м и больше при толщине стебля 2—4 см.
Такие лианы могут подниматься на самые высокие деревья, вынося
свои листья ближе к свету.
На поперечном разрезе в большинстве случаев стеблевая часть
побегов имеет форму более или менее правильного круга, реже
стебли бывают трехгранные (у осок) или четырехгранные (у гу-
боцветных) . Иногда стебель бывает сплюснутый или даже плоский
(некоторые виды мятлика — Роа compressa, опунций — Opuntia
и др.). Поверхность стебля может быть ребристой или иметь вдоль
106
междоузлий выросты в виде пластинок; в последнем случае стебель
называется крылатым.
Размер и возраст стеблей. Побег, возникший в текущем году
из почки, может просуществовать один, два, несколько или много
лет. Стебли однолетних побегов обычно не древеснеют и сохраня-
ют зеленый цвет до конца своей жизни- По наступлении неблаго-
приятного времени года (зимы или жаркого сухого лета) они отми-
рают, утрачивают зеленую окраску и быстро разрушаются.
Есть много однолетних травянистых растений, обычно мелких,
которые, образовавшись из семени, быстро отцветают, дают плоды
Рис. 49. Формы стеблей. А -• прямостоячий, Б — цепляющийся,
В — вьющийся, Г — ползучий
и отмирают. Цветут эти эфемеры обычно ранней весной и, следо-
вательно, побеги у них живут всего несколько недель и размеры
побегов редко превышают 10—20 см. Однако однолетние не дре-
веснеющие стебли могут достигать и весьма значительных разме-
ров, например стебель главного побега кукурузы достигает высоты
4 м и более.
107
Стебель, живущий несколько лет, обычно древеснеет, у деревьев
он может достигать огромных размеров; например, ствол монопо-
диально ветвящегося дерева (ели, кипариса) есть не что иное, как
стебель главного побега этого растения, который может существо-
вать несколько сот лет и достигать размера 50 м и более. Наибо-
лее крупные деревья (эвкалипты, секвойя) достигают высоты
140 м, а диаметр их 10—12 м. Наиболее долговечные (секвойи,
тисс, кипарисы) живут до 3000—5000 лет.
АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЯ
В процессе развития из семени почечка зародыша развивается
в побег, а верхушечная ее часть становится конусом нарастания,
или апексом. Анатомическое строение апекса различных растений
в общем сходно (рис. 50). Весь он состоит из образовательной
ткани — протомеристемы. Один или несколько наружных слоев
клеток апекса отличается от остальной массы клеток способом
Рис. 50. Продольный разрез через вер-
хушку стебля:
/—туника; 2 — зачатки листьев; 3 — апекс;
4 — прокамбий; 5 — зачатки пазушных почек;
6 — проводящий пучок листа; 7 — слияние про-
водящего пучка листа с пучком стебля; 8 —
сердцевина
ную меристему и прокамбий. В
деления: клетки этих слоев
делятся перегородками, ори-
ентированными перпенди-
кулярно к поверхности ко-
нуса (антиклинально).
Клетки, лежащие ib глубине
конуса нарастания, делятся
в различных направлениях.
Наружный слой (или
слои) клеток, делящихся
антиклинально, т. е. перпен-
дикулярно поверхности ко-
нуса, составляет тунику
апекса; внутренняя часть
апекса, клетки которой де-
лятся в различных направ-
лениях, называется корпу-
сом.
Антиклинальное направ-
ление плоскостей деления
клеток туники обусловлива-
ет рост ее в поверхность;
следовательно, туника диф-
ференцируется в протодер-
му, из которой затем разо-
вьется эпидермис. Корпус
дифференцируется на основ-
быстроты
результате неодинаковой
деления поверхностных клеток в отдельных пунктах апекса в неко-
тором отдалении от его кончика вырастают листовые бугорки —
зачатки будущих листьев. Уже в зоне зачаточных листьев можно
видеть, что в результате дифференциации промеристемы заклады-
108
ваются пучки первичной образовательной ткани — прокамбия.
Клетки промеристемы, из которых образуется прокамбий, делятся
перегородками, направленными преимущественно по длине органа,
поэтому они отличаются от соседных клеток остальной массы об-
разовательной ткани вытянутой формой.
Прокамбиальные тяжи закладываются внутрь апекса, по его
оси и во внутренней части оснований будущих листьев. Прокамби-
альные тяжи листьев удлиняются в двух направлениях: кверху,
в будущую пластинку листа, и книзу, по направлению к прокамби-
альным пучкам будущего стебля. Сделав поперечный разрез апек-
са в его основании, мы можем видеть на поверхности среза эти
прокамбиальные тяжи, из которых затем будут формироваться
проводящие пучки стебля.
За счет деятельности прокамбия и остальной меристемы апекса
формируется первичное строение стебля. Структура, возникающая
в результате деятельности камбия, называется вторичной, она
сложнее первичной.
Стебли однодольных, особенно травянистых злаков, имеют бо-
лее простое строение, чем стебли двудольных растений, им свойст-
венно, за редким исключением, первичное строение, в то время как
для двудольных растений типично вторичное строение.
Целесообразно рассмотреть вначале первичное строение стеб-
ля однодольных, а затем первичное и вторичное строение стебля
двудольных. Первичное строение стебля однодольных мы рассмот-
рим на примере кукурузы и ржи.
Строение стебля злаков. На поперечном разрезе стебля кукуру-
зы (рис. 51) можно видеть, что с поверхности стебель покрыт одно-
слойным эпидермисом, стенки клеток эпидермиса утолщены. Сна-
ружи эпидермис покрыт слоем кутикулы. Под эпидермисом распо-
лагается сплошное кольцо мелкоклеточной склерифицированной
паренхимы, постепенно переходящей в обычную крупноклеточную
паренхиму. Ширина склерифицированного кольца (фактически —
толщина цилиндра) неодинакова; в молодых стеблях хлорофилло-
носная паренхима подходит непосредственно к эпидермису, и в этих
местах можно заметить устьица-
В молодом стебле стенки клеток субэпидермального цилиндра
тонки и слабо одревеснены; процесс одревеснения клеточных стенок
только начинается. С возрастом происходят такие изменения: уве-
личивается толщина склерифицированного цилиндра, стенки кле-
ток все более древеснеют и становятся толще, различия между
этими клетками и клетками обычной паренхимы становятся все
более резкими.
Основную массу стебля составляет паренхима. Клетки парен-
химы крупные, с тонкими стенками; между ними — межклетники.
В молодых стеблях паренхима сочная, клетки сильно вакуолизи-
рованы, вакуоли заполнены сладким клеточным соком. К концу
вегетации большая часть паренхимных клеток заполняется возду-
хом, благодаря чему вся сердцевина становится белой, очень мяг-
кой, почти как вата.
109
В паренхимной ткани без заметной закономерности разбросано^
большое количество сосудисто-волокнистых пучков. Те из них, ко-
торые находятся ближе к периферии, размещены более густо,—
они непосредственно примыкают к склерифицированному цилинд-
Рис. 51. Поперечный разрез стебля кукурузы:
/ — эпидермис; 2 — склерифицированная паренхима; 3 —основная паренхима; 4 —
проводящий пучок
ру; ближе к центру стебля пучки размещены реже, но каждый
пучок становится крупнее. Пучков очень много: даже в молодых
стеблях их можно насчитать свыше сотни.
Проводящие пучки злаков коллатерального типа, они не со-
держат камбия, следовательно, это закрытые пучки (см. рис. 36)..
Такие пучки формируются в результате дифференциации прокам-
биальных ^яжей, закладываемых в меристеме в процессе формиро-
вания стебля и листьев.
Первоначальные проводящие элементы ксилемы представлены
несколькими узкими кольчатыми и спиральными сосудами прото-
110
ксилемы, окруженными мелкоклетной паренхимой. К моменту
окончательного формирования пучка эта паренхима разрывается,
и под сосудами образуется воздухоносная полость. Позднее обра-
зуются два широких пористых сосуда метаксилемы, расположен-
ных симметрично по бокам, ближе к флоэме, которая занимает
часть пучка, ориентирован-
ную к периферии поперечно^
го разреза стебля.
Каждый сосудисто-во^
локнистый пучок у злаков
окружен склеренхимной об-
кладкой, или чехлом, в виде
сплошной трубочки, внутри
которой находятся проводя-
щие и паренхимные элемен-
ты пучка. Мощность скле-
ренхимной обкладки пучков
возрастает по направлению
к периферии стебля, так что
механически наиболее осна-
щенные пучки занимают
периферийное положение.
Здесь наружный ряд пучков
сливается своей склеренхим
ной обкладкой с общим на
р у жны м скл ер и фицир ов ан
ным кольцом, и в результа-
те такого размещения меха-
нических элементов в стебле
обеспечивается наибольшая
его механическая прочность
при минимальном расходо-
вании строительного мате-
риала.
Стебель ржи, -и вообще
большинства злаков, полый
(рис. 52); такой стебель на-
зывается соломиной. По-
лость образуется на ранних
этапах формирования стеб-
ля в результате отмирания
сердцевинной паренхимы. В
оставшемся периферическом
слое паренхимы размещены
закрытые коллатеральные
пучки, так же, как мы это
видели в стебле кукурузы;
однако пучков значительно
.меньше.
Рис. 52. Строение стебля ржи на по-
перечном разрезе:
1 — эпидермис; 2 — склерифицированная
паренхима; 3 — хлорофиллоносная парен-
хима; 4, 5 — проводящие пучки; 6 — основ-
ная паренхима
111
Так же, как и у кукурузы, пучки одеты склеренхимным чехлом.
Наиболее периферийные из них размещаются в общем склерифи-
цированном цилиндре, примыкающем к эпидермису. Стенки клеток
эпидермиса и склеренхимы с возрастом древеснеют.
Непосредственно под эпидермисом в толще склерифицирован-
ного цилиндра располагаются парные тяжи хлорофиллоносной па-
ренхимы, на поперечном разрезе они имеют вид двух продолгова-
тых участков, прилегающих к эпидермису длинной стороной.
В эпидермисе стебля над хлорофиллоносными участками имеются
устьица. Поверхностные тяжи хлорофиллоносной паренхимы хоро-
шо заметны при рассмотрении еще не пожелтевшей соломины; по-
верхность стебля полосатая, темно-зеленые полосы чередуются
с более светлыми.
В узлах стебель злаков несколько утолщен. У растений с полым
стеблем в этом месте перегородка. Утолщения в узлах связаны
с тем, что здесь находятся обильные сплетения мелких стеблевых
Рис. 53. Строение стебля подсолнечника:
/ — эпидермис; 2 — колленхима; 3 —основная паренхима; 4 — крахмалоносный слой
(эндодерма); 5 — склеренхима; 5 —флоэма; 7 — камбий; 8 — ксилема
пучков с пучками, выходящими из листьев; между этими пучками
имеются многочисленные анастомозы. Стебель на узлах оснащен
добавочным количеством склеренхимной ткани.
Строение стебля двудольных. Одним из наиболее удобных объ-
ектов для изучения структуры стебля служит подсолнечник
(рис. 53). Рассматривая поперечные срезы стебля в различном воз*
расте, мы можем проследить изменения в анатомической структуре
112
с момента ее возникновения вблизи апекса до окончательного
оформлёния в закончившем рост стебле.
В анатомическом строении двудольных и голосеменных расте-
ний принято различать первичную и вторичную структуру. Первич-
ная структура возникает в результате дифференциации апикаль-
ной меристемы, переход ко вторичной структуре начинается с мо-
мента деятельности камбия.
Строение стебля удобнее рассматривать, начиная с поверхности,,
переходя последовательно к центральной части органа. В такой
последовательности мы рассмотрим на поперечном разрезе строе-
ние молодого стебля подсолнечника, сохранившего еще первичное
строение.
С поверхности стебель, как обычно, покрыт эпидермисом. На
небольших выростах эпидермиса, состоящих из группы мелких
клеток, сидят жесткие волоски, состоящие из одного ряда клеток.
Всю остальную массу стебля на поперечном разрезе разделяют на
два слоя: первичную кору и центральный осевой цилиндр.
Первичная кора на нашем объекте состоит из разнообразных
элементов. Непосредственно под эпидермисом несколько слоев пла-
стинчатой колленхимы, клетки которой содержат хлорофилловые
зерна; затем идет тонкостенная паренхима, в ее клетках количество
хлоропластов постепенно уменьшается по направлению к центру;
переход между колленхимой и паренхимой не резкий.
Последний, внутренний слой клеток первичной коры называет-
ся эндодермой, или крахмалоносным влагалищем; в клетках этого
слоя имеются крахмальные зерна, роль которых недостаточно вы-
яснена. Крахмальные зерна эндодермы не расходуются даже при
голодании растений. В коровой паренхиме разбросаны смоляные
ходы — узенькие канальцы, окруженные мелкими клетками основ-
ной ткани: у других видов травянистых растений могут быть раз-
личные отклонения: колленхима или отсутствует вовсе, или, если
стебель имеет ребристую поверхность, располагается тяжами,
в таком случае колленхима обычно уголковая или переходного
характера. Выдающиеся ребрышки, в основном, и состоят из такой
колленхимы. Вместо колленхимы может быть сплошной цилиндр
или отдельные прослойки склеренхимы; последняя размещается
всегда глубже колленхимы, между склеренхимой и эпидермисом
остается слой хлорофиллоносной паренхимы.
Первый слой (или несколько слоев) паренхимных клеток цент-
рального осевого цилиндра, или стелы, непосредственно примыка-
ющий к эндодерме, относится к перициклу, клетки которого, как
уже сказано, способны к возобновлению меристематической дея-
тельности. Клетки перицикла могут превращаться в лубяные волок-
на, которые называются первичными, в отличие от вторичных,
образованных камбием. Перицикл дает начало придаточным кор-
ням. Глубже перицикла располагается основная паренхима, в ко-
торой размещены коллатеральные пучки, расположенные здесь
закономерно, по кругу. Пучки возникают из прокамбиальных тя-
жей, закладываемых еще апикальной меристемой.
113
В отличие от однодольных, у которых весь прокамбий расходу-
ется на образование постоянных элементов пучка, у двудольных
прокамбий полностью не дифференцируется; клетки узенькой про-
слойки прокамбия между сформировавшимися первичными ксиле-
мой и флоэмой сохраняют способность делиться и превращаются
в постоянную действующую прослойку первичного камбия.
Камбий продолжает формировать элементы ксилемы и флоэмы.
В отличие от элементов, образованных прокамбием, ксилема
и флоэма, возникающие в результате действия камбия, называют-
ся вторичными. С момента деятельности камбия начинается фор-,
мирование вторичной структуры стебля. На ней мы остановимся
позже, а сейчас вернемся к первичному строению стебля.
Между пучками, образованными прокамбием, остаются про-
слойки сердцевинной паренхимы, играющие важную роль в горизон-
тальном перемещении растворов. Эти прослойки называют сердце-
винными, радиальными лучами или межпучковыми зонами. Цент-
ральный участок стебля, состоящий из паренхимы, отграниченный
внутренними концами пучков, называется сердцевиной. Сердце-
винные пучки соединяют сердцевину с перициклом.
В случае хорошо развитой сердцевины у древесных наружная
часть ее с более мелкими клетками паренхимы называется пери-
медуллярной зоной. При отмирании сердцевины клетки этой
зоны сохраняются живыми и служат вместилищем запасов крах-
мала.
У некоторых растений (лен, сурепка, мыльнянка и др.) отдель-
ные тяжи прокамбия еще в конусе нарастания соединяются и дают
сплошное кольцо камбия, который продуцирует кнаружи элементы
вторичной флоэмы, внутрь — вторичной ксилемы. Такое строение
стебля называется беспучковым.
Вторичные изменения в стебле пучкового строения не ограни-
чиваются деятельностью пучкового камбия. Уже в самом начале
вторичных изменений в том участке первичного сердцевинного лу-
ча, который расположен между камбиальными прослойками двух
соседних пучков, клетки основной ткани приобретают свойство
энергично делиться. В результате этого деления появляется уча-
сток образовательной ткани, соединяющейся в конце концов с со-
седними участками пучкового камбия.
Вновь образованные прослойки образовательной ткани называ-
ются межпучковым камбием. Таким образом, межпучковый кам-
бий по своему происхождению является вторичным камбием. Вто-
ричный камбий соединяется с первичным пучковым камбием, обра-
зуя общее камбиальное кольцо (цилиндр), клетки которого и
продолжают функционировать согласованно, подстилая вторичную
флоэму под первичную и наслаивая вторичную ксилему на первич-
ную. Переход от первичного строения стебля ко вторичному схе-
матично изображен на рис. 54.
Межпучковый камбий обычно вначале продуцирует паренхиму,
наращивая таким образом первичные сердцевинные лучи; у неко-
торых растений (например, у клевера лугового) он формирует ме<
114
ханическую ткань — склеренхиму. При существенном утолщении-
стебля, как, например, у подсолнечника, межпучковый камбий фор-
мирует добавочные пучки вторичного происхождения; у этих пуч-
ков нет первичных элементов, а ксилема и флоэма у них только
вторичные. В результате разрастания старых и вновь возникающих
пучков сердцевинные лучи настолько стискиваются, что ксилемы
соседних пучков почти сливаются, образуя сплошной древесинный
цилиндр.
В стеблях беспучкового строения сплошное кольцо камбия об-
разуется прямо из прокамбия и, следовательно, на всем протяже-
нии имеет первичное происхождение. Деятельность этого кольца
камбия ничем не отличается от г
деятельности камбиального
кольца, возникающего более
сложным путем при пучковом
типе строения стебля. Таким
образом, независимо от того,
какова была первичная струк-
тура стебля растений, в ре-
зультате вторичных изменений
у них мюжет образоваться оди-
наковая структура: общее кам-
биальное кольцо, внутрь от не-
го— ксилема, кнаружи — фло-
эма. Так формируется вторич-
ная структура у большинства
травянистых растений, у хвой-
Рис. 54. Образование общего камбиаль-
ного кольца (схема поперечного среза)_
А — первичное строение, Б — образова-
ние кольца камбия:
1 — кора; 2 — сердцевина; <3 — первичная флоэ-
ма; 4 — пучковый камбий; 5 — первичная кси»
лема; 6 — межпучковый камбий
ных и многих древесных дву-
дольных. У некоторых древесных лиан (например, у винограда)
межпучковый камбий формирует только паренхиму лучей и поэ-
тому при вторичных изменениях сохраняется пучковое строение.
У большинства древесных растений (например, у липы) сплош-
ное камбиальное кольцо образуется в самом начале формирования
стебля. Межпучковые зоны очень узкие, образуется сплошной ци-
линдр ксилемы (древесины) с очень тонкими прослойками основ-
ной ткани; во флоэме эти прослойки обычно шире, особенно в пе-
риферийной части.
Благодаря деятельности сплошного кольца камбия первона-
чальный осевой цилиндр, свойственный первичному строению, раз-
деляется на два слоя: внутренний — вторичную древесину и на-
ружный — вторичную кору. Древесина — это ксилемная часть стеб-
ля, вторичная кора — флоэмная; вторичную флоэму называют так-
же вторичным лубом.
В результате деятельности камбия в стебле накапливается все
большая и большая масса элементов вторичного происхождения,
а роль и значимость в общей массе первичных элементов структуры
быстро сходит на нет. Следовательно, основная роль в формиро-
вании стебля двудольных, особенно древесных, принадлежит кам-
бию.
115
Формирование камбием элементов ксилемы и флоэмы происхо-
дит с неодинаковой быстротой; на одну клетку флоэмы камбий
отделяет несколько клеток ксилемы; следовательно, ксилема нара-
стает гораздо быстрее, чем флоэма. У многолетних древесных ра-
Рис. 55. Строение стебля липы (Т ilia cor data)'.
/ — эпидермис; 2 — пробка; 3 — паренхима коры; 4 — сердцевинный луч;
5 — лубяные волокна; 6 — флоэма; 7 — камбий; 8 — весенняя древесина;
9 — осенняя древесина; 10 — сердцевина
стений к тому же происходит постепенное отмирание элементов
флоэмы снаружи, поэтому в них на долю ксилемы приходится поч-
ти вся масса ствола и ветвей; флоэма же составляет сравнительно
тонкий слой коры.
Клетки камбия делятся преимущественно тангентально, благо-
даря чему эта ткань приобретает характерное строение: клетки
располагаются правильными стопками, радиально направленными
рядами. В связи с непрерывным утолщением ксилемного цилиндра
окружность камбиального цилиндра также увеличивается, что про-
исходит в результате радиального деления клеток камбия.
116
За счет деятельности камбия в составе вторичных ксилемы
и флоэмы формируются разнообразные элементы. Наибольшее
разнообразие их достигается в многолетних стеблях древесных ра-
стений (рис. 55).
Вследствие периодической деятельности камбия в ксилеме дре-
весных растений возникают годичные кольца. Весной, благодаря
обилию воды и питательных веществ, камбий формирует крупные
элементы древесины; к концу вегетационного периода деятель-
ность камбия затухает — в древесине все больше преобладают ме-
ханические элементы и узкие сосуды.
Мы рассмотрели онтогенез и законченное строение стебля,
свойственное высшим цветковым растениям. Анатомическая струк-
тура стебля у этих растений достигла наибольшего совершенства
лишь в процессе эволюции, длившейся сотни миллионов лет. У пер-
вых высших растений — псилофитов, мхов, плаунообразных
и др. — стебель имел более простое строение; в частности, его
внутренняя осевая часть — стела — у первых высших растений
была устроена более примитивно: ксилема и флоэма не были рас-
членены на участки; сплошной цилиндр из ксилемы был окружен
слоем флоэмы, без радиальных прослоек паренхимной ткани. Та-
кое строение создавало худшие условия для снабжения всех частей
растений водой и питательными веществами, по сравнению со
строением, свойственным семенным растениям.
ЛИСТ И ЕГО ВИДОИЗМЕНЕНИЯ
Настоящие листья, в отличие от энаций (поверхностных выро-
стов, свойственных первым высшим растениям) возникли в процес-
се эволюции боковых веточек главного побега в результате их
уплощения. Превращение обычных веточек в плоские органы резко
увеличило поверхность надземной части растения и явилось сле-
дующим этапом развития растений на пути приспособления к наи-
лучшему выполнению двух функций: фотосинтеза и испарения. Эти
две функции — основные для листьев. Следы стеблевого происхож-
дения листьев хорошо сохранились у папоротников; в отличие от
всех остальных растений, листья у них, подобно стеблю, растут
своей верхушкой.
Листья имеют самые разнообразные формы, но тем не менее
можно установить некоторые общие и основные черты строения
(рис. 56). Плоская и обычно расширенная часть листа называется
пластинкой. У большинства растений пластинка листа прикрепле-
на к стеблю посредством черешка, который кажется ответвлением
побега. На самом деле это нижняя сильно суженная часть листа,
спадает лист вместе с черешком.
Нередко пластинка листа не имеет черешка, тогда лист называ-
ется сидячим. Листья с черешками называют черешковыми. Нали-
чие черешка — прогрессивный признак; черешковые листья могут
•свободнее располагать свою пластинку по отношению к падающе-
му свету.
117
Рис. 56. Различные формы листьев. А — игольчатый, Б — мечевидный, В — линей*
ный, Г — ланцетный, Д — яйцевидный, Е — овальный, Ж—округлый, 3 — обрат-
нояйцевидный, И — сердцевидный, К — почковидный, Л — щитовидный, М — стре-
ловидный, Н — копьевидный, О — неравнобокий, П — перистолопастный, Р —
тройчатолопастный, С — пятилопастный, Т — перистораздельный, У — пальчато-
лопастный, Ф — перисторассеченный (лировидный), X — прерывистоперисторассе-
ченный, Ц — пальчаторассеченный, Ч — двоякоперисторассеченный, Ш — тройча-
тосложный, Щ — пальчатосложный, Э — непарноперистосложный, Ю — двояко-
перистосложный
У злаков и некоторых других растений (осок, орхидных, многих
зонтичных) нижняя часть листа расширена и охватывает стебель
в виде желобка пли трубки; эта часть называется влагалищем
листа. У злаков влагалища листьев играют особенно важную роль:
они обхватывают нежную, растущую часть стебля, расположенную
над узлом, и тем самым предохраняют ее от повреждения. У неко-
торых злаков часть листьев не имеет пластинок и состоит из одних
влагалищ. Влагалища листа выполняют и общие функции, свойст-
венные листу,— фотосинтез и испарение, но менее интенсивно.
У многих злаков на границе влагалища и пластинки имеется
более или менее длинный пластинчатый вырост — язычок. Он со-
ставляет как бы продолжение влагалища и плотно прилегает к
стеблю. У некоторых видов язычок заменен пучочком волосков.
Верхние уголки влагалища имеют клиновидные выросты, охваты-
вающие стебель; они называются ушками. При изучении злаков
эти части (язычок и ушки) имеют большое диагностическое значе-
ние, биологическая роль их, по-видимому, сводится к лучшему вы-
полнению влагалищем защитной функции.
У основания листа у многих растений можно видеть особые ли-
стовидные выросты обычно меньшего размера, чем сам лист. Эти
выросты называются прилистниками. В процессе эволюции при-
листники отделились от нижней части листовой пластинки, следо-
вательно, они имеют более позднее листовое происхождение. При-
листники выполняют те же функции, что и листья. Редко они быва-
ют крупнее самих листьев (как у некоторых видов чины). Так же
как листья, прилистники могут превращаться в чешуйки и колючки.
Срастаясь, прилистники образуют раструб, расположенный выше
прикрепления листа (например, у гречишных).
В пастбищном корме и сене листья — наиболее ценная часть,
они лучше перевариваются и содержат больше протеина по срав-
нению со стеблями.
Жилкование листа.. Пластинка листа обладает достаточной ме-
ханической прочностью и при обычном воздействии ветра или воды
•не разрывается. Прочность листа в значительной степени зависит
от разветвляющихся в нем жилок, или «нервов», состоящих из ме-
ханических волокон и тканей, проводящих воду и растворенные в
ней вещества. Следовательно, жилки не только механические, но
и проводящие элементы листа, благодаря им лист равномерно
снабжается водой и питательными веществами, которые корень
поглощает из почвы и подает в стебель. Вместе с тем посредством
тех же жилок органические вещества, образующиеся в листе в ре-
зультате фотосинтеза, направляются в стебель и корни растения.
Отходя от черешка или непосредственно от стебля, если лист
сидячий, жилки различным образом распределяются и разветвля-
ются в пластинке. Они могут идти более или менее параллельно
друг другу (у злаков, осок); в таком случае обычно нельзя из об-
щей массы жилок выделить центральную, главную. Чаще же име-
ется одна главная жилка, служащая как бы продолжением череш-
ка, а от нее отходят боковые. В одних случаях они образуют поло-
119
гие дуги и в конце листа опять сближаются; в других случаях жил-
ки, отходя от главной жилки под более или менее острым углом,
доходят до края листа. Наконец, встречаются листья, у которых от
черешка сразу отходят в виде лучей несколько крупных жилок,
расходящихся к краям листа; от них в обе стороны отходят более
мелкие жилки; мелкие разветвления жилок обычно образуют гу-
стую анастомозирующую сеть. Общая длина жилок в среднем по
размеру листа составляет около 100 см, а среднее расстояние меж-
ду двумя соседними жилками — около 100 мкм.
По характеру жилкования различают листья параллельно-
нервные (у злаков), дугонервные (у ландыша, подорожника), пе-
ристонервные (у вишни, груши) и пальчато- или дланевиднонерв-
ные (у клена, клещевины).
Форма и размер листовой пластинки. Листья имеют самую раз-
нообразную форму- Они делятся на простые, если у них листовая
пластинка цельная или расчлененная на лопасти, и сложные, если
лист состоит из нескольких пластинок (листочков). Сложные ли-
стья филогенетически возникли из простых.
Пластинка простого цельного листа может иметь различное
очертание. Она может быть более или менее правильно округлой,
продолговатой, яйцевидной, обратнояйцевидной, ланцетной, обрат-
ноланцетной, ромбической, лопатчатой, сердцевидной, почковидной,
стреловидной, копьевидной, линейной, игольчатой.
По степени рассеченное™ различают листья лопастные, раз-
дельные, рассеченные, а по расположению лопастей их называют
перистолопастными и пальчатолопастными, перистораздельными
и пальчатораздельными, перисторассеченными и пальчаторассечен-
ными. Перистораздельный лист с крупной верхней долей называ-
ется лировидным (например, у сурепки, редьки, брюквы). Лопаст-
ные или рассеченные листья меньше затеняют друг друга и легче
образуют мозаику.
Сложные листья подразделяются на пальчатосложные, тройча-
тосложные и перистосложные. Перистосложные листья, имеющие
нечетную верхнюю долю, называются непарноперистосложными,
с четным числом листочков — парноперистосложными. Встречают-
ся листья еще более сложного строения: дваждыперистосложные,
дваждытройчатосложные, пальчатоперистосложные и пр. Общая
ось сложного листа называется рахисом.
Нередко можно встретить растения, у которых листья имеют
резко различную форму, причем такие неодинаковые листья рас-
полагаются даже на одном и том же побеге. Это явление называ-
ется гетерофиллией, что означает разнолистностъ (рис. 57). Осо-
бенно характерно это явление для водных растений (например,
у стрелолиста) и здесь оно легко объяснимо: листья, находящиеся
в воде, имеют узкую лентовидную форму, наиболее устойчивую
против разрыва текучей водой; надводные листья, наоборот, имеют
широкую пластинку, которая лучше улавливает солнечный свет.
Различают также листья по характеру края пластинки. Редко
края пластинки бывают ровными, такие листья называются цель-
120
Рис. 57. Гетерофиллия. А — подводные листья водяного лютика, Б —
надводный лист того же лютика, В — тесьмовидные подводные листья
и стреловидные надводные листья стрелолиста
нокрайними. Обычно они более или менее зазубрены, причем форма
зазубринок бывает разнообразной. Различают такие края пластин-
ки: зубчатые, пильчатые, городчатые, выемчатые, двоякопильчатые,
извилистые и др.
Размеры листа могут быть самые разнообразные. Особенно
крупные листья у некоторых тропических пальм; например, у ра-
фии они достигают 20 м и более в длину и до 12 м в ширину. Очень
крупные листья у бананов. У наших растений тоже встречаются
довольно крупные листья, например, у лопушника, подбела, бор-
щевика; крупные листья у кукурузы. С другой стороны, многие
травянистые растения имеют очень мелкие листья, иногда едва за-
метные простым глазом.
Независимо от размера пластинки, благодаря многоярусному
расположению листьев, общая поверхность их может во много раз
превышать площадь земли, на которой растение произрастает. Так,
например, общая листовая поверхность растений, произрастающих
на 1 га, составляет (приблизительно) у кукурузы 12 га, у пшеницы
25 га, у картофеля 40 га. Еще больше она у древесных растений.
Видоизменения листьев. Помимо настоящих листьев, хотя и
имеющих самые разнообразные формы, но выполняющих обычные
для них функции — фотосинтез и транспирацию, на растениях ча-
сто можно видеть различные их видоизменения. Эти видоизменен-
ные листья подчас бывают настолько непохожи на обычные, что
для доказательства их листовой природы приходится прибегать
к онтогенетическому методу исследования, т. е. изучать эти органы
в процессе их возникновения, развития.
Обычные видоизменения листа — чешуи, лишены хлорофилла^
Ими одеты, например, почки почти всех древесных и кустарнико-
121
вых растений. В почках можно наблюдать постепенный переход
от чешуй к нормальным, хотя и неразвившимся еще листочкам.
Биологический смысл данного видоизменения ясен — чешуи играют
защитную роль. Такую же роль играют крупные и тонкие чешуи,
покрывающие луковицы, например, обычного репчатого лука. Но
основная масса луковицы состоит из сочных чешуй, богатых саха-
ром и другими питательными веществами; здесь чешуи служат
органами запаса.
Своеобразные видоизменения листьев — семядоли. Форма
и размер их резко отличаются от нормальных листьев данного ра-
стения. Семядоли служат органами запаса (у двудольных) или
с их помощью питательные вещества извлекаются из эндосперма
и передаются развивающемуся растению (у злаков). У большинст-
ва двудольных растений семядоли при прорастании семени выно-
сятся наверх, зеленеют и служат первыми фотосинтезирующими
органами. После распускания настоящих листьев они отмирают.
Листья, подобно стеблям, могут видоизменяться в колючки
и усики. В природе можно наблюдать различную степень превра-
щения листа в колючки. В одних случаях в колючки превращаются
кончики жилок, доходящих до края листа, тогда получается колю-
чий лист, свойственный, например, различным чертополохам и осо-
там. У барбариса (Berberis vulgaris) часть листьев превращена
в 3—7-раздельные колючки, в пазухах которых сидят укороченные”
побеги с несколькими нормальными листьями. Все формы перехо-
да от нормальных листьев к колючкам можно наблюдать у так на-
зываемой верблюжьей колючки (Alhagi camelorum). У некоторых
астрагалов в колючку превращается рахис перистосложного листа.
В колючки могут превращаться также и прилистники, например
у белой акации- Колючки играют защитную роль, предохраняя
растения от поедания животными, хотя уже само название одного-
из таких растений — «верблюжья колючка» говорит о том, что не
во всех случаях это приспособление действует. С другой стороны,
можно заметить, что превращение листьев в колючки чаще бывает
в странах с жарким и сухим климатом, где сокращение листовой
поверхности уменьшает испарение.
Превращение листьев в усики широко распространено в семей-
стве бобовых: у видов гороха, вики, чины и других в усики превра-
щены центральные жилки только верхних листочков (простые уси-
ки) или несколько листочков (ветвистые усики). У некоторых чин
в ветвистый усик превращен весь лист, а роль листа выполняют
прилистники (например, у чины прилистниковой). Иногда роль
усика выполняет черешок листа — у ломоноса (Clematis), настур-
ции (Tropaeolum) и др.
Таким образом, колючки и усики могут быть и листового, и
стеблевого происхождения. Органы различного происхождения,
но имеющие сходное строение и выполняющие одинаковую функ-
цию, называются аналогичными.
С другой стороны, один и тот же орган (например, лист) в про-
цессе приспособительной эволюции может резко видоизменяться
122
Рис. 58. Насекомоядное растение венерина
мухоловка (Dionaea muscipula)
л превратиться по существу в новый орган с совершенно иными
функциями, например лист-колючка. Такие органы, имеющие оди-
наковое происхождение, но видоизменившиеся в различные формы,
выполняющие неодинаковые функции, называются гомологичными.
Органом, гомологичным листу, является также филлодий — листо-
образное разращение че-
решка у австралийской ака-
ции, а из растений нашей
флоры — у одного из южных
видов чин (Lathyrus nisso-
lid).
Явление гомологии ха-
рактеризует большую пла-
стичность вегетативных ор-
ганов высших растений в
процессе эволюции. Ярким
примером такой пластично-
сти могут быть ловчие орга-
ны насекомоядных расте-
ний.
Известно свыше 400 ви-
дов, преимущественно тро-
пических растений, у кото-
рых листья превратились в
особые улавливающие аппа-
раты, иногда довольно слож-
ного строения, с помощью
которых они ловят насеко-
мых (рис. 58). Растения эти
чаще живут на почвах, бед-
ных усвояемым азотом и
минеральными веществами,
в связи с чем они приспосо-
бились пользоваться допол-
нительным питанием за счет улавливаемых насекомых. Перевари-
вая их, насекомоядные растения получают добавочные источники
азотистых и минеральных веществ. Лишенные такого дополни-'
тельного питания, они растут значительно хуже, что было точно
установлено Ч. Дарвином, который весьма обстоятельно изучал
биологию насекомоядных растений.
Строение улавливающих аппаратов и способы ловли насекомых
очень разнообразны. В одних случаях насекомое садится на ли-
сток, прилипает и затем крепко удерживается специальными вор-
синками растения, выделяющими липкую жидкость; в других —
растение активно ловит насекомое с помощью специальной хло-
пушки, в которую превратилась листовая пластинка; в-третьих
случаях насекомое попадает в специальный кувшинчик, пузырек,
из которого уже не может выбраться. Во всех этих случаях насеко-
мое переваривается при помощи кислот и пищеварительного фер-
123
мента, подобного пепсину желудочного сока. Пищеварительная
жидкость выделяется в ответ на раздражение, производимое насе-
комым на липкой поверхности улавливающего листочка, или скап-
ляется на дне кувшинчика, пузырька. Такого же эффекта можно
достичь, если в улавливающий аппарат класть маленькие кусочки
мяса или яичного белка.
Анатомическое строение листа. Анатомическая структура листа
формируется в апексе одновременно с формированием стебля; по-
кровы листа являются продолжением покрова молодого стебля,
проводящая система листа вливается в проводящую систему стеб-
ля, пучки, выходящие из листа, образуют в стебле так называемые
листовые следы — часть пучков, обнаруживаемых на поперечном
срезе стебля. Щель, образуемая над тем местом, где листовой след
отходит в лист, и заполненная паренхимой, называется листовым
прорывом или лакуной. В строении черешка листа и стебля можно
обнаружить много сходного, что подчеркивает общность их фило-
генеза. Вместе с тем наблюдаются и существенные различия, кото-
рые определяются иной функцией листа и его недолговечностью
по сравнению со стеблем.
В начале формирования лист растет своей верхней частью. За-
тем верхушечный рост затухает, растет более или менее равномер-
но вся пластинка и постепенно зона роста сохраняется лишь в ниж-
ней части листа. Позже всего формируется черешок путем наращи-
вания верхней части.
Вообще же форма листа складывается довольно рано, обычно
сразу же по развертывании почки, а затем идет лишь увеличение
пластинки за счет разрастания клеток. Листья злаков и некоторых
других однолетних растений могут, однако, длительное время
удлиняться благодаря деятельности зоны меристемных клеток в
нижней части пластинки — подобно вставочному, интеркалярному
росту стебля злаков.
Пучки в листьях однодольных растений закрытые, коллатераль-
ного типа, подобны пучкам стебля. В листьях двудольных растений
только в крупных жилках имеется камбий- Деятельность его по-
степенно затухает по направлении? к периферии листа. Мелкие
жилки имеют только первичную структуру, сформированную про-
камбием. Вторичные изменения, следовательно, не играют суще-
ственной роли в структуре листьев. Это объясняется, прежде всего,
небольшой продолжительностью жизни листьев.
С обеих сторон лист покрыт кутинизированным эпидермисом,
защищающим внутренние ткани листа от высыхания и неблаго-
приятных воздействий факторов внешней среды. Защитная функ-
ция эпидермиса усиливается наличием кутикулярного слоя, воско-
вым налетом и различного рода выростами эпидермиса. Газообмен
между фотосинтезирующими клетками и атмосферой осуществля-
ется через устьица. Строение эпидермиса, строение и работа устьиц
были уже рассмотрены в разделе о тканях.
Между двумя слоями эпидермиса находится мезофилл, или хло-
ренхима, составляющая основную массу листа (рис. 59).
124
В большинстве случаев лист ориентирован в пространстве бо-
лее или менее горизонтально, и в нем можно различать верхнюю,
брюшную, или вентральную, сторону и нижнюю, спинную, или дор-
зальную, В любом листе, независимо от его расположения, разли-
3 8 7
Рис. 59. Строение листа подсолнечника:
/ — кутикула; 2 — верхний эпидермис; 3 — палисадная паренхима; 4— парен-
химная обкладка жилки листа; 5 — ксилема; 6 — флоэма; 7 — устьице; 8 — за-
мыкающие клетки; 9 — нижний эпидермис; 10 — воздухоносная полость; 11 —
губчатая паренхима
чают сторону адаксиальную, направленную к стеблю, и абаксиаль-
ную— противоположную. В листьях такого дорзовентрального ти-
па мезофилл обычно бывает дифференцирован на палисадную, или
столбчатую, паренхиму и губчатую ткань.
Один или несколько слоев столбчатых клеток примыкают
к верхней стороне листа. Такая ориентировка клеток связана с ос-
новной функцией палисадной ткани — фотосинтезом. Клетки па-
лисадной ткани направлены перпендикулярно к поверхности листа.
В условиях чрезмерного освещения, при котором усиливается раз*
рушение хлорофилла, хлоропласты размещаются преимущественно
на вертикальных стенках клеток и тем самым взаимно затеняют
друг друга; при недостаточном освещении хлоропласты перемеща-
ются на горизонтальные стенки клеток, параллельные поверхности,
листа.
Наиболее хорошо выражена палисадная ткань в листьях расте-
ний, обитающих в условиях обильного освещения, например,,
у хлопчатника. Наоборот, у растений теневых, где условия освеще-
ния более мягкие, палисадная ткань менее выражена, длина клеток,
этой ткани не намного превышает их ширину.
125
Губчатая ткань, примыкающая к нижней стороне листа дорзо-
вентрального строения, состоит из более или менее изодиаметриче-
ских, рыхло расположенных клеток. Между ними большие меж-
клеточные пространства, сообщающиеся с устьицами. Это ткань
преимущественно испаряющая, транспирационная, хотя функция
фотосинтеза свойственна и ей.
Рыхлое расположение клеток губчатой паренхимы в огромной
степени увеличивает испаряющую поверхность. Внутренняя поверх-
ность листьев при этом в 6—30 раз превышает наружную, достигая,
например, у деревьев средней величины 5000 м2. В мезофилле вод-
ных и болотных растений образуются воздухоносные пространства,
превращающие мезофилл листа в аэренхиму.
У листьев, ориентированных вертикально или под острым углом
к стеблю, резкого различия между палисадной и губчатой тканью
может не быть. В условиях засушливого климата палисадная ткань
примыкает у некоторых растений не только к верхней, но и к ниж-
ней стороне листа (например, у лебеды). В отличие от листьев
дорзовентралъного строения, листья, у которых в анатомическом
строении не заметна дифференциация на нижнюю и верхнюю сто-
роны, называются изолатералъными.
Механическая прочность листа обеспечивается, в первую оче-
редь, сосудисто-волокнистыми пучками, образующими нервацию
листа. Наиболее крупные жилки хорошо оснащены механической
тканью — склеренхимой. По мере утончения жилок они утрачивают
склеренхимную обкладку, остаются только проводящие элементы
пучка. У очень тоненьких жилок исчезают элементы флоэмы, а со-
суды ксилемы заменяются трахеидами.
Заканчиваются жилки в мезофилле узенькими одиночными тра-
хеидами, по которым подводится вода и растворенные в ней веще-
ства непосредственно к фотосинтезирующим клеткам. Отток асси-
милятов вначале происходит по обычным клеткам мезофилла, затем
по специальным вытянутым обкладочным клеткам хлорофиллонос-
ной паренхимы, образующих обкладку вокруг трахеи. По этим
клеткам продукты ассимиляции передаются уже ситовидным труб-
кам флоэмы. Клетки флоэмы жилок часто имеют не совсем типич-
ный вид: они слабо вытянуты и иногда не имеют ситовидных пла-
стинок. В общем они приближаются к обычным вытянутым парен-
химным клеткам.
Помимо пучковой склеренхимы, механическая ткань в листьях
может размещаться и самостоятельными прослойками; непосредст-
венно под эпидермисом встречается и слой колленхимы.
Механическая прочность краев листа иногда усиливается раз-
ветвленными склереидами, в которые дифференцируются конечные
трахеиды тонких жилок.
Листья злаков по анатомическому строению значительно отли-
чаются от типичных листьев дорзовентрального типа (рис. 60).
Различия видны уже в строении эпидермиса, клетки которого здесь
имеют сильно удлиненную форму и на. поперечном разрезе неоди-
наковы по величине. В том месте, где к эпидермису примыкает
126
механическая ткань, клетки мельче, чем над паренхимной тканью.
Благодаря неравномерному размещению механических тканей-
(склеренхимы) по длине листа, поверхность его становится волни-
стой и даже ребристой.
На верхней стороне листа, между ребрами, на поперечном раз-
резе можно заметить группы веерообразно расположенных круп-
ных клеток. Это так называемые двигательные клетки, они сильно-
вакуолизированы и при достаточном количестве влаги находятся^
Рис. 60. Детали внутреннего строения листа кукурузы:
1 — шарнирные (двигательные) клетки эпидермиса; 2 — обкладочные клетки
в состоянии тургора. При недостатке влаги тургор в клетках пада-
ет, и листья свертываются в трубку, благодаря чему предупрежда-
ется излишнее испарение. Такое целесообразное приспособление
особенно хорошо развито в листьях узколистных злаков типа ко-
выля или типчака, свойственных степям и полупустыням.
Своеобразным строением отличается и мезофилл в листьях
злаков. Он не дифференцирован, как обычно, на палисадную
и губчатую ткань. Пучки окружены слоем клеток, несколько вытя-
нутых по длине пучка и образующих вокруг него обкладку. На по-
перечном разрезе эти клетки расположены в виде розетки. Об-
кладки двух соседних пучков, идущих параллельно в листе, могут
контактировать между собой.
Обкладочные клетки не у всех злаков выражены одинаково.
Особенно хорошо они развиты у злаков из группы просовидных
(просо, кукуруза, сорго и др.). Хорошее развитие обкладок свя-
зывают с лучшим использованием воды злаками этой группы; на
единицу создаваемого органического вещества они расходуют
меньшее количество влаги. Характерные особенности строения ли-
ста злаков наиболее отчетливо выражены в крупных листьях ку-
курузы.
127
Анатомическое строение прилистников зависит от величины их.
Крупные прилистники имеют такое же строение, что и листья.
У мелких прилистников проводящая система сильно редуцирована.
Самостоятельных следов в стебле прилистники, как правило, не
дают; проводящая система их сливается с таковой же листа еще
до поступления ее в центральный цилиндр стебля.
Своеобразное строение имеют листья хвойных (рис. 61). На
рисунке видно, что под эпидермисом хвои располагается особый
склеренхимный слой гиподермы. Мезофилл — складчатая паренхи-
Рис. 61. Хвоя сосны (Pinus silvestris), поперечный срез:
/ — эпидерма; 2 — складчатая паренхима; 3 — смоляные ходы; 4 — устьица; 5 — эндодер-
ма; 6 — паренхимные клетки с окаймленными порами; 7 — флоэма проводящего пучка;
8 — склеренхима; 9 — гиподерма
•ма, в ней смоляные ходы. Центральная часть хвои отделена от
мезофилла эндодермой; внутрь от эндодермы — проводящие пучки.
В анатомическом строении листового черешка есть много об-
щего с первичным строением стебля: вместе с тем здесь наблюда-
ется большое видовое разнообразие, благодаря чему анатомиче-
ская картина поперечного среза черешка используется в система-
тике покрытосеменных.
128
Лист недолговечен. Обычно он живет не дольше одного вегета-
ционного периода, у так называемых «вечнозеленых» деревьев
и кустарников — до 10 и более лет. Опадение листьев подготавли-
вается заранее: в основании черешка в результате растворения
срединной пластинки или слоя клеток поперек черешка возникает
отделительный слой; после опадения листа под образовавшимся
рубцом очень быстро возникает перидерма, и обнажившиеся живые
ткани затягиваются защитным слоем феллемы (пробки).
Процесс фотосинтеза. Лучистая энергия солнца и другого ис-
точника света, падая на поверхность листа, частично отражается
и рассеивается,* частично проходит сквозь лист и лишь около 50%
ее поглощается хлорофиллом. Еще классическими исследованиями
К. А. Тимирязева было установлено, что не все части спектра сол-
нечного света одинаково поглощаются хлорофиллом. Наиболее ин-
тенсивно поглощаются лучи красной и несколькр слабее сине-фио-
летовой части спектра. Лишь несколько процентов (до 5 и в ис-
ключительных случаях до 10) поглощенной хлорофиллом энергии
используется в процессе фотосинтеза, т. е. превращается в конеч-
ном счете в скрытую химическую энергию синтезированных орга-
нических веществ. Свыше 90% поглощенной энергии переходит
в тепловую или расходуется в процессе транспирации для превра-
щения воды в парообразное состояние. Энергия света, поглощен-
ная хлорофиллом, превращается им в энергию химическую, кото-
рая и аккумулируется в ходе реакций фотосинтеза. О сущности
этого процесса было сказано при описании клетки.
Материалом для фотосинтеза служит углекислый газ СО2, ко-
торый должен попасть внутрь листа. Поступив внутрь листа через
устьица, СО2 растворяется в воде и в форме угольной кислоты —
Н2СО3 проникает в клетки сквозь клеточные оболочки, задержи-
вается в цитоплазме, а затем поглощается хлоропластами.
Количество хлоропластов в листе огромно, и общая поверх-
ность их составляет в одном травянистом растении несколько квад-
ратных метров. Не удивительно, что при очень слабой концентра-
ции СО2 в воздухе (около 0,5 мг/л), процесс поглощения ее идет
весьма интенсивно.
Углеводы — не единственные продукты фотосинтеза; помимо
них могут синтезироваться и другие органические вещества, в том
числе аминокислоты и белки. Однако углеводам принадлежит пер-
вое место, поэтому об интенсивности фотосинтетического процесса
судят обычно по количеству отложенного в растении крахмала.
Интенсивность фотосинтетического процесса поддается регули-
рованию, и это имеет большое производственное значение. В на-
стоящее время широко практикуется выращивание овощей с ис-
пользованием дополнительного искусственного освещения в осен-
не-зимний период. Для этого используются как обычные лампы
накаливания, так и люминесцентные.
Положительные результаты получены также в опытах выращи-
вания растений в атмосфере с повышенным содержанием углекис*
лого газа. Увеличивая в несколько раз концентрацию СО2 в воз-
5—326
129
духе,, добивались повышения урожая оранжерейных культур более
чем вдвое.
Процесс транспирации. Во время фотосинтеза через открытые
устьица одновременно с выделением кислорода происходит испа-
рение воды поверхностью клеток мезофилла. Этот неизбежный для
сухопутных растений процесс называется транспирацией.
Интенсивность транспирации, особенно в сухом и жарком кли-
мате, обычно превышает физиологическую необходимость, т. е. ис-
паряется воды больше, чем нужно для нормального существования
растения. Это одно из противоречий в жизни растений: для осуще-
ствления фотосинтеза нужно, чтобы устьица были открыты, но
через устьица испаряется вода, а чрезмерное испарение может
даже вызвать гибель растения в результате высыхания клеток.
Но в процессе транспирации имеется и положительная сторона:
испарение воды с поверхности листьев обеспечивает постоянный
приток к ним воды с растворенными в ней веществами; ток про-
хладной воды, двигающейся по растению, и сам процесс испарения
охлаждают растение. В условиях высоких температур такое охлаж-
дение играет большую положительную роль, предохраняя растение
от перегрева и ожога солнечными лучами. Не удивительно, что
даже в самые жаркие дни поверхность деревьев никогда' не нагре-
вается, как, например, может нагреваться деревянный столб. На
интенсивность испарения влияют многие факторы.
1. Величина испаряющей поверхности. Одна из форм приспо-
собления растений к сокращению испарения — уменьшение испа-
ряющей поверхности. У типичных степных злаков — типчака и ко-
выля— листья узкие и продольно свернуты, почти волосовидные.
2. Подвижность воздуха. Ветер сдувает с поверхности листьев
слой влажного воздуха и тем самым повышает транспирацию.
Густой покров из отмерших волосков, свойственный многим
растениям степей и полупустынь, смягчает влияние этого
фактора.
3. Влажность воздуха. Чем меньше недостаток насыщения ат-
мосферного воздуха водяными парами^ тем слабее транспирация.
Поэтому в естественных условиях обитания растений умеренной
и засушливой зоны достаточно высокий уровень относительной
влажности рассматривается как явление положительное. В оран-
жереях, где воздух насыщен водяными парами почти до предела,
транспирация падает до минимума, имеющего уже. отрицательное
значение; растения не охлаждаются и нередко подвергаются сол-
нечным ожогам.
4. Уровень барометрического давления. Разрежение атмосферы
усиливает транспирацию.
5. Сила освещения. Свыше 80% поглощенной листом солнечной
энергии расходуется на испарение воды с поверхности листа. Бла-
годаря большому испарению даже в самые жаркие дни поверх-
ность листьев на ощупь прохладная. В числе прочих факторов,
повышающих транспирацию, солнечному свету принадлежит пер-
вое место.
130
6. Строение самого листа. Рыхлое расположение клеток губча-
той ткани, мезофилла увеличивает внутрилистовую испаряющую
поверхность, межклеточные пространства образуют систему про-
ветривания листа. Устьица являются отдушинами этой системы.
Вместе с тем слабое испарение с листа идет и помимо устьиц
через слой кутикулы; это так называемая кутикулярная транспи-
рация. Чем моложе лист и чем тоньше слой кутикулы, тем боль-
шую роль играет эта форма транспирации. Тем не менее основная
масса испаряющейся с поверхности растений воды идет через
устьица.
Существует закономерность (закон Стефана), согласно которой
испарение с малых поверхностей идет пропорционально не площа-
ди их, а диаметру отверстий. В силу этой закономерности при боль-
шом числе устьиц и малом их размере с эпидермиса листа вода
может испаряться почти с такой же скоростью, как с равновеликой
площади открытой водной поверхности.
В физиологических и агрономических исследованиях отношение
растений к потреблению воды характеризуют транспирационным
коэффициентом, который показывает число граммов воды, израс-
ходованной растением за вегетационный период при накоплении
одного грамма сухого вещества. Величина эта колеблется в широ-
ких пределах — от 125 до 1000, т. е. для того-чтобы накопить 1 г
сухого вещества, растению, в зависимости от вида, сорта и усло-
вий выращивания, надо израсходовать от 125 до 1000 г воды.
Обратная величина называется продуктивностью транспирации.
Ее выражают в граммах накопленного сухого вещества на каж-
дый килограмм израсходованной воды. Эта величина колеблется
в пределах от 1 до 8; другими словами, расходуя 1 кг воды, ра-
стение может накопить от 1 до 8 г сухого вещества.
При выращивании большинства культур в средних условиях
обеспечения влагой транспирационный коэффициент обычно бы-
вает близок к 300, а продуктивность транспирации — к 3.
КОРЕНЬ И ЕГО ВИДОИЗМЕНЕНИЯ
Первые высшие растения настоящих корней не имели. Роль
корней как всасывающих органов играли ризоиды — тонкие ните-
видные нежные выросты из нижней части стебля. Такие ризоиды
мы можем видеть у ныне живущих зеленых мхов. Эти предшест-
венники корней слабо прикрепляли растение к субстрату и, следо-
вательно, механической роли почти не играли. Настоящие корни
возникли из нижней части стебля, и появление их знаменовало
дальнейший этап приспособления растений к жизни на суше.
МОРФОЛОГИЯ КОРНЯ
Корень, развившийся из зародышевого корешка, называется
главным. При прорастании семени он направляется вниз и зани-
мает более или менее вертикальное положение. Очевидно, это
реакция на земное притяжение. Свойство частей растения расти
5*
131
или перемещаться в определенном направлении под влиянием од-
носторонне действующей причины называется тропизмом. В зави-
симости от действующего фактора (свет, вода, земное притяжение
и др.) различают фототропизм, гидротропизм, геотропизм и т. д.
В данном случае мы имеем дело с положительным геотропизмом,
так как корень направляется в сторону действующего фактора.
Примером отрицательного тропизма может служить рост главного
побега.
От главного корня отходят боковые корни, от крупных боковых
корней — более мелкие, от них — мельчайшие и т. д. Образуется
более или менее густая сеть корней, которая называется корневой
системой данного растения;
Корневая система растения выполняет очень важные функции:
1) закрепляет растение в почве~'2) поглощает из почвы воду.и ра-
створенные в ней вещества и' снабжает ими все части растения;
всасывающая способность корней значительно увеличивается бла-
годаря корневым волоскам; 3) синтезирует важнейшие органиче-
ские соединения, из которых затем образуются белки и другие
вещества растений. Это три основные функции корня. Очень часто
корень служит вместилищем запасных питательных веществ. Не-
редко из части корня восстанавливается целое растение, т. е. ко-
рень служит в таком случае и для вегетативного размножения рас-
тений.
Новые корни могут возникать не только из главного или боко-
вых корней; они могут образовываться также из стебля и даже из
листа. Такие корни называются придаточными. У однодольных
растений, например у зерновых злаков, лука, чеснока, вся корневая
система состоит из большого количества придаточных корней.
Способность растений образовывать придаточные корни имеет
большое биологическое значение: 1) создаются дополнительные
возможности для развития корневой системы, 2) в случае гибели
корневой системы, возникшей из главного корня, растение не по-
гибает, образуя корневую систему заново, 3) возникает возмож-
ность вегетативного размножения. Последнее широко используется
при размножении растений клубнями, луковицами, черенками, от-
водками и другими частями.
Если у растения хорошо развит главный корень, то в таком слу-
чае корневая система называется стержневой. Такая система свой-
ственна большинству двудольных растений. Однодольным же
растениям, например злакам, свойственна мочковатая корневая
система, состоящая из пучка придаточных корней.- Придаточные
корни у злаков, развившиеся близ зерновки и заложенные еще
в зародыше — «зародышевые корни», образуют их первичную кор-
невую систему. Более мощно развиваются придаточные корни на
нижних частях побегов, от так называемого узла кущения. Они
образуют вторичную корневую систему злаков, играющую обычно
гораздо большую роль в жизни растения.
Корневая система растений может быть развита по-разному
(рис. 62). У злаков, например, основная масса корней в виде гу-
132
Рис. 62. Формы корней. А — стержневой, Б — веретеновидный, В — реповид-
ный, Г — мочковатый
*стого пучка размещена преимущественно в верхних слоях почвы.
У травянистых и древесных растений, обладающих стержневой си-
стемой, основная масса корней может быть размещена на различ-
ных, иногда на больших глубинах. Если главный корень рано пре-
кращает свое развитие или он искусственно удален, корневая си-
стема формируется в основном за счет боковых корней, разраста-
ющихся горизонтально. В таком случае основная масса корней
расположена на сравнительно небольшой глубине, но зато сильно
распространена вширь. В других случаях не только главный, но
и боковые корни направлены вглубь, и тогда вся корневая система
расположена на большой глубине^ занимая в ширину меньшее
пространство. Наибольший объем почвы используется тогда, когда
равномерно развит и главный корень, и идущие в стороны боковые
корни.
Различные типы корневых систем позволяют большому числу
видов произрастать в- одной группировке, что очень часто бывает
при естественном произрастании растений. ’Корни разных видов
растений распределяются в различных горизонтах почвы, и объем
почвы, освоенный корнями, таким образом используется наиболее
полно.
На формировании корневой системы в большей степени сказы-
вается воздействие внешних условий, состояние и характер почвы,
обеспеченность ее водой, а также различного рода повреждения
корней. Изменяя эти условия сознательно, можно регулировать
процесс формирования корневой системы. Например, обрезая
главный корень, можно заставить растение формировать корневую
133
систему в основном за счет боковых корней; развивается более
мощная и широко разветвленная корневая система. Этот прием
используется в садоводстве, овощеводстве, цветоводстве.
Разрыхляя почву, мы тем самым стимулируем развитие корне-
вой системы именно в этом пахотном слое, наиболее богатом пита-
тельными веществами. Обрабатывая стеблевые черенки растений
так называемыми ростовыми веществами, можно усилить образо-
вание придаточных корней, а у тех древесных пород, которые не
обладают такой способностью, вызвать их образование искусствен-
но. Этот прием используется в садоводстве и лесоводстве. Корне-
вые системы растений развиты более мощно, чем нам может по-
казаться. Такое ошибочное представление объясняется тем, что
при извлечении растений из почвы мелкие разветвления корней
обычно обрываются и мы их не замечаем.
В связи с приобретением некоторых дополнительных функций
корни более или менее резко видоизменяются. Такие видоизмене-
ния называются метаморфозами корня. Наиболее широко распро-
страненными метаморфозами являются мясистые корни или так
называемые корнеплоды, у
таких растении, как свекла,
морковь, турнепс, брюква,
редька и др. Нижняя часть
корнеплода по своему про-
исхождению — главный ко-
рень, верхняя относится к
стеблю, между ними нет за-
метной разницы. Корнепло-
ды служат вместилищем за-
пасных питательных веществ
и обычно используются в ка-
честве сочных кормов и как
пищевой продукт. Иногда
Рис. 63. Опорные придаточные корни куку- питательных веществ на-
РУЗЫ * столько много, что их спе-
циально извлекают из кор-
неплодов. Так, например, корнеплоды сахарной свеклы используют
для получения сахара.
Корни с большим запасом питательных веществ образуются
обычно у двулетних растений; на второй год жизни у таких расте-
ний из корнеплода развивается цветочный стебель, растение плодо-
носит и затем отмирает.
Корнеплоды развиваются у растений, имеющих хорошо выра-
женную стержневую 'Корневую систему. Реже утолщаются прида-
точные корни, тогда образуются корневые клубнщ их не следует
смешивать с более широко известными клубнями стеблевого про-
исхождения, например клубнями картофеля. Корневые клубйи
образуют цветочное растение георгина, ранневесеннее растение
чистяк и др. Особенно крупные корневые клубни образуются
у тропических растений — батата (из семейства вьюнковых) и ям-
134
са (семейство диоскорей-
ных). В тропических странах
клубни этих растений широ-
ко используются в пищу ме-
стным населением.
Придаточные корни, вы-
растающие из надземной ча-
сти стебля, могут играть роль
подпорок, например у куку-
рузы (рис. 63). В тропиче-
ских странах такие корни-
подпорки, или ходульные
корни, широко распростра-
нены у деревьев, растущих в
топких местах с зыбким
грунтом, в зоне морских при-
ливов и отливов. Такие кор-
ни придают необходимую
устойчивость деревьям и
вместе с тем выполняют и
всасывающую функцию. За-
росли деревьев с ходульными
корнями называют мангро-
вой растительностью,
АНАТОМИЧЕСКОЕ
СТРОЕНИЕ КОРНЯ
Анатомическое строение
корня может быть первич-
ным и вторичным. Первич-
ное строение возникает в ре-
зультате дифференциации
меристемы апекса; вторичное
строение — результат дея-
тельности камбия.
Первичное строение кор-
ня свойственноюднодольным
растениям на всем протяже-
нии их жшзни; у двудольных
первичное строение корня
довольно быстро сменяется
вторичным, в результате че-
го корень у них может с воз-
растом значительно утол-
щаться.
Рис. 64. Первичное строение корня
ириса:
1 — центральный осевой цилиндр; 2 — остатки
эпиблемы; 3 — экзодерма; 4 — основная парен-
хима коры корня — мезодерма; 5 —эндодерма;
6 — перицикл; 7 — флоэма; 8 — ксилема
Первичное строение корня. В первичной структуре корня, вбли-
зи его кончика, различают три слоя: наружный — эпиблему (эпи-
дермис), средний — первичную кору и центральный осевой ци-
линдр— стелу (рис. 64 и 65).
135
можно установить по длине
Рис. 65. Поперечный срез центрального ци-
линдра корня лютика (первичное строение):
1 — эндодерма; 2 — пятна Каспари; 8 — перицикл;
4 — флоэма; 5 — ксилема
Разделение корня на три слоя намечается уже в апексе корня,
в меристематической ткани. Здесь соответственно различают: дер-
матоген, который затем дифференцируется в эпиблему, периблему,
превращающуюся в первичную кору, и плерому, из которой фор-
мируется центральный цилиндр — стела.
При рассмотрении^ кончика, корня на продольном разрезе
его следующие части: корне-
вой чехлик, зону роста и рас-
тяжения, зону корневых во-
лосков, или всасывания, и
зону заложения боковых
корней.
Корневой чехлик с по*
верхности постепенно стира-
ется, клетки его 'Ослизняют-
ся и слущиваются. Ослизне-
ние клеток облегчает про-
никновение корешка вглубь.
Изнутри клетки чехлика на-’
ращиваются за счет деления
инициальных клеток. В зоне
роста и размножения клетки
меристемы энергично делят-
ся, а выше этого места вытя-
гиваются в длину, здесь на-
блюдается наибольшая ско-
рость роста. Длина корне-
вых волосков колеблется в
широких пределах— 190—
1500 мкм, а общая длина
кончика корня, включая зо-
ну всасывания, составляет обычно 3—4 мм. На 1 мм2 поверхности
всасывающей зоны корня приходится до нескольких сот корневых
волосков.
Чем больше поверхность эпиблемы, тем лучше корень будет
выполнять свою основную, всасывающую функцию. Именно в свя-
зи с этой функцией часть клеток эпиблемы вытянута в корневые
волоски, суммарная поверхность которых во много раз увеличи-
вает всасывающую поверхность корня (см. рис. 28). Подсчитано,
что суммарная длина корневых волосков одного растения достига-
ет огромных величин, измеряемых километрами; соответственно»
общая поверхность их достигает*сотен квадратных метров.
Степень развития корневых волосков в большЪй мере зависит
от состояния окружающей среды. Корни водных растений не име-
ют корневых волосков, но и обычные наземные растения, если
корни их поместить в .роду, чаще всего не образуют корневых во-
лосков. Наоборот, некоторые водные растения (желтая кубышка,
аир и др.), растущие по краям водоема, на корнях, проникающих
в почву, волоски образуют. Вырастает волосок в течение 1—3 дней.,
136
рост его на^йнается с выпя-
чивания бугорка наружной
стенки клетки эпиблемы; буг
горок растет, и в него пере-
ходит ядро данной клетки.
И клетка, и волосок, выра-
стающий из нее, сильно ва-
куолизированы. Продолжи-
тельность жизни волоска в
большинстве случаев нахо-
дится в пределах нескольких
дней, но имеются растения,
у которых волоски гораздо
долговечнее, например воло-
ски у растений из семейства
злаковых, сложноцветных.
Так, у некоторых сложно-
цветных корневые волоски
могут жить в течение 1—2
вегетационных периодов;
при этом стенки волоска
утолщаются и древеснеют.
Вслед за отмиранием во-
лосков отмирает обычно и
соответствующий участок
эпиблемы. Наружным слоем
корня становится верхний
слой клеток первичной ко-
ры — экзодерма. На смену
отмершим волоскам в более
нижней части корня разви-
ваются новые волоски, и та-
ким образом наиболее дея-
тельная, всасывающая зона
корней все время перемеща-
ется вглубь и в стороны
вслед^за растущими кончика-
Рйс. 66; Продольный срез кор-
ня (/) и возникновение вторичного
строения корня (II). А — корневой
чехлик, Б — зона роста и растяже-
ния, В — зона корневых волосков,
'или всасывающая, Г — начало зо-
ны боковых корней:
1— начало роста бокового корня; 2 --
корневые волоски на эпиблеме (3); 4~
кора корня; 5 —эндодерма; 6 — пери-
цикл; 7 — центральный цилиндр; 8 —
клетки первичной коры и эндодерма;
9 — перицикл; 10 — камбий; // — пер-
вичная флоэма; 12 <— первичная кси-
лема
137
ми разветвленной корневой системы. При этом общая всасывающая
поверхность корней, если этому не препятствуют изменившиеся
внешние условия, все время увеличивается.
Стенки клеток экзодермы утолщаются и к моменту отмирания
эпиблемы в большинстве случаев пробковеют, а у некоторых ра-
стений одновременно и древеснеют (например, у некоторых гера-
ниевых). Однако пробковеют не все клетки, среди опробковевших
клеток остаются клетки с целлюлозными или слегка одревеснев-
шими оболочками — это пропускные клетки, через которые почвен-
ные растворы могут свободно проникать в глубь-коры.
Не у всех растений экзодерма хорошо выражена; здесь мы
наблюдаем широкое* варьирование от почти полного отсутствия
какой-либо дифференциации этих слоев до хорошо выраженного
слоя экзодермы с сильно утолщенными и опробковевшими стенка-
ми клеток. В многослойной экзодерме иногда развиваются волокна
склеренхимы. Можно заметить такую закономерность: чем дольше
функционирует первичная кора, тем лучше выражена экзодерма.
У растений, не имеющих экзодермы (например конопля, горох, гре-
чиха и др.), первичная кора очень быстро отмирает и сбрасы-
вается.
Под экзодермой располагается основная паренхима коры — ме-
зодерма. Клетки крупные, особенно в средних слоях, стенки их
целлюлозные, между ними многочисленные межклетники. Здесь
же, в мезодерме, у некоторых растений находятся вместилища вы-
делений, смоляные ходы, кристаллические включения, а у одно-
дольных и волокна склеренхимы. При затрудненности аэрации
в основной паренхиме бывают развиты (например, у осок, некото-
рых злаков, живущих в затопленной почве) крупные воздухонос-
ные ходы.
Эндодерма у большинства растений резко отличается от осталь-
ных слоев первичной коры формой клеток и толщиной клеточных
стенок. На поперечном разрезе клетки эндодермы имеют форму
прямоугольника с закругленными углами. Вначале оболочки кле-
ток эндодермы тонкие, целлюлозные; с возрастом корня стенки
клеток утолщаются и пропитываются лигнином и суберином; этот
процесс начинается в радиальных стенках и затем распространя-
ется на внутренние тангентальные; наружные тангентальные стенки
обычно остаются менее утолщенными. Хорошо заметные на попе-
речном разрезе утолщения радиальных стенок получили название
«пятна Каспари» (см, рис. 65). Особенно сильно утолщаются, дре-
веснеют и пробковеют стенки клеток эндодермы у однодольных
растений. Это связано с тем, что у однодольных растений эндодер-
ма сохраняется на всем протяжении жизни растения. Процесс
пробковения захватывает не все клетки эндодермы. Часть клеток
сохраняет оболочки тонкими и неопробковевшими, эти клетки на-
зываются пропускными. Через них вода с растворенными в ней
веществами свободно проникает в зоне всасывания из первичной
коры внутрь центрального осевого цилиндра, а выше по корню —
в обратном направлении, т. е. из цилиндра в первичную кору;
138
другими словами, через пропускные клетки осуществляется связь
между корой и центральным осевым цилиндром.
Наружный слой клеток центрального цилиндра представляет
перицикл. Реже перицикл состоит из нескольких слоев клеток.
Радиальные стенки клеток перицикла не совпадают с радиальны-
ми стенками клеток эндодермы. Перицикл — тонкостенная парен-
хима, способная к меристематической деятельности. О функции
этой ткани уже было сказано; в частности, здесь закладываются
боковые корни; перицикл участвует также в формировании вто-
ричной структуры корня у двудольных, клетки его, размножаясь,
способны превращаться в камбий, феллоген.
Внутрь от перицикла идет основная паренхимная ткань цент-
рального цилиндра, в которой размещается сложный проводящий
пучок радиального строения; радиально расположенные участки
ксилемы чередуются с 'участками флоэмы, количество радиусов
ксилемы обычно бывает небольшим (2—5), но у некоторых одно-
дольных может достигнуть двадцати и больше. В ксилемных ра-
диусах ближе к периферии цилиндра формируется протоксилема,
ближе к центру — метаксилема; сосуды последней крупнее. В за-
висимости от числа ксилемных радиусов различают корни моно-
архные, диархные, триархные и т. д.
Вторичное строение корня. Вторичные изменения структуры
корня, связанные с деятельностью камбия, свойственны двудольным
растениям. У однодольных это явление встречается очень редко,
лишь у некоторых древовидных форм (у драцены, например).
Процесс вторичных изменений начинается с появления прослоек
камбия под участками первичной флоэмы, внутрь от нее (рис. 66).
Камбий возникает из слабо дифференцированной паренхимы цен-
трального цилиндра. Возникнув в виде ограниченных очагов (при
взгляде на поперечном разрезе), прослойки вторичного камбия
разрастаются по направлению к внешним концам ксилемных ра-
диусов. Здесь они смыкаются с перициклом, который также при-
нимает участие в формировании камбиального кольца (фактиче-
ски цилиндра).
Кольцо это вначале имеет извилистую форму, прогибаясь
внутрь в участках под' флоэмой. Однако довольно быстро кольцо
камбия расправляется и принимает правильную форму; это про-
исходит потому, что возникший камбий начинает энергично откла-
дывать элементы вторичной ксилемы напротив участков первичной
флоэмы. Одновременно под первичную флоэму подслаиваются
(изнутри) элементы вторичной флоэмы, но нарастание флоэмы,
как обычно, происходит медленнее, чем ксилемы.
Вся флоэма, и первичная и вторичная, отодвигается к перифе-
рии центрального цилиндра. В результате деятельности вторич-
ного камбия, таким образом, в промежутках между лучами пер-
вичной ксилемы формируются типичные коллатеральные пучки от-
крытого типа.
Вторичные изменения в структуре корня затрагивают также
периферическую часть центрального цилиндра и первичную кору.
139
Прежде всего изменения происходят в эндодерме; пробковеют тан-
гентальные стенки не только обычных, но и пропускных клеток
эндодермы, в результате чего она превращается в сплошной опроб-
ковевший цилиндр, изолирующий первичную кору от центрального
цилиндра.
В это же время от перицикла отчленяется слой феллогена, из
которого начинает формироваться перидерма, подслаивающаяся
под эндодерму. В результате вся первичная кора отмирает, ссыха-
ется и постепенно сбрасывается; наружным слоем корня становит-
ся перидерма, постоянно возобновляемая1 деятельностью феллоге-
на. Флоэмная часть цилиндра между камбием и перидермой ста-
новится вторичной корой корня.
Видоизменения корней. У корнеплодов верхняя часть главного
корня и гипокотиль разрастаются и становятся мясистыми. Утол-
щение происходит за счет увеличения массы запасающей парен-
химы, так что корнеплод состоит в большей своей массе из основ-
ной. ткани. Механические ткани развиты слабо.
У представителей различных семейств утолщение корней про-
исходит по-разному. У крестоцветных (редька, редиска, турнепс,
репа и др.) сильно разрастается ксилемная часть корня, флоэма
образует лишь тонкий периферический слой (рис. 67).
У зонтичных (морковь, петрушка и др.) утолщение происходит
в основном за счет флоэмы: ксилема образует внутренний более
твердый и волокнистый стержень корня (рис. 68).
Своеобразно происходит утолщение корня у свеклы (рис. 69).
В центральном цилиндре первичного строения у свеклы имеется
два луча ксилемы и два участка флоэмы. Вторичная перестройка
происходит обычным способом: возникает сплошное камбиальное
кольцо, которое и отодвигает участки первичной флоэмы на пери-
ферию центрального цилиндра. Центральная часть цилиндра за-
полняется элементами вторичной ксилемы, затем идет камбий
и кнаружи от него слой вторичной флоэмы; непосредственно к по-
следней примыкают продолговатые участки первичной флоэмы.
Первое камбиальное кольцо существенной роли во вторичном
утолщении корня не играет, деятельность его вскоре прекращает-
ся. Дальнейшее утолщение корня свеклы связано с деятельностью
перицикла. Внутрь перицикл отделяет новый’слой камбия,-который
откладывает паренхиму и в ней сосудисто-волокнистые пучки, рас-
положенные по окружности. Как только между новым слоем кам-
бия и перициклом отложится несколько слоев паренхимы, за счет
деятельности перицикла возникает еще один слой камбия и т. д.
Количество слоев камбия перициклического происхождения может
достичь 10 и даже более, в зависимости от условий питания
корня.
Увеличение числа камбиальных колец происходит быстро и без
существенного утолщения корня. Затем продолжается согласован-
ная работа всех камбиальных слоев, и в результате этой работы
происходит утолщение корня, характерное для свеклы. Степень
утолщений опять-таки зависит от условий питания и состояния
140
Рис. 67. Вторичное строение корня редьки
Рис. 68. Вторичное строение корня моркови
почвы; плотная глинистая почва, например, задерживает разра-
стание корня в толщину.
Установлено, что между деятельностью камбия и развитием
листьев имеется связь; число листьев в розетке кратно числу кам-
биальных колец.
Своеобразные изменения происходят в корнях, вступающих
в сожительство с низшими гетеротрофными организмами — бакте-
риями и грибами.
Клубеньковые бактерии {Bacterium radicicola'), поселяющиеся
на корнях бобовых растений и способные усваивать молекулярный
азот, проникая в коровую паренхиму корня, производят раздраже-
ния, стимулирующие размножение клеток перицикла. В результате
деятельности перицикла и интенсивного деления клеток флоэмной
паренхимы происходит мощное разрастание вторичной коры; об-
разуется опухоль—клубенек, состоящий в основном из паренхи-
мы, богатой крахмалом (рис. 70).
142
Рис. 69. Строение корня свеклы
Внутри клубенька разрастается бактероидная ткань — скопле-
ние большого количества азотусваивающих бактерий. Снаружи
он, как и сам корень, покрыт перидермой. Питание клубенька во-
дой и растворёнными в ней веществами осуществляется посредст-
вом ответвлений проводящих пучков, отходящих от проводящей
системы корня.
Бактерии получают от высшего растения органическое и мине-
ральное питание; в свою очередь высшее растение использует сое-
динения азота, синтезируемые бактериями. Такое обоюдополезное
сожительство двух организмов, как известно, называется сим-
биозом.
443
Корни многих высших растений находятся в симбиозе с гриба-
ми, образуя микоризу (рис. 71). Анатомическая картина такого
сожительства может быть различной. В одних случаях гифы (ни-
ти) грибов оплетают с поверхности тоненькие корешки высшего
растения, образуя более или менее плотный чехлик. Кончики гифов
проникают лишь в самые поверхностные слои корня. Такая мико-
риза называется наружной, или эктотрофной. Гифы эктотрофной
микоризы заменяют корневые волоски, поэтому в большинстве
Рис. 70. Клубенек на корне люпина
случаев последние не образуются. Гифы грибов могут проникать
также и в глубокие слои корневой коры, образуя в клетках парен-
химы древовидные разветвления или сплетения в виде клубочков.
Такая микориза называется внутренней, или эндотрофной. Встре-
чается также микориза и смешанная, наружно-внутренняя или
экто-эндотрофная. В том случае, когда гифы гриба только окру-
жают корень, совершенно не проникая внутрь его, микоризу назы-
вают перитрофной.
Корни, участвующие в образовании микоризы, обычно более
усиленно ветвятся, и кончики их несколько утолщаются. Гифы
гриба, проникшие внутрь живых клеток корня, постепенно перева-
риваются в них.
Микориза — весьма распространенное в природе явление. На-
пример, эктотрофная, а иногда и экто-эндотрофная микориза име-
ется у березы, дуба, клена, орешника, липы, осины, сливы, груши
и других деревьев и кустарников. Эндотрофная микориза встреча-
ется у таких древесных, как грецкий орех, виноград, шелковица
и др. Не менее широко распространена микориза у травянистых
растений. Эндотрофная микориза найдена у лисохвоста, костра,
144
овсяницы, орхидеи, клевера, донника, люцерны и у многих других
растений. Эктотрофная микориза имеется у пшеницы, проса, льна-
кудряша и др.
Микориза — явление древнего происхождения. На протяжении
сотен миллионов лет многие высшие растения эволюционировали
в тесном симбиозе с грибами. Древность этого явления говорит,
о весьма важной роли каждого симбионта в совместном существо-
вании. Однако до сих пор сущность микоризы, роль одного симби-
онта в жизни другого недостаточно выяснена. По-видимому, гриб
и
Рис. 71. Эктотрофная микориза дуба
с его ферментами помогает высшему растению превращать в более
легко усвояемую форму как вещества почвы, так и запасные ве-
щества самого растения. Установлено, что гриб снабжает высшие
растения витаминами (Bi и др.), стимулирующими рост корней;
некоторые грибы могут фиксировать молекулярный азот, и соеди-
нения его затем становятся доступными для высшего растения.
О роли эктотрофной микоризы, заменяющей корневые волоски,
уже говорилось. Некоторые растения, например береза, вовсе не
имеют корневых волосков и, следовательно, не могут существовать
без сожительства с грибом. Высшие растения в свою очередь
снабжают гриб органической пищей, освобождающейся в процессе
изнашивания тканей корня.
Известны и другие метаморфозы корня: он может превращаться
в колючки (у некоторых лиан), воздушные корни эпифитов содер-
жат аэренхиму и могут впитывать атмосферную влагу; в тропиках,
на заболоченных почвах на концах корней образуются особые пнев-
145
матофоры, которые, вырастая наружу, снабжают корни воздухом;
очень широко распространены корни-присоски и т. д.
Всасывающая деятельность корня. Посредством корня растения
получают из почвы воду и элементы почвенного питания. В течение
своей жизни даже сравнительно небольшое растение потребляет
огромное количество воды. Для того чтобы создать только один
грамм сухого вещества, растение должно испарить несколько сот
граммов воды. Одно растение кукурузы, например, испаряет за
вегетационный период до 200 кг воды. И вот эту воду корневая
система должна отнять у почвы. Для того чтобы иметь правильное
представление об этой работе, надо знать, в каком состоянии вода
находится в почве.
При достаточном насыщении почвы водой большая часть ее на-
ходится в промежутках между частицами мелкозема. Та часть ее,
которая заполняет крупные почвенные капилляры и подчиняется
силе тяжести, называется гравитационной водой; мелкие капилля-
ры, где уже действуют силы поверхностного натяжения, заполне-
ны капиллярной водой. И гравитационная, и капиллярная вода
легко забираются корнями растений.
С большим трудом растение может отнять пленочную воду,
облекающую тонким слоем каждую частицу почвы. Чем тоньше
становится ^та пленка, тем сильнее она удерживается на поверх-
ности частиц почвы силами молекулярного притяжения. И совер-
шенно недоступна для растений гигроскопическая вода, которая
удерживается почвой даже при длительном ее высушивании. Чем
мельче частицы почвы, тем больше удерживается в ней такой воды.
Так, например, в песчаной почве гигроскопической воды остается
около 1%, а в тяжелых глинистых почвах количество ее возраста-
ет до 10% и более.
Таким образом, в почве остается мертвый запас воды, не до-
ступный растениям. Эта величина резко изменяется в зависимости
от механического состава почвы и всегда превышает гигроскопиче-
скую влажность (в 1,2—2,0 раза).
Одна из причин, обусловливающих постоянный ток воды в ра-
стениях снизу вверх, это убыль воды в процессе испарения —
транспирации. Создается недостаток насыщения, влекущий за со-
бой пассивное засасывание воды корнями растений. Однако это
не единственная причина. Можно сделать следующий простой опыт.
Срезать всю надземную часть растения и на оставшийся пенек
надеть трубку с манометром. Поступающий из корня водный раст-
вор заполнит трубку и поднимет на определенную высоту ртуть
манометра. По разности в уровнях ртутного столба можно судить
о силе так называемого корневого давления, т. е. силы, с которой
корень активно отбирает воду от почвы и проталкивает ее вверх
к стеблю; она может достигать величины 1—2 атм и более.
Выделение с кончиков листьев капельно-жидкой воды можно
наблюдать и в естественных условиях у многих растений (карто-
феля, гречихи, декоративного растения настурции, ивы ломкой,
у комнатного растения Arum и др.). Это явление, как уже было
146
сказано, получило название гуттации. Выделение воды из перере-
занных сосудов растения называется плачем растения.
Следовательно, корень активно забирает воду из почвы и про-
талкивает ее вверх по сосудам. Это сложный процесс, связанный
с жизнедеятельностью клеток и свойствами цитоплазменных мем-
бран. Необходимым условием всасывания воды корнем является
разность концентрации раствора в почве и сосудах растения; ра-
стение может забирать воду из почвы, если концентрация почвен-
ного раствора ниже, чем в сосудах. Поэтому-то большинство ра-
стений не может существовать на засоленных почвах; такие почвы
становятся физиологически сухими, так как находящаяся в них
вода недоступна растению.
Физиологически сухой может быть также и холодная почва.
Для каждого растения существует минимальная температура, при
которой прекращается поступление воды в корень, даже если она
имеется в избытке. Лишь немногие растения приспособились жить
на почвах с низкими температурами; это растения Крайнего Севера
и высоких гор. Явлением физиологической сухости объясняется
морфологическое сходство растений, обитающих на Крайнем Се-
вере и в альпийской зоне с растениями сухих степей и полупу-
стынь.
Поскольку процесс активного поглощения воды корнями связан
с обменом веществ в клетках корня, постольку он может протекать
только при доступе кислорода, необходимого для дыхания. Поэто-
му длительное затопление почвы вызывает угнетение и даже ги-
бель растений. Нарушение процесса дыхания тормозит поглощение
воды корнями растений. Если, например, подействовать на корень
растения наркотическими веществами, угнетающими дыхание,
плач растения прекращается.
Большие'трудности представляет изучение механизма переме-
щения воды и органических веществ по растению. Согласно разра-
ботанным гипотезам (Сабинин, Курсанов и др.), одностороннее пе-
ремещение воды по живым клеткам паренхимы и ситовидных тру-
бок может быть объяснено различиями в проницаемости и харак-
тере обмена веществ в клетках. Если проницаемость цитоплазмы
выше у стенок, ориентированных к центру корня, это должно по-
влечь перемещение растворов от периферии корня к центру, т. е.
к сосудам. Неодинаковость в типе обмена веществ в массе прото-
пласта может привести к полярным различиям свойств цитоплаз-
мы, а это вызовет одностороннее перемещение растворов мине-
ральных и органических веществ. При этом следует иметь в виду,
что хотя проводящие элементы ксилемы предназначены для про-
движения растворов из корней в листья, а флоэмы — в обратном
направлении, в отдельные моменты жизни растения возможны
перемещения и в противоположных направлениях; детали этих
перемещений исследованы еще далеко не полно.
Перемещение воды по сосудам связывают (Вотчал, Уршпрунг
и др.) с силой сцепления молекул воды, которая может достигать
200—300 атм и более. Эта сила способна обеспечить подъем воды
147
по сосудам на большую высоту при наличии непрерывных водных
нитей от кбрней до листьев. Ток воды по растению обеспечивается
совокупным действием всех указанных сил. Скорость перемещения
воды может достигать 1—2 и более метров в час. Перемещение
органических веществ связано с комплексом процессов обмена ве-
ществ в растении.
Корневое питание растений. Через корень идет снабжение расте-
ний элементами азотистого и минерального питания.
Азот входит в состав органического вещества и улетучивается
при сжигании растений, элементы минерального питания остаются
в несгоревшей золе. Анализ золы многих растений показывает, что
в общем счете в золе могут встретиться почти все элементы, кото-,
рые мы находим в земной коре. Это, однако, еще не означает, что
все найденные элементы действительно нужны растению. В необ-
ходимости для растения того или иного элемента можно убедиться
на специально поставленных опытах.
.Все элементы, входящие в состав растения, принято делить,
исходя из степени их массового участия, на две группы: макро-
элементы и микроэлементы.
Роль того или иного элемента не определяется количеством его
в составе растения; например, железо встречается в растениях
в ничтожных количествах и все же отсутствие его ведет к гибели
растения; наоборот, натрия может быть в золе растения очень
много, тем не менее полное исключение его из раствора не вредит
растению.
Вопрос о том, какие элементы почвенного питания необходимы
растению, изучается уже около ста лет методом водных культур.
Сущность его заключается в том, что растения выращивают на
растворах смеси солей и, судя по тому, какой эффект вызывает
отсутствие в растворе того или иного элемента, решают вопрос
о его роли в жизни растений.
В подобных опытах еще в конце прошлого столетия было ус-
тановлено, что для нормального роста и развития растений необ-
ходимы следующие семь элементов, получаемых растением из поч-
вы: К, Са, Mg, Fe, S, Р и N. Вместе с С, получаемым из воздуха,
Н и О, входящими в состав воды, это составит десять элементов,
которые длительное .время считались необходимыми и достаточ-
ными для нормального существования растений.
Начало использованию метода водных культур было положено
в середине прошлого столетия немецким ученым Кнопом. Раствор,
предложенный им, и в настоящее время используется, под названи-
ем кноповского раствора. Этот раствор на 1 л воды содержит 1 г
Ca(NO3)2; 0,25 г КН2РО4; 0,25 г MgSO4; 0,125 г КС1 и следы
Fe2O3.
Позднее были предложены и другие рецепты. Один из них был
составлен советским ученым Д. Н. Прянишниковым, длительное
время изучавшим питание растений методом водных культур и усо-
вершенствовавшим этот метод. Д. Н. Прянишников предложил
смесь следующего состава: на 1 л воды —0,24 г NH4NO3, 0,172 г
148
СаНРО4-2Н2О; 0,16 г КС1; 0,344 г CaSO4-2H2O; 0,025 г Fe2Cl6<
•6Н2О; 0,06 г MgSO4.
Подбирая состав солей для питательных растворов, приходится-
учитывать избирательную способность корня, который может
в большем количестве выбирать из данного раствора одни катионы
и анионы и в меньшей степени — другие. В результате может изме-
ниться реакция раствора и из нейтральной сделаться кислой или
щелочной, вредной для растения.
Так, например, из раствора соли (NH4)2SO4 растение будет
энергично забирать катионы аммония и в гораздо меньшем коли-
честве анионы серной кислоты, которые, накопляясь в растворе^
будут подкислять его. Наличие в растворе соли NaNO3 приводит
к подщелачиванию его, так как растения будут забирать анион
ЫОз, почти не трогая катион натрия. Первая соль в данном случае
будет выступать в роли физиологически кислой, вторая — физиоло-
гически щелочной. Чтобы избежать вредного влияния избытка ки-
слоты или щелочи, следует время от времени менять раствор,,
в котором выращиваются растения.
Постепенно, в меру усовершенствования методики исследования
и в связи с применением химически чистых солей, стали накапли-
ваться факты, указывающие на необходимость для жизни растений
все новых элементов, нужных растениям в ничтожно малых коли-
чествах, так называемых микроэлементов. В настоящее время уже-
доказана необходимость для нормального роста растений марган-
ца, цинка, меди, бора, алюминия, йода, фтора, молибдена. В связи
с этим в питательные растворы вводят наряду с макроэлементами
соли, содержащие в своем составе и микроэлементы. Выявляется-
необходимость и ряда других микроэлементов. Потребность в мик-
роэлементах бывает настолько малой, что она удовлетворяется
запасами, отложенными растением в семени.
Некоторые элементы, например кремний, хлор, хотя и встре-
чаются в составе растений в больших количествах, однако для
жизни растений они необходимы лишь- в ничтожных количествах;
поэтому их тоже следует отнести к микроэлементам.
Остановимся на роли отдельных элементов почвенного пита-
ния.
Азот необходим растению как составная часть белка — основы
живого вещества.
Сера и фосфор также входят в состав некоторых белков. Ог-
ромную роль соединения фосфора играют в обмене белков, жиров
и углеводов, в процессах фотосинтеза и дыхания. Сера входит-
в состав витамина Bi, играющего большую роль в процессах обме-
на веществ растений и животных.
Калий в виде катиона принимает активное участие в жизне-
деятельности цитоплазмы, способствует большей ее гидратации
и подвижности. Он также, по-видимому, играет существенную роль,
в деятельности ферментов.
Магний играет роль, сходную с калием; кроме того, он входит
в состав молекулы хлорофилла.
149
Кальций уравновешивает действие калия, выступая в роли ан-
тагониста. В противоположность калию кальций уплотняет цито-
плазму, уменьшает ее гидратацию и делает менее подвижцой, уве-
личивает вязкость. С возрастом клеток в них увеличивается
содержание кальция за счет уменьшения калия. Кальций нейтра-
лизует ядовитую щавелевую кислоту, образуя в отмирающих ча-
стях растений кристаллы щавелевокислого кальция.
Железо входит в состав окислительных ферментов, играющих
•существенную роль в процессах дыхания; оно участвует в обра-
зовании хлорофилла. В отсутствие железа растение заболевает
хлорозом/, желтеет и умирает от истощения ввиду отсутствия хло-
рофилла.
Менее выяснена специфическая роль микроэлементов.
Марганец, входящий в состав некоторых ферментов, также
необходим для нормального роста растений. Марганцевые удобре-
ния повышают урожай и содержание сахара в корнях сахарной
свеклы.
Бор благоприятно влияет на поступление и передвижение мине-
ральных веществ в растениях. Установлено положительное влияние
его на цветение и плодоношение, поэтому борные удобрения при-
меняют, например, для увеличения урожая семян многолетних
трав. В последние годы установлена разносторонняя роль бора
в процессе обмена веществ у растения: бор необходим для нор-
мальной активности апикальной меристемы и усвоения кальция,
регулирует поступление кислорода в корни и др.
Цинк совершенно необходим при создании главнейших фер-
ментных систем, и недостаток его резко сказывается на росте.
Медь необходима для правильного развития растений. Дока-
зано положительное влияние медных удобрений на торфянистых
почвах.
Молибден необходим для нормальной работы листового аппа-
рата и развития меристемы. Отсутствие его нарушает также азот-
ный и водный обмен в растениях. Молибден также необходим для
работы бактерий — азотфиксаторов.
В науке накапливается все больше фактов, говорящих об ог-
ромной и всесторонней роли микроэлементов в жизни растений.
Особое место в питании растений занимает азот — обязатель-
ная составная часть белка, основы живого вещества. Азота в при-
роде очень много: около 80% атмосферного воздуха составляет
молекулярный азот, но, за исключением небольшой группы микро-
организмов, он не может непосредственно использоваться расте-
ниями.
В атмосфере имеется также небольшое количество связанного
азота. Это главным образом окислы азота, образующиеся в ре-
зультате электрических разрядов, и аммиак, выделяющийся при
разложении остатков растений и животных. Эти соединения могут
усваиваться листьями и корнями растений, но содержание их
в атмосфере настолько ничтожно, что практического значения в пи-
тании растений они не имеют. Поэтому основным источником азо-
150
тистого питания, непосредственно используемым растениями, слу-
жат соединения азота, находящиеся в почве. Азотистые вещества,
находящиеся в почве, делят на три группы: 1) соли азотной и азо-
тистой кислот, 2) соли аммиака и 3) органические соединения.
Общее количество этих веществ по отношению к сухой массе поч-
вы обычно измеряется долями процента.
Многочисленными исследованиями, среди которых следует осо-
бенно отметить долголетние опыты Д. Н. Прянишникова, доказа-
но, что азот нитратов и солей аммиака высшие растения усваива-
ют одинаково хорошо, т. е. корни растений поглощают и анионы
ЫОз, и катионы NH4. Из органических соединений азота растения-
ми усваиваются только те, которые растворимы в воде: аспарагин,
мочевина, очень слабо — аминокислоты.
Вместе с тем известно, что в почве основная масса азота на-
ходится именно в составе органических веществ, и о богатстве поч-
вы обычно судят по количеству в ней гумуса — сложного вещества,
накопляющегося в почве в результате разложения отмерших ра-
стительных и животных организмов.
Высшие растения используют эти соединения лишь при посред-
стве бактерий, переводящих органические соединения азота в соли
аммиака и азотной кислоты. Эта работа осуществляется несколь-
кими группами бактерий. Гнилостные бактерии расщепляют белки
до аммиака, а нитрифицирующие окисляют аммиак до азотной кис-
лоты, которая, соединяясь со щелочами, дает соли азотной кисло-
ты— нитраты. Нитрифицирующие бактерии, в отличие от боль-
шинства представителей этой группы организмов, могут самостоя-
тельно создавать органическое вещество, используя энергию, осво-
бождающуюся в ходе химических реакций. Этот процесс, в отли-
чие от фотосинтеза, называется хемосинтезом.
В природе имеются также бактерии, усваивающие свободный
молекулярный азот воздуха. Одна такая группа живет в симбиозе
с высшими растениями: это клубеньковые бактерии, обитающие
в корнях бобовых растений, о чем говорилось выше. Синтезируе-
мые клубеньковыми бактериями азотистые вещества используются
высшими растениями. Отмирая, бактерии обогащают почву сое-
динениями азота, которые, после их минерализации, также всасы-
ваются корнями растений.
В почве имеются также и свободноживущие бактерии, усваи-
вающие молекулярный азот, так называемые бактерии-азотсоби-
ратели. Это, во-первых, широко распространенный Azotobacter
chroococcum и, во-вторых, Clostridium pasteurianum. Азотобактер
усваивает азот в присутствии кислорода, т. е. это аэробная бакте-
рия. Клостридиум относится' к группе анаэробных бактерий, раз-
вивающихся в бескислородной среде.
Польза, приносимая азотусваивающими бактериями, огромна:
они могут ежегодно накапливать в почве на каждый гектар до
10—20 кг связанного азота. Особенно велика их роль в жарких
странах, где на почвах, бедных азотом, в течение многих лет мож-
но получать высокий урожай сельскохозяйственных культур без
151
внесения азотистых удобрений. Практикуется внесение в почву
препаратов азотобактера — азотобактерина, фосфоробактерина,
а также препаратов силикатных бактерий. Фосфоробактерин со-
держит бактерии, минерализующие органические соединения фос-
форной кислоты. Силикатные бактерии разрушают содержащиеся
в почве калийные силикаты и тем самым улучшают условия ка-
лийного питания. Рекомендуется также внесение гуминовых удоб-
рений, оказывающих стимулирующее влияние на многие стороны
•обмена веществ в растении.
Помимо бактерий, играющих специфическую роль в снабжении
растений необходимыми питательными веществами, огромное зна-
чение для корневого питания растений имеет колоссальная масса
микроорганизмов различных систематических групп, населяющих
почву: бактерий, грибов, актиномицетов, водорослей. Общая био-
масса этих организмов, накапливающаяся в пахотном слое почвы,
достигает 10—20 т на 1 га. В результате жизнедеятельности мик-
роорганизмов в почве накапливается большое количество различ-
ных витаминов (группы В, пантотеновой, никотиновой, фолиевой
кислот и др.) и других биологически активных веществ, которые
могут благотворно влиять на жизнь высших растений.
Роль корней не ограничивается всасыванием готовых органиче-
ских соединений. В последние годы все более накапливается фак-
тов, указывающих на важную синтезирующую роль корней. Ис-
следованиями Курсанова, Колосова, Литвинова, Потапова и др.
доказано, что используя углекислоту, имеющуюся в почве, и амми-
ак, корни могут синтезировать аминокислоты, а поглощая мине-
ральный фосфор, корни могут синтезировать нуклеопротеиды
и другие органические соединения фосфора. По-видимому, корни
способны синтезировать многие жизненно важные органические
соединения. Подобно листьям корни, экспонированные на свету,
.могут синтезировать хлорофилл (Рубин и др.).
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ
ВЫСШИХ РАСТЕНИИ
Высшие семенные растения обычно размножаются при помощи
семян. Однако новое растение может возникнуть также из любого
вегетативного органа или его части. Такое размножение называет-
ся вегетативным, оно основано на регенерации части растения
в новый индивид.
Вегетативное размножение, как мы уже знаем, широко рас-
пространено в природе и свойственно не только высшим, но и низ-t
шим растениям. Однако для нас наибольший интерес представля-
ют высшие семенные растения.
В естественных условиях и в сельскохозяйственной практике
вегетативное размножение имеет большое значение. При вегета-
тивном размножении растение может сохраниться в тех случаях,
.когда условия препятствуют семенному возобновлению. Кроме то-
152
го, особи, полученные вегетативно, более точно воспроизводят-
свойства и признаки материнского растения, чем это бывает при
размножении семенами.' Поэтому в практике разведения растений
нередко прибегают к вегетативному размножению.
И. В. Мичурин широко использовал прививки для продвижения-
на север ценных пород плодовых. Он изучал явления взаимодей-
ствия подвоя и привоя, использовал установленные закономерно-^
сти в практических целях для создания новых сортов плодовых де-
ревьев и кустарников.
В природе существуют разнообразные формы вегетативного'
размножения. В практике разведения растений человек использует
не только эти формы, но и разрабатывает новые приемы.
Размножение выводковыми почками. Некоторые растения обла5'
дают способностью развивать почки, которые, опадая на почву или
в воду, дают начало новому растению. Почка представляет собой'
зачаточный побег; следовательно, размножение при помощи вы-
водковых почек мы можем рассматривать как одну из форм раз-
множения при помощи побегов.
Выводковыми почками размножаются мятлик луковичный,,
у которого почки образуются в соцветии, некоторые камнеломки,
(Saxifraga nivalis, S. сегпиа), очиток (Sedum villosum) и другие
дикорастущие растения. В комнатных условиях часто разводят два,
вида тропического растения бриофиллума (Bryophyllum), образу-
ющего выводковые почки на листьях. Этим способом их размно-
жают и у нас, и притом очень быстро, так как каждое растение-
может дать сотни выводковых почек. Как уже сказано, выводко-
выми почками ,могут размножаться и споровые растения — мхи..
Растения, размножающиеся при помощи выводковых почек,,
называются живородящими, так как почка может сразу же, без
периода покоя, прорасти в новое растение.
Размножение при помощи укореняющихся побегов. Многие ра-
стения образуют побеги, предназначенные для размножения. На-
пример, земляника и клубника, наряду с укороченными побегами,
образуют длинные ползучие побеги, лишенные развитых листьев,
так называемые усы.
На их }гзлах при соприкосновении с влажной почвой образу-
ются побеги, развивающие собственную корневую систему. Каж-
дый ус может дать несколько таких побегов, расположенных на
значительном расстоянии друг от друга (до 10 см и больше).
После того как дочерние побеги укоренились, плети, соединяющие
их, перегнивают и таким образом количество индивидов увеличи-
вается.
В практике садоводства Этот процесс размножения ускоряют,
рассаживая растения пореже и разделяя укоренившиеся дочерние
побеги искусственно, перерезая усы. Укореняться могут не только
удлиненные ползучие побеги (плети, усы), но и обычные. На этом-
свойстве основано размножение растений отводками. Некоторые
деревья и кустарники (например, пихта, скумпия, рододендрон
и др.) образуют ветви, часто лежащие прямо на земле. При доста-
153
точной влажности на узлах этих ветвей образуются придаточные
корни, а из почек развиваются побеги.
В условиях культуры такой укоренившийся побег разрезают на
части по числу сформировавшихся новых побегов и рассаживают
как самостоятельные растения — отводки. Отводками размножают
крыжовник, шелковицу (тутовое дерево), смородину и другие ра-
стения.
Размножение при помощи корневищ. От идущих под землей
корневищ на некотором расстоянии от материнского растения от-
ходят надземные побеги (см. рис. 45). Рано или поздно старые
корневища перегнивают, и тогда возникает несколько вполне са-
мостоятельных индивидов.
9 При благоприятных условиях растения, имеющие длинные кор-
невища, могут очень быстро расселяться, захватывая все новые
и новые площади. Размножающийся таким способом пырей ползу-
чий при возделывании почвы может стать очень опасным, трудно
искоренимым сорняком. В культуре при помощи корневищ раз-
множают ландыш, ирисы и некоторые другие растения.
Размножение корневой порослью, или корневыми отпрысками.
Многие деревянистые и травянистые растения могут давать над-
земные побеги из горизонтально распространяющихся под землей
корней. Таким свойством обладают, например, из древесных —
вишня, слива, малина, ежевика, осина, тополь, ивы, сирень, белая
акация и другие, из травянистых — осот желтый (Sonchus) и лило-
вый (Cirsium arvense), горчак (Acroptilon picris), вьюнок и др.
С сорняками, размножающимися таким образом, очень трудно
бороться, так как после уничтожения надземной части растения из
его корней возникают новые побеги. При разрезании корней на ча-
сти процесс размножения сорняка только усиливается, из каждой
части возникает новое растение.
Корневой порослью часто размножаются деревья и кустарники
в практике лесоводства и садоводства. Например, это обычный
способ размножения сирени, малины, золотистой смородины.
Подобно корневой может возникать пневая поросль. Это проис-
ходит, обычно, когда дерево срезано или сильно повреждено. У ос-
нования ствола из спящих или придаточных почек «появляются
новые побеги. Вначале они живут за счет материнского растения,
а потом развивают и собственную корневую систему, а корневая
система материнского растения отмирает.
При помощи пневой поросли возобновляют дубовые леса, а так-
же восстанавливают в культуре айлант {Ailanthus altissima), гле-
дичию, каталъпу (Catalpa bignonioides) и другие породы деревьев.
Размножение клубнями и луковицами. Клубни — это сильно ме-
таморфизированные стебли (у картофеля, топинамбура) или корни
(георгина, батата), луковица — сильно видоизмененный побег.
Клубнями стеблевого происхождения размножают картофель.
Можно использовать и часть клубня, лишь бы на ней был хотя бы
один, глазок. Экономически выгодно иногда размножать картофель
верхушками клубня, т. е. морфологически верхней его частью, на
154
которой больше глазков. Остальная большая часть клубня исполь-
зуется на продовольствие.
Размножение луковицами основано на том, что взамен отмер-
шей материнской луковицы обычно образуется несколько дочерних.
В природе дочерние луковицы сидят сближенно, в культуре моло-
дые луковицы выкапывают и рассаживают. Луковицами размно-
жают лук, чеснок, лилии, тюльпаны, гиацинты, нарциссы и некото-
рые другие растения.
Размножение черенками. Побеги очень многих растений, отре-
занные и отломленные от материнского растения, могут укоре-
няться и давать начало новому растению. У некоторых растений
это происходит без вмешательства человека. Например, ветви ивы
ломкой (Salix fragilis) легко обламываются ветром, падая на
влажный песчаный субстрат (ива растет обычно вблизи рек, озер,
прудов и т. д.), обломленные ветви на морфологически нижнем
конце образуют корни и продолжают расти, образуя вокруг дерева
молодую поросль.
Вегетативное размножение растений отрезками побега очень-
распространено в культуре. Части побегов, специально заготовлен-
ные для размножения, называются черенками, черенки винограда
называют также чубуками. Черенки могут быть не только стебле-
вого происхождения. Некоторые растения размножают отрезками
корня, н-апример малину, сливу, вишню, айву, розы и др. Отрезки
корня называют корневыми черенками. И сравнительно немногие
растения (некоторые виды бегонии, колеусы, глоксинии и др.)
могут восстанавливаться из отрезанных листьев, листовых че-
ренков.
Все же размножение отрезками корня или листьями встречается
редко, поэтому черенком обычно называют отрезок стебля, несу-
щий пазушные или спящие почки и образующий придаточные кор-
ни. Черенками размножают очень многие деревья и кустарники.
Черенки, взятые после опадания листьев, называются зимними,
а черенки облиственные называются летними.
В черенке различают морфологически нижний, или базальный,
конец, и верхний — апикальный. Физиологически эти концы раз-
личны: на нижнем конце черенка образуются корни, на верхнем —
побеги. Поэтому нижний конец черенка называют корнеродным,
верхний — побегообразующим. Явление физиологической неравно-
значности черенка по длине называется полярностью. Полярность
наиболее отчетливо проявляется у молодых побегов, с возрастом
она затухает, ослабевает. Причина полярности, по-видимому, за-
ключается в неравномерном распределении ростовых веществ по
длине побега. Подвергая черенок центрифугированию или одно-
стороннему освещению, можно изменить его полярность. При по-
садке черенков свойство полярности учитывают и сажают черенки
базальным концом вниз.
Стеблевыми черенками размножают не только многолетние де-
ревянистые растения, но и травянистые однолетние и многолетние
растения (томаты, картофель, баклажаны, тыквенные, а также
155
цветочные многолетние культуры: пеларгонию, гвоздику и др.).
Размножение черенками — один из, наиболее эффективных спо-
собов вегетативного размножения, позволяющий быстро увеличи-
вать количество растений, однако до недавнего времени примене-
ние его ограничивалось неспособностью многих растений давать
придаточные корни. В настоящее время в качестве стимуляторов
роста применяют гетероауксин (бета-индолил-уксусная кислота)
и другие ростовые вещества, которые позволяют укоренить черенок
почти любого растения.
Прежде чем произойдет укоренение черенка, на концах его при
благоприятных условиях температуры и влажности образуются за-
метные на глаз наросты — мозолистое тело, или каллюс. Каллюс
состоит из живых паренхимных клеток, богатых питательными ве-
ществами, преимущественно углеводами. Питательные вещества
расходуются образующимися в каллюсе придаточными корнями
и почками до тех пор, пока в них не разовьется своя проводящая
система, соединяющаяся с питательной системой самого черенка.
Каллюс возникает из клеток меристемы и паренхимной ткани,
«в какой бы части черенка эти ткани ни находились. Легче всего
каллюс образуется из меристемы. Являясь по структуре своей ос-
новной запасающей тканью, каллюс легко производит вторичную
меристему, из которой и развиваются корни и почки. На поверх-
ности наплыв каллюса бывает покрыт несколькими слоями мелких
клеток, играющих роль покровной ткани, стенки этих клеток даже
могут пробковеть.
Прививка растений. В практике садоводства широко использу-
'етс’я метод размножения растений путем прививок. Этот метод
заключается в пересадке частей одного или нескольких растений
на другое растение, имеющее корневую систему. Обычно в этой
операции участвуют два растения:* одно из них, имеющее корневую
систему,1 называется подвоем, второе, которое сращивают с подво-
ем,— привоем. В результате трансплантации и сращивания воз-
никает единое растение, части которого имеют различное проис-
хождение. В садоводческой практике прививка делается-для того,
чтобы в одном растении сочетать лучшую выносливость и приспо-
собленность к данным условиям подвоя с ценными качествами
прививаемого растения. Поэтому в качестве подвоя чаще берут
дичок, в качестве привоя — какой-нибудь культурный сорт.
Существуют различные способы прививок. Наиболее распрост-
ранена прививка почкой, или глазком,— окулировка. Нередко для
прививки используют черенки, прививка черенками называется
копулировкой. Сравнительно редко применяют метод сближения
или аблактировку. Окулировку производят в конце лета с тем,
чтобы до конца вегетационного периода глазок (почка) сросся
с подвоем. Перед окулировкой заготовляют отрезки одревесневших
побегов, на которых листья срезают, оставляя лишь черешок, иног-
да с небольшим кусочком листовой пластинки. Эти побеги назы-
ваются окулянтами. Окулировку проделывают специальным оку-
лировочным ножом с острым и тупым лезвиями. Острым лезвием
156
на небольшом расстоянии от почвы делают Т-образный разрез на
стволике подвоя, тупым лезвием отодвигают кору, срезают глазок
вместе с плоским кусочком древесины и вставляют его под кору
Т-образного разреза. Затем на место операции накладывают плот-
ную повязку. Через 10—15 дней глазок с подвоем срастаются. При-
вивку черенков, или копулировку, можно производить по-разному.
При одинаковой толщине подвоя и привоя на них делают косые
срезы, прикладывают их друг к другу поверхностями среза и на-
кладывают повязку. Для более прочного соединения на срезанной
поверхности делают острый выступ — язычок: заклиниваясь, язы-
чок хорошо скрепляет подвой с привоем. Если приходится приви-
вать черенок на ветвь большего диаметра, прививают в расщеп
или под кору. Широко применяемые в последнее время прививки
травянистых растений (томаты, паслен, табак, тыквенные и др.)
делают в расщеп. Злаки обычно прививают зародышем зерновки
привоя в эндосперм подвоя, а также в корневую шейку. (
Аблактировкой пользуются чаще в экспериментальной обста-
новке. Для этого нужно, чтобы соединяемые растения росли рядом.
На сближаемых поверхностях делают одинаковые по длине срезы
коры, срезанные поверхности взаимно прикладывают и место при-
вивки плотно забинтовывают мочалом. В таком состоянии расте-
ния находятся все лето и зиму. Делая прививку, необходимо об-
ратить внимание на то, чтобы у привоя на стороне, противополож-
ной срезу, обязательно находилась почка. На следующий грд по-
вязку снимают и часть растения-привоя ниже места сращения сре-
зают; у подвоя, наоборот, срезают верхнюю часть выше того места,
где произошло срастание. Аблактировкой пользуются, когда не-
обходимо срастить растение, с трудом поддающееся прививке.
Культура растительных клеток и тканей. Вегетативное воспроиз-
ведение высшего растения может начаться с одной клетки, взятой из
любой уже специализированной ткани. Выращивание делящихся
клеток вне организма показывает, что таким путем можно получить
растения со всеми присущими им органами: корнями, стеблями,
листьями и цветками. Такое воспроизведение растений связано
с образованием каллюса, причем началу каллюса может дать лю-
бая клетка растения, даже такая сугубо специализированная, как
клетка пыльцы и клетка механической ткани — колленхимы. Кал-
люсные клетки можно выращивать на питательных средах, вне
организма, неопределенно долго, периодически освежая, «меняя»
среду. Выросшие из каллюса ткани и целые растения обладают
в полной мере видовой специфичностью; ткани лекарственных ра-
стений, выращенные таким путем, синтезируют и выделяют в пи-
тательную среду соответствующие полезные вещества, причем да-
же £ большей концентрации, чем это делает целое растение в ес-
тественных условиях. Следовательно, каждая клетка растительно-
го организма в потенциальном состоянии содержит все наследст-
венные задатки (геном), необходимые для воспроизведения целого
организма. На этом, собственно, и основаны все приемы вегетатив-
ного размножения растений.
ГЛАВА III
СИСТЕМАТИКА РАСТЕНИИ
Потребность в классификации растений возникла в связи
с практикой их использования. В наиболее древних описаниях
растений мы уже видим попытку сгруппировать их по характеру
использования. Отец ботаники Феофраст описал около 500 видов
растений, среди которых он выделял деревья, кустарники, кустар-
нички, травы. Описывая растения, Феофрцст обращал внимание
на связь растений с окружающей средой: почвой, климатом, место-
расположением и указывал на характер использования растений,
подразделяя их на лекарственные/ пищевые, медоносные, плодо-
вые, овощные и т. д. Эта работа была продолжена в трудах рим-
ских писателей: Плиния Старшего, который описал уже до 1000
растений, Диоскорида, Варрона, Колумеллы и др.
Во время Средневековья большой вклад в развитие ботаники
внес последователь Аристотеля Альберт Великий (1193—1280). Он
описал большое количество растений, подразделив их по морфо-
логическим признакам на деревья, кустарники, полукустарники
и травы; впервые отметил различия между однодольными и дву-
дольными.
В эпоху Возрождения ботаническая наука обогатилась боль-
шим количеством описательного материала, были, собраны богатые
гербарии как в Европе, так и в заморских странах. Количество из-
вестных и описанных растений возросло до нескольких тысяч, по-
явилась необходимость классифицировать этот богатый материал.
В этот период итальянский врач и ботаник Чезальпино (1519—
1603) создал первую систему растений, основанную на строении
семян и плодов. Из 16 классов его системы 15 классов включали
цветковые растения и лишь один класс был отведен для всех ос-
тальных растений. Английский ученый Рей (1623—1705) разделил
цветковые растения на однодольные и двудольные и в своей клас-
сификации использовал не только описания плодов и семян, но
и форму цветка. Он дал оцределение вида у растений и сделал по-
пытки применить бинарную номенклатуру. Несколько позже фран-
цузский ботаник Турнефор (1656—1708) создал свою систему ра-
стений, основанную на изучении форм цветка, и ввел понятие рода.
Системы Чезальпино, Рея и Турнефора были искусственными
системами; в одну группу объединялись растения, не имеющие ни-
чего общего, кроме сходства в каком-либо одном признаке. Такой
же искусственной системой была система Линнея (1753), в осно-
ву которой также было положено строение цветка, число и разме-
щение частей андроцея и гинецея. Однако линнеевская система
158
была разработана более основательно; одновременно с ее разра-
боткой Линней ввел систему двойных обозначений, так называе-
мую бинарную номенклатуру. Эта система была принята не только
в ботанике, но и в зоологии.
Весь растительный мир был разделен Линнеем на 24 класса.
В первые 23 класса вошли цветковые растения, названные Линне-
ем явнобрачными, все остальные растения, названные им тайно-
брачными, вошли в один 24-й класс.'Такое соотношение говорит
о слабой изученности во времена Линнея нецветковых растений.
Работы Линнея сыграли огромную положительную роль в раз-
витии ботанической науки. Достаточно сказать, что Линней наиме-
новал и систематизировал около 10 000 видов растений, известных
в- то время. Все описания растений были приведены в стройную
систему, в которой легко было разобраться. Применение единой
системы наименований на латинском языке устранило путаницу,
связанную с разноязычными названиями, и позволило обобщить
усилия ученых многих стран, направленные на изучение быстро
растущего описательного и гербарного материала. В конце своей
жизни Линней, сознавая несовершенство и ограниченность искусст-
венной системы, начал работу по объединению растений в естест-
венные группы, которые позже получили названия семейств.
На смену искусственной системе Линнея в конце XVIII — нача-
ле XIX в. пришли естественные Системы растений. В этих систе-
мах- растения объединялись по сходству^многих признаков, а такое
сходство бывает связано с общностью происхождения, следова-
тельно, в систематические группы объединялись родственные меж-
ду собой растения.
Первой такой системой была естественная система А. Жюссье
(1748—1836). Руководствуясь сходством по ряду основных призна-
ков, Жюссье разбил все растения на 100 внутриродственных групп,
введя для этих групп понятие семейства. Семейства объединялись
им в 15 классов, классы были распределены между шестью ряда-
ми: 1) бессемядольные, 2) односемядольные, 3) двусемядольные
безлепестные, 4) двусемядольные, однолепестные (спайнолепест-
ные), 5) двусемядольные многолепестные и 6) раздельнополые.
Как и в системе Линнея, все нецветковые растения, включая гри-
бы и водоросли, объединялись в один ряд — бессемядольные. Все-
го в системе А. Жюссье значилось уже около 20 000 растений.
В начале XIX в. создал естественную систему швейцарский бо-
таник О. Декандоль (1778—1841). При построении системы он
пользовался не только морфологическим, но и анатомическим ме-
тодом. Это дало ему основание разделить весь растительный мир
на четыре группы: 1) явнобрачные, 2) тайнобрачные сосудистые,
3) тайнобрачные бессосудистые лиственные и 4) тайнобрачные
бессосудистые безлистные.
Систему Декандоля усовершенствовал Р. Браун (1786—1858);
он выделил из группы сосудистых тайнобрачных голосеменные
растения. Система Р. Брауна была опубликована в 1827 г. К этому
времени число видов растений возросло уже до.75 тысяч.
159
В 1859 г. была опубликована книга Ч. Дарвина «Происхожде-
ние .видов путем естественного отбора», утвердившая в науке идею
эволюции.
Теория Дарвина явилась теоретической основой для разработ-
ки филогенетической системы растений, которая отражает картину
последовательного развития растительного мира и взаимное родст-
во отдельных групп растений.
Большое значение имели для построения филогенетических си-
стем работы Гофмейстера и Беляева в области онтогенеза расте-
ний. Изучая половой процесс у различных групп растений, они
установили общность происхождения мохообразных, папоротников
и голосеменных растений. Таким образом, была установлена не-
прерывная филогенетическая связь между водорослями, с одной
стороны, и цветковыми растениями — с другой.
В XX столетии почти в каждой стране с развитой ботанической
наукой создаются филогенетические системы. Особое внимание уде-
ляется разработке филогенетических систем покрытосеменных —
группы, наиболее богатой видами и представляющей огромный ин-
терес в связи с всесторонним использованием цветковых растений
в жизни людей.
Современная филогенетическая систематика пользуется разно-
образными методами для установления родственных связей и ме-
ста в системе отдельных групп растений.
Сравнительно-морфологический метод был и остается основным
методом в систематике растений. Пользуясь им, о родственной бли-
зости судят на основании сходства и различия в структуре изучае-
мых растений. При этом слово «морфология» понимается в широ-
ком смысле, имея в виду, что исследуется структура на всех уров-
нях: от макроскопического изучения внешних форм до микроско-
пического исследования микро- и ультраструктуры растительного
организма. Для изучения микроскопической и субмикроскопиче-
ской структуры (ультраструктуры) используются оптический и элек-
тронный микроскопы. Применение современных оптических и элек-
тронных микроскопов позволяет широко использовать для целей
систематики данные цитологии, кариологии, палинологии.
Огромное значение для современной филогенетической систе-
матики имеет палеонтологический метод. Изучение ископаемых
форм дает возможность решать вопросы происхождения крупных
систематических групп растений, восстанавливать недостающие
звенья в системе растений, решать вопрос о геологическом возра-
сте той или иной группы. Наравне с морфологическим — это ос-
новной метод в современной филогении.
Эмбриологический метод также широко применяется в наше
время. Родственная близость растений зачастую легче выявляется
в зародышевом состоянии, чем во взрослом. Прослеживая детали
в онтогенетическом развитии, удается установить связи между
группами растений, во взрослом состоянии весьма несхожих.
Успехи современной биохимии позволяют привлечь для реше-
ния вопросов систематики растений данные химического состава
160
продуктов жизнедеятельности растений, а также информацию о мо-
лекулярной структуре белков, рибонуклеиновых кислот и других
жизненно важных соединений. Эти возможности создают перспек-
тивы для все более широкого использования в систематике расте-
ний биохимических методов.
Существенную роль в современной филогенетической науке
играют ' также эколого-морфологический, морфолого-географиче-
ский методы исследования. Эти методы бывают в’есьма эффектив-
ны при установлении родства мелких систематических единиц от
вида и ниже.
Применение данных всех, по существу, биологических, и биоло-
го-географических наук делает систематику растений синтетичес-
кой наукой, в которой интегрируются обшйрные знания всесторон-
него изучения растительного мира. Систематика, в силу этого, ста-
новится необходимой не только для глубокого понимания эволю-
ции жизни на Земле, но и для практического использования ра-
стительных богатств для нужд человека. Для решения вопросов
практического использования того или иного растения необходимо
иметь точные данные о таксономической принадлежности рас-
тения.
Основной таксономической, или классификационной, единицей
в филогенетической систематике является вид. Виды объединяют-
ся в роды, роды в семейства, семейства в порядки, далее идут
классы и отделы. Это основные таксоны. Существуют и промежу-
точные, например подкласс, подрод, подсемейство и т. д. С другой
стороны, основной таксон — вид — подразделяется на подвиды,
разновидности, биоморфы. Какое же содержание надо вкладывать
в понятие «вид»?
Впервые понятие «вид» было введено в науку, как уже сказа-
но, в конце XVII в. Джоном Реем. Под видом Д. Рей понимал
группу растений, настолько сходных между собой, насколько сход-
ны особи, происходящие из семян одного растения; формы,
принадлежащие к данному виду, постоянно сохраняют свою
природу. Подобных же взглядов на вид придерживался и
Линней.
Ламарк первый ввел динамическое, историческое понимание
вида: постоянных, неизменяющихся видов не существует; виды из-
меняются вслед за изменением условий среды. Дарвин закрепил
в науке такое историческое понимание вида своей классической
формулой: «разновидность есть зачинающийся вид, вид — обосо-
бившаяся, определившаяся разновидность».
Крупнейшие русские ботаники-дарвинисты К. А. Тимирязев
и В. Л. Комаров в своих определениях -вида исходили из дарви-
новской формулы. «На вопрос: естественно-исторический вид —
отвлеченное понятие или факт, — мы должны отвечать двояко, со-
ответственно двоякому смыслу, который, очевидно, связан с этим
словом. Вида, как категории, строго определенной, всегда себе рав-
ной и неизменной, в природе не существует. Но рядом с этим и сот
вершенно независимо от этого вывода мы должны признать* что
6—326 161
виды в наблюдаемый нами момент имеют реальное существование,
и это факт, ожидающий объяснения»
«Вид представляет настоящее явление природы. Не неподвиж-
ный факт, а именно явление, т. е. нечто развертывающееся, так
сказать, текущее мимо нас и понятное лишь постольку, поскольку
мы принимаем во внимание, что охватить все явления невозможно,
а исследуется лишь один из периодов в его развитии, случайно
оказавшийся современным нам самим». «Виды, как особи, имеют
свою молодость, свой зрелый возраст, когда они размножаются
особенно интенсивно, и свою старость, когда они постепенно вы-
мирают» 1 2.
«Вид есть совокупность поколений, происходящих от общего
предка и под влиянием среды и борьбы за существование обособ-
ленных отбором от остального мира живых существ; вместе с тем
вид есть определенный этап в процессе эволюции»3.
Особи данного вида приспособлены к определенным условиям
существования; для различных видов диапазон этих условий мо-
жет быть то более широким, то более узким. Вид имеет также
свою область распространения, или ареал. В процессе естественной
эволюции и под влиянием условий, создаваемых' человеком, вид
может приспосабливаться к новым условиям существования и' из-
менять свой ареал.
Каждый вид имеет научное латинское название, составленное
из двух слов, из которых первое, означающее род данного вида,
пишется с прописной буквы, например Ranunculus repens L. — лю-
тик ползучий, Trifolium pratense L. — клевер луговой, Betula ulba
L. — береза белая. Рядом с латинским названием растения в науч-
ной литературе всегда ставят начальные буквы фамилии, а иногда
и имени ботаника, впервые назвавшего или описавшего данное
растение. Буква L., стоящая справа от видового названия расте-
ния, говорит о том, что это название дано Линнеем.
Сходные виды, связанные между собой родственными отноше-
ниями, объединяются в род (genus); роды, так же как и виды, ре-
ально существуют в природе. Следующей более крупной таксоно-
мической категорией является семейство (familia), которое вклю-
чает один, несколько или много родов, имеющих общее происхож-
дение. Семейство — важнейший таксон, наряду с видовым широко
употребляемый; латинские названия семейств в своем окончании
имеют суффикс -асеае> например Ranunculaceae, Fabaceae и т. д.
Семейство — самый низший из высших таксонов. Следующий бо-
лее высокий по рангу таксон — порядок (ordo); в порядке может
быть одно и несколько филогенетически тесно связанных семейств.
Следующие еще более обширные таксоны — класс (classis) и от-
дел (divisio). Отделы объединяют несколько классов; это наиболее
крупные подразделения растительного мира.
1 Тимирязев К. А. Собр. соч., т. III. М., Огиз-Сельхозгиз, 1949, с. 458.
2 Комаров В. Л. Избр. соч., т. 1. Л. — М., Изд-во АН СССР, 1945, с. 32—34.
3 Там же, с. 373.
162
Хотя в настоящее время существует и более современная клас-
сификация органической природы, однако общеупотребительно по-
ка разделение всех живых существ на два царства живой приро-
ды: царство растений (Vegetabilla, Plantae) и царство животных
(Animalia). Мы будем придерживаться этого подразделения.
Общепринято весь растительный мир делить на низшие и выс-
шие растения. Низшие растения характеризуются относительной
простотой строения, многие из них имеют весьма малый, микро-
скопический размер и состоят из одной или немногих клеток.
В тех случаях, когда тело низшего растения состоит из многих
клеток, его строение все же значительно проще, чем высших рас-
тений. Многоклеточное тело низшего растения называется слоеви-
щем или талломом, и такие растения объединяются под общим
названием слоевищных или талломных растений (Thallobionta).
Среди низших растений имеются как автотрофные, так и гетеро-
трофные организмы. Некоторые авторы относят гетеротрофные не-
животные организмы к особому царству природы, к микоидам,
ввиду глубокого различия их в обмене веществ.
Наиболее характерная черта высших растений — сложная рас-
члененность" их тела. За исключением небольшого числа парази-
тов, вторично утративших хлорофилл, все высшие растения авто-
трофны. Этим они сходны с водорослями, от которых и произошли.
Но, будучи сходны по окраске с зелеными водорослями, высшие
растения резко отличаются от них сложностью строения тела.
Большинство высших растений имеет стебель, корень, листья и бо-
лее сложные, чем у низших растений, органы размножения. Слож-
ности внешнего расчленения соответствует совершенство внутрен-
него и большая сложность индивидуального развития — онтогене-
за, связанная с появлением зародыша.
По наличию зародыша все высшие растения получили назва-
ние Embryobionta, а в связи с тем, что тело их обычно дифференци-
ровано на стебель, корень и лист, всю группу высших растений на-
зывают кормофитами (Kormobionta) от слова «кормос» — ствол.
В современной классификации в подцарстве низших растений
насчитывают 14 отделов (типов), в подцарстве высших растений
8 отделов. Названия отделов оканчиваются суффиксом -phyta.
I. Низшие растения (Thallobionta):
1. Бактерии (Bacteriophyta).
2. Синезеленыё водоросли, или Цианеи (Cyanophyta).
3. Пиррофитовые водоросли (Pyrrophyta).
4. Золотистые водоросли (Chrysophyta).
5. Диатомовые водоросли (Bacillariophyta).
6. Бурые водоросли (Phaeophyta).
7. Красные водоросли (Rhodophyta).
8. Желтозеленые водоросли (Xanthophytd).
9. Эвгленовые водоросли (Euglenophyta).
10. Зеленые водоросли (Chlorophyta).
11. Харовые водоросли (Charophyta).
6*
163
12. Слизевики (Myxophyla).
13. Грибы (Mycophyta').
14. Лишайники (Lichenophytd).
II. Высшие растения (Kormobionta, Embryobionta):
1. Риниевые, или Псилофиты (Rhyniophytd).
2. Моховидные (Bryophyid).
3. Псилотовые (Psilotophytd).
4. Плауновидные (Lycopodiophyta).
5. Хвощевидные (Equisetophyta).
6. Папоротниковидные (Polypodiophyta').
7. Голосеменные (Pinophytd).
8. Цветковые, шли Покрытосеменные (Magnoliophyta).
НИЗШИЕ РАСТЕНИЯ (Thai lob ion ta)
ОТДЕЛ 1. БАКТЕРИИ (BACTERIOPHYTA)
Это мельчайшие прокариотные организмы, имеющие клеточное
строение: величина большинства бактерий не превышает несколь-
ких микрометров, многие имеют размер нескольких десятых долей
микрометра, такие бактерии с трудом можно рассмотреть в обыч-
ный микроскоп даже при самом сильном увеличении.
Бактерии в большинстве имеют форму микроскопических ша-
риков, прямых или изогнутых палочек или спирально изогнутых
ниточек. Шаровидные — называются кокками, палочковидные —
бациллами, или бактериями, изогнутые — вибрионами, спирально
извитые — спириллами, или спирохетами. Бациллы внешне сходны
с бактериями, но в отличие от них образуют споры. Кокки, сцеп-
ленные попарно, называются диплококками, соединенные в виде
цепочки — стрептококками. Реже встречаются нитчатые формы
бактерий (рис. 72).
Несмотря на ничтожные размеры, многие бактерии имеют один,
несколько и очень много протоплазменных жгутиков, при помощи
которых активно передвигаются.
Оболочка бактериальной клетки не содержит целлюлозы и со-
стоит из особого вещества — муреина (аминосахара в соединении
с несколькими аминокислотами).
Поверхность оболочки способна ослизняться. Встречаются бак-
терии и'без плотных оболочек, они называются микоплазмами.
Морфологически оформленного ядра в клетках бактерий нет. Ос-
новная масса наследственного вещества представлена огромной
циркулярно замкнутой хромосомой, состоящей из одной молекулы
ДНК и занимающей более или менее центральное положение
в клетке. Кроме этой огромной молекулы ДНК, в которой закоди-
рованы основные видовые свойства бактерий, в протопласте име-
ется еще несколько молекул мелких, тоже циркулярно построен-
ных молекул ДНК. Эти внехромосомные молекулы ДНК, получив-»
164
шие название «плазмиды», также способны к репликации авто-
номно от основной хромосомы. Плазмиды могут перемещаться
в’другие, даже систематически отдаленные (до семейства) бакте-
риальные клетки, изменяя их наследственность, объединяясь
иногда с хромосомой новой бактерии. Такое перемещение плазмид
совершается в момент конъюгации бактерий, причем бактерия-до-
нор может рассматриваться как мужская клетка, бактерия-реципи-
А б В Г Д Е
Рис. 72. Формы бактерий: А — бактерии, Б — бациллы, В —
спирохеты, Г — кокки, Д — спириллы, Е — вибрионы, Ж —
стрептококки, 3 — сарцины, И — нитевидные формы, К — жгу-
тиковые формы, Л — реснитчатые формы
ент — женская. Плазмиды можно рассматривать как агенты пере-
дачи наследственной информации, в том числе и такой генной
информации, которая сообщает патогенные свойства бактерии-реци-
пиенту. С переносом плазмид связано также возникновение у пато-
генных бактерий устойчивости против антибиотиков.
Ядрышки у бактерий не обнаружены, однако в цитоплазме
имеется огромное количество рибосом, доходящее у некоторых ви-
дов до 20 000.
В матриксе широко развиты мембранные структуры в виде ка-
нальцев, ламелл, пузырьков, тилакоидов, содержащих иногда пиг-
менты, у автотрофных структур — фотосинтезирующие. Среди мем-
бранных структур — аналоги митохондрий и аппарата Гольджи.
Выпячивания наружной мембраны внутрь клетки образуют склад-
чатые структуры — мезосомы.
165
За немногим исключением, бактерии бесцветны. Лишь некото-
рые, пурпурные и зеленые, содержат в протоплазме пигменты, по-
добные зеленому хлорофиллу и красному фикоэритрину (хлорин
и пурпурин).
Размножаются бактерии путем деления одной клетки надвое.
Быстрота размножения зависит от внешних условий; в потенции
она огромна: в благоприятных условиях деление может происхо-
дить каждые 20—30 мин. В последнее время у некоторых бакте-
рий обнаружены упрощенные формы полового процесса.
Споры у бактерий являются формой переживания неблагопри-
ятных условий. Перейдя в состояние споры, бактерия выдерживает
длительное высушивание, нагревание свыше 100° С и охлаждение
почти до абсолютного нуля. В обычном же состоянии бактерии
неустойчивы к неблагоприятным внешним воздействиям и быстро
погибают при высушивании, под воздействием солнечного света,
при нагревании до температуры 65—80° С, под действием спирта
и других дезинфицирующих веществ.
Огромное большинство бактерий гетеротрофно, и этим опреде-
ляется их роль в природе; они минерализуют отмершие организмы
животных и растений и возвращают в круговорот минеральные ве-
щества, использованные автотрофными растениями в процессе пи-
тания. Процесс распада белков в результате деятельности бакте-
рий называется гниением, а процесс распада углеродсодержащих
небелковых веществ — брожением.
Немногие бактерии автотрофны, некоторые из них, подобно зе-
леным растениям, используют для создания органического веще-
ства солнечную энергию; к таким фотосинтезирующим бактериям
относятся, например, зеленые бактерии, содержащие пигмент бак-
териохлорин, и пурпурные бактерии, у которых имеется фотосинте-
зирующий пигмент — бактериопурпурин.
Большинство автотрофных бактерий создает органическое ве-
щество, используя химическую энергию окисления других веществ.
Этот процесс называется хемосинтезом. К хемотрофным бактери-
ям относятся, например, нитрифицирующие бактерии, переводящие
аммонийные соединения в нитратные, серные бактерии и железо-
бактерии.
Особую роль в природе играют бактерии, обладающие свойст-
вами усваивать свободный молекулярный азот; населяя почву, та-
кие бактерии обогащают ее азотом. К этой группе относится сво-
бодноживущий азотобактер и клубеньковые бактерии; поселяю-
щиеся на корнях бобовых растений. Большинство же бактерий пи-
тается азотом органических, аммонийных и нитратных соединений.
По отношению к кислороду бактерии делят на аэробных, нуж-
дающихся в свободном молекулярном кислороде, и анаэробных,
живущих в отсутствие воздуха и извлекающих кислород из его
соединений, минеральных и органических.
Значение бактерий в жизни людей огромно. Некоторые бакте-
рии весьма полезны и широко используются человеком в хозяйст-
ве. Это, например, молочно-кислые бактерии, играющие основную
166
роль в процессах силосования кормов, при квашении овощей и ис-
пользуемые для приготовления разнообразных пищевых продуктов
из молока; бактерии маслянокислого брожения, наоборот, попадая
в молочные продукты, портят их.
Для приготовления уксуса используют бактерии уксуснокисло-
го брожения.
Бактерии пектинового брожения вызывают мацерацию расти-
тельных тканей; этим свойством бактерий пользуются в практике
мочки волокнистых растений: льна, конопли и др.
Огромный ущерб здоровью человека и животных наносят мно-
гочисленные виды бактерий, вызывающие болезни (холера*, чума,
желудочно-кишечные заболевания, сибирская язва, бруцеллез, ту-
беркулез и многие другие).
Такие патогенные бактерии относятся к группе паразитов. Но
и бактерии-сапрофиты наносят большой ущерб человеку, вызывая
порчу продуктов питания, кормов и других материалов раститель-
ного и животного происхождения.
Подобно патогенным бактериям действуют вирусы — ультра--
микроскопические, видимые только в электронный микроскоп, не-
клеточные организмы. Вирусы не имеют собственного обмена ве-
ществ и живут только внутри клеток растений, животных и чело-
века, вызывая многие опасные болезни. Вирусы, поселяющиеся
в клетках бактерий и вызывающие их гибель, называются бакте-
риофагами или просто фагами. Фаги размножаются с огромной
быстротой.
Подробное изучение бактерий входит в задачу курса микробио-
логии, а изучением вирусов занимается специальная наука вирусо-
логия.
К бактериям относятся так называемые лучистые грибки или
актиномицеты (Actinomyces). Это мельчайшие нитчатые формы,
по внутренней организации сходные с бактериями. Многочислен-
ные почвенные сапрофиты, а также паразиты на растениях и жи-
вотных (например, возбудители актиномикоза челюстей рогатого
скота). Многие актиномицеты выделяют антибиотики; например,
актиномицет Streptomyces griseus выделяет в процессе жизнедея-
тельности важнейший антибиотик — стрептомицин. Из актиноми-
цетов получают также тетрациклиновые антибиотики, эритроми-
цин, нистатин, олеандомицин и многие другие.
ОТДЕЛ 2. СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA)
Простейшие и наиболее древние автотрофные прокариотные ор-
ганизмы. Протопласт клеток слабо дифференцирован. Фотосинте-
зирующий пигмент диффузно распределен в периферических сло-
ях цитоплазмы, а ядерное вещество (ДНК) сконцентрировано
в центральной части клетки.
Наиболее характерен для синезеленых водорослей синий пиг-
мент фикоциан, который в сочетании с хлорофиллом и -создает
сине-зеленую окраску, свойственную большинству водорослей -это-
167
го типа. Однако присутствие каротина и фикоэритрина придает
некоторым представителям синезеленых водорослей желтоватый,
красноватый и другие оттенки.
В результате фотосинтетического процесса, образуются полиса-
харид гликоген и некоторые другие продукты обмена, играющие
роль запасных питательных веществ.
Примитивность внутреннего строения клеток, склонность к мик-
сотрофному питанию, особенности обмена веществ, сближающие
синезеленые водоросли с животным миром (образование гликоге-
на), способность их выдерживать нагревание до 90° С (в воде го-
Рис. 73. Синезеленые водоросли — Cyartophyta. А —
осциллатория — Oscillatoria, Б — носток — Nostoc, В —
анабена — Anabaena, Г — лингбия — Lyngbya, Д — риву-
лярия — Rivularia, Е — глеокапса — Gloeocapsa, Ж —
хроококк — Chroococcus:
1 — общий вид; 2 — вид при малом увеличении микроскопа;
3 — отдельная нить при большом увеличении; 4 —> гетероцисты
рячих источников), отсутствие полового процесса — все это гово-
рит о древности синезеленых водорослей. Об этом свидетельствуют
и палеонтологические данные: ископаемые формы синезеленых во-
дорослей находят в древнейших пластах земной коры.
В процессе эволюции синезеленые водоросли достигли сравни-
тельно небольшой величины и сложности: большинство видов име-
ет одноклеточное строение, есть также нитчатые и колониальные
формы. Размер одноклеточных и нитчатых форм микроскопический;
лишь скопления клеток или нитей в виде слизистых колоний до-
стигают значительной величины (рис. 73).
168
Клетки синезеленых водорослей разнообразны по форме, от
шаровидных до палочковидных. Оболочки клеток состоят из пекти-
новых веществ, способных ослизняться. Очень часто на поверхно-
сти клеток выделяется слизь из разбухшей гемицеллюлозы, слизь
окутывает группы делящихся клеток (например, у Microcystis) или
клубки нитчатых водорослей (у Nostoc).
Ниточки многоклеточных форм бывают прямые (например,
у Oscillatoria) или извилистые и даже спирально изогнутые (Spf-
rulina). Чаще нити не ветвятся, но встречаются и ветвящиеся фор-
мы, причем ветвление у них ложное: боковые ветви вырастают
в местах разрыва нити.
Одноклеточные синезеленые водоросли размножаются путем
деления клеток, нитчатые и колониальные — путем разрыва и рас-
падения колонии на части или- на одиночные клетки, реже нитча-
тые водоросли образуют споры, служащие для размножения. Ку-
сочки нитей называются гормогониями. Разрыв нитей у многих во-
дорослей происходит через особые, несколько увеличенные клет-
ки — гетероцисты. Половой процесс, как уже сказано, у синезеле-
ных водорослей неизвестен, нет у них также подвижных жгутико-
вых форм. У некоторых, преимущественно морских, видов внутри
клеток образуется помногу неподвижных* эндоспор, служащих
и для размножения. У многих видов клетки водоросли в неблаго-
приятных условиях превращаются в неподвижные споры пережи-
вания.
Синезеленые водоросли обитают преимущественно в пресных
водах (реках, озерах, лужах, колодцах и пр.), весьма широко рас-
пространены на поверхности и в пахотном слое почвы. Здесь они
играют существенную роль в накоплении органического вещества.
Будучи азотфиксаторами, они могут накопить на 1 га почвы за ве-
гетационный период до 20 кг азота. Поселяясь на субстрате, лишен-
ном органического вещества (скалы, пески), синезеленые водо-
росли подготавливают почву для заселения ее более высокоорга-
низованными растениями.
В морях живет сравнительно немного видов синезеленых водо-
рослей, но размножаться они здесь могут в таких огромных ко-
личествах, что меняют цвет морской воды. Так, например, Красное
море получило свое название из-за того, что иногда здесь размно-
жается в массе один из видов водорослей (Trichodesmium eryth-
raeum), имеющих красноватую окраску.
Еще чаще можно наблюдать массовое размножение синезеле-
ных водорослей в пресных водах, где они вызывают «цветение во-
ды» {Microcystis, Andbaena и др.). Такая вода становится непри-
годной для питья и даже может вызвать отравление из-за присут-
ствия некоторых ядовитых представителей (Microcystis). Загрязне-
ние речной воды органическими веществами, а также тепловое за-
грязнение часто приводят к массовому размножению синезеленых
водорослей даже в крупных реках (Днепр и др.). Отмирая, массы
водорослей отравляют воду ядовитыми продуктами распада и мо-
гут вызывать в прудах и озерах массовую гибель рыбы — так на-
169
зываемый замор; попадая в водопроводные системы, синезеленые
водоросли наносят иногда ущерб, закупоривая фильтры насосных
станций.
Филогенетических связей синезеленых водорослей с другими
группами растений не установлено.
Известно около 1400 видов синезеленых водорослей.
ОТДЕЛ 3. ПИРРОФИТОВЫЕ ВОДОРОСЛИ (PYRROPHYTA)
Одноклеточные водоросли (за редким исключением), как и все
последующие в систематике растения, — эукариотные, т. е. имеют
морфологически обособленное ядро; в цитоплазме клеток много-
численные дисковидные хроматофоры, содержащие хлорофилл,
желтый пигмент ксантин и у большинства еще бурый пигмент —
перидинин; запасные вещества — крахмал и масло.
Лишь немногие, более примитивные не имеют твердой оболоч-
ки, у большинства же видов клетка одета панцирем из целлюлоз-
ных пластинок, придающих телу водоросли своеобразную форму
(рис. 74).
Из 1100 видов водорослей этого отдела свыше 1000 принадле-
жат к классу периДиней (Peridineae). Тело водоросли имеет дор-
зовентральное строение и по форме или близкое к шаровидной
(Peridinium, Noctilucd), или вытянутое (Ceratium); в последнем
случае форма водоросли асимметрична и весьма своеобразна,
с длинным рогом в передней части и 2—3 рогами — в задней ча-
Рис. 74. ’Представители пиррофитовых водорослей: А — перидиниум,
Б — церациум, В — ноктилюка
170
сти. Тело водорослей, одетых в панцирь, опоясано поперечной
и продольной бороздками; на вентральной стороне два жгутика,
один из них в поперечной бороздке, второй — в продольной борозд-
ке, кончик его выступает за пределы панциря. Вегетативное деле-
ние пиррофитов — путем деления клетки надвое, при этом Peridi-
nium сбрасывает створки, Ceratiuin делит створку между двумя
дочерними клетками, вторая половина створки доращивается; бес-
панцирные формы (Noctiluca) размножаются путем почкования,
а также образуют зооспоры. Известен и половой процесс — путем
слияния двух протопластов. Форма переживания неблагоприятных
условий — толстостенная циста, в которую превращается прото-
пласт водоросли.
Перидинеи обитают в морских и пресных водах. В морях и оке-
анах они (вместе с диатомовыми) составляют основу растительно-
го планктона, где вызывают ночное свечение моря. Ночесветка
(Noctiluca) и некоторые другие виды.
ОТДЕЛ 4. ЗОЛОТИСТЫЕ ВОДОРОСЛИ (CHRYSOPHYTA)
В этом отделе преобладают одноклеточные и колониальные
подвижные формы и лишь как исключение — одноклеточные и нит-
чатые формы неподвижные. В цитоплазме обычно два крупных зо-
лотисто-желтых хроматофора, в которых помимо хлорофилла,
еще два каротиноида: желтый лютеин и буроватый фукоксантин;
запасные питательные веще-
ства— масло и углевод лей-
козин. Оболочки — целлю-
лозно-пектиновые или клет-
ки голые. В оболочках часто
известковые или кремниевые
пластинки, иногда с иглами.
Количество жгутиков 1—2,
но бывает и 3—4, равной и
неравной длины.
Золотистые водоросли —
обычные обитатели достаточ-
но чистых пресных вод, ред-
ко встречаются в морях.
Размножаются путем деле-
ния клетки надвое, причем
в колониальных формах
одна дочерняя клетка оста-
ется в материнском жестком
бокальчике, а вторая при-
крепляется рядом, строит
свой домик и обычно прира-
щивается. Многие виды об-
разуют также зооспоры, а в
неблагоприятных условиях
Б
Рис. 75. Представители золотистых водорос-
лей: А — синура, Б — динобрион
171
подвижные клетки могут превращаться в эндогенные цисты. Новые
колонии могут возникать также путем распада старых.
Золотистые водоросли весьма разнообразны: от микроскопиче-
ских одноклеточных форм до колоний, достигающих иногда длины
20—30 см (Hydrurus). Общее количество видов достигает 400.
В качестве представителя широко распространенных одноклеточ-
ных форм можно назвать хромулину (Chromulina) и малломонаду
(Mallomonas). Характерны и внешне очень красивы колонии сину-
ры (Synura) и динобриона (Dinobryori) (рис. 75). Те и другие
весьма энергично двигаются в воде, синура в виде вертящегося
золотистого волчка; у динобриона обращают на себя внимание
прозрачные целлюлозные бокальчики, в которых сидят толые про-
топласты клеток с двумя неравными жгутиками.
ОТДЕЛ 5. ДИАТОМОВЫЕ ВОДОРОСЛИ (BACILLARIOPHYTA)
Преимущественно одноклеточные, реже колониальные орга-
низмы (рис. 76). Протопласт клеток дифференцирован на цито-
плазму, ядро и хроматофоры. Большую
Рис. 76. Диатомовая водоросль
пиннулярия (Pinnularia). А —
вид со стороны пояска, Б — со
стороны створок
часть объема клеток занимают вакуо-
ли; ядро находится в тяже цитоплазмы
и занимает центральное или (реже)
пристенное положение. Хроматофоров
один или несколько; кроме хлорофил-
ла в них содержится несколько разно-
видностей ксантинов желто-бурого
цвета. Различное соотношение этих
двух пигментов дает общую окраску
водорослей — зеленовато-желто-бурую.
Крахмал не образуется, вместо него
в цитоплазме и вакуолях накаплива-
ются капельки масла.
Оболочка пектиновая, наслоенная
снаружи кремнеземом. Это наиболее
характерная черта диатомовых водоро-
слей, благодаря которой они называ-
ются также кремнистыми водорослями.
Окремнелая оболочка состоит из двух
створок, надетых одна на другую напо-
добие коробочки и крышечки. Меныпая
створка (коробочка) называется гипо-
текой, большая — эпитекой. Окремне-
лые оболочки не растут, поэтому даль-
нейший рост клетки внутри створок не-
возможен.
Панцири клеток имеют различного
рода утолщения и выросты. Эти скуль-
птурные детали обычно очень тонки и
красивы; каждому виду водоросли
172
свойствен свой рисунок, поэтому скульптура створок — основной
систематический признак.
Живут диатомовые водоросли в морях,* пресных водах и на
влажной почве. Незаметны простым, невооруженным глазом. Игра-
ют большую роль в природе как создатели органического вещест-
ва в гидросфере. Особенно велика роль их в морях и океанах, где
они составляют основную массу растительной части планктона
(организмы, находящиеся во взвешенном состоянии в толще во-
ды). Толща слоя, населенного планктоном, может достигать 100 м.
В пресных водоемах большую роль играют бентосные диатомеи,
т.е. водоросли, чья жизнь связана с дном водоема.
Некоторые представители диатомовых водорослей (класс Cen-
tricae) образуют жгутиковые формы, которые обнаруживаются во
время полового процесса. Предполагают-, что это сперматозоиды,,
которые участвуют в, изогамном, гетерогамном и оогамном поло-
вом процессе.
По строению и форме створок отдел диатомовых делят на два
класса: Centricae и Pennatae. Представители класса Centricae име-
ют радиальное строение; -это преимущественно планктонные водо-
росли морей и океанов. Водоросли из класса Pennatae имеют дву-
сторонне-симметричную, вытянутую форму; по образу жизни это
бентосные и напочвенные водоросли.
С типичными особенностями водорослей класса Pennatae обычно
знакомятся на примере широко распространенной бентосной водо-
росли пресных водоемов пиннулярии (Pinnularia). Тело; пиннудя-
рии вытянуто, длина в.несколько раз превышает ширину. При рас-
смотрении со стороны эпитеки или снизу, со стороны гипотеки,
водоросль имеет форму сильно вытянутого эллипса, сбоку, со сто-
роны так называемого пояска, — форму вытянутого прямоугольни-
ка. На внутренней стороне нижней створки можно видеть три бу-
горка— узелка: два по концам, один в центре. Между узелками
узкая щель, так называемый шов. Через шов цитоплазма сообща-
ется с внешней средой. Внутри клетки два больших хроматофора
корытообразной формы, по одному в каждой створке водоросли.
Пиннулярия способна передвигаться. Механизм перемещения
подобен гусеничному ходу трактора.; цитоплазма, имеющая враща-
тельное движение, продавливается сквозь щель шва; покрывающий
цитоплазму слой слизи соприкасается с субстратом и, в силу сце-
пления .с его поверхностью, проталкивает водоросль в сторону, про-
тивоположную движению цитоплазмы. Многие представители клас-
са Pennatae не имеют шва*и не способны передвигаться. *
Зооспор пиннулярия и другие представители класса не имеют;
вегетативное размножение — типичное для диатомей: путем деле-
ния надвое с последующим доращиванием меньшей половины
кремниевого панциря. Половой процесс происходит путем попарно-
го слияния клеток двух водорослей. Слиянию предшествует редук-
ционное деление в каждой водоросли, в результате чего образуют-
ся тетрады. Но две клетки из каждой тетрады отмирают, остальные
две попарно сливаются, образуя диплоидные зиготы, зигота одева-
173
ется в оболочку и превращается в ауксоспору, которая, вырастая,
становится новой водорослью (у водорослей класса Pennatae).
Ядра диатомовых водорослей всегда диплоидны, т. е. содержат
двойной набор хромосом, гаплоидны лишь клетки, подготовившие-
ся к слиянию в половом процессе.
Неподвижные представители класса Pennatae часто образуют
колонии. Они возникают в процессе вегетативного деления водо-
росли и склеивания образующихся новых особей. Колонии имеют
форму цепочек (Tabellaria), лент (Fragillaria), кустиков или нитей
(Navicula), У некоторых видов клетки склеены своими концами
и колония имеет форму звездочки (Asterionella).
Всего известно до 6000 видов диатомовых водорослей.
ОТДЕЛ 6. БУРЫЕ ВОДОРОСЛИ (РЙАЕОРНУТА)
Сюда относятся около 800 видов почти исключительно морских
водорослей. Все бурые водоросли — бентосные, т. е. они населяют
Дно водоемов и, следовательно, обитают вблизи берегов. Таллом
их многоклеточный и достигает размера нескольких десятков и да-
же ста метров. При таком огромном размере и внутреннее стро-
ение таллома бурых водорослей довольно сложно; здесь намеча->
ется уже дифференциация клеток на ткани. Нитчатое строение
таллома свойственно только наиболее примитивным формам; при-
митивность строения связана с небольшими размерами водо-
росли.
Клетки, из которых состоит таллом бурых водорослей, покры-
ты целлюлозной оболочкой, пропитанной пектиновыми вещества-
ми. В клетке одно ядро и зернистые хроматофоры бурых оттен-
ков. Цвет хроматофоров зависит от примешивания к хлорофиллу
бурого пигмента фукоксантина, каротина и ксантофилла. Фуко-
ксантин специфичен для бурых водорослей и в основном опреде-
ляет их окраску. Примесь остальных пигментов снижает интенсив-
ность окраски до зеленовато-оливковой. По химической природе
фукоксантин близок к каротину и также нерастворим в воде; по-
этому, отмирая, бурые водоросли не сразу теряют свойственную
им окраску.
В качестве конечного продукта ассимиляции вместо крахмала
образуется специфичный для бурых водорослей полисахарид лами-
нарии,- растворимый в воде. Бурые водоросли прошли длинный
путь эволюции от примитивных нитчатых форм почти микроскопи-
ческого размера до огромных водорослей с расчлененным пластин-
чатым талломом сложного анатомического строения.
Почти всем бурым водорослям свойственно чередование поко-
лений. Это явление заключается в том, что в онтогенезе этих во-
дорослей бесполое,поколение (спорофит) дает споры, из которых
развивается половое поколение (гаметофит), дающее половые
клетки (гаметы). В результате слияния гамет образуется зигота,
из которой развивается бесполое поколение (спорофит).
174
Рис. 77. Ectocarpus. А — внешни* вид (видны интеркалярные зоны рос-
та, зооспорангии), Б — зооспорангий (сильно увеличен), В — гаметан-
гий
Талломы спорофита и гаметофита могут быть одинаковыми (изо-
генератные формы) и резко различными (гетерогенератные фор-
мы). У некоторых водорослей (фукусовые) бесполого размноже-
ния нет.
Изогенератные формы в наших морях обильно представлены
эктокарпусом {Ectocarpus). Это небольшие кустики из одноряд-
ных нитчатых веточек, прикрепленных к камням или к более круп-
ным водорослям (рис. 77). В первой половине лета развиваются
обычно диплоидные спорофиты. Зооспорангии развиваются сбоку
ветвей, в них гаплоидные зооспоры с двумя жгутиками. Из зоо-
спор развиваются такие же кустики, но гаплоидные. Это гамето-
фиты. На них развиваются многоклеточные гаметангии. В гаме-
тангиях развиваются гаплоидные подвижные гаметы, они морфо-
логически одинаковы, но физиологически раздельнополые. Слива-
ясь попарно, гаметы дают зиготу (изогамия), которая сразу же,
без периода покоя, прорастает в спорофит. Иногда гаплоидное по-
коление выпадает, тогда зооспоры диплоидные и сразу прорастают
в спорофит.
Подобно эктокарпусу происходит чередование поколений у рас-
пространенной в Черном море диктиоты {Dictyota), которая в от-
личие от эктокарпуса имеет пластинчатый таллом и вместо под-
вижных зооспор образует лишенные жгутиков шаровидные тетра-
175
Рис. 78. Бурые водоросли. А — ламинария
сахарная (Laminaria saccharina), Б —.лами-
нария пальчатая (L. digitata)
щих из небольшого числа клеток.
споры (по 4 в одном тетраспорангии). Диктиота — растение дву-
домное: из внешне одинаковых тетраспор развиваются однополые
женские гаметофиты с оогониями и мужские с антеридиями. Поло-
вой процесс оогамный.
В качестве представителя гетерогенератных бурых водорослей
может быть рассмотрена ламинария (Laminaria).
Два ее вида — ламинария сахарная (L. saccharina) и пальча-
тая (L. digitata) — широко распространены в наших северных мо-
рях (рис. 78). Таллом их расчленен на ризоиды, стеблевую и ли-
стовую части. У ламинарии сахарной листовая часть в виде
длинной широкой ленты, у
пальчатой листовая пластин-
ка пальчатораздельная. Об-
щая длина черешка и листо-
вой части достигает несколь-
- ких метров. Внутреннее
строение таллома ламина-
рий также довольно сложно,
заметна дифференциация на
ткани.
Описанный таллом лами-
нарии представляет собой
диплоидный спорофит. На
поверхности листовой пла-
стинки спорофита группами
образуются одногнездные
зооспорангии, в них много-
численные зооспоры. Зооспо-
ры гаплоидны и физиологи-
чески неравнозначны. Про-
растая, зооспоры дают мик-
роскопического размера га-
метофиты— заростки в виде
ветвящихся ниточек, состоя-
На заростках образуются органы полового размножения: на
одних (женских) — оогонии, на других (мужских) — антеридии,
и в тех и в других по одной половой клетке; в антеридии — спер-
матозоид, в оогонии — яйцеклетка. После оплодотворения из
зиготы без периода покоя вырастает новый диплоидный
таллом.
У ламинарий наших северных морей зооспоры образуются ле-
том; половой процесс — весной следующего года. В течение осени
и зимы развитие гаметофитов очень медленное, причем процесс
этот происходит только при пониженной температуре.
Из фукусовых водорослей, не имеющих бесполого поколения,
в северных морях (Белое и Баренцево) широко распространены
заросли фукуса (Fucus vesiculosus), в Черном море — цистозейры
(Cystoseira barbata).
476
ОТДЕЛ 7. КРАСНЫЕ, ИЛИ БАГРЯНЫЕ,
ВОДОРОСЛИ (RHODOPHYTA)
Около 4000 видов, почти все они — обитатели морей. Преобла-
дающая окраска малиновая, но бывают водоросли и других оттен-
ков: бледно-розовогр и даже желтовато-зеленого и синеватого.
Рис. 79. Красные водоросли. А — делессерия (Delesseria), Б — хондрус
(Chondrus), В — полисифония (Polysiphonia)
Различная окраска зависит от наличия в клетке нескольких пиг-
ментов — красного фикоэритрина, синего- фикоциана и зеленого
хлорофилла. Первые два пигмента белковой природы и легко рас-
творяются в холодной воде после гибели клеток.
177
Все красные водоросли многоклеточны, таллом их пластинча-
тый и нитчатый, разветвленный, размером от нескольких санти-
метров до нескольких десятков сантиметров (рис. 79). Целлюлоз;
ные оболочки клеток обычно пропитаны пектиновыми веществами,
благодаря чему поверхность водорослей ослизняется. У некоторых
видов оболочки клеток пропитаны углекислым кальцием и маг-
нием.
Протопласт обычно одноядерный, с многочисленными зерни-
стыми хроматофорами, без пиреноидов. В хроматофорах находят-
ся названные выше пигменты. Продуктом фотосинтеза является
особый полисахарид — багрянковый крахмал, близкий по своим
свойствам к обычному крахмалу.
Бесполое размножение — посредством неподвижных моноспор
и тетраспор (по количеству спор, образующихся в каждом споран-
гии). Жгутиковых форм совершенно нет. Моно- и тетраспоры гап-
лоидны. У моноспоровых водорослей чередования поколений нет,
из споры развивается гаплоидное растение, на котором развива-
ются и моноспорангии, и. органы полового размножения. У тетра-
споровых изоморфное чередование поколений. Из тетраспор раз-
виваются гаплоидные гаметофиты с органами полового размноже-
Рис. 80. Органы размножения красных водорослей.
А — тетраспорангий с тетраспорами, Б — выпадение
гетраспор, В — ветвь с карпогоном (о), трихогиной
(тр) и антеридиями (а),' Г — цистокарпий, развив-
шийся из оплодотворенного карпогона, вверху трихо-
гина (тр)
178
ния. После оплодотворения образуются диплоидные спорофиты,
внешне не отличающиеся от гаметофитов, но с тетраспорангиями.
Половой процесс оогамный. Женский половой аппарат в виде
одной колбообразной клетки с сильно вытянутым горлышком. На-
зывается этот аппарат карпогоном, вытянутый конец — трихогиной
(рис. 8Q). Антеридии — маленькие бесцветные клеточки, вырастаю-
щие на талломе. В каждом антеридии находится одна неподвиж-
ная голая клетка — спермаций. Спермации пассивно, током воды,
переносятся на кончик трихогины и переливаются внутрь карпо-
гона. После оплодотворения зигота многократно делится, в ре-
зультате чего образуются гаплоидные карпоспоры.
Большинство красных водорослей образует тетраспоры, наибо-
лее характерные формы из них показаны на рис. 80. К этой же
группе относится распространенная в Черном море, имеющая про-
мысловое значение филлофора (Phyllophora nervosa). Из моно-
споровых следует назвать тоже характерные формы: порфиру
(Porphyra), немалион (Nemalion) и одну из немногих пресновод-
ных водорослей — батрахоспермум (Batrachospermum).
ОТДЕЛ 8. ЖЕЛТОЗЕЛЕНЫЕ ВОДОРОСЛИ (XANTHOPHYTA)
Наиболее характерная черта этих водорослей — наличие у веге-
тативных форм и у зооспор двух жгутиков разной длины и строе-
ния, за что их называют также Разножгутиковыми (Heterocontae).
Один из жгутиков длинный, перистый и направлен вперед, второй
короткий, гладкий (назад) и направлен чуть в сторону.
Рис. 81. Желтозеленые водоросли — Xanthophyta,
А — хлорамеба — Chloramqeba, Б — трибонема — Tri-
bonema, В — ботридий — Botrydium:
1 — распад цитн; 2 — зооспора трибонемы; 3 — зооспора бот-
рндня
179
Сюда принадлежат одноклеточные, колониальные, многоклеточ-
ные (нитчатые), а также водоросли неклеточного строения. Клет-
ки этих водорослей имеют обычно пластинчатый или зернистый
хроматофор, содержащий хлорофилл и большое количество жел-
тых и красных каротиноидов, почему и водоросли имеют желтова-
то-зеленый цвет. Запасные питательные вещества (масло, крах-
5
2 1
Рис. 82. Жизненный цикл вошерии (Vaucheria). А — бесполое размноже-
ние, Б — половое воспроизведение:
1— материнское растение; 2 — зооспорангий; 3 — выход зооспоры; 4 — зооспора; 5 —
прорастающая зооспора; 6 — антеридий; 7 — оогоний; 8 — сперматозоиды; 9.— яйце-
клетка; 10 — зигота
мал) не образуются. Оболочки клеток пектиновые, реже целлю-
лозные. Размножаются преимущественно вегетативно или бесполо
при помощи зооспор, реже половым путем — изогамно или оогам-
но. Распространены в пресных и соленых водах, а также на поч-
ве, стенах домов, камнях и т. д.
Известно около 250 видов. Представитель одноклеточных хлор*
амеба (Chloramaeba), живущая в пресных и морских водах; из
нитчатых в пресных водоемах распространена трибонема (Tribo-
пета); на почве вблизи водоемов или в колеях иногда очень обиль-
но размножается неклеточного строения водоросль ботрйдиум
180
(Botridium) — грушевидной формы пузырьки диаметром до 2 мм;
нижняя часть таллома расчленена в ризоиды. При избытке влаги
образует большое количество зооспор, при цысыхании субстрата
многочисленные мелкие ядра переходят в ризоиды, которые рас-
падаются на цисты (рис. 81).
К желтозеленым водорослям неклеточного строения относят
также и вошерию (Vaucheria), широко распространенную бентос-
ную водоросль преимущественно пресных водоемов (рис. 82). Тело
ее представляет собой ветвящуюся нить неклеточного строения.
В протопласте многоклеточные ядра и хлорофилловые зерна; за-
пасное вещество — масло, оболочка целлюлозная’; бесполое раз-
множение при помощи многоядерных зооспор или апланоспор, от-
деляемых перегородкой на концах- разветвленного таллома. Зоо-
споры многожгутиковые; жгутики отходят попарно от каждого из
ядер, расположенных в периферической части зооспоры. Споры не-
посредственно прорастают в новую особь. Половой процесс оогам-
ный. В антеридии образуются многочисленные сперматозоиды гру-
шевидной формы с двумя неравными жгутиками. После оплодот-
ворения в оогонии зигота развивается в диплоидную ооспору; при
прорастании ее происходит редукция хромосом, поэтому ядра в ни-
ти вошерии гаплоидны.
ОТДЕЛ 9. ЭВГЛЕНОВЫЕ ВОДОРОСЛИ (EUGLENOPHYTA)
Одноклеточные водоросли веретеновидной или эллипсоидной
формы (рис. 83). Целлюлозной оболочки нет. Снаружи клетки уп-
лотненный слой — перипласт. У типичного представителя эвглены
(Euglena) перипласт тонкий, позволяющий клетке менять форму;
у факуса (Phacus) перипласт более плотный и форма клетки по-
стоянная, слегка сплюснутая; трахеломонас (Traclielomonas) по-
верх протопласта имеет жесткий панцирь, пропитанный слоями же-
леза. Хроматофоры зеленые, содержат хлорофилл, каротин и ксан-
тофилл. Запасное вещество — углевод близкий к крахмалу — пара-
милон.
На переднем конце жгутик (иногда два), выходящий из углуб-
ления и прикрепленный к базальному тельцу. В протопласте ядро,
сократительные вакуоли и (у большинства) красный глазок.
Размножаются вегетативно, путем продольного деления надвое.
У трахеломонас, имеющих твердый домик, одна из дочерних кле-
ток выскальзывает через жгутиковое отверстие и создает собст-
венный домик.
Эвгленовые питаются миксотрофно; в сильно загрязненных
пресных водоемах широко распространенная эвглена зеленая
(Ей. viridis) размножается в огромных количествах, а эвглена крас-
ная (Ей. rubra), имеющая красный пигмент гематохром и размно-
жаясь массово в водоемах, придает воде кроваво-красный цвет.
Наличие эвглен — показатель сильной загрязненности воды орга-
ническими веществами.
181
в
Рис. 83. Представители эвгленовых водорослей. А — Euglena viridis, Б —
Phacus, В — Trachelomonas
ОТДЕЛ 10. ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA)
Это' наиболее широко известная группа низших автотрофных
организмов, насчитывающая около 6000 видов. Обитают зеленые
водоросли преимущественно в пресных водах, реже в морях. До-
вольно часто их можно встретить на почве, на мокрой поверхности
камней и даже на арктическом снегу.
Зеленые водоросли очень разнообразны по величине и строе-
нию: от одноклеточных, не видимых простым глазом, до крупных
нитчатых или пластинчатых форм, достигающих размера 10 см
и более. Общее для всех водорослей этого отдела — наличие
в клетках.зеленого пигмента, не маскируемого пигментами других
окрасок.
В отличие .от представителей предыдущего отдела клетки зе-
леных водорослей (за редким исключением) вырабатывают в ка-
честве запасного питательного вещества крахмал, а стенки их по-
строены обычно из целлюлозы с примесью пектиновых веществ,
реже из пектина и гемицеллюлозы. Это типичные зеленые расте-
ния.
Отдел зеленых водорослей подразделяют на два класса: 1. Рав-
ножгутиковые зеленые водоросли (Isocontae); 2. Сцеплянки, или
конъюгаты (Conjugatae).
182
Первый класс представляет собой многочисленную и разнооб-
разную группу водорослей .Объединяющий их признак — наличие
подвижных зооспор, снабженных двумя одинаковыми жгутиками.
В половом процессе также принимают участие подвижные жгути-
ковые формы. При этом наблюдаются: изогамия — слияние двух
одинаковых по размеру подвижных половых клеток — гамет; гете-
рогамия — слияние двух разных по величине подвижных гамет
и оогамия — слияние неподвижной женской гаметы и подвижной
мужской.
Во второй класс объединяются организмы, представляющие
слепую и слабо подвинувшуюся ветвь эволюции зеленых водорос-
лей. Здесь утрачены подвижные жгутиковые формы; половой про-
цесс приобрел своеобразную форму конъюгации — слияние двух
вегетативных клеток водорослей.
Класс равножгутиковые (isocontae)
Класс равножгутиковые делят на четыре порядка, отражающих
ступени и направления эволюции: вольвоксовые (Volvocales); хло-
рококковые (Chlorococcales); улотриксовые (Ulotrichales); сифо-
новые (Siphonales).
Порядок вольвоксовые. Представлен одноклеточными и коло-
ниальными формами (см. рис. 84 и 85); те и другие в обычных ус-
ловиях существования подвижны; клетки их снабжены парой жгу-,
тиков (одинаковых), штопорообразное движение которых передви-
гает водоросль.
Порядок хлорококковые. В своем происхождении хлорококко-
вые непосредственно связаны с вольвоксовыми; представлены они
также одноклеточными и колониальными формами (см. рис. 87).
Эта группа эволюционировала в направлении утраты подвижно-
сти. Жгутики или совершенно утрачены, или сохранились только
у клеток, служащих для размножения, т. е. зооспор и гамет.
Порядок улотриксовые. Наиболее прогрессивная ветвь эволю-
ции собственно зеленых водорослей. Этот порядок объединяет мно-
гоклеточные формы, имеющие нитчатый и, реже, пластинчатый
таллом. Нитчатый таллом может быть неразветвленный и ветвя-
щийся (см. рис. 88).
Порядок сифоновые. Наиболее своеобразные водоросли имеют
относительно крупный таллом неклеточного строения (см. рис. 90).
Для характеристики класса приведем описание нескольких
представителей из каждого порядка; из порядка вольвоксовых опи-
шем хламидомонаду гониум, вольвокс; из порядка хлорококко-
вых — хлорококк, хлореллу и сценедесмус; из порядка улотриксо-
вых — улотрикс, кладофору, у льву, плеурококк; из порядка сифоно-
вых — каулерпу.
Многочисленные представители рода хламидомонада (Chlami-
domonas) широко распространены в пресных водоемах и особенно
часто встречаются в лужах, колеях и других временных скоплени-
ях воды.
183
Обильно размножаясь, они придают воде ярко-зеленый цвет.
Одноклеточное тело водоросли покрыто оболочкой из пектина
и гемицеллюлозы. Клетки имеют шаровидную или овальную форму
(рис. 84). От их переднего, несколько вытянутого конца отходят
два жгутика равной длины..
Протопласт состоит из цитоплазмы,
большого чашевидного хроматофора,
ядра и так называемого глазка, пятныш-
ка из пигмента, близкого к каротину,—
гематохрома; глазок размещен в перед-
ней половине клетки. В нижней части
чашевидного хроматофора расположен
пиреноид — тельце, богатое белками, во-
круг которого образуется первичный
крахмал; в углублении хроматофора, в
окружении цитоплазмы, находится ядро.
В толще цитоплазмы — несколько пуль-
сирующих вакуолей.
Такое строение клеток типично для
всех вольвоксовых.
Хламидомонады быстро размножают-
ся; материнская клетка теряет жгутики,
протопласт ее делится на 2—4—8 частей,
каждая часть оформляется в самостоя-
тельную особь, одевается оболочкой, раз-
вивает жгутики и в таком виде покидает
материнскую оболочку. Вышедшие особи
быстро подрастают, и обычно уже через
сутки каждая из них, в свою очередь,
приступает к делению.
В неблагоприятных условиях (недо-
статок воды или кислорода) молодые
особи не приобретают жгутиков и не рас-
ходятся; на поверхности их выделяется
Рис. 84. Хламидомонада
(Chlamidomonas):
I — зеленый хроматофор; 2 — пи-
реноид; 3 — ядро; 4 — жгутики;
5 — красный глаз; 6 — пульси-
рующие вакуоли; 7 — цитоплаз-
ма
Рис. 85. Колониальная водо-
росль гониум (Goniutn)
слизь, покрывающая скопление клеток
общим слоем. Внутри слизистого комоч-
ка клетки продолжают делиться. Такая
форма переживания неблагоприятных ус-
ловий называется пальмеллевидным.со-
стоянием. В пальмеллевидном состоянии
хламидомонады могут находиться дли-
тельное время, при попадании их в све-
жую чистую воду клетки приобретают
жгутики и расходятся во все стороны.
Ядра хламидомонад, как и большинст-
ва зеленых водорослей, гаплоидны; следо-
вательно, в процессе деления клеток не
происходит редукции хромосом. Диплоид-
ны лишь зиготы, возникающие в резуль-
184
тате полового процесса. У большинства хламидомонад половой
процесс изогамный, но встречается и гетерогамный.
Гаметы образуются внутри материнской клетки подобно беспо-
лому размножению, но только в большем количестве, в числе 32—
64. Каждая гамета имеет два жгутика и во всем похожа на зоо-
спору, только меньшего размера. По выходе из материнской обо-
лочки гаметы некоторое время движутся, а затем попарно копули-
руют. Как правило, копулируют лишь гаметы, происшедшие от
различных материнских особей. Это явление называется гетеро-
таллизмом в противоположность гомоталли$му, при котором могут
копулировать гаметы, происшедшие из одного материнского орга-
низма.
Возникшая зигота одевается толстой оболочкой, протопласт ее
обогащается запасными питательными веществами, и в таком со-
стоянии зигота-циста может длительное время выдерживать небла-
гоприятные условия. В благоприятных условиях внутри зиготы
образуются четыре новые хламидомонады, размножающиеся затем
бесполым путем. Одно из делений внутри зиготы редукционное,
после которого восстанавливается гаплоидность хламидомонад.
Гониум (Goniurn)—колониальная водоросль, состоящая из
16 клеток, соединенных в округлую пластинку путем склеивания
оболочек. Клетки подобны хламидомонадам; жгутики Обращены
наружу и загнуты в одну сторону, благодаря чему пластинка дви-
жется перпендикулярно своей плоскости (рис. 85).
В результате бесполого размножения внутри каждой из 16 кле-
ток колонии образуется тоже по 16 клеток, составляющих новую
'колонию. После ослизнения оболочки материнской клетки такая
колония выходит в воду, где и дорастает до нормального размера.
При половом воспроизведении (изогамия, реже гетерогамия)
молодая колония распадается на отдельные клетки — гаметы. Ко-
пулировать могут только гаметы, происходящие из разных колоний
(гетероталлизм). Из зиготы после периода покоя образуются 4
гаплоидные зооспоры, каждая из которых дает начало колонии из
16 клеток.
У более высокоорганизованного представителя вольвоксовых —
вольвокса (Volvox) половой процесс оогамный.
Вольвокс — наиболее сложная в этом порядке колониальная
водоросль. Колония имеет форму шарика ярко-зеленого цвета диа-
метром 0,5—2 мм; в одной колонии от нескольких сот до несколь-
ких десятков тысяч клеток (рис. 86); каждая клетка такой коло-
нии подобно хламидомонаде, снабжена двумя одинаковыми жгути-
ками, направленными вовне; клетки располагаются в один слой,
внутри шарика слизь. Цитоплазмы соседних клеток соединены
плазмодесмами. При помощи жгутиков вольвокс относительно
быстро перемещается, и в это время можно заметить, что шарик
всегда движется вперед какой-то одной стороной. На противопо-
ложной, задней по движению стороне располагаются клетки, кото-
рые, делясь, образуют впячивание стенки внутрь. В конце концов
это впячивание выворачивается наизнанку, отшнуровывается,
185
и внутри шара оказывается маленькая дочерняя колония; таких
дочерних колоний может сформироваться до 10.
Внутри дочерних колоний таким же способом могут сформи-
роваться внучатые. После отмирания клеток материнской коло-
нии дочерние колонии становятся свободными и продолжают са-
мостоятельное существование. Так, у вольвокса осуществляется
вегетативное размножение. Для полового процесса небольшая часть
клеток (5—15) превращается в
2 к®
Рис. 86. Жизненный цикл вольвок-
са — Volvox. А — бесполое размноже-
ние, Б — половое воспроизведение:
1 — материнская особь; 2 — партеногони-
дия; 3 — начало формирования дочернего
организма; 4 — дочерний организм; 5 —
оогоний; 6 — пучок сперматозоидов; 7 —
сперматозоид; 8 — яйцеклетка; 9 — зигота
антеридии и оогонии (до 30).
В антеридиях образуется по 32Д-
64 двужгутиковых сперматозои-
дов; клетка оогония сильно уве-
личивается и превращается в
яйцеклетку. После оплодотворе-
ния зигота превращается в ооспо-
ру, которая после периода покоя
превращается в результате много-
кратного деления в новый воль-
вокс. Клетки вольвокса гаплоид-
ны, диплоидна только зигота;
вольвоксы бывают однодомные и
двудомные.
Представитель порядка хлоро-
кокковых — хлорококк ( Chloro-
соссит) —одна из наиболее рас-
пространенных одноклеточных зе-
леных водорослей. Обитает в
пресной воде, а также на влажной
почве, на коре деревьев. Шаро-
видные клетки этой водоросли
покрыты целлюлозной оболочкой.
Внутреннее строение такое же,
как и у хламидомонады, только
нет пульсирующих вакуолей и
глазка.
Бесполое размножение посред-
ством двужгутиковых зооспор,
которые образуются из протопласта материнской клетки в числе 32.
Половой процесс изогамный — копуляция двух подвижных гамет,
образующихся подобно зооспорам.
Хлорелла (Chlorella) встречается в тех же условиях, что и хло-
рококк. Отличается от последнего отсутствием подвижных спор;
взамен зооспор у хлореллы неподвижные апланоспоры, образую-
щиеся в числе нескольких из протопласта материнской клетки.
Половой процесс неизвестен (рис. 87).
Так же как и хлорококк, хлорелла — обычный симбионт ли-
шайников, но часто поселяется и в теле гидр, инфузорий.
Сценедесмус (Scenedesmus) — представитель простейших коло-
ниальных водорослей из порядка хлорококковых, обычный в планк-
тоне наших пресных водоемов. Колония состоит из четырех про-
186
долговатых клеток, соединенных в один ряд; реже восемь клеток
в два ряда. Колония снабжена длинными шипиками, помогающи-
ми ей держаться в толще воды. Размножается апланоспорами,
которые образуются в числе 4 в каждой клетке ценобия.
Представитель порядка улотриксовых улотрикс (Ulotrix) —
многоклеточная нитчатая зеленая водоросль (рис. 88). Нить невет-
Рис. 87. Хлорококковые водоросли. А — хлорелла (Chlorella), Б — рафидиум
(Raphidium), В — сценедесмус (Scenedestnus)
вящаяся, самая нижняя клетка нити
вытянута в длинный бесцветный ри-
зоид, которым водоросль прикрепля-
ется 'к субстрату. Остальные клетки
одинаковы, каждая имеет один хро-
матофор с пиреноидами и ядро. За
счет поперечного деления клеток
нить удлиняется.
Бесполое размножение посредст-
вом четырехжгутиковых зооспор, ко-
торые образуются в числе несколь-
ких из .протопласта вегетативных
клеток. Зооспоры двух сортов: круп-
ные и мелкие четырехжгутиковые.
Половой процесс изогамный. Га-
меты двужгутиковые образуются по-
добно зооспорам, но в большем чис-
ле. Зигота после периода покоя дает
четыре неподвижные гаплоидные
клетки, из которых вырастают новые
нити.
Нити (талломы) улотрикса гете-
роталличны.
Кладофора (Cladophora) — нит-
чатая ветвящаяся водоросль, рас-
пространенная в пресных и морских
водах. Нити достигают длины одно-
го метра. Как и улотрикс, кладофора
Рис. 88. Улотрикс (Ulothrix).
А — нить с зооспорами и гаме-
тами, Б — зооспоры, В — гаме-
ты и их копуляция
187
прикреплена к субстрату, но в связи с крупной величиной Цасто от-
рывается и образует плавающие скопления, так называемые тины,
темно-зеленого цвета, слизистые на ощупь. В отличие от улотрикса
клетки кладофоры многоядерны (рис. 89).
Бесполое размножение при помощи зооспор подобно улотриксу.
Половой процесс изогамный. У большинства видов кладофоры на-
блюдается чередование поколений, смена генераций. Бесполое по-
коление, дающее зооспоры, диплоидно; зооспоры гаплоидны, из них
развивается половое поколение, дающее гаметы; из зиготы разви-
вается диплоидное бесполое поколение. Следовательно, редукция
хромосом происходит при образовании зооспор, восстановлении
Рис. 89. Многоклеточная зеле-
ная водоросль кладофора (С1а-
dophora)
диплоидности — в половом процессе.
Бесполое диплоидное поколение на-
зывается спорофитом, половое (гапло-
идное) — гаметофитом. Внешне они
одинаковы.
У льва (Ulva) имеет пластинчатый
таллом; пластинка из двух слоев кле-
ток. Широко распространена по побе-
режью наших южных и северных мо-
рей. Крупная пластинка ее и по цвету,
и по форме напоминает листья салата,
отсюда и название ее — морской салат
(Ulva lactuca). Пластинка ульвы вна-
чале прикрепляется к субстрату, но
легко отрывается и продолжает жизнь
в таком состоянии.
Примером улотриксовой водоросли,
приспособленной к наземному образу
жизни, может быть плеврококк (Pleu-
rococcus). Таллом плеврококка реду-
цирован до одиночных клеток, иногда
непрочно соединенных в короткую нить,
размножение только вегетативное, пу-
тем деления клеток надвое. Обитают
плеврококковые на стволах деревьев,
заборов, образуя налет. В таких же ус-
ловиях можно обнаружить похожую
на плеврококк трентеполию (Trentepoh-
Па)\ клетки ее коричнево-красного цве-
та: хлорофилл маскируется красным
пигментом гематохромом, растворен-
ным в масле, капельки которого дис*
пергированы в цитоплазме.
Сифоновые водоросли морфоло-
гически весьма разнообразны, но все
они объединяются по признаку отсут-
ствия внутренних перегородок внутри
таллома, что дает основание говорить
188
Рис. 90. Водоросль неклеточного строения каулерпа (Caulerpa). А — общий вид,
Б — продольный разрез таллома
о неклеточном строении. Сюда относится крупная, до 1 м длины,
морфологически сложно дифференцированная морская водоросль
каулерпа (Caulerpa). Механическая прочность таллома усиливается
внутренними целлюлозными тяжами, пересекающими полость тал-
лома в различных направлениях (рис. 90). Таллом сифоновых дип-
лоидный, размножение обычно половое: изогамия или гетерогамия,
гаметы двужгутиковые; зооспоры редко, как исключение.
По внутреннему строению таллома к сифоновым близки воше-
риевые, это тоже водоросли неклеточного строения. В качестве
примера была рассмотрена широко распространенная в пресных
водах вошерия. Некоторые систематики в виду особенностей обме-
на веществ вошерии относят ее к отделу желтозеленых водорослей,
о чем уже сказано выше.
Класс сцеплянки, или конъюгаты (Conjugatae)
Одноклеточные и нитчатые водоросли, живущие в пресной воде,
реже на влажной почве. Характерной чертой этого класса является
отсутствие зооспор и апланоспор. Размножение путем вегетатив-
ного деления одноклеточных водорослей надвое или путем разрыва
нити. Половой процесс в виде слияния протопластов двух вегета-
тивных клеток с последующим редукционным делением диплоидно-
го ядра; вегетативные особи гаплоидны.
Большое количество видов конъюгат (около 4500) разделяют
на три порядка: 1) небольшой порядок наиболее примитивных ме-
зотениевых (Mesotaeniales), представленных одиночными клетка-
ми, обычно одетыми слизистым чехлом; 2) наиболее обширный по-
рядок десмидиевых (Desmidiales); в этом порядке около 3900 ви-
дов, в основном одноклеточных и лишь в небольшом количестве
многоклеточных нитчатых; третий порядок зигнемовых (Zygnema-
tales) также представлен небольшим количеством видов: здесь
только многоклеточные нитчатые водоросли.
189
Рис. 91. Spiro tae-
nia condensata.
Строение клетки.
Виден слизистый
чехол
вают оболочку,
У мезотениевых все четыре гаплоидные клетки, на которые де-
лится зигота, прорастают в новые особи. Это обитатели пресных
вод, бедных минеральными веществами (торфяные болота и т. п.).
Довольно типична для этого порядка спиротения
{Spirotaenia) —овальная клетка, покрытая
слизью; единственный крупный спиральный хро-
матофор с многочисленными периноидамн
(рис. 91).
У десмидиевых только две из четырех клеток
зиготы прорастают, что можно рассматривать как
шаг вперед в процессе эволюции. Огромное боль-
шинство десмидиевых — одноклеточные водорос-
ли, форма которых имеет резко выраженную дву-
стороннюю симметрию. Рисунки 92, 93 дают
представление о внешнем и внутреннем устрой-
стве типичных представителей этого порядка:
клостериума (Closterium) и космариума (Cos-
marium). У обоих родов в каждой из полу-
клеток продольновытянутые хроматофоры, у
клостериума с радиально расходящимися лопа-
стями. Ядро клетки расположено в срединной
части клетки, у космариума в этом месте пере-
тяжка. В оболочке поры, через которые выдавли-
вается слизь; клостериум, отталкиваясь от суб*'
страта посредством выделяемой через концевые
поры слизи, может активно перемещаться (рис.
92, 93).
При вегетативном размножении клетка рас*
падается на две симметричные части, и каждая
половинка достраивает вторую половину клетки.
При половом размножении две сблизившиеся
клетки одеваются общим слоем слизи, сбрасы-
протопласты их сливаются. После двукратного
деления ядра два из четырех ядер отмирают, а оставшиеся два
ядра, разделив между собой цитоплазму, превращаются в новые
особи. Обитают десмидиевые водоросли в topi же среде, что и мезо-
тениевые.
Из зигнемовых рассмотрим спирогиру (Spirogyra). Это одна из
наиболее широко распространенных зеленых водорослей (рис. 94).
Ярко-зеленые скользкие на ощупь нити ее образуют основную массу
тины, взвешенной в толще воды медленно текущих рек, озер и пру-
дов. Нить спирогиры состоит из цилиндрических клеток. Полость
клетки заполнена крупной вакуолей. Ядро висит на протоплазмен-
ных тяжах в центре клетки.
В пристенном слое цитоплазмы лентовидные спирально скру-
ченные хроматофоры. Края хроматофоров узорчатые; вдоль них,,
по средней линии, расположены пиреноиды, обычно окруженные
зернышками крахмала. Удлинение нити происходит за счет деле-
ния и роста составляющих ее клеток.
190
Оторвавшиеся -кусочки нити продолжают расти — так происхо-
дит вегетативное размножение спирогиры.
Половой процесс зигогамный: две нити сближаются, между
рядом лежащими клетками образуются канальцы, по которым со-
держимое клеток одной нити (ее можно считать мужской особью)
Рис. 93. Космариум. А — клётка
сверху, Б — спереди, В — деление
Рис. 92. Клостериум. А — вид клетки сбоку,
Б — поперечный разрез клетки
клетки
переливается в клетки другой особи (женской). Две рядом лежа-
щие конъюгирующие водоросли с поперечными перегородками
между ними напоминают своим видом лестницу, поэтому такая
конъюгация называется лестничной конъюгацией. Реже наблюда-
ется так называемая продольная конъюгация; при этом половой
процесс происходит внутри одной нити; протопласт одной клетки
переливается в соседнюю клетку той же нити. После слияния
протопласта двух клеток образуется зигоспора, покрытая тремя
оболочками, из которых средняя сильно утолщена и имеет бурую
окраску.
После периода покоя делится диплоидное ядро. В результате
двух делений образуются четыре гаплоидных ядра; три из них
отмирают, остающееся ядро получает весь протопласт зиготы и
прорастает в новую нить спирогиры.
191
Рис. 94. Спирогира (Spirogyra). А —
клетка спирогиры с тремя спираль-
ными хроматофорами, Б — нить спи-
рогиры, В — конъюгация спирогиры
В тех же условиях, что и спи-»
рогира, довольно часто встречают-
ся тоже нитчатые водоросли —
зигнема (Zygnema), клетки кото-
рой имеют’по два звездчатых хро-
матофора, и мужоция (Mougeo-
tid), у которой в клетках по
одному пластинчатому хромато-
фору.
ОТДЕЛ 11. ХАРОВЫЕ ВОДОРОСЛИ
(CHAROPHYTA)
Наиболее сложно устроенные
из пресноводных водорослей. Цел-
люлозные стенки клеток сильно
инкрустированы солями кальция.
Хроматофоры в виде мелких зе-
рен, без пиреноидов. Таллом мно-
гоклеточный, мутовчато-развет-
вленный, разделенный на междо-
узлия, высотой до 50 см. Из двух
широко распространенных родов
хара {Chard) и нителла {Nitelld)
наиболее известны хары, напоминающие по внешнему виду хвощи.
Обычная среда распространения харовых — пресноводные водое-
мы с жесткой водой, насыщенной солями кальция. Таллом хары
внутренне дифференцирован на коровую и сердцевидную части.
Вегетативное размножение — укореняющимися нижними ответ-
влениями таллома или специальными клубеньками, образующи-
мися на нижних узлах «стебля» или на ризоидах, которыми хара
закрепляется в субстрате. Спор у харовых нет. Половой процесс
оогамный. Органы полового размножения, оогоний и антеридий,
многоклеточны и довольно сложного строения (рис. 95), они обра-
зуются на узлах таллома, оогоний расположен над антеридием-.
Стенку оогония образуют пять спирально закрученных нитей, об-
разующих на верхушке коронку из пяти клеток. Внутри оогония
единственная яйцеклетка. Антеридий из 8 плоских клеток — щит-
ков, при созревании оранжевого цвета. С внутренней стороны свер-
нутые в клубок сперматогенные нити, каждая из 100—300 клеток,
превращающихся в сперматозоиды. Созревший антеридий распа-
дается на отдельные щитки, и сперматозоиды разъединяются. Оп-
лодотворенная сперматозоидом яйцеклетка превращается в ооспо-
ру, которая после периода покоя прорастает в новую хару. Клетки
хары гаплоидные, диплоидна только ооспора.
Рис. 95. Хара (Chara). А — общий вид, Б — предросток:
/-«ризоиды; 2 — клубеньки; 3 — боковая ветвь; 4 — оогоний; 5 — антеридий;
6 — щиток антеридия; 7 —* сперматогенные нити; 8 —* сперматозоиды
РОЛЬ ВОДОРОСЛЕЙ В ПРИРОДЕ И ИСПОЛЬЗОВАНИЕ
ИХ ЧЕЛОВЕКОМ
Водоросли широко распространены во всех естественных и ис-
кусственных водоемах земного шара. По количеству производимой
живой массы в реках и озерах большую роль играют бентосные
водоросли, в морях и океанах — планктонные.
Общее количество биомассы, создаваемое водорослями, особен-
но планктонными, колоссально. Достаточно привести несколько
цифр ежегодного «урожая» водорослей. Так, например, один гек-
тар открытой поверхности Баренцева моря дает растительного
планктона 30—50 т, а общая годовая продукция фитопланктона
этого моря составляет 5,6 млрд, т, фитобентоса — несколько мил-
лионов тонн; годовая продукция 1 га растительного планктона
пресноводных озер составляет также несколько десятков тонн сы-
рого вещества. Еще больший урожай могут давать заросли бен-
тосных морских водорослей (преимущественно бурых) —100—
200 т и больше с 1 га. В сравнительно небольшом Каспийском
7—326
193
море ежегодная продукция фитопланктона составляет около
1 млрд, т, фитобентоса — около 3 млн. т.
В процессе фотосинтеза водорослей освобождается огромное
количество кислорода, который частично растворяется в воде, час-
тично выделяется в атмосферу, поддерживая более или менее по-
стоянный уровень этой составной части воздуха. За счет органиче-
ского вещества, создаваемого водорослями, живет богатейший
и разнообразный мир рек, морей и океанов.
Немало водорослей живет и в почве; подсчитано, что общая
масса их составляет на 1 га почвы свыше 100 кг.
Растительный бентос и планктон — основная кормовая база
для рыбы и крупных морских млекопитающих, добываемых чело-
веком.
Диатомовые водоросли, отмирая, оставляют кремневые оболоч-
ки, которые могут образовать мощные залежи на дне моря. По
выходе морского дна на поверхность эти запасы оказываются до-
ступными для человека. Под названием горной муки, трепела или
диатомита этот материал широко используется в промышленности
как термический изолятор, нри изготовлении строительных мате-
риалов, в качестве фильтрационного слоя и для многих других
целей.
Вблизи морей бурые и отчасти красные водоросли, вернее пе-
регной, который из них образуется, широко используется для удоб-
рения полей.
В последние годы начаты опыты по выращиванию некоторых
хлорококковых водорослей (преимущественно хлореллы) в специ-
ально построенных водоемах. Органическое вещество этих водо-
рослей весьма богато белками, жирами и витаминами. Предпола-
гают, что выращиваемые водоросли могут быть использованы для
получения ценного пищевого концентрата и для людей. Многие ви-
ды красных водорослей давно служат обычной пищей в Японии
и на некоторых островах Тихого океана. В Китае употребляют
в пищу синезеленую водоросль носток. Широко используются в ка-
честве лечебного средства (йод) некоторые виды бурых водо-
рослей, преимущественно ламинарий, под названием морской
капусты.
Морские водоросли используются также и как промышленное
сырье. Из золы бурых водорослей добывают йод, калийные и на-
триевые соли, а из органической части — клеящие вещества.
Вываривая некоторые виды красных водорослей, получают сли-
зистое вещество — агар-агар. Большое количество агар-агара рас-
ходуется в лабораторной бактериологической практике для изго-
товления питательной среды. Агар-агар также широко использует-
ся в пищевой промышленности при изготовлении мармелада, па-
стилы и других изделий. Применяют его в текстильном и бумаж-
ном производстве.
Филлофора, добываемая в Черном море (Phyllophora nervosa),
используется для добывания агар-агара и йода.
194
ФИЛОГЕНЕЗ ВОДОРОСЛЕЙ
Древнейшие из всех водорослей — синезеленые, как уже было
сказано в начале курса; они возникли около 2 миллиардов лет то-
му назад, позже, чем первые гетеротрофные организмы, с которых
началась жизнь на Земле. Первые организмы нашей планеты —
эобионты, очевидно, были устроены значительно проще, чем про-
исшедшие от них современные бактерии, наиболее древние из ны-
не живущих прокариотных организмов. Синезеленые водоросли
возникли, по-видимому, не от бактерий, а от более примитивных,
древнейших амебоидных прокариотных организмов — эобионтов.
Согласно гипотезе симбиогенеза возникновение первых эука-
риотных организмов связано с проникновением в прокариотную
клетку спирохетоподобной бактерии. Так возникли простейшие жгу-
тиковые формы, обладающие оформленным ядром. Проникновение
в клетки таких организмов синезеленых водорослей положило на-
чало первым автотрофным эукарибтным организмам — водорослям.
Все девять отделов эукариотных водорослей, по-видимому, пред-
ставляют самостоятельные эволюционные ветви (филы), берущие
начало от группы первичных эукариотных организмов.
ОТДЕЛ 12. МИКСОМИЦЕТЫ (MYXOPHYTA)
Тело миксомицета представляет собой скопление голой прото-
плазмы с большим количеством ядер (рис. 96). Ядра диплоидные.
Слизистые тела, или плазмодии, миксомицетов можно обнаружить
среди гниющего корья, в щелях старых пней или досок, под опав-
шими листьями в лесу. Живущие
здесь миксомицеты питаются сап-
рофитно. Окраска плазмодия раз-
личная: беловатая, желтая, крас-
новатая и др. Размер от несколь-
ких миллиметров до нескольких
десятков сантиметров.
Миксомицеты могут амебооб-
разно передвигаться и избегают
света. К моменту размножения,
наоборот, плазмодий выползает
на свет, и тело его превращается
в скопление спорангиев, заполнен-
ных спорами вперемешку с тон-
кими упругими волоконцами —
капиллицием. Волоконца эти раз-
рыхляют массу из спор и способ-
ствуют лучшему распылению их.
Спорангии иногда сливаются, об-
разуя общие вместилища — эта-
лии.
Споры гаплоидны. Попадая во
влажные места, споры прораста-
Рис. 96. Миксомицет. А — часть плаз-
модия миксомицета, Б — прорастания
споры миксомицета, В — зооспора
195
ют, при этом содержимое их превращается в две одножгутиковые
зооспоры, способные делиться. Спустя некоторое время зооспоры
втягивают жгутики и превращаются в амебы, которые также могут
размножаться делением. Передвигаясь, амебы попарно копулиру-
ют, и при этом их гаплоидные ядра сливаются.
Диплоидные амебы, сливаясь по нескольку, образуют тело плаз-
модия, при этом ядра уже не сливаются. Плазмодий питается, вса-
сывая раствор органических веществ всей своей поверхностью. Мо-
жет питаться также и твердыми частицами, например бактериями,
поглощая их и переваривая внутри своего тела. В этом он обнару-
живает сходство с животным организмом.
Существует несколько видов паразитных миксомицетов; наибо-
лее широко известен Plasmodiophora brassicae — паразит, вызыва-
ющий болезнь «капустную килу». Голый плазмодий этого паразита
можно обнаружить в коровой паренхиме на корнях капустных рас-
тений. Поселяясь здесь, этот паразит вызывает образование опу-
холей; заболевшее растение хиреет, а то и вовсе погибает.
Пораженные клетки, содержащие одноядерные голые протоплаз-
матические тела паразитов, не сразу погибают, они продолжают
делиться; при этом тело плазмодия распределяется между двумя до-
черними клетками. Так происходит размножение паразита и увели-
чение его массы. В результате слияния амеб ядра копулируют и
диплоидный плазмодий продолжает дальнейший рост за счет пита*
тельных веществ, создаваемых растением.
После отмирания клеток хозяина голые клетки плазмодия прев-
ращаются в споры, в состоянии которых миксомицет переживает
неблагоприятные условия внешней среды; спорангии эти слизевики
не образуют. Споры превращаются в двужгутиковые зооспоры или
амебы, которые, по-видимому, попарно копулируют, и диплоидный
плазмодий проникает в нежные ткани корня растения, скорее всегр
через корневые волоски. Возможно также, что копуляция происхо-
дит внутри клеток хозяина.
Паразит, вызывающий болезнь капусты, может поражать и дру-
гих представителей семейства крестоцветных, например брюкву.
В северо-западных областях нашей страны он наносит значитель-
ный ущерб сельскому хозяйству. Серьезный ущерб запасам картофе-
ля наносит также паразитический миксомицет спонгоспора (Spon-
gospora solant), вызывающий порошистую паршу клубней карто-
феля.
Что касается сапрофитных представителей этого отдела (их боль-
шинство), то они никакого практического значения не имеют. При-
мером их могут быть Lycogala — оранжево-красный плазмодий или
Trichia, имеющий чаще желтоватую окраску. Оба миксомицета жи-
вут на мертвой, гниющей древесине и коре.
Всего известно около 500 видов миксомицетов, большинство из
них сапрофиты.
Слизевики по своей биологии близки к низшим грибам, однако
произошли они, по-видимому, самостоятельно, непосредственно от
первичной группы бесхлорофильных эукариотных организмов.
196
ОТДЕЛ 13. ГРИБЫ (FUNGI, MYCOPHYTA)
К грибам относится обширная группа организмов исключитель-
но гетеротрофного питания. Способ питания, а также формы раз-
множения — это основное, что объединяет все грибы в один отдел,
в остальном они весьма разнообразны. Насчитывается около 100 000
видов грибов.
Размер относящихся к этому отделу организмов колеблется в
широких пределах. Тело очень многих видов микроскопически ма-
ло, и в этом отношении они сближаются с бактериями. С другой
стороны, среди грибов, например из группы трутовиков, живущих
на древесине, имеются экземпляры, тело которых достигает полу-
метра и более.
Протопласт клеток гриба не содержит пластид; ядра мелкие,
одно или несколько. Крахмал не свойствен грибам, вместо него
в качестве запасного питательного вещества отлагается гликоген.
Это сближает грибы с животным миром. Имеются капельки масла.
Клеточные оболочки лишь у немногих, примитивных низших гри-
бов целлюлозные, у большинства же видов они имеют более слож-
ный состав: углевод, близкий к
целлюлозе, соединен с азотсодер-
жащим веществом, сходным с хи-
тином насекомых. Наличие при-
митивных форм с целлюлозными
оболочками указывает на общ-
ность происхождения грибов и во-
дорослей.
Таллом гриба состоит из тон-
ких нитей—гиф. Лишь некото-
рые, наиболее примитивные или
упростившиеся грибы имеют од-
ноклеточное строение. Совокуп-
ность гиф называется мицелием
или грибницей (рис. 97). Мицелии
могут быть гомоталличными —
физиологически однозначны и ге-
тероталличными — физиологиче-
ски разнозначны, или разнополые,
сплетаются, образуя нечто, напоминающее ткань; такое образова-
ние называется ложной тканью или плектенхимой. Тугое сплетение
гиф, на поверхности или внутри которого образуются споры, служа-
щие для размножения, называется плодовым телом. Таким плодо-
вым телом обладают, например, обычные шляпочные грибы.
Плотные сплетения гиф, заполненные запасными питательными
веществами, называются склероциями. В форме склероциев неко-
торые грибы переживают неблагоприятное время года.
Питается гриб путем всасывания растворенной пищи всей по-
верхностью мицелия. Мицелий обычно бывает сильно разветвлен,
это увеличивает всасывающую поверхность и, следовательно, об-
Рис. 97. Развитие мицелия гриба
из споры
Иногда нити мицелия очень туго
197
легчает процесс питания. Иногда пучки гиф образуют длинные
плотные шнуры — ризоморфы (у опенка, домового гриба и др.).
Эволюция мицелия началась с неклеточного строения, и у мно-
гих грибов при очень большой длине гиф в мицелии совершенно
отсутствуют клеточные перегородки. Низшие грибы имеют нерас-
члененный мицелий с большим количеством ядер или вовсе его не
имеют; высшие грибы имеют клеточный мицелий и филогенетически
находятся на более высокой ступени развития. Клетки многих гри-
бов многоядерны, а клеточные поперечные стенки имеют отверстия.
На мицелии развиваются органы спороношения. Поэтому мице-
лий иногда называют вегетативным телом гриба, противопостав-
ляя его плодовым телам и спороносящей части гриба.
Мицелий имеет различную продолжительность жизни, от не-
скольких дней (например, у плесени) до многих лет (например,
у шляпочных грибов). Долговечностью мицелия объясняется на-
хождение некоторых грибов из года в год в одном и том же месте.
Любители собирать съедобные грибы знают такие грибные места
и каждый год их посещают.
По способу питания грибы можно разделить на сапрофитов
и паразитов. Сапрофиты питаются мертвыми органическими веще-
ствами, паразиты нуждаются в живом белке. Многие грибы-пара-
зиты могут питаться и сапрофитно, например, трутовик вначале
поселяется на живом организме, а после его смерти продолжает
питаться за счет отмерших тканей.
Меньшая часть грибов-паразитов может жить только на живых
организмах, со смертью хозяина погибает и паразит. Такие пара-
зиты называются облигатными в отличие от частичных, или факуль-
тативных, которые могут жить и на мертвом органическом веще-
стве.
Характерной чертой грибов является разнообразие форм раз-
множения. Грибы могут размножаться вегетативно — кусочками
мицелия, почкованием или клетками, на которые иногда распада-
ется мицелий; такие клетки носят название оидий или хламидо-
спор, в последнем случае они одеты толстой темной оболочкой.
Бесполое размножение осуществляется при помощи специальных
спор; с разнообразием спор мы познакомимся при рассмотрении
отдельных групп грибов. Наконец, грибам свойствен и половой
процесс.
У низших грибов встречается хологамия, изогамия, гетерогамия
и зигогамия. После некоторого периода покоя зигота у низших гри-
бов делится редукционно. Гаплоидные клетки дают начало новому
мицелию, спорам или зооспорам, которые, как и мицелий, гаплоид-
ны; следовательно, диплоидна только зигота.
У высших грибов половой процесс упрощен: содержимое жен-
ского полового устройства оплодотворяется переливающимся про-
топластом мужского устройства или мелкими клетками — спермия-
ми, сливаются протопласты двух вегетативных клеток, сливаются
попарно ядра в одной клетке. У высших грибов широко распростра-
нен незаконченный половой процесс (плазмогамия), когда слива-
198
ется только цитоплазма, а ядра только сближаются, образуя дика-
рионы. Дикарионы могут размножаться. Половой процесс завер-
шается слиянием парных ядер (кариогамия), после чего сразу же
происходит редукционное деление.
Отдел грибы подразделяется на шесть классов.
1. Хитридиомицеты (Chytridiomycetes). Тело гриба — плазмодий
или очень слабо развитый мицелий. Одножгутиковые зооспоры.
Половой процесс в форме изогамии, гетерогамии, оогамии или хо-
логамии (слияние двух вегетативных особей).
2. Оомицеты (Oomycetes). Мицелий хорошо развит, но некле-
точный. Двужгутиковые зооспоры. Половой процесс оогамный.
3. Зигомицеты (Zygomycetes). Мицелий неклеточный, много-
ядерный. Половой процесс зигогамный.
4. Аскомицеты, или сумчатые грибы (Ascomycetes). Мицелий
клеточный- Бесполое размножение при помощи спор — конидий.
Споры, возникшие в результате полового процесса, развиваются
в особых вместилищах — сумках (эндогенно).
5. Базидиомицеты (Basidiomycetes). Мицелий клеточный. Бес-
полое размножение спорами. Споры полового размножения разви-
ваются на особых органах — базидиях (экзогенно).
6. Несовершенные грибы (Fungi imperfecti). Мицелий клеточ-
ный. Размножение спорами. Половой процесс неизвестен. Первые
три класса относят к низшим грибам, их свыше 1000 видов, аско-
мицеты и базидиомицеты — к высшим. Несовершенные грибы по
строению мицелия примыкают к высшим грибам.
Класс хатридиевые (Chytridiomycetes)
Этот класс непосредственно примыкает к бесцветным жгутико-
вым, которые рассматриваются как простейшие представители
животного мира. К хитридиевым относятся наиболее древние при-
митивные формы, не утратившие еще связи с водной средой. Боль-
шинство из них паразитирует на водорослях, некоторые — на выс-
ших растениях. Всего известно около 300 видов хитридиевых.
Из этого класса подробно рассмотрим только одного предста-
вителя, паразитирующего на капусте и приносящего ущерб сель-
скому хозяйству,— олышдиум (Olpidium brassicae).
Голые комочки протоплазмы, представляющие тело гриба, об-
наруживаются в клетках коровой паренхимы корневой шейки рас-
тений капустной рассады. Все тело паразита превращается в ша-
ровидный зооспорангий (рис. 98). Из трубчатого отростка
спорангия выходят одножгутиковые зооспоры. При наличии воды
зооспоры некоторое время двигаются, затем прилипают к поверх-
ности стебелька капусты, теряют жгутики и переливают свое со-
держимое в клетку эпидермиса. Отсюда паразит перебирается
в более глубокие слои коры и вновь превращается в зооспорангии.
Зооспоры могут, как гаметы, сливаться попарно, давая подвижную
двужгутиковую зиготу, которая и заражает здоровые растения.
199
Весь цикл развития обычно заканчивается в 2—3 дня, и, следо-
вательно, при благоприятных условиях перезаражение происходит
очень быстро. Зимует гриб в виде споры — цисты, снабженной
толстой оболочкой. Весной спора развивается в зооспорангий.-
Поврежденное паразитом растение гибнет, при этом стебелек
рассады чернеет; отсюда название болезни — «черная ножка ка-
пустной рассады». Меры борьбы с болезнью — умеренный полив
и полная дезинфекция парников перед посевом новой рассады.
Из паразитических представителей грибов этого класса боль-
шой вред народному хозяйству способен наносить гриб синхитриум
(Synchytrium endob ioticum), вызывающий болезнь клубней карто-
феля — рак картофеля.
Подобно ольпидиуму, зооспоры синхитриума также имеют один
жгутик, но в отличие от ольпидиума этот гриб образует группу зоо-
Рис. 98. Ольпидиум (Olpidium brassicae). А — заболевание
«черпая ножка» капустной рассады, Б — зооспорангий в клет-
ках ткани капусты, В — зооспоры, Г — спорангии в состоянии
покоя
спорагиев (сорус); помимо зооспор синхитриум образует и одно-
жгутиковые гаметы; половой процесс изогамный. Зигота зимует
в клубне в виде цисты, которая весной прорастает в новое поколе-
ние паразита. Пораженные ткани клубня разрастаются в темные
наросты.
Класс оомицеты (Oomycetes)
В качестве представителя этого класса рассмотрим паразитиче-
ский гриб фитофтору (Phytophthora infestans), относящуюся к по-
рядку переноспоровых (Peronosporales). Этот гриб вызывает
болезнь картофеля: поражает ботву и клубни; оставаясь в клубнях
200
и попадая вместе с ними в хранилища, вызывает массовое загни-
вание и гибель картофеля.
Заболевание растения обнаруживается в появлении темнык
пятен на листьях картофеля (рис. 99). В мезофилле такого листа
находится мицелий гриба. Разветвленные кончики гиф — споран-
гиеносцы высовываются из устьиц пораженного листа; разветвле-
ния заканчиваются спорангиями округло-удлиненной формы. По-
раженным грибом листья быстро отмирают.
Осыпающиеся спорангии, попадая на капельки воды, прора-
стают, и из них выходит около десятка двужгутиковых зооспор.
Подвигавшись некоторое время, зо-
оспоры одеваются оболочкой и за-
тем прорастают в гифу. Гифа прони-
кает сквозь эпидермис внутрь листа
и там развивается в мицелий. Таким
же путем происходит заражение и
клубней. При отсутствии капельно-
жидкой воды спорангий непосредст-
венно прорастает в гифу, энергия за-
ражения при этом, следовательно,
уменьшается.
Опадающие на почву спорангии
также могут прорастать; возникаю-
щий из них мицелий питается сап-
рофитио. Следовательно, фитофто-
ра — факультативный паразит.
Фитофтора — гетероталличный
гриб, однако половое воспроизведе-
ние известно только на родине это-
го вида — в Мексике.
Источник первичного зараже-
ния— высаженные весной поражен-
ные болезнью клубни.
В северо-западных районах на-
шей страны этот гриб в годы массо-
Рис. 99. Фитофтора (Phythophtho-
га) — разрез листа, на котором
видны гифы гриба и спорангиенос-
вого размножения наносит огром- цы со спорангиями
ный ущерб, вызывая гибель большо-
го количества продовольственного и кормового картофеля. Ради-
кальная мера борьбы — выведение сортов, устойчивых против фи-*
тофторы.
Сходно с фитофторой развивается плазмопара (Plasmopara
viticola), вызывающая болезнь винограда — мильдыо.
К этому классу относятся сапролегниевые грибы (Saprolegnia),
относящиеся к порядку Saprolegniales, для которого характерно
образование в оогонии нескольких яйцеклеток. Чаще это сапро-
фиты, живущие в водной среде на трупах насекомых (например,
мух) п других мелких животных. Но сапролегния может также
паразитировать на икре и мальках рыб, на пораненных рыбах.
Этим она наносит ущерб рыбным заводам и аквариумам.
201
Класс загомицеты (Zygomycetes)
Преимущественно сапрофиты, но имеются и паразиты, живущие
на позвоночных животных (рыбах, лягушках, ящерицах) и насеко-
мых. Наибольший интерес представляет сапрофитный гриб—• го-
ловная плесень (несколько, видов рода Мисог — М. mucedo,
М. alba-ater и др.).
Широко распространенный сапрофит, живущий на разнообраз-
ных продуктах растительного происхождения, а также на навозе
Рис. 100. Гриб мукор (Mucor mucedo). А—общин вид мицелия
со спорангием, Б, В, Г — развитие спорангия, Д, Е, Ж — половой
процесс и развитие зиготы, 3 — зрелая зигота и ее прорастание
(рис. 100). Белые гифы мицелия мукора густо оплетают поверх-
ность субстрата и проникают внутрь него.
На мицелии развиваются толстые спорангиеносцы, заканчиваю-
щиеся шаровидными спорангиями. В спорангиях большое количе-
ство гаплоидных спор; к моменту созревания споры покрываются
темной оболочкой, поэтому спорангии с созревшими спорами ста-
новятся почти черными. Такие споры, развивающиеся внутри спо-
рангия, называются спорангиеспорами или эндоспорами. Из спор
202
вырастают новые мицелии, и, таким ооразом, происходит очень
быстрое распространение плесени.
Половой процесс зигогамный. Кончики гиф двух мицелиев —
гаметангии (Мисог— гриб гетероталличный) сближаются и отделя-
ются перегородками. Стенки соприкоснувшихся кончиков мицелия
растворяются, два многоядерных протопласта сливаются в зиготу,
при этом гаплоидные ядра соединяются попарно. Зигота одевается
толстой оболочкой и превращается в зигоспору. После непродол-
жительного покоя зигоспора прорастает в гифу с зародышевым
спорангием.
В зародышевом спорангии в результате редукционного деления
образуются гаплоидные споры. Споры эти физиологически нерав-
нозначны. Возникающие из них гаплоидные мицелии также физио-
логически различны (явление гетероталлизма) и производят спо-
ры только одного знака. Диплоидность восстанавливается только
в зиготе.
Мукоровая плесень, поселяясь на сочных плодах и овощах,
портит их при хранении. С другой стороны, размножаясь в огром-
ных количествах в почве, грибы этой группы приносят существен-
ную пользу, разлагая органические вещества и вовлекая их в даль-
нейший круговорот.
Мукор относится к наиболее богатому видами порядку мукоро-
вых (Mucorales). Почти все мукоровые — сапрофиты, однако име-
ются и паразиты, например виды рода абсидии (Absidia), пора-
жающие дыхательную систему человека. У представителей поряд-
ка энтомофторовых (Entomophthorales), представленных парази-
тами, бесполое размножение экзоспорами — конидиями. Энтомо-
фторовые паразитируют на насекомых и представляют интерес для
биологического метода борьбы с вредителями. Представитель этого
порядка Etnpusa muscae паразитирует на комнатных мухах.
Класс аскомицеты (Ascomycetes)
Высшие грибы с расчлененным, многоклеточным мицелием.
Споры бесполого'размножения не заключены в спорангии, они
образуются свободно на кончиках и разветвлениях гиф. Такие
споры называются конидиями. Для грибов этого класса характерно
также образование особых вместилищ — сумок, в которых нахо-
дится всегда определенное количество спор, обычно восемь; споры
эти носят название аскоспор или сумчатых спор.
Сумки развиваются на дикарионном мицелии, который являет-
ся продуктом незаконченного полового процесса. Лишь у наиболее
примитивных представителей сумка развивается непосредственно
из зиготы.
В наиболее простой форме у самых примитивных аскомицетов
половой процесс выражается в слиянии одноядерных протопластов
из выростов двух соседних клеток мицелия.
В типичном случае женский половой орган, называемый архи-
карпом, состоит из нескольких клеток. Нижняя часть его вздута
203
и называется аскогоном, вытянутая вверх часть — трихогиной. Ря-
дом с архикарпом развивается антеридий, имеющий форму удли-
ненной цилиндрической клетки. Аскогон и антеридий многоядерны.
К моменту оплодотворения кончик антеридия срастается с кон-
цом трихогины, и содержимое антеридия переливается через три-
хогину в аскогон. Ядра антеридиального протопласта не сливают-
ся с ядрами аскогона (плазмогамия), а только сближаются с ними,.
Рис. 101. Схема развития сумки со спорами (А) и базидии (Б)
образуя так называемые дикарионы. В дальнейшем из аскогона
развивается целый клубок гиф, состоящих из дикарионных клеток.
В каждой клетке может быть по нескольку пар ядер, из которых
одно материнское, второе отцовское.
Гифы, содержащие дикарионы, называются аскогенными-, на
концах этих гиф образуются сумки, или аски.
Несколько упрощенно можно представить процесс образования
сумки и сумчатых спор (аскоспор) следующим образом (рис. 101).
К началу формирования сумки два ядра дикариона конечной клет-
ки аскогенной гифы сливаются (кариогамия), этим завершается
половой процесс, предварительным результатом которого (плазмо-
гамия) были дикарионы. Диплоидное ядро, полученное в результа-
204
те слияния ядер дикариона, делится редукционно, в результате
чего опять восстанавливается гаплоидность ядер. Гаплоидные ядра
делятся еще дважды, в результате чего в разросшейся клетке ока-
зывается уже восемь гаплоидных ядер. Цитоплазма клетки рас-
пределяется между восемью ядрами; образуется восемь клеток,
они одеваются твердой оболочкой и превращаются, таким образом,
в аскоспоры. Оболочка материнской клетки становится вмести-
лищем спор — сумкой.
Биологический смысл своеобразной формы полового процесса,
приводящего к образованию аскогенных гиф, заключается в том,
что один половой процесс дает не одну зиготу, как обычно, а не-
определенно большое количество их, поскольку каждая клетка
аскогенной гифы в потенции может после слияния ядер дикариона
превратиться в сумку.
Лишь у некоторых наиболее примитивных аскомицетов сумки
расположены непосредственно на мицелии. У большинства же
сумчатых грибов сумки, оплетенные аскогенными гифами, распо-
лагаются на поверхности или внутри особых плодовых тел, или
аскокарпов. Основа плодовых тел состоит из плотного сплетения
обычных вегетативных гаплоидных гиф; по характеру строения
плодовые тела бывают трех типов.
Клейстокарпии — замкнутые плодовые тела. Сумки освобожда-
ются в результате разрыва или сгнивания плодового тела.
Перитеции — полузамкнутые плодовые тела, имеющие форму
кувшинчика. Сумки высовываются через горлышко перитеция,
и споры активно выбрасываются из сумок.
Апотеции — открытые плодовые тела в форме блюдца. Сумки
располагаются на поверхности апотеция вертикально, вперемежку
с кончиками бесплодных гиф — парафизами. Слой из сумок и па-
рафизов называется гимением или гимениальным слоем.
У многих аскомицетов сумки встречаются очень редко, и у них
преобладает бесполое размножение при помощи спор — конидий.
Всего имеется свыше 25 000 видов сумчатых грибов. Класс
аскомицетов делится на два подкласса.
1. Подкласс голосумчатые (Protoascomycetidae), не имеющие
плодовых тел.
2. Подкласс плодосумчатые (Euascomycetidae), имеющие пло-
довые тела.
Первый подкласс содержит два порядка: Protascales и ТарЬгн
nales; во втором подклассе рассмотрим пять порядков: Plectascales,
Erysiphales, Pyrenomycetales, Discomycetales, Tuberales. Порядки
пиреномицетов и дискомицетов в современной классификации де-
лятся на несколько более мелких порядков.
Подкласс голосумчатые (Protoascomycetidae, или
Gymnoascomycetidae)
Порядок протасковые (Protascales). Наиболее примитивные
формы, преимущественно сапрофиты, но некоторые паразитируют
на животных и человеке. По происхождению граничат с зигомице-
205
тами, что находит выражение в многоспоровости сумок у некото-
рых древнейших форм; такая сумка аналогична зародышевому
спорангию зигомицетов.
Наибольший интерес в этом порядке представляют пивные
дрожжи (Saccharomyces cerevisiae) (рис. 102). Этотвид дрожжей,
известный только в культуре, сбраживает глюкозу с образованием
этилового спирта; выделяются углекислый газ и тепло. Пивные
дрожжи применяются в пивоварении, хлебопечении и в производ-
стве спирта из углеводов. Винные дрожжи используются в вино-
делии.
Под влиянием жизни в благоприятной жидкой среде дрожжи
сильно упростились. Мицелий гриба состоит из почкующихся кле-
ток овальной или округлой формы.
Половой процесс в форме слияния двух гаплоидных клеток.
Из зиготы образуется сумка, обычно с четырьмя аскоспорами, ко-
торые многократно почкуются. После слияния аскоспор диплоид-
ные клетки могут или сразу формировать сумки, или (у большин-
ства) могут длительное время почковаться в состоянии диплоид-
ной фазы. Немногие виды первичносумчатых грибов образуют
споры бесполого размножения — конидии.
Рис. 102. Пивные дрожжи (Saccharomyces cerevisiae). А — отдель-
ная клетка, Б — редуцированный мицелий из нескольких клеток,
В, Г, Д — почкование клеток, Е — сумка с четырьмя аскоспорами
Порядок тафриновые (Taphrinales). Сюда относятся исключи-
тельно паразитические виды, вызывающие болезни косточковых
плодовых деревьев. Болезнь выражается в появлении уродливых
листьев (курчавость листьев) или плодов. Представитель Т. defor-
mans вызывает курчавость листьев персика- В утолщенных местах
обычно деформированных листьев под кутикулой или глубже обра-
зуются сумки с 8 аскоспорами. Зрелые сумки прорывают кутикулу,
а находящиеся в них аскоспоры выходят наружу; аскоспоры могут
почковаться. При слиянии аскоспор образуются дикарионы, кото-
рые многократно делятся; половой процесс заканчивается лишь
перед образованием сумок.
206
Подкласс плодосумчатые (Euascomycetidae, или
Carpoascomycetidae)
Порядок плектасковые—Plectascales. Бесполое размножение
конидиями, сумки в беспорядке и в неопределенном количестве
расположены в клейстокарпиях, обнаруженных лишь у немногих
видов; сумки освобождаются при сгнивании оболочки клейстокар-
пия.
Интерес представляют дца рода из этого порядка: Penicillium
п Aspergillus (рис. 103)..
Конидии пеницилла образуются на кончиках пальчаторазвет-
вленных конндиеносцев. Этим пеницилл морфологически отличает-
□ о о
Рис. 103. Пеницилл (Penicillium) и аспергилл (Aspergillus). А — об-
щий вид пеницилла, Б — конидиеносец с отделяющимися конидиями
аспергилла
ся от аспергилла, у которого конидии формируются на слегка взду-
том кончике конидиеносца.
Грибы этих двух родов в остальном внешне сходны между со-
бой и образуют так называемую сизую или зеленую плесень, кото-
рая, поселяясь на влажных кормах (например, в плохо подсушен-
ном сене или на овощах, плодах), делает их негодными к употреб-
лению. Скармливание животным сена или других грубых кормов,
пораженных плесенью, может вызвать тяжелые заболевания ды-
хательных путей и пищеварительных органов.
Некоторые виды грибов живут в почве и выполняют здесь по-
лезную роль разрушителей органических остатков. Виды пеницил-
ла, например Р. roqueforti, принимают участие в созревании ценных
207
сортов сыра. Два вида пеницилла — Р. notation и Р. chrysogenum—
в настоящее время используются для приготовления пенициллина,
ценного и широко применяемого антибиотика. С этой же целью
используются и некоторые виды аспергилла-
Порядок мучнисторосые (Erysiphales). Сюда относятся парази-
тические грибы, вызывающие болезнь — мучнистую росу (рис. 104).
Это название связано с появлением белого мучнистого налета на
пораженных органах, состоящего из большого числа конидий. Ко-
нидии отшнуровываются с кончиков нитей мицелия, который в виде
паутинки заплетает поверхность пораженного органа; посредством
присосок мицелий проникает в клетки эпидермиса, паразитируя
на них.
Рис. 104. Мучнисторосые грибы. А — мицелий с конидиями на поверхно-
сти листа, Б — треснувший клейстокарпий мучнистой росы злаков
(Erysiphae)
С помощью конидий происходит массовое заражение растений.
Истощив пораженный орган, гриб образует клейстокарпии, в кото-
рых одна или небольшое число сумок, расположенных правильно,
в форме пучка. В сумках — аскоспоры. На клейстокарпиях выра-
стают различного вида придатки, способствующие удержанию
клейстокарпия на мицелии до полного созревания.
Сумки зимуют в клейстокарпиях, опавших с пораженными ор-
ганами. Весной сумки разбухают, разрывают клейстокарпий, и вы-
шедшие из сумок аскоспоры производят первичное заражение.
Пример — мучнистая роса злаков (Erysiphae graminis), сферотека
крыжовника (Sphaerotheca mors uvae), «пепелица» винограда
(Uncinula necator), микросфера дуба (Microsphaera alphitoides).
208
Порядок пиреномицеты (Pyrenomycetales). Включает свыше
10 000 видов, среди них паразиты и сапрофиты. Плодовое тело —
перитеций вытянутой формы, с отверстием на верхнем конце. Соз-
ревшие сумки высовываются через отверстия в перитеции и актив-
но разбрасывают споры — способ более совершенный, чем у грибов
с замкнутым плодовым телом, клейстокарпием. Перитеции у неко-
торых представителей собраны группами в особом сплетении гиф —
строме.
Среди паразитических видов грибов этого порядка много воз-
будителей заболеваний травянистых растений и плодовых деревьев.
Широко известен гриб, паразитирующий на ржи и некоторых
других злаках,— спорынья (Claviceps purpurea). Поражение ржи
Рис. 105. Спорынья (Claviceps purpurea). А — колос ржи, пораженный спорыньей,
Б — проросший склероций со стромами, В — разрез через строму, Г — перитеций
с сумками, Д — сумка с восемью нитевидными спорами, Е — медвяная роса
на цветущем колосе, Ж — конидии
спорыньей обнаруживается к моменту налива зерна. В это время
в колосе ржи можно заметить темно-фиолетовые рожки длиной
1—3 см, развивающиеся вместо зерен (рис. 105). Опадая на почву
или попадая туда вместе с посеянным зерном, склероции лежат до
весны.
Весной на склероции вырастает около десятка красноватых
головок на ножках диаметром в 1—2 мм. Эти головки называются
стромами. В периферической зоне стромы в ее мякоть погружены
перитеции, заполненные сумками. В каждой сумке восемь аско-
спор нитевидной формы. Споры выталкиваются из сумок и токами
воздуха разносятся по полю.
Попадая на рыльца цветков ржи, споры прорастают, и мицелий
внедряется в завязь. Вскоре здесь развивается конидиальное спо-
209
роношение. Одновременно с образованием конидий на пораженной
завязи выделяются капельки сладкой жидкости — медвяной росы.
Насекомые, потребляющие медвяную росу, переносят конидии на
здоровые цветки, вызывая массовое перезаражение ржи. К мо-
менту созревания зерна в колосе на месте пораженной завязи обра-
зуется упомянутый склероций. К этому же порядку относятся воз-
будители широко распространенной болезни плодовых: парша
яблони, груши и др. Болезнь вызывается грибом из рода вентурия
(Venturia). Конидиальная форма развивается на поверхности ли-
стьев и плодов в виде темно-оливкового налета- С помощью кони-
дий происходит перезаражение растений. Сумки образуются в опав-
ших листьях; аскоспоры весной разносятся токами воздуха и про-
изводят первичное заражение молодых листьев плодовых деревьев.
Порядок дискомицеты (Discomycetales). Грибы имеют плодовое
тело блюдцевидной формы — апотеций, на котором сумки со спо-
рами сидят открыто, к этому порядку относится много почвенных
сапрофитов, среди которых съедобные грибы — сморчки и строчки.
Из паразитов наибольший интерес представляют виды Sclerotinia,
поражающие плоды яблони, сливы, персика, абрикоса, вишни.
Пораженные плоды буреют, а на поверхности их появляются
сероватые подушечки из скопления конидиеносцев, отчленяющих
конидии. Затем плоды твердеют, мумифицируются и превращаются
в склероции. Весной на склероции развиваются конидии, которые
и заражают деревья. На овощах склеротиния вызывает болезнь
«белая гниль». Апотеции образуются очень редко.
Порядок трюфелевые (Tuberales). К этому порядку относятся
съедобные грибы —трюфели (Tuber). Плодовое тело трюфеля раз-
вивается в поверхностном слое почвы; вначале это обычный апо-
теций, затем края апотеция завертываются и образуют замкнутое
плодовое тело, внутри которого находятся сумки.
Класс базидиомицеты (Basidiomycetes)
По уровню развития и богатству видов этот класс занимает
в системе такое же положение, как и класс аскомицетов. Это две
параллельные ветви эволюции.
Подобно сумчатым грибам, половой процесс вначале ограничи-
вается плазмогамией, т. е. слиянием протоплазмы; образуются ди-
карионы. После окончания полового процесса (кариогамия) из
зиготы в результате ее разрастания в длину образуется базидия
(см. рис. 101). Базидиоспоры формируются на базидии обычно
в виде овальных выпячиваний, сидящих на ножках — стеригмах;
базидиоспор обычно 4.
Сама базидия при этом может оставаться одноклеточной (холо-
базпдия), но может и разделиться на четыре клетки (фрагмобази-
дия), каждая из которых дает одну базидиоспору.
В отличие от аскомицетов базидиомицеты не имеют ни половых
органов, ни половых клеток. Половой процесс по форме упрощен
и заключается в слиянии протопласта мицелиев, развивающихся
210
из гаплоидных базидиоспор. В случае гомоталлизма половой про-
цесс совершается и в пределах одного мицелия. Гаплоидный мице-
лий в отличие от аскомицетов не развит и живет недолго. Преобла-
дает дикарионный или диплоидный мицелий, из него же (дикари-
онного мицелия) развиваются плодовые тела, которые также могут
быть открытыми и закрытыми. На плодовых телах или внутри них
развиваются базидии; иногда же базидии могут развиваться просто
на дикариопнОхМ мицелии, подобно голосумчатым грибам.
Бесполое размножение у базидиомицетов встречается значи-
тельно реже, чем у аскомицетов; осуществляется оно или посред-
ством конидий, или, чаще, спорами, представляющими собой клет-
ки, на которые распадается мицелий. По существу это вегетативное
размножение. Споры вегетативного размножения называются
оидиямп или хламидоспорами; в последнем случае они одеты плот-
ной темноокрашенной оболочкой.
Базидиомицетов известно свыше 25 000 видов. По строению
базидий их делят на два подкласса: холобазидиомицеты (Holoba-
sidiomycctidae)—с одноклеточными базидиями и фрагмобазидио-
мицеты (Phragmobasidiomycetidae) —с многоклеточными (четы-
рехклеточными) базидиями.
Подклассы делятся на порядки- Мы рассмотрим по два порядка
в каждом подклассе: порядки гименомицетов и гастеромицетов
в первом подклассе и порядки головневых и ржавчинных во вто-
ром подклассе.
Подкласс холобазидиомицеты (Holobasidiomycetidae)
Порядок гименомицеты (Hymenomycetales). Насчитывает около
16 000 видов, среди которых наиболее широко известны шляпоч-
ные грибы. Эти грибы имеют плодовые тела, на поверхности кото-
рых сплошным слоем расположены базидии вперемежку с бесплод-
ными парафизами. Этот слой называется гимением, откуда и на-
звание порядка.
Парафизы придают плотность гимениальному слою и одновре-
менно препятствуют слипанию базидиоспор, развивающихся на
концах базидий. Поверхность, несущая гимениальный слой, назы-
вается гименофором. Гименофор может быть трубчатый (рис. 106)
и пластинчатый (рис. 107). У наиболее примитивных гименофор
гладкий, без выростов.
Плодовые тела, на поверхности которых образуется гимениаль-
ный слой, чаще однолетние, но могут быть и многолетними (напри-
мер, у трутовиков). Многолетние плодовые тела из-за сильного
утолщения и уплотнения стенок гиф имеют деревянистую консис-
тенцию; живут до 80 и более лет. У большинства грибов с много-
летним твердым плодовым телом гимениальный слой ежегодно
отмирает и затем нарастает заново. Возникает слоистость плодо-
вого тела, по числу слоев можно судить о его возрасте.
К грибам с трубчатым гименофором относят домовый гриб
(Merulius lacrimans), трутовики (Forties), белый гриб (Boletus
211
Рис. 106. Базидиальные грибы с трубчатым гименофором. А— плодовое
тело трутовика (Fotnes). Б — поперечный разрез через трубчатый гимено-
фор, В — белый гриб (Boletus edulis)
edulis) и др.; к грибам с пластинчатым гименофором — шампиньон
(Psalliota campestris), сильно ядовитую бледную поганку (Ama-
nita phalloides), рыжик (Lactarius deliciosus) и др.
Порядок гастеромицеты (Gasterbmycetales). Почвенные грибы
с замкнутым плодовым телом обычно круглой формы. Плотная обо-
лочка плодового тела называется перидием, рыхлая ткань внутри —
глебой. Созревший гриб заполнен огромным количеством темных
базидиоспор вперемешку с капиллицием.
Представители — так называемые дождевики — растут обычно
на выгонах (из рода Bovista и др.). Крупные плодовые тела дож-
девиков содержат несколько триллионов спор. Молодые плодовые
тела гастеромицетов съедобны. Гастеромицетов известно свыше
1000 видов.
Подкласс фрагмобазидиомицеты
(Phr agmobasidiomycetidae)
Порядок головневые (Ustilaginales). Насчитывает около 900 ви-
дов, паразитирующих на высших, преимущественно злаковых ра-
стениях. Наибольший интерес представляют виды, паразитирующие
на хлебных злаках (рис. 108) и тем самым приносящие большой
вред народному хозяйству.
212
Рис. 107. Базидиальные грибы с пластинчатым гименофором. А — шам-
пиньон (Psalliota canipestris), Б — разрез через пластинчатый гименофор,
В — опенок (Armillaria tnellea)
Соцветия, плоды или другие части растений, пораженные го-
ловней, полностью или частично превращаются в черную массу
спор; растение производит впечатление обугленного, отсюда и на-
звание— головневые грибы.
Черные споры образуются из клеток мицелия гриба; оболочки
клеток, на которые распадаются гифы мицелия, ослизняются, клет-
ки округляются, и на них образуется плотная темная оболочка.
Такие споры вегетативного происхождения называются хламидо-
спорами.
Хламидоспоры диплоидны; из них развиваются базидии, на ба-
зидиях— гаплоидные базидиальные споры, физиологически разно-
значные— плюс и минус базидиоспоры. Из базидиоспор развива-
ются гаплоидные мицелии. Копулируют только мицелии, развив-
шиеся из разнозначных спор; могут копулировать и сами споры
и даже клетки фрагмобазидии. Из зиготы развивается диплоидный
мицелий, который и поражает растение. Гаплоидный мицелий, если
он возникает, имеет кратковременное существование, микроскопи-
чески мал и непосредственно не способен заражать растение. По
способу заражения головневые грибы хлебных злаков можно раз-
делить на три биологические группы:
1) заражение происходит во время цветения злаков; диплоид-
ный мицелий проникает в завязь цветка;
2) заражение происходит во время прорастания зерна; дипло-
идный мицелий проникает в развивающийся зародыш или проро-
сток растения;
213
Рис. 108. Головневые грибы (Ustilaginales). А — пыльная головня овса (Ustilago avenae), Б — пыльная
головня проса (Ustilago panici miliacei), В — пыльная головня пшеницы (Ustilago tritici), Г — твердая
головня пшеницы (Tilletia tritici), Д— пузырчатая головня кукурузы (Ustilago zeae):
1 — спора пыльной головни пшеницы; 2 — ее прорастание; 3 — спора твердой головни пшеницы; 4 — опа же с конидией;
5 — зерновка со спорами; 6 — прорастание споры пузырчатой головни кукурузы
3) заражение происходит в любом возрасте злакового растения;,
диплоидный мицелий проникает в молодые нежные ткани цветка,
листа, стебля и даже корня.
Из каждой группы мы рассмотрим по одному типичному пред-
ставителю.
Пыльная головня пшеницы (Ustilago tritici). К моменту цвете-
ния пшеницы соцветия растения, пораженного головней, превра-
щаются в черную пыль — скопление головневых спор. Споры,
попадая на рыльце цветка, прорастают, из них вырастают базидии;
базидиоспоры не образуются, копулируют соседние клетки фраг-
мобазидии.
Из зиготы вырастает диплоидный мицелий, который внедряется
в завязь, проникает в семяпочку и сохраняется в зародыше разви-
вающегося семени. Здесь в состоянии покоя микроскопический ми-
целий пребывает до момента прорастания семени; семя внешне-
совершенно здорово; мицелий трогается в рост одновременно с
прорастанием семени и развивается внутри хозяина, удлиняясь-
в меру его роста.
К моменту выхода колоса из трубки все его части, кроме цент-
ральной оси, уже полностью разрушены паразитом, вместо них мы
обнаруживаем скопление спор, на которые распадается мицелий
гриба. Споры при помощи ветра переносятся на рыльца цветущих
растений, и с этого момента начинается новый цикл развития-
гриба.
Подобно пыльной головне пшеницы, происходит развитие пыль-
ной головни ячменя (Ustilago nuda) и др. От зачаточного мицелия)
пыльной головни избавляются путем термической обработки по-
севного материала, в результате которой мицелий погибает.
Твердая, или вонючая, головня пшеницы (Tilletia tritici). Пора-
женное зерно отличается более темным цветом. Оно оказывается
состоящим из массы темных спор; при смачивании этой массы от
нее исходит отвратительный запах, откуда и название головни.
Вместе с посевным зерном споры попадают в почву и прорастают
одновременно с пшеницей. Из споры вырастает базидия, на ней
удлиненные базидиоспоры; базидиоспоры попарно копулируют,
и возникающий диплоидный мицелий проникает в проросток пше-
ницы. В дальнейшем мицелий развивается так же, как и у пыльной
головни; зерновка превращается в массу черных склеенных в комо-
чек головневых спор.
Подобно твердой головне, развивается большинство головневых,
поражающих хлебные злаки: головня проса (Ustilago panicimilia-
cei); пыльная головня овса (U. avenae), твердая головня ячменя
(U. hordei) и др.
Меры борьбы: протравливание посевного материала.
Головня кукурузы (Ustilago zeae). Пораженная ткань разра-
стается, образуя характерные опухоли, заполненные мажущейся
темной массой, состоящей из спор. При подсыхании споры разле-
таются и, попадая в почву, прорастают. На базидии образуются
базидиоспоры, способные почковаться, давая конидии. Гаплоидные
215
конидии физиологически разнозначны. Попадая на растение, кони-
дии прорастают, давая гаплоидный мицелий; при встрече с мицели-
ем противоположного знака происходит копуляция их, и возникший
диплоидный мицелий поражает ткань, вызывая на ней опухоли.
Пока на растении кукурузы имеются молодые нежные ткани,
перезаражение таким образом может происходить несколько раз.
Меры борьбы: срезание и уничтожение опухолей; глубокая вспаш-
ка, при которой пораженные части растений закапываются на глу-
бину, исключающую дальнейшее распространение болезни, сево-
оборот.
Порядок ржавчинные грибы (Uredinales). Весьма распростра-
ненные паразиты, живущие на листьях и стеблях высших растений.
Всего насчитывается около 7000 видов ржавчинных грибов.
В отличие от головневых, патогенностью обладает не только
диплоидный, но и гаплоидный мицелий, причем эти мицелии не-
редко живут на различных растениях (разнохозяйственность) •
Характерно разнообразие спороношений, связанное со сменой хо-
зяина и фаз развития. Споры содержат масло, окрашенное в оран-
жевый цвет липохромом; отсюда и название грибов.
В качестве типичного примера ,разнохозяйственного ржавчин-
ного гриба, обладающего всеми типами спороношений, рассмотрим
линейную ржавчину хлебов (Puccinia graminis) (рис. 109). Этот
гриб имеет двух хозяев; барбарис и один из злаков. На барбарисе
развивается гаплоидный мицелий и образуются дикарионы, на зла-
ке живет дпкарионный мицелий. Развитие на злаках заканчивает-
ся образованием базидиоспор, которые заражают листья барба-
риса.
Весной на листьях барбариса появляются ржавого цвета пятна,
очень мелкие на верхней стороне листа и более крупные на ниж-
ней. Мелкие пятнышки на верхней стороне листа представляют
собой отверстия углубленных в мезофилл колбочковидных вмести-
лищ. Крупные пятна на нижней стороне листа — раструбы урно-
чек — также погружены в мезофилл. Те и другие — вместилища
спор; на верхней стороне пикниды, или спермогонии, с пикноспо-
рами, или спермациями, на нижней стороне листа — эцидии с эци-
диоспорами; последние по времени появления называются весенни-
ми спорами.
Пикниды представляют собой сплетения одноядерных гаплоид-
ных гиф, с кончиков которых и отчленяются очень мелкие пикно-
споры. По мере созревания они выдавливаются кверху через узень-
кое отверстие, окруженное кончиками бесплодных гиф. Из пикни-
ды к моменту созревания пикноспор выступает пахучая сахаристая
жидкость.
Зрелая эцидия заполнена сплетением дикарионных двуядерных
гиф, отчленяющих двуядерные эцидиоспоры. Эцидиоспоры попа-
дают на листья злаков, здесь они прорастают; дикарионный мице-
лий проникает внутрь листа, разрастается там и образует большое
количество двуядерных уредоспор, или летних спор. Скопляясь
под эпидермисом, уредоспоры прорывают его и, разносясь ветром,
216
Рис. 109. Линейная ржавчина хлебов (Puccinia graminis). А — листья
барбариса, пораженные ржавчиной, Б — эцидий и пикнида на листе
барбариса, В — стебель и влагалище листа, пораженные ржавчиной,
Г — кучка уредоспор, Д — кучка телейтоспор, Е — проросшая телей-
тоспора с двумя фрагмобазидиями и восемью базидиоспорами
перезаражают здоровые растения подобно эцидиоспорам. Такое
перезаражение может в течение лета происходить несколько раз;
для развития каждой новой генерации уредоспор нужна одна не-
деля. За лето может развиться 5—6 поколений уредоспор, в резуль-
тате из одной эцидиоспоры могут образоваться многие миллиарды
уредоспор — происходит массовое заражение поля ржавчиной.
К концу лета образуются более темные двуклеточные телейто-
споры, или зимние споры. Весной в телейтоспорах заканчивается
половой процесс — два гаплоидных ядра сливаются в одно дипло-
идное.
217
Телейтоспоры зимуют в почве или на остатках отмершего расте-
ния. Весной из каждой клетки телейтоспоры вырастает типичная
фрагмобазидия с четырьмя базидиоспорами. Базидиоспоры зара-
жают листья барбариса, и в результате этого заражения образуют-
ся пикниды и эцидии.
Будучи гаплоидными, базидиоспоры не могут дать начало ди-
карионным гифам эцидия. Дикарионность создается в результате
попадания в эцидий гаплоидного мицелия, развившегося из пикно-
споры, причем пикноспора должна быть по знаку противоположна
базидиоспоре, из которой начал развиваться мицелий, формирую-
щий эцидию. Пикноспоры переносят насекомые, пользующиеся
сладкой жидкостью, выделяемой пикнидами. Пути проникновения
в эцидию пикноспоры или развивающегося из нее гаплоидного
.мицелия недостаточно выяснены. Дикарионные гифы могут образо-
ваться также и в том случае, если в эцидии развились по крайней
мере два мицелия, возникших из базидиоспор с противоположными
знаками (+ или —).
Кроме линейной ржавчины злаков, широко распространены ко-
рончатая ржавчина овса (Puccinia coronifera), ржавчина подсол-
нечника (Р. helianihi), ржавчина кукурузы (Р. maydis) и многие
другие виды, поражающие культурные растения.
Меры борьбы с ржавчиной хлебов: выведение сортов, устойчи-
вых против этого заболевания, а также хорошая агротехника.
Класс несовершенные грибы (Fungi imperfecti)
Обширная группа грибов, насчитывающая свыше 25 000 видов
сапрофитного и паразитного образа жизни- По наличию клеточно-
го мицелия виды этого класса должны быть отнесены к высшим
грибам, но они не имеют ни сумчатого, ни базидиального спороно-
шения, и представлены либо совершенно стерильными грибницами,
либо размножаются при помощи оидий или конидий.
Класс несовершенных грибов — искусственная группа. В про-
цессе изучения выясняется, что многие формы, описанные как не-
совершенные грибы, представляют собой конидиальные стадии уже
известного сумчатого гриба. Одна из искусственных систем этого
класса — система итальянского миколога Саккарда, согласно кото-
рой этот класс грибов подразделяется на три порядка: гифомице-
ты, меляпкониевые и сферопсидные (рис. НО).
Порядок гифомицеты (Hyphomycetales). Конндиеносцы одиноч-
ные или в небольших пучочках коремиях. К этому порядку отно-
сятся: Monilia— конидиальная стадия Sclerotinia, поражают пло-
довые деревья; Verticillium — некоторые виды которого паразити-
руют в сосудах хлопчатника, вызывая его увядание — болезнь
вилт; Fusarium — сапрофитные формы которого широко распро-
странены в почве, паразитные вызывают заболевание многих куль-
турных растений (фузариозы): хлебных злаков, льна, хлопка и т. д.
К этому же порядку относится грибок стахиботрис (Stachybotrys
218
alternans), вызывающий опасную
болезнь лошадей — стахиботрио-
токсикоз.
Порядок мелянкониевые (Ме-
lanconiales). Конидиеносцы соба
раны плотным слоем. Сюда отно-
сится род Gleosporium, паразити-
ческие виды которого поражают
надземные части многих культур-
ных растений (клевер, фасоль
тыквенные), вызывая заболев^
ние под названием антракнозы
на пораженных растениях появ
ляются язвы, затем ткани отми-
рают.
Порядок сферопсидные
(Sphaeropsidales). Конидиеносцы
внутри шаровидных вместилищ —
пикнид. Сюда относится род Pho-
та, многочисленные виды которо-
го поражают листья свеклы, ви-
нограда и других, вызывая чер-
ную гниль. На винограде же пара-
зитируют виды филлостикта —
Phyllosticta.
Рис. ПО. Спороношения несовершен-
ных грибов (Fungi irnperfecti). А —
фузариум (Fusarium), Б — глеоспори-
УхМ (Glaeosporium), В — филлостик’га
(Phyllosticta)
ФИЛОГЕНЕЗ ГРИБОВ
Существуют различные точки зрения на происхождение грибов.
Сторонники полифилетического происхождения выводят грибы из
различных групп водорослей (зеленых и красных), утративших
хлорофилл, сторонники монофилетического происхождения вы-
водят грибы из группы первичных организмов, обладавших
жгутиками; вторая точка зрения признается более вероят-
ной.
Простейшая группа грибов представлена классом хитридиевых.
С хитридиевыми непосредственно связаны оомицеты, имеющие зоо-
споры, что указывает на приспособленность к водной среде. Зиго-
мицеты, не имеющие уже подвижных зооспор, явились выражением
приспособления грибов к наземному существованию; по-видимому,
они также произошли от хитридиевых. Дальнейшее приспособле-
ние к жизни на суше—конидиальное спороношение, свойственное
высшим грибам. Через первичносумчатых с зигомицетами связаны
аскомицеты. У плодосумчатых эволюция шла от закрытых плодо-
вых тел к совершенно открытым апотециям. Базидиальные также,
по-видимому, произошли от зигомицетов, а затем линия эволюцио-
нировала параллельно аскомицетам. Эволюция шла от холобази-
диомицетов к фрагмобазидиомицетам.
219
РОЛЬ ГРИБОВ В ПРИРОДЕ И ИХ ХОЗЯЙСТВЕННОЕ
ЗНАЧЕНИЕ
Роль грибов в природе определяется способом их питания. Раз-
рушая остатки отмерших растений, грибы тем самым возвращают
содержащиеся в них вещества в круговорот веществ, обеспечивая
постоянный поток жизни на Земле. Находясь в почве в огромном
количестве, они наряду с бактериями восстанавливают ее плодоро-
дие, разрушая и минерализуя растительные остатки.
Грибы играют большую роль в жизни человека, в его хозяйстве.
Многие грибы съедобны; они составляют заметное подспорье в пи-
тании людей и особенно ценятся за их специфический вкус и запах.
Дрожжевые грибы используются для получения спирта, в хлебо-
печении, при изготовлении пива, вина, кумыса, кефира и др.; из
аспергилла и пеницилла получают лимонную кислоту, а также
применяют эти грибы для созревания некоторых сортов сыра. В по-
следнее время многие грибы 1используются для получения
антибиотиков. Ценнейшее лекарственное сырье — склероции
спорыньи.
С другой стороны, грибы наносят большой вред человеку; они
являются возбудителями большинства болезней культурных расте-
ний; многие грибы вызывают заболевание животных и чело-
века; грибы губят огромное количество древесины, разрушают
деревянные сооружения, вызывают порчу продуктов и кормов.
Развиваясь на хлебных злаках и попадая в муку, грибы могут
вызвать опасные заболевания человека и животных. Наконец, мно-
гие шляпочные грибы ядовиты.
ОТДЕЛ 14. ЛИШАЙНИКИ (LICHENOPHYTA)
Лишайник — своеобразный растительный организм; тело его
состоит из гиф гриба и синезеленых или зеленых водорослей.
Большинство лишайников имеет вид более или менее шерохо-
ватой пленки на камнях, коре деревьев или другом твердом суб-
страте (рис. 111). Окрашена такая пленка (и вообще лишайники)
чаще в неяркие тона: сероватый, зеленовато-серый, белый, бурый
или почти черный. Лишь желтые.и оранжевые лишайники выделя-
ются яркостью окраски.
Лишайники в виде тонкой пленки, плотно соединенной с суб-
стратом, называются накипными или корковыми. Накипной ли-
шайник не может быть отделен от твердого субстрата без разру-
шения его тела.
Таллом многих лишайников имеет форму измятой пластинки,
более или менее плотно прилегающей к субстрату; такие лишайни-
ки называются листоватыми. Листоватые лишайники часто обита-
ют на коре деревьев и деревянных заборах.
Наконец, таллом лишайника может иметь форму ветвящегося
«стволика» или длинных разветвленных нитей, свисающих с'дере-
ва. Лишайник с такой морфологией называется кустистым. Посе-
220
Рис. 111. Лишайники (Lichenes). А — письменный лишайник на камне (Graphis
scripta), Б — листоватый лишайник пармелия (Parmelid), В — бородатый лишай-
ник (Usnea barbaia), Г и Д — олений лишайник ягель (Cladonia rangiferina),
Е — разрез таллома пармелии, Ж — соредия
ляясь на деревьях во влажных лесах, кустистые лишайники обра-
зуют длинные космы — «бородатые лишайники».
Двойственную природу лишайника можно установить при рас-
смотрении под микроскопом внутреннего строения таллома. Уже
при небольшом увеличении заметно, что таллом лишайника состо-
ит в основном из гиф гриба; между сплетениями гиф включены од-
ноклеточные или нитчатые водоросли.
221
По характеру анатомического строения все лишайники делят
на гомеомерные и гетеромерные-
У гомеомерных лишайников водоросли более или менее равно-
мерно распределены по всей толще таллома. Так устроено тело
всех накипных и небольшой части листоватых и кустистых лишай-
ников.
Большинству листоватых и кустистых лишайников свойственно
гетеромерное строение. У гетеромерных лишайников верхняя сто-
рона таллома состоит из плотного сплетения гиф, это верхняя кора
таллома. Под верхней корой, между более рыхло сплетенными
гифами, размещаются шаровидные клетки зеленой водоросли. Это
так называемый гонидиальный слой. Название дано давно, когда
еще не была известна истинная природа лишайников, и клетки во-
доросли относили к органам размножения.
Под гонидиальным слоем находится сердцевина таллома, со-
стоящая из рыхло соединенных гиф. Нижняя поверхность таллома
сходна с верхней и называется нижней корой, от нее отходят пучки
гиф, которыми таллом лишайника соединяется с субстратом.
Водоросль, входящая в симбиотический организм лишайника,
будучи отделена от гриба, может существовать самостоятельно.
Гриб, наоборот, отдельно от водоросли существовать не может.
В качестве автотрофного симбионта в составе лишайников
встречаются синезеленые водоросли Nostoc, Chroococcus, Glaeocap-
sa и др., из зеленых обычно Chlorella, Chlorococcunz, Pleurococcus,
реже нитчатая Cladophora, Trentepholia и др.
Гетеротрофными симбионтами лишайника являются сумчатые
грибы и лишь у немногих тропических лишайников базидиальные
грибы. Сумчатые грибы относятся к порядкам пирено- и дискоми-
цетов. Принадлежность гриба к тому или иному порядку хорошо
устанавливается по форме грибных плодовых тел, развивающихся
на талломе лишайников: перитециев или апотециев. В этих плодо-
вых телах находятся сумки, в сумках аскоспоры, способные к про-
растанию.
Из аскоспор могут даже развиться микроскопические мицелии,
но этот мицелий без водоросли существовать не может, и если он
случайно не натолкнется на клетки соответствующей водоросли,
то быстро погибнет. В положительном случае гифы гриба окутыва-
ют встретившиеся клетки «своей» водоросли, заново образуя ли-
шайник. Клетки водорослей, заключенные в таллом лишайника,
могут размножаться там только вегетативно.
Размножение лишайника как единого организма происходит
различными способами: обломившимися кусочками таллома и по-
средством соредий и изидий. Соредия, — специальное устройство,
состоящее из одной или нескольких клеток зеленой водоросли, опле-
тенных гифой гриба. Образуются соредии в гонвдиалыном слое
и через разрывы в верхней коре выдуваются наружу.
Изидии встречаются реже, чем соредии. Они образуются в виде
выростов на периферии таллома и также состоят из нескольких
клеток водоросли, оплетенных гифами гриба. В отличие от соре-
222
дий изидии снаружи одеты уплотненным, корковым слоем лишай-
ника. Попав в благоприятные условия, и соредии и изидии дают на-
чало новому таллому лишайника.
Характер взаимоотношения водоросли и гриба в лишайнике—•
не вполне гармоничное симбиотическое сосуществование. Эти два
симбионта, конечно, приносят друг другу взаимную выгоду: водо-
росль снабжает гриб созданными ею углеводами и получает от него
воду с растворенными в ней неорганическими веществами. Но гри-
бу недостаточно сапрофитного питания. Установлено, что гифы
гриба дают отростки — гаустории, с помощью которых он проника-
ет в клетки водоросли и питается их протопластом, а водоросль
погибает. Это носит уже паразитический характер. Правда, пара-
зитизм в данном случае умеренный; погибает лишь незначительная
часть водорослевых клеток. Зависимое положение грибов проявля-
ется также и в неспособности их к самостоятельному существова-
нию.
Характерная особенность биологии лишайников — медленный
их рост. Накипные лишайники ежегодно нарастают в диаметре
всего на несколько миллиметров; листоватые и кустистые — на
1—30 мм. Возраст лишайников достигает 50—100 лет. Известно
около 20 000 видов лишайников.
Распространены лишайники в природе довольно широко, за ис-
ключением мест, где воздух загрязнен вредными газами. Лишайни-
ки очень чувствительны к загрязнению воздуха и поэтому быстро
погибают в крупных городах и вблизи заводов и фабрик.
Поселяются лишайники на самом разнообразном субстрате: на
песках, скалах, деревьях, деревянных и каменных зданиях и т. д.,
немногие лишайники живут в воде. Лишайники очень нетребова-
тельны к субстрату и, выделяя угольную и другие кислоты, могут
растворять многие минералы. Они поселяются даже на таких не-
обычных субстратах, как оконные стекла, старое железо, фарфор,
фаянс, кожаные и матерчатые изделия и др. Поселяясь на оконных
стеклах, лишайники делают их мутными и непрозрачными, поверх-
ность хорошо отполированного гранита — шероховатой.
Лишайники длительное время переносят полное высыхание;
смоченные водой они оживают и продолжают жизнедеятельность.
Существенная роль лишайников в природе заключается в том,
что, поселяясь па совершенно бесплодных субстратах, они тем са-
мым создают условия для более требовательных к условиям суще-
ствования растений. Так происходит постепенное заселение расте-
ниями совершенно голых скал.
В хозяйстве человека важную роль играют лишайники из рода
Cladonia, так называемый олений мох — ягель, широко распростра-
ненный в зоне тундры и лесотундры и являющийся здесь основным
кормом для северного оленя. Наибольшее значение здесь имеют
такие виды: Cladonia rangiferina, С. rnitis, С. alpestris, С. silvatica.
Другие виды этого рода обитают на почве наших южных степей,
где также иногда, при выгорании травянистой растительности, мо-
гут быть кормовыми растениями. Северные олени поедают также
223
лишайники и некоторых других видов (Cetraria, Alectoria, Usnea
и др.). Лишайники некоторых видов рода Cetraria используются
в медицине для приготовления лечебных слизистых отваров; ли-
шайники других видов этого рода содержат большое количество
витамина С.
Лишайники некоторых видов рода Lecanora дают так называе-
мую лишайниковую манну — комочки накипного лишайника, легко
переносимые ветром. В странах западной Азии и северо-восточ-
ной Африки лишайниковая масса употребляется в пищу. Лишай-
ники из родов Roccellia и Ochrolechia используются для получения
лакмуса и других сине-фиолетовых и пурпурных красок. В парфю-
мерии используются лишайники рода Evernia,
Поселяющиеся на живых деревьях лишайники не являются па-
разитами, но вредны растению тем, что закупоривают поры и слу-
жат убежищем вредным насекомым. Поэтому в садоводческой
практике принято стволы деревьев очищать от лишайников. Оби-
лие лишайников на стволе и ветвях дерева свидетельствует о пло-
хом росте его, что может быть связано с неудовлетворительным
уходом за насаждениями или болезнью деревьев.
ВЫСШИЕ РАСТЕНИЯ (Kormobionta, Embryobionta)
Около полумиллиарда лет назад наступил такой момент в исто-
рии Земли, когда растения начали заселять и сушу. Выход на су-
шу положил начало высшим растениям. Их так называют прежде
всего за сложность строения, которая явилась следствием приспо-
собления к новым, более жестким наземным условиям существо-
вания. Из первых сухопутных растений развился в конечном счете
весь многообразный и сложный мир современных высших растений.
Какая из групп водорослей дала начало первым наземным ра-
стениям, наука пока не дает единого ответа. По условиям жизни
ближе всего к высшим растениям стоят зеленые водоросли; неред-
ко они живут вне воды, на влажной почве, коре деревьев и на дру-
гих предметах, находящихся в условиях обильного увлажнения.
Однако по сложности строения бурые и красные водоросли стоят
выше зеленых. Отсутствие ископаемых остатков первых сухопут-
ных растений, непосредственных потомков водорослей, затрудняет
решение этого вопроса.
Первые высшие растения, отделившиеся около полумиллиарда
лет тому назад от своих предков — водорослей, имели очень про-
стое строение. Сохранились ископаемые остатки древнейших из
известных сухопутных растений — псилофитов (рис. 112). Самые
древние из них с наиболее простым строением имели вид неболь-
шого, высотой до 30 см и толщиной до 5 мм, вильчато разветвлен-
ного стебелька без листьев и корней.
Нижняя, подземная, часть стебельков была горизонтально за-
гнута, и на ней вместо корней вырастали тоненькие одноклеточ-
ные ниточки-ризоиды. Стебельки были вначале совершенно голые
224
и ассимилировали углерод всей своей зеленой поверхностью. Пред-
ставителями этой наиболее примитивной формы высших растений
могут быть риния (Rhynid) и горнея {Horned).
Позднее в процессе эволюции на стебельках появились тонень-
кие чешуйчатые выросты, которые значительно увеличили ассими-
лирующую поверхность. Примером такого растения может быть
Рис. 112. Псилофиты. А — горнея (Ногпеа), Б — риния
(Rhynia), В — астероксилон (Asteroxylon)
псилофит астероксилон (Asteroxylon). Среди современных нам ра-
стений подобные неразвитые листья в виде тоненьких чешуек или
пластинок находим у мхов, плаунов.
Позднее развились настоящие корни, как подземные разветвле-
ния нижней части стебля, и настоящие листья, состоящие из многих
слоев клеток. Филогенетически, т. е. с точки зрения исторического
развития, листья возникли из боковых ответвлений стебля, приняв-
ших плоскую форму,— наилучшее приспособление к более полному
улавливанию солнечного света.
Из ныне живущих наиболее близки к псилофитам — этим ис-
ходным формам высших растений, плауны, хвощи, папоротники
(рис. 113—115) и мхи. Мхи, плауны и хвощи — небольшие травя-
нистые растения; папоротники — более крупные (в тропических
лесах встречаются даже их древовидные формы).
Выход растений на сушу был очень важным, как говорят, пере-
ломным моментом в эволюции растительного мира. От этого мо-
мента нас отделяет свыше 450 млн. лет, и за этот длительный
8—326 225
период мир растений все время изменялся, изменялись условия су-
ществования на Земле. С выходом растений на сушу их эволюция
пошла гораздо быстрее, так как условия существования здесь го-
раздо сложнее и разнообразнее, чем в водной среде. Растения
постепенно изменялись соответственно новым условиям обитания.
Рис. 113. Плаун булавовидный (Lycopodium clavatum).
А — растение-спорофит со спороносными колосками, Б —
спорофилл со спорангием, В — споры, Г — заросток
Для биологии высших растений характерно правильное, рит-
мичное чередование поколений, или фаз развития, возникшее уже
у водорослей. Бесполое поколение, или спорофит, сменяется поло-
вым поколением, или гаметофитом. Воспроизводимые гаметофитом
половые клетки, сливаясь, дают зиготу, из которой развивается
спорофит. Спорофит дает споры, из которых развивается гамето-
фитное поколение.
У высших растений спорофит всегда представлен диплоидной
фазой. Редукция хромосом происходит при образовании спор. По-
этому гаметофиты гаплоидны. Диплоидность восстанавливается
при слиянии гамет.
226
Высшие растения не образуют зооспор. Споры их кутинизиро-
ваны и приспособлены к пассивному перемещению вне водной сре-
ды. Половое воспроизведение нового поколения с участием по-
движных гамет, свойственное водорослям, оказалось в несоответ-
Рис. 114. Хвощ полевой (Equisetum arvense). А — растение-спорофит
с вегетативными и спороносными побегами, Б, В — спорофилл со
спорангиями, Г, Д, Е — споры
ствии с новой, воздушной средой обитания высших растений.
Однако освобождение полового процесса от водной среды длилось
многие десятки миллионов лет, и до наших дней сохранились еще
группы высших растений, половое поколение которых нуждается
в капельножидкой воде.
8*
227
Рис. 115. Мужской папоротник (Dryopterls filix mas). А — расте-
ние-спорофит, Б — часть листа с сорусами на нижней поверхно-
сти, В — поперечный разрез соруса с покрывальцем, Г — споран-
гий с высыпающимися спорами
Необходимость для полового процесса водной среды, по-види-
мому,— основная причина прогрессирующей редукции гаметофита,
приведшая к тому, что у семенных растений это поколение полно-
стью лишилось самостоятельности и существует за счет хорошо
развитого спорофита.
Уже через несколько десятков миллионов лет после массового
выхода растений на сушу (срок небольшой в истории растительно-
го мира) из первых небольших сухопутных растений — псилофи-
тов — развился богатый и разнообразный мир крупных древовид-
ных растений: лепидодендронов (рис. 116), сигиллярий (от которых
228
Рис. 116. Ископаемое споровое растение — лепидодендрон
(Lepidodendron)
произошли современные плауны), каламитов — родоначальников
хвощей и огромных древовидных папоротников.
Климат в то время был влажный и теплый, отмирающие расте-
ния падали на дно многочисленных водоемов или погружались в
болотистую почву. Здесь они из-за недостатка кислорода не разла-
гались и скоплялись большими массами. Из этих залежей отмер-
ших растений впоследствии образовался каменный уголь, поэтому
и время, когда iHa Земле преобладали такие влажные леса, назы-
вается каменноугольным периодом.
Первые вышедшие на сушу растения размножались и распро-
странялись посредством специальных клеток — спор. Во влажных
условиях споры хорошо выполняют это свое назначение.
К концу каменноугольного периода споровые растения начина-
ют уступать место растениям семенным. Семена имеют более слож-
ное строение, чем споры, и несравненно крупнее их. Это уже гото-
вые многоклеточные зародыши будущих растений, снабженные
запасом питательных веществ и защищенные оболочкой от небла-
гоприятных воздействий внешней среды.
Распространение семенных растений связано с изменением кли-
мата в сторону большей сухости. В условиях сухого климата семе-
на гораздо успешней выполняют задачу размножения и распро-
странения растений, по сравнению.со спорами.
Первыми растениями, размножавшимися при помощи семян,
были семенные папоротники, от них произошли и другие группы
семенных растений. Вначале семенные растения не имели плодов,
семена у них образовывались открыто, не были замкнуты в плодо-
вые оболочки, поэтому такие семенные растения называются голо-
семенными. Из голосеменных наиболее широко распространены
хвойные растения: сосна, ель, пихта и др.
Появление семенных растений можно считать вторым перелом-
ным моментом в эволюции растительного мира. Это произошло
около 350 млн. лет назад. При помощи семян растения быстрее
осваивали сушу и успешней, легче приспосабливались к изменяю-
щимся условиям наземного существования.
Господство голосеменных растений длилось около 100 млн. лет.
До этого они около 150 млн. лет сопутствовали споровой флоре,
занимавшей господствующее положение. Позднее появились высо-
коорганизованные цветковые растения. С момента возникновения
первых цветковых растений прошло, по-видимому, около 200 млн.
лет, и за это время они распространились по всему земному шару.
Цветковые растения, таким образом,— самая молодая группа ра-
стений, вместе с тем это самые совершенные растения.
Как показывает само название — цветковые, эти растения име-
ют особые органы — цветки, возникшие как приспособление к пе-
рекрестному опылению при помощи насекомых. Участие насекомых
в процессе опыления у цветковых растений сыграло большую по-
ложительную роль в эволюции растительного мира. Из части цвет-
ка— завязи — образуется плод, в котором находятся семена; отсю-
да происходит и другое название цветковых растений — покрыто-
230
семенные; семена у них находятся внутри особых органов растения,
в плодах.
К цветковым, или покрытосеменным, растениям относятся, все
сельскохозяйственные растения. Жизнь человека и современных
сухопутных животных, включая и птиц, связана прежде всего с этой
группой растений; от них человек получает продукты питания, кор-
ма для животных, сырье для изготовления одежды, многочисленные
лекарственные и различные химические вещества, используемые
в промышленности. Без цветковых растений жизнь человека была
бы невозможна.
Появление цветковых, или покрытосеменных, растений мы мо-
жем считать третьим переломным моментом в эволюции раститель-
ного мира. Наступил период бурного расцвета цветковых растений,
быстрого увеличения численности и разнообразия видов этой груп-
пы. Период господства цветковых растений длится уже около
100 млн. лет.
В настоящее время насчитывается до 250 тыс. видов цветковых
растений. Из этого богатства еще первобытный человек начал от-
бирать наиболее полезные для него растения, положив тем самым
начало новому этапу в эволюции растительного мира, связанному
вначале с бессознательным, а затем и сознательным вмешательст-
BOxM человека в жизнь природы. При непосредственном участии че-
ловека начался процесс создания культурных растений.
История культурных растений сравнительно непродолжитель-
на, всего 10—12 тыс. лет. Однако и за этот сравнительно короткий
промежуток времени человек настолько изменил облик и качество
возделываемых им растений, что для многих из них сейчас уже
трудно найти дикого, первобытного предка. Возможности для даль-
нейшей деятельности человека в этом направлении безграничны;
достаточно сказать, что из общего числа 250 тыс. видов цвет-
ковых 'растений окультурено пока не более 3000 видов, т. е.
около 1,2%.
Рассмотрение высших растений начнем с древнейших — псило-
фитов.
ОТДЕЛ 1. ПСИЛОФИТОВЫЕ, ИЛИ
РИНИЕВЫЕ (PSILOPHYTA, RHYNIOPHYTA)
Древнейшие и самые примитивные из всех известных высших
растений. Возникновение первых псилофитов относят к началу си-
лурийского периода; к концу девона псилофиты вымерли. Следо-
вательно, период существования псилофитов длился примерно свы-
ше 70 млн. лет из 450 млн. лет, которые насчитывают высшие
растения вообще.
В течение такого длительного периода псилофиты, естественно,
не оставались неизменными; простейшие их формы эволюциониро-
вали в более сложные.
Первые, самые примитивные псилофиты, как уже было сказано
в начале этой главы, представляли собой голые, дихотомически
231
ветвящиеся стебельки; росли они в непосредственной близости
к воде, вместо корней у них были ризоиды. На кончиках дихото-
мически разветвленного стебелька сидели примитивные спорангии
в виде простых мешочков, без особых приспособлений для откры-
вания. Высота псилофитов не превышала 50 см, толщина — 5—
6 мм.
Внутреннее устройство стебля весьма примитивно. Стела без
сердцевины, в центре ее расположена ксилема, состоящая из тра-
хеид с кольчатыми и отчасти лестничными утолщениями. Ксилему
окружают элементы флоэмы; стела такого простого устройства
называется протостелой. Камбия у псилофитовых не было, мери-
стемы только верхушечные.
У наиболее примитивных риниофитов протостела была пред-
ставлена гаплостелой с округлым очертанием ксилемы на попереч-
ном разрезе, у более продвинутых {Asteroxylori) — актиностелой
с лопастно расчлененной ксилемой. В процессе дальнейшей эволю-
ции высших растений строение стелы совершенствовалось.
Наличие устьиц на поверхности стебельков и сильная кутиниза-
ция споровых оболочек у первых псилофитов говорят о достаточно
продвинувшейся уже приспособленности к сухопутному образу
жизни. Достоверные остатки гаметофитов у псилофитов не най-
дены.
Представителями наиболее примитивных псилофитов могут
быть горнея {Horned) и ршшя {Rhynid) (см. рис. 112) , а наиболее
специализированных — астероксилон (Asteroxylori), остатки кото-
рого найдены в Казахстане, Западной Сибири. Подземная часть
астероксйлона более мощно развита и дихотомически разветвлена
до тонких веточек, приближающихся по своей функции к настоя-
щим корням. Надземная часть стебля также дихотомически раз-
ветвлена.
Стебель покрыт мелкими листочками, имеющими шиловидную
или коническую форму. Такие листья представляют собой выросты
(энации) поверхностных слоев стебля; в отличие от настоящих
листьев их называют филлоидами. Внутри филлоидов отсутствуют
проводящие пучки, они заходят лишь в их основания, благодаря
чему на поперечном срезе стебля обнаруживаются листовые следы.
Псилофиты давно вымерли, но от них берут начало ветви эволю-
ции, представленные мохообразными, плауновидными, членистыми
и папоротникообразными. Папоротникообразные, в свою очередь,
дали голосеменные и покрытосеменные.
Многие ученые считают, что мохообразные ведут начало непо-
средственно от водорослей. Кроме основных четырех ветвей сохра-
нилось несколько видов псилотов, по-видимому, прямых потомков
псилофитов.
ОТДЕЛ 2. МОХОВИДНЫЕ (BRYOPHYTA)
Относящиеся к этому отделу растения довольно разнообразны
по величине, внешнему и внутреннему строению, но имеют ряд об-
щих черт. Гаметофит всегда по своему размеру и развитию преоб-
232
ладает над спорофитом, который вырастает на гаметофите и пол-
ностью или частично питается за его счет. Этим мохообразные
принципиально отличаются от остальных отделов высших расте-
ний, у которых спорофит преобладает над гаметофитом. Мохооб-
разные не имеют корней, тело их представлено или небольшим
травянистым стебельком с тонкими упрощенными листьями, или
имеет вид листовидного таллома, прилегающего к земле. Спора
прорастает в зеленую нить или пластинку — протонему, из которой
затем развивается собственно гаметофит. Развитие гаметофитов
начинается из почек, которые закладываются на протонеме. Гаме-
тофиты могут быть обоеполыми и однополыми. У моховидных ши-
роко распространено вегетативное размножение посредством вы-
водковых почек.
Отдел моховидные разделяется на три класса: 1. Антоцеротовые
(Anthocerotae). 2. Печеночники (Hepaticae) и 3. Лиственные мхи
(Musci).
Класс антоцеротовые (Anthocerotae)
Небольшая группа (300 видов) наиболее просто устроенных
моховидных, распространенных преимущественно в тропической
зоне. В СССР всего несколько видов. Общий вид антоцероса пред*
ставлен на рис. 117. Небольшое лопастное слоевище распростерто
на почве и прикреплено к пей при помощи ризоидов. В ткань тал*
Рис. 117. Примитивные архегониаты. А — антоцерос — Anthoceros laevis (общий
вид со зрелыми спорангиями), Б — женский орган — архегоний, В — мужской
орган — антеридий (внутри развивающиеся сперматозоиды):
/ — брюшко архегония (в центре яйцеклетка); 2 — шейка (внутри шейковые канальцевые
клетки)
233
лома погружены антеридии и архегонии. Спорогон в форме вытя-
нутого до нескольких сантиметров стручочка с тонкой колонкой
внутри раскрывается двумя створками. Спорогон может удлинять-
ся за счет меристемы, расположенной в его основании. Внутрен-
нее строение таллома слабо дифференцировано; в клетках немно-
гочисленные хроматофоры, снабженные пиреноидами.
Сторонники самостоятельного происхождения мохообразных
от водорослей рассматривают антоцеротовых как наиболее прими-*
тивных высших растений, наиболее близких филогенетически к зе-*
леным водорослям.
Класс печеночника (Hepaticae, Marchantiatae)
Тело печеночника (гаметофит) имеет в большинстве случаев
дорзовентральное строение, т. е. в нем можно различать верхнюю,
или спинную {дорзальную'), сторону и нижнюю, или брюшную
{вентральную), сторону. От вентральной стороны отходят однокле*
точные ризоиды, соединяющие растение с субстратом.
Рис. 118. Маршанция (Marchantia polymorpha). А—женский таллом, Б —
мужской таллом
Всего известно свыше 8500 видов печеночников, среди них име*
ются листостебельные и талломные формы.
Класс подразделяют на 3 порядка: Маршанциевые (Marchan-
tiales), Юнгерманиевые (Jungermaniales) и Сферокарповые
(Sphaerocarpales). Наиболее богат видами (7000) порядок Юнгер-
маниевых, но виды этого порядка распространены преимуществен-
но в тропических странах. Небольшой порядок Сферокарповых за-
нимает промежуточное положение между Юнгерманиевыми и Мар-
шанциевыми, среди которых наиболее известна в нашей флоре
маршанция обыкновенная (Marchantia polymorpha), обитающая
преимущественно в северных и умеренных широтах нашей страны.
234
Это напочвенная талломная форма в виде дихотомически ветвя-
щейся пластинки размером до 10 см дорзовентрального строения
(рис. 118). Талломы раздельнополые, следовательно, растение это
двудомное. Органы полового размножения размещаются на особых
подставках. На мужских талломах подставки лопастные, и на верх-
ней стороне их сидят антеридии; на женских талломах подставки
звездчатые; между лучами подставки группами расположены
архегонии шейкой вниз. После оплодотворения из зиготы развива-
ется спорангий в виде коробочки, сидящей на короткой ножке.
Споры, выпадающие из коробочки (спорангия), дают сначала
пластинчатую протонему, из которой уже развивается гаметофит.
Споры перемешаны с пружинками — элатерами, которые разрыхля-
ют массу спор, способствуя их распространению. Для вегетативного
размножения служат двулопастпые выводковые почки, которые об-
разуются в открытых вместилищах — корзинках, вырастающих
на верхней стороне таллома. Выводковые почки, естественно, про-
растают в гаметофиты.
Класс лиственные мхи (Musci, Bryatae)
Имеется свыше 14 000 видов лиственных мхов, распространен-
ных преимущественно в тропическом поясе. При этой многочислен-
ности видов лиственные мхи отличаются сравнительным однообра-
зием строения. Из 14 000 видов 13 500 относятся к обширному по-
рядку зеленых мхов (Bryales), типичных для данного класса.
Около 300 видов объединяются в своеобразную группу белых,
или сфагновых, мхов—лорядок Sphagnales.
Третий порядок — андреевые мхи (Andreales) представлен не-
большим количеством видов, занимающих промежуточное место
между зелеными и сфагновыми мхами: внешний облик их сходен
с зелеными мхами, внутреннее строение ближе к сфагновым.
Порядок сфагновые мхи (Sphagnales). Единственный относя-
щийся сюда род Sphagnum (рис. 119) содержит свыше 300 сходных
между собой видов. Сфагновые мхи особенно широко распростра-
нены в северной лесной зоне Европы и Северной Америки, где они
покрывают огромные заболоченные пространства. Нередко можно
встретить сфагнум и в горах. Здесь он произрастает в местах с по-
вышенной влажностью и с бедным каменистым субстратом.
Из споры сфагнового мха развивается пластинчатая протонема,
из которой вырастает сильно ветвящийся стебелек гаметофита, гус-
то усаженный листочками типа филлоидов. Филлоиды имеют
своеобразное внутреннее строение — большую часть их объема
составляют крупные мертвые клетки с кольчатыми и спиральными
утолщениями, между ними сетка из тонких живых хлорофиллонос-
ных клеток. Преобладание в листе мертвых, заполненных воздухом
клеток обусловливает белесоватый цвет мха, почему сфагновые
мхи называют также белыми мхами. Такие же мертвые клетки со-
ставляют главную массу коры стебля. Преобладание в ткани
листьев и стеблей пустых воздухоносных клеток определяет огром-
235
Рис. 119. Торфяной мох сфагнум (Sphagnum). А—общий вид,
Б — микроскопическое строение листа
ную гигроскопичность сфагнового мха; одна часть сухого мха мо-»
жет впитать 25—30 частей воды.
Биология развития сфагновых мхов такая же, как и зеленых.
Сфагновые болота образовали мощные толщи торфа. Торф име-
ет огромное народнохозяйственное значение; его используют как
топливо, а также в качестве сырья для химического производства.
Сфагновый мох может служить прекрасной подстилкой для скота,
а также используется при строительстве домов в качестве прокла-
док между бревнами.
Порядок зеленые мхи (Bryales). Эти лиственные мхи распростра-
нены преимущественно в местах с избыточным или хотя бы доста-
точным увлажнением. Спора дает хорошо развитую протонему
обычно в форме ветвистой нити. На протонеме закладываются поч-
236
Рис. 120. Зеленый мох «кукушкин лен» (Polytrichurn commune). А — общий
вид, Б — поперечный разрез стебля, В — лист, Г — верхушка мужского га-
метофита, Д — верхушка женского гаметофита, Е — коробочка спорангия:
/ — мужской гаметофит; 2 — женский гаметофит со спорофитом; 3 — общий вид лис-
та; 4 — поперечный разрез листа; 5 —антеридий; 6, 8 — парафизы; 7 — архегоннй;
9 —спорогоний с колпачком; 10 — он же без колпачка; // — продольный разрез спо-
рогония
ки, из которых развивается листостебельное тело гаметофита.
Ризоиды многоклеточные, хорошо развитые. Спорофит развивается
на гаметофите и представляет собой спорогоний, состоящий из нож-
ки и коробочки, внутри находится спорангий. Основание ножки
играет роль гаустория, с помощью которого спорофит извлекает
питательные вещества из гаметофита.
Более детально с лиственными мхами познакомимся на приме-
ре широко распространенного зеленого мха кукушкина льна
(рис. 120).
237
Кукушкин лен (несколько видов рода Polytrichuni) образует
густые дерновины в хвойных лесах, на лугах, болотах. Это один
из наиболее рослых мхов, стебель его достигает высоты 20 см
и более. Стебель мха покрыт жесткими листьями — филлоидами —
клиновидноланцетной формы. Подземная часть стебля представ-
лена корневищем, от которого отходят ризоиды. В центральной
части стебля проходит примитивный проводящий пучок, снаружи
стебель одет эпидермисом.
Стебелек заканчивается группой антеридиев или архегониев,
окруженных пучком листьев.
Листья, окружающие антеридии, имеют красноватую или жел-
товатую окраску. Антеридий представляет собой удлиненный мешо-
чек, заполненный сперматозоидами; стенка антеридия из одного
слоя клеток.
Между антеридиямм имеются выросты в виде однослойной пла-
стинки — парафизы.
Архегоний имеет форму колбочки; на дне колбочки яйцеклетка,
над ней брюшковая клетка, а в горлышке архегония цепочка шейко-
вых клеток. К моменту оплодотворения брюшковая и шейковые
клетки расплываются в слизь.
Оплодотворение совершается при помощи капельки воды. Из зи-
готы развивается спорогоний, который состоит из коробочки, сидя-
щей на длинной ножке. Коробочка прикрыта волосистым колпач-
ком, развивающимся из брюшка архегония; в коробочке спорангий
со спорами. Коробочка имеет довольно сложное строение. Она сос-
тоит из урны и крышечки. В центре урны возвышается колонка,
наверху колонка шляпкообразно расширяется и закрывает урночку
эпифрагмой — тонкой эластичной пленкой. Внутри урночки цилин-
дрический мешочек, заполненный спорами,— спорангий.
К моменту созревания спор вначале слетает волосистый колпа-
чок, затем отваливается крышечка. Через отверстия, расположен-
ные по краям эпифрагмы, споры высыпаются наружу. Споры очень
мелки и легко развеиваются ветром; попадая в благоприятную
среду, они прорастают в протонему, из которой развивается листо-
стебельный гаметофит.
Сходно с кукушкиным льном размножаются и другие многочис-
ленные виды зеленых мхов. Среди них встречаются формы и очень
мелкие, всего несколько миллиметров высотой, и очень крупные,
достигающие 0,5 м высоты.
Помимо обычного способа размножения спорами зеленым мхам
свойственно также вегетативное размножение при помощи вывод-
ковых почек, образующихся на листьях, стебле или в особых вмес-
тилищах. Форма почек разнообразна.
Филогенетические связи между тремя классами мохообразных
не ясны; по-видимому, антоцеротовые, печеночники и лиственные
мхи представляют самостоятельные ветви эволюции, отходящие
от общей родоначальной формы. Относится ли этот родоначальник
к псилофитовым или был непосредственно связан с зелеными водо-
рослями— на этот вопрос пока нет точного ответа.
238
ОТДЕЛ 3. ПСИЛОТОВЫЕ (PSILOTOPHYTA)
Отдел представлен немногими видами, относящимися к двум
родам: псилот (Psilotum) и тмезиптер (Тmesipt&ris). Это неболь*
шие растения, обитающие в тропической и субтропической зонах
обоих полушарий. Ведут наземный или эпифитный образ жизни.
Стебли, достигающие длины 100 см, покрыты чешуевидными
Рис. 121. Псилот — Psilotum triquetrum. А — общий вид спорофита,
Б — часть таллома, В — спорофилл, Г — гаметофит, Д — разрез гамето-
фита:
1 — синангий; 2 — ризоид; 3 — архегоний; 4 — антеридий; 5 — микориза
листьями, имеющими, однако, проводящие пучки. Нижняя часть
стебля переходит в подземные «корневища» — ризомоиды длиной
свыше 1 м.
Проводящие элементы стебля образуют актиностелу. Споран*
гии срослись в двух- или трехгнездные синангии. Из споры выра*
стает подземный гаметофит, внешне похожий на ризомоид и живу*
щий в симбиозе с грибами (микориза): по всей поверхности гаме*
тофита рассеяны антеридии и археогонии. Сперматозоиды
многожгутиковые. Как уже сказано, псилоты, по-видимому, могут
являться прямыми потомками псилофитов. Рисунок 121 дает пред*
ставление о внешнем и внутреннем строении псилотовых.
239
ОТДЕЛ 4. ПЛАУНОВИДНЫЕ (LYCOPHYTA, LYCOPODIOPHYTA)
Древнейшие представители этого отдела произошли от псило-
фитов еще в силуре, но наиболее пышного расцвета плаунообраз-
ные достигли в каменноугольном периоде; влажные леса этого
периода в большей степени состояли из плаунообразных, достигав-
ших размеров крупных деревьев (см. рис. 116).
Современные представители плаунообразных лепидодендроны,
сигиллярии — крупные травянистые растения, снабженные слабо-
развитыми корнями и небольшими тонкими листьями типа фил-
лоидов. Среди них имеются и равноспоровые, и разноспоровые
формы.
Мы познакомимся с представителями обеих форм, относящи-
мися к двум порядкам: Lycopodiales и Selaginellales. Сохранились
также представители третьего порядка — Isoetales (полушнико-
вые)—водные растения; два вида этого порядка встречаются
и на территории СССР.
Плаун булавовидный (Lycopodium clavatum (см. рис. 113) от-
носится к семейству плауновых (Lycopodiaceae). Распространен
в хвойных лесах. Стебель стелющийся, дихотомически разветвлен-
ный, покрытый мелкими линейно-шиловидными листьями. Он до-
вольно прочный, достигает длины 6 м и более. Проводящие эле-
менты оформлены в актиностелу или плектостелу. К почве стебель
прикрепляется тонкими, слабо развитыми придаточными корнями.
Верхушечные побеги заканчиваются спороносными колосками.
Колоски обычно парные; верхняя прямостоячая часть веточки,
на которой они сидят, дихотомически разветвлена и покрыта более
редко расположенными листочками. Спороносный колосок состоит
из стержня, покрытого тонкими листочками,— спорофиллами. В па-
зухе спорофиллов сидят одиночные почковидные спорангии. В спо-
рангиях большое количество спор тетраэдрической формы, снабжен-
ных толстой сетчатой оболочкой. Морфологически и физиологи-
чески споры одинаковы. В благоприятных условиях из споры вы-
растает маленький (2—3 мм) кубаревидный заросток — двуполый
гаметофит плауна. Гаметофит лишен хлорофилла, он оплетен гифа-
ми гриба, образующего микоризу. На верхней стороне гаметофита
имеются органы полового размножения — архегонии и антеридии,
и те и другие типичной формы. Сперматозоиды с двумя жгутиками,
овальной формы. После оплодотворения из зиготы развивается за-
родыш, из зародыша — взрослое растение спорофитпого поколения.
На образование из споры гаметофита и развитие из пего спорофита
требуется свыше 20 лет.
Распространены плауны преимущественно в тропической зоне,
всего насчитывается их свыше 100 видов; в СССР— 14 видов.
Споры плаунов имеют практическое применение, они исполь-
зуются в медицине в качестве детской присыпки, для обсыпки
пилюль, в металлургии при тонком литье, а также при изготовле-
нии фейерверков. Как корм плауны никакой ценности не пред-
ставляют; они практически не поедаются животными.
240
Селагинелла (Selaginella) относится к семейству селагинелло-
вых (Selaginellaceae). Стебли селагинелл нежнее, чем у плаунов;
внутреннее строение их бывает сложное: встречается сифоносте-
ла, как производное протостелы.
Встречающиеся в нашей стране немногочисленные виды пред-
ставлены небольшими, дихотомически ветвящимися кустиками
до 10 см длиной.
Тропические вьющиеся формы достигают длины 20 м. Колоски
селагинелл имеют спорангии двух типов: микроспораигии с боль-
Рис. 122. Селагинелла (Selaginella). А — общий вид растения, Б — коло-
сок, В — микроспорангий, Г — мегаспорангий, Д — проросшая мегаспора
с женским заростком (гаметофитом)
шим количеством микроспор, и мегаспорангии, в каждом из кото-
рых находится по 4 большие мегаспоры (рис. 122).
Микро- и мегаспорангии сидят в пазухах спорофиллов на одном
и том же спороносном колоске.
В отличие от плаунов селагинеллы, следовательно, разноспо-
ровые растения, и это является выражением прогресса. Гаметофи-
ты, развивающиеся из спор, сильно редуцированы, особенно муж-
ские. Заростки не покидают оболочки споры. Мужской заросток
состоит из антеридия и одной вегетативной клетки; на почти мик-
роскопическом женском заростке несколько архегониев типичного
241
вида. После оплодотворения образуется зародыш, а из него разви*
вается взрослый спорофит.
Всего известно около 600 видов селагинелл, преимущественно
в тропиках. В Советском Союзе найдено только восемь видов —
в горах Кавказа, Урала, Сибири, Дальнего Востока и в Арктике.
Хозяйственного значения селагинеллы не имеют, но тропические
виды часто разводятся в оранжереях как красивые декоративные
растения.
ОТДЕЛ 5. ЧЛЕНИСТЫЕ, ХВОЩЕВИДНЫЕ
(ARTICULATAE, ИЛИ EQISETOPHYTA)
Название свое растения этого отдела получили по строению
стебля, разделенного на членики, и по форме листьев у некоторых
ископаемых форм. В отличие от плауновых листья членистых —
это настоящие листья, филломы-, филогенетически они имеют стеб-*
левое происхождение: это сильно видоизмененные боковые веточ-
ки. Членистые возникли почти одновременно с плауновидными
(в начале девона) и древовидные формы их — каламиты — вместе
с лепидодендронами, сигилляриями и папоротниками образовали
мощные леса каменноугольного периода. Подобно плауновидным,
членистые сохранились лишь в форме невысоких травянистых рас-
тений— хвощей, играющих сейчас, как и плауны, ничтожную роль
в формировании растительного покрова.
Современные хвощи, их около 30 видов, широко распростране-
ны преимущественно во влажных местах. В нашей стране распрос-
транены хвощи: полевой, луговой, болотный, лесной, топяной, вет-
вистый и другие виды. Мы рассмотрим биологию хвоща на при-
мере наиболее широко распространенного вида — хвоща полевого
(см. рис. 114).
Хвощ полевой (Equisetum arvense) распространен на полях
в условиях избыточного увлажнения. Многолетнее растение. Зиму-
ющая часть — мощно развитые подземные корневища с запасами
питательных веществ. Органами запаса являются также неболь-
шие клубни, сидящие на корневищах. От узлов корневища в ниж-
ней части надземного стебля отходят придаточные корни. Прово-
дящие элементы расчленены <на участки, образующие артростелу;
в середине стебля полость.
Ранней весной на корневищах вырастают бледно-розовые спо-
роносные побеги высотой около 20 см. На узлах побегов сидят му-
товки листьев; заканчиваются побеги спороносными колосками.
Позднее развиваются вегетативные побеги с мутовчато располо-
женными боковыми веточками. Спороносный колосок состоит
из стержня, перпендикулярно которому на ножках сидят спорофил-
лы в виде шестиугольной пластинки — щитка. Вокруг ножки споро-
филла, под щитком, находится 6—10 спорангиев, в них многочислен-
ные округлые споры зеленоватого цвета.
Вначале щитки спорофилла сидят плотно, без зазоров; позд-
нее, к моменту созревания спор, стержень колоска удлиняется,
242
между щитками образуются промежутки, через которые и высыпа-
ются споры из созревших спорангиев.
Шаровидная спора обмотана четырьмя пружинками — элатера-
ми. Элатеры во влажном воздухе прилегают плотно к споре, в су-
хом воздухе концы их раскручиваются. Сцепляясь между собой
элатерами, споры комочками, по (нескольку штук вместе, разно-
сятся токами воздуха и, попадая во влажную среду, прорастают.
Гаметофиты, развивающиеся из спор хвоща полевого, двуполые,
причем архегонии и антеридии у них развиваются не одновременно,
а чередуясь во времени. Замечено, что лучшие условия питания
способствуют развитию архегониев. При загущенности гаметофи-
тов и на очень бедном субстрате развиваются преимущественно ан-
теридии. У других видов гаметофиты бывают и раздельнополые,
и, следовательно, споры у них физиологически разнозначны.
Архегонии и антеридии погружены в ткань заростка. В антери-
диях образуется свыше 200 сперматозоидов в каждом. Спермато-
зоиды спирально закручены, с большим числом жгутиков. Опло-
дотворение происходит при помощи воды. Из оплодотворенной
яйцеклетки развивается зародыш, из него — взрослый спорофит.
После рассеивания спор у хвоща полевого спороносные побеги
отмирают и на смену им вырастают зеленые ассимилирующие му-
товчато разветвленные побеги. То же происходит у хвоща лесного
(Е. silvaMcum) и лугового (Е. pratense). У других видов (болотный,
топяной, ветвистый) спороносные колоски образуются на верхуш-
ках обычных ассимилирующих стеблей.
Хвощи достигают в высоту 0,5—1 м. Лишь один из тропических
видов вырастает до 12 м.
Значение хвощей чаще отрицательное. Поселяясь на луговых
и полевых землях, они, будучи в большинстве своем несъедобными,
засоряют их; некоторые виды хвощей (топяной, болотный, полевой)
ядовиты. Причина ядовитости, по-видимому, в содержании у хво-
щей алкалоидов. Однако хвощ ветвистый поедается животными
без видимых признаков отравления. В таежной зоне имеются виды
хвощей, ценные в кормовом отношении (Е. variegatum, Е. scirpoi-
des), а хвощ зимующий (Е. hiemale) неплохо поедается лошадьми,
крупным рогатым скотом и другими видами сельскохозяйственных
животных.
ОТДЕЛ 6. ПАПОРОТНИКОВИДНЫЕ (PTEROPHYTA, POLYPODIOPHYTA)
Наиболее сохранившийся среди высших споровых растений от-
дел. Древнейшие ископаемые формы известны с девона. В карбоне
крупные древовидные папоротники наравне с другими споровыми
растениями составляли обширные влажные леса, остатки которых
дают теперь залежи каменного угля. К нашему времени сохрани-
лось свыше 10 000 видов папоротников, многие из них (в Австра-
лии и на Новой Гвинее) достигают размеров крупного дерева.
Как и предыдущие отделы высших растений, папоротниковидные
произошли от псилофитов; сохранились ископаемые формы папо-
ротников, 'Непосредственно примыкавшие к псилофитовым.
243
В отдел папоротниковидных входит несколько порядков, в ко-
торых представлены и равноспоровые и разноспоровые. Из равно-
споровых наиболее богат видами порядок настоящие папоротники
(Polypodiales). В этом порядке свыше 10 семейств, большинство
видов относится к семейству Polypodiaceae.
Разноспоровые водные представлены двумя порядками: Сальви-
ниевые (Salviniales) и Марсилиевые (Marsiliales).
В качестве типичного представителя настоящих папоротников
рассмотрим обычный в лесах папоротник Aspidium, Dryopteris
filix mas (см. рис. 115), называемый щитовником мужским, широко
распространенный в лесах умеренной зоны. Из корневища, покры-
того бурыми чешуями, на поверхности почвы вырастает пучок
листьев. У основания взрослых листьев молодые листья свернуты
в улитку. Новый лист начинает развертываться под поверхностью
почвы и лишь на третий год жизни окончательно развивается, до-
стигая в длину 1 м. Лист двоякоперистораздельный, растет верхуш-
кой. Все взрослые листья каждый год поздней осенью отмирают.
В корневищах многочисленные пучки концентрического типа, в со-
вокупности образующие диктиостелу.
К середине лета на нижней поверхности листа развиваются спо-
рангии. Спорангии сидят группами (сорусами) на толстом вырос*
те — плаценте, из которой вырастает зонтиковидное покрывальце —
индузий. Спорангий в форме овального плоского мешочка; стенки
его состоят из одного слоя клеток. На гребне мешочка находится
кольцо из толстостенных клеток, охватывающих 3/4 окружности.
Наружная тангентальная стенка клеток кольца не утолщена, бла-
годаря этому при подсыхании спорангия радиальные стенки клеток
сближаются, раскручивая кольцо в обратную сторону, спорангий
раскрывается, и споры разбрасываются.
В благоприятных условиях спора прорастает. Зеленая пластин-
ка заростка имеет сердцевидную форму, размер ее до 1 см (рис.
123). Посредством ризоидов заросток прикрепляется к почве.
На нижней поверхности его развиваются, ближе к почве, антери-
дии, выше них — архегонии. Те и другие типичной формы. Спермато-
зоид штопоровидный, на переднем конце его пучок жгутиков,
на заднем — пузырь из цитоплазмы. Оплодотворение происходит
под каплей воды, осевшей на заросток. Из зиготы развивается за-
родыш, соединенный с заростком; после укоренения зародыша
заросток отмирает. Зародыш постепенно развивается во взрослое
растение — спорофит. Спорофит диплоидный, споры и гаметофит
гаплоидные. Папоротники могут размножаться и вегетативно,
при помощи особых выводковых почек.
В Советском Союзе насчитывается около 100 видов папоротни-
ков. Большинство из них практически не поедается животными.
Наоборот, среди них имеются ядовитые папоротники. Экстракт
из корневища щитовника мужского используется как сильнодейст-
вующее глистогонное средство. Вместе с тем имеется многолетняя
практика (за рубежом и у нас) скармливания скоту корневищ
папоротников без видимых ’вредных последствий. Такие наблюде-
244
ния имеются, например, в отношении широко распространенного
в лесной зоне папоротника орляка (Pteridiurn aquilinutn). По-види-
мому, ядовитость папоротников сильно меняется в связи с условия-
ми произрастания, фазой вегетации и др. На пастбищах и полях
папоротники — злостные сорняки, подлежащие искоренению.
Рис. 123. Заросток (гаметофит) папоротника Dryopteris filix mas
Из водных папоротников рассмотрим на юге СССР салъвинию
(Salvinia natans). Это плавающее однолетнее растение встречает-
ся на озерах или >в протоках в большом количестве, например
в низовьях Днепра. Тонкий стебелек сальвинии длиной до 15 см
имеет на каждом узле по 3 листа: 2 обычного вида плавающих
и один, погруженный в воду, рассеченный на нитевидные доли, по-
хожие на тонкие корни (рис. 124). У основания подводных листьев
гроздьями сидят шаровидные замкнутые сорусы — спорокарпии.
В сорусах — спорангии различного размера, в одних в небольшом
числе крупные — мегаспорангии, в других многочисленные мелкие —
микроспорангии. В мегаспорангиях полного развития достигает
лишь одна мегаспора; в микроспорангии обычно 64 споры. Сорусы
со спорангиями зимуют на дне водоемов. К весне оболочки споро-
карпиев сгнивают и спорангии всплывают. Споры прорастают
внутри спорангиев. Многочисленные антеридии прорастают сквозь
245
Рис. 124. Сальвиния пла-
вающая — Salvinia па-
tans.
А — спорофит; Б — лист
(поперечный разрез), В—
спорокарпий (попереч-
ный разрез), Г — микро-
спорангий с мужским га-
метофитом, Д — завер-
шающие стадии разви-
тия мужского гаметофи-
та с восемью спермато-
зоидами, Е — мегаспо-
рангий, Ж — мегаспо-
рангий с женским гамето-
фитом, 3 — архегоний,
И — мегаспорангий с
женским заростком и
развивающимся зароды-
шем спорофита, Л" — мо-
лодой спорофит:
1 — спорокарпий; 2 — подвод-
ные рассеченные листья; 5 —
микроспорангий; 4 — мега-
спорангий; 5 спермато-
зоид; 6 — мегаспора; 7 — ар-
хегоний; 8 — яйцеклетка; Р —
тело гаметофита; 10 — заро-
дыш спорофита; 11 — обо-
лочка мегаспоры; 12 — оста-
ток стенки мегаспорангия
стенку микроспорангия, мегаспора прорастает в небольшой зеленый
веерообразный заросток, который прорывает стенку мегаспорангия;
большая часть его остается в оболочке мегаспорангия. После опло-
дотворения из зиготы развивается зародыш, из которого формиру-
ется взрослое растение. Первое время зародыш живет, питаясь
за счет заростка — женского гаметофита.
ОТДЕЛ 7. ГОЛОСЕМЕННЫЕ (GYMNOSPERMAE, PINOPHYTA)
В отличие от высших споровых голосеменные растения размно-
жаются и расселяются при посредстве семян. В условиях сухопут-
ного образа ’жизни семя выполняет эту функцию несравненно
успешней, нежели спора, поэтому возникновение семени явилось
важным этапом в эволюции высших растений. В типичном выраже-
нии семя — это зародыш спорофита, снабженный запасом пита-
тельных веществ для дальнейшего развития и защищенный от не-
благоприятных воздействий внешней среды более или менее мощ-
ной оболочкой.
Первые голосеменные растения возникли еще в конце девона,
около 350 млн. лет назад. По-видимому, они произошли от первич-
ных папоротников из порядка Archaeopteridales, вымерших в на-
чале каменноугольного периода. Все голосеменные представлены
деревьями или кустарниками; травянистых растений среди них нет.
Количество видов голосеменных невелико, всего около 700, но рас-
пространены они по всему земному шару, и в холодной зоне север-
ного полушария занимают огромные территории, образуя хвойные
леса.
Расцветом флоры голосеменных был конец палеозоя и мезозой.
К концу мезозоя сушей овладели покрытосеменные, а многие груп-
пы голосеменных почти или полностью вымерли. Филогенетиче*
ская система голосеменных растений строится с учетом не только
ныне живущих групп, но и вымерших форм.
Отдел голосеменных разделяют на три класса: 1. Саговниковые
(Cycadatae). 2. Шишконосные (Pinatae). 3. Оболочкосеменные
(Chlamydospermatae, Gnetatae).
Класс саговниковые (Cycadatae)
Этот класс включает три порядка: семенные папоротники
(Pteridospermales); саговники (Cycadales) и беннеттиты (Bennetti-
tales). Из трех порядков сохранилось только небольшое число ви-
дов саговников. Другие два порядка представлены вымершими фор-
мами.
Порядок семенные папоротники (Pteridospermales)» Появились
в конце девона, вымерли в триасовом периоде; роль их в филоге-
незе аналогична псилофитам; от них, вероятно, произошли все ос-
тальные семенные растения. Это были деревья, похожие на совре-
менные древовидные папоротники. В отличие от современных па-
поротников они были разнополы и размножались посредством
семян.
247
На крупных двоякоперистых листьях формировались группы
микроспорангиев (синангии) и мегаспорангии, одетые дополнитель-
ными оболочками — интегументами. Такие мегаспорангии называ-
ются семенными зачатками, поскольку из них развиваются семена.
С семенными папоротниками обычно сближают небольшую груп-
пу ископаемых растений неясного происхождения — кейтониевые
(Caytoniales) прибрежно-водного обитания. Семезачатки кейтоние-
вых в числе 6—30 обрастали частью мегаспорофилла, и возникало
подобие завязи, что внешне сближало кейтониевые с покрытосемен-
ными. Кейтониевые жили в первой половине мезозойской эры (три-
ас и юра).
Порядок саговники (Cycadales). Возникли в конце палеозоя,
наибольшего расцвета достигли в юрском периоде. К нашему вре-
мени сохранилось около 100 видов саговников. Распространены
в тропических странах Старого и Нового Света. Это крупные де-
ревья, напоминающие пальмы; большие перисторассеченные
листья расположены и а вершине ствола. Наиболее крупные
представители порядка достигают высоты 25 м, а ствол — тол-
щины 1 м.
Саговники — растения двудомные. Мегаспорофиллы или похо-
жи на обычные листья (у рода Cycas), или собраны в шишки —
стробилы. На каждом спорофилле по 1неокольку мегаспорангиев —
семезачатков. Микроспорофиллы всегда собраны в стробилы —
шишки, вырастающие на верхушке стебля. Женские стробилы
с семезачатками у некоторых саговниковых достигают огромных
размеров и массы в несколько десятков килограммов; семезачат-
ки достигают размера 5 см. Оплодотворение при помощи подвиж-
ных сперматозоидов.
Сердцевина некоторых видов саговников съедобна, из нее де-
лают крупу под названием «саго». В Советском Союзе саговники
выращиваются как декоративные растения.
Порядок беннеттитовые (Bennettitales). Наиболее древние остат-
ки найдены в отложениях конца пермского периода. В середине
мезозоя беннеттиты достигли наивысшего расцвета, но в верхнем
мелу уже вымерли. По внешнему и внутреннему строению они похо-
дили на современных саговниковых; ровный и чаще толстый ствол
заканчивается на верхушке пучком крупных перисторассеченных
листьев. Существенное различие было в органах размножения.
Большинство беннеттитовых имели двуполый стробил, напоминаю-
щий по типу строения цветок современных покрытосеменных (рис.
125). Микроспорофиллы с большим количеством микроспорангиев
располагались на периферии стробилов; редуцированные мегаспо-
рофиллы размещались в центральной части стробила, каждый
из них имел по одному семезачатку; интегументы семезачатка
вытянуты в характерную микропилярную трубку. Спорофиллы бы-
ли окружены покроволистиками, аналогичными околоцветнику
покрытосеменных. Опыление у беннеттитовых осуществлялось
при помощи насекомых. Беннеттиты, так же как и саговники, про-
изошли от семенных папоротников.
248
Рис. 125. Продольный разрез двуполых стробилов беннеттитов
Класс шишконосые (Coniferatae, Pinatae)
Относящиеся сюда растения имеют разветвленный стебель, срав-
нительно не крупные листья игольчатой или пластинчатой, линей-
ной формы. Этим они отличаются от саговниковых, для которых
характерен одиночный неразветвленный ствол с крупными перисты-
ми листьями на верхушке.
К этому классу относятся три порядка: кордаитовые (Cordaita-
les)—полностью вымерший; гинкговые (Ginkgoales)—представ-
ленный одним видом, и хвойные (Coniferales)—наиболее богатая
видами группа голосеменных растений.
Порядок кордаитовые (Cordaitales). Был представлен многочис-
ленными видами древовидных форм, начиная с карбона и до начала
мезозойского периода; вымерли в триасе.
249
Заросли кордаитов походили на современные хвойные леса, вы-
сота деревьев достигала 30 м, толщина ствола — 1 м. Стебель имел
сильно развитую сердцевину — сходство с саговниками. Линейные
листья достигали длины 1 м при ширине от нескольких сантимет-
ров до 20 см. Стробилы однополые; растения однодомные и дву-
домные.
Кордаитовые произошли от семенных папоротников, а сами, дали
гинкговые и хвойные.
Порядок гинкговые (Ginkgoales). Остатки гинкговых известны
еще с верхнего карбона, но наибольшего расцвета эта группа рас-
тений достигла в середине мезозоя. К нашему времени сохранился
всего один вид Ginkgo biloba. Его называют живым ископаемым, так
как ближайшие родичи этого растения десятки миллионов лет на-
зад вымерли. Ginkgo biloba в культурном состоянии широко рас-
пространен в Японии, а оттуда вывезен в Европу и Северную Аме-
рику. Это крупное красивое дерево, достигающее высоты 70 м;
очень долговечное, доживает до 2000 лет. Листья имеют вееровид-
ную лопастную пластинку на тонком черешке; жилкование дихо-
томическое. На зиму листья опадают.
Гинкго — двудомное растение; оплодотворение осуществляется
подвижными сперматозоидами. Разводят как декоративное дерево.
Семена съедобные.
Порядок хвойные (Coniferales, Pinales). Наиболее сохранившая-
ся и самая многочисленная группа голосеменных. Возникнув в кон-
це каменноугольного периода, хвойные, как и другие голосеменные,
наибольшего расцвета достигли в мезозое; но и в наше время име-
ется около 600 видов хвойных. Огромные массивы хвойных распро-
странены на севере Евразии и Северной Америки; наибольшее
многообразие их видов представлено в бассейне Тихого
океана.
В палеогене и неогене хвойные были распространены севернее
теперешней своей границы и заселяли территории, покрытые в на-
стоящее время вечными арктическими снегами и льдами.
Почти все виды хвойных представлены деревьями, достигающи-
ми у некоторых видов более 100 м и возраста свыше 4000 лет. Не-
многие виды имеют кустарниковую форму; травянистых растений
среди хвойных совсем нет. У большинства хвойных листья игольча-
тые (хвоя), реже чешуевидные и еще реже с широкой пластинкой,
напоминающей листья ископаемых кордаитовых. Кроме зеленых
листьев, у хвойных обычно коричневые чешуйки — рудименты на-
стоящих листьев. За исключением лиственницы и нескольких дру-
гих родов, у всех хвойных листья не опадают в неблагоприятное
время года.
Древесина хвойных устроена гораздо проще, чем у деревьев
цветковых растений, и состоит в основном из трахеид.
Стробилы у хвойных исключительно однополые, а растения од-
нодомные, реже двудомные. Форма и величина стробилов широко
варьируют. Жесткие мегастробилы обычно называют женской
шишкой, мягкие микростробилы — мужской шишкой.
250
4
Рис. 126. Сосна обыкновенная (Pinus silvestris):
/ — женская шишка первого года; 2 — женская шишка прошлого года; 3 — раскрывшаяся
(старая) шишка; 4— мужская шишка (стробил); 5 — то же (увеличено); 6 — женская шиш-
ка; 7 — семенная (плодущая) чешуя (мегаспорофилл) и два семезачатка; 8 — микроспора
Чередование поколений у хвойных, как и вообще у голосемен-
ных, в связи с сильной редукцией гаметофитов весьма затушевано.
Для расшифровки этого явления мы детально рассмотрим все фазы
индивидуального развития (онтогенеза) на примере наиболее ши-
роко распространенного у нас представителя хвойных — сосны
обыкновенной (Pinus silvestris).
В мае у основания молодых побегов сосны образуются пучки
серовато-желтых мужских шишек — стробилов (рис. 126). Каждая
шишка состоит из оси, густо одетой микроспорофиллами. На ниж-
ней стороне микроспорофилла два микроспорангия. Внутри микро-
спорангия многочисленные микроспоры, устроенные очень своеоб-
разно. Микроспора представляет собой одну клетку, одетую двумя
оболочками: внутренней — интиной и наружной — экзиной. Экзина
с двух противоположных сторон вздута в виде двух пузырей, увели-
чивающих летучесть споры.
Женские стробилы-шишки возникают на концах побегов теку-
щего года в числе одной или нескольких; молодая, только что сфор-
мировавшаяся шишка красноватого оттенка, состоит из стержня
и плотно прилегающих к нему чешуй — мегаспорофиллов. Чешуи
двух родов: наружная маленькая кроющая и внутренняя — круп-
ная семенная. У основания семенной чешуи, на ее верхней стороне,
расположены два беловатых семенных зачатка.
251
Рис. 127. Семезачаток сосны:
1—4 — последовательные стадии развития женского гаметофита
<4
В процессе формирования стробила каждый семенной зачаток
возникает в виде бугорка, обрастающего покровом — инте гумен*
том; интегумент вверху не срастается, оставляя пыльцевход —
микропиле. Бугорок превращается в нуцеллус, или ядро семенного
зачатка., Нуцеллус — гомолог мегаспорангия споровых растений.
Внутри нуцеллуса обособляется одна клетка, отличающаяся вели-
чиной и более густой цитоплазмой. Это археспориальная клетка.
Она двукратно делится, из образующихся четырех клеток три от-
мирают, одна остается и увеличивается в размерах. Это единствен-
ная мегаспора внутри нуцеллуса — мегаспорангия. Мегаспора про-
растает внутри мегаспорангия и дает женский гаметофит — зарос-
ток с двумя типичными архегониями (рис. 127). Заросток богат
питательными веществами и является эндоспермом будущего
семени.
Мужской гаметофит начинает развиваться внутри микроспоры,
когда та еще находится внутри пыльника — микроспорангия. Раз-
витие начинается с того, что ядро микроспоры делится и от клетки
микроспоры отделяется.маленькая проталлиалъная клетка, которая
затем делится на две: возникают две проталлиальные клетки (го-
молог мужского заростка), они вскоре рассасываются. Затем ядро
микроспоры делится еще раз, в результате чего образуется малень-
кая антеридиальная и болыПая вегетативная клетка — клетка
пыльцевой трубки. В таком состоянии микроспоры высыпаются
из раскрывшегося пыльника и разносятся ветром.
Попав в пыльцевход семенного зачатка, микроспора прилипает
к нуцеллусу и тут остается в состоянии покоя до следующего года,
при этом края интегумента сближаются и пыльцевход закрывается.
Развитие семени заканчивается на следующий год. Этим сосна от-
личается от большинства остальных хвойных, у которых все разви-
тие заканчивается в течение одного года.
У сосны развитие мужского гаметофита продолжается через год.
Вегетативная клетка разрывает экзину и прорастает в пыльцевую
трубку, которая проникает через стенку нуцеллуса до архегониев.
В это время антеридиальная клетка разделяется на две: генератив-
252
ную и базальную. Содержимое базальной клетки переливается
в пыльцевую трубку. Генеративная клетка делится и дает два спер-
мия; один из спермиев через разрыв в пыльцевой трубке проникает
в архегоний и сливается здесь с яйцеклеткой. Второй спермий мо-
жет проникнуть в другой архегоний и тоже оплодотворить яйце*
клетку.
В результате дробления зиготы развивается зародыш семени,
женский заросток становится эндоспермом семени, нуцеллус и ин-
тегумент образуют оболочку семени. Одновременно с формирова-
нием и ростом семени увеличивается .и древеснеет шишка. Семена
высыпаются зимой, на второй год после образования женской
шишки и переноса на нее микроспор.
Таким образом, у хвойных, как и у других голосеменных, муж-
ской гаметофит редуцировался до размера нескольких клеток; за-
канчивает свое развитие мужской гаметофит на поверхности мега-
спорангия. Женский гаметофит развивается внутри мегаспорангия
и остается соединенным с материнским растением — спорофитом —
вплоть до сформирования внутри и за счет его зародыша следую-
щего спорофитного поколения, который в качестве основной части
семени начинает следующий цикл развития.
Порядок хвойные разделяют на девять семейств. Из ныне жи-
вущих наиболее богаты видами и широко распространены в тропи-
ческой и субтропической зонах семейства араукариевые (Агаиса-
fiaceae) и подокарповые (Podocarpaceae), в умеренной зоне — тис-
совые (Тахасеае), сосновые (Pinaceae), кипарисовые (Cupressa-
сеае) и таксодиевые (Taxodiaceae). Мы познакомимся лишь с неко-
торыми представителями последних четырех семейств.
Семейство тиссовые (Тахасеае). К этому семейству относятся
несколько родов, распространенных в северном полушарии. Для нас
представляет интерес род тисс (Taxus), наиболее крупный в семей-
стве.
Тисс ягодный (Taxus baccata) широко распространен в среди-
земноморских странах, в Западной Европе и у нас на Кавказе,
в Западной Белоруссии и Западной Украине (Беловежская пуща
и Буковина) и в Южном Крыму. Тисс ягодный — большое дерево,
достигающее 35 м высоты и свыше 1 м в диаметре, живет
до 3000 лет. Древесина тисса тяжелее воды, очень прочная и кра-
сивая, без смоляных ходов. Хвоя плоская, широкая. Растение дву-
домное. Древесина, кора и хвоя тисса ядовиты. Тисс очень теневы-
носливое растение.
Семейство сосновые — Pinaceae. Самое крупное и наиболее ин-
тересное в хозяйственном отношении семейство в порядке хвойных.
Виды этого семейства распространены в северном полушарии,
преимущественно в умеренной приполярной зоне, где они образуют
обширнейшие хвойные леса. Деревья, редко кустарники, за неболь-
шим исключением вечнозеленые, с жесткой хвоей. Однодомные.
В семействе 10 родов и свыше 200 видов. Для нас пред-
ставляют интерес виды пяти родов: сосна, ель, пихта, листвен-
ница, кедр.
253
Сосна (Pinus). Деревья или стланиковые формы. Побеги двух
типов: обычные и укороченные. Обычные побеги покрыты незеле-
ными чешуевидными листьями; укороченные — неразвитые побеги
и заканчиваются пучком листьев (хвои) обычно в числе 2—5. Жен-
ская шишка деревянистая. Наибольший хозяйственный интерес
представляет сосна обыкновенная (Р. silvestris) —основная лесооб-
разующая порода в европейской и азиатской частях Советского
Союза. Растет на разнообразных почвах, но предпочитает пески;
нередко ее высевают в целях лесомелиорации. На укороченных по-
бегах по два листа — хвои. Шишки небольшого размера. Светолю-
бивое растение. Дает хорошую строевую и .поделочную древе-
сину; из живицы получают скипидар, канифоль и другие про-
дукты.
Сосна кедровая, или сибирская (Р. sibirica). На укороченных по-
бегах пучок из 5 листьев; кроме ценной древесины, дает крупные
съедобные семена — кедровые орешки, из них добывают ценное
кедровое масло. Дерево теневыносливое.
Ель (Picea). Крупные деревья, достигающие высоты 50 м ха-
рактерной пирамидальной формы. Укороченных побегов нет. Оди-
ночные хвои обычно четырехгранные. Шишки кожистые, повислые.
Древесина более ценная, чем у сосны. Дерево теневынос-
ливое.
В Советском Союзе наиболее широко распространена в европей-
ской части ель обыкновенная {Picea excelsa), в Сибири ель сибир-
ская {Р. obovata).
Пихта {Abies). Внешне похожа на ель и также не имеет укоро-
ченных побегов. Хвоя одиночная, плоская. Шишки прямостоячие,
при созревании рассыпающиеся. Так же как и ель, теневыносливое
дерево, образующее темные леса. В Советском Союзе наиболее ши-
роко распространена пихта сибирская (Л. sibirica). Ареал ее охва-
тывает Сибирь и северо-восток европейской части СССР. На край-
нем западе СССР распространена белая пихта (A. alba), на Кавказе
пихта кавказская (A. nordmanniana). Древесина пихты менее цен-
на, чем сосны и ели, и используется преимущественно в производ-
стве бумаги; из коры добывают ценный пихтовый бальзам. Из всех
хвойных, встречающихся в СССР, пихта кавказская достигает
наибольшей высоты — 75 м.
Лиственница {Larix). Светолюбивое дерево, образующее свет-
лые леса. Древесина твердая, смолистая, долговечная. В отличие
от остальных хвойных, обитающих в СССР, лиственница сбрасыва-
ет на зиму хвою. Подобно сосне, образует обычные и укороченные
побеги; на концах укороченных побегов пучки мягкой хвои. Шишки
мелкие, созревают в течение года, но держатся на ветвях 2—3 го-
да. В СССР широко распространены лиственница сибирская
(L. sibirica) на Урале и в Западной Сибири и лиственница
даурская (L. dahurica) в Восточной Сибири, на Дальнем
Востоке.
Кедр {Cedrus). Красивейшее декоративное дерево; побеги
обычные и укороченные, на укороченных побегах пучки хвои. Шиш-
254
ки очень плотные, распадающиеся при созревании семян. У нас
растет только в культуре по Черноморскому -побережью Кавказа
и на южном берегу Крыма. Представлен тремя видами: кедр ливан-
ский (С. libanii), атласский (С. atlantica), гималайский (С. deodara).
Семейство кипарисовые — Cupressaceae. Представлено некруп-
ными деревьями или кустарниками, а также стланиковыми фор-
мами. Большей частью однодомные, но бывают и двудомные.
Листья игольчатые или чешуевидные, супротивные или мутовча-
тые. У можжевельников чешуи женского стробила к моменту со-
зревания семени становятся сочньши и превращаются в «ягоду».
В Советском Союзе распространены представители четырех ро-
дов: кипарис, можжевельник, биота, туя.
Кипарис (Cupressus). Очень долговечные деревья с плотной
красивой древесиной. Листья чешуйчатые. Женские шишки некруп-
ные, деревянистые. В Крыму и на Черноморском побережье Кав-
каза широко культивируется красивый пирамидальный вечнозеле-
ный кипарис (С. sempervirens). В этих же районах произрастают
в культуре и другие, непирамидальные формы кипариса.
Можжевельник (Juniperus). Наиболее богатый видами род. Пре-
обладают кустарниковые формы, но есть и деревья. Листья игло-
видные и чешуевидные. Женские шишки более или менее сочные,
образуют «ягоду». В горах Средней Азии распространены светлые
леса из можжевельника туркестанского (J. turkestanica); здесь же,
на границе растительности, характерным ландшафтным растением
является стланиковая форма можжевельника зеравшанского
(J. seravschanica). В европейской части СССР распространены
можжевельник обыкновенный (J. communis) с игольчатыми листья-
ми, можжевельник казацкий (J. sabina) с чешуевидной хвоей.
Заросли кустарникового можжевельника широко распространены
на каменистых склонах южного берега Крыма.
Биота (Biota) и туя (Thuja)—декоративные растения с харак-
терными побегами, веточки которых распространены в одной плос-
кости. Листья чешуевидные, зрелые шишечки деревянистые.
Семейство таксодиевые — Taxodiaceae. В этом семействе пред-
ставлены самые крупные хвойные деревья.
Секвойя (Sequoja). Единственный вид этого рода S. sempervie-
rens произрастает на тихоокеанском побережье Америки, в Кали-
форнии. Это огромные деревья, достигающие высоты 100 и более
метров и до 10 м в диаметре. Возраст достигает 4000 лет. Весьма
ценная, не поддающаяся гниению древесина. Широко культивиру-
ется у нас в Крыму и на Кавказе в декоративных целях. Мексикан-
ский болотный кипарис (Taxodium mucronatum) —дерево, достига-
ющее высоты 50 м и до 30 м в обхвате. Мягкая, но очень ценная
негниющая древесина. Живет до 6000 лет. Культивируется в Кры-
му и на Кавказе как декоративное дерево. Криптомерия японская
(Cryptomeria japonica) —также очень декоративное дерево с цен-
ной древесиной, высотой до 60 м, диаметром 1—2 м. Известна так-
же под названием японский кедр. Культивируется в Крыму
и на Кавказе.
255
Класс оболочкосеменные (Chlatnydospermatae,
Gnetatae)
К этому классу также относятся три порядка: эфедровые
(Ephedrales), вельвичиевые (Welwitschiales) и гнетовые (Gnetales).
В каждом порядке по одному монотипному семейству с единствен-
ным родом в каждом семействе: Ephedra, Welwitschia, Gnetum.
Эти три рода сближаются по наличию общих признаков: супротив-
ные листья, трахеи во вторичной ксилеме (на смену трахеидам),
дихазиальное ветвление собраний однополых стробилов, наличие
покровов вокруг стробилов, ПОХО-
ЖИХ на околоцветник покрытосе-
менных, длинные микропилярные
трубки у семезачатков, двудоль-
ные зародыши, двудомность.
Сходство с покрытосеменными в
свое время дало даже основание
приблизить оболочкосеменные не-
посредственно к цветковым рас-
тениям, в качестве их ближайших
предков; однако сейчас эта точка
зрения оставлена.
Род Ephedra имеет около 40
видов, распространенных в пу-
стынных областях Европы, Азии
и Америки. У нас несколько видов
эфедры, из них наиболее распро-
странена Ephedra distachya. Это
небольшой полукустарник с силь-
но редуцированными листьями,
похожий на хвощ или веточку
хвойного дерева (рис. 128). Эфед-
ру часто называют хвойником.
Внутреннее строение ее также
сходно с хвойным.
Эфедра — растение двудомное,
редко однодомное; стробилы, сле-
довательно, однополы. Однако не-
Рис. 128. Эфедра двуколосковая
(Ephedra dlstachya). А — общий вид,
Б — побег с микроспорангиями:
/ — женский побег; 2 — мужской побег;
3 — микроспорангий
редко в однополых стробилах находят рудименты — органы проти-
воположного пола. Мужские стробилы состоят из небольшого чис-
ла чешуй, в пазухе верхних чешуй микроспорофиллы, на концах
которых по 2—8 микроспорангиев. Ось женского стробила одета
несколькими бесплодными чешуями и заканчивается единственным
семезачатком, одетым двумя интегументами. Внутренний интегу-
мент вытянут в микропилярную трубку, улавливающую микроспо-
ры. Под интегументами нуцеллус, а в нем женский заросток (гаме-
тофит). Как и у хвойных, женский гаметофит представлен эндоспер-
мом, в который погружены архегонии; обычно их два, но иногда
бывает один, редко три.
256
Архегоний имеет длинную шейку, заполненную клетками (их
до 32). Оплодотворяется яйцеклетка в одном из архегониев. После
оплодотворения появляется семя, окруженное мясистым «около-
плодником», образовавшимся из сочного покрова семезачатка
и кроющих чешуй. Этот «плодик» обычно краснеет, напоминая
ягоду.
Некоторые виды эфедры дают ценнейшее лекарственное сырье —
алкалоид эфедрин. Для сбора сырья используются обширные зарос-
Рис. 129. Вельвичия удивительная — Welwitschia mirabilis. А — молодое женское
растение, Б — собрание мужских шишек,- В —г собрание женских шишек, Г — раз-
рез женских шишек, Д — разрез мужских шишек:
/ — покров; 2 — интегумент; 3 —нуцеллус семезачатка; 4 — рудиментарный семезачаток;
5 — микроспорофилл
ли хвойника в южном Казахстане. Хвойник неплохо поедается вер-
блюдами, овцами, козами, но содержащийся в нем алкалоид ино-
гда вызывает отравление.
Род Welwitschia представлен единственным видом — велъви-
чией удивительной (W7. mirabilis), обитающей в прибрежных пус-
9—326 257
тынях юго-западной Африки (рис. 129). Это действительно удиви-
тельное по своему внешнему виду растение имеет длинный корень,
толстый и короткий ствол до 1 м толщиной и до 50 см высотой
и два крупных листа, сохраняющихся в течение всей жизни
(до 100 лет). Листья достигают 2—3 м длины и постоянно нара-
стают у основания, отмирая с конца. Разветвленные стробилы од-
нополые, однако в микростробилах имеются рудименты семезачат-
Рйс. 130. Гнетум — Gnetutn gnemonoides. А — репродуктивный
побег мужского растения, Б — мужская шишка (элементарная),
В — репродуктивный побег женского растения, Г — продольный
разрез женской шишки, Д — семя на укороченном побеге:
1 — собрание мужских шишек; 2 — собрание женских шишек; 3 — наруж*
ный интегумент; 4 — средний интегумент; 5 — внутренний интегумент
с микропилярной трубкой; 6 — нуцеллус семезачатка
ков. Растение двудомное. Архегония нет, образуется несколько
женских половых клеток, одна из которых участвует в оплодотво-
рении, в результате которого и образуется двудольное семя.
Pod Gnetutn содержит, как и Ephedra, около 40 видов. Это лиа-
ны, деревья и кустарники, обитающие во влажных тропических
лесах Южной Азии, Африки и Южной Америки. У гнетумов боль-
шие, цельные кожистые супротивные листья на коротких черешках.
Растения двудомные. В мужских стробилах рудименты мегаспо-
рангиев (рис. 130). Архегония нет, оплодотворяется одна7 из нес-
кольких яйцеклеток, образующихся в женском гаметофите. В этом
отношении гнетумы похожи на вельвичию. Семена некоторых ви-
дов гнетума в Южной Азии употребляются в пищу.
Филогенетически оболочкосеменные связаны с беннеттитами; это
подтверждается многими сходными признаками, в частности нали-
чием микропилярной трубки у семезачатков, дихазиальным ветвле-
нием стробилов и др. Наличие рудиментов мегаспорангиев в муж-
ских стробилах говорит о происхождении их от двуполых строби-
лов, которые свойственны беннеттитам.
258
ОТДЕЛ 8. ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ (ANGIOSPERMAE,
MAGNOLIOPHYTA)
Покрытосеменные — наиболее совершенные из современных рас-
тений. Вместе с тем это и наиболее многочисленная группа, насчи-
тывающая до 250 000 видов. На большей части суши покрытосемен-
ные дают основную массу растительности; даже в хвойных лесах,
где по количеству производимой массы преобладают голосеменные,
видовое разнообразие остается все же за цветковыми растениями.
Это говорит о наибольшей приспособленности покрытосеменных
к современным условиям существования на Земле.
Наиболее характерный признак, отличающий цветковые рас-
тения от голосеменных,— наличие у них специального органа —
цветка. Одна из существенных частей цветка — завязь — превра-
щается в плод, внутри которого находятся семена. Отсюда и второе
название этого типа растений — покрытосеменные.
Естественно будет начать знакомство с цветковыми растениями
изучением специфического для этой группы растений органа —
цветка. '
ЦВЕТОК, СОЦВЕТИЯ, ПЛОДЫ
Достигнув определенного возраста, растения начинают образо-
вывать органы, специально предназначенные для размножения,
такие органы называются генеративными. У покрытосеменных —
это цветки и развивающиеся из них плоды с семенами. Цветки
могут быть одиночными или сидят группами, образуя соцветия.
Однолетние и двулетние растения зацветают и воспроизводят
плоды один раз в течение своей жизни и этим заканчивают ее. ААно-
голетние растения цветут и дают плоды не один, а много раз в те-
чение жизни; по этой особенности многократно давать плоды они
называются поликарпическими растениями, или поликар пиками,
Очень немногие многолетние растения зацветают только один раз
в жизни, например агавы, бамбуки, некоторые крупные зонтичные
(из рода Ferula), растущие у нас в Средней Азии, немногие пальмы
(например, Corypha, Metroxylori). Растения, дающие «плоды один
раз в жизни, (Называются монокарпическими, или монокарпи-
ками.
Онтогенетически цветок развивается из цветочной почки, похо-
жей на обычную почку будущего побега. Это наводит на мысль, что
листовая и цветочная почки имеют одинаковое происхождение.
И действительно, в настоящее время почти все ботаники считают,
что филогенетически цветок не что иное, как сильно видоизменен-
ный укороченный побег, а все его части, за исключением цветоложа,
имеют филломную, т. е. листовую природу. Более детально этот
вопрос будет рассмотрен позже; здесь же мы ограничимся лишь
рассмотрением строения и функции генеративных органов цветко-
вых растений.
9*
259
Строение цветка и его частей
Цветок занимает на растении верхушечное или’пазушное поло*
жение и обычно сидит на цветоножке. Но нередко встречаются цвет*
ки без цветоножки, они называются сидячими. Верхнюю, расширен*
ную часть цветоножки, к которой присоединяются все части цветка,
называют цветоложем. Цветоложе может иметь различную форму:
плоскую, выпуклую или вогнутую. У основания цветоножки или не*
Рис. 131. Схема цветка. А — общий вид цветка сверху, Б — продольный
разрез:
1 — чашелистики; 2 — лепестки; 3 — тычинки; 4—пестик; 5 — цветоножка; 6 — цвето-
ложе
посредственно под цветоложем у некоторых растений расположены
один или несколько прицветников, имеющих форму небольших,
несколько видоизмененных листьев.
У типичного цветка на цветоложе расположены такие части:
чашечка, состоящая из чашелистиков, венчик — из лепестков, ты-
чинки и один или несколько пестиков (рис. 131). Чашечка и венчик
составляют околоцветник; этим названием подчеркивается, что
чашечка и венчик — не существенные части цветка. И действитель-
но, есть растения, у которых цветки имеют или чашечку, или вен-
чик, или не имеют ни того, ни другого, и это не мешает цветкам
выполнять их функцию. Цветки, имеющие и чашечку, и венчик,
называются двупокровными', при наличии чего-либо одного, чашеч-
ки или венчика, цветок называется однопокровным, редко встре-
чаются цветки беспокровные, или голые (например, у ясеня).
Таким образом, чашечка и венчик называются еще покровами
цветка, и это опять-таки подчеркивает второстепенность этих час-
тей, по сравнению с тычинками и пестиками, их вспомогательную
роль.
Чашечка» Чашечка — самая наружная часть цветка; она обычно
зеленого цвета и состоит из нескольких, чаще одинаковых по раз-
меру и форме частей — чашелистиков. Чашелистики могут быть
совершенно свободными, не сросшимися, в таком случае чашечка
называется раздельнолистной. Если же чашелистики на большем
или меньшем протяжении срастаются, чашечка называется срост-
нолистной.
На цветках с раздельнолистной чашечкой нетрудно видеть, что
чашелистики — органы листового происхождения.
260
Функция типичной чашечки, прежде всего, защитная; она обе-
регает внутренние, более нежные и важные части цветка в то время,
когда цветок еще не расцвел и находится в состоянии бутона.
Но и у распустившегося цветка чешел-истики обычно не утрачивают
своей защитной роли, поддерживая лепестки и оберегая их от не-
благоприятных внешних воздействий. Но иногда, например у мака,
чашечка отпадает в момент цветения.
Чашечка способна видоизменяться и приобретать совершенно
новую функцию. Например, у сложноцветных (одуванчик, осот
и др.) чашечки цветков превращены в хохолки, которые способст-
вуют разносу плодов; у других растений (например, у череды,
лопушника) чашелистики превращаются в крючочки, зазубренные
щетинки, при помощи которых плоды разносятся животными.
Иногда чашечка приобретает яркую окраску и тогда она играет
роль венчика (например, в сем. лютиковых).
Венчик. Венчик — наиболее заметная часть цветка. ОбычноЧэн
ярко окрашен и почти всегда крупнее чашечки. При ближайшем
рассмотрении венчик оказывается состоящим из нескольких одно-
образных или различных по форме и величине частей — лепестков.
Лепестки вырастают на цветоложе или совершенно разобщенно,
или бывают в большей или меньшей степени сросшимися; в первом
случае венчик называется раздельнолепестным, во втором — спайно-
лепестным. У некоторых растений, например у вьюнка, петунии,
табака, лепестки настолько полно срастаются, что граница между
ними едва заметна, а венчик имеет форму правильной воронки.
Части раздельнолепестного венчика могут иметь самую различ-
ную форму, причем нижняя его часть бывает резко суженной;
у лепестка различают широкую верхнюю часть — пластинку и ниж-
нюю, суженную, — ноготок, который обычно бывает светлее окра-
шен, чем пластинка (например, у гвоздики).
У спайнолепестного венчика различают нижнюю, суженную,
часть — трубочку и верхнюю, расширенную,— отгиб; место пере-
хода трубочки в отгиб называют зевом. Не всегда эти детали вен-
чика бывают хорошо выражены. В зеве могут быть разнообразные
выросты венчика; такие выросты широко распространены у расте-
ний семейства бурачниковых и являются существенным системати-
ческим признаком, т. е. характеризуют собой принадлежность рас-
тения к тому или иному роду и виду. У нарциссов и некоторых
других растений эти выросты очень крупны, более ярко окрашены
и образуют как бы второй венчик. В таком случае эти выросты
называются привенчиком или коронкой; их присутствие делает цве-
ток более красивым. Хорошее развитие таких добавлений к вен-
чику— в большей мере' результат длительного искусственного
отбора.
Спайнолепестность — вторичный признак; древнейшие цветковые
растения (например, магнолия, лютики) были раздельнолепестко-
выми и лишь в процессе последующей эволюции возникли спайно-
лепестные цветки. Следовательно, филогенетически это более моло-
дой признак.
261
Форма цветка бывает самой разнообразной (рис. 132). Чаще
венчик полисимметричен, т. е. его можно рассечь на симметричные
части в нескольких направлениях; таковы, например, цветки лю-
тика, гвоздики, первоцвета, шиповника и др. Цветки такого строе-
ния называют актиноморфными, или правильными. Очень часто
встречаются растения, у которых венчик может быть разрезан
на симметричные части только в одном направлении (например,
Рис. 132. Формы цветка. А — актиноморфный, Б — зигоморфный, В — асиммет-
ричный
цветки шалфея, гороха, львиного зева и др.). Такие цветки называ-
ются зигоморфными или неправильными. Наконец, очень редко
встречаются венчики совершенно асимметричные, например у де-
коративного растения канны.
Актиноморфные цветки филогенетически более древние, из них
в процессе эволюции возникли зигоморфные, как лучше приспо-
собленные к перекрестному опылению при помощи насекомых.
Ярко окрашенный венчик возник как. приспособление к лучшему
опылению цветка насекомыми. Лепестки цветка произошли или
из тычинок, утративших пыльники, или, реже, из верхушечных
видоизмененных листьев; в последнем случае чашечка и венчик име-
ют совершенно одинаковое происхождение.
Количество лепестков в цветке бывает чаще небольшим и опре-
деленным для данного вида растений. Но нередко в природе можно
встретить цветки с ненормально увеличенным числом лепестков
(например, у лютика, сирени и др.). Число лепестков может быть
значительно увеличенным, и тогда цветок называют махровым.
Источники махровости могут быть различными: удвоение числа
лепестков у растений с простым венчиковидным околоцветником
(тюльпан, лилия), превращение в лепестки тычинок (пионы, маки,
шиповник, розы), расщепление лепестков (фуксия — Fuchsia)
и др. Сильно выраженная махровость у декоративных цветочных
растений (пионы, розы, маки, гвоздики — Dianthus и др.) —резуль-
тат искусственного,отбора и с биологической точки зрения рассмат-
ривается как явление уродливости.
Тычинки. Тычинки в совокупности составляют мужскую часть
цветка, или андроцей. Чаще количество тычинок в цветке невели-
262
ко и определенно, но нередко встречаются растения, цветки кото*
рых имеют большое и неопределенное число тычинок, например
лютики, шиповник, мак.
Каждая тычинка, как правило, состоит из тычиночной нити
и пыльника (рис. 133). Тычиночная нить может быть очень длин-
Рис. 133. Строение тычинок. А, Б — внешний вид, В — поперечный разрез
через гнезда пыльников:
/ — тычиночная нить; 2 — пыльник; 3 — связник; 4 — высыпающаяся пыльца; 5 — гнез-
да пыльника еще не вскрылись; 6 — гнезда пыльника вскрылись; 7 — проводящий
пучок в связнике
ной, так что пыльники далеко выдаются за пределы венчика,
или очень короткой, и тогда пыльники почти сидячие. На попереч-
ном разрезе тычиночная нить чаще бывает округлой или овальной,
но встречаются и такие растения, у которых нити тычинок плоские,
листоватые (например, у птицемлечника — Ornithogalum), что ука-
зывает на листовое происхождение этого органа растения. Эволю-
ция формы тычинок изображена на рис. 134.
Рис. 134. Стадии эволюции микроспорофилла от примитивного
микроспорофилла до высокоспециализированного типа (по Тах-
таджяну)
263
Рис. 135. Строение гине-
цея. А — ценокарпный,
Б — апокарпный:
1 — завязь; 2 — столбик; 3 —
рыльце
Пыльник в большинстве случаев состоит из двух половинок,
каждая из двух гнезд. Окончание тычиночной нити, связывающее
две половинки пыльника, называется связником. Обычно пыльник
прикреплен к тычиночной нити неподвижно, но встречаются тычин-
ки (злаки, лилии,, некоторые губоцветные
и др.), у которых пыльник прикреплен к
. связнику подвижно, как бы с помощью шар-
'нира. При опылении ветром такое приспо-
собление имеет определенный биологический
смысл, помогая рассеиванию пыльцы.
У некоторых растений (аистник — Его-
dium из сем. гераниевых) часть тычинок
утрачивается, в процессе редукции пыль-
ники остаются недоразвитыми; такие бес-
плодные тычинки называются стаминодия-
ми. Стаминодии часто приобретают но-
вую функцию, превращаясь в нектарники,
т. е. железки, выделяющие сладкую жид-
кость, которой питаются насекомые. Пре-
вратиться в нектарники могут также и ле-
пестки. Вообще же нектарники имеют самую
разнообразную форму, помещаются в глуби-
не цветка на различных его частях, выделя-
ются своей блестящей поверхностью.
Созревший пыльник вскрывается, и пыль-
ца, находящаяся в нем, высыпается наружу.
Способы вскрытия пыльников различны: трещины, дырочки, кла-
паны и т. д.; отверстия расположены также по-разному.
Пестик. В центре цветка находятся один или несколько пести-
ков, или плодников. Они составляют женскую часть цветка, кото-
рая называется гинецеем (рис. 135). У некоторых растений из се-
мейства мотыльковых или лютиковых легко можно видеть, что'
гинецей также имеет листовую природу. Например, у водосбора
или калужницы из семейства лютиковых пестик представляет со-
бой просто свернутый вдвое зеленый листок. Рис. 136 прекрасно
иллюстрирует постепенные стадии превращения примитивного пе-
стика, состоящего из вдвое сложенного плодолистика, в типичный
пестик.
Таким образом, все основные части цветка филогенетически
имеют листовое происхождение, и цветок действительно представ-
ляет собой сильно метаморфизированный, укороченный побег.
Видоизмененные листочки, из которых формируется пестик,
называются плодолистиками (или карпеллами), так как из пести-
ка, его нижней части, в конечном счете образуется плод. Пестик
может быть построен из одного или нескольких плодолистиков.
В нервом случае гинецей называется апокарпным, во втором —
ценокарпным.
Пестики могут иметь самую разнообразную форму, но при всем
многообразии их почти в каждом пестике можно различать ниж-
264
нюю, расширенную, наиболее массивную часть — завязь, над за-
вязью возвышается тонкая часть пестика — столбик, заканчиваю-
щийся рыльцем. Рыльце может иметь самое разнообразное строе-
ние: в виде булавы, диска, головки, перышка и т. д. Во всех
случаях рыльце приспособлено к лучшему улавливанию и удержа-
нию пыльцы.
На внутренней поверхности завязи сидят одно, несколько или
много беловатых телец овальной формы, это семенные зачатки
Рис. 136. Стадии эволюции плодолистика от примитивного к высокоспециа-
лизированному типу (по Тахтаджяну)
{семяпочки). Из них после опыления и оплодотворения, цветка
развиваются семена, в то время как из завязи в целом развивает-
ся плод, причем стенки -завязи превращаются в околоплодник.
Реже в образовании плода принимает участие столбик и очень
редко (у мака) —рыльце.
При апокарпном гинецее свободные плодолистики образуют
камеры, на внутренней поверхности которых, обычно вдоль шва,
сидят семяпочки. При ценокарпном гинецее каждый плодолистик
может сформировать самостоятельную камеру, или гнездо, завязи.
Такие многогнездиые ценокарпные гинецеи называются синкарп-
ными.
Обычно происхождение синкарпного гинецея из нескольких
плодолистиков довольно легко устанавливается. Однако ценокарп-
ные гинецеи могут быть и однокамерными, причем семенные за-
чатки в них размещаются по-разному. В одних случаях они сидят
на центральной колонке, поднимающейся со дна завязи; при та-
ком размещении семяпочек гинецей называется лизиксСрпным.
В других случаях семезачатки сидят по внутренней поверхности
одногнездной завязи; тогда гинецей называется паракарпным.
Следовательно, ценокарпный гинецей может быть синкарпным,
лизикарпнькм и паракарпным. Наиболее же древний тип гине-
цея— апокарпный, состоящий из одного или отдельных плодоли-
стиков. Срастание .плодолистиков — результат последующего эво-
люционного процесса. На рис. 137 представлена схема эволюции
основных типов гинецея.
265
Рис. 137. Схема эволюции основных типов ги-
нецея (по Тахтаджяну)
Завязь может различно располагаться по отношению к осталь-
ным частям цветка. Если завязь свободно сидит на цветоложе
внутри цветка, она называется верхней. Под околоцветником на-
ходится только расширенная верхняя часть цветоножки. При вы-
пуклом или плоском цветоложе, в случае верхней завязи, осталь-
ные части цветка прикреплены под пестиком, поэтому и цветок
называется подпестичным. Но бывает цветоложе вогнутое, кувши-
нообразное (например, у
шиповника, вишни, сливы
и др.). В таком случае ос-
тальные части цветка при-
креплены к верхнему краю
кувшинчика, на уровне
вершины завязи; цветки
такого строения называ-
ются околопестичными.
Если околоцветник и
тычинки сидят на самой
вершине завязи, ее назы-
вают нижней, а цветок —
надпестичным. Происхож-
дение нижней завязи мо-
жет быть различно; в од-
них случаях завязь охва-
тывается вогнутым цвето-
ложем и срастается с ним,
в других—завязь сидит на
цветоложе и охватывается
нижними частями около-
цветника и тычинок, так-
же срастаясь с ними. Вто-
рой способ распространен
шире, и его следует счи-
тать основным. Устано-
вить истинное происхож-
дение нижней завязи бы-
вает нелегко.
Различают еще полунижнюю завязь — у бузины (Sambucus),
жимолости (Lbnicera), камнеломки (Saxifraga). При такой завязи
не вся, а только нижняя ее половина срастается с цветоложем или
околоцветником и тычинками. Такой цветок называется полунад-
пестичным.
Семенной зачаток (семяпочка) прикрепляется к внутренней по-
верхности завязи посредством семеножки, или фуникулуса. Место
прикрепления семеножки к стенке завязи называется плацентой,
а место прикрепления семеножки к семенному зачатку — рубчи-
ком. Если семезачаток в своем росте ориентирован в том же на-
правлении, что и семеножка, и составляет как бы продолжение
последней, он называется прямым, или атропным. Такие семеза-
266
чатки встречаются редко. Чаще семезачаток бывает по отношению
к своей семеножке повернут на 180° и растет по направлению
к плаценте. Такой семезачаток называется обратным, или ана-
тропным, Наконец, в более редких случаях сам семезачаток изог-
нут, тогда он называется кампилотропным (рис. 138).
Выше было описано строение двуполого /цветка; у которого
имеется и андроцей, и гинецей. Растений с такими цветками при-
мерно 75% от общего количества
видов покрытосеменных, у ос-
тальных 25% растений цветки
однополые, мужские (тычиноч-
ные) или женские (пестичные).
Растения, у которых на одном
и том же экземпляре имеются и
мужские, и женские цветки, на-
зываются однодомными (кукуру-
за, тыква, огурцы, дуб <и др.);
если же однополые цветки сидят
на различных особях, растения
называются двудомными (коно-
пля, тутовник — Morus, ива
и др.).
Однополые цветки возникли
из двуполых в результате полной
редукции органов противополож-
ного пола. Во многих случаях у
однополых растений можно обна-
ружить рудименты органов дру-
гого пола (например, у некоторых
кленов).
Число и расположение частей
цветка. Число частей околоцветни-
ка, количество тычинок и пести-
ков может быть различно*. Чаще
оно небольшое и определенно для
данного вида. Но имеются цветки,
у которых количество лепестков,
Рис. 138. Форма семезачатков. А —
прямой, или атропный, Б — согнутый,
или кампилотропный, В — обратный,
или анатропный
Рис. 139. Построение диаграм-
мы цветка
и пестиков велико и не-
определенно, например в семействе лютиковых.
При большом числе частей цветка они располагаются спираль-
но, п такие цветки называют ациклическими. При небольшом и оп-
ределенном числе частей цветка они располагаются кругами, по-
этому их называют циклическими. Большое число частей цветка
и ациклическое (спиральное) их расположение — первичный, при-
митивный признак.
Первый круг составляют чашелистики, иногда чашелистики
располагаются в два круга (у некоторых розоцветных и мальво-
вых), тогда нижняя чашечка обычно называется подчашием. Ле-
пестки обычно располагаются в один круг, но бывают цветки
и с двумя кругами лепестков, например у касатика, лилии. Тычин-
267
ки довольно часто располагаются в несколько кругов, например
у лилейных, гвоздичных, гераниевых, розоцветных и др.
Части гинецея образуют самый внутренний круг; число частей
(плодолистиков) здесь бывает'меньшим по сравнению с количе-
ством тычинок и лепестков; число тычинок может быть равным
или кратным числу лепестков и чашелистиков.
.Цветки, таким образом, бывают построены как бы по опреде-
ленному плану, который может быть выражен формулой и диа-
граммой. В формуле цветка каждый круг обозначается начальной
буквой латинского названия данной части цветка, а число частей
цветка — цифрой; используются также и другие условные обозна-
чения.
Простой околоцветник обозначается буквой Р, чашечка — К>
венчик — С, андроцей — А, гинецей — G.
Значком оо обозначается большое и неопределенное число ча-
стей цветка, значком f —зигоморфность, звездочкой —акти-
номорфность, значком $ — женский цветок, значком *о — муж-
ской цветок.
Если части цветка сросшиеся, цифру, обозначающую их число,
берут в скобки. В гинецее цифрой обозначают число плодолисти-
ков, причем, если завязь верхняя, эту цифру подчеркивают снизу,
если нижняя — черту ставят над числом. Для пояснения можно
привести формулы нескольких цветков:
Лютик K5C5A00G00
Сурепка K2+2C4A2+4G (2)
Касатик P3+3A3+0G (3)
Наперстянка K5C5A4G (2)
Диаграмма дает более полное и наглядное представление
о плане строения цветка (рис. 139). Диаграмма — это проекция
частей цветка на плоскость, перпендикулярную цветоножке. Если
цветок сидит в пазухе листа, то диаграмму ориентируют таким об-
разом,-чтобы кроющий лист был внизу. Условные фигуры, обозна-
чающие части цветка, в случае срастания их, соединяются линия-
ми. Диаграмма и формула цветка взаимно дополняют друг друга.
Типы соцветий. Как уже сказано, формирование соцветий в про-
цессе эволюции следует рассматривать как одну из форм приспо-'
собления к перекрестному опыдению. Формирование соцветий
в процессе эволюции происходило по-разному, и не всегда легко
определить их границы. Все разнообразие форм соцветий можно,
по способу формирования их, разбить на две группы: соцветия
монрподиальные, или рацемозные (неопределенные), и со~цветия
симподиальные, или цимозные (определенные). Моноподиальные
соцветия формируются за счет развития центральной или верху-
шечной его части (акропетально). У таких соцветий самые моло-
дые цветки находятся в центре или на верхушке соцветий. Наобо-
рот, у симподиальных соцветий первый верхушечный цветок за-
канчивает собой главную ось соцветия, и дальнейшее развитие
соцветия идет за счет развития боковых осей первого порядка,
268
Рис. 140. Простые моноподиальные соцветия. А — колос; Б — сережка,
В, Г — початок, Д — кисть, Е — щиток, Ж — зонтик, 3, И — головка,
Л, Л — корзинка
затем второго порядка и т. д. (базипетально). Однако на практике
иногда не легко бывает установить тип соцветий, переходы п про-
межуточные типы.
Моноподиальные соцветия могут быть простыми, у которых
цветки сидят непосредственно на главной оси соцветий, или слож-
ными, у которых цветки прикреплены к разветвлениям главной
оси соцветий. К простым моноподиальным соцветиям относятся
кисть, простой колос, початок, щиток, зонтик, головка, корзинка
(рис. 140). К сложным моноподиальным соцветиям относятся ме-
269
телка, или сложная кисть, сложный колос, сложный зонтик, слож-
ный щиток (рис. 141). Сережка может быть простой или сложной;
это та же простая или сложная кисть, но со слабой повисшей
осью — центральным стебельком соцветия.
К симподиальным соцветиям от-
носятся монохазий, подразделяемый
еще на извилину и завиток, дихазий,
или развилка, и плейохазий, или
ложзный зонтик (рис. 142).
Рис. 142. Симподиальные соцветия. 4 —
монохазий-завиток, Б — монохазий-изви-
лина, В — дихазий, Г — плейохазий
Рис. 141. Сложные моноподиальные
соцветия. А — сложный колос, Б —
сложный зонтик, В — метелка
Опыление
Для того чтобы цветок выполнил свою функцию как орган раз-
множения, должно произойти оплодотворение, т. е. слияние жен-
ской гаметы — яйцеклетки, и мужской половой клетки (гаметы) —
спермия\ из оплодотворенной яйцеклетки — зиготы — развивается
зародыш будущего растения.
Мужские гаметы — спермин — образуются в результате деления
пыльцевой клетки, яйцеклетка — внутри семенного зачатка. Следо-
вательно, для того чтобы произошло оплодотворение, пыльца
должна быть перенесена на женскую часть цветка — пестик; про-
цесс такого переноса и называется опылением.
Перенос пыльцы на рыльце пестика может осуществляться раз-
личными способами. Пыльца данного цветка может попасть на
рыльце пестика этого же цветка; такой способ опыления называ-
270
ется самоопылением. Перенос пыльцы на рыльце другого цветка
называется перекрестным опылением. Перекрестное опыление мо-
жет осуществляться посредством различных агентов; при помощи
насекомых, ветра и, в более редких случаях, при, помощи воды
и даже небольших птичек (в тропических лесах). Соответственно
способу переноса пыльцы различают энтомофилию — опыление
с участием насекомых, анемофилию — опыление при помощи вет-
ра, гидрофилию, или опыление при помощи воды, и, наконец, ор-
нитофилию— опыление при участии птиц.
Первые покрытосеменные растения опылялись, по-видимому,
при помощи насекомых. Эти функции они могли выполнять еще
в эпоху господства голосеменных растений, питаясь их пыльцой,
но наибольшего-совершенства насекомоопыление достигло лишь
в процессе эволюции цветковых растений.
Ветроопыление у покрытосеменных возникло в процессе эволю-
ции позже, и группы растений, опыляемых при помощи ветра, фи-
логенетически более молодые, чем примитивные насекомоопыляе-
мые. Гидрофилия принципиально не отличается от анемофилии,
орнитофилия — от энтомофилии.
Самоопыление также вторичное явление; к самоопылению ра-
стения перешли, по-видимому, вынужденно, в условиях, когда ка-
кие-то обстоятельства препятствовали перекрестному опылению.
Самоопыление биологически менее полноценно, чем перекрестное
опыление, при котором начало новому организму кладется поло-
выми клетками (гаметами), более разнокачественными. При таком
сочетании у потомства создается более обогащенная наследствен-
ность, позволяющая новому растению лучше приспосабливаться
к условиям среды.
С этой точки зрения перекрестное опыление между различными
растениями (ксеногамия) биологически более полноценно, чем
перекрестное опыление в пределах данного растения (гейтонога-
мия). Поэтому двудомность, при которой возможна только ксено-
гамия,— более прогрессивное явление, чем однодомность.
Несмотря на то что большинство растений имеет двуполые
цветки, это необязательно влечет за собой самоопыление; само-
опылению препятствуют различные приспособления, к числу кото-
рых относится дихогамия.
При дихогамии в данном цветке происходит не одновременное
созревание пыльников и подготовка рылец к приему пыльцы.
У одних растений раньше созревают пыльники (у губоцветных,
бобовых, свеклы, кукурузы и др.); это явление называют проте-
рандрией. У других растений (крестоцветных, розоцветных, пасле-
новых, душистого колоска, ситников и др.) раньще созревает рыль-
це; это явление называется протерогинией.
Растения обладают многобразпыми приспособлениями, способ-
ствующими перекрестному опылению; особенно многочисленны
такие приспособления у насекомоопыляемых растений. К этим
приспособлениям относится, прежде всего, яркая окраска венчика,
что делает цветок заметным’для опыляющего насекомого. Часто
271
окраска не бывает однородной, и от этого цветок становится еще
более заметным. Цветки нередко собраны в различного рода со-
цветия— более заметные, чем одиночный цветок. Примером может
служить корзинка подсолнечника, ромашки, маргаритки и другие
представители семейства сложноцветных, у которых соцветия име-
ют двойную окраску: краевые цветки или их рудименты по окрас-
ке резко отличаются от цветков центральной части соцветия.
В зависимости от способа опыления приспособления носят со-
вершенно различный характер.
Энтомофилия» Цветки, опыляемые насекомыми, как правило,
ярко окрашены, о чем уже говорилось. Иногда бывают окрашены
не только венчик, но и чашечка. Почти во всех случаях насекомо-
опыляемые цветки имеют нектар. Нектарники обычно расположены
в глубине цветка, так что опыляющее насекомое при посещении
цветка неминуемо касается пыльников и рылец.
По ярко окрашенному цветку насекомые могут ориентировать-
ся лишь на близких расстояниях; более чувствительный сигнал —
запах цветков, который зависит от выделения легколетучих эфир-
ных масел. В большинстве случаев это приятные запахи, реже
встречаются дурно пахнущие растения. По запаху насекомое отыс-
кивает нужное ему растение с большого расстояния; приблизив-
шись, оно ориентируется и по окраске цветка.
Помимо нектара, насекомое использует пыльцу (например,
у мака) и различного рода мясистые выросты на частях цветка
в виде волосков, чешуек, бородавок и т. п. (например, у некоторых
растений из сем. бурачниковых).
Характерным показателем энтомофилии является также каче-
ство пыльцы. У насекомоопыляемых растений она, как-правило,
более крупная, обычно с неровной поверхностью, часто клейкая.
Эти качества облегчают захват и перенос пыльцы насекомыми.
Орнитофилия. Этот способ опыления встречается лишь в тропи-
ческих лесах, где распространены мелкие птички колибри, медосо-
сы и др.- Для. цветков, опыляемых птичками, характерны яркая
окраска и обилие нектара. Крупные цветки тропических растений
могут опыляться даже летучими мышами.
Анемофилия. Ветром опыляется сравнительно небольшая часть
видов покрытосеменных, примерно около Vio всех видов. Цветки
ветроопыляемых растений резко отличаются от насекомоопыля-
емых. Они невзрачные, почти такой же окраски, как и вегетатив-
ные части растения. Типичным примером этой группы растений
могут быть злаки, а из древесных — тополь, дуб, тутовник и др.
Околоцветник ветроопыляемых растений сильно редуцирован,
иногда представлен небольшой чешуйкой, что не препятствует
свободному поступлению пыльцы внутрь цветка. К_этому следует
добавить, что пыльца у ветроопыляемых растений мелкая, сухая
и, следовательно, легкая, свободно разносимая ветром. Пыльца
образуется в огромном количестве.
Гидрофилия. Этот способ опыления, естественно, встречается
лишь у водных растений (зостера— Zostera, наяда — Najas, рого*
272
листник—Ceratophyllum и др.). Зерна пыльцы у них лишены эк-
зины, обычно имеют вытянутую форму, в виде ниточек: наиболее
удобная форма для переноса их водой. Опыление подводное, что
облегчается плотностью пыльцы, которая примерно равна плот-
ности воды; поэтому пыльна находится не на поверхности, а в тол-
ще воды.
Самоопыление. Биологически этот способ опыления менее пол-
ноценен, чем перекрестное опыление, при котором в одной особи
объединяются наследственные^ задатки более различающихся
между собой родительских форм^ В природе самоопыление встре-
чается реже, чем перекрестное опыление, тем не менее оно может
быть биологически необходимым; чередуясь с перекрестным опы-
лением, самоопыление способствует закреплению генотипа в по-
томстве.
В том случае, если в силу каких-либо условий не может совер-
шиться перекрестное опыление, многие перекрестноопылители са-
моопыляются. При этом происходит изменение во взаимном распо-
ложении рылец и пыльников: они или соприкасаются, или сбли-
жаются настолько, что собственная пыльца легко попадает на
рыльце, например у грушанок \Pyrola), арники (Arnica),
Самоопыление широко распространено у культурных растений
(пшеница, ячмень, овес, просо, рис, горох, лен, томаты и др.). Са-
моопыляемые растения могут иметь яркий двугубый венчик (на-
пример, у шалфея), явно приспособленный к перекрестному опыле-
нию, да и цветки злаков-самоопылителей по своему виду не отли-
чаются от цветков большинства злаков, которые опыляются пере-
крестно, при помощи ветра. Такое несоответствие объясняется вто-
ричностыо приспособления этих растений к самоопылению; предки
их, по-видпмому, опылялись перекрестно.
Наблюдения показывают, что у огромного большинства само-
опылителей, в том числе и у названных культурных растений, не
исключено и перекрестное опыление, которое, видимо, время от
времени и ' происходит. Но у* некоторых растений (фиалка —
Viola, недотрога — Impatiens и др.) наряду с обычными яркими пе-
рекрестно опыляемыми цветками имеются цветки мелкие, невзрач-
ные и никогда пе раскрывающиеся — клейстогамные. В них может
произойти только самоопыление. В таких цветках пыльца может
прорастать даже в закрытом пыльнике; пыльцевые трубочки .прора-
стают стенку пыльника и таким путем достигают рыльца.
Опылению предшествует образование микроспор и созревание
женской половой клетки — яйцеклетки. Процесс образования ми-
кроспор называется микроспорогенезом.
Яйцеклетка образуется внутри женского гаметофита, который
развивается из мегаспоры. Процесс формирования мегаспоры на-
зывается мегаспорогенезом. Прежде чем перейти к рассмотрению
оплодотворения, мы должны познакомиться с этими двумя анало-
гичными процессами и с последующими явлениями, приводящими
к возникновению двух родов половых клеток: спермия и яйце-
клеток.
273
6
Рис.
разрез
4 — незрелый
пыльник, j
щийся зрелый
143. Поперечный
через пыльник.
"[ закрытый
Б — вскрыв-
пыльник
Микроспорогенез и развитие
мужского гаметофита
Микроспорогенез, т. е. процесс образования микроспор, можно
наблюдать внутри пыльцевых гнезд развивающихся тычинок. Раз-
витие каждой тычинки (начинается с появления .на апексе цветонос-
ного побега бугорка из образовательной ткани. Постепенно ткань
бугорка дифференцируется в эпидермис и паренхиму, в центре бу-
горка закладывается проводящий пучок, который у развитой ты-
чинки пройдет через связник в тычиночную нить.
В однородной ткани бугорка формируются четыре пыльниковых
гнезда (микроспорангии). Гнезда расположены попарно и при
окончательном формировании превращают-
ся в два двойных пыльцевых мешка
(рис. 143). Процесс формирования пыльце-
вого .гнезда начинается с обособления в
паренхиме бугорка нескольких более -круп-
ных клеток — археспория. Каждая клетка
археспория делится тангентально на две
клетки: внутрь — спорогенная, к перифе-
рии — париэтальная клетка. Спорогенные
клетки многократно делятся и в конечном
счете дают материнские клетки микроспор.
Каждая из материнских клеток затем дваж-
ды делится, давая тетраду микроспор; одно
из этих делений редукционное, поэтому мик-
роспоры получаются гаплоидные. Тетрады
распадаются на отдельные микроспоры к
моменту .созревания и открытия пыльника.
У некоторых растений, например у рогоза,
тетрады не распадаются на отдельные мик-
роспоры.
Париэтальные клетки делятся танген-
тально и образуют вокруг спорогенной тка-
ни несколько дифференцированных слоев:
1) наружный субэпидермальный фиброзный, входящий в состав
стенки пыльника; 2) промежуточный, клетки которого в процессе
созревания пыльника дегенерируют и рассасываются; 3) внутрен-
ний выстилающий слой — тапетум (рис. 144). Этот слой непосред-
ственно прилегает ж спорогенной ткани. Клетки тапетума богаты
питательными-веществами (жирами и каротиноидами). Это по су-
ществу питательная ткань, которая постепенно используется архе-
спориальными клетками в процессе их размножения. К моменту
формирования тетрад тапетум полностью разрушается и от него
ничего не остается. Перед раскрытием пыльника перегородки меж-
ду двумя парными микроспорангиями разрушаются и весь пыльник
оказывается состоящим из двух-пыльцевых мешков.
Микроспора при своем возникновении представляет собой одно-
ядерную клетку, покрытую двумя оболочками: внутренней, тонкой
и нежной,— интиной, и наружной, толстой и плотной,— экзиной.
274
В состав интины входит целлюлоза, экзина состоит из липоидного
вещества спорополленина. Экзина обычно имеет различного рода
выросты, шероховатости, шипики, помогающие микроспоре за-
крепиться на рыльце пестика. Утонченные места в экзине в форме
пор или борозд, через которые происходит прорастание пыльцы,
называются апертурами. ФормьГ и размеры микроспоры различны
и индивидуальны для каждого вида. Протопласт микроспор (пыль-
цевых зерен) имеет развитую эндоплазматическую сеть, диктиосо-
мы и многочисленные митохондрии.
Рис. 144. Поперечный разрез одного из гнезд пыль-
ника:
/ — эпидермис; 2 — субэпидермальпый слой; 3 — выстилаю-
щий слой (тапетум); 4 — материнские клетки микроспор
Развитие мужского гаметофита из микроспоры начинается еще
в пыльнике. Ядро микроспоры кариокинетически делится надвое,
вокруг каждого из ядер обособляется протоплазма и внутри обо-
лочки микроспоры образуются две кДетки, большая — вегетативная
и меньшая — генеративная (рис. 145). Вегетативную клетку можно
рассматривать, как гомолог мужского заростка, генеративную —
как гомолог антеридия. Таким образом, у покрытосеменных расте-
ний происходит крайняя редукция мужского гаметофита, который
и представляет собой содержимое пыльцевого зерна.
„Дальнейшее развитие мужского гаметофита происходит обыч-
но после перенесения пыльцевого зерна «на рыльце пестика. Здесь
происходит прорастание пыльцевого зерна. Содержимое его, одетое
интиной, выпячивается в виде трубочки через утончения или пору
275
Рис. 145. Развитие мужского гаметофита покрытосеменных. А — прорастание
пыльцы, рост пыльцевой трубки и образование спермиев (вегетативное ядро
в копчике трубки), Б—сильно увеличенное пыльцевое зерно, в нем округлое
ядро вегетативной клетки и вытянутая генеративная клетка
в экзине; трубочка прорастает внутрь пестика, достигает завязи,
а затем и семенного, зачатка. Протопласт вегетативной и генера-
тивной клеток опускается в пыльцевую трубку. Ядро генеративной
клетки делится надвое, образуется два спермия. Спермин без жгу-
тиков и имеют различную форму: вытянутую, шарообразную, спи-
ральную и др.
Мегаспоронегез и развитие
женского гаметофита
Мегаспорангий, или семезачаток, так же как и микроспорангий,
начинает свое развитие с образования на внутренней поверхности
плодолистика меристематического бугорка. Бугорок окружен ва-
ликом, который в будущем развивается в покров семезачатка, или
интегумент. Сам бугорок превращается в нуцеллус. Покров (инте-
гумент) не полностью обрастает нуцеллус, оставляя свободный
проход к нему — пыльцевход, или микропиле. Интегумент часто
бывает двойной.
Одна из клеток нуцеллуса, расположенная поближе к пыльце-
входу, увеличивается в размере и становится археспориальнойу
или материнской, клеткой мегаспоры. Археспориальная клетка
дважды делится, образуя тетраду. Одно из делений (первое) ре-'
276
A
о
Рис. 146. Схема развития семезачатков. А — прямой семезачаток, Б —
обратный, В — согнутый
Аукционное. Клетки тетрады располагаются обычно цепочкой?
в глубь нуцеллуса. Одна из клеток тетрады, обычно самая внут-
ренняя, начинает увеличиваться в размере и становится единствен-
ной мегаспорой. Разрастаясь, мегаспора сдавливает остальные три*
клетки тетрады, и те дегенерируют. Затем начинается развитие
женского гаметофита из мегаспоры. Развитие это сопровождается
увеличением в размерах всего мегаспорангия — семезачатка
(рис. 146).
Развитие женского гаметофита происходит следующим обра-
зом. Ядро мегаспоры делится трижды без цитокинеза. Образуется
восемь ядер, четыре из которых отходят ближе к микропиле и че-
тыре— в противоположную сторону. Клетка при этом удлиняется,,
а цитоплазма вакуолизируется. Затем от каждого полюса к центру
клетки перемещаются по одному из гаплоидных ядер и здесь сли-
ваются, образуя вторичное диплоидное ядро. Это ядро одевается
собственным слоем цитоплазмы; точно так же одевается цитоплаз-
мой каждое из шести остальных ядер (рис. 147).
277
Рис. 147. Мегаспорогенез и развитие зародышевого мешка. Л — образо-
вание мегаспоры, Б — развитие из мегаспоры зародышевого мешка
(женского гаметофита)
Таким образом, из' протопласта мегаспоры образуется семь
клеток: по три на полюсах и одна в центре. Все это образование*,
гомологичное женскому гаметофиту, называется зародышевым
мешком. Три клетки, расположенные ближе к микропиле, состав-
ляют яйцевой аппарат; одна из них, более крупная,— яйцеклетка,
две остальныет— синергиды (клетки-помощницы). Клетки, распо-
ложенные на противоположном полюсе зародышевого мешка, на-
зываются антиподами.
Оплодотворение и развитие семени
Образовавшиеся в результате деления генеративной клетки два
спермня направляются по пыльцевой трубочке внутрь семезачатка,
к зародышевому мешку. Здесь оболочка пыльцевой трубочки раз-
рывается, и один из проникших сюда спермиев сливается с яйце-
клеткой, а другой — с вторичным диплоидным ядром; происходит,
таким образом, двойное оплодотворение. В результате такого оп-
лодотворения из оплодотворенной яйцеклетки развивается зародыш
будущего семени, а из оплодотворенного вторичного ядра — эндо-
сперм-— питательная ткань семени.
Эндосперм может образоваться двумя способами: 1) оплодо-
творенное ядро вначале многократно делится, образуя многоядер-
ный протопласт, и лишь в конце формирования эндосперма одно-
временно возникают клеточные перегородки; такой тип развития
называется ядерным, или нуклеарным*, 2) клеточные оболочки об-
разуются сразу же после каждого деления ядра; этот тип называ-
ется клеточным или целлюлярным. Эндосперм расходуется на пи-
тание зародыша в процессе его формирования и дальнейшего раз-
вития в новое растение при прорастании семени.
278
Таким образом, эндосперм покрытосеменных не гомологичен эн-
досперму голосеменных. У голосеменных — это женский заросток,
гаметофит, обогащенный за счет спорофита питательным ве-
ществом; у покрытосеменных — своеобразный продукт полового
процесса, лишь внешне сходный с эндоспермом голосемен-
ных.
Яйцеклетка сразу же после оплодотворения одевается оболоч-
кой и начинает делиться. В результате многократного деления фор-
мируется тело зародыша, которое постепенно врастает в массу эн-
досперма.
Нуцеллус под давлением разрастающихся эндосперма и заро-
дыша обычно сильно истончается и превращается в тонкую плен-
ку, одевающую семя, а иногда вовсе рассасывается. У некоторых
растений нуцеллус, наоборот, разрастается, клетки обогащаются
питательным веществом и образуется особая ткань — перисперм,
играющий ту же роль, что и эндосперм.
Интегументы семенного зачатка превращаются в оболочки
семени.
В конкретных случаях у отдельных групп растений дальнейшее
развитие семени может пдйти по-разному.
1. В процессе формирования семени весь эндосперм может быть
израсходован на развитие зародыша, при этом нуцеллус не разра-
стается. В таком случае получаются семена без эндосперма, со-
стоящие из одного зародыша и кожуры; питательные вещества при
этом отлагаются в семядолях семени. Семена такого типа свой-
ственны бобовым, сложноцветным, тыквенным, розоцветным
и др.
2. В процессе развития семени эндосперм сохраняется, но пери-
сперм не получает развития. Такие семена, состоящие из зароды-
ша, эндосперма и кожуры, свойственны злакам, пасленовым, зон-
тичным и др.
3. Развиваясь, зародыш полностью расходует весь эндосперм,
но одновременно в семени из нуцеллуса формируется перисперм,
семя состоит из зародыша, перисперма и кожуры. Семена такого
типа свойственны растениям из семейства маревых, гвоздичных,
кувшинковых и др.
4. Наконец, очень редко в семени сохраняется эндосперм и раз-
вивается перисперм (черный горошковый перец).
Размер семени может быть различным, от почти микроскопиче-
ского (например, у орхидей) до огромных семян некоторых пальм;
например, масса семени сейшельской пальмы достигает 15—25 кг,
а длина — 50 см. На семенах могут развиваться мясистые выросты,
у основания семезачатка—ариллусы, вблизи микропиле — карун-
кулы, Эти выросты способствуют распространению семян живот-
ными.
Как правило, после оплодотворения, одновременно с развитием
семян, завязь цветка разрастается и превращается в плод.
279
Отклонения от нормального процесса формирования
семян и плодов %
У некоторых растений зародыш семени может развиваться из
•яйцеклетки без оплодотворения. Такое явление называется парте-
ногенезом. Яйцеклетка при этом может быть диплоидной (у ман-
жетки, одуванчика и др.) или- гаплоидной (у дурмана, табака
и др.). Но бывает и так. ядро яйцеклетки дегенерирует и замеща-
ется ядром спермия; набор хромосом остается одинарным, отцов-
ским; развивается гаплоидное растение. Такое явление называется
-андрогенезом.
В иных случаях зародыш семени может развиваться не из яйце-
клетки, а из неоплодотворенных клеток — синергид или антипод.
Такое отклонение от нормы называется апогамией.
Могут быть также случаи, тогда зародыш развивается хотя
и в зародышевом мешке, но без оплодотворения, из диплоидных
клеток нуцеллуса или интегумента, т. е. из элементов, относящих-
ся к спорофиту. Такое явление называется апоспорией.
Все эти случаи ненормального развития зародыша без слияния
половых клеток, объединяются под общим названием апомиксиса.
Апомиксису противопоставляется нормальный, процесс двойного
оплодотворения — амфимиксис.
Плоды могут образоваться без оплодотворения и последующего
развития семян; получаются плоды бессемянные. Такое явление
называется партенокарпией.
Свойство растений давать плоды без семян человек широко ис-
пользовал, и теперь выведено много сортов плодовых растений,
дающих партенокарпические плоды (апельсины, мандарины, ли-
моны, виноград, бананы и др.). Размножают-такие сорта вегета-
тивным способом.
Строение и классификация плодов
Как уже ранее было сказано, плод развивается из нижней ча-
сти пестика — завязи; реже в образовании плода принимает уча-
стие столбик и очень редко — рыльце. Если в образовании плода
принимает участие не только пестик, но и другие части цветка,
чаще всего основания околоцветника и тычинок, реже цветоложе,
то плод называется ложным.
Плод может иметь самые разнообразные формы и величину,
а околоплодник — различную консистенцию. У сочных плодов хо-
рошо выражены три слоя околоплодника: наружный — экзокар-
пий, обычно более плотный, кожистый; средний — мезокарпий —
наиболее развитый и мясистый; и внутренний — эндокарпий,—
имеющий различную ‘консистенцию; иногда он очень твердый,
например у вишни, сливы. У плодов с с^им околоплодником эти
слои менее выражены.
Плоды могут быть простыми, образовавшимися из единствен-
ного пестика в цветке, и сложными—из нескольких или многих
пестиков одного цветка. Группы плодов, образовавшихся из со-
280
цветня, но расположенных так тесно, что в целом создается впе-
чатление одного плода, называются соплодиями, например сопло-
дие шелковицы (Morus), инжира (Ficus carica).
Перечисленные различия в строении плодов, положены в основу
их классификации. Тогда все разнообразие плодов можно будет
подразделить на две группы: плоды с сухим околоплодником^
вскрывающимся (рис. 148) и невскрывающимся (см. рис. 149)
и плоды с сочным околоплодником (см. рис. 150 и 151). Эти груп-
пы в свою очередь подразделяются на более мелкие, конкретные
формы.
Плоды с сухим околоплодником
Вскрывающиеся, обычно многосеменные (рис. 148). Листов-
ка— одногнездный плод из одного плодолистика, вскрывается по
одному, брюшному шву; пример — плод живокости {Delphinium
consolida). Может быть и сложная листовка, например у водо-
сбора (Aquilegia), калужницы (Caltha).
Боб — также одногнездный плод, образованный одним плодо-
листиком, но раскрывается по двум швам:- брюшному и спинному;
примеры — горох (Pisum), вика (Vicia). Бывают и односеменные
плоды, например у эспарцета (Onobrychis).
Стручок — удлиненный двугнездиый плод, между двумя створ-
ками продольная перегородка, образуется из двух плодолистиков;
пример — белая горчица .{Sinapis alba).
Рис. 148. Сухие вскрывающиеся плоды. А — листовка, Б — сложная лис-
товка, В — боб, Г — стручок, Д — стручочек, Е, Ж — коробочка
281
Рис. 149. Сухие невскрывающиеся плоды. А — орех, Б — орешек,
В — двукрылатка, Г — сложная (сборная) семянка на сочном
цветоложе, Д — зерновка, Е — крылатка, Ж — семянка, 3 — дроб-
ный стручок, И — желудь
Стручочек — то же, что и стручок, но укороченной формы, дли-
на его не более чем втрое превышает ширину; пример — пастушья
сумка (Capsella bursa-pastoris).
Коробочка — одногнездный или многогнездный плод, образует-
ся из нескольких плодолистиков, вскрывается дырочками, продоль-
ными или поперечными трещинами; примеры — мак (Papaver),
белена (Hyosciamus), хлопчатник (Gossypium).
Невскрывающиеся односеменные (рис. 149). Зерновка — сем^
плотно срастается с тонким пленчатым околоплодником; приме-
ры— пшеница (Triticum), рожь (Secale).
Семянка — околоплодник кожистый, не срастающийся с семе-
нем; часто снабжена хохолком или летучкой; примеры — подсол-
нечник (Helianthus annuus), одуванчик (Taraxacum). Семянки
могут быть и сложные (сборные), на общем сухом (лютик) и соч-
ном (земляника) цветоложе. Своеобразный плод у шиповника—•
это сложная семянка на вогнутом мясистом цветоложе. В семейст-
ве зонтичных — двусемянки, например у моркови.
282
Крылатка — семянка с крыловидным придатком; пример —
ясень (Fraxinus)} у кленов {Acer)—двукрылатка, из двух срос-
шихся крылаток.
Орех — околоплодник жесткий, деревянистый; пример — лес-
ной орех {Corylus avellana). Маленький орех — орешек, например
у конопли (Cannabis).
Желудь — тот же орех, нижняя часть которого погружена
в особую плюску, образованную из сросшихся прицветников; при-
мер— дуб {Quercus robur).
Плоды с сочным околоплодником
Ягода — многосемянный плод; эндокарпий и мезокарпий соч-
ные, экзокарпий кожистый; примеры —гвиноград (Vitis vinifera),
томат (Lycopersicum esculentuni) (рис. 150).
Яблоко — многосемянный ложный плод, в образовании кото-
рого, кроме завязи, принимает участие разросщееся основание око-
лоплодника; экзокарпий и мезокарпий сочные, эндокарпий жестко-
кожистый или окостеневший; примеры — яблоня. (Malus), груша
(Ругus), айва (Cydonia), боярышник (Crataegus).
Тыквина — многосемянный ложный плод, в образовании ко-
торого принимает участие и цветоложе; экзокарпий твердый, иног-
да деревянистый, мезокарпий и эндокарпий сочные; пример — ар-
буз {Citrullus), тыква (Cucurbita) (рис. 150).
Рис. 150. Сочные многосемянные плоды. А и Б — ягоды, В — тык-
вина, Г — померанец
283
Рис. 151. Сочные односемянные плоды —
костянки
Померанец — плод цитрусовых, многосемянный; экзокарпий
мягкокожистый, богатый эфирными маслами, мезокарпий сухой,
губчатый, эндокарпий сочный; пример—лимон {Citrus limonium)
(рис. 150).
Костянка — односемянный плод; экзокарпий тонкий, кожистый,
мезокарпий сочный, эндокарпий каменистый; в процессе созрева-
яия плода мезокарпий может высыхать (у миндаля — Amygdalus)\
примеры — вишня (Cerasus),
слива (Prunus). Сложная ко-
стянка у малины и ежевики
(Rubus) (рис. 151).
Приспособления для распространения
плодов и семян
У многих растений плоды
пли семена имеют специальные
приспособления, -способствую-
щие их распространению. При-
способления эти очень разнооб-
разны. Прежде всего-сюда сле-
дует отнести наличие сочного
околоплодника. Представители
животного мира: птицы, млеко-
питающие и насекомые, поль-
зуясь такими плодами, как пищей, растаскивают их на большие
или меньшие расстояния от места произрастания. Семена, попадаю-
щие в пищеварительный тракт, не теряют всхожести и даже, наобо-
рот, получают лучшие условия для прорастания.
Весьма широко распространены различного рода выросты на
плодах, при помощи которых плоды прицепляются к проходящим
животным (или человеку). Так, например, распространяются плоды
дурнишника (Xanthium), череды (Bidens), подмаренника (Gali-
um), лопушника (Arctium), якорцев (Tribulus) и многих других.
Не менее широко распространены приспособления к рассеива-
нию плодов ветром; это различного рода хохолки, парашютики,
крылышки, пушинки. Примером могут служить одуванчик, бодяк
{Cirsium), козлобородник (Tragopogori), вяз, клен, ясень и др.
Приспособлением к распространению семян ветром можно счи-
тать также и чрезвычайно малую массу их у некоторых растений,
например, у тропических орхидей масса полмиллиона семян все-
го 1 г.
Особая форма приспособления для разбрасывания семян в ус-
ловиях степного простора — формирование надземной части расте-
ния в виде «перекати-поле». Такие растения приобретают в очер-
тании форму шара, корень их в конце лета быстро перегнивает
и они легко перекатываются по полю. В это время плоды посте-
пенно подсыхают, раскрываются, и семена рассеиваются (кер-*
мек — Statice).
284
У некоторых растений плоды вскрываются с такой силой, что
семена разлетаются на значительное расстояние, например у жел-
той акации, вики, недотроги, бешеного огурца.
Наконец, следует упомянуть о растениях, плоды которых обла-
дают специальным приспособлением для самозарывания; таким
свойством обладают плоды ковыля, аистника, овсюга и- др.
Происхождение цветковых растений
Даже из того краткого обзора, который мы сделали, можно
было убедиться, насколько сложнее устроены органы размноже-
ния у покрытосеменных по сравнению с голосеменными. Цветки
и плоды у цветковых растений представлены бесконечным разно-
образием форм, расцветок и размеров. Однако совершенство по-
крытосеменных выражается не только в наличии цветка и плода.
Они лучше, полнее голосеменных используют солнечный свет, что
связано с большой пластичностью листовых органов и более вы-
сокой организованностью ассимиляционного процесса. У покрыто-
семенных более совершенна проводящая система и лучше органи-
зованы механические, покровные и запасающие ткани. Проводя-
щая система организована, как у хвойных, в эвстелу и атакостелу
(у однодольных).
Исключительного разнообразия достигли органы размножения
растений — цветки; в обеспечении их функции, в процессах опы-
ления и оплодотворения, наблюдается большое количество самых
разнообразных приспособлений.
Способность покрытосеменных растений создавать огромное
количество разнообразных органических веществ расширяла мно-
госторонние связи этой высшей группы растений с представителями
животного мира; наличие среди синтезируемых веществ сильнодей-
ствующих ядовитых соединений (алкалоиды, глюкозиды и др.)
обеспечивало лучшую охрану растений от поедания животными.
Ни одна группа растений не может сравниться с покрытосемен-
ными в отношении приспособляемости к самым разнообразным
условиям существования; холодные арктические тундры, высочай-
шие горы, жаркие и сухие пустыни, солончаки и болота, пресные
и морские водоемы — все эти резко различающиеся по совокупно-
сти жизненных факторов места в той или иной мере обжиты цвет-
ковыми растениями.
Соответственно условиям обитания покрытосеменные обладают
бесконечным разнообразием форм; гигантские деревья и мельчай-
шие, едва различимые глазом, плавающие в воде растения; сочные
широколистные травы и сухие колючие кустарники. Особенно
большого разнообразия и пышности форм достигают растения во
влажных тропических лесах, где все факторы жизни имеются
в избытке.
За исключением, пожалуй, лишайниковых тундр и хвойных
лесов, облик растительного покрова Земли определяется цветко-
выми растениями; все остальные растения в этом отношении игра-
ют второстепенную роль.
285
Стремительное завоевание суши цветковыми растениями про-
изошло в течение сравнительно короткого времени. Первые по-
крытосеменные появились, по-видимому, около 200 млн. лет назад
в начале юрского или, возможно, даже-в конце триасового перио-
да. Но многие миллионы лет, до середины мелового периода, пер-
венцы этой группы растений заметно не выделяются среди господ-
ствующей флоры папоротников и мезозойских голосеменных: гинк-
говых, саговниковых, беннеттитовых и хвойных. Период такого не-
заметного существования покрытосеменных длился около 100 млн.
лет, первые достоверные ископаемые остатки цветковых растений
известны только из нижнего мела.
Существуя в этом периоде небольшими по числу особей группа-
ми, покрытосеменные растения, по-видимому, достигли большого
разнообразия; внезапное и быстрое распространение покрытосе-
менных с середины мелового периода началось сразу с большого
числа форм. К концу мезозоя’флора покрытосеменных сформиро-
валась уже почти в том составе, в котором мы ее сейчас наблюда-
ем; остатки меловых покрытосеменных принадлежат современным
семействам и даже родам.
Благодаря большой пластичности покрытосеменные растения
могут формировать сложные, иногда многоярусные сообщества
и в составе таких внутренне отлаженных формирований мигриро-
вать по земному шару.
Причины такого быстрого распространения покрытосеменных
в середине мелового периода объясняются следующим. В конце
юры в конфигурации суши северного полушария произошли боль-
шие изменения; в связи с отступлением моря значительно увели-
чилась поверхность суши, климат изменился в сторону большей
сухости, уменьшилась облачность. Происшедшие изменения оказа-
лись неблагоприятными для влаголюбивой мезозойской раститель-
ности, она начала вымирать или изменяться в сторону большей
ксероморфности. С другой стороны, представители флоры покрыто-
семенных, длительное время живя среди господствующих мезозой-
ских споровых и голосеменных растений, имели время и возмож-
ность приспособиться к разнообразным условиям существования,
выработать различные приспособительные формы. Такое разнооб-
разие п’риспособительных качеств и было использовано покрытосе-
менными в момент вытеснения ими отживающей мезозойской фло-
ры. Немаловажную роль сыграли при этом те сложные и обоюдо-
полезные взаимоотношения с представителями животного мира —
насекомыми, в которые вступили ближайшие предки покрытосе-
менных растений.
Питаясь пыльцой, насекомые переносили ее с одного стробила
на другой и, таким образом, были первыми агентами перекрестно-
го опыления. Пыльцы производилось очень много, и поедание'части
ее насекомыми не наносило заметного ущерба, в то время ’"’как
польза от перекрестного опыления была несомненная. Но, не огра-
ничиваясь пыльцой, насекомые приспособились поедать и семеза-
чатки. Это уже наносило существенный вред размножению расте-
286
ний, так как семезачатков было ограниченное количество. Реак-
цией на такое отрицательное влияние насекомых явился отбор
приспособительных форм с закрытыми семезачатками; мегоспоро-
филлы замкнулись и образовали пестик, гинецей, превратившись
тем самым в плодолистики. В дальнейшем в связи с обогащением
мира насекомых процесс опыления усложнялся и разнообразился.
Семезачатки, оказавшись внутри замкнутых мегаспорофиллов,
получили защиту не только от насекомых, но и от неблагоприят-
ных воздействий внешней среды, особенно от вреднего влияния
сухого воздуха. Вместе с тем хорошее укрытие привело к упроще-
нию строения семезачатков *и к увеличению числа их, создало бла-
гоприятные условия для редукции и ускоренного развития женско-
го гаметофита.
Образование плодолистиков (карпелл) явилось предпосылкой
для возникновения ценокарпии и возможности последующего об-
растания ценокарпного гинецея цветоложем и другими частями
растения. С другой стороны, образование рыльца из верхних ча-
стей плодолистиков способствовало избирательности оплодотворе-
ния, а приспособление цветков к определенной группе насекомых —
опылителей способствовало биологической изоляции <и ускорило
процесс дивергенции и образования новых видов.
Отмечая ведущую роль насекомоопыления на первых этапах
эволюции покрытосеменных, не следует упускать из виду, что поч-
ти во всех случаях энтомофилия не исключала анемофилию и, ког-
да благоприятствовали условия, это являлось предпосылкой для
возникновения и эволюции ветроопыляемых линий покрытосемен-
ных растений. Вместе с тем двуполость цветка первичных покрыто-
семенных не исключала также возможность запасного акта — са-
моопыления, когда обстоятельства мешали использованию приспо-
соблений к перекрестному опылению. Самоопыление, чередуясь
с перекрестным опылением, могло сыграть положительную роль
в эволюции, способствуя закреплению и изоляции новых генотипов,
которые при последующем перекрестном опылении создавали но-
вые комбинации наследственных задатков.
Важную роль в эволюции покрытосеменных сыграло, по-види-
мому, двойное оплодотворение, которое было открыто в 1898 г.
русским ученым С. Г Навашиным. В результате сильной редукции
женского гаметофита и возникновения двойного оплодотворения,
эндосперм голосеменных, имеющий только материнское происхож-
дение, заменился эндоспермом двойственной природы — продуктом
полового процесса. Этот второй акт оплодотворения, надо пола-
гать, оказывает стимулирующее влияние на развитие зародыша
и является главным фактором значительно более быстрого форми-
рования семени у цветковых растений по сравнению с голосемен-
ными. С другой стороны, положительную роль сыграло и более
экономное расходование питательных веществ, идущих на образо-
вание эндосперма: у покрытосеменных эндосперм возникает и раз-
растается одновременно с развитием зародыша — потребителя это-
го материала.
287-
Какую группу растений следует считать ближайшими предка-
ми покрытосеменных? Эту группу, очевидно, нужно искать среди
растений, уже имевших семена, т. е. среди голосеменных. Наибо-
лее древние представители покрытосеменных имели двуполые цвет-
ки, следовательно, предков покрытосеменных нужно искать среди
голосеменных, имевших двуполые стробилы. Такие стробилы свой-
ственны только беннеттитовым (см. рис. 125) . Поэтому беннеттито-
вые можно было бы считать наиболее вероятными предками по-
крытосеменных. Однако признанию беннеттитовых ближайшими
предками цветковых растений препятствуют существенные обстоя-
тельства: мегаспорофилл беннеттитовых сильно редуцирован и ут-
ратил листовую форму, в то время как плодолистики покрытосе-
менных имеют явно листовое происхождение. Кроме того, ни у од-
ного из представителей покрытосеменных нет свойственной беннет-
титовым мпкропилярной трубки семезачатков. Роль улавливающе-
го пыльцу аппарата у цветковых выполняет рыльце, т. е. верхняя
часть сросшихся в пестик плодолистиков. По-видимому, покрыто-
семенные произошли от одной из примитивных групп семенных
папоротников, и, возможно, они в этой группе имели общих пред-
ков с беннеттитовыми.
Такова одна из наиболее убедительных теорий происхождения
покрытосеменных.
Эта теория, как видно, исходит из предположения, что цветок
покрытосеменных произошел из стробила голосеменных и, .следова-
тельно, все части цветка, кроме цветоложа, имеют листовое про-
исхождение. Теория эта называется стробилярной или эуантовой,
ее разделяет большинство ботаников, и она убедительно подтверж-
дается многими фактами. В противоположность эуантовой теории
существует и псевдантовая теория, которая считает, что цветок —
это видоизмененное редуцированное соцветие из однополых цвет-
ков, в центре — женских, на периферии — мужских.
Соответственно этим двум теориям сторонники псевдантовой
теории считают, что древнейшей, первичной формой цветка были
однополые цветки и что цозже возникли двуполые цветки. Наобо-
рот, сторонники эуантовой теории считают первичной формой дву-
полый цветок, однополые же цветки возникли в результате редук-
ции или гинецея (у мужских цветков) или андроцея (у женских
цветков).
Многочисленные факты доказывают правильность эуантовой
теории. Убедительным является совпадение у наиболее древних
цветковых растений примитивности в анатомическом строении
древесины с наличием двуполого цветка с большим количеством
тычинок и пестиков, по типу сходного с двуполым стробилом. Та-
ков примитивный цветок магнолии, все части которого, исключая
цветоложе, имеют явно листовое происхождение. На этом цветке
отчетливо видно, что цветок есть не что иное, как сильно укоро-
ченный спороносный побег.
Листовая природа частей чашечки и венчика легко прослежива-
ется у большинства цветковых растений. Сложней обстоит дело
-288
с тычинками и плодолистиками, которые подверглись, в связи со
специфическими функциями, наиболее глубокому метаморфозу.
Однако, если мы сопоставим формы этих органов, свойственные,
наиболее примитивным группам цветковых растений (Magnolia-
сеае и др.), с формами у более поздних групп, мы вполне сможем
себе представить, в каком направлении шла эволюция.
Итак, цветок — двуполый стробил, на котором собраны микро-
и мегаспорангии. Четырехгнездный пыльник есть не что иное, как
четыре микроспорангия, сросшиеся вместе; такие группы споран-
гиев называются синангиями. В микроспорангиях находятся мик-
роспоры, из которых в будущем будут развиваться мужские гаме-
тофиты.
Семенной зачаток представляет собой мегаспорангий, одетый
дополнительными покровами — интегументами. Единственная ме-
гаспора его развивается в сильно редуцированный женский гаме-
тофит— зародышевый мешок.
СИСТЕМАТИКА ПОКРЫТОСЕМЕННЫХ
В начале XX в. ученые различных стран создают первые фило-
генетические системы покрытосеменных. За рубежом наибольшей
известностью пользуются филогенетические системы покрытосемен-
ных, разработанные Энглером, Веттштейном, Бэсси, Халлиром
и др.
Немецкий ученый А. Энглер и Р. Веттштейн (Австрия) в пост-
роении систем исходили из основных положений псевдантовой
теории, поэтому наиболее примитивные группы у них однопокров-
ные и беспокровные. Систёма Энглера наиболее подробно разра-
ботана, поэтому ею пользуются и в настоящее время.
Бэсси (США), Халл-ир (Германия), Хатчисон (Англия) и дру-
гие-сторонники эуантовой (стробилярной) теории, и в качестве
наиболее примитивной группы они рассматривают многоплоднико-
вые (раналиевые, магнолиевые).
Бэсси можно считать основоположником эуантовой теории
(1915). Графическое изображение его системы настолько ориги-
нально, что его стоит воспроизвести (рис. 152). Рисунок напоми-
нает 1кактус-опунцию (отсюда «Опунция Бэсси»), Площадь каждой
фигуры в рисунке соответствует (приблизительно) объему порядка
по количеству видов, взаимное расположение фигур отражает род-
ственные связи между порядками.
Халлир создавал свою систему одновременно с Бэсси.
Оригинальной чертой системы Хатчисона является разделение
им филогенетического древа на три ствола: два ствола двудольных,
отходящих от гипотических первичных покрытосеменных. Один
ствол представлен первично древесными формами и берет начало от
деревянистых Magnoliales, второй ствол берет начало от травянистых
Ranales и представлен травянистыми формами. Третий ствол, также
берущий начало от гипотических первичных покрытосеменных,
представлен однодольными. Система Хатчисона вполне современна*
10—326
289
Рис. 152. Схема происхождения и родства порядков цветковых растений
согласно Charles Е. Bessey (по Жуковскому)
В нашей стране филогенетические системы разрабатывались
X. Я. Гоби, Н. И. Кузнецовым, Н. А. Бушем, Б. М. Козо-Полянским
и др. Н. И. Кузнецов придерживался взгляда о дифилетическом
происхождении цветковых растений, поэтому в качестве исходных
форм он принимает и группу многоплодниковых, и однопокровные
растения. Взгляды Гоби, Буша и Козо-Полянского близки к совре-
менным, причем Гоби опубликовал свою систему одновременно
с Бэсси.
Наиболее разработанными и современными системами покрыто-
семенных в СССР являются системы А. А. Гроссгейма и А. Л. Тах-
таджяна. Оба они последовательно придерживаются эуантовой те-
ории происхождения покрытосеменных.
290
В системе А. А. Гроссгейма (1945) покрытосеменные лодраз^
деляются на три эволюционные ступени, отражающие различный
уровень совершенства органов размножения; от многочленных цвет-
ков с неопределенным' строением к цветкам с меньшим количеством
частей и более строгим и определенным их размещением, от апокар-
пного к ценокарпному гинецею и от верхней завязи к нижней.
По.таким ступеням идет развитие большинства ветвей (стволов)
филогенетического древа покрытосеменных.
Обычно принято филогенетическое древо графически изображать
в профиль. На таком профильном изображении можно видеть, что
от основания древа отходит несколько расходящихся стволов; более
крупные из них разделяются на ветви, отражающие линии эволюци-
онного процесса. А. А. Гроссгейм изобразил филогенетическое древо
иначе — в виде проекции кроны этого древа на плоскость, перпен-
дикулярную основанию древа. При таком изображении стволы и
ветви будут представлены расходящимися от центра к периферии
лучами. Лучи заканчиваются кругами, символизирующими поряд-
ки растений.
Объем каждого порядка находит свое отражение в размере кру-
гов, являющихся проекциями шара на плоскость; объем шара про-
порционален количеству видов, объединяемых данным порядком.
В центре схемы помещена'исходная группа растений, от которой от-
ходят все линии эволюции.
Для разграничения трех ступеней эволюции все поле схемы раз-
деляют на три части двумя окружностями; внутри первой окружно-
сти сосредоточены порядки, в которых преобладают виды с прими-
тивными признаками; на периферии схемы, за пределами второй
окружности, находятся порядки, наиболее филогенетически продви-
нувшиеся. Между первой и второй окружностями размещаются по-
рядки, занимающие промежуточное место в филогенетической систе-
ме; такие порядки составляют большинство.
Графическое изображение филогенетической схемы в оригинале
у Гроссгейма очень сложно из-за большого числа порядков, отража-
ющих флору всего земного шара; их около 70. Около половины всех
порядков объединяют тропические виды, которые не представляют
для нас практического интереса.
М. В. Культиасовым разработана сокращенная схема филогене-
за, построенная по тому же принципу, что и полная схема Гросс-
гейма. Схема Культиасова включает только важнейшие для нас по-
рядки, имеющие своих представителей на территории СССР. В этой
схеме только 7 стволов эволюции, в то время как оригинальная схе-
ма Гроссгейма включает 11 стволов. В отличие от схемы Гроссгей-
ма, схема Культиасова имеет контур полукруга. Рисунок 153 дает
наглядное представление о принципе графического изображения
системы А. А. Гроссгейма?? В настоящем учебнике систематика покры-
тосеменных изложена соответственно теоретическим взглядам и
системе А. Л. Тахтаджяна. Система Тахтаджяна наиболее совре-
менна. На протяжении длительного периода, начиная с первой ее
публикации (1954), эта система уточняется и совершенствуется ав-
10*
291
Fagales
Juglandales ( ) [Trie a co ae
у' ( Rubiales 1 \Microspermae\ (Cgper ales j тпа Ceranlales у \ \ / [Lilli florae С ) Thyphales \ ХХ|y^yPrlnci pes Spathi -lorae_ штЬешЬууУ--—\ 1 -les J yy X / /Хх/ ( Tubiflorae j / X^iales ) XX^K * \ \ J S'—\Ericales\ RnacardtaA ^У(Maly \ у X. у les 1 \ \poles // X r X / \ \УУ{ Myrtales ) М^®0\ \Compo5itales\ "X X/ \ \ \ rales j у / Legumlnopales\ [Resales \ 'K \ tales ) r~~^\ \ ь / г x^» Cucurbl-y ) x—x \ ^^hcales f V^^xX /~xtQles У-У( j \ \ Z/X_—J ^XXT i Primulalesy^^ ) \ < Г yi \ Zx***^**^ X \ jRhoeadoies У / niP"j УхУРопа1ео^^^\У^У У~У \ у / Hltsmales ^^^\yoiucarolcae)^^^^^Pntrny^<Z/^-^ \ / Упо11о1ез\ \lV^\spermoe) J d. y0lycarpi-f ,(O / V// \ttzf,/ у У \ Polggonales 10 J /у 4 J 1
Рис. 153. Сокращенная схема филогенетического древа цветковых растений (составлена М. В. Культпасовым
по А. А. Гроссгейму).
Римскими цифрами обозначены стволы развития, окружности — ступени развития
тором, и в настоящее время она получила широкое международное
признание. В последней редакции эта система опубликована
в 1978 г., и в ней использованы достижения филогенетических сис-
тем XX в., а также результаты новейших работ в области сравни-
тельной морфологии и систематики цветковых растений.
Каковы были первичные, исходные формы покрытосеменных?
Это первый и основной вопрос, который возникает при построении
филогенетической системы. От решения этого вопроса будет зави-
сеть, какие признаки растения считать первичными, примитив-
ными и какие вторичными, более совершенными, прогрессив-
ными.
Современная филогенетическая систематика, используя все мето-
ды исследования, дает нам возможность получить ответ на постав-
ленный вопрос. Тем самым мы получаем критерии, необходимые для
построения филогенетического древа покрытосеменных.
С точки зрения стробилярной, эуантовой теории происхождения
покрытосеменных, следующие признаки необходимо считать первич-
ными, примитивными: деревянистый стебель; простые, вечнозеленые,
спирально расположенные листья; одиночные насекомоопыляемы-е
актиноморфные двуполые цветки с двойным околоцветником,
с большим числом спирально расположенных тычинок и пестиков;
свободные, несросшиеся части цветка; апокарпный гинецей; верхняя
завязь; двудольное семя.
Все эти признаки примитивности-, сближающие покрытосеменные
с голосеменными предками, наиболее полно сосредоточены в поряд-
ке магнолиевых. Поэтому растения этого порядка рассматриваются
в свете стробилярной теории как самые древние и наиболее прими-
тивные представители покрытосеменных.
Признаки, противоположные перечисленным выше,— вторичные
и характеризуют растения, стоящие на более высокой ступени эво-
люции.
Особый интерес представляет вопрос о происхождении одно-
дольных растений. Большинство ботаников считает, что однодоль-
ные произошли от двудольных растений. Данные палеоботаники
и другие факты говорят о том, что однодольные произошли на са-
мых ранних этапах эволюции покрытосеменных.
Какая же группа первичных двудольных наиболее близка к од-
нодольным? Это должны быть травянистые растения, ибо самые
примитивные однодольные — травянистые - формы. Искомые фор-
мы— представители порядка Пастуховых — Alismales. Из всех
групп однодольных растений частуховые ближе всего стоят к ним-
фейным (Nymphaeales)—тоже водным растениям. По-видимому,
ближайшие предки однодольных произошли от какой-то группы
водных растений, близких к современным нимфейным, следователь-
но, вся линия эволюции однодольных растений имеет первично вод-
ное или земноводное происхождение. Этим, вероятно, объясняется
упрощенность в строении вегетативных органов у однодольных
и, в частности, мочковатый тип корневой системы. Древесные формы
однодольных — вторичного происхождения; это находит свое выра-
293
жение и в совершенно ином способе утолщения ствола однодольных
древесных.
Монофилетичность происхождения однодольных растений дает
основание сохранить традиционное деление отдела покрытосемен-
ных на два класса: двудольные и однодольные.
Двудольные отличаются от однодольных по ряду признаков:
Двудольные Однодольные
1. Семя с одной семядолей
Корневая система мочковатая, из
придаточных корней
Листья с параллельным или дуго-
нервным жилкованием
Стебель с закрытыми пучками, рас-
положенными без видимой законо-
мерности
Цветки преимущественно трехчлен-
ные
6. Часто образуют заросли, состоя-
щие преимущественно из одного
вида
1. Семя с двумя семядолями
2. Корень у подавляющего большин-
ства видов стержневой
3. Листья с
4. Стебель с
женными
пучками
5. Цветки
членные
6. Редко образуют сплошные зарос-
ли, состоящие из одного вида
сетчатым жилкованием
концентрически располо-
открытымн проводящими
преимущественно пяти-
2.
3.
4.
5.
Следует, однако, отметить, что не всегда различие между дву-
дольными и однодольными резко выражено, и некоторые призна-
ки однодольных (например, мочковатый корень) могут встретиться
и у двудольных.
В системе А. Л. Тахтаджяна, опубликованной в 1970 г., 440 се-
мейств, из них 373 двудольных и 67 однодольных. Количество по-
рядков соответственно 94, 74 и 20. В классе двудольных около
176 000 видов, однодольных — около 64 000, всёго приблизительно
240 000 видов.
В системе Тахтаджяна введены два промежуточных таксона:
подкласс и надпорядок. Введение в систему таксона, объединяюще-
го несколько близких порядков, следует признать весьма удачным
и с методической точки зрения; это облегчает изучение филогенети-
ческих отношений порядков, указанных в схеме (рис. 154). Каждый
надпорядок включает от одного до 15 порядков, выражающих на-
правление эволюции и ступени филогенетической продвинутости.
Например, к первому надпорядку отнесены 6 наиболее примитивных
порядков, среди них самый примитивный первый порядок Magnolia-
les, наиболее близкий к гипотетической исходной группе, положив-
шей начало всем ныне живущим примитивным порядкам; к послед-
нему 15-му надпорядку класса двудольных отнесены три порядка,
составляющих вершину филогенетического древа, среди них поря-
док Asterales с одним семейством сложноцветных (Asteraceae, или
Compositae), наиболее высокоорганизованным из двудольных.
В принятой в учебнике системе покрытосеменных названия всех
высших таксонов, от семейства до класса, основаны на названии
типового рода; это также облегчает запоминание научных названий
таксонов и делает всю номенклатуру более стройной.
В добавление к графическому изображению системы (рис. 154)
ниже дана табл. 2, в которой указано соподчинение таксонов выс-
294
* 74Asterales 71.Lamiales
75.Calycerales
72.0ampanulales
7O.Scrophulariale$
I 82. Orchidales
89.Poales
25Juglandales
24.Myrlcales
ISMymelales JS^mamelidales
13. Cercidiphy Hales
12. Trochodendrales
36.CuCurbiialeS ьоМШк
26.Leitnerialcs ,, suegomalis . I
WassM's ilJh laeaUs
W. Primula les 7or \ | /
MCapparales \ii6.Euphorbiales
59.Polygalale$
66.Proteles
65.ElQcagnales
25.Balaaopaley
22Mulales
21. Fagales
20.0asuarinales
19.Barbeyates\
18.Urttcales 4
ITEucommlales
92.Diapensiales
41. Erl cates
9.Plumbaginales
\jo.Thellgonales
45. Ma Iva les
/ 58.Ceraniqles
55.Hippuridal.es
ct u . . I 57$apindales
^^1
KjJ.Podostemai
'52.Nepenthales'
69.Posales?y^.Y .
„n 3i:vikaiss
33. Theales
6gPolemontales
83.9uncales
84.Cyperales /
fdd. Pestionales
87. Erlocaulales
51. Connaralcs
94. Typhales
90. Ar ecalcs
'81.7lnglberales
пк / 68.0entianales
62.Rh am nates
'53. Oleates
8O.lridales
61. CelastraLes /67Dipsacales
OO.Cornales
78. Triuridales
50. Fabales \
77,Najadales \
76. Hydrocharltale\
86. Commc'linales
85.Bromeliales
03. Panda nates
Oi.Cyclanthales
SRaffleslales
9.Ranun^ulales 7O n ^Aristolochiates
' J2-PaeotlLaLes3i:Dlllcnialcs 7n. . 75. Alternates 79Llt,‘,aies
3.Piperalcs
7.1lh.ctalcs
LMagnoliales
Рис. 154. Схема вероятных взаимоотношений порядков цветковых растений (по Тахтаджяну)
8.Hclumbonalcs
n , 6.Numphacales
2. Laurates
Таблица 2
Система покрытосеменных А. Л. Тахтаджяна (1970) (классы, подклассы,
надпоря^ки и порядки)
Подклассы Надпорядки Порядки
Отдел Magnoliophyta (Angiospermae)
Класс Magnoliatae (Dicotyledones)
Magnoliidae I. Magnolialiae
Ranunculidae II. Ranunculanae
Hamamelididae III. Hamamelidanae
Caryophyllidae IV. Caryophyllanae
Dilleniidae V. Dillenianae
VI. Ericanae
VII. Malvanae
Rosidae VIII. Rosanae
IX. Myrtanae X. Rutanae
XI. Aralianae XII. Celastranae
XIII. Proteanae
Asteridae XIV. Lamianae
XV. Asteranae
Magnoliales, Laurales, Piperales,
Aristolochiales, Rafflesiales, Nym-
phaeales.
Illiciales, Nelumbonalcs, Ranuncula-
les, Papaverales, Sarraceniales.
Trochodendrales, Cercidiphylales
Eupteleales, Didymelales, Hama-
melidales, Eucomiales, Urticales,
Barbeyales, Casuarinales, Faga-
les, Betulales, Bolanopales, Myri-
cales, Juglandales, Leitneriales.
Caryophyllales, Polygonales, Plum-
baginales, Theligonales.
Dilleniales, Paeoniales, Theales, Vio-
lates, Passiflorales, Cucurbitales,
Begoniales, Capparales, Tamari-
cales, Salicales.
Ericales, Diapensiales, Ebenales,
Primulales.
Malvales, Euphorbiales, Thymelae-
ales.
Saxifragales, Rosales, Fabales, Con-
narales, Nepenthates, Podostema-
les.
Myrtales, Hippuridales
Rutales, Sapindales, Geraniales,
Polygalales.
Cornales (Araliales)
Celastrales, Rhamnales, Oleates,
Santalales
Elaeagnales, Proteales
Dipsacales, Gentianales, Polemoni-
ales, Scrophulariales, Lamiales.
Campanulales, Calycerales, Astera-
les.
Класс Liliatae (Monocotyledones)
Alysmidae
Liliidae
Commelinidae
Arecidae
I. Alismanae
II. Lilianae
III. Juncanae
IV. Commelinanae
V. Arecanae
Alismales, Hydrocharitales, Najada^
les. >
Triuridales, Liliales, Iridales, Zingi-
berales, Orchidales.
Juncales, Cyperales.
Bromeliales, Commelinales, Eriocau-
lales, Restionales, Poales.
Arecales, Cyclanthales, Arales, Pan-
danales, Typhales.
296
ших рангов (до порядка) в системе Тахтаджяна. Эта таблица дает
возможность найти место в филогенетической системе любого расте-
ния, в том числе описанного в учебнике, если известно, к какому по-
рядку растение относится. С этой целью в учебнике всегда указы-
вается порядок, к которому относятся изучаемые семейства.
Класс двудольные (Dicotyledones —\Magnoliatae)
Подкласс 1. Магнолиидные (Magnoliidae)
Надпорядок I. Магнолиевидные (Magnolianae)
К этому надпорядку отнесено 6 наиболее примитивных порядков,
объединяющих 27 семейств. Каждый порядок характеризуется боль-
шим или меньшим комплексом примитивных, первичных признаков,
но не все порядки равнозначны. Наибольшее количество первич-
ных, примитивных признаков сосредоточено у растений порядка
Magnoliales — это исходная группа растений для всего филогенети-
ческого дрёва.
Однако и в этОхМ порядке нет ныне живущих растений, у кото-
рых сочетались бы все примитивные признаки. Начало филогенети-
ческих линий, которые исходят от порядка Magnoliales, могли дать
какие-то еще более примитивные, вымершие формы, близкие к сов-
ременным растениям, относимым к этому порядку. Очень близко
к магнолиевым стоит порядок Laurales, в котором преобладают дре-
весные формы; магнолиевые же представлены только древесными
формами: деревьями, кустарниками и лианами. В порядке Piperales,
который близок к Laurales, преобладают уже травы. Еще больше
травянистых растений в порядке Aristolochiales; порядок же Raf-
flesiales представляет особую линию эволюции: это исключительно
паразиты, живущие на корнях и стеблях преимущественно тропичес-
ких растений. Особое место занимает порядок Nymphaeales; как
уже об этом сказано, нимфейные — группа двудольных, наиболее
близкая к формам, давшим начало всему классу однодольных.
Порядок магнолиецветные (Magnoliales)
Семейство магнолиевые (Magnoliaceae). Тропические или суб-
тропические деревья, кустарники и лианы, преимущественно вечно-
зеленые. Листья плотные, кожистые, богатые эфирными маслами,
чаще цельнокрайние. Цветки с хорошо выраженными признаками
примитивности; на удлиненном цветоложе большое количество спи-
рально расположенных тычинок и пестиков (рис. 155). Завязи верх-
ние. Листья очередные. Признаки примитивности особенно хорошо
выражены у магнолии.
Магнолия (Magnolia) распространена в Юго-Восточной Азии
и Северной Америке. В Советском Союзе на ЧерноморскохМ лобе-
.297
Рис. 155. Цветок магнолии (Magnolia)
режье Кавказа и на Южном
берегу Крыма широко распро-
странена как декоративное де-
рево магнолия крупноцветная
(М. grandiflora). У нее крупные
белые цветки с сильным прият-
ным запахом, большие жесткие
с блестящей поверхностью про-
стые эллиптические листья, не
опадающие на зиму. В мезо-
филле листа желёзки с аромат-
ными эфирными маслами и
алкалоиды. Лекарственное.
Порядок лавроцветные (Laurales)
Семейство лавровые (Lauraceae). Большое семейство тропиков
и субтропиков. За немногим исключением, деревья и кустарники
с кожистыми листьями, очередно расположенными. Листья с эфир-
ными желёзками. Цветки одно- и двуполые, околоцветник простой,
число частей околоцветника и количество тычинок обычно кратно 3.
Плод костянка или ягода.
Лавр благородный ( Lauras nobilis)—дерево с вечнозелеными
листьями. Разводится в приморских районах Кавказа и Крыма. Вы-
сушенные листья используются как пряность.
К этому же семейству относится лавр камфорный (Cinnamomum
camphora), из древесины которого добывают ценнейшее лекарствен-
ное средство камфору; авокадо (Persea gratissima)—тропическое
дерево, дает богатые жиром плоды, возделывается в тропической
Америке.
Порядок кирказоноцветные (Aristolochiales)
Семейство кирказоновые (Aristolochiaceae). Преимущественно
тропические и субтропические многолетние травы, кустарники и лиа-
ны. Через примитивный род Saruma (Китай) семейство непосредст-
венно связано с магнолиевыми.
Кирказон ломоносовидный (Aristolochia clematitis). Довольно
крупное многолетнее травянистое растение с прямым или извили-
стым стеблем и ползучим корневищем. Листья сердцевидно-яйце-
видные. Цветки трубчатые, зигоморфные, желтоватые. Тычинок 6,
гинецей ценокарпный, плод — висячая грушевидная коробочка.
Ядовитое и лекарственное растение. Поражает центральную нерв-
ную систему, желудочно-кишечный тракт, почки.
Порядок кувшинкоцветные (Nymphaeales)
Семейство нимфейные (Nymphaeaceae). К этому семейству от-
носятся исключительно водные и болотные растения. Среди них
имеются древние формы с совокупностью многих примитивных приз-
298
каков, но есть и формы, значи-
тельно продвинувшиеся в про-
цессе эволюции. Как уже ука-
зывалось выше, в свете совре-
менных представлений первые
однодольные растения в своем
происхождении -связаны ю ним-
фейными, в связи с чем утверж-
дается теория водного проис-
хождения однодольных. Семей-
ство нимфейных представлено
преимущественно тропическими
видами, но несколько видов
встречается и в нашей стране.
Кувшинка белая (Nymphaea
alba). Под названием «водя-
ной лилии» широко распрост-
ранена в СССР в медлейно те-
кущих пресных водах. Круп-
ные, лежащие на воде листья;
многочисленные спирально рас-
положенные лепестки перехо-
дят в тычинки, которых также
много. Гинецей, однако, цено-
карпный и завязь полунижняя,
обросшая выпуклым цветоло-
жем (рис. 156). Содержит ал-
калоид.
К этому же роду относится
настоящий египетский лотос
Рис. 156. Кувшинка белая (Nymphaea
alba) — плавающий лист и цветок; внизу
слева — тычинки и постепенный переход
их к лепесткам, справа — плод
(М lotos), г к близкому роду — знаменитая Victoria regia из бас-»
сейна Амазонки; листья ее, лежащие в виде огромной сковороды
на воде, имеют диаметр до 2 м и удерживают груз до 50 кг, цветки,
очень похожие на цветки нашей кувшинки, достигают 35 см в по-
перечнике.
Кубышка желтая (Nuphar luteum). Встречается, в тех же мес-
тах, что -и белая кувшинка. Листья такие же крупные, но цветки
мельче, с большим числом желтых лепестков. Содержит алкалоиды.
Подкласс 2. Ранункулидные (Ranunculidae)
Надпорядок И. Лютиковидные (Ranunculanae)
В этом надпорядке сосредоточены также преимущественно при-
митивные, но все же более продвинувшиеся группы растений, о чем,
в числе прочих признаков, говорит 3-бороздность микроспор, в то
время как в первом надпорядке споры почти исключительно 1-бо-
роздные или безаппертурные. Порядок Illiciales, хотя и связан
в своем происхождении с Magnoliales, однако отличается от магно-
299
лиевых и палинологическими по основным числам хромосом ( = 14,
у магнолиевых =19). Порядок Nelumboales-представлен двумя ви-
дами водных растений (род Nelumbo — лотос). Порядок Ranuncula-
les — характеризует направление эволюции в сторону выработки
травянистых форм. Порядок Papaverales очень близок к Ranuncula-
les, в то время как растения, объединяемые порядком Sarraceniales,
эволюционировали в совершенно своеобразном направлении—•
в сторону насекомоядное™ (плотоядности).
Порядок иллициецветные (Illiciales)
Семейство лимонниковые (Schizandraceae). Заметно продвинув-
шееся семейство, о чем говорит наличие однополости цветков.
Лимонник (Schizandra)—деревянистая лиана. Двудомное. На
вытянутом цветоложе многочисленные пестики, превращающиеся
в красные ягоды. Сильно удлиненное цветоложе несет гроздь таких
ягод. На Дальнем Востоке распространен лимонник китайский
(Sch. chinensis). Кора, корни и плоды лимонника содержат-тонизи-
рующие, укрепляющие вещества и используются в качестве лекар-
ственного сырья.
К близкому семейству иллициевых (Illiciaceae) относится цен-
ное пряное растение бадьян, или звездчатый анис (Illicium verum).
Вечнозеленое дерево. Используются плоды — сложные листовки.
Порядок лютикоцветные (Ranunculales)
Семейство лютиковые (Ranunculaceae). Весьма полиморфное
семейство, в котором можно проследить различные ступени эволю-
ции цветка, начиная с наиболее примитивной; однако завязь всегда
верхняя. Листья, за-редким исключением, очередные. Преимущест-
венно травянистые растения, распространенные в основном в уме-
ренной зоне северного полушария. Большинство видов семейства от-
носится к непоедаемым или плохо поедаемым растениям, так что
в кормовом отношении это семейство большого интереса не пред-
ставляет. Много растений ядовитых и лекарственных.
Ядовитость растения зависит от наличия в клеточном соке силь-
нодействующих ядовитых веществ. В клеточном соке лютиковых со-
держатся различные алкалоиды, гликозиды, сапонины, лактоны,
синильная кислота.
Алкалоиды — сложные органические соединения, содержащие
углерод, водород, азот; большинство алкалоидов содержит также
и кислород. Алкалоиды представляют собой щелочи, образующие
с органическими кислотами легко растворимые в воде соли. В фор-
ме раствора таких солей алкалоиды и присутствуют в клеточном
соке.
Гликозиды — органические соединения, распадающиеся на угле-
вод и аглюконы. Аглюконы имеют различный химический состав,
они и определяют токсичность гликозидов. Гликозиды горькие на
300
Рис. 157. Адонис весенний (Adonis
v emails)', отдельно внизу — цве-
ток; вверху — сборный плод из се-
мянок (односемянных листовок)
Рис. 158. Купальница европейская
(Trollius europeus);' вверху —
сборный плод, внизу — отдельная
листовка
вкус. Одну из групп гликозидов представляют сапонины; сапони-
ны при взбалтывании с водой дают стойкую пену — мылятся.
Лактоны — также органические соединения, по химической при-
роде это ангидриды оксикислот.
Многие ядовитые вещества в небольших дозах обладают лекар-
ственными свойствами; увеличение дозы вызывает отравление, ко-
торое нередко заканчивается гибелью животного.
Описания отдельных представителей семейства расположены по
филогенетическому принципу.
Адонис (Adonis) — многолетние или однолетние растения с пе-
ристо- или пальчаторассеченными листьями. Цветки актиноморф-
ные. Чашелистиков 5—8, лепестков 5—много, тычинок и пестиков
много, плоды сборные из большого количества семянок (односемен-
ных листовок). Наиболее известен адонис весенний (A. vernalis) —
ценнейшее лекарственное растение (рис. 157).
Купальница (Trollius) — многолетнее травянистое растение Арк-
тики, лесной и высокогорной зон северного полушария. Листья
лальчато рассечены. Чашелистиков 5—много, большое количество
мелких лепестков; тычинок и пестиков много. Плод состоит из 5—
301
многих листовок. Окраска оранжево-желтая. В европейской части
СССР наиболее широко распространена купальница европейская
(Т. europeus) ('рис. 158). В культуре — декоративные формы. Расте-
ния подозрительные на ядовитость.
Калужница (Caltha) — весеннецветущее, многолетнее травяни-
стое растение влажных мест. Листья цельные. Околоцветник венчи-
ковидный из 5 — многих лепестков; тычинок и пестиков много. Плод
состоит из 5—12 многосеменных листовок. В северном полушарии
широко распространена калужница болотная (С. palustris) с круп-
ными золотисто-желтыми цвет-
ками (рис. 159). Растение ядо-
витое. При поедании у живот-
ных поражаются центральная
нервная система, пищевари-
тельный тракт и почки. Отрав-
ления редки, так как растение
имеет острый, жгучий вкус.
Рис. 160. Ветреница лютичная (Апе-
топе ranunculoides)
Рис. 159. Калужница болотная (Caltha
palustris) — часть растения, плоды и се-
мена
Ветреница (Anemone) — многолетнее растение с тропчато- или
пальчатораздельными листьями. Околоцветник простой, из 5—10
лепестков; тычинок и пестиков много; плод — сборная семянка. Пре-
имущественно лесные растения; наиболее широко распространены
ветреница лютичная (A. ranunculOides) и дубравная (A. nemorosa)
(рис. 160). Многие виды ядовиты, но не настолько, чтобы представ-
лять большую опасность. Ядовиты также близкие к ветреницам
прострелы (Pulsatilla).
302
Чистяк (Ficaria)— многолетнее растение с цельными листьями.
Чашелистиков 3. Лепестков 8—12 желтой окраски; тычинок и пес-
тиков много. ‘Плод из большого числа семянок. Наиболее известен
чистяк лютичный (F. rahunculoides) (рис. 161). Лекарственное рас-
тение. При поедании его скотом в больших количествах ядовито.
Рис. 1G1. Чистяк лютичный (Ficaria гапип-
culoides)
Лютик (Ranunculus) — многолетние, реже однолетние растения
(рис. 162). Листья перисто- или пальчатораздельные, реже цельные.
Чашелистиков и лепестков 5, тычинок и пестиков много. Плод соб-
ран из семянок. Цветки, за редким исключением, желтые. Широко
распространенные растения — засорители пастбищ и сенокосов.
Несъедобные, а некоторые и ядовитые растения. Особенно ядовит
лютик ядовитый (R. sceleratus). Лютик едкий (R. асег), ползучий (R.
303
repens) и др. менее ядовиты. Ядовитые начала (лактоны) поража-
ют центральную нервную систему, пищеварительный тракт, почки.
Ядовит также близкий к лютикам рогоглавник (Ceratocephalus) —
известны случаи отравления овец.'
Аконит (Aconitum)—многолетнее растение с пальчатораздель-
ны}ли листьями. Цветки зигоморфные, собранные в кисть, 5 лепест-
Рис. 162. Лютик едкий (Ranunculus
асег); вверху — отдельный цветок,
внизу — лепесток с нектарником
в основании
ковидных чашелистиков, -из которых один шлемовидный; два ле-
пестка срослись, много тычинок, 3—5 пестиков. Плод состоит из
3—5 листовок. Преимущественно горные растения; ядовиты, при
поедании у животных поражается центральная нербная система
и пищеварительный тракт. Используются как лекарственное сырье.
Один из видов — A. napellus «культивируется как декоративное рас-
тение (рис. 163).
304
Живок&сть, шпорник (Delphinium) — многолетние и однолетние'
растения с перисторассеченными листьями. Цветки зигоморфные;
5 окрашенных чашелистиков, один из них со шпорцем; 2—4 срос-
шихся лепестка. Тычинок много, плод из 1—5 листовок. Наиболее
распространено сорное растение — живокость полевая (D. consolida)
(рис. 164). Ядовито. Симп-
томы заболевания такие-
же, как и при отравлении
аконитом, но степень ядо-
витости меньшая. Не-
сколько видов культиви-
руются как декоративные-
растения.
Рис. 163. Аконит джунгарский Асо-
nitum soongoricum); справа — сбор-
ный плод из трех листовок
Рис. 164. Живокость полевая (Delphi-
nium consolida); отдельно — цветок и
группа плодов-листовок
Василистники (Thalictrum) подозрительны на ядовитость.
К этому порядку относится семейство барбарисовых (Berberida-
свае), в котором колючий кустарник барбарис (Berberis) со съедоб-
ными плодами. В листьях барбариса амурского (В. amurensis) л
обыкновенного (В. vulgaris) алкалоиды кровеостанавливающего
действия.-В этом же семействе лекарственное растение Leontice —
травянистое, с желтыми цветками, с подземными клубнями, из ко-
торых и добывают лекарственные вещества — алкалоиды.
305
Порядок макоцветные (Papaverales)
Рис. 165. Мак снотворный (Papa-
ver somniferum) (А), коробочка,
разрезанная поперек (5), семена,
наружный вид и продольный раз-
рез (В)
Семейство маковые (Papaveraceae). Наличие у представителей
этого семейства большого количества тычинок сближает маковые
с примитивным семейством лютиковых. Лепестков 4, чашелистиков
2. Завязь верхняя, плод — коробочка. Растения имеют членистые
млечники, млечный сок оранжевый, желтый или беловатый. Боль-
шинство относящихся к этому семей-
ству растений представлены травя-
нистыми формами; опыляются насе-
комыми.
Чистотел (Chelidonium majus) —
довольно крупное многолетнее рас-
тение, произрастающее в тени. Жел-
тые цветки собраны зонтиковидны-
ми соцветиями. Тычинок много,
плод — одногнездная коробочка.
Оранжевый млечный сок этого рас-
тения ядовит, однако случаи отрав-
ления редки, так как растение имеет
неприятный и жгучий вкус.
Мак (Papaver) представлен у
нас большим числом видов. Травя-
нистые растения с белым млечным
соком, содержащим ценные алка-
лоиды (морфин, кодеин, папаверин
и др.), используемые в медицине
(рис. 165). Особенно много целебных
алкалоидов в млечном соке мака
снотворного (Р. somniferum), кото-
рый специально высевается для по-
лучения опия. Алкалоиды имеются
также и в млечном соке обычного
мака-самосейки (Р. rhoeas), поэто-
му этот мак относится к группе ядо-
витых растений; содержащиеся в
нем ядовитые вещества поражают
центральную нервную систему.
Цветки мака актиноморфны,<име-
!от 2 чашелистика, которые отпадают в начале цветения, 4 лепест-
ка, большое число тычинок. Плод коробочка, с большим количе-
ством семян. Из семян мака добывают ценное масло, имеющее
пищевое и техническое применение. Сами семена используются
в кондитерском производстве.
Семейство дымянковые (Fumariaceae). По сравнению с преды-
дущим семейством более продвинувшееся: число частей гинецея
и андроцея невелико и определенно, цветки зигоморфные.
Хохлатка (Corydalis). Один из лепестков зигоморфного венчика
со шпорцем. Чашелистиков может совсем не быть. Тычинки 2, каж-
306
дая на верхушке трехраздельна. Цветки собраны в соцветие —
кисть; плод — удлиненная коробочка. Листья дважды или трижды
тройчатораздельные. Многолетние виды образуют клубень. В лесах
распространена хохлатка Галлера — С. halleri и др. Ядовиты. Со-
держат алкалоиды.
Дымянка (Futnaria) — небольшое растение с мелкорассеченны-
ми сизоватыми листьями. Цветки также зигоморфные, сходные
с цветками хохлатки, но мельче. Плод — округлый орешек. Обыч-
ные сорные растения; в европейской части СССР распространены
F. officinalis и другие виды. Ядовиты, содержат алкалоиды.
Подкласс 3. Гамамелидидные (Hamamelididae)
Надпорядок III. Гамамелидовидные (Hamamelidanae)
В этот.надпорядок автор системы (А. Л. Тахтаджян) объединя-
ет 15 порядков с 22 семействами. На схеме филогенетического дре-
ва это самостоятельная большая ветвь, берущая начало от Magno-
liales, порядок Hamamelidales в этой ветви занимает особое место,
от него отходит веер ответвлений — все более и более продвинув-
шихся порядков: Eucomiales, Urticales, Barbeyales, Casuarinales,
Fagales, Betulales, Balanoporales, Myricales, Juglandales, Leitneria-
les.
Основное направление эволюции здесь — приспособление к вет-
роопылению; отсюда и утрата околоцветника, и раздельнополость,
и возникновение соцветия — сережки. Однако, в виде исключения,
в этой группе имеются и вторичные приспособления к насекомо-
опылению, например у инжира (Ficus caria), относящегося к семей-
ству тутовых (Могасеае), порядку Urticales.
Исходный порядок—Hamamelidales объединяет очень древние
растения, ископаемые остатки их находят в пластах мелового пери-
ода; по-видимому, это вымирающее звено, связующее уже вымер-
шие, переходные к Magnoliales еще более примитивные формы
с современными группами растений, приспособившимися к ветро-
опылению. Представителем этого порядка в нашей флоре может
быть платан (Platanus), относящийся’к семейству платановых.
Несколько особняком стоит порядок Trochodendrales. В нем
всего два семейства с несколькими видами. Виды, близкие к маг-
нолиевым, но давно уже обособившиеся в сторону анемофилии (ут-
рата околоцветника, сережковидные соцветия). Очень близок к не-
му монотипный порядок Cercidiphyllales.
Порядок Eupteleales, состоящий из одного семейства и рода, то-
же занимает изолированное положение в этой ветви. Такое же мес-
то, но несколько выше, занимает порядок Didymelales, также пред-
ставленный одним родом.
Порядок гамамелидоцветные (Hamamelidales)
Семейство платановые (Platanaceae) представлено одним ро-
дом (Platanus) с несколькими видами. Платан, или чинар,— боль-
шие деревья с пальчатолопастными крупными листьями, располо-
307
женными поочередно. Растения однодомные, цветки однопокров-
ные, невзрачные. Околоцветник 3—8-членный, тычинок обычно 3—4,
гинецей апокарпный из 3—8 плодолистиков. Плод — многоорешек,
В СССР в естественном состоянии встречается два вида платана'—•
в Закавказье (Р. digitata) и Средней Азии' (Р. orientalis).
К порядку Eucomiales относится семейство эйкомиевых (Еисо-
fniaceae), в котором ценнейшее дерево эйкомия вязолистная (Enco-
mia ulmoides). Разводят на Кавказе для получения гуттаперчи.
Порядок крапивоцветные (Urticales)
Семейство крапивные (Urticaceae). В нашей стране это семейст-
во представлено лишь травянистыми формами. Листья очередные
и супротивные. Околоцветник простой, чашечковидный, из свобод-
ных или сросшихся листочков в числе 4—5. Тычинок также 4—5;
завязь верхняя, одногнездная. Плод — семянка. Растения двудом-
ные и однодомные. В кормовом отношении наибольший интерес
представляют крапивы (Urtica).
Крапива двудомная (U. dioica) давно используется в корм жи-
вотным в свежем и запаренном виде, а также в виде сенной муки.
Это крупное корневищное растение, покрытое жгучими волосками;
вегетировать начинает ранней весной. Скармливание крапивы по-
вышает молочную продуктивность коров и яйценоскость птицы
(рис. 166). Вместе с тем это лекарственное растение (листья).
Порядок букоцветные (Fagales)
Семейство буковые (Fagaceae). Деревья с простыми листьями.
Мужские цветки в сережчатых или головчатых симподиальных со-
цветиях. Женские цветки одиночные или в неразвитых дихазиях;
околоцветник четырех-восьмичленный. Соцветие окружено при-
цветниками, срастающимися и образующими при плодах обверт-
ку — плюску.
Бук (Fagus)—теневыносливое дерево с овальными яйцевидны-
ми листьями. Мужские соцветия короткие, головчатые. Из цветков
женского соцветия развиваются два трехгранных плода — ореха, ок-
руженных общей обверткой, снабженной мягкими игловидными вы-
ростами. Бук — ценнейшее дерево, дающее прекрасную поделочную
древесину. Буковые орехи богаты маслом и очень питательны.
В Юго-Западной Украине распространен бук европейский (F. silva-
tica), на Кавказе и в Крыму — восточный (F. orientalis). Из буково-
го дегтя добывают лекарственные средства: креозот, гваякол и др.
Дуб (Quercus); наиболее широко распространенная у нас лесо-
образующая порода — дуб обыкновенный, или черешчатый (Qu.
pedunculata), с перистолопастными листьями. Мужские сережки
длинные, висячие. Женские цветки одиночные или в немногоцветко-
вых дихазиях. Плод — желудь с плюской, образованной из срос-
шихся прицветников. Крепкая и красивая древесина дуба находит
широкое применение в деревообрабатывающей промышленности.
Дубовая кора — ценное дубильное сырье (техническое и лекарст-
венное).
308
Рис. 166. Крапива двудомная (Urtica dioica):
1 — ветвь мужского растения; 2 — тычиночный цветок до раскрывания; 3 — он же
в раскрытом состоянии после выбрасывания пыльцы; 4 — веточка с женским соцве*
тием; 5 — пестичный цветок
Каштан настоящий (Castanea sativa) — крупное дерево, распро-
страненное в естественном состоянии и в культуре на Кавказе.
Листья широколанцетные, остропильчатые. В общей плюске 1—3
ореха. Орехи каштана настоящего вкусные, питательные. Древесина
весьма ценная. Кора богата дубильными веществами.
Порядок березоцветные (Betulales)
Семейство березовые (Betulaceae). Деревья или кустарники. Ли-
стья простые, очередно расположенные. Мужские цветки собраны
в сережки, женские — в сережки или пучки. Плод — односемянный
орешек.
309
Береза (Betula)—деревья или кустарники; наиболее широко
распространена береза белая, или бородавчатая (В. alba), реже бе-
реза пушистая (В. pubescens). Мужские соцветия—сережки — зак-
ладываются с осени, женские формируются весной. У женских цвет-
ков'околоцветник отсутствует, у мужских он сильно редуцирован.
Плод — односемянный, крылатый орешек. Береза — ценная лесооб-
разующая порода, широко распространенная в лесной зоне и в го-
рах Кавказа и Средней Азии. Из коры ее получают березовый де-
готь. Для сухой перегонки используют также и древесину. В меди-
цине применяют отвар из березовых почек, а также весенний
березовый сок, деготь.
Ольха (Alnus)—деревья и кустарники. Наиболее широко рас-
пространена ольха клейкая (A. glutinosa). Ольховые насаждения
приурочены к заболоченным, сильно увлажненным почвам. На кор-
нях ольхи обитают азотусваивающие 'бактерии, образующие
клубеньки. Кора ольхи используется как дубильное сырье. Почки
и листья — лекарственное сырье.
Семейство лещинные (Corylaceae). Деревья и крупные кустар-
ники. Листья простые, очередно расположенные. В отличие от-пре-
дыдущего семейства мужские цветки вовсе не имеют околоцветни-
ка. Плод — орех или орешек в листовой обвертке/
Орешник, или лещина (Corylus), — крупный кустарник, реже де-
рево. Мужские цветки в длинных висячих сережках. Околоцветника
нет. Женские цветки попарно сидят в почке из чешуй. Плод — орех
в обвертке из разросшихся прицветников. В СССР произрастает не-
сколько видов лещины, из них наиболее широко распространена
лещина обыкновенная (С. avellana)..Плоды орешника съедобны,
питательны, из них добывают ценное масло.
Граб (Carpinus). Наиболее распространен у нас граб обыкно-
венный (С. betulus) в европейской части СССР и на Кавказе и граб
восточный (С. orientalis) —на Кавказе и в Крыму. Листья простые,
очередно расположенные. Цветение граба начинается одновремен-
но с развертыванием листьев. Мужские, цветки без околоцветников,
из 4—12 раздвоенных тычинок, собраны в сережку. Женские цвет-
ки также собраны в сережку, попарно сидят в пазухе мелких кро-
ющих чешуй, позднее опадающих. Плод — орешек в листовой'ло-
пастной обвертке.
Порядок орехоцветные (Juglandales)
Семейство ореховые (Juglandaceae). Наибольший интерес для
•нас представляет орех грецкий (Juglans regia). Крупное и весьма
декоративное дерево с большими непарноперистыми листьями, об-
ладающими приятным запахом. Растение однодомное. Плод — кос-
тянка; выпадающая из нее косточка обычно называется орехом.
Мякоть ореха очень питательна, богата маслом и обладает высоки-
ми вкусовыми качествами. Листья, свежие несозревшие плоды
и околоплодники — лекарственное сырье.
310
Подкласс 4. Кариофиллидные (Caryophyllidae)
Надпорядок IV. Гвоздиковидные (Caryophyllanae)
В этом надпорядке всего 4 порядка. Центральный из них —
гвоздикоцветные (Caryophyllales), он непосредственно связан с
Rapunculales и характеризуется большой полиморфностью (16 се-
мейств). Здесь имеются и более примитивные формы, с двойным
околоцветником, приспособленным к насекомоопылению; но есть
семейства, значительно продвинувшиеся по пути приспособления
к ветроопылению, утратившие яркий венчик (.семейства Amarantha-
ceae, Chenopodiaceae и др.). Следующие два порядка (Polygonales
и Plumbaginales) монотипные и содержат лишь по одному семейст-
ву. Наиболее высокое место в системе занимает порядок Plumbagi-
nales; растения этого порядка обладают уже спайнолепестным вен-
чиком — показатель более высокой ступени эволюции. Неясно поло-
жение порядка Theligonales.
Порядок гвоздикоцветные (Caryophyllales)
Семейство гвоздичные (Caryophyllaceae). Травянистые, реже по-
лукустарниковые растения. Листья супротивные, иногда срастаю-
щиеся своими основаниями. Цветки правильные, с двойным около-
цветником (иногда венчик редуцирован). Чашечка свободнолист-
ная и спайнолистная; венчик из 5 свободных лепестков. Тычинок
5 или 10. Завязь верхняя, одногнездная, из 2—5 плодолистиков, со-
ответственно чему и столбиков бывает от двух до пяти. Плод — ко-
робочка или орешек.
Большинство видов этого семейства плохо поедается скотом или
вовсе не поедается, поэтому как корм большого значения не имеет.
Имеются растения, в которых содержатся ядовитые вещества са-
понин-гликозиды. Водные растворы сапонинов при взбалтывании
дают мыльную пену, поэтому толстые корни некоторых растений
этого семейства (из рода Saponaria) используются для получения
мГоющих средств.
Кукушкин цвет (Coronaria flos-cuculi)—лекарственное рас-
тение.
Большой ядовитостью обладают семена куколя (Agrostema
githago); довольно крупное растение, засоряющее озимые и яровые
посевы. Цветки темно-розовые. Чашечка спайнолистная, тычинок
10, столбиков 5, плод — коробочка. Отравляются животные обычно
при поедании зерновых отходов, засоренных семенами этого расте-
ния.
Несколько видов гвоздик (Dianthus barbata, D. sinensis, D: ca-
ryophyllus) возделываются как декоративные растения.
К порядку гвоздикоцветных относится также семейство какту-
совые (Cactaceae) с большим количеством морфологически разнооб-
разных видов (Южная и Центральная Америка).
311
Семейство маревые (Chenopodiaceae). Травы, полукустарники,
редко деревья. Листья очередные, иногда супротивные. Цветки ма-
лозаметные; околоцветник чашечковидный, зеленый или пленчатый,
иногда сочный. Частей околоцветника 1—5, столько же и тычинок.
Завязь верхняя или полунижняя; рылец 2—3. Плод односеменный
с пленчатым или сочным околоплодником.
Распространены маревые преимущественно в зоне сухих степей,
на солонцеватых почвах полупустынь и пустынь, где играют суще-
ственную роль в кормообеопечении животных, в основном как паст-
бищные растения.
Рис. 167. Свекла (Beta vulgaris). А — растение первого года жизшц
Б — растение второго года жизни, В — колосовидное соцветие, Г —
цветок, Д — цветок в разрезе, Е — образование соплодий, Ж—со-
плодие в разрезе, 3 — семя в разрезе, И — прорастающее соплодие
312
К этому семейству относится свекла. Наибольшее кормовое зна-
чение после свеклы имеют кохия, несколько видов соляной (Salso-
la), саксаул, терескен — все это растения сухих мест. Как сорные
распространены: многочисленные виды мари (Chenopodium), лебе-
ды (Atriplex), курай (Salsola pestifer). Некоторые виды этих родов
имеют и кормо1вое значение.
Свекла (Beta vulgaris) — двулетнее растение, дающее в первый
год жизни корнеплоды (рис. 167). Цветки двуполые, пятичленные.
Завязь полунижняя с 2—3 рыльцами. Плодики собраны в клубочки.
В последние годы советскими селекционерами выведены сорта с-од-
носемянными клубочками, что имеет большое значение при обработ-
ке посевов. Имеются разновидности сахарной, кормовой и столовой
свеклы.
Кохия, или прутняк (Kochia prostrata),— многолетнее полукус-
тарниковое растение с очередными листьями. Цветок пятичленный,
при плодах части околоцветника разрастаются, на них образуются
крыловидные придатки.
Одно из основных кормовых растений засушливой солонцеватой
степи юго-востока и в песчаной полупустыне; используется для соз-
дания искусственных пастбищ. Хорошо выдерживает вытаптывание,
Терескен (Eurotia ceratoides) распространен в зоне пустынь и по-
лупустынь. Полукустарник. Цветки однополые, четырехчленные;
растение однодомное. На Памире — основное кормовое растение, за-
готовляемое также на топливо. Введен в культуру.
Саксаул (Haloxylon) — несколько видов кустарников и деревь-
ев, распространенных в пустынях Средней Азии. Зеленые веточки
саксаула в условиях пустыни — ценный корм.
Анабазис (Anabasis)—несколько видов многолетних трав или
полукустарников с-членистыми стеблями и редуцированными листь-
ями. Распространен в полупустынных районах Средней Азии. Ядо-
витые растения с алкалоидом анабазином, обладающим сильным
инсектицидным действием. Некоторые виды мари (Chenopodium am-
brosioides и др.) —ценное лекарственное сырье для получения про-
тивоглистных препаратов.
К этому же порядку относится семейство щирицевых (Amaran-
thaceae) с широко распространенным родом щирицы (Amarantr
hus) —сорные, декоративные и др.
Порядок гречихдцветные (Polygonales)
Семейство гречишные (Polygonaceae). Травянистые, реже дере-
вянистые растения. Листья очередные, два сросшихся прилистника
образуют характерный для этого семейства раструб. Цветки пра-
вильные, однопокровные; околоцветник чашечковидный или венчи-
ковидный. Частей околоцветника 4—6, тычинок 4—8, завязь верх-
няя, плод — семянка.
К этому семейству относится ценная продовольственная и медо-
носная культура — гречиха (Fagopyrum sdgittatum)^
313
Горец (Polygonum) — этот род представлен многочисленными
видами, среди которых имеются ценные кормовые растения. Широко
распространен спорыш (Р. aviculare), или птичья гречка. Это не-
крупное стелющееся растение, обильно произрастающее вдоль
дорог, на выгонах; питательно, поедаемое всеми видами сельскохо-
зяйственных животных. Горец змеиный (Р. bistorta) — ценное расте-
ние в составе растительности субальпийских лугов. Горец вьюнко-
вый (Р convolvulus), известный больше под названием гречихи
вьюнковой,— весьма распространенный полевой сорняк. Ядовит для
лошадей.
Щавель (Rumex)— менее интересен в кормовом отношении;
большинство видов почти не поедается; сорняки пастбищ и сеноко-
сов. Листья щавеля кислого (R. acetosa), конского (R. confertus) и
несколько других видов охотно поедаются животными. Щавель кис-
лый используется как овощное растение. Известны случаи отравле-
ния животных щавелем воробьиным (R. acetosella).
Подкласс 5. Диллениидные (Dilleniidae)
Надпорядок V. Диллениевидные (Dillenianae)
Это крупный надпорядок, насчитывающий 10 порядков, а в них
51 семейство, преимущественно тропические и субтропические. По-
рядком Dilleniales, включающим наиболее примитивные формы,
начинается крупная ветвь филогенетического древа, большая часть
которой состоит из разветвлений — порядков, входящих в настоящий
надпорядок. Это прежде всего крупнейший порядок чаецветных
(Theales), включающий 20 семейств, преимущественно тропических.
Из нашей флоры в этом порядке наиболее известно семейство зве-
робойных, а из завезенных и возделываемых — чай (Thea sinensis)
из семейства чайных (Theaceae). От порядка чаецветных идут фило-
генетические линии к четырем порядкам, входящим уже в следую-'
щий надпорядок — Ericanae, объединяющий более высоко продви-
нувшиеся формы.
Вторым важнейшим в филогенетическом отношении порядком
изучаемого надпорядка является порядок Violates; от него отходят
пять ответвлений, характеризующихся различным уровнехМ и на-
правлением развития. Это порядки Passiflorales, Cucurbitales, Ве-
goniales, Capparales. В Capparales важнейшее семейство кресто-
цветных Tamaricales и порядок Salicales с единственным семейст-
вом ивовых.
Особое место занимает порядок Paeoniales. Он близок к дилле-
ниевым, но отличается от них по ряду морфологических признаков:
мясистый лопастный нектарный диск, толстостенные йлодолисти-
ки, сильно развитые наружные интегументы и др.
Некоторые из названных порядков и семейств мы рассмотрим
более подробно.
314
Порядок пионоцветные (Paeoniales)
Семейство пионовые (Paeoniaceae). Включает один примитив-
ный род, близкий к Dilleniales, — пион, ранее включавшийся в се-
мейство лютиковых.
Пион (Paeonia) — многолетнее травянистое растение, реже не-
большое деревце (-рис. 168). Листья перисторассеченные, цветки
одиночные, крупные. Чашечка пя-
тилистная, лепестков пять и боль-
ше: многочисленные тычинки рас-
положены по спирали. Гинецей
апокарпный, плод из 2—8 много-
семянных листовок. Распростра-
нены в умеренной зоне северного
полушария. Большое количество
культурных разновидностей. В ди-
ком виде в СССР широко распро-
странен пион узколистный (Р te-
nuifolia). Один из видов пиона —
Р. anomala, широко распростра-
ненный в Сибири, — ценное ле-
карственное растение.
Порядок чаецветные (Theales)
Семейство зверобойные (Ну-
pericaceae). Относящийся сюда
род Зверобой .(Hypericum) Пред- рис 168. Пион кавказский (Paeonium
ставлен в флоре СССР несколь- caucasica) — лист, цветок и плоды
кими видами, из которых зверобой
обыкновенный (Н. perforatum) известен как ценное лекарственное
растение. Это травянистое растение с супротивными ''листьями.
Цветок с 5 лепестками и многочисленными тычинками, сросшимися
у основания в 3—5 пучков. Завязь верхняя, плод коробочка.
Порядок фиалкоцветные (Violates)
Семейство фиалковые (Violaceae). Типичные представители это-,
го семейства относятся к роду фиалка (Viola), многочисленные ви-
ды которого широко распространены в СССР. Растения травяни-
стые, с очередными и супротивными листьями. Цветки зигоморфные,
с пятью лепестками и тычинками; плод — коробочка, раскрывающа-
яся тремя створками. Наиболее известны фиалка душистая (V odo-
rata) и анютины глазки (V tricolor). Лекарственное растение.
Порядок тыквоцвеДные (Cucurbitales)
Семейство тыквенные (Cucurbitaceae). Преимущественно тропи-
ческие растения. В нашей флоре однолетние, травянистые, лазящие
растения с усиками, простыми или разветвленными. Цветки обычно
315
одиночные, тычинок 5, более или менее сросшихся .между собой; ги-
нецей ценокарпный, как правило, из трех плодолистиков. За редким
исключением растения однодомные. Завязь срастается с цветоло-
жем, образуя ложный сочный плод — тыквину. Наибольшее значе-
ние имеют следующие роды.
Огурец (Cucumis). Этот род включает несколько видов. Огурец
(С. sativus) — широко распространенное овощное растение родом из
Юго-Восточной Азии (рис. 169). В пищу употребляются незрелые
разросшиеся завязи. Имеется очень много сортов. В диком виде не-
известен.
Дыня (С. melo) имеет плоды различной формы, богатые саха-
ром. С древнейших времен культивируется в Средней Азии и стра-
нах Ближнего Востока. Теплолюбивое и солнцелюбивое растение.
Арбуз (Citrullus) родом из Южной Африки (пустыня Калахари),
где он и сейчас встречается в диком ваде. В отличие от остальных
Рис. 169. Огурец (Cucumis sativus). А — репродуктивный по-
бег, Б — пестичный и тычиночный цветки, В — тычинки, Г —
плод в поперечном разрезе
316
тыквенных, -имеет глубокораздельные, лопастные листья. Плоды^
шаровидные или удлиненные. Как и дыня, растение теплолюбивое,
требующее для вызревания много солнца. Разводится арбуз в двух
видах. Арбуз съедобный (С. edulus) имеет розовую или красную
сладкую мякоть; арбуз кормовой (С. colocynthoides) имеет крупный
плод, удлиненный, с желтоватой несладкой мякотью. Плоды выдер-
живают более длительное хранение, нежели плоды съедобного ар-
буза.
Колоцинт (С. colocynthus)—лекарственное растение из Север-
ной Африки, у нас встречается в Туркестане. Экстракт из плодов —
сильное слабительное.
ТыАра (Cucurbita). Растения, относящиеся к этому роду, имеют
плоды самой разнообразной формы, различных размеров и окрасок.
Плоды широко используются в пищу и корм окоту. У тыквы круп-
ноплодной (С. maxima) плоды обычно округлые, очень крупные, на-
гладких, без граней плодоножках, а у тыквы мускатной (С. moscha-
ta) плоды удлиненные, на четырехгранных плодоножках. Последняя
более требовательна к теплу, разводится на юге. Тыква обыкновен-
ная (С. реро), культивируется несколько разновидностей: обычная,
столовая тыква, кабачки, патиссоны.
Кабачки в небольшом количестве можно использовать как соч-
ный корм для скота с середины и до конца лета. Тыква дает пло-
ды, хорошо сохраняемые до конца зимы, широко используется на
корм.
Переступень белый (Bryonia alba) — однодомное растение и пе-
реступень двудомный'(В. dioica) — ядовитые и лекарственные рас-
тения. Многолетники. Поражают желудочно-кишечный тракт с по-
следующим параличом центральной нервной системы.
Порядок каперсоцветные (Capparales)
Семейство крестоцветные (Brassicaceae, Cruciferae). Очень мо-
нотипное семейство. Растения травянистые или полукустарники. Ли-
стья очередные. Цветки правильные: 4 свободных чашелистика-
и столько же лепестков; тычинок 6, из них 2 наружные, более .ко-
роткие; пестик из двух плодолистиков. Плод— стручок, стручочек
#или, реже, односемянный орешек. Ввиду большого однообразия в.
’строении цветка и вегетативных органов основное значение при оп-
ределении растений этого семейства имеют плоды.
В этом семействе много ценных продовольственных и кормовых-
растений, но много также и сорняков. Многие крестоцветные содер-
жат горчичные гликозиды, поражающие при поедании растений ор-
ганы дыхания и пищеварительного тракта (горчицы, гулявники,
желтушники, резушка, жерушник и др.).
Род Brassica—наиболее важный в хозяйственном отношении
Сюда относятся следующие виды.
Капуста (В. oleracea) —двулетнее растение (рис. 170), представ-
ленное несколькими разновидностями. Капуста кормовая (В. olera-
cea v. acephala), с большими толстыми листьями, дающими хороший
317
Рис. 170. Капуста (Brassica oleracea). А — соцветие, Б — не-
зрелый плод, В — зрелый плод, Г — семя, Д — зародыш семе-
ни, Е — поперечный разрез семени, Ж — андроцей ‘ и гинецей
цветка капусты
сочный корм. Капуста кочанная (В. oleracea v. capitata) — основное
овощное растение в нашей стране. Употребляется в свежем, кваше-
ном и вареном виде. Возделывается во всех зонах СССР. Капуста
цветная (В. oleracea v. botrytis) —диетический овощ, деликатес. В
пищу употребляются метаморфизированные сочные соцветия. В ди-
ком виде кочанная капуста произрастает по атлантическому побе-
режью Франции.
Репа (Brassica гара) — культурное растение в двух формах:
столовая репа, с небольшим округлосплющенным корнеплодом, и
кормовая репа, называемая также турнепсом, — с удлиненными
крупными корнеплодами. Двулетники. Возделываются преимуществ
венно в северной и умеренной зоне.
Брюква (В. napus v. esculenta) —двулетнее растение, дающее
крупный съедобный корнеплод с желтоватой'мякотью.
Рапс (В. napus v. oleifera) — однолетнее кормовое масличное
растение, возделываемое в озимой и яровой форме.
Горчица полевая (В. campestris). Озимый и яровой однолетник.
Возделывается как масличная культура.
Сарептская горчица (В. juncea) — однолетняя масличная куль-
тура. Из жмыхов, остающихся после выжимки масла, изготовляют
горчицу — приправу к пище и средство, применяемое с лечебными
целями. Горчица также хороший медонос.
318
Горчица ( Sinapis)
представлена нескольки-
ми дикорастущими вида-
ми. Один из них — горчи-
ца белая (S. alba), иног-
да выращивается в куль-
туре как хороший медоьГос
и масличное растение
(рис. 171).
Редька (Raphanus) —
имеется несколько видов
сорных растений; один
вид — R. sativus — культи-
вируется в двух разновид-
ностях: собственно редька
(R. sativus v. niger) и
редис (R. sativus v. radi-
cula). Редька полевая
(R. raphanistrum) счита-
ется ядовитым растением.
Гулявник (Sisymbri-
um) объединяет несколько
видов сорных растений.
Особенно широко распро-
странены виды: S. altissi-
albd), спра-
Рис. 171. Горчица белая (Sinapis
ва — плод стручок
mum под названием ро-
гачки, гулявник Лозеля
(S. loeselii), гулявник
Софья (S. sophia, или Des-
curainia sophia). При поедании в большом количестве все эти рас-
тения могут вызвать воспаление дыхательных путей и пищевари-
тельного тракта (ядовитые семена).
Порядок ивоцветные (Salicales)
Семейство ивовые (Salicaceae). К этому семейству относятся
два важных рода — ива (Salix) и тополь (Populus). Ива представ-
лена многочисленными видами деревьев и кустарников, распростра-
ненных во всех зонах СССР, тополь — деревьями. Листья очеред-
ные; соцветия — сережки; растения двудомные. Тычинок две—-
много, плод — коробочка. У тополей ветроопыление; ивы имеют
нектарники и опыляются насекомыми. Околоцветник у обоих родов
сильно редуцирован.
Наиболее распространенные виды ивы: ива белая (S. alba), лом-
кая (S. fragilis), прутовидная, или корзиночная (S. viminalis).
Из рода тополь наиболее распространена осина (Р. tremula),
а также тополь: белый (Р. alba), черный (Р. nigra) и пирамидаль-
ный (Р. pyramidalis).
319
Надпорядок VI. Вересковидные (Ericanae)
Небольшой надпорядок, состоящий из 4 порядков, в которых 19
семейств*; из них 10 в порядке верескоцветные (Ericales), непосред-
ственно связанном с порядком чаецветных. Порядок Ebenales пред-
ставлен в основном тропическими и субтропическими деревьями
и кустарниками, среди которых для нас наибольший интерес пред-
ставляет хурма (Diospyros), возделываемая на Кавказе как плодо-
вое дерево. Третий, -наиболее продвинувшийся порядок — Primu-
lales. У примуловых тычинки расположены напротив частей спай-
нолепестного венчика, что рассматривается ,как признак более
высокой организации. Этот порядок представлен в основном холо-
достойкими растениями. Небольшой порядок Diapensiales, вероят-
но, имеет общее происхождение с Ericales.
Порядок верескоцветные (Ericales)
Семейство вересковые (Ericaceae) — богатое видами семейство,
^широко распространенное от (субтропиков до полярных зон обоих
полушарий. Кустарники и полукустарники, обычно вечнозеленые
листья, очередно расположенные. Цветки насекомоопыляемые,
актиноморфные, с двойным околоцветником, тычинок.10, 8, 4. За-
вязь верхняя. Плод коробочка, костянка или ягода. Ъ Советском
‘Союзе растения этого семейства распространены преимущественно
в лесной и приполярной зонах, а также в субальпийском и альпий-
ском поясах гор. Наиболее известны: вереск обыкновенный (Calluna
vulgaris), багульник (Ledum palustre), несколько видов рододенд-
рона (Rhododendron), брусника (Vactinium vitis idaea), черника
'(V. myrtyllus), голубика (V. uliginosum), клюква (Oxycoccus quad-
rip etalus).
Порядок первоцветные (Primulales)
Семейство первоцветные (Primulaceae). 'Травянистые растения
<с прикорневыми листьями. В этом семействе наиболее известен род
первоцвет (Primula). Цветки правильные, венчик спайнолепестный,
тычинок 5, сросшихся с венчиком. Плод — коробочка. Первоцвет ле-
карственный (Р. officinalis) — ценное лекарственное растение. К
этому же семейству относятся цикламен (Cyclamen), проломник
.(Andros асае), вербейник (Lysimachia).
Надпорядок VII. Мальвовидные (Malvanae)
В этом надпорядке 3 порядка, семейств 18. Начинается он по*
рядком Malvales, у представителей которого примитивные признаки,
сближающие их с Violates. Остальные два порядка Euphorbiales
и Thymelaeales занимают значительно более высокое место в систе-
ме; хотя и у Euphorbiales имеются примитивные представители,
•близкие к Violates.
320
Порядок мальвоцветные (Malvales)
Семейство липовые (Tiliaceae). Древесные и травянистые расте-
ния с очередными листьями. Цвет-ки с многочисленными тычинками.
В СССР два рода: липа, джут.
Липа (Tilia) — крупное дерево. Листья сердцевидные, цветки
в дихазиях, сидящие на видоизмененном прицветнике. Плод — од-
ногнездный орешек. Цветки богаты нектаром, благодаря чему ли-
па — хороший медонос. Широко распространена липа мелколистная
(Т, cordata), очень декоративна липа войлочная (Т. tomentosa)<
Древесина липы мягкая, широко использу-
ется для бытовых изделий. Соцветия липы
«Липовый цвет» — лекарственное сред-
ство.
Джут (Corchorus)—волокнистое одно-
летнее растение тропического происхожде-
ния. Дает ценное негигроскопическое волок-
но, используемое для изготовления упако-
вочной ткани, шпагата и др. В среднеазиат-
ских республиках разводят джут длинно-
плодный (С. olitorius).
Семейство мальвовые (Malvaceae). Обла-
дает рядом примитивных признаков: боль-
шое количество тычинок, верхняя завязь,
очередное расположение листьев. В этом се-
мействе несколько ценных в практическом
отношении родов (рис. 172).
Растения этого семейства преимущест-
венно теплолюбивые, распространены по
всем материкам. Травянистые и древесные формы. Многочисленные
тычинки срослись нитями в трубочку, обнимающую столбики. Цвет-
ки двуполые, лепестков 5. Плод —сложная листовка, сложная се-
мянка или коробочка.
Канатник (Abutilon theophrastii) — однолетнее травянистое рас-
Рис. 172. Строение цвет-
ка у мальвовых:
I — наружная чашечка (под*
чашне); 2—внутренняя ча-
шечка; 3 — венчик; 4 — ан-
дроцей; 5 — пестик; 6 — семе-
зачаток
тение. Листья цельные, пальчатонервные. В отличие от остальных
мальвовых подчашия нет. Плод — сборная коробочка. Культивиру-
ют; из стебля добывают грубое волокно, используемое для производ-
ства мешковины, веревок, шпагата и др. В семенах до 20% масла,
пригодного для технических целей.
Алтей (Althaea) — однолетние и многолетние травянистые рас-
тения. Цветки с подчашием. Плод — сборная семянка. Алтей лекар-
ственный (A. officinalis) — дикорастущее лекарственное растение;
используется корень. Штокроза (A. rosea) — широко распростра-
ненное декоративное двулетнее растение. Цветки различных расцве-
ток.
Мальва (Malva) — близкий к алтею род травянистых, преиму-
щественно однолетних растений. Виды этого рода под названием
калачиков — довольно распространенные сорняки и вместе с тем
удовлетворительно поедаемые питательные растения. Мальва лес-
11—326
321
ная (М. silvestris) —лекарственное растение; используются цветки,
реже листья. Мальва курчавая (М. crispa) и др. виды вводятся
в культуру как кормовые.
Хлопчатник (Gossypium) — важнейшая культура в этом семей-
стве (рис. 173). Стебель деревянистый, хотя растение и однолетнее
(в наших условиях). Листья пальчатонервные, цветки крупные,
Рис. 173. Хлопчатник (Gossypium hirsutum)
с подчашием. Плод 3—5-гнездная коробочка. Семена покрыты одно-
клеточными выростами — волокнами, ради которых хлопчатник
и разводят. Тепло- и светолюбивая культура, выращиваемая у нас
в среднеазиатских республиках и в Азербайджане. Возделываемые
в СССР сорта хлопчатника относятся к видам упланд (G. hirsu-
tum) и египетский хлопок (G. barbadense). Оба вида американского
происхождения. Из семян хлопчатника добывают масло; жмых идет
на корм скоту; содержит ядовитое вещество госсипол, оказывающее
вредное влияние при длительном скармливании и в больших дозах.
Особенно чувствительны к госсиполу свиньи.
322
Порядок молочаецветные (Euphorbiales)
Семейство молочайные (Euphorbiaceae). Полиморфное семейст-
во, особенно по строению и размеру вегетативных органов; в этом
семействе имеются деревья, кустарники и травянистые растения.
Листья и стебли у многих растений сочные. Суккулентные виды
с такими признаками свойственны жарким пустыням. При большом
разнообразии относимых сюда видов имеются и общие признаки:
наличие млечного -сока и своеобразное строение завязи, состоящей
из трех гнезд, каждое из которых дает орешек. Кроме наиболее ти-
пичного рода молочая, в этом семействе имеется ряд ценных расте-
ний: клещевина, хевея бразильская и др., относящиеся к самостоя-
тельным родам.
Молочай (Euphorbia) — древесные и кустарниковые виды этого
семейства, распространены в тропической и субтропической зонах
(рис. 174). В нашей стране— многолетние и однолетние травы, ред-
Рис. 174. Соцветие молочая (Euphorbia lathyris).
А — с покрывалом, Б — без покрывала:
1 — тычиночный цветок соцветия; 2 — пестичный цветок
соцветия
ко полукустарники. Листья очередные, редко супротивные. Цветки
молочая с редуцированным околоцветником собраны в своеобраз-
ные соцветия — циации, которые легко принять за цветки. Циаций
состоит из одного женского беспокровного цветка и большого чис-
ла (10—20) однотычиночных мужских цветков; при основании каж-
дой тычиночной нити небольшой чашевидный прицветничек. Жен-
ский цветок состоит из пестика с трехгнездной завязью, сидящей на
длинной ножке; столбиков 3, каждый из них имеет двухлопастное
рыльце. Женские и мужские цветки окружены общей колокольча-
той обверткой, по краям которой 5 тонких лопастей; между лопас-
тями своеобразные утолщения — пластинки-нектарники. Молочай,
таким образом, растение однодомное.
11* 323
Молочаи часто являются вредной примесью в растительности
естественных пастбищ и сенокосов, многие из них ядовиты. Немно-
гие виды (Е. canescens, Е. cheirolepis) в условиях песчаной пустыни
хорошо поедаемы и безвредны. Молочай лозный (Е. virgata) —
частый сорняк полей.
Клещевина, или рицина (Ricinus communis), — многолетнее рас-
тение из тропической Африки. Вне тропической области выращива-
ется как однолетняя культура. Очередные пальчатолопастные ли-
стья. Растения однодомные. Околоцветник однопокровный, чашеч-
колистный. Мужские многотычинковые цветки в нижней части кис-
тевидного соцветия, женские однопестичные — в верхней. Плод —
трехсемянная коробочка, покрытая шипиками. Семена ядовиты.
Из семян клещевины добывают цепное невысыхающее масло, ис-
пользуемое в технических и лекарственных целях.
Хевея бразильская (Hevea brasiliensis). Каучуконосное дерево.
Родина — бассейн р. Амазонки. Листья — тройчатые. Растение од-
нодомное. Плод — трехсемянная коробочка. Хевея дает свыше 90%
мировой добычи натурального каучука.
Пролесник (Mercuriales annua) — двудомное растение с невзрач-
ными цветками. Ядовит.
Порядок тимелеецветные (Thymelaeales)
Семейство волчеягодниковые (Thymelaeaceae). В этом семейст-
ве интересен род волчьи ягоды (Daphne), Это маловетвистый кус-
тарник высотой до 1,5 м, листья очередные, собранные на концах
ветвей, цельные, обратноланцетные. Цветки двуполые с простым
четырехраздельным околоцветником, распускаются до появления
листьев; плоды — ярко-красные костянки, образуемые прямо на
деревянистых побегах. Наиболее широко распространены волчьи
ягоды обыкновенные (D. mesereum). Все части растения, особенно
кора, сильно ядовиты для животных и человека. Действующие на-
чала — смолоподобное вещество мезерин и гликозид дафнии.
Подкласс 6. Розидные (Rosidae)
Надпорядок VIII. Розовидные (Rosanae)
Большая и полиморфная группа в составе 39 семейств, распре-
деленных между 6 порядками. Порядки Rosales и Saxifragales свя-
заны с Dilleniales, а через него с Magnoliales. В этих порядках со-
средоточены наиболее примитивные формы в пределах рассматри-
ваемого надпорядка.
От Saxifragales отходит целый -веер линий развития, некоторые
из них составляют порядки, входящие в данный надпорядок. Моно-
типный порядок Nepenthales включает лишь один род узко спе-
циализированных в сторону насекомоядности растений. Небольшой
порядок Podostemales представляет гидрофильную линию эволю-
ции с сильно редуцированными и метаморфизированными вегета-
324
тивными органами. Самостоятельную ветвь образуют порядки Fa-
bales и Connarales. В этом большом надпорядке мы рассмотрим
лишь семейства, имеющие практическое значение.
Порядок розоцветные (Rosales)
Семейство розанные (Rosaceae). Деревья, кустарники и травя-
нистые растения, распространенные преимущественно в умеренной
зоне северного полушария. Листья цельные и непарноперистослож-
ные, обычно с прилистниками; за редким исключением расположены
поочередно. Цветки актиноморфные, с раздельнолепестным венчи-
ком. За немногим исключением чашелистиков и лепестков 5, число
тычинок кратно числу лепестков
ный и ценокарпный. Завязь
верхняя, средняя и нижняя.
Для цветков характерно нали-
чие подчашия, образованного
уменьшенными листьями вер-
хушки цветообразующего по-
бега.
Семейство полиморфное и
разделяется на 4 подсемейства,
отражающие различные ступе-
ни и направления эволюции
внутри семейства (рис. 175).
К семейству розанных отно-
сится большинство плодовых и
ягодных культур СССР. В кор-
мовом отношении это семейст-
во не имеет большого значения.
1. Подсемейство спирейные
(Spiraeoideae). Представлено
кустарниками. Плодолистики
образуют пять и больше листо-
вок, срастающихся иногда в
коробочку. Цветки подпестич-
ные. Наиболее широко извест-
на спирея (Spiraea), неболь-
шие обильно цветущие кустар-
ники.
2. Подсемейство розовые
(Rosoideae). Цветки околопес-
тичные. Гинецей апокарпный;
многочисленные плодолистики
образуют односеменные плоди-
ки—семянки или (костянки,
сидящие на выпуклом или во-
гнутом цветоложе. Кустарники
и травы.
и достигает 50. Гинецей апокарп-
Рис. 175. Цветки представителей различ-
ных подсемейств семейства розанных
(Rosaceae). А — таволга (Filipendula)—
подсемейство спирейных, Б — малина —
(Rubus idaeus), В — шиповник (Rosa
majalis)—подсемейство розовых, Г —
миндаль степной (Prutnus tenella) — под-
семейство сливовых, Д — яблоня (Matus
silvestris) — подсемейство яблоневых
325
К роду Rubus 'относится несколько видов: малина, ежевика, ко-
стяника, морошка, поленика. Это небольшие кустарники или травя-
нистые растения, у которых плод сочный, состоит из нескольких
или многих мелких костянок.
К роду Fragaria относятся земляника (F. vesca), клубника
(F. collina). Травянистые растения с ползучими укореняющимися
побегами — усами. Многочисленные плодики-семянки расположены
на сильно разросшемся сочном цветоложе. Мякоть цветоложа слад-
кая, ароматичная, окраска от кремовой до темно-красной.
Лапчатка (Potentilla) — многолетние, реже однолетние травы
с желтыми цветками. Многие виды этого рода широко распростране-
ны в различных природных условиях северного полушария. Кормо-
вые достоинства растений невысоки.
Шиповник (Rosa) — различного размера кустарники, широко
распространенные в СССР, особенно в горных районах. Побеги
покрыты шипами. Цветки различных расцветок—белые, желтые,
розовые, оранжевые. Лепестков 5, тычинок и пестиков много.
У махровых форм часть тычинок превращена в лепестки. Многочис-
ленные волосистые семянки сидят внутри разросшегося цветоложа
(гипантия), которое к моменту созревания плодов становится соч-
ным и приобретает оранжевую или красную окраску. «Плоды» ши-
повника богаты витамином С. Виды многочисленны, наиболее ши-
роко распространены: R. canina, R. lutea, /?. cinnamomea .и др.
Культурные, декоративные и эфиромасличные сорта роз выведены
из диких шиповников. Размножают розы вегетативно, прививая на
дикорастущий шиповник.
3. Подсемейство яблоневые — Pomoideae. Цветки надпестичные,
завязь нижняя, сросшаяся с цветоложем, поэтому плод ложный.
Лепестков 5, тычинок до 25 (у яблони иногда до 50), плодолистики
в числе 2—5 образуют цеиокарпный гинецей. Деревья и кустарники.
Яблоня (Malus) в числе нескольких видов распространена в го-
рах и в лесной зоне. Дерево, реже кустарник. Листья опушены, ле-
пестки розовые. Завязь пятигнездная, в каждом гнезде по 4—6 се-
мян; столбики в основании срослись. Цветки собраны в зонтики.
Большинство культурных сортов, преимущественно гибридного про-
исхождения, объединяются в вид яблоня домашняя. Свыше 40 сор-
тов яблонь с прекрасными потребительскими качествами создано
И. В. Мичуриным.
Груша (Pyrus)—цветки в отличие от цветков яблони собраны
в щитки, лепестки всегда белые, столбики свободны, плоды своеоб-
разной грушевидной формы или шаровидные, мякоть плода с каме-
нистыми клетками. Деревья. В дикорастущем состоянии широко
распространена в европейской и азиатской частях СССР груша
обыкновенная (Р. communis), в Крыму — груша лоховидная (Р. elae-
agnifolia). Культурные сорта выведены из груши обыкновенной или
имеют гибридное происхождение. Большая часть сортов приспособ-
лена к теплому климату. Пользуясь методом скрещивания и направ-
ленного воспитания, И. В. Мичурин вывел ряд зимостойких сортов,
что дало возможность продвинуть грушу дальше па север.
326
к подсемейству ябленевых относится также еще несколько рас-
тений менее ценных в хозяйственном отношении. Рябина обыкновен-
ная (Sorbus aucuparia) и боярышник (Crataegus) отличаются от
яблони, груши и айвы твердым, костянистым внутриплодником.
Пользуясь методом гибридизации и отбора, И. В. Мичурин вывел не*
сколько сортов; сорт Гранатная получен им в результате межродо*
вого скрещивания рябины и боярышника.
4. Подсемейство сливовые (Prunoideae). Деревья и кустарники.
Цветоложе, как и у яблоневых, вогнутое, но завязь с цветоложем не
срастается, цветок околопестичный. Тычинок много, гинецей реду-
цирован до одного плодолистика с 1—2 семяпочками, плод кос-»
тянка.
Миндаль (Amygdalus) в отличие от остальных представителей
этого подсемейства имеет плод с кожистым мезокарпием; съедобно
лишь семя. Небольшое дерево или кустарник с розовыми или бе-
лыми цветками, распускающимися ранней весной, до появления лис*
тьев. Семена миндаля используются в кондитерской промышлен-
ности, в парфюмерии и медицине. Вместе с тем дикий миндаль — бо-
бовник (А. папа) иногда накапливает в семенах ядовитые вещества,
которые могут вызвать отравление животных.
Слива (Prunus) — небольшие деревья и кустарники. Широко
распространены дикорастущие виды этого рода: терн (Р. spinosa),
алыча (Р. divaricata), слива уссурийская (Р. ussuriensis) и др. Боль-
шинство культурных сортов относится к виду слива домашняя
(Р. domestica). Это, по-видимому, гибридный вид. Чернослив (тер*
нослив) относится к виду Р. insititia. И. В. Мичуриным выведены
ценные сорта слив и тернослива, получившие широкое распростра-
нение; им также выведены десертные сорта терна.
Абрикос (Armeniaca). В диком состоянии встречается несколько
видов; культурные формы происходят от абрикоса обыкновенного
(A. vulgaris), произрастающего в диком виде в горах Тянь-Шаня.
Персик (Persica) — более теплолюбивая культура, чем абрикос.
Большинство культурных сортов относится к виду персика обыкно-
венного (Р. vulgaris). Основные районы выращивания персиков —
Средняя Азия, Кавказ, Крым.
Вишня (Cerasus) — род, наиболее богатый дикорастущими фор-
мами, деревьями и кустарниками. Культурные сорта относятся
к вишне обыкновенной (С. vulgaris), черешне (С. avium) и к гибри-
дам между этими двумя видами. В Советском Союзе разводится
много сортов вишен и черешен, в том числе около 20 сортов, выве-
денных И. В. Мичуриным. Черешня — южная культура, но И. В. Ми-
чуриным выведены сорта, распространяющиеся далеко на север, до
Москвы и Ленинграда. Плоды вишни более ароматные, чем череш-
ни, и содержат больше кислоты.
К подсемейству сливовых относится также черемуха (Padus га-
cetnosa) с малосъедобными плодами, широко распространенная
в лесах СССР, и лавровишня лекарственная (Laurocerasus officina-
lis) — вечнозеленый кустарник, произрастающий на Кавказе: ле-
карственное растение.
327
Порядок камнеломкоцветные (Saxifragales)
Семейство крыжовниковые (Grossulariaceae). Смородина черная
(Ribes nigrum). Цветки серовато-розовые. Плоды черные со специ-
фическим запахом, листья тоже пахучие. Ягоды богаты витамином
С и широко используются в пищевой промышленности.
Смородина красная (R. vulgare). Плоды красные или беловатые
без запаха. Цветки зеленоватые или желтоватые.
Смородина золотистая (R. аигеит). Цветки желто-золотистые.
Плоды буровато-красные до черных, съедобные, но малоценные.
Растение из Северной Америки, культивируют как декоративный
кустарник.
Крыжовник (Grossularia reclinata). Обычен в культуре, но часто
дичает. Стебли покрыты шипами, цветки одиночные или по два-три
в пучках. Ягоды покрыты железистыми волосками, зеленые или
слегка пурпуровые. Имеются многочисленные сорта.
Порядок бобоцветные (Fabales, Leguminosales)
Обширный порядок, состоящий из трех семейств: мимозовые, це-
зальпиниевые, бобовые. Наиболее богато видами семейство бобовые.
Первые два семейства в нашей стране имеют небольшое значение.
Из мимозовых в Закавказье и по Южному берегу Крыма разводит-
ся шелковая акация, или альбицция (Albizzia julibrissin), — неболь-
шое, очень декоративное дерево. На Черноморском побережье Кав-
каза раннецветущая желтая акация «мимоза» (Acacia dealbata). Из
цезальпиниевых на юге широко распространена гледичия (Gledi-
tschia triacanthos) — крупное дерево с красивой ажурной кроной;
ветви гледичии обычно покрыты огромными колючками. Растение
двудомное и однодомное; происходит из Северной Америки. В тро-
пической Африке кассия (Cassia). Некоторые виды ее (С. angu-
stifolia и др.) — ценные лекарственные растения.
Семейство бобовые (Fabaceae). Растения этого семейства рас-
пространены по всему земному шару. Травянистые, кустарники, ре-
же деревья. Очень много лиан, травянистых и деревянистых.
Листья очередные, с прилистниками, перисто- или пальчатослож-
ные, реже простые. Цветки двуполые, обычно в соцветиях, реже оди-
ночные. Чашечка зубчатая или двугубая, венчик пятилепестный,
своеобразной «мотыльковой» формы; верхний лепесток—парус,
боковые — крылья, или весла, два нижних срослись в лодочку. Ты-
чинок 10.
Лишь у очень немногих видов все тычинки свободны, это наибо-
лее примитивный признак. Чаще наблюдается срастание в трубочку
всех десяти тычинок, такие тычинки называются од нобр ат ст венны-
ми. У огромного большинства -В1ИДОВ 9 тычинок срослись нитями, де-
сятая остается свободной — двубратственные (рис. 176). Трубочка
из сросшихся тычинок охватывает пестик. Пестик образуется из од-
ного плодолистика; плод — боб, обычно многосемянный, но у неко-
торых видов односемянный, невскрывающийся. Семя без эндоспер-
328
Рис. 176. Горох посевной (Pisum sativum). А — общий вид растения, Б —
цветок, В*—продольный разрез цветка, Г—андроцей и гинецей цветка,
Д — пестик, Е — части цветка (вверху парус, по бокам весла, внизу лодоч-
ка), Ж — зародыш семени
ма, из одного зародыша и кожуры: зародыш имеет две толстые се-
мядоли.
Интересная особенность бобовых —симбиоз их корней с бакте-
риями, усваивающими атмосферный азот. Поселяясь в тканях корня,
бактерии вызывают их разрастание, образуя клубеньки; отсюда
и название бактерий — клубеньковые.
Бобовые имеют большое народнохозяйственное значение; среди
них много ценных продовольственных и кормовых растений, отли-
329
Рис. 177. Люпин узколистный синий (Lupinus
angustfolius)
чающихся высоким содер-
жанием белка. Семена не-
которых бобовых наряду
с высоким процентом бел-
ка содержат много жира.
В кормовом отношении
это семейство занимает
второе место после зла-
ков.
Большую роль играют
бобовые растения в обра-
зовании травостоя сухо-
дольных и заливных лугов
лесной зоны, немного
меньшую — в лесостепной.
В условиях степи бобовые
бывают обильны лишь в
травостое поемных лугов.
Горные луга, так же как
и луга лесной зоны, бога-
ты бобовыми.
При создании культур-
ных пастбищ и сенокосов
бобовым, наравне со зла-
ками, принадлежит основ-
ная роль. Весьма сущест-
венна роль бобовых так-
же и в полевом кормопро-
изводстве. Вообще без бо-
бовых очень трудно обес-
печить животных полно-
ценными кормами. Трава
молодых растений из се-
мейства бобовых бедна
клетчаткой и хорошо пере-
варивается.
Люпин (Lupinus) —
травянистое растение с
пальчатосложными листь-
ями (рис. 177), культурные сорта выращиваются на зеленое удоб-
рение и корм. Семена обычно содержат горькие алкалоиды, которые
могут вызвать отравление животных. Культивируются многолетние
и однолетние виды. Расцветка венчика различная. Из-за красивого
соцветия нередко разводят как декоративное растение. Происхож-
дение— Средиземноморье и Северная Америка. Из многочисленных
видов чаще высевают люпин узколистный (L. angustifolius), жел-
тый (L. luteus), белый (L. albus), многолетний (L. perennis), мно-
голистный (L. polyphyllus) и др. Кормовое значение имеют только
безалкалоидные и малоалкалоидные сорта.
ззо
Рис. 178. Люцерна (Medicago sativa), внизу,— пло-
дики и цветок в разрезе
Люцерна (Medicago) — многолетние и однолетние травы с трой-
чатыми листьями (рис. 178); дают траву и сено отличного качества.
Мелкие цветки собраны в короткие головчатые 'кисти. Лепестки
желтые, голубые, фиолетовые, синие. Тычинки двубратственные.
В культуре многолетние: люцерна посевная (М. sativa), серповид-
ная (М. falcata) и гибриды между ними.
Люцерна посевная — рослое хорошо облиственное растение, до-
статочно требовательное к почвам, засухоустойчивое. Цветки мелкие,
синие или фиолетово-синие, собранные в соцветия — кисть. Плод —
спирально закрученный боб. Хорошо отрастает и на поливе дает до
4—5 укосов. Возделывается преимущественно в южных районах
страны, на орошаемых землях. В остальных районах возделывается
обычно синегибридная люцерна — гибриды между люцерной посев-
ной и серповидной; гибридные сорта более засухо- и морозоустой-
чивы.
Люцерна серповидная имеет более мелкие, узкие листья и ме-
нее облиственна по сравнению с люцерной посевной. Плоды серпо-
видные. Засухоустойчива, солевынослива и зимостойка. Эти качест-
ва как раз и ценятся при скрещивании люцерны серповидной и по-
севной.
Для посева в пастбищных травосмесях перспективна широко
распространенная в диком виде люцерна хмелевидная (М. lupulina)
331
одно-двулетнее, нежное, стелющееся растение с мелкими желтыми
цветками, собранными в головчатое соцветие.
Рекомендуется высев люцерны в смеси со злаками: житняком,
райграсами, костром безостым и др.
Донник (Melilotus) — двулетние или однолетние крупные травя-
нистые растения (рис. 179). Листья тройчатые. Мелкие цветки соб-
раны в соцветия — кисти. Боб продолговатый, с 1—3 семенами. Наи-
большее кормовое значение имеет донник белый (М. albus) и ле-
карственный (М. officinalis).
Растения с сильным запахом
кумарина; используются
обычно как пастбищные и
идут на силос.
Клевер (Trifolium) —
многолетние, однолетние и
двулетние травянистые рас-
тения с тройчатыми листья-
ми (рис. 180). Мелкие цвет-
ки собраны в головки. После
цветения венчики не отвали-
ваются, остаются в головке.
Большинство клеверов —
прекрасные кормовые расте-
ния. Наиболее широко рас-
пространены и возделывают-
ся клевер луговой, розовый,
ползучий и др.
Клевер луговой (Т. рга-
tense) —мощное растение,
многочисленные боковые по-
беги которого образуют боль-
шой куст. Цветки лилово-
красные, редко белые; боб
Рис. 179. Донник лекарственный (Melilotus мелкий, односемянный. Ши-
officiualis) роко распространен в диком
состоянии. В культуре возде-
лывается в двух формах: позднеспелый (одноукосный) и ран-
неспелый (двуукосный). Возделывается в лесной и лесостепной
зоне.
Клевер розовый (Т. hybridum) .внешне похож на луговой, но со-
цветия обычно мельче, цветки розовые, или розово-белые. Распрост-
ранен в диком виде и возделывается преимущественно в лесной
зоне. В отличие от лугового клевера хорошо переносит .кислые
почвы.
Клевер ползучий, или белый (Т. repens), имеет стелющийся сте-
бель, побеги легко укореняются. Отличное пастбищное растение,
прекрасно отрастает, выдерживает вытаптывание. Долговечен. Рас-
тет повсюду, но возделывается без полива лишь в лесной и лесостеп-
ной зонах.
332
В северных районах нашей страны клевера высеваются в смеси
с тимофеевкой и другими злаками. В засушливых районах клевер
уступает по урожайности люцерне.
Лядвенец (Lotus) —- многолетнее травянистое растение (рис.
181). Листья тройчатые, с двумя прилистниками. Цветки желтые,
крупнее, чем у люцерны. Собраны в зонтиковидные головки. Боб
удлиненный, цилиндрический, слегка согнутый. Высокой кормовой
ценностью отличается ши-
роко распространенный
лядвенец рогатый (L. cor-
niculatus). Хорошо выдер-
живает вытаптывание, од-
нако в цветущем состоя-
нии не поедается из-за
ядовитых горьких веще-
ств, накапливающихся в
соцветии. Введен в куль-
туру.
Рис. 180. Клевер луговой (Trifolium pratense)
181. Лядвенец рогатый
Рис.
(Lotus corniculatus), внизу —
плодик боб
Астрагал (Astragalus) — многолетние, редко однолетние травы,
полукустарники и кустарники с непарноперисто-сложными листья-
ми. Род, наиболее богатый видами; в СССР известно свыше 800 ви-
дов. У деревянистых форм центральные черешки часто превращены
в колючки — трагакантовые астрагалы; стебли таких астрагалов со-
держат камедь, широко используемую в промышленности (текстиль-
ной, бумажной, спичечной и др.). Многие, особенно горные, виды —
прекрасные и хорошие кормовые травы пастбищного использования.
333
Рис. 182. Эспарцет виколистный (Onobrychis
viciifolia), справа — плодик
Цветки разных расцветок собраны в кисти, иногда в форме головки.
Некоторые виды, ценные в кормовом отношении, испытываются
в культуре, например астрагал прутяной (A. virgatus) на юге Ук-
раины. Есть виды, вызывающие при поедании заболевания. Особен-
но широко распространены астрагалы в Средней Азии; в субальпий-
ских и альпийских пастбищах ценен в кормовом отношении астра-
гал альпийский (A, alpinus). Ценны также астрагал бороздчатый
(A. sulcatus), сладколистный (A. glycyphyllus), эспарцетовидный
(A. onobrychis) и др.
Эспарцет (Onobrychis) — многолетнее травянистое растение
с непарноперистыми листьями (рис. 182). Цветки розовые и пурпур-
ные. Собраны в большую кисть. Плод — односемянный боб, окру-
гло-сплюснутый, морщинистый, часто покрытый шипами. Большин-
ство видов обладает хорошими кормовыми качествами. Возделы-
вают эспарцет виколистный, или посевной (О. Viciifolia), песчаный
(О. arenaria) и некоторые др. По сравнению с люцерной эспарцет
менее требователен к почве. Высевается в смеси с костром безос-
334
Рис. 183. Арахис, земляной орех (Arachis hypogaea)
тым и другими многолетними травами, преимущественно в лесостеп-
ной и степной зонах. Хороший медонос.
Арахис, или земляной орех (Arachis hypogaea), — однолетнее
культурное растение, родина — Бразилия (рис. 183). Листья пар-
ноперистые. Чашечка двугубая. Цветки мелкие, желтые; верхние
бесплодные, нижние — клейстогамные, плодущие; после оплодотво-
рения они на сильно удлиняющейся цветоножке (вернее, трубочке
чашечки) — гинофоре — внедряются в почву, где и происходит раз-
витие плода. Плод двух-, четырехсемянный, вздутый, невскрываю-
щийся. Семена богаты маслом, используются в прокаленном виде
335
как лакомство, а также для добывания ценного масла и приготовле-
ния лечебных эмульсий. Жмыхи используются в кондитерском про-
изводстве. Арахис дает много зеленой массы и высевается также для
получения сена и зеленой подкормки.
Нут, или турецкий орех (Cicer orietinum),— однолетнее куль-
турное растение, завезенное из Южной Азии. Листья непарнопе-
ристые, стебель прямостоячий, цветки одиночные, .сидящие, венчик
белый, розоватый, реже голубоватый; плод вздутый, обычно двусе-
мянный. Все растение железисто-пушистое. Семена округло углова-
тые, белые и темноокрашенные. Засухоустойчивая зернобобовая
культура. Семена идут в пищу человеку и в корм скоту, обычно
в размолотом виде. *
Вика, или горошек (Vicia),— широко распространенные много-
летние, реже однолетние травянистые растения с перистосложными
листьями, которые заканчиваются усиком или острием. Бобы много-
семянные, реже двухсемянные, обычно мелкие. Цветки различной
величины и расцветки. Среди многосемянных видов есть ценные
кормовые травы, введенные в культуру для получения зеленого кор-
ма и сена: вика посевная (V. sativa), озимая (V. villosa) и др. Один
из видов (V. faba) под названием русских или кормовых бобов куль-
тивируется как зернобобовая продовольственная и кормовая куль-
тура.
Чечевица (Lens esculenta) — однолетняя продовольственная
и кормовая культура. Перистосложный лист заканчивается усиком
или острием. Стебель слабый, лежачий. Цветки мелкие, бобы тоже
мелкие, сплюснутые, с 1—3 семенами. Имеется ряд разновидностей,
отличающихся формой и размером плодов. Растение, по-видимому,
азиатского происхождения.
Чина (Lathyrus) — многолетние и однолетние растения, цепные
в кормовом отношении. Перистосложные листья имеют крупные
прилистники и заканчиваются усиком или острием. Стебель слабый,
цепляющийся. Морфологически чины сходны с виками. Для получе-
ния сена высеваются чина луговая (L. pratensis), чина посевная
(L. sativus) в смеси с ячменем и овсом на зеленый корм, а также
как зерновая культура. Семена клиновидные и округлоугловатые;
используются в размолотом виде преимущественно на корм скоту.
Горох (Pisum) — однолетние и многолетние культурные и дико-
растущие растения. Перистосложные листья заканчиваются усиком?
Цветки обычно крупные (см. рис. 176). Боб многосемянный, плос-
кий. Как продовольственная культура высевается обычно горох по-
севной (Р. sativum); имеются многочисленные сорта. В корм скоту
выращивают горох полевой, или пелюшку (Р. arvense), с угловато-
круглым.и тем1ноо(крашенными семенами. Для получения травы го-
рох, как и вику, выращивают в смеси с овсом, пшеницей и рожью.
Фасоль (Phaseolus)—однолетнее культурное растение. Листья
тройчатые, с прилистниками. Бобы крупные. Растения вьющиеся.
Под различными названиями возделывается несколько видов. Наи-
более широко распространена фасоль обыкновенная (Ph. vulgaris)
в большом количестве сортов.
336
Лобия, или вигна, коровий горох, лагута (Vigna sinensis) сходна
с фасолью, но стебель не вьющийся, прямостоячий или стелющийся.
Бобы длинные, саблевидные. Теплолюбивое южное растение. Воз-
делывается как продовольственная культура. В зеленом виде ска-
шивается также на корм скоту. Рекомендуется для смешаных по-
севов с кукурузой.
Соя (Glycine hispida) — продовольственная, кормовая и техни-
ческая культура. Однолетние растения; листья тройчатые. Все рас-
тение щетинисто-волосистое. Стебель прямостоячий. Семена бога-
ты белком и маслами. Возделывается главным образом как зернобо-
бовая культура. Используется также для получения зеленой массы
и как зеленое удобрение. Родина — Восточная Азия. Рекомендуется
высевать сою в смеси с кукурузой и суданской травой на зеленый
корм.
Среди бобовых есть и ядовитые растения: термопсис (Т her mop-
sis), распространенный в Средней Азии, ракитник (Cytisus), софора
(Sophora), пажитник (Trigonella arcuta), козлятник (Galega)
и др.
Надпорядок IX Миртовидные (Myrtanae)
Небольшая группа, состоящая всего из двух порядков: Myrtales
и Hippuridales. Порядок Myrtales близок к порядку Saxifragales.
Для нас представляют интерес лишь немногие растения из порядка
Myrtales: кипрей (Epilobium) и энотера из семейства Onagraceae,
дербенник (Lythrum) из семейства дербенниковых (Lythraceae), во-
дяной орех (Тгара) из семейства Тгарасеае. Из семейства миртовых
(Myrtaceae) интересны эвкалипты (Eucalyptus), крупнейшие де-
ревья из Австралии, культивируемые у нас на Кавказе, и декоратив-
ное дерево — мирт (Myrtis communis). Всего в этом падпорядке
16 семейств.
Надпорядок X. Рутовидные (Rutanae)
Второе ответвление от центрального порядка Saxifragales начи-
нается порядком Rutales, где преобладают древесные формы, затем
порядок Sapindales, также в основном из деревьев и кустарников,
и через порядок Geraniales эта ветвь заканчивается далеко про-
двинувшимся порядком Polygalales. В двух последних порядках пре-
валируют травянистые формы. Всего, следовательно, в этом надпо-
рядке 4 порядка, семейств в этой группе очень много — 47, но боль-
шинство из них не представлено в нашей стране. Мы остановимся
лишь на тех, которые представляют интерес для нас.
Порядок рутоцветные (Rutales)
Семейство рутовые (Rutaceae). К важнейшему роду этого семей-
ства (Citrus) относятся вечнозеленые деревья и кустарники. Листья
овальные, сверху глянцевые. В мезофилле листьев находятся желез-
337
ки с эфирными маслами. Расположение листьев очередное. Цветки
белые или розовые, пахучие, двуполые, реже однополые. Чашечка
с 3—5 сросшимися долями; лепестки в числе 4—8, также иногда
сросшиеся в основании. Тычинок много, в нескольких пучках. Гине-
цей синкарпный, плод многогнездиый.
К этому роду относится несколько видов, родина которых Южная
и Юго-Восточная Азия. Район промышленного разведения цитрусо-
вых у нас — Грузия.
Наибольшее распространение имеют 3 вида: лимон (С. Ито-
nium), мандарин (С. reticulata), апельсин (С. sinensis). Многие
сорта мандарина имеют бессемейные плоды (партенокарпия).
Семейство сумаховые (Anacardiaceae). Ценнейшее тропическое
растение кэшью (Anacardium occidental). Дает разнообразную про-
дукцию: орехи, богатые маслом, инсектицид, ценную древесину и др.;
скорлупа ореха ядовита.
Порядок сапиндоцветные (Sapindales)
Семейство кленовые (Асегасеае). Более или менее крупные де-
ревья с супротивными пальчато-лопастными реже перистослож-
ными листьями. Цветки двуполые или растения однодомные и дву-
домные (A. negundo)\ число частей цветка изменчиво за исключени-
ем пестика, который всегда -с двугнездной верхней завяью <и с двумя
столбиками. Плод во всех случаях — характерная двукрылатка.
Соцветие кистевидное.
В СССР семейство кленовых представлено родом клен (Acer) со
значительным количеством видов, распространенных в лесах евро-
пейской части СССР, Дальнего Востока, Кавказа, Средней Азии.
Л^ногие виды широко используются для озеленения населенных
пунктов и при создании парков. Среди них наиболее известны клен
остролистный (A. platanoides), клен татарский (A. tataricum),
клен явор (A. pseudoplatanus) > клен полевой (A. campestre), клен
ясенелистный) или американский (A. negundo), и др. Древесина
большинства видов клена отличается высокими качествами.
Порядок гераниецветные (Geraniales)
Семейство леновые (Linaceae). По ряду признаков это семейство
близко к рутовым; цветки актиноморфные, тычинки срослись осно-
ваниями нитей, завязь верхняя. Шагом вперед по сравнению с руто-
выми является сокращение числа тычинок до 10, а плодолистиков
до 5. Гинецей ценокарпный, но столбики свободные.
Лен (Linum) является важнейшим и типичным представителем
этого семейства в нашей флоре. Этот род представлен однолетними
и многолетними травянистыми растениями. Листья преимуществен-
но очередные; цветок актиноморфный, пятерного типа: 5 чашелисти-
ков, 5 свободных лепестков, 10 членов андроцея, их низ 5 стамино-
дий; 5 плодолистиков, сросшихся в ценокарпный гинецей; плод —
коробочка. Цветок самоопыляется.
338
Рис. 184. Лен-долгунец (Linum usita-
tissimum)
Среди большого числа видов
льна наибольшее значение имеют
лен обыкновенный, долгунец
(L. usitatissimum) (рис. 184), вы-
ращиваемый для получения во-
локна, и лен-кудряш (L. humile),
выращиваемый для получения из
семян масла. Лен-долгунец имеет
длинный стебель, ветвящийся
только на вершине. Лен-кудряш,
наоборот, низкорослый и сильно
ветвится уже от основания стебля.
Лен — одна из древнейших
культур, распространившаяся по
всему миру, по-видимому, из Ин-
дии. Египетские мумии, насчиты-
вающие 4—5 тыс. лет, обмотаны
тканями, сделанными из льна.
Наибольшее количество льна-дол-
гунца выращивается в нашей
стране, наш лен издавна славится
высоким качеством волокна. Льня-
ной жмых — ценнейший белковый
корм в рационе сельскохозяйст-
венных животных.
Семейство гераниевые (Gera-
niaceae). Типичный представитель
этого семейства — род герань (Ge-
ranium) с многочисленными вида-
ми. Растения травянистые, одно-
летние и многолетние. Листья
супротивные, пальчатонервные.
Цветок сходен с цветком льна,
пятерного типа: 5 чашелистиков,
5 лепестков, 10 тычинок, 5 плодо-
листиков, сросшихся в синкарп-
ный гинецей. На высокогорных
пастбищах Киргизии некоторые
виды герани хорошо поедаются
скотом и являются нажировоч-
ным кормом.
В семействе парнолистниковых
ственное растение гармала (Peganum harmala); при поедании жи-
(Zygophyllaceae) ценное лекар-
вотными может вызвать тяжелое отравление.
Надпорядок XI. Аралиевидные (Aralianae)
Надпорядок состоит всего из одного порядка Araliales (Сог->
nales), 14 семейств и представляет своеобразную линию в эволюции
двудольных, которая заканчивается далеко продвинувшимся боль-
339
шим семействохм зонтичных — Apiaceae (Umbelliferae). Порядок
Cornales связан с исходным порядком Saxifragales. Наибольший
интерес для нас представляет семейство зонтичных, на котором
мы и остановимся.
Порядок аралиецветные (Araliales, Cornales)
Семейство зонтичные (Apiaceae, Umbelliferae). Травянистые
растения, очень широко распространенные в умеренной зоне север*
иого полушария. Название свое семейство получило по наиболее
характерному признаку — строению соцветия, имеющего форму
сложного, реже простого зонтика. Характерным признаком явля*
ется также наличие в тканях растений пахучих эфирных масел
и смол. Листья чаще сильно рассечены, реже простые, с влагали*
щами.
Цветок актиноморфный, реже слегка зигоморфный, в основа*
нии его нектароносный диск, под ним нижняя двугнездная завязь,
превращающаяся при созревании в плод — двусемянку. На завязи
два столбика, тычинок 5. Лепестков и чашелистиков также по 5.
Чашечки сильно редуцированы, часто совершенно незаметны.
При большом однообразии цветков плоды имеют различный размер
и форму.
В семействе несколько ценных овощных и пряных культур
и много сильно ядовитых растений, нередко засоряющих естествен*
иые сенокосы и пастбища в лесной зоне.
Морковь (Daucus carota) в диком виде встречается повсеместно
как сорняк. Двулетнее растение с мелко перисто рассеченными
листьями, покрытое жесткими волосками. Культурные формы мор*
кови, отличающиеся сильным развитием запасающей ткани в кор*
не, выделяются в самостоятельный вид (Daucus sativus). Ценней*
шая продовольственная и кормовая культура. Корень богат каро-
тином и витамином С.
Петрушка (Petroselinum sativum) —двулетнее растение с утол*
щенным корнем. Листья блестящие, дважды перистые. Культиви*
руется, корни используются в кулинарии как пряность.
Пастернак (Pastinaca sativa)—двулетнее сорное растение.
Культурные формы с утолщенным корнем, который используется
как овощ и пряность. Стебли и листья, как у петрушки, блестя-
щие; листья перисторассеченные.
Сельдерей (Apium graveolens) —двулетнее растение с веретено-
видным корнем. Блестящие стебель и перисторассеченные листья
голые. В южных приморских районах растет в диком виде. В куль*
туре разводится как пряное растение; используются все его части:
корень, стебель, листья и плоды.
Укроп (Anetum graveolens)—культурное растение, в южных
районах часто дичает. Листья мелко рассеченные на нитевидные
доли. Вся надземная часть растения используется как ароматиче*
ская приправа. Однолетник.
340
Анис (Pimpinella ani-
sum) — однолетнее расте-
ние. Тройчатоперисторас-
сеченные листья. Плоды
используются как пряно*
сти и с лекарственными
целями. Легко дичает.
Тмин {Сarum carvi) —
двулетнее растение с
триждыперисторассеч е.н-
ными листьями. Дикора-
стущее и культивируемое
как эфиромасличное и
пряное растение. Исполь-
зуются плоды.
Кориандр (Coriandrum
sativum') — однолетнее
культурное растение, лег-
ко дичает. Листья пери-
стораздельные. В отличие
от большинства растений
этого семейства, плод ша-
ровидный. Эфиромаслич-
ное и пряное растение.
Болиголов (Conium
maculatum). Очень ядови-
тое растение, обитающее
в затененных местах
(рис. 185). Крупный дву-
летник. Все растение го-
лое. Листья триждыперисторассеченные. Распространен повсе-
местно. Содержащиеся в болиголове алкалоиды (кониин,
конпцеин и др.) вызывают паралич центральной нервной
системы.
Вех ядовитый (Cicuta virosa)—также очень ядовитое широко
распространенное растение. Крупный многолетник (рис. 186).
Предпочитает влажные места. Листья дважды, трижды перисто-*
раздельные с остропильчатыми линейноланцетными долями. Тол-
стое корневище. Как и предыдущее растение, может быть причи-
ной опасного отравления животных. Смерть при отравлении вехом
ядовитым наступает от паралича органов дыхания.
К этому же семейству относится еще несколько ядовитых рас-
тений: бутенъ опьяняющий (Chaerophyllum temulum), омежник
водяной (Oenanthe aquatica) и другие виды этого рода, поручейник
(Slum), собачья петрушка (Aethusa cynapium) и др. Многие
растения используются как лекарственные: амми зубная (Ammi
visnaga), айован душистый (Trachyspermum ammi), дягиль
лекарственный (Archangelica officinalis), а также болиголов,
вех и др.
341
Рис. 186. Цикута, или вех ядовитый (Cicata virosa), справа
внизу — разрезанное вдоль корневище
Надпорядок XII. Бересклетовидные (Celastranae)
В этом надпорядке 4 порядка и 27 семейств, представляющих
одну из ветвей эволюционного древа, отходящую от Saxifragales.
Филогенетически, с точки зрения уровня развития, порядки, вхо-
дящие сюда, располагаются так: Celastrales, Rhamnales, Oleales,
Santalales. Большинство семейств, относимых к этому надпорядку,
не представлено в нашей флоре. Мы имеем возможность остано-
виться лишь на немногих.
342
Порядок бересклетоцветные (Celastrales)
Семейство бересклетовые (Celastraceae). Деревья и кустарники
обычно с супротивно расположенными простыми листьями. Цветки
актиноморфные, как правило, двуполые, насекомоопыляемые. За^
вязь верхняя, всех частей цветка по 4—5. Плод — 4—5-гнездная
коробочка.
Бересклет бородавчатый (Evonimus verrucosa) — кустарник
с бородавчатыми ветвями, кора содержит до 10% гуттаперчи, ра*
ди которой этот вид у нас разводится; в диком виде встречается
в лесах умеренной зоны европейской территории СССР. Более ши^
роко в нашей стране распространен бересклет европейский, с мень*
шим содержанием гуттаперчи (до 4%).
Порядок крушиноцветные (Rhamnales)
Семейство крушинные (Rhamnaceae). Кустарники с очередно
или супротивно расположенными листьями. Цветки двуполые или
растения двудомные. Плод — костянка. Сюда относятся два широко
распространенных кустарника.
Крушина ломкая (Rhamnus frangula) —растение с очередными
листьями, с обоеполыми цветками пятерного типа. Отвар коры
дает хорошее слабительное и рвотное лекарство.
Крушина слабительная, жостер (Rh. cathartica). Двудомное
растение. Частей цветка по 4, листья супротивные. Плоды — хоро-
шее слабительное средство.
Семейство виноградные (Vitaceae). Деревянистые лианы с оче-
редно расположенными пальчатолопастными или пальчатосложны-
ми листьями. Цветки невзрачные; обычно двуполые, собранные
в кистевидные или щитковидные соцветия, превращающиеся затем
в гроздь винограда. Лепестков 4—5; тычинки расположены против
лепестков в том же числе. Завязь верхняя, плод — ягода.
Виноград (Vitis) —деревянистые лианы с лопастными листья-
ми. Цветки двуполые, реже растения двудомные. Двуполые цвет-
ки часто бывают физиологически однополыми ввиду недоразвития
пыльцы. Имеются хорошо развитые нектарники — растения насе-
комоопыляемые. Лепестки срастаются верхними концами, и при от-
цветании венчик опадает целиком. Соцветие — кисть. Различают
дикорастущий виноград (V. Silvestris), встречающийся на юге Ук-
раины, включая Крым, в Средней Азии и на Кавказе, и культур-
ный виноград (V. vinifera), представленный огромным количеством
сортов.
Порядок маслиноцветные (Oleales)
Семейство маслинные (Oleaceae). Деревья и кустарники обычно
с супротивными листьями. Околоцветник двойной или простой
из 4 частей, иногда цветки без околоцветника, тычинок 2, завязь
верхняя, плоды различного типа из двух плодолистиков. Самое
343
ценное растение в этом семействе — оливковое дерево, маслина
(Olea europea), широко возделываемая в присредиземноморских
странах для получения оливкового масла и маслин. В нашей фло-
ре многочисленные виды ясеня (Fraxinus), сирень (Syringa vulga-
ris и др.), широко распространенный кустарник — бирючина
(Ligustrum vulgaris).
Надпорядок XIII. Протейновидные (Proteanae)
Всего два монотипных порядка; Elaeagnaceae, в котором широко
распространенный кустарник лох (Elaeagnus), и Proteales, не име-
ющий представителей в СССР.
Порядок лохоцветные (Elaeagnales)
Семейство лоховые (Elaeagnaceae) широко распространено на
территории нашей страны и представлено двумя родами: лохом
и облепихой.
Лох (Elaeagnus)—деревья и кустарники с колючими ветвями
и очередными серебристыми листьями. Цветки двуполые, актино-
морфные с простым околоцветником (4) и четырьмя тычинками.
Гинецей апокарпный, завязь верхняя, плод — костянковидный орех.
В естественном состоянии распространен на Кавказе и в Средней
Азии лох узколистный (Е. angustifolia). Из Северной Америки за-
везен лох серебристый (Е. argentea). Оба вида широко возделыва-
ются у нас как хорошие закрепители почвы, а также ради съедоб-
ных плодов.
Облепиха (Hyppophae rhatnnoides). Дерево с мощными колюч-
ками. Листья линейноланцетные, серебристые. Растение двудом-
ное, околоцветник простой из 2 или 4 частей, тычинок 4, гинецей
апокарпный, плод — костянка. Хороший закрепитель почвы. Плоды
очень ценные, обладают целебными свойствами. На территории
СССР широко распространена по песчаным -и галечниковым бере-
гам рек.
Подкласс 7. Астеридные (Asteridae)
Надпорядок XIV. Яснотковидные (Lamianae)
К этому и следующему надпорядку относятся семейства, увен-
чивающие ветви филогенетического древа, отражающего эволюцию
двудольных растений. Здесь собраны наиболее продвинувшиеся се-
мейства с наиболее богатым комплексом прогрессивных признаков:
сростнолепестковость, немногочисленность частей цветка, совершен-
ство приспособлений к перекрестному опылению насекомыми и др.
В надпорядке 46 семейств, распределенных между 5 порядками.
Наименее продвинувшиеся семейства собраны в порядки Dipsa-
cales и Gentianales. Из порядка Dipsacales в нашей флоре пред-
ставлены семейства Caprifoliaceae, Valerianaceae, Dipsacaceae.
344
Из порядка Gentianales семейство горечавковые: (Gentianaceae)
с большим родом преимущественно альпийских растений —
Gentiana. От Gentianales отходит ветвь порядков, отражающих
процесс все более совершенного приспособления цветка к пере-
крестному опылению насекомыми: порядки Polemoniales, Scrophula-
riales, Lamiales.
Порядок ворсянкоцветные (Dipsacales)
Семейство валерьяновые (Valerianaceae). К этому семейству
относится важнейшее лекарственное растение — валерьяна лекар-
ственная (Valeriana officinalis). Это крупный многолетник с супро-
тивными непарноперистыми листьями. Мелкие неправильные цвет-
ки собраны в соцветия — полузонтики. Венчик асимметричный
трубчато-ворончатый, розовато-белый. Развитых тычинок 3. Завязь
нижняя, трехгнездная, но лишь одно гнездо развивается в плод —
орешек. Растение обитает во влажных, преимущественно затенен-
ных местах. Целебное эфирное масло добывают из подземной час-
ти растения. В лесостепной зоне культивируется.
Порядок горечавкоцветные (Gentianales)
Семейство мареновые (Rubiaceae). Крупное семейство, виды ко-
торого распространены преимущественно в тропической зоне; здесь
они представлены чаще древесными формами, в умеренных облас-
тях— травянистыми растениями. Листья супротивные, с прилист-
никами, иногда листья рассечены на узкие доли, которые вместе
с такими же прилистниками производят впечатление мутовки.
Нижняя завязь у большинства видов двугнездная, реже одно-
и многогнездная. Плод — ягодоподобиая костянка, коробочка или
распадается на несколько орешков. Частей околоцветника и тычи-
нок 4—6.
Из растений флоры умеренных областей в этом семействе широ-
ко распространены виды подмаренника (Galium). Некоторые из них
(подмаренник настоящий—G. verum) имеют кормовую ценность.
Кормовую ценность имеют также некоторые виды ясменника
(Asperula).
Из тропических растений этого семейства наибольший интерес
представляют кофейное дерево (Coffea arabica) и хинное дерево
(Chinchona lancifolia). Семена кофейного дерева используются
для получения напитка кофе. Из коры и других частей хинного
дерева добывают хинин и другие ценные алкалоиды. Родина ко-
фейного дерева — Африка, хинного — Южная Америка.
Порядок полемониецветные (Polemoniales)
Семейство вьюнковые (Convolvulaceae). Стебель травянистый,
чаще вьющийся. Листорасположение очередное. Цветок правиль-
ный, венчик ворончатый, сросшийся из 5 лепестков. Тычинок 5,
345
Рис. 187. Повилика европейская(Cuscuia
еигораеа), вверху — продольный разрез
цветка, внизу — поперечный разрез завязи
гинецей ценокарпный, из 5—2 плодолистиков; завязь — 5—2-гнезд-
ная. Плод — коробочка, реже ягода.
Вьюнок полевой (Convolvulus arvense)—широко распростра-
ненный многолетний сорняк с млечным соком. Стебель вьющийся.
Остатки корня дают корневые отпрыски, что затрудняет борьбу
с ним. Обладает хорошими кормовыми качествами, однако для ло-
шадей ядовит.
Семейство повиликовые (Cuscutaceae). К этому семейству отно-
сится повилика (Cuscuta), морфологически близкая к вьюнку.
Представлена несколькими
сходными видами, паразити-
рующими на дикорастущих и
культурных растениях (рис.
187). Повилика — однолет-
нее, незеленое растение с ре-
дуцированными листьями;
мелкие, обычно пятичленные
цветки собраны в короткие
кисти, плод — коробочка, се-
мена очень мелкие. Особен-
но большой вред наносят
повилики, живущие на лю-
церне (С. racemose), клеве-
рах (С. trifolii), конопле и
других культурных растени-
ях (С. australis, С. arvensis).
Крупные виды повилики
(С. lupuliformis) паразити-
руют на деревьях и кустар-
никах, часто вызывая их
гибель.
Семейство бурачниковые (Boraginaceae). Травянистые растения,
обычно с жесткими волосками. Листья чаще цельные, поочередные.
Соцветие — завиток. Цветки правильные, редко слегка зигоморф-
ные, пятичленные; завязь распадается на 4—2 орешка. Тычинки
срастаются с венчиком. В зеве цветка у многих растений — чешуй-
ки, также в числе 5.
В этом семействе преобладают травы, малоценные в кормовом
отношении, или сорняки; есть лекарственные и декоративные рас-
тения.
С лекарственными целями используются чернокорень лекарст-
венный (Cynoglosutn officinale), окопник лекарственный (Sym-
phytum officinale), медуница лекарственная (Pulmonaria officinalis)
и др. Немногие растения имеют кормовое значение; к ним отно-
сится, например, окопник жесткий (S. asperum). Большинство рас-
тений из семейства бурачниковых — хорошие медоносы. Из ядови-
тых— гелиотропы (Heliotropium), триходесма (Trichodesma inca-
пит)—весьма ядовитое растение, распространенное в Средней
Азии.
346
Порядок норичникоцветные (Scrophulariales)
Рис. 188. Дурман обыкновен-
ный. (Datura stramonium), спра-
ва — зрелая коробочка
Семейство пасленовые (Solanaceae). Травы, реже кустарники
с очередными листьями. Цветки правильные, редко слегка зиго-
морфные, обычно пятичленные. Плод — коробочка или ягода.
Для большинства растений этого семейства характерно содержание
алкалоидов.
К растениям, у которых плод — коробочка, относятся табак, бе-
лена, дурман, петуния. Ягоду имеют кустарник дереза, или лиций
(Lycium), помидоры, стручковый перец, белладонна.
Табак {Nicotina). Этот род представлен несколькими видами.
Наибольшее значение имеет табак настоящий {N. tabacum), про-
исходящий из тропической Америки.
Это крупное, с большими листьями од-
нолетнее растение. Разводится преиму-
щественно для получения курительного
табака. Действующее начало — алка-
лоид никотин. Меньшее значение имеет
табак махорка (АЛ rustled) из Северной
Америки. Из листьев и стеблей получа-
ют экстракт, используемый для
борьбы с накожными паразитами
животных. Значительное количество
махорки используется также для
курения.
Белена {Hyosciamus niger) и дур-
ман {Datura stramonium) — сорные
ядовитые растения, используемые так-
же, как лекарственные (рис. 188).
Алкалоиды, содержащиеся в белене и
дурмане (гиосциамин, атропин, скопо-
ламин), вызывают поражение цент-
ральной нервной системы.
Паслен {Solatium)—к этому роду
помимо сорняковых пасленов относятся
ценные культурные растения. Картофель {Solanum tuberosum) —
разводят большое количество сортов. Различают сорта столовые,
кормовые и технические, служащие для получения крахмала и спир-
та. Родом из Южной Америки. Синие баклажаны (S. melongena) —
южная овощная .культура. Плоды темно-фиолетовые, крупные. Из
Индии. На полевых, особенно орошаемых землях, широко распро-
странен однолетний сорняк — паслен черный. {S. nigrum). Паслен
сладкогорький (S. dulcamara) ядовит.
Помидор (Lycopersicum esculentum) — ценнейшая однолетняя
культура с красными или желтыми плодами. Происходит из Юж-
ной Америки.
Белладонна (Atropa belladonna)—ядовитое травянистое рас-
тение с черными ягодами. Содержащиеся в растениях алкалоиды
(атропин и др.) используются в медицине.
347
Семейство норичниковые (Scrophulariaceae). В умеренной зоне
это семейство представлено в основном травянистыми растениями.
В субтропиках и тропиках преобладают деревянистые формы, пре-
Рис. 189. Льнянка обыкновен-
ная (Linaria vulgaris), спра-
ва — цветок
имущественно кустарники. Внут*
ри семейства можно наблюдать
редукцию лепестков от 5 до 4, а
тычинок — от 5 до 4 in 2. Завязь
всегда верхняя, двухгнездная,
плод — коробочка. Форма венчика
в меньшей или большей степени
зигоморфная, до двугубой. Распо-
Рис. 190. Заразиха
ветвистая (Orobonche
ramosa). Растение за-
разихи с корнями,
приросшими к главно-
му корню растения-
хозяина (конопли)
ложение листьев очередное и супротивное. Для семейства харак-
терно наличие полупаразитных и паразитических растений. Есть
ядовитые и лекарственные растения. К семейству норичниковых
близко семейство заразиховых.
348
Коровяк (Verbascum)—крупные, преимущественно двулетние
растения с очередными листьями. Венчик пятилопастный, слегка
зигоморфный, тычинок 5. Коровяка шерстистый (V phlomoides)
и тапсовидный (V. thapsiforme) имеют лекарственное значение.
Наперстянка (Digitalis)—также довольно крупное растение,
двулетнее, с очередными листьями. Венчик с четырехлопастпым
отгибом, тычинок 4. Наиболее широко известна наперстянка пур-
пурная (D. purpurea)—одно из ценнейших лекарственных расте-*
ний; культивируется. Вместе с тем при поедании животными на-*
перстянки возникают тяжелые отравления, вызывающие расстрой-
ство сердечной деятельности.
Льнянка (Linaria) —растение с очередными и супротивными ли-*
стьями (рис. 189). Венчик двугубый, со шпорцем. Тычинок 4.
Льнянка обыкновенная (L. vulgaris) ядовита, вызывает поражение
центральной нервной системы и пищеварительного тракта. Расте-
ние содержит гликозид, отщепляющий синильную кислоту.
Вероника (Veronica) имеет супротивные листья. Венчик четы-
рехраздельный, слегка зигоморфный. Тычинок 2. Некоторые виды
вероники (вероника лекарственная и др.) имеют лечебное значение.
Семейство заразиховые (Orobanchaceae). Представлено исклю-
чительно паразитными растениями. Из нескольких родов, относя-
щихся к этому семейству, наиболее широко распространен и богат
видами род заразихи (Orobanche).
Виды очень изменчивы, сходны между собой. Листья редуциро-
ваны до чешуек. Цветки с двугубым венчиком и 4 тычинками. За-
вязь верхняя, плод — одногнездная коробочка. Растения однолет-
ние. Паразитирует заразиха на различных растениях, из культур-
ных чаще поражает подсолнечник, капусту, коноплю, табак, клевер,
бахчевые культуры и др. Наибольший вред культурным растениям
приносят заразиха ветвистая (О. ramosa) (рис. 190), подсолнечни-
ковая (О. ситапа).
По строению цветка заразиховые ближе всего стоят к норични-
ковым, среди которых также широко распространен паразитизм.
Порядок ясноткоцветные (Lamiales)
Семейство губоцветные (Lamiaceae, Labiatae). Растения облада-
ют рядом характерных признаков; стебель четырехгранный, листья
супротивные, цветки обычно двугубые, тычинок 4, реже 2, завязь
верхняя, четырехлопастная; плод распадается на 4 орешка. Травы
или полукустарники. Растения с сильным запахом, часто приятным.
Среди растений этого семейства много эфиромасличных, лекарст-
венных, встречаются ядовитые. Кормовое значение сравнительно
невелико.
Мята (Mentha) — многолетние, редко однолетние травянистые
растения с сильным характерным запахом. Венчик с четырехлопаст-
ным отгибом, почти актиноморфный, маленький. Большинство видов
обитает в местах с избыточным увлажнением, по берегам рек
349
Рис. 191. Шалфей луговой (Salvia pratensis), А, Б — общий
вид, В — цветок в продольном разрезе, Г — тычинка с шар-
нирным приспособлениехМ, Д — чашечка
и озер. Как лекарственные и ароматические растения разводят
мяту перечную (М. piperita), курчавую (М. orispa) и некоторые
другие виды.
Шалфей (Salvia) —широко распространенные многолетние тра*
вянистые растения, реже полукустарники. Венчик двугубый, ты-*
чинок 2 (рис. 191). Несколько видов культивируются как эфиронос-*
ные и лекарственные растения: шалфей мускатный (S. sclarea),
лекарственный (S. officinalis).
Пикульник (Galeopsis)—довольно широко распространенные
сорные растения, преимущественно однолетние. Некоторые виды
пикулъника (G. ladanum и др.) ядовиты; наиболее богаты ядовиты-*
ми веществами семена и соцветия.
Кроме названных в этом семействе имеется еще много лекарст
венных растений: шлемник байкальский (Scutellaria baicalensis),
яснотка белая (Lamium album), пустырник волосистый (Leonurus
villosus), мелисса лекарственная (Melissa officinalis) и др.
350
Надпорядок XV. Астровидные (Asteranae)
Включает три порядка (Campanulales, Calycerales, Asterales),
заканчивающих собой одну из ветвей филогенетического древа.
Семейств тут немного, всего 9, но зато единственное семейство
порядка Asterales — Asteraceae — самое богатое видами: в -нем до
20 000 видов; вместе с тем это самое высокоорганизованное семей-
ство среди двудольных, и находится это семейство в .состоянии бур-
ного развития.
Порядок колокольчикоцветные (Campanulales)
Семейство колокольчиковые (Campanulaceae). Наиболее богат
видами, которые широко распространены и в нашей флоре, род
колокольчик (Campanula). Многолетние и двулетние травянистые
растения с очередными листьями. Венчик пятилепестковый, коло-
кольчатый, тычинок 5, плод — 3—5-гнездная коробочка. Широко
распространены колокольчик сибирский (С. sibirica), персиколист-
ный (С. persicifolium) и др. Колокольчик скученный (С. glomerata)
применяется в народной медицине.
Порядок астроцветные (Asterales)
Семейство сложноцветные (Asteraceae, Compositae). Наиболее
прогрессивное, утратившее почти все примитивные признаки. Цве-
ток спайнолепестный, пятерного типа: 5 лепестков, 5 сросшихся ты-
чинок. Плодолистиков 2, образуют одну нижнюю завязь, столбик
заканчивается двухлопастным рыльцем. Цветки обычно очень
мелкие, собранные в плотное соцветие — корзинку, часто внешне
похожее на один цветок. В связи с таким строением соцветия ча-
шечка сильно редуцирована, а то и вовсе отсутствует. У большин-
ства растений энтомофилия; соцветие приспособлено именно к это-
му способу опыления.
Редуцированная чашечка нередко превращена в хохолок, игра*
ющий роль парашюта при разносе плодов ветром.
Группа цветков в корзинке окружена обверткой из видоизме-
ненных листьев. Листочки обвертки могут быть зелеными, могут
внешне походить на чешуи, лишенные зеленого пигмента; часто
они окрашены и производят впечатление лепестков. Нередко роль
обвертки играют бесплодные воронковидные цветки.
Венчики цветков — различных категорий (рис. 192). Чаще всего
встречаются трубчатые венчики, заканчивающиеся на верхушке
пятью зубчиками. Из такого рода цветков состоит внутренняя часть
корзинки большинства видов.
Меньшая часть видов имеет корзинки, состоящие из язычковых
цветков. Примером растений с таким соцветием может служить
одуванчик.
351
Рис. 192. Цветки сложноцветных. А — трубчатый,
Б — язычковый, В — двугубый, Г — ложноязычко-
вый
По фор-ме основных цветков, составляющих соцветие, семейство
сложноцветных делят на два подсемейства: трубкоцветные
(Tubiflorae) и языкоцветные (Liguliflorae).
В семействе сложноцветных мало продовольственных и хоро-
ших кормовых растений, много декоративных, лекарственных и сор-
няков; имеются технические растения (например, каучуконосы
гвайюла, кок-сагыз).
1. Подсемейство трубкоцветные (Tubiflorae). Сюда относятся
следующие культурные и дикорастущие растения.
Подсолнечник (Н elianthus annuus) — самая ценная масличная
культура. Жмыхи — прекрасный корм, богатый белком. Топинам-
бур, или земляная груша (Н. tuberosus), — ценное кормовое расте-
ние. Высокоурожайный клубнеплод. Клубни — хороший углеводи-
стый корм; обычно на корню они поедаются свиньями; стебли си-
лосуют.
Ромашка (Matricaria chamomilla)—лекарственное растение;
широко распространена в естественных условиях как сорное расте-
ние; культивируется.
Тысячелистник (Achillea) —несколько близких видов этого рода
широко распространены на естественных пастбищах и сенокосах
и являются кормовыми растениями удовлетворительного качества.
Наиболее известны тысячелистник обыкновенный (A. millefolium)
(рис. 193) —лекарственное растение.
Полынь (Artemisia) представляет интерес как кормовое расте-
ние. Несколько видов многолетних полыней (A maritima и др.)
являются основными кормовыми растениями полупустынь. Исполь-
зуются поздней осенью и зимой. Полынь A. taurica — ядовитое
растение. Поедание ее вызывает заболевание центральной нервной
системы, сердца, пищеварительного тракта и почек. Полынь цитвар-
352
Рис. 193. Тысячелистник обыкновен- Рис* 194. Осот полевой (Sonchus
ный (Achillea millefolium), справа— arvensis), справа — отдельный цветок
средний трубчатый цветок и краевой
ложноязычковый
пая (A. cina)—ценнейшее лекарственное растение, содержащее
сантонин, — эффективное антигельминтное средство.
Осот розовый (Cirsium arvense)—один из наиболее вредных,
трудноискоренимых корнеотпрысковых сорняков. Засоряет поля
часто на межах, вдоль дорог.
Крестовник (Senecio) — представлен большим числом видов,
многие из которых ядовиты.
Сушеница (Gnaphalium uliginosum)—ценное- лекарственное
растение.
Горчак (Acroptilon picris)—еще более опасный сорняк из той
же группы корнеотпрысковых. Распространен на юге Украины.
Ядовит.
Васильки (Centaurea) —многочисленные виды, засоряющие
пастбища, сенокосы и полевые земли. В кормовом отношении ма-
лоценные растения. Есть декоративные формы. Наиболее известен
василек синий (С. су anus).
2. Подсемейство языкоцветные (Liguliflorae). В качестве приме-
ра остановимся на следующих его представителях.
12—326
353
Одуванчик обыкновенный (Taraxacum officinale) — широко рас-
пространенное луговое растение. Распространению его способствует
интенсивная пастьба. Лекарственное.
Цикорий (Cichorium inthybus) растет вдоль дорог как сорное
растение; цветки голубые. Лекарственное.
Осот полевой (Sonchus arvensis) — сорное растение поле-й и ого-
родов (рис. 194).
Класс однодольные (Monocotyledones — Liliatae)
Подкласс 1. Алисмидные (Alismidae)
Надпорядок I. Частухо видные (Alismanae)
Это линия эволюции, выражающая процесс перехода цветковых
растений к водному образу жизни. 18 семейств, объединенных
в 3 порядка, и всё это — растения, в большей или меньшей степени
приспособленные жить в воде. В основании этой ветви эволюции
порядок Alismales. Здесь сосредоточены виды земноводные,
Рис. 195. Частуха подорожниковая
(Alisma plantagoaquatica), слева вни-
зу — цветок
не вполне еще оторвавшиеся от
суши. Остальные два порядка —
Hydrocharitales и Najadales —
почти исключительно водные рас-
тения. Наибольший интерес пред-
ставляет первый порядок —
Alismales.
Порядок частухоцветные
(Alismales)
Семейство частуховые (Alisma-
сеае). Не имеет кормового значе-
ния. Тем не менёе его следует
рассмотреть, так как это семей-
ство интересно в филогенетиче-
ском отношении. По-видимому,
частуховые — наиболее древние
из сохранившихся однодольных,
примыкающих к двудольным рас-
тениям, а именно к семейству
нимфейных — водяная лилия, или
белая кувшинка (Nymphaea
alba).
Оба семейства представлены
формами, произрастающими в
водной среде или живущими в ус-
ловиях избытка влаги в почве.
Частуховые непосредственно свя-
заны с формами, из которых про-
354
изошли лилейные, — центральный порядок в классе однодольных.
К лилейным примыкают многие другие семейства однодольных.
Цветки Пастуховых тройного типа: частей околоцветника 3 + 3,
тычинок тоже 3 + 3; гинецей апокарпный, пестики многочисленные
(примитивный признак).
В СССР наиболее широко распространены типичные представи-
тели этого семейства; част у ха (Alisma plantagoaquatica) и стрело-
лист (Sagittaria sagitifolia). Оба растения — гигрофиты (рис. 195).
В порядке Najadales интересны многочисленные виды рдестов
(Potamogeton), обитающих в пресных водах, и два вида взморника^
морской травы (Zostera), обитающих в морях.
Подкласс 2. Лилиидные (Liliidae)
Надпорядок II. Лилиевидные (Lilianae)
Это самый крупный надпорядок в классе однодольных, в нем
почти половина всех семейств (32) и пять порядков. Исходный
для всей группы порядок Liliales, от него отходит веер из шести
филогенетических ответвлений, четыре из которых представлены
порядками, входящими в надпорядок лилиевидных. По уровню
развития они располагаются так: Triuridales, Iridales, Zingiberales,
Orchidales.
В последнем порядке однодольные растения достигли наиболее
тонкого приспособления формы и биологии цветка к перекрестному
опылению насекомыми. В нашей флоре наиболее полно представ-
лен порядок лилиецветных (Liliales). Из порядка ирисоцветных
(Iridales) у нас широко распространены в естественном состоянии
и в культуре ирисы (Iris) и гладиолусы (Gladiolus), из раннецве-
тущих весенних растений очень красивы крокусы (Crocus).
Ирисовые в отличие от лилейных имеют нижнюю завязь.
Порядок лилиецветные (Liliales)
Семейство лилейные (Liliaceae). Многолетние растения; травя-
нистые, за редким исключением. Подземная часть растений очень
часто имеет луковицу или корневище. Листья чаще линейнонервные
или дугопервные. Околоцветник простой, чашечковидный или вен-
чиковидный, состоит, как правило, из 6 сросшихся или свободных
частей. Тычинок столько же, сколько лепестков. Завязь верхняя,
обычно трехгнездная; плод — коробочка, реже ягода.
В семействе лилейных мало хороших кормовых растений, име-
ются ядовитые растения. Наиболее часто на пастбищах и сеноко-
сах встречаются виды следующих родов: лук и чеснок (Allium),
гусиный лук (Gagea), спаржа (Asparagus), чемерица (Veratrum).
Интерес представляют и такие ядовитые растения, как ландыш
(Convalaria), вороний глаз (Paris) и купена (Polygonatuni), кото-
рые также могут встретиться на естественных пастбищах.
Лук, чеснок (Allium) — к этому роду относится .несколько видов
культурных овощных растений и большое число дикорастущих рас*
12*
355
тений, широко распространенных во всех зонах Советского Союза.
Подземная часть растений этого рода^сьшбжена^уковицей./Листья
дрлыеил и .плоские. Мелкие цветки собра’ньГТТонтик, который^ мо-
жет^ быйъ различной .формы; Околоцветник простой, венчиковид-
ньшГсостоит^из шести свободных или сросшихся"в основании'1Ге-
пестков. Тычинок 6. Завязь верх-
няя с нитчатым столбиком, закан-
чивающимся головчатым трехло-
пастным рыльцем. Плод — тонко
стенная коробочка (рис. 196).
Рис. 197. Гусиный лук желтый
(Gagea lutea), справа внизу —
продольный разрез цветка
Рис. 196. Лук репчатый (Allium сера).
А — общий вид, Б — соцветие простой
зонтик, В — с удаленным околоцветни-
ком, Г — поперечный разрез завязи
Все растения этого рода имеют специфический запах, который
при поедании передается молоку. Поэтому луки на пастбищах
и в сене оцениваются, в основном, как вредные растения; некоторые
виды лука, например A. caspium, ядовиты. Тем не менее многие
виды лука поедаются животными.
Гусиный лук (Gagea) — мелкие весеннецветущие луковичные
растения, распространенные преимущественно в зоне степей и полу-
пустынь. Лцстья узкие, нитевидные, плоские или полые. Цветки
одиночные или собранные в немногоцветковые зонтики или щитки
(рис. 197). Венчик из шести свободных лепестков желтой окраски.
Завязь трехпнездная, коробочка трехгранная. Гусиный лук охотно
поедается животными на весенних пастбищах, особенно овцами
и козами.
356
Чемерица (Veratrum)—крупные многолетние растения, рас-
пространенные в травянистом ярусе лесов и на горных лугах. Под-
земная часть имеет корневище. Стебель мощный, листья крупные,
с влагалищами. Околоцветник раздельнолистный, зеленоватый,
белый или пурпуровый. Плод — коробочка. Все части растения со-
держат алкалоиды (протовератрии, протовератридин и др.), пора-
жающие центральную нервную систему. Поэтому чемерица оцени-
вается как ядовитое растение, хотя и наблюдаются случаи поедае-
мости ее на пастбищах. Наиболее распространенный вид—чемери-
ца Лобеля (V. lobelianum).
В лесах и средн кустарников лесной и лесостепной зон, а также
в горных районах довольно часто встречаются ядовитые, а вместе
с тем и лекарственные растения: ландыш майский (Convalaria
majalis), купена аптечная (Polygonatum officinale) и вороний глаз
(Paris quadrifolia). Все три растения содержат гликозиды (конваля-
марип, паридин и др.), действующие на сердце.
В семействе лилейных имеется большое количество видов с кра-
сивыми цветками. Многие виды этого семейства давно введены
в культуру и разводятся как декоративные: лилии, тюльпаны, гиа-
цинты, юкка и др.
В этот же порядок входит семейство амариллисовые (АтагуШ-
daceae), с нижней завязью. Из этого семейства широко известны
подснежники (Galanthus), нарциссы (Narcissus).
Порядок орхидноцветные (Orchidales)
Семейство орхидные (Orchidaceae). Наравне с сем. сложноцвет-
ных, орхидные — крупнейшее семейство на земном шаре, насчитьк
вающее до 30 000 видов. Это преимущественно тропические расте-
ния, все травянистые; очень многие виды представлены эпифитами.
Растения этого семейства характеризуются многочисленными, ино-
гда очень сложными приспособлениями к перекрестному опылению
с помощью насекомых. Цветки двуполые, зигоморфные; околоцвет-
ник из двух-, трехчленных кругов лепестковидных листочков, ино-
гда очень своеобразного строения. Тычинка чаще одна, нить ее
срастается со столбиком, образуя так называемую колонку; ре-
же— тычинок 2; пыльца слипается в один комочек — поллиний,
который при опылении целиком переносится насекомыми на рыль-
це другого цветка. Завязь нижняя, плод — коробочка. Семена
у орхидных очень мелкие, зародыш почти не дифференцирован
на зачаточные органы. В Советском Союзе наиболее широко рас-
пространены виды рода ятрышник (Orchis); ятрышник шлемонос-
ный (О. militaris), пятнистый (О. maculata), широколистный
(О. latifolia) и др.
Большинство видов этого единственного в своем порядке се-
мейства— тропические растения. Среди них лиана ваниль (Vanilla
fragrans)—пряное растение мирового значения. Виды, входящие
в остальные порядки этой группы, также обитают преимуществен-
но в тропиках и субтропиках. Много декоративных.
12*—326
357
Подкласс 3. Коммелинидные (Commelinidae)
Надпорядок III. Ситниковидные (J ипсапае)
Сюда относится всего 2 порядка (Juncales и Cyperales) с тремя
семействами. Это линия развития, отходящая от лилейных, но пол-
ностью приспособившаяся к опылению ветром. Отсюда редукция
околоцветника, превращение яркого венчика в малозаметные че-
шуи; нередко околоцветник и вовсе редуцирован. Представители
обоих порядков обитают в сходных экологических условиях.
Порядок ситникоцветные (Juncales)
Семейство ситниковые (Juncaceae). Филогенетически непосред-
ственно примыкает к лилейным; тип цветка остается тем же,
но внешность его резко изменилась под влиянием приспособления
к ветроопылению. Вместо яркоокрашенного венчика здесь малоза-
метные чешуи, однако число чешуй 6 (3 + 3), околоцветник акти-
номорфный. Тычинок также 6, и расположены они в два круга,
иногда один круг редуцирован. Плод—коробочка, трехгнездная
или одногнездная, завязь верхняя, с тремя рыльцами.
Рис. 198. Представители семейства ситниковых. А — ситник расходящийся (Jun-
cus effusus), Б — ожика волосистая (Luziila pilosa)
358
Внешне растения этого семейства похожи на злаки (рис. 198).,
у них такие же узкие листья с влагалищем. В хозяйственной клас-
сификации ситниковые (вместе с осоками) относились некоторыми
авторами к группе «кислых злаков». Этим подчеркивается их сход-
ство с подлинными злаками. Соцветие ситниковых также сходно
с соцветиями злаков; растения чаще многолетние, редко
однолетние.
В Советском Союзе распространены два рода этого семейства:
ситник (Juncus) и ожика (Luzula).
Многочисленные виды рода Juncus широко распространены пре-
имущественно на влажных заболоченных лугах. Многие виды вы-
держивают засоление почвы и произрастают по берегам морей
и соленых озер. Нередко здесь можно встретить сплошные заросли
ситника. В кормовом отношении ситники не представляют большой
ценности.
Один из наиболее распространенных видов этого рода — ситник
Жерарда (J. gerardi). Он же наиболее ценен в кормовом отноше-
нии; выносит сильное засоление. Широко распространены также
ситник сплюснутый (J. compressus), блестящеплодный (J. larnpro-
carpus) и др.
Другой род этого семейства — ожика (Luzula)—имеет еще
меньшее кормовое значение. Зарослей не образует,, изредка встре-
чается в лесах и на заболоченных лугах. В качестве примера можпе
указать ожику волосистую (L. pilosa).
Порядок осокоцветные (Cyperales)
Семейство осоковые (Сурегасеае). В эволюционном отношении
осоковые продолжают линию развития, начатую ситниковыми,
В результате приспособления к ветроопылению околоцветник под-
вергся еще большей редукции; чешуи превратились в щетинки,
а у большинства родов полностью редуцировались; однако есть
виды, у которых они имеют листоватую форму и тем самым мало
отличаются от околоцветника ситниковых.
В кормовом отношении преобладают растения среднего досто-
инства. Наибольший интерес представляют только осоки (Carex)
и кобрезни (Cobresia). Остальные роды этого семейства имеют не-
сравненно меньшее значение; они представлены растениями, плохо
или вовсе непоедаемыми.
Осока (Carex) — самый крупный род семейства и один из наибо-
лее богатых видами. К тому же осоки образуют часто сплошные
заросли почти без примеси растений из других семейств. В СССР
произрастает около 400 видов осок, большинство из них — растения
избыточно увлажненных мест.
Внешне, по форме листьев и соцветий, осоки похожи на злаки,
но строение цветков у них иное (рис. 199). Цветки однополые,
собранные в колосья. Колосья могут состоять из цветков одного
пола, но бывают и смешанные. Преобладают растения однодомные
Цветки сидят в пазухах кроющих чешуй. А1ужские имеют 3 ты-
12**
359
чинки, женские имеют пестик, образованный из 2—3 плодолистиков,
соответственно чему и рылец бывает 2 или 3. Пестик заключен'
в особый мешочек, образованный из прицветничка. Плод — орешек;:
если он образован тремя плодолистиками, то имеет трехграннук>
форму; образованный двумя плодолистиками — двояковыпуклый
или плосковыпуклый.
Из огромного количества видов лишь сравнительно немногие
ценны как кормовые растения. Это мелкие осоки степи, полупусты^
.ни и пустыни: осока толстостолбиковая (С. pachystylis), пустынная
(С, physodes), ранняя (С., ргаесох) и др.; а также некоторые осоки
горных лугов и степей: осока низкая (С. humilis), траурная
(С, tristis) и др. Большое хозяйственное значение имеют некоторы^
Рис. 200. Кобрезия (Cobresia). А — общий
вид С. capillifolia, Б — часть колоса с двумя
цветками (вверху слева), В — обоеполый
цветок, Г — колос кобрезия Болларда
(С. bellardii)
Рис. 199. Осока (Carex inf la-
ta), справа вверху — тычи-
ночный цветок, ниже — пес-
тичный цветок и его про-
дольный разрез
360
осоки тундр. Они поедаются ранней весной северным оленем. Цен-
ные виды имеются также в лесах Дальнего Востока.
Большинство осок в кормовом отношении оценивается низко
ввиду плохой поедаемости, бедности белком, солями кальция
и фосфорной кислоты. Нередко листья осок имеют так много крем-
незема, что даже ранят слизистую оболочку пищеварительного
тракта животных.
Кобрезии (Cobresia) очень сходны с осоками (рис. 200), однако
в отличие от последних у них плодик не заключен в мешочек,
Кобрезии — растения высоких гор. Здесь они создают целые гор®
пые ландшафты, прекрасные овечьи и конские пастбища. В горах
Средней Азии особенно широко распространена кобрезия волоса*
листная (С. capillifolia). Широко распространена и кобрезия Бел*
ларда (С. bellardi).
К этому же семейству относится цепное пищевое растение —
чуфа (Cyperus esculentus)у съедобные клубни, содержат 10—23%
масла.
Надпорядок IV. Коммелиновидные (Commelinanae)
В этом надпорядке всего 5 порядков, 11 семейств. Для нас пред-
ставляет интерес лишь последний порядок Poales с единственным
семейством злаковых Poaceae (Gramineae). Растения четырех ос-
тальных порядков (Commelinales, Ericaulales, Restionales, Bromel-
liales) в нашей флоре не встречаются, за исключением одного вида
коммелиевых. Порядки этой группы также представляют ветвь,
идущую от лилейных. Заканчивается эта ветвь формами, приспо-
собленными к ветроопылению, причем эта приспособленность наи-
высшего своего выражения достигает в семействе злаковых.
Порядок мятликоцветные (Poales)
Семейство злаковые (Poaceae, Gramineae). Имеет важное хо-
зяйственное значение; к злаковым относятся основные продоволь-
ственные и кормовые растения.
Огромное большинство злаков — многолетние и однолетние тра-
вы. Лишь одна группа злаков — бамбуки — представлена древес-
ными и кустарниковыми формами. У большинства злаков стебель
внутри полый, в местах прикрепления листьев перегородки — узлы.
Листья линейные, часто очень узкие, почти волосовидные.
Цветки злаков собраны в колоски, из колосков формируются
сложные соцветия: метелка, колос или колосовидная метелка.
Цветок злаков устроен очень своеобразно (рис. 201). Части
околоцветника имеют форму чешуй и пленок, бесцветных или окра-
шенных хлорофиллом в зеленоватые тона. Цветок состоит из двух
цветковых чешуй, верхней и нижней, нижняя часто бывает с остью,
двух маленьких пленок — лодикул, трех тычинок и одной верхней
завязи с двумя столбиками с перистыми рыльцами.
361
Ркс. 201. Колосок и цветок ржи (Secale сегеа-
fes). А — двуцветковый колосок, Б — цветок
5ез цветковых чешуй, В — лодикулы и пестик
Колосок может состо-
ять из одного, нескольких
или многих цветков. У ос-
нования колоска обычно
находятся две чешуйки —
колосковые. Плод у зла-
ков— зерновка, редко (у
бамбука) ягодовидный.
От этого типичного
строения бывают отклоне-
ния, характерные для от-
дельных родов семейства
злаков.
У большинства злаков
цветки двуполые, реже од-
нополые.
В кормлении сельско-
хозяйственных животных
злаки играют первосте-
пенную роль. Основу кон-
центрированных кормов
составляют зерна злаков
или отходы их переработ-
ки. Зерновая культура —
кукуруза, используемая
также в качестве зеленого
корма или для приготов-
ления силоса, принадле-
жит к этому семейству.
На естественных сенокосах и пастбищах основу травостоя чаще
составляют злаки. В смешанном травостое злаки играют сущест-
венную роль при высушивании травы на сено, способствуя лучше-
му просыханию травы и сохранению листьев двудольных растений.
Будучи примешаны к травам бобовых, злаки улучшают гигиениче-
ские качества зеленого корма. Лишь в зоне тундр, а также в полу-
пустынях и пустынях злаки в естественных кормовых угодьях усту-
пают главенствующее место представителям других групп расте-
ний.
В Советском Союзе произрастает почти 150 родов злаковых
растений; существенное сельскохозяйственное значение имеют ра-
стения, принадлежащие примерно к 30 родам, которым и будет
дана краткая характеристика.
Кукуруза (Zea mays) —однолетнее однодомное растение. Роди-
на— Мексика. В последние столетия распространилась по всему
земному шару.
Один из крупнейших травянистых злаков, стебель достигает вы-
соты 5 и более метров, листья крупные, шириной до 12 см; мужские
цветки собраны в верхушечное соцветие — метелку, состоящую из
колосовидных веточек (рис. 202). Колоски расположены на веточ-
362
Рис. 202. Кукуруза (Zea mays). А — общий вид растения, Б —
пестичный колосок, В— тычиночный колосок, Г — початок с зер-
новками
ках парами; один на ножке, второй почти сидячий. Каждый колосок
состоит из 2 колосковых чешуй, между которыми 2 мужских цветка;
цветковых чешуй 2. Тычинок, как и у подавляющего большинства
злаков, 3.
Женские цветки собраны в пазушные соцветия — початки. Каж-
дый початок имеет многослойную обвертку, образованную влага-
лищами листьев. Одноцветковые женские колоски с пленчатыми
колосковыми и цветковыми чешуями расположены попарно вдоль
початка параллельными рядами. Из двух парных колосков разви-
вается один, цветок в другом колоске остается бесплодным. Стол-
бик длинный, нитевидный с раздвоенным рыльцем. Пучок столби-
ков к моменту цветения высовывается из початка.
363
Мужские цветки у данного растения созревают на несколько
дней раньше женских, чем и достигается перекрестное опыление,
осуществляемое посредством ветра.
Кукуруза — кормовая, продовольственная и техническая куль-,
тура. По занимаемой площади на земном шаре она уступает только
пшенице. Огромное количество кукурузы выращивается для приго-
товления силоса. Стебли, листья и початки кукурузы богаты саха-
ром и прекрасно силосуются. Зерно используется в корм скоту,
а также для производства разнообразных пищевых продуктов. Из
стеблей кукурузы и кукурузных кочерыжек химическая промыш-
ленность изготовляет целый ряд синтетических продуктов. Сухие
листья и стебли кукурузы используются для изготовления бумаги.
Цветочные столбики заготовляются как лекарственное сырье (жел-
чегонное средство). Существует «несколько разновидностей кукуру-
зы и сотни сортов. Высокой урожайностью отличаются гибриды.
Сорго (Sorghum) — в Советском Союзе произрастает 8 видов, из
них лишь один — многолетнее растение. Все однолетние виды воз-
делываются в культуре. Это крупные растения с метельчатым со-
цветием.
Сорго сахарное (S. caccharatum) — возделывается на Украине,
на Северном Кавказе и в Нижнем Поволжье. Достигает высоты
свыше 2 м. Стебель заполнен паренхимой, богатой сахаром. Ис-
пользуется на сено, зеленый корм, как пастбищное растение и на
силос. В молодых листьях иногда при условиях, ухудшающих рост,
накапливается синильная кислота, вызывающая отравление живот-
ных. Зерно также используется на корм скоту. Засухоустойчиво.
Большое хозяйственное значение имеет сорго-гумаевый гибрид —
многолетнее кормовое растение, полученное в результате скрещи-
вания сорго с многолетним корневищным сорняком — гумаем
(S. halepense). Рекомендуется к возделыванию на песчаных землях
засушливой зоны СССР.
Сорго поникшее, или джугара (S. сегпиит), возделывается
в Средней Азии как зерновая культура. Зерно употребляется в пи-
щу и как концентрированный корм.
Суданская трава (S. sudanence) распространена в культуре на
черноземных и каштановых почвах. Интенсивно кустится и хорошо
отрастает. Дает пастбищный корм, зеленую подкормку и сено.
Очень засухоустойчива.
Из многих видов рода просо (Panicuni) широко распространено
просо посевное (Л miliaceum)—продовольственная и кормовая
культура. Соцветие у проса — метелка. Возделывают просо для по-
лучения зерна, зеленой массы и как пастбищное растение. Зерно
используется преимущественно для изготовления крупы — пшена,
а также на корм птице. Пастбищная трава проса иногда вызывает
отравление животных, поэтому просо используется как зерновая
культура.
В засушливых районах СССР возделывается однолетний вид
африканского проса, относящийся к роду Pennisetum (Р. typhoide-
иш). Это крупное растение с широкими темными листьями. Соцве-
364
тие в виде плотной метелки, похожей на початок. Возделывается
для получения зеленого корма и зерна, употребляемого в пищу
человеком, и как концентрированный корм скоту, а также на
зеленый корм.
Рис посевной (Oryza sativa) — вторая по мировому значению
после пшеницы продовольственная культура. Крупный гигрофиль-
ный злак размером до 150 см.
Соцветие — метелка. Колоски од-
ноцветковые. Колосковых чешуй
4. Цветковые чешуи плотно при-
легают к зерновке, нижняя обыч-
но с остыо. Тычинок 6 (рис. 203).
В СССР возделывается на Даль-
нем Востоке, в республиках Сред-
ней Азии, в Закавказье, на Кубани
и в южных районах Украины. Ро-
дина риса — Юго-Восточная Азия.
При возделывании риса планта-
ции обычно заливают водой; со-
ветскими учеными выведены сор-
та, дающие урожаи и при перио-
дических поливах. Отходы от пе-
реработки зерновок риса исполь-
зуют на корм скоту.
Ковыль (Stipa) — многолетний
ксерофитный злак с узкими щети-
новидными листьями. Образует
плотные дерновины. Соцветие —
Рис. 203. Рис посевной (Oryza saliva).
4 — общий вид растения, Б — цветок,.
В — цветок без цветковых чешуй
(6 тычинок), Г — метелка безостой
формы, Д — метелка остистой формы
метелка. Колоски одноцветковые.
Нижняя цветковая чешуя с длин-
ной остью, перистой или волосо-
видной. Многочисленные виды
ковыля распространены на целин-
ных землях степей и полупус-
тынь. На пастбищах п в сене
ранних фазах вегетации.
ковыли хорошо предаются лишь в
Из перистых ковылей в европейской части СССР наиболее ши-
роко распространен ковыль Лессинга (S. lessingiana)-, менее широ-
ко— ковыль-волосатик, или тырса (S. capillata).
На-ковыльных пастбищах овец можно пасти только до момента
выколашивания ковыля. Плоды ковыля имеют острое и очень твер-
дое заострение и легко втыкаются в нежную кожу овцы. Ость ковы-
ля в основании спирально закручена, при смачивании спираль рас-
кручивается и зерновка все глубже проникает в тело овцы. Так она
может пройти сквозь мышцы и достигнуть внутренних органов.
Овца истощается и может погибнуть. Особенно опасны зерновки
ковыля-волосатика.
Молодая трава ковыля — высокопптательпый молокогонный
корм, особенно ценный для лошадей.
365
Тимофеевка (Phleuni) представлена в Советском Союзе не-
сколькими видами. Наибольшее значение имеют два: тимофеевка
луговая (Ph. pratense) и степная (Ph. phleoides)—рыхлокустовые
многолетние травы (рис. 204). У обоих видов соцветие — плотная
колосовидная метелка. Тимофеевка луговая — более рослая и с ши-
рокими листьями; тимофеевка степная мельче, с более узкими ли-
стьями, засухоустойчивая. Оба вида растений хорошо поедаются
скотом.
Лисохвост (Alopecurus) —злак, похожий на тимофеевку, но бо-
лее влаголюбивый. Соцветие — тоже колосовидная плотная метел-
ка. Листья нежные, широколинейные. За небольшим исключением
все относящиеся к этому роду виды хорошо поедаются животными.
Наибольшее значение имеют два вида: лисохвост луговой (А. рга-
iensis) и вздутый (A. venlricosus). Оба—многолетние короткокор-
невищные злаки. Лисохвост луговой распространен преимуществен-
но в лесной зоне, лисохвост вздутый — в лесостепной и степной,
хорошо выносит значительное засоление почвы.
К роду полевица (Agrostis) относятся за немногими исключе-
ниями многолетние корневищные злаки с тонкими стеблями и не-
Рис. 204. Тимофеевка луговая (Phleuni pratense'}
(4) и лисохвост луговой (Alopecurus pratensis)
(S):
1 — колосовидная метелка; 2 — пестик; 3 — открытый
4 — закрытый колосок
366
широкими листьями. Преобладают гигрофиты. Соцветие — рыхлая
метелка. Среди довольно многочисленных видов этого рода имеются
виды высокого и среднего кормового достоинства. В культуру вве-
ден наиболее ценный, полиморфный вид — полевица белая
(A. alba).
Вейник (Calamagrostis)—это крупные, хорошо облиственные
многолетние корневищные злаки. Нередко образуют сплошные
обширные заросли. Соцветие — метелка. В европейской части
СССР наиболее широко распространен по песчаным наносам вей-
ник обыкновенный (С. epigeios)-, в Сибири — вейник Ландсдорфа
(С. landsdorfii).
Рис. 205. Овес посевной (Avena sativa) (Л), овсюг (Avena faiua) (Б):
1 — метелка; 2 — отдельные колоски; 3 — колосок.с цветками; 4 — цветок с цветочны-
ми чешуями; 5 — цветок без чешуй; 6 — общий вид овсюга; 7 — отдельный колосок;
8 — нижняя цветковая чешуя с остью
367
Вейники дают большую кормовую массу, однако листья их бы-
стро грубеют и становятся дочти недоедаемыми. Вейниковое сено
грубое, малопитательное.
Овес (Avena)—преимущественно однолетние злаки. Листья
широкие, соцветие — метелка. Колоски крупные, 2—4-цветковые
(рис. 205). Колосковые чешуи крупнее цветковых. Нижняя цветко-
вая чешуя обычно с остью. В различных зонах возделывается кор-
мовая и продовольственная культура — овес посевной (Л. sativa).
Выращивается как для получения зерна, так и для зеленого корма;
в последнехМ случае овес обычно высевается в смеси с бобовыми
культурами — викой и горохом.
Посевы зерновых бывают засорены овсюгом пустым (A. fatua)
и овсюгом Людовика (A. ludoviciana)—растениями, в молодом
возрасте похожими на овес. При правильной агротехнике овсюг,
как однолетний сорняк, сравнительно легко уничтожается, однако
при плохой агротехнике становится опасным сорняком и может
полностью заглушить посевы хлебов.
К роду райграс (Arrhenatherum) относится райграс высокий
(A. elatius)—многолетний рыхлокустовый злак, ценный в кормо-
вом отношении. Давно .введен в культуру; в диком состоя-
нии распространен преимущественно в лесной и лесостепной
зонах.
Свинорой (Cynodon) —род представлен единственным видом —
свинорой, или пальчатая трава (С. dactylon), распространенным
в зоне сухих степей, полупустынь и пустынь. Растение многолетнее
с длинными ползучими корневищами. Прекрасно выносит вытап-
тывание, является хорошим пастбищным растением. Отзывчив на
полив. Злостный сорняк орошаемых земель Средней Азии; солевы-
носливый и засухоустойчивый.
Один из видов рода бекмания (Beckmania) — бекмания обыкно-
венная (В. eruciformis)—ценное в кормовом отношении влаголю-
бивое многолетнее растение. Имеет подземные корневища с ползу-
чими побегами. Широко распространена в местах с избыточным
увлажнением в степной, лесостепной и лесной зонах. Бекмания
введена в культуру, выносит засоленные почвы.
Тростник (Phragmites communis)—широко распространенный,
очень крупный гигрофитный злак. Вдоль берегов южных рек обра-
зует заросли, характерные для плавней. Встречается и вне водое-
мов. Растение многолетнее, с мощными корневищами. Соцветие —
метелка. Лишь молодые побеги пригодны в качестве зеленого кор-
ма и в силос. Взрослые сухие стебли — топливо; строительный
материал и сырье для производства картона и бумаги.
Виды рода тонконог (Koeleria) встречаются во всех зонах
СССР, но преимущественно в степной и в горных районах. Дерни-
стые многолетние злаки. Соцветие — плотная колосовидная метел-
ка. Колоски 2—5-цветковые. Наиболее широко распространен
тонконог стройный (К. gracilis)—хорошая кормовая трава паст-
бищного использования, хорошо выносит вытаптывание; сплошных
зарослей не образует.
368
К роду прибрежница (Aeluropus) относится прибрежница со-
лончаковая (A. littoralis); имеет большое значение в районах с со-
лончаковыми почвами. Один из немногих злаков, выносящих край-
нюю степень засоления. Многолетнее растение с длинными ползу-
чими побегами. Стебли и листья жесткие, богатые минеральными
солями. Соцветие — плотная метелка с колосовидными веточками.
Судя по химическому составу, прибрежница солончаковая может
быть отнесена к злакам довольно высокого кормового достоин-
ства.
Среди нескольких видов рода ежа (Dactylis) наиболее широко
распространена ежа сборная (D. glomeratd) — многолетний рыхло-
кустовый злак с широкими листьями. Соцветие — односторонняя
плотная метелка с многоцветковыми колосками, собранными в гу-
стые пучки. Она широко распространена в лесной и лесостепной
зонах, а также в горных лугах Киргизии; здесь она образует об-
ширные сплошные заросли.
Сенокосная и пастбищная трава; хорошо п многократно отра-
стает после стравливания и скашивания,
Мятлик (Роа)—род с большим количеством видов, широко
распространенных во всех зонах и в самых разнообразных эколо-
гических условиях. Растения нередко образуют сплошные заросли.
Преобладают многолетние корневищные и рыхлокустовые формы.
Стебель тонкий; листья преимущественно неширокие или узкие.
Соцветие — метелка, колоски 2—10-цветковые.
Несколько видов мятлика представляют кормовую ценность.
Мятлик луговой (Р. pratensis) —луговое корневищное и корне-
вищно-рыхлокустовое растение, дающее хороший урожай при до-
статочном увлажнении. Растение преимущественно пастбищного
использования (рис. 206). Более теневынослив и влаголюбив мят-
лик обыкновенный — Р. trivialis. В кормовом отношении оценива-
ется ниже, чем мятлик луговой. Еще требовательнее к влаге мятлик
болотный — Р. palustris, также корневищный злак с ползучими по-
бегами.
Мятлик луковичный, живородящий (Р. bulbosa v. vivipara) ши-
роко распространен в степях, полупустынях и пустынях. Начинает
вегетировать очень рано. В условиях засухи — растеньице с низень-
кой дерновиной и тоненьким стебельком. При достатке влаги раз-
растается в мощное растение сенокосного использования. Обычно
же это типичное пастбищное растение, хорошо переносящее даже
интенсивную пастьбу.
К роду бескилъница (Atropis) относятся многолетние дернистые
растения, приспособленные к сильно засоленным почвам. Соцве-
тие— развесистая нежная метелка с многоцветковыми колосками.
Листья обычно узкие, даже щетиновидные. Наиболее широко рас-
пространена бескилъница расставленная (A. distans), которая час-
то образует на засоленных почвах сплошные заросли. Как и дру-
гие виды этого рода, растение пастбищного и сенокосного исполь-
зования, однако после цветения быстро желтеет, грубеет и
поедается значительно хуже, чем в молодом возрасте.
369
Рис. 206. Мятлик луговой
(Роа pratensis).
А — корневище с побегами, Б — метелка, В —
колосок без колосковых чешуй (вверху) и ко-
лосковые чешуи (внизу), Г — цельный коло-
сок
Многочисленные виды
рода овсяница (Festuca)
наиболее широко распро-
странены в нашей флоре.
Встречаются во всех рас-
тительных зонах Совет-
ского Союза и в самых
разнообразных экологиче-
ских условиях. Рыхло- или
плотнокустовые многолет-
ние злаки. Соцветия — ме-
телка, колоски 5—10-
цветковые с заостренны-
ми или остистыми цветко-
выми чешуями. Наиболь-
шей известностью пользу-
ется овсяница луговая и
бороздчатая, или типчак
(рис. 207).
Овсяница луговая
(F. pratensis) — растение
крупное, с широкими
листьями, хорошо облист-
венное, влаголюбивое.
Распространено в лесной
и лесостепной зонах. Хо-
рошо поедается скотом.
Овсяница бороздчатая,
или типчак (F. sulcata),—
основное растение южных
дернистых степей. Наряду
с ковылямп составляет ос-
нову травостоя сухих сте-
пей. Типично пастбищное
растение, хорошо перено-
сящее пастьбу; на богатых и хорошо увлажненных почвах достигает
высоты более 50 см и вполне может быть использовано под сенокос.
Плевел (Loliurri)—многолетние и однолетние рыхлокустовые
злаки. Соцветия — сложный колос. Колоски, за исключением вер-
хушечного, имеют лишь одну колосковую чешую и обращены
к главной оси колоса ребром (рис. 208).
Райграс многоукосный (Л. multiflorum)—одно- и двулетнее
растение; нижняя цветковая чешуя у него с остью. Райграс паст-
бищный (Л. регеппе) — многолетний низовой злак, нижняя цветко-
вая чешуя без ости. Растения обоих видов райграса хорошо поеда-
ются скотом.
Костер (Broinus)—многолетние и однолетние растения, рас-
пространенные во всех зонах Советского Союза; нередко образуют
сплошные заросли. Соцветие костра — развесистая или сжатая ме-
370
Рис. 207. Овсяница луговая (Festuca pratensis)—А (метелка, колосок),
овсяница бороздчатая — типчак (Festuca sulcata) —Б
телка. Колоски многоцветковые, что очень характерно для видов
этого рода. Почти у всех видов нижняя цветковая чешуя с остыо.
Кормовое значение имеет костер безостый {В. inermis) —многолет-
нее корневищное растение (рис. 209). Среди большого числа форм
371
и сортов этого вида имеются как влаго-
любивые, так и хорошо переносящие
засушливый климат. Костер безос-
тый — высокоурожайный верховой
злак, дающий хорошее сено и пастбищ-
ный корм, особенно в смеои с бобовы-
ми (например, эспарцетом, люцерной).
Из однолетних костров широко рас-
пространен костер кровельный (В. tec-
tor ит). Как и большинство других од-
нолетних костров, это эфемер, дает
удовлетворительного качества зеленую
массу лишь весной; затем он быстро
высыхает и, по существу, играет роль
сорного растения. Плоды костра кро-
вельного легко обламываются и сильно
засоряют шерсть овец.
Род пырей (Agropyrum) в настоя-
щее время обычно подразделяют на
два самостоятельных рода: собственно
пырей (Elytrigia) и житняк (Agropy-
Son). У пырея колосковые чешуи без
киля, у житняка — с килем.
У обоих видов соцветие — сложный
колос, колосок многоцветковый. Все от-
носящиеся сюда растения — многолет-
ники. Хорошо поедаются скотом.
Из собственно пыреев широко рас-
пространен пырей ползучий (A. re-
pens). Это верховой корневищный злак,
встречающийся и в луговых и в степ-
ных растительных формациях (см. рис
208). Хорошо выносит засоление поч-
вы. Дает хорошее сено и пастбищный
корм. Пырей ползучий может быть
злостным сорняком полей. Житняк
гребневидный (A. pectiniforme) — наи-
более широко распространенный вил
и хорошо переносит засоление почвы.
Рпс. 208. Пырей ползучий
(Agropyrum repens)—Д, рай-
грас пастбищный {Loliuni ре-
геппе) — Б
житняка. Засухоустойчив
Очень полиморфный вид. Различным экологическим условиям свой-
ственны формы, отличающиеся мощностью куста, шириной листьев,
толщиной стебля и формой колоса. Кормовые качества житняков
хорошие.
Род рожь {Secale) — представлен в Советском Союзе несколь-
кими дикорастущими видами; на больших площадях культивиру-
ется рожь посевная (S. cereale). Ближайшим предком посевной
ржи является сорнополевая рожь (S. segetale), в настоящее время
распространенная на Кавказе и в Средней Азии, где засоряет посе-
вы зерновых культур. Стебель посевной ржи достигает при благо-
372
Рис. 209. Костер безостый (Bromus inermis), слева —
колосок, справа — корневище
приятных условиях высоты 2 м, листья широкие, соцветие — слож-
ный колос. Колоски двухцветковые расположены на оси в два ряда.
Нижняя цветковая чешуя с длинной остью (см. рис. 201). Ржаная
дерть — обычный вид концентрированного корма.
Пшеница (Triticum)—однолетние, за исключением нескольких
видов, возделываемые растения. Всего насчитывается около 20 ви-
дов шпеницы, (из которых мировое значение как продовольственные
культуры имеют два вида: пшеница мягкая, или обыкновенная
(Г. vulgare) и твердая (71. durum).
373
Мягкая' пшеница — наиболее широко распространенный вид.
Колос этой пшеницы более или менее плотный или рыхлый. Колос-
ки 4—5-цветковые, нижние цветковые чешуи с остью или безостые
(рис. 210); соломина полая. Зерновка мучнистая или мучнисто-
стекловидная. Возделываются озимые и яровые формы.
Твердая пшеница менее широко распространена. Колос плот-
ный. Колоски 3—4-цветковые, два нижних цветка в каждом колонке
Рис. 210. Колосок и цветок (с чешуями и без них)
пшеницы
обязательно с длинной остью. Соломина выполнена белой парен-
химой или имеет лишь узкий просвет. Зерновки стекловидные.
Оба вида возделываемых пшениц имеют большое количество
разновидностей. Виды и разновидности пшениц легко скрещивают-
ся между собой.
Пшеницы введены в культуру с древнейших времен. Дикие
предки их не найдены.
Пшеница — продовольственная культура. Отходы от переработ-
ки зерна пшеницы (отруби и др.) представляют собой ценный кон-
центрированный корм.
Ячмень (Hordeurri)—однолетние «и многолетние растения. Со-
цветие— сложный колос. Характерная морфологическая особен-
ность: колоски сидят на оси не в одиночку, а группами по три ко-
лоска. Колоски одноцветковые. Колосковые чешуи щетиновидные
или ланцетошиловидные. Нижняя цветковая чешуя с длинной
остью. Зерновка обычно срастается с цветковыми чешуями, но бы-
вают и голозерные ячмени.
Существенное хозяйственное значение имеют два вида возделы-
ваемых ячменей: ячмень обыкновенный ( Н. vulgare) и двурядный
(Н. distichum). Ячмень возделывается с древнейших времен. Дикий
предок ячменя двурядного — ячмень дикорастущий (Н. spontane-
um}, в настоящее время произрастает на Кавказе и в Средней
Азии.
Ячмень — кормовая, продовольственная и техническая культу-
ра. Зерно ячменя идет на приготовление крупы, используется в пи-
воварении, а также идет на корм скоту, ячменная солома — широко
используемый грубый корм. Ячмень высевают также для получения
зеленого корма.
374
Подкласс 4. Арецидные (Arecidae)
Надпорядок V. Пальмовидные (Агесапае)
Последняя ветвь родословного древа покрытосеменных, вклю-
чающая 5 порядков, 7 семейств. Группа эта имеет общее происхож-
дение с лилейными, но вместе с тем относящиеся сюда растения
имеют и своеобразные черты. Первый порядок (Arecales, или Pal-
males), представляющий самостоятельное ответвление в эволюции,
характеризуется древесными формами, это пальма. Четыре осталь-
ных порядка (Cyclanthales, Arales, Pandalales, Thyphales)—фор-
мы, которые эволюционировали в сторону вторичного приспособле-
ния к обитанию в условиях избытка влаги.
Порядок пальмоцветные (Arecales, Palmales)
Семейство пальмовые (Агесасеае). Деревянистые растения,
обычно с колонновидным стволом, заканчивающимся пучком круп-
ных листьев. Цветки мелкие, однополые (редко двуполые), собран-
ные в метельчатые соцветия, прикрытые до цветения общим покры-
валом. Растения однодомные и двудомные; плод — ягода, костянка
или орех.
Наиболее широко известны пальма финиковая (Phaenix dactyli-
fera, Ph. canariensis), кокосовая (Cocos nucifera) и др. На Черно-
морском побережье Кавказа хорошо растет декоративная паль-
ма— хамеропс низкий (Chamerops humilis).
Порядок ароидноцветные (Arales)
Семейство ароидные (Агасеае). В этом семействе преобладают
тропические растения. В Советском Союзе из этого семейства на
влажных местах распространен белокрыльник болотный (Calla
palustris). Цветки этого растения собраны в початок, окруженный
общим белым покрывалом. У двуполых цветков полностью редуци-
рован околоцветник, тычинок 6, плоды ягодообразные, красные.
Листья сердцевидные. Все растение сильно ядовито и нередко вы-
зывает массовое отравление животных. В условиях леса распрост-
ранены растения рода аронник (Arum), также обладающие ядови-
тыми свойствами. У растений этого рода мелкие однополые цветки,
собранные в початок; женские — в нижней части початка, муж-
ские— в верхней, широкое большое покрывало охватывает соцве-
тие. Листья стреловидные. Наиболее широко распространены
аронник пятнистый (A. maculata), удлиненный (A. elongata)
и др.
Семейство рясковые '(Lemnaceae). К семейству относятся широ-
ко распространенные в стоячих водах мелкие водоплавающие рас-
тения— ряска (Lemna). Растения многолетние, однодомные, ссиль-
375
но редуцированными вегетативными органами. У каждого растения
всего 1— 3 очень мелких цветка: один женский и 1—2 мужских
с одной тычинкой. Чаще встречается ряска малая (L. minor), ред-
ко — трехраздельная (L. trisulca). Ряска — ценный корм для рыбы
и водоплавающей птицы.
Порядок рогозоцветные (Thyphales)
Семейство рогозовые (Thyphaceae). Многолетние травянистые
растения, широко распространенные по берегам рек и озер. Расте-
ния однодомные, цветки собраны в длинное цилиндрическое соцве-
тие, коричневое к моменту созревания плодов. Плод — маленький
орешек. Корневища у рогоза (Thypha)' толстые, содержат до 40%
крахмала, который отличается высокими техническими качествами.
Наиболее широко в нашей стране, распространен рогоз широколи-
стный (Т. latifolla), реже узколистный (Т. angustifolia).
376
ГЛАВА IV
ЭЛЕМЕНТЫ ЭКОЛОГИИ И ГЕОГРАФИИ РАСТЕНИЙ
Условия жизни растений на Земле в течение многих миллионов
лет изменялись и продолжают изменяться в наше время. Большие
изменения в условия существования растений вносит деятельность
человека, более того, человек во многом сам создает эти условия.
Современный лик Земли, включая и растительные ландшафты, на
больших пространствах носит следы бессознательной и сознатель-
ной деятельности человека. В условиях социалистического общест-
ва становится возможным планомерное, полезное для человека
преобразование природы.
Изменение природы, подчас коренное, не лишает нас интереса
к истории естественной растительности на Земле, к исследованию
ее современного состояния. Более того, это необходимо для рацио-
нального использования природных ресурсов. Многие миллионы
квадратных километров земной поверхности, покрытой естествен-
ной растительностью, представляют и в своем первобытном состоя-
нии большой научный и производственный интерес. С другой сто-
роны, вмешательство в природу, изменение растительности только
тогда может принести пользу человеку, когда с достаточной глуби-
ной изучены закономерности существования растений и развития
растительного покрова.
На протяжении всей истории своего развития растительный мир
находится в неразрывном единстве с изменяющимися условиями
среды, образно выражаясь, он является продуктом этой среды,
в этой среде растения находили и находят необходимые условия
своего существования и развития. Однако если мы сопоставим
особенности внешнего и внутреннего строения ныне живущих рас-
тений, их биологию с современными условиями существования, то
не всегда найдем полную согласованность; будучи в общем приспо-
собленными к нынешним условиям, многие растения проявляют
в своей жизни некоторые черты, отражающие какие-то иные, не
современные нам условия существования. Например, всем извест-
ные растения, так называемые подснежники, начинают вегетацию
очень рано, еще под снегом, быстро отцветают и на протяжении
всего теплого периода года остаются в состоянии скрытой жизни.
Что мешает им использовать теплый и в общем более благоприят-
ный период года для своей активной жизни? Мешает консерватизм
их природы, сохранившаяся приспособленность к каким-то иным,
ранее существовавшим условиям жизни. В изменившихся условиях
они выхватывают для своей жизни небольшой отрезок года с ком-
13—326
377
плексом факторов, при которых формировались их отдаленные
предки, и с наибольшей полнотой используют этот отрезок. Под-
снежники— это, очевидно, растения, весь уклад жизни которых
сложился еще в отдаленную от нас эпоху ледникового периода, в
эпоху с очень коротким холодным и влажным летом.
Жизнь и строение растений, таким образом, зависят не только
от современных условий жизни, но и от условий, при которых раз-
вивался длинный ряд предков ныне живущих растений, уходящих
в глубь веков.
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Наука, изучающая строение и жизнь растений в связи и во взаи-
модействии с конкретными сочетаниями внешних условий, называ-
ется экологией. Отсюда и элементы внешней среды, небезразлич-
ные для жизни растения и формирования его структуры, получили
название экологических факторов. Свет, температура, вода, воздух,
ветер, почва называются абиотическими факторами. Неровности
земной поверхности в большей степени перераспределяют эти фак-
торы, обусловливая степень и характер их воздействия на расте-
ния. Поэтому в элементы экологии входит также рассмотрение эко-
логической роли рельефа.
Из всей совокупности элементов внешней среды выделяют под
названием биотических факторов влияние на растение представи-
телей мира животных и растений. Особо рассматривают влияние
человека — антропогенные факторы. Как уже сказано выше, строе-
ние и жизнь растений определяются также всей предшествующей
историей предков данного растения, т. е. историческими фактора-
ми, о чем не следует забывать, исследуя влияние экологических
факторов.
В целях методического удобства рассмотрим роль каждого
фактора в отдельности. Однако это лишь искусственное выделение.
В природе на растение действует весь комплекс внешней среды,
и влияние отдельных факторов всегда тесно переплетается. Вместе
с тем в данный момент жизни растения какой-либо один фактор
может оказаться решающим. Это, например, могут быть крайние
температуры, дефицит влаги, недостаток света и т. д. Чрезмерное
выражение или, наоборот, недостаток какого-либо одного фактора
может свести на нет благоприятное сочетание остальных факторов.
АБИОТИЧЕСКИЕ ФАКТОРЫ
Роль климатических факторов. Коротко остановимся на каж-
дом факторе в отдельности.
Свет. Свет необходим зеленым растениям как источник энер-
гии, используемой в фотосинтезе. Этим, однако, не исчерпывается
роль света в жизни растений. Каждому пункту земной поверхности
свойствен свой ритм светового режима, и этот ритм отражается
в биологии развития произрастающих здесь растений. Эта зако-
378
номерность получила название фотопериодизма. Сущность его за-
ключается в том, что растения, принадлежащие к различным ви-
дам, по-разному реагируют на соотношение продолжительности
дня и ночи в вегетационный период, а этим определяется возмож-
ность нормального развития растений на различных географических
широтах. Свет также влияет на всю обстановку, окружающую рас-
тение: температуру воздуха и почвы, влажность, деятельность мик-
роорганизмов и т. д. В этом сказывается косвенная роль света.
Свет доходит до растения в самых разнообразных дозировках
как в количественном, так и в качественном отношении. Например,
в северных районах из-за большой облачности преобладает рассе-
янный свет; на юге, особенно в зоне пустынь и полупустынь,— пря-
мые солнечные лучи. В свете, который приходится на долю высо-
когорных растений, относительно больше ультрафиолетовых лучей.
Под пологом леса свет иной, чем рядом, на открытом месте. Про-
ходя сквозь крону деревьев, свет резко меняет свой качественный
состав, так как не все части спектра одинаково задерживаются
листьями. Вблизи предметов, отражающих свет, увеличивается
содержание невидимых инфракрасных лучей и т. д.
В процессе эволюции растения приспособились к различному
световому режиму; внешняя и внутренняя структура растения но-
сит на себе отпечаток различных условий освещения. Даже части
одного и того же растения, попадая в неодинаковые условия осве-
щения, меняют свое строение.
Различают светолюбивые и теневыносливые растения. Светолю-
бивые растения характерны для открытых мест: степей, полупу-
стынь, пустынь, высокогорных лугов. Под пологом леса произра-
стают теневыносливые растения. Эти растения настолько приспосо-
бились жить в условиях слабого освещения, что яркий свет для них
губителен. Поэтому после вырубки леса большинство произрастав-
ших в лесу травянистых растений погибает.
Различная теневыносливость хорошо выражена у древесных по-
род. Теневыносливые породы (например, тисс, бук, граб, пихта,
ель) имеют густую крону, и под пологом этих деревьев очень мало
света. Наоборот, светолюбивые деревья (лиственница, береза, сос-
на) образуют светлые леса; кроны этих деревьев пропускают мно-
го света. Ажурность кроны светолюбивых деревьев формируется
в результате того, что побеги этих деревьев, попадая в тень, быстро
отмирают. Остаются только те веточки, к которым попадает доста-
точно много света.
Почти все культивируемые растения, в том числе и кормовые,—
светолюбивы.
Растения, выросшие на свету или в тени, имеют характерные
черты внешнего и внутреннего строения. По сравнению с теневыми
растениями у растений, выросших на свету, более приземистый
и прочный стебель, более густое расположение листьев; хорошее
развитие механических тканей; листья более толстые, с хорошо
дифференцированным мезофиллом; эпидермис покрыт более тол-
стым слоем кутикулы.
13*
379
Для теневого растения характерны как раз противоположные
признаки; стебель тонкий, с длинными междоузлиями, слабо осна-
щенный механическими тканями; листья тонкие, палисадная ткань
не выражена; слой кутикулы очень тонкий или его совершенно нет.
Весь облик растения меняется, если его выращивают в условиях
недостатка света (рис. 211).
Говоря о реакции растений на различный световой режим, есте-
ственно возникает вопрос: не может ли световой режим какого-ли-
бо географического района, например нашего Севера, ограничивать
Рис. 211. Фасоль, выросшая при недостаточном освеще-
нии (А) и при достаточном освещении (Б)
возможность получения максимального урожая выращиваемых
культур. Специальные исследования, проведенные в этом направ-
лении (Л. А. Иванов и др.), показали, что света достаточно на лю-
бой широте и что световой режим в любой зоне не может быть
ограничивающим фактором для получения достаточного урожая
растительной массы.
Тепло. Каждое растение может нормально существовать в оп-
ределенных температурных границах. Это наследственное свойство
организмов, выработанное растениями в процессе эволюции.
Тепловой режим закономерно изменяется по земной поверхно-
сти в направлении от полюсов к экватору. Поэтому различают
последовательно холодную полярную зону Земли, умеренно-холод-
ную, умеренно-теплую, субтропическую и тропическую зоны.
Каждой зоне свойственна растительность, состоящая из расте-
ний, приспособленных к колебаниям температуры в определенных
границах. Из этого, конечно, не следует, что в пределах данной зо-
ны растительность однообразна.
На характере растительного покрова сказываются и другие
экологические факторы: влажность почвы и др. Да и сам темпера-
турный режим в пределах одной зоны варьирует в зависимости от
380
многих обстоятельств; на южном склоне больше тепла, чем на
северном, в глубине растительной группировки дневные колебания
температуры менее резки, чем вне ее; вдали от морей и океанов
температура колеблется в более широких пределах в течение суток
и на протяжении года; лето здесь более жаркое, а зима более хо-
лодная, чем вблизи океанов и морей. Сказывается также и влияние
господствующих ветров, наличие горных хребтов и т. д.
При всем различии растительности в пределах одной зоны тем-
пературный режим все же является основным ограничивающим
фактором, препятствующим переносу растений из одной зоны в дру-
гую. Поэтому для каждого культурного растения существуют оп-
ределенные географические границы возделывания. В результате
акклиматизации растений границы эти могут быть передвинуты.
При подъеме в горы температурный режим изменяется анало-
гично тому, как мы это наблюдаем при перемещении с юга на се-
вер; чем выше, тем становится холоднее. Соответственно изменяет-
ся и растительность: наиболее приспособлены к резким колебаниям
температуры растения высокогорного альпийского пояса.
Приспособленность растения к определенному тепловому режи-
му выражается не только в перенесении крайних температур. Каж-
дый этап в развитии растения приспособлен к определенной выра-
женности температурного режима.
В общем, растения гораздо легче переносят значительные от-
клонения от оптимальной температуры в сторону холода, чем в сто-
рону более высокой температуры. Однако в сельскохозяйственной
практике мы чаще сталкиваемся со случаями гибели или повреж-
дения растений от низких температур. Для борьбы с этими явле-
ниями важно знать о факторах, повышающих холодостойкость
растений. Вопросы эти исследованы акад. Н. А. Максимовым.
Под действием температур ниже нуля происходит обезвожива-
ние цитоплазмы, и это одна из причин гибели клеток. Обезвожива-
ние происходит потому, что кристаллизация воды начинается
в межклеточниках. Нарастая, кристаллы отнимают воду от цито-
плазмы и одновременно механически ранят клетку. Разрывается
клеточная оболочка, поверхностный слой цитоплазмы, и клетка по-
гибает.
Чем труднее клетка отдает воду, тем устойчивей она против
действия низких температур. Водоотдача затрудняется повышени-
ем осмотического давления клеточного сока; в этом отношении
большую роль играет накопление в клетке сахаров; концентрация
их повышается осенью, при постепенном понижении температуры.
Наоборот, внезапное похолодание может застать растения неподго-
товленными, и тогда опасность гибели возрастает.
Опасны также оттепели, при которых, в результате жизнедея-
тельности клеток, количество сахара в них снижается. Наступаю-
щие затем холода губят растения. Морозоустойчивость клеток по-
вышают и другие вещества; установлено, что к таким веществам
относятся антоциан, жирные масла. Большую роль, по-видимому»
играют индивидуальные свойства самой цитоплазмы.
381
Морозоустойчивость клеток повышается при обеднении их во-
дой. Сухие семена и споры выдерживают охлаждение, близкое к
абсолютному нулю. Набухшие семена погибают даже при неболь-
ших морозах.
Интересно отметить, что теплолюбивые растения могут погиб-
нуть от охлаждения даже при положительной температуре (тропи-
ческие— уже при 10° С). Гибель наступает, по-видимому, от рас-
стройства обычно согласованной деятельности ферментов цито-
плазмы; в результате накапливаются продукты, не свойственные
цитоплазме, которые и вызывают отравление клетки.
В борьбе растений с вредным влиянием высоких температур
выработалось такое важное приспособление, как поглощение хло-
рофиллом наиболее горячих лучей. В результате накопления в
клетках солей повышается температура свертывания цитоплазмы,
что позволяет растениям до известного предела выдерживать на-
гревание живых тканей. Транспирация, отнимая от живых тканей
тепло, также помогает растению противостоять губительному дей-
ствию чрезмерно высоких температур.
Формирующая роль теплового фактора недостаточно выяснена.
Заметного влияния на морфологию растений тепловой режим как
таковой не оказывает.
Растения одного вида, выросшие в разных, климатических зонах,
практически неразличимы.
Очень велико и многообразно косвенное влияние теплового
фактора. В конечном счете все физиологические процессы зависят
от степени выраженности (уровня) этого фактора. Степенью на-
грева определяется интенсивность транспирации; охлаждение поч-
вы может быть причиной физиологической сухости ее; от теплового
режима зависят рост и развитие растений и т. д.
Вода неравномерно распределена по земной поверхности. Не-
равномерность эта — основная причина большого разнообразия
растительности, произрастающей на одной и той же широте. Жар-
кие сухие пустыни Африки расположены на одной параллели
с влажными тропическими лесами. Среди пустынь встречаются
оазисы — островки роскошной растительности; экологический фак-
тор, определяющий такую разительную перемену,— вода.
Экологическое значение имеет и атмосферная, и почвенная во-
да, но главную роль в жизни сухопутных растений играет вода, по-
лучаемая растением посредством корней из почвы.
Для водных растений вода — необходимая среда обитания, вне
ее эти растения очень быстро погибают.
Осадки, выпадающие из атмосферы, играют различную эколо-
гическую роль.
Снежный покров предохраняет зимующие растения от вымерза-
ния. Недостаточность снегового покрова в степных районах нашей
страны часто является ограничивающим фактором для возделыва-
ния озимых культур. Большие массы снега часто обламывают де-
ревья и кустарники. Стаивая, снег образует ледяную корку, под
которой растения могут погибнуть.
382
На Крайнем Севере и высоко в горах снеговой покров лежит
большую часть года; продолжительность вегетации резко сокра-
щается, и в таких условиях отбираются растительные формы, начи-
нающие развитие при низких температурах и быстро заканчиваю-
щие вегетацию.
Выпадающий обычно в теплое время град может сильно повре-
дить растения.
В условиях жарких песчаных пустынь большое значение в жиз-
ни растений имеет роса. Капельки росы проникают в толщу песча-
ного субстрата и перехватываются корнями растений. В летний
период здесь — это основной источник водоснабжения растений.
Вода, в большей или меньшей степени насыщающая атмосферу,
оказывает существенное влияние на водный режим растений.
При сухом воздухе, особенно если еще дует ветер, резко повыша-
ется транспирация, и растение начинает страдать даже при нали-
чии влаги в почве. Однако чаще атмосферная засуха сопровожда-
ется почвенной; в условиях сухой и ветреной погоды почва также
быстро высыхает.
Вместе с тем растения сами являются существенным источни-
ком атмосферной влаги. Под пологом леса воздух всегда более
влажный, чем в открытом месте. Внутри травостоя относительная
влажность воздуха также значительно выше, чем на голой пло-
щадке.
Атмосферная влага в большинстве случаев играет лишь косвен*
ную роль, уменьшая транспирацию и испарение воды из почвы.
Лишь немногие растения способны поглощать воду из атмосферы.
В нашей флоре таким свойством обладают мхи и лишайники.
В тропических лесах это свойственно эпифитам — растениям, посе-
ляющимся на стволах деревьев. Воздушные корни этих растений
могут поглощать влагу прямо из воздуха. Следует иметь в виду,
'что во влажных тропических лесах воздух сильно насыщен водя-
ными парами.
Дождь — основной источник воды, поступающий в почву. Уже
в процессе выпадения капельножидкая вода играет некоторую эко-
логическую роль. В этой воде содержатся окислы азота, аммиак;
стекая по растению, вода обмывает его. Все это имеет положитель-
ное значение. С другой стороны, сильный дождь механически по-
вреждает растения и нередко наносит им существенный вред.
Для водоснабжения растений очень существенно распределение
осадков в течение вегетационного периода. Например, для полу-
чения хорошего урожая зерновых хлебов в условиях средней поло-
сы особенно ценны майские и июньские дожди. Если же весна
и начало лета были сухими, то последующие июльские и августов-
ские дожди в данном случае уже не могут исправить положение.
Для степных пастбищ очень важны весенние и раннеосенние
дожди. Такое распределение осадков полезно для весеннего от-
растания степных трав и для осеннего их возобновления.
Способность растений использовать почвенную воду зависит
от многих обстоятельств. Прежде всего большое значение имеет
383
механический состав почвы: чем крупнее частицы почвы, тем пол*
нее растения могут использовать воду. Например, на песчаных
почвах недоступны растению всего лишь 1—2% воды; вся вода
сверх этого количества свободно используется корнями растений.
На глинистых же почвах даже 10—15% влаги недоступны расте-
нию. Этим объясняется то обстоятельство, что песчаные пустыни,
даже при меньшем количестве осадков, гораздо богаче жизнью,
чем глинистые пустыни. Избыток солей, низкая температура также
ограничивают способность растений забирать из почвы влагу.
Это состояние называется физиологической! сухостью.
Разнообразие условий водообеспечения на земной поверхности
привело к выработке различных биоморфологических типов рас-
тений, приспособленных к тому или иному водному режиму. С этой
точки зрения различают три основных вида сухопутных растений:
гигрофиты, мезофиты и ксерофиты.
К гигрофитам относятся растения, приспособленные жить в ус-
ловиях избыточного увлажнения; во влажных тенистых лесах или
на открытых местах, но на почве, насыщенной влагой. 1\ гигрофи-
там следует, например, отнести такие злаки, как тростник обыкно-
венный (Phragmites communis), манник водяной (Glyceria aquati-
са), канареечник тростниковидный (Phalaris arundinacea). Из раз-
нотравья к гигрофитам относится калужница болотная (Caltha
palustris), некоторые лютики; из культурных растений — рис (Oryza
sativa).
Гигрофитам обычно свойственна своеобразная внутренняя
и внешняя структура.
От гигрофитов следует отличать гидрофиты — растения, обита-
ющие в водной среде, например белая кувшинка (Nymphaea alba),
рдесты (Potamogeton), элодея и др. Эти растения, вынутые из во-
ды, быстро погибают.
Гигрофиты переходными формами 'соединяются с мезофи-
тами.
Мезофиты — растения, произрастающие в условиях достаточно-
го, но не избыточного увлажнения. К мезофитам относится боль-
шинство ценных дикорастущих кормовых и возделываемых куль-
турных растений. Можно сказать, что это растения средних усло-
вий жизни; соответственно структура их не имеет черт
приспособленности к крайнему выражению какого-либо фактора.
Типичный мезофит страдает и при переходе к длительному избы-
точному увлажнению, и в условиях крайней сухости.
В природе можно найти формы, промежуточные между типич-
ными мезофитами и гигрофитами. Так же трудно провести резкую
границу между мезофитами и ксерофитами.
К формам, переходным от мезофитов к гигрофитам, можно от»
нести лисохвост луговой (Alopecurus pratensis), бекманию (Beck-
mania eruciformis), мятлик болотный (Роа palustris). К нетипич-
ным мезофитам, приближающимся к ксерофитам, можно отнести
житняки, волоснец сибирский (Elymus sibiricus), мятлик узколист-
ный (Р. angustifolia).
384
Рис. 212. Поперечный разрез листа ковыля:
/ — пластинка листа свернулась в трубку; 2—пластинка листа развернутая
Ксерофиты — растения засушливых местообитаний, наиболее
богаты приспособительными формами. Ими богаты естественные
кормовые угодья степей, полупустынь и пустынь. По-разному рас-
тения приспособились к экономному расходованию влаги, к суще-
ствованию в условиях засушливого климата; соответственно этому
выработались и совершенно различные формы ксерофитов. На тер-
ритории Советского Союза преобладают ксерофиты, у которых
ксерофитная структура характеризуется более сухими, жесткими
листьями и стеблями. Такие ксерофиты получили особое назва-
ние— склерофиты. Типичные склерофиты— ковыль, типчак. Листья
у этих злаков узкие и к тому же бывают свернуты в трубочку,
стороной с устьицами внутрь (рис. 212). У некоторых двудольных
склерофитов листовая пластинка резко сокращается, например
у верблюжьей колючки (Alhagi camelorum), а то и совсем редуци-
руется— у испанского дрока (Sparcium junceum), саксаула
(Haloxylon) и др. Представители этой группы ксерофитов — боль-
шинство полыней, у которых листовая пластинка сильно рассечена
и листья покрыты густым слоем (волосков, препятствующих испаре-
нию, например Artemisia maritima.
Менее распространены в СССР ксерофиты типа суккулентов.
Это растения, ткани которых сильно обогащены водой. У одних
растений, например у кактусов, сочным становится стебель, листья
при этом редуцированы в колючки (рис. 213). У других, напротив,
сочными становятся листья, например у очитка (Sedurn). Суккулен-
там свойствен очень медленный обмен веществ, в процессе которого
вода расходуется постепенно в малых количествах. Зато и рост
385
Рис. 213. Суккулент опунция (Opuntia)
суккулентов в период засухи также очень медленный. Вода в тка*
нях суккулентов удерживается с большой силой даже в условиях
очень жаркой и сухой погоды.
Таким образом, способность ксерофитов противостоять засухе
зависит не только от анатомо-морфологической структуры, но и фи-’
энологических свойств клеток, от стойкости самой цитоплазмы
и особенностей биохимических процессов, протекающих в расте*
ниях.
Ксероморфную структуру может вызвать не только физическая,,
но и физиологическая сухость. Высоко в горах или на севере, в ус-
ловиях холодных, хотя и обе-
спеченных влагой почв, можно
встретить растительные формы,
очень похожие на ксерофитов
жаркой степи. Такие растения
относят к особой экологиче-
ской группе психрофитов.
К психрофитам можно отнести,
например, злаки: белоус (Nar-
dus stricta), овсяницу пеструю
(Festuca varia), встречающую-
ся в горах Кавказа, кобрезию
(Cobresia capillifolia), высокогорное растение из семейства осо-
ковых.
Избыток солей, как известно, тоже может сделать почвы фи-
зиологически сухими. К таким почвам можно отнести мокрые со-
лончаки. На них произрастают растения с явными признаками
ксероморфной структуры. Тут и сочные суккуленты, например
солерос (Salicornia herbacea) (рис. 214), сведа (Suaeda maritima)
и др., и сухие склерофиты — бескилъница (Atropis distans) и др.
Есть еще своеобразная биологическая форма, возникшая в про-
цессе приспособления к ограниченному водному режиму. Это рас-
тения, использующие для своей вегетации кратковременный период
весны и начала лета. За краткость существования они получили
название эфемеров. Озимые эфемеры начинают развитие из семе-
ни осенью, яровые — ранней весной. И те и другие быстро разви-
ваются, зацветают, плодоносят и отмирают. Особенно богата эфе-
мерами растительность степей и пустынь. Вся жизнь яровых
эфемеров здесь укладывается в несколько недель. Несмотря
на кратковременное существование, эфемеры — ценный в кормовом
отношении компонент растительного покрова полупустынных
и пустынных пастбищ. К наступлению летней жары от них оста-
ются только семена.
К наиболее часто встречающимся эфемерам относится веснянка
(Erophila verna), некоторые вероники (V. verna, V ргаесох), зон-
тичник (Holosteum umbellatum), репяшок (Ceratocephalus orthoce-
ras) и многие другие.
Сходную биологию развития могут иметь и многолетние расте-
ния. В отличие от эфемеров, их называют эфемероидами. К ним от-
386
носятся гусиные луки (Gagea), крокусы (Crocus), безвременники
(Colchicum), тюльпаны (Tulipa), гадючий лук (Muscari), мятлик
луковичный (Роа bulbosa), некоторые пустынные и степные осоки
(Carex) и др.
Воздух. В состав воздуха, как известно, входят: азот (78%
по объему), кислород (около 21%), аргон (0,94%), углекислый газ
(0,03%), следы водорода и различные газо-
образные примеси. Кроме этого, в атмосфер-
ном воздухе всегда содержатся водяные
пары, а в нижних слоях, над сушей, и раз-
личного происхождения пыль.
Из основных газов, входящих в состав
воздуха, наибольшим колебаниям подверга-
ются кислород и углекислый газ, и колеба-
ния эти во многом связаны с жизнедеятель-
ностью организмов. Источники поступления
в атмосферу кислорода и углекислого газа
распределены по земной поверхности нерав-
номерно. Например, с поверхности морей и
океанов поступает огромное количество кис-
лорода, выделяемого водорослями. Наобо-
рот, основная масса углекислого газа выде-
ляется из почвы, богатой органическим ве-
ществом. Тем не менее, благодаря явлениям
диффузии, постоянному движению и пере-
мешиванию воздуха, состав его, как прави-
ло, отличается постоянством.
Гораздо большее экологическое значение
имеют случайные примеси к обычному со-
ставу воздуха. Чаще всего это двуокись се-
ры, в большом количестве выделяемая в
воздух при сжигании угля. Вблизи химиче-
ских предприятий в воздух могут выделять-
ся и другие ядовитые газы, повреждающие
растения. Ядовитость сернистого ангидрида
настолько велика, что даже ничтожная при-
месь его (тысячные доли процента) отрав-
ляет растения.
Чувствительность к ядовитым газам у
различных растений неодинакова. Очень
чувствительна, например, обыкновенная ель
(Picea excelsa) и очень стойки туи, можже-
Рис. 214. Суккулент соле-
рос (Salicornia herbacea),
вверху веточка увеличе-
на
велъник, тополь, клен американский. При озеленении городов это
свойство деревьев и кустарников обязательно учитывают.
Особой чувствительностью к загрязнению воздуха отличаются
лишайники. Поэтому в крупных городах и вблизи металлургических
заводов лишайники встречаются очень редко.
В атмосферном воздухе, как правило, имеется большее или мень-
шее количество пыли. Экологическое влияние небольшой примеси
387
Рис. 215. Флагообразная ветровая форма кроны дерева
пыли трудно уловимо. Но бывают моменты, когда в воздух под-
нимается огромное количество пыли, так называемые пыльные или
черные бури. Вред от таких бурь разносторонний. Затемнение,
вызываемое частицами пыли, резко снижает фотосинтез, листья
покрываются слоем пыли, затрудняющим газообмен. Травянистая
растительность иногда засыпается толстым слоем пыли, под кото-
рым растения погибают. В других местах, откуда выдувается мел-
козем, обнажаются корни растений, и они гибнут от высыхания.
Прошедшая, особенно продолжительная пыльная буря может за-
метно изменить облик растительного покрова. Особенно сильно
страдают от пыльных бурь озимые и яровые посевы хлебных куль-
тур. После таких бурь нередко приходится значительные площади,
занятые этими культурами, заново пересевать.
Движение воздуха оказывает разностороннее влияние на расте-
ния. Положительная роль ветра и более слабых перемещений воз-
духа сказывается в процессе опыления и разноса плодов и семян.
Вредное влияние ветра может быть прямым и косвенным. Пря-
мое действие ветра сильнее сказывается на деревьях и кустарни-
ках. Сильные ветры часто ломают деревья (бурелом) или вывора-
чивают их с корнем (ветровал). Под действием ветров, дующих
преимущественно в одну сторону, вырабатываются особые, флаго-
образные формы крон у деревьев и высоких кустарников (рис. 215).
На границе леса с тундрой иссушающие ветры являются причи-
ной гибели древесной растительности. Безлесье тундры объясняют
именно этой причиной. Лишь под покровом леса и под защитой
388
травянистого покрова сохраняются мелкие, стелющиеся деревца
полярной ивы (Salix polaris) и карликовой березы (Betula папа).
Прямое вредное влияние ветра на травянистую растительность
редко приходится наблюдать. Лишь при очень сильном ветре могут
обламываться и полегать высокие травянистые растения. Зато очень
часто сказывается косвенное влияние ветра на травянистые расте-
ния. Ветер резко повышает испарение с поверхности листьев, часто
вызывает повреждения, а то и гибель растений при суховеях.
Если сухой и жаркий ветер дует во время налива зерна злаков,
зерно становится щуплым и за несколько дней может погибнуть
до половины ожидаемого урожая; к тому же резко снижается и ка-
чество зерна. Испарение во время суховея настолько резко возрас-
тает, что растения страдают даже при вполне достаточном количе-
стве воды в почве; корни не справляются с подачей воды.
Косвенное влияние ветра сказывается также и в ветровой
эрозии почвы, в переносе песка в песчаных пустынях и полупус-
тынях.
Создаваемые на полях лесополосы замедляют движение ветра
и тем самым смягчают его вредоносное действие. Хорошо устроен-
ная ажурная полоса в зимнее время способствует равномерному
распределению снега на полях.
Экологическая роль почвы. Подавляющее большинство растений
произрастает на почве, из которой они берут воду и элементы поч-
венного питания. Поэтому почва имеет не меньшее экологическое
значение, чем климатические факторы. Чаще, пожалуй, выступает
косвенная роль почвы как среды, определяющей действие других
климатических факторов.
Но почва может выступать и как непосредственно действующий
фактор, и тогда связь характера растительности с качеством почвы
становится настолько выразительной, что говорят даже о растени-
ях-индикаторах, присутствие которых прямо указывает на опреде-
ленные качества почвы. Растения могут указывать на бедность
или богатство почвы питательными веществами.
Характерную структуру имеют растения, произрастающие
на сильно засоленных почвах. Растения этой экологической группы
называют галофитами. Внешне они похожи на типичных ксерофи-
тов. Среди галофитов имеется большая группа сочных солянок-
суккулентов. Из-за избытка солей галофиты в свежем виде практи-
чески непоедаемы; лишь поздней осенью, после отмирания клеток
и выщелачивания солей, сочные галофиты становятся съедобными.
Состав солей в клеточном соке галофитов в общем отражает
солевой состав почвы.
Физические свойства почвенного субстрата также не остаются
безразличными для растений. Экологическая роль механического
состава почвы особенно отчетливо видна на растениях, приспособ-
ленных к жизни на песках, — псаммофитах, или песколюбах. Ха-
рактерное свойство песков — подвижность их, накладывает особый
отпечаток на морфологическую структуру и биологические свойства
псаммофитов.
389
Растения, живущие на песках, в процессе приспособительной
эволюции выработали ценное свойство образовывать придаточные
корни на засыпаемой части стебля и, наоборот, закладывать почки
на обнажаемых корнях. При постоянном перемещении песков такие
растения не погибают. Типичными растениями-песколюбами явля-
ются виды селина (Aristida) в Средней Азии, на южно-украинских
песках — пырей пушистоцветковый (Agropyrum dazyanthum).
Экологическая роль рельефа. Рельеф следует отнести к косвенно
действующим экологическим факторам. Под влиянием рельефа по-
разному складывается комплекс климатических и эдафических
(почвенных) факторов. Особенно 'резко выступает экологическая
роль рельефа в горных странах. Например, в условиях Средней
Азии, где горы возвышаются в окружении пустынь, можно наблю-
дать такую смену растительности: полупустыня, степь, лугостепь,
субальпийский пояс, альпийский пояс. В субальпийском поясе —
леса и субальпийские высокорослые луга, альпийский пояс обычно
лишен древесной растительности, здесь низкорослые луга и степи.
В условиях гор очень резко сказываются на характере расти-
тельности также экспозиция и крутизна склонов.
В условиях сухого климата даже незначительные неровности
почвы заметно сказываются на растительности. Причина этого яв-
ления— изменение в степени засоления почвы.
Комплексность растительного покрова, особенно хорошо выра-
женная в условиях гор, весьма ценится в пастбищном хозяйстве,
повышая поедаемость травы и увеличивая в целом эффективность
использования естественных кормовых угодий.
БИОТИЧЕСКИЕ ФАКТОРЫ
Распределение растений по земной поверхности и их существо-
вание неразрывно связаны с жизнью бесчисленного множества
живых организмов. Даже в местах, наиболее бедных жизнью, на-
пример в арктических или высокогорных пустынях, растения всегда
находятся во взаимодействии не только с факторами неживой
природы, но и с организмами, не всегда видимыми невооруженным
глазом.
Достаточно рассмотреть субстрат, в котором закреплены расте-
ния, и поверхность подземных и надземных частей растений, «и мы
убедимся, что всюду находятся микроорганизмы, составляющие
микрофауну и флору данного местообитания. Растения, следова-
тельно, всегда существуют в сообществе с другими растениями,
животными и микроорганизмами. Такие сообщества называются
биоценозами.
Можно привести бесчисленное множество примеров взаимо-
влияния растений и животных, они могут быть более или менее
нейтральными, взаимополезными или односторонне ущербными.
Мы остановимся на нескольких примерах, представляющих
не только общебиологический, но и производственный интерес.
390
Влияние животных. 1. Пастьба животных оказывает существен-
ное влияние на растительный покров пастбища. Наблюдения по-
казывают, что полное отсутствие пастьбы, например в степи, ведет
к ее ухудшению. Такое, на первый взгляд, парадоксальное явление
объясняется, очевидно, тем, что естественная травянистая расти-
тельность исторически сложилась во взаимодействии с пасущимися
животными. В природе всегда, еще до появления человека, травя-
нистые растения служили источником питания для травоядных
животных. Непосредственная причина деградации растительности
в отсутствие пастьбы — прогрессирующее накопление отмерших
частей растений и закупорка почвы этим материалом. С другой
стороны, чрезмерная пастьба неминуемо ведет к деградации расти-
тельного покрова. Уменьшается общая масса растительности,
а главное, растения, ценные в кормовом отношении, заменяются
несъедобными и плохопоедаемыми. Умеренная пастьба, наоборот,
является фактором, необходимым для поддержания травостоя
на определенном, не снижающемся уровне.
2. Насекомые, реже птицы, опыляют растения. Отсутствие соот-
ветствующих опылителей может быть причиной бесплодия рас-
тений.
3. Огромную роль играют представители животного мира
и в процессе разноса плодов и семян. Такое распространение может
быть и пассивным, и активным со стороны животных.
4. Велика экологическая роль землероев. Делая бесчисленные
ходы в почве и перерывая ее, они могут улучшать ее воздушно-
водный режим. В этом может сказаться положительная роль их.
Вместе с тем, выворачивая на поверхность подпочву и образуя
кочки, землерои отрицательно влияют на растительный покров,
изменяя его состав в нежелательную сторону и ухудшая условия
использования кормового угодья.
5. Общеизвестна положительная роль беспозвоночных, населя-
ющих почву. Особенно велика роль земляных червей, отмеченная
еще Ч. Дарвином. Достаточно сказать, что каждые несколько лет
весь плодородный слой почвы пропускается через пищеваритель-
ный тракт червей. Это способствует обогащению почвы элемен-
тами питания растений и улучшению физической структуры почвы.
6. Насекомые и их личинки нередко наносят огромный вред
растениям, поедая корни или уничтожая листья. Например, свекло-
вичный долгоносик повреждает посевы сахарной свеклы. Большой
вред может причинить саранча.
Взаимовлияние растений. 1. Произрастающие совместно расте-
ния затеняют друг друга. Поэтому в результате приспособления
в естественных группировках сожительствуют растения с различ-
ной требовательностью к свету; под пологом более светолюбивых
растений обитают растения тенелюбивые. Приспособления эти
настолько глубокие, что после вырубки леса гибнет большая часть
видов травянистых растений, обитавших под пологом деревьев,
и возникает травянистая растительность иного состава. Деревья,
выросшие в лесу, в окружении других деревьев, имеют совершенно
391
Рис. 216. Сосна, выросшая в лесу (слева) и на открытом
месте
иной вид по сравнению с деревьями, выросшими вне леса, на от-
крытом месте (рис. 216).
2. Некоторые растения служат опорой для других растений,
имеющих слабый стебель. Особенно распространено это в темных
тропических лесах.
3. Совершенно односторонним и часто губительным для одной
из сторон является паразитизм растений на растениях. К числу
'наиболее известных растений-паразитов относятся многочисленные
виды повилик и заразих.
4. Примерами обоюдополезного сожительства могут служить
симбиоз бобовых растений и клубеньковых бактерий, корней выс-
ших растений и грибов (микориза) и, наконец, тесный симбиоз
между водорослями и грибами в едином организме лишайников.
392
5. Совместное обитание нескольких видов высших растений
позволяет более экономно использовать факторы жизни растения:
свет, влагу, элементы почвенного питания. На этом основаны
преимущества возделывания смеси трав перед чистыми посевами
тех же видов.
АНТРОПОГЕННЫЕ ФАКТОРЫ
Влияние человека на растительный мир растет вслед за разви-
тием цивилизации и научно-технического прогресса. Во второй по-
ловине XX в. антропогенный фактор стал самым мощным среди
всех сил, влияющих на живую природу. Создавая огромное коли-
чество новых форм растений, более продуктивных и более полез-
ных, человек явился причиной переноса вредных сорняков и пара-
зитных растений с одного континента на другой и внутри конти-
нентов. Перевозя и акклиматизируя полезные растения, человек
нередко вместе с ними переносит и возбудителей болезней этих
растений.
Пастуший образ жизни древних народов был причиной превра-
щений когда-то цветущих областей в пустыни (Сахара, пустыни
стран Ближнего Востока, Центральной Азии); непосредственная
причина опустынивания — чрезмерный, нерегулируемый выпас
животных. Миллионы гектаров пастбищ и в умеренной, влагообес-
печенной зоне по той же причине превращены в низкопродуктив-
ные пустоши. Нерациональное земледелие нередко приводит к мас-
совому размножению и расселению полевых сорняков.
Человек проводит каналы и миллионы гектаров пустынных зе-
мель превращает в цветущие оазисы. Но вместе с тем часто в ре-
зультате неправильной эксплуатации орошаемых земель они пре-
вращаются в бесплодные солончаки. Человек уничтожил свыше
двух третей всех когда-то бывших лесов — мощных накопителей
влаги и регуляторов водного режима на нашей планете. Работы
по посадкам и восстановлению лесов далеко не восполняют потерь
в лесе, возрастающих с каждым десятилетием. Но пожалуй, наи-
больший вред растительному миру наносят негативные стороны
научно-технического пропресса, выражающиеся в прогрессирующем
загрязнении биосферы ядовитыми веществами, которые извлека-
ются человеком из земных недр и выбрасываются как продукты
отхода производственных процессов. Участи суши, обжитой чело-
веком, не избегли реки и Мировой океан: мало сохранилось рек
с чистой водой, а прогрессирующее загрязнение морей и океанов
угрожает уже самому существованию растительного планктона
и бентоса — необходимого элемента биосферы, в большей мере
определяющего условия существования человека па Земле. Охра-
на природы — это прежде всего охрана растительного мира — ос-
новы существования человека. Современный уровень науки и тех-
ники, однако, дает возможность нейтрализовать негативные
последствия научно-технического прогресса и совместными усилия-
ми всех стран мира сохранить растительный мир нашей планеты
и приумножить его богатство для нужд человека.
393
ПОНЯТИЕ О ФЛОРЕ И РАСТИТЕЛЬНОСТИ
Эти два понятия не являются синонимами. Современная бота*
ническая наука в понятие «флора» и «растительность» вкладывает
совершенно различное содержание.
Под флорой следует понимать совокупность видов, произра-
стающих на данной территории. Можно говорить о флоре в целом
земного шара, какой-либо страны, государства или природного
района. Каждому периоду в истории Земли соответствует своя
флора, потому что в процессе эволюции одни виды вымирают, дру-
гие возникают и, таким образом, современная флора сложилась
на протяжении всей истории Земли. Растительность — совокупность
естественных сочетаний, группировок растений, покрывающих зем-
ную поверхность.
Подобно флоре можно говорить о растительности земного ша-
ра или какой-либо части земной поверхности. Но если флора опе-
рирует отдельными видами, родами, семействами растений, то объ-
ектами исследования науки о растительности являются такие ка-
тегории, как лес, степь, тундра, луг и другие естественные группи-
ровки растений. Чем разнообразнее и благоприятнее условия жизни
растений в данной стране, тем богаче флора и растительность ее.
Например, на огромном протяжении тундровой зоны насчитывает-
ся не более 300 видов цветковых растений, а в одной лишь Москов-
ской области, занимающей около Vio площади всей тундровой зоны,
произрастает 1100 видов. В Курской области произрастает уже
1300 видов, а в Крыму — свыше 2000. В горной части Средней Азии
произрастает около 6000 видов, а в пустынной части Средней Азии
на территории, в несколько раз большей,— всего 600 видов. На Кав-
казе, отличающемся большим разнообразием природно-климатиче-
ских условий, произрастает более 6000 видов.
Во всем Советском Союзе насчитывается около 18 000 видов
цветковых растений, в Бразилии же, площадь которой в 2,5 раза
меньше площади СССР, — 40 000 видов. На сравнительно неболь-
шом острове Калимантан известно 11 000 видов.
Флора каждой страны слагается из всех видов, ее населяющих.
Вместе с тем каждый вид имеет свою область распространения.
Область распространения данного вида называется его ареалом.
Можно также говорить об ареале рода, семейства, порядка. Неко-
торые растения имеют огромный ареал, иногда охватывающий
несколько материков. Такие виды называют растениями-космопо-
литами. К ним относится, например, тростник (Ph. communis).
Некоторые сорные растения (одуванчик — Taraxacum officinalis,
крапива— Urtica dioica, осот — Sonchus oleraceus) благодаря чело-
веку стали космополитами. Другие виды, наоборот, обитают
на очень ограниченной площади. Такие растения называют энде-
миками. Особенно много эндемиков произрастает в горных райо-
нах, на пустынных островах. Например, около 20% видов всех рас-
тений Кавказа — эндемики. Причина широкого распространения
394
эндемизма в горных странах и на отдаленных островах заключа-
ется в том, что затруднено распространение семян и других зачат-
ков (диаспор).
ЖИЗНЕННЫЕ ФОРМЫ РАСТЕНИЙ
В процессе естественного отбора у растений вырабатывается
определенный внешний облик (габитус), отражающий их приспо-
собленность к условиям среды; растения, по-одинаковому приспо-
собленные к условиям существования, объединяются в одну жиз-
ненную форму, иначе — биологическую форму, биоформу. На од-
ном и том же участке суши могут одновременно обитать,
существовать несколько различных жизненных форм, по-разному
использующих данный комплекс внешней среды. К.одной и той же
жизненной форме могут принадлежать растения совершенно раз-
личных, даже систематически весьма отдаленных семейств.
Наиболее распространена и популярна классификация жизнен-
ных форм, предложенная в начале XX в. датским ботаником
К. Раункером. Эта классификация основана на различном положе-
ни почек возобновления на растении по отношению к уровню суб-
страта (почвы) и снегового покрова. Этот признак характеризует
большую или меньшую защищаемость почек в неблагоприятное
время года.
Раункер выделил следующие пять главных типов жизненных
форм: фанерофиты (Ph)—почки возобновления расположены вы-
соко над землей (деревья, кустарники, деревянистые лианы, эпи-
фиты); хамериты (Ch) — почки расположены не выше 20—30 см
над землей и обычно зимуют под снегом (кустарнички, полукустар-
нички, стелющиеся растения); гемикриптофиты (НК) — многолет-
ние травы, почки которых расположены на уровне почвы и прикры-
ты снегом и отмершими листьями прошлых лет; криптофиты (К) —
почки скрыты под землей (корневища, клубни, луковицы) —геофи-
ты или под водой — гидрофиты; терофиты (Th) — однолетники, пе-
реносящие неблагоприятное время года в виде семян.
Установлено, что раункеровский признак — расположение пере-
живающих почек, хорошо коррелирует с другими признаками, ха-
рактеризующими внешний облик растений.
Классификация Раункера используется для построения биоло-
гического спектра растительности различных зон и других регионов
земного шара. Каждой почвенно-климатической зоне свойственно
характерное количественное соотношение видов, отнесенных к раз-
личным раункеровским типам жизненных форм.
ПОНЯТИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ
В условиях крайнего недостатка факторов жизни растения рас-
тут в одиночку. Так бывает в арктической или, наоборот, сухой
пустыне; на скалах или среди песков; в горах на большой высоте,
где вообще прекращается возможность произрастания высших рас-
395
тений. Обычно в естественной обстановке растения живут сообща;
на одном участке, даже небольшом, произрастают многие десятки,
сотни и тысячи растений. Лишь в исключительных случаях это
бывают растения одного вида; как правило, даже на совсем не-
большом участке суши встречаются растения, принадлежащие
к разным видам.
Произрастающие совместно растения разносторонне воздейст-
вуют друг на друга и на окружающую среду. Нередко они всту-
пают в антагонистические взаимоотношения, борясь за влагу в поч-
ве, свет и даже пространство. Конкретные группировки растений,
однородные по составу и строению, называются растительными
сообществами, или фитоценозами.
Естественные растительные сообщества — не случайные объеди-
нения растений, они сложились исторически, в процессе длительной
эволюции. Поэтому растительные сообщества отличаются извест-
ным постоянством и приуроченностью к данному комплексу эколо-
гических факторов. Если внимательно присмотреться к любому рас-
тительному сообществу, почти всегда можно заметить, что на од-
ном месте произрастают растения, по-разному использующие
факторы жизни, по-своему приспособленные к местным условиям
существования. Например, даже в сравнительно простом травянис-
том сообществе можно найти растения, корневая система которых
не идет глубоко и черпает воду и минеральную пищу из верхних
горизонтов почвы; рядом с такими растениями произрастают виды
со стержневыми корнями, идущими глубоко в почву. То же самое
можно сказать и об использовании света и других факторов;
под пологом высокорослых растений произрастают более мелкие,
которые мирятся с худшим освещением.
Растения, по-одинаковому приспособленные к условиям сущест-
вования, объединяются в одну жизненную форму. К одной и той же
жизненной форме могут- быть отнесены растения совершенно раз-
личных, даже значительно отдаленных семейств. Например, к жиз-
ненной форме мелких эфемеров южной степи можно отнести и вес-
нянку (семейство крестоцветных), и зонтичник обыкновенный (се-
мейство гвоздичных), и веронику раннюю (семейство норични-
ковых) .
Растительные сообщества не остаются неизменными, можно го-
ворить лишь об относительном постоянстве. Даже без вмешатель-
ства человека растительный покров любой страны изменяется. Из-
менения эти могут быть очень медленны, но они неизбежны, так
как на протяжении веков меняется и климат, и почва; да и сами
растения, их естественные сообщества, длительно произрастая
на одном месте, меняют среду обитания и тем самым принимают
участие в эволюции растительного покрова.
В науке, изучающей фитоценозы, из которых состоит раститель-
ный покров Земли, — геоботанике (фитоценологии) основной систе-
матической (таксономической) единицей для классификации рас-
тительных сообществ принята растительная ассоциация, аналогично
тому как в систематике растений основной таксономической еди-
396
ницей принят вид растения. Подобно тому как каждое растение-
относится к определенному виду, каждое растительное сообщество,,
или фитоценоз, относится к определенной растительной ассоциации.
Так же, как в систематике, изучая данный вид растений, мы имеем
дело с конкретными особями, так и в фитоценологии, изучая ассо-
циации, мы изучаем конкретные растительные сообщества, фитоце-
нозы.
В практической работе зоотехника, при исследовании естествен-
ных пастбищ и сенокосов, возникает необходимость описания рас-
тительности естественного кормового угодья, его сельскохозяйст-
венной, зоотехнической оценки.
Приступая к геоботаническому описанию растительного покро-
ва, его прежде всего расчленяют на основные ассоциации и уста-
навливают количественное (по занимаемой площади) участие каж-
дой ассоциации в растительности данного участка. Затем возникает
задача — дать название каждой ассоциации. Название ассоциации
удобней всего составлять из названий растений — эдификаторо&
данного фитоценоза.
Эдификаторами называют один или несколько (обычно не боль-
ше трех) основных видов из данного сообщества, определяющих
весь его строй; это те растения, которые в основном создают внут-
реннюю биотическую среду фитоценоза. Например, в ковыльной
степи это будет ковыль; в ковыльно-типчаковой степи — ковыль
и типчак и т. д.
Для составления названий обычно пользуются латинскими*
названиями, соответственно чему названия отмеченных выше ассо-
циаций будут выглядеть так: в первом случае — асе. Stipa lessin-
giana, во втором— асе. Stipa lessingiana-\-Fesiuca sulcata.
Если второе (и третье) доминирующее растение играет заметно-
меньшую роль по сравнению с главным эдификатором, говорят
иногда о субэдификаторах.
Ассоциации, имеющие общего эдификатора, объединяются
в группы ассоциаций. Сходные по экологическим условиям группы
ассоциаций объединяются в формации. Например, можно говорить
о формации целинной дерновинно-злаковой степи, она будет
объединять ассоциации со сходными эдификаторами: ковылями
и типчаком.
Описание фитоценозов. Каждый фитоценоз при описании его
характеризуют, как минимум, следующими признаками: видовой
состав, ярусность, обилие, покрытие. Для хозяйственной характе-
ристики необходимо также определить производительность фитоце-
ноза, т. е. установить количество растительной массы, произраста-
ющей на единице площади данного фитоценоза.
Видовой состав. Видовой состав растительности описывае-
мого фитоценоза определяет все его признаки и хозяйственные
качества. В списке растений в отношении каждого растения указы-
вают, в какой фазе вегетации оно находится, а также рост растения
в сантиметрах.
397
Рис. 217. Ярусность в степи Аскания-Нова
Яр у сност ь. Как правило, естественный растительный покров
расчленен на ярусы — по относительной высоте растений, образу-
ющих фитоценоз (рис. 217, 218). В травянистых фитоценозах час-
то бывает 2—3 яруса, в лесных больше. Например, в южно-украин-
ской степи можно различать три яруса: 1-й ярус — ковыль,
2-й ярус — типчак и тонконог, 3-й ярус — мелкие однолетники.
В сосновом лесу — четыре яруса: 1-й ярус — сосна, 2-й ярус —
орешник, 3-й ярус — черника, 4-й ярус — зеленые мхи.
В дубовом лесу иногда можно насчитать до семи ярусов.
К первому ярусу относятся наиболее высокорослые растения.
Расчленение растительного покрова на ярусы имеет большое эко-
логическое значение, оно позволяет сочетать в одном фитоценозе
растения с различными требованиями к свету, влаге.
Обилие. Для характеристики фитоценоза важно установить,
в каком количественном отношении находятся виды растений, вхо-
дящие в данное сообщество. Обилие одних видов и, наоборот, не-
большое количество экземпляров других зависит от многих обстоя-
тельств, в том числе от размера растений, большей или меньшей
их приспособленности к наличному экологическому комплексу и др.
Существуют различные шкалы для обозначения обилия видов.
Наиболее употребительна шестибалльная шкала Друдэ. Эта шкала
пользуется следующими условными обозначениями.
Soc. (sociales)—растения смыкаются своими надземными час-
тями и образуют фон, на котором незаметны другие растения.
398
Рис. 218. Схема ярусности в лесу. Слева — трехъярусный, справа — пятиярусный1
фитоценоз (римскими цифрами обозначен порядок ярусов)
Сор.з (copiosae)—растения встречаются весьма обильно,
в очень большом количестве.
Сор.2— растения встречаются обильно, в большом количестве.
Cop.i — растения встречаются довольно обильно, в немалом ко*
личестве экземпляров.
Sp. (sparsae) —растения встречаются изредка, рассеянно, в не*
большом количестве экземпляров.
Sol. (solitariae)—растения встречаются очень редко, единич*
ними экземплярами.
Хотя определение обилия в известной мере субъективно, тем
не менее при достаточном навыке между описаниями двух исследо-
вателей обычно баллы по данному растению различаются не более
чем на одну ступень.
Покрытие. Этот показатель более точно, чем обилие, отража*
ет степень участия данного вида в формировании травяного покро-
ва. Степень покрытия определяют при взгляде на растительный
покров сверху вниз и выражают этот показатель в процентах.
Существует несколько методов определения покрытия. В про*
стейшем случае мы мысленно фиксируем небольшой участок данно*
го фитоценоза и на глаз прикидываем, какую часть почвы (в про-
центах) покрывают надземные части данного растения. Более точ*
но этот показатель можно определить посредством квадратной
метровой рамки, разделенной проволокой на клетки. Наиболее
удобно разделить метр на 25 клеток, тогда каждая клетка будет
соответствовать 4% покрытия. Покрытие — более объективный по-
казатель, чем обилие, и, если нет очень большой разницы в высоте
растений, этот показатель довольно близко отражает степень учас-
тия каждого вида в формировании фитоценоза.
Для более тонкой кормовой оценки растительного покрова паст*
бищ и сенокосов применяют еще некоторые приемы исследования:
изучают структуру травостоев и структуру урожая многолетних,
трав.
399
При изучении структуры травостоев определяют массу надзем-
ных органов, их объем, поверхность, особенно листовую, в наиболь-
шей степени отражающую кормовую ценность растений. Исследу-
ют также размещение органов растений в пространстве: в течение
вегетации и от года к году.
При изучении структуры урожая многолетних трав подсчитыва-
ют число генеративных, вегетативных удлиненных и вегетативных
укороченных побегов; определяют массу каждой категории побегов,
причем генеративные побеги разделяют на соцветия, стебли
и листья, вегетативные удлиненные побеги — на стебли и листья.
Весовой анализ. При геоботаническом исследовании пастбищ
и сенокосов для хозяйственной оценки их необходимо сделать весо-
вой анализ растительного покрова, т. е. определить весовое участие
каждого вида в создании общей растительной массы на момент
обследования.
Для правильного решения этой задачи необходимы повторные
определения по каждой ассоциации. Практически наиболее удоб-
ны квадратные метры, накладываемые не менее чем в четырехкрат-
ной (лучше десятикратной) повторности на каждой ассоциации
изучаемого кормового угодья. Ограниченную рамками метра расти-
тельность срезают овечьими ножницами у самой поверхности поч-
вы, а затем срезанные растения разбирают по видам и взвешивают.
Лучше разбирать сразу же на месте и определять массу в сы-
ром, а затем, после высушивания, и в сухом состоянии. При таком
срезе мы определяем валовую производительность данного участка.
Для кормовой оценки растительного покрова пастбищ и сеноко-
сов при весовом анализе виды растений объединяют в кормобота-
нические группы. В лесной, лесостепной и степной зонах обычно
составляют следующие 5 групп: злаки, бобовые, осоки, разнотравье,
ядовитые. В группу «разнотравье» объединяют все двудольные
растения за вычетом бобовых и ядовитых.
Растительность полупустынь и пустынь более разнообразна
в кормовом отношении. Здесь обычно растения распределяют
на 10 групп: злаки, бобовые, осоки, эфемеры, полыни, солянки су-
хие, солянки сочные, прочие травы и полукустарники, кустарники,
ядовитые.
Хозяйство интересуется не валовой, а кормовой производитель-
ностью массы, которая может быть отчуждена при сенокосном
или пастбищном использовании. В первом случае задача решается
проще: растения срезают на уровне среза сенокосилкой (5—10 см)
и срезанную траву анализируют. Сложнее обстоит дело на паст-
бищах. Для правильного учета кормовой производительности здесь
необходимо знать степень стравливания каждого вида в нормаль-
ных условиях пастьбы (в процентах). Срезав траву у поверхности
почвы и разобрав ее по видам, массу каждой фракции затем ум-
ножают на соответствующий коэффициент и полученные величины
складывают.
Пасущееся животное в течение пастбищного сезона один и тот
же участок пастбища стравливает несколько раз. Поэтому и опре«
400
деление производительности пастбищ приходится делать не один,
а несколько раз за сезон, имитируя пастьбу. Гораздо точнее эта
задача решается в условиях загонной пастьбы, когда учитывают
запас травы до использования и остаток после стравливания. Де*
тально об этом говорится в соответствующих методических посо*
биях.
Геоботанические исследования растительного покрова являются
обязательным элементом в системе мероприятий по культурному
использованию пастбищ и сенокосов.
ОСНОВНЫЕ ТИПЫ РАСТИТЕЛЬНОГО
ПОКРОВА СССР
Растительность СССР имеет хорошо выраженный зональный
характер. Рассматривая карту растительности нашей страны, легко
заметить, что изменение растительного покрова следует определен*
ной закономерности; обширные пространства тундры, простираю-
щейся сплошной полосой от границы с Норвегией на западе
и до Чукотки на востоке, сменяются полосой леса, также вытяну*
той с запада на восток; южнее леса сменяются степями, еще далее
на юг степи замещаются пустынями. Эти обширные полосы в об-
щем однородной растительности, называются растительными зо-
нами.
Растительные зоны совпадают с соответствующими почвенными
зонами. Изучая почвенный покров нашей страны, В. В. Докучаев
впервые сформулировал свое учение о зонах природы.
Зональность в распределении растительности (и почв) отража*
ет общую закономерность распределения на земном шаре клима*
тических факторов, в частности таких факторов, как солнечная
радиация, количество тепла и насыщенность почвы и атмосферы
влагой. Как раз эти последние два фактора в основном и опреде-
ляют характер растительности той или иной страны.
Отчетливому проявлению зональных закономерностей благо*
приятствуют обширность и относительная равнинность нашей стра-
ны. От Балтийского моря на западе и до Охотского моря на вос-
токе простирается равнина, почти не нарушаемая крупными возвы-<
шенностями. Уральские горы настолько сглажены, что не вносят
существенных изменений в общую закономерность. Растительные
зоны, дойдя до Урала, почти неизменно продолжаются дальше
на восток, за Уральским хребтом.
Конечно, речь идет о растительности, покрывающей основные
элементы рельефа, т. е. относительно ровные водораздельные про-
странства, которые получили название плакорных. На общем фоне
плакорной зональной растительности, в любой зоне можно встре-
тить растительность, приуроченную к понижениям, возвышен-
ностям или склонам различной крутизны. Эта растительность хотя
и .подчиняется зональным влияниям, однако может резко отличать-
ся от основной, зональной растительности; ее называют интразо-
401
нальной растительностью. Интразональна, например, раститель-
ность лугов и болот.
Зональность, о которой идет речь, получила название горизон-
тальной. Эта смена растительности следует за географической
широтой. В условиях гор, в связи с поднятием над уровнем моря,
можно также наблюдать изменение растительности по зонам.
Это выражение вертикальной зональности.
По мере поднятия в горы климатические факторы изменяются
подобно тому, как это можно наблюдать при перемещении от эк-
ватора к полюсу. Однако здесь эти изменения происходят гораздо
быстрее. Достаточно бывает подняться на несколько сот метров,
чтобы попасть из одной вертикальной зоны в другую, в то время
как ширина горизонтальных зон измеряется сотнями километров.
Вертикальные зоны часто называют растительными поясами. Этим
оттеняется нетождественность этих двух сходных явлений.
Вертикальная зональность еще более резко нарушается в связи
с характером рельефа, нежели горизонтальная зональность. В го-
рах, нередко в непосредственной близости, можно встретить луго-
вую, степную и полупустынную растительность. Все зависит
от экспозиции и крутизны склонов, определяющих качество почвы
и степень обеспечения ее влагой.
В кратком обзоре растительного покрова СССР мы остановим-
ся лишь на характеристике основных растительных зон, на интра-
зональной растительности лугов и болот и на основных закономер-
ностях распределения растительности горных районов нашей
страны.
Арктическая пустыня. К этой зоне относятся северная оконеч-
ность Таймырского полуострова и северные острова Ледовитого
океана: Шпицберген, Земля Франца Иосифа, северный остров Но-
вой Земли, Северная Земля, Новосибирские острова и остров
Врангеля.
В арктической пустыне сплошного растительного покрова нет.
Почвенный покров лишь в первоначальной стадии образования,
преобладают голые скалы и каменные россыпи. Растительность
представлена в основном накипными лишайниками и куртинками
мелких зеленых мхов.
Лишь в местах, защищенных от ветра, можно встретить одиноч-
ные растения или куртинки цветковых растений.
Зона тундры. Характерная черта растительности тундры — без-
лесие. Немногие деревянистые растения — мелкие стелющиеся де-
ревца и кустарники. Травянистые растения почти все многолетники.
Хотя осадков в тундре выпадает очень мало, не больше, чем
в степи, растения произрастают здесь при избытке влаги в воздухе
и почве. Это объясняется большой облачностью, низкой испаряе-
мостью и наличием вечной мерзлоты, препятствующей проникнове-
нию влаги в глубь почвы..
Пространства, занимаемые тундрой, огромны, свыше 3 млн.
км2, что составляет 15% всей территории СССР. Естественно, что
на таком огромном пространстве тундры не могут быть однородны-
402
ми: сказываются различия в рельефе, почвенном субстрате, боль-
шей или меньшей суровости климата.
Различают лишайниковую тундру на песчаных почвах. Как по-
казывает название, здесь преобладают лишайники (Cladonia,
Cetraria и др.). В кормовом отношении — это лучший тип тундр,
так как лишайники — хороший корм для северного оленя. На такой
же сухой песчанистой почве бывают развиты кустарниково-лишай-
никовые тундры. Наряду с лишайниками здесь произрастают кус-
тарники: карликовая береза (Betula папа), голубика (Vactinium
uliginosum), водяника (Empetrum nigrum) и др.
На суглинистом субстрате преобладают моховые тундры с гос-
подством зеленых мхов; моховые тундры в кормовом отношении
представляют меньшую ценность.
По низинам в зоне тундр очень много болот, преимущественно
осоковых, торфяников (сфагновых) немного. В комплексе с боло-
тами участки бугристых тундр. На буграх растут осоки (С. rariflo-
ra, С. rotundata и др.), пушица (Eriophorum angustifolia) и др.
Здесь же можно встретить карликовую березу (Betula папа).
Флора тундры крайне бедна цветковыми растениями. Это ис-
ключительно многолетние формы: травы, полукустарники и кустар-
ники.
Невысокая продуктивность растительного покрова тундры ком-
пенсируется огромными площадями, поэтому в кормовом отноше-
нии тундра представляет большую ценность; здесь выпасаются
многочисленные стада северного оленя — основного сельскохозяй-
ственного животного в тундровой зоне.
Зона лесов. Эта обширнейшая зона широкой полосой простира-
ется от западной до восточной границы СССР. С севера к ней при-
мыкает зона тундр, с юга — зона степей. Между тундрой и лесами
лежит переходная область — лесотундра; между лесом и степями —
лесостепь. В этих переходных областях тундра и степь занимают
плакорные места, леса — понижения, балки, долины рек. Леса за-
нимают около V2 всей территории СССР.
В среднем в зоне лесов выпадает около 500 мм осадков; это ко-
личество обеспечивает устойчивость земледелия. Значительные пло-
щади в лесной зоне распаханы, особенно в европейской части Со-
ветского Союза.
В СССР преобладают хвойные леса, площадь лиственных лесов
составляет не более !/ю от общей площади лесной зоны. Леса с пре-
обладанием хвойных пород получили название тайги. Слово «тай-
га» обычно применяется по отношению к хвойным лесам Сибири.
Основные массивы лиственных лесов расположены на юго-за-
паде европейской части Советского Союза, в районе Карпат, на юге
Западной Сибири и на Дальнем Востоке, в Приморье и южном
Приамурье. В европейской части Советского Союза между подзо-
нами хвойных и лиственных пород широко распространены сме-
шанные леса, состоящие из хвойных и лиственных пород.
Хвойные леса подразделяются на темнохвойные и светлохвой-
ные, в зависимости от господствующей породы. Темнохвойные
403
леса образуют ели, пихты и сибирский кедр. Светлохвойные леса —
сосны и лиственницы. Все эти хвойные породы, кроме лиственницы,
сохраняют хвою круглый год. У лиственниц хвоя нежная, опадаю-
щая на зиму. Темнохвойные леса преобладают в европейской части
лесной зоны, в Западной Сибири и на Дальнем Востоке. В Восточ-
ной Сибири преобладают светлые, преимущественно лиственнич-
ные леса.
В хвойных смешанных лесах европейской части Союза распро-
странены два вида ели: в западной половине ель обыкновенная
(Picea excelsa), в восточной половине ель сибирская (Р. obovata).
Эта же ель распространена и в Сибири. К обоим видам ели бывает
примешана сосна обыкновенная (Pinus silvestris), которая нередко
образует и чистые насаждения — боры.
Так же как и ель, сосна входит в состав смешанных лесов. На-
ряду с елью сибирской в темнохвойной западносибирской тайге
распространен кедр сибирский (Pinus sibirica) и пихта сибирская
(Abies sibirica).
Основными лесообразующими породами светлохвойных лесов
восточной Сибири являются лиственницы: на западе сибирская
(Larix sibirica), на востоке даурская (L. dahurica). Нередко в сооб-
ществе с лиственницей сибирской растет и сосна обыкновенная.
Хвойным породам нередко сопутствуют лиственные деревья
и кустарники: дуб (Quercus), липа (Tilia), орешник (Corylus) и др.
На почвах, хорошо увлажненных, но не заболоченных, в хвой-
ном лесу бывает хороший травянистый покров, растут кустарники
(смородина — Ribes nigrum, шиповник — Rosa, жимолость — Loni-
сега) и лиственные деревья (черемуха, черная ольха — Alnus glu-
tinosa, липа и др.). В светлых сосновых лесах, растущих по песча-
ным дюнам, напочвенный растительный покров нередко состоит
из лишайника — оленьего «мха» (Cladonia).
Лиственные леса подразделяются на широколиственные и мел-
колиственные. Такое подразделение основано на размере листовой
пластинки. К деревьям с крупной листовой пластинкой относятся
такие породы, как дуб, клен (Acer), ясень (Fraxinus), бук (Fagus),
граб (Carpinus), вяз (Ultnus) и др.
Крупные листья создают больше тени, поэтому леса, где преоб-
ладают широколиственные породы, более тенисты, составляющие
их деревья характеризуются большей теневыносливостью.
К мелколиственным породам относятся осина (Populus tremula)
и береза (Betula).
Крупные массивы широколиственных лесов сохранились в Кар-
патах и в прикарпатских районах. Основная порода здесь бук
(Fagus silvatica).
Буковые леса широко распространены также в горах Кавказа
и Крыма, но здесь другие виды бука: на Кавказе бук восточный
(Fagus orientalis), в Крыму — таврический (F. taurica).
В южной части лесной зоны на европейской территории Совет-
ского Союза широко распространены дубовые леса — дубравы.
Эдификатор этих лесов — дуб черешчатый (Quercus robur). Сопро-
404
вождающие породы: клен остролистный (Acer platanoides), ясень
(Fraxinus excelsior), липа (Tilia cordata), ильм (Ulmus). В под-
леске обычен орешник (Corylus avellana), а также бересклеты
(Evonimus), свидина (Cornus sanguinea), жостер (Rhamnus cathar-
tica). Летом в дубравах бывает довольно обильный травянистый
покров из широколистных злаков и двудольных.
На Дальнем Востоке в широколиственных лесах основная по-*
рода — дуб монгольский (Q. mongolica). К нему примешивается
береза даурская (В. dahurica). В подлеске: орешник разнолистный
(Corylus heterophyНа), леспедеца (Lespedeza bicolor) и др.
В Западной Сибири на южной границе лесной зоны имеются
большие массивы березово-осиновых лесов, по-видимому, первич-
ного происхождения.
Зона степей. Как уже сказано, между зоной лесов и степями
лежит область лесостепи. По водораздельным, плакорным элемен-
там рельефа здесь распространены степи (в северных районах —
лугостепи), а в понижениях — леса. Среди лесных пород лесостепи
на европейской территории СССР преобладает дуб, за Уралом дуб
замещается березой. Пространства лесостепи, свободные от леса,
почти полностью распаханы. Целинная растительность, сильно из-
мененная чрезмерным выпасом, сохранилась лишь на более крутых
склонах.
В зоне степей первобытная целинная растительность на значи-
тельной территории сохранилась лишь к востоку от Урала, хотя
здесь и распаханы в последние годы десятки миллионов гектаров.
В европейской части Советского Союза степи полностью распаха-
ны, если не считать небольшие участки, охраняемые в заповедни-
ках. Эти заповедные участки дают возможность составить представ-
ление о степях, когда-то покрывавших огромные пространства степ-
ной зоны.
В степях преобладают пространства с ровным рельефом. Расти-
тельность в общем хотя и ксерофитного характера, однако расти-
тельный покров более или менее сомкнут. В отличие от лугов степи
в середине лета испытывают недостаток влаги, и это заметно ска-
зывается на летнем состоянии растительности, в южной части
степной зоны она даже выгорает.
При большой протяженности степной зоны с севера на юг
и с запада на восток растительный покров ее не может быть оди-
наковым. По направлению от северной границы к югу и с запада
на восток степи становятся суше и беднее. Юго-восточный край
степной зоны граничит уже с переходной областью полупустыни.
В северной части степной зоны выпадает 400—500 мм осадков,
и при благоприятном распределении их в году это количество до-
статочно для получения высоких урожаев полевых культур. К югу
годовое количество осадков уменьшается до 300 мм, весна и лето
здесь часто бывают засушливые. Однако и в этой части степной
зоны получают достаточно высокие урожаи зерновых культур.
Вместе с тем это зона, наиболее подходящая для развития
пастбищного скотоводства и овцеводства.
405
В связи с большими различиями в растительном покрове при-
нято зону степей делить на две подзоны: северные степи — с боль-
шим содержанием в травостое разнотравья, и южные степи — с пре-
обладанием в травостое ковыля и типчака. Растительный покров
северных степей почти полностью сомкнут, в южной степи расти-
тельность более разреженная, между дерновинами ковыля и тип-
чака здесь нередко проглядывает голая почва. Северные степи
полностью распаханы, сохранились лишь небольшие участки в за-
поведниках.
Характеристику южных степей, еще имеющих производственное
значение, можно дать на примере асканийской степи (И. К. Пачо-
ский, М. С. Шалыт).
Для южной степи характерно обилие весенних эфемеров и эфе-
мероидов. Из эфемеров в это время обильно цветут крупка
(Erophila verna), вероника (Veronica verna), незабудка (Myosotis
arenaria), ясколка (Cerastium ucrainicum) и др.
Из эфемероидов — мятлик луковичный, живородящий (Роа bul-
bosa v. vivipara), несколько видов гусиного лука (Gagea pusilia,
G. bulbifera, G. schovitsii) и др. Несколько позже зацветают тюль-
паны (Tulipa schrenkii) и касатики (Iris pumila). К концу весны
зацветают перистые ковыли (Stipa lessingiana, S. ucrainica), тип-
чак, а из разнотравья — будяк (Carduus uncinatus) и коровяк
(Verbascum phoeniceum). В разгар лета цветут фломис (Phlomis
pungens), шалфей (S. nemorosa, S. aephiopis), васильки (Centaurea
rhutenica), ромашник (Pyrethrum millefoliatum) и др.
К концу лета зацветает ковыль тырса (Stipa capillata), а из раз-
нотравья кермек (Statice tatarica), луки (A, guttatum, A. paczoskia*
num), грудница (Linosyrus villosa) и др. К середине лета количест-
во цветущих растений резко падает и степь начинает засыхать,
общий фон ее после отцветания перистых ковылей становится блек-
ло-желтым. Если благоприятствует погода, осенью степи зеленеют;
отрастают дерновинки ковылей и типчака, возобновляет рост мят-
лик луковичный, появляются обильные всходы озимых однолетни-
ков (вероника, незабудка) и других весеннецветущих эфемеров.
Южные степи, благодаря преобладанию дерновинных злаков,
хорошо выдерживают вытаптывание и являются прекрасным весен-
нераннелетним пастбищем. Однако чрезмерная и бессистемная
пастьба приводит к деградации и этих степей. Ценные дерновинные
злаки уступают место мятлику луковичному, полыни австрийской
(Artemisia austriaca), молочаю (Euphorbia seguieriana), и, наконец,
при очень сильном сбое, на пастбищах преобладает спорыш
(Polygonum aviculare) и устели-поле (Ceratocarpus arenarius) —
колючий стелющийся однолетник.
При такой степени деградации степь уже трудно вернуть к ее пер-
вобытному, естественному состоянию, обеспечивающему, хотя
и не высокий, но стабильный урожай прекрасного пастбищного
корма.
Зона пустыни. От северо-восточных берегов Каспийского моря
на западе и до границы с Китаем на востоке, в пределах Средней
406
Азии, простираются пустыни, прерываемые лишь руслами рек,
оазисами и хребтами высоких гор. На севере пустынная зона через
переходную область полупустынь граничит с зоной степей, на юге
пустыни доходят до государственных границ СССР.
Переходная область полупустыни характеризуется комплекс-
ностью почв и растительности. В растительном покрове участки
разреженной сухой степи комплексируются с участками, на кото-
рых пробладают ксероморфные полукустарники типа полыни мор-
ской (Artemisia maritima) и кохии распростертой (Kochia pros-
trata.)
Такая мозаичность растительного покрова связана с засолен-
ностью почвы; в условиях избытка солей даже незначительные по-
нижения рельефа резко сказываются на характере растительности.
Полупустыня является переходной зоной и в климатическом
отношении. Континентальность климата, свойственная уже южным
степям, наивысшего выражения достигает в зоне пустынь.
Климат полупустынь более сухой и жаркий, чем в зоне степей,
но здесь все же удается иногда в благоприятные годы получить
урожай зерновых без полива. В зоне пустынь возможно только
поливное земледелие, настолько климат здесь засушлив. Количе-
ство осадков в .пустыне не превышает 200 мм в год. К этому при-
соединяются очень высокие летние температуры (до 40° С) при до-
вольно холодной для данной параллели, хотя и короткой зиме.
Характерная черта климата пустынь — та же сухость воздуха и ог-
ромная испаряемость, в 5—7 раз превышающая количество выпа-
дающих осадков.
По характеру почвенного субстрата различают 4 типа пустынь:
1) глинистые, 2) песчаные, 3) солончаковые, 4) каменистые (гип-
совые).
Основной северный массив глинистой пустыни простирается
широкой полосой от Каспийского моря до гор Тянь-Шаня и Алтая
на востоке, занимая северную часть советской Средней Азии. Лишь
в районе Аральского моря и озера Балхаш глинистые пустыни пре-
рываются крупными участками песков. Этот массив глинистой
пустыни на севере граничит с областью полынно-злаковой полу-
пустыни и по площади составляет свыше !/3 всей территории Сред-
ней Азии.
Для этого основного участка глинистых пустынь характерно
преобладание в растительном покрове ксероморфных полукустар-
ников: полыни (A. terrae-albae), биюргуна (Anabasis alba), кокпека
(Atriplex alba), итсегека (Anabasis aphylla). Здесь же заросли чер-
ного саксаула (Haloxylon aphyllum). Это растения в форме неболь-
ших корявых деревьев с редуцированными листьями. Ассимилиру-
ют зеленые веточки.
Глинистые пустыни южного типа занимают меньшую площадь.
Они расположены на крайнем юге Средней Азии, в предгорьях
западного Тянь-Шаня, Памиро-Алая и Копетдага. Основу траво-
стоя глинистой пустыни южных частей Средней Азии составляют
два растения: осока пустынная (Carex pachystylis) *и мятлик луко-
шт
винный (Роа bulbosa). К ним в большом количестве видов приме-
шиваются мелкие однолетники, преимущественно из семейства
крестоцветных, гвоздичных, бобовых и злаковых.
Весной, когда в почве имеется достаточно влаги, растительный
покров этой пустыни напоминает луг. Он густым ковром покрывает
почву. Это очень ценная в кормовом отношении растительность.
Однако в таком состоянии растительность находится недолго; на-
ступает лето, и все растения засыхают, активный период жизни
их продолжается всего около трех месяцев в году.
Солончаковые пустыни и территориально и экологически при-
мыкают к глинистым пустыням северного типа. Почвы солончако-
вых пустынь насыщены солями. В местах наиболее засоленных
растительный покров вообще отсутствует. Участки менее засолен-
ные покрыты редкой растительностью из экологической группы
галофитов. Это прежде всего многочисленные виды солянок из ро-
дов Salsola, Sueda, Anabasis, Salicornia, Halocnemum и др., относя-
щиеся к группе суккулентных растений. Галофиты-несуккуленты
относятся преимущественно к семействам Plumbaginaceae, Tama-
ricaceae, Caryophyllaceae. Характерен также галофитный злак
прибрежница солончаковая (Aeluropus littoralis). Высокую степень
засоленности выносит упомянутый выше черный саксаул.
Совершенно иначе выглядят песчаные пустыни. Растительность
их богаче, разнообразней. Это объясняется лучшей влагообеспе-
ченностыо: песок водопроницаем, и под верхним сухим слоем его
скопляется значительное количество влаги; песок конденсирует во-
дяные пары из воздуха; ночью вода по широким капиллярам опу-
скается вглубь и затем может быть использована корнями растений.
В песчаной пустыне произрастают деревья белого саксаула
(Haloxylon persicum), многочисленные кустарники (виды Calligo-
num, Halimodendron, Ammodendron, Tamarix, Elaeagnus, Astraga-
lus) и др., из полукустарников Ephedra виды астрагалов. Из травя-
нистых растений наиболее характерны злаки из рода селин (Aristi-
da) и особенно песчаная осока (Carex physodes). Роль этой осоки
аналогична роли пустынной осоки в глинистой пустыне. Здесь
ей также сопутствует мятлик луковичный, несколько однолетних
видов костра (Bromus tectorum, В. oxydon) и др.
Песчаные пустыни занимают огромные пространства к юго-за-
паду от р. Сырдарья (пустыни Каракумы, Кызылкумы и др.). Это
круглогодичные овечьи пастбища, широко используемые в караку-
леводстве.
Каменистые пустыни развиваются на почвах, богатых гипсом,
в комплексе с глинистыми пустынями. Обилие гипса обусловливает
каменистость почвенного субстрата. К растениям-полукустарникам,
указанным для глинистых пустынь, здесь присоединяются виды,
специфичные для данного типа пустынь: полукустарничек тас-би-
юргун (Nanophyton erinaceum), растения-колючки (Zygophyllum)
и другие пахучие растения — эфироносы из семейства Lamiaceae,
Rutaceae, Apiaceae; растения, выделяющие из поверхности гипс
(Statice, Tamarix, Frankenia и др.). Вообще в связи с особенностью
408
почвенного субстрата здесь большой процент своеобразных энде-<
мичных видов.
На пустынях мы заканчиваем краткий обзор зональных типов
растительности Советского Союза.
Луга. Лугами называют пространства, покрытые сплошным,
сомкнутым ковром мезофитной травянистой растительности. Такая
растительность может быть только в условиях достаточного,
но не чрезмерного увлажнения.
Больше всего настоящих мезофитных лугов встречается в лес-
ной зоне, обычно на месте сведенных лесов, в поймах больших
и средних рек. Луга, однако, имеются и в лесостепной области,
и в степной зоне, но для этих зон они не типичны. Широко и раз-
нообразно представлены луга в горных условиях. Об этих лугах
мы скажем при описании растительности горных районов СССР;
здесь же речь будет идти только о равнинных лугах.
В зависимости от того, какие геоморфологические элементы
земной поверхности занимает луговая растительность, различают
луга материковые, развитые на водораздельных пространствах,
и не заливаемые полыми водами, и луга заливные, или поемные,
расположенные в поймах рек и заливаемые полыми водами.
Материковые луга подразделяются на суходольные, располо-
женные на возвышенных элементах рельефа, и низинные, располо-
женные в понижениях рельефа. Суходольные луга получают воду
только за счет осадков, да и то часть выпадающих здесь осадков
скатывается в понижения рельефа. Низинные луга обеспечиваются
не только водой осадков, но почвенно-грунтовой и натечной.
Суходольные луга, особенно в наиболее возвышенных местах,
имеют небогатую и нпзкопроизводительную растительность. Из зла-
ков здесь встречаются овсяница красная (F. rubra), мятлик луговой
(Р. pratensis), полевица обыкновенная (Agrostis vulgaris), белоус
(Nardus stricta), душистый колосок (Anthoxanthum odoratum),
трясунка (Briza media) и др. Из семейства бобовых здесь могут
быть клевер луговой (Trifolium pratense), ползучий (Т. repens), чи-
на луговая (Lathy г us pratesis), лядвенец рогатый (Lotus cornicu-
latus) и др. Из других семейств василек луговой (Centaurea jaceae),
тысячелистник обыкновенный (Achillea millefolium), подорожник
средний (Plantago media), лапчатка серебристая (Potentilla ar gen-
tea), лютик едкий (R. асег), подмаренник северный (Galium
boreale) и многие другие. В связи с небольшой производитель-
ностью и изобилием разнотравья луга этого типа в кормовом отно-
шении расцениваются невысоко .и, как правило, нуждаются в ко-
ренном улучшении.
Низинные луга более производительны, но травостой здесь за-
частую содержит малоценных растений больше, чем на суходоль-
ном лугу. Очень часто и в большом количестве встречаются мало-
ценные злаки: щучка дернистая (Deschampsia caespitosa), полеви-
ца обыкновенная и собачья (A. vulgaris, A. canina), душистый
колосок. Но есть и ценные злаки: тимофеевка луговая, лисохвост
луговой, овсяница луговая; из бобовых — клевер ползучий; из дру-
14—326
409
гих семейств: купальница (Trollius europeus), горец, или раковые
шейки (Polygonum bistorta), гравилат речной (Geutn rivale).
Потенциально низинные луга богаче суходольных; после удале-
ния избыточной влаги здесь можно создавать ценные травостои
с большим количеством кормовых растений.
Заливные луга ежегодно или почти ежегодно заливаются полы-
ми водами, которые несут с собой большое количество мелкоземис-
тых частиц, смытых с водосборной площади реки. Оседая, эти
частицы образуют весьма плодородные аллювиальные (наносные)
почвы.
Пойму по ее ширине можно разделить на три части: прирусло-
вую, примыкающую к руслу реки, центральную и притеррасную,
прилегающую к незаливаемой возвышенной полосе берега.
В прирусловой зоне поймы преобладает разнотравно-крупно-
злаковая растительность. Очень характерен для песчаных наносов
этой зоны подбел (Petasites spurius) с большими лопухообразными
листьями. Обладая мощными корневищами, это растение не боит-
ся ежегодных изменений в уровне песчаных наносов. Тут же за-
росли ив (S. triandra, S. viminalis) и др. Рыхлый субстрат благо-
приятен для развития зарослей корневищных злаков: пырея ползу-
чего, вейника наземного, костра безостого. Из бобовых встречаются
люцерна желтая, клевер горный (Т. montana) и др. Из разнотравья:
порезник горный (Libanotis montana), тысячелистник обыкновен-
ный, подмаренник (Galium verum), лапчатка серебристая и др.
В общем преобладают растения, приспособленные к рыхлому
субстрату с довольно глубоким расположением водоносного слоя.
В притеррасной зоне, наоборот, обитают растения, приспособ-
ленные к близкому стоянию грунтовых вод: много осок, ситников,
тростник, камыш. Из злаков — щучка дернистая. Из древесных
растений характерна ольха клейкая (Alnus glutinosa)—в местах
с поверхностным скоплением воды. Нередко луга переходят в коч-
карные и моховые болота.
Центральная часть поймы наиболее богата ценными кормовы-
ми растениями. Среди них тимофеевка луговая, лисохвост луговой,
овсяница луговая, мятлик луговой, пырей ползучий, полевица бе-
лая; из бобовых — клевера, вики, чины. Разнотравья сравнительно
немного. В условиях рационального использования в центральной
пойме можно накашивать много сена. Это самые ценные естествен-
ные кормовые площади, подлежащие всемерной охране.
Болота. Наиболее крупные площади заболоченных земель распо-
ложены в лесной зоне, особенно в северной части этой зоны. Очень
большой процент заболоченных земель в северо-западных районах
европейской части Советского Союза и Западной Сибири.
Для болота характерны два основных признака: 1) избыточное
увлажнение и 2) накопление неразложившегося мертвого органи-
ческого вещества.
Болото может возникнуть различными путями. В северных рай-
онах основные площади болот образовались в результате заболачи-
вания лесов, лесных вырубок или гарей и лугов. В лесостепных
410
и степных районах болота образуются обычно в результате зара-
стания и заторфовывания водоемов.
Заболачивание лесов, лесосек и пожарищ (гарей) начинается
с поселения мхов. Мхи, особенно белый мох (Sphagnum), облада-
ют большой гигроскопичностью. В условиях слабой испаряемости,
свойственной лесной зоне, происходит прогрессирующее наращи-
вание слоя мха, напитанного водой.
Заболачивание лугов может происходить в результате скопле-
ния застойных* почвенных вод и развития плотнокустовых злаков
и осок. Дерновни-ки плотнокустовых растений, отмирая, закупори-
вают почву, на поверхности ее и в толще отмершей растительной
массы скапливается избыточная влага; поселяются мхи, вначале
зеленые, а затем и сфагновые, — на месте луга возникает болото.
Заболачиванию лугов способствует также чрезмерная нерегули-
руемая пастьба.
Заболачивание (зарастание) водоемов начинается с накопления
на дне водоема (например, озера) ила. Вначале ил в основном
минерального происхождения, затем, по мере обогащения водоема
живыми организмами (планктон и бентос), накапливается органи-
ческий ил — сапропель, состоящий из отмерших остатков микроско-
пических организмов. Слой сапропеля постепенно нарастает, и во-
доем мелеет.
Глубоководные растения (хары, рдесты, роголистники), цели-
ком погруженные в воду, заменяются растениями, листья которых
лежат на поверхности воды (белая кувшинка, желтая кубышка).
В более мелких местах поселяются камыш, рогоз, тростник, отно-
симые к группе гигрофитов. Выше располагаются осоки и другие
сопутствующие им растения. Отмершие остатки этих растений,
скопляясь на дне водоемов, превращаются в торф.
Кормовое значение растительности болот невелико. Пастьба
животных здесь затруднена труднопроходимостью, да и хороших
пастбищных растений здесь почти нет. Уборка трав на сено или си-
лос также связана со значительными трудностями. Сено из болот-
ных трав невысокого качества.
РАСТИТЕЛЬНОСТЬ ГОРНЫХ РАЙОНОВ СССР
Растительность гор, с точки зрения ее кормового использова-
ния, представляет особый интерес. Здесь располагаются лучшие
естественные пастбища. Разнообразие в рельефе и абсолютной вы-
соте позволяет иметь на сравнительно небольшом протяжении
пастбища различных экологических типов и всех или почти всех
сезонов использования.
В связи с этим в горных странах издавна получила распростра-
нение своеобразная форма отгонного животноводства. Особенно
богаты прекрасными альпийскими и субальпийскими пастбищами
горы Средней Азии. Мы дадим здесь лишь самую краткую харак-
теристику растительности Карпат, Крыма, Кавказа, Средней Азии
и Алтая.
14*
411
Растительность Карпат. В Карпатских горах различают следую-
щие основные пояса растительности: 1) пояс хвойношироколист-
венных лесов, где основные растения пихта (Abies alba), ель (Picea
excelsa), бук (Fagus silvatica), граб (Carpinus betulus), дуб (Quer-
cus robur) и др. (абсолютная высота 600—1200 м); 2) пояс елово-
пихтовых лесов с примесью бука (1200—1600 м); 3) пояс субаль-
пийской и альпийской растительности. Для животноводов пред-
ставляет интерес именно этот последний, наиболее высокогорный
пояс растительности (1600—2000 м); здесь расположены основные
пастбища Карпатских гор — полонины.
Для субальпийских луговых ассоциаций характерно преоблада-
ние в травостое малоценного в кормовом отношении злака белоуса
(Nardus stricta). Обилие этого злака говорит о сильной деграда-
ции пастбищ, наступившей в результате чрезмерного и бессистем-
ного стравливания, практиковавшегося многие годы. К белоусу
примешаны другие, более ценные злаки: овсяница красная (F. rub-
ra), овсяница рисованная (F, picta), полевица скальная (А. ги-
pestris), а также второй малоценный злак — шучка дернистая
(Deschampsia caespitosa). Из разнотравья — лапчатка прямая
(Р. erecta), арника горная (Arnica niontana), чабрец альпийский
(Thymus alpestris), ястребинка альпийская (Hieraceum alpinum),
осоки (Carex pilulifera и др.), ожика (Luzula sudetica, L. nemorosa
и др.). Из полукустарников — черника (V actinium myrtilis), из дре-
весных— стланиковая сосна (Pinus mughus), можжевельник (J. si-
birica). Эти формы можно видеть на границе леса и безлесных
пространств — полонии.
Для альпийской растительности, расположенной обычно на вы-
соте свыше 1800 м, характерны ассоциации с господством осоки
вечнозеленой (С. sempervirens) или овсяницы лежачей (F. supina)
и рисованной (F. picta). Из разнотравья обильны эдельвейс (Leon-
topodium alpinum), скабиоза блестящая (Scabiosa lucicla), сон-тра-
ва (Pulsatilla alba), колокольчик альпийский (Campanula alpina),
примула маленькая (Primula minima) и др. Из полукустарников —
голубика (Vactinium uliginosum). Кустарники представлены родо-
дендроном (Rhododendron kotschyi).
Растительность Горного Крыма. Для нижнего пояса характерны
ксерофитные можжевело-дубовые леса с господством можжевель-
ника благородного (Juniperus excelsa) и дуба пушистого (Quercus
pubescens). Выше располагается зона господства крымской сосны
(Pinus pallasiana) в сообществе с дубом пушистым растет граб
(Carpinus betulus), кизил (Cornus mas).
Еще выше располагается пояс с господством бука (F. silvatica
и F. orientalis). Вторые места занимают сосна крымская и обыкно-
венная, клен (A. hyrcanum), редко тисс (Taxus baccata), из кус-
тарников— бересклет (Evonimus latifolius). На границе с яйлой —
стелющийся можже в ельник (J. depressa).
Верхняя безлесная часть Крымских гор — яйла — представляет
для пас наибольший интерес: здесь расположены основные масси-
вы крымских горных пастбищ. Растительность на яйле луго-степ-
412
ная; преобладают ксерофитные злаки: типчак (Festuca sulcata), ке-
лерия (Koeleria gracilis)} несколько видов костра. Очень характе-
рен «крымский эдельвейс» (Cerastium biebersteinii). Часто встре-
чается манжетка (Alchernilla pubescens), клевер непостоянный
(Т. ambiguum), проломник пушистый (Androsacae villosa), крупка
(Draba cuspidata), альпийская фиалка (Viola altaica), осока низ-
кая (Carex humilis) и др.
Растительный покров крымской яйлы сильно изменен чрезмерно
интенсивной пастьбой, во многих местах он сильно изрежен и даже
полностью разрушен. В настоящее время выпас на яйле огра-
ничен.
Растительность Кавказа» Флора и растительность горного Кавка-
за отличаются богатством и разнообразием. Здесь можно хорошо
наблюдать, как в зависимости от высоты над уровнем моря и экс-
позиции склонов меняется характер растительности. Смену расти-
тельности по поясам лучше всего рассмотреть, пересекая северный
скат Главного Кавказского хребта, например в направлении Наль-
чик — Теберда — Клухор.
До предгорий северного ската хребта доходит зона южных сте-
пей. Выше степи сменяются лесостепью. На фоне степной и луго-
степной растительности все большую площадь занимают островки
и более крупные массивы леса довольно сложного видового соста-
ва: здесь можно встретить дуб, ясень, берест (Ulmus foliacea),
несколько видов клена, груши, яблони. В подлеске — лещина, алыча,
боярышник (Crataegus monogyna), желтый рододендрон (Rhodo-
dendron flavum), кизил, бересклет, бирючина (Ligustrum vulga-
ге) и др.
Выше располагается лесная зона Предкавказья. Примерно
до высоты 1500 м горы покрыты лиственными породами. На не-
высоких горах произрастает дуб (Q. petraea), бук, граб, ясень,
клены, вязы, липа, яблони, груши. Выше, где количество осадков,
увеличивается, появляется самшит (Buxus colchica), тисс (Taxus
baccata), клекачка (Staphylea colchica), иглица (Ruscus hypophyl-
lum) и другие чисто кавказские виды.
В верхней полосе лесной зоны располагаются хвойные леса
с господством ели кавказской (Р. orientalis) и пихты кавказской
(A. nordmanniana). В подлеске лесов — падуб (Ilex aquifolium)
и понтийский рододендрон (Rhododendron ponticum). В наиболее
высоких горизонтах лесной зоны — сосна крючковатая (Р. hamata),
береза Литвинова (В. litwinovii), горный клен (A. trautvetteri)
и стланиковый можжевельник (I. depressa).
На высоте 1400—2400 м начинается субальпийская зона.
На Кавказе она характеризуется, прежде всего, обилием высоко-
рослых трав, так называемым высокотравьем. Здесь растут гигант-
ские колокольчики (С. lactiflora), борщевик (Heracleum pubescens)
высотой до 3 м, аконит (A. orientate), водосбор (Aquilegia olympi-
са), живокость (D. speciosus), чемерица (Veratrum lobelianum),
валерьяна (V alliariaefolia), девясил (Inula magnifica), розовая
гречиха (Polygonum carneum), кавказский лютик (R. caucasica),
413
цефалярия (Cephalaria tatarica), скабиоза (Scabiosa caucasica),
телекия (Telekia speciosa). Большинство из этих растений имеет
крупные яркие цветки или соцветия, поэтому во время цветения
субальпийского луга зелень листьев часто скрывается за сплошным
разноцветным ковром цветков.
Из древесных в этой зоне обычна береза (В. raddeana), клен
(A. irautvetteri), понтийский дуб (Q. pontica), крушина (Rh. iniere-
tina), рододендрон (Rh. caucasicum).
За субальпийской зоной следует зона альпийских лугов, подни-
мающаяся до 3000 м и выше. Для альпийских лугов характерна
мизкотравность и плотная задернованность почвы. Высота растений
обычно не превышает 30 см. Основу дерна образуют осоки (С. tris-
tis, С. oreophila) и кобрезия (С. capillifolia) \ злаки представлены
мятликом альпийским (Р. alpina), тимофеевкой альпийской
(Ph. alpinum). Из ярко цветущего разнотравья — низкорослые ко-
локольчики (С. ciliata, С. collina), генцианы (Gentiana pontica,
С. caucasica), примулы (P. algida), незабудки (Myosotis alpestris),
фиалки (V caucasica), хохлатки (Corydalis alpestris), куропаточья
трава (Dryas caucasica), кавказская живокость (D. caucasicum),
астрагал (A. alpinus) и др. В этой зоне изредка встречаются не-
большие болотца с покровом из осок и сфагновых мхов.
В альпийской зоне расположены лучшие высокогорные пастби-
ща Кавказа.
Растительность гор Средней Азии. Флора гор Средней Азии
не менее богата, чем флора Кавказа. Не менее разнообразна и рас-
пгтельность. По сравнению с Кавказом она отличается в общем
большей кеероморфностью и преобладанием травянистых форма-
ций. Здесь нет темных влажных лесов, свойственных горам Кав-
каза;. Преобладание травянистых формаций, высокая кормовая цен-
ность многих растений делает горы Средней Азии краем лучших
и наиболее обширных в Советском Союзе горных пастбищ. Особен-
но в этом отношении выделяются высокогорные пастбища Кир-
гизии.
Горы Средней Азии представляют собой сложный комплекс
хребтов и замкнутых высокогорных долин. Взаиморасположение
хребтов и размещение их по отношению к огромным пустыням,
окружающим горные сооружения Средней Азии, определяют слож-
ную и пеструю картину в распределении осадков. К этому еще при-
соединяется резкое различие в тепловом режиме южных и северных
склонов гор. В силу этих обстоятельств одни и те же растительные
пояса могут располагаться на различных абсолютных высотах.
Наоборот, на одной и той же высоте, но на разных склонах можно
встретить и мезофитный субальпийский луг, и сухую степь или по-
лупустыню.
Обобщая, можно -сказать, что в горах Средней Азии можно
встретить следующие пояса растительности: 1) низкогорные полу-
пустыни; 2) низкогорные сухие степи, 3) низкогорные лугостепи
и луга, 4) пояс субальпийской растительности, 5) пояс альпийской
растительности.
414
В Средней Азии можно различать два основных типа низкогор-
ных полупустынь: эфемеровые и полынные.
Эфемеровые низкогорные полупустыни распространены преиму-
щественно по предгорьям хребтов, на высоте 400—1000 м.
Основные растения этих полупустынь — мятлик луковичный
(Р. bulbosa) и осока пустынная (С. pachystylis). К ним присоеди-
няются злаки-эфемеры: костры (В. japonicus, В. danthonia), ячмени
(Н. tnurinum, Н. crinitum), из бобовых — астрагалы (A. filicaulis
Л. sesamoides), эспарцеты (О. micrantha, О. pulchella), псоралея
(Psoralea drupacea) и др. Из разнотравья — зопник бухарский
(Phlomis bucharica), вьюнки (С. subhirsutus, С. pseudocanthabrica)^
солодка (Glycyrhiza as река).
Растительность эфемеровых пастбищ используется в основном
как весеннее, а иногда и осеннее пастбище.
Полынные полупустыни распространены на высотах от 700
до 2600 м. Обычное их месторасположение — сухие части замкнутых
горных долин, склоны южной экспозиции, шлейфы и предгорья
невысоких хребтов. Растительность редкая и не богатая видами.
Основные растения — несколько местных разновидностей полы-
ни морской (A. sublessingiana, A. incana), к ней присоединяется
кохия (Kochia prostrata), из злаков — ковыли (S. caucasica, S. orien-
talis), житняк (A. cristatum), мятлик оттянутый (Р. attenuate)^
Нередко — полукустарник эфедра (Ephedra distachya). Из разно-
травья характерен многолетний колючий вьюнок (С. spinifera)..
В общем растительный покров низкогорной полынной полупусты-
ни очень похож «а соответствующий тип зональной полупустыни,
для (Него также характерна комплексность, связанная с засо-
ленностью почвы.
Полынные полупустыни используются в основном как осенне-
зимние пастбища, меньше — как весенние.
Выше полупустынь расположены низкогорные сухие степи. Они
представлены несколькими типами. Наиболее широко распростра-
нены дерновинно-злаковые степи с преобладанием в травостое
типчака (F. sulcata), ковыля (S. kirghisorum) и др. К ним присоеди-
няются овсец пустынный (Avenastrum desertorum), тонконог
(Koeleria gracilis), костер безостый (В. inermis), пырей волосо-
носный (A. trichophorum), из бобовых — люцерна серповидная
(М. falcata); разнотравье представлено полынью-эстрагоном
(A. dracunculus), тысячелистником обыкновенным, лапчаткой дву-
лопастной (Р. bifurca), скабиозой (Scabiosa alpestris).
Менее широко распространены степи с преобладанием в траво-
стое корневищного злака — пырея волосоносного (A. trichophorum)~
Этот пырей часто сопровождают ячмень луковичный (И. bulbosa)^
костер безостый, свинорой (Cynodon dactylon)\ из бобовых — чины
(L. tuberosus, L. mulkak), из разнотравья — алтей бедноцветный
(Althea nudiflora).
Низкогорные сухие степи используются как весенние и ранне-
летние пастбища. Степи с преобладанием пырея волосоносного
дают хороший сенокос.
415
На высоте 1600—1800 м степи сменяются луго-степями и низко-
горными лугами. Эта зона простирается примерно до высоты 2400—
2600 м, откуда уже начинается субальпийский пояс гор. Нередко
лугостепи и луга образуют комплексы; на южных склонах — луго-
степи, на северных, теневых — луга.
Ботанический состав этих типов растительности весьма разно-
образен. В луго-степях, естественно, степные формы встречаются
рядом с луговыми. В лугах широко распространен пырей ползучий,
костер безостый, мятлик луговой, люцерна серповидная, горечавки,
подмаренники, эстрагон, характерные растения из семейства зон-
тичных: прангос (Prangos pabularia) и ферула (Ferula ovina). По-
следние характерны для южных хребтов. Для северных хребтов
характерно крупное растение из семейства сложноцветных (Ligu-
laria altaica), крупное лилейное (Eremurus), скабиоза (Scabiosa
soongorica) и др.
И в северных, и в южных лугах широко распространена ежа
сборная (Dactylis glomerata). Нередко этот ценный злак образует
почти чистые заросли, простирающиеся на сотни и тысячи гектаров.
В местах интенсивной пастьбы луга из ежи сборной бывают сильно
засорены конским щавелем.
Низкогорные луга с преобладанием разнотравья — удовлетвори-
тельные летние пастбища; с преобладанием злаков — прекрасные
летние пастбища и сенокосы.
В зоне ннзкогорных лугов встречаются также небольшие остро-
ва древесной растительности. Для ферганского хребта характерны
леса из ореха (Juglans regia, J. f allax). Здесь же дикие яблони, гру-
ши (Pyrus heterophyНа), алыча (Prunus divaricata), клен (A. tur-
kestanicum), экзохорда (Exochorda alberti), дикая вишня (Cerasus
ulmifolia). Широко распространены шиповник, боярышник и дру-
гие крупные и мелкие кустарники.
Растительность субальпийского пояса гор представлена субаль-
пийскими разнотравно-злаковыми лугами и хвойными лесами из ели
(Р. schrenkiana, Р. tianschanica), пихты (A. semenovii) и древовид-
ного можжевельника арчи (J. tallasica, J. serawschanica, J. semiglo-
bosa). В качестве подлеска в этих лесах — тянъшанъская рябина
(Sorbus tianschanica), жимолость, бересклет, шиповник и др.
В травостое субальпийских лугов’ преобладает разнотравье:
зопник (Phloinis oreophila), герань (G. ferganense), манжетка
(Alchemilla vulgaris), купальница (Trollius altaicus), незабудка
(M. suaveolens), водосбор (Aquiligia karelini) и др.
Злаки играют второстепенную роль. Наиболее часто встречают-
ся рэгнерия тяньшаньская (Raegneria tianschanica), лисохвост луго-
вой (A. pratensis), овсец (Avenastrum pubescens, A. asiaticum),
тризетум (Trisetum sibiricum, Т. flavescens), овсяница красная и др.
Из осок чаще наиболее широко распространена осока вечнозеленая
(С. senipervirens). Бобовых мало.
В комплексе с лугами иногда встречаются участки высокогорных
степей с преобладанием ковыля киргизского. Вообще субальпий-
ские луга в горах Средней Азии почти всегда в той или иной мере
416
остепнены и никогда не достигают такого пышного развития, какое
наблюдается на Кавказе.
Субальпийские луга в горах Средней Азии — прекрасные летние
пастбища; отдельные небольшие участки, где в травостое преобла-
дают злаки, пригодны для сенокоса.
Расположены субальпийские луга на высоте 2400—3000 м. Даль-*
ше, до высоты 4000 м, располагается альпийская зона.
Растительность альпийской зоны представлена почти исключи-
тельно травянистыми ассоциациями. Из древесных форм встреча-
ются только стланиковая форма можжевельника арчи (J. turkesta-
nica) и кое-где низкорослая жимолость (Lonicera olgae, L. hispid а).
Травянистая растительность представлена не только альпийски-
ми лугами, но и ксерофитными высокогорными степями, полупусты-
нями и пустынями. Ксерофитные формации представлены в замкну-
тых долинах и на высокогорных плато. Примером такой высоко-
горной долины с преобладанием степного ландшафта может быть
известная Алайская долина. В Центральном Тянь-Шане имеются
обширные пространства высокогорных плато, так называемые
сырты, на которых преобладает степная и полупустынная расти-
тельность. Степи — аналоги низкогорных ковыльных, ковыльно-
типчаковых и типчаковых степей. Полупустыни полынные и полын-
но-злаковые имеют типичный вид зональных полупустынь.
Растительность Памира, особенно его восточной части, представ-
лена пустынями. Это своеобразная холодная высокогорная пусты-
ня. Основное растение здесь — полукустарник терескен (Eurotia
ceratoides). Второстепенную роль играют полынь (A. skornjakovii),
ковыль (S. orientalis), подушковидные акантолимоны (Acantholi-
mon diapensoides) и остролодка (О. ponsinsii). Из бобовых также
пажитник Эмода (Trigonella emodi) и др. Все эти растения состав-
ляют скудное пастбище памирской пустыни.
В альпийских лугах Средней Азии основную роль играет кобре-
зия волосолистная (Cobresia capillifolia). Она составляет основу
травостоя обширных пастбищ Киргизии. Меньшую роль играют
другие виды кобрезии (С. stenocarpa, С. humilis и др.).
Совместно с кобрезией часто произрастает птилагростис мон-
гольский (Ptilagrostis mongolica), мятлик альпийский (Р. alpina).
Разнотравья обычно немного: первоцвет (Primula algida), альпий-
ский василистник (Thalictrum alpinum), астра альпийская, неза-
будка, несколько видов мелколепестника (Erigeron), горечавок
(Gentiana) и др. На сухих местах обилен типчак, на влажных —
осока вечнозеленая.
В большинстве случаев растительный покров альпийских лугов
монотонный из-за преобладающей роли кобрезии. Пестрых альпий-
ских лужаек немного.
Очень характерно для высокогорья Средней Азии комплексиро-
вание в одном массиве альпийских и субальпийских лугов. Такая
комплексная растительность представляет особую ценность как са-
мое лучшее высокогорное пастбище. Используются луга альпийской
зоны только как летнее пастбище.
417
Растительность горного Алтая. В горах Алтая различают нес-
колько растительных поясов:. 1) субальпийский, представленный
высокотравными субальпийскими лугами, кедровыми (Pinus sibi-
rica) и лиственничными (Larix sibirica) лесами; 2) альпийский,
представленный альпийскими лугами, низкорослыми, но очень кра-
сочными из-за обилия ярко цветущего разнотравья; 3) горнотунд-
ровый, занимающий здесь наибольшие пространства; 4) мохово-ли-
шайниковый, где преобладающие растения — мхи и лишайники;
вместе с тем для этого пояса характерно широкое распространение
карликовой березы (Betula папа) и нескольких видов ив (S. arbus-
cula, S. glauca, S. krylovii, S. arctica и др.).
Таким образом, обширные территории Алтайского высокогорья
покрыты тундровой растительностью, а в составе этой растительно-
сти много видов, свойственных Крайнему Северу. Эта особенность
объясняется северным расположением Алтайских гор, а также на-
личием здесь обширных бессточных плато.
Растительность горного Алтая используется как хорошее летнее
пастбище. Сенокосов, ввиду обилия крупного разнотравья, сравни-
тельно немного.
ЛИТЕРАТУРА
Алехин В. В. География растений. — М.: Учпедгиз, 1950.
Базилевская Н. А., Белоконь М. П., Щербакова А. А. Краткая история бота-
ники. — М.: Наука, 1968.
Боген Г Современная биология. — М.: Мир, 1970.
Ботанический атлас.—М.; Л.: Изд-во сельскохозяйственной литературы жур-
налов и плакатов, 1963.
Вальтер Г., Алехин В. В. Основы ботанической географии. — М.; Л.: Биомел
гиз, 1936.
Вилли К., Детье В. Биология. — М.: Мир, 1975.
Воронов А. Г. Геоботаника. — М.: Высшая школа, 1963.
Вульф Е. В., Малеева О. Ф. Мировые ресурсы полезных растений. — Л.: Нау-
ка, 1969.
Голенкин М. И. Победители в борьбе за существование. — М.: ГИЗ, 1927.
Горбунова Н. П. и др. Малый практикум по низшим растениям. — М.: Выс-
шая школа, 1976.
Жизнь растений. Т. I—III. — М.: Просвещение, 1974—1977.
Жуковский П. М. Ботаника. — М.: Высшая школа, 1964.
Жуковский П. М. Культурные растения и их сородичи. — Л.: Колос, 1971.
Зеров Д. К. Очерк филогении бессосудистых растений. — Киев: Наукова дуж-
ка, 1972.
Имс А. Морфология цветковых растений. — М.: Мир, 1964.
Пост X. Физиология клетки. — М.: Мир, 1975.
Комарницкий Н. А., Кудряшов А. В:, Уранов А. А. Систематика растений.—
М.: Учпедгиз, 1975.
Кронквист А. Г., Тахтаджян А. А. и Циммерман В. Высшие таксона
Enibryobionta. — Бот. журн., 1966. Т. 51, вып. 5.
Купцов А. И. Введение в географию культурных растений. — М.: Наукд,
1975.
Марков М. В. Общая геоботаника. — М.: Высшая школа, 1962.
Мейер К. И. Морфология высших растений. — М.: Изд-во Московского ун-та,
1958.
Павлов Н. В. Ботаническая география зарубежных стран. — М.: Высшая
школа, 1965.
Первухина И. В. Проблема морфологии и биологии цветка. — Л.: Наука,
1970.
Попов М. Г. Основы флорогенетики. — М.: Изд-во АН СССР, 1963.
Приступа А. А. Основные сырьевые растения и их использование. — Лл
Наука, 1973.
Робертис де Э., Новицкий В., Саэс Ф. Биология клетки. — М.: Мир, 1971
Рубин Б. А. Курс физиологии растений. — М.: Высшая школа, 1977.
Сукачев В. Н. Растительные сообщества.— М.; Л.: ГИЗ, 1928.
Тахтаджян А. Л. Высшие растения. Т. I.— М.; Л.: Изд-во АН СССР, 1956.
Тахтаджян А. Л. Основы эволюционной морфологии покрытосеменных.—
М.; Л.: Наука, 1964.
Тахтаджян А. Л. Происхождение и расселение цветковых растений. — Л.:
Наука, 1970.
Тахтаджян А. Л. Происхождение покрытосеменных растений. — М.: Высшая
школа, 1961.
Тахтаджян А. Л. Систематика и филогения цветковых растений. — Л.: Нау-1
ка, 1966.
419
Тимирязев К. А. Жизнь растений. Избр. соч. Т. III. —ОГИЗ— Сельхозгиз,
1949.
Тимирязев К. А. Чарльз Дарвин и его учение. Избр. соч. Т. IV. — ОГИЗ —
Сельхозгиз, 1949.
Тутаюк В. X. Анатомия и морфология растений. — М.: Высшая школа, 1972.
Хржановский В. Г Курс общей ботаники. Систематика, элементы экологии
и география растений. — М.: Высшая школа, 1976.
Хржановский В. Г Курс общей ботаники. Цитология, гистология, органо-
графия, размножение. — М.: Высшая школа, 1976.
Шенников А. П. Введение в геоботанику. — Л.: Изд-во Ленинградского ун-та,
1964.
Шостаковский С. А. Систематика высших растений. — М.: Высшая школа,
1971.
Эсау К. Анатомия растений. — М.: Мир, 1969.
Ярошенко П. Л. Геоботаника. — М.; Л.: Изд-во АН СССР, 1961.
Яценко-Хмелевский А. А. Краткий курс анатомии растений. — М.: Высшая
школа, 1961.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ
Abies 254
— alba 254, 412
— nordmanniana 254, 413
— semenovii 416
— sibirica 254, 404
Absidia 203
Abutilon theophrastii 321
Acacia dealbata 328
Acantholimon diapensoides 417
Acer 283, 338, 404
— campestre 338
— hyrcanum 412
— negundo 338
— platanoides 338, 405
— pseudoplatanus 338
— tataricum 338
— trautvetteri 413, 414
— turkestanicum 416
Aceraceae 338
Achillea millefolium 352, 353*, 409
Aconitum napellus 304
— orientale 413
— soongaricum 305*
Acroptilon picris 154
Actinomyces 167
Adonis vernalis 42, 301*
Aeluropus littoralis 369, 408
Aethusa cynapium 341
Agropyron 372
— cristatum 415
— pectiniforme 372
Agropyrum 372
— dazyanthum 390
— repens 372*
— trichophorum 415
Agrostema githago 311
Agrostis alba 367
1 Звездочкой указаны страницы, на
— canina 409
— rupestris 412
— vulgaris 409
Ailanthus altissima 154
Albizzia julibrissin 328
Alchemilia pubescens 413
— vulgaris 416
Alectoria 224
Alhagi camelorum 122, 385
Alisma plantagoaquatica 354, 355
Alismales 293, 296
Alismanae 296, 354
Alismatales 354
Alismidae 296, 354
Allium 355
— сера 356*
— guttatum 406
— paczoskianum 406
Alnus glutinosa 310, 404, 410
Alopecurus pratensis 366*, 384, 416
— ventiricosus 366
Althaea 321
— nudiflora 415
— officinalis 321
— rosea 321
Amanita phalloides 212
Amaranthaceae 311, 313
Amaranthus 313
Amaryllidaceae 357
Ammi visnaga 341
Ammodendron 408
Amygdalus 284, 327
— nana 327
Anabaena 168 *, 169
Anabasis 313, 408
которых помещены рисунки.
421
— alba 407 — taurica 352
— aphylla 407 — terrae-albae 407
Anacardiaceae 338 Articulatae 242
Anacardium occidentale 338 Arum 146, 375
Andreales 235 — elongata 375
Androsacae 320 — maculata 375
— villosa 413 Ascomycetes 199, 203
Anemone nemorosa 302 Asparagus 355
— ranunculoides 302* Asperula 345
Anetum graveolens 340 Aspidium 244
Angiospermae 259, 296 Asteraceae 351
Anthoceros laevis 233* Asterales 296, 351
Anthoxanthum odoratum 409 Asteranae 296, 351
Apiaceae 340, 408 Asteridae 296, 344
Apium graveolens 340 Asterionella 174
Aquileqia 281 Asteroxylon 225*, 232
— karelini 416 Astragalus 333, 408
— olympica 413 — alpinus 334, 414
Araceae 375 — filicaulis 415
Arachis hypogaea 335* — glycyphyllus 334
Arales 296, 375 — onobrychis 334
Araliales 296, 339, 340 — sesamoides 415
Aralianae 296, 339 — sulcatus 334
Araucariaceae 253 — virgatus 334
Archaeopteridales 247 Atriplex 313
Archangelica officinalis 341 — alba 407
Arctium 284 Atropa belladonna 347
Arecaceae 375 Atropis distans 369, 386
Arecales 296, 375 Avena 368
Arecanae 296, 375 — fatua 367*, 368
Arecidae 296, 375 — ludoviciana 368
Aristida 390 — sativa 367*, 368
Aristolochia clematilis 298 Avenastrum asiaticum 416
Aristolochiaceae 298 — desertorum 415
Aristolochiales 296, 297, 298 — pubescens 416
Armeniaca 327 Azotobacter chroococcum 151
— vulgaris 327
Armillaria mellea 213* Bacillariophyta 163, 172
Arnica 273 Bacteriophyta 163, 164
— montana 412 Bacterium radicicola 142
Arrhenatherum elatius 368 Balanoporales 307
Artemisia 352 Barbeyales 296, 307
— austriaca 406 Basidiomycetes 199, 210
— cina 353 Batrachospermum 179
— dracunculus 415 Beckmania eruciformis 368, 384
— incana 415 Begoniales 296, 314
— maritima 352, 385, 407 Bennettitales 247, 248
— skornjakovi 417 Berberidaceae 305
— sublessingiana 415 Berberis amurensis 305
422
— vulgaris 122, 305
Beta vulgaris 312*, 313
Betula alba 162, 310
— dahurica 405
— litwinovii 413
— nana 389, 403, 418
— pubescens 310
— raddeana 414
Betulaceae 309
Betulales 296, 307, 309
Bidens 284
Biota 255
Bolanopales 296
Boletus edulis 212*
Beraginaceae 346
Botridium 179, 181
Brassica campestris 318
— juncea 318
— napus v. esculenta 318
— napus v. oleifera 318
— oleracea 317, 318*
— oleracea v. capitata 318
----v. botrytis 318
----v. acephala 317
— rapa 318
Briza media 409
Bromeliales 296, 361
Bromus 370
— danthonia 415
— inermis 371, 373*, 415
— japonicus 415
— oxydon 408
— tectorum 372, 408
Bryales 235, 236
Bryatae 235
Bryonia alba 317
— dioica 317
Bryophyllum 153
Bryophyta 164, 232
Buxus colchia 413
Cactaceae 311
Calamagrotis 367
— epigeios 368
— landsdorfii 368
Calla palustris 375
Calligonium 408
Calluna vulgaris 320
Caltha 281, 302
— palustris 302*, 384
Calycerales 296, 384
Cammelinidae 358
Campanula alpina 351, 412
— ciliata 414
— collina 414
— glomerata 351
— lactiflora 413
— persicifolium 351
— sibirica 351
Campanulaceae 351
Campanulales 296, 351
Cannabis 283
Capparales 296, 314, 317
Caprifoliaceae 344
Capsella bursa-pastoris 282
Caragana arborescens 105
Carduus uncinatus 406
Carex 359, 387
— humilis 360, 413
— inflata 360*
— oreophila 414
— pachystylis 360, 407, 414
— physodes 104, 360, 408
— pilulifera 412
— praecox 360
— rariflora 403
— rotundata 403
— sempervirens 412, 416
— tristis 360, 414
Caricaceae 87
Carpinus 310, 404
— betulus 310, 412
— orientalis 310
Carpoascomycetidae 207
Caryophyllaceae 311, 408
Caryophyllales 296, 311
Caryophyllanae 296, 311
Caryophyllidae 296, 311
Carum carvi 341
Cassia angustifolia 328
Castanea sativa 309
Casuarinales 296, 307
Catalpa bignonioides 154
Caytoniales 248
Caulerpa 189*
Cedrus 254
— atlantica 255
— deodara 255
423
— libanii 255
Celastraceae 342
Celastrales 296, 342, 343
Celastranae 296, 342
Centaurea 353
— jaceae 409
— cyanus 353
— ruthenica 406
Centricae 173
Cephalaria tatarica 413
Cerastium biebersteinii 413
— ucrainicum 406
Cerasus 284
— ulmifolia 416
Ceratium 170, 171
Ceratocarpus arenarius 406
Ceratocephalus 304
— orthoceras 386
Ceratophyllum 273
Cercidiphyllales 296, 307
Cetraria 224, 403
Chaerophyllum temulum 341
Chamerops humilis 375
Chara 192, 193*
Charophyta 163, 192
Chelidonium majus 306
Chenopodiaceae 311, 312
Chenopodium ambrosioides 313
Chinchona lancifolia 345
Chlamidomonas 183, 184*
Chlamydospermatae 247, 257
Chloramaeda 179*, 180*
Chlorella 186, 187*, 222
Chlorococcales 183
Chlorococcum 186, 222
Chlorophyta 163
Chondrus 177*
Chromulina 172
Chroococcus 168* 222
Chrysophyta 163, 171
Chytridiomycetes 199
Cicer orietinum 336
Cichorium inthybus 354
Cicuta virosa 341, 342
Cinnamomum camphora 298
Cilia cordata 405
Cirsium 284
— arvense 154, 353
Citrullus 283, 316
— colocynthus 317
— colocynthoides 317
— edulus 317
Citrus 337
— limonium 282, 338
— reticulata 338
— sinensis 338
Cladonia 223, 403, 404
— alpestris 223
— mitis 223
— rangiferina 221*, 223
— silvatica 223
Cladophora 187, 188*, 221
Claviceps purpurea 209*
Clematis 122
Closterium 190
Clostridium pasteurianum 151
Cnetatae 256
Cobresia 359, 360*, 361
— bellardii 360*, 361
— capillifolia 360*, 361, 386, 414, 417
— humilis 417
— stenocarpa 417
Cocos nucifera 375
Coffea arabica 345
Colchicum 387
Commelinales 296, 361
Commelinanae 296, 361
Commelinidae 296
Coniferales 249, 250
Coniferatae 249
Conium maculatum 341*
Conjugatae 182, 189
Connarales 296
Convallaria 355
— majalis 357
Convolvulaceae 345
Convolvulus arvense 346
— pseudocanthabrica 415
— spinifera 415
— subhirsutus 415
Cordaites 249
Corchorus olitorius 321
Cariandrum sativum 341
Cornales 296, 339, 340
Cornus mas 412
— sanguinea 405
Coronaria flos-cuculi 311
Corydalis alpestris 306, 414
— halleri 307
Corylaceae 310
424
Corylus 310, 404
— avellana 283, 310, 405
— heterophylla 405
Corypha 259
Cosmarium 190
Cotinus 42
Crataegus 283, 327
— monogyna 413
Crocus 355, 387
Cruciferae 317
Cryptomeria japonica 255
Cucumis melo 316
— sativus 316*
Cucurbita 283, 317
— maxima 317
— moschata 317
— pepo 317
Cucurbitaceae 315
Cucurbitales 296, 314, 315
Cupressaceae 253, 255
Cupressus sempervirens 255
Cuscuta arvensis 346
— australis 346
— europaea 346
— lupuliformis 346
— racemosa 346
— trifolli 346
Cuscutaceae 346
Cyanophyta 163, 167, 168*
Cycadales 247, 248
Cycadatae 247
Cycas 248
Cyclamen 320
Cyclanthales 296
Cydonia 283
Cynodon dactylon 368, 415
Cynoglossum officinale 346
Cyperaceae 359
Cyperales 296, 358, 359
Cyperus esculentus 361
Cystoseira barbata 176
Cytisus 337
Dactylus gromerata 369, 416
Daphne mesereum 324
Datura stramonium 347*
Daucus carota 340
— sativus 340
Delesseria 177*
Delphinium 305
— caucasicum 414
— consolida 281, 305*
— speciosus 413
Deschampsia caespitosa 409, 412
Descurainia sophia 319
Desmidiales 189
Dianthus 262, 311
— barbata 311
— caryophyllus' 311
— sinensis 311
Diapensiales 320
Dicotyledones 296, 297
Dictyota 175
Didymelales 296, 307
Dagitalis 42, 349
— purpurea 349
Dillenianae 314
Dilleniales 296, 314, 315, 324
Dilleniidae 296, 314
Dinobryon 172
Dionaea muscipula 123*
Diospyros 320
Dipsacaceae 344, 345
Dipsacales 296, 344
Discomycetales 205, 210
Draba cuspidata 413
Dryas caucasica 414
Dryopteris filix mas 228*, 244
Ebenales 296, 320
Ectocarpus 175*
Elaeagnales 296, 344
Elaeagnus 104, 344, 408
— angustifolia 344
— argentea 344
Elymus sibiricus 384
Elytrigia 372
— repens 102
Embryobionta 163, 164
Empetrum nigrum 403
Empusa muscae 203
Entomophthorales 203
Ephedra 256, 258
— distachya 256*, 415
Ephedrales 256
Epilobium 337
Equisetophyta 164, 242
Equisetum 227
— arvense 242*
— hiemale 243
425
— pratense 243
— scirpoides 243
— silvaticum 243
— variegatum 243
Eremurus 416
Ericaceae 320
Ericales 296, 320
Ericanae 296, 314, 320
Ericaulales 361
Erigeron 417
Eriocaulales 296
Eriophorum angustifolia 403
Erodium 264
Erophila verna 386, 406
Erysiphae 208*
— graminis 208
Euascomycetidae 205, 207
Eucalyptus 337
Eucomiaceae 308
Eucomiales 307
Eucommia 44
— ulmoides 308
Euglena 181
— rubra 181
— viridis 182*
Euglenophyta 163, 181
Eucomiales 296
Euphorbia 323
— canescens 324
— cheirolepis 324
— lathyris 323*
— seguieriana 406
— virgata 324
Euphorbiaceae 323
Euphorbiales 296, 320, 323
Eupteleales 296, 307
Eurotia ceratoides 105, 313, 417
Evernia 224
Evonimus 405
— latifolius 412
— verrucosa 44, 343
Exochorda alberti 416
Fabaceae 162
Fabales 296, 328
Fagaceae 308, 328
Fagales 296, 307, 308
Fagopyrum sagittatum 313
Fagus 308, 404
— orientalis 308, 404, 412
— silvatica 308, 404, 412
— taurica 404
Ferula 80, 259
— ovina 415
Festuca picta 412
— pratensis 370, 371*
— rubra 409, 412
— sulcata 99, 370, 371* 397, 413, 415
— supina 412
— varia 386
Ficaria 303
— ranunculoides 303*
Ficus carica 281, 307
Filipendula 325*
Fomes 211, 212*
Fragaria collina 326
— vesca 326
Fragillaria 174
Frankenia 408
Fraxinus 283, 344, 404
— excelsior 405
Fuchsia 262
Fucus vesiculosus 176
Fumaria officinalis 307
Fumariaceae 306
Fungi 197
— imperfecti 199, 218, 219*
Fusarium 218, 219*
Gagea 355, 387
— bulbifera 406
— lutea 356*
— pusilia 406
— schovitsii 406
Gamianae 296
Galanthus 357
Galega 337
Galeopsis ladanum 350
Galium 284, 345
— boreale 409
— verum 345, 410
Gasteromycetales 212
Gentiana 345, 417
— caucasica 414
— pontica 414
Gentianales 296, 344, 34Б
Geraniaceae 339, 345
Geraniales 296, 337, 338
426
Geranium 339
— ferganense 416
Geum rivale 410
Giliatae 296
Ginkgo biloba 250
Ginkgoales 249, 250
Gladiolus 355
Glaeocapsa 168*, 222
Glaeosporium 219*
Gleditschia triacanthos 328
Glyceria aquatica 384
Glycine hispida 337
Glycyrrhiza aspera 415
Gnaphalium uliginosum 353
Gnetales 256
Gnetatae 247
Gnetum 256, 258
— gnemonoides 258*
Ganium 184*, 185
Gossypium 322
— barbadense 322
— hirsutum 322*
Gramineae 361
Graphis scripta 221*
Grossularia reclinata 328
Gymnoascomycetidae 205
Gymnospermae 247
Halimodendron 408
Halocnemum 408
Haloxylon 313, 385
— aphyllum 407
— persicum 408
Hamamelidales 296, 307
Hamamelidanae 296, 307
Hamamelididae 296, 307
Helianthus annuus 282, 352
— tuberosus 352
Heliotropium 346
Hepaticae 234
Heterocontae 179
Hevea 44, 87
— brasiliensis 324
Hieraceum alpinum 412
Hippuridales 296, 337
Holobasidiomycetidae 211
Holosteum umbellatum 386
Hordeum bulbosa 415
— crinitum 415
— distichum 374
— murinum 415
— spontaneum 374
— vulgare 374
Hornea 225*, 232
Hydrocharitales 296, 354
Hydrurus 172
Hymenomycetales 211
— niger 347
Hypericaceae 315
Hypericum perforatum 315
Hyphomycetales 218
Hyppophae rhamnoides 344
Ilex aquifolium 413
Illiciaceae 300
Illiciales 296, 299, 300
Illicium verum 300
Impatiens 59, 273
Inula magnifica 413
Iridales 296, 355
Iris 90*, 102, 355
— pumila 406
Isocontae 182, 183
Isoetales 240
Juglandaceae 310
Juglandales 296, 307, 310
Juglans fallax 416
— regia 310, 416
Juncaceae 358
Juncales 296, 358
Juncanae 296, 357
Juncus 359
— effusus 358*
— gerardi 359
Jungermaniales 234
Juniperus communis 255
— depressa 412, 413
— excelsa 412
— sabina 255
— semiglobosa 416
— seravschanica 255, 416
— sibirica 412
— tallasica 416
— turkestanica 255, 417
427
Kochia prostrata 313, 407, 415 Koelera gracilis 368, 413, 415 Kormobionta 163, 164 Lilianae 296, 355 Liliatae 296, 354 Liliidae 296, 355 Linaceae 338
Labiatae 349 Lactarius deliciosus 212 Lamiaceae 349, 408 Lamiales 296, 345, 349 Lamianae 344 Laminaria 176 — digitata 176* — saccharina 176* Lamium album 350 Larix 42, 254 — dahurica 254, 404 — sibirica 254, 404, 418 Lathyrus 336 — mulkak 415 — nissolia 123 — pratensis 336, 409 — sativus 336 — tuberosus 415 Lauraceae 298 Laurales 296, 297, 298 Laurocerasus officinalis 327 Laurus nobilis 298 Lavandula vera 41 Lecanora 224 Ledum palustre 320 Leguminosales 328 Leitneriales 296, 307 Lemna 375 — minor 376 — trisulca 376 Lemnaceae 375 Lens esculenta 336 Leontice 305 Leontopodium alpinum 412 Leonurus villosus 350 Lepidodendron 229* Lespedeza bicolor 405 Libantotis montana 410 Lichenes 227* Lichenophyta 164, 220 Ligularia altaica 415 Liguliflorae 352, 353 Ligustrum vulgaris 344, 413 Liliaceae 355 Liliales 296, 355 Linaria vulgaris 348*, 349 Linosyrus villosa 406 Linum 338 — humile 338 — usitatissimum 339* Lolium multiflorum 370 — perenne 370, 372* Lonicera 266, 404 — hispidae 417 — olgae 417 Lotus corniculatus 333*, 409 Lupinus albus 330 — angustifolius 330* — luteus 330 — perennis 330 — polyphyllus 330 — angustifolius 330 Luzula 359 — nemorosa 412 — pilosa 358*, 359 — sudetica 412 Lycium 347 Lycogala 196 Lycopersicum esculentum 283, 347 Lycophyta 240 Lycopodiaceae 240 Lycopodiales 240 Lycopodiophyta 164, 240 Lycopodium clavatum 226, 240 Lyngbya 168* Lysimachia 320 Lythraceae 337 Lythrum 337 Magnolia 297, 298* — grandiflora 298 Magnoliaceae 289, 297 Magnoliales 297, 299, 307 Magnalianae 296, 297 Magnoliatae 297 A'lagnoliidae 296, 297 Magnoliophyta 164, 259, 296 Malvanae 320 AAallomonas 172 Malus 283
428
— silvestris 322, 325*
Malva 321
— crisp a 322
Malvaceae 321
Malvales 296, 320, 321
Marchantia polymorpha 234*
Marchantiales 234
Marchantiatae 234
Matricaria chamomilla 352
Marsiliales 244
Medicago falcata 331, 415
— lupulina 331
— sativa 331*
Melanconiales 219
Melillotus albus 332
— officinalis 332*
Melissa officinalis 350
Mentha 349
— crispa 350
— piperita 41, 350
Mercuriales annua 324
Merulus lacrimans 211
Mesotaeniales 189
Metroxylon 259
Microcystis 169
Microsphaera alphitoides 208
Monilia 218
Monocotyledones 296, 354
Moraceae 307
Morus 267, 281
Mougeotia 192
Mucorales 203
Mucor 202, 203
— alba-ater 202
— mucedo 202*
Muscari 387
Musci 235
Mycophyta 164, 197
Myosotis alpestris 414
— arenaria 406
— snaveolens 416
Myricales 296, 307
Myrtaceae 337
Myrtales 296, 337
Myrtanae 296, 337
Myrtis communis 337
Myxophyta 164, 195
Najadales 354, 355
Najas 272
Nanophyton erinaceum 408
Narcissus 357
Nardus stricta 386, 409, 412
Navicula 174
Nelumbo 300
Nelumbonales 296
Nemalion 179
Nepenthales 296
Nerium oleander 101
Noctiluca 170, 171
Nicotina rustica 347
— tabacum 347
Nitella 192
Nostoc 168*, 169
Nuphar luteum 299
Nymphaea alba 104, 299*, 354, 384
— lotos 299
Nymphaeaceae 298
Nymphaeales 293, 296, 297, 298
Ochrolechia 224
Oenanthe aquatica 341
Olea europea 344
Oleaceae 343
Oleales 296, 342, 343
Olpidium brassicae 199, 200*
Onagraceae 337
Onobrychis 281, 334
— arenaria 324
— micrantha 415
— pulchella 415
— viciifolia 334*
Oomycetes 199, 200
Opuntia 166, 386
Orchidaceae 357
Orchidales 296, 355, 357
Orchis latifolia 357
— maculata 357
— militaris 357
Ornithogalum 263
Orobanchaceae 349
Orobanche cumana 349
— ramosa 348,* 349
Oryza sativa 365*, 384
Oscillatoria 168*, 169
Oxycoccus quadripealus 320
429
Padus racemosa 327 Phytophthora infestans 200, 201
Paeonia anomala 315 Picea excelsa 254, 387, 404, 412
— caucasica 315* — obovata 254, 404
— tenuifolia 315 — orientalis 413
Paeoniaceae 315 — schrenkiana 416
Paeoniales 314, 315 — tianschanica 416
Palmales 375 Pimpinella anisum 341
Panicum millaceum 364 Pinaceae 253
Pap aver 282, 306 Pinales 250
— rhoeas 306 Pinatae 247, 249
— somniferum 306* Pinnularia 172*, 173
Papaveraceae 306 Pinophyta 164, 247
Papaverales 296, 306 Pinus 254
Paris 355 — hamata 413
— quadrifolia 101, 357 — mughus 412
Parmelia 221* — pallasiana 412
Passiflorales 296, 314 — sibirica 254, 404, 418
Pastinaca sativa 340 — silvestris 128*, 251*, 404
Peganum harmala 339 Piperales 296, 297
Pelagonium roseum 41 Pisum 281, 336
Penicillium 207* — arvense 336
— chrysogenum 208 — sativum 336
— notatum 208 Plantago media 409
— roqueforti 207 Plasmodiophora brassicae 196
Pennatae 173, 174 Plasmopara viticola 201
Pennisetum typhoideum 364 Platanus digitata 307, 308
Peridineae 170, 171 — orientales 308
Peridinium 170 Plectascales 205, 207
Peronosporales 200 Pleurococcus 188, 222
Persea gratissima 298 Plumbaginaceae 408
Persica 327 Plurrtbaginales 296, 311
— vulgaris 327 Poa alpina 414, 417
Petasites spurius 410 — angustifolia 384
Phacus 182* — attenuata 415
Phaeophyta 163, 174 — bulbosa 387, 408, 415
Phalaris arundinacea 384 v. vivipara 369, 406
Phaseolus 336 — compressa 106
— vulgaris 336 — palustris 369, 384
Phleum 366 — pratensis 369, 370*, 409
— alpinum 414 — triviais 369
— phleoides 366 Poaceae 361
— pratense 366* Poales 296, 361
Phlomis bucharica 415 Podocarpaceae 253
— oreophila 416 Podostemales 296, 324
— pungens 406 Polemoniales 296, 345
Phoma 219 Polygalales 296, 337
Phragmites communis 368, 384 Polygonaceae 313
Phragmobasidiomycetidae 212 Polygonales 296, 311, 313
Phyllophora nervosa 179, 194 Polygonatum 102, 355
Phyllosticta 219 — officinale 357
430
Polygonum aviculare 314, 406
— bistorta 314, 410
— corneum 413
— convolvulus 313
Polypodiaceae 244
Polypodiales 244
Polypodiophyta 164, 243
Polysiphonia 177
Polytrichum 238
— commune 237*
Pomoideae' 326
Populus alba 319
— nigra 319
— pyramidalis 319
— tremula 31.9, 404
Porphyra 179
Potamogeton 355, 384
Potentilla 326
— argentea 409
— bifurca 415
— erecta 412
Prangos pabularia 416
Primula algida 414, 417
— minima 412
— officinalis 320
Primulales 296, 320
Proteales 296, 344
Proteanae 296, 344
Protoascales 205
Protoascomycetidae 205
Prunoideae 327
Prunus 284, 327
— divarticata 416
— domestica 327
— insititia 327
— spinosa 327
— tenella 325*
— ussuriensis 327
Psalliota campestris 212, 213*
Psilotophyta 164, 239
Psilotum triquetrum 239*
Pteridium aquilinum 90*, 245
Pterophyta 243
Ptilagrostis mongolica 417
Puccinia coronifera 218
— graminis 216, 217*
— helianthi 218
— maydis 218
Pulsatilla 302
— alba 412
Pyrenomycetales 205, 209
Pyrethum millifoliatum 406
Pyrola 273
Pyrrophyta 170
Pyrus 283, 326
— communis 326
— elaeagnifolia 326
— heterophylla 416
Quercus mongolica 405
— pedunculata 308
— petraea 413
— pontica 414
— pubescens 412
— robur 283, 404, 412
Raegneria tianschanica 416
Rafflesiales 296, 297
Ranales 289
Ranunculaceae 162, 300
Ranunculus 303
— acer 303, 304*, 409
— caucasica 413
— repens 162, 304
— sceleratus 303
Raphanus sativus 319
---v. niger 319
---v. radicula 319
---v. raphanistrum 319
Raphidium 187*
Restionales 296, 361
Rhamnaceae 343
Rhamnales 296, 342, 343
Rhamnus catharctica 343, 405
— frangula 343
— imeretina 414
Rhododendron 320
— caucasicum 414
— flavum 413
— kotschyi 412
— ponticum 412
Rhodophyta 163, *177
Rhynia 225*, 232
Rhyniophyta 164, 231
Ribes aureum 328
— nigrum 328, 404
— vulgare 328
Ricinus communis 324
Rivularia 168*
431
Roccelia 224
Rosa 326, 404
— canina 326
— cinnamomea 326
— - lutea 326
— majalis 325*
Rosaceae 325*
Rosales 295, 324, 325
Rosanae 296, 324
Rosidae 296, 324
Rosoideae 325
Rubiaceae 345
Rubus 284
Rumex acetosa 314
— acetosella 314
— confertus 314
Ruscus 104
— hypophyllum 413
Rutaceae 337, 408
Rutales 296, 337
Rutanae 296, 337
Saccharomyces cerevisiae 206*
Sagittaria sagittifolia 355
Salicaceae 319
Salicales 296, 314, 319
Salicornia herbacea 104, 386, 387*
Salix alba 319
— albuscula 418
— artica 418
— fragilis 155, 319
— glauca 418
— krylovii 418
— polaris 410
— triandra 410
— viminalis 319, 410
Salsola 313, 408
— pestifer 313
Salvia 350
— aephiopis 406
— nemorosa 406
— officinalis 350
— pratensis 350*
— sclarea 41, 350
Salvinia natans 245, 246*
Salviniales 244
Sambucus 266
Santalales 296, 342
Sapindales 296, 337, 338
Saponaria 311
Saprolegnia 201
Saprolegniales 201
Saxifraga 266
— cernua 153
— nivalis 153
Saxifragales 296, 324, 328, 337, 342
Scabiosa alpestris 415
— caucasica 413
— lucida 412
— soongorica 416
Scenedesmus 186, 187*
Schizandra chinensis 300
Sclerotinia 210, 218
Scrophulariales 296, 345, 347
Scutellaria baicalensis 350
Secale 282, 372
— cereale 362*, 372
— segetale 372
Sedum 385
— villosum 153
Selaginella 241*
Selaginellaceae 241
Selaginellales 240
Sequoja sempervierens 255
Sinapis 319
— alba 281, 319*
Siphonales 183
Sisymbrium altissimum 319
— loeselii 319
— sophia 319
Sium 341
Solanaceae 347
Solanum dulcamara 347
— melongena 347
— nigrum 347
— tuberosum 347
Sonchus 154
— arvensis 353*, 354
Sorbus aucuparia 327
— tianshanica 416
Sorgum caccharatum 364
— ccrnuum 364
— helepense 364
— sudanense 364
Sparcium junceum 385
Sphaerocarpales 234
Sphaeropsidales 219
Sphaerotheca mors uvae 208
Sphagnales 235
Sphagnum 235, 236*, 411
432
Spiraea 325
Spirogyra 190, 192*
Spirotaenia condensata 190*
Spirulina 169
Spongospora solani 196
Stachybotrys alternans 219
Staphylea colchica 413
Statice 284, 408
— tatarica 406
Stipa 99
— capillata 365, 406
— caucasica 415
— kirghisorum 415
— lessingiana 365, 397, 406
— orientalis 415, 417
— ucrainica 406
Streptomyces griseus 167
Suaeda 408
— maritima 386
Symphytum asperum 346
— officinale 346
Synchytrium endobioticum 200
Syringa vulgaris 344
Tabellaria 174
Tamaricaceae 408
Tamaricales 296, 314
Tamarix 408
Taphrinales deformans 206
Taraxacum 282
— officinale 354
Taxaceae 253
Taxodiaceae 253, 255
Taxodium mucronatum 255
Taxus 253
— baccata 253, 412, 413
Telekia speciosa 414
Thalictrum 305
— alpinum 417
Thallobionta 163
Thea sinensis 314
Theaceae 314
Theales 296, 314, 315
Theligonales 296, 311
Thermopsis 337
Thuja 255
Thymelaeceae 324
Thymelaeles 296, 320, 324
Thymus alpestris 412
Thypha angustifolia 376
— latifolia 376
Thyphaceae 376
Thyphales 296, 376
Tilia 321, 404
— cordata 116, 321
— tomentosa 321
Tiliaceae 321
Tilletia tritici 214*, 215
Tmesipteris 239
Trachelomonas 181, 182*
Trachyspermum ammi 341
Tragopogon 284
Trapa 337
Trapaceae 337
Trentepholia 188, 222
Tribonema 179*, 180
Tribulus 284
Trichia 196
Trichodcsma incanum 346
Trichodesmium erythraeum 169
Trifolium ambiguum 332, 413
— hybridum 332
— montana 410
— pratense 162, 332, 333*, 409
— repens 409
Trigonella arcuta 337
— emodi 417
Trisetum flavecens 416
— sibiricum 416
Triticum durum 373
— vulgare 373
Triuridales 296, 355
Trochodendrales 296, 307
Trollius altaicus 416
— europeus 301*, 302, 410
Tropaeolum 122
Tuber 210
Tuberales 205, 210
Tubiflorae 352
Tulipa 387
— schrenkii 406
Ulmus 404, 405
— foliacea 413
Ulotrix 187*
Ulva 188
— lactuca 188
Uncinula necator 208
Uredinales 216
433
Urtica dioica 308, 309*
Urticaceae 308
Urticales 296, 307, 308
Usnea 224
— barbata 221*
Ustilaginales 212, 214*
Ustilago avenae 214*, 215
— hordei 215
— nuda 215
— panici miliacei 214*, 215
— tritici 214*, 215
— zeae 214*, 215
Vaccinium myrtyllus 320
— uliginosum 320, 403, 412
— vitis idaea 320
Valeriana alliariaefolia 413
— officinalis 345
Valerianaceae 344, 345
Vanilla fragna 357
Vaucheria 180*
Venturia 210
Veratrum 355, 357
— lobelianum 357, 413
Verbascum phlomoides 349
— phloeniceum 406
— thapsiforme 349
Veronica praecox 386
— verna 386, 406
Verticillium 218
Vicia 281, 336
— faba 43, 336
— sativa 336
— villosa 336
Victoria regia 299
Vigna sinensis 337
Viola 273, 315
— altaica 413
— caucasica 414
— odorata 315
— tricolor 315
Violaceae 315
Vitis silvestris 343
— vinifera 283, 343
Volvocales 183
Volvox 185, 186*
Welwitshia 256, 257
— mirabilis 257*
Xanthium 284
Xanthophyceae 179*
Xanthophyta 163, 179
Zea mays 362, 363*
Zingiberales 296, 355
Zostera 272, 355
Zygnema 191
Zygnematales 189
Zygomycetes 199, 202
Zygophyllaceae 339
Zygophyllum 408
ОГЛАВЛЕНИЕ
Стр.
Введение
Определение науки и объект ее изучения . 3
Возникновение и эволюция растительного мира 0
Растения — основной компонент биосферы . 9
Разделы ботаники И
Развитие ботанической науки 13
Глава I. Растительная клетка 18
Форма и величина клеток 18
Молекулярные компоненты клетки 20
Строение клетки, ее составные части . 21
Протопласт 22
Включения в протопласт 38
Клеточная оболочка 46
Основные функции клетки 51
Размножение клеток 59
Г лава II. Ткани и вегетативные органы растений . 66
Ткани 57
Образовательные ткани — меристемы 67
Покровные ткани 69
Основные, или питающие, ткани . 76
Выделительные ткани 80
Механические ткани 80
Проводящие ткани 84
Сосудисто-волокнистые проводящие пучки 88
Вегетативные органы высших растений, их онтогенез и функции 91
Развитие вегетативных органов из зародыша семени 91
Побег и его видоизменения 95
Морфология стебля 106
Анатомическое строение стебля 108
Лист и его видоизменения 117
Корень и его видоизменения 131
Морфология корня 131
Анатомическое строение корня . 135
Вегетативное размножение высших растений 152
Г лава III. Систематика растений 158
Низшие растения (Thallobionta) 164
Отдел 1. Бактерии (Bacteriophyta) . 164
Отдел 2. Синезеленые водоросли (Cyanophyta) 167
435
Стр.
Отдел 3. Пиррофитовые водоросли (Pyrrophyta) 170
Отдел 4. Золотистые водоросли (Chrysophyta) 171
Отдел 5. Диатомовые водоросли (Bacillariophyta) 172
Отдел 6. Бурые водоросли (Phaeophyta) 174
Отдел 7. Красные, или багряные, водоросли (Rhodophyta) 177
Отдел 8. Желтозеленые водоросли (Xanthophyta) 179
Отдел 9. Эвгленовые водоросли (Euglenophyta) 181
Отдел 10. Зеленые водоросли (Chlorophyta) 182
Класс равножгутиковые (Isocontae) 183
Класс сцеплянки, или конъюгаты (Conjugatae) 189
Отдел 11. Харовые водоросли (Charophyta) 192
Роль водорослей в природе и использование их человеком 193
Филогенез водорослей 195
Отдел 12. Миксомицеты (Myxophyta) 195
Отдел 13. Грибы (Fungi, Mycophyta) 197
Класс хитридиевые (Chytridiomycetes) 199
Класс оомицеты (Oomycetes) 200
Класс зигомицеты (Zygomycetes) 202
Класс аскомицеты (Ascomycetes) 203
Подкласс голосумчатые (Protoascomycetidae, или Gymnoascomycetidae) 205
Подкласс плодосумчатые (Euascomycetidae, или Carpoascomycetidae) 207
Класс базидиомицеты (Basidiomycetes) 210
Подкласс холобазидиомицеты (Holobasidiomycetidae) . 211
Подкласс фрагмобазидиомицеты (Phragmobasidiomycetidae) 212
Класс несовершенные грибы (Fungi imperfecti) 218
Филогенез грибов 219
Роль грибов в природе и их хозяйственное значение 220
Отдел 14. Лишайники (Lichenophyta) 220
Высшие растения (Kormobionta, Embryobionta) 224
Отдел 1. Псилофитовые, или риниевые (Psilophyta, Rhyniophyta) 231
Отдел 2. Моховидные (Rryophyta) 232
Класс антоцеротовые (Anthocerotae) 233
Класс печеночники (Hepaticae, Marchantiatae) 234
Класс лиственные мхи (Musci, Bryatae) 235
Отдел 3. Псилотовые (Psilotophyta) 239
Отдел 4. Плауновидные (Lycophyta, Lycopodiphyta) 240
Отдел 5. Членистые, хвощевидные (Articulatae, или Eqisetophyta) 242
Отдел 6. Папоротниковидные (Pterophyta, Poiypodiophyta) 243
Отдел 7. Голосеменные (Gymnospermae, Pinophyta) 247
Класс саговниковые (Cycadatae) 247
Класс шишконосные (Coniferatae, Pinatae) 249
Класс оболочкосеменные (Chlamydospermatae, Gnetatae) 250
Отдел 8. Покрытосеменные, или цветковые (Angiospermae, Magnoliop-
hvta) 259
Цветок, соцветия, плоды 259
Строение цветка и его частей 269
Опыление 279
436
Стр-
Микроспорогенез * и развитие мужского гаметофита 274
Мегаспорогенез и развитие женского гаметофита . 276
Оплодотворение и развитие семени 278
Отклонения от нормального процесса формирования семян и плодов 280
Строение и классификация плодов 280
Плоды с сухим околоплодником 281
Плоды с сочным околоплодником 283
Приспособления для распространения плодов и семян . 284
Происхождение цветковых растений 285-
Систематика покрытосеменных 289
Класс двудольные (Dicotyledones — Magnoliatae) 297
Подкласс 1. Магнолиидные (Magnoliidae) 297
Надпорядок I. Магнолиевидные (Magnolianae) 297
Порядок магнолиецветные (Magnoliales) 297
Порядок лавроцветные (Laurales) . 298
Порядок кирказоноцветные (Aristolochiales) 298-
Порядок кувшинкоцветные (Nymphaeales) 298
Подкласс 2. Ранункулидные (Ranunculidae) 299
Надпорядок II. Лютиковидные (Ranunculanac) 299
Порядок иллициецветные (Illiciales) 300
Порядок лютикоцветные (Ranunculales) 300
Порядок макоцветные (Papaverales) 306-
Подкласс 3. Гамамелядидные (Hamamelididae) 307
Надпорядок III. Гамамелидовидные (Hamarnelidanae) 307
Порядок гамамелидоцветные (Hamamelidales) 307
Порядок крапивоцветные (Urticales) 308
Порядок букоцветные (Fagales) 308
Порядок березоцветные (Betulales) 309
Порядок орехоцветные (Juglandales) 310
Подкласс 4. Кариофиллидные (Caryophyllidae) 311
Надпорядок IV Гвоздиковидные (Caryophyllanae) 311
Порядок гвоздикоцветные ( Caryophyllales) 311
Порядок гречихоцветные (Polygonales) 313
Подкласс 5. Диллениидные (Dilleniidae) 314
Надпорядок V. Диллениевидные (Dillenianae) 314
Порядок пионоцветные (Paeoniales) 315
Порядок чаецветные (Theales) 315
Порядок фиалкоцветные (Violales) 315
Порядок тыквоцветные (Cucurbitales) 315
Порядок каперсоцветные (Capparales) 317
Порядок ивоцветные (Salicales) 319
Надпорядок VI. Вересковидные (Ericanae) 320
Порядок верескоцветные (Ericales) 320
Порядок первоцветные (Primulales) 320
Надпорядок VII. Мальвовидные (Malvanae) 320
Порядок мальвоцветные (Malvales) . 321
Порядок молочаецветные (Euphorbiales) 323
Порядок тимелеецветные (Thymelaeales) 324
437
Стр.
Подкласс 6. Розидные (Rosidae) 324
Надпорядок VIII. Розовидные (Rosanae) 324
Порядок розоцветные (Rosales) 325
Порядок камнеломкоцветные (Saxifragales) 328
Порядок бобоцветные (Fabales, Leguminosales) 328
Надпорядок IX. Миртовидные (Myrtanae) 337
Надпорядок X. Рутовидные (Rutanae) 337
Порядок рутоцветные (Rutales) 337
Порядок сапиндоцветные (Sapindales) 338
Порядок гераниецветные (Geraniales) 338
Надпорядок XL Аралиевидные (Aralianae) 339
Порядок аралиецветные (Araliales, Cornales) 340
Надпорядок XII. Бересклетовидные (Celastranae) 342
Порядок бересклетоцветные (Celastrales) 343
Порядок крушиноцветные (Rhamnales) 343
Порядок маслиноцветные (Oleales) 343
Надпорядок XIII. Протейновидные (Proteanae) 344
Порядок лохоцветные (Elaeagnales) 344
Подкласс 7. Астеридные (Asteridae) 344
Надпорядок XIV. Яснотковидные (Lamianae) 344
Порядок ворсянкоцветные (Dipsacales) 345
Порядок горечавкоцветные (Gentianales) 345
Порядок полемониецветные (Polemoniales) 345
Порядок норичникоцветные (Scrophulariales) 347
Порядок ясноткоцветные (Lamiales) 349
Надпорядок XV. Астровидные (Asteranae) 351
Порядок колокольчикоцветные (Campanulales) 351
Порядок астроцветные (Asterales) 351
Класс однодольные (Monocotyledones — Liliatae) 354
Подкласс 1. Алисмидные (Alismidae) 354
Надпорядок I. Частуховидные (Alismanae) 354
Порядок частухоцветные (Alismales) 354
Подкласс 2. Лилиидные (Liliidae) 355
Надпорядок II. Лилиевидные (Lilianae) 355
Порядок лилиецветные (Liliales) 355
Порядок орхидноцветные (Orchidales) 357
Подкласс 3. Коммелинидные (Commelinidae) 358
Надпорядок III. Ситниковидные (Juncanae) 358
Порядок ситникоцветные (Juncales) 358
Порядок осокоцветные (Cyperales) 359
Надпорядок IV. Коммелиновидные (Commelinanae) 361
Порядок мятликоцветные (Poales) 361
Подкласс 4. Арецидные (Arecidae) 375
Надпорядок V. Пальмовидные (Агесапае) 375
Порядок пальмоцветные (Arecales, Palmales) 375
Порядок ароидноцветные (Arales) 375
Порядок рогозоцветные (Thyphales) 376
438
Стр.
Глава IV. Элементы экологии и географии растений . 377
Экологические факторы 378
Абиотические факторы 378
Биотические факторы 390
Антропогенные факторы 393
Понятие о флоре и растительности 394
Жизненные формы растений 395
Понятие о растительных сообществах 395
Основные типы растительного покрова СССР 401
Растительность горных районов СССР 411
Литература 419
Указатель латинских названий 421
Тихомиров Федор Константинович
БОТАНИКА
ИБ № 1694
Редактор Т. А. Рыкова
Художник Ю. С. Шлепер
Художественный редактор Т. А. Коленкова
Технический редактор Т. Д. Гарина
Корректор С. К. Марченко
Изд. № Е-324. Сдано в набор 16.03.78. Подп. в печать 04.09.78.
Формат 60X90718. Бум. тип. № 2. Гарнитура литературная. Печать высокая. Объем
27,5 усл. печ. л. 30,33 уч.-изд. л. Тираж 30 000 экз. Зак. № 326 Цена 1 р. 40 к.
Издательство <Высшая школа»,
Москва, К-51, Неглинная ул., д. 29/14
Московская типография № 8 Союзполиграфпрома
при Государственном комитете Совета Министров СССР
по делам издательств, полиграфии и книжной торговли,
Хохловский пер., 7